/
























































































































































































































































































































































































































































Текст
у....
ЛЕНИНГРАДСКИЙ ОРДЕНА ЛЕНИНА государственный
УНИВЕРСИТЕТ имечп А. А. ЖДАНОВА
В. А. ДОГЕЛЬ
ОБЩАЯ ПАРАЗИТОЛОГИЯ
Переработано и дополнено
Ю И Полянским к Е М Хеисиным
ИЗДАТЕЛЬСТВО
ЛЕНИНГРАДСКОГО УНИВЕ РСИТЕТА
19Ь2
ПРЕДИСЛОВИЕ
Предлагаемая вниманию читателей книга «Общая паразитология»
представляет собою дополненное п переработанное издание книги про
фессора Ленинградского университета В. А. Догеля «Курс общей пара-
зитологии», вышедшей в 1911 г и переизданной в 1947 г. Необходимость
переработки книги возникла в связи с тем, что с 1917 г. по настоящее
время как в Советском Союзе, так и за границей появилось множество
новых работ но вопросам паразитологии, которые необходимо было от-
разить В результате около половины текста написано заново и
часть устаревшего материала, а также имеющего лишь косвенное отно-
шение к вопросам общей паразитологии, изъята. Несмотря на
значительные дополнения и исправления, основные мысли и идеи
В А. Догеля, изложенные им в «Курсе общей паразитологии», не только
полностью сохранены в данной книге, но и являлись руководящими для
нас при привлечении нового материала Поэтому мы считали необходи-
мым и правильным сохранить авторство за нашим учителем — профес-
сором В А. Догелем, тем более что п книге одно из самых главных мест
занимает раздел «Парлзптофауиа и среда», отражающий основное на-
правление эколого-паразитологических исследований догеленской шко-
лы, зародившейся в стенах Ленинградского университета. Задачей эко-
логической паразитологии является «изучение зависимости паразнто-
фаупы, взятой в целом, от изменения внешних условий, окружающих
хозяина, и от изменения физиологического состояния самого хозяина»
(Догель, 1947). Работы, аналогичной «Общей паразитологии» В А. До-
геля, нет пи в советской, ни в зарубежной литературе.
Книга «Общая паразитология» состоит из четырех частей. В первой
рассматриваются общие черты паразитизма как биологического явле-
ния и вопрос о происхождении паразитизма в животном мире. Вторая
часть посвящена формам адаптации к паразитизму и проблеме жизнен
них циклов. В третьей излагаются вопросы отношений между парази-
тами и средой. В четвертой рассматриваются формы взаимоотношений
между паразитом и хозяином, проблема вида и видообразования у па-
разитов.
1*
3
Рассмотренные в книге вопросы отражают значительную часть про-
блем общей паразитологии, но, конечно, не исчерпывают их полностью.
В частности, в книге лишь в малой степени затронуты вопросы биохи-
мии и физиологии паразитов, представляющие самостоятельную область
исследования.
При переработке книги часть глав и некоторые разделы были напи-
саны заново, а именно «Приспособление эмбриональных и ларвальных
стадий паразитов», «Зависимость одних членов паразнтофауны от на-
личия других», «Зависимость паразнтофауны от географических факто-
ров», «Опыт зоогеографического районирования по паразитологическим
данным», «Паразиты как компоненты биоценоза», «Влияние на па-
разитофауну животных хозяйственной деятельности человека», разделы,
посвященные вопросам иммунитета и специфичности, и ряд других Че-
рез всю книгу более четко, чем в предыдущих изданиях, проведен эко-
логический аспект понятий «паразитизм» и «симбиоз». Значительно пе-
реработана и дополнена 3-я часть книги («Паразитофауна и среда»),
главным образом за счет многих новых данных, полученных советски-
ми паразитологами.
Значительное место уделено библиографии, которой мы придаем
очень большое значение.
При подготовке книги мы использовали советы и консультации ря-
да наших коллег. В особенности мы благодарны проф. Л С. Мопчадско-
му, старш. науч, сотр С. С. Шульману, доп. М М. Белопольской, доц.
Т. А. Гинсцинской. Огромную техническую помощь нам оказали сотруд-
ники кафедр зоологии беспозвоночных ЯГУ и лаборатории протистоло-
гии Института цитологии АН СССР И. В. Кулемина, Г. А. Штейн, Р Е.
Шульман, М Н Голикова. Всем нм мы выражаем нашу признатель-
ность. Рисунки для книги заново сделаны Н. Г Коробовой, которую мы
просим принять нашу искреннюю благодарность за прекрасное выпол-
нение столь сложной работы.
Профессора Ленинградского университета
Ю. И. Полянский, Е. М. Хейсин
Часть I
ПАРАЗИТИЗМ. ЕГО РАСПРОСТРАНЕНИЕ
И ПРОИСХОЖДЕНИЕ
-----------------------------—— —v-
ГЛАВА I
ПОНЯТИЕ О ПАРАЗИТИЗМЕ И ПАРАЗИТАХ
Явление паразитизма есть один из видов взаимоотношений между
организмами — животными или растительными. В целях ограничения
предмета мы будем рассматривать паразитизм в животном царстве, ш
касаясь многочисленных примеров его, имеющихся в мире растении.
Слово «паразит» греческое и дословно обозначает того, кто питает-
ся за счет другого или других (рага—около, sitos — питание); таким
образом, по-русски можно было бы передать этот термин словом «при
хлебатель». В древнем Риме, куда слово «паразит» перешло из Греции,
оно получило специальный оттенок. Здесь так стали называть особую
профессию люден, живших тунеядцами исключительно за чужой счет
В этом своем значении слово «паразит» перешло в старинную медицину,
где под этим названием понимался всякий организм, живущий внутри'
другого организма и питающийся за его счет.
По мерс развития наук, а именно в XVII в., стала все более и более
выясняться общность паразитов (как животных организмов) и свободно
живущих животных. До того времени паразитов выделяли в совершен
но изолированную группу и самому их появлению иногда придавали за
гадочиый смысл, считая, что паразиты возникают в теле хозяина посрел
ством самозарождения. С включением паразитов в общую систему жн
вотного царства потребовался пересмотр и более точное определение
понятия паразита
Первая такая попытка характеристики животных паразитов и точ
ной формулировки особенностей, отличающих их от свободно живущих
организмов, принадлежит Ленкарту (Leuckart, 1879) «Паразита-
ми,— говорит Лейкарт, — в широком и настоящем значении этого
слова называются все те существа, которые находят пищу и жили
ше на другом живом организме... К паразитам принадлежат нс только
внутренностные глисты и сродные с ними формы, ио и такие существа,
которые сродни некоторым свободно живущим животным, за исключе-
нием рода их пищи».
Таким образом, согласно Лейкарту, главным отличительным при-
знаком паразита следует считать способ питания: всякий организм, пи
тающийся за счет другого живого организма, не убивая его, как это де-
5
лает хищник, является паразитом. То обстоятельство, что паразит нс
убивает своей жерты, расценивается как единственное отличие его от
хищника. «Границы паразитизма, — пишет Лейкарт,— нарушаются,
однако, такими животными, которые, как, например, пиявки, являются
паразитами лишь в отношении некоторых животных, а именно: лишь
тогда, когда добывают нишевые вещества от более крупных и сильных
организмов, но в отношении равных себе и слабейших они ведут се-
бя как настоящие хищники. Паразит всегда меньше и слабее животно-
го-хозяина, нс будучи в состоянии одолеть последнего, он довольствует-
ся тем,, что питается его соками или твердыми составными частями,
смотря но потребностям»
Из этих слов вытекает, что паразитизм представляет как бы одну
из форм хищничества, обусловленную главным образом взаииоотноше
нием размеров тела хищника и его жертвы Образ жизни — на теле или
внутри тела др’ того животного организма — является для Лейкарта вто-
ростепенным признаком, отнюдь не обязательным для всякого пара
зита.
Это oiipi де пенне понятия «паразит» Лейкартом, крупнейшим биоло-
гом своего времени и основоположником современной паразитологии,
прочно вошло в биологи» >, и его в той или иной степени придерживается
большинство современных паразитологов. Например, Коллери (Caulk-
гу, 1922) харамеризмет паразитизм как «нормальное и необходимое ус-
ловие жизни организма, который питается за счет другого организма
хозяина, — не разрушая этого последнего подобно тому, как делают
хищники со своей добычей».
В свете совпеменных знаний лейкар^овское определение, отразив-
шее в себе очень важную черту паразитизма, тем не менее является не-
сколько односторонним, ибо один критерий питания недостаточен для
определения паразитизма.
Из такого понимания следует, чго никакой естественной группы па-
разитов быть не может, как не может быть и никаких общетеоретических
проблем, касающихся паразитарных отношений. В категорию паразитов
искусственно придется объединить разнообразнейшие организмы, hi
имеющие ничего общего, кроме сходства в способе питания. К парази-
там придется отнести вампиров — летучих мышеи рода Desmodus, ко
торые по ночам нападаю! на люден и животных с целью кровососания,
комаров и постельного клопа, которые «временно посещают тело хозяи-
на с целью питания», и новозеландского попугая Nestor notabilis, на-
падающего на овец и выклевывающею у них кусочки мяса для пропи-
тания.
С другой стороны, поскольку решающим для определения является
характер питания, совершенно вне сферы понятия «паразитизм» должны
оказаться многие кишечные простейшие (Entamoeba coli. Trichomonas)
н даже черви (как Trichosirongylidae), которые питаются не за счет
тканей и соков хозяина, а поедают кишечных бактерий или простейших.
В связи с недостаточностью определения паразита по признаку пи-
тания некоторые авторы пытались уточнить это понятие путем присоеди
нения к данному определению признака вредности паразита для хозяи
на f'Vlin. 1ни. 1412: Dobell, 1919; Якимов, 1931; Холодковский, 1933,
и др.). Скрябин (1923), например, считает, что паразитизм есть «такой
тип биологических взаимоотношений между организмами, когда одил
Из них — „паразит”— обитает временно иди постоянно на поверхности
тела или в глубине органов и тканей другого — „хозяина”, — питаясь
за счет последнего и нс оказывая ему взамен никакой услуги». Здесь
учтены пространственные (но нс экологические) отношения, характер
литания и одностороннее использование одного члена сожительства дру-
гим. По определению Холодковского, «паразитизм состоит не только в
том, что гость (паразит) находит себе приют и пищу, но н в том, что он
питается за счет самого тела и соков хозяина, нанося ему прямой физи
ческий вред». Непригодность такого дополнения к определению особен-
но хороню видна из того, что по отношению к большинству паразитов,
в особенности встречающихся в диких животных, наличие вреда, прино-
симого хозяину паразитом, не доказано, а иногда и недоказуемо.
Кроме того, как правильно отметил Л. Фплипчепко (1937), нель-
зя определять объект (паразита) по признаку, принадлежащему исклю-
чительно Другому объекту (хозяину), в данном случае признак вреда
относится к хозяину? а не к паразиту. Это понятие является к тому же
весьма относительным и во многих случаях субъективным. Один и тот
же паразит \ разных хозяев, так же как и у одного хозяина, может вы-
бывать различные последствия. В силу различных физио логических со-
стоянии пар; зиг в одних случаях вызывает значительный патогенный
эффект, в других же — оказывается совершенно безвредным. Патоген-
ность паразита определяется не свойствами самого по себе паразита, а
соотношением его свойств со свойствами организма хозяина.
Неправильность определения паразитизма по принципу вреда —
пользы вытекает еще из того, «то паразит сам испытывает часто вред-
ное влияние со> стороны хозяина, что лучше всего иллюстрируется яв-
лением иммунитета. В частности, антитела, вырабатываемые хозяи-
ном, ограничивают жизнеспособность паразита, его плодовитость и т. д.
Наконец, если главной особенностью паразита является вред, ко-
торый он приносит хозяину, то при таком толковании паразитология
приобретает узкий меди ко ветеринарный характер.
Третья точка зрения на сущность паразитизма впервые была выска-
зана еще Ьпаупом (Braun, 1883). Последний считал, что «для парази-
тов характерен образ жизни, а нс организация... паразиты не система-
тическая I руина, а биологическая, и о них можно говорить только в том
же смысле, как о наземных или водных животных и т. д., не принимая
это сопоставление за классификацию». Следовательно, Браун считал,
что паразитизм есть понятие не систематическое и не Физиологическое,
а чисто экологическое
Эта правильная характеристика нс получила тогда дальнейшего
развития, и последующие паразитологи продолжали рассматривать па-
разитизм как один из видов сожительства между организмами. Между
тем при паразитизме одни из партнеров — хозяин — является для пара
зпта не просто сожителем, а той внешней средой, которая в первую оче-
редь определяет возможность существования паразита и к которой все-
цело адаптирован паразит. Павловский (1935), определяя паразитизм,
отмечает, что «паразиты используют организм хозяина не только как
источник питания, но н как место своего постоянного пли временного
обитания». Эту мысль Павловский развил особенно в своей статье
«Организм как среда обитания» (1931). Хозяин есть не один из компо-
Ht-нтов сожительства двух животных организмов — паразита и хозяи-
на,— а та среда, в которой живет и к которой приспособляется паразит.
Свободно живущий организм непосредственно сталкивается и взаи-
модействует с абиотическими и биотическими факторами внешней сре-
ды окружающего биотопа. В то же время отношения паразита со средой
биотопа обычно опосредуются через организм хозяина. Организм, ве-
дущий паразитический образ жизни, как пишет Мошковскпй (1'116),
«перекладывает бремя регуляции отношений с внешней средой на орга-
низм хозяина», через него же регулируется и получение паразитом пн-
7
щи, являющейся нажнейшим фактором внешней среды. В этом, но его
мнению, и лежит принципиальное отличие паразитического и свободно-
го образов жизни. Это не значит, однако, что паразит не связан с внеш
ней средой, в которой обитает хозяин. Напротив, многочисленные дан-
ные, которые будут приведены ниже, показывают, что паразиты в той
или иной степени связаны со средой, окружающей хозяина, н подвер-
гаются в большей или меньшей степени влиянию факторов этой внешней
среды, действующих, как правило, через организм хозяина пли в ряде
случаев непосредственно на паразита.
Говоря о паразитах, приходится по отношению к ним различать
двоякую среду обитания (Павловский, 1931): среду первого порядка —
непосредственное место обитания, т. с. организм хозяина, и среду вто-
рого порядка — условия, окружающие хозяина (среда биотопа).
Жизнь паразита, все его процессы и изменения, какие в нем про-
исходят. в первую очередь обусловлены и зависят от организма хозян
на. Эта особенность и является наиболее характерной и общей чертой
всех истинных паразитов, отличающая пх от всех других организмов н Г/
позволяющая объединить их в одну естественную группу. При такой
точке зрения на паразита он может быть определен, как правильно от-
метил А. А. Филиппенко (1937), как ^организм, средой обитания кото-
рого является другой живой организм». Из этого определения вытека-
ет, что паразиты являются такой же экологической группой организмов,
как, например, наземные или водные организмы.
Однако явление паразитизма весьма разнообразно, и не всегда воз-
можно провести резкую грань между категорией паразитических орга-
низмов и организмов, ведущих иной образ жизни. Между различными
типами приспособлений к окружающей среде и взаимоотношений орга-
низмов имеются всевозможны! переходы, а налйчие переходов, облегчая
нам понимание возникновения паразитов из свободно живущих форм,
вместе с тем чрезвычайно затрудняет формулировку самого понятия
«паразитизм».
По-виЙнмому, для того чтобы охватить все необычайное разнооб-
разие паразитических форм, необходимо как-то расширить определе-
ние паразитизма. Для выработки этого определения в свою очередь не-
обходим исторический подхоз к явлению паразитизма, а именно: изуче-
ние происхождения данного явления. Отдельные случаи паразитизма
могут иметь, как мы увидим далее, весьма различное происхождение,
но под всей совокупностью путей, ведущих к паразитизму, лежит одна
общая база, а именно: тенденция наилучшего и наиболее полного и эко-
номного использования пространства и пищевых ресурсов окружающей
природы со стороны бесчислен кого множества живых существ, ее на-
селяющих,— борьба за пространство и борьба за пищу.
Мы предпочитаем принимать для явления паразитизма определение,
наиболее широко охватывающее, хотя бы и менее конкретное. С пашей
точки зрения, паразиты—это такие организмы, которые используют
другие живые организмы в качестве среды обитания и источника пищи,
возлагая ппи этом (частично или полностью) на своих хозяев задачу ре-
гуляции своих взаимоотношений с окружающей внешней средой.
При этом мы ограничиваем применение термина «паразитизм» лишь
теми случаями, где хозяин и паразит относятся к разным видам, и ос-
тавляем в стороне внутриутробное развитие плода.
Конечно, расширение формулировки всегда идет несколько во пред
ее определенности, но увеличивает ее правильность. В природе обычно
нет тех резких границ между биологическими явлениями, которые мы
вводим своими формулировками.
8
Идя в противоположном направлении, можно детализировать оп-
ределение паразитизма и паразитов, стремясь охватить в расширенном
определении как можно большее количество сторон многообразного яв-
ления паразитизма. Это удачно провел Павловский в своем «Руковод-
стве по паразитологии человека», изданном в 1946 г Приведем опреде-
ление Павловского полностью: «Паразитами называют животных, ко-
торые живут за счет особен другого вида, будучи биологически н эко-
логически тесно связаны с ними в своем жизненном цикле на большем
или меньшем его протяжении. Паразиты питаются соками тела, тканями
или переваренной пищей своих хозяев, причем такой паразитический
образ жизни является специфическим видовым признаком данного па-
разита, многократно (в противоположность хищникам) пользующегося
для питания своим хозяином. Кроме того, паразиты постоянно или вре-
менно используют организм хозяина как территорию своего обитания
Паразит и хозяин являются повседневными партнерами в жизненных
соотношениях; разнообразие их противоречивых интересов определяет-
ся видовыми свойствами этой биологической системы организмов, ин-
дивидуальными качествами и состоянием данной пары партнеров и кон-
кретными особенностями среды обитания паразита и хозяина па фоне
их филогенетического развития».
Итак, паразитизм является преимущественно экологическим поня-
тием. В соответствии с этим паразитология дЬлжШГ изучать не только
самих паразитов и их хозяев, но особенно те соотношения и приспособ-
ления, которые возникают как последствие поселения одного организма
в другом или на другом организме.
Эту точку зрения па сущность явлении паразитизма впервые стали
детально развивать и обосновывать американские паразитологи лет
около 30 тому назад. По словам одного из крупнейших их представите-
лен Хегнера (Hegner, 1927), задача паразитологии состоит нс в изуче-
нии паразита—это задача зоологии или ботаники, в зависимости от
природы паразита; равным образом и изучение вреда, приносимого пара
литом хозяину, по Хегнеру, не может быть предметом паразитологии: это
объект изучения медицины, ветеринарии или сельскохозяйственных наук.
Объектом же изучения паразитологии как самостоятельной дисциплины
может быть только изучение взаимоотношений двух организмов — хо-
зяина и паразита, т. е. тех изменений в них, которые развиваются
как результат жизни одного из них в другом. «Отношения, — говорит
он, — между паразитом и хозяином как биологическое явление должны
изучаться совершенно так же, как мы имеем обыкновение изучать отно
шения между свободно живущими организмами и их средой».
Нужно отметить, что эта оценка задач общей паразитологии яв-
ляется необходимой базой для выработки практических мер борьбы с
различными паразитами. Общая паразитология имеет капитальное зна-
чение для правильного понимания и развития вопросов частной парази-
тологии, а частная паразитология находится в теснейшей связи с прак
тической жизнью и служит се интересам.
Весьма характерно, что экологический подход к явлениям парази-
тизма в последние годы все шире проявляется в исследованиях многих
паразитологов (Ваег, 1952; Noble, 1958; Theodorides, 1954; Stammer
1955, и др ).
Изучение отношений между организмом н средой является объектом
специальной дисциплины — экологии. Имеются ли в таком случае ка-
кие-нибудь основания для выделения изучения экологических отноше-
ний паразитов из обшей экологии в особую дисциплину — паразитоло-
гию? Несомненно, имеются.
$
Прежде всего в пользу этого говорит своеобразие отношений «па-
разит— хозяин», нс встречающееся в других ассоциациях. Дело в том.
что экология изучает поведение организма в отношении факторов внеш-
ней среды, т. е. те реакции (изменения функционального и морфологи
ческого характера), посредством которых организм приспособляется к
изменяющимся условиям своего существования При исследовании этих
приспособлений изучаются, конечно, и среда, и те изменения, которые
в ней происходят. Последние, однако, лишь в незначительной степени
бывают обусловлены особенностями самого организма, и поэтому для
экологии они представляют интерес не сами по себе, а только по их воз-
действию на организм. Иное дело в отношениях паразита и хозяина. Не
говоря уже о том, что с практической точки зрения главный интерес в
явлениях паразитизма представляет в большинстве случаев не организм
(паразит), а среда его обитания (хозяин), и с теоретической точки зре-
ния эти отношения представляют значительный интерес. Среда парази-
та, будучи таким же живым организмом, как и ее обитатель, реагирует
на присутствие последнего иногда очень сильно. При этом реакция сре-
ды по своему существу оказывается такой же приспособительной реак-
цией, как и реакция самого паразита Взаимодействие между организ-
мом и средой приобретает в этом случае совсем иной характер, чем
обычное взаимодействие свободно живущего организма и его среды
обитания. Отношения в смысле обратных ответов и реакций здесь не
только значительно более сложны, но нередко и резко отличны по своей
природе от аналогичных процессов, разыгрывающихся между неорга-
нической средой и обитающими в ней свободными организмами. Таким
образом, если в экологии вообще изучаются преимущественно односто-
ронние влияния среды на организм, то в отношениях паразитарных при-
ходится главным образом иметь дело с двухсторонними влияниями ор-
ганизма на среду и среды на организм в нх постоянном взаимодействии.
Вторым существенным моментом является метод изучения этих
взаимоотношении. Обычные методы экологического исследования среды
оказываются недостаточными для изучения явлений паразитизма. По-
мимо них здесь приходится пользоваться целым рядом специальных ме-
тодов. Поскольку главное место в изменениях среды, вызываемых па-
разитом, занимают явления иммунитета, постольку иммунобиологиче-
ская методика приобретает здесь чрезвычайно важное значение. Резкая
физиологическая изменчивость паразита под влиянием воздействия со
стороны среды (т. е. хозяина), проявляющаяся в виде изменений виру-
лентности и патогенности паразита, тоже требует для своего исследова-
ния особых методов, не известных экологии.
Таким образом, с точки зрения объекта изучения и метода, выде-
ление изучения всех явлений паразитизма в самостоятельную дисцип-
лину можно считать вполне обоснованным, законным и рациональным.
В процессах и явлениях, возникающих в результате взаимоотноше-
ний паразита и хозяина, отнюдь не следует видеть что-либо особенное,
нс встречающееся в жизни свободно живущих организмов. В основном
биологические процессы и явления в обоих случаях качественно тож-
дественны. Но наряду с этим в них имеются значительные различия, что
делает явления паразитизма особенно удобными и ценными для изуче-
ния целого ряда обшебпо. югпческих проблем В свяш с такими особен-
ностями среды обитания паразитов, как ограниченность во времени и
пространстве, многие биологические процессы приобретают у паразитов
крайне резко выраженный характер. Процессы (медленно — на протя-
жении веков и тысячелетий — и широко — на протяжении огромных тер-
риторий), протекающие у свободно живущих организмов, у паразитов
10
как бы конденсируются во времени н пространстве. Сильное изменение
окружающей среды свободно живущего животного (климат, почва, ра
сгительный покров, биотическое окружение) требует огромных проме-
тку тков времени, а отсюда и процессы изменчивости, состоящие в при-
способлении организма к этим переменам во внешней среде, протекают
весьма медленно, неуловимо для глаза исследователя.
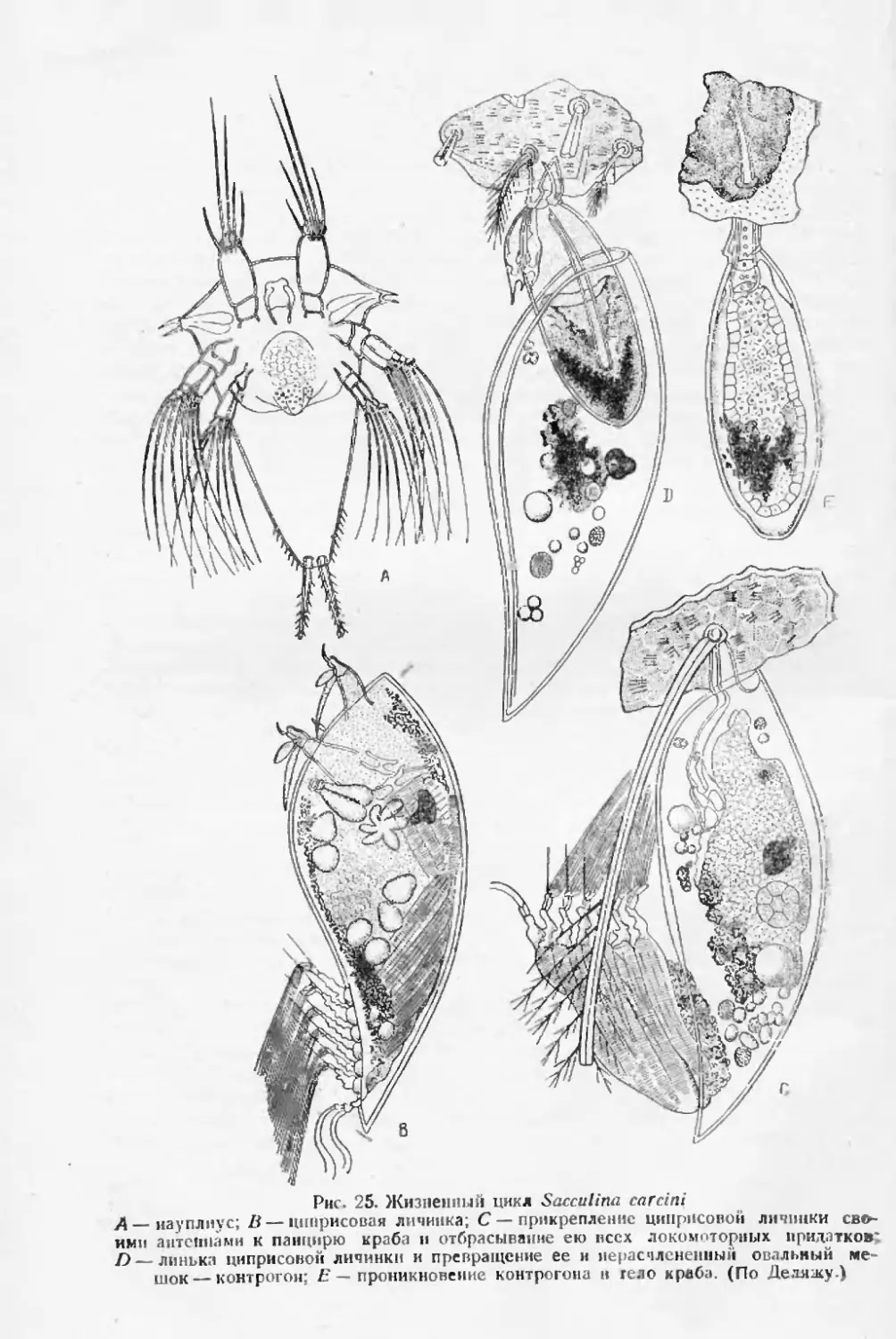
Vue. 1. Изменения инфузории Eniudlnitint сатШит от формы, снабженной хвостовы-
ми шинами (Л), к форме невооруженной (G) пол влиянием изменения пищевого ре
жима. (По Полянскому и Стрелкову.)
У паразита окружающая его среда может резко меняться на протя-
жении очень короткого срока, исчисляемого иногда неделями и даже
днями (заболевание хозяина, выработка нм иммунитета и т. д.), в связи
с чем и процессы изменчивости паразита ускоряются до размеров, не
встречающихся у свободно ж ивущих организмов Особенно ясно это по
отношению к некоторым паразитическим простейшим, которые могу г
претерпевать изменения при каждом бесполом размножении. Для Oph-
ryoscolecidae из желудка жвачных Полянский и Стрелков (1ОД8) пока-
зали, что изменение диеты хозяина может уже в течение двух недель
привести к превращению снабженной сильными хвостовыми нишами
(Ьормы Entodiniutn caudaiurn в форму вполне бесхвостую (рис 1).
Возникновение географических разновидностей и рас у свободно
живущих организмов связано с преодолением больших пространств или
и
с возникновением непреодолимых географических барьеров между об-
ластями, которые ранее составляли одно целое. Аналогичные до извест-
ной степени процессы у паразитов, состоящие либо в смене места обыч-
ной локализации паразита на новую локализацию в организме того же
хозяина (переход от кишечного паразитирования к тканевому или к
кровяному), либо в смене хозяина (переход кишечных паразитов рога-
того скота к лошадям и т. д.), могут происходить на самом ограничен-
ном пространстве Именно в силу этих моментов конденсации процессов
во времени и пространстве Шимксвич (1898) й говорил, что изучение яв-
юний паразитизма, помимо практического, представляет громадный
теоретический интерес: «Ничто не доказывает нам с такой убедитель-
ностью изменяемость форм и их способность приспособляться к усло-
виям существования, как изучение форм паразитических».
Мы поставили задачу в последующих главах проследит
общую естественную историю взаимоотношений паразита и хозяина и
вскрыть те закономерности, которые лежат в основе этих взаимоотно-
шений и касаются главным образом процессов взаимной адаптации ор
ганизмов паразита и хозяина.
ГЛАВА II
ЯВЛЕНИЯ СИМБИОЗА И ИХ ВЗАИМООТНОШЕНИЯ
С ПАРАЗИТИЗМОМ
Паразитизм не представляет собой явления, резко обособленного в
природе. К нему близки некоторые другие виды сожительства между
организмами, и норою бывает трудно провести грянь между явлением
паразитизма и другими формами взаимоотношений.
Если сожительство двух организмов рассматривается с точки зрС'
ния вреда или пользы членам ассоциации, то обычно различают три ос-
новных формы взаимоотношений мутуализм, комменсализм и парази-
тизм. При мутуализме оба члена сожительства являются полезными
друг другу, при комменсализме один получает пользу, а другому не на-
носится никакого вреда, при паразитизме один из сожителей получает
пользу, а другой вред. Однако мы уже видели в предыдущей главе, «то
критерий пользы — вреда не может быть положен в основу определения
паразитизма По этому признаку нельзя определять и другие формы
взаимоотношении организмов, что заставляет нас полностью отказаться
о г приведенной выше и пользующейся широким распространением клас-
сификации форм сожительства организмов.
Выясняя основные черты, характеризующие паразитизм, мы отме-
тили, что основное различие между паразитическими и свободно живу-
щими организмами лежит в их неодинаковом отношении к внешней сре-
де. Свободно живущий организм непосредственно сталкивается и взаи
моденствуст с внешней средой, подвергается прямому действию клима-
тических. эдафнчсских, биотических элементов своего биотопа, тогда как
существование паразитических организмов в значительной степени за-
висит от хозяина, устанавливающего свои отношения с внешней средой
самостоятельно без участия партнера (паразита) и обеспечивающего
возможность существования паразита.
Экологический принцип, положенный в основу определения пара-
зитизма, может быть применен п для характеристики основных черт дру-
гого, близкого к нему круга явлений взаимоотношения организмов —
к явлениям симбиоза. Существенным отличием этих явлений от парази-
тизма является иное отношение партнеров к внешней среде, включая и
пищевые связи Для разных форм симбиоза характерно, что оба чл< на
ассоциации совместно участвуют в регуляции отношений с внешней
13
средой. В одних случаях, как мы увидим на приводимых далее приме-
рах, ни одни из партнеров не может самостоятельно свободно сущест-
вовать, ибо каждый из них в отдельности менее жизнеспособен, чем в-
сочетании друг с другом (взаимно облигатный симбиоз, по Мошков-
ско.му). В других случаях идин нз партнеров использует другого для
своего приспособления к внешней среде, т. е. совместное существование
жизненно необходимо только одному партнеру, которым частично со
храняет самостоятельность своих связей с внешней средой. В обоих слу-
чаях организм как бы пополняет отсутствующие у него особенности, не-
обходимые для приспособления к данным условиям существования, пу-
тем объединения с другим организмом, у которого эти особенности
имеются.
Такне виды сожительства объединяются понятием «симбиоз»
(sin — вместе, bios — жизнь). Мошковскмн (1946) достаточно удачно
определил это явление, как такой тип сожительства организмов двух ви-
дов. при котором «оба партнера вступают в непосредственное взаимо-
действие с внешней средой, регуляция отношений с которой осущест-
вляется совместными усилиями, сочетанной деятельностью обоих орга-
низмов». При этом следует отметить существенную черту такого совмест-
ного существования: отсутствие в нем случайности. Этим симбиоз отли-
чается от случайного поселения каких-нибудь сидячих животных на дру-
гих, более крупных организмах. На какой-нибудь раковине можно найти
богатое сборище гидроидов, мшанок и т. д. Но те же гидроиды и мшан-
ки могут быть обнаружены и на сваях пристаней, на прибрежных кам-
нях и т. п. Значит, здесь перед нами случайное поселение одного жи-
вотного на другом без наличия между ними каких-либо тесных взаимо
отношений. Если совместное сожительство не носит такого случайного
характера и сочетание двух организмов регулярно повторяется, то мы
имеем перед собой пример симбиоза.
Сожительство организмов имеет важное значение для их приспо
соблення к передвижению, защите от неблагоприятных факторов внеш-
ней среды и хищников и, наконец, к улучшению своего питания, дыха-
ния и т. п.
Многочисленные явления симбиоза можно рассматривать, с одной
стороны, но принципу пространственных взаимоотношений между парт
верами, с другой — по большей или меньшей степени связи, установив-
шейся между симбионтами. Остановимся более подробно на рассмотре-
нии разных форм симбиоза.
Весьма разнообразны случаи сожительства типа «квартирант-
ства» — с и и о и к и я (sin — вместе, oikos— дом). При этом один из парт-
неров по своим меньшим размерам, а часто и вследствие своей боль-
шей подвижности помещается в виде спутника на другом или в жилище
другого организма. Использование одним из сожителей другого исклю-
чительно в качестве места поселения можно было бы в свою очередь
разделить па несколько групп в зависимости от характера предостав-
ляемой одним животным другому квартиры.
Простое квартирантство состоит в поселении квартиранта только в
жилище квартирохозяина или даже только в ареале его действия. По-
следний случай иллюстрируется следующим примером рыбка Amphi
prion держится среди щупалец крупных актиний (Crambactis и Dis-
cosonta), содействуя, по-видимому, своими движениями циркуляции
свежей воды, необходимой для дыхания актиний. При втягивании акти-
нией щупалец рыбка оказывается закрытой щушальпамн н на время
проникает при этом в гастральную полость партнера, пользуясь не
только защитой его стрекательных клеток, но и частью пищи. Всроят-
14
но, Amphiprion обладает известным иммунитетом по отношению к яду
актинии, защитное значение актиний для Amphiprion доказал Слюитер
(Sluiter, 1888). Посадив в аквариум с актинией и Amphiprion несколько
хищных рыб, исследователь держал там эту рыбку целыми месяцами.
Но стоило удалить актинию из аквариума, и Amphiprion уже через не-
сколько часов был уничтожен хищными рыбами. Сходно! отношение
квартирантства к медузам обнаруживают и плавающие рачки Нуре-
ridae.
Десятиногий рак Arete dorsalis держится рядом с морским ежом
Heterocentrotus в дырах, которые последний образует в полипняке мад-
Рис. 2. Примеры синойкин (кварти-
плнтст-а)
А и Л— двустворчатый моллюск
Leda яр. нл Японского мооя с окру-
каюти'.н его выходной сифон мел-
кими моллюсками из рода Modiola,
играющими роль комменсалов;
(' — краб Hmiixa, живущий в труб-
ках черня Chaeioptcrus. (Оригинал.)
P'lft 3. Эииойки и.। отряда усопших раков
(Ciriipedia)
/I — Anctax’>a sijitiilicrda, живущий па коже
акул; /•’ — Cornnula baliienurum, глубоко
внедряющийся в кожу китов, вид сверху;
в центре видно отверстие, ведущее в по-
лость мантии, содержащей мощные извест-
ковые пластинки. (Я — но P*fi Ланкастеру,
В - но Бриану.)
I — мясистый стебелек. ннодреииыП п кожу хоипиш.
'2 голиппой отдел паразита; J - рисщснленные
грухние иажъи; -/ — мясистая лиустнорчзтая мхтин,
охшииошая тело дш клише.
репоровых кораллов; при этом рачок имеет одинаковую с ежом окраску
тела. Некоторые раки из родов Polyonyx и Pinnixa (рис. 2) регулярно
встречаются в трубках живых сидячих полихет, например Loimia. C.hae-
loptenis.
Энной ки я включает такие случаи квартирантства, когда кварти-
рант поседштся на поверхности тела животного-хозяина. Характерный
пример такого рода представляют различные Sirripedia, питающиеся
планктоном, поселяющиеся па коже китов и акул. Таковы Anelasma.
sQuaiicoia. Curotiula (рис. 3) и др Очень много примеров эпиойкни сре-
ди простейших. Так, среди сосущих инфузорий есть много видов узко
специфичных в выборе животного, на котором они поселяются: Dendro-
cometes paradoxus на Gammarus, Stylocomeies digitatus на Asellus. Эти
эпнойки питаются ресничными инфузориями и, таким образом, не имеют
никаких пищевых связей с хозяином. Однако они гибнут, бу.чхчи сняты
с рачков: вероятно, движения рачка облегчают газовый обм^н сидящих
иа них простейших.
Штаммер (Stammer, 1955) приводит интересные факты поселения
инфузории Prntridia на различных водных насекомых. Причем одни
инфузории (Epistylis rotans, Vorticella campanula и др.) могут прикрсп-
15
ляться и жить не только на насекомых, но и на частичках детрита, тог-
да как для других субстратом всегда служит поверхность тела различ-
ных водных организмов. В одних случаях инфузории поселяются лишь
на определенных представителях водных животных (Orbopercularia
atninosa— на Gyrinus, Epistylis anophelis— на личинках комаров),
в других нс наблюдается такой приуроченности (например, Epistylis
nympharum, Vorticella crassicaulis).
Иногда квартиранты приходят в более интимную связь с хозяином,
заселяя впадины на его теле или его систему открытых полостей. Сюда,
Рис 1. Рыба — квартирант голоту-
рии (/ lulolhuria tubulosa)
А — рыба Fierasfcr unheris, вы-
лезающая из клоаки Holothuria
tubttlosn, в которой она живет
в качестве квартиранта: Н—Fie-
rasfer imberbis, вынутый на голо-
турии. (Оригинал.)
например, принадлежат относящиеся к
разным группам животные, которые посе-
ляются в мантийной полости моллюсков,
по периодически покидают последнюю
для добывания пищи. Такова рыбка 3—
6 см длины Apogonichtys strombi, живу-
щая в мантийной полости Strombns pi-
gas, крабы Pinnotheres — в мантийной
полости пластинчатожаберных моллю-
сков, олигохета Chaetogasler limnaei— у
Limnaea stagnalis и др.
Рыбки Fierasjer (рис. 4) из сем. Ат-
niodytidae., живущие в водяных легких
голотурий, периодически выходят через
клоаку наружу и питаются мелкими рач-
ками, получая от голотурии только квар-
тиру. В эту же группу эппойков можно
отнести любопытное сожительство червя
Aspidosiphon с актинией И ete.ropsammia
(Bouvier, 1895). Aspidosiphon в молодом
возрасте прячет конец тела в маленькую
пустую раковину Gastropoda. Па ту же
раковину потом салится личинка акти-
нии, которая разрастается и плотно охва-
тывает самое тело растущего червя как
муфтой. Червь высовывается из хода в
теле актинии, которой он доставляет сред-
ство передвижения, тогда как актиния
оказывает червю защиту своими стрека-
тельными клетками (рис. 5, А).
Классический случай симбиоза по типу эппойкии представляет со-
жительство раков отшельников с различными актиниями. При этом Sa-
gartia parasitica живет на раковинах, занятых несколькими различны-
ми видами раков, a Adamsia palliata обнаруживает специфичность по
отношению к Enpagurus prideauxi. В этих симбиозах, особенно у Adam-
sia, оба партнера вступают друг с другом в очень интимную связь. Pa-
gurus asper и другие, меняя свою жилую раковину, пересаживают сво-
их симбионтов, а именно Sagartia, со старой раковины на новую. Adam-
sia (рис. 5, С) обрастает Eupagurus своей подошвой, так что брюшко
последнего отчасти заключено в пустой раковине моллюска, отчасти же
в футляре, образованном телом актинии. Adamsia обращена ртом кни-
зу, причем ее ротовое отверстие приходится несколько позади рта рака,
так что часть пищи последнего всегда поглощается актинией. Вне сим-
биоза с раком Adamsia palliata жить неспособна.
Наконец, допустимо существование симбионтов-квартирантов, ко-
торые проникают внутрь хозяина. Это явление энтойкии
J6
Ifo существу оно представляет собой переход к явлению паразитиз
ма н трудно отгранииимо от последнего, Обычно энтойкия иллюстри
руется на примере жгутиконосцев из кишечника термитов. В задней ча
стн кишечника очень многих термитов живут в большом количестве
многожгутиковые жгутиконосцы отряда 1 lyperniast'mda (рис. 6). На
холя себе приют и пшцу в кишечнике термита, эти простейшие, несом
ионно, извлекают из этого сожительства определенную пользу. Но в то
же время, как ясно следует из работ Кливлэнда и других (Cleveland,
РнС 5. Различные примеры симбиоза
А — Aspidosiphon, охваченный подошвой актинии Heteropsammia cochlea
которую он raci ает за собой Н — освобождении ft от актинии Aspidosl-
phon, С—рак-отшельннк Eapagurus prldeauxi в симбиозе с актинией
Adamsui palliuia. Актиния охватывает раковину отшельника сноси подош-
вой, края которой сходятся на его спнннон стороне; снизу из-под брюшка
рака вы гл я цы пл ет часть щупалец актинии. (Но Коллери)
1925а, б, с, 1926, 1928; Hungate, 1939; Grasse, 1952а и ар.), термиты так-
же получают пемхпьшук) пользу or своих еообитг гелей. Взаимоотноше
пне между этими животными становится столь тесным, что у термитов
вырабатываются па его почве даже некоторые специальные инстинкты,
и ле толькз жгутиконосцы не могут су цествовать без термитов, но и
термиты—без жгутиконосцев. Пищей некоторых термитов с |ужит ис-
ключительно клетчатка, которую они добывают, грызя древесину су-
хого дерева. Переваривание пищи у них происходит в заднем отделе ки-
шечника, где находятся жгутиконосцы. Как это ни странно, но в кишеч
пике самих термитов нс обнаружено никаких ферментов, растворяю-
щих клетчатку. Между тем, тело Hyperniastigida сплошь набито ог-
рызками древесины, которые они заглатывают и, несомненно, нсрсва
ривают. Отсюда возникла мысль, не играют лн Hyperniastigida какой
пибудь роли в пищеварении их хозяев. В пользу того же иредположе
ния говорило и наличие жгутиконосцев решительно во всех особях тех
видов термитов, где они вообще имеются. Для разрешения этого вопроса
Кливлэнд прежде всего выработал технику дефаунирования термитов,
т. е. освобождения их от жгутиконосцев. При содержании термитов в
кислороде под давлением в 3 агм все жгутиконосцы в скором времени
2 В А. Догель
17
погибают, тогда как термиты остаются живыми. После дефаунировання
Кливлэнд помещал в одинаковые нормальные условия с обильным на
шчием нищи, т. е древесины, как дофах ннрованных насекомых, так и в
качестве контроля обыкновенных особен, г. е. содержащих I-lypermasti
gid? Термиты сс жгутиконосцами жили в своих опытных сосудах не-
Рис. 6. Различные Hypcrmastigkla, живущие в кишечнике термитов
А — Trichonympha tiirkesfanica-, И - MiTcspironympha porfer,, C Calony-
mpha grassii; D Kofoidia loriculata. (Д но Вериштейн. H. C— по Яшщ-
кому. Г) — но Лайт.)
определенно долгое время, иногда до 1—1V? лет, тогда как дефауниривап-
ные особи выдерживали не более 10—14 дней п потом неизбежно поги-
бали. При этом они все время питались, но пища, очевидно, нс могла
ими утилизироваться. Стоило искусственно заразить дефаунироваиных
термитов жгутиконосцами, как такие насекомые приобретали способ-
ность продолжать свое существование не погибая
18
Исследованиями Тангеита (IInngate, 1939, 1943) было доказано на-
личие у симбиотических жгутиконосцев фермента, расщепляющего
клетчатку, — целлюлазы. По-впднмому, одним из продуктов расщепле-
ния клетчатки в теле термитов наряду с глюкозой является уксусная
кислота. Возможно, что она в первую очередь и используется термитами
(Grassi. 1962а).
Опыты Кливлэнда показали, до какой тесной органической связи
доходит симбиоз термитов и ilypermusligicla По-видимому, жгутико-
носцы переводят клетчатку в растворимое состояние, используя при этом
сами лишь часть пере работанного вещества. Вполне естественно, что
при этом у обоих членов сожительства выработались черты, способ-
ствующие постоянному заражению термитов их столь полезными сим-
бионтами. Р« шлющее значение в этом смысле имеет инстинкт слизывания
друг у друга жидких экскрементов, выступающих из заднепроходного
отверстия термитов. У молодых личинок, воспитываемых взрослыми
рабочими термитами, такой способ питания является даже единствен-
ным. Легко видеть, что слизыванием экскрементов достигается двоя-
кая цель. С одной стороны, термит проглатывает содержимое задней
кишки своих собратьев, где находится в некотором количестве
переведенная в ,свояемое состояние клетчатка, с другой — он получает
возможность заражения жгутиконосцами, что особенно важно для ли-
чинок, которые при выходе из яйца свободны от симбионтов.
Казалось бы, что возможность заражения затруднена тем обстоя-
тельством, что большинство Hypermastigida не обладает способностью
и.нцнстироваться, и потому проглоченные термитом жгутиконосцы мо-
гут быть переварены пищеварительным соком хозяина во время своего
следования через кишечник от рта до задней кишки, где имеются усло-
вия, благоприятные для их существования. Однако Догель (1916—
1922) обнаружил, что в шдней кишке термитов наряду с нормальными,
активными жгутиконосцами, всегда имеется небольшое количество му-
мифицированных I lyperrnastigida. Такие особи лишены пшщвых ча-
сгип, эктоплазма их сильно уплотнена, длинные жгутики их сплетены и
образуют вокруг тела род войлока. В таком неипцистированном (ибо
никакой оболочки животные вокруг себя не выделяют), ио мумифици-
рованном состоянии жгутиконосцы, по-видпмому, приобретают способ-
ность без вреда для себя проходить через весь кишечник термита до
задней кишки и обеспечивать заражение хозяина.
Большое значение Ilyperrnastigida в жизни термитов видно уже из
того. что. поданным Капни н Кирби (Katzin а Kirby 1939), вес жгути-
коносцев, отфильтрованных из кишки термитов, достигает 36% веса те-
ла хозяина. Вес этот так велик, что невольно рождается мысль, не слу-
жит ли часть жгутиконосцев, проходящих через кишку термитов в сли-
занных экскрементах, дополнительным источником питания этих насе-
комых.
Аналогичный характер отношений обычно принимался и для жи-
вущих в громадном количестве в рубце и сетке жвачных инфузорий
отряда Entodiniomorpha сем Ophrx oscolec idae, и для их хозяев. Й здесь
обычно все особи животного-хозяина бывают заражены инфузориями,
число которых доходит до 1 000000 на 1 с.«3 содержимого желудка. При
этом Ophryoscolec.dae, наподобие llypermastigida, всегда набиты ог-
рызками травы, растительными волокнами и т. д. Одни из них — Diplo-
dinium и Eudiplodinium— переваривают целлюлозу, так как вырабаты-
вают энзимы — целлюлазу и целлобиазу, тогда как представители дру-
гих родов (например, Efitodiniuni и т. и.) не обладают этими энзимами
и пе могут перерабатывать целлюлозу. Положительное значение инфу
2*
19
зорий для хозяина может заключаться в том. что они. попадая в сычуг,
п массе гибнут и тем самым служат некоторым дополни тельным источ
ником снабжения хозяина углеводами и животными белками. Цефауни-
рованные жвачные, т. е. те, которые освобождены от инфузорий, однако,
также способны использовать целлюлозу, но только в том случае, если
опн содержат бактерии, расщепляющих целлюлозу Опыты Беккера и
Эверетта (Becker a. Everett, 1930), Полянского и Стрелкова (1933) го-
ворят о том, что молодые десЬаунироваииые ягнята развиваются вполне
нормально и с нормальной быстротой растут и увеличиваются в весе.
Поэтому вряд ли правильно считать взаимоотношения инфузорий Oph
ryoscolueidaa с хозяином симбиотическими; эти инфузории с большим
нрадом должны быть отнесены к паразитам, для которых рубец жвач-
ного является их средой обитания.
То же необходимо сказать и о взаимоотношениях между термита-
ми и жгутиконосцами. Хотя они часто н именуются симбиотическими,
но в свете принятого нами выше определения паразитизма их можно
рассматривать и как отношения паразита и хозяина. Кишечник термита
служит для жгутиконосцев средой псового порядка, и их связь с внеш-
ней средой второго порядка регулируется через организм хозяина.
Чрезвычайно тесные взаимоотношения, вероятно симбиотического
характера, наблюдаются между животными и организмами растнт<-ль
ной природы, водорослями, грибками, бактериями.
В тканях некоторых беспозвоночных с давних нор известно нали-
чие мелких, округлых включений желтого или зеленого цвета. Еще I Leu
конский (1871) предложил толковать эти включения как одноклегоч
иые водоросли, причем у морских животных в тканях встречаются жеч
гые зооксантеллы, а у пресноводных — зеленые зоохлореллы. В настоя-
щее время эти симбионты констатированы у многих простейших (осо-
бенно у радиолярий и инфузории), у некоторых губок (Spon^illidae) и
кишечнополостных (Chlorohydra viridistdma, некоторые меду пя, многие
Anlhozoa) и у отдельных представителей других типов (турбелляоии
Convnluta, Vortex и др., коловратка Ascomorpha Helvetica, мшанка Zoo
hothryum, моллюски Tridacna и Elyna viridis и т. д.).
Самостоятельность и растительная природа зоохлорелл были не-
опровержимо доказаны опытами выделения их из тела хошина (напри-
мер, раздавливанием тела инфузории, содержащей водоросли) н куль-
те рой их в искусственных условиях. Зоохлореллы в таких условиях мо-
гут жить и размножаться неопределенно долгое время. Равным образом
в некоторых случаях (например, с Chlorohydra viridissirna) удается
освободить и хозяина от симбионтов, без которых он также в состоянии
нормально сушсс гвовать.
Особенно тесным, по крайней мере для одного из сожителей, являет-
ся симбиоз некоторых грибков и бактерий со многими прсимсщественно
кровососущими, ио также и иными насекомыми. Правильнее, однако, и
эти взаимоотношения называть скорее паразитарными, так как для бак-
терий и грибков насекомое служит средой обитания, а не просто сожи-
телем.
В различных местах тела насекомых (и желудочном диске вшей, в
трахеальном органе желудочного овода, в третьем брюшном сегменте
клопов, в стенках средней кишки мухи цеце, в брюшке тлен и червецов,
в жировом теле тараканов) имеются специальные образования, так на-
зываемые мицетомы, состоящие нз скопления клеток, населенных бакте-
риями или одноклеточными грибками (рис. 7). Мицетомы часто окра-
шены в зеленоватый или буроватый цвет. Очень часто мицетомы распо-
ложены вблизи яичников, так что они могут попадать в яйцевые клетки
20
и таким образом передаются из поколения и поколение. В иных случа
ях Бухнер (Buchner, 1912, 1918. 1921) и другие авторы описывают ряд
чрезвычайно оригинальных и разнообразных приспособлений, обеспечи-
вающих передачу грибков или бактероидов потомству насекомого-хо-
зяина. В указанных организмах различные авторы распознали дрожже-
вые грибки (Schizosaccharomyces aphidis у некоторых тлей, Sdccliaromy-
ces, Oospora. Coccidbmyces и т д.). Изредка, по Бухнеру, у одного вида
насекомых имеется даже два или три сорта мицетомов с различными
симбионтами или паразитами.
Рис. 7 Внутриклеточные симбионты насекомых
А— личинка тли Drepanosiphon platanoldes с мицетомом.
или зеленым телом; Н — зародыш чернена Iteryn puchasi с .ми-
цетомой; С—отдельная клетка из мицетома I)actylopius ci tri
с бактериоидными серповидными тельцами и ядром. (Но
Коллери.)
1 — мицетом; 2 — Сиктсриоилиое тельце; 3 — ядро.
Вопрос о значении так называемых симбионтов для организма хо-
зяина не может считаться окончательно выясненным По-видимому,
внутриклеточные симбионты приносят своим хозяевам определенную
пользу, и некоторые из них, безусловно, необходимы для жизни насе-
комых. .V тлен и червецов, но Пьерантони (Pierantoni. 1910), симбион
ты выделяют энзимы, способствующие перевариванию сахара, в н.зобм
лии получаемого этими насекомыми в соке растений
Бухнер (I960) приводит интересные примеры значения симбионтов
в развитии некоторых насекомых. Так, например, личинки хлебного то-
чильщика Sitodrepa paniceum не росли нормально н не были способны
к линьке, если у них не было симбионтов (Fraenkel a. Blewett, 1943)
В то же время личинки с симбионтами росли и проделывали линьку
нормально. Если же стерильных личинок кормить дрожжами, т. е. да-
вать им витамин В, то личинки растут и развиваются так же, как имею
тис симбионтов. Поэтому можно думать, что симбионты выделякл ни
тамин В, необходимый для роста хозяина
До известной степени особый тип симбиоза, а именно п ара си м-
биоза (симбиоз бок о бок), дают многие мирмикофилы и термито-
филы, т. е. симфиты муравьев и термитов (рис. 8). Обычный тип симби-
оза заключается в попарном парасимбиотическом сожительстве; даже
21
еслисимбионтов много, то во всяком случае каждый мелкий симбионт
составляет со своим хозяином одну пару индивидуумов. Но возможны
случаи парасимбиоза «общественного», когда ни один из симбионтов не
вступает в прочную связь с определенным индивидом другого вида, но
Рис. 8. Ра «личные муки— мнрмикофилы
А — Lomechusa slrumosa; Н — Onussus turciciis; С — Mimcciton pulex
D— (’laviger testareus. (I Io Коллери.)
с целой общиной особей. Жук Paussus, снабжающий муравьев прият
ными для них выделениями особых желез, обслуживает любого члена из
гнезда муравьев и получает услуги также от любой особи. Точно так
же и тли пользуются защитой муравьиной колонии, взятой в целом, и
дают свои сладкие выделения всякому проходящему ччпавью
Рис. 9. Общин
ннд Udoni Ila.
(По Иванову.)
Рис. 10. I идроиды Hydrichthys, cyclothori'S, пара
актирующие на рыбе Cyclothone signata
А — рыба с двумн колониями гидроидов (отмече-
ны двумя стрелками); Н — колония Hydrichthys
при большом увеличении; черные пятна вокруг
колонии—светящиеся органы рыбы. (Ио Д-ма.)
Итак, вы видели, что комплекс отношений, именуемый термином
симбиоз, содержит в себе самые разнообразные переходы от отношений
более или менее индифферентных до таких, когда оба члена сожитель
ггва нс могут существовать один без другого
Особо надо остановигься па явлениях комменсализма, которые
нередко рассматриваются, наряду с симбиозом и паразитизмом, как осо-
бая категория взаимоотношений между организмами. Мы уже говорили
выше, что, с нашей точки трения, подобный взгляд несостоятелен. Под
комменсализмом мы понимаем лишь способ питания одного организма
остатками пищи другого. Комменсализм может возникать при разных
формах сожительства как симбиотических, так и паразитарных. Ком
менсализм может иметь место и средн свободно живущих организмов,
нс образующих ассоциаций
Рис. 11 Hllobiophrya ttonacis на жабрах Donax vittutiis
^По Lllai гону.)
1 инфузория. отдслnioiiidH от себя дочернюю особь, 2 дочернин
особь. рсшишная хтдннй венчик ресниц; .7 перекладина жабср
нпй решетки; J — ресницы жаберного эпителия.
Комменсализм средн паразитических организмов наблюдаетоя }
многих кишечных простейших, как, например, у жгутиконосцев Tricho-
mastix и C.hitnrnastix, поедающих бактерий, грибков и т. п. в кишечнике
позвоночных хозяев, или инфузории Balantidium elongatum и др. из тон-
кой кишки амфибий и других животных. К этой же категории можно
отнести и некоторых паразитов кишечника, питающихся осмотическим
путем, например опалин, заселяющих заднюю кишку амфибий.
Среди эктопаразитов комменсализм наблюдается у червя Udonella
который прикрепляется к яйцевым мешкам Ca'igidae и питается отбро-
сами пищи хозяина (рис. 9)
Много примеров комменсализма среди симбионтов
23
Около порошицы роющего морского ежа Echinocariiiuni очень ча
сто держится пластинчатожаберный моллюск Montacuta lerruginosa;
вокруг отверстий сифонов дальневосточного пластинчатожаберного
моллюска Leda sp. целыми гроздьями прикрепляются значительно бо
лее мелкие Modiola (см рис. 2)
Интересный случай представляет поселение некоторых морских
гидроидов (Hydrichthys cyclothonis) на коже рыб (Cyclothone signata).
обыкновенно неподалеку от порошицы хозяина (рнс. 10) Очевидно, гид-
ранты используют в качестве пищи экскременты хозяина.
В кожных ^давлениях по бокам основания хвоста у головастиков
травяной лягушки очень часто поселяются колониальные Peritricha.
Другие Peritricha, как, например, Ellobiophrya donacis, поселяются на
жабрах пластинчатожаберного моллюска Donax, причем инфузория
охватывает одну из жаберных нитей хозяина своим выступающим в двух
направлениях основанием наподобие кольца и свободно болтается на
нити, как висячий замок на скобке (рнс. 11)
Небольшой краб Pinnixa живет в полости раковины дальневосточ-
ного пластинчатожаберного Pecten jessoends, причем краб становится
слепым, что говорит о потере его связи с внешним миром. Все эти орга-
низмы питаются остатками пищи хозяина, т с. являются типичными
комменсал иста ми.
Как ясно видно нз приведенных выше примеров, иллюстрирующих
некоторый формы сожительства организмов, имеются разные степени свя-
зи между обоими членами сожительства. Наименьшая связь наблю
дается при некоторых формах синонкни Дальнейшее упрочение связи
идет в двух направлениях: с одной стороны, по линии усиления взаим
ноги использования обоими партнерами друг друга, с другой — ио ли-
нии использования одним членом сожительства другого члена в каче
-стве среды обитания, что приводит к настоящему паразитизму.
ГЛАВА III
РАЗЛИЧНЫЕ ФОРМЫ СВЯЗИ ПАРАЗИТА И ХОЗЯИНА
1. ФАКУЛЬТАТИВНЫЙ И ЛОЖНЫЙ ПАРАЗИТИЗМ
Между паразитическим и свободным образом жизни можно найти
всевозможные промежуточные звйкья и иевеходы Наличие в природе
переходных состояний, представляющих собой постепенные адаптации
к паразитическому образу жизни, иллюстрируется очень хорошо явле-
нием факультативного па раз и г и з м а Гак называют примеры
паразити>ма животного, которое нормально живет в свободном состоя
нии, но при случайном попадании на подходящих хозяев или в н< которых
из них непосредственно переходит к паразитизму. Следова гельпо, в жй-з
пенном цикле таких животных стадия паразитизма является не обяза
тельной
Среди пиявок (рис. 12). например, европейская Glossosiphonia com
planala является свободно живущим хищником, который питается мел
кими беспозвоночными. Ложноконская пиявка Hnqmopis в отношении
мелких животных ведет себя так же. Limnalis nilotica, нападая на круп^
ных позвоночных животных, ограничивается временным сосанием крови
Среди клопов виды рода Reduvius являются настоящими свободно
живущими хищниками, поедающими различных насекомых однако Re-
duvius personaius, наряду с таким же хищничеством, может иногда на
падать на человека и сосать у него кровь. Особенно типичны случаи фа
культативного паразитизма у некоторых круглых ч> рвей. Скрябин при
обследовании челове ческих кише шых паразитов в Донбассе нашел в ки
шечнике человека яйца, личинок и взрослых червей двух видов Rhabdili.\
которые обычно живут в почве.
Типичным факультативным паразитизмом обладает круглый червь
Aloionema appendiiulatum. Обыкновенно он является свободным обита-
телем почвы. Однако иногда личинки Aloionema приникают в слизня
Arion ater, где и живут некоторое время в кишечнике, продолжая расти
и достигая вдвое большей величины по сравнению с нормой. Впрочем по-
ловозрелого состояния достигнуть в слизне они не могут и должны для
этого выйти наружу, где размножаются. Их потомство может одинаково
хорошо существовать как на воле, так и в слизнях.
Факультативный паразитизм представляет интерес с точки зрения
вопроса о происхождении паразитизма. Во-первых, переход к парази-
тизму у свободно живущих организмов можеу осуществляться лишь в
том случае, если в их строении и функциях или образе жизни имеются
какие-либо особенности, позволяющие вести паразитический образ жиз
ни.1 Это может выражаться, например, в особой прочности покровов,
спасающей животное от действия пищеварительных соков хозяина, пли
в особенностях дыхательного обмена, например животные, встречаю-
щиеся в
гниющих веществах, могут легко жить в анаэроиных условиях.
Пиявки с различными способам» пн-
тания
4 - Glossoslphoma complanata (Rhynchobdi l-
lid.i); В— Hhemopis sanguisuga. поедающая
деждепого черня; С — Hacmadipsa ceylonica,
цейлонская наземная пиянкл на древесном
листе. (По Кюкенталю.)
Рис 12.
а следовательно, способны лег-
ко адаптироваться к жизни в
кишечнике хозяина, где кисло-
род отсутствует
Указанный пример с Aloi-
ипегпа свидетельствует о чрез-
вычайной быстроте изменении,
вызываем ых паразитизмом
сильное увеличение размеров
паразитических личинок в пре-
делах одного и того же поко
Ленин. Во вторых, пример с
Rhabditis показывает нам не
которые из обстоятельств, спо-
собствующих переходу свобод-
но живущих организмов в па-
разитическое состояние. Преж-
де всего этому благоприят-
ствует усиление контакта меж
ду факультативным паразитом
и будущим хозяином. Находка
Скрябиным Rhabditis именно в
Донбассе ш случайна. Там. в
глубоких подземных шахтах.
тысячи люден ежедневно и на
небольшом сравнительно пространстве приходят в самое тесное сопри
косновение с землей, в которой обитают свободные Rhabditis; таким об-
разом, этому организму открывается путь к переходу в паразитически*
«стояние. То обстоятельство, что тысячи рабочих находятся в самом
тесном контакте друг с другом, способствует дальнейшему распростри
нению зарождающегося паразитического вида. Наконец, равномерная
температура и достаточная влажность в шахтах являются благоприят-
ными факторами для существования здесь Rhabditis. В данном случае
сочетается целый ряд условий, благоприятствующих тому, чтобы парази-
тофауна человека обогатилась еще одним паразитом
С понятием факультативного паразитизма более или менее совпа-
тает выросшее главным образом на медицинской почве понятие о «л о ж
н о п а р а з и т и з м е». Под этим названием объединяют таких свободно
живущих организмов которые в состоянии некоторое время жить внутри
тела другого животного при чисто случайном попадании в последнее
Таковы, например, личинки ряда видов мух (комнатной мухи, серой
мясной мухи, сырной мухи Ру oph.il а), которые нормально живут в навозе
или в падали, или в сыре, но обладают такой жизненной стойкостью, что
способны противостоять даже действию пищеварительных соков кишеч
ника н могут известное время жить в такой чуждой для них среде и даже
вредят при этом своему случайному и временному хозяину.
Аналогичное явление наблюдается у тнроглифондных клещей, вре-
дителей зерна, сыра и других продуктов. Они могут попадать с пищей в
• Вопрос о преадаптациях см Osche. (1957) н Stammer (1955).
26
пищеварительный тракт челов! ка 11 задерживаться там некоторое время,
вызывая при этом иногда даже кишечные расстройства Возможно, что
ложноиара.йпи м отличается несколько от факультативного лишь своим
еще более редким и случайным характером.
Из вышеуказанного видно, чю факультативный паразитизм в неко-
торых случаях может служить переходным этапом к типичному парази
тнзму, который обозначают как паразитизм обязательный, или облигат
ный. К данной категории принадлежат все формы, у которых в жизш н
ный цикл обязательно входит паразитическая фаза Разбираясь в пеобы
чайном разнообразии отношений паразитов к их хозяевам, можно раз-
делять облигатных паразитов на группы, руководствуясь различными
критериями. Можно взять в качестве критерия пространстветшые отноше-
ния паразитов к хошину или отношения паразита к хозяину во времени,
если учитывать длительность контакта паразита с хозяином в течение
жизненного цикла паразита
2. ПРОСТРАНСТВЕННОЕ ОТНОШЕНИЕ ПАРАЗИТОВ К ХОЗЯЕВАМ
Подобно тому как любой биотоп суши или моря может быть пасе
лен живыми существами. так и каждая живая ткань и каждый орган
животного могут служить местом обитания для паразитов. Соответствен-
но с локализацией паразита в организме хозяина их разбивают на две
категории: эктонара даты — обитают на внешних покровах хозяина,
на коже, на жабрах; э и д о пара з и т ы — живут во внутренних полостях
тканях и клетках хозяина.
Эндопаразиты некоторыми авторами разделяются еще на полост
п ы х, т к а п е в ы х, в н у т р и к л е т о ч н ы х и т. д.
Имеются целые группы паразитов, характеризующиеся или эктопа
разитичсски.м, или эндопаразитическим образом жизни. Так, среди при
стейших класс Sporozoa состоит целиком из эндопаразитов, тогда как »
числе паразитических Mastigophora и Infusoria встречаются и экто-, и
эндопаразиты. Класс Monogencfidca характеризуется эктопаразитическим
образом жизни, класс Trematodes — эндопаразитическим Однако среди
Monogenoidea мы находим несколько интересных переходов к эндопа-
разитизму. Первый пример дают представители рода Polystoma. Родичи
Pulysloma живут на жабрах морских, реже пресноводных рыб. Такой жг
образ жизни следует приписывать и предкам этого рода. Однако соврс
хинные виды. как. например, Polystoma mtegerrtmuni и некоторые дру-
гие, лишь в личиночной своей стадии держатся пй жабрах головастиков
в виде э(<топаразнтув. С превращением головастика в молодую лягушку
паразит, в связи с атрофией иабгр и замыканием жаберных щелей у
хозяина, переходит в его кишечник, спускается в клоаку, а из последней
проникает в мочевой пузырь, где достигает половой зрелости и на всю
жизнь уже остается эндопаразитом
Целлер (Zeller, 1872), а позднее Галлион (Gallion, 1935) показали,
что в некоторых случаях раннего проникновения личинок Poluslotna в
головастиков личинки успевают еще до метаморфоза головастика, т. е
Оставаясь на жабрах', достигнуть половой зрелости и начать откладывать
яйца. Этот факт проливает свет на прошлое рода: предки его обладали
чисто эктопаразитическим (на жабрах) образом жизни. В том же роде
Pnlystoma имеются виды, паразитирующие па черепахах, например Poly-
yioma (Polystomoidcs) ocellalum, которые поселяются в ротовой полости
хозяина иод его языком, или Р. rugosum (рис. 13) в ноздрях черепахи
Trionyx.
Попытки к переходу в эндопаразитическое состояние делались неза
27
висим» представителями разных групп mohoiенетнчсскнх сосальщиков
Так, средн отряда Monopisttiodisciiica,паразитирующего на жабрах iihi
коже карповых рыб, род Acolpenteron (рис. 14) ушел с покровов хо
зянна н проник, очевидно через наружные мочевые отверстия, в моче
точники почек Одни из видов этого рода описан Гвоздевым (1945) ич
пределов СССР (в мочеточни-
ках небольшой рыбки Diplo-
physa)
1 ipHMep факультативного
перехода к эндопаразитическо-
му состоянию одного и < Мопо-
genoidca, обычно паразитирую
|цею па жабрах рыб, констати-
ровал Рушковскпн (Ruszkou
Рис. 15.’' Монотеистически»
сосальщик Amphibdella tor
pediitis. Взрослый червь с
жабер Torpedo nntrmoraia
(По Нихонскому ।
Рис. 13- Poly stoma rugosiim и i
ноздрей черепахи Trionyx fe-
гох. (Но Мак- Каллюму.)
I глотка. 2 яйцо; J семенник;
4 кишечник; 5 - нрнаккн; 6
галнщс; 7 — яичник; 8 ниделнгель-
1ЫМ нора.
Рпс М. Монотеис-
тический сосаль-
щик Acolpenteron
ru phriticuni из мо-
четочников рыбы
Diplophysa sp. (По
1 возлеву )
ski, 1931) у Amphibdella 'torpedinis (рис. 15). Этот представитель уж.
давно описан с жабр скатов Средиземного моря (Torpedo ocellata п
Г. rnarrnorala) Исследовав в Неаполе 27 скатов, Рушковский
убедился в том, что очень часто, а именно у 50% рыб, в ссрд
не имелись характерные яйца Amphibdella lorpedintb, а в 30% слу
чаев и сами черви в половозрелом состоянии. Способ проник
новения A. torpedinis в сердце остался невыясненным, но несомненно,
что, наряду с эктопаразити.«мом на жабрах, этот вид способен перехо
дить к глубокому внутренностному паразитизму. По-видимому, уход в
кровеносную систему является для него тупиком, не дающим возмож
пости дальнейшему распространению, но этот уход знаменателен как по
казатель широкой потенции паразитов к перемене своей локализации в
2«
геле хозяина. Возможно, что в других случаях указанная потенция могла
привести к полному переходу паразита к паразитированию в крови. Так.
быть может, следует трактовать случайный переход некоторых кишеч
пых жгутиконосцев через 'стенку кишки в кровь хозяина
Все цестоды всегда являются типичными эндопаразитами, равно как
и Acanthoccphala, и нематоды. Напротив, в тех редких случаях, когда
аннелиды переходят к паразитизму, они почти всегда становятся Элтона
разитами. Таковы паразитирующий на коже рыб Ichthyoionius sanguida
ritis среди Polychaeta и 2 рода из сем I listribde.Iidae, паразитирующие
на раках. Только некоторые роды сем. Euniciciae паразитируют в полости
тела других полнхет и являются эндопаразитами. Среди Oligochaeta
имеется лишь небольшое число эктопаразитов (Branchiobdellidac на прес-
новодных раках и немногие Encliytraciilae на дождевых червях). Этим,
вероятно, обьясняется то, что генетически связанные с Oligochaeta
пиявки содержат в своем числе тоже исключительно эктопаразитов.
Немногочисленные паразитические моллюски принадлежат к обеим
категориям Наконец, среди Arthrdpoda. особенно среди раков и насе-
комых, имеются многочисленные как эктопаразиты, так и эндопаразиты.
Для каждого паразита характерно определенное место обитания на
хозяине или в нем Одни паразиты па всех стадиях своего цикла локалн
зуются только в одном органе хозяина, как эго наблюдается у многих
кишечных амеб, жгутиконосцев, грегарин, кокцидпи, некоторых нематод,
эктопаразнтических раков, клещей и насекомых. Изменение локализации
этих паразитов во вр^мя их жизненного цикла в данном хозяине проис-
ходит только в пределах пораженного органа.
У других паразитов в процессе их индивидуального развития проис
ходит смена мост обитания. Например, преэритроцита рные шизонты
Plasmodium vivax разоиваются в печеночных клетках, а их последующие
стадии—шизонты и гамонты—локализуются в эритроцитах. Взрослые
грихинеллы минут н стенке кишечника, а личинки обитают в мышцах,
взрослые Delafondia (Slrongylrdae) паразитируют в книичпике лошади,
а личинки проходят развитие в брыжжссчиых артериях Такая смена
мест обитания па протяжении индивидуального развития паразита за-
висит от разных причин и часто от способности личиночных или других
стадий развития г миграции по телу хозяина. Нередко такая миграция
связана с тем, что окончательное место локализации не совпадает про-
странственно с местом внедрения паразита в хозяина Локализация пара-
зитов на г уз и к своему постоянному месту обитания может быть назва-
на временной. Так, например, локализация личинок анкилостом или
аскарид в кровяном р еле, шчсии, легких, дыхательных путях является
времегпои, а обитание взрослых паразитов в кишечнике считается по-
стоянной локалн шцней.
Помимо тех органов, где паразиты встречаются постоянно и находят
паилучшие условия для своего развития, питания и размножения, они
могут иногда локализоваться и в других органах, где условия, хотя и
менее благоприятны, все же не ирг лятствугот их развитию, например до-
стижению половой зрелости у взрослых паразитов. При дальнейшем
отклонении от нормальной локализации паразит попадает в такие усло-
вия, при которых уже не может развиваться и обычно погибает, йнкан-
сулируясь или рассасываясь в тканях хозяина.
3. ВРЕМЕННЫЙ ПАРАЗИТИЗМ
Уже со времени Лейкарта стало общепринятым деление паразитов,
смотря по времени, проводимому ими в непосредственном телесном кон-
такте с хозяином, на временных и стационарных. Под названием вре-
29
мснпых понимаются такие паразиты, которые недолго остаются на хозяи-
не и не привязаны к нему прочно; они обычно не размножаются и не
развиваются на хозяине, но процесс принятия ниши сочетается у них с
переходом к жизни на хозяине. Иногда временными паразитами считают
такие организмы, которые только на короткий срок посещают хозяина
для приема пищи. В остальной период жизни они не связаны с ним.
Временный паразитизм характеризуется сравнительно слабым кон-
тактом паразита с хозяином, что в свою очередь сказывается в слабо
выраженных адаптациях животного к паразитической жизни. В предо
лах этой категории можно наметить ряд ступеней усиления контакта
между обоими членами ассоциации (паразит —хозяин); от короткой
по срокам связи с хозяином, когда ещ( не наступает истинных парази-
тарный отношений, до стационарною Паразитизма. Наименьший контакт
имеется в тех случаях, когда один организм использует другой только
для получения пиши, как это, например, наб подается у комаров и многих
других кровососов. Дтя них животное, на котором они получают пищу,
не является средой обитания. Всю жизнь они проводят свободно, вне хо-
зяина. Такие фактически свободно живу щи * организмы обычно нс свя-
заны ни с определенной особью животного, ни даже с одним видом
хозяина Комары, например, нападают без особого разбора на человека
и различных животных. Причем прием пищи связан с животным только
у самок комаров, однако их существование в течение долгого периода
можно поддерживать подкармливанием сахарной водой, т. с. без всякого
контакта с животным.
Далее, контакт с хозяином у комаров в течение приема пищи ограни-
чивается часто I—2 минусами. Соответственно со слабостью контакта
между комарами и их хозяевами мы находим у кровососущих комаров
морфологические ад штанин исключительно в их ротовом аппарате, тог-
да как в прочих от ношениях они существенно нс отличаются от других
комаров, не питающихся кровью.
Такой же характер связи с хозяином и \ цейлонских пиявок Паета-
dipsa ceylonica, которые живут па кустах и время от времени нападают
на свои жертвы для сосания крови, не npni't гая к иным способам пита-
ния. Норовые клещи Ornithodorus сосут кровь на животных и человеке
очень короткое время, в течение от нескольких минут до 2—3 часов,
после чего покидают хозяина и возвращаются на обычные места своего
обитания в Природе, где н проводят всю свою жизнь. Как показали ис-
следования Павловского с сотрудниками (1946—I95U), нимфы и взрос-
лые клещи Ornithodorus papillipes могут оставаться, без пищи во внеш-
ней среде, сохраняя жизнеспособность до 9- 14 лет Постельный клоп
(Cimex lectulur'iis) питается кровью человека, домашних птиц и других
животных; взрослые особи клопа сосут кровь лишь около 15 минут, а
личинки всего 2- 3 мсгнуты. В ю же время вне хозяина клопы могут
находиться длительное время: они иногда около года сохраняют жизне-
способность, по получая при этом нищи. У аргазовых клещей и у клопов
наблюдается некоторое приближение в пространственном отношении к
хозяину, так как они поселяются в помещении человека, в гнездах птиц
или норах диких животных.
У иксодовых клещей намечается сближение с хозяином в отношении
удлинения срока пребывания паразита на хозяине во время приема пи-
щи. Особи каждой фазы развития Ixodes riciims — треххозяйного кле-
ща— после кратковременного питания на хозяине отпадают от него и
проделывают дальнейший метаморфоз во внешней среде. Личинки сосут
кровь 3—5 дней, нимфы — 4—6 дней и взрослые (самки) —7- 8 дней.
'Иногда этот срок у самок может удлиняться до 14--20 дней в зависи-
30
мости от физиологического состояния хозяина. Следовательно, общая
продолжительность пребывания клещей на хозяине составляет от 14 до
20 дней и лишь в некоторых случаях около месяца, тогда как ио внешней
среде метаморфоз каждой фазы длится много месяцев, и весь цикл раз
вития клеша растянут от 3 до 6 чет. У двуххозяипого клеша Нyalomma
delritum личинка после насыщения кровью нс отпадает о г хозяина и на
нем же превращав гея в нимфе, которая продолжает сосать кровь и толь-
ко затем отпадает Общая продолжительность сосания кропи личинками
и нимфами составляет 20—22 дня. Самка питается на другом животном
7—25 дней. Весь цикл развития клеща, включая пребывание во внепньи
среде, занимает один год.
У однохозяйного клеща Boophilus calcaratus личинки нападают па
животное, присасываются к нему и, не переменяя места, превращаются
в нимфу, а затем во взрослого клеща. Самки насасываются крови и ос-
тавляют хозяина, чтобы на почве осуществить яйцекладку. На хозяине
клещ проводит в общей сложности 22—30 дней, а полное развитие одно-
го поколения продолжается около двух месяцев. Другой однохозяйный
клеш Hyalomma scupense еще дольше пребывает па хозяине. Личинки
сосут кровь 5—7 дней, затем через несколько дней превращаются в нимф,
не покидая хозяина. Нимфы начинают сосать кровь приблизительно
через 4—7 дней после отрождепия и продолжают сосание 3—6 дней.
Дней через 1U, в*течение которых нимфы находятся в состоянии покоя,
появляются половозрелые клещи. Таким образом, продолжительность
стадии нимфы равна 17—22 дням. На юге СССР личинки нападают на
крупный рогатый скот с конца лета до начала ноября, а их нимфы встре-
чаются всю зиму. Вышедшие из нимф зимой взрослые клещи остаются
на хозяине и в противоположность Boophilus начинают питаться не сра-
зу, а покидают место линьки и ползают в шерсти хозяина, приса-
сываясь лишь спустя 1—2 месяца. Фактически, все фазы развития
клеща пребывают па крупном рогатом скоте, не отпадая с хозяина при-
близительно с октября по апрель.
Блохи гнезда животных (beopsylla и др.) нападают на хозяина
только для кровососаиня на короткий срок и большую часть жизни
проводят в гнезде, а блохи шерсти животных (Clenocephabdes и др)
многократно и часто принимают пищу п, насосавшись крови, не поки-
дают тела хозяина, хотя длительное время могут существовать вне хо-
зяина. Развитие блох (стадии личинки и куколки) протекает во внешней
среде.
Временный паразитизм характерен для некоторых паразитических
раков, а именно: для Argulus и других Branchiura. Карпосды течение
длительного времени держатся на рыбе для приема пищи. Argulus очень
легко покидают рыб*, кровь которой они сосут, после чего в течение
некоторого времени свободно плаваю! в воде, а затем снова могут са-
диться па другую рыбу для продолжения питанья
Рыбья пиявка Piscicola geomet га в общем большую часть жизни
проводит на тело рыбы, легко перебираясь с одной рыбы на другую, а
в период размножения вообще покидает хозяина и откладывает коконы
с яйцами на дно водоемов.
4. СТАЦИОНАРНЫЙ паразитизм
Под стационарным паразитизмом разумеют те случаи, когда пара-
зит на долгое время, иногда на всю жизнь поселяется на или в хозяине,
так что последний является постоянным носителем паразита. Стационар-
ный паразитизм в свою очередь распадается на периодический, когда пара-
31-
зит проводит известную часть своей жизни вне свя щ с хозяином, и на
постоянный, т. с. продолжающийся в течение веси жизни паразита.
Периодический паразитизм
Периодический паразитизм может варьировать в «движимости от
ого, к; кой характер носит периодичность. В общем периодический пара
зитизм встречается значительно чаще постоянного.
Рис. 1<>. Rhabdias hu fonts
Л — паразитическое поколение; В и С--самец и самка свободно гиву-
ШС10 поколении. (По Клаусу.)
1 рот; 2 нррииос кольцо: 3 — ж, пгкпе nojnimc nriivpmic; 4 полония железа;
5 кишка; 6 порошица; 7 яичник; S - семенник; ° спикулы.
Чередование паразитических и о к о л е и и й со сво-
бодно живущими. Этот род паразитизма до известной степени
связан с факультативным паразитизмом, описанным ранее для Alnione-
та appendiculaturn. Rhabdias bujunis (рнс. 16), живущие в легких ля-
гушки, предъявляют собой «ермафродитпос поколение (гермафродн гных
самок) данного вида. Яйца откладываются этими особями внутри хо-
зяина и выводятся наружу через рот или кишечник. Па земли из них
выходя г личинки, дающие свободно живущее раздельнополое поколение.
Самцы оплодотворяют самок, которые производят личинок, снова про-
никающих в хозяина (лягушку). Свободное поколение Rhabdias сильно
походит по строению на свободно живущих нематод, которые принадле-
жат к роду Rhabditis.
Таким образом, у Rhabdias паразитизм является обязательным обра-
зом жизни не для каждого поколения, а через поколение Паразитизм
в данном случае еще не вполне завладел всем жизненным циклом пара-
зита. Нечто аналогичное наблюдается у интересных гидроидных медуз
из группы Narcomedusae. В наиболее простых случаях у Cunoctantha
32
jctonaria (рис 17) свободные половозрелые медузы дают планулообраз
иых личинок, которые забираются в субумбреллярпую полость других
медуз и там паразитируют. Планула превращается в маленькую медузу,
которая вводит свои ротовой хоботок внутрь хоботка хозяина и питается
за счет последнего. Затем медуэка отпочковываст на себе новых медуз,
которые отрываются от нее, становятся
свободными и развивают в себе половые
продукты. Личинки, происходящие от
этих медуз, снова становятся паразитами.
Здесь, следовательно, имеется правиль-
ное чередование полового свободного по
колония с бесполым паразитическим
Паразитический образ жиз-
ни на определенных фаза х
развития организма. При этом
могут быть два случая, которые мы назо-
вем паразитизмом личиночным и парази-
тизм ом и маги иа л ы iы м
Л и ч и п о ч н ы й п а р а з и т и з м
встречается в целом ряде групп. Немато-
ды из сем. Mermithidac во взрослом со-
стоянии живут свободно в воде, куда и
откладывают яйца. Выходящие из яиц
личинки проникают внутрь моллюсков
или насекомых, где проходят в паразити-
ческом состоянии все стадии роста. Дли-
тельность периода свободной жизни силь-
но варьирует: иногда животные копули-
Рис. 17. Медуза Cunortantha octo-
— nar'ta
А — свободная медуза с сильно
редуцированным ротовым хобот-
ком, Л — три личинки С. octonaria
разных возрастов, прикрепившиеся
своими длинными ротовыми хобот-
ками к хоботку медузы Turrltop-
sis nutricola. (TIo Бруксу.)
I, г, 3 — личинки; 4 — ротовой хойоток
Т. nutricvla.
руют вскоре после покидания ими хозяи-
на, иногда же (Mermis nigrescens) самки
достигают половой зрелости лишь после
двух лет свободной жизни. Такой же, при-
близительно, цикл развития имеют и во-
лосатики (Gordiacea).
Среди Copcpoda имеется своеобраз-
ное сем. Monstrillidae (рис. 18 и 19).
Во взрослом состоянии они быстро
плавают в морском планктоне при
помощи сильных гребных ног, ориентируясь своим науплиусовым
глазом Кишечник взрослых Monstrillidae атрофирован настолько, что
они совсем не питаются. Из яиц оплодотворенных самок вылупляются
личинки, пауплпусы, лишенные пищеварительного канала. Они плавают,
отыскивают определенные виды Polychaeta (например, Polydora, Sabna-
cina) и проникают в них. В теле червя они забираются не известным
еще способом в спинной кровеносный сосуд, где, слиняв, приобретают
сначала вид недифференцированного многоклеточного овального тельца.
Затем па будущем переднем конце зародыша постепенно развиваются
два длинных щупальцевпдиых выроста, которые служат животному в
качестве органов абсорбирования пищи. Далее зародыш еще раз линяет,
и под хитиновой кутикулой следующей стадии постепенно дифференци-
руется взрослый рачок со всеми его конечностями и с половыми железа-
ми. Сформированный рачок прорывает кутикулу, покидает хозяина и пе-
реходит к свободной жизни.
Другой пример, представляет сем. Gnathiidae из отряда Isopoda
(рис. 20). Виды этого семейства обладают столь сильным половым ди
3 В А Догель
33
морфизмом, что самцов и самок их долгое время считали за разные
роды. Взрослые Gnalliiidae живут свободно, но не питаются, п поэтому
большая часть их ротовых органов атрофирована. Их личинки, извест-
Рис. 18. Развитие Haemocera danae, паразитического веслоногого рачка
из сем. Monstrillidae
А — полихета Salmacina dysteri с двумя сформированными коиепочит-
нымн личинками в целомической полости; В — науплиус, приникающий
в тело хозяина; С. F. — развитие личиночной стадии и хозяине с образо-
ванием выростов, служащих для увеличении поверхности всасывания;
D— свободно плавающий науплиус. (По Малакену.)
ные под названием Praniza, паразитируют на коже рыб, Они прокалы
вают кожу хозяина своими игольчатыми челюстями и сосут его кровь.
Итак, в обоих приведенных случаях ясно выступает биологическое
значение ларвального паразитизма. Паразитическое питание изобильной
жидкой нищей дает возможность личинкам накопить в себе достаточное
34
количество питательного материала, чтобы обеспечить на всю жизнь не
только существование взрослой стадии, но и возможность развития в ней
половых пролу ктов. Между
стадиями жизненного цикла
Monstrillidae и Gnathiidae на-
ступает разделение жизненных
функций: питание сосредоточи-
вается на стадии личинки, раз-
множение—на стадии взрос-
лою животного.
Регулярным личиночным
паразитизмом обладает гро-
мадное большинство Panlo-
poda. причем личинки разных
представителей этого класса
обнаруживают переходы от
экто- к эндопаразитнзму
(рис. 21 и 22). Мирские пауки
выходят из яйца в состоянии
шестиногой личинки, снабжен-
ной некоторыми провизорными
органами, которые облегчают
личинке нападение на хозяина.
Таковы особые прядильные
железы передних конечностей,
выделяющие две прочные шел-
ковистые нити, которые слу-
жат для прикрепления к хо-
зяину. Сильное вытягивание
когтей конечностей у личинок
Anoplodactylus и шипов на пе-
редних конечностях у личинок
Pycnogonum является приспо-
соблением. которое позволяет
им находиться некоторое вре-
мя во взвешенном состоянии в
воде, пока они не осядут на хо-
зяина. Хозяевами громадного
большинства личинок Pantopo-
da явпяются колонии гидроид-
ных полипов. При этом у одних
из них (Ammothea, Pycnogo-
num) личинки просто прицеп-
ляются к основанию одного из
гидрантов, надкалывают своим
хоботком оболочку ствола ко-
Рис. 19. Самка Нарпю.гга (Mnnstrillid те)
А — самка в последней личвнпчнон и:ктр-
ке, вцнрепаровакпая и > спинного сосуда
погихеты; Н — взрослая свободно живущая
самка. (По Малакену.)
лонин (тэку) и сосут соки и ткани хозяина (в кишечнике личинок можно
заметить проглоченные стрекательные клетки гидранта). У других
видов (Endeis) личинка забирается на гидранта и, погружая свой хобо-
ток в рот полипа, пользуется содержимым его кишечника.
Ряд других видов переходит к еще более глубокому эндопаразитиз-
му. Так, личинки Anoplodactylus линяют, сбрасывают две пары конеч-
ностей и забираются внутрь гидранта, где и проделывают остальной ме-
таморфо’ Личинки рода Phoxichil'diuni проникают еще глубже, а имен-
но: переходят из желудка гидранта в ствол колонии; там они сильно
3*
35
разрастаются, причем вокруг них ствол образует шаровидное вздутие,
напоминающее галлы на растениях. Закончив метаморфоз и сформиро-
вав все свои конечности, животные переходят к свободному образу жиз-
ни. Они ползают по колониям гидроидных полипов и целиком поедают
гидрантов, т. е ведут себя как хищники
Рис. 20. Различные стадии жизненно! о цикла
равноногого рачка Gnathiamaxillarls, в личи-
ночном возрасте паразитирующего на коже
рыб
Я — взрослый самец; Л — молодая свободная
личинка; С — паразитическая личиночная ста-
дия, или так называемая Praniztr. D — взрос-
лая самка. (По Кюкенгалю.)
Рис. 21 Личинка Pycnogo-
пит litorale, сидящая у ос-
нования си драя га Clava mil-
ticornis и сосущая его.
(По Догелю.)
J - гонофоры гидранта; г гид-
рант J личинка.
Рис. 22. Личиночный паразитизм Pantopoda
.4 — шестиногая личинка Ammothea laevis, сосущая гидранта ОЬеЦа,
прицепившаяся к его основанию; Л — личинка Anoplodactylus petio-
latus о гастральной полости гидранта Obelia; С — личинка Endeix
\spinosus, высасывающая гидранта Obelia. (По Догелю.)
Особенно много случаев ларвального паразитизма у насекомых, где
им обладают целые большие группы Так, все наездники (Ichneumoni-
dae) откладывают свои яйца в яйца, личинок и куколок других насеко-
мых Личинки, развивающиеся из яиц, живут в теле своего хозяина до
периода окукления, питаясь его соками и тканями. Вышедшие из куколок
взрослые наездники ведут, однако, свободный образ жизни. То же на-
блюдается у мух (Tachinidae), у оводов (Ocstridae) и у многих других.
Среди моллюсков прекрасный пример личиночного паразитизма
дают Unionidae (рис. 23) Эти пластинчатожаберные моллюски откла-
36
дывают яйца в полость наружной жабры; там на них развиваются осо-
бые, снабженные прикрепительной биссусовой нитью и зазубренной по
краю раковиной личинки, глохидии. Когда мимо моллюска проплывает
близком расстоянии рыба, моллюск выприскнвает через свое сифо-
налъвое отверстие множество глохидиев, часть которых врипепляетси к
Рис- 23. Глохидии Uniom’dae
А—жабра рыбы, зараженная глохидиями; Л— три жаберных лспс
стка рыбы с шестью обросшими эпителием глохидиями на них
С — глохидий Symphynota costata с зазубренными краями створок
(По Кокеру.)
плавникам или к жабрам рыбы. Раздражая эпителий хозяина сомкнув-
шимися крючками сгворок своей раковины, глохидии вызывают его про-
лиферацию Вокруг личинки происходит опухание ткани хозяина, н она
обрастает личинку. Птходясь внутри опухоли, глохидий усиленно питает-
ся соками хозяина, проделывает метаморфоз, принимает окончательную
форму и затем прорывает стенкх опухоли и падает на дно водоема в ви-
де вполне сформированного маленького моллюска. Метаморфоз зани-
мает 1—2 месяца Всю остальною многолетнюю свою жизнь моллюск
проводит свободно.
В рассмотренных нами явлениях ларвального паразитизма обращает
внимание то обстоятельство. что в одних случаях, например у раков и
оводов, личинка накапливает в ое.бе такое количество питательною мате-
риала, которое обеспечивает у взрослых особей развитие половых про-
дуктов б<? дополнительного приема пищи (редукция пищеварительного
аппарата). В других же случаях ларвальный паразитизм не имеет зиа
чення непосредственной подготовки животного к половому размножению.
В связи с этим постларвальный свободный период жизни таких форм
длителен, и взрослые стадии интенсивно питаются, имея вполне разви-
тый пищеварительный аппарат.
Паразитизм и м а г и и а л ьн ы й встречается чаще личиночного.
Он характеризуется тем, что паразитический образ жизни свойствен по-
ловозрелым особям паразита, а личинки являются свободно живущими
организмами. Имагннальный паразитизм встречается у многих нематод,
например подотряда Strongylata. Так, Ancyiostotna duodenale (рис. 24)
в половозрелом состоянии живет в кишечнике человека. Яйца выводятся
37
с экскрементами во внешнюю среду, где из них вылупляется личинка
первой стадии, которая проделывает две линьки и превращается в ли-
чинку третьей стадии Эта последняя является инвазионной для хозяина.
Личинки живут длительно,- вр^мя во влажной почве и обладают значи
тельной подвижностью. Они проникают в хозяина через кожу и продол
Рис 21. Жизненный цикл Ancylostoma duodenate
1 —.елмец; /Т — са мка; С, D, И — первая, вторая и третья стадии личинок (послед-
няя стадия Е инвазионная). (По Чендлеру.)
жают далее свое развитие, претерпевая две линьки во время миграции
по крове носиой системе через легкие в пищеварительный тракт, где и
вырастают в половозрелые особи.
Имагинальный паразитизм очень распространен у паразитических
Copepoda. Из яиц прикрепленного к коже рыбы эктопар.азитического
рачка выходят шестиногие, плавающие в планктоне личинки — науплну
сы, которые, проделав две или более линек, переходят в так называемую
копеподитную стадию и только тогда оседают на рыбу. То Же самое
имеет место и у Saccuana и ее родичей. Выходящий из отложенного сак
кулинон яйца науплиус плавает, растет, линяет, превращается в гак
38
называемую циприсовндную стадию с двустворчатом мантией и только
на этой стадии оседает на краба, который служит саккулине хозяином
(рнс. 25). Типичным нмагинальпым паразитизмом характеризуются и
блохи, личинки которых и куколки живут на свободе и только имаго по-
селяются на хозяине. Среди них многие являются временными паразита-
ми, периодически нападающими на хозяина; но ряд видов может считать-
ся С1ационарны.ми паразитами, так как они приспособлены к длитель-
ному пребыванию на хозяине. Так, например, самки блох Vermipsylla
alacurt и V. dorcadia, паразитирующие па овцах и других копытных в
тирах Тянь Шаня, осенью присасываются к хозяину и остаются на нем
в течение всей зимы (5—6 месяцев), питаясь кровью. По мере созре-
вания яиц самка увеличивается в длину от 2 до 16 лл. Япца отклады
ваются в шерсти животного и затем осыпаются на землю. Весной самки
отмирают, и в течение всего теплого пеонода во внешней среде происхо-
дит метаморфоз блох. Песчаная блоха Tutiga (Sarcopsylla) penetrans,
живущая в тропиках, не только присасывается после спаривания на дли-
тельный срок к коже человека и животных, но и целиком погружается
в псе, ьбуравливаясь при помощи верхних челюстей. Наружу остается
торчать лишь кончик брюшка. При этом брюшко разрастается до раз-
меров горошины, и в нем созревает огромное количество яиц Яйца по-
надают па землю, где и происходит дальнейший метаморфоз блохи.
К категории имагннальпого паразитизма непосредственно примы-
кают и тс многочисленные случай, когда во внешнюю среду выводятся
только япца паразита пли первые личиночные стадии, тогда как осталь-
ные стадии метаморфоза и взрослые ос-обл ведут паразитический образ
жизни. При этом Bipoc iue н личинки могут обитать в одном пли разных
хозяевах Примером могут служить многие Ascaridata, у которых в яй-
цах, выведенных во внешнюю среду, развивается личинка второй стадии,
а последующие личиночные стадии формируются уже в теле хозяина, где
они превращаются во взрослые формы, У цестод во внешней среде нахо-
дится яйца с онкосферой или корацидпй {Diphyllobothrium и др ). а
взрослые и личинки паразитируют обычно в разных хозяевах У нематод
Protostrongylus и др. во внешней среде развиваются только личинки
первой стадии, а затем они проникают в промежуточного хозяина. Во
всех этих случаях свободная стадия паразита обеспечивает возможность
ЗараиКння новых особей хозяина.
Паразитизм, повторяющийся на разных фазах
развития в течение одного поколения. Такого рода яв-
ления наблюдаются, например, у иксодовых клещей, которые имеют три
пост эмбриональных фазы развития: личинка, нимфа и имаго (рнс. 26).
Как было отмечено выше, паразитизм каждой фазы прерывается свобод
нымн периодами.
Повторность паразитирования в течение одного
жизненного цикла у р а з и ы х п о к о л е н и й. Этот тин наблю-
даемся у паразитов, обладающих наиболее сложным жизненным циклом,
а именно у Trematodes.Жизненный цикл трематоды представляет собой
очень сложную гетерогонию. У них в один и тот же жизненный цикл
входит очень часто не. менее трех разных поколении: одно гермафродит
ное дззрослые двуустки) и два состоящих из партеногенетических самок
(спороцисты, редин).
Соответственно этому в жизненном цикле трематоды мы различаем
несколько паразитических периодов, прерываемых периодами свободной
жизни. Взрослые особи гермафродитного поколения паразитируют в поз
воночном хозяине Отложенные ими яйца дают в воде мнрацидиев
(1-я свободная фаза). Мираииднн проникают в моллюсков, где превра
39
Рис. 25. Жизненный цикл Sacculina carcini
А — науплиус; В — цнприсовая личинка; С — прикрепление циирнсовой личинки сво-
ими антеннами к панцирю краба н отбрасывание ею всех локомоторных придатков;
D— линька циприсовон личинки и превращение ее н нерасчлененный овальный ме-
шок— контрогон; Е — проникновение контрогона к тело краба. (По Де.тяасу )
таится в сппроциоы и редин. В редиях формируются цсркарии (т. « бу
дущне гермафродиты). Цсркарни покидают моллюсков и обычно выхо-
дят в воду (2 я свободная
фаза), где отыскивают вто-
рого промежуточного хозяи
на, чаще всего какое-нибудь
членистоногое или рыбу. По-
ступая во второго промежу
точного хозяина, церкария
сбрасывает хвост и стано-
вится мстацеркарисн. Про
межуточный хозяин боль
шей частью проглатывается
Рис. 26. Ixodes ricinus, сталии развитии
А - шест niiot ая личинка; И — нимфа; С — взрос-
лая самка; в А следу#т отмстить отсутствие
стигм, в Н — наличие стигм, но отсутствие по
левого отверстия. (Л и В — по Брумпту. С — по
Павловскому.)
I [.niajii.iioe отнврстн*; 2 - стигмы; 3 — половое отверстие.
позвоночным и перевари-
вается. причем метацерка
рия освобождается, доби -
раезся до места своей окон-
чательной локализации и
вырастает там в половозре-
лою гермафродита. Следо-
вательно, переход в окончательного хозяина совершается в большинстве
случаев без прерывания паразитического образа жизни
Постоянный паразитизм
Постоянный паразитизм кара.ктеризуется тем, что паразит во внеш
ней среде совершенно не может существовать.
Постоянный паразитизм в одном хозяине. Хорошим
примером являются вши. живмцие всю свою жизнь на хозяине и питаю-
щиеся на всех стадиях развития исключительно его кровью. Тот же ха-
рактер носит паразитизм у пухоедов и власоедов, которые всю жизнь
держат* я на теле хозяина и даже япца свои, подобно вшам, приклеивают
к сто перьям или шерсти. К постоянным паразитам относятся и чесоточ-
ные зудни (Acarus stro, рис. 27), паразитирующие в коже человека и
некоторых млекопитающих. Самки живут в толще кожи, прогрызая в пен
ходы в которых откладывают яйца. Развившиеся нз яиц, клещи но дости
женин половой зрелости (что происходит на стадии телеоннмфы, пред
шёетвующен последней линьке) выползают на поверхность кожи и копу-
лируют. Самцы вскоре умирают, а телеоннмфы снова внедряются в кожу,
линяют еще один раз, превращаясь во взрослых самок, и вскоре начина-
ют откладывать яйца. Жизнь такой самки от момента начала откладки
яиц продолжается 6 8 недель, после чего самка умирает.
Постоянные паразиты имеются и среди простейших. Так, Trypano-
soma equiperdum передается от одного хозяина к другому того же вида
во время полового акта и вне своего хозяина (лошади) ни в какой ста-
дии существовать не может.
Постоянный паразитизм, сопровождаемый сме
ной хозяев Эга интересная категория заключает в себе паразитов,
которые ни одного .момента свой жизни не проводят на свободе, а вместе
стом, параллельно с чередованием поколений, регулярно сменяют своих
хозяев. Особенно много примеров подобного рода среди простейших, где
к данному типу принадлежат все Haemospor.dia. Так, у малярийного
паразита (Plasmodium) бесполое размножение протекает в человеке, по-
ловое—в комаре, причем передача от одного хозяина к другому север
шается без прохождения свободной фазы
41
Сходным образом происходит передача трипанозом через мух цене
(Glossina) л слепней от одного хозяина к другому или лей шмаиий через
москитов. В этих случаях паразит передается при питании одного хозяи-
на— переносчика кровью другого — позвоночного хозяина. Schellackia
boliuari—паразит ящерицы или Hepalozoon inuris— паразит мышей пе-
редаются от хозяина к хозяину чере- га тазовых клещей. При кровососа-
нии клеит получают паразита от позвоночного хозяина, а последний
заражается, поедая таких клеццей.
Рис. 27. Жизненный цикл ч| соточнЛ о зудня (Acartis siro)
А— самка с брюшной стороны; В— сямкл в ходе, проделан-
ном ею п коже человека, и отложенные яйца; С -самец
с брюшной стороны; D— личинка с тремя парами ног.
(11о Бнльдингу.)
Здесь огромной лабораторией природы производится каком постоян-
ный, не прерывающийся в течение тысячелетий опыт передачи паразитов
от одного организма другом’.
Из паразитических червей постоянный паразитизм наблюдается у
трихины (TriMnella spiralis), которая передается от одного хозяина
другому в виде личинки, инцистированной в мышцах хозяина во время
поедания новым хозяином этого последнего. Попавшая в кишечник ли-
чинка достигает там половой зрелости и копулирует. Самцы после копу
ляцпн гибнут, самки же внедряются в стенки кишечника п отрождают
в капиллярах субмукоэы множество личинок, которые разносятся током
крови и постепенно проникают в мышечные клетки хозяина, несколько
вырастают и инкапсулируются, переходя в инфекционную стадию покоя.
Постоянный и а р а з и т и з м, с о п р о в о ж д а е м ы й смело и
хозяев и передачей паразита в последующих поколе-
ниях одного н того же хозяина. Такой паразитизм, назы-
ваемый обычно трансовариальным, может, очевидно, иметь место
лишь в случае передачи нарашта от самок хозяина непосредственно их
потомству через зараженные яйцевые клетки. Примером подобного рода
могут служить кровепаразиты домашних животных сем. Piroplasmidae
(Babesiidac), которые распространяются иксодовыми клещами. Самки
клещей заражаются пироплазмами во время кровисосаиня на больных
животных и передают возбудителя своему потомству. По данным ряда
авторов (Regendanz u. Rcichcnow, 1933; Dennis, 1932; Полянский и Хей-
син 1959, Муратов и Хсйснн 1959 и др ), из кишечника клеща паразиты
приникают в гемоцсль, где приобретают булавовидную форму и оттуда
проникают в развивающиеся ооциты клеща. В дальнейшем булавовид-
ные формы пироплазм оказываются в отложенных самкой яйцах и далее
-12
в развившихся из них личинках и последующих фазах метаморфоза
клеща (рис. 28). Следовательно, поколение личинок, нимф и взрослых
клещей, разнившихся из яиц, отложенных зараженной самкой, может
быть инвазионным и при нападении иа хозяина для кровососания эти
формы заражают его пироплазмами.
Трансовариальная передача известна для Piroplasma bigeminum у
Boophilus calcaratus, для Babesiella bovis у Ixodes ricinus, для Piroplas-
ma caballi и P. canis из Dermacentor п других клещей, Piroplasma ovis
и Babesiella ovis у Rhipicepfialus bursa и т. д. Работами Брумнта
(Brumpt, 1937), а также Маркова А. А. и его сотрудников (1952—
1957 гг.), была доказана повторная трансовариальная передача пиро-
плазм на протяжении нескольких поколении клещей. Например, Р. canis
Рис. 28. Булавовилвпя стадии Piroplasma bigeminum
nt яйца Boophilus calcaratus. (Оригинал.)
в клещах Rhipicephalus sanguineus передается па протяжении 5* поколе
ний, Р. caballi в клещах Hyalomtna plumbeum— 11 поколений, В. ovis
в клещах Rhipicephalus bursa—1-1 поколений (Марков, и др., 1957;
Абрамов, 1955, 1957).
Сходный пример, по факультативной трансовариальной передачи
наблюдается у Nosema bombycis. Когда в 60-х годах XIX в. обеспокоен-
ные эпидемическим характером пебрины шелковичного червя хозяева
шелководческих предприятий Франции обратились за помощью к вели-
кому Пастеру, последний выяснил следующую любопытную особенность
этой болезни. Сильно зараженные паразитом черви погибают, но слабо
инфицированные выживают, окукляются и дают бабочек, у которых во
всех внутренностях, между прочим и в яичниках, имеются споры Nosema.
Вследствие этого бабочки откладывают зараженные яйца, н после вы
лупления из них гусениц болезнь разгорается в следующем поколении
заново. Основываясь на этой особенности, Пастер предложил проводить
отбор (или «гренаж») яиц шелкопряда, что спасло шелководство от
громадных потерь. Следовательно, \ Nosema, кроме обычного способа
заражения новых особей хозяина через посредство спор, вываливающих
ся наружу из погибших червей, имеется еще иной способ распростране-
ния— непосредственная передача спор следующему поколению хозяина.
43
ГЛАВА IV
РАСПРОСТРАНЕНИЕ ПАРАЗИТИЗМА В ЖИВОТНОМ МИР!
Паразитизм есть явление, необычайно широко распространенное
в природе, причем распределение паразитов в разных группах живот-
ного мира представляет некоторые интересные особенности (Кнорре,
1937). Веглый обзор данных по разным группам показываетследукнцук
картину.
В типе простейших (Protozoa) все пять классов имеют паразити-
ческих представителей, а два из них состоят целиком из паразитов
В прочих классах имеется несколько чисто паразитических отрядов
или подклассов (Hyperniastigida, Opalinida, F.ntodiniomorpha). Самые
крупные группировки паразитических простейших имеют еле д ующий
состав.
В классе Sarcodina крупную паразитическую группу составляю”
амебы. Так, в составе отряда АпюеЫпа Мёллер (Nxiller, 1922) насчиты-
вает около 70 паразитических видов. В других отрядах Sarcodina пара- .
зиты составляют редкость [Pseudospora средн Heliozoa и некоторые
другие). Принимая во внимание, что после сводки Мёллера появился
ряд описаний новых видов пара штнчсских амеб, например из термитов,
можно считать, что пока известно около 100 видов параштнчсских
Sarcodina.
Гораздо труднее даже приблизительно подсчитать количество па ра
знтических Mastigophora. Паразитические жгутиконосцы имеются,
прежде всего, в отряде Pcridinea (рис. 29). Шаттоп (Chatton, 1919)
дает список 43 типичных паразитических Per-dinea и представителей
рода Haplozuon, описанного Догелем л принадлежащего к тому ж<
отряду.
В настоящее время можно считать 'iik.io известных паразит
чсских Peridinea достигшим 60 видов. В отряде Euglenoidida копстати
ровано паразитирование в кишечнике головастиков одного вида Eug
lena, Euglenamorpha hegneri (Wenrich, 1924), а также Hegneria Icpto-
dactyli. В общем паразитический образ жизни в этом отряде представ
ляет редкое исключение.
Напротив, в отряде Protomonadina паразитизм широко распро
странен Венной (Wcnyon, 1926) в своем большом руководстве приводит
около 300 видов, в том числе свыше 70 видов Trypanosoma. С то пор
44
количество описанных форм значительно увеличилось, так что для дан
ного отряда можно принять не менее 350 видон паразитов.
Гораздо лучше изучены представители Polymastigida и Hypermas
igida, встречающиеся в термитах и тараканах, вследствие наличия
Рис. 29. Паразитические Peridinca
Л — ClausocaJantis arcuicornis с тремя особями Blastodinium
pruvoti. паразитирующими в желудке: В — бесполое размно-
жение Blastodmium, внутри его оболочки видны крупная
клетка и множество мелких, из которых произойдут жгути-
ковые зооспоры; С н D — одноклеточная и многоклеточная
стадии Haplozoon Птшге; Е— Н аг та turn; Е — El delicatu-
lum. (Д и В — но Шаттону, C — F—ио Догелю.)
I — стилет; 2 — ллпасиыс стилеты: 3 — клетки ратйоягсинн; 4 - осоСн Bla-
stodinium pruvuti
сводки Кирби (Kirby, 1937). До сих пор изучено лишь около трети
общего числа термитов, содержащих Mastigophora. причем в 140 видах
термитов обнаружено 140 разных видов Polymastigida и 85 видов
Hypermastigida.
45
Целиком паразитической группой являются Opalinida, живущие
исключительно в кишечнике амфибий. К ним относится не менее 150
зидов.
Таким образом, в общем, в классе Mastigopl юга можно насчитать
не менее 750—800 паразитических представителей.
Рис. 30- Различнее представители инфузорий из отряда Fntodinio-
morpha, паразитирующие в кишечнике травоядных млекопитающих
А — DiplO'iinium maggii из рубца .кнлчных; С — (.alo scolex came-
linns из желудка верблюда; С—Ophryoseolex caudatus из рубца
жвачных; D — Elephanthophilus из слепой кишки индийского слона.
(Л — С—но Догелю, D — по Кофонду.)
I — околоротовэп ч-чна мемйрапелл; 2-сял пин зона мембранелл; 3 -мнкроиуклеус;
4 - лояолиипмы1ые зоны иембранелл: 5 уок|л«1н1слы1ые вакуоли: 6 — анальное
огиерстна, 7 — находящиеся в хвйетоооЯ лонасгн нк.чоченш неизвестною назиачешн;
S — макронуклсус; 9 — скелетные пластинки.
В классе In.'usoria крупной, целиком паразитической группой яв-
ляются Епtodiniomorpha с 200 видов (рис. 30). В отряде Holotricha
почти все паразитические формы сосредоточены в трех группах. Одна
из этих групп паразитирует в кишечнике млекопитающих и охватывает
около 190 видов, вторая состоит из морских форм (Apostomata),живу-
щих частью на различных морских беспозвоночных, частью внутри их;
сюда по сводке Шаттона и Львова (Chattori et Lwoff, 1935) относится
до 40 паразитических видов. Третья группа, а именно: подотряд без-
ротых (Astomata), содержит около 150 видов. Инфузории Thigmotricha
46
содержат по крайней мере 80—100 видов, живущих на жабрях различ-
ных моллюсков. В отряде Ik-temtricha в настоящее время отмечено
свыше 80 паразитических видов, тогда как отряд Hypotricha содержит
очень мало паразитов, всего около 10 видов (Кггопа с Hydra и не-
сколько инфузорий из кишечника морских ежей). Из кишечника морских
ежей известно не менее 50 видов инфузорий, относящихся к отрядам
Holotricha и Oligotricha.
Среди Peritricha паразитируют только некоторые (около 50 видов)
представители семейства Urceolaridae. Наконец, в подклассе Suctoria
известно всего около трех десятков видов, паразитирующих на иле
в ресничных инфузориях, а также на рыбах. Всего среди инфузорий
имеется не менее 900 паразитических видов.
Большое количество видов включает чисто паразитический класс
Sporozoa. Отряд Gregarinida содержит около 500 видов, отряд
Coccidia —350—400. отряд Haemosporidia и другие группы кровепара ш-
тов неизвестной природы (Piroplasmidea, Theileriidca) имеют около
200 видов. Большое количество видов включает также чисто па-
разитический класс Cnidosporidia: к отряду Microsporidia относится
свыше 200 видов, а к отряду Му cosporidia принадлежит больше
700 видов.
В общем же весь тип простейших на 20000 видов содержит при-
мерно 3500 видов паразитов.
Своеобразные, относящиеся к классу Mesozoa (отряды Dicyemida
и Heterocyem’da) паразиты почек головоногих моллюсков включают
40 видов. Группа Orthoneclida (положение которой в системе остается
неясным), паразитирующая преимущественно в моллюсках, а также
в иглокожих и некоторых других беспозвоночных, включает
15 видов. 1 • 1 < г / I1
Тип губок (Spongia) образован исключительно свободно живу-
щими видами.
В типе кишечнополостных (Coelenterata) паразитические формы
встречаются разбросано и единично. Можно указать на Polypodium
hydrirjorme из яиц стерляди, на два десятка Narcomedusae (Cunina,
Cunocantha и др.), из гастроваскулярной системы других медуз и па
паразитическую ктенофору Gastrodes, живущую в полости тела сальп.
Следовательно, весь тип кишечнополостных едва насчитывает около
25 паразитических видов.
Средн 24 классов червей 13 имеют паразитических пред-
ставителей, а 6 классов целиком состоят из паразитов (Tremato-
des, Monogenoidea, Ccstodes, Acanthocephala, Nematomorpha, Myzo-
stomida).
В классе Turbellaria из 81 семейства 21 имеет паразитических
представителей или же 8 семейств содержат исключительно парази-
тов; общее число паразитических видов этого класса превышает 80.
Дигенетические сосальщики (Trematodes) представлены
примерно 4000 видов. К Monogenoidea относится пе менее
1100 видов.
Класс Cestodes. по Фурману' (Fuhrmann, 1908), состоит из
1500 видов. Класс Acanthoccphata имеет около 500 представителей
(Meyer, 1933; Петроченко, 193*5) Из 10 000 Nematodes около 5000
являются паразитами, a Nematomorpha охватывают менее 100 видов.
Класс Nemertini содержит всего около десятка паразитических форм
(Malacoodella и Carcinonemerteg:').
Очень малочисленны паразиты и среди Polychacta, где мы имеем
опять-таки около 20 паразитических форм (Ichthyotomus, Histriobdella
47
и некоторые Eunicidae). Ь группе Oliguchaeta имеется всего около
40 паразитов (главным образом принадлежащих к сем Branchiobdel
lidae, которые живу г на жабрах раков). Переход к эк i опара штизму
произошел в классе пиявок (Hirudiiiea), так. как из 300 пиявок около
250 ведут паразитический образ жизни. Группа Myzostomida, пред-
ставляющая собой продукт видоизменения Polychaeta под влиянием
паразитизма, содержит III видов. Последним классом червей, имею-
щим паразитических представителей, являются коловратки (Rotato-
ria), среди которых насчитывается около 20 паразитов.
Крайне беден паразитическими формами тин Mollusca, содержа-
щий 3 паразитных Lamellibranchia и около 100 ведущих паразитиче-
ский образ жизни Gastropoda.
Необычайно богатый видами (больше 1000 000) тип Artliropoda
дает в общем довольно значительное число паразитических форм,
хотя в нем имеется всего один небольшой (75 видов) класс Pvntasto-
mida, целиком состоящий из паразитов. Представители другого
класса, Pantopoda, за редкими исключениями характеризуются личи
ночным паразитизмом на гидроидных полипах или внутри их. P«nto
poda имеется около 330 видов. Рассматривая но порядку прочие
классы, мы видим, что в классе раков (Crustacea) имеется одни цели
ком паралитический отряд Bcanchiura (около 70 видов;, а в отрядах
Copepoda, Cirripedia. и Isopoda процент паразитических форм весьма
значителен. Так, например, из 10 подотрядов Copepoda — 8 чисто
паразитических. Два подотряда Cirripcdia (Rhjz.ocephala н \scotliora-
cida) тоже сплошь состоят из паразитов. В общем можно считать, что
класс раков имеет около 2000 паразитических представителей
Большое количество паразитов насчитывает класс Arachnoidea
в своем отряде клещей (Acarina). Лишь в одном подотряде (Or.ycho
palpida) паразитов нет К подотрядам Mesostiginata и Ixodides при-
надлежат наиболее важные в медико-ветеринарном отношении клещи,
В этих подотрядах насчитывается до 1500 паразитических видов, среди
которых около 400 принадлежит к подотряду Ixodides. В подотряде
Sarcop'iformes, куда относятся чесоточные зудни и перьевые клещи
птиц, насчитывается свыше 900 видов. Подотряд Troinbidiforrm s
включает паразитов растений (Eriophyidae, Tetranychidac), число
видов которых достигает 800, и паразитов животных (личинки боль-
шинства Hydracarina и Trombiculidae) числом около 1000 видов.
Таким образом в отряде клещей всего можно насчитать около
3500 паразитов животных и до 800 паразитов растений.
Мелкие классы Palaeostraca, Tardigrada и Protracheata, а из
более крупных — класс Myriapoda паразитов не содержат Что касает-
ся громадного по объему класса насекомых (около 1000000 видов),
то в нем имеются как полностью паразитические отряды (вши, пухо-
еды, блохи, Strepsiptera), так и отдельно разбросанные в разных
отрядах паразитические формы или группы таковых (наездники,
тахины, оводы) При этом к эндоьаразитизму способны лишь личиноч-
ные стадии насекомых, тогда как эктопаразитнческий образ жизни
могут вести и взрослые насекомые (вши, блохи и пухоеды). Больше
всего паразитов в группе наездников, где в настоящее время насчиты-
вается до 40 000 видов. Кроме того, в том же отряде перепончатокры-
лых, но в других его группах (Clirysidoidea, Mutilloidea), имеется еще
5400 паразитических видов. Пухоедов известно не менее 1500 видов,
блох — около 800, Среди двукрылых большую группу составляют
комары (CuJicidae), которых мы решили с оговорками относить
к паразитам. Имеется 1400 видов кровососущих комаров, в том числе
48
164 из группы Anophelini, куда относятся переносчики малярии
Следовать тын», всего насчитывается не менее 50 00U видов паразитиче-
ских насекомых.
В типе иглокожих (Echinoderniata) паразитических форм ист,
если не считать нескольких офпур, которые являются квартирантами
.морских е,кей. Эго очень мелкие формы (диаметр диска от 1 до
5 ,и.и), оживленно ползающие среди игл ежа, по выражению Мортен-
сена (Mortensen, 1933), наподобие обезьян в тропическом лесу. Спе-
цифическое приспособление этих офиур к эшии'кному образу жизни
сказывается в том, что иглы на последних члениках их лхчеп пре-
вращаются в крючки (рнс. 31, В), помогающие мм при лазании по
иглам ежа, а копны л* чей приобрели особую гибкость, напоминая
ценим хвост обезьян. Кроме того, почти все эти виды офиур (3 вида
из 1; обладают совершенно несвойственной офнурам, но встречаю-
щейся иногда у паразитов особенностью длительного спаривания
полов. При этом карликовый самец постоянно держится па диске
самки, прижавшись ртом к ротовому отверстию последней (рис. 31,71).
Рис. -31. Офиурл Ntiniiophiiirn lagani, живущая на иглах
морских ежей
.4—самка офиуры с прицепившимся к ротовой поверхности
диска карликовым самцом И—конец луча офиуры с превра-
тившимися в крючк* иглами. Jlo Мортенсену).
Вся совокупность этих признаков показывает, что офиуры (\ иши-
phhiru lagani, Amphilucus androphorus) представляют собой живот-
ных, специально приспособившихся к эпиойкин, з отсюда лишь один
шаг до паразитизма. То же отсутствие паразитов наблюдается и среди
Chordata. Мелкие рыбки родов Fieraster, Oxibelus и Euriialyopliis,
встречающиеся в клоаке и водяных легких голотурии, являются
квартирантами, а не паразитами. Миксины же, вбуравливающиеся
в волость тела рыб, скорее хищники, чем паразиты.
Наконец, среди рыб различные горчаки (Rhodeus, Acanthorhif-
deus) откладывают свою икру внутрь раковин пресноводных дву
створчатых моллюсков, на их жаберные пластинки. Выходящие из
икры личинки горчаков доволы+р долго живут па жабрах моллюска
и ра (вивают ряд приспособлений, которые заставляют считать их
комменсалами, или, ио меньшей перс, хорошо адаптированными
квартирантами своих хозяев I ак, у обыкновенного горчака (Rhodens
sericeus) развиваются позади головы два уховидных придатка, кото-
рыми он зацепляется за жаберные нити хозяина. У Aianthorhodeus
asrnussi па теле личинки развиваются особые «первичные чешуйки»,
торчащие вершинами наружу и помогающие передвижению личинки
4 В. Л. Догель
49
но поверхности жабр, наподобие шнпнков, усаживающих поверхность
тела многих сосальщиков.
Хищничество, приближающееся к паразитизму, имеет место
у летучих мышей Южной Америки из рода Desmodus (Acosta u-
Romana, 1938). Они нападают ночью на крупных млекопитающих,
прорезают их кожу острыми верхними резцами и сосут выступающею
кровь. Передняя часть кишечника этих летучих мышей специально
приспособлена к такому образу питания: острые резцы, узкий нише
вод, особый придаток, служащий для накопления крови. Сходство
с другими кровосос, щнм и паразитами, например из насекомых,
выражается в том, чго Desmoaus являются переносчиками некоторых:
трипанозом лошадей (Trypanosoma equimun и др.), наподобие того
как слепни переносят Т. evansi — возбуди геля сурры.
Итак, общее, число известных в настоящее время паразитических
животных равно не менее 65—70 тыс. Принимая общее число видов,
известных для науки животных, близким к 1200 000, мы видим, что
паразиты составляют средн них не менее. 6 7%, Арндт (Arndt, 1940)
дат сравнение числа паразитов и свободно живущих животных для
фауны Германии, показав при этом, что на самом деле процент пара"
зитов еще более велик. Арндт насчитал для всей фауны Германии
приблизительно 40 000 видов. Относя к паразитам также комменсалов
(500 видов) и кровососов (комары, мошки и др. — всего 250 видов),
он получил 10 000 паразитических видов, т. е 25% фауны. Из этого
числа около 60% падает на долю перепончатокрылых (наездники)
Отсюда как нельзя лучше видно, какую огромную роль играют пара
зиты во всем животном царстве, взятом в целом.
Сведения по наличию паразитических видов в разных системати
ческн.х гр1 плах 1В<дсны к; мн в схему (рис. 32).
Такова общая картина распространения паразитизма в животном
мире. Мы видим что к паразитическому образу жизни переходят
самые различные группы, не имеющие между собой ничего общего.
Однако, несмотря на такую кажущуюся случайность распределения
паразитов по филогенетическому древу, в этом распределении все же
намечаются некоторые закономерности.
Прежде всего бросается в глаза резкое различие в отношении
к паразитизму между Deuterostomia и Protostoinia. Среди 17 классов
Dcuterostomia не паразитирует ни одни. Между тем, пз 37 классов
Protostomia 20 классов, т. е. более половины, имеют паразитических
представителей, а 7 — целиком паразитические (Trematodes. Mono-
genoidea, Cestodes, А< aninoucphala, Nematomorpha, Myzostmnida,
Pcntastomida). •
Далее интересно, что совокупность тех групп среди Protostomia.
которые обнаруживают ряд черт, сближающих их с Dcuterostomia.
а именно: классы Bryozoa Brachiopoda, Phoronidea, Kamptozoa, пара-
зитов не содержат и в этом смысле опять-такп приближаются
к Dcuterostomia. Итак, признак отсутствия в их числе паразитических
представителей можно включить в общую характеристику большой
группы животного царства, а именно: Dcuterostomia.
С известной осторожностью можно сделать вывод, что наряду со
многими другими факторами переход к паразитизму обусловливается
и высотой организации той пли иной группы. Среди билатеральных
животных больше всего паразитов в низшей группе— в типе плоских
червей. Такая же тенденция наблюдается нередко и внутри отдель-
ных классов. Так среди Gastropoda паразиты имеются только в низ-
50
Рис. 32. Распространение паразитизма в разных группах животных
На генеалогическом древе черным цветом изображены классы, целиком состоящие
из паразитов; серым — классы, содержащие наряду' со свободно живущими и пара-
зитические формы; белым — классы, образованные исключительно свободно живущими
видами. (По Кнорре, с изменениями.)
4*
51
тем отряде — Prosobranchia. В классе Oligochaeta есть два отряда:
Arckioligochaeta — низшие и Neoligochaeta— высшие. Среди Archi-
oligochaeta из 5 семейств паразиты содержатся в 4, а из 18 семейств
Neoligochaeta — только в 2, и притом самых низших. Далее, в классе
раков из 5 отрядов низших раков (Tntomostraca) паразитические
формы имеются в 3 (Copepoda, Branihiura. Cirripedia). а из 10 отря-
дов высших раков (Malacostraca)—только в 2 (Isopoda и Amphi-
poda). Не будем, однако, закрывать глаза на то, что среди насекомых
наибольшее число паразитов дают высшие отряды.
Конечно, простота организации есть лишь один из способствую-
щих переходу к паразитизму факторов. В самом деле, средн низших
многоклеточных (Spongia, Coelenterata) паразитов почти пет, а часто-
та паразитизма среди Protozoa объясняется не только простотой их
организации, но и малыми размерами.
Некоторые закономерности наблюдаются и в распределении % но
филогенетическому древу тех животных групп, которые преимуще-
ственно служат хозяевами для различных паразитов.
Рнс. 33. бокоплавы Gammarux locustu и Cyamus
А—бокоплав Canimurus lociisfa и вынутый из такою же бокоплава
плероцеркоид цесюды Diplocotyle при одинаково*! увеличении; В — бо-
коплав Cyamus, паразитирующий на коже китов. (4 по Догелю и Вол-
ковой, Н— по Кюкенгалю.)
Так, Protozoa вследствие своих малых размеров лишь в очень
редких случаях являю! ся хозяевами для паразитов (некоторые грега-
рины для Metchnikove.llidae, некоторые ресничные инфузории — для
сосущих инфузорий). По тем же причинам япца, т. е. одноклеточные
стадии Metazoa, лини. редко служат пристанищем для паразитов,
и иритом главным образом из типа простейших: некоторые кокцнднп
паразитируют в яйцах червей (у T/ialassenia) и клещ* й, некоторые
Myxosporidia найдены в икре- рыб. Из многоклеточных животных толькд
наездники дают длинный ряд паразитов, уничтожающих яйца насеко-
мых и пауков.
Почти столь же редко попадаются паразиты у Coelenterata (кроме
ларвальных стадий Pantopoda в гидроидных полипах). Черви служат хо-
.ясвамп почти исключительно для простейших, очень редко для Crustacea
и нередко для личинок черве.й (мстацеркарнн сосальщиков, личинки
нематод). Моллюски имеют в общем мало паразитов (Coccidia,
не лногие виды инфузории, изредка раки или клещи), если нс считать
партеногенетических поколений трематоды, для которых моллюски
служат обязательным первым промежуточным хозяином. Водные
Arthropoda, т. с. главным образом раки, явтяются хозяевами для
простейших (грегарины, микроспоридии, инфузории), а также для
паразитических раков (для Cirripedia и Isopoda); кроме того, в раках
паразитируют личинки сосальщиков и ленточных глистов, причем
в некоторых случаях паразиты (плероцеркоиды Diplocotyle) в
6—10 раз превышают в длину размеры хозяина (Gammarus locusta,
рис. 33). Наземные Arthropoda и в том числе насекомые используются
простейшими, личинками плоских червей и нематод, а главным обра-
зом личиночными стадиями паразитических насекомых.
Иглокожие характерны гем, что почти исключительно к ним при-
урочены все паразитические Gastropoda и Myzostomidae; кроме того,
в них встречаются простейшие (грегарпны, инфузории и др.) и изредка
раки и ресничные черви (из Rhabdocoela). Среди хордовых мы имеем
две резко разграниченные группы. Низшие хордовые ведут себя
относительно паразитопоси гельства подобно беспозвоночным. В них
есть грегарпны (в Ascidiae), на них (подобно тому как на Copepoda)
паразитируют некоторые Peridinea, а из многоклеточных они дают
убежище комменсальным рачкам (Copepoda). Напротив, высшие хор-
довые, т. е. позвоночные, показывают совсем иную картину. Эта
группа наиболее щедро населена паразитами. Помимо самых разно-
образных простейших (кроме грсгарип), позвоночные служат оконча-
тельными хозяевами для всех (за редчайшим исключением) Tremato-
des, Monogenoidea, Cestodes, Acanlhocephala и для огромного боль
шинства паразитических Nematodes. Кроме того, Vertebrata являются
исключительными окончательными хозяевами всех Pentastomida.
Наконец, водные позвоночные, а именно рыбы, очень часто несут ла
себе раков (Copepoda, Branchiura, Isopoda), а наземные — насекомых
(вши, блохи, клопы, пухоеды) и клещей (перьевые клещи, чесоточные
зудни, нксодовые клещи и др.).
Таким образом, из приведенного перечня мы видим, что Protozoa
и Vertebrata представляют собой две противоположности в смысле
использования их паразитами. Малые размеры и малое количество
питательного материала, которое может представить отдельная особь
Protozoa, тормозят заселение * простейших паразитами. Напротив,
крупные размеры и большие запасы живой пищи, предоставило
мые позвоночными, привлекают к ним большое разнообразие па-
разитов.
Далее, просмотр того же перечня обнаруживает известную связь
некоторых свободно живущих и паразитических групп: некоторые
группы паразитов как бы специально приспособлены к определенным
группам хозяев. Такая связь замечается, например, между грегари
нами и всеми беспозвоночными; между гидроидными полипами
н пантоподами; между моллюсками и партеногенетическими поколе-
ниями сосальщиков; между некоторыми группами свободных и парази-
тических раков; между Echinodermata, с одной стороны, и паразити-
ческими Myzostomida и Gastropoda — с другой.
В чем лежат причины подобной связи — сказать большей частью
трудно. Личиночный паразитизм Pantopoda в гидроидных полипах,
быть может, связан с тем, что взрослые пантоподы питаются псклю
чительно гидроидами и как бы пасутся на целых лугах колониальных
гидроидов Вероятно, именно вследствие медлительности своих дви-
жений Echinodermata являются подходящими хозяевами для таких
медленно движущихся паразитов, как Gastropoda. Все эти толкования
имеют, однако, лишь предварительный характер.
Интересно, что головоногие моллюски, отличающиеся и сравни-
тельно большими размерами и образом жизни, напоминающим рыб.
обладают паразитофауной, по-видимому, более разнообразной, чем
53
гие. S4. Инфузория /inllodora dimorpha
с брюшных ножек мокрицы. I руипл инфу-
зорий, сидящих на стебельках. Мелкие
особи готовящиеся оторваться — бродя-
жки. (По Догелю и Фурсенко.)
прочие моллюски. Так. помимо кокцидпй и инфузории, а также личи-
нок цестод (Corallubothrium) п Dicycriiida, мы находим па них в каче-
стве эктопаразитов моногенетическнх сосальщиков (Jsajiasfrurn loligi-
nis) и веслоногих раков (Archicaligus на Nautilus# из групп, обычно
паразитирующих па рыбах.
Кроме размера, играет известную роль и высота организации
животных-хозяев (Киршенблат. 1941). Действительно ра .личные
органы хозяина представтяют для паразитов различные условия
обитания, а чем разнообразнее последние, тем большее количество
видов паразитов может существовать одновременно в данном хозяине.
()1частп поэтому крупные, по низко органи ованпые животные (на-
пример актинии) имеют малора.люобразиый контингент пара-
зитов.
Многие группы паразитов одинаково широко распространены
и видных животных, п у наземных, др\чие имеют хозяев только сред!
гьцветных, ведущих определенный образ жизни. Наконец, осп. и та-
кие систематические группы паразитов, в которых наряду с образом
жизни, характерным для большинства представителей, имеются
отдельные «выходцы», перешед-
шие к паразитированию в несвой-
ственной для данной группы сти-
хни. Обычно такой переход на-
земных паразитов в воду и вод-
ных па сушу связан с изменением
образа жизни их исконных хо-
зяев.
Переход водных паразитов
на сушу иллюстрируют некоторые
кругорссничные ннфезорин (Pe-
rithelia). Так, инфузория Ballo-
dora (Догель и Фурсенко. 1920)
(рнс. 34) с жабр мокриц пред-
ставляет собой близкого родича
пресноводных сувоек и вышла па
сушу вместе со своим хозяином,
предки и близкие родичи которо-
го ведут водный образ жизни.
Точно так же инфузория Trichodi-
nopsis (очень близкий родич жп
вущей на гидрах, водных моллю-
сках и рыбах Trichodina) сохра-
нилась в вышедшем на сушу брю-
хоногом моллюску (Cyclosloma — Pomatias). Многоустка (Polystoma
integerrimum) обыкновенной травяной лягушки есть потомок жаберных
сосальщиков рыб, и самая локализация ее в мочевом пузыре хозяина
есть резу тътат приспособления к частично наземному' образу жизни хо-
зяина и к утрате последним жабр.
С другой стороны, тюлени взяли с собой в воду целый ряд вшей
и клещей, морские змеи—клеща Atnblyomtna nitidum (один из нксо-
довых клещей). Перепончатокрылые, в частности наездники, — типично
наземные животные; однако, среди 27 000 наземных наездников имеет-
ся 4 или 5 родов (Prestwichta, Agriotypus), спускающихся временами
в воду и кладущих свои яйца в япца жуков-плавунцов и других вод-
ных насекомых.
Однако при переходе хозяев в иную стихию наблюдается и дру-
51
joe явление: переход id них чужеродных, т. е. принадлежащих дру-
гой стихии, паразитов. Особенно инструктивны в этом отношении
китообра иые как формы сухопутного происхождения, ио глубоко
адаптировавшиеся к водному образу жизни Ча китах, и только на
китах, поселилось несколько родов усоногих раков (Coronula.
Tul'icinella), а также паре шгическне бокойлавы Cydmus (см. рис. 33».
выедающие па коже китов язвы в виде ямок Паразиты из ракообраз-
ных имелись и па коже вымершей стеллеровой короны (Rhytina
stelle'i) из отряда енреповы.х (Sirenia), которых считают \шедшими
вводу отдаленными потомками копыТнцх. В глазу бегемота поселил-
ся монотеистический с.осалыцнк Oculotrema — потомок жаберных
сосальщиков рыб.
Из паразитов водного происхождения, вышедших на землю,
можно вспомнить цейлонскую пиявку (Haetnadipsa beylonica). живу
тую в лесах азиатских тропиков и нападающую на крупных млеко-
питающих, па которых опа сваливается с придорожных деревьев
я КУСТОВ
ГЛАВА V
ЯВЛЕНИЕ ГИПЕРПАРАЗИТИЗМА
Мы уже сидели, как широко распространено в природе использова-
ние одними животными других и в качестве местообитания, и в качестве
Рис. 35 Сосущие инфузории (Siictoria) из кишечника лошади, илрази
тирующие на кишечных ресничных инфузориях
А — Allantosoma intestinalis; Н — Al. cucutnix, присосавшаяся к хвосто-
вым пучкам Cycloposthium ede.ntatunr, С—Al. dicorniger, D — Al. bre-
vicorniger. ирнсо'савшаяся к lilepharozoum. (По Стрелкову.)
1 - мзкроиуклсус 2 — микронуклсус: 3 — сос;пслы1ыс шуиазниа: 4 - naxiurj полопнно iiir-
фумрий Cyclopusthium 3- инфузория Blepharozoum 6 п ~ — нршосаншпсс-и Allitntosotna
источника пиши. Примерами этого служат все паразиты. Однако в ряде
случаев паразиты, хорошо упитанные и богатые запасными питательны
мп веществами, в свою очередь становятся источником получения нищи
для более мелких паразитов второго порядка: паразитизм делается двух-
•56
степенным: таким образом, возникает особое явление называемое гппер-
паразитпзмом. Явление это, в общем редкое, но исследования послед-
них лет раскрыли нам достаточное количество примеров гнперпарази-
тизма, часть которых оказалась имеющем даже, известное практическое
значение.
Интересно, что рассматриваемое нами явление изредка наблюдается
уже по отношению к самим мелким представителям паразитов, в лите-
ратуре имеется ряд указаний, что
жить паразитические простейшие.
Вполне естественно, что вслед-
ствие малых размеров простей-
ших их гиперпаразптамн могут
быть лишь другие одноклеточные,
а именно: простейшее же или
грибки.
Очень хороший пример дает
одна сосущая инфузория, Allan-
tosotna (рис 35), которая поселя-
ется на ресничных инфузориях из
группы Entodiniomorpha, парази-
тирующих в толстой кишке ло-
шади. В настоящее время извест-
но уже несколько видов Allanto-
sotv.a, впервые описанной Тассов-
ским (1918). Все они имеют кол-
басовидное тело с двумя пучками
сосательных щупа тьцев или про-
сто с двумя сосательными щу-
пальцами на противоположных
концах животного. Allantosoma
можно наблюдать присосавшей-
ся одним из этих пучков к телу
какой-нибудь крупной ресничной
инфузории (Cycloposthium и др.),
там, где кутшЛ’ла инфузории
хозяевами гиперпаразнтов могут слу-
Рис. 36. Паразитические амебы (Entamoeba)
в теле опалины Ci‘p.‘dca tonga. (По Стэб
леру и Чену.)
1 ядра опалины: 2 паразитические амебы.
яаляется более нежной
В последнее время (начиная
с 1933 г.) появились указания о
паразитизме амеб в теле различ-
ных Opalinida, а именно: в Zelle-
riela, Cepedea и Opahna (Stabler, 1933; Carini und Reichenow, 1935; Stabler
and Chen, 1936; Эпштейн, 1941). В плаз е этих инфузорий иногда в очень
больших количествах встречаются типичные мелкие амебы 5—14 р в диа-
метре, принадлежащие к роду Entamoeba (рис. 36 и 37). Карнин и Рейхе-
пов (Carini und Reichenow, 1935) предполагают,что это,возможно,Е.га-
пагшп, паразитирующая в кишечнике амфибий и переходящая к иарази
тпзму в опалинах. В теле опалин находятся не только вегетативные ста-
дии, но и цисты амеб. Кроме того, Стэблер и Чен (Stabler and Chen,
1936) обнаружили амеб п внутри цист, образованных опалинами. Это
обстоятельство объясняет легкость распространения амеб и большую
экстенсивность заражения, доходящую до 160%.
Особенно часто паразитируют в паразитических простейших Micro-
sporidia и некоторые близкие к ним организмы. Для микроспоридий из-
вестно паразитирование в грегарппах (Perezia в I.ankesteria ascidiae
и др.), в инфузориях (Nosema в Balantidium). в мпксоспорпднях (Е'о.ас-
ma marionis i Certilomyxa coris). Частыми паразитами грегарнн являют-
ся представители семейства Mctchnikovcllirtac (рис. 38)—одноклеточны :
организмы, систематическое положение которых недостаточно хороню
выяснено, по, по-видимому, близко к Microsporidia.
Особенно часто в грегарина.х наблюдаются продолговатые инеты
Metchnikovellidae с нескольким!) отдельностями внутри. Наконец, во мно-
гих эндопаразитических амебах, инфузориях (X'yctolherus. Entodinio-
morpha; констатированы грибки из рода Spha.-riia.
Piic. 37. Гипсрчаразп гпзм у Protozoa
А — Entamoeba гапагит, паразитирующая п опаншах. с яд-
ром и заглоченными бактериями; В - амеба, паразитирующая
внутри опалины /.elleriella, с ядром и паразитирующим
в амебе грибком Sphaerita\ С — цис га Cepedva longa с ядром
и четырьмя а.мебами. (По Стэблеру и Чену.)
1 — ят]ю Cepgiiea lunga: 2 пара «iitii'ici'kiic амсби. J грибок Sphairifa
Микроскоподпн, по-вндимому, мало разборчивы в выборе хозях в,
так как отдельные прел ставите ли их, кроме паразитических Protozoa,
были найдены в паренхиме или желт очниках сосальщиков (Nosema Irgf’i
в Gytnnophallus somaleriae и др.), ленточных глистов (в паренхиме и го-
надах Taenia bacillpris, Moniezia expansa и др.), в нематодах (Nosema
mystacis в iопадах Ascaris myslux из кошки, Thelohania remformis в
Prolospirtira muris из мыши) (pin 39). Микроспоридии из рода Nosema
обнаружены в тканях церкарий (Cort cl al., 1960). Пои ,том интересно,
что в этих примерах дело пс может идти о случайном заражении глиста
микроспоридиямч, встречающимися у его хозяина, так как, например,
млекопитающие (кошка, мышь) никогда не с. г жат хозяевами микро-
споридий. Средн эктопаразитов микроспоридии (Nosema pullets. .V. cteno-
cephali) встречены в некоторых блохах.
Иногда гнперпаразитамн паразитических червей Moryi быть и жгу
пгконосцы. Так, Гюншшен и Впхтермап (Ilunninen and Wichterinann,
193«) нашли, что кишечный жгутиконосец морских угрей Hexamitw;
очень часто (в 20 из 35 вскрытых рыб) может быть встречен в матке,
яйцеводах и яйцах сосальщика Deroprislis Uiflata, иногда до 20 штук в
одном яйце.
Особенно интересный случай представляет жгутиконосец Hisiomo-
nas meleagridis, паразитирующий в кишечнике кур и индеек, где совмест-
но с пим нередко попадается нематода Heterakis gallinae. Грейбилль п
Смит (Graybill and Smith, 1920) указывают, что данный жгутиконосец
58
нс только может паразитировать в нематоде, но и нередко проникает
в яида Hpterakis. Выводимые вместе с экскрементами яйца могчт слу-
жить источником заражения нс только нематодами, по и жг\ тнкопосца-
чп. .Межд\ том инвазия Hislotnonas приводит к тяжелым и даже смер-
тельным заболеваниям птицы.
Рис. 38. Me chnikt \eilidac.
лара штирующке и грегарн-
иа.х
.1 — споры Metcltuikovt Hit
spionis в rpei арине /’o-
lyrhabdina spionis; fi — ве-
гетативные стадии в те же
грегарпне (живой обьеьт).
С — споры в грегарине (ок-
рашенный объект); [) - изо-
лированная спора Metcfini-
кгчч-На spioitis. (По Грассе.)
Гипсрпараз!1 гизм на теле сравнительно круп-
ных Monogenoidi а обнаружен недавно у инфу-
зорий Trichodina. Так, например, Т. stre/kovi
обычно вс гр-чается на Aucyroccphalus niogu-
rundae и Ancylodiscotdes asoti, живущих,
в свою очередь, н i
жабр; ч амурских
рыб Simperca cfiu-tsi
п ParasUlurits aso-
I us.
Рис. ЗУ. Микроспоридия The-
lohania reniformis - пшер
паразит нематоды Pryfospi-
rura mttris. iFlo Кудо и
Гедеринггоп.)
Ila.ieiin четыре клетки kiihic’iiio-
го эпи гелия Р. marls с ]мг»пыин
стадиями развитии Thelohnnia re-
ntfor mis вн угри них. Направо -
три спори Th reniformis, нижняя
с выстреленной стрекательной
питью.
Рис. 40. Паразитирующий
1>а рыбах рачок Lepaoph-
theiriis sp. с сидящими
на не и гиисрпаразига ин.
(Оригинал.)
/ Чернь Udunella calllgoritm:
2 — инфузории Conidophrys
sp.; 3 — яичные мешки рачка,
на которых частью сидят пшер-
нарзлнтм.
Гораздо реже, чем олнок юточные, гииерпаразитами являются черви.
Имеются немно) очпе ченные сведения, что в некоторых сосальщиках
и ленточных глистах были обнаружены круглые черви или личинки тре-
матод п нестод. Метацср1Гарин /. trucoiyle сем. Strigeidae нередко пара-
знтириот в реднях э.хнностомнд (Итог, 1955), а один вид Т. variegate
1ЛПЧИНКП Cotylurtis pileutus) —в теле плероцеркоидов Ligu'a cfdumhi из
яолости тела шиповки fCobitis taenia) п пескаря {Gobio goido) (Дубн-
59
инна, 1956). Мета перкарин Telracotyle variegata обычно локализуются в
субкутнкулярном слое пли в паренхиме плероцеркоида.
К гиперпаразитам следует отнести н своеобразных червей Udonella.
включенных Ивановым (1952) в класс Udonellida (рнс. 40).
Трематода Cyclobothrium живет в ранноногом рачке Meinertia—
эктопаразите морских рыб, a Derogenes— в паразитических Copepoda
(Dollfus, 1951). Кроме того, имеется довольно много случаев паразити-
рования личинок в различных эктопаразитах. Нередко эти эктопаразиты
даже являются
окончательным
Рнс. 41. Гнперпардзнтнзм веслоногого рачка
Aspidoecia normani. (По Жнару.)
1 рачок Erythrnps sp. (Scliiznpocta); ! — паразитическое
ракообразное Aspidophryxus sp. (Isopoda) па рачке;
! — ruiicpiiapMHTliMecKiiit рачок Aspidoecia normani na
Aspidophryxus.
связующим передаточным звеном личинок глистов их
хозяевам. В качестве примера напомним тыквовндного-
цеиня (Dipyiidium caninuni) со-
баки, цнегнцеркопды которого
живут в собачьей блохе (Cte-
nocephalus cards) и во власо-
еде (Trichodectes cards).
Довольно много гпперна-
разптов среди раков, причем
они живут на других раках, па-
разитирующих часто, в свою
очередь, на более крупных
ракообразных; в таких случаях
получается как бы трехэтаж
ное наслоение трех видов раз-
личных Crustacea. Такого рода комплекс образуют упоминавшиеся ра
нее представители Liriopsidae (Isopoda), например Danalta curvata, со-
сущие саккулину (Sacculina), которая сидит на брюшке краба Inachus
Некоторые Cabiropsidae из Isopoda поселяются на других Isopoda, а
именно: на Bopyridae (Cabirops— па Bopyrus, Gnomoniscus— на Po-
dascon), паразитирующих в жаберной полости Crustacea (Decapoda).
Жиар (Giard, 1911 —1913) указывает, что па пелагическом рачке
Erythrops (рис. 41) живет паразитическая ичопода Aspidophryxus, а под
брюшком последней нередко можно видеть приценившегося веслоногого
р^чка Aspidoecia normani.
Однако больше всего сверхпаразитов, а также и хозяев сверхпаразп
тов в классе насекомых. Очень многие наездники, откладывающие своп
яйца или личинки в других насекомых, сами становятся жертвой наезд-
ников или та хин. Вопрос о гпперпаразнтах приобрел в прикладной энто-
мологии большое значение. Дело в том, что особый биологический метод
борьбы с вредителями сельскохозяйственных растений заключается в
массовом искусственном разведении паразитов вредителей и в выпуска-
нии этих паразитов (главным образом наездников) на волю, где они своим
усиленным размножением ставят преграду чрезмерному размножению
вредителя. В обширной имеющейся но этому вопросу литературе приво-
дится длинный ряд данных об удачной борьбе при помощи наездников
с насекомыми — вредителями сельскохозяйственных культур.
Однако в этой борьбе с вредителями сверхпаразиты могут приво-
дить к самым нежелательным последствиям, сводя к нулю эффект меро-
приятий по борьбе с вредителями. Насколько полезен в этих случаях пер-
вичный паразит, истребляющий вредителя, настолько вредоносен вторич-
ный паразит, уничтожающий первичного. Поэтому, вводя для борьбы с
каким-нибудь вредителем культур его паразитов, энтомологи в настоя
щес время обращают усиленное внимание на то- чтобы вместе с парази-
тами не пустить в круговорот природы и их гиперпаразитов. Между тем
отношения паразитов п гиперпаразитов порой столь сложны, что могут
приводить к самым серьезным в практическом смысле ошибкам.
60
Многие из наездников, паразитирующих в насекомых-вредителях,
подвергаются нападению со стороны нескольких различных видов вто-
ричных паразитов. Так, известный паразит капустной белянки, наездник
Apanteles glonier-atus, страдает приблизительно от 20 гиперпаразптов,
гамымн важными из которых, но данным Фор (Faure, 1926), являются
Tetrastichus rupo, Dibrachys bouclieanus н J'emifeles fitivipes. Насколько
сильно бывает влияние этих вторичных паразитов, можно видеть на при-
мере Apanteles fulbipes — паразита непарного шелкопряда. Будучи в
Европе очень эффективным истребителем шелкопряда, в Хмерика . ’ ful-
vipes не лает желаемых результатов. I Iomiimo других причин, мешающих
сасп рост ранению этого наездника в США- важным препятствием к его
разведению оказались вторичные паразиты. Действительно, из 5456 ко-
конов Apanieles при обследовании 2573 оказались заражены гпнерпара-
зитамп.
Рис. 42 Некоторые чэ паразитов кукурузного мотылька (Pyraitsid nuhi-
lalis) н их гипериаразиты
А — Pyraustti nubilalis; /< — наездник / imtwria, один вид которою пара-
зитирует в /’уг«п.х/<г; С— наездник Ihmiteles, гпперп.1разнтирую1И1,п и
Limnvria; D — иаезлинк Pimpla, некоторые виды которого гппёрнар.вп-
nipyior и Limmria; — наездник рода /lesoc’torus, некоторые ниты ко-
торого гппернарлзптнруют в Limn -ria; Р — н >ездннк /Sopitin' pun^toria,
п.тразнгнрую1цнн в Pyrausta nubilalis; G— наездник Pimpta, rinicpii.ipa-пт
АпцШа; H — наездник I upteromains nidulans, niiiepnapa iiir Angitia.
(Рисунок составлен по ряду авторов.)
Для иллюстрации гиперпара.чптнзма мы лаем прежде всего два ,
(из приблизительно двадцати) паразитов мкхрхзпого мотылька {Ру-
rajusia nubilalis), а именно: Angiiia puncioria и Litntieua (рис 42'
С каждым из них связано по поскольку гиперпаразптов. Па схематичным
рис. 43 представлены круг паразитов луишого мотылька и часть их
гиперпаразптов (по .Мейеру, 1937).
Гиперпаразиты могут жить п в паразитах, относящихся к другому
отряду насекомых: наездник lapinogalus pulchellu развивается в парази-
тической мухе-тахпне Exorista lobeliae, а наездник Ixodiphagus—даж-
в клеще Ixodes ricinus (рис 44).
11 t\ । '"r9?As^fA I \ \ \ t- -7 \ affinia i 1 1 \fl,i/nneria\ У. J\ 1 UrruxM^) I I tz" “X I CMoms \ I \ (Liberia \\yr*X'&s I \\<}eaicvtator'\b y- 1 1 (Jietpus у 1 X I robcratcr J 1 jLimfiana \ x. 1 \yu(i(trrwj~^ ' X .у'"'! Limntria V n„ n r- /I/" "XT \pleuralis) Рис <13. Схема, выясняющая / {1_;тпепд\ \. взаимоотношения между хо- Салили] У зяином, его паризитами и / >А. у Z^, их I itnepnapaiHT.iMii. (По / iMeitepv.) I Mesocbarus J/lngdia A В центре почетен кружок, «знача- tuberculiger J*\d>rys<istide} копии хозяина. а именно лугоногп ^х. / у шпилька (Loxostegti sticticilt*). К нему схоиися стрелки, гнущие f >. от его пара <и гон; расположенные fjfrtgitto \ (Xlnoitia \ ни периферии прямоугольники и» \armiUdta ) \fenestrZis] начагиг « опой rnnepnapii Итон, соя X. J Vе 7 запных стрелками с частью пара- X.. у X. _ опои лугоного мотылька, и кото- рых рин Нарвойтируют. H^WxAz->f (/lpani?tes\ 1.4ра~!е!н\ f.^ontvlti \ ICeZn ..fc\ Iflsrdxbilny f/Masta \/Ц1 1 rujierus j \sctanarJu-: t yse.'tirefiB 1 l кй'мп 1 I j ( ehtntnu jl 11 *\ z^" ^\l Z' Z' /' /Xiulaita \ ill LTmtsliA I Гг,</аГ \\l Pl^» A fX A n'otata J/// И t/ Z^\ \^y / / / 'S--<zxx^-o i —< /xL fA /// ^^x/Xv4 x\ \' i / ! / ' /<3 } S’" / / ^x^\xXx*x* it/У/у^\зн«г/»1л]I I ' '/Л 7i [f/У W> ti.ubr-»i/rha\ / / .J?7 //ffZara.WgM -ydebelis I 1 1 ~~~UA St/ctlCiH/5 Ifjl ~ \ J У x / — , - > Ж J}l< ~. ~ f Ofmrga A XtiQubrinall y^ /// / / \ ' \ W X I L/mnert,3\ / у // / / 1 \ ' \ \ 4'- ^X yilyriMlj 1 Z Z / / / 1 \ \ \ x.'\^X \ >z / / / / ' \ \ \ ''''x XyxoMa j (^а/уугзУ f Clemehs\ f Vona \ I£xonsb\ f ThyfUa ( [^ZodtfXX t civilis 1 \pu//ah j yura/is j I mlt<o J \b/orabs j ypZ^anx 1 {hygadeaon ^iygadeuon Phygadetion grandiceps зас/юпп loxastoginis
Вопрос о гиперпаразитизме у насекомых усложняется еще тем, что
на гиперпаразите, или вторичном паразите, иногда паразитирует тре-
тичный паразит, деятельность которого является полезной, так как он.
