









































































































































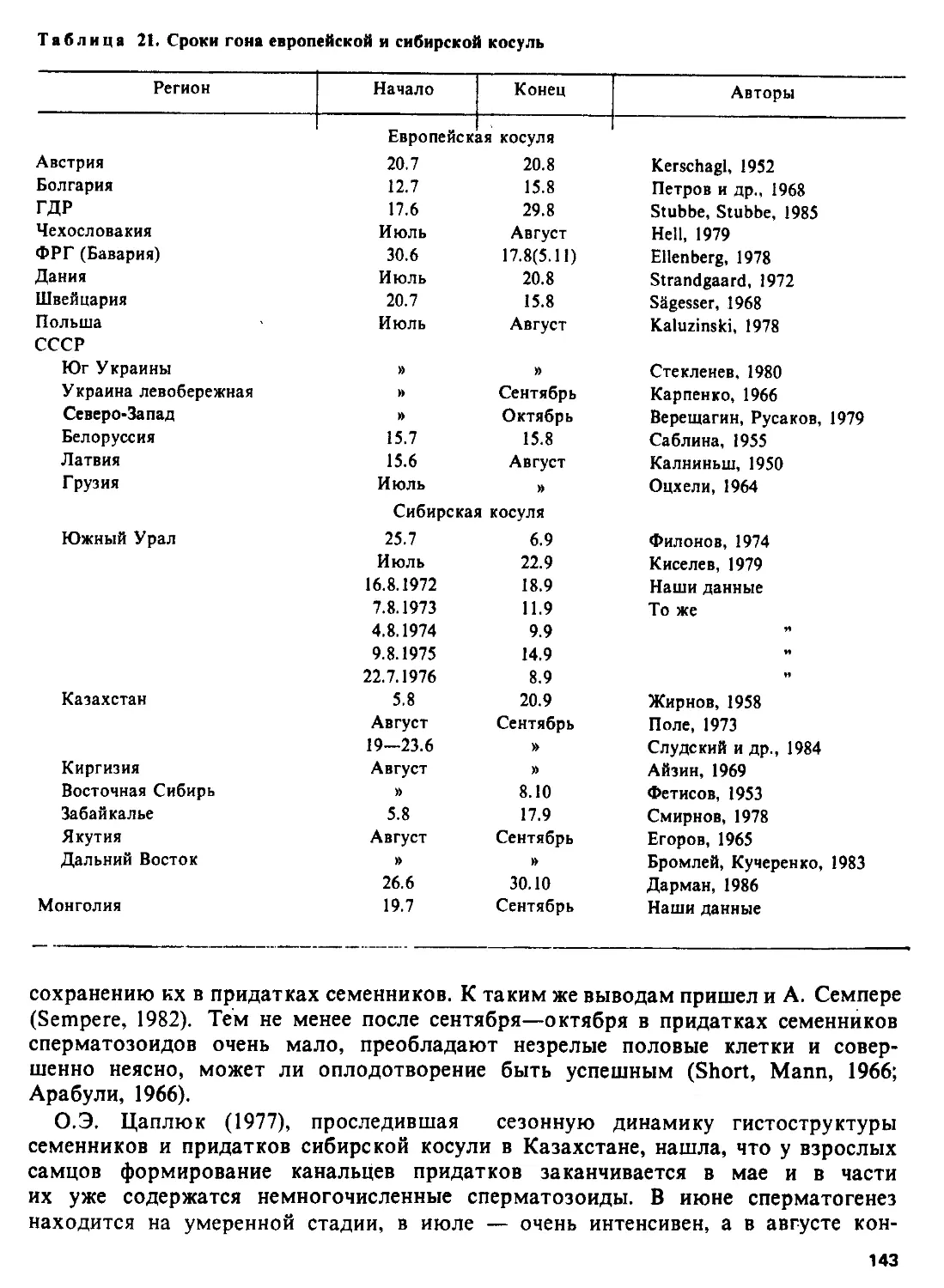

















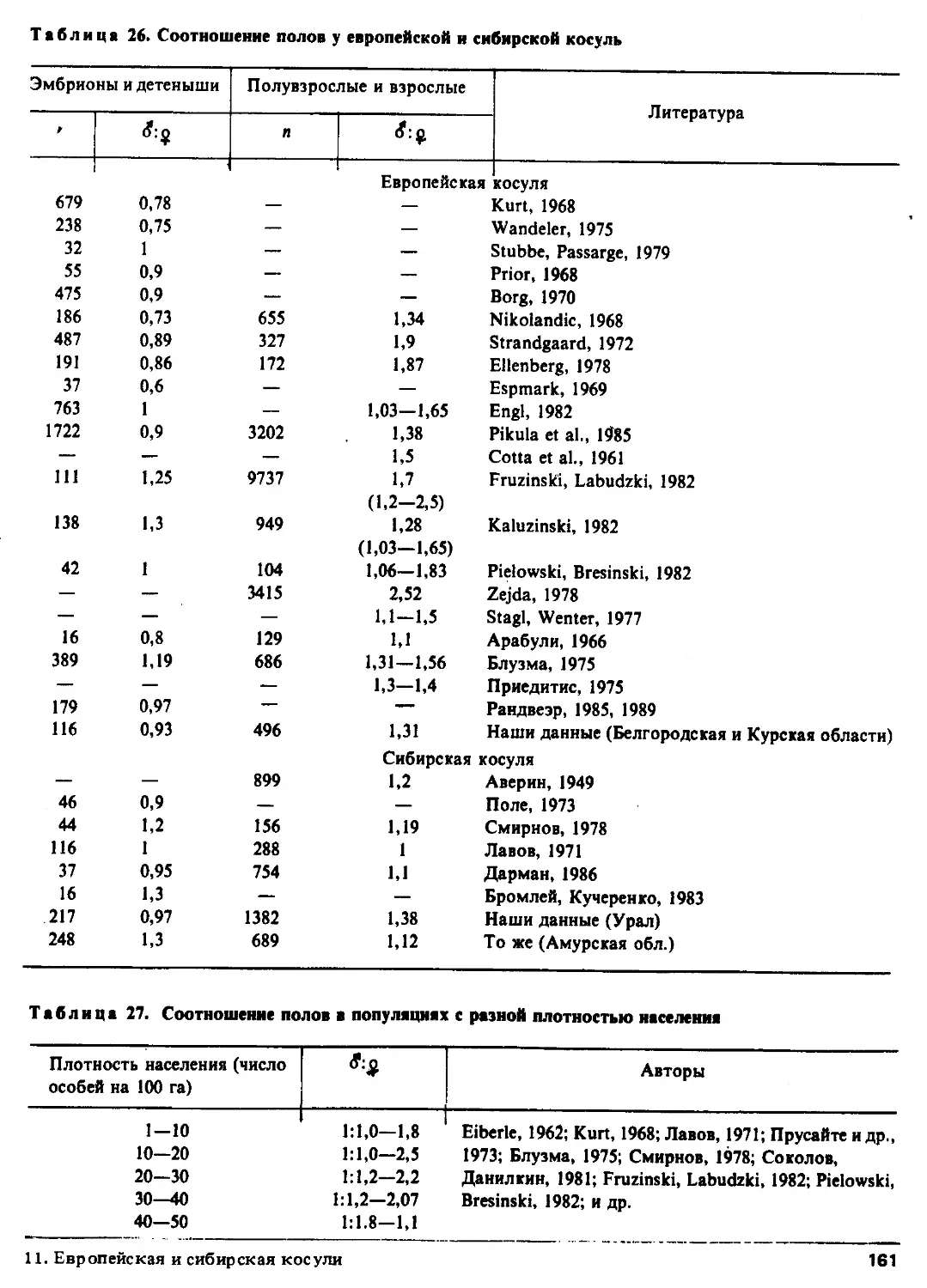























































































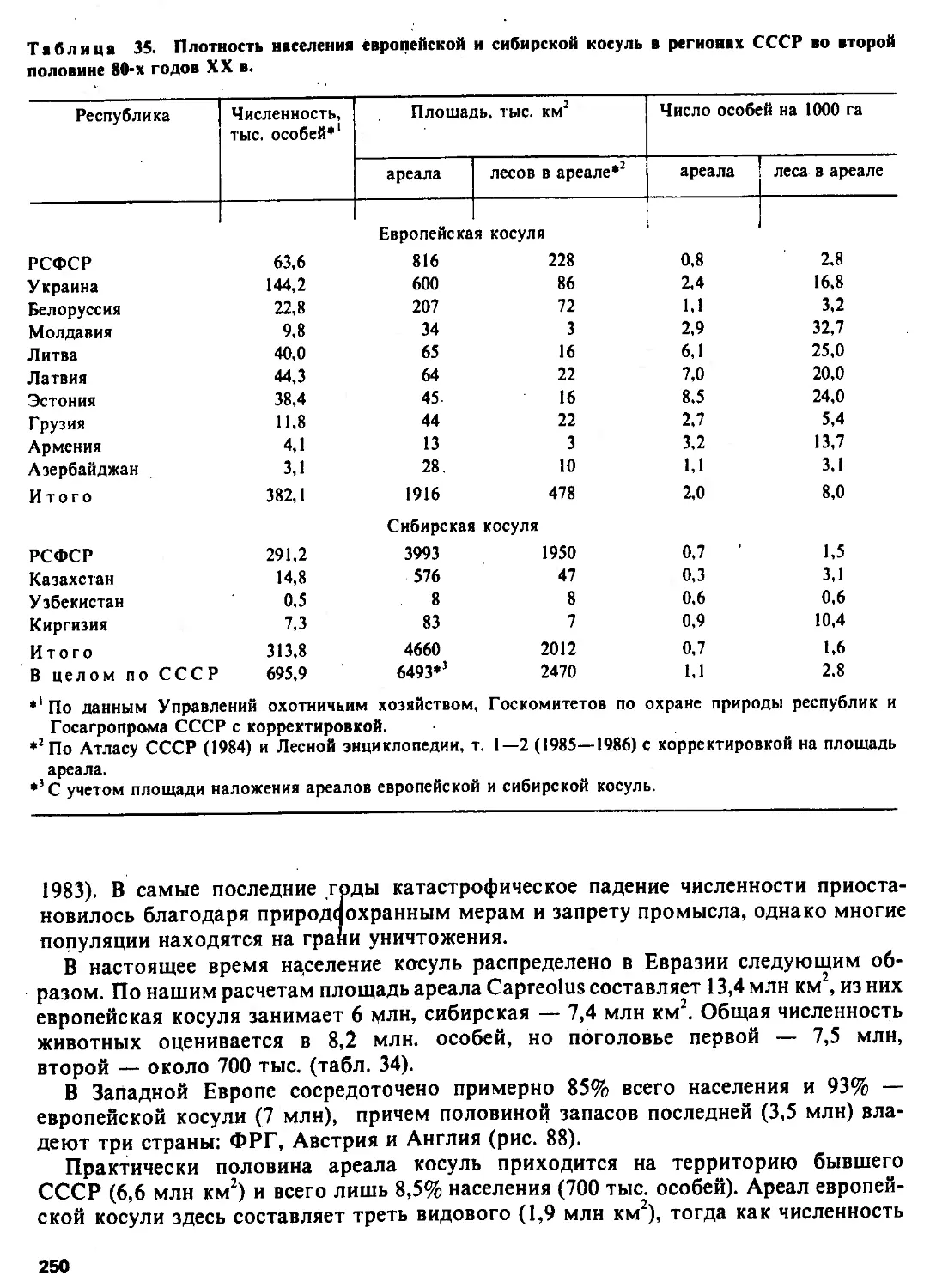





















































































































































Автор: Соколов В.Е.
Теги: ruminantia (ruminants) биология зоология эволюция охотоведение москва наука исследование систематики косули филогенетика
ISBN: 5-02-005417-8
Год: 1992




Российская академия наук
Институт
эволюционной морфологии
и экологии животных
им. А.Н. Северцова
Научный совет по программе
"Животный мир: изучение,
охрана и рациональное
использование"
I&B
Виды
фауны России
и сопредельных
стран
Российский комитет
по программе ЮНЕСКО
"Человек и биосфера"
Европейская
и сибирская
косули
Систематика, экология, поведение,
рациональное использование
и охрана
С
Издательство «Наука»
~7
Russian Academy of Science
A.N.Severtzov Institute
of Evolutionary Animal
Morphology and Ecology
Scientific Council
for the Programme
"Wildlife: Investigation,
Protection and Management"
\J Species
» "jp4Q °^ the ^auna
lyfcftj of the Russia
and the Contiguous
Countries
Russian Committee
for the UNESCO Programme
"Man and Biosphere"
European
and Siberian
Roe Deer
Systematics, Ecology,
Behaviour,
Rational Utilization,
Conservation
Editor-in-Chief
V.E. SOKOLOV
Moskow
"Nauka"
1992
Российская академия наук
Институт
эволюционной морфологии
и экологии животных
им. А.Н. Северцова
Научный совет по программе
"Животный мир: изучение,
охрана и рациональное
использование"
N$ffi
Виды
фауны России
и сопредельных
стран
Российский комитет
по программе ЮНЕСКО
"Человек и биосфера"
Европейская
и сибирская
косули
Систематика, экология,
поведение,
рациональное использование
и охрана
Ответственный редактор тома
академик В.Е. СОКОЛОВ
Москва
«Наука»
1992
УДК 599.735.3
Европейская и сибирская косули: Систематика, экология, поведение,
рациональное использование и охрана. М.: Наука, 1992. 399 с.— ISBN 5-02-005417-8
Книга представляет собой первое достаточно полное сравнительное исследование систематики
и биологии косуль в пределах всего ареала. Рассматриваются филогения, Эволюция, систематика,
изменчивость, ареал, поведение, численность и факторы, ее определяющие. Даются рекомендации по
охране и рациональному использованию ресурсов этих животных. Для зоологов, экологов, этологов,
охотоведов и охотников.
Табл. 48. Ил. 111. Библиогр. 26 с.
European and Siberian Roe Deer: Systematics, Ecology, Behaviour, Rational
Utilization, Conservation. Moscow: Nauka, 1992. 399 p. —ISBN 5-02-005417-8
The book is the first comprehensive comparative research into the systemftics and biology of roe deer throughout
their entire range. The phylogeny, evolution, systematics, variability, range, behaviour, numbers and its underlying
factors are considered. Recommendations for the conservation and rational utilization of their resources are
given. For zoologists, ecologists, ethologists, game biologists and hunters.
Tabl. 48. Fig. 111. Ref. 26 p.
Редакционная коллегия
В. Б. СОКОЛОВ
(отв. редактор)
В.Н. БОЛЬШАКОВ
Р.С. ВОЛЬСКИС
И.М. ГРОМОВ
И.С. ДАРЕВСКИЙ
П.А. ЗАЯНЧКАУСКАС
А.В. ИВАНОВ
В.Д. ИЛЬИЧЕВ
Н.С. ЛЕБЕДКИНА
Е.И. ЛУКИН
Б.П. МАНТЕЙФЕЛЬ
В.М. НЕРОНОВ
Т.С. РАСС
А.Н. СВЕТОВИДОВ
В.А. СВЕШНИКОВ
Н.Н. СМИРНОВ
Я. И. СТАРОБОГАТОВ
В.Б. СУХАНОВ
Л.С. СТЕПАНЯН
Х.М. ХАБЕРМАН
М.И. ШАТУНОВСКИЙ
Н.Н. ЩЕРБАК
А.В. ЯБЛОКОВ
Editorial Board
V.E. SOKOLOV
(editor-in-Chief)
V.N. BOLCHAKOV
R.S. VOLSKIS
I.M. OROMOV
I.S. DAREVSKY
P.A. ZAJANCHKAUSKAS
A.V. IVANOV
V.D. ILYCHEV
N.S. LEBEDKINA
E.I. LUKIN
B.P. MANTEIFEL
V.M. NERONOV
T.S. RASS
A.N. SVETOVIDOV
V.A. SVESHNIKOV
N.N. SMIRNOV
Ja.I. STAROBOGATOV
V.B. SUKHANOV
L.S. STEPANYAN
Kh.M. KHABERMAN
M.I. SHATUNOVSKY
N.N. SCHERBAK
A.V. YABLOKOV
F Ш7Ю0М-237 М1тПг, полугадне
042(02)-92
ISBN 5-02-005417-8
© Коллектив авторов, 1992
® Российская академия наук, 1992
От редактора
Объект настоящего исследования — косули — заслуживают самого
пристального внимания ученых и специалистов охотничьего хозяйства.
Велика народнохозяйственная роль этих животных, но с каждым годом
она будет еще усиливаться вследствие большой экологической
пластичности и приспособленности зверей к жизни в культурном ландшафте,
жизни рядом с человеком, что делает их наиболее перспективными
видами для современного охотничьего хозяйства. Состояние
популяций в пределах обширного ареала неодинаково. Европейские косули
в настоящее время — самые многочисленные животные среди диких
копытных, и во многих районах требуется искусственное снижение их
населения. Напротив, в Азии, где сибирские косули еще в прошлом веке
составляли основу промысла и добыча их исчислялась сотнями тысяч
особей, многие популяции находятся на грани исчезновения, и здесь
необходимы серьезные меры по охране и восстановлению поголовья.
Несмотря на хозяйственное значение косуль, до сих пор не было
глубокого исследования их биологии в пределах всего ареала, весьма
отрывочными были знания о систематике, распространении, популя-
ционной биологии. Настоящая монография в значительной мере
восполняет этот пробел и, по сути, представляет собой биологические
основы рационального использования и охраны животных, научные
основы управления популяциями. Настоящая фундаментальная сводка,
входящая в серию "Виды фауны России и сопредельных стран" и
являющаяся итогом многолетних исследований .ведущих
специалистов, несомненно будет интересна как для зоологов, экологов
и этологов, так и для практиков-охотоведов и охотников.
Академик В.Е. Соколов
5
Предисловие
Идея подготовки монографии о косулях принадлежит академику В.Е. Соколову,
который всячески способствовал осуществлению программы исследований,
принятой на Координационном совещании специалистов по косуле
(Москва—Вильнюс, 1979 г.).
К сожалению, не все задуманное осуществлено. В частности, по ряду причин
не удалось одновременно с первым томом монографии подготовить второй,
в который планировалось включить разделы о развитии и росте косуль,
морфологии, биохимии, физиологии, главу о болезнях и паразитах. В связи с этим далеко
не весь материал, имеющийся у нас, отражен в настоящей монографии.
Большие сложности возникли в организации совместных исследований и в сборе
стандартизированной информации по регионам, некоторые популяции остались
практически неизученными. По-прежнему остается много вопросов, требующих
рассмотрения. Далеко не все ясно в происхождении косуль и видообразовании.
Необходимы более тщательные сравнительные исследования современными
методами европейской и сибирской косуль и подвидов последней, более
пристальное изучение популяций в Центральном Китае, Передней Азии, в Якутии и на
Алтае, на востоке европейской части России и на Кавказе. Большой интерес
вызывает ситуация в "зоне гибридизации" европейской и сибирской косуль на
правобережье Волги.
Не все главы монографии равноценны по объему и однотипны по стилю
изложения. Учитывая важное народнохозяйственное значение косуль и то
обстоятельство, что в числе читателей, помимо специалистов, будут и охотники, было
решено расширить объем главы "Хозяйственное значение, рациональное
использование ресурсов и охрана" и по возможности уменьшить ее насыщенность
литературными ссылками. В других разделах авторы, напротив, стремились привести
литературные источники как можно более полно, чтобы отразить разнообразие
точек зрения исследователей, т.е. дать "пищу для размышлений" нашим
последователям.
Обработка собранного материала и первоначальное редактирование
осуществлены А. А. Данил киным, им же написано большинство разделов. Большую
помощь в сборе данных для настоящей монографии оказали специалисты
Управлений охотничьего хозяйства республик Советского Союза и других стран,
сотрудники заповедников и охотничьих хозяйств, коллеги из многих зоологических
и биологических институтов и музеев страны и охотники. Нет возможности
перечислить всех помогавших нам. Мы благодарны всем и особенно В. Бресинскому
(Польша), К. Матраи, Л. Шугару, И. Хелтаи (Венгрия), С. Дуламцэрэну (Монголия),
Ю.А. Дарману, В.Б. Поле, Г.Г. Собанскому, А.Б. Арабули, А.А. Киселеву, А.А.
Гусеву, В.Г. Швецу, СП. Кучеренко, К.А. Морозову и другим зоологам и охотоведам.
б
приславшим первоисточники своих публикаций, предоставившим свои коллекции
или давшим ценные советы. Искреннюю признательность хочется выразить
К.П. Филонову, Л.В. Жирнову, Л.М. Баскину, Г.В. Кузнецову, В.А. Кузякину,
И.Я. Павлинову, Н.Ш. Булатовой, В.П. Сухову, прочитавшим рукопись или
отдельные главы и высказавшим полезные замечания, а также А.Ю. Стручкову,
О.В. Чекалиной, М.Н. Воронцовой и В.М. Гудкову за помощь при подготовке
монографии к печати.
Рукопись монографии, подготовленная к публикации в 1989 г., около трех
лет находилась в печати. За это время в мире произошли существенные изменения:
Советский Союз распался на ряд независимых государств, не стало на
политической карте ГДР, в Европе образовались новые страны, меняются названия
городов и других населенных пунктов, и процесс этот продолжается. Такие
изменения влекут за собой необходимость уточнения географических названий и
коренной переделки рукописи. Однако авторы и редакционная коллегия решили
не изменять текст, отобразив тем самым историю исследований биологии
косули и состояние популяций животных в мире на конец 80-х годов текущего
столетия.
Глава первая. Филогения, эволюция, систематика
Классификация семейства оленьих Cervidae Gray, 1821, как и многих других
парнокопытных, разработана недостаточно четко. В полной мере это касается косуль
и близких к ним форм — современных Capreolus Gray, 1821 и древних Ргосар-
reolus Schlosser, 1924. Во многом не решены также вопросы их эволюции и
родственных взаимоотношений. Это объясняется рядом причин, и в первую очередь
малочисленностью и фрагментарностью костных остатков ископаемых форм и
отсутствием точных определений их геологического возраста.
Современных косуль рода Capreolus рассматривают либо в составе подсемейства
Cervinae Baird, 1857 (Флеров, 1952; Соколов И.И., 1959; Вислобокова, 1981), либо
относят к подсемейству Odocoileinae Pocock, 1923 (Simpson, 1945; Гептнер и др.,
1961; Viret, 1961; Haltenorth, 1963; Соколов В.Е., 1979; Барышников и др., 1981; Ко-
роткевич, 1984). Симпсон (Simpson, 1945) выделяет в составе последнего трибу
Capreolini Simpson, 1945, включив сюда, кроме Capreolus, род вымерших косуль
Procapreolus. И.Я. Павлинов и О.Л. Россолимо (1987) предлагают за трибой
Capreolini сохранить авторство Брукса (Brookes, 1828) и заменить наименование
подсемейства Odocoileinae Pocock, 1923 на Alcinae Brookes, 1828 (Павлинов,
Россолимо, 1987).
Место древних косуль в системе оленьих также долгое время не было точно
определено. Их относили и к подсемейству Pliocervinae (Zdansky, 1925), и к Cervinae
(Czyzewska, 1968; Вислобокова, 1983), и к группе Capreoli вместе с древними
цервулинами (Dawkins, 1878). Procapreolus рассматривали в составе рода Cervus
(Bravard, 1828; Croizet, Jobert, 1828; Deperet, 1854; Pomel, 1853; Gervais, 1859; Daw-
kins, 1878; Boule, 1905; Macarovici, 1936; Heintz, 1970), относили к роду Cervavus
(Deperet, Croizet, 1884; Schlosser, 1903), роду Capreolus (Pavlow, 1896; Венюков,
1902) и к Cervocerus (Czyzewska, 1960).
В последние годы появились новые исследования, отражающие филогенетические
отношения в семействе Cervidae. На основании детального анализа эволюции
и распространения цервид Р. Харрингтон (Harrington, 1985a) пришел к выводу
о полифилетическом происхождении представителей телеметакарпальной и пле-
зиометакарпальной групп оленей (рис. 1).
Основания для этого, помимо морфологических, следующие. Первые имеют
высокий репродуктивный потецциал, относительно рано созревают, питаются
преимущественно молодыми побегами, менее устойчивы к заболеваниям и др.
Вторые отличаются сравнительно низким репродуктивным потенциалом, самки
поздно становятся половозрелыми, животные могут существовать на грубой
пище, они более устойчивы к паразитарным заболеваниям. У телеметакар-
пальных форм полиморфизм по определенным белкам и ферментам крови
невысок, хромосомные наборы в целом сходны (2л = 70), но степень репродуктивной
изоляции между видами явственно больше, чем у плезиометакарпальных оленей,
8
Moschus Capreolus
Htidropotes Alces
0
Telemeiacarpi
Rangifer Blasiocerus
Odocoileirs Ozotocerus
Hippacamelus
i Mazama Pudu
J-
->•: 30 миллионов лет
Munijocus
Elaphodus
Lama Terras
Axis Elaphurus
f'lesiomeiacarpi
Рис.4. Филогенетическая диаграмма Cervidae (из: Harrington, 1985a)
у которых диплоидный набор хромосом различен, но гибридизация между
видами идет сравнительно легко.
По своим параметрам косули явно относятся к группе телеметакарпальных
оленей, однако по некоторым показателям не вписываются в нее (табл. 1).
Сбрасывание рогов у них, как у большинства представителей этой группы,
происходит осенью или в начале зимы, но рост рогов начинается практически сразу же
после сбрасывания, тогда как у других — только весной. Отличаются они в этом
отношении и от плезиометакарпальных оленей, у которых возобновление роста
рогов происходит сразу же после сбрасывания, однако они сбрасывают рога
весной, а не осенью. К тому же только у Capreolus активация гонад,происходит
зимой, тогда как у других оленьих — в начале лета, (Sempere, 1982).
Есть и еще очень существенное отличие от остальных цервид. Период
спаривания у Capreolus начинается летом, у других оленей — осенью (см. табл. 1). Это
единственные парнокопытные с латентным периодом беременности. Явно, что
в семействе Cervidae косули занимают особое место.
Это подтверждается также гельминтологическими данными. Большинство
представителей семейства, включая Cervinae и Odocoileinae являются хозяевами одной
или нескольких форм нематод подсемейства Elaphostrongylinae, к которым косули
не восприимчивы ни спонтанно, ни в эксперименте. Личинки этих гельминтов
погибают, достигнув их легких. В данном отношении они оказались" сходными
с зайцеобразными, грызунами и полорогими. Возможны два объяснения насто-
9
Таблица 1. Некоторые отличительные признаки отдельных представителей Cervidae, имеющие
систематическое значение
Вид
Capreolus pygargus
С. capreolus
Odocoileus hemionus
О. virginianus
Rangifer tarandus
Alces alces
Cervus elaphus
C. nippon
С dama
C. axis
Группа*
T
Т
Т
Т
Т
Т
П
П
П
П
Хромосомный
набор, 2л
70+0" 14)
70
70
70
70
70—68
68
68—64
68
66
Сроки (месяцы)
сбрасывания
рогов
XI
XI
I—II
I—II
XI—XII
XI—XII
III—IV
IV—V
IV
IV
*T — телеметакарпальные, П — плезиометакарпальные.
начала
роста
рогов
XII
XI
IV—V
IV—V
III—IV
IV—V
IV
IV—V
IV—V
IV—V
гона
VII—IX
VI—VIII
IX—XI
X—XI
IX—X
IX—X
IX—X
IX—XI
IX—X
IX—X
Пятни-
раски у
взрослых
_
-
-
-
-
-
- +
+
+
+
Наличие
предглаэ-
ничных
желез
_
-
+
+
+
+
+
+ •
+
+
ящему феномену — или косуля была хозяином элафостронгилин в прошлом,
а потом стала невосприимчивой к инвазии (что маловероятно), или филогенетическая
ветвь рода Capreolus дифференцировалась от общего ствола оленей до того, как
у последних исходная форма протостронгилид эволюционировала до
элафостронгилин, т.е. косули обособились раньше, чем произошло разделение Cervinae
и Odocoileinae. Таким образом, в свете гельминтологических данных косуль
следовало бы выделить в самостоятельную систематическую группу, видимо на
уровне подсемейства (Прядко, 1972).
Поддерживая эту точку зрения, мы тем не менее считаем, что для окончательных
выводов необходимы более тщательные сравнительные исследования представи-
Таблица 2. Генетическое сходство (I, над диагональю) и различие (D(;, под диагональю) (по: Nei,
1972; из: Baccus et al„ 1983)
Вид
A. americana
В. bison
В. taurus
A. alces
Antilocapra americana
Bos bison
B. taurus
Alces alces
Capreolus capreolus
Cervus elaphus canadensis
C. elaphus
Odocoileus hemionus
O. virginianus
Rangifer tarandus caribou
R. tarandus
—
1,422
1,670
1,321
1,164
1,133
0,925
1,127
1,164
1,330
1,294
0,241
—
0,085
1,221
1,415
0,902
0,656
1,406
1,491
1,059
1,030
0,188
0,918
—
1,143
1,123
0,843
0,661
1,313
1,392
0,993
0,975
0,267
0,295
0,319
—
0,619
0,822
0,809
0,771
0,759
0,650
0,469
10
телей семейства Cervidae всеми современными методами, и в первую очередь
иммунологическими и биохимическими. В генетическом отношении косули
оказываются более близкими к Odocoileus, Alces и Rangifer, чем к Cervus (табл. 2),
что подтверждает правомерность классификации оленьих, предложенной Симпсо-
ном (Baccus et al., 1983).
Включение древних Procapreolus и современных Capreolus в состав трибы Сар-
reolini подсемейства Odocoileinae нам пока что представляется более оправданным.
Основанием для отнесения косуль к трибе Capreolini служат прежде всего
особенности строения черепа, метакарпальных костей (степень редукции боковых
метаподий), последовательность развития отростков на рогах, отсутствие
надглазничного отростка. Последним признакам придается особое значение, так как рога
вымерших оленей лучше сохраняются в ископаемом состоянии, чем другие части
черепа.
Онтогенетическое развитие рогов Procapreolus прослежено на серии различных
возрастных стадий этих образований у P. florovi Korotkevich, 1974 из раскопок
у с. Черевичное Одесской области. На начальной стадии рог имеет вид одно-
конечной "спицы" без розетки, несколько уплощенной, особенно в верхней части
(рис. 2, А). На следующей возрастной стадии рог округляется, удлиняется и
разветвляется вверху (рис. 2, Б). При этом передний отросток вилочного
разветвления является продолжением ствола рога, а задний представляет собой вторичное
образование. Затем происходит закладка третьего по порядку появления отростка
за счет небольшого выпячивания передней стенки ствола (рис. 2, В). На
последующих стадиях (рис. 2, Г—Е) этот отросток удлиняется и рог приобретает
типичную для косуль форму (Короткевич, 1974).
Одновременно с Procapreolus существовали близкие к ним по размерам и
имеющие аналогичное строение рогов и зубов олени — плиоцервусы. Однако у них в
отличие от Procapreolus после одноконечной стадии рога и последующего
дихотомического разветвления его верхушки продолжением ствола служит задний
отросток, а передний (второй по порядку появленя) является надглазничным.
Последующее развитие рогов происходит за счет дихотомического разветвления
основного ствола (рис. 3). Рога плиоцервус и цервавитус (рис. 3, III) на двухконечной
С. capreolus
0,312
0,243
0,325
0,539
—
0,719
0,824
0,811
0,718
0,832
0,512
Се. canadensis
0,322
0,406
0,430
0,440
0,487
—
0,290
0,989
0,896
0,850
0,706
С. elaphus
0,396
0,519
0,517
0,445
0,439
0,749
—
0,889
0,936
0,841
0,791
О. hemionus
0,324
0,245
0,269
0,460
0,445
0,372
0,411
—
0,149
0,783
0,665
О. virginianus
0,283
0,225
0,249
0,468
0,488
0,408
0,392
0,862
—
0,766
0,548
R.t. tcaribou
0,264
0,347
0,371
0,522
0,435
0,427
0,431
0,457
0,465
—
0,174
R. tarandus
0,274
0,357
0,377
0,626
0,599
0,494
0,454
0,514
0,578
0,840
11
Рис. 2. Возрастная изменчивость рогов (А—Е) у Procapreolus florovi Korotkevich. Черевичяое, Одесская
обл., конец мэотиса
/—3 — последовательность развития отростков рогов; А—Е — см. в тексте
Л
ш
Рис. 3. Последовательность развития отростков (/—3) рогов у Procapreolus (/), Pliocervus (II) и Cervavitus
стадии развития по форме и особенностям строения крайне близки к рогам цер-
вулйн типа эупрокса, отражая их филогенетические связи.
Отсутствие надглазничного отростка и последовательность развития остальных,
с одной стороны, служит четким отличительным признаком косуль от настоящих
оленей подсемейства Cervinae, а с другой — свидетельствует об очень раннем
ответвлении Capreolini от филогенетического ствола оленьих.
У древних оленей и косуль было различным и эволюционное направление
изменения зубов (рис. 4, 1—30). У древних цервулин — эупроксов, как и у церва-
витусов (первые, возможно, стоят в основе развития настоящих оленей), строение Р4
сохраняет примитивные особенности: внутренние долинки открыты, метаконид
в той или иной степени вздут (см. рис. 4, 26, 29, 30). Такой же тип строения Р< и
у Cervidae gen. из понта Одессы (10). У оленеобразных из района Кучургана (рис. 4,
5—13) (представленных, судя по строению рогов, оленем плиоцервусом, древней
косулей прокапреолюс и древними цервулинами, близкими по общим размерам)
зубы характеризуются разнообразным типом строения. У мэотических (рис. 4,
14—25) и сарматских (рис. 4, 27, 28) видов прокапреолюс эти зубы менее моляри-
зованы, чем у современных косуль: намечается слияние переднего конца метаконида
с параконидом, как и на зубах современных и субфоссильных Cervus elaphus,
у которых прослеживается тенденция к замыканию передней долинки в результате
слияния переднего конца метаконида с параконидом (рис. 4, 3, 4). У современных
же видов Capreolus (рис. 4, /, 2) Р4 моляризован в значительно большей степени,
у С. pygargus (рис. 4, 1) замкнута даже задняя долинка (при одинаковой стертости
зуба).
В целом у оленей изменение Р4 сопровождалось постепенным его усложнением
от первичной примитивной формы с открытыми внутренними долинками до
сложной, отражающей степень моляризации этого зуба — с уплощенным мета-
конидом, образующим внутреннюю стенку зуба за счет замыкания внутренних
долинок. У косуль моляризация Р4 происходила за счет разъединения прото-
конида и метаконида и соединения последнего с энтоконидом (рис. 4).
Полное разделение хоан выступающим далеко назад сошником, задний край
которого достигает и касается основной клиновидной кости, считается основным
отличительным признаком Cervinae от Odocoileinae (Флеров, 1952; Соколов И.И.,
1959). У косуль задний край сошника достигает основной клиновидной кости.
Он имеет вид тончайшей пластинки, следы которой прослеживаются на
субфоссильных черепах.
В основу филогенетической ветви, идущей к Procapreolus и Pliocervus,
развивавшихся некоторое время параллельно, К.К. Флеров (1950) ставил прокапреолюс,
а цервавитуса относил к другой филогенетической ветви, но происходящей от
общего с ним ствола древних цервулин — дикроцерин, которых он вел от верхне-
олигоценовых—раннемиоценовых Palaeomericinae. И.А. Вислобокова (1983)
выводит косуль, лосей, большерогих и настоящих оленей из плиоцервин, отходящих
от ветви Dicrocerini, связывая последних с Dremotheriini.
На наш взгляд, исходя из приведенных выше данных, Pliocervini как и Dicrocerini
не могут лежать в основе развития представителей трибы Capreolini. Последние,
скорее всего, отделились от основного ствола оленьих на более ранних этапах
эволюции.
Гипотетическим предком косуль, возможно, могли быть представители рода
13
^в^ && G& из5
и
№цъ
/у
75
Л?
27
27
6 /2 V 21 ' 28
23
•TV>
Л?
20
1см
Z*f
25
2S J"
Рис.4.Строение жевательной поверхности предкоренных зубов (Р4)
/ — Capreolus pygargus Pall.; 2 — С. capreolus L.\ 3, 4 — Cervus elaphus (сильно- и слабостертые);
5—9, 11—13 — оленеобразные из Кучургана (Одесская обл., конец раннего—начало среднего плиоцена);
10 — Cervidae gen. (Шкодова гора, окр. Одессы, ранний плиоцен); 14—15 — Procapreolus cf. florovi
(Андреевка, Николаевской обл., вторая половна мэотиса); 16—20 — P. florovi (Черевичное, Одесская обл.,
конец мэотиса); 21—25 — P. cf. ucrainicus; 26 — Cervavitus variabilis Alexjev (Новоелизаветовка, Одесская
обл., начало мэотиса); 27—28 — P.' ucrainicus (Новоукраинка, Одесская обл., конец позднего сармата);
29 — Cervavitus sp. (Гребеники, Одесская обл., вторая половина позднего сармата); 30 — Euprox sp.
(Климентовичи, Хмельницкая обл., начало среднего сармата)
Procervulus Gaudry из нижнего-среднего миоцена Евразии. Процервулюсы
рядом исследователей включены в подсемейство Lagomerycinae Pilgrim, другие же
относят их к подсемейству Palaeomericinae Matthew (см.: Вислобокова, 1983), что,
вероятно, правильнее. Рога процервулюса характеризуются двураздельным
разветвлением верхушки довольно высокого и тонкого ствола, слабо овальной
формой его сечения, бороздчатой структурой основания ствола в нижней части,
отсутствием розетки (Thenius, 1948). В определенной мере рога процервулюс
по строению и размерам близки к начальным стадиям развития рогов прокапреолюс.
Последующие филогенетические связи представителей трибы Capreolini и их
эволюция прослеживаются следующим образом. В раннем-среднем миоцене, скорее
всего, появились косулеподобные формы со слаборазвитым средним отростком
14
Рис. 5. Форма рогов и строение коренных зубов Procapreolus ucrainicus Korotkevich из Новоукраинки
А — левый рог, Б — правый рог молодых особей; В — рог взрослой особи; Г — верхние коренные
М2—М3; Д — нижние коренные Мг—Мз
на рогах, которые и стали в основе развития древних косуль рода Procapreolus.
Позднемиоценовые виды этого рода уже имели полностью сформированные
лировидные рога с тремя отростками, как у современных косуль. О форме рогов
у предковых форм прокапреолюс можно судить на основании строения рогов
молодой особи наиболее древнего вида из известных в настоящее время P. ucrainicus.
Для него характерно сравнительно слабое развитие среднего отростка и
небольшие размеры рогов, а также примитивный тип строения зубов (сильное развитие
палеомериксовой складки, воротничка, дополнительных элементов эмали)
(рис. 5).
Время существования P. ucrainicus в Восточной Европе определяется концом
позднего сармата—началом мэотиса (Короткевич, 1965). Возможно, что он близок
типовому виду рода P. latifrons из Монголии (Schlosser, 1924) и Китая (Zdansky, 1925),
геологический возраст которых трудно сопоставим с таковым европейских видов.
Из числа последних можно назвать P. florovi, обитавшего в конце мэотиса
в Восточной Европе, и P. cusanus, остатки которого отмечены здесь
преимущественно в отложениях нижнего плиоцена (ранний виллафранк). P. wenzensis
описан из среднего плиоцена Польши (Czyzewska, 1960). Этим возрастом ограничено
распространение рода Procapreolus в Восточной Европе (Стратиграфия СССР.
Четвертичная система, 1982, Т. 1), вероятно на основании включения Кучургана
15
в состав молдавского русильена. Нам же возраст кучурганского фаунисти-
ческого подразделения представляется более древним.
Самый поздний представитель рода — Procapreolus sp. отмечен в квабебской
фауне (Восточная Грузия), возраст которой определен низами верхнего плиоцена
(Векуа, 1972). По многим признакам он близок к P. cusanus из Этуэра (Франция,
нижний виллафранк). Этот вид и близкие к нему формы известны также с
территории севера Италии, Венгрии невероятно, Румынии.
Таким образом, в составе рода Procapreolus насчитывается не менее пяти
видов, время существования которых определяется концом сармата—началом
позднего плиоцена. Их обитание в основном связано с прибрежными районами
Восточного и Центрального Паратетиса — древнего Средиземноморья. Для
территории Восточной Паратетиды известно три вида: P. ucrainicus — P. ftorovi —
P. cusanus, характеризующие последовательные по времени фаунистические
подразделения гиппарионовой фауны Северного Причерноморья. У этих видов
происходит постепенная моляризация предкоренных зубов, особенно четко
проявившаяся на Р4, уменьшение базальных образований и дополнительных элементов
(складок) эмали на коренных зубах, уменьшение палеомериксовой складки вплоть
до ее исчезновения у более молодых видов, укорочение длины роговых пеньков,
относительное понижение среднего отростка, увеличение общих размеров.
Плиоценовые виды рода Procapreolus, вероятно, стояли в основе рода Capreolus.
Последние по морфологическим особенностям строения черепа (см. ниже диагнозы
родов) и рогов (рис. 6) представляют собой как бы последующую эволюционную
стадию их развития. Признаки, сближающие оба рода, наиболее четко проявляются
у сибирской косули Capreolus pygargus Pall. Кости конечностей Procapreolus
(метаподии, большая берцовая, копытные фаланги и др.) по морфологическим
признакам и пропорциям близки к таковым у современных косуль. Строение
резцов у них практически такое же — самый большой Ii, остальные
последовательно уменьшаются в направлении Ь; клык (С) по длине коронки равен Ь;
коронки резцов симметричной формы, умеренно широкие, высокие, на их внутренней
поверхности имеется одно продольное ребро. Резцы, как и у Capreolus, не
приспособлены для подрезания коры, но обеспечивают относительно широкий захват
растительной пищи.
Морфологические особенности строения черепа и зубов у Procapreolus отражают
их адаптации больше к питанию мягкой сочной древесно-кустарниковой, чем
травянистой растительностью. Животные, видимо, обитали в условиях
субтропического климата, характерного для начальных этапов развития гиппарионовой
фауны, среди густых зарослей кустарника, невысоких деревьев и высокотравья,
окаймлявших многочисленные водоемы. Во второй половине плиоцена площади
таких биотопов сокращаются, поскольку климат становится более сухим и
постепенно происходит остепнение ландшафта. Скорее всего, именно эти изменения
способствовали эволюционным преобразованиям у Procapreolus.
Первое появление представителей рода Capreolus в Восточной Европе указано
в котловинской фаунистической группировке, возраст которой сопоставляется
с нижним виллафранком. Присутствие их прослеживается на протяжении
существования таманского фаунистического комплекса, однако они не найдены
в тираспольском и сингильском, но четко обнаруживаются в хазарском комплексе
(Вангенгейм, Флеров, 1982). По последним данным костеносные отложения Котло-
16
Рис. б. Форма рогов Procapreolus (A—В) и
Capreolus (Г, Д)
А—Б — P. florovi из Черевичного; В —
P. cf. florovi из Андреевки; Г — С. pygargus;
Д — С. capreolus
вины разновозрастны: остатки косули, вероятно, происходят из отложений хапров-
ского фаунистического комплекса, возраст которого отвечает среднему вилла-
франку или позднему плиоцену. Остатки косуль из нижнеантропогеновых
отложений известны с юга Украины, Молдавии, Передней Азии, Закавказья
(Громов В.И., 1948; Пидопличко, 1951а,б; Татаринов К.А., 1970; Алексеева Л.И., 1977;
и др.).
В Западной Европе описана С. suessenbornensis Kahlke, 1956, которая по
основным признакам строения близка к европейской косуле С. capreolus L.(Kahlke, 1969).
Возраст фауны Зюссенборна определяется низами среднего плейстоцена (миндель),
что сопоставимо с фауной тираспольского комплекса, возраст которого по
стратиграфической схеме, принятой в СССР, соответствует нижнему плейстоцену
(Кальке, 1971).
Формы Capreolus, близкие к современным, в Восточной Европе отмечены в
позднем плиоцене, а в Центральной — в начале антропогена. В Азии наиболее древняя
Capreolus, сопоставляемая с таманским фаунистическим комплексом, обнаружена
в Павлодарском Прииртышье (Вислобокова, 1973). Э.А. Вангенгеим (1977) эту
Capreolus sp. относит к кизихскому комплексу, характерному для первой половины
позднего эоплейстоцена Западной Сибири, и отмечает близость его одесскому
комплексу и верхневиллафранкским фаунам Центральной и Западной Европы.
2. Европейская и сибирская косули
17
Ряс. 7. Места находок остатков Procapreolus (1) и Capreolus (2—7)
1 — плиоцен; 2 — поздний плиоцен — ранний плейстоцен; 3 — плейстоцен; 4.5 — голоцен; 6,7 — 1 —2-е
тысячелетие н.э. (б — сибирская и 7 — европейская косули)
Основная же масса находок косуль в Азии датируется верхним плейстоценом и
голоценом (рис. 7)1.
Следует особо подчеркнуть, что из-за скудности ископаемых остатков и
отсутствия точных определений их геологического возраста нельзя однозначно
говорить о более раннем или более позднем появлении косуль в том или другом
регионе. По совокупности имеющихся сведений о плейстоценовых млекопитающих
Европы Б. Куртен (Kurten, 1986) пришел к выводу, что косуля иммигрировала
сюда из Азии вместе с другими, азиатскими по происхождению, млекопитающими.
Эоплейстоценовые Capreolus sp. из Павлодарского Прииртышья, судя по
остаткам рогов, хранящихся в Палеонтологическом институте РАН, были очень
крупными животными. На большие размеры ископаемых косуль в Азии и мелкие
в Европе указывают почти все исследователи. Особенно четко эти различия
прослеживаются в голоцене. Костные остатки "крупной" Capreolus, найденные
в Европе (Boessneck, 1956), по размерам не выходят за пределы варьирования
признаков, свойственных европейской косуле (Паавер, 1965), но достоверно
отличаются от таковых ископаемых сибирских косуль (Косинцев, 1981).
На основании данных по истории становления группы, а также исходя из того,
что в плейстоцене существовали естественные водные и ледниковые барьеры на
границе Европы и Азии, можно заключить, что развитие европейской и сибирской
косуль длительный период времени происходило обособленно, и это привело
к существенным генетическим различиям и частичной репродуктивной изоляции
(см. ниже).
В общем виде систематическое положение Procapreolus и Capreolus представляется
следующим образом.
СЕМ. CERVIDAE GRAY, 1821 — ОЛЕНЬИ
ПОДСЕМ. ODOCOILEINAE РОСОСК, 1923 — ЛОСИНЫЕ
Триба Capreolini Simpson, 1945
Род Procapreolus Schlosser, 1924 — Прокапреолюсы
Cervus: Croizet, Jobert, 1828, p. 226; Bravard, 1828, p. 136; Pomel, 1853, p. 103—112; Gervais, 1859, p. 149;
Dawkins, 1878, p. 402—420; Deperet, 1884, p. 247—284; Boule, 1905, p. 32—33; Macarovici, 1936, p. 349—367;
Heintz, 1970, p. 67—91.
Cervavus: Deperet, Croizet, 1884, p. 269—274; Schlosser, 1903, p. 118.
Capreolus: Pavlow, 1896, p. 1—12; Венюков, 1901, с. 33; 1902, с. 123—133.
Procapreolus: Schlosser, 1924, p. 634—640; Zdansky, 1925, p. 24—26; Gaal, 1943, p. 87—88; Верещагин,
' Места находок остатков Capreolus приводятся на основании следующих литературных
источников: Горбачев, 1915; Бируля, 1929; Young, 1932; Герасимов, 1940; Громов, 1948; Громова, 1948, 1965;
Толстое, 1948; Хороших, 1949; Окладников, 1950; Пидопличко, 1951а,б, 1956; Корнеев, 1952;
Верещагин, 1954, 1956, 1959, 1971; Kahlke, 1955; Boessneck, 1956; Цалкин, 1956, 1960, 1962а,б, 1963,1966; Бара-
баш-Никифоров, 1957; Lehmann, 1958, 1960; Марвин, 1959; Никифорова, 1960; Бажанов, Костенко, 1962;
Thenius, 1962; Бибикова, 1963; Ермолова, 1963, 1978; Равский и др., 1964; Марков и др., 1965; Паавер,
1965; Успенский, Лозаи, 1965; Верещагин, Батыров, 1967; Марков, Величко, 1967; Верещагин, Оводов,
1968; Татаринов, 1970; Кузьмина, 1971; Иванова, 1972; Маркович-Марьянович, 1972; Шарапов, 1972а,б;
Тимченко, 1972; Вислобокова, 1973; Смирнов, 1975; Смирнов Н„ 1976а,б; Алексеева, 1977, 1980; Ван-
генгейм, 1961, 1977; Кожамкулова, 1977, 1981; Алиев, 1978; Верещагин, Русаков, 1979; Stubbe, Passarge,
1979; Верещагин, Барышников, 1980а,б; Huang, 1980; Косинцев, 1981, 1986; Давид, 1982; Li et a!., 1984;
Savage, Russell, 1983; Косинцев, Ражев, 1985; Бурчак-Абрамович, 1986; Динесман и др., 1986; Каспаров,
1986; Nobis, 1986; и др. (см. текст).
19
1959, с. 52—55; Короткевич, 1963, с. 1390—1393; 1964, с. ш—336; 1965, с. 60—67; 1970, с. 62—72; 1974,
с. 68—77; 1968, с. 549—572; Векуа, 1972, с. 209—211.
Поздний миоцен—средний плиоцен Европы и Азии.
Procapreolus cusanus Croizet, Jobert, 1828.
Средний плиоцен Украины, Молдавии, Франции, Венгрии, Румынии.
Procapreolus latifrons Schlosser, 1924.
"Плиоцен" Китая, Монголии. Тип рода (? ранний плиоцен Монголии).
Procapreolus wensensis Czyzewska, I960. Плиоцен Польши.
Procapreolus ucrainicus Korotkevich,1965.
Поздний миоцен Украины, Молдавии, Кавказа.
Procapreolus florovi Korotkevich, 1974.
Конец миоцена—начало плиоцена Украины.
Характеристика рода Procapreolus. Вымершие представители оленьих средних
размеров. Излом мозго-лицевой оси черепа выражен. Мозговой отдел относительно
длинный, а дентальная часть лицевого — короткая, передний край глазницы
находится на уровне середины длины М2. Слуховые пузыри большие, округлые.
Рога лировидные с тремя отростками. Надглазничный отросток отсутствует,
а аналогичный ему средний расположен высоко от розетки. Розетка небольшая,
жемчужевидная. Пеньки высокие, широко расставлены, расстояние между ними
не менее их поперечника. Лобные кости широкие без продольных валиков.
Слезные ямки глубокие, четкие. Верхнечелюстная кость высокая, превышает
длину ряда М1—М3. В верхней челюсти имеются клыки. Коренные зубы брахио-
донтные, их боковые стенки скошены от основания коронки к жевательной
поверхности. Базальные образования и выросты эмали во внутренних лунках, как
и палеомериксовая складка, уменьшаются, а моляризация, удлинение ряда пред-
коренных зубов увеличивается у геологически более молодых видов по сравнению
с более древними. На Р4 намечается замыкание передней долинки.
Род Capreolus Gray, 1821 — Косули
1755. Capreolus. Frisch, Natur-Syst. vierfiiss. Thiere, p. 3.
1821. Capreolus. Gray, London Med. Reposit., v. 15, p. 307.
1837. Caprea. Ogilby, Proc. Zool. Soc. London, 1836, p. 135.
Поздний плиоцен, плейстоцен — современность Европы и Азии.
Capreolus capreolus Linnaeus, 1758 — Европейская косуля.
Linnaeus С, Systema Naturae, ed. 10, Т. 1, p. 68. Средний плейстоцен — современность Европы
и Малой Азии. Тип рода (Швеция).
Capreolus pygargus Pallas, 1771 —Сибирская косуля.
Pallas P.S., Reise Russl., Bd 1, S. 453. Средний плейстоцен — современность Азии и Восточной Европы.
Характеристика рода Capreolus. Телеметакарпальные олени мелкого и среднего
размеров. Длина тела особей в среднем от 107 до 145 см, масса — от 20 до 50 кг.
Хвост рудиментарный (2—3 см), скрыт в волосах. Передние ноги немного короче
задних. Шея длинная, грива отсутствует. Уши большие (13—16 см). Предглазничные
железы нефункционирующие. Рога в норме только в самцов, чаще с тремя
выростами. Надглазничного отростка нет. Розетки рогов хорошо выражены.
Поверхность рогов у основания бугристая.
Окраска зимой серо-бурых тонов, летом красновато-рыжая, у телят пятнистая;
околохвостовое "зеркало" ярко выражено зимой, но летом у части особей
отсутствует. Линяют два раза в году — осенью и весной.
Череп удлиненный, мелких и средних размеров (в среднем 190—245 мм), его
наибольшая ширина (84—106 мм) составляет менее половины длины. Слезные кости
20
короткие, короче поперечника глазницы. Предглазничная ямка на слезной кости
широкая, но в норме мелкая, слабо выражена и расположена близко к краю
глазницы. Этмоидальное отверстие изменчиво.по величине. Межчелюстные кости
равномерно суживаются вперед, не образуя лопастеобразных выступов на наружном
крае. Передние концы носовых костей вильчато раздвоены и соприкасаются с пред-
челюстными. Дентальная часть лицевого отдела сравнительно короткая, передний
край глазниц находится на уровне М2—М3. Глазницы средних размеров, их
вертикальный диаметр равен или несколько превышает длину ряда коренных зубов
(М1—М3). Верхнечелюстная кость сравнительно высокая и равна длине ряда
коренных зубов. Зубная формула:
lfCfP|M| = 32.
Резцы с расширенными, несимметричными длинными коронками; Ii в полтора
раза больше Ь, который во столько же раз больше 1з. Нижние клыки по величине
равны Ь, верхние, как правило, отсутствуют. Р4 с двураздельной средней лопастью
(передняя ветвь очень мала). Коренные зубы с коронкой средней высоты, мало
суживающейся от основания к жевательной поверхности; их наружная стенка почти
отвесная; М3 с простой не замкнутой вперед задней складкой наружной поверхности.
Роговые выросты лобной кости направлены назад и вверх, их высота с внутренней
стороны не превышает ширину; они сближены (у европейской косули) или
сравнительно широко расставлены (у сибирской косули), расстояние между внутренними
сторонами пеньков не более чем в 1,5 раза превышает их ширину.
Кариотип (2л = 70—84) состоит из 70 основных и у сибирской косули от 1 до 14
добавочных В-хромосом. Y-хромосома акроцентрическая и самая маленькая,
Х-хромосома субметацентрическая и самая крупная в наборе.
Сравнение. Capreolus отличается от Procapreolus слабо развитой предглазничной
ямкой; относительно меньшим размером слуховых пузырей; более короткими
роговыми пеньками, относительно уже расположенными и слабее отклоненными
назад к крыше черепа; присутствием выростов в основании ствола рога;
относительно меньшей шириной лобной кости; отсутствием палеомериксовой складки
и дополнительных элементов складок эмали на зубах; более высокой коронкой
и слабым развитием на них базальных образований; более моляризованными
предкоренными, особенно Р4; отсутствием в норме верхних клыков.
Сравниваемые роды сближают такие признаки, как общий тип строения рогов,
последовательность формирования на них отростков, отсутствие предглазничного
отростка, относительные размеры глазниц и основных отделов черепа.
Глава вторая. Изменчивость и таксономия
История исследований
Capreolus названа так, вероятно, в связи с некоторой схожестью с Сарга — козлами,
отсюда "дикая коза", "козуля", "косуля". Сибирская косуля получила название
pygargus при следующих обстоятельствах. П.С. Паллас (1809), путешествуя
по разным провинциям Российской Империи, в октябре 1768 г. за Волгой на правом
берегу р. Сок увидел.животных, у которых "зад до самой спины покрыт большим
белым пятном, по чему можно было сего зверя почесть за Пигарга древних",
и посчитал, что "сии дикие козы совсем особливого рода и разнятся от всех
Европейских родов оленей", так как "величиной более обыкновенной дикой K03bi(Dama),
а цветом сходствует с серною (Capreolus)".
Фактически на примерно таком же натуралистическом подходе зиждется вся
таксономия косуль. Виды, подвиды и расы описывались на основании одного-двух
экземпляров рогов или черепов, по окраске одной или нескольких шкур или
по визуальным показателям — одни животные больше, другие меньше. Поскольку
амплитуда географической изменчивости Capreolus весьма велика, это давало
повод для выделения в пределах ареала от одного до нескольких видов и
множества рас и подвидовых форм. Только их простое перечисление заняло бы
несколько страниц текста (см., например: Ellerman, Morrison-Scott, 1951). Приведем
лишь основную синонимику, в определенной степени отражающую историю
исследований (по: Каталог млекопитающих СССР, 1981).
1758. capreolus. Linnaeus, Systema Naturae, ed. 10, t. 1: 68.
1771. pygargus (Cervus). Pallas, Reise Russl., Bd 1: 453.
1780.ahu (Cervus). Gmelin, Reise Russl., Bd 3: 496.
1830.dorcas. Burnett, Quart. J. Sic. Lit. Art. 1829, v. 2: 353.
1832. vulgaris. Fitzinger, Beitr. Landesk. Osterreichs., Bd 1: 317.
1832.+fossilis. Meyer, Palaentologica: 95.
1843. caprea. Gray, List Spec. Mamm. Brit. Mus.: 176.
1846. europaeus. Sundevall, Svenska Vet. Ak. Handl. 1844: 184.
1889. pygargus mantschuricus. Noack, Humboldt, Bd 8: 9.
1906. tianschanicus. Satunin, Zool. Anz., Bd 30: 527.
1907. transsylvanicus. Matschie, Weidwerk Wort, Bild, Bd 16: 224.
1908. bedfordi. Thomas, Proc. Zool. Soc: 645.
1910. rhenanus. Matschie, Weidwerk Wort, Bild, Bd 19: 263.
1911. melanotis. Miller, Proc. Biol. Soc. Washington, v. 24: 231.
1916. zedlitzi. Matschie, Sitzungsber. Ges. naturforsch. Freude Berlin: 272.
1923. coxi. Cheesman et Hinton, Ann. Mag. Natur. Hist., v. 12: 608.
Большинство зоологов XIX в. принимали одновидовую концепцию рода
Capreolus. Ее поддерживал и сам П.С. Паллас (1811), отказавшийся признать за pygargus
статус самостоятельной видовой единицы (по: Пастернак, 1955). В первой половине
XX в. многие исследователи выделяли все же два-три вида (чаще С. capreolus,
22
С. pygargus и С. bedfordi) с разным числом подвидов, правда без достаточного
обоснования (Satunin, 1906; Динник, 1910; Браунер, 1915; Флеров, 1928,1929; Лавров,
1929;'Даль, 1930; Jacobi, 1932; и др.). Позднее снова возобладала "видообъедини-
тельная" тенденция, господствовавшая до настоящего времени (Allen, 1930, 1940;
Barclay, 1933; Соколов И.И., 1959; Соколов В.Е., 1979, Honacki et al., 1982; и др.).
В середине века наибольшее распространение получила классификация Capreolus,
предложенная К.К. Флеровым (1952):
С.с. capreolus L. — Европа до восточных районов европейской части СССР и Малая Азия;
С.с. pygargus Pall. — восточные районы европейской части СССР, Северный Кавказ, азиатская часть
СССР до Хабаровского края, Северная и Северо-Западная Монголия;
С.с. bedfordi Thomas — Хабаровский и Приморский края, Северный и Северо-Восточный Китай,
Корея;
С.с. melanotis Miller — Китай.
Эта классификация в основном совпадала с таковой Эллермана и Моррисон-
Скотта (Ellerman, Morrison-Scott, 1951), за исключением подвида С.с. melanotis,
который включался в состав С.с. bedfordi, что, вероятно, было более правильным.
Еще Аллеи (1940) по исследованиям косуль в Китае и Монголии заключил, что
С. bedfordi и С. melanotis идентичны. Однако упомянутые выше и другие ученые
в силу ограниченности материала не смогли провести сравнительного
систематического анализа косуль в пределах их ареала. К тому же во многих работах
допускались серьезные методические просчеты.
Наиболее масштабное изучение предпринято Ф.А. Пастернаком (1952, 1955),
который нашел, что по морфометрическим показателям географические расы
связаны рядом переходных форм, находящихся между собой в состоянии строгого
географического викариата, и укрепился во мнении о монотипическом составе
рода Capreolus. К сожалению, как справедливо заметил еще В.Г. Гептнер (1961),
представленный материал был недостаточно обработан статистически.
В последние десятилетия в основном общепринята систематика косуль,
предложенная В.Г. Гептнером и др. (1961). В.Г. Гептнер считал воззрения К.К. Флерова
(1952) и Эллермана, Моррисон-Скотта (1951) слишком упрощенными, полагая, что
расы единственного вида Capreolus capreolus явственно разбиваются на две группы:
европейскую и сибирскую. К европейской группе он относил только европейскую
косулю С.с. capreolus L., 1758 (Малая Азия и Европа, включая Украину и
Белоруссию, Крым и Закавказье), хотя и подчеркивал, что закавказской форме,
вероятно, должно принадлежать имя armenius Bladder, 1916. К сибирской группе
причислялись:
Сибирская косуля, С.с. pygargus Pall., 1771 — от восточных областей европейской части СССР на
восток до Амура, включая Северную Монголию;
Тянь-шанская косуля, С.с. tianschanicus Satunin, 1906 (syn. ferganicus) — Тянь-Шань, Китай;
Дальневосточная (маньчжурская) косуля, С.с. bedfordi Thomas, 1908 (syn. mantschuricus, ochracea) —
Уссурийский край и Приамурье, Северо-Восточный Китай, п-ов Корея;
Северокавказская косуля, С.с. caucasicus Dinnik, 1910 — северные склоны Главного Кавказского
хребта.
Как видим, В.Г. Гептнер относил северокавказскую косулю к сибирской группе,
хотя в противоположность К.К. Флерову не считал ее идентичной С.с. pygargus
и прямо указывал на малую изученность этого подвида. Эта же классификация
была принята Г.Ф. Барышниковым и др. (Каталог млекопитающих СССР, 1981).
Корбет (Corbet, 1978), а также Штуббе и Пассарге (Stubbe, Passarge, 1979) к этому
добавляли все же С.с. melanotis.
23
Таким образом, несмотря на большие разногласия по вопросу таксономии
Capreolus, большинство ученых считали их единственным видом рода, выделяя
европейскую и сибирскую косуль тем не менее в качестве групп, подвидов или форм
и отмечая тем самым существующие между ними различия. Сомнения в
монотипичности рода стали появляться вновь лишь в последнее десятилетие, после
установления отличий в хромосомном наборе европейской и сибирской косуль
(Соколов и др., 1975, 1978; Данилкин, 1978; Sokolov, Danilkin, 1980; Zernahle, 1980),
обнаружения репродуктивных барьеров при их гибридизации (Штуббе, Брухгольц,
1979, 1980) и разницы в общем уровне метаболизма (Граевская и др., 1980).
Существование генетической и морфометрической разнородности наряду с
репродуктивными барьерами и географической изоляцией явно указывали на видовую
самостоятельность европейской и сибирской косуль. Становятся очевидными
также неправомерность выделения некоторых подвидов в указанных границах
ареала и отнесение их к той или иной группе. В частности выяснилось, что
северокавказские косули имеют одинаковый набор хромосом с европейской формой и,
следовательно, являются европейскими, а не сибирскими (Соколов и др., 1980;
Соколов, Данилкин, 1981; Данилкин и др., 1983).
На основании последних исследований зоологи снова начинают принимать
точку зрения о разновидном составе рода Capreolus. Семпере (Sempere, 1982),
не вдаваясь в детали, показывает в пределах ареала три вида: С. capreolus, С. pygargus
и С. bedfordi. Леман и Зегессер (Lehmann, Sagesser, 1986) рассматривают
европейскую и сибирскую косуль как два разных вида, при этом принадлежность
caucasicus к тому или другому виду не определена. Указывая на наличие
значительной внутривидовой изменчивости, они выделяют только у европейской
косули 12 подвидовых названий, большую часть которых считают все же
недействительными:
capreolus (Linnaeus, 17S8) — Скандинавский полуостров, Дания, Германия к востоку от Рейна;
transsylvanicus Matschie, 1907 — Восточные и Южные Карпаты;
thotti Lonnberg, 1910 — Англия, Шотландия;
canus Miller, 1910 — Испания;
cistaunicus Matschie, 1913 — северо-запад Германии, Нидерланды, Бельгия, Франция;
decorus Cabrera, 1916 — северо-западная Испания;
joffrei Bladder, 1916 — Франция;
zedlitzi Matschie, 1916 — Восточная Пруссия, восточная Польша, европейская часть СССР до Днепра;
italicus Festa, 1925 — Италия;
grandis Bolkay, 1925 — Югославия;
whittalli Barklay, 1936 — европейская Турция;
illyricus Car, 1967 — Югославия (Кроатия).
Как видим, таксономия косуль к настоящему времени еще более усложнилась
и весьма далека от ясности.
Возникшие проблемы можно решить лишь при детальном разноплановом
сравнительном изучении животных в пределах всего ареала морфологическими,
цитогенетическими, биохимическими, этологическими и прочими методами.
Учитывая важную практическую значимость этих животных для народного хозяйства,
на координационном совещании специалистов по косуле стран — членов СЭВ
(Москва—Вильнюс, 1979 г.) было решено провести ревизию систематики Capreolus
и более тщательно изучить биологию зверей. Результаты этих исследований
приводятся ниже.
24
Морфометрический анализ
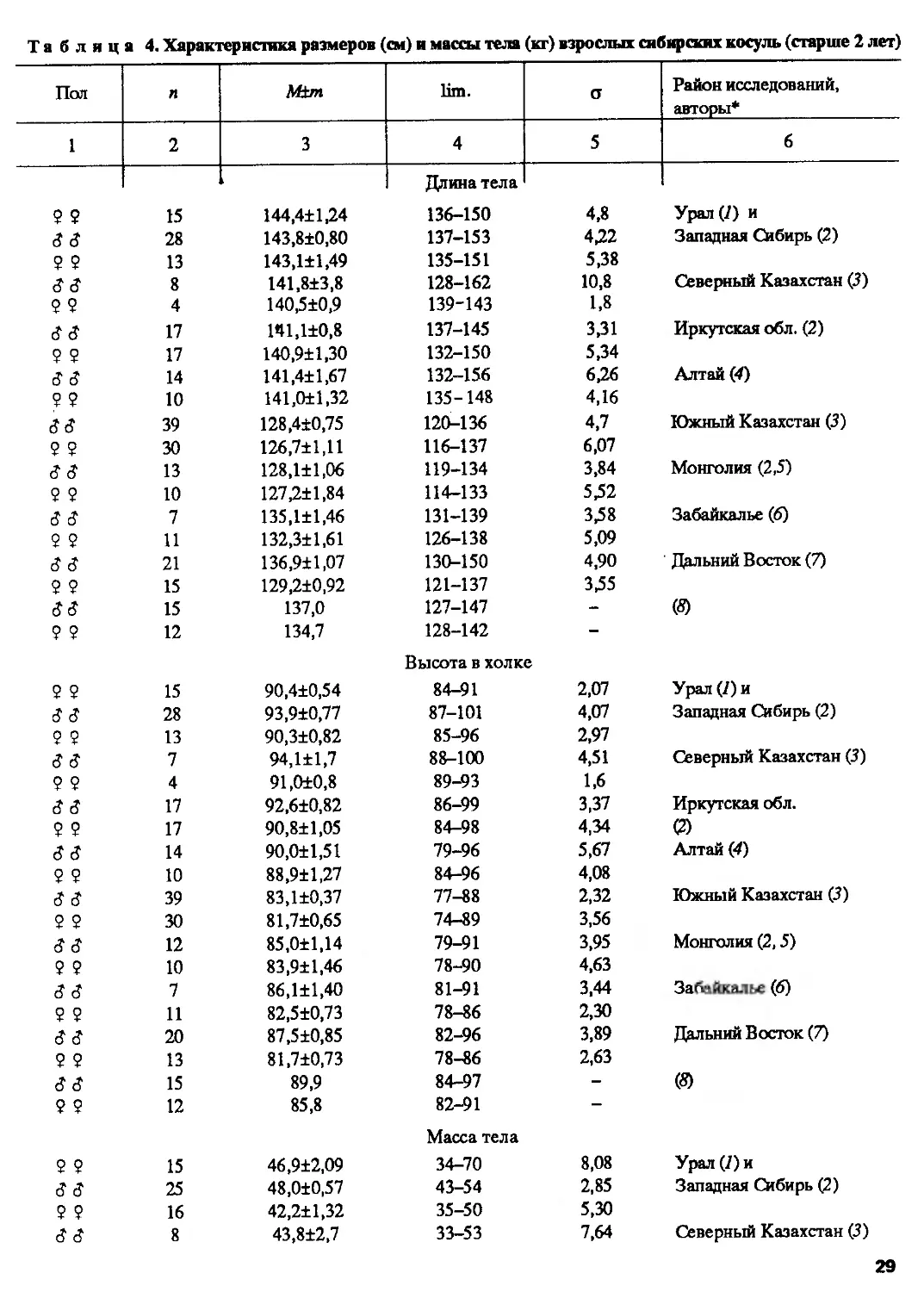
Размеры и масса тела1. Морфометрические показатели, приведенные в литературе,
к сожалению, не всегда сопоставимы из-за нестандартности измерений, и к тому же
весьма значительны погрешности при промерах тела. К примеру, в эксперименте
ошибка при измерениях длины тела одной и той же косули шестью специалистами
составила ±2,5 см. Масса тела животных значительно колеблется по сезонам
и даже в течение суток может изменяться в пределах 1—3 кг в зависимости от
количества потребляемой пищи и воды. Сравнение массы потрошеных особей
также затруднено, поскольку звери взвешиваются или без всех внутренностей,
или без желудка и кишечника, с головой или без нее. К тому же размеры и масса
животных меняются с возрастом и во многом зависят от конкретных условий
обитания, и в первую очередь от пищевых ресурсов (Sagesser, 1966; Wandeler,
Huber, 1969; Ellenberg, 1978; Stubbe, Passarge, 1979; HanuS, Fiser, 1981).
И все же в пределах ареала выявляются определенные закономерности
географической изменчивости популяций (табл. 3 и 4). Прежде всего видно, что
европейские косули в целом мельче, чем сибирские, и существенно отличаются по
средним показателям, но максимальные показатели особей первых перекрываются
с минимальными параметрами вторых (рис. 8). В географическом аспекте
изменчивость размеров и массы тела косуль в целом укладывается в рамки известного
правила Бергмана: на севере и на востоке в областях с более холодным климатом
животные значительно крупнее, чем на юге и на западе, где климат мягче. Ф.А.