истребляя вторичного паразита, тем самым способствует сохранению
первичного паразита, уничтожающего вредителей. Бывают случаи, koi да
Лело доходит до гиисрпаразитпзма в четвертей степени. Впрочем, пара-
зптнзм третьей или четвертой стадии не носит такого правильного харак-
тера, как простой гинерпарази гизм. Так, например, в совке Prays oleellus
Сильвесгри (Silvestri, 1906) на-
шел 20 видов паразитов, нз кото-
рых только один являлся исклю-
чительно первичным паразитом
(Ageniaspis juscicollis). Семь
дру гих видов были нсключнтель
ио вторичными паразитами. Про-
чие могли быть как первичными,
так и вторичными паразитами,
без разбора, а многие из видов
при случае могли становиться и
паразитами третьего и четверто-
го порядков. Вообще у третичных
паразитов наблюдается малая
разборчивость по отношению к
объекту их нападений.
Среди других групп живот-
ных паразитизм третьего порядка
встречается редко. У амеб, кото-
рые, как указывалось ранее, па-
разитируют в опалинах лягушек,
иногда наблюдается заражение
Рис. 41. 1 lae ith ih /.v<> fiphagus utucurtei,
откладывающий яйцо н лимфу Ixodes
ricintis
грибком Spiiacrita.
Помимо гиперпаразитизма, у паразитических насекомых часто встр<
чается явление, называемое с и н и а р а з л т н з м о м, или соиарази-
тнзмом, когда в одну особь хозяина помещают свои япца или личин-
ки несколько разных видов паразитов. В таком случае эти паразиты
конкурируют друг с другом и могут оказывать Друг на др\ га более пли
менее вредоносное влияние. Разбирая отношения между такими одновре-
менными паразитами одного хозяина, Штельваг (Stellvxaag, 1921) при-
знает возможность одного из следующих случаев:
I Один из паразитов остается в живых, другой погибает: 1) раньше
вылупившаяся личинка уничтожает свою соперницу; 2) один из маразм-
тов вызывает гибель хозяина, а следовательно, и другого паразита.
II. Как паразит, так и сона разит остаются в живых.
III Оба паразита гибнут в результате смерти хозяина, вызванной
чрезм* рным истощением
Как примеры сопаразнтизма, можно привести личинок наездников
Aphidius и Aphelinus, попадающихся в одном и том же экземпляре тли.
При этом присутствие личинок ‘phelirtus тормозит развитие личинок
Aphidius. Точно так же в случае одновременного паразитирования ли-
чинок Zygobothria nidicola и Meteorus в гусенице златогузки развитие
Zygobothriu нарушается. Интересно отметить гот факс, что если одни из
паразитов является наезднике -м. а дру гой тахиной, то в б льшинствс слу
чаев погибает личинка наездника.
Вредоносными по отношению друг к другу сопаразнтамн могут ока-
заться и личинки одного и того же вида Томпсон (Thompson- 191-3) ука-
63
зывает, что тахины Euphoracea в таком числе нападают на колонии пи-
лильщиков Lophyrtts, что большинство личинок пилильщика становятся
перезараженными. В результате отродившиеся паразиты имеют карли-
ковый рост и являются нежизнеспособными. Нильсен (Nielsen, 1917) на-
шел 150 гусениц пестрянки е отложенными на них 767 яйцами тахины
Larvaevora larvarum. Из этого количества яиц Нильсену удалось по-
лучить всего 1-11 тахины, остальные погибли.
Рассматривая явления паразитизма и гипсрпаразптизма у насеко-
мых, мы наталкиваемся иногда на весьма оригинальные случаи паразп
•тпческих отношений. Так, например, такие чисто наземные животные.
Рис. -15. Водные наездники
Л — Prestwirhia aquaiica, крылатая самка; Н — тс же. самка с рудиментар-
ными крыльями; С — кокон ручейника с выдающейся наружу дыхатель-
ной лептой наездника Agiiotvpus sp. (Л п Л — по Римскому-Корсакову,
С—по Бишофу.)
как наездники, иногда приспосабливаются к паразитированию в водных
насекомых. Так, Hemifeles biannulatus нападает на личинок ручейников
сначала в качестве эктопаразита, а позднее внедряется в тело хозяина.
В ручейниках же паразитирует Agrioiypus armafus, спускающийся при
откладке яиц на 10 и более минут в воду. Некоторые Braconidao пара-
зитируют в Hydrellia. минирующих водные растения личинках. Разные
виды Prestwichia (рис. 45) живут в яйцах разнообразных водных насеко-
мых. Наконец. Anagrus и Caraphraaus развиваются в яйцах стрекоз,
плавунцов и водяных клопов. При откладке яиц и при вылуплении из ко-
кона эти наездники приходят в непосредственное соприкосновение с водой.
Несмотря па приспособление к новой среде обитания (воде), все
эти паразиты водных насекомых испытывают крайне мало специальных
изменении в строении. По отношению к эмдопаразнтарным стадиям их
развития это вполне понятно, ибо они находятся в таких же условиях су-
ществования, как н паразиты наземных животных, т. е. внутри хозяина.
Только личинка Agriotypus (рис. 45) при окуклении на крышечке чехли-
ка своего хозяина — ручейника — изготовляет из пряжи длинную тонкую
ленту, в ячеях которой содержатся пузыри воздуха. Эта лента имеет ка-
кое-то важное значение для дыхания, так как при отрезании се наездник
не заканчивает развития. Зато у взрослых водных наездников не обнару-
жено особых изменений, хотя, например, крылья у них находят совер-
шенно особое употребление, а именно: при погружении в воду эти наезд-
ники работают крыльями, как веслами.
61
5 В А. Догель
ГЛАВА VI
ПРОИСХОЖДЕНИЕ ПАРАЗИТИЗМА
I ПРОИСХОЖДЕНИЕ ЭКТОПАРАЗИТИЗМА
Явление паразитизма имеет многообразное происхождение. Даже
один и тот же тип паразитизма, например эктопа ра.зитнзм, у разных
грх ип животных мог возникнуть разными путями. Во всех случаях про
похож [енис эктопаразитпзма связано со сменой прежнего способа лига
пня. свойственного свободно живущим предкам, и переходом к питанию
за счет позвоночного хозяина, его тканями, кровью или выделениями.
\ эго в свою очередь и вводило к усилению топической связи между
двумя организмами и возникновению системы паразит — хозяин.
Большая чисть эктопаразитов происходит из хищников. Уже приво-
дившиеся выше примеры с пиявками и клопами показывают, как из
хищничества мог развиться настоящий эктопаразитизм. Особенно много
примеров такого постепенного перехода мы имеем среди членистоногих,
комары, москиты и т. д. .мало чем отличаются от насекомых .хищников.
Клопы представляют дальнейший переход к паразитизму. Чтобы стать
паразитами, им не хватает только пребывания и питания па хозяине в
течение более или менее длительного времени. У некоторых клещей (на-
пример, Ixodes) мы встречаемся с выработкой длительного контакта с
хозяином. Контакт в еще более сильной степени развит у не которых блох;
у них только личинки и куколки живут на свободе, половозрелые же
особи большую часы, жизни проводят па хозяине. От этих длительных
но все же периодических паразитов остается лишь один шаг до постоян-
ных, обитающих в течение всей жизни, во всех ее стадиях на хозяине.
У подобных форм, например у вшей, япца нс выбрасываются во внеш-
нюю среду, а откладываются и развиваются здесь же на хозяине. Такой
переход к эктопа разитиз.му на наземных позвоночных мог осуществиться
в условиях максимального контакта членистоногих с позвоночными жи-
вотными. Это прежде всего и с наибольшим успехом могло происходить
в норах, гнездах н других убежищах позвоночных, где обитают также и
различные представители членистоногих, Контакт с позвоночным этих
норовых или гнездовых обитателей приводит сначала к питанию некото-
рых представителей на легко доступном хозяине, а затем и к длитель-
ному существованию на нем. Так мог возникнуть паразитизм у гамазо-
вых клещей в норах грызунов, у клещей Argasidae, вероятно у перьевых
65
ii чесоточных клещей, \ клопов, пухоедов, блох, некоторых мух-кровосо-
сок и многих других членистоногих (Беклемишев, 1951).
В отношении пухоедов (Mallophaga) можно думать, что эти эктопа-
разиты происходят от предков, питавшихся сначала различными
(частью животными частью растительными) остатками, вроде Oribatidae
среди клешей и Psocidae среди насекомых. Быть может, сначала они
поселялись в гнездах птиц, на дне которых всегда скопляется значитель-
ное количество органических остатков. Впоследствии они стали с под-
стилки гнезда перебираться на его обитателей и питаться их омерт-
вевшими. т. с. ороговевшими, вы-
ростами покровов. Интересно, что
некоторые из пухоедов, например
Physoslontutn, вторично перехо-
дят к питанию кровью хозяина.
Пухоед пеликана Tetrophthalmus
(рис. 16) мигрировал с перьев
и гиды в ее громадный подклюв-
Рис. 46. Аберрантная локализация пухоеда Tetrophthalmus
/1 — пухоед Tetrophthalmus titan с пеликана: Н — голова пеликана с рас-
крытым клювом для показа молодых пухоедов у края клюва и взрослых
впереди п позади дыхательной щели. (По Дубинину.)
Слсп.1 пт головы_ показано в упсличеинпм мэсшгшТе т-;>о пеликана с приклеенным к нему
нйцом пухоеда. Стрелки иоказыинют пути миграции пухоедин и разные моменты нх цикла'.
Часть основания клюка вырезана д.ы показа дыхательного отнерстия-
1.ып мешок и вместо перьев стал питаться кровью. Для откладки
яиц Tetrophthalmus выходит из ротовой полости на оперение хозяина
(Дубинин. 1917).
Некоторые членистоногие, как, например, иксодовые клещи, личинки
краснотелок, многие двукрылые и др., не обитающие в норах и гнездах,
могли перейти к эктопара.зптизму в каком-либо открытом биотопе (на
пастбище) при нападении на животных, не пользующихся убежищами
(рептилии, копытные, некоторые хищники)
И ряие случаев путь х возникновению эктопаразитпзма ведет от
сидячего образа жизни Такого происхождения, вероятно, паразитизм
у кругореспичиых инфузорий Trichodina. все родичи которых ведут при-
крепленный образ жизни, а многие из них при этом прикрепляются не
ко дну водоема, л к живым организмам. Так же поселяются на теле
водных животных усоногие раки (Cirripedia), причем некоторые из них
глубоко внедряются в кожу хозяина, как это делает Coronula на теле
китов (см. рис. 3).
об
Or хищных прямокишечных турбеллярпн (Rliabdoeoela) произошли
очевидно, моногенетвческщ сосальщики (AVjiiogenoidea), большая часть
которых является эктопаразитами рыб, паразитирующими прен.муще
ственно на жабрах, плавниках, коже. Некоторые Mbnogenoidea, как, на-
пример. виды рода Gyrodactylus, питаются слизью, выделяемой покрова
мн хозяина. Другие виды, наряду с эпителием, начинают питаться
кроьью (Dcrctf iogyrus uustaior). Наконец, некоторые виды питаются
Исключительно кровью хозяина (большинство высших Monogenoidea
Diclybotbriidae, Hexabothriidae и др.).
Известное представление о том. как мог воз-
никнуть эктопарлзнгизм Monogenoidiа дает и*,
большая группа ресничных червей Ternnocephala
(рис. 17). Эти мелкие черви поселяются на теле
крабов и мечехвостов. На переднем конце их име-
ются пальцевидные выросты — щупальца, а на
заднем присоска. Они получают от хозяина
только квартиру, питаются же различными мел-
ки in животными пол тающими но телу их хозяев,
т. е- ведут себя, как настоящие мелкие хищники
Нужно иметь в виду, однако, что 1 einnocephala
нельзя рассмагривагь. как переходную группу
от Rhabdocoela к \\onogenoidea. Это — самостоя-
тельная небольшая слепая филогенетическая
вётвь
В выработке эктопаразнтнт.ма несомненную
роль сыграл постепенный переход приспособь <по-
щихсч к порази! и зму мелких хищников от поли-
фагии к монофагии. г. с все большее епецналнзп-
Рис. 47. Организация
Temnocephal.i с покри-
вив Limulus pot themu*
‘ 1 Io Бресслау п Райзин-
геру )
/ ш у 11.1.1 та: и*Л1Н и..ь-
цые капали; 3 — мичсысж ну-
iupt> it инделительнзн пора
i глотка; 5 ~ матка: 6 — се
мспняки; 7 - яичник; Л - п<>-
-Шюое от игр< тис; 9 — присоска;
Ю peiits; П кишечник;
/'? - мозг.
рование их в смысле характера пищи Так, на-
пример, среди колющих Diptcra есть такие (мно-
гие комары), которые питаются не исключитель-
но кровью, Другие (каковы мухи цеце) сохраня-
ют еще все повадки двукрылых, но питаются уже
исключительно кровью, которую они высасывают.
Гретьи (каковы Ornithomya, H'ppobosca среди
Pupinara) уже поселяются па хозяине, ио легко
оставтяют его и нер-ходят на другие особи того
же или близких видов, так как еще сохранили
функционирующие крылья (рис. 18). Наконец,
такие представители кровососок, как Stenopteryx hirundinis лас-
точек. Linopl^na оленей, Melophugus ovums овец, Nycteribia летучих
ышей, испытывают в различной степени редукцию крыльев или обл.т
мывают их во время индивидуальной жизни {Lipoptena) и становятся
длительными эктопаразитами. Таким путем устанавливается прекрасный
постепенный переход от свободно живущих двукрылых к типичным экто-
паразитам
Эктопаразитами наземных позвоночных могут быть личинки иски
торых членистоногих, и такой личиночный паразитизм возник, вероятно.
путем перехода некрофагов (питающихся мертвыми тканями животною,
трупами) к питании» тканями живого организма и длительному прерыва-
нию на его теле. Дичинки Calliphora и Lnciha питаются падалью пли
мясом животного, но могут быть факультативными паразитами в язвах
и ранах человека, выедая отмершие ткани хозяина. В Австралия и Индии
l.icilia откладывает япца в шерсть здоровых овец, и личинки ее внедря-
ются лаже в неповреж тенную кожу, причиняя большой уцон овцеводству
и кожевенной промышленности (.Martini, 1952). Личинки ряда тропиче-
ских мух Cochliomyia hominivorax. Cordylobia anthropophaga н дп. ак-
тивно вбуравливаются в кожу человека и животных, питаются живой
тканью и вызывают кожные миазы. Личинки африканской мухи Auch-
meroniyia lyieola перешли даже к кровососани ю, нападая для этого
ночью на спящего человека.
Рис. 48. Различные мухи-кровососки (Hippoboscidae), иллюстрирующие
пр >цесс [сд; кцпи крыльев
/1 — l.ynchia тайга', Н — hipoptena cerci с остатками крыльев; С ~ Melupha-
gus ovinus—овечий рунец. (IIo Брумпту.)
2. ПРОИСХОЖДЕНИЕ ЭНДОПАРЛЗИГИЗМА
Что касается паразитизма внутреннего, пли эндопаразитизма, ю в
некоторых отдельных случаях происхождение его можно связать с пред-
шествующим ему эктопаразитическим образом жизни. Примером этого
могут служить инфузории Trichodina. которые, как правило, являются
эктопаразита мп жабр, кожи н плавников рыб, но небольшая часть
видов (Tr. urinaria из окуня, Tr. renicola из черной щуки. Tr. alburni
из \клеи и др.) перешла к эндопаразитизму в мочевом пузыре, мочеточ-
никах и в яйцеводах. Весьма вероятно, что такой переход в мочеполовую
систему мог осуществиться именно у инфузории, паразитирующих на
жабрах. Это связано с тем, что жабры и органы выделения рыб имеют
некоторое функциональное сходство. Через жабры, так же как и через
ночки рыб выводятся аммиак и мочевина (Smith, 1929). Из выделитель-
ной системы трихолимы проникли уже в яйцеводы. Свидетельством это-
му является тот факт, что Tr. oviducti из яйцевода ската не утратила еще
способности существовать и в мочевом пузыре (Полянский, 1955).
Другой, более известный пример подобного рода дает лягушечья
многоустка (Polystoma integerrimum) (см. стр. 27). Интересно, что в од-
ном из приведенных случаев (Trichodina urinaria и др.) имеет Место
активное заселение, паразитом новых территорий па теле хозяина; в дру-
гом случае (Polystoma) сам хозяин как бы заставляет паразита своим
метаморфозом и редукцией того органа, па котором ранее паразитировал
сосальщик, искать себе новое местожительство в тело хозяина.
В некоторых специальных случаях путь к эндопаразитизму лежал
через изменение инстинкта откладки яиц. Взамен откладки яиц па раз-
личном гниющем органическом материале яйца могли начать отклады
ваться па язвах и ранах живых животных, где условия для их развития
оказались более благоприятными, что и привело к преимущественному
выживанию особей, обладавших этой привычкой. А отсюда не далек был
68
путь к откладке тех же ниц в сообщающиеся с впешпеп средой полосги
(рот, задний проход, нос, '.ши и т. д.). Вольфартова муха (IVohtfahrtia
magnifiaa) (рис. 49) так и делает, откладывая своих личинок то на раз-
личные язвы и раны, то в рот, уши и нос различных живогны.х. Ес личин-
ки могу г проникать затем дальше, даже в лобные и челюстные пазухи
и ведут там образ жизни настоящих эндопаразитов Те же отношения
можно проследить \ некоторых оводов.
Однако большинство случаев внутреннего паразитизма, а имении:
паразитизм в кишечнике, представляет собой первичное явление, разви-
вавшееся в результате случайного заноса в пищеварительную систему
яиц пли покоящихся стадии различные сво-
бодно живущих организмов. Так, Добелль
(Dobell, 1919), специально занимавшийся этим
вопросом в отношении человека, описывает
под названием «коп розой пых простейших че-
ловеческих экскрементов» 12 видов амеб и
жгутиконосцев, как панболте часто встречаю-
щихся в фекалиях простейших. Как правило,
цисты всех этих видов проходят транзитным
порядком пищеварительный тракт человека и
только по выходе из него развиваются в актив-
ные формы, живущие в качестве сапрофитов в
отложенных экскрементах. Не раз описыва-
лось нахождение этих организмов в только что
отложенный: экскре.мснтал уже не в виде цист,
а в виде активно подвижных форм. Хотя по-
добные описания в большинстве слу чаев ста-
вятся под большое сомнение, трудно отрицать
a priori возможность того, что в очень редких
случаях эксннстпрованш происходит уже в
нижних отделах кишечника В процессе же
эволюции такие р» лкж ненормальности могут
навсегда закрепиться и превратиться в норму.
Кишечная среда выгодна дня простейших по
Рнс. И1. Вильфлртоп..
муха ( Wahl/ahriia та-
gnifica)
А — взрослая муха; Н —
личинка последней ста-
дии, вид с бритвой
стероны. (По Павлов-
скому. )
обнтию содержащихся в пей питательных веществ. Средн простейших, к
сожалению, нам почти неизвестны случаи несомненного переход
ного состояния от свободной жизни к кишечному паразитизму,
когда бы организм был cinq способен к с\ шествованию и в той и в дру
гой среде. Лишь отчасти это наблюдается уг некоторых жгутиконосцев
рода Trichomonas, паразитирующих в кишечнике различных животных
и в то же время способных длительно выживать (днями, а иногда и не-
дслями) в отложенных экскрементах В редких случаях описывались слу
чаи нахождения живых Trichomonas даже прямо в различных водоема',
куда они, очевидно, случайно попали со стоками нечистот.
В сточных водах Москвы Чалая (1941) обнаружила свободно
живущего представителя рода Entamoeba, названного ею Е. moshkovskii
Морфологически эти формы не отличимы от паразитирующих в кишеч
нике человека Е histolytica. Возможно, что здесь имеет место редкий
случай перехода паразитической амебы (род Entamoeba состоит исклю-
чительно из паразитов) к свободному существованию в среде, очень бо-
гатой органическими веществами.
Значительно больше примеров, говорящих о происхождении парази
тнзм'а путем случайного проглатывания, мы встречаем среди нематод.
Значительное число представителей этого класса ведет свободный образ
жизни в воде или в почве. Яйца и личинки таких свободно живущих чер-
69
Рпс. 3'1. Кшпечнля утрнц.т бУ'/гф/геу-
loieics stercoral is). ( По Павловском».)
Сленл фслэи гаикл in кишечника че-шкека:
посредине rnvp.\v личинка; по-
средине Miiwjy |]>11ли|»1к*мн,1пая личинка; cups-
«а наверху елмсн. иин <у самка спосолии
ЖИ1Л-IHCI п IIUKCMCHIIU.
вей. отличающихся и большинстве своем очень м<. жими размерами Hie
более I—2 с,ч), несомненно, нередко заносятся случайна в кишечник раз-
. нчных животных, где в большинстве с. п чаев гибнут, либо остаются без
всяких изменений- пока не буд\т выведены п^ужу Что из таких случай-
ных заносов может ра питаться факультативней паразитизм, доказывя-
кч нам некоторые представители сем. Rhabditidae. '1 ак, Aloionema
appendiculinum нормально ведет свободный образ жизни. Однако личин
i n ее могут случайно попадать в кишечник слизня Arion liter и, иста-
каясь там в течение известного времен:!, могут расти и достигать вдвое
большей величины по сравнению с свободно живущими особями. Поло-
возрелыми, однако, эти нематоды становятся, лишь покинув слизня, при
чем взамен развивающихся у свобод-
но живущих особей 30—40 яиц, у та
кпх случайных паразитов насчитывает-
ся до 500--600 яиц. Потомство же этих
особей, в случае возвращения к сво-
бодному образу жизни, имеет снов1
обычные размеры и дает малое число
яиц.
Дру гоп род того же семейства
Sirongyloidcs (рис. 50) продвинулся в
своей эволюции в направлении к обли-
гаторному кишечному паразитизмуеш-
дальше. Здесь паразитизм стал уже
обя штелькым в тон или иной стадии.
У Strongyloides siercorahs паразити-
рующей в кишечнике обезьяны Aides
geofjroyi (Strongyloides sinrcoralis ти-
па Ateles). пичиика, попав в хозяина не
только растет, но и развивается в иар-
тепогеиегичсскую самку, принимая, та-
ким образом, на себя функцию размно-
жения. Яйца такой самки выносятся
наружу, из них развиваются рабдито-
видные личинки, вырастающие в сво-
Гн дно живущих самцов и самок. По-
томство этих свободных особей в виде
фнлириевндиых личинок снова возвра-
щается к пара титивескому образу жиз-
ни. Таки I образом, здесь о.тновр( менно с паразитизмом ра ввивается и
чередование поколений (См, схему).
Другой подвид того же Strongyloides, паразитирующий у обезьяны
Pitheeus rasas (Strongyloides типа I'lt/iecus), проделывает тот же цикл, с
той, однако- ра пищей, что личинки, развившиеся из яиц партеногенети-
ческой самки, могут, минуя развитие в половозрелых самцов и самок,
вернуться прямо к паразитированию и превратиться снова в партеногене-
тических самок Здесь отсутствует чередование дв\х поколений, так как
выпадает сталия свободно живущих половозрелых особей.
X третьего типа Strongyloides (тип Ното), пара.штир\ юшего у че-
ювека. прежде всего исчезает партеногенетическое поколение самок и
замещается образованием половозрелых паразитических самцов и самок.
В зависимоегп or внешних условий цикл развития Str. stercoralis может
протекать различно. В жарком и влажном климате личинки, вышедшие
из яиц еще в кишечнике, выводятся во внешнюю среду п превращаются
г свободно живущих самцов и самок. Это поколение даст начало рабди-
Схема циклов развития разных форм Strongylofdes stercoralis
озяин
Внеш-
няя
с. eia
Тип Atclcs
Тип Pitherus
Гцп Ното
Партеногенетическая
самка *
Яйцо
I
Рабднтовплная
личинка
I
Рабдитовнднля
личинка
I
2____ Фнляриешп-
| пая личинка
Рабдите видная *
личинка-----------
Партеногенетическая
самка
1
Яйцо
I
Рнбдпто- фплярне-
еидпая —* видная
личинка личинка
1
Яйцо
4
Рабдито-
видная
личинка
Рабдито-
видпая
личинка
I
Ян цо
Филярие-
— видная
личинка
t
филярнс-
— витая
личинка
f
аблито-
нидная
личинка
товмдным п филяршвидным личинкам, которые и инвазируют человека,
проникая в него через рот или через кожу. Здесь сохраняется чередова-
ние поколений. В условиях более низкой температуры (в зоне умеренно
го климата) свободное поколение самцов и самок выпадает и остается
только паразитическое раздельнополое поколение, а во внешней среде
рвбдитовидные личинки превращаются в филяриевидные, которые инва-
зируют человека. Па этой стадии эволюции- когда свободный образ жиз-
ни ведут только личинки, находятся сейчас многие паразитические не-
матоды (подотряд Strongyl ala). У Sir. stercoralis эта свободно живущая
стадия в некоторых случаях может выключаться из цикла. Рабднтовпд-
ная личинка в кишечнике человека превращается в филяриевидную ста-
дию, которая не выходит во внешнюю среду, а внедряется в слизистую
кишечника, проделывает миграцию по теле хозяина и превращается в
половозрелую форму (Faust, 1932; Faust and Groat, 1940).
Дальнейшая эволюция многих нематод приводит различными путя-
ми к постепенному сокращению свободно живущей личиночной стадии.
Во многих случаях личинки не покидают яйца и совсем не ведут свобод-
ной жизни.
У пологе) ряда нематод (Ascaridae. Trichoceohalidae) в яйце, вышед-
шем из хозяина наружу, получается личинка, длительно сохраняющаяся
в нем дез тех пор, пока такое яйцо нс будет снова занесено в организм
подходящ* го хозяина, где только и происходит выход личинки из яйца
п ее дальне йшее развитие. Здесь, однако, пребывание яйца во внешней
среде оказывается необходимым условием, потом что личинки получа-
ются только и выведенных наружу яйцах при свободном доступе кисло-
рода. Яйцо же- задержавшееся в хозяине или попавшее тотчас ке по
выходе из хозяина в другого подходящего .хозяина, оказывается неспо-
собным к развитию н гибнет. Можно допустить, однако, существование
кишечных червей, для развития личинок которых в япце срок пребывания
во внешней среде сокращается в силх того, что часть развития оказы-
вается осуществимой в кишечнике хозяина.
С исчезновением необходимости пребывания яиц во внешней среде
они заканчивают развитие в организме .хозяина и образовавшиеся личин-
ки нс выходят из него. Паразит превращается в постоянного эндопара-
зита (Filariata, Trichinella). Такой шреход связан с изменением биоло-
гии паразита и прежде всего с изменением способа его передачи от
отиого хозяина к другому, без которого не может продолжаться суще-
ствование ВИ,д<1
71
3. ПРОИСХОЖДЕНИЕ КРОВЕПЛРЛЗИТИЗ.МЛ
Особую форму эндопаразитизма представляет паразитизм в крови.
Рассмотрим его на примере простейших. Относительно того, что эта фор-
ма паразитизма есть явление вторичного порядка, возникающее из ки-
шечного паразитизма, разногласии нет. Одидко, признавая это обшее
положение, разные авторы в отношении вопроса п первичных хозяевах
кровепаразитов придерживаются двух различных точек трения.
По мнению одних исследователей, кровепаразнты позвоночных были
первоначально исключительно кишечными паразитами различных бес
позвоночных и лишь с переходом их хозяев к кровососущему образу
жизни па позвоночных вторично постепенно приспособились к новой
среде обитания в кровяном русле этих последних, куда случайно стали
попадать при акте кревососания от их первых хозяев. Постоянно попадая
обратно в кишечник беспозвоночных благодаря тому же акту кровосо-
сания. они не утеряли способности обитания п в первоначальной среде
и оказываются в настоящее время одинаково хорошо приспособленными
цля жизни в обоих хозяевах: в кишечнике кровососущих беспозвоночных
и в кровяном русле позвоночных.
В противоположность этому взгляду Мнпчнй (MincWih, 1912) выска-
зал гипотезу, согласно которой кровепаразнты позвоночных первона-
чально были кишечными паразитами тех же хозяев, откуда постепенно
перешли к паразитированию в кровяном русле. и, найдя здесь более
благоприятные условия и для существования, и для распространения,
постепенно совершенно утеряли способность жить в первоначальном
месте их обитания, приобретя взамен этого вторично способность па-
разлтнрования в кишечнике беспозвоночных.
По-видпмому, оба взгляда являются правильными, и разные группы
кровенаразитическнх простейших имеют различное происхождение.
Рассмотрим прежде всего возможный путь возникновения кровспа-
рлзитнз.ма у различных представителей Sporozoa. Этот путь, по-види-
мому, начинается от представителей кишечных Sporozoa, паразитирую-
щих в позвоночных хозяевах.
Представители подотряда Eimeriidca (отр. Coccidiida) являются,
как правя ю, чисто кишечными и иритом кишечноэпптелпальпыми пара-
зитами. Многие виды кокцидий, как, например, Eimeria inteslinaiis.
Е. perforans, Е. / irijormis из кролика, Е. aceruulina,. Е. mills из кур н др ,
проходят все стадии развития (шизогония, гаметогопия, оплодотворение,
образование ооцист) в эпителии кишечника. Однако в никоторых слу-
чаях наблюдается проникновение различных стадии развития кокцидий
в более глубокие слои стенки кишечника, а именно: в соединительную
ткань. Например, гамонты Е. media, Е irresidua, Е cnecicola в кишечни-
ке кролика начинают развитие в эпителии, а затем погружаются в сое-
динительную ткань ворсинки. У Е. magna не только гамонты, но и ши-
зонты развиваются субэпителиально. Аналогичное явление наблюдается
также у Е. subcpiihcliulis из кишечника карпа, у isospora bigemina из
кишечника собак и кошек и у других кокцидий С другой стороны, у
Eimeria tenella из кур только шизонты 2-й генерации развиваются суб-
эпигелиалыю, а гамонты локализуются в эпитетии. Локализация разных
стадий развития кокцидий в соединительной ткани связана с тем. что по-
раженные паразитом эпителиальные клетки выталкиваются „з эпнте
лиального пласта и погружаются в подлежащую ткань, где паразит про-
должает свое развитие. Здесь же образуются ооцисты, споруляция ко-
торых обычно не происходит до тех пор, пока они не будут выведены во
внешнюю среду (Е magna. Е. tenelfa и др.). В то же время имеются виды
72
кокциднй, как, например, Е. subepithelialis (рыбы). Isospora bigemitia
(собака) и / dirutnpens (кишечник змеи Bills arietans), ооцисты которых
созревают (т. е. спорулируют), находясь еще в соединительной ткани
кишечника, а затем уже выводятся во внешнюю среду и служат для
заражения нового хозяина.
Следовательно, у разных кокцндии намечается постепенный процесс
погружения шизонтов, га-
монтов и ооцист в соеди-
нительную ткань стенки
кишечника; при этом, од-
нако, сохраняется способ-
ность ооцист к существо-
ванию вне организма хо-
зяина и нс возникает еще
приспособления к переда-
че спорозоитов в другого
хозяина иным путем, не
через кишечник.
Дальнейшая эволю-
ция кишечных паразитов
в сторону кровепарази-
тизма могла произойти
только с включенном в их
жизненный цикл нового
звена, а именно: перенос-
чика из числа кровососу-
щих беспозвоночных Пер
вын этап в этом направ-
лении даст нам развитие
кокцндии Schellackia bo-
livari (рис. 51), парази-
тирующей у ящериц
(Acanthodactylus. Psam-
modromus). Согласно ис-
следованию Реихенова
(Reichenow. 1919) жиз-
ненный цикл этой кокци-
дии начинается с пассив-
ного заноса спорозоита в
кишечник ящерицы, где
он внедряется в эпителий
и Начинает размножаться
в нем посредством шизо-
гонии.
После нескольких
бесполых поколений фор-
мируются мужские и жен-
ские первичные половые
клетки (микро- и макро-
гамонты). Первые раз-
Рнс. 51. Жизненный цикл кокцндии Schellackia 1м>-
livari в ящерице и в клеще Liponyssus saurarum.
(По Рейхенову.)
1 — вхождение спорозоитом в клетку кишечника ящерицы:
а-.; - сталии ишююнии н стенке кишки; 6—9 — рост микро-
гамстоиита и образование микрогзмет н эпителии кишечника;
10. 11 — рост макрогаметы всубэпителнлльной ткани 1£ — опло-
дотворение макрогаметы; 13 /о рост зиготы н образование н
получающейся из исе ооцисте спорозоитов; 17 — проникновение
спорозоита и кровеносный сосуд ящерицы; IS — спорозоиты
внутри эритроцитов; /9 — красные кроенные тельца со споро-
зоитами фагоцитируются клетками кишечного эпителия клеша:
20 — красные кропимые тельца внутри кишечных клеток клеша;
21 — 1 руина спорозоитов, скопившихся и кишечной клетке клеша
после того, как содержавшие их эритроциты ящерицы подверг-
лись перевариванию. При послания клеша 119—21] ящерицей
(1-1S) спорозоиты проникают и кишечный эпителий яшериин.
и пик.т начинается снова.
виваются в тех же эпителиальных клетках, где шло бесполое размноже-
ние, вторые же погружаются в субэпителиальпую соединительную ткань
и уже там созревают в макрогаметы. Образовавшиеся из микрогаметоци-
та мужские гаметы затем тоже проникают в субэпителиальную ткань и
оплодотворяют макрогамету. Получившаяся зигота развивается
73
здесь же, но не образует спор, а лопается, и освобождающиеся из нее
спорозоиты попадают в капилляры кровеносной системы, ветвящиеся в
соединительной ткани, и внедряются в красные кровяные клетки, В по-
следних они не развиваются н остаются до тех пор, пока не попадут
вместе с кровью в кишечник клеша Liponyssus saurarum — временною
паразита ящериц. Клещ сосет кровь, после чего эритроциты вместе
с заключенными в них спорозоитами захватываются эпителиальными
клетками кишечника. Внутри клеток эритроциты перевариваются, а спо-
розоиты освобождаются, по не перевариваются, а пассивно остаются в
клетках кишечника клеща до тех пор, пока он не будет случайно съеден
ящерицей, как это часто бывает с Liponyssus. В кишечнике ящерицы
клещ переваривается, а спорозоиты активно внедряются в эпи гелий
кишки и возобновляют только что описанное нами развитие.
Итак, у Schellackia мы встречаем ряд серьезных изменений п в жиз-
ненном цикле, и в морфологии. Прежде всего происходит временный
отрыв паразита от стенок кишки и переход к жизни в эритроцитах.
Однако эта часть цикла еще сравнительно ничтожна (всего одна ста-
дия), не активна и не сопровождается ростом; в обоих хозяевах пара-'
шт еще длительно живет в клетках кишки, и эритроцитная стадия есть
лить необходимый передаточный пункт от кишечной клетки ящерицы к
кишечной клетке клеща. Далее, переход в кровь неизбежно влечет за
собой появление промежуточного хозяина (клеща). Наконец, измене-
нием в морфологии паразита является утрата оболочек спор. Она
вполне объяснима биологически, ибо Schellackia ни одного момента
жизни не проводит вне тела одного из хозяев, а потому спо-
розоиты и не нуждаются в защите, и оболочки спор становятся излиш-
ними.
Примером дальнейшего развития кровепаразитизма может слу-
жить кокцидия Lankeslerella minima, паразитирующая в лягушке. Как
и у Schellackia. заражение здесь происходит через кишечник. Однако, но
исследованиям Цёллере (NoIler, 1920). спорозоит Lankeslerella (рис. 52)
сразу проникает через эпителиальный и субэпителиальный слон кишки
в капилляры и затем внедряется в эндотелиальные клетки сосудов раз
пых органов. В этих клетках происходит бесполое размножение пара-
зита. Освобождающиеся из клеток мерозоиты попадают в кровь и зара-
жают новые эндотелиальные клетки, где повторяют процесс бесполого
размножения. В результате ряда шизогоний возникают гамонты, созре-
вающие н совершающие процесс оплодотворения в тех же эндотелиаль-
ных клетках. Здесь образуется зигота, которая покрывается оболочкой
и становится ооцистой. В ней формируется без образования споробластов
п спор множество спорозоитов. Последние выходят из ооцисты, снова
попадают в кровь, где и внедряются в эритроциты. В них спорозоиты
остаются без всяких изменений до тех пор, пока не попадут в кишечник
пиявки Hemiclepsis marginala. где проникают в клетки эпителия кпшеч
пика. Как спорозоиты Schellackia в клеще, так и спорозоиты Lankesfe-
rella в пиявке никакого развития в организме этих хозяев не пределы
вают, а просто сохраняются до тех пор, пока нс попадут в кишечник ля-
гушки, нередко пожирающей пиявок. Здесь они снова начинают свой
жизненный цикл.
Следовательно, у Lankeslerella весь жизненный цикл проходит
внутри эндотелиальных клеток, и только спорозоиты проникают в эри-
троциты. Дальнейшая эволюция в направлении кровеиаразитизма со-
стоит в том. что паразиты начинают внедряться в эритроциты нс на
стадии спорозоитов, а на более ранних этапах развития, в связи с чем
развитие в эндотелии сосудов все более сокращается
71
нкгхкохоЛон.' o.> ехлнпоо bki-'oiIe — t/r
:эмхэ1Г.ч ионч.тнга loiri.e 11 L<t>t(iiPfl'xliiK(fu<li.i -ffijinio» - л :«лчо omx-H-m иончь-р
-нглхоине и нике., иояэпэж эмпаЛояли.огио A .hOi.ijo,' XBMiairM хнн-цггнгэхогп*.
x.iKv.iod.xint >HHBiiot'i>*d >0 и рхипох.и-ех оюилилк ijod - > 9 :XEMIM'M•хннчивш'пхог
-П€ Я ПОХНЦЛ 1ЭКГ..1 НГ«1оф ЭНГиИПн < !u.4tU4l!t I Ц ЛНИ <1НГ И 3HHI4SU0H •HllKlCOdOIl.»
j м»1 iiiiiioxorнт огпьен ’aoi лпрУ хвнхагн xriicirviili'nioLiie п hhiiojoeiihi ннггхх - /
слс!о|ггл[] о]р JMm.UKV я uuniuui inf.u.ns^i/ui^ fiiunati;c[ 3££ ’.’•Щ]
У некоторых Haeniosporidia (Haemoproteus) развитие в эндотелиаль-
ных клетках сосудов внутренних органов доходит лишь до стадии гаме-
тоцитов (спорозоит — шизогония — мерозоит — гаметоцит), которые вне-
Рис. 53. Жизненный цикл'.//aenioproteus; columbite,
схема (Но Веньону.Ч
in, 2а рост женский гаметы в крленнм кровяном tl-.ii.uc;
Ю, 20 — рост мужского гаметоцита; ,Т«, In округление
женской гаметы н выход ее из красного кроимного тельца
голубя: ЛЬ, 40 — округление мужского гаметоцита и образо-
вание мнкроглмет; 5 — онлиьотворепне; 6. 7 «jopaaoiwuiiH*
оокинеты. которая проникает через стенку кишки кровососки
l.ynrhla тайга'. 8 - молодая ооциста на стенке кишки кро-
вососки; 11 - зрелая ооциста со еппрпзоптачп. которые про-
никают и слюнные железы I. тачга и затем попадают и крон),
голубя; 10 спорозиш' проникает и клетку эидоте.шя сосуда
голубя; it - рост спорозоита н giiiore-iiia.o.nort клетке;
12—15 — после лона тельное размножение паразита путем шизо-
гонии; 18 — 17 — распаление продуктов niiixoroiiiiii на мно-
жество мелких молодых гам питии, проникающих в красные
кротиные клетки.
дряются уже в эритроциты
(рис. 53). А у представите-
лей рода Plasmodium в эрп-
ipomrrax развиваются не
только гамонты, но также и
шизонты, размножающиеся
многократно путем шизого-
нии. Однако процесс шизо-
гонии у малярийных парази-
тов происходит как в эри-
троцитах, так и экзоэрптро-
цнтарно. подобно Haemopro-
teus. Спорозоиты малярий-
ных плазмодиев птиц Plas-
modium ctithemerium или Pl.
gallinucciini проникают, no
данным Хаффа с сотр. (Huff
a. al., 1944—1946) и Каул-
стопа. Кантрелла и Хаффа
(Collision, Cantrell a. Hull,
1945), в клетки ретикуло-
эндотелиальной системы,
преимущественно в эндоте-
лий кровеносных сосудов, в
макрофаги, моноциты, рети-
кулярные клетки различных
внутренних органов, где пре-
терпеваю г преэрнтроцитар-
иую шизогонию; образовав-
шиеся при этом мерозоиты
проникают уже в эритроци-
ты. Экзоэритроцитарная ши-
зогония PI. mexicanum в
ящерицах может протекать
также и в молодых клетках
эритроцитарного ряда (эри-
тробласты и нормоб.тасты).
Прсэритроци гарна я шизого-
ния } Pt. vivax, PI. falcina-
rutn человека и Pl- knowlcsi.
Pl. cynomolgy, Pl. gouderi
обезьян осуществляется в
клетках печени (рис. 54).
Приведенные приме ры
дают ясную картину (рис
55) того, как первоначально
чисто эпителиальные кишеч-
ные формы кокцидий (Eimc-
ria intestinalis, Е. perforans)
становятся субэпителиальными (Е. magna, Е. irresidua). Далее,
еще в пределах группы кокцидий, происходит (Schellackia) сна-
чала кратковременный отрыв от эпителия кишечника позвоночного и
76
Pile. 54. Цикл развития малярийного царазнта Plasmodium viuax. (Оригинал.)
/ — снороз’оигн: 2—4 — лре^рпгроиитариые стмим развития, шизогония в клетках печени; 5— 10 — эрн-
ipouiirapiiue сталии развития;.»—s> - шизогония; 9 — сегментация шизонта и образование мерозоигои; К) —
иерозоиты. вышедшие на эритроцитов; 11—14 — гамонты; 11—13 — .макрогачонты; 14 -14 — янкрогяаюнты:
jj _ макрогамета; 16 — микрогамегацнт; 17 — образование микрогамет; 18 — копуляция; 19-20 - зигота
и ш оокинета; SI — проникновение оокинеты через эпителий стенки желудка комара; 22 26 - ооцисты.»
которых происходит рязнитис спорозоитов; 27 — спорозоиты в клетках слюнных желез комара.
появляется короткая (всего одна фаза — спорозоит) внугрнэрн
троцнтная стадия. Еще далее (l.ankesterella) отрыв от эпителия киш
ки позвоночного делается постоянным (эпителий служит лишь
для прохождения через него спорозоитов), но паразит проходит все свое
развитие в эндотелии сосудов, т. е. все же в эпителиеподобной ткани-
внутри эритроцитов паразит по-нрежнемх проходит лишь стадию споро-
зоита.
Наконец, при переходе к отряду Haeinosporidia совершается они
больший отрыв от эндотелия и переход гамонтов (Haemoproteus) изн
шизонтов и гамонтов (Plasmodium) к локализации внутри эритроцитов;
наступает типичный кповепаразитнзм (см. рнс 55) *
ХОЗЯи 4 (nC3t)ilh04ht>U) ieuinn9 cpela .Zww*
Кишеччик Hsfrrnw )ри"'рЗЦ1/-ПЦ
3nu,neruu -V
Г tinert а -&ilt salts c 0@
Г HWO | По 0 o? о e. (®? 0 0@
Isosрога oi^emtna 0 ©V 0 О/
Schellacfi’a bit Ivar i я V
Lanhesicrtllj ятю >J08 0
Чаеп/oprateus columba о
PiZS medium gsHinaceurn o
Р Стряют □ б Оо^та
@ S gj g Мг&сгЯет.) А Оош-т
ipumpouuire Г*ъ/ ъе C7S
О " .?{£?. - w 0« зосаюь " f'. СТ Ооцигти со
Qj ".Г-ЕГШЗТ ‘ {У V/ гчерок.-г ЗУ
Рис. 55 Схима. иллюстрирующая перезол у Sporozoa ог кишечного паразитизм;! к
кроненар.ичтизму. (Орнгии; и
Однако с того .момента, как в эритроцитах оказываются не споро-
оиты, а более ранние стадии, например гаметоциты, один перенос их ст
одного хозяина к другому (как эго имеет место у Sc/iellackia и Lan
kesterella) ока ывается недостаточным. Для возможности сражения
нового позвоночного хозяина требуется предварительное окончание раз-
витая до стадии спорозоита, которое в этих случаях и перемешается и,
организм переносчика кровососущего членистоногого. Вместе с перено-
сом части жизненного цпкза во второго хозяина меняется и способ рас-
пространения паразита, прежний способ—пожимание нромежхточного
хозяина основным —оказывается нс вполне выгодным для паразита, ибо
нередко в этих случаях в основного хозяина будут попадать паразиты, пи-
закончившие еще своего развития и потому не способные прижиться
у него. Поэтому данным способ будет вытесняться другим — непосред-
ственным возвращением спорозоитов из переносчика в позвоночного во-
время акта кровососания. В дальнейшем такой способ передачи стано-
вится единственным (Haemosporidia).
Таким образом, можно думать, что среди Sporozoa кровепаразиты
позвоночных имеют в качестве своих отдаленных предков кишечных па-
разитов тех же позвоночных Однако но отношению к кровяным жгутике -
носцам, каковы многочисленные виды Trypanosoma, правильнее всею
предполагать, что их первоначальными хозяевами были различные бес-
позвоночные, главных» образом насекомые, не имевшие специализирован
ного кровососущего способа питания Предки трппанозом жплщв кишем
нике таких хозяев. В пользу этого очень важ
ным свидетельством является то обстоятель
ство, что в настоящее время имеется много на-
секомых (блохи, мухи и др.) как кровососу
тих. так даже и не кровососущих, в кишечни-
ке которых живут ближайшие родичи трипа-
нозом. а именно: жгутиконосцы Leptomonas и
Crithidia. Эти жгутиконосцы (рис. 56) весь
свой цикл проделывают в кишке членистоного-
го, а заражение ими совершается через рот,
т с. они не имеют никакой связи с кровепара-
зитнзмом. Когда впоследствии эти насекомые
стали питаться кровью позвоночных, строго
специализируясь в смысле питания, у кишеч-
ных жгутиконосцев получилась возможность
контакта с позвоночными и увеличились шан-
Рис 5G. Схема сгроенн'
трш1.1нозомид. (По Гоару.Х
I — рол Leptomonas: /<
pot Crithidiu: С— род
Trypanosoma.
сы попадания во время акта сосания в кровь
Кровь оказалась более питательной средой, чем содержимое кишечника.
Поэтому случайно попадающие в кровь жгутиконосцы оставались в ней
подолгу жить и даже продолжали размножаться. В свою очередь, дли-
тельность пребывания жгутиконосцев в крови давала им шансы па воз-
врат в кишечный тракт насекомого при одном из последующих актов кро-
вососания. Не утратив способности жить в кишечнике насекомого, такие
попавшие из позвоночного в насекомое паразиты сохранялись в послед-
нем, нс погибая. Позднее жизненный цикл таких жгутиконосцев диффе-
ренцировался на ряд стадий, часть которых регулярно проводится пара
знгом в позвоночном, другая же часть столь же регулярно — в кишечни-
ке кровососущего насекомого (Ноаге, 1960)
Произведенный обзор ясно показывает, что путей к паразитиче-
скому образу жизни от свободного имелось несколько, причем в извест-
ной связи с этим обстоятельством находится и необычайное разнообра-
зие отношений между паразитами и хозяевами.
4. ДРЕВНОСТЬ ПАРАЗИТИЗМА И УСЛОВИЯ ЕГО ВОЗНИКНОВЕНИЯ
Закончим эту главу рассмотрением вопроса о времени возникнове-
ния паразитизма в ходе эволюции жизни в условиях, способствующих его
возникновению Вопрос о том. когда в ходе эволюции жизни на земле
мог впервые возникнуть паразитизм, естественно, не поддается точному
определению, по позволяет высказать но этому поводу некоторые косвен
ные соображения
Пространственные и пищевые взаимоотношения (хозяин как место-
обитания н источник пиши паразита) в паре паразит — хозяин таковы.
что паразиты обыкновенно значительно мельче хозяина. .Чиненные раз-
меры паразита, как правило, не менее чем в десятки, а объемные, сле-
довательно, в сотни рл меньше размеров хозяина. Отсюда напраши-
вается вывод, что паразитизм мог развиться в животном царстве в ши
роком .масштабе лишь после того, как животные организмы успели
аволюцпонировать в значительной степени и дать целый ряд Форм, су
щеетвенно различающихся своими размерами. Только тогда меньшие
формы получили возможность поселиться на больших или внутри их.
Несколько более конкретно можно подойти к вопросу о том, ни
когда появился паразитизм, а когда он впервые получил более или ме-
нее широкое распространение. Прежде всего начали, вероятно, приспо-
собляться к паразитизму простейшие, в Ч( м им способствовали их микро-
скопические размеры. Однако и среди них паразитизм как болте или
менее широко распространенное явление мог развиться не ранее, чем
появились различные группы свободно живущих червей В самом деле,
средн простейших отсутствуют формы, паразитирующие у губок, и лишь
в виде редчайших исключений имеются простейшие, наразнтипгющие у
кишечнополостных (жгутиконосцы Trypanophis в гастральной полости
епфонофор, Amoeba hydroxena, инфузории Trichoclina и Кегопа на шдре,
инфузории Foettingeriidae в актиниях). Только начиная с червей, а в
частности с кольчатых червей, мы наталкиваемся на широкое приспособ
лбине простейших к паразитизму. В аннелпдах п других высших червях
имеются многочисленные инфузории, сотни видов грегарии, кокцидин-
микроспоридии, Actinoniyxidia, многие виды Peridinea (Haplozoon и д|
в Polychaeta). Итак, время более широкого развития паразитизма среди
одноклеточных мы можем относительно определить как совпадающие
или последующее за эволюцией кольчатых червей.
Развитие паразитизма у паразитических червей относится уже, ве-
роятно, к топ части палеонтологической летописи, которая нам известна,
а именно: сопутствует или следует за появлением позвоночных. В самом
деле, все представители современных паразитических гельминтов в поло-
возрелом состоянии, как правило (кроме очень немногих сосальщиков и
нематод), живут в позвоночных. В беспозвоночных встречаются только
личиночные стадии их, дтя которых моллюски и членистоногие служат
промежуточными хозяевами. Но самое явление развития с промежуточ-
ными хозяевами есть явление вторичное, которому предшествовало раз-
витие простое, без смены хозяев. Таким образом, беспозвоночные вклю-
чились в никл гольмии job лишь позже, чем они стали паразитами позво-
ночных.
Путем этих рассуждений мы приходим к выводу, что паразитические
черви появились не ранее силура—девона. Правда, среди нематод имеет-
ся несколько десятков видов, паразитирующих в перепончатокрылых,
жуках и др., но развитие их должно быть отнесено к гораздо более позд-
ним эпохам, к мезозою, так как сами хозяева их начинают появляться не
ранее верхних горизонтов палеозоя. В.ттьшая группа паразитов (до
40 000 видов) образуется в'среде двукрылых и перепончатокрылых (та-
хины. наездники), но эволюцию этих паразитических форм можно сг-
нести не ранее как к юрской эпохе, так как самые отряды двукрылых и
перепончатокрылых ведут свое начало с юры. Возраст некоторых пара-
зитических раков труднее поддается определению, так как среди них
иногда целые группы (Bopyridac, Entoniscidae. многие Cirripediа) при-
способились к жизни на или внутри других ракообр иных или гаже коль-
чатых червей (Xenocoeloma, Herpyllobius и др. из Copepoda), а также
в игло. ежих и в клоакалыюн полости асцидий.
В последнее время относительно отдельных видов паразитов како-
во
пились ьолее конкретные данные, которые мы приводим, так как они
ие лишены интереса. Наиболее древние несомненные находки паразитов
в ископаемом состоянии относятся к видоизмененным кольчатым червям,
Vlyzostomidae, образующим галлы, или так называемые зооцецидии, на
теле морских лилий. Такие галлы найдены нс только у современных мор-
ских лилии, по и у ископаемых вплоть до юрского периода.
В балтийском янтаре, г. е. в олигоцене, не раз были найдены насеко-
мые с выходящими из их тела нематодами. В том же янтаре была обна-
ружена блоха Palaeopsylla debsiana. Отсутствие у нее глаз, указывает
па то, что она прошла уже долгий предварительный путь эволюции в
паразитическом состоянии.
Павловский (1916 а) приводит целый ряд любопытных данных о
наиболее давних сведениях относительно некоторых паразитов человека.
Наиболее древней находкой является обнаружение объизвесгвленных
яиц кровяной двуусукп (Schistosoma haemainbium) в почках мумии
Х\ династии фараонов Египта (1200—1090 до н. э.). О глистных заболе-
ваниях упоминается в вавилонско-ассирийских клинописных таблицах.
PnCiHKii на фигурных глиняных сосудах древних перуанцев гово-
рят о том, что еще за много столетий до открытия Америки европейцами,
население этого материка страдало от песчаной блохи (Sarcopsylla
penetrans) и от лейшманиозов (болезнь Ута), Новые доказательства
древности челове icckhx паразитов дает исследование кишечника двух
трупов, найденных в торфяниках бывшей Восточной Пруссии. Шил ат
(Szidat, 1911) сообщает, например, о нахождении в кишечнике обоих
трупов яиц аскариды, власоглава, а также, .может быть, и лептона широ-
кого. Между тем обе означенные находки датируются ранним леднико-
вым периодом.
Наконец, в вопросе о происхождении паразитизма немалую роль
играют и морфологические, и физиологические, и экологические особен
пости живо! пых, становящихся хозяевами паразитов, Паразнтофауна
любого животного в се современном составе отнюдь не представ <яет
чего-нибудь строго постоянного и устойчивого: она может меняться как
в сторону уменьшения, так и в сторону увеличения се разнообразия.
Один п тот же хозяин в разных местах и при разных условиях может со-
держать в значительной мере разную паразитофауну, становится хозяи-
ном ряда новых паразитов.
Этот процесс становления организма хозяином паразитов представ-
тяет большой интерес и рассматривался Догелем (1927 6), а затем под-
вергся специальному анализу со стороны Павловского (1946 а). Целый
ряд экспериментальных данных показывает, что многие организмы лими-
тируются в приобретении ими новых паразитов не только какими-нибудь
эндогенными ииршпдмн (специфичность), но в начителыюй мере ря-
дом вшшпих условии, устранение которых ведет к тому, что организм
становится хозяином новых для него видов паразитов. Таким образом,
помимо действительного, или фактического, набора паразитов опреде-
ленного хозяина, можно еще говорить о его потенции я ь п о й п-а р а-
3 н то фа у н с.
Понятие о потенциальных паразитах имеет большой теоретический и
практический интерес Теоретически оно затрагивает вопрос о приобре-
тении организмом свойственного им.у ныне набора паразитов в процессе
эволюции. Практически потенциальны! паразиты организма являются
угрозой, всегда нависающей над организмом их возможного хозяина.
Разбирая ф а к >ор га, способствующие заражению хо
з я и и а о и р е д г л е и и ы м паразита м, Павловский относит к числу
их следующие условия.
Ь В. А. Догель
81
Во-первых, факторы, пре драсполагающие организм к заражению его
теми пли иными паразитами. Такими являются, с од.юн стороны, строе
нпс и функции организма, при которых возможно лроникнг»в1 ипе пара-
зита в организм. С дру гой стороны, сюда относится .характер биохнмпзма
опосредуемого паразитом организма, при котором возможно не только
переживание паразита в новом хозяине, но и дальнейшее прохождение
в нем жизненного цикла.
Во-вторы.х необходимо действие факторов, устанавливающие биоц>
нотичсскне связи между организмом и его потенциальным паразитом
(через посредство промежуточных хозяев пли через иные способы инва-
зии).
Первую категорию факторов мы можем называть эндогенными,
зависящими лишь от самого организма хозяина, вторую — экзоген-
н ы м и, или зависящими от окружающих условии.
Наконец, третьим моментом становления животного хо .яипом пара-
зитов является наступление условий, при которых факторы внешней
среды, действующие и на восприимчивого хозяина и на вирулентного
нара.шта, в конечном счете ос’ Ществляют возможность приживания ви-
рулентного паразита, проникшего в организм чувствительного хозяина
(Павловский и Гнездилов, 1939).
Сочетание всех этих разнообразных условий и их воздействие очень
хорошо иллюстрируется экспериментами Павловского, Гнездилова и Та-
лызина (1936, 1939) над плероцеркоидами Diphyllobcrthrium latum. Они
паразитируют во внутренних органах хищных рыб (окунь, щука, налим
и др.). При поедании одной более крупной рыбой другой, Солее мелкой,
зараженной плероцеркоидами, проглоченные вместе с добычей плероцер-
коиды проходя г сквозь стенку желудка проглотившей хищной рыбы н
поселяются в полости тела. В то же время в мирных рыбах, как, напри
мер, в карпе, плероцеркоиды никогда не обнаруживаются. Однако при
искусственном введении живых плероцеркоидов в желудок карпа они
пролезают через стенки желудка в полость тела н там приживаются.
Отсюда ясно, что в данном случае кр\г промежуточных хозяев
плероцеркоидов среди рыб определяется не устройством рта и кишечника
определенных видов рыб (рот карпа способен проглотить плероцеркоида)
и не биохпмпз.мом разных видов рыб (ибо плероцеркоиды, введенные
в кишечник карпа экспериментальным путем, удачно приживаются в
этой рыбе), tipufcnibi нсзаражениостп карпа (и других мирных рыб)
плероцеркоидами, очевидно, экзогенного характера и зависят от отсут
с гния пищевой связи между карпом и содержащими плероцеркоидов
рыбами (щука, налим и др.), т. е.. вернее, односторонностью этой связи
хищные рыбы поедают карма, но не поедаются им
Столь же легко, как у карпа, приживаются экспериментально введен
ные плероцеркоиды и у миноги, которая нормально никогда их не со-
держит. По причина этого отсутствия, по-видимому, иная, чем у карпа,
и зависит от анатомо-физиологических особенностей пищеварительного
аппарата миноги. Миноги нападают на разных рыб. причем своим круг-
тым ртом присасываются к телу рыбы и перетирают ее ткани множе-
ством роговых 31 бон ротовой воронки. Такому' перетиранию должны
подвергаться и плероцеркоиды, если они имеются в поедаемой миногой
рыбе.
Однако возможности заражения плероцеркоида ми новых хозяев го-
раздо более широки и далеко выводят за пределы класса рыб. Аналогич-
ные вышеприведенным успешные опыты введения плероцеркоидов были
проведены и с травяной лягушкой, жерлянкой, разными видами ящериц,
ужами, гадюкой и даже черепахой.
42
Таким образом, и здесь причиной отсутствия плероцеркоидов в пере-
численных животных служит не действительная невозможность этих
животных заражаться плероцеркоидами, а отсутствие пищевых связей
между ними и нормальными хозяевами плероцеркоидов. I включение со-
ставляет только черепаха, у которой отсутствие заражений может зави-
сеть не только от отсутствия пищевых связен (экзогенная причина), но
и от строения желудка взрослых черепах (эндогенная причина), а имен-
но: заражение черепах удается лишь у молодых, годовалых черепах,
тогда как хрящеватая плотность стопок желудка взрослой черепахи ме-
шает заражению плероцеркоидами.
Кроме того, опыты с ящерицами выявили наличие еще одного фак-
тора, а именно: температурного. Дело в том, что опыты заражения яще-
риц плероцеркоидами удавались лишь при комнатной температуре, т е.
при температуре, которая приблизительно отвечает таковой нормального
местообитания плероцеркоидов (температура пресноводных водоемов в
летнее время). При помещении же ящериц в термостат с температурой
37° С в опытах миграции плероцеркоидов не происходило, и они поги-
бали.
Другой пример влияния температуры, хотя и в обратном направле-
нии, даю г опыты .Матова (Matofi, 194-4) по заражению трихинами обычно
не свойственных им хозятв, а именно: различных амфибий, т. е. холодно-
кровных позвоночных. Заражая амфибий (жаб, саламандр) мускуль-
ными трихинами при температурах 18—25°С, Матов не добился никаких
ясных результатов. Однако при содержании подопытных животных в
термостате при 31—37° С личинки трихин выходили в их кишечнике из
капсул, развивались в самцов и самок, а у саламандр дело доходило
даже до откладки самками личиной и до развития мускульных трихин.
Правда, в последнем случае постановка опытов была искусственной, так
как самки трихин вводились автором прямо в полость тела саламандры.
Все эти примеры показывают нам, что потенциально весьма многие
организмы могли бы стать хозяевами для гораздо более многочислен-
ного контингента паразитов, чем тот, которым они в действительности
обладают, и, наоборот, многие паразиты могли бы иметь потенциально
гораздо более широкий круг хозяев. Кроме того, разобранные случаи
говорят, в какой сильной мере приобретение паразитом новых хозяев
зависит от комбинации факторов разного порядка и частью эндогенного,
частью экзогенного характера: строение и функции организма хозяина,
биоценотнческие связи хозяина с проспективным паразитом, наконец,
условия среды, способствующие или не препятствующие проникновению
паразита в организм хозяина (температура и др.) и приживанию его в
последнем.
Отсюда Павловский (1940а) приходит к выводу, что установление
связи «паразит и хозяин» возникает на базе случайных благоприятных^
признаков и особенностей, физиологически развившихся вполне незави-
симо у разных организмов, но в процессе дальнейшей эволюции эти
случайно благоприятные признаки и соотношения становятся исходной
базой для естественного отбора. При этом создаются и закрепляются но-
вые особенности и связи, приводящие к возникновению биологических
систем — «хозяева и их паразиты».
6*
ГЛАВА VII
РАЗЛИЧНЫЕ ПУТИ ПРОНИКНОВЕНИЯ ПАРАЗИТОВ
В ОРГАНИЗМ ХОЗЯИНА
Разобрав различные способы происхождения паразитизма целе-
сообразно подвергнуть рассмотрению вопрос о путях проникновения
паразита в хозяина, так как оба эти вопроса весьма тесно связаны.
Соответственно особенностям локализации в теле и на теле
хозяина паразиты, как мы уже видели, могут быть разделены на
эктопаразитов и эндопаразитов, причем среди последних,
в свою очередь, удобно различать кишечных и внутренностных.
Под кишечными мы подразумеваем тс формы, которые живут
либо в самом кишечнике, либо в непосредственных его выпячива-
ниях, как-то: желчный пузырь, печень, мочевой пузырь амфибий,
клоака и ее выросты, как, например, фабрпцпева су мка птиц.
Внутренностными (не внутренними) мы называем всех
паразитов, локализующихся между наружными покровами и стенками
кишечника; таким образом, сюда будут принадлежать и полостные,
и кровяные паразиты, и формы, живущие в мускулатуре, соедини-
тельной ткани и т. д. Последних часто определяют специальным тер-
мином «тканевые паразиты».
Теоретически рассуждая, для проникновения паразита в организм
имеются два главных пути — через покровы или через рот и далее
мере? пищеварительный канал, соответственно с чем мы и будем раз-
личать экзогенный и эндогенный пути проникновения. Вполне
естественно, что установление связи эктопаразитов с хозяевами всегда
носит экзогенный характер, и, как правило, эктопаразиты проходят
весь своп цикл развития в покровах хозяина или на них.
Однако все же есть случаи, когда эктопаразптической стадии
предшествует внутренностная Это имеет место в одном из классиче- »
ских примеров паразитизма. а именно: у SaccuLna. Взрослая Sacculina
живет в виде мешковидного нароста на нижней стороне брюшка
у различных крабов, например у Inachus. Этому ее состоянию пред-
шествует длительная внутренностная стадия, когда паразит целиком
находится в полости тела краба и только после многинедельной
скрытой жизни образует опухоль на брюшной стороне тела; потом
эта опухоль прорывается, Sacculina высовывается наружу и становит-
ся эктопаразитом (см. рис. 104).
S4
Нечто аналогичное представляют и личинки бычьею оьода
llypoderma lineahun (рис 57), которые вылупляются сначала на
коже хозяина из яиц. прикле-
енных I- шерсти, и вбуравли
каются в кожу. Далее личинки
проделывают внутри животно-
го длительную миграцию. Они
поднимаются по соединитель-
ной ткани в дорсальном на-
правлении, проходят тангенцп
альио через стенки нитевода
(не попадая в его просвет), по-
том мигрир\Ю1, часто Через
спинномозговой канал позво-
ночника, под кожные покровы
спины. Здесь они задержпва
ются в подкожной соединитель-
ной ткани, устанавливая не-
посредственную связь с вн< ai-
неи средой при помощи дыха-
тельного отверстия в коже хо-
шина. Таким образом, тесная
Рис. 57. Силин разлития кижного оно да Ну-
pudi-rma liii'-atum (/-/. bovis)
Л — яйца опола, прикрепленные к полосу;
И— личинки первой сталии; С—кусок пище-
вода быка с личинками оволч; D — выросшая
личинка овола. вынутая из кожи быка. (По
Скрябину.)
связь подрастающих личинок с покровами хозяина является вторичной
и следующей за долгим периодом внутренней миграции.
Для кишечных паразитов эндогенное иро-
Рпс. 58. Схема миграции
личинок Ascaris Itimbri-
coides в теле человека.
[По Павловскому.)
I — рог; 2 — глотка. 3 — пи-
meiioj; 4 — иылуплспис личи-
нок в кишечнике: 5 - миграции
их в пени кишки; о - печеш.;
/ — пцанос гердпе; S — легкие;
9 — ко нрат .iii'iiiiiKK в пище-
вод: ТУ — икЕвксиие .TI14HUKU-
ми подовом зрелости и кишеч-
нике.
iHiKHOLi’itne в хозяина через рот является
совершенно естественным и потому наиболее рас-
пространенным Однако и от этого правила име-
ются отступления, например у Anculostoma duo-
denal? При контакте обнаженных покровов ч<
ловека с землей находящиеся в ней личинки Ап-
cyloatoma вб\ ра вливаются в кожу, попадают в
кровеносные сосуды и кровью заносятся сначала
в сердце, потом в легкие. В легких личинки вы-
нол ают в полость бронхов, потом отсюда мигри-
руют в трахею и глотку, а из глотки спускаются
по пищеварительному каналу вплоть до двена-
дцатиперстной кишки, т. е к месту своей оконча-
тельной локализации. Здесь, следовательно, ки
щечному паразитизму предшествует внутренпост
ный и притом экзогенного происхождения. Впро-
чем заражение может происходить и при загла-
тывании тичинок хозяином, но тогда миграции
не происходит. В природе этот путь осущест-
вляется весьма редко.
Внутренностный пара з и т и з м,
предшествующий кишечному, наблю-
дается. например, у Ascaris, проникающих в .хо-
зяина, в противоположность анкилостоме, эндо-
генным путем.
Яйца аскарид, выброшенные с экскремента-
ми и развивши*- в себе личпцку, заглатываются
через рот, попадают в кишечник и достав-
тяются таким образом к месту своей дефинитивной локализации. Но
на самом деле личинки немедленно покидают кишку, вбуравливаясь
85
в ее стейку, и попадают в капилляры, а затем гем же способом, как
и личинки Ancylostoma, достигают легких. Там личинки живут в тече-
ние нескольких дней и лишь после этого через трахею, глотку и пище-
вод вторично проникают в кишечник и там прочно поселяются и дости-
гают половозрелости (рис. 58).
В дополнение к указанным аберрантным случаям проникновения
паразитов в кишечник можно указать еще па один паразитологический
парадокс, а именно: на веслоногого рачка Xenocoeloma brumpti
(рис. 59), паразитирующего па черве Polycirrus (Polychaeta) и
описанного Коллери и Мениль (Caullerj et Mesnil, 1919). Xenocoeloma
Рис. 59. Рачок Xcnocucloma brumpti (Copepoda), паразпгирую-
tiitiit ил по.шхете /’olycirrttx
Л — два рачка с яйцевыми мешками, прикрепившиеся к
хозяину: В — пара.шт, си iniiptii иа участке тела хозяина, при
большом увеличении. (Но Коллери и .Мениль.)
1 — семенники Xenocoeloma . 2 — яичник; 3 — псекэн полость рачкэ; 4
сонрериклиие ооинги: 5 — кишечник Pi tvcirrus, наполнении» песчинками.
имеет вид бесформенного колбасовндпого образования с двумя яйце
выми мошками — типичный признак Copepoda—на заднем конце
тела. Он внедряется через кожу хозяина в полость тела полихеты,
прорастает через весь целом и врастает своим передним концом
в стопку кишки хозяина Между тем задний конец рачка с яйцевыми
мешками торчит далеко наружу из тела Polycirrus. Итак следователь-
но. часть тела Xenocoeloma находится в целоме хозяина, большая часть
паразита и яйца торчат наружу, так что вылупляющиеся личинки
могут выхолить прямо в воду, по пищ’’ свою рачок пол\чает из кишем
пика хозяина, напоминая в этом отношении кишечных паразитов.
Наибольшее разнообразие путей проникновения в хозяина обнару-
живают внутренностные эндопаразиты.
Часть внутренностных паразитов проникает к месту своей лока-
лизации через стенки кишки. Это наблюдается, например,
у гемигрсгарлны KaryoLysus lacertaruin. ра увивающейся в эндотелии
86
сосудов и в эритроцитах ящериц, или у Hepatozoon muris, локализую
щейся в клетках печени и моиопуклеарах крыс и мышей. Спорозоиты
лэ переносчика (тамазовые клещи) попадают в кишечник хозяина
и оттуда через эпителий ворсинок в капилляры кровеносных сосудов
и далее в места окончательной локализации. Через кишечник в кровь
проникают и трпланозомы с контамипативьым способом заражения
(Trypanosoma lewisi, Т melophagium)
Многочисленные полостные грегарины встречаются в целоме,
кров( поеных сосудах, семенных мешках, мальпигиевых соеудах, жиро-
вом теле и других органах червей, иглокожих и насекомых. Таковы,
например, различные виды Urospora. Monocystis Diplocystis и др.
Рис ИО. Полостные iрегзрины
/1 — пара полостных грегарин I’l'ospnra iiiahtaneoriini из
пплихеты Atahttine', li—— грегарип.з Ггоарога chiriiioiae н i
i плоту pun Chiriilotn (/> — совсем молодая особь из кровемос-
пого сосуда голот рнп; С взрослая вегетативная особь с
Hpo.io.ibMoii борптдог из кровеносного сосун голотурии; /)—
дне спарившиеся особи, выдающиеся из сосуда в полость
тела хозяина; Е— то ;ке. В правоп особи я гро разделил ась на
мною дочерних ядер, левая з;иь<лнен.< нарляппрующчми и
iperapi.'iv .челками снсровикаме Нуа ’’/sp/ta га). (Но Догелю )
Ооцисты этих грегарпн тем или иным путем выходят наружу из тела
животного н потом случайно проглатываются подходящим для них
хозяине и В кишечнике пос 'едиего из ооцист выходят спорозоиты,
которые пооходят через стенку кишки и проникают в ткани пли
целом хозяина. Притом даже у разных представителей одного и того
же рода можно наблюдать в различной мере глубокое проникновение
внутрь хозяина. Так, в роде Urospora вид U. chiridotae <рис. 60).
паразитирующий в го. ioT\pini Chiridoia pellucida. ведет себя так, что
часть особей, проходящих сквозь стенки кишки, застревает уже
в спинном кишечном кровеносном сосуде, т е. не достигает целома:
другая часть пронизывает перитонеальный эпителий и выдается
в целом, по своим задним концом остается погруженной в стейку
кровеносного сосуда (рис. 60, Д,Е). Наконец, особи Urospora rnalda-
37
neoruni из поли.хеты Maidane совершенно у солят в целом и плавают
в заполняющей сю жидкости
Таким же путем проникает и большое количество различных
червей, обладающих сменой хозяев; личинки этих червей имеют
покоющуюся стадию в мышцах или в полости тела промежуточной
хозяина. Громадное большинство личйпочйых стадии нематод
и пузырчатых стадий цестод проникает в ткани хозяина именно
через стенки кишки. Чере > кишку же вносится в мускулатуру хозяина
и Trichinella spiralis в своей личиночной стадии. Toi же путь проник-
новения характеризует собой некоторых паразитических Gastropod.
(Entocolax, Enteroxenos): подробнее об этом будет сказано дальше.
Среди насекомых тоже имеются внутренностные паразиты (например,
личинки многих мух-тахин), проникающие в ткани хозяина через
кишку.
Иногда путь попадания паразита впугрь хозяина косвенно опре-
деляется местом его локализации. Так, некоторые почечные паразиты
(Xephrotrenta iruncatmn крота, свайник-гигант, пли Dioctophijnn’
renale, собаки) встречаются исключительно в правой ночке. Отсюда
Эисмопт (Ejsmont, 1936) делает вывод, что личинки или молодые
паразиты должны проникать в почку через печень: печень лежит
целиком на правой стороне тела и тесно прилегает к правой почке
В пользу этого говорит и неоднократное нахождение молодых свай-
ников на печени или ее мезентерии. Эйсмонт полагает, что молодь
свайника понадает в печень с током крови и оттуда, с кровью же или
активно, попадает в правую почку.
Дрхгая большая категория внутренностных паразитов проникает
в тело хошина через покровы. При ртом некоторые формы
сохраняют до известной степени связь с внешней средой, другие
утрачивают ее совершенно; один проникают в тело хозяина само-
стоятельно и активно, другие — через посредничество либо пере-
носчиков, либо при откладке яиц родительскими формами, как это
имеет место у наездников.
Примеры сохранения связи внутренностного паразита с внешней
средой, равно как ряд интересных переходов к эндопаразитизму,
показывают паразитические Gastropoda, живущие в иглокожих
(рис. 61).
Так. свободно живущий род брюхоногих Melanella. обладающий
типичными для Gastropoda строением и спиральной раковиной, со-
держит. кроме свободно живущих, несколько эктопаразптпческих
видов, которые живут на хозяине, погружая свой несколько увели-
ченный хобот в его кожу. У других родов (Mucronalia. Stilii'cr)
прочнее оседающих на хозяине, нога постепенно редуцируется
а вокруг основания хобота разрастается откидывающаяся иг .ад, на
все тело, кольцевая кожная складка — ложная мантия (psiudopal-
liuni), обрастающая в виде мешка всего паразита. Парнзйт погру-
жается всем телом в толщу стенок тела хозяина, однако задний
конец раковины паразита и края ложной мантии еще выдаются
наружу.
Дальнейший этап изменений иллюстрирует род Gaslerosiplion,
который помещается целиком в полости тела голотурий Deiiiui. Он
имеет вид мешка с двумя мускулистыми трубками на концах. Одна
из трубок есть обращенный внутр!, тела хозяина хоботок, которым
моллюск внедряется в спинной кишечный сосуд голотурии и питается
ее кровью Другая трубка, или сифон, возникает за счет удлинившихся
краев ложной мантии, идет к коже и через отверстие в покровах
88
AUJMJ lid tUUUUU ТГГГ. С7Ю71i UUSlL'f ,____
че.тает совершенно. Все выше приведенные формы в личиночном воз-
расте внедряются в тело хозяина извне, т. с. относятся к паразитам
экзогенного происхождения. Еще сильнее изменены и совершенно
внедрены внутрь хозяина Entocolax Entoioncha. Parenteroxenos
Enteroxenos. Оли живут в полости тела, имеют вид длинного мешка
и своим сифоном прикреплены уже нс к покровам, а к кишке хозяина.
Рнс. 61. Приспособление Gastropoda к паразитизму
А — Mehiiu Uu eqtiestris\ Н — Mucronalidpalmipedes’, С — Stilifer linckidae.;
D — Gastcrosiphon deimatis (продольным разрез); l — самка Entocolax
ludwigi’, F—Enteroxenos fistergreni (ироюльнып разрез!. (Из Иванова.)
Интересно, что эти, по общему мнению, происходящие от эктопарази-
тов, но наиболее измененные формы модифицировали не только
ориентировку своего тела (сифоном к кишке), но и характер проник
новепия в тело хозяина. Именно, по даннгям Боцпеви (Boimexic, 1902»
относительно Enteroxenos. по данным Швапвнча (1917) относительно
Entocolax и Иванова (1947) относительно Purenleroxenos, личинки этих
моллюсков поступают в кишечник голотурии и проникают в ее полость
тела через стенку кишки, представляя собой, паразитов эндогенного
характера проникновения.
Через покровы проникает в кровь хозяина громадное большинство
кровепаразптов: Haemosporidia. Piroplasmoidea, T-ypanosomidae средн
простейших, различные филярии среди нематод Так, например.
wuchereria bancrofti проникает в тело человека через кожу при укусе
89
комаров. Личинки филярии собираются в нижней губе комара и, про-
рывая тонкую хитиновою пленку, попадают отг\да на кожу; здесь,
по-видимому, даже не использхя места укуса, личинки филярии про-
буравливают эпителий и проникают в кровь.
Как видно из вышесказанного, большинство кровепаразитон
попадает в кровь экзогенным путем, но через посредство переносчи-
ков. Такими переносчиками служат различные кровососущие живот-
ные. Это правило имеет, однако, ряд исключений. Так, среди трипано-
зом возбудитель «дурины» Trypanosoma equiperdum проникает при
случке лошадей самостоятельно через слизистую оболочку половых
путей из одной особи в другую. Церкарни кровяных двуусток (Schisto-
sornatidac) активно вбуравливаюгся в покровы хозяина.
Рис. H2. МВтаг'ерклрнп Neascas culicola
А — красноперка из дельты Дуная с многочисленными пнгмептиров.'ншымг
мстацсркарнями Ncascus Ctific6la\ If — метпцеркарчя Л'сдлти*-. окруженная
черным пигментом. (По Петрушевскому и Шульману.)
Много примеров экзогенного проникновения в хозяина дают со-
сальшики. Так. у двуусток мирацидни проникают через нежные
покровы моллюсков в их печень или половый железы, где протекает
жизнь партеногенетических поколений. Таков /ке путь к р;. «личны й
внутренним органам хозяина, прилагаемый церкарпямп многих дву-
усток. Через покровы забираются в тело хозяина под кожу рыб церкя-
рии kcascus culieola (рис. 62), N. tnusculicola — в мышцы, Hemistomum
spathaceum, Tylodelphys clavata— в глаза рыб. Tetracotyle ardeae—
под кожу цапель (Дубинина, 1937) и многие дрхгие.
У мнрацидиев и в особенности у некоторых церкарий описаны
специальные органы проникновения в виде стилетов или усаженного
шипикамн мускулистого «фронтального органа» (фуркоцеркарии) и
всегда сопутствующих им «стнлетных желез» или «желез проникнове-
ния», которые исчезают после ннцпетнровани-я в тканях хозяина. Мира-
цидип и церкарии обычно очень быстро проникают через неповреж-
денные покровы животного. Кроме них, также и миог не кишечные
простейшие личинки нематод и других гельминтов, не обладающие
часто специальными органами проникновения, легко внедряются в
стенку кишечника или совершают миграции по всему телу хозяина,
проходя при этом через эпи гелий кишечника, эндотелии кровеноСкых
сосудов и другие разнообразные клеточные преграды.
90
Такая способность легко преодолевать клеточные и тканевые
барьеры связана с тем, что эти организмы выделяют особый энзим —
гиалуронидазу,— который оказывает специфическое деполимеризую-
щее действие на мукополисахарид—гиалуроновую кислоту, входя
тую в состав основного вещества соединительной ткани и в вещество.
цементирующее эпителиальные клетки кожи, стенки кишечника, эндо
телий сосудов и т. п. Выделяемая паразитами гиалуронидаза раз-
рушает межклеточное вещество (гиалуроновую кислоту) и тем самым
способствует проникновению паразита
или его токсинов в глубь тканей хо-
зяина. С гиалуронидазой связана ин-
вазионность многих патогенных бакте-
рий, быстро распространяющихся по
тканям пораженного животного Гиа-
луронидаза обнаружена также у Ba-
lantidium coh и Entamoeba histolytica,
трофозоиты которых могут проникать
из просвета кишки в соединительно-
тканный слой ее стоики. Е- histolytica
проникает оттуда в печень, где
образует абсцессы.
Гинецинская (1950) обнаружила
гиалуронидазу в организме стилстных
церкарнй из моллюсков Limnaea stag-
nates и высказала предположение, что
она продуцируется железами проник-
новения. Вероятно, церкария разрезает
с помощью стилета хитин насекомых
(вторые промежуточные хозяева) а
выделяющийся секрет желез, содержа-
щих гиалуронидазу, способствует
быстрому внедрению перкарин в тело
хозяина. Гиалуронидаза выявлена
и у перкарин Schistosoma. Наличие
гиалуронидазы установлено у ин-
Рис. 63. Откладка яиц некоторыми
наездниками
/1 — наездник из сем Iclineumo-
mdae с чрезвычайно удлиненным
яйцекладом (длина тела -2 с.н,
длина яйцеклада — 15 с.и); В—
наездник F.phie.lhs, приготовляющий-
ся воткнуть свой яйцек'ад в кору
дерева; С — наездник Epfilaltes, вты-
кающий яйцеклад в дерево при оты-
СКИПЛН1П1 сверлящей дерево личинки
жука. (Пл Бишофу.)
вазионных личинок Ancylostoma canimim Staphanurus denlatus, Stron-
gyloides ransotni, Ascaris sutim. Ascaridici gal It. Strongylus sp. (Oshio
and Furuta, 1955).
У личинок Strongylus equinus. Sir. edentatus гиалуронидаза скон-
центрирована в области пищевода и, вероятно, является продуктом
(орзаль.чых пищеводных желез (Deiana, 1957) У взрослых червей
гиалуронидаза не обнаружена.
Такой механизм внедрения во внутренние органы свойствен,
вероятно, большинству паразитов как с эндогенным, так и с экзоген-
ным способом проникновения в хозяина.
Большой материал по экзогенному проникновению в хозяина
дают насекомые, в особенности наездники (рис. 63). При этом одни
наездники (Ryssa, Ephialtes) прицепляют свои яйца к покровам дру-
гих насекомых, например гусениц, предоставляя личинкам самим
нбуравлинаться в тело хозяина после своего вылупления из яйца.
Другие наездники при помощи своего длинного яйцеклада вносят
яйца непосредственно внутрь тела своей жертвы (Aphclinus, и др.).
Естественно, что экзогенный характер носит большей частью и зара
жение насекомых тахпнами; делают они это приблизительно таким же
способом, как наездники, но способы заражения ими хозяев особенно
91
разнообразны. Таунзенд (Townsend, 1908) различает тахин следую-
щие пути заражения хозяина:
1. Откладка яиц на листья кормового растения хозяина, где они
случайно поедаются последним. Так заражает гусениц шелковичного
червя муха Crossocostnia sericariae, личинки которой живут в полости
тела червя.
2. Откладка яиц на тело хозяина. Молодая личинка проникает
через кожу в полость тела хозяина (гусеницы), К этой категории от-
носится Parexorista cheloniae — паразит гусениц I.iparis chrysorrlioea.
3. Откладка живых личинок на кожу хозяина. Такой способ имеет
место у мух Dexidae.
4. Откладка личинок под кожу хозяина при помощи колющего
яйцеклада. Это наблюдается, например, • Dexodes nigripes — у пара-
зита гусениц Liparus chrysorrlioea.
5. Откладка живых личинок на листья и ветки, находящиеся из
пути движения хозяина (гусеницы). Личинки прицепляются к про-
ползающей мимо гусенице и проникает внутрь се тела. Таков способ
заражения у мухи Eupeleteria magnicornis.
Вольфартова муха (Wtohljarthia tnagnifica) рождает своих личинок-
па кожу животных и человека, после чего личинки забираются под
кожу, в раны, в носовую и гайморов полости.
Некоторые иные пути проникновения паразитов в
хозяина. Известен ряд случаев, когда паразит попадает в хозяина
не через кожные покровы или рот, а через какие-нибудь другие от
верстия внутренних органов. Например, полостные оводы (Rhineestriis
purpureas, Oestrus ovis) на лету выпрыскивают из полового отверстия
струю живых личинок, стараясь попасть ею в ноздри лошади, овцы
или в глаз человека. Попавшие в тело личинки пробираются в глубь
носовой полости, доходят до лобных па зух, разруппсют нередко решет
чатую кость и даже могут проникать в полость черепа.
Некоторые виды инфузорий Trichodina (Т. renicola, Т. urinaria)
живущие в мочевом пузыре рыб (см. рнс. 86), проникают к месту
своей локализации, несомненно, через наружное мочевое отверстие
Личинки мухи Pollenia rudis проникают в тело дождевых червей
через отверстия семеприемннков, из которых они впоследствии внед-
ряются в глубь тела хозяина (Keilin, 1925). Такие случаи, в общей,
крайне редки.
Соответственно со способом проникновения в хозяина Скрябин и
Шульц (1931) предложили делить паразитических червей на гео-'
гельминтов и б и о г е л ь м и и то в. Первые получаются хозяином
прямо с субстрата и отвечают паразитам, не имеющим промежуточ-
ных хозяев Вторые передаются всегда через какого-нибудь промежу
точного хозяина — носителя инвазии.
В некоторых редких случаях паразиты доставляются на тело
хозяина совершенно особым способом, носящим название форе» пи.
При этом паразиты используют для заражения новых особей хозгапа
обычно подвижных животных, не имеющих никакого отношения к их
нормальному жизненному' циклу. Прекрасный пример форезии даю г ио
которые нухоады птиц, например Philopteius sLurni (рнс. 64) скворца
(Марков, 1938). Не обладая крыльями, пухоеды могут заражать новых
хозяев только во время гнездового периода, когда они переползаю'
с родителей на оперяющихся птенцов. В качестве дополнительного
средства распространения им служат, кроме того, мухп-кровососки
Ornithotnyia, живущие на скворцах и других птицах. О/ mthoniyia
обладают хорошо развитыми крыльями и могут перелетать с одного
92
хозяина на другого. Но окончании гнездового периода Марков наблю-
дал, что к ножкам и к телу Ornithomyia, находимых им на скворцах,
нередко прицепляются своими жвалами пухоеды Philoplcrtts. Иногда
на одной мухе сидело до восьми пухоедов. Нет
никакого сомнения в том, что таким способом
малоподвижные пухоеды доставляются с од-
ной птицы на другую при перелетах мух-но-
сительниц, Статистические данные Маркова
показывают, что форезия может играть в рас-
пространении некоторых пухоедов немаловаж-
ную роль, а именно: он нашел пухоедов на
25% из снятых им со скворцов мух, причем
общее число добытых таким способом пухо-
едов равнялось 30.
Удивительный случай форезии показывает
южноамериканский овод Dertnatobia cyani-
ventris (рнс 65). Этот овод приклеивает пач-
ку своих яиц к брюшку мух и комаров. Через
несколько дней сформировавшиеся внутри яиц
личинки приподнимают своими головными
концами крышечку яйцевой скорлупы. Они
ожидают того момента, когда их носитель ся-
дет на какое-нибудь млекопитающее (бык, со-
бака, агути или человек). Тогда личинки Der-
Рис. 64. Кровосос кд Огш-
thomyia со скворца с
прикрепившимися к ней
пухоедами скворца- Phi-
lopterus sttirni — пример
форезии. (По Маркову.)
1 пухоеды «ч-ворна.
matobia вываливаются из яйцевых скорлупок на кожу своего подлинно-
го хозяина, внедряются в волосяные фолликулы и своим дальнейшим
развитием вызывают образование подкожной гнойной опухоли.
Рис. 65. Явление форезии
/1 — овод Dertnatobia cyantv&itris (увеличено в 2 раза); В —
кладка яиц De.rmatobia не брюшке мухи, служащей перенос-
чиком молодых личинок овода; С — выросшая личинка Der-
matobia. (По Мартини.)
1 — кладка яиц.
Различные способы, которыми паразиты покидают
тело хозяина. Рассмотрим, какими путями различные стадии раз-
вития паразитов покидают тело хозяина для передачи или распростра-
нения инвазии, В очень многих случаях, в особенности в слу-
чаях ларвального паразитизма, паразит вообще не оставляет тело
93
хозяина и освобождается из него только тогда, когда данный хо*янн
(нромсжуточпЧзГй) съедается другим подходящим хозяином (оконча-
тельным). Некоторые простейшие (Schellackia), многие адолескарии
сосальщиков, финки ленточных глистов, личинки скребней и нематод
ведут себя именно таким образом.
Способом выведения наружу, наиболее естественным для кишеч-
ных паразитов, является выход через заднепроходное отверстие
вместе с экскрементами.
Столь же естественно, что эктопаразиты, покидая хозяина, просто
отрываются от его тела, как это делают пиявки, карпоеды (Argulus),
клещи и многие другие. Нередко, однако, эктопаразиты столь прочно
приживаются к хозяину, что сами не покидают его вплоть до своей
смерти, но дают подвижное потомство, которое отрывается от хозяина
в поисках новых хозяев. Таковы личинки Monogenoidea, науплиусы
паразитических Copepoda и Cirripedia (Coronula, Sac с ulina и др.).
Многие подкожные и мышечные, а иногда и более глубоко лока-
лизованные паразиты большей частью для выхода прорывают стенку
тела хозяина Иногда это происходит пассивно. Так, например, цисты
Myxobolus pfeifferi образуют в мускулатуре усачей (Barbus) большие
опухоли; опухоли прорываются наружу и превращаются в открытые
нарывы, из которых высыпаются наружу бесчисленные споры Myxo-
bolus Таким же путем освобождаются и споры многочисленных Мухо-
sporidia, образующих цисты в жабрах рыб. Ришта (Dracunculus
medinensis) получает сообщение с внешней средой путем прорыва
кожи человека на разных местах тела, в которых помещаются самки
этой нематоды. Самка остается в подкожной клетчатке, но получает
таким способом возможность отрождения наружу личинок, которые
для дальнейшего развития должны попасть в виду, а оттуда в тело
своего промежуточного хозяина — рачка Cyclops.
Пассивным же путем освобождается из опухолей на жабрах рыб
развивающаяся в них из глохидш-в молодь пластинчатожаберные
моллюсков Umomdae.
Другие внутренностные паразиты прорывают стенки тела хозяина
активно, как, например, Mermithidae среди нематод, Gordiacea, личин-
ки многих наездников и многих паразитических мух и т. д. У сосаль-
щиков Sanguinicola—паразитов кровеносной системы рыб, яйца от-
кладываются прямо в кровь хозяина (Ejsmont, 1925). Они попадают
через жаберные артерии и капилляры жабр, где и застревают
вследствие своей угловатой формы. Здесь из них прямо в кровь вы-
ходят мирацидии. Они активно прободают стенки капилляров н эпи-
гелий жаберных лепестков рыб и попадают в воду.
Особый способ выхода из хозяев имеет большинство кровепара-
зитов, передающихся через кровососущих переносчиков. Позвоноч-
ное— хозяин — покидается этими паразитами всегда пассивно, так
как они втягиваются вместе с кровью в кишечник переносчика при
сосании им крови позвочкого животного. Кровепаразнты покидают
тело про.межуточного хозяина значительно более разнообразными
способами. Например, попавшие в кишечник кровососущего живот
ного кровепаразнты могут покидать его обычным для кишечных пара-
зитов способом, т. е. через заднепроходное отверг i не. Так дело обстоит
с Trypanosoma lewisi из крыс (рис. 66,А). Беспозвоночным хозяином
этой формы служит крысиная блоха CeratophyUus fasciatus. По Пел-
леру (Ndller. 1912), попавшие в кишечник блохи грипаноземы усилен-
но размножаются и превращаются в мелкие, так называемые мета
циклические формы, которые занимают «заднюю позицию» в кишеч-
94
нике (рис. 66, А, В) и затем выводятся наружу с экскрементам:!. При
чистке шерсти зубами крыса загрязняет рот частицами экскрементов
блох и получает трипанозом в свой пищеварительный тракт, через
стенки которого жгутиконосцы и попадают в кровь крысы. Это та*
называемый к о н т а м и н а т и в н ы и
Кроме проглатывания содержащих
может поедать самих блох и зара-
жаться тем же коптаминативным
способом (быть может, этот способ
заражения даже доминирует в слу-
чае с Trypanosoma lewisi). При
этом паразиту вовсе не надо поки-
дать тело беспозвоночного хозяина
для того, чтобы попасть в позвоноч-
ное животное.
Таким же путем поедания овцой
промежуточного хозяина, а именно:
рунца (Mclophagus ovinus), проис-
ходит заражение овен Trypanosoma
melophagiurn. Конта ммнативным пу-
тем человек заражается Т. cruzi от
клопов Tnatoma, когда их фецес,
содержащий метациклинически х
трипанозом, попадает на слизистую
оболочку рта или носа.
При другом, пнокулятнв-
пом способе заражения
(рис. 66, С) позвоночного хозяина,
кровенаразигы, не выходя за пре-
делы кишечника беспозвоночного
хозяина занимают «переднюю пози-
цию» в кишечнике, накопляясь иног-
да в его придаточных, а именно:
слюнных железах; отсюда инвазнон-
тнп заражения у трипа новом
трипанозом экскрементов, крыса
Рис. 66. Развитие гршшиозом (Trypa-
nosoma) в теле переносчиков
.4-В-при коитамннативио.м заражении;
С —D — при ннокулятпвиом заражении.
A—Trypanosoma lewisi в крысиной блохе
(Cciatophyllns fasciatusV, li ~ Т. melo-
phagium в овечьей кровососке (Ме/о-
phagux ovinus'r. С — Т gamhiensc и Т.
brucei в мухе цеце ((ilossina palpalis)-.
D — Т- rotarorium в пиявке (/lemtclepsis
margina(a). (Но Гоару.)
1 iiuiiicno.i; 2 средняя кишка; 3 мальпи-
гиевы сосуды; 4 - задняя кишка: 5 — прямая
кишка; ь хоботок; 7~ глотка; S — u.iaia.-Hiuc
хо'ютка; сплошным черным инетом л юбражепы
ные стадии паразитов поступают в места локалнзшиш мета11нк.1ических трин шотом
„ • (инманопиыс формы), точками — места нреоблада
КрОВЬ ХОЗЯИНа при укусе его Пере- НИЯ крит>с11алм1ьх и TpuuauoioMHi.x форм
носчиком. Таким путем осуществ-
ляется заражение у большинства трипанозом (Г. gambiense сонной
болезни. Т. brucei «наганы» рогатого скота, Т. rotatorium лягушки
и т. д.) и у лейшмапнй (Leishmania) Иногда заражению инокулятив
ним путем предшествует внутренностная фаза развития кровепаразпта
в промежуточном хозяине. Так, малярийные паразиты (Plasmodium),
попав в кишечник комсра-переносчика, претерпевают половой про
цесс; образовавшаяся при этом оокинета прободает кишечную стенку
и инцистируется (становится ооцистой). Образующиеся в ооцисте
спорозоиты проникают в полость тела, откуда током крови приносятся
к слюнным железам и активно забираются в них. Здесь, следователь-
но, паразиты достигают слюнных желез кружным путем, но способ,
каким они покидают переносчика, от этого не меняется: это проис-
ходит инокулятпвным путем, во время укуса.
Таким же приблизительно путем личинки (микрофилярии)
Wuchereria bancrofti, вызывающей слоновую болезнь человека, поки-
дают тело различных комаров-переносчиков (Culex fatigans и др.).
Когда эти комары сосут кровь носителей филярий, то плавающие в
крови личинки попадают в желудок комара. Оттуда они вбуравли-
95
ваются в полость тела, проделывают фазу роста и затем скопляются
у основания ротовых органов насекомого. Когда комар сосет кровь
человека, то вонзает в его кожу хоботок. При этом происходит раз-
рыв тонкой сочленовой перепонки между концевыми дольками хобот-
ка, из полости которого личинки Wuchereria выходят наружу и вбура-
вливаются в кожу человека (рис 67). Здесь, таким образом, выход
Рис. 67. Проникновение филярпевияиоп
личинки IV 'uchereria hancrofti через кожу
но время ак1.| кроиососяиия комаром. (По
Мартини.)
J — филярневилнап личинка.
паразитов из тела комара проис-
ходит, собственно говоря, через
разрыв стенки тела, а самое зара-
жение человека представляет как
бы смесь мноку.тятивного с кон-
тактным
В некоторых отдельных слу
чаях паразит покидает хозяина
еще каким-нибудь менее обычным
способом. Так, иногда паразиты
покидают хозяина через посред-
ство его мочевых путей.
Прежде всего этим способом по-
кидают хозяина обитатели моче-
вого пузыря и почек: Trichodi.no
renicola и Т. urinaria рыб, Phyllo-
distomum из мочеточника рыб,
Renicolidae и др. из почек птиц.
Dioctophyme renale из почечной
лохани собаки, тюленя, выдры.
Trichosomoides crassicouda из мо-
чевого пузыря крыс. Таким путем
выводятся покоящиеся стадии
т. е. цисты или яйца вышеназванных паразитов. Таким же путем выходят и
яйца кровяной двуустки (Schistosoma haematobium), которые при помо-
щи особого колющего шипа их оболочки пронизывают стенки вен моче-
вого пузыря, проникают внутрь последнего п затем выводятся наружу с
мочой.
Паразиты, имеющие связь с половой системой, выходят через
стенки половых органов (Trypanosoma equiperdum. случной
болезни) или через половое отверстие (Trichomonas vaginalis из влага-
лища человека, грегарина Urospora gonadipertha. паразитирующая в
половых железах голотурии Cucumaria frondosa: эти грегарнны обра-
зуют в гонадах голотурии многочисленные цисты со спорами, причем
споры выводятся через половые протоки).
Личинки полостных оводов (Rhinoestrus, Oestrus ovis) готовые к
окуклению, выходят наружу через ноздри хозяина; иногда это
облегчается сильным чиханием хозяина, вследствие раздражения
личинками слизистой оболочки носовой полости.
Часть II
ХДАПТАЦИИ В СТРОЕНИИ И ЖИЗНЕННЫХ ЦИКЛАХ
ПАРАЗИТОВ
ГЛАВА VIII
МОРФОЛОГИЧЕСКИЕ АДАПТАЦИИ ПАРАЗИТОВ
К ИХ ОБРАЗУ ЖИЗНИ
Нигде в животном царстве по сказываются с такой силой приспосо-
бительные потенции организмов, как у паразитов Приспособления па-
разитов к их специфическому образу жизни настолько многочисленны
и разнообразны, что мы остановимся лишь на главных категориях их,
сгруппировав изменения по тем системам органов, которые ими затра-
гиваются. При этом сплошь и рядом приспособления одной и той же
системы у эктопаразитов и эндопаразитов носят совершенно различный
характер.
1. ФОРМА ТЕЛА
Эктона разнты. Общей чертой самых различных эктопаразитов
является сплющивание тела в лорзовентральном направлении, способ-
ствующее лучшему прикреплению к телу хозяина. Для доказательства
этого можно привести такой длинный ряд форм, как Trichodina, Chilo-
don, Kerona (с гидры) из инфузории, все Monogenoidea, Malacobdella
из немертин, Myzostomida, Hirudinea, Branchiura, клещи, клопы, вши
и т. д.
Присматриваясь к этим животным (рис. 68), нетрудно заметить, что
их уплощенная форма тела имеет в разных группах разное происхож-
дение. У одних паразитов плоская форма тела унаследована от свобод-
но живущих предков и явилась одним из факторов, облегчивших дан-
ному организму переход к паразитическому образу жизни. Таков, на-
пример, Chilodon cypririi с кожи карпов: свободно живущие виды того
же рода имеют такую же уплощенную форму тела. Точно так же и ин-
фузория Kerona уплощена подобно всем прочим брюхоресничным ин-
фузориям (HypotrfcKa). Значит, здесь сплющенность тела имеет допа-
разитарное происхождение. Такого же происхождения и листовидная
форма тела Monogenoidea, так как они ведут свое происхождение от
плоских Turbellaria. Отсутствие влияния образа жизни на происхожде-
ние плоской формы тела у паразитических плоских червей зидно, меж-
ду прочим, и из того, что плоскими среди них являются не только экто-
паразиты, но и эндопаразиты, хотя они нередко живут в таких усло-
виях, например в гонких протоках желез (Fasciola hepatica), где ци-
7 В. А Догель
97
липдрическая форма тела, казалось бы, выгоднее плоской Следова-
тельно, и у Trematodes мы имеем лишь наследование сп пощснн>ч1
формы тела от свободно живущих предков.
Инос положение имеет место у Malacobdella. Myzostomida, Hirudi-
пса, Branchilira, ибо их ближайшие свободно живчщие родичи не обна-
руживают сплющивания тела.
Рис. 68. Приспособление к паразитизму у пемертин и
мизостом.и
А— немертина Malacobdella grossa. паразьтирующая в но-
jocth мантии лиуствоочатого моллюска Cyprine.ua заднем
конце имеется хорошо рз_ъптая присоска; В и С — Муго-
stoma pulvinar в своей женской < В) и мужской (C"i фазе.
У В по краям тела видны измененные иараподпн; малень-
кий самец (мужская фаза) сидит позади на теле женской
особи. Фигура С изображена при большем увеличении,
чем В. । По Целлеру.)
В громадном большинстве случаев, как сказано, сплющивание про-
исходит в дорзовентральном направлении, что и наиболее выгодно в
смысле плотного прилегания к телу хозяина. Латеральное сплющивание
наблюдается лишь у блох и связано, вероятно, с выгодностью сжатой
с боков формы тела для животных, движущихся в узких промежутках
между волосами, которые густо покрывают тело млекопитающих, этих
преимущественных хозяев блох
Что касается укороченное™ или вытянутости тела у эктопаразитов,
то чаще наблюдается тенденция к укорачиванию животного. Нрав за,
пиявки, Ichthyotomus среди Polychaeta, Penella и некоторые, другие па-
разитические Copepoda не короче своих свободных сородичей. Однако
Malacobdella и Myzostomida определенно короче, свободных немертин
и полихет. а клещи и Branchilira тоже имеют короткое, широкое тело.
В гармонии с укорачиванием тела находится нередко наблюдаемое
у эктопаразитов (а также и у эндопаразитов) сглаживание метамерии
у сегментированных паразитов. Так, все Mvzostumida в сильной степени
утратили внешнюю членнстость, резко отличаясь от своих богато рас-
члененных предков — Polychaeta. Процесс слияния сегментов еще боль
те выражен у паразитических раков. Свободно живущие Copepoda. как
известно, имеют ясно расчлененную грудь (кроме первою, слитого с го-
ловой сегмента) и брюшко Между тем, у паразитических Copepoda су-
ществует целый ряд переходов в направлении к полной редукции чле-
нис гости. Начало этого ряда составляют формы, вроде паразитирую,
щего на рыбах Ergasilus. тело которого так же богато расчленено, как
у свободных циклопов. У Caiigidae (см. рис. 40) с головой сливается
уже четыре сегмента, генитальный сегмент брюшка сильно разрастает-
ся, а остальное брюшко вообще не расчленено. Кроме того, основание
каждой аитеннулы образует по присосочке (luntgh). а все тело сильно
95
уплощается и расширяется (признак подвижного чктопаразититма).
Наконец, у Lcrnaeidae и Lernaeopodidne (рнс. 69 и 70) все брюшко
расчленено или не расчленено, генитальный сегмент его крайне разра-
стается, а все остальное тело \ взрослых самок вполне утрачивает чле
нттость и становится
мешковидным. Самцы и
неполовозрелые самки
обоих последних семейств
сохраняют еще облик сво-
бодных Copepoda, оста-
ваясь долгое время на
коиеподитной стадии раз-
вития. В таком виде про-
исходит оплодотворение,
после которого самцы
умираю!* а самки претер-
певают характерные из-
менения и редукцию.
Иногда тело паразити-
рующих на рыбе Сореро-
Рис. 70. Паразитирующие па рыбах рачки сем.
I ernae.idae
I — самка Lrrnaea esocina; В—F — Li'rnaeoura
briiiichialis; В— самец с жабр камбалы С—ие-
оилодотиорепная самка, с брюшной стороны; D —
коиеиолитная свободно плавающая личинка; I
взрослая сачка с вполне развитыми яйцевыми
мешками. (По Клаусу.)
1 — ГО.1О11ИЫ1- П|>ИКрС||||Те.И.||ЫС orpin'г ки. которыми ч.тр»шт
нН стряс гея и и-ло рыпы; 2 - яйцевые чешки;./ антенны, служл-
nine ЫЯ iiphKperi.teiiiiH к пок[>о|>ам рыбы. / - uiai;-5 Kilnui.i
6 г|1С(>М1>т<>||юрпые карманы, 7 челки шыг.ножки.
Рис. 69. Паразитические
Copepoda
А — самка\l.ernaeocera Ьгап-
tfiialis па жабрах наваги;
Я— изолированный рачок
со спирально закрученными
яйцевыми мешками и ветви-
стым прикрепительным ан
паратом. । Но Петрушевско-
му и Шульману.)
da сохраняет вьпянутую форму (Pliilichthus,. рис. 71, С), но конечности
его сильно редуцируются, да и членистость становится менее ясной.
Крайнюю слепень влияния паразитизма обнаруживает Xcnocoelmua
(см. рис. 59), у коюрого тело совсем не расчленено и имеет вид ци-
линдра с закругленными концами. Параллельно с исчезновением пле-
чо
нистостп у ряда вышеперечисленных Copepoda происходит прогресси-
рующая редукция конечностей, которые совсем исчезают у Xenococloma.
Еще более резкую картину утраты членнстости дают паразитиче-
ские усоногие Peltogaster и Sacculina, тело которых имеет вид округло
го или овального .мешка. Это не удивительно, ибо во время развития
ципрнсовидная личинка этих раков отбрасывает все брюшко и всю
грудь, сохраняя только головной отдел, из которого формируется тело
половозрелого паразита.
Рис 71. Паразитические ракообразные
А—Cyproniscus cypridina? (hop иа), паразн гирующин на Cypridina
sp. (Osiracoda). самка; Li — самка Cyprtmiscus, сидящая на Cypridina sp.
в ракомине последней; (. — PhiLichthys xiphia (Copepoda), паразити-
рующий в полостях лобных костей меч-рыбы. (По Кюкснталю.)
1 — самка Cyproniscus.
Такой же ряд форм можно наметить и среди паразитических Iso-
poda (см.рис 71 и 76),начиная с еще сильно расчлененных Cyinothoidae
с жабр рыб и кончая паразитами из жаберной полости десятиногих ра-
ков. Самки этих паразитов, относящихся к семействам Bopyridae и Еп-
toniscidae, испытывают сильную деформацию тела и слияние члеников
его между собой. Самцы же этих паразитов в значительно большей сте-
пени сохраняют расчлененность тела, хотя и паразитируют вместе с
самками, иногда прицепляясь к телу последних. По-видимому, большая
деформация самок зависит от переполнения их тела многочисленными
яйцами. Интересно, что некоторые Isopoda (Cyproniscus cypridinae,
рис. 71) приспосабливаются при этом к паразитированию на очень
мелких рачках более примитивного, чем они сами, отряда Ostracoda.
Утрата членнстости видна и па видоизмененных паразитизмом по-
томках Polychaeta, а именно: на Myzostomida (см. рис. 68). Поселяясь
на теле морских лилий и других иглокожих или эндопаразитируя в них,
Myzostomida теряет главную характерную внешнюю черту кольчецов:
длинное, по зеленное насечками на многочисленные сегменты тело. Те-
ло Myzostomida укорачивается, но расширяется, становится плоским
и почти дисковидным. Следы членнстости сохраняются лишь в виде пя-
ти пар сильно измененных параподнй и в некоторых чертах внутренней
организации (несколько пар метанефридиев, иногда несколько гангли-
ев брюшной нервной цепочки). Пара подии Myzostomida сильно укоро-
чены п превращены в крючки, спрятанные в особых влагалищах, т. е. и
здесь переход к паразитизму сопровождается редукцией конечностей.
Гораздо труднее найти закономерности, управляющие изменением
формы тела эндопаразитов, тем более что эти изменения следуют раз-
ными путями у кишечных паразитов и у внутренностных.
ЮТ
Кишечные эндопаразиты. В общем, кишечные обитатели
скорее имеют тенденцию к удлинению, чем к укорачиванию тела. В этом
отношении мы имеем явную конвергенцию у самых различных групп ки-
шечных паразитов. Сильная вытянутость тела наблюдается у многих
кишечных грегарин (Porospora gigantea, Grebnickiclla gracilis, Seleni-
dium),y многих инфузорий Automata (Hoplitophryu, Mesnilclla и др.). Це
столы ясно выражают эту особенность, да и у скребней она нередко хо-
рошо выражена (Macracanthorhynchus. .Moniliformis. Echinorhynchus
gaili и др.).
Рис. 72. Кишечные паразиты, имеющие членистую форму тела
A—HapldzMn (Peridinca): if — Cychirtyw ha strobila 0lyperniasti-
gida); C—Radiophrya (Asiomata); D — raeniocystix mira; E — C.< s-
todes; /•'— Oligacanihorhynchiis I Acantbocephala). (Ориншал.)
Это вытягивание тела сопровождается нередко другим явлением,
довольно распространенным у кишечных паразитов, а именно: расчле-
нением тела в поперечном направлении (рис. 72). Расчленение на про
глоттиды составляет характернейшую черту цестод, оно внекоторойсп-
пени намечено в виде перетяжек у скребней типа Moniliformis и у боль-
шинства Pentaetomida. Кишечные грегарпны также обнаруживают разде-
ление тела на протомернт и дейтомсрит, а у некоторых форм, например
\ описанной Леже (Leger, 1906) Taeniocystis mira и у некоторых грега-
рин из кишечника крабов (Merocystis), дейтомсрит опять-таки подраз-
деляется на участки тонкими, нежными плазматическими прослойками.
.У интересного рода Ilaplozoon (представитель Peridinea, паразитирую-
щий в кишечнике лолихет) то же свойство сказывается в делении тела
на поперечные ряды клеток. Наконец, среди 1 lypermastigiqa из кишеч-
ника термитов удивительная Cyclonympha sirobila подразделяется на
участки опоясывающими ее жгутиковыми кольцами. Значение этого рас-
членения в разных случаях, по-видимому, несколько различно. С одной
стороны, оно предохраняет тело от разрыва: длинное узкое тело должно
менее сопротивляться разрыву, чем тело, поделенное на участки перего-
родками, которые, быть может, играют роль скреплений. В других слу-
чаях. однако, а именно: у Haplozoon и Cestodes, имеется ясная связь
между членистостыо тела и размножением У цестод периодически от-
деляются членики и группы члеников, наполненные зрелыми яйцами,
у Haplozoon отделяются поперечные ряды клеток размножения, кото
рые по выходе из тела хозяина превращаются в маленьких жгутико-
носцев. I
Другая, сравнительно небольшая категория кишечных паразитов
ведет себя по отношению к стенкам кишки так же, как эктопаразиты
J01
по отношению к нархжной стенке юла. Такова, например, Latnblia
сплющенное тело которой вогнуто на одной стороне, причем этой во-
гн\'остью паразит садится на свободные концы эпителиальны’: клеток
Рис. 73. Форма тела тканевых и полостных паразитов
.1 и В — самец и самка нематоды Tel га meres из стенок желудка п пт; С—
самка нематоды Heterodi-ra thachti из сахарной свеклы: !> — грегарина Diplor-
ysO’x phrwsanaie 11:1 полости тепл Phryganea grandis. (По Догелю )
Рнс 71. Конвергентное развитие ветвистой формы тела
Л у грегарпн Aikinrtocyxtis нз дождевого черня; В — Uruspora malda-
tieorum нз морских полижет. (Оригинал.)
кишечника. Сплющенное и расширенное тело многих трематод, быть
ложет, имеет такое же значение.
В н \ г р с н н о с т н ы с паразиты подчиняются совершенно иным
принципам формообразования. Полостные паразиты отрезаны от внеш-
него мира, гарантированы от отзыва, от хозяина и в некотором смысле
походят на яйцевые клетки хозяина по своих* отношениям к окружаю-
102
Рис. 75. Паразитические ракообразные Ascothoracida
А и В— две самки видов Dendrogaster. D. astropecteni и D.
dichotomtis (Dendrogasteridae нз полости тела иглокожих);
С* — самец D. dichotonuis; D — личинка I). astericola, облада-
ющая членистым, характерным для раков телом и конечно-
стями. (А—С—по Вагину, D — по Книповичу.)
103
щей среде. Соответственно с этим, многие полостные паразиты обладают
приблизительно шаровидной формой тела (рис. 73). В отличие от вы-
тянутых кишечных грегарин многие полостные гретарины (например,
Diplocystis из тараканов и ручейников) округлы; это же следует сказать
о многих Tetracolyle, личинках сем. Strigddae; теми же причинами мож-
но, вероятно, объяснить шаровидную или овальную форму финок це
стод, округло-овальную форму тела у нематоды Allantonema из полости
тела шмелей. Особенно ясна зависимость формы тела от локализации
паразита у нематоды Tetratneres (рис. 73), самка которой внедряется
в стенку желудка птиц, тогда как самец сохраняет первичный способ па-
разитирования в просвете желудка. Самец остается вытянутым в дли-
ну, подобно прочим нематодам, тогда как у самки середина тела взду-
вается шаровидно.
Рис. 76. Равноногий рачок Porfunion, паразитирующий в жаберной
полости десятиной х раков
А — самец: В— самка ( 1о Лангу и Коллери.)
1 — парные торакальные пластинки, прикрываю пне выводковую камеру; 2 челю-
стные ножки; 3 — (штейны и ншенпулы. 4 — гол он но а отдел; 5 — Срюшко с сто
п.1.1СТ11Н'игыми конечностями.
При внутренностном паразите есть, однако, случаи когда на-
блюдается тенденция к увеличению поверхности тела, очевидно, в це-
пях усиления всасывания пиши. Среди полостных грегарин такова Uros-
pura maldaneorum (см. рис. 60, 74), один из концов тела которой много-
кратно дихотомирует. Интересен и доказателен в смысле конвергирую-
щего влияния одинаковых экологических условий — жизнь в полости те-
ла кольчатых червей — другой пример среди грегарин, а именно: с Aiki-
netocystis singularis (рис. 74). Если Urospora живет в полости тела мор-
ской полихеты Maldane, то Aikinetocystis— в целоме дождевого червя
Eutyphaeus и не имеет с предыдущей ничего общего, относясь к совер-
шенно другому семейству Сдиако у обоих один конец тела закруглен,
другой же многократно дихотомически ветвится. При этом в обоих сл>
чаях ветвление происходит в одной плоскости и совершается в одном
случае три, в другом — четыре раза. Dendroguster (рис. 75) —аберрант-
ная форма низших раков (отряд Ascothoracida) из полости тела мор
ских звезд, а также некоторые Entoniscidae (Portunion, рис. 76), обла-
J04
дают сильно разветвленным телом. Сильно ветвится н пузырчатая ста-
дия некоторых цестод (Sparganum некоторые виды Multiceps). Впро-
чем, здесь неправильная ветвистость, быть может, способствчет внедре-
нию паразита в ткани хозяина, так как эти паразиты локализуются в
мускулатуре, печени и т. п.
2. РАЗМЕРЫ ТЕЛА
Общего для всех паразитов влияния их образа жизни на размеры
животных не замечается. По в тех случаях, когда такое влияние есть,
оно сказывается в увеличении размеров тела при переходе к паразити-
ческому образу жизни
Особенно ясна эта связь в классе нематод. Длина большинства сво
бодных нематод колеблется в пределах 1 л.м, средн наземных и пресно
водных лишь немногие достигают 5—10 мм длины: среди морских сво-
бодных форм насчитывается несколько видов длиной до 3—5 с.и. Пара-
зитические нематоды достигают значительно больших размеров как в
среднем, так и в отношении отдельных своих представителен Полово-
зрелые Mermithidae достигают 15 см. Среди паразитов крупных живот-
ных известны настоящие гиганты. Так, лошадиная аскарида (Parascaris
equorum) имеет до 30 см длины, a Dioctopkytne renale из почечных ло-
ханок собаки— 1 м.
Подкожный паразит человека — ришта (Dracunculus medinensis)
достигает длины 1,2 м при толщине 1,5 лс.и. Описанная Губановым
(1951) гигантская нематода Placentonemagigantissima. паразитирующая
в плаценте кашалота, достигает в длину 8—8,4 и и в толщину — 1,5—
2,5 си
Влияние паразитического образа жизни i:a размеры лучше всего до-
казывают факультативные паразиты Aloionema. Личинки этого червя
из слизня Arion достигают сивое больших размеров, чем личинки, остав
шиеся на свободе. Gordiacea. паразитирующие, в течение всей личиноч-
ной жизни в насекомых, тоже иногда бывают до 160 с.я длины.
Крупными размерами отличаются и некоторые скребни, достигаю-
щие 70 см длины при довольно большой толщине. Трематоды, в общем, не
крупнее свободно живущих Ttirbellaria, но цесгоды, ведущие свое про-
исхождение от того же корня, достигают в некоторых случаях гигант
скоп длины в 10 м (Diphyllobothrium latum), тогда как наибольшая
длина Turbellaria измеряется €0 см.
Очень хороший пример паразитического гигантизма дают веслоно-
гие рачки (Copepoda). Свободно живущие Copepoda большей частью не
превышают 1 леи и только в редких случаях бывают до 1 си длиной
(Valdiviella). Между тем среди веслоногих рачков, паразитирую-
щих на рыбах, длина в -1—5 см не редкость (Tracheliastes stellutu.i
с осетров), а самки некоторых видов Penella, паразитирующих на
акулах, достигают даже общей длины в 25—30 си. принимая червеоб-
разную форму.
Большинство паразитических инфузорий естественно не отличает-
ся по своим размерам от свободно живущих видов. Однако и здесь ерй
ди паразитов наблюдаются отдельные случаи гигантизма Так, живу-
щая в кишечнике даманов паразитическая инфузория Pycnothrix достн
гает длины до 3 лби. В теле этой инфузории находят иногда проглочен
ных мелких личинок нематод. Крупными размерами, достигающими до
2 мм длины, характеризуются также некоторые инфузории группы Asto
mata (например, из рода Radiophrya), паразитирующие в кишечник
Oligochaeta.
Последний пример в интересующем нас вопросе дают живущие в
кишечнике термитов представители жгутиконосцев из отрядов Polymas-
tigida и Hyperniastigida.
Среди большого разнообразия жгутиконосцев, которые живут в ки-
шечнике термитов, встречаются виды, напоминающие по форме и раз-
мерам кишечных Mastigophora позвоночных. Таковы трихомонады, на-
пример Trichomonas termitis: размеры их — в пределах величин, кото-
рые известны для немногих свободно живущих Polymastigida,— 15—20 ц.
Судя по всему, от подобных исходных форм, широко распространенных
у разных групп животных (> амфибий, рептилий, птиц, млекопитающих),
и ведут начало все разнообразные Mastigophora, для эволюции кото-
рых столь подходящим местом оказалась задняя кишка термитов.
В благоприятных для паразита условиях паразитического образа
жизни произошло прежде всего сильное увеличение размеров живот-
ных, а кроме того, и чрезвычайное усложнение их строения. Некоторые
паразиты термитов (Gigantomonas) вполне сохраняют еще структуру
Trichomonas (три свободных жгутика, волнообразная перепонка, аксо-
стпль), но увеличиваются в размерах до величин (70н). не известных
среди прочих трихомопад. Другие претерпели в течение длительной эво-
люции сложные изменения структуры и приобрели еще большую вели-
чину. Так, представители рода Joenia достигают 250ц, а некоторые Tri-
chonympha — даже 450ц длины. Какова была в данном частном случае
причина особого увеличения размеров, решить трудно, но, по-видимо-
му, в этом играла какую-то роль чисто клетчатковая диета большинства
термитов. По крайней мерс, в термитах той группы Prototerniitidac, ко-
торые питаются грибками или смешанной пищей, жгутиконосцев очень
мало или они отсутствуют вовсе, а размеры их очень малы
Итак, мы приходим к выводу, что в общем неограниченное количе-
ство хорошо усваиваемой пищи, всегда находящейся в распоряжении
паразитов, очень часто приводит к увеличению размеров тела парази-
тических животных.
Интересно сделать некоторые сопоставления между размерами са-
мих паразитов и величиной их яиц и личинок, т е. их стадий размноже-
ния. Оказывается, что у всех эндопаразитов размеры тела не только не
отражаются на размерах яиц и личинок, но, напротив, яйца эндопарази-
тов всегда характеризуются малыми размерами, даже по сравнению с
яйцами свободно живущих организмов. Это зависит, с одной стороны,
вероятно, от тенденции к произведению возможно большего количества
яиц, с другой — от того обстоятельства, что мелкие яйца с большей лег
костью осуществляют инвазию нового хозяина, с большей легкостью
попадают в его организм. Эктопаразиты, у которых заражение происхо-
дит большей частью посредством активного отыскивания молодью па-
разита нового хозяина, не обнаруживают тенденции к уменьшению раз-
меров яиц, как это можно видеть на монотеистических сосальщиках.
3. ОКРАСКА ПАРАЗИТОВ
Вопрос об окраске паразитов надо рассматривать по отдельности
для эндо- и эктопаразитов.
Эндопаразиты. Общим правилом для внутренних паразитов
является отсутствие пигментации, беловатый или слегка желтоватый
цвет тела. Отсутствие пигментации есть следствие нахождения парази
тов в темноте, аналогичная картина наблюдается у многих пещерных
или подземных животных. В тех случаях, когда эндопаразиты все же
так или иначе окрашены, цвет их зависит не от пигментации покровов,
106
как это бывает большей частью у форм, живущих на свету, но or окрас-
ки кишечник», запасных питательных материалов, яиц или от наличия
дыха!ельиых пигментов.
Окраску кишечника в красный или черный цвет от свежей или пере-
вариваемой крови обнаруживают некоторые сосальщики (Polystoma из
мочевого пузыря лягушки, ддолескарии Clino tlomuin с жабр рыб) н не-
которые нематоды. У большинства сосальщиков сквозь стенки тела про-
свечивают в виде, желтоватых, рыжеваты' или буроватых пятен жел-
точки ки, причем красящее вещество носит жировой характер. Жировыми
запасами в виде капель разных размеров бывают окрашены некоторые
скребни, например Acanlhocephalus anguillae из карповых рыб красно-
вато-оранжевого цвета. У Echinorhynchus salmonis из кишечника сигов
наблюдается даже известное разнообразие в окраске: одни экземпляры
обычного для паразитов беловатого цвета, другие — оранжевого (от
жира), третьи — темного, почти черноватого (судя по форме окрашен-
ных образований, тоже от жира, только чем то видоизмененного).
Оплодотворенные и окруженные оболочкой яйца многих гельминтов
(сосальщиков, цестод, нематод) приобретают желтую, буроватую и да-
же чернею окраску в зависимости от того, насколько оболочка успела
отвердеть. Вследствие этого наполненная яйцами матка выделяется на
фоне тела в виде окрашенного канала. Наконец, некоторые нематоды
имеют розовую 114111 даже ярко-красную окраску в связи с дыхательны-
ми функциями полостной жидкости. Розоватый цвет от присутствия ге-
моглобина имеет паразит кишечника рыб Carnallanus, ярко красным
цветом обладают некоторые полостные и тканевые нематоды рыб, ка-
кова Ft/aria sanguinea- кроваво-красный цвет имеет громадная (до I л)
нематода из почки собаки и других хищников — Dioctophyme renule.\ та-
ким же цветом обладают и личинки близких к Dioctophyme нематод
Euslrongylides, инкаису шрованные в мышцах рыб. Во всех последних
случаях пигмент зависит, по-вндпмому, от присутствия гемоглобина в
полостной жидкости или тканях паразита и имеет дыхательное значение.
Интересное явление окраски спороцист наблюдается у трематоды
Leucochlorid ит (Urogonimus). В наземном брюхоногом мочлюске Sir-
cinea спороцисты проникают в щупальца, где в них развиваются мета-
церка рин (о цикле Leucochloridium см. гл. X). При этом спороцисты
приобретают яркую окраску и своеобразный кольчатый рисунок, что
связано, по-видимому, с воздействием на паразита света, легко прони-
кающего сквозь тонкие покровы щупалец (Wesenberg-Lund, 1931). Эта
окраска и рисунок постоянны для отдельных видов Leucochloridium (Ги
нсцннская, 1951 б) и имеют, вероятно, биологическое значение. Ярко
окрашенные спороцисты несколько походят на мелких гусениц и склевы-
ваются птицами, которые таким путем заражаются сосальщиком.
Эктопаразиты. С точки зренья окраски эктопаразиты не пред
ставляют каких-нибудь интересных закономерностей. Моногеиетическйе
сосальщики водных животных скрыты большей частью под жаберными
крышками хозяев и не отличаются по цвету от эндопаразитов. Пиявки
паразитические ничем особенным не отличаются по цвету от свободно
живущих Паразитические ракообразные или полупрозрачны, или серо-
ватого цвета с просвечивающими гонадами. У паразитирующего на
жабрах рыб веслоногого рачка Ergasilus яйцевые мешки окрашены в
синеватый цвет и внутри тела также имеются синие пигментные пятна.
Эктопаразитнчсскнс насекомые, живущие на теле хозяина, не обна-
руживают каких-либо особенностей в окраске. В общем можно только
сказать, что у них всегда отсутствуют яркие и пестрые тона, часто свой-
ственные свободно живущим насекомым Имеются некоторые сведения.
107
что пухоеды, паразитирующие на белого цвета птицах, светлее окра-
шены, чем птхоеды с хозяев темных цветов.
Все те насекомые, которые паразитируют лишь в личиночном воз-
расте (наездники, паразитические пчелы и осы, мухи-тахнны, оводы
и т. д.), нс отличаются особенно по окраске от родственных им свободно
живущих форм. Оно п не удивительно, так как вся имагипалиная стадия
этих паразитов протекает на свету в обычных для всех насекомых ус-
ловиях.
4. СТРОЕНИЕ ТЕЛА
Органы прикрепления
Рнс. 77. Прикрепитель-
ные аппараты у Astu-
inata
Д н С hitoschcllina
poljanskyi; Н — Radioph-
rya iitrnbriculi; L) - A*.
i oplitas. (По XciicHiiy.)
Органы прикрепления составляют одну из самых характерных осо
ценностей большинства паразитов. Для поддержания существования
паразит должен быть неразрывно связан с хозяином, п это обстоятель-
ство дает толчок к развитию и усовершенство-
ванию прикрепительных аппаратов. При этом
любопытно, что, несмотря на необычайно ши-
рокое распространение органов прикрепления
в животном мире, устроены они сравнительно
однообразно. В строении органов прикрепле-
ния наблюдаются мпогочпе пенные конверген-
ции. В самом деле, два основных мотива, на-
блюдаемых в выработке органов прикрепле-
ния,— это образование крючков или присосок
Оба эти типа органов прикрепления одинаково
богато представлены как средн эктопаразитов,
гак и среди внутренних паразитов. Между тем
казалось бы, что набор органов прикрепления
мог быть значит! льно разнообразнее. Мало
использованы паразитами в качестве прикре-
пительных аппаратов приспособления тичч
клещей, ножниц или распорок.
В общем весь комплекс паразитических
животных дает следующие категории органов, служащих для прдкре.п-
чеиия к хозяину.
Крючки — одна нз самых распространенных форм органов при-
крепления, встречающаяся начиная с простейших вплоть до насекомых
Среди простейших крючками обладают грегарины и инфузооип. Многие
из кишечных грегарнп имеют венчик крючьев на эпимерите или иначе
устроенные крючкообразные выросты. У некоторых инфузорий (рис. 77)
группы Astomata (Hoplitophrya, Radiophrya и др.) имеется на брюшной
стороне переднего конца тела один подвижной крючок, служащий и для
прикрепления, и для внедрения в эпителий. МонойтИетическш» сосали
щики очень часто имеют крючки на заднем конце тела. Большей частые
крючки сидят венчиком на заднем конце тела, причем основным числом
часто является 16 мелких крючочков. Однако чаще 2—4 крючка при-
обретают особенно сильное развитие, тогда как остальные остаются
мелкими или частично редуцируются. Так обстоит дело у Gyrodactylus
и Dactylogyrus с одной парой крупных, а у Tetraonchus с двумя парами
крупных крючьев.
У кишечных трематод настоящие крючки большей частью отсут-
ствуют, и только у некоторых семейств, например Echinostomidae. на пе-
реднем конце тела развивается воротничок из крупных шипов, функцио-
нирующих как крючки. Эти шипы вырабатываются из мелких шипиков.
108
которые у довольно многих трематод в большом количестве рассеяны в
кутикуле. В классе цестод (рис. 78) намечается в отношении крючков
следующая группировка форм. Одни лентецы (Pseudoph\ Ilidea, часть
Cyclophyllidea) обходятся в качестве органов прикрепления только при-
сосками, прочие же, кроме присосок, имеют еще разным образом рас-
положенные крючья. Изредка крючки лежат в небольшом числе прямо
Рис. 78. Крючки на сколексах Ceslodes
А — головка Triaenophorus nod'tlosus (P>ei dophyllinea)
с четырьмя сложными крючками (видны дна) и двумя
присосками (видна одна); it — головка Telrarhynchus
(Tehailbnchklea). паразитирующего в скатах ив го-
ловке имеются четыре длинных, усаженных мпточис-
jieiiHi ми крючьями, выдвижных хоботка, могущих втя-
I ввиться в особые мускулистые влагалища; (.'—уча-
сток хоботка Telrarhynchus. »саженный крючками раз-
ной формы и размеров; D—।оливка Cysticercus pisi-
formis(Cycloph; llldta) с апикальной стороны для показа
двойного венчика крючк в. (По Кюкеиталю.)
I — хоботок; 2 мускулистое влагали нс.
на поверхности головки (Т riaenophotus, Tetraphyllidea). Гораздо чаще
крючки сидят в виде простого или двойного венчика на особом хоботке,
занимающем центр передней поверхности головки (Cyclophyllidea): из-
редка хоботок усажен несколькими венчиками мелких крючьев (Dipyli-
dium caninum). Наконец, у Tetrarhvnchidea из кишечника акул и скатов
сколекс снабжен четырьмя очень длинными хоботками, которые могут
вворачиваться в мускулистые влагалища. Каждый хоботок снабжен
сложной системой многочисленных крючков, вследствие чего хоботок
Tetrarhynchidta является очень эффективным органом прикрепления.
Наконец иногда (Davainea, Proteocephalus osculatus — передняя при-
соска) внутренняя поверхность присосок покрыта .многочисленны ми
мелкими крючочками.
Интересно, что лаже у тех форм, которые во взрослом состоянии
являются невооруженными, личинка онкосфера обладает 6—10 крюч-
109
ками (Diphyllobothriufn. Amphilina). У вооруженных форм эти личиноч-
ные крючки так же, как и у невооруженных, имеют провизорный харак-
тер, и окончательное вооружение возникает вне связи с ними.
У скребней (Acanthocephala, рис. 79) крючки служат в качестве
единственных органов прикрепления Крючки расположены на торча-
щем вперед хоботке в виде нескольких венчиков; в самом простом слу-
Рис. 79. Крючки Acanthocepliala
Л — Pallixentis umbellatus, передняя половина животного с
крючками на хоботке, шейке и на теле; И — Serruscntis social ix,
самец; кроме хоботковых и шейных крючков, имеются попереч-
ные ряды крючков на одной (брюшной?) стороне п ллшпца; С
Rhadinofhynchus horridus, справа — передний конец тела при
большом увеличении; I)— Согупихпта sfrumosum, самец; кроме
передних крючков имеются крючки на заднем конце тела, вокруг
отверстия полопой сумки. (Но Мейеру.)
чае таких венчиков три ( Xeoechinorhynchus к др.). Только у одного вида
скребней (Apororhynchus hemignathi) крючков совершенно нет. У неко-
торых родов (Corynosoma в некоторых других), кроме хоботка, крюч-
ки, хотя и проще устроенные, имеются и на большей пли .меньшей ча-
сти остального тела Весьма возможно, что хоботковые крючья пред-
ставляют результат сильного развития мелких кутикулярных крючков,
1Н>
или шипов, некогда покрывавших все тело скребней, но исчезнувших
повсюду, кроме хоботка. Сильнее всего развито вооружение у рода Rha-
dinerhynchus.
У нематод (рис. ЯО) крючковидные органы прикрепления разви-
ваются сравнительно редко. Мц находим хорошо выраженные крюч-
ковидные шипы у некоторых Dioctophyinata, например у Hystrichis.rM
они покрывают переднюю
треть тела. Пара трехзубых
крючков имеется у Seuralia
из Aet'ariidac; передний вен
чик головных сосочков полу-
чает крючковидный харак-
1 ср у Eustrongyhdes (Do-
ctopliymata). У Hirudinea
крючков нет. У паразитиче-
ских моллюсков крючки Ik
известны.
Рнс. 81. Способы прикрепления
к хозяину у грегарин
.-1 и В — молодая и взрослая
грегарииа Апсчга sagittald', С—
пар । грегарин /'nterocystis ен~
sis в состоянии сизигия. (Я
и В—ио Рейхевовг. С. — по
I (веткову.)
1 — ядро: 2 ипутрнялерипс тельце:
.1 — 'ТСрСШЯИ wool,.
Рнс. 80. Органы прикрепления круглых червей
(Nematodes)
А — передний конец Hystric/iis varispinosus
(Dioctophyinata) с многочисленными крючками
из кишки 4f. rpus serrator, В — передний ко-
нец S<urafia[shipleyi (Acuaiiidac) сбоку. (По
Кюкенталю.)
/ — ппропшчок пколсротпвых крючкол: 2 — пищевод; .3
1ЫПН нз >вух мощных трехзубчжтых боковых крючьев;
4 — аил риал коротких iiiHiinn; Л — грелимп кишка; 6 —
выделительный канал с вы целительным отверстием.
Среди Arthropoda мы довольно часто встречаем крючковидные ап-
параты, ио здесь они в громадном большинстве случаев являются не
простыми выростами тела, а более или менее сильно измененными ко-
нечностями. Таковы антенны Ergasilus, антенну л ы Argulus, коротки?
крючковидные изогю тые торакальные ножки некоторых Cymdlhoid^c,
мощные, на вид совершенно простые, две пары крючков Pentastomida,
крючковидные мандибулы личинок многих мух. Такое же значение при-
цепок или крючков имеют и разнообразные коготки конечностей у вшей
и других мелких паразитов из членистоногих.
С известной оговоркой к типу органов прикрепления посредством
крючков могут быть отнесены приспособления некоторых кишечньг
грегарин, тело которых образует распорки, помогающие паразиту удер
жпватьс» в просвете кишки, хотя он и не обнаруживает непосредствен-
ного прикрепления к самим стенкам кишечника. Примером этого слу-
жат гр era р и вы An enra sagiltata из кишечника Capirella и Enierocystis cri-
sis из личинок Caerijs (Цветков, 1926). Интересно, что у Enterocystis еп-
III
sis лишь передний член пары конъюгирующих грегарин, или примят,
образует по бокам тела два торчащих вбок выроста, тогда как задний,
сателлит, сохраняет обычную для грегарин червеобразную форму. Та-
ким образом, распорки, образованные примятом, служат аппаратом, за-
держивающим в кишечнике не только данною особь, но образование
более сложною порядка, а именно конъюгирующую пару (рис. 81).
Присоски представляют собой более или менее глубокие ямки
на геле, обведенные валиком особо дифференцированной мускулатуры,
которая состоит из нескольких систем мускульных волокон: радиальных,
Рис. 82. Органы прикрепления ленточных червей
(Cesiodes)
А— Diphyllobothrium latum (Pseuduphyllide<i) — го-
ловка с дгумя присасывательными ямками по бокам;
Н — liothridium pythonis (Dsendupliyllid<a) — свободные
края обеих присасывательных ямок на известном про-
тяжении спаялись, так чго ямкн превратились в две
открытых на ьпнпах и расширенных кпереди трубки;
С — Lecanicephalum abruption (TeiraphyII dea) из ки-
шечника скатов с мощной теменной и четырьмя боко-
выми присосками; D - MyzoihJlobothrium rubrum из
кишечника скатов: позади четырех присосок головки
имеются четыре дополнительных присасывательных
аппарата: Е — головка Thysaniezia ovilla с четырьмя
мощными присосками. (Но Кюкенталю.)
кольцевых. Среди волокон разбросаны ганглиозные клетки. Присоски
могут иметь различный влд и обнаруживать большую или меньшую
дифференцировку. Самыми примитивными являются присасывательные
ямкн, встречающиеся у PseudophvIlidea (рис. 82) Они имеют вид вы-
тянутых в продольном направлении ямок, или щелей, не обладающих
ясно обособленной мускульной обкладкой, так что мускулатура ямок
постепенно переходит в мышцы окружающей парен имы. Присасыва-
тельных ямок обычно бывает две, расположены они по бокам сколекса
(Diphynobothrium, Ligula и др ). У некоторых Pseudophyllidea обе ям-
ки смещаются сильно вперед и обращены своими вдавлеииями кпереди,
но еще ясно разделены друг от друга перегородкой (Bothrimonus из
112
осетровых рыб Каспия). Наконец, иногда (Cyathocephalus из кишечника
окуня, форели и др.) обе направленных вперед присасывательные ямки
вполне сливаются друг с другом, образуя общую воронку.
Типичные присоски — чаще всего округлой, реже овальной формы,
с хорошо обособленной мускульной обкладкой. Такого рода присоски
(рис. 82—84) широко распространены, встречаясь у некоторых Turbclla-
ria (мощная задняя присоска Bdelloura. Syncoelidiurn). почти у всех со
сальщиков. у цестод, Malacobdella из немертин (одна задняя присоска),
у пиявок, Oligochaeta (Branchiobdellidae) и др.
Рис. S3. Присоски Turbeilaru
Л — Dldymorrhig paranephropis (Rhabdocoela) — эктопаразит
npeciiOBO.moi о рака Гагат phrops яр ; В и С — молодая
и взрослая Fecampia xanthocephala-, D — Feeampia xanthuce-
phala внутри грудного отдела рачка fdothea rfsclectai E и
P—Bdelloura condlda (Tiiclada) c Limulus polymorphus, при
виде сверху и сбоку. (Из Догеля )
/—глотка; 2 — желточннкн; 3 — кишка; 4 — ничпик: .5 — матка; 6н8 —
д,.ч семенника; 7 — присоскиобра пипс расширение заднего конца тела;
u — Л'л ampin xanthocephaia.
Чрезвычайно разнообразны органы прикрепления моногенетичс
ских сосальщиков, у которых имеются разного типа присоски, крючья и
прикрепительные клапаны в самых различных сочетаниях. Образования
типа присосок у моногснетических сосальщиков могут быть расположе-
ны как на переднем, так и на заднем конце тела. На переднем конце
можно различить два типа присоскоподобных образований. Одни из них
не связаны с ротовым отверстием, вторые же формируются в связи с ро-
товым отверстием Первые представлены обычно двумя присасыватель-
ными ямками или присосками, расположенными по сторонам ротового
отверстия (например, Nitzschia, Tristomuni. см. рис. 37). Второй тип
присосок на переднем конце моногснетических сосальщиков — это ро-
8 В А. Догели | |3
товая присоска, окружающая ротовое отверстие (пример — Polystoma
см. рис. 13). Наиболее разнообразны прикрепительные аппараты, рас-
полагающиеся па заднем копие тела монотеистических сосальщиков. Ос
нову их представляет прикрепительный диск, па котором располагают-
ся различные прикрепительные образования. У некоторых монотеисти-
ческих сосальщиков сам прикрепительный диск приобретает форму при
соски, внутренняя поверхность которой может быть поделена па камеры
радиальными перегородками (Nitzschia, Trochopus, Tristomum, см. рис
87, Л), образуя сложные присоски. Обычно на таком прпсоскообразном
диске располагаются, кроме того,
прикрепительные крючья. Иногда у
Monogenoidea не сам диск превра-
щается в присоску, а на Нем возни-
кают вторичные более мелкие при
соски. Хороший пример такого рода
имеет место у обычного паразита
мочевого пузыря лягушки Polylsto-
tiia integerrimum.
У трематод обычно развиты две
присоски — ротовая и брюшная
(рис. 84). Чаще всего брюшная при
соска располагается на некотором
расстоянии и от переднего и от зад
него конца тела, но иногда брюшная
присоска оказывается сдвинутой на
самый задний конец (например у
Diplodiscus subclavatus из кишечни
ка лягушки). У Monostomidae зад
няя присоска исчезает совершенно.
У Cestodes чаще всего разви-
ваются четыре присоски по краям
сколекса, но у паразита рыб Proteo-
cephalus на переднем конце головки,
где у других представителей отряда
имеется хоботок, лежит маленькая
пятая, или теменная, присоска.
Присоски пиявок имеются в
числе двух: ротовой и задней. Зад-
няя присоска пиявок интересна тем,
что представляет собой не простой
мускулистый вырост тела, но ре-
зультат видоизменения последних
4—7 сегментов животного. У трема-
тод (рис 84) и у пиявок дно перед-
ней присоски пронизано ротовым от-
верстием.
Типичные присоски встречаются,
хотя сравнительно редко, и у чле-
нистоногих. где края их ограничены
мощной хитиновой рамкой. Очень ти
пнчны две присоски Branchilira (рис.
85). образующиеся за счет мак-
силл На переднем краю головогр\дпого щита Caligidae (рис. 85) за
счет основания их антепнул тоже образуются две небольшие, присоски,
или лунки (lunula) Небольшие стебельчатые прнсосочки на концах ног
Рис. 84 Присоски Trematodes
А — Syncoi’lium katuwo и < тунца,
со стебельчатой брюшной присоской;
Н — Jiunoderu luciopi-rcae и < окуня;
передняя треть тела с ротовой ш и-
соской, окруженной венчиком со-
сочков; С — Parainphistoinum gi-
gantocofyli1 из гиппопотама с громад-
ной брюшной присоской; L)—(iaatra-
discus aegyptiaeus in кишечника
лошади; вогнутая брюшная сторона
тела, покрытая сосочками, образует
присасывательный диск, тогда как
маленькая брюшная присоска отсту-
пает ил второй план. (Д — но Яма-
гутн, И—Г)—по Кюкешалю.)
114
многих клешем (например, у зудней Sarcoptes) отличаются от типичных
присосок отсутствием в них мускулатуры, ибо они являются чисто хи-
тиновыми образованиях!;!.
Уместно сказать несколько слон о наличии присосок или присоско-
образных приспособлений у некоторых простейших (рис. Ь6). Среди
жгутиконосцев подобным приспособлением обладает кишечный паразит
Рис. 85. Крючки и присоски у раков
л Л — Brgasilus sieboldi, вид сбоку; В — антеннула
Lrpr&pirrfu’ii US' с мощной прпсасынатсльн' й ямкой
* v основания; С —передний конец тела hepefipktfmrtis
с анюипулами и присасывательными ямками; L) —
Argiilus fcliaceus (Branchiura). (Л и D - но Марко-
вичу. В и С - по Ямагути.)
1 — крючков н in не 1кнтешп>|; 2 — ротовое отверстие; 3 — нйнешо*
мешки; 4 — присасывательные ямки; 5 — измененные и крючки
антгмиулы; 6 — сложные гляэл; / -вторая пара мякснлл. шио
измсшнинПхся в мощные присоски.
многих позвоночных — Larnblia. Брюшная сторона его тела вогнута, об-
разуя обведенную тонким скелетным волокном ямку. Этой ямкой пара
зит садится на обращенные к просвету кишки концы кишечных клеток
и может временно плотно приставать к ним.
Кроме Larnblia, присоски встречаются еше у некоторых инфузорий,
возникая на самых различных местах тела. Так, у некоторых Astomata
(Radiophryu, Mrazekiella) в качестве присоски функционирует одна из
сторон тела, несколько вогнутая в виде ложки. Вдоль этой стороны при-
ходит в наружном слое инфузории ряд продольных кутикулярных ребер.
117
Рнс. 86. Присоски у Protozoa
А и В — Haptophrya michigancnsis, передний конец снизу
н сбоку лая показа присоски, С и D - Trichodina renirola
из почек Esgc niger (С—разрез через мочеточник щуки
с присосавшимися к его станке ।риходшими, D — отдельная
особь TrFhodina, несколько снизу н сбоку); Е—Lamblia
intestinalis с брюшной стороны. видна обведенная опорной
нитью присоска; F—L. intestinalis в профиль; G— Polycycla
discophora. (Я и В — но Бушу, С н D — по Мюллеру, Е н
F — по Эпштейну, G — но Полянскому.)
I — мусктльиаи сЛо.ючкл мочеточника; 2 — эпителий мочеточника; 3 — трн-
ходины; 4 — адоральная спираль ресничек; .5—тело животного: Н —край
нрнсзсывзтелыюго лиска. Z — венчик ресничек; 8 — венчик крючьев; 9—ядро
10 — аксостилп.
116
которые, очевидно, способствуют прикреплению. У Plyihoslumuin при-
соской служит передний юнец тела. Сложнее всего дифференцирован
прикрепительный аппарат у I rccolannav, например у инфузорий, пара-
зитирующих на гидрах, циклопах, а главным образом на рыбах. Эти
простейшие (рис. 8G) проводят свою жизнь в подвижно-сидячем состоя-
нии, скользя своим базальным полюсом, на котором расположена при-
соска, по телу хозяина и от времени до времени прикрепляясь к п> му
Противоположный свободный конец тела несет ротовое отверстие. Ба-
зальный конец тела образует вогнутый слегка кутикуляризованный диск,
на дне которого помещается венчик кутикулярных крючков, содействую-
щих прикреплению. Диск окаймлен венцом ресничек. Благодаря такому
устройству Trichodina может произвольно то прочно приставать к телу
хозяина, то плавно скользить по нему на своем диске. Они отрываются
от тела хозяина лишь для временного плавания.
Настоящая нрисЛжа имеется у паразитической инфузории Polycycla •
(рис. 86), живущей в кишечнике плклрорьи голотурий. При помощи ее
инф'зория присасывается к эпителиальным клеткам хозяина (Полян-
ский, 1951).
Присоски сложные (рис. 87) являются результатом дальней-
шей дифференцировки простых присосок. Опп обычно происходят пу-
тем подразделения первоначально простых присосок валиками или пе-
регородками на несколько участков. Так, у Tetraphyllidea (Cestode?)
образуется сначала одна продолговатая присоска, а затем она подразде-
ляется поперечными валиками на 2. 3. 4 и значительно большее количе-
ство отдельных участков, пли ареол. Такие сложные присоски называют
бот р иди я м и. причем они принимают иногда чрезвычайно сложную,
складчатую форму, сидят на особых стебельках, и г. д. Кроме Tetraphy-
Пнка, сложные присоски имеются у монотеистических юсалыииков. как
это уже отмечалось выше. Среди трематод чрезвычайное развитие по-
лучает брюшная присоска Aspidogastridae (рис. 87, В и С). Она зани-
мает ВчЮ брюшную поверхность тела и дробится на вторичные
ячеи пли даже разделяется на несколько рядов лежащих друг за другом
присосочек.
Клапан ы представляют собой аппараты, основанные на принципе
захлопывания двух створок и ущемления между ними известного участ-
ка тела хозяина. К этому типу принадлежат прежие всего клешни чле-
нистоногих, которые, однако, лишь очень редко развиваются у парази-
тических Arthropoda. Единственный несомненный пример дают парази-
тирующие* на гидроидных иолипах или внутри их личинки Pantopoda
(см. рис. 21 и 94). обладающие клешневидными передними конечно-
стями.
В высшей степени разнообразны по своей структура прикрепитель-
ные к. шпаны Monogenoidea. Они представляют собою видоизменения
присосок. Однако принцип их действия иной: они действуют, как за-
щемляющие, а нс присасывательные аппараты. В простей ше м случае кла-
пан очень напоминает присоску (рис. 88) и снабжен лишь тремя хши-
нондными частями, из которых одна располагается в перегородке, раз-
деляющей клапан на 2 половинки, а две остальных — по правому и ле-
вому краю (Hexastoniatklae). Такой клапан — присоска складывается
по своей поперечной оси. У большинства моногенетических сосальщиков,
обладающих прикрепительными клапанами, последние устроены истее
сложно, причем усложнение идет по линии увеличения числа и разме-
ров хитиноидных частей (рис. 88). Каждый клапан состоит из двух смы-
кающихся половинок. По продольной оси клапана лежит крупная хи-
тиновая пластинка, играющая роль пружинки. К нон присоединяется
117
Рис. 87. Сложные присоски Monogenoidea, Trem«tod<s и Ccslodee
А — Tristoma iMo-ogenoidcaj с жабр скатов, 13 н С — Lophn-
taspis valid (сбоку н с брюшной стороны) со сложным присасы-
вательным диском, занимающим нею брюшную сторону тела: D —
сколекс Acanthohothrium ijimai (Tetraphy 11 de<1) с четырьмя богри-
днями, разделенными на три камеры каждый; Е — сколекс Echcnei-
hothrium flexile с четырьмя сложными ботридиямп. (По Кюкенталю.)
/ — передние нригасыплтелынде имин; 2 — но.юпое отверстие; 3 - penis; 4 — семс-
провоя; Л - влагали не; 6 - матка: 7 — гемсирнемник; Ь’ — жетточнпкь; 9 — семе-
проиоя: 10 — желточныД проток; 11 — сложяпч ирисоск<; н - ларилльпне крючки;
I.i - ceniH. леляжпе присоску к» камеры; II семенники. 15 - ветви кишки;
16 - кишка; 17 яичник; Ш — иредстате’ьитч железа; 19 — пищевод; ГО — глотка;
21 — рот.
118
еще ряд хитшюндпых элементов, лежащих по краю клапана. В результа-
те такой клапан действует, как зажим или щипцы. Число клапанов на
прикрепительном диске у разных Monogenoidea различно: от шести
(Didybothrium, 1 lexabothriidae) до нескольких десятков (Heteraxine,
Axine, Microcotylidae). Иногда клапаны располагаются на довольно
длинных стебельках (Microcotyle)
Рис. 88. Прикрепительный аппарат Monogenoidea
А — MicroaAyle mugiiis взрослый червь с жабр M'igU
auratus-, В и С — схема строения примитивного прикрепи-
тельного клапана Hexostomatidae (В — клапан развернут
полностью. С—coiHyT); D — типичный прикрепительный
клапан. Черным обозначены хитиновые части. Мускулатура
заштрихована. (По Быковскому)
Интересный прикрепительный аппарат этого рода вырабатывается у
некоторых трематод сем. Ecliinostomidae. Обычно передний конец тела
этих двуусток окаймлен воротничком из крупных шипов, котсрые функ-
ционируют в качестве венчика крючьев Воротничок на брюшной стороне,
т. о. позади ротовой присоски, прерван. У Patagtfer (рис.89) воротничок
сильно развит, и ряд его крючьев прерван не только на брюшной сто-
роне, но и па спинной, т. с. впереди ротовой присоски. Вследствие этого
воротничок становится двулопастным, и обе его лопасти получают спо
собносгь захлопываться, образуя аппарат типа клешей. Интересно, что
такого же рода физиологическую функцию выполняют и присоски у не-
которых пиявок; так \ Notostomum laeve (рис. 89) с кожи рыб присоски
перегнуты на две половины, правею и левую, и усажены по свободному
краю особыми роговыми сосочками. Обе половины присоски захлопы-
ваются. действх я подобно присасывательным клапанам Polyopisfhocoty-
linea ,
119
Наконец, по типу клещей захлопываются створки глохидиев, т. е.
личинок LJnionidae. после того, как они пристанут к телу рыбы при по
мощи своей биссусовой нити.
Тип обхватывающего прикрепления приближается к
типу клещей, но отличается от него следующим существенным обстоя-
тельством. В органах прикрепления, устроенных по типу клещей, обе
створки клещей защемляют между собой какой-нибудь участок тела
хозяина, будь то волосок, конечность или просто лоскуток кожи. При
обхватывающем типе прикрепления сходящиеся друг с другом выросты
Рис. 80. Различные типы органов прикрепления
А и В — Patagifer bilobus, очень молодой и взрослый
экземпляры (Д нарисован при значительно большем увеличе-
нии чем В, при одинаковом увеличении А имеет размеры,
показанные па Л1. Видно сильное развитие и дифференци-
ровка воротничка крючьев в днуств рчатый клапан, тогда
как значение ротовой присоски отступает на второй план);
С — Acanthocotyle lohiancoi — жаберный сосальщик с прови-
зорным личиночным прикрепительным диском, который хотя
и сохраняется, но у взрослого животного функционально
заменяется другим, вышележащим диском; Г) н Е — задний
конец тела пиявки Notostonium laeuc с брюшной стороны и
сбоку; присоска приняла характер двустворчатого клапана.
(Д и В — по Дубинину, С — ио Кюкенталю, D и Е— по
Васильеву.)
1 — ротовая присоска; 2 — воротничок крючков; 3 — глотка; 1 — брюшная
присоска; 5 — лиунетвнетая кишка; 6—семенники; 7—две околиpoioisux
присасывательных ямки Acanthocotyle; 6 — дефинитивный присасывательный
диск с радиальными рядами мелких крючьев; 9 провизорный, личиночный
присасывательный диск; 10 — присоска Notostomum.
тела паразита лишь обхватывают какой-нибудь участок тела хозяина,
смыкаясь вокруг него и виде кольца, не ущемляя тела своего носителя.
Мы уже видели пример такого прикрепления на инфузории Ellobiophrya,
обхватывающей двумя выростами своего основания жаберную нить мол-
люска Donax (см. ряс. 11). Аналогичное прикрепление нам удалюсь об-
наружить у одного веслоногого рачка (Ergusilus arnplectens) с жабр ке-
120
фали на Дальним Востоке. Своими необычайно вытянутыми крючковлд
ними аптепп'а’Ми этот рачок обхватывает жаберный лепесток рыбы. при-
чем оба крючка захотят друг за друга, образуя вокруг лепестка сплош-
ное юльцо (рис! 90)- Прикрепленные экземпляры
Е. amplectens можно иголкой свободно поворачивать
вокруг лепестка, как вокруг осп, а при попытках про-
сто отодрать паразита от места прикрепления сто ан-
ichhli не раскрываются, а где-нибудь обрываются.
Это обстоятельство говорит о том, что скрепление
обеих антенн друг с другом нагнется весьма прочным.
Пи некоторым препаратам нам кажется, что антенны
не только зацепляются Друг за Друга концевыми ког-
тями, но в склеиваются этими концами при помощи
какого-то цемента.
Быть может, таков же тип прикрепления и у свое-
образного жаберного сосилыцика Dajrielius (см.
рис. 97).
Р а с п о р к и. Этот редкий и своеобразный тип ор-
ганов прикрепления встречается только у эктонарази
тнческой полихеты I chthyotomus sanguinarius (рис.
91). На переднем конце тела этого червя имеется
острый двойной стилет, работающий до известной сте-
Рис- 90. Веслоно-
гий рачок Ergasi-
lus amplectens с
жабр дальнево-
сточной кефали;
длинные антенны
пени наподобие ножниц. При прикреплении к хозяину
стилет вонзается в кожу рыбы сложенным и затем
раскрывается в ране наподобие распорки или рас-
крытых ножниц, разрывая при этом кровеносные со-
суды. /chthyotomus прикрепляется очень прочно, так
что его трудно оторвать от рыбы, не повредив послед-
ней.
Стилеты. Под таким названием можно выде-
лить плотные, прямые, отходящие от переднего конца
тела выросты, вонзающиеся в тело хозяина. В одних
случаях стилеты не втяжные и служат для постоянно-
го прикрепления. Такой характер носит эпи.мерит ки-
охиатывают жабер-
ный лепесток в
виде кольца, пл
котором паразит
болтается, как
замок на скобе
(Оригинал.)
Рис. 91. Паразитическая полпхета Ichthyotomus sanguinarius
/1 - IcMhyottnnus sanguinarius, прикрепившийся к плав-
нику рыбы; Н—передний конец/, sanguinarius-, С—стилеты
или челюсти /. sangu-'narius. (По Эйзшу.)
1 — стилеты: 2 — медшпнмя аплеты; 3 — боковые рудиментарные антенны;
! — моет. 5 — г 133; 6— гемофильные желс/ы, выле.-’янлнме ьешегтии. кото-
рое метле г епертыпанню кропи; z — пищевод; S — гтотка.
121
шечной грегарины Stylorhynchus. Жгутиконосцы Oxyinonas из кишечни-
ка термитов и Pytsonytnpha (из отряда Polymastigida, рис. 92) имеют
залегающую внутри тела осевую палочку аксостиль; на переднем конце
тела аксостнль продолжается в длинное копье, которое вонзается в стен-
Рис. 92. Жгутиконосец
Proboscidiclla kofoidi
(Polymastigida), парази-
тирующий в кишечнике
т рмитов, прикрепляю-
щийся к стенке кишки
при помощи особого
стебельковилиого вы-
роста на переднем конце
тела. Близ узкого конца
тела мины ядра, часть
жгутиков нарисована
полностью, часть обреза-
на на конце; телт прони-
зано в юль лентовнднымн
зксос! илями и наполнено
комочками нищи. (Но
Кирби.)
I сге6ел1.кон1гли14*| вырост;
2 идрл: J и 4 — жгутики;
о — дкеости.ти.
ку кишечника хозяина- Подвижные стилеты
имеются у церкарий Trematodes, но они при-
обретают здесь иное назначение, помогая цер-
кариям внедряться в тело их хозяина. Такое
же значение имеет и копье некоторых нематод,
паразитирующих в растениях.
Стрекательные нити. Эти образо-
вания представляют собой частный случай вре-
менного прикрепительного аппарата, встре-
чающийся у Myxosporidia- и Microsporidia.
Споры этих простейших содержат от 1 до 4
овальных хитиноидных мешочков — полярных
капсул, внутри которых скручена в виде упру-
гой спирали длинная тонкая нить. Длина этой
нити у iMicrosporidia (рис. 93) может превы-
шать 500р, тогда как размеры самой споры
равняются 5 .6р. В общем, эти капсулы
устроены вполне по образцу стрекательных
капсул кишечнополостных и способ действия
их приблизительно одинаков. При действии не-
которых раздражителей (йод, азотная кислота
и др.) или даже просто при механическом на-
давливании стрекательня пять с силой вывора-
чивается из полярной капсулы, выстреливаясь
наружу. По одним представлениям, эта нить
служит лишь для временного прикрепления
споры к стейке кишечника хозяина, в которого
случайно попадают вместе с пищей и споры.
Задерживая спору в кишечнике, нить дает
время амебоидному зародышу, находящемуся
в споре, выйти из нее и внедриться в стенку
кишки.
В 1937 г. появилась небольшая работа
Ошима (Ohshima), который несколько иначе
толкует значение «стрекательных» нитей. В
спорах Microsporidia он мог наблюдать, что
при выбрасывании нити амебоидный зародыш
выходит на конце нити наружу через се про-
свет. Таким образом, стрекательная нить слу-
жит на самом деле трубкой для выведения
содержимого споры, напоминая этим вывора-
чивающиеся спородукгы цист некоторых гре-
гарин. Выть может, нить имеет и то значение,
что через ее посредство зародыш из споры по-
падает прямо в стенки кишки, не подвергаясь
вредном) воздействию пищеварительных соков хозяина в просвете его
кишки. Недавно эти наблюдения Ошима были подтверждены исследова-
ниями Гиббса (Gibbs, 1956) и Крамера (Kramer, 1960) и чешских прото-
зоологов Лома и Вавры (Lorn and Vavra).1
1 Доклад на 1 Международной конференции по протозоологии, август 1961.
122
Липкие, или прядильные, нити. Эти инти представляют
собой продукт выделения особых желез: они помогают паразиту при-
клеиваться к телу хозяина или закручиваются вокруг выростов тела хо-
зяина. Таким способом прикрепления пользуются лишь мелкие личиноч-
ные стадии развития, ведущие паразитический образ жизни. Прекрас-
ный пример таких органов прикрепления дает биссусова нить личинок
L’nionidae— глохидиев. На брюшной стороне тела личинки, где впослед-
ствии развивается нога, имеется небольшая биссусова железа, выделе-
ние которой выходит из протока в виде шелковистой, желтоватой, длин-
ной нити Это биссусова нить, или биссус. Она выдастся далеко наружу
Рнс. 93. /Vbsezna bombycis < Microsporidia), диаграмма
жизненного цикла, протекающего в кишечном эпителии
шелковичного червя. Налево, внутри кишечной клетки,
изображено деление вегетативных стадий и формиро-
вание спор, направо I- споры с выстреленной стрека-
тельной нитью, из которой выходит дзуядерпыи аме-
боидный зародыш. (По Штемпелю.)
из раковины глохидия и служит для его первоначального прикрепления
к телу хозяина (рыбы). Уже после прикрепления нитью личинка вцеп-
ляется в эпителий хозяина захлопыванием своих зазубренных на краю
створок раковины.
У целого ряда паразитических Copepoda (Caligidae, Lernaeidae, Lcr-
naeopodidae) имеется ларвальный железистый орган на переднем конце
головогруди — фронтальная железа. Эта железа выделяет плотную, до-
вольно длинн.ю нить, при помощи которой конеподитиая личинка Са-
ligidae прикрепляется к коже рыбы, на которой она паразитирует. По-
зднее оба пола Calirjidae отделяются от тела хозяина и получают спо
собность плавать свободно, лишь периодически прикрепляясь к телу
рыбы для сосания крови. Самки Lernaeidae и Lernauvpodidae остаются
прикрепленными к рыбе и но взрослом состоянии.
Еще один пример выделения биссусовой нити дает класс Pantopoda,
а именно: личинки этих животных (рнс. 94). Правда, биссусова нить ли-
чинки елхжит, главным образом, для подвешивания к телу- родителей,
123
но также и для временного прикрепления к гидроидам, на которых ли-
чинки паразитируют.
Для jioi'i цели в основном членике передней пары конечностей ли-
чинок имеется довольно крупная двуклеточная или многоклеточная пря-
дильная железа, тонкий проток которой открывается на вершине особо-
го длинного прядильного шипа. Из шипов торчат наружу две очень
Рис. 9-1. Личинки различных Pantopoda
4 — Nymplum strOmii; В - fyenogbnum litoral?-, С — Anoplu-
dactylus petiolatus; I)—Phuxichilidittm femoratum. (По Догели.)
/—хоботок: 2 — железы клешней; 3 — прядильная жс1сла; 4 — основной членик
клешней; и 6 — вторая и третья пары кот; 7 — средняя кишки; .V — ганглии. У —
кожные железки; 10— конисиые членики ног; 11 прядильный шип.
длинные нити, во много раз превышающие длину самой личинки. При
помощи нитей личинки некоторых Pantupoda (Chaeionymphon, Pallene)
замечательно прочно приклеиваются к ножкам родителя. У других ви-
дов прикрепление к родительскому телу отсутствует, и здесь, по всей вс
роятности, нити получают функцию прикрепления к хозяину.
Нитевидные отростки тела. Это образования, аналогич-
ные биссусовым нитям по функции, ио совершенно иного происхожде-
ния Они имеются у молодых личинок Pantopoda и иногда, ио-види.миму,
заменяют собой прядильные нити. Так, у личинок Anoplodactylus
(рис. 94) последний когте образный членик конечностей 2-й и 3-й пары
вытягивается в своеобразный крайне длинный усик, при хот шео изогну*
12-1
нрикре-
Рнс. 95. Стебельки, служащие для
пления паразитических Peridinea
А — Qodlnium fritillariae с кожи O>ko-
pleura-, В — Apodinium mycetouies с кожи
Fritilljria', С —часть тела и стебелек с кор-
невыми отростками Apodinium mycctoides
с кожи аииенднкуляриь (По Шаттону.)
1 — ядро паразита; 2 — прикрепительный диск; 3 —
паразит; 4 — стебелек; исеилоиолии или корневые
отростки; б — основное вздутие стебелька.
гый При помощи этих усиков личинка запутывается в колониях гидро-
идных починов. У личинок рода Pycnogonum для той же цели служил не
померно развитый прядильный шип на первой паре конечностей. Таким
образом, у Pycnagonutn полу 1ается весьма своеобразная замена про
дукта выделения шипа, т. е. биссусовой нити, самим шипом при его
сильном вытягивавши
Стебельки. Это очень редкий тип прикрепления, встречаемый
прежде всего у некоторых паразитических Peridinea. С гебсльком обла
дает род Oodi'uwn (рис. 95).
паразитирующий на различных
планктонных животных (саль-
пах. аппечдикуляриях). (Оваль-
ное тело паразита прикреп-
ляется одним из концов к по-
кровам хозяина. Из прикре-
пленного полюса тела выходит
довольно толстый, волокнисты и
стебелек, проникающий внутрь
тела хозяина и рассыпающий-
ся там на сложную систему
корневидных ра ^ветвлений
Стебелек OodiniiuQ очевидно,
соединяет в собе две функ-
ции— прикрепления и осмоти-
ческого питания за счет соков
хозяина. Гакпм же стебельком,
ио гораздо более длинным п
тонким, обладает и Apodinium
rnyceioides, живущий на теле
аппендикулярпй Нечто подоб-
ное мы находим и у своеобраз-
ной г.ру шы Catenate (род Нар-
lozoon), которые представляют
собой аберрантных Peridinea,
приспособившихся к жизни в
кишечнике некоторых Poly-
chaeta. На переднем конце те-
ла Haplozoon имеется, по дан-
ным Догеля (1919), небольшое
отверстие, через которое высо-
вывается наружу пучок до-
вольно нежных длинных нитей,
напоминающих псевдоподии,
которые глубоко внедряются в
эпителий кишки хозяина.
Строение этого нежного сте-
белька Haplozoon заставляет
препслагать. что стебельки всех
паразитических Р. iidinea представляют собой пучок измененных и уплот-
ненных псевдоподий, внедряющихся в тело хозяина. В пользу этого
свидетельствует, между прочим, и способность некоторых свободных
Peridinea выпускать из отверстия, имеющегося в их панцире для вы-
хода жгутиков, типичные ветвящиеся псевдоподии.
Дрхтая группа паразитов, обладающих стебельком, принадлежит к
отряду уеоногих раков (Cirt.pedia); это подотряд Rhizoccphala, а имен-
125
Рис. 96. Вторичные органы прикрепления некою
рых Cesiodes
.1 — головка Priapocephalus granais (Cyclophyl-
lidea) из кишечника полосатика с исчезнувшими
присоска ..и; В—метасколекс Goezella siluri (Те-
traphylhdea) позади сохранившегося сколекса с
присосками; С — сколекс и сильно складчатый
in евдосколекс у Fimbriaria jasciolaris из кишеч-
ника утки. (Пи Кюкенталю.)
но: Saccutina, Peltogaster и др. Взрослая Sacculina, сидящая на брюш-
ке краба, внедрена в тело хозяина при пимиши короткого стебелька,
который дает внутри кпаба сложнейшую сеть разветвлении. Разветвле-
ния пронизывают все тело хозяина, заходя вплоть до концевых члени-
ков ног. Стебелек Rhizocephala выполняет две функции: прикрепления
к хозяину и высасывания последнего.
11 р и к р е в л е и и я пот и п у якоря, к данной категории мы от-
носим паразитов, у которых служащий для прикрепления конец тела
вздувается, внедряясь в тело хозяина, и таким способом обеспечивает
весьма прочное и, если можно так выразиться, необратимое прикрепле-
ние к хозяину. Органы та-
кого типа встречаются как
у эндо-, так и у эктопарази-
тов
Среди кишечных пара-
зитов прекрасным примером
может служить ленточный
глист Priapocephalus (рис.
96 Д) с жсл у девидно взду-
тым сколексом. Среди экто-
паразитов у многих Lernaeo
podiflae майсиллы сильно
вытягиваются вперед, срас-
таются вместе и образуют на
конце непарный прикрепи-
тельный аппарат в виде бу
завы, груши или колокола,
который глубоко внедряется
в тело рыбы-хозяина. У не-
которых Lernaeopodidae
(Traclieliasies st elicit us) этот
аппарат образует пять ради-
альных выростов и прини-
мает вид пятиконечной звез
ды. .Аппарат такого рода на
поминает снабженный крюч-
ками эпнмерлт некоторых
грегарин. т. и. приближается уже к другому типу органов прикрепления.
Такой же приблизительно вид имеет и разветвленный передний конец те-
ла некоторых Lernaeidae. К описываемой нами группе относятся и лен-
точные глисты Pnylloboihrium, Tryzanocephalum, Goezella и Др., у кото-
рых хотя и сохраняются присоски, но они уменьшаются и принимают
рудиментарный характер, напротив, следующая за головкой шейка рас-
ширяется и образует массивный метасколекс (рис. 96, В).
Наконец, ленточный червь Finbriaria прикрепляется по головкой,
а послесколексовым отделом цепочки, который принимает складчатый
вид и несколько внедряется в выемки стенки кишечника. Этот участок
цепочки обозначают особым термином ^псевдосколскс» (рис. 96. С).
Общие замечания об органах прикрепления. В ре-
зультате обзора органов прикрепления мы видим, что у громадного
большинства паразитов они построены по типу крючков или присосок,
и только в отдельных случаях таковые заменяются какими-нибудь
другими прикрепительными аппаратами. Крючки распространены во
всем животном царстве, присоски встречаются у форм с сильно хи ти-
пизированными покровами (Arthropoda) сравнительно редко. Большей
126
частью описанные нами приспособления служат лишь для прикрепле
ния к хозяину, но иногда они, кроме того, применяются для раздраже-
ния или разрывания тканей хозяина (стилеты Ichthyotomus. эпи мерит
некоторых грегарин), а в отдельных группах применяются и для вса-
сывания пищи (стебелек Peridinea и Sacculina).
Заслуживает особого внимания вопрос о ларвальных и нмагиналь-
ных органах прикрепления в тех случаях, когда парашты прикреп-
ляются к хозяину на разных стадиях своего жизненного цикла.
У таких паразитов органы прикрепления могут быть одинаковы ли
в течение всей жизни (органы прикрепления большинства трематод,
а также эктопаразитическич насекомых и т. п.) и только увеличивают
в размерах с возрастом паразита или же при переходе животного во
взрослое состояние они могут заменяться или дополняться новыми
прикрепительными приспособлениями. Некоторые паразиты дают пре-
красные примеры субституций, совершающихся в течение индивиду-
альной жизни животного. В этом смысле показательные картины
наблюдаются у многих Monogenoidea. Плавающая личинка этих
животных часто бывает снабжена иными органами прикрепления, чем
взрослое животное. Так, например, личинка лягушечьей многоустки
(Polystopia integerrimurn) вначале имеет на >адием конце тела округ
ль’й прикрепительный диск, на котором расположено 16 мелких крюч-
ков. По мерс метаморфоза личинки на диске развиваются шесть
мощных присосок, большая часть крючков сохраняет эмбриональный
характер и перестает функционировать, а задняя пара разрастается
в пару крупных, функционирующих и у взрослого организма крючьев.
У молодых стадий Dtjrfpzoon paradoxiun, или так называемых днпорп,
можно прекрасно наблюдать, как, параллельно с ростом животного,
усиливается его прикрепительный аппарат: сначала дипорпа имеет
всего два присасывательных клапана, к которым постепенно прибав-
ляются еще шасть, пока /кивотное не приобретет признаков, типичных
для Octocoiylidae.
Молодые стадии Dactylogyrus характеризуются наличием 16 мел-
ких крючков, тогда как взрослая обладает 14 мелкими и двумя очень
крупными, которые иногда раз в десять крупнее первичных крючков.
Описанный Быховским (1936) соседний с Dactylogyrus род Dogielius
(рис. 97) иллюстрирует способ образования крупных крючков: к паре
сначала мелких крючков прирастает резко обособленное от них круп-
ное и массивное основание. Форма крупных крючков Dogieliiis легко
показывает, что рост их протекает в два ясно отделенных друг от
друга паузой периода и что основная, большая часть крючков пред-
ставляет собой вторичную надставку к первичным крючкам Чрез-
вычайно любопытный сравнительно-анатомический объект представ-
ляет собой Acanthocofyle (см рис. 89). В молодом возрасте животное
имеет на заднем конце тела очень небольшие присоску. У взрослых
Acanthocotyle непосредственно впереди от этой присоски развивается
крупный присасывательный диск, усаженный многочисленными рядами
радиально расходящихся от его центра крючков. Личиночная присоска
сохраняется позади диска в виде маленького р\димента, показы-
вающего, что здесь имела место типичная субсттуция органов.
Среди трематод печ-о подобное наблюдается у паразитирующего
в кишечнике каравайки Palagifer bilobus (Echinostomidae). Попавшие
в кишечник хозяина аюлескарии Patagii.er (см. рис 89) обладают от-
носительно крупной ротовой присоской и воротничком из шипов,
характерным для Echinostomidae. При дальнейшем росте червя
диаметр присоски увеличивается сравнительно мало, ио зато по бокам
127
головного конца гела разрастается очень широкий двулопасгын мус-
кулистый воротник с мощными шинами. Обе лопасти воротника.
правая и левая, захлопываются на брюшную сторону, действуя как
клещевидный клапан и совершенно заменяя стегающую в росте
ротовую присоску.
Было бы неправильным, но нашему мнению, относить к данной
категории замену эмбриональных крючков дефинитивными, совершаю-
щуюся у большинства Цсстод, так как оба ро-
да крючков имеют здесь различное физиологи-
ческое значение. Ь взрослых цеетод крючки яв-
ляются настоящими органами прикрепления к
стенке кишечника хозяина, тогда как 6—10
крючков личинки образуют (подобно стилету
перкарин сосальщиков) сверлящий аппарат,
облегчающий проникновение в хозяина.
Еще один пример возрастной замены ор-
ганов прикрепления можно найти у Caligidae.
Личиночное прикрепление нитью секрета лоб-
ной железы заменяется у взрослых рачков
прикреплением при помощи крючковидных но-
жек и посредством присосковидно вогнутой
брюшной стороны тела
Нередко у одного и того же животного
комбинируются двоякого рода органы прикре-
пления, чаще всего присоски и крючки: моно-
генетические сосальщики, многие цестоды,
Branchiura и др. Кроме тою, известно доволь-
но много случаев, когда обе функции, приса-
сывания и припенлення, соединяются в одном
и том же образовании. Это мы наблюдаем в
вооруженных многочисленными мелкими крю
мочками присосках цеетод Davaineidac и Gun-
gesia, в покрытых крючками присасыватель-
ных дисках многих Monogt noidea.
Насколько специальными и узко адапти-
рованными для определенной цели могут иног-
да быть органы прикрепления, показывает
пример перьевого клеща Michaelichus hetero-
pus с бакланов (рис. 98). Ноги этого клеща
заканчиваются сидящими па последнем члени-
ке круглыми присосками. При этом самый по-
следний членик на второй паре ног с правой
стороны имеет нормальную коническую фор
му. На левой же стороне он сильно вытянут на
конце в виде кочерги (рис. 99). Это полезная
для клеща адаптация прикрепительного свойства. Значение ее но Дуби
нину (1951), описывающему этот случай, выясняется при наблюдении
положения клеша на пере. Оказывается, что Michaelichus сидит на бо-
родке лера, ппижавшись к ее стержню своей правой, нормально постро-
енной стороной и вытянув в продольном направлении все ноги, кроме
второй левой ноги. Но вторая левая нога его. обладающая крючковидно
и -огнутым концевым члеником, отгибается сильно влево и зацепляется
своим крючком за веточки соседней бородки Таким образом, мы имеем
здесь дело с тончайшим и чрезвычайно частным приспособлением всего
одного членика одной моги к узко специальной цели.
?ис. 97. Жаберный со-
сальщик Dogielitis for-
ceps
’/I — целое животное; /?—
одни из двух главных
крючков. (По Быховгко-
му.)
I — глаза; 2 - глотка; .4 —
желточинкн; 4 — кольцеобраз-
ный кишечник; .5 — отверстии
ииагашнц*; б — влагали .це;
7 яичник; .У — дополнитель-
ные крючки; 9 — гл а ни не
крючки; iO — семенник; Н -
семелровод; /2 - pars prostatl-
ci; 73 — матка; 74 — пилинис
отверстие.
126
Но пример с Michaelichus становится особенно интересным, если
мы проследим, как отражается это приспособление на всей остальной
Рис. 98. Перьевой клеш Michaelichus heteropns с
маховых перьев баклана. Справа — самец клеша
с брюшной стороны, слепа — характер локализации
клещей на опахале пера. (По Дубинину.)
морфологии животного. Усиленное функционирование левой второй
йоги отражается прежде всего па некотором увеличении ее размеров
Рнс. 99. Концевые членики второй пары ног
Michaelichus; последний члеппк на левой (на ри-
сунке— справа) ноге сильно удлинен, согнут под
углом н служит для закрепления на проксимальных
лучах соседней бороздки (см. рис. 98). Членики
правой ноги развиты нормально. (По Дубинину.)
и мощности ио сравнению с правой (рис. 99). Таким образом, Michae-
lichus является своего рода левшой, подобно тому как у человека
сильнее развита правая рука. Но здесь дело идет дальше. Большая
9 И- А Догель
J 29
развитость леьой второ i йоги влечет за собой и соответственную
пшсртрофию отвечающей ей эпимералыюй пластинки, а это, в свою
очередь, ведет к несколько асимметричному строению всего комплек-
са эпимералъных пластинок. Наконец, известное «.псреразвнтие» левой
стороны тела компенсируется некоторым недоразвитием левого хвос-
тового придатка, который явно короче своего правого партнера.
В результате одно мелкое изменение адаптивного свойства (измене-
ние членика ноги) приводит к целому ряду далеко иду щпх корреля-
тивных изменений (смещение брюшною эпи.меоального комплекса
пластинок, уменьшение левого хвостового отростка) и к ясно выра-
женной асимметрии всего тела. Анализ данного частного случая пока-
зывает, какого пристального внимания заслуживают даже мелкие мор-
фологические изменения, когда удается связать их с биологическими
особенностями и выяснить их причинные зависимости.
Пищеварительная система
Вполне естественно, что переход к паразитическому образу жизни
не мог не отразиться на пищеварительной системе, так как у многих
паразитов способы питания в значительной мере отличаются от спо-
собов питания свободно живущих организмов.
У пазличпых групп паразитов изменения пищеварительной систе-
мы могли идти в различных направлениях.
Во-первых, у многих форм, питающихся кровью, а особенно у
таких, которые питаются периодически, с длинными паузами между
отдельными трапезами, наблюдается известная гипертрофия некото-
рых частей пищеварительной системы В одних случаях при этом мор-
•фологня кишечника не испытывает особых изменений, кроме того, что
стенки кишки становятся особенно растяжимы, так что кишечник раз-
дувается ст пищи, деформируя все тело животного. Это имеет место
у насосавшегося клопа, комара или у мухи цеце (рис. 100); у насосав-
шейся мухи брюшко чрезвычайно выпячивается на брюшную сторону,
так как на нижней стороне абдомена хитин более нежны» , на верх-
ней— сравнительно грубый. В других случаях кишечник кровососущих
животных образует специальные выпячивания, увеличивающие его
вместимость. У пиявок имеется значительное количество боковых
карманов кишки, у Леда (Isopoda) средний отдел кишечника снабжен
двумя громадными дивертикулами, наполняемыми кровью, у многих
кровососущих клещей средняя кишка снабжена системой боковых
выростов Сильное раздувание клещей во время питания — явление
общеизвестное; размеры голодной самки Ixodes равны 0,4 см ал’цпы,
тогда как длина насосавшейся самки—1 си. Вес голодной самки —
2—3 мг, насосавшейся — 254—100 мг. Увеличение объема кишечника
есть приспособление к редкому, но обильному приему пищи. Известно,
что клопы, насосавшиеся крови, могут голодать несколько лет, не
погибая, а некоторые иксодовые клещи живут без пищи до
2—3 лет.
Другим приспособлением кровососущих форм является развитие
желез, выделяющих секрет, препятствующий свертыванию крови.
Пиявки имеют многочисленные глоточные слюнные железки, выделяю-
щие гемофилии, или гирудин. Действие последнего легко может
быть доказано экспериментальным путем. Впрыскивание в вены
собаки вытяжки из растертых глоток медицинских пиявок лишает
кровь собаки на известное время способности свертываться. То же
наблюдается у /chthyotomus и др.
130
Во-вторых, у других групп паразитов вырабатываются противо-
положные только что рассмотренным отношения, а именно: редукция
кишечника и переход к осмотическому питанию через поверхность
тела Такие изменения' встречаются как у кишечных, так и у полостных
паразитов. Среди простейших все грегарины питаются осмотическим
путем, полное отсутствие рта и пищеварительных вакуолей характе-
ризует также всех Opalinina. паразитирующих в кишечнике амфибии.
Целая группа равиоресничных инфузорий получила название Ast-o-
inata, т. е. безрогых, ибо они лишены пищеварительного аппарата
Рис. 100. Растяжимость стенок кишки у кровососущих паразитов
Л—рачок Aega (Isopodn), сосущий кровь рыб, имеет пару вмести-
тельных боковых дивертикулов кишечника; В— насосавшаяся личинка
клона Triatoma megista, передающая трип ано юмпую болезнь Чагаса
человеку; С н D—Glossina palpalis сбоку в голодном п насосавшемся
состс яппи; /Г н В— самка клеща Zxodes riainus в голодном и насосавшем-
ся сос гояппи. (4 — но Рэп Ланкастеру. В — но Брэму, С и D — по Брумпту,
Е и В—на .Большого практикума по зоологии беспозвоночных" под ред.
Полянского.)
— глотка; 2 — сосательный жслулок. но боком его две’пэры печёночных нн; ен тои; Л —
слепые лиисртикулы кишки 4— та дня н кишка.
Сюда относятся Anoplophrya, Hoplitophrya, Mesnilella и другие роды,
паразитирующие главным образом в Oligochaeia. Вторичный
характер отсутствия рта у этих форм доказывается данными
Мияшита и Раабе (Miyaschita, 1933; Raabe, 1933), которые у Protu-
anoplophrya (рис. 101) нашли миниатюрный рудимент рта. Россолимо
(1926), по-видимому, правильно подчеркивает сборный характер
группы Astomata, происходящей из различных групп свободно живу-
щих равноресничных инфузории, причем отсутствие рта у инх пред-
ставляет собой результат конвергентной адаптации к одинаковому,
а именно— паразитическому образу жизни.
8*
131
Рис. 101. Ptotoanoploph-
rya bylhiniae с рудимен-
тарным ртом
А — целое животное; И —
4,11 ть околоро ГОВОГО НОЛЯ
с ротовой щелью- (По
Раабе.)
/ |кпоиая ше.и.; 2 — м.чкро-
Hjb'.ioyc; 3 — мнкронуклсус; 4
— сокрзтигсльныс вакуоли: 5 —
6a.u.ibiiuc зерна ресниц.
На сборном характере Astoniata настаивает и Пнйторак tPuvto-
ас, 1954) в своей большой работе, посвященной этим безротым инфу-
ориям. Этот автор предлагает даже уничтожить Astoniata как само-
юятельную систематическую категорию я относит большинство
kstomata к группе Thigmotricha. Наконец, многие паразитические
Mastigophora, а особенно кровяные и тканевые — трппано юмы,
фйшмании— совершенно лишены пищеварительного аппарата.
Среди многоклеточных той же чертой
обладают целые классы животных, а именно
все Atesozua, все Cestodes и Acanthocephala.
В общем, различные степени редукции кишеч-
ника имеются у многих групп паразитов.
Дигенетнческие сосальщики у целого
ряда видов обнаруживают хкорачивание
обеих ветвей кишечника, а такая разветвлен-
ная пищеварительная система, какой обладает
Fasciola luipatica, является у сосальщиков
большой редкостью. Наибольшее укорачива-
ние ветвей кишки наблюдается у Sanguinicola
из крови карповых рыб. У Sanguinicola остает-
ся лишь маленькая розетка полых выростов
на конце пищевода, которую Некоторые даже
не считают за гомолог кишки. Еще более упро-
щен кишечник у партеногенетических поколе-
ний Digcnea. У спороцист вовсе пет кишечни-
ка, у редин кишечник имеет вид простого ме-
шочка, нередко непропорционально малого по
сравнению с размерами самой редин. У лен-
точных глистов и скребней на всех стадиях
развития нет даже никаких рудиментов ки-
шечника
Среди нематод Ю/ке имеются примеры редукции кишечника, но
полная его атрофия наблюдается у Muopicea borelli (Mcrmitliida) из
мыши, у Robertdollfusa paradoxa (Spirurala) из глаз Corxidae.
(Chabaud et Campana, 1950), а также у аберрантной Buddenbrockia.
паразитирующей в полости тела пресноводных мшанок Plum atelia
(рис. 102). Buddenbrockia имеют вид мелких (до 3,5 мм) червячков,
?т< нка тела которых состоит из наружного эпителия, из четырех
рядов мышечных клеток и из слоя генеративного эпителия, выстилаю-
щего довольно обширную центральную полость и дающего начало
половым клеткам обоих полов. Всякие следы рта, порошицы и кишеч-
ника отсутствуют. У других нематод если и бывает, то юлько частич-
ная атрофия пищеварительного канала. Так, у Trichinella и Мег-
Tiithidae пищевод сводится до степени чрезвычайно узкой внутри-
клеточной кутикулярной трубочки, кот. рая у Mermithidae даже
замыкается сзади слепо. Средняя кишка у iWermitliidae совсем
утратила свой просвет и состоит из одного ряда клеток, набитых
опасными питательными веществами. У некоторых Filariata редуди-
ювана задняя кишка, так что средняя заканчивается слепо. У Gor-
Касса личинка обладает полным кишечником, у взрослого же червя
гсчезает пищевод; остаются средняя и задняя части кишечника,
шшепныс возможности функционировать.
Ряд интересных изменений испытывает пищеварительная система
шразнтических Gastropoda (рис. 103). Прежде всего, хоботок их
утрачивает способность вворачиваться и становится органом прикреп-
32
лення к тел} хозяина и высасывания его соков. У эндопаразитических
Gastropoda хоботок может совершенно исчезать (Asterophilidae). По
мере приспособления к паразитиз-
му исчезают радула, челюсти и слюн-
ные железы, а иногда и вся глотка
(S Pilfer celeb crisis, Paedophoropus).
Пищевод паразитов развит всегда
хорошо. Наиболее характерен про-
цесс редукции, связанный с упроще-
нием печени. Печень принимает вид
простого мешка, который непосред-
ственно сообщается, с одной сторо-
ны, с пищеводом, с другой — с тон-
кой кишкой (у Megadenus urrhyn-
chus)- Таким образом, печень ста-
новится на место желудка и его
заменяет. У Asterophi'idae вся часть
кишечника позади печени атрофи-
руется, так что пищевод открывает-
ся в замкнутый печеночный мешок.
Наконец, у Enteroxenos osiergreni
весь кишечник исчезает, и питание
становится сапрофитным.
Хорошие примеры редукции ки-
Рис. 102. Buddenbrnckia plumaiellae силь-
чоупрощенная нематода из полости тела
мшанки Plumatella
А — целое животное: внутри видны яйца,
на одном конце виден очаг образования
живчиков; В— передняя часть тела при
большем увеличении. (По Шредеру.)
шечиика имеются у некоторых пара-
зитических раков. Мы уже видели
что веслоногие сем. Monstritlidac
лишены кишечника, начиная с ли-
чиночных стадии и вплоть до поло-
возрелого состояния. Пот пост ыо от-
сутствует кишечник у взрослых Rhi-
zocephaJa (Saffculina и др.), хотя Ди-
1 — живчнкн; 2 — генсрятнввые клетки; 3 —
мускульные клетки; 4 — наружный эпителий; S —
1ЙЦЛ.
чинки их и снабжены функционирующим кишечником. Все питание
взрослой Saccubna (рис. 104) совершается осмотическим путем через
Рис- 103. Изменения пищеварительной системы у паразити-
ческих Gastropoda
А — свободно живущие Fitlimaiae-, В — Megadenus holoihtt-
r:rnla\ С—Af. arrhynchus\ I) — Ast< rnphiia japoniar, E—
Paedophoropus dieoeb>bius\ F—Entocolax ludwigi; G—
I ntocuntiia mirabilis. (По А. Иванову.)
1 и 2 — печень и полость печени; —желудок; 4 — пншепол; 5 — глотка:
6 рот; 7 — гонкая кишка; Ь' —nnpoiniitta; 9 - выпячивания глотки.
посредство ветвистого стебелька, внедренного в тело краба хозяина.
Отсутствием кишечника характеризуется и не раз упоминавшаяся, пара-
1-33
зь i приобщая в нолихетах копепода Л еnocoeluma. л некоторых других па-
разитических раков отсутствует задняя кишка, так чю средняя кишка
замкнута слепо (Dendrogaster, некоторые Entuniscidae).
R противоположность ракам паразити-
Рнс. 104. Краб, несущий на
важней стороне брюшка поло-
возрелую мешконндную Sac-
cuhna; внутри краба видны
сложные разветвления соса-
тельного стебелька Sacctilina.
(По Коллери.)
/ гели Sacculina; 2 рдипстилсиня
«'tcne.ii.Mi.
ческие личиночные стадии насекомых обык-
новенно имеют хорошо развитый пищева-
рительный канал.
Полная редукция кишечника у доволь-
но многих 'внутренних паразитов объясняет-
ся тем. что они живут в жидкостях (пище-
вая кашица, кровь, лимфа), содержащих
вещества, которые животный организм
в состоянии использовать почти без даль-
нейшее подготовительной обработки. Вслед-
ствие этого пищеварительный аппарат де-
лается для паразита до известной степени
излишним, и паразит начинает всасывать
питательные вещества стенками своего тела
dap и чрезвычайно разветвленному стс
бельку Sacculina (рис. 104).
Совершенно аберрантные приспо-
собления для всасывания пищи имеют-
ся у Polypodiutn и у паразитических
личинок Unionidac. Polypodiutn hydri-
inrme—маленький полип, который на
молодых стадиях развития нараантн-
рурт в икринках стерляди (рис. 105).
Во время паразитической стадии заро-
дыш Potypodium выворачивается наиз-
нанку— своей эндодермой по направ-
лению к желтку хозяина, и в таком ви-
дг вырастает в длинный колбасовид-
пый мешок, столон (Липин, 1925; Рай
кова, 1958)- От стенок столона внутрь
него впячиваются около двух десятков
• юлипоидных почек. Впоследствии сто-
лон разрывается, полипы выворачива-
ются эктодермой наружу и покидают
икринку, переходя к свободному обра-
зу жизни. Способ заражения икринок
остается до сих пор невыясненным. В
биологии Polypoditim особенно инте-
ресно выворачивание эндодермы нару-
по образцу клеток хозяина.
Параллельно с отмеченной нами редук-
цией кишечника у некоторых паразитов про-
является тенденция к увеличению наруж-
ной поверхности тела, с.Ц/кащей для всасы-
вания пищи. li'ieiiHo 1 акои значение следует
приписывать паре длинных тонкое генных
выростов паразитических стадий Monsfrllli-
Рнс. 105. Polypodium hxdriforme
А — икринка стерляди со вкрученным
внутри нее столоном Polyrodium’, Н
— столон, выходящий из икринки; С
— участок столица г тремя почками,
щупальца которых ввернуты внутрь
столона, а энтодерма обращена кна-
ружи, т. е. к желтку икринки; D -
огориавшийся н принявшим нормаль-
ное положение, т. с. эктодермой па-
рт жу. полип Р. hydriforme (Л — ио
Усову, С — по Липину. Ви D — по
Райковой.)
Жу, причем паразит питается салро-
фитно, но не через покровы, а через изменившую свое первоначальное
положение эндодерму, т. с. как бы через вывернутые наружу стенки
своего кишечника.
134
Личинки Unionidae—глохидии — паразитируют под кожей рыб
При этом глохидий совершенно лишен пищеварительной системы,
а снаружи одет двустворчатой раковиной. В связи с этим, для всасы-
вания пищи служат внутренние стенки ого мантии, клетки которой
сильно разрастаются и становятся высокими, цилиндрическими
(рис. 106). При окончании метамор-
фоза эпителий мантии глохидия сме-
няется новым эктодермальным эпи-
телием.
Нервная система
Вопрос о нервной системе и ор-
ганах чувств не требует долгого об-
суждения. У всех паразитов, а в
особенности у эндопаразитов, нерв-
ная система более или менее упро-
щается.
Это упрощение прежде всего
сказывается в утрате органов
чувств, которые могут присутство-
вать . свободных стадий паразитов
и исчезают по мере перехода живот-
ного к паразитической жизни. Инте-
ресную картину дают в этом отно-
шении, с одной стороны. Trematodes,
с другой — Monstrillidae. Молодая
личинка сосальщиков, мпрацидип,
плавающая на воле и отыскиваю-
щая промежуточного хозяина, снаб-
жена непарным глазком; следующие
Рис. 106. I лохи uni Unto
Л — с раскрытыми створками; В—в
поперечном разрезе; глохидий иахо-
, штся в подкожной соединительной
ткани рыбы. (Но Гар.му.)
1 — блсохмвэ нить; 2 - мускул — замыкатель
рэкоиипы; 3 — край рэкьинпи; 4 — кишечник;
5 - педальные гинглин; 6 — минтнм илроглого
жннипнэго; 7 — личиночная mjhtiii; 8 — май
тнйиая полость.
стадии развития — спороциста и ро-
дня— живуг внутри моллюсков и
лишены органон чувств. Из редин
выходят церкарин, которые покида-
ют моллюска и проходят плавающую стадию жизни в поисках нового хо-
зяина. Довольно многие цепкарии в связи с этим обладают двумя или
тремя главками (рис. 107). Когда, наконец, церкария попадает
в окончательного хозяина и превращается в половозрелую двуустку,
эта эндопаращтцчсская стадия вновь теряет глаза. У Monstrillidae
свободно плавающая личинка науплиус снабжена пазком; когда
личинка забирается внутрь червя Polycirrus, она теряет глазок; перед
выходом из червя взрослые Monstrillidae снова приобретают непар-
ный глаз.
Утрата органов чувств настолько характерна для паразитов,
что в некоторые случаях по отсутствию глаз у животных, например
у раков, обычно обладающих таковыми, .можно предполагать
о вгроятнэм эндопаразитическом характере таких видов. Лишь в очень
редких случаях переход к эидопаразитизму не сопровождает-
ся полной редукцией глаз, как, например, у взрослых Polystoma
iniegerrimum, сохраняющих две пары рудиментарных, лишенных
пигмента глазков. Напротив, при полном переходе к внутреннему
паразитизму (Cestodes, Acanthocephala, Saccultna) исчезают все
органы чувств и сохраняются лишь отдельные, разбросанные в коже
чувствительные клетки или нервные окончания.
135
Известное исключение из этого правила составляют лишь не-
которые равнорссничные инфузории из кишечника копытных. У них
близ переднего конца тела развивается особая, сложно устроенная
«вакуоль с конкрециями», которую Догель (1929) рассматривает как
своеобразный органоид чувства равновесия.
Центральная нервная система подвергается меньшему воздеи
ствию условий паразитического образа жизни. Однако нередко
и в отношении се наблюдаются известные упрощения, выражаю-
щиеся как в концентрации ее ганглиозных составных чаете.. так
Рис. 107. Церклрня с глазами. (По Гинсцинскон.)
А— церкария Notocotylus sp.; ii — поперечны^ разрез перка-
рин Nvturotylas sp.
I — глазные пятна; 2 — шлелительный канал; .4 — ветип кишечника; 3 —
мозговой ганглий; 5 — шнцев-. г.
и в ослаблении системы периферических нервов. Так, в отличие от
Polychaeta, брюшная нервная цепочка Myzostomida испытывает силь-
ное укорачивание и концентрацию ганглиев (рис. 108). Только
у Protomyzostomum polynephr-s, по Федотову (1915), нервная систе-
ма сохранила вид брюшной нервной лестницы. Средн Cirripedia
группа эндопаразитических Rhizocephala показывает слияние всей
нервной цепочки в одни ганглий. Однако у паразитических личинок
различных насекомых никаких особых изменений в нервной системе
не замечается. Зависит последнее обстоятельство, вероятно, от того,
что эндопаразитические и даже внутренностные личинки, например
Hynienoptera. ведут нс только нс менее, но порой более подвижный
136
образ жизни, чем свободно живущие личинки перепончатокрылых
(например, ос, пчел, шмелей, муравьев и др.)
Выделительная и осморегуляторная система
Выделительная система паразитов не показывает каких-либо
специальных изменении. В одних случаях выделительные органы
паразитов подвергаются частичной редук-
ции, как, например, у Myzostomida, В
противоположность мета мерному строе-
нию выделительных органов у Polvchacta,
у Myzostomida сохраняется обыкновенно
всего одна пара очень мелких иефридиев.
Лишь у Protoirvzoslomum имеется пять
пар иефридиев, причем Федотов отмечает
наклонность части этих нефрид'иальных
каналов атрофироваться в течение инди-
видуальной жизни животного. Совершен-
но обратные отношения имеются у Ne-
matodes, где боковые выделительные ка-
налы и фагоцитарные клетки получают
у паразитических форм сильное раз-
витие.
Очень распространенной чертой эн-
допаразитических простейших являемся
исчезновение пульсирующих вакуолей.
Их нет у всех Sporozoa, у паразитических
амеб и жгутиконосцев. Там, где они име-
ются (Entodiniomorpha, Balantidium.
Nyctolherus, Astomata и др.), пульсация
их сильно замедлена (Хейсии, 1952), но
не всегда, ifc сравнению с таковой у сво
бодно живущих инфузорий. Тенденцию к
атрофии выделительной системы парази-
тические простейшие разделяют с мор-
скими Protozoa, что зависит от сходных
осмотических условий окружающей сре-
ды. Как паразитические, так и морские
простейшие живут в среде, значительно
более богатой солями, чем среда, в кото-
рой живут пресноводные. Вследствие это-
го разница в осмотическом давлении
междуг содержимым их тела и окружаю-
щей средой меньше, чем у пресноводных
форм, а это, в свою очередь, уменьшат г
необходимость наличия особых алпара*
Рнс. 108. Концентрация нервной
системы у Myzostomida
А—схема нервной системы
Polycliaeta, от которых произо-
шли Myzostomida; И нервная
система Proiom vzostomumpoly-
nepftris, паразитирующего в
офпурах (нервная система еще
сохранила до известном степени
вытянутый характер, но мозго-
вые ганглии сильно редуциро-
вались); между обеими поло-
винками нервной цепочки
залегает тонкий непарный про-
межуточный нерв; С—схема
нервной системы типичных
Myzostomirm- D — парочка
Myzostomam cysticolum внутри
кожной цисты, или зооцецн тя
тон, откачивающих поступающую в тело
извне воду, каковыми являют ся пульси-
рующие вакуоли.
на амбулакральной стороне
луча морской лилии Anthomclra
adriani. (По Кюкенталю.)
Дыхательная система
Дыхание эктопаразитов подчиняется тем же законам, что
и дыхание свободно живущих организмов. Полостпые и тканевые
паразиты находятся, очевидно, в таких же условиях дыхания, как
и соседние ткани самого хозяина, т. с кислород доставляется им
137
кровью хозяина пли (у Arthropod;! Trachea(а и у паразитов, живу-
щих в дыхательных иугях позвоночных) непосредственно органами
дыхания хозяина. Совершенно особые условия дыхания окружают
кишечных паразитов они живу г в почти бескислородной среде.
В течение долгого времени считалось, что кишечные паразиты
обладают чисто анаэробных! типом обмена веществ, т. с. что источни-
ком энергии у них являются процессы распада (брожения) углеводов
и жиров, протекающие без использования свободного кислорода. Так,
Вейнлаид (\\ inland, 1901) давно уже показал, что у аскариды
в тканях в большом количестве накапливается гликоген и источником
энергии является его расщепление, происходящее с выделением угле-
кислоты в валериановой кислоты. Однако в настоящее время можно
считать доказанным (см., напрммер, Брандт, 1951; Brandt. 1952), что
V кишечных паразитов наряду' с анаэробными процессами брожения
(расщепления) имеют место и аэробные процессы, связанные с исполь-
зованием свободного кислорода. Такого рода смешанный тип обмена
носит название аиоксибио за. У кишечных паразитов при анокенбиозе
благодаря низкому парциальному давлению кислорода преобладают
расщепптельные процессы. Известный знаток этих вопросов Брандт
в цитированной выше монографии 1951 г., посвященной вопросам
анаэробиоза у беспозвоночных, пишет: «...по нашему мнению, ста-
рую альтернативу „аэробиоз” или „анаэробиоз” в отношении кишеч-
ных паразитических червей надо признать несостоятельной, ибо
в зависимости от своей организации одно и тоже живогпоз может
вести преимущественно анаэробное существование в той самой среде,
в которой другое способно жить в основном аэробно».
Анаэробиоз благодаря отсутствию респнтеза продуктов броже-
ния характеризуется значительно меньшей энергетической эффектив-
ностью, чем оксибноз. При анокенбиозе в зависимости от большей
или меньшой степени преобладания анаэробных процессов над
аэробными энергетическая эффективность может быть различной.
Например, у аскариды в процессе обмена веществ освобождается
примерно 9°о тон энергии, которая освободилась бы при полном
окислении углеводов. Тем нс менее по сравнению с анаэробными
формами брожения у микроорганизмов (бактерий, дрожжей), аиакси-
биоз паразитических червей характеризуется все же относительно
высокой энергетической эффективностью. Так, освобождение энергии
при маслянокислом брожении равно всего 2.2%, а при спиртовом
брожении — 4% (но сравнению с полным окислением).
В теле паразнгов-аноксибнонтов почти всегда имеются значи-
тельные запасы гликогена, который и является основным энергетиче-
ским источником.
Экспериментально доказано (Марков, 19396, 1943, 1958), что при
перс л.иванни паразитов вне тела хозяина в растворах, лишенных
питательных веществ (например, раствор Рингера), происходит трата
гликогена. Чем выше температура и чем активнее движение парази-
та. тем быстрее потребляются запасы гликогена. Кроме гликогена
в тканях паразитов-анокепбнонтов обычно отлагаются значительные
количества жира, который является конечным продуктом при броже-
нии гликогена (Brardt und Weinland, 1924, Лутта, 1939; Марков,
1950. 1958).
В некоторых случаях наблюдаются интересные биологические
приспособления внутренностных паразитов, облегчающие им полу-
чение атмосферного воздуха. Так, нами уже говорилось, что личинки
кожного овода, располагаясь под кожей рогатого скота, устанавли-
138
вают связь с внешней средой прн помощи прободающего кожу
отверстия. Другой интересный пример дает представитель паразити-
ческих .мех Crnssoeosmia (UgintyiaJ sencariae (рис. 109). Яйца этой
мухи откладываются сю на листья тутового дерева и поедаются
вместе с нищей гусеницами тутового шелкопряда. .Личинки мухи
вылупляются из яиц в кишечнике гусеницы, ппободают стенку кишки
и сначала внедряются в ганглии брюшной нервной цепочки хозяина.
Вскоре они, однако, покидают ганглий и выходят в полость тела
гусеницы, где располагаются так, чтобы их находящиеся на заднем
конце тела дыхальца соприкасались с одним из дыхалец хозяина.
Рис. Ю9. Личинки мухи Crossucnxmia s-ricariae
пара иппрую'щне в тутовом шелкопряде
Д — личинки мухи, проникающие в брюшной ганглий гусеницы
тутового шелкопряда; Л‘ — личинка, вылезающая из кокона
шелкопряда; С—гусеница шелкопряда с тг иным пятном около
одной и 1 перс шнх С.ТШ м — признак зар гжепня гусеницы личин-
ками мух. (По Сасаки.)
2— гангдпн: 2—трахейные стшмы; .3 .iiviiitiiui мухн; J — опт из отхо-
дя 1И11Х ОТ raiir.'IIIM IICJ1IMHI.
Подобное положение паразитов настолько постоянно, что о присут-
ствии их внутри гуссипц шелкопряда узнают по наличию около
одного из дыхалец хозяина буроватого пятна, вызываемого парази-
том. Очевидно, в этом тесном контакте со стигмой хозяина мы имеем
дыхательное приспособление личинок Crossocosnva (Uginiyia).
Очень интересные приспособления для дыхания обнаруживают
некоторые вши морских млекопитающих, а именно: ластоногих. Не-
которые виды вшей ластоногих не имеют специальных морфологиче-
ских адаптаций и располагаются преимущественно на морде своего
хозяина, г. е. па том месте тела, которое чаще всего выставляется
хозяином из воды на воздух. Здесь, следовательно, мы имеем биоло-
гическое приспособление паразита, в смысле его локализации. У дру-
гих вшей, относящихся к роду Antarctophtliirius. на теле развиваются
особые плоские чешуйки (рис. 110). При погружении хозяина в воду
эти чешуйки прижимаются насекомыми к его телу, зажимая в про
страистве между чешуйкой и телом маленький пузырек воздуха. Таким
путем паразит получает на время погружения хозяина некоторый запас
уносимого иод воду воздуха. Подобное морфологическое лриспособле
пне позволяет Antaectophtiurius жить на всей поверхности тела
хозяина.
В настоящее время \ довольно многих беспозвоночных в крови
пли в полостной жидкости установлено вещество, близкое к гемогло-
бину и имеющее, подобно последнему, значение в дыхательном обме-
не. Ввиду ею некоюрою химического отличия от гемоглобина
Лнндро-т (Undrql.li, 1943) предлагает называть это вещество более
139
Рнс. 110. Сямка тип Antareto-
phthirius mierwhir с тюленя
Zalophus calijurnianus
Л — общий пил насекомого; И
— группа чешуек, которыми к
телу прижимаются nv <ырькп
воздуха. (По Феррису.)
индифферентным термином «эритрокруорин». Эритрокруорин найден
и у некоторых эндопаразитов, которым он может придавать розова-
тую пли даже красную окраску. Таковы, например, некоторые не-
ма годы; паразит кишечных рыб Gamallauus, паразит почек собаки
громадный Dioctophyme renale, тканевый паразит рыб Philometra
sanguinea и некоторые другие. Трахейные клетки личинок желудоч-
ного овода (Gastrophilus equi) крупны и окрашены в красноватый
цвет — они содержат эритрокруорин, ио Кемницу, (Keinnitz, 1916).
Некоторые веслоногие рачки, прячущие
большую часть своего тела внутрь орга-
низма .хозяина (таков Herpyllobiiis), име-
ют полостную жидкость розоватого цвета.
Интересующим нас в данном случае
обстоятельством является го, что боль-
шей частью у свободно живущих и сво-
бодно дышащих воздухом родичей таких
паразитов эритрокруорин отсутствует,
т. е. приобретение его паразитами одна
чает собою одну из адаптаций к иарази-
ческому образу жизни. Очень хорошо
обосновывает эту мысль Кемниц сравне-
нием личинок желудочного овода с тако-
выми бычьего {Hypodenna lineatum).
Личинки бычьего овода живут в подкож-
ных желваках, сообщающихся с внешней
средой, и дышат наружным воздухом; их
трахейные клетки лишены красного пиг-
мента, тогда как личинки желудочного
овода отрезаны от свободного доступа
воздуха и имеют эритрокруорин. Таким
же образом свободно живущие нематоды
не обнаруживают красной окраски, а у
некоторых эидоиара штиых круглых чер-
вей последняя имеется. По-видимому, эритрокруорин некоторых эндопа-
разитов содействует им в поглощении кислорода из бедной этим газом
ок ру ж а юте й среды.
Органы движения
Органы движения паразитов, в особенности внутренних, при
приспособлении к паразитизму подвергаются большей или меньшей
редукции. Мы уже видели, что параподии Myzostoinida по сравнению
с таковыми Polychaeta испытывают уменьшение н упрощение. Очень
сильной редукции или полной атрофии подвергаются конечности мно-
гих паразитических раков. На свободных личиночных стадиях разви-
тия (копеподитная стадия у Copepoda. цпирнсовпдная у Rhizocephala)
эти раки обладают нормально развитыми конечностями, но по мере
перехода к паразитическому образу жизни ноги у них атрофируются.
Полная атрофия конечностей имеет место у Rhizocephala и у таких
Copepoda. как Lcrnaeocera, Penella, Xctiocoeloma; частичную атрофию
в возрастающей степени иллюстрирует целый ряд эктопара литиче-
ских веслоногих раков. Эндопаразитические личинки некоторых
Pantopoda сбрасывают вторую и третью пары конечностей (Phoxichi-
lidium, Anoplodactyliis) и развивают их вновь лишь перед превраще-
нием во взрослое свободно живущее животное. Две нары заметных
ни
бугорков с хитиновыми крючьями, имеющиеся ла переднем конце
тела PentastoniKia. рассматривают как рудимент более развитых
к он еч и остей. Эндопаразитические зудни (Sarcopt idae) отличаются
iильным укорачиванием ног.
Конечности паразитических
насекомых нс испытывают
особых изменений, но зато
чрезвычайно распространен-
ным \ них явлением оказы-
вается полная атрофия
крылы в.
Целые отряды парази-
тических насекомых вторич-
но утратили крылья: вши,
блохи, пухоеды,, Особенно
показательны те груипы эк-
топара •нгнческих насеко-
мых, из которых можно со-
став! |ть морфологический
ряд постепенной редакции
крыльев Это, как мы видели
(стр. р8). легко сделать по
отношению к мухам-крово-
соскам. где о г снабженных
нормальными крыльями Ог-
nithumyia мы через Stenop-
teryx имеем переход к впол-
не бескрылым Meloohapus.
Кровососки интересны и в
несколько ином отношении:
Рис. 111. Изменение строения некоторых
насекомых в связи с паразитизмом
А нН — муха Ascodiptcron spaiseritututn,
паразитирующая в коже iilfio-малаЯских ле-
тучих Msiini fl (Л — самка м ухи до погружения
в кожу; />’—самка, погруженная до копна
брюшка в кожу хозяина) (по Мюир); С — in -
сочная б.юха Sarropxylla penetrans. сильно
вздувшаяся самка; L> — самец 5' penetrans
(Из Шимкевича.)
средн НИХ имеются формы, /-грудь
которые теряют крылья в то-
чечно индивидуальной жиз-
ни, по мере установления более тесного
Ascudiptcron speiserianum (рис. Ill)
ii голова 1111Я-1*11вншс<*« в ее брюшко;
2 — TipiuiitKO.
контакта с хозяином. Взрослые
обладают хорошо развитыми
Рпс. 112. Редукция крыльев в связи с
пара, нтнзмом
А -самка Sfylnps sp. (отр. Strcpsipiera) в
брюшке пчелы Andrena vaga-. Н — самка
I'lenchus (Strepji,ptent), вил сбоку; С—самец
Delphacixertos (Sirepsiptera) (11о Веберу.)
/ передние j рудиментарные крылья.
крыльями и конечностями, а
также торчащим вперед хо-
ботком. Оплодотворенная сам-
ка садится па хозяина (одна
из летучих мышей Малайского
архипелага! и прикрепляется у
основания ущеп. Она постепен-
но внедряется в кожу хозяина,
пока снаружи нс останется
торчать в виде бугорка лишь
задний конец тела кровососки.
При этом муха испытывает
глубокие изменения. Ес крылья
отваливаются, ноги отрывают-
ся на уровне вертлужного чле-
ника, голова и грудь впячиваются в основание брюшка, становящегося
огромным. Самец остается свободным и сохраняет соответственно с этим
нормальное строение.
Очень сильную редукцию крыльев и конечностей испытывают
самки Sirepsiptera (рнс. 112), паразитирующие в перепончатокрылых.
1 И
Самка Strepsiplera, живущая внутри хозяина (например, Xenos
rossi в личинке, а позднее во взрослом Policies), не имеет ни сяж-
ков. пи глаз, ни ног. ни крыльев и своим видом напоминает безногих
личинок некоторых насекомых.
Редукция органов движения происходит и у паразитических мол-
люсков где можно отметить сильное уменьшение ноги у Mucronalia,
низведение се до степени р\ гимента у Stilifer и полное исчезновение
ноги у эндопаразитических Etitocolax и Enteroxpnos.
Половая система
Функция размножения у большинства паразитов становится осо-
бенно доммнируюшеп: кроме обычных способов размножения, часто
появляются еще иные, усиливающие способность животного к размно-
жению. В жизни паразитов все решительно функции вполне подчи-
нены одной, а именно — функции размножения, что далеко не так ясно
выражено у свободно живущих организмов.
Паразитический образ жизни, прикованность паразитов к их
хозяевам затрудняют встречу между особями разного пола. Это
характерный фактор паразитической жизни, приводящий к тому, что
паразиты обнаруживают несомненную наклонность к гермафроди
тизму. Средн паразитов имеется очень большое количество гермафро-
дитов Самый характер гермафродитизма, а именно его способ
происхождения у папазитов может быть двояки .к Часто гермафроди-
тизм паразитов объясняется историческим моментом, а именно- про-
исхождением от гермафродитных свободных предков. Достаточно
вспомнить о классах Trematodes и Cestodes, ведущих начало от
гермафродитных TurbeJIaria; точно так же гермафродитизм Rliizocepha-
1а может быть объяснен гермафродитизмом прочих усоногих, т. е. сво-
бодно живущих предков Khizocepliala. Такого же происхождения и гер-
мафродитизм у пиявок (потомков гермафродитных Oligochaeta).
Иной характер носи г гермафродитизм у некоторых нематод, па-
пример. у паразитического поколения Rhabdias bufonis (гл X), ибо все
свободно жив'щие нематоды раздельнополы. В таких случаях можно
говорить о влиянии на половую систему паразитического образа жизни.
К этой же категории относится гермафродитизм Myzostomida, ибо гро-
мадное большинство Polychaeta раздельнопото, а также гермафродит-
ное состояние многих паразитических Isopoda (Cymothoidac. Entoni-
scidae). Очень ясен переход к гермафродитизму, связанный с парази-
тизмом. у Prosobranchia. Свободно живущие Prosobranchia, в том числе
и род Eulinici раздельнополы. Несколько видов Etilima становятся экто-
паразитами морских лилий и звезд, сохраняя раздельнополость. Род
AEicronalia, имеющий ту же родословную, ио сильнее адаптированный
к паразитическому образу жизни, все еще, по-видимому, является раз-
дельнополым, равно как и еще сильнее измененный род Sliltjer. Однако
средн представителей последнего рода имеется два несомненно герма-
фродитных вида. Эти виды иллюстрируют раыын способ происхождения
гермафродитов у данного рода. Нирштрасс и Шен.мап (Nierstrass urid
Sclirpinan. 1909) и Джоикер (Jonker, 1916) описали у них неотепиче-
ских. личнпкообразных карликовых самцов, которые, ио-видимому,
отвечают дополнительным самцам Cirripedia. Поэтому можно думать,
что гермафродитные особи возникали путем изменения самок и превра-
щения их в гермафродитов.
У паразитических Gastropoda в семействе Entoconchidae, а именно
у родов Enteroxenos (из полости тела голотурии Stidwpus tremulus) и
142
Parentctoxcnos (полость тола голотурии Cuciitnaria jupotiiaa), также
наблюдается типичный гермафродитизм. Поскольку у некоторых пара-
зитических Gastropoda имеются карликовые самцы (например, род
Entocolax), то представляется вероятным, что гермафродитизм пара
зитических Gastropoda так же, как и среди некоторых ракообразных,
возник путем изменения самок (приобретения ими семенника) и утери
самцов (Bornevie, 1902; Иванов, 1947; Шванвпч, 1917).
Наконец, гермафродитом является и чрезвычайно сильно адапти
роваиный к паразитизму веслоногий рачок Xenocoeloma.
Рис. 113. Организация Nematobothrium fiiarina. (По Ямагути.)
1 — рот; 2 — глотки; 3 — по.кппя клоака: J — семенник; 5—семепровод;
О' — яичник; 7 — желсочник; i S — кишечник.
В каж щемся противоречии с намеченной линией изменений нахо-
дится обратный процесс возникновения в пределах гермафродитных
групп отдельных раздельнополых видов с резким половым диморфиз-
мом Повторяем, что указанное противоречие только мнимое, так как
в каждом отдельном случае рука об руку с выработкой раздельно-
полости идет адаптация, обеспечивающая каким-нибудь способом пере-
крестное оплодотворение. Чаще всего это приспособление заключается
в том. что самец и самка более или менее прочно связываются друг с
другом на всю свою жизнь.
Среди сосальщиков есть краппе интересное семейство Didymozoo-
nidae. которые живут иод кожей морских рыб, на их жабрах пли в
ротовой полости. Они встречаются всегда попарно, в особых подкожных
кансхлах. Некоторые из представителей этого семейства, например
Dtdimozoon scombri со скумбрии, типичные гермафродиты. Здесь, следо-
вательно, мы имеем приспособление к перекрестному оплодотворению
и только. Однако у других Didymozounidae, а именно у Netnafo-
buthriiim fiiarina (рис. 113) из жаберной полости Sciaena, две особи
в одной и той же капсуле хотя и гермафродитны, но различаются раз-
мерами и способностью к продукции яиц. В одной особи их мало, и они
143
остаются светлыми и. по-видимому, неспособными к развитию, у ару
гон жл многочисленные япца имеют нормальный вит. Наконец, у треть-
его члена семейства, Wedlia bipartiia (рис, 114} из тунца (Thynnus). обе
особи каждой пары различаются не только по размерам (4 и 9 мм), но
и но строению полового аппарата. N меньшей особи i яеется отлично
развитый мужской половой аппарат, а женский хотя и имеется, но
миниатюрных размеров и не способен функционировать Таким обра-
зом эта особь представляет собой, собственно говоря, самца. Большая
особь имеет полностью развитую женскую половую систему, а от муж-
ской у нее сохраняется лишь рудимент ссмепровода.
Рис. 114. Сосальщик Wedlia bipartite «Didy niozoonidae) из
тунца
A — самец н сачка, выделенные из общей капсулы (увеличено
и 5 раз); В — изолированная самка при увеличении в 15 раз;
С — самец (Х20). (По Одперу.)
1 — половое отверстие; 2 — poron.ni присоска; 3 — ссмспровот; J — истин
кишечника; 5 — мап.а, б — семенник. 7,—рудимент Ленской ио-аоиой
системы.
Ямагути (Yamaguti, 1938) описал еще одно интересное изменение
организации у Di lyniozoonidae. связанное с их попарным нахождением
в одной цисте. У Diplotrema обе особи, находящиеся в цисте, сра-
стаются своими задних и половинами воедино, тогда как тонкие перед-
ние половины их остаются раздельными (рис. 115).
Кроме Didynnozoonidae, раздельнополостью и постоянной спарен-
ностыо полов среди сосальщиков обладает еще семейство кровяных
двуусток (Schistosomatidae). Широкий, ио короткий самец St hisiosoma
обхватывает узкую и длинную самку боковыми краями своего тела.
В таком виде пара этих червей держится в кровеносных сосудах чело-
века и млекопитающих.
144
Строгая спаренпость индивидов обоих полов но всех указанных
случаях представляет собой крайне интересную загадку. Для раз-
делыюполы.х Trematodes вероятнее всего простое отыскивание молодыми
особями одного пола особен другого, так как именно таким способом
дело обстоит у гермафродитных, но спаривающихся на всю жизнь осо-
бей Diplozoon parda'bxtrtn (из Monogenoidea). Diplozoon (рис. 116)
паразитирует па жабрах карповых рыб. Молодые
особи этого вида, известные под названием «дч-
порпа», живут на жабрах поодиночке, но но мере
приближения половой зрелости отыскивают друг
друга и сначала скрепляются одна с другой крест-
накрест пйзарно при помощи специальной Неболь-
шой присоски, находящейся приблизительно посе-
редине тела, а затем и срастаются друг с другом в
этой области. Особенно интересно при этом, что
мужские половые протоки одной особи вступают в
связь с женскими протоками другой, чем обеспечи-
вается перекрестное оплодотворение. Дипорпы, не
нашедшие себе партнера, не достигают половой зре-
лости и погибают.
Нечто аналогичное наблюдается и у раздельно-
полых сосальщиков. У Schistosoma mansoni самки,
не соединенные с самцами, не развиваются до поло-
вой зрелости. У самки половые продукты развива-
ются только тогда, когда она понадает в гипеко-
форчый канал самца. У Sch. taponicum наблюдает-
ся взаимное влияние полов. Ни самка, ни самец
не достигают половой зрелости без спаривания
(Vogel, 19-11).
Отмечается также влияние половых гормонов
хозяина па развптш Sch. mansoni определенного
пола. Например, у кастрированных самцов белой
мыши задерживается развитие мужских особен па-
разита (Berg, 1957). Интересно, что Sch. mansoni в
Рнс. 115. DipWrema
pelamydis (Didvinozo-
nrndae'l; сросшаяся па-
ра сосал ымгкпв к ।
цисты с жабр тупца
। Euthynnns peSamysl
(По Ямагучи. изме-
нено ।
/ ccMviiiiiikii; ? яични-
ки: Л передние! конец тс-
.1.1 <<> р1пм п гло;кой,'
J — ccNciipriticn; 5 - жел-
HiMinti н. Mdih'j, чаиплнмю-
ния все тело животного,
опущена.
морских свинках или кроликах может развиваться
.аже в гер маерродпт ные особи (Vogel, 1947).
С другой стороны, для Didymozoonidac (см.
рис. 115) можно предположить и другие отношения,
напоминающие тс, что имеют место при определении
пола у ВореШа. а именно Йс жпо думать, что кап-
сула сначала образуется вокруг одной особи, кото-
рая получает женскую ориентировку. Если позднее
в рыб' попадают другие мета перкарин того же ви-
да. то они отыскивают одиночные капсулы, прони-
кают в них н становятся особями, орпеитированны-
МИ В MV/KCKVH) сторону.
Единичные случаи перехода к раздельнополости имеются и среди
Cestodes. Так, прежде всего известную тенденцию к разделению полов
обнаруживает удивительная Gynandrotaenia stammeri (рис. И7),
описанная Фурманом (Fuhrmann, 1936). Она замечательна правильным
чередованием в стробиле мужских и женских члеников, причем первые
меньше вторых. Таким образом, здесь каждый членик содержит одно-
полый половой аппарат. У Dioicocestus из кишечника некоторых водо
плавающих птиц дело идет дальше. Эти цсстоды (рис. Н8) имеют
стробилы или цепочки двоякого рода: один — целиком мужские, дру-
|f> в Л. Догель
145
lie. I б. Dip'nioon рагаИчхит
Л — две сросшиеся нзрис.и ' особи; Н — яиио с нитевидным
pVpucTKOM скорлупы С - свободно плаваюта личинка. (По
Быховскиму )
Рнс. 117. Строение Ciythituira-
taertin stammgri
zl - стробила (iviuindi'otaeitia
sfainmt'fi. Видно правильное
чередование мужских ( екого-
рыс с выпяче ним цпррусом) и
женских iipoi.ior hi h гою-
в .а с nepeiii'1 ныро-гом
(нроскилекг)и BiHiiyniM Хобот-
ком (По Фурману.)
Рнс. 1 8. Дне
стробилы
раздельно-
по лоб це с то-
ды I ioico-
cestus
А — же н’кая
(?j и 13-
мужская (з).
(По <жрма-
ну.’ I
гие— чисто женские. При этом мужские и женские цепочки встре-
чаются всегда попарно. Такое удивительное совпадение предположи
телыю можно объяснить тем, что в финке Dioicocestus развиваются две
головки одна из которых дает мужскую, другая — женскую цепочки
при подобном устройстве окончательный хозяин всегда должен иметь
в своем кишечнике две цепочки разного пола
Пример Dioicocestus прекрасно пока-
зывает, как разделение полов у парази-
тов сопровождается приспособлением,
гарантирующим нахождение одного по-
ла другим.
Попарное прочное соединение обоих
полов в тех же целях наблюдается иног
да и у раздельнополых паразитов, напри-
мер у Syngamus trachea (рис. 119) из
трахеи птиц. У этой нематоды самец проч-
но прикрепляется, почти прирастает к
тел) самки в области ее полового отвер-
стия, так что животные всегда встречают-
ся парами, состоящими из маленького
самца и втрое более крупной самки. По-
добные же отношения мы встречаем у
нематоды Hedruris (рис. 119), паразити-
рующей в пищеводе и желудке водных
холоднокровных позвоночных. Самка
Hedruris. снабженная особым хвостовым
клапаном, держится за слизистую обо-
лочку пищевода хозяина, самец же не-
сколько раз штопорообразно обертывает-
ся вокруг самки
Сна ревность особен разного пола
принимает парадоксальный характер у
нематоды Tridiosomoides crassicaudd
tpne. 120), паразитирующей в мочевом
пузыре крыс. Самцы этого вида проника-
ют в половые протоки самки и ведут там
паразитарный образ жизни, выполняя в
то же время свою функцию оплодотворе-
ние. 119. Попарное соединение
полов у паразитических нематод
Л — самка Hedruris andmphora,
прикреплениям прпсосковидчым
задним концом к стенке кишки
хозяина н обпитая в неско.ижо
оборотов самцом./{— парочка
Sytigamtis trachea-. маленький
самец прирастает к телу самки
в области ее полового отвер-
стия; внутри самки просвечи-
вают нзвшые трубки половой
системы. (По Кюкенталю.)
НИЯ янц.
Наряду с приспособлениями, гарантирующими оплодотворение
яиц, изредка у паразитов наблюдаются и особенности, препятствующие
преждевременной копуляции или повторному совокуплению с уже
оплодотворенной другим самцом самкой. Случай первого рода демон-
стрирует нематода Stenurus minor из дельфина; уг незрелой самки
этого круглого червя половое отверстие покрыто двумя направлен-
ными навстречу Друг другу кутикулярными выростами. У вполне со-
зревших самок эти выросты отведены в сторону п половое отверстие
открыто.
Пример второго рода мы находим у некоторых скребней. Самец
в конце копуляции выпускает на задний конец самки, несущий па себе
половое отверстие, секрет своих цементных желез. Секрет обволакивает
задний полюс самки, застывает на нем в виде плотного колпачка и
таким образом закупоривает входное отверстие к женской половой
системе самки
к
117
1 тигельная спаренноегь полов установлена у некоторых клешей,
например, у Schizocoptes conjugatus (сем. Listrophoridae) с африкан-
ского золотого крота (Chrysochloris, или Bematiscus). Самцы этого вида
находятся в состоянии длительного спаривания с женскими нимфами.
При этом оба партнера соединяются задними концами, будучи направ-
лены головами в противоположные стороны (рис. 121). У самца имеются
два крючковидных выроста на заднем полюсе, служащие для обхваты
вашгя конца тела
а кроме того, две мощных брюшных присоски,
которым у самки соответствуют два бугра, или
запонки. Соединенные животные двигаются и
ведут, себя, как один организм.
В общем можно еде 1ать вывод, что у па-
разитических, а особенно у эндопаразитиче-
ских животных имеется некоторая тенденция
к выработке гермафродитизма. Сплошь н ря-
дом у гермафродитов развитие мужских и жен
скпх г<шад происходит разновременно, причем
самки,
Рис. 120 Самка Ti'.ho-
somoides cnissieamta из
мочевого пузыря крысы
с самцом в ее половых
притоках. < П:» Кюкеп-
та.по.)
/ — самец.
Рнс. 121. Клещ Schtzocopre4-c»njuffaiiis, живу-
щий па шерсти африканскою крота Chryso-
rliloris sp.
А — самец с брюшной стороны: В — самец в
.uiiTe.n.noii копуляции с женской нимфой (на
рисунке нимфа обращена голАниыя концом
КПП .у). <Г10 .'I l.VfKI'Cy )
обычно сперва развиваются мужские половые железы, позднее женские —
гак называемый протсрапдрнческни гермафродитизм Это явление при-
водит к том*. чго животное сначала фу акционирует как самец, потом как
Самка, что новело к некоторым ошибкам в исследовании паразитов. Так,
Myzostoniida сначала считали рездельнополыми, потому что они часто
встречаются попарно в особых вздутиях на теле морских лилий, причем
одна особь имеет хорошо развитую мужскую, другая — женскую поло-
вую систему. «Мужская» особь значительно мельче «женской» и могла
рассматриваться как карликовый самец. На гамом деле оказалось, что
обе особи суть гермафродиты в разных фазах их существования и
что «самец.» постепенно вырастает, развивает женские гонады и стано-
вится «самкой».
Еще яснее выражен такой последовательный гермафродитизм у пе-
кпгорых паразитических Isopoda (Linopsidae из Cryptoniscidac). Так, у
DanaUa curvata (рис. 122) из яйца выходит ясно расчлененная личинки
11Х
на так называемой эппкаридной стадии; она превращается в самца
(стадия CryptoniSKiis), который тоже имеет ясное метамерное строение
и вполне развитые семенники. В это время личинка прицепляется к
пелагическим веслоногим рачкам, надкалывает их и сосет. Затем такой
самец отыскивает самку, оплодотворяет ее и переходит после копуля-
ции на coin ршеппо иного хо $янна, а именно на Sacculina.
Самец Diaialia прикрепляется к саккули-
не, линяет, сбрасывает все конечности и теряет
все следы членнстости, превращаясь в мешок,
снабженный хоботком, который вонзается в
тело хозяина саккулины, т. е. краба Inachus. В
теле краба хоботок Danalia внедряется в сте-
белек саккулины п таким образом начинает пи-
таться за счет паразита краба, вырастает, раз-
вивает яичники и постепенно превращается в
самку. На стадии самца Danalia имеет всего
1 ял длины, на стадии же самки — до 13 .«.и.
Протерогинический гермафродитизм, ко-
гда женская половая система развивается
раньше мужской, встречается редко. Приме-
ром его может служить один ленточный червь
Progynopilidiwn nd Iler i.
J раздельнополых паразитов количествен-
ные соотношения особей обоих полов большей
частью не представляют каких-либо интерес-
ных особенностей, кромт тех случаев, когда
одинаковое количество спаренных самцов л
самок гарантируется, по-видимому, самым
способом их происхождения. Это мы видели,
например, у сосальщиков Didymozoonidae, у
цестоды Dioiсосе sfиs.
Самый акт оплодотворения в одних слу-
чаях строго ограничивается попарным сочета-
нием самки с самцом, тогда как в других слу-
чаях мы сталкиваемся с полигамией. Карину
первого рода дает, например, нематода Syn-
gunifis trachea, где самих как бы прирастает
к ио 'оному отверсгш<1 самки, а также скребни,
i них копуляция заканчивается тем, что поло-
вое отверстие самки наглухо заклеивается.
Случаи полигамии известны для упоми-
навшейся ранее Trichosomcides crassicauda, у
которой половая система самки содержит
шихся в нее карликовых самцов (полиандрия). Редкую комбинацию
обратного характера (полигинии) обнаруживает раздетьнопо.тый
питии сосальшик н.з кровяных двуусток — Macrobdharzia тасго-
biiha'riia. Самы этого вида гораздо многочисленнее самцов, но зато
и значительно мельче их. При этом в брюшном желибе самца (ги-
некофорный канал), где происходит оплодотворение, можно встретить
иногда до 6 самок (Me Intosh, 1933). Своеобразие этой формы допил
нястся еще тем, что у нее, по-видимому самки отыскивают самца, а не
самец самок.
Помимо тенденции к гермафродпгнзлу, имеется еще один широко
распространенный у паразитов признак половой системы: громадная
плодовитость, огромное количество продуцируемых паразитами яиц.
Рис. 122. Различные половые
фазы Danalia curvata
(isopoda)
А — молодая личинка с
брюшной стороны, X 100:
Н — зрелый самец (стадия
Cryptoaiscut} со спинной
стороны, X 40 С и D — пере
.ходпые стадии (С — во время
линьки. /> — прикрепления).
Е — молодая самка. X 20;
/•’ — взрослая самка, X t
(По Коллери.)
/ антенны; 2 торакальные к->
нечншпи; 3 - абдоминальные ко
нсчнистп; I 'емепникп; о — янч-
1.11X11; 6 с шияишаа шкурка; 7 —
копоток; .5’ — «|йцсподы.
иногда до 6 забрав-
-(то свойство, называемое законом большого числа яиц, является при-
способлением, которое помогает паразитам в борьбе с тем множеством
неблагоприятных условий, которые приходится паразиту преодолевать
во время его сложного жизненного цикла.
Конечно, большая плодовитость паразитов не представляет чего
шбо свойственного исключительно паразитическим организмам. Среди
свободно живущих видов тоже встречаются примеры чрезвычайной
плодовитости «Царица» термитов Termes bellicosus, по наблюдениям
'Ипериха (1909), откладывает до 30 тыс. яиц в день, т. е. до 10 млн. в
год, а за 10 лет своей жизни—до 100 млн. Сельдь ежегодно мечет до
’.О тыс. икринок, треска до 9 млн., а у луны-рыбы в яичнике насчиты-
вается до 300 млн. яиц.
Крайнее увеличение плодовитости паразитов главным образом обу-
словливается, вероятно, двумя моментами. С одной стороны, усиленное
пиIанис паразита, осуществляемое без всяких перерывов н перебоев,
может, ^несомненно, способствовать более сильному развитию полового
аппарата и половых функций На примере Aloionema (гл. VI) мы уже
видели, как временное паразитирование ее личинки увеличивает в
десять раз число яиц, развивающихся у выросшей из такой личинки
половой особи. Вторым, не менее важным моментом является непре-
рывно действующий естественный отбор наиболее плодовитых особей.
Отбор этот находится в прямом соотношении с темп трудностями, кото-
рые встречаются на пути распространения паразитов и которые, как
правило приводят к тому, что из миллионов зародышей достигнуть
половой зрелости удается в лучшем случае лишь единицам. Чг м боль-
шее число зародышей будет гибнуть при преодолении этих трудное ген.
тем меньше шансов будет для малоплодовитой особи длительно нод-
- держивать существование вида. В процессе эволюцииалоплоповитые
индивиды будут отметаться, и только способные давать максимально
большое число потомства окажутся в состоянии распространяться в
лице свои к потомков на все новые и новые поколения хозяев
Наши сведения о половой продукции паразитов, к сожалению,
очень ограничены, но все же в отношении некоторых червей эти данные
настолько уточнены, что позволяют судить о закономерности этого яв-
ления.
Свободно живущие нематоды продуцируют обычно небольшое
чис . ) яиц. исчисляемое десятками и сотнями. TncluneUa spiralis отрож-
iaer, несмр’Лря на своп малые размеры, до 10 тыс. личинок, Xecator
amcricanus производит в день около 9 тыс. яиц. Ancylostoma duo-
dena le --около 20 тыс. Поскольку срок жизни Ancylostoma исчисляется
приблизительно в 4—5 лет, одна самка в течение своей жизни может
отложить около 25—30 млн. яиц. У анкилостомы, личинка которой
может двигаться во внешней среде, нападать па нового хозяина и
активно в него вбуравливагься, возможность попасть в нового хозяина
не так мала, как у какой-нибудь аскариды, пассивно и чисто случайно
заносимой в нового хозяина на стадии яйца. Быть может, этим обстоя-
тельством н объясняется то, что Ascaris lumbricoides, например, за 5—6
месяцев своей половозрелой жизни продуцирует уже до 50—60 млн.
яиц (в среднем 250 тыс. за одни сутки), т. с. гораздо больше, чем аикп-
чостома.
Относительно близкие между' собой сосальщики Clonorchis sinensis
и Fasciolopsis buski продуцируют в сутки первый — 2.5, а второй —
25 тыс. яиц. Clonorchis в своем жизненном цикле имеет окончательного
и двух промежуточных хозяев и передается от одного хозяина другому
путем хищничества, т. е. пожирания одного хозяина другим. Fasciolop-
I-S11
sis в стадии церкарии инцистирустся на водяных растениях и в виде
цисты пассивно и случайно заносится в организм хозяина, обычно при
питье воды.
Шансов попасть в организм окончательного хозяина при выше-
описанных циклах развития будет у Clonorchis конечно значительно
больше, чем у Fasciolopsis. В связи, быть может, с этой особенностью
продуктивность первого оказывается в 10 раз меньше таковой второго.
Иногда половая продукция паразитов поднимается до поистине
гигантских размеров. Так, например, свиной солитер Taenia solium за
свою долгую жизнь (до 10--15 лет) производит сотни (200 -300) мил-
лионов яиц. Шапиро (Scbapiro, 1937) вычислил для продукции янц
Taenia saginata еще более астрономические цифры. По ei о подсчету
этот крупный ленточный глист человека дает в год до 600 млн. яиц.
что при средней продолжительности жизни данного глиста в 18 лет
(по Лейкарту, 1863), говорит о продукции им за всю его жизнь до
10 млрд. 700 млн. яиц (10700 000000!). По приблизительному подсчету
вес отложенных самкой Ascnris lumbticoides за год ее жизни янц и
1700 раз превышает собственный вес животного. Одна из самых плодо-
витых из свободно живущих форм — матка пчелиного улья—продуци-
рует в год количество яиц. лишь в 100 раз превышающее ее собствен
ный вес. .
Не меньшие цифры дает размножение некоторых паразшпческих
простейших, у которых роль янц в распространении вида играю!
цисты, т. е. покоящиеся стадии. По данным Кофоида (Kofoid. 1925),
человек, зараженный дизентерийной амебой (Entamoeba histolytica).
выделяет в своих экскрементах в день по 50 млн. цист этих амеб, в
среднем при обследовании нескольких человек в течение двух месяцев
Получилась цифра в 15 млн. цист, выводимых человеком за день.
В связи с увеличением половой продукции паразита, у последнего
вырабатываются и соответственные морфологические изменения, со-
стоящие главным образом в значительном увеличении размеров поло
вых органов. Увеличение это иногда может быть выражено в чрез-
вычайно сильной степени. У трема!од, например, половые органы за-
нимают большую часть тела. У ленточных червей половые органы не
только заполняют собой весь члепнй', но и соответственно с разделе •
ином тела па членики сильно увеличиваются в числе, повторяясь в каж-
дом членике. Повторение половых аппаратов по продольной осп тела
наблюдается даже у таких форм, тело которых не расчленено, напри-
мер у Ligula, Triaenophorus Иногда же у членистых Cestodcs насту-
пает умножение числа половых аппаратов н в поперечном направле-
нии: в каждом членике развивается два набора половых органов
(Dipylid'um caninum. Djplogonoporus, рис. 123).
Процесс гипертрофии половых органов может заходить так далеко,
что весь организм пр< вращается как бы в едва заметный придаток
своей собственной половой системы. У нематод рода Atractonema (над-
сем. Aphelenchoidea) (рис. 124) самки, развивающиеся из личинок на
свободе, после оплодотворения проникают в своего хозяина (личинка
комарика Cciidomyia pini) и продолжают там расти главным образом
за счет выворачивания наружу своего влагалища, в котором помещают-
ся все половые органы и скопляются оплодотворенные я iiiia; такое выво-
роченное влагалище достигает размеров, равных всему остальному
телу самки. У другой нематоды того же семейства, Sphaerutaria bombt
паразитирующей в полости тела шмелей, разрастающаяся матка выпа-
дает через половое отверстие наружу, принимает в себя половые
оркшы и часть кишечника, достигает размеров в 15 ллъ пРе
151
Рис. 123. Членики ленточных глистов (Cestodes) с у двоен-
ным половым аппаратом
А — DiplogqtiopArus grandis (Pseudophyllidea). дна зрелых
членика; В — Moni"zia expansa (Cyclopliyllidea); С— Dipy-
hdium caninuni (Cyctophyllidea). (По Кюкснтллю I
/ —совокупите льны и орган; 2 влагалище: 3 — ннчмик; J — семени н mi ;
5 — межч.1сник<>1'ыс железки: 6 жг.почиик.
ь Ito,
Рис. 124.'Гипертрофия мл тки у некоторых паразитических Ncni.il»»d.-s
Д—Atractimema giblntsiim взрослая салка с вывернутой маткой;
Н — U — Spltaeriilaria bnmhi (И — моле тая самка с ничнигиошичся
выпячиванием матки; С—матка выпятилась п Bifliiy.ia в г- г pt. себя
гонады), D — взрослая самка па нпжйем конце г ромадноп, разрос-
шейся матки. (По Филпиьену.)
восходя по объему все остальное тело в 13 — 20 тыс. раз. При таком
соотношении сам организм паразита выглядит едва заметным придат
ком, прикрепленным к как бы самоегоятетыто существующей маткс-
(рпс 124).
Однако паразиты не ограничиваются указанным приснособлснн'-м
для умножения числа особей и продления жизни вида; многие из них
изменяются в сторону развития различных дополнительных способов
размножения, о чем будет говориться в одной из следующих глав.
Морфологические особенности яиц пара .итов не даю' какой-нибудь
нслыюй картины (см. рис. 125 и 126). Средн приспособлений, встречае-
мых в яйцах различных групп паразитов, отметим несколько разных
примеров
Рис, 125. Янцл рлзличиых мовогеиетнчсскдох сосальщиков
А — Diplectanum aciilcflfunt (под янцом концы ножек двух яиц при большом
увеличении); /? — МапнГаех alosae-. С — nenedenia derzkavini; D -кла ik.<
яиц SUrrticotyh' gotoi\ / —группа яиц (с общим основанием; Ac uithncotyle
verr'lli; /- — Prbtoancyrocephalus streikowi; (i — Dtplozoon partidoxum\
H — Microcotyte gotoi. (По Быховскому.)
У некоторых Monogeaoidca яйца имеют разнообразные придатки
(рис. 125), среди которых Быховский (1957) различает ножку и фила-
менты. Ножка формируется на нижнем полюсе яйца, филамент—на
верхнем. Последний отличается наличием особой крышечки. Биологи-
ческое значение этих придатков может быть различно. У некоторых
видов (Mtzschia Mazocraes) ножка служит для прикрепления (вероятно
приклеивания) яиц к покровам хозяина. В других случаях ножка и
филаменты являются, по-вндимому, приспособлениями для «'парения»
в воле ана огично планктонным организмам Интересное приспособле-
ние описывает Быховский у яиц моноп нетпч» ского сосальщика А1ьги
cotyle gotoi (рис. 125), паразитирующего на жабрах придонных мор-
ских рыб рода Hexagrammus. V этого червя яйца откладываются не по
одиночке, а пачками по 75- Г25 яиц за один раз. Эти яйца оказываются
скрепленными друг с другом благодаря скручиванию очень длинных
филаментов. Когда такой пакет попадает в воду, то он пе.реворачн
153
чается вниз клубком филаментов, яйца же расходятся веером во все
стороны, образуя нечто вроде парашюта. Такой клубок очень медленно
опускается вниз и обычно не достигает дна, запутываясь филаментами
в водорослях и т. п. Это способствует заражению рыбок Hexagrammus,
которые ведут придонный образ жизни средн зарослей водорослей, вы-
ходящими из яиц личинками. Белопольская (1938) обнаружила у неко-
торых видов цестод отряда Cyclophyllidea приспособления для флоти-
рования яиц в воде. Промежуточными хозяевами этих цестод являются,
очевидно, планктонные Copepoda и флотирование яиц является биоло-
гическим приспособлением, облегчающим возможность поедания яиц
Рис. I21-. Приспособление ниц цсстот к нарепню н вще
А — fliimi Anmtujlacnia plalyrhynchtv. Is яйип Anontvtaenta paramirro-
rltyiiena (Ho Be.iuiio.ibi Kuii.)
рачками J1 одних (Anotnotacnia stentorea, Dilepis glareula, Diorchys
nyrocae) япца снабжены щумя палыми филаментами, во много раз
превышающими длину яйца (рнс. 126). Такие яйца держатся в воде
в вертикальном положении и очень медленно iiorpj жаются. ' других
nu:n)B(Anoinotaenia platyrhyncha, Dilepis ochropodis) яйца, имеющие
тоже по 2 ффламента, соединяются в цепочки (рис. 126), которые могут
длительное время флотировать в воде. У Anomotaenia puramicrorhyni ha
яйца обладают одним филаментом. Они соединяются концами, лишен-
ными филаментов, в пакеты, которые также парят в воде.
Длинные филаменты имеются и у якц некоторых паразитических
нематод из различных рыб. Примером могут служить яйца видов рода
Ascarophis.
Очень сложное строение имеет оболочка яиц некоторых Mermi-
thidae, например Mermis xulmigrescens. На обоих полюсах яйца нахо-
дится по очень сложно ветвящемуся отростку. Эти отростки сложены во
круг яйца, пока оно находится внутри матки, но у отложенных яиц от-
ростки расправляются в капле дождя и прилипают к листьям после
высыхания. Травоядные насекомые, являющиеся хозяевами Mermis,
поедают листья с приставшими к ним яйцами и таким образом зара-
жаются.
Яйиа многих постоянных эктопаразитов, например вшей и пухо-
едов, прочно приклеиваются к волосам и перьям (или у платяной вши
15 J
к бслыо) хозяина при помощи особого цемента, выделяемого железами
половых путей самки.
У яиц некоторых паразитов наблюдаются приспособления, способ-
ствующие выходу их или из тела хозяина, или же из тела самого пара
зита, т е. материнского организма. Первый случай встречается у неко
торых кровепаразиюв. Отложенные в кровь яйца различных Schistose
matidae не имеют непосредс гвенного открытого сообщения с внешней
средой. Весьма вероятно, что имеющийся на оболочке яиц некоторых
видов червей острый шип способствует разрыву капилляров и выход\
из них яиц. Кроме того, продвижение яиц по сосудам и их выход вызы-
вается сокращением стенки сосуда.
Яйца большинства скребней (Acanthocephala) имеют сильно вытя-
нутую, веретеновидную форму. Можно думать, что такая форма яиц
помогает им в неповрежденном виде проскальзывать в кишечник раков
и насекомых, которые служат промежуточными хозяевами для личинок
скребней.
ГЛАВА IX
ПРИСПОСОБЛЕНИЯ ЭМБРИОНАЛЬНЫХ И ЛАРВАЛЬНЫХ
СТАДИЙ ПАРАЗИТОВ
В большинстве случаев эмбриональные стадии развития паразитов
обнаруживают относительно мало специальных черт приспособлена;
к паразитизму и нередко бывают относительно незначительно изменены
но сравнению с близкими свободно живущими формами. Это объяс-
няется, вероятно, тем, что во многих случаях развитие яиц паразитов
-происходит во внешней среде и не испытывает влияния со стороны
хозяина. Например, эмбриональное развитие аскариды ничем специ
альным не отличается от развития свободно живущих нематод. В тех
же случаях, когда яйцо парашта развивается в организме хозяина,
обычно имеется система оболочек, которые изолируют яйцо ог окру-
жающей среды, и эмбриональное развитие его протекает относительно
автономно. Примером может служи! ь развитие яиц многих трематод
н цеетод.
Случаи настоящею эмбрионального паразитизма, когда
устанавливается более или менее тесная связь между развивающимся
эмбрионом и организмом хозяина, относительно редки. Наиболее много-
численные примеры этою рода дают паразитические перепончатокры-
лые (наездники и некоторые другие группы), личиночные стадии кото-
рых паразитируют в других насекомых. Здесь имеет место настоящий
‘эмбриональный паразитизм, тесная связь эмбриона с хозяином и обычно
питание развивающегося зародыша за счет хозяина Взрослые же насе-
комые являются здесь типичными свободно живущими перепончато
крылыми, без каких-либо следов влияния паразитического образа
жизни.
Зародыши и личинки обнаруживают весьма разнообразные при-
способления к паразитизму, причем можно наметить различные ступени
этих приспособлений.
О. М. Нвапова-Казас (1948, 1954, 1962) в ряде работ подробно ра
смотрела вопрос об эмбриональной паразитизме перепончатокрылых
Она отмечает ряд глубоких изменений эмбриогенеза, обусловленных
паразитизмом. Эти изменения в основном связаны с приспособлениями
позволяющими па все более ранней стадии онтогенеза питаться за счет
хозяина. При этом происходит обеднение янц желтком и уменьшеиш
150
их размеров. Можно наметить два основных пуп и в обусловленной
паразитизмом эволюции эмбрионального развития наездников.
Первый путь характеризуется тем. чго питание за счет .хозяина
становится возможным лишь после выхода личинки из яйца. Янко
обладает нормально развитым хорионом. Все эмбриональные процессы
при этом упрощаются, зародышевые оболочки, утратившие свою защит-
ную функцию, редуцируются. Личинка выходит недоразвитой и сразу
же начинает питаться за счет тканей насекомого хозяина. Окопчапш
ее онтогенеза происходит уже на личиночной фазе. Наиболее ярко
выражен этот путь приспособления у Prestwichia. Вылупляющаяся из
яйца личинка (рис. 127) не сегментирована, лишена конечностей. нерв-
ной. дыхательной и кровеносной систем и мускулатуры. Хорошо развита
и функционирует лишь пищеварительная система. Все остальные
органы развиваются в процессе ностэмбриопального развития.
Рис. 127. Только что вышедшая из япца личинка Prestwickia aqiui-
rica. (По Иванозой-Казас.)
I — ротопое отнерггис; S — проток слюнной железы; 3 — пищевод; 1 - с чоннзт
железа; 5 — средняя кишка; 6 — задняя кишка; 7 — amuibjioe отверстие.
Второй путь адаптивной эволюции эмбриогенеза наездников харак-
теризуется способностью зародыша начиная с самых ранних стадий
извлекать питательные вещества из организма хозяина. Этот путь
также связан с глубокими изменениями эмбриогенеза (роды Litjio
mastix, Plulygaster. Macrocentrus и др.). Хорион становится очень гон-
ким и легко пропинаемым, а иногда исчезает вовсе. Яйца лишаются
желтка и резко уменьшаются в размерах (по сравнению со свободно
живущими видами перепончатокрылых). Образуются специальные при-
способления в виде трофамниопа (трофических оболочек), через кото-
рые и осуществляется питание эмбриона. Эти оболочки возникают
часто па самых ранних стадиях развития и образуются или за счет
одного из первых двух бластомеров дробления или даже за счет ядер
направительных телец, которые как бы отторгают от яйцевой клетки
часть протоплазмы (рис. 128).
Вокруг зародыша образуется обычно соединительнотканная кап-
сула хозяина, которая используется благодаря наличию амниона для
питания зародыша. Вследствие питающей функции амниона происхо-
дит быстрый и энергичный рост зародыша, который резко увеличи-
вается в размерах. По подсчетам Пваповоп-Казас (1948) в некоторых
случаях масса зародыша увеличивается при этом до 2U тыс. раз, а
иногда и больше того. Питание эмбриона за счет хозяина открывает
новые эволюционные возможности — возникает полиэмбриония. При
15"
этом масса зародыша распадается па множество групп клеток
(рнс. 128), из которых каждая в дальнейшем- дает личинку. Таким
путем из одного яйца может развиваться до 3000 личинок (у наездника
Lithomastix truncatellus). В других случаях число личинок, развиваю-
щихся из одного япца, бывает менее велико и измеряется десятками
или сотнями. Полиэмбриония — это бесспорное следствие паразитизма
и вместе с тем приспособление к паразитизму, значительно повышаю-
щее число особей вида. Это — особая форма проявления обшей законо
мерности, выражающейся в повышении плодовитости паразитов по
сравнению со свободно живущими формами, о чем нам уже приходи-
лось подробно говорить в другом месте (гл. VIII).
Риг. 528. Начлльпыс стадии ра жития Lithornastix floridanus
Д — яйцо в момент оплодотворения; В—стадия двух бла-
стомеров; С — образование трофачшюна; D — разделение
эмбриональных клеток на группы, каждая нз которых пре-
вратится впоследствии в одного зародыша. (По Петерсону.)
1 — редукционные ядра; 2 — проиуклсусы; 3 — трофамшюп; 4 - зародыш;
5 — эмбрионjльные клетки.
Интересно отмстить, что имеются несомненные черты сходства
в эмбриональном развитии, вызванные, с одной сторо-
ны, паразитизмом, а с д р' г о й — ж п в о р о ж д е и и г м. Эгот во-
прос был давно \же поставлен и рассмотрен Фаусеком (1913) и дру
гимн авторами. Фаусек по поводу этого сходства писал, что живорож-
дение— это «частный случай паразитизма, временный паразитизм в
пределах одного вида каждого последующего поколения в предыду-
щем». Среди насекомых имеются относительно немногочисленные слу-
чаи живорождения. Примерами могут служить виды семейства Poh
ctenidac (отряд полужесткокрылых) и своеобразный отряд Sirepsiptera.
У Pol} ctenidac развитие происходит в яйцеводах матери. Яйца очень
бедны желтком. Зародышевые оболочки образуются, при этом сероза
приобретает трофическую функцию. Оча тесно прикладывается к стен-
кам яйцевода, и через нее осуществляется питание зародыша.
158
Большие черты сходства с наездниками имеются в эмбриональном
развитии Strepsiplera, где развитие зародышей происходит в жировом
теле матери. Здесь яйца тоже бедны желтком, у некоторых видов
наблюдается тотальное дробление, образуется трофамииоп, и питание
осуществляется за счет тела матери, что приводит к большому росту
зародыша на эмбриональных стадиях. Наконец, у некоторых видов
Strepsiptera дело доходит до полиэмбрионии (например, у Huliioxenox
simpiitis), Многие черты в эмбриональном развитии живородящих
позвоночных — млекопитающих — также обнаруживают, как известно,
далеко идущие черты сходства с гем. чго имеет место у паразитических
Hymenoptcra. Здесь также наблюдается обеднение яйца желтком,
тотальное дробление, образование трофического аппарата питания за
счет матери, рост эмбриона и т. п. Наконец, в редких, правда, случаях
и у млекопитающих дело доходит до полиэмбрионии, примером чему
является американский броненосец (Tathusia) из отряда неполно
зубых (Edentata).
В тех случаях, когда мы не имеем дела с ларвальным паразитиз-
мом, который может влиять модифицирующе на соответственные
стадии, ранние свободные фазы развития паразитов часто не несут на
себе никакого отпечатка влияния паразитизма и могут служить цеп-
ными документами для суждения о филогении сильно измененных па-
разитизмом взрослых стадий паразитов. Можно сказать, что уг многих
паразитов по мере приближения к половой зрелости изменение тела
иод влиянием паразитизма все время прогрессирует.
Филогенетическое значение личиночных форм паразитов пли п.х
ранних стадий развития доказывается целым рядом примеров. В поло-
сти тела пли в кишечнике Copepoda пара штпруют замечательно
своеобразные Blastodinidae. Они представляют собой аберрантную
группу' Peridinea, но во взрослом состоянии ничем не обнаруживают
своей истинной природы. Свою жизнь Blastodinidae начинают, однако,
со стадии подвижкой «диноспоры», которая по строению вполне
походит на некоторых голых Peridinea (Gymnodinium), что дало воз
можность Шаттону (Chatton, 1919) выяснить происхождение Blasto-
dinidae. На том же основании можно говорить о происхождении от
Pc-rid'nea группы Catcnata, паразитирующей в кишечнике Polychaeta
и описанной Догелем. Мужские гаметы кокцидий и грегарин даюг
повод предполагать, что эти споровики произошли от жгутиконосцев
типа Bodo. Мерцательные личинки трематод и некоторых цестод
< Diphyllobothrium) рассматриваются иногда как рекапитуляция
этими группами турбеллярнеобразных, т. е. покрытых ресничками,
предков. Личинки Myzostorida напоминают некоторых измененных
трохофор Polychaeta. а наличие рабдиговидных личиночных стадий
у многих паразитических нематод говорит о связи этих форм
с свободно живущими нематодами из группы Rhabditidae.
Мы видели, каким громадным изменениям подвергаются парази-
тические стадии некоторых брюхоногих моллюсков (Entocolax. Ente-
roxenos. Entoconcha). низводимые эндопаразитпзмом до степени бес-
форменного мешка. Однако получающиеся из яиц личинки этих пара-
зитов имеют вид типичного маленького переднежаберного моллюска
со спирально завитой раковиной н даже с крышечкой, столь харак-
терной для Gastropoda Prosobranchia. Особенно ясно выражен
анцсстральпый характер у личинок паразитических веслоногих раков,
которые во взрослом состоянии нередко теряют конечности и де-
формируются до неузнаваемости. Однако личинки Copepoda, вылуп-
ляясь из яйца, имеют строение типичных пауплнусов, свойственны \
1-59
свободно живущим веслоногим. То же наблюдается и у Rhizoccphala,
которые сходны со свободными Cirripcdia не только в с гадин
пауплпуса, но и в последующей цмприсовидной стадии своего раз-
вития.
Все эти многочисленные примеры показывают, что личинки боль-
шей частью избегают модифицирующего влияния паразитизма
и сохраняют облик соответственных стадии развития предков.
Таким образом, личинки лишь в тех случаях приобретают специ-
фичные черты «паразитарного склада», когда сама личиночная ста-
дия, так сказать, рассчитана на паразитизм j\ это обстоятельство
чаще всего отражается па двух категориях признаков: на призна-
ках защитного характера и на органах, помогающих отыски-
ванию хозяина, т. е. главным образом на органах движения.
Ведь главное назначение личинок очень многих паразитов состоит
в широком распространении вида, а потому н выработка приспособле-
ний, содействующих активному или пассивному передвижению их,
вполне естественна. Однако во многих случаях потребность в выра-
ботке таких приспособлений уже /довлегворяегся совокупностью
ларвальных признаков, унаследованных личинками от предков. Это
те же признаки, которые служили для распространения свободно
живущим предкам (реснички турбеллярим, плавательные конечное ги
нахпллуса и цпнрисовидпой личинки раков, реснички парусинка
моллюсков и т. п.). В таких случаях специальных двигательных
приспособлений «паразитарного склада» не развивается. В других
случаях старых приспособлений к новому, паразитическому образу
жизни оказывается недостаточно, и тогда вырабатываются адаптации
движения, обязанные своим происхождением паразитическому образу
жизни. Они-то иас в настоящий момент и интересуют.
К числу активных ларвальных органов передвижения такого
происхождения, несомненно, относится хвост церкарнй трематод.
Если мы отбросим недостаточно обоснованную гипотезу Синицы-
на (1905) относительно происхождения сосальщиков от коловраток,
го хвост церкарнй можно охарактеризовать как вполне оригинальное
образование, не имеющее себе гомолога у' свободных предков сосаль-
щиков и выработавшееся под влиянием паразитизма взрослых
трематод.
Своеобразный двигательный аппарат чисто паразитарного про-
исхождения представляет собой так нaзывac'’ы,, фороцпт зародышей
паразитических медуз Carina и Cunoctaniha (рис 129). Оплодо-
творенное яйцо этих медуз делится на две клетки, из которых толы^о
одна Дает зародыш. Другая разрастается, выпускает псевдоподии
н сначала поглощает зародыша, а позднее носи г его па себе, рас-
пластанного в виде шапочки мелких клеток. Эта вторая клетка,
или фороцпт, питает зародыша и переносит его в один из радиаль-
ных каналов материнской медузы. В кишечнике матери зародыш
продолжает развиваться до стадии молодой медузкп. которая поки-
дает материнский организм и после короткого периода свободной
жизни проникает в гастральную полость других медуз (Geryonia.
I.iriope и др.), в которых паразитирует. Хотя образование и функцио-
нирование фороцпта предшествует свободному’ периоду жизни моло-
дой Cunina, тем не менее приспособление это, встречающееся исклю-
чительно у паразитических медуз, явно сложилось иод влиянием
паразитического образа жизни Cunina в теле материнской особи.
У науллиусов веслоногих рачков Monstrillidae для плавания
служат три пары конечностей, как и у свободно живущих весло-
160
ногнх, но третья пара получает крючковидную форму и служит,
очевидно, не столько для движения, сколько для временного при-
крепления к телу хозяина или для ползанья на нем.
Чрезвычайно разнообразны приспособления ларвальных свободно
живущих стадий дигенетических сосальщиков, направленные на
нахождение н проникновение в окончательного пли второго иромежу
точного хозяина. Можно указать два основных способа нронпкпове
пня перкарин в хозяина — активный, путем вб\равливания в ткани
хозяина и пассивный, осуществляемый обычно в результате заглаты-
вания перкарин хозяином.
Церкарии. как известно, обладают специальным органом движе-
ния— хвостом, который во всех
беппо важную роль. Рассмо-
трим несколько конкретных
примеров. Перкарин рода San-
guinicala (рис. 130) снабжены
длинным раздвоенным на кон-
• цс хвостом (фуркоцеркарии) и
плавательной мембраной в ви-
де гребня. Благодаря этим при-
способлениям они могут дли-
тельно «парить» в толще воды.
В случае контакта с рыбами
церкарии активно оседают на
их покровы, а затем вбуравлн-
наются в жабры, проникая в
сосуды, где и превращаются
в половозрелого сосальщика.
Носкблько иной характер но-
сят приспособления церкарий
Diplosjpmum spathaceiun, пара-
штирующей в половозрелом
Рис. 129. Фороциты паразитических медуз
Д—дробящееся яйцо медузы Cunina рго-
boscitiea вну ц и амевсм.щой клетки форо-
цпта, J3 — молодая мерца тельная личинка
медузу Cunoctantha parasitica, распластан-
ная па своем фориннте. (11g (Мечникову.)
1 — ядро фщо^нта; S лнчинкэ; 3 - i|iO|ioiuit.
состоянии в кишечнике птиц. Вторым промежуточным хозяином здесь
являются различные рыбы, у которых метацеркарип локализуются
в хрусталике и стекловидном теле глаза. Церкарии обладают
(рис. 130) длинным и разветвленным на конце хвостом, снабженным
волосками. Этот хвост распластывается в воде горизонтально, что
позволяет церкарии подолгу неподвижно «парить» в воде. Периоди-
чески активными движениями они всплывают вверх, а затем вновь
«парят» в воде, очень медленно при этом опускаясь вниз. При сопри-
косновении с рыбой церкарии быстро, активно присасываются к ней.
отбрасывая хвост, и активно внедряются в покровы. У некоторых
других пелагических церкарий, например Lepocreadiutn album, кроме
жуточиым хозяином которой служит планктонная полнхета Alcyope.
по обеим сторонам хвоста развиваются два ряда длинных щетинок,
способствующих «парению» в воде. Среди пассивно заглатываемых
перкарин также можно отмстить ряд приспособлений, способствующих
проникновению в хозяина. Вторым промежуточным хозяином морских
сосальщиков семейства Hemiuridae являются некоторые планктоииьн
организмы (Copepoda, Sagitta). В соответствии с этим церкарии
(рис. 130) обладают разнообразными приспособлениями, обеспечи-
вающими их длительное «парение» в толще воды. Тело церкарий
<аключено в особую прозрачную капсулу и снабжено разнообраз-
ными длинными филаментами, иногда щетпнковидными выростами,
имеющими вид зонтика и т. п. (рис. 130). Очень своеобразно также
[ I В. Л. Догель
НИ
строение перкарин сосальщика Azygia lucii, паразитирующей во
взрослом состоянии в кишечнике иг к и некоторых других хищных
рыб. Здесь имеет .место явление мимикрии: неркария имитирует
строение личинок комаров сем. Culicirlae (рис. 130). Движения пер-
карин также напоминают движение личинок. Мальки рыб, активно
поедающие двигающихся личинок насекомых, поедают вместе с тем
и церкарий Azygia, заражаясь таким путем сосальщиками.
Pric. 130. Церкарии различных Trematodes с разного рода приспособле-
ниями для заражения хозяев
Л — церкарня рода Sangitinieola, живущая в половозрелом состоянии
в крови рыб; тело и хвост снабжены плавательной оторочкой; В — церка-
ргя рола Azygia, имитирующая личинку комара, тело заключено в камеру,
предохраняющую церкарню от повреждения в момент заглатывания рыбой;
С — церкарня рода Heogurus, придатки хвоста обеспечивают планктонный
образ жизни перкарин; D — церкарня рода Diplostcmum, расставленные
ветви хвоста служат для поддержания тела во извещенном состоянии в
воде; Е. Л* — положение тела перкарин Diplostomum и Co/ylurns во время
флотирования. (Л. D. Е. Е— оригинальные рисунки Гниецииской, В—по
Вез* иберг-Лупду, С — по Чубрик.)
У некоторых церкарий хвост гипертрофируется, становится гро-
мадным. а в переднем основном конце его имеется кармашек, в кото-
рый может втягиваться сама церкарня (Cercaria calli'istomae, цер-
карня Azygia). Хвост подобных церкарий имеет, вероятно, много-
образное значение. Так, для церкарий Azygia предполагается, чтс
увеличение хвоста делает церкарню более заметной для молодых
щучек, которые, по наблюдениям Шндата (Szidat, 1932), охотно
хватают относительно громадных церкарий Azygia. Кроме того,
прячась в кармашке хвоста в момент заглатывания рыбой, церкарня-
предохраняется от повреждений.
J62
Приспособлениями для пассивного плавания в воде обладают
и пропагативные стадии других паразитов. Такова, частично, роль
длинных отростков тела (прядильного шипа, конечностей) у некото-
рых личинок иантопод (стр. 126). Безусловно для увеличения поверх-
ности служат крайне длинные огростки спор некоторых Myxosporidia
и Actinomyxidia (Cerutumyxa longispina, Triactiuomyxon н др.)
(рис. 131).
Рис. 131. Споры Myxosporidia' п Aclinomyxidia, облада-
ющш длинными oi рос псами, которые способствуют фло-
тнрочаипю их и воле
,4 — Ctirahimyxa xpinosa: li — Mitraspora caudata-. C. —
lb nncgufa D — SphatsracfiriAmyxoii i h — Triactinn
myxon ignottim F—Hexactinomyxon pxammoryctes.t (Io
Beriboiiy.)
Совершенно особый тип ларйалвных приспособлений вырабаты-
вают некоторые наездники, например Perilampidae (рис. 132),
Eucharidae. Обычно выходящая из яйца личинка наездников и-меет
вид беловатого червячка; она лишена ног и всяких других придатков
и обладает сравнительно слабой подвижностью. Такой характер
личинок естественно вытекает из того, что яйцз откладываются
паездпикамч в яйца или личинки других насекомых, т. е. в живой
резервуар готовой пищи. Perilampidae и Eucharidae, однако отклады-
вают своп мелкие яйца на различные части растений, так что выходя-
щая личинка должна сама отыскивать себе подходящего хозяина.
В связи с этим у данных наездников из яиц вылупляются вытянутые
в длину, темно окрашенные, сильно хитпнизироваппые личинки —
планидии, или бродяжки. Они снабжены хорошо развитой головой
и двумя придатками (cerci) на заднем конце тела. Личинки эти
мелки и крайне подвижны. Вот как описывает Смит (Smith, 1917)
поведение плапиднев Perilatnpus chrysopae. Вылупившийся нз яйца
планидии сначала очень подвижен, но затем прикрепляется задним
концом к поверхности листа, передний вытягивает вертикально кверху
и остается неподвижным, пока поблизости его нс окажется подходя
щее насекомое (личинка Chrysopa). Тогда планидии проявляет беше-
ную активность, ощупывает своим головным концом ближайшие
окрестности и при соприкосновении со своим будущим хозяином
молниеносным движением прикрепляется к нему. Взобравшись па ли-
чинку Chrysopa, планидии прочно прикрепляется к пей своими роговы-
ми крючками. Таким образом, у этих наездников в связи с иным спосо-
бом заражения хозяев, при котором активная роль переходит от
матери к потомству, т. с. вылупляющейся личинке, в самое начало
постэмбрнонального развития вставляется особая личиночная стадия
II*
163
иланлдия, физиологически отличающаяся большой подвижностью,
а морфологически — вышеупомянутыми признакам^. Развитие Peri
iampidae получает особую надставку, или, правильнее, хнредставкх».
Рис. 132. Развитие Pertlampus sp.
Д — сильно увеличенный плапидий с брюшной стороны; видны
облегчающие движение поперечные ряды шнннков, хороню
развитая голова и придатки заднею конца тела; И — то же,
в меньшем масш.-абе; видна 1с’ильная хппшизацпя покровов;
С — нерехо i из состояния планндия к стадии взрослой личин-
ки: D — взрослая личинка; Е — кукочка Pertlampus сбоку;
F — куколка мухи (Varidiaeta). на которой п .разитиреют/’сп-
lampus-, па куколке видны два плаиидпя разных возрастов.
(По Уилеру,)
Подобного рода удлиненный метаморфоз получил особое наименова-
ние г и пер метаморфоз а.
ГЛАВ.А X
РАЗМНОЖЕНИЕ, ДЛИТЕЛЬНОСТЬ ЖИЗНИ И ЦИКЛЫ РАЗВИТИЯ
ПАРАЗИТОВ
1. ИЗМЕНЕНИЕ ОСНОВНЫХ ЖИЗН1 ИНЫХ ФУНКЦИИ ОРГАНИЗМА В СВЯЗИ
С ПАРАЗИТИЧЕСКИМ ОБРАЗОМ ЖИЗНИ
Основными биологическими моментами жизни всякого организма
являются приспособления, обусловливающие сохранение особи
и вида. Сохранение жизни особи прежде всего зависит от питания,
сохранение жизни вида — от размножения.
. В зависимости от различных условии, времени и места эти две
стероны в жизни вида могут приобретать различное значение. Иногда
опп оказываются даже в прямом противоречии друг другу. Сохране-
ние вида в некоторых случаях находится в прямой зависимости от
числа составляющих его особе'. Ио увеличение числа последних,
благоприятное для сохранения вида, может приводить к перенаселе-
нию и связанному с ним недостатку в пище, что оказывается неблаго-
приятным для особи.
Разрешается это противоречие путем расселения вида и завоева-
ния им новых мест обитания. В определенных границах организм
путем приспособления к условиям обитания преодолевает трудности,
стоящие на пути сохранения вида или особи, поддерживая нужные
для этого функции на должной высоте. Однако способность ппнепо
собленпя всякого организма не безгранична, и, что самое главное,
в процессе эволюции могут вырабатываться такие приспособления,
которые оказываются трудно обратимыми. Благодаря последнему
обстоятельству жизнь такого организма оказывается всецело обуслов-
ленной определенными условиями среды обитания, и ои не способен
к существованию в иной среде.
Среда обитания паразита чрезвычайно отлична от любой среды
обитания свободно живущих организмов, и у паразитов вырабаты-
ваем я целый ряд почти необратимых приспособлений. В основе и.х
кет ничего специфически присущего только паразитам, так как
любое отдельно взятое паразитарное приспособление можно встре-
тить н у свободно живущих организмов, особенно у ведущих сидячий
образ жизни; но комплекс этих приспособлений качественно отли-
чает паразита от свободно живущего организма.
В основе всех таких приспособлений паразита лежат особенности
двух вышеназванных основных его функций, обусловленные особен-
165
костями среды обитания, Для свободно живущих организмов пробле
ма питания оказывается очень часто оанон из наиболее сложных
Разрешая ее, организму приходится преодолевать много трудностей,
в связи с чем у него вырабатывается целый ряд особенностей, спе-
циально приспособленных для осуществления задачи питания (раз-
витие органов чувств и органов движения, выработка сложной системы
органов пищеварения и т. д.) Между тем второе основное биологи
ческое требование, направленное к сохранению вида и связанное
с размножением, оказывается относительно менее сложным.
V паразитов мы сталкиваемся как раз с обратным отношением
По мерс того как организм все больше переходит к паразитическому
образу жизни, проблема питания для него все больше упрощается.
С разрешением этой проблемы, с получением непрерывного усилен-
ного питания начинает увеличиваться репродуктивная деятельность
организма. Но это усиленное размножение паразита, в связи с про-
странственной и временной ограниченностью места ого обитания, при-
водит к относительно быстрому перенаселению этого места и связан-
ной с этим необходимости расселения вида для его сохранения
Проблема расселения, относительно легко разрешимая для сво-
бодно живущих организмов, для паразита оказывается чрезвычайно
сложной и трудной. Главная трудность состоит в том, что для
«аселення новых особен хозяина пара :пт часто Должен преодолеть
среду, к жизни в которой он не приспособлен. В связи с этим
у паразитов вырабатывается большое число очень сложных адапта-
ции 'обеспечивающих им возможность расселения через внешнюю
среду на новые особи хозяина и тем гарантирующих сохранение
вида
Прежде чем перейти к рассмотрению этих приспособлений,
остановимся на тех изменениях, которые претерпевают у паразитов
две основные физиологические функции организма—питание и раз-
множение.
Паразитизм, как мы видели, начинается со случайного заноса
в организм какого-либо животного зародышей будущего паразита.
В тех случаях, когда зародыш одерживается в новом месте обитания,
он находит там усиленное питание, которое быстро отражается на
общем состоянии такой особи (500—600 яиц, отгладываемых
Aioionama, выросшими в кишечнике слизня, и 20—30 яиц, которые
кладутся особями Aloionema, развивавшимися из свободно живущих
личинок).
Функция питания у паразитического организма может быть все-
цело перенесена на паразитическую личиночную стадию и полностью
ликвидирована у свободно живущей фазы, как это и имеет место
у мермисов (Mermithidae), волосатиков (Gordiacea) и др. В свобод-
ной фазе, обнимающей часть личиночной и всю половозрелую стадию,
эти животные не питаются вовсе. Таким образом, за свободной
фазой остается лишь фу акция размножения, т е. питание и размно-
жение оказываются строю распределенными между двумя стадиями
жизненного цикла паразита, приспособительно к особенностям тех
сред обитания, в которых они живут.
Усиленное питание в личиночной стадии начинает приводить по
только к усилению репродуктивной деятельности особи вообще
(у некоторых мермитпд, например у Agarnermis ecaudate, разви
вается уже свыше 5600 яиц), но и к преждевременному созреванию
половых продуктов. В связи с этим постепенно возникает способность
к партеногенетическому развитию яиц, как мы это видим у Rhabdiax
166
nyfonis и Strongyloides slercoralis. Здесь на паразитическую личипоч
ную стадию переносится нс только функция питания, по частично
и фупкния размножения. При дальнейшем ходе эволюционного про-
цесса функция размножения полностью перепоен гея на паразитар
ную фа у, свободная же фаза, необходимая для распространения
вида, ограничивается той или иной частью личиночной стадии разви-
тия. Такое сокращение стадии свободной жизни до известной степени
обусловливается тем обстоятельством, что ио мере удлинения парази-
тарной фазы у паразита развивается ряд специальных адаптаций
к существованию в хозяине, делающих его совершенно неспособным
к жизни в иной среде. Свободные фазы у части паразитов в конце
концов совершенно выпадают из жизненного цикла.
2. ПРИСПОСОБЛЕНИЕ ПАРАЗИТОВ h РАСПРОСТРАНЕНИЮ ВИДА
Глубокое приспособление к паразитическому образу жизни,
являясь крайне выгодным для сохранения индивидуальной жизни
особи, оказывается недостаточным для сохранения вида. Это обстоя-
тельство и влечет за собой целый ряд специальных приспособлений
к распространению вида, когоро< осуществляется в большинстве
случаен через внешнюю среду. У всякого паразита во время пребыва-
ния в последней имеются три биологические задачи I) сопротивление
вредному воздействию различных факторов наружной среды;
2) доведение своего развития до стадии, способной к существованию
в организме хозяина; 3) нахождение хозяина и проникновение в него.
Разрешение первых дв' х задач обеспечивается главным образом
приспособлениями, увеличивающими устойчивость свободных фаз
паразита (яиц, цист, личинок) в отношении различных факторов
снешней среды (температура, влажность, действие солнечных лучей
и т. и.).
Для разрешения последней задачи служат такие специальные
приспособления паразита, как увеличение плодовитости со связанны-
ми с этим изменениями в половой системе, усложнение циклов разви-
тия. введение в них чередования поколений и смены хозяев. Такое
разделение приспособлений, конечно, чисто схсматическо, В деьствн-
гельности все они тесно связаны друг с другом, нередко дополняют
н координируют друг друга, Так, недостаточная устойчивость в отно-
шении факторов внешней среды пли необходимость относительно
длительного в иен пребывания, связанная с гибелью части зародышей,
нередко компенсируется резким усилением репродуктивной деятель-
ности паразита. Повышение устойчивости свободно живущих стадий
служит косвенно разрешению задачи нахождения паразитом своего
хозяина, а система промежуточных хозяев — задаче развития парази-
та до инвазионной стадии и т. д. Все это лишь различные моменты
одного процесса — процесса передачи паразита от одного индивида—
хозяина — к другому,— моменты, взаимно обусловленные.
В тех случаях, когда паразит распространяется во внешней среде
на стадии личинки, в зависимости от особенностей среды обитания
последней, а отчасти и от способа проникновения се в хозяина, у него
могут встречаться различные приспособления. У некоторых паразп
тов, личинки которых живут исключительно в воде, приспособление
к длите.- ыюй жизни на воле почти созсем отсутствует. Эти лпчипки
(Mupavii.iiiH и церкарнй трематод, корацидии лентецов, личинки
ришты), если нм не удается проникнуть в хозяина, быстро гибнуI.
Причиной гибели, судя по всему, является истощение небольшого.
IG7
имеющегося в личинках запаса питательных веществ при отсутствии
у них приспособлении для самостоятельного питания на воле. По
крайней мере Лутта (1939) наблюдала, как имеющиеся сначала
в мпранидпи печеночюй двуустки п в кора индии широкого лентеца
запасы гликогена уже через 48—70 часов после выхода из яйца
оказываются израсходованными, движение личинок параллельно
с этим замедляется, н они вскоре гибнут. Мирацидии печеночной
двуустки расходуют гликоген п гибнут через 6—8 часов. То же отме-
чает Аксмэн (Ахшапп, 1947) для мирацидиев шистозом.
Динамика гликогена в период свободной жизни церкарий был
подробно изучена Гинсцинскон (1960). У церкарий гликоген со
держится в паренхиме тела и главным образом хвоста, где он
сосредоточен в особых клетках — резервуарах (каудальные тела).
В период свободной жизни церкарий в первую очередь расходуете»
гликоген в хвосте, являющемся органом движения личинок. Продол
жигелыюсть жизни, различная у разных видов церкарий, находится
в прямой зависимости от количества содержащегося в их тканях
гликогена По мере расходования гликогена активность и инвазионная
способность церкарий падает, и после полного истощения его запасов
наступает гибель организма. Темп расходования гликогена и дли-
тельность жизни церкарий находятся в непосредственной зависимости
от температуры среды.
У других паразитов (многие нематоды) личинки ведут наземны.
обра ? жизни. Проникновение их в хозяина часто обусловлено прслва
рнтелыгым развитием личинки во внешней среде, в связи с чем врем»
пребывания в этой среде значительно удлиняется — иногда до мнопь
месяцев н даже лет.
Так, например, личинки нематод Sirongylidae, Trichostrongylidae.
Ancylostomidae и др. должны проделать во внешней среде не меньше
1вух линек и лишь после этого становятся способными к внедрению
в хозяина. Поскольку в последней свободно живущей ларвальной ста-
дии личинкам иногда довольно долго приходится выжидать случая
внедриться в хозяина, подвергаясь в это время неблагоприятным
климатическим условием, покровы их приобретают ряд особенностей,
благодаря которым личинки становятся устойчивыми к высыханию
и проникновению в них различных вредных веществ.
Так, по исследованиям Завидовского (1929), личинки трихострон
ШЛ11Д (Trichostrongvlidae) кишечника копытных в возрасте пяти
дней способны оставаться живыми при полном высыхании при
температуре 22° более двух месяцев. Они выносят без всякого вреда
для себя действие насыщенного раствора медного купороса,
1%-ного раствора сулемы, 4°/о-пого раствора формалина и т. д.
в течение многих часов. Замораживание до —8П - 10° переносится
личинками более 6 месяцев, кратковременное нагревание (в сухом
состоянии) до —| 60—80 тоже не убивает личинок.
Яйца и цисты обладают еще более мощными зач’нтнымн при-
способлениями. В отношении длительности пребывания яиц во внеш-
ней среде все паразиты, выносимые из хозяина во внешнюю среду
в состоянии яйца.'могут быть схематично разделены па две группы.
У паразитов первой группы (например, Ancylosloma. Diphyllohothriuni
latum, Fasciola hepatu-р и др ) личинки выходят из яиц во внешней
среде, у паразитов второй группы (например, Dicrotoetium
dendriticum. Taenia solium и др,)—в организме полого хозяина. Яйца,
из которых личинки выходят во внешней среде, имеют назначение
лишь защитить зародыш во время его эмбрионального развития,
its
Яйца же второй категории предназначены защищать зародыш ш
только во время развития, по и после его окончания, вплоть до того
момента, когда такое яйцо будет случайно занесено в нового хозяи-
на. В связи с этим и Сроки их пребывания во внешней среде значи-
тельно удлиняются. Совершенно очевидно, что устойчивость таки?
ниц будет иная, чем яиц первой категории. Например, яйца Апсу
lostoma duodenale быстро погибают в 3—6%-ном растворе N tCl
или 0,25%-ной H2SOi. тогда как яйца Parascaris equinum длительно
сохраняют жизнеспособность в насыщенном растворе Nad или
в 5%-пой II2SO4.
Яйца обеих категорий могут выноситься наружу либо совсем
незрелыми (Ascaris. Diphyllobothrium latum, fasciola hepatica и др.),
либо частично развившимися (Ancylostoma, Enterobius и др.), либо,
наконец, уже закончившими эмбриональное развитие н содержа щи
ми зародыш или личинку (Dictyocuulus, Hynieiiolepis папа, Schisto-
soma и др.). Вполне вероятно, что яйца, все эмбриональное развитие
которых проходит во внешней среде, должны быть, при прочих рав-
ных условиях, более устойчивы, чем яйца, выносимые наружу уже
после прохождения ими части развития в организме хозяина.
Свойства оболочек яиц и цист определяются главным образом
необходимостью приспособления последних к особенностям среды,
в которой они развиваются. Например, у аскарид из наземных позво-
ночных яйца развиваются па суше н имеют плотную скорлупу, состоя-
щую > из пяти отдельных оболочек: наружная, или так называемая
эпителиальная, выделяется эпителием поповых протоков; за ьсй еле
дуст трехслойиая глянцевитая оболочка. Все они выделяются самим
Я1.дом и имеют белковую природу. Внутренняя, или волокнистая,
оболочка полупроницаема; опа очень слабо пропускает воду, а соли
i многие органические вещества через нее совсем не проникают. Она
пропинаема лишь для веществ, растворяющих ее или растворяющихся
в иен. .Внутренняя оболочка состоит из липидов и служит для защиты
яйца от химических агентов. Благодаря наличию липидной оболочки
яйца аскариды продолжают развиваться и сохраняют месяцами
жизнеспособность даже в таких средах, как насыщенный раствор
сулемы. C.uSO.1. ZnSO(. п./8 КОН, и./50 HCI и др. (Завадовскнй, 1915,
1925). Иное наблюдается у аскаридат пз водных хозяев (Anisakis, Соп-
tracoccum). Их яйца попадают всегда в воду и пмгют в связи с этим
тонкую оболочку, легко проницаемую для воды. Яйца Dicrocoeliutn
detidriticum, приспособленные. к существованию па суше, обладают
значительно более высокой стойкостью к пониженной влажности
и действию некоторых химических веществ, чем яйца Fasciola hepa-
tica, которые развиваются в воде. Ооцисты кокцидий кролика снабже-
ны двумя белковыми оболочками, слабо пропускающими воду. Полу-
проницаемой является внутренняя оболочка, так же как и у аскарид,
хотя она и иного строения. Ооцисты обладают значительной устои
чпвостыо к действию химических веществ Насыщенные растворы
солей, неорганических кислот и щелочей ие убивают их в течение
многих дней, а г насыщенном растворе сулемы ооцисты гибнут через
30 минут. В то же время оописты Eimeria carpelli из карпов (Хенсин
и Заика, 1957) имеют только одну топкую оболочку, легко проницае-
мую для воды. Эти ооцисты в течение нескольких .минут погибают при
недостатке влаги, а ооцисты кокцидий кролика могут выдерживать
высушивание много часов.
Устойчивость к воздействию факторов внешни! среды крайне
необходима для этих покоящихся стадий, так как они проходят
169
значительную часть развитий (образование личинки \ аскариды,
образование спорозоитов у кокцидин) во внешней среде, после чего
еще долго ожидают случайного заноса в организм новой особи
хозяина.
Защитные приспособления яиц и цвет могут обеспечиваться
и иным строением оболочек. Так, яйца Diphylloboihrium latum имеют
толстую скорлупу, легко проницаемую для химических веществ,
и студенистую оболочку (вероятно, мукопротеиновую), трудно прони-
цаемую для различных растворенных в воде веществ.
Цисты Entamoeba histolytica имеют только одну белковую, полу-
проницаемую оболочку, и по сравнению с ооцистами кокцидин кро-
лика они менее устойчивы к химическому воздействию. Некоторые
простейшие даже в иеипцистированиом состоянии могут выживать вне
хозянца. Трофозоиты Trichomonas horninis несколько дней могут жить
в воде при комнатной температуре и выживают в течение часа при
температуре 28
3. ДЛИТЕЛЬНОСТЬ ОТДЕЛЬНЫХ СТАДИЙ РАЗВИТИЯ
В предыдущем изложении мы видели, что жизнь паразитов сла-
гается нз разных периодов, в пей чередуются свободная п паразити-
ческая, личиночная и взрослая, активная и покоящаяся фазы. Дли-
тельность отдельных фаз у разных паразитов весьма различна, да и
общая длительность всего жизненного цикла (от япца до яйца сле-
дующего поколения) тоже сильно варьирует. Вариации могут зависеть
либо от специфичных свойств самих паразитов, либо от длительности
жизни их хозяев, либо, наконец, от условий внешней среды, в которой
протекает часть жизни паразита, п колеблются в весьма широких пре-
делах: от нескольких дней и до многих лет.
В некоторых случаях срок жизни паразита ограничивается дли-
тельностью жизни хозяина. Так, у паразитов коловраток (споровики
рода Eertramla) длительность жизни хозяина часто нс превышает
10—12 дней, а следовательно, и длительность активной, вегетативной,
стадии паразитов нс может быть большей. Еще короче, вероятно,
жизнь сосущей инфузории Endosphaera (рис. 133), паразитирующей
внутри других инфузорий (Paramecium, Opisthonecta).
Срок жизни паразитов у позвоночных хозяев может значительно
удлиняться в соответствии с жизнью хозяина. Яркий пример дает в
этом отношении Polystoma integer rimtun из мочевого пузыря травяной
лягушки. Она заражает лягушку в состоянии головастика; многоустка
проникает в мочевой пузырь еще до окончания метаморфоза и остается
в нем около пяти лет, т е. в течение всей жизни лягушки. Некоторые
данные свидетельствуют о том, что ленточные глисты человека (Taenia.
Diphylloboihrium) в отдельных случаях остаются в организме хозяина
свыше 25 лет. Clonorchis sinensis (из сосальщиков) живет в человеке
более 25 лет, Schistosoma haematobium — от 5 до 18 лет, Sch. japoni-
сит — до 28 лет. Gastrodiscus aegyptiacus зебры живет в хозяине до
9 лет. Нематоды Loa loa держатся в человеке более 15 лот, Wuchere-
ria bancrofti—17 лет, Ancylostoma duodenal?.— до 5—7 лет, Necator
arnericanus — до 12 лет. Для кровяной двуустки (Schistosoma haemato-
bium) срок жизни определяется в 5—18 лет, но иногда заражение
затягивается до 24 лет.
Таким образом, некоторые паразиты обладают до известной сте-
пени неопределенным сроком жизни, зависящим ог длительности
жизни хозяина. Интересные цифры имеются относительно способности
ГО
некоторых нематод долгое время сохраняться живыми в апабпотичс
ском состоянии. Tylenchus tritici остается в подсушенных зернах пше-
ницы, не погибая, в течение 9 лет fQpodey, 1923), а раиыпе этот срок
считали еще более длительным — до 27 лег. После вымачивания таких
зерен находящиеся внутри них личинки Tylenchus оживают.
Рис. 133 Opisthatv’cta henmguyi. и которой паразити-
руют сосущие и 'фузорнн / ndosphaera engetmanni.
(По Ленчу и Ноблю.)
1 мионемы ре. ракте.ры ргл Э1Ю1 о пеичнка ре нии; « — сфинктер:
Л MiiKpiiiiVK-ieyc О. hem eguyi'- < л uhiiii пснчнк ресниц; а -
млкропеклеус 6. henneguyr. 6 сократиre.iиные h.ikvg.ih: 7 мер-
цл1е.1КныЛ лродын! Е. engilnitinni. iiokha«ioihh7i хоЗДшы; н — Е.
engelmunni с мерцательный зародышем кнутри: 9 Е. engeEranni
(стадТг.1 росте)
Полную противоположность подобным паразитам составляют
такие, жизнь которых в половозрелом состоянии более или менее
ограничена. Так, относительно Schistosoma japonicum известно, что
самоизлечение от заболевания этим паразитом у лошади происходит
4Q рез 270 дней, чему, очевидно, и отвечает длительность жизни Schi-
stosoma в этом окончательном хозяине. Быховскнн (1929), исследуя
зремаюд волжских рыб, заметил, чго зимой заражение ими чрезвы-
чайно снижается, что говорит в пользу одногодичное™ их цикла.
Позднейшие исследования (Ляйман, 1940; Комарова, 1950, 1951 а, б;
Маркова, 195В) подтверждают, что значительная часть эндопаразити-
ческих червей (дигснстические сосальщики, скребни, ленточные черви)
пресноводных рыб имеют одногодичные жизненные циклы.
13 последнее время особенно Догелем и его школой выяснена дли-
тельность жизни ряда птичьих сосальщиков и ленточных глистов. При
этом у сосальщиков выяснились очень интересные различия в дли-
171
дельности половозрелой стапгн, с одной стороны, печеночных, с дру-
гой— кишечных видов..Для целого ряда печеночных паразитов стрижа
п ласточки (Platynosommn clalhraium, Lyparpsbimun olssoni L. salebro-
sum) длительность жизни оказалась сравнительно велика, равняясь
минимум нескольким месяцам. Наоборот, среди кишечных трематод
преобладают краткосрочные инвазии окончательного хозяина. Так
для Plagiorclus maculosiis и особенно для Plianeropsolus micrococcus
установлено, что срок их половозрелой жизни в кишечнике птицы не
превышает месяца. Хорошо может быть определен срок жизни тре
матод, обитающих в фабрнцисвоп сумке птиц Prosihogommus ovatots
живет у стрижа менее года. Молодые стрижи ннвазнруюгся этим чер-
вем в конце июля, а весной следующего года (в мае) оказываются
уже свободными от паразитов. Интересно, что у паразитов фабрицие
вой сумки предел жизни, помимо всего прочего, определяется сроком
существования самого органа, в котором они обитают: у большинства
птиц фабрицпева сумка атрофируется еще на первом году жизни.
Кратковременность половозрелой стадии кишечных сосальщиков
из семейства Strigeidae известна уже давно, они живут всего 7—10
дней. Mefagonimus katsurcidai — маленький сосальщик кишечника
млекопитающих — живо г в экспериментально зараженной мыши не
более 32 дней. Длительность жизни большинства Dactylogyndae и
G rodaotylidac (Monogenoidea) не превышает года, а некоторые, на-
пример Dactylogyrus vastmor (Изюмова. 1956), живут только 25--40
дней, и лишь отдельные экземпляры этого паразита перезимовывают
и гибнут в возрасте 6—7 месяцев.
Ленточный червь Hymenolepis fraferna держится в кпшечнию
крыс от 6 до 20 недель, a Moniezia expunsa в овцах — о г 2 до 6 меся
цев, достигая за это время 1,5 м. Среди нематод самки трихинеллы
живут 1з кишечнике крысы не более 3 недель, а в кишечнике человека
около 2—3 месяцев, между тем самцы удаляются сразу после спари-
вания. Enterobius vermicularis живет в кишечнике человека около
1 месяца, a Aschris lutnbricoides — до 9—11 месяцев. Кратковремен-
ность жизни многих паразитов в организме хозяина может быть
объяснена развитием у хозяина иммунитета, препятствующего даль-
нейшему существованию паразита (см. гл. XXI).
Половозрелая фаза эктопаразитов варьирует в своей длительности
очень сильно. Например, рачок Caligus lacustus с южного Каспия в
мае-нюне принадлежит к числу наиболее обычных паразитов на коже
очень многих пород рыб, между тем как исследования в сентябре-октя-
бре в том же районе не дали ни одной находки (Догель и Быховскпй.
1938).
Столяров (1936) подвергал круглогодичным наблюдениям Lttrnaea
cypriiiacea, паразитирующую на коже карасей, и выяснил следующее.
Осенью караси в Ропшинскнх прудах показали обильное заражений
взрослыми, но еще не имеющими яйцевых мешков самками Lernaca
(рис. 134). В течение зимы прежде всего наблюдалось отмирание
очень значительного процента еще не достигших стадии откладки яиц
самок. С ноября и по конец мая из 223 рачков, находившихся на 13
карасях, умерло 68; в другой партии рыбы из I э 1 рачков погибло 197.
(От умерших рачков под чешуей рыб остаются передние прикрепи-
тельные аппараты, что позволяет вести точный учет паразитов). Снль
пая смертность обнаружилась и в июне, по одновременно были най-
дены (16 июня) самки с яйцевыми мешками. К 23 июня плодоноСЙщне
самки отмерли и после этого срока вообще больше не встречались на
карасях, а из их яиц вылупились личинки--науплиусы. Очевидно.
172
как это следует и из данных Вильсона (Wilson, 1916) относительно
Lcrnaea variabilis, пбловая зрелость достигается рачками в конце лета
(август-сентябрь) на четвертой конеподитноп стадии развития, после
чего самка садится на карася и остается на нем до июня, постепенно
вырастая. За зиму и вплоть до июня около половины самок погибает,
ио эта гибель не представляет собой нормального отмирания, а яв-
ляется результатом неблагоприятных зимних условий существования.
В июне происходит откладка яиц, вылупление личинок и массовая
гибель самок, закончивших процесс размножения.
Рис. 134. Развитие Lernaea cyprinacea, кожного эктопа-
разита карповых рыб
А — науплизс после выхода из яйца; Н — конеподитная
свободно плавающая стадия, напоминающая циклопов.
1 la этой стадии происходит оплодотворение и прикреп-
ление к хозяину; С — молодая прикрепленная самка с
рудиментарными грудными ножками. (По Столярову.)
Из вышесказанного следует, что Lernaea обладает одногодичным
•циклом, причем половозрелое ее состояние длится с осени по июнь,
т. е. около 10 месяцев.
Длительность половозрелой стадии зудня Acaris stro равна 40
50 дням,' самки платяной вши живут не более 46 дней, а самцы — 32
дня; головная вошь менее долговечна: самка живет не более 38 дней,
а самка Phlhirius pubis—17 н более дней. Как видно из примера
платяной вши, долговечность у обоих полов неодинакова. По отноше-
нию ко многим видам замечено, что самцы отмирают значительно
раньше самок. Важнейшие функции самцов закапчиваются оплодотво-
рением самок, тогда как у самок за оплодотворением следует иногда
весьма длительный срок откладки яиц или зародышей или вынашива-
ния потомства (Copepoda). Такие же отношения наблюдаются и у
свободно живущих организмов.
Самки некоторых клещей (Ixodes ricinus) способны выживать до
1 — Р/г лет, а для разных видов блох длина этого периода колеблется
от 100 (Xenopsylla cheopis) до 513 (Pulex irritans) дней, в зависимости
от условш питания, температуры п влажности окружающей среды.
Приведенные нами примеры показывают, что длина половозрелой
стадии определяется не филогенетическими связями паразитов, а
скорее какими-то условиями их биологии: зудни и вши стоят ближе
друг к другу в смысле сроков жизни, чем зудни к клещам, а вши к
блохам.
173
На длнтелоность жизни некоторых штонаразитов влияет темпе-
ратура и частота корлтлопня. Взрослые самки вшей, например, при
температуре 28' и разовом каждодневном питании живут около
30 дней, при двукратном принятии пищи в сутки и при температурах
25э днем и 35е ночью жизнь их удлиняется до 45 дней.
Стадии яйца (у многоклеточных) или цисты (у простейших)
в отличие от половозрелой стадии представляют собой пассивное со
стояние паразита, в течение которого он не нуждается в питании и яв
ляется совершенно неподвижным. , (лителыюсть этих состояний варьи-
рует чрезвычайно широко, в значительной мерс в зависимости от той
категории, к которой относятся яйца, и в зависимости от условий
внешней среды.
Очень часто срок жизни в яйце определяется лишь промежутком
между развитием в нем зародыша и попаданием яйца в хозяина, а
этот промежуток, как легко можно понять, зависит от счастливой
случая н может затягиваться неопределенно долго При этом при-
ходится говорить скорее не о нормальной длительности яйцевого
периода, а о степени выживаемости яиц и цист. Вот некоторые из от
носящихся сюда фактов. Цисты Entamoeba histolytica выживают в экс-
крементах, разбавленных водой, до 35 дней. По видимому, на жише
способность цист вредно влияют бактериальные процессы, ибо те ж*
цисты, тщательно отмытые от экскрементов и помещенные в дистил-
лированную воду, выживают при темпера type 12—22° до 7 месяцев
(Boeck, 1921). Цисты Balantidium coli, по данным Оги (Ohi, 1924).
при комнатной температуре в воде остаются жизнеспособными в тече-
ние по меныпей мере двух месяцев. Ооцисты Fimeria rnagna сохра
н-яют жизнеспособность при нормальном доступе кислорода до 1,5 лет,
а в анаэробных условиях погибают через 3—5 недель.
Среди паразитических червей особенной жизненной Стойкостью
отличаются яйца Ascaris, которые при благоприятных условиях могут
оставаться живыми 5—6 лет.
Более определенные сроки отмечаются главным образом для яиц
некоторых’ эктопаразитов, особенно для таких, яйца которых не
нуждаются в переносе на другого хозяина. У сусликового клеща
Rhipicephalus schulzei личинки вылупляются в лаборатории через 17
35 дней после, откладки. Яйцз часоточного зудня (Sarcoptes) разви-
ваются в 2,5—3,5 дня. Скорость развития зародышей в яйцах вшей,
в зависимости от температуры, колеблется от 16 дней (при 25°) до
7—14 дней (при 30°) и до 6-8 дней (при 35 ). У различных блох
фаза яйца длится от 5 до 14 дней.
Личиночный период, следующий за эмбриогенезом, тоже от-
личается большим разнообразием сроков У многих паразитов, обла
дающих свободными ларвальными стадиями, этот период жизни очень
мал, ограничиваясь одними сутками или даже то'о менее. Таковы ми-
рани inн трематод, личинки монотеистических сосальщиков, онкосферы
широкого лентеца (Diphylloboihrium), личинки ришты и др. Личинки
быстро гибнут, если не встретят промежуточного хозяина. Подобные
личинки обыкновенно очень мелки и снабжены малым количеством
пнш( вых запасов.
Полную противоположность личинкам такого рода составляют
личинки некоторых нематод, например представители Strongylata.
У Ancylostoma duodenal? из яйца выходит рабдитовидная личинка,
способная к самостоятельному питанию. Опа живет во влажной почве
2—3 дня, затем дважды линяет и превращается в стронгнлоидную
личинку. При второй линьке старая кутикула не сбрасывается, а
174
остается в виде чехлпка, внутри которого находится инвазионная ли
чинка 3-н стадии. Такая ппцистированпая личинка может длительное
время жить за счет потребления запасных питательных веществ, отло-
женных в ее теле, и проявлять значительную устойчивость к неблаго-
приятным воздействиям. В природных условиях личинки в некоторых
случаях могут сохранять жизнеспособность до 6 месяцев. Личинки не-
которых трнхостронгилцд (например, Nematodirns filicollis и др.) спо
собны выдерживать зимовку, не погибая.
К числу паразитов с потенциально крайне долго живущими на
свободе личиночными стадиями относятся и клещи Ixodidae, личинки
которых при известных, особенно благоприятных, условиях могут
существовать около года после выхода из яйца, не принимая пищи.
Перейдем теперь к срокам жизни личинок с момента проникнове-
ния их в хозяина или поселения на хозяине и до окончания ими
метаморфоза. Когда личинки попадают не сразу в своего окончатель-
ного хозяина, а сперва в промежуточного, то для них сначала обыкно-
венно наступает период роста, а затем период временного покоя, кото-
рый нередко является чрезвычайно длительным. Оно и понятно, так
как увеличение длительности этого периода соответственно повышает
шансы личинки попасть в окончательного хозяина путем поедания
последним промежуточного. Мнрацпдин сосальщиков, попав в мол-
люска, уже на 2—3-й день превращаются в спороцисту и сразу же
начинают развивать в себе реднй и затем церкарий — здесь, следо-
вательно, период покоя отсутствует. Напротив, метацеркарии испы-
тывают во втором промежуточном хозяине более или менее длитель-
ный период покоя, определяемый моментом попадания промежуточ-
ного хозяина в окончательного
Очень любопытный и даже кажущийся почти невероятным факт
сообщает Ротшильд (Rotschild, 1942) относительно длины срока жизни
реднй сосальщика Cryptocotyle lingua в моллюске Littorina litiorea.
Ротшильд содержала крупный экземпляр этого брюхоногого моллюска
в аквариуме с фильтрованной морской водой в течение 7 лет. И за все
это время инвазия не только держалась в моллюске, ио и продолжа-
лось оживленное размножение паразитов. По приблизительному под-
счету за это время из моллюска вышло более 5 млн. церкарий. Этот
факт говорит нам, с одной стороны, о большой длительности инвазий,
с другой — о той огромной потенции к размножению, которой нередко
обладают паразиты.
Ряд последовательно сменяющих друг дрхта стадий обнаруживает
развитие Diphyllobothrium latum (рис. 135). Развитие подвижной мерца-
тельной личинки (корацидия) в яйце занимает около месяца; далее, сле-
дует короткий (около суток) период плавания корацидия в воде до мо-
мента проглатывания его первым промежуточным хозяином. В полости
тела циклопа корацидий за 2—3 недели превращается в следующую ста-
дию— процеркоида. При проглатывании зараженного циклопа некото-
рыми породами рыб (щукой, палимом, окунем, ершом) процеркоид про-
никает через стенку желудка в полость тела второго промежуточного хо
зянна н там вырастает в следующую личиночную стадию — плероцерко-
ида. Длительность жизни процеркоида определяется сроком жизни ци-
клопа и потому коротка, в лучшем случае циклоп живет не более 1—1,5
месяца. Напротив, плероцеркоиды, находясь в многолетних хозяевах, ка-
ковыми являются рыбы, обладают возможностью неопределенно долгого
существования. И действительно, плероцеркоиды остаются в полости тела
рыбы очень долгое время, не изменяясь. Мало того, эксперименты Гнез-
дилова и Талызина (1936) с прижизненно окрашенными плероцеркои-
175
да .ми доказали, чго в случае поедания мелкой зараженной рыбы более
крупными щуками плероцеркоиды мелкой рыбы не перевариваются в же-
лудке крупной щуки, а пробираются через стенки желудка в полость
тела щуки и. таким образом, продолжают свое существование во вто-
рой особи своего промежуточного хозяина. Последующие опыты Павлов-
ского и Гнездилова (1939) показали, что плероцеркоиды могут проделы-
вать миграцию п еще раз, если будут введены в желу док четвертой особи
озянна (т. е. рыбы). Все это показывает, что стадия плероцеркоида
Рнс. 135. Развитие Diphyllobothrium latum
4. В— стадии формирования корацнлии; С — корацитий;
Г) — процеркоид в полости тела циклона;/ —плероцеркоид;
F—процеркоид. (По Розен.)
очень дл.ггелыш п тянется месяцы, может быть годы. Тем не менее, не-
редко плероцеркоиды погибают в рыбе. В крупных щуках часто можно
наблюдать, наряду с нормальными живыми плероцеркоидами, плероцер-
коидов сморщенных, пожелтевших и мертвых; они постепенно ликвиди-
р- ются деятельностью тканей хозяина.
Возникновение покоящейся стадии (плероцеркоида) в жизненном
иклс сильно удлиняет ларвальный период. У других ленточных глистов
длительность этой затяжной стадии во втором промежуточном хозяине
возрастает в связи с тем. что на нее переносится процесс роста, который
в большинстве случаев, как, например, н у Diphyllobolhrium, отклады-
вается до момента попадания личинки в окончательного хозяина. Этим
достигается двоякая выгода. Во-пергых, используется для роста период
пассивной жизни в промежуточном хозяине; во-вторых, ускоряется про-
цесс образования половых органов и откладки яиц после попадания вы-
176
росшей личинки в окончательного хозяина. Прекрасный пример такого
vO.ua дает ремнец (f.igula). У широкого лентеца, достигающего в полово-
зрелом cocioHitHH почти 10 .и длины, плероцеркоид в рыбе имеет не бо-
лее 2—3 см длины. Между гем у Ligula взрослая стадия из кишечника
тины дистгает максимально 1 я, тогда как плероцеркоиды, добытые из
крупных Лешей, могут быть до 80 см.
I.'ikhm образом, личинка приобретает в промежуточном хозяине раз-
меры. свойственные взрослому червю. Следовательно, съеденной окон-
чательным хозяином личинке предоставляется возможность немедленно
приступить к формированию половых продуктов.
1 акую жг картине' даст Triaenophorus nodulostis — кишечнып пара-
зит щуки, жив’mini в полости тела окуня, ерша и других своих проме-
жуточных хозяев. Личинки Triaenophorus находятся в этих рыбах внутри
округлых капсул п имеют вид длинной ленты — до 20 см дппцой, в то
время как взрослый червь бывает до 30 см длиной. Относительно пле-
роцеркоидов Triaenophorus известно, что они могут оставаться в полости
гела промежуточного хозяина, не погибая, от 1 до 3 лет
Довольно сильный подготовительный рост проделывают личинки не-
матод Eustrongylides в мускулатуре рыб Взрослые Eustrongylides из
кишечника водяной птицы имеют от 35 до 90 длины, тогда как в раз-
личных каспийских рыбах мы находили личинок этих червей, имевших
не менее 50 м и длины Если мы учтем, что личинки ришты (Dracunculus).
имеющей в человеке до 120 си длины, у своего промежуточного хозяина
(циклопа) достигают размеров лишь в I лью ю Судет вполне ясно, ка-
кую подготовку в смысле роста проходит личинка Eustrongylides в теле
рыбы.
Особенно долгое время могут сохраняться личинки в теле много-
летних хозяев. Так, мускульные трихины в некоторых Случаях обнару-
живались живыми в теле человека через 31 год после инвазии.
Сроки полового созревания. Иначе обстоит дело с личин-
ками после того, когда они наконец попадут в своего окончательного
хозяина. В громадном большинстве случаев метаморфоз ими быстро за-
канчивается, н животное скоро переходит во взрослое состояние. Пере-
ломным моментом в этом периоде жизни является начало откладки яиц
как переход к следующему поколению, к следующему жизненному
циклу. Момент этот у разных паразитов наступает в различной степени
быстро.
Особенно короткий период созревания наблюдается у некоторых со-
салыцчков ”3 семейства Microphallidae (рис. 136). Метацеркарнп М>сго~
phallus pigmaeus в окончательном хозяине уже через 36 часов становят-
ся половозрелыми особями, приступают ими к откладке яиц (Бстополь-
ская, 1949) Microphathis nicolli становится половозрелым в кишечнике
птицы через 12 чзеов, a Pseudolevinsiella client достигает половозрелости
даже через 6 часов (Tsai Shang-ta, 1955). Microphallus minus при экспе-
риментальном заражении белых мышей уже через 3 часа имели в матке
3—4 яйца и через 5 часов — до 115 яиц (Yeh a. Wu. 1951). Это, несо-
мненно, связано с тем, что метацеркария во втором промежуточном хо-
зяине обладает уже развитой половой системой, не требующей доразви-
тня при попадании в окончательного хозяина, и имеет размеры, сходные
Со взрос пым червем. Здесь проявляется явная тенденция к прогенезу
(см. стр. 207).
С "р1 гой стороны, адолескарии Fasciola hepatica или метацеркарии
Dicrocoeiium dendritiaun не имеют развитой половой системы, и поэтому
половозрелого состояния первый вид в крупном иогртом скоте достигает
только через 3—4 месяца после заражения, а второй — в овцах — через
[2 В А. Догель
177
78—85 дней. То же наблюдается п у ленточных червей Для тех из них.
у которых личинки уже в промежуточном хозяине приобретают значи
тельную величину и имеют хорошо выраженный комплекс половых орга
нов (Ligula. Schistocephalus). достаточно каких-нибудь одного-трех
дней, проведенных в окончательном хозяине, чтобы паразиты начали про
Аудировать яйца. В то же время Diphyllobothrium latum в окончатель
ном хозяине (человек) достигают половой зрелости и начинают выделять
яйца только через 3—5 недель после заражения. Это, несомненно, свя
зано с тем, что плероцеркоид D latum из рыб морфологически соответ
стоуст самым ранним неиивазпоппым стадиям развития плероцеркоида
Рис- 136. Microp/iatlus papillorobitstus
А — взрослый сосальщик; Г. — метацеркария из Gammaru.\
locusta. (По Белопольской.)