Пастернак (1955) эти обстоятельства в свое время приводил как доказательство
невозможности видового разграничения косуль по морфометрическим признакам.
Действительно, наиболее крупные европейские косули регистрируются на севере
ареала — в Швеции (Essen, 1966), к востоку от Днепра и в горах Северного Кавказа.
Однако совершенно очевидно также, что самые крупные особи из популяций
европейской косули, обитающих на востоке ареала, по своим параметрам достоверно
отличаются от граничащих с ними в районе Волги популяций сибирской косули,
т.е. налицо прерывание постепенности изменения морфологических признаков
(см. табл. 3 и 4).
Кроме того, обнаруживается, что очень крупные по морфометрическим
показателям популяции, населяющие огромный регион от Волги до Байкала к северу
от Алтайско-Саянской горной системы, весьма существенно отличаются от
популяций Южного Казахстана, Монголии, Забайкалья и Дальнего Востока (см. табл.4).
Более четко эти различия проявляются при краниометрическом анализе.
Размеры и масса тела животных, их рост и развитие, как известно, зависят
не только от условий обитания, но и в значительной мере определяются
генотипом. Темпы роста и развития и обмен веществ неодинаковы даже у
близкородственных видов. Следовательно, если европейские и сибирские косули — разные
формы, то при содержании в одинаковых условиях они должны различаться по этим
параметрам.
В эксперименте (в вольере под Москвой) масса тела новорожденных телят
европейской косули составляла в среднем 1650 г., сибирской — 1850, однако у
последних при рождении троен она была меньше, чем у первых, — 1530 г. В период роста
европейские косули прибавляли в весе в среднем на 4 кг в месяц, а сибирские — на 6 кг,
'Схему измерений см. на рис. ПО.
25
160
140
^ izo
%
<ъ100
I
180
40
20 -
H'
m
ф«*«
_l_
<* о
;
7/7
ffl?
!
1
20
10
3 4 S В 7 В
Признаки
Рис. 8. Изменчивость размеров и массы тела европейской (/) и сибирской (II) косуль
Точкой обозначены средние значения, прямоугольником — пределы средних для популяций, линией —
предельные значения для особей
/ — длина тела; 2 — высота в холке; 3 — обхват туловища; 4 — длина туловища; 5 — длина ступни;
б — длина уха; 7 — масса тела; 8 — масса туши. Схему измерений см. на рис. ПО
и, кроме того, значительно превосходили по размерным показателям (обхват
груди). В возрасте 4—5 мес максимальная масса европейских косуль не превышала
22 кг, тогда как у сибирских достигала 28 кг, а у взрослых соответственно 33 и 42 кг.
Разница во все сезоны года была примерно одинаковой — в среднем 8—9 кг, или
около 20% от массы сибирской косули. Таким образом, при одинаковых условиях
обитания и продолжительности постнатального развития темпы роста сибирской
косули явно выше, т.е. очевидны различия на генетическом уровне (Громов, 19886).
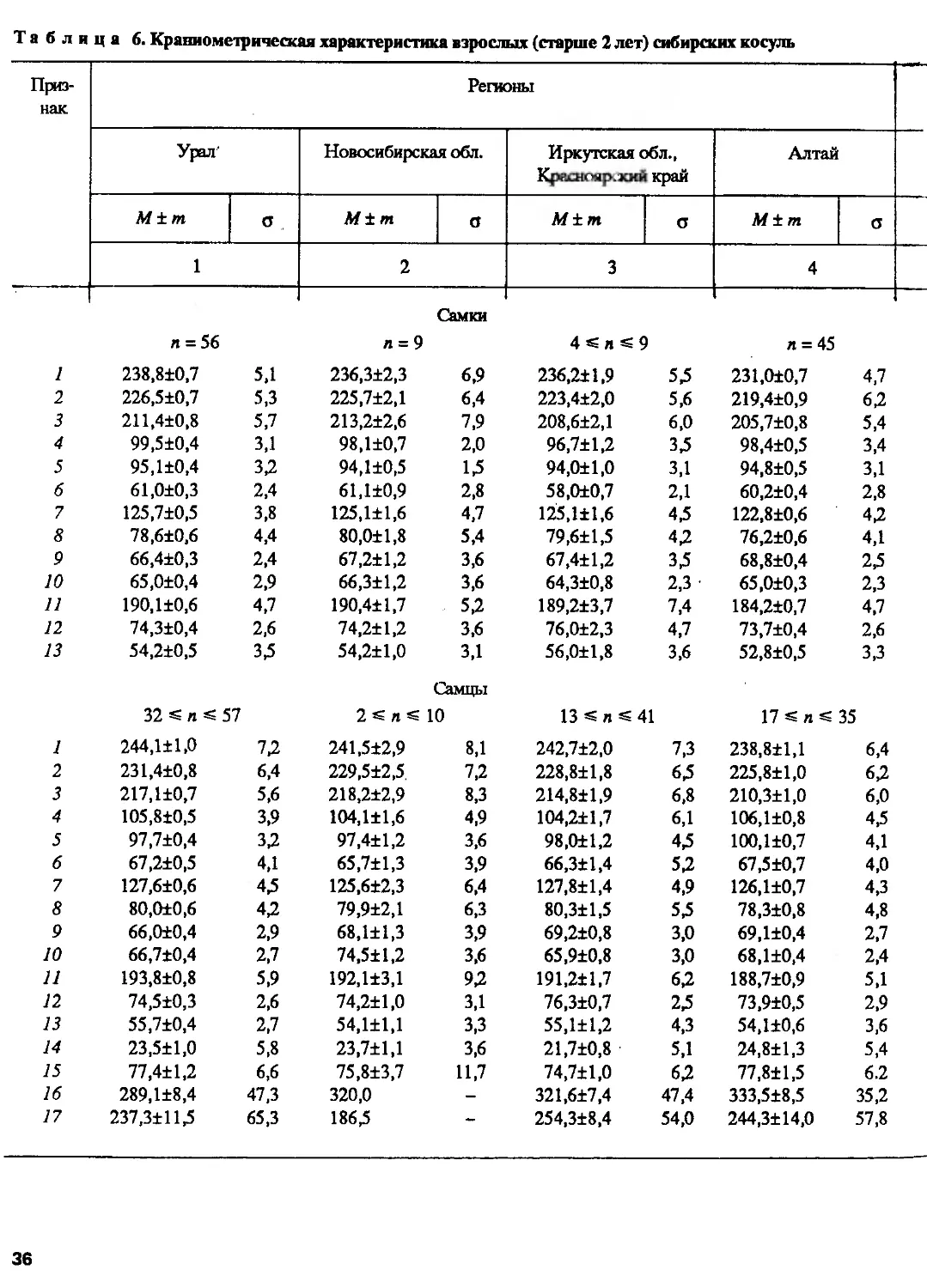
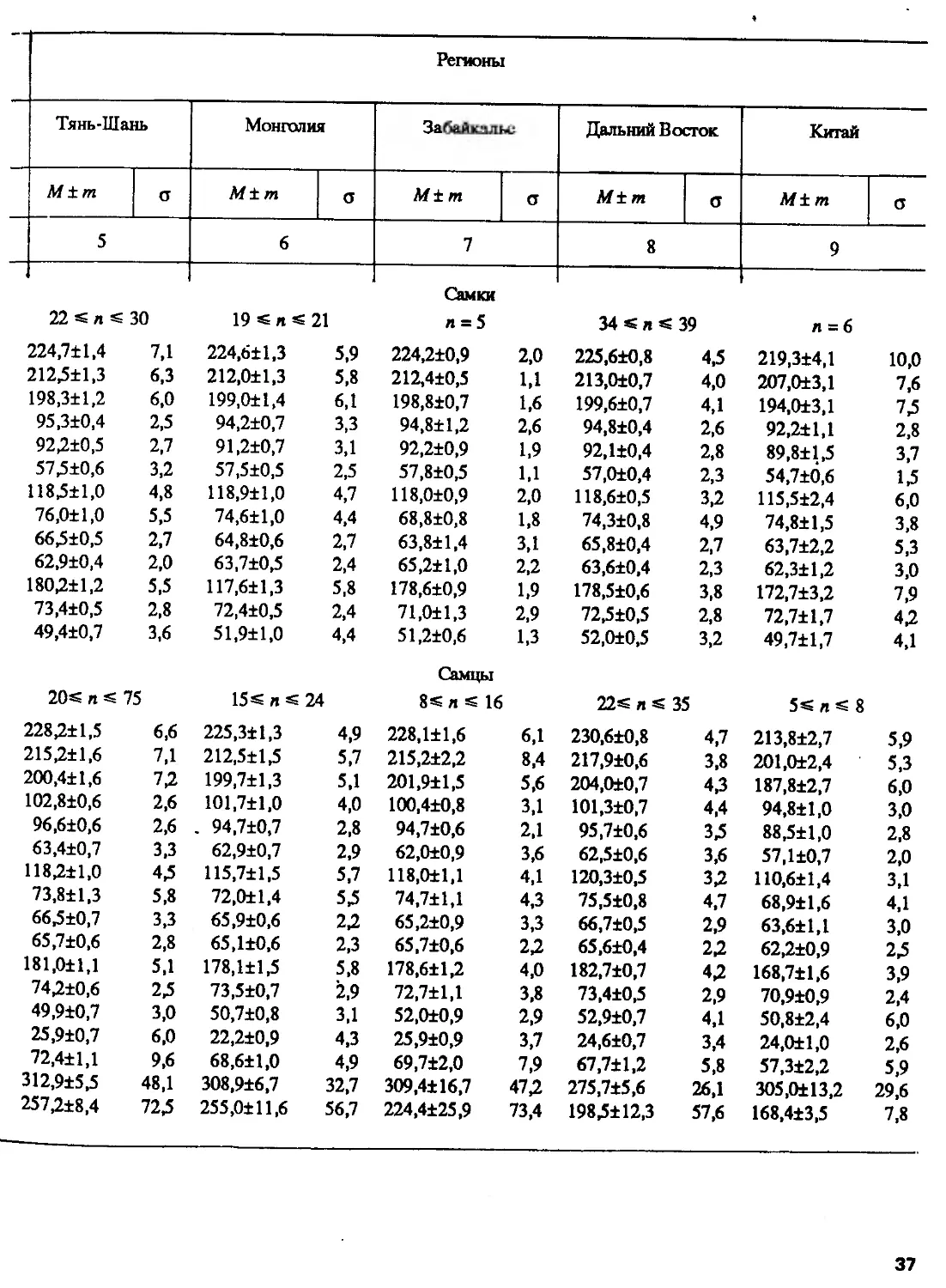
Краниометрические исследования. В соответствии с программой изучения
систематики и биологии косуль две группы сотрудников ИЭМЭЖ РАН и Института
зоологии Болгарской АН провели измерения черепов косуль, хранящихся в
основном в музеях и институтах СССР и некоторых других стран и в частных коллекциях,
по схеме, приведенной на рис. 111. Выборки представляли популяции с большей
части ареала. Статистическая обработка проводилась разными методами на ЭВМ
отдельно для самцов и для самок из-за четко выраженного полового диморфизма.
Одна группа ученых, измерив около 500 черепов косуль старше 2 лет, пришла
к выводу, что по размерам и пропорциям черепа они достоверно разделяются
на четыре группы: 1) европейские косули, имеющие наименьшие размеры и
наибольшую относительную ширину черепа; 2) косули Урала и Северо-Восточного
Казахстана, имеющие самый крупный череп с относительно коротким верхним
рядом зубов и узкой мозговой капсулой; 3) популяции Восточной Сибири и
Дальнего Востока с относительно более длинным рядом зубов и широкой мозговой
капсулой и 4) популяции Тянь-Шаня и Монгольского Алтая, отличающиеся отно-
26
Та блица 3. Характеристика размеров (см) и массы тела (кг) взрослых (старше 2 лет) европейских
косуль
Пол
1
зз
9 9
33
9 9
33
33
9 9
33
9 9
33
9 9
33
9 9
33
9 9
33
9 9
33
9 9
33
9 9
33
9 9
33
9 9
33
33
9 9
33
99
33
9 9
33
99
33
99
33
99
33
■9 9
33
п
2
547
379
50
17
125
53
47
29
36
43
55
52
51
118
157
333
300
112
176
>50
>50
б
б
9
7
4
15
16
555
379
50
17
54
44
28
36
56
89
52
51
118
М±т
3
112±0,2
Ш±0,3
115,б±0,93
114,4±2,07
118,6
112,4
111,0
118,5
116,5
117,1±0,87
115,0±0,20
115,7±О,80
114,4±0,70
108,2-111,9
107,1-114,1
111,7-115,2
111,0-114,0
119,6±0,3б
118,9±0,28
112,3-119,8
111,9-119,7
124,8±1,77
124,2±1,10
126,5±2,35
125,7±1,90
123,7±1,38
116,0
117,6
66±0,2
66±0,2
74,8—0,90
70,7±0,93
76,4
75,6
81,5
78,8
73,5±0,6
72,1±0,36
74,3±0,51
74,0±0,60
69,2-74,9
lim.
4
Длина тела
93-129
96-125
96-135
88-140
98-139
-
-
-
-
108-128
105—128
102-127
102-125
86-131
75-129
98-128
101-123
-
-
-
-
121-132
119-127
121-139
119-132
121-127
110-129
о
5
5,23
5,05
6,60
8,53
-
6,0
5,1
64
5,5
5,71
1,48
5,8
5,0
5,9-8,5
5,4-10,4
3.9-5,7
3,6-4,6
3,7
3,65
-
-
4,35
2,69
7,06
5,03
2,75
-
Высота в холке
56-77
55-77
54-96
59-82
-
-
-
-
65-84
63-84
64-83
63-86
56-96
3,55
3,56
6,99
3,85
44
4,1
2,2
3,0
4,49
3,39
3,7
4,3
34-6,0
Район исследований,
авторы*
6
ГДР (7)
Чехословакия (2)
Венгрия (3)
Лес (4)
Поле (4)
Болгария (5)
(б)
Польша (7), Лес
Поле
Литва (8)
Эстония (9)
Украина (70)
Курская, Белгородская
области (70)
Северный Кавказ (70)
Закавказье (77)
ГДР (7)
Чехословакия (2)
Венгрия (4). Лес
Поле
Болгария (5)
(б)
Польша (7). Лес
27
Таблица 3 (окончание)
12 3 4 5 6
9 9
SS
99
ss
9 9
ss
9 9
S 3
99
S3
99
<?<?
9 9
S3
SS
99
157
333
300
112
176
75
48
>50
>50
6
6
9
6
4
15
16
70,4-74,6
71,6-72,6
72,3-72,8
78,4±0,42
75,4±0,70
77,0±1,1
76,0±1,3
73,7-80,5
73,1-77^
80,6±1,36
83,3±0,80
80,7±0,88
81,2±1,13
78,5±1,25
74,0
73,2
58-80
64-81
62-86
-
-
-
-
-
75-84
81-86
77-84
77-84
76-82
63-79
4,6-7,2
2,6-3,3
2,9-3,5
4,46
9,25
9,53
9,01
-
3,33
1,97
2,63
2,77
2,52
-
Поле
Литва (8)
Латвия (12)
Эстония (9)
Украина (10)
Курская, Белгородская
области (10)
Северный Кавказ (10)
Закавказье (11)
Масса тела
SS
99
SS
99
SS
9 9
SS
99
SS
99
SS
99
SS
99
SS
9 9
SS
9 9
SS
9 9
SS
SS
9 9
50
17
30
27
12
15
52
73
26
20
131
213
74
49
>50
' >50
5
4
9
8
2
15
16
24,0±0,30
22,6±0,90
23,7
22,9
28^
23,7
26,3±0,35
23,9±0,26
24,7±0,78
22,6±1,04
29,2
26,1
28,8±0,37
26,7±0,90
27,4-28,5
24,6-29,0
29,6±1,60
29,0±1,29
32,0±0,82
30,0±0,66
29,3
25,2
24,3
15-32
21-34
-
-
-
-
20-31
20^29
15-32
15-32
-34,6
-
-
-37,4
-35,2
24-33,3
26-32
28-36
25-34
27-31,6
18-30
2,82
3,72
2,6
22
3,1
2,3
2,52
2,22
3,96
4,66
-
3,18
6,30
-
-
3,58
2,58
2,45
2,44
—
-
Чехословакия (2)
Венгрия (4). Лес
Поле
Болгария (5)
(б)
Литва (8)
Латвия (/2)
Эстония (9)
Украина (10)
Курская, Белгородская обл.
СИ)
Сев. Кавказ (10)
Закавказье (11)
* Данные; 1 - Stubbe, Smimov, 1972; 2 - Hell, Herz, 1968; 3 - Galamb,. Tusnadi, 1973; 4 - K. Matrai, L.
Sugar, I. Heltay (личное сообщение); 5 - Петров и др., 1968; б - Г.Г. Марков; 7 - Frazinski et al., 1982;
8 - Блузма, 1974, 1975; 9 - Рандвеэр, 1985; 10 - А. А. Данилкин; // - Арабули, 1966; 12 - Янсон, 1975.
28
Таблица 4. Характеристика размеров (см) и массы тела (кг) взрослых сибирских косуль (старше 2 лет)
Пол
1
я
2
М±т
3
lim.
4
Длина тела
о
5
Район исследований,
авторы*
6
99
SS
99
SS
99
с?<?
9 9
SS
99
SS
9 9
SS
9 9
SS
9 9
SS
9 9
SS
9 9
99
с? с?
9 9
ss
9 9
С? (J
99
6*3
9 9
SS
9 9
SS
9 9
SS
9 9
33
9 9
SS
99
99
33
9 9
SS
15
28
13
8
4
17
17
14
10
39
30
13
10
7
11
21
15
15
12
15
28
13
7
4
17
17
14
10
39
30
12
10
7
11
20
13
15
12
15
25
16
8
144,4±1,24
143,8±0,80
143,1±1,49
141,8±3,8
140,5±0,9
1«U,1±0,8
140,9±1,30
141,4±1,67
141,0±1,32
128,4±0,75
126,7±1,11
128,1±1,06
127Д±1,84
135,1±1,46
132,3±1,61
136,9±1,07
129,2±0,92
137,0
134,7
90,4±0,54
93,9±0,77
90,3±0,82
94,1±1,7
91,0±0,8
92,6±0,82
90,8±1,05
90,0±1,51
88,9± 1,27
83,1 ±0,37
81,7±0,65
85,0±1,14
83,9± 1,46
86.Ш.40
82,5±0,73
87,5±0,85
81,7±0,73
89,9
85,8
46,9±2,09
48,0±0,57
42,2±1,32
43,8±2,7
136-150
137-153
135-151
128-162
139-143
137-145
132-150
132-156
135-148
120-136
116-137
119-134
114-133
131-139
126-138
130-150
121-137
127-147
128-142
Высота в холке
84-91
87-101
85-96
88-100
89-93
86-99
84-98
79-96
84-96
77-88
74-89
79-91
78-90
81-91
78-86
82-96
78-86
84-97
82-91
Масса тела
34-70
43-54
35-50
33-53
4,8
4,22
5,38
10,8
1,8
3,31
5,34
6,26
4,16
4,7
6,07
3,84
5,52
348
5,09
4,90
3,55
-
—
2,07
4,07
2,97
4,51
1,6
3,37
4,34
5,67
4,08
2,32
3,56
3,95
4,63
3,44
2,30
3,89
2,63
-
"
8,08
2,85
5,30
7,64
Урал (/) и
Западная Сибирь (2)
Северный Казахстан (3)
Иркутская обл. (2)
Алтай (4)
Южный Казахстан (3)
Монголия (2,5)
Забайкалье (б)
Дальний Восток (7)
®
Урал (/) и
Западная Сибирь (2)
Северный Казахстан (3)
Иркутская обл.
(2)
Алтай (4)
Южный Казахстан (3)
Монголия (2,5)
За.. • .- (6)
Дальний Восток (7)
Ю
Урал (/) и
Западная Сибирь (2)
Северный Казахстан (3)
29
Таблица 4 (окончание)
1
9 9
33
9 9
33
9 9
33
9 9
33
99
33
9 9
33
у 9
33
9 9
2
4
16
17
14
6
32
24
9
8
7
11
12
И
15
12
3
41,5±2,0
45,3±1,06
43,0±1,03
48,6±1,59
44,5±1,61
35,0±0,55
32,0±0,66
34,9±1,20
32,3±1,05
36,6±1,16
33,4±0,96
39,7±1,33
33,9±0,99
39,2
34,3
4
38-47
36-54
36-55
41-60
38^t7
3(МЗ
28-39
3(М0
28-37
32-40
30-90
36-45
31—41
32^19
ЗСМО
5
4,0
4,21
4,23
5,93
3,94
3,1
3,21
3,59
2,97
2,83
3,02
4,60
3,30
-
-
6
Иркутская обл. (2)
Алтай (4)
Южный Казахстан (?)
Монголия (2,5)
Забайкалье (б)
Дальний Восток
(7)
(5)
* Данные: J - Киселев, 1975; 2 - А.А. Данилкин; 3 - Поле, 1974; 4 - Г.Г. Собанский (личное
сообщение); 5 - С. Дуламцэрэн; 6 - Смирнов, 1978; 7 - Дарман, 1986; 8 - Бромлей, Кучеренко, 1983.
сительно широким черепом среди косуль Азии. Соответственно выделены 4
подвида: 1)европейский — вся европейская часть ареала и Кавказ; 2)сибирский —
Предуралье, Урал, Северо-Восточный Казахстан и Западная Сибирь; 3)
дальневосточный — Восточная Сибирь (Красноярский край, Иркутская область и
Забайкалье) и Дальний Восток; 4) тянь-шаньский — Тянь-Шань и Монгольский Алтай
(Соколов, Громов, 1985а).
Однако этот анализ строился на традиционном сравнении выборок по регионам
без детального сопоставления каждого черепа со всеми остальными, что
методически не вполне корректно. Поэтому ошибок не удалось избежать;
разнокачественные популяции Предбайкалья и Забайкалья рассматривались в одной
выборке, а однотипные популяции животных из Монголии — в разных, причем
совершенно необоснованно выделялись косули Монгольского Алтая — ни в СССР,
ни в Монголии материала из этого района практически нет. В конечном счете
методические в основном просчеты привели и в этой и в других работах по морфо-
метрии косуль к некоторым сомнительным выводам (Соколов, Громов и др., 1986;
Громов, 1986а; Громов, Скулкин, 1986).
Другая группа исследователей, обработав около 1300 экз. черепов и рогов и
применив шаговый дискриминантный анализ, пришла к другому заключению: по
краниометрическим показателям европейских и сибирских косуль можно считать
хорошо дифференцированными видами, что подтверждается также достоверными
различиями в темпах роста их черепов (рис. 9) (Соколов, Марков и др., 1985а,б).
Сибирские косули, как выяснилось, разбиваются на две большие группы,
заслуживающие подвидового ранга — Ср. pygargus Pall, (от Волги до Байкала)
и Ср. tianschanicus Satunin, 1906 (= Ср. bedfordi, Thomas, 1908), включающую
популяции Тянь-Шаня, Монголии, Забайкалья, Дальнего Востока и Китая, хотя
30
L
60
40
9
i i
Ф
i
6 12 18
Возраст, мес
Рис. 9. Абсолютный рост черепа сибирской (а, б) и европейской (в, г) косуль
а, в — самцы; б, г — самки. Цифры на графиках — краниометрические признаки (обозначения
признаков 1—13 см. в табл. 5 и на рис. 111)
последние уклонялись по некоторым признакам, что позволяло при
ограниченном материале подразумевать существование в районе Тибета Ср. melanotis
(Данилкин и др., 1985; Соколов, Данилкин и др., 1986; Данилкин, Марков, 1987).
Географической границей между сибирским и тянь-шаньским подвидами косуль
явно служит барьер из горных хребтов Алтая, Западного и Восточного Саян
и оз. Байкал (Данилкин, 1986в). В Казахстане границу между ними ранее
относительно точно определил В.Б. Поле (1974): популяции первых обитают к северу
от оз. Балхаш и к северо-востоку от Зайсанской котловины, популяции вторых —
южнее этих районов. В Якутию могли проникнуть особи того и другого подвидов.
Более детальный анализ европейской косули показал ее неоднородность и
обособление популяций в две группы (Марков и др., 1985), однако для выделения
таксонов требовалось сравнение с популяциями из Швеции, где описан вид,
и привлечение дополнительного материала. По краниометрическим показателям,
так же как и по кариотипу, косули Северного Кавказа оказались сходными
с популяциями, обитающими на востоке Европы (на Левобережной Украине,
в Курской и Белгородской областях), т.е. европейскими, а не сибирскими, как
принято считать. По размерам они гораздо крупнее косуль из Закавказья, что
31
It
i'
J*
ю
О
s<
в
Э
о
* Ш
Я£
§ 8
В к
§ в
я о\
_ а
- • о\
•ь. я
к * I
й Р "Е
S * е
2 S S
g 2 Д
' ё §
1 е S р
и 5 и
s £ те "
я в "о —
i»ss^
я 8 и "■
s о - • I
* ? "К
5 i
К -
S* 9Й " О
KJ
141
It
til!
ПН
*40v^A.U»N>b^-O^OOo>s)0\^i-^UiKj
W ">J W W W W О *W О 4 "6 W О W "*J "-^ "-J
h-h-i+h-i+h-h-h-h-h-h-i+h-h-i+i+h-
ЧО C\ W *-*0 О _*- О p j^i ^ J— P P .Г* ^-* ,Г~
lo "^ "о ^о "o ~4*. 1л Vi Ъч 1л "о Ъо "о Vi Vj Vi Va
ОЛ i—' W W^ О JO i-» ^- 0\ J— Ы О ^ Ы Ы Ы
"vo b "w M "w o\ "os b b ^ Vj m b w "о о b
OO i-i »—■ i—' t—* I—*
КЛ\Л^Л>—'4^СЧ(-АО\(-Л(ЛО(-ЛОООО--ЛОО
vq'0^0\IjJN)AOOOU(OIj}UJs]UOOO
1л Ыо u In oo о О "м 1л 1л 1л К) О О
и-н-н-н-и-яГн-н-н-н-н-н-н-н-
^Оу»^— jD^-j— j^^j^ ►—J-- ^ J-»* l_-
Vj ^-« ^o "vo "1л 1л "^ "to b In "-J \o 1o w
A W О si О SO OO 1л О О W О '
I Ъ\ *tO
VO \D ^ i- ^0\U)CNUlU\DlACi0V0sl0uO
W lA V> W О W jU N) jU Д OO "-• Л W t- S) Ы o^
1л ^ 7э ^ 1л 1л 1л Tjj To "La Vi "Vj **-• V> "I—,,^o1*J о
_w w p p p p p p p p p p p p p о р /Л
^^1л"^1>л1^Ъ\^1^1л'4ь1>л1>л'^"а\,~--о.'*-'4 5i
/л
^ЮЮ(ОД>-(ОДЫЮМыи1ли1
^1оЪ\4^ЬчЪо1^К»^1'Ъ\1лТо'^-*1л'*-о.
Ю oo
1л "н-
н- н-
J*^ °°
Ъ\ "о
I—• !-• I-* I— (О
1Лм^и\1лЛУ10\ОУ<ООО^ооО
^ ^л ^- 1л oo to ^ о to и» ф >-* -о *о >—
1л'о1лЪо1л'»--»Ъ\,^о'оо'о*--'*^4'чо'\о"о
Н-Н-Н-Н-Н-Н-Н-Н-Н-Н-Н-Н-Н-Н-Н-
р р р р ^ р р ООро Он-^
оо^1лЬ\^Ъ\Ъ.ю1о.Ъч^\о''~о1о"'.рь
yjtototo-uioto^k
Ъ\ "о "^ "to 1o Vj lo "to
N3 W W
1л t- \о '
I OS ОЧ
UjM»N.04)OoNJO\Oi-^U)IOk-
^51СлОч^олчо^оооослоочо
Ыа N 0< ю ° w * ^ °Г
1л w ° < *w lo "w w Vi b b
н- н-н-н-н-н-н-н-н-
О и>'-*0>--0^'--и-
\о
to oo "jk to a "to b 1л
1Л W О M О JO
vi "^ Ъ\ 1— bs "o '
to
i Ъ\
^0\1Л1ЛУ<О\ОА0000"О.00\О
-w^oa\otooowo\wo\-vj
Ъо^оТоЬч'Ъч'ач'оо'о'о'Ьо'оо'о'Ь?
H-H-H-H-H-H-H-H-H-H-H-H-ff
j— P ^ p „O W f* .*— О P ,f* ~ f°
"o't^'w'oo1o'"-o":-j'oo1ftVi1obo'to
NJ О W ^ JsJ OO W J^ p- j- Д 4^
"w oo b "со ь- *w "-J "*-» "м 1л ы н- '
^a\o>uiuiui«AMco^siio
Oto>—\owwooootoa\-j40»-»
lAlftlAbN'tO'^loVl'tO'uj'tO'ib^--* ^.
г+н-н-н-н-н-н-н-н-н-н-н-и- а
p О р О О О р О О О О О О |f
1о1о^1о1о^%1о1о'Ьо1л1л1л ^
(Л
jojoytjojo^j&.jotojoyiyijTs
^]1л1о_4^.'и>^3,4^Ь\Ъч1о1о1о,и-'
да\1Л1л^10\о-^оооо^оо^
_и- ^ Оч ЧО СГ\ p W Ю Ы у» ^ vl ^
1л'о'чо1о*^о'^о'й-*1л*й-'\о1лл"^ °°
н-н-н-н-н-и-н-н-н-н-н-н-Н-
р р р о р р р р р р р р —
1л 1>j "(Ж 7>j "oJ \0 "(7\ ~^. "^ 1л "чО "чо о
S3 ^о .рь. ^-> I— y\ oj jo to w у» у» ел
Ъо "о "J^ "^О Ъо 1о ~С7\ "ji. "и- "О "^ N3 Ъ\
Сам
5_
—
W
ы
*-
S;
н-
3
Q
а
н-
3
a
а
и-
3
а
5:
н-
3
a
>
нглия
Франция
S?
I
1
£ 1
(восточ
сть)
я
и
§
гария
я а
11
Регионы
1
!
макс
к
£
»
!э
о-
ное рас
о
ч
о
Е
s
т
Я
К
Та
•5
1
s
"я*
т
2
т*
расст
Г)
»
в
S
л
между
Е
X
1
X
о
ч
орон
диаст
п
X
(Г
X
ижней
л
п
а
S
_Х
>-«
л.
1
расе
N
8
я
Е
ч
п>
т>
жне
рог
о
_ш
•**
1
X
X
а
X
о
><
X
"3
'»
X
ним
X
3
о
"8
X
3 8
£ я
ё 3
§"3
II
р о
_ X"
Ъъ
§1
- • *
►«ч t
1 1
(-ntJU»^-^-LftH-OOjIi.vON3<-fti--j>jyt(«n
Ъл^Ъ\>оЪч^^о\и*Ъ\^1^ТльоЪо'1о'^
н-н-н-н-н-н-н-н-н-н-н-н-н-н-н-н-н-
1л"очю1лЪ\1л^Ъ»Ъч^^,сл1л,оо'1о'и>1л
1л "х*. ^° И°° *■* Ъ "н- "-о "к- "Ьо Ь> 1о 1л "to "** ">- V
to — i— ^- и- СО
\0(01Лн-.ио\0\0\1ЛО\ОСПОООООЧОО
1»л1>лЪоЪоЪо1лЪо1л1л^о1л1я'о1л'?оЪо1л
н-н-н-н-н-н-н-н-н-н-н-н-н-н-н-н-н- ,,
>— ОО .— О _>— ~ tO р ^ _Ю ^ О р ^— N> _S> JO W
ToVi^o\^^to^\,o'ik.*oo'b%Vjl>jo'a4'vo 'Л
w J^J to i— ^w *- 4* j- *- ^ w и-» иь- to ^ у» u»
to^lol^boo^woVilrtu>'4*'a\'b^--'oo
to [л
-О OO
"оч "стч
и- н-
to w
i-* и— r- i— (у
Lrtt— ■Ск0Ч0чСТ\^ЛС7\О(-Л00Ч000\ОО
Н-Н-Н-Н-Н-Н-1+Н-Н-Н-Н-Н-Н-Н-Н-
OOOO^-OOOOOOOOt— О
Ъ\ V "o\ V "о "i^ ~4*. "oo "о\ *. "*ь Ъч so "о so
-^j Хо "** "i»> "*" 1> w "^ to Ъ\ "to "j* ".u "w Ъо 1о Ъо
££ -
H- '0|ЧОоо'Ь\Ъ\'оо'Ьо'К>"Ь\1»лмэ,^-'оо'Ь\'1о
H-H-H-H-H-H-I+H-H-H-I+H-I+H-H-I+
yi -o ~ p p p ^- p p p ^— p p p _^- ^и-
'охо'Ьо'^о'стч'оо'о'сл'^^о^'-оЪч'оо'^'^
i± 3
oo /Л
3
/Л
to is> ы д s> ы
U Ш M U c7t (Tt <^
) 4* ~o 1^ In
Vj To "»-* "Ьо Ъ\ "bo "»— Ъо Ъ\ "ль. Ъ\ Ъо tc*.
H-H-H-H-H-H-H-H-I+H-H-I+.+
P P Г- P P P P P ро.ии Г-
IrtVj'obo'bo'oo'vo'ValA'cTN'ui'to'to
to'^-'to'bolnwso'vovo'ji.'bs'to^o
"oo tpk "o "oo "to Ъ\ "o\ ".Рь "oo "to "o\ "oo "4^
н-н-н-н-и-н-н-н-н-н-н-н-н-
Г ,и .м Г* и ,M w ° ° и м ы ы
loVjoo^Vi'UjVi'bN'ii.'wVj'to'bo
N)l>lO\ts)OJUUli-'OW0N-400
^1То1о1л"--о'Ьо'чо'1>»"оо'(э'»-*1о*Ь\
и- »— »— и-.. (О
yj JO j» _00 1Л p (>j p 4^ OO (7\ о *•*
*to Vi чо'Ъо Vjln'bo'bo,N—'o4'"-4l-»,to
H-H-H-H-H-H-H-H-H-H-H-H-H-
P P P P -° P -° P О О wO О p
"oo Vi "vo 1л *w "^ w 1л "ь- oo oo Vi о
и- *-* »— t— to
yijb.4b.*+OP\4b.0Q**y\ О *J 0\ OO
Vj'to'w'uj'vo'o'oolftXo'o'cAwVi
н-н-н-н-н-н-1+H-H-hPH-H-H-
иО jD p О p p p p p О ^ *~ *~
1п1л^?"4^1л'чоЪо*4^,1лЪч1о,Ьо^О
JOtOj(i-lON>^U>N>tO Ы1Л У1 W
^t/»"bN'4i.ln'4uVj"l>j'00'ol«V]'bN
$
X
l-ft
ОЧ
-J
oo
S:
H-
3
a
5:
H-
3
Q
s:
H-
3
a
H-
3
a
Польша
В
се
еция
$
х
бал тик
Ч
?!
рай
на,
Бело{
■Z
8
2
Регионы
c^t^^C^^Nnir^co^v^c^NOc^o
ел" сп сп гТ с4* ^~ сп en сч" г4^ сп~ --" ~-~
я
со
S
5-
Ол Ом ON V\ Щ ^t ON С\ \С «Л оо ^ **"1
—Г —Г о" о" о" сГ о" о" о" о" —Г о" о"
+Н-Н-Н-Н-Н-Н-Н-Н-Н-Н-Н-Н-Н
OONTTf-ONr-ONrSnrtMMN
1П_ Ов Ов ^ Г^ Tj-^ тГ_ С^ О^ 00^ \Ои On_ Г-^
о6\С\ОСНСЪСНтТ*Л-*С$1Г)-*СН
X
Е
-н
5
°\ **"! *0. ***! ""i ^1 ^1 ^ ^"- ~*~ еГ) °° *0.
■п-" го" со" ^ г-н" ^н" cs сз о -* с4" о" —<" *■
-н-н-н-н-н-н-н-н-н-н-н-н-н =
OONOOOOOOIOOVOVIVOVOVO^- С.
fNj ^- -Ч — ,-,
"1 ^ Ч °. №. Ч Ч ^ °i 'l. "^ ^ °i
vn"v£vT'co*c^r<fcTeo"^c'4'<ro'co"c-4
i
§
s
5
с _," _" —<" o" o" o" o" —" o" o" —Г —<" о"
-н-н-н-н-н-н-н-н-н-н-н-н-н
Mwnvi>ooo-'(Soooiwo^
C-? ON* V~T ^h" r~-" »-Г >-ч" -чг" Г--" ** со" ГГ*
•-1 ON CO ON CO NV)
<N -» ~-
NO «O NO NO \D
a
в
Ю
G x
'^Г^ГП'^'Л^ОКОсО.О'ЧГ^Ргч
Г""! "1. ^ **! Q. Q. °1 ^ ^ ""1. ^T ^ ^ °\ ^ °°~ ^4
rf m" ri № № m r-< г-}- ts г-f со со r^c^»oi—« о
со со
\D vfi «Л VC VO VO •^■-ОЧ1»Пц'^- 0^0^40,^0^
o" o" o* o" o" o" o" o" o" o" о o" o" o" —Г
•н -н -н
f- f-
1-4- oC Г-" «-Г \C~ ^ "З" 0\ ОО О С\1Л CS \b" ТГ NO О
ooohacct/'Oviin^irivo-Nf'-iniSH
С^го^О^и^^о^^О^со^с^оо-^со^т^г^е^г^
\o"io',^:'r^ co"c^fco"u^c4'co"c^c4'c-i'cn' со"~ч-г о"
СО CS
ОО П ОО № О М и
ri •-" ~* о" о" о" ---ч cq о" о" о* о" о" -^ ~ in оС
-H-H-H+t-H-H-H+4-H-H-H-H-H+l-H—'+)
*-^ ча^ Т"£ Г^ ^ i~1 00^ N£>_ СО_ f^ С^ О^ ^ О^ О0_ +1 00_
iHO\ooc»cemOvo«nvovovo^'--<'/imw
tS *-н -н ^ .-. О -н
c^t^^v^^oo^t^o^o^vc^^c^^oo^r^c^cn^
у^т^^со'с^'с^со'со'сТ^со'со'со'.^т-г'с^О'С
^-Г —Г *-Г ^н" о" о" —Г *-ч" -ч" о" -ч ^" о" о* —Г оГ г-"
-Н-Н-Н-Н-Н-Н-Н-Н-Н-Н-Н-Н-Н-Н-Н-Н-Н
^Я ■""!. *Ч 'Ч ^*"_ ^1 **\ "~t СЯ **! *"\ ^ "1 'Ч ^ ^Ч °°-
г^ о" nc" in" оо" чо^ о" тг" оо" c*f vT v^T \о^ wi" »n" cs" »/i*
^-«O00C?\00»n»-'4DW-)40VDV0,4j''-4inr^r-
Cn-J OJ ^н ^н ^ r-.
*>CNjfn4lO45N00O\O,>iCNlr^",3-lO,ON
I
250
2 tO
220
200
180
160
140
120
100
80
ВО
40
20
V iM
_L
J_
12
dqdqdqdq dqdqdqdqdq d о с? о
/ 2 3 4 5 6 7 в З 10 11
Признаки
Рис. 10. Изменчивость размеров черепа взрослых европейских и сибирских косуль
Обозначения см. на рис. 8; признаки 1—13 — в табл. 5 и на рис. 111
d $
13
позволяло рассматривать их на уровне подвидов (Данилкин, Марков, 1985). Тем
не менее, учитывая постепенность изменения признаков с востока на запад и на юг,
европейская косуля на данном этапе исследований была признана монотипическим
видом (Данилкин, 1986а).
В итоговой публикации В.Е. Соколов и B.C. Громов (1988) значительно меняют
свои взгляды на таксономию косуль и приходят практически к тем же основным
выводам, что и вторая группа исследователей (Данилкин и др., 1985; Данилкин,
1986а), за исключением подвидового деления сибирской косули: они выделяют все же
три подвида (сибирский, тянь-шаньский и дальневосточный), но уже в новых
границах. В качестве основного критерия различия ими используется С/)-коэффи-
циент, предложенный Майром (Майр и др., 1956). Однако при этом весьма нечетко
обосновывается выделение тянь-шаньского и дальневосточного подвидов по
параметрам рогов: сравнивается не вся выборка, а лишь наиболее крупные по размерам
особи из горной популяции Центрального Тянь-Шаня (см.: Поле, 1974) с наиболее
мелкими — из Приморского края. Границу между тянь-шаньским и
дальневосточным подвидами авторы проводят уже не по Монгольскому Алтаю, а "в районе
35
NJO\^4U)tO^OVOCoNO\OiAUltv}K
to
w to и- ►-. и- to to
-w VO -O. W 1Л jfb _l>i ^OV ^CTv О jO. -O -O y\ jO. ^н-
t+ '►-''^.'Lft^o'i-ft'boVio'o'bN'toVi'bo'e-'ii.
— н-н-н-н-н-н-н-н-1+н-н-н-н-н-н-
*"' .°° .""* -^ -° -° -° -° -° -° -° -° -° -° -° -°
V»^'tJo"-4^,u»'oo'iik'4^'CT\'b\'Lft'^'lAV]'oo
to
■-■ to
£ /Л
"о а
/Л
OO tO
a\ о
1л о
*— и- м tO tO (J
sJtO^<«OslO\NJMO\«OOMtJ^
U Ы ^^tO*iOO\OUy«vl^oo\0 >—'
^oVi^bJ^-'Oi'I-*'vo'b\Vj>h^-'io,lAUi
H-H-H-H-H-H-H-H-H-H-H-H-H-H-H-
Vj *►-» "►-• о "J-1 'bj'w,»—I>j1>i*to"c4^ov»^o
tO Ы >-» »— м tO tJ M
Uit0^tO^-^,00\ff\OOtOO\,OOb'(OA
^. и- JSw ►-* 1лр\^-«^Л^Ор^0Ч QOJb.Jb.00 tO
T^^ViVi^T^Va^Vil^bow'o'ioooboVi
н-н-н-н-н-н-н-н-н-н-н-н-н-н-н-н-н-
■^ -^ 0 ^ -^J -
lOOOOVl^-^tO'sl^OOO
/Л
at
/Л
1Л 4^
^^1о^1^Хл>оо^\лчо'Ьо'1л'^-Ъо'Сл'1>>
и- м н- N) M ts)
ts*u» t—» i- м i-> cj [g n
jNoj^J^^wpo^oo^opo^j^i © 5K p J-л po
"wIa'oo'oo'*—'^oVi'*--'!-» 1>*,ь-»Ъ»'»—T—'w'oo'bo
H-H-H-H-H-H-H-H-H-H-H-H-H-H-H-H-H-
H- j» _^- _^- О О О p p p p p О p ,►— _н- j—
.^ 1л 1л "w o\ Irt о V "*. oo ">j Vj "o 00 о о "►-'
о
ил w
NlU)0\LnigN)^tOM^-b>^^^^9\C7\
"00 to ^tpk^^^'tPkVi'bo'wo'^-tAo'So'i^
UjKi^O'OOsnO^OiA.UjM^.
■—' >-» tO tO (O
Ul-J^^^NJtOONOOi-tOW
4i.jC^P^rtO40OtyiH- 1Л *0 и ft OO
Ы W "ь- О V 0\ V] О "^ 1л Д Irt OO
p P p p p p p p о p p p p 3
Ъ1^Ъ\'*к'и*Ъч,1лТ>л"1рк.'^'оо"--о'^4 II
ft
5:
H-
3
II
01
я
я
в
4?
Xл'ст\Vi'^o'^'t^Ъo"*k'ЬJ'*-•,"■-4'^>A,^--
«— >— ю to to
О Л j*J О Vi _►- ^. OO yi yi р\
V W "tO О н> н- м м to "nJ W
f+H-H-H-H-1+H-H-H-H-H-
Г* ~ ^ Г- Г- -° ° ° J° J° J°
"-^I'to'to'oo'os'vo'i^Vi'bv'H-'w
3:
3
£
s
ы и (л ы ы ia .
"►-^ ft "tO "ft "ft "^
to м to < л л
oo 1л о "ю "^ "ю
1
>-- *- to to to
i^-o.ooftft-ototA*©*ootow
0\9\<ОА^Юи1'00^№00и)0\
OOtOW^O\MQO<o\«,tO
Г- J° -w -° Г- Г* Г* ° ^ Г- i^ J° "-
00 w Vj 00 to 1л <л *"*j "о Ъ "*-> о vo
*. _-g nj w *» a to ы w o\ w yi
, "^1^1>*'1л'1о'1л'н-'»--'1ло'Ь\,1л
/Л
a
/Л
ЧО
S:
H-
3
w
я
"Я
■1
1
H- H- tO tO tO
^■0000M7\>4t00\>OOO>-'W
м w л и» oo ft м о -t^p°y»^o^—
OO Vl "tO О OO "tJ OO N OO V "^ "*» О
H-H-H-H-H-H-H-H-H-H-H-H-H-
p p p p p p p p о p p p p
Ъ»^^Т>л^ЪчЪч'4ь'1лТл'оо'ю'^о
a:
H-
3
1
WtO^N)N)^^WWWUlO\*.
"oj "ft ^ "w 1л Т— Ъо Ъо "J-- "^ *^. "bo *Vj
K£
*J
к>
н-
оо
*.
-J
S)
U
w
"NO
H-
u>
1Л
*.
00
■u
^- ■— ^ ts) (О tO
^Xo^|o^VjVioo'(oV,b\'bo'jil.'to,bj
H-H-H-H-H-H-H-H-H-H-H-l+H-H-H-
r- P P P г* P P r- -^ P P P r* j- r*
Ъч^^1л^—'ооЪа'оо'1л'ила\,<7\,Ьо,ь-Ъ\
S) Ы >-» *-* и- t—• tsJ tO
1Л OO OO M О W ОО (Л Lft W Wl W ^ и- Ю N) W>
b Ъ o\ to 'nj 1л "»- ">- "vo b "sj *\o ">j Vj Vj 1л w
H-H-H-H-H-H-H-H-H-HhH-H-H-hfH-H-H-
■—» 0\ е-» О О О •—•00»-*»-,0©t~i>—■ i—' ■—*
^Vi^^boVa^ososV'OiVi'Vjo'w'Lrt'w
LA I»
Vj^^l>j^^bot^ToViVj'\o'ooo^-e^-jTo
to oi
к- tO Ю Ю
fOQCT\lOLA«0-J040S^li-»0\vOQ&^-tO
Ю и- N» О О ^- •— © О ►— *- О О О и- tO *- ^
У* <* о To To "►— "to Ъ\ \o w ^- no "o\ "оо 1л "to Ъ\
vO -J /Л
-J 4^
^Ь*То^1\оЪоо1о'и>'со'>--Ъ\'^*'*-ЪчЪ.'1-»
»Г N>
м- tO W tO
ОО СЛ -O Ji Ы Ы W 1Л Л U» О N> W Г *
w
_ _ _ e _ JO. ©
н-(+н-н-н-н-н-н-н-н-н-н-н-|+нГн-(+
*- w* ~ о p о p © о p © p о © p © ©
1°Ъч^э^л^1л^л^Ъ|'оо'1лЪ\Ъч'^л'^,Ь\'оо
La
к
-4^CrtW4^tO^tOt4>Jk.(>*U»0J^J^(>i-^
"£л 1— оо "jk "и- To To to To Vi To Ъч Хл V "w оо "-о
s > p p _°° -M -w -w p >• -°° .* ,-J г ,w
"Ъо
OOOvO-JtOasvOOS^-LAOOOOQ
ffHrHrH-t+H-H-H-t+H-H-H:*
.. H-H-H-H-H-H-H-H-H-H-.
ЮОи-Ои-^-н-О*— t—tOtOtO
1л w w о V b 'л "vo 1- "ел V Vi b b "-o. V Vi
-J 45 y» N) ON Ю W Ю W _A W W Ю U) 0\ У1 У»
Ъо^ТоЪчо^То1л©'1--,^-ооо©©'1>аТо
N- *~ H- tO tO
JSh^J00CT\0\-J<—LAvO\OvO«—tO
40L0Ot00\CS00—JNJLAOOtOJi.
Ъ>^1о\оЪ»оЪ»1лТо"1>л'иоТл'*.о.
H-H-H-H-M-H-hfH-H-H-H-H-H-
P p _^- p О ^- ^ь- p p p ^ J- и-
^Tvi'to'i*'lA©ol7\"Lft'!^"to'w'!f*.
У* JO ^Л JO JO Lrt jU j>J JO JO рч, _СЛ -4
ЪчЪоЪ*©Т^'1л'Ьо'ь>~'-о'1ло'1»*Т—
.— i— i— tO tO
UlN]h-0\0\vjM.Ui^O'0'0^tJ
To^"c>Vibob\To'0»'to'too©'o".
н-н-н-н-н-н-н-н-н-н-н-и-н-
I- p ^ p p ^" ^ p p p ¥^ I-» »-»
©Ъ^Ъл^лЪчОоХлХдТо.'^к'Ьо'иа
■^ JO Ul W W Д _
"j*. "jb. oo "j*. *%j "ль.'
i— H- .— tO Ю
^ *~ OQ y* y* CO OO -4JO .fb. j» JO J».
s^o^w'oo'bo'o'ooto'oo'bo'ikto
H-H-H-H-H-H-H-H-H-H-H-H-H-
p J— p J- J— p p p p J- p p p
Ъч1>лТо©'^,ЬоТоТлТо'1о'-о'1лТо
ь- JO ^- JO y» и- JO *- ^— to^p-и- tO
Ip To To "to ^— Ъо'о^-чоЪч'Ьч'^-'Ъ
H- ■— H-* tO tO
1Л-*Л^10ЧО\-Лн-1ЛЧОЧО*0*-10
JOJOpoyiyijtbpoj-JJOjUvOyjpi
1р1л'1лЪч'оо'1>л,С7ч'о'и-'оо,Ьч'рЪч
i+H-hfi+H-H-H-i+H-H-H-ffH:
p p p p p p p p p p p p о
W W w S) M ^ Ы Ы N) S) «> -^ -U
tjboooT^Vjvobj'Lo'oo'bs^-OLrt
и- _ и- и-* tO tO
^OJObJbOyijfk.yij&.sOJOjfr.jO. \0
"sj *ч4 Vj w Vl CO "1Л "nJ OO W О Q W
►-'^w.T-to^-too-^-^-ww.**
Т^^^^оТоХл^'слТлХ— V-T— "^-
"m Is) « b w "oo b "u> Vi оо 1л b\ b
JO ул ул р\ ух yi
1л ^-* to "»— Ъо То
t/l
<7\
—1
OO
\o
5:
H-
3
9
5:
H-
3
Q
a:
H-
3
0
£
H-
3
а
5:
H-
3
о
H
^
1
S
1
1
w
№
Дал
Br"
й Восток
i?
i
1
J?
ГИ0НЫ
Таблица 7. Распределение косуль из разных регионов по краниометрическим показателям (по
результатам кластерного анализа)
Регион
Англия
Франция
ГДР
Болгария
Польша
Швеция
Прибалтика
Украина, Белоруссия
Курская, Белгородская области
Северный Кавказ
Закавказье
Урал'3
Западная Сибирь
Предбайкалье
Алтай
Тянь-Шань
Монголия
Забайкалье
Дальний Восток
Китай
л
4
9
220
66
46
9
75
41
19
12
13
113
17
16
80
38
32
15
67
10
Распределение
по 13 признакам
1
100
100
100
100
100
100
100
100
100
100
100
—
—
—
—
5
3
7
—
50
2
—
—
—
—
—
—
—
—
—
—
100
100
100
100
95
97
93
100
50
по кластерам, %
по 5 (/,
приэна
4, 8, 9, 12)
сам
1
100
100
100
100
100
100
100
98
100
100
100
—
—
—
—
3
—
—
_
10
2
_
—
—
—
—
—
—
2
—
—
100
100
100
100
97
100
100
100
90
Обозначения признаков см. в табл. 5.
Промеры черепов (явно сибирской косули) из района великокняжеской охоты на Кубани не включены
в анализ, поскольку не исключена возможность завоза зверей из Сибири.
*3Урал—Предуралье, Зауралье, Северный Казахстан.
*4Предбайкалье — Красноярский край и Иркутская обл.
Средней Монголии, продолжив на север до Байкала", хотя по нашим сведениям
здесь не существует сколько-нибудь заметной географической преграды для
косуль, как нет и достоверных различий по морфометрическим параметрам
между особями из Западной и Восточной Монголии (см. табл. 9). Несостоятельно
и утверждение об отсутствии существенных географических барьеров в ареале
сибирской косули (см. главу "Ареал").
Учитывая возникшие разногласия, было решено еще раз произвести
статистическую обработку дополненного материала (см. табл. 5—10)'.
1 Использован собственный материал и коллекции зоомузеев МГУ (Москва), ЗИН (Ленинград),
Институтов зоологии (Киев, Алма-Ата, Баку, Ереван, Тбилиси), Иркутского сельскохозяйственного
института, Института биологии СО АН СССР (Новосибирск), Института зоологии и паразитологии
(Вильнюс), Центрально-Черноземного заповедника (Курск), Института общей и экспериментальной
биологии Монгольской АН (Улан-Батор), Института леса Болгарской АН (София), Национального
музея природы (Париж), Музея зоологии и энтомологии университета г. Лунд (Швеция), Британского
музея естественной истории (Лондон, кол. V.P.W. Lowe). Свои коллекции или некоторые данные по мор-
38
Таблица 8. Распределение европейской и сибирсхой косуль из разных регионов по 13
краниометрическим показателям (по результатам кластерного анализа)
Регион
Западная Европа (разные регионы)
ГДР
Болгария
Польша
Швеция
Прибалтика
Украина, Белоруссия
Курская, Белгородская области
Северный Кавказ
Закавказье
Урал, Северный Казахстан
Западная Сибирь
Предбайкалье
Алтай
Тянь-Шань
Монголия
Забайкалье
Дальний Восток
Китай
п
Европейская косуля
Сибирская
18
220
66
46
9
75
41
19
12
13
косуля
111
16
16
78
36
31
15
65
10
Распределение по кластерам, %
1
72
92
67
72
44
40
10
—
8
62
77
69
50
47
6
3
7
6
10
2
28
8
33
28
56
60
90
100
92
38
23
31
50
53
94
97
93
94
90
Табл. 5, 6 и рис. 10 наглядно демонстрируют, что по многим средним
краниометрическим показателям, как и по средним значениям размеров и массы тела,
европейские и сибирские косули существенно различаются, но максимальные
показатели у отдельных особей первых перекрываются с минимальными
размерами вторых. Среди европейской косули самыми крупными по
краниометрическим показателям являются популяции Восточной Европы, включая Северный
Кавказ, самыми мелкими — популяции Западной Европы и Закавказья.
Граничащие на востоке Европы популяции наиболее крупной европейской косули
достоверно отличаются по размерам черепа от уральской популяции сибирской косули,
т.е. снова налицо разрыв постепенности изменения морфометрических
показателей, указывающий на генетические различия. Популяции сибирской косули,
обитающие по обе стороны от Алтайско-Саянской горной системы, явно
различаются по средним размерам черепа: уральско-предбайкальские популяции гораздо
фометрии косуль любезно предоставили В. Бресинский (Польша), М. Бек (Англия), Ю.А. Дарман,
В.Г. Швец, К.А. Морозов, А.А. Киселев, П.П. Блузма, А. Б. А рабу л и. В.Е. Локтионов, В.Б. Поле, Г.Г. Со-
банский и многие охотоведы и охотники Советского Союза и других стран. Всем коллегам,
предоставившим материал, приносим глубокую благодарность и считаем их полноправными соавторами
настоящего раздела монографии. Мы очень признательны также А.В. Абрамову и Б.Н. Науменко за помощь
в статистической обработке материала на ЭВМ.
39
Таблица 9. Краниометрические характеристики и значения коэффициента различия CD (по: Майр
и др., 1956) популяции Западной и Восточной Монголии
Признак*
/
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
•Обозначени
Западная Монголия
самки
л
9
9
9
9
9
10
9
8
10
10
8
10
8
М±т
222,8±1,4
210,4±1,6
197,0±1,4
94,6±0,7
92,2±0,8
57,1±0,9
118,1±1,4
74,0±1,5
64,4±0,6
62,7±0,7
174,8±1,5
72,9±0,7
49,8±1,1
о
4,3
4,8
4,1
2,2
2,5
3,0
4,1
4,2
2,1
2,2
4,1
2,2
3,2
самцы
п
9
9
8
12
12
12
8
10
И
12
8
7
8
16
15
17
14
я краниометрических признаков см. в табл. S.
М±т
224,8±1,3
212,4±1,7
199,0±2,4
101,9±1,2
93,8±0,9
60,8±0,7
П4,9±2,3
70,8±1,3
65,7±0,6
65,2±0,6
180,4±2,2
74,4±0,8
51,5±1,3
22,2±1,1
64,5±2,4
30,1±9,6
240±13
о
4,3
5,2
6,7
4,0
3,0
2,5
6,4
3,8
2,2
2,2
6,3
2,1
3,6
4,3
9,4
40
47
крупнее тянь-шаньско-дальневосточных, среди которых наиболее маленькие черепа
имеют косули Центрального Китая.
Обработка массива "полных" данных (черепа 417 самцов и 488 самок косуль
из разных географических районов, измеренных по всем 13 признакам) на ЭВМ
методом кластерного анализа (авторы программы А.В. Абрамов и Д.С. Авиндор),
при котором каждый отдельный череп в общем массиве сравнивается по всем
признакам со всеми остальными, дает следующие результаты (табл. 7).
При анализе по 13 признакам (отдельно по полу) массив данных разделяется
на две группы: в первую включаются все экземпляры из Европы, единичные
черепа из некоторых регионов Азии и половина выборки из Центрального Китая,
во вторую — только сибирские косули. Еще более четко разделение на
европейскую и сибирскую группы проявляется при анализе массива по 5 признакам,
наиболее часто употребляемым в таксономии (длина и ширина черепа, длина
носовых костей и верхнего и нижнего ряда зубов). Лишь одна особь из "гибридной"
популяции Днепропетровской области попадает в группу сибирских, и, напротив,
только один экземпляр из популяции Тянь-Шаня и один из Китая оказываются
в европейской группе. Особенно наглядны различия европейской и сибирской
косуль по длине зубных рядов (см. табл. 5 и 6).
Этот результат, на наш взгляд, убедительно свидетельствует о существенной
40
Восточная Монголия
самки
я
6
6
6
6
6
6
6
5
6
6
6
6
5
М±т
227,3±3,1
213,8±3,2
200,7±2,9
92,8±1,8
90,0±1,5
57,5±0,8
121,3±2,3
77,6±2,6
65,3±1,2
65,2±0,9
180,5±3,1
71,8±0,9
53,0±2,1
■
с
7,6
7,8
7,2
4,4
3,6
2,1
5,6
5,7
3,0
2,1
7,8
2,3
4,7
самцы
1
л 1
8 1
7
8
10
10
10
7
9
10
11
7
7
7
9
9
8
7
'
М±т
225,4±1,8
212,3±2,2
199,5±1,7
99,6±1,4
94,1±1,1
62,2±0,8
116,7±1,8
74,3±2,5
65,8±0,6
64,0±0,8
177,4±1,9
72,1±1,3
50,9±0,8
22,2±1,5
66,1±1,8
304±13
251±28
°
5,3
5,7
4,7
4,5
3,3
2,5
4,6
7,4
1,9
2,6
5,0
3,5
2,0
4,5
5,6
37
74
CD
самки
0,38
0,27
0,33
0,27
0,36
0,08
0,33
0,36
0,18
0,58
0,48
0,24
0,41
самцы
0,06
0,01
0,04
0,27
0,05
0,28
0,16
0,31
0,02
0,25
0,27
0,41
0,11
0
0,11
0,04
0,09
морфологической дифференциации европейской и сибирской косуль и полностью
подтверждает выводы, полученные нами ранее другими методами. В то же время
относительное сходство особей из Центрального Китая с европейскими в
определенной степени говорит о близком филогенетическом родстве и монофилети-
ческом происхождении разных форм косуль.
Выделенные группы европейской и сибирской косуль оказались
неоднородными по составу, внутри каждой как бы обособляются две "морфы" (табл. 8).
Восточноевропейские популяции в целом дифференцируются от западноевропейских,
однако обозначить четкую географическую границу между ними не представляется
возможным, как и невозможно достаточно аргументированно выделить таксо-
номически отличающиеся подвидовые формы. Сибирские косули, напротив,
достаточно четко разделяются разными методами обработки материала на ЭВМ
только на две крупные, сравнительно обособленные и таксономические
отличающиеся подгруппы: уральско-предбайкальскую и тянь-шаньско-дальневосточ-
ную, включающую популяции Китая. Косули Алтая, Восточного Казахстана и
юго-востока Иркутской области имеют по некоторым признакам "переходные
черты", что позволяет говорить о смешанном составе этих популяций.
Ни по С/)-коэффициенту Майра* соответствующему величине 1,28, ни другими
методами не выявляются таксономически значимые отличия косуль Тянь-Шаня
41
500
wo
% 300
К
kzoo
1
1±
14
I.) Hi
Признаки
17
Рис. И. Изменчивость размеров рогов взрослых европейских
и сибирских косуль
Обозначения см. на рис. 8; признаки 14—17 — в табл. 5
и на рис. 111
и юга Казахстана от популяций Монголии,
Забайкалья и Дальнего Востока РФ, как нет
существенных различий между популяциями
животных Западной и Восточной Монголии (табл. 9),
относимых В.Е. Соколовым и B.C. Громовым
(1988) к разным подвидам — тянь-шаньскому
и дальневосточному.
Рога как таксономический показатель.
Изменчивость рогов косуль настолько велика и так
зависит от конституции, возраста и
экологических условий, что весьма проблематично
таксономического показателя. Тем не менее
использовать их в качестве
практически во всех руководствах рога европейских и сибирских косуль
приводятся в качестве одного из основных отличительных признаков.
Действительно, по средним показателям и индексам выборки рогов достоверно
различаются (Соколов, Марков и др., 1985), но индивидуальное перекрывание
показателей настолько значительно, что безошибочно можно распознать лишь
крупные рога сибирской косули, длина которых превышает 30 см, а размах 25 см
(см. табл. 5, 6 и рис. 11). Однако на востоке ареала европейской косули нельзя
исключить появление (или существование) особей с рогами, несколько
превышающими эти размеры.
Сравнительный анализ массива рогов (л = 460) методом кластерного анализа
(табл. 10) показывает, что по выделенным признакам нет четкого разделения
Таблица 10. Распределение самцов косуль из разных регионов по параметрам рогов* (по результатам
кластерного анализа)
Регион
Англия
Франция
ГДР
Болгария
Польша
Швеция
Прибалтика
Украина,
Белоруссия
Курская,
Белгородская области
я
3
2
60
33
30
3
61
17
6
Распределение
кластерам, %
1
100
100
92
91
87
100
21
18
100
2
—
8
9
13
—
79
71
—
по
3
—
—
—
—
—
—
11
—
Регион
Северный Кавказ
Закавказье
Урал
Западная Сибирь
Предбайкалье
Алтай
Тянь-Шань
Монголия
Забайкалье
Дальний Восток
Китай
п
4
15
32
2
41
17
75
24
8
22
5
Распределение
класте
1
25
47
—
—
—
—
—
—
—
—
—
рам, %
2
75
53
41
50
17
18
23
12
38
50
100
по
3
—
59
50
83
82
77
88
62
50
42
на европейскую и сибирскую группы, как это выявляется по признакам черепа.
Массив рогов разделяется на три группы, не имеющие ясной географической
дифференциации, хотя по среднестатическим показателям популяции европейской
и сибирской косуль достоверно различаются (см. табл. 5, 6 и рис. 11). Однако
ни у одних, ни и у других по признакам рогов невозможно выделить применяемыми
методами какие-либо значимые на таксономическом уровне группы, хотя такие
попытки предпринимались (Громов, 1986а; Соколов, Громов, 1988). В то же время
следует отметить, что у сибирской косули наиболее крупные рога имеют особи из
Алтая, Тянь-Шаня и Предбайкалья (из горных районов), у европейской
относительно крупными размерами рогов отличаются популяции Прибалтики, Украины
и Кавказа.
Окраска как таксономический признак
Окраска как таксономический признак фигурирует во многих работах. Наиболее
типичное описание окраски косуль дал зоолог и художник К.К. Флеров (1952,
с. 93—94): "Окраска взрослых одноцветная. Зимой общий цвет серый или серо-
бурый, иногда серовато-рыжий, постепенно темнеющий кзади, где переходит в
бурый, иногда в темно-коричнево-бурый (особенно на верхней стороне зада).
Вся окраска постепенно светлеет вниз. Зеркало небольшое, белое или светло-
рыжевато-белое, не заходящее на бока и верх тазовой области, но резко
ограниченное темной окраской зада. Конечности книзу постепенно рыжеют и ниже
пяточного сустава имеют рыжую окраску. Голова или одноцветна с туловищем
или несколько бурее и рыжее, иногда более серая, особенно в верхней части. Уши
одноцветные с туловищем с наружной стороны или немного бурее, постепенно
темнеющие от основания к вершине; внутренняя сторона их беловатая или
рыжеватая. На нижней губе большое темное пятно. На подбородке белое пятно,
изменчивое по размерам. Окружность носа и верхняя губа черно-бурые или серо-бурые,
иногда край верхней губы белый. Брюхо и внутренняя сторона конечностей
желтовато-кремовые. Летний мех рыжий, иногда на хребте буровато-рыжий. Вся окраска
светлее книзу, но в общем более ровная, чем в зимнем меху. Зеркало почти
отсутствует и лишь слабо намечено небольшим посветлением общего рыжего тона.
Подмышки и пах беловатые. Брюхо беловато-рыжее или рыжее, окраска головы
часто более серая или бурая, чем туловище, переносье и лоб иногда более темные
и бурые. Уши снаружи рыжие, в верхней части темнее, чем при основании, внутри —
-грязно-белые или рыжевато-белые. Вокруг носа черно-бурое пятно, передний край
верхней губы около голого пространства и подбородок белые или буровато-
белые. Конечности рыжие, иногда светло-рыжие.
Окраска молодых пятнистая. Общий тон окраски верхней части тела
новорожденных бледный, рыжевато-желтый; брюхо в передней части и внутренняя
сторона конечностей светло-охристые, конечности снаружи светло-рыжие, задняя
часть брюха беловатая. Голова сверху близка по тону к общей окраске спины,
но более серая, внешняя сторона ушей, брови и щеки сероватые. Кольцо вокруг
глаз темное, буроватое, переносье серовато-охристое. Нижняя часть головы
светло-охристая со слабым сероватым оттенком. Пятна на спине расположены
тремя продольными полосами с каждой стороны, четвертая полоса едва
намечается. Две средние, наиболее яркие, начинаются за ушами и идут по верхней
стороне шеи и спине до зада. Спереди эти полосы сероватые, однотонные с ушами,
43
но кзади постепенно белеют, принимая слабый желтоватый оттенок. Следующие
полосы начинаются на плечах и постепенно бледнеют книзу, так что третья (нижняя)
очень слабо выражена. Сзади в области таза появляются еще полосы, одна ниже
третьей, другая между первой и второй".
Как видим, цветовая гамма окраски животных, меняющаяся и по сезонам, и с
возрастом, настолько велика, что весьма проблематично ее точное описание,
поскольку каждый исследователь может трактовать цветовые оттенки, зависящие
к тому же от освещения, по-разному. Ф.А. Пастернак (1955), наиболее детально
изучивший окраску косуль в географическом аспекте, пришел к выводу о
невозможности выделения по этому признаку систематических групп в пределах ареала.
С запада на восток и с юга на север явно наблюдается постепенное осветление
общей окраски меха косуль и изменение цветовой тональности. Вместе с тем
по краям ареала при повышении влажности климата происходит увеличение числа
феомеланинов, обусловливающих более темную и яркую окраску, что характерно
и для других оленей.
Леман (Lehmann, 1971) подтвердил эту точку зрения, считая различия в структуре
и цвете волосяного покрова связанными с климатом. По мере продвижения на
восток и север окраска летнего наряда европейской косули становится менее яркой,
насыщенный красный цвет постепенно переходит в желтоватые охристые тона,,
волос становится длиннее и толще, окраска основания волос несколько меняется.
К.К. Флеров (1952), признавая эти закономерности распределения окраски
в пределах ареала, тем не менее дал весьма точные, на наш взгляд, отличительные
признаки европейской и сибирской косуль. У европейских косуль окраска в зимнем
меху серовато-бурая, иногда темная, переходящая в коричнево-бурую, а на боках —
в серую. В летнем наряде верхняя сторона головы серая или бурая, резко
отличающаяся от рыжей окраски спины и боков. Основания волос серо-бурые или
темно-бурые, эпидермальный слой кожи светлый, не пигментированный. У
сибирских косуль зимой цвет меха серый с охристым оттенком, на спине буроватый,
брюхо и внутренняя сторона конечностей кремовые. Летом окраска головы рыжая,
однотонная со спиной и боками. Основания волос светлые, эпидермальный слой
кожи буровато-серый, пигментированный. К тому же у европейской косули явно
выделяются светлое носовое (муфлоново) пятно и бурый или темно-бурый цвет
метатарзальных желез (Meyer, 1968; Stubbe, Passarge, 1979).
Позднее эти отличительные признаки приводили в своих работах И.И. Соколов
(1952) и В.Г. Гептнер (1961). К таким же выводам пришел и B.C. Громов (1986а,б),
показав, что они появляются уже у телят.
Просмотрев во всех перечисленных выше музеях СССР и некоторых других стран
шкуры косуль и сравнив по цвету более сотни добытых и отловленных в природе
зверей разного пола и возраста (в общей сложности более 600 особей), мы пришли
к убеждению, что в целом в пределах ареала окраска этих копытных имеет те же
закономерности, о которых говорил Ф.А. Пастернак. Однако европейская и
сибирская косули явно различаются между собой по большинству из перечисленных
признаков, т.е. окраска имеет таксономическое значение.
Вместе с тем из-за очень большой индивидуальной, возрастной и сезонной
изменчивости далеко не всегда можно по этим признакам отличить европейскую
косулю от сибирской. К тому же во многих музеях из-за длительного и
небрежного хранения цвет шкур стал весьма далеким от естественного.
Особенно изменчива окраска головы. В летнем наряде у отдельных европейских
44
косуль она бывает одного цвета с верхом туловища, что характерно для сибирской
косули, и, напротив, у некоторых сибирских особей, особенно молодых, окраска
переносья и лба серовато-бурая, как и у части европейских. У некоторых старых
сибирских косуль на носу выделяется светлое пятно, как и у европейских. Нередко
по цвету оснований волос невозможно отличить одну косулю от другой.
Достоверных различий в окраске между подвидами сибирской косули мы не нашли.
У значительного числа сибирских косуль, так же как и у европейских, в летнем
наряде имеется четко выраженный, а иногда и очень большой "каудальный диск",
или "хвостовое зеркало" (см., например: Соколов, Данилкин, 1981, рис. 4, 15, 39,40).
Напротив, у значительного числа европейских косуль, как и у многих сибирских,
оно отсутствует (см., например, фото: Honak, 1970; Stubbe, Passarge, 1979). В природе
из 93 встреченных летом 1986 г. в Белгородской области европейских косуль четкий
"каудальный диск" был лишь у 59 животных, у остальных он едва прослеживался
или отсутствовал. Следующим летом (по сообщению охотоведа О.В. Чекалиной)-
после затянувшейся на месяц линьки из-за поздней весны почти все звери этой же
популяции не имели "хвостового зеркала". В результате многолетних наблюдений
за одними и теми же сибирскими косулями на Южном Урале мы установили также,
что "хвостовое зеркало" в летнем наряде является признаком, передаваемым по
наследству. Все дети самок, имевших летом большие белые "каудальные диски",
имели их в отличие от большинства детенышей других самок, у которых "хвостовое
зеркало" отсутствовало.
Таким образом, "каудальный диск", выделенный В. Громовым (1986а,б) как
отличительный признак европейской и сибирской косуль, не может использоваться
в качестве таксономического показателя.
Пожалуй, наиболее существенным отличительным признаком является цвет
метатарзальных желез, не выделяющийся из окраски волос ноги у сибирской косули
и более темного цвета у европейской. Однако и в этом случае из 34 европейских
особей, осмотренных в Белгородской области, у 6 окраска желез была "сибирского
типа" и, напротив, среди сибирских косуль на Южном Урале и на Дальнем Востоке
встречались животные с темной ("европейского типа") окраской.
Предпринятое нами исследование окраски и расположения губных пятен нижней
зоны головы и шеи у различных популяций в ареале также не дало однозначного
результата. Практически в каждой из изученных популяций европейской и
сибирской косуль мы находили особей, сходных с типами, описанными М.Н.
Смирновым (1978) у животных Западного Забайкалья. Вместе с тем в каждой популяции
тот или иной признак имел большее или меньшее количественное выражение.
Особи с аномальной окраской (альбиносы, меланисты и пятнистые) иногда
встречаются как среди сибирских, так и среди европейских косуль, однако
описаны они в основном у последних из-за более пристального внимания к ним
Очень большой интерес представляет популяция "черных" косуль, населяющая
низменные заболоченные районы Нижней Саксонии в Германии (рис. 12).
Здесь же живут косули и с нормальной "красной" окраской, среди которых черные
особи составляют от 2 до 80% в отдельных стациях. Эта популяция
документально известна с 1591 г. Взрослые особи после весенней линьки имеют блестящую
черную окраску; зимний мех тоже черный, но матовый, к животу свинцово-серый,
нередко с серебристой остью. "Зеркало" меньше, чем у "красной" формы,
рыжеватое, бурое или черное. Детеныши рождаются черными, реже пегими. В популяции
у самцов повышена доля черепов с клыками в верхней челюсти: у красной формы 7%,
45
Рис. 12. Изменчивость окраски у
европейской косули
А — нормальная (слева) и черная
(фото В. Горгасс); Б — пятнистая; В —
альбинос (фото К. Штуббе)
у черной — 33% и более длинные хвосты. По размерам и массе тела они не
отличаются, поведение их сходно, они образуют смешанные пары, однако черные
косули явно более, чем красные, предпочитают равнину горной местности и на
равнине — влажные лесистые биотопы. Черная окраска, вероятно, является рецессивной
мутацией, которая вызвана обитанием в относительно холодном и влажном климате
низменности. С другой стороны, учитывая наличие клыков, более длинного хвоста
и отличной окраски, нельзя не задуматься о возможности сохранения у этой
популяции генов предкового типа (Meyer-Brenken, 1986; Stubbe, Passarge, 1979).
Как бы то ни было, ясно, что особи одной и той же популяции европейской
косули, живущие вместе, по окраске отличаются гораздо больше, чем европейские
от сибирских. Этот пример еще раз говорит о том, что к окраске косуль как к
таксономическому признаку следует относиться с осторожностью.
46
Цитогенетическая изменчивость
Все исследованные популяции европейской косули имеют одинаковый набор
хромосом 2л = 70 (Gustavsson, 1965; Amrud, Nes, 1966; Wurster, Benirscke, 1967;
Gustavsson, Sundt, 1968; Zivkovic*, Isakovic-, 1972; Соколов идр., 1978; Данилкинидр.,
1983; Susa, Kaluzinski, 1984; Марков, Добриянов, 1985; Zima, Havrankova, 1987).
Все популяции сибирской косули характеризуются присутствием "микрохромосом",
относящихся к группе добавочных, или В-хромосом, — 2л = 70 + (1—14) (Соколов
и др., 1975, 1978; Zernahle, 1980; Данилкин, 1978, 19856; Соколов, Данилкин, 1980,
1981; Данилкин идр., 1983; Графодатский и др., 1989). Аутосомы как у европейской,
так и у сибирской косуль акроцентрические. Половые хромосомы самок XX,
самцов — XY. X — хромосома самая крупная (двуплечая) субметацентрическая,
Y-хромосома самая маленькая, акроцентрическая. В-хромосомы метацентрические
и акроцентрические. Внешне морфологически сходные их кариотипы тем не менее
разнятся по морфометрическим параметрам: 11 хромосомных пар из 34 основного
набора аутосом, а также Y-хромосомы статистически достоверно различаются
(Соколов и др., 1986).
Кариотипы европейской косули стабильные (с одинаковым числом хромосом
на всех метафазных пластинках), в то время как у сибирской они двух типов —
стабильные и мозаичные (с разным числом В-хромосом у одного и того же
животного и у разных особей в одной и той же популяции). Явление мозаицизма наиболее
отчетливо выражено у косуль Дальнего Востока, где обнаружены 4—5 клоновые
мозаики. К примеру^ у одного из самцов число добавочных хромосом варьировало
от 5 до 10 (2л = 75—80) с преобладанием клонов с 6 и 8 В-хромосомами (75%);
15% клонов были с 5, по 25% — с 6, 7, 8 и 10% — с 10 "микрохромосомами". У самок
их количество колебалось от 6 до 10 (диплоидные числа соответственно
изменялись от 76 до 80) и преобладали клоны с 8 (40%) и 10 (30%) добавочными
хромосомами. Трехклоновым мозаиком (2л = 78, 80, 82) с преобладанием клона с 12
добавочными хромосомами (60%) оказался самец косули, поставленный в Москву
Восточносибирским зоокомбинатом. К сожалению, точный район отлова этого
зверя неизвестен (предположительно Забайкалье). Клоны с 5,6,7 и 8 В-хромосомами
были найдены у двух самок косули из Восточной Монголии. Кариотипы косуль
из Предуралья, Зауралья и Предбайкалья отличались большей стабильностью,
при этом число, "микрохромосом" в них было минимальным — от 1 до 4 (Данилкин,
Баскевич, 1987). Наибольшая вариабельность кариотипа обнаружена в популяции
косуль Алтая — 2л = 72—84 (Графодатский и др., 1989).
В-хромосомы наследуются гибридами, как было установлено в экспериментах
по скрещиванию европейской и сибирской косуль, проведенных в ГДР (Zernahle,
1980). Самцы сибирской косули, участвующие в опыте, имели в хромосомном
наборе по 10 В-хромосом. Хромосомный набор у гибридов Fi оказался равным
70 + 3 (т.е. меньше теоретически предполагаемого диплоидного числа 2л = 70 + 5).
Возвратные скрещивания гибридных самок с самцом сибирской косули привели
к повышению числа добавочных хромосом у потомства до 6—7, что также ниже
теоретически ожидаемого. Этот результат говорит как об относительно случайном
распределении "микрохромосом" в мейозе, так и об "обогащении" сибирского
генотипа при возвратном скрещивании.
Гибриды в опытах наследуют фенотип сибирского родителя. Означает ли это,
что фенотип косуль определяется добавочными хромосомами и их числом? Вряд ли,
47
поскольку в одной и той же популяции могут существовать особи с разным числом
В-хромосом и особи-мозаики. К тому же животные без добавочных хромосом
(2л = 70) и имеющие их (2л = 71, 72) из одних и тех же популяций на левобережье
Украины и на Северном Кавказе, по нашим данным, не различаются достоверно
по размерам тела и окраске. Последние, имея одинаковый кариотип с сибирскими
косулями Предуралья, отличаются от них очень существенно.
С другой стороны, нельзя не обратить внимания на отчетливое морфометри-
ческое сходство 1—4 В-хромосомных популяций сибирской косули из Заволжья,
Зауралья и Предбайкалья и их значительное отличие от 5(6)—10(12)
В-хромосомных популяций Южного Казахстана, Киргизии, Монголии и Дальнего Востока,
также морфометрически сходных между собой. Эти крупные и относительно
изолированные географические популяции косуль оказались генотипически сходными
на очень значительном протяжении ареала, а изменение числа добавочных хромосом
в значительной степени совпадает с географическими барьерами.
В целом же видно, что с востока на запад ареала число В-хромосом у косуль
редуцируется, исчезая полностью в Центральной и Западной Европе, и это явление
явно носит характер клинальной изменчивости (рис. 13).
Генетическая роль добавочных хромосом, их природа и полиморфизм по ним
и у косуль, и у других млекопитающих во многом неясны. Применение этой
кариологической характеристики в качестве таксономического критерия
оказывается полезным на межвидовом уровне, но встречает определенные трудности
на уровне внутривидовом. Большая вариабельность числа хромосом у особей-
мозаиков и у разных особей в одной и той же популяции все же не позволяет четко
дифференцировать подвиды и популяции сибирской косули, как ожидалось. В»то же
время изучение цитогенетическои изменчивости по В-хромосомам открывает
возможности для обсуждения происхождения тех или иных популяций косуль и
зоогеографических реконструкций.
Присутствие в диплоидном наборе сибирской косули добавочных хромосом
наряду с палеонтологическими данными, приведенными выше, может
свидетельствовать о большей ее древности по сравнению с европейской. Если это принять
за основу, то, учитывая дискретность хромосомных различий, можно
предположительно вычленить возможные центры формообразования у Capreolus и проследить
эволюционные связи. Расселение косули, вероятно, могло идти из центра
формообразования в Центральной Азии в основном южнее Алтайско-Саянской горной
системы. Западнее Алтая (не исключены и другие районы) косули проникли на север,
откуда расселение шло на восток — к Байкалу и на запад — в Европу. Вместе с тем,
судя по исключительной вариабельности числа В-хромосом, таким центром вполне
мог быть и Алтай (см. рис. 13).
Постепенная редукция числа добавочных хромосом наводит на мысль о
существовании географической зоны "переходного" генотипа от сибирской к европейской
косуле, где особи имели бы в кариотипе минимальное количество В-хромосом
или не имели бы их совсем. Такая зона теоретически должна была бы находиться
западнее Волги, и именно такой кариотип 2л = 70; 70 + 1 и 70 + 2 мы нашли у неясных
в систематическом отношении популяций косуль в Самарском лесу
Днепропетровской области и на Северном Кавказе. В популяции косуль Самарского леса из
9 исследованных животных 2 не имели добавочных хромосом, 3 содержали по одной,
а 4 особи — по две В-хромосомы. На Северном Кавкае добавочные хромосомы (2)
были обнаружены только у одной особи, добытой вблизи г. Апшеронска Красно-
48
Рис. 13. Изменчивость числа хромосом косуль
дарского края. Во всех других районах от Волги до Днепра в начале XX в. косули
были полностью уничтожены, практически исчез и "переходный генотип".
Возможна и другая гипотеза, которая в свете последних исследований нам
кажется более правильной. Не исключено, что в становлении современного
кариотипа европейской косули существенную роль сыграли ледники,
способствовавшие географической изоляции популяций, содержащих и не имеющих в кариотипе
добавочных хромосом. Европейские популяции, географически изолированные от
сибирских, накопили генетические различия и со временем приобрели признаки,
обеспечивающие частичную репродуктивную изоляцию с популяциями предкового
типа. Из одного родительского возникли два хорошо интегрированных генных
комплекса.
В послеледниковый период сибирская косуля проникла на запад по меньшей мере
до Днепра, в чем нет сомнения, и, может быть, на Северный Кавказ, где обитала
европейская косуля. В результате образовывались "зоны гибридизации", в которых
особи имели генотип как европейской, так и сибирской косули.
Предлагаемая гипотеза аллопатрического видообразования у косуль, на наш
взгляд, хорошо согласуется со всеми известными до сих пор фактами
генетической и фенотипической изменчивости и дает объяснение происхождению
неясных в систематическом отношении популяций. Конечно же, при накоплении
данных предполагаемый путь расселения косуль будет уточнен, однако
магистральное направление их географического видообразования в целом нам кажется
достаточно ясным.
Нет сомнения также в том, что необходимо дальнейшее цитогенетическое
исследование популяций, представляющий особый интерес в свете изложенных
гипотез. Таковыми прежде всего являются популяции Кавказа, Поволжья, Алтая,
Тянь-Шаня, Якутии и Китая (район Тибета), где возможны новые интересные
находки, которые могут весьма изменить существующие представления о цито-
генетической изменчивости и видообразовании у косуль.
Биохимический и иммунологический анализ
Методом зонального электрофореза установлено (Соколов, Шурхал и др., 1986),
что среди 11 ферментов, кодируемых 14 локусами, в выборке из популяции
европейской косули 3 локуса (эстераза Д, ИДГ и Г6ФДГ) полиморфны. У сибирской
косули обнаружены только два полиморфных локуса: ЭСТ-Д и Г6ФДГ. Ни в одном
из 14 аллозимных локусов не выявлено различий по электрофоретической
подвижности фракций ферментов и только для одного локуса ИДГ найдены различия
по частоте встречаемости генотипов. По распределению аллелей исследованных
систем ферментов отличий между уральскими и дальневосточными популяциями
сибирской косули не обнаружено.
Методом изоэлектрического фокусирования белков плазмы крови выявлены
к тому же четкие различия между европейскими и сибирскими косулями по
отдельным фракциям в преальбуминовой зоне (ПРА) спектра в области значений
рН 5,2 (рис. 14). Отличия по этим фракциям фиксированы, каждая из
европейских косуль отличается от каждой из сибирских.
Различия такого же уровня установлены и по ИЭФ-спектрам растворимых
белков мышечной ткани, изменчивость показана для фракций ИЭФ-спектра,
лежащих в области значений рН 6,6 и 7,8 (рис. 15). Анализ данных по оптической
so
*
*
t I I
/ г з ч 5 6 7 1 z з ч s e
Рис. 14. Спектр изоэлектрического фокусирования белков сыворотки крови сибирской (1—4) и
европейской (5—7) косуль
Рис. 15. Спектр изоэлектрического фокусирования растворимых белков мышечной ткани
сибирской (1—3) и европейской (4—6) косуль
плотности и характеру распределений значений pi этих фракций позволяет
предположить, что в каждом из случаев различающиеся белковые фракции являются
продуктами альтернативных аллелей трех разных локусов у сравниваемых форм
жосуль. В спектре ИЭФ белков плазмы по распределению генотипов локуса ПРА
обнаружена дифференциация между правобережными и левобережными амурскими
популяциями сибирской косули; в левобережной популяции распространено три
аллеля этого локуса, а в правобережной — только один. ИЭФ-спектры мембранных
белков мышечной ткани европейской и сибирской косуль сходны.
Таким образом, из 14 локусов, кодирующих системы ферментов, и 8 локусов,
кодирующих синтез избыточных белков ИЭФ-спектра, у европейской косули
полиморфны 3 локуса, относящиеся к системам ферментов. У сибирской косули
полиморфны также три локуса, но один из них является локусом ИЭФ-спектра
белков плазмы — ПРА. Уровень полиморфизма или доля полиморфных локусов
как для европейской, так и для сибирской косули оценивается равной величиной
в 13,5%. Величина средней гетерозиготности для европейской косули составляет
Н = 0,0123 и 0,0111. Относительно низкие значения Н хорошо согласуются с
данными других исследователей, также отметивших низкий уровень Н у копытных
животных.
Какова же степень генетических различий между исследуемыми формами?
Анализ данных показал, что применение традиционных методов разделения белков
посредством зонального электрофореза не позволяет выявить фиксированных
различий между ними. Лишь по одному локусу показана гетерогенность по частоте
встречаемости аллелей, а обобщенная оценка различий в единицах генетического
расстояния по Нею составляет всего D = 0,0044. Такие значения D чаще отражают
Дифференциацию между слабоизолированными географическими популяциями.
51
Применение более тонких методов разделения сложных смесей белков, в
частности изоэлектрическое фокусирование, позволило выявить диагностические
признаки, представляющие собой фракции спектров ИЭФ, распределение которых
имеет характер неперекрывающейся изменчивости — альтернативные фракции
закреплены соответственно у разных форм косуль.
В целом установлены различия по трем альтернативным признакам,
представленным в виде фракции изоэлектрических спектров белков плазмы крови
и растворимых белков мышц. С высокой вероятностью можно предполагать,
что эти фракции являются белковыми продуктами альтернативных аллелей трех
разных локусов и отражают достаточно высокую степень генетической изоляции
европейской и сибирской косуль. Если объединить данные по всем ферментным
системам и по ясно различаемым фракциям ИЭФ-спектров растворимых белков
плазмы и мышц, то значение генетического расстояния, по Нею, в этом случае уже
будет составлять величину D = 0,7183, которая обычно характеризует достаточно
надежные виды (см. табл. 2). Тем не менее следует отметить, что ключевые для
такого вывода данные были получены на относительно ограниченном материале
(13 проб от европейской косули и 15 — от сибирской) и для окончательного решения
вопроса требуются дополнительные исследования.
Обнаруженные незначительные генетические различия между правобережными
и левобережными амурскими популяциями сибирской косули, вероятнее всего,
отражают их внутривидовую экологическую специализацию. Неясные в
систематическом отношении косули Днепропетрвоской области (Самарский лес), часть из
которых имеет В-хромосомы, по биохимическим показателям относятся к
европейской косуле (Шурхал и др., 1985; Соколов, Шурхал и др., 1986).
Сходные данные были получены и при сравнительном иммунохимическом
исследовании (Марков, 1985; Марков, Кехаиовидр., 1985). Тестирование нативных
и абсорбированных (путем насыщения сывороткой от другой группы косуль)
антисывороток посредством двойной иммунодиффузии (по методу Ouchteriog)
и иммуноэлектрофореза (по микрометоду Scheideger) показало, что антисыворотка
против белков сыворотки сибирской косули (анти-С.р.) реагирует с гомологичной
сывороткой, образуя в агаре многочисленные преципитационные полосы.
Аналогичные числа и расположение полос найдены и при реакции антисыворотки
против сыворотки европейской косули (анти-С.с). Этот многократно полученный
результат дает основание предположить, что сыворотка крови сибирской косули
содержит все антигены, присутствующие и у европейской косули. Подтверждением
этому служит также то обстоятельство, что после абсорбции анти-С.р. с
сывороткой крови европейской косули антисыворотка перестает реагировать как с
сывороткой европейской, так и сибирской косули. Полное сходство антигенов
устанавливается и при исследовании анти-С.р. в условиях иммуноэлектрофореза.
Тестирование на анти-С.с. выявило наличие антигенов, которые характерны
только для сыворотки крови европейской косули. После абсорбции анти-С.с.
сывороткой крови сибирской косули эта антисыворотка перестает реагировать
с сывороткой крови сибирской косули, но образует при иммунодиффузии и при
иммуноэлектрофорезе две преципитационные полосы при реакции с сывороткой
европейской косули (рис. 16).
Дополнительные данные об отличиях антигенного состава сывороток крови
европейской и сибирской косуль были получены и при их сравнительном
исследовании в условиях полиакриламидного электрофореза в градиентном геле (рис. 17).
52
Рис. 16. Иммуноэлектрофореграмма сывороток и
антисывороток крови европейской (Capreolus capreolus —
С.с.) и сибирской (Capreolus pygargus — Ср.) косуль.
В лунках А 3 и 5 — сыворотки крови С.с, в лунках 2
и 4 — сыворотки крови Ср., в канавках — анти-С.с-
сыворотка, абсорбированная полной сывороткой Ср.
° 2
о *
6 7 8 9 10 11 12
я
ft
л
Рис. 17. Электрофореграмма в полиакриламидном градиентном геле белков сыворотки крови
европейской (1—8) и сибирской (9—12) косуль
Маркеры F\ и Fi расположены в зонах рН 7—7,5 и 5—5,5 соответственно
53
°лт
200 300 V т
Рис. 18. Гельная хроматография сыворотки крови европейской косули
F\ и Fi — специфические маркеры сыворотки крови
Рис. 19. Иммунодиффузия специфических маркеров сыворотки крови европейской косули
В лун ке / — анти-С.с.-сыворотка, абсорбированная сывороткой крови Ср.; в лунке 2 — антисыворотка
против ft; в лунке 3 — Лив лунке 4 — Fi
Рис. 20. Иммуноэлектрофорез специфических маркеров сыворотки крови европейской косули
В канавке — анти-С.с.-сыворотка, абсорбированная сывороткой крови Ср. В лунках — хроматогра-
фические фракции Ft (Р-подвижность) (/) и Fi (аг-подвижность) (2)
Спектр белков сыворотки крови европейской косули отличается по двум фракциям
в области средне- и быстромигрирующих его частей, что, очевидно, соответствует
двум добавочным антигенам.
Для получения некоторых ориентировочных биохимических и
физико-химических характеристик маркеров сыворотки европейской косули были применены
опыты по изоляции этих белков в относительно гомогенный вид. При хроматогра-
фическом разделении (рис. 18) методом колоночной гель-хроматографии оба
маркера (обозначены как Fi и F2) идентифицированы как разные фракции. Эти
данные, показывающие значительные различия в размерах массы молекул обоих
антигенов, совпадают с результатами полиакриламидного электрофореза и изо-
электрического фокусирования. Кроме того, оба антигена различаются и по своей
антигенной специфичности (рис. 19) и электрофоретической подвижности: у Fi
Р-подвижность, у F2 аг-подвижность (рис. 20).
Таким образом, в результате проведенного сравнительно-иммунохимического
исследования установлено существование двух антигенов, специфичных для
54
сыворотки крови европейской косули, что дает нам основание дифференцировать
европейских и сибирских косуль на видовом уровне. С другой стороны,
идентичный набор остальных антигенов сыворотки крови позволяет говорить об их
близком филогенетическом родстве.
Гибридологический анализ
Одним из основных критериев вида, который выработан систематикой в своем
длительном историческом развитии, является критерий репродуктивной или
генетической изоляции (нескрещиваемости). В связи с этим большой интерес
представляют опыты по гибридизации европейской и сибирской косуль.
Наиболее тщательный эксперимент проведен в ГДР Г. Штуббе и 3. Брухголь-
цем (1979, 1980). Двух самцов сибирской косули 32 раза спаривали с самками
европейской косули, из них 13 раз безуспешно. Из 19 случаев родов кесарево сечение
пришлось делать 9 самкам и в 3 случаях крупный плод был извлечен ручным
способом. Двух гибридных самцов Fi 10 раз ссаживали с гибридными самками Fi,
но приплод не был получен. Напротив, возвратные скрещивания гибридных самок
с самцами как европейской, так и сибирской косули оказались возможными.
В этом эксперименте была установлена стерильность гибридных самцов и
показано, что самки европейской косули, покрытые самцами сибирской, не могут
нормально родить более крупный плод. В природе это привело бы к их гибели.
Аналогичные опыты, проведенные на экспериментальной базе Института
эволюционной морфологии и экологии животных АН СССР под Москвой,
подтвердили наличие трудностей при гибридизации косуль (Соколов, Громов, 1Я856;
Громов, 1986а). Из шести ссаживаний двух самцов сибирской косули с тремя
самками европейской успешными были только три, но одна самка из них погибла
при родах, другая — перед родами, и лишь одна принесла гибридного самца.
Все попытки скрещивания двух самцов европейской косули с девятью самками
сибирской (12 ссаживаний) не дали результата. Однако возвратное скрещивание
гибридного самца Fi с самками европейской и сибирской косуль оказалось
успешным, в результате чего получены четыре детеныша.
Таким образом, выяснились следующие новые обстоятельства: не все гибридные
самцы косуль стерильны; скрещивание самцов европейской косули с самками
сибирской практически невозможно, как считают авторы, из-за отсутствия
нормальной реакции у самцов на самок в период течки. Эти ученые предположили,
что «в выделениях самок европейской косули присутствует специфический запа-
ховый компонент, обладающий свойствами аттрактанта для самцов азиатской
косули, а у самок азиатской косули такого аттрактанта для самцов европейской
косули нет»1. B.C. Громов (1986а) нашел также различия в половом поведении
европейской и сибирской косуль, которые рассматривает как таксономический
признак: «Для европейских косуль в период гона характерен специфический ритуал
полового поведения, способствующий образованию кольцевых "гонных" троп.
У азиатских косуль такая особенность поведения отсутствует».
Наши наблюдения в этой же вольере и в природе в корне противоречат такому
утверждению. Неоднократно гонные пары сибирской косули на наших глазах
В этих работах (Соколов, Громов, 19856; Громов, 1986а) сибирская косуля названа азиатской.
55
проявляли типичное половое поведение (многократные пробежки вокруг кустов,
деревьев, пней, кочек и т.п.), приводящее к образованию кольцевидных или в виде
восьмерки троп (см. рис. 83). Эта особенность полового поведения у сибирских
косуль уже была описана ранее (Поле, 1973, с. 139; Слудский и др., 1984, с. 71;
Соколов, Данилкин, 1981, с. 95; Дарман, 1986).
Более того, ранее уже сообщалось о получении гибридов при скрещивании
самцов европейской косули с самками сибирской косули (Intern. Zoo Yerbook, 1970,
vol. 10, p. 249; 1971, vol. 11, p. 259). Эксперименты в Ленинградском зоопарке
наглядно показали, что гон у этих пар протекает обычным образом, самец четко
реагирует на запах самки в течке, плодовитость не снижена, однако обращает на себя
внимание относительно большее число мертворожденных (Данилкин, 19866).
Скрещивание гибридных самцов с самками здесь специально не проводили,
поэтому их репродуктивные возможности остались не выясненными. Однако
замечено, что гибридные самцы реагируют на самок сибирской косули в течке и
пытаются крыть их.
При возвратном скрещивании гибридной самки с самцом европейской косули
потомства не было получено, хотя гон и спаривание имели место. Та же гибридная
самка в 1971 г. была снова подсажена к европейскому самцу, но это произошло
после обычных сроков спаривания — в декабре. В марте 1982 г. наблюдался гон
(вероятно, "ложный"), спаривание было бечуспешным.
Следует отметить, что гибридная самка имела аномальные рога. Рост рога
начался в годовалом возрасте, и к осени 1970 г. он был высотой 6—7 см. Рог рос
не только вверх с периодическим сезонным сбрасыванием верхней части, но и во все
стороны от основания. Второй рог появился у самки в апреле 1973 г., незадолго
до забоя, и вырос на несколько сантиметров. Появление такого рода аномалий,
вероятно, может быть следствием гибридизации. Масса тела этой самки-гибрида,
забитой из-за чрезмерного разрастания рогов 11 мая 1973 г., оказался равным 35 кг,
что больше массы самок европейской косули.
Масса трех новорожденных гибридов составила в среднем 1660 г, что близко
к массе телят сибирской косули (Громов, Данилкин, 1984). В десятимесячном
возрасте один из гибридов весил 21,5 кг.
Из приведенных выше опытов по гибридизации европейской и сибирской косуль,
итоги которых приведены в табл. 11, видно, что некоторые результаты оказались
прямо противоположными. Если в ГДР гибридные самцы не дали потомства,
то в вольере под Москвой оно получено. В то же время здесь, как утверждают
авторы, самцы европейской косули не реагировали на самок сибирской косули
в течке и от них не был получен приплод, хотя в зоопарке Ленинграда размножение
косуль в таком сочетании протекало весьма успешно.
В чем прич'ина таких разнородных результатов — в разном происхождении
производителей и их генотипе, физиологическом состоянии, условиях содержания
и выращивания, в климате или в чем-то другом? Пока на этот вопрос однозначно
ответить нельзя, но некоторые предположения сделать все же можно.
Воспроизводство косуль в вольерах может отличаться от размножения в
естественных условиях. Почти безуспешное спаривание в Ленинградском зоопарке
на протяжении ряда лет одного из самцов европейской косули с самками того же
вида наводит на мысль либо о недостаточной половой способности отдельных
самцов, либо о существовании репродуктивных трудностей даже при скрещивании
европейских косуль, что маловероятно. В других опытах (Соколов, Громов, 1985а)
56
Таблица 11. Обобщенные результаты опытов по гибридизации европейской и сибирской косуль (по: Штуббе, Брухгольц, 1979,1980;
Соколов, Громов, 19866; I . i >-. 19866)
Ссаживания
самка
л
самец
л
Результат
число
спариваний
успешные
*1
спаривания
благополучные
*3
роды J
Новорожденные
п
масса, кг
л
3:2
Сиб.
F,
Евр.
F,
Евр.
Сиб.
F,
Ft
BCi сиб.
ВС, евр.
12
1
13
7
11
2
1
3
2
2
Евр.
Евр.
Сиб.
F,
F,
F,
Евр.
Сиб.
Сиб.
Евр.
4
1
4
2
3
2
2
2
1
1
20
1
38
10
11
2
3
3
2
2
4(20%)
1.7
12
8 (21%)
25
2.0-2,2
25
1 (9%)
1
2
3
1
2
2
4
4
3
1,9
1,45
1.45
2.2
2,1
5:1
1:2,1
7 (35%)
Нет
22(58%)
Нет
1(9%)
1
3
3
2
2 _____
Условные обозначения: Сиб. - сибирская косуля; Евр. - европейская; Fi — гибрид первого поколения; BCi сиб. - первые потомки от пары самка
F\ х самец сибирской косули;ВС| евр. - от самца европейской косули.
* Спаривания, приведшие к • • • • эмбрионов.
* Нормальные роды (без помощи человека) живых телят
некоторые самки, безуспешно спариваясь с одним самцом, могли принести
гибридное потомство при спаривании с другим; либо одна из двух самок (обе из Литвы)
могла успешно, а другая безуспешно спариваться с одним и тем же самцом.
Очевидно, что о результатах гибридизации косуль можно судить лишь тогда,
когда эксперименты проведены в разных местах, в разных условиях и при большом
числе особей в опытах. Здесь же в каждом отдельном эксперименте было не более
двух самцов-производителей (к тому же выращенных в неволе, что могло заметно
изменить их поведение), и этого явно недостаточно для уверенных выводов по
каждому отдельному опыту.
По итогам же всех экспериментов можно считать доказанным, что
гибридизация европейской и сибирской косуль может быть относительно успешной как
в случае, если самец сибирской косули спаривается с самкой европейской, так
и при скрещивании самца европейской косули с самкой сибирской. Также можно
считать очевидным факт относительной фертильности гибридов.
В то же время нет сомнения в том, что в целом гибридизация европейской
косули с сибирской значительно затруднена (см. табл. 11). Только около 20% самок
из общего числа ссаживаемых могли нормально (без помощи человека) родить
живых телят, т.е. налицо определенные репродуктивные барьеры. В пользу их
существования свидетельствует также почти вековой и в целом неудачный опыт
интродукции сибирской косули в ареале европейской. Уже давно замечено (Тигбек,
1951), что мелкие самки европейской косули, покрытые выпущенными крупными
самцами сибирской косули, погибали при родах крупного плода.
Все это, несмотря на возможность гибридизации и относительную фертиль-
ность гибридов, говорит о дифференциации европейской и сибирской косуль
на уровне видов.
В настоящем анализе, вероятно, следует упомянуть и об опытах по скрещиванию
особей разных подвидов сибирской косули (сибирского, тянь-шаньского и
дальневосточного) между собой и с европейской косулей (Соколов, Громов, 19856;
Громов, 1986а). Животные для этого эксперимента были доставлены в Москву
Дальневосточным зоокомбинатом (Хабаровск), зообазой г. Фрунзе и
Восточносибирским зоокомбинатом (Иркутск). Точный район отлова зверей в
сопроводительных документах не указывался. Последней организацией они могли быть
отловлены в обширном регионе: в Иркутской или Читинской областях или в Бурятии.
За исключением Иркутской области, все остальные (включая Туву, Дальний Восток
и Киргизию) входят в зону обитания тянь-шаньского подвида сибирской косули.
Самец, доставленный из Иркутска, имел хромосомный набор 2л = 78—82, явно
более сходный с кариотипом популяций тянь-шаньского подвида, чем сибирского.
Специально исследованный нами хромосомный набор косуль (3 66 и 3 9?) из
Иркутской области оказался иным — 2л = 74. Кроме того, перед началом опытов
по гибридизации намеченные сибирские косули из разных регионов содержались
нами вместе (для изучения поведения) и несколько самцов покрывали самок
поочередно (Соколов, Данилкин, 1981), а приплод, также неясного происхождения,
впоследствии использовался для гибридизации.
Следовательно, сообщение о скрещивании особей разных подвидов
сибирской косули между собой следует принимать с осторожностью. По этой причине
желательно критически отнестись и к данным хромосомного анализа, приведенным
в работе Соколова и Громова (1988).
Очевидно, что необходимо в расширенном варианте повторить эксперименты,
58
обратив особое внимание на скрещивание европейской косули с особями сибирского
подвида и последних — с особями тянь-шаньского подвида. Крайне важно эти
опыты проводить при тщательном цитогенетическом, биохимическом и иммунном
контроле и при обязательной паспортизации скрещиваемых особей.
Поскольку в эксперименте можно считать доказанной возможность частичной
гибридизации европейской и сибирской косуль, возникает вопрос о гибридизации
в природных популяциях. Гибридные особи могли вполне появляться на Северном
Кавказе и на Русской равнине, где еще в начале нашего века сохранялись
небольшие популяции сибирской косули (см. главу "Ареал"). "Зона гибридизации"
образовалась в настоящее время на стыке ареалов европейской и сибирской косуль
на правом берегу Волги. Гибриды, вероятно, существовали в Подмосковье и
в других районах, где производились искусственные выпуски сибирской косули
в ареале европейской.
Какова судьба популяций европейской и сибирской косуль в "зоне гибридизации"?
Судя по приведенным выше результатам экспериментов, здесь следует ожидать
повышенную смертность самок европейских косуль при родах крупного гибридного
плода; низкую плодовитость самок, еще более низкую у гибридов; значительное
количество мертворожденных, т.е. смешанная популяция будет иметь очень низкий
уровень воспроизводства. При численном преобладании особей сибирской косули,
фенотип популяции будет "сибирским", при большем числе особей европейской
косули — "европейским", но хромосомный набор — "сибирского" типа.
Моделирование динамики численности "смешанной" популяции европейской и
сибирской косули дает следующий результат. Интродукция нескольких десятков
сибирских особей в ареале европейской косули приводит со временем к полному
исчезновению первых, что подтверждается неудачной в целом практикой расселения.
Локальные популяции сибирской косули, оказавшиеся в окружении популяций
европейского вида, в течение нескольких десятилетий обречены на замещение их
европейскими. Такая ситуация, по-видимому, имела место в Са'марском лесу
в Днепропетровской области. Через 30—40 лет после иммиграции сюда европейской
косули локальная популяция сибирских косуль фактически утратила свои типичные
черты и по морфометрическим и биохимическим параметрам стала сходной с
европейской, хотя генотип этой популяции все еще несет признаки сибирской формы—
В-хромосомы (Соколов, Данилкин, 1981; Данилкин, 19856; Соколов, Шурхал и др.,
1986).
В "зоне гибридизации" при наложении естественных ареалов европейской и
сибирской косуль модель предсказывает процесс "самоуничтожения" смешанной
популяции (при тех условиях обитания, в которых одновидная популяция заведомо
не депрессируется), причем более быстрая депрессия численности проявляется
в популяции европейской косули. При высоких коэффициентах выживания следует
ожидать смещения границ ареалов европейской и сибирской косуль на запад, т.е.
сибирская форма при прочих равных условиях может вытеснять европейскую.
Однако процесс этот очень медленный из-за постоянного притока мигрантов
с той и другой стороны, да и высокие коэффициенты выживания здесь вряд ли
реальны, что, вероятно, будет "удерживать" зону гибридизации на месте. Это
явление известно и для других видов животных (Майр, 1974).
59
Таксономия Capreolus
в свете современных исследований
Что представляет собой тот или иной вид животных, в особенности с позиций
эволюционной экологии? Абсолютного критерия вида не существует, поэтому
в каждом конкретном случае обосновывают доказательства или опровергают
дискретность ранга группировки (Майр, 1968, 1974; Тимофеев-Ресовский и др.,
1977; Haffer, 1986). Из классической систематики известно, что вид должен
отличаться от других категорий биологической самостоятельностью и морфофизиоло-
гической уникальностью, а также отвечать тройному критерию: 1) генетическая
изолированность(нескрещиваемость), 2) хиатус (прерыв постепенности изменения
признаков) между самыми близкими видами и 3)самостоятельность ареала.
С. С. Шварц (1980) справедливо считал, что четвертый критерий вида — его
экологическая специфичность, приспособленность к определенным условиям среды.
Новый вид отличается от исходной популяции предкового вида более совершенной
приспособленностью, которая определяется не только ее функциональной
эффективностью, но и энергетической стоимостью. Ведущую роль в приспособлении
видов к определенным условиям играют глубокие биохимические изменения.
Основу различий между видами животных он видел в биохимических отличиях,
в тканевых перестройках, определяющих специфику их взаимосвязи с окружающей
средой. Некоторые из его положений нам представляется необходимым привести
дословно: «...нет буквально ни одного вида (по крайней мере среди высших
позвоночных), морфофизиологические особенности которого не распространялись
бы и на тканевый уровень. Все формы, удовлетворяющие тем требованиям, которые
предъявляются современной систематикой к видам (важнейшее из них —
репродуктивная изоляция), специфичны уже на тканевом уровне. В этом отношении между
видом и резко выраженным подвидом различие принципиальное...
Вид — это подвид, существенно отличающийся на тканевом уровне; мера
существенности — тканевая несовместимость».
В последние десятилетия большинство специалистов принимают биологическую
концепцию вида, важнейшим критерием которого является репродуктивная
изоляция. Из этой концепции следует, что невозможность получения гибридов при
скрещивании двух форм или их стерильность могут рассматриваться как
безусловные доказательства их дифференциации на уровне вида. Однако плодовитость
гибридов не' может служить доказательством принадлежности родительских форм
к одному виду из-за существования многих других изолирующих механизмов,
поддерживающих чистоту вида в природе (Майр, 1968, 1974; Мейер, Г986).
По уровню репродуктивной изоляции в какой-то мере можно оценить степень
филогенетической близости видов. При скрещивании представителей разных родов
гибриды обычно стерильны, если они могут получиться (Gelder, 1977). В ранг
"подрода" чаще возводят те формы, которые при скрещивании дают гибриды
бесплодные или с низкой продуктивностью. В случаях, когда гибриды фер-
тильны, но отличаются низкой интенсивностью размножения и повышенной
смертностью, родительские формы могут рассматриваться как "хорошие виды" (Хавесон,
1960).
Европейская и сибирская косули, как показано выше, по этому критерию
относятся более чем к "хорошим" видам. Отметим для сравнения, что у других оленей
межвидовая гибридизация — явление не редкое. Описаны гибриды лани и благо-
60
родного оленя, пятнистого и благородного, белохвостого и чернохвостого оленей
и другие, причем многие гибриды получаются легко, вполне плодовиты и образуют
гибридные популяции, существующие в природе (Стекленев, 1975, 1986; Wishart,
1980; Barto's et al., 1981; Harrington, 1985b; Чегорка, 1986).
Однако, по выражению Шварца (1980), «виды не потому виды, что они не
скрещиваются, а они потому не скрещиваются, что они виды». Что же обособляет
европейскую и сибирскую косуль как виды? Бесспорно, что они имеют самостоятельные
ареалы (см. рис. 22, 25) и морфофизиологическую уникальность. По морфо-
метрическим параметрам они достаточно четко дифференцированы: с высокой
достоверностью можно отличить каждую особь европейской косули от каждой
сибирской по краниометрическим показателям. Четко выражен прерыв
постепенности изменения признаков у граничащих популяций, что также указывает на
видовые различия.
Как показано выше, они значительно (на уровне видов) обособлены и по
цитогенетическим, иммунохимическим и биохимическим показателям, т.е.
существенно отличаются на тканевом уровне и, видимо, уже существует
определенная тканевая несовместимость. Сибирская косуля явно лучше европейской
приспособлена к более суровым условиям жизни, особенно к глубокоснежью и
низким длительным температурам, и, вероятно, отличается и на энергетическом
уровне, определяемом биохимическими изменениями.
Это убедительно подтверждается найденным разным общим уровнем
метаболизма у них (Граевская и др., 1980, 1988). Известно, что основные функции
метаболизма у организмов направлены на образование высокоэнергетических
соединений, необходимых для обеспечения клетки энергией. Одним из регуляторов,
способствующих приспособлению организма к меняющимся условиям среды,
являются гормоны-катехоламины. Их уровень в надпочечниках европейской
косули в 3 раза выше, чем у сибирской, но в крови наблюдается обратное
соотношение. Концентрация глюкозы в крови достоверно различается. Содержание
гликогена в печени европейской косули в 2 раза ниже, чем у сибирской, но
активность ключевого фермента углеводного обмена глюкозо-6-фосфатазы у первой
в 7 раз выше, а щелочной фосфатазы — в 3 раза ниже. Индекс регуляции углеводного
обмена и индекс энергетического резерва у европейской косули примерно в 5 раз
выше, но индекс энергетического уровня оказался ниже в 4 раза. Для поддержания
равновесия между внутренней и внешней средой ей, видимо, требуется меньше
энергозатрат по сравнению с сибирской.
В то же время по многим морфометрическим и морфологическим показателям
европейская косуля является как бы уменьшенной копией сибирской. На близкое
филогенетическое родство указывает также в целом сходный набор хромосом,
антигенов и белков и значительное сходство в поведении и экологии этих форм
(см. ниже).
Таким образом, практически по всем критериям классической систематики,
так и по критерию экологической специфичности европейская и сибирская косули
дифференцируются как разные, но филогенетически весьма близкие виды —
Capreolus capreolus L. и Capreolus pygargus Pall.
Подвидовое деление косуль, принимавшееся ранее, также явно не соответствует
действительности и требует пересмотра. Четкого критерия подвида, как и вида,
к сожалению, пока не существует. По определению Майра и др. (1956), подвиды —
это географически обособленные группы местных популяций, таксономически
61
отличающиеся от других таких же подразделений вида. В последнее время подвид
чаще трактуется как группа особей, занимающих определенный экологический
или географический ареал и отличающихся от других особей этого вида по одному
или нескольким диагностическим признакам, присущим всем или большинству
особей. В качестве подвидов выделяют в основном "крупные" и "ясные" группы
особей, занимающих обширные ареалы (Тимофеев-Ресовский и др., 1977).
Выделение таких групп в ареале европейской косули проблематично. Сибирская
косуля, напротив, четко разделяется на две "крупные" и "ясные" таксономически
отличающиеся группы особей, занимающих обширные ареалы, разделенные
географической преградой — горными хребтами Алтая, Западного и Восточного Саян,
оз. Байкал, Становым нагорьем и Становым хребтом (см. главу "Ареал"). Под-
видового ранга заслуживают идентичные по многим показателям популяции
Предуралья, Зауралья, Северного Казахстана, Западной и Восточной Сибири
(до Байкала) и, скорее всего, Якутии и сходные между собой популяции Южного
Казахстана, Средней Азии, Монголии, Забайкалья, Дальнего Востока и.Китая.
Популяции Алтая, Восточного Казахстана и, видимо, юго-востока Иркутской
области находятся на стыке ареалов, и здесь вполне вероятна гибридизация части
особей сибирского и тянь-шаньского подвидов.
Подводя итог изучению систематики косуль, можно констатировать, что
комплексные исследования, проведенные нами в последние годы, значительно
расширили знания о видообразовании у этих животных и изменили представления
об их таксономическом статусе. На данном этапе работы таксономия косуль
представляется следующим образом:
Capreolus capreolus Linnaeus, 1758 — европейская косуля
Диагноз. Размеры мелкие — средняя длина тела в разных популяциях 107—126 см,
масса тела 22—32 кг, кондилобазальная длина черепа 180—200 мм, длина нижнего
зубного ряда 58—66 мм. Рога обычно не длиннее 27—30 см, их размах чаще не более
10—15 см. Слуховые пузыри на черепе небольшие и не выступают из барабанной
ямки. Окраска головы в летней шерсти и метатарзальных желез обычно
значительно темнее окраски туловища. В-хромосомы отсутствуют. В отличие от
сибирской косули имеются два дополнительных антигена сыворотки крови.
Распространение. Европа до Волги и Передняя Азия (см. рис. 25).
Состав. Вид монотипичен.
Capreolus pygargus Pallas, 1771 — сибирская косуля
Диагноз. Размеры средние и крупные — средняя длина тела в разных популяциях
126—144 см, масса тела 32—48 кг, кондилобазальная длина черепа 201—231 мм,
длина нижнего зубного ряда 71—76 мм, рога обычно длиннее 27—30 см, их размах
превышает 15—20 см. Слуховые пузыри на черепе крупные и заметно выступают
из барабанной ямки. Окраска головы в летней шерсти и метатарзальных желез
в основном не отличается от цвета туловища. В хромосомном наборе присутствуют
В-хромосомы.
Распространение. Восток Европы, Азия.
Состав. Включает два подвида.
62
Capreolus pygargus pygargus Pall., 1771 — сибирская косуля
Диагноз. Размеры крупные — средняя длина тела в разных популяциях 140—
144 см, масса тела 41—48 кг, кондилобазальная длина черепа 223—231 мм, длина
лицевой части 125—127 мм, длина носовых костей 78—80 мм, длина нижней челюсти
189—194 мм, расстояние между внешними сторонами осевых стержней рогов чаще
превышает 74 мм. В хромосомном наборе обычно не более 4 В-хромосом: 2и =
= 70+ (1-4).
Распространение. Поволжье, Урал, Северный Казахстан, Западная и Восточная
Сибирь (до Байкала), Алтай и Якутия (см. рис. 25).
Capreolus pygargus tianschanicus Satunin, 1906 —
тянь-шаньская косуля
Диагноз. Размеры средние — средняя длина тела для разных популяций 126—
137 см, масса тела 32—40 кг, кондилобазальная длина черепа 201—218 мм, длина
лицевой части 110—120 мм, длина носовых костей 68—76 мм, длина нижней челюсти
168—183 мм, расстояние между внешними сторонами осевых стержней рогов
обычно менее 74 мм. Хромосомный набор в основном содержит более 4
В-хромосом — 2и = 70 + (5—14). Окраска более ярких тонов, чем у сибирской косули.
Распространение. Юг Казахстана, Тянь-Шань, Южный Алтай, Монголия,
Забайкалье, Дальневосточные области России, Корея, Северный и Центральный
Китай (см. рис. 25). Не исключено проникновение в Якутию и Предбайкалье.
Глава третья. Ареал
История ареала рода Capreolus
На раннем этапе своего существования в конце плиоцена—начале плейстоцена
косули, близкие к современным формам, судя по ископаемым находкам (см. главу
"Филогения..."), обитали в Центральной и Западной Европе, включая
Великобританию (но не смогли заселить острова: Ирландию, Корсику и Сардинию, уже
отделившихся к тому времени от материка), занимали Переднюю Азию, юг Сибири
и Дальний Восток. Малочисленность костных остатков Capreolus этого периода,
к сожалению, не дает полной картины их распространения, и о ней можно судить
лишь весьма приблизительно (рис. 21).
В плейстоцене ареал рода Capreolus явно значительно расширился (рис. 21).
Косули современного типа заселили Крым, Приазовье, Нижний Дон, Северный
Кавказ (Громов, 1948; Верещагин, 1959). На Урале и в Сибири они продвинулись
далеко на север, достигнув в Предуралье верховьев Печоры, а в Сибири — Якутии
(Ермолова, 1963, 1978; Кузьмина, 1971; Вангенгейм, 1977; Алексеева, 1980; и др.).
В голоцене (см. рис. 21) по мере потепления и отступления ледников начинается
расширение зон лесов, что, в свою очередь, влечет за собой вспышку численности
лесных копытных и экспансию их ареалов (Бибикова, 1975; Верещагин,
Барышников, 1980а). Европейские косули проникают в Скандинавию через существующую
в послеледниковый период перемычку из Дании в Южную Швецию (Stubbe,
Passarge, 1979), расселяются по Русской равнине. Сюда же с востока
распространяются и сибирские косули. Последние в послеледниковое время гораздо чаще
стали встречаться среди добычи древних охотников Азии, а на Урале, в Пред-
байкалье и в Якутии они становятся основными или одними из основных объектов
охоты (Окладников, 1950; Пидопличко, 1956; Цалкин, 19626; Паавёр, 1965;
Ермолова, 1978; Косинцев, 1981).
Ареал рода в историческое время вплоть до второй половины XX в.
практически полностью совпадает с их распространением в голоцене. Существенное
приращение ареала происходит лишь в Скандинавии в самые последние
десятилетия (см. ниже).
Накопленный, фактический материал с учетом современного распространения
косуль позволяет достаточно точно провести контур естественного максимального
(восстановленного) ареала рода Capreolus (рис. 22).
На северо-западе он охватывает Англию и север Скандинавского полуострова,
выходит на востоке его к Кандалакше, огибает с юга Онежское озеро и по линии
Кострома, Нижний Новгород, Вятка доходит до западных склонов Уральского
хребта, поднимается к верховьям Печоры и Северной Сосьвы, идет по Оби, Кети,
Ангаре, верховьям Нижней Тунгуски, левобережью Лены и Вилюю до Верхоянского
хребта и отсюда спускается на юг к низовьям Амура, захватывает Приморье
и Корею. Южная линия восстановленного ареала с востока на запад проходит
64
по центральным районам Китая к Тибету, поднимается по его склонам на север и,
огибая пустынные районы, уходит круто на северо-восток к западным отрогам
Большого Хингана (Гептнер и др., 1961); затем по Керулену, югу Хэнтэя и Хангая
выходит к Монгольскому Алтаю, огибает его и, минуя с севера пустынную
Джунгарию, подходит к Тянь-Шаню1.
На Тянь-Шане линия восстановленного ареала идет по Алайскому хребту,
захватывает Вахшскую долину, по границе с Афганистаном и самыми северными
районами Ирана (севернее Эльбурса) и Ирака через Сирию и Израиль выходит
к Средиземному морю и далее охватывает Переднюю Азию и Европу.
За пределами естественного восстановленного ареала европейская косуля в XX в.
была выпущена в Северной Америке в штате Техас (Bump, 1970).
Движение ареала и численность косуль в плейстоцене и голоцене всецело
определялись природными факторами. Антропогенный фактор начинает оказывать
всевозрастающее воздействие лишь вж втором тысячелетии нашей эры, особенно
в конце его.
Вплоть до XIX столетия н.э. исторический ареал рода, скорее всего, был
сплошным. Еще в XVI—XVIII вв. по свидетельству очевидцев леса в Молдавии
"кишели" косулями, а на Украине добывали их "неимоверное количество" (Кири-
ков, 1959, 1966, 1983; Аверин, 1960а,б). С XVIII в. из-за сведения лесов, распашки
степей, скотоводства и интенсивной охоты численность и ареал косуль начинает
сокращаться во многих районах. Впервые наиболее ощутимо эти явления
проявились в Западной Европе: к началу века звери уцелели лишь в лесах на севере
Шотландии, в Швеции их осталось около 100 особей в самой южной провинции,
в Швейцарии они были истреблены полностью (Шарфф, 1918; Meikle, 1975; Stubbe,
Passarge, 1979).
В Восточной Европе и в Азии тенденция к сокращению ареала проявляется
несколько позже — в XIX и начале XX в. (Кириков, 1959). В середине XIX столетия
косули были уничтожены в Центральночерноземных областях (Барабаш-Ники-
форов, 1957), в Поволжье, на юге Украины, что привело к разрыву ареала. К концу
века их не стало во многих районах на северо-востоке Европы, однако в Псковской
области даже в начале XX столетия косуля составляла предмет значительного
промысла (Соловьев, 1922). Последняя косуля в Ярославской области была
убита в 90-х годах прошлого века (Лавров, 1929). Из-за неумеренного промысла
и обилия хищников исчезла она и в большинстве районов на севере Казахстана
и в Западной Сибири (Шухов, 1929).
В это время становится очевидным и обратный процесс — восстановление
численности и ареала европейской косули в Западной Европе, и в первую очередь
в Шотландии (Meikle, 1975).
' Учитывая, что в четвертичном периоде климатичес кие и ландшафтные условия претерпевали сложные
и неоднократные изменения и существенно отличались от теперешних (Марков и др., 1965), а также
принимая во внимание потенциальные возможности расселения косуль (см. гл. 5), южная граница
восстановленного ареала в Китае, возможно, шла несколько иначе: по северным склонам Тибета сразу же
на Тянь-Шань.
S. Европейская и сибирская косули
65
ф
О)
Рис. 21. Распространение рода Capreolus и близких к нему форм в четвертичном периоде (реконструкция)
/ — верхний плиоцен—нижний плейстоцен; 2 — плейстоцен; 3 — голоцен
Рис. 22. Максимальный ареал рода Capreolus Gray и максимальные видовые ареалы С. capreolus L. (/) и С. ру-
gargus Pall. (2)
Ареал в XX веке
Наибольшие изменения исторического ареала рода Capreolus произошло в XX в.
в основном на территории б. СССР и на Скандинавском полуострове (рис. 23). В
самом начале его из-за хищнического уничтожения (см. гл. 11) и все увеличивающегося
антропогенного воздействия на биотопы ареал настолько сократился, что
оказался разбитым на несколько частей (западную, крымскую, кавказскую, уральско-
западносибирскую и сибирско-дальневосточную) практически полностью
изолированных одна от другой (Гептнер и др., 1961)2. Между ними сохранилось лишь
несколько отдельных мелких очагов обитания: в Самарском лесу Днепропетровской
области, в Черном лесу Кировоградской области, близ г. Ставрополя, в
Воронежской, Липецкой, Тульской областях и, вероятно, в, некоторых районах Западной
Сибири и Северного Казахстана (Лавров, 1929; Залесский, 1934; Белышев, 1934;
Кузнецов, 1948; Насимович, 1955; Барабаш-Никифоров, 1957; Гептнер и др., 1961).
Косули фактически были истреблены на огромном пространстве от Днепра
до Урала, в Западной Сибири, в Северном и Центральном Казахстане. Значительно
отодвинулась на юг северная граница их распространения в Восточной Сибири.
Своей кульминации депрессия ареала достигла в 20-е годы. Во многом этому
способствовали войны 1914—1920 гг., вооружившие массы людей. Очень много
дальнобойного оружия оставалось у населения и помеле войны.
Однако уже с 30-х годов становится отчетливо заметной тенденция к
расширению ареала, что связано с уменьшением промысла, потеплением климата
и малоснежностью зим, интенсивным освоением темнохвойных лесов и заменой
их на лиственные породы. Возможно, существовали и другие стимулы
естественного расселения, выделить которые не представляется возможным. Расселение
косуль шло очень быстро во всех направлениях. Уже к середине 50-х годов
наблюдается соединение изолированных частей ареала в Сибири. Однако западный,
крымский и кавказский участки по-прежнему остаются отделенными один от
другого и от области распространения сибирской косули.
История ареала в первой половине XX в. очень подробно описана в сводке
В.Г. Гептнера и др. (1961) и в целом в несколько измененном и дополненном
виде отражена на рис. 23. Со второй половины XX в. его движение проходило
следующим образом.
Европейская косуля. В 60—70-е годы благодаря охранным и биотехническим
мероприятиям и отсутствию крупных хищников численность европейской косули
в Западной и Центральной Европе сильно возросла, что привело к занятию ею
всех пригодных биотопов (в том числе полевых) и к очень существенному
продвижению на север, особенно на Скандинавском полуострове. В начале века она
обитала только на юге Швеции примерно до 60° с.ш. (Шарфф, 1918), к середине его
достигла 65° (Гептнер и др., 1961), а к настоящему времени заходы ее отмечены до
70—71° с.ш., т.е. на северной оконечности полуострова, далеко за пределами
исторического ареала. В 1970 г. косуля впервые проникла на территорию
Финляндии. Расселение шло по трем направлениям: в южную часть Финляндии из
СССР, в северную — из северной Норвегии и вдоль побережья Ботнического
залива из Швеции (Pulliainen, 1980) (рис. 24).
2 Проанализировав первоисточники, мы пришли к выводу, что для выделения среднеазиатской
части нет достаточных оснований.
68
Рис. 23. Изменения ареала косуль в XX в. на территории б. СССР и на севере Европы
Распространение: / — в период максимальной депрессии ареала в начале столетия; 2 — в середине века
(по: Гептнер и др., 1961; с изменениями и добавлениями); 3 — во второй половине 80-х годов
О)
СО
Рис.24. Современный ареал косули в Западной Европе (по: Lemann, Sagesser, 1986; Prior, 1987, с добаВ'
лениями; см. текст)
Заходы обозначены кружками
В эти десятилетия расширение исторического ареала европейской косули
наблюдалось и на Северо-Западе Европы. В 60-е годы благополучные зимовки и дальние
заходы стали обычны в Карелии, где звери регистрировались в окрестностях
Петрозаводска, у оз. Пюхеярви, в Сортавальском районе (Данилов, 1974). В 1970 г.
несколько особей благополучно перезимовали в Пудожском районе, в период
с 1971 по 1975 г. животных неоднократно видели в Кондопожском и Прионежском
районах. В 1974 г. косули появились (вероятно, из Скандинавии) в Кандалакшском
70
районе Мурманской области (см. рис. 24). Однако основная граница их
распространения на Северо-Западе СССР в эти годы шла по южному побережью Финского
залива на г. Пушкин и далее на Тосно — Кириши — Бокситогорск в
Новгородскую область (Верещагин, Русаков, 1979). Следует отметить, что северная граница
ареала в этом регионе носит нестабильный пульсирующий характер и во многом
поддерживается за счет иммигрантов из Прибалтики.
Практически после векового отсутствия были отмечены заходы косуль в
Костромскую область из Ярославской (Сапоженков, 1981).
Такими же быстрыми темпами, как и на севере, расширение ареала европейской
косули шло на юге Украины, что привело к воссоединению его западной
и крымской частей. В 60-е годы животные заняли практически все лесные
районы Украины и к концу их широко распространились по Причерноморским
степям, достигнув берегов Черного моря (Гурский, Назаренко, 1969; Гурский,
1975). В 1973 и 1974 гг. зверей неоднократно отмечали в северных районах Крыма,
возможно зашедших сюда вдоль лесополос по трассе Северо-Крымского канала
из Херсонской области (Дулицкий и др., 1975).
Особенно интенсивно европейская косуля рассеялась в восточном направлении.
В конце 50-х годов она стала заселять Тульскую, Орловскую, Липецкую,
Ростовскую и Волгоградскую области, единичные заходы отмечены в Тамбовской
и Саратовской областях. В 60-е годы на Востоке Европы происходило медленное
нарастание численности, в 70-е —уже более быстрое (табл. 12), чему способствовало
улучшение охраны зверей. Естественным путем животные заняли практически
все пригодные для жизни биотопы в этих областях. Проводимое здесь же
искусственное расселение косуль за небольшим исключением не дало ожидаемого результата
(Фадеев, 1981).
В 80-е годы европейская косуля с низовьев Дона проникает в Предкавказье
на Ставропольскую возвышенность в район обитания локальной популяции
сибирской косули, образовавшейся в результате искусственного расселения
(Емельянов, Рыбалов, 1968) и граничащей с областью распространения европейской
косули на Северном Кавказе, т.е. кавказская часть ареала снова сомкнулась с
основным ареалом рода (см. рис. 27). Однако связь эта пока непрочная и
поддерживается в основном заходом отдельных особей. Соединения ареала европейской
косули в Предкавказье пока не произошло.
Сибирская косуля. С середины века заметно интенсивное продвижение сибирских
косуль с Урала на запад. В самом конце 50-х и начале 60-х годов единичные заходы их
с востока отмечены в Куйбышевскую (Белянин, 19806) и Ульяновскую области
и Татарию. В 1960 г. 4 косули зашли в Кировскую область, где прожили полгода
у с. Константиновка Малмыжского района. Зимой 1971 г. в центре области (у с. Ады-
шево) был пойман самец, в 1972 г. одна косуля провела всю зиму в пойме р. Воя
у впадения ее в Вятку, в 1974 г. животные появились на юге области в Верхне-
Полянском районе (Злобин, 1975).
Началом массового заселения косулями левобережных областей Поволжья
следует считать конец 60-х годов. Зимой 1967/68 г. несколько сот особей
мигрировали с западных склонов Уральских гор в Альметьевскии и Бугульминский
районы Татарии. Часть их, возможно, вернулась назад или была истреблена, а часть
осталась здесь жить. В эти же годы заметное увеличение численности сибирских
косуль отмечено и в других левобережных областях Поволжья (см. табл.12).
В конце 60-х годов сибирские косули, расселяясь на запад, достигли Волги;
71
Таблица 12. Динамика численности косуль (особей) в центральных и восточных областях
европейской части СССР в 1960—1970-е годы*
Область, республика
1961
1962
1963
1964
1965
1966
1967
1968
Белгородская
Волгоградская
Воронежская
Калужская
Куйбышевская
Курская
Липецкая
Московская
Орловская
Пензенская
Ростовская
Рязанская
Саратовская
Смоленская
Тамбовская
Татария
Тульская
Ульяновская
Итого
700
50
413
100
Ед.
70
50
300
25
25
40
—
20
300
—
20
67
Ед.
2180
700
50
400
60
30
90
45
420
40
25
100
—
26
400
—
Ед.
100
Ед.
2486
800
180
400
70
60
300
80
330
50
50
175
—
10
400
Ед.
20
200
Ед.
3125
1500
200
400
70
60
600
55
380
50
50
150
—
10
200
Ед.
20
200
Ед.
3945
1800
200
800
130
600
600
65
460
40
50
175
—
10
180
Ед.
20
40
Ед.
5170
1800
200
900
200
600
600
70
510
40
33
200
20
10
300
Ед.
20
20
Ед.
5503
2000
200
1200
260
150
600
45
512
40
ПО
220
15
10
300
Ед.
20
30
Ед.
5710
2100
200
1400
150
230
1200
60
55
100
250
Ед.
23
290
Ед.
35
30
5
6640
причем в районе Жигулевского заповедника на правом берегу они появились в 1960 г.
(Белянин, 1975). В Саратовской области на правый берег Волги косули вышли
в 1971 г„ в Ульяновской — в 1972 г. В эти же годы они стали заселять
Волгоградскую область и заняли практически все крупные острова по Волге: Колтовской,
Подлесный, Орловский, Семеновский и др. Во время весеннего разлива звери
с островов переплывали в основном на правый берег. В середине 70-х годов
в Красноармейском районе Саратовской области отмечены переходы их по льду
как с левого берега на правый, так и в обратном направлении3.
К концу 70-х годов сибирские косули продвинулись еще дальше на запад и
достигли восточных районов Мордовии и Пензенской области и вышли к рекам
Хопер и Дон. Здесь, по Хопру и Дону в Волгоградской области, западная и урало-
сибирская части ареала рода косуль соединились, т.е. снова произошло частичное
наложение видовых ареалов европейской и сибирской косуль (см. рис. .25).
В Зауралье, по данным Л.П. Сабанеева (1872, 1875), в середине прошлого века
северная граница ареала сибирской косули простиралась до 58°30' с.ш. Из-за
интенсивного промысла и многоснежных зим в начале XX в. численность зверей здесь
резко сократилась. В первую очередь прекратили существование наиболее
малочисленные северные популяции, и граница ареала снова сдвинулась южнее примерно
на 2° (Демидов, 1940). В 20-е годы становится заметным новое продвижение их
к северу (Лавров, 1929), чему способствовало ограничение промысла, интенсивное
3 В конце 60-х — начале 70-х годов в Приволжских областях практиковалось также искусственное
расселение косуль.
72
1969
2100
90
1800
250
290
1450
40
560
60
200
320
Ед.
27
250
Ед.
80
30
5
7552
1970
1170
120
1800
250
690
1850
44
530
55
300
450
Ед.
58
500
Ед.
130
120
15
8082
1971
1830
180
3020
500
1530
2200
42
430
55
300
580
Ед.
210
530
Ед.
190
100
60
11757
1972
1120
300
1930
670
2800
1300
46
520
30
350
675
Ед.
230
360
Ед.
200
100
122
11463
1973
1700
450
2400
600
2900
1200
60
620
40
450
1000
Ед.
600
400
Ед.
300
300
200
12640
1974
1750
600
2200
1000
3500
1450
100
800
50
570
1200
Ед.
800
700
Ед.
1100
500
500
16770
1975
2200
700
2400
2000
3600
1500
150
900
100
100
1800
Ед.
610
700
60
500
640
600
18110
1976
2500
900
1800
2100
4400
1500
300
1100
850
100
1900
Ед.
800
800
100
700
800
700
21050
1977
1700
1100
2000
2000
4000
1450
280
1600
820
100
2200
Ед.
600
800
200
1200
810
750
22410
1978
1300
2100
3500
4000
1820
300
2300
900
200
2650
Ед.
1500
900
130
1700
610
400
26010
освоение темнохвойных лесов и ряд малоснежных зим. В первой половине 30-х годов
животные вновь появились к северо-востоку от р. Сосьва, в 1933 г. замечены на
р. Тавда (Демидов, 1940), а чуть позднее они проникают в бассейн Конды,
практически до южной границы Кондо-Сосьвинского заповедника и на северо-восток
к р. Обь примерно до 61° с.ш. (Скалой, 1946). С 1918 г. за 20 лет ареал косули
расширился на север вдоль восточных склонов Уральского хребта более чем на 300 км
(см. рис. 23), а одиночные заходы зверей наблюдались вплоть до 63—64° с.ш.
(Гептнер и др., 1961). По западным склонам Урала северная граница ареала также
продвинулась на север до бассейна Камы, а отдельные заходы наблюдались вплоть
до 62°30'с.ш. В многоснежные зимы 1940/41 и затем 1945—1947 гг. косули исчезли
из Висимского района и бассейна Чусовой (Насимович, 1955).
Таким образом, на Северном Урале к середине века исторический ареал
практически полностью восстановился, но позднее из-за интенсивной добычи,
многоснежных зим его северная граница снова заметно отодвинулась к югу. В конце
70-х годов она стала такой же, какой была в конце прошлого века (Киселев, 1979).
Аналогичная картина движения ареала наблюдалась и в Западной Сибири.
В прошлом веке северная граница ареала в этом районе, вероятно, проходила
между 56 и 57° с.ш. В середине 80-х годов большие стада косуль встречались
у Тюмени, к востоку от р. Тобол на территории бывших Юргинского и
Ялуторовского округов, но их не отмечали на правобережье Иртыша (Словцов, 1892). В
Омской области звери обитали до 1905 г., затем были полностью истреблены. Лишь
в середине 20-х годов они появились к северо-западу от Омска в Крутинском районе
и в южной части бывшего Тарского округа (Залесский, 1934; Корш, 1941).
73
Однако в 20-х и начале 30-х годов в Прииртышье косули были не так редки, как
принято считать. В это время они жили в Курганских борах, на западе и северо-
западе Тарского и Омского округов, в Большереченском районе как по правому,
так и по левому берегу Иртыша, встречались в Омском районе и боровых массивах
между нижними течениями Томи и Оби (Шухов, 1929; Белышев, 1934), с середины
30-х годов стали обитать в Ёарабинской лесостепи (Зверев, 1937). В начале 50-х годов
косули встречались уже севернее Новосибирска и Кемерово, южнее Томска
(Янушевич, Благовещенский, 1952; Насимович, 1955).
Незамеченными остались сведения И.П. Лаптева (1955, 1958). Тем не менее он
приводит наиболее дальние заходы косуль на север Западной Сибири, что в корне
меняет представление о границах восстановленного ареала в этом регионе. Зимние
заходы косуль в конце 40-х — начале 50-х годов зарегистрированы в район дельты
р. Яя, на р. Чулым возле с. Куяново, в 30 км севернее с. Ново-Мариинка, в пойму
Оби в 20 км севернее с. Каргасок, к с. Усть-Балык Сургутского района, в нижнем
течении р. Юган и по р. Вогулка в 20 км выше с. Березово, т.е. на 300—500 км
севернее обычно проводимой границы ее распространения. В этих же районах косули
отмечены и в летнее время: в бассейне р. Салым, окрестностях сел Белогорье,
Троицкое и Леушки (примерно у 62° с.ш.) и в долине р. Вогулка (см. рис. 26). Позднее
были зарегистрированы встречи косуль в Самаровском и Кондинском районах
Ханты-Мансийского автономного округа в пределах 61—62° с.ш. (Шаргаев, 1966).
Расширение ареала в Западной Сибири достигло своей кульминации к началу
60-х годов, и затем из-за браконьерства, распашки земель и перевыпаса скота
(Белов и др., 1980) он стал быстро сокращаться. Численность косуль к 1980 г. по
сравнению с началом 60-х годов в регионе сократилась почти в 10 раз, а площадь
ареала — в 2 раза (Царев, 1969; Николаев, 1982). Лишь в последние годы снова
стал заметен процесс увеличения их населения и восстановления ареала.
В Северном Казахстане в период наибольшей депрессии южную границу
распространения сибирской косули проводят от южной оконечности Урала на север
по восточным его склонам до Троицка, по правобережью Тобола до широты Кус-
таная и от Кургана на юг к Кокчетаву. Подробно ареал в началу века описан
Н.П. Лавровым (1929). С 30-х годов здесь явно становится заметным процесс
расселения зверей на юг (Антипин, 1941), к 50-м годам южная граница ее ареала
проходила уже от Уральска через бассейны рек Илека и Хобды, южнее
Актюбинска, между городами Иргиз и Тургай, охватывала с юга горы Улытау, Актау
и Ортау вдоль северного побережья Балхаша, огибая его, выходила на Джун-
гарский Алатау (Гептнер и др., 1961).
В начале 60-х годов численность косуль в Северном Казахстане заметно
возросла и звери начали распространяться на юг от Уральска в пойменные леса
и особенно далеко по пойме Урала. В начале 70-х годов звери были отмечены
на севере Гурьевской области (Павлов и др., 1974). К середине 80-х годов на
западе Казахстана они заселили Урдинский сосновый бор, расположенный южнее
Джаныбека, верховья Чижа, заходы ее отмечены повсеместно по р. Кутум на юг до
оз. Жалтырколь. В настоящее время косуля обитает в лесной даче Кара-Агаш
по р. Булдырты и других пригодных местах к востоку от р. Урал.
В Актюбинской области она занимает бассейн р. Хобда, по р. Илек распространена
на юг почти до Октябрьска и лишь немного не доходит до г. Хромтау. Отдельные
очаги обитания имеются по р. Уил (у пос. Уил), по р. Эмба северо-восточнее поселка
Эмба и рядом с поселком Иргиз. В Тургайской области самый южный локальный
74
участок обитания косуль — пески Тосынкум (Тусум) по левому берегу р. Тургай
(Лобачев, 1972; Чельцов-Бебутов, 1976). В целом в Центральном Казахстане южная
граница распространения сибирской косули в настоящее время совпадает с ее линией
в 50-е годы и лишь в районе к северу от Балхаша сдвинулась еще дальше на юг,
проходя теперь всего лишь в 50—100 км от его северного побережья (Капитонов,
1980). К северу от Балхаша по всему Центральному и Северному Казахстану
распространение косуль носит очаговый характер: они живут здесь в поросших
кустарником горах, по мелкосопочнику, долинам рек и озер и практически по всему
побережью Иртыша.
В Средней Азии изменения ареала в XX в. не были такими значительными, как
в Европе, на Урале, в Казахстане и Сибири. В настоящее время он почти такой же,
каким был, вероятно, в конце прошлого века, за исключением исчезновения очага
их обитания по Амударье, очевидно существовавшего до 30-х годов (Разевиг, 1909;
Зарудный, 1915; Лавров, 1929; Кашкаров, 1931; Флеров, 1935а, 1952; Гладков,
Никольский, 1935; Антипин, 1941; Кузнецов, 1948; Толстое, 1948; Корелов, 1956;
Ишунин, 1961; Айзин, 1969; Янушевич и др., 1972; Закиров, 1974; Слудский и др.,
1974). По сравнению с серединой века в последние десятилетия ареал несколько
расширился на юге региона (см. рис. 23).