ремнецов с неразвитой половой системой. Поэтому, когда такие плеро
неркоиды попадают в окончательного хозяина, требуется длительный
срок, чтобы в процессе метаморфоза во взрослого червя произошло фор
мирование всего комплекса половых органов и началось выделение яиц
(Дубинина, 1957 а). Развитие половой системы у многих других цеетод
в окончательном хозяине также протекает медленно Например. Monie-
zta expansa в телятах достигает половой зрелости через 37—40 дней, а
Taeniarhynchus saginatus у человека — через 3 месяца.
Среди нематод, с одной стороны, имеются вилы созревающие в
окончательном хозяине очень быстро. Например, мускульной трихине,
попавшей в кишечник хозяина, требуется для достижения половой зре-
лости 6—7 дней, a Strongyloides ratii созревает даже в 4 дня. С другой
стороны, с момента выхода личинки Ascons lumbricoides из яйца в ки
шечппке зараженного субъекта и вплоть до появления первых янц в его
экскрементах проходит 2—3 месяца, нз которых 10—12 щей уходит на
миграцию личинки.
178
Длительные сроки достижения половой зрелости наблюдаются у
некоторых представителен монотеистических сосальщиков. По данным
Быхоиского (1957), Dactylogyrus iwanovi заражает молодых особен
Lcuciscus brandii, скатывающихся в море в нюне-июле; черви достигаю!
половой зрелости через 2 года в морской период жизни хозяина и начи
нают кладку яиц при подходе хозяина в р'екн, в марте-апреле.
Более резко очерченные границы имеет ларвальный период у экто-
паразитов, проводящих всю свою жизнь на геле хозяина. У зудней (Sar-
coptes), например, оз вылупления личинки из яйца и до ее превраще-
ния во взрослого клеща проходит 8—10 дней, у Phthirus pubis личиноч-
ный период длится 15—17 дней, у Pediculus— от 10 до 14 дней.
Подводя итоги рассмотрению относительной длительности отдель-
ных стадий, мы замечаем, что в цикле паразитических животных прояв-
ляются некоторые более или менее определенные тенденции. Во-первых,
нередко наблюдается тенденция к удлинению стадий покоящихся яиц.
покоящихся личинок в промежуточном хозяине и свободных личинок
предназначенных к проникновению в окончательного хозяина. Это удлн-
ш нис вполне объяснимо, так как оно способствует встрече паразита с
хозяином, и таким образом дальнейшему сохранению вида. Далее, иног-
да имеется тенденция к подготовке развития полового аппарата <ше
в личиночном возрасте, чем ускоряется начало размно?кения при
попадании личинки в окончательного хозяина. В некоторых слу
чаях замечается наклонность к удлинению пмагннальиой стадии, ибо
этим удлиняется не только период размножения, но и увеличивается
продз кция яиц.
Другой путь, ведущий к тем же результатам, имеет диаметрально
пВэтивоположный характер и заключается в укорачивании как взрослой
стадии, так и прочих стадий цикла, но в увеличении на том же отрезке
времени числа поколений животного. Однако это наблюдается лишь в
исключительных случаях, ибо при кратковременности всех стадий умень-
шаются гарантии встречи их с соответственными хозяевами, что, конеч-
но, должно неблагоприятно отразиться на сохранении вида. Такое уско-
рение цикла возможно лишь при аутоинфекции (пример — Hytnenolepis
папа) и тогда, когда передача паразита от одного хозяина к другому со-
вершается путем непосредственного контакта. Поэтому мы находим,
например, сильное ускорение цикла с соответственным уменьшением чис-
ла откладываемых яиц у таких форм, как чесоточные зудни. Тогда как
непривязанные постоянно к телу хозяина Ixodidae имеют длительный
жизненный цикл (полгода и более) и продуцируют многие тысячи яиц.
мелкие зудни Acarus siro, передающиеся посредством контакта и раз-
множающиеся на самом теле хозяина, имеют жизненный цикл длиной
около двух месяцев и дают всего 40—50 яиц. И. несмотря на малую пло-
довитое гь, частота смены поколений \ зудней приводит к очень интен-
сивному заражению хозяина.
В отдельных случаях укорачивание взрослой стадии не сопровож
дается умножением числа поколений, что мы имеем, например, у кишеч-
ных трихин, которые созревают уже па 7-й день пребывания их в кишеч-
нике, хотя получающимся от них личинкам иногда предстоит целые годы
оставаться в мускуллтуре той же особи хозяина до их передачи другому
хозяину. Быть может удлинение срока жизни кишечных трихин было бы
бесцельным или даже вредным для вида, так как чрезмерное увеличение
потомства, сопутствующее удлинению половозрелой стадии, могло бы
привести к быстрой гибели хозяина и затруднило бы передачу инвазии
на других хозяев. Заражение организма слишком большим количеством
мускульных трихин заканчивается для нею смертельным исходом.
17*2
12-
4. ПРИСПОСОБЛЕННОСТЬ ЖИЗНЕННЫХ ЦИКЛОВ ПАРАЗИТОВ
К ЖИЗНЕННЫМ ЦИКЛАМ ХОЗЯЕВ
Помимо отдельных морфологических и физиологических адаптаций
к паразитическому образу жизни, весь жизненный цикл паразита неред-
ко обнарх.кивает очень детальное ирпспсссбление к жизненному циклу
хозяина. Примеры такого совпадения имеются в большом количестве в
самых различных группах паразитов
Рис. 137. Житпепныи цикл лягушки и се кишечных
паразитов; наружный круг — Nyctotherus cordiformis,
средний круг — Opalina гапагит. (Но Боевой-Нетру-
шевскр^ )
1 — вегетативная стадии; 2 4 — Гесиолос размножение делением;
5 — HHHHCTiipoBaiiiie; о — iihxui из пнет; 7 образов, пне нос| ед*
ством ледени 4 ii|iexoiibiurain(.n j ДГугТо/й^'ш eorthformis пли
гамету Opalina гапагит; Я коны .гл ни к у Д’, tortli form т и.ш
копуляция гзмет у О. rar-arum; 9 — эксконъюгант у Д . cordijor-
mis или liiimicTiipouaHiLin зигота у О. ranu'um.
Полное совпадение имеется в цикле инфузорий и жгутиконосцев,
населяющих кишечник лягушки, с циклом хозяина. Нересгеймер (Ne-
resheimer, 1907) и Суханова (1953) показали это для Opalina, а Воева-
Петрушевская (1933) для Nyctotherus лягушек (рис. 137). В течение
всего года опалины и Nyclotherus размножаются в кишечнике лягушки
только бесполым способом, посредством деления надвое Когда весной
лягушки приступают к половому размножению, наступает и период под-
готовки к половому процессу у инфузории и опалин. Они инициируются,
цисты попадают в воду вместе с экскрементами и проглатываются ма
ленькнми головастиками. Здесь опалины многократно делятся, образуя
очень мелкие гаметы, которые потом копулируют; Nyclotherns соединя-
ются попарно и конъюгируют. У Opalina в головастиках образуются в
конечном счете цисты, которые выходят снова в воду и проглатываются
крупными головастиь амп. Образование цист в лягушках приурочено ко
времени икромет iпня и выхода головастиков из икры, вследствие чего
стадии распространения паразитов получают возможность стыка с новым
поколением хозяина, которое и заражается цистами.
По Кодрсану (Codreanu, 1930) развитие Ophryogletia, паразитирую
щнх в личинках поденок, тесно согласовано с развитием хозяина. В мо
лодых личинках инфузории остаются в ннц.чстпровапном состоянии, со-
храняя вместе с тем способность делиться. Зараженные мужские личин
кн поденок превращаются в нормальное имаго, последующа^ гибель ко
торого влечет за собой и смерть паразитов. Инфузории Женских личинок
выходят из своих цист и проникают в яичник самки поденки, содержимое
которого они и пожирают. Кастрированная самка тем не менее участвует
в брачном полете и опускается на воду, но вместо янц откладывает в воду
многочисленных инфузорий. Инфузории расплываются в воде и отыски-
вают личинок поденок.
t Щ|1|1Г1г-1|
2 CJ12J
Рнс. *ЗЯ. Цикл развит ня грегарипы Diplocystls phryganeat
из полости тела Phrvganea grmuiis. (Но Федоровой-
Виноградовой.)
1, 3 и 5 — годичны г цикл мстаыпрфтз Ph. gnindis; 2. 4. б — стаит
l>a»niiru>i D. phryguneuC, J —ЛНЧИПСИ; 2 — minuet иргноипыс парочки
(ен-лигин); 3 — куколки; 4 — цисты си спорами S — нзрослые ручсЛпнкн;
6 — одиночные ucrcTniiiuirue сидни гре1д| ии.
4 Сходное, но несколько иное совпадение циклов имеется у грегарш-
Diplocystis plirygaueae (рис. 138), паразитирующих в полости тела ру
чейника Phryganca grandis. Федорова-Виноградова (1924) нашла, чго
в личинках ручейника грегарнны остаются в вегетативном состоянии,
но когда наступает время окукления, большая часть грегарин соедини
ется попарно, пнцнстнруется и проделывает половое размножение и
образует ооцисты. Таким образом, в вылетающих имаго преобладают
ооцисты. После откладки яиц в воду самками Phryganea последние здесг
же на воде вскоре погибают и служат, таким образом, источником зара
женпя личинок.
Другие грегарипы, Schneideria metumorphosa нз личинок мухи
Sciara coprophila, тоже начинают половой процесс при превращении ли
чинки в куколку. Ооцисты выталкиваются наружу вместе с кишечным
содержимым во время линьки, предшествующей окуклению.
Коллери и Мениль (Caullcry et Aicsnil, 1899) наблюдали, что гре
гарины Gonospora longissima нз полости тела лолихет спорулнруют в
период половой зрелости червя, причем ооцисты выводятся вместе с по-
ловыми продуктами хозяина. Высказывается предположение, что червь
во время своего гаметогенеза образует в себе вещества, которые стн.му
лиру ют грегарин к половому процессу.
181
Очень четко выражено совпадение жизненного цикла жгутиконосцев,
населяющих кишечник тараканов Cryptocercus, с циклическими измене
киями хозяина. Исследованиями Кливленда (Cleveland, 1957) было пока
за но, что у представителен Polymastigida и Hyperniastigida, размножаю
1ЦПХС.Я обычно агамным путем, в определенный момент их жизни на-
ступает половой процесс, характеризующийся образованием гамет и их
копуляцией. При этом было установлено в отношении представителей
10 родов жгутиконосцев, что переход к половому процессу у них всегда
связан с периодом линьки хозяина. Этот процесс у самых молодых нимф
Cryptocercus совершается два раза в году, а у более старых только один
раз и все развитие длится несколько лет.
Рис. В9. соотношение гаметогенеза, on.iojoi впрепия п мейоза у простейших из
Cryptocercus к линьке хозяина. Покачана продолжительность этих процессов у раз-
ных видов простейших. (По Кливленду.)
Z гаметогенез; ” — гшд о лот nopei Hie; о — мейоз, О — момент линьки.
Период подготовки к линьке наступает вскоре после начала выде-
И'ння гормона линьки, задолго до самого момента сбрасывания кути
кулы (ecdysis). У разных видов жгутиконосцев образование гамет и ко-
пуляция происходят в разные сроки подготовительного периода или
вскоре (через 1—2 дня) после самого процесса линьки (рис. 139). На-
пример, у Trichonyntplia гаметогенез начинается за 5 диен до линьки, а
копуляция гамет происходит на второй день после линьки; у Leptospiro-
nynipha wachula весь половой процесс осуществляется за 2G—30 дней до
линьки; у Oxytnonas эти процессы протекают за 32—1*1 дня, а у Barbulo-
nympha гаметогенез и копуляция происходят даже за 40 50 дней до
линьки. Если задержать период линьки, то задерживается соответственно
и наступление полового процесса у жгутиконосцев. Это связано с тем,
что гормон, вызывающий линьку у хозяина, влияет вместе с тем и на по-
ловой процесс его паразитических жгутиконосцев. Поэтому циклические
изменения паразитов так тесно связаны с циклическими изменениями
хозяина.
182
Детальные совпадения жизненного цикла Polystoma integerrimurn
с жизнью лягушки выяснены Ьыховскнм (1957) (рис. 140). Личинки
многоустки, пока у головастиков имеются жабры, сидят на них в виде
эктопаразитов, по после зарастания жаберных шелеп проходят через
кишечник в мочевой ну !ырь, где и проделывают остальное развитие
Гаким образом, каждая лягушка заражается многоустками лишь раз
в жн.знн, на стадии головастика. В связи с особенностями жизненного
чикла лягушки жизненный цикл многоустки краппе замедлен; вероятно.
Рис. 1Ю. Polvsinma integerrimum из мочевого пузыря лягушки. Цикловые
изменения половых желез (в частности жслточнпков) в связи с биологией
хозяина. (По Ьыховскому.)
Желточннки (сплошной черны3 цист ii.hi < срные точки! логинают полного рззг-итин н октябре и
естаютс» н т ком визе ло икромет'Пня л iryuieK. когта нронсхолнт и р зчножеипе Potystomi' после
чтого запасы и желточннках летом столяг и нет по с ai-rycr* начин ются новое увеличение желточ-
iiiiKOU. нрололжаюшееся до октябри.
это един из наиболее медленно растущих сосальщиков. Многоустка до-
стигает половой зрелости лишь на третьем году жизни, весной, т. е. к той
же поре, когда впервые начинает размножаться и лягушка. К этому
времен ч желточннки. степень развития которых — хороший показатель
половой зрелости многоустки, проходят через ряд ступеней созревания,
достигая окончательного размера в октябре второго года жизни парази-
та. В таком виде желточннки остаются до апреля; в начале мая много-
устка опоражнивается от янц и желточных клеток во время икрометания
лягушки в воде. Вылупление подвижных личинок из янц «подогнано»
таким образом, что оно происходит в то время, когда в водоеме уже
имеются вышедшие н? икры головастики лягушек. Получается смыкание
цикла паразита и хозяина, и головастики заражаются личинками. Между
тем взрослые многоустки остаются в мочевом пузыре лягушек, и у них
прежним темпом идет нарастание желточников и всей половой системы
183
до весны следующего года, когда следует новое половое размножение
хозяина и паразита. Многоустка остается в .мочевом пузыре лягушки
по-видимому. вплоть до смерти лягушки. Как видно, каждый этап жизни
многоустки приурочен к отдельным этапам жизни хозяина.
Много хороших совпадений в циклах развития обнаруживают раз
личные наездники и насекомые, которые служат для них хозяевами. Так.
например, для успешного развития наездников, которые откладывают
свои яйца в яйца или личинки другого насекомого, абсолютно необходи
мо, чтобы самки паразита появились в то время или даже несколько
раньше, чем появится половозрелая стадия хозяина. И действительно,
такое совпадение наблюдается у многих видов Ageniaspis и Anastatus
с одногодичной генерацией. В других случаях, когда хозяин имеет не
сколько генераций в году, соответственное число поколений развивается
и у паразита; три поколения наездника Ageniaspis [uscicollis praysincola
совпадают с соответственными поколениями бабочек Prays oleeHtii.
Иногда, видимо, совпадения циклов нет. но это обстоятельство иахс
дит себе объяснение в тех или иных особенностях жизни паразита или
хозяина. Так, например, полное совпадение циклов может отсутствовать
без вреда для паразита в том случае, если последний способен заражать
разные стадии развития хозяина. Так. паразит непарного шелкопряд;
(Apanteles melanoscelus) способен откладывать свои яйцт в гусениц
разного возраста. Другой случай кажущегося расхождения циклов даст
наездник Kaltenbachia bimaculata, имеющий три генерации, тогда как
ею хозяин, оса Odynerus laevipes. имеет лишь одну. Однако в данном
случае покоящаяся стадия хозяина, в которой паразитирует личинка
Kaltenbachia, так длительна, а срок развития паразита столь короток,
что три поколения наездника имеют возможность провести свою парази
тическую стадию в куколках одного поколения Odynerus.
Особенно любопытный пример представляет наездник Perilampus
hyalinus. Его первая личиночная стадия, или планидий, проникает
внутрь гусеницы бабочки Hypantria, но питается не за ее счет, а отыски
вает паразитирующих внутри гусеницы личинок мухи-тахины Varichaeta
пли же наездника Limnesiiun. Планид ий забирается внутрь этих личинок
и остается в них без изменения до .момента их окукления. Тогда плани
дий вылезает, питается содержимым куколки и проделывает преврати
ннс. При этом личинка Varichaeta окукляется осенью, а личинка Limne-
sium зимует и окукляется лишь весной. Соответственно с этим и жизнен
«ный цикл у личинок Perilampus. попавших в муху или в наездник.,
складывается различно в отношении сроков развития. Помимо гиперпа-
разитизма в этом случае важное значение имеет то обстоятельство, что
мы наблюдаем в пределах одного вида Perilampus hyalinus начал*
внутривидовой дивергенции, как бы намечающийся путь к формирова
ншо двух биологических или физиологических видов (см. главу XXII)
Такие отношения мог\т быть даже у паразитов яиц. Шопар (Cho
pard, 1923) приводит интересный пример с яйцеедом наездник;
Podagridn pachymerutn, имеющим две генерации в год;. тогда как хо
зяин его, богомол Mantis rcligiosa, имеет всего одну генерацию. Двои
пая генерация этого паразита делается возможной благодаря тому, чг<
не все яйца в оотеке богомола развиваются сразу, но часть из них
сильно запаздывает и служит добычен второму поколению наездника.
Отмеченная согласованность нс является, однако, абсолютным пра
внлом. и иногда бывают случаи тупиков развития. Мы уже видели, что
Ophryoglena в самцах подонок умирают, не оставив потомства. Парази
ты водяных личинок насекомых, которые во взрослом состоянии имеют
лишь слабый контакт с водой (например, стрекозы), имеют очень мало
184
шансов снова попасть в воду, если нс успеют покинуть .хозяина до его
превращения. Так, иногда можно наблюдать стрекоз, к телу которых
прикреплены нимфы водяных клещей (Hydracarina). Тупиком развития
служит для трихины человек, так как она способна проделывать в нем
все свое развитие, но дальнейшего распространения получить не может;
для осуществления его необходимо было бы наличие каннибализма.
В огромном большинстве случаев, как мы видели, различные фазы
жизни паразитов удивительно подогнаны к биологии хозяина. Однако
изредка встречаются примеры явных нецелесообразностей в этом отноше-
нии. В полости тела волжских лягушек встречаются в большом числе
крупные личинки трематоды Codonocephalus, дальнейшее развитие ко-
торой заканчивается в малой выпи. Заражение этими личинками лягушек
начиная с ранней весны растет и у младших возрастов продолжает
расти до зимы (Дубинина, 1950в). Но у половозрелых лягушек, идущих
на икрометание, паразиты выметываются вместе с икрой в воду. Вслед-
ствие этого процент заражения лягушек летом сильно снижается, снова
несколько повышаясь лишь к осени. Так как выброшенные с икрой на-
ружу личинки Codonocephalus обречены на гибель, то цикл этой тремато-
ды, гармонично согласованный с ходом жизни молодых и подрастающих
лягушек (где личинки этой трематоды не выбрасываются частично на-
ружу). очевидно, нс вполне приспособлен к биологии взрослых лягушек
вследствие отсутствия выработки какого-то приспособления, задержи-
вающего паразитов внутри хозяина во время икрометания.
Интересное совпадение не с жизненным циклом, а с инстинктами
промежуточных хозяев обнаруживают некоторые нематоды. Личинки
(микрофилярии) паразитирующей в человеке и вызывающей слоновую
болезнь Wuchereria bancrofti, передаваемые другому хозяину через по-
средство ночных кровососущих насекомых, а именно комаров, появля-
ются в периферической крови только в ночные часы.
Обратные отношения представляет другой паразит человека, нит-
чатка Loa loa, личинки которой циркулируют в крови днем; ее перенос-
чик слепень Chrysops нападает на человека в самое жаркое время дня
5. ЧЕРЕДОВАНИЕ ПОКОЛЕНИИ И ЖИЗНЕННЫЕ ЦИКЛЫ. КЛАССИФИКАЦИЯ
ЖИЗНЕННЫХ ЦИКЛОВ
Мы уже видели, что затруднения, испытываемые паразитами в из-
вестный период жизни, а именно при переходе от одного хозяина к дру
грму, могут быть с успехом преодолены лишь при условии усиления спо-
собности паразитов к размножению. Эта способность и выражается в
разобранном нами ранее законе большого числа яиц. Однако усиление
продукции яиц является далеко не единственным средством компенсации
большого процента погибающих во время процессов распространения
паразита яиц или личинок. У многих групп паразитов в жизненный цикл
животного вставляется особая фаза размножения посредством партено-
генеза, или бесполого размножения, н, таким образом, жизненней цикл
приобретает .характер чередования типичного полового поколения с по-
колениями партеногенетическими, или бесполыми.
В одних систематических группах паразитов (например, дигсистнче
ских сосальщиков и кокцидин) чередование поколений является прави
лом, но среди других (например, у нематод, цестод и грегарин) одни
представители обладают сменой поколений, другие нс обладают.
Вопрос усложняется тем обстоятельством, что у многих паразитов
чередование поколений комбинируется со сменой двух или более хозяев
причем жизнь одного нз поколений протекает в одном хозяине, другого —
в другом.
18">
Присматриваясь к различным циклам и к их постепенному усложне-
нию. можно заметить, что в циклах паразитов н известном порядке сме-
няются дна процесса. В одном из поколений, обыкновенно в бесполом,
или партеногенетическом, наблюдается накопление паразитов в одном
из хозяев; в другом поколении, именно половом, пропагативные (т е. слу-
жащие для распространения) стадии паразита, покидая хозяина, рассеи-
ваются во внешней среде, служа для заражения новых хозяев. Эгп
две фазы мы будем обозначать терминами накопления, или
а г гл ом е р а ц и и, и р а с се я н и я, или д и с п с р с и и, соответствен-
но отмечая их на нижеприведенных схемах буквами Л и I). В наиболее
сложных циклах может быть несколько фаз А или D
Фаза аггломерацнн выражается в накоплении значительного числа
особен паразита в одном индивиде хозяина, что, обыкновенно, происходит
путем усиленной э н л о тени о й и н фекц и и. С такой аггломерацией,
являющейся следствием жизненного цикла паразита, не следует смеши-
вать накопления паразитов, происходящего в одной и той же особи хо-
зяина посредством ее повторных инфекций. Всякий хозяин, например,
может неоднократно проглатывать яйца паразита и, таким образом,
будет происходить накопление паразитов в данной особи хозяина. По
такое экзогенное накопление не имеет ничего общего с эндо
генной аггломерацией паразитов в крови малярика, которая получается
в результате бесполого размножения паразитов. Экзогенное накопление
может в таких случаях прибавляться к эндогенному, если человек будет
укушен зараженными комарами дважды. Экзогенная и эндогенная аггло-
мерации отсутствуют лишь в тех редких случаях, когда размеры хозяина
позволяют паразитировать в нем одной особи паразита (личинки некото-
рых наездников, паразитирующие в яйцах или личинках других насе-
комых).
Классифицировать жизненные циклы довольно сложно, так как иног-
да кажущийся простым жизненный цикл па самом деле произошел в
результате упрощения гораздо более сложного цикла. У представителей
одной и той же группы имеют место иногда различные циклы, а один и
тот же тин цикла может проявляться в разных группах паразитов.
При рассмотрении жизненных циклов по мерс их постепенного услож-
нения видно, что сходный характер могут приобретать циклы несколько
различного происхождения и характера.
Существенное затруднение проявляется при сравнении циклов про-
стейших и многоклеточных. У многоклеточных зигота, как правило, обра-
зует (кроме редких случаев полиэмбрионии) один взрослый индивид,
тогда как у простейших часто зигота в виде покоящейся цисты пронзко-
дит в себе посредством деления (т. е. бесполым способом) несколько
зародышей. Кроме того, среди простейших есть много представителей
(все кишечные и кровяные жгутиконосцы, все кишечные амебы), у кото-
рых до спх пор известно лишь бесполое размножение, — особенность, не
встречающаяся совсем средн многоклеточных. Вес эти обстоятельства
настолько затрудняют классификацию жизненных циклов паразитов, что,
например, Коллери (Caullcry, 1922) и поздп-е Грассэ (Grasse, 1935)
вовсе от нее отказались, просто давая ряд описании нескольких наиболее
типичных циклов.
Действительно, расположить циклы столь разнохарактерного кон-
гломерата форм, и притом руководствуясь одновременно несколькими
критериями (наличие или отсутствие полового размножения, наличие
плз отсутствие смены поколений и хозяев, количество и расположение
фаз аггломерацнн и дисперсии), чрезвычайно трудно. Поэтому, делая
в дальнейшем попытку классификации, мы решаемся на это лишь вслед-
IK6
етвие необходимости когда-нибудь сделать такую попытку, хотя бы и
весьма несовершенную. При этом нам думается, что с точки зрения па-
разитологии более важное значение имеют количество н распределение
фаз эндогенной аггломерацни и дисперсии, чем самый путь, при помощи
которого совершаются эти процессы, т. с., например, половой или беспо-
лый характер размножения и т. п.
Ниже мы приводим графическое изображение ряда жизненных инк
лов паразитов. При этом буквами A, У н Z обозначены хозяева паразитов.
13 случае наличия всего одного хозяина он отмечается буквой Л, при на
личин смены хозяев добавляются буквы У в Z. Процесс дисперсии выра-
жается буквой D и указывается расходящимися от хозяина стрелками,
тогда как на экзогенную аггломера-
цию указывают стрелки, сходящиеся
к хозяину. Буквы enA. стоящие под
буквой, обозначающей хозяина, вы-
ражают эндогенную аггломерацию.
Знак ~ указывает на неопределен-
но большое количество паразитов
при эндогенной аггломерацни. Знак
W обозначает наличие полового
размножения у паразита, знак $ —
партеногенетическое поколение.
Паразиты без чередования
поколений и без смены хозяев.
А Без эндогенной а г г ло-
ме р а ц и И. Этот тип довольно ши-
роко распространен среди многокле-
точных, для которых он обыкновен-
но является наиболее примитивным
и наиболее простым.
В хозяине происходит рост и
размножение паразита, формируют
ся его яйца или личинки, которые вы-
водятся наружу п заражают других
особей того же самого вида хозяина.
По этому типу складывается жизненный цикл у всех монотеистических
сосальщиков, у многих нематод (рнс. 141), у пиявок, многих раков и у
других животных. У всех этих форм имеется лишь половое размножение
Схема подобного жизненного цикла имеет следующий вид:
Рис. 141. Ascaris suum
/1 — слепа самец, справа самка; В —
пеицвазионное (нсразшшшеесн) яйцо;
С—инвазионное яйцо с личинкой. (По
Анишину.)
В отдельных случаях как у многоклеточных, так и у простейших цикл
подобного типа получается посредством упрощения циклов более слож
иого характера.
Гаков никл человеческого liymenolepis папа (рис. 142). Из яии
паразита, попивших в кишку человека, вылупляются зародыши, которые
проникают в ворсинки тонкой кишки и превращаются в маленькую финку
иистнцеркоидпого строения. Через 4—5 дней цнетнцеркоид вываливается
187
в просвет кишки, присасывается к се стенке и превращается в полово-
зрелого ленточного червя. Чернь производит яйца, которые выводятся
с экскрементами и служат источником заражения других людей. Сравне-
ние с другими цестодами показывает. что у Н. папа, несомненно, имеет
место вторичное у прощение цикла, выражающееся в выпадении проме-
жуточного хозяина: у этого вида и финка, и ленточная стадия разви-
ваются в о зной и тон же особи хозяина.
Результатом х прощения является, судя по всему, и жизненный цикл
настоящих грегарин (Eugregarinae), по всем своим деталям отвечающий
циклам данного тина. Здесь беспо-
Рпс. U2. Цикл разв гия Ihmenolepis
папа из кишечника человека. (По
Догелю.)
1 — изроелзя строСнлл: £ — яЗш». вышедшие из
erpnt илы н длю цпе икни пкрюч них э *|:одыш<* i;
J - ШфГТИКрЮЧНЫ Л 3:>ролыи1 В МОрСИНКС КН1ПСЧ-
пню; -Л - прелр.ч.цепне зародыша в шюицеркондз;
Л х HHCiinicpKdiu иыяора’щи. етси it посдупдсг в
прогнет кн печника; г» — получиипынсн головка
Н| нк[ vii-ъктся к стенке кишки и пырни уст и
ст।де илу. Снизу изображены при солыпим увели-
чении HKCfiiKpioniiu I jдродию, иистицерконд !!
ГО-10ВК9 стро( II.ты.
лое размножение отсутствует вооб
щс, накопление паразитов в одной и
той же особи хозяина совершается
только путем поступления новых па-
разитов извне, а половое размноже-
ние приводит к образованию ннцис-
тирующихся зигот — ооцист в кото-
рых формируются спорозоиты.
Ооцисты служат для дисперсии во
внешней среде По-видимому, цикл
грегарин произошел от цикла, в ко-
тором имелось и бесполое размноже-
ние. как у Scilizogregarinae. Впо-
следствии бесполое поколение у
предков грегарин выпало, и осталась
одна половая часть цикла, вслед-
ствие чего он оказался принадлежа
щим к первому типу.
Б. С эндогенной а г г л о м е-
рацией. Цикл данного типа встре-
чается у некоторых кишечных про-
стейшие (амебы, жгутиконосцы),
для которых до сих нор известно од
но бесполое размножение. Цикл Еп
tanioeba или Lainblia протекает так
вышедшие из инет в кишечник<
хозяина паразиты усиленно размно-
жаются посредством деления. Позд
нее, наряду с делением в свободно
подвижном состо иши, часть особей
инцнст||руется и продолжает делить-
ся в покоящейся стадии, а сами
цисты выводятся наружу и слу кат
для рассеяния паразитов. Значит,
отличие от предыдущего цикла заключается в том, что в хозяине носред
ством размножения паразитов наступает эндогенная аггломерадзя, а
дисперсия паразитов происходит при помощи пнет, образовавшихся бес
полым способом. Получается схема, выражаемая так:
188
Отсутствие полового процесса, столь важного для сохранения вида.
заставляет думать, что циклы
видимому, представляют собой
размножения.
По наличию стадии эндо-
генной аггломсрацнн эта раз-
новидность I типа прибзижд-
ется ко II типу, который итли
чается однако, тем, что, поми-
мо бесполого, характеризуется
и половым размножением
Паразиты с чередованием
поколений и без смены хозяев
А. С эндогенной а г -
г л о м е р а ц и е й. Среди про-
стейших таким циклом обла-
дает большинство кокциднй.
например Eitneria (рис. 143)
кроликов и кур, Isospora собак
и кошек и др Ооцисты этих ки-
шечных кокцидпй попадают в
кишку хозяина и освобождают
находящиеся в них спорозои-
тов. Спорозоиты проникают в
кишечный эпителий и присту-
пают к бесполому размноже
нию посредством шизогонии.
Бесполые формы (шизон-
ты) распадаются на множе-
ство мерозоитов, которые про-
никают в клетки кишечника и
вновь претерпевают шизого-
нию. Этот процесс может по-
вторяться несколько раз. В
конце концов, мерозоиты пре-
вращаются в дифференциро-
ванные в половом отношении
гамонты. Последние си тьно
растут. У микрогамонта ядро
во время роста многократно
делится, и в результате из каж-
дого микрогамоита формирует-
ся множество одноядерных
микрЛамет с двумя жгутика-
ми. Из каждого макрогамонта
образуется только одна круп-
ная ' макрогамета, снабженная
запасными питательными ве-
ществами. Микрогамета прони-
кает в макрогамету, и их ядра
сливаются. Образовавшаяся в
результате этого процесса .зи-
гота инцистпрустся (становит-
некоторых кишечных простейших, по-
рсзультат вторичной утраты полового
Рнс. 1 *3. Пикл развития Fimeria magna из
кишечника кролика. (По Хейсипу.)
1 — cirnj он нт; 2 5 - шизогония ц? — милело» шизонт в
эпителии кишечника кролика, л / рдзинл-ио ни ст
шизонт. 5 — ссгмситнроиаипыл шиэоп): о — мерозиит;
7—/У- panfiiniK* те.юмеризингоп (7 р пняя стлил раз-
пития шизонта. Я-9 - расту.цис шидпнгы, 10 - ceriicnni-
ровмшый шизонт): II — телпмерозонгы; 12 td — р»з-
niiriic мнкроымег (J2 рдпнти ст j ли и развитие мик|юги-
метоццтз. /3—J-5 — рог г мнкрогаметоипта и деление ялир.
Гл — образование мпкригзмст); 17 — мнкрог«мета; 18 -
20 развитие мэкрлглметы: 2/ мисрогАмста о белко-
выми гранулами in перифефш: 22 - оплодотворение;
23 - сформпроиапшэяся к кишечниас ооциста; at 2-*1
развитие лопнет (спорогония) (2/ — ооциста с округлив-
шимся протоплазматическим шаром. 25 27 - обр зоианис
снороблаетоп. в центре — остаточное тело 28— рдеи"тие
сплропкст, £9—ооциста с 4 с пирам и и которых содер-
жится по 2 спорозоита).
189
ч-я ооцистой) и выносится во внешнюю ередх. Внутри ооцисты прсйк.хо-
дит деление зиготы па споробласты; последние выделяют на поверхности
оболочку н становятся спорами, внутри которых последовательным деле-
нием обратится спорозоиты. В цикле развития Einieriidea преобладают,
как видно, бесполые поколения, служащие для расселения в кишечнике
хозяина. В результате полового процесса образуются такт- стадии раз-
вития (ооцисты), которые, предназначены для дисперсии и заражения
новых особен хозяина.
Схематично этот тип никла может быть изображен так.
Б. Без эндогенной аггломерацни Таким циклом раз
вития обладают некоторые нематоды, у которых паразитическое поколе
ние представлено партеногенетической самкой, а другое, свободно живу-
щее,— раздельнополыми особями. Это наблюдается, например, у Rhab
dias bujoiiis и Strongyloides stercoralis (формы нз обезьян Ateles).
В связи с отсутствием бесполого размножения здесь нет эндогенной аг
гломерацнн. Схематически такой цикл может быть изображен так:
X
Паразиты без чередования поколений с однократной < меной) хозяев
А. Без эндогенной аггломерацни. Сюда принадлежат
формы, которые проделывают в споем развитии метаморфоз, причем
ларвальная стадия проводится паразитом в одном хозяине, нмагиналь-
ная — в друюм Как видно из определения, к данной группе принадле-
жит прежде всего большое количество нестод, а именно все ленточные
глисты, которые в стадии финки не имеют бесполого размножения.
Taenia, Taeniurkynchus, Dipylidiurn и др. Далее, мы находим в данной
категории многих нематод, как. например, Protoslrongylus, Muellerius —
представители Spirurala, Dracunculus inedinensis. Wuchereria ban
crojti в др.
К этой же группе относятся почти все скребни и .многие паразити
ческне раки как средн Copepoda, так и среди Isopoda. Так, у Lernaeocera
branehiulis живо!ное выходит из яйца на стадии шестиногой личинки —
науплнуса. Науилнус достигает первой копеподитной стадии и садится
на жабры рыб из группы камбаловых, прикрепляясь при помощи секре
та лобнпй железы. Затем паразит теряет подвижность, испытывает силь-
ный регресс плавательных торакальных ножек и переходит в стадию
«куколки». Далее «куколка» линяет, к животному возвращай тся подвиж
пость, и оно оставляет хозяина на стадии, отвечающей взрослому состоя
нню свободно живущих Copepoda. Паразиты копулируют, находясь на
жабрах камбаловых, причем развитие самца на этом заканчивается,
самка же отыскивает второго хозяина нз семейства тресковых, прикре-
IW
иляется к его жабрам, врастает головным концом глубоко в ткани рыбы
и подвергается глубокому регрессивному метаморфозу. Ножки паразита
редуцируются, половой сегмент брюшка чрезвычайно разрастается и ста-
новится червееобразным, так что он делается совершенно непохожим на
ракообразное. В таком состоянии самка откладывает два длинных яйце
вых шнура, приклеивая их к основанию своего брюшка. Равноногие Epi
carida паразитируют в одной из личиночных фаз на пелагических Cope-
poda. позднее же переходят на других ракообразных, где и приступают
к половому размножению.
Графическое изображение такого цикла принимает следующую
форму:
К этому же третьему типу можно условно отнести и своеобразный
и упрошенный цикл развития Trichinella spiralis. Одна и та же особь по-
звоночного хозяина играет для трихинеллы роль и окончательного, и про-
межуточного хозяина. При заражении животное становится сначала
окончательным хозяином, когда в его кишечнике образуются половозре
лые трихинеллы и самки отрождают личинок Затем эти личинки разно
сятся по телу и проникают в поперечнополосатые мышцы, где инкапсу-
лируются, тогда хозяин становится промежуточным. В данном цикле в
результате полового размножения происходит как бы внутренняя дис-
персия, ведущая не к заражению нового хозяина, а к накоплению личи
нок в тканях хозяина. При этом наблюдается своеобразная эндогенная
аггломерация совершенно иного происхождения, чем у паразитов с бос
полым размножением. Заражение нового хозяина происходит при по
едании им животного, в .мускулатуре которого имеются личинки трихи
неллы.
Схема такого цикла может быть графически изображена так
В. С эндогенной а г г л о м е р а ц и е н. Этот тип цикла иаблю
дается, например, в семействе Trypanosomidae (рис 144 и 145). Громад
ное большинство трипанозом передается посредством инокуляции от кро
Bocjcyiuero беспозвоночного позвоночному во время акта сосания крови
и таким же образом попадает из крови позвоночного в кишечник крово-
соса. Новым моментом в цикле трипанозом является то обстоятельство,
что как в беспозвоночном, так и в позвоночном хозяине имеет место бес-
полое размножение трипанозом, т е. имеются в двух местах цикла фазы
эндогенной аггломсрацпп. Напротив, у всех Metazoa, относящихся к
третьему типу, эндогенная аггломерация нс наблюдается вовсе.
Аналогичный тип цикла наблюдается также и у представителей пн
роплазмид (Piroplasniidae)—паразитов крови различных млекопитаю-
щих, передающихся посредством инокуляции через пкеодовых клешей.
191
Рис. 141. Диаграмма развития Trypano-
soma gatnbieixe в кропи человека и в
кишке мухи цепе Glnssina palpalis. (По
Веньону.)
1,2 — обыкновенные и делящиеся формы трипя-
нозим п крови челонек*; J — трипвюэомы попада-
ют в хосогок мухи ия тоботка в кишку (Т|, из
кишки в слюнные железы мухи (о); о — яличные
трипзплзомы. нопллюдие из кишки в слюнные
железы через оепонание хоботкз; 7- критнлилль-
пые формы, получающиеся из длинных и слюн-
ных железах; 8 — мстшиклические формы трнна-
нозпм. получаю неся в слюнных железах из
критнли.1ЛЫ|'ых. Эти формы вызывают инфекцию,
попадая нз слюнных желез в хоСоточ н оттуда
при укусе в кровь человека.
Рис. I45. Диаграмма развития Т<ypanosoma
lewisi в крови крысы и в кишечнике крысиной
блохи. (По Веньону.)
/ — трпппцозомы в крови Крысы; 2 — триплкаюмы п
9!1нтели.|.11>1Юй клетке ккшечгнжл блохи: 3 одни
трип тиолом, прикрспимппич'я к стенке лилией кишки;
ЯВ1МЮЦИ1 крпгиформы в мстлииклпчсскую;
4 — свободные мета шили четкие трнпзпоюмы из з.!.пней
кишки блохи, вызынмопше инфекцию крыс; 5 —трипзнозо-
мы и примой кишке; 6 - трппаиозоми н средней кишке
блохи.
В эритроцитах позвоночного
юзяина пироплазмы размно-
жаются бесполым путем и бы-
стро увеличиваются в количе-
стве. Попадая в организм кле-
ща при сосании крови послед-
ним, паразиты продолжают
размножаться бесполым путем
(деление надвое или множе-
ственным делением), что при-
водит к накоплению паразита
в теле клеща-переносчика и
далее при трансовариальной
передаче (т. е. через яйцевые
клетки)—в новом поколении
клещей. В данном случае дис-
персии паразитов во внешней
среде нс происходит, а эндоген-
ная аггломерания паразита в
хозяине служит одновременно
и для их распространения.
Графически этот тип цикла
изображается так:
Некоторая вариация этого
цикла наблюдается у Trypano-
soma lewisi, Т. cruzi и Т. те-
lophagium- В этих случаях
имеет место дисперсия мета-
инк тческих трипанозом во
внешней среде, которая, одна-
ко, может быть не обязатель-
ной. Позвоночный хозяин зара-
жается либо поедая экскре-
менты беспозвоночного, в кото-
рых имеются метациклические
трипанозомы, либо путем по-
едания самого беспозвоночного
хозяина (так называемый кдн-
тамвнативнын путь зараже-
ния).
Поразить! без чередования
поколений с двукратной сме-
ной хозяев, без эндогенной аг-
гломерации. 1< данной катего-
рии относится сравнительно не-
большое количество отдельных
паразитов, принадлежащих
192
• лавпым образом к ленточным глистам и скребням. Так, один из обыч-
ных скребней Финского залива и Каспийского моря, Corynosoma slrumo-
<ит, во взрослом состоянии живет в кишечнике тюленя. Яйца Согупо-
sorna проглатываются рачьим-бокоплавом Pontoporcia ap'inis, после чего
молоденькие личинки прободают стенку кишки рачка и поселяются в по-
лости его тела. Когда зараженные рачки проглатываются рыбой самых
разнообразных пород (миноги, сельди, налимы, бычки, угри и некоторые
другие),'несколько подросшие личинки скребня опять-таки прободают
кишку и инкапсулируются в по пости тела рыбы Наконец, поедание ин-
фицированной рыбы ведет к заражению тюленя (рнс. 146).
Рис. 146. Жизненный цикл скрг-бня Corynosmna strumosum.
(Орш пиал.)
t - n-рослыЗ скребень на кишечник i тн л«их. г — яйца скребня, выходя цис
нз кишечникр тюленя н вод} и поедаемые рачкпм р<>„» рогеш (3). в полость
которого проник-ет личинкг- (б). ьы- >дн ипя из нйн». Р..чок поедается |ыбой
(5>. причем личинка скребни, пмешпаисн и рлчке. проходит Через стенку
кишки в полость те п рыбы и гам инкапсулируется (в). Инвазии тюленя
происхишт при поедании им зараженпол put ы.
Подобный же цикл развития подозревается и для другого скребня,
Pomphorhynchus laevis. причем первым хозяином служит бокоплав Gam-
marus, вторым — мелкие рыбы (линь, гольян, пескарь), окончательного
же развития паразит достигает в кишечнике хищных рыб. Впрочем, на-
личие второго обязательного хозяина для Pomphorhynchus не вполне до-
казано, и, быть может, единственным промежуточным хозяином его слу-
жит бокоплав.
Также и в отношении цикла развития Corynosoma высказывается
мнение (Петроченко. 195о; Рыжиков. 1954;. что рыба — необязательный
хозяин, и тюлень может заражаться, непосредственно поедая бокопла-
вов. Другими словами, возможно, что здесь пет второго промежуточного
хозяина, а рыба является резервуарным хозяином (см. гл. X). Вопрос
этот, однако, требует специального экспериментального исследования.
Примеры данного типа также наблюдаются у представителей Pseu-
dophyllidea: широкий лентец (Diphyllobothr,um latum) с vro последова-
тельным паразитированием в циклопах, в рыбе и в окончательном хо-
зяине— человеке, того же типа и паразитизм ремнеца (Ligula), полово-
зрелые стадии которого живут в кишечнике водяной птицы, главным
13 В. Л. Догель
193
образом чаек. Яйца ремнецов попадают в воду, из них вылупляются
корацидпи, которые проглатываются рачками рода Diaptomus. Здесь они
превращаются в процеркоидов. Рачки поедаются одним из видов карпо-
вых рыб, в которых личинки Ligula проходят через стенку кишки в по-
лость тела и там дают громадных (до 80 с.«) плероцеркоидов; наконец,
птицы заражаются, поедая содержащую плероцеркоидов рыб^. Такой
же. в общем, цикл развития у ряда других цестод: Schislocephalus и
Triaenophorus.
Среди Cyclophyllidea развитие с двумя промежуточными хозяевами
наблюдается у Mesucestoides linceaius' (сем. Mesocestoididae)—пара-
зита собак, кошек, лисиц, соболей и др> гих плотоядных. Членики пара-
зита с экскрементами выбрасываются во внешнюю среду и из них осво
бождаются яйца, которые заглатываются первым промежуточным хозяи-
ном. Таковыми являются панцирные клещи Oribatei. В fix теле онко
сфера превращается в стадию цистнцеркоида (Солдатова, 19-14). Зара-
женные клещи вместе с пищей заглатываются вторым промежуточным
хозяином (грызуны, ящерицы, птицы), в полости тела которых цисти
церкоид становится тетратцридшгм, си' ыю вытянутой червеобразной ли-
чинкой с головкой на одном конце. Плотоядные животные заражаются
при поедании этих хозяев. Тетратпридин были найдены также и в по-
лости тела собак, кошек, соболей и куниц.Следовательно, они становн
лись вторым промежуточным хозяином, если случайно с пищей съедали
клещей, зараженных цистнцеркоидами.
Схематическое изображение циклов этого типа имеет следующий
вид:
Паразиты с чередованием поколений,
эндогенной аггяомерацией и сменой хозяев
А. Однократная смена хозяев Ср* ди многоклеточных
этот тип встречается у тех цестод (Mulliceps, Echinococcus), у которых
пузырчатая стадия обнаруживает процесс бесполого размножения по-
средством выпочковываиия многочисленных головок.
Дисперсия у этих паразитов происходит только в окончательном хо-
зяине, а в промежуточном наблюдается эндогенная аггломерация, значи-
тельно увеличивающая возможность попадания паразита в окончатель-
ного хозяина при поетаиип последним промежуточного Схема цикла
приобретает здесь следующий вид:
-><? Б
Сходный цикл наблюдается и у ряда дигенетических трематод (Fas-
ciola, Schistosoma и др j с той разницей, что в промежуточном хозяине
они размножаются, вероятно, не бесполым путем, а партеногенетнчески
194
В результате этого процесса происходит эндогенная аггломерация пара
зита, обеспечивающая в большинстве случаев дисперсию церкарий во
внешней среде. Размножение партеногенетических поколений (партенит)
в моллюсках бывает иногда весьма интенсивным: \ Cotylophoron coiy
lophoruni образуется около 200 церкарий, у Parorchis acanthus — Солее
тысячи, у Paragonimus— от 10 до 20 тысяч, у фасциолид— до 25 тысяч
и из одного мирацидия Schistosoma mansoni может образоваться до
Рис. 147 Сгони жизненного цикла Fasciola hepatica
Д — половозрелая особь; Н — яйцо; С" — у иранидий; D — молодая снороцн
ста;/:' — взрослая спороциста; /•'—редия с развивающимися церкарнями;
G — церкарня;//—нпцисгированиая метацеркарня (адолескария). (/1 — по
• Чендлеру. В— //—по Бильдннгу.)
200 тысяч церкарий (Faust and Hoffman, 1934). Fasciola hepaticu
(рис 147) в промежуточном хозяине (Galba truncatula) образует обычш
два партеногенетических поколения (спороцисты и редни), а в некото
рых случаях в редиях может формироваться еще одно поколение редин
и затем уже образуются церкарин. Последние покидают промежуточ
ного хозяина, подплывают к берегу или поверхности водоема и ннцисти
руютдя на прибрежной траве или под поверхностным слоем воды. Пас\
щнйся у берегов водоема скот, съедает цисты печеночной двуустки вме
сте с травой и таким образом заражается, У трех видов Schistosoma, в<
взрослом состоянии живущих в крупных венозных сосудах брюшной по
лостн человека, промежуточным хозяином служат моллюски Planorbis
13’ 195
(подроды Biotnnhalaria, Ausfralorbis), Bulinu-., Phtsopsis, Oncomelania
Образовавшиеся в них церкарии с длинным вилообразным хвостом по-
кидают моллюска н активно проникают в тело ку шюшихся пли берущих
вод} людей, вбуравливаясь в кожные покровы Схема такого цикла со-
сальщиков выглядит так:
Циклы многих простейших, а именно ряда Eimeridea (Aygregata
(рнс. 148), Schellackia и др.), а также Haemosporidia в принципе при-
надлежат к этому же четвертому типу, но имеют некоторые особенности
У Schellackia половое и бесполое размножение имеет место в одном хо-
зяине, а именно в позвоночном, тогда как беспозвоночное содержит в
С(бе лишь покоящуюся стадию паразита и является простым перенос-
чиком. Схема такого цикла имеет вид:
ос впД
♦
Бесполое размножение приводит к эндогенной аггломсрацнн, а в
результате полового процесса и последующей спорогонии в крови позво-
ночного хозяина накапливаются спорозоиты, имеющие значение для
распространения паразита. Здесь происходит как бы внутренняя диспер-
сия паразита, обеспечивающая попадание его в переносчика. Цикл
Aggregate отличается от ScheUackia тем, что половое размножение и
спорогония происходят в одном хозяине (Sepia), а бесполое в другом —
крабе (Portunus). Дисперсия ооцист происходит в первом хозяине, а
эндогенная аггломерация — во втором. Схема цикла вполне совпадает
с тем, что изображено выше для эхинококка
Для Haemosporidia характерно, что эндогенная аггломерация проис-
ходит в позвоночном (промежуточном) хозяине, так же как н в цикле
Echinococcus, Fasciola и Aggregate. В отличие от них у Haemosporidia
этот процесс носит более интенсивный характер. Бесполое размножение
путем шизогонии приводит к образованию огромного количества пара-
зитов в хозяине В окончательного хозяина Haemosporidia проникают
при кровососании, и таким же п}тем спорозоиты возвращаются обратно
в промежзточного позвоночного хозяина. В окончательном хозяине (кро-
вососущие двукрылые) происходит половой процесс и спорогония, при-
водящая к накоплению спорозоитов в слюнных железах насекомого. Это
обеспечивает передач} паразита восприимчивому хозяину. Особенности
цикла Haemosporidia выражены в следующей схеме:
1%
Рис. 14ч. Жизненный цикл Aggregate cberthi
Л — недиффсгснилровапнля стадия паразита, развивающаяся нз мсро
зонта; 13 — />-рост мпкроглчетош'та и развитие микрон мст; Е—G —
развитие макрс!аметы и оплодотворение; Н—зигота, н тало первою
( едуьционпою) деления ядра з*ноты; /—анафаза редукционного деле
ния ядра; J К — телепне ядер в сио| оцисте и образование споробластов;
L — спора с тремя спорозоиты и н с таточным телом Л1 — в *хсд . ло-
розоитов в кишечнике краба; N-Q — ро^т шизонта н шизогония; Н —
жрозоит, проникающим в Sepia. ^По Добеллю-)
14?
Б. Двукратная смена хозяев. Этот наиболее сложный тип
развития характеризует собой большинство трематод. Приведем прежде
всего несколько примеров циклов этого типа. Дйууст.ка Paragonimus
wcslertnanni (рис. 119) живет в легких кошки, собаки, свиньи, человека
Яйца выводятся или прямо при отхаркивании с мокротой наружу или
попадают из гортани в глотку и пищевод и поступают наружу с экскре
Рнс. 14П. Стадии жизненного цикла Paragonimus westermannl
А—половозрелая особь; Л? — зрелая редпя (второго поколения);
С — церкария. вышедшая из моллюска; D — метанеркария из
краба (оболочка не нарисована); Е— G— крабы, служащие про-
межуточными хозяевами (£—Potamon dehaanr, F- Stsarma
t chaani; G — Eriocheir japonirus}’, fl — моллюск Pomiiiopsis
laphlaria — первый промежуточный хозяин Paragonimus ki-lli-
cotli (Л, H - по Чандлеру, В. С* — но Эмилю, D — по Чену,
£ — 6 — iio bpjMiny.)
ментами. В яйцах имеется мнрацпдий, который вылупляется при попа
цапни яиц в воду и вбуравлпвается в первого промежуточного хозяина —
улитку Melania. В печени моллюска мнрацпдий превращается в бесфор-
менный мешок — спороцисту, — наполненный крупными клетками — за-
родышевыми шарами. Спороцисты представляют собой партеногенетиче-
скую самку, т. с. особое поколение в цикле двуустки, а зародышевые
шары — развивающиеся партеногенетические яйца. Каждый зародыше
вын шар путем развития дает одну особь следующего поколения, а
(98
именно редию, которая тоже является партеногенетической самкой с
зародышевыми шарами внутри.
Редин остаются в печени моллюска: от спороцист они отличаются
наличием кишечника и особого «родильного отверстия». В редиях из
зародышевых шаров образуются или снова редин или же прямо заро-
дыши другой формы — церкарин, которые представляют собой, собствен-
но говоря, молодую двуустку, но с провизорным, личиночным придат-
ком, хвостом. Церкарин покидают первого промежуточного хозяина,
г. е. моллюска, свободно плавают в воде и проникают во второго проме-
жуточного хозяина, которым являются раки (Astacus japonicus) и прес-
новодные крабы, например Eriocheir japonicus и др. Церкарин пробурав-
ливают имеющимся у них стилетом покровы краба и инцистпруются в
его мышцах, печени и т. д., образуя стадию метацеркарии. При поедании
зараженных крабов метацеркарии пронизывают стенку кишки оконча-
тельного хозяина и выходят в брюшную полость, а оттуда активно до-
бираются до легких, где и начинают питаться, а затем размножаются
половым путем
Таким образом, в выбранном нами случае наиболее сложного раз-
вития имеется чередование, как и у многих других двуусток, по меньшей
мере трех различных поколении. Типичное половое гермафродитное поко-
ление, или, иными словами, поколение взрослых двуусток, находящееся
в окончательном хозяине, сменяется двумя поколениями партеногенети-
ческих самок (спороцисты и редин); последнее поколение этих самок
дает начало поколению личинок двуусток, т. с. церкариям, которые пона-
дают в окончатезьного хозяина через организм второго промежуточного
хозяина (краба) В цикле имеется эндогенная аггломерация, протекаю
шая в промежуточном хозяине (моллюске), и двукратная дисперсия —
мнрацидиев и церкарий, так что цикл принимает следующий вид:
По типу Paragonimus развиваются очень многие другие трематоды
(Opisthorchidae, Plagiorchidae, Heteropliy idae, Hemiuridae, Microphal-
lidae и пр.). Например, у Gorgoderina vitelliloba из мочевого пузыря ля-
гушки партеногенетические поколения паразитируют в двустворчатых
моллюсках шаровках (Sphaerium), а церкарий инцистпруются в теле
личинок стрекоз.
Вторым промежуточным хозяином двуусток могут, кроме членисто-
ногих (насекомые и ракообразные), служить другие беспозвоночные
(пиявки, полихеты моллюски) или же позвоночные. Так, кошачья дву-
устка (Opislhorchis jclincus, рис 150) во взрослом состоянии живет в
печени кошки, собаки, человека; первым промежуточным хозяином эта
двуустка имеет улитку Bythinia. а вторым — различных карповых рыб
(Rutilus г util us. Leuciscus leuciscus).
Следовательно, у трематод наблюдается один тип цикла, в котором
чередуются гермафродитное поколение с поколениями партеногенетиче-
ских самок и эндогенная аггломерация происходит в промежуточном
хозяине. У одних представителей может быть одни промежуточный хо-
зяин, у других — два (а у Alaria muslella. вероятно, и три), что по суще-
ству представляет лишь вариацию одного типа цикла. У некоторых со-
199
сальшиков из сем. Lchinostoinidae и др. цикл с однократной сменой хо
зясв может рассматриваться как результат упрощения цикла с двукрат
нов сменой хозяев, из которого выпал второй промежуточный хозяин
В других случаях (Fasciola п др.), однако, наличие одного промежуточ-
ного хозяина является, вероятно, первичным в цикле трематод, у третьих
же второй промежуточный хозяин включился в цикл дополнительно, как
важное приспособление, способствующее достижению инвазионной сти
диен паразита окончательного хозяина.
Рис. 150. Ст;.Tin жизненного цкь.ы Oplsthorr^is febn^us
A iin.inBup лая особь;/? мнрацндий в яйнсн/ й 16 -то-
чке; С — ciiop'»iin/та с ра )пнпаю111нмнся ре.тиями; D
.молотая рент с рл видающимися церклриямн; Г—пер-
карин, I - тело церкаре и с брюшной стороны; 6 — моло-
дая метаиеркарна и:« рыбы; Н— .моллюск Bylht'iia leachl
с редня.мн внутри тела. (Д по < тилел н Хасаль, В— (i —
ио Фогелю, Н — по Брумпту.)
Рассмотренные выше два наиболее типичных цикла со сменой
одного (Fasciola) и друх хозяев (Paragonimus) могут испытывал! у раз
ных сосальщиков многочисленные изменения. Они касаются прежде всего
выпадения отдельных стадий цикла, притекающих в первом промежуточ
ном хозяине, хотя вообще без первого промежуточного хозяина трема-
тоды обойтись не могут. Пример выпадения спороцист мы встречаем у
Mctaqorumus yokaga&'at (рис. 151), паразитирующих в кишечнике чело
200
века, кошки, собаки и пеликана на Дальнем Востоке и в Японии. Их пер-
вым промежуточным хозяином являются моллюски Melania. Btanlordia
и др. В моллюске из мирацидия формипуется прямо редия, которая сна-
чала дает тоже редни, а в последних формируются хвостатые церкарии.
Церкарии покидают моллюсков и внедряются в чешую или в толщу по-
кровов жабр и плавников целого ряда рыб (Coregnnus ussuriensis
Рис. 151. Жизненный цикл Metagonimus vokagawal
А - взросл.!» двуустка, В — яйпо Л/ yokaganai с ип-
раиндием внутри; С рс^ия со еле lyioimiM поколением
реднй внутри; D — церкария с гребным хвост ом и па-
рой глазных пятен: Е цисты с метацеркариями в
чешуе р.абы. (Чо Павловскому.)
Plecoglossus altivelis и др.), которые служат червю вторым промежуточ
ным хозяином. Человек заражается Metagonimus, поедая недостаточно
прожаренную или проваренную рыбу.
Пример выпадения реднй дает Dicrocoelium dendriticum. Яйца с ми-
рацчдием внутри заглатываются наземными моллюсками (роды Helicella
Zebrina и Др.), в которых мирациднй превращается в спороцисту. В по-
следней образуются дочерние спороцисты, а в них уже—церкарии
Последние выходят в мантипную полость, где обволакиваются с лзыо и
образуют «сборные цисты», попадающие па траву или почву. ЛАуравьи
(Fortrica [usca) поедают их, затаскивая в Муравейник, и в полости тела
201
муравья, как второго промежуточного хозяина, развиваются метацер-
карии (рис. 152).
Выпадение редни наблюдается также у Leucochloridiiim paradoxum
(рис. 153), в половозрелом состоянии живущей в кишечнике певчих птиц
Их ягца выходят вместе с экскрементами птицы наружу и падают на
листву. Там их вместе с листьями поедают улитки Succinea. В кишечнике
Рис. 152. Сталин жизненною цикла Dicrocoelium lanceatum (D. d^ndrlticum)
А- половозрелая особь; В — мнрацндий в оболочке яйца; С — церкарня;
D - слизистые комочки из цист метацеркарии; Е - инцистировапная ме-
тацеркария; F — восьмидневный неполовозрелый червь из окончательного
хозяина; G - муравей Formica ru/iburbus. ннватрлвапный метаиерклрия-
ми; Н—К—моллюски — промежуточные хозяева (Н - Chondrtila triduns;
I — Н‘ ticdla cr‘'nimargn\ Д' ~H. dirbi-ntina'i, (Л. F — по Heiiraycy. В —
по Бнльтишу, С — по Фогелю. D, Е. G. Н К — по Сваджапу.)
улитки из яйца Leucochioridium выходит мирацидий, который проникает
через стенку кишки и забирается в печень. Там он превращается в спо-
роцисту, которая постепенно чрезвычайно разрастается и разветвляется,
заходя своими отростками в самые различные уголки тела моллюска.
Внутри разветвлении спороцисты образуются многочисленные, вполне
сформированные молодые двуустки, окруженные каждая прозрачной
толстой оболочкой. Окончательный хозяин заражается этими молодыми
двуустками, откусывая у улитки щупальца, в которые заходят Ветви спо-
роцисты Как видно из описания, Leucochioridium может служить иллю-
страцией редукции нс только стадии редий, но н исчезновения всех сво-
бодно живущих стадий развития, т. е. свободного мнрацидия и евпбод-
202
ной перкарин. Все эти стадии проходятся паразитом в теле первого про
межуточного хозяина, тогда как второй промежуточный хозяин вына
даст, будучи заменен первым. Получается сильное укорачивание и упро-
щение цикла.
Рис. 153 Развитие Leucochlori Чит ((Arogonimus)
А — моллюск Sucriwa г двумя выростами спороцисты сосаль-
щика в сильно вздувшихся голевных щупальцах;& — ветви-
стая спороцис га Leticochloridium paradoxum. один из выростов
которой сильно разросся и приобрел сходВво с личинкой
насекомого; С — о ща из молодых двуусток Leycochloridium
paradoxum, которыми переполнены выросты ciiopon.ic >ы;
Г)—взрослая лвуустка нз чдоэки певчих птиц. (Из Догеля.)
1 — ротон <я r.pncoi'K.i « - глотка: 3 - протоне<|>рилнальные каналы: 4 — лве
веши киш и; 5 — Сркшш га прксоскл; 6— семенники; 7 — яичных;£ — мочевое
огверсгис; 9 - яичник; 10 — лазферов къ<1эл; 11 — семенник.
Очень полную картину последовательного сокращения цикла у тре-
матод семейства Microphalli Jae проследила Белопольская (1962). Исход-
ным для этого семейства является цикл с дв' мя промежуточными хозяе-
вами (моллюски и ракообразные). Вторичным следует считать выпаде-
ние второго промежуточного хозяина и последовательные стадии упро-
щения в строении церкарнй (редукция хвоста и стилета). Последнее
203
связано с тем, что церкарии тертлит активно подвижные функции и инци
стируются в первом промежуточном хозяине, который поедается окон
нательным хозяином (птицами).
Еще большее упрощение цикла наблюдается у Ctjclocoelutn micro-
stomum и Tracheophylus sisowi из дыхательной трубки и воздушных меш-
ков водоплавающих птиц. В их яйцах развивается мнрацидий, содержа-
щий в себе вполне развитую редию (рис. 154). Мирацидий выходит в
Рис. 154. Стадии жизненного цикл, Cyclocoelum microstomuin
А — MHpamutiii с редней внутри; В—редия с церкариями;
С — взрослая особь из полости тела лысухи (РиПса аг га).
(По Гннецинской)
воду и активно внедряется в покровы моллюсков Limnea и Pianorbis.
Сам он не входит глубоко в ткани улитки, а только пробивает путь для
родни, которая и проникает внутрь тела хозяина. Там в родии форми-
руются церкарии, которые инцнстируются, не выходя из моллюска. Пти-
цы заражаются, поедая моллюсков с ннцпетированнымн метацеркариями
(Szidat, 1933; Гинецпиская, 1954а).
Наконец, в очень редких случаях у сосальщиков может встретиться
еще более прямое развитие. Один из таких примеров Иеген (Jegen, 1917)
видит в развитии Co'dyriclum faba (рис. 155). Этот сосальщик живет
парами в подкожных капсулах, располагающихся преимущественно по
соседству с отверстием клоаки некоторых птиц (воробей и др.). Птица,
ухаживая за своими перьями, вскрывает клювом капсулы и случайно
проглатывает яйца паразита. В экскрементах из яиц вылупляются
204
Рчс. 155. Collyrictum faha,
паразитирующий пот кожей
воробья. (По Скрябину, с
изменениями.)
1 — ротимя присоска: 2 - глотка;
J — кишка: 4 - жсаточники: 5 — семен-
ники 6 — метка; 7 - цнррус: 8 — пмле-
лнтсльное отверстие; у — яичник.
эмбрионы, лишенные ресничек и соответствующие мирацндням, которые
уже содержат в себе зачаток обоих членов будущей пары, и выходят
наружу с экскрементами. Выходящие из клоаки мирацидии попадают на
гнездовых птенцов и активно внедряются в перьевые мешочки, в том
числе и в области анального отверстия, там зачатки пары сосальщиков
созревают и кладут яйца. Если наблюдения Иегена правильные, а они
нуждаются в некотором подтверждении, то мы имеем у Collyriclum по-
следний возможный этап сокращения жизненного цикла сосальщиков.
Большой интерес представляет цикл
семейства Aspidogastridae, обладающих
в отличие от прочих трематод вместо
простой брюшной присоски сложным
брюшным щитом, разделенным на боль-
шое количество мелких присосок. Неко-
торые представители этого семейства
имеют, по-видимому, простое развитие с
одним хозяином. Их только что вылупив-
шаяся из яйца личинка уже походит на
взрослую стадию, отличаясь от нее лишь
наличием двух головных желез, содей-
ствующих проникновению в ткани хозяи-
на. В этом цикле нет каких-либо призна-
ков его вторичного характера, так что
простота цикла в данном случае, един-
ственном среди трематод, по-видимому,
является примитивной.
Мы нарочно остановились подробнее
на циклах сосальщиков, так как эта груп-
па отлично демонстрирует, как типичный
цикл группы может подвергаться необычайно разнообразным измене-
ниям.
При толковании цикла развития трематод мы придерживаемся ши-
роко распространенной точки зрения, что процесс образования дочерних
спороцист или реднй внутри спороцисты и церкарий внутри реднй пред-
ставляет собой партеногенетическое размножение, и признаем споро-
цисты и редин за поколения партеногенетических самок, образующих
яйца, претерпевающие процесс созревания. Сторонники этой точки зре-
ния называют поэтому спороцисты и редин партенитами.
По мнению Вудхеда (Woodhead, 1931, 1954, 1957). исследовавшего
представителей Bucephalidae, Paragonimus, Schistosomatium и некото-
рых других трематод, в материнских и дочерних спороцистах образуются
мужские и женские половые клетки; при нх образовании происходит
редукционное деление и наблюдается процесс оплодотворения. Оплодо-
творенные яйца, например у Schistosomatium douthitti. дают начало
эмбрионам, которые развиваются в церкарий. Эти наблюдения не под-
тверждаются другими исследователями (Ciordia. 1956 и др.), и поэтому
вопрос о наличии полового процесса пока следует оставить открытым.
Существует еще одно мнение (Brooks, 1930; Cort et al., 1954; Guilford,
1958, ii др.), что размножение спороцист и реднй происходит путем поли-
эмбрионии. Заключенные в мирацидии (материнская спороциста) и рано
обосабливающиеся при дроблении половые клетки делятся мнтотнческн
и дают начало последующей стадии развития. В спороцистах и реднях
половые клетки, образовавшиеся непосредственно от предыдущих, также
размножаются без процесса созревания и без оплодотворения, давая на'
чало зародышам церкарин, в которых формируются половые зачатки
205
Чаблюлаемая у раздельнополых кровяных двуусток однополость
всех церкарнй, происходящих при экспериментальном заражении из
одного яйца (миряцидия), находит наиболее вероятное объяснение в
процессе полиэмбрионии.
Совершенно ясно, что для окончи тельного решения вопроса о сио
собе размножения ди генетических трематод в моллюсках необходимы
дальнейшие и весьма тщательные цитологические исследования.
6. ОБЩИЕ ЗАМЕЧАНИЯ О ЖИЗНЕННЫХ ЦИКЛАХ
Подводя итоги сказанному о жизненных циклах паразитов, прихо-
дится еще раз отметить необычайное их многообразие, которому трудно
найти параллель у свободно живущих организмов. Сильная затруднен-
ность распространения паразитов и передачи их от одного хозяина к дру-
гому вызвала и самого различного рода приспособления для преодоле-
ния трудностей развития. Мы могли привести лишь главные типы жиз-
ненных циклов, но есть не мало циклов, которые нс вошли ни в один из
вышеуказанных четырех типов. Такие циклы развития встречаются
главным образом среди паразитов с непосредственной передачей зара-
жения от одной особи хозяина к другой и представляют собой резуль
таты упрощения циклов более сложных
Очень большая простота цикла имеет место, например, у Trypano-
soma equiperdum — возбудительницы случной болезни лошадей Т. equi-
perdum размножается всегда лишь при помощи простого и дающего по-
добных материнскому организму особен деления надвое. У нее пет ни-
каких специальных мстациклнческих форм, появляющихся у дрхгих три
панозом при переходе их в беспозвоночного промежуточного хозяина.
Передача совершается непосредственно от одной лошади к другой через
слизистые оболочки половых органов при случке. Один и тот же процесс
деления надвое в подвижном состоянии у этого паразита служит как для
эндогенной аггломерацни его. так и для дисперсии. Нет никакого сомне-
ния в том, чю этот цикл выработался путем упрощения из циклов, со
державших в себе промежуточного хозяина кровососа.
При наличии большего разнообразия жизненных циклов у парази
тов, нередко довольно трудно бывает решить вопрос, которые из них
являются более п р и м и т и в п ы м н, которые произвол и ы м и.
. Нам думается, однако, что в общих чертах можно установить следую-
щую закономерность. Жизненные циклы простейших, с одной стороны, и
многоклеточных, с другой — испытали разную эволюцию. Циклы многих
паразитических простейших (Sporozoa) в основе своей имеют цикл с че-
редованием поко тений, и если такового нет в рассматриваемом частном
случае, то этот случаи (например, цикл i регарип) представляет собой
результат упрощения. Лишь у паразитических амеб, так же как у сво-
бодно живущих, размножающихся только агамным нутом, отсутствие че-
редования поколений можно рассматривать как первичное явление.
Напротив, у многоклеточных исходной точкой циклов различных па
разитов .1в шется простое половое размножение без всякого чередования
поколений; последнее возникает лишь впоследствии, как нопезная адап-
тация в жизненном цикле перешедших к паразитическому образу жизни
животных.
Такой взгляд поддерживается рассмотрением жизненных циклов сво-
бодно живущих простейших и многоклеточных. Свободно живущие клас-
сы простейших обладают в своем жизненном цикле так или иначе выра-
женным чередованием поколений. Иногда это явление носит типичный
характер правильной смены полового и бесполого размножения, как \
20Г
корненожек. В других случаях в длинную серию бесполых поколении
периодически вставляется половое размножение: конъюгация, или авто-
гамия у инфузорий, образование зооспор — гамет — у лхчевиков, обра-
зование половых колоний у Volvocidae. Таким образом, для большин-
ства паразитических простейших чередование поколений есть такое яв-
ление, которое уже было свойственно их свободно живущим предкам и
сохраняется ими при переходе к паразитизму.
Рис. 156, Прогеиетнческие метанеркарин
А — метацеркарня Dinnrus noiatus из к реветкн Cera/aspis1 В — лрогенетн -
ческая метацеркарня Coitocoecum test:obliq urn in Potitogamtnarus bosnicus;
C — прогенстичсская метацеркарня из Syndesmya. (Л - по Дольфусу,
И — ио Вишневскому, С—по Латышеву.)
I — ( отсоая присоска; 2 - fipioiHHai присоска 3 маткэ, пзГига« яйцами, 4 — оылсли-
тепьный канал; 5 — кишечник; 6 - Гформир01чи1иие яйц 1.
У многоклеточных дело обстоит иначе. Приспособление к парази-
тизму возникает впервые среди червей (мы отбрасываем в данном случае
те два десятка примеров паразитизма, которые известны для Coelente-
rata), т. е. в пределах тон группы, где у свободно жив1 тих форм чере-
дование поколений или даже вообще наличие бесполого размножения
составляет большую редкость. Бесполое размножение известно лишь у
очень немногих RhaLdococla, Triclada и у очень небольшого количества
аннелид, да и то оно лишено правильной периодичности. Следовательно,
у многоклеточных все изменения никла в сторону выработки чср1дова-
гия поколении являются новоприобретенпем.
В связи с изложением вопроса о жизненных циклах уместно будет
затронуть еще некоторые изменения этих циклов, имеющие биологиче-
ский интерес.
Про генез и п р 0 г е и с т и ч е с к и е формы. В последнее вре
мя списан ряд случаев, когда паразит приступает к половому размно-
жению и к продукции янц ранее, чем ему полагалось бы при нормальном
коде жизненного цикла, а именно: оставаясь в своем промежуточном хо-
зяине и еще не достигнув окончательного, в котором нормально только
207
ii начинаются половые процессы Это явление и получило название про-
генеза. Целый ряд лрогенстических форм описан главным образом Доль-
фусом (Dcllfus, 1927а, б, 1934) средн сосальщиков (рис. 156). Большей
частью дело идет о метацеркариях трематод, которые достигают половой
зрелости в различных раках, служащих для них вторым промежуточным
хозяином. Таковы, например, метацеркарии в Gammarus pulex, взрослой
стадией которых является, no-видимому, Allocreadium transversals из
кишечника рыб. У Coilocoecum tesliobliquum, паразитир) ющей в поло-
возрелом состоянии в кишечнике форели, метацеркарии начинают проду-
цировать яйца, еще находясь в ннцнетированном состоянии в полости
тела рака Ponlogatntnurus bosntcus (Wisniewski, 1932—1933). Обыкно-
венно в таких случаях в метацеркарии образуется л.лшь очень неболь-
шое число яиц, но Дольфус (1927а) описал метацеркарию Dinaras по-
tatus (Hemiuridae), найденную однажды в креветке Cerataspis, у кото
рой (метацеркарии) вся матка была заполнена тысячами вполне сфор-
мированных яиц. Взрослый Dinurus notatus встречается в кишечнике
рыбы Coryphaena, питающейся пелагическими животными, в том числе
креветками.
Прогеисз обнаруживает и метацеркария Lecithodenrium chtlostomum
(из кишечника летучих мышей), покоящаяся в полости тела ручейников
(Phryganea grandis). Наконец, известно несколько случаев прогенсза у
метацеркарии, вторым промежуточным хозяином для которых служат
моллюски. Так, Серкова н Быховский (1940) нашли прогенетическую
метацеркарию в улитке Byihinia tentaculala; во взрослом состоянии эта
церкарня, принадлежащая Asymphylodora progenetica, живет в кишке
карповых рыб. Латышева (1939) обнаружила прогенетическую метацер-
карию в пластинчатожаберном моллюске Syndesmya из Азовского моря.
Особенно интересный пример представляет сосальщик Ratzia parva.
Взрослый сосальщик живет в кишечнике змеи Zamenis, метацеркарии
же его инцистпруются в лягушке (Rana esculenta). Эти метацеркарии
откладывают яйца, внутри которых образуются вполне сформированные
мирацидии. Заразить этими мирацидиими моллюсков, однако, не удает-
ся. На основании изучения Ratzia Дольфус (1927 6) высказал гипотезу:
не имеется ли у некоторых сосальщиков, помимо нормального жизнен-
ного цикла, циклов укороченных, из которых выпадает взрослая стадия.
Подобное изменение цикла можно представить себе для Ratzia следую-
щим образом:
мирацидии
спороциста £Т
ретия о =
церкарня 2
меганеркярня *
взрослый червь
— свободная стадия
— в моллюске
— ti моллюске
— свободная стадия
— в лягушке
— в змее
Если мы допустим, что укорачивание цикла войдет в норму, то из
цикла выпадут и взрослое состояние червя и его окончательный хозяин.
Паразит примет неотеннческпн характер и станет дв\ ххозяйным. Раз-
личные примеры явления прогенсза мы находим в работе Буттпера
(Buttner, 1951).
Представление об укороченных циклах и об их значении в толкова-
нии некоторых загадочных форм цестод возникло впервые у Яннцкого
(Janicki, 1930), который применил его к толкованию Amphilina Joliacea
(рнс. 157). В отличие от всех других ленточных глистов (кроме Archi-
getes), Amphilina живет не в кишечнике, а в полости тела хозяина
(осетр, севрюга, белуга, стерлядь). Далее, тело Amphilina не сегменти-
208
ровано н содержит всего один половой аппарат. Наконец, вместо двух
промежуточных хозяев, имеющихся j родственной амфилине группы
Pseudopbyl lidea (широкий лентец и др.), этот вид обладает всего одним
(рачки Dikerogatntnariis, Gammarus platycheir, Corophiurn, Metamysis
strauchi).
Все эти обстоятельства заставили Яницкого предположить, что
Arriphilina является потомком какого-то типичного членистого ленточного
глиста и представляет собой неотсническую плероцеркоидную личинку,
Рис. 157. Цикл ра жития Amphilina foliacea в нзстиЯ|Цем и в прошлом.
(По Яинцкому.)
1 — «1цп с зародышем. оиоряжиипаемое и коду стерлядью черед се .п6доми113.11.Н|4к
поры; 2 - стлан» iipoiiepKniii.i к перпчы промежуточном xo«iiiine И. atnmurus. Af tamy-
sitY. 3 • neoreiiiviei-iia i сопрсченнап чле|юцсркои шин егздпн Amphilina н ппдчети тела
второго проме «уточного xo'oinir т. е. стерли-н; < — гииогсгпческан пэловозрелая
строки та н KKinviHiiKe мслоаопской рептилии.
приобревшую полную видовую самостоятельность. Яиицкпн предпола-
гает. чго в прежние времена современная половозрелая стадия Amphilina
была плероцеркоидом типичной членистой цестоды, которая жила в ки-
шечнике какого-то хищника, а осетровые были лишь вторым промежуточ-
ным хозяином цикла. Этомх о пи чают и печленпстый, цельный характер
амфилины и сс нахождение в полости тела хозяина — обычное месго-
чребыв-чше плероцеркоидов цестод. Впоследствии окончательные хозяева
амфилины вымерли, а плероцеркоилная стадия се приобрела способ-
ность размножаться половым способом.
Принимая во внимание случаи прогенеза у сосальщиков, можно
тумать, что предварительно плероцеркоиды Af'iphilina приобрели проге
нетический характер и начали продуцировать яйца еще во втором проме-
жуточном хозяине, т е. в осетровых.
Кто же мог быть окончательным хозяином Amphilina в прошедшие
времена, раз се промежуточные хозяева (осетровые рыбы) имеют такие
крупные размеры, а между тем заражршс окончательного хозяина, по
всей вероятности, должно было происходить при пожирании им промс-
|| В. А Догель
209
Рис. 158. Неотеиические цестоды
.1—Archigetes crypfpbolJiriits из I.unnyitrllus
hofnieisferi; /i — Caryophytlaeus taticeps. личинка
ил Tubifcx tubifex; C.—взрослый Caryophyllaejix
hitireps из кишки кариа.(/1 — no BitiiinciicKOxiy.
/Г и C— но Секутовичу.)
I голонка; J семенники. ,i желточннки; / совокупи.
Tv.iLiiNM npi .in; 5 млтка; 6 iiw.KJiiacTiiofi инчник; 7 xiwi-
< птпй при laroK.
ж\ точного? Яницкий высказывает для объяснения этого смелую и остро-
умную догадку. Окончательным хозяином Amphilina в незапамятные 14ч
мена были какие-то мезозойские крупные рептилии, которые в изобилен
населяли моря мезозойской эры. Современный никл амфилины предстаг
лист собой «осколок» более полного мезозойского цикла этого червя
В таком же смысле толкуют в настоящее время развитие напоми-
нающего по своемч строению процеркоида Archigeres и похожего на п.к -
роцерконда Caryophyllin
us. Archigeies паразити-
рует в полости тела Oligo-
eliaeta (Limnodrilus. Ти-
bifex). a Caryophyllae-
us — в кишечнике разных
видов рыб (рис. 158)
Особенно говорпг за нео-
теннческий характер этих
форм нахождение полово
зрелых стадий Archigetes
в беспозвоночном, чего Hr
бывает ни у одной из всех
прочих цеетод. Вероятно,
и здесь мы имеем дело с
неогеинчеекнми формами,
получившими способность
размножаться половых;
способом.
Наконец. Гинецпн
ская (1949) обнаружила
своеобразный случай про-
генеза у цеетод Cyclophyl-
lidea. В 1933 г. Фурманн
(Fuhrmann) установи.,
новый отряд цеетод Аро
riilea в составе двух видов
рода Мет а i ори г и taen in.
паразитирующих в ки-
шечнике лебедей. Они от-
личаются отсутствием
многих характерных для
цеетод признаков (отсут
стнпем членпстости, по-
ловых отверстий и копулятивных органов, обособленных жел
точкикон. двух из четырех стволов выделительной системы) и наличием
совершенно оригинального устройства половой системы. Гннецинская
нашла под роговой выстилкой мускульного желудка уток еще одни вид
Gastrotaenia dogieli. обладавший такими же признаками, но ины i
устройством сколекса, и путем сравнения его с Nematopara taenia уста
повила, что оба рода вовсе не образуют особого отряда в классе цеетод,
а являются иеогеннческимп личиночными стадиями каких-то птичьих
цеетод, относящихся к двум разным семействам: Nematoparataenia к Da-
vaieneidae Gastrotaenia к Hymenolepididae.
Если вышеприводнмые нами толкования справедливы, то явление-
нрогенеза, постепенно закрепляясь, могло в известных случаях приводить
к образованию новых видов. Этот процесс аналогичен явлению неотенин
у свободно живущих видов.
210
П ол я э ,м б р и о н н я Грис. 159) есть одно из приспособлении к ум» о-
zKt нию потомства, которые столь разнообразны \ паразитов. Это явле-
ние открыто Маршалом (Marshal, 1‘>0<5) у пара литических перепончато-
крылых (Chalcididae и Proct jtrypidae). Процесс этот оы 1 подробно изу-
чен па Encyrtus (A^eruas[ji4) fusdeuilis—наезднике, паразитируют» м
в г сен.шах бабочки НуропетелИа. Яйцо этого представителя Clinic 11
dae откладывается н.м в яйцо бабочки осенью н до зимы проделывает
ишь начальные стадии дробления. Зимой развитие останавливается и
возобновляется весной. Яйцо паразита с самого начала окружено -иш
гелнальным мешочком, прянадлежашнм тканям хозяина В дробящемся
Рис 1аЧ. Полиэмбрионии в развитии наезтника Ptilygnorus
ininutiis
1 — яйцо н периоде дробления, распадающееся на отдель
чые клетки,; Н—из отдельных бластомеров возникают дробя
in неся зародыши; С— дальнейшие стадии развития злродынк-ii,
получившихся из одного яйца. (По Маршалю.)
1 — ла[ниЫ111И: 2 питательные ядра iipoHCiiic.uiiiie на я пне вой клетки. ио
не у>|.1стиею1цие й оГ'рл илмнчн эмйриииип.
япце вскоре выделяется крупное трофическое ядро — парануклеус— и
много мелких шродышевых ядер. Последние группируются в мелкш
морулообразные клеточные скопления, каждое из которых впоследствии
дифференцируется в отдельного зародыша. Таким образом, из бласто-
меров первоначально единственного зародыша получается до сотни за-
родышей Позднее* полиэмбриония была найдена у целого ряда других
наездников, а именно: у Polygnotus rne/iutus из Cecidomyiq destructor
у Liihomitlstix truncatidlus из lycennu Piitsia gamma и т. д. .V Lithonbdstix
Сильвестри (Sil\e>tri, 1906) наши, что из одного япца формируется до
5п00 личинок, не считая известного количества абортивных, или. по тер
микологии Сильвестри, бесполых, личинок. Эги личинки лишены полового
зачатка и дегенерируют, не заканчивая метаморфоза.
Легко видеть, каким мощным средством размножения является по
лнчмбриония и насколько ина экономит силы откладывающей япца еам
кп паразита При небольших размерах вышеназванных наездников каж
дый из них может продуцировать за свои собственный счет лишь не-
большое. число яиц. Но попадая в среду, крайне богатую питательными
веществами, яйцо наездника дает начало множеству зародышей, уве-
личивая интенсивность продукции каждой особи наездника в 100
1000 раз.
К 211
Рис. 160. Gyroductvlus
eJectins с жабр золотой
рыбки. । "'а Фурману.)
/ — головные железы; 2 — гло-
тка; .? — Г1НМ1КуП1174*.1ЬНЫЙ
оргии' 4 — 7— ис гл пленные ниш
в други л ли роды in и четырех
пендедок пел иных поколение:
Л — мн nt- и ля клетка; 9 - сними;
10 ~ яичник; П — семенник;
/2 и t3 — сланные и побоч-
ные крючки ПрНКрС1]|1ТеЛЫ1ПГО
диска.
Как известный переход от нормального развития к полиэмбрионии
можно рассматривать онтогенез наездника Platy paster hiemalis, парази-
тирующего и гессенской мухе. По данным Хилла (Hill, 1923), яйца Pta-
tygaster то нормальны, то имеют полнэмбрионный характер, причем в
таких случаях нз яйца формируется только два паразита, т. е. полиэм-
бриония выражена слабо.
По-видимому, к категории явлений полиэмбрионии должно быть
отнесено и чрезвычайно своеобразное размно
жеиие сосалыциков Gyrodactylus (рнс. 160),
встречающихся на коже и на жабрах пресно-
водных рыб. Gyrodaclylus— живородящ; каж-
дая его особь развивает в себе единовременно
лишь одного зародыша следующего поколе-
ния, а тот, в свою очередь, зародыша третьего
поколения. При этом яйцо дробится в матке
материнской особи, образуя слой мелких кле-
ток. расположенных вокруг одного крупного
макромера. За счет мелких клеток формирует-
ся весь зародыш, тогда как микромер снова
начинает делиться и дает серию мелких кле-
ток, представляющих собой зародыша следую-
щего поколения. Далее макромер еще дважды
испытывает деления, давая начало еще двум
зародышам В общем, из первичного яйца та-
ким способом происходят четыре личинки,
представляющие собой четыре последователь-
ных поколения, . включенных друг в друге. Ка
таринер (K.-thariner, 1904), исследовавший
эти явления, толкует их как один из случаев
нолпэмбрнопин. Разница пи сравнению с ранее
рассмотренными случаям» заключается в том,
что у наездников все зародыш.к возникающие
нз одп эго яйца, развиваются одновременно, у
Gyrodaclylus же они, будучи но существу
сестринскими индивидами, тем не менее раз-
виваются последовательно, в виде четырех
следующих друг за другом поколений.
Процесс бесполого размноже-
ния как составная часть ци к л о в
п а р а з н т и ч ских живот и ы х. Одним из
средств усиления интенсивности инфекции и
повышения продуктивности паразитов служит
вставка в их жизненный цикл фаз бесполого
размножения, которое особенно часто встре-
чается у простейших и реже у многоклеточных
паразитов. Из вышеприведенной общей оцен-
ки жизненных циклов следует, что у простей-
ших это бесполое размножение не представ-
ляет каких-либо отличий от процессов, наблю-
даемых у свободно живущих простейших, а потому мы и не будем на них
останавливаться. Напротив, у многоклеточных бесполое размножение в
циклах является новойрнобретенкем, вызванным адаптацией к парази-
тическому образу жизни. Поэтому-следует ознакомиться с разными ти-
пами бесполого размножения, которые, по-видимому, совершенно неза-
висимо возникли у разных групп паразитических животных.
212
Рис. 161. I rocystiiHum gtvnrni-
ptiruni из Желчных ходив он
датры (Drtrfrt/л/ zibethicul; от
цепочки отпичкнвми.тютси ио
вые молодые стробилы. (11ч
Беддардл)
Среди немногочисленных паразитических кишечнополостных широ-
ко распространено размножение почкованием. Единственный паразити-
ческий полип Polypodiutn hydriiortne во время своей жизни в икринки
стерляди имеет вид длинного колбасовыдного сто юна, выночковываю-
щего на себе до 30 почек, которые впоследствии превращаются в само-
стоятельных полипов. Паразитические личинки медуз Cunina Cunocantha
и др., живущие в гастральной системе других медуз (Geryonidae), на
очень ранней стадии развития дают начало целому ряду почек.
Очень много и притом разнообразных примеров бесполого размно-
жения представляют цестоды. Во-первых, самый процесс образования
членистых цепочек, или стробпляиня, многими авторами рассматривался
как бесполое размножение, а каждый чле-
ник— как отдельная особь. 11 если мы счи-
таем особые всю цепочку, или стробилу, це-
ликом, а членик—небольшой частью инди-
вида, то все же образование многочислен-
ных члеников с самостоятельными половы
ми аппаратами есть адаптация, ведущая
к умножению потомства и по своему про-
спективному значению сходная с бесполым
размножением. Но у некоторых цестод, как,
например. Urocysiidiuin gemntiparum (рис.
161), живущей в половозрелом состоянии в
желчных протоках печени ондатры, мы
встречаем па стадии пеночки настоящее
почкование новых стробил от боковых краев
члеников. Является ли это нормальным
процессом, пока не выяснено.
Несравненно чаще бесполое размноже-
ние встречается у и,дырчатых стадий ленточ-
ных глистов, пребывающих в различных вну-
тренних органах промежуточных хозяев.
Плотные массивные финки типа плероцеркоидов почкуются редко
По данным Ивата (Iwata, 1934) плероцеркоиды «Sparganuin» (Diphylio-
bothriuni erinacei-europaci), встречающиеся в Восточной Азии у лягушек,
змей, свиней, крыс, а также у человека и имеющие удлиненное тело
(около 30 ли длины) с одной головкой, способны к бесполому размиожс
нию. По-вндимрму. такой процесс наблюдается лишь у искусственно ио-
врежденных плероцеркоидов, а в норме он не происходит.
Гораздо чаще бесполое размножение вставляется в цикл у Cyclu-
phyllidea, имеющих пузырчатую стадию в виде типичного пузыря, папол
ж иного жидкостью, в которую вдается от стенки пузыря зачаток головки
будущей стробилы. В некоторых случаях эта пузырчатая стадия вместо
одного зачатка головки образует разными способами ж сколько ско-
лексов.
Так, финка Taenia niloiica из кишечника птицы Си г sori its europucus,
живущая в дождевых чернях, образует на своих стенках несколько го
ловок. У мозговика (Multiceps), половозрелая стадия которою живет в
кишечнике собак, а пузырчатая, или так называемый цепур,— в мозгу
овец и крупного рогатого скота, пузырь достигает гораздо больших раз-
меров, пог его стенок вдаются внутрь около сотни головок. Еще одно
усложнение встречается у Taenia serialis собаки, цепур которой разви
вается в соединительной ткани зайцев и кроликов. На внутренней стенке
ценура головки располагаются рядами, а на одной из сторон ценура
отшпуровиваются дочерние п\ зырьки, тоже способные к образовании
головок. Наконец. у Lduwcorctis granulosus (рис. 162) бесполое размно-
жение достигает своего максимума. Пупырчатая стадия эхинококка, до-
Рис 162. Цепень эхинококк. (' ehiiuxvo wx
granulosus)
.1 — 1 зрослып червь, состоянии! из сколекса
и ipex члеников; I)— пупырчатая стадия
jxiiiioKOKK.i. пре тс гав. нпо и ,.тя собой громадный
материнский пузырь, в котором на внутренних
стенках образуются выводковые кЬпсулы, а
па стенках их — молодые сколексы. От
наружных стенок материнского пузыря могут
"тделятыя ра твнвлклцисея самостоятельно
дочерние пузыри
; i-b№ivKru; з - кянсу-м; ючерпие- irsupti.
стигающая иногда размеров
детской головы, образует на
своей внутренней стенде много-
численные выводковые капсу-
лы € большим количеством го-
ловок и дочерние id зыри, каж-
дый нв которых по своем'
строении» отвечает цсиуру моз
говпка В средней величины
эхинококке образуются таким
способом многие тысячи голо
вок. Кроме того, при случай-
ных разрывах эхинококка
часть сколексов может превра-
титься в небольшие вторичны'
пузыри, отче! ) заражение дан-
,ной особи хозяина еще усили-
вается.
Все перечисленные измене-
ния пузырчатой стадии ленточ-
ных глистов представляют со
бон процесс почкования перво-
начальною простого одиночно-
го зародыша. Особенно инте-
ресно то обстоятельство, что у
цеетод процесс бесполого раз-
множения вырабатывается, ви-
димо, в виде компенсации у
тех видов паразитов, которые
обладают сравнительно малой продуктивностью
в vBOi.m половозрелом состоянии. Так, Taenia so-
lium, Т. saginalus. Dipuiidium caninuni и другие
чс-рвн, развивающиеся по типу простого цисгн
церка образуют обыкновенно сотни члеников,
продуцирующих огромные количества яиц.
Mutticeps. развивающийся в промежуточном
хозяине по типу цепура с его сотней головок
имеет в половозрелом состоянии всего несколько
десятков члеников, a f-'chinococcus с его бесчислен-
ным множеством головок в (Ьи.чознон стадии
имеет всего 3—1 членика в состоянии стробилы.
Чрезвычайно любопытны й факт прелставляст
бесполое размножение некоторых Rhizocephala.
т. е паразитических усоногих рачков. 3 дан-
ном случае мы имеем перед собой пример проти-
воречия ыижду характерным для всех Arthropoda
свойством, а именно неспособностью к бесполо
му разложению, и между влиянием паразити-
ческого образа жизни, при котором вставка в
жизненный цикл бесполого размножения являет-
ся особенно выгодной. У ряда Rlr yiccpihala новые
•кологические условия одерживают верх над у наследованными
тенденииями животного, и в цикле таких паразитических раков возни-
2(1
кает бесполое размножение. Thompsonia (рнс. 163), паразитирующие на
тропических десятиногих раках, всегда встречаются в большом количе-
стве на одной особи хозяина. Однако вес эти мешковидные паразиты
развиваются нз одном личинки, проникающей в хозяина. Внутри деся-
тиногого рака имеется одна общая система корневых нитей, от которой
ырастают и выпячиваются нз тела хозяина многочисленные мешковид-
ные особи, наполненные, половыми продуктами. Во время линьки хо-
шина эти мешки отваливаются, после чего по наблюдениям Поттса
• Potts, 1915) от корневой системы нитей выпочковывается новая ге-
нерация мешковидных паразитов, вроде последовательных нарастаний
шляпных грибов из грибницы. У другого рода [Chlorogastcr), парази-
тирующего на раках-отшельниках, Перез (Perez, 1928) констатировал
четырехкратное отпадение мешков п замену их новыми. Такие же сведе-
ния относительно бесполого размножения получены для родов Pelto-
gaster и Peliogusterella.
Весьма вероятно, что у Rhizocephala развитие бесполого размноже-
ния становится возможным в связи с необычайной морфологической
дифференцировкой этих животных, которые утратили во взрослом со-
стоянии всякие признаки ракообразных и превратились внутри хозяина
в бесформенное ветвистое слоевище. Мешковидные выросты этого слое-
ища. наполненные половыми клетками, даже не вполне могут отвечать
понятию о резко обособленном индивиде.
Сделанный памп обзор показывает необычайное распространение и
разнообразие, процессов, при помощи которых у паразитов повышается
продукция потомства и обеспечивается сохранение вида.
ГЛАВА XI
ПРОМЕЖУТОЧНЫЕ И РЕЗЕРВУАРНЫЕ ХОЗЯЕВА
И ВОПРОС ОБ ИХ ПРОИСХОЖДЕНИИ
Теперь нам остается подробнее остановиться на одном явлении,
которое, наряду с увеличением плодовитости параанта, имеет целью
обеспечение распространения паразита и более успешное проникно-
вение его в новых хозяев. Это приспособление состоит, как мы
видели из рассмотрения жизненных циклов, в усложнении цикла пара-
зита путем введения в него системы так называемых промежуточных
хозяев. Как мы увидим дальше, это явление увеличивает шансы
паразита в борьбе за существование, доставляя паразиту максималь-
ные возможности для его распространения.
Сущность системы промежуточных хозяев состоит в том, что
зародыш паразита, покинувший организм хозяина, в котором он обра-
зовался, прежде чем попасть в хозяина, где он достигнет половой
зрелости и осуществит функцию полового размножения, должен
активно или пассивно попасть в другое животное; в этом втором
животном зародыш претерпевает некоторые, иногда очень сложные
изменения, лишь после, чего он оказывается способным к дальней-
шему развитию в своем основном хозяине.
То животное, в котором паразит достигает половой зрелости
и размножается половым путем, называется окончательным (дефини-
тивным) хозяином. Тот же организм, в котором паразит живет
в молодом, личиночном состоянии или размножается бесполым путем
и нартеногенетически, называется промежуточным хозяином.
Последнее определение нуждается, однако, в некотором уточне-
нии Лело в том, что ire всякое животное, в котором молодой паразит
проводит часть своего развития, может быть с одинаковым основанием
названо промежуточным хозяином. Промежуточным хозяином можно
считать только то животное, прохождение через организм которого
является необходимым и обязательным условием для дальнейшего
развития паразита в организме окончательного хозяина.
В определении промежуточного хозяина мы сталкиваемся иногда
с некоторыми трудностями, это касается тех случаев, когда паразит
не обладает половым процессом как это наблюдается, например,
у триианозомид. В позвоночном и беспозвоночном хозяине (муха цеце.
2IG
пиявка и т. п.) происходит бесполое размножение паразита, и по
этому в данном случае правильнее нс употреблять понятия оконча-
тельного и промежуточного хозяина, а говорить о беспозвоночном
хозяине как о переносчике (vector) трипанозомнд. Терминологиче-
ские неудобства возникают и в отношении наименования хозяев
малярийного плазмодия и других гемосиоридий. Комары для маля-
рийного паразита являются окончательным хозяином, а человек —
промежуточным. Здесь также комаров лучше именовать переносчи-
ками.
Поведение паразита в промежуточном хозяине может быть
очень разнообразным Поэтому и та роль, которую играет промежу-
точный хозяин в жизненном цикле паразита, может быть различна.
Дать исчерпывающую картину рассматриваемого нами явления очень
трудно, однако все же можно наметить несколько главных типов
пром е ж у точи ы х х о з я t в, по их взаимоотношению с паразитом.
К перво м у т и п у будут относиться те редкие случаи, когда
паразит не проделывает в организме хозяина какого-либо заметного
развития и вся роль хозяина сводится к какой-то подготовке пара-
зита для внедрения в его окончательного хозяина л для дальнейшего
развития в нем. Примером может служить кокцидия Adelea ouata.
Ооцисты Adelea проходят все свое развитие вплоть до образова
ния в пнх спорозоитов в кишечнике своего окончательного хозяина —
многоножки Lithobius jorjicalus и выходят наружу с экскрементами
многоножки в совершенно, казалось бы, готовом для дальнейшего
развития виде. Однако в случае заноса такой ооцисты в кишечник
другой многоножки заражения последнего не происходит, и ооцисты
без всяких изменений проходят через его кишечник и выносятся
с экскрементами снова наружу. Для того же, чтобы произошло
заражение новой многоножки, необходимо предварительное заглаты
ванне ооцисты мокрицей Oniscus marariits. Ооцисты остаются
в кишечнике мокрицы без всяких видимых изменений до тех нор, пока
многоножка не съест мокрицу. Попав в кишечник многоножки,
ооциста лопается, и вышедшие из нее спорозоиты начинают свое
дальнейшее развитие в кишечном эпителии.
Сходные взаимоотношения с хозяином-переносчиком наблюдаются
у Schellackia. п Lankeslerella (см. гл X). 1\ тому же типу следует
отнести п случаи механической передачи некоторыми переносчиками,
например слепнями (Tabantis), трипанозом (Trypanosoma evansi)
лошадей и верблюдов. Слепни принимают пищу, т. е. кровь, малыми
порциями, часто прерывая акт кровососанпя и перелетая с одного
животного на другое. При сосании зараженных трипанозомамп особей
слепень мажет свой хоботок кровью, содержащей жгутиконосцев
Если в течение 15 минут, следующих за первым сосанием, т. е, пока
хоботок не обсохнет и трппанозомы на нем не погибнут, слепень
успеет укусить новое животное, живые трппанозомы попадают
с хоботка в ранку на теле укушенного животного. Укус того же
слепня, но произведенный через более длинный срок, не вызывает
инфекции. Отсюда ясно, что хоботок слепня играет в данном случае
такую же роль, какую мог бы сыграть шприц или скальпель при
искусственном заражении.
Ко второму типу можно отнести всех тех промежуточных
хозяев, в организме которых паразит претерпевает значительные
изменения в виде метаморфоза, эмбрионального или постэмбриональ-
ного развития, ио без какого-либо размножения этих личиночных
стадий. В этих случаях число выходящих нз промежуточного хозяина
2Г
паразитов не превышает числа попавших в него зародышей паразита.
Средн простейших можно привести, например, Schellmkia и Lan-
kesterella. Примеры подобного рода изобилуют среди нематод
(Drncunnilus, \i uchereria, l.va, Metastrongylus, Protosirongylus,
НыЬгопета и т, д.). Среди цестод так же себя ведут личинки семей
ства Tavniidae, I ly rnenolepid'dau, Anoplocep halidac, имеющие форму
цистпиерка (пу зирчагая личинка с одной головкой) или цистн-
церкоида (финка с вздуты передней частью и с хвостообразпым задним
придатком, в котором расположи ны эмбриональные крючья (Taenia
sntinin. Та. niarhyrichus saginatus. Dipyli^iwn. Moniezia и др.).
Скребни проделывают такое же развитие в своих промежуточных
хозяева, — пазлнчиых ракообразных или насекомых для скребней,
паразитирующих у наземных позвоночных. Личинка скребня, вышед-
шая из оболочек янпа в кишечнике промежуточного хозяина и назьт
ваемая акантором, проникает в полос п> тела, где претерпевает ярко
выраженный процесс метаморфоз. Она теряет свои личиночные
органы (крючки, шины, мышцы и пр.) и окружается оболочкой.
Внутри тела появляются гигантские клетки являющиеся зачатками
внутренних органов. Акаитор превращается в следующую стадию
развития — ироакаителлу, для которой характерно постепенное увели-
чеине размеров, сопровождаемое закладкой и формированием всех
внутренних органов скребня. Наконец, эта личиночная стадия npi
вращается в акаителлу — личинку, находящуюся в неактивном и нн-
ийртироваимим состоянии и представляющую гобой почти сформиро-
ванного скребня (Moore, t946: \ ап Cleave, 1947; Петроченко, 1у56)
Накопеп, третий тин промеж уточных хозяев составляют такие,
в организме которых личинка паразита не только сохраняется и раз
вивается, ио и размножается
В этих случаях и з одного зародыша, пинавшего в промежуточ-
ного хозяина, развивается иногда огромное количество, исчисляемое
тысячами зародышей, которые способны инвазировать новых оконча-
тельных хозяев. Тип этот, распространен главным образом среди
простейших (Haemosporidia, Haejnogregarina). а из червей — у тре-
матод (Fasciota. Schistохота и др.) и некоторых цестод (Multiccps,
Echinococcus и др.)
Нередко паразит в своем жизненном цикле не ограничивается
одним промежуточным хозяином, а проходит через двух промежуточ-
ных хозяев. Последний в литературе обозначается иногда как
дополнительный хозяин («1 lil fswirt») (Скрябин и Шульц.
1940). Как мы видели из предыдущей главы, двукратГгфя смена хозяев
происходит у большинства тнгенегическпх трематод, у многих це»тод
(Diphyilobothrium. Triaenophorus и др.), у некоторых скребней
(Curynosomu) и у немногих нематод (Gnathostoma spinigerum).
Последняя в половозрелом состоянии паразитирует в стенках желуд
ка п.чстеялных. Первый промежуточный хозяин — циклоп, второй
рыбы, лягушки, змеи.
Во втором промежуточном хозяине личиночные стадии не прз
терисвают размножения, ио проходят некоторые этапы онтогенеза или
даже совершают метаморфоз, как, например, у некоторых трематод
происходит превращение церкарий в метацеркарии. Обычно во втором
промежуточном хозяине личинки находятся в ипцпетнроваином
состоянии. По своим взаимоотношениям с организмом паразита этих
промежуточных хозяев можно отнести ко второму типу.
Существ ет еще одна категория хозяев. Это гак называемы.-
резервуарные хозяева, т. е. такие, которые в отличие от про-
21*
межуточных не являются обязательными в жизненном цикле пара-
зита. по накапливают в себе инвазионных личинок паразита и спо-
собствуют передаче их окончи тельному хозяину I Рыжиков, 155-4;
Шульц и Лаитян, 19556, и др.).
Как правило, в резервуарном хошине паразиты не претерпевают
развития, а просто переживают (иногда в инкапсулированном состоя-
пип) где-либо в его внутренних органах в течение длительного вре-
мени, сохраняясь при этом в инвазионном состоянии.
Это явление обнаружено у разных гельминтов, имеющих не
1Олько прямое развитие, но и развитие с промежуточными хозяевами
Примером резервуарного паразитизма у re. bmihitob с прямым разви-
тием может служить нематода Syngamus trachea — паразит трахеи
кур, фазанов, индеек и других куриных, а также воробьиных птии
Птица может заражаться паразитом, склевывая с почвы яйца с инва-
зионными личинками. Однако заражещн возможно и путем сплевыв; -
ния дождевых червей, наземных моллюсков, многоножек или насеко-
мых, которые предварительно про; ютили яйца паразита и сами стали
носителями инвазионных личинок. Эти хозяева не обязательны в цикле
развития нематоды, ц в них не происходит развития личинок. В дожде-
вых чернях при этом личинки могут сохранять инвазионное и» до
I лет Следовательно, ли беспозвоночные играют роль резервуарные
хозяев, накапливающих и транспортирующих инвазионное начало
к окончательному хозяину. Резервуарный хозяин часто облегчает сам
процесс заражения. Так, птицы скорее имеют возможность заразиться
нематодой Sypgamus через посредство дождевых червей, которых они
во множестве поедают, чем непосредственно склевывая яйца паразита
с почвы. Такое же явление наблюдается н у Capillaria ptitori— пара-
зита хорьков, соболей и других куньих.
Такой никл развития с резервуарным хозяином можно изобра-
зить следующей схемой:
V гельминтов со сложным циклом развития резервуарный пара
штнзм обнаруживается у целого ряда видов нематод, цесгод, скреб-
ней и сосальщиков, паразитирующих главным образом у хищных
животных. Имеется ряд примеров включения в цикл развития
резервуарного хозяина при наличии одного промежуточного хозяина.
Так, например, нематода Spirocerca lupi в половозрелом состоянии
является паразитом собак, волков и других хищников. Япца паразита
попадают с экскрементами хозяина во внешнюю среде, где они
поедаются жукамн-копрофагамп (промежуточные хозяева). В теле
жука из яиц вылупляется личинка, которая развивается до ипвазион
нон стадии. Если жук бхдет съеден окончательных! хозяином, то
в организме последнего формируется половозрелая стадия червя
Однако жук может быть съеден ящерицами, птицами, насекомоядны-
ми млекопитающими и другими животными. В их организме личинки
219
паразита не развиваются до половозрелого состояния, а проникают
из пищеварительного тракта в другие внутренние органы, где
инкапсулируются и могут длительно сохраняться в инвазионном
состоянии. Эти животные, резервуарные хозяева паразита, легко могут
стать пищей окончательного хозяина, и тогда цикл паразита замы-
кается. В данном случае резервуарные хозяева хотя и нс являются
обязательными в жиг пенно i цикле паразита, но представляют собой
важное звено, облегчающее проникновение личинки Spirocerca
в окончательного хозяина, так как они составляют существенную
часть его пищевого спектра.
Сходное явление наблюдается у другой нематоды Physocephaln^
sexalntus — паразита желудка свиней, этих всеядных животных, кото
рые могут заражаться не только при заглатывании промежуточных
хозяев — жуков-копрофагов, но и при случайном noi данин резервуарных
хозяев (крыс, мышей, некоторых птиц. »мей. лягушек) и ряда других
животных, ннвазировапных личинками паразита. При поедании одних ре-
зервуарных хозяев другими животными — резервуарными хозяевами,
личинки переходят в организм последних и сохраняются в них дли-
тельное время способными для превращения в половозрелую форму.
Круг резервуарных хозяев обычно бывает весьма широким, что
является важным приспособлением паразита, обеспечивающим достиже
мне нм окончательно!о хозяина.
Можно предполагать, что в никл развития скребней также вклю
чаются резервуарные хозяева. Ио эго остается пока предположением,
потому' чю экспериментально не установлено, являются ли они обяла
тельным или не обязательным звеном в цикле различных скребней.
В них, однако, личинка паразита пе претерпевает какого-либо разви-
тия и остается на тон стадии, на какой она проникла в этого хозяина.
Предполагаемые резервуарные хозяева — это в основном позвонтчньн
животные, в пищу которых входят членистоногие, являющиеся про-
межуточными хозяевами скребней. В кчшке таких позвопочпьг
животных личинка скребня (акаптелла) пе в состоянии достигнуть
половозрелости. Она проходит через стенку желудка или кишечника
в полость тела или в мускулатуру, где инкапсулируется. В таки'
хозяевах происходит накопление паразита. Например, у Ро!у-
inorphus— паразита водоплавающих птиц, личинки ннвазируют раз-
личные виды рыб. личинки Macracantliorhynchus catuliniis— паразита
плотоядных (лисица, корсак и др.) — накапливаются в тех же хозяе
ва.х, а также в грызунах и мелких хищниках, которые могут съесть
промежуточных хозяев — жуков (Дубинин, 19186).
Вероятно, окончательный хозяин может заразиться и пспосрсл
ствышо от промежуточного хозяина и при ноеда«ш второго хо яиии
(резервуарного), в котором имеются личинки скребней Количество
их в таком хозяине может быть весьма значительным Так, в хищных
рыбах в дельте Волги личинки Corynnsoma были найдены в 39.6%
случаев, причем в некоторых рыбах встречалось до 89 -100 парази
тов (Дубинин, 1949, 1952). Принимая во внимание, что рыба служит
основным источником питания тюленя (окончательный хозяин), их
роль в доведении личинок Corynosuma до окончательного хозяина
оказывается весьма существенной. Рыбы являются в данном случш
тем важным звеном, может быть и не обязательным, между промежу-
точным и окончательным хозяином, которое дает возможность паразит •
замкнуть свой никл развития.
Схематически приведенные циклы с включением резервуарного
хозяина могут быть изображены так:
220
Окончательный
хозяин
—» Внешняя
Резервуарные хо лева могут включаться также н в цикл ра «вития
паразита с дуумя промежуточными 'озяевами. Это наблюдается
у . ситец в и среди трематод у некоторых представителей SLrigeata
(Ala ria).
Схематическое изображение такого цикла имеет следующий вид
Окоичатель-
iti й хозяин
Внешняя
среда
.У Diphylloboihrium latum первым промежуточным хозяином
являются L)i. ptotiitis и Cyclops, а вторым — некоторые рыбы (налим,
щука, окунь, ерш и др.), которые заражаются. поедая веслоногих
рачю в. Кроме того, хищные рыбы могут быть и резервуарными хозяе-
вами, если они заражаются путем поедания рыб (вторых промежу-
точных хозяев), зараженных плероцеркоидами. Последние, попав
в желудок лнцщ и рыбы, не перевариваются, а проходят через его
стенку и инкапсулируются где-либо в тканях. В этих рыбах проис-
ходит накопление паразита. Так, например, во взрослых щуках Онеж-
ского озера было обнаружено в среднем до 100 плероцеркоидов па
рыбу. Интересно, что передача плероцеркоидов возможна не только от
рыбы к рыбе, но (в экспериментальных условиях) п от рыбы к ам-
фибиям п рептилиям (Павловский и Гнездилов. 1939). В этих хозяе-
вах п ’ире.иеркоид нс терял инвазионной способности длительное время.
Таким образом, амфибий и рептилий можно рассматривать как
иогедгпальиых резервуарных хозяев, пс имеющих, однако, значения
в распространении паразита в природных условиях.
Основная роль в передаче Diphyllobothrium latum к окончатель-
ному хозяину принадлежит, несомненно. щ.ке и налиму как резер-
вуарным хозяевам, накапливающим плероцеркоидов. Для D. erinacei-
europaci — паразита собак, кошек и других хищников — резервуарными
хозяевами служат представители разных классов позвоночных (уж,
полоз, лягушка, ящерица, некоторые грызуны, ласка, куница, некото-
рые птицы, обезьяна и даже человек). Плероцеркоид вместе со вто-
рым промежуточным хозяином (лягушка, уж, поло i) может попадать
как в окончательного, так и в резервуарного хозяина. При наличии
пищевых связей у этих хозяев цикл развития паразитов может
завершиться столь же успешно, как и при заражении от второго про-
межуточного хозяина (Дубинина. 1951)
221
Таким обрядом, резервуарный хозяева, накапливающие инвазион-
ных личинок, в значительной степени облегчают процесс заражения
окончательного хозяина, если последний имеет с ними постоянные
пищевые связи. Теиствигелыю. плотоядные млекопитающие или хищ-
ные рыбы имеют больше шансов заразиться через мелких жнвотйых
(растительноядных или питающихся членистоногими), которые обычно
служат нм добычен, чем от промежуточных хозяев беспозвоночных
животных. которые не всегда доступны для них при питании.
С другой стороны, роль резервуарного хозяина мо/кет быть менее
значительной в распространении паразита, если окончательный
хозяин имеет более теш/ю пищевхю связь с промежуточным хозяи-
ном н от него в основпо .1 заражается. Это, например, имеет место
у свиней, которые иолхчают Physocephalus. как правило, от жуков.
Резерьх арпые хозяева, таким образом, пе все и не всегда могу|
реально передавать инвазию. Так, личинки Physocephalus накапли-
ваются в таких позвоночных, которые практически никогда не попадаю!
в пишу свиньям; это, например. сова, чайка и другие организмы,
имеющие пищевые связи с промежуточным хозяином, по не с оконча-
тельным. Следовательно, средн круга потенциальных резервуарных
.хозяев можно выделить, кроме реальных, еще н «фиктивных»
(Шульц и Давтян. 19556), т. е таких из которых личинка пе имеет
выхода в окончательного хозяина.
Резервуарными хо яевами. по мнению Шульца н [лвтяпа
(19556), становятся те животные, которые могут в процессе пйтания
получить инвазионную личинку по последняя не находит в этом
хозяине подходящих условий для своего дальнейшего развития
в кишечнике, вследствие наличия у него первичного ичмхнитета. Сле-
довательно, резервуарные хозяева это ни специфичные для половозре-
лая стадии паразита , нвотные. В ^том от ношении между ними и про-
межуточными хозяевами имеется большое сходство В ряде случаев
резервуарные хозяева могут становиться промежуточными, т. е. обяза
тельными в цикле развития паразита
Относительно происхождения промежуточных хозяев
теоретически допустимы две возможности, подобные двум выше
разобранных! возможностях! происхождения кровенаразиГизма^ либо
промежуточный хозяин являлся первичным хозяином, в котором пара-
зит первоначально достигал половой зрелости, либо он лини, позже
вошел в жн .пенный цикл паразита, который сначала паразитировал
лишь у окончательного хозяина. Согласно твум возможностям имеются
и две главные гипотезы о происхождении промежуточных хозяев.
Точка зрения, согласно которой пром е ж уточни н х о з я н и
является первичным, высказана первоначально Пенкартом
(I euckart, 1К63). «Промен уточные хозяева,— говорит он. или лишь
позднее были вставлены в жизненную историю гельминтов, или они -
первоначальные истинные хозяева, которые с самого начала доводили
1 воих внутренностных червей до половой зрелости, ио позднее были
низведены до степени промежуточных хозяев потому именно, что исто-
рия их паразитов растянулась на более шачнтсльное число стадий».
Главным аргументом Леннарта было то соображение, что дефп
ннтнвпые хозяева состоят почти исключительно из позвоночных, кото
рыс появились на земле значительно позже беспозвоночных, являю-
щихся. как правило, промежуточными хозяевами. «Если я высказы-
ваюсь,— говорит далее Лейкарт, — безусловно за второй случай
(т. е. первичность промежуточного хозяина), то вследствие того, что
взрослы* н зрелые в половом отношении эндопаразиты в настоящее
время, за немногими исключениями, находятся только у позвоноч
пых. .. Если мы нс будем объяснять этого обстоятельства тем, что
глисты возникли вообще лишь одновременно с позвоночными или что
древнейшие представители их все погибли, а то и другое при бес
пристрастном обсуждении кажется маловероятным, то остается только
допустить, что глисты беспозвоночных со временем изменили свои харак-
тер и вследствие дальнейшего развития в позвоночных животных
из зрелых в нолевом отношении особей стали молодыми формами»
Кроме этого общего соображения, Лейкарт считает, что извест-
ным доказательством первичности промежуточного хозяина служит
наличие определенных особенностей у тех немногих червей, которьь
паразитируют только в беспозвоночных. Большинство нематод бес
позвоночных оказывается очень близким но своей морфологии к сво-
бодно живущим Rhabditidae, от которых они, несомненно, произошли
Ленточный червь Archige.ics, паразитирующий в полости тела мало-
щетинковых червей (l.imnodrilus. Tubifex), «в морфологическом отно
шепни есть нс что иное, как процеркоид, т. е. ленточный глист, окап
чивающий свое развитие на такой ступени, которая у обыкновенных
ленточных глистов представляет переходную форму, прпнадлежашук
промежуточному" хозяину». «Наконец, сосальщик Aspidogaster conchi-
eola, паразитирующий в двустворчатых моллюсках IJnionidae, — пишет
Лейкарт,— в смысле развития и устройства кишечного аппарата,
настолько похож на так называемую редню, что я нисколько не
колеблюсь соединять его с ре.диеп, несмотря на его половую зрелость».
На основании всех этих данных Лейкарт считает, что редин
(и спороцисты) и цнетицерки являются древнейшими паразитарными
формами червей, некогда достигавшими в своих беспозвоночных
хозяевах половозрелого состояния, как это имеет место у Archigefes
н Aspidogaster. С появлением нового хозяина, возникшего, вероятно,
в результате хищничества, паразит приобрел способность достигать
половой зрелости лишь в нем, сохранившись в первом хозяине только
в личиночном состоянии, которое удержало следы прежнего половозре-
лого состояния в виде партеногенетического размножения.
Приблизительно так же представляет себе происхождение кроме
жуточны.х хозяев и Шимкевич (1898). По его мнению, первичное
состояние, паразитизма было таково, что в половозрелом виде паразит
жил на свободе, паразитическое же поколение его размножалось
иартеиогеиетичсски. как это имеет место, например, у Rhabdias bufonis.
С появлением хищника, пожирающего этого первичного хозяина пара
знта. последний перенес в хищника свою половозрелую стадию, т. е.
как бы заменил свою свободно живущую фазу паразитированием во
втором хозяине. Leucochioridium paradoxum является, по Шимкевичу.
примером такого наиболее, простого и древнего цикла: из яйца,
съеденного моллюском янтарной (Succiuea), выходит зародыш, внутри
которого в моллюске партеногенетическим способом развиваются
зародыши будущего поколения. Когда-то они выходили во внешнююсрс
дун развивались там в половозрелое поколение. С появлением птиц,
пожирающих моллюсков, эти зародыши стали развиваться в половозре-
лую форму- не во внешней среде, а в организме птицы.
Синицын по своим взглядам примыкает к Лейка рту и Шимкевичу
Противоположной точки зрения согласно которой по.рвнчны-
м и х о зя е в а мн паразитов б ы л и и х сов р е м е и и ы е око и
нательные хозяева, а промежуточные лишь впоследствии вошли
в жизненный цикл паразита, придерживаются Лоос (Looss, 1892),
Клаус (Claus, 1889), Моньг (Monioz. 1880), Мордвилко (1908) и др.
223
Лоос высказывается но этому вопросу следующим образом: «Если
мы обратим внимание на взаимные отношения, в которых состоят
хозяева и промежуточные хозяева ныне существующих паразитов, то
скоро выясняется тот мало обращающий на себя внимание факт, что
промежуточный хозяин принадлежит к животным, преимущественно
составляющим пищу истинного хозяина» Первоначально свободные
личиночные стадии паразитов, обитавших только у позвоночных, слу-
чайно внедрялись в различных низших животных. В тех случаях
когда последние служили нишей окончательных хозяев паразитов,
такой способ распространения оказывался более выгодным для пара-
та, как более обеспечивающий запое его в окончательного хозяина,
и в связи с этим закреплялся естественным отбором. Другие проме-
жуточные хозяева, не служивши* пищей окончательному, постепенно
отпали в процессе естественного отбора
Более детально эту точку зрения Лооса развил Мордвилко, счи-
тающий, что «явление промежуточных хозяев могло возникнуть
первоначально лишь в жизни хищных или плотоядных животных,
и притом исключительно на пути к более успешной инфекции дефи-
нитивных хозяев». Процесс этот Мордвилко в общем представляет
себе так. Первичной формой эндопаразитизма является кишечный пара-
ми» sv Последний в большинстве случаев возникает из пассивного
заноса паразита в кишечник хозяина, Это первоначальный способ проис-
хождения кишечного паразитизма, сохранившийся и до сего времени у
большинства растительноядных животных. При усилении н'одовитостл
паразитов зародыши последних могш быть занесены нс только в их
истинных хозяев, но случайно и в различных других животных.
Если истинным хозяином был хищник, то для пего открывался и
второй способ заражения, а именно через поедание других животных,
в которых случайно попали зародыши паразитов хищника.
Огромным преимуществом для паразита в таких случаях было
то, что сам хозяин (хищник), разыскивая себе пищу (т. е. других
животных), как бы разыскивал и своих паразитов. К тому же служа-
щие пищей хищникам растительноядные животные по самому образу
питания приходят в более тесный контакт с почвой и поточу легче
могут проглотить яйца паразита хищника, чем сам хищник. Следова-
тельно, добыча хищника была крайне подходящем передаточным
пунктом для его паразитов. При этом особенно хорошо обеспечива-
лась передача у таких паразитов, которые не ограничивались кратко-
временным пребыванием в кишечнике растительноядного, по прони-
кали в его внутренние органы, окружаясь пистон или капсулой. Этим
удлинялся срок пребывания в растительноядном, и вместе с тем воз-
растало число шансов паразита попасть в дефинитивного хозяина,
т. е. хищника. Кроме того, при попадании пот< мства паразитов
в растительноядных животных исчезала необходимость самостоятель-
ного добывания ими пищи до попадания в дефинитивного хозяина.
Отсутствие необходимости накопления в яйце запасных питательных
материалов для долгого выживания потомства на воле дало паразиту
возможность производить более мелкие яйца, а это повлияло на уве-
личение продукции яиц. По мере того как потомство паразитов хищ-
ных хозяев стало проделывать часть своего развития в растительно-
ядных животных, оно утратило способность непосредственного
заражения дефинитивного хозяина, т. е никл паразита прочно стал
двуххозяйным.
Высказанные Л^ордвилко общие соображения о происхождении
промежуточных хозяев у паразитов из хищных животных могут быть
224 ,
вполне приложимы и к паразитам из хозяев с иным характером пита-
ния (Anoplocephalidac из травоядных млекопитающих, Proteocephalus
и другие из различных мирных рыб)
Промежуточными хозяевами в цикле развития таких паразитов
могли стать те беспозвоночные животные, жизни которых связана либо с
почвой, либо с дном и толщей воды водоема, где обычно может осу-
ществиться контакт с инвазионными стадиями развития паразита (яйца,
личинки). Эти животные, кроме того, должны попадать в качестве
пищи в организм позвоночного хозяина (например, циклопы с личинками
Proteocephalus заглатываются рыбой) или вместе с пищей в его
'рганизм (клещи Onbaiidae с цистицеркоиламн Moniezia exptinsa
г.опадают к крупному рогатому скоту с травой).
Процесс формирования промежуточного хозяина, несомненно, во
многих случаях начинался путем включения в никл развития сначала
резервуарного хозяина. Так, например, хищники могут заражаться
Capillaria putori как прямым путем, так и через дождевых червей,
являющихся резервуарными хозяевами. В то же время ряд смежных
видов капнллярпй (С. aerophila, С. plica, С. mucronata) развивается
только при участии тех же дождевых червей, играющих уже. роль
промежуточных хозяев (Скарбилович, 1950). Резервуарный хозяин
в данном случае в процессе длительного отбора закрепился в цикле
развития капиллярий и превратился в промежуточного.
Включение в цикл развития второго промежуточного хозяина
наблюдается в основном у паразитов плотоядных и всеядных живот-
ных. Вторым промежуточным хозяином могут стать животные, являю-
щиеся главным звеном в н пшеном спектре окончательного хозяина
н экологически тесно связанные с первым промежуточным хозяином
(общие условия обитания) Это может произойти в том случае, если
личинка паразита пассивно или активно проникает в животное, в кото-
ром она не находит подходящих условий для достижения ноловозре-
лости В таком организме при полном несоответствии условий среды
личинка, проникшая через рот, в кишечнике не приживается и погибает.
Если же личинка преодолевает сопротивление хозяина, гибели ее не
происходит, но тогда она уходит из желудка или кишечника через
стенку этих органов в полость тела, откуда может уже проникнуть
во внутренние органы, где обычно инкапсулируется, претерпев
иногда развитие или метаморфоз Эта реакция «бегствам пара-
зитов из. пищеварительного тракта, наблюдаемая в хозяевах, чужды-,
тля взрослого паразита (плероцеркоид в хищных рыбах, амфибиях,
рептилиях; личинки анкилостомы в кроликах, хомяках; метацеркарня
Alaria alata в мышах или в мелких хищниках, и т. п.), является одним
из проявлении естественного иммунитета (Шульц и Давтян, 1955а).
В формировании второго промежуточного хозяина (так же как
и первого) с\ шественную роль мог т иногда играть ре юрвуарные
хозяева, которые сначала включаются в цикл развития паразита как
необязательное звено, а затем они становятся обязательными, т. с.
промежузочными хозяевами.
Возможность включения в качестве второго промежуточного
хозяина в цикле развития паразита определяется еще и степенью
развитья самой личиночной стадии паразита, попадающей в организм
.хозяина. Так, например, с лребенъ Leplorhynchoides thecaius, живущий
во взрослом состоянии в морском окупе и во многих других морских
и пресноводных рыбах Северной Америки, имеет промежуточного
хозяина — бокоплава Hyalela uzteca.
i Эксперименты Де Гиусти (De Giusti, 1949) показали, что если
15 В. Дсгсл» 2’23
бокоплав, в котором личинка скребня развивалась 31 день, будет
съеден какой-то рыбой, то в ней личинка превратится в половозрелого
червя, а если личинка имела возраст 27—28 дней, то, попав в рыбу,
она проходит через стенку желудка и инкапсулируется в брыжейке,
где доразвивается до инвазионного состояния. Следовательно, рыба
в данном случае становится вторым промежуточным (или резервуар
ным) хозяином, поедание которого другой хищной рыбой приводит
к заражению последней половозрелой стадией. Если в дальнейшем
такой путь развития закрепится, то в цикле развития данного пара-
зита будет не один, а два промежуточных хозяина.
В отношении возникновения промежуточных хозяев у простей-
ших, вернее только у кокцидий, Рейхенов (Reichenow, 1913) развивает
приблизительно ту же точку зрения, что и Мордвилко, представляя
себе ход эволюции следующим образом.
Первоначальной формой паразитирования кокцидий является,
несомненно, кишечный паразитизм у копрофагов и травоядных, рас-
пространявшийся исключительно путем пассивного заноса паразита
в кишечник хозяина. Дальнейшая эволюция паразитизма происходила
при участии хищников, причем к старому типу паразитизма стали
присоединяться и другие, развивающиеся, вероятно, в такой последо-
вательности
I. Взаимное поедание, встречаемое сейчас, среди многоножек
Lithobius. С хозяином поедаются и паразиты на различных стадиях
развития, продолжающие цикл в новой особи хозяина.
2. Поедание копрофагов, в которых случайно заглоченные ооцисты
оставались без изменения
3. То же самое, но уже с некоторым изменением в состоянии
ооцисты в виде подготовки ее для раскрытия и освобождения споро-
зоитов в основном хозяине, как это имеет .место у мокрицы в отноше
нии Adelea
4 Выход спорозоитов в промежуточном хозяине и начало их раз-
вития в нем же. Здесь жизненный цикл паразита уже закономерно
распределялся между двумя хозяевами, и, следовательно, промежу-
точный хозяин оказывается абсолютно необходимым для развития
паразита Так дело обстоит у кокцидии Aggregata eberthi, половая
часть цикла которой протекает в кишечнике головоногих, а бес-
полая— в кишечнике крабов, которыми головоногие питаются
5. Полное развитие системы промежуточных хозяев наступает
у кокцидий с переходом кишечных паразитов к кровепаразитизму.
где часть развития, проходящая во внешней среде, совсем упразд-
няется и в отношениях паразита и хозяина достигается такое состоя-
ние, при котором перенос и распространение паразита обеспечиваются
без гибели хозяина.
Таковы вкратце гипотезы о происхождении промежуточных
хозяев. Несомненно, недостатком всех их является стремление
уложить развитие всех существующих форм взаимоотношений пара-
зита и хозяина в одну общеобязательную для всех паразитов схему.
В действительности, поскольку явление промежуточных хозяев раз-
вивалось в виде приспособления к паразитическому образу жизни,
одни и те же отношения могли возникать в результате различных
путей развития В одних случаях первичным хозяином мог быть
современный позвоночный хозяин, и таких случаев гораздо больше;
в других — современный беспозвоночный. И задача исследователя
состоит в том, чтобы в каждом отдельном случае проследить возник-
новение исследуемого явления
Часть III
ПАРАЗИТОФАУНА И СРЕДА
ВВЕДЕНИЕ
В настоящей части нашей книги мы намерены рассмотреть зани
симость паразнтофауны животною, взятой в целом, от окружающей
ее среды. Как в экологии можно различать изучение экологических
отношений одного какого-нибудь вида и синэкологию, рассматриваю-
щую сложные и запутанные взаимоотношения со средой членов целого
биоценоза, так и в паразитологии можно по отдельности разбирать
закономерности отношений между каким-нибудь видом паразита и его
хозяином и условия, управляющие развитием всей паразнтофауны
того или иного животного, взятой в целом. Это вытекает из самого
понятия о паразитах, как о животных, средой обитания которых
являются другие организмы. Раз это так, то совокупность паразитов,
обитающих в одном каком-либо хозяине, представляет собой свое-
образный биоценоз, имеющий свои закономерности развития и свою
динамику. Изучая эту динамику паразитоценозов, по терминологии
Павловского (1937), мы должны, однако, учитывать одно весьма
существенное обстоятельство. Средой обитания паразитов является не
только сам хозяин, но и внешняя среда, окружающая этого хозяина.
Особенно ясно это может быть показано на эктопаразитах водных
животных, которые непосредственно зависят и от температуры и от
гидрохимических особенностей внешней среды. Вот почему, говоря
о паразитах, приходится по отношению к ним различать двоякую
среду обитания (Павловский, 1934) Средой первого порядка является
их непосредственное место обитания, т. е. хозяин. Но, кроме того,
паразиты многими прочными нитями связаны и с внешней средой,
окружающей самого хозяина, которую можно назвать средой второго
порядка. Эту двойную зависимость паразитов от внешней среды
Теодоридес (Theodorides, 1954) называет «законом двойного биотопа»
(«loi du double biotope»). Это необычайно ярко сказывается, например,
па особенностях паразнтофауны хозяев, ведущих наземный или вод-
ный образ жизни Очень многие особенности распространения парази-
тов водных животных совершенно иные, чем паразитов животных
сухопутных, целый ряд паразитов встречается либо только у водных,
либо лишь у наземных хозяев и т д.
15*
227
Вот почему в содержаний этой части будет рассматриваться
зависимость паразитофауиы как от среды первого, так и от среды вто-
рого порядка. Иными словами, ту же задачу можно определить как
изучение зависимости паразитофаины взятой в цс юм, от изменений
внешних условий, окружающих хозяина, и от изменений физиологи-
ческого состояния самого хозяина, т. е. среды первого порядка
(Догель, 19276, 1933, 1935, 1938а. 1948а, 1958).
ГЛАВА XII
ЗАВИСИМОСТЬ ПАРАЗНТОФАУНЫ ОТ ВОЗРАСТА
ЖИВОТНОГО ХОЗЯИНА И ОТ СЕЗОНА ГОДА
I ЗАВИСИМОСТЬ ПАРАЗНТОФАУНЫ ОТ ВОЗРАСТА ЖИВОТНОГО-ХОЗЯИНА
Первым и весьма важным фактором, действующим на состав
паразнтофауны животного, является возраст хозяина Изучение пара-
зитологии человека ясно показывает, что появление многих паразитов
при.рочено к определенному возрасту. Новорожденные обыкновенно
свободны о г паразитов, младенцы, питающиеся исключительно моло-
ком, пе имеют кишечных паразитов и т. д. По отношению к животным
в последнее время в этом отношении накопилось тоже значительное
количество фактов.
Прежде, всего мы имеем по данном) вопросу ряд отдельных,
эмпирически накопленных данных. Так. известно, что у всех млеко-
питающих весь внутриутробный период жизни остается, как правило,
забронированным от большинства паразитов (всех червей, всех экто
паразитов и т. д.). Мало того, для многих паразитов млекопитающих,
особенно для паразитов кишечника, хозяева остаются недоступными
н во весь период питания детеныша исключительно молоком матери
Это прекрасно видно на целой группе инфузорий Ophryoscolecidafc,
которые в огромных количествах населяют рубец п сетку взрослых
жвачных животных, между тем молодые, телята и ягнята за все то
время, пока кормятся только молоком матери, совершенно свободны
от этих паразитов. Стоит нм начать питаться травой или сеном, и в li.x
желудке уже через несколько дней показываются Ophryoscolecidae.
Количе -тво паразитов, проникающих в тело млекопитающего во
внутриутробном периоде его существования, невели о. Лишь очень
немногие паразиты преодолевают барьер между телом матери и за-
родышем, создаваемый плацентой. Таким способом может происходить
инфекция плода малярийным плазмодием, трипапозомами и пиро
плазмами (описаны случаи заражения как человека, так и живот-
ных), а также спирохетами сифилиса и возвратного тифа. Помимо
простейших, и некоторые черви могут проникать через плацент)
в зародыша, как это установлено для Ancylostoma duodenalc. собачьих
аскаркд, Schistosoma и др. Известны случаи находок печеночных дву-
усток в желчных хотах печ. ни породившихся ягнят, пузырен
эхинококка у зародышей человека и т. д. Такого рода заражение (через
229
плаценту) называется плацентарным. Другого рода зароженнс,
осуществляемо? через половой аппарат, представляют случаи герми-
нативной. или т р а п с о в а р и а л ь н о й, инфекции, когда паразит
проникает в теле хозяина в оплодотворенное яйцо и таким образом
сопровождает зародыш уже с самых первых стадий дробления, начи-
нающего развитие яйца. Подобный способ заражения известен, как мы
уже ранее говорили, для пироплазм, например для Pirnplasma bige-
minum рогатого скота в клеще Boophilus calcaralus
Итак, имеется несколько видов паразитов, которые проникают
в тело хозяина очень рано Далее известны многие паразиты, инвазия
которыми характерна лишь для молодых стадий хозяина и которые,
подобно «детским болезням», должны считаться паразитами дет-
ского возраста хозяина. Так, Ascaridia lineata может паразити-
ровать только в кишечнике цыплят. То же, до известной степени,
наблюдается и в отношении паразита дыхательных путей птиц
Syngamus trachea, a Trypanosoma laewisi встречается почти всегда
у молодых крыс.
С другой стороны, имеются инфекции, характерные для более
или менее зрелого возраста Trypanosoma equiperdum, передаю-
щаяся во время полового акта, нс .может, конечно, проникать в орга
ннзм, не достигающий половой зрелости или уже вышедший из периода
функционирования полового аппарата. Грегарина Diplodma gone
dipertha начинает заражать голотурию Cucumaria, в гонадах которой
она живет, по-видимому, начиная с наступления половой зрелости
1олот}рии. Наконец, мы увидим. что и некоторые мнксосиоридин
начинают заражать рыб также лишь при достижении последними
половой зрелости
Таковы отдельные данные, касающиеся повозрастного распростра-
нения отдельных видов паразитов. Посмотрим теперь, какие ре: уль
тэты дает повозрастный анализ всей паразитофауиы того или иною
животного-хозяина
Для того чтобы судить о влиянии на паразигофауну именно
возраста, а не какого-либо иного фактора, удобнее всего пользоваться
такими хозяевами, которые живут достаточно долго и притом все
время в одинаковых условиях и возраст которых можно легко опре-
делить. Этим требованиям отвечают, например, пресноводные, пепро-
ходные рыбы, особенно живущие в хорошо обособленных небольших
водоемах.
В настоящее время имеется ряд исследований в этом направле-
нии, позволяющих установить общую картину возрастных изме-
нений паразитофауиы пресноводных рыб- работа Горб}
новой по плотве и щуке (1936), Дубинина — по хариусу (1936) и осет-
ровым (1952), Быховской-Павловской — но окуню (1940), Столярова
(1934), Ляймана (1916) Малевиикой (1952), Изасик (1953) — по
карпу и карасю. Богдановой (1957)—по карповым рыбам дельты
Волги, Полянского и Шульмана (1956)—по рыбам некоторых озер
Карелии, Бауера и Никольской (I957) по ладожскому сигу и др.
Рассмотрим несколько примеров изменения паразитофауиы
пресноводных рыб в зависимости от возраста.
Горбунова при исследовании возрастных изменений паразите
фауны щуки и плотвы установила следующие группировки их пара-
зитов.
1. Паразиты, не зависящие в своем распространении ог возраста
хозяина. Из 18 часто встречающихся у щуки паразитов к этой катего-
рий относятся 6. Это были 2 вида миксоспори дни— Myxosoma dujar-
230
dint и Henneguyu lobosu, сосальщик Gasierosieum fibriatutn, скребни —
uiiithocephalus lucii и Neoechechindrhynchus rutili, рачок Ar galas
joliaceum
2. Паразиты, убывающие в числе с возрастом хозяина Сюда
относится у щуки всего 2 вида. Типичным представителем данной
группы является лишь ленточный глист Proteocephalus сегрпие, распре-
деление которого по щукам разного возраста таково
Возраст щуки (в годах) Сеголетки 12 3 4 5—11
Заражение (в процентах) 70,6 26,4 13.2 6,6 14.7 19
Такая закономерность распространения объясняется, по-видимому,
тем, что в первые месяцы своей жизни щука питается преимуществен
но планктоном, в котором находятся промежуточные хозяева Proteo-
ceptudus— веслоногие рачки.
Другой парашт данном категории — слизистый споровик Неппе-
guya oviperda — часто встречается в трехлетием возрасте, после чего
сражение нм постепенно снижается.
3. Паразиты, возрастающие в числе с возрастом хозяина. Таких
фирм у щуки оказалось 10. Из подсчета видно, что данная категория
паразитов, безусловно, доминирует над прочими. Усиление инвазии
прекрасно выражается не только в экстенсивности, но н в интенсив-
ности инвазии, как это можно видеть для ряда форм дайной катего-
рии из прилагаемой табл. I. В таблице для Myxidium lieberkiihni
интенсивность не показана, так как у простейших число особей пара-
зитов, имеющихся в хозяине, установить невозможно. Кроме отдель-
ных случайных hi согласованностей (например, у трехлетних щук
инвазия Rhaplnduscaris, очевидно, имелась, но не была почему-либо
обнаружена), цифры таблицы прекрасно отражают постепенность
процесса заселения хозяина паразитами параллельно увеличению его
возраста.
Имеется еще один показатель, иллюстрирующий усиление зара-
жении с возрастом хозяина: учет среднего числа видов рйразптов.
приходящихся на одну рыбу определенного возраста. Для щуки
и плотвы в этом отношении получаются следующие данные;
Возраст (в годах) Сеголетки 1 2 3 4 5 11
Щука (число видов паразитов) . 3.4 4,8 5.8 7,6 8 9,4
Не
Плотва (число видов паразитов) 0,13 1.85 4 1.3 5,3 было
Совершенно аналогичные данные получаются в общем и для дру-
гих исследованных рыб. Так, у хариуса, по Дубинину, число видов
паразитов, встречающихся в среднем в одной рыбе, меняется с возра-
стом следующим образом: сеголетков на I рыбу приходится 1,4 вида,
у годовиков — 2.G, у двухлеток— 2.5. у трехлеток — 2.1 и у четырех-
леток— 2,7. У исследованной в 1933 г. Догелем и Петрушевским
молоди семги из Сев. Двины те же возрасты имеют соответ-
ственно в среднем 1 вид, 1,3 вида, 2.3 вида и 2,7 вида паразитов
на I рыбу.
То же очень хорошо показал Петрушевский (1937) у культурной
форели в прудах, а Столяров (1934) у карпов. Так, по Петрушев-
скому, у сеголетков форели в среднем приходится 0,06 видов парази-
тов па одну особь (т. е. большинство сеголетков совершенно свободно
от паразитов), у годовых форелей эта цифра поднимается до 0,4.
231
О ; п
у двухлетних—до 1.4. а у трехлет-
них— до 2,8 видов паразитов на
1 рыбу.
Интересно проследить динами-
ку заражений паразитами на самых
ранних стадиях жизни рыб — в воз-
расте до одного года. Ряд подобных
примеров, касающихся различных
пресноводны’ рыб из озер Карелин
(леща, плотвы, уклеи, щуки, окуня,
ерша), находим в работе Полянско
го и (Пульмана (1936). В качестве
примера приведем данные об нзме
нении паразнтофауны окуня в теч»
пне первого года жизни (табл 2).
Из приведенных в таблице дан-
пых видно, что самые молодые из
исследованных окуней (1— 1,5 меся-
ца) были уже сильно заражены двт
мя видами трпходин (Trichodina те-
gamicronucleata и Т. percarum) л
также еще двумя паразитами, раз-
вивающимися без промежуточны*
хозяев' (Trichodina urinaria и Zschok-
kella nova), и одним пара гигом (Тс
tracoiyle uariegata). активно прони-
кающим в хозяина. Кроме того, и у
самых молодых окуней имеется епн
цестода Proleocephaltis регсае, зара-
жение которой происходит при по-
едании планктонных Copepoda. В
дальнейшем паб.подается общее на-
растание числа видов паразитов, эк
степсивиости и интенсивности зара-
жения. Однако некоторые виды ока-
зываются свойственными лишь са-
мым молодым возрастам (пример:
Trichodina megamicronuclecda) и по
мере роста рыбы в дальнейшем вы-
падают.
Таким образом, по отношению
к пресноводным рыбам т дается со-
вершенно отчетливо вывести прави-
ло, говорящее о том, что интенсив-
ность и экстенсивность инвазии в,
общем увеличиваются с возрастом
хозяина.
Причины этого явления для от-
дельных видов паразитов различны,
но сама закономерность несомненна
В качестве некоторых факторов,
влияющих па повышение интенсив-
ности и экстенсивности заражения,
намечаются, например, усиление с
возрастом прожорливости хозяина и
Таблица 2
Изменения паразитофауиы окуня в зависимости от возраста (в %
Зара же ,ир» (по Полянскому и Шульманх)
Название паразита Органы 13 зраст рыб и количество вскрытых экземпляров
1 — 1,5 мес. (24 экз.) 2—2,5 мес. (15 экз.) 1 -ь (14 экз.)
Myxosporidia Zschokkclla поел .... Желчный птзы) ь 4,2 7.1
Mvxobolus pse.tidodispar Почки — 6.5 —
1 lenncguya psorosperm.i ca Жабры 6.6 —
Infusoria
Trichodina percarum . . . Жабры, плавники 70,8 46,6' 92,8
Trichodi na me gam i cron uele-
ata Жабры, плавники 37,5 — —
Trtchodida urinaria .... Мочевой пузырь 4,2 — 78,5
Monogenoidea
Ancyrocephaltts paradoxus . Жабры — — 21,4
Trematodes
Diplostomulum spathaceum Хрусталик — — 4'2.8
Diplostomulum ciavatum Стекловидное тело — 6,6 —
Tetracotyle percae .... Почки — — 7.1
Tetracotyle variegata . Ткани внуiренних органов 8.3 2:5.6 14,3
iiunodcra hiciopcrcae . . . Кишечник 26,6 —
Cestodes TrfacHuphorus itodulosus
Печень 20.0 7.1
Protcocephalus percae . . . Кишечник 4.2 — •28,5
Mcllusca
Glochidium Жабры 1 — 7.1
увеличение размеров добычи (которая может служить в качестве проме-
жуточного хозяина паразитов), пожираемой животным. Для эктопарази-
тов, вероятно, имеет также значение увеличение площади, пригодной для
заселения. Например, жабры молодых щук никогда не содержат столько
Ergasilus и Telraonchus, как жабры старых, крупных рыб. Совершенно
естественно увеличение интенсивности заражения такими паразитами, ко-
торые живут в хозяине в теченщ нескольких лет: с каждым годом про-
исходит аккумуляция их в хозяине. Для щуки это легко демонстрируется
заражением личинками широкого лентеца. Конечно, при оценке подоб-
ных случаев надо учитывать и возможность влияния возрастного имму-
нитета.
Для большинства пресноводных рыб. кроме вышеуказанного правя
ла, выясняется и другая отчетливо выраженная закономерность. Оказы-
вается, что раньше всего хозяин большей частью заражается такими
паразитами, которые не имеют смены хозяев или которые активно про
пикают в тело хозяина (как Tetracotyle пли Diplostomulum). Кроме при-
веденных выше примеров, в настоящее время накопился значительный
фактический материал, подтверждающий справедливость этого правила.
Интересно, что у пресноводных рыб заражение паразитами, не имеющи
ми промежуточных хозяев, начинается в самые первые дни после выкле-
233
на из икринки мальков или личинок Так, Шульман наблюдал у личинок
леща к плотвы заражение Trichndina megandcronucleata на 3-и день,
у щуки тот же паразит был найден на 7-й день и т. п. Мальки леща в
возрасте 2 месяцев имели \же 8 видов паразитов, но все они относились
к формам, развитие которых протекает без промежуточны.^ хозяев, либо
к активно проникающим в рыбу видам (Diplostoimilum)
В значительной мере иной характер носят возрастные изменения
паразнтофауны морских видов рыб, У которых отсутствует непосредствен-
ный контакт между молодью и взрослыми особями.
Рис. 164. Кривые возрастных изменении парззптофаун ы
трески. (По Полянскому.)
I — Podocotylc alomon; 2 — Lepidapedon gadi-, J — Hemiurus levinteni;
Л — Derngrncs viiricus: 3 Abaft num gadi; 6 — Pyrumicocephalus
phocarum; 7-Contracpecum aduncam <u кишечнике); 8 - то жен
орижсикс; 9 — то же и печени; 16 At.lsakls (и брыжейке); 11 то же
в печени; 12 — Pchincrhynclius gadl.
Хороший и наглядный пример такого рода отношищй дают треско-
вые северных морен и в том числе трбска (Ciadas morhua morhua) (По-
лянский, 1955), которые характеризуются резкими различиями в эколо
гни мальков и взрослой рыбы. У баренцевоморскон трески после икро-
метания в марте-апреле \ северных берегов Норвегии пелагическая икра
и личинки Нордкапским течением подносятся к берегам западного и
восточного Мурмана. В конце июня — начале июля мальки трески мас-
сами заходят в губы и заливы м\рмапского побережья, где проходят
так называемую литоральную стадию.1 В это время они держатся в са-
1 Нужно иметь в виду, что не все популяции мальков трески проходят нитораль-
иую стадию. Часть мальков остается в открытом морс.
234
ной прибрежной зоне и во время прилива питаются на литорали. I [низ-
кого контакта со взрослой треской, находящейся в открытом море, маль-
ки на этой стадии нс имеют. В годовалом возрасте молодь постепенно
отходит от берегов, опа все дачьше и дальше проникает в открытое море
и начинает в области своего распространения контактировать со взрос-
лой треской.
Таблица 3
Изменение паразитофауны баренцевоморской трески в зависимости от возраста (в % заражения) (по Полянскому)
1(азвание паразита Орган Возраст рыб м количество вскрытых экземпляров
4- -5 мес. (39 экз.) 10—11 мес. (29 экз) 1 + (50 экз.)
Protozoa
Trichodina sp Жабры плавники 10,3 —
Myxidium bergense . . Желчный пузырь 2.6 — —
Pilstophora sp Мышцы 2,6
M^nogenoidea
Gyrodactylus mariaus Жабры . — 3.4 —
Trematodes
Podocctyle atonion . . Кишечник 23,6 34,Л 40.0
Podocotyle r&flexa . — 20,7 —
Lepidapedon gudi . . . — 62,1 .6,0
Hemiurus levinst ni . . Желудок 7,7 3,4 16,0
Derogenes varicus . . . 18,0 3.4 ю.с
Lecithaster sp Кишечник 2,6 —
Cestodes
Scolex poiymorphtis . . 28.2 20.0 16.0
Abothrium gadi .... — — 2,0
Pseudophyilidea gen. sp.,
larvae .Мезентерий 2,6 — 2,0
Nematodes
Contramectirn aduncum Кишечник, печень.
juv. брыжейка 28,2 79.3 42,0
Xnisakis sp., larvae . . Мезетсрий, печень 2,6 6.8 10,0
Acanthocephala
Echtnorhynchus gadi . . Кишечник — 10.3 42,0
Copepoda
Caligus curtus .... Поверхность тела 2,6 — —
Lernaeocera branchia its Жабры — 3,4 —
Clavella uncinata . . . - — 3,1 10,0
Как отражается эта сложная карч ина возрастных изменений эколо-
гии хозяина па у го паразнтофагне? Ответ па этот вопрос дают данные,
суммированные в табл. 3 и кривые (рис. 1G4).
В период пассивного дрейфа вдоль берегов Мурмана личинки трески
совершенно лишены паразитов. По как только они заходят в губы и на-
чинают питаться, паразиты сразу же появляются. В возрасте 4—5 меся
цев (т. е. через 1—3 месяца после подхода к берегам) число видов пара-
зитов уже довольно значительно (табл 3, рис. 164). Однако в отличие
от рассмотренных выше пресноводных рыб это почти исключительно
паразиты, развивающиеся с промежуточными хозяевами и попавши * в
235
мальков через промежуточных хозяев. Паразиты с прямым развитием
совершенно единичны (Trichodina sp., .X-lyxidium bergense, Caligus cur-
ias) и относятся к видам с широкой специфичностью, которые могут
переходить на мальков трески с прибрежных, например камбаловых, рыб.
Основная же масса паразитов трески на ее «литоральной» стадии — это
черви со сложными циклами развития, проникающие в треску вместе
с пищей. Например, сосальщик Podocolyle alomon приобретен от лито-
ральных Gamrnaridae. в которых находятся метацеркарня этого вида
(Успенская, 1950). Ряд других видов (Ilemiurus levinseni, Derogcnes vu-
ricus, Lecithaster sp.. Scolex polymorphus, нематоды Anisakis и Contru-
coecutn) получен от пелагических Copepoda. составляющих в этот период
жизни трески ее основную пищу.
Интересно отметить, что некоторые паразиты трески, появляющиеся
у них в течение первого года жизни, носят ясно выраженный «детский»
характер. Это, во-первых, Podocolyle alomon и, во-вторых, Lepidapedon
gadi. Первый характерен для литоральных рыб (заражение через лито
ральиых Amphipoda), у взрослой трески он не встречается, вторым
треска заражается при поедании червей Nereis pelagica, в которых пара-
зитируют мстацеркарпп (Амосова, 1955) Отходя от берегов, треска
теряет н этого паразита.
Совершенно сходная с треской возрастная цшамика паразитофау-
ны наблюдается и у баренцевоморской сайды (Pollachius virens). Био-
логия этой рыбы очень близка к таковой грес.кн.
Глухова (1956) изучила возрастную динамику паразитофауны рав-
ных видов камбал Белого моря. Она наблюдала также (как это имеет
место у трески) увеличение числа видов, экстенсивности и интенсивно-
сти заражения с возрастом. При этом в первую очередь происходит за-
ражение паразитами, развивающимися с промежуточными хозяевами
Например, мальки Pleuronecles flesus раньше всего заражаются скреб
нем Ne.oechinorhynchus rutili, сосальщиком Podocolyle alomon п пемато
доп Contracoecum aduncam (larva). Паразиты с прямым циклом появ-
ляются значительно позже.
Возрастная динамика паразитофауны некоторых черноморских мор-
ских рыб подробно изучена Решетниковой (1955а, б) на примере трех
видов кефалей (Mugile aural us. М. cephalus, М. saliens). У 51. auralus
мальки в возрасте примерно до 8 месяцев питаются планктоном, затем
постепенно происходит переход их к питанию бептодстритом Стайки
мальков держатся обособленно от взрослых рыб. У М auralus с возрас-
том происходит обогащение видового состава паразитов, повышается
экстенсивность и интенсивность заражения. Но дело этим нс ограничи-
вается. У мальков М. auralus, так же как у баренцевоморской трески,
имеются «детские паразиты», которыми мальки заражаются в период пи-
тания планктоном. Таковыми являются некоторые сосальщики сем. Не-
tniuridae. личиночные стадии цеетод Scolex polymorphic, нематоды Con-
tracoecum. После перехода к питанию бентодеТрптом «детские паразиты»
постепенно исчезают и заменяются видами, характерными для взрослых
кефалей. Некоторое время паразитофауна сеголетков носит смешанный
характер — в ней сочетаются «детские паразиты» с видами, типичными
для взрослой кефали. Характерно, что раньше всего Mugile auralus зара-
жаются паразитами, развивающимися с промежуточными хозяевами.
Заражение паразитами с прямыми циклами осуществляется значительно
позже, когда становится возможным контакт молоди со взрослыми ры-
бами.
Таким образом, для многих морских рыб сформулированное выше. и
справедливое для пресноводных рыб правило о первоочередном зараже-
236
нии паразитами, не имеющими смены хозяев, оказывается не примени-
мым. Причина этих различий всецело определяется особенностями эко-
логии хозяев — характером питания и степенью контакта молоди со
взрослыми рыбами. Особенно интересно отметить, что у тех морских рыб,
у которых контакт взрослых форм с мо.тодыо сохраняется на всех этапах
жизни, возрастная динамика паразитпфауны вновь приобретает черты,
характерные для пресноводных рыб. Например, у трехпглоп колюшки
[Gasterosteus acuteatus), живущей в губах Мурманского побережья Ба-
ренцева моря, где контакт между рыбами разных возрастов все время
остается тесным, мальки в первую очередь заражаются паразитами, раз-
вивающимися без промежуточных хозяев,—триходинамн и моногепетп-
ческими сосальщика ми.
Правильность сделанного нами ранее заключения о постепенном
усилении инвазии с возрастом хозяина доказывается нс только на рыбах,
по и на других группах хозяев, а именно: на птицах и млекопитающих —
это есть не частная, а общая закономерность.