На Алтае до 30-х годов сибирская косуля обитала практически по всей территории
автомной области, за исключением высокогорий и темнохвойных лесов (Дмитриев,
1938). К началу 40-х годов ее численность из-за неумеренной охоты сократилась,
однако за период войны восстановилась повсеместно, и в 50-е годы косуля снова
распространилась повсюду. В 60-е годы из-за перепромысла и непродуманных мер
по химизации сельского и лесного хозяйства численность зверей резко сокращается
и соответственно существенно уменьшается площадь ареала. Животные были
уничтожены на северо-востоке Алтая, в окрестностях Горно-Алтайска, в нижнем
и среднем течении р. Чуя и в других районах. Лишь в начале 70-х годов становится
заметным рост численности мигрирующих популяций и восстановление ареала
(Собанский, 1986).
В Восточной Сибири в начале века также из-за перепромысла и ряда снежных
зим северная линия ареала значительно отодвинулась к югу (Троицкий, 1930).
В 30-х годах она пересекала Енисей у 57° с.ш., далее на восток поднималась
до 58°, шла вдоль р. Чуна к Нижне-Илимску, затем поворачивалана юг к верховьям
Лены и Киренги и опять поднималась к северу, обходя северную оконечность
Байкала. К началу 50-х годов граница ареала на левом берегу Енисея продвинулась
к 58° с.ш., а на правобережье косуля практически полностью восстановила свой
исторический ареал. В 70-х годах она заселила правобережье Ангары, проникнув
в летнее время на 58° с.ш. (Ельский, 1983).
К северу от Байкала в плейстоцене косули обитали по р. Лене (Равский и др.,
1964), во времена неолита и бронзы они были широко распространены в Якутии;
многочисленные костные остатки обнаружены в районе г. Якутска (Окладников,
1950). Еще в XIX в. звери жили в верховьях Лены, забегая отсюда на Нижнюю
Тунгуску до вершины р. Непа (Маак, 1886). В конце XIX — начале XX в. северную
границу их распространения обычно проводили по 60—62° с.ш. (Лавров, 1927,
1929; Дементьев, 1933; Флеров, 19356; Подаревский, 1936), лишь изредка отмечая
заходы севернее в Сунтарскую излучину Вилюя и восточнее нижнего течения рек
Чары и Олёкмы. В 40-е годы косуль находят на Вилюе севернее 63° с.ш. и в юго-
восточной Якутии в Тимптонском районе (Скалой, 1946). Позднее более подробные
75
сведения дал В. И. Белык (1953), в основном по ним была составлена карта
максимального ареала косули в этом регионе в середине века (Гептнер и др., 1961).
В последующие годы были получены новые данные (Егоров, 1965; Попов, 1970;
Тавровский и др., 1971; Попов и др., 1980), которые вместе с материалами
Управления охотничье-промыслового хозяйства Якутии включены в карту современного
ареала.
В Забайкалье, на Дальнем Востоке и в Монголии изменения ареала сибирской
косули в историческое время не были существенными. Движение его в Китае,
к сожалению, не представляется возможным проследить.
Таким образом, в XX в. мы стали свидетелями максимальной депрессии ареала
рода Capreolus, разрыва его на отдельные части, затем практически полного
восстановления и даже расширения границ на севере Европы (см. рис. 23).
Видовые ареалы европейской
и сибирской косуль
Проанализировав литературные источники, в которых приводятся данные об
ископаемых находках косуль (см. гл. 1), мы пришли к убеждению, что с плейстоцена
отчетливо прослеживается существования по меньшей мере двух форм: мелкой
Capreolus capreolus в Европе и крупной Capreolus pygargus в Азии. Сведения о
распространении сибирской косули в Центральной Европе в плейстоцене и голоцене
вплоть до исторического времени (Kahlke, 1955; Raesfeld, 1956; Lehmann, 1958,1960),
вероятно, не соответствуют действительности. К.Л. Паавер (1965) на основании
остеометрических исследований показал, что размеры даже очень крупных костей,
найденных в Центральной Европе, все же уступают наибольшим показателям
позднеголоценовой европейской косули из Восточной Прибалтики. В то же время
сравнение последней популяции с голоценовыми остатками сибирской косули из
Зауралья показывает достоверные различия между ними (Косинцев, 1981).
Не исключено, что крупная форма aff. pygargus, найденная в пещерах Палестины,
Сирии и Ливана, обитавшая здесь со среднего плейстоцена до неолита (Верещагин,
1959), на самом деле представляет собой крупную форму европейской косули.
Плейстоценовые ледники на Русской равнине и трангрёссии Каспийского моря,
простиравшиеся по Волге далеко на север (Бадер, 1950; Верещагин, 1959; Марков
и др., 1965), вероятно длительное время служили существенной преградой для
расселения сибирской косули на запад. Лишь по мере отступления этих преград
в послеледниковое время она заселяет с востока Русскую равнину вплоть до
Днепра (Флеров, 1952) и, видимо, в позднем средневековье достигает Северного
Кавказа (Верещагин, 1959).
На Северном Кавказе две формы косуль могли жить вместе вплоть до начала
XX в. (Динник, 1910), до недавнего времени локальные популяции сибирской косули
обитали в Днепропетровской и Кировоградской областях (Браунер, 1915; 1923;
Мигулин, 1927, 1929; Шарлемань, 1937; Корнеев, 1952; Соколов, Данилкин, 1981).
Остатки сибирских косуль обнаружены в Орловской области (Горбачев, 1915;
Флеров, 1952); в низовьях Дона в городище Саркел, датируемые VIII—XIII вв. н.э.
(Верещагин, 1959); при раскопках славянских городищ IX—X вв. н.э. вблизи
Воронежа (Громова, 1948); в слоях торфа вблизи станции Заворич Киевской области
(Корнеев, 1952); в среднем Поднепровье (Тимченко, 1972).
Специальными сравнительными исследованиями находок в городищах конца
76
1-го — начала 2-го тысячелетия н.э. в Орловской, Курской и Воронежской областях
В.И. Цалкин (1963), на наш взгляд, достаточно убедительно показал их
принадлежность к сибирской форме. К ней он отнес и костные остатки из Троицкого
городища вблизи с. Бородино Можайского района Московской области и, видимо,
находки из археологических памятников Полтавской и Харьковской областей.
На правобережье Днепра костные остатки примерно этого же времени принадлежат
европейской косуле.
На левобережной Украине в голоцене, видимо, существовала "зона гибридизации",
что усложняет определение костных остатков на местах раскопок и отнесение их
к тому или иному виду. В этот период отдельные особи или группы сибирской
косули, вероятно, заходили на запад от Днепра вплоть до Центральной Европы
и на север — в Прибалтику, где также могли образовываться смешанные с
европейской косулей популяции.и происходила гибридизация, вследствие которой
и сейчас иногда появляются особи с рогами "сибирского типа"(Соиа, 1969; Lehmann,
1976). Однако репродуктивные барьеры и многочисленность европейской косули
не благоприятствовали сохранению их генофонда.
В XIX— начале XX в. сибирская косуля была практически уничтожена человеком
на территории Восточной Европы (см. рис. 23), за исключением нескольких
локальных популяций по Днепру, которые в последние десятилетия гибридизи-
ровали с европейской косулей. Возможность обитания сибирской косули на Русской
равнине в историческое время наглядно подтверждается быстрым современным
расселением ее из Предуралья на запад в район рек Хопра и Дона.
Современный ареал европейской косули (рис. 24, 25) с запада охватывает Европу,
включая Англию и Скандинавский полуостров. Далее на восток его северная
граница идет на Кандалакшу, Петрозаводск, Тихвин, Бологое, Вышний Волочек,
Тверь, Конаково, Талдом, Загорск. Коломну, Рязань (редкие заходы наблюдаются
к северо-востоку от этой линии), включает юго-запад Мордовии, а также западные
районы Пензенской, Саратовской и Волгоградской областей (восточнее р. Хопер)
и по правобережью Дона круто поворачивает на запад, где от Волгодонска идет
уже по левому берегу Дона вплоть до Азовского моря. С низовьев Дона европейская
косуля проникает на юг до г. Сальска и по р. Егорлык на Ставропольскую
возвышенность.
Современная граница ареала европейской косули в Предкавказье проходит
южнее р. Кубань на Невинномысск, Черкесск, Минеральные Воды, Грозный,
Махачкалу и Баку. Не доходя до Баку, круто поворачивает на запад по
предгорным районам Тбилиси, затем на юго-восток — на Кировабад, Степанакерт до
государственной границы с Ираном, идет по его территории на восток, снова
выходя в райо'не Ленкорани.
Современное распространение европейской косули в Передней Азии
недостаточно ясно, однако на основании имеющихся сведений (Harrison, 1968; Kumerloeve,
1975) можно полагать, что она обитает в Турции, вероятно еще сохранилась
в горах на северо-западе Сирии, северо-востоке Ирака и на западе Ирана (в горах
Загрос и в его северной части в провинциях, примыкающих к Каспийскому морю).
Участники XVIII конгресса биологов-охотоведов (Краков, 1987 г.) сообщили,
что с 1970 г. не стало косуль в Израиле (в Хайфе) и, вероятно, нет их теперь в Ливане
(Y. Yom-Tov); нет в настоящее время косуль на о-ве Сицилия, а по территории
Италии они распространены отдельными очагами преимущественно в северной
части страны (G. Ferrario by M.Pavan, 1983); в Испании на юге и в центральных
77
с. 25. Современные ареалы европейской и сибирской косуль
/ — Capreolus capreolus L., 2 — С. pygargus pygargus Pall., J — С pygargus tianschanicus Satunin
районах их ареал также имеет очаговый характер, и лишь на северо-западе страны
они обитают повсеместно (L. Costa, E.J. Purroy).
Современный ареал сибирской косули (рис. 25) на западе ограничен линией
Киров, Чебоксары, Саранск, Пенза, Балашов, Поворино, реками Хопер и Дон до
Волгодонйка. Северная граница идет с запада на восток южнее Кирова, по рекам
Вятка и Кама, по границе Башкирии и Пермской области на Нижний Тагил,
Тюмень, Тару, отсюда к устьям Чулыма и Ангары, по правому берегу которой
и р. Бирюсе достигает г. Братска, огибает с севера Братское водохранилище
и через Жигалово поднимается к северу до Улькана, затем резко уходит на юг
вдоль западных склонов Байкальского и Приморского хребтов, упираясь в
Восточный Саян.
Северная граница распространения косули на северо-восток от Байкала
проходит от устья Витима по левому его берегу до Ленска, далее на север к Мирному,
восточнее его по левобережью Вилюя к устью Мархи, откуда по правому берегу
Вилюя достигает Лены ниже впадения в нее Алдана и по ее долине поднимается
на север примерно до устья Линде. На юг по Алдану выходит к устью р. Учур,
по долине которой, а также по долине р. Мая узкими языками заходит из Якутии
в Аяно-Майский район Хабаровского края. От Учура линия ареала круто
поворачивает на северо-запад к среднему течению р. Амги, отсюда на юг по
правобережному бассейну Буатамы и верховьям Амги к среднему течению Олёкмы,
а от предгорий Станового хребта на северо-запад к Чаре и далее на север к Лене.
Относительно изолированный участок обитания имеется в верховьях Витима.
Ареал косуль в Якутии совпадает с очертаниями Приленского плато. На севере их
распространение ограничено многоснежными районами Центральноякутской
низменности, с востока и юга крупными горными массивами, как бы разрезающими
ареал за Байкалом на две части — северную(описанную выше) и южную.
Южная часть ограничена с севера системой хребтов Станового нагорья и
Становым хребтом, по южным отрогам которого граница ареала доходит на востоке
до верхнего течения р. Уда, огибает с запада хребет Джагды до истоков Селемджи,
с юга хребет Турана до верховьев Бурей и Буреинский хребет, затем по восточным
склонам Баджальского хребта выходит в долину Амгуни, однако Амура по ней
не достигает. Огибая с запада хребты, разделяющие Амгунь и Амур, линия
распространения косули в нижнем течении Амура доходит до с. Богородское и по
западным склонам Сихотэ-Алиня уходит на юг, пересекая хребет примерно у
р. Великая Кема, откуда по восточным склонам поднимается на север до р. Ботчи .
Современное распространение косуль в Корее и Китае не совсем ясно. Известно,
что в настоящее время животные обитают на северо-востоке Китая, включая
северо-западную часть Великой Китайской равнины (Zhang, 1984), особенно много
их в провинции Шаньси (Shen et al., 1982), но нет их южнее, в провинции Сиянь
(Zhejiang) (Zhuge, 1982), отсутствуют они в списке млекопитающих Тибета (Feng
et al., 1983).
Современный ареал косуль в Китае и Корее (см. рис. 25) обозначен в основном
по личным сообщениям профессора Ю. Ma (Ma Yiging) и других членов китайской
делегации на XVIII конгрессе биологов-охотоведов.
Ареал сибирской косули в Монголии изучен нами достаточно подробно (Соко-
4 Более подробное описание ареала в Забайкалье и на Дальнем Востоке см. в работах (Смирнов, 197S,
1978; Кучеренко, 1976; Бромлей, Кучеренко, 1983).
79
лов и др., 1982). Его южная граница выходит сюда из Китая на 114° восточной
долготы, идет на восток по государственной границе, снова заходит на территорию
Монголии в районе сомона Эрдэнэ-Цаган и включает левобережный бассейн
р. Халхин-Гол. Зимой нередко мигрирующие стада из Китая проникают глубоко
в степи Монголии, достигая к западу пос. Матад и горы Их-Чулут. От оз. Буйр-Нур
южная линия ареала идет к востоку от государственной границы, входя на
территорию Монголии в районе оз. Шаварт-Нур, спускается круто на юг по линии
железной дороги до сомона Хавирга, поворачивает на запад к Улан-Батору. Отдельный
очаг обитания косули находится южнее р. Карулен возле пос. Тумэн-Цогт. От
Улан-Батора по правобережью Толы, огибая Хангай по границе распространения
лесов и по хребтам Хан-Хухийн-Ула и Турген-Ула выходит к границе с СССР,
захватывая Туву, и упирается в Западный Саян5. На самом северо-западе Монголии
в последние два-три десятилетия из-за неумеренного промысла образовался
своеобразный разрыв ареала к востоку от Монгольского Алтая. Несколько очагов
обитания косуль на Монгольском Алтае сохранились до наших дней возле самой
границы с Китаем, самый южный из них находится примерно на 46° с.ш. Отсюда
граница ареала идет по западным склонам Монгольского Алтая, огибает с севера
пустынные районы и пески Дзосотын-Элисун и по северо-восточным склонам Тянь-
Шаня выходит в Среднюю Азию.
Распространение косуль в Средней Азии наиболее мозаично: среди высоких
горных хребтов, полупустынь и пустынь относительно редки пригодные для них
биотопы. Долины рек, пожалуй, являются здесь основными биотопами косуль,
и по ним они пронидсают далеко в глубь Тянь-Шаня и в пустынные районы к северу
от него. Южная граница ареала в Тянь-Шане с востока на запад проходит по долине
р. Нарын, хр. Ат-Баши, Алайскому и Туркестанскому хребтам, захватывает
Ферганскую долину, огибает с запада отроги Чаткальского, Пскемского, Угамского
хребтов и хр. Каржантау, где многочисленна, и чуть восточнее Чимкента, по
западным склонам хребта Каратау поднимается на север примерно до пос. Жанатас
(Ишунин, 1961; Айзин, 1969; Закиров, 1974; Янушевич и др., 1972; Слудский и др.,
1984). Отсюда линия ареала поворачивает на северо-восток к оз. Большой Камкалы,
включает долину р. Чу, огибает с юга Балхаш и севернее его через горы Котанэмель,
Бектаута (Капитонов, 1980) почти по прямой уходит на запад к Джесказгану и
через горы Улытау, поселки Иргиз и Шабаркудук Актюбинской области, север
Гурьевской области южнее Джаныбека выходит к Волге в месте впадения в нее
р. Еруслан.
Отдельный очаг обитания сибирской косули, образованный в результате
искусственного расселения, находится в Предкавказье на Ставропольской
возвышенности.
Видовые ареалы европейской и сибирской косуль в начале 80-х годов нало-
жились один на другой на правобережье Волги в Волгоградской области (Данилкин,
1985в) и в районе к востоку от Хопра образовали "зону гибридизации" (см. рис. 25).
В целом ареал рода Capreolus находится в умеренном поясе Евразии, совпадает
с лесостепной зоной, захватывает южную часть лесной и северную — степной.
По берегам рек звери проникают в глубь полупустынь и пустынь, в высокогорные
районы поднимаются до высоты 3000 над ур.м.
Распространение косуль, как показано выше, определяется природными и антро-
5 Распространение косуль в Туве см. в работах (Никифоров, 1969; Смирнов, 1980, 1985).
80
>> /
л/.
.■~^-
■< ■■' '
-■ Ч'"
i~; / '.'
'''V -'
Ул:Л\'
т~*^
s ' ■
:~~г.
^utr.
3
1
Рис. 26. Факторы, определяющие границы распространения косуль
/ — основной ареал рода Capreolus; 2 — заходы за пределы основного ареала; 3 — изолиния высоты
снежного покрова в 50 см; 4 — районы с высотой снежного покрова свыше 60 см; 5 — территории с
продолжительностью снежного периода более 230—240 дней; 6 — зона темнохвойных таежных и горнотаежных лесов, горных
и арктических тундр и пустынь; 7 — основные горные системы (осевые части); 8 — зона пустынь
09
24 30 36 42 48 64 60 66 72 78 84
24 30 36 42 48> 64 60 v^' 66 72 78 84
Рис. 27. Структура современных ареалов европейской (/) и сибирской (2) косуль в России и сопредельных
странах
3 — зона наложения ареалов; 4 — заходы
погенными факторами. Последние оказывают решающее воздействие лишь в
последние столетия, когда человеком оказались прямо или косвенно уничтожены
многие популяции, что привело к резкому сокращению и разрыву ареала. При
снижении антропогенного пресса ареал животных быстро восстанавливается.
Колебания численности явно отражаются и на площади распространения косуль:
при высокой численности они занимают больший ареал, при низкой — он
соответственно сокращается.
Границы ареала не являются постоянными даже в течение одного года, они
изменяются (пульсируют) по сезонам, особенно в Азии. Здесь летние стации зверей
по многих районах находятся гораздо севернее, в горах выше, чем зимой, что
обусловлено сезонными миграциями (см. главу "Перемещения..."). В малоснежные
зимы граница ареала проходит севернее, чем в многоснежные.
Высота снежного покрова является, пожалуй, одним из самых существенных
факторов, лимитирующих распространение косуль. Весовая нагрузка на след косули
составляет 300—360 г на 1 см опорной площади конечностей, что больше, чем
82
90 96 102 108 114 1 20 126 132 138
i fC . .. ^ i— 1- f I
90 96 ,02 ~108 " П4 120 126 132 138
у кабарги, северного оленя, серны, рыси, волка, но меньше, чем у благородного
и пятнистого оленей и лося. При плотности снежного покрова 0,15—0,25, типичного
для лесных и лесостепных районов, критическая его высота для взрослых особей
сибирской косули 40—50 см, для более мелкой европейской косули 30—40 см
(Насимович, 1955). Однако снег не залегает равномерно и плотность его различна,
что позволяет животным жить в районах, где высота его превышает эти цифры.
Среднюю высоту снежного покрова в 50 см обычно считали предельной для
расселения как европейской, так и сибирской косуль (Формозов, 1946), лишь
изредка отмечая выдвижение животных за эту линию (Насимович, 1955; Гептнер
и др.,. 1961). Действительно, северная граница ареала европейской косули
практически полностью совпадает с линией высоты снежного покрова в 50 см (рис. 26),
но для более крупной сибирской косули она не является пределом; только
высота в 60 см в известной мере ограничивает распространение этого вида.
В то же время в Восточной Сибири есть много районов, где высота снежного
покрова менее 60 см, но звери здесь не живут. При внимательном рассмотрении
83
оказывается, что на распространение сибирской косули наряду с высотой
существенно влияет еще и продолжительность залегания снежного покрова: животных
нет там, где снег лежит свыше 230—240 дней в году. Европейская косуля же
практически отсутствует в областях с продолжительностью его залегания свыше 140—
150 дней.
Третий лимитирующий фактор, помимо высоты и продолжительности снежного
покрова, — растительность. Косули избегают горных тундр, темнохвойных лесов.
Проникновение их в зону темнохвойной тайги стало возможным лишь после
сведения ее на больших площадях, что особенно заметно в Скандинавии, на
Северном Урале и в Сибири.
Четвертый фактор, ограничивающий распространение косуль на юге, — зона
пустынь. В Казахстане и Средней Азии (см. рис. 26), в Монголии и Китае граница
ареала практически полностью совпадает с границей зоны пустынь.
Каждый из перечисленных факторов в своем крайнем выражении является
лимитирующим, однако первые три нередко действуют вместе, особенно в горах.
Оптимум ареала европейской косули находится в Западной и Центральной
Европе в регионах со снежным покровом менее 20 см, со средней месячной
температурой+10 не менее 150—160 дней в году и с годовой совокупностью осадков
450—650(800) мм. В высотном отношении наиболее благоприятна для нее зона от
300 до 600 м над уровнем моря; появление на больших высотах является сезонным
(Hell.Herz, 1970; Novakova, Hanzl, 1974; Hell, 1979; Pedroli et al., 1981; Hromas, 1983).
Сибирская косуля явно приспособлена к более суровым климатическим условиям
и живет в областях, где зимняя температура достигает -60° (в Якутии), а летняя
превышает +40° (в Средней Азии).
Наиболее оптимальны для нее регионы, где высота снежного покрова,
сохраняющегося не более 160 дней, не превышает 30 см, т.е. отдельные районы Средней
Азии, Забайкалья, Монголии и юга Дальнего Востока.
Структура ареалов европейской и сибирской косуль несколько отличается:
у первых область распространения практически сплошная, у вторых — носит более
очаговый характер (рис. 27). Очевидно, это объясняется неоднородностью
ландшафтов в Азии и большей миграционной способностью сибирской косули,
имеющей более высокие экологические возможности для расселения.
Анализ особенностей распространения косуль показывает, что горные массивы
являются более существенными барьерами для них, нежели равнинные степи
и полупустынные участки. На рис. 27 отчетливо просматривается разрыв ареала
сибирской косули по горным хребтам Алтая, Западного и Восточного Саян,
оз. Байкал, Становому нагорью и Становому хребту. Эта линия как бы разделяет его
на две части: северную и южную. Наличие этого географического барьера
выявляется и при морфометрическом исследовании косуль — северные популяции
отличаются от южных на уровне подвидов (см. выше).
Однако горные хребты сами по себе не могут служить существенной преградой
для косуль. Их изолирующая роль определяется экологическими факторами,
особенно на азиатской части ареала. Снег, выпадающий в высоких поясах гор
раньше, чем в предгорьях и на равнине, и особенно глубокий в зоне тайги,
вынуждает зверей ежегодно мигрировать в предгорья и на малоснежные равнинные
участки. Из-за этого к югу от горного барьера северная граница распространения
животных сдвигается еще южнее, а с северных склонов горных хребтов косули
отступают к северу, увеличивая тем самым зону разрыва ареала до нескольких
84
сот километров, Весной звери возвращаются обратно, но проникновению их
через рубеж мешает снежный покров, который в горах тает позднее, чем начинается
занятие самцами территорий и отел у самок.
Система горных хребтов Азии сложилась очень давно и несомненно оказала
влияние на формообразование у косуль, обитающих по разные стороны этой
преграды, простирающейся от Чукотки до Памира. Из центра формообразования,
где бы он ни был, звери могли расселяться, вероятно, вдоль данного рубежа.
Разрыв в цепи горных хребтов (так называемые Джунгарские Ворота) позволил
форме, близкой к современному тянь-шаньскому подвиду, распространиться по обе
стороны от барьера. Морфометрическая идентичность популяций Дальнего Востока,
Забайкалья, Монголии и Средней Азии убедительно подтверждает это.
Главный Кавказский хребет также явно является барьером, разделяющим две
разнокачественные географические популяции европейской косули.
Установленные выше закономерности распространения косуль позволяют
прогнозировать движение границ ареалов. Очевидно (см. рис. 26), европейская
косуля практически исчерпала свои видовые возможности, заселив пригодные
биотопы, за исключением Предкавказья, куда следует ожидать ее продвижение.
Сибирская косуля, напротив, при увеличении численности имеет реальные
возможности занять огромную территорию • на Среднесибирском плоскогорье,
расширить свои границы в некоторых северных районах Сибири и европейской части
России, потеснить европейскую косулю с Русской равнины и заселить Северный
Кавказ.
Глава четвертая. Среда обитания
Ареал косуль находится в умеренном и субтропическом поясах, включающих
области с климатом от влажного морского до сухого континентального. Они
живут почти во всех природных зонах: в широколиственных и смешанных лесах,
в тайге, в лесостепи, степи, полупустынях, по рекам проникают в пустыни;
обитают на равнинах и в высокогорье (см. главу "Ареал"). В то же время в пределах
ареала выявляются определенные закономерности их биотопического
распределения и отчетливо прослеживаются факторы, оказывающие влияние на их
размещение.
Основные биотопы
Европейская косуля. На западе ареала она населяет практически все леса,
предпочитая их мозаику в открытом ландшафте (Southern, 1964; Prior, 1968;
Stagl, Margl, 1977; Mayer, 1977; Pedroli et al., 1981; Reimoser, 1982). Наиболее
типичные биотопы — разновозрастные леса с богатым кустарниковым ярусом,
находящиеся в окружении лугов и полей1. В последние десятилетия одними из
основных стаций стали сельскохозяйственные поля (Mottl, 1954; NeCas, 1963;
Raesfeld, 1965; Pielowski, 1970; Szederjei, Szederjei, 1971; Strandgaard, 1978; Stubbe,
Passarge, 1979; Casanova, 1981) (рис. 28).
На северо-западе Европы косуля преимущественно живет в небольших
смешанных лесах, встречается в зарослях тростника, на лугах и полях (Cederluhd, 1982).
Животные избегают больших лесных массивов, но обычны на опушках лесов.
Зимой они предпочитают смешанные насаждения, поляны, разреженные сосняки
и зарастающие вырубки; весной роль лесных стаций снижается; летом большая
часть животных обитает в сравнительно открытых высокотравных биотопах,
поросших кустарником; осенью по мере увязания травянистой растительности
снова наблюдается переход в лесные угодья (Верещагин, Русаков, 1979).
В Литве в лесных районах очевидно тяготение их к сравнительно открытому
ландшафту (Балейшис, Прусайте, 1980). К оптимальным стациям относится
смешанный елово-лиственный лес, к худшим — чистые лишайниковые боры (Па-
дайга, 1984).
В Белоруссии в жизненно необходимые биотопы зверей входят смешанные
сосново-дубово-еловые леса, сосновые молодняки, спелые боры, болотистые
луга с кустарником и грабово-дубовые насаждения; дубравы, лиственные
молодняки, ольхово-ясеневые леса и сфагновые болота они используют лишь
периодически (Саблина, 1955; Гатих, 1975). Южнее на Украине косули
распространены практически во всех лесах, проникают по лесополосам далеко в степные
' В тексте биотоп — синоним местообитания и стации (Реймерс, 1980).
86
Рис.28.Сельскохозяйственные поля — одна нз основных стаций европейской косули в Западной Европе
районы, часто встречаются в садах, на полянах и на целинных степных участках
(Гречушкин и др., 1975; Гурский, 1975).
В Крыму для них наиболее характерны пять стаций: буковые насаждения
на северных склонах на высотах 700—1000 м над уровнем моря; дубравы на
южных склонах; смешанные двухъярусные дубово-буково-ясеневые леса; небольшие
сосновые массивы по склонам гор и молодые смешанные насаждения,
образованные дубом, грабом, ясенем, кленом, осиной, липой и ивой. По сезонам
картина местонахождения животных существенно меняется (Лавров, 1929;
Пастернак, 1955).
На Востоке Европы косуля заселяет прежде всего лесостепные дубравы
с густым подлеском, сосновые боры, пойменные леса, заросшие кустарником
овраги и заливные луга, но их распределение по стациям подвижно и зависит
от ряда факторов, основные из которых кормность угодий и доступность
кормовых участков (Новиков, Тимофеева, 1965; Ватолин, 1975; Простаков,
Обтемперанский,- 1986).
На Кавказе косули держатся преимущественно в разряженных дубовых лесах,
поднимаясь летом до высоты 3500 м над уровнем моря (Динник, 1910; Шахмар-
данов, Рамазанов, 1980). В Закавказье область их распространения совпадает
с зоной грабово-дубовых, буковых и каштановых лесов и достигает нижней
границы субальпийской зоны (2200 м над уровнем моря). Зимой, как и на
Северном Кавказе, звери здесь мигрируют вниз и совершают горизонтальные пере-
87
мещения, переходя с северных заснеженных склонов на южные и восточные.
Осенью многие из них перекочевывают в дубовые и каштановые леса (На-
симович, 1939; Арабули, 1963, 1967).
Сибирская косуля. От Волги до западных склонов Уральских гор она населяет
практически все островные леса; по лесополосам и поймам проникает далеко
в степные и полупустынные районы. На Южном Урале ее основными стациями
являются березовые колки вблизи водоемов и кустарниковые заросли на
высокотравных лугах, смешанный лес с большой долей осины в древостое и зарастающие
гари и вырубки (рис. 29). На Среднем Урале зимой косуля занимает 90% площади
основных типов лесов, характерных для северной лесостепи и сосново-березовый
подзоны. Наибольшая плотность (4—12 особей на 100 га) наблюдается в
высокотравных и разнотравных березняках и сосняках, перемежающихся с лугами,
на вырубках и гарях. В темнохвойной тайге звери отсутствуют. В горах наиболее
плотно заселяются пологие склоны, кустарниковые долины, средней густоты
молодняки и (летом) высокотравные луга вблизи водоемов (Сабанеев, 1875;
Ушков, 1947; Соколов, Данилкин, 1981; Дворников, 1984).
В Западной Сибири распространение сибирской косули приурочено к южным
лесостепным и степным районам, где она обитает в березово-осиновых колках,
сосновых борах и кустарниковых зарослях по берегам рек и озер и в лесополосах.
Зимой основными местами кормежки зверей раньше служили высокотравные
и полынные степи. В связи с распашкой целинных земель площадь зимних
пастбищ косуль в этом регионе значительно сократилась, что отрицательно
сказалось на их численности. В предгорных и горных районах Кузнецкого
Алатау животные встречаются практически во всех типах леса, исключая темно-
хвойную тайгу. С выпадением снега они мигрируют в малоснежные районы
(Янушевич, Благовещенский, 1952; Метельский, 1976).
На Алтае косуля живет повсеместно в ленточных борах на равнине, в
кустарниках и островных хвойных лесах, преимущественно лиственничных, по склонам
гор, летом обычна в высокотравных долинах (Дмитриев, 1938; Савинов, 1961).
На Южном Алтае звери предпочитают лиственный лес с густым подлеском
(Слудский и др., 1984). Темнохвойной захламленной тайги, болот и открытых
низкотравных степей они избегают.
В западной части Казахстана животные обитают в основном в пойменных
лесах с густым подлеском и многочисленными полянами. На севере Казахстана
они занимают осиново-березовые колки с подлеском из ив, шиповника,
боярышника, степной вишни и др., пойменные леса и искусственные посадки, зимой
выходят на открытые участки степи и на зарастающие полынью пашни. В
степной зоне основные их стации представлены пойменными лесами и
кустарниками в островных и ленточных борах. В равнинной части юго-востока
Казахстана косули встречаются по тугаям, перемежающихся полянами и участками
тростника по берегам водоемов.
В горах Северного Тянь-Шаня и Джунгарского Алатау косули живут повсеместно
от предгорий до субальпийского пояса включительно. Лучшие биотопы —
заросли из диких яблонь, урюка и осины, еловые и елово-пихтовые леса по северным
склонам и березовые островные леса (Слудский и др., 1984). Смена стаций
в вертикальном отношении выражено очень четко (рис. 30). На Центральном
Тянь-Шане звери обычны в тугаях по поймам рек, в островных еловых лесах
по северным и северо-западным склонам гор, изобилующих высокотравными
88
-ж-
**V ц .,* ■"' 'и,-
*^ВД1
■jt*:i:
■:*■■■£■
43;iw№ti;t
Г -I?..
^K;:r#^
'■ '%>
:<>&•
if'
я:/
& ^
sm.v
Рис. 29. Основные кормовые стации сибирской косули на Урале — высокотравные луга (А) и зарастающие
вырубки и гари (Б)
I ЛШШТШШШТХХЛШ
Месяцы
Рис.30.Биотопическое (вертикальное) распределение косуль в Таласском Алатау (в процентах от числа
особей, встреченных за месяц) (по: Грачев, Смирнова, 1980)
/ — полусаванны (1300—J800 м над ур. м.); } — арчевникй (1800—2300 м); 3 — субальпийская зона
(2300—2900 м)
лугами, а в летний перибд встречаются и на высокотравных субальпийских
лугах. Зимой в субальпийской зоне и в нижней части гор из-за глубоких
снегов они отсутствуют, хотя выше, в поясе арчевника (на высоте около 3000 м
над уровнем моря) обитают во все сезоны года (Антипин, 1941; Жирное, 1958).
В горах Киргизии наиболее типичные стации — островные леса с
разнотравными полянами и кустарниковые долины, тополево-ивовые насаждения по
долинам рек. В арчевых зарослях и на альпийских лугах косули встречаются
реже (Айзин,1969). На равнине животные заселяют кустарниковые и тростниковые
заросли по берегам рек и ручьев.
В Восточной Сибири сибирская косуля особенно многочисленна в лесостепи,
в высокотравных березовых и осиновых колках, в разреженных лиственничных
лесах с поддеском, в ерниковых зарослях по берегам водоемов,
представленных кустарниковой березой и низкорослой ивой, на зарастающих гарях, богатых
кипреем! и в разнотравных сосняках. Зимой из горных многоснежных районов
она мигрирует в злаково-разнотравные степи. Косуля практически не живет
в темнохвойной тайге, избегает моховых болот, каменистых россыпей и гольцов
(Фетисов, 1953; Зырянов, 1975; Ельский, 1975, 1978).
В Западном Забайкалье к основным стациям косули М.Н. Смирнов (1978)
относит леса с преобладанием сосны и лиственницы, березово-осиновые колки,
вырубки, зарастающие гари, степные участки среди горного леса, среднеувлажнен-
ные луга, осоковые болота, залежи, речную урему. Животные явно тяготеют
здесь к открытым угодьям с хорошо развитой кустарниковой и травянистой
растительностью. По сезонам стации изменяются, что связано прежде всего с их
меняющейся кормностью (Ельский, 1979). В глубокоснежных районах обычны
миграции.
В Якутии сибирские косули обитают главным образом в биотопе, имеющем
90
облик лугостепи с небольшой высотой снежного покрова. Наиболее
предпочитаемые угодья — опушки, ерниковые поляны, высокотравные долины рек,
зарастающие вырубки и гари. Летние и зимние пастбища из-за неглубоких
снегов совпадают, и миграций в этом регионе не отмечается (Тавровский и др.,
1971).
В Амурской области Ю.А. Дарман (1983, 1986) выделяет шесть основных
биотопов: 1) разреженные дубовые леса с густым подлеском из леспедецы
и лещины и подростом осины, дуба, липы, березы даурской; 2) горные
мелколиственные леса; 3) заросли лещины на южных склонах; 4) широкие безлесные
луговые долины рек и ручьев (пади); 5) густые заросли ивняков с примесью
осины и березы маньчжурской, образующие вместе с лугами прирусловый комплекс;
6) влажную лесостепь с заболоченными лугами и участками черноберезниковых
и лещинных релочных лесов, характерную для всей Зейско-Буреинской равнины.
В течение года биотопическое распределение косуль меняется, но не так резко,
как в горах. В бесснежный период большинство животных предпочитают
влажную лесостепь, зимой они концентрируются в дубняках и дубово-чернобере-
зовых лесах.
Наиболее характерное местообитание сибирской косули в Амуро-Уссурийском
крае — низкогорные ландшафты с мягкими формами рельефа, лесостепь с
лиственными разнотравными лесами с густым подлеском и многочисленными
лугами. Наиболее высока ее численность в дубняках близ сельскохозяйственных
полей, на слабо заболоченных лугах с обилием водно-болотной
растительности, на лесосеках и гарях с осиново-березовым подростом. Лучшие стации
в этом регионе — леспедециево-лещинные дубняки, разнотравно-кустарниковые
лесистые участки в окружении лугов, колки в полях, молодняки на гарях
(Кучеренко, 1976; Бромлей, Кучеренко, 1983). В последнее время
предпочитаемыми становятся соевые поля. При условии улучшения охраны зверей и
увеличения их численности здесь можно ожидать формирования полевых
популяций.
Стации косули в Монголии представляют особый интерес (Данилкин, Ду-
ламцэрэн, 1981; Соколов и др., 1982). Южная граница ее ареала совпадает
с южной границей распространения лесов и выходит за ее пределы лишь при
появлении в степи островков кустарниковой растительности, представленных
различными видами ив и карликовой березы. Животные явно предпочитают
для жизни лесостепь, разреженные островные лиственничные леса (рис. 31,А)
по пологим склонам гор и особенно многочисленны в высокотравных долинах
рек и ручьев, заросших кустарником. На Хангае и Хэнтее во многих местах
плотность их населения достигает 2—10 особей на 100 га.
На востоке Монголии звери обитают в тростниках и кустарниках по берегам
озер и рек, в островках низкорослого кустарника по понижениям в степи (рис. 31, Г),
в высокотравных безлесных долинах среди невысоких и низкотравных
возвышенностей, на посевах зерновых культур. В предгорьях Большого Хингана в
бассейне р. Халхин-Гол косуля живет в песчаных барханах, поросших кустарником,
в высокотравных кустарниковых долинах рек и ручьев (рис. 31, В), в тростниковых
зарослях и в высокотравной холмистой степи (рис. 31, Б). Обращает на себя
внимание тот факт, что в степных биотопах нередко на десятки километров
вокруг нет источников воды и животные обходятся той влагой, которая
содержится в растениях и на них в виде обильной росы, а также пьют воду из луж
91
:■■ ч ■■■'
Ч^..**1 •■ -St.*. ,y*
'■■■■ fc
f*
&
Щ
'$!»:<
».
> > '^M*1**
i? и '';*"«»
< ^^й
*.
■urn.
■:*'
Г \
tajjl^*
■■f
ЭЙ
■*4fc ':■*:
>4,
,;^-. :»ww
■■**ч.
««•"•"IfS"
:-йк. *:' «\ F KpAjtor *№V ■■*■ ■■
:. *
[if%i
* ■%. "*f^
5'
.,.,,., ь|^ it:
Щ&*0 Ч& ;
':*:■■'■ "$?'■■■ *-Ъ ■■ ■::i*:-'/-"
'■'■-'JV*
■<x' '' 'S.r-.
!'' «■
.'„ ;) >; s:vs
^rt»1*
,#■■
.■■:^,.^l>,
* ?:..
Рис. 31. Типичные стации сибирской косули в Монголии
А — разреженные высокотравные лиственничные леса; Б — высокотравная холмистая степь; В —
кустарниковые долины рек и ручьев; Г — островки низкорослого кустарника по понижениям в степи; Д —
низкотравная холмистая степь
92
«м *.< ^??т
1 j£"aSs.■ ■ *'■ .\ "Я? .
Р"у
^.6i< Г*.; .(-„
гть:
2V*
!мжм ,„■■,;*■'>*#
Д?»^!:^1
ИЦ*1
► &■■■.;" <•
i' *
*:Ш>»
,,,,%, ^# Ifei-;?*^ #' '■,
*"' $* itw .ц
1, 41 .,
^Iffi
'-.1
.vW
«те^.-ц
>,:-*,»*'*
93
после дождей. Плотность их населения в степных высокотравных биотопах
летом весьма велика — до 3—8 особей на 100 га.
На юго-востоке по государственной границе с Китаем они живут в
низкотравной холмистой степи, изрезанной оврагами (рис. 31, Д). Зимой мигрирующие
стада косуль заходят на десятки и сотни километров в глубь равнинных степей
в типичные стации дзерена. Некоторые группы остаются на редких здесь
возвышенностях, однако из-за отсутствия укрытий звери становятся легкой добычей
браконьеров.
На востоке Монголии, таким образом, существуют степные популяции косуль.
Они есть, очевидно, и в сопредельных районах севера Китая.
Факторы, влияющие
на биотопическое размещение
На биотопическое размещение косуль оказывает существенное влияние весь
комплекс биотических и абиотических факторов. Наиболее значимы среди
них следующие.
Как подробно анализируется в главе "Питание", косули имеют очень широкий
спектр кормов, но склонны к потреблению прежде всего легко усваиваемой
растительной пищи, богатой питательными веществами и водой. С этим в тесной
связи и находится выбор местообитания.
Их пищевым требованиям лучше всего отвечает стации сравнительно открытого
мозаичного ландшафта — небольшие островные леса среди
сельскохозяйственных полей и высокотравные луга с вкраплениями кустарников. В лесных районах
особенно благоприятные условия звери находят на зарастающих гарях и
вырубках, где, помимо богатого веточного корма, хорошо развивается травяной
покров. Средневозрастные и перестойные леса без подлеска и травостоя,
а также лесные массивы таежного типа малокормны, и животные их
избегают.
Предпочтение определенных стаций приводит к неравномерному распределению
косуль в угодьях, в лучших плотность населения бывает наивысшей, несмотря
на жесткую конкуренцию за территорию. Поэтому между качеством биотопа,
плотностью населения и продуктивностью популяций существует высокая
корреляция.
Наиболее оптимальны для их жизни, как показывают исследования, леса
площадью от 100 до 500 га, имеющие сравнительно большую длину опушечной
линии (Ueckerman, 1957; Sagesser, 1966; Юргенсон, 1968; Herz, 1971; Блузма, 1975;
Hell, I979; Балейшис, Прусайте, 1980; Прусайте и др., 1983). Лиственные и прежде
всего широколиственные леса для косуль имеют большее значение, чем хвойные
(Novakova, Hanzl, 1974; Strandgaard, 1978). Смешанные леса охотнее заселяются
ими по сравнению с монокультурами, особенно хвойных пород. В
разновозрастных древостоях косули выбирают молодые посадки, имеющие максимальный
запас веточных кормов (Тимофеева, 1985). При оценке качества лесного биотопа
кустарниковый и кустарничковый ярусы имеют принципиальное значение;
наиболее ценны для косуль те леса, где доля полян и лугов превышает 20%
(Ueckermann, 1952; Mottl, Janda, 1955; Vincent et al., 1979).
Для животных, обитающих в полях, наибольшее значение имеет состав
культур. В межвегетационный период кормовая база здесь представлена в основном
94
остатками урожая кукурузы, сахарной свеклы и картофеля, озимой пшеницей,
люцерной и клевером. В вегетационный период выбор кормов и соответственно
стаций гораздо шире — почти все выращиваемые на полях культуры и
растительность на пустошах (Zejda, Homolka, 1980; Zejda, Bauerova, 1985).
Убежища необходимы для косуль не менее, чем пища, особенно в открытых
степных и полевых ландшафтах. Если нет укрытий, звери не живут здесь даже
при изобилии пищи. Особенно четко это видно в степных районах Монголии.
В равнинной степи они обитают лишь там, где есть островки карликового
кустарника, высокой травы по понижениям или посевы зерновых
сельскохозяйственных культур. В безлесной, но высокотравной и холмистой степи
животные обычны и плотность их населения сравнительно высока, поскольку
здесь и высокая трава, и сама холмистая местность обеспечивают укрытие;
в низкотравной же холмистой степи они встречаются преимущественно там,
где есть овраги. Мигранты остаются жить в степных районах только при наличии
убежищ.
В лесу укрытий для косуль достаточно, если около 10% его площади
представлено густыми зарослями (Strandgaard, 1978). В период отела наличие мало-
просматриваемых заросших участков жизненно необходимо для самок, которые
занимают их перед родами и защищают от других особей (см. ниже).
Существенным фактором, влияющим на биотопическое размещение, следует
считать беспокойство, вызванное деятельностью человека (туристы, сборщики
грибов и ягод и т.п.), выпасом скота и присутствием других видов диких
копытных. Плотность населения косуль в часто посещаемых людьми стациях может
быть ниже на 30% по сравнению с однотипными угодьями, где деятельность
человека низка или отсутствует (Novakova, Hanzl, 1974; Strandgaard, 1978; Hell, 1979).
Места с интенсивным выпасом скота или высокой плотностью населения
других видов копытных косули покидают (Владышевский, 1968; Hell, 1979;
Pulliainen, 1980; Pedroli et al., 1981; Данилкин, Дуламцэрэн, 1981).
Среди абиотических факторов, как указывалось выше, наиболее важен
снежный покров. К примеру, европейская косуля в северной части Брянской области,
где высота снежного покрова более 50 см, отсутствует или очень редка,
тогда как в южной части при средней его высоте в 29 см плотность ее населения
достигает 8,3 особей на 10 км2 (Ватолин, 1975).
На сильное увеличение снежного покрова животные реагируют уменьшением
размеров участка обитания или откочевывают в малоснежные биотопы. Тем не
менее при наличии троп и высококачественного естественного корма или
подкормки и отсутствия фактора беспокойства (хищников в первую очередь),
косули могут пережить зиму даже при очень глубоком (свыше 1—1,5 м), но
плотном снежном покрове. Уплотнение снега позволяет не только относительно
свободное передвижение по нему, но и расширяет кормовую нишу зверей,
способных при таких условиях доставать корм, находящийся на высоте до
3 м от земли, и объедать прежде недоступные им верхушки кустарников и
небольших деревьев (рис. 32). Фактор беспокойства в этих экстремальных
условиях вызывает увеличение локомоторной активности косуль, что ведет к большим
затратам энергии и значительно снижает их шансы на выживание.
В горных биотопах размещение косуль в большей степени, чем на равнине,
обусловливается комплексом факторов, из которых решающую роль играют
кормовая база (состав и структура растительности), состояние (высота, плот-
95
/.г
•S к*
Ч- { • in
Рис.32.При высоком, но плотном снежном покрове косулям становятся доступны верхушки
кустарников
ность) снежного покрова и крутизна склонов (Метельский, 1975). Поэтому
смена стаций (вертикальные кочевки) здесь часто выражены более четко.
Относительно объективным показателем предпочтения косулями определенных
биотопов в пределах ареала может быть плотность их населения, хотя
отсутствие унифицированных и точных методов учета численности существенно
затрудняет сравнение результатов. Проанализировав собственные данные и
пригодные для анализа литературные сведения (и = 139), мы определили среднюю
плотность населения косуль в типичных биотопах Европы. Она, как выяснилось,
изменяется в пределах от 0,1 до 65 особей на 100 га.
В лиственных и смешанных лесах с преобладанием лиственных пород
плотность зверей довольно часто достигает 13—18 особей на 100 га и лишь изредка
выше; в смешанных лесах, где в древостое преобладают хвойные породы, —
10—15; в хвойных лесах плотность обыкновенно ниже — от 2 до 6 особей на 100 га
(Mottl, 1954; Bobek et al., 1974; Bobek, 1977; Блузма, 1975; Reichholf, 1980;
Vincent, 1982; Blant et al., 1982; Reimoser, 1982; Падайга, 1984). В лесных угодьях
средняя плотность населения косуль прямо зависит от плодородия
преобладающего типа почв, доли лиственных пород в составе лесонасаждений и находится
в обратной тесной связи с лесистостью отдельных районов (Блузма, 1975;
Падайга, 1975).
В полевых биотопах плотность населения косуль довольно разнообразна:
в безлесном ландшафте Южной Словакии 1,1 (Hell, 1979), в Венгрии 3 (Szederjei,
Szederjei, 1971), в ГДР 4—13 (Stubbe, 1987), на западе Польши 7 (Kaluzinski,
96
1982),в Южной Моравии в Чехословакии 12 особей на 100 га (Zejda, 1985).
Она определяется прежде всего круглогодичным наличием корма и убежищ
в виде колков, оврагов, полезащитных лесонасаждений и высокотравной
растительности. В открытых полях плотность населения животных минимальна (до 1 особи
на 100 га), а при наличии укрытий может достигать 20 особей.
Мозаика полей и лесов считается лучшим из биотопов косули, но с юга
ареала на север плотность в них падает с 18—20 до 3—4 особей на 100 га
(Sedlaczek, 1964; Балеишис, Прусайте, 1980; Cederlund, 1982), что указывает
на возрастающее негативное влияние на популяции комплекса факторов среды.
Максимальные плотности населения (40—60 особей на 100 га) отмечаются
лишь на сравнительно небольших площадях и во многих случаях сезонно
(Muller, 1973; Stagl, Margl, 1977; Mayer, 1977; Reichholf, 1980; и др.). При пересчете
на большую территорию эти цифры заметно снижаются. Например, в Литве
на площади 27,8 км2 (83% — по^я) плотность популяции определена в 6—7 особей
на 100 га, на участке в 24,3 км (поля — 55% площади) учтено в среднем от 10 до 12
косуль и в маленьких лесах площадью от 200 до 1000 га плотность составила
уже 15—20 особей (Прусайте и др., 1977, 1983).
С севера на юг ареала по широтному градиенту плотность населения
косуль увеличивается: в подтаежной зоне она не превышает 0,1 особи на 100 га,
в зоне смешанных и лиственных лесов достигает 3—6, а в лесостепи и зоне
широколиственных лесов — 5—12 особей.
Из приведенных выше данных отчетливо видно, что уровень плотности
населения животных определяется качеством биотопов и, в свою очередь,
часто служит показателем экологической емкости угодий. Плотность населения'
косуль существенно зависит и от общей численности зверей в регионе (см. главу
"Численность"). Для малочисленной сибирской косули она почти на порядок
ниже, чем для европейской, и лишь в наиболее излюбленных ими стациях и
при зимних концентрациях достигает 10—20 особей на 100 га на сравнительно
небольшой площади (Смирнов, 1978; Соколов, Данилкин, 1981; Данилкин,
1985; Дарман, 1986).
Приспособленность популяций
к определенным биотопам и их изменениям
Локальные популяции в процессе эволюции, как известно, отбираются по
приспособленности к специфическим условиям среды, в которой они обитают
(Майр, 1974). Существуют ли "экотипы" косуль и какова первичная среда
их обитания — лес или открытые пространства?
На этот счет нет единого мнения. Большинство исследователей относят
их к лесным видам животных, однако этому противоречит тот факт, что в
загущенных естественных лесах численность зверей низка или же они отсутствуют
вовсе. К тому же косули могут круглогодично обитать в совершенно
безлесных степных районах и на полях, что свидетельствует в пользу гипотезы
об их "степном" происхождении. С другой стороны, в большинстве регионов
животные явно избегают открытых биотопов, лишенных убежищ.
При рассмотрении этих вопросов из поля зрения зоологов почему-то выпадает
одно очень немаловажное обстоятельство. В Азии многие популяции сибирской
косули осенью мигрируют из лесных биотопов в степные (реже наоборот) и
7. Европейская и сибирская косули ду
весной кочуют в противоположном направлении (см. главу "Перемещения"),
т.е. в течение года ведут себя то как "лесные",то как "степные". Кроме того,
многие особи, живущие в лесу, регулярно выходят кормиться на поля или
в степь, а самки предпочитают рожать детенышей в более открытом, но имеющем
убежища ландшафте. Эти факторы указывают на очень большую
экологическую валентность косуль в пределах ареала и характеризуют их как
животных лесостепного типа, более приспособленных к жизни в густой высокой
траве и кустарниках, чем в густом древостое или в открытой степи. Очевидно,
что отнесение их к обитателям мозаичного ландшафта более обосновано
(Соколов, 1959; Raesfeld, 1956; NeSas, 1963; Szederjei, Szederjei, 1971; и др.).
Морфологические признаки косуль — некоторая укороченность передних
конечностей, сравнительно длинная шея, присутствие хорошо развитых боковых
пальцев и сокращенная мягкая пяточная часть копыта (Флеров, 1952), узкое
в плечевом поясе туловище — также свидетельствуют в большей степени о
приспособленности их к обитанию в более открытых, сравнительно влажных,
высокотравных и кустарниковых биотопах, нежели в лесных стациях.
Передвижение зверей при опасности прыжками длиной до 4—7 м с
периодическим подпрыгиванием вверх на 1,5—2 м больше роднит их с обитателями
саванн, нежели с жителями леса.
Действительно, как показано выше, косули обычны прежде всего в лесостепи,
а в лесном биотопе предпочитают разреженный древесный ярус с обилием
полян и богато развитым кустарниковым подлеском и травостоем (рис. 33).
Поразительно, но факт, что своими маркировочными действиями (см. главу
"Поведение") самцы производят разреживание подроста. Каждый из них на своем
участке обитания серьезно повреждает сотни деревьев, способствуя образованию
световых островов в лесу, где животные находят более, высококачественный
корм, чем под густым пологом леса. Частичное сведение человеком лесов,
особенно темнохвойных, привело к значительному расширению ареала и
способствовало быстрому росту численности этих животных. Поскольку косуля
часто концентрируется на постепенно зарастающих вырубках и гарях, Госсов
(Gossow, I977) справедливо назвал ее "сукцессионным типом".
В последние десятилетия во многих странах Европы появились
многочисленные полевые популяции косуль. Этому явлению способствовали высокая
плотность их населения в лесах и конкуренция за участки обитания,
интенсификация лесного хозяйства и всевозрастающее беспокойство со стороны человека,
увеличение площадей агроценозов и наличие здесь корма в течение всего
года, а также уменьшение беспокойства в полях в связи с механизацией
сельскохозяйственных работ.
Можно ли в связи с этим считать, что возник новый "полевой" экотип косуль?
Наблюдения показывают, что, проводя в открытом ландшафте практически
весь год, полевая косуля все-таки имеет определенную связь с древесными
насаждениями, которые служат дополнительным источником корма, местом
отдыха и убежищами при неблагоприятных погодных условиях (HoliSova et al.,
1982; Zejda, Bauerova, 1985). Многочисленна она лишь там, где крупные поля
имеют сеть полезащитных полос или мозаику маленьких лесов: в агрикультурном
безлесном ландшафте звери отсутствуют или плотность их популяций очень
низка (Hell, Herz, 1970; Hell, 1979; Stubbe, Passarge, 1979; Pielowski, 1970).
В экспериментах (Graczyk, 1978) взрослые особи полевой популяции после
98
Рис. 33: Наиболее предпочитаемый биотоп у косуль — лесостепь
многомесячного содержания в лесу, в котором они дали потомство, возвращаются
в полевую экосистему, а взрослые особи из лесной популяции, содержавшиеся
в полевых условиях, уходят в лес. Детеныши "лесной" косули, рожденные
в поле, тоже возвращаются в лес; здесь же остаются и детеныши "полевой",
рожденные и выращенные в загороженном лесном массиве. Следовательно,
"полевые" косули пока еще не обладают экологически обусловленными
наследуемыми признаками приспособленности к специфическим условиям обитания,
что обычно характеризует разные экотипы.
Приведенные выше данные, указывающие на значительную экотипическую
изменчивость, не позволяют выделить у Capreolus четко ограниченные "экотипы"
("лесной", "полевой" или "степной"). Само определение косуль в качестве
животных лесостепного типа при отсутствии у них узкой морфоэкологической
специализации указывает на возможность обитания как в открытых, так и в закрытых
ландшафтах.
Следует отметить, что термины "полевой экотип", "лесной экотип" и "степной
экотип" уже прочно вошли в зоологический лексикон для обозначения
популяций, обитающих преимущественно в полях, в лесу или в степи, и не стоит
отказываться от них.
Тем не менее биотопы оказывают существенное влияние на образ жизни
и поведение. Отличительная черта экологии зверей, круглогодично или сезонно
живущих в закрытом биотопе, — более тесная связь с убежищами (лесом,
кустарником, тростниковыми зарослями): при опасности они бегут почти всегда
из открытого биотопа в закрытый, а не наоборот; у них короткая дистанция
99
бегства; затаивание взрослых особей наблюдается в сравнительно редких случаях
и в основном в положении стоя; зимой животные живут небольшими группами.
Косулям, круглогодично обитающим в степи и на сельскохозяйственных
полях, закрытый биотоп служит лишь кратковременным убежищем — при
опасности они бегут на значительно большее расстояние ("полевые" косули
минуют при этом лесной массив). В степи, где травяная растительность ниже
их роста, звери ложатся в "островках" высокой травы или в складках оврагов
без ориентировочной позы (см. главу "Поведение"), мгновенно исчезая из виду,
затаиваются и не убегают, даже если человек или автомашина проходит в 2—10 м
от них. Зимой в открытых ландшафтах образуют большие стада.
При рассмотрении вопроса об изменении среды обитания в историческом
плане и влиянии этого процесса на популяции становится совершенно
очевидным, что в целом на большей части ареала человек постепенно и постоянно
улучшал их биотопы: вырубал сплошные леса, создавая мозаику
разновозрастных древостоев с островами лугов и полей, а в последнее два века благодаря
лесополосам в степях и изменению агроценозов возникли предпосылки для
заселения открытых стаций и образования полевых популяций. По сравнению
с другими дикими копытными европейская и сибирская косули оказались наиболее
приспособленными к измененным человеком биотопам, и их без сомнения
можно характеризовать как виды будущего в антропогенном ландшафте.
Однако антропогенное изменение среды обитания далеко не всегда
положительным образом сказывалось и продолжает сказываться на популяциях этих
животных. Полное сведение лесов и распашка целинных высокотравных степей,
которые служили основными кормовыми стациями зверей, наряду с другими
факторами способствовали снижению их численности в ряде регионов.
Мелиорация земель, осуществляемая на огромных территориях, приводит к уменьшению
площади мозаичных угодий и прямому уничтожению лучших биотопов —
высокотравных лугов, поросших кустарником.
В малоснежных районах потеря целинных степей и лугов, к счастью,
компенсируется возможностью обитания животных в агроценозах, но в областях
с высоким снежным покровом замена их на полевые угодья означает практически
полную утерю лучших, а часто и единственно пригодных биотопов.
В Приамурье, например, в результате распашки лугостепных массивов
уничтожено более 50% зимовочных стаций сибирской косули (Швец, 1980).
Аналогичная ситуация в Западной Сибири, где к тому же значительно
увеличилась площадь пастбищ домашнего скота и усилился фактор беспокойства
(Белов и др., 1980). Ареал европейской косули на Западном Кавказе в результате
хозяйственного освоения территории в последние 20 лет сократился, и его
границы сдвинулись в сторону Главного Кавказского хребта на 10 км в
черноморской и до 50 км в кубанской части (Дуров, 1980). Значительные площади
основных пойменных стаций этих копытных, исчисляемые миллионами гектаров,
безвозвратно уходят под воду при заполнении водохранилищ многочисленных
гидростанций. Не благоприятствует им и тенденция к монокультурному
выращиванию хвойных пород в лесном хозяйстве.
При высокой плотности населения популяции косуль и сами могут существенно
влиять на биотопы, угнетая подрост и изменяя прежде всего состав
растительных ассоциаций. Подробнее эта проблема, имеющая практическое значение,
рассматривается в главе "Хозяйственное значение...".
Глава пятая. Перемещения
«
Оседлость и миграции
Косули — очень пластичные в экологическом отношении животные. Они весьма
"привязаны" к своему участку обитания, но на неблагоприятные факторы
среды реагируют временным перемещением в другие районы. Особенно отчетливо
эта особенность проявляется у сибирской косули, обитающей во многих
регионах в таких условиях, в которых лишь при своевременных миграциях ее
популяции могут выжить. И только такие популяции, у которых в процессе
эволюции выработалось сезонное миграционное поведение, обеспечивают
относительное процветание вида в областях, где отрицательные факторы среды,
казалось бы, превышают его экологические возможности1.
Одним из первых ученых, которые обратили пристальное внимание на
перекочевки животных и причины их вызывающие, был А.Ф. Миддендорф
(1869), который писал: "Если вся пища съедена или истреблена суровым временем
года, либо укрылась в недоступные места, или покрыта глубоким слоем снега,
то животное, которое питается этой пищей и не хочет умереть с голоду, должно
отыскивать ее в более отдаленных местах... Страны света тут ничего не значат;
нередко животные на зиму уходят к северу. Богатые снегом местности или,
особенно, снежные горы обыкновенно явно усиливают кочевание с гор". В
дальнейшем явление миграций у косуль подробно рассматривалось А.Н.
Формозовым (1948), А.А. Насимовичем (1955) и другими исследователями.
В настоящей монографии сделана попытка свести воедино разрозненные
сведения о перемещениях косуль в разных регионах и составить карту их
миграций. Наиболее интересные сообщения, отражающие процесс миграции, мы сочли
необходимым цитировать.
Европа. Массовое мечение косуль в. Европе показало, что большинство
особей регистрируются повторно в пределах 1—3 км от места мечения.
В Дании из 146 животных 102 были замечены на расстоянии менее 3 км,
44 — в 3 км и только 3 зверя ушли за 10 км. Наиболее дальний заход (25 км)
отмечен в Западной Ютландии (Strandgaard, 1972). Максимальная дистанция
перемещения косуль в Швейцарии и Франции — 8—9 км (Blankenhorn, 1975;
Sempere, 1979). В лесном районе в Польше из 84 особей 84% самцов и 54% самок
наблюдались не далее 1 км от места мечения и лишь 17% самок ушло на расстояние,
превышающее 3 км. Максимальная дистанция составила для самцов 2,3 км, для
самок 5 км (Fruzinski et al., 1983). Сходные данные получены и для полевой
популяции (Pielowski, Bresinski, 1982).
' Миграция у косуль — перемещение животных за пределы сезонного или годового (многолетнего)
участка обитания.
101
По сведениям В. Рика (Rieck, 1956, цит. по: Stubbe, Passarge, 1979), который
проанализировал возвраты 1361 метки, 74% телят и 41% взрослых косуль
найдены в радиусе 1 км от места мечения; 95% сеголеток и 76% взрослых —
в радиусе 3 км; 96 и 86% соответственно — не далее 5 км; на расстояние более 10 км
переместились только 2 и 7% животных, более 50 км — 0,3 и 0,6%. Такие же
результаты приводит Элленберг (Ellenberg, 1978), обработавший сведения о 1058
меченых особях из нескольких стран Средней Европы; большая часть зверей
обнаружена повторно в пределах 2,5 км от места мечения, и лишь отдельные
мигрировали далее 10 км, максимально на 70 км. В Австрии по данным мечения
2734 детенышей косуль среднее расстояние повторных встреч самцов не
превышает 2,4 км, самок — 2 км, сеголеток — 0,6 км, более половины взрослых и 83%
сеголеток обнаружены в пределах 0,5 км, 3 молодых самца найдены в 25—32 км
и одна самка — в 20 км (Reimoser, Zandl, 1987). Наиболее значительны перемещения
отдельных зверей на севере Европы: в Норвегии на 90 км (Myrberget, 1965) и
в Швеции на 205 км (по: Strandgaard, 1972).
Эти и другие исследования указывают на ярко выраженную оседлость
большинства особей европейской косули (см. главу "Структура популяций"). С другой
стороны, очевидно, что в оседлых популяциях есть животные, предпринимающие
дальние перемещения, которые на севере и востоке ее ареала, а также в горах
могут носить характер регулярных сезонных миграций.
Отмечены сезонные вертикальные кочевки в восточной части Грузии (Арабули,
1963а) и в предальпийских областях Швейцарии (Robin, 1975]; осенние перемещения
в Центрально-Черноземных областях (Простаков, Обтемперанский, 1986);
спорадические летние передвижки животных в Молдавии, связанные с усыханием
водоемов (Успенский, Лозан, 1965). Миграционный путь косуль зарегистрирован
в Прибалтике (см. рис. 34). "Почти ежегодно с завидным постоянством в мае—
июне косуля движется здесь с запада на восток, переплывая Нарву небольшими
группами (по 2—3 особи) в одном и том же ее участке протяженностью
около 15 км, расположенном между деревнями Кукин Берег и Пустой Конец.
В несколько меньших масштабах такие миграции прослеживаются в верхнем
течении Нарвы между побережьем Финского залива и Ивангородом, а также
на р. Луге и в районе Сабск-Ивановское" (Верещагин, Русаков, 1979; Русаков,
1978).
Протяженные миграции европейской косули зафиксированы на территории
Финляндии, где на севере природные условия не совсем благоприятны для вида.
Мигранты, одиночные самцы и самки с телятами придерживаются здесь морских
побережий, речных долин и берегов озер, где высота снега меньше и есть
корм в виде стогов сена. Отдельные особи удалялись на 700 км от границ
основного ареала (Pulliainen, 1980). Животные-мигранты прослежены и в Швеции.
Одна из меченых самок вернулась вместе со своим теленком на прошлогодний
зимний участок, находившийся примерно в 40 км от летнего участка, и в мае снова
мигрировала отсюда. Другая совершала миграции за 6 км от летнего участка
(Cederlund, 1982). Мигрирующие звери зарегистрированы и в других оседлых
популяциях в Западной и Центральной Европе, однако они составляют здесь,
вероятно, не более 1—2% (Strandgaard, 1972; Ellenberg, 1978).
С помощью радиопрослеживания достоверно выявлена регулярная сезонная
осенняя и весенняя миграция части особей в заповеднике "Лес на Ворскле"
Белгородской области на расстояние, превышающее 10 км (Соколов, Данилкин
102
и др., 1986а). Осенне-зимние перекочевки здесь весьма растянуты по срокам в
зависимости от высоты снежнего покрова, весенние — гораздо более заметны.
Первыми в конце марта — начале апреля в разгар таяния снега начинают
целенаправленно перемещаться к своим территориям взрослые самцы. Один из них,
форсировав в период половодья реку шириной 200 м, прошел расстояние около
13 км за 5 дней. Взрослые самки начинают передвигаться к местам отела позднее
самцов уже после полного таяния снега и появления в массе первоцветов2.
Одна из взрослых самок, снабженная радиопередатчиком, ежегодно покидала
заповедник в апреле: в 1984 г. 13-го, в 1985 — 16-го, в 1986 — 10-го числа, преодолевая
расстояния в 10—12 км за один день. Осенью в октябре—ноябре она вместе
с детьми возвращалась в заповедник на один и тот же участок. Весна в 1987 г.
запоздала в этом районе примерно на месяц, соответственно на месяц сдвинулись
сроки миграции.
В заповеднике совершенно четко выявилась картина движения особей
популяции в пространстве в зависимости от высоты снежного покрова (см. рис. 38).
При высоте снега менее 20—30 см большая часть популяции оставалась на летней
территории. При снежном покрове от 30 до 40 см на своих летних участках
оставались лишь отдельные особи, да и то только там, где на их участке имелись
малоснежные зоны или участки с большим запасом корма. При высоте рыхлого
снега более 40 см буквально все косули покидали свои летние участки и
перемещались на окраину заповедника или за его пределы, концентрируясь в
малоснежных местах по долине реки или во фруктовых садах, богатых веточным
кормом. Сюда же стекались особи со всех окрестных лесов, что приводило
к увеличению численности животных в районе наблюдений минимум в 3—4 раза.
В этих же стациях звери обитали и в особенно многоснежную зиму 1986/87 г.,
когда снег достигал 1,5 м. В этой экстремальной ситуации большая часть
популяции все-таки выжила благодаря тропам кабана, по которым шло
перемещение, и относительно быстро установившемуся плотному насту на открытых
местах (при спокойной ходьбе пальцы у них широко раздвигаются, что
увеличивает площадь опоры и позволяет передвижение). Во фруктовых садах и ивняках
косулям стали доступны практически все ветви на высоте до 2,5—3 м, и это резко
улучшило их кормовую базу. При преследовании звери пытались передвигаться
прыжками, глубоко проваливались в снег и легко становились жертвами
браконьеров и собак. На лыжах не составляло особого труда через 10—15 мин
преследования догнать косулю.
У европейской косули, вероятно, все-таки нет таких массовых сезонных
миграций, как у сибирской, хотя сообщения о них имеются. На Украине в
середине XVI в. во время сезонных перекочевок зверей убивали тысячами.
В 30—40-х годах XVIII в. зимой животные собирались в лесостепи в стада
до 100—300 голов, что также может косвенно говорить о возможности миграций
(Кириков, 1959, 1966). На Северном Кавказе в начале века отмечалась
миграция косуль из Темного леса на многоснежной Ставропольской возвышенности
в долину р. Кубани (Динник, 1910). Однако все эти факты вполне могли
относиться к сибирской косуле, скорее всего, обитавшей в этих районах ранее (см. главу
"Ареал").
Урал. Миграции косуль (рис. 34) здесь в прошлом веке наблюдал и описал
1 Аналогичные наблюдения приводит для Швеции Седерлунд (Cederlund, 1982).
103
Л.П. Сабанеев (1872, 1875). Осенью «...они в огромном количестве, большими
или меньшими стадами, до 20—50 голов, переходят через Урал и
сосредоточиваются исключительно в Каслинском Урале, где остаются до наступления весны,
когда начинается обратное переселение... Сюда стекаются козули с севера,
запада и юга из дач Полевского, Сысертского, Нязе-Петровского, Златоустов-
ского заводов; здесь как бы сборный пункт и центр их распределения; дальше
на юг и запад они малочисленнее и встречаются исключительно в летнее время...
Приходящие с запада в одних и тех же местах переходят Карабайскую гору,
а приходящие с Каштымского, Златоустовского и Миасского Урала — р. Вязовку.
Большинство козлов собирается около Татоша, Ташкуля и Семикуля, Козлиной,
Карабайской и Вишенных гор... Один охотник встретил раз осенью в 1850 году
стадо в 500 голов, шедшее из Златоуста через Кыштымскую дачу, как стадо
баранов...».
О причинах, вызывающих миграции косуль на Урале Л.П. Сабанеев (1875)
пишет следующее: "В Каслинской даче не бывает снегов, притом леса ее редки,
вершины увалов более или менее обнажены, а нередко и эти снега уносятся
в лога северо-западными ветрами. Совсем другое видим мы к северу, югу и
особенно на западном склоне Урала, снега там глубже и в Уфалейских, особенно
Нязе-Петровских дачах нередко достигают полутора-двух аршин. При таких
условиях козулям становится затруднительным как добывание пищи, так и
само передвижение".
Позднее, с уменьшением численности косуль на Урале, характер миграций
зверей в этом районе сильно изменился. Вот что по этому поводу говорит
Аверин (1949): «Регулярный уход козуль осенью из Миасского и
Златоустовского Урала в Каслинский и возвращение их весной, как писал Сабанеев,
сейчас места не имеет. Современные кочевки не носят характера массовых
регулярных передвижек... По словам старых промышленников только раз на
их памяти была массовая перекочевка козули с севера на юг. Ход животных
начался поздней осенью 1911 г. и продолжался с перерывами до зимы.
Встречались стада по 50 голов. Козуль всюду было так много, что некоторые
охотники добыли за зиму 1911/12 г. до 300 козуль каждый. Дневная добыча
в 2—3 штуки считалась небольшой; обычно убивалось больше — до 9 штук.
Обратного хода весной не наблюдалось».
К середине XX в. на Урале весьма значительно изменилась экологическая
обстановка. На путях миграций зверей были построены железные дороги, заводы
и города. Основная масса косуль из высокогорной части Урала стала кочевать
на восток и северо-восток в Ильменские горы, откуда часть их уходила дальше
в восточном и юго-восточном направлении в лесостепь, доходя до Казахстана.