Для проверки такого же правила на птицах требуется брать приме-
ры среди оседлых птиц, для того чтобы к влиянию возраста не примеши-
валось влияние миграций (молодые перелетные птицы еще ни разу нс
проделали миграции, тогда как взрослые уже бывали па зимовке в юж-
ных широтах). Кроме того, возраст птиц определяется гораздо труднее
возраста рыб, а потому у них в большинстве случаев удается различать
только три возрастных категории: гнездовые птенцы, взлетыши, или мо-
лодые птицы, и взрослые птицы (выведшиеся в прошлом годх или ранее).
Очень показательную картину дают эндопаразиты воробья (Побе
доносит в, 1940), где заражение эндопаразитами наблюдается
у 14% птенцов, 54% молодых и 80% взрослых птиц.
Наличие у птиц чисто «детских» паразитов, накладывающих совер-
шенно особый отпечаток на паразнтофауну птенцов, лучше всего пока-
зывают некоторые сосальщики (Proslhoffonimus, Episthmium bursicola
различных цапель, рис. 165 и др.), паразитирующие в фабрициевой сум-
ке, т. е. в органе, который исчезает у большинства птиц еще до конца
первого года жизни. Вместе с органом птица теряет и соответственных
паразитов
Довольно подробно возрастная динамика паразитофауиы была изу-
чена Зерновым (19*196) для некоторых видов врановы.х птиц (Corvidae).
В отношении эндопаразитов грача (Corpus frugilegus) Зехпов обнару-
жил общее повышение экстенсивности заражения с возрастом с 40% до
98,6%. Увеличивается также число видов гельминтов: у молодых птенцов
он обнаружил 2 вида, у старших птенцов—6 видов, у летных птенцов —
10 видов, у взрослых птиц— 12 видов. Однако увеличения интенсивности
заражения (т. е. числа особей паразитов на одного хозяина) не наблю-
далось. Наоборот, с возрастом наблюдалось снижение интенсивности
инвазии, что автор ставит в связь с изменением характера питания.
Эндопара Лой грача Зехнпв по характеру распространения и чис-
ленности их в птицах различного возраста подразделяет на пять групп.
К первой группе он относит гельминтов, встречающихся только у птен-
цов (Plagiorchis brauni, Prostliogonitnus ovatus, P. cuneaius, Porrocoecun
cnsicaudatuni). Среди этих гельминтов виды рода Prosthngoniinus пара-
зитируют в фабрициевой сумке, поэтому, естественно, могут встречаться
только у птенцов. Ко второй группе эндопаразитов грача относятся
Hymenolepis serpenlulus, Syngamus trachea, Capillaria reseda. Они встре-
чаются как у птенцов, так н у взрослых пгнн, ио преобладают у птсидов.
Третью группу составляют Anoinofaenia constrida, Dilepis undula и Ca-
pillaria contorta, встречающиеся у взрослых птиц, по более мпогочислсн-
237
иые у летных птенцов Четвертую группу образуют Acuaria anthuris и
Microiilaiia sp., которые появляются у летных птенцов, но экстенсивность
и интенсивность заражения которыми увеличивается с возрастом Нако-
нец. к пятой группе относятся Tamerlania zarudnyi, Agamospirura sp.
Dinlotacnia tricuspis и Microtetrunit-res inertuis, обнаруженные только у
взрослых птиц. Возрастные изменения паразнтофауны грача Зехнов свя
6
Рис. 165. Сосальщики. живущие в фабрпциевой
сумке птиц
А — Prosthog^nimus ovatus; В — Episthmlum
bursicola. (4—по Кюкенталю, Л—по Дубинину.)
I — ротоиэн присоска; 2 — половое отверстие; .3 — брюш-
ная присоска; I — желточпикн; .5 — семенники; 6 — матка;
7 - выделительное отнерстне.
зывает с двумя категориями
причин: изменение состава пи-
щи и возрастание невосприим-
чивости (иммунитета) к неко-
торым паразитам с возрастом.
Последний вопрос подробнее
рассматривается нами ниже
(стр. 404).
Быховская - Павловская
(1953, 1962) на большом мате-
риале птиц, относящихся к раз-
ным отрядам, устанавливает
следующие закономерности
возрастной динамики фауны
гельминтов. В первую очередь
птенцы заражаются цестодами,
несколько позже — скребнями
и круглыми червями и, нако-
нец, сосальщиками. Все это
происходит быстро и полутора-
двухнедельные птенцы, напри-
мер у водоплавающих птиц,
уже имеют паразитических
червей, относящихся ко всем
группам
Интересно отметить, что
согласно Быховской Павлов-
ской и ряду других авторов у
большинства видов водопла-
вающих и воробьиных птиц
молодые заражены значитель-
но сильнее (как в отношении
экстенсивности, так и интен-
сивности), чем взрослые. Be
роятно, это
большим количеством и составом потребляемой
птицы в отношении закономерностей возрастной динамики эндопара ш-
тов несколько отличаются от рыб. У последних, как мы видели выше,
обычно с возрастом увеличиваются интенсивность и экстенсивность за-
ражения.
Особенно отчетливо возрастные изменения паразитофауны птиц вы
ражены у видов со смешанным питанием, когда птенцы питаются пре-
имущественно насекомыми, а взрослые птицы — преимущественно расти-
тельной пищей. Примером может служить зяблик (Frinp'tlla coelebs)
(Соколова, 1959). Сосальщиками заражены преимущественно птенцы.
У взрослых трематоды отсутствующ или встречаются редко.
По-видимЬ.му, существуют различия в возрастных изменениях пара-
знтофауны между видами птиц, имеющих гнездовых и выводковых птен-
цов. Эти различия затрагивают самые молодые возрасты и связаны с
стоит в связи с
пищи. Следовательно.
238
различиями н биологии хозяев. Гнездовые птенцы уже в возрасте
1—4 дней заражаются эктопаразитами — кровососами Это обнаружено
у воробья (Passer domesticus) (Победоносцев, 1940), каравайки (Plegu-
dis jascinellus) (Дубинин, 1938), скворца (Sturnus vulgaris) (Марков,
1939а), пеликанов (Дубинин, 195-16). Заражение эндопаразитическими
червями у них происходит несколько позже, хотя тоже очень рано
(у скворца кишечные гельминты были найдены впервые на 5—6-й день,
у всробья — на 11-й день, у канарейки — на 5—6-н день и т. и.). Однако
иногда, при особом типе питания, заражение гнездовых птиц гельминта-
ми может происходить почти сразу после выхода из яйца. Это имеет
место у пеликанов, когда родители отрыгивают полуиерсваренную пинг
(рыбу), захватывая при этом в рот голову птенца (Дубинин, 19546).
У выводковых птиц (пока имеются некоторые данные только по
водоплавающим) первыми появляются эндопаразиты со сложными цик-
лами, источником заражения которыми является пища. Эго обнаружено
у гаги (Somateria molissima) (Белопольская, 1952; Кулачкова, 1958) и
поганки (Poaiceps crisiatus) (Гипсцинская, 19526). Эктопаразиты перье
вого покрова появляются у выводковых птиц несколько позже, что зави-
сит от меныпей степени контакта их с родителями, чем у гнездовых птиц.
Вопрос о возрастной динамике эктопаразитов у птиц, в особенности у
птенцов, требует дальнейших исследований.
Очень удобным объектом изучения возрастных изменений
паразитофауны у млекопитающих являются, как указывает
Киршенблат (1938), мышевидные грызуны, которых он исследовал в
Закавказье. На основании вскрытия значительного количества особей
разного возраста Microtus sociahs, Apodemus sylvaticus и Mus musculus
(на воле) Киршенблат приходит к следующим выводам по отношению
к грызунам. Количество видов гельминтов в грызунах увеличивается с
возрастом, хотя самые старые полевки (Microtus) заражены меньшим
количеством глистов, чем полевки среднего возраста. Раньше всего в
лесных и домашних мышах появляются гельминты, развивающиеся без
посредства промежуточных хозяев, а в полевках представители ленточ-
ных глистов Anoplocephalidae. С возрастом особенно увеличивается эк-
стенсивность заражения такими гельминтами, продолжительность жизни
которых в хозяине сравнительно велика, а также теми видами, для кото-
рых сами грызуны являются промежуточными хозяевами, Таким обра-
зом, главные положения Киршенблата, касающиеся гельминтов, во мно-
гом подтверждают нарисованную нами ранее картину возрастной дина-
мики паразитофауны пресноводных рыб. Однако возрастные изменения
в количестве и в видовом составе эктопаразитов у грызунов отсутствуют.
Богатый материал по вопросу о возрастных изменениях пара.знто-
фауны грызунов Таджикистана имеется в работе Сосниной (1957), Она
обнаружила (в согласии с данными Киршенблата). что количество ви-
дов паразит ическнх червей увеличивается с возрастом грызунов, при этом
особенно возрастает количество видов, развивающихся с промежуточны-
ми хозяевами и заражающих грызунов позже, по сравнению с гельмин-
тами, заражение которыми осуществляется без промежуточных хозяев
Еще более сильные возрастные изменения паразитофауны должны
быть у хозяев, развитие которых сопровождается сложным метаморфо
ю.м, например у амфибий. У травяной лягушки (Rana temporaria).
как и у большинства амфибий, длительность жизни равна нескольким
годам, развитие же связано с метаморфозам, со сменой местообитания
и характера пищи. Водный образ жизни и вегетарианское питание голо-
вастика заменяется у молодой лягушки наземной жизнью и животным
питанием (насекомыми). В дальнейшем лето обычно лягушки проводят
22$
на суше вблизи водоемов, а на зиму уходят в воду и зимуют подо льдом.
Весной третьего года жизни лягушка в воде мечет икру.
Это разнообразно жизненных условий, несомненно, должно сильно
отражаться на паразитофауне в различные периоды жизни. Не следует,
конечно, забывать, что в данном случае на наразитофауну влияет не воз-
раст сам по себе, а именно периодическая смена условии существования.
Но эти смены неизбежно отражаются на исследовательском экране в
виде возрастных изменении паразитофауиы
Рис. 166. Диаграмма головастика, окруженного комплексом его наиболее
характерных паразитов
Д — [fhabdias bufonis (в легких); В — Diplodiscus subclavatus (в кишечни-
ке); С — Polystoma integerrimum (в жабрах);/? — Amphileptus branchiarum
(в жлбрах); Е — Trichodlna sp. (к жабрах); F — Nydotherus cordiformls
(в кпш. чинке); G — Balantidium elongatum (в кишечнике); Н — Opaline
ranarttm (в кишечнике); J — Mastigina (в коже); К — Peritrieha gen. sp.
(в коже у основания хвоста). (Оригинал;)
Возрастное исследование паразитов травяной лягушки, предприня-
тое Марковым и Рогозой (1953, 1955) в Петергофе, показало, что голо-
вастики лягушки несут богатую фауну эктопаразитов, состоящую глав-
ным образом из простейших- Trichodlna. прикрепленные Peritricha,
Amphileptus branclriarum, Mastigina hylae н др., а также из личинок
Polystoma integerrimum на жабрах головастика (рис. 166), Эта экто-
фауна постепенно уменьшается и исчезает перед концом метаморфоза,
т. е. пока животное находится еще в воде, но приняло уже вид молодой
лягушки с частично редуцированным хвостом.
Эндопаразиты начинают проникать в головастика только перед
концом метаморфоза, кроме простейших (Opalina, Xyctofherns и др.),
которые заселяют кишечник головастика значительно раньше. Первыми
многоклеточными эндопаразитами головастиков являются церкарин со-
сальщиков Strigeidae. Они внедряются в головастика, образуя подкож-
ные пли целомические цисты с личинкой типа Tetracotyle. Почти в то же
21П
время в кишечнике головастиков начинают изредка появляться молодые
сосальщики Dipfodiscus subclavalus, а немногим позже молодые Rhab-
dias bujonis. Отличие паразитофауны лягушки от головастика видно из
того, что у взрослой лягушки в Петергофе имеется 7 видов сосальщиков
и 5 видов нематод, т. е. паразитофаупа взрослого животного несравненно
богаче и резко отличается от паразитофауны головастиков по видовому
составу.
Очень ясно н более подробно показаны возрастные отличия гельмин-
тофауны Дубининой (1950в) у озерной лягушки (Rana ridibunda) в
дельте Волги, как видно из табл. 4.
Таблица 4
Возрастные изменения гельминтофзуны Rana ridibunda дельты Волги
(по Дубининой)
Возраст лягушек Число видов паразитов Всего видов паразитов
iretnatodes Cestodes Nematodes
Гол<>влстикн 3 1 4
Сеголетки И — 4 15
1 год ..... • . . . 11 — 4 15
2 тола 15 — 5 20
3 I 1 — 5 1b
4 17 1 7 25
5 лет 20 1 7 28
*
Волгарь-Пасту.ховл (1959) подробно изучила возрастную динамику
паразитофауны шести видов бесхвостых амфибий дельты Дуная. У голо-
вастиков опа обнаружила лишь паразитов, развивающихся без промежу-
точных хозяев или активно проникающих из воды личинок (/1/оло alata.
Distomum acervocalciferuni). Резкая смена состава паразитофауны про-
исходила во время метаморфоза.
Вопрос о зависимости паразитофауны от возраста (и размеров) хо-
зяина затрагивается в некоторы.' американских работах. Брандт (Brandt,
19361 приводит некоторые данные о годичном цикле паразитофауны
шести видов североамериканских лягушек. При этом, однако, ларваль-
ный период лягушек автором не исследовался. Для Rana catesbiana
автор разбил исследованных особей на две группы: больше 100 лл дли-
ны и меньше 100 лл. В первой группе заражение паразитами рав-
но 100%, а среднее число особей паразитов на одну лягушку равно 47.3;
у более мелких тягушек эти числа равны соответственно: 89,4% и 10,12.
Взрослые пестоды встречались только в крупных лягушках. Нематоды
были значительно более многочисленны в крупных лягушках как в
смысле числа видов (II и 8), так и в смысле процента заражения (100 и
60.6) и среднего числа и-разнтов па одну особь хозяина (16.6 и 1.6).
Личинки скребней (Centrorhynchus) имелись в лягушках обеих групп,
но у больших лягушек они встречались ч-чце. чем у мелких (78 п 5.25%)
и были многочисленнее (12,6 п 0.3 экземпляра на одну лягушку)
Ранкин (Rankin, 1937) произвел аналогичное исследование 19 видов
Salamandrina из штата Сев. Каролины, вскрыв в общем 1000 экземпля-
ров. Исследование Ранкина распространяется и на ларвальные стадии
амфибий. Данные по влиянию возраста, однако, немногочисленны и ка-
саются лишь трех видов амфибий. Так, для тритонов (Triturns) указы-
вается, что сосальщики Plagifura встречаются в личинках значительно
чаще (88%), чем во взрослых (52%). По-вндпмому, эти сосальщики с
16 В. А. Догель
241
возрастом постепенно элиминируются хозяином С другой стороны, со-
сальщики мочевого пузыря (Gurgoderina) встречаются только во взрос-
лых тритонах. Взрослые нематоды более обычны во взрослых (88.6%).
чем в личинках (4%), главным образом потому, что Capillaria inaequaHs
отсутствует в личинках, находясь у 81 % взрослых тритон,ув.
У амблистомы (Amblystonia орасигп) кишечные сосалыцикл и со
салыцики мочевого пузыря, а также нематоды и клещи встречаются
только во взрослых особях, а скребни ирису гствуют обычно в личинках
В гораздо ме.чыией степени изученным является вопрос о возрас т-
I ы х и з м с и е н и я х п а р а з и т о ф а у и ы • у р а з л и ч н ы х б е с п о-
з в о и о ч н ы х ж и в о т и ы х. Однако все же имеются дан пые, говорящие
о закономерностях возрастной динамики пара итофауны близких к та
ковьгм у позвоночных. Очень отчетливо нарастание экстенсивности (про-
цента) и интенсивности заражения паразитами (главным образом пар-
тенитами трематод) наблюдается как у морских, так и у пресноводных
моллюсков. Так, по наблюдениям Чубрнк (1954 195“) у широко распро-
страненною брюхоногого моллюска Мурманского побережья Баренцева
моря Purpura lapiilus, достигающего половозрел ости на 3 4-м годе
жизни, зараженность партенитами трематод в первые два года жизни
равна пулю, в 3 года— 0,1%, в 4—1,3, в 5 — 3,0, в 6 — 24,0, в 7 — 32.5
и в 8 лет- 43,5%.
Очень сходную картину нарастания зараженности трематодами t
возрастом дают, согласно наблюдениям ряда авторов, и пресноводные
моллюски (Чорногореико-Б1дулл1на, 1958; Фролова, 1958>. Аналогии
нь1е данные имеются и в отношении некоторых морских ракообразных
Успенская (1950, 1955) наблюдала повышение с возрастом процента за
ражгппя Gammarus метацеркариями Podocotyle utomvn и краба Ilyas
агапеи.ч личинками скребней.
Особый характер приобретают возрастные изменения паразитофах
ны у тех беспозвоночных хозяев (в первую очередь насекомых), у которых
имеется ясно выраженный метаморфоз. Здесь, аналогично тому как это
мы видели выше у амфибии, паразиты личиночной фазы резко отлп
чаются ог паразитов имаго, что связано с различиями в экологии (r.iai
ным образом в инщс) личинок и имаго: чем значительнее эти различия,
тем больше разницы в составе паразитов. .
В качестве крайнего случая подобных различии приведем пример,
который дает Теодоридес (Theodor ides, 1955) в большом исследовании
ио паразитам жуков. Личинки жука-бронзовки (Celoruа)—сапрофаги
Они живут в богатой перегноем почве или даже в перепревшем навозе
Теодоридес приводит для них следующий список паразитов: грегарпна
Gregorian cetoniae, амеба Hartmunella zograli. жгутиконосец Polymastix
nielolonthae. нематоды Thelastoma macrainyhidium, Cephalobellus brevi-
caudatu4. Spirura talpae. скребни A'uiracarithorhynchus furudinaceus (ли-
чинки), личинки насекомых Scolia hirta и Billaea peefinata. Взрослые
C.etorr.a питаются на цветах и совершенно лишены паразитов. Аналогии
пых примеров, касающихся насекомых, можно было бы привести мио
жество.
Таким образом, совершенно ясно, что паразитофауна большинства
хозяев испытывает ряд закона мерных изменений параллельно с возра-
стом хозяина Эти изменения могут касаться только отдельных чтонов
паразитарного комплекса или накладывать отпечаток па общий харак-
тер паразитофауны. Изменения мог\т зависеть от весьма мпогообраз
пых причин В одних случаях фактором, вызывающим изменение, являют-
ся просто меньшие разме ры молоди, не позволяющие селиться в ш .
кронным паразитам, в других случаях морфологические особенности
242
молодых животных (фабрициева сумка птенцов птиц), в третьих — ка-
кие-нибудь экологические особенности молодого возраста (длительное
пребывание в норе или гнезде, иной характер пищи, иная внешняя сре-
да— у головастиков травяной лягушки — и т. д.). Но все эти биологи-
ческие особенности молодых животных, отличающие их от животных бо
лее старых возрастов, делают состав паразитофауны в течение жизни
животного из стабильного подвижным, постепенно изменяющимся, что
мы и постарались показать в данном разделе пашей книги.
2. СЕЗОННЫЕ ИЗМЕНЕНИЯ ПАРАЗИТОФАУНЫ
Сезонные изменения природы необычайно сильно сказываются в
первую очередь на органическом мире. Поэтому естественно ожидать,
что паразиты, хотя и не имеют (кроме эктопаразитов) непосредственной
связи с внешней средой, не должны оставаться безучастными к клима-
тическим изменениям времени года. Действительно, сезонные различия
в частоте нахождения отдельных видов паразитов не представляют со-
бой редкости и неоднократно констатированы в литературе; однако об-
следований, касающихся сезонных изменений всей паразитофауны ка-
кого-нибудь животного вообще или хотя бы значительной ее части, пока
еще очень мало.
Первые попытки к выяснению качественных и количественных се-
зонных вариаций целой паразитофауны, как и вообще первые шаги к
широкому биологическому изучению целой паразитофауны, принадлежат
английским ученым Эльтону, Форду и Бекеру (Elton, Ford, Baker. 1931)
В довольно большой работе они пытаются выяснить зависимость перио-
дических изменений смертности у некоторых грызунов, в особенно
сти у лесных мышей (Apodemus sylvaticus) от различных факторов, в
том числе и от влияния паразитов. При этом ими в течение ряда лет про-
слеживался ход различных протозойных, гельминтозных и артроподных
инвазий у трех видов грызунов. Наряду со своей основной задачей авто-
ры касаются и таких вопросов, как сезонное распределение паразитов
и заражение ими хозяев разной величины, а следовательно, и возраста
К сожалению, авторы не делают попытки как-нибудь суммировать свои
исследования по паразитофауне, так что мы принуждены сделать свод-
ку из отдельных данных их работы, останавливаясь главным образом на
паразитах лесных мышей как наиболее подробно изученном объекте
Было исследовано около 700 лесных мышей, причем в них найдено
три вида сосальщиков, три вида цестод и четыре вида нематод. Зна-
чит, в смысле видового состава указанные классы глистов представлены
довольно одинаково, однако в смысле частоты встречаемости дело об-
стоит не так. Сосальщики, взятые все вместе, дают около 2,5% зара-
жения, цестоды— 8,5%, а нематоды — около 78%. Такое различие хо-
рошо объясняется преимущественно вегетарианской диетой хозяина. Со
салъщики и цестоды, попадающие в мышь при поедании ими промежу-
точных хозяев (вероятно насекомых), представлены очень слабо как в
смысле экстенсивности, так и, ио-видимому, в смысле интенсивности ин-
вазии. По крайней мере, в отношении наиболее часто встречающегося
у лесных мышей сосальщика Harmostomum recurvum сообщается, что
обычно его находили в числе 1—2 экземпляров. Напротив, нематоды
часто находят себе путь в хозяина или через кожу (как это авторы пред-
полагают для самого частого паразита лесной мыши (Heli^mosotnum
dubium). имеющегося у 60% мышей), или через загрязнение пищи’
яйцами (как это имеет место для второго наиболее частого паразита
мышей (Syphacea obvelata), встречающегося у 32% Apodemus). Поэто-
му частое нахождение нематод у мышей является вполне естественным..
В качестве эктопаразитов на лесных мышах попадается девять ви-
дов блох, из которых, однако, лишь один (Ctenophihalimis agyrtes)
встречается на 38 о мышеи и один (Сет atophy tins lurbidus)— на 4%.
Все прочие — лишь случайные гости лесной мыши и попадались не бо-
т<_е чем у 1% особен. Далее, на лесных мышах имелся одни вид вшей
(Polyplax serrata), один вид клещей Ixodidae (/xodes tenuiroslris) и
10 видов Laelapidae и Tmnibidiidae-
Рнс. 167. Кривил динамики заражеитя л чнэй мыши Ар ntemtis
sylvaticus неилгодами MeligHiosotuum tffbium
Д — экст»чЬ1ин->С1Ь ннчмии мышеи нематодами по кварталам
года (начиная с 4-ги т. е. октябрь декабрь) за тра после-
дующих года; В - р icnpe слепне н.-магод по кошевам раАпч-о
возраста, критерием которого взят и-с мышей в грамма(По
Элыс-пу)
Нз рис. В сплошная линия — распределенн- .|этод в самцах, лрернинстая — п
самках ми tn ей.
Что кесастся сезонного распределения паразитов лесной мыши
(рнс. 167), то картина для разных видов паразитов меняется. Так, для
Heligmostynum diibitun частота нахождения, если считывать только
взрослых мышей, приблизительно одинаков! в течение целее года. Дру-
гая нематода (Sypliacea obvcluta) не показала сколько-нибудь опреде-
ленных результатов, так как первый год исследования дал по каким-то
причинам чрезвычайно пониженные цифры заражения; такчо же кар-
тину, но с повышенным заражением в течение пе рвой зимы рок >зал лен-
точный инет Catenotaenia. Прочие внутренние парашты попадались
столь редко, что пе могли быть использованы для решения данного во-
проса. Таким образом, сезонность распределения внутренних паразитов
у лесных мышей остается неясной.
Среди эктопаразитов блохи и вши дают картину повыше пня инва-
зии весной (II квартал) п летом (III квартал). Так, для Polyplax сред-
няя зараженность мышей в I квартале равна 9%, во II —15, в III —
16,4 и в IV —8,6%. У клешей некоторая сезонная изменчивость могла
быть подмечена только по отношению к клещу Aotoedres, который в
наибольшем колнчсствг попадался в первую половин! года. Паразити-
ческие простейшие (Lamblia. Entamoeba, Etmeria и др.) не показали ни-
каких сеюнных колебаний.
244
Интересно сравнить с данными Эльтона и его сотрудников сведения
о паразитах некоторых грызунов, собранные Харке.моп (Нагкеша, 1936)
в Сев. Каролине. Следует только при сравнении учитывать, что резуль-
таты Харкемы получены на значительно менее обильном материале:
Тогда как Эльтон имел возможность в течение трех последующих лет
исследовать ио нескольку сот грызунов трех различных видов, Харкема
работал приблизительно год п за это время всесторонне исследовал
II американского дикого кролика (Sylvilagus ftoridanus maliurus)>
53 белки (Sciurus curoiineiisis), 54 белоногих мыши (Peromyscus leuco-
piis), 84 домашних мыши и 55 серых крыс (Rattus not ucgicus). Харкема
сделал нарочно выбор хозяев, сильно различающихся по образу жизни.
Белка выбрана как древесное животное, домашние мыши и крысы —
как домашние и притом всеядные грызуны, Peromyscus и кролик — как
дикие растительноядные формы.
Сезонных изменении паразитофауны белки автор не обсуждает, так
как материал по белке отсутствовал у пего за тип летних месяца (июнь-
август). Однако приводимые Харкемон таблицы показывают резкое уве-
личение зараженности белки вшами в течение трех зимних месяцев
(ноябрь—январь).
Харкема говорит о сезонных изменениях фауны паразитов у кро-
лика и крысы, которые были в его распоряжении в достаточном количе-
стве в течение всего года. Для большинства эндопаразитов этих живот-
ных он не мог усмотреть заметных сезонных вариации, но Citlotaenia
pectinata явно превалирует в апреле — июне, a Trichostrongylus calca-
ratus кролика дает резкий скачок зараженности (90% и 2000 нематод
на одну особь хозяина) в весенние месяцы и в начале лета (март
июнь). Автор объясняет эту особенность и падение заражения к разгару
лета тем обстоятельством, что после усиленного весеннего заражения
животное приобретает по отношению к этому паразиту частичный имму-
нитет. Слабая инвазия Trichostrongylus в зимние месяцы толкуется, как
результат влияния низких температур па свободно живущие личиночные
стадии этой нематоды. Среди эктопаразитов сезонность обнаруживают
клещи — Ixodoidca и блохи. Блохи кролика (Cediopsylla simplex и
Odotitopsyllus multispinosus) превалируют в течение февраля — апреля,
когда кролики, по мнению автора, долгие периоды времени проводят в
своих порах. К сожалению, выводы Харкемы базируются на малом ко-
личестве исследований (2—5 кроликов в месяц), что заставляет отно-
ситься к ним с осторожностью.
Очень Детальный анализ сезонной изменчивости паразитофауны
(как экто-, так и эндопаразитов) для многих видов грызунов Средней
Азии (Гнссарской долины п южного склона Гпссарского хребта в Тад-
жикистане) приводится в работе Сосниной (1957). В районе Душанбе
максимальная зараженность грызунов почти всеми видами паразитов
имеет место весною. Летом происходит снижение зараженности. Осенью
наблюдается второй максимум с последующим ослаблением заражен
пости зимою. Это отмечается для ленточных и круглых червей, гамазо
вых клещей, вшей и блох. Интересно, что в другом районе Таджикиста-
на— в ущелье Кондара— Соснина не наблюдала летнего снижения за-
раженности. Для разных видов паразитов в этом районе была выявлена
довольно пестрая картина возрастной динамики, но в общем у болышш
ства паразитов обнаружилось возрастание зараженности от весны к
осени с минимумом заражения зимою. Указанные различия в сезон-
ной динамике паразитофауны между двумя районами Таджикистана
Соснина ставит в связь с различиями климатических условий. В районе
Душанбе летом наблюдается очень жаркая и сухая погода, вызываю-
245
щая снижение зараженности паразитами. 3 ущелье Кондара такой лет-
ней жары не бывает и влажность летом здесь значительно выше, чем
в Душанбе. Таким образом, сезонная динамик паразитофауны сказы
вается тесно связанной с климатическими факторами.
Иногда у птиц довольно ясно выраженными оказываются се-
юнчые изменения паразитофауиы в пределах одного и того же вида
хозяина. Так, Зехпов (1919а) отмечает, что у галки в несение-летний
период параллельно с
пищи
та ми,
видов
параллельно с увеличение л в пищевом рапноне животной
(в июне, июле, августе) увеличивается зараженность гельмин-
иолучаемыми через поедание промежчточных хозяев. У отдельных
паразитов галки (например, у
Anomotaeiiia constricia) частота
заражения в летние месяцы по-
вышается до 50- 60%, тогда
как в зимние не превышает
12.5%.
Аналогичную картину се-
зонной динамики паразнтофау-
иы Зехпов наблюдал и у грача.
Довольно сложную картн-
ну представляет сезонная дина-
мика эктопаразитов галки, так
же подробно изученная Зехно-
вым (1919а). Например, в от-
ношении перьевых клещей пай
большая экстенсивноегь и ин-
тенсивность заражения наблю-
дается в летние месяцы. Она
снижается зимою, чго, вероят-
но зависит от непосредствен-
ного влияния па клещей
Эктопаразитов низкой температуры. Большинство видов клещей зимою
не размножается. Заметное уменьшение количества к leineii имеет место
также в период
стенное влияние на количество ж товара .итов, как показал
оказывает купание галок, осуществлщ.401 наиболее интенсивно
л в начале лета. При этом часть клицен и их яиц с
удаляется.
Паши знания о сезонных изменениях па р а з и то ф а) пы
•чес>щы
Рис. 168. Сезонное расирстеюние общего
заражения всеми многоюкл очными паразитами
аламандры DasmognaUA ftiscus jusctis. (По
Ранкину.)
шньки хозяина — в сентябре-октябре. Наконец, суще-
Зсхнов,
вест но
перьев
амфи-
бий пока еще очень фрагментарны. Довольно многочисленные, хотя и
недостаточно систематизированные снедения о сезонности распростране-
ния паразитов различных Sjlainandrina (рис, 168) из Сев. Каролины
зает Ранкин (Rankin, 1937). Различные группы паразитов здссо, по его
наблюдениям, Ьедут себя по-разному. Гак, кишечные нросгсннше дают
высокий процент заражения в течение всего года Целый ряд сосальщи-
ков (Brachycdtyium, Plapitura, Amphistomum) имеет тенденцию к усп
леплю инвазии в мае—августе) тогда как зимой зараженность падает.
Автор правильно лредшмагагт. что это зависит от характера образа
жизни промежуточных хозяев, которыми служат какие-нибудь насеко-
мые, распространенные в начале лета. Паразиты, полученные хозяином
в этот период, потом постепенно вылирьюг. Скребни встречаются в
саламандрах почти исключительно в гечсние зимних месяцев Очень
ясную картину сезонности дают личинки клеща Иаппетапта dumii. за-
ражение которым держится с августам и достигает максимума в ноябре.
Личинки почти не встречаются с февраля по июль По-виличому. весну
и лето клещ проводит в половозрелом состоянии, откладывая в копие
246
лета яйца, личинки выходят из яиц осенью, попадают на саламандр и
держатся на них до весны.
Помимо собственных, Ранкин приводит также некоторые лнтера-
турнвй данные о сезонных различиях фауны у саламандр. По Холлю
iHoll, 1932), Triturus имеет максимальное заражение паразитами в те-
чение зимы. Брандт (Brandt, 1936) находит, что лягушки удерживают
зимой своих кишечных паразитов. Ранкин считает, что период инвазии у
саламандр согласован с периодом размножения, когда они уходят в
воду, т. е. с весной. Вследствие этого максимум зараженности (в особен-
ности сосальщиками) падает на лето. Саламандра Amblyslomd орасит
в отлично от прочих размножается осенью, и па этот период ее жизни
падает инвазия одной формой Diploslomuliun.
В общем, Ранкин даст такие картину распространения инвазии у
североамериканских саламандр:
Весна — кропят.ie 1 'rotozoa, Capillaria, С osrnoepreoides. Hexamitus.
Лето — Amphiftomum, Brachycoelftfni. метаиеркарнн Corgoderidac. Ptagitura.
Осень — Allocreadium, взрослые Cestodes, различные формы Diplostomulum,
Oxyuris.
Зима — Acanthocephala, кровяные Protozoa, Capillaria, цисты цесто.т. llannetnan-
ilia, Trifrichoniusiix.
Во все сезоны гола: Camatlanus, Oswaldocruzia, Physaloptera. большинство ки-
шечных Protozoa.
Некоторые данные о сезонной динамике паразнтофауны бесхвостых
амфибий приводятся в работах Д бннппон (1950в) и Мазурмовича
(1951). Наиболее подробно Мазурмовичем изучены сезонные изменения
гельминтов двух видов лягушек — озерной (Rana ridibunda) и зеленой
(R. esciilenta) — в OKpvciплетях Киева. Результаты <гго исследований в
отношении гельминтов зеленой лягушки приведены в табл. 5.
I* а б л и ц а 3
Изменения процента заражения Rana escutenta
гельминтами в окрестностях Киева по временам года
(по Мазурмовнчу)
Название группы гельминтов Осень Зима Веспа Лето
Все группы гсльмнн- TOII 100,0 94.1 81,5 100,0
Tremaindcs -4- Moiioge- noidru 92,1 82,4 63.0 85.7
Nematodes .... 46 1 5.9 H.l •12,9
Acanliiocephala . . . 7.7 17 I 17.1 0
Мазурхе вич указывает, что причины, обусловливающие сезонное
изменение, мнлюобразпы и сложны Искать их следует в сезонных пз.ме
нениях словпн среды н характера питания, а также в особенностях жиз-
ненных циклов самих паразитов. Во время Ьимовки половозрелые нема-
тоды, вероятии, завершают свой казненный цикл и гибнут. Поэтому про-
цент заражения ими зимой резко снижается. Начиная с весны и летом
происходит заражение лягушек нематодами, развивающимися без про-
межеточны.х хозяев. Опп достигают по.товозрелостн летом н осенью.
Сосальщики амфибий развиваются с промежуточными хозяевами,
каковыми являются моллюски, водные насекомые и их личинки. После
м'мовкп большинство половозре 1ых сосальщиков, завершив свой цикл,
погибает, а их личинки заражают промежуточных хозяев. Массовое за-
247
ряжение лягушек сосальщиками происходит во второй половине лета
и осенью.
Сосальщики удерживаются в хозяине па зиме. Дубинина объясняет
это явление тем, что в зимние месяцы лягушки впадают в спячку, при
этом развитие сосальщиков замедляется, что и приводи^ к их задгржк»
в хозяине. Паразитирующий в зеленой лягушке скребень Acauthocepha
his гапае имеет промежуточным хозяином рачка Asellus aquaticus, ко-
торый достигает массового развития в районе Киева летом. Веспой пе-
резимовавшие в лягушках скребни гибнут, и происходит очищение хо
зяев от этого паразита. Летом личиночные стадии Acanihocephalus гапае
развиваются в Asellus Поедая их в копне лета и в первой половине
осени, лягушки заражаются скребнями, достигающими половой зрелости
во время зимовки. В результате всех этих сложных взаимоотношений
слагается определенная закономерная сезонная динамика паразнтофау
ны лягушки.
Сезонные изменения паразитофауны рыб исследованы относи
только слабо. Большинство относящихся сюда работ касается изучения
в сезонном аспекте жизненных циклов отдельных видов паразитов, и
лишь в немногих исследованиях рассматриваются сезонные изменения
всей совокупности паразитов того пли иного вида рыб.
Изюмова (1958. 19596) изучала сезонную динамику паразитофауны
некоторых рыб (лещ, чехонь, судак, окунь, плотва, ерш) Рыбинского
водохранилища (верхнее течение Волги). В отношении некоторых групп
или отдельных видов паразитов она обнаружила вполне закономерны»
сезонные изменения. Большинство видов мнксосиорндий развивается в
значительных количествах в зимние месяцы, тогда как летом они почти
отсутствуют. Автор ставит это в связь с тем, что зимой рыбы мало
активны и собираются в глубоких частях (ямах) водоема группами, чтс
облегчает заражение развивающимися в это время спррамн мнксоспо
| 'ИДИН.
Интересную н довольно сложи’ю картину дают mohoiенетмчсскш
сосальщики. Значительная часть видов этой группы обнаруживает ясно
выраженный летний максимум развития и зимой в активном состоянии
отсутствует (Dactylogyr^as auricula!us, D. falcaitis, D. nanus н др.)
К другой гр'ппе можно отнести монотонен, дающих летний максимум,
но не исчезающих полностью зимой (D plozoon paradoxum, Ancyrocc-
phalus paradoxus). Наконец, третья группа — это моногспси, встречаю
щпсся круглогодично и нс обнаруживающие катгон-либо цикличности
(Dactylopyrus simplii imallcata). Столь различный хара тер сезонной
цикличности в пределах одной группы зависит, вероятно, в какой-те
мере от отношения к температуре. Изюмова (1953) экспериментально
показала, что два близкие вида рода Dactylogyrus (Г) vastator и D so-
lidus). паразитирующие па карпе, имеют весьма различный температур
нып оптимум развития и яйцекладки. Оптимальная температура яйце
кладки D. vastator—18—2 Г, D. suldus гораздо более холодолюбив,
температурный оптимум его яйцекладки—10—14 Глубокие различия
в отношении этих двух видов к условиям среды (и в частности к темпе-
ратуре) опредетяют и различный характер сезонного цикла. D. vastator
может размножаться только летом, D. solidus — осенью и даже зимой.
Этот пример показывает, что изучение сезонной динамики паразите
фауны необходимо сопровождать экспериментальными исследованиям и.
вскрывающими отношения разных видов паразитов к отдельным фак-
торам внешней среды.
Многие другие паразиты рыб Рыбинского водохранилища имеют
летний максимум развития (некоторые виты Trichudinu. рачки Ergasi'us
248
и др.). Характерный одногодичный цикл развития с зимним максимумом
обнаруживает сосальщик Bunodera luciopercae (.Пяйман, 1940; Кома-
рова, 1944). Заражение ими происходит осенью, к концу зимы они до-
стигают половозрелости, весной рыба очищается or них почти пол-
ностью. По-видимому, годичный цикл, захватывающий, однако, различ-
ные сезоны, характерен для многих паразитических червей рыб. Кома-
рова (1950) доказала его для Acanthocephalus lucii, Маркова (1958)
для Azygia lucii. Малахова (1959) для Allocreadiuni isoporuni и др.
Еще менее, чем для пресноводных рыб, изучен вопрос о сезонных
изменениях паразитофауны морских рыб. Шульман и Шульман-Альбова
(1953) для скребня Echinorhynchus gadi из Белого моря описывают ти
личный одногодичный цикл. Заражение рыб происходит в сентябре
К концу зимы паразит достигает половозрелостн, весной и летом проду-
цирует яйца, а затем погибает. Определенную зависимость от сезона
года эги же авторы обнаружили для нематоды Cucullaiius keterochrous
и рачка Lernaeocera branchialis из морских камбал. Здесь максимум
развития падает на лето и начало осени. Нобль (Noble, 1957) изучал
сезонные изменения паразитических простейших некоторых морских рыб
(Microstomas kitt, Callionytnus lyra, Cadus merlangus) в окрестностях
Плимута (Англия), Он приходит к выводу, что миксосгюрндин желчного
пузыря наиболее многочисленны летом, а гемогрегарины в крови —
зимой.
Таким образом, зимний максимум развития миксоспоридий, уста-
новленный рядом авторов для пресноводных рыб, по-видимому, не рас-
пространяется на морских рыб. Полянский (1955), исследовавший пара-
зитов рыб Баренцева моря в различные сезоны, не обнаружил значи-
тельных сезонных различии паразитофауиы. Возможно, что это связано
с относительно меньшими сезонными колебаниями гидрологического ре-
жима (благодаря теплому Нордкапскому течению), по сравнению с кон
тннентальным арктическим водоемом, каковым является Белое море.
В еще меньшей степени, чем в отношении паразитов рыб, изучена
сезонная динамика паразитофауны разных беспозвоночных Одна-
ко даже имеющийся в этом направлении материал показывает, что здесь
также имеется определенная сезонная динамика паразитофауиы. Пожа-
луй, полнее всего этот вопрос изучен в отношении партеногенетических
поколений трематод в пресноводных и морских моллюсках. Для тех и
для других многие авторы отмечают быстрое возрастание процента и
интенсивности заражения партеногенетическими поколениями сосаль-
щиков от весны к концу лета и затем падение зараженности к осени и
почти полное отсутствие зараженности зимой (Чорногоренко-Б1дулл|’на.
1958; Фролова, 1958; Суханова, 1958; Чубрик, 1957; Гинецинская, 1959).
Вероятно, что основными факторами здесь являются температура в
водоеме и увеличение активности и количества окончательных .хозяев по-
звоночных. Существуют закономерные сезонные соотношения разных
фаз развития паразита (Чубрик, Фролова). Зимой, если трематоды и
встречаются в моллюсках, то развиты только спороцисты и редии. Вес-
ной же и летом появляются активные стадия — церкарин.
Ранкин (Rankin, 1939) для партеногенетических поколений сосаль-
щиков, паразитирующих в моллюске Pseudosuccinea columella (США,
штат Массачузст), наблюдал отчетливо выраженную сезонность в жиз-
ненном цикле. При этом разные виды сосальщиков обнаруживали мак-
симум развития в разные сезоны года. Эхиносто.мндные церкарин да
вали резко выраженный максимум развития в зимние месяцы (декабрь,
январь, февраль), кенфидиоцеркарии (xiphidiocercariae)— летом (июнь,
июль, август).
2-4'»
В отношении некоторых паразитов пресноводных ракообразных
также намечаются закономерности сезонной динамики паразитофауны.
Штеп? (1957) в некоторых озерах Карелин наблюдала наличие се-
<онпых изменений в заражении некоторых членистоногих сосальщика
мн. Ойо отчетливо было выражено у амфипод и личинок вислокрылок
(Sialis). У вислокрылок Штейн наблюдала два максимума заражения
зимой и летом. У бокоплавов наблюдается летний максимум заражения
(в июле). Вероятно, в летнем увеличении зараженности существенную
роль играет массовый выход церкарнй, нападающих на вторых проме-
жуточных хозяев. Одним из факторов снижения процента заражения к
осени является выход молодого незараженного поколения бокоплавов,
влияющего своим появлением на общий процент зараженности по-
пуляций
Отчетливую картину сезонных изменений зараженности наблюдала
также Успенская (1955) на некоторых бентических ракообразных Барен-
цева моря.
Необходимо иметь в виду, что сезонные изменения паразитофауны,
лишь кратко рассмотренные выше, и еще во многом недостаточно изу-
ченные, представляют собою очень сложные и комплексные явления,
определяемые множеством абиотических и биотических факторов среды.
Среди них большое значение имеют климатические факторы и в первую
очередь температура.
У паразитов понкилотермных животных наблюдаются отчетливо
выраженные изменения теплоустойчивости в разные сезоны года. В этом
отношении большой интерес представляют наблюдения Сухановой (1959,
1962) над простейшими — паразитами кишечника амфибий. Суханова
изучала стойкость паразитов к высокой летальной температуре (обычно
около 38°) и определяла теплоустойчивость по быстроте гибели пара-
зитов. Она потучита для Opalina ranarutn нз травяной лягушки следую
щис данные относительно быстроты гибели при 38°:
Зима (январь)
6,32 (J (18 мин
7 5O-U.17 .
11.49^0,(18 -
Веспа (апрель)
7,83 ±0,06 мин
9,22 ±0.10 .
10,39 ±0,02 ,
Лето (июль)
17,7-1 ±0,21 мин
18,96 ±0.17 .
22.00±0(18 .
Осечь (сентябрь)
И, 45 ±0.17 мни
14,67 ±0,21 .
1-7,62 ±0 05
Эти наблюдения показывают, насколько значительно меняется теп
лоусгончнвость паразитов в разные сезоны года В данном случае это
изменение теплоустойчивости зависит от температуры и представляет
собою яркий пример зависимости физиологического состояния паразита
от внешней среды (т. е. от среды второго порядка) Сухановой удалось
очень убедительно доказать зависимость теплоустойчивости паразитов от
температуры внешней среды экспериментальным путем. Она помешала
лягушек в разные температурные условия и при этом наблюдала такие
же изменения теплоустойчивости паразитов, которые в естественной об
становке происходят в разные сезоны года (Суханова, 1959).
С сезонными климатическими изменениями тесно связаны измене
ния характера питания и образа жизни хозяев, влияющие на возмож
преть и вероятность заражения паразитами. Существенным моментом
для паразитов являются сезонные изменения активности хозяев—их
подвижность, образование стай и т. п. Важное значение имеет также эко-
логия промежуточных хозяев и ряд других факторов. Изучение сезон-
ных изменений паразитофауны должно сводиться нс только к конста-
тированию самого факта в его различных проявлениях, но такжр к ана-
25U
лизу отдельных факторов, влияющих на тот иди иной ход сезонных изме-
нений паразитофауны. Именно в этом направлении сделано еще, как
видим, очень немногое.
3. ВАРИАЦИИ ПАРАЗИТОФАУНЫ В РАЗЛИЧНЫЕ ГОДЫ
По отношению к отдельным видам паразитов (в особенности у че-
ловека и домашних животных) имеются довольно многочисленные дан
ные о том, что в различные годы, отличающиеся один от другого в кли-
матическом отношении, зараженность данным паразитом может варьи-
ровать в очень широких пределах. Достаточно вспомнить о значитель-
ном усилении зараженности рогатого скота печеночной двуусткой в
дождливые, сырые годы.
Однако во отношению к паразитофауне какого-нибудь животного,
взятой в целом, такие наблюдения немногочисленны, ибо требуют чрез-
вычайно долгой п трудоемкой работы.
Ряд данных по этому вопросу имеется у Элтона, Форда и Бэкера
< 1931) относительно паразитов грызунов в Англин. Так, например, если
мы возьмем зараженность лесной мыши (Apodemus sylvaiicus) в одни
и те же месяцы трех последовательных лет, то получим следующую кар-
тину (табл. С):
Таблица 6
Процент заражения Apodemus разными паразитами в октябре — декабре
1925—1927 гг. (по Элтону и др.)
Годы наблюдений iieligmo- sornum Syphacia Cate notae- nia Cn-noph- thalmus Polyp- lax Notoed res
1925 100 97 16 15 16 Данных iiei
1926 74 22 4 31 11.5 13
1927 79 26 10 9 1 6,5
Данные табл. 6 свидетельству ют пока лишь о том, что у многих па-
разитов имеются очень резкие годичные колебания экстенсивности за-
ражения. К сожалению, каких-нибудь причинных связей и зависимости
от климатических условий разных годов авторам подметить не удалось.
При оценке годовых различий в паразитофауне необходимо обра-
щать внимание лишь па случаи крупных отличий, тогда как мелкие рас-
хождения в 10 и даже 20% не выходят за пределы возможной ошибки
Подходя с подобным ограничением к данным Зсхпова (1949а) о
паразитофауне галки в Вологде, мы получаем у некоторых членов эндо-
фауны этой птицы вполне ощутимые годовые колебания.
Зехпов изучал экто- я эндопаразитов галки в течение двух следую
щих друг за другом лет (1938 и 1939 гг), довольно существенно отли
чавшихся друг от друга по метеорологическим условиям. В 1937—
1938 гг. была продолжительная и холодная зима, весна — теплая, лето
жаркое и сухое. В 1938—1939 гг. зима была значительно теплее, весна,
напротив, была холоднее и с гораздо меныпим количеством осадков,
чем в предыдущем году. Лето 1939 г. было менее жарким, чем в 1938 г.,
и с еще меньшим количеством осадков.
Вероятно, в связи с этими различиями во внешних факторах зара-
женность некоторыми паразитами в эти два следующие друг за другом
года существенно различалась. Так. в 1938 г. птенцы были заражены
251
Prosthogonimus ovatus на 70,6% при средней интенсивности в 15,2 эк-
зсмкляра на одного хозяина. В 1939 г. зараженных птенцов было всего
25% при средней интенсивности 2,3 экземпляра. Цестода Anomotaeniu
в птенцах в 1938 г.— 17,6% при высокой интенсивности (13,5 экземпля-
ра), тогда как в 1939 г.—18,4%, но с меньшей интенсивностью (4,7 эк-
земпляра). Нематода Capillaria в птенцах дата в 1938 г. 31,7% с Интел
снвностыо 19,0 экземпляра, а в 1939 г. — 51,5% при интенсивности 22,Ь
экземпляра. У взрослых птпц различия в зараженности паразитами
в 1938 и 1939 гг. были менее резкими, хотя в отношении некоторых ви-
дов довольно отчетливо выражены.
В отношении же некоторых других видов паразитов (например, Syn-
gamus trachea, Tamerlania zarudniji) существенных различий в зара
женносги галки в 1938 и 1939 гг. не наблюдалось.
Интересные данные о годичных изменениях паразитофауны сквор
па приводит Марков (1910), сравнивший в этом отношении 1935 п
1936 гг. в окрестностях Ленинграда (в Петергофе). В 1935 г. была за
тяжная, холодная и дождливая весна и прохладное лето с большим ко-
личеством осадков. 1936 год характеризовался ранней и теплой вес-
ной, теплым и очень засушливым июнем (засуха, пересыхание мелких
водоемов). В июле и августе — вновь много осадков и тепло. Как эти
метеорологические особенности двух соседних .чет отразились на пара'
знюфауне скворцаэ
Весной 1936 г. (по сравнению с 1935 г.) условия для развития яиц
и промежуточных хозяев были гораздо более благоприятными. В связи
с этим наблюдалось значительно большее развитие всех групп гельмии
тов как в отношении процента, так и интенсивности заражения. Напри
мер, для Plagiorchis brauni эти различия представлены в табл 7
Таблица 7
Распространение Plagiorchis brauni у птенцов скворца в 1935 н 193о гг.
в Петергофе (по Маркову)
Г оды наблюдении Процент зараже- ния Всего червей Средняя интенсив- ность зара- жения Максималь- ная интен- сивность Впервые появились Возраст птенцов впервые за- раженных (в днях)
1935 31 НН5 5 14 31 V 10-15
1936 56 1253 52 230 19 V 5
Аналшичная картина в мае наблюдалась и в отношении других па-
разитических червей скворца, как видно из следующих данных:
1935 г 193'' г.
Plagiorchis maculosus ........... 25% 82%
Prbsthogqriwius ovatus P. cuneatus ... 16 43
Lait’rotrt'nia ve'fans ............ 1 40
Примерно в два раза сильнее в 1936 г. были заражены птенцы
скворца по сравнению с 1935 г. также ленточными червями (Нутепо-
lepis, Choanolaenia, Paricterotaenia).
Интересные изменения произошли в июне 1936 г. в связи с засухой.
Capillaria contorta совершенно исчезла, тогда как в 1935 г. ею были
заражены 50% птиц. Syngamus trachea в 1936 г. в июне дали лишь 5%
252
заражения взрослых скворцов, тогда как в 1935 г.—47%. В июле и авгу-
сте после окончания засухи, но при сохранении теплой погоды вновь
наблюдалось быстрое увеличение зараженности скворцов гельминтами
Засушливый июнь явился как бы биологическим барьером для развития
многих паразитов.
Эти данные Маркова представляют интерес в том отношении, чго
они позволяют связать годичные колебания в паразитофауне с опреде-
ленными годичными колебаниями метеорологических условий.
Сведения о годовых различиях паразитофауны амфибий крайне не-
значительны. Наибольший интерес в этом направлении представляет
работа Маркова и Рогоза (1955), касающаяся паразитов травяной ля-
гушки (Rana teinporaria). Работа была выполнена в те же годы и в том
же месте, чго и рассмотренное выше исследование по паразитам сквор-
ца. Так же как и для скворца, авторы отмечают для лягушки значитель-
ное пс вышенпе зараженности в теплом 1936 г. по «рйвпейию с 1935 г
Различия сводятся к двум моментам. 1) повышение зараженности в
1936 г. по сравнению с 1935 г. у лягушки было менее отчетливым, чем
у сльорца, и 2) июньская засуха 1936 г. не вызвала заметного снижения
:аражениссти лягушек. Последнее вполне понятно, так как лягушки
держались в сырых местах у берегов более крупных, непересы.хающих
водоемов. Сравнение зараженности травяной лягушки в 1935 и 1936 гг
по месяцам представлено в табл. 8.
Таблица 8
Зараженность Rana temporaria из Петергофа
дигенетическими счсальщиками в 1935 и 1936 гг.
(по Маркову и Рогоза)
Месяцы Экстенсивность заражения в процентах
1935 г. 1936
Апрель 60 92
Май — июнь .... 70 Si.)
Июль август . . . R1
Сентябрь — оыябрь. 70 95
Как пйдно. метеорологические различия 1935 и 1936 гг. оказали глубо-
кое влияние на паразнтсфау ву разных животных.
Очень мало игвестно об изменениях паразитов рыб одного водоема
в разные годы Имеются, правда, исследования по изучению изменений
паразитоф-г ны рыб при строительстве водохранилищ. Однако эти дан-
ные не могут быть использованы в связи с рассматриваемым нами во-
просом. так как при <| ормнрованни водохранилищ резко меняются в ре-
зультате деятельности человека ус тория жизни рыб н их паразитов, а
потому' эти явления представляют собой самостоятельною подлежащую
изучению проблему, которая и обсуждается нами ниже (гл. XIX). В дан-
ном же разделе речь идет об изменении паразнтофауны рыб в водое-
мах, не подвергающихся целенаправленному и резкому воздействию со
стороны человека н результате его хозяйственной деятельности.
1 !екоторые существенные изменения в паразитофауне За ряд лет
произошли, по-видимому, в ряде озер Карелии Так, по данным Петру-
шевского и Быховс»',ой-Павлпвскоп, в 1935 г рачок Ergasilus siebo'di
в Кончозере был очень редким паразитом. При исследовании Шульма-
ном и Рыбак (1961) тех же озер в 1953—1951 гг. оказалось, «тл Е. sie-
bold- — обычнейший паразит, дающий у щук, например, почти 100%-ное
253
заражение. Существенные изменения были констатированы и в отно-
шении некоторых других паразитов. Чем они вызваны — остается не-
выясненным. Шульманом высказывается предположение (личное сооб-
щение), что эти изменения паразитофауны озер Карелин, возможно,
связаны с процессами постепенной эутрофикаини водоемов. Однако
имеются некоторые данные, указывающие на то, что в других случаях
состав паразитофауны и зараженность рыб в течение длительных про-
межутков времени почти не изменяются.
Догель и Петрушевский в 1933 г. изучили состав паразитофауны
Невской губы В 1958—1959 гг., т. е через четверть века, эти же работы
на том же водоеме и с таким же количеством материала были повто-
рены в нашей лаборатории У Бао-Хуа (1961). Получилось почти полное
совпадение результатов. Никаких существенных изменении в паразито-
фауне рыб Невской губы за 25 лет не произошло. Можно думать, что и
гидрологический режим этого эстуарного водоема не претерпел сколько-
нибудь существенных изменении.
Имеются только совершенно единичные данные, касающиеся го-
дичных изменений паразитов беспозвоночных. Например, Штейн (1957.
I960), изучавшая паразитов членистоногих в озерах Карелии, отмечает
довольно значительные колебания процента заражения личинок насеко-
мых и Amphipoda метацеркарпями трематод в разные годы. Например
личинки ручейника Stenophylax из Сямозера в 1954 г. были заражены
метацеркарпями на 17,2%, а и 1955 г —на 66.1%. Подобного рода коле
бання отмечены ею и для других паразитов членистоногих Очевидно.
>ти различия определяются сложным комплексом биотических отноше
нпй и метеорологических факторов, природа которых не ясна.
Широко известно, что у насекомых, дающих вспышки массового
размножения, обычно вслед за увеличением численности насекомого-хо-
зяина происходит массовое развитие паразитов, резко снижающих чис-
ленность насекомого. Здесь, таким образом, численность паразитов, так
же как и хозяев, в разные годы оказывается очень различной.
Как ясно из изложенного, вопрос о годичных колебаниях паразите
фауны и факторах, их обусловливающих, находится на первоначальной
ступени накопления фактического материала. Дальнейшее изучение
его представляется важным и с практической стороны, так как оно
будет способствовать прогнозированию численного колебания парази-
тов, многие из которых имеют важное практическое значение. f
Долговременные наблюдения над годовыми колебаниями парази-
юфауны были проведены Васильевым (1939) в нашей лаборатории в ок-
рестностях Ленинграда (в Петергофе) и касаются эндопаразитов зем-
лероек. Данные эти но ряду видов паразитов сведены нами в табл. 9.
Таблица 9
Изменение зараженности землероек разными видами паразитов в различные
годы в Петергофе (в %) (по наблюдениям Васильева)
Годы на- блюдений I.ongistria- ta dept fssa Plagiorrhis exasperates Harrnn- stemum Choanotae- nia Ilymenolepis singularis Hymenalepis scutigera
1934 I ОН 40 45 94 25 44
1935 60 _»n 40 40 20 0
1936 Наблюдений не были
1937 89 46 ? 48 o 0
1938 83 29 21 96 45 II
1939 87 62 12 6? 37.5 50
254
Из таблицы, в которую включены лишь наиболее распространенные
у землероек виды гельминтов, видно, что почти все они в разные годы
могут обнаруживать весьма различное заражение в одной и той же
местности и в одно и то же время года (все данные взяты в июле и авгу-
сте и выражены в процентах заражения).
Годичная разность сказывается не только на частоте нахождения,
но и на времени появления определенных паразитов. Так, Дубинин (19158)
констатирует, что в дельте Волги в прохладное лето 1936 г. некоторые
перьевые клещи каравапкн (Megninia) появились на птенцах лишь
29 июня, тогда как в жаркое и сухое лето 1935 г. они перешли на птен-
цов уже 20 июня. Аналогичное запоздание дали и клеши ночной цапли,
или кваквы
ГЛАВА XIII
ЗАВИСИМОСТЬ ПАРАЗИТОФАУНЫ ОТ ПИЩИ
И ОБРАЗА ЖИЗНИ ХОЗЯИНА
I. ПАРАЗИТОФАУНА И ПИЩА ХОЗЯИНА
Наличие известной зависимости состава паразитофауны животного
от характера его нищи не подлежит никакому сомнению и ясно следует
из того, что многие паразиты, как говорилось ранее, самый доступ в тело
хозяина получают через пищеварительный канал при поглощении пищи
Однако легко можно видеть, что характер пищи должен оказывать на
иаразитофауну более частное воздействие, чем возраст, ибо от влияния
нищи хозяина совершенно ускользают все эктопаразиты, а также те
внхтреннсстные паразиты, которые проникают в тело хозяина экзоген-
ным путем, например кровепаразнты. Таким образом, характер пищи
хозяина оказывает несомненное влияние на кишечных паразитов и на
часть внутренностных паразитов. При этом одинаковость пищи влияет,
судя по всему, в двух направлениях С одной стороны, одинаковость пи-
щи, например подспрание с земли трапы травоядными млекопитаю-
щими. влечет за собой заглатывание покоящихся стадий одних и тех же
паразитов, которые вследствие этого получают шансы приспособиться
к жизни в кишечнике различных хозяев, объединяемых общностью од-
ного фактора — характером пиши. С другой стороны, можно думать,
что одинаковость пищи приводит к сходному химизму кишечной среды,
а это в свою очередь создаст условия, благоприятствующие заселению
одинаково питающихся хозяев сходными паразитами.
Рассматривая вопрос о зависимости состава паразитофауны от ха-
рактера пищи и других экологических и физиологических факторов, не-
обходимо иметь в виду, что заражение хозяина тем или иным видом па-
разитов определяется не только возможностью его проникновения, но
также п возможностью развития и завершения цикла. Здесь мы сталки-
ваемся с явлением специфичности как приспособленностью паразита к
определенному кругу хозяев (о специфичности см. главу XXII). Парази-
ты узкосИенпфичные приурочены к одному или немногим видам хозяев
и в случае попадания в других хозяев в них не развиваются. Паразиты
широкоспеипфпчные могут развиваться во многих хозяевах. Именно по-
следняя группа н обнаруживает особенно отчетливо выраженную завн
симость от экологических факторов среды Видовой состав паразитофау-
ны определяется как бы равнодействующей двух сил: с одной стороны.
256
факторами исторического порядка — прошлой эволюционной историей
хозяина и паразита, а с другой — многочисленными ныне действующими
экологическими факторами, среди которых способ питания и характер
пищи игрист очень существенную роль.
Отдельные факты, говорящие о влиянии пищи на состав паразито-
фауны, известны в достаточном количестве. Так, для объяснения значи-
тельного сходства к плечной и внутренностной паразитофауны свиньи
с таковой человека указывают на всеядность обоих этих организмов.
Действительно, у человека и свиньи имеется значительное число общих
или чрезвычайно близкородственных паразитов. Средн простейших ука-
жем на инфузорию nalanlidium coll из толстой кишки, среди ленточных
глистов — taenia solium, которая в своей пузырчатой стадии может
развиваться не только у свиньи, но и у человека. Круглые черви лают
нам пример общности паразитов свиньи и человека в лице трихины.
Кроме того, Ascaris lurnbricoides человека морфологически не отличима
от Л. suurn свиньи, так что если это и различные виды, то дифференциро-
вавшиеся от одной исходной формы. Скребс нь-велнкан Macraranthor-
hynchus hirudinaceus кишечника свиньи может. но-види.мому, в отдель-
ных редких случаях ока заться паразитом человека.
Прекрасный пример влияния одинаковой пн щи тают с
одной стороны, копытные, с другой — грызуны. Животные эти принадле-
жат к различным отрядам млекопитающих, но содержат ряд общих или
близких кишечных паразитов, в особенности кишечных инфузорий Для
рубца жвачных и для толстой кишки однокопытных необычайно ха-
рактерна пышная фауна инфузорий подотряда Entodiniomorpha, при-
чем в жвачных встречаются исключительно представители семейства
Ophryoseolccidae (до 150 видов), а в однокопытных — члены сем. Cyclo-
posthiidae (около 50 видов). Кроме того, в толстой кишке однокопытных
очень богато представлен отряд равиоресиичных инфузорий (Holotri-
cha), а именно: многочисленные представители семейств Biitscliliidae и
Paraisotrichidae.
Да К\нья (da Cunha) в целом ряде работ, начинающихся с 1914 г.,
нашел, что в некоторых бразильских грызунах (морская свинка, водо-
свинка), несомненно, имеются инфузории, принадлежащие к несколь-
ким родам из семейств Ophryoscolectdae (Entodnvum) и Cycloposthiidac
(несколько видов типичных Ct/cloposlhium в водосвинке). В водосвинке
констатированы п представители рода Paraisotricha из сем. Paraisotri-
chidae, а в морской свинке—Blepharocorys — род, характерный для од-
нокопытных.
Не менее интересен факт нахождения близких между собой инфузо-
рий из сем. Pycnothrichidae (рис. 169) в кишечнике грызунов и даман»'В,
г. е. разных отрядов .млекопитающих. Все эти инфузории отличается
очень характерной общей чертой, а именно: глубокой продольной бо-
роздкой, пробегающей вдоль одной стороны тела животного. Сначала
был найден род Pycnothrix в кишечнике даманов, потом Шаттон п Пе-
pap (Chaiton el Pcrard, 1919, 1921) описали двы новых рота — Nicnllella
и CollineUa— из кишечника африканского грызуна гундн (Ctenodaclylus
ttondi). Все эти ролы относятся к семейству Pycnothrichidae и нахожде-
ние их у столь различных групп животных, по всей вероятности, вызы-
вается одинаковостью питания их хозяев.
Еше один пример, взятый из типа простейших, дают Hypermastrgida,
паразитирующие в кишечнике насекомых, главным образом термитов.
Этот отряд жгутиконосцев насчитывает свыше 100 видов, большин-
ство которых встречается лишь в питающихся клетчаткой видах терми-
тов. Кроме того, дпл вида, относящихся к особому роду Lophornonas,
1“ В Л. Догг-i
257
' же. давно известны из кишечника тараканов (Blaila urienimis) К.пн.
лэнд (Cleveland. 1934) с сотрудниками нашел богатую фауну Hup- rrtia-
stigida и притом отчасти ге же рплы (Trichonympha), которые живут в
термитах, н кишечнике одного североамериканского таракана (Сгурю
cercus). При этом оказалось, что данный род таракана, подобно терми-
там, питается исключите тьно клетчаткой. Пищевые связи между фауна
mi: Cryptocercus и термитов очевидны
Гораздо меньше имеется по этому
ся всей паразитофауны сравниваемых
вопросу наблюдений, касающих-
жи потных. При этом правильнее
всего сравнивать двух живот-
ных. не имеющих между собой
родственных связей, вед1 щих
одинаковый образ жизни и от
л ичаюшнхся друг от други
только в одном отношении, а
именно по характеру пи
ши; таким нутом выпячивает
ся значение именно одной
фактора питания. Хороший ма
тернал такого рода могут пред-
ставить мелкие Insectivora и
Rodentia. Это животные при
б.шзнтельно одинаковых раз
моров, живущие в норах и т. д .
Рис 169. Инфу зерни из кишечника различных трапоялпых
животных, сходные по наличию у них па теле глубскон про
дол i.iioii борозды
.4—PycnnthriK monocystoides из даманов; Н — Collinella
gundi из африканского ipuayiia: ('—Paraisotrichopsis coni
posita из лошади. (1 и Л — ио Шагтопу, С —по Стрел-
кову.)
/ язкрпнуклсус: 2 продольна и Гнцшддл.
но одни из них насекомоядны, тогда как др\гие зерноядны
Некоторый материал для сравнения мы находим в статье Жиикииа
(1931) о паразитах млекопитающих Узбекистана. Среди других млеко-
питающих Жннкин изучал ежей (Hemiechinus) и больших песчанок
(Rltombomys opitnus). При этом оказалось, что у данной нары живот-
ных фауна эктопаразитов в отношении встречающихся на них групп
(блохи, клещи) была в общем сходной, тогда как кишечные паразиты
показали резкую разницу: песчанка оказалась почти незаряженной,
тогда как \ ежа '•!' нг ходим в значительном количестве тенточных гли-
стов. кругл!.:- червей и скребне:).
Многочисленные и убедительные примеры зависимости состава на-
।*азпге.фауиы. в первую очередь эндопаразитов, от характера питания
25к
даю г ниты. Зернояднм шпцы, как правило,: обладают гораздо более
бедной фауной эндопаразитов' чем насекомоядные и хищные. Это в осо-
бенности касается тех паразитов, промежуточными хозяевами которых
являются беспозвоночные и в том числе насекомые. И пример, по дан-
ным Победоносцева (1949) у воробья (Passer dorneslicus) в окрестно-
стях Саратова было обнаружено всего два вида гельминтов — Choano-
taeniu passe ппа и Acttaria skrjabini. Зараженность первым из них весь
ма низка, он был обнаружен всего у 6% вскрытых птиц (общее колнче
ство вскрытий— 200). Воробел— зерноядная птица, и только птенцы
частично питаются насекомыми. В противоположность этому насекомо-
ядные нгицы. живущие, так же как и воробьи, в близком соседстве с чело
веком, обладают гораздо белее богатей фауной эндопаразитов. Догель
и Навпсвпч (1936), например, у ласточки (Hirundn rustica) в окрестно-
стях Ленинграда нашли 12 видов гельминтов, в том числе 4 вида трема
год, 3 вида цестод, н 5 видов круглых червей, многие из которых давали,
высокий процент заражения. Дубинин и Дубинина (1910) показали на
птицах дельты Волги, что наибольшее число паразитов, развивающихся
при участии промежуточных хозяев, наблюдается > птиц со смешанным
питанием (водные беспозвоночные, рыбы, амфибии). У чисто рыбоядных
птиц (бакланы, пеликан) число этих видов паразитов .шачителыи*
меньше.
Зависимость состава наразитифауны птиц иг.харакгеиа пищи q'ijqhl
убедительно показана Белопольской (1952) в работе, посвященной изу-
чению паразитофауны птиц восточной части Баренцева моря. По харат
п ру питания Бе 1ипольск„я разделяет птиц Мурманского побережья н;
3 группы*
1. Птицы, питающиеся только беспозвоночными; к этой группе огни
сигся обыкновенная гага (Somateria mollisshuaj. гага i ребенушка
(S. speciabelis), кулик-песочник {Calidris martlima).
2. Птицы, питающиеся рыбой и беспозвоночными, - морская чайка
(Larus mariuus), сизая чайка (L. canus), серебристая чайка (L. агреп
talus), моевка (Rissa iridatlyla}, поморник (Slercoraritts parasiticus}.
крачка (Starna paradisea) и чистик (Ceppiius pi pile).
3. Птицы с чисти рыбным питанием—длинпоклювая кайра (L'ria
uulpe), корпткиклювая кайра (U. lomuia), гагарка (Alert tarda), тупик
\Fratercula arcticu), краснозобая raiapa iCiauia stellaia], баклан (Pha-
lacrocorax aristotelis).
Зараженность, перечисленных выше птиц сосальщиками и лепгоч
ними червями представлена в табл. 10
Беспозвоночные являются промежуточными хозяевами большинства
трематод в цестод. чем и объясняется прямая зависимое 1ь между коли-
чеством беспозвоночных в нище (в особенности моллюсков) и числом
видов н интенсивностью зараженности сосальщиками и ленточными чер
вями. Особенно большое количество беспозвоночных поедает гага. Этим
определяется ее исключительно сильная зараженность гельминтами.
Из данных таблицы отчетливо видно постепенное ослабление зара
женности от 1-й группы к 3-й. Птицы, питающиеся исключительно ры-
бой, имеют очень небольшой как но составу видов, так и по интенсивно-
сти j.ipazKLHHH набор гельминтов. Аналогичную зависимость заражен
нгстн птиц гельминтами от характера питания устанавливает Кураш-
ни.ь; Т1957) для птиц Грузни.
Интересные примеры зависимости паразитофауны от характера ни-
щи хозяина приводит Гипсцинская (19-19) в работе, посвященной пзуче
шли паразитов утиных птиц дельты Волги. Здесь речь идет о птицах
близких как в систематическом. так и в экологическом отношениях. Од-
259
Т .ч б л п ца 10
Интенсивность и экстенсивность заражен тя разшчных птиц трем толами и
цесгодами в зависимости от ха(акгсра пищ» (по Белопольской)
1HIJ.II rillllAll ] Вид Tnmeiodv» | Cesiodrs
ЧИСЛО ВИДОВ % зара- жения NBKCil- МИЛЫ1. ш!тен- О1ВП. ЧИСЛО видов Н заря- жен! я и а к си- МЛ.1Ы1 нитей- V 1П и.
1 1 ' Somatcrra inollissima . 10 1 >п.<» 21'04 5 88.0 195(10
‘ 1 Calidris tnariiima . . . 11 lud и 487 П 91.1 161
I.anis argentaius . . . 9 с 0,0 56 7 Н'0,0 51
Larus cantts 9 3J.it 46 7 88.0 163
/tissti tndartvla .... 4 21.1 К) 5 65.9 61
Stercontrius parasiticus. 2 26.6 Я 4 30.0 41
Sterna paradisea . . 1 2 .<» 13 4-л0 5
Cepphus grylle . . . 1 18.6 27 .4 4л 15
Uria aalge ...... 1 2.0 4 Л 36 it 9
Uria lotnvia 1 7.1 1 2 64 1 14
> Fraterrula tirtica . . . 9 11,1 4 2\1 1
и Aiea tarda - - 9 1К.2 2
/ 'ha lac row a x a ris tote Ii s I 44 4 7 1 3 5,3 3
(la via stellain .... 2 3 >.3 2 1 к.0,0 10b
паки некоторые виды уток довольно значительно различаются но ха-
рактеру пищи. Например, серая утка f.lnus streperu) преимущественно
растительноядна, тогда как в питании кряквы (Л. plaihyrhyncha) суще-
ственную роль играет животная пища, в том числе моллюски и личин-
ки насекомых. Этими различиями в питании обусловливается значитель-
ная разница в гсль.минтофауне этих птиц. В серой утке 12 видов гель-
минтов, средн них паразитов, заражение которыми происходит при по
едапии моллюсков, всего 2 вида. В крякве найдено 34 вида паразитиче-
ских червей, из которых 13 видов передаются через моллюсков. Этот
пример поучителен: если близкие виды одного рода, живущие бок о бок,
различаются по характеру питании, то и различие в паразитофаупе ока
зывается весьма существенным.
Киршенблат (1938) замечает, что у некоторых грызунов, регулярно
питающихся, помимо растительной пищи, еще и насекомыми, количество
гельминтов, развивающихся при посредстве промежуточных хозяев, зна-
чительно больше, чем у чисто растительноядных грызунов.
Аналогичным примером, также почерпнутым в среде млекопитаю-
щих, может служить сравнение паразитофауны грызунов п насекомояд-
ных в окрестностях Ленинграда, по материалам, собранным в нашей
лаборатории В. Васильевым (1949) н еще полностью не опублико-
ванным
Сравниваемыми хозяевами служили в данном с.тсчае полевка (['.со-
tomys glureolus) и лесная мышь (Apodernus jlauicoHis), с одной сторо-
ны, и землеройки (Sorex araneus и Neomys fodiens)— с другой. При
этом оказалось, что чисто или почти чисто растительноядные полевки и
лесная мышь были необычайно слабо (па 2,5—4%) заражены сосаль-
щиками всего одного вида (Ptagiorchis tnuris). так как трематоды мо-
гут попасть в окончательного хозяина только через поедание промежу-
точных хозяев (моллюски, насекомые). В отличие от мышей и полевок
насекомоядная землеройка оказалась обладательницей 4 видов сосаль-
щиков, которыми были заражены 50% исследованных землероек
260
lie менее яркую картину дает и заражение сравниваемых живот-
ных .инточными глистами. В условиях окрестностей Ленинграда в по-
левках ( казалось 2 вида цеетод, давших 30% заражения. У лесной мы-
ши найдено тоже 2 вида цеетод, заразивших 20% мышей Напротив то
го, \ землероек число видов цеетод достигает 9, а экстенсивность зара-
жения ими--77%. Картина заражения поделок, мышей и землероек
нематодами менее ясна вследствие того, что пути заражения хозяина
многими из нематод еще не установлены. Землеройки наиболее богаты
и этой гр’ пней паразитов. В результате общее число видов кишечных
гельминтов, обнаруженных у полевки, равнялось 7, у лесной мыши — 5
н \ землеройки -19. Приведенные цифры достаточно ясно свидетель
ствуют о влиянии животного характера пищи на увеличение разнообра-
зия кишечной паразитофауны гельминтов.
Некоторые данные о зависимости паразитофауны от пищи ветре
чаются п в работе Пирса (Pearse, 1930) над грызунами Нигерии. Пирс
сравнивает хозяев, пища которых содержит более 50% рас тигельного
корми, с такими, у которых вегетарианская лита составляет менее 50%
(таб.л II).
Таблица II
Зависимо!.ть паразитофауны грызунов Нигерии от состава нищи (в •?«)
(по Пирсу)
Состав гнчци х- «зяпна Protozoa Cesiodcs. половозре- лые Ceslodes, финки Nematodes, проникаю- щие через ро Хеша lodes, проникаю- щие через кожу
Болес 5о% раститель- ной и ниш .... 51 33 5 53 63
Meiive .50% раститель- ной нищи , , . 7 7а 3 45 2
Среди данных таблицы обращают внимание два обстоятельства: во-
первых, скудность простейших в хозяевах, менее 50% пищи которых со
ставляют растения; во-вторых, преобладание у них кишечных стадий
цестгд Скудность простейших вызывается, вероятно, тем, что, судя по
Хегнеру (Hegner, 1921), кишечные простейшие грызунов плохо перено-
сят мясную диету хозяина. Богатство данных хозяев-грызунов кишечны-
ми нестидамн есть результат \ потребления ими в пишу насекомых, ко-
торые служат промежуточными хозяевами для многих цеетод. К сожа-
лению, сведения о нематодах, приведенные в табл 11, в особенности >
проникающих через кожу формах, дают неожиданный и неестественный
результат. Следовало бы думать, что не зависящие от нищи в смысле
пути своего проникновения в хозяина формы круглых червей будут оди-
наково богато представлены как у вегетарианцев, так и у невегетарнан-
цев. Между тем проникающие через кожу нематоды у вегетарианцев за-
ражают гЗ% особей, а у видов менее вегетарианских — всего 2%.
Применение возрастного анализа вскрывает иногда очень
интересные детали заражения некоторых животных через пищу, связан-
ные с экологией этих животных. Так, например, Горбунова (1936) на-
шла, что щука в Кончсзере на первом же году своего существования,
т. е. будучи еще сеголетком, сильно (47%) заражается Bucephalus ро-
lymorpllus Л\ежду тем, поданным Быховской-Павловской (1940), окунь
на том же озере начинает заражаться Bucephalus лишь на третьем год\
своей жизни. Как объяснить эти различия? Нам кажется несомненным,
что различие зависит от характера питания обоих видов рыб. Щука —
261
это столь быстро растущий хищник, что она уже трех месяцев от роду
начинает питаться молодые карповых рыб, содержащей личинок Buce-
phalus. Между том окунь переходит к типичному хищническому образу
питания значительно позже, чем объясняется и более позднее заражение
его Bucephalus.
Среди рыб как пресноводных, так и морских можно привести мно-
го весьма показательных примеров влияния на паразигофауну харак
тора пищи. Большинство карповых рыб питается, роясь и грунте и до-
бывая различных мелких беспозвоночных, которые могут явиться про-
межуточными хозяевами различных паразитических червей. По подуст
(Chondrostoma nasus) характеризуется своим растительноядным пита
пнем, в связи с чем он испытал довольно сильное изменение устройства
рта, обращенного книзу и приспособленного к срыванию растении.
Следовательно, подуст лишь случайно может поглощать промежуточ-
ных хозяев. Эти особенности питания подусга очень резко сказываются
на его паразитофауне.
Чрезвычайно характерным оказалось при этом почти полное отсут-
ствие у подуста кишечных паразитов, попадающих в организм рыбы пу
тем поедания промежуточных хозяев. Среди 65 ппдустов только у че-
тырех оказался в небольшом количестве Proteocephalus torulosus. Ila
против, если мы возьмем для сравнения сходное количество других кар
новых рыб, исследованных в тех же районах, то получается совершенно
иная картина. Так, например, исследование 75 лещей дало 44 находки
кишечных червей, относящихся к пяти разным видам трематод, пг.стод
и AcantlioCcphala. Исследование 45 штук плотвы дало 30 находок чер-
нен, относящихся к четырем видам Trematodes. Из этих цифр ясно вид
но сильное подавляющее влияние растительной листы на заражение кар
новых рыб кишечными глистами, передающимися через промежуточ
пых хозяев.
Агапова (1945) и Кузьменко (1945) подтвердили это же наблюде-
ние на некоторых растительноядных рыбах из Сыр-Дарьи, а именно: на
остролучке (Capdefobrama kuschakezeitschi) и на Сырдарьинскоп быст-
рины (Squalalburnus faenialus). В данном водоеме эти рыбы, питаю
щнсся почти исключительно спирогирой и мелкими растительными
остатками и оказались свободными от кишечных паразитов.
Интересный пример зависимости паразитофауны некоторых сиговых
иг характера пищи приводит Бауер (1948) для сигов Енисея. Пелядь
(Coregoniis peled) ведет пелагический образ жизни н питается планкто-
ном; чир (С. nasus)—придонная рыба, питающаяся бентосом и в том
числе моллюсками, В отношении эктопаразитов обе эти рыбы имеют
очень сходную паразнтофауиу. Зато их кишечные паразиты сильно раз-
нятся. У бентофага С. nasus многочисленны трематоды Crcpidostomum
(заражение через моллюсков) п малочисленны цестоды. Планктоиофаг
Coregonus peled, напротив, беден трематодами и богат постолами, ко
горые он получает, поедая планктонных ракообразных.
Многочисленные примеры зависимости паразитофауны от характера
питания можно было бы привести и в отношении морских рыб. Мы
ограничимся лишь немногими из них.
Питание разнообразным бентосом и рыбой приводит к обогащению
морских рыб эндопаразитами. Примером может служить барепцевомор-
ская треска (Gadits morhua), обладающая очень разнообразным спект-
ром питания. У нес обнаружено 27 видов паразитов (Полянский, 1955).
среди которых 7 видов являются эктопаразитами, а 20 видов — эндопа-
разитами; из них 16 видов развиваются с промежуточными хозяевами,
входящими в число пищевых объектов трески. При этом интенсивность
262
сражения многими видами паразитов у трески бывает очень велика.
Чапрнмер, в печени и полости тела нередко находятся сотни кр’.глых'
червей из родов Contracoecuni и Anisakis, в кишечнике — десятки и сот-
ни скребней Echinorhynchus gadi, сосальщики llemiurus и др Совершен-
но другую картину зараженное гп да юг мурманские иланкгоноядиые
морские рыбы, напри иер такой массовый вид, как сельдь (Clupea haren-
gus}. Число видов паразитов, обнаруженных у них, нс превышает 9.
Средн них некоторые гельминты встречаются очень редко (1—2% зара-
жения) (пример Hemiurus levitiseni, Bracliyphaltus crenatus). Несколъ
ко более часто вст печаются некоторые трематоды (Lecithosier) и нема-
тоды (Anisakis, Contracoccum), промежуточными хозяевами которых
являются планктонные рачки.
В тех случаях, когда у животных-хозяез, далеких в систематическом
отношении, наблюдается сходство в питании, и паразитофауна приобре-
тает сходные 4i (ты. Это явление отмечено для разных групп позвоноч-
ных Быховская-Павловская (1955) отмечает, что птицы, далекие в си-
.•тематиноском отношении, но питающиеся сходной пищей, обнаружи-
вают черты сходства в фауне сосальщиков. Опа отмечает некоторые об-
щие виды трематод для некоторых хищников, рыбоядных яистообраз-
ных и всеядных враиовых. Значительное сходство фауны эндопаразитов
обнаруживается среди морских рыб Баренцева и Белого морей между
(алеко стоящими в системе рыб камбаловыми (Pleuronectidac) и зу-
батками (Апarhichadidac) (Шульман-Альбова 19526. Полянский 1955).
•У баренцевоморскнх зубаток и камбал имеются 9 общих паразитов, что
составляет половину всех паразитов нбаток. Среди этих общих парази-
тов имеется несколько видов (днгепстическпе сосальщики Diphterosto-
mutii tnicroat eiabult'tn. Steringophorus [urcigrr, Zoogot’-tides uiviparus),
icTpi чающихся в Бар< ивовом море только в камбаловых и зубатках.
Чем объясняется это, на первый взгляд малопонятное сходство? Оче-
видно, что близость видового состава паразитов камбаловых и зубаток
есть результат сходства условий жизни и в первую очередь питания. П
те, и другие—типичные беитосоядные рыбы, в пищевом рационе кото-
рых большую роль играют моллюски, нгтокожие и в меньшей степени —
бентические ракообразные. Пищевой спи гр зубаток и камбаловых об-
наруживает значительные черты с.'одства. В этом п заключается при-
чина большого сходства их паразитофауны.
Заслуживают внимания, хотя еще недостаточно проверены, указа-
ние некоторых авторов на детель м и н т и з иру ющее в л и я н и е
некоторых пищевых веществ, потребляемых различными жи-
вотными. Так, имеются указания, что суслики, питаясь перед залеганием
на зимнюю спячку полынью, освобождаются в результате такого пище-
вого режима от кишечных гельминтов. Боутон (Boughton, 1932) отме-
чает, что в Канаде дикие кролики (Syivilagus) почти совершенно теря-
ют к зиме цеетод Cttioiaenia по мере того, как переходят от зеленой пи-
щи к питанию корой. О.чигер (1940) полагает, что аналогичными причи-
нами вызвано исчезновенье кокцндцП (Eimeria) из кишечника рябчп
ков поздней осенью. В течение всего лета рябчики питаются смешанной
нищей (ягоды, семена, изредка мелкие беспозвоночные). С половины
сентября рябчики переходят на питание почти исключительно смолисты-
ми сережками ольхи и березы, и с этого времени в их кишечнике исче-
зают кокнидлп.
[Ц\льман-Альбова (1954) у фазана (Phasianus colchicus) наблюда-
ia во вторую половину- лета при переходе от животного питания к расти-
тельному постепенное полное очищение от гельминтов. Это ставится ею
в связь с дегельмнн гизнрующим действием некоторых растений.
263
Из приведенных выше .материалов и примеров явствует, что харак
тер питания и состав пищи в значительной мере определяют состав па
разитпфауны животного. Ио, с другой стороны, наличие определенных
видов паразит св может служить прекрасным показателем некоторых
фракции пищи животного. Т< кв.м образом, возникает понятие о порази
голо ‘ических индикаторах характера питания (Догель, 1958). Так, на
пример, по нахождению скребня Ecliinorhynclitis salmonts в кншечпикг
рыбы можно ' твержлать, что спа питается реликтовыми ракообразными
(промежуточные хозяева этого червя). По нахождению у рыбы большой
количества кишечных пестод можно заключить о питании ее планктон
ними рачками и т. д. Паразитологические индикаторы питания являют
ся одним из показателей, который позволяет по характеру паразнтофах
иы делать заключения о разных сторонах экологии хозяина.
2. ПАРАЗИТОФАУНЛ И ОБРАЗ ЖИЗНИ ХОЗЯИН,А
Вопрос, которого касается данный раздел, еще мало затронут в ли-
тературе, хотя интерес к пому, по-видимому, начинает пробуждаться.
Под образом жизни мы разумеем в данном случае общий характер жиз
недеятельное! и животного, связанный с особенностями окружающей его
среды пли с его собственными повадками и инстинктами. При этом, ра-
зумеется, изменение образа жизни бывает связано и с изменением ха
рактера нищи, что было специально рассмотрено нами выше.
В каждой систематической группе животных-хозяев имеются от-
дельные представители, ведущие образ жизни, в чем-нибудь существен-
ном отличный от тягового большинства их родичей.
Интересные наблюдения проведены Сосниной (1949) над древесным
грызуном— сеней (G/t.s gds) в Кавказском заповедник^ Сравнение с
лесными же, но наземными грыЙлша.мп того же района, намрумер лесной
мышью ( Apodcnius sylvaltcus), выявило несравненно менынсе 'кгзнооб-
разне паразит офах ны сони. У нее было найдено всего три mm эндопа-
разитов (Krachylaernus recurvus. Hyrnenolepis tnyoxi, Heligmosomum
gracile). Между тем у исследованной одновременно с сеней в т^м же
заповеднике лесной мыши обнаружено 10 видов глистов (2 сослришка,
3 цестоды и 5 нематод). Разница в богатств? энтофахчы, завис щая от
древесного или наземного образа жизни хозяина, вполне очсв" ’’Г1.
Харкема (l larkema. I93C) нашел у амернка! ской белки fS -iurus
carolinensis) только одного кишечного паразита (нематода Longistriuta
hassalli). Эт\ бедность мы объясняем. как и у сопи, отрывом ст г.гчвы.
с которой связано заражение многими паразитами либо непосредствен-
но, т. е. заглатыванием яиц и личинок вместе с пищей, либо 11 ‘-»сз по-
средство промежуточных хозяев, живущих в почвенном растит тьном
покрове.
Не лишним будет объяснить, в каком смысле мы понимаем термины
«бел н а и «б о г а т а я» п а рази тофа т н ы. Было бы совершенно
ошибочно определять степень богатства паразитофауны по исчислении
общего количества видов паразитов, которые мот встречаться в дан
пом животном. Термин «богатство» паразнтофауны в нашем ст чае дол
жен иметь относительнее значение и притом значение двойственное,
а именно в разобранном выше примере следует сравнивать паразите
фа ну не с< ни воебще с таковой других грызх лов вообще, а соню Кав-
кавейорф заповедника с другими грызунами той же местности. Кроме то
го, следует учитывать не только количество видов, встречающихся в бел-
ке и в других грызунах, но также экстенсивность и интенсивность зара
ження. Если при одном н том же количестве видов экстенсивность и
2о1
интенсивность их распространения у белки будет значительно слабее
зараженности других грызунов их паразитами, то мы будем расценивать
на рази: гофауну белки как более бедную.
Другой иллюстрацией аберрантного образа жизни среди грызунов
может служить слепец (Spalax leucodon) — крупный грызун с сильно
редуцированными глазами, всю свою жизнь проводящий под землей и
не выходящий на се поверхность. Такой полный отрыв oi поверхности
почвы опять-таки создаст условия, способствующие обеднению паразит-
ной фауны. Киршснблаi (1917). исследовавший паразитов слепца в Ар
мепип. считает фа'гу последнего как бы сильно обиднейшей наразнто-
фауион суслика (Cylelltis xanthoprymnus), живущего в большом коли
честве. в той же месгпосгн, н находит у слепца лишь четыре вида гель-
минтов (одну личиночную стадию цестод, два вида нематод п один вид
скребнем — Moniliformis moniliformis). В то же время у суслика той же
местности имеется 9 видов гельминтов: ланцетовидная двуустка, три
вида цестод (в том числе две личиночных формы), четыре вида нематод
и один вид скребней (Кирше.нблат, 1939). Помимо меньшего разнообра-
зия видов, слепец обнаруживает и меньшую экстенсивность распростра-
нения паразитов. На 16 вскрытых слепцов скребень Moniliformis был
найден лишь один раз, т. е. встретился у 6,2% особей, тогда как среди
-10 взрослых сусликов нм были заражены 33%. Кроме (iongylonema. ко-
торая была констатирована пять раз, все остальные черви попадались
редко. В числе четырех гельминтов слепца три были идентичны с пара-
зитами зуслика (личиночная стадия Cestodes, (Jongylonema, Monilifor-
mis). а четвертый (Ascaris sp.) был представлен неполовозрелыми эк-
земплярами. Ro всяком случае отсутствие специфичности гельминтов
слепня ясно выражено, что делает высказанное Киршеиблатом предпо
ложеннс о равдоподобпым.
Изменения не во всем образе жизни, а только в некоторых его чер
тах соответственно с этим оказывают лишь частичное влияние на пара-
зитофауну. Насколько даже изменения одной какой-нибудь бпологнче
ской особенности могут отразиться на паразитофауне животного, пока-
тывает пример кукушки, разработанный Догелем (1936ет) в отношении
пухоедов этой птицы. Кукушка отличается от всех прочих птиц отс т-
ствием инстинкта гпездбстроптельства и выкармливания птенцов. Та-
ким образом, у этой птицы родители не имеют никакого контакта со сво-
ими собственными птенцами, г. е. с шинами следующего за ними поко-
ления. Между тем среди эктопаразитов нашей европейской кукушки
(Cuculus canorus) имеется целый ряд пухоедов, строго специфичных по
отношению к своему хозяину. Это Docophorus latifrons, Nirmtts fenestra
ttis, Mcnopon phanerostigma (рис. 170, 171).
Возникает вопрос о том, каким способом происходит заражение ку
кушек специфичными для них пухоедами. У других птиц молодой птенец
обычно получает своих пухоедов еще в гнезде от своих родителей, когда
оперение птенца достигает определенной стадии развития. Но молодые
кукушки вырастают в чужих гнездах, где они развивают и свое опере-
ние. Если рассуждать теоретически, птенцы кукушки могли Сы заря-
жаться от своих приемных родителей, с которыми находятся в тесном
контакте. Однако против этого говорит узкая специфичность пухбёдоц
кукушки, которые никогда не встречаются у певчих птиц, дающих яйцам
кукушки невольный приют. Мы просмотрели 11 свежпбптых, же выле-
тевших нз гнезда молодых кукушек и 25 шкурок молодых кукушек в Зоо-
логическом музее Академии наук и только два раза обнаружили на них
по одному пухоеду из рода Nirmus. но из вида не свойственной) кукуш
ке. Отсюда можно сделать вывод, что молодые кукушки не заражаются
263
or приемных родителей, а если отдельные экземпляры пухоедов и пере-
ходят па пих с певчих птиц, то не размножаются и не удерживаются на
кукушках. На взрослых кукушках (как на свежеубитых, так и на их
шкурках) в изобилии были найдены нами специфичные для кукушки
Docophdrus lutijrons и Nirmus fenestratus.
Итак, с одной стороны, несомненно, что кукушки по первому году
оставляют северные широты, будучи еще свободными от пухоедов.
С другой стороны, взрослые кукушки заражены специфичными пухоеда-
ми. Для объяснения заражения остается лишь одна возможность: пе-
редача пухоедов от одной типы к другой совершается во время спари-
вания, т. е. иначе, чем у прочих птиц. Спаривание, которое повторяется
v кукушки' несколько раз в лето, доставляет пухоедам кукушки возмож-
ность переходи еь со старых птиц на птиц более молодых поколений.
Рис. 170. Docophorus.
latifrtms пухоед ку-
кушки. (Оригинал.)
Рис. 171. Мспчрг/П phane-
rttsligmit пухоед кукуш-
ки. (Оригинал.)
Итак, мы получаем следующее представление о происхождении
Mallopfriaga кукушки. Вероятнее всего, предки современных пухоедов,
подобно некоторым Psocidae, были сначала просто обитателями птичьих
гнезд п лишь позднее поселились на самих птицах. Так же поселились
пухоеды и на кукушках в те отдаленные времена, когда эти птицы еще
не утратили инстинкта гЕЕездостронтельства и имели собственные гнезда.
Поселившиеся на кукушках предки пухоедов специализировались и по-
степенно развились в современные специфичные виды.
. В прежние времена они переходили с одного поколения кукушек па
следующее тем же способом, что и у др\ гих птиц, т. с еще в гнезде, и
притом с родителей на птенцов. Когда, однако, половые инстинкты к)
кушки изменились и старый способ инвазии сделался невозможным, за-
ражение пухоедами стало осуществляться лишь путем использования
акта спаривания .хозяина. Весьма возможно, что и прежде пухоеды мог-
ли случайно переходить с одной кукушки на другую при спаривании, но
теперь этот случайный способ заражения сделался единственным и обя-
зательным. Узкая специфичность Mallophaga не позволила пухоедам
приемных родителей переселиться на птенцов кукушки и приспособиться
к ним. Вследствие этого кукушка удержала лишь своих стародавних
Mallophaga. Пример с пухоедами кукушки показывает нам, что измене-
2Ы1
ним инстинкти хозяина могут оказывать существенное влияние на те или
иные части его паразитофауны. Правда, имеется еще одна возможность
заражения кукушек их пухоедами, а именно через крылатые формы
кровососок вроде Ornithomyia, которые могут перелетать с одной ку-
кушки на другую и переносить пухоедов. Но и в таком случае мы имели
бы замену одного доминирующего у прочих птиц способа заражения
другим, т. е. адаптацию к специфическим чертам биологии кукушки.
Дубинин (1951) подтверждает данные Догеля, указывая, что спе-
цифические клещи и пухоеды кукушки появляются на ней лишь во вто-
рую половину лета после первого прилета .молодой птицы на север (при
чет 12 мая, первые перьевые клещи — в июле). До тех нор молодые ку-
кушки совсем свободны от вышеназванных паразитов. Во время жизни
в гнезде приемных родителей птенец кукушки получает некоторое число
чужих клещей и пухоедов, по которым можно даже судить о виде при-
емных родителей. Так, на птенце, вылетевшем нз гнезда серой вороны,
были, например, найдены Analgopsis corvinus и пухоед Philopterus сог-
vinus, а на кукушке из гнезда камышовки — два экземпляра часто встре-
чающегося у камышовки клеща Proctophyllodes utnpelidis. Однако эти
чуждые кукушке паразиты вскоре после вылета совершенно утрачивают-
ся ею.
Выше памп был рассмотрен специальный случай — заражение ку-
кушки Mallophaga в связи со своеобразным образом жизни этой птицы.
Можно привести много других примеров, показывающих зависимость
паразитофауны птиц от их образа жизни. Быховская-Павлоиекая в ряде
работ, посвященных всестороннему исследованию трематод птиц фауны
СССР (1952, 1953, 1955, 1962), приходит к выводу, что степень за-
раженности р а з и ы х экологических г р у пи пт и ц в ес ь-
м а различна. Изучая птиц Западной Сибири, она нашла, что наи-
больший процент заражения обнаруживают птицы, непосредственно
связанные с добыванием пиши в воде (пастушки, чайки, гусиные). Чис-
ю видов трематод у них также велико. Болотные птицы (кулики) обна-
руживают несколько меньший процент заражения, но по богатству ви-
дов сосальщиков они занимают первое место. Слабее всего заражен-
ными оказались наземные воробьиные итппы— это в одинаковой сте-
пени относится как к общему проценту заражения, так и к числу видов.
Таблица 12
Зараженность сосальщиками пгиц разных экологических групп
(по Быховской-Павлопской)
Группа птиц Виды % заражен. Число ви- дов Место добывания нищи
I Пастушки . 70.3 20 Поверхность и толща воды, дно
водоемов
Водяные ‘laiiK’.i . . 60.1 22
Гусиные 48,6 34
Болотные Кулики . . 37,5 37 Заболоченные водоемы, Соло-
тпетля почва, грязь, заросли
гростникл и г. д
Наземные Воробьиные 17.1 12 Вне воды (земля, воздух и т. д )
13 табл. 12 суммированы данные но зараженности птиц разных эколо-
гических гр\пп Западной Сибири. Очевидно, эти различия, связанные
с различным образом жизни, в первую очередь определяются характе-
ром питания, обилием в пище промежуточных хозяев.
257
Существенное влияние на паразитофауну птиц и в особенности на
богатство эктопаразитами оказывает способ гнездования — о д и -
ночи ы li и л и колон и я м и. Для птиц, гнездящихся в дельте Волги,
этот вопрос был подробно изучен Дубининым и Дубининой (19'0) в Аст-
раханском государственном заповеднике. У птиц, гнездящихся и живу-
щих колониями (разные виды цапель, кваква—Nycticorax nyciicurax. ка-
равайка Plegadis falcinellus. баклан Phalacrocorax carbo и мн. др.), на
блюдается сильное заражение эктопаразитами, в особенности пухоеда
мп, достигающее 80—100% при высокой интенсивности заражения.
В противоположность этому птицы, которые держатся чаще всего в оди
почку (поганки— Podiceps, водяные пастушки — Pallas и др.), зараже-
ны Mallophaga значительно слабее.
Очень интересную зависимость от образа жизни показывают пара-
зиты хозяев, отличающихся амфпбиотическимп повадками с попере-
менными переходами от наземного образа жизни к водному и обратно
Таковы, например, бесхвостые амфибии (Anura).
Еще Лоос (Looss. 1894) указал, что степень заражения разных ля
гушек сосальщиками стоит в связи с длительностью их пребывания в во-
де или около воды, так как в воде живут промежуточные хозяева этих
червей.
Пауль (Paul. 1934) для амфибий Силезии пришел к тем же выводам
относительно сосальщиков, а кроме того, выяснил, что заражение круг-
лыми червями идет в обратной последовательности. Сильнее заражен-
ными нематодами оказываются виды амфибии не в воде и не на де-
ревьях (Hyla), а на влажной земле, т. с. преимущественно наземные ви-
ды. Это зависит от того, что яйца и личинки большинства круглых чер
вей развиваются во влажной почве и прямо из нее попадают в хозяина.
Этот же вопрос изучала Дубинина (1950в) в дельте Волги и Волгарь-
Пастухова (1959) в дельте Дуная.
Суммарные данные о зараженности взрослых амфибий дельты Ду-
ная различными группами паразитических червей сведены в табл. 13
Т а б л и и л 13
Зараженность амфибий делыы Дуная различными классами паразитических
червей (по Волгарь-Пасгуховой)
Вилы зеинов >д111..х Trematodes Nematodes Ccsrodes Acamho- cephala Итш о:
ЧИСЛО ВИДОВ % зара- жения число ВИДОВ * зара- жения ЧИ ЛО ни до в 3 за pa- жен г я число НИЛОВ % за ра- же пн я ЧИСЛИ ВИД|)П % л ри- жски»
Rana rifhbtttuta . ?l 82,77 7 57,76 2 6,85 1 1.3 31 87.92
Rana .'srii.'enta 24 81,41 5 54.33 о 6.09 1 2.52 88.9
Вот'чм bornbina 11 г>4.1о 4 Г8.4) 2 6.28 — — 17 83.51
Ji via a borca . . 13 48,12 1 3.48 1 2.5 1 2.08 Ч) 51,61
Uujn tiriiHs 12 43,37 9 80,39 1 1,75 1 22,97 3 HI,99
Pelobates fusctis . 9 ico.ou 3 16.b5 1 6,66 1 З.ьб 11 Ь ю,оо
Как видно из приведенных данных, наибольшую зараженность трема
тодами имеем у двух лягушек, ведущих водный образ жизни — озер
пая лягушка (Rana ridibunda) и зеленая лягушка (/?. esculenla)
Меньше всего видов паразитов у чесночницы (Pelobates fuscus),
относительно мало связанной с водой и ведущей полуподземный o6pai
жизни в дюнах, вызывающий общее обеднение паразитофауны.
268
Больше всего нематод имеет зеленая жаба (В и jo viridis), тесно
связанная с влажной землей и живущая в воде лишь в период икро-
метания. Кажущимся исключением из общего правила является жер-
лянка (Bornbitia bnmbina). Хо .я большую часть времени она про-
водит в воде, пара гитофауна ее относительно бедна. Вероятно, это
зависит от некоторых экологических особенностей указанною вида.
Дубинина (1950в) по этому поводу справедливо указывает, что в то
время как лягушки населяют постоянные, чаще большие водоемы
(озера, пру.ды, реки) с постоянной н богатой фауной беспозвоночных,
жерлянки поселяются преимущественно в небольших, мелких, пере-
сыхающих водоемах временного характера, в которых фауна бес-
позвоночных (т. е. промежуточных хозяев) носит случайный характер
н обычно бедна.
Интересные данные приводит Дубинина в отношении озерной
шгушкл, обладающей широким географическим ареалом распростра-
нения и жир/щей в довольно различных экологических условиях. Эти
различные ’гловия обитания в пределах даже одного вида наклады
вают onpi (е.ннпый отпечаток на ее иаразитофах ну. Например, озер-
ная лягушка из водоемов Средней Азин слабо заражена сосальщика-
ми. сравнительно сильно нематодами и редко жребиями. Гак, для
Старой Бухары наблюдалось всего 2,7% заражения одним видом тре-
матод и <51.3% — нематодами, скребни не были найдены совсем
(Масснпо, 1927). В Кулябе было обнаружено заражение трематодами
V 37.7% дягмчек (5 видов), 79,2% были сражены нематодами
(Быховскнй. 1935). Аналогичные результаты были получены н для
других районов Средней Азии. В противоположность этому озерная
лягушка из районов Европейской части СССР и Кавказа очень сильно
заражена сосальщиками (обычно около 100%) при большом их видо-
вом разнообразии (10- 20 видов). Наряду с этим зараженность круг
л. мн червями была слабее, чем в Средней Азии (30—б4%). Причиной
указанных различии являются особенности условий обитания хозяина.
В Средней Азии, бедной водой, лягушки живут обычно в замкнутых,
небольших, мелких и частично пересыхающих в жаркое время во we-
мах. которые содержат очень одпообра ив ю. бедную фауну беспозво-
ночных и в особенности моллюсков — пром< жуточных хозяев трема-
тод. При этих условиях нематоды становятся преобладающими пара
зитамн. Последнее связано, вероятно, и с тем, что лягушки значитель-
ною часть времени проводят не в воде, а во влажных местах. Совер-
шенно иную картину представляют водо< лы Европейской части СССР
и Кавказа с их богатой фауной беспозвоночных н с преимуществен-
ным пребыванием озерной лягушки в воде.
Для паразитов морских животных, например рыб, несомненное
значение должна иметь р а з л и ч пая г л у бин а во л ы, как среды,
окр/жающен их хозяина. Первудо попытку к разрешению этого
вопроса сделал в ряде статен Мантер (Manter, 1931), работавший
в биологической лаборатории на островах Тортугас. Сначала он
исследовал фауну сосальщиков у 1500 экземпляров различных мелко-
водных рыб, после чего произвел аналогичное исследование 720 глу-
боководных рыб, принадлежащих к НО различным видам. До Мантера
свечения о паразитах глубоководных рыб бычн чрезвычайно скудны.
Укажем в этом отношении находку Distomlitn lialosawi н« мочеточни-
ков Haloscuirus mucrochir с глубины 2000 .и <ВсП. 1.887) и скребня
Echinn'hifiuiius abi/ssicola, которого описал Дольфус (Dollius, 1931).
из Pachifcura obesa с глубины 47F3 м. 14а глубоководной рыбе Масги-
rus Бриан | Brian, 1931) наанАл веслоногого рачка Rebalula edivardsi.
2ii9
Мантер не имел дела с рыбами очень больших глубин, гак как го
материал происходил с глубин от 80 до 1060 м. Ему удалось обнару-
жить в этих глубоководных рыбах около 50 вп юв трематод. При этом
ам(рпкаискии автор подчеркивает, что глубоководны^ трематоды
явственно отличны от сосальщиков мелководья. Средн сосальщиков,
найденных глубже 200 только 3 вида встречались и в мелких
водах, все же прочие были глубоководными. Можно было бы думать,
чго такое распределение ганнеит исключительно от характера рас
пространення рыб-хозяев. Однако Мантер указывает, чго .многие из
паразитов глубоководных рыб пе особенно разборчивы в выборе
хозяев. Кроме того, имеются сосальщики, которые огратаВшваются
вполне определенными глубинами, например сосальщик Megenteruni
crasstmi был найден лишь на глубинах 360—550 .и, Xpotturus inter
ntf’dius— 360 150 м н т д.
Интересно указать, что некоторые сосальщики, которых Мантер
встречай только на начнтельных глубинах, в других географических
широтах пе являются глубоководными. Хороший пример — Derogates
various. Man гор находил этого сосальщика на глубинах свыше 200 .и
в Баренцевом же море он встречается па любых г копнах, в том
числе у литоральных рыб. Вероятно, в данном случае распространи
нле Derogates связано с термическим фактором: он является север
ным видом и в тропиках опускается на глубины, г ie температура
в течение круглого го да сохраняется ни экой.
Среди пресноводных водоемов лишь немногие обладают боль
шнми глубинами Особый интерес в этом отношений представляет
озеро Байкал, максимальные глубины которого превышают 1200 м.
Изучение паразитофауны рыб этого своеобразнейшего водоема
(Догель и Боголепова, ’1957; Догель, Боголепова, Смирнова, 1949)
представило большой юогеографическнн интерес и вместе с тем
показало. чго па больших глубинах встречается ряд видов паразитов
бычков, которых можно считать настоящими глубинными формами
этого о тора. К числу их относятся мпксоспорплии Myxidiutn p Tni-
cjosum, Henneguiiu hatcalensis. моногенетнческне сосальщики Dactylo-
gyrus colonut G if redact tins comephori н некоторые другие. Вопрос об
экологических особенностях паразитов абнссальны животных еще мачо
изучен и требует дальнейших исследований.
На морских рыбах было показно, что возможно установить законе
мерную связь характера паразитофауны (богатство видового состава
паразитов, наличие определенных характерных видов, экстенсивность
и интенсивность «араженпя) с определенными экологическими груп-
пировками рыб.
Речь идет об э к о л о г и ч е с к и х к о м и л е к с а х и а р а з и г о в
рыб. Для паразитов костистых рыб Баренцева моря, например» По
янским (1955, 1958) были установлены следующие комплексы:
1. Паразиты придонных рыб открытого моря, питающиеся бентосом
и мелкой рыбой. Очень богатый в видбйс м отношении комплекс, сл •
гающипся нз многочисленных видов. Особенно разнообразны мнксоспо-
рндин н сосальщики. Весьма характерны виды родов Rbodoirenta. Sle-
ringophorus. Обычны Echiiwrltynchus gadi.
2. Паразиты литоральных и прибрежных рыб Значительно беднее
ио составу видов, чем предыдущий комплекс. Имеется ряд характерных
видов, связанных в своем развитии с литоральными беспозвоночными
как-то: сосалыцикн Podocolyle atomon и Lfipidapedon gadi.
3. Паразиты нланктоноядных рыб. Этот комл< кс беден как ни раз-
нообразию видового состава, так и но интенсивности и экстенсивноеп.
заражения. Сюда относится ряд характерных сосальщиков, которыми
рыбы заражаются через пелагических Copepoda.
4 Паразиты проходных рыб. Комп текс характерен сочетанием,
пресноводных и морсЙнх паразитов, что связано с явление м мнгпгцнн
подробно рассматриваемым нами ниже.
Разумеется намеченные выше комплексы не резко разграничены.
Имеется ряд видов, общих для всех или для некоторых из них. П отно-
шении некоторых видов паразитов различие касается г 1авпым образом
количественных показателен (интенсивность заражения). Тем не mchci
каждым из указанных выше комплексов имеет свой фахнистпческий и
экологический облик в такой же примерно степени реальный и объек-
тивный, как реальны н объективны понятия — бентосоядная рыба,
планктоноядная нектонная рыба и т. и. Экологические комплексы па-
разитов морских рыб, очень близкие к таковым для Баренцева моря,
были установлены и для некоторых других морей СССР. Например,
подобную работу осуществил 1О. Стрелков (I960} для рыб Берингова
моря у побережья Камчатки
Несомненно, что наметить экологические комплексы паразитов воз-
можно и для пресноводных рыб. Однако здесь они будут мепее отчет
лпво очерчены, так как разграничение экологических группировок их
хозяев в море выражено зля большинства рыб более резко, чем в прес-
новодных водоемах.
3. ВЛИЯНИЕ СПЯЧКИ ХОЗЯИНА НА ПАРАЗИТОФАУНУ
Мы выделяем этот частный вопрос в особый раздел ввиду тех
чрезвычайно резких сезонных изменений внешних условии существова
ния п внутреннего состояния хозяина, которые явление спячки вносиi
в жизнь целого ряда животных. Действительно, мы знаем, что, напри
мер, летучие мыши в условиях северо-западной области СССР 7 -8 не
сяцев в году проводят в полном оцепенении, Из акгивш-го состояния
животное переходит в совершенно пассивное: прием пищи и дефекация
прекращаются, все жизненные процессы замедляются, и даже темпе
ратура тела падает до 7°, и летучие мыши становятся на это время хо
лоднокровнымн, У сусликов, в особенности у некоторых ?еркесгапских
видов (Citelius fulvus. по Кашкаровх 1944), спячка длится до 9 меся
цев и захватывает не только зимний период, но и наиболее жаркую и
сухую часть лета. Летняя спячка наблюдается у целого ряда тропиче-
ских позвоночных: некоторых пресноводных рыб (Protopterus), кроко-
дилов и др.
Спрашивается, как ведет себя паразитофаупа всех >тнх животных
во время длительного голодания и полного покоя? Вопрос этот
изучен еще далеко не достаточно, однако за последние годы накопился
все же некоторый материал, представляющий значительный экологи-
ческий интерес. Исследование летучих мышей показывает, как много
неожиданностей готовит разработка этого вопроса. Заранее можно
ожидать у перезимовавших мышей крайне ослабленную в смысле
числа фауну кишечных паразитов, состоящую из немногих вполне
половозрелых особей. На самом деле паразитофаупа мышей uoc.ni
зимовки носит совсем иной характер.
Еще Ван Бенедеп (Van Beiieden. 1873), исследовавший паразитов
летучих мышей Бельгии, заметил, что зимние особи мышей содержали
обильную фауну паразитов В 1938 г. Марковой в окрестностях Лсигч-
града было произведено планомерное с.с юфное обследование ияра
зитофауны двух видов мышей: Epsetici' nilssonl п Plecofus auritus
кив, принадлежащих к трем видам, ко
Рис. 17'2. Трематоды из кншивиикв летучих
мышей
А — Phigiurrhis uesperiilionis-, U Lccilhotldn-
drium asci Ча. (Л — по Штрочу, /< оригинал 1
I — рот<ш;и1 iif игоск:'; 2 глогка; 3 шгтии кишки; f
penis: о — яичники; ft* семенники; 7 — ччю; Л — жсл-
TiriiiMkii; 9 f.рюнпшм iq нгпека.
При этом летучие мыши разного возраста вскрывались в течение
лета, затем вскоре после их отлета на спячку в пещеры, далее во вто
рой половине апреля, перед вылетом из пещеры на летйпп период.
Вопреки всем ожиданиям чистый и совершенно свободный от нищи
кишечник кончающих зимовку мышей был набит сотнями сосальщн-
орые встречаются у взрослых
мышей и в течение всею лета
(ряс. 1721. Особенно интересно
было ю Обстоятельства, что
сосальщика находились на раз-
личных стадиях роста и зре
лостн. начиная с самых моло-
дых. Как это могло случиться
при отсутствии питания, а сле-
довательно, и новых заражений
в тсчише всей долгой зимы?
Очевидно, сосальщики были
получены мытами в конце лет-
него сезона предыдущей о года,
причем момент перехода хо-
зяина к зимовке оказался для
них моментом полной останов-
ки в развитии—дичнаузой.
Паразиты одновременно с хо-
зяином погрузились в глубокий
зимний покой. Интересно, что
у летучих мышей зимняя спяч-
ка распространяется и на неко-
торые особенности процесса
размножения, а именно: сово-
купление п[ онсхотпту летучих
мышей перед началом зимней
Спячки, ио развитие зародыша
начинается лишь с весны. Вес-
пой и хозяин, и паразиты при-
ступают к по ювому р.чзмноже-
ипо, а кроме того, начинаются
инвазии летучих мышей новы-
ми личиночными стадиями па-
разитов через их промежуточ-
ных хозяев. Кроме сосальщи-
ков, в кишечнике ленинград-
ских мышей лишь изредка по-
падаются нематоды H'stiosf-
rongultis tipula, которые тоже, сохраняются в течение всей зимовки.
Помимо эндопаразитов, летучие мыши имеют под Ленинградом
два вида кровососущих клещей н муху Nycterihia (рис. 173). Один из
видов, относящийся к Laelapirlae, ведет себя так же. как сосальщики,
и на крыльях зимующих мышей находились без различия личинки,
нимфы и взрослые клещи обоих полов. Другой клещ (Spiniurnix)
представлен на зимующих мышах исключительно самками, внутри
которых имеются эмбрионы на определенной стадии развития. Очевид-
но, у Spinttirnix жизненный цикл построен так, что ко времени зчмов
ки хозяина паразиты достигают определенной стадии, и лишь на этой
стадии их застает период покоя. Итак, две близких между собой
272
Рис. 173. Некпторы. эктопаразиты летучих
мышей
А — Nycteribia «п. бескрылая муха; В —
клещ Spinturnix. (Ориги- ал.)
I — «пОпок: ЗиЛ- гребешки.
формы (клещи) обнаруживают существенные различия в жизненном
цикле, несмотря на одинаковые внешние условия.
По-видимому, иной, чем у летучих мышеи во время зимней спяч-
ки, является судьба паразитов у млекопитающих, впадающих в лет-
нюю спячку. Имеются еще старые наблюдения Бланшара (Blan-
chard, 1903) о том, что сурки (Marmota), впадающие в спячку, осво-
бождаются полностью от ки-
шечных паразитов. Аналогич-
ное явление наблюдали Дуби-
нин и Лешкевнч (1945) у тар-
баганов (Martnota sibirica) в
Забайкалье, длительная спячка
которых захватывает несколь-
ко сезонов. Эти зверьки впада-
ют в спячку осенью и просыпа-
ются лишь в начале лета. Пе-
ред началом спячки их кишеч-
ник полностью освобождается
от паразитов, основную массу
которых составляют Ascons
larbagani.
Влияние спячки хозяина
на паразитов кишечника под-
робно изучено Дубииниой
(19496) у степной черепахи
(Testudo horsfieldi) в окрест-
ностях Дмнанбе. В условиях
Средней Азин черепаха прово-
дит в состоянии спячки 9 меся-
цев в году. Ее активная жизнь
длится всего 3 месяца (апрель,
май, июнь). Летняя спячка не-
посредственно переходит в зим-
нюю. За это зремя в кишечнике
черепах развивается очень
большое количество нематод,
основную массу которых со-
ставляют \iractis dactylurus и
вилы рода Тiichygonetria С по-
гружением хозяина в спячку
развитие нематод полностью
не прекращается, но сильно за-
медляется. Благодаря этому в
течение первых 4—5 месяцев
интенсивность заражения па-
дает незначительно, и боль-
шинство особен достигает по-
ловой зрелости. Во время
нослъ заражения нематодами
времени все молодые черви становятся
лыс стареют и вымирают. К концу спячки остаются почти нсключи-
чительно только половозрелые особи червей. Таким образом, пара-
зиты черепахи в период спячкн ведут себя совершенно иначе, чем
паразиты летучих мышей Они не впадают в спячку, а продолжают
свое развитие.
bi opoii
сильно
половины спячки интенсив-
падает, так как к этом\
половозрелыми, а половозре
R В. Л. Догель
273
Согласно исследованию Дубининой (1950в), проведенному над
озерной лягушкой (Rana ridibunda) в дельте Волги, зимняя спячка
оказывает различное влияние на разные группы паразитов Продол
жителыюсть зимней спячки озерной лягушки 4—5 месяцев (с середн
ны октября до начала марта). В это время лягушки собираются
большими массами на глубине 1 —1,5 м под вымоинами берегов, где
они находятся в неподвижном состоянии и не принимают нищи
Паразитические простейшие (инфузории, Opalinina) остаются в актив-
ном состоянии и процент заражения ими сохраняется без изменений
Личинки сосальщиков (Neodiplostomulurn, Tetracotyle), заражающш
лягушек путем активного внедрения церкарий через покровы, зимой
их не заражают. Часть церкарий дегенерирует, часть остается живыми
Кишечные сосальщики (Pietirogenes claviger, Pleurogenoides medians.
Opisthioglyphe ranae) ведут себя так же, как сосальщики летучих
мышей, т. е. вместе с хозяином погружаются в спячку. Нематода
кишечника Cosrnocerca ornata и легочный сосальщик Haematoloechus
variegatus не погружаются в спячку, а продолжают медленно разви
ваться вплоть до достижения половой зрелости. Но выходе из спячки
половозрелые особи Cosrnocerca и Haematoloechus вскоре погибают
а затем заражение указанными паразитами начинается заново. Кишем
пая нематода Oswaldocruzia filiformis имеет короткий срок жизни
В зимующих лягушках она сначала достигает половозрелостн. а затеи
отмирает Веспою у выходящих из спячки лягушек не остается пи
одного экземпляра этого вида нематод. Таким образом, разные виды
паразитов лягушек по-разному реагируют на весьма своеобразные
условия зимней спячки.
Разнообразна также судьба паразитов у рыб, впадающих в зим
нее оцепенение. Этот вопрос тоже подробно изучен Дубининой (1949а»
иа нескольких вилах рыб дельты Волги. Некоторые виды пресиовод
пых рыб залегают здесь на зиму в углублениях дна (зимовальные
ямы), где и скапливаются большими массами. Дубининой была изу
чека в условиях зимовки пара.зитофауна четырех видов рыб: леща
(Abratnis brarna), сазана (Cyprinus carpio), сома (Silurus glatiis)
и судака (Lucioperca lucioperca). Из них два первых являются
«мирными» рыбами, два последних — хищными. Лещ, сазан и сом во
время зимовки не питаются, судак же полностью не теряет активно
сти и продолжает питаться, хотя и в ограниченных размерах. Именно
в силу этих различий эндопаразиты судака зимой ведут себя иначе,
чем у трех других, впадающих в спячку рыб.
У леща, сазана и сома зимой не происходит новых заражений
эндопаразитическими червями, а к январю исчезают все кишечные
сосальщики и нематоды, заражение которыми произошло в активный
период жизни хозяев. У этих рыб трематоды кишечника в отличие от
озерной лягушки или летучих мышей не впадают вместе с хозяином
в состояние спячки, а продолжают развиваться, достигают полово»
зрелости и погибают. Своеобразные изменения претерпевают лепточ
ные черви, У них происходит отбрасывание и отмирание всеГ1 стро
билы (процесс дестробиляции), так что сохраняется лишь сколекс
Веспой, при переходе рыб в активное состояние, сколексы вновь
регенерируют стробилу. В отличие от леша, сазана и сома у судака
питание которого зимой нс прекращается, заражение эндопаразит»
чески ми червями происходит и зимой, и очищение кишечника от пара-
зитов не наступает Таким образом, потерю некоторыми рыбами
паразитов кишечника во время спячки следует связывать не с пони
жением температуры, а прекращением питания У зимующих рыб при
274
низкой температуре прекращается размножение почти всех паразитов.
Большинство видов Monogenoidea сохраняется в виде единичных
шмующих половозрелых особей.
Совершенно иначе ведут себя зимой в дельте Волги рачки Агци
lus и пиявки Piscicola geomefra и Cystobranchus fasciatus. Они скапли-
ваются массами на рыбах в зимовальных ямах; экстенсивность
и интенсивность заражения ими зимой оказываются во много раз
больше, чем летом. Они активно питаютр! кровью и наносят зимую
ишм рыбам большой вред, повреждая их покровы.
Приведенные факты показывают, что разные группы и виды
паразитов впадающих в зимнюю спячку рыб выработали самые раз
личные приспособления для перенесения зимнего режима.
Сведения о паразитах беспозвоночных, впадающих в спячку,
в литературе почти отсутствуют. Лишь недавно были опубликованы
интересные данные Барбозы (Barbosa, 1958) о судьбе спороцист
сосальщика Schistosoma rnansuni во время спячки моллюсков
Australarbis glabratus, вызванной пересыханием водоемов в летнее
время (наблюдения проводились в Бразилии). При этим спороцисты
сохраняли жизнеспособность, но развитие их прекращалось и зрелые
церкарнй в спороцистах не образовывались. Через 10—13 дней после
помещения зараженных моллюсков в воду наблюдалось выделени<
перкарин. В связи с летней спячкой моллюска-хозяина развитие
спороцист прекращается, а затем при наступлении благоприятных
условий вновь возобновляется.
ГЛАВА XIV
ЗАВИСИМОСТЬ ПАРАЗИТОФАУНЫ ОТ МИГРАЦИЙ ХОЗЯИНА
Одним из факторов, наиболее мощно изменяющих наразнтофаупу
животного, являются его миграции. Дальние миграции ставят живот-
ное в столь различные условия существования, что паразиты не могут
не реагировать на эти изменения внешней среды. Кроме того, мигра-
ции сплошь и рядом сопровождаются существенными изменениями
физиологии хозяина, которые присоединяют свое действие на пара-
зитофауну к влиянию миграций.
Прежде всего влияние миграций хорошо иллюстрируется изме
нениями паразитофауны проходных рыб. Классическим
примером проходных рыб служит логосы До трех-четырехлстнего
возраста лосось живет в верховьях северных рек. плохо питается
и слабо растет. Затем молодь спускается в море, где начинается
период нагула, сопровождающийся бурным ростом Последний осо-
бенно сказывается на размерах чешуи, которая является как бы
паспортом, где отмечаются важнейшие события в жизни лосося.
После двух- или трехлегиего периода морской жизни лосось в пер-
вый раз поднимается в реки для нереста. При этом он проводит
в пресной воде несколько месяцев, иногда почти год. Во все время
пребывания в реке для нереста лосось ничего не ест и между тем
тратит массу энергии как на поднятие против течения (иногда на
тысячи километров), так и на развитие половых продуктов. После
нереста лосось, как говорят, скатывается в море и только через год
или два морской жизни снова поднимается для нереста в реки. Вся
эта гамма разных местообитаний и физиологических состояний не
может не отразиться на паразитофауне лосося.
Сначала была описана фауна паразитов поднимающегося в реки
взрослого лосося, причем Гейц (Heitz. 1918) в прекрасной работе,
посвященной рейнскому лососю, подробно разобрал обеднение этой
рыбы паразитами по мере поднятия ее к верховьям рек. Догелю
и Петрушевскому (1934. 1935) удалось получить полно»' прсдставле
ние о перемг нах фауны паразитов лосося в течение всей его жизни,
собрав материал не только ио взрослому лососю, но и по молоди,
проживающей в верховьях наших северных рек до своего первого
ската в море. Материал был получен из ряда рек: Онеги, Выга.
221i
а главным образом из притоков Северной Двины и состоял и i значи-
тельного числа (около 150 экземпляров) молода разных возрастов
а также взрослых лососей (50 экземпляров). Среди иссйедованиой
молоди были сеголетки, имевшие всего 3—4 месяца от роду, рыбки
одно-, двух-, трех- и четырехлетнего возрастов. Первый скат в море
совершается молодые, как сказано уже выше, в
летнем возрасте, так что имелся и материал,
собой предельный период пребывания молодого
воде. Взрослые исследованные лососи имели от 4 до
10, по чаще всего 5 лет, т. е. получилась полная воз-
растная цепь лососей на разных периодах жизни.
Первым результатом этих исследований явилось
установление в молоди лосося до тех пор неизвест-
ной фауны пресноводных паразитов (рис. 174). По-
следняя ие представляет собой чего-либо специфич-
ного, но отвечает сильно обедненной паразитофауне
родичей лосося из того же района. Фауна эта у ис-
следованных мальков состояла из 12 видов, в том
трех- или четырех-
харакгеризовавший
лосося в пресной
Рис. 174. Пресноводные паразиты молпди лосося (Salrno safar)
Л—почечный сосальщик Phyllotf'.siomaiii cenostomum; В — личинка со-
сальщика Diplostomulum spatbaccum. (По Догелю i; С — D. spathaceum
в хруста ике глаза рыбы: D - Neoechinorhynchus rutlii.
1 — хоботок; 2 — нлагалмше хоботка; 3 — лемниски; з — гигантские я яра н коже: « < v
мен ники; (7 — цементные железы; 7 — полопчя клоэк.ч.
числе из самых типичных пресноводных паразитов: Crepidoslomum [а-
rionis, Phyllodistotnum conoslomum, Diplostomulum spathaceum из глаз,
Trimnophorus nodulosus, молодых Proteocephalus, Neoechinorhynchas ru-
lili, Spiropt >ra tenuissima, Rhabdochona, Rhaphidascaris acus.
Эта фауна появляется в мальках не вся сразу, а постепенно, по
возрастам, чем подтверждается правило о характере возрастных изме-
нений паразитофауны пресноводных рыб, высказанное нами ранее. Как
показывают наши данные, значительный процент сеголетков вообще
свободен от паразитов, у остальных же встречаются в небольшом
количестве лишь две нематоды (Spiroptera и Rhabdochona) и один раз
был найден в глазу Diplostomulum. У однолетних мальков, или голо
виков, заражены почти все особи, и к списку параяитов прибавляются
Phyllodisfomum мочевого пузыря и Neoechinorhynchas. У двух-
и трехгодовалых рыб появляются как бо_ее или менее частые
компоненты паразитофауиы Diplostomulum и Crepidoslomum, а также
в виде отдельных находок и некоторые другие новые виды; наконец,
у четырехлеток инвазия, сохраняя тот же видовой состав, обогащается
в смысле интенсивности и экстенсивности заражения.
Этот процесс постепенного обогащения фауны мальков может
быть иллюстрирован также в цифровом отношении средним количе-
ством видов паразитов, приходящихся па одного зараженного малька
в разных возрастах. Для сеголетков мы находим в среднем 1 вид
паразитов па 1 зараженную рыбу, для годовиков—1,3 (т. е. имеются
уже отдельные, рыбки, несущие два вида паразитов), для двух-
и трехлеток — 2,3 вида и для четырехлеток--2 7 вида паразитов.
Усиление интенсивности инвазии демонстрирует Spiroptera; заражен-
ные сеголетки имеют в среднем по 2 червя па рыбу, годовики — по 3.3.
а двух- и трехлетки — по 9 червей.
Далее те же исследования показали, что в море лосось нацело
освобождается от своих пресноводных паразитов и получает новый
набор паразитов—морского происхождения. Действительно, иарази-
тофауна возвращающихся в реки и пойманных в реках взрослых
лососей, состоящая из 15 видов, содержала всего 3 пресноводных
вида: Camallanus iacuslris (найден I раз в числе 1 особи). Cupilia
ria sp. (I раз) и Diplostomulum spaihaceum (довольно часто, но
в очень небольшом количестве особей). Однако и эти немногочислен
яые пресноводные паразиты нс сохранены лососем от малькового
возраста, а благоприобретены им во время поднятия в реки, как
показывает D. spathaceum. У мальков эта личинка сосальщиков
встречается в своем нормальном местообитании, а именно: в хруста-
лике глаза. Напротив, у взрослых лососей ее находили исключительно
в стекловидном теле глаза. Очевидно, мальковая инвазия этими
личинками со временем ликвидировалась (паразиты рассосались),
а в глаза взрослой рыбы проникли новые личинки, которые, однако,
вследствие большой плотности хрусталика взрослого лосося не могли
вбуравнться в пего и остались в стекловидном теле.
Все остальные паразиты взрослого лосося чисто морские. Таковы
представители семейства сосальщиков Heiniuridae (Brachyphallus
crenaius, Hemiurus. два вида Lecithaster), а также Deiogenes varicus.
сосалыник, распространенный у весьма многих морских рыб. Мор-
ского происхождения н личинки цестод Telrabolhrium, а вероятно,
также и Eubothrium crassum. Единственный вид скребней, имевшийся
во взрослых лососях (Echiuorhynhus gadi) обычно встречается
в треске., а личинки нематоды Porrococcum capsularia констатированы
для 90 различных морских рыб. Наконец, типичным морским парази
гом является и рачок Lepeophtheirus stromii (рнс. 175), попадающий-
ся па лососях, входящих в реки.
Все эти морские виды сопровождают лосося в его ападромальнон
миграции в реки, но в пресной воде, как уже наблюдалось многими
авторами, происходит постепенная утрата лососем его морской пара-
зптофаупы. Прежде всего рыба освобождается от эктопаразитов,
которые подвергаются непосредственному воздействию пресной воды.
Так. в пункте Сорока на Белом море все лососи поголовно были
заражены рачками Lcpeophiheirus. Между тем лососи, поднявшиеся от
этого пункта па 10 км вверх по реке Выг, показали экстенсивность
инвазии уже всего на 25%, а рыбы, пойманные, еще выше по течению
реки, были вообще свободны от Lepeophtheirus.
За потерей эктопаразитов следует постепенная утрата кишечной
паразитофауны, причем падение видового состава и абсолютного
числа кишечных морских паразитов находится в прямой зависимости
от срока, проведенного рыбой в пресной воде. Средн речных лососей
можно благодаря некоторым признакам отличать «осеннюю» рыбу,
которая недавно вошла в реку, от так называемых «лохов», которые
278
провели в пресной воде уже много месяцев и вполне готовы к нере-
сту. Сравнивая ггаразигофауну обеих этих гр^чп лосося, мы находим,
что у лохов морские паразиты представлены несравненно беднее,
чем у осенних лососей. Так, Derogenes various встречается
у 93,7% осенней рыбы и только у 53,3% лохов. Параллель
но с этим падает и интенсивность инвазии. У осенней рыбы
на одного лосося приходится на р. Ниве 12'J Derogenes varicus,
а у лохов—всего 12, т. е. в десять раз меньше. Аналогичную
к. ртину наблюдал уже раньше Гейц (Heitz, 1918), сравнивая лососей
нижнего и верхнего течения Ренна. Несколько Чтаче ве-
дет себя ленточный червь Eubothrium cras^tim-. Число го-
товок его в кишке осенних лососей и лохов приблизи-
тельно одинаково, но у лохов стробилы Eubothrium чрез-
вычайно укорочены, вследствие отрывания и утраты поч-
ти всех своих члеников. Некоторые из кишечных морских
паразитов вовсе исчезают у лохов — таков Echiuorhy'n-
chus gadi.
Меньше всего отражается влияние миграций па по-
достных морских паразитах, которые не подвергаются
непосредственному действию пресной волы н не могу г
быть выведены из полости гста наружу. Поэтому, на-
пример, личинки Porrocoetum capsularia приблизительно
равномерно распространены у осенней рыбы н лохов.
Можно задать себе вопрос, почему лосось захваты-
чает во вр< мя своей анадромальпой миграции столь ни-
чтожное число пресноводных паразитов. Ответ на это
дает нам физиология анадромалыгого лосося. Во время
странствования он не питается, а потому главный путь
инфекции — кишечник — остается закрытым. Сравни-
тельно частое заражение анадромальны.х лососей одним
пресноводным паразитом, а именно личинками Diplosto-
muluin, не ослабляет, а поддерживает наше предполо-
жение. Церкарии большинства Strigeida, к которым при-
надлежит Diplostomulurn, активно проникают в тело про-
межуточного хогяина через покровы, т. с. не зависят в
отногшпин инвазии or принятия хозяином пищи.
Мы нарочно остановились на паразитах лосося под-
робнее, чтобы иллюстрировать положительные стороны
грименепня возрастного анализа фауны при иаразнто
логических исследованиях, а также потому, что здесь
ся динамик? п?ряцЛтпфауНи живо проходит перед нами
га примере одной рыбы, жизнь которой состоит из ряда
глубоких контрастов. Работы над тососем показали целый ряд законо-
мерностей в изменениях паразитофаулы: постепенное нарастание пресно-
водной паразитофауны у мальков, ее утрата и замена морской; правиль-
ная последовательная утрата части последней у анадромальны.х лососей
и восстановление морской паразитофауны при каждом возвращении
лосося в море. Изменения фауны паразитов имеют вид несколько раз
•годинчающейся и падающей кривой (рис. 176).
Если мы рассмотри.’.! для сравнения паразитофауну одного из
родичей лососевых, а именно чисто пресноводного хариуса (Дубинин,
1936) из тс - же самых водоемов (притоки Сев. Двины), то увидим,
что здесь с возрастая идет постепенное поднятие первой (мальковой)
части кривой паразитофауны лосося, и на этом дело оканчивается
(рис. 177).
Рнс. I 75.' Пара-
зи тирующие на
лососе вес юно-
гис рачки t.epe-
ophtheirux strfi-
mit
A — с брюшной
стороны-. /- co
спинной. (Пл
Солдатову.)
череду гощи хся
570
Другим примером влияния миграций на иаразнтофаупу рыб mojkci
служить голец (Salvelinus alpinas) с Новой Земли (Догель и Марков,
1937). Эта рыба известна в двух разновидностях: одна живет
в пресноводных озерах северной Европы, другая—в Ледовитом
оксане. Эта морская разновидность проделывает правильные анадро
мальные миграции для нереста в верховья рек. Голец с Новой! Земли
Изменение паразитофауны
Рис. 176. Диаграмма изменений паразитофауиы лосося, происходящих в зависимости
от его миграции (Оригинал Т
Из осп абсцисс от.зожепм голы жизни лосося, речно-1 л.ш морем* 1 ие-Изж скапынает па iiihTiimhiiii
лосося в соответственные голы п пресной или соленой воле. I - подъем и тление пресноводной ii.ip.-i *ито-
<||зуны; 2 — изменение морскол пзразитофзМы. Левы.1 рисунок пзпбрзжлет малька, прапы.3 — взрослого
лосося.
нерестится в речках и озерах этого арктического острова, там он раз
вивается и там же проводит большую часть года даже во взрослом
состоянии, спускаясь для нагула в море лишь на 2—3 месяца в году
Мальки гольца развивают в себе несколько более бедную (6 видов)
н иную пресноводную паразитофауну, чем мальки лосося. Однако
изредка (одна находка у 54 мальков) у мальков встречаются и чисто
морские черви (Derogcnes varicus). Причиной этого явления служит,
вероятно, с одной стороны, короткость речных потоков острова, с дру-
гой— влияние морских приливов, загоняющих морскую воду в устье
рек. Таким путем морские паразиты получают доступ к малькам
Взрослые гольцы, пойманные в различных заливах Нозой Земли,
содержат, как и следовало ожидать, более или менее морскую пара-
зитофауну, к которой примешивается, однако, целый ряд пресновод
пых форм, указывающих на длительное пребывание взрослых гольцов
в пресных водоемах. Около 65% паразитов взрослого гольца имени
чисто морской, прочие же 35% более или менее пресноводный харак-
тер. Вольшая примесь пресноводных паразитов явственно отличает
280
гольца от лосося и говорит за то, то голец, вероятно, не вполне
прекращает питаться в пресной воде. Ниже мы приводим список
паразитов взрослых гольцов, в котором звездочками отмечены пресно
водные виды, а цифры показывают экстенсивность инвазии в про
центах:
* Tetraonchus arcticus 2
Dproirrnes various . 43
F.uhothrium salvcluti 70
Hemiurtif levinseni 40
Brachyphallus c enatus 7
Diplocott/le olriki . 53
Te(raboihrium sp. 23
Echinorhynchus gadi 16
*Echinnt hynchus salrnonis 23
Porrocoecum sp. . 43
* Cyslidicola farionis IB
Conlracoecum si). . . 12
Poclocotylc at от on 2
Lecituiister gibbosus *
• Salmincola al pitta 7
*Saltnincola extamescens . b
Интересные экологические
взрослых гольцов, а именно:
недавно спустилась из озер Но-
вой Земли и идет в морс, и осенней,
направляющейся из моря в реки.
Летние гольцы содержали боль-
шее количество видов и больший
процент пресноводных паразитов,
чем осенние. Так, например, у лет-
них гольцов в изобилии имелись
рачки Salmincola exlumescens и
S. alpina, отсутствовавший у осен-
них. Морской скребень Echinorhyti-
chus gadi был найден у 4% летних
и 28% осенних гольцов, а пресновод-
ный скребень Е. salrnonis— у 32%
летних и 14% осенних гольцов.
Нахождение у осенней, т. о. воз-
вращающейся в пресные воды,
рыбы некоторых потамофнлььых
паразитов (Cyslidicola jartonis,
Echinorhynchus salrnonis) объяс-
няется отчасти, быть может, тем, что
промежуточные хозяева этих пара-
зитов имеются и в заливах Новой
Земли (что вероятно для Е salrno-
nis), отчасти же сравнительно
коротким пребыванием (2 — 3 месяца)
штрихи дает сравнение двхх категории
так называемой летней рыбы, которая
6С
с. гтпiiiiiim।вциптп
I 2 1 4 5 6 г в s го и ч и н ts
Годь> жизни
Рис. 177. Три кривых, показывающих
изменения паразитофауны в зависи-
мости от образа жизни хозяина
А - пресноводная рыба (хариус —
Thymallus thymallus) В—рыба, про-
водящая долгие сроки в море(лосось
Salmo salary, С - рыба, прово-
дящая во взрослом состоянии еже> од-
но три месяца и море, остальное же
время • озерах (новозсмсльскни го
леи - Salvelinus alpinus). (Оригинал.)
1 подъемн и падения прсспопппюи napa.iit
тофвуиы. В wopiKiM ip.i штофцупл. Замер
пены гроки ьрлиилимые рибои и морс.
гольца в море, что делает для
части пресноводных паразитов возможным удержаться в хозяине до
срока его возвращения в реки.
Не менее интересные картины изменений получаются и при изу
чеинн паразитофауны катадромальных рыб, примером которых
является угорь (Anguilla anguilla). Взрослые угри оставляют евроиен
ские реки для того чтобы нереститься в глубинах Атлантического
океана, где они погибают, уже не возвращаясь более в пресные воды
Личинки угря в течение трех лет ведут пелагический образ жизни,
Постепенно мигрируя от середины Атлантического океана к европей-
ским берегам. Достигнув берегов, личинки входят в реки, совершенно
изменяют свой образ жизни и становятся донными животными. В ре-
ках молодые угри растут и остаются до периода нереста, когда при-
ступают к своей катадромальной миграции.
Парэзитофауна взрослого угря изучена в Невской губе Догелем
2а 1
и Петрушевским (1933). тогда как личинки угря (Lcptocephalus)
и молодые угри, готовые войти в реки, исследованы Догелем (19366)
Личинки угря в течение всего трехлетнего периода своей пелагической
жизни совершенно свободны от паразитов. То же самое следует
сказать и о периоде его превращения в молодого угря, которое зани-
мает около года. Таким образом, угорь представляет полную противо-
положность лососю, будучи свободен от паразитов в течение первых
3 4 лет своей жизни. Это различие отчасти обусловливается весьма
слабым развитием пищеварительного капала у личинок угря, питание
которых до сьх лор представляет собой загадку, так как в их кишеч-
нике не находили никакой нищи. Как только молодой угорь, закончив
метаморфоз, опускается на дно и начинает питаться речным бентосом
(личинками Simuhum п т. д.), наступает инвазия его различными
паразитами. В исследованном Догелем случае первые появляющиеся
у молодого угря паразиты принадлежали к категории паразитов, не
имеющих промежуточного хозяина. Так, у шести молодых угрей
(около 70 j!.u длиной), пойманных близ Казерты под Неаполе м, инна-
шя СЛ1ННСТЫМ споровиком Myxidiurn giardi встретилась шесть раз,
(jyrodactylus обнаружен на жабрах два раза, тогда как из числа
паразитов, нуждающихся в промежуточных хозяевах, встретились по
одному разу только какая-то личинка сосальщиков и скребень
]canthocephalus nnguillae Этими данными подтверждается высказан-
ное ранее (гл. \11) правило о порядке заражения пресноводных рыб
в первою очередь паразитами, не имеющими промежуточных хозяев
или активно проникающими в хозяина.
Угорь интересен еще в одном отношении: у него наблюдается
известный половой диморфизм в смысле паразитофауны, совпадающий
с подом животного-хозяина. Дело в том, что самки проникают далеко
в реки и другие пресные водоемы, где и протекает многолетний
период их нагула. Громадное большинство самцов (и очень неболь-
шой процент самок) задерживается, однако, сразу после окончания
метаморфоза в устьях рек. остается там во все время нагула и еще
до приобретения брачного наряда начинает свое откочевыванпе в море
тля переста Поэтому взрослые самки, исследованные в Невской губе
при их скате в море, имеют чисто пресноводную паразвтофаупу из
11 видов многоклеточных паразитов. Однако для других место-
нахождений угря некоторыми авторами отмечаются находки у пего
чисто морских паразитов, каковы сосальщики Deropristis. Heniiurus.
/.ecithasier, Podocotyle, Sterrhurtis, скребень Corynosotnu и др. Эти
морские формы, по нашему предположению, должны встречаться
почти исключительно у самцов угря, держащихся в солоноватоводных
г.'туариях рек.
Влияние миграций на паразвтофаупу можно видеть также на
примере миног. Шульман (1957) показал, что Невская минога (Lam
petra /luviatilis) во время пребывания в море заражается кишечными
паразитами Proteocephalus sp., Eubothrium sp. и Dacn.tis slelmioides.
Во время периода жизни в реке, куда минога входит для икрометания
и где она не питается, все кишечные паразиты утрачиваются и про-
исходит лишь заражение мстацеркарнями Dlplostomulum spathaceum.
локализующимися в глазу.
Значительный интерес для паразитологии представляют также
случаи «миграций» рыб нпого характера. Речь идет в данном случае
нс о периодических перемещениях рыб в течение их индивидуального
жизненного цикла, а о переходе того или иного вида рыб
к жизни в необычной для дайной группы рыб среде.
Пресноводные по своему происхождению рыбы могут выселяться
в море, и, наоборот, рыбы морские по происхождению могут пере-
ходить в пресную воду.
Прекрасный пример этому дает р. Амур и Японское море. Здесь
типичная карповая, т. е. пресноводная, рыба краепоперка-\ган
(Leuciscus brandli) эмигрировала в море и только во время нереста
посещает приустьевые пространства рек. С другой стороны, китайский
окунь (Sinipcrca chua-tsi). принадлежащий к морскому семействе
Serraiiiciac, полностью приспособился к жизни в реках, которые оп
вообще не покидает.
Возникает вопрос, какие изменения могли произойти в паразита
фауне этих обеих «аномальных* рыб. Оказывается, каждая и.з них
увлекла за собой в свое новое местообитание часть своих исконных
паразитов и, с другой стороны, приобрела комн пеке паразитов, свой-
ственных новому местожительству Таким образом, в результате этого
река, с одной стороны, и морс — с другой, обогатились некоторыми,’
ио существу7 чуждыми им паразитологическими элементами. Чтобы не
быть голословными, приведем следующие данные Китайский окунь
па Амуре имеет целый набор паразитов пресноводного облика п про-
исхождения. Таковы жаберный споровик Leniospora chua-tsi, сосаль-
щики Tetraonchus dispar, Bucephalus polymorphus. Isuparorchis hypse-
iobugrt, цестод. Tetracampus, скребень Paracanthocephalus tenuirostris
нематода Rhabdochona denudata. рачки Ergasilus jlavimanus, Pseuder-
gusilus major., l\ undulaius. t. e. 10 видов паразитов. Однако, кроме
них, китайский окунь обладает двумя паразитами, говорящими о дав-
них морем 'lx связях этой рыбы. Во-первых, это специфичная для него
кипи чная нематода Contracoecu.ni sinipercae. N чисто пресноводных
рыб нематоды рода Contracoecum встречаются лишь в личиночном
состоянии и приюм в полости тела. Напротив, у морских и проход-
ных рыб они известны, помимо того, и из кишечника, где живут
в половозрелой ста тип. Следовательно, п этом отношении китайский
окунь приближается к рыбам морского происхождения. Во-вторых,
в его мочевом пузыре живет споровик Henneguya sinipercae. У чисто
пресноводных рыб Huineguya встречаются на жабрах, под кожей,
в мускулатуре. У некоторых морских и солонОватоводных рыб Нсппе-
,иуа констатированы в мочевом пузыре и почках.
С другой стороны, в паразитофауне краспоперкп-утая обнар’ кн-
наются вполне ясные следы пресноводного происхождения. Кроме
ряда морских паразитов, как-то: Podocotyle. Lecohaster, Zoogonoidcs
скребень Rhadinorhynchus, Scolex pub morphus и некоторые другГЛе.
зтот вид имеет пять паразиюв пресноводного характера. Эго рачки
Tracheliastes sacchalinensis и Ergasilus wilsoni, споровик Myxobolus
marinus и сосальщик Dactylogyrus iwanovi, нематода Rhabdo< honu
denudata. Роды Tracheliastes н Myxptfblus свойственны рыбам прес
nJx вод. Однако Исакова-Кео (19Г>2) нашла Мул fbolus на жабрах \тая—
дипствсиный пример жаберных Myxobolus у рыб, живущих в море.
Еще более интересен Dactylogyrus iwanovi, открытый на жабрах
того же у гая Быховскпм (рис. 178). Помимо того, что род
Dactylogyrus вообще свойствен пресноводным рыбам, вид. найденный
им из угас, показывает ряд изменений, вызванных в нем переменой
среды, окружающей хозяина. А именно, все пресноводные Ductylogy-
rus (а их мною десятков видов) живут свободно па поверхности
жаберных лепестков. Один лишь Dactylogyrus iwanovi угая прочно
прицепляется к определенному месту лепестка, после чего жаберный
эпителий разрастается вокруг основания паразита в виде высокого
2ЯЗ
Рис. 178. Dactvlogvrus
'Wtinovi с жабр дальне-
восточной краснонерьн-
уган (Lritcisctis brandli)
Я D. iuanovi на жа-
берном лепестке, п гру-
женный задним концом
глубоко в ямку разрос-
шегося вокруг него эпи-
телия хозяина; В то же.
снятый с хозяина; на зад-
нем копне К1Щ1Ы тва
крюч-а прикрепительно-
го диска 'По Быховскому )
воротничка или валика. Под защиту этого воротничка паразит может,
вероятно, временно втягиваться. Мало того, эта пресноводная
группа (Ьорм, по-видимому, вполне приспособилась к морской среде
Rhubdochona deruidala, кроме своего первоначального хозяина, ветре
чепа уже у типичного морского бычка Alcichlfiys. Таким образом,
утаи, переселившись в море, начинает как бы «навязывать» часть
своей пресноводной паразигофауны морским рыбам.
Еще более общее значение и общий интерес представляю1
после,юваиия паразитофауны перелетных пт и ц.
По отношению к перелетным птицам Дог.
.чем п его сотрудниками был применен возраст
посезонный анализ паразитофауны, состоящий
в том, что в определенной местности детально
изу |астся паразитофауна какого-нибудь вида
птицы, начиная с первых дней ее прилета и
вплоть до самого отлега; в то же время изучает
ся и паразитофауна птенцов от момента их вы
лупления и до первого отлета на юг. Применение
этого метода дало очень важные результаты
(Догель, 19496) Таким методом исследовано уж<
значительное количество птиц, а именно: стриж
(Догель п Каролинская, 1936), ласточка (Догель
и Навцевич, 1936», каравайка (Дубинин, 1938)
кваква, пли ночная цапля Nyctic&rax (Дубини
на, 193"), скворец (Марков, 1939), пеликан (Д.
бинчн, 195-16), утиные птицы и кулики (Гинг
цинская, 1949, 1953; Гпиецпиская и Наумов. 1958-
Белэпольская, 1956, 1959). Ряд работ выполнен
по разным группам перелетных птиц Быховской
Павловской (1953, 1955, 1962), Догелем и Быхов
ской-ПавловсКой (1955), Дубининой (19506.
19536) и другими.1 Для сравнения с перелетными
теми же методами исследователями изучались
некоторые оседлыс или кочующие птицы серая
куропатка—Барышевой (1939); рябчик—Олигер
(1940); домашний воробей — Победоносцевым
(1940); галка — Зсхиовым (1949а), и др. Благо
даря применению возрастно-сезонного анализ i
в том конгломерате паразитических форм, кото
рын как нечто статическое признавался до сих пор дпя каждой пере-
летной птицы, удалось различить несколько естественных групп, обла-
дающих каждая своей собственной особого рода динамикой.
Начнем с характеристики паразитофауны птиц, только что при
летевших на места гнездования. Эти птицы имеют известный набор
паразитов, которых они приносят с собой с юга. ио по своему гене
псу этот комплекс паразитов не является однородным. Количество
и разнообразие приносимых с собой паразитов у ра ных видов птиц
подвержено довольно сильным вариациям Этих паразитов, принесен
ньгх с юга. постигает различная судьба. Одни из них, как, например,
пухоеды и перьевые клещи, не только остаются на птице во время е<
пребывания па севере, но и переходят с родителей на птенцов и затем
уносятся как птенцами, так и родителями на юг. Другие принесенные
паразиты постепенно утрачиваются взрослыми птицами па севере,
а птенцы не обнаруживают зараженности этими паразитами в первое
лето своего существования. Третья группа паразитов, а именно; часть
284
юг. наконец, имеются эндо-
Рис 179. Схема распре слепня на
взрослом стриже повсеместных на ра-
зи гон — убиквистов (пппрамСр, Mal-
1орИака, Sarcoptiformesi. (По До-
гелю ) На взрослых стрижах убикви-
сты встречаются в течение вей о гида,
на птенцах — начиная с изнссгного
мом пта развит и з оперения.
Сплошные линии ибозилч шот грипицы меси пен.
мупк1ирные липни гпасчзют моменту прилета
и отлета птиц; пункт при..ш линия, дсляшзи
irio.ii. в секторе птеипсн*. отмечает период mi-
луплепня iircimoib Штрихиин» плначзет на-
личие лампой группы паризитов.
>ндопаразптов, держится у взрослых в течение всего лета, но не пере-
ходит на птенцов вплоть до момента отлета.
Однако, помимо паразитов, занесенных с юга, на местах гнездо-
нания паразитофауна птицы обогащается целым рядом новых форм.
Прежде всего сюда относятся эктопаразиты, связанные с гнездом
птицы, как-то: блохи, некоторые кровососущие мухи и др Далее, сюда
принадлежат эндопаразиты, которыми на севере заражаются как
взрослые, так и птенцы и которые, однако, ко времени отлета выми-
рают, так что птицы не уносят их па
паразиты, которые появляются у птиц
на севере, уносятся ими на юг, но от-
сутств\ ют у птиц, возвращающихся на
места гнездования.
Несмотря на кажущуюся калейдо-
скопичность такого состара паразито-
фа\ны, в ней уже на основании сделан-
ного нами только что беглого обзора
удается по происхождению паразитов
различать три главных группы:
I) убиквисты, встречающиеся в хозяи-
не в течение круглого года, безразлич-
но как на севере, так и на юге; 2) юж-
ные формы, заражение которыми про-
исходит исключительно на местах зи-
мовий; 3)‘северные формы, заражение
которыми совершается опять-таки ис-
ключительно на местах гнездования. 1
К этим трем основным группам на
основании недавно опубликованных
исследований Белопольской (1956)
следует добавить еще одну группу па-
разитов: 4) группа паразитов, зараже-
ние которыми происходит во время
пролета птиц по миграционным путям.
Каждая из этих групп может
делиться, как мы увидим далс
на подгруппы, к описанию состава
и особенностей которых мы и пере-
ходим.
1. Убиквисты (рис. 1791 перелетных птиц. Это, прежде всего,
все ~е паразиты, которые живут на самом теле птицы. Они тесно
связаны с телом хозяина во всем своем жизненном цикле и сопро-
вождают птицу во всех ее странствованиях. Эту естественную и одно-
родную первую подгруппу убиквистов составляют пухоеды
и перьевые клещи (Sarcopliformcs).
Пухоеды (М tllopha^a) всю свою жизнь проводят на птице и на пти-
це же откладывают свои яйца, приклеивая их к перьям (см. рис. 180).
Поэтому Mallopha^a можно встретить и на только что при летевших
птицах, и в течение всего пребывания птицы па севере, и при отлете ее
на юг; есть полное основание утверждать, чго они держатся па хозяине
и во время всей его зимовки. Таким образом, это типичные и притом
кр\ 'логодичные паразиты — биквнеты Однако .можно все же выска-
зать предположение о первичном происхождении пухоедов и перьевых
клетей. Как те, так и другие могли стать паразитами птиц лишь в таких
условиях, когда птица имеет наиболее теспое и длительное сопри-
28т
косновение с субстратом. Между тим на местах зимовки птицы не имею!
строго постоянного местожительства, и точкой наиболее длительно! о
соприкосновения их с субстратом является гнездо, где к тому же облег
чсна и передача эктопаразитов от одной особи хозяина к другой. Вот
почему мы думаем, что первоначальной родиной эктопаразитов перелет
ных птиц являются места их исконных гнездовании.
В распространении пухоедов у перелетных птчц есть много любо
пытных моментов, представляющих общий биологический интерес.
Прежде всего, пухоеды дают очень хороший материал по вопросу
о возрастных изменениях паразитофауны. Дело в том, что первое зара
жнше птицы пухоедами совершается во время птенцового периода i
гнезде, где птенцы получают пухоедов от своих родителей. Наблюдение
над гнездом и измерение птенцов показывает, что это первоначально!
заражение птенцов пухоедами у каждого вида пищ строю приурочен^
к определенной стадии развития оперения, а значит и к определенному
возрасту птен на
Так, у стрижа пухоеды (и перьевые клещи) впервые появляются
на птенцах в то время, когда крыло их (от места сгиба до конца наибо-
лее длинного махового пера) достигает 120—125 длины, тогда как
крыло взрослой птицы имеет 170- 180 мм длины. Длина в 120 лиг при-
близительно отвечает 14—15-дневному возрасту. Таков нижний порог
заражаемости пухоедами стрижа. У городской ласточки крыло взрослой
птицы имеет 112—114 мм длины, а пухоеды начинают появляться на
птенцах с крыльями, достигающими 50—60 лл длины. У скворца крнти
ческим моментом, с которого начинается массовая миграция пухоедов на
птенцов, является возраст около 10 дней, соответствующий длине крыла
в 50 .ил (длина крыла взрослого скворца равняется 126—127,ил).
У некоторых птиц срок заражения пухоедами сдвигается либо к бо-
лее раннему, либо к более позднему возрасту Так, у кваквы (Nuclicorax)
(Дубинина, 1937) первые заражения птенцов пухоедами Eslhioplerum
ardcae наблюдались на II, 15 и 22-м дне их жизни, а мас-
совое заражение наступает лишь перед самым отлетом из гнездовых
колоний.
С другой стороны, у иовоземельскоп кайры (Uria lomvia) Марков
(1937) наблюдал, что пухоедами были згражены птенцы уже 1—5-днев
ного возраста. Быть может, это обстоятельство вызывается в дашюл
случае влиянием северных шпрот когда иерывон покров птенца растет
быстрее.
Наконец, время перехода пухоедов па птицу зависит не только от
вида хозяев, ио п от видовых особенностей самих пухоедов, как показал
Дубинин (19381 на каравайке, причем у этой птины решающую роль
играют абсолютные размеры пухоедов. Чем меньше пухоед, тем раньш<
переходит он на хозяина.
Птхоедами взрослые птицы большей частью заражены поголовно,
причем таким же стопроцентным становится и заражение птенцов ко
времени осеннего перелета.
Как известно, на одном виде птиц может быть несколько видов
пухоедов, причем в таких случаях иногда можно отметить явную регио-
нальность распространения паразитов на теле хозяина. Дубинин, напри-
мер- различает три категории пухоедов, населяющих строю определен-
ные части оперения каравайкп (рис. 180). Он пишет по этому повод'
(1947): «Это позволяет нам на теле птицы начертить своеобразную
географическую карту ареалов обитания отдельных родов п видов пу-
хоедов». Аналогичную закономерность удается установить и в отноше-
нии перьевых клещей, о чем подробнее будет сказано ниже.
286
По отношению к некото
рым птицам, а именно: к кара-
вайке, удалось подметить зави-
симость известных моментов
жизненного цикла пухоедов от
миграции хозяина. Дубинин
(1947) пишет, что длительный
перелет ведет к потере части
эктопаразитов. У каравайкп
пухоеды Ferribia и частично
ibidoecus и Esthioplerum при-
способились к сопутствованию
мигрирующей птице в виде ио
коящнхся стадий — яиц, кото-
рые приклеены к перьям и бо-
лее гарантированы от потери
во время пути. Ferribia и клещ
Megninia мигрируют исключи-
тельно па стадии яиц, о чем
свидетельствует накопление
громадного количества яиц пе-
ред отлетом н резкое уменьше-
ние числа взрослых и молодых
особей. Точно так же и весной
развитие эмбриона в яйцах
большинства пухоедов кара-
вайки приурочено ко времени
прилета каравайкп в дельту
Волги. Вторичная откладка
яиц пухоедами и появление мо-
лоди первой северной генера-
ции совпадает со временем по-
явления птенцов. Другие виды
Mallophaga пе обнаруживают
подобных зависимостей и мо-
гут быть найдены на приле-
тающей птице в виде имаги-
нальных стадий.
Перьевые клещи Sarcopti-
formes (рис. 181) представляют
много параллелен в своем рас-
пространении с пухоедами с
которыми они сходны и по ха-
рактеру питания и по характе-
ру всего жизненного цикла,
Клещи тоже приносятся в виде
имагииальных стадий или в ви-
де яиц (Megninia у каравайкп)
взрослыми птицами, а во вре-
мя гнездового периода перехо-
дят с родителей на птенцов.
Время заражения птенцов кле-
щами нередко совпадает со
временем перехода на них
пухоедов. Так дело обстоит
Рис. 180. Распределение различных видон
пухоедов на карасайке (ibis falcinellus)
(По Дубинину.)
I — Jbidoecus bislgnutus (с перьев головы и пс-н);
2 — Metiopon plegadts [с иеры-u грули, 6рюх« н бо-
ков); в — пилы ColpOcephalum и Irrrtbla (с нерьси
крыльев и хниств); 4 — b.striioptrrum rhaphiamm
(с кроющих перьев спины).
Рис. 181. Analgopsis passerintis (Sarcoptifor-
iiies, Analgesidae) с перьев воробьев
A — самка; /?—самец. (По Кюкенталю.)
Третья пара ног самца чризкычанни yio.iinciw и служш
для oCxdjtuujiihm симки при копуляция.
287
у ласточки и стрижа, а также у скворца. У зяблика клещи появляются
иа птенцах с длиной крыла в 36—45 льи, у каравайки переход на птенцов
клеща Megninia совершается позже (на 18-й день), чем пухоедов.
У клещей тоже наблюдаются в жизненном цикле приспособления
к миграциям хозяина. Мы уже видели, что Megninia каравайки проде-
лывает перелет на стадии яйца. Интересные отношения, по данным Д)
бининон (1937), намечаются у кваквы (\'ycticorax) и других цапель.
Прилетающая в дельту Волги в первой половине апреля кваква не
имеет на себе ни одного из свойственных ей клещей Pterolichus ardeae,
которые начинают появлять
ся в небольшом количестве
лишь с 18 апреля. В это Же
время весь подкожный слой
только что прилетевшей птн
цы переполнен дейтопимфа-
мн клеща, которые являют-
ся стадией развития Pteroli-
chus. В следующие три меся-
ца идет постепенное увели-
чение интенсивности зара-
жения накожными формами
Pterolichus и, наоборот
уменьшение заражения дей-
тонимфами. В августе же
(предотлетный месяц) идет
обратный процесс умень-
шения как экстенсивности,
так и интенсивности зараже-
ния накожными Pterolichus
ardeae и увеличение интен
снвности заражения дейто
нимфами. Таким образом,
суля по всему, перелет со-
вершается Pterolichus ar-
deae на стадии подкожной
деитоннмфы, для которой
риска отпасть от птицы во
время миграции совершен
но нс г.
Pacnpete-npHUP клещей
Рис. 182. Диаграмма распр’делсиия неры-вых
клещей (Sarcopliionnes) на .маховых перьях у
ласточки. (Оригинал.)
1 расппслелеп11е клечней у пяроглых ласточек: " — распре-
деление Sarcoptifnrnws у летних iitljiikju; 3 — w же у i нез-
юных игениоп.
В расселении клещей но телу птицы нередко имеются вполне опре-
делеиные закономерности (рис. 182). Их излюбленным местом поселения
служат маховые перья крыла, причем иногда клещи селятся на них не
равномерно, по предпочитая определенные номера перьев. Так, у стрижа
таким предпочтением со стороны Eusthatia cultrifer и Chauliacia
securiger пользуется 2-е маховое перо, как это видно из следующих цифр
средней зараженности различных маховых перьев, полученных на осно-
вании подсчета клещей на 30 взрослых стрижах. На 1-м маховом пере
встречаются в среднем 3, 4 клеща, на 2-м — 194, на З-.м —82, на 4-м —37.
на 5-м — 7, 5, на 6-м —3, на 7-м и прочих— 1. То же различие наблю-
дается и у птенцов стрижа, но выражено несколько слабее.
У ласточки имеется аналогичная неравномерность, выраженная
сильнее всего у взрослых птиц, слабее у летных птенцов и еще слабее у
птенцов гнездовых (табл. 14).
Скворец, по Маркову (1939), обнаруживает аналогичные отноше-
ния, только особенно густо заселенными, кроме 2то, оказываются еще
288
3-е и 4-е маховые перья. 11 здесь у гнездовых птенцов клещи распростра-
нены на перьях более равномерно.
Т а б л it it а 14
Средняя густота населения клещей на маховых перьях
ласточки (по Догелю и Навцечич)
Возраст ласточек Маховые перья
1-е 2-е ье 4-е 5-е | б-е 7-е 8-е
Взрослые птицы .... 6 83 40 31 19 17 17 13
Летные птенцы .... 0 24 10 2 5 2,3 1.3 2,8 0
1 нездоные птенцы . . . 1 11 11 11 7 3 » 0,0
У каравайкп Дубинин (1936) обнаружил более или менее равномер-
ное расселение клещей на первых 12 маховых перьях (кроме 1-го, которое
всегда почти свободно от клещей). 1\ моменту переползания клещей на
птенцов указанная выше закономерность нарушается и клещи располза-
ются не только на все 27 маховых перьев взрослой птицы, но и на крою-
щие. перья крыла.
Несколько иной характер имеет закономерность распространения
клещей на маховых перьях баклана по данным Никольской (1939) л
Дубинина (1951), На баклане имеется три вида клещей, которые распре-
деляются на маховых перьях крыла следующим образом. Allopies sub-
crassipes расселяются на 6—20-м маховых перьях баклана с максимумом
у 10-го пера; Megninia phalcicrocoraci.s— на 4—8-м перьях, с максимумом
у 5-го пера; наконец. Midiae/ichus heieropus заселяют 1—7-е маховые
перья, давая максимум густоты на 3—4-м перьях.
Закономерности распределения перьевых клешей по перьевому по-
кровх птиц особенно тщательно изучены Дубининым п изложены в пер-
вом томе его монографии Analgesoidea (1951). Автор подробно рассма-
тривает нс только самый характер распределения клещей на разных
группах перьев, по даст также анализ причины этого явления. Мы не
имеем здесь возможности подробно рассмотреть результаты исследова-
ний Дубинина и ограничимся лишь кратким изложением его основных
выводов.
Дубинин считает, что закономерное распределение клещей по перье-
вому покрову птиц является прежде всего результатом исторически сло-
жившихся в процессе эволюции взаимных приспособлений паразита и
хозяина, адаптации определенных родов и видов клещей к определенным
условиям обитания. Существенную роль пои этом играют особенности
пространственного положения и функции отдельных групп перьев, в том
числе общность макро- и микроскопического строения перьев внутри
определенных групп (например, маховые перья 1-го порядка, 2-го по-
рядка и т. п.) и резкое различие микроструктур перьев в разных груп-
пах. Очень большое значение имеют аэродинамические особенности
(быстрота, направленно и сила воздушного потока при полете) разных
групп перьев, которые связаны с общей конфигурацией крыла и харак-
тером полета птицы. Все эти особенности находят свое выражение в
строго закономерном для каждого рода и вида клещей распределении
по перьевому покрову.
Вторую биологическую подгруппу среди убиквистов
состав тяют те эндопаразиты птиц, заражение которыми происходит оди-
наково как на юге, на местах зимовки, так и у нас на севере. В обоих
районах жительства хозяина протекает полностью весь цикл развития
в промежуточных и окончательных хозяевах и в обоих же местах могут
1Q В Л Догель
2Я9
наступать новые, заражений пара ином. 11 ан большим количеством эндо-
паразитов-убиквистоЕ обладают, ио-видимому, птицы, не меняющие зимой
характера своей пищи и отлетающие на зиму сравнительно недалеко,
например в южную Европу Таков скворец, летящий но Францию, в Бель
ппо н во всяком случае не дальше южного побережья Средиземного МО
ря. Среди 21 вида гельминтов скворца, обнаруженных Марковым в
Петергофе, несомненными убиквистами являются 8 (1 сосальщик, 2 цс
стоды и 5 нематод). Такие виды, с одной стероны, имеются весной у
прилетных скворцов, с другой стороны, они же попадаются и летом у
птенцов, которые могли заразиться только на севере. Кроме того, три
вида {Leucochloridium paradoxuni, Puriclerotaenia par’та it Choanotaenia
masculoaa), по всей вероятности, тоже убиквисты, т. е. к убиквистам
принадлежит 55% гельминтов скворца, а также коКцндня Isospora la-
cazei. Попять такое обилие убиквистов нетуудно, если мы учтем, что
в условиях зимнего сезона южной Европы шпорец питается теми же
пли очень близкими видами дождевых червей, моллюсков и даже пасеки
мых (особенно роющихся в земле личинок), которыми он питался и
у нас на севере.
Вторым примером могут служить утки, паразитофауна которы
изучалась Гинсцинской (1949).в дельте Волги. Утки, гнездящиеся в ниж-
нем течении Волги, зимуют ла южном побережье Каспия п совершают,
следовательно, лишь относительно небольшой по протяженности пере-
лет Основная масса паразитов ток согласно 1’ннецпнекой является
убиквистами н лишь единичные виды могут быть отнесены к южным
(Echinuria uncinata) и северным (Tetracotyle ardeae, Apora dogieli)
формам.
Иначе обстоит дело у таких птиц (стриж, ласточка), которые осеныс
летяг далеко иг» экватор и таким образом резко меняют не только вс>
свои условия существования, но и свои кормовые базы. Трудно ожидать
найти в Центральной Африке и в северной Европе идентичные формы
насекомых, которые могли бы служить промежуточными хозяевами для
одних и тех же паразитов зимой и летом. Вит почему среди 15 видов
гельминтов стрижа, только 4 быть может относя гея к убиквистам и
встречаются у взрослых стрижей уже в первые дни после прилета. У лас-
точки средн 12 видов гельминтов, встречающихся в ней в Петергофе,
Догель и Навцевнч не могут указать нн одного несомненного убиквиста,
имея известные сомнения только относительно трех видов цестод. Точно
так же у кваквы (ЛусИсбгих) Дубинина средн 52 видов червей лишь
одного считает несомненным убиквистом, а Дубинин находит лпшь двух
убиквистов х каравайкп, насчитывающей в дельте Волги 29 видов гель-
минтов. Даже если мы отнесем к убиквистам у кваквы и каравайкп
несколько видов гельминтов сомнительного происхождения, все же
у биквисты составляют ничтожный процент в эндофауне эти к птиц.
2. Южные формы, т. е. паразиты, заражение которыми проис-
ходит исключительно на местах зимовки. Теоретически рассуждая, дан-
ную группу паразитов можно прежде всего разделить па две большие
подгруппы. К перво й п о д г р у п н е ю ж к ы х фор м относятся па-
разиты, весь цикл развития которых протекает па местах зимовий, так
что в нашей северной обстановке мы пх вообще не находим. О существо-
вания таких паразитов цока мы можем то4п.ко догадываться по анало-
гии с Некоторыми северными паразитами, которые утрачиваются птицей
еще до оглега ее на юг.
Вторую подгруппу южных паразитов образуют ви-
ды, цикл развития которых проделывается ими па юге, но па одной нз
стадии цикла эти виды заносятся типами па север, где мы их и обна
wo
руживаем. Однако существенным отличием их от убиквистов является
отсутствие у них возможности размножаться на севере. Это временные,
заносные элементы, которые размножаются лишь на юге. Подобные па-
разиты в свою очередь могут быть двух сортов: один из них (катего-
рия А) умирают ио время пребывания птицы на севере, и таким обра-
зом птица заражается ими заново по возвращении на места зимовий
(рис. 183); другие (категория В) переживают срок пребывания птиц
на севере и снова уносятся ими на юг. где, возможно, продолжают раз-
множаться (рис. 18-1).
Рнс. 783. Схема распределения в
стрижах южных паразитов пл ка-
тегории А (например, Agamonema sp ).
Приобретенные птицей в Африке па-
разиты имеются только у взрослых
птиц и постепенно утрачиваются ими
па севере. (По Догелю.)
Обозначения тс же, что на рнс. 179.
Рис. 18-1. Схема распределения в
стрижах южных паразитов пл кате
горни В (например, PhiiynOsomum
ctaihratum, Lyperosomum olssoni).
Приобретенные птицей в Африке па-
разиты имеются лишь во взрослых
стрижах и унос ятся ими обратно на
юг. (По Догелю.)
Обозначения Те же, 'по па рис. 179.
К южной категории Л относится, например, Tanaisia fedtehenkoi из
почек каравайки (дельта Волги». Tanaisia попадалась только но взрос-
лых каравайках и притом лишь в первую половину пребывания хозяина
на севере (до 15 нюня). При этом средн найденных червей было много
отмирающих, а в июне из 47 найденных только 2 были живыми
Гораздо быстрее освобождается каравайки от другого южного пара
зита, относящегося к категории Л. а именно: от нематоды Echinuria соп-
torta, которая живет под кутикулой мышечного желудка. Вс1речаясь
лишь у взрослых птиц и сначала будучи довольно обычным паразитом
(у 36,8% особей), эта нематода позже 15 мая Дубинину вообще не по-
падалась. .\ кваквы (Xycticorax) к этой категории паразитов относятся
Opisthorchis longissirna из желчных протоков печени н скребень Echinor-
hynchus spiralis. Оба паразита найдены только во взрослой птице, и
большинство их было в умирающем или в умершем состоянии. Находки
скребня относились лишь к апрелю—июню, Opisthorchis встречалась до
начала августа. У стрижа категория А содержит только одну форму, а
именно: личиночную форму нематод /1 / атопета (см. рис. 183). обра-
зующую округлые цисты в стенках кишечника. Они были констатированы
лишь в первую полонии' лета (до половины июля) и только у взрослых
птиц. Таким образом, получается небольшая, но резко очерченная груп-
па южных форм, гибнущих в птицах до периода их отлета па юг.
19’
291
Категория В тоже содержит целую серию паразитических форм
Сюда относятся, например, почечный сосальщик каравайкп — Renicola
В стрижи- имеется группа сосалыцикив (Platynvsonuuii clathratum, Lype-
rosomum salebro^um и L. olssoni)- живущих в печени и желчном пузыре
взрослых птиц, причем все они принадлежат к данной категории. В про-
тивоположность членам предыдущей категории эти сосальщики могли
быть констатированы во взрослых стрижах вплоть до самого отлета,
хотя часть этих паразитов отмирала в течение лета и попадалась в полу-
разложпвше.мся состоянии. У ласточки, кроме Plalynosomum (см
рис. 184), к этому же разряду, по-видимому, относится и полостная не
матода Diplutriuena obtusa, а у каравайкп — небольшие цистицеркп
Cysticercus sp. и почечный сосальщик Renicola, ннцистнрованные в стен-
ках кишечника. У кваквы наряду с теми же цпетпцерками мы имеем
еще одного паразита дайной категории — Роггосоесшп reticulatum. Этот
вид до сих пер был встречей в пределах Африки и Индии, и заражение
нм взрослых квакв, очевидно, происходит на юге. Заразившись в Афри-
ке, кваква приносит половозрелых червей на места гнездовья, где они
продолжают существовать вплоть до отлета птицы назад. Частота зара-
жения этим паразитом не меняется в течение всего лета, что говорит
о полной выживаемости его в наших северных условиях.
В пределах обеих категорий южных эндопаразитов, мы замечаем
щну особенность: в них очень редки кнщрчные паразиты и преобладают
печеночные, почечные и полостные гельминты. В самом деле, среди
13 форм, взятых нами в качестве примера, лишь четыре относятся к ки-
шечным паразитам (Echinuria, два вида скребней и Porrocoeciun]. Не-
видимому, на север заносятся главным образом такие паразиты, которые
нс логут быть легко и быстро выведены наружу с экскрементами.
В пользу этого говорит п то обстоятельство, что среди четырех кишечных
форм Echinuria живет под кутикулой желудка, т. е. трудно поддается
выведению, а скребни глубоко внедряются в стенку кишки своим воору-
женным крючьями хобот.юм. Такое малое количество кишечных пара-
зитов наводит на мысль о двух моментах: во-первых, оно говорит в поль-
зу того, что на юге у перелетных птнц имеется кишечная фауна пара-
зитов, ио она вымирает до прилета тпц па север, подобно тому как
многие северные формы (см. далее) вымирают в птицах еще до их от-
лета на юг; во-вторых, печеночные и почечные сосальщики обладают
сравнительно с кишечными трематодами значительно большей длитель-
ностью жизни.
3. Северные формы проделывают свой цикл развития на
местах гнездовий. Если они выпадают из паразитофауны птицы еще
до ее отлета на юг, то мы относим их к к а т е г о р и и С (рис. 183); если
же они \ носятся птицей на юг, но обратно следующей весной при прилете
не возвращаются, то мы включаем их в к а т е го р и ю D (рис. 186 и 187).
К категории С, естественно, относятся прежде всего все эктопара-
зиты перелетных птиц, связанные не столько с телом хозяина, сколько
с его гнездом. Эти эктопаразиты ведут крайне активный образ жизни
во время короткого гнездового периода птицы; вылупляются из покоя-
щихся стадий, усиленно питаются, копулируют и размножаются. Однако
с вылетом птицы из гнезд все эти формы утрачивают свой источник пи-
тания, а потом) н цикл их к этому времени заканчивается, и паразиты
вплоть до следующей весны уходят в том или другом виде на покой
Число видов гнездовых паразитов у разных видов птиц сильно варьи-
рует, ио иногда бывает очень значительным. К гнездовым паразитам
принадлежат прежде всего кровососущие клещи Laelapidae. Они в общг м
мало связаны с телом хозяина и попадаются главным образом при изу-
292
Рис. iJ‘5. Схема распрехелеипн паразитов ласточки
/1 — схема распределения северных паразитов ласточки из категории (
(наир) мер, /‘fuiAcropsolfis micrococcus). Паразиты приобретаются на севере
н там же вымираю!; В — схема распределения паразитов гнезд ласточки.
Паразиты появляются на птице тотчас же после заселения ею гнезд и
па птенцах тотчас же после их вылупления. Момент вылета ласточек
из гнезд обозначает время освобождения птиц от гнездовых паразитов
(По Догелю п Н вценич.’) ►>
Обозначения те же. чти из рис. 179.
Рис. 187. Схема распределения север-
ных паразитов стрижа hi категории
Р (например, Pltigiorchis nmcilosus)-
Паразнты приобретаются лишь ра
севере как в ipoc.ion птицей, так и
птенцами. Некоторая часть их уносит
ся птицей при отлет. па юг, где
постепенно отмирает. (По Догелю.)
Обиаизчеиим тс же. что из рис. 179.
Рнс. 186. Схема ра .'нре деления
северных нарлзнюн стрижа пз кате-
гории D (например, Prosthogonimus
ovatus). Исключительно пярашпы
птенцов, уносимые ими па юг, где
паразиты утрачиваются еще до пер-
вою возвращения птицы па север.
(По Догелю.)
Обоаначеппя тс же, чю пл рис. 179.
24
ченип гнезд последнего. Лишь иногда часть этих клешей удерживается на
птенцах, взятых из гнезда для осмотра.
В гнездах птиц встречается также целый ряд кровососущих насеко-
мых, принадлежащих к разным отрядам. У стрижен (Догель и Каролин-
ская, 1936) изредка попадались постельные клопы (Cirnex lectularius).
а у ласточек—особый род клонов (Oeciacds hirundinis, рнс. 188) У не-
которых птиц гнездо обильно заселено блохами. Так. например, в гнез-
дах скворца паразитируют Ceralophyllus gallinae. Зимой они не гибнут.
। переходят на домашних птиц или перезимовывают в пмагииально.м
состоянии в скворешниках. Особенно многочисленны блохи (Cerato-
phyllus) у ласточек, где иногда мы в одном гнезде находили до 100 взрос-
1ых блох и до 150 личинок их
Рис. 188. Насекомые-эктопаразиты из гнезд ласточки
А— Oeciacus hirundinis — клон ласточки, В — блоха нз рода
Ccratophyllus представители которого встречаются в гнездах
ласточек. (/1 — по Брумпту. В — по Павловскому.)
Всего разнообразнее же в гнездах фауна двукрылых. В гнездах на
птиц, в особенности па птенцов, нападают многочисленные Ilippobosci’
dae, кровососущая муха Camus hernapterus, а в дельте Волги и Simulii
dae (Simuliurn niaculatiun). Все эти мухи тесно связаны с гнездовым
периодом, в особенности Camus, которая утрачивает крылья почти сразу
после выхода из куколки, а на взрослой птице никогда не попадается.
Это типичный паразит птенцов. Дубинин (1938) очень ясно проследил
динамику Camus в гнездах каравайкп в разное время года: в феврале -
шреле в пустых еще гнездах каравайкп можно найти лишь 1 —10 куко-
лок, в мае—июне около 35% гнезд содержит взрослых насекомых, иногда
то 80 экземпляров на гнездо. В июле зараженность гнезд взрослыми
мухами резко падает, а в августе—сентябре в гнездах опять остаются
чдни коконы (1 — К) экземпляров на гнездо). Таким образом, Camus
проводит зиму в состоянии куколки. Некоторые нз Hippoboscidae, на-
ipiiMCp Ornithoniyia. благодаря хорошему развитию крыльев менее тес
но связаны с гнездом. Марков находил Ornithomyia и на убитых в июле
скворцах. В Петергофе муха О. chloropus позже 1 сентября иа птицах
пе попадалась, т. е. видимо из юг муха не уносится. Чисто северный
характер совершенно ясен у кровососки Stenopteryx hirundinis ласточки,
которая обладает узкими, ис способными к полету крыльями и вслед-
ствие этого тесно связана с гнездом. Под осень (с начала августа) в
брошенных ласточками гнездах остаются зимовать куколки этих мух.
Столь же несомненен северный характер некоторых мух из семейства
тахнн (Protocalliphora sordida). паразитирующих на ласточках в личи
ночном состоянии. Личинки этих мух сос_\т кровь птенцов ласточки (и не-
которых других птиц) н оюкляюгся в гнездах. Часть куколок остается
шмонать в гнездах.
Итак мы видим, что целый комплекс эктопаразитов приурочен к
гнездам птиц и вследствие этого связан с северным местопребыванием
птицы. Особенно богат этот комплекс у ласточки, где гнездо содержит
I aekipidae, клопов Oeciacus. блох Ceratophyllus и мух Sh nopleryx и
Pralocalliphnra.
Дрхгая часть паразитов категории С не имеет никакого отношения
к гнезду и состоит из эндопаразитов, пре имущественно кишечных'. Се-
верный характер этих форм доказывается, с одной стороны, тем, что они
не встречаются i тн очень pi дко встречаются у убитых вскоре после
Рис. 189. Сосальщик Phuneropsolus micrococcus ив кишечника ласточ-
ки и кривая его распространения в ласточках Петергофа в разные
месяцы. (!!•> Догелю и Навцевич.)
Нэ сии абсцисс отложены месяцы, на оси ординат нокллэпа частота нахождении паразита
п разнос время сезона. Крппэи пока эынзет, что заражение Phaneropxolns, начинаясь
сразу после прилета ласточек, постепенно попытается ли конца июля, по затем но как им-то
причинам стреми rc.ii.no падает и прекращается п л мал питых числах am уста, цесиитры ня
то, что ласточки остлштси и Петергофе еще ДО десятых ’iiice.i сентября.
прилета взрослых птиц, обнаруживая разгорание инвазии лишь в более
поздний период лета. С другой стороны, эти паразищ обычно заражают
и птенцов, чем прекрасно доказывается местный характер источника ин-
вазии. Они начинают исчезать из птицы еще до ее отлета ил юг, чго
показывает принадлежность >тпх форм, в частности, к категории С
Ряд видов данной грхины имеется среди сосальщиков, причем осо-
бенно показателен пример Phaneropsolus micrococcus мелкого сосаль-
щика (см. рис. 185 и 189). живущего в кишечнике ласточки. По
Догелю и Навцевич (1936), ласточки заражаются в Петергофе этим со-
сальщиком сразу после прилета. В первый период после прилета (до
I нюня) заражено 30% взрослых птиц с интенсивностью заражения
в 3 паразита. В июне и июле (до 20 июля) экстенсивность и интенсив-
ность инвазии достигают максимума (70% и 17 паразитов на птицу).
Третий период (с 21 июля но 10 августа) характеризуется той же экстен-
сивностью инвазии (70%), по сильным падением интенсивности зара-
ь аия (G паразитов на итипу). Наконец, с 10 августа и вплоть до отлета.
29".
который происходи.) но второй половине сентября, Phaneropsolus был
найден всего у одной из 30 вскрытых ласточек и то один экземпляр.
Такчч образом, совокупность данных заставляет нас принять сле-
дующую картину динамики этого вида у ласточек. Прилетающие зеспой
ласточки заражаются Phaneropsolus и заражают им своих птенцов, кормя
их насекомыми, содержащими метацеркарнн. Инвазия подновляется вс<
время, пока продолжается лет насекомых — промежуточных хозяев. Од-
нако за некоторый срок до 10 августа (в 1934 г.) лет этих насекомых
прекращается, и тогда паразиты, обладающие, подобно многим другим
сосальщикам (например. Slrigeidac), короткой длительностью жизни,
вымирают, чем и объясняется полное отсутствие Phaneropsolus во вторую
половину августа. Отлет ласточек из Петергофа происходит лить во
второй половине сентября, п тем самым получается биологический барьер
распространению данного паразита на месте зимовий ласточек.
Судя но всему, такой же динамикой обладает и сосальщик Р1а-
giorchis rnaculosus ласточки и стрижа, а также, но-видпмому, и больший
ство цесгод этих птиц Дубинин 11938) относит той же категории Pla-
giorchis uhlaorini и четыре вида цеетод каравайки, а Дубинина (1937) —
некоторых сосальщиков и одну цестоду (Velipora rmitahiiis) кваквы.
Марков (1939) для скворца причисляет к категории С ряд сосальщиков
(Plagiorchis brauni, Р. rnaculosus. а также, возможно, и I .ateroiremu
uexans). Следовательно, к данноп группировке пока удается несом пенно
отнести только часть сосальщиков и ленточных глистов перелетных птиц.
Наконец, следует остановить еще одну категорию 13 северных форм
куда относятся виды, которые проделывают своп цикл развития на се-
вере, но задерживаются в организме хозяина до осени, вследствие чего
и захватываются им на юг. Судя по всему, они вымирают па юте, н
новое заражение ими происходит опять на севере.
Среди паразитов птиц, исследованных в нашей лаборатории в Пе-
тергофе (ласточка, стриж, скворец, зяблик), безусловно к этой группи-
ровке принадлежат лишь сосальщики рода Piosthogownus (см.
рис. 186), хотя, быть может, сюда относятся и некоторые виды катего-
рии С, по крайней мерс у птиц, отличающихся ранним отлетом (стриж).
Так, например, у ласточки и скворца сосальщики рода Plagiorchis
icm. рис. 187) вымирают сто до отлета птиц, тогда как у стрижей,
улетающих уже в половине августа, Plagiorchis попадаются вплоть до
отлета н уносятся птицей на юг..
Напротив, в дельте Волги паразиты категории D оказались у кваквы
самыми распространенными, к ним относится 12 видов (5 сосальщиков.
2 цестоды н 5 нематод), а у каравайки — 6 видов (3 сосальщика и 3 не-
матоды). Некоторые из этих в\дов, принадлежащие к сем. Stripeidae,
судя по краткости срока половозрелой стадии представителей этого се-
мейства, вымирают уже на пути в Африку, другие, вероятно, держатся
значительно дольше. К таким типичным представителям категории В
принадлежат виды рода Prosthogonimus (Р. ovaitis, Р. cuneatus). Этот
сосальщик встречается только у птенцов очень многих видов птиц и
живет в фабрицчевой ехмке. Это обстоятельство, с одной стороны, го-
ворит о северном происхождении этих видов Prosihogonitnus, с другой —
объясняет, почему птицы не могут принести данного сосальщика с юг;
снова на север Фабрициева сумка у большинства птиц редуцируется
еще до конца первого года жизни, а потому и паразиты, живущие в ней
обречены па гибель в течение первой зимы, следующей за отлетом птен-
цов на юг. Молодые птицы в первый раз возвращаются на север, уж*
освободившись о г Prosthogonimus.
4. Особую экологическую группу паразитов птиц представляют так
296
называемые миграционные формы, реальность с^'цесгвования
которых была убедите 1ьно обоснована в работах Белопольской (1956
1959).
Особенно"отчетливо наличие этом груп ы паразитов 'дается обна-
ружить у видов птиц с длинными миграционными путями. К ЧПСЛ5 такн>
птиц относятся, например, нулики, проделывающие во время перелетов
очень длинный пун. от северных шпрбт до тропиков, а некоторые даже
до берегов Австралии. При этом многие виды куликов летят вдоль мор
скпх побережий (на востоке преимущественно вдоль Тихоокеанского по-
бережья). Во время этих длительных перелетов (особенно при отлете
который совершается менее стремите .ыю, чем прптет, п занимает го
раздо больше времени) птицы делаю г. ос ганпвкй. питаются и, поедая
различных беспозвоночных, заражаются паразитами. Таким образом
возникает особая категория — миграционные паразиты, заражение ко
горымд связано с самим процессом пролета. При этом, наряда с приобре
гением новых видов, происходит постепенное очищение птиц от ранее
полеченных паразитов — утра га северных визов (в том случае, если речь
идет об отлете). Для куликов, летящих вдоль морских побережии,
«миграционными паразитами», как показала Белопольская, являются
главным образом сосальщики сем. Microphallidac. Их личиночные ста
дни — церкарнй и метацеркарии—уже вполне сформированы к этому
времени в промежуточных хозяевах. Проникая в птиц, они оч<нь быстро
приступают к откладке яиц, рассеивая их в окружающей среде.
Кроме мнкрофаллнд, к «миграционным паразитам» могут относиться
также некоторые виды цестод, нематод п скребней. Например, Дубини
на (19536) у называет, что птицы, гнездящиеся в Западной Сибири, при-
обретают некоторых цестод на пролете.
В Германии в 1910 г. по вопросу об экологии паразитов перелетных
птиц появилась интересная статья Шидата (Szidati о паразитах белого
аиста и об их отношении к экологии и филогении аисгов. Сравиение
иаразптофаупы белого аиста с таковой черного (Ciconta nigra) поражает
их однотипностью, н в го же время паразиты аистов резко отличаются
от паразитов серой цапли. Эго дает автору основание заключать, что
сем. Ardcidae не находится в близком родстве с остальными семействами
Gressores и что цапли должны были весьма рано отделиться от аистов
н весьма Длительно развиваться в условиях другой прародины.
Далее, на основании литературных данных о распространении дру-
гих аистовых, а также и их паразитов автор приходит к выводу о том,
что и те и другие имеют определенно южный отпечаток, .хотя Шидату
и удалось найти цикл развития ряда специфичных сосальщиков аиста
(Chaunocephahts ferox, Cathueniasia liians, Tylodelphys excavata) в на
ших европейских моллюсках (первый промежуточный хозяин) и в зеле
ион лягушке (Rana escuh'nfa—второй промежуточный хозяин). Указы
вая, что все ближайшие родичи Chaunocephalus и Cathaemasia найдены
на Пиле, в Ин.ши и в Южной Америке вместе со своими хозяевами —
другими представителями семейства аистовых, — Шидат высказывает
мысль, что основная родина и места гнездования наших аистов должны
были ранее находиться гораздо южнее, вероятнее всего приблизительно
у истоков Пила (рнс. 190), откуда места гнездовий постепенно пере
двинулись на север, чему могло содействовать отст vr.тонне на север
льдов в Европе (после ледникового периода). При этом к нам па север
попали и первоначальные южные виды сосальщиков. Этому способствова
ло широкое распространен!!! их второго промежуточного хозяина — зе
Лс-яой лягушки (Rana esculenta), которая и сейчас доходит на юг до
30' северной шпроты, т. е. до низовьев Нила.
297
Рис. 190. Карш пе>релетои белого листа Ciconia alba (По Шпдату.)
Пиистироиапна» полоса — путь псрслепш; точки и Европе и Северной Африке
—’места гнездований, в Южной Африке - места янмешкн; пунктирным китч ром
очерчена ирежиылгземан рилина сем. Clcoiilidae; пунктирные стрелки — н>ти
ри сселеним предсгашпсмем атагн гемейстпа: fiiJinimio.i контур — прслно-ип аемая
рилина белого аиста, енлнншыс стрелки пути расселения.
Таким образом, южный ни своему происхождению паразиты при
расселении аиста на север должны были начать развиваться здесь в
новых промежуточных хозяевах (моллюсках), которые не встречаются
на юге.
Обозначения «северный» и.«южный» в применении к паразитам пе-
релетных птиц относительны: они правильны только для паразитов опре-
деленного вида птицы и в определенных ареалах ее распространения.
Нас не удивит, если, например, некоторые «южные» паразиты петергоф-
ских стрижей и ласточек где-нибудь в Крыму, у крымских особей тех же
птиц или в Италии окажутся принадлежащими к убиквистам.
Быховская Павловская |1955, 1962). анализируя с эколого-парази-
тологической точки зрения фауну трематод птиц Советского Союза, при-
ходит к выводе- что огромное болыпнпст.чо их (не менее 300 видов)
приобретается птицами на гнездовьях. Одной из главных причин этого
является, вероятно, совпадение гнездового периода со временем паи
большею разнообразия потребляемой птицами пиши. Значительно мень-
шая часть паразитов приносится птицами с зимовок (62 вида), i биквп
стами среди трематод I йЛовская-11ав.товская считает 43 вида. Не
настаивая на абсолютной точности приведенных цифр, можно считать,
что они все же дают известное представление о количественном соотно-
шении разных экологических категорий трематод. Имеются основания
думать, что примерно такие же соотношения имеют место и в пределах
других грунн паразитических червей— нематод, цесгод, скребней. Чго же
касается эктопаразитов, то здесь отношения, как мы частично указывали
выше, иные. Среди эктопаразитов, с одной стороны, имеется большое
количество убиквистов (пухоеды, перьевые клещи), а с другой — чисто
северных форм (паразиты гнезд).
Чем отличается п а р а з и i о ф а у и а птиц на гнез-
довье от таковой на пролете и .на чимовке?
1 (аразитпфауна птиц на зимовках в тропической зоне у видов, гнез-
1ЯЩИХСЯ на территории Советского Союза, почти не изучена. Имеющиеся
отельные фрагментарные данные не позволяют дать характеристику
паразитофауны этих птиц в зимний период. Здесь необходимы дальней-
шие планомерные исследования. Иная картина имеет место в отношении
птиц, у которых зимовка нротекаш в южных районах Советского Союза.
За последние годы этот вопрос изучался рядим исследователей. Многие
виды птиц, гнездящихся в Западной Сибири, шмуют в южных частях
Гаджикистана, где они н были подвергнуты паразитологическому изу-
чению: цестоды изучались Дубининой (19506), круглые черви — Дуби-
ниной и Серковой (1951), сосальщики — Быховскон-Павловской (1355,
1962). Паразпгифауна этих же видов птиц в отношении эндопаразитов
была этими же авторами изучена и на местах гнездовий (в Западной
Сибири) и частично па пролете в дельте Волги (Быховская Павловская,
1953) Все это дало возможность в отнонк Mini многих групп паразитов
составить представление об их динамике в течение всего годичного никла
жизни хозянна-птицы.
В отношении всех групп гельминтов констатировано значительное
обеднение видового состава н интенсивности заражения во время пролета
и па зимовках. Например, Дубинина указывает, что на гнездовье в За
падкой Сибири у гусиных птиц найдено 36 видов цеетод. У этих же птиц
па пролете их в дельтах Волги и Дуная Найдено только 18 видов, из ко-
торых 11 (Вносятся к категории убиквистов, 6 северных видон н 1 —юж-
ный. Таким образом, утеря большого количества видов, относящихся к
категории северных, происходит во время пролета очень быстро. Весьма
.... данные, полученные Быховскон-Павловской при сравне
29 I
пни зараженности птиц, относящихся к разным отрядам, сосальщиками
Западной Сибири (места гнездованья) и в Таджикистане (места знмов
ки). Некоторые из этих данных суммированы в табл. 15.
Таблица 15
Заражение птиц сосальщиками в Таджикистане и Западной Сибири
(по Быховской-Павлоиской)
Отнят Заладили Сибирь Южный Таджикистан
колич. видов нгиц К заражен, сосал ыц. колич. видов СООЛЫЦ. колич. видон птиц % заряжен, сосальщ. КОЛИЧ видов сисальш.
Пастгш*и . . 2 70.3 20 3 78,5 10
Гусиные 14 48.1 34 11 5*9 15
Хищники 5 54 Я 17 5 62.5 8
Воробьиные .... 21 17.1 12 34 7 2 10
Как видно из этих данных, число видов сосальщиков на месте зи
монкп резко \бывает во всех отрядах птиц. При этом процент щраженин
сосальщиками не снижается, а даже, наоборот, на месте зимовки яь
ляется для большинства отрядов птиц (кроме воробьиных) несколько
более высоким, чем на месте гнездования. В отношении видового соста
ва сосальщиков, сравнивая птиц Годил.х и тех же видов) из Западней'!
Сибири и Таджикистана, Быховская-1кнзловекая уназывает па выпаде
ине на юге шачительного количества северных видов (в частности, от
сутствнс видов рода PhigierohiS) н на появление ряда новых, отсутство-
вавших в Сибири, южных видов [СИ похитит coniplahatiipi, Oszvaldoiu
mosyuensis, Cyclocoeluni rnutabile и др.). В общем аналогичную картину
изменений обнаруживают круглые черви. .У некоторых ленточных червей
во время пролета происходит явление дг сгробп тяшш (потеря и выход
из хозяина всех члеников, кроме сколекса), аналогичное тому. которо<
наблюдается у цестод от некоторых рыб, впадающих в спячку (Дуби-
нина. 1919а, 1950а). Вероятно, этот процесс Дестробнляции представляет
собой своеобразное приспособление цестод к неблагоприятным условиям
которые создаются как на пролете птиц (недостаток нищи), так и на
зимов! е рыб в зимовальных ямах. Это подтверждается эксперимеиталь
ными данными Рейда (R< id, 1910), которому удалось у ленточного червя
Raillieiina cesticillus из кур наблюдать дестробпляцию после голодания
хозяина в течение всего I—2 суток.
Значительное обеднение утиных птиц гельминтами на местах зимо
вок (окрестности г. Поти, Кавказ) по сравнению с местами гнездовии
обиархжпл также Рыжиков (1956).
Из сделанного выпи* описания видно, чго динамика паразитофауны
перелетных птиц, в связи с миграциями хозяина, имеет очень сложный и
интересный характер. Н этой динамике переплетается влияние различ-
ных факторов, а именно: миграций, сезона года, характера пищи, жиз-
ненного цикла промежуточных хозяев, образа жизни окончательных
хозяев и т. д.
Очень часто комбинируется действие нескольких разных (Ьактороь
Напримгр. при сравнении паразитоф уны ласточки и скворца (рис. 191)
выясняется, что состав пара штов, кроме ла ьиостп -миграционных путей
в значите тыюй мере зависит от того, что ласточка хватает свою добычу
на лету, в воздухе, скворец же ловит ее на земле или роясь в почы
Вследствие этого ласточки прежде всего всегда свободны от Syngamu.'-
300
rachea. который для заражения должен быть подхвачен с земли (в вид»
лиц или личинок) и нередко имеется у скворца. Другим отличием кишеч-
ной паразитофауны скворца от таковой ласточки является длительная
однородность ее состава. Эю зависит от того, что ласточки ловят насе-
комых, т. е. промежуточных хозяев своих паразитов, только в период
их лета. т. с. на сравнительно короткой имягиналыюй их стадии. Вслед-
ствие этого при прекращении лёта одних насекомых и начале лёта дру-
гих у ласточек сменяется
паразитов. Скворцы же
собирают с земли как
взрослых, так н личинок
насекомых, а также дож-
девых червей н моллюс-
ков. Таким образом, пи-
щевой режим скворца в
течение всего сезона ос-
тается более или менее
одним и гем же, а зара-
/кенный личинкой какого-
нибудь скворцового пара-
шта промежуточный хо-
зяин, например насеко-
мое, имеет гораздо более
долгий срок контакта со
квориом, чем это имеет
место у ласточки. В са-
мом деле, скворец ловит
'Начала личинок этого
laccKOMoro. например ли-
чинок жука, потом выка-
пывает его куколок и, на-
П1йца, а значит, может измениться и состав
Рнс. 191. Диаграмма, иллюстрирующая влияние
дальних (ласточка) и ближних (скв рсц) нерелсов
иг- ц на их паразитофауну. При дальних перелетах
паразиты-убиквисты отсутствуют, з число южных
H.ipa »птов относительно велико. Напротив, у
скворца убиквисты сильно преобладают над
северными и южными формами (Оригин.'Л.)
/ — число гепгрпых форм паразитов; 2 — число пзрззнгов-
уЛмКПИСТПВ; J — ЧИСЛО ЮЖНЫХ форм liapfl iHTOB-
конец, может питаться
имаго во время его лёта.
Следовательно, зараже-
ние определенным пара-
зитом может держаться
у скворца гораздо доль-
ше, чем у ласточки.
Совершенно особый
характер имеют .мпгра-
цин некоторых птиц в высокогорных местностях, где они
совершают вертикальные переселения — к лету нз долин в горы, и к осс-
ни — в обратном направлении. Резкие изменения окружающих условий
(растительность, состав фауны беспозвоночных как промежуточных хозя-
ев многих червей ) должны создавать в па разитосрауне вертикально миг-
рирующих птиц изменения, аналогичные тем. какие наблюдаются у птице
горизонт.1ЛЫ1ЫМН миграциями. Бауер (1939) проверил это предположе-
ние па горной трясогузке 1 Molacilla cinerea) и горной овсянке (F.mbe-
riza cia) на Кавказе, под Тбилиси. Ч^сть материала собиралась нм в
апреле под Тбилиси (высота 400 .и), часть в мае—августе на высоте
2000 лг Исследование показало зависимость некоторых компонентов па-
разитофауны от вертикальных миграций. Так, заражение птиц малярией
(Plasmodium cathemerium) происходит исключительно в долинах. Жи-
вущие в высокогорной обстановке птенцы и птицы (дятлы, клесты), пе
пускающиеся в долины, совершенно свободны от этих паразитов.
301
Напротив, Haemoproteus, другой кровяной споровик, передастся
через мух кровососок, связанных с хозяином преимущественно во время
гнездового периода. Соответственно с этим птицы, убитые в долинах, н<
имели данного паразита, а у нагорных особей птиц заражены были оди-
наково как взрослые, так и птенцы
Наконец, у некоторых группировок паразитов, к которым относится,
например, Anomoiaeiiia borealis, заражение происходит независимо от
места обитания, по-видимому, в течение круглого года.
В действительности имеется еще немало привходящих иных, боле»
частных факторов, которые могут действовать не на всю паразитофауиу.
а па отдельные се компоненты.
Рис. 192 Freyana anatina — перьевой клещ уток и его поведение
во время линьки хозяина
/1 — взрослый клещ; Н — нормальная локализация клещей на
опахале пера — зачерненные места; С — клещи, спустившиеся и
перьевую сумку старого, выпавшею пера и взобравшиеся па очин
ноиого пера, где они остаются до вырастания опахала: D — часть
тою же рисунка при большем увеличении. (Но Гннеципской.)
Таково, например, влияние, которое оказывает линька птиц и.
с-с эктопаразитов. По этому вопросу накопился уже довольно большой
материал. Особенно подробно он освещен в работах Дубинина (1917,
1918а, I951). Линька птицы прежде всего в.тцяет задерживающе на про-
цесс размножения эктопаразитов. Например, на пухоедах ряда птпи
Дубининым было подмечено, что перед линькой число копулирующих нар
паразитов очень невелико. Напротив, резкое усиление копуляции (соответ-
ственно откладка яиц) наблюдается после окончания линьки. Иногда
во время линьки вымирает и теряется значительная часть некоторых ви
дов пухоедов, как это наблюдал, например, Марков для скворца. Иптен
сивность заражений скворца пухоедами Philopterus и Degeericlla в июле
по сравнению с июнем снижается в 2—2.5 раза. Это же явление имеет
место и в отношении перьевых клещей. Па скворце, например, в июле,
в разгар линьки, интенсивность заражения взрослых птиц клещами
уменьшается в G раз по сравнению с маем.
Линьки влияют на распределение клешей но перьям птицы. У мио
ги.х птиц в период линьки наблюдается расползание клешей по перьям
всего крыла, тогда как до и после линьки они локализуются лишь на
определенных маховых перьях, как это было показано выше. Располза-
ние клещей тю время линьки go всему крылу предохраняет их от гибели
или удаления с птицы вместе с выпавшими перьями. Вновь вырастающее
302
пери, как только оно достигнет размера I 13 125 лиг, снова заселяется
клещам и.
Особенный па торсе должна представлять судьба пухоедов и поры
вых клещей у таких птиц, линька которых протекает бурно, с одповрс
менным выпаданием всех маховых перьев (утиные). Этому вопросу уде
. ила внимание Гннсшшская (1949), исследовавшая паразитофаупу уток
дельты Волги При этом оказалось, что и разгар линьки (пюль1 август)
экстенсивность заражения падает со 100% до 16% для пухоедов и до
50% для перьевых клещей. У последних и возрастной состав приобретает
особый характер: исчезают все стадии цикла, кроме готовых к метамор
фозу нимф
Наконец, у самого распространенного клеща уток (Freyana pnatina)
Гинецинская обнаружила очень любопытное специальное биологическое
приспособление, направленное к сохранению ла хозяине во время линьки
Оказывается, что незадолго до выпадения старых маховых перьев эти
клеши, покидая обычные места своей локализации на опахале пера,
опускаются но его стержню и заползают в перьевую сумку, где затем
располагаются па новом, растущем пере. Они сидят правильным венчи-
ком на месте выхода нового пера и по мере роста.его сохраняют то же
положение, оставаясь таким образом все время, пока не вырастет опа
хало пера, иод защитой его перьевой сумки (рис. 192).
Итак, F. лиаНпа обнаруживает своеобразные и вполне определенные
адаптации в своих инстинктах, направленные к сохранению клещей па
птице в критический для паразитов период линьки хозяина.
Если с периодическими явлениями в жизни паразитов перелетных
птиц, вызванных миграциями хозяина, сравнить сезонные изменения па-
разитофауны оседлых птиц, то они посят иной характер. У оседлых птиц.
как это указывалось памп выше, происходит закономерное изменение
паразитофауны по сезонам, но оно не носит характера тех глубоких пре-
образований всей паразитофауны птиц, которые обусловливают я пере-
летами
ГЛАВА XV
ЗАВИСИМОСТЬ ПАРАЗИТОФАУНЫ ОТ ЧАСТОТЫ
ВСТРЕЧАЕМОСТИ И ОБЩЕСТВЕННОГО ОБРАЗА ЖИЗНИ
ХОЗЯЕВ. ОБМЕН ПАРАЗИТОФАУНАМИ И САМООЧИЩЕНИЕ
ОТ ПАРАЗИТОВ
Существенное влияние на состав паразитофауны оказывает часто
га встречаемости хозяина в его ареале. Предположим, что
на какой-то территории сущее гв’ ют -два хозяина, которые по своим осо-
бенностям (размеры, образ жизни, характер питания и т. д.) могут да-
вать пристанище одинаковому контингенту паразитов, по один нз них
принадлежит к животным редким, встречающимся спорадически, тогда
как другой представляет собой одно нз самых обыкновенных животных
дайной местности. Если при этом «заражающая способность» паразитов
и восприимчивость обоих хозяев одинаковы, то ясно, что реже встречаю-
щийся хозяин будет обладать Солее бедной паразитофауной и меньшей
интенсивностью заражения, так как его паразиты будут иметь меньше
шансов передачи от одной особи хозяина к другой. Весьма вероятно, что
этим обстоятельством объясняется, почему различные рыбы у границы
ареала своего распространения, где они встречаются реже, чем внутри
ареала, обладают бедной паразигофауной. Гаковы ерш и лосось в Араль-
ском море, чехонь и со.м в Невской губе и т. д.
Помимо только что сказанного, весьма важный момент представляет
во многих случаях степень оседлости пли отсутствие се у хозяев.
Дело в том. что у хозяев, имеющих различного рода гнезда, норы и то-
му подобные убежища, в которых они выводят детенышей, передача
паразитов от родителей к потомству нередко связана именно с этими
местами приюта для следующего поколения. Здесь переходят с родителем
на детей различные эктопаразиты (пухоеды, мухи кровососки и т. л.)
В норах и гнездах облегчается и заражение эндопаразитами, цисты и
яйца которых скопляются здесь в Со тыном количестве.
Если же взять, как другую крайность, животных.оставляющих свое
потомство па произвол судьбы с самого момента рождения, например
рыб, выметывающих икру, то в этом случае возможность непосредствен-
ного заражения потомства от родителем сходит почти па нет. Особенно
заметно это выступает у таких животных, у которых вывод молоди про-
исходит в ином месте, чем место жительств г взрослых животных (лосось
и т. и.). Естественно, что таких животных фауна мололи и взрослого
животного может сильно различаться. Мы видели это выше в отношении
лосося, совершающего длительные миграции из моря в реки для нереста.
304
Но семья является первым зачатком «общественного образа жизни
Это приводит нас к весьма нитересиозп вопрос'- о паразитах о я и-
юч пых и «общественных» животных. В общем, следует
сказать, что по сношению < легкости заражения паразитами одиночные
и «общественные» животные находятся в таком же положении, как фор-
мы, редко встречающиеся и обыкновенный. Общественный образ жизни
Ви много раз увеличивает возможность заражения пара штамп.
Это повышение заражаемости ска-
зывается даже не на типичных обще-
ственных животных, а уже на таких,
которые ведут групповой inn стадный
образ жизпи. Несомненно, тесный кон-
такт между стадными животными
имеет большое значение для распро-
странения их паразитов. Лучшим при-
мером в тайном случае служат инфу-
зории в желудке пли в толстой кишке
разных копытных животных. Разнооб-
разие и оои тис кишечных инфу юрпй
(Entodiniomorpha и др.) у копытных
поразительно: они насчитывают не ме-
нее 200 видов.
У копытных же можно хорошо об-
наружить зависимость процветания
паразитов от большей или меньшей ча-
стоты нахождения хозяев: инфузории
из кишечника диких копытных (неко-
торых антилоп и оленей, не образую-
щих больших стад) гораздо менее раз-
нообразны. чем таковые быка, овцы
или козы. Для быка известно около
70 видов и вариететов, для овцы — 35,
для козы — 20, между гем для антилоп
(Trtigelaphus scriptus, Bubalis cokei
и др.) это число колеблется от 1 до 17.
У северного оленя описан 21 вид инфу-
зорий. v ипочнх же оленей и v лося —
Рис. l')3. Troglodytel/a gorillae
(Entodiniomorpha j — инфузория из
кишечника i ориллм. (Но Ренхе-
нову.)
I — енолиротинл и .-юн- мембрэмели; 2 —
бсльшлп, сооТошизч нз мелких яч«ек«
ckc.ktii.iu ii.iaci цикл; .¥ — четыре зоны
мембрацслл.опоясыпзюпких тело; /- поро-
шиц); 5 — согрэштельныс вакуоли.
I до 8. Это обстоятельство по-
казывает, что многочисленность особен рогатого скота и тесное сожи-
тельство н.ч гарантируют перекрестную передачу паразитов, а потому
фауна их инфузорий получает возможность и как бы толчок к более
пышному своему развитию.
Очень близкая к инфузориям копытных инфузория нз того же се-
мейства Opliryoseolccickie, а именно Troglodyiella (рис. 193), живет в ки-
шечнике шимпанзе и гориллы. Это совпадение с копытными в паразитах
объясняется, с одной стороны, одинаковостью пищевого режима, который
у шимпанзе и гориллы является преимуществ! ино растительным, с дру-
гой стороны, общественным образом жизни этой обезьяны. Рей хеков
(Reichenow, 1920), наблюдавший шимпанзе в Камеруне и нашедший
у mix Trogjodytclla. говорит, что шимпанзе живут небольшими стаями
в 12—15 особен и устраивают общие логовища.
Не mi нее резко выступлюг такие же отношения у некоторых настоя-
щих «общественных» животных, например у термитов. В таком же раз-
нообразии и пышности, как инфузории у копытных, в кишечнике терми-
тов развились жгутиконосцы нз отрядов Polyniasti^ida п Hypermastigida.
Их 6iviee 200 видов, причем кишечник одного вида термитов, несмотря па
2П I А. Догель
305
малые размеры хозяина, может содержать до 20 видов жгутиконосцев
а кроме того, еще грегарин, инфузорий к амеб. Все термиты одного и
того же нита заражены жгутиконосцами поголовно. Итак, термиты даю-1
прекрасную иллюстрацию обогащения своей паразитофауны под влпя
пнем общественного образа жизни.
Другие «общественные» насекомые—пчелы — тоже богаты парази
та.мя, хотя их число значительно меньше, чем у термитов. У пчел отме
чено 7—8 видов простейших, в числе которых есть .микроспоридии А >.<₽-
та apis. вызывающие тяжелое кишечное заболевание—нозематоз. II
паразитических червей в пчелах встречаются представители нематод.
Кроме того, имеется целый ряд паразитических к шщей (например
Pediculoides ventricosus, Acarapis woodi) На пчелах паразитирует такжя
бескрылая слепая муха firaula caeca, а ее личинки развиваются в сотах
В ульях пчел часто встречаются в большом количестве личинки иасеко
мых (жука Meloe, восковой моли Galleria и др.), которые поселяются в
ячейках сот и являются опасными врагами пчел, разрушающими н.\
постройки.
Однако нельзя утверждать, что общественная жизнь у животных
все! да без исключений сопровождается особым изобилием паразитарных
заболеваний. Муравьи, живущие всегда самыми тесными сообществами
в общем бедны паразитами. Быть может, причиной этого является му-
равьиная кислота, содержащаяся и теле Муравьев и щедро орошающая
как их самих, так и муравейник Кислота представляет собой естествен
ный дезинф* кгор. гибельный для многих паразитов, в особенности для
простейших.
Во {действие общественной ж и з н и человека на распро
странениг паразитарных заболеваний весьма многосторонне. Присма-
триваясь к различным заболеваниям человека, как паразитарным, так
и бактериальным, можно видеть, что значительный пропет их зависит
от общественного склада человеческой жизни.
Самая важная сторона общественного быта, связанная с вопросом
о паразитизме, — это скученность населения. В первую очередь это от
носится к городам. Нам представляется, что именно в недрах городе»
древнего Востока с большой скученностью населения, особенностями его
бы га и теплым климатом с.щпались и крепли связи между различными
паразитарными и бактериальными организмами и человеком, вырабагы
вались и укоренялись болезни с тем, чтобы потом постепенно распро
страняться во все стороны.
Уже в средине века значение Востока как рассадника болезней
ясно проявляется хотя бы в таком факте, как периодические чумны<
эпидемии, беру щие начало всегда на Востоке. Мы думаем, чго параллель
но могло происходить и обогащение фауны паразитов человека в Европе.
Что такое обогащение происходит именно по указанному' направлению,
видно из примеров новейшего времени. В самом деле, свайник двенадца
тнперстной кишки (Ancylostoma duodenale} проник в южную Европ'
лишь в 70-х годах XIX в. и притом, ио-видимому, из Египта.
Эти примеры по только показывают, что каждый паразит имеет из-
вестный очаг возникновения, но отчасти характеризуют нам и пути рас-
пространения паразитарных заболеваний. Это, прежде всего, усиление
связей между различными странами.
Следует сказать, что факторы, особенно способствующие распростра-
нению и размножению паразитов, будут несколько отличными для экто-
и эндопаразитов. Для эктопаразитов единственным достаточным уело
висм являете я скученность населения при одновременном наличии анти-
санитарных условий Один из примеров этого явления мы видим в чрез
306
мерном распространении вшей в лагерном и траншейной обстановке во
время прежних войн. Для эндопаразитов к этому часто прибавляется
необходимость наличия промежуточных хозяев и особых условий для
развития личиночных стадий.
Дивольно многие паразиты человека встречаются преим>шественн<
у лиц, занимающихся о п р е д с л е и н ы м н профессия м и, что объ-
ясняется какими-нибудь особенностями профессии, ставящими человека
в более тесный контакт с тем или иным пара нтом. Так, относительна
балан гидиоза установлено, что Balantidium coii нередко обнаруживает-
ся у мясников, режущих свиней, т. с. дричнной заражения является ь
данном случае контакт со свиньями. Ancylostoma duodenale наиболее
обыкновенен у людей, занимающихся земляными работами, но не у кре-
стьян, а у работающих в угольных копях, роющих туннели и т. д. Извест
ная эпидемия анкилостомоза, давшая около 1000 заболеваний, произо-
шла в 70-х годах XIX в. средн рабочих, пробивавших Сен Готардский
туннель. Скопление людей при таких работах и постоянное соприкосно-
вение их с землей позволяет личинкам анкилостомы легко проникать
через кожу в тело рабочих.
Нередко заражению паразитами способствует соблюдение извест
них обычаев, особенностей жизненного уклада люден в
дайной местности или данной стране. Так объясняется, например, чрез-
вычайно широкое распространение человеческой аскариды в Японии
Во многих местностях Японии до 80% населения заражено этими гли
стами, инвазия которыми происходит посредством проматывания выве
денных вместе с экскрементами яиц /Iscor/s. Сильное заражение аскари
дами находи гея в прямой связи со способом удобрения огородов. Ввиду
недостатка удобрений японские земледельцы пользуются в качестве та
ковых фекалиями. Таким путем и экскременты зараженных аскаридам^
объектов, содержащие в себе яйца аскарид, попадают на огороды и
смешиваются с землей. Вследствие этого яйца аскарид очень часто при
стают к поверхности разных овощей и затем попадают в кишечник чело
века.
Широкое распространение трихиноза в прежнее время в Германии
объясняют употреблением там в пищу полусырой «вестфальской» вет
чины. Сильная инвазия населения Карелии широким лентецом вызы
валась гем, что там часто употребляли в пищу сырую или плохо нрожа-
реннмо рыбу, а потребление в пищу замороженной строганой сырой
]!ыбы в Сибири имеет результатом учащение инвазии кошачьей двууст
кой (Opisthorchis [еlinens).
Тесиое совместное обитание на одной территории нескольких разных
видов животных влияет на их паразитофау ну. D результате такого сов-
местного обитания возникает еще очень мало подвергшееся оценке яв.ц
ине обмена паразитарными фаунами. Зак .ючается оно в
следующем. Всякое животное обладает известным набором своих сне
цифичиых паразитов. Ввиду того, что специфичность многих паразитов
далеко не абсолютна, а приспособляемость их весьма значительна, из
вестная часть паразитов данного вида животного при переходе к сов-
местному обитанию и контакту с другими видами животных получает
возможность распространяться на всех или некоторых особей других ви-
дов животных. С другой стороны, их паразиты могу г переходить на пер
вый вид животного. Последствием этого будет обогащение паразитарной
фауны отдельных видов животных.
Судя по ряду данных, обмен паразитарными фаунами имеет месте
в более широком масштабе, чем это можно было бы думать, так что у
многих животных мы должны отличать первичных паразитов, или, так
20- 30”
сказать. аоорнгенов, от вторичных, которые повали в животное в резуль-
тате обмена паразитами, па ёймом деле являющихся аборигенами дру-
гих животных-хозяев. Для иллюстрации этого приведем несколько при-
меров относительно фатны инфузории, населяющих кишечник копытных
животных.
В желудке коров и овец встречается множество видов инфуЗорин из
сем. Ophryoscolecidac, большинство которых относится к родам Ento-
diniunu Diplodinium и Ophrtjoscolex. Это семейство вообще характерно
для желудка жвачных. Одни из видов офриосколецнд живут только в
крупном рогатом скоте (например: Eniodiaium пйп'пиип, Е. rosiratum,
Diplodinium d ’tiliailaiuin, Opiinjoscolcx purkinjei и др.), другие — только
в овнах (Entodinium vorax bispinosum, Ophryoscolex caudcnus quadricinc-
tus и др.). Большинство же видов MOzKerбыть встречено как в тех,так и в
других. Среди этой обширной категории имеются виды, одинаково часто
встречаемые у коров и у овен (Eniodinium caudatiun, Е. simplex, Diplo-
dinium multivesicidatum и др.), и.такие, которые, встречаясь предпочти-
тельно в одном нз поименованных животных, изредка попадаются и в
другом. Так, Eniodinium longinucleatum, Diplodinium mgleciutn bovis.
D. gracile селятся чаще в желудке коровы, тогда как D. triloricatiun и
Ophryoscolex caudalus fricincfus — в желудке овны.
При оценю происхождения всей этой паразитарной фаупы можно
с определенностью сказать, что имеются две группы видов, из которых
одна специфична для коровы, другая— для овны. Происхождение же
видов, одинаково часто встречающихся у коров и овец, не поддается
учету.
Среди инфузорий сем. Ophryoscolecidac обнаруживается явление об-
мена фаунами. Формы, предпочитающие коров и лишь изредка встре-
чакчцшея у овец, были сначала исключительно паразитами крупного
рогатого скота и перелили к овцам лишь в результате совместной жизни
в одомашненном состоянии. То же следует сказать и о видах Ophryos-
colccidae, предпочитающих овец, только здесь переход осуществляется
в обратном направлении,
Сделанное нами предположение подтверждается ознакомлением с
фауной инфузорий диких баранов с Конет Дата н с Камчатки (Ovis
orianlalis cijcloccrns, О. nivalis). Эта фауна состоит из таких видов, ко-
торые либо вовсе не встречаются у коровы, либо попадаются у псе
лишь в редких случаях, являясь) частыми обиталлями домашних овец.
Гак как домашние овцы произошли от диких праро цгге-тей, то выше-
указанное обстоятельство подтверждает первичную специфичность для
овец видов Ophryoscolecidac, которые в настоящее время бывают лишь
редкими гостями коровы. Корова и онпы испытали обмен наразитофау-
намп
При известных условиях обмен паразптофау нами может стать одно-
сторонним. Так, у верблюдов в зоологических садах Европы нередко
имеется фауна ннф’ зорпй, тождественная с таковой желудка коровы.
Между тем у верблюдов в Туркестане, живущих, так сказать, в своей
естсстшнной обстановке, имеется целый ряд специфичных видов и даже
родов (Caloscole*). Этот факт может быть объяснен только одним спо-
собом. Вырванные нз естественной обстановки н тесного общения с себе
подобными и поставленные в условия контакта с европейским рогатым
скотом верб поды постепенно утрачивают свою оригинальную фауну ин-
фузорий, но заражаются чуждыми нм инфузория ми коровы и овцы.
В сказанных до сих пор примерах обмен фаунами касается инфузо-
рий из желудка различных жвачных, т е. в данном случае паразит,
пенял своего хозяина, не менял своей локализации. Но некоторые пара-
308
зиты обнаруживают, по-видимому, еще более широкую приспособляе-
мость: переселяясь в другого хозяина, они вместе с тем изменяют свое
место жительства в кишечнике последнего. Так, представители ct мейства
инфузории Blcpharccoridae обычно встречаются в слепой толстой кишке
лошади. Таковы многочисленные виды рода Bleyharoiorys (Догель, 1926;
Стрелков, 1939). Однако вид Blepharocorys ventriculi (рис. 194), очень
похожий iki прочие виды, ио меньших размеров, найден в желудке ко-
ровы. Вне всякого сомнения, этот вид представляет собой видоизменив-
шегося под влиянием новых жизненных условий переселенца из толстой
кишки лошади.
Несколько примеров такого рода мож-
но найти в паразитофауне каспийского тю-
леня, как об этом говорится в разделе, по-
священном вопросу об особенностях парази-
тов реликтовых животных.
Направление обмена паразитами нахо
дится в зависимости от качественного набо-
ра и количественного состава животных,
служащих хозяевами паразитам в том пли
ином биоценозе, например в водоеме, лесу
и т. п. Исследования последних лет пока-
зали. что в таких случаях преобладающая
в данном биоценозе группа хозяев, оказы-
вает своего рода руководящее влияние на
состав паразитофауны ряда других хозяев,
не принадлежащих непосредственно к до
минируют й группе.
Очень хорошую иллюстрацию этому
даег паразитофаупа рыб Ладожского озера
ио сведениям Барышевой (1949), Барыше-
вой п Бауера 11957). Ихтиофауна озера со-
держит весьма большой процент лососевых,
дающих, например, до 25% промысловых
уловов в озере. В видовом отношении лосо-
севые тоже составляют свыше 25% всей их-
Рис. 191. Близкие виды пардон
гпческих ии.ручорип из лошати
и ।оровы
.1 Btephurorarys ven ricitli из
желудки быка; В — lilepbaroev-
г ys jitbata нз толстой кишки
.Ю1И.11 . (.1 — но Догелю, 1> —
по Стрелкову )
1 — г.ютк.1. 2 - чикронуклсус; J—мак
[jOiiyK.itfyc / - сократительная из-
куо.ис о — мдпий лучок ресничек.
тисфауны. В связи с этим обстоятельством паразиты, обычно специфичны»
для лос.авых. в Л иожском озере распространяются па ряд других ви-
дов рыб, которым они вообще по свойственны. 1ак, например, специфич-
ный для лососевых в >д миксоспориднй (!} enneguya zachokkei) встре-
ченный в 33% особей ладожских сигов, наблюдался, однако, кроме по,
у таких рыб. как и длим, щука, сом, и даже у ряда карповых, как плотва,
лещ, сырть, уклейка;- чехонь. Такой же переход па иеспсцафлчпых хо-
зяев наблюдался для ленточного глиста Eiiboihriuiii crassuni. Пом <ми
лососе вых, зараженных им поголовно, он папдг-и у таких не свойствен-
ных даипому паразиту хозяев, как сом, ерш, колюшка, и даже у язя и
чехони. Характернейший скребень сигов (Echi nor hynchus saimonis), за-
ражающий 100% этих рыб па Ладоге, оказался, кроме того, у сома
(33%), угря (22%), окуня (26%), осетра, колюшки (20%), налима
(66%) и даже у карповых (сырть — 13%, B.’icca bjorkna —13%). Очс-
видио, здесь играет роль главным образом количественное обилие зараз-
ного начала (спор для Henneguya. зараженных промежуточных хозяев
для гельминтов). Большое изобилие заразных стадий какого-нибудь
вида паразитов в данном водоеме способствует переход) их со своих
первичных хозяев па хозяев, им первоначально ио свойственных. Иначе
говоря, мы видим здесь пример обмена паразитофаунами в определен
30ч
пом направлении (от численно преобладающих форм хозяев к более под
чиненным). Само собой разумеется, что подобные сдвиги могут служить
первым шагом к более прочному завоеванию некоторым паразитами
новых для них хозяев.
Обмен паразитами описан и у птиц. В особенно широких масштабах
он осуществляется в местах массовых скоплений птиц. Это имеет место,
например в дельте Волги, где гнездятся многочисленные виды птиц, от-
носящиеся к различным отрядам. Вопрос этот рассмотрен в специаль-
ной статье Гипецннскон (1952а). Анализируя под этим углом зрения
гельминтов птиц Астраханского заповедника, расположенного в дельте
Волги. Гипецниская приходит к выводу, чго 32 вида паразитических
червей встречаются в хозяевах, по существу чуждых нм. При этом наи-
большее число видов, способных паразитировать в неспецнфнчных хо-
зяевах, приходится на сосальщиков, где эти виды составляют 23% от
числа найденных.
Рассматривая судьбу паразитов, развивающихся в
чуждых им хозяевах, Гииецпнская различает несколько случаев.
Во-первых, некоторые паразиты достигают в чуждых хозяевах нормаль
них размеров и половой зрелости и дают высокий процент и интенсив-
ность заражения (пример — паразит желудка утиных Tetrameres fis-
sispina, который в дельте Волги переходит па цапель).
Другую группу составляют вилы, тоже достигающие половозрелости,
но встречающиеся в чуждых хозяевах редко и в единичных экземплярах
Таких оказывается большинство (около 20 видов)*
Третью группу составляют паразиты, хотя н достигающие в чуждом
хозяине половозрелости, по характеризующиеся при этом небольшими
размерами. Подобное уменьшение размеров указывает на то, что новая
среда обитания оказывается для них неблагоприятной. Примером может
служить развитие скребней Corynosoma strumosum (паразит тюленей)
г; птицах.
Наконец, четвертый случай — это паразиты, не достигающие в чуж-
дом хозяине половозрелости. К их числу относится характерный паразит
баклана Eustiongylides raergorum, встречающийся во многих рыбоядных
птицах, но становящийся половозрелым только в бакланах. Здесь мы
встречаемся с явлениями специфичности и врожденного иммунитета, ко-
торые будут рассмотрены ниже в главах XXI и XXII
Широкий обмен паразитами птиц дельты Волги наблюдал также
Дубинин (1951). Например, на некоторых гусиных птиц здесь перехо-
дят перьевой клещ Frcyarta anulina, характерный для уток, и три вида
llloptes, характерные для куликов и чаек.
Вполне естественно, что обмен паразитами наблюдается также меж-
ду животными ч человеком. Далеко не все паразиты, встречающиеся в
настоящее время у человека, являются его первичными паразитами. При
анализе паразитофауны человека выясняется, что очень многие ее ком-
поненты вторичного, пришлого происхождения и проникли в человека
лишь после привлечения в человеческую общину ряда других животных,
одомашненных пли нахлебников. Различные одомашненные и приручен-
ные животные внесли в паразнтофауну человека разное количество пара-
зитов в зависимости от тесноты и характера контакта данного животного
с человеком или от каких-нибудь других обстоятельств.
Так, значительное участие в Обогащении паразнтофауны человека
принимала, по-видимому, собака, как давнишний и близкий сожитель
человека. DipyHdiutn caninum, собачья блоха (Ctenocaphalus canis) и
многие другие случайные или редкие паразиты человека являются пер-
вичными паразитами собаки. Таково же, вероятно, и происхождение за
310
эажения человека эхинококком. Эхинококк живет в состоянии ленточной
глисты в .лппечнике собаки, тогда как пузырчатая стадия ею развивает-
ся в раз. 1ИЧНЫХ внутренних органах человека, свиньи, лошади, коровы н
некоторых других животных, которые заражаются, случайно прогла-
тывая выходящие нз кишечника собаки яйца. Нам думается, что перво-
тачальный никл развития эхинококка распределялся между дикими
предками собаки и ди. ими травоядными копытными. Нахождение тех и
других в обиходе человека привело к распространению эхинококка и на
.воден.
Свинья внесла в паразптофауну человека очень много компонентов,
чо главным образом благодаря другому обстоятельству, а именно: своей
всеядности. Всеядный характер пи гания человека и свиньи сближает
sth организмы между собой, делает, очевидно, их кишечную среду сход-
ной и таким путем облегчает обмен паразитами. Вероятно вследствие
этих причин человек приобрел в свою паразитофа’ ну Balantidium coll,
а человеческая аскарида (Jsturis lutnbricoides) произошла, вероятно, от
свиной (Ascaris suum). Мы уже видели ранее, что для трихины человек
[рсдставляет собой побочный тупик развития. Несомненно, чго этого
паразита человек приобрел путем потребления в пищу мяса свиньи.
Точно так же и многие другие паразиты человека могут быть в
своем происхождении прослежены вплоть до какого-нибудь другого, Со-
лее древнего и первичного их хозяина.
С другой стороны, и некоторъи паразиты самого человека могут
переходить ни животных, поселяющихся в человеческом жилье и т. п.
Достаточно напомнить, что человеческий постельный клоп (Citnex lectula-
rius) поселяется в гнездах стрижа, нападает на летучих мышей, селя-
щихся па чердаках.
На интенсивность паразитарной инвазии в ряде случаев влияет фак-
тор активного самоочищения хозяина от паразитов (глав-
ным образом эктопаразитов). При внимательном анализе данного во-
проса можно убедиться в том. что не так редки случаи, когда животные,
зараженные известными паразитами, пытаются тем или иным способом
избавиться от них. Способы освобождения могут быть весьма разнооб-
разными, то более или менее пассивными, то даже активными, особенно
у наиболее высоко развитых животных — птиц, млекопитающих. Обыч-
но попытки освобождения хозяев oi паразитов являются ответной реак-
цией хозяина на раздражение или болезненное ощущение, вызываемое
паразитом.
Наиболее бросаются в глаза активные попытки многих птиц и млеко-
питающих избавиться от эктопаразитов посредством удаления их со
своих покровов. Так, многие млекопитающие освобождаются от блох и
вшей, выбирая их из шерсти или волос зубами, чесанием при помощи
конечностей. Наиболее совершенно проделывается это обезьянами, ко-
торые выбирают паразитов из шерсти пальцами и уничтожают их.
Птицы при помощи клюва производят двоякого рода работу: с
одной стороны смазывание оперения секретом копчиковой железы, с
дрсгой — выбирание пухоедов и других паразитов. Имеются любопыт-
ные сведения о том, что многие птицы пользуются муравьями для целей
дезинсекции своих перьев при помощи муравьиной кислоты До настоя-
щего времени подобные наблюдения сделаны ' же над многими десят-
ками видов птиц, принадлежащих к 13 разным семействам
(Ivor, 1943).
Итаны садятся на участки земли, изобилующие муравьями (напри-
мер, при роении последних), хватают муравьев клювом, расправляют
крылья и проводят мхравьямн по нижней поверхности .маховых перьев.
311
как бы смазывают муравьем перья, т, е. как раз там, где сидят в боль-
шом количестве перьевые клещи (Analgina). Для обозначения этого яв-
ления в последние годы выработался даже особый глагол «муразеини-
чать», или «муравьить» (to ant), который применяют для птиц, занятых
процессом дезинсекции.
Аналогичные явления наблюдал Дубинин (1951) у разных птиц. По
сто данным, многие перьевые клещи после, воздействия на них муравья
ми начинали интенсивно переползать по крылу и вскоре многие из них
погибали. После «муравьения» численность клещей Pterodectes bilobu
tus на степном коньке (Anlhus campestris), например, в течение 24 часов
уменьшилась на 30,5%. К сожалению, детали всего процесса, а также
сам процесс воздействия муравьев на клещей пока не выяснены.
Пассивную защиту от паразитов осуществляют те млекопитающие
(буйволы, носороги), которые позволяют некоторым птицам собирать
на своем теле паразитических насекомых. Правда, привычка эга выра-
боталась, по-виднмому, па основе использования птиц в к..честве преду-
преждения животного о приближающейся опасности, но она вместе с
тем приобрела и дезинсекционное значение.
Очень интересные приемы освобождения от эктопаразитов наблю-
даются у многих птиц из семейства- куриных. Летом, в жаркое время,
куры охотно принимают песочные ванны, садясь в песок или в пыль,
взъерошивают перья, пропускают через них песок и этим частично осво-
бождаются от пухоедов (а может быть и от перьевых клещей).
Такие же явления констатированы и для диких куриных, причем у
рябчика Олигер (1940) нашел прямую связь между началом периода
песочных ванн и значительным уменьшением числа пухоедов: к концу
июля активные стадии пухоедов Lagopccus и Goniocotes на рябчике ис-
чезают. Дубинин (1951) подтверждает это просеиванием пыли нз пе-
сочных ванн, в которых только что «купались» птицы. Он находил после
просеивания значительное количество перьевых клещей, свалившихся с
птиц. Впрочем, с другой стороны, пылевые ванны служат и местом пере-
хода клещей с одной пгнцы на другую при повторных их купаньях в
одной и той же ванне.
Мы склонны считать мерой освобождения от паразитов, или, вернее,
спасения от них, одно явление в жизни ласточек. Гнезда деревенской
ласточки часто являются рассадником всевозможных кровососу-
щих паразитов: клещей Gainasidae. блох, клопов, личинок мух Pseudo-
caliiphora. Трудно собе представить, как птенцы ласточки могут выдер-
живать существование в таких гнездах. Немедленно после того, как осен-
ний выводок ласточек научится летать, родители вместе с ним бросают
гнезда и переселяются на последние две-три недели своего прелотлет
кого периода на ночевки в камыши или другие укромные места. У стри-
жей же, гнезда которых обычно совсем свободны от паразитов, летные
птепны вылетают из гнезд незадолго до отлета.
1 Л Л В A XVI
ЗАВИСИМОСТЬ ПАРАЗИТОФАУНЫ ОТ ГЕОГРАФИЧЕСКИХ
ФАКТОРОВ И ЗНАЧЕНИЕ ПАРАЗИТОЛОГИИ ДЛЯ РЕШЕНИЯ
НЕКОТОРЫХ ВОПРОСОВ ЗООГЕОГРАФИИ И ФИЛОГЕНИИ
В предыдущих главах нами рассматривалось главным образом влия-
ние факторов, так или иначе тесно связанных с самим хозяином, как-то:
возраст хозяина, его пища, образ жизни, миграции, а также влияние
факторов климатического характера: зависимость состава паразнто-
фауиы от сезона и т н Несомненно, что распространение паразитов, по-
добно распространению свободно живущих организмов, должно обусло-
вливаться современным ареалом хозяина, а также и историческим прош-
лым этого ареала. Все зоогеографические понятия, деления и т. д. в
такой же мере приложимы к паразитам, как и к их хозяевам. Мало того,
сплошь и рядом изучение состава паразитофауны различных животных
дает чрезвычайно ценные сведения для зоогеографической характери-
стики хозяев.
Общее число паразитов, встречающихся в каком-нибудь организ-
ме, может быть весьма различным. Так, например, в человеке одних
только гельминтов известно свыше 150 различных видов. В рубце быка
насчитывается около 70 вадов и разновидностей инфузорий Ophryosco-
keidae. Для лосося описано около 65 видов разных паразитов (llcitz,
1918). Дольфус в монографии по паразитам трески (Dolll'us, 1953) при-
водит список 71 вида паразитов, найденных в различных частях север-
ного полушария. Однако в Баренцевом море из их числа встречаются
всего 27 видов (Полянский. 1955). а в Белом море — 24 (Шульман и
Шульман-Альбова, 1953).
Не следует, однако, думать, что все это количество видов можно
встретить в одном каком-нибудь районе, а тем более в одной особи жи-
вотного-хозяина. Так, у человека редко можно встретить более 2—3 ви-
дов червей одновременно, а одни индивид лосося редко содержит более
5 видев паразитов. Точно так же н в полом для вида животного-хозяина
в каждом районе его области распространения характерен известный,
более или менее ограниченный контингент паразитов. Мы напрасно
стали бы искать в коренных обитателях . книнграда таких паразитов,
как ришта, свайник двенадцатиперстной кишки, леишманпя и т. д., ко-
торые характерны для более южных широт. Помимо такой общей зави-
симости от географических условий, имеются более специальные зоогео
313
графические особенности известных ареалов местообитания, наклады
вающие свой отпечаток на паразптофауну. Вопрос об этих зависимостях
в настоящее время выяснен ио отношению к паразитам еще далеко не
юсгаточно. На большое теоретическое и практическое значение изуче-
ния зоогеографии паразитов (в частности гельминтов) неоднократно
указывал Скрябин (1946, 1958).
1 ВЛИЯНИЕ ГЕОГРАФИЧЕСКИХ ФАКТОРОВ НА ПАРАЗПТОФАУНУ
Значение ландшафтно-климатических зон
Общая зависимость паразитофауны от географических факторов вы
ражастся в том, что она испытывает влияние не одного какого-нибудь
фактора, а целой совокупности таковых, как-то: различные климатиче
ские условия, на личие или отсутствие промежуточных хозяев, различный
состав почвы (для наземных) или воды (для водных животных) и т д.
Поэтому подобного рода зависимость носит наиболее неопределенный и
наименее в каузальном отношении выясненный характер. Рассмотрим
кратко два примера, из которых один касается трематод птиц на террп
горни СССР, а второй — паразитов рыб рек Сибири.
Зоогеографический анализ фауны трематод птиц, проведенный Бы-
ховской-Павловской (1955, 1962) на очень большом материале, охваты
вающем все основные семейства птиц во всех ландшафтно-географиче-
ских зонах Советского Союза, показал наличие существенных различий
между разными зонами.
В арктической зоне фауна трематод птиц характеризуется относи
тельной бедностью видового состава (найдено 53 вида), причем экстен
сниность и интенсивность заражения некоторыми видами бывает очень
высокой. Преобладающими семействами являются Microphallidae и
Gymnophallidae— типично морские паразиты, промежуточными хозяе-
вами которых в большинстве случаев являются морские литоральные
беспозвоночные.
Зова лесов умеренного климата характеризуется чрезвычайным
разнообразием видового состава трематод (211 вида, что составляет
50,3% всех известных трематод от птиц СССР). Наиболее широко рас
иространенными семействами здесь являются Plagiorchidae, Eucotylidae,
Bracliylaemidae, Leciihodendriidae. В качестве промежуточных хозяев
особенное значение приобретают насекомые. В этой зоне особый харак
тер носит фауна трематод Дальнего Востока, включающая 47 видов со
сальщнков, нигде более не встречающихся и паразитирующих в дальне-
восточных видах птиц.
Зона лесостепи является переходной. Она характеризуется весьма
разнообразным и гетерогенным составом фауны трематод, из которых
около 80% встречаются в смежных зонах (лесной и степной).
В степной зоне, обладающей пониженной влажностью, наблюдается
меньшее, чем в лесной, разнообразие трематод, хотя общее число видов
продолжает оставаться значительным (157 видов).
Зона пустынь умеренного климата (в се типичных ландшафтах) ха-
рактеризуется значительной бедностью видового состава трематод (око-
ло 30 видов). Это связано в основном с условиями, неблагоприятными
для существования их промежуточных хозяев. Преобладающими семей-
ствами здесь являются Dicrocoellidae и Plagiorchidae, связанные в своем
развитии с насекомыми и наземными моллюсками. Весьма обедненной
является также фауна сосальщиков птиц горных ландшафтов, где также
преобладают Dicrocoellidae и Brachylaemidae.
Детальное сравнение видового состава сосальщиков птиц отдельных
зон, с одной стороны, показывает налично различий между зонами и,
с другой — постепенность перехода от одной зоны к другой при взаим-
ном проникновении отдельных элементов фауны сосальщиков из одной
зоны в другую.
Наряду с общими закономерными изменениями фауны сосальщи-
ков по ландшафтно-климатическим зонам, в пределах различных зон
отдельными пятнами могут возникать биотопы, носящие азональный ха-
рактер. Обычно такие биотопы бывают связаны с водоемами озерного
пли речного типа. Характерный пример — дельта Волги. Она лежит в
зоне пх стынь умеренного пояса и тем не менее характеризуется исклю-
чительным богатством фауны трематод, что непосредственно обуслов-
лено богатством в этом районе орнитофауны и фауны водных беспозво-
ночных.
Второй хороший пример зависимости паразитофауны от географиче-
ского фактора дает изучение паразитов рыб Сибири (Петрушевский и
Бауер, 1948). Протяженность этих рек очень велика. По ходу своего
течения они пересекают различные климатические ландшафтные зоны,
начиная с пустынь и кончая арктической сибирской тундрой.
Анализируя шпротное распределение паразитов рыб сибирских рек
Петрушевский и Бауер различают три группы паразитов.
К первой группе они относят паразитов, распространенных по всему
течению рек, независимо от широты и климатической зоны: например
мнксоспоридин Myxidium lieberkuhni, монотеистический сосальщик
Tetradnchus monenteron, трематода Cotylurus variegatus, нестода Triae-
nophorus nodulosus. нематода Rhaphidascaris acus и др. Число таких
форм от общего количества паразитов достигает 30—45% (в разных ре-
ках). Более половины этой группы принадлежит к видам, развиваю-
щимся без промежуточных хозяев.
Ко второй группе относятся северные формы (на Оби — 30%, на
Енисее — 34%), которые распространены в среднем п главным образом
в нижнем течении. Это мнксоспоридин Heuneguya zschokkei, Chloro-
тухит coregoni, монотеистические сосальщики Tetraonchus arcticus.
Discocoiyle sagittatwn, многие трематоды — Crepidosiomum farionis.
Phyllodistomum conostornum, ленточные черви Triaenophorus crassus.
Eubolhrium crassum, Cyathocephalus truncatus, Diphylloboihrium minus,
скребни Echinorhynchus salrnonis, E. truttae, нематоды Cyslidicola fario-
nis, Coregonema sibirica и ряд других.
Третья группа — это южные паразиты. На Оби она охватывает око-
ло 25%, на Енисее — лишь 11% видов паразитов. Характерными форма-
ми здесь являются инфузории Ichtyophthirius muitifiliis, трематоды
N'eascus cuticula, ленточный червь Caryophyllaeus laticeps и ряд других.
Средн северных н южных форм преобладают виды, развивающиеся с
промежуточными хозяевами.
Приведенные примеры ясно показывают зависимость паразитофауны
от ландшафтно-климатической зоны. В этом отношении паразиты, как
и все другие животные, подчиняются общим зоогеографическим законо-
мерностям
Влияние географического фактора выступает также очень отчетливо
прп изучении некоторых более частных географических зависимостей.
Закономерности распределения гельминтов от различных групп позвоноч-
ных животных по ландшафтно-географическим зонам па территории
СССР рассматривает в ряде статен Левашов (1950, 1953, 1959). Перей-
дем теперь к нескольким случаям более частных географических зави-
симостей.
315
Зависимость паразитофауны от размеров площади распространения
хозяина и от степени ее изолированности
Речь идет о паразитах животных, обитающих в небольших, более
п in пенсе замкнутых водоемах и на островах По отношению к свободно
живущим организмам фактор изоляции играет весьма важную роль в
их распространении. Относительно паразитов мы имеем пока лишь раз-
розненные данные, наибольший интерес средн которых представляют
снедения Быховской-Лавловскон (19366, в) о паразитофауне рыб в замк
иутых маленьких озерах Карелин. Опа сравнивает паразитофаунх' одних
и тех же видов рыб в крупных озерных водоемах Карелин, например в
Коичозере, и нескольких небольших «ламб», т. е. глухих лесных озер
того же района. Автор исследовал четыре очень мелких ламбы, в двух из
которых имелся только окунь, в одной — плотва и окунь, а в одной,
кроме того, и щука. Эти оверкн имели в среднем 100—21Ю л в диаметре.
Сравнительные данные о паразитофауне рыб этих ламб с паразитофау-
ной рыб в Коичозере приведены в табл. 16.
Таблица 16
Число видов паразитов некоторых рыб в речных водоемах Каре чин
<по Быховской Павловской)
Наименование рыб Число паразитов в волоемаi
Кругла i л а мба Лам ба Ч» рновского Кинламба Внзаламба К он ч озеро
RlthluS rtililus . . —- - 5 •1 17
Perea f aviatllis . . 1 1 5 4 2Э
Esox Indus — — — 5 21
Итак, первый факт, который поражает при сравнении мелких озер-
ков с крупным озером —эю резкое обеднеПпе фауны в ламбах. Для
большинства паразитов интенсивность заражения ими рыб в ламбах
тоже меньше, чем в Коичозере.
В оинОшеннн качественного с( става сравнение разных ламб тоже
дает очень интересный результат, который лу4и*е всего иллюстрируется
на окуне. Табл. 17 показывает, что паразитофаупа рыб в ламбах отлн-
Т л б л и и .1 1“
Видовой состав паразитов окуня в ламбах Карелин (по Быховской Птнлэвгкоп)
Количество паразитов и (Ч) в но темах
Название паразитов Круглая лайба .Намба Чертовского Кпйл'ймба Ви’,.- ламба Пол.лзмба
Lentospoi a tiermatobia . — -— 33.1 — —
Муха olus CartlKsii —- —. 26.7 6 7
Ancyroivphalus parade-
XUS ...... — — — 1 раз
Tylodelphis clavata . . — — 6.7 — I раз
liunodej a iuciapercae — 26.7 —-
Pr.)te/>cephatu\ errnuae . — — 13.4 13.1 —
Tr'aenophorus ruxfutosus — — — 33 3 ——
Acanthoa’/iftalus lurii . . — — — Щ.2 93,3
Canal lartus lac us tris 93 3 93,3 — — —
Ergasilus sp — — 6,7 — —
316
чается не только своей бедностью, но и своим случайным характером.
Параз:пы. весьма обычные для окуня в одной дамбе, могут совершенно
отсутствовать в окунях, взятых нз соседней дамбы.
В дамбах, т. с. в водоемах с сильно обедненным населением беспоз-
воночных, которые играют роль промежуточных хозяев, можно предпо-
лагать преобладание числа видов «прямых» паразитов над паразитами,
в цикле которых обязательны промежуточные хозяева. II эта гипотеза
подтверждается на деле, ибо, например, в Польламбе 62% всех пара-
зитов. найденных у рыб этой дамбы (плотва, красноперка, окунь и ту-
ка), являются прямыми, тогда как среди паразитов тех же рыб в Конч-
озере только -40% относятся к прямым Насколько качественный состав
паразитов в дамбах вообще случ ie.H, настолько же он постоянен во вре-
мени, ибо исследования дамб велись 3 года подряд, давая все время
одни п те же результаты.
Мы остановились на примере дамб несколько подробнее, так как он
дает очень красивую паразитологическую характеристику целон кате-
гории пресноводных водоемов
Аналогичные результаты должны, вероятно, получиться при изуче-
нии паразитофауны каких-нибудь животных па островах, отделенных
от остальной суши водным барьером. Афанасьев (1941), работая на Ко-
мандорских островах, прекрасно подтвердил правильность сделанных
нами выводов и для островных паразнтофауп. На двух близко располо-
женных островах эгон группы (острове Медном и острове Беринга) па-
разнтофауна песца, например, разнится вследствие отсутствия ряда
форм, завезенных на остров Беринга вместе с собакой, тогда как на
Медный остров собака не завозилась. Кроме того, на Медном острове
отсутствие полевок, имеющихся на острове. Беринга, влечет за собой
опять-таки выпадение нз паразитофауны песцов некоторых видов, для
которых полевки служат промежуточными хозяевами.
К сожалению, соответственные наблюдения немногочисленны
Влияние на паразитофауну близости местонахождения хозяина
к границам своего ареала
Оказывается, что помимо всего прочего, паразнтофауна животного
иосит несколько различный характер, в зависимости от того, исследуем
ли мы хозяина, взятого из района с оптимальными для пего условиями
существования или же добытого откуда-нибудь с границ ареала вида,
где этот хозяин постепенно сходит па нет. очевидно вследствие каких-
нибудь неблагоприятных окружающих условий. Обыкновенно в послед-
нем случае мы наталкиваемся или на общее обеднение паразитофауны
животного, или на обеднение ее специфическими для данного хозяина
видами.
Такую картин\ дают, например, в Невской губе чехонь н сом, т. е.
рыбы южного происхождения. На Каспии чехонь на 100% заражена
специфичным жаберным сосальщиком Dactylogyrus sbnplicimalleata;
в Белом озере этот сосальщик у чехони тоже имеется, но в Невской губе
обнаружен не был. То же следует сказать о сосальщиках Aucyrocephalus
siluri сома, Отсутствуя в Невской губе, Л. siluri поражает 100% сомов
во всех южных водоемах. Чрезвычайно характерный для сома ленточ-
ный глист Proteocephalus osculalus на юге (Арал, Каспий) встречается
у всех сомов поголовно, в Невской же губе встречался лишь у 40% их,
п притом в очень малом количестве экземпляров.
К гой же категории фактов следует отнести и бедность паразито-
фауиы у волжской белорыбицы. Все родичи белорыбицы, как, например,
317
нельма н другие, тяготею! к северным bua<jM, тогда как белорыбица
продвинулась в своем распространении далеко на юг и нагуливается в
Каспии. Левашов (1924), исследовавший гелцминтофауну белорыбицы,
нашел у нее, вскрыв 50 белорыбиц, лишь два вида червей: сосальшика
Crepidostomum farionis и нестоду Eubolhrium crassum.
Аналогичная картина обнаруживается х окуней {Perea fluviatdis}
Ахтаринскнх лиманов (А ювскос море), где эта рыба находится у южных
пределов своею распространения. Еыховская-Павлпвская и Быховский
(19 10) нашли, что паразитофауна окуня в этом водоеме, по сравнению
с другими, является очень обедненной и включает всего 5 видов парази-
тов. Значительное обеднение паразитофауны по сравнению с более юж-
ными районами констатирует Марков (1952) для паразитофауна репти-
лий Ленинградской области, близкой здесь к северной границе их
ареала.
Описанные особенности паразитофауны животных, находящихся
<у предела», объясняются, вероятно, двумя причинами. С одной стиро-
ны, не вс" промежуточные хозяева различных паразитов доходят в своем
распространении до границ вода окончательного хозяина. Как только
наступает разрыв в распространении промежуточных н окончательных
хозяев, так существование паразита в данном ареале становится невоз-
моя- ным
С другой! стороны, у пределов своего распростДпення животное ста
новится обыкновенно белее, редким, а мы видели, что паразиты редко
встречающихся животных имеют меньше шансов нз сохранение, чем па
разиты животных, изобилующих в данной местности.
Интересны некоторые исключения нз изложенного выше правила.
Они наблюдаются тогда, когда в силу тех или иных причин данный вид
хозяина у границы своего ареала испытывает массовое размножение и
образует попхляцип с большой плотностью населения. Это наблюдала,
например, Шульман-Альбова в Карелии (!п litteris) в отношении неко-
торых грызунов (мышевкя северная Soc’sia belulina. и мышь-малютк;
Micromys minulus), давших летом 195^ г. значительную вспышку раз
множежия у самой северной границы ареала. Эта вспышка сипровожда
лась усиленным развитием фауны их эктопаразитов— клещей Наето-
laelaps dogieli, Laelaps micromydis и др. В данном случае можно гово
рить, что исключение подтверждает правило, ибо обычно на границ»
ареала плотность популяции бывает невелика.
Влияние реликтовых условий существования хозяина
на его паразитофауну
Паразитофауна реликтовых животных до сих нор не служила пред
устои специальных исследований. Между тем она. несомненно, пред
ставляет большой биологический интерес
Реликты — это формы, сохранившиеся в данном ареале нередко с
весьма давних времен, несмотря на изменение окружающих условий
существования. Это животные, принадлежащие по существу или другой
геологической эпохе жизни земли, или совсем иным географическим
широтам, но сумевшие сохраниться в не свойственных данному виду
окружающих условиях. Вполне естественно задать себе вопрос: какими
особенностями отличается паразитофауна реликтов?
Первой совершенно естественной закономерностью, выяснившейся
при исследовании этой фауны, оказалось обеднение паразитофауны ре-
ликтов. Реликты живут в местах, где первобытная фауна данного района
в значительной .мерс вымерла, а соответственно с этим вымерли и мно
31Я
rue промежуточные хозяева паразитов реликтового животного. Отсюда
следствие: выпадение соответственных паразитов. С другой стороны, в
новых условиях существования к паразитированию в реликтах могли
приспособиться совсем новые формы паразитов.
Хорошим объектом для выяснения iiiirepi сующего нас вопроса яв
ляется каспийский тюлень (Phoca caspica), исследованный Шулаковым
(1936). Для сравнения с ним ел1 жат северные тюлени Phoca vituluia.
Ph hispida и Histrionhoca groentandica. к ж формы близкие к каспий
скомх тюленю.
Иаразитофауна северных тюленей довольно богата. Она содержит
прежде всего эктопаразитов в виде вшей (3 вида Echinophthtrius) и со
гласно Делима Ре i!95o) 21 вид гельминтов.
Рис- 195. Схема реликтового озера Могильного ни острове
Кил .дине, отделенного от моря барьером. (Оригинал.)
Г — верхние, опресненные горизонты озера; 2 — средние горизонты озера с
приблклителтп нормально.! морской водемкв них сохранилась треска и другие
морские ж шм иные; J — ипжпне горизонты, огра ил сивые серит иородом.
У каспийского тюленя список паразитов очень мал: всего 6 видов.
Целые группы паразитов северных тюленей у него отсутствуют: вши.
ленточные глисты. Причина отсутствия цестод, очевидно, лежит в вы-
падении из фауны Каспия каких-то морских ракообразных, вероятнее
всего Copepoda, служащих для цестод в качестве первого промежуточного
хозяина. Но и среди остальных эндопаразитов идентичными с паразита-
ми севепных тюленей являются только три формы: Corynosoma strumo
sum, Dioctophyme и Crijptoiaiyle lingua. Кроме них, каспийский тюлень
имеет в своем кишечнике еще сосальщика Pseudechinosiomum advena
и нематод Eustrongylides excisus и Anisakis.
Таким образом, каспийский тюлень, потеряв почти всех паразитов,
свойственных северным его родичам, испытал некоторое обогащение
паразитофауны за счет 2—3 новых видов. Присматриваясь к этим новым
паразитам тюленя, мы невольно приходим к убеждению, что в приобре-
тении их тюленем играли большую роль птицы, вернее их паразиты.
Eusirongyhdes excisus. безусловно, представляет собой птичий вид не
матод, свойственный бакланам и лишь в условиях Каспийского моря
включившийся в паразнтофауну тюленя. Сосальщик же Pseudechino-
stomum чрезвычайно близок к птичьим Echinostonjidae и тоже, по всей
вероятности, перешел на тюленя с пгнц.
Итак, па поимере тюленя мы видим, как лиризитофауна реликто-
вого животного оскудевает его исконными паразитами и пополняется
немногими новыми видами имеющими чуждое для тюленя пронсхож
дение
Второй интересный елхчай — 5то реликтовая треска из озера Мо-
гильного на острове Кгльдипе (рис. 195). В этом водоеме, отделенном
от моря довольно высоким перешейком, верхние 5 м воды вполне прес
пые, самые глубокие стон озера с 12 до 16 .« насыщены сероводородом
и непригодны для жизни, а промежуточная толща воды в 7 .« состоит
31'
из морской воды. в которой и держится треска. Озеро обособилось от
моря давно (о нем имеются сведения от XV! в.), столь давно, что треска
уже успела образовать в нем особую морфу. Кроме трески, в озере со-
хранилось и много морских беспозвоночных.
Оригинальные условия существования в реликтовом водоеме сильно
отразились на паразитах трески. .Морская треска нз соседних с Кильдн
ном районов Баренцева моря, по данным Полянского (1955), содержит
27 видов, тогда как в могильиекскон треске найдено всего 7 видов пара-
зитов, идентичных с таковыми морской трески (Догель, 1936а). Итак,
правило об обеднении паразитофауны реликтов подтверждается и на
примере могильнсиской трески.
Очень любопытный факт обнаружен относительно одного вида со-
сальщиков. На жабрах трески оз. Могильного нашлись в значительном
количестве ипцнетнровапные молодые сосальщики, которые во взрослом
состоянии встречаются, по всей вероятности, в кишечнике каких-нибудь
рыбоядных птиц. Между тем тресКа озера отрезана 5 и пресной воды
от всякого сообщения с птицами. Поэтому остается лишь одно правдо-
подобное объяснение наличия сосальщиков в озере: птицы, пролетая над
озером, сбрасывают туда свои экскременты с яйцами сосальщиков. Из
яиц выходят личинки и проникают в моллюсков, где нз них в конце кон-
цов получаются церкарии, заражающие треску. Но последнего этапа раз-
вития, а именно поедания трески птицей, не происходит, и потому жа-
берные сосальщики озерной «трески не могут никогда развиться до
половой зрелост и и в конечном счете обречены на гибель. Не будь по-
стоянного подновления заражения извне, эти сосальщики должны были
бы выпасть из паразитофауны оз. Могильного. Следовательно, обеднение
паразитофауны реликтов может обусловливаться выпадением не только
промежуточных, по и окончательных хозяев.
Наиболее сильное влияние оказал реликтовый образ жизни на пара-
штов бычка Myoxocephalus quadricornis в Онежском озере (Догель и
Розова, 1911). М. quadricornis — морская форма, в настоящее время
встречающаяся в Балтийском море, в Белом море и в Ледовитом океане.
Однако в Онежском озере, а также в некоторых более мелких озерах
Швеции и Финляндии сохранились реликтовые формы этой рыбы, кото-
рые нам удалось получить для исследования (около 20 экземпляров из
Онежского озера). Из морских Сычков описан целый ряд типичных мор-
ских паразитов, в особенности сосальщиков. Al. quadricornis Онежского
озера не только утрат.:.! без остатка всю свою морскую паразптофауну,
но и не приобрел взамен почти никаких пресноводных паразитов.
.V Онежских четырехрогпх бычков были найдены лишь некоторые ин-
фузории на жабрах (Trichodina, сосущая инфузория Trichophrya), а нз
многоклеточных паразитов только мелкие личинки нематод (по-видимому
Rhaphiduscaris), инкапсулированные в печени н в стоиках кишки. Онеж-
ский четырехрогий бычок не приобрел, следовательно, ни одного пресно-
водного паразита, достигающего в нем половой зрелости, прежняя же
паразитофауна его исчезла бесследно.
Большой интерес в отношении реликтовых форм представляет озеро
Байкал. Около половины видов рыб Байк.гта (свыше. 20) составляют
бычки (семейства Cottidae, Cottoconicphoridae, Comeplioriciae), которых
рассматривают как морские реликты более птл менее древнего происхож-
дения.
Изучая паразптофауну бычковых рыб Байкала (Догель и Боголе-
пова, 1957), датось убедиться, что она слагается из нескольких групп
различного происхождения Одну небольшую группу составляют так
называемые «палеоэидемнки», т. е. очень древние и, вероятно, морские
320
по своему происхождению виды. Таковых имеется всего 2 вида—это
мнксоспоридин Myxidiurn perniciosurn и Myxobilatus baicalensis. Вторую
большую группу паразитов представляют собою псоэндемики в количе-
стве 8—9 видов. Они имеют совершенно иное происхождение. Это (по
сравнению с палеоэндемиками) относительно недавнее приобретение.
Они перешли на бычков с других чисто пресноводных байкальских рыб,
но успели на бычках уже дать начало новым видам. Относительно этих
видов паразитов можно с уверенностью сказать, что часть их произошла
от паразитов карповых рыб (Myxobolus talievi, М. spatulatus, Daclylo-
gyrus colonus, Allocreadium polymorphum), другие же — от паразитов
лососевых и хариусовых (Crepidcstomum baicalense, Соте phorone та,
Salmincola cottidarum). Эта уверенность основывается на большой си
стемятическон и .морфологической близости одной части паразитов
к соответствующим паразитам карповых, а другой — к паразитам лосо-
севых и хариусовых.
Резюмируя сказанное выше о паразитах байкальских бычков, мы
видим, что переход первоначально морских бычков в чисто пресновод-
ные условия ознаменовался, с одной стороны, вспышкой видообразования
паразитов у самих бычков, с*другой — резкам изменением их паразито-
фауны. При этом старая .морская паразитофауна почти вымерла (но
не полностью!) и заменилась новой, которая принадлежала другим
пресноводным рыбам. Однако переход этих чуждых бычкам паразитов
на бычков привел к видоизменению и образованию группы новых, спе-
цифичных для бычков эндемичных паразитов. Таким образом, здесь на
бычках Байкала, наряду с обычным для реликтов обедненном паразито-
фауны, произошло ее дополнение и обогащение из новых источников,
сопровождавшееся, кроме того, вспышкой видообразования. Вероятно,
это связано с. тем, что сами хозяева — байкальские бычки — не представ-
ляют собой вымирающую или находящуюся в состоянии депрессии груп-
пу, но, наоборот, испытали в-Байкале вспышку видообразования с силь-
ным увеличением числа родов, видов, а также и особей.
Пример байкальских бычков показывает, что паразитофауна релик-
тов может- испытывать изменения различного рода в зависимости от
дальнейшей судьбы самого хозяина.
Необходимо подчеркнуть желательность дальнейших исследований
паразитов всякого рода реликтовых животных, что представляет боль
шой зоогеографический и эколого-паразитологический интерес.
2. ОПЫТ ЗООГЕОГРАФИЧЕСКОГО РАЙОНИРОВАНИЯ
ПО ПАРАЗИТОЛОГИЧЕСКИМ ДАННЫМ
На основе рассмотренных нами выше разнохарактерных влияний
внешней среды и эволюционной истории фауны возникли большие
группировки паразитов, населяющих различные зоогеографические об-
ласти, подобласти, провинции и т. д., па которые разбивается поверхность
нашей планеты.
Несмотря па го, что достаточно подробных данных о распределении
паразитов вообще еше сравнительно мало, некоторые группировки их
уже поддаются зоогеографическому р а й о и и р о в а и и ю вроде
того, которое применяется для свободно живущих организмов. Подобное
районирование может быть произведено, исходя из двух разных отправ-
ных точек. С одной стороны, можно построить картину зоогеографичс
ского распространения отдельных систематических групп
паразитов, например сосальщиков, цестод н т. п. С другой стороны,
можно дать отображение зоогеографической характеристики всей пара-
21 в. а. Догель
321
зитифауны определенной группы животных-хозяев i
местах распространения последних — распространение паразитов репти
лпи, птиц и т. п. Этот второй тип зоогеографического анализа с точю
зрения паразитологии представляет большой интерес.
В настоящее время попыток произвести такой июгеографический
анализ для всех паразитов какой-нибудь группы хозяев в .мировом мас-
штабе еще очень мало. Наибольший интерес в этом плане представляет
зоогеографический раздел монографии Деляму ре (1955), посвященный
гельминтам морских млекопитающих (китообразные и ластоногие)
Однако эта работа касается не всей паразитофауны, а лишь паразите
ческих червей.
Первый вопрос, который ставит Делямуре, касается распределения
гельминтов в северном и южном полушариях. В какой мере они подчи-
няются тем общим закономерностям распределения в мировом океане,
которые установлены в отношении свободно живущих организмов? Ос-
новные результаты анализа, дающие ответ на этого вопрос, представле-
ны в табл 18.
• Таблиц а1ь
*
Распространение гельминтов ластоногих и китообразных в северном и южном
полушариях (по Делямуре)
I руплы гельминтов Количество видов гельминтов, зарегистрированных Bcei<i
исключительно в северном полу- шарии исключительно в южном полушарии в обоих полу- шариях (общие виды)
Tremalodes 35 1 2 38
Cestodes . 24 10 4 38
Nematodes . 41 14 У 64
Acamhiicephola .... Я 5 4 17
Итого 108 30 19 157
Как видно из приведенных данных, из общего количества 157 е.идое
паразитических червей исключительно в северном полушарии найдено
108, исключительно в южном — 30. а 19 видов встречено в обоцх полу-
шариях. Таким образом, оказывается, что распределение гельминтов
между акваториями обоих полушарий является неравномерным — число
видов в северном полушарии значительно больше, чем в Южном. Это
хорошо согласуется с распределением свободно живущей морской фауны
в водах северного и южного полушарий. Зенкевич (1951) в своей извест
ной сводке по гидробиологии морей указывает, что северное полуша-
рие, более однообразное в отношении наземной зоогеографии, дает паи
большее зоогеографическое разнообразие морской фауны. Совершенно
обратное соотношение имеет место в южном полушарии. Приведенные
выше данные о распространении гельминтов морских млекопитающих
подтверждают эту общую зоогеографическую закономерность.
Представляет интерес рассмотреть распределение гельминтов по по-
лушариям для ластоногих и китообразных отдельно, учитывая большие
различия в их экологии. Эти данные представлены в табл. 19.
Приведенные в табл. 19 данные показывают, что установленные выше
закономерности справедливы как для ластоногих, так и для китообраз-
ных. У обеих групп хозяев богатство видами гельминтов в северном по-
32'2
Таблица 19
Распространение гельминтов морских млекопитающих в севеоном и южном
полушариял. отдельно для ластоногих и китообразных (по Делямуре)
Количество зарегистрированных видов гельминтов
Систематические группы гельминтов исключительно в северыом полу- шария исключительно в южном полушарии в обоих полу- шариях
ластоно- гие китооб- разные ласто- ногие китооб- разные ласто- ногие китооб- разные
Trematodes , 13 23 1 1 Нет 2
Cestodes . 14 J2 8 2 Нет 4
Nematodes 14 27 7 7 3 7
Acanthocepliala 7 4 3 2 Нет 4
Ито! о и 66 19 V 3 17
л; шарни значительно больше, чем в южном. Однако между ластоногими
и китообразными в этом отношении наблюдаются существенные разли-
чия, которые выражаются в том, что у китообразных имеется гораздо
больше видов гельминтов (притом во всех группах), общих для север-
ного и южного полушарий. У ластоногих таких видов имеется всего 3 и
все они относятся к нематодам Эти существенные различия между груп-
пами хозяев Делямуре ставит в связь с особенностями их экологии.
Китообразные совершают очень большие миграции, во время которых
они частично переходят из одного полушария в другое.
Среди ластоногих, хотя и имеются виды, совершающие значитель-
ные миграции (гренландский тюлень — Pagophoca groenlandica, котик —
Callorhinus ursinus), таких видов немного, и протяженность миграций
щачительно меньше, чем у китообразных.
Отчетливо удается также установить закономерности рас-
пространения гельминтов ластоногих и китообраз-
ных по зоогеографическим областям мирового
океана—Арктической, Бореальной, Тропической, Анти бореальной и
Хнтарктической. Распределение гельминтов по этим зоогеографическим
областям представлено на рис. 196 (суммарно для всех морских млеко-
питающих и отдельно для ластоногих и китообразных). Из этих схем
видно, что на первом месте но числу видов гельминтов стоит Бореальная
область (94 вида), на втором — Антибсреальная (38'видов), на
третьем—Тропическая (31 вид), на четвертом — Арктическая (25 видов):
и на последнем, пятом — Антарктическая (14 видов) Преобладание ви-
дов гельминтов в умеренных областях закономерно повторяется для
цестод, нематод, скребней. Единственное исключение составляют трема-
тоды Антпбореальной области.1
Сопоставляя приведенные данные о распределении гельминтов с
данными о биологической структуре океана, Делямуре подчеркивает,
что гельминты обнаруживают принципиально то же симметричное рас-
колом (нее явлений жизни в океане, которое установлено для свободно
живущих организмов (Зенкевич, 1951). Однако существует различие, ко-
торое сводится к тому, что Тропическая зона океана, как известно, ха-
рактеризуется большим качественным разнообразием свободно живущих
1 Причины этого исключения пока не ясны Пам представляется возможным до-
пустить, что отсутствие трематод связано с отсутствием необходимых для завершения
цикла промежуточных хозяев.
21е
323
организмов — обилием родов и видов, что, как мы видим, не имеет места
у гельминтов морских млекопитающих.
Это различие и его объяснение Делямуре представляется нам вполне
правдоподобным. Оно определяется некоторыми биологическими особен-
ностями паразитических червей, по сравнению со свободно живущими
водными организмами. Тропическая зона океана характеризуется боль-
шим качественным многообразием, однако в количественном отношении
(плотность популяций отдельных видов) она относительно бедна. Напро-
тив, в умеренных областях оксана создаются условия для массового
т с
4 7
р>тичесд£
PKTWQL-
Тропическая
4 0 0 2 14 3 2 11 2 • 10
А
о > е
N А Т С N
6 3-20 ? 0 4
бор едльна я
7 II 7 4 • 29 24 15 25 0 • 72
Антисорсальная
0 6 9 3-18 О 3 15 5* 21
1 4 Ю 1 3 2 0-6
Т С N А Т с N А
Рнс. 196. Распределение гельминтов водных млекопитающих по зоогеографическим
, областям
Д — обшая картина распространения гельминт в ластоногих н китообразных, Н— рас-
пространение гельминтов ластон i их и китообразных раздельно Т — Trematodes; С —
Cestodes; N — Nematodes; Л — Acanilrocephala. (Но Делямуре.)
Арлбскис цифры укллыияют iu числи виши.
размножения видов, хотя видовой состав организмов менее разнообразен.
Плотность популяций животных-хозяев (в особенности ластоногих) в
Тропической зоне значительно меньше, чем в умеренной и арктической.
Таким образом в умеренной зоне наиболее благоприятно сочетаются
важные положительные для развития гельминтов факторы: наличие не-
обходимых дефинитивных и промежуточных хозяев, значительная плот-
ность их популяций, облегчающая возможность заражения, и наличие
благоприятных абиотических условий. Этими решающими причинами,
вероятно, и определяется богатство гельминтофауны водных млекопи-
тающих умеренных областей. Чго касается Арктической и Антарктиче-
ской областей, то уменьшение числа видов гельминтов, очевидно, связа-
но с общим ухудшением условии жизни в связи с менее благоприятными
абиотическими условиями и с уменьшением числа видов промежуточных
хозяев.
Широкие миграции некоторых морских млекопитающих (особенно
китообразных), очевидно, несколько нарушают картину распределения
гельминтов по географическим областям океана, но тем не менее общие
закономерности этого распределения, как мы видели, выступают вполне
отчетливо.
По паразитам других групп позвоночных не существует пока дан-
ных, которые позволили бы провести зоогеографическое районирование
324
в пределах земного шара. В отношении пресноводных рыб это возможно
сделать для более ограниченной территории, а именно для территории
Советского Союза, т. е. для одной шестой суши земного шара. Зоогео-
графическое районирование паразитов пресновод-
ных рыб СССР проводилось с учетом всех групп жнвогных-парази-
юв. Впервые это было осуществлено Догелем (1947). а затем расширено
и уточнено Шхльманом (19546, 19586). Зоогеографическому райониро-
ванию паразитов рыб предшествовало планомерное фаунистическое и
экологическое изучение их, которое осуществлялось большим коллекти-
вом советских паразитологов по единому плану на всей территории
СССР в течение 25 тет. В результате этих работ предстояло прежде
всего ответить на вопрос, в какой мере на паразитологическом материа-
ле можно уловить различие нс только крупных зоогеографических еди-
ниц— областей и подобластей, но также и более мелких подразделе-
ний— зоогсографнческн.'. провинции и окру] он. Второй вопрос общего
характера сводился к тому, в какой мере зоогеографическое райониро-
вание паразитов соответствует районированию их хозяев-рыб.
Зоогеографическое .районирование, пресноводных рыб СССР было
с большой полнотой осуществлено Л. С. Бергом (1949). Принимаемые
км подразделеипядерриторнн СССР и при'ежаишх стран представлены
на прилагаемой карте (рис. 197). Паразитологические исследования не
с одинаковой полпотАг охватили все географические районы, в част-
ности еще недостаточно нзхчена в зоогеографическом отношении Нагор-
но-Азиатская подобласть, Охотско-камчатский и Анадырский округа и
некоторые другие. Тем не менее общая картина распространения парази-
тов рыб на территории СССР достаточно ясна для общих выводов зоо-
географического порядка. Прежде всего можно утверждать, что суще-
ствует полный параллелизм в зоогеографическом районировании рыб и
их паразитов. Отдельные зоогеографические подразделения но видовому
составу паразитов рыб характеризуются главным образом различны и
соотношением групп разного географического происхождения и в неко-
торых случаях наличием эндемичных для данного зоогеографического
района видов.
Мы не имеем возможности в рамках данной книги дать подробную
характеристика и списки видов паразитов рыб. встречающихся в отдель-
ных зоогеографических районах СССР Эги данные приведены в упомя-
нутой выше работе. Шульмана < 19586). Мы ограничимся тем, что сравним
в качестве примера йаразитофауиу двух округов —европейского и си-
бирского. относящихся к лсдовнтоморской провинции Голарктической
области. Это сравнение вместе с тем покажет нам реальность различий
в паразитофачне рыб между самыми мелкими единицами зоогеографиче-
ского районирования —округами.
Состав паразитофауны европейского округа Шульман выражает сле-
дующими данными:
Голарктические 78 питов. т. е. 50,3 ч
Циркумполярные 19 . . 12,3
Ле.товитоморскнс Североевропейские, характерны! i-тя 22 . 14.2
европейского округа 3 . 1.9
Понто-арало-каспийские 17 , . ю.з
С невыясненным ареалом 16 . . и,о
Всею................155 10U
Соотношение разных групп паразитов сибирского округа харакге
ризуется след чшшми данными:
32'
Рис. 197. Карга зоогеографических областей для пресноводных рыб. (По Бергу.)
Д - Голарктическля область: / — Циркумполярная подобласть, 1 — ле ювитоморская провинция, ЬО — евро;-
псйский округ, СО — сибирский округ, 2 — тихоокеанская провинции, ЛнО — анадырский округ О-КО — охотско-
камчатский округ; II — Байкальская’ подобласть; III -Средизехп* .морская подобласть, 8 — балтийская провинция,
РО — рейнский'округ, НО — невский округ, 4 — понто-арало-касппйская провинция. 40 —черноморский округ,
КО- каспийский округ, ЛрО — аральский округ, 5—иранская провинция, 6 — туркестанская провинция;
IV—Нагорпо Азиатская подобласть, 7—западпом'оигольская провинция, 8 — балхашская провинция, 9 — тарим-
ская провинция, 10 — тибетская провинция. В — Амурская переходная область; II — амурская провинция,
— амурский округ, СахО — сахалинский округ, 12 — приморская провинция,ПрО— приморский округ,
ОИ — округ Иезо, или Хоккайдо, 13 — корейски-маньчжурская провинция, // — японская провинция, С — Си
ноивдийская область: Г—Китайская пс.юбластг (По Бергу.)
Рис. 198. Пути проникновения южных форм паразитов рыб в водоемы балтийской
провинции и ев олейского округа ледовитоморскон пропннцнк. (По Шулымаку.)
1 - южные виды па, алитов (числами обозначено количество, видов); 3 — в том числе черноморские виды
j - пути проникновения видов и северные водоемы.
327
Г олзрктические 38 видов, г. е. 50,6 %,
Циркумполярные 23 . 13,2
Ледпннтоморскнс 23 . 13,!
Эндемики Сибири Характерные для европейского 7 . 4,0
округа 1 . 0,6
Понто-арало-кзспийскис . , 3 . 1,7
Байкальские 1 . 0.6
Амурские 1 0.6
Морские 3 . 1.7
С невыясненным ареалом 24 . . 13.8
Всего 174 1П0
Сравнивая приведенные цифры, между обоими округами можно об-
наружить некоторые существенные различия. Прежде всего для сибир-
ского округа характерно наличие некоторых эндемичных видов (Согс-
“onerna sibirita, Achtheres strigalus, Coregonicola producta, C. orienlalis.
Salmincola jacutica. Basunistes woskoboinikowi), при этом роды нематод
Coregorierna и Coregonicola эндемичны для Сибири. Во-вторых, в сибпр
ском округе по сравнению с европейским значительно слабее проникно-
вение южных иопто-арало-каснийских элементов паразитофауны В си-
бирском округе имеются элементы, характерные для Байкальской под-
области н Амурской области, полностью отсутствующие в европейском
округе. Все эти различии, взятые в целом, создают определенный фауни-
стический облик паразитов рыб европейского н сибирского округов.
Если взять для сравнения зоогеографические районы более высокого
ранга (провинции и области), то различия между ними в cocthiu
паразитофауны рыб окажутся еще значительно более резко выражен-
ными •
Зоогеографическое районирование и сравнительное изучение, пара-
зптофаупы рыб различных районов Советского Союза позволяют в ряде
случаев решать вопросы и сто р и ч е с к о й зоогео гр а ф и и
происхождение фаун. Хорошим примером, разработанным
Шульманом, может служить проникновение южных видов паразитов на
Север, имевшее место в послеледниковую эпоху. Сравнительным изуче-
нием видового состава паразитов рыб на территории Европейской части
СССР показано (рис. 19S), что проникновение южных видов на Север
осуществлялось через водные бассейны ДнщГра, Дона и Западной Дви-
ны, через балтийскую провинцию. Выясняется, что Волга, как путь
расселения, нс играла существенной роли. Это связано, очевидно, с тем
что до рисского оледенения ее началом были истоки Камы и опа приня-
ла свой современный характер сравнительно поздно, лишь с вюрмского
оледенения. Приведенные соображения о путях расселения паразитов,
рыб хороню согласуются и с ихтиолого-географическими и геологически-
ми данными.
Аналогично тому как это было сделано для паразитов пресноводны
рыб, зоогеографическое районирование было осуществлено и для паря-
зптов морских рыб морей Советского Союза (Полянский, 1938).
Пе останавливаясь на рассмотрении этих работ, укажем лишь, что
они показали полную возможность использования паразитологического
материала при разработке и обсуждении вопросов морской зоогеогра-
фии. Зависимость распространения паразитов от целого сложного ком-
плекса факторов исторического и экологического порядка делает их
особенно «.чу вствительными индикаторами» изменений условий сущест-
вования. Это в свою очередь особенно пенно для зоогеографии, строя
щенся на экологической основе.
:;28
3. РОЛЬ ПАРАЗИТОВ ДЛЯ РЕШЕНИЯ НЕКОТОРЫХ ВОПРОСОВ
ЗООГЕОГРАФИИ И ФИЛОГЕНИИ
До сих пор при оценке различных общих вопросов зоогеографии
(о происхождении тех ити иных фаун, о значении областей и под-
областей) принимались во внимание почти исключительно свободно
живущие организмы. Между тем паразитические животные для эти*
целей представляют собой объект не только не менее, но порой даже
более удобный и ценный. Поэтому учитывать паразитические группы
при зоогеографических рассуждениях совершенно необходимо.
Отчасти значение паразитологии для зоогеографии было показано
нами в начале данной главы. В дополнение к сказанному' остановимся
еще на некоторых аспектах этой проблемы.
Существенные данные может дать паразитология при решении
вопросов происхождения фаун зоогеографических областей
Первым на путь широкого использования данных паразитологии для
решения вопросов исторической зоогеографии стал Иернпг (lhering
1891 и 190'2), занимавшийся вопросом о происхождении южноамери
канской фауны позвоночных животных. При этом Исринг полагает,
что в случае общего происхождения двух хозяев они содержат одина-
ковые или близкие друг другу виды паразитов. Опираясь па рас
смотрение различных гельминтов (сосальщиков, цеетод, нематод
и скребней) у южноамериканских и североамериканских млекопитаю-
щих, Иернпг приходит к ряду важнейших зоогеографических выводов
По его мнению, соединение обеих ХмериЛ в его современном виде про
изошло не раньше плиоцена. Далее; фауна Южной Америки слагается
из двоякою рода элементов. Одни из них — древние автохтоны, дру
гие— более новые поселенцы, пришедшие после плиоцена из Север
нон Америки. Долгая изоляция автохтонов в течение третичной эпохи
должна была привести н привела к развитию в них особых, само
стоятельных видов гельминтов, тогда как гетерохтонные пришельцы
содержат в себе паразитов, сходных с таковыми голарктических
млекопитающих. И. действительно, только южноамериканские автох-
тоны содержат, например, особые, своеобразные виды скребней.
Хотя часть фактических данных Ht ринга при последующих пара
штологнческнх работах оказалась не. вполне правильной, все его
общие рассуждения вполне подтвердились; попытку этого автора
использовать для зоогеографии отношения между паразитами и хозяе
сами можно называть методом Неринга.
Почти одновременно с Перин том, а также в ряде более поздних
статей Келлог (Kellog, 1896, 1908, 1913, 191-4) обсуждал значение
птичьих пухоедов для опенки филогении их хозяев. Так, отмечая
наличие обших видов Mallophaga у разных видов европейских
и американских птиц (iiaripnMi. р, двух разных видов корольков,
у двух разных видов ворон, европейского и американского), Келлбг
толкует этот факт, как признак происхождения такой пары хозяев
от одного общего предка: этот вид дивергпровал и дал два самостоя
тельных вида, тогда как паразиты его остались неизменными. Тот жт
путь суждений Келлог применяет и для оценки родственных отноше-
ний между бескилевыми птицами: Struthiones Африки, Rheiformes
Южной Америки и Casnariiformes Австралии имеют несколько обших
видов Mallophaga что. по мнению Келлога, говорит о генетической
близости этих групп птиц.
На том же материале, т. е. на пухоедах, разбирал генетически*
отношения их хозяев и Гаррисон (Ilarrison 1912 1914, 1922) в цс-
32е
лом ряде своих работ, пользуясь одинаковым с Ксллогом ходом рас
суждений. Таким путем он старается доказать, что наличие общего
сем. Mallophaga на австралийских сумчатых и южноамериканских
днкообразах свидетельствует о былой южной связи между материка
мн Австралии и Америки.
Он же указывает, что характер AXallophaga пингвинов говорит
против всякого их родства с антарктическими водоплавающими пти-
цами. а скорее за связь их с типаму, курами и голубями; пухоеды
новозеландского киви (Apteryx) родственны Mallophaga ржанковых,
чем доказывается отсутствие связен между киви и страусами.
Хорошим показателем геологического распространения южных
речных раков служат их эктопаразиты из группы Temnocephala.
Наличие Temnocephala па раках из Австралии, Новой Зеландии,
Мадагаскара и Южной Америки, при отсутствии их на раках север
него полушария, говорит как о бывшей сухопутной связи между
указанными странами, так и об известной независимости северных
раков от южных.
Позднее, в 1928 г., Гаррисов особенно ясно высказывается отно-
сительно важности использования сочетаний «хозяин и паразит» для
зоогеографических выводов.
Наконец, очень большое внимание аналогичным паразитологиче
•ским данным уделяет в ряде своих работ по простейшим нз кишечнп
ка амфибий американский ученый Меткаф (Metcalf, 1923, 1929, 1940).
Он широко пользуется распространением этих паразитических простей
ших для оценки не только современного распространения их хозяев,
но и для прослеживания путей их расселения в прошлом.
Исследовав Opalinida из амфибий всего мира, Меткаф применяет
для зоогеографической характеристики отдельных амфибий следую-
щие данные: 1) вид встречающихся в хозяине Opalinida, 2) вид
самого хозяина, 3) семейство, к которому относится хозяин, 4) гео-
графическое распространение данного простейшего в хозяине, 5) гео-
графическое распространение вида н рода хозяина. Такне сведения,
в совокупности с картиной распределения суши и моря в настоящую
и в предшествовавшие эпохи жизни земли, приносят большую пользу
при изучении как современного, так и прошлого распространения
паразита и его хозяина, а также и их путей и направлений рас-
селения.
Как далеко решается идти Меткаф в своих зоогеографических
рассуждениях, базируясь на комбинации вышеуказанных данных,
видно из нижеследующих примеров. В амфибиях встречаются четыре
рода Opalinida (рис. 199) Protoopalina, Zelleriella, Cepedea и Opalina.
Самый древний род—Protoopalina— был в Экваторни (Австралия
вместе с Африкой и Южной Америкой) еще в триасе, и наиболее
архаический подрод этого рода—подрод I1—до сих пор сохранился
на названных трех континентах. Прочие семь подродов Protoopalina
возникли так: подрод II возник в Австралии; подрод 111 дифферен-
цировался еще до изоляции Австралии в ней и в юго восточной Азии
в юрскую эпоху; затем в мелу он распространился па север в Европу,
а позднее — в третичную эпоху проник с северо-востока в Африку;
подрод IV появился в юрскую эпоху в Австралии или юго-восточной
Азии и т. д.
Род Zelleriella возник в Патагонии до отделения ее от Антарк
гики, которое совершилось, вероятно, в середине миоцена. Род это!
1 Metcalf разбивает род Protoopalina на 8 подродов, которым он не дает, однако,
самостоятельных латинских названии, а обозначает римскими цифрами от I до VIII.
<30
Рис. 199. Представители четырех родов, образующих группу Opalinida
4 Protoopalina inteslinalis', Р—zellefielta maironucleaia\ C Cepedea
pkrynomantidls; D — Opalina rauarum.
Рис. 2'Ю. Распространение рола Zellericlla. (Но Меткафу ) Этот род.
встречающийся преимущественно в лягушках сем. Lepiodaciyfidae, имеется
в Австралии и особенно в Америке, отсутствуя в Африке, хотя там живут
чва или три вида Leptodactylidae. Зачерненные участки указывают места
нахождения Zelieriella.
331
появился лишь после потери связи Патагонии с Африкой, так как
в Африке он не встречается. Его первоначальными хозяевами были
южные лягушки (Leptodactylidae). Присутствие этого рода в Южной
Америке и Австралии (рис. 200), наряду с отсутствием в Евразии,
говорит об имевшейся некогда южной сухопутной связи между Амери
кой и Австралией. Если бы, как думают некоторые, различны*
Leptodactylidae развились независимо и параллельно на разных
материках в их современных местах распространения, то совершенно
непонятно было бы нахождение в них па разных материках весьма
близких видов рода Zelleriella.
С другой стороны, если считать, что лягушки Leptodactylidae
были сначала распространены повсюду на севере, но затем были
оттеснены на юг и оказались лишь на южных континентах, то они
успели бы передать своих опалин Zelleriella хотя бы каким-нибудь
видам современных северных лягушек, тем более чго у себя на юге
Leptodactylidae передают инвазию Zelleriella другим бесхвостым
амфибиям, например Bufo, Hylidue. Поэтому полное отсутствие Zelle-
riella у лягушек па севере говорит о том, что ни Zelleriella, ни ее
хозяева, т. е. Leptodactylidae, на севере никогда не были. Таким
образом, единственным объяснением, охватывающим все имеющиеся
налицо факты, является допущение в прошлом сухонугной связи
Южной Америки с Австралией. В таких же деталях обсуждаются
Меткафом и роды С.еребеа и Opalina.
Далее, Меткаф устанавливает место и время возникновения
нескольких семейств лягушек и жаб и пути, приведшие к заселению
ими современного ареала. *
Часто имеют вес пе только положительные, но и отрицательные
паразитологические данные. Отсутствие рода Opalina в Южной
Америке доказывает, что настоящие лягушки (Raninae) никогда не
распространялись из Северной в Южную Америку. С другой стороны,
единственная евразийская древесница (Htyla arborea) с ее песколь
кимп подвидами есть иммигрант из Северной Америки, так как опа
содержит североамериканский вид Opalina. Приведенных примеров
достаточно, чтобы видеть, к каким важным обобщениям, пусть даже
иногда еще гипотетичным, приводи г Меткафа комбинированный спо-
соб использования для зоогеографии филогении хозяев совместно
с их паразитами.
Очень интересны соображения одного из ведущих американских
паразитологов Хегнера (Hegiier, 1928) о взаимоотношениях одно-
клеточных паразитов человека и обезьян. Большинство простейших
человека и обезьян, говорит Хегиер, принадлежит к одним и тем же
видам или к видам, морфологически нс отличимым друг от друга. Это
обстоятельство особенно поразительно, если мы сравним простейших
нз обезьян с простейшими нз различных животных, тесно сопри-
касающихся с человеком. Здесь с домашними животными почти
никакого сходства нс наблюдается. Два приведенных сопоставления
дают еще одно доказательство общему происхождению человека
и обезьяны.
Итак, вы видим, что для решения ряда .зоогеографических
и филогенетических вопросов все более привлекаются на помощь
паразитологические данные.
Однако, чтобы правильно оценить их значение, необходимо учи
тывать все многообразие самых различных факторов — историки- ео-
графичсски.х. экологических, физиологических и др., т. е. всех факто-
ров, определяющих распространение того или иного вида паразитов
332
Прежде всего необходимо рассмотреть, всегда ли совпадает рас-
пространение паразитов с таковым их хозяев, и от каких факторов
могут зависеть наблюдаемые случаи несовпадения. 11е разобрав эти
факторы- легко можно сделать в своих зоогеографических и фило-
генетических выводах ряд ошибок.
Мы уже видели в начале главы, что паразнтофауна одного
и того же животного может различаться в различных районах ареала
его распространения. Из этого примера следует и обратное положе-
ние, а именно, очень многие паразиты далеко не всегда следуют за
своим хозяином во все места его распространения, но имеют преры-
Рнс. 2 И Очаговый характер распространения широкого лентеца
Diphyllobotkrium latum в Европе и Азии. (Ор чгинад ) Показаны линь
главные очаги распространения — в Швейцарии, па Дунае, в Прибал-
тике, па Оби и на Енисее.
Достаточно припомнить, что песочная блоха (Sarcopsylla penetrans)
человека заселяет тропическую зону Южной Америки, что малярий-
ный плазмодии, свайник двенадцатиперстной кишки и многие другие
паразиты человека имеют ясную северную границу распространения,
далеко не совпадающую с распространением хозяина. Припомним
только, что Dracunculus medinensis, например, в пределах СССР имел
явственно очаговое распространение: в недавнем прошлом 1 этот пара-
зит встречался в трех городах Средней Азии, а именно: в Самаркан-
де. Джизаке и Бухаре. Широкий лентец (Diphyllobothrittm latum)
известен из Швейцарии, Италии, Румынии, Прибалтики, Карелин, на
Оби, занесен человеком в США (рис 201).
Такими же фактами изобилует, конечно, и паразитология диких
животных, несмотря на до, что сведений о них имеется значительно
меньше. Укажем два-три примера. Azygia lucii имеется в щуках
Ленинградской области, встречается на Волге, но отсутствует в Арале
и на юге Каспия (под Ленкоранью). Neascus cuticola разных карпо-
вых рыб встречается только в южных водоемах, отсутствуя в Нев-
ской губе и озерах Карелии. Achieves percaruni. распространенный
с севера вплоть до Каспия, отсутствует у аральских окуней и суда
ков Mxxosporidia имеются в устье Аму-Дарьи, но отсутствуют в мор
ском районе Арала.
• В настоящее время заболевание человека Dracunculus medinensis на территории
Советского Союза ликвидировано.
ЗЭЗ
Рис. 202. Инфузории из желудка сайги (Saiga
tafarica)
,1 — Entodinium kazahstaiticunr, В - E. saigac\
C—E. exigaum; D—E. bicornutum (А и В— спе-
цифичные для caiii и. Си D — общие для сайги
и северного оленя.) (По Догелю.)
Очень интересный случай прерывистого распространения парази
тов лают некоторые кишечные инфузории копытных из сем. Ophryos
colecidae. Исследуя инфузорий из рубца сайги. Догель (1946а)
нашел, что два из имеющихся у нее видов, а именно: Entodinium
exiguum и Е. bicornutum (рнс. 202). помимо сайги были находимы
до сих пор только у северного оленя. На первый взгляд такое
совпадение кажется совершенно непонятным, так как ареалы обита-
ния обоих хозяев (сайги и северного оленя) разделены огромными
пространствами. Однако находки черепов сайги в четвертичных
отложениях далекого севера
(вплоть до 73 с. ш.) и данные
по встречному распростране-
нию северного оленя на юг го-
ворят о том, что было время,
когда ареал обоих копытных
совпадал. Мензбир (1931) ри
сует яркую картину того, как
во вторую межледниковую эпо
ху на территории теперешнего
Казахстана «стада северных
оленей, сайгаков, джейранов,
лошадей и куланов пасутся ря
дом друг с другом». При таких
уелозиях сайга и северный
олень легко могли в это время
обменяться некоторыми компо-
нентами своей инфузорной
фауны и таким образом иметь
ряд обших видов Ophryoscole-
cidae. Прошли тысячелетия: из-
менение климатических уело
вин и истребительная деятель-
ность человека отодвинули оле-
ня далеко на север, а сайгу —
на юг, и некоторые общие им
инфузории оказались теперь
встречающимися в отдаленных
одна от другой областях.
Аналогичный случай опи
сали Полянский и Соловьева
I1959) в отношении паразитических инфузорий горала (Nemorhaedus саи-
datus). ,У этого дальневосточного горного копытного, ареал которого
тянется от Амура до Бирмы, наряду с некоторыми другими видами
инфузорий сем. Ophryoscolecidac найден, как и у сайги. Entodinium
exiguum— типичный вид для северного оленя. В настоящее время
ареалы северного оленя и горала не совпадают, однако в прошлом
ареал горала (Мензбир, 1934; Соколов, 1959) был шире и частично
совпадал с ареалом северного оленя
Эти случаи дают нам, с. одной стороны, примеры прерывистого
распространения паразитов и его объяснение; с другой стороны, опи
еще раз четко иллюстрируют то важное значение, которое приобре-
тают многие паразитологические данные для оценки зоогеографиче-
ского распространения животных-хозяев.
Причины только что отмеченного несоответствия в распростране-
нии паразитов и хозяев чрезвычайно многообразны и отчасти затро
334
нугы в предыдущих главах. Благополучное существование паразита
в ванной местности возможно лишь при сочетании всего комплекса
условий, необходимых для него на разных стадиях его жизненного
цикла. Недостаток хотя бы одного из этих условии при наличии всех
прочих может повлечь за собой отсутствие паразита в данном месте
Иллюстрировать эти отношения лучше’ всего на каком-нибудь
частном примере, в качестве которого мы возьмем парази гофауну
щуки (рнс. 2U3). Эта паразнтофауна, во-первых, зависит от ряда
факторов, находящихся в связи с самим хозяином, и, во-вторых, от
группы факторов, связанных с водоемом в его настоящем и прошлом
Прежде всего в щуке находятся
виды, например Henneguya lobosa.
Myxiduum lieberkilhni, которые
встречаются только в данном виде
рыбы, но по-вцдимому, повсюду, где
бы он ни имелся (от* Мурманска
до Лрала и от Франции до Северной
Америки). Здесь, следовательно, мы
имеем зависимость паразита глав-
ным образом лишь от одного фа к-
гора — его специфичности к дан-
ному хозяину. Такой же характер,
по-видимому, носит и распростране-
ние жаберного сосальщика Tetraon-
chus monenteron.
У других паразитов щуки, на-
пример у глохидиев, наблюдается
зависимость от времени года, т. е.
сезонный паразитизм. Глохидии
встречаются па жабрах щуки лишь
в период размножения ракушек
Unionidae.
Очень велика зависимость пара-
зитофауны от гидробиологии во-
доема. Так, например, ясно, чго
Рис. 203- Схема зависимости паразита
фауны водного животного (например,
щуки) от различных факторов (июбра-
и.енных на схеме в виде секторов)
внешней среды и от факторов, связан-
ных с особенностями хозяина. (Пг>
Догелю.)
глохидиев можно ожидать лишь
гам, где имеются взрослые Unionidae. Или очень часто паразиты
щуки связаны с распространением их промежуточных хозяев, дающих
приют их молодым стадиям развития. Таковы Azygia (промежуточ-
ный хозяин — моллюск), Camallanus (рачки), Proteocephalus (цик-
лопы). В Финском заливе, а также в большинстве озер Ленинградской
области щуку нередко заражает скребень Acanthocephalus lucii, про-
межуточным хозяином которого служит водяной ослик (Asellus
aquaticus). В Арале все высшие раки представлены лишь одним
видом ((jonirnarus at alensis), а потому и скребни в Арале совершенно ог
сутствуют. Для других паразитов щуки с более сложной сменой хозяев
необходимо присутствие в водоеме не только щуки (которая служит
для них вторым промежуточным хозяином), но и первого промежу-
точного хошина и окончательного хозяина. Таков скребень Согупо-
sonia strumosum. для полного цикла которого нужны бокоплав Ponto-
poreia affinis. рыба и тюлень. Вот почему паразит этот не может
находиться в щуках Арала и Волги, но может встретиться в щуках
Каспия и Финского залива. В Арале и Волге пет ни Pontoporcia.
ни тюленя, тогда как в Финском заливе и в Каспии присутствую!
все три необходимых компонента.
335
Однако нас не должно удивлять, если мы тем не менее встретим
Corynosotna strumosum в Волге у сельдей (Caspialosa volgensis).
Хотя в Волге тюленей и нет, но сельди приходят в Волгу из Каспия,
где имеются и тюлени и рыба в качестве второго промежуточного
хозяина (Corynosotna не специфичен по отношению к щуке и может
жить во многих ьидах рыб). Таким образом, важное значение полу-
чает и стационарный или мигрирующий характер жизни хозяина:
если бы щука была проходной рыбой, вроде сельди, то в Волге
можно было бы ожидать присутствия в щуках скребня Corynosorna.
Мейсе заметно, хотя, вероятно, не
Рис 204. Распространение сосаль-
щика Neascus cuticola (Sirigeidae)
в Касипи и па Волге (обозначено
косыми штрихами). (Оригинал.)
Будучи обнаружен во всех местах
Каспия, где пр 'изводились пссле-
довамня (дельта Вол» н. устье Ура-
ла, Кзыл Агач, Га» сан-Кулид на
Волге Neascus не доходит до го-
рода Куйбышева.
менее велико, влияние гидрологии во-
доема на паразитофауну. В качестве
примера укажем, что отсутствие в мор-
ском районе Арала мнксоспоридин
(Myxosporidia) мы приписываем боль
шому количеству сульфатов в араль-
ской воде.
Даже размеры водоема отражаются
на составе паразитофауны, как это
можно было видеть на примере очень
маленьких глухих лесных озер Каре-
лии, или ламб.
Ист сомнения в том, что и геогра-
фическое положение водоема (южное
или северное, горное или низинное),
отражаясь на температуре воды, рас
тительности н т. д., играет роль суще-
ственного фактора: прямого (влияние
на самого паразита) или косвенного
(влияние на хозяина). Этим объяс-
ияется, быть может, незахожденпе не-
которых южных паразитов щуки (Ne
ascus culicola) далеко на север
(рис. 204).
Примером влияния северных ши-
рот на распространение паразитов мо-
жет служить уже упоминавшийся на-
ми жаберный монотеистический со-
сальщик осетровых рыб Mlzschia stu
rionis. Он известей с европейского и се-
вероамериканского побережий Атлан-
тики, имеется в Средиземном и Черном морях и удержался в качествг
морского реликта в Каспии Однако многочисленные вскрытия Acipenser
baeri в низовьях Оби и в Обской губе (Волкова. 1941; Петрушевский.
Мосевич и Шулаков, 1948) дали отрицательный результат, равно как
и работы Бауера (1948) па Енисее, Лене и Амуре. Таким образом.
Nitzschia sturionis представляет собой чисто атлантическую и притом
не заходящую дальше бореальных вод форму.
Далее, при оценке паразитофауны водоема надо учитывать и его
историческое прошлое, его бывшую связь с другими водоема
ми или отсутствие этой связи и т. д. Так, например, Каспийское морс
по своему характеру в настоящее время представляет собой водоем
скорее озерного типа, что сказывается и на его ихтиофауне, и на
паразитофауне. Однако еше в третичную эпоху Каспий входил в со-
став гораздо более крупного морского бассейна. Следы этого мор-
336
скоро прошлого сохранились ь паразнтофауие бычков Каспия, в поч-
ках и мочевом пузыре которых имеются Myxosporidici из рода Сега-
tomyxa, обыкновенно встречающегося только у морских рыб
(рис. 205).
Наконец. в некоторых случаях при оценке зоогеографического
распространения паразита надо принимать во внимание даже повад-
ки хозяина в разных районах
его обитания, например в раз-
ных водоемах. Так, в Невской
губе, наряду со щукой и нали-
мом, крайне частым хозяином
(около 90% заражения) плеро-
церкоидов широкого лентеца
служит срш. Между тем в
Кончозере. у большого числа
'ершей удалось обнаружить
лишь одного плероцеркоида.
В кончозерской группе озер ер-
,ши ловятся на значительной
глубине, в ямах, тогда как щу-
ка и налим — повсюду. Между
Рнс. 205. Некоторые паразитические простейшие- рыб
А — Ceratomyxa caspia — морской слизистый споровик, нс речающпнен
н почках каспийских* бычков; В и С—спора и плазмодий слизистого
споровика Chlorотулит leydigi из желчного пузыря акул и скатов;
D— ооциста Eimeria sardinne (кокцндии из семенников сельдевых рыб;
с четырьмя всретенонндиы чн спорами, внутри которых видно ио два
спор «опта; Е — участок семенника иваси, зараженной множеством ооцист
Е. sardinac. (Л — но Догелю. В н С — по Дофлейну. /? —по Веньону.
Е оригинал.)]
тем в Невской губе с ровным и плоским дном все три видя рыб живут
на одном уровне. Весьма вероятно, что наблюдаемые различия в зара-
женности ерша широким лентецом зависят от разности местообитания,
причем в озерах вследствие глубоководного образа жизни ерша уничто-
жается стык между окончательным хозяином лентеца, т. е. человеком, и
его промежуточными хозяевами — ершом и циклопом.
Разобранные главным образом на одном частном примере
(щука) чрезвычайно сложные отношения показывают, насколько
22 В- ' Догеяь
337
осторожным надо быть при оценке географического распространения
нара hi гоп,
В русской литературе распространение паразитов тоже и некото-
рых случаях привлекалось при решении зоогеографических вопросов.
Сивинский (1904) в своем капитальном труде о развитии фауны
Попто-Арало-Каспийского ’ бассейна для доказательства северного
характера некоторых компонентов фауны Каспия привлекает и их
паразитов (а именно Corynosotna— паразита каспийского тюленя)
Догель и Быховский (1938) в своем подробном описании паразитов
рыб Каспийского моря неоднократно затрагивают вопросы проис-
хождения свободно живущих обитателей Каспия, базируясь на дан-
ных их паразитофауны. Так, они на основании паразитологических
данных подтверждают наличие в Каспии двух самостоятельных стад
осетровых рыб — северного и южного. Далее, учитывая распростране
ние как самих паразитов, так и их хозяев, вышеназванные авторы
делят паразитов рыб Каспия на несколько групп разного происхожде-
ния: космополиты, северные формы, южные и юго-восточные. Кроме
того, ими указываются морские (сосальщики Nilzschid и Mazocraes
споровик Ceratomyxa. скребень Corynosotna) и пресноводные элементы
в паразитофауне рыб Каспия; последние составляют громадное боль-
шинство.
Попытки дифференцировки отдельных элементов паразнтофауны
рыб Черного моря делаются и Османовым (1940), хотя паразиты рыб
в данном водоеме еще недостаточно исследованы для окончательных
выводов. Интересные наблюдения сделаны Османовым над акуловы-
ми рыбами Черного моря, паразитофауна которых по сравнению
с теми же рыбами Средиземного моря значительно беднее. У акуло-
вых рыб Черного моря отсутствуют в желчном пузыре характерные
и широко распространенные мнксоспоридин акул и скатов, Chloro-
tnyxum leydigi (рис. 205) и совершенно отсутствуют моногенетиче-
скце и днгенетическпе сосальщики. Вообще, паразитофауна многих
средиземноморских рыб в Черном море окажется, по-внднмому,
потерявшей часть своих компонентов
Любопытно, что зоогеографический характер Черного моря опре
деляется в значительной мере особенностями его гидрохимии. Это сле-
дует из того, что выпадение средиземноморских паразитов падает на
долю форм, не имеющих в своем цикле промежуточных хозяев
(Myxosporidia, iMonogenoidea), т. е. зависящих лишь от характера окру-
жающей среды. Догель исследовал дополнительно в Керчи около ста
экземпляров черноморских рыб (Belone, Umbrina, Mullus и др.) и под-
твердил чрезвычайную бедность их миксоспорндиями. Догель же
(1940, 19486) делает попытку объяснить некоторые особенности рас-
пространения Eimeria в семенниках сельдевых рыб, прибегая к мето
ду Неринга.
В семенниках большинства Clupeidae встречаются очень свое-
образные представители рода Eimeria (Е. sardinae) с очень крупными
ооцистами и очень длинными веретеновидными спорами. Эта или блнз^
кие ей формы Eimeria известны нз атлантических и средиземномор-
ских сельдевых (Clupea harengus membras, сардинка и др ), а Фужита
(Fujita, 1934) описал их (Eimeria nishin) из дальневосточной сельди.
Кроме того, Догель нашел их на Дальнем Востоке у иваси (Sard<-
nella melanostica) и Etrumeus micropus, а в Белом море — у Clupea
h'arengus tnembras На Каспии Eimeria sardinae отсутствует у всех
сельдевых, т. е. у Caspialosa и Cltipeonella.
Очень интересные результаты дает Черное море. У Caspialosa
338
и Clupeonclla кокцндии здесь тоже не были найдены, ио у 25% сам-
цов Engraulis encrassicholus Догель нашел типичных Eimeria sardinae
На этом основании Догель предполагает, чго Eimeria или отсутство-
вали, или вымерли у Clupeidae Понто-Арало-Каспийскот о бассейна
_io его окончательного соединения со Средиземным морем. Когда это
соединение совершилось, в Черное море проник нз Средиземного
Engraulis, который и «импортировал» с собой Eimeria sardinae.
В дальнейшем можно ожидать перехода Е. sardinae с хамсы
(Engraulis) на других сельдевых Черного моря. Вероятно, в прежние
эпохи Eimeria имелась у понто-каспийских эндемиков (Caspialosa
Ciupeonella), но вымерла под влиянием временного сильного опресне-
ния черноморского бассейна.
Применяя паразитологический анализ, Быховский и Полянский
(1953) пытались подойти к решению вопроса о соотношении некото-
рых элементов ихтиофауны сем рной Атлантики и северной Пани
фнки. Они изучали моногенетических сосальщиков сем. Gyrodactylida»
на разных рыбах Баренцева моря и северной части Тихого океана.
Эти черви характеризуются весьма узкой специфичностью и отдельные
виды их встречаются лишь на одном или на немногих близких видах
рыб. Изучение фауны Gy rodactylidae показало чрезвычайную бли-
зость их в северной Атлантике и Паиифике. У атлантической трески
(Gadus morhua morhua) в Баренцевом море, у тихоокеанской трески
(G. morhua macrocephaltis), а также у минтая [Theragra chalcogrammu
chalcogramma (Pall)] на жабрах живет одни и тот же вид червей-
Gyrodactylus marinus. В настоящее время известные ареалы баренцевс
морской трески и дальневосточных тресковых не только не сопри-
касаются, ио разорваны на всем протяжении морей северного
побережья Азии. Сходство моногенетических сосальщиков Баренцева
моря с тихоокеанскими видами не ограничивается паразитами треско-
вых, но распространяется п на ряд других видов рыб и их моногене-
тических сосальщиков.
На беломорской наваге (Eleginus navaga Pall.) и на дальневосточ
нон наваге — вахне (Eleginus gracilis) живет один и тот же вид
Gyrodactylus gerdi (иа вахне он образует особый подвид—G gerd>
crientalis). Эга общность фауны моногенетических сосальщиков
затрагивает и ряд других видов рыб и их паразитов — моногенетнчс-
ских сосальщиков, перечислять которых мы здесь нс будем. О чем
говорит эта общность моногенетических сосальщиков северной Атлан
тики и Северной Пацнфнки? Очевидно, о широких связях, которые
относительно недавно существовали между этими акваториями.