Численность особей в группах мигрирующих животных осенью и зимой составляла
2—6 особей, редко больше, весной в обратном направлении мигрировали
группы от 5—6 до 30—120 животных. В продолжительную многоснежную
зиму 1941 г. наблюдалась массовая гибель зверей, в основном молодых и
беременных самок от истощения и воспаления легких (Ушаков, 1954).
В последние десятилетия, когда депрессия численности косуль на Урале
достигла своего апогея, а также из-за сильной урбанизации районов, по которым
проходили пути миграций, массовые перемещения зверей на востоке хребта
практически прекратились (Соколов, Данилкин, 1981), однако при увеличении
численности их следует ожидать.
104
г
Q
<£?=>
ш^
/л,
I ^^3?М£Ш А/
<v^/
■Si
5' V ,
£ЙУ
Рис. 34. Схема основных путей сезонных миграций популяций косуль
/ — границы ареала; 2 — основные горные системы (осевые части); 3 — основные направления осенне-
зимних перемещений; 4 — зоны зимней концентрации животных
8
С западных склонов хребта, по сообщению охотинспекций, звери
мигрируют небольшими группами на запад, доходя до Татарии и Куйбышевской
области.
Северный Казахстан и Западная Сибирь. О массовых миграциях косуль
в Кустанайской области в 1951 г. известно из сообщения Ф.Т. Герасимова
(Куражсковскии, 1952). Одиночками и стадами до 50 голов они шли с севера
со стороны бора Аман-Карагай (возможно, из Троицко-Челябинского района)
в сторону Аман-Тельды. Животные были сильно истощены. Ббльшую часть
их истребили люди и собаки, но часть уцелела и осталась жить в песках
близ Тургая.
Сюда же, в Северный Казахстан, мигрируют звери из соседних областей:
Тюменской, Курганской и Омской. В Павлодарскую область они заходят
из Новосибирской области и, вероятно, из Алтайского края (Слудский и др.,
1984). В Новосибирской области откочевки косуль на юг отмечены Ю.С.
Царевым (1969).
Южный Казахстан. В Заилийском Алатау отмечены вертикальные миграции
(Белоусов, 1978). По данным А.А. Слудского и др. (1984), косули, живущие
в пойме р. Или, оседлы. В Терскей Алатау с верховьев р. Каркары они уходят
осенью в долины рек Большой Кокпак и Баянкол. В Джунгарском Алатау звери
кочуют в западном и северо-западном направлении в предгорья и равнинные
тугаи долины р. Лепсы, в пески Каракум, Таш-Каракум, Аралкум и Алаколь-
скую котловину, с горного массива Текели — в пойму р. Тентек и в пески
Сарыкум.
С северных склонов хребта Тарбагатай животные мигрируют на север в Зай-
санскую котловину. Западная часть Зайсанской котловины и пески Кызылкум
служат зимовкой косуль, обитающих на Колбинском, Нарымском и в западной
части Курчумского хребтов (Поле, 1974). В восточную часть Зайсанской .
котловины (пески Айгыркум) кочуют звери с восточных склонов Курчумского
хребта и горных массивов, расположенных близ оз. Маркаколь.
Восточный Казахстан и Алтай. На Алтае косули зимой скапливаются в
предгорьях в Майминском, Онгудайском, Алтайском, Солонешенском и Чарышском
районах (Савинов, 1967). Часть их уходит в более южные районы (Лавров, 1929),
куда, возможно, мигрируют звери из Восточно-Казахстанской области с Убин-
ского и Ивановского хребтов (Слудский и др., 1984). С верховьев Катуни,
где высота снежного покрова более 1 м, они кочуют вниз по течению в долину
р. Бухтарма. Оставшихся на зиму особей добывают соболевщики или росомахи
(Насимович, 1955). В конце прошлого века были известны миграции животных
через Иртыш в районе между Бухтарминским и Александровским редутами
(Никольский, 1883).
В настоящее время на Алтае существуют три основных потока мигрантов
(Собанский, 1987). Основной из них — с северо-запада из предгорий и
равнинных участков на юго-восток в горные районы по долинам рек Ануй и Песчаная
через Ануйский, Чергинский и Семинский хребты, где часть животных остается
на зимовку. Наиболее мобильные стада пересекают Чуйский тракт в районе
сел Калмак и Черга, переплывают через Катунь выше устья Семы и задерживаются
на ее правом берегу. Фронт этого миграционного потока достигает 50 км,
протяженность его от 70 до 200 км. Второй, менее многочисленный поток мигрантов
из западных предгорий направляется по долине Чарыша в ее верховья, оста-
106
навливаясь в районе сел Усть-Кумир и Талица, а в многоснежные годы звери
достигают села Усть-Кан и распространяются восточнее. Некоторые стада из
предгорий в начале зимы уходят на север, на равнину в сравнительно
малоснежные сельскохозяйственные угодья. Сроки миграции на этих двух направлениях
зависят от начала обильных снегопадов и растянуты с конца октября до середины
января. Третий миграционный поток отчетливо выражен на юго-западе Алтая.
В конце октября — начале ноября животные кочуют из многоснежных районов
Восточного Казахстана и с хребтов Коксуйского, Холзуна и других вдоль
р. Кокса до ее правого притока — р. Карагай, распределяясь на зимовку
в ее нижнем течении и на левобережье Коксы. При сильных снегопадах
происходит откочевка косуль в восточном направлении вниз по долине реки.
Из-за резкого снижения численности зверей в 60-е годы практически "иссякли"
два других миграционных потока: с севера на юг по долине Катуни в восточную
часть Шебалинского района и с востока из бассейнов р. Лебедь, верхних
притоков Бии, из Чуйского и Турачакского районов вдоль р. Сара-Кокши на
запад в малоснежные районы по правобережью Катуни. В 80-е годы с
увеличением поголовья намечается восстановление этих миграционных путей.
Мигранты на Алтае движутся чаще всего в светлое время суток небольшими
группами в 3—5 особей. Число животных, приходящих в горные районы, в
отдельные годы существенно колеблется (на 70—100%) в зависимости от высоты снежного
покрова. В малоснежные зимы большая часть косуль не мигрирует и зиму
они проводят в летних стациях (Собанский, 1987).
В Горном Алтае наряду с мигрирующими популяциями обитают и оседлые.
Большинство районов здесь отличаются малоснежьем и животным нет надобности
покидать их. Наблюдается лишь незначительная передвижка в более s
благоприятные места: летом большая часть их поднимается в верхний пояс горных
лесов, в сентябре спускается в предгорные районы (Дмитриев и др., 1937; Дмитриев,
1938).
Восточная Сибирь. Здесь миграции выражены очень отчетливо. В
Красноярском крае выделяются две крупные группировки популяций (географические
популяции): одна ограничена с запада Кузнецким Алатау и Абаканским хребтом,
с юга — Западным Саяном, с востока — западными отрогами Восточного
Саяна; другая находится восточнее Енисейского кряжа и с юга и юго-востока
ограничена Восточным Саяном и его отрогами. На севере границы популяций
определяются зоной темнохвойной тайги (см. рис. 34).
Косули первой географической популяции осенью мигрируют с Кузнецкого
Алатау и Абаканского хребта на восток, с Западного Саяна — на север в
Хакасские степи. С западных склонов Восточного Саяна звери направляются
к Енисею. Они движутся широким фронтом по одиночке и стадами до 60 и более
особей по пяти миграционным путям, перемещаясь в среднем на 50—60 км.
Осенняя миграция начинается здесь в конце октября — начале ноября при
высоте снежного покрова 15—30 см, заканчивается в конце ноября — начале
декабря. Весенняя миграция в обратном направлении идет с начала апреля
до конца первой декады мая (Лапсин, Соколов, 1971; Субботин, 1973;
Мельников, 1978; Ельский, 1983).
С образованием водохранилища Красноярской ГЭС традиционные пути
миграций косуль нарушились. Ниже плотины Енисей не замерзает на
протяжении 100 км и становится существенной преградой для животных, переправы
107
чаще всего оканчиваются гибелью зверей (Зырянов, 1975). До образования
Красноярского моря до тысячи косуль ежегодно переходили Енисей по льду
ниже Сарагаша, теперь путь миграции сдвинулся южнее на 70—100 км в район
Ербы, где на западный берег Енисея кочуют всего 150—200 особей. Основная
часть популяции остается на восточном берегу в менее благоприятных для
зимовки местах. Переправе косуль на другой берег берег мешает значительная
ширина скользкого льда. Раньше лед, образовавшийся после шуги, не был
таким гладким и не препятствовал передвижению (Лапсин, Соколов, 1971).
Отрицательное воздействие на миграционные пути косуль в Хакассии
оказала также сеть новых железных дорог, но особенно пагубно Для популяций
сказалась распашка целинных земель — основных зимних кормовых стаций
животных и браконьерство (Байкалов, Семенов, 1971). Миграции здесь из-за комплекса
отрицательных факторов и снижения численности стали затухать.
Косули второй популяции из долин рек Тасеева и Бирюсы со второй декады
октября мигрируют на юг на расстояние от 40 до 100 км по трем путям. Южнее
нет четко выраженных путей кочевок: с гор Восточного Саяна звери постепенно
(с ноября до февраля) по мере увеличения снегового покрова передвигаются
в малоснежные предгорные и лесостепные угодья в район Канска. Отдельные
стада остаются в местах летнего обитания, там, где им легкодоступны зимние
корма (Субботин, 1973; Лавов, 1975).
Третья крупная географическая популяция довольно четко определяется в
Иркутской области: с севера она также ограничена таежной зоной, с запада и
юга — восточными склонами Восточного Саяна и с востока — Приморским и
Предбайкальским хребтами. С горных хребтов и из северной тайги звери
мигрируют осенью в разреженные леса и малоснежные пади в Приангарье, в
Ордынские степи и другие степные и лесостепные районы (Троицкий, 1930; Подарев-
ский, 1936; Фетисов, 1953). Свои впечатления о миграциях в этом районе А.С.
Фетисов описывает так: «Козули из многочисленных мест Саянских гор идут
сюда как одиночно, так и группами в 3—5 и более голов. Внезапно выпавший
глубокий снег заставляет их передвигаться более быстро. В Черемховском
районе Иркутской области после выпавшего такого снега 7, 8 и 9 ноября 1943 г.
мне пришлось наблюдать ход козуль на расстоянии по меньшей мере в 8—10 км
шириной. Снеговой покров был буквально испещрен следами этого копытного.
Миграция проходила кратковременно, интенсивно. Глубина снегового покрова
в местах наблюдения достигала 35—40 см. В горах, южнее места наблюдений,
он был, конечно, еще более глубоким. "Ходовая" козуля мало
останавливается во время миграции на одном месте. Внезапно выпавший снег буквально
гонит ее к малоснежным местам. Основной причиной таких миграций является,
несомненно, боязнь козули глубокого снега, покрывающего кормовую ветошь,
не позволяющего козуле свободно передвигаться и лучше спасаться от почти
постоянно преследующих ее волков и других хищных зверей».
Забайкалье. Горный рельеф местности с обширными высокогорными гольцами
и глубокоснежьем, значительные открытые степные пространства отчетливо
расчленяют этот регион на пригодные и непригодные для косули биотопы.
Горные леса и здесь являются временными стациями косули. Ежегодно осенью
животные вынуждены из-за многоснежья спускаться с горных хребтов в предгорья
и лесостепь, где высота снега не превышает 30—40 см.
С xf>. Хамар-Дабан косули с выпадением снегов мигрируют на восток и юго-
108
восток, с северных склонов Джидинского хребта — на север; с южных — на юг
в Монголию; с Малханского, Заганского и Кударинского хребтов — на их
северные и южные предгорья; с хр. Улан-Бургасы — в верхне-среднее течение
р. Оны, левобережную часть Курбинской котловины в лесостепи, прилегающие
к долине р. Уды; с хребтов Цаган-Дабан и Цаган-Хуртей — в долины рек Худун,
Кижинга, Илька (Смирнов, 1978).
В этом регионе.известны миграции косуль от Байкала вверх по р. Баргузин
(Доппельмаир, 1926). С Черского, Яблоневого и других близлежащих хребтов
животные уходят в долины рек Джиды, Чикоя, Ингоды, Нерчи, Онона, Шилки,
Ульдурги, Уды и Кижинги. Во время миграций эти и другие крупные реки не служат
им заметным препятствием (Кузнецов, 1929; Фетисов, 1953; Скалой, 1960).
В многоснежные годы в южную часть Забайкалья могут перемещаться звери
с северо-западных отрогов Большого Хингана. По описанию Н. Кирилова (1902),
«зимой и в начале весны 1894/95 г. в Дучарской волости избито до 2000 козуль,
бежавших к нам с Маньчжхрской стороны Аргуни».
Расстояния, на которые мигрируют косули в Западном Забайкалье, невелики —
20—60 км (Смирнов, 1978). А.А. Насимович (1955) считает, что на Витимском
плоскогорье они имеют протяженность от 100 до 200 км.
Первые перемещения стад косуль в районы зимовок начинаются в сентябре.
Группы мигрантов обычно состоят из 2—7 особей, редко из 40—50 особей
(Лавов, 1978). Весенняя миграция менее выражена и более растянута по срокам:
начало ее отмечено в апреле, конец — в начале июня.
Расчлененность рельефа и постоянные из года в год пути перемещений
зверей приводят к образованию локальных группировок. Тем не менее
популяции не совсем изолированы одна от другой и между ними возможен обмен
особями (Смирнов, 1970а, 1978).
Якутия. Здесь регулярных сезонных миграций не установлено, что, по-видимому,
объясняется довольно сходными климатическими и кормовыми условиями,
совпадением летних и зимних стаций (Егоров, 1965) и относительно малой
численностью.
Дальний Восток. Очень интересные описания миграций косуль в
Уссурийском крае сделаны Н.М. Пржевальским (1870), который также считал причиной
переселений косуль глубокие снега, «при которых козе весьма трудно ходить,
а еще труднее добывать пищу. В береговой полосе, где снегу выпадает меньше,
нежели внутри Уссурийского края, дикие козы остаются круглый год на одних
и тех же местах... Зимой козы до того исчезают из уссурийских лесов, что редко-
редко где попадаются одиночки, вероятно отсталые или, по какой-либо
особенной причине, не успевшие уйти вместе с другими. Впрочем, близость или дальность
переселения зависит от количества выпавшего снега, так что в малоснежные
зимы множество коз остается в горах западной части Ханкайского бассейна».
Осенние миграции косуль в этом регионе приурочены к октябрю—ноябрю,
весенние — к концу марта — апрелю. По наблюдениям П.И. Ветлицина (1902)
продвижение косуль заметно уже в сентябре. Звери скапливаются у рек в
определенных местах и здесь выжидают ледостава. "Но лишь только станут реки,
а то бывает в конце октября, дикие козы трогаются в путь, но только в том случае,
если выпал уже большой снег, что обыкновенно в это время случается. В это
время козы идут большими стадами и не торопясь, жируя по пути на еланях
и по небольшим увалам, покрытым мелким кустарником и дубняком... Ход
109
козы не всегда бывает большой; если снега мало, коза идет в незначительном
количестве. Последние четыре зимы почти не было снега и коза почти не шла.
Обыкновенно каждая снежная метель, или как здесь говорят "пурга", вызывает
ход козы».
В XIX в. миграции были столь интенсивны, что по описанию Н.М.
Пржевальского и П.И. Ветлицина во время массового хода охотник мог добыть до сотни
особей. Из "Приложения к отчету военного губернатора Амурс/ой области
за 1888 г." известно, что многоснежной зимой этого года был "неслыханный
ход козы с северных плоскостей по направлению Маньчжурии", во время которого
охотники добыли около 150000 штук. В "Охотничьей газете" за тот же год
(N 16, с. 184) сообщалось, что в декабре вследствие глубоких снегов
наблюдалось массовое передвижение косуль с севера на юг. В течение месяца только
возле Благовещенска было убито около 10000 косуль.
Такие же массовые миграции, как в 1865 и в 1888 гг. отмечались в разных районах
Дальнего Востока в 1912, 1917, 1920, 1927, 1932, 1942, 1950, 1956, 1959 и 1972 гг.
(Баранчеев, 1962; Раков, 1965; Кучеренко, Швец, 1977).
Очень подробное описание миграций косуль на Дальнем Востоке составил
Н.В. Раков (1965), который пришел к заключению, что сезонные перемещения
(осенние — с конца октября до декабря, весенние — с конца марта до июня)
звери совершают в пределах локальных районов. Соседним крупным
группировкам свойствен очень слабый обмен населением. На территории Амуро-Уссурий-
ского края, вероятно, существует четыре географические популяции: зейско-
буреинская, нижнеамурская, приханкайская и приморская, каждая из которых
образована более мелкими экологическими популяциями, имеющими свои
постоянные миграционные пути и районы зимовок.
Популяции подразделяются на два типа: зимующие за пределами летних
областей обитания (преимущественно кочующие) и зимующие в пределах
области летнего обитания (преимущественно оседлые). При неблагоприятном
снежном режиме северные популяции быстрее и полнее смещаются с места-зимовок,
вызывая "массовые ходы" животных в поисках благоприятных малоснежных
кормовых биотопов.
Таким образом, Н.В. Раков считает, что "массовый ход" косуль является
лишь концентрированным выражением обычной осенне-зимней миграции,
развивающейся в обстановке недостаточной урожайности кормов и высокой
снежности. В случае дружных и повсеместных снегопадов, особенно сырых,
ускоренно двигающиеся к местам зимовок животные скапливаются в большом
количестве в локальных районах. Перенаселение, в свою очередь, служит толчком
к смещению местной популяции. Поэтому именно в районах зимовок, где
существует больше возможностей для создания перенаселения, и отмечаются
массовые миграции, проходящие по не совсем обычным направлениям, что нередко
заканчивается гибелью почти всех особей из какой-либо локальной популяции,
надолго исчезающей затем из районов летнего обитания. Это случалось
неоднократно с удской, верхнезейской, урканской, амгунь-горийской, хунгарийской,
коппинской, ботчинской и нельминской популяциями.
Массовую миграцию косуль в Амурской области, причиной которой
послужило резкое похолодание и обильные снегопады на севере области в октябре
1959 г., описал Л.М. Баранчеев (1962). Пожалуй, это наиболее подробный
фактический материал о массовой миграции, поэтому некоторые выдержки из его
110
работы мы также приводим дословно. «Раньше всего "ходовые" косули появились
на севере Мазановского района несколько севернее Усть-Норска, передвигаясь
с севера на юг. Отдельные табуны косуль стали замечаться уже 5 октября. С 15 по
25 октября наблюдалась массовая миграция. В это время численность табунов
достигала 20—30 и больше голов. Двигались они тремя основными потоками
с северо-востока на юго-запад. Первый поток переходил р. Нору в ее устье,
второй — в 20 км выше устья и третий — в 35 км. Переправлялись через реку
вплавь, вначале мелкими группами по 2—4 штуки, а затем от 10 до 15 и более.
В один из таких дней егерь B.C. Самокрутов насчитал 127 косуль, переплывших
р. Нору. После ледостава вторая волна массовой миграции приходится на 7—8
ноября. Шли косули ночью и, судя по следам на снегу, очень большими
табунами. К 20 ноября миграция закончилась... Река Зея не представляла для них
непреодолимого препятствия, хотя в это время шла уже шуга, а позже большие
льдины. Подходившие к берегу табуны косуль, не задерживаясь, прыгали в воду
и плыли на противоположную сторону. Переплывшие реку в течение первых
40—50 мин отдыхали, после чего продолжали путь на юг... В период хода сплошной
шуги по р. Зее с 7 по 10 ноября отмечался наиболее интенсивный переход
косуль. Большие табуны, численностью до 70 голов, подходя к реке, бросались
в воду и плыли. На правом берегу у кромки льда преобладающее большинство
косуль тонуло из-за невозможности выбраться на гладкий скользкий лед.
От табуна в 70 голов выходило на берег не более 14—16. В районе с. Буссе,
расположенного на левом берегу Амура, наибольшая концентрация животных
отмечалась в конце октября. Косули шли с запада на восток. Пытаясь преодолеть
водный рубеж, животные прыгали на проплывающие льдины. Многие из них сразу
же тонули среди идущей по реке плотной массы шуги. После ледостава
наблюдалось немало случаев, когда косули забегали в сараи и на скотные дворы
с. Буссе. При этом животные почти не боялись людей. Кормились они в основном
на соевых полях. Закончилась миграция 17 ноября 1959 г. Весной на маршруте
массового перехода было обнаружено большое количество трупов косуль».
Отдельные стада во время этой миграции насчитывали 200, 350 и даже 600
особей3. Зимовка зверей проходила на левобережье Амура в районе Благовещенска,
излюбленнымм местам жировок служили соевые поля. Весной они шли по тем
же путям в обратном направлении с середины апреля по 1 июля. Во время
ледохода также отмечалась гибель косуль в реках.
На территории Верхнего Приамурья выделены два основных миграционных
потока: северный и южный. Северный поток включает 4 миграционных пути:
первый идет от верховьев рек Ига и Альдикон двумя ветвями на Зейско-
Буреинскую равнину и Амуро-Зейское плато; второй — из котловины оз. Огорон
и бассейна р. Деп через долину р. Гарь, где пополняется особями из бассейна
р. Нора, до правобережья р. Селемджи и затем к долине р. Зея, которую звери
преодолевают вблизи сел Ураловка и Практич. Часть животных, переправившись
через Селемджу, уходят на Зейско-Буреинскую равнину. По третьему
миграционному пути животные из Верхнезейской котловины через Зейское ущелье
направляются в бассейн р. Уркан, по четвертому — от хребтов Тукурингра и Янкан,
которые они преодолевают в сентябре еще до снега, до бассейнов рек Уркан
'По сведениям старожилов, в ноябре—декабре 1912 г. в Буреинском районе встречали стада
численностью до 1000 голов в каждом (Баранчеев, 1962).
111
и Ольдой. Южный миграционный поток с правобережья Амура имеет два основных
направления к зимовке: между реками Буриндой и Осежиной и по долине р. Уль-
мин, причем мигранты с правого берега Амура появляются на левобережье
ранее северных популяций4. Осенью в этом регионе первыми перемещаются
группы взрослых самцов, состоящие из 10—15 особей, а позднее идут смешанные
стада, насчитывающие до 100 особей5. На интенсивность миграции здесь также
оказывает существенное влияние высота снежного покрова (Дымин, 19756).
С образованием Зейского водохранилища пути миграций косуль оказались
перекрытыми широкими заливами. Переходы зверей через Зейское ущелье
прекратились, а численность верхнезейской популяции резко уменьшилась.
Сейчас наметились новые миграционные пути через Огоронскую низменность
и понижения хребта Тукурингра западнее пос. Золотая гора (Щетинин, 1975; Бром-
лей, 1984; Ильяшенко, 1984).
На Дальнем Востоке очень четко видна разница (до 400—500 км) между
летней и зимней областями распространения косуль, т.е. ежегодно происходит
сезонная пульсация ареала (Раков, 1965; Кучеренко, 1976; Бромлей, Кучеренко,
1983), вызванная протяженными миграциями. Районы зимовок косуль приурочены
в основном к долине Амура и его притоков.
В 60—70-е годы из-за распашки целинных земель площадь лучших зимних
биотопов косуль на Зейско-Буреинской и Уссурийско-Ханкайской равнинах
существенно сократилась, что наряду с другими факторами отрицательно
сказалось на численности зверей. Некогда грандиозные их миграции стали не столь
заметными и в некоторых районах прекратились совсем. Кроме того, для
многих мигрирующих популяций оказалась катастрофической многоснежная зима
1972/73 г. Снег высотой до 1—1,5 м, выпавший уже в конце октября, отрезал
их от района зимовок. Много зверей утонуло при переправах через реки,
превратившиеся в ловушки из-за тонкого льда, покрытого пухлым слоем снега.
Оставшиеся в живых бедствующие животные становились легкой добычей браконьеров
и хищников, гибли от бескормицы, замерзая целыми стадами до 20—30 особей.
К весне их численность сократилась в 3—4 раза (Швец, 1975; Кучеренко, Швец,
1977).
В последние годы поголовье косуль на Дальнем Востоке увеличилось,
особенно в Амурской области, где миграции снова стали ярко выраженными.
Поскольку сведения о характере миграций скудны и порой противоречивы,
а структура мигрирующих популяций практически не изучена, мы провели
в 1989—1991 гг. специальные исследования в Мазановском районе Амурской
области на водной переправе через р. Нора примерно в 5 км от устья6.
Непрерывные наблюдения в сентябре и октябре в светлое время суток с помощью
биноклей и зрительной трубы (усиление 30X60) дали возможность определить
пол, возраст и число особей, пересекающих реку (рис. 35) на двухкилометровом
4 В настоящее время этот миграционный поток практически угас из-за пограничных сооружений
по р. Амур.
Г. Ельский (1983, 1986) считает, что первыми мигрируют самки с молодыми, а затем взрослые
самцы.
6 В работе принимали участие зоологи Ю.А. Дарман и Р. Прайор, инженеры А.Н. Минаев-,
Ю.Д. Вулис и сотрудники Норского республиканского зоологического заказника, которым автор
приносит глубокую благодарность за помощь в работе.
112
"„.Тш£-:,,-;Л» '■
"'^.■''J':''*'■•'"■
Щ.' ч ... J'^
■ -ц^Гд irr'liS^^j|Haj<t"f
„»Ч»>11,..
*,.,; .**r... .a.- rf ,.',: „.*£"■
.,».■■« -J г-* *
^Lr^&ffr*
JpSb»"
'^v.wsw'*'^
:;ml
m щи
й**
■■■*
<*
"j «^*
'a
i
' <;>"!
f,
r *vV:-
J*-
• •?"* -':
- 'як . •'■". •■ *■ ilttf ни»».... «^
. Рис. 35.Сибирские косули — мигранты (р. Нора, сентябрь 1989 г.)
8. Европейская и сибирская косули
20.
/00%
ВО
60
W
20-
9 22.9 21.9 263 28.9 30.9 2.10 4.10 6.10
Б Латы
-j=
-
I
ll'l
|Д
I
1
§
1
7 в 9 101112 13141516 171819 20
Времягч
j c*9 Juir
Mdd gad
^dd ad
20 22 24 26 28 30
Сентябрь
20-30.09
Рис. Зб. Половая и возрастная
структура (Б) и миграционная активность (А, В)
популяции сибирской косули (Амурская
обл., р. Нора, сентябрь—октябрь 1989 г.)
А — дневная, В —почасовая
активность
участке, а также зарегистрировать их поведение и последующий путь благодаря
мечению радиопередатчиками и слежению с самолета АН-2. При ночных
наблюдениях фиксировалось только количество переплывающих зверей.
Миграция косуль из северных районов Амурской области начинается не в октябре,
как полагал Л.М. Баранчеев (1962), а в начале сентября. Судя по сообщениям
егерей Норского республиканского зоологического заказника, первые особи,
пересекающие р. Нора и ее притоки, появляются ежегодно примерно с 10 сентября,
однако в массе — в последней декаде этого месяца.
Следует особо подчеркнуть, что в исследуемом районе осенняя миграция
сибирских косуль идет в бесснежный период года, когда дневная температура
воздуха находится в пределах +10...+20°С, а ночью лишь иногда опускается
чуть ниже 0°. Следовательно, несмотря на очевидную в целом связь сезонных
перемещений косуль с режимом снежного покрова, этот фактор не всегда служит
"пусковым механизмом" осенних миграций.
Поток мигрантов неравномерен (рис. 36, А) и явно образует "волну миграции",
пик которой осенью 1989 г. пришелся на 27 сентября. В этот день только
на двухкилометровом участке реки зарегистрировано 318 плывущих зверей,
а всего за 11 дней в светлое время суток реку форсировали здесь 1352 косули,
преимущественно утром и вечером, т.е. во время своей обычной активности
(рис. 36, В). Плывущие животные отмечены наблюдателями в любой час суток,
ночью миграция не прекращается. По приблизительным расчетам в темное время
суток реку переплывают от 20 до 50% особей по отношению к дневному потоку
114
мигрантов. В общем через р. Нора и р. Селемджа на протяжении миграционного
пути в 60 км переправляется более 20 тыс. особей.
Первыми начинают перемещаться семейные группы — мать с детенышами,
однако в пик миграции соотношение животных по полу и возрасту примерно
одинаково и соответствует структуре популяции (рис. 36, Б).
Водные преграды косули преодолевают, как правило, в одних и тех же местах.
По данным радиопрослеживания с самолета 16 косуль, оснащенных
радиопередатчиками, протяженность их миграционного пути 108—178 км, средняя скорость
перемещения осенью 6,5 км (максимальная 26 км), весной — 3,8 км в сутки;
расчетный период осенней миграции составляет 17—27 дней, весенней — 30—47 дней.
Район зимовки популяции находится на левобережье Зеи (рис. 37). В обратном
направлении животные мигрируют в конце марта—начале апреля по тому же
маршруту, что и осенью. Летние и зимние участки обитания особей одни и те же
из года в год.
Интересно поведение мигрирующих косуль на переправах через реки.
Большинство взрослых самцов форсируют их в одиночку или же в группе,
преимущественно состоящей из самцов. Максимальный размер такой группы 17 особей,
в которой находились 12 взрослых самцов. Попытки больших "самцовых" групп
пересечь реку в большинстве случаев безуспешны. Самец-лидер, проплыв несколько
метров, возвращается назад, уступая дорогу следующему за ним, второй — третьему
и т.д. Они долго кружатся в воде и, наконец, возвращаются на берег. Успешными
были переправы больших (до 14 особей) групп, если лидером становились
взрослая самка, имеющая телят. Именно они лидировали в большинстве случаев,
причем семьи косуль (мать с одним, двумя или тремя детенышами) чаше
переправлялись отдельно. Иногда к ним присоединялись полувзрослые самки,
обычно следующие поодиночке или в паре с полувзрослым самцом. Большие
группы образовывались чаще по утрам. Складывается впечатление, что некоторые
косули ждут рассвета, не рискуя плыть ночью.
На переправах через реки матери, несмотря на звуковой контакт, нередко
теряют своих детенышей, отставших на воде и вернувшихся назад вместе
с другими взрослыми косулями, которые испугались плыть дальше. Телята,
потерявшие мать, громко пищат, пытаются присоединиться к другим группам
косуль, но те обычно прогоняют их. На писк детенышей отзывается мать,
ожидающая их на противоположном берегу, и через некоторое время телята
плывут к ней. Иногда матери возвращаются за ними, поэтому ежедневно
наблюдается несколько животных, плывущих против основного потока мигрантов.
При опасности мать, потерявшая детей, удаляется от берега, а детеныши,
иногда объединившись в группу из 3—5 особей, переплывают реку. Не найдя
матерей на противоположной стороне, некоторые плывут назад, выбиваясь
из сил. В пик миграции оба берега оглашаются писком детенышей. Очевидно,
что они представляют легкую добычу для хищников.
Во время миграции мы неоднократно регистрировали половое поведение
косуль. Концентрация животных на путях миграций несомненно способствует
успешному размножению животных. Неоплодотворенные самки, пришедшие
в течку, гарантированно покрываются самцами.
Монголия. Сезонные миграции косуль отмечены на севере и на востоке
страны (Банников, 1954; Соколов и др., 1982). Из Китая с хр. Большой Хинган
они большими стадами и мелкими группами уходят на территорию Монголии
115
v-\^ ) -г1
л \ ^ f
Гарь ^^/V_ >^ ;^^^^\ Ч
-^~N^ ^ '^~J/^?P^-^--- _-^^-— -—--^^ffl ^
0*X——-7^~Z^S JP?I——J2^>*^~-^
s*—^"T__»---"^^ "/N^^-^^^^ ( ^-^—^
—^•±^~J£r/ / ^ifr^^'^*' _А--^ у
rJ^^^^^/^%^^A /
L ^^ /^ / /~^s*s^T" sl-—' 7*-—' o^-Z\ Норсм /
Vivl 7*3) (J4) p£7 ^/£<7у'Л'1' ) /\
\ 1* W/7if/v^/W \ P /fcr^L-^'^
V-ЛЧагоян . X^:'-':^\ 1 1 V':\ •jCsjLlJ ^< ri£x/~
4 (^W^/ 3 iH>
1 ( / / ^''"'•ЧЗ-^^—
) ^~^ - <v / /
—) ,^S\D P
С А %)""^
I Норокиенекий Увал ( Мазанопо1
Г
Vv
/
/
Ч
V
i С с л е м д щи чек
~^' у^^"^
-/^—-—\ г\ Jyi fDenpant.cn
^^T^-^^wV
^^^^^^ \ с^а\
^^/ \ Л
„ilKOW jS—Ч У
V/ 1 Я
IX]»L©_,
-з. [z:
Л Г x
*l« 1 x
mil ?
0 0 10 km
j i
' 1 ■ ■ * 1
Д
С
*
з
5^^-
Y^ Чагоян
10 H
h-T-r-r-= - -
Г
£]
Гарь ^v/~\
(Тз/
"Щ^^^2У У/ 2
-^ ^i^ -^ • /-~-f 1
ЩЪх-^ -^ -S" ' УЛ ] ^
10 км
-1
if »^_
^ 7
1 "* I У
( д ^
С J3>.X
#,з/ \
^^S/ xi*^^^^^-^---*®l3
-/pN^Tli) ^?i£?—^—-- г-г-" '
/JJAIAL/^—^^^^^1-:.J ^ -^
УС^г- "^jli^Z'/i. VlP4^—•'"" n-^ ^V^v
s'/j ^"^4 / ^/-^"~"~"^
/ ( /^ч Sjj
у / >s | ^w^S
1 Нойон и «некий Упал (.Мазаноно) ^
Сел'емджинси /
^^У s®
jg) Шевральск
\ ^~
4
„Л^.
Рис. 37. Сезонные пути миграции и участки обитания косуль, оснащенных радиопередатчиками
/4 — осенняя, £ — весенняя миграции; а — пункты отлова и мечения; б, в — пути миграции косуль,
помеченных в сентябре 1989 и 1990 гг., соответственно; г — зимние, д — летние участки обитания; е — пункты
метеонаблюдений; ж — места гибели косуль; з — исчезновение сигнала радиопередатчика. Цифры — номера
косуль
s C.cJCp.
\ 50160
4 ,
% 40/50
\ 30/10
I 20/30
* 10120
Рис.38.Схема пространственной
стратегии популяций косуль в зависимости
от высоты снежного покрова
/ — популяция оседла; 2 — популяция
мигрирует; Се. — европейская косуля;
Ср. — сибирская косуля
*§ 5 10 20 500
Расстояние миграции км
в равнинные степи за десятки и сотни километров от летних участков обитания,
туда, где снега меньше. 'Наиболее заметными и протяженными были
перемещения косуль в особенно многоснежные зимы 1977 и 1978 гг., когда мигрирующие
косули достигли районов Тамсаг-Булака и Матада — типичных мест обитания
тысячных стад дзерена.
Наряду с перемещениями, вызванными неблагоприятными факторами среды,
у европейских и сибирских косуль отчетливо выражено выселение молодых
особей, которые также можно отнести к разряду сезонных миграций.
Ежегодно весной значительная часть животных в возрасте одного года,
преимущественно самцы, покидают материнские участки обитания и эмигрируют
на значительные расстояния от них. Причиной выселения являются
социальные факторы, в первую очередь агрессивность взрослых особей, изгоняющих
полувзрослых зверей со своих территорий.
В целом эти миграции не такие массовые и протяженные, как описанные
выше. Обычно они не превышают нескольких километров, лишь как исключения
бывают перемещения на десятки километров от места маркировки (Bramley,
1970; Strandgaard, 1972; Myrberget, 1973; Ellenberg, 1978; Соколов, Данилкин, 1981;
Fruzinski et al., 1983; Reimoser, Zandl, 1986, 1987). Пик эмиграции молодых
особей в Западной Европе приходится на конец апреля — начало мая, на востоке
Европы и в Азии — на июнь.
Осенью многие годовалые особи, как показали наши наблюдения с помощью
радиопеленгации, возвращаются к месту рождения и зиму проводят в тех
районах, где они прошлой зимой находились с матерью. Весной, уже в
двухлетнем возрасте, они снова эмигрируют или занимают свободные участки
(если такие имеются) в районе своего рождения.
Подводя итог сказанному выше, можно констатировать, что сезонные
миграции, вызванные неблагоприятными факторами среды, присущи как сибирской,
так и европейской косуле. Там, где экологические условия не позволяют
круглодичного оседлого обитания в одном и том же локальном районе, у
популяций вырабатывается сезонный миграционный стереотип поведения, проявляющийся
тем ярче, чем больше высота снежного покрова. В схематизированном виде
это заключение иллюстрируется рис. 38. Снежный покров, затрудняющий или
делающий невозможным передвижение животных и кормодобывание, является
причиной зимних миграций. Им же в основном и определяются сроки миграций:
118
в горах и в северных районах, где снег выпадает раньше, раньше всего (иногда
до снега) начинается перекочевка. Направление перемещения зверей в основном
перпендикулярны горным хребтам, а на равнинной местности они идут чаще
в южном направлении к малоснежным зонам.
Пути миграции косуль постоянны из года в год, но при изменении
экологической ситуации (распашка зимних пастбищ, образование водохранилищ)
могут меняться. При снижении численности миграции "затухают", при высокой
численности, напротив, наиболее выражены.
Миграционное поведение несомненно благоприятствует сохранению популяций.
Вероятно, оно может передаваться последующим поколениям путем "обучения"
и "закрепляться" в памяти потомков, что проявляется при переселении особей
из таких популяций в другие многоснежные районы, из которых они также
стремятся мигрировать (Юргенсон, 1969). Однако оседлость многих популяций
в малоснежных районах не дает оснований считать миграционное поведение
отличительной чертой рода Capreolus.
Эмиграция молодых животных, происходящая весной и летом, обусловлена
социальным фактором и более выражена при высокой плотности населения.
Выселение части особей популяции непосредственно перед появлением нового
поколения также весьма благоприятно для вида, так как обеспечивает большие
возможности выживания детенышей из-за снижения пищевой конкуренции и
способствует расселению зверей.
Естественное и искусственное расселение
При всей своей кажущейся оседлости косули способны расселяться с
относительно большой скоростью. В Швеции менее чем за 100 лет границы
ареала продвинулись на север примерно на 1000 км, в Восточной Европе за 30 лет
с середины века как европейская, так и сибирская косуля расселились
естественным путем почти на 500 км, т.е. ежегодная скорость движения границ
основного ареала превышает в среднем 10—15 км (см. главу "Ареал").
Нет сомнений, что основным способом расселения косуль, особенно
сибирских, являются сезонные миграции. Заходы отдельных особей, групп и стад
регистрируются за сотни километров от основного ареала. Отмечено много
случаев, когда мигранты задерживались в новых районах, приносили потомство
и, если позволили условия, оставались здесь жить навсегда. Так, в Монголии
в 1957 г. около десятка косуль, кочуя зимой с севера на юг, перешли р. Керулен
и обосновались на горе Тумэн-Цогт, где их потомки живут до сих пор в отрыве
от основного ареала в нескольких небольших островках карликовой березы
и осины общей площадью не более 10 га. Такая же ситуация имела место в
безлесных горах в окрестностях сомона Матад на Востоке Монголии, где примерно
из сотни мигрировавших сюда зверей зимой 1977 г. остались 13 особей,
которые принесли здесь потомство, но из-за отсутствия укрытий позднее были
уничтожены. Очевидно, что именно благодаря миграциям с Урала сибирская
косуля быстро восстановила свой ареал на левом берегу Волги и вышла на его
правобережье (см. главу "Ареал").
Возникшие на новом месте очаги обитания, в свою очередь, становятся
источником материала для расселения. По мере нарастания плотности населе-
119
Рис.39.Схема процесса естественного
расселения косуль
/ — многочисленная популяция; 2 —
сезонные миграции; 3 — популяция
мигрантов; 4 — расселение молодых особей
ния и учащения социальных конфликтов все в большей степени происходит
процесс выселения молодых особей за пределы родительского участка,
завершающийся занятием собственных участков обитания на еще свободных
территориях поблизости от родительских владений, т.е. таким образом в основном
происходит "насыщение" территорий. Тем не менее дальние перемещения
молодых особей, как показано выше, не редкость, и образовавшиеся семьи также
могут служить ядром новой локальной популяции. В схематичном виде процесс
естественного расселения косуль отражен на рис. 39. ..
Естественному расселению косуль может в какой-то степени способствовать
фактор беспокойства. Нами достоверно с помощью радиопередатчиков
зарегистрированы два случая, когда по этой причине беременные самки европейской
косули непосредственно перед родами покинули свои участки и эмигрировали:
одна из них прошла примерно 23 км, а другая — более 10 км. Поскольку фактор
беспокойства в наши дни имеет весьма существенное место, можно полагать,
что такие перемещения косуль происходят довольно часто.
Характеристика перемещений косуль будет далеко не полной без данных
об их искусственном расселении, которое велось в больших масштабах. В
основном эта работа преследовала две цели: расширение существующего ареала и
вселение более крупной сибирской косули в область обитания мелкой
европейской с целью "освежения крови", повышения массы тела и улучшения качества
трофеев последней.
Известно, что за пределами исторического ареала XIX в. европейская
косуля была интродуцирована в Ирландию, где хорошо размножалась в течение
70 лет, но впоследствии, после решения о восстановлении лесов, популяция
была полностью уничтожена. В XX в. ее акклиматизировали в штате Техас
в США. Сибирскую косулю начиная с прошлого века неоднократно выпускали
в Европе: в Англии, Германии, Бельгии, Словакии и на западе России (Север-
цов, 1941; Тигбек, 1951; Bump, 1970; Stubbe, Passarge, 1979).
В пределах естественного ареала их расселяли во многих европейских странах.
В частности, только во Франции с 1952 по 1979 гг. выпущено 4780 европейских
косуль (Boisaubert, 1980). В целом по экспертной оценке искусственно были
перемещены не менее 10 тыс. особей.
С большим размахом эти работы велись в СССР. С 1925 г. по настоящее
время здесь расселено более 3 тыс. особей, преимущественно в европейской
части. На Украине выпустили более полутысячи зверей (из них 60 сибирских особей),
в Московскую и Калининскую области их завезли около 1,5 тыс. (Павлов и др.,
1974; Вершинин, 1969; Фадеев, 1969а, б; Насимович, Шубникова, 1976; Денисов,
1978). От общего количества выпущенных животных сибирские косули состав-
120
ляли более половины. Только Зоообъединением Главохоты РСФСР с 1950
по 1980 гг. их передано в охотничье хозяйство 1,1 тыс. (Шурупов, 1982).
По данным перечисленных выше авторов, а также по материалам Главохоты
РСФСР и РосохОтрыболовсоюза, республиканских и областных госохотин-
спекцией составлена карта искусственного расселения косуль (рис. 40), на которой
достаточно наглядно видна роль человека в перемещении животных и в смешении
генофонда популяций.
Итог этой многолетней и дорогостоящей работы в общем за небольшим
исключением оказался почти безрезультатным из-за неудовлетворительной
организации, полного отсутствия научных исследований и многочисленных ошибок
при выпусках. Зверей расселяли мелкими партиями, редко превышавшими
20 особей, в половом соотношении от 1:9 до 6:1, более половины животных
выпущены в многоснежных районах на самой окраине их ареала, подкормка
зверей и охрана от хищников и браконьеров не были должным образом налажены.
Подавляющее большинство животных погибли в местах выпусков в первые же
годы.
Кроме того, одна из основных причин неудач работ по расселению — смешение
в одних и тех же районах особей разных подвидов и видов. Расселение
сибирского косули в современном ареале европейской неизбежно влекло за собой
их гибридизацию со всеми вытекающими отсюда негативными последствиями
(см. главу "Изменчивость и таксономия"). Процесс эволюции и
видообразования привел к возникновению репродуктивных барьеров и, вероятно, к
определенной тканевой несовместимости между европейской и сибирской косулями.
Как показали исследования (см. выше), многие гибридные самцы оказываются
стерильными, европейские самки часто не могут родить более крупный плод
и гибнут вместе с потомством от сибирских самцов. Все это наряду с
естественной смертностью ведет к быстрой элиминации небольшого числа интродуцентов
и к "поглощению" сибирского генотипа.
Полученные все же "гибриды европейской и сибирской косуль по массе тела
и величине рогов действительно оказываются крупнее, чем европейские косули.
Казалось бы, одна из целей (повышение качества трофеев), ради которой
проводилась интродукция, достижима. Однако результаты этой грандиозной
по замыслу и трудоемкой по содержанию работы сводятся на нет из-за очень
низкого уровня воспроизводства косуль в смешанных парах.
При анализе результатов интродукции сибирского вида становится ясно,
что в Европе успешными были лишь два выпуска и оба в тех районах, где не было
европейской косули или численность ее была ничтожно мала. В 1886 г. в Гатчинскую
охоту царского двора (Ленинградская обл.) было выпущено 59 сибирских косуль,
которые успешно размножались. В 1909 году их насчитывалось уже 1068 (Север-
цов, 1941), но впоследствии все звери были уничтожены. В 1956 г. 15 сибирских
косуль из Приморского края и Киргизии выпустили в Темном лесу на территории
Шпаковского района Ставропольской области (Емельянов, Рыбалов, 1956).
Через 10 лет численность их выросла до 200 голов, они начали расселяться
и заняли практически всю Ставропольскую возвышенность.
В настоящее время эта локальная популяция сибирской косули оказалась
в окружении европейской косули (см. рис. 25), что в дальнейшем неизбежно
приведет к их гибридизации.
121
Рис.40. Места выпуска европейской (/) и сибирской (2) косуль
Линия указывает на район отлова
Таким образом, очевидно, что искусственное смешение разных видов косуль
недопустимо. Пользу, вероятно, может принести только внутриобластное (или
из соседних областей) переселение животных в те районы, откуда они по каким-
либо причинам исчезли или численность их резко снизилась. Успешным в этом
плане можно считать перемещение европейской косули из Воронежской области
в соседнюю Липецкую (Побединский, 1987) и в Ростовскую область в
низовья Северского Донца, где были искусственно созданы мощные очаги ее
обитания.
Практика показала, что естественный путь расселения косуль при
возрастающей численности оказывается более эффективным. Сейчас европейские и
сибирские косули уже полностью восстановили свой исторический ареал и не имеет
смысла продолжать расселение зверей.
Глава шестая. Питание
Общая характеристика питания
Пищевая стратегия копытных определяется четырьмя параметрами: размером
тела, типом пищеварительного тракта, соотношением между объемом желудка
и массой тела, размерами рта (Hanley, 1982). Европейская и сибирская косули —
одни из самых мелких видов оленьих, по способу питания относящиеся к
обкусывающим животным. Соотношение между объемом желудка и массой тела
у них ниже, чем у других жвачных такого же размера (Necas, 1975; Dreschner-
Kaden, 1974, 1976; Perzanowski, 1978; Henry, 1978a). Набор кормовых растений,
емкость пищеварительного тракта и строение стенок рубца свидетельствуют
о том, что они склонны к потреблению легко перевариваемой и богатой
питательными веществами пищи (Esser, 1958; Bubenfk, 1959; K16tzli, 1965; Dreschner-
Kaden, 1974; Dreschner-Kaden, Seifelnasr, 1977; Hofmann et al., 1976; Koniget al., 1976;
Gttnther, 1978; Reichholf, 1980; HoliSova et al., 1982).
В отличие от других жвачных косули очень избирательны в питании и
поедают в основном наиболее предпочитаемые виды растений или их части (Саб-
лина, 1959; Klotzli, 1965). Из-за этой особенности их считают "разборчивыми"
(Kurt, 1970; Nec"as, 1975; Hell, 1979), "требовательными к питанию" (Stubbe,
Passarge, 1979) и им приписывают какую-то "пищевую мудрость" (Lockie, 1967;
Rusterholz, Turner, 1978). Состав пищи косуль очень разнообразный и
отличается как в отдельных частях ареала, так и в разных биотопах. Он может
изменяться в течение суток и сезона года, зависит от пола, возраста и состояния
здоровья особи, обилия и доступности корма и других факторов.
В пределах ареала косули употребляют в пищу более 1289 видов растений.
Тем не менее представленный ниже список кормовых растений (см. приложение)
не является исчерпывающим. Самую большую долю в питании занимает
двудольные травянистые растения (55,2%), на втором месте древесные породы:
деревья, кустарники и полукустарники (24,5%). Однодольные травянистые
растения поедаться хуже (15,8%) (табл. 13). Эти три категории вместе составляют
более 95,5% общего числа видов растений, найденных в пище животных. Другие
группы растений относятся к второстепенным источникам пищи.
В течение года доля древесных пород в пище почти не изменяется.
Двудольные травянистые растения в основном поедаются летом, а однодольные зимой.
Количество принимаемой пищи и ритм пищевой активности у жвачных
животных во многом определяется емкостью рубца. У косуль масса рубца и сетки
с содержимым по отношению к общей массе тела составляет 6—8,5% (Nagy,
Regelin, 1975; Stubbe, Passarge, 1979), а к массе туши — 13—19% (Szaniawski,
1973). Общий объем рубца и сетки у взрослых особей достигает 4—5,8 л (Ne6as,
1975).
124
Таблиц* 13. Корма европейской и сибирской косуль
Корм
Древесные породы
Травянистые растения
Двудольные
Однодольные
Папоротники
Мхи
Лишайники
Грибы
Всего
Число видов
Европейская
косуля
226
497
146
17
12
6
11
915
24,7
54,3
16,0
1,9
1,3
0,7
1,2
растений и
Сибирская
косуля
131
350
97
7
—
4
10
599
21,9
58,4
16,2
1,2
—
0,7
1,7
их доля
Общие
42
135
40
1
—
3
4
225
(%) в питании
виды
18,7
60,0
17,8
0,4
—
1,3
1,8
Всего
315
712
203
23
12
7
17
1289
видов
24,5
55,2
15,8
1,8
0,9
0,5
1,3
Относительно небольшой объем рубца косуль говорит о том, что пища
не может долго оставаться в нем (Hofmann, 1978a). Скорость ее прохождения
через пищеварительный тракт зависит от времени усвояемости (Perzanowski,
1978; Drozdz, 1979) и колеблется в зависимости от строения отдельных частей
растений и, возможно, от содержания клетчатки (Bubenik, 1959; Lochman, Barth,
1965). Клетчатка разлагается в преджелудках жвачных животных посредством
воздействия симбиотической микрофлоры (Baldwin, 1984). Для деятельности
микрофлоры необходима совершенная фрагментация принятой пищи (Pond et al.,
1984; HoliSova et al., 1985).
Небольшой объем желудка и относительно быстрый процесс пищеварения
требует частого питания. В течение суток в нормальных условиях косули
питаются 5—11 раз (Bubenik, 1962; Lochman, 1965; Klotzli, 1965; Соколов, Данил-
кин, 1981). Сразу или через непродолжительное время после приема пищи следуют
короткие периоды пережевывания. При оптимальной трофической ситуации
(в вегетационный период) животные принимают пищу приблизительно через
каждый час (Lochman, 1965; Hofmann, 1978b).
В популяциях, неумеренно преследуемых человеком, ритм питания особей
резко нарушается; звери выходят на кормежку в сумерки или ночью, днем
практически не кормятся. На ритм пищедобывательной активности в
значительной степени влияют также погодные факторы, количество и доступность
корма, сезон года. Пасутся звери в любое время суток, наиболее
продолжительные кормежки-утром и вечером. В жаркие летние дни кормятся реже, чем в
прохладные. Зимой кормежки продолжительнее, чем летом (Соколов, Данилкин,
1981).
Кормовая зона у европейской косули находится на уровне 0—120 см от земли,
у сибирской — 0—150 см, однако при высоком плотном снеге она увеличивается
до.2—3 м. Диаметр скусываемых побегов достигает 6 мм, а в среднем 2—4 мм.
Косулям присуща максимальная оптимизация питания по сезонам,
проявляющаяся в разной степени утилизацией растений. Пасущиеся животные постоянно
перемещаются по участку. Летом они никогда не объедают все растения вокруг
себя, а срывают лишь одно или часть его и идут дальше. С многолистных
125
растений косули срывают только 1—2 листочка или обкусывают соцветие,
после чего растение может куститься и биомасса его в целом нередко
увеличивается, что дает возможность кормиться им еще несколько раз. Такая
особенность питания в сочетании с одиночным образом жизни особенно ценна
с точки зрения ведения сельского и охотничьего хозяйства, поскольку эти
копытные не причиняют значительного вреда сельскохозяйственным посевам.
Зимой в поисках корма они раскапывают снег передними-ногами иногда
на глубину 30—50 см. Причем сибирские косули нередко раскапывают снег
даже вокруг высокостебельных растений, откусывают растение у самого
основания и полностью поедают его. Летом, как отмечено выше, они этого никогда
не делают. И в этом тоже проявляется оптимизация питания, позволяющая
животным максимально использовать в неблагоприятных зимних условиях
скудные запасы корма.
Выбор корма и избирательность питания
При выборе подходящего корма косули руководствуются обонянием и вкусом.
Опытами показано, что их привлекает запах некоторых растений, например
пихты и бересклета. Сухие, старые и сильно деревянистые части растений,
твердые злаки и осоки, растения, содержащие токсичные вещества (алкалоиды,
сапонин, фенолы и глюкозиды), не поедаются или поедаются плохо (Esser,
1958; Klotzli, 1965; Stubbe, Passarge, 1979). Наиболее предпочитаются самые
молодые органы с самым низким содержанием клетчатки (Wagner, 1961).
Помимо этого, значительную роль играет и присутствие в растениях
дубильных экстрактов (Klotzli, 1965), определенных минеральных веществ, витаминов,
воды и т.п. в зависимости от физиологических потребностей особи (Voser-
Huber, Nievergelt, 1975).
В списке растений, приведенных в приложении, все же находятся виды,
которые обыкновенно считаются ядовитыми для человека и домашних
животных: Daphne mezerum, Rhus typhina, Aconitum lycoctonum, Atropa belladonna,
Rannunculus acer, Solanum dulcamara, S. nigrum, S. tuberosum, Paris quadrifolia
и др. Косули, видимо, сравнительно малочувствительны к влиянию
токсичных веществ, содержащихся в них (Liepmann, 1972; Prior, 1968). Вероятно,
растительные яды в их пищеварительном тракте частично нейтрализуются или разлагаются
благодаря биохимическим процессам. Не исключено, что противоядием могут
служить и дубильные вещества, содержащиеся в пище (Тимофеева, 1985).
Многие исследователи указывают на избирательное питание особей
различными видами полыней, которые звери, скорее всего, используют в качестве
лекарственного противопаразитного средства (Дементьев, 1933; Фетисов, 1947,
1953).
Выбор определенных растений проявляется даже в пределах одного биотопа
(Klotzli, 1965). Хвоя пихты, например, привлекает косулю только тогда, когда
дерево растет на солнечных участках, так как содержит больше эфирных масел
(Wagner, 1961). Еловая и сосновая хвоя одиночно стоящих деревьев
предпочтительнее хвои молодых растений, растущих в подлеске, поскольку имеет более
высокую пищевую ценность (Prieditis, 1984).
Предпочтение определенного источника пищи может быть перенесено
"традицией" или "научением" на остальных особей (Ellenberg, 1974; Mtiri, 1983). За-
126
мечено, что особой "популярностью" у зверей пользуются новые виды древесных
пород (часто экзотических), которые были высажены в лесную поросль (Miiri,
1978; Riiedi, 1956), и корм, наиболее дефицитный в данном биотопе: лиственные
породы в хвойных лесах и, напротив, хвойные породы в лиственных лесах
(Падайга, 1965).
Установлено также, что косули предпочитают не отдельные виды растений,
а целые "блоки видов", причем интенсивность потребления одного из видов
в блоке сопряжена с интенсивностью потребления остальных. Пищевые блоки
характерны для разных экосистем и зависят от структуры фитоценозов (Kossak,
1976; Рубис, 1982).
Разумеется, избирательность в питании животные проявляют лишь при
большом разнообразии доступного корма. Такая ситуация обычна в течение
вегетационного периода и в естественных ненарушенных биотопах. Вне
вегетационного периода при лимите кормов это проблематично (Nudds, 1980).
Благодаря выбору отдельных частей растения пища в рубце косуль
оказывается более богатой питательными веществами, чем содержится их в целом
растении (Соколова, Данилкин, 1977). Очень высокая избирательность в питании
приводит к заметному изменению суммарного биохимического состава травостоя
в биотопах этих животных. По данным А. Бубеника (Bubenik, 1959), после их
пастьбы общая кормовая ценность травостоя может снижаться по протеину
на 54%, по кальцию на 46,5% и по фосфору на 23,2%.
В зависимости от того, как часто и в каком количестве отдельные виды
растений встречаются в рационе, различают две категории кормов — основные
и второстепенные.
Основные и второстепенные корма
К основным источникам пищи косуль, как отмечалось выше, относятся
двудольные травянистые растения, древесные породы и в меньшей степени
однодольные травянистые растения. В эту же категорию входят растения,
которые в определенном биотопе и в определенный период года
предпочитаются большинством особей из популяции.
В содержимом рубцов находят от 3 до 42 видов растений (Siuda et al., 1969; Се-
derlund et al., 1980). Однако число видов, представленных в пище в большом
количестве (более 5% от общего содержимого рубца), обычно колеблется от 1 до 6
в течение всего года. В Венгрии животные, живущие в лесу, зимой чаще всего
потребляют 7—10 видов (в среднем 8,6), а особи, обитающие в полях, — 5—9 видов
(в среднем 6,8) (Matrai et al., 1983). В содержимом пищеварительного тракта
косуль в южной Моравии (Чехословакия) летом присутствовало 3—16 видов
растений (в среднем 13,6), зимой от 4 до 27 (в среднем 14,9) (HoliSova et al., 1982,
1984). В лесостепных дубравах Белгородской области в пище
европейской косули в снежный период зарегистрировано около 90 видов растений,
но только 17 из них, преимущественно древесно-кустарниковые, принадлежали
к основным кормам (Тимофеева, 1985).
В пище косуль преобладают листья двудольных травянистых растений и
листья и молодые побеги древесных пород (см. приложение). Этот корм может
составлять 51—58% от общего объема рациона (Bubenik, 1959), а в течение
вегетационного периода — около 90% (Hell, 1979). Относительная доля листьев
127
и побегов древесных пород и листьев травянистых растений в пище сильно
изменяется в зависимости от типа биотопа. Косули, живущие в лесу, предпочитают
листья и побеги древесных пород, в отличие от полевых, которые питаются
в основном травянистыми растениями (HoliSova et al., 1982, 1984, 1986а, b).
В их рационе часто встречаются некоторые лекарственные виды,
используемые в медицине (Stubbe, ZSrner, 1977). Возможно, что вещества,
содержащиеся в них, имеют отрицательное влияние на эндопаразитов животных. То же
самое, вероятно, касается и надземных частей папоротников с высоким
содержанием дубильных экстрактов (K16tzli, 1965).
Гораздо меньшую составную часть их корма представляют надземные части
злаков и другие однодольные растения как по видовому разнообразию, так
и по объему (Hell, 1979; Schafer, 1974; Melichar, 1960). Однако злаковые
культурные растения весьма предпочитаемы животными, обитающими в полях.
Косули охотно потребляют сочные плоды деревьев, кустарников и
полукустарников (Vaccinium sp., Rubus sp., Malus sp.,Pirus communis, Rosa sp., Sorbus
aucuparia, Prunus sp., Crataegus oxyacantha и др.). Очень любят они яблоки,
буковые орешки, желуди, каштаны. Животные собирают их с поверхности почвы,
а зимой выкапывают из-под снега (M6hring, 1963; NeJas, 1975; HoliSova et al., 1984,
1985a). Плоды и семена растений способствуют образованию запасов
подкожного жира (Ne6as, 1975; Stubbe, Passarge, 1979) — важного источника энергии
в зимний период. Довольно часто в их рационе присутствуют цветы, соцветия
и целые вершинные части растений (K16tzli, 1965; Borowski, Kossak, 1975; Kossak,
1983; HoliSova et al., 1984, 1986a, b). Подземные части растений служат основным
источником пищи там, где они случайно становятся доступны в большом
количестве. Это касается корнеплодов сахарной свеклы и клубней картофеля,
остающихся после сбора урожая.
Особого упоминания заслуживает потребление косулями хвои (Gunther, 1978;
Kampmann, 1983). Одни исследователи считают ее основным, другие —
второстепенным кормом, поедаемым животными зимой лишь при недостатке более
качественной и в другое время более предпочитаемой пищи (Oszlanyi, 1982).
Экспериментально установлено, что звери употребляют хвою даже при наличии
богатого питательными веществами корма (зерен злаков, желудей, лиственного
корма, гранулированных сушеных осиновых листьев и клеверного сена) (Kuen,
Bubenik, 1980; Prieditis, 1984). Можно предположить, что она необходима
животным в качестве источника воды (Juon, 1963; Jackson, 1974) или
витаминов. Это заключение Подтверждается наблюдениями за пятнистыми оленями
(Obrtel et al., 1985).
В категорию второстепенных кормов входят растения, встречающиеся
в пище с малой частотой (у небольшого числа особей в популяции) или в очень
незначительном количестве. В большинстве случаев они попадают в желудок
при "опробовании" животными новых видов или качества растений и их частей.
Во многих районах европейской и сибирской косулями в большом
количестве поедаются карпофоры грибов (см. приложение). Однако в целом они
составляют незначительную долю в питании и являются все же второстепенным
кормом, так же как мхи, лишайники, папоротники, хвощи и плавуны.
Полевые косули потребляют иногда кукурузу, пораженную головней. В
исключительных случаях в содержимом рубца и в экскрементах зверей находят
остатки мелких млекопитающих и насекомых, вероятно, проглоченных вместе
128
с растительной пищей (Velich, 1925; Holiiova et al., 1982,1984,1986). Потребление
животной пищи косулями и другими видами Оленьих объясняют как один из способов
удовлетворения их потребностей в редких элементах и белковых веществах (Bubenik,
1966) и в кальции (Prior, 1968; Raesfeld, 1965).
Вместе с растениями и при посещении солонцов в пищеварительный тракт
зверей попадают разные примеси: камни, щебенка, пластмасса, кусочки
металла, песок, глина и помет мелких млекопитающих. Очень часто встречается
шерсть, причем зимой в больших количествах, проглоченная, видимо, при
вылизывании тела (Cederlund et al., 1980; HoliSova et al., 1982, 1984).
Видовые, региональные
и биотопические аспекты питания
Питание косуль в пределах обширного ареала имеет свои видовые и
региональные особенности. В пище европейской косули выявлено 915 видов растений,
у сибирской — 599. Однако как для одного, так и для другого вида животных
этот список (см. приложение) далеко не полный. В целом пищевая стратегия
европейских и сибирских косуль сходна, что видно хотя бы по одинаковому
соотношению групп растений в питании, но по составу их корм существенно
различается — общими оказались всего лишь 17,4% видов растений, в основном
двудольные (см. табл. 13). Тем не менее это отличие в большей степени
отражает разницу в видовом составе растений в Европе и в Азии, нежели
видовые особенности животных в питании.
В разных регионах количество поедаемых видов растений колеблется от 64 до
320. В пределах локальных районов пища косуль состоит в среднем из 80—130,
в крупных регионах — из 130—250 видов растений (табл. 14).
В каждом регионе у косуль есть свои излюбленные виды растений. На Южном
Урале, например, у одной из популяций летом наиболее предпочитаемый корм —
иван-чай, обнаруженный в содержимом 80% исследованных рубцов, причем
в большинстве их он составлял от 40 до 90% объема растительной массы.
У двух сеголеток, добытых в августе, желудок, помимо свернувшегося молока
(266 и 218 г.), был заполнен листьями исключительно этого растения. Здесь же
звери охотно поедали порезник сибирский, клевер луговой, вейник наземный.
В другой популяции наиболее предпочитаемыми были молодые побеги осины
во все сезоны года (Соколов, Данилкин, 1981). В северной части Южного Урала
в Ильменском заповеднике основной корм сибирской косули — сосна, осина
и береза; весной — ивы, лютик и иван-чай, осина; летом — клевер, иван-чай,
некоторые виды зонтичных, вахта, ястребинка и др.; осенью — иван-чай, горец,
зонтичные и сложноцветные (Аверин, 1949).
В Восточной Сибири сибирские косули больше всего любят осинник, ивы,
троелистку, охотно едят полыни (Фетисов, 1953); в Западном Забайкалье —
астрагалы, чины, вики, клевер, люцерну (Смирнов, 1978); на Дальнем Востоке —
листья и побеги осины, ив, леспедецы, липы, рябины, берез, ясеня, дуба, жасмина,
винограда, лещины, малины, а также прострела и сои (Бромлей, Кучеренко,
1983; Дарман, 1986).
В западных районах Белоруссии основу зимнего питания (84,6%) европейской
косули составляют крушина, граб и дуб, в восточных — дуб и ива (70,7%) (Рубис,
1982).
9. Европейская и сибирская косули
129
Таблица 14. Количество видов растений в пище косуль в разных част» ареала
Регион
Швейцария
Польша
Попильно
Чемпинь
Беловежская Пуща
Чехословакия
Южная Моравия
Румыния
Болгария
Лудогоре
Стара Планина
Рило, Пирин, Родопите
ГДР
СССР
Литва
Калининградская обл.
Беловежская Пуща
Левобережная Украина
Белгородская обл.
Курская обл.
Крым
Восточная Грузия
Южный Урал
Алтай
Восточная Сибирь
Забайкалье
Амурская обл.
Юг Дальнего Востока
Число видов
растений
Европейска i косуля
95
178
85
97
252
130
129
148
76
81
70
64
628
118
113
90
320
122
165
145
120
Сибирская косуля
93
ПО
60
130
140
129
92
Авторы
Voser-Huber, Nievergelt, 1975
Siuda et al., 1969
Kaluzinski, 1982c
Kossak, 1983
Mottl, 1957
HoliSova et al., 1984
Manolache, Babulia, 1969
Петров и др., 1968
Knorr, Briedermann, 1976
См. приложение
Прусайте и др., 1983
Романов, Ромашин, 1982
Саблина, 1955
Карпенко, 1966
Тимофеева, 1985
Гусев, 19846
Лавров, 1929; Даль, 1930 v
Арабули, 1966
Соколов, Данилкин, 1981
Аверин, 1949
Пастернак, 1955
Фетисов, 1947, 1953
Смирнов, 1978
Дарман, 1986
Бромлей, Кучеренко, 1983
Однако несмотря на определенные региональные различия в питании,
достаточно четко видно (см. приложение), что в пределах близких географических
регионов набор основных кормов косуль совпадает. Яркий контраст наблюдается
лишь там, где резко меняется видовой состав растений в целом.
Пища косуль отличается не только в разных частях ареала, но и в зависимости
от типа биотопа, занимаемого популяцией или отдельными особями (K16tzli,
1965; Bubenik, 1970; Jackson, 1974; Владышевский, Ельский, 1974; Strandgaard,
1977; Necas, 1975). В частности, в Литве в лиственных лесах с примесью ели
их основной зимний корм — ясень и дуб; в смешанных елово-лиственных
лесах — рябина, крушина, осина и дуб; в сосновых лесах с примесью
лиственных пород — крушина, рябина, дуб и осина (Падайга, 1965). В содержимом
262 рубцов косуль из Латвии относительная доля полукустарников (прежде
130
всего Calluna vulgaris и Vaccinium sp.) колебалась с 2 до 57% в сосновых лесах,
с 0,3 до 9% в еловых и с 0,01 до 0,4% в лиственных лесах; доля древесных пород
(деревьев и кустарников) изменялась с 9 до 21% в сосновых, с 3 до 15% в еловых
и с 1 до 8% в лиственных лесах. Эти отличия согласуются с различной степенью
встречаемости основных источников пищи в сравниваемых биотопах (Яисон,
Падайга, 1975).
Косули, живущие в лесу, чаще используют в пищу древесные породы, полевые —
травянистую растительность (Stubbe, Passarge, 1979; Hell, 1979; HoliSova et al.,
1986b). В содержимом рубца европейских косуль, отстрелянных в лесу,
окруженном полями, преобладают травянистые виды (57,6%, из них 16,2% — культурные)
(Балейшис, Прусайте, 1980). Пища сибирских косуль, населяющих леса вблизи
соевых полей на Дальнем Востоке, осенью и зимой на 70—75% состоит из
бобов и стеблей соли (Дарман, 1986).
Различия в составе рациона популяций косуль, обитающих в одной и той же
географической области (в Литве), но в биотопах с разной долей лесных
угодий (7 и 24,8%) установили Прусайте и др. (1983). У первой популяции
в общем объеме пищи нашли 53,2% культурных, 30,2% травянистых растений
и только 16,5% древесных пород, во второй — 10,2% культурных растений,
желудей, полукустарников, грибов и лишайников, 50,1% травянистых растений
и 39,7% древесных пород. Эти данные свидетельствуют о высокой степени
приспособления животных к местным экологическим условиям.
Сезонные особенности питания
Доля отдельных видов растений в пище косуль значительно отличается по сезонам
года (см. приложение), что связано, во-первых, с периодичностью вегетации
(Jackson, 1974, 1980) и, во-вторых, с различной степенью использования кормов
в зависимости от изменяющегося физиологического состояния животных
(HoliSova et al., 1984, 1986 а, b). В зимний период из-за снежного покрова существенно
уменьшается ассортимент корма и постепенно истощаются его запасы. Однако
в это время заметно снижается общий уровень метаболизма и потребность
в пище (Ellenberg, 1978; Холодова, 1986). Весной, с началом вегетации корм
появляется в изобилии, повышается обмен веществ и ускоряется процесс
пищеварения благодаря мягкой сочной растительности. Летом в их распоряжении
самое высокое разнообразие и обилие корма. Осенью прекращается вегетация
растений, но созревают семена и плоды, которые становятся доступными
для животных.
Сезонную изменчивость питания косуль отмечают почти все исследователи
(см. приложение). Зимой состав пищи у них наименее разнообразный. Если
в период вегатации в содержимом желудков сибирских косуль на Южном Урале
находили от 10 до 26 видов растений (в среднем 16), то зимой значительно
меньше — от 5 до 11 видов (в среднем 8) (Соколов, Данилкин, 1981). Однако
в Западном Забайкалье сибирские косули очень полно используют набор кормов
в течение всего года; из 140 видов потребляемых растений зимой зарегистрировано
93,5%, весной — 86,3, летом — 84,9, осенью — 87,7% (Смирнов, 1978).
Объемное соотношение древесно-кустарниковых и травянистых кормов по
сезонам может меняться очень значительно. В польской части Беловежской Пущи
оно было весной и летом 20:80%, осенью и зимой — 35:65 (Gebczynska, 1980),
в Амурской области весной составляло 1:99%, летом — 0:100, осенью — 65:35, зи-
131
мой — 75:25% (Дарман, 1986), в Западном Забайкалье весной и летом 0:100,
осенью — 7:93%, зимой — 80:20% (Смирнов, 1978).
Зимой в хвойных лесах основная пища косуль — побеги ели, пихты, хвоя
сосны, вегетативные части черники, ежевики, брусники и вереска в подлеске,
побеги с почками разных видов лиственных древесных пород. В лиственных
лесах важную составную часть пищи представляют однолетние побеги и почки
и травяная ветошь (Саблина, 1955, 1959; Новиков, Тимофеева, 1964; Esser, 1958;
Siuda et al., 1969; Henry, 1978b; Ельский; 1978; Helle, 1980; и др.). Тропления
взрослых животных показывают, что они за сутки зимой скусывают 1700—2100
концевых древесных кустарниковых побегов, весящих около 1 г каждый
(Тимофеева, 1985). В годы урожая семян бука и дуба звери в значительном количестве
поедают эту концентрированную высококалорийную пищу (Komarek, 1954; Романов,
Ромашин, 1982).
В полях самое большое значение в рационе косуль зимой имеют
сельскохозяйственные культуры (табл. 15): початки и листья кукурузы, люцерна, соя,
листья и колоски пшеницы, свекла, а также листья и побеги Rubus sp., Ligustrum
vulgare и др. (HoliSova et al., 1982; Matrai et al., 1983; Дарман, 1986).
Зимой различий в составе пищи самцов и самок, обитающих в полях не найдено
(HoliSova et al., 1984), нет их и в отношении времени питания (Turner, 1979).
У косуль, живущих в лесу, напротив, они установлены: в одном и том же биотопе
хвоя ели и сосны содержалась в пище 22,2% самок и только у 0,12% самцов
(Григоров, 1976).
Состав и степень использования зимних кормов в лесах находится в прямой
зависимости от высоты снега (табл. 16), которая, в свою очередь, v связана
с сомкнутостью крон. Наиболее используются запасы корма в высокололнот-
ных древостоях (Ельский, 1978).
При глубоком снеге из-за недостатка пищи косули вынуждены переходить
на питание хвоей. Их зимовка проходит нормально, если хвоя составляет
менее 50% рациона. При более высоком уровне ее потребления звери ежемесячно
теряют 7—8% массы (Prieditis, 1984). В критических ситуациях они могут
глодать кору деревьев (Киселев, 1979).
Весной основным кормом в лесу становятся распускающиеся листья и
доступные соцветия древесных пород и травянистые виды растений (Siuda et al., 1969;
Фетисов, 1953; и др.). В этот период наиболее охотно поедаются проростки
первоцветов, относительно часто используются злаки и другие однодольные
растения (K16tzli, 1965; Тимофеева, 1985). В полях они кормятся прежде всего
листьями озимых зерновых культур, люцерной, клевером и рядом видов сорных
растений, растущих здесь же. При случае косули обкусывают также поросль
древесных пород в ветрозащитных полосах и ремизах (HoliSova et al., 1986).
Летом самое большое значение в их питании в лесных биотопах имеют
двудольные травянистые растения (Gebzcynska, 1980; Suida et al., 1969; Смирнов,
1978; Соколов, Данилкин, 1981), меньшую роль играют листься древесных
пород и относительно малозначимы однодольные растения (Klotzli, 1965; Kuen,
Bubenek, 1980; Kioroglanidis, 1981).
Полевые косули в начале лета чаще всего потребляют молодые листья
кукурузы, листья и цветочные части люцерны, незрелые колоски пшеницы, листья
сахарной свеклы. Поздним летом их пища состоит главным образом из листьев
132
Таблица 15. Состав пищи (%) полевой косули в Южной Моравии (HoliJova et a)., 1986)
Корм
сентябрь-
ноябрь
январь
февраль
март
апрель
май—июль
июль—
сентябрь
Древесные части растений
Листья деревьев и
кустарников
Листья и стебли травянистых
растений
Листья и стебли злаков
Семена и плоды
Цветочные части
Корнеплоды сахарной свеклы
14,4
17,4
18,1
9,3
35,9
5,7
3,6
11,8
4,4
10,5
4,8
31,2
5,3
39,1
10,3
7,6
11,3
11,3
28,3
1,8
32,6
13,6
3,0
10,5
23,3
18,8
0,7
37,1
7,8
3,5
33,3
23,3
14,0
10,3
2,0
6,0
9,8
32,0
15,3
23,1
13,8
—
3,5
6,5
27,4
1,1
36,4
20,8
—
Таблица 16. Состав корма (дола от остатков растений в рубце, %) косуль, обитающих в смешанных
лесах (Приедитис, 198S)
Корм
Июнь—октябрь
(я = 68)
Ноябрь—апрель
без снега (и = 10) снег 15 см (л=28) снег 30 см (я=48)
Февраль—март
Хвоя ели и сосны
Ветки лиственных пород, летом листья
деревьев
Вечнозеленые кустарнички
Разнотравье
Злаки
Грибы и лишайники
Корни папоротника
Другие виды кормов, включая
корнеплоды
•Обычно сено.
—
23,4
3,7
54,1
12,7
2,9
—
0,1
10,6
28,4
26,0
23,1
1,1
—
2,4
4,8
75,3
4,7
5,9
0,6
6,3
60,7
13,1
7,1
3,6*
15,5*
—
—
3,2
10,8
и цветов люцерны, недозрелых початков кукурузы и колосков пшеницы (Но-
lisovd et al., 1984).
Осенью косули предпочитают концентрированную пищу — семена в плоды
древесных пород и травянистых растений. Это связано с образованием жировых
запасов, которые позволяют им пережить возможный критический период во второй
половине зимы и ранней весной (Hofmann, 1979b). В пище лесной косули осенью
возрастает доля однолетних побегов (Саблина, 1959; Melichar, 1961). Полевая
косуля в это время использует послеуборочные остатки сахарной свеклы,
картофеля, кукурузы, семена сорных растений и зерновки дикорастущих злаков
(HoliSova et al., 1982, 1986b; Stubbe, Passarge, 1979).
Значение отдельных составных частей пищи косуль изменяется не только
в разные периоды года, но также и в течение того же самого периода в разные
годы (Jackson, 1974; Kuen, Bubenik, 1980), что связано с обилием доступной
133
Таблица 17. Относительное значение (%) некоторых видов растений в пище косули в зимний период
в разные годы (Григоров, 1976)
Растение
1973/74
Год
1974/75
1975/76
Rubus sp.
Pinus silvestris
Picea excelsa
Fungi
Fagus silvatica
Salix caprea
Травянистые растения
Sambucus nigra
Carpinus betulus
Другие древесные породы
84,64
0,01
6,09
0,33
8,92
97,80
2,14
0,03
0,03
26,49
1,95
0,08
69,52
1,47
0,17
0,15
0,09
0,04
0,04
Таблица 18. Состав пищи (%) полевой косули в течение двух сезонов разных лет в Южной Моравии
(HolKovaetal., 1986b)
Корм
Древесные части растений
Листья деревьев и кустарников
Листья и стебли травянистых
растений
Листья и стебли злаков
Семена и плоды
Цветочные части
Корнеплоды сахарной свеклы
Январь, февраль
1979
9,5
5,7
8,9
7,2
29,9
0,7
43,7
| 1980
|
1
13,1
4,7
8,3
4,5
39,4
9,8
24,4
1984
9,9
5,7
14'. 1
8,2
27,8
2,7
38,8
Март, апрель
1979
11,9
7,3
30,3
11,3
10,3
0,1
17,8
1980
11,6
0,5
9,5
24,4
25,5
2,3
36,8
1984
8,0
1,7
22,1
37,1
18,3
15,8
3,6
пищи. Их вариабельность более высокая в марте и апреле (т.е. в начале весны),
чем в январе и феврале (табл. 17 и 18).
Нельзя не отметить оптимальное использование косулей "сезонных" кормов,
быстроисчезающих или имеющих кормовую ценность сравнительно короткий
срок (грибы,, ягоды, плоды, цветы). Животные поедают их в возможно большом
количестве, как бы собирая урожай. Осенью их желудки бывают заполнены
некоторыми видами грибов, фруктами, ягодами и листьями брусники, черники
и др., весной — цветами первоцветов, летом — листьями наиболее излюбленных
растений (Фетисов, 1953; Смирнов, 1978; Соколов, Данилкин, 1981).
134
Количество потребляемой пищи
Количество потребляемой пищи связано с питательностью кормового рациона,
зависит от периода года, доступности корма, возраста, пола и,
физиологического состояния (Necas, 1975; Ellenberg, 1974, 1978).
Масса содержимого рубца значительно изменяется в зависимости от того,
когда животное было отстреляно: в начале или в конце периода пищевой
активности, поэтому приводимые исследователями данные могут заметно
отличаться. У европейской косули вес содержимого рубца составляет в среднем
1,6—1,9 кг(МоШ, 1957;Siudaetal., 1969; Балейшис, Прусайте, 1980; Knorr, Briedermann,
1974; HoliSova et al., 1982). У косуль в Литве средняя масса содержимого
рубца в январе—феврале у однолетних была существенно ниже (700 г), чем
у взрослых особей (1137,5 г). Зимой она гораздо выше (1230 и 1530 г,), чем весной
(700 и 680 г) (Балейшис, Прусайте, 1980; Прусайте и др., 1983).
Масса содержимого желудков взрослых сибирских косуль на Южном Урале
в среднем составила 2,5 кг (от 1,0 до 4,7 кг), причем в период вегетации — 2,2 кг
(от 1,0 до 3,9 кг), в снежный период — 3,2 кг (от 2,1 до 4,7 кг) (Соколов, Данил-
кин, 1981). Близкие к этим значения найдены у зверей в Забайкалье (Смирнов, 1978).
Суточный рацион европейской косули находится в среднем в пределах от 1,5—
2,5 кг до 4 кг зеленой растительной массы (Mottl, 1957; Briedermann, 1974; NeCas,
1975). В вольерах дневное потребление побегов древесных пород, кустарников
и полукустарников достигает 1,1 кг у однолетних, 1,7 кг у 1—3-летних и до 2,3 кг
у 5—9-летних особей (Padaiga, 1975).
Примерно такие же данные приводит А. Приедитис (1985). Полевые
косули потребляют до 2,15 кг зеленой массы озимых зерновых в день (что
соответствует 390 г вещества в сухом виде) или до 3,5 кг люцерны (615 г
сухого вещества) (Pielowski, 1970). Общее дневное потребление пищи
коррелирует с массой особи. Максимальный дневной прием пищи составляет 650 г
(в пересчете на сухое вещество), минимальный — 350—400 г. Максимальное
потребление сухих веществ из расчета на 1 кг 0,75 живой массы:
весной — 80 г, летом около 60—70 г и зимой 40—30 г в течение 24 ч. В
естественных условиях эти показатели выше примерно на 40%, так что средний
суточный рацион может оцениваться в 753 г сухих веществ, или 84,5 г/кг 0,75.
Потребность в перевариваемом белке в зависимости от возраста,
физиологического состояния, времени года составляет 70—150 г в день. Энергетические
затраты косули в сутки в среднем 2000 ккал (летом — 2800, зимой — 1300)
(Ueckermann, 1964, 1986; Droidi, Osiecki, 1973; Briedermann, 1974; Papageorgiou et
al., 1980; Prieditis, 1984; Strothman, Oslage, 1986).
Следует учитывать, что экспериментальные исследования питания косуль
в неволе могут приводить к ошибкам, поскольку пища, предлагаемая опытным
животным, как правило, значительно отличается от избираемой ими в природе
и по качественному составу и по разнообразию (Droidi, Osiecki, 1973; Perzanowski,
1978; Droidi, 1979).
Годовое потребление растительной массы взрослой особью равняется в
среднем 750 кг (Weiner, 1975). За это время косуля обкусывает около 3/4 млн побегов
(Leibundgut, 1976).
Количество поедаемого корма прямо пропорционально степени его
перевариваемое™ и содержанию белка и обратно пропорционально содержанию
135
Таблица 19. Состав молока (%) косуль и домашних животных(по: Stubbe, Passarge,1979, с изменениями
и добавлениями)
Компонент
Вода
Жир
Белок (протеин)
Молочный сахар (лактоза)
Минеральные вещества (зола)
Питательные вещества
Европейская
косуля
83,8
9,8
9,2
3,5
1.4
20,0
Сибирская
косуля
—
9,6
8,9
3,3
1,2
19,0
Корова
87,5
3,7
3,3
4,8
0,7
13,0 .
Овца
6,2
5,4
4,3
0,9
16,0
Коза
87,1
3,9
3,8
4,4
0,9
13,0
клетчатки. Наибольшее количество растительной массы употребляется весной (до 3 кг
в сутки, что соответствует 250 ккал на 1 кг массы), когда перевариваемость
достигает в среднем 73,5%. Зимой она резко сокращается, соответственно
уменьшается и количество поедаемой пищи (Droldz, 1979). Особенно низко
потребление корма при снежном покрове. Максимумы потребления падают на раннюю
весну, первую половину лета и осень, что связано у самок с периодом
выкармливания телят, у самцов с территориальным поведением (Ellenberg, 1978).
Детеныши с первых минут после рождения и обычно до августа—октября
питаются молоком, и лишь в редких случаях выкармливание длится дольше
(Borg, 1970). В неволе европейские косули кормят детей молоком в течение
162—194 дней и за это время каждый косуленок получает 128—184 л (Kossak, 1981b).
В каждый из нескольких приемов кормления теленок высасывает от 30 до 80 мл
(Espmark, 1969), а за сутки — до 1200 мл (Bubenik, 1965). Молоко косуль
отличается ббльшим содержанием жира и питательных веществ по сравнению
с молоком домашних животных (табл. 19).
Растительную пищу детеныши начинают пробовать через 5—10 дней после
рождения (Bubenik, 1965; Соколов и др., 1985). С возрастом в их рационе
быстро увеличивается количество видов растений: в 11—15 дней косулята поедают
уже около 13 видов, на 40-й день жизни — 43, и на 90-й — 66 (Kossak, 1981b).
С трехмесячного возраста пища телят как европейской, так и сибирской косуль
сходна с пищей взрослых животных.
Каждый из обоих полов имеет отличающуюся жизненную функцию. Из этого
следуют неодинаковые проявления активности и поведения, неадекватность
энергетических требований и метаболических процессов, удовлетворение которых
ведет к различиям в потреблении корма. Самки более консервативны в
потреблении энергии, чем самцы, и в отличие от них имеют тенденцию снижать
энергетические затраты в периоды повышенных энергетических требований
(Ellenberg, 1978; Turner, 1979; Eisfeld, 1985).
Существуют различия в питании самцов и самок в период гона: самцы
принимают значительно меньшее количество пищи (Lochman, 1961; Соколов,
Данилкин, 1981), и скорость ее переваривания у них более длительна, чем у самок
(Lochman et al., 1974).
Минимальное дневное потребление пищи у самца косули весом около 25 кг
составляет 600 г сухой массы, которая содержит 35 г переваримых белков
и около 300 г крахмальных единиц (Becker-Dillingen, 1945). В период лактации
136
Таблица 20. Потребление энергии у косуль(ккал/кг массы тела 0,75/сутки)(no: Ellenberg, 1978)
Самцы
а
Небеременные
самки
Лактирующие
самки
Время
159
145
127
195
210
135
176
168
190
218
—
190
173
128
ПО
131
—
—
143
150
160
150
221
173
171
162
—
—
—
—
—
—
—
275
370
—
—
—
—
Январь—февраль
Начало марта
Начало апреля (самцы спокойны)
Конец марта (самцы соскабливают кожу с рогов)
Начало мая (самцы территориальные)
Конец апреля—май (самцы спокойны)
Начало июня
Конец июня
Начало июля
Гон
1-я течка у самок
Сентябрь—октябрь
Ноябрь—декабрь
самки потребляют больше пищи, чем самцы в это время (Lochman, Barth, 1965).
У кормящей самки дневное потребление пищи достигает 1300 г сухой массы,
содержащей 120 г переваримых белков и 200 г крахмальных единиц (Esser, 1958).
А. Приедитис (1985) приводит такие данные. Зимой в неволе сеголетки съедают
в среднем 1 кг корма в сутки (0,4—0,6 кг сухого вещества), взрослые особи
соответственно 1,3—1,6 кг (0,6—0,9 кг). Летом эти величины у взрослых косуль
достигают 4,3—4,6 ki (1—1,3 кг), у лактирующих самок — 7,3—7,5 кг (2,0—2,3 кг
сухого вещества).
Потребность косуль в энергии тесно связана с физиологическими функциями
полов и имеет четкие сезонные аспекты. Наиболее основательные
исследования в этом отношении проведены Г. Элленбергом (Ellenberg, 1978). Некоторые
результаты его работы мы приводим в табл. 20.
Элленберг установил, что летом потребность в энергии у косуль примерно
на 50% больше, чем зимой. У самок она во все сезоны меньше, чем у самцов.
У беременных самок потребление энергии возрастает по сравнению с холостыми
лишь за 4 недели до отела, что по времени совпадает с началом вегетации.
Энергетические потребности лактирующих самок в среднем на 150% больше,
чем самок без детенышей. Причем, чем больше телят у самки, тем больше
ее затраты и, соответственно, потребности в энергии. Особенно велики затраты
косуль в период лактации и гона.
Минеральное питание и потребление воды
Для нормального развития и обмена веществ косули, как и другие копытные,
нуждаются в минеральном питании и в воде, которые получают обычно с
растительным кормом. В период беременности и лактации у самок и роста рогов
у самцов потребность в минеральных веществах увеличивается в 1,5—2 раза
(Becker-Dillingen, 1945; Bubenik, 1959; Esser, 1958; Drescher-Kaden, 1974; Briedermann,
1974). Подробный обзор литературы по минеральному питанию европейской
косули в норме содержится в монографии К. Штуббе и X. Пассарге (1979).
137
Рис.41. Естественный солонец, посещаемый косулями и маралами (Монголия, Хангай, 1979 г.)
В отдельных регионах косулям явно требуется дополнительное минеральное
питание. Например, в Зейском районе Амурской области, которая находится
в геохимической провинции с недостатком фосфора, меди, кобальта, йода и
частично кальция, местная популяция обладает низкой продуктивностью, у
самцов слабые рога необычно черного цвета с неочищенной кожей даже у взрослых
особей. Благодаря искусственным солонцам с добавлением микроэлементов
(см. ниже) удалось резко повысить ее продуктивность и качество, рога у самцов
стали нормальными и симметричными (Морозов, 1982).
В таких провинциях особенно заметна тяга зверей к естественным и
искусственным солонцам. Эту особенность поведения отмечали многие
исследователи (Туркин, Сатунин, 1902; Даль, 1930; Насимович, 1938; Аверин, 1949; Фетисов,
1953; Арабули, 1966; Смирнов, 1978; и др.). Естественные солонцы известны
многим поколениям косуль в округе, так же как и многим поколениям охотников,
устраивающим здесь засады. Часто размер сухих естественных солонцов
достигает нескольких десятков квадратных метров. Поедая почву, звери выгрызают
настолько большие ямы, что порой скрываются в них (рис. 41). Другой способ
получения минеральных веществ — водно-солевой и водно-грязевой: косули
пьют воду из источников, богатых минеральными солями, и поедают грязь
на берегах (Фетисов, 1953).
Химический анализ естественных солонцов показывает, что они богаты
солями натрия, серы, кальция, магния и другими элементами и соединениями,
но в разных регионах их химический состав отличается.
В настоящее время существует несколько гипотез, объясняющих причины
138
солонцевания диких копытных. Помимо рассмотрения этого явления, исходя
из необходимости минерального питания и потребления микроэлементов,
указывалось на привлекательные вкусовые свойства солонцов, на необходимость
механического фактора в пищеварении и на благотворное влияние солонцевания
на желудочно-кишечные микроорганизмы животных.
П.В. Елпатьевский и A.M. Паничев (1980), проведя геохимический анализ
зверовых солонцов Сихотэ-Алиня, считают, что причиной посещения животными
природных солонцов в первую очередь является потребность в натрии, в меньшей
мере — в кальции, в железе, марганце и, может быть, фторе. Большая
потребность в натрии объясняет причину охотного посещения копытными искусственных
солонцов, содержащих поваренную соль. Кроме того, не исключено, что солон-
цевание вызвано нарушениями минерального обмена, периодически возникающего
у копытных в связи с их физиологическими ритмами и воздействием факторов
среды, и необходимо им для минерально-лечебного питания. Использование
в пищу минеральных грунтов, обладающих ионообменными свойствами,
позволяет животным регулировать состав и концентрацию ионов в пищеварительной
полости, особенно в весеннее время в связи с резким преобладанием в составе
зеленого корма калия над натрием (Паничев, 1987).
Свободная вода, безусловно, нужна косулям, как и всем другим
животным. Однако по сравнению с другими оленями необходимость в ней меньше
выражена. Их суточная потребность в воде при естественном корме составляет
около 1350 мл/10 кг живой массы (Bubenik, 1959). При кормлении сухим
сеном потребность в воде возрастает до 400 мл (Becker-Dillingen, 1945). Бридерман
(Briedermann, 1974) указывает, что для полноценного кормового рациона косулям
необходимо 1500 г свободной воды в день.
В летнее время косули получают достаточное количество воды с
растительной пищей, богатой влагой, особенно в утренние и вечерние часы и после
дождей. Поэтому они могут жить за десятки километров от естественных
источников, как, например, в степях Монголии. Однако при наличии поблизости
водоема животные регулярно посещают его.
Лактирующие самки испытывают ббльшую, чем обычно, потребность в воде
и могут ежедневно ходить на водопой на большое расстояние. У воды чаще всего
разнообразнее и обильнее корм, растительность гуще и выше, что в
совокупности создает предпосылки для повышенной плотности их населения. Образование
искусственных водоемов в местах обитания косуль поэтому должно
приветствоваться.
Зимой большая часть водного баланса также покрывается за счет содержания
ее в естественной растительной пище и очень незначительная — за счет поедания
снега.
Глава седьмая. Размножение
Половая зрелость
Половая активность впервые проявляется у самцов: косуль в возрасте 4—6 мес.
В это время происходит незначительное увеличение массы семенников и диаметра
семенных канальцев (рис. 42), повышение гормонального уровня (рис. 43),
зафиксированы первые стадии сперматогенеза вплоть до образования
сперматоцитов первого порядка и в редких случаях сперматозоидов. Однако уже с конца
декабря сперматоцитов в канальцах придатка семенника не видно, и они появляются
вновь лишь в мае—июне. Поскольку половые продукты полностью практически
не созревают, этот первый активный половой период у подавляющего
большинства полугодовалых самцов нерепродуктивен. Тем не менее первые стадии
сперматогенеза способствуют относительно раннему для копытных животных
наступлению половой зрелости. Единичные собщения об успешном покрытии
детенышами своих матерей или других самок в вольерах представляются
исключительными (Stieve, 1950; Wandeler, 1975; Цаплюк, 1977; Sempere, 1982).
Половая зрелость у самцов косуль наступает в начале 2-го года жизни. Это
подтверждается как наблюдениями за их поведением, так и исследованиями
гистологического строения семенников и придатков. У однолетних самцов
к началу гона резко повышается уровень содержания тестостерона (см. рис. 43).
Семенники и диаметр семенных канальцев значительно увеличиваются в размерах
(см. рис. 42). Канальца придатков могут наполняться сперматозоидами почти
с такой же высокой плотностью расположения, как и у взрослых особей,
что характеризует годовалых особей как полноценных производителей. Однако
семенники их развиты все же слабее, а диаметр семенных канальцев у них меньше,
чем у взрослых (Stieve, 1950; Оцхели, 1964; Bramley, 1970; Поле, 1973; Sempere, 1982),
что указывает на более высокую половую потенцию последних. Молодые самцы
уступают взрослым в физическим развитии и изгоняются ими с территорий
(см. главу "Структура популяций"). Не имея к периоду гона собственной территории,
большинство однолетних особей в естественных условиях не могут участвовать
в размножении (хотя в вольерах успешное покрытие ими самок обычное явление)
и представляют собой некий репродуктивный резерв популяции, включающийся
в размножение лишь при недостатке территориальных самцов.
Половое созревание самок, напротив, наступает уже на первом году жизни.
В августе в яичниках самок-детенышей появляется множество мелких фолликул,
что, видимо, связано с приближением эстрального цикла. К октябрю они атре-
зируются, но часть сохраняется, созревает, и в это время возможно
оплодотворение (Цаплюк, 1977). Однако в размножении участвуют лишь немногие сеголетки
в силу социальной незрелости или из-за невозможности оплодотворения
самцами, у большинства из которых осенью уже нет сперматозоидов. Желтые
140
7 W Ж Л I Ш 1 Ш Ж Ш I Ж 7 Ш Ж Ш I Месяцы
I 3 5 7 9 11 13 15 17 19 21 > Возраст,мес
Рис. 42. Возрастные и сезонные изменения массы семенников (7) и диаметра семенных канальцев (2)
у сибирской косули (по: Цаплюк, 1977; с изменениями)
3 — период гона
/ 3 5 7 9 11 13 15 17 19 21 > возраст, же
Рис.43. Возрастные и сезонные изменения концентрации тестостерона (/) и Л Г (2) в плазме крови у самцов
европейской косули (no:Sempere, 1982; с изменениями)
3 — период гона
тела беременности у сеголеток европейской косули регистрировались
неоднократно: Георги (Georgii, 1974) обнаружил их у 4 из 67 телят, 9 из 39 годовалых
косуль овулировали, будучи детенышами; Ванделер (Wandeler, 1975) нашел
желтые тела у 5 из 60, а Чепмен (Chapman, Chapman, 1971) — у 2 из 5 осмотренных
молодых животных. Желтые тела присутствовали у детенышей сибирской косули
в Казахстане (Цаплюк, 1977), мы нашли их у 1 самки из 7 осмотреных на Южном
141
Урале в возрасте до одного года. Однолетние особи уже могут приносить
потомство, однако такие случаи редки (Neubert, 1970; Borg, 1970; Ellenberg, 1978).
Таким образом, большинство самок становятся половозрелыми, как и самцы,
на 2-м году жизни и приносят первое потомство в возрасте 2 лет.
Спаривание
Гон (см. главу "Поведение") у европейской косули в основном проходит
в июле—августе, у сибирской — в августе—сентябре, т.е. сроки спаривания
у них перекрываются (табл. 21). Начало гона у Capreolus имеет тенденцию
к временному смещению с запада на восток и с юга на север, изменяется
в одних и тех же районах по годам и в зависимости от высоты местности
над уровнем моря. В Австрии, например, на низменных участках спаривание
идет чаще в период с 20 июля по 7 августа, на холмистой местности с 25 июля
по 15 августа, а в горах с 3 по 20 августа (Kerschagl, 1952).
Предположение о том, что начало гона зависит от погодных условий, до
сих пор не удалось подтвердить. Тем не менее после мягкой зимы вероятны
более ранние сроки гона. Элленберг (1978) нашел у европейской косули
сравнительно постоянный интервал между сроками отела и течки — 67,3±1,9 дней.
У однолетних самок течка начинается несколько раньше, нежели у взрослых
(Kaluzinski, 1978; Eberle, 1986). Ее начало, возможно, в какой-то мере
синхронизируется посредством "датчиков времени", из которых важнейшим является
длительность дня, равная 15J/« ч (Lehmann, 1960).
В литературе упоминается о проявлениях полового поведения косуль
практически в любом месяце, однако случайные эпизоды не могут отражать реальную
картину размножения этих животных. Спаривание у них происходит со второй
половины июня по конец сентября, и лишь малая часть взрослых самок,
оставшихся непокрытыми, и очень незначительная часть самок-сеголеток приходят
в течку позднее вплоть до декабря. Успешное оплодотворение самок после
этого срока и до мая практически невозможно по физиологическим причинам.
Этот вывод основан на следующих данных. Масса и объем семенников,
а так же диаметр семенных канальцев взрослых самцов в течении года изменяются
с четкой цикличностью (см. рис. 42): зимой показатели в среднем в 5—10 раз
меньше, чем летом. Максимальных значений масса семенников достигает к июлю,
т.е. к сезону размножения, минимальных — в период с декабря по март, однако
рост гонад заметен уже с февраля. На Урале, где обитает самый крупный
подвид сибирской косули, масса семенников взрослых особей изменялась с 3,5 до
54,3 г (Соколов, Данилкин, 1981), в Казахстане у особей тянь-шаньского
подвида — с 2,9 до 44 г (Поле, 1973). Аналогичные данные получены по европейской
косуле (Stieve, 1950; Short, Mann, 1965, 1966; Bramley, 1970; Стекленев, 1980;
Sempere, 1982).
Циглер (Ziegler, 1848) установил, что в ноябре—декабре сперматозоидов
в семенниках уже нет и, следовательно, спаривание у косуль поздней осенью
и зимой не может быть продуктивным. Позднее эту точку зрения подтвердили
Бишоф, Кейбел (Bischoff, 1854; Keibel, 1899) и другие исследователи. Однако
Штиве (Stieve, 1950), изучив гистоструктуру половых органов европейской
косули, показал, что, несмотря на быструю редукцию семенников после гона
и прекращение продуцирования семенных клеток, оплодотворение живыми
сперматозоидами теоретически возможно вплоть до конца декабря благодаря
142
Таблица 21. Сроки гона европейской и сибирской косуль
Регион
Австрия
Болгария
ГДР
Чехословакия
ФРГ (Бавария)
Дания
Швейцария
Польша
СССР
Юг Украины
Украина левобережная
Северо-Запад
Белоруссия
Латвия
Грузия
Южный Урал
Казахстан
Киргизия
Восточная Сибирь
Забайкалье
Якутия
Дальний Восток
Монголия
Начало
Конец
Европейская косуля
20.7
12.7
17.6
Июль
30.6
Июль
20.7
Июль
»
»
»
15.7
15.6
Июль
20.8
15.8
29.8
Август
17.8(5.11)
20.8
15.8
Август
»
Сентябрь
Октябрь
15.8
Август
»
Сибирская косуля
25.7
Июль
16.8.1972
7.8.1973
4.8.1974
9.8.1975
22.7.1976
5.8
Август
19—23.6
Август
»
5.8
Август
»
26.6
19.7
6.9
22.9
18.9
11.9
9.9
14.9
8.9
20.9
Сентябрь
»
»
8.10
17.9
Сентябрь
»
30.10
Сентябрь
Авторы
Kerschagl, 1952
Петров и др., 1968
Stubbe, Stubbe, 1985
Hell, 1979
Ellenberg, 1978
Strandgaard, 1972
Sagesser, 1968
Kaluzinski, 1978
Стекленев, 1980
Карпенко, 1966
Верещагин, Русаков, 1979
Саблина, 1955
Калниньш, 1950
Оцхели, 1964
Филонов, 1974
Киселев, 1979
Наши данные
То же
"
"
"
Жирное, 1958
Поле, 1973
Слудский и др., 1984
Айзин, 1969
Фетисов, 1953
Смирнов, 1978
Егоров, 1965
Бромлей, Кучеренко, 1983
Дарман, 1986
Наши данные
сохранению кх в придатках семенников. К таким же выводам пришел и А. Семпере
(Sempere, 1982). Тем не менее после сентября—октября в придатках семенников
сперматозоидов очень мало, преобладают незрелые половые клетки и
совершенно неясно, может ли оплодотворение быть успешным (Short, Mann, 1966;
Арабули, 1966).
О.Э. Цаплюк (1977), проследившая сезонную динамику гистоструктуры
семенников и придатков сибирской косули в Казахстане, нашла, что у взрослых
самцов формирование канальцев придатков заканчивается в мае и в части
их уже содержатся немногочисленные сперматозоиды. В июне сперматогенез
находится на умеренной стадии, в июле — очень интенсивен, а в августе кон-
143
центрация сперматозоидов в придатках особенно велика. Однако уже с августа
сперматогенез идет на убыль и преобладает спермиогенез. К концу сентября
сперматогенез прекращается и эпителий на отдельных участках канальцев
придатка разрушается. К концу ноября все сперматогенные клетки и оставшиеся
в канальцах единичные сперматозоиды дегенерируют, и к концу декабря живых
сперматозоидов в половых органов самцов сибирской косули нет. В январе
процессы резорбции в семенниках и придатках усиливаются, в феврале они
продолжаются в семенниках, но в придатках уже начинается смена эпителия. В марте
в семенных канальцах идет митоз сперматогоний и в апреле наблюдается
образование сперматогенных клеток. У самцов европейской косули (во Франции)
свободные сперматозоиды присутствуют в половых органах с марта (Sempere,
1982).
Следует отметить, что у Capreolus активация гонад происходит зимой и
сочетается с увеличением светового дня, тогда как у других оленьих
возобновляется в начале лета и их репродуктивный цикл совпадает с уменьшением
светого дня. Каковы же в таком случае гормональные изменения?
После периода спаривания концентрация тестостерона в крови косуль резко
уменьшается, зимой находится на минимальном уровне и повышается весной,
достигая максимума к периоду гона. Изменения концентрации гормона гипофиза
(ЛГ) идентично профилю годового цикла гормона семенников (тестостерона),
но опережает его примерно на месяц (см. рис. 43). Годовой цикл ЛГ
возобновляется между декабрем и январем, что также отличает косулю от других
млекопитающих. Кастрация не изменяет существенным образом профиль цикла ЛГ в
плазме. Сезонные изменения активности семенников и пролактинемии строго
синхронны. Реактивация экзокринной и эндокринной функций семенников
происходит почти одновременно, но процессы, участвующие в образовании
сперматозоидов, развертываются весьма быстро, а эндокринные имеют замедленную
активность в течение весны. Поэтому трудно допустить, что андрогенные
и сперматогенные функции полностью коррелируют между собой. Тем не менее
возрастные и сезонные изменения их циклов весьма сходны (см. рис. 42, 43),
т.е. активность семенников явно находится под гипоталамо-гипофизарным
контролем (Short, Mann, 1965; Sempere, 1982).
Гистологические исследования гипофиза подтверждают это предположение
(Cadariu et al., 1977). Активность гонадотропных (j-клеток проявляется в мае,
усиливается в июне к началу сперматогенеза и, достигнув в июле максимума,
снижается. В А-клетках найдены такие же изменения. Максимальная
активность n-клеток зарегистрирована с начала до конца августа. Авторы прямо
связывают эти явления с репродуктивным циклом.
Таким образом, самцы косуль физиологически способны к размножению
только в течение одного полового цикла в году — с мая (марта) по декабрь,
хотя успешное покрытие ими самок в самом начале и конце этого срока
проблематично (рис. 44).
Указания Штиве (Stieve, 1950) на два брачных периода в году у самок: в июле—
августе и в ноябре—декабре — вызывают сомнения (Short, Mann, 1966; Aitken,
1974). Тем не менее убеждение о наличие основного и дополнительного гона
у косуль прочно утвердилось, хотя и не совсем соответствует
действительности.
О.Э. Цаплю к достаточно убедительно показала на примере сибирских косуль
144
Рис.44.Репродуктивный цикл у Capreolus
из Казахстана, что самки являются сезонно-полиэстральными животными с одним
длительным брачным периодом. В их гонадах в январе присутствуют только
атретические фолликулы; в феврале фолликулярная зона оживляется,
развиваются первичные и вторичные фолликулы, а полостные в отдельных случаях
достигают в диаметре 5 мм. В марте картина гистоструктуры остается без
существенных изменений, но уже с мая преобладают большие вторичные
фолликулы. В августе у самок обнаруживается множество полостных фолликулов,
появление и атрезия которых предшествует созреванию овуляторных
фолликулов и, возможно, благодаря выделению биостимуляторов способствует
обеспечению последних стадий их роста и созревания. В сентябре и октябре в
яичниках самок, помимо формирующихся желтых тел, находятся 1—2 крупных
нормальных фолликула, которые атрезируются в ноябре и уже глубоко атретичны
в декабре. Желтые тела в начальных стадиях формирования, видимо, способствуют
быстрому развитию полостных фолликул последующей генерации, обеспечивая
условия полиэстральности.
Отметим и еще одно немаловажное обстоятельство. Яичники самок в одно
и то же время могут находиться на разных функциональных стадиях (Popovici
et al., 1978). Отдельные фолликулы имеют по две яйцеклетки, а в последних
могут быть по два ядра (Цаплюк, 1977). Распределение желтых тел,
формирующихся после овуляции из клеточных компонентов фолликула, в обоих
яичниках возможно в следующих вариантах: 0+1, 0+2,0+3,0+4,1+1,1+2,1+3,2+2
(Borg, 1970; Прусайте и др., 1977; Blant, 1987). Это косвенно говорит о том, что
яйцеклетки могут созревать неодновременно, вызывая иногда повторную течку.
Г. Элленберг (Ellenberg, 1978) наблюдал ее у разных особей через 9, 14, 17, 27,
32 и 84 дня. X. Страндгард (Strandgaard, 1972) также зарегистрировал
повторные проявления полового поведения у двух самок через 20 и 22 дня. Неодина-
10. Европейская и сибирская косули _ 145
ковые промежутки времени между повторными течками однозначно говорят
в пользу высказанного предположения. Тем не менее у большинства самок
беременность является результатом единичного короткого процесса гона (см. главу
"Поведение").
Следовательно, оплодотворение самок косуль в принципе реально с мая
(апреля?) до декабря (см. рис. 44), но в начале этого срока (в мае—июне)
покрытие большинства из них практически невозможно из-за продолжающейся
беременности и последующей интенсивной лактации. В конце срока спаривания
(в октябре—декабре) оплодотворение затруднено из-за прекращения процесса
сперматогенеза у самцов.
Осенний всплеск половой активности у этих копытных в основном вызван
проявлением полового поведения у части рано созревших сеголеток и
адекватной реакцией взрослых особей. Позднее в декабре—январе у беременных самок
происходит перестройка в половых органах при имплантации бластулы, вероятно,
сопровождаемая усилением деятельности желез. В результате самцы иногда
приходят в половое возбуждение и даже совершают половой акт, однако не
приводящий к оплодотворению, так как самки уже были покрыты ранее (Schumacher,
1939; Bettmann, 1961).
В целом гон в осенний период вряд ли имеет большое значение для
популяций. Во всяком случае эксперименты Г. Элленберга (1978) ясно показали,
что взрослые самки европейской косули, выпущенные к самцам в октябре
и содержавшиеся с ними до лета, не забеременели. Наличие этого периода
половой активности у Capreolus, скорее всего, является отголоском осенних
сроков гона у древних косуль.
Беременность
Косули — единственные из копытных, имеющие латентный период беременности,
в связи с чем их репродуктивный цикл (см. рис. 44) заметно отличается даже
от близкородственных животных1.
Оплодотворенная яйцеклетка на стадии морулы проникает в матку где быстро
делится, достигая стадии бластоцисты диаметром 0,1 мм. После этого
наступает 4,5—4-месячный инкубационный период с минимальной митотической
активностью и медленным затяжным развитием, в результате которого бластула
вырастает всего лишь до 5 мм в диаметре. Во время так называемого периода
предбеременности с июля—августа по декабрь—январь бластула лежит в матке
свободно и ее жизнь поддерживается "маточным молоком" (Short, Hay, 1966;
Aitken, 1974, 1981). В этот момент часты случаи миграции бластоцист из одного
рога матки в другой, но в конечном счете у самок с двумя зародышами в каждом
роге всегда находится по одному эмбриону, независимо от того,
выталкиваются ли яйцеклетки из одного и того же или по одной из каждого яичника
(Strandgaard, 1972). Появление однояйцовых близнецов — большая редкость
(Hamilton et al., 1960; Borg, 1970).
1 Принято считать, что впервые явление задержки эмбрионального развития и существование
отсроченной овоимплантации описал Циглер (Ziegler, 1843), но объяснил его значение Бишоф (Bischoff,
1854). Однако О.Э. Цаплюк (1977), ссылаясь на Амораэо и Финна (Amoraso, Finn, 1962), уточняет,
что явление диапаузы в эмбриональном развитии было установлено более 300 лет назад Гарвеем
(Harvey, 1651).
146
Задержка имплантации у косуль в отличие от куньих (Mustelidae), вероятно,
не зависит от светового периода. В экспериментах при ускоренном сокращении
светового дня, а затем удлинении фотопериода (что на 2 мес опережало
естественную сезонную картину) у них не удалось вызвать изменения латентного
периода беременности, хотя звери отчетливо реагировали на опыты
преждевременной (на 2 мес) линькой (Lincoln, Guinness, 1972). Кроме того, в
противоположность другим млекопитающим с эмбриональной диапаузой, у косуль в течение
всего периода беременности отмечена весьма значительная активность яичников
(Aitken et al., 1973; Цаплюк, 1977). Уровень прогестерона в ходе эмбриональной
диапаузы у беременных самок сравним с таковым у небеременных, но
значительно выше, чем у сеголеток, что хорошо подтверждает секреторную активность
желтого тела. Однако в ходе собственно беременности прогестеронемия
существенно увеличивается, тогда как у небеременных взрослых самок, вероятно
из-за дегенерации желтого тела, быстро снижается, достигая очень низкого
уровня, наблюдаемого у сеголеток (Hoffmann et al., 1978; Sempere, 1982).
Эмбриональная диапауза у Capreolus, скорее всего, обусловлена недостатком
каких-то веществ, вызывающих и поддерживающих процесс эмбрионального
развития. С начала января они находятся в секрете, выделяемом железами
матки и содержащем специфические белки, белки сыворотки крови, около
20 свободных аминокислот, глюкопротеиды, глюкозу и свободную кетозу.
Внезапное удлинение бластоцисты связано с повышением концентрации
эстрогена, видимо, Стимулирующего секреторную активность маточных желез. Однако
в этот период функциональная активность яичников не изменяется, поэтому
повышенное содержание эстрогена скорее является следствием эмбрионального
роста, чем его причиной (Aitken et al., 1973; Aitken, 1974, 1981). Выражение
"задержанная имплантация" в свете этих исследований не отражает
действительную ситуацию, поскольку сдерживается не образование анатомической связи
между бластоцистой и маткой, а развитие самой бластоцисты.
В Центральной Европе период предбеременности у половины самок
завершается к середине января, т.е. эмбриональная диапауза длится в среднем
около 168 дней (Wandeler, 1975). В областях с более суровым климатом
этот процесс заканчивается раньше (Hubner, 1938). У косуль, покрытых поздней
осенью, период предбеременности практически выпадает и сразу начинается
собственно беременность. В январе происходит плацентарное прикрепление
(имплантация) эмбриона, масса которого не превышает 1—2 г (Aitken, 1974;
Wandeler, 1975). В то же время есть наблюдения, что внедрение зародышевого
пузыря в матку в период предбеременности может происходить за много дней
до его развития (Short, Hay, 1966).
Период донашивания характеризуется бурным ростом эмбриона и
сопровождается повышенным выделением секрета желез рогов матки, в результате
чего в промывных водах содержание углеводов (особенно фруктозы) возрастает
по сравнению с латентным периодом в 15—30 раз (Aitken, 1976).
Эмбрионы у впервые рожающих самок развиваются медленнее, и рожают они
обычно на неделю позднее, чем взрослые (Ellenberg, 1978). В более северных
областях и высокогорных районах беременность явно продолжительнее. В Швеции,
например, эмбрионы развиваются почти на месяц дольше, чем в Германии.
На западном побережье Ютландии беременность длительнее на 12—14 дней
по сравнению с восточным, отличающимся более мягким климатом (Strandgaard,
147
1972). Сходные наблюдения приводят и другие исследователи (Rieck, 1955;
Sagesser, Kurt, 1966; Kurt, 1968; Wandeler, 1975). Даже в одной и той же местности
сроки беременности могут различаться по годам, что видно по сдвигам во
времени отела (см. ниже).
Не исключено, что длительность беременности у косуль может варьировать
в зависимости от погоды. При более высоких температурах воздуха весной
происходит более ранняя и быстрая вегетация растительности, соответственно
имеется больше корма для матерей и он лучшего качества. При качественном
питании развитие эмбриона ускоряется, при плохом — замедляется (Sagesser,
1968). В ФРГ поздней весной 1973 г. беременность косуль была продолжительнее
на 10 дней (Ellenberg, 1978). В Белгородской области весной 1987 г. в результате
запоздавшего на месяц начала вегетации первые косулята появились лишь в конце
июня, а массовый отел шел в июле, т.е. период беременности (при обычных
сроках гона) удлинился по сравнению с предыдущими годами не менее чем
на 10—15 дней.
В целом период беременности у европейской косули, судя по литературным
данным, может колебаться от 264 до 318 дней, при позднеосеннем гоне эти сроки
могут сокращаться до 5 мес. В среднем они составляют в разных регионах
Европы 294—300 дней (Prior, 1968; Wandeler, 1975; Ellenberg, 1978; и др.). У
сибирской косули в связи с более поздним временем начала течки сроки
беременности несколько короче. На Урале по нашим наблюдениям они составили в среднем
287 дней (от 279 до 296 дней), на Дальнем Востоке — 284 дня (Дарман, 1986).
Срок беременности европейской и сибирской косуль, содержащихся в одной
и той же вольере под Москвой также оказался неодинаковым: у первых 306 дней,
у вторых в среднем 280 дней (Громов, 1986а).
Эмбриональная диапауза в период беременности играет огромную роль,
при ее отсутствии рождение потомства пришлось бы на самое неблагоприятное
время — на зиму. Явно, что задержка эмбрионального развития является
вторичной, выработавшейся в процессе эволюции адаптацией. Однако до сих
пор не ясно, почему эта особенность размножения смогла появиться именно
у косуль и почему ее нет у других жвачных, известных с плиоцена и раннего
плейстоцена и обитавших вместе с ними в сходных климатических зонах и
биотопах?
Возможность беременности самок как с длительной эмбриональной диапаузой,
так и без нее, а также проявление первого цикла половой активности сеголеток
(в возрасте 4—6 мес) осенью (см. рис. 42,43) наводят на мысль о "подстройке"
функции размножения самок к изменившимся с осенних на летние сроки наивысшей
половой потенции взрослых самцов, что, очевидно, связано с территориальностью
(см. главу "Структура популяций"). На наш взгляд, именно из-за явления
территориальности, приуроченной к началу вегетации растительности, происходит
раннее достижение самцами пика гормональной и сопряженной с ней половой
активности. Это приводит к преждевременному и энергичному преследованию
самок в летний период (см. главу "Поведение"), что, в свою очередь,
стимулирует их и способствует успешному оплодотворению. Однако преждевременно
оплодотворенная яйцеклетка не может развиваться (по пока еще неясной причине)
вплоть до начала "нормального" зимнего срока беременности, характерного
для Cervidae, обусловливая возникновение эмбриональной диапаузы.
148
Отел
Детеныши европейской косули обычно рождаются с конца апреля до середины
июня, у сибирской — чаще со второй половины мая до середины июля (табл. 22).
Родовой период длится в каждом году примерно одинаковое время, смещаются
лишь его кульминационные моменты, что обусловлено влиянием погодного
фактора на срок беременности.
Рик (Rieck, 1955), использовав сведения о 1660 маркированных детенышах,
показал, что в Европе в мае и июне появляется на свет 96% телят. Половина
из них рождается до 1 июня, наибольшее количество родов приходится на 2 июня.
Средний срок рождения смещается все более на поздние числа июня в направлении
с юго-запада на северо-восток и от равнины к высокогорью. В Швейцарии
массовые роды в 1965 г. пришлись на 3 июня, в 1966 г., когда была наиболее
высокой температура, — на 26 мая, в 1967 г. — на 30 мая (Sagesser, 1968), в ФРГ
Таблица 22. Сроки отела у европейской и сибирской косуль
Район исследования
Швейцария
Швеция
Дания
ФРГ (Бавария)
ГДР
Венгрия
Польша
Болгария
СССР
Белоруссия
Грузия
Северо-запад
Белгородская область
Урал
Киргизия
Казахстан
Восточная Сибирь
Забайкалье
Якутия
Дальний Восток
Московская обл. (вольера)
Монголия
Начало
Европейская
24.4
21.5
5.5
1.5
3.5
14.4
15.5
24.4
Май
»
Апрель
22.5
Сибирская
17.5
23.5
10.5
22.5
31.5
Май
20.5
Май
»
»
1.5
Апрель
2.6
Май
Конец
косуля
16.6
3.7
11.7
3.6
15.6
17.5
20.6
5.6
Июль
Июнь
»
11.7
косуля
20.6
Июнь
Июль
23.6
—
—
Июль
»
Июнь
»
25.7
Июнь
17.6
17.7
Авторы
SSgesser, Kurt, 1966
Espmark. 1969
Strandgaard, 1972
Ellenberg, 1978
Stubbe, Stubbe, 1985
Szederjei, 1971
Kaluzinski, 1982
Драгоев, 1965
Сержанин, 1955
Арабули, 1966
Верещагин, Русаков, 1979
Наши данные
Дворников, 1984
Киселев, 1979
Аверин, 1949
Наши данные
Филонов, 1974
Айзин, 1969
Поле, 1973
Фетисов, 1953
Смирнов, 1978
Егоров, 1965
Дарман, 1986
Бромлей, Кучеренко, 1983
Громов, Данилкин, 1984
Наши данные
149
Рис. 45. Роды у косуль
Пояснения в тексте
в 1972 г. — на 19, в 1973 — на 29, в 1975 — на 25 и в 1976 — на 17 мая (Ellenberg,
1978). На Дальнем Востоке среднерасчетная дата отела сибирской косули 29 мая
(Дарман, 1986), на Урале — 9 июня (и = 41). Очевидно, что косули рожают
детей в наиболее благопрятное теплое время, когда уже развит травяной покров,
обеспечивающий укрытие и обильный корм.
Примерно за месяц до отела взрослая самка отделяется от группы и занимает
один и тот же из года в год небольшой по площади родовой участок, защищая
его от других самок (Kurt, 1966; Соколов, Данилкин, 1981). При выборе родового
участка беременная самка руководствуется прежде всего тремя факторами:
защитными и кормовыми условиями и близостью водоема в заслушливых областях.
"Просматриваемость" родовых участков оказывается значительно меньше, чем
на окружающей их местности. Близость жилища человека не влияет существенно
на выбор родового участка там, где нет чрезмерного преследования,
некоторые самки рожают в 100—200 м от строений. Однако фактор беспокойства
может привести к эмиграции беременных самок непосредственно перед родами
(Соколов, Данилкин и др., 1986а).
Наиболее предпочитаются для отела опушки леса и луговые поймы, где
имеется густой высокий травяной покров. В лесостепных биотопах большая часть
косуль рожает на лугах. В Швейцарии здесь появляются на свет 74% детенышей,
в лесу — 23%, а в полях всего 3%. В траве высотой до 20 см самки рожают лишь
17% детенышей, при высоте от 20 до 50 см — 51% и свыше 50 см — 32%(Blankenhorn,
1975). В горах и холмистой местности наиболее привлекательны южные склоны,
при высокой плотности населения дичи возрастает конкуренция за места отелов
на них (Sagesser, Kurt, 1966).
Мы неоднократно регистрировали случаи отелов одних и тех же самок на одном
месте в течение нескольких лет. Есть наблюдения, что, став половозрелыми
и заняв участок матери (в случае ее гибели или эмиграции), молодые особи
могут рожать точно там же, где родились сами (Aylmer, 1977).
Большинство случаев отелов приходится на светлое время суток. Самка
рожает детеныша лежа (Bubenik, 1965; Espmark, 1969) или стоя (Holler, Prilkofer,
1980), при рождении нескольких телят эти две позы могут чередоваться
(Соколов, Данилкин, 1981; Громов, Данилкин, 1984). Начало родов характеризуется
наступлением схваток, во время которых поведение самки становится
беспокойным: она периодически встает, облизывает область влагалища и появившиеся
части околоплодных оболочек. Во время схваток, которые следуют через 2—9 мин,
косуля стремится лечь на бок, при потугах поворачивает голову назад и издает
звук, напоминающий сдавленный выдох. Примерно через полчаса после начала
схваток из влагалища появляются копытца и ножки теленка, которые вставшая
самка может, вылизывать. Первый детеныш рождается на свет примерно
через 8—40 мин после появления из родовых путей его конечностей. Плод
выходит передними ногами и головой вперед. Второй и третий теленок рождаются
с интервалами от 10 до 40 мин. Послед отделяется через 12—50 мин после
рождения последнего детеныша. В целом нормальные роды протекают за 1,5—2 ч,
реже требуются 4—5 часов (рис. 45).
Материнское и детское поведение в послеродовой период описано в главе
"Поведение".
152
Воспроизводство
Скорость воспроизводства является одним из ключевых факторов динамики
популяций. Она определяется числом участвующих в размножении животных,
процентом беременных самок, числом родившихся и выживших детенышей и
длительностью их полового созревания. Косули отличаются от других оленьих
высокой плодовитостью, которая обусловлена ранним созреванием и участием
части самок в размножении уже на первом году жизни, а самцов на втором,
длительным периодом спаривания (возможен с мая по декабрь) и полиэстраль-
ностью и многояйцовостью самок.
В размножении у косуль в нормальных условиях участвует большинство
половозрелых самок. В Литве из 206 исследованных особей яловыми оказались
только семь животных (3,4%) из всех возрастных групп (Прусайте и др., 1977),
но в отдельные годы не размножались до 9% самок (Прусайте, Балейшис, 1975).
В ГДР без признаков беременности были 10 из 174 самок (5,7%) европейской
косули, в Польше — 6,5% (Kaluzinski, 1982a) и 14% (Fruzinski, Labudzki, 1982),
в Дании в зависимости от плотности популяции — от 0 до 5% (Strandgaard, 1972).
Все 55 самок сибирской косули старше 1,5 лет, добытые в разные годы в различных
районах Казахстана, оказались беременными (Поле, 1973). Беременными были
все 28 половозрелых самок, осмотренных нами на Урале, и, кроме того, каждый
год успешно рожали еще 23 меченые в природе или содержащиеся в вольерах.
В Забайкалье М.А. Лавов (1971) среди 31 косули не нашел ни одной яловой.
Лишь Ю.А. Дарман (1986) обнаружил на Дальнем Востоке 3 прохолоставших
самок из 18, однако он исследовал в основном трупный материал.
Самки как европейской, так и сибирской косуль в большинстве своем рожают
двух детенышей, реже одного и трех. Четыре детеныша в помете встречаются
в виде исключения, хотя четыре эмбриона в матке находят нередко (табл. 23),
иногда их число доходит до 5 (Аверин, 1949; Саблина, 1955; Сержанин, 1955;
Егоров, 1965; Myrberget, Milton, 1971; Верещагин, Русаков, 1979), и в
исключительны случаях — до 6 (Черкасов, 1884, по изд. 1962).
Исследования показывают, что в пределах ареала в среднем на одну самку
приходится от 1,7 до 2,5 желтых тел и от 1,6 до 2,3 эмбрионов (см. табл. 23).
В процессе беременности возможна потеря яйцеклеток и резорбция части
эмбрионов: у взрослых здоровых особей потери не превышают 1—4% (Short,
Hay, 1966; Chaplin et al., 1966; Strandgaard, 1972; Wandeler, 1975; Слуцкий и др.,
1984; Дарман, 1986), у больных или найденных мертвыми они гораздо
существеннее — от 9 до 22% (Borg, 1970; Wandeler, 1975). В эксплуатируемой популяции
в ГДР у 164 беременных самок резорбция эмбрионов составила 1,7%, что совпадает
с данными, приведенными выше.
Число желтых тел и соответственно эмбрионов зависит от возраста
животных. Полувзрослые самки имеют меньшее количество желтых тел, чем взрослые;
у последних чаще встречаются по 3 и более желтых тела (табл. 24), однако
смертность эмбрионов у старых особей выше, чем у средневозрастных.
Убедительные результаты в этом отношении получил Борг (Borg, 1970),
изучив матки 362 самок. Среднее число эмбрионов составило у двухлетних
зверей 2,23, у трех-пятилетних — 2,35, у шести-семилетних — 2,14, у
восьмилетних и старше — 1,88. В ГДР среднее число детенышей на самку увеличивается
с возрастом с 1,76 до 1,94 до 4 лет и после пика снижается с семилетнего
153
Таблица 23. Плодовитость европейской и сибирской косуль
Регион
Швейцария
Швеция
Дания
Англия
ФРГ
ГДР
Польша
СССР
Литва
Латвия
Казахстан
р. Или, Терскей-
Алатау
Урал, Зап. Сибирь
Предбайкалье
Забайкалье
Дальний Восток
Якутия
Авторы
Kurt, 1968
Wandeler, 1975
Blant, 1987
Essen, 1966
Borg, 1970
Andersen, 1953
Standgaard, 1972
Там же
и
Hamilton et al., 1960
Chaplin et al., 1966
Prior, 1968
Short, Hay, 1966
Chapman, Chapman, 1971
Ellenberg, 1978
Stubbe u.a., 1982
Fnizinsta', Labudzki, 1982
Kaluzinski, 1982
Прусайте и др., 1977
Приедитис, 1975
Слудский и др., 1984
Наши данные
Устинов, Лобанов, 1983
Лавов, 1971
Дарман,1986
Тавровский и др., 1971
Число
самок, л
желтых тел
на самку
Европейская косуля
114
99
179
46
41
8
32
35
17
152
2,12
2,00
2,51
1,98
1.9
2,4
1,77
1,80
1,71
2,16
Сибирская косуля
12
20
31
2,21
2,0-2,3
1J
самок, л
573
438
47
362
24
26
29
38
14
54
56
149
71
138
25
21
28
13
8
эмбрионов на
самку
1,69
1,90
2,30
2,19
1.9
1.»
1,9
1,9
1,8
1,9
1,67-2,15
1,85
1,56
1,88
1,71
1,9
2,3
2,21
1,92
2,25
Из них(%) с
1
17
10
15
16
9
5
-
15
17
13
14
17
21
12
13
28
25
14
29
15
—
2
76
77
47
50
85
93
63
85
79
82
86
78
72
68
62
56
42
64
71
77
75
3
7
10
32
33
6
-
37
_
4
5
-
4
7
12
22
16
33
18
_
8
25
4
1
1
6
1
-
-
-
_
_
_
_ -
2
_
1
3
_
—
4
_
_
_
Таблица 24. Число желтых тел (и эмбрионов) у европейской косули ■ разных возрастных группах
Авторы
Прусайте и др., 1977
Borg, 1970
Georgii, 1974
Wandeler, 1975
Strandgaard, 1972
Blant, 1987
Kaluzinski, 1982
Fruzinski, Labudzki, 1982
1-2
1,94
2,38
(2,23)
1,56—1,93
1,94
1,57
1,87—1,82
(2,00)
(1,40)
Возрастные группы, лет
3-5(6)
2,17
2,54
(2,35)
1,92-2,0
2,08-2,22
2,08
2,36—2,00
(2,00-2,54)
(1,67—2,00)
6(7) и более
2,54
2,54
(2,14-1,88)
2,20
2,32
2,08
2,00
(1,78-1,60)
(2,00)
Таблица 25. Успешность воспроизводства в зависимости от массы тела самок (по: Ellenberg, 1978)
Масса тела в декабре, кг
Результаты размножения
7
8
9
22
20
11
10
30
Возраст
12,4
13,1
15,2
15,9—17,6
Возраст
19,7
22,7
Возраст
22,7
24,8—25,1
0+
1+
2+
Умерли зимой
Умерли их детеныши
Вырастили по 1 детенышу
Вырастили по 2 детеныша
Яловые или умерли их детеныши
Вырастили по 2 детеныша
Вырастили "плохих" детенышей
Вырастили "хороших" детенышей
возраста до 1,73 (Stubbe et al., 1982). Примерно та же закономерность установлена
в Польше (Kaluzinski, 1982; Fruzinski, Labudzki, 1982) и в Чехословакии (Pi-
kula et al., 1985b). Показатель рождаемости у полувзрослых особей может
быть на 14—27%, а у взрослых — на 5—10% меньше, чем показатель
беременности (Wandeler, 1975; Ellenberg, 1978).
Приведенные выше данные свидетельствуют о том, что в популяциях косуль
средневозрастные самки представляют наиболее продуктивную ее часть, и это
обстоятельство следует учитывать при рациональном использовании ресурсов
животных.
Следует отметить, также сравнительно четкую тенденцию к увеличению
плодовитости в зависимости от массы тела. По данным Блента (Blant, 1987), средняя
масса самок старше 2 лет с 0—1 желтым телом составляла 16,2 кг, с 2 — 18,2 кг и
с 3 — 21,3 кг. Чем крупнее косули-матери, тем больше и более полноценных
детенышей они могут вырастить. Этот вывод очень хорошо аргументирован
Элленбергом (табл. 25).
155
Масса самок и успешность размножения зверей, вероятно, являются функцией
плотности популяции и качества биотопа. В Швеции, например, скорость
воспроизводства малочисленных популяций выше, чем в Средней Европе (Essen, 1966;
Borg, 1970). В Дании в перенаселенной популяции на самку приходится в среднем
1,6 желтых тела, тогда как в популяции с нормальной для этого района
плотностью (25 особей на 100 га) — 1,9—2,4 (Strandgaard, 1972).
Еккерман (Ueckermann, 1964) обнаружил, что при повышении плотности дичи
с 17 до 40 особей на 100 га прирост популяции снизился с 129 до 74% от числа
самок. На другом участке при удвоении численности с 150 до 324 особей
он уменьшился с 68 до 57%. В Северной Литве рост неэксплуатируемой популяции
прекратился, как только плотность населения достигла 19—20 особей на 100 га,
хотя условия обитания не изменились; к зиме на 1 взрослую самку приходилось
в этот период лишь от 0,76—0,54 сеголетка (Прусайте и др., 1973). На
увеличение прироста с 67 до 93% при снижении плотности населения популяции с 21 до
10 особей на 100 га указывает Хайн (Hein, 1966). Четкую отрицательную
зависимость между числом самок в популяции и размером прироста определил
Курт (Kurt, 1968) в Швейцарии. Он объясняет это явление запаздыванием
на год и более половой зрелости, усиливающейся резорбцией эмбрионов,
повышенной смертностью детенышей в первые недели после рождения.
Можно привести много других примеров, подтверждающих данную точку
зрений, однако при сравнении цифр плотности дичи и величины прироста
убеждаешься, что при одних и тех же плотностях уровень воспроизводства в разных
популяциях различен или, наоборот, при разных уровнях плотности населения
прирост одинаков, т.е. здесь явно действуют и другие факторы. В частности,
анализ размножения косуль в популяциях с разной плотностью не показал
статистически достоверных отличий (Bobek, 1977).
Элленберг (Ellenberg, 1974, 1978) на основании тщательных опытов в вольере
приходит к выводу, что плотность популяции оказывает несравненно меньшее
влияние на темп размножения и выживаемость молодняка, чем обеспеченность
кормом. Действительно, в вольерах даже при очень высокой плотности, но
при качественном кормлении продуктивность бывает максимальной (от 150
до 212%). На результат воспроизводства влияют резервы пищи в биотопе,
пол и количество телят в приплоде и социальный статус матери. Благодаря
лучшему питанию в лучших по качеству биотопах телята растут и развиваются
быстрее, особенно если они у матерей единственные. У доминантных самок,
занимающих оптимальные биотопы, детеныши более крупные, чем у
иерархически подчиненных. Более развитые сеголетки легче переживают зиму, отставшие
в росте погибают гораздо чаще. Кроме того, слабое их развитие означает,
как минимум, замедление оборота популяции из-за того, что молодые самки
вступают в размножение на год позже. Там, где в популяциях много мелких
по размерам сеголеток, с полной уверенностью можно говорить о
перегруженности биотопа из-за слишком высокой плотности населения.
На скорость воспроизводства в популяции косуль очень большое влияние
оказывает доля самок, участвующих в размножении, и доля самок в приплоде.
В естественной популяции среди взрослых животных преобладают самки.
Соотношение полов у детенышей может варьировать в зависимости от условий
питания самок перед оплодотворением. Подробнее об этом говорится в главе
"Структура популяций".
156
Все приведенные выше факты свидетельствуют о том, что популяция,
находящаяся в хороших кормовых условиях при умеренной плотности населения,
обладает наибольшим воспроизводительным потенциалом: самки при хорошей
кондиции более плодовиты, рожают больше детенышей женского пола, которые
лучше развиваются и выживают, раньше созревают и эффективнее участвуют
в размножении.
Успешность размножения во многом зависит и от характера биотопа. На
южных склонах гор величина приплода может быть выше, чем на других (Kurt,
1968). В лесу прирост популяций выше, чем в поле. Причиной высокой
смертности детенышей в открытых биотопах наряду с погодными факторами
является высокий уровень механизации и химизации сельского хозяйства. В
популяциях полевых косуль ежегодный прирост колеблется от 0 (после суровых
зим) до 100% (Pielowski, 1977). Тем не менее в Болгарии прирост популяции
в открытых биотопах оказался выше (170—180%), чем в лесу (140—150%) (Колев,
1980).
Потенциальная плодовитость европейской и сибирской косуль в среднем
составляет примерно 2 детеныша на одну половозрелую самку (см. табл. 23).
Реальный прирост популяции оказывается меньше этой цифры. В Шотландии,
например, при продуктивности популяций, достигающей в некоторые годы 210
телят на 100 самок, смертность детенышей составляет 55—65% (Ratcliffe, Rowe, 1985).
Часть детенышей гибнет уже вскоре после рождения, особенно у молодых,
впервые рожающих матерей. По этой причине практически не оказывают влияния
на воспроизводство однолетние самки, забеременевшие будучи детенышами.
Гибель самок вместе с детенышами при родах невелика, тем не менее в некоторых
регионах эти потери могут достигать нескольких процентов. Элленберг приводит
данные по вольере с высокой плотностью населения, в которой при родах
погибли 5 из 109 беременных (4,6%). В популяции на научном стационаре Хакель
(ГДР) смертность составляла 4,1%.
Особенно существенны потери детенышей в первый месяц жизни (Reimoser,
1984). В ГДР только в первую неделю их отход составляет 10% (Stubbe, Stubbe,
1985). Значительный ущерб причиняют хищники, и в первую очередь лисицы
(Рухлядев, 1948; Владышевский, 1968; Strandgaard, 1972; Wandeler, 1975; Слудский
и др., 1984).
Добывать затаившихся и не убегающих при опасности в первые дни жизни
телят могут также шакалы, енотовидные собаки, волки, бродячие собаки,
дикие кабаны. В Сибири сеголетки могут быть одним из .основных
компонентов пищи рыси (Смирнов, 1978)2.
Однако в агроценозах потери детенышей от хищников все же меньше, чем
урон от сельскохозяйственных машин, который может составлять 26—35%
прироста популяции (Драгоев, 1965; Beklova et al., 1982; Reimoser, Zandl, 1986).
До 40% телят погибает весной в результате неблагоприятных погодных условий
(Stubbe, Stubbe, 1985).
С момента рождения и до осени в Казахстане гибнет до 16% сеголеток
сибирской косули (Поле, 1973), на Дальнем Востоке — от 13,7 до 32,3% (Дарман, 1986).
На Урале в заповедной популяции по расчетным данным отход составлял от 11,5
2 О смертности косуль см. подробнее в главе "Численность".
157
до 19,3%, однако ни одна из 23 находящихся под нашим наблюдением самок
не потеряла до осени ни одного детеныша; в Ильменском заповеднике
смертность телят к этому времени достигает 25% (Филонов, 1974). В популяциях
европейской косули потери детенышей к осени могут быть еще выше. В Польше
с мая по ноябрь в среднем гибнет 37,6% (33,6—44,6%) (Fruzinski, Labudzki, 1982),
в Литве и в Швейцарии смертность их в отдельные годы достигает 50% (Прусайте
и др., 1974; Blant, 1S87).
Величина детской смертности в конечном счете определяет прирост
популяции к осени (к промысловому сезону)3. В ГДР на одну самку приходится
в среднем 1,35 теленка (Stubbe, Stubbe, 1985); в Швейцарии — 1,46 (1,33—1,60)
(Blant, 1987), в Дании на одном и том же участке количество детенышей на
одну половозрелую особь колебалось от 1,6 до 0,5 (Anderson, 1953; Strandgaard,
1972); в Англии каждая самка имеет в среднем 1,56 детныша (Loudon, 1978);
в Эстонии — от 0,36 до 0,95, но в пересчете на отелившихся животных — от
1,27 до 1,87 детей (Рандвеэр, 1985).
В Забайкалье в эксплуатируемой популяции сибирской косули прирост к осени
достигает 125% (Лавов, 1971), по другим данным в этом же регионе молодняк
к ноябрю составляет всего лишь 46% от количества самок (Смирнов, 1978).
В Казахстане на одну рожавшую самку приходится в период с 20 сентября
по 20 октября 1,6 косуленка (Поле, 1973). На Урале в неэксплуатируемой
популяции и при отсутствии хищников прирост достигал 180% от числа самок при
плотности популяции 2—5 особей на 100 га, но с ростом численности волко-
собачьих гибридов и рысей снизился до 132%.
Зимой происходит дальнейший существенный отход детенышей в результате
промысла и из-за естественной смертности. Убыль сеголеток за год может
быть различной в разных районах и колеблется от 20 до 90%, соответственно
отражаясь на цифрах прироста популяции (Саблина, 1955; Петров и др., 1968;
Лавов, 1971; Падайга, 1961; Поле, 1973; Филонов, 1974; Прусайте и др., 1973, 1974;
Bobek et al., 1974; Рандвеэр, 1985; Приедитис, 1985; Stubbe, Stubbe, 1985; Ratcliffe,
Rowe, 1985; и др.).
К весне из двух телят чаще всего остается один, но в разных
популяциях в пределах ареала цифры реального прироста колеблются от 0,1 до 1,5 (Поле,
1973, Соколов, Данилкин, 1981; Дарман, 1986; Fruzinski, Labudzki, 1982; Kaluzinski,
1982а; Blant, 1987; и др.).
Приведенные выше данные показывают, что прирост у косуль — величина
непостоянная, подверженная значительным колебаниям в зависимости от
внешних факторов среды. Потенциальная биологическая плодовитость, близкая
к 200% от числа участвующих в размножении самок, может реализоваться
'Следует отметить,- что приводимые во многих работах данные о воспроизводстве популяций,
основанные на визуальных регистрациях самок с телятами или без них в летние месяцы, приводят
к грубым ошибкам в сторону занижения реальных цифр прироста и не анализируются в настоящем
исследовании. Ошибки возникают из-за незнания или недоучета особенностей материнского и детского
поведения косуль: родившие самки часто пасутся без телят, телята затаиваются, кормление их в первые
две-три недели происходит поочередно, во время гона матери держатся отдельно от детенышей
(см. главу "Поведение").
С наименьшей погрешностью прирост у популяций европейской косули можно подсчитать лишь
с конца августа, у сибирской — с середины сентября, когда происходит синхронизация
пространственно-временной активности у членов семьи.
158
только в очень благоприятных климатических условиях, при полноценном
питании, отсутствии естественных врагов и болезней, низком уровне механизации
и химизации сельского хозяйства. В природе к осени прирост популяции
обычно снижается в лучшем случае на 10—20%, в худшем — более чем на половину.
К следующему сезону размножения при особенно тяжелых условиях зимовки
цифра прироста стремится к нулевым значениям, определяя депрессию
численности популяции, при хороших условиях она может превышать 150% от числа самок.
Устраняя или смягчая неолагоприятные факторы среды и избирательно
воздействуя на половой и возрастной состав, на практике возможно
поддерживать воспроизводство популяции на уровне, близком к максимальной
плодовитости вида. Основные принципы управления популяциями подробно
рассматриваются в главе "Хозяйственное значение, рациональное использование ресурсов
и охрана".
Глава восьмая. Структура популяций
Половая и возрастная структура
В популяциях европейской и сибирской косуль соотношение полов у
детенышей близко 1:1 (табл. 26), но оно, вероятно, может сдвигаться в ту или
другую сторону адекватно изменившимся условиям среды. Елленберг (EUenberg,
1978) довольно убедительно показал, что при качественном питании самок
в летнее время и хорошей их упитанности к началу гона в следующем
году рождается в 3 раза больше детенышей-самок, при неблагоприятных
условиях соотношение становится 3:1 в пользу детенышей мужского пола. К тому
же при плохих кормовых условиях в период отелов повышается смертность
телят-самок, что увеличивает преобладание детенышей-самцов. Молодые и
старые самки чаще приносят телят мужского пола.
С возрастом происходит сдвиг соотношения полов в пользу самок. Это
вызывается следующими обстоятельствами, связанными с авторегуляторными
процессами в популяции. Большинство годовалых самцов из-за преследования
их взрослыми территориальными особями вынуждены эмигрировать из мест
рождения, в то время как многие молодые самки остаются вблизи участка
матери. Изгнанные молодые самцы чаще, чем самки, подвергаются опасности
в незнакомой местности и нередко погибают, не дожив до двухлетнего
возраста. В многочисленных популяциях двухлетние самцы по-прежнему не могут
занять постоянную территорию и в отличие от самок снова вынуждены
скитаться. Во многих районах Европы промысловый пресс на самцов выше,
чем на самок. Добавим к этому меньшую природную осторожность самцов
и большую их агрессивность, приводящую к дракам, травмам и гибели
некоторых из них в территориальный период.
Четких доказательств зависимости соотношения полов от плотности населения
популяций пока не найдено: даже при 40—50-кратной ее разнице цифры
соотношения полов примерно одинаковы (табл. 27).
Половая и возрастная структура популяций европейской и сибирской косуль,
как показывает анализ, не имеет существенных различий. Обобщив имеющиеся
данные по их возрастному составу в разных регионах (Anderson, 1953;
Strandgaard, 1972; Pielowski, Bresinski, 1982; Pikula et al., 1985; Лавов, 1971;
Смирнов, 1978), мы получим на графике некую кривую, отражающую
возрастную структуру популяций рода Capreolus и ее закономерные изменения
(рис. 46). Отсюда следует, что доля животных старшего возраста с каждым годом
становится все меньше и меньше и после 7—10 лет их насчитываются
единицы, т.е. предельный возраст естественной популяции косуль равен в среднем
10±2 года. Тем не менее известны случаи, когда продолжительность жизни
160
Таблица 26. Соотношение полов у европейской и сибирской косуль
Эмбрионы и детеныши
«?:<
Полувзрослые и взрослые
Литература
679
238
32
55
475
186
487
191
37
763
1722
111
138
42
16
389
179
116
46
44
116
37
16
217
248
0,78
0,75
1
0,9
0,9
0,73
0,89
0,86
0,6
1
0,9
1,25
1,3
1
0,8
1,19
0,97
0,93
0,9
1,2
1
0,95
1,3
0,97
1,3
655
327
172
3202
9737
949
104
3415
129
686
496
899
156
288
754
1382
689
Европейская косуля
— Kurt, 1968
— Wandeler, 1975
— Stubbe, Passarge, 1979
— Prior, 1968
— Borg, 1970
1,34 Nikolandic, 1968
1,9 Strandgaard, 1972
1,87 Ellenberg, 1978
Espmark, 1969
Engl, 1982
Pikula et al., 1985
Cotta et al., 1961
Fruzinski, Labudzki, 1982
1,03—1,65
1,38
1,5
1,7
(1,2-2,5)
1,28
(1,03-1,65)
1,06—1,83
2,52
1,1-1,5
1,1
1,31—1,56
1,3-1,4
1,31
Сибирская
1,2
1,19
1
1,1
1,38
1,12
Kaluzinski, 1982
Pielowski, Bresinski, 1982
Zejda, 1978
Stagl, Wenter, 1977
Арабули, 1966
Блузма, 1975
Приедитис, 1975
Рандвеэр, 1985, 1989
Наши данные (Белгородская и Курская области)
косуля
Аверин, 1949
Поле, 1973
Смирнов, 1978
Лавов, 1971
Дарман, 1986
Бромлей, Кучеренко, 1983
Наши данные (Урал)
То же (Амурская обл.)
Таблица 27. Соотношение полов в популяциях с разной плотностью населения
Плотность населения (число
особей на 100 га)
6":<
Авторы
1-10
10—20
20-30
30—40
40—50
1:1,0—1,8
1:1,0—2,5
1:1,2—2,2
1:1,2-2,07
1:1.8-1,1
Eiberle, 1962; Kurt, 1968; Лавов, 1971; Прусайте и др.,
1973; Блузма, 1975; Смирнов, 1978; Соколов,
Данилгин, 1981; Fruzinski, Labudzki, 1982; Pielowski,
Bresinski, 1982; и др.
11. Европейская и сибирская косули
161
АО
40
}
15 20
I
1 2 3 Ч 5 6 7 в 3 10 11 12 13 14
Возраст t лет
Рис,46. Возрастная структура популяций европейской и сибирской косуль
Штриховкой обозначена область варьирования средних значений для разных популяций
отдельных маркированных особей была в природе 15—17 лет, а в неволе
19—25 лет (Strandgaard, 1972; Stubbe, Passarge, 1979).
Доля сеголеток в популяции максимальна и сразу же после периода
отела может достигать 50%. Однако из-за большой смертности число
детенышей снижается с каждым месяцем и тем заметнее, чем в худших
экологических условиях находится популяция (см. главу "Размножение"). К примеру,
осенью доля сеголеток в популяции европейской косули колеблется по годам
в Литве от 27,1 до 36,1% (Прусайте и др., 1974; Блузма, 1975; Padaiga, 1975),
в разных районах Эстонии от 18,9 до 39,1% (Рандвеэр, 1985), в Польше
от 28,7 до 48,5% (Fruzinski, Labudzki, 1982; Pielowski, Bresinski, 1982).
У сибирской косули доля сеголеток в осенней популяции примерно такая
же: в Забайкалье 20—42,4% (Лавов, 1966, 1971; Смирнов, 1978), на Дальнем
Востоке 30,6—41,6% (Дарман, 1986), на Южном Урале 35,1—48,1% (Киселев,
1979; Дворников, 1984; Соколов, Данилкин, 1981; неопубликованные данные).
К весне их становится еще меньше, особенно при суровых условиях
зимовки:, на Дальнем Востоке 14,6—34,1%, на Южном Урале 18,3—36,2%.
Аналогичные показатели характерны для полевых популяций европейской . косули
(Kaluzinski, 1982).
Динамика полового и возрастного состава популяции европейской косули
наиболее тщательно исследована в Дании Страндгардом (Strandgaard, 1972).
В 1950 г. при тотальном отстреле косуль на участке леса площадью 165 га
были добыты 107 особей (Anderson, 1953). Осенью 1965 г. здесь оказались
96 животных, в 1966 г. — 112, в 1967 г. — 92. Соотношение полов у
косуль до 9-месячного возраста было приблизительно 1:1 (по годам
соответственно 1:1, 1:1, 1:0,6; 1:1), среди половозрелых особей на одного самца
приходилось в среднем 2 самки (1:1,7; 1:2,2; 1:2,2; 1:1,8). Доля сеголеток в
осенней популяции в эти годы составляла 47, 41, 35 и 18% (рис. 47), хотя
продуктивность самок все время была примерно одинакова — в среднем
162
SO
50
40
30
го
w
1
л
11 •.
" •■'* \
;1\\
- \\\
- >L.
уй^
&
1 1 1 1 Г
// 1Z3456783
Возраст, лет
ГЛ 1 1 Г-
11
-i—г
Z 3 1 5 Б
Возраст t лет
Рис. 47. Половая и возрастная
структура популяции европейской косули
в разные годы (по: Strandgaard, 1972)
/ — 1950, 2 — 1965, 3 — 1966,
4 — 1967
2,0 Corpora lutea, что говорит о потере части детенышей уже после
рождения.
В течение 2,5 лет в этой неэксплуатируемой популяции произошло
стремительное обновление: в ее состав, насчитывающий 57 животных старше одного
года, вошли 99 новых особей, в то же время из нее убыли 77 косуль.
Прирост этой популяции происходил исключительно из родившегося и
выросшего в этом районе молодняка. Наиболее существенной причиной убыли самцов
одно- и двухлетнего возраста и однолетних самок была эмиграция; взрослые
животные погибали в результате несчастных случаев й болезней. Оборот
популяции самцов был быстрее, чем самок: у первых в возрасте от 0,5 до 2,5 лет он
достигал ежегодно 50%, у более взрослых — 25%, тогда как у самок в
возрасте от 6 до 18 мес — 33%, а после этого — около 20%, т.е. в
неэксплуатируемой популяции косуль теоретически число самцов, достигших
шестимесячного возраста, в первые два года ежегодно уменьшается вдвое и затем на
четверть, в то время как у самок уменьшаестя в первый год на треть, а в
163
*<*:& 52:38
2-3 З-Ч 4-5 5-6 6-7 7-8 8-10 >10
Рис.48.Возрастная структура популяции ев
результатам отстрела в сезон охоты (Б) (из
1<- 1-2 2-3 3-1 1-5 5-6 6-7 7-8 8-Ю ->W
Возраст, лет
ропейской косули по данным тотального отстрела (А) и по
Pielowski, Bresinski, 1982b)
последующем ежегодно на 1/5 часть. Соответственно одно- и двухлетние
самцы имеют "запас" продолжительности жизни в 1,4 года, старше 2 лет — 3,3,
однолетние самки — 2,6 и более взрослые — 3,3 года. Быстрый оборот
особей приводит к омоложению популяции: средний возраст самцов в период
с 1965 по 1967 г. составлял здесь 2,1; 1,8, и 2,3 года (в среднем 2,1 года),
возраст самок — 3,2; 3,6 и 3,8 года (в среднем 3,5 года), средняя
продолжительность жизни этой популяции была 2,9 года (в 1950 г. — 2 года).
В других регионах Европы цифры среднего возраста популяций весьма
изменчивы, что, скорее всего, связано с разным уровнем их эксплуатации:
в Чехословакии — 2,85 года (у самцов — 2,7, у самок — 2,98) (Pikula
et al., 1985а); в ФРГ — 2,72 года (Ellenberg, 1978); в Польше они
колеблются от 2,4 до 3,5 лет (Bobek et al., 1974; Kaluzinski, 1982a; Pielowski et al.,
1982; Fruzinski, Labudzki, 1982); в Литве средний возраст популяции самцов
3,8, самок 4 года (Padaiga, 1975); в Забайкалье — 27—38 мес (Лавов,
1971; Смирнов, 1978).
Средний возраст популяции существенно меняется в течение года. К
примеру, в ГДР вскоре после отела он составляет 1,8 года, а перед рождением
нового поколения достигает 2,4 года (Stubbe, Stubbe, 1985).
Половой и возрастной состав эксплуатируемых популяций может в
значительной мере изменяться в зависимости от степени пресса на те или иные
половозрастные группы. Практика показывает, что даже при селективном остреле
под выстрел наиболее часто попадают средневозрастные животные, наиболее
продуктивные в популяции (Fruzinski, Labudzki, 1982; Pielowski, Bresinski,
1982). Это положение весьма показательно иллюстрируется данными польских
экологов (рис. 48).
При загонных способах охоты, практикуемых в осенне-зимний период во многих
164
областях России, отстреливают преимущественно самок (до 74%), из которых около
70% составляют половозрелые особи, имеющие телят. Такое воздействие
охотничьего пресса на половозрастную структуру необходимо обязательно
учитывать при управлении популяциями косуль.
Социальная организация
Социальная организация популяции косуль существенно изменяется в течение
года. В летний период большинство животных ведет одиночный и семейный
образ жизни, в зимний — преимущественно семейно-групповой и стадный.
Летом на одиночных особей приходится 60—90% всех встреч, а группы из
четырех и более животных практически отсутствуют. Пары наиболее часто
представлены самкой с детенышем, двумя детенышами, взрослым самцом и
самкой во время гона; реже пары образуют годовалые особи и иногда
взрослый самец имеет спутника — молодого самца. Группы из трех особей
являются преимущественно семейными: мать и два детеныша. В начале осени
семейные группы из двух и трех членов составляют почти половину от
всех встреченных косуль. Зимой наибольший удельный вес в популяции имеют
смешанные группы, состоящие из четырех и более особей (рис. 49, 50).
Размер группировок в популяциях европейской и сибирской косуль,
обитающих в сходных биотопах, одинаков (рис. 50). Некоторые отличия заметны
лишь весной и осенью, что связано с более поздними сроками начала
вегетации, более ранним и резким наступлением холодов и становлением
снежного покрова в Азии.
В открытых биотопах число животных в группах значительно выше
(нередко до 40—90 особей), чем в лесу, где лишь иногда можно встретить
вместе более 10—15 косуль. Средняя численность особей в группах в одной
и той же местности увеличивается при повышении плотности населения,
снижении температуры и увеличении высоты снежного покрова и при
уменьшении площади кормовых участков. Следовательно, социальная организация
популяции косуль во многом зависит от характеристик среды, среди которых
основное значение имеют обилие и распределение ресурсов и укрытий (Graczyk,
Bereszynski, 1978; Zejda, 1978; Reichholf, 1980; Bresinski, 1982; Maublanc et al.,
1987; Соколов, Данилкин, 1981; Дарман, 1986). В совокупности все эти
факторы проявляются в некоторых регионах на азиатской части ареала,
вызывая массовые миграции косульг во время которых животные нередко
объединяются в стада численностью в несколько сотен особей (см. главу
"Перемещения").
Группа формируется ранней осенью из одной или двух семей (нередко
родственных "и живущих по соседству) и присоединившихся к ним годовалых
самок. Позднее в их состав могут входить другие косули, и в частности
полувзрослые и взрослые самцы. Однако последние часто живут отдельно
не только летом, но и всю зиму. Среди одиночных зверей их
насчитывается около 70%, в маленьких группах они, как правило, бывают по
одному (Bramley, 1972), в больших группах и стадах взрослые особи
составляют около 30% (Zejda, 1978). Во время зимних миграций в одном стаде
могут находиться несколько десятков взрослых территориальных и агрессивных
165
^%%%%%V*% 4%%%%%%%V*%VVVV'
Состав групп--
ш
1
• •
смешанный
66
сеголетка
99+*^
i cf+9+*^
=
1
И
cf+9
9+oteroff
1
9
с*
-i 1 1 г
7ШШШШ1ШИ11 Л Ш Ш Y
Месяцы
Рис.49. Сезонная динамика социального состава полевой популяции европейской косули (из: Bresinski,
1982)
о • о •, о»о»о»о»о»о«о»о
1ИШШЖ1ИМ1 Ж Ш Ш
Месяцы
Число „
особей
В группе
Рис. 50. Динамика количественного состава группировок лесостепных популяций европейской (/) и
сибирской (2) косуль в течение года
/ — по 2470 встречам 5586 особей в Белгородской области (1982—1986 гг.); 2 — по 2366 встречам
5172 особей на Южном Урале (1971 — 1978 гг.)
166
Рис.51. Лидеры в группах сибирской(Л и европейской (2)
косуль
в летний период друг к другу самцов. Иногда
в январе—марте (апреле) наблюдаются группы,
состоящие исключительно из них (до 7 особей).
Большие группы чаще всего формируются на
местах кормежки при опасности: обеспокоенные
чем-либо животные стараются сблизиться со
спокойно стоящими или лежащими. Вид бегущей
особи вызывает следование за ней других, и
образовавшаяся группа может сохраниться в таком
составе на протяжении нескольких часов, дней
или, реже, недель.
Состав небольших групп наиболее стабилен.
Члены семейной группы обычно держатся
вместе на протяжении всей зимы (Bideau et al., 1983a; Данилкин,
Минаев, 1988). В больших по численности группах беспрестанно происходит
смена особей, хотя их состав и кажется стабильным (Strandgaard, 1972).
Наши наблюдения за европейскими и сибирскими косулями показали, что при
опасности стадо легко разбивается на мелкие субъединицы (чаще семьи) и
формируется затем уже в другом составе. Рассеянные при внезапном испуге члены
одной семьи могут оказаться в разных группах, но через несколько дней они вновь
находят друг друга, встречаясь на местах кормежки. Иногда отставшие животные по
следам разыскивают свою группу.
Оставшись без матери, сеголетки могут стать постоянными членами другой семьи
или группы. Тем не менее они принимаются неохотно и во многих случаях косули
агрессивны к ним. Поэтому некоторые телята всю зиму вынуждены держаться
обособленно, лишь изредка при опасности присоединяясь к разным группам.
Отдельно или парами всю зиму могут жить и полувзрослые животные. Одиночные
взрослые самцы особенно легко входят в состав различных групп и покидают их.
Например, один из них, помеченный радиопередатчиком в Белгородской области,
только в марте 1984 г. был членом 9 разных групп, а за весь зимний период его
регистрировали в 17 группах, причем в один из дней — в 5.
Такая структура популяции (одиночки—семьи—группы—стада) характерна для
всего зимнего периода. К концу марта (в апреле) заметно увеличивается число
одиночных особей, однако окончательный распад групп происходит на месяц
позднее (см. рис. 50). Семьи сохраняются дольше всего; в наших исследованиях
последняя дата их встреч у сибирской косули 28 мая, у европейской —19 мая.
Таким образом, семья у косуль служит ядром большинства группировок и в
конечном счете составляет основу всей социальной организации популяций.
Взрослые самки-матери к тому же обычно являются лидерами — вожаками групп. У
сибирских косуль в лесостепной популяции они оказывались ими в 86% случаев,
взрослые самцы — в 9,5%, полувзрослые самцы — в 1,5% и телята — в 3% (Соколов,
Данилкин, 1981). Примерно такую же картину мы зарегистрировали в лесостепой
популяции европейской косули на востоке ареала (рис. 51). В полевых популяциях
вожаки в большинстве случаев также представлены взрослыми самками, но
2 п*200
Взрослые
самки
Самцы Сеголетки
1В?
процентное соотношение их ниже (Bresinski, 1982) и здесь чаще лидирует особь,
первой среагировавшая на опасность (Graczyk, Bereszynski, 1978).
Анализируя приведенные выше и другие литературные сведения по европейской
(Саблина, 1955; Raesfeld, 1956; NeSas, 1960,1963; Kurt, 1968;Nikolandic, 1968; Bramley,
1970, 1972; Zejda, 1978; Reichholf, 1980; Bresinski, 1982; Тимофеева, 1985; Maublancet
al., 1987; и др.) и сибирской (Аверин, 1949; Филонов, 1974, 1977; Смирнов, 1978;
Соколов, Данилкин, 1981; Слудский и др., 1984; Дарман, 1986) косулям, мы приходим
к твердому убеждению, что социальная организация их популяций в этом плане
сходна; разница заключается лишь в сроках сезонного изменения образа жизни,
который у сибирской косули из-за климатических факторов запаздывает примерно
на месяц весной, а осенью проявляется чуть раньше.
Приведенная выше характеристика социальной организации популяций косуль
будет далеко не полной без учета взаимоотношений особей в пространстве.
Пространственно-этологическая структура
Структуру популяций млекопитающих можно рассматривать с различных точек
зрения (Наумов, 1963; Petrusewicz,1966; Шварц, 1967; Флинт, 1977; и др.), но
наибольшее значение имеют пространственная (статическая система закономерного
размещения особей по территории) и этологическая структура (динамическая
система взаимоотношений между особями) (Wynne-Edwards, 1962, 1964; Наумов,
1967, 1971, 1972, 1977; Шилов, 1972, 1977).
Для характеристики пространственно-этологической структуры популяций
животных в экологической и этологической литературе применяют многочисленные
термины: пространство жизнедеятельности (action area), пастбищное пространство,
сезонное пастбище, индивидуальный ареал, индивидуальный участок, участок
обитания (home-range), охраняемый участок, охотничий участок,
материнский участок, территория (terrytory) и др. Существует различная трактовка
этих терминов (Burt, 1943; Hennig, 1962; Барабаш-Никифоров, Формозов,
1963; Наумов, 1963; Jewell, 1966; Kurt, 1966,1968a,b; Strandgaard, 1972; Walther,
1972; Баскин, 1976; Шилов, 1977; и др.), поэтому необходимо уточнить
понимание некоторых из них по отношению к косулям.
Под индивидуальным участком подразумевается совокупность всех участков
обитания особи, включая пути ее кочевок, т.е. всех мест, где животное
бывает в течение всей жизни.
Участок обитания — пространство, на котором особь (или группа)
находится в течение определенного времени (сутки, сезон, период, год). Участки
обитания животных могут широко перекрываться.
Родовой участок — пространство, защищаемое беременной самкой от других
особей перед родами и некоторое время после них.
Территория — охраняемая самцами часть участка обитания с определенными
для данной особи границами, которые известны соседним животным и
поддерживаются чаще всего взаимным избеганием заходов. на чужие участки или
же активной защитой их. В это понятие сознательно не включается нередко
употребляемое выражение "маркируемая часть участка обитания", поскольку
у косуль маркироваться может не только своя, но и чужая территория
(см. главу "Поведение"). Взрослых самцов, защищающих свою территорию,
называем, как общепринято, территориальными; молодых, у которых она
отсутствует, — нетерриториальными.
168
Благодаря массовому мечению и радиопрослеживанию в последние годы
удалось получить весьма подробную картину пространственной структуры
нескольких популяций европейской (Kurt, Л968а,Ь, 1970; Bramley, 1970, 1972;
Strandgaard, 1972; Ellenberg, 1978; Sempere, 1982; Pielowski, Bresinski, 1982;
Fruzinski et al., 1983; Zejda, Bauerova, 1985; Cederlund, 1982, 1983; Bideau
et al., 1983b; Vincent et al., 1983; Vincent, Bideau, 1987; Соколов, Данилкин и
др., 1986а; Данилкин, Минаев, 1988) и сибирской косуль (Соколов, Данилкин,
1981). В качестве модели ниже приводится пространственно-этологическая
структура лесостепной популяции европейской косули, состоящей из оседлых и
сезонно мигрирующих особей и имеющей типичные черты как оседлой, так
и мигрирующей популяции (Данилкин, Минаев, 1988) . Этот пример дает
хорошую возможность сравнительного анализа и способствует выявлению
закономерностей внутрипопуляционной организации Capreolus.
Пространственная структура популяции косуль, как и вся их социальная
организация, существенно изменяется в течение года, но относительно сходна в
два больших периода: летний (репродуктивный и территориальный), когда
животные, ведущие преимущественно одиночный образ жизни, диффузно
распределены по площади; и зимний (атерриториальный), когда происходит
концентрация преимущественно объединенных -в группы зверей на кормных участках,
а распространение их по площади имеет агрегированный характер. Кроме того,
в летний период территориальное поведение зверей различно в зависимости
от пола « возраста.
Внутрипопуляционная структура в летний период
Территории у взрослых самцов. Весной, сразу же после таяния снега
взрослые самцы европейской и сибирской косуль из года в год занимают в
основном одни и те же территории, которые охраняют от других самцов и
маркируют ольфакторно-оптическими метками (см. главу "Поведение").
Захватив однажды территорию, большинство из них остаются на ней в
течение всей жизни. Во всяком случае, в модельной популяции ни один из пяти
взрослых самцов не сменил ее в течение 4—5 лет. Аналогичные данные
приводит Лоудон (Loudon, 1978). Страндгард (Strandgaard, 1972)
зарегистрировал эмиграцию только одного взрослого самца, Бремли (Bramley, 1970) —
двух. Право на владение территорией самцам каждый год приходится
отстаивать в борьбе с другими особями, особенно молодыми, на что указывают
бои между ними и шрамы на голове и шее, имеющиеся практически у
каждого территориального животного.
В модельной популяции европейской косули в Белгородской области
размеры территорий самцов (при летней плотности населения косуль в
заповеднике около 5 особей на 100 га) в среднем 114 га (от 62 до 157 га,
л=9), что значительно больше, нежели в других частях ареала. Однако,
самая большая по площади территория самца (204 га) была за пределами
заповедника, где плотность животных была минимум втрое ниже. С учетом
' Подробнее об экологии этой популяции косуль, обитающей на востоке ареала в
заповеднике "Лес на Ворскле" в Белгородской области, см. в работе (Новиков, Тимофеева, 1965).
169
(№10 /№2B /№43
Рис. 52. Пространственная структура мужского населения популяции европейской косули летом (15
апреля—сентябрь) (по данным радиопрослеживания и визуальным наблюдениям в Белгородской обл.
в 1982—1986 гг.)
/ — территории взрослых самцов; 2 — изменение границ территории самца N 3; 3 — эмиграция
годовалых особей; 4 — участок обитания двухлетнего самца N 26; 5 — лес; 6 — фруктовые сады; 7 — строения
поселков; 8 — участок обитания годовалого самца N 1; 9 — пустошь; 10 — луг с кустарником; /; — поля
этого участка размеры территорий самцов в районе исследований в среднем
равнялись 122 га («=-10) (рис. 52). .
Анализ размеров территорий самцов в пределах ареала показывает, что их
площадь обратно пропорциональна плотности населения популяции и находится
в пределах от 2 до 200 га (табл. 28).
Размеры территорий самцов, скорер всего, определяются качеством летнего
биотопа: в лучших выше плотность их населения и меньше размеры территорий
(Mottl, 1962;Bobek, 1977) (табл. 29).
Действительно, несмотря на относительно диффузное пространственное
размещение косуль в летний период, повышенная плотность их населения
наблюдается на опушках леса, высокотравных лугах, вблизи водоемов, где изобилие
пищевых ресурсов и хорошие защитные условия. В таких биотопах четко
проявляется конкуренция за территории, тогда как в соседних, худших по качеству
угодьях, зверей может совсем не быть или их плотность значительно ниже.
Однако структура фитоценоза одного и того же биотопа мало влияет на
170
Таблица 28. Размеры территорий самцов косуль при разной плотности населенна популяций
Число особей
на 100 га
Размеры территорий, га
Европейская
косуля
Сибирская
косуля
Авторы
1—2
3—10
10—20
20—30
30—40
40—50
60
126—204
38—157
6—100
12—39
7—20
5-10
2—9
100—170
35-120
18—37
—
—
—
—
Hennig, 1962; Mottl, 1957; 1962; Kurt, 1968; Prior,
1968; Bramley, 1970, 1972; Strandgaard, 1972;
Bobek, 1977; Ellenberg, 1978; Janeau et al., 1981;
Sempere, 1982; Fruzinski et al., 1983; Cederlund,
1983; Bideau et al., 1983; Соколов, Данилкин,
1981; Соколов и др., 1982, 1986; Дарман, 1986
Таблица 29. Средний размер территорий и участков обитания косуль в зависимости от летней
и зимней кормовой емкости некоторых европейских лесов (по: Bobek, 1977)
Лесные экосистемы
Хвойные
Хвойно-лиственные
Влажные хвойно-лиственные
Лиственно-хвойные
Горные лиственные
Лиственные
Ольховые
Сухая масса пищи;
т/100 га
лето
40,4
83,3
64,3
70,4
30,0
72,9
112,4
зима
13,9
8,6
6,6
3,4
6,1
1,5
2,0
Размер
территорий самцов,
га
69,6
15,7
26,7
22,2
128,5
20,6
8,5
Размер
участКормовая емкость ле-
ков самок, п са, число особей на
35,4
8,0
13,6
11,3
65,3
10,5
4,3
100 га
лето
4,3
18,9
11,1
13,3
2,3
14,4
32,5
зима
246,9
152,7
117,2
60,4
108,3
26,6
35,5
размеры территорий самцов, и площади их здесь примерно одного порядка
(Strandgaard, 1972; Sempere, 1982).
Зависимости размера территории от возраста животных, на что указывал Бо-
бек (Bobek, 1977), скорее всего нет, как это наглядно показал Лоудон
(Loudon, 1978). В нашем исследовании за 4 года ни у одного из взрослых
самцов также не произошло приращения или заметного уменьшения
территории, за исключением временного захвата одним из них части территории,
принадлежавшей соседнему самцу, погибшему летом (см. рис. 52). К такому же
выводу пришел и Элленберг (Ellenberg, 1978), изучавший косуль в вольерах.
Конфигурация территорий при неизменной плотности населения, очевидно,
почти не меняется. При высокой плотности некоторые молодые самцы могут
захватить и удержать за собой часть территории или изгнать с нее старого
самца (Sempere, 1982). Территорию самца, погибшего зимой, занимает весной
молодой самец. При гибели хозяина летом часть его территории могут занять
соседние самцы (Bramley, 1972; Strandgaard, 1972; Ellenberg, 1978; Данилкин,
Минаев, 1988).
171
В естественных условиях территории соседних самцов при умеренной плотности
населения практически не перекрываются (см. рис. 52) или перекрываются очень
незначительно (Kurt, 1968b; Prior, 1968; Bramley, 1972; Соколов, Данилкин,
1981). Границы территорий поддерживаются в основном взаимным избеганием
заходов на чужой участок. Однако у большинства самцов наблюдаются
отдельные кратковременные "рейды" по чужим территориям и возврат на свой
участок в тот же или на следующий день.
Границами территорий служат заметные ориентиры: опушки леса, дороги,
проселки, тропинки, склоны оврагов, ручьи и реки. У некоторых зверей они
проходят в однородном массиве леса, и принцип их разграничения в данном
случае остается неясным.
При очень высокой плотности населения косуль в природе и в вольерах,
где жизненное пространство ограничено, участки взрослых самцов могут
частично накладываться один на другой за счет савместного использования
кормовых площадей (Соколов и др., 1982), но наиболее посещаемые зоны обычно
четко разделены (Sempere, 1982; Maublanc et al., 1987). Частичное
наложение территорий самцов наряду со значительными сокращениями их площади
позволяет популяции образовывать высокую плотность населения даже при ярко
выраженной территориальности самцов.
Размеры суточных участков взрослых самцов зависят от площади их
территорий и поведения животных. В Белгородской области они равнялись в
среднем 49 га (от 4 до 97 га) и составляли около 40% от площади летнего
участка. В Швеции (Cederlund, 1983) суточные участки были меньше (20,3 га) и
занимали лишь 21% территории. Эти цифры означают, что ежедневно каждый
зверь охраняет (осваивает) лишь часть своей территории. До 70% точек
радиопеленгации приходится на центральную зону (рис. S3), т.е. для многих
самцов она, очевидно, представляет наибольшую ценность. Однако при маленьких
территориях каждый день осваивается практически вся площадь.
Во время гона территориальная организация популяции косуль существенно не
изменяется (Bramley, 1972; Sempere, 1982; Соколов, Данилкин, 1981). В
начале гона самцы обычно спариваются с самками, обитающими на их
территории. Очевидно, что при такой пространственной структуре преимущество
в размножении получают особи, на территориях которых имеется больше участков
обитания самок (см. ниже). В конце гона учащаются "рейды" самцов за
пределы своей территории в поисках самок в течке, тем не менее
территориальность еще сохраняется, хотя с каждым днем заметно уменьшается, особенно
в мигрирующих популяциях.
Участки обитания у одно-двухлетних самцов. В модельной популяции,
обитавшей в островном лесу площадью около 1000 га, было отчетливо заметно
значительное сокращение числа молодых самцов с апреля по июль. К периоду
гона в лесу оставались только территориальные самцы и не более 5—6 широко
кочующих одно-двухлетних особей. Напротив, за пределами лесного массива
по долине реки и во фруктовых садах встречались преимущественно годовалые
самцы, иногда в парах с самкой того же возраста.
Эти наблюдения полностью совпадают с данными радиопрослеживания и
визуальными регистрациями животных. Из пяти меченых однолетних самцов
весной эмигрировали четыре, а один занял участок размером около 2 га между
территориями взрослых самцов, разделенных рекой, и лишь изредка заходил
172
/
+
+
+
III
+
+
:•:
+
+
+
in
/
+
V
III
;■:
III
III
+
/
+
V
V
III
III
III
X
+
.
Ill
III
Ш
ffl
V
ffl
X
+
+
■
ffl
Ш
Ш
+
+
+
+
+
III
III
Ш
■
■
+
+
+
+
+
III
■
■
■
a
in
+
.
•
+
v
Ш
■
Ш
+
+
+
i^*
III
III
+
■
■
■
ffi
ffl
+
^
+
+
Ш
в
■
III
III
+
Ш
+
^
Ill
III
V
ffi
III
III
■+
+
'.*
+
+
III
III
•
+
III
•
yt
7J
+
в
L.
IKM
,
Рис. 53. Структура участка обитания взрослого самца европейской косули (N 5)
«AT, ™™ZyZCrZ\7s ST^Ll Г™ °бИТаНИЯ - Lb^д на6лк>де„ия (03.84-
.-^астоквокт^Г-в'ио^ГГ^
J^ST™ С * П° " ЗВГУСТа 1Ш - В ~ — P-oneSr4e„'„9(B S2&'LS%.
в лес (см. рис. 52). Осенью в заповедник возвратились только три звеоя
из них один погиб зимой, другой снова эмигрировал в начале лета Т„ет„й
зимой и следующим летом широко кочевал по" всему заповеднику те по
территориям почти всех самцов, но территориальным он стал только наrvl
лого самцаЗНВсегоНЯиВ УЧаСТ°К ИСЧСЗНувШеГ0 ^°»™* всего, погибшего)
«зрелого самца. Всего, из четырех маркированных двухлетних особей из лесйот
массива эмигрировали три. Некоторые молодые самцы, покинув лес несколько
недель жили поблизости, но в июле ушли за пределы ^килом^таовой зоны
ГоХшихТоГйПлощади летних участков обит™ ^7нт:;ро„во1Роко
кочующих особей в зоне радиопрослеживания превышали 1000-2000 га что
несравненно больше, чем у взрослых животных в этот период.
Причиной выселения молодых зверей из места рождения служит
агрессивность к ним взрослых самцов, изгоняющих их со своих территорий
Энергичное преследование годовалых особей начинается сразу же'Зе распада
173
семейной группы, как только они остаются без матери. Большинство их (до 58—
90%) вынуждены эмигрировать из района своего рождения (Bramley, 1970;
Strandgaard, 1972; Loudon, 1978; Соколов, Данилкин, 1981; Дарман, 1986;
Данилкин, Минаев, 1988), и выселение молодых самцов носит характер
сезонной миграции. Осенью значительная часть эмигрантов возвращается в район
своего рождения, обычно покидая его вновь следующей весной.
В популяциях с очень высокой плотностью и в вольерах не все трехлетние
самцы могут стать территориальными, тогда как в разреженных некоторые
годовалые ведут себя как территориальные (Ellenberg, 1978). В плотно
заселенных биотопах на место изгнанных молодых косуль приходят другие из
соседних угодий и взрослым самцам приходится вновь и вновь защищать
свою территорию. К тому же далеко не все одно-двухлетние самцы покидают
район, где они родились и выросли. Часть их широко кочует летом по
территориям нескольких соседних самцов, а некоторые из них становятся
"спутниками" территориальных самцов, сопровождая их повсюду по меньшей мере
до периода гона (Соколов, Данилкин, 1981; Дарман, 1986). В редких случаях
хозяин территории не проявляет к молодому зверю явной агрессивности,
позволяя находиться на его участке.
В целом молодые самцы косуль в отличие от взрослых показывают более
разнообразный спектр пространственного поведения (см. рис. 55):
эмигрируют из района рождения;
занимают свободные участки за пределами территориальной зоны;
кочуют по территориям нескольких самцов;
занимают маленький участок в пределах "буферной зоны" между территориями
соседних самцов;
обитают на территории одного из самцов-"пацифистов", не проявляющих к ним
явного агрессивного поведения;
становятся "спутниками" территориального самца, всюду еледуя за ним.
Молодые самцы представляют собой явный экологический резерв популяции.
Благодаря такому пространственному поведению популяция быстро расселяется
и заполняет образовавшиеся в результате смертности взрослых самцов бреши в
территориальной структуре, что способствует успешному размножению и
выживанию.
Участки обитания у взрослых самок. Пространственное распределение у самок
в летний период имеет другой характер, нежели у самцов (рис. 54). В
модельной популяции в конце апреля — начале мая беременные самки
отделялись от группы и занимали примерно за 3—4 недели до отела одни и те же на
протяжении нескольких лет родовые участки размером в несколько гектаров.
Почти половина из числа меченых самок (л=14) каждый год в апреле
мигрировала за пределы заповедника на расстояние более 7—10 км. Там они
приносили телят и в октябре—декабре вместе с ними ежегодно возвращались
в район заповедника. Две оседлые прежде самки при беспокойстве
эмигрировали со своих родовых участков за 1—2 недели перед родами и,
благополучно отелившись на новом месте, вернулись осенью на свой зимний
участок. Следующей весной они ушли на новый родовой участок и снова
осенью вместе с детьми пришли в район заповедника.
Беременная самка защищает свой родовой участок быстрыми наскоками
с опущенной головой и явно агрессивными намерениями, а иногда и при помощи
174
Рис. 54. Пространственная структура женского населения модельной популяции европейской косули
летом (май—август)
/ — участки обитания взрослых самок; 2 — участки полувзрослых самок; 3 — места отелов; 4 —
эмиграция беременных самок, вызванная фактором беспокойства; другие обозначения см. на рис. 52
передних конечностей, вынуждая этим других самок и своих детей из
прошлогоднего помета покинуть его. Такое же поведение самок зарегистрировано и в
других популяциях (Kurt, 1968b; Bramley, 1972).
В биотопе далеко не все участки пригодны в качестве родовых. Как
показывают наблюдения, они должны отвечать следующим требованиям: быть
поблизости от источника высококачественного корма, обладать хорошими
защитными условиями и, желательно, находиться неподалеку от воды. Таким
условиям чаще всего отвечают опушки леса, высокотравные луга, долины
ручьев и рек, поросшие кустарником. Плотность населения беременных самок в
этих биотопах максимальна. В некоторых случаях ими защищается каждая
отдельно стоящая группа кустов на лугу, что нам довелось видеть в
Белгородской области, в то время как на большей части лесного массива,
видимо, не отвечающего необходимым требованиям, самок не было (см. рис. 54).
Таким образом, в отдельных биотопах даже при относительно низкой
плотности населения косуль (4—5 особей на 100 га) наблюдается дефицит родовых
участков.
Спустя одну-две недели после отела участки обитания самок начинают
заметно расширяться и с увеличением возраста детенышей все более наклады-
175
ваются один на другой. Большинство матерей остаются поблизости от места
родов вплоть до осени, т.е. до конца сезона размножения. Концентрация их в
определенных биотопах приводит к тому, что в период гона на территориях
одних самцов находятся несколько самок, на территориях других они могут
отсутствовать. Однако в модельной популяции некоторые самки в конце июня —
в июле полностью покидали свои родовые участки и вместе с телятами
переходили на несколько сотен метров на другие, вероятно более защищенные,
участки, попадая таким образом на территорию другого самца, который
покрывал их.
Эти обстоятельства (расширение и смена участков у части самок) позволяют
участвовать в размножении и тем самцам, на чьих территориях нет родовых
участков. Более того, самец, имевший на своей территории до 10 достигших
половой зрелости самок, на следующий год может остаться без них при
изменении структуры фитоценоза (Strandgaard, 1972).
Площади суточных участков обитания лактирующих самок в наших
исследованиях в Белгородской области были в среднем 24,1 га (от 7 до 34 га,
л=16), сезонных (с конца апреля по август включительно) — 73 га (от
39 до 107 га, и=7), что близко к данным Седерлунда (Cederlund, 1983) для
косуль в Швеции (суточные участки 12,3 га, сезонные — 94,6 га). Размер
суточных участков обитания детенышей летом в несколько раз меньше, чем у
матери: в июне и июле он минимален — 1—8 га, в августе — от 3 до 14 га.
С середины сентября площадь участков матерей и их детей становится
одинаковой, что говорит о синхронизации активности и пространственного поведения
семьи (см. главу "Поведение").
Как и у самцов, размеры сезонных участков обитания самок, очевидно,
определяются качеством биотопа: чем он лучше, тем выше плотность их
'населения и тем меньше пространство, занимаемое ими (см. табл. 29). В
пределах ареала оно колеблется от 1—7 га .в период отела до 180 га к концу
летнего сезона; в популяциях с высокой плотность^ населения в среднем
от 7 до 60 га (Mottl, 1957, 1962; Kurt, 1968b, 1970; Nikolandic, 1968; Bramley,
1972; Strandgaard, 1972; Ellenberg, 1978; Janeau et al., 1981; Fruzinski et al.,
1983; Cederlund, 1983).
Участки обитания у одно-двухлетних самок. В модельной популяции из трех ра-
диомеченных в заповеднике однолетних самок эмигрировали в апреле две
(одна из них вместе с братом). Осенью вернулась сюда лишь одна из них,
но следующей весной она снова покинула район исследований. Из 6 двухлетних
особей в общей сложности весной эмигрировали 4; две из них осенью
вернулись с телятами и следующей весной снова ушли из заповедника.
Примечательно, что все молодые особи-эмигранты зимой находились в группах
вместе со взрослыми ежегодно мигрирующими самками и, вероятно, являлись
их дочерьми, перенявшими их поведение.
Две молодые самки после распада семьи держались поблизости от
родительских участков, иногда контактируя с матерями на местах кормежки.
Территориальные самцы не проявляли к ним агрессивности. В начале сентября одна
из них влилась в семейную группу, другая всю зиму вела одиночный образ
жизни. Площади их летних участников обитания равнялись 72 и 133 га. На
следующий год весной обе заняли прошлогодние участки и отелились здесь.
В популяциях с умеренной плотностью населения большинство молодых
176
Рис. 55. Схема пространственной структуры популяции косуль в репродуктивный период
/ — территории взрослых самцов; 2 — участки обитания половозрелых самок с родовым участком;
3 — участки обитания годовалых самок; 4 — эмиграция однолетних особей; 5 — различные варианты
участков обитания одно-двухлетних самцов (пояснения см. в тексте)
животных занимают свободные площади, прилегающие к материнским или
перекрывающие их. После гибели или при искусственном удалении взрослой самки
(Bramley, 1972) на ее участке обитания сразу же обосновывается молодая
самка, обычно ее дочь, которая может телиться на том же самом родовом
участке, что и мать (Aylmer, 1977).
При высокой плотности населения значительная часть молодых самок
покидает в конце весны родительский участок (Bramley, 1972; Strandgaard,
1972; Loudon, 1978). Пока что известны два момента, способствующие их
миграции: агрессивность к ним беременных самок непосредственно перед родами и
спустя некоторое время после отела (Kurt, 1968) и агрессивность территориальных
самцов, направленная конкретно не на молодую самку, а на ее брата, если
пара прошлогодних детенышей держится вместе после распада семьи. Такое
поведение мы наблюдали в модельной популяции.
Это обстоятельство способствует тому, что новые участки заселяются не
только молодыми самцами-эмигрантами, но и молодыми самками, а в общем
этот процесс способствует расселению вида. Механизм расселения косуль
показан в главе "Перемещения" (см. рис. 39).
В целом молодые самки демонстрируют меньшую вариабильность
пространственного поведения, нежели самцы-одногодки (см. рис. 55):
эмигрируют из места рождения;
занимают участки поблизости от участка обитания матери;
в редких случаях широко кочуют по территориям нескольких взрослых самцов
в паре с однолетним самцом, скорее всего братом.
Сложные семьи. Большинство взрослых косуль, как показано выше, занимают
12. Европейская и сибирская косули 177
из года в год одни и те же участки обитания. Зоны выращивания
детенышей находятся в тесном соседстве или накладываются одна на другую прежде
всего среди близкородственных особей — матерей и их прошлогодних детей —
самок, обосновавшихся на участках, граничащих с материнским. Эти зоны
чаще всего находятся в пределах территории одного и того же самца,
который менее агрессивен к своим повзрослевшим детям, нежели к другим. Гон
проходит обычно на территории самца, спаривающегося с одними с теми же
самками, причем нередко с дочерьми.
Таким образом, в репродуктивный период основу пространственной структуры
популяций косуль составляют участки обитания "сложной семьи" (состоящей из
нескольких близкородственных самок и их детенышей), "привязанные" к
территориям определенных самцов (Kurt, 1968, 1970; Strandgaard, 1972; bllenberg, 1978;
Stubbe, Passarge, 1979; Cederlund,1983; Vincent et al., 1983; Maublanc et al.,
1987; Данилкин, Минаев, 1988). Зимой эти самцы могут входить в состав
семейной группы, "сложной семьи" или объединения, в которое входит их
семья. Однако последнее не является правилом, и многие из них вне
репродуктивного периода живут отдельно.
Следовательно, несмотря на кажущийся одиночный образ жизни, популяция
косуль в репродуктивный период в основном состоит из совокупности "сложных
семей". Члены семей очень хорошо знают друг друга, что сокращает
количество взаимных столкновений, облегчает спаривание, объединение в группы
в зимний период и способствует лучшему сохранению нового поколения.
В общем виде картина пространственной структуры стабильной популяции
косуль в летний период схематично изображена на рис. 55.
Внутрипопуляционная структура в зимний период
В зимний период (октябрь—апрель) внутрипопуляционная пространственная
структура существенно отличается от летней. Она к тому же различна в разных
регионах и является отражением общей пространственной стратегии популяций
косуль, в основном определяемой неблагоприятными экологическими факторами и
в первую очередь высотой снежного покрова. Есть популяции, сезонно
мигрирующие и полностью покидающие летние участки обитания, есть оседлые и
найдены популяции, в которых имеются как оседлые, так и сезонно мигрирующие
особи, причем число тех и других зависит от высоты снежного покрова
(см. рис. 38).
Картина их внутрипопуляционной структуры усложняется и тем, что ранней
осенью и весной одни животные уже изменяют свое территориальное поведение,
другие еще нет и в этот момент происходит как бы наложение одного
на другой двух периодов — территориального и атерриториального.
Участки обитания отдельных особей и разных групп в зимний период
широко перекрываются (рис. 56). В модельной популяции их размеры с
октября до апреля составили в среднем 508 га (от 124 до 1002 га, л=24) и
были примерно одинаковыми у животных разного пола и возраста. В пределах
сезонного участка отчетливо выделялись одна или несколько кормовых зон,
на которых звери проводили большую часть суток.
Приведенные выше площади участков обитания в зимний период являются
максимальными для европейской косули. Относительно близкие к ним значения
178
1 ' i i i i i 26 -v —v- J'i
-"—77 - 35
7, Iff » ч " " id —— 29 —l — ! 'Ю
11111 10 *- — ?0,7t i-Hb-MMj/ ++.+.+■ чг, -'/5
Рис. 56. Пространственная структура модельной популяции европейской косули в зимний период
(октябрь—март)
2—43 — участки обитания животных, снабженных радиопередатчиками и другими метками (1982—
1986 гг.)
179
Рис. 57. Суточные участки обитания европейских косуль зимой по данным одновременного радиопро-
слеживания (5 декабря 1984 г.)
Обозначения см. на рис. 56
дают Седерлунд (Cederlund, 1982, 1983) для косуль Швеции и Зейда, Бауерова
(Zejda, Bauerova, 1985) для полевой популяции в Чехословакии (от 40 до 812 га,
в среднем 200 га). Во всех других популяциях в Западной и Центральной
Европе они меньше в 3—10 раз (Mottl, 1962; Kurt, 1970; Ellenberg, 1978;
Bideau et al., 1983b; Vincent et al., 1983; и др.). Эти отличия, скорее всего,
объяснимы условиями обитания исследуемых популяций: чем хуже
экологическая ситуация, тем шире животные вынуждены перемещаться в поисках
источников корма, что особенно характерно для мигрирующих и полевых
(Zejda, Homolka, 1980) популяций.
180
Соответственно при ухудшении экологической ситуации гораздо большими
становятся и суточные участки обитания. В модельной популяции, например,
при высоте снежнего покрова до 25 см они были в среднем 103 га (от
30 до 243 га, л=24) (рис. 57). Однако увеличение кормовцй площади при
снижении запасов корма продолжается до известных пределов. С резким
повышением уровня снежного покрова суточные участки обитания зверей заметно
снижались, и при его высоте от 50 до 150 см некоторые особи и группы
при отсутствии фактора беспокойства в течение одной-двух недель оставались
на площади 2—7 га.
Значительное уменьшение участков обитания в особенно суровые и
многоснежные зимы наблюдалось и в других популяциях (Cederlund, 1983; Maublanc et al.,
1987), что, видимо, связано с экономией энергетических затрат на
передвижение и выгодно с точки зрения биоэнергетики. В это время также
существенно снижается метаболизм животных и потребление корма.
На протяжении зимнего периода участки обитания даже у оседлых косуль
могут смещаться, меняются в конфигурации и в размерах по месяцам (см. рис. 53)
и сезонам года: весной они нередко больше чем зимой или осенью (Sempere, 1979;
Janeau et al., 1981). В Швеции, к примеру, средние площади сезонных
участков обитания косуль в разные годы были следующими: зимой — у самцов
91 и 216 га (суточные в среднем 17,5 га), у самок 108 и 122 га (15 га);
осенью соответственно 95 га (23 га) и 128 га (27 га); весной — у самцов
132 и 254 га (37 га), у самок 100 и 176 га (Cederlund, 1983). Весной
у зверей резко увеличивается метаболическая активность, вызывающая большее
потребление пищи. С уменьшением и сходом снежного покрова они начинают
широко передвигаться в поисках появившейся зеленой растительности и
перемещаются к своим летним участкам. У самцов в период занятия территорий
возрастает локомоторная активность. Увеличение размеров участка обитания в
осенние месяцы (по сравнению с летним периодом) объясняется снижением
территориальности при большой свободе передвижения в бесснежный (или
малоснежный) период. Крайнее выражение таких перемещений — сезонные
осенние и весенние миграции.
Годовые и индивидуальные участки обитания
Годовой участок обитания у косуль складывается из их участков в летний
и зимний периоды и путей перехода от одних к другим, если они различны.
В оседлых популяциях у особей, живущих в благоприятных биотопах, сезонные
участки обитания совпадают; при худших условиях участок обитания в
зимний период обычно шире летнего и включает часть его или иной. В
мигрирующих популяциях, как уже отмечалось, они полностью отличаются у всех
особей.
В схематичном виде структура годовых участков обитания с учетом
изложенных обстоятельств отражена на рис. 58. В модельной популяции были
представлены все эти образцы, причем у многих особей в течение
нескольких лет наблюдений встретились все их вариации при неизменной летней
площади. В особенно многоснежные зимы участок обитания отдельных косуль
с осени до весны имел примерно такие же изменения, как показано на схеме.
Размеры годовых участков в пределах ареала различны. На Южном Урале
181
© а о *—►
• Ск I 1
Рис. S8. Схема структуры годового участка обитания косуль в зависимости от экологических условий
Участки обитания: / — летом, 2 — зимой
у четырех меченых сибирских косуль они составили в среднем 520 га (от 332
до 828 га). Близкими к ним по площади оказались годовые участки
обитания европейских косуль на востоке ареала: в модельной популяции (без учета
однолетних самцов-эмигрантов) — 471 га (от 189 до 845 га, л=26), у взрослых
самок — 459 га (от 237 до 701 га, и = 11), у полувзрослых самок — 598 га (от
257 до 825 га, я=5), у взрослых самцов они были меньше, чем у самок, — 385 га
(от 189 до 845 га, и=9), у двухлетнего самца — 733 га. Телята в
возрасте до одного года имели примерно такой же участок, как и их матери.
У всех самцов-эмигрантов в возрасте от 1 до 2 лет, возвратившихся осенью
на место первой зимовки (на родительский участок), их площадь превышала
2000 га.
Индивидуальные (многолетние) участки взрослых особей этой популяции широко
перекрывались (рис. 59) и по размерам были близки к годовым, хотя и больше
их — в среднем 583 га (от 304 до 1132 га, л = 10). У обоих полов они
оказались примерно одинаковы: у самцов 595 га (от 304 до 1132 га, и = 5),
у самок 590 га (от 342 до 716 га, л=5).
Сходные данные получены для полевой популяции косуль в Чехословакии
(Zejda, Bauerova, 1985), где площади перекрывающихся индивидуальных участков
равнялись в среднем 546 га (от 440 до 755 га), у самцов 447 га (от 148 до
574 га), у самок 592 га (от 160 до 755 га). Несколько меньшего размера
они были в Швеции (Cederlund, 1982, 1983). В Польше у полевой популяции
индивидуальные участки составили: у самцов 168 га, у самок 219 га, годовые
соответственно 151 и 141 га (Pielowski, 1984). У лесной популяции эти
показатели не превышали 52 га у самцов и 77 га у самок (Fruzinski et al.,
1983). В других районах Европы размеры годовых и индивидуальных участков,
как правило, находятся в пределах 20—100 га (Mottl, 1962; Strandgaard, 1972;
Bramley, 1972; Janeau et al., 1981; Bideau et al., 1983; Vincent et al., 1979,
1983), причем они могут быть примерно одинаковыми у самцов и самок или же
большими или меньшими у одного из полов2.
Интересные результаты получил в просторной вольере "полезной" площадью
около 130 га Элленберг (Ellenberg, 1978). Индивидуальные участки самцов
здесь оказались в среднем на треть больше, чем у самок (35—40 и 25—30 га
соответственно); у территориальных самцов они составили около 21 га и были
существенно меньше в лучшем биотопе (12,5 га), нежели в худшем по ка-
2 Более детальное сравнение размеров участков обитания косуль затруднено из-за
использования различных методик обработки данных.
182
Рис. 59. Многолетние участки обитания
взрослых европейских косуль в
модельной популяции (1982—1986 гг.)
честву (от 24 до 36 га). С возрастом участок обитания у самок уменьшается
в среднем с 23,5 до 14 га, у самцов он имел наибольшие значения в
первые три года (29—24 га), а в последующие был примерно одинаковым (18—
20 га).
При увеличении плотности населения косуль в вольере вдвое размер
годовых участков обитания самок остался примерно таким же, но у самцов
сократился почти вполовину. Эти показатели прямо говорят о том, что животные
по-разному реагируют на увеличение численности: у самок участки обитания
накладываются один на другой во всевозрастающей мере, самцы, напротив,
обычно не допускают свободного доступа на свои участки, что при
учащающихся столкновениях приводит к уменьшению их площади. В обоих
случаях пищевая база для каждой особи уменьшается.
Следует отметить, что пространство участков обитания используется
косулями неравномерно. В оседлых популяциях звери отдают предпочтение
локальным маленьким площадям, близким к центру годового участка. В Швеции
по меньшей мере 80% всех точек радиопеленгации каждого животного во все
сезоны были найдены в пределах 400—500 м от него, причем сезонные
геометрические центры активности и самца и самки, живущих на одной территории,
находятся близко один от другого (Cederlund, 1983).
Из приведенных в этом разделе данных видно, что у полевых косуль и
популяций, находящихся на севере и востоке ареала в худших экологических
условиях, годовые и индивидуальные участки обитания больше (при меньшей
плотности населения!), чем у популяций, живущих в более благоприятных
биотопах. К тому же у первых индивидуальные участки весьма значительно
накладываются один за другой, хотя и происходит это в основном за счет
зимнего периода.
Анализ показывает, что внутри популяций выделяются группировки особей,
гораздо более тесно связанных друг с другом, чем с особями из других
аналогичных группировок. Чем ближе расположены их участки обитания, тем
теснее эта связь и тем чаще животные находятся вместе.
Существование таких группировок хорошо известно для многих видов
животных. Н.П. Наумов (1967, 1971) назвал их парцеллами, или "парцеллярными
группировками", и объяснил биологическое значение этих внутрипопуляционных
структурных единиц. И.А. Шилов (1977), исследовавший роль
внутрипопуляционных группировок с других позиции, нашел, что именно их существование
обеспечивает устойчивость популяции в целом.
Описанные выше своеобразные пространственные отношения у косуль имеют
глубокий экологический смысл, выполняя важную роль в регуляции численности
популяции непосредственно перед появлением нового поколения. Подробнее
социальные механизмы регуляции численности освещаются в главе "Поведение".
Сопоставляя пространственную структуру и социальную организацию популяций
европейской и сибирской косуль, мы убеждаемся в их практически полной
идентичности. Разница проявляется лишь в большей миграционной способности
многих популяций сибирской косули. Однако не следует забывать, что здесь
же в Азии в районах с благоприятными экологическими условиями имеются
и оседлые популяции.
184
Глава девятая. Поведение
Онтогенез и материнское поведение
Развитите поведения детенышей европейской и сибирской косуль находится в тесной
связи с материнским поведением. Поэтому нам представляется более правильным
в монографии рассматривать ту и другую формы в комплексе. Этим
вопросам были посвящены специальные исследования (Bubenik, 1965; Dathe, 1966; Kurt,
1968b; Espmark, 1969; Соколов, Данилкин, 1981; Громов, Данилкин, 1984;
Соколов и др., 1985), результаты которых приводятся ниже.
Детское и материнское поведение
в послеродовой период
Сразу же после появления на свет теленок пытается выбраться из
околоплодной оболочки (рис. 60,о) и тихо пищит. Это первый коммуникативный
сигнал у детеныша, на который мать отвечает иногда приглушенным шипением.
Мать освобождает его от пленки (рис. 60,6), ложится рядом и почти
непрерывно в течение 5—20 мин вылизывает (рис. 60,в). Вылизывание, скорее
всего, способствует высушиванию кожного покрова и нормализации теплообмена,
а также стимулирует локомоторную активность детеныша. Материнское
поведение, вероятно, в свою очередь, усиливается под воздействием тактильных и
запаховых ощущений (Blauvelt, 1955; Lent, 1971; Баскин, 1976).
При рождении нескольких детенышей самка уделяет больше внимания
первому. Наблюдались случаи, когда второго теленка самка начинала
вылизывать только через 5 мин, а третьего — спустя 1,5 ч после рождения
(Громов, Данилкин, 1984).
Околоплодные оболочки и траву в том месте, где лежали детеныши, мать
сразу же съедает, а затем съедает и послед, который выходит через 30—40 мин
после рождения последнего теленка (см. рис. 45).
Детеныш, еще не обсохший, ползет к вымени, тыкаясь носом в разные
части тела матери и, найдя его, сосет молоко в течение 2—3 мин в несколько
приемов (рис. 60,г). Насытившись, теленок расслабляется (рис. 60,е)), а мать
в это время рожает (см. главу "Размножение") второго или третьего теленка.
Если мать после первого кормления не меняет позы, детеныш сосет молоко
в несколько приемов (до 6—7 раз). Суммарное время сосания в этот период
может достигать 12 мин.
Отдохнув, малыш пытается подняться: выпрямляет задние ноги, встает на
широко расставленные передние ноги или на согнутые фаланги пальцев (рис. 60,е),
делает 1—2 шага и падает, вновь встает и опять падает, но все-таки
продвигается на 1—3 м до высокой травы, где ложится в типичной позе "ка-
185
a
6
Рис. 60. Детское и материнское поведение в послеродовой период
Пояснения в тексте
f
ш.
Таблица 30. Позы и реакции новорожденных (л = 7) сибирской косули (по: Соколов и др., 1985)
Позы и реакции
Начало появления
реакции с момента рождения,
мин
среднее мин.-макс.
Позы и реакции
Начало появления
реакции с момента рождения,
мин
среднее мин.-макс.
Поднимание головы
Распрямление ушей
Первая попытка встать
Ползанье
Писк
Первое сосание (лежа)
6,5
11,3
12,4
17,8
23,5
27,3
2—12
10—12
2-33
6—34
1—51
7-102
Первое вставание
Хождение на фалангах
пальцев
Хождение на прямых ногах
Первое сосание стоя
Отдых вне места родов
29,7
29,3
40,2
69,5
94,0
20—47
21—47
28-51
45—94
79—113
лачиком" (рис. 60,ж). Через несколько минут некоторые косулята поднимаются,
подходят к матери и снова отходят на несколько метров от нее.
Второе кормление, лежа или стоя (рис. 60,з), происходит через 1—5 ч после
первого по инициативе матери: она подходит к теленку, обнюхивает, лижет его
и ложится рядом или стоит над ним. В последнем случае детеныш встает,
ищет вымя в области груди и передних ног, в подмышечной впадине и,
тыкаясь носом, постепенно передвигается к задней части тела самки. Найдя
вымя, захватывает сосок и принимает типичную позу сосания (рис. 60,з);
мать же в это время вылизывает его аногенитальную область.
Насытившись, детеныш может самостоятельно отойти от матери на 3—30 м и
лечь в высокой траве. Некоторые следуют за матерью, которая явно стремится
увести их от места родов в разные стороны (рис. 60,ы), но, устав, ложатся
в густую траву или в тень под деревом или кустом. Максимальное
перемещение теленка в естественных условиях на Южном Урале в первые пять
часов жизни составило около 170 м от места родов.
Следует отметить, что в первые часы жизни телята могут следовать за лю-
бцми объектами, близкими по размерам к матери (импринтинг), и лишь к
концу первой недели жизни уверенно отличают ее от других косуль, однако
человека они воспринимают как опасный объект уже через 1—2 сут. Самка
узнает своего детеныша, вероятно, по запаху, уже в конце послеродового
периода, т.е. запечатление у нее, видимо, происходит в первые 2—5 ч после
родов, но в эксперименте (Bubenik, 1965; Kurt, 1968) подмена телят возможна
до трехнедельного возраста. По голосу и внешнему виду она не способна
различить своего детеныша, поэтому и реагирует на писк любого теленка или
имитацию его.
На этом послеродовой период, при котором мать с детьми довольно
длительное время находятся вместе, заканчивается. Последовательность развития
и продолжительность поз и реакций новорожденных показаны на примере
сибирской косули в табл. 30.
189
Развитие поведения в период затаивания
У копытных есть два основных типа семейных взаимоотношений: неотступное
следование детеныша за матерью или затаивание его в пределах небольшого
по размерам участка в перйые недели и месяцы жизни (Walther, 1968).
Европейская и сибирская косули явно относятся к последним. Период
затаивания у детенышей косуль длится примерно 2,5—3 мес, но за это время
происходит значительное изменение поведения от максимально возможной
разобщенности членов семьи и асинхронности их активности до полной
синхронизации поведения во времени и пространстве. Для большей четкости изложения
рассмотрим развитие основных форм поведения в этот период раздельно
(Соколов и др., 1985).
Оборонительное поведение. Стереотип оборонительного поведения семьи после
появления потомства направлен на рассредоточение ее членов по участку
и на ограничение контактов друг с другом. Мать в течение 2,5—3 мес
отдыхает отдельно от телят и кормит их в первые недели по очереди, а детеныши
в первый месяц жизни ложатся отдельно друг от друга на расстоянии
5—200 м. После кормления косулята меняют место лежки, но все перемещения
их происходят в пределах очень маленького участка (см. главу "Структура
популяций").
При опасности мать подает телятам сигнал "тревоги": она с шумом
прыгает и убегает, громко стуча копытами о землю. Детеныши стремглав бегут
к ближайшему укрытию (высокой траве, дереву, кусту), где с разбегу
ложатся и затаиваются в характерной позе, поджав ноги к туловищу, вытянув
и прижав голову к земле (рис. 61).
В первые 2—5 дней жизни затаившийся косуленок при приближении любого
объекта не делает ни малейшей попытки к бегству, даже если до него
дотронуться, но при проведении манипуляций с ним пронзительно пищит.
С 3—7-дневного возраста он убегает с этим звуком, если объект, не похожий
на мать, подходит на 1—2 м. В возрасте 1—3 недель теленок не подпускает
к себе ближе 10 м, затем эта дистанция увеличивается до 15—30 м, убегает
он уже без звука, но будучи пойманным, громко пищит.
На этот сигнал и на приближение человека или животных к теленку мать
в первые 1—2 недели их жизни реагирует агрессивным поведением (см. ниже).
Позднее у нее начинает доминировать оборонительная реакция, но к другим
самкам косуль она остается агрессивной около двух месяцев.
Уничтожение матерью околоплодных оболочек, последа и травы на месте
родов; пятнистая маскирующая окраска телят (рис. 60) и неразвитость их кожных
желез, не оставляющих сильного запаха; жизнь на маленьком участке наряду
с рассредоточением и ограничением контактов членов семьи (см. рис. 62, А),
затаиванием, сигналами тревоги и активной защитой детей — все это
позволяет косулям относительно эффективно противодействовать прессу
многочисленных хищников при фактически полной беззащитности потомства в первые
дни жизни.
Затаивание при опасности наблюдается у косулят до сентября. После линьки
матерей и появления у них большого белого "зеркала", которое служит
морфологической "поддержкой" следования особей за лидером, оборонительное
поведение членов семьи синхронизуется и на опасность они реагируют сов-
190
Рис. 61. Поза затаивания
местным бегством. К этому" времени телята путем подражания усваивают
и демонстрируют и другие оборонительные позы и реакции: позу "тревоги",
"шараханье", сближение со спокойно стоящими особями и т.п. (см. ниже),
причем оборонительное поведение и особенно бегство у них проявляется чаще,
чем у взрослых особей, имеющих жизненный опыт и способных оценить
степень опасности.
Ориентировочно-исследовательское поведение. Первые поведенческие реакции
на различные стимулы внешней среды (осматривание, обнюхивание,
прислушивание) проявляется у косулят с первого часа жизни. С трехдневного возраста они
подолгу обследуют различные предметы, движение объектов сопровождается
четкой ориентировочной реакцией. При встрече с матерью или другими ко-
сулятами становится обязательным назо-назальный контакт. Со второй недели
они начинают реагировать на следы жизнедеятельности: мочу, фекалии, метки
самцов на земле и на деревьях, подолгу обнюхивая их. С двухнедельного
возраста ориентировочно-исследовательские реакции детенышей практически не
отличаются от таковых у взрослых особей.
Пищевое поведение. В первые 2- -3 недели самка кормит своих детей
преимущественно поодиночке в определенной очередности, нарушаемой лишь
изредка. Если теленок не голоден и не встает с лежки при ее
приближении, самка уходит, но через некоторое время снова возвращается, и так
несколько раз в течение часа. Позднее мать в большинстве случаев кормит
своих детенышей вместе (рис. 62, Б), однако периодичность кормления
сохраняется.
Контакты с телятами в первые дни их жизни происходят в основном по
инициативе матери. Начало кормления, вероятно, стимулируется наполнением
вымени молоком. Мать прекращает пастись, ее походка становится беспокойной
и переходит в кратковременный бег. Она целенаправленно идет к месту пред-
191
20 40 60 ВО 100
Возраст; дни
20 40 ВО
Возраст гдни
20 40 60 80 WO
Возраст г дни
мин
70
50
30"
В
/
~"г г—^
"/ ч "
t i t 1 i t
мин
12
■уо
1
8§8
Ъ
6*6
•8
- ч
'цг
М
Г
Г
-\
- \
\
\
\
- * \
^^^
.^>^^
1 1 1 1 1 1
20 W
Возраст, дни
60
Возраст дна
Е
Возраст, Пни
Рис.62. Характеристика материнского и детского поведения косуль (на примере европейской косули, по:
Espmark, 1969; с изменениями)
А — доля встреч матери без детенышей (/) или вместе с ними (2), Б — с одним (/) или двумя (2)
детенышами; В — средняя продолжительность (мин) периода кормления (1) и их число в сутки (2); Г— средняя
продолжительность сосания за период кормления; Д — доля прекращений кормления по инициативе
матери (/) или детеныша (■?); £ — доля различных форм игрового поведения у косулят: / —
самостоятельные "исследовательские" прогулки, 2 — взбрыкивания и прыжки, 3 — пробежки, 4 —
преследования, 5 — игровая борьба, 6 — сексуальные игры
192
полагаемой лежки детеныша, часто останавливаясь и пристально осматривая
это место, и, найдя теленка, обнюхивает и вылизывает, после чего тот
встает и начинает сосать. Через несколько дней поведение самки меняется.
Она останавливается за несколько метров от лежащего косуленка, вытягивает
к нему шею, иногда 2—3 раза кивая головой; он встает и после назо-
назального контакта сосет мать.
Контакты по инициативе детеныша также имеют свою специфичность во времени.
В первые 10 дней жизни проголодавшийся теленок не уходя от места
лежки настойчиво пищит, подзывая мать (см. раздел "Звуковая сигнализация").
Позднее он пытается самостоятельно искать ее вокруг места отдыха, издавая
время от времени звуки, и реагирует на всех проходящих мимо косуль,
подбегая к ним. С 1,5—2-месячного возраста косулята утоляют голод
растительной пищей и подбегают к самке лишь завидев ее.
Чужих телят самки обычно не кормят, прогоняя их после обнюхивания или
убегая от них. Взрослые самцы чаще демонстрируют исследовательское
поведение к подбежавшему детенышу, нежели агрессивное.
Научившись находить вымя, телята через несколько подходов обучаются
принимать правильную позу при кормлении (рис. 60,з), при которой самке
доступна их задняя часть. Сосание в первые дни начинается сразу же при
прикосновении к вымени, позже они несколько раз тыкают носом в вымя.
Во время кормления детеныши пищат от нетерпения, если очень голодны, и
часто меняют соски. Сосут они обычно в несколько приемов, прерываясь
для отдыха, во время которого могут отходить от матери, а затем снова
припадают к вымени. Число таких подходов в первую декаду в среднем
4—5, максимально 11, но к 1,5—2 месяцам сокращается до одного.
Соответственно общее время сосания уменьшается с 10—12 до 1 мин (рис. 62, Г).
В первые 10—15 дней, насытившись, теленок обычно сам прерывает кормление
(рис. 62, Д), позднее кормление прерывает самка; она перешагивает через
теленка и уходит или убегает, не допуская его к вымени. Количество периодов
кормления молоком в зависимости от количества детенышей в первый месяц
достигает 5—9 раз в сутки, во второй — 2—4 раза, позднее 1—2 раза
(рис. 62, В), однако время совместной активности членов семьи увеличивается
с 30—40 до 70 мин. Лактация в основном прекращается в августе и лишь
изредка длится дольше (до октября—ноября). С этого момента телята
полностью переходят на питание растительной пищей, которую начинают
понемногу употреблять уже с конца первой декады жизни (см. главу "Питание").
Комфортное поведение. У новорожденных практически полностью отсутствуют
реакции комфортного поведения. К ним обычно относят уход за волосяным
покровом, почесывание, взаимные чистки и подготовку места лежки.
Реакция почесывания задней ногой головы и шеи формируется на 2—3-й день,
однако координация движений еще несовершенна и не всегда эти действия
достигают цели. Первые попытки чистки тела вылизыванием регистрируются
на 4—6-й день, в конце декады косуленок достает аногенитальную область,
а в возрасте 3—4 недель уже может самостоятельно ухаживать за волосяным
покровом. Эту функцию до достижения ими 2—2,5-месячного возраста в основном
выполняет мать в периоды кормления, особенно продолжительно и
тщательно в первые две недели их жизни, когда телята испытывают трудности с
самостоятельным испражнением (рис. 60,в,з). Взаимные чистки зарегистрированы
13. Европейская и сибирская косули ^gg
с конца первой декады; помимо гигиенической, они, скорее всего, несут и
социальную нагрузку.
Место лежки косулята выбирают самостоятельно с первого дня, явно
предпочитая укрытия в виде густой высокой травы и кустарников и избегая
прямого солнечного света. С месячного ьозраста они нередко ложатся в течение
нескольких дней на одно и то же место, возвращаясь сюда после кормления.
Оборудование лежки (царапание земли передними ногами и отгребание верхнего
слоя почвы) наблюдают у них впервые в возрасте 2,5 мес. Зимой они точно
так же отбрасывают снег, докапываясь до земли, а при глубоком снежном
покрове делают лунку овальной формы.
Игровое поведение (рис. 62, Е). Прыжки на месте и взбрыкивание наблюдаются
у детенышей с трехдневного возраста. К концу первой недели становятся
регулярными пробежки: быстрый беспричинный бег от матери или вокруг нее
и внезапные остановки. Наибольшую интенсивность эти формы игрового поведения
получают в период совместного кормления детенышей и дополняются
взаимными преследованиями, иногда в этой игре принимает участие и мать. С
1,5-месячного возраста преследования часто прерываются позами угрозы, атаками и
боданием.
Игровое половое поведение у телят-самцов регистрируется с 1,5—2-месячного
возраста. В этот период появляются также действия, напоминающие
маркирование: трение головой и шеей о деревья и кусты. С 4-месячного возраста
они демонстрируют при игре полный набор поз, свойственных агонистическому
поведению взрослых самцов. Интересно, что всплеск агонистического поведения
у детенышей коррелирует с увеличением половых желез и повышением уровня
гормональной активности (см. главу "Размножение", рис. 42, 43).
Активность. Периоды активности у детенышей косуль в первую декаду
жизни совпадают с периодами кормления1; каждый из них длится около
получаса. Детеныши встают только при подходе матери и сразу же ложатся при
ее уходе, но уже со второй недели они в течение нескольких минут до и
после кормления ходят одни. С месячного возраста время каждого периода
активности увеличивается до 1—1,5 ч, с двух месяцев — до 1,5—2 ч, причем
половину его косулята проводят без матери, затрачивая до 30-40% времени
на пастьбу вокруг лежки. Несмотря на постепенное сокращение числа периодов
кормления молоком, число периодов активности телят остается примерно
одинаковым — 5—7 раз в сутки, но общее время активности существенно
увеличивается с возрастом, тогда как у самок оно остается одинаково
высоким в течение всего периода лактации (рис. 63).