Этот вывод, который диктуется паразитологическим материалом,
хорошо согласуется как с ихтиологическими, гак и с геологическими
данными. Интересно отметить, что внимательный анализ Gyrodacty
lidae с близких видов рыб Атлантики и Пацнфнки позволил Быхов
скому и Полянскому высказать предположение о том, в каком именно
направлении происходили перемещения тех или иных тресковых рыб
в период потепления вдоль северного побережья Евразии — с запада
на восток или с востока на запад. Не вдаваясь в детали этого апа
лиза. укажем, например, что треска, вероятно, проникла на восток
с запада. Об этом говорит тог факт, что в Баренцевом .море паразити-
рующий на ней Gyrodactylus mariiiys на других рыбах (на пикше —
Melanogrammus aeglefinus) дает специальные подвиды, тогда как на
востоке его внутривидовая изменчивость значительно меньше. Это
говорит об относительно недавнем нахождении его иа востоке и более
продолжительной истории вида на западе.
22*
339
Обратные отношения имеем у наваги, которая, как указывают
паразитологические факты (согласующиеся со взглядами некоторых
ихтиологов на рассмотрении которых мы уже не будем останавливать-
ся, отсылая читателя к специальной литературе), имеет дальневосгоч
иое происхождение и проникла в отлитие от трески в Белое и Барен
иево моря с Востока.
Мы привели этот пример с Gyrodactylidae для того, чтобы показать,
что паразитологический материал даст возможность осуществить до-
вольно детальный анализ далеки нс простых зоогеографических и фило-
генетических вопросов.
Очень широкие выводы зоогеографическою и филогенетического
характера в отношении тресковых рыб и камбал (Gadidae и Hetcros-
lomafa) делает в недавно вышедшей большой работе Шидат (Szidat.
I960). Изучая паразптофауну указанных групп рыб у побережья
Аргентины (Южная Атлантика), он устанавливает среди встречаю-
щихся здесь Gadidae два разных комплекса паразитов. Один из них
характерен для Merlticcius hubbsi, Macruronus magellatilcus и некото-
рых других видов, второй — для Urophycis brasiliensis. Первая группа
обладает комплексом паразитов, которых Шидат считает происходя-
щими из южной Пацпфики, в то время как Urophycis является
носителем североатлантической паразитарной фауны. Шидат делает
отсюда вывод о том, что Gadidae, живущие в настоящее время
у побережья Аргентины, двоякого происхождения: одни из них
пришли нз северной Атлантики, другие—из южной Пацпфики.
Паразитофа} на' последних слагается нз видов более примитивных,
чем у атлантических. На этом основании Шигат считает, что одним нз
центров происхождения Gadidae является южная часть Тихого океана.
Эта гипотеза противоречит взглядам многих ихтиологов, кажется
еще спорной и мы не будем обсуждать ее здесь в деталях. Во
всяком случае, следует подчеркнуть, что работа Шндата представляет
большой интерес как пример широкого использования паразитологи-
ческих данных для решения зоогеографических и филогенетических
проблем
ГЛАВА XVII
ЗАВИСИМОСТЬ ОДНИХ ЧЛЕНОВ ПАРАЗИТОФАУНЫ
ОТ НАЛИЧИЯ ДРУГИХ
Это явление, па которое раньше мало обращали внимание, весьма
вероятно, играет известную роль в формировании паразитофауны
животного. В одном хозяине часто обитает помногу разных парази-
тов, и в особенности богатой и разнообразной оказывается иаразито-
фауна кишечника. Наряду с животными-паразитами в кишечнике
обитаю! cine бактерии, грибки и другие микроорганизмы. Совокуп-
ность всего паразитарного населения организма хозяина, включая не
только животных-паразитов, но и микроорганизмы, Павловским
(1937) была обозначена термином паразитоценоз. При этом в даль-
нейших его исследованиях (Павловский 1948а, 19556) было отмечено,
что между отдельными членами паразнтоценоза, относящимися
к одному или разным видам, существуют определенные взаимо-
отношения, изучение которых имеет немаловажное значение для пони-
мания структуры паразнтоцеиоза в целом и некоторых сторон взапмо
действия паразита и хозяина Павловский и Гнездилов (1953)
обратили внимание па то, что качественный и количественный состав
популяции паразитов различных хозяев определяется не только эколо-
гическими факторами и особенностями состояния самого микроорга-
низма, как среды обитания, но и характером взаимоотноше-
ний между отдельными видами, входящими в состав
п а р а з и т о и е н о з а.
В одних случаях эти взаимоотношения выражаются в антагони-
стических отношениях, когда при наличии особей одного вида исклю-
чается возможность существования другого или других видов. В иных
случаях отношения могут носить синергетический характер, когда
одни паразиты способствуют существованию других паразитов.
В настоящий момент мы еще далеко не всегда можем точно
выявить характер этих отношений в пределах того или иного пара-
знтоиенсза. Одним из путей, позволяющих подойти к выяснению таких
взаимоотношений, является, по мнению Павловского и Гнездилова
(1953), «наблюдение и анализ статистических данных по сочетанным
заражениям особен хозяина различными видами паразитов». Другой
путь, непосредственно связанный с первым. это экспериментальным
3-1)
анализ предполагаемых синергетических или а и 1 а гоп нети
ч е с к и х отнош они й между паразитами.
Любопытный факт наблюдал Вильсон, (Wilson, 1916) при изучении
зараженности рыб глохидиями и веслоногими рачками. Рыбы,
зараженные глохидиями, были обычно свободны от рачков и. обратно,
рыбы, зараженные рачками, были лишены глохидиев. Возможно, что
здесь имеет место антагонизм паразитов. Для паразитов рыб отме-
чены и другие аналогичные примеры. Так, ШеЯерипг iScheuring.
1923) наблюдал, что при сильном заражении щук Triaenophorus coda-
losus в кишечнике обычно отсутствуют скребни. По наблюдениям
Петрушевского (1940, 1955) во многих водоемах у 96,3% зараженных
щук Muxidium. Heber kiihni в мочевом пузыре встречались всегда раз-
дельно от сосальщиков Phyllodistomum folium. Только у 3,7% рыб
наблюдалось совместное заражение этими паразитами. Существует
предположение, подтвержденное экспериментально что сосальщик
питается слизью со стенок мочевого пузыря и вместе с ней заглаты-
вает плазмодии и споры Myxidium Heberkiihni (Бауер. 1959). В кишеч-
нике ряпушки из Северной Америки, по данным Кросса (Cross, 1933),
при наличии большого числа (42,09 экземпляра на рыбу) ленточных
червей Proteocephalus exiguus. скребней Neoechinorhynchus sp. в этих
же рыбах было очень мало (1.09 экземпляра). С другой стороны, при
большом количестве скребней (27 экземпляров) число ленточных
червей было равно всего в среднем 1,33 экземпляра. В сибирской
ряпушке (Coregonus sardinella) ленточные черви и скребни всегда
встречались в разных экземплярах рыб (Петрушевский, 1955).
Интересный пример антагонизма между близкими формами
обнаружила Банина (1952) у Opalina из кишечника лягушек. На
очень большом материале она показала, что в тех случаях, когда
какой-нибудь вид лягушек бывает в известной местности заражен
двумя разными родами и даже видами опалин, то никогда при этом
не наблюдается смешанных инвазии. Каждая 1араженная лягушка
содержит всегда лишь один из воз ьолчных двгх видов опалин.
Для паразитов лягушки Маэурмоьич (1957) отмечают антагонисти-
ческие отношения между легочными трематодами (Haematoloei has)
и нематодами (Rhabdias bufonis). Совместное заражение этими пара
знтами не было обнаружено. Для кишечных паразитов он отмечает,
что скребни Acanthocephalus гапае очень редко встречались изолиро-
ванно от трематод.
С другой стороны, исследования Маркова (1955) и наши данные
Un litteiis) показали, что в зегких Rana temporaria сосальщик Haplu
rnetra cylindracea и Rh. bufonis встречаются совместно достаточно
часто (от 20 до 60% случаев в разной местности) и антагонизм
между ними не наблюдается. Однако было отмечено некоторое сни-
жение интенсивности заражения грома годами при наличии большого
количества нематод в легких.
В отношении эктопаразитов наземных животных Уотерсов
(Waterston, 1914) отмечает, что пухоеды редко встречаются на птицах,
сильно зараженных перьевыми клещами (Analgina).
Труассар (Trouessart, 1885) уже давно наблюдал, что при поселе-
нии клещей рода Syringnphil’is (сем. Cheyletidae) в очинах перьев
птиц, зараженных перьевыми клещами нз рода Syringobia, последние
полностью уничтожаются первыми. Дачные эти подтверждаются
Дубининым (1951) на паразитах кулька Totanus caltdris. Если кулики
заражаются клещом Syringophilus bipeciinatus. то эти клещи
. ничтежают Syringobia и поселяются в очинах всех перьев,
312
потребляя в пищу чешуйки эпидермиса сосочка „ера. Последующее
сражение таких куликов Syringobia становится невозможным, так
как они тотчас же поедаются Syringophilus.
Ряд примеров совместной или раздельной встречаемости парази-
тов можно привести в отношении эндопаразитических видов, встре-
чающихся у млекопитающих и человека.
Гнездилов (1951) проанализировал зараженность человека в раз-
ных районах СССР гельминтами и Lamblia и пришел к заключению,
«то, как правило, средн людей, зараженных анкилостомой, ин в одном
с.ь iae ни было найдено лямблий. Случаи совместной встречаемости
этих паразитов наблюдались только при крайне слабой интенсивности
«аражеипя анкилостомой. В одной нз обследованных групп населения
отмечалась более редкая встречаемость Lamblia (6,1%) у лиц.
аражопных аскаридами (2170 человек), нежели встречаемость их
(10,7%) у лиц, не зараженных аскаридами (5570 человек). Иная
картина наблюдалась при сопоставлении зараженности Hyrnenolepis
папа и Lamblia При наличии ленточного червя лямблии встречались
в два раза чаще (25%), чем у пезараженпых этим червем (10%).
Гнездилов высказывает мигнис, что круглые черви действуют на
лямблий отрицательно, а ленточные благоприятствуют их жизни.
Павловский и Гнездилов (1953) отметили, что у собак, заражен
дых Toxocara canis, значительно меньший процент плероцеркоидов
Diphyllobothrium latum приживался при экспериментальном зараже-
нии, чем у собак, свободных от аскариды. Кроме того, наблюдя юсь, что
Toxocara canis задерживает рост и развитие лентеца широкого. Так же
влияет и Taenia nydaligena. При наличии этого паразита только
‘0 60% плероцеркоидов приживалось в кишечнике собаки п давало
гробилу. а без Т. hydatigeiiu приживалось 77—80% плероцеркоидов.
В некоторых случаях замечается определенная связь между рас-
пространением Lamblia и различной патогенной флорой кишечника.
У детей, больных дизентерией Кр* зо-Зоние, зараженность лямблией
оказалась равной 8,4%, т. е. в 5 раз ниже, чем у обследованных здоро-
вы' детей (40,3%) той же местности Такая же низкая зараженность
была обнаружена у гетей, больных паратифом. С другой стороны,
дети, больные дизентерией Флекснера, оказались зараженными лямб-
лией иа 77,3% (Полянская, 1954). Это явление не имеет, вероятно,
общего значения, так как исследования Чалой и Тарасовой (1958) не
выявили какой-либо зависимости паразитирования Lamblia от пали*
чня того или иного возбудителя дизентерии.
Особый интерес представляют данные Карапетяна (1960) о синер-
гетических отношениях Lamblia с дрожжеподобными организмами
Candida guilliermondi, всегда встречающимися совместно с Lamblia.
Кроме того, культирование Lamblia оказалось возможным только при
наличии Candida, а применение антибиотика нистатина, действующею
на Candida, ускоряло излечение и от лямблиоза. Механизм этих
взаимоотношений остается пока еще не ясным.
Весьма интересны взаимоотношения между Entamoeba histolytica
и кишечными бактериями. Многие исследователи склоняются к мысли,
чго основным^ фак гопом, определяющим развитие патогенного процес-
са при амебиазе, является сопутствующая бактериальная флора
кишечника (Stewart a. Jones, 1948; Weslfal, 1942; Phillips et а!.. 1955,
и др.). Escherichia coli и бактерии группы paracoli усиливали пораже-
ние кишечника крыс, зараженных Entamoeba histolytica, тогда как
сами по себе они не вызывали никаких поражений. С другой сто-
роны, заражение крыс культурой Е. histolytica и культсроп Strepto-
313
coccus faecalis ослабляло патологический процесс (Slewart, Jones,
Rogers, Г>18). Вероятно, это связано с тем, что продуктом метаболиз-
ма Str. faecalis является молочная кислота, которая в малых кон-
центрациях неблагоприятна для амеб и вызывает их гибель.
Эксперименты Филлипса с сотр. (Phillips et al., 1955) особенно
. бедителыю показали значение сопутствующей бактериальной флоры в
развитии острого амебиаза у экспериментальных животных. Штамм
амеб, культивируемый без бактерии (с Schizotrypanum cruzi), вызывая
у обычных морских ейинок (имеющих бактериальную флору в кишеч-
нике) типичные поражения кишечника. В то же время у стерильно
выращенных морских свинок тот же штамм не вызвал никаких пора
женин. и амебы находились только в просвете кишечника. Если же
стерильным морским свинкам перед заражением амебами вводилась
через рот культура бактерий Escherichia coll или Aerobacter аегопе-
nes, то у них развивались типичные амебные поражения. Косвенным
подтверждением рели бактериального фактора в развитии амебной
дизентерии является факт усиления положительного эффекта действия
амебопидных препаратов при добавлении некоторых антибиотиков
(пенициллин, грамицидин). Они подавляют сопутствующую бакте-
риальную (|).iop> и изменяют среду обитания Entamoeba histolytica
в неблагоприятную для нее сторону.
Этими примерами не исчерпываются все случаи взаимоотношений
между разными видами в паразитоцеиозс, и можно думать, чго даль-
нейшие. главным образом экспериментальные исследования в этом
направлении принесут новые сведения о влиянии одних сочленов
паразнтоцеиоза иа др'гих.
ГЛАВА XVIII
ПАРАЗИТЫ КАК КОМПОНЕНТЫ БИОЦЕНОЗА
Выше мы рассмотрели различные условия, которые определяют
состав паразитофауны отдельных видов животных, и главное внимание
обратили на факторы внешней среды, влияющие па формирование
системы паразит — хозяин. Кроме того, были рассмотрены различные
типы циклов развития паразитов и выяснены некоторые стороны
взаимоотношений между отдельными членами паразитоценозов и ря-1
;ругпх вопросов Однако мы почти не капа тись общих закономерностей
существования паразитов в каком-либо биоценозе и циркуляции их
средн членов биоценоза, а также факторов, которые в одних случаях
способствуют*, в других— Препятствуют осуществлению кругбворода
паразитов. Изучение этих вопросов имеет немаловажное значение для
понимания причин формирования определенного комплекса паразито-
фауны, свойственного какому-либо хозяину, входящему в состав опре-
деленного биоценоза. Как отмечает Павловский (1937, 1948а), паразиты,
входящие в состав паразнтоцеиоза хозяина, так же как и сам хозяин,
являются частями биоценоза того или иного биотопа. Это касается не
только тех паразитов, которые проводят часть своей жизни вне хозяина
(периодический паразитизм), но и постоянных паразитов, потерявших
связь с внешней средой. Они связаны с пси через хозяина, будучи его
обитателями, и циркуляция их осуществляется при пищевых связях
биоценоза.
В природных условиях с\шествование паразитов в биоценозе обес-
печивается многими и весьма разнообразными )словиями: наличием
подходящих окончательных и промежуточных хозяев, наличием между
ними биоцеиотичсских связей (главным образом пищевых), делающих
возможным проникновение паразита в хозяина, наличием факторов
внешней среды, действующих прямо на фазы развития паразита, нахо-
дящиеся вис хозяина, или косвенно на паразита через хозяина (Павлов-
ский, 194Са).
В каждом конкретном биоценозе возможности циркуляции пара-
зитов мог^т оказаться и обычно оказываются, но одинаковыми в смяли
с тем, что видовой состав его будет различен, так же как различны бу-
дут и биоценотпческие связи между отдельными его членами.
г •.
315
I. Экологические основы распространения трансмиссивных
ЗХБОЛЕВАНИЙ ЧЕЛОВЕКА И ЖИВОТНЫХ И УЧЕНИЕ ОЬ ИХ ПРИРОДНОЙ
ОЧАГОВОСТИ
Для изучения сложных процессов циркуляции паразитов н биоце-
нозу Павловским (1935, 1948а,б, 1955а) и его ученицами в СССР были
предприняты широкие комплексные эколого-паразитологические иссле-
дования различных переносчиков и возбудителей многих заболеваний
человека и домашних животных. Эти исследования позволили Павлов-
скому разработать учение о природной очаговости трансмиссивных за-
болеваний. Такие исследования проводились над различными клеща-
ми— переносчиками пироплазмозов и тейлериозов, клещевого энцефа-
лита— н над другими объектами.
Цля примера опишем одну из схем комплексного экологического
исследования клещей Ixodes ricinus — переносчиков бабезиеллоза
крупного рогатого скота на северо-западе СССР, осуществленного с
целью практической разработки мер борьбы с этим заболеванием. Эги
исследования проводились в 1923—1927 гг. Первым условием в иссле-
довании была констатация вида клеща-переносчика в составе фауны
биоценоза и ; становление действительной возможности передачи им
возбудителя скоту в природных условиях используемой территории.
Второй вопрос — это выяснение количества и колебаний численное!i
переносчика на данной территории. Третьим вопросом являлось нэучг-
пие биоцрнотнческих соотношений между клещами п их хозяевами в
разные сезоны года. Чс-твсргый вопрос — выяснение круга хозяев, кото-
рые служат источником прокормления различных фаз метаморфоза
клеща и установления ведущих сочленов биоценоза, обеспечивающих
циркуляцию всех фаз развития клеща на данной территории. Изучение
биоценотииескнх отношений клешей с разнообразными хозяевами и
условиями внешней срё^ы, позволило поставить вопрос о рациональных
клешеистребитсльных мероприятиях с целью выведения /. ricinus из
состава биоценоза в данной местности.
Сходным образом проводились исследования других переносчиков
в других биоценозах с учетом конкретных особенностей местности и спе-
цифики биологии переносчиков, а также возбудителей заболеваний
(11авловскнн, 1948а).
Рассмотрим несколько более подробно роль переносчиков в различ-
ных биоценозах при формировании системы паразит — хозяин.
У многих эктопаразитов и в особенности у временных, фаза сво-
бодной жизни из-за своей длительности нередко имеет первенствуют» е
шачение в цикле их развития. Такне эктопаразиты, кратковременно
связанные с организмом хозяина, являются членами различных биоце-
нозов, в которые могут включаться в качестве одного из сочлеш в также
и их потенциальные хозяева. Между тем. будучП кровососами позво-
ночных животных, эти организмы (клещи, комары, мошки, мокрецы,
слепни, мухи и пр.) переносят от одного животного к другому возб'
дителей вирхеных, бактериальных, риккетсиозных, (Жпрохетозных, про-
гозонных и других заболеваний, играя тем самым весьма существенную
роль в циркуляции соответствующих возбуди гелей в природе. Переход
возбудителей заболевания от одного хозяина к другому через посредство
переносчиков обеспечивает существование их видовой жизни.
Такие заболевания, которые передаются с помощью переносчики,
называются Трансмиссивными. Примером могут служить малярия чело
века, распространяемая комарами рода Anopheles, или пироптазмозы
домашних животных, переносимые иксодовымл клещами и многие
310
другие. К настоящему времени известно около 50 трансмиссивных забо-
леваний человека, распространение которых происходит только через
переносчиков (облигатно трансмиссивные болезни), и несколько забо-
леваний (например, чума, туляремия, сибирская язва), заражение кото-
рыми через переносчика является лишь одним из путей их распростра-
нения (факультативно трансмиссивные болезни). Роль кровососущих
членистоногих в распространении этих заболеваний неодинакова. Наи-
более многочисленными и всесторонними связями с возбудителями за-
болеваний обладают иксодовые клеши и комары, являющиеся перенос-
чиками многих болезней (табл 20). Другие кровососы специализи-
руются на передаче лишь отдельных групп возбудителей.
Таблица 20
Установление связей между систематическими группами переносчиков и
возбудителей трансмиссивных заболеваний позвоночных животных и человека
(по Беклемишеву, 1949 Г., с изменениями)
Возбуди•ели
Ixodidae................
Argasidae...............
Trombidiidac............
Gatnasoidca ......
Culicidae...............
Simuliidac...............
Helcidae................
Phlebotornus............
Tabanidae ..............
Glossina................
Reduvitdae..............
Pediculus...............
Aphanipiera
Передача возбудителен от животных к животным или к человеку
требует для своего осуществления ряд биотических и абиотических
условий. Для биотических условий, определяющих циркуляцию
возбудителя в природе, характерно наличие в данном биотопе в составе
определенного биоценоза следующих основных компонентов: 1) возбу-
дителя данного заболевания, 2) животных (или человека) —носителей
возбудителя (донор), 3) переносчика, черпающего от них инвазивное
начало, и 4) животных (или человека), восприимчивых к данному забо-
леванию (реципиент). Последний в свою очередь может стать донором
для переносчиков и т. д. (Павловский, 1948а). Следовательно, поддер-
жание видовой жизни возбудителя болезни любой природы связано с
его циркуляцией среди сочленов биоценоза.
Однако процесс циркуляции не изолирован от прямого пли косвен-
ного влияния факторов внешней среды, которые то благопрп
ятствуют осуществлению этого процесса, то, наоборот, в корне его пре
секают. В первую очередь это влияние сказывается на переносчиках',
как пойкилотермных организмах. Возбудитель в переносчике весьма
чувствителен к колебаниям температуры окружающей среды. При ее
повышении до определенного предела возбудитель в переносчике может
быстро закончить свое развитие, образовав инвазионную стадию; с дру-
ЗГ
гой стороны, при низкой температуре развитие возбудителя может за-
держаться па длительный срок и он просто погибает в переносчике, не
достигнув инвазионного состояния.
Хорошо известно, что Plasmodium uiuax в Anopheles maculipennis
может развиваться только при температуре нс ниже 16°. При 16—17
развитие до образования спорозоитов протекает 45 дней, при 25° -
10 дней, а при 30°—6,5 дней (Николаев, 1935). При повышении темпе-
ратуры завершения спорогонии в более короткий срок нс происходит.
Установлено, чго при температуре 32° цисты в желудке комара обра-
зуются, по спорозоиты при этом в его слюнных железах пе появляются
(Stratrnan-Thomas, 1940); при 37° плазмодии в желудке комара быстро
погибают.
У комаров An. quudrimaculatus, содержащихся при температуре
15° в течение 17 дней, не образовывались ни цисты, ни спорозоиты
Plasmodium vivax, даже после того как эти комары были переведены
на два месяца в условия оптимальной температуры (22°). Следователь-
но, распространение малярии возможно только в той местности, где
в летний сезон температура превышает 15—16°, в течение срока, необ-
ходимого для созревания в комаре спорозоитов. В местности, где низ-
кая температура препятствует развитию паразита в комаре, несмотря
даже на присутствие людей-гаметоноентелей и восприимчивых контин-
гентов, распространение заболевания не произойдет.
Появление заболеваний японским энцефалитом на Дальнем Во-
стоке во вторую половину лета и в начале осени связано отчасти с тем,
что низкие температуры со второй половины апреля до конца июня
в Приморье препятствуют реактивации вируса японского энцефалита
в переносчиках Culex pipiens и С. tritaeniorhynchus после зимовки.
В июле и августе вирус в комаре активируется и в конце августа — на-
чале сентября комары становятся способными заражать человека
(Павловский, 19176). Комары Aedes acgypli при температуре 37° пере-
дают вирус желтой лихорадки через I дня после заражения, а при 18*
остаются безвредными до 30 дней. При более низкой температуре ко-
мары не способны передавать вирус, но сам вирус при этом не разру-
шается, и его передача комаром восстанавливается, как только прои-
зойдет достаточное повышение температуры (Павловский, 19476).
Важным моментом для осуществления циркуляции возбудителя
заболевания среди людей или животных является возможность
контактирования переносчика с хозяином — носителем
соответствующего инвазионного начала и с восприимчивым организмом
Контактирование переносчика с человеком или с животными осу-
ществляется как в условиях естественного ландшафта в открытых био-
гопах или в природных убежищах (норы, пещеры и т. п.), так и в насе-
ленных пунктах, в условиях помещений, посещаемых или заселенных
человеком или домашними животными. По способам нападения на поз-
воночного хозяина п контактирования с ним можно различить несколь-
ко экологических групп кровососущих членистоногих: активно нападаю-
щие кровососы, подстерегающие в убежищах пли па пастбищах, и
постоянные эктопаразиты (Беклемишев, 1945, 1951) В каждой группе
возможность контактирования с позвоночным хозяином зависит от це-
лого ряда причин, определяемых и контролируемых факторами внеш-
ней среды.
Нападению на хозяина предшествует его отыскание. Активно напа-
дающие и подстерегающие кровососы иногда на большом расстоянии
определяют положение позвоночного хозяина по его запаху благодаря
наличию у них специальных обонятельных хеморецепторпых аиппара-
348
7ов. Это даст возможность отыскал, хозяина, приблизиться к нему и
напаси» на него. Большой тонкостью обоняния отличаются многие иксо-
ювые н аргазовыс клещи. Достаточно, например, человеку попасть
н помещение, населенное- клещами Orntthodorus, как последние привле-
каемые его запахом, начинают выползать из своих укрытий и пере-
двигаться в сторону добычи.
Хорошее обоняние таежного клеща Ixodes persulcatus приводит
к тому что голодные самки и самцы обычно распределяются в лесном
биотопе нс равномерно а в больших количествах скапливаются на лес-
ных тропах. Это происходит в результате того, что на них остается
запах от животных и человека, которые обычно передвигаются но этим
тропам. В одном из участков таежного леса за один и тот же отрезок
времени вдоль трон было поймано 327 клещей, а в стороне от них, в гу-
ще леса, всего 65 клещей (Померанцев и Сердюкова, 1948).
Но непосредственное контактирование голодных клещей с хозяином
в лесу обеспечивается также некоторыми особенностями поведения
самих клешей. При оптимальной температуре н влажности голодные
клещи lx. persulcatus проявляют активность, выражающуюся в том,
что они поднимаются с поверхности почвы па вершины листьев травя-
нистых растений, примерно на высоту 20 50 см и принимают харак-
терную позу «ожидания». Тремя парами ног клещ удерживается за
растение, а передняя пара, снабженная хеморецепторным апппаратом
Галлера, выставляется вперед н вверх и с.т жнт, с одной стороны, как
бы сигнальным аппаратом, определяющим приближение хозяина, с дру-
гой— как бы «якорем», цепляющимся своими когогкамн за проходящих
кивотных или человека. Легкое задевание сидящего в такой позе кле-
ща, например рукой или тканью, ведет за собой быстрое его прицепле-
ние. Or характера одежды во многом зависит возможность попадания
клещей на человека. Особенно легко клеши прицепляются к ворсистой
ткани. Совершенно не прицепляется к резиновым сапогам.
Соответствующее поведение клеща, облегчающее его попадание на
хозяина, зависит от физиологического состояния самого
ьлыца и определяется в значительной степени внешними факторами.
Появившиеся после линьки самцы н самки lx. persulcatus во второй
половине лета длительное вр-хы остаются насснвн.мп. Это значит, что
при сохранении подвижности они не проявляют чувства голода, не обла-
дают положительным хемотаксисом, т. е. не привлекаются запахом
хозяина (видимо вследствие нефхнкцпонирования органа Галлера), и
не взбираются на растения для ожидания хозяина. В таком физиологи-
ческом состоянии они не могут в естественных условиях контактировать
с хозяином. Активность приобретается только после некоторого периода
«физиологического доразвития» (Сердюкова, 1952; Балашов, 19571
В начале этого периода (в течение 5—10 дней) происходит затверде-
вание наружных покровов клеща и идет процесс переваривания нише-
вых включений, оставшихся от питания в нимфатыюй фазе, сопровож
дающийся частой дефекацией буро черных кристаллов гематина, кото
рыс смешаны с выделениями мальпигиевых сосудов. Одноврсменн
происходит дифференцировка клеток кишечника (обособление клеточ-
ного эпителия) и слюнных желез. Когда клещ готов к нападению,
в просвете его кишечника остаются одиночные клетки с пищевыми
включениями. Основная их масса разрушается (Балашов, 1957)
Ixodes persulcatus, так же как п lx. nciruis. на cent ре своего ареала
приобретают активность обычно посте зимовки, в теплое время года.
В это время их поведение и, соответственно, возможность нападения
на хозяина находя гея в зависимости от колебания температуры воздуха.
349
относительной влажности и количества выпавших осадков. В лесах
Карелин lx. persulcatus делается активным, т. е. вползает на вершины
растений при температуре от 5 до 20г и относительной влажности oi
99 до 1О7о. Температура выше 20—25° особенно при низкой влажности
Iоколо 40%) действует неблагоприятно на клещей, они уходят на по-
верхность почвы в подститку и тем самым их контакт сохозяином
затрудняется (Херин, 1953). Во время сильных дождей активность
клещей также резко снижается, и нападение на человека в такую по-
году, как правило, не происходит (Померанцев и Сердюкова, 1948).
Взрослые клещи Dennacentor pictu-s и D. inarginatus нападают
на животных только ранней весной, когда температура воздуха еще
низка, а летом при высоких температурах они полностью утрачивают
свою активность. Будучи выпущенными па траву, клещи тотчас направ-
ляются к земле и, достигнув ее поверхности, впадают в неподвижное
состояние (Алфеев. 1939. 1951).
Комары н другие летающие кровососы привлекаются запахом чело-
века и животных, но, кроме того, в их поисковом лете особенно па
близких расстояниях немалую роль играет зрение. Интенсивность напа-
дения летающих кровососов зависит от сочетания действия различных
факторов внешней среды н физиологического состояния переносчика
Подробный анализ факторов внешней среды, влияющих на актив-
ность нападения различных видов комаров родов Aedes, Т heobaidia и
Anopheles, произвел Мончадский (1946, 1949, 1950 1956) в различных
географических зонах Советского Союза: на Дальнем Востоке, в юж
нем Таджикистане, в дельте Волги и в районах Субарктики. Как пока
зали эти исследования, некоторые из факторов, в частности относитель
пая влажность воздуха, не влияют на изменение численности нападаю-
щих комаров Слабый или средней силы ветер снижает активность
щ падения, а ветер в 2,5—3 м/сек. практически прекращает нападение
комаров.
Суточный ритм нападения комаров, имеющий существенное при
способительное значение для жизни отдельной особи и для вида в це-
лом, определяется в основном взаимодействием факторов
света и температуры, имеющим строго закономерную суточную
периодичность.
Наиболее мощное влияние на активность и нападение комаров
оказывает свет, его интенсивность и в особенности быстрые изменения
освещенности, связанные прежде всего с периодами захода и восхода
солнца. Так как изменения силы света очень быстро проникают во все
микростацип любого наземного биотопа, реакция на эти изменения
наступает одновреме нно у всех физиологически подготовле тых к ней
особей комаров данной популяции. Наступающее при заходе солнца
быстрое уменьшение освещенности, переходящее в сумерки, оказывает
резко стимулирующее воздействие на сидящих спокойно в течение дня
в местах природных дневок голодных самок комаров. Вечерний макси-
мум нападения комаров обычно приурочен к поздним сумеркам.
Массовое нападение комаров разных видов может наступать при
неодинаковой силе освещенности. Так, например. Anopheles tnaculipennis
rnesseae в дельте Волги в открытых стациях нападет особенно активно
при освещении от 1 до 100 лк. При более сильном освещении наступает
резкое снижение нападения, а при освещении свыше 9000 лк нападение
прекращается. У Andes vexans лёт и нападение прекращаются пол-
ностью при освещенности свыше 20 тыс. лк, освещенность в 10 тыс. лк
еще неблагоприятна для нападения, при такой силе света ксмары мо-
гут нзпадать лишь под влиянием непосредственного раздражающего
350
воздействия близости человека пли животных. При более низкой ост
[ценности активность нападения усиливается и нгквыешая численность
комаров наблюдается при сумеречной освещай пости (около I лк).
Наступление полной темноты определяет коней вечернего макси
.чума нападения. Ночной минимум длится в течение всего периода пол-
ной темноты. Первые проблески рассвета вызывают наступление утрен-
него максимума нападения, который обычно приходится па начало рас-
света. По мере усиления освещения постепенно прекращают активный
лет сначала менее (Anopheles), а потом и более световыноелнвые
(rkdes) виды комаров. Днем активные поиски добычи и нападение
практически отсутствуй'!. Закономерные изменения чнсленпиосгп на
падающих комаров в период ночного мнипм ма могут происходит!,
в соответствии с фазами луны (Мончадский, 1956).
Суточный ритм нападения комаров в том виде, в каком он сло-
жился под влиянием периодические колебаний освещенности, может
претерпевать существенные изменения в зависимости от окружающей
температуры. В типичном виде суточный ритм (вечерний и утрешни
чакешмумы, ночной и дневной минимумы) пров.чяется только в пределах
температур, допускающих нормальную активность комаров Па севере,
например, комары Лё4<?$ нападали в температурных пределах от 3 до
29°. Зона оптимума лежала между 9 и 24°. Если температура прибли-
жается к нижней границе пороговой зоны или заходит за нее, происхо-
дит резкое изменение суточного ритма нападения. Осенью- и весной при
низких утренних температурах утренний максимум на рассвете отсут-
ствует; комары нападают в поздние утренние часы: ион высокой осве
[ценности. При низких температурах сильное освещение в меньшей
степени угнетает активность комаров, чем в жаркий период сезона.
Поэтому при низких температурах ранней весной и поздней осенью на-
падение комаров наблюдается только в наиболее1 теплые дневные часы,
а утрОхМ и вечером происходит снижение численности нападения, cooi-
встственно ходу кривой температуры. В условиях Севера, когда солнце
летом нс заходит за горизонт, суточный ритм нападения комаров це-
ликом регулируется колебаниями температуры, за исключением начала
вечернего максимума в период наиболее низкого стояния солнца. Таким
образом, оптимальная температура является фактором, изменяющим
пинь количественную сторону нападения, но не нарушающим суточный
ритм нападения комаров связанный с изменением освещения. Однако
низкие, угнетающие комаров температуры, вызывают уже качественные
изменения их суточного ритма.
При этом было установлено, что температурные границы актив-
ности нападения комаров разных видов не являются неизменными, а
смещаются в зависимости от изменения температурного режима в дан-
ной местности в данное время. Летом в более жаркий период эти гра-
ницы смещаются в сторону более высоких температур; наоборот, весной
или осенью при более низком температурном режиме они лежат значи-
тельно ниже. По наблюдениям Моцч-адского (1946) в Таджикистане
кпмары Mansonia richiardii летом нападали в пределах температуры
16—3x3°, а максимум их нападения наблюдался при 25- 27°, тогда как
осенью нападение происходило при 14—29° с резко выраженным макси-
мумом между 22- 24°. Не менее значительные смещения активности
наблюдались у Aedes caspius caspius в тех же местах. Летом макси-
мальная активность была при температуре 25—30', а осенью — при
17,5 25,5°. Аналогичное явление наблюдалось у мошек (Sirnuliidac)
п мокрецов (Heleniae), нападающих на добычу в природе: первые
только днем, а вторые — днем и в сумерки.
351
Такая способность к смешению температурных границ активности
несомненно, носит адаптивный характер и дает возможность комарам
н другим кровососам в широки к пределах естественно меняющихся
температурных условий окр\ жающен среды найти объект кровососания
и на нем напитаться. Тем самым обеспечивается получение перенос-
чиком инвазионного начала и его распространение среди людей или
животных, населяющих данную местность.
Нападение па хозяина летающих кровососов определяется, кроме
того н физиологическим состоянием самого кровососа.
Взрослые комары, мокрецы, мошки, Ирскнты в течение первых дней
окрыления еще не активны, и поиски хозяина у инк начинаются лишь
тогда, когда развитие яичников у самок достигнет определенной степени
У wimok Anopheles rnaculipennis, изученных в этом плане весьма нод-
робно (Беклемишев, 1911, 1949), ihicthhkt жажды крови проявляется
с того момента, когда в цитоплазме яйцевых клеток появляются мелкие
желточные зерна. В это время самки становятся зрелыми и вступают
в период активной гонотрофнческой жизни. У An. rnaculipennis rnesseae
период дозревания самки длится при тгмиерат}ре ‘20 около 10 дней,
при 25е срок сокращается до 4 дней.
Вся жизнь зрелой самки комара, так же как и у других кровососов,
изученных в этом плане, распадается на ряд последовательных гонотро-
фпческих циклов, связанных с периодическим принятием крови и
откладкой яиц. Совокупность процессов приема кровн, ее п<реварива-
ния, созревания и откладки яиц составляет такой гонот рофпческнп
цикл. У малярийного комара он складывается из трех периодов:
1) поиски тобычи и нападение, 2J переваривание крови и созреваний
яиц, 3) поиски водоема и откладка яиц. После этого самка снова ока-
зывается голодной, ищущей добычу, т. е. вступает в первый период
следующего гонотрифического цикла. Одного приема крови обычно
достаточно для поиюю развития яичников и откладки одной порции
яиц (гоиотрофнческая гармония). В течение своей жи.нн комар про-
делывает несколько гонотрофнческих циклов, т. е. он несколько раз
может приступать к кровососанию и откладке яиц.
В первый период гонотрофнческого цикла комары приходят в воз-
бужденное состояние, у них появляется положительный фототаксис к
к слабому свету, н с наступлением сумерек они вылетают из убежищ
в поисках добычи. По исследованиям Беклемишева (1944, 1949) и его
учеников, Anopheles rnaculipennis. так же как и ряд других видов
Anopheles, совершают длительный полет от их места выплода по на-
правлению к крупны.»! скоплениям добычи в населенных пунктах, где
они и нападают на человека „лн на животных в помещениях, па улице
в самом писетке или на его окраине. При отыскивании добычи на боль-
шом расстоянии (до 3 км) комары руководствуются преимущественно
обонянием и летят против ветра, приносящего запахи. Вблизи, при
нападении па самую добычу, им помогает не только обоняние, но и
зрение, и особенно большое значение имеет тепловое чувство. Голодный
комар нападает па любой теплый предмет. 1£с.тк поверхность тела
имеет температуру 28е, то комар нс сосет кровь, но он охотно нападает
при температуре поверхности тела не ниже 32°.
Каждое место, где ио вечерам скапливаются люди или животные,
является центром притяжения для голодных самок Anopheles. В необ-
житой местности это могут быть стада диких копытных, в обжитой
местности—главным образом люди или скопления домашних живот-
ных. Комары, как и все свободно нападающие кровососы, редко специа-
лизируются на каком-либо одном виде добычи. Но, однако, например.
352
< ulex pipiens обычно нападает на птиц; наобопот, Anopheles maciilipea-
nis питается в основном за счет крупных млекопитающих, а на птиц
почти нс нападает. Некоторые подвили, как, например. An. niaculipennis
ulroparvus, предпочитаю! копытных человеку, некоторые (An. niaculi-
nennis sachnrovi) одинаково часто нападают на тех и других.
На выбор добычи влияет относительная численность каждого вида
добычи в данной местности При наличии в поселке большого количества
домашних животных (на один двор 4 головы скота) 98% нз исследован-
ных I/ь niaculipennis niaculipennis содержали кровь животных, а у того
у д' подвида нз другого пос- тка, где было мало скота, 80% исследован
ntyx комаров содержали кровь человека (Беклемишев. 1949).
Связь комаров с человеком или домашними животными в зиачи-
г чьной степени определяется еще и т о н о г р а ф пческ им распре-
делением в поселках тех и других. Если па окраине поселка на
пути полета комаров ог водо.ма имеется скопление скота (в скотнь
дворах), то большинство комаров насасывается крови на животных и
далее в поселок нс летит. J аким образом, при наличии такого шелона
з скотпых дворов между очагом выплода комаров и человеческим
жильем значительно снижается возможность контактирования малярш!
пых комаров с человеком и тем самим предотвращается pacnpociране-
ние малярии в данной местности. На этом принципе основана система
«оопрофилактики малярил (Беклемишев, 1949).
После принятия крови начинается второй период гонотрофического
цикла кровососа. Из состояния активноеги самка комара переходит
в состояние покоя, когда происходит переваривание крови и одновре-
менно с этим идет процесс созревания яичников При температуре 10
. 2,5V переваривание полной порции крови длится 247- 291 час, а при
.HJ--35°—4]—43 часа (Шленопа, 1938). За этот срок у самки созре-
вает и среднем от 130 до 244 яиц. An niaculipennis переваривает кровь
находясь внутри помещения, тогда как у An. hyrcanus, An. bifurctiius
и др. это совершается в природных убежищах. Самки с соревшнмп
яичниками в вечернее время вылетают из помещения или других убе-
жищ и направляются к ближайшему водоему для кладки янц.
После этого, третьего, периода гонотрофического цикла комары
вновь приступают к поискам добычи. Возвращаются они не обязательно
в тот же дом, откуда улетели к водоему. Куда они попадут— дело слу-
чая. Чем больше гонотпофическнх циклов приделает самка, тем больше
разных помещений она может посетить. А это имеет существенное зна-
чение для распространения малярии в данной местности. Комар, аара-
швшись в одном доме, должен, прежде чем в нем созреют спорозоиты,
проделать по крайни) мере (при температуре 25—30 ) 4 гонотрофиче-
еких цикла, т. е. столько же раз может сменить иомешение. Поэтом)
он вероятнее всего заразит человека совсем в др гом доме, нежели
в гом, где он нолхчил возбудителя.
В конце лета вновь окрыляющиеся самки ко?.; ,ров дплпахзпруют,
что проявляется у них в торможении развития яичников. Они больше
не подчиняются правилу гонотрофическоЙ гармонии. Если лаже самки
и пьют кровь, то яичники у них теперь не рашиваются Диапаузирую
щне самки могут питаться сладкими растительными соками, в резуль-
тате чего у них происходит значительное ожирение за счет сильного
увеличения жирового тела. Только такие днапаузирующне самки спо-
собны Зимовать, тогда как летние аь гивные самки погибают к осени.
С наступлением ожирения самки /4«. niaculipennis niesseae уже нс
отыскивают добычу, а выбирают прохладные помещения или природные
убежища (норы, пещеры, дупла и т. п.), где они остаются в состоянии
В. А Дсгсть
353
покоя на всю зиму. Весной, в зависимости от широты местности и уело
вин погоды, перезимовавшие самки, вышедшие из состояния диапаузы,
вылетают из мост зимовок н начинают период активной жизни.
Наступление диапаузы в условиях средней полосы Европейской
части СССР начинается у комаров разных поколений уже в начале
июля, при высоких температурах, и принимает массовый характер во
второй половине июля—начале августа. В пределах ареала An. macuti
pctmis messeae сроки наступления массовой диапаузы изменяются oi
конца июля до 3-й декады «вгуста (юг Европейской части СССР, Сев
Кавказ). В соответствии с этими сроками прекращается и массово)
питание комаров кровью. ‘
Циклические изменения развития малярийного комара и переход
его к состоянию диапаузы регулирются, по мнению ряда исследовате-
лей (Данилевский. 1948, 1961; Данилевский и Глиняная, 1958; Виногра-
дова, 1958; ШиппцынЕ 1957, 1959), изменением режима освещения,
т. е. явлением фотопериодизма. В эксперименте развитие в течение ли-
чиночной и имапшальнои фазы в условиях короткого дня (12—15 ча-
сов), приводит к полной диапаузе даже при сравнительно высоких тем
iupa-турах. Самки, развившиеся при коротком дне, не обнаруживают
следов развития яичников, и, наоборот, длинный день (20—22 часа)
обеспечивает у самок нормальное течение гонотрофического цикла
В природе сроки начала диапаузы определяются степенью уменьшения
продолжительности дня, при которой происходило развитие водных фаз
Например, в условиях средней полосы Европейской части СССР массо-
вая диапау :а наступала при уменьшении длины дня до 18 часов 30 ми
нут, в то время как на юге она начиналась при длине дня 12—13 часов
Приведенные данные показывают, сколь сложны и разнообразны
словня, определяющие контактирование переносчика с позвоночным
хозяином и возможность распространения возбудителей заболевания
от одного хозяина к другому.
Во время акта кровососання переносчик получает возбудителя
от позвоночного хозяина. Однако ис каждая особь данного спецнфиче
ского вида переносчика способна заразиться при питании на хозяине
Это обусловливается состоянием как возбудителя, так и самого пере-
носчика. Например, заражаемость малярийного комара отчасти
зависит от количества гаметоцитов в крови человека. Так, при пали
чин от 1 до 100 гамонтов Plasmodium vivax в 1 леи3 крови зарази
лось только 50% комаров, тогда как при числе гамонтов от 1000 до
2000 заразилось 100% комаров (Mackerras a. Ercole, 1948) Заражае-
мость комаров на одном и том же больном в отдельные дни то
повышалась, то понижалась, что, вероятно, объясняется колебанием
в периферической крови человека количества гамонтов, способных
к развитию в комаре. Инфекционность гамонтов Pl. falciparum была
высокой с 4-го по 10-й день их появления, а после этого срока
(иногда даже и ранге) инфекционность гамонтов падает и даже
исчезает совсем.
Заражаемость мух пене трнпанозомами Trypanosoma gambiense
и Т. rhodosiense сравнительно низка; в эксперименте она нс превы-
шает 10%. а в природных условиях в эндемической зоне обычно
можно найти не более 1% зараженных мух. Многие мухи обладают,
видимо, врожденной невосприимчивостью. Однако незарэжаемость
зависит и от трипанозом. У разных штаммов трипанозом наблюдают
ся вариации в отношении способности развития их в переносчике.
Одни трппанозомы развиваются в желудке мухи, но пе проникают
в ее слюнные железы, другие не развиваются даже в желудке. Такие
354
штаммы нс способны к передаче через мух (non-transmissible strains;
Ноаге, 1949). Кроме того, инфекционность трипанозом для мух выше
в первый период болезни и постепенно снижается в последующие
периоды развития заболевания.
Степень заражаемости переносчика зависит еще от условии
внешней среды и главным образом температуры в момент попадания
возбудителя в переносчика. Если, например, малярийный комар сразу
после кормления будет помешен на 3 часа при температуре 37,7°, то
его заражаемость снизится в 25 раз, а после 4 часового пребывания
в таких условиях комары совсем не заражаются.
Выше было показано, что развитие возбудителя в переносчике
може,т происходить лишь при определенной температуре внешней
среды. Кроме того, и ряд других факторов оказывает влияние на
созревание возбудителя в переносчике Так, например, удалось уста
повить любопытный факт влияния дополнительного углеводного или
солевого питания на развитие леншманин в москитах. Многие годы
не удавалось получить заражения экспериментальных животных или
людей-добровольцев кожным лейшманиозом через укус москитов
Лишь единичные опыты были удачны
Однако, после того как стали применять подкормку москитов
углеводной пищей (изюмный сок), в их кишечнике происходило осо-
бенно обильное развитие жгутиконосцев, которые поступали в перед
пий отдел кишечника и закупоривали пищевод. При таких условиях
во время сосания москитом крови ленгомонады в большом количестве
попадали в тело позвоночного хозяина и вызывали заражение.
(Smith, Holder, Ahmed, 1940, 1941; Swaminath, Shortt. Anderson. 1942)
Вероятно, фруктовый сок увеличивает вирулентность паразитов. Адлер
и Бэр (Adler и Вег, 1941) кормили москитов Phlebotomus papatasii
через перепонку взвесью культуры Le.ishmania tropica с 3 частями
2,7%-ного раствора поваренной соли и 1 части дефибрированной кро-
личьей крови. Через некоторое время укусами этих москитов удалось
заразить кожным лейшманиозом 5 человек. Авторы приписывают этот
эффект активирующему действию солевого раствора. Вероятно, это
вещество, так же как и фруктовый сок, влияет на передвижение и на-
копление лептомонадных стадий в дистальной части глотки москита,
что является предпосылкой для успешного заражения человека.
При контактировании переносчика с хозяином создаются условия,
при которых инвазионное начало вводится в восприимчивый организм
позвоночного животного. Способы заражения хозяина через
переносчиков зависят от того, где в переносчике локализуются инва-
зионные стадии возбудителя, т. е. каковы пути их выхода нз переносчика
В одних случаях это связано с самим актом сосания, причем воз-
будитель вводится путем инокуляции либо со слюной переносчика
(малярийный плазмодий, Trypanosoma gambiense, пироплазмы, тепле
рии, спирохеты клещевого рекурреиса, риккетсии сыпнотифозных лихо-
радок, вирусы желтой лихорадки и клетевого энцефалита и др ), либо
непосредственно из переднего отдела пищеварительного канала
(Leishmania, Trypanosoma evansi, Pasteurella pestis), либо даже из
самих ротовых органов (личинки филярий). Возбудитель в перносчике
иногда должен, прежде чем достигнет инвазионной стадии, претерпеть
развитие или размножение. Поэтому переносчик не может сразу же
при повторных кровососапиях заразить восприимчивого хозяина. Для
этого требуется некоторое время, контролируемое условиями внешней
среды. Например, при разных условиях температуры Leishmania
tropica в москитах становится инвазионной через 6 или 8 дней, Тгура-
'23*
355
nosoma gambiense н мухе цеце — через 15—35 дней, Plasmodium
falciparum в комаре — через 8—35 дней.
Но возбудитель может передаваться, как мы видели выше, и меха-
ническим путем при повторных актах кровососания, (например, через
слепней возбудители сибирской язвы и туляремии, Trypanosoma
evansi и др ). В других случаях заражение происходит контамипатпв-
но, т. е. переносчик (часто во время кровососания) загрязняет кож-'
ные. покровы или слизистые оболочки хозяина своими экскрементами
или другими выделениями, заключающими в себе возбудителей
заболеваний. Примером могут служить Picket sia сыпного тифа,
выделяющиеся с экскрементами вшей на кожу человека и при ее расчесы
ванин проникающие в кровь. Так же проникает и Trypanosoma cruzi
из клопов Triatoma. Через коксальную жидкость выделяются на
поверхность кожи спирохеты африканского рекурреиса во время крово-
сосання клещей Ornithodorus moubata. Если возбудитель проникает
в гсмоцель переносчика и там остается все время, то заражение в та-
ком случае возможно только при его раздавливании-. Это наблюдает-
ся, например, при заражении человека эпидемическим рекурренсом
через вшей. К категории коптамннатпвиого заражения можно отнести
случаи проглатывания переносчика. Так заражаются крысы, прогла-
тывая блох с' Trypanosoma lewisi. или ящерицы, проглотив клеща
Liponyssus saararum с Karyolysus lacerfttm.
В отдельных случаях один и тот же возбудитель может пере-
даваться как ипокулятивно, так и коигамииатнвно. Так, передача Bacil-
lus lularense возможна при укусе, комара (механически) и при попада-
нии на кожу экскрементов зараженного комара (Олсуфьев, 1938).
Заражение восприимчивого, не иммунного, позвоночного хозяина
происходит большей частью при однократном введении переносчиком
тем или иным путем инвазионного начала. Достаточно, например,
одного укуса комара чтобы вызвать заражение человека малярией
пли желтой лихорадкой. Уже одноминугное сосание крови клешем
Ornithodorus papillipes может заразить морскую свинку возвратным
тифом, тогда как обычно клещ сосет 30—40 минут (Павловский,
1947а). В то же гремя заражение человека лейшманиозом происходит
в результате лишь нескольких повторных укусов москита. Для зараже
яия лошадей Piroplasma caballi необходимо, чтобы самка клеща сосала
кровь не менее 2,5—3 дней. В течение первых дней питания клещ не мо
жет пнокулировать заражающую стадию; се формирование в слюнных
железах клеща происходит только после принятия первых порций
крови (Марков А., 1952). Клещи Ixodes perstilcalus вводят в первые,
дни кровососания столь немного вируса энцефалита, что он не вызы-
вает заболевания человека, а лишь приводит к его иммунизации
(Павловский, 1947а).
Осуществление контакта переносчика с позвоночным хозяином
обеспечивает, таким образом, попадание возбудителей в переносчика
н возврат инвазионного начала к позвоночному хозяину. В тех слу-
чаях, когда возбудитель относится к категории мопоксениых нарази
тов, г е. обитает в одном виде хозяина, переносчик получает пива
знойное начало только от него и передает возбудителя другим особям
того же вида хозяина. Это имеет место, например, при передаче
заражения Plasmodium vivax, риккетсий сыпного тнфя и лихорадки
паиатачи от человека к человеку соответственно через комара, вошь
и москита, пироплазм домашних животных — через клещей и т. п.
Во многих случаях, однако, возбудитель заболевания является
поликсенпым организмом, н тогда переносчик может воспринять воз-
356
будптеля от разных видов хозяев в зависимости от имеющихся эколо-
гических связей между ним и позвоночными хозяевами. В отношении
человека это имеет большое значение, так как возбудители петого
ряда заболеваний могут передаваться ему через переносчиков от раз
личных позвоночных животных, которые в одних случаях являются
синатрипамп, а в других— обитателями дикой природы. Такне транс
мисснвные заболевания являются зоонозами. От домашних животных
к человеку передаются сибирская язва (слепнями, мухами жигалка-
ми). средиземноморская форма ката-азара (москитами) и другие,
заболевания. Кроме того, было выяснено, что человек может заражаться
через переносчиков от диких животных, которые подвержены множеству
болезней Такие заболевания возникли и возникают у человека нере«
ко тогда, когда он проникает в безлюдную местность, где подвергаете,
нападению раг-шчных кровососов. Так, tianpi.мер, возникли а боле
вання таежным клещевым энцефалитом в связи с освоенном тайги
Дальнего Востока или появились заболевания остронекротизирунлцен
формой кожного лепшмациоза у людей, приехавших в необжиты
районы пустыни Туркмении, и многие другие.
Болезням подобного рода, получаемым человег ом от диких живо
пых иерез переносчиков, свойственна, следовательно, природная
очаговость, как это установит Павловский сщ в 1938 г В даль-
нейшем это явление подробно исследовалось нм самим и сю школой
и ныне разрослось в целое учение о природной очаговости трансмис-
сивных заболеваний. Этому понятию Павловский (1948а) дает сле-
дующее определение: природная очаговость —это явление, когда
«возбудитель, специфический его переносчик и животные-—резервуа-
ры возбудителя в течение смены своих поколений неограниченно дол
гое время существуют в природных условиях вне зависимости от чело-
века, как по ходу своей уже прошедшей эволюции, так и в настоящий
се период». Если в такойеючаг, где возбудитель с помощью перенос
чика циркулирует между дикими животными, попадает человек, то
переносчик, почерпнувший возбудителя у диких животных, можс
передать его человеку, и последний, если он восприимчив к данному
возбудителю, заражаетея. Однако здесь человек является побочным
веном, не обязательным для существования возбудителя как вида
В общей форме схему распространения природно-очагового
габолеванпя можно изобразить так:
4ле||||сти1ЮП1ь
переиоС'ччси
. 1ик!И- ни шоночные
Дикие позвоночные
4t доеск
Благодаря исследованиям Павловского н его учеников (1939.
1947а,б, 1948а, б, 1957) на территории СССР выявлен целый ряд при
родных очагов трансмиссивных заболеваний человека и всесторонне
изучены условия их существования. Эго оказалось возможным
потому, что в основе исследования был положен эколого-паразитоло-
гический метод, позволяющий вскрыть и установить всестороннш
связи в пределах биоценоза определенною биотопа между itg сочле-
нами (возбудитель—членистоногие—позвоночные животные) и выяс-
нить путч циркуляции возбудителей.
В настоящее время можно считать, что природная очаговость свои
ствеи а нескольким десяткам трансмиссивных боле шей разной этио-
логии. Некоторые из них мы назовем в качестве примерз. Клещевой
357
энцефалит—вирусная болезнь многих диких млекопитающих и птиц.
Вирус был выделен из мозга бурундука (Tamias sibiricus), красносерой
полевки (Evoibniys rufocanus), ежа амурского (Erinaceus amurensis),
крота уссурийского (Mogera robusta), дрозда сизого [Tardus hortulo-
гит). синего соловья (Larvivora суапеа) и многих других животных
(Павловский, 1947 а). Переносчиком является Ixodes persulcatus и ряд
других клешей. Сыпнотифозная лихорадка Востока, выявленная у чело-
века, оказалась заболеванием диких крыс, сусликов, бурундуков’ поле-
вых мышей и других грызунов. К человеку она передавалась через
клещей Derniacentor niittalli п ряда других видов этого рода. Возбуди-
тель клещевого возвратного тифа (Treponema sogdianum) в природе
выявлен среди тушканчиков, мышей, ежей, летучих мышей, песчанок
н других животных. Переносчиком является Ornithodorus papdlipes.
Остропекротизирующая форма кожного лейшманиоза (Leishmanta
tropica) является болезнью диких грызунов — песчанок (Rhombomys
qpimus, Merinnes.erythrourus), суслика (Spermophylopsis leptodactylus)
и др. Передача осуществляется москитами.
Вне пределов СССР природная очаговость свойственна, вероятно,
желтой лихорадке. В Африке и Южной Америке отмечено, что резер-
вуаром вируса этого заболевания являются обезьяны. Болезнь Чагаса
(возбудитель— Trypanosoma cruzi), распространяемая клопами Reciu-
viidae. имеет очагн в природе. Резервуарами этих трипанозом являются
многие сумчатые, неполнозубые, грызуны, рукокрылые и другие млеко
питающие. Есть предположение, что в малайских джунглях возбудитель
малярии обезьян Plasmodium knowlesi передается к че ювеку (Garnharn.
1958). Это тем более вероятно, что человек заражается этим паразитом
при введении их с кровью (Ciuca, 1955). Можно говорить о природной
очаговости и некоторых гельминтов. Например, резервуаром в природе
для Loa loa в Камеруне служат обезьяны (Gordon, Kershaw, Crewe.
Oldroyd, 1950), а резервуаром элефантиазиса (Wuchereria tnalayi) в
Малайе также являются обезьяны (Presbytis melalophos) и лори (МурИ-
cebtis coucand) (Nelson a. Heiscli, 1957). Дифиллоботрноз. по-видпмому.
также имеет природные очаги, так как паразиты были обнаружены у
тюленя, медведя, лисиц и некоторых других рыбоядных хишников.
Природные очаги трансмиссивных заболеваний характеризуются
приуроченностью к определенным ландшафтам той или иной геогра
фической зоны. Напри мер, лесному ландшафту свойственны очаги кле-
щевого энцефалита, степному ландшафту — очаги клещевого сыпного
тифа и чумы, ландшафту поименных а также суходольных лугов на
местах вырубленных лесов на Востоке свойственны очагн японского
энцефалита. С ландшафтом жарких пустынь и полупустынь связаны
очаги кожного лейшманиоза.
В пределах данного ландшафта в зависимости от условий обитания
и характера мест нахождения переносчика природные очаги транемне
енвных болезней могут иметь более пли менее ограниченное распростра-
ненно. Например, не вся территория таежного леса или вторично воз-
никшего смешанного леса представляет одинаковую опасность в отно-
шении инфицирования клещевым энцефалитом. Заражение происходит
там, где возможно контактирование с Ixodes persulcatus, а места их
наибольшего обилия приурочены к участкам леса с преобладанием
лиственных пород и с хорошо развитым травяным покровом. Но и здесь
клещи распределены неравномерно. Больше всего их скапливается
вдоль троп. Именно здесь создаются наиболее благоприятные условия
(ля нападения клещей на человека и его заражения.
В однородном на первый взгляд ландшафте песчаной пустыни или
‘58
юлупустыпн можно выделить микроландшафты, где сосредоточиваются
различные животные, как позвоночные, так и кровососущие членисто-
ногие, и где возможна циркуляция возбудителя заболевания и, следо-
вательно, развитие природного очага той или иной болезни человека.
Очень часто таким очагом является пора животного или пещеры, в ко
торых создаются сложные биоцснотпчеекие комплексы, сочленами ко-
торых являются пе только позвоночные, членистоногие, постоянные и
временные паразиты, но и возбудители ряда заболеваний различной
природы.
Так, в пещерах и норах дикообразов были обнаружены клещи
Ornithodorus, являющиеся носителями спирохет рекурреиса. При ночев-
ке в такой пещере люди заболевали возвратным тифом, будучи под-
вергнуты укусам клещей. Норы большой песчанки (Rhombomys opitnus)
являются типичными микроочагами кожного лейшманиоза. Здесь вы-
плаживаются и живут москиты, которые распространяют среди обита-
телей нор лейшманиоз. Песчанки, например, до 60—80% в некоторых
районах Туркмении заражены лейшманиозом. Зараженность жгутико-
носцами Phlebotomus papatasii, выловленных из таких нор, оказалась
равной 6% (Латышев и Крюкова, 1941). Москиты могут вылетать из
пор и разлетаться па расстояние до 1,5 клт в зависимости от рельефа
местности. Человек заражается при попадании в зону такого очага.
Переносчик, получив возбудителя от диких животных, передаст его
разным животным. Одни из них оказываются «тупиками» на пути цир-
куляции возбудителя, так как не представляют собой среду, подходя
щую для его развития. Такова, например, судьба спирохет клещевого
рекурреиса в крови кур. Другие животные заболевают или становятся
явными или латентными носителями возбудителя заболевания. Они
могут служить источником заражения переносчика. При видовом
обилии восприимчивых животных пе все они могут играть одинаковую
роль в циркуляции возбудителя и поддержании очага. Это зависит от
их численности в данном биотопе, от возможных условий их контакти-
рования с переносчиками и ряда других условий.
Среди многих видов кровососущих насекомых и клещей в данном
биотопе не все могут сделаться переносчиками возбудителей, являю-
щихся сочленами того же биоценоза. Некоторые нз них получают воз-
будителя, но последний в них не способен развиться до инвазионного
состояния. Для ряда заболеваний установлено по нескольку видов
переносчиков, роль которых в передаче заболевания человеку
может быть, однако, далеко не одинаковой. Например, в пределах
очага клетевого сыпного тифа возбудителя этого заболевания человеку
передают только иксодовые клещи, а среди диких животных передача
осуществляется также и через гамазовых клешей, не нападающих па
человека. Вирус клещевого энцефалита передается тремя видами кле-
щей: Ixodes persulcatus, Haemaphysalis concinna и Dermacentor silvarum.
Однако в Ixodes persulcatus вирус развивается значительно интенсив
нее, чем в Haemaphysalis. Кроме того, количественное преобладание в
тайге Дальнего востока Ixodes persulcatus по сравнению с Haemaphy-
salis позволяет выделить первый вид как наиболее важного перенос-
чика этого заболевания (Павловский, 1947 а).
Во многих случаях члснистоногие-перспосчнкн играют существен
ную роль в развитии и поддержании природных очагов трансмиссивных
болезнен. В них длительно сохраняется возбудитель: не только па про
тяжепин эпидемического периода, ио и в межэпидемнческин период,
который при явно выраженной сезонности многих природно-очаговых
щболсвапий бывает весьма продолжительным. Гак, вирус клещевого
359
энцефалт а в Ixodes persulcatus мои с г перезимовывать (Ч\маков, 194!)
Принимая во внимание трансоварпалсиую передачу вируса при дли
тельном развитии одной генерации клеща от 2 до 4 лет в разных по
графических зонах, можно считать, что клещи являются подлинными
резервуарами вирусе! в природе, сохраняющими его в течение многих
лет в ряду последовательных поколении.
Orniihodorus papillipes при содержании в лаборатории в тсчешн
многих лет сохранял способность заражать восприимчивого хозяина
спирохетами рекурренса. Одна самка этого вида в 1е115 г. заразила
морскую свинку после 13 лет пребывания в лаборатории (Павловский
и Скрынник, 1946). Вероятно, п з природе, они уогут так же Долго сохра-
нять спирохет, оставаясь иногда многие годы в голодном состоянии.
С другой стооопы, резервуаром кожного лейшманиоза служат пес-
чанки, болеющие 7—9 месяцев и сохраняющие возбудителя в знмшч
время, а москиты не играют роль резервентов. так как взрослые насе-
комые не доживают до следующею эпидемического периода (Латышев.
Кожевников. Поваливший. 1953). Таким образом, в каждом конкретном
случае сохранение и поддержание природного очага , ibhcht от разных
условии и главным образом от тех сложных отношений, которые возни
кают между сочленами биоценоза.
При вмешательстве человека, нарушающего биоценотичеч кие снял'
в данном ландшафте, свойственный ему природный очаг заболевания
может постепенно затухать. Так, при хозяйственном освоении тайги
человеком очаги клещевого энцефалита ослабевают, так как создаются
н благоприятные условия для существования и развития как перепое
чиков, так и диких животных — носителей вируса. Закапывание нор
песчанок в одном из очагов кожного лейшманиоза привело к резком1
снижению заболеваний в ближайших поселках с 700 до 1 человек па
тысячу населения (Латышев, Кожевников, 11овалшнииа, 1953).
Некоторые трансмиссивные болезни с природной очаговостью могут
одновременно распространяться п в условиях жилья человека, если
переносчик может жить в этих условиях. Клещевым рекурренсом можно
заразиться и непосредственно в жилых помещениях, которые заселены
клешами Orntihodorus. При наличии в помещениях до левых грызунов
клеши могу г от них черпать спирохет и передавать их человеку. Вдаль
нейшем сам человек сможет стать донором для клещей, если они бул т
пить кровь больного.
Анализ факторов природной очаговости трансмиссивных болезней
необходим для понимания их возникновения и условий развития
вспышек соответствующих заболеваний. Равным образом он является
основой для построения системы радикальных мероприятий по обез
вреживанию и полной ликвидации очагов болезней.
2. ЭКОЛОГИЧЕСКИЕ ОСНОВЫ РАСПРОСТРАНЕНИЯ ПАРАЗИТОВ
В ВОДОЕМАХ
Сравнительно недавно изучением крт Говорова паразитов в биоиено
зах различных типов пресных водоемов занялся Вишневский со своими
сотрудниками (Wisniewski, 1955. 1958) в Польше. Была поставлена
задача изучить весь комплекс паразитов, входящих в состав какого
либо конкретного биоценоза водоема, выявить наиболее тннп«пыс
доминантные виды паразитов и установить их циркуляцию среди видов,
составляют!г: биоценоз. Исследования в этом плане были начаня 1 а
эутрофном озере Дружно и продолжены для сравнения на мезотрофном
озере Голдаинво. Анализ полученных данных был проведен путем изу-
Зш
чеиня качественного состава паразитов всех членов биоценоза и коли
явственных показателем экстенсивности и интенсивности заражения
хозяев на разных этапах развития паразита.
Следует отметить, что имеющиеся в настоящее время данные не
являются полными, так как исследования, к сожалению, нс могут счп-
гагься завершенными. Однако они представляют определенный интерес
в связи с новым подходом к изучению паразитофауны, входящей в
состав конкретного биоценоза.
Началом исследования явилось установлен^ основного состава
населения ценоза и выявление «ведущих его сочленов», которые пре
обладают количественно и могут пграть наиболее су шестсенпую роль
в круговороте паразитов. Паралелльно с этим устанавливались и все
существующие Сиопенотическис связи между его н letta.ni.
В озере Дружно было выявлено около 170 видов возможных хозяев
паразитов, относящихся к беспозвоночным и позвоночным животным.
Подробными исследованиями на основании большого материала в этих
хозяевах выявлено .значительное количество паразитов. У окопчатель
пых хозяев констатировано 92 вида паразитов, у первых промежугочп i.x
хозяев — 78 видов и вторых промежу точных — 51 вид. На основании
данных по экстенсивности и интенсивности заражения были установ-
лены наиболее типичные виды паразитов для данного биоценоза, а также
выявлены основные п второстепенные виды хозяев для каждого вида
паразитов. Главное внимание было обращено на сосальщиков, цеетод
и скребней. В результате были изучены жизненные циклы около 60 ви
дов сосальщиков и многих видов ленточных червей. Кроьте того, им
основе фаунистического исследования был произведен анализ путей
круговорота паразитов в биоценозе. Для сосальщиков
установлено 12 таких путей, а для цеЛгод—6.
Эти путл для сосальщиков следующие: 1) легочные моллюски —
рыбы — тицы. Этот путь используют 11 видов (например, Diplosto-
niutn spathaceuiu. Tylt delphis clavata и др.); 2) легочные моллюски —
амфибии — птицы (Tylodelphis excavate., Codonocephalus urntgerus.
2 вида Echinostomatidae и др.); 3) легочные моллюски — пиявки,
олигохеты — птицы (Apatemon gracilis, Cyaiocoiyle prussica); 4) легоч-
ные моллюски—моллюски — птицы (несколько видов сем. Ecliinosto-
matidae), 5) легочные моллюски — моллюски — рыбы (Asymphylodoru
tincae)\ 6) легочные моллюски — раковины моллюсков или предметы
на дне водоема — ипшы. Этот путь свойствен Nolocolilus, Catatropis
и другим видам.
Все эти пути, весьма характерные для эутро'фного водоема, могут
осуществляться и в других водоемах, но лишь в ограниченных пре-
делах. Так, в мезотрофпых водоемах 4-й и 6-й пути могут иметь место
только в условиях литорали.
7) моллюски — планктонные ракообразные — рыбы (характерно
только для Bunodera luciopercae и Corgodera vitelliloba)'. 8) мол-
люски— рыбы — рыбы (хпшпые). Этот путь свойствен Bucephalus,
piiipidi/cofylc (мссссвые виды) и ряду’ других видов. Осуществлена
этого пути возможно только в водоемах богатых рыбой; 9) мол-
люски—рыбы (кровеносная система). Этот путь характерен только
для видов рода Sanguinicola; 10) моллюски — птицы (кровеносная
система). Этим путем развиваются Trichobilharzia и Bilfu rziella.
9-й и 10-й пути осуществляются при активном проникновении цер
карий и могут иметь место в водоемах разного типа.
Следующие пути характерны для паразитов амфибий: II) мол-
люски— насекомые — лягушки (Pleuragenes claviger, Prosotocus, Gor-
361
godera и др.), и 12) легочные моллюски — раковины моллюсков
лягушки (только Diplodiscus subclavatus).
В отношении цестод в озере Дружно выявлены следующие нуги —
циркуляции Наиболее часто используется путь 1) планктон (Cope-
poda, Ostracoda)—птицы (18 видов сем. Hymenolepididae); другие
нети оказываются более редкими: 2) легочные моллюски — птицы
(2 вида сем. Hymenolepididae), 3) планктон (Copepoda)—рыбы —
рыбы (Triaenophorus lucii), 4) планктон (Copepoda)—рыбы — птицы
(Ligula), 5) планктон — рыбы — млекопитающие (Diphylloboihrium
latum) н 6) олигохеты — рыбы (Caryophyllaeus, Monobothriurn)
Для скребней отмечен один путь через Asellus aquations к амфи-
бням, рыбам п птицам. Это. конечно, не все пути циркуляции, но, ве-
роятно, наиболее существенные и ясно выраженные.
Характерным для круговорота подавляющего большинства паразн
тов является то, что он осуществляется но путям пищевых связей
между членами биоценоза, за исключением случаев активного про-
никновения в хозяина (9 н и 10-й пути сосальщиков) или пассивного,
не через пищу (6-й путь у Notocotylus и др.). В общем можно отме-
тить, что в круговороте паразитов птицы имеют доминирующее значе-
ние, как окончательные хозяева. Действительно, для 90% ленточных
червей и для 64,7% сосальщиков они являются таковыми. С другой
стороны, рыбы, гораздо реже используются как окончательные хозяе-
ва. Только 10% ленточных червей п 17,6% сосальщиков заканчивают
развитие в рыбах. Так же и амфибии для 17,6% сосальщиков служа
окончательными хозяевами. Главная маг та рыб в озере Дружно вклю-
чается в круговорот сосальщиков в качестве промежуточных хозяев.
Характер паразитофауны в биоценозе озера Дружно в значитель
ной степени определяется присутствием специфичных видов хозяев
и их количественным показателем. Отсутствие или редкое появление
какого-либо одного из хозяев исключает или резко уменьшает коли-
чество опредленного вида паразита в биоценозе. Циркуляция пара-
зита легко может осуществляться при наличии основных хозяев пара-
пета; однако, если в водоеме имеются только второстепенные хозяева,
то циркуляция паразита оказывается резко ограниченной. Например,
Triaenophorus lucii является редким паразитом рыб в озере Дружно,
что обусловлено наличном в озере только второстепенных первых
промежуточных хозяев (Cyclops albidus и др.), зараженность которых
крайне низка (0.038%).
Вообще распределение паразитов у окончательных и промежуточ-
ных хозяев в количественном отношении оказывается весьма неравно-
мерным. Обычно окончательные хозяева заражены более часто и бо-
лее интенсивно, чем промежуточные хозяева. Это особенно заметно
в тех случаях, когда заражение окончательных хозяев происходит
через беспозвоночных животных. Например, для Runodera luciopercae
в озере Дружно первым промежуточным хозяином является Sphae-
rium corneum, который был заражен на 1%; второй хозяин — Масго-
eyclops oithonoides — заражен еще более слабо (на 0,22%). В то же
время окунь и другие рыбы, как окончательные хозяева, заражены
приблизительно на 45%.
Кома циркуляция паразита осуществляется от беспозвоночного
через позвоночною к другому позвоночному, количественные соотпо
шеппя паразитов на родных стадиях цикла оказываются несколько
нш мп. Наименьшее количество паразитов и наиболее низкая заражен-
ность наблюдаются всегда у первого промежуточного хозяина; эти
показатели возрастают у второго промежуточного хозяина, если он
имеет нишевые связи с первым («нтанпе беспозвоночными). Заражен-
ность окончательного хозяина иногда может еще повыситься или
часто остается на уровне заражения второго промежуточного хозяина
и зависимости от особенностей нх питания и условий контактирования.
Вишневский иллюстрирует это явление на примере Ligula intesti-
tialis. Первым промежуточным хозяином служит Cyclops olbidus,
заражение которого процеркоидами не превышает 0,06%. Второй
промежуточный хозяин — рыбы ffu/ilus ruttlus — заражен плероцер-
коидами па 7.4%, a Rhodens rhodeus даже на 72%, и наконец, птицы
заражены так же часто, как н рыбы. Аналогичное явление наблюдается
у Piphyllobothrium latum. Циклоны, очень редко заражены процеркои-
(ами, а рыбы заражены часто и интенсивно плероцеркоидами,
иногда даже в большей степени, чем заражены половозрелыми
стадиями окончательные хозяева.
Объяснить это явление можно тем, что окончательные хозяева
поедают в большом количестве промежуточных хозяев, численность
которых в водоеме иногда очень велика (ракообразные или насеко-
мые) и таким образом аккумулируют в себе паразитов. В тех слу
чаях, когда второй промежуточный хозяин, например рыба, питается
беспозвоночными (первыми промежуточными хозяевами), то у него
также наблюдается значительное накопление личинок паразита. Это
особенно ясно выражено в циклах развития ленточных червей. Сильное
заражение вторых промежуточных хозяев создает максимальные воз-
можности для широкого заражения окончательных хозяев.
Между тем в ряде случаев усиление зараженности вторых про-
межуточных хозяев может наблюдаться и при активном проникнове-
нии личинок в хозяина, в случае отсутствия пищевых связей у про-
межуточных хозяев. Это отмечено Гннецннской (1959) для некоторых
сосальщиков. Гак, например, в Рыбинском водохранилище заражен-
ность прудовиков партенитами Diplostornum (Cercaria chromalophora)
была равна 0,62%, а зараженность рыб личинками Diplostornum,
локализующимися в хрусталике глаза, достигала 33,3% (у судака)
ХП% (у окуня).
В данном случае, как и в некоторых других, степень аккумуля-
ции личинок во втором промежуточном хозяине (например, в рыбах)
обусловлена в большей мере их подвижностью и продолжитель-
ностью жизни. Перемещаясь с места на место, рыбы соприкасаются
с источником инвазии — моллюсками. Последние, зараженные парте-
нитами сосальщиков, выделяют в течение своей жизни, как правило,
огромное количество церкарий, и тем самым создается возможность
нх проникновения во многие особи второго промежуточного хозяина.
На протяжении своей жизни рыбы накапливают значительное количе
ство метацеркарий, и поэтому их зараженность оказывайся более
высокой, чем моллюсков
В мелких водоемах с большой плотностью населения аккумуля-
ция паразитов вторым промежуточным хозяином проявляются еще
более резко. Так, в одном нз прудов 0,4% Limnaea stagnalis были
заражены партенитами Cofylurus. Вторым промежуточным хозяином
а.ля них служит тот же вид моллюска, который оказался зараженным
метацеркарпями (Tetracotyle lypica) па 98% при интенсивности до
600 паразитов па одного моллюска (Гинецинская, 1959).
Отмеченные выше различия в экстенсивности заражения оконча-
тельного, первого и втооого промежуточных хозяев, кроме указанных
причин, зависят вероятно от численности популяций хозяев. Количе-
ство особей первых промежуточных хозяев обычно бывает во много
363
раз больше, чем окончательных (например, количество циклонов
и диаптомусов, с одной стороны, и рыб — с другой, в случае с Diphyllo-
boihriuni la/шн). Таким образом, инвазионные стадии выдстяемые
окончательным хозяином (яйца, личинки и т. и.) распределяются на
огромное число особей популяции црбмежуточного хозяина,
В осуществлении круговорота паразитов, как отмечает Епишев
скин (1955), важным фактором является локальность пли оч а го-
тов ость заражения многих беспозвоночных животных
в ратных стадиях водоема. В тех местах, где концетрируются птицы
и рыбы, наблюдается и значительное ч;;сло беспозвоночны1 заражен-
ных личиночными стадиям» паразита. Следовательно, нс во всех
местах обитания в водоеме беспозвоночный хозяин в одинаковой сте-
пени заражается паразитам^. Б одних местах собирается большое
количество видов хозяев, более сильно зараженных, чем в други',
местах. Это в особенности касается тех форм, которые сравнительно
мало передвигаются в водоеме (моллюски, некоторые личинки асеко-
мых и др.). В результате не каждая популяция промежуточного
хозяина в водоеме может в одинаковой степени обеспечить заражение
окончательного хозяина. Эго необходимо иметь в виду пои анализе
зараженности окончательных хозяев. Сходные данные были получены
Фроловой (195MJ по ih ряжению моллюсков партенитами сосальщиков
в водоемах Кар».тин, Гпненипскоп (1959) и другими в Рыбинском воде-
храни, ище.
Обюр паразитофауны эутрофного озера Др,жио показывает
значительное количественное преобладание в его биоценозе цестод
и трематод, представленных главным образом личиночными форма
мн. Нематоды, скребни и паразитические раки представлены сравни-
тельно незначительным числом видов и особей.
Необходимо отметить, что некоторые паразиты, встречающиеся
особенно часто и в больших количествах, играют определенную рол:,
в регуляции численности хозяев. Это наблюдается, например, при
заражении глаз рыб личинками Diplostomum spaihaceutn или при
поражении мышц личинками Posthodiplostonmm nulicola и в ряд>
других случаев. Однако такое явление в биоценозе озера Дружин
наблюдается не часто и не носит закономерный характер.
Исследование; посвященное выяснению путей циркуляции пара-
зитов в биоценозе, выполненное на небольшом озере в Калининград-
ской области, опубликовала недавно Голикова (1961). Установленные
ею закономерности весьма близки к таковым, обнаруженным в рас-
смотренных выше работах польских авторов. Голикова устанавливаем
30 путей циркуляции паразитов в изученном ею водоеме.
Совпадение результатов в двух независимо выполненных исследо-
ваниях указывает на общий характер установленных закономерностей
На морском материале изучение циркуляции паразитов в биоце-
нозе, по ограниченное только ди генетическими сосальщиками, прово-
дила в СССР Зелнкман (1950, 1955). Она осуществила эколого-пара-
зитологпческпй анализ биоценоза литорали Кандалакшского залива
и выяснила круговорот сосальщиков между всеми сочленами этого
биоценоза. Было установлено, что параллельно пищевым связям, кото
рые осуществляются меж у птицами и рыбами, с одной стироны, и лип
ра ьными беспозвоио шы.мн—с другой, создаются и связи паразитарные
Не останавливаясь более па этих вопросах, отметим, что дальней
шее изучение циркуляции паразитов и их взаимоотношений в различ-
ных биоценозах представляет не только общепаразитологнческни н<
и большой практический интерес.
ГЛАВА XIX
ВЛИЯНИЕ ИД ПАРАЗИТОФАУНУ ЖИВОТНЫХ
хозяйственной деятельности человека
Хозяйственная деятельность человека является мощным факто-
ром, воздействующим на природу и изменяющим ее. Естественно, что
эти воздействия затрагивают и паразптофауну животных, так или
иначе связанных с хозяйственной деятельностью человека. В настоя-
щей главе мы кратко рассмотрим относящиеся к этой проблеме,
представляющей большей нс только теоретический, но и практиче-
ский интерес, три вопроса: I) особенности паразитофауны домашних
животных, 2) влияние на паразптофауну акклиматизации и интродук-
ции н 3) изменение паразитофауны рыб при строительстве водо-
хранилищ.
1. ОСОБГЧНОСГИ ПАРАЗИТОФАУНЫ ДОМАШНИХ ЖИВОТНЫХ
Видовой состав паразитофауны домашних животных и птиц
прежде всего определяется тем «наследством», которое они получают
от своих диких предков. Многие из встречающихся в настоящее время
паразитов домашних животных являются именно такими «ан пе-
стра льны мн» формами. К числу их у лошади и осла относятся,
например, роды Anoploceplvala, Habronema, Trichonetna, Cylindropha-
rynx, Gyalocephalus, Gastrophdus; у крупного рогатого скота
b'iscliacderius Carmyeriu s, Hypodernia; у свиней — Giobocephalus.
Ilyastrongylus. Mac.racanUwrhynchus; v собак — Spirocerca. Dirojilaria:
у кур—Davainea, Oxyspirura. Lipeurus (Маркевич. 1944). Бесспорно,
что значительная часть (но не все) видов инфузорий Entodinioiiiorpha.
живущих в рубце рогатого скота, овец и коз и в толстом кишечнике
лошади п осла, также является их исконными паразитами, унаследо-
ванными от диких предков.
В дополнение к аипестральным формам пара.штофа' ка домашних
животных претерпела значительное обогащение за счет обмена
паразитами между разными видами до я а-иних- и синантропных
кнвотпых Этот вопрос был уже рассмотрен выше на примере пара н-
гпческ < инфузории кишечного канала копытных. Дополним его не-
колькими примерами, касающимися гельминтов Касимов (1936)
Зк ">
указывает, что иестода Amuebotueiiiu sphenoiaes относится к числу
анцестральных паразитов кур (род Gallus). Она встречается и у ди
ких кур в Индии, Бирме и иа Цейлоне. Блеете с домашними курами
она распространялась по всему свету, а затем, вторично (в условиях
одомашнения), перешла иа индеек (птиц другою семейства). Нема-
тода Capillaria colutnbue— паразит голубей, — по данным Касимова,
наблюдается иногда у домашней' курицы и индейки.
Между разными домашними 'животными установилась ест<
ст вен на я циркуляция некоторых видов их лара ihtob, причем
одни оказались окончательными хозяевам^» другие— промежуточны-
ми. Общеизвестными примерами подобных отношений являются
Echinococcus granulosus (окончательный хозяин — собака, промежу-
точный— крупный рогатый скот), Multiceps multiceps (окончательный
хозяин — собака, промежуточный — овца).
Паразитофаупа домашних животных обогащается так ж
за счет диких видов, более или менее близких к ним
в систематическом отношении. Хороший пример в этом
направлении дают домашние копытные. Почти все домашние живот-
ные Америки завезены туда из стран Старого Света. В Америке мно-
гие из них получили новых паразитов от представителей местной
фауны. Например, в печени рогатого скота в Северной Америке встре-
чается Fascioloides тирпа. Этот сосальщик перешел от североамери-
канских оленей. В Южной Америке подобным же новоприобрегенным
от местных копытных паразитом является Balanorchis anastrophus,
а также еще некоторые гельминты (Thysanosoma actinoides.
Haernonchus cunatus) Относительно часто домашние пгицы зара
жаются паразитами от диких родичей. Хорошо известно, что нематода
Sungamus trachea, паразитирующая в трахеях многих диких воробьи
ных и куриных птиц, легко переходит на домашних кур, передке
вызывая среди последних эпизоотии.
Ряд интересных примеров, иллюстрирующих роль диких птиц
в распространении гслъмннтоюв среди домашних куриных приводит
Гвоздев (1958), изучавший этот вопрос в Казахстане. Он указывает
на эпизоотии, возникающие у юмашпих кур и вызванные заражением
их от диких птиц трематодами Piosfhogonimus ovatus и Р. cuneatus,
паразитирующими в фабрицневой сумке и в яйцеводах кур. На
домашних кур в Казахстане переходит также сосальщик Echino-
paryphtum westsibiricum на домашних индеек — сосальщик Postliar
mostoinum gallinum (от диких перепелов) От диких птиц домашние
куры и индейки получают цестод Choanolaenia infundibulum и Rail
lietma cesticillus. Аналогичных примеров можно было бы привести еще
очень много. Все они ясно показывают, что дикая фауна является
резервуаром, постоянно обогащающим домашних животных парази
тами.
Вероятно, паразиты при переходе их на домашних животных
(а также и на человека) и длительном существовании в новых для
них условиях сами меняются. Происходит процесс расо- и видообразо-
вания. Сначала он затрагивает преимущественно физиологические,
а затем и морфологические особенности. Можно думать, что Ascaris
suum свиньи и /1. lumbricoides человека имеют обшее происхождение
Это же справедливо для человеческого Tbichocephalus trichiurus и сви-
ного Т. suis.
Стадный образ жизни и обычно благоприятные условия питания
являются факторами, способствующими распространению паразитов
среди домашних и синантропных животных Это приводит к тому
366
что пр» отсутствии должного ветеринарного вмешательства и конт
роля средн домашних животных возникают паразитарные эпизоотии,
наносящие значительный экономический ущерб. Примеры иаразнтар
пых эпизоотий домашних животных настолько общеизвестны (пиро
плазмозы и тейлериозы рогатого скота, трихостронгплезы лошадей,
фасциолезы рогатого скота, и т. д ), что нет необходимости здесь на
них специально останавливаться (см., например, Д. П. Антипин,
В С. Ершов, II Л. Золотарсв и В. А. Саляев, «Паразитология и ннва
знойные болезни сельскохозяйственных животных», 1959).
Поскольку многие причины экологического характера способ
ствуют широкому распространению паразитов среди домашних
и синантропных животных, особенно серьезным в практическом отно-
шении является вопрос о борьбе с паразитарными заболеваниями. Опа
может быть эффективной лишь в том случае, если в основу разработки
мер борьбы с паразитами будут положены глубокие знания жизненных
циклов паразитов, а также всей системы экологических факторов, спо-
собствующих их распространению. В Советском Союзе эти мероприя-
тия носят плановый характер. Особенно много для их теоретического
обоснования и практической разработки (в отношении гсльмиитозов)
сделано работами акад. Скрябина (1946, 1958 и др.) и его школы,
наметившими пути полной ликвидации некоторых глистных инвазий
человека н домашних животных.
Поголовную дегельминтизацию акад К И. Скрябин (1945) обозна
чает термином «девастация». Под девастацией он понимает наступа-
тельную. активную профилактику, направленную на истребление воз-
будителей заболеваний па всех фазах их жизненного цикла всеми
доступными способами механического, химического, физического или
биологического воздействия
2. ВЛИЯНИЕ НА ПАРАЗПТОФАУНУ АККЛИМАТИЗАЦИИ И ИНТРОДУКЦИИ
Природа подвергается со стороны человека многочисленным пере
стройкам, к числу которых относятся разнообразные опыты по акклн
матнзацни и интродукции новых полезных животных в места, где их
до тех пор не было. Многие из этих опытов, проводившихся главным
образом с рыбами и млекопитающими, увенчались полным успехом,
и акклиматизируемые животные стали размножаться, иногда в очень
больших количествах, в новых местах обитания.
Особенно широко работы по акклиматизации проводились за
последние 25—30 лет в Советском Союзе. Рассмотрим, как отражается
искусственное переселение животных на их паразитофаупе отдельно
для рыб и для млекопитающих.
В СССР акклиматизация рыб осуществлялась и осуще-
ствляется очень широко и преследует хозяйственные цели — увеличе-
ние рыбопродуктивности водоемов за счет ценных пород рыб. В каче-
стве примеров можно указать успешную пересадку чудского сига
(Coregonus lavaretus maraenoides) во многие озера Урала и в озеро
Севан на Кавказе, рипуса (Coregonus albula ladogensis) — нз Ладож-
ского озера в озера Урала, сазана (Cyprinits carpio) — в озеро
Балхаш и во многие другие озера Средней Азии, севрюги (Acipenser
sfellalus) — из Каспийского моря в Аральское, кефали (Mugil аига-
tus) — из Черного мор» в Каспийское и т. д.
В некоторых случаях для акклиматизации использовались рыбы
не только нз отечественной ихтиофауны, но и нз других стран и мате-
риков. Из Америки и Канады, например, в ряде водоемов СССР
367
акклнматирована радужная форель fSalmo irideus). Карликовый
сомик (Amiurtis n^bulosus) из Ссв<риой Америки успешно прижился
во м nonig озерах Западной Украины и Белоруссии. Все эти л многие
другие случаи успешной акклиматизации рыб сопровождались пара
читологпчегкнми исследованиями, которые проводились как на
местах, откуда брались рыбы для пересадки, так и на местах их
нового обитания. Не давая подробного описания всех этих отдельных
опытов, остановимся лишь на общих закономерностях изме-
нения паразитофауны, которые при этом были обнаружены
(Догель, 19386. 19,» 1954; Петрушевский, 1954, 195®, Петрушевский
и Бауер. 1953) и могу г быть сведены к следующему.
1 Наблюдается общее обеднение паразитофауны, полная или
частичная утрата паразитов, свойственных рыбе в материнском
водоеме. В особенности это касается видов, развивающихся с проме-
жуточными хозяевами. Последнее совершенно понятно, так как в но-
вом водоеме часто не оказывается необходимых промежуточных
хозяев.
Приведем несколько конкретных примеров. Ринус в Ладожском
озер? имел 11 видов паразитов, в озерах Урала в нем было обнару-
жено лишь 5 видов; сиг в Чудском озере имел 6 видов, а в озере
Севан — только 2 вида, форель в озере Севан имела 10 видов, она же
в озере Иссык-Куль — 3 вида и т. д.
2. В новых водоемах рыбы приобретают и некоторых новых, не
свойственных им паразитов за счет паразитофауны местных рыб.
Обычно эти вновь приобретаемые паразиты относятся к числу видон
с широкой специфичностью. Например, сиги, пересаженные из
Ладожского и Чудского озер в озеро Севан на Кавказе, утеряли
почти всех своих паразитов, но приобрели скребней Po.nphorliyncliu^
laevis и Echinorhynchus baeri, которые перешли на них от сегаискн'
форелей. Серебряный амурский карась (Cvassius auralus tn. gibclio)
неоднократно перевозился с Дальнего Востока в европейскую часп
Советского Союза Па его родине, на Амуре, в нем было найдено
19 видов паразитов. После акклиматизации в прудовых хозяйствах
он утерял всех своих паразитов, кроме дв> х, по приобрел ряд новых,
шцроко распространенных, таких как Ichtyopkihirius niultijiliis.
CliHodon cyprini, Dactylogyrus anchoraius и ряд других. В некоторых
случаях при этом было отмечено, что на акклиматизируемых рыб
могут переходить паразиты от очень далеких хозяев. Например,
Богатова (1936) описала интересный факт перехода i а акклиматизи-
рованного в озере Тургояк (на Урале) сига паразитических клешей
рода Unionicola. обычно паразитирующего в моллюсках.
3. При акклиматизации из числа старых паразитов преимуще-
ственно сохраняются виды с прямым развитием, в частности мопс
генетические сосальщики. Интересный и поучительный пример в этом
отношении дает так называемая солнечная рыба (Leponiis gibbosus).
Эта маленькая декоративная рыбка была привезена в Европу из
Северной Америки еше в конце прошлого века и сначала разводилась
в аквариумах. Затем она проникла в пруды и водоемы Западной
Езропы Распространяясь далее на восток, она проникла в Дунай.
1де была впервые обнаружена в 1918 г. Изучая паразнтофауну этой
рыбки в Дунае, Роман (1953), наряду с европейскими паразитами
(Diplostonumi spaikaceurn, Cbitpcoecum. skrjabini, Hepaticola petru-
schewskii). обнаружила двух моногенегическпх coca тыцикор
Urocleidus dispar u U simili’,. — которые были описаны с этой рыбки
в Америке и которые до сих пор были известны только для амери
канской фахпы. Таким образом, эти паразиты сохранились на Lepotnis
после акклиматизации ее в Европе в течение более полустолетия.
4. При акклиматизации рыб иногда наблюдается Переход пари-
зитов с вселяемой рыбы ни аборигенов. При этом йцюгда случается,
что местные рыбы, нс будучи приспособленными к новому паразиту,
очень сильно от него страдают. В некоторых случаях в результате
такой пересадки возникает эпизоотия средн местных рыб. Яркин при-
мер такого рода имел место при пересадке севрюги (Acipenser stel-
kit us) нз Каспийского в Аральское море в тридцатых годах. Вместе
с севрюгой был перенесен монотеистический сосчлыцик iVitxsc/iia
В Аральском море этот парази перешел иа местную очень
цепную осетровую рыбу—шипа (Acipenser nudiveJitris)— п вызвал
массовую гибель последнего, размножившись па нем в огромных
количествах (Догель и Лупа. 1937: Нута, 1910).
Вторым примером заноса паразита в новые условия при акклима-
тизации рыб может служить Myxosotna cerebraiis— паразит, локали-
зующийся в хрящевой ткани лососевых рыб и вызывающий тяжелое
аболсванне, которое обычно кончается смертью. Эго заболевание
тавно известно в Германии, где возбудитель его был описан Гофером
(Hofer, 1903, 1906), а затем подробно изучен Шенерклаусом
(Schaperclaus. 1931, 1954). В Германии этот паразит наносил очень
большой ущерб форелевому хозяйству В настоящее вр< мя в ГДР
и ФРГ в основном благодаря исследованиям Шеперклауса разрабо-
тана система мероприятий по борьбе с этим заболеванием, в резуль-
тате ее применения заболевание форелей в ГДР и ФРГ почти не
наблюдается. В начале 50-х юдов этот паразит вместе с посадочным
материалом был завезен в некоторые рыбные хозяйства в СССР, где
и был в 1952 г. впервые констатирован и изучен Успенской (1957).
В настоящее время в Советском Союзе также приходится проводить
ряд мероприятий для борьбы с этим заболеванием.
Недавно (Малевнцкая, 1958) был обнаружен случай заноса пара
шта со сложным никлом ра звития — Bolliriocephalus goivkongcnsis.
Это—цестода китайского происхождения, которая проникла в Амур,
а затем с акклиматизируемым белым амуром (Ctenophyngodon idella)
была занесена в прудовые хозяйства европейской части СССР Пара-
зит перепил на карпов и в первый же год нанес значительный эконо-
мический ущерб.
Приведенный выше случай эпизоотии шина в Аральском море,
вызванной h'ilzsclua sitiriotiis. и занос Myxosoma cerebraiis и Bothrio
i ephalus gowkungensis в СССР — это поучительные примеры, пока-
зывающие необходимость строгого паразитологического контроля при
акклиматизационных мероприятиях.
Хорошо изученный в паразитологическом отношении пример
н п т р о д у к ц п и м л е к о п и т а ю щ и х представляют собою акклима-
тизация ондатры (Ondatra xjbethica] в Советском Союзе. Этот крупный
грызун представляет значительную хозяйственную ценность, так как
обладает хорошим мехом. Он легко приживается в различных геогра-
фических зинах и быстро размножается. По данным .Лаврова (1916),
за период с 1928 по" 1944 год в ЗЯ областях Советского Союза было
выпуин но более 48 тыс. шерьков, причем во многих областях акклп-
мат’лзяцня прошла успешно. В частности, ондатра хорошо прижилась
на севере СССР в Карелии в Бурятской АССР и ча юге
в Казахстане.
На своей родине (Северная Америка) ондатра имела довольно
богатую парадитофаупу, которая слагалась из 40 видов: 2 вида про-
21 В. Л. Догель Зи9
стейших. 23 вида сосальщиков, 4 вида цеетод, 7 видов нематод, 4 вида
эктопаразитов (Barker. 1913, 1915, 1916; Price, 1931; Warwick, 1936).
Из них 26 видов можно считать специфичными для ондатры.
При акклиматизации ондатры в Карелии в первые годы Василье-
вым (1939) было обнаружено всего пять видов паразитов (рис. 206).
из которых два вида трематод (Echinostoma armigerurn и Notocoty-
lus quinqueserialis) оказа^сь представителями ее исконной амери-
канской фауны, остальные три — новоприобретеннямн за счет нового
" Рис. 206. Четыре сосальщика из кишки карельской он.инры
.4 — Plagiorchis eatamiatus zihethicas\ И — lichinostoma armige.runr, С — Niilo-
cotylus quinqueserialis; D—Psoioirenia marki (.4 и L>— приобретены и Европе,
Li н С—завезены из Америки), (По Васильеву.)
места обитания. Поскольку у ондатры Карелии сохранились амери-
канские трематоды, то можно утверждать, что они нашли себе среди
местной фауны беспозвоночных новых промежуточных хозяев. Более
того, имеются некоторые наблюдения, показывающие, что трематоды
американской фауны начинают переходить на местных грызунов. Так,
например, Скворцовым (1934) Notocotylus quinqueserialis были обна
ружены в водяных крысах (Arvicola terrestris).
В дальнейшем в Карелин продолжалось обогащение ондатры
местными паразитами. Серкова (1948). изучавшая паразитов ондатры
в Карелии через несколько лет после Васильева, нашла у нее 9 видов
паразитов. Из американских форм сохранился лишь Notocotylus
quinqueserialis, все остальные представляли собою яовоприобретеиие.
Среди них интересно отметить цестоду Paranoplocephala omphalodes —
паразита, характерного для водяной крысы, скребня Polyrnorphus
tninulus — вид, паразитирующий в водяных птицах. Таким образом
ондатра получила некоторых паразитов от весьма отдаленных видов
370
хозяев, связанных с нею, идиако. единством среды обитания
(водоемы).
Догель и Раппопорт (1944) изучили паразптофауну ондатры,
акклиматизированной в Казахстане, в низовьях реки Или, где наблю-
далось очень своеобразное сочетание -паразитов. Гельмиптофауна
ондатры состояла здесь из четырех видов: Notocotylus quinqueserialis,
Plagiorchis eutumlatus, Trichocephalus siifs и Macracanthorhyuchus
hirudinaceus. Первые два оказались общими с паразитами карельских
ондатр, но два остальных представляют собою паразитов свиней,
а вовсе не грызунов. Это странное на первый взгляд обстоятельство
легко объясняется тем, что дельта реки изобилует кабанами, от кото-
рых ондатра и позаимствовала некоторых паразитов.
Аналогично протекало изменение паразитофауны и в других
районах акклиматизации ондатры в СССР. Терялись почти полностью
ее исконные американские паразиты и приобреталась более или
менее случайная паразитофауна от хозяев, связанных с ондатрой
экологически. Среди этих вновь приобретенных паразитов оказалось
несколько видов от очень далеких хозяев, как-то: Trichocephalus suis
(от свиньи), Coniracoecum spiculigerum (от утиных птиц). Общин
список паразитов, приобретенных ондатрой в различных районах ее
интродукции в СССР, довольно значителен. Спасский (1951) приводит
список в 24 вида гельминтов и, кроме того, еще несколько видов
эктопаразитов, из которых только три вида имеют американское
происхождение. Однако эти паразиты разбросаны по различным райо
нам, так что в каждом отдельно взятом месте обитания паразите
фауна акклиматизируемой ондатры бедна и более или менее слу
чайна.
Совершенно аналогично изменялась паразитофауна ондатры и при
акклиматизации ее в Англин (Warwick, 1936), где она сохранила
5 видов паразитов американского происхождения (2 вида гельминтов
и 3 вида эктопаразитов) и приобрела ряд новых видов от местных
животных.
3. ДИНАМИКА ИЗМЕНЕНИЯ ПАРАЗИТОФАУНЫ РЫБ В ВОДОХРАНИЛИЩАХ
Строительство крупных гидроэлектростанций на больших реках
и улучшение условий судоходства на них осуществляется в Советском
Союзе в очень широких масштабах. В результате создаются большие
водохранилища — пресноводные моря, — условия существования в ко-
торых для водных организмов существенно отличаются от условий
в реках, давших начало водохранилищам. В 1941 г. было создано
гигантское Рыбинское водохранлище в верховьях Волгл, в 1952 г.—
Цимлянское водохранилище на Допу, в 1955 г. — Горьковское и Куй-
бышевское на Волге, в 1958 г.— Волгоградское на Волге, в 1959 г.—
Братское на Ангаре—в Сибири и множество других крупных и более
мелких водохранилищ.
При строительстве многих водохранилищ проводились гидробио-
логические, ихтиологические и паразитологические исследования,
которые позволили изучить динамику биологических процессов при
этих глубоких изменениях природы, осуществляемых человеком в про-
цессе целенаправленной хозяйственной деятельности.
Изменение паразитофауны рыб при формировании Рыбинского
водохранилища в течение 12 лет (1942—1954) подробно изучалось
Столяровым (1954, 1955), а затем эти работы были продолжены
Изюмовой (1959а). Имеются паразитологические исследования и по
24
371
другим водохранилищам Цимлянскому (Смирнова, 1$>59|, Выгозср-
скому (Нагибина. 1957) и по ппоторым другим
Остановимся на рассмотрении лишь основных изменений, кото-
рые претерпевает паразитофауна рыб в процессе формирования водо-
хранилища. Эти изменения тесно связаны с теми 1,','бокнмн гидро
логическими и гидробиологическими преобразованиями, которые при
лоч происходят. Л
Фауна паразитических простейших (инфузории, мкксоснорплип.
кокпндпи) и моногенетических сосальщиков — паразитов, развиваю
ИЧ1ХСЯ без смелы хозяев, при образовании водохранилища не пре-
терпевает существенных изменений но сравнению с исходной. Выпа-
дают лишь некоторые виды, характерные для реофильны.х рыб,
исчезающих из водоема В некоторых случаях наблюдается даже
усиленное ра витие этих паразитов, особенно в приилотинной зоне,
где создаются условия слабой проточное га. иапомпающие таковы,
озера. Несколько по особенному ведут себя лишь паразитические
рачки. В первый год число их резко сокращается. Это св тано.
вероятно, с неблагоприятными условиями, создающимися для свобод-
но живущих копеподитню стадий, о чем можно прел полагать, так
как в первый год существования водохранилищ копсподный планктон
их бывает очень беден, Уже начиная ср второго года количество
паразитичесюгх рачков увеличивается и через 3—4 года достигает
большой экстенсивности и интенсивности. На 4 5-й годы существова-
ния водохранилищ в них отмечались даже эпизоотии среди рыб, вызы-
ваемые массовым развитием паразитических рачков ErguSilus fieboldi
и ArgiUus foliaceus. Особенно быстро нарастание их происходит
в водохранилищах южной и уме репной зоны и гораздо медленнее на
Севере.
Иначе веауг себя паразиты, развивающиеся со сменой хозяев.
Среди них мы можем различить две группы: у одних промежуточ-
ными хозяевами служат планктонные рачки, у других — различные
бенз нческие беспозвоночные
Для первой группы, куда относится большинство ленточных чер
вей (многие Psendophyllidea. все Tctraplivllidu-a)» характерно резкое
снижение численности в первый год существования водохранилища
(слабое развитие конеподиою планктона!) к быстрое нарастание
в последующие годы. Последнее обусловлено массовым развитием
планктона, среди которого начиная со второго года видная роль вновь
принадлежит свободно живущим Copepoda. Особенно хорошо это
можно проследить на примере Ligula На четвертый год существова-
ния Рыбинского водохранилища ligula заражала уклею па 30%.
плотву — па 60%. Та же картина наблюдалась и в других, в особен-
ности южных видохраии тпщах. Цестодозы, а особенно лпгулсз.
оказываются важнейшими паре итариыми заболеваниями рыб в но-
вых водохранилищах, приносящими нередко значительный экономиче-
ский ущерб.
Обратимся теперь к рассмотрению второй и.; намеченных выше
групп. С бентосом в своем развпгпн связано небольшое количество
ленточных червей, многие нема готы, поч ти все скребни и все сосаль-
щики. Для цсстод сем CaryophyIlacidiie промежуточными хозяевами
явтяются олигохеты, для цистод сем. Л mphil inidae. Cyalhocephaliidae.
для сем. Rhabdoclionidac и большинства скребней — бентические рако-
образные из Amphipoda и Isopoda, для всех трематод — моллюски.
Судьба всех перечисленных групп промежуточных хозяев в создаю-
щихся водохранилищах несколько разится между собой, что отра-
372
жаегся и на динамике нх паразитов Для ГвоЗлнчнивов (Caryophyl
laeiJae) характерно резкое снижение численности. иногда почти пол-
ное исчешовение в первый гид существования водохранилища, но со
второю и в последующие годы начинается быстрое увеличение нх
количества. Например, лещ в Рыбинском водохранилище в 1942 г.
(второй год его существования) был заражен Caryuphyllaeus latteeps
на 2G%, в 1945—i:a 40% и в 1947— на 80%. Совершенно та же кар-
тина наблюдалась и в Цимлянском 'водохранилище Очевидно, это
явление связано с быстрым накопленьем нсЯдне водоема олигохет,
в особенности Tubifex.
В отношении паразитов, промежуточными хозяевами которых
являются высшие раки, в первые годы происходит снижение заражен-
ности. которая затем начинает из года в год мс тленно возрастать. Это
связано с относительно медленным увеличением численности Amphi
poda н Isopoda.
Очень характерна и типична для вновь создающихся водохранп
лнщ судьба ди генетических сосальщиков. Эта группа паразитов почти
полностью выпадает нз состава паразитофауны рыб сразу же после
образования водоема. Восстановление ее затягивается на многие
годы. В Рыбинском водохранилище, наиболее детально изученном
в этом направлении Столяровым, процесс формирования фауны тре-
матод занял свыше 10 лет. Такое постепенное увеличение численности
грома гит в течении целого ряда лет связано с промежуточными
хозяевами —моллюсками, фауна которых в водохранилищах форма
рустся чрезвычайно медленно.
В крупных водохранилищах (вроде Рыбинского) удается иод
летнть некоторые различия в дппамкк. паразитофауны и разных
частях водоема, что также связано с особенностями протекающих
в них гидробиологических процессов. Па этой стороне вопроса мы
не будем, однако, специально oi ганавливаться (сводку этих данных
см. в работе Бауера и Столярова, i95<S).
Необходимо отметить, что из массовых заболеваний рыб в водо-
хранилище наибольшую опасность представляют лнгулез (южная
и умеренная климатическая зона), поражение паразитическими рач-
ками (главным образом в южной зоне), чернонят пиетая болеть
(в южной зоне), трнэнофороз (в умеренной и северной зонах).
Следует подчеркну гь, что познание закономерностей формирова-
ния паразитофауиы рыб в водолраин .пшах, кроме большого эколого-
паразнтологического интереса, имеет существенное практическое зна
чепие. Необходимо предвидеть, какие эпнзооищ могут возникнуть
среди рыб. населяющих водохранилище, и своевременно принять •
профилактические меры, рассмотрение которых не входит, однако
в гадачу этой книги.
Часть IV
ВЗАИМООТНОШЕНИЯ МЕЖДУ ПАРАЗИТОМ И
ХОЗЯИНОМ. ПРОБЛЕМА ВИЛА У ПАРАЗИТОВ
ВВЕДЕНИЕ
Эта последняя часть нашего труда посвящена более детальному
разбору взаимоотношении между паразитом и хозяином. Рассматривай
сочетание «паразит—хозяин», можно видеть, что каждый член пары
не только воздействует в различных направлениях па своего партнера,
но и сам испытывает воздействие партнера. В данной части книги нас
будут интересовать уже не вопросы, касающиеся всей паразитофауны
животного в целом, не динамика паразн гпфаены, не циклы развития
паразитов и т. д., а характер взаимоотношений, который устанавли-
вается в пределах определенной исторически сложившейся системы
«паразит—хозяин». Мы рассмотрим нормальный характер этих взаимо-
отношений, а также различные сдвиги их, которые могут происходить
в результате различных экспериментальных воздействий.
Важность изучения этих взаимоотношений впервые подчеркнул
Хегнср (Hefner, 1927). Позднее вопросу о функциональном взаимодей-
ствии организмов паразитов и хозяев специальное внимание уделил
MpiHKOBCKHH (1916). Последний предлагает введенный Хегнером гро-
моздкий термин «host parasit relationship» (взаимоотношение между
паразитом и хозяином) заменить термином «функциональная паразито-
’огия». которая составляет особый раздел паразитологической науки.
Особенно существенной стороной (руикцпопальиых взаимодействии
является взаимное воздействие по линии антигенно-иммунологических
отношений.
В понимании Мошковского, несколько разнящемся от такового
Хегнера, предмет функциональной паразитологии составляют лишь те
взаимоотношения между паразитом и хозяином, которые возникают
с момента проникновения паразита в хозяина и кончаются моментом
покидания нм организма хозяина. Между тем Хегиер был склонен
включать в область host parasit relationship все фазы жизненного цикла
паразита, вне зависимости от того, где они протекают.
Разрабатывая предмет функциональной паразитологии более де-
тально, Мошковсклй разбивает жизненный цикл паразита с точки зре-
ния его отношения к данному хозяине на четыре, как он называет,
«такта»: 1) внедрение паразита в организм хозяина; 2) развитие в орга-
374
низме хозяина; 3) выход паразита (в виде пропагативной стадии) из
хозяина; -1) период пребывания или развития паразита во внешней сре-
де или (при смене хозяев) в организме других видов хозяев.
В рамки функциональной паразитологии данной системы «пара-
зит—хозяин» входят лишь первые три такта.
Как следует из этого, в жизни многих паразитов, развивающихся
с промежуточными хозяевами, имеется, если можно так выразиться,
несколько раздельных и иногда весьма рнзличпых «функциональных
паразитологии». Поясним эту мысль примерами. Так, для процеркоида
широкого лентеца функциональные отношения существуют только с ци
клопом, т. е. первым промежуточным хозяином, а все прочие фазы
цикла относятся к четвертому такту; для плероцеркоида к четвертому
такту относится почти весь цикл лентеца (в том числе и пребывание
в циклопе), кроме стадии паразитирования в рыбе; у взрослой стадии
лентеца функционально-паразитологические взаимоотношения имеются
тшь по отношению к теплокровному окончательному хозяину (чело-
век и др.), все же остальные фазы составляют чуждый этой физиологии
четвертый такт.
Совокупность этих тактов можно назвать функционально-паразито-
логическим звеном, причем весь жизненный путь паразита слагается из
ряда звеньев двоякого рода. Одни нз звеньев характеризуются на-
личием функциональных отношений с организмом хозяина, другие —
отношениями с внешней средой (свободные стадии паразитов); эти
двоякого рола звенья обычно чередуются.
В последующих главах мы ограничиваемся лишь вопросами соб-
ственно функциональной паразитологии, т. е. отношениями между пара-
зитом и хозяином во время пребывания первого в последнем.
ГЛАВА XX
ВОЗДЕЙСТВИЕ ПАРАЗИТОВ НА ХОЗЯИНА
Как мы видели выше, паразит и хозяин находятся в тесных взанмо-
отношеннях и взаимодействии друг с другом. Если паразит оказывает
на организм хозяина весьма многостороннее влияние, то и хозяин в свою
очередь, реагируя на поселяющегося в нем или па нем паразита, ока-
зывает на него те или иные воздействия, приводящие иногда к сущест-
венным нарушениям жнзисдсятельноети паразита.
Характер воздействия паразита на организм хозяина зависит от
ряда условий: с одной стороны, от вида паразита, его морфо-физиоло-
гических особен нос геи и вирулентности, а с ди гоп стороны, от видов.и
специфики хозяина, высоты его организации, локализации паразита
в определенных клетках, тканях и органа а также от того, насколько
быстро хозяин реагирует на воздействие паразита.
Рассмотрим прежде всего, в чем выражаются эти воздействия
паразита на хозяина. Их можно вести к следующим моментам.
I) механические воздействия', 2) отнятие у хозяина веществ, необходи-
мых для ею нормальной Лчизнсдеяте.тыюсти; 3) выделение токсических
в» ществ, действующих локально н >п на весь организм хозяина В целом;
4) открытие путей для проникновения внутрь организма хозяина паю
генных микробов или вирусов, .й) общее ослабление организма хозяина,
в результате чего повышение его чувствительности к другим заболева-
ниям.
Необходимо огмегпн., что эти разнообразные способы воздействия
паразита на хозяина в большинстве случаев взаимно связаны, и какие-
либо одно воздействие, как правило, мотке i привести к воздействию
иного характера; в результате затрагивается весь организм в целом
1. МЕХАНИЧЕСКОЕ ВОЗДЕЙСТВИЕ
Присутствие в организме чужеродного тела само но себе механиче-
ски действует на организм, а кроме того, паразиты часто наносят хо-
зяину механические повреждения при помощи своих органов прикреп-
ления u ni во время принятия нищи. Раздражение органов и тканей хо-
зяина .может причиняться даже просто движением паразитов по те.чх
хозяина. Так действует, например, движение бесчисленных инфузории
376
Рис. 207. Клубок гигантских ги.тнпиков
(Dioi-tuphyiiie rrnale'l внутри вскрытой почки
соваки: ипутрс-ппость почки совершенно
уничтожена параutravu. illo Эйсмопту.)
Trichodina н .н Chilodon на покровы рыб. Результатом раздражении
1 рыб является помутнение кожи, усиленное выделение слизи к «ahum.i
железами и т. д.
Некоторые кишечные паразиты достаточно крупных размеров могут
обусловливать закупорку и плохую проходимость кишечника. Это на-
блюдается, например, при наличии большого количества экземпляров
Asians iumbricoides в кишечнике человека, при заражении ленточными
червями Eubolhrium crassutn лосося или при заражении форели скреб
нем Echinorhynchus trutlac и т. и.
Еще легче закупориваются паразитами тонкие протоки ш которых
органов, например желчные npoToijji — печеночными двуустками. Такое
же воздействие могут оказы-
вать н легочные иематоты
(Dictyocaulus Muetterius, Ме-
taslrongylus и др )• От лечены
случаи закупорки бронхов пли
гортани большим количеством
нематод, свернувшихся в клу-
бок у телят, ягнят, поросят
Кроме такого во {действия,
некоторые паразиты вызывают
своим присутствием механиче-
ское давление на ткани, орга-
ны и соответственно приводят
к нарушению их функций. Так.
вайпик-гнгант (Dioctophymr
renalc), достигающий 1 .if дли-
ны, своим присутствием в иоч
ке собаки вызывает атрофию
большей части ткани почки, от
которой сохраняется только
тонкостенный мешок, содержа-
щий паразитов (рис. 207).
11 резвы чай но сильные из-
менения печени трески вызы-
вают поселяющиеся в ней ли
чинки нематод Conlracoecum и
Роггосоесит. причем ясно за-
метит! зависимость степени по-
.врежденип печени oi числа пива шровавшпх ее паразитов. У рыб едина
новой длины вег печени, пораженной I- 2 личинками, был равен 175 .
а при наличии |5—63 эк емпляров ее вес снижался до 5 Такая силь-
но пораженная печень резко уменьшается в размерах, съеживается, при
обретает неровные контуры (рнс. 208). а содержание жира в ней очень
сильно падает, Давление на ткани и их атрофию вызывают растущие п’
ырп Echinococcus granulosus в промежуточном хозяине. В случае чока-
лизапин эхинококка ио соседству с важными сосудами или нервами раз
растающинся пузырь своим давлением па эти органы вызывает иногда
тяжелые болезненные симптомы: били, нарушения кровообраще-
ния и т. д.
Еще сильнее ска зевается влияние локализации паразита на силе
его механического воздействия у мнзговнкя (Mulliccps muhiieps). ко-
торый паразитирует в мозгу ягнят. Давление на мозг и разрушение
мозгового вещества приводят к нарушению регу ininiii движений овны
(болезнь «вертеж») и даже к смерти ее.
377
Значительные механические повреждения тканей могут вызывать
те паразиты, которые снабжены специальными органами прикрепления
Среди простейших это особенно ясно видно у грегарин, отростки
эпнмернта которых внедряются или между клетками кишечного эпите-
лия или даже внутрь этих клеток. Моногенетическне сосальщики на-
носят покровам хозяина повреждения своими крючками (Dactylogyrus)
или присосками (Nitzschia и др ), а дигенетнчсские сосальщики—при-
сосками. Очень крупные желваки вызывает па стенках желчного пу-
зыря стрижа Platynosomiun dathratum своим присасыванием. Некото-
рые церкарии наносят повреждения покровам своим стилетом при про-
никновении в тело хозяина (Shistosomatidae). Даже яйца сосальщиков
Рнс. 208. Печет- oi грех экземплярец трески, имеющих
одинаковые размеры (67 г.ч длины) но в разной степени
зараженных нематодами Степень заражения возрастает от
верхнего изображения к нижнему. (По (Пульману и
Петрушевскому.)
являются иногда причиной механических повреждений. Снабженные
шипом яйца кровяной двуустки (Schistosoma haematobium), проходя
через стенки сосудов мочевого пузыря, раняг их, так что в моче появ-
ляется примесь крови.
Ленточные черви, обладающие сходными с трематодами и моногс-
кетпческнмн сосальщиками органами прикрепления, вызывают анало-
гичные повреждения у хозяина. На местах прикрепления Тriaenophortis
nodulosus к стенкам кишечника щуки получаются кратерообразные
углубления стенок. Еще более сильное внедрение в стенку кишки на-
блюдается у паразита кишечника китов Priapocephalus (см. рис. 96) и
некоторых других.
Сходные повреждения кишечника получаются и от паразитирова-
ния скребней. Большей частью особых видимых изменений скребни не
378
вызывают, но при массовом заражении (Echfnorhynchus sainionis \
сигов) стенки больших участков кишки покрываются грубокими вдав-
ленпями, а на вынутых из ямок хоботках паразитов всегда имеются
обрывки тканей хозяина. Всего сильнее воздействие иа хозяина таких
видов скребней, у которых шейка хоботка вздута в луковицу, например
у паразита рыб Pomphorhynchus laevis и паразита водоплавающих
птиц Filicollis anatis. Они вызывают глубокие впячцванцй стенки кишки
в полость тела. Среди нематод чаще всего причиняют повреждения
формы, снабженные шипами, зубами и крючками. Так, например, пара-
зиты рыб Goezia, птиц Acuaria н другие своими шипами переднего
конца тела вызывают повреждения кишки. Многие Dioctophymata
(Euslrongylides) прошивают своим телом стенку желудка птиц, вда-
ваясь в его просвет обоими концами тела. Человеческая аскарида
иногда пробуравливает стенку кишки и выходит в брюшную полость.
Власоглав глубоко внедряется сильно вытянутым нитевидным перед-
ним концом в стенку кишки хозяина.
Очень заметные повреждения у рыб своими органами прикрепле-
ния вызывают нередко паразитические рачки. Таковы, например,
Lernaeopodidac, у которых челюстные ножки сливаются концами и об-
разуют мощную луковицу или звезду, которой паразит глубоко вне-
дряется в мускулатуру хозяина.
Важно отметить, что механические повреждения нередко соче-
таются с принятием паразитом пищи. Это наблюдается при паразити-
ровании многих организмов, имеющих вооружение в ротовом аппарате.
Инфузории Chilodon cyprini вонзают в кожу рыбы залегающий
у них в стейках глотки палочковый аппарат. Dacfylogyridae питаются,
по-видимому, отчасти эпителием жабр рыб, пиявки проделывают в коже
хозяина глубокие простые или трехгранные (соответственно трем че-
люстям) ранки.
Некоторые кишечные нематоды могут активно внедряться в ткань
органа, вызывая нарушение его целостности. Например, Ancylostoma
duodenale повреждает слизистую оболочку кишечника своими зубами,
расположенными в ротовой капсуле. Другие нематоды, например Syn-
gamas merulae, фиксируются ротовой капсулой, имеющей форму при-
соски, к трахеям дроздов и вызывают образование небольших бугор-
ков на поверхности трахей, соответствующих по форме присоске черня.
Некоторые раки (Lernaeocera branchiahs) внедряются своим головным
отделом вплоть до брюшной аорты рыбы, служащей этому паразиту
хозяином. Artliropoda, как правило, нарушают целость покровов своими
ротовыми частями при укусах, а некоторые формы идут глубже и вы-
грызают подкожные ходы ( личинки И ohlfartia, клещи Acarus и др.).
Механические воздействия обыкновенно носят локальный характер,
но нередко повреждение какого-либо одного жизненно важного органа
сказывается на нарушении функции других органов и всего орга-
низма.
2. ОТНЯТИЕ У ХОЗЯИНА ВЕЩЕСТВ, НЕОБХОДИМЫХ ДЛЯ ЕГО НОРМАЛЬНОЙ
ЖИЗНЕДЕЯТЕЛЬНОСТИ
Эта форма воздействия связана, во-первых, с использование м
паразитом некоторых ингредиентов пиши хозяина и, во-вторых, с
питанием кровью и выделением а н т и к о а г у л и и о в, в резуль-
тате чего поисходит значительная потеря крови и уменьшение содер-
жания гемоглобина, что является причиной серьезных нарушений здо-
ровья организма хозяина.
3?ч
Частичное лишение хозяина отдельных ингредиентов пиши произ-
водится всеми кишечными паразитами, по результат этого воздействия
не всегда отчетливо выявляется и отражается на организме хозяина.
Можно отметить, что крупные паразиты или множественные скопления
более мелких кишечных паразитов оказывают наиболее значительное
влияние па организм хозяина, чем мелкие паразиты или встречающиеся
в малом числе.
Действи le.'ibiio, многие крупные ленточные черви (например.
Diphyllobothrium latum. Taeniarhynchus sagina^fs, Moniezia expansa
и др.) вызывают v хозяина тяжелые заболевания, обусловленные от-
части отнятием пищи. Эти черви обладают весьма быстрым ростом.
Например, но данным Штрома (1939), скорость роста Taeniarhynclius
saginatus в сутки составляет 7—10 см в молодом возрасте и 6—7 с.п
в зрелом. Естественно, что для этого требуется потребление большого
количества питательного материала хозяина, который ленточные черви
поглощают осмотическим путем. 4
Большое скопление паразитов в кишечнике может иногда вызы-
вать общее истощение организма. Так, например, Eubothrium crassum
собирается в кишечнике лосося в таком количестве, что вес червей в
одной рыбе достигает 200 ?; при весе лосося в 6 кг вес паразитов
составляет, таким образом, 3% веса хозяина. В результате наблюдается
задержка роста хозяина. То же наблюдается п при сильных заражениях
цестодами Moniezia expansa; шестимесячные ягнята недодают в живом
весе в среднем II кг. Эндрыос, Кауфман и Девис (Andrews, Kaufmann
and Davis, 1942) нашли, что количество пищи, необходимой для проду-
цирования ягнятами 1000 фунтов живого веса, приблизительно вдвое
более велико у искусственно зараженных нематодой Trichostrongylus
eolubrijormis особен, чем у ягнят, свободных от паразитов
Паразиты могут не только непосредственно использовать пищевые
материалы, полученные хозяином, но могут также выделять вещества,
являющиеся ингибиторами ферментов, вследствие чего нарушается спо-
собность хозяина перерабатывать полученные нишевые продукты в тон
мере, как это происходит у незаряженного организма. Интересно, что
некоторые кишечные паразиты могут отнимать v хозяина не только
пищевые вещества, но и необходимые для него витамины. Так, напри-
мер, Diphyllobothrium latum может накапливать в своих тканях боль-
шое количество витамина В, ., который синтезируется в кишечнике глав-
ным образом бактериями Escherichia coli. Лишая хозяина этою аитп-
апемнчеекого витамина, широкий тептец вызывает тем самым развитии
у человека тяжелой формы анемии.
Некоторые нематоды и отчасти трематоды, так же как и кровосо-
сущие насекомые и клещи, в процессе питания пог дотают у хозяина
значительные порции крови, что оказывает па него иногда весьма па-
губное воздействие. Кишечные гематофаги Ancylostoma duodenale я
/Vecalor americanus повреждают слизистую кишечника н вызывают
значительные потерн крови не только за счет заглатывания ее в свой
кишечник, но и за счет кровотечения из ран, наносимых паразитами
вследствие действия антпкоагулшюв поступающих из пищеварительных
желез червя. Каждая собачья анкилостома в сутки поглощает 0,7
0.8 лл3 крови, а человеческая анкилостома — 0,37 ллР. В связи с интен-
сивным заражением анкилостомами может развиться, анемия, при ко-
торой число эритроцитов падает иногда до 800 000 и гемоглобин до 12
15%. Гематофагия с соответствующими последствиями свойственна
также различным другим Slrongylata и, вероятно, аскаридам, власо-
главу и острицам.
380
Воздействие паразита, связанное с отнятием различных веществ,
необходимых для нормальной жизнедеятельности организма хозяина,
сказывается в сильной степени на половых железах хозяина. Голодание
хозяина, вызванное паразитом, который использует пищевые решествз
хозяина, сравнительно быстро отражается на строении и функции его
гонад, позже наступают дистрофические изменения в других органах
и, наконец, общее истощение. Более чунеч витсльными при этом являются
яичники, нежели семенники. Под влиянием паразитизма у хозяина на-
блюдается разрушение, дегенерация и, наконец, полная атрофия поло-
вых желез, а также изменение других частей полового аппарата и вто-
рично половых признаков. Для обозначения этих изменений широкое
распространение получил термин «паразитарная кастрация»,
введенный Жиаром (Giard, 1911- 1913).
Рис. _’О9. Грсгариаи Dlplodina goiutilipt-rihn, паразитирующая в
половых железах Cmwiiarie frandosa
/1—участок iinjoiuni трубки Cucumaria с пыл ляющимися на се
фоне светлыми грегарииа.мй; Н u С — поперечные разрезы семенника
Cticumuria с днуяирными грегаринамп </). вытеснившими большую
часть ткани семсиннкл; I) — две расположенных в ряд споры
Diplodiaa. (По Дьяконову.)
Следует отметить, что угнетающее влияние паразитов на гонады
хозяина нередко является для них выгодным биологическим приспособ
лением. Большое количество питательных веществ, которые в незаря-
женном хозяине используются для роста гонад и созревания половых
продуктов, паразит как бы переключает па себя, употребляя их дтл
своего роста и развития своих половых продуктов.
Во многих случаях паразитарная кастрация может обусловли-
ваться непосредственным воздействием паразита на половые железы.
При этом паразит может частично или полностью разрушать половую
/железу или вызывать лишь некоторые нарушения ее функции Так,
например, грсгарппы в семейных метках дождевых чернен, кокцндии
Eirneria sardiuae в семенниках сельдей, инфузория Orchitoplirya
riellanun в семенниках морских звезд, грегарииа Diplodina gonadiperihu
(рис. 209) в половых железах Cucumaria и целый ряд других паразитов
ч 1СТНЧ1Ю вытесняют и разрушают 1кань половых желез, ш вызывая
3Й1
полного нарушения половых функций, а лишь снижая плодовитость
В других случаях действие паразитов может быть более сильным.
По наблюдениям Кузнецова и Чубрик (1950) па побережье восточ-
ного Мурмана (Баренцево море) брюхоногие моллюски Littorina saxci-
tills сильно заражены личиночными стадиями трематод родов Podoco-
tyle и Spelotrema. Эти паразиты вызывают полную кастрацию моллюс-
ков. Процент зараженных литторин на некоторых участках побережья
восточного Мурмана достигал 52. Таким образом, паразиты оказываю)
большое влияние на размножение и -количественный состав популяции
моллюсков.
Личинки овода Cuterebra emasculator, поселяясь в мошонке амери-
канского бурундука Tatnias, вызывают разрушение семенников. Пред-
ставитель Orthoncctidae Rhopalura ophiocomae поселяется ио соседству
с гермафродитными железами Amphiura squamata, причем яичники
останавливаются в развитии, семенники же продолжают функциониро-
вать.
Разветвленные спороцисты Bucephalus polytnorphus, поселяясь в
гонадах пластинчатожаберных моллюсков сем. Unionidae, кастрируют
последних. ft
Степень влияния паразитов на половые железы хозяина в значи-
тельной мере зависит от соотношений размеров паразита и хозяина и
интенсивности заражения. Если хозяин велик, а паразит мал и числен
пость его не велика, то заражение обычно не вызывает заметного изме-
нения гонад. При малой величине хозяина наличие даже единичных
особей паразита уже может вызвать атрофию половых желез. Напри-
мер, личинки волосатика (Gordius), паразитирующие в крупных насе-
комых (плавунцы), не оказывают заметного влияния на гонады а в
мелких жужелицах эти личинки используют для своего питания пище-
вые резервы жирового тела, вследствие чего гонады хозяина не разви-
ваются.
Весьма часто наблюдается непрямое действие паразита на гонады
хозяина, причем паразит находится от гонад на известном расстоянии,
но тем нс менее вызывает кастрацию хозяина. Чаше всего примеры
подобного рода встречаются у членистоногих.
Так, на раках-отшельниках встречается паразитический усоногий
рак Peltopaster, а к нему присасывается равноногий рачок I.iriopsis
руртаеа (из Eptcarida). В течение всего времени, пока Liriopsis дер-
жится на Pellopaster, яичник последнего содержит лишь дегенерирую-
щие половые клетки.
При поражении шмелей самками нематоды Sphaerularia bornbi
созревание яиц у шмелиных самок задерживается и ослабляется их
инстинкт заботы о потомстве: зараженные шмели остаются праздными
в то время года, когда нормальные самки уже ревностно занимаются
сбором пыльцы и т. д.
Паразитарная кастрация может иногда сопровождаться различны-
ми внешними изменениями хозяина, как это показал сна-
чала Перез (Perez, 1886), а потом и другие на одиночных пчелах, за-
раженных насекомыми из отряда Sirepsiptera (Stylops или Xenos).
Подвижные личинки Sirepsiptera внедряются в личинок цикад или
перепончатокрылых (Andrena, Polisies, Sphex, Odynerus и мн. др.) и
проделывают в них весь метаморфоз (рис. 210) параллельно с развити-
ем хозяина, пе вызывая его смерти. Когда личинка перепончатокрылого
превращается в куколку, личинка паразита выставляет передни!'! копен
своего тела между двумя соседними брюшными сегментами хозяина
и остается в таком положении до окончания метаморфоза. Вскоре
382
после окрыления хозяина самцы паразитов выходят наружу в виде
очень мелких и черезвычайно подвижных летучих насекомых. Женские
личинки Strepsiptera испытывают мало изменении, так как самка этих
насекомых имеет вид неподвижного, личинкообразного мешка, внутри
которого развивается громадное число личинок. Вследствие этого Насо-
нов (1892) полагает, что размножение самки близко напоминает педо-
генез. В одном хозяине встречается иногда до 15, но чаще всего 1—2
паразита.
Рис. 210. Паразитические насекомые из отряда
SUepsiptera
Л — схема жизненного цикла Siresrplcra, паразитирующих
в перепончатокрылых; Н—самец A'enos vesparum
вылезающий из брюшка Poiistes gallicus на границе
между двумя брюшными сегментами. (По Ульриху.)
I — пчела с самкой Sircpsipiera, -горчащей наружу из границе двух
члеников; 2 - выползшие ил самки лимитен; .3 ' кэп зление лимит»:
Sirepsiplera па личинок пчелы: 4 — эндпгыразитпая стадия личинок
SlrepAfptcfti »дичинках пчелы; 5 то же и ее куколках; 6 — вылете-
вшие пчелы с паразитами; 7 паузит одних пчел остастсн в них
н лает нзрослую самку Strepsiptera. riapa.usr других мокихает
хозяина в виде карликового крылатого самца Slrepsiptera.
Некоторые виды поражаемых перепончатокрылых не показывают
никаких внешних изменений. Напротив, зараженные Strepsiptera особи
Andrena испытывают такие изменения, что их называют «стилоииро-
ванными» (от названия рода Stylops). При этом модификации самцов
н самок хозяина в общем сводятся к изменению в сторону противопо-
ложного пола, вследствие чего стнлопнроваппые особи принимают по
внешнему виду до известной степени характер интерсексов (рис. 211).
У самок Andrena наблюдается некоторая редукция аппаратов для соби-
рания пыльцы, потеря анальной бахромы, приобретение угловатых щек
(характерных для самца), появление желтого цвета на головном щитке,
редукция жала и ядовитого аппарата. У самца появляются длинные
щетинки, характерные для самки, задние лапки расширяются, образуя
зачаточный аппарат для собирания пыльцы, угловатость шск сглажи-
вается, головной щиток развивает черную окраску, вокруг порошицы
развивается бахрома, совокупительный аппарат уменьшается в разме-
рах. Получается как бы известный обмен признаками между обоими
полами. Стилопнрованные самки Andrena становятся менее активны и
нс собирают пыльцы.
3S3
Наиболее всесторонне изучено кастрнру юшес действие Saccuhna
на крабов Carcinus и Jnaclius, \ которых прежде всего изменяется под
влипшим паразитов весь обмен веществ. У самок Carcinus кровь нор-
мально бесцветна, но во время созревания яичников она желтеет, гак
Рис. 211 Пчели Andrena xolidaginis
I голова норма итого самца; В голова сти.гоннро-
ва л ого самца; С — гопнормальной самки. (!<>
Сольту.)
зараженных Sacculina, очень
Рнс. 212. .МодифнкацЬ] брюшка
крабов /nachtix mauritanuus над
влиянием плразнтнроиания в них
Sarrtilina
/I и D — нормальные самец и
сам» а краба Н. С п £— самцы и
самка зараженные паразитом
(По Сингу.)
как содержит до 0,2% жира, а также гетронзритрин и лютеин. У сам-
цов кровь бесцветна и содержит всего 0,06% жира, Содержание жира
в печени варьирует от 4 до 12%, причем больше всего жира у самок
с созревшими яичниками.
Так дело обстоит У нормальных особей. У крабов обои, полов
много жира в печени, а кровь даже у
самцов близка по содержанию жира к
таковой половозрелых самок. Следова-
тельно, паразитирующая на крабах Sac-
cuiina поворачивает обмен веществ хо-
зяина в сторону женского, оказывая на
животное такое же действие, как созре-
вающий яичник. Это изменение обмена
веществ отражается и на внешней мор
фолзгии крабов, особенно у Inachus.
Строение самок, обладающих широ-
ким брюшком с пятью парами дву
волосатых ножек, не ме-
самцы подвергаются уднвн-
измг-нению (рнс. 212). Hop-
имеют сильно
ветвистых
ияется. но
тельному
малыше самцы fnncliiis
суженное брюшко с одной только парой
палочковидных копу лятнвпых конечно-
стей па первом брюшном цементе. У за-
ражг иных саккулиной самцов имеются
Bci градации перехода к строению
брюшка, характерному для самки. КояуйяНшньТе ножки уменьшаются,
а на прочих сегментах брюшка вырастают ЛвуветПнстые перистые
ножки; иногда превращение становится столь полным, что различить
нол животного сдается лишь при вскрытии. Изменения сказываются
и ца некоторых других признаках, как. например, па клешнях, которые
v нормальных самцов бывают значительно более мощными 1аже после
отпадения саккулины Смит I Smith, 1910) находил при реконструк-
ции гонады самца и его половой железе молодые оноцнты, что показы-
вает, какой сильной перестройке подвергалась гонада во вр мя пара-
зи'1 ировання саккулины.
3S4
Те же явления, но в более слабой степени наблюдаются и у оаков-
птшельников, на которых паразиг.чрус! Peltogaster.
Можно привести еШе целый ряд нрнмцр'ов кастрирующего дечштвпя
н’р,'питон на хозяина. Таковы, например, веслоногие рачки Рсгоаегта
ulindricum, кастрирующие сардинок, или рачки Enterocoia из кишеч-
ника асцидий.
Встает вопрос, чем вызывается изменение хозяина в сторону дру-
гоп пола (или известная его интерсексуальность) иод влиянием пара-
рта. Ответить па этот вопрос трудно; некоторые данные говорят за ю.
что такие изменения могут иногда
Рлс 213. Нлпянпс паразитических не.ча-
то i на мураи ьск
2) мермпгэргат Pheidole cammutata со
спермуluiiHMi я и клууЬк AhTriiis sp.
iiayipii брюшка; /» типичный риёгЬчиИ
мураисн /’Л. coiiimuliulti. Д1и Иммсу).
происходить и без кастрации гонад
товялиа В насекомых thelia bitnas-
atlotu (Ilomoptera Mernbracidae)
паразитируют личинки наездника
Aphelopus ihtdiae. Под влиянием па
р.тлнтов Thelia би тыией частью ка-
стрируются и приобретают интерсек-
су а тьные признаки. Зараженные
самцы, достигая взрослого состоя
пня, принимают некоторые черты
самки: увеличиваются в размерах,
приобретают характерное распреде-
ление брюшных шипов и характер-
ную <рорх у брюшных сегментов, у
них происходит ум( ныпенне муж-
ского полового аппарата. Самка из-
меняется меньше, испытывая лишь
меньшение частей своего полового
аппарата. Однако в одном случае
корп.хаузер (Kornliauser, 1919) наблюдал, что «афелонпзировапиый» и
сильно измененный в женском направлении самец содержат семенники
нормл ibnoii величины с большим количеством енермиев, т. е. в данном
частном случае внешняя интерсексу атьность по зависела от кастрации
самих гонад.
Нечто аналогичное интерсексам ранее рассмотренных форм пред-
ставляют собой «интеркасты» известных видов муравьев, возникающие
под влиянием паразитирования в них нематод Mertnis. Они обнаружи-
вают смесь признаков пт двух разных полов, но различных каст этих
общественных насекомых. У этих интеркастов получается смещение
признаков половозрелых самок н рабочих, т. е. самок бесплодных.
Средн интеркастов можно различать мермитогпнов и мермитэр-
гатов (рис. 21-3). ЛЧсрмитогпна Xi и называются плодущие самки, зара-
женные Mermis. Они очень походят на нормальных «цариц», но обла-
дают редуцированными крыльями, а голова и-грудь их меньше нормы.
Их жировое тело не содержит жира, а яичники их недоразвиты. Ин-
стинкты их тоже несколько изменены, так как мермипчниы чч*чн. ак-
тивны н выполняют некоторые работы, свойственные рабочим. Уиллер
(Wheeler, 1928) полагает, что они происходят из шчинок, которые
должны были дать половозрелых самок.
Мермитэргаты — рабочие, измененные под влиянием Mering. Опи
крхпнес нормальных рабочих, могут развивать лобные глазки, имею-
щиеся, как правило, лишь у половозрелых самок, обладают окраской,
напоминающей цариц, и т. д.
Гёсвальд (Gosswald, 1938) обнаружил и модификации сахщов
муравьев под влиянием заражения мермитидамп, причем нередко зара-
25 в- Л. Догель
385
женне сказывается нс только в разбухании брюшка, но и и зпачпте.н
ном укорочении крыльев (v Lasius jlavas)
Видоизмененные под воздействием Meanis особи встречаются у
Lasius, Pheidole, Myrtntca, Camponotus и других Муравьев. По-види-
мому, вначале личинки муравьев педпффврепцнровалы и развиваются
затем в сторону той или другой касты, в зависимости от ко шчеств;
доставляемой нм пиши. Паразит вызывает у личинок нарушении обмена
веществ, а разные степени изменений зависят от того мом<9ита, ко; да
Merntis проникает в личинку. Чем рапыне происходит проникновении,
тем более значительны претерпеваемы'- муравьем изменения.
3 ТОКСИЧ1(ког воздействие паразита на хозяина
Дня хошина пс безразличны те продукты обмена веществ паразита
или нх секреты, которые поступают в его организм. Эти вещества прс»-
никают в кровь или лимфу, разносятся по всему телу и вредно денет
вуют на организм хозяина, вызывая различные нарушения в деятель-
ности его отдельных органов. Все эти веще/ тва. выделяемы* парази-
тами и влияющие в той или иной степени на организм хозяина, обычно
обозначаются наименованием «тексты». Выделение паразитом toi
сипов объясняют часто наблюдаемое при заражении явление, когд
паразит оказывает на хозяина не местное, а общее болезнетворное
в ши пне. Это наблюдается не только у эндопаразитов, но н у эктопара
знтов, питающихся кровью хозяина Они часто выделяют в кровь секрет
своих слюнных желез, нередко обладающий более или менее ядовиты
ми свойствами. Действие этого секрета может быть не только местным,
но и общим. Секреты и экскреты, выделяемые паразитом и поступаю
шие в кровь, могут действовать и как антигены, вызывая у хозяин
образование особых защитных веществ — антител. С ними связано раз-
витие у хозяина резнслентцости к соответствующему napa-girtTv* (см
гл XXI).
Остановимся кратко на различных формах токсическом
воздействия паразитов на хозяина, принимая во вп-м.
ние, что этот вопрос имеет в болинен степени медицинский и ветери-
нарный интерес.
Во многих случаях поступление в организм токсических вещеив
сопровождается изменением состава ф-о р м е н н ы х
элементов крови и в частности увеличенном количества эозшю
фнлов, что связано с нарушением нормальной деятельности клетного
мозга. Это наблюдается при заражении многими ленточными червями
Талызин (1949) отметил, что при самозаражении Taeniarhynchus sagi
natug эозинофилия достигла 16,5% и только после изгнания паразита
1 Недавно Киршцнб-яат 095»j высказал мнение, что термин «токсин» в отиоше
вин веществ, образующихся в теле паразитов и оказывающих разнообразные воздей-
ствия на организм хозяина, не яи тяется рацион» ’ьным Некоторые, так называемые
токсины, являются просто зкекретамн паразита, другие образуются после смерти парэ-
«нта в качестве продуктов его распада и не имеют непосредственного биологического
начення для паразита, третьи являются специфическими веществами, которые выраб;
тываются живым паразитом и воздействуют на организм хозяина, Нот для таких вс
щрстн Кчршеиблат предлагает новый термин — ксегаг’оны. Характер этих веществ
крайне разнообразен, и все они могут быть объединены в несколько полгрупп: (—гн-
‘-толпзгны, вызывающие нарушение целости тканей и имеющие важное значите для
паразита при внедрении его в тело хозяина и во время миграций; 2— ^антиферменты,
ирепятствулрщне действию ферментов, вырабатываемых организмом хохшпа; 3 — тро-
фогоиы, воздействующие иа организм, вызывая приток пищевых веществ к месту лока-
лизации паразита 4 — тнлакогеиы, вещества, которые вызывают реактивнее разраста-
ния тк анен :озяниа в области локализации паразита.
3S6
число эозинофилов снизилось ди нормы. Анкилостома, аскарида и не-
которые другие круглые черви выделяют гсмолитическн действующие
вещества. У Ancylosioma они выделяются головными н шейными желе-
зами. В результате при заражении этими червями часто наблюдается
значительное снижение числа эритроцитов. Это отмечается не только
у высших позвоночных, но и v рыб при паразитировании на них моно-
генетических сосальщиков и цестод. Например, Gyrodactylus elegant
вызывает у линя снижение числа эритроцитов в 2—3 раза в гемогло-
бина до 20 27% (Казадаев, 1954),
Наблюдаемые у человека, зараженного аскаридами, лептецем ши-
роким, гименолеписом и другими паразитами, различные нервные яв-
ления могут быть связаны с общей интоксикацией, вызванной наразн
тами.
Имеется много наблюдении, показывающих, что экстракты из па
разитов оказывают иногда сильное действие на организм хозяина
Полостная жидкость Ascaris вызывает сильное раздражение слизистых
оболочек человека, лошади и др. Еще более сильное воздействие полу-
чается при впрыскивании этой жидкости под кожу. Так, у собак при
впрыскивании 8—10 си3 на 1 кг веса наблюдается усталость, рвота,
пониженное кровяное давление, удушье, паралич задних конечностей
и даже смерть.
Экстракт, полученный из Taeniarhynchus saginaius в разведении
1 : 1000 и выше, вызывал на изолированном сердце кошки первона-
чально кратковременное возбуждение и последующее длительное
угнетение (Павловский и Дунаева, 1936) Этот же экстракт в меньшем
разведении действует угнетающе на моторную функцию кишечника,
а .затем вызывает тонизирующее действие (Талызин, 1949). Алфеева
(1941) установила сильную интоксикацию морских свинок при кормле-
нии их аскаридами. Интоксикация выражалась в дистрофическом ожн
рении подкожной клетчатки шейной части, области половых желез н
щитовидной железы, кровоизлияниях в мозговых оболочках, атрофии
селезенки, увеличении количества лимфоцитов, снижении числа эршро-
дитов, нарушении сперматогенеза и сокращении плодовитости.
Для членистоногих тоже известно выделение токсинов, например
для личинок кожного овода (Hypoderma). В Дании известно заболева
нне коров, которое называют розовой лихорадкой. Веспой у животных
распухают и отекают веки, морда, губы, вымя. Бродерсеп ( Bro<ierso.n.
1919) поставил эту болезнь в связь с искусственным выдавливанием
шчинок овода из кожи коров, которое производится в Данни весной,
а Йенсен (Jensen, 1919) проверил это предположение инъекцией гелей
кх вытяжки нз личинок Hypoderma. Уже через полчаса после инъекции
у теленка обнаружились все признаки розовой лихорадки.
Впрочем здесь острота реакций зависит, но-вндимому, не столько
от особой токсичности вытяжки из личинок, сколько от особо чувстви-
тельного, или анафилактического, по отношению к этой вытяжке состоя
ния, в которое приходят зараженные личинками кожного овода особи
скота.
Ядовитость секретов многих эктопаразитов также совершенно ш
сомнения, Кожные покровы специфически реагируют на поступле-
ние слюйы различных эктопаразитов в организм хозяшш. В одних
случаях реакция ограничивается лишь появлением воспалительного
пятна, в других—па месте укуса развиваются крупные папулы или
некротические везикулы. Сильное воздействие на кожные покровы ока-
зывает слюна клещей. На месте укола роговыми органами Orniihodorus
pupillipes на коже появляется сначала темно-красный узелок и затем
25* 387
развивается остры!! воспалительный процесс с образованием мощных
инфильтратов но сосудам в отеке дермы (Павловский и Штейн, 1936).,
Среди насекомых ядовитыми свойствами обладает секрет бобовид-
ных ( но не подковообразных) слюнных желез вшей (Pediculus). Через
rS—10 часов после инъекции человеку эмульсин из бобовидных желез
вшей на месте впрыскивания начинается сильный зуд и появляется ма
леиькая папула синеватого цвета, держащаяся до 4 дней. Если эмуль-
сию предварительно прокипятить, то эффекта не получается (Павлов-
ский и Штейн, 1923).
У постельного клопа слюнные железы содержат как ядовитое ве-
щество, так и аптикоагулпн. Секрет слюнных желез блох действует
в том же роде, как у вшей, но несколько слабее. Такие же ядовитые
секреты обнаруживаются в слюнных железах комаров, москитов, мошек.
Особенно это выражено у Simulium colunibaczense, укусы которого вы-
зывают образование припухлостей, отеков, При массовых укусах скота
этой мошкой самочувствие искусанного животного ухудшается, пульс
ускоряется, наступает удушье, а иногда и смерть ( на б—7-й день).
Среди клешей группы Ileterosligniata несомненное явление отрав-
ления вызывает у человека Pediculoides venlricosus, паразитирующий
на личинках некоторых амбарных вредителей (Tinea granella, Calandra
granaria и др.). Когда человек спит на зараженном субстрате (рогоже
и т. и.), эти клещики переходят на него и вызывают своими укусами
сильный зуд, крапивную лихорадку, а в тяжелых случаях даже настоя-
щую лихорадку и боли в суставах.
4. ОТКРЫТИЕ ПУТЕЙ ДЛЯ ПРОНИКНОВЕНИЯ ВНУТРЬ ОРГАНИЗМА
ХОЗЯИНА ПАТОГЕННЫХ МИКРОБОВ ИЛИ ВИРУСОВ
Мы уже. отмечали, что многие наружные и кишечные паразиты
производят своими ротовыми или прикрепительными органами мелкие
поранения на теле хозяина. Эти поранения могут являться воротами
инфекции для заболеваний, не имеющих ничего общего с данными па-
разитами На коже рыб. поврежденной карповой вошью и другими
рачками, поселяется паразитический грибок сапролегпия. Через ранки,
нанесенные этими рачками, в кровь карпов проникает, вероятно, воз-
будитель красну хи. Высказывается мнение (Kocylowski, 1952), что
повреждения слизистой кишечника карпа, вызываемые Caryophyllaeus
и кокипднямн, также способствуют возникновению \ рыб краснухи.
С прикреплением к слизистой червеобразного отростка человека
власоглава или остриц часто связывают некоторые случаи бациллярного
воспаления этого отростка (аппендицит). Весьма вероятно, чго такие
простейшие, как Entamoeba, Balantidium, Eimeria, вызывающие силь-
ные повреждения стенки кишечника, .могут способствовать проникнове-
нию патогенных бактерии внутрь организма.
Кроме того отмечается, что некоторые мигрирующие из кишечника
ио Te.iv хозяина взрослые паразиты или их личинки могут транспорти-
ровать патогенных микробов, способных вызывать обшес заболевание
хозяина или воспалительные очаги во внутренних органах. Николаев-
ский (1944) описывает своеобразный путь проникновения инфекции,
осуществляемый инфузориями нз рубца северного оленя при известных
условиях неформального его питания в летнюю жару, когда, с одной
стороны, подкисляется содержимое рубца и живые инфузории уходят
из его просвета, внедряясь в сгенку рубца и попадая в кровь; с дру-
гой— нарушается сокоотделение в сычуге (возникает ахилия) и инфу-
зории проскакивают в кишечник, но, не находя там благоприятных
388
условии существования, внедряю ica г> сошку кишечника и тоже nas'iiQ
сятся кровью по всему организму. В обоих случаях инн могут захва-
тывать с собой болезнетворных микробов. С этна явлением связывается
но ihhkhobciiiic у оленей заболевани i некрибацн.мезом.
Личинки Delafondta у тошадей при миграции заносят в oftranifcvfl
возбудителен паратифа, мыта и др., в рсзу.т .тате чего развиваете;! сеп-
тическое воспаление корпя брыжейки. Случаи гнойного воспаления,
с лизанного с проникновением патогенной флоры, наблюдаются при
внедрении личинок Oesopliagostonium в слизистую тол ?той кишки
Возможна также инокуляция микробов лччнпктмп паразитов (напри-
мер. Ancylostoma), проникающих через кожу. Косвенно об этпм евп
детельетву ет факт нахождения множества микробов па поверхности
личинок анкилостом Как предполагает Каваииши (Kawanishi, 19311
приставшие к личинкам бактерии могул быть занесены при перкутан-
ном проникновении личинок Внутрь организма хозяина.
Ц| лый ряд исследователей (Стефанский. 19551. однако, отмечает. чго
далеко Не так часто паразиты, повреждающие слизистую кишечника
или мигрирующие во телу хозяина, способствую! проникновению ми-
кробов внутрь организма хозяина. Действительно, при частом зараже-
нии сельскохозяйственных животных многочисленными паразитами,
повреждающими слизистую оболочку кнйГечинка, следовало бы ожи
дать множество случаев развития еешицпмип. как следствие инвазии
паразитами. Однако на самом деле такие случаи наблюдаются сравни-
тельно редко. Tai же и некоторые экспериментальные данные показы
вают, что паразиты пе всегда открывают путь для проникновения бак-
терий. Например, заражение кроликов личинками rricl!ostrongulu,>
retortaeformi ч и (iraphid'um strigosuni повреждающими соответственно
слизистую топкой кишки и желудка, при одновременном сведении kv.ii
туры Bacillus suipestijer н< вызывало Заболевания, несмотря на большое
количество нематод (Taylor, 1935) Одновременное заражение мышей
яйцами Ascaris suum и ку тьтурой возбудителя рожи свиней не привело
к повышению процента смертности этих животных но сравнению с
контр > тем (Stejanski, 1955). Можно, наконец, привести много фактоь
отсутствия случаев нагноения слизистой кишечника, поврежденной при-
крепительными аппаратами различных червей.
Таким образом, паразиты открывают «ворота инфекции» далеко не
всегда, а только при каких-то определенных условиях взаимоотношения
паразита и хозяина.
Паразит, воздействуя на хозяина, может вызывать у пего повыше-
ние чувствительности к другим заболеваниям как паразитарного, так
и иного проис'ождеиня. Отмечается, например, что мигрирующие
в паренхиме печени молодые печеночные двуустки снижают pt шетент-
ность хозяина. Попавшие в печень и находящиеся в латентном состоя
пни споры анаэробной бактерии Clostridium в результате повреждения
двуусткой печечт! начинают развиваться, н у овец наступает острая фор-
ма поноса (blacft diesease).
ГЛАВА XXI
РЕАКЦИЯ ХОЗЯИНА НА ПАРАЗИТА, ИММУНИТЕТ
Вполне естественно, что при тех разносторонних «агрессивных»
воздействиях паразитов па хозяина, которые имеют место, организм хо-
шина нс остается индифферентным к паразитам и даст па них ответные
реакции. Часть этих реакции не поддастся точному определению в смыс
те их биологического значения, другие реакции имеют определенную за
щнтную функцию. Эти реакции можно разделить на кзеточпые и ткане-
вые, и гуморальные.
I. КЛЕТОЧНЫЙ и тканевые реакции
Очень частым ответом па внедрение паразитов, в особенности мел-
ких. протозойного характера является гипертрофия одной или не-
скольких прилежащих клеток хозяина. Причины паразитарной ги-
пертрофии, вероятии, связаны с механическими и химическими воздей-
ствиями внутриклеточного паразита, в результате чего наступают функ-
циональные изменения инвазнрованпоп клетки. Происходит активация
клеточного метаболизма, что приводит к гигантизму зараженной клегки.