С 2,5—3-месячного возраста наблюдается синхронизация активности у членов
семьи как в пространственном, так и во временном отношении. Они вместе
пасутся^' в одно и то же время ложатся и вместе спасаются бегством от
опасности. К этому времени детеныши перенимают от родителей все основные
формы поведения, однако территориальное и половое поведение молодых косуль
еще весьма отличается от поведения взрослых особей (см. главу "Структура
популяций").
' Период активности — промежуток времени, в течение которого косуля не находится на лежке.
Период кормления — промежуток времени, в течение которого мать и детеныш находятся
вместе и мать обязательно кормит его молоком.
194
30 50 70 30
Возраст, дни
Рис. 63. Суточная а ктивность матерей (/, 2)и детенышей (3, 4) европейской (1,3) к сибирской (2, 4) косуль
(по: Espmark, 1969; Соколов и др., 1985)
Анализируя и сравнивая онтогенез поведения и материнское поведение
европейской и сибирской косуль, мы не смогли уловить какой-либо заметной
разницы между ними: развитие поведения имеет одни и те же закономерности,
абсолютно одинаково поведение матерей в период выращивания нового потомства,
сходна в целом и активность тех и других, что еще раз указывает на близкое
их родство.
Агрессивное поведение
Агрессивное поведение в той или иной степени свойственно большинству
(если не всем) млекопитающих и играет немаловажную роль в их жизни.
Интерес к этому явлению огромен и носит дискуссионный характер как в
определении понятия, так и причин, вызывающих агрессию. Общепринятого
термина агрессивного поведения нет. В этологии этот термин используют в
узком смысле, имея в виду поведение одной особи по отношению к другой,
связанное с установлением определенного иерархического статуса, предъявлением
прав на территорию, пищу, какой-либо объект или их защиту, приводящее
к физическому столкновению или к демонстрации нападения. Этой трактовки
придерживаемся и мы. Во многих случаях выделяют собственно
демонстративное поведение, а проявление агрессивности включает в территориальное
поведение, оборонительное, пищевое и т.д. Агрессивное поведение входит в
сложный комплекс, называемый "агонистическим поведением", образуемый
демонстрацией "уверенности в себе", угрозой, нападением, подчинением и бегством, и
выделить из этого комплекса агрессивное поведение как таковое достаточно
трудно. Нет сомнения, что агрессия возникает в первую очередь вследствие
нарушения другой особью определенной дистанции и вызывается зрительными,
слуховыми и обонятельными раздражителями* а также может возникнуть в
результате "конфликта" между различными видами активности. Уровень
агрессивности зависит от наследственности (например, высокой степенью агрессивности
наследственно обладают фокстерьеры и боевые петухи), гормонального состоя-
195
ния и врожденного типа нервной деятельности (Крушинский, 1960; Lorenz, 1967;
Шовен, 1972; Тинберген 1974; Хайнд, 1975; Баскин, 1976; Шилов, 1977;
Панов, 1983).
Следует заметить, что косули агрессивны не только к особям своего вида,
но и к другим животным и к человеку. Агрессивность у них проявляется
в основном через территориальные отношения в репродуктивный период (см. главу
"Структура популяций").
Социальные связи самок со своим подросшим потомством нарушаются за 2—4
недели до рождения нового поколения. Причиной этому служит возросшая
агрессивность матерей, занявших родовые участки, которые они защищают перед
отелом от других особей.
Наиболее отчетливо агрессивное поведение выражено в первые 1—2 недели
после родов: самки демонстрируют его практически к любому объекту,
приблизившемуся к ее детенышу, особенно при подаче ими звукового сигнала
(Kurt, 1968b; Espmark, 1969; Соколов, Данилкин, 1981). Мать принимает позу
угрозы, прижимает уши, низко наклоняет голову к земле, ударяет передними
конечностями о землю, иногда издает щипящие звуки и внезапно устремляется
к нарушителю, вынуждая его к бегству и преследуя 50—100 м, ил»
пробегает с шумом рядом и разворачивается, чтобы повторить атаку. Если другая
самка косули не убегает, то противники сходятся в позе "импонирования"
с вертикально поднятыми шеями и головами, направленными чуть в сторону,
в 4—5 м останавливаются, принимают позы угрозы и бросаются навстречу,
ударяясь лбами. После столкновения они расходятся и через несколько секунд
снова нападают друг на друга. Победителем почти всегда оказывается та особь,
телята которой находятся поблизости. В этот же период самки иногда
демонстрируют угрозу по отношению к взрослым самцам, хотя в другое время
они боятся их и отступают или убегают при встрече.
По мере подрастания детей агрессивное поведение лактирующих особей
затухает, их участки обитания начинают перекрываться и с конца августа
соседние (родственные) семейные группы могут объединяться, что свидетельствует о
практически полном исчезновении антагонизма после сезона размножения.
Учитывая, что агрессивность самок заметно повышается в последние недели
беременности и максимальна сразу же после родов, можно предположить ее
гормональную обусловленность.
Социальная нетерпимость между самками, состоящими в родстве, очевидно,
все же меньше, чем между неродственными. Это позволяет неэмигрировавшим
полувзрослым дочерям оставаться вблизи материнского участка обитания и рожать
здесь же через год, что приводит к наложению участков и ведет к
образованию "сложной" семьи (см. главу "Структура популяций"). Члены "сложной"
семьи нередко проявляют "семейный" антагонизм к животным и особенно к
сеголеткам, стремящимся присоединиться к их группе. С другой стороны,
сеголетки, потерявшие мать, и полувзрослые особи без затруднений
принимаются "родственной" самкой.
Система доминирования в зимнее время проявляется у самок косуль лишь
при высокой плотности населения и при ограниченных пищевых ресурсах (Geiger,
Kramer, 1974a,b; Espmark, 1974); и в этих экстремальных условиях она в целом
благоприятна для потомства особей, занимающих более высокое иерархическое
положение. Дети доминантных матерей, как установил Элленберг (Ellenberg, 1978),
196
—! 1 1 1 1—4—
1 1 3 4 5 В 7
<33
8 9
51^
о — о
—I—I—I—I—I—I—I—I
10 11 12 1 2 3 4 S В 7
<HL
о о
О о
в 9
ЕЕ>
+—н—I—i
10 11 12
Месяцы
1
ШШ з
%ч
_-_ в
Рис. 64. Соотношение периодов территориального (агрессивного, маркировочного) (/) и полового (б)
поведения, развития рогов (2—4) и образования "кожного щита" (5) у самцов европейской (А) и
сибирской (Б) косуль
2 — начало роста рогов у особей в разных частях ареала; 3 — сроки очищения их от "бархата"; 4 —
твердые рога и сбрасывание
лучше развиваются, имеют больше шансов выжить и чаще становятся
"акселератами" — будущими мигрантами.
Самцы косуль более агрессивны, чем самки. Начало их агрессивного
поведения совпадает, (в вольерах слегка опережает) со временем очищения рогов
от кожи и маркировочными действиями (рис. 64). На западе ареала такое
поведение регистрируется на 1—2 мес раньше, чем на севере и востоке,
и приуроченно оно к сходу снежного покрова и началу вегетации
растительности. Синхронность агрессивного и маркировочного поведения и получение к
территориальному периоду "оружия" (рогов)(см. ниже) свидетельствует о том, что
все эти атрибуты территориальности имеют сдвиг в сторону максимально
раннего их проявления, насколько это возможно при больших энергетических
затратах. Эволюция у самцов явно шла по пути отбора особей, наиболее
рано демонстрирующих агрессивное поведение, что в конечном итоге
способствует захвату и удержанию за собой территорий.
Максимум агрессивных взаимодействий наблюдается во время занятия тер-
197
л и ш ш г
Л Ш Ш Ш X
Масяцы
Рис. 65. Проявления агрессивного (А) и маркировочного (Б и В) поведения самцов европейской косули
в вольере (в % к общему числу наблюдений, из: Ellenberg, 1978)
А: 1 — всего столкновений, 2 — импонирование (угозы), 3 — погоня, схватки; Б: 1 — маркировка.
2 — царапанье земли, 3 — лай, 4 — бодание предметов; В: 1 — у взрослых территориальных, 2 — у впервые
территориальных, 3 — у нетерриториальных, 4 — у годовалых
198
риторий и передгоном (рис. 65,А), причем между взрослыми и молодыми !
особями агрессивные контакты наиболее часты в момент очищения рогов у
последних и в период спаривания, а между взрослыми — в начале территори- ;
ального периода (Sempere, 1982). Осенний всплеск агрессивности самцов,
обнаруженный Елленбергом в условиях вольеры (рис. 65), пока не находит"'
достаточно убедительного объяснения. Автор проводит аналогию с осенним
пением птиц на своих участках и считает, что это беспокойное поведение
перед началом зимы может приводить к миграциям на зимние участки. На
наш взгляд, корни этого явления следует искать глубже. Скорее всего, у
предков косуль период размножения приходился, как и у оленей, на осенние
месяцы, и такое поведение есть не что иное, как отголосок изменившихся в
процессе эволюции сроков размножения, что особенно очетливо видно по
осеннему всплеску половой активности сеголетков (см. главу "Размножение").
Взрослые самцы особенно нетерпимы к нетерриториальным особям, которых
они преследует визуально или по запаху следов сразу же, как только обнаружат их.
При приближении хозяина те обычно покидают его территорию, и лишь иногда
такие встречи заканчиваются столкновениями. Конфликты между соседними
территориальными самцами, которые хорошо знают друг друга на протяжении
многих лет, сравнительно редки и, несмотря на взаимные угрозы, чаще всего
ограничиваются демонстрациями силы. Бои происходят в основном на спорных
участках или во время "рейдов" по чужим владениям в тех случаях, когда
нарушитель не заметил вовремя приближение хозяина и не успел скрыться.
Демонстрации и бои европейской и сибирской косуль (рис. 66) протекают
сходным образом (Hennig, 1962a,b; Kurt, 1966, 1968; Meyer, 1968; Смирнов,
1978; Соколов, Дьнилкин, 1981). Заметив друг друга, противники сближаются,
примерно в 30—40 м они останавливаются и принимают ориентировочную
позу и, если хозяин территории сразу не набрасывается на пришельца, то
медленно, твердо ставя передние конечности на землю, сходятся еще ближе.
В 4—5 м они снова останавливаются и затем, став в профиль к противнику,
идут параллельно один другому2. Шея у них при этом поднята вертикально
вверх, голова направлена чуть в сторону от соперника, но глаза фиксируют
его. Внезапно звери разворачиваются и идут обратным курсом. Эта своего
рода демонстрация "уверенности в себе" или "силы" (рис. 66, А) длится от
2—5 до 40 мин, чередуясь с трением головой и шеей о деревья и
кустарники, боданием их (рис. 66, В и Г) и скребующими землю движениями
передних конечностей (рис. 66, Д). Иногда они срывают траву и листочки с
деревьев, как бы кормясь, но, по-видимому, не проглатывают их.
Демонстрация "силы" все чаще переходит в угрозу (рис. 66, Б), при
которой спина выгнута, шея опущена, уши прижаты, рога направлены на
противника, шерсть вздыблена, видны белки глаз, у отдельных особей (в вольере)
появляется пена у рта. Следует резкий бросок на противника одного из
зверей или одновременно обоих, однако в начале боя они не всегда
сталкиваются — расстояние между ними остается от 0,5 до 2,5 м, после чего
животные взаимно отступают, внимательно следя друг за другом. В
промежутках между ложными выпадами и непосредственными столкновениями самцы
2Этот ритуал у оленей получил название "параллельный марш" (Vos et al„ 1967; Федосенко, 1977).
199
Рис. бб. Агонистическое поведение самцов косуль
Обозначения см. в тексте
200
демонстрируют описанные выше элементы поведения, но в этой ситуации
преобладают бодание кустов и небольших деревьев и скребущие землю движения
ногами (см. рис. 66, Г, Д). -•
Очень интересным моментом является нападение обоих самцов не на
противника, а на одно и то же дерево или куст, который они с
ожесточением избивают, причем рога их почти соприкасаются (рис. 66, £).
Возбужденные звери временами издают "шипящие" звуки, на которые другой самец
реагирует броском вперед. Наконец, после очередной угрозы самцы, иногда
поднимаясь на задние ноги (рис. 66, Ж), устремляются навстречу и сталкиваются
рогами. Столкнувшись, каждый из бойцов пытается оттеснить другого (рис. 66, J) и,
если силы равны, несколько минут топчутся на одном месте. У них напряжены
все мышцы, особенно шейные. Сцепившись рогами, противники пытаются
"вывернуть" друг другу голову, не упуская случая освободить рога и быстро
нанести удар в голову или шею. После столкновения они медленно
расходятся и, если не выявлен сильнейший, вновь следует демонстрационное
поведение, описанное выше, и снова звери бросаются друг на друга.
Количество столкновений в одном бою от 1 до 8, а количество ложных нападений
доходит до 22.
Побежденный самец спасается бегством (рис. 66, К). Победитель преследует
его чаще всего до границ своей территории, а иногда и дальше. После
боя он высоко держит голову (рис. 66, Л), нередко лает и еще какое-то
время продолжает бодать деревья и кусты и "копытить" землю.
Побежденный" зверь находится в типичной позе подчинения (рис. 66, Я), деревья не
бодает, но касается их головой и шеей. Если победитель не выявлен, то оба
самца, медленно пятясь и делая вид, что поглощены кормежкой (но фиксируя
глазами противника), расходятся каждый в свою сторону, затем с высоко
поднятой головой лают и быстро уходят в позе победителя, бодая по пути
деревья и кусты.
Поведение косуль при конфликтах, таким образом, в значительной мере
носит ритуализированный характер. Их демонстрационное поведение, как и у
многих других животных, представляет собой определенную систему оптической
сигнализации, состоящую из отдельных стереотипных элементов — поз, несущих,
с одной стороны, информацию о мотивационном состоянии демонстрирующей
особи, а с другой — соответствует тому или иному комплексу внешних
раздражителей. В демонстрационном поведении противников во время стычек
можно выделить конфликтные тенденции к нападению (агрессивность) и к бегству
(страх); изменение "относительной силы" этих тенденций и вызывает
определенный тип поведения, что известно и для других животных (Панов, 1969, 1983;
Тинберген, 1969, 1970, 1974; Хайнд, 1975; и др.).
У самцов косуль очень часто наблюдается изменение направления атаки, ]
т.е. переадресованная реакция: вместо Нападения на противника они бодают >
кусты и деревья. Изредка встречается и "смещенная активность" — срывание I
листьев с деревьев и травы во время конфликта — поведение, явно не ■
соответствующее ситуации и похожее на поведение дерущихся петухов, ко-1
торые время от времени начинают клевать землю, словно подбирая корм./
Отступление противников, равных по силам, после боя также похоже на отступ-'
ление петухов или гусей после драки. Трение самцами* головой и шеей ol
201
I деревья и кусты, на наш взгляд, тоже легко объяснимо — это не что
; иное, как маркировочные действия.
""Отметим также, что приведенные выше элементы агонистического поведения
у косуль не строго фиксированы и могут идти в различной последовательности.
Самцы при сближении иногда сразу же начинают угрожать один другому
или сначала трутся о деревья и бодают их и только потом проявляют
демонстрацию "силы". Некоторые элементы поведения могут выпадать
совсем (например, маркировочные действия, если самцы встретились на лугу
или в поле, где деревьев нет). Кроме того, различно и расстояние, с которого
они производят демонстрации, — от метра до десятков метров.
При увеличении плотности населения самцов и соответственно с уменьшением
площади их территорий частота встреч и конфликтов между ними, очевидно,
возрастает. Судя по тому, что каждый взрослый самец, отловленный или
отстрелянный в природе, имеет шрамы от ран, можно заключить, что
бои между ними имеют место чаще, чем удается фиксировать их при
наблюдениях.
Выше отмечалось, что все взрослые самцы за редким исключением нетерпимы
к молодым особям, которых активно преследуют и изгоняют со своих
территорий. Однако обнаруживается любопытная особенность: взрослые звери
особенно агрессивны к ним в утренние и вечерние часы, т.е. во время своей
наивысшей активности. Днем нетерриториальные самцы иногда могут кормиться
и даже ложиться в нескольких метрах от хозяина, который изгонял их со своего
участка утром и непременно будет преследовать вечером. Кроме того,
наблюдаются индивидуальные различия в уровне проявления агрессии. Одни
взрослые животные, видя нетерриториального самца, становятся крайне
возбужденными и сразу же атакуют и энергично преследуют его, другие довольно
медленно идут за ним, изредка маркируя по пути деревья, до тех пор,
пока не выгонят со своего участка.
Проявление агрессивности у самцов легко объяснить исходя из
территориальности косуль. Труднее осмыслить тот факт, что некоторые взрослые особи в
территориальный период имеют постоянного "спутника" — молодого самца и не
проявляют к нему агрессивности, хотя в то же время не терпят на своем
участке других самцов.
Довольно убедительные аргументы на этот счет представил Страндгард
(Strandgaard, 1972), который нашел четкую взаимосвязь между степенью
физического развития и частотой миграций годовалых особей. Наиболее развитые
молодые животные весной начинают проявлять стереотип территориального
поведения, включая агрессивные и маркировочные действия, что вызывает
адекватное поведение территориальных самцов. Маленькие звери со слабо
развитыми рогами, напротив, не имеют территориальных претензий, и некоторые
взрослые самцы относятся к ним вполне терпимо, изгоняя со своего участка
лишь на следующий год.
Сходную зависимость между физическим развитием и тенденцией к миграциям
у молодых особей обнаружил и Элленберг (Ellenberg, 1978), выявив к тому же,
что территориальные самцы менее агрессивны к потомству самок, живущих
на их участках, чем на территориях соседних самцов. Первых они терпят
до тех пор, пока о*ни не слишком явно демонстрируют территориальное
поведение, вторых изгоняют без компромиссов.
202
Таким образом, хорошо развитые молодые особи в результате давления
на них со стороны территориальных самцов оказываются мигрантами, что
вызывает разобщение членов семьи и ограничивает возможность
близкородственного скрещивания по линии сын—мать и брат—сестра, а также способствует
регуляции численности и расселению вида (см.. главу "Перемещения").
Наиболее развитые годовалые особи — "акселераты" как бы рождаются для
эмиграции и колонизации новых областей, где образуют новые центры
развития популяций. В свете этих данных становится понятным "эффект
иммиграции", при котором косули, заселившие новую область обитания, отличаются
необычно крупными размерами.
После периода размножения агрессивность косуль заметно снижается, вплоть
до почти полного исчезновения. В естественных условиях зимой все члены
группы могут кормиться рядом, не проявляя заметного антагонизма. Более
того, в конце зимы иногда наблюдается временное объединение взрослых
территориальных летом самцов в однополые или преимущественно однополые
группы. Причем у сибирской косули агрессивность в таких группах не
регистрировалась даже в конце апреля, хотя все взрослые самцы уже имели
полностью развитые (но еще в бархате) рога (Соколов, Данилкин, 1981).
Тем не менее зимой антагонизм животных наблюдается в местах их
концентрации у кормушек и на подкормочных площадках. Еспмарк (Espmark, 1974)
в Швеции у одной из кормушек зарегистрировал 1048 агрессивных контактов
и нашел прямую зависимость между их числом и количеством особей в группе.
Гейгер и Крамар (Geiger, Kramer, 1974a,b) в Швейцарии по наблюдениям
за мечеными косулями на месте зимней подкормки установили, что
агрессивные взаимодействия более часты между самцами, чем между самками, причем
ранг животных определяется расстоянием до их территории. Иерархия почти
линейна и довольно стабильна на протяжении зимы. Взрослые самцы
доминируют над всеми другими особями, сбрасывание рогов понижает их ранг,
и, напротив, очищение рогов от кожи сразу же повышает социальный статус.
Поведение самок у кормовых точек определяется принципом приоритета;
раннее появление особи в начале зимы у места кормежки повышает ранг.
Детеныши мужского пола занимают более высокое положение, чем их сестры,
и в конце зимы иногда могут доминировать над всеми самками группы,
включая своих матерей.
В вольерах, где в силу ограниченного пространства невозможно территориальное
разделение, отношения косуль носят ярко выраженный линейный иерархический
характер на протяжении всего года, что можно рассматривать как средство
адаптации к измененной окружающей среде. Один из взрослых самцов,
наиболее мощный и с хорошо развитыми рогами, доминирует над всеми
другими особями. Далее в иерархической цепочке идут другие взрослые и
полувзрослые самцы, взрослые и полувзрослые самки и сеголетки. Если нет
раздражителей извне, доминант не проявляет особой агрессивности к ним; легкого
наклона головы с прижатием ушей ему обычно достаточно, чтобы вызвать
у подчиненных особей оборонительную реакцию — отступление или бегство.
Тем не менее проявление агрессии здесь регистрируется практически
ежедневно и очень мешает нормальной жизни, нередко приводя к гибели
субдоминантных животных, что следует учитывать при разведении косуль в неволе.
В экспериментах ранг самца-доминанта сразу же понижается после спиливания
203
у него рогов, а осенью, после сбрасывания их остатков, доминировать над
ним могут даже самки. С появлением новых твердых рогов (с момента
очищения их от кожи) между взрослыми самцами в вольере на протяжении
нескольких дней снова происходят многократные ожесточенные сражения.
Победитель становится доминантом и ежедневно подолгу преследует побежденного.
Последний временами принимает позу угрозы, но при приближении доминанта
почти всегда отступает, проявляя при этом демонстрацию "уверенности в себе"
и маркировочное поведение в противоположном углу вольеры. Постепенно
маркировочное и агрессивное поведение субдоминанта затухают, и на
приближение доминанта он реагирует только отступлением и бегством. При
разъединении самцов в этой же вольере перегородкой (сетчатой) оба проявляют
стереотип поведения доминанта и подолгу демонстрируют соответствующие
агрессивному поведению позы и бодают друг друга через сетку, т.е. в данной
ситуации зверь, бывший субдоминантом, обретя свою территорию, получает
какой-то дополнительный стимул, способствующий усилению агрессивного и
маркировочного поведения.
Аналогичная ситуация наблюдается в естественных условиях. При вступлении
на чужую территорию у нарушителя резко изменяется поведение от
агрессивного на своем участке к продолжительному
ориентировочно-исследовательскому на чужом и к оборонительному (поспешному бегству) при появлении
хозяина. Объяснение этому феномену уже найдено при исследовании других
млекопитающих: у зверя, попавшего на чужую территорию, доминирует
ориентировочная реакция — рефлекс, временно подавляющий другие формы поведения
и легко переходящий в пассивно-оборонительную реакцию при виде хозяина
территории. Напротив, у хозяина доминирует агрессивное поведение,
подкрепляемое "чувством дома". Исход встречи решает различная "настройка" их высшей
нервной деятельности (Шилов, 1977).
В зависимости от условий окружающей среды поведение косуль, как показано
выше, может значительно варьировать от жесткой линейной иерархии до
четкой территориальной структурированности в период размножения или
практически полного отсутствия антагонизма в нетерриториальный период.
Эволюция социального поведения у Capreolus, видимо, шла именно таким путем —
от стадного к одиночному образу жизни.
Отличительной чертой поведения диких копытных и косуль, в частности,
является природная осторожность и боязнь хищника и человека. При
встречах с ними у косуль доминирует пассивно-оборонительное поведение — бегство
и затаивание. Контактов с крупными копытными (лосями, оленями, ланями и
кабанами) они явно избегают и обычно уходят при их приближении, к
мелким млекопитающим и птицам относятся, как правило, индифферентно.
Межвидовая агрессивность в природе проявляется весьма редко. Самки
демонстрируют ее только в первые одну-две недели после отела при защите телят,
у которых в это время отсутствует реакция бегства. Самцы в территориальный
период при высоком уровне мотивации проявляют позы угрозы по отношению
к лисицам, к некрупным собакам, зайцам, большим птицам, изредка к другим
видам копытных и к человеку.
Б.А. Кузнецова (1929) свидетельствует: «Мне несколько раз приходилось
в период "гоньбы" встречаться в тайге с диким козлом, который вместо
того, чтобы спасаться бегством, начинал меня, по выражению местных промыш-
204
ленников, "пугать". То он начнет стучать копытом о землю, то делает вид,
что сейчас бросится на человека, как бы собираясь бодаться». Иногда
угрозы их даже в естественной обстановке кончаются нападением на людей
(Holmes, 1973).
В неволе поведение косуль изменяется значительно. Детеныши,
выкормленные человеком, весьма ласковы и привязаны к своей "приемной матери",
следуют за ней повсюду, воспринимают окружающую обстановку как
"нормальную" и не боятся людей и домашних животных. У самок дружелюбное
отношение к человеку сохраняется при неизменных условиях на всю жизнь.
Самцы, напротив, со второго года жизни становятся агрессивными к нему,
практически теряют чувство страха и нападают на него, демонстрируя при
этом весь набор специфических поз и реакций, как при борьбе с особями
своего вида.
В особое возбуждение зверей приводят реакции и звуки человека, хотя
бы отдаленно напоминающие действия противника. Например, царапающие
движения ногой при высоком уровне мотивации самца в большинстве случаев
вызывают немедленное его нападение. Причем, увидев однажды
пассивно-оборонительную реакцию человека (бегство), возбужденный зверь будет преследовать
его, отличая от других людей, все время.
Межвидовое агрессивное поведение самцов косуль, так же как и
внутривидовое, имеет ярко выраженный сезонный и временной характер: сильнейший в
весенне-летние месяцы антагонизм к человеку осенью заметно снижается, зимой
большинство из них очень дружелюбны, но с весны, с момента очищения
рогов от кожи, агрессивность проявляется с новой силой, особенно утром и
вечером (в периоды наивысшей активности).
Опыты показывают, что в неволе самцы косуль реагируют гораздо сильнее
на неспецифический раздражитель (человека), нежели на особь своего вида,
и особенно агрессивны они к наиболее знакомым им людям. Звери
отличают их от других по голосу, одежде и распознают, очевидно, по запаху
или по лицам, даже если они в необычной одежде. После ухода человека,
к которому доминант только что проявлял агрессивное поведение, реакция
переадресуется на субдоминантных животных. При появлении человека
агрессивность снова направляется на него, или, при высоком уровне мотивации,
на любой движущийся объект независимо от формы и цвета, причем более
крупные предметы вызывают у него большее возбуждение. Немотивированный
предварительно самец в ответ на предъявление тех же самых предметов
проявляет только исследовательское поведение. При провоцировании его и
вызове агрессивной реакции возбужденный зверь снова демонстрирует ее по
отношению к любому движущемуся предмету (Соколов, Данилкин, 1981).
Таким образом, в целом агрессивное поведение косуль существенно зависит от
сезона года и физиологического состояния особей, времени суток, "типа нервной
системы", внешних раздражителей и наличия или отсутствия территории и "чувства
хозяина", оно в значительной мере определяется гормональными факторами (Barth
et al., 1976; Schams, Barth, 1982; Sempere, 1982), четко коррелируя с уровнем
концентрации в крови тестостерона (см. главу "Размножение"). Между самцами в
репродуктивный (территориальный) период идет напряженная борьба, в которой
большую роль играют некоторые морфологические адаптации к поединкам.
205
Морфологические адаптации
к борьбе у самцов
Эволюция млекопитающих, как сейчас уже хорошо известно, шла и по пути
выработки особой формы внутривидового антагонизма, сопряженного с
минимумом причинения физического вреда. У многих видов животных для
выявления доминанта обычно достаточно демонстраций. У косуль, видимо, как и у
оленей, роль визуального показателя "силы" особи, определяющего
доминирование без физического насилия, прежде всего выполняют рога и мощные
туловище и шея. Тем не менее драки1 между взрослыми самцами, которые
часто связывают с размножением, нередки, и даже известны случаи их гибели
в поединках (Дементьев, 1933; Аверин, 1949; Северцов, 19516; Кириков, 19S2;
Соколов, 1959; Черкасов, 1962; Holmes, 1973; Stubbe, Passarge, 1979; и др.).
Однако, как показано выше, борьба между ними идет не за самку
непосредственно, а носит территориальный характер и определяется агрессивностью ,-к себе
подобным.
Успех борьбы между самцами, как это отмечал еще Ч. Дарвин (1935),
нередко зависит от наличия особого "оружия" или средств защиты, и даже
слабое преимущество в этом может привести к победе. Вопрос о
биологическом смысле таких органов в дальнейшем был разработан С.А. Северцо-
вым (1951а), подчеркнувшим их принадлежность к категории внутривидовых
приспособлений, и в отличие от других приспособлений, названных конгруэнтными.
У косуль к ним относятся рога, кожный "щит" и сезонные утолщения мышц
шеи.
В настоящее время принято считать, что рога копытных являются
турнирным оружием, служащим для нападения и защиты в конфликтах между
самцами (Северцов, 1951а,б; Bubenik, 1966b, 1968; Geist, 1968; Lincoln, 1972).
Их развитие контролируется половыми гормонами (Wislocki et al., 1947;
Chapman, 1975; Bubenik u.a., 1976; Jaczewski, 1976; Jaczewski et al., 1976). Роль
рогов в защите от хищников, добывании пищи и терморегуляции, очевидно,
вторична (Lincoln, 1972). Существует также мнение, что у высокогорных
копытных рога являются дополнительным очагом кроветворения —
адаптацией к пониженному парциальному давлению кислорода (Коржуев и др., 1978).
Эксперименты со спиливанием рогов (см. выше) ясно демонстрируют, что
они служат у самцов косуль важнейшим инструментом во внутривидовой
борьбе, определяя их иерархический ранг. Особенно необходимы рога в
качестве "оружия" при защите или захвате территорий и развиваются они именно
к территориальному периоду (рис. 64).
Рога взрослых животных быстрее растут, достигают большего размера, чем у
полувзрослых особей, и на 2—3 недели раньше очищаются от кожи (рис. 67).
Процесс очищения рогов от покрывающей их кожи длится недолго — от 2 до
5 дней; самец многократно трется рогами о стволы и ветви деревьев,
оставляя на них куски кожи, которые тут же съедает, а ветки облизывает
(рис. 68). Рога имеют в это время красный цвет от крови из лопнувших
кровеносных сосудов и, по-видимому, сильно зудят, что вынуждает .самца
подолгу, иногда по 40—50 мин подряд чесать их. Существенная разница
в сроках приобретения "оружия" дает значительное преимущество именно
взрослым самцам, позволяя им первыми захватывать территории, получая
206
Рис. 67. Уровень развития рогов у самцов косуль разных возрастных групп
А — сеголетки, Б — полувзрослые, В — взрослые; I—V — месяцы
одновременно и психологический выигрыш — "чувство хозяина", подкрепляемое
маркировочными действиями..
Наличие мощных рогов во многом способствует успеху в борьбе. В бою
отростки рогов, задевая один за другой, ослабляют дгдар по черепу и снижают
возможность проникающих ранений. Тем не менее Иногда рога одного
противника проскакивают, не задев рогов другого, и удар приходится по голове,
Рис. 68. Процесс очищения рогов от кожи у самцов сибирской косули. Лоскуты кожи, оставленные на
дереве, самец съедает
208
УЛ
Рис.69.Шрамы от ударов рогов противников у взрослых самцов
чаще всего в затылок, в лоб, щеки. У взрослых самцов, осмотренных
летом, поврежденной оказывается в основном кожа на голове и шее (рис. 69),
причем чем меньше по размерам рога зверя, тем больше ран на е,го теле
(рис. 70). Максимальное количество ран, обнаруженное у одного животного,
27, минимальное — 2, наибольшая глубина раны 8 мм, длина 7 см.
Проникающих ранений почти не бывает исключительно благодаря очень толстой
коже в этих местах.
Измерения показывают отчетливый половой диморфизм, возрастные и
сезонные изменения толщины кожи у самцов в области головы и передней части
туловища, особенно значительные у более крупной сибирской косули. Кожа
самок (рис. 71,в, 72), напротив, не имеет резких сезонных различий на
разных участках туловища: максимальная толщина у них не превышает 2,5 мм.
У взрослых самцов кожа на голове и шее
летом в 2—5 раз толще, чем на спине и на боках,
и представляет собой как бы "щит",
покрывающий переднюю часть животного до лопаток,
а по хребту — несколько дальше назад (см.
рис. 71,в, 72). Максимально утолщена она
(до 10,5 мм) за рогами. Осенью кожа в 2—3 раза,
а поздней зимой в 3—5 раз тоньше, чем летом,
и становится почти той же толщины, что и
у самок.
Кожный "щит" самцов косуль начинает
Рис. 70. Соотношение между длиной рогов и числом ран у
взрослых самцов сибирской косули
20 30 40
Длина рогов\ см
210
•13
щ а 12 in is
7 в 91011121213
ib*'
;h*z
II
It
a
a
n,
m
13
is
•>s\
• w
■ 20
•o ,
•20-
•10
. 10
77888 '10
•10
•10
•10
•10
798877788 '.%
•9
ws' 10-
S' 9-
7t>
я":
i9i
w I
Wcm
Рис. 71. Толщина кожи взрослых
сибирских косуль
а — самка (июль): б — самец (март);
в — самец (июль); единица измерении —
0,1 мм. Точками указаны места
измерений. Пунктиром ограничен участок с
наиболее утолщенной кожей
развиваться с раннего возраста. У 2—3-месячных самцов в июле—августе уже
намечается его утолщение, но у 9—10-месячных зверей кожа здесь почти в 2 раза тоньше.
У 13—14-месячных самцов он выражен уже очень четко: на голове и шее кожа
примерно в 3—5 раз толще, чем у 9—10-месячных особей (рис. 73).
Процесс утолщения кожи у самцов косуль совпадает по времени с
усилением их агрессивного поведения и соответственно с ослаблением агрессив-
211
5
if
О
*:
О
а;
3
3"
10
150
30 50 70 90 110 130
Полоса кожи пт г.убы до ануса по хребту, см
Рис. 72. Сезонные изменения толщины кожи у взрослых косуль
Самцы, добытые: / — в июне—августе; 2 — в ноябре; 3 — в марте—апреле; 4 — самки, добытые
в июле—августе, ноябре и марте
10 30 51) W 911 110 130 1511
Полоса коти от губы Но ануса по хребту, см
Рис. 73. Возрастные изменения толшины кожи у самцов
Самцы в возрасте: / — 2—3 мес (добытые в июле—августе); 2 — 9—10 мес (март); 3 — 13—14 мес (июль);
4 — старше 3 лет (июль)
212
Рис. 74. Изменение толщины шеи сибирских косуль по
сезонам года
/ — самки во все сезоны; 2 — самцы зимой; 3 — самцы
летом
Рис. 7S. Толщина одноименных мышц шеи (m. sterno—masto-
ideus)
1, 4 — мышцы самцов; 2, 3 — мышцы самок
■4
- у^«.»•*■■...
"■* '.^ч'Л'
WI
ч '■",
ности кожный щит редуцируется. Очевидно, кожный "щит" расположен в местах,
наиболее подверженных ударам рогами в бою, и максимального развития он
достигает к периоду боев, т.е. имеет несомненное адаптивное значение,
предохраняя жизненно важные органы от повреждения и уменьшая смертность
самцов в драках. С другой стороны, значительное увеличение размера кожных
желез в этом утолщенном участке кожи позволяет самцам эффективно
использовать его для ольфакторной маркировки территории.
Другая замечательная морфологическая адаптация у самцов косуль —
утолщение шеи в летний период. Окружность шеи самок летом и зимой
одинакова; у самцов же летом она на несколько сантиметров больше, чем в другие
сезоны года (рис. 74). Происходит это в основном за счет утолщения ко-
213
жи, как показано выше, и отдельных мышц шеи. Так, мышцы шеи (m. sterno-
mastoideus) у самцов сибирской косули летом в 3,5—5 раз толще (3,5—4,6 см),
чем у самок (0,8—1,0 см) (рис. 75). Зимой у самцов толщина этой мышцы
уменьшается примерно в 2 раза (1,3—2,3 см); у самок она одинакова во все
сезоны года.
Биологический смысл этих морфологических приспособлений, на наш взгляд,
легко объясним. Эволюция у самцов косуль, видимо, шла по пути
индивидуального отбора на высокую агрессивность в репродуктивный период, дающую
им преимущество в борьбе за территорию. Победителями оказывались самцы
наиболее развитые физически, с большими рогами и мощными мышцами
шеи, несущими основную нагрузку при борьбе с противниками, и наиболее
защищенные ("кожный щит"). С другой стороны, мощная шея при
демонстрационном поведении самцов, скорее всего, служит одним из визуальных
показателей силы противника. Уменьшение после сезона размножения
толщины кожи на голове, шее и средней части туловища, уменьшение
толщины мышц шеи, сбрасывание рогов и снижение агрессивности говорят о том,
что в эволюции у косуль имеется и противоположная тенденция — отбор
на снижение агрессивности, не препятствующий объединению особей в
определенный (зимний) период года.
Близкие предки Capreolus исходно, видимо, не были такими агрессивными и
вели преимущественно стадный образ жизни. Необходимость защиты территорий,
очевидно, оказавшейся для них более выгодной формой пространственных
отношений, привела к развитию специфического поведения, отличного от
поведения других оленей. Примечательно, что и эти морфологические адаптации
у косуль появляются совершенно в другие сроки, нежели у оленей. Годовой
цикл их функционирования, сопряженный с проявлением территориального
поведения, показан на рис. 64.
Маркировочное поведение
и химическая коммуникация
Самцы косуль с весны и до осени маркируют свои участки ольфакторными
и~ ольфакторно-оптическими метками (Raesfeld, 1956; Hennig, 1962a,b; Тигбек,
1962; Kurt, 1966, 1968; Stark, 1966; Meyer, 1968; Prior, 1968; Смирнов, 1978;
Соколов, Данилкин, 1981). J Ольфакторной меткой служит секрет сильно
увеличенных летом кожных желез головы и шеи, оставляемый зверем при трении
о деревья, кустарники и высокостебельные травы лбом, щеками и шеей
(рис. 76). Оптической меткой являются поврежденные рогами деревья (рис. 77),
но при повреждении их самец случайно или "преднамеренно" касается кожей
головы ободранных участков ствола или ветвей, оставляя здесь же и оль-
факторную метку. Замечено, что такие ольфакторно-оптические метки издали
привлекают самцов.
-— Интересен метод маркировки. Прежде чем поставить метку, зверь
обнюхивает дерево, на котором, видимо, остается запах секрета от предыдущего
мечения, трется лбом о дерево, пропущенное между рогов, и сдирает с него
кору, обнюхивает это место, снова трется лбом или щекой, вновь
обнюхивает и так многократно.
Т" Косули предпочитают метить (повреждать) таким образом небольшие де-
^4
-г if"
j т..
w^
i :i
* "''Ы^\Я -
* ' V-1-.:
I 1
wr
1
1
1
;. ■'*■" I i .
fev'
■*..:■■
"»,
f
Л7 .7
St
,;■,.;■ .;■■■: лЛ«
■v. {■!■■■
V.
"*.? .. Ц!,_
■*
1
Рис.76. Маркировка деревьев самцами косуль
лбом, щекой и шеей
215
**■
4'^ ..
X
■;..■«*
i» "I
«У *'
V
'&•*!
Г
*&
R*
'*
ЪЁ&
к-г
*.*'
ревца диаметром до 8—12 см на
высоте от 15 до 100—120 см. При
большом возбуждении самец не только
трется о них, но и бодает, сгибает
до земли, ломает дерево или ветви.
В таких случаях особенно отчетливо
заметно, что маркировочное поведение
переходит в агрессивное и
маркируемые предметы начинают служить
как бы "условным противником", с
которым сражается этот зверь
(рис. 78).
Ольфакторно-оптические метки
представлены также хорошо заметными
"пятачками" оцарапанной копытами
земли, достигающими 1 м в длину и
0,5 м в ширину. На такой площадке
остается запах секрета межпальцевых
желез (рис. 77).
В редких случаях при ольфакторно-
оптическом мечении территории
возбужденный самец лает, однако вряд
ли этот звуковой сигнал служит
специально для маркировки,скорее всего
он является выражением "избытка
мотивации".
На западе ареала самцы
европейской косули начинают маркировку
территории посредством трения о
деревья уже с февраля, оптические
метки на деревьях появляются после
очистки рогов от кожи, а метки
на земле — только в марте. На севере
и востоке ареала ольфакторное мечение
наблюдается с конца марта, а ольфак-
торно-оптическое — с апреля.
Сибирские косули на Урале маркируют
участки лишь с мая. В районах с высоким
и продолжительным снежным
покровом маркировочные действия
совпадают с началом вегетации.
Маркировочное поведение взрослые
животные проявляют на две-три
недели раньше, чем молодые (см. рис. 65, В).
Это дает им возможность раньше
обозначить свою территорию и
получить несомненное "психологическое"
216
?■■;
■ ■> •:.■■
<t j Л *^
1 * -;-V'ki^**- .
Рис.77.Ольфакторно-оптические метки самцов косуль на земле и на деревьях
217
преимущество. С другой стороны, немаркированный участок явно свидетельствует
о его незанятости, что, очевидно, служит указателем для молодых особей,
позволяя им с наименьшими затратами энергии найти и утвердиться на
нем.
Маркировочная активность самцов имеет два пика — в начале
территориального периода и непосредственно перед гоном, что совпадает с проявлениями
агрессивного поведения. Снижение маркировочной деятельности в начале лета
обусловлено тем, что большинство молодых самцов изгнано с занятых
территорий, границы их установлены, размечены и поддерживаются незначительными
усилиями. Это дает возможность взрослым самцам, основным производителям,
снизить энергетические затраты непосредственно перед гоном. После гона
маркировочное поведение затухает и с сентября—октября свежие метки в природе
практически не встречаются (EUenberg, 1978; Sempere, 1982; Соколов, Данилкин,
1981).
Самцы маркируют в,сю занимаемую ими площадь территории (рис. 79),
но наиболее часто некоторые участки по границам, тропы (иногда каждое
второе рядом стоящее дерево) и отдельно растущие или редкие в данном
биотопе деревья. Наличие высокого показателя мечения периферийной зоны
территории при преимущественном освоении ее центра (см. главу "Структура
i.l:Jmmm:..'; ■ Ш1 ■'■■ ■:!;!V.i.,
Рис. 77 (окончание)
218
&* -■>
''■* *f#t
■•за»: f
■* ■.?
•* *'J
•*"#
-У¥г
Рис.78. Маркировочное поведение самцов часто переходит в агрессивное и маркируемые предметы
начинают служить "условным противником"
219
.■'Г
s
■а*
№ .>&.
;*■*
f ' &V&. Hi'" ' л>
Рис. 78 (окончание)
220
100м
Рис.79.Метки самцов сибирской косули на одном из участков леса на Южном Урале
Пунктиром выделена территория самца с радиопередатчиком
популяций") указывает на посещение границ в основном с целью их мар- ■
кировки.
Взрослые самцы маркируют не только свои владения, но и территории
соседних самцов во время "рейдов"; они метят те же или рядом стоящие
деревья, которые незадолго до этого маркировал хозяин, но в наибольшее
возбуждение приходят не от его меток на деревьях, а от "пятачков" земли,
выбитых копытами и мест уринации." .. -
Маркировочные действия на чужих территориях могут производить и одно-
двухлетние самцы, иногда в присутствии хозяина. При этом молодой зверь
маркирует сигнальное дерево, на котором есть метка, и идет и следующему.
Взрослый самец следует за ним и метит те же деревья, предварительно их
обнюхав, а после этого пытается догнать нарушителя. Тот при его
приближении становится в позу угрозы, но боя не принимает и быстро убегает
до следующего дерева с оптической меткой, и так многократно. ^
В период с весны до осени каждый из территориальных самцов остав-
224-j
Таблица 31. :. е по повреждению (мечению) деревьев самцами сибирских косуль на контрольном участке на Южном Урале (1972 г.)
Показатель
Площадка (50 х 50 м), Mi
Всего
%
Высота деревьев, м
Всего деревьев
Из них повреждено
В том числе за одно лето
Из них
слабые повреждения
сильные повреждения
погибли
новые повреждения по старым
меткам
0.5-2.0
84
31
27
11
11
5
8
1.0-2.5
29
22
13
8
2
3
8
2,0-4,5
160
51
6
5
1
-
3
0,3-U
173
32
И
6
4
1
2 •
1,0-2,5
70
23
18
8
7
3
1
-
516
159
75
33
25
12
22
-
100
30,8
14,5
50,7
33,3
16,0
20,8
Таблица 32. Данные по повреждению (мечению) деревьев самцами сибирских косуль в листве:
в Монголии (1975 г.)
лесах на Восточном Хангае
Показатель
Всего деревьев
d 1-5 см
d5-70 см
Из них помечено
Из них сильно
повреждены
Новые помеченные
деревья
Из них повреждены
1
179
152
74
41
23
10
2
1286
444
116
49
11
7
Площадка (100 х
3
525
2104
116
39
9
5
4
5
409 127
518 511
123 77
52 33
7 6
,
1
100м),№
6
738
753
121
66
5
7
657
,673
56
21
4
2
8
729
1251
63
35
8
3
9
1670
1583
97
45
6
1
Всего
6320
7989
843
38
79
30
Доля от
общего
количества
деревьев
100
5,9
2,7
0,6
од
,%
подроста
100
-
13,3
6.0
U
0,5
На 1га.
шт.
1589,9
93,6
42,3
8,8
33
ляет от 200 до 700 ольфакторно-оптических меток на земле и маркирует
(повреждает) сотни деревьев (см. рис. 77, табл. 31, 32). Многие из этих деревьев
засыхают, но еще много лет служат предметом мечения.
Интенсивность маркировки во многом зависит от сезона, времени суток,
погоды и ситуации на участках. В начале территориального периода самцы
могут наносить до 500—600 пахучих меток в день, в середине — 40—150 и в
конце его — до 10 меток (Ellenberg, 1978). В дождливую и холодную погоду
и в дневные часы звери или совсем не маркируют участки, или такие
действия очень редки.
Ольфакторные метки регулярно обновляются, одни из них чаще, другие
реже. Наши наблюдения показали, что в августе на протяжении 6 дней
самец сибирской косули пометил 457 деревьев, из них: одно дерево — 8 раз,
50 — по 4 раза, 132 — по 3, 93 — по 2 и 181 — 1 раз. Наиболее
часто в этом случае маркировались деревья, стоящие отдельно и на
границах территории.
При внешних раздражителях, и особенно при нарушении территорий, частота
маркировки резко увеличивается. Например, у самцов сибирской косули при
отсутствии внешнего раздражителя она может составлять около 100 касаний
предметов за день, при нахождении на участке знакомого им человека число
соприкосновений с деревьями увеличивается почти в 4 раза, а при
обнаружении свежего следа нарушителя (молодого самца) интенсивность маркировки
достигает 400 соприкосновений за час. В состоянии возбуждения самец
косули только за 3 ч может пометить более 370 деревьев. Причем, если на
территории нет нарушителя, мечёние чередуется с кормлением. В противном
случае самцы заняты только маркировкой (Соколов, Данилкин, 1981).
Плотность меток существенно зависит и от структуры растительности:
в лесной чаще их меньше, чем в других типах леса; на вырубках, в степных
и полевых биотопах преобладают метки на земле.
В вольерах маркировочные действия самцов косуль почти всегда сопутствуют
агрессивному поведению. Здесь они метят не только деревья, но и сетку,
различные сооружения. Доминант может проявлять маркировочное поведение
даже зимой, чего, в естественных условиях не наблюдается, но в это время
маркируемые предметы не обнюхиваются им. При агрессивном поведении
самец за 1 мин касается в среднем 4—5 предметов и делает при этом
3—4 гребущих землю движений ногой, но не всегда касается земли копытом.
Маркировочное поведение косуль в пределах ареала, таким образом, имеет
четко выраженный сезонный характер, совпадает с началом вегетации
растительности, связано с агрессивными действиями (см. рис. 65), защитой
территорий и коррелирует с уровнем гормонов в крови (см. рис. 43) (Barth u.a., 1976;
Sempere, 1982). Метки явно выполняют сигнальную функцию, указывая другим
особям на занятость участка, что способствует избеганию прямых столкновений.
Однако, несмотря на большое число ольфакторно-оптических меток, оставляемых
самцами на своих территориях, их границы нарушаются весьма часто. Тем не
менее, как хорошо известно для других животных, "психологический" выигрыш
маркировавшая особь все же получает. Пахучая метка не задерживает
нарушителя, но, вступив на чужую территорию, он теряет уверенность, и хозяин
при столкновении имеет больше шансов на победу.
У самок в отличие от самцов специфического маркировочного поведения
223
не наблюдается, хотя отдельные элементы, вероятно, могут иметь место (Kurt,
1968).
Другими сигнальными веществами, несущими определенную информацию о
присутствии на данной местности косуль и, возможно, информацию о возрасте,
поле и физиологическом состоянии зверей, являются моча, кал, по-видимому
слюна, попадающая на землю и растительность при кормлении и облизывании,
а также пахучие выделения половых органов, метатарзальных и
межпальцевых желез и железистых комплексов в коже дистальных отделов плюсны и
пясти.
Косули могут отыскивать друг друга по запаху на следах. Идя по ним,
они низко опускают голову к земле и постоянно принюхиваются. В данном
случае животные реагируют в основном на запах выделений межпальцевых
желез, оставленный на уровне земли, и на запах выделений расположенных
выше на ногах желез в коже плюсны и пясти и метатарзальных желез,
который остается на уровне растительности и снега.
В период гона самец, преследующий самку в течке, держит голову и шею
вытянутыми параллельно земле на уровне ее гениталий, что свидетельствует
о том, что в данной ситуации он реагирует на запах выделений половых
органов самки. Однако стоит самке скрыться от преследователя, как он
начинает искать ее по следам, а найдя и сблизившись, обнюхивает половые
органы. Косули отчетливо реагируют и на мочу друг друга, особенно самцы
на мочу самок в течке. Это видно по характерному поднятию ими верхней губы
и оголению зубов ("флемен"). Летом они приходят в сильное возбуждение,
обнюхав место, где мочился другой самец. При встрече незнакомых особей
после назо-назального контакта (вероятно, животные реагируют на запах слюны
или кожных носовых желез) следует обнюхивание тела: головы, туловища,
метатарзальных желез, гениталий (более продолжительно). Очевидно, что
химическая коммуникация играет важную роль в жизни косуль.
Гистологическое исследование кожного покрова европейской и сибирской косуль
убедительно свидетельствует о возможности оставления пахучего секрета желез
на земле и растительности (Meyer, 1968; Соколов, Данилкин, 1981).
Наибольшую роль в химической коммуникации и поведении косуль играют сильно
увеличенные летом сальные и потовые железы кожи головы, шеи и передней
части туловища и специфические железы: метатарзальная, межпальцевая передних
конечностей и межпальцевая задних конечностей, а также железистые комплексы
в коже дистальных отделов плюсны и пясти, которые по своему строению
могут быть приравнены к специфическим железам (рис. 80).
Экспериментами в ольфакторных камерах выявлено, что косули способны по
запаху секрета метатарзальных желез отличать половозрелых особей от
неполовозрелых, а при предъявлении запаха секрета кожных желез лба взрослые
самцы демонстрируют агрессивное и маркировочное поведение. Эти данные
подтверждают возможность индивидуального распознавания по запаху секрета
метатарзальной железы и указывают на информативную роль кожных желез
лба в территориальном поведении (Broom, Johnson, 1980). По сообщению
Элленберга (Ellenberg, 1978), газохроматографический анализ секрета желез
показывает, что территориальные самцы производят гораздо больше пахучих
веществ, чем нетерриториальные, а лактирующие самки больше по сравнению с
полувзрослыми и самками без детенышей. Состав секрета межпальцевых желез
224
Рис. 80. Зоны с увеличенными сальными и потовыми
железами
1 — метатарзальная железа; 2 — передняя
межпальцевая железа и железистый комплекс дистального отдела
пясти; 3 — задняя межпальцевая железа и железистый
комплекс дистального отдела плюсны; 4 — кожные железы
головы, шеи и передней части туловища (увеличены
только у самцов летом)
у разных особей отличается, что также может
косвенно указывать на возможность
индивидуального различения по запаху. ^
Особо следует рассмотреть вопрос о
"наличии" у самцов косуль на голове
специфической кожной железы. Многие авторы
указывают, что эта железа расположена на лбу
перед рогами или между ними (Schumacher,
1939, по: Meyer, 1968; Schaffer, 1940; Raesfeld,
1956; Nec"as, 1963; Pielowski, 1970). Однако Мейер
(Meyer, 1968), сделавший тщательный
гистологический анализ кожи головы европейской
косули, специфической кожной железы здесь
не обнаружил, как не нашли и мы ее у си-2
бирской косули (Соколов, Данилкин, 1977,
1981).
При исследовании не выявляется каких-либо
обособленных участков с утолщенной кожей, указывающей не наличие
гипертрофированного четко оконтуренного жележистого комплекса, как, например, в мата-
тарзальной и в межпальцевой областях. Зимой кожа на голове и шее самцов
повсеместно тонкая, к лету в отличие от других участков она равномерно утолщается
по направлению от носа к темени (см. рис. 71). Соответственно летом сильно
увеличиваются в размерах сальные и потовые железы, которые осенью и
зимой снова уменьшаются в несколько раз, и их спавшиеся секреторные
отделы, по-видимому, находятся в это время в нефункционирующем
состоянии.
У самок косуль разницы в степени развития потовых и сальных желез в
коже головы и шеи, а также остальных участков тела почти не наблюдается.
Однако летом все железы у них чуть больше, чем зимой. У сеголеток они
развиты очень слабо, но у годовалых самцов уже близки по размерам к
таковым взрослых животных.
Мы считаем, что у самцов косуль нет на голове "лобного органа",
предназначенного для маркировки территорий. В коже их головы, шеи и передней
части туловища имеется лишь множество сильно увеличенных летом сальных
и потовых желез, что тесно связано с сезонным изменением толщины кожного
покрова в этих местах.
В связи с этим вполне вероятно следующее направление эволюционного
развития маркировочного поведения у косуль. Увеличение кожных желез,
скорее всего, может происходить при сезонном утолщении кожи любого участка
15. Европейская и сибирская косули 225
тела животных. Первично "кожный щит" служил у Capreolus (или Procapreolus)
исключительно для защиты во внутривидовой борьбе. Однако в процессе
эволюции (при переходе от стадного образа жизни к территориальности) это
сезонное увеличение размеров кожных >::елез "щита", соответственно
приводящее к большему продуцированию пахучего секрета, получило новое
функциональное назначение как органа маркировки. Отсюда маркировочное поведение,
видимо, следует считать вторичным явлением, выработавшимся уже на основе
имеющегося органа — кожного "щита".
Звуковая сигнализация
Звуковые сигналы играют значительную роль в социальной жизни европейской
и сибирской косуль. Наиболее подробное их описание и попытка
классификации содержится в исследованиях Майра и др. (Meyer, 1968; Stubbe, Passarge,
1979; Смирнов, 19756, 1977; Данилкин,1977; Соколов, Данилкин, 1981; Соколов и др.,
1987). Основываясь на анализе этих и других работ, а также на наблюдениях в
Таблица 33. Звуковые сигналы европейской (I) и сибирской (II) косуль
Мп/п
Тип сигнала и транскрипция
Вид сигнала; длительность, мс;
частотная характеристика,
мГц*
Пол, возраст
1 Писки "пии, пии..."
2 Шипение "чуф-ш-ш-ш"
Гармонический;
I. 144-175; 1,4-2,5
П. 165-190; 0,7-2,1
I. 365-489; 1,0-2,1
П. 237-583; 0,8-2,1
Негармонический;
I, П. 397-510; 1-2,7
Самки и телята, очень редко
самцы
Тоже
Самцы, реже самки
Лай "бооу, бооу..
"бо-бо-бо"
Негармонический;
I. 145-185; 1,4-2,5
П. 216-292; 0,7(1,0Ь2,3
Взрослые самцы и самки, реже
неполовозрелые особи
Скуление
Гармонический;
П. 151 ±9; 0,5-0,6
I, П. Гармонический
1,П
Самцы
5 Верещание
6 Звуки невокалыюго
происхождения:
1) при "топаний"
2) при гребущих землю I, П
движениях ногой
*По: Соколов и др. (1987); длительность сигнала — средние цифры, частотная характеристика
—максимум энергии.
Оба пола, все возрасты
Тоже
Тоже
226
природе и в вольерах, у косуль можно выделить 6 основных типов сигналов
(см. табл. 33).
Писки (или свисты) — гармонические сигналы двух разновидностей: тихие,
короткие (140—200 мс) и громкие, длинные (230—590 мс), имеющие разную
социальную значимость (табл. 33, рис. 81, А—Е).
Тихий писк, служащий сигналом взаимодействия особей на близком
расстоянии, наиболее обычен при контакте матери и детеныша и в разных ситуациях
является или призывным звуком, или выражением беспокойства. Чаще всего
его издают новорожденные, потерявшие визуальный контакт с матерью и
проголодавшиеся телята, подзывая мать. Реже такие звуки слышны у самок,
ищущих своих детенышей.
Сходным образом пищат субдоминантные особи при контакте с доминантом
(или с человеком), что выражает состояние беспокойства и дискомфорта и,
может быть, подчинения. Этот звук обычен у молодых зверей и самок, реже
у безрогих взрослых самцов (зимой). При приближении доминанта подчиненная
особь приседает, вытягивает шею и наклоняет к земле голову, нередко мо-
Годовойцикл
Причины и значения
Май-сентябрь, редко в другое
время
Тоже
1. Социальный контакт мать - дитя, призывные звуки
2. Выражение беспокбйства, дискомфорта и подчинения
3. В период гона способствует поиску и преследованию самки
1. Сильное смятение, испуг
Май-октябрь, реже в другое
время
Весь год, наиболее часто с
весны до осени
Июль-август
1. Сильное возбуждение: у самцов - во время гона, при агрессивном и
маркировочном поведении; у самок - при агрессивном поведении;
у сеголеток — при игровой борьбе
2. Сигнал контакта мать-дитя, доминант-субдоминант
1. Выражение беспокойства
2. Способствует исследовательскому поведению
3. Средство настораживания соседних особей
4. У самцов (предположительно) "выражение избытка мотивации"
1. Предположительно выражение дискомфорта
1. Возможно, выражение дискомфорта во время гона
Весь год
Тоже
Тоже
1. Раненая или пойманная косуля; страх, боль
1. Выражение беспокойства, нерешительности
2. Средство настораживания соседних особей, запугивания врага или
противника
1. Сигнальное значение только при демонстративном поведении самцов
во время конфликтов
чится и от 1 до 10 раз тихо пищит, после чего следует назо-назальный
контакт или бегство.
Точно так же, как и подчиненные особи,- ведут себя самки в течке при
приближении к ним взрослых самцов. Чуть громче пищат они в начале
гона при преследовании партнерами, особенно когда те ведут себя агрессивно.
После садок самка уже меньше боится самца, сигнал слышится реже или же
совсем перестает звучать. На заключительной стадии ухаживания и во время
садок самки сибирской косули издают тихие короткие (55±3 мс) сдвоенные писки,
следущие короткой серией из двух-трех звуков с интервалом 462±14 мс
(Соколов и др., 1987).
В редких случаях короткие писки регистрируются и у самцов как
европейской, так и сибирской косуль, преследующих самок по следу.
Сильно встревоженные звери издают очень громкий, пронзительный и
продолжительный свист, который можно охарактеризовать как сигнал испуга
или сильного смятения. Скорее всего, он развивается на основе тихого писка:
с возрастом длительность сигнала увеличивается в 2—3 раза, а основная
частота снижается в 1,5—2 раза и максимальная энергия иногда переносится на
вторую гармонику (рис. 81, £).
Наиболее типичен этот звук у телят при явной опасности. На него или его
имитацию особенно бурно реагируют лактирующие самки в первые недели после
родов и в редких случаях самцы, находящиеся поблизости (они устремляются
к источнику звука и демонстрируют по отношению к раздражителю агрессивное
поведение). У взрослых особей сигнал испуга иногда сменяется лаем.
Шипение (пыхтение или сипение) выражает сильное возбуждение
животных. В зависимости от ситуации этот сигнал может быть тихим и коротким,
громким и пронзительным, прерывистым (рис. 81, Ж—К).
Тихий сигнал наиболее обычен при подходе самца к самке в течке, но
становится громким при ее энергичном преследовании. Иногда при периодическом
прикрывании рта звук приглушается, в результате чего получается
продолжительный (до 1000 мс) прерывистый сигнал, состоящий из нескольких
коротких звуков (рис. 81, Ж, И).
Шипят косули и при агрессивном поведении (рис. 81, А): самцы — во
время борьбы друг с другом или "условным" противником (кустом или деревом),
при преследовании другого самца визуально или по следу, перед нападением
на человека и при учащенном мечении территории; лактирующие самки —
при защите недавно родившегося теленка от хищника, человека или другой
самки. Впервые этот звук регистрируется у 3—4-месячных телят при игровой
борьбе.
Очень тихое шипение, слышимое человеком не далее 20 м, издают матери
при подходе к своим детенышам и доминантные особи при приближении к
субдоминантам. Скорее всего, такой звук служит сигналом желаемого контакта.
Рис.81.Сонограммы звуков европейской (А, £, Ж, 3, И, К, М) и сибирской {Б, В, Г, Д, Л) косуль (по:
Соколов и др., 1987; с изменениями)
Писки: А —В — детенышей при потере контакта с матерью, Б— новорожденного, ГиД (сдвоенный) —
самки во время гона, Е — при испуге; шипение самцов: Ж — при преследовании самки, 3 — переходящее
в писк, И — прерывистое, К — при агрессивном поведении: Л — скуление; М — лай
228
кГц
\
т
\
> 1
"i'1'i
i^*rtii
. 4г-
J
2
1 -
4ft
*
При повышении возбуждения тихое шипение переходит в громкое, а в редких
случаях в другой тип сигнала — писк (рис. 81, J) или лай. Такой переход
мы регистрировали в вольерах и природе как у европейской, так и
сибирской косуль.
Лай — наиболее типичный и интенсивный негармонический сигнал,
состоящий чаще всего из серий звуков (от 3 до 30, в исключительных случаях
до 140), из которых первый отличается максимальной интенсивностью и
длительностью (рис. 81, АО- В среднем длительность отдельных звуков находится
в пределах от 140 до 330 мс с интервалами от 0,3—1,4 с, максимум
энергии сосредоточен в области от 1 до 2,5 кГц. Лают в основном стоящие
животные (бооу, бооу), при движении сигнал становится очень коротким
(бо-бо-бо). Динамический спектр лая имеет большое сходство с аналогичным
у Cervinae, особенно у бухарского оленя (Никольский, 1976).
Детеныши косуль этот звук не воспроизводят. Впервые он проявляется
в возрасте 6—8 мес, что наводит на мысль о постепенном его развитии
из другого типа сигналов. '
Звук слышен на расстоянии до 3 км. Лают обычно потревоженные или
обеспокоенные чем-то особи в сумерках или ночью, реже днем. Они
реагируют так не только на подозрительный шорох и запах, но и на видимый
раздражитель, например на человека, приблизившегося неожиданно вплотную
к ним, или волка, проходящего невдалеке. Если раздражитель не опознан,
то нередко, лая и стуча копытами (возможно, провоцируя), зверь огибает
подозрительное место и заходит под ветер, чтобы с помощью обоняния
установить причину беспокойства.
Весной на неопознанный раздражитель нередко лают одновременно несколько
животных из группы и им могут вторить другие особи из групп, находящихся
неподалеку. В это время особенно четко заметно, что звук настораживает
зверей и служит составной частью ориентировочно-исследовательского поведения.
Летом на лай одной встревоженной косули также отзывается несколько (до 11)
особей в округе, но чаще происходит обратное — другие животные
реагируют на сигнал настораживанием лишь в первый момент, а затем, если нет
явной опасности, несмотря на повторяющийся звук, продолжают кормиться или
отдыхать.
Самцы иногда лают после боя (победитель или оба противника, если
борьба закончилась ничейным исходом) и при высоком уровне мотивации во
время маркировочных действий.
Некоторые исследователи считают, что косули лают и без видимой причины,
расценивая такие звуки и самцов как акустическую маркировку территории
(Meyer, 1968; Смирнов, 19756, 1977). По нашим многолетним наблюдениям
лай всегда имеет четкие причины связи, но не всегда человек имеет
возможность их установить. Специального сигнала, предназначенного для мечения
территории, у них, скорее всего, нет, но лай (наряду с "шипением" и
звуками невокального происхождения) может играть определенную роль в
территориальных отношениях по принципу мультифункциональности.
При сравнительном изучении в природе обнаруживается, что сибирские
косули лают чаще европейских и самцы чаще, чем самки. Летом на Южном
Урале этот звук зарегистрирован в 24,5% встреч с животными, из них в 70%
случаев у самцов. Европейские косули в Белгородской и Курской областях при
230
тех же ситуациях лаяли лишь в 7% встреч, но и здесь самцы издавали
этот сигнал чаще (67,8%) самок. В зимний период с ноября по март как те,
так и другие более "молчаливы" — звук отмечен соответственно в 4 и 1%
встреч. Наиболее интенсивен лай (самцов и самок в равной мере) весной,
когда большинство из них еще ведут групповой образ жизни: у сибирской
косули в 29% случаев, у европейской — в 9%. Однако большая
"молчаливость" европейской косули вызвана, видимо, не столько видовыми
особенностями, сколько разным уровнем антропогенного пресса, гораздо более
высоким на европейской части ареала, в то время как сибирские косули
менее привычны к данному раздражителю.
Скуление — гармонический сигнал, состоящий из серии звуков (от 3 до 20)
длительностью около 150 мс с основной частотой 0,5—0,6 кГц (рис. 81, Л),
очень похожий на скуление собак. Он регистрируется у отдельных самцов
сибирской косули в период гона: во время остановки после длительного
преследования самки (когда зверь стоит или лежит), при потере контакта с
самкой и при невозможности сближения с ней (если звери находятся в разных
вольерах). Значение его неясно, видимо, звук является смешанным сигналом,
отражающим дискомфорт (писк) и половое возбуждение (шипение).
Верещание (стон, рев) — гармонический сигнал (вероятно, смешанного
типа) сильной интенсивности, издаваемый раненым или пойманным животным и,
скорее всего, выражающий страх, боль.
Звуки невокального происхождения производятся косулями при
топаний ногами (при этом животное, медленно передвигаясь, высоко поднимает
поочередно передние ноги и с силой опускает их на землю), когда они
обеспокоены неопознанными или явными раздражителями. Эти звуки являются
составным элементом исследовательского, оборонительного и агонистического
поведения и, скорее всего, выражают нерешительность зверя перед принятием
определенного решения к дальнейшему действию, служат одним из средств
предупреждения находящихся рядом особей об опасности и, возможно,
способствуют запугиванию врага или противника. Такие удары передними ногами
зарегистрированы при наступлении косуль на врага — лисицу, собаку,
человека и во время демонстрационного поведения конфликтующих взрослых
самцов.
Сюда же мы относим звуки при гребущих землю движениях передних
ног, приводящих к образованию "пятачков" на земле. Такого рода движения
обычны при раскопке снега (для добывания корма и изготовления лежек),
однако сигнальное значение они имеют, видимо, только летом в качестве
составной части демонстрационного поведения при конфликте самцов — такие
сигналы одного из противников вызывают рывок к нему другого самца.
Итак, у косуль в раннем возрасте усматривается только один тип
акустических сигналов — писк (тихий и громкий), вероятно, трансформируемый
затем в другие звуки. Все сигналы, за исключением скуления, у самцов
и самок совпадают. В целом у европейской и сибирской косуль они сходны
и имеют одно и то же функциональное значение, что говорит об их близком
филогенетическом родстве. Однако у самцов европейской косули пока не
отмечено аналогов скулению (рис. 81, Л), обнаруженному | у отдельных особей
сибирской косули (Соколов, Данилкин, 1981), у самок — сдвоенных писков
(рис. 81, Д), у детенышей несколько отличаются характеристики писков при
231
потере контакта с матерью (рис. %\,А,В) (хотя на слух сигналы
неотличимы и с возрастом становятся сходными?), и к тому же у европейской
косули более высокая частота издаваемых звуков (Соколов и др., 1987).
Тем не менее делать окончательный вывод об отличиях акустических
сигналов у европейской и сибирской косуль пока преждевременно, поскольку
сравнительное исследование характерных звуков проведено на очень
ограниченном числе особей, а отличающиеся сигналы регистрируются далеко не у всех
животных. Скорее всего, аналоги их будут найдены. Масса европейской и
сибирской косуль различается, что, вероятно, может в какой-то степени влиять
на частотную характеристику и длительность звуков. Эта особенность
наглядно проявляется с возрастом у сеголеток.
Половое поведение
Поведение диких копытных в период гона разнообразно (Адольф, 1954;
Слудский, 1956; Walther, 1958, 1963; Савинов, 1962; Устинов, 1967; и др.),
но именно для косуль слово "гон" наиболее полно соответствует этому
понятию. Самец на большой скорости с шипящими звуками преследует самку
в течке, стремящуюся убежать от него, и у наблюдателя складывается
впечатление, что он или гонит самку или же гонится за ней (рис. 82 £, В).
Одни исследователи считают, что самцы косуль во время гона очень
агрессивны и избивают самок (и даже убивают в некоторых случаях), тем самым
вынуждая их убегать (Туркин, Сатунин, 1902; Даль, 1930; Флеров, 1952;
Аверин, 1949; Гептнер и др., 1961; Черкасов, 1962). Другие полагают, что он не
гонит самку, а бежит за ней, стараясь не упустить из вида, и самка не
боится партнера (Смирнов, 19756, 1978).
По-своему правы как те, так и другие ученые. По нашим многочисленным
наблюдениям в первый день гона самцы ведут себя с самками, особенно
молодыми, довольно агрессивно, вплоть до ударов рогами. Взрослый самец,
приближаясь к самке (как и к любой особи субдоминанту), наклоняет голову,
выставляет рога вперед и нередко "шипит", что означает угрозу и вызывает
оборонительное поведение; она поворачивает голову к нему, приседает,
мочится, издает несколько свистящих звуков и, когда тот приближается
вплотную, срывается с места и быстро убегает. Самец начинает преследовать ее
незамедлительно или, если самка мочилась, после обнюхивания мочи на земле и
"флемена"(рис. 82, А, Б).,
После непрерывной продолжительной погони большими кругами уставшая
самка начинает бегать вокруг деревьев, кустарников, высоких кочек, ям, и,
наконец, обессиленная останавливается и позволяет не менее уставшему самцу
сделать садку, после чегЬ оба ложатся отдыхать. Многократно пробегая по
одному месту, косули выбивают характерные для гона кольцевидные или в виде
восьмерки тропы (рис. 83). Иногда уставшая самка еще до садки самца
ложится прямо на тропе, но возбужденный самец бодает ее, заставляет
подняться и делает садку.
После садок поведение животных заметно изменяется. Самка уже меньше
боится самца, реже стремится скрыться от него, и повторные спаривания
нередко происходят сразу же при сближении зверей: после назо-назального
контакта он обнюхивает и быстрым движением языка облизывает голову,
232
Рис. 82. Гон
А — самка боится
приближающегося самца и убегает от
него; Б — преследование иа
большой скорости в начале гоиа; В —
следование за уставшей самкой
непосредственно перед
спариванием
в
Рис. 83. Характерные тропы европейской и сибирской косуль во время гона
бок, спину и гениталии самки и делает садку (рис. 84). У молодых самцов
из-за отсутствия опыта садки не всегда