В качестве примеров достаточно указать хотя бы на кокцидню Са-
rtfotropiia mesiiili, паразитирующую в сперматогониях нолихеты Polym-
nia. и па ряд грегарин. Caryotropha (рис. 211) вызывает гипертрофию
клетки, и которой развивается, а также сильные изменения в соседних
сперматогониях. Грегарпна Clepsidrina davini, прикрепляясь энпмерн-
том к стежке кишки хозяина, приводит к слиянию в синцитий кишечных
клеток, которые соприкасаются с эпнмернтом; другая грегарпна, Slylor-
hyncfuis. прикрепляется к одной кишечной клетке, которая сильно гипер-
трофируется. Вокруг паразитирующих в субмукозе задней кишки Cepha-
lopoda кокпнднй Aggregate! дифференцируются особые гигантские клет-
ки, обособляющие паразита от окружающей ткани.
У грегарин Lithocystis schneideri, паразитирующих в полости тела
морского ежа (Echinocardiiun^ordatuin), подвижные стадии не вызыва-
ют никаких реакции со стороны тканей хозяина. Но, как только грсга-
рины соединяются попарно и инцистпруются, переходя к половому раз
множеиию, они окружаются сплошной массой оседающих на цисте тей-
з«и
тоцитов. Эти лейкоциты сливаются в общий синцитий и образуют торча-
щие кнаружи волосковидиые отростки.
Реакция на паразита часто выражается в воспалительных
процесс а х, развивающихся на месте его внедрения или в местах вре-
менной или постоянной локализации. Эта реакция со стороны хозяина
носит защитным характер и может иметь различное проявление. Очень
часто вокруг паразита образуется соединительнотканная капсула, кото-
рая более или менее изолирует паразита от окружающих тканей. Обра-
зование подобных капсул является правилом для большинства покоя-
щихся стадий паразитов, проводимых ими в промежуточных хозяевах;
Рис. 214. Клеточные реакции хозяина по отношению
к rpernpiiii.iM и кокцидиям
zl — Caryotropha nwsnili. внутри гигантском клетки
хозяина; вокруг более пли менее измененные снер.чато-
гоннн хозяина; — Clepxidrtna <! acini. вы «ыв.тюшля
своим эпимер >им образование синцн ия в эпителии
кишки хозяина; С грегарппп Siyfi>rhyn> has longicoilis.
вызывающая споим эпнмернтом гипертрофию ki щечной
клетка хозяина. (По Коллери.)
I гмрдзпт; 2 — эпимер', г Vy.'rjr/iynr/r«y; J - гпн е|прифн>к>п;| пи ан
клетка хозяина.
метацеркарии сосальщиков, личинки нематод, скребней. Обыкновенно
вокруг проникшего в известную ткань ii.ni орган паразита происходит
расширение сосудов, образуется эксудат, накапливаются лейкоциты.
В некоторых случаях, например у Trichinella spiralis, образующаяся во-
круг паразита капсула имеет сложную структуру п снабжается со сто-
роны хозяина сетью капилляров и нервными окончаниями (Березаицсв
1957, 1961). Личинки кожного овода, внедряясь в ткани, вызывают вос-
паление, приводящее к некрозу.
Пногд^ в стенке соединительнотканной капсулы отлагаются соли
извести (Trichinella). Образование вокруг паразита капсулы не мешает
его жизни н даже росту. Впрочем при длительном пребывании внутри
капсулы паразиты нередко в конце концов погибают, как это наблюдает-
ся у плероцеркоидов DiphyUobolhrium latum в полости тела щуки. Кап-
сулы образуются в каких угодно органах, ибо соединительная ткань
имеется повсюду.
Особый гни реакции тканей хозяина на паразита представляет обра-
зование вокруг активных стадий паразита особых разрастаний, напоми-
нающих галлы, вызываемые насекомыми на растениях. У животных по-
добные образования получая^ название тилаций (термин Жиара
(Giard, 1913)) или з о о п е ц п л и е в. На месте локализации паразита
ткань хозяина образует вокруг последнего большое вздутие или разра-
стание, полое внутри; в полости помещается паразит. От опухолей или
391
капсуч зооцецидии отличаются постоянством формы и некоторыми осо-
бенностями строения, придающими образованию известную полярность.
Чаще всего зооцецпдин вызываю ня некоторыми эктопара штамп.
Оседая на поверхность тела хозяина, они заставляют эту поверхность
податься вглубь, образовать глубокое впячнвание, карман, сообщаю-
щийся с внешней средой более или менее узким отверстием. Это впячи
ванне и есть зооцеццдин Очень хороший пример таких отношений лает
равноногий рачок из семейства Cymothoidae—Livoneca anturensis, пера-'
зитнрмощий на некоторых карповых рыбах реки Амура (А- миров, 1939).
Другие представите!и С\muthoidac живут в качестве типичных эктопа
разнтов на жабрах ii.au па коже многих морских рыб. Livoneca атигеп
sis (рве. 215) прикрепляется к телу рыбы на уровне грудных плавников
Рис. '.’15. Нар.тпическш ранионогн-fi рачок Livoneca amnrensis
А — амурская рыба Leiicisrus walcckii с равноногим парази-
тическим рачком Livoneca arnnrensis, торчащим нз глубокое»
ко;кного ииячивання нот группам плавником /Т— самка
Livoneca, nuiiyiaH in ииячивання. [По Ахмерову.)
/. L.amurensls.
и вызывает образование глубоко!о, тонкое генного впячивання. далеко
вдающегося в полость тела. Внутри впячивання обыкновенно помещаю!
ся самец и самка паразита, причем самка (до 3 с.ч длины) па-атолькл
раздувается от переполняющих ее янц, что уже не может покинуть зоо-
цеиндий через узкое отв< рстне впячивання. Аналогичную картину даю г
равноногие рачки Bopyridae. поселяющиеся в жаберной полости десяти-
ногих раков. Бесформенное тело паразита вдается в полость тела хо [чи-
на, но на самом деле повсюду отделено от нее тонкой стенкой кармано
образного зооцецидия Веслоногий рачок Pionodesmotes phormosomae
(рис. 216) производит подобие настоящих галлов на мягкотелом мор-
ском еже Phormosoma. Вокруг Pionodesmotes образуется вдающееся в
полость тела ежа толстостенное1 шаровидное* впячнвание, в стенках ко-
торой откладывается большое количество углекислой извести: полу
чается шаровидный галл. Входное отверстие этого юощ-цндия так узко,
что сидящая1 внутри крупная самец Pionodesmotes оказывается замуро
ванной в нем. Подобным же образом копепода Astrochordeuma живет в
офнуре Aslrochuris gracilis (рис. 217).
Яичники некоторых мух та.хпн (Tachinidae), внедряющиеся в тело
насекомых, образуют на теле последних мубокое впячнвание. в котором
личинка проделывает первые линьки, будучи обращена дыхальцами к
выходу из впячивання.
Зооиспндни образуются и некоторыми из брюхоногих мол носков,
паразитирующими в игтокожих. Таков Megadentis arrhynchns, парази-
тирующий в стенке тела звезды Antedonoides rugulosus; вокруг паразита
стенка тела сильно утолщается .<1 счет увеличения скелетных пластинок
392
Megadenus делает больниц галлы в иглах одного морского ежа; вздутш
только узким отверстие и сообщается с внешним миром, а внутри впячп-
вания сидят самец п самка Megadenus.
Наконец, очень многие представители
Myzostomida образуют на пннну чах, или
на рукдх, морских лилий камеры с утолщен-
ными стенками, сообщающиеся с внешней
средой узким каилдом. По наличию таких
зооцециднев на ископаемых Ciiiioidca юр-
ского периода можно уверенно говорить о
существования MyzostonBcki по меньшей
мере с юрского периода.
Приведенные многочисленные примеры
пока’ывают одну любопытную черту целый
ряд паразитов нз различных групп живот-
ного царства образует юоцецндин па пред-
ставителях одного типа, а именно: па игло-
кожих. Поэтому мы можем говорить, что у
разных хозяев имеется в разной степени вы-
раженная предрасположенность к образо-
ванию зооцециднев. С другой стороны, при-
веденные случаи показывают, что формиро-
вание зооцециднев нс есть специфическая
Рис. 2;ь. Гi,ipa3iiTif<iecKini вес-
лоногий рачок J'ionodcsmolrs
phorinuxomac
Л - участок панциря морского
ежа I’hormosama uranus (пил
изнутри) с Apiiciore.icHijM фона-
рем и miioiчисленными iiiapo-
iii 1Ш.1МИ галлами рячка Hiniu>
tiesmoles pin>rm<fsumae’, В —
внутренность отдельного галла
с самкой рачка Pivnodesmvit’x.
(По Бочнье.)
/ — 111*фО|ШДШ4С ГЛЛЛН, е — ApllCWTC-
леи фонарь.
черта определенных паразитов, но скорее
специфическим ответ онредс icinibix хозяев
на раздражение какими угодно паразитами.
Несколько реже зооцецидпн возникают
под влиянием эндопаразитов. Так, тремато-
ды Hallouria monogatna (рнс. 218) нз ки-
шечника марабу образуют шаровидные вы-
пячивания стенки кишки, сообщающиеся с
кишечником лишь тонкий порой; внутри вы-
пячивания помещается всегда пара парази-
тов. Аналогичные дивертикулы в кишке гры-
зу нов образуют нематоды Heligtnosomides.
а нематода niarioides bronclnalis —
в бронхах хорьков.
Запо. >ающие внутрь i асгра ть-
мой полости колониальных гидрои-
дов личинки некоторых пантопод,
например PluixichVidiuin, разраста-
ясь, вызывают в местах своей лока-
лизации сильные вздутия ствола ко-
лонии.
О причинах образования зооце-
цндиев мы еще плохо осведомлены,
по. судя по аналогии с растительны-
ми галлами, они являются резуль-
татом реакции тканей хозяина на
химические и механические раздра-
Рпс. 217. Вгслоно! nil рачок Asirochor
deuma appendiculoxtHn
галл н лучах офнуры Astrocharix
gracilis’ В — вскрытии галл с паразитом
внутри. (Но Стефенсону и Мортенсону.)
жения со стороны паразита.
Наряду с зооцецидиями можно упомянуть о жемчуге, как резуль-
тате во [действия некоторых паразитических червей на мантию Lamelli
branchia. У целого ряда пластинчатожаберных моллюсков мелкие яи-
чники пюских червей, [[поникающие в промежуток .между наружным
ЗУ’.
жемчужина. Личинка,
Рис. 218. Сосальщики Hal-
fouria monogaitia, лежащие
попарно в цисте кишки
optilas eminent fe г. (Ио
Кюкенталю.)
Drascheia megastoma
слоем эпителия мантии и створками раковины, окружаются отложениями
перламутра и замуровываются в блестящий перламутровый шарик.
В большинстве случаев, например у Mytilus и других, слой перламутра,
окружающий паразита, очень тонок и прозрачен; однако у некоторых
морских пластинчатожаберных, например Meleagrina. личинка паразита
окружается многочисленными концентрическими слоями перламутра
так, что на месте инкапсулированной личинки получается настоящая
давшая толчок к образованию жемчужины, в
конце концов погибает и рассасывается, так
что на месте ее остается едва различимое цен-
тральное зерно. Жемчужины образуются во
круг метацеркарии трематод, а также вокруг
личинок некоторых ленточных червей (Tetrar-
liynchidca) Tylocephalum margaritiferum.
Взрослый Tylocephalum живет в кишечнике
скатов, которые в значительной степени пита-
ются моллюсками.
Представляет интерес неопластиче-
ская реакция тканей х о з я и н а, воз-
никающая в результате воздействия паразита.
Отмечено, что некоторые паразитические черви
могут вызывать новообразования как в резуль-
тате токсического, так и механическо-
го воздействия. Примером может служить
(подотряд Spirurata), паразитирующая в же-
лезистом отделе желудка лошади и вызывающая в нем опу-
холевидные образования величиной до куриного яйца. Нематоды
Gonyylonema neoplasiicum локализуются под слизистой оболочкой пи-
щевода или желудка крыс и вызывают иногда образование палиломы
или типичных раковых опухолей желудка, способных давать метастазы
Злокачественные опухоли образуются также и в печени грызунов, зара-
женных цнетннеркамн Hydafigera tacniaeformis (Cysticercus fasciolaris).
При шистозо.матозах человека нередко образуются карциноматозные
разрастания в мочевом пузыре и прямой кишке на почве раздражения
тканей огромным количеством яиц этих паразитов. Имеются данные, что
у лиц, зараженных Clonorahis sinensis, раковые опухоли встречаются в
3 раза чаще, чем у людей не нивазнроваиных.
2 ГУМОРАЛЬНЫЕ РЕАКЦИИ
Гуморальные реакции со стороны хозяина — явление широко распро-
страненное. Эти оеакцни вначале были выявлены по отношению к бак-
териям, позднее по отношению к паразитическим простейшим, а в на-
стоящее время доказано, что такие же реакции вызываются и паразити-
ческими червями. Эти реакции тесно связаны с явлением иммунитета н
заключаются в образовании в крови хозяина особых, специфичных по
отношению к различным паразитам защитных веществ — антител. Вы-
работка таких антител в организме хозяина доказывается многочислен-
ными опытными данными. Так. ft. :и простейших' весьма показательна
реакция агглютинации и преципитации. Если кролику впрыскивают в
кровь культуру Leishmania. то его кровь" в дальнейшем приобретает
свойство убивать этих жгутиконосцев. Если к жгутнконосным культур-
ным стадиям Leishmania прибавить кровяной сыворотки такого кролика,
то жгутиконосцы агглютинируются, т. е. слипаются в комки, и прецлпи-
тируются, г. е. осаждаются иа дно стекла; в крови кролика, следова-
тельно, выработались антитела.
394
Некоторые паразитарные инвазии Делают Животное сверхчувстви-
тельным ио отношению к антигену, производимому данным паразитом.
Так, если под кожу подозреваемого па зараженность известным па-
разитом животного впрыскивают очень небольшое количество антигена,
приготовленного из этого паразита, го в случае зараженности живот-
ного на месте инъекции появляется краснота, отек и образование вол-
дырей. Если животное не заражен-. то никакого внешнего эффекта от
ш.рыскивапня не получается. Путем такой аллергической реакции диаг-
ностируются, например, эхинококкоз и аскаридоз.
С формированием антител в организме хозяина связано развитие
резистентности в отношении паразитов; при помощи этой защитной реак-
ции хозяин либо полностью избавляется от паразита, либо препятствует
его вредному влиянию
3. ИММУНИТЕТ
В настоящее время различают две основных формы иммунитета:
врожденный (или естественный) и приобретенный. Врожденный иммуни-
тет включает отношения между паразитом и хозяином не связанные с
предшествующим взаимодействие:! организма хозяина с данным видом
паразита. П риобретенный же иммунитет развивается как результат реак-
ции микроорганизма на внедрение соответствующего паразита. Поэтому
совершенно естественно, что при врожденном иммунитете хозяином при-
водятся в действие совершенно другие механизмы защиты нежели в
случае приобретенного иммунитета. Если в приобретенном иммунитете
главную роль в защите игратот специальные иммунологические факторы
(развитие специфических антител, фагоцитоз), то в случае врожденного
иммунигета ведущее значение приобретает морфо-физиологическое и
биохимическое состояние организма. К ним может быть отнесено дей-
ствие* желудочных и кишечных энзимов, уничтожающих большую часть
инородных организмов, проходящих Ч( рез пищеварительный тракт, сте-
пень кислот постя желудочного сока п соков в кишечнике, неблагоприят-
ная для развития паразита, биохимическая, не пригодность тканей для
существования паразитов, гормональное состояние организма, не спо-
собствующее* развпти!о|паразнтй, структура кожных покровов, препят-
ствующая проникновению партзнтои, и т. п. Немалую роль играет в
развитии врожденного иммунитета также* и активность клеточных реак-
ций, проявляющаяся в фагоцитозе.
П p:i наличии а б с о л ют но й врожденной н е в о с п р н и м ч и-
в ост и невозможность развития паразита в том или ином животом
обусловливался полным несиогветсгвш м условий Для его существова-
ния, н это явление не связано с выработкой антител. Тот факт, что ни
один вид паразита не может заражать всех хозяев, а встречается только
у определенных видов, всегда отсутствуя у других, зависит именно от
того, чго паразит не в каждом животном, в которое проникает или
может проникнуть его инвазионнаястадия, находит необходимые усло-
вия для своего развития. У одних животных их морфо-фпзиологиче-
скпе н биохимические особенности могут оказываться вполне подходя-
щими для осуществления жизнедеятельности паразита, и тогда происхо-
дит формирование системы паразит—хозяин, у других эти особенности
не обеспечивают существования паразита, ir поэтому- данное животное
не становится хозяином соответствующего паразита. В таком случае
мы скажем, что это связано с врожденной невосприимчивостью, или
врожденным иммунитет ом.
При этом следу с г иметь в виду, что не всякое отсутствие зараже-
ния хозяина данным паразитом свидетельствует о наличии у Него врож-
ЗЭ>
денной нс восприимчивости. Это может быть связано с определенными
условиями, которые препятствуют встрече данного вполне восприимчи-
вого животного с нивазнонны ш стадиями развития паразита. Напри
мер, трихинеллез распространен средн всеядных животных, грызунов п
плотоядных, i травоядных трихинелла не встречается, чти определяется
способом питания этих животных. В эксперименте же травоядные легко
заражаются трихинами. Поэтому наличие врожденnoil невоенриимчи
востн .может быть доказано только после попытки экспериментального
заражения животного.
Абсолютная врожденная невосприимчивость свойственна обычно
всем особям какого-либо вида животного, вследствие чего ее можно
называть видовой невосприимчивостью. Одним из ее ирояв-
Ленин является специфичность паразитов в отношении определенных
хозяев, что более подробно будет рассмотрено в следующей главе.
Врожденная невосприимчивость может наблюдаться как в отнонн-
пин всех стадий развития паразита, так и в отношении только какой-
либо одной стадии. Например, ни одна стадия развития бычьего соли-
тера никогда не развивается в организме собаки, а в человеке парази-
тирует только половозрелая стадия, но не личиночная. Крупный рога
тын скот невосприимчив к половозрелой стадии эхинококка, и в то же
время легко заражается личинкой.
Такая абсолютная невосприимчивость в отдельных случаях при нс
которых условиях может быть «прорвана», т. е. преодолена паразитом.
В таком случае животное может оказаться зараженным паразитом, не
свойственным с му в обычном состоянии. I с..и жаб или саламандр со
держать при температуре 35”, то при экспериментальном заражении нх
личинками трихинелл можно подучить нс 1ово»релы.х червей и в даль-
нейшем мышечные формы трпхнпел i (Matoil, 1911). В то же время при
низкой температуре никакого заражения получить нс удается.
Наряду с полным несоответствием условий в организме животного
для существования партита могут иметь место такие взаимоотноше-
ния паразита и хозяина, когда паразит хотя и преодолевает соиротпн
леине хозяина, но развивается в нем только до известных пределов, и
в этих случаях происходит большее и ш меньшее ограничение; его жизне-
деятельности. Это явление относительной врожденной не-
восприимчивости может выражаться в том, что развитие пара-
зита в той итн иной степени задерживается, хотя некоторые особи все
же могут достигать иоловозрслости. В других случаях паразит не до-
стигает половой зрелости и проделывает в нммх ином хозяине лишь на-
чальные стадии развития. При этом не развившиеся окончательно пара
знт либо удаляется из хозяина, либо инк^су тируется в его тканях.
Примером может сложить новедс нш личинок Ascari.x liinibricoules
итн Parascaris equorum и др. в лабораторных животных, которые в
природе являются относительно иммунными в отношении этих парази-
тов. Эти личинки мш рнрхют через печень в легкие, по разщгтня до поло-
возрелого состояния не происходит, и часть личинок инкапсулируется
главным образом в печени, другие же выводятся наружу при мшрации
через легкое. Аналогичное явление наблюдается и в отношении Ancylosto-
ma cLivniun при искусственном заражении крыс. /I caninum у своего
нормального хозяина — собаки — при заражении через рот не мигри-
рует. но в организме крысы или морской свинки, или даже человека
личинки ее проделывают полную миграцию, во (вращаются в кишечник
и остаются там некоторое время (у крыс до 21 дня), не обнаруживая
дальнейшего роста, а затем личинки выводятся с испражнениями во
внешнюю среду.
396
Весьма вероятно, что именно с явлением относительного врожден-
ного иммупнюга можно связать феномен м и г р а ц и н л и ч н и о к мно-
гих гельминтов внутри организма хозяина. Во многих i пчаях. если не
во всех, причиной такой миграции является «стремление» личинок- из-
бегнуть гибельного действия кишечного содержимого на сше очень мел-
кие и нежные личинки, находящиеся па ранней фазе своего эндопара-
игичискчго периода жизни. Следовательно, переход к .миграции личинок
многих паразитических черве.i. есть не что иное-, как «реакция бегства»
паразита нз неподходящей среды. Такая миграционная способность у
личинок некоторых нематод проявляется лишь в том хозяине, который
шляется по отношению к ним относительно иммунным, тогда как в
об штатном .хозяине, т. е. ирётостав.iflioiilCM все условия для нормального
развития паразита, они не мигрируют (анкилостома в человеке пли со-
баке при заражеимм через рот) и пеноср< тстенно в кишке достигают
полосой зрелое!и.
Относительный врожденный иммунитет имеет, несомненно, наиболь-
шее значение в возникновении ренрвуарных хозяев для некоторых па-
разитов (см. главу XI). Наличием такого пммунитега объясняется ни
капе лироваиие личиночных стадий в ре^?рвуарном хозяине. Иогтсд-
ние не обладают необходимыми условиями для развития в них кишеч-
ных паразитов до половозрелого состояния. в( .едствне чею личинки,
попавшие в такой организм, нс задерживается в кпшечнихе, а ухолят
из него в полости, юла и далее инкапсулируются где-либо во виутрсн-
И1.х органах, что можно рассматривать как проявление защитной ре-к-
ним организм,) хошина. При этом личинки длительный срок сохраняют
жизнеспособность и пива iiioiinoeib. Нанрпхкр, личинки Phy^oceplialus
и таки ре п рвуавиых хозяевах выживают до 6—7 месяцев. Если эти ли-
чинки скармливать свинье (окончательный хозяин), то в ее кишечнике
сии превратятся в половозрелые стадии, а при скармливании относи-
тельно имму тому озяпну, лп‘.\1ки вновь инкапсулируются.
Быть может, и вторых промежуточных хозяев можно до некоторой
стопеич рассма грива ;ь как организмы с резко выражнтым врожден-
ным иммунитетом, препятствующим достижению паразитом потовой зре-
locT.i. Это гем более вероятно, что \1Ы знаем целый ряд примеров про
। епеза (см. главу X), когда в промежуточном хозяине метацеркарии
дос гига in половой зрелости. В этих случаях ослаблялось тормозящее
влиянии на иар.’зпга со стороны хозяина, и гем самым развитие паразита
полностью завершалось. Наличие врожденной невосприимчивоегн у про-
межуточного хозяина к личиночным стадиям паразита выражается еще
н в том, что далеко не каждое животное способно заражаться соответ-
ствующим паразитом и при заражении иногда наблюдается небольшой
процент развития личинок. Кроме того, пе в!^ личинки достигают в
таком хозяине инвазионного состояния. Так, например, при заражении
36 ягнят онкосферами Multi,eggs miilticeps у I I нс было обнаружено пу-
зырчатых стадий, кроме того, при введении животному 500 онкосфер
образоватись всего лишь единичные пузыри и некоторые из них оказа-
лись мертвыми (111 льц н Бондарева. 1955).
Рассмотрим теперь с т у ч а и о т и о с и т с л ь и о г и в р о ж д с li-
no г о и м м и итога, при котором паразит ы, несмотря на за-
труднения и ограничения в развитии, все же достигают полово-
зрелого состояния. Хозяин при этом но предоставляет паразиту
необходимых для его нормального существования условий и оказывает
в большей или меньшей степени неблагоприятное влияние на проникаю-
щего в пего паразита. Это в зияние, которое испытывает паразит си сто-
роны хозяина, выражается в следующих проявлениях.
397
1. Понижение Звр^жа'йыостн животных как в отношеййи экстенсив-
ности, так и нип мощности. Относи гслыю имму.шыо животные зара-
жай тся в меььиш-м проценте и число развивающихся паразитов у них
сравнительно невелико Гак, кОнЯн оказались менее восприимчивыми к
A’icylosioma dtiodeuGle, чем собаки iScoit, 1928), мыши менее, чем
крысы, к Nippostrongylus tnuris и т. и.
2 Уменьшение размеров паразита одного вида как проявлении отно-
свтельного иммунитета хозяина может быть иллюстрировано на пример!
Diphyllobothrium latum из человека, где паразит Достигает 6 -9 .« и нз
кошки где он, как правило, не превышает 1.5 .и. Opisthorchis fclirteus
в желчных протоках кошки имеет меньшие размеры, чем тог же вид из
печени человека.
3. Удлинение сроков, необходимых для развития паразита до полов»,
зрелого состояния, наблюдается, например, у Aucylnslmna caniumn в
кошках где этот срок равен 17 дням, в то время как у собаки нормаль
пая скорость развития 11 дней.
4. Сократите срок жизни паразита наблюдается, например, у
Diphyllobothi чин latum и организме кошки, где они живут только 3
1 недели ГЦ 1 н.х об а лов а. 19501, в то время как у человека этот паразит
живет голами. Длительность жи ши Slroagyluides sierajrplis в кошках
2- 7 недель, в собаках 3- 7 месяцев и в человеке много лет,
5. Понижение плодовитости паразита очень ясно выражено у Апсу-
lostoma CMiiruim. паразитирующей у к>иш к. В этом отчине каждая
самка в среднем чткладывлег в суптЯ 2300 яиц, тогда как в собаке про-
дуктивность достигает 16 000 янц в сутки
6. Огранит; пне жизнеспособности яиц и личинок паразитов. Так;
например, у Diphyllobothrium latum, паразнтир' ющего в собаках и кош-
ках. всего лишь около 10% яиц способно образовать корацнднн, тогда
как 90% яиц этого же паразита из человека заканчивает развитие. Тот,
ко 28% яиц. отложенных Aniylostoma cuninum. нз кошки, образуют ли
чинки. Яйца, полсчегзтцс от этого же паразита из собаки, были жпзнс
> пособным‘1 на 89%.
Приведенные примеры, число которых можно .значительно умни-
жить, свидетельствуют о видовой относительной резистентности. Между
тем имеются наблюдения, указывающие на различную степень устончн
вости к одному н тому же виду паразита у разных иоро i или лаже раз-
ных особей в пределах одного вида животного. Так, например, норе гы
овец архаромериносов в Казахстане значпжлыю б„тее устойчивы ь
ценурозу, чем г] хбошеретные местные породы. Рида ила иды, плимутроке
более усчийчнвы к Asccuidia lineata. чем леггорны и минорки. Иидквп
дуальная резистентность отдельных особей мышей была обнаружена П!н
'обаловоп (19491 при заражении Trichocephalus muri ? Давтян (19491
наблюдал повышенную устойчивость у отдельных особей овец при искус-
ственном заражении их Dictyoealllus i&uria. В таких овцах развитие па
разнта до прловозрелостн длилось 18—73 дня. тогда как обычно он»
развивались 36 - 41 дня. Личинки у них выделялись только 21 33 дня по
сравнению с обычными сроками в 86 180 диен.
Индивидуальный иммунитет в общем менее прочен, чем видо-
вой. Под влиянием внешних условий (голодание, утомленно, сопут-
ствующая инфекция и т. и.), так ши иначе ослабчяюших общее состоя
ине организма, наблюдается снижение индивидуальной ре шстенгиостн и
в некоторых с ту чаях полное ее снятие. Это имеет место и при относи
тельном видовом иммунитете, о че.м будет также сказано ниже.
Важным фактором, влияющим на врожденную невосприимчивость,
является вснрасг хозяина. Как правило, молодые животные более вое
39*
нринмчнвы к паразитам но сравнению с» старыми н, кроме того, моло-
дые болеют часто более сильно, чем взрослые хозяева. Это объясняется
дем, что но мере роста и развития организма у животного повышается
естественная невосприимчивость к паразитам и их токсинам, не связан-
ная с перенесенной ранее инвазией, но зависящая от возрастных изме-
нении общего физиологического состояния организма. Эта повышенная
резистентность взрослых животных определяется как возрастной
иммунитет. У молодых животных защитные механизмы либо отсут-
ствуют, либо действие их ослаблено, по ср ibiichhio с тем как они про
являются х взрослых организмов.
Возрастной иммунитет проявляется в ограничении заражаемости па-
разитами. замедленном развитии, сокращении срока пребывания пара-
зита в хозяине и в ослаблении патогенного эффекта. Среди кишечных
простейших возрастной иммунитет известен по отношению к Lamblia и
многим представителям кокцидий родов Eimeria и Jsospora. Папболег
восприимчивыми к заражению Eimeria tenellu являются цыплята в воз
расти до 3 месяцев, так же как Е. magna и Е intesfinalis развиваются
обычно у 3 1-месячных крольчат.
Взрослые куры и кролики если и заражаются кокцидиямп, то раз-
множение последних в них происходит в ограниченной степени, и ника-
кого заболевания при этом ие вызывает. Так же и овцы, крупный рога-
тый скот и другие животные, как правило, наиболее восприимчивы к
кокцидням в молодом возрасте. 11моются, однако, случаи, когда кокцн-
дип чаще встречаются у взрослых, нежели у молодых животных. Это
относится к Е. necalrix нз кур в Е. stiedae из кролика и др. В отношении
кронсиаразитов отмечается, что молодые крысы более чувствительны к
заражению Trypanosoma le&isi, чем взрослые.
Много примеров возрастного иммунитета можно продемонстриро-
вать в отношении паразитических червей. Ограниченная заражаемость
Нymenolepis fraterna наблюдается, цапрнмер, у крыс в возрасте свыше
трех месяцев В опытах Данскер и Пик (1937) крысы до трех месяцев
заражались на 80%. а более старшие-—на 12,1%. В то Же время имму-
нитет к 11. пипа у крыс развивается уже в возрасте I месяца. Крысы
старшего возраста совершенно не заражались Н. папа. Взрослые крысы
с трудом заражаются также и Helerakis spumosa, причем только из
малого процента янц развивались взрослые черви (Winfield, 1933).
У взрослых собак и кошек до половозрелого состояния развивается
3.96—5,53% введенных личинок Ancylostoma braziliense, а у котят н
щенят в 10 раз больше (32,31- 14,42%) (Sarles, I92t). Паразит трахей
птиц Syngamus trachea сильнее всего поражает молодых птиц. Подан
ным Маркова (1939а), Syngamus встречался в Петергофе у 50% птенцов
скворца и лишь у 30% взрослых птиц.
Возрастной иммунитет проявляется также в ограничении роста, раз-
вития, плодовитости и сроков жизни паразита Это было констатпро
вапо у многих паразитов. Например, Ancylostoma braziliense у котят
достигает зрелости на 14—16 день, а у взрослых кошек— на 18—25 день,
при этом длительноси, жизни паразита у котят около II—32 педель, а
у взрослых кошек они живут менее 2 недель (Sarles, 1929) В 3-месяч
ны.х щенках самки .1. caninum дают больше яиц, чем паразитирующие в
6-месячных щенках. Срок развития личинок Dictyocaulus filaria до поло-
возрелого состояния у овен н коз в возрасте II—12 месяцев равен 51
72 дням, в то время как \ 1 6-месячных ягнят, как правило. — 36
(2 дням Количество выделенных личинок половозрелыми червями у
старых овец было значительно Меньше, чем х молодых (Давтян, 1949).
Ascaridia lineala во взрослых особях растет медленнее и достигает мет.
зчч
ших размеров, чем в молодых (ЛскеП, Pert® a. Beach, 1935), Так. в
цыплятах 5-дш.вного возраста прирост длины паразита был равен 5,3 ял,
я 30-дпевиых цыплятах — 3,6 льи, в 2-х месячных — 2.6 л.и и 3-х месяч-
ных 0.-1—0.1 ,iMi (Herrick, 1925). Сходное явление были отмечено для
Hymennlepis fralerna и Heterakis spumosa из крыс (Shorb, 1933:
Winfield, 1933) н для других паразитов.
Болес высокая резистентность взрослых животных выражается и в
более легком течении инвазии. Взрослые животные, как правило, не бо
леют кокцидиозом, и заражение протекает бессимптомно, тогда как у
молодняка кокцидиоз является весьма тяжелым заболеванием. Анкило-
стомоз тяжелее протекает у Детей. нежели у взрослых людей и 1. д.
Создается впечатление, чго возрастной иммунитет развивается
обычно у млекопитающих и ПТИЦ, НО не у низших ПОЗВОНОЧНЫХ II OCCIKi-
гвоночных животных. Примеры возрастного иммунитета у рыб в отноше-
нпн к некоторым монотеистическим сосальщикам (Ляйман, 1948; Ляйман
и Садковская 1952) нуждаются. несомненно, в экспериментальной про-
верке, чтобы устранить влияние экологических факторов, определяющих
иногда большую зараженность молодняка по сравнению со взрослыми
(см. гл. XII). Следует быть осторожным в этом вопросе при опенке дай
пых различной заражаемое!и разных возрастов некоторых беспозвоноч-
ных (моллюски) партенитами сосальщиков.
Представляет несомненный интерес тот факт, что у одного животно-
го-хочяпна с возрастом повышается резистентность пе ко всем видам
паразитов. II (ихобалова, (1949) демонстрир овала у белых мышей одина-
ковую зараженность Trichocephalus muris в любом возрасте, тогда как
у того же хозяина ясно выраженный возрастной иммунитет развивается
к f/t/menolepis fraterna и Nippostrongylas muris. Целый ряд гельминтов
н простейших в одинаковой степени заражает как взрослых, так н мото-
дых животных (Entamoeba histolytica. Ascaris luntbricotdes). Имеются,
кроме того, примеры обратной возрастной резистентности, когда моло-
дые особи оказываются менее чувствительными к заражению, чем взрос-
лые Например, при заваженпи круного рогатого CKoia Piroplasma
bigeminum или Babesiet.a bovis телята то I года более стопки к зара-
жению. и пироплазмоз протекает у них бессимптомно, тогда как взрос-
лые живчтные болеют очень тяжело Наблюдаемые случаи более часто-
го и и нт< неявного заражения взрослых рыб нт и амфибий и других
животных по сравнению с молодью объясняются в перь ю очередь эколо-
гическими моментами (см. главу XII) н, вероятно, не связаны с явте-
ниями возрастной невосприимчивости.
Во шикает вонрф. чем обусловливается возрастное
повышение резистентности хозяев. По этому поводу суще-
ствуют разные точки зрения, каждая из которых в тон или иной степени
дает рациональное объяснение разнообразным случаям возрастного им-
мунитета. Некоторые авторы высказывают предположение. что возраст-
ной иммунитет зависит от нзмДОння общего физиологического состоя-
ния организма и его реактивности. Эккерт и Портер (Ackert a. Wrier,
1931) считают, что в крови кур с возрастом появляются активные ве-
щества, препятствующие росту паразитов (Ascaridta lineataj. После кро-
вопускания, т. с. уменьшения указанных веществ в 1>рови. у кур снижа-
лась резистентность к этойу паразиту. Фостер (Foster, 1936} нашел, что
понижение с возрастом шраженности собак анкилостомами зависит от
постепенного повышения количества гемоглобина в крови. Молочная
диета и связанное с ней у мсныпение железа в пище, а также кровопуска-
ние вызывают анемию у собак, в связи с чем у них снижается возраст-
ная ре ыстептногть к анкилостомам. Можно думать, что по мерс роста
400
животное приобретает качества, повышающие его общую физиологиче-
скую устойчивость п одновременно с этим устойчивость к паразитарным
инвазиям. Возрастным изменением реактивности можно объяснить тот
факт, что при перкутанном заражении собак анкилостомой сразу на-
ступает клеточная воспалительная реакция, в результате чего движение
личинок в коже быстро останавливается, У молодых собак такой реак-
ции не наблюдается, следствием чего является быстрая миграция личи-
нок через южу внутрь тела, т. е. успешное заражение.
Резкое снижение восприимчивости взрослых кур к Ascaridia lineata
находит объяснение в относительном увеличении количества бокаловид-
ных клеток в эпнтель.т двенадцатиперстной кишки (Ackert и Edgar, 1938;
Ackert 1942, и др.). У цыплят по мере их роста с повышением числа
бокаловидных клеток возрастает и количество слизи, ими выделяемой.
Эксперименты Эккерта показали, что слизь бокаловидных клеток яв-
ляется фактором, препятствующим развитию аскарндпн в организме хо-
зяина. Это особенно наглядно было демонстрировано в опытах in \iiro.
Молодых аскарндпн помещали в питательный раствор, содержащий се-
крет бокаловидных клеток, вгятый от птиц разного возраста. При этом
у них наблюдя юсь замедление роста по сравнению с паразитами, поме-
щенными и контрольную среду. Слизь, взягая от Цыплят в возрасте 3 ме-
сяцев, вызывала значительно большую задержку роста, чем секрет бо-
тсе молодых птнн.Это вещество, тормозящс! рос г червей, является термо-
стабильны и легко проходит через фильтр Беркефельда. Слизь, взя-
тая ' птнп в Ti сроки, когда наступает ясно выраженная возрастная
резистентность к /1 scandia, оказывает in vitro резко тормозящее нВ истине
на р( сг червей. .Аналогичные данные получены также п в отношении
крыс (Ackert, 1942).
В некоторых случаях возрастная незс^прнимчнвость имеет морфо-
логическую осиовх н связана со структур н ы м и и з м с и с и и я м и
хозяина. Так, иногда повышение резистентности с возрастом связано
с изменением толщины кожного покрова, в результате чего у старых
животных затрудняется перкутанное проникновение личинок паразита
(Ancylostoma caninum и др.). Тем же, а также появлением большого ко-
личества кожных желез, вероятно, можно объяснить более легкое про-
никновение церкарии Diplostomum flexicaudiim через кожу головасти-
ков, нежели через кожу лягушки. То же можно сказать и в отношении
птичьего сосальщика Prosthogonimus, паразитирующего в фабрицпевой
еумке только на первом году жизни птиц. Ко второму году, когда этот
орган исчезает, отпадает и возможность заражения этим паразитом.
Другой аналогичный пример дают клеши Acarapis, вызывающие у
пчет массовые и тяжелые заболевания. Эти клеши живут в трахеях, при-
чем заражают обычно лишь молодых пчел, до 9 дневного возраста. Этот
«возрастной иммунитет», по мнению Оросн-Паль (Orosi-Pai. 1936), ве-
роятнее всего, вызывается гем. что у старых пчел вход через дыхатель
ныс стигмы затрудняется вследствие большого затвердения мптша по
краям стигм и венчика волосков, окружающих стигмы. Он эксиеримен-
татыго удалят у старых пчел околостнгмальные волоски и получил под-
держивающие его гипотезу результаты. В его опытах нормальные моло-
дые пчелы заразились па 32,7%, нормальные старые.—на 1,8%. Старые
пчелы с удаленным венчиком стигма тьных волосков оказались заражен-
ными на 13.5%. т. е. в 6 раз сильнее, чем контрольные
Во многих случаях различная зараженность хозяев разных возрас-
тов не определяется изменениями их морфо-физнологнческих особенно-
стей. т. е. нмммгологнче.скым состоянием, а объясняется экологическими
факторами, о чем было сказано в главе XII.
26 В А. Догель
401
Одной из первых попыток объяснения явлений возрастного иммуни
тета может считаться высказывание Сэндграхнда (Sandground, 1929).
сделанное им на основе опытов с анкилостома мн собак и кошек и Si/nga
mus trachea кур и индеек. По мнению этого автора, возрастная невос
приимчивость развивается лишь у тех хозяев, которые являются не нор
малышм н (или второстепенны м и) хозяевами для данного па-
разита, т. е. такими, к которым паразиты приспособлены несовершенно.
У таких хозяев только молодые особи не обладают еще специфическими
биохимическими и физиологическими особенностями, свойственным!*
взрослому организму, и поэтому лишь в молодом организме могу г пара-
зитировать не свойственные данному хозяину паразиты, находящие в
не .1 бтагоприятные условия для развития. Но мере роста среда хозяина
становится вес менее подходящей для паразитов вследствие повышения
ес биохимической специфичности, и заражение таких хозяев становится
затруднительным, а приобретенные ранее паразиты выводятся из хо-
зяина. Сэндграунд считает, что развитие у кур возрастного иммунитета
к Syngamus и у собаки к -1. duodenale объясняется именно тем, что это
хозяева, не нормальные для паразитов, тогда как у индеек и человека —
нормальных хозяев этих паразитов — возрастной иммунитет соответ
ственно не развивается.
Между тем многие факты показывают, что возрастной иммунитет
может развиваться не только у «ненормальных» хозяев в отношении не-
достаточно приспособленных к нему паразитов, но и в «нормальных»
хозяевах. Так. у крыс имеется ясно выраженный возрастной иммунитет
к Heterakis spumosa и Nipposlrongylus marts, хотя крысы являются нор
мзльнымн хозяевами дтя них ^Africa, 1931; Winfield, 1933). Го же отме-
чается и в отношении Hymenolepis fraterna (крысиного штамма). Одна»
к этому виду иммунитет развивается значительно позже, чем к нс специ-
фичному для крыс Н. папа (от человека). Несомненно, гипотеза Сэнд-
граунда проливает некоторый свет па явление возрастного иммунитета,
ио не имеет общего значения, которое ей склонен был приписывать сам
автор. Вполне вероятно, чт0> и в облигатных хозяевах это явление мо-
жет иметь место так же, как и в ненормальном хозяине, по опо будет \
них выражено тем определеннее, чем мрнсс взаимоприспособлспы друг
«.другу паразит и хозяин в процессе эволюции.
Весьма врроятно, что. кроме отмеченных объяснений причин воз-
растного иммунитета, немаловажную роть в этих явлениях должна
играть гормональная деятельность животного, изменяющаяся с возра
стом. Было, например, отвечено, что молодые крысы становятся значи-
тельно более устойчивыми к Trypanosoma lewisi при введении нм внут-
римышечно гипофиза от взрослых крыс (Herrick a. Cross. 1936). Таких
данных еще мапо, чтобы делать каКне-лнбо выводы, но исследования в
этом паправле ши следовало бы продолжить.
Приобретенный иммунитет развивается в результате воздействия
паразита на хозяина. Он может быть а к т и в и ы м нл и пассивны м.
Первый в свою очередь может быть приобретен или в результате сстест
венной (нормальной) инвзши, или в результате вакцинации. т. е. искус-
ственного введения антигена. Пассивный иммунитет вырабатывается у
животного после введения ему сыворотки крови от и чму иного живот-
ного. содержащего в своей крови антитела. Существенная черта имму-
нитета— его специфичность. Рассмотрим лишь некоторые примеры при
обретенного иммунитета, нс останавливаясь на этом явлении подробно,
принимая эо внимание, что более или менее потные сводки по этому
вопросу при паразитарных заболеваниях имеются на русском и аиглпй
ском языках (Ши.хобалова, 1950, Кулбертсон, 1948; Taliaferro, 1929).
402
Резистентность к повторному заражению развивается нередко при
наличии в организме хозяина паразитов, оставшихся от предшествую-
щей инвазии. Эту форму иммунитета называют термином «пре му ин
ция» (Sergent, 1935), или нестерильным иммунитетом (су-
перипвазионным, ио Шнхобаловой, 1950) В данном случае \стойчиво<
состояние существует одновременно с латентной инфекцией и исчезает
после гибели паразита. Наряду с нестерильным иммунитетом имеется еще
и так называемый стер н л ь н ы н и м м у ните т, сохрани юшпйся пос-
ле окончания инвазии. Его нередки обозначают постинвазионным нмму
нитстом, или иммунитетом к рс инвазии. Следует отметить, чго обе эти
формы иммунитета пе всегда легко можно разграничить, и но существу
принципиальной разницы между ними нет. Различия между ними пост
лишь количественный характер: в од|г м случае иммунитет оказывается
слабым и прекращается после удаления антигенного начала из организ-
ма хозяина, в других случаях развитие защитных механизмов удержи-
вается более продолжительное врефя, уже после того как паразит вы-
веден из хозяина.
Приобретенный иммунитет в отношении простейших обнаружен,
например, у человека в отношении Leishmania tropica — паразита, вызы-
вающего болезнь Боровского (Латышев и др.. 1953). То же можно ска-
зать п в отношении сонной болезни. Крупный рогатый скот, перенесший
заболевание наганов (Tri/panosomctfbriicei), не подвергается повторному
заражению. Крысы приобретают иммунитет к Т. fewisi и Т. cruzi. В тО
же время лягушек можно многократно заражать Т. rotaioriiitn, а овец —
Т. melophagium (Кулбертсон, 1948).
Пироплазмоз и тейлериоз крупного рогатого скота вызывают срав-
нительно стойкий приобретенный иммунитет к повторным заболевания а
При пироплазмозе развивается иммунитет типа прсмуннпин, а при тей
лериозе он, вероятно, является стерильным.'
Приобретенный иммунитет при малярии v человека выражается
прежде всего в том, что по мере развития острой фазы заболевания у
аиц, оставленных без лечения (при экспериментальной малярий), при-
ступы могут прекращаться спонтанно при высокой степени паразитемни.
и количество паразитов после этого постепенно уменьшается (Boyd М..
1947; Blackburn. 1948, и др.) Часто при этом происходит исчезновение
их из периферической крови, но иногда они обнаруживаются в крови
длительное время после прекращения приступов. Следовательно, сна-
чала вырабатывается устойчивость организма хозяина к клиническим
проявлениям инфекции, а в дальнейшем развивается сопротивляемость
по отношению к паразитам, сдерживающая их размножение. Обпаружн
ваегся гибель мерозоитов, так что только 1 мерозоит доходит до стадии
меруляции. Приобретенный иммунитет проявляется еще и в том, что при-
ступы при рецидивах начинаются тогда, котла в крови имеется около
3000 паразитов на 1 лл3, между тем как первичный приступ происходи
при наличии 10- 20 паразитов в 1 лиг3 в крови.
В основе иммунитета при малярии лежит явление премуниции (Во
yd, 1947; Taliaferro, 1948). При кокцидиозе кур. кроликов, крупного ро-
гатого скота п других животных вырабатывается иммунитет, главным
образом препятствующий развитию заболевания. Повторные заражения
могут иметь место, но симптомы заболевания при этом и смертность от
кокцидиоза в значительной степени снижаются. Это выявлено при зара-
жении цыплят ооцистами Eimeria tenella или кроликов при заражении
их Е. magna и Е. neol. poris. Чем сильнее первичное заражение, тем от-
четливее развивается реакция иммунитета. У крупного рогатого скота
иммунитет развивается через 14 дней после первого заражения и сохра-
2ГД 403
няется от 2—3 ю 7 месяцев. Доза в 1000 ооцист вызывала менее дли-
тельный иммунитет, чем доза в 30 ОСО ооцист (Sengcr i. др., 1959). Доза
в 0000 ооцист Е. neoteporis потностыо предохраняла кроликов от забо-
левания, тогда как меныпис дозы лишь частично снимали симптомы.
Известные намеки на приобретенный иммунитет можно найти пост
ношению к миксоспорндням Ilenneguya oviperda, паразитирующим в
яичниках щуки. Заражение Henneguua наблюдается главным образом у
3—4-летних щук. т. с. вскоре после достижения рыбой половой зрелости.
Более старые рыбы заражены значительно реже. Возможно, что послед-
нее обстоятельство вызывается тем, что большая часть старых щук уже
успела перенести заболевание в 3—4-летнем возрасте и потому больше
не. заражается.
Иммунитет, приобретенный в результате инвазии гельминтами, в
большинстве случаев оказывается не абсолютным, а относительным
Стучась полной приобретенной невосприимчивости к повторным зара-
жениям крайне мало, и они не вттолне достоверны. Можно указать, на-
пример, что суперинвазия человека, зараженного Taenia solium, как
правило, не происходит, или крысы, имевшие естественную инвазию цис-
стицеркамн (Cysticercus [asciolaris). вторично не заражались (Баку-
лева, 1940). Чаще наблюдается относительная невосприимчивость, когда
происходит торможение в развитии паразита у иммунного хозяина. Эт
проявляется прежде всего в виде ограничения зараЛюмости при по-
вторных введениях паразита в хозяина При заражении собак шнстозо-
ма.ми вторично у них развивается меньше паразитов, чем у зараженных
впервые То же наблюдалось у крыс при заражении Hymenolepis frater
па. Имеются данные, что у человека развивается приобретенный имму-
нитет к Diphyllobothritun latum. Тарасов (1935) заражал себя в первый
раз 7 плероцеркоидами, и у него развилось 7 червей, во второй раз через
год из 6 раувилось 2 и в третий раз из 7 — ин одного. Более .многочмелен-
ные данные имеются по приобретенному иммунитету в отношении не-
мал од.
Так, у мышей при повторных заражениях Trichocephalus muris па
20—30-й день развивается в 8—10 раз меньше червей, чем при первич-
ном заражении! одинаковым количеством яиц. При этом часть червей вы-
делялась, не достигнув половозрелое™ При ренпвазии через 10—15
дней наблюдалось снижение интенсивности заражения в 3—4 раза. Им-
мунитет сохранялся в течение I —1.5 месяцев после отхождения парази-
тов, а затем крысы вновь воспринимали инвазию в одинаковой степени
с контрольными (Шихобалова, 1950) Напряженность иммунитета во
многом зависит от интенсивноегн первичного заражения. У мышей, по-
лучивших первично 50 янц Т. muris, при суперпнвазнн топ же дозой
развилось в 4 раза меньше паразитов, чем у контрольных, не подвергав-
шихся первичному шражепию. У другой группы мышеи, инзазпрован-
пой первично 400 янц, в результате вторичного заражения (50 яиц) раз-
bh.tcci» в 22,4 ра ta леныие власоглавов, чем у мышей, не иммунизиро-
ванных (Шнхобаловл, i960). Маккой (Me Соу. 1931) наблюдат, что у
крыс после повторного заражения трихинеллами развивалось 0,01—
2,8%, тогда как у контрольных 2.”—35.2% личинок.
Приобретенный iimmj нитст проявляется также и в том, что пара-
зиты в иммунном хозяине претерпевают замедленное развитие умень-
шаются в размерах, понижается плодовитость самок и сокращаются
сроки пребывания паразита в хозяине. Это наблюдается, например, у
Dictyocauhts и CystocauJus из овец (Давтян, 1949; Давтян н Паносян,
1946), у Trichocephalus muris из мышей (Шн.хобалова, 19-19), Heterakis
spttmosa у крыс (Winfield, 1933), Xypposlrongylus muris (Chandler,
40-1
1932}, Ancylostoma caninum (Scott, 1928; Herrick, 1928) н других гель-
минтов, гласным образом нематод.
Таким образом, в отношениях между паразитом и хозяином паб. пи
даются ясные взаимовлияния, которые выражаются в том, что в ре-
зультате антигенного воздействия паризита на хозяина происходят так-
же и нарушения нормальной жизнедеятельности самих паразитов. Эти
можно еще Демонстрировать и некочоЦымп особенностями развития па
ралиов в случаях интенсивного и неиитепсивноги заражения хозяина
Примером могут служить наблюдении над Trichocephalus muris (Шихо-.
балсьа, 1949, 1950), Trichinelltt spiralis (Шихобалоза 1952), Ascaridia
lineafa (Шихобалова и Кустова, 1950). Syngamus trachea (Ши юба.това,
1956), Cystocaulus у овен (Давтян и Шульц, i9-19), f)ipiiylb>boiltriuni la
turn -(Натовский и Гнездилов, 1949) и др.
Исследования Шчхобаловой над 7richocephalus muris показали, что
у интенсивно зараженных .мышей .Лчинки гибнут, не достигнуз полово-
!релости; в большем количестве, чем при слабых заражениях. При до.и
1009 яиц на одну мышь до половозрелого состояния развилось 8,58%, при
дозе 500 яиц — 20,4%, при дозе 50 яиц — 78,8%. Далее подмечено, чго
чем больше власоглавов развивается в хозние одновременно, гем мед
ленкее происходи г это развитие и юм позд нес самки приступают к яйце-
кладке. У мышей, получивших 5U яиц, размеры самок на 10-ii день бы
ли равны 23.57 мм. а при заражении дозой 4500 яиц их длина в среднем
равнялась 16,75 jui. Половое созревание самок происходит на 2 дня
быстрее при заражении дозой в 50 яиц, чем при дозе 400 яиц. Чем боль-
ше самок одновременно имеется в хозяине, тем меньше становится су-
точное количество яиц, откладываемых одной самкой. Так, если имеется
1—10 самок, то выделяется 7880—1117 яиц в сутки, а при 40—55 сам-
ках — от 820 до 580 яиц.
Шихобалова трактует эти явтения нс как результат перенаселения
или недостатка пищи, а как следствие приобретенного иммунитета,
быстро развивающегося при заражении большими дозами паразита. То1
же взгляд поддерживают Давтян и Шульц (1949) изучавшие взапмоот
ношения между Cystocaulus и организмом овец. Они пишут, что «чем нн
тснсивисе инвазия, тем сильнее создается иммунологическая напряжен
посты и тем больше ограничивается развитие гельминтов в процессе того
же первичного заражения». Наблюдаемое нередко самопроизвольное от-
хождение гельминтов при интенсивном или даже слабом заражении,
также можно рассматривать как одно нз проявлений приобретенного
иммунитета.
При повторных заражениях нередко наблюдается развитие пмм'ни-
тета, направленного против действия токсинов, выделяемых паразита-
ми. Например, овцы, переболевшие днктнокаулезом, при последующа
заражениях не дают клинических явлении, свойственных заболеванию
при первичном заражении (Пухов, 1940). Крысы легко переносят пи
вторные заражения летальными дозами трихинелл. У морских свинок
повторное заражение личинками аскарид не вызывает кровоизлияний ь
легких, как это наблюдается при первичной инвазии, и т. д.
Активный иммунитет, приобретенный в результате вакцинации,
был демонстрирован на большом материале. Наиболее ясные данные
были полечены с тканевыми паразитами (Cysticercus (asciolaris. С. pi
stformis, Echinococcus granulosus) или с теми паразитами, которые про-
ходят миграцию в организме хозяина (Trichinella spiralis, Ascaris и др.).
В меньшей степени возможна вакцинация животных против кишечных
паразитов, не совершающих миграции. Вакцинация экспериментальных
животных проводилась, кроме вышеуказанных, еще против следующих
405
гельминтов: Schistosoma, japonicutn, Fasciola hepatica, Hymenolepis
fraterna, Moniezia expansa, Trichocephaius niuris, Toxocara mistax, Ancy-
lostoma caninum, Cyslticaulus nigrescens и др. (Кулбертсон, 1948; Ши.хо-
балова, 1950). Для вакцинации использовались различным способом
приготовленные антигены и нх активные фракции. Например, телята,
иммунизированные полисахаридным антигеном, приготовленным из чле-
ников бычьего солитера, оказались резистентными к последующему за-
ражению их финнозом (Бабаджанов, 1949). Полисахаридные антигены
были приготовлены 1>гд.1я многих других гельминтов В отношении Cys-
ticercus fasciolaris наиболее активными оката пись глобулиновая и эу-
глобулиновая фракции (Campbell, 1937).
Возможность пассивной иммунизации была установлена
для некоторых гельминтов: Hymenolepis папа, Cysticercus bovis, Ancy-
lostoma caninum, Ascaris suum, Ascaridia Uneaia. Trichinella spiralis
и других (см. Шихобалова и Ленкина. 1948).
Пассивная иммунизация вызывает более слабое проявление имму-
нитета, чем активная,.и се действие обычно сравнительно кратковре-
менно.
Не вдаваясь в летали механизмов, при посредстве которых прояв-
ляется приобретенный иммунитет, мы отмстим лишь, что специфический
ответ животного на действие антигена выражается в выработке ан-
тител и повышенной активности фагоцитирующих
клеток. Существенное значение в выработке и осуществлении имму-
нитета принадлежит р с т и к у л о - э и д о т е л и а л ь н о й системе
т к а и с и, которая является поставщиком антигенов. Поэтому всякая
стимуляция этих тканей ведет к сильной ответной реакции, и. наоборот,
когда функция ретикуло-эндотелиальной системы нарушается, иммуни-
тет ослабевает. Действительно, в ряде случаев, например у крупного ро-
гатого скота, спленэктомия приводит к резкому снижению резистентно
стн к пироплазмозу. Даже в тех случаях, когда животное было абсолютно
невосприимчиво к определенному паразиту, удаление селезенки сни-
мало естественный иммунитет. В этом отношении исключительный инте-
рес представляют наблюдения Скрабало и Деяновича (Skrabalo a. Dea-
novic, 1957) над заражением человека нс свойственным для пего пара-
зитом Babesia bovis и эксперименты Гэрнем и Брэй (Garnham а. Вгау,
1959) над заражением сплснэктомированных шимпанзе паразитом В. di-
vergens. Аналогичные данные имеются и в отношении малярии. С дру-
гой стороны, резистентность к лейшмапиям у животных, лишенных се-
лезенки, не снижалась.
Роль гуморального и клеточного факторов в развитии приобретен
ного иммунитета нс одинакова для разных паразитов. Активность кле-
точных реакций, фагоцитоз, в значительной степени определяет развитие
резистентности при многих протозойных заболеваниях (малярия, пиро-
плазмоз и др.). Фагоциты захватывают паразитов переваривают их.
Впрочем, иногда паразит способен развиваться в фагоцитах, как это
наблюдается, например у Leishmania или у Plasmodium calhemerium.
Но наряду с клеточной реакцией происходит и появление в крови анти-
тел, и часто во времени эти реакции совпадают.
По отношению к многоклеточным паразитам фагоцитарная реакция
развивается менее часто и главным образом к тканевым паразитам.
Мигрирующие личинки гельминтов (в особенности нематод) всегда за-
держиваются в разных органах. Первым барьером оказывается стенка
кишечника пли кожа, вторым — печень, лимфатические железы, треть-
им— легочная ткань и пр. Быстрая мобилизация подвижных клеток рс-
гикуло-эндотелнальной системы приводит к иммобилизации паразита и
406
в конце концов к инкапсуляции его в соответствующих органах. Этот
фактор обычно действует наряду с гуморальным и тесно с ним связан.
Менор (1937) отмечает ясную фагоцитарную реакцию у гусениц ка-
пустницы, зараженных яйцами наездника Apanleles glomerulus. Часть
яиц постепенно обволакивается кровяныхп^клеткамп хозяина, и в конце
гонцов они распадаются и погибают, окруженные плотным синцитием
из слившихся клеток.
Врожденный и приобретенный иммунитет не является стабильным
у животного и может в значительной степени усиливаться или ослаб-
ляться под влиянием факторов, влияюща* на организм хозяине. Эти
влияния могут быть обусловлены различной диетой, сопутствующими
заболеваниями и другими факторами.
Полноценный пищевой режим с достаточным количеством витами-
нов, белков, солей и т. п. значнтелыЙГповышает врожденный и приобре-
тенный иммунитет животных к паразитам. С другой стороны, недоста-
ток витаминов некоторых минеральных солей снижает резистентность
организма и создаст условия, содействующие «пробиванию» однажды
/же приобретенного иммунитета.
Так, например, в опытах Эккерта (.Ackert, 1935) при коамленин цып-
лят авитаминозной пншей и заражении их Ascartdia в них развивалось
в среднем по 58 червей, тогда как в цыплятах, воспитывающихся на нор-
мальной. витаминозной пище, развивалось в среднем 4.4 червя. Авита-
минозные кошки при заражении их большой дозой яиц Тохосага саН со-
держали в себе от 75 до 231 паразита, тогда как нормальные — всего 50.
Фостер и Корт (Foster a. Cort, 1932, 1935) содержали подопытных собак
на диете без солей и витаминов и нашли, что неполноценная диета не
только снижает устойчивость собак, но «пробивает» возрастной и приоб-
ретенный иммунитет. Яйцепродукция паразитов в авитаминозных хозяе-
вах значительно повышалась.
Клэфеи (Claphani, 1931) изучал влияние отсутствия солей кальция
на паразитирование Heierakis gallinae у цыплят. При бескальциевой ди-
ете заражение было более сильным (81 червь па цыпленка), чем при нор-
мальной диете (50 червей на цыпленка) Портер (Porter, 1935) исследо-
вал влияние молочной диеты, сопряженной с отсутствием железа, па
устойчивость крыс к ftippostrongyhes muris. Он содержал крыс на мо-
локе 45—172 дня и в этот период заражал их личинками Nippostrogylus.
При молочной диете инвазия получалась вдвое сильней, чем при нор-
мальной.
Крысы, находящиеся на диете, лишенные витамина А, снижают ес-
тественную и приобретенную резистентность к h'ippostrongylus muris и
к Slrongyloides raiti (Spindler, 1933; Lawler. 1941, и др.). Это, вероятно,
связано с нарушением функции кишечника. Недостаток в рационе крыс
комплекса витамина В снижает резистентность к Hymenolepis fraterna
(Вавилова, 19-16 и др.). Цыплята, получающие недостаточное количе-
ство витамина В, более чувствительны к заражению Ascaridia (Ackert,
1942; Watt, 1944). Недостаток биотина приводит к тому, что цыплята
очень тяжело переносили заражение Plasmodium lophurae. На высоте
ннфедппш среднее количество паразитов на 10000 эритроцитов состав-
ляло 9524. в то время как • контрольных, при нормальном питании, оно
составляло 3550 (Trager, 1947). Мыши, содержавшиеся на диеге без ка-
зенна, не вырабатывали им.м\тштгта к Plasmodium berghei (Prakash,
1957). Крысы, получающие углеводную диету значительно легче зара-
жаются Bulaniidium, Trichomonas, Larnblia, чем находящиеся на белко-
вой диете (Ilcgner, 1937, и др.). Крысы, получавшие мясо или дрожжи,
497
не только не заражались лямблиями, но и освобождались от них, если
до этого были заражены (Глуховцев, 1935).
Введение в пищевой рацион небольших котичеств аткоголя также
снижает резистентность хозяина. Это показано, нагример, на крысах
при заражении их Hymenolepis fraterna /'Largh, 19-15) пли на мышах,
зараженных Plasmodium berqhei | Sauted, Pastacaldi, Vuillet, 1956).
В целом ряде случаев можно наблюдать и обратные соотношения,
когда резкое истощение организма хозяина под влиянием голодания
или гипоавптгмнноза приводит к гибели паразитов, так как сильно из-
мененная cpejfi обитания оказывается неблагоприятной и для самих па
разитов. Так, например. Tuxocara cati покидает чрезмерно рахитичных
котят (Винницкий, 1937 ft Рейд (Reid, 1940, 1942) наблюдал, что це-
стода RailUctina cesticillus. паразит кур, отбрасывает всю стробиле пос
ле голодания хозяина в точите 24—48 часов, теряя при этом свыше
90% своего гликогена. Рейд именно этой потери! гликогена объясняет
н отрывание от юловк.н стробил, лишающихся мышечной энергии, до-
статочной для сонротивлен!-. перистальтике кишок. Сколексы, однако,
остаются в стенках кишки •даже после 20 дней голодания курицы.
Ra.llietina настолько чувствительны к пищевому режиму, что содержа-
ние в них гликогена обнаруживает правильные суточные колебания от
7,15% в 6 часов вечера (т. е. после целого дня кормежки) и до 3,68%
в 6 часов утра, т. е. после проведенной хозяином без питания ночи.
Имеются данные, что сопутствующие заболевания снижают резис-
тентность к заражению паразитами. Так, у мышей, ослаб пенных пара-
тифозной инфекцией наблюдается аутоинвазия Hymenolepis fraterna
(Hiinninpn, 193G). В отношении этих же паразитов имеются данные, что
у беременных мышей развивается вдвое больше цнетицеркондоз при за
раженнн одинаковой дозой, чем у нсбсремеиных.
Таких примеров, иллюстрирующих влияние внешних факторов на
взаимоотношения паразита и хозяина, можно привести еще множество,
и все они свидетельствую г о подвижности системы «паразит—хозяин».
Важно еще и то, что при ослаблении резистентности хозяина различны-
ми факторами, в особенности диетой, может теряться его видовая спе
цифнчность и тем самым у ш го могут прижиться такие паразиты, ко-
торые при обычных условиях не развиваются. Повторение таких случаев
и отбор довершают дело, способствуя образованию новых видов пара-
зитов, удаляющихся от исходной формы.
I
Г/IABA XXII
СПЕЦИФИЧНОСТЬ ПАРАЗИТОВ К ХОЗЯЕВАМ И ПРОБЛЕМА
ВИДА У ПАРАЗИТИЧЕСКИХ ЖИВОТНЫХ
1. СПЕЦИФИЧНОСТЬ ОТНОШЕНИИ ПАРАЗИТОВ К ХОЗЯЕВАМ
Одной из характерных особенностей паразитизма является извест-
ная специфичность, т. с. постоянство сочетания системы «паразнт-хозя-
нн» или известная приуроченность определенных видов паразитов к on
ределепным же видам хозяев.
Однако, как мы увидим далее, такое определение специфичности не
может считаться полным и в достаточной степени точным. Следует ска-
зать, что вообще единого взгляда на сущность явления специфичности
до сих нор нет. Многие исследователи высказываются по этому вопро-
су, и за последние 10—15 лот накопилась большая литература. Значи-
тельнее внимание проблеме специфичности паразито-хозяйных отноше-
ний уделяют советские паразитологи, опубликовавшие немало статей
обобщающего характера (Кнорре, 1937; Киршепблат, 1911; Дубинин,
1950 Орлов. 1948, 1957; Дубинина, 1953а; Шульман, 1954а 1958а,
Шульц и Давтян. 1955а; Гусев, 1955; Полянский, 1955; Быховскпй. 1957,
и др.). Этот вопрос дебатируется также и паразитологами других стран
(Stammer, 1957; Baer. 1952; Szidat, 1956: Michajlow, 1959; Stuncard IL.
1957. и др.), свидетельством чего межет служить состоявшийся в апре-
ле 1957 г. в Ныошателе первый интернациональный симпозиум по спе-
цифичности, в котором приняли участие Ж- Бэр (J. Васг). Мантер (Man-
ter), Шабо (Chabaud), Дюбуа (Dubois), Гольван (Golvan). Лыоэллин
(Llevellyn), Дольфус (Dollfus) и др. Несомненно, возросший интерес к
проблеме специфичности обусловлен тем, что она представляет большое
значение для решения некоторых вопросов эволюции и филогении как
самих паразитов, так и нх хозяев. Мы рассмотрим только некоторые ос-
новные аспекты проблемы.
Мы уже отмечали, что хозяин представляет собой для паразита сре-
ду обитания. Свободно живущие организмы всегда приспособлены к оп-
ределенным условиям окружающей среды и для всякого организма есть
какие-то оптимальные условия существования,’ при которых он лучше
всего живет и размножается. Так же и паразиты адаптированы только к
определенной среде, т. е. к какому-то одному определенному виду хозяи-
на или к группе видов. Однако адаптция паразитов носит своеобразный
характер в связи с тем, что среда, к которой они приспособлены, являет -
404
ся живым организмом, реагирующим в свою очередь на паразита путем
выработки различных приспособлений защитного порядка. Поэтому при-
способ пенноегь паразита и возможность его существования в хозяине
определяются не только морфр),физиологически ми особенностями само-
го паразита, но и его способностью преодолевать защитные механизмы
хозяина, противопоставляемые паразиту и выраженные в специфиче-
ских морфо-фнзнологпчсских и биохимических свойствах организма хо-
зяина.
В ге^ случаях, когда паразит находит у хозяина подходящие усло-
вия для своего существования, тогда возможно формирование систсчы
«паразит-хозяин». Следовательно, специфичность можно понимать как
приспособленность паразитов к определенном!/ виду или группе видов
хозяев (среда ц^нтания), которая проявляется в виде приуроченности
паразита к хозяину. Паразит специфичен (приспособлен) к хозяину, так
же как к хозяин специфичен по отношению к паразиту.
Для каждого организма имеготся оптимальные условия существо-
вания, при логорьииоп лучше всего живет и размножается. Всякий орга-
низм способен существовать и при некоторых отклонениях от этого оп-
тимума. Для различных животных максимум такого отклонения весьма
различен; можно говорить о формах с узкой и широкой нормой реак
иии по отношению к различным факторам внешней среды. Виды, выно-
сящие большие колебания внешней среды, называются эурибионт-
н ы м и, тогда как требу юшие определенных и узкоограниченных условий
носят название сто цобионтн ых. Можно, например, говорить об
эури- и стенотермных (талинных и т. п.) организмах. То же относится и
к паразитам, одни из них обитают только в одном хозяине н не могут
существовать в друч их. а иные приспособлены к жизни в нескольких или
многих видах хозяев. Первых можно считать стенадаптирны ми
(моноксснпымн) паразитами, вторых эу р и а д а пт и в н ы м и (олиго-
ксенпымн и полнксенными) (Sandground, 1929).
Поэтому в наиболее общей форме специфичность может быть опре-
делена как норма реакции на среду, принимая во внимание, однако, что
в формировании взаимоотношений между паразитами и хозяином ак-
тивную роль играет также и организм .хозяина.
Адаптация паразита к организму хозяина, так же как и хозяина
к паразиту, складывается не сразу, а в течение длительного срока, и по-
этому специфичность, как функция времени, отражает определенный
этап эволюции паразмго -хозяйпых отношений. В приспособлении к хо-
зяину большую и существенную роль играют не только условия сущест-
вования паразита в организме хозяина, по и те экологические условия,
которые окружают самого хозяина н определяют возможность форми-
рования системы «паразит—хозяин». Таким образом, специфичность яв-
ляется экологически обусловленным явлением.
Рассмотрим теперь различные степени специфичности па-
разитов к н х хозяева м. Здесь следует оговорить, что. рассмат-
ривая распределение паразитов по хозяевам, мы обычно говорим об их
встречаемости в том или ином хозяине и на основании этого судим о
приуроченности паразита к хозяин' или хозяевам. Однако в целом ряде
случаев встречаемость паразита нс отражает всех возможностей его су-
ществования уг хозяев. Обычно потенциальная специфичность шире ре-
альной (Киршенблат, 1941), г. е., отмечая встречаемость паразита, мы
нс определяем еще весь круг его потенциальных хозяев, который может
быть шире фактического (Павловский, 1946а). Быховский (1957) счита-
ет, чю специфичность реализуется в фирме встречаемости. Для спреде'
лення потеница.тыюн специфичности паразита нужны эксперименты или
410
широкие обследования животных, предполагаемых хозяев данного па-
1а₽тся и
ифнчпость которых
для кокцндии кур, коз, овец и некоторых дру-
проверена экспериментально.
Parrnieroxenos dogieli
длинным из всех брюхо-
плрлнпгнрует в
Рис. 219.
— наиболее
но тих моллюсков,
волосгп телп дальневосточной голо-
турии Cucurnaria japonica-, уменьше-
но вдвое. (По Иванову.)
разита. «ч
Узкая специфичность (м о н о к с е н и я), или приурочен
ность к одному виду хозяина, наблюдается, например, у кокциднй рода
Eimeria, каждый вид которого специфичен в отношении определенного
вида хозяина. Кокцндии кролика нс .могут существовать в зайце и на-
оборот. 'I о же отмечается
тих животных, спет
Сходное явление наблюдается у Piro-
plasmida и Tlieileriida Bobesiella
bovis крупного рогатого скота не
может развиваться в других животных,
хотя такую строгую специфичность
можно преодолеть путем спленэктомии
нормально невосприимчивого организ-
ма (см.стр. 406).
11екоторые Myxosporidia, как
Chloromyxum esocinum, встречаются
только у щуки (Шульман, 1958а).
По данным Быховского (1957),
74,1% известных моногснетических со-
сальщиков встречется только' на одном
виде хозяина. Многие трематоды встре-
чаются также только у одного хозяина.
Например, поданным Быховскоп-Пав-
ловскон (1935, 1957), из 112 видов со-
сальщиков, найденных в 72 видах птиц
Барабйнской степи, 57 являются моио-
ксеинымн. Мантер (Manter, 1957) от-
мечает, что нз 296 видов трематод, мор-
ских рыб Японии 226 (76,6%) встреча-
ются только у одного хозяина. У цеетод
это характерно для Teania solium и
многих других видов. Средн нематод
известно, например, что Cystoopsis aci-
penseris встречается только у стерля-
ди, у которой по бокам тела образуют-
ся кожные опухоли. Wuchereria bim-
crofii обнаружена только у человека,
гнй моллюск Purenteroxenos dogieli найден до сих пор лишь на дальне-
восточной голотурии Cucurnaria japonica (рнс. 219). Средн паразитиче-
ских раков Ergasilus gibbus встречается только у угря, a Tracheliastes
sfellijer—у сома. Строгая специфичность свойственна многим перьевым
клещам (Дубинин, 1950) и Mallophaga (Clay, 1957). У наездников, с
другой стороны, такое явление наблюдается очень редко. Можно отме-
тить Sycosoler lauagnei в личинках жука Hypoborus ficus и несколько
других примеров.
Гораздо чаще наблюдаются случаи более ши рок ой специфич-
ности. когда паразиты способны существовать более чем в одном или
на одном виде хозяев (олигоксены пли поликсены). Один пара-
зиты специфичны для видов одного и того же рода хозяев. Так, грега-
рины рода Pterocephalus встречаются у разных видов Scolopendra, ин-
фузория Trichodinopsis paradoxa живет только в представителях рода
Pomatias (Gastropoda). 95 видов, или 9,9% моногснетических сосальщи-
ков паразитируют па представителях одного рода рыб. Среди трематод
ПаразитнческнЛ брюхоно-
411
таким примером может служить Phyllodistomum felts, характерный
для рода Anarichas.
Другие парази гы могут развиваться у различных родов одного и то-
го же семейства или у разных представителей одного отряда. Так, напри-
мер. инфузория Meturadiophryttt litmbrici (Astomaia) встречается только
в представителях семейства Luinbricidac. Hewieguya zschokkei характер-
на для лососевых. Isospora felis— для кошек и собак. Dactylogyrus го
buslus паразитирует толы® у представителей родственных родов Leiicis
cus и /Isp/us, а паразит жабр Diplozoon paiadoxum— на всех Карповы'
рыбах. Средн трематод, например, Leucochioridium phrag/nitophila
встречается у 3 видов воробьиных, a Plagiorchis nanus— у 12 видов ку-
ликов (Быховская-1 (авловская. 1957). У ленточных червей Echinococcus
granulosus и Taenia hydafigena найдены у разных представителей ср
бачьих. Paranoplocephalus amphilodes — в подсемействе Microtinae
Andrya cuniculi— у кроликов и зайцев. Разные виды цестод из птиц,
как правило, встречаются только у представителей одного отряда.
Много примеров приуроченности видов к представителям одного отряда
(Дубинин, I960) дают пухоеды и перьевые клеши. Наездник Trichogram-
та eoanescens заражает разных представителей бабочек и т. д.
Есть еще более широкоспецпфичные паразиты, которые ветре
чаются не только в пределах разных отрядов одного класса, но
в в разных классах. Так, Entamoeba histolytica паразитирует в кишечни-
ке человека, ио может развиваться у кошек, собак, кроликов, крыс ’ хо
мячков. Balaniid urn colt известен у свиней, крыс и человека. Fasciola he
patica. Opisthorchis felineus, Diphyllobothrium latum, Trichinella spiralis
и ряд других червей паразитируют у разных млекопитающих. Три шел-
ла, как показывают экспсриметпы, может развиваться как в представи-
телях класса амфибии, так и птиц (Matofi, 1936. 1944). В млекопитаю
щих и птипах паразитируют Corynosnma strumosum, Meiagontmus joka-
guwai (человек, собака, пиликан). Cryptocotyle lingua (чайка, баклан,
тюлень, собака) в др
Шульман (1954а, 1958а) отмечает, что иногда специфичность про-
является не по отношению к какой-нибудь систематической группе. а к
определенному исторически сложившемуся фаунистическому комплек
су. Так. Echinorhynchiis salmonis в личиночном состоянии встречается
только у рачкпв-бокоплавов Pontoporeia afftnis и Pallasea quadri-spinosa,
которые являются представителями так называемого «иольднев.т > ( мп
лекса», сохранившимися в некоторых озерах в качестве реликта фауны
бывшего 11ольХ.нева моря.
Бывают случаи, когда средн паразитов какого-нибудь рода, специ
фнчных по отношению к одной вполне определенной группе хозяев, выде-
ляются виды, которые приспосабливаются к паразитированию иа совер-
шенно иных хозяевах и таким образом, резко отличаются от своих ро
дичай. Так. например, род Protoopalina, характерный для кишечника зм
фибий, заключает в себе один вид (Р. saturnalis), паразитнрушип в киш-
ке морской рыбы (Box boops); род опалил Zelleriella, тоже свойственный
амфибиям, имеет одного представителя в рыбах (Z. piscicola) и
одного в змеях (Z. boipevae). Веслоногие рачки из рода Lernaeu
обычно паразитируют на рыбах, кроме L. гапае, встречающегося
на головастиках лягушки (Rana clamitans), а также кроме редких
случаев, когда L. cyprinacea вместо паразитирования на карповых
рыбах поселяется на головастиках амфибий Рачки сем. Са-
ligidae живут, как правило, на рыбах, по Archicaligus живет и? готово
йогом моллюске Nautilus. Клещи из сем. Arga'sidae сосут кровь т« пло
кровных позвоночных, но Ornithodorus furicata присасывается к амсри
412
канской субтропической черепахе Gopherus polyphemus Весьма веооят-
но, что мы имеем здесь дело с более поздними, вторичными, приспособ
лениями паразитов к повой группе животных-хозяев. **
1.ЮГДЗ целые паразитарные фауны гшеют загадочное происхожде-
ние, какова фауна кишечных простейших у морской рыбы Вос boops.
Она состоит из опалин Protoopalina salurnalis, из трихомонад Oclomitus
и нз амеб. т. е. представляет полную аналогию с кишечным населением
лягушек. Между тс м прочие морские рыбы вообще ие содержат в ки-
шечнике почти никаких простейших.
В общем половозрелые стадии одного и того же вила паразитов
лишь редко переходят в своем распространении за границы одного оп-
ределенного класса хозяев (Киршеиилат, 1911). Притом подобные крап-
ине случаи объясняя тся обычно ограниченностью и полным совпаде-
нием в составе нищи у чуждых между гобой хозяев. Так, coca циник
Crijpiucoiyle lingua живет в кишке у водоплавающих птиц и у тюленей,
нематоды Porrocoecum decipiens и Р. anfarclica в кишке акул и тюленей,
т с. рыбоядных животных.
Почти во всех случаях, когда паразит обнаруживается у нескольких
хозяев, удастся заметить, что в одном из них он встречается более часто,
вырастает до больших размеров, бы.трее достигает половой зредости,
продуцирует больше яиц и вообще оказывается оптимально приспособ-
ленным к условиям существования, и его развитие ие испытывает задер-
живающего влияния со стороны хозяина. Таких хозяев можно назвать
главны м и, или основ и ы м п (облигатными, по Скрябину и Шуль-
цу, 19-10), или нормальными (американские авторы). В других хо-
зяевах паразит если даже и встречается, то менее часто и в меньшем ко-
личестве, отстает и росте и испытывает со стороны хозяина значительное
сопротивление. Другими словами, паразит слабо приспособлен к такому
хозяину вследствие иевотможностн преодолеть его относительную невос-
приимчивость (см. главу XXI). Эти хозяева могут быть названы вто-
ростепенными (вспомогательными, по Михайлову, 1955). или fie-
ri о р м а л ь н ы м и (но терминологии американских паразитоло! ов). 11а-
коисц. еще в некоторых хозяевах паразит встречается совсем редко и,
как правило, с трудом в них развивается. Их следует считать с л уча й-
н ы м и, и ш не специфическими хозяевами (факультативны-
ми. по Скрябину и Шульцу, 1940).
Второстепенные хозяева нередко бывают филогенетически близки-
ми к главному, но могут быть сходными с ним только экологически и не
иметь близкородственных отношений.
Diphylloboihrium latum встречается у многих рыбоядных хозяев, нов
человеке достигает наибольших размеров, развивается быстрее всего и
95% отложенных червем яиц заканчивают развитие. Это главный хозяин.
В то же Bpiмя в кошке, собаке, свинье паразит имеет меньшие размеры,
медленнее развивается, живет немного времени и его яйца в малом про-
полте дают кораннднн. Эю. несомненно, второстепенные хозяева. Их
роль в распространении паразита меньше, чем человека. Сходные отно-
шения наблюдаются v OpistliWcliis felineus из человека, кошка,
собаки.
Крупный сосальщик Azygia lucii встречается в Ленинградской обла-
сти у §()'!(> щук и только у 10% судаков н ершей. При этом в ерше co-
ca тьщпки имеют небольшие размеры, часто бывают неполовозрелыми
или с зауГетпо уменьшенным числом яиц. Fasciola hepa.'ica и Dicrocoeli
urn dertdtfiicum встречается обычно у овец и крупного рогатого скота, ио
перед-о паразитирует у других животных {грызуны) а также у чело-
века. В этих случаях главный и второстепенный хозяева определяются
413
не только различной невосприимчивостью, но н экологическими уело
виямн, в основном возможноегью встречи инвазионного начала с хозяи
ном. ,
В зависимости от конкретных эюлогнческнх условий разные виды
могут играть роль го главного, то второстепенного хозяина. Так, для
Xzygia lucii в одних водоемах щука— главный хозяин, а многие другие
рыбы являются в горостепенными, но на Балхаше, где щука отсутствует,
гласим хозяином становится эндемический окунь Perea schrenki (До-
гель, 1-947).
Нередко наблюдается, что второстепенные хозяева бывают зараже
ны только в молодом возрасте, вследствие того что у взрослых хозяев
развивается невосприимчивость. Например, Trypanosoma abratnidis в
1лавном хозяине — леще—встречается в любом возрасте, тогда как у
Pntilus rttfilus caspicus она найдена только в крови молодых рыб (Дуби
пни, 1932). Для H'unastia rnilitaris lEchinostoniaticlae) кулики являются
главными хозяевами, но этот паразит заражает и гагу (Somateria molis-
sitna}, причем встречается только у птенцов, и к тому же сосальщики в
этом хозяине не образуют яиц (Белопольская. 1952).
Главные и второстепенные хозяева могут быть выявлены нс только
у окончательных хозяев, но и среди промежуточных. В этом отношении
интересны данные Дубининой (1953а) по личинкам ремнецов и Чихай
лова (1955) по процеркоидам Triaenophorus nodulosus. Дубинина иссле-
довала развитие Ligula intestinalis и Digrairnna interrupts, Ола выясни
ла, что плероцеркоиды могут хорошо развиваться у леща, тогда как в
уклее и пескаре плероцеркоиды D. interrupta хотя и развивались, но ш
достигали инвазионного состояния, и многие ш них (до 14,1%) дегенс
рировали.
Михаилов в своих опытах по заражению веслоногих рачков корацл
днями Triaenophorus noduiosus выяснил, что имеется 5 групп видов про
межуточных хозяев с различной восприимчивостью для паразита. В 1-й
группе, куда относятся Cyclops viridis. Diaptomus amhlyodon и др., онко
сфгры погибают в пищеварительном тракте. Это песпсцифпчпые хозяева
для данного паразита. Во 2-й группе (Cyclops laucarti) немногие онко-
сферы выживают в кишечнике, проникающие же в полость тола медлен-
но растут, но не превращаются в процеркоиды. В 3-н группе^Сус/орх ser
rulaiusj большинство онкосфер выживает, ио, попав в полость тела, так
же не превращается в процеркоиды. В 4-н группе (Cyclops (uscus, D<ap
ronins gracilis} часть онкосфер погибает, но те, которые проникают в
полость тела, развиваются в процеркоиды. Наконец в 5-й группе (Cyc-
lops strenuus и С. vicinus и др.) онкосферы в огромном большинстве вы-
живают и в полости тела дают процеркоидов. Таким образом, хозяев
5-н группы Михайлов считает главными, а 4-й группы — второстепеп
ними. Не исключена возможность, что при изменении условий хозяева
2-й и 3-й ipynn могут также играть роль второстепепнпых хозяев.
Но данным Гпнецниской (1954а), оказалось, что продуктивность
реднй Cyclocoelum niicrostoniutn из моллюска Limnaea равна 30—40цер-
кариям, тогда как в Planorbis родии формировали только 13—14 церка-
рий при одинаковой интенсивности заражения. В Planorbis многие родни
дегенерировал1! Очевидно. Limnaea является главным хозяином, а Р1а
norbis— второстепенным, где развитие протекает в значительной степени
заторможенно.
Возможность существования паразита в разных хозяевах имеет
важное практическое значение. При борьбе с патогенными паразитами
необходимо поэтому проводить уничтожение их не только в главном хо
зяине. но и во второстепенном; в противном случае сохранившиеся во
414
второстепенных хозяевах паразиты могут привести к возобновлении
инвазии у главного хозяина.
Взаимоотношения пара шта с главным хозяином нередко рассмат-
риваются как наиболее усзачовивишсся в протесе эволюции, тогда как
отношения со второстепенными хозяевами считаются сто нс устоявши
мися и в эволюционном с числе возникшими, вероятно, сравнительно не-
давно (Chandler, 1940).
Некоторые паразитологи высказывают также мнение, что высокая
степень патогенности паразита проявляется именно в относительно но-
вых для данного паразита хозяевах, быстро реагирующих иа воздей-
ствие паразита. Trypaifosqma brucei — возбудитель «наганы» рогатого
скота н лошади, приводящий больных животных к смертельному исхо-
ду,— не должен считаться основным для данных животных паразитом
Основными же хозяевами Т. brucei являются африканские антилопы,
для которых заражение этими паразитами протекает совершенно безбо
лсзнспно. Антилопы здесь служат, как говорят, резервуаром инвазии.
Очевидно, антилопы, с незапамятных времен заражающиеся Т. bru-
cei, успели приобрести известный иммунитет к данному паразиту,
тогда как завезенные нз Европы домашние животные иммунитета еще не
выработали.
Однако такие взаимоотношения паразита и хозяина нс являются
общими и могут иметь место обратные случаи, когда паразит тотькл в
главном хозяине вызывает патогенный эффект, как, например, Diphyl-
lobothrium latum \ человека и т. п. (Ball. 1943).
У одного и то! о же вида паразитов специфичность может быть в раз-
личной степени выражена на разных этапах развития ив
разных хозяевах — промежуточном резервуарном
и окончательном. Это очень хорошо иллюстрируется многими при-
мерами. При этом обращает на себя внимание следующий факт. Специ-
фичность паразитов в отношении хозяев оказывается наиболее узкой на
тон стадии, которая претерпевает в организме хозяина значительное раз
витке органов размножения, осуществляет половой процесс или парте-
ногенетическое размножение, т. е. претерпевает значительные морфо-фи-
зиологические перестройки. Наоборот, более широкая специфичность
свойственна тем стадиям, которые не развиваются в хозяине или претер-
певают лишь рост с некоторой дифференцировкой органов и не размно-
жаются половым путем Поэтому для многих паразитов широкая специ-
фичность проявляется часто в отношении второго промежуточного хо-
зяина и. как правило, в отношении резервуарных хозяев, что довольно
ясно выражено среди трематод и некоторых цестод.
Например, партениты дпгенетпчсскнх сосальщиков, как правило,
развиваются всего в одном-двух вилах моллюсков. Напротив, покидаю
щие моллюсков церкарин проникают во многих хозяев и метацерка-
рин. которые сравнительно мало развиваются, оказываются наиболее-
широко специфичными (метацеркарии (Jpislhorchis felineus встре-
чаются более чем в 10 видах рыб. Alaria alata — во многих млекопитаю-
щих и т. и.) Взрослые сосальщики опять-таки iimcioi менее широкий
круг хозяев, чем их метацеркарии. В тех с.и чаях, как, например, у Mic
rophallidae, когда метацеркарии в теле промеж^ точного хозяина пре-
терпевают .значительное развитие (формирую!ся все полозые органы и
могут образовываться даже яйца), нх специфичность ко второму про-
межуточному хозяину сказывается весьма узкой; в то же время взрослые
мнкрсфаллнды могут жить в разнообразных позвоночных хозяевах, так
как фактически в кишечнике последних дальнейшего развития нс претер-
певают У них происходит только продуцирование яиц. и черви очень
415
быстро покидают .хозяина. По наблюдениям Белопольской (1952, 1957).
метацеркарии Microphallus papillorobustum развиваются только в Gam-
marus locusia, а взрослые — у 14 видов куликов, 2 видов чаек, 2 видов
уток и I вида воробьиных.
Сходные закономерности наблюдаются у некоторых цестод, в осо
бспностп у ремнец в и Diphyllobothriuni. Ремпсцы (Ligula. Digramma)
в фазе плероцеркоида сравнительно узкоспецифнчиы и встречаются толь-
ко у разных представителен карповых рыб (Дубинина. 1953а, 1957а).
Это, вероятно, объясняется тем, что их плероцеркоиды претерпевают су-
щественные изменения, связанные с органогенезом половой системы.
Плероцерхоид до инвазионной стадии растет 12—14 месяцев, причем в
это время у личинки формируются все органы взрослого червя, но ие
происходит образования яиц Взрослые ремпсцы, развитие которых сво-
дится только к образованию яиц, являются широкоспецнфичпыми и мо-
гут развиваться не только во многих птицах (до 7 отрядов), но. в экспе-
рименте, и у млекопитающих. В io время как стадия процеркоида, на-
пример у Liquid intestinalis. приурочена лишь к циклопам и диаптому-
сам (Diaptomus gracilis, Cyclops strenuus, Accnthocyclops viridis, Meso-
cyclops dy bowski, Eucyclops serraltilus).
Сходные отношения наблюдаются у Schistocephalus solidus, встре-
чающегося в фазе плероцеркоида в колюшках и редко в бычках; взрос
лые формы развиваются нс только у рыбоядных птиц, fro и у зернояд-
ных (воробьи, куры) и млекопитающих при экспериментальном зараже-
нии (Дубинина, 1953а, Joyeux et Ватт 1942).
Однако данные ио DiphyllobolMum latum, Triaenophorus и ряду
других цестод стоят в некотором противоречии с вышеотмечейцой зако-
номерностью. У [Diphylloboihrium latum фаза плероцеркоида пе претерпе-
вает органогенеза головой системы и значительного роста, и потому его
развитие мон ет обеспечиваться менее специфическими условиями среды
(организм хозяина); плсрощ рконды встречаются у представителен раз
пых семейств и отрядов рыб (щука, нашим, судак, окнуь, ерш. угорь и
др.). В эксперименте они могут развиваться у амфибий и рептилий (Пав-
ловский и Гнрздилов 1939). В то же время половозрелая фаза D. latum,
требующая роста и значительного развития, паразитирует у 22 видов
млекопитающих.
У Triaenophorus nodulosus взрослый чернь встречается в кишечнике
щуки, процеркоиды — у одного-дву < видов циклопов, а плероцеркоиды,
достигающие большой длины н инкапсулирующиеся — у 30 видов прес-
новодных рыб. У других цестод финки большей частью узкосненифич-
ни, так же как и взрослые стадии или даже еще более. Впрочем из этого
правила имеются нередкие исключения. Финки Echinococcus granulosus
распространены у рогатого скота, свиней, лошадей, оленей, человека и
др., тогда как взрослая форма имеется у собак, реже у волка и лисицы.
У скребней личинки сплошь и рядом встречаются у одного какого-ни-
бгдь вида рачков (например Echinorhynchus salrnonis нз Pontoporeia af-
finis), тогда как в зрослые скребни встречаются у целого ряда рыб (раз-
ные сиги, лосось, форель, угорь и др ).
Особенно широкоспецифнчры личинки червей, встречающиеся в ин-
капсулированном состоянии в полости геза рыб. Так. например, личин-
ки скребня Gorynosoma 'slrumostim, первым промежуточным хозяином
которого в Каспии является только рачок Pontoporeia affinis, а окопча
тельным — только каспийской тю снь. имеют в качестве второго проме-
жуточного (или резерв’арпого) хозяина до 20 видов каспийских [ тб, в
том числе столь ра дюрол иы< формы, как минога, осетровые карповые,
судак и др Почти такой же широкий размах в смыс ie выбора прохкжу-
416
точных хозяев обнаруживает Contracoecum squalii, личиночная форма
которой встречена у 11 видов каспийских рыб.
У простейших, обладающих сменой хозяев, мы тоже нередко имеем
сильные различия в специфичности ио отношению к окончательным п
промежуточным хозяевам. Так. промежуточным хозяином Plasmodium
t'ivax является человек, а окончательным (переносчиком) служат многие
ниды Anopheles. Поданным Йонга и Бургоса (Jong и Burgess, 1948), наи-
более восприимчивы к паразиту A. maculipetmis, в меныисн степени
1. punctipennis и Л. quadrimaculatus и очень слабо Л. albimanus. Plasmo-
dium vivax развивается также в Anopheles bijurcatus и Л. ptilcherimus.
С другой стороны, возбудитель малярии птиц Plasmodium gallina-
сешп встречается у многих видов птиц, а также может распростра-
няться различными переносчиками, относящимися к родам .4ct/es, Arni-
geres, Culex, Mansonia, Theobaldia, Anopheles. Более узкая специфич-
ность характерна для половых стадий Plasmodium berghei, которые раз-
виваются только у Anopheles durani (Garnham, 1957), хотя не лишено
вероятности, чти переносчиками могут служить и некоторые другие ви-
лы комаров.
Trypanosoma brucei переносится очень ограниченным числом видов
рода Glossina на много различных видов и родов антилоп и па несколь-
ко родов домашних животных.
Специфичность паразита к определенному хозяину может быть не-
одинаково выражена в различных географических зонах
их ареала.
Так, например, промежуточным хозяином для печеночной двуустки
(Fasciola hepatica) в Европе служит почти исключительно Limnaea trun-
lalula. В других европейских брюхоногих моллюсках, например в Lim-
naea stagnalis, мнрацидпп двуустки проделывает только начало разви-
тия. Но занесенная на другие материки, где нет L. trunca-
tula. печеночная двуустка приспосабливается к паразитированию в
других видах, Limnaea: в Северной Америке — в L. humilis, в Южной
Америке — в L. viator, па Сандвичевых островах — в L. oaltnensis. в Ав-
стралии— в Balinas tenusfriatus и т. д. Таким образом, в разных странах
Fasciola может приспосабливаться к разным промежуточным хозяевам.
То же наблюдается у Diphyllobothrium latum. Первым промежуточ-
ным хозяином в Европе является Cyclops strenuus и Diaplomus gracilis,
.а в США — Cyclops brevispinostis, С. prasinus, Diaplomus oregonensis,
D. silis, D silicoides. Этим отчасти объясняется такое широкое распрост-
ранение указанных паразитов. С другой стороны, партениты Opisthor-
chis felineus могут развиваться только у Bithynia leachi и в тех случаях,
когда инвазионное начало распространяется с окончательным хозяином
«а пределы ареала этого моллюска, цикл развития паразита там не мо-
жет осуществиться вследствие неспособности партенит развиваться в ка-
ком-либо другом хозяине, кроме В. leachi.
Как уже отмечалось. взаимная приспособленность паразита и хо-
зяина, выражающаяся в строгой специфичности, вырабатывалась. как
правило, в течение длительного времени при тесной связи филогенеза хо-
зяина и паразита. С другой стороны, под влиянием факторов, влияющих
на паразита и хозяина, а также экологических условий, возникает воз-
можность формирования новых связей паразита и хозяина,т е. переход
паразита, специфичного в отношении какого либо одного или несколь-
ких хозяев, на новых хозяев, не свойственных данному паразиту
Одним из факторов, необходимых для выработки специфичности, яв-
ляется возможность контактирования паразита и хо-
зяин а.
4Г
27 В. А. Догель
Очень ботьшое значение имеют при выборе и отыскивании хозяин.,
г рои из мы паразита. Так, например, мирацидии сосальщиков в.
время своей свободной жизни подчиняются положительному фототак-
сису и хемотаксису по отношению к определенным видам моллюсков
При активном проникновении личинок нематод известную роль играет
термотаксис личинок, которые стремятся ко всякому теплому предмету
находящ< муся с ними по соседству.
Пикар (Picard, 1921) в ряде опытов выяснил факторы, руководящие
выбором хозяина со стороны одного наездника Pimpla instigator. В при-
сутствии куколок бабочек самка Pimpla приходит в возбуждение и гот
час же начинает откладывать в них яйца, тогда как в присутствии гусе-
ниц она остается вполне индифферентной. Бумажная трубочка, смочен
пая кровью куколки капустной белянки, усердно прока 1ывается самкой
Pimpla, пытающейся отложить в нее япца. Ясно, что здесь самкой пара
знта pj ководит хемотаксис по отношению к куколкам.
У наездника Torynus nigricornis (Chalcididae) характер хемотакси
са несколько иной. Этот наездник вводит свои япца только в галлы Dip
lolepis quercus, причем па него хемотаксически влияет. по-впдимому, вс
щество самого галла, а не его хозяина. В пользу этого свидетельствует
то обстоятельство, что личинка Torynus нападает без различия на личн
нок Diplolepis, его куколок пли даже па других паразитов, ранее посе-
лившихся в том же галле. Точно так же другой наездник, Brucon viator
испытывает известное притяжение к головкам чертополоха и огктады
вает на них свои яйца, вис зависимости от того, кого эти головки содср
жат — личинок мух или жуков.
Возможность смены хозяина может зависеть от широты нормы
реакции паразита на хозяина. Если при столкновении с новым
хозяином у паразита будут достаточно лабильны его морфо-физиологн
ческис механизмы, преодолевающие сопротивление хозяина, то станет
реальным переход паразита в новые условия существования.
Наконец, возможность перехода на нового хозяина зависит от сте-
пени его врожденной невосприимчивости и факторов
ослабляющих се (см. гл. XXI). В эксперименте, например, при кормле-
нии пищей с дефицитом витаминов частично снимается врожденная не
восприимчивость. Свиньи па диете без витамина А заражаются челове-
ческий аскаридой и т. п. Так же и в природе, при попадании организма
в измененные условия (акклиматизация, одомашнивание, искусе гневное
разведение и т. п.) наблюдается часто заражение их неспецнфпчпымг.
паразитами (см. г i. XIX).
Переход может осуществляться как иа родственных хозяев, так и не
родственных, но обладающих сходными условиями существования и
сходной пищей. Для паразитов водных хозяев Быховскпй (1957) отме-
чает, что первый случай характерен для форм с активным способом за-
ражения (Aionofrnoiclea), второй—дтя форм с пассивным способом
(Myxpsporidia). Вообще возможность перехода на новых хозяев связана
отчасти с древностью отношений между паразитом и хозяином. Чем эти
отношения древнее, тем труднее смена условий обитания паразита
Шульман (195$а) отмечает, что паразиты таких древних рыб. ка:;
акуловые и осетровые, строго к ним приурочены и не переходят на но-
вых хозяев, а специфичные паразиты молодых групп рыб легко могут
перейти из представителей других родов и семейств, п лаже на фьлоге-
гетнцески более древних рыб Кроме того чаще переходят в новые ус-
ловия обитания кишечные паразиты и некоторые эктопаразиты, в то
время как взрослые паразиты кровеносной, лимфатической системы и
почек более строго специфичны и обычно не способны к смене хозяина
4И
Таким образом, специфичность паращ|то-хозяйных отношении не
является стабильной и может изменяться в разных направлениях в про-
цессе эволюции под влиянием различных факторов внешней среды. С яв-
лением специфичности, несоменно, тесно связан и вопрос о видообразо-
вании у парази гов.
2. ВИД У ПАРАЗИТОВ
Несмотря на то, что влд представляв г собой, казалось бы, основное
и строго определенное понятие в биологии, подход к этому понятию в
разных отраслях биологии может быть не вполне одинаковым. Ботаник
понимает вид не вполне так, как зоолог, а палеонтологический вид носит
опять-таки несколько иной характер, чем виды, входящие в состав совре-
менной фауэы. Зависит это обстоятельство и от характера объекта н от
различия методов, применяемых разными дисциплинами для определе-
ния понятия о виде.
Естественно поэтому возникает вопрос о том, как обстоит дело с по
питием о виде в паразитологии.
Понятие о виде и о его подразделениях особенно детально разраба-
тывалось на насекомых в зоологии н на цветковых растениях в бота-
нике. В результате и в зоологии, и в ботанике установилось представ
ление о виде как о некотором сложном комплексе, который состоит нз
ряда более дробных катеЬернй. Так, Семенов-Тян Шанский (1910) при-
нимает следующую категори >ацию дроблений вида:
1. Вид.
2. Раса, или подвид (dibspc cies) —категория, базирующаяся в зна
читальной мере на географической изоляции.
3. Племя (nalio) —более мелкое, чем раса, ио основанное тоже на
географическом распространении деления вида.
4. Морфа (rnorpha) —зависящее не от географической изоляции, а
от резкого изменения внешних условий, неустойчивое уклонение основ-
ного вида или подвида: нередко может возвращаться к Первоначальному
состоянию при изменении условий.
5. Аберрация (abermtio)—индивидуальная вариация.
С некоторыми изменениями аналогичные градации принимаются и
многими другими систематиками.
Все вышеупомянутые подразделения характеризуются известными
морфологическими отличия .т. Но нс следует забывать, что в настоящее
время в упомянутую систему вклинивается еще введенное в науку Хо
лолковскнм (1910) понятие о биологических видах, т. е. неразличимых
морфологически, но разнящихся друг от друга в физиологическом или
экологическом отношениях.
Обращаясь к паразитическим животным, мы прежде всего, находим,
что понятие о виде по отношению к ним (особенно к эндопаразитам) раз-
работано еще явно недостаточно.
В громадном большинстве случаев паразитолог оперирует с видом
как с конечной систематической единицей. Более низкие категории уста-
навливаются исключительно редко. Между тем более глубокий анализ
стрч ктуры вида показывает, что у паразитических организмов можно
найти в некоторых о/ношениях сходные со свободно живущими формами
внутривидовые категории.
Когда вид паразита обладает широкой специфичностью и живет в
несктьких различных хозяевах, принадлежащих иногда к разным се-
мействам или отрядам, то в разных его популяциях могут возникнуть от-
дельньц расы, или штаммы, более приспособленные к паразитированию
27*
119
в одном или немногих из этих хозяов. Эти различия в одних случаях
носят морфологический характер, в других — чисто физиологический или
биологический и представляют собой типичные внутривидовые подраз-
деления.
Прекрасный пример возможной начинающейся внутривидовой ди-
вергенции вила дает, по Миккелю (Miekei, 1928), представитеть парази-
тических ос Mutillidae— Dasymutilla bioculata. Личинки этого вида па-
разитируют на куколках ос Bembicidac. Собирая Dasymutilla bioculata
в больших количествах в окрестностях Миннеаполиса. Мпкксль нашел,
что кривая размеров длины взрослых Dasymutilla имеет явный бимо-
дальный характер (рис. 220) Одна н вершин кривой, составтенной для
самок D. bioculata, лежит при 7,5 мм, тогда как другая—при 13 жле.
Аналогичные, по несколько иные в цифровом отношении (10 и 14 /<.«)
результаты дает и кривая для самцов.
Рис. 220 Дпувегипипная кривая размеров самок Dasymutilla
bioculata. зависящая от того, чго личинки данном популяции
паразитировали на двух различных хозяевах. (.По Миккелю.)
Исследуя причины этого явления, Миккель выяснил, что малорослые
особи Dasymutilla происходят из личинок, которые воспитывались на ку-
колках iMicrobembex rnonodonta, тогда как крупные особи паразита рос-
ли за счет куколок Bernbex pruinosa. Промеры обоих видов хозяев дали
для Microbembex размах вариаций длины от 8 до М мм, а для Bcmbex—
от 16 до 19 мм. Таким образом, более крупным хозяевам отвечала н
группировка белее крупных особен паразитов. Миккель делает отсюда
правильный вывод, что размеры взрослых Dasymutilla bioculata нахо-
дятся в прямой зависимости от количества пищи, которую получают их
личинки, паразитируя в мелких или крупных хозяевах. Отсутствие дру-
гих экологических и климатических влияний на Dasymutilla было устра-
нено тем, что все наблюдения велись на одном и том же небольшом
участке песчаной дюны.
Для нас в данном примере особенно интересна явная двувершин-
ность кривой, которая может привести к постепенной прочной диверген-
ции вида. В самом деле, теоретически рассуждая, если мпкросамиы в
состоянии легко оплодотворять макросамок, то для обратных отношений
может наступить момент, когда слишком большой копу зя гивный аппа-
рат макросамцов не будет в состоянии оплодотворять ми кросамок. Та-
ким образом, возможна физиологическая изоляция двух рас D. bioculata
420
исключительно на базе "<гостальных» различий и даже в условиях одном
и той же местности.
Отчетливо выраженная внутривидовая изменчивость имеет место
и у некоторых сосальщиков. Еще Дольфус (Dollfus, 1922) указывал
на изменчивость формы тела и строения полового аппарата у Dicro-
coeUum dendriticum {= D. lanceatum). Белопольская (1954) также обна-
ружила вариации размеров, формы тела
и положения семенников у D. detulrili-
сит, порученных из разных хозяев и из
разных географических пунктов. Напри-
мер, в Cervus clapiuis в Приморье разме-
ры червя равны б,4Х2,3 лиг, а в Cilellus
xaiithoprymnus из Армении 4,5X0,87 м.п.
У зайца, изюбря и оленя иа Дальнем Во-
стоке преобладают двуустки с семенни-
ками, расположенными симметрично по
бокам тела, у гималайского медведя и у
оленя, акклиматизированного в Горьков-
ской области, встречаются сосальщики с
семенниками, лежащими наискось по от-
ношению продольной оси тела, а у сусли
ка преобладают формы с продольным
расположением Семянников (рис. 221).
Некоторые различия в форме семен-
ников и яичников, размерах тела и поло-
жении желточников обнаружила Шуль-
ман-Альбова (1952а) у Podocotyle а/о-
топ из разных рыб. Феррис (Ferris, 1932,
1933, 1935) обнаружила морфологические
различия у вшей Haematopnius от свиней,
коз и лошадей, взятых из разных геогра-
' фичсских пунктов.
Средн простейших у Trypanosoma
eoansi Гоар (Ноагс, 1956) констатировал
наличие mhoi их рас из разных млекопи-
тающих, отличающихся, в частности, друг
от друга размерами тела Различия меж-
ду ними, однако, нс достигают степени,
которая позволила бы рассматривать нх
как самостоятельные виды или подвиды.
У Т. evansi, паразитирующей в лошадях,
наблюдается постепенное увеличение
размеров в разных географических
районах по направлению с запада на
восток.
Существуют, кроме того, внутривидо-
вые группировки, морфологически тож-
дественные. ко отличающиеся биологиче-
скими, экологическими иля физиологическими особенностями. Они пред-
ставляют собой биологические расы, связанные с паразитированием в
различных хозяевах с разным географическим распространением и в не-
которых случаях также с различным отношением к промежуточным хо-
зяевам. Среди гельминтов можно отметить Ancylostoma caninum, кото-
рая образует две биологические расы, отличающиеся своими взаимоот-
ношениями с opi анизмом хозяина (Scott, 1929).
Рис. 221. Измен’ anoci ь Dlcroco-
eliurn dendriticum из различных
хозяев
/1, В — нз Cervus elaphus (При-
морье); С — нз l.'rsuS tibeia-
ntts (11рнморьец D — из Citellus
xanlhoprymnus (Армения). (По
Бе.юпольсьон.)
421
Раса кошачья даст высоки» процент заражения у котяг
(45% искусственно введенных личинок) и весьма низкий у щенят
(1%). Наоборот, собачья раса давала большую заражаемость у щенят
(50% вызревания личинок). При этом также удалось показать, что
размеры получающихся анкилостом определяются нс столько взятой
расой, сколько характером хозяина. Анкилостомы как собачьего, так
и кошачьего происхождения достигали меньшего размера в кошках,
чем в собаках, и обратно. Таким же способом Шорб (Sborb, 1933)
установил наличие двух рас у Hymenolepis fralerna из крыс и мышей.
Расы Strongyioides stercoralis из человека и обезьян отличаются
характером своих жизненных циклов (см. гл. X).
Некоторые виды паразитических простейших также образуют
отчетливые биологические расы, отличающиеся нище выми потребно-
стями, местом обитания, патогенностью и т. п. Весьма вероятно, что
эти расовые отличия, не достигающие степени подвидовых, при морфо-
тогическом сходстве организмов, зависят от различий в антигенной
структуре. Гоар (1956) предлагает именовать эти внутривидовые
группировки термином «дем» (о г греческого слова — demos — населе
ние). В зависимости от характера различий к слову «дем» делается
соответствующая приставка: экодем (различия в условиях обита
ния), нозодем— для клинических вариантов, серодем —для
штаммов, отличающихся иммунологическими свойствами, ксено-
1ем — различия по специфичности для различных хозяев, клино-
дем — географические разности. В само понятие «Дем» Гоар ш
вкладывает таксономического содержания, и по существу оно близко
к понятию популяция. Ксенодемы, например, образуют Trypanosoma
evansi, формы, приспособленные к верблюдам, лошадям и рогатому ско-
ту. Некоторые группировки Т. evansi можно рассматривать как клине
демы и нозодемы, встречающиеся на западе и востоке у домашних
животных и вызывающие при продвижении на госток более тяжелое
заболевание. Ксенодемы образует Babesia canis, приспособленная
в Европе к клещам Dermacentor, а в Азин — к Rhipicephalus. Разны'
штаммы Plasmodium falciparum адаптированы к разным видам
\nopheles {Hodre, 1956).
Весьма вероятно, что в процессе эволюции такие демы могли
стабилизироваться путем отбора в новом хозяине и превратиться
в наследственно закрепленные подвиды и виды, потерявшие способ-
ность к существованию в исходном хозяине. Такне виды, сформн
юзанные на основе физио, югнческнх и экологических отличий, пред-
ставляют собой биологические виды, приспособленные к раз
ным хозяевам и не отличающиеся морфологически друг от друга.
Гаковыми являются, например. Entamoeba histolytica (человек).
Е гапагит (лягушка), Е. invadens (рептилии) и Е. mushkowski
(своботно живущая форма); Trypanosoma brucei, вызывающая пата-
ну домашних животных, и Т. gambiense— возбудитель сонной боле ши
человека—тоже относятся к этой категории и их Игличает только
гостальная специфичность (Н >аге. 1956).
Биологическими видами могут считаться Eimeria arloingi
и Е. faurei из коз и овец. Ооцисты этих видов нз разных хозяев
совершенно сходны между собой, однако перекрестного заражения
ими не происходит (Крылов, 1961), что свидетельствует о фи-
:шологйческо$ и экологическом различии между ними. Boot ще решить
вопрос, действительно ли мы имеем дело с биологическим видом,
можно только путем опыта перекрестного заражения разных видор
хозяев
122
Плазмодии малярии человека и шимпанзе нс отличимы морфо-
логически друг от друга. Plasmodium falciparum соответствует
Pl. reichenowi шимпанзе, но перекрестное заражение ими не удается.
Человеческий штамм Pl. vivax у обезьяны вызывает скрытое зараже-
ние. так же как и Pl. malariae дает у них бессимптомное заражение.
Следовательно, Pl. falciparum человека и Pl. reicheno&i являются з же
[обособившимися биологическими видами, a Pl. vivax и PI. malariae
представлены каждый двумя расами, нз которых одна адаптирована
лучше к человеку, а другая к обезьяне (Мозге, 1949).
Не останавливаясь на других примерах, можно сказать, что био-
логические виды отражают начальные стадии дивергентного процесса
видообразования. Они являются как бы «молодыми видами», видами
.in slatu nascendi» морфологические различия между которыми не
всегда могут быть обнаружены. Эго хорошо иллюстрируется примера-
4И происхождения некоторых видов трипанозом. весьма подробно
разобранными в работах Гоара (Ниаге, 1956, 1957). Гоар считает, что нет
оснований сомневаться в том, что вид Trypanosoma evansi, вызываю-
щий болезнь сурра у домашних животных и распространяемый слеп-
нями. произошел от Т- brucei, вызывающей нагану н распространяе-
мой мухой цеш* Это могло случиться, когда верблюды проникли
в ареал Glossina и заражались там наганов, а затем возвращались
ча территорию, свободну ю от пене. Здесь Trypanosoma brucei стала
переноситься слепнями и с течением времени превратилась в моно-
морфный вид Т. evansi. Наблюдаемые иногда в некоторых расах
вспышки полиморфизма у Т. evansi можно рассматривать как атавизм
(Т. brucei является полиморфным видом, морфологически сходным
с Т. evansi). Кроме верблюда, Т. evansi приспособилась к другим
животным в странах востока. С лошадьми она проникла в Южную
Америку, где образовала новый вид, лишенный кииегонласта, пато-
генный для местных лошадей (Т equinum, вызывающая Mai de Gade
ras (Ноаге, 1957, 1960в).
Внутривидовые разности, определяемые как демы, могут
возникать и в пределах одного хозяина. Так. например.
Leishmania donovani, вызывающая висцеральный лейшманиоз
(Кала Азар), представлена индийским, средиземноморским и Судан
ским типами (Ноаге, 1949), различающимися но клиническим, пмму-
нологическнм и эпидемиологическим признакам, хотя во всех случаях
паразиты морфологически сходны. Кожный лейшманиоз (болезнь
Воровского). вызываемый L tropica, в Старом Свете представлен двумя
различающимися по эпидемиологии нозодемами— сельским, остронс-
ХрОТНДПруЯщИ-М, И городским, Г1ОЗ,1НОНЗЬЯВ.1Я1ОЩИ'!СЯ. — нс дающими
перекосе,гного иммунитета.
Plasmodium vivax и Pl. falciparum образуют географические расы,
отличающиеся вирулентностью, длительностью лттентпого периода
и приспособленностью к разным переносчикам (Boyd, 1949). Для
PI. vivax установлено, что он обладает двумя расами с длинной
(7—12 месяцев) и короткой (8—23 дня) инкубацией (Николаев, 1949).
Гnian.oe'ta Histolytica образует несколько рас, отличающихся по
своей вирулентности, представляющих собой нозо- или ссродсмы
(Ноаге a. Nacl. 1955: Матевосян, 1951; Ноаге, 1960а).
Наряду с биологическими расами в пределах одного хозяина
может наблюдаться у паразитов внутривидовая диффер*п
ц н р о в к а морфологического характера Добелл и Джене
(Dobell a Jepps 1918) описали 5 рас Entamoeba histolytica, различаю
щихся по размерам цист Среди жгутиконосцев это наблюдается
4‘Л
Рис. 222. Дне расы Entodinium
caudatum из рубна козы. (По
Полянскому и Стрелкову.)
у Trypanosoma brucei Этот вил образует не только иозодемы п серо-
делил, но, вероятно, и морфологически различимые расы. Городской
клинической форме кожного лейшманиоза соответствует возбудитель
Leischmania tropica f. minor, а сельск iff форме— L. tropica f. major
Обе эти формы лейшманий несколько отличаются друг от друга раз
мерами
Наиболее сложная картина внутривидовой дифференцировки
наблюдается у инфузорий и в особенности в сем. Ophryoscolccidae.
населяющих рубец и сетку жвачных (Догель, 1927а; Полянский
и Стрелков, 1938; Киль, 1940). Было установлено, что многообразие
морфологических форм у этих инфу :орий частично обусловливается
модификационной изменчивостью, размах которой у разных видов
оказывается не одинаковым. Например, у
Entodiniiini caudatum (см. рис. I) наблю-
дается широкий । азма.х изменчивости, а
Eudiplodinium magii отличается необычной
константностью своих признаков. Эти раз-
нообразные морфологические изменения,
определяемые в основном условиями среды,
< бычно именуются термином «форма» (До-
гель, 1927а).
Наряду с модификационной изменчи-
востью, вызывающей полиморфизм вида,
имеет место еще наличие генотипически не-
однородных групп в пределах вида, которые
представляют соб'*й р а с ы и биотипы
Наличие нх с несомненностью удалось уста-
новить у Entodinium caudatum (рис. 222)
Одна раса (А) заметно более крупная, чем
вторая раса (Bj. Они отличаются, кроме то-
го, и нормой реакции. Хотя каждая из них
и дает все формы от caudatum до simplex (см рис. I), тем не менее ра
са В легче, чем раса Л «превращается» в бссшнпное состояние.
Наконец, в пределах каждой расы удается обнаружить еще
более мелкий наследственно констангные формы, соответствую-
щие биотипам. Они также отличаются друг от друга размерами
и характером реагирования на изменения факторов окружающей
среды (разный порядок и характер редукции шипов). У Diplodinium
dentatum один из биотипов легко превращается в бесшипиую форму,
тогда как другой никогда не удается целиком перевести в такое
состояние.
В некоторых случаях внутри вида могут образовываться геогра-
фические и экологические расы, в пределах которых ветре
чаются различные формы. Примером географических рас может слу-
жить раса Epidiniuni ecaudatum нз южного рогатого скота, которая
отличается небольшими размерами тела, короткими и не очень мощ-
ными шипами от другой расы Ер. ecaudatum из рогатого скота
северных районов СССР. В пределах обеих рас имеются разные
формы {t. caudatum, bicaudatum, tricaudatum). Особую мелкую эколо-
гическую расу этот же вид дает в северном олене (Догель, 1927а).
Имеются, наконец, и некоторые намеки на наличие у эндопарази-
тов известных топографических вариаций. Так, м икспспопо-
дня Mi/xidium giardi, паразит жабр угря, может с одинаковой
легкостью образовывать цисты и в почках той же рыбы. При этом
споры, взятые из разных .мест локализации, отличаются небольшими.
424
но определенными различиями в размерах и в пропорциях длины
и ширины. Споры из почек имеют размеры 9—10 X 5 -7. а из жабр
13Хб,5р. Таким образом, здесь местонахождение паразита внутри
хозяина отражается на морфологических особенностях паразита
Точно так же при исследовании способ разных многоклеточных пара
лггнческих динофлагеллят Haplozoon armatum из кишечника мпого-
щетинкового червя Travisia forbesi Догель (1910) отметил, что
экземпляры, взятые из самой кишечной трубки, имеют более плот
нос. компактное строение, чем особи, добытые нз слюнных желез
хозяина; эти индивиды были тонки и сильно вытянуты в продольном
направлении.
Рис. 2'23. Prosptocus confiisus из зеаснлй лягушки {Rana csculenta}
А — из кишечника; Н — из желудка. (По Догелю.)
Сосзльщнк Prosolocits confusus обычно паразитирует в верхнем
отделе тонкой кишки зеленой лягушки, но иногда попадается н в жг
лудке При этом особи из желудка, как правило, отличаются от
кишечных размерами, строением яичника, формой желточных фолли
кулов и соотношением размеров брюшной и роговой присосок
(рис. 223). В том же разрезе можно трактовать и явление возраст
кого диморфизма у Poliistoma integerrimum (Быховскнй. 1957)
Жаберные «взрослые» многоустки отличаются от настоящих взрослых
достигающих половой зрелости в мочевом пузыре, размерами, подо
развитием прикрепительного аппарата и продукцией малого количс
ства янц.
В- шикает вопрос, могут ли такие различия играть известную рол>
в процессе дивергенции и формообразования паразитов в пределах
одного хозяина В пользу этого говорят некоторые предпосылки
и факты. Прежде всего в процессе топологической дифференциации
у паразитов происходит в известной степени пространственная изоля-
ция тих особен популяции от других— обстоятельство, способ
ствующее дивергенции. Кроме того, в пользу высказанного предполо-
жения свидетельствует далеко по редкое у паразитов явтеине так
называемых сопряженных видов (Догель, 1949а). Это такт
виды, относящиеся к одному роду, которые в одном хозяине про
427
странственно отграничены друг от друга, при условии, что между
ними удается установить непосредственную генетическую связь. Хоро
шим примером сопряженных видов могут служить две формы челове-
ческой вши (Pediculus), относительно которых даже еще окончи
ли они обособились как различные
Р. vestimenti) или они представляют
приспособленные к ра ным условиям
было, оба вида пли подвида имеют
тельно не установлено, вполне
виды рода Pediatlus (Р. capitis,
собой еще подвиды Р. humanis,
существования. Как бы то ни
общее происхождение, и
Нис. 224 Схема образования
«сопряженных внасп* у перье-
вых клешей рода Freyana на
перс ут> них птиц
Л — исходный пид F. largijolia-,
ft — F. anatina—vw 1. происшед-
ший из предыдущего. (Но
Дубинину.)
исходным видом для них, несомненно, был
Р. humanis capitis, а причиной его дивер-
генции явилась редукция волосяного покро-
ва на веем теле человека. Р humanis vesfi-
nienti возникла после появления на человеке
ОД1 ж ты.
Дру гим примером уже обособленных
видов могут служить перьевые клеши, изу-
ченные Дубининым (1950, 1954а). Парази
ты пластинчатоклювых, рода Freyana, раз-
множаются на маховых перьях в той части
опахала, где расширенные брюшные края
бородок первого порядка образуют так на-
зываемые воздушные «коридорчики». Они
защищают клешей от вымывания дождем и
охлаждения. У F. anatina (рис. 224) с кряк-
вы, обитающих в «коридорчиках», прикре
ни тельный аппарат на заднем конце тела
ратвит слабо. С другой стороны, у близкого
вида F. largijolia (рис. 221). живущего на
более открытых частях опахал тех же
маховых перьев, более сильно развиты
различны! аппараты прикрепления к перу
(хвостовые щетинки, гребни на голенях
и др.). У всех утиных, обладающих выра-
женными «коридорчиками». наблюдается
одновременное существование двух группи-
ровок клещей рода Freyana типа anatina и типа largijolia. У тех утиных.
где маховые крылья лишены «коридорчик >в» и потому пред-
ставляют однородную среду обитания, всегда находят по одному-
виду клещей из сем. Freyanidac. На этих птицах дивергенция в пре-
делах одного хозяина не произошла. Там, где дивергенция имела
место, исходным типом являются формы, живущие вне «коридорчи-
ков». а скрывшиеся в них виды типа anatina испытывают упрощение
прикрепительного аппарата в результате переселения в более защи-
щенш ’С пространства.
Аналогичные примеры дивергенции дают Trichodina dotnerguei
и Т. carassii — паразиты карася. Первая расселена на повео'лости
тела и иногда заходит на жабры, вторая, более мелкая, живет исклю-
чительно на жаберных лепестках. Этот вид. вероятно, является
дериватом п< рвого. Примеры сопряженных видов можно найти среди
монотеистических сосальщиков (Гусев. 1955). Гак, па жабрах карпо-
вой рыбы Erythrorulter oxycephalic из о.ера Ханка встречается
Dactylogyrus branchialis, а на тлавпнках тон же рыбы обнаруживает-
ся другой вид — D. pterigialis, очень похожий па предыдущий, ио
вместе с гем отличный от него более мелкими крючьями прикрепи-
тельного аппарата и деталями строения поддерживающей пластинки
126
последнего. Так как жабры являются обычным местом локализации
Dactylogyrus, то поселение их на плавниках можно считать новым
местом обитания, куда перешли паразиты с жабр, и в результате
естественного отбора здесь образовался сопряженный вид с более
слабым прикрепительным аппаратом, обеспечивающим удержание
ч менее сильных токах воды, обтекающих плавники (Гусев, 1955г
Рис. 225. Схема лок-ллпз «цин эндогенных стадий двух
сопряженных видов кокцидий в тонкой юпике коолнкл
,4 — Eimeria intesiinalis; Ji.-E. magna. (По Хейспну.)
Среди эндопаразитов хороший пример сопряженных видов дают
кокцидий рода Eimeria из кроликов и кур У кролика один вид
Е. stiedae локализуется в печени, а в кишечнике паразитируют 7 ясно
различимых видов Eimeria. Все они дифференцируются топологиче-
ски, не только по длине кишечника, по и по расположению в различ-
ных участках слизистой, Например, Е. media занимает верхний отдел
кишечника, Е. magna — нижний, Е. piriformis— только толстую киш-
ку и т. д. Если локализация видов совпадает по длине кишки, то
всегда наблюдается пространственное разобщение эндогенных стадий
в толще слизистой. Так, Е. magna и Е. inlestinatis встречаются в ниж-
нем отделе тонкой кишки, по все стадии первого вида развиваются
субэпителналыю, а второго—только в эпителии ворсинок н т. д.
|рис. 225). Все кокцндии кролика узко специфичны н не могли
перейти в него от других хозяев, поэтому, несомненно, они представ-
ляют собой сопряженные виды, возникающие в пределах данного
хозяина (Хснсин. 1947,1957).
Два близких вида грегарин L'rospora iuiestinalis и U. pitlmonalis
паразитируют в голотурии Cucumaria japonicat. первая — в толще эпи-
телия средней кишки, вторая — в водяных легких. Эти грегарпны
сходны своими вегетативными стадиями, но различаются формой
спор.
Вероятно, три вида Trichomonas у человека (Т. hominis— кишка.
Т. vaginalis — половая система. Т. fenax— ротовая полость) могут
быть также признаны за сопряженные виды, где два последних про-
изошли, вероятно. 01 первого—кишечного. Средн цестод явление
сопряженных видов обнаружено у Anuplocephala. A. magna встречается
в тонкой кишке лошади, а Д. perjoliata — в толстой. У овец и коз
один вид Stiaiesia globipunclata локализуется в кишечнике, другой
вид—5. hepalica — в желчных протоках.
Совершенно очевидно, что явление сопряженных видов прекрасно
иллюстрирует процесс видообразования, связанный с освоением новых
мест обитания в одном хозяине и изоляцией популяций при контрол»
естественного отбора и независящий от дивергенции хозяина. Следует
иметь в виду, что не всякая пара или несколько видов одного рода,
обитающих в одном хозяине, являются сопряженными. Действительно,
мы знаем случаи, когда рядом друг с другом обитает несколько видон
одного рода. Например, на жабрах амурских сомов локализуется
несколько видов Ancijlodiscoides (Monogenoidea), а на lleinibarbus —
ряд видов Dcictylogyrus (Гусев, 1955). Трудно сказать, чем обуслов
лево такое явление, ро в некоторых случаях это может быть связано,
при широкой специфичности паразита, с происхождением из разных
хозяев — близкородственных или живущих в одинаковых экологии»-
скнх условиях. Для решения этих вопросов в каждом случае следует
тщательно анализировать генезис исследуемых видов паразитов.
Резюмируя все сказанное, приходится прийти к выводу, что
вопрос о внутривидовых группировках паразитов, в особенное?;
у эндопаразитов, нуждается в дальнейшей разработке. Именно это
обстоятельство и побудило нас подробнее остановиться па низших
систематических категориях паразитов, ставя данный вопрос до
известной степени в дискуссионном порядке.
3. ОСОБЕННОСТИ ЭВОЛЮЦИИ ПАРАЗИТИЧЕСКИХ ВИДОВ
При эволюции свободно живущих организмов ведущую рол
играют изменения окружающей среды. Эти изменения могут пропс
ходить на тех же самых местах, где данный вид сформировался, или
же организм наталкивается на них, расширяя площадь своего распро
странснпя. У паразитических форм этот вопрос осложняется тем, что,
помимо окружающей природы с различными ее изменениями, которая
для паразитов играет ограниченную роль (среда второго порядка),
средой обитания паразитов в первую очередь является организм
хозяина (среда первого порядка).
Отсюда вытекает одно существенное различие в эволюции св»
бодно живущих и паразитических животных. Свободно живущщ
виды эволюционируют в основном самостоятельно, завися от других
видов лишь постольку, поскольку те являются одним нз многих био
тических факторов окружающей их среды. Паразитические виды
в своей эволюции обнаруживают зависимость от эволюции вида их
хозяина, вследствие чего их развитие в некотором отношении носит
определенно направленный характер, а именно: эволюция паразитов
особенно специфичных и специализованных, протекает сопряженно
с хозяином во времени и пространстве. Дивергенция паразитов
обычно следует за дивергенцией вида хозяина. «Отсюда следует, что
родственные хозяева населены и родственными паразитами, а схема
филогенетических отношений между спецнализованными паразитами,
как правило, соответствует схеме филогенетических отношений между
их хозяевами, т. е. наблюдается филогенетический параллелизм
(Рубиоз, 1940).
Этот вопрос привлекал п привлекает внимание многих паразито-
логов. Значительный интерес представляют высказывания Фурмана
(Fuhrman. 1909, 1928. 1932) ио этому поводу. На основании изучения
ленточных червей птиц он пришел к выводу, что определенные виды
нестод всегда встречаются только в определенной группе птиц
и характерны для нее (1908). Фактически «каждый отряд птиц обла-
428
дает своей особой фауной цестод» (1932). Это положение часто
фигурирует в литературе как «правило Фурмана» (Асе. 1933, и др.)
Оно, несомненно, отражает те реальные эколого-физиологические
взаимоотношения паразита и .хозяина, которые складывались в тече-
ние длительного исторического периода. Хотя это «правило» и тре-
бует ряда оговорок, но все же до известной степени оно и сейчас
удерживает свое значение.
Более подробно вопрос о соотношении эволюции паразитов и их
хозяев рассматривается в ряде работ Эйхлера (Eichler, 1940а, 19406,
1942, 1948, 1955), Шидата (Szidat, 1956, I960) и Шгаммера (Stammer,
1957). Эйхлср формулирует и принимает три основных «паразпто-
фнлетических правила», сущность которых сводится к следующему:
1, Хозяева и паразиты развиваются параллельно, и поэтому фило-
генетические взаимоотношения паразитов отражают Taimnup тпч«рв
(«правило Фсгерхольца»).
2. В более примитивных в филогенетическом о-
встречаются более примитивные паразиты, в более в
ых хозяевах — более высокоорганизованные naj
11ицш-Ксллога»).
3. Группы хозяев, богато расчлененные в систематическом отноше-
нии, обладают большим богатством видов паразитов, чем группы хо-
<яев, менее расчлененные. По поводу этих «правил» Штаммер совершен-
но справедливо указывает: «Очевидно, что первое правило параллель
ной эволюции является основным, из которого два других о высотг
организации и расчленения легко можно вывести». Поэтому в дальней-
шем мы и остановимся главным образом на рассмотрении вопроса о
параллелизме в развитии паразитов н хозяев.
Паразитология знает целый ряд прекрасных примеров таких
весьма далеко идущих параллелизмов в развитии паразитов и их
хозяев, из которых мы ограничимся лишь некоторыми примерами.
При этом большей частью эволюция систематических единиц
паразитов несколько отстает от таковой их хозяев. Если
хозяин успел днвергировать на два самостоятельных вида, то его
специфичные паразиты остаются иногда еще па положении одного
прежнего вида или дифференцируются на два подвида. Если из
первоначального вида хозяина создались уже два самостоятельных
рода, то этому отвечает разделение первоначального вида паразита
на два вида, и т. д.
Процесс этот прекрасно иллюстрирует эволюция желудочных
оводов, которые ограничены в своем распространении двумя отряда
ми млекопитающих: хоботными (Proboscidea) и непарнокопытными
(Perissodactyla). Принадлежа к одному надсемейству Oestroidea,
оводы хоботных и непарнокопытных относятся, по Рубцову (1939),
к двум семействам: Cnbboldiidac и Gastrophilidae. Соответственно
с делением современных хоботных па два рода (африканский слон —
Loxodon и индийский — Elephas) единственный род сем. Cobboldiidae
распадается на два вида: Cobboldia loxodontis и С. elephanfis. У двух
семейств отряда непарнокопытных (носорогов и лошадиных)
Gastrophilidae образуют два рода: Gyrosligma у носорогов
и Gastrophilus у лошадиных. Далее, сем. лошадиных имеет ряд видов
Gastrophilus: на лошади (Equus caballus), зебре (Е. zebra и др.)
и па осле (Asians asinus), а современные виды носорогов дают приют
трем видам Gyrostigma. Прилагаемый рис. 226 лучше всяких слов
иллюстрирует полный параллелизм в развитии хоботных и непарно-
копытных и их оводов.
420
Не менее, если не более интересные примеры, дают вши чело-
века и обезьян (Ferris, 1932, 1933, 1935). Вши известны для всех
групп обезьян, начиная с широконосых и кончая человекообразными
Отряд обезьян распадается на два подотряда: более примитивных
широконосых, или обезьян Нового Света, и более высокоорганизован
ных узконосых, куда принадлежат семейства мартышек, гиббонов
Рис. 2'26. Схема развития желу:точных оводов в сняли с эволюцией
их хозяев — хоботных и пспрр;1О..опытиых. (По Рубцову.)
человекообразных и людей. Соответственно подразделению подотряда
узконосых паразитирующие на них вши обнаруживают меныпую
степень дивергенции, чем хозяева, и принадлежат целиком к одному
семейству (PedicuIdae). виды которого группируются в два под-
семейства: Prdtcinae (несколько видов Pedicinus) и Pcdiculinae (раз-
ные вилы рода Pediculus и 2 вида Phthirius). Pcdicinae свойственны
сем. мартышек. Pediculinae— остальным трем семействам. Вши
последних дпвергироналн очень .медленно, образован у трех семейств
хозяев лишь разные виды рола Pediculus (Р. oblongus— у гиббона.
Р. scliajfi— у шимпанзе и Р. humanis — у человека). До сих нор
параллельная эволюция вшей и их хозяев прослеживается очень
хорошо и просто.
Своего рода загадку представляет собою, однако, открытие чело-
веческой вши н особых (хотя н чрезвычайно близких к человеческой
вши — Peliculus т/obergi) видов из рода Pedt'tilus ча южноамери-
канских обезьянах из подотряда широконосых. Трудность обьясн?
.130
ния заключается в гом, что широконосые с эоцена живут гео
графически изолированно От узконосых, тогда как сам р эд Pediculus
сложился у узконосых лишь в олнгоцене. Таким образом, единствен-
ным объяснением этого язляется допущение вторичного перехода
в Южной Америке человеческой вши с человека на местных обезьян
В таком случае дальнейшая выработка особых видов Pediculus на
широконосых обезьянах из первоначального материала, т. е. челове
ческой вши, должна была идти весьма быстро, так как человек
появился в Южной Америке, вероятно, лишь после палеолита.
Вышеупомянутый автор, Феррис, находит доказательства парал
лелнзма эволюции вшей и их хозяев в значительном соьпадении
числа разных филогенетических ветвей паразитов и хозяев. При пали
чип всего 220 видов вшей они распределены на 215 родах хозяев.
Вши объединены Феррисом в 31 рода, а хозяева принадлежат
к 30 семействам. Вшн группируются, в 7 подсемейств, а их основные
хозяева относятся к 7 отрядам. Этот подсчет говорит и о пггратле-
лнзме эволюции и о более медленной дивергенции паразитов по срав
нению с хозяевами, как это видно из более высокого ранга системати
чсских категорий хозяев.
Подобно вшам отчетливую картину параллельной эволюции
с хозяевами обнаруживают паразитирующие на птицах Alallophaga
(Eichler, 1940а, б; Clay, 1957).
Некоторые примеры параллельной эволюции паразита н хозяина
показывают нам монотеистические сосальщики (Ьыховский, 1957
Гусев, 1955). Гусев отмечает, чго на амурских рыбах Gnathopogon
chankaensis встречаются два вила Dactylogyrus и па Cliilogobio
szerskii еще одни близкий к ним вид. Их взаимная связь во времени
представляется так: на предках G. chankaensis паразитировала близ
кая к одному из его современных видов форма; когда от липин раз-
вития пескарей обособились виды рода Chilogobio, то исходная форма
Dactylogyrus дала начало виду D. latjmanianus. За эго же время
исходная форма днвергнровала на близком к предкам Gnathopogon,
образовав два сопряженных вида Dactylogyrus zachvalkiiii и D. du-
bius.
Наряду с примерами отставания или совпадения эволюции пара
зитов и их хозяев можно привести пебозыпое число примеров опере-
жения темпов эволюции хозяев паразитами. Один из
примеров касается инфузорий из сем. Ophryoscolecidac, встречаю-
щегося в желудке жвачных (Догель, 19166). Ophryoscolecid ае принад-
лежат к отряду инфузорий Spirotriclia. подо гряду Enlodiniomorpha,
куда, кроме того, относится др} гое крупное сем. Cycloposthiidac, из
кишечника непарнокопытных
Сем Ophryoscolecidac содержит приблизительно десяток родов
с сотней видов, распределенных между разными жвачными; часть
указанных родов и видов узкоспецпфнчна. другие широко распро-
странены у разных хозяев, особенно у домашних животных.
В морфологическом отношении различные формы Ophryoscolecidac
располагаются в следующий ряд этапов постепенно усложняющегося
строения:
1. Низшая ступень организации: род Entodinium—одна зона рес-
ниц, отна сократительная вакуоль, скелета пет, ядро простой колбасо-
видной формы, хвостовые шипы обычно слабо развиты.
2. След)пгшая ступень: род Anoplodiniitnt— две зоны ресниц. две
вакуоли, скелета нет, ядро иногда сильно вытягивается и сгибается
шипы слабо развиты.
43!
3. Третья ступень: род Eudiploatnium — две зоны ресниц, дне на
куоли, сколет слабо развит, шипы также.
4. Четвертая ступень: роды Epidinium, Opisihotriihum, Ostraco-
dinium— две зоны ресниц, две-четыре вакуоли, скелет в виде одной-
трех широких пластин, шипы сильнее развиты
Anaplodi.i'um
EntoJiTiiunt
Рнс. 227. Схема градации морфологического
усложнения инфузорий сем. Ophryoscolecidae.
(Оригинал.)
5. Пятая ступень: роды
Polyplaslron, Caloscolex, осо-
бенно Oplv.yoscnlex— дю
зоны ресниц, много вакчо-
лен, часто образующих во-
круг тела венчики, скепет в
виде трех-пяти пластинок,
хвостовые шипы ра звиты
сильно, иногда образуя не-
сколько концентрических
венцов.
Этапы эти, как видно,
выражаются в увеличении
числа ресничных дут и со-
кратительных вакуолей, в
усложнении формы ядра, в
появлении и усилении ске-
летных пластин, наконец, в
появлении и усиленном раз-
витии хвостовых шипов
(рис. 227).
На основании добычах
главным образом Догелем
данных сейчас возможно со
ставить представление о раз-
витии Ophryoscolecidae в
следующих' главных ветвях
парнопалых копытных: в се-
мействе верблюдов из полот
ряда Tylopoda и в семей-
ствах оленей (Cervidae) и
быков (Bocidae—быки, ба-
раны, козы и антилопы) нз
подотряда Ргсога. Инфузо-
рии семейств оленьков, ви
.порогов и жирафов остаются
еще неизученными.
Рассматривая составленное по Ромеру («Палеонтология позво-
ночных», 1933) филогенетическое древо исследованных нами трех
семейств, можно определить положение в истории земли точек рас-
хождения друг от друга различных
Учитывая далее распространение у
Ophryoscolecidae, специфичных для
систематических групп жвачных,
разных жвачных родов и видов
отдельных групп хозяев, можно
получить довольно ясное представление о времени появления и дивер-
генции соответственных паразитов (рис. 228).
Так, у верблюдов специфичными являются род Caloscolex, один
вид рода Lntodinium (Е. ouum-rajae) и о тин вид Anoplodiniuri
(A. Cfimeli). Из этого вытекают два обстоятельства. Во-первых, роды
Etdudintum и Anoplodinium. общие Tylopoda и всем Pccoia, должны
были залсжиться еще до того, как произошло расхождение этих
432
двух ветвей жвачных, т с. в эоцене. Интересно, что с этим согласует-
ся примитивность данных родов — они относятся к первому и вто-
рому этапам вышеприведенной нами шкалы. Другие роды Ophryoscolc-
cidae в эоцене еще не сформировались. Что касается рода Cuioscolex,
специфичного для верблюдов, то точно определить время его
возникновения невозможно, но, судя по тому, чго одинаковые с ним
по сложности строения роды Opisthotrichuin и Ophryoscolex разви-
лись, по-вндимому, в плиоцене, сюда можно отнести и обособление
данного рода.
В пределах подотряда Ресога, помимо уже упомянутых Ento-
dinium и Anoplodinium. видами, специфичными, с одной стороны, лишь
для Corvidae, с другой — только для Bovidae обладают еще роды
Eudiplodinium и Epidinium. Судя по этому, данные два рода должны
были сформироваться еще до того, как разделились указанные
два семейства, т. е. ^см. рис. 228), вероятно, в олигоцене. Специфич-
ные виды рода Ostracodinium отсутствуют в сем. оленей, но имеются
у всех групп Bovidae. Рассуждая уже примененным нами ранее
способом, мы должны считать, что род Ostracodinium дифференциро-
вался уже после отделения ствола оленей от ствола Bovidae, но еще
то распадения последних па антилоп и на козо-барано-быков, т. е.,
с\дя по всему, в миоцене.
Наконец, род Opisthoirichuni констатирован лишь у некоторых
антилоп, тогда как роды Polyplastron п Ophryoscolex лишь у козо-
барано-быков; поэтому все эти роды должны были во шикнуть лишь
после разделения Bovidae на антилоп и прочие группы. т. е. в копне
шоцеиа — начале плиоцена. Наиболее позднее появление этих трех
родов опять-такп очень хорошо увязано со значительной сложностью
их организации (4-й и 5-н этаны морфологической эволюции). В пре-
делах козо-барано-быков род Ophryoscolex дал виды, специфичные
для быков (О. purkinjet) и для козо-баранов (О. caudatus)-, очевидно,
эти виды возникли лишь в конце плиоцена пли в плейстоцене. К тому
же времени следует относить и ряд видов и подвидов других, более
просто организованных родов Ophryosculecidue, а именно таких видов,
который узкоспецифичны для определенных видов Ресога. Таковы,
например, Entodinium anieronucleatum, Е. bicornutum, Anoplodinium
rangijeri, Lpidinium ecaudatum gigas у северного оленя; Entodinium
saigae и E. kazahstanicutn у сайги; Entodinium ovinum, E. vprax.
'noplodinium crsta-galli. Ostracodinium triloricatum у козо-баранов
н свыше десятка видов других разных родов у быка. Все они
возникли в плиоцене или плейстоцене.
Пример с инфузориями жвачных представляет интерес в том
отношении. что, по сравнению с предыдущимн, обладает некоторыми
особыми чертами, В примерах с вшами человека мы имели дело
с эволюцией одного-двух близких родов. В случае с Ophryocc olecidac
нам удастся до известной степени проследить развитие целой фауны,
слагающемся нз десятка родов с сотней видов и двумя сотнями разно-
видностей. Далее, этот пример показывает, что сопряженность
и параллелизм развития паразитов с эволюцией хозяев нельзя пони-
мать слишком узко. Дело в том, что у Ophryoscolecidae видообразова
ние идет в весьма больших размерах даже без тон стимуляции этого
явления, которую даст дивергенция вида хозяина на новые виды.
У жвачных в пределах одного вида хозяина наблюдается образова
ние нелых -^букетов» видов и разновнлно'теп иифезорпй. Так, напри-
мер. у быка встречается свыше 20 хороших видов рода Entodinium
в том числе 5 видов узкосиеиифичиых, т. е. по всей видимости возник-
26 В Л. Дыии.
431
I
I
I
Celosr. ее met (bus
Pnopl.cairteli
Ent. ovum-rajae
Лама ВерЕллд Оленьи Се&овень Олень Носове Днгимпа Сайга
Catoscotex
Ostracodiniuni
flnopfodiniu/n
Entod/mum
Qphryoscolex
Polyplastfpn
Epid/n/um
Eudiptodi/Hum
Ep id eeaudalum p/pas
Pnopi -onpi/eri
Ent bicornutum
Ent anteron<Jcl.faSum
ОрП‘.саи(РьП,гг>
Pstn Irdoricatu-)
Eudtpi affirta
EntsasaPst finmt.enala-gaUt
fntsaigae Entovinum
Ent varan
faiU ОРца
Ophr.purfa'njei
Ostr abtusum
Ostr. dentatum
Eudtpi mtratum
dtnopt. dent,
бак denticbtatum
£nt. rostratum
Ent. bimast.’us
Ent bicarinatum
OpiMotritbum
Рис. 228. Схема эволюции инфузорий ссм. Ophryoscolecidac в желудке различных
жвачных. (Оригинал.)
ших 1з пределах рода Bos у северного оленя — 3 специфичных вида
Entudiniurn и т. д. Таким образам, эволюция паразитов, по крайней
мере в пределах одного рода, может идти не только вверх по фило-
генетическому древ} их хозяев (т. е. по мере дивергентной эволюции
последних), но и вширь, т. е. у одного и того же вида хозяина, чем
в дальнейшем увеличиваются возможности и родовой дивергенции
паразитов.
При этом отмечается, что филогенетическая дифференцировка
крупных систематических единиц (родов) у Ophryoscolecidae не-
сколько отстает от эволюции хозяев. Жвачные распались на ряд
семейств и подсемейств, тогда как у инфузории дело ограничилось
дифференциацией родов. Но с другой стороны, процесс видообразова-
ния у нпфуюрий протекал более быстрым темпом, чем у хозяев.
Другим примером могут служить жгутиконосцы отрядов Ро \-
mastigida и Hyperiiiastigida паразитирующие в задней кишке терми-
тов и некоторых тараканов (Cryptocercus). По данным Кливленда
с сотрудниками (Cleveland et al., 1934), Кирбп (Kirby, 19з7), Грассе
(Grasse. 1952а) и других, взаимоотношения этих паразитов с хозяева
.ми являются весьма древними и эволюция жгутиконосцев протекала
в значительной мере параллельно с происхождением отряда Isoptera
от предков Protoblattoidea, давших, с одной стороны, термитов, с дру-
гой— современных тараканов. Отряды Polymastigida и Hypermasti
gida обраювали значительное число семейств, распадающихся на
несколько десятков родов. Большинство родэв имеет по 1—3 вида,
и сравнительно немногие (Truhunymphu и др.) имеют большое коли
чество видов. Представители Ilypermastigida и Polymastigida встре-
чаются как в термитах, так и тараканах Cryptocercus, в связи с чем
можно предположить, что эти группы сформировались еще до дивер
гепнии Вlatloidea и Isoptera. Эволюция Polymastigida осуществлялась
в осповно .1 в термитах, где образовалось 30 родов. Обмен жгутико-
носцами между хозяевами, вероятно, сильно ограничен в связи со
спецификой заражения и биологией хозяев. Поэтому распределение
паразитов по хозяевам в основном отражает их филогенетические
отношения.
В пределах отряда термитов некоторые жгутиконосцы, как, напри-
мер, виды рода Devestovina дифференцировались в основном парал-
лельно с дивергенцией хозяина (род Caloterrnes). Более часто наблю-
далось некоторое ускорение эволюции паразитов по сравнению
с хозяевами. Так, например, эволюция сем. Oxymonadidae связана
с термитами рода Caloterrnes и тараканом Cryptocercus. В первом
образовались роды Oxymonas, Microrhopalodina и Barroella, а во
вором—Saccinobaculus и Notila. Сем. Pyrsonyniphidae связано
с термитами рода Reticulotermes, в которых образовалось два рода
(Pyrsonympha и Dinenympha). Сем. Calonymphidat- (около 50 видов)
распадается на роды Coronympha, Metacoronympha, Stephanonympha.
Calonympha, Shyderella, и его эволюция сопряжена с полиморфным
родом Caloterrnes.
В отличие от инфузорий Ophryoscolecidae дивергенция в преде-
лах очного хозяина у этих жгутиконосцев, видимо, происходит в весьма
слабой степени. Случаев одновременного нахождения нескольких
видов одного рода в одном хозяине относительно мало. Можно
отмстить, например, что в Reticulotermes flaviceps встречается 4 вида
Dinenympha или в Termopsis angusticollis 3 вида Trichonympha.
Гораздо чаще разьые виды одного рода встречаются в разных видах
хозяев.
28*
Вообще следует отметить, что филогенетический параллелизм
выражен и доказуем далеко не всегда, так как часто экологические
факторы доогут затушевывать параллелизм изменений паразитов
и хозяев на протяжении нх длительной эволюции, а в некоторых слу-
чаях и значительно искажать его.
С правилом параллелизма обстоит в этом отношении дело так
же. как с биогенетическим законом. Общая распространенность этого
закона признается в топ или иной мере всеми, а ярких и далеко
прослеживаемых случаев его вовсе не гак много.
Филогенетическая близость паразитов у родственных хозяев дает
возможность, в ряде случаев, использовать паразитологический кри-
терий для установления систематической близости хозяев.
Прекрасную иллюстрацию этому дает Быховский (1933) в своем
исследовании видов Dactylogyrus. паразитирующих па разных пред-
ставителях рода Abramis. Три вида рыб рода Abramis, а именно лещ
(Л. hrama). синец (A. balle.rus) и белоглазка (A. sapa), несут па себе
совершенно различные виды Dactylogyrus, не переходящие с одной
из этих рыб на другую. Между тем лещ имеет некоторые виды
Dactylogyrus, которые, кроме того, могут попадаться на плотве
(Ruiilus) н на сырти (Vimba), т. е. на карповых, относящихся к другим
родам. Это разъясняется тем фактом, что три вида рода Abramis
хотя и живут и нерестятся в одинаковых условиях, тем не менее
помесен между собой не дают. Между тем для леща известны
помеси с плотвой и сыртыо. Таким образом, получается впечатление,
что. несмотря на родовые различия, между этими рыбами имеется
большее хромозом альное или физиологическое сходство, чем между
тремя видами рода Abramis. Нибелин (Nybelin, 1936) считает, что
по специфическим паразитам можно даже точно устанавливать роди-
тельские формы гибридов.
Интересно, что эти соображения, возникающие на основании
биологических и паразитологических данных, находят себе косвенное
подтверждение в данных систематики. Некоторые ихтиологи высказы-
вали мнение, что лещ. синец и белоглазка должны быть отнесены
к разным подродам пли даже родам. Судя но распределению видов
Dactylogyrus, именно такая классификация и является наиболее пра-
вильной.
Наряду с фактами, свидетельствующими о филогенетическом
параллелизме эволюции паразита и хозяина, встречается немало при-
меров, которые не могут быть объяснены этой закономерностью. Мы
уже отмечали, что условия обитания хозяина, специфика его питания
и другие экологические факторы, а также географическая изоляция
накладывают значительный отпечаток на состав паразитофауны в це-
лом и встречаемость отдельных видов, се составляющих. Организмы,
относящиеся к разным отрядам, но живущие в одном биотопе, питаю-
щиеся сходной пищей, имеют нередко больше общих паразитов, чем
филогенетически близкие хозяева, по обитающие в разных условиях.
Примером могут служить человек н человекообразные обезьяны,
имеющие меньше сходных видов паразитов, чем человек и некоторые
животные, имеющие с ним сходство в питании и часто контактирую
щие с ним. Например, с крысами общими являются Hyrnenolepis
папа, Trichinella, Balantidium coli, Entamoeba histolytica, с собака-
ми— Dipylidiutn caninum, свиньями—Balantidium coli, Trichinella
i! др., рыбоядными млекопитающими — Opisthorchis. Clonorchis и др.,
травоядными — Fasciola hepatica, Trichostrongylidae и т. д. Diphyl-
lobothriiun latum, Opisthorchis felineus встречаются у рыбоядных
136
животных, не имеющих близкородственных отношении между собой.
Представители Anoplocephalidae, передающиеся через панцирных клс
щей Oribatidae, встречаются в разных млекопитающих, поедающих тра
ву (жвачные, грызуны, -нитеобразные)
Обстоятельный обзор и анализ вопроса о параллелизме в эволю-
ции паразитов и хозяев н ее закономерностях, базирующийся па
многолетних исследованиях своей школы, опубликовал недавно Штам-
мер (Stammer, 1957). Он рассматривает под э-тим углом зрения
несколько групп паразитических организмов. У грегарин имеет песто
известная при роченность некоторых семейств к определенным груп-
пам хозяев (Monocystidae у Ohgochaeta, Dactylophoridae у Chilopoda
н т. п.). однако параллель .ма в эволюции установить не удается
Точно так же и у богатых видами Infusoria Astoinata, большинстве,
которых паразитирует в Otigochaera параллелизм эволюции парази
тов и хозяев не выражен. Это справедливо и в отношении большей
части инфузории эктокомменсалов (Peritricha, Suctoria), живущих
на различных морских и пресноводных беспозвоночных.
Совершенно другую картину обнаруживают некоторые паразити-
ческие нематоды, паразитирующие в жуках-коросдах (Ipidaej
и подробно изученные в лаборатории ЁУтаммера Рюмом (Riihm,
1955, 1956). Здесь имеет место исключительно отчетливо выраженный
параллелизм в эволюции паразитов и хозяев, позволяющий но фило
генетическим взаимоотношениям паразитов предсказывать и опреде
лять соответствующие отношения у хозяев. Иную картину дали пара-
зитирующие в полости тела различных насекомых нематоды отрядов
RhabdiLida и Tylenchida (Wachek, 1955). Большое количество видов
черве» этой группы распределяется по хозяевам совершенно бес
порядочно, без какой бы го ни было связи с систематическими н фило-
генетическими вза пмоо гпопн нпямн хозяев. Очевидно, в данном слу-
чае определяет мн являются экологические факторы и в том числе
принадлежность паразитов и хозяев к одному биотопу. На при мер-
нематод семейства Acuariidae, паразитирующих в птицах, Ошг
(Osche. 1957) дает обстоятельный анализ факторов, определяющих’
их эволюцию п распространение в хозяевах. При этом наряду с фак-
торами филогенетического порядка очень большую роль играют
экологические факторы, а также особенности промежуточных н резер-
вуарных хозяев. Не удалось также Штаммеру и его сотрудникам оонару
жи ъ параллелизм эволюции между насекомыми и паразитирующими и .
них клешами. Подводя итог рассмотренному мате опалу, Ш гаммер ninuei
«.Обобщая, можно сказать, что между паразитическими простейшими,
клещами и нематодами н их хозяевами в отдельных случаях имеет
место параллелизм в эволюции п таким образом реализуется первое
паразнтофилстнческое правило».
Резюмируя все сказанное об эволюции видов паразитов, можно
прийти к следующему выводу. Теоретически рассуждая, эволюция
паразитов, а особенно паразитов более или менее специфичных но
отношению к каким-нибудь определенным хозщвам. должна идти
в общем параллельно эволюции хозяев, часто с некоторым запоз.ц
пнем в дивергенции систематических единиц со стороны паразигог.
Однако этп общие законе мерности часто нарушаются различными
экологическими факторами, в особенности у паразитов, обладают!! ,
широкой специфичностью и сложными циклами развития.
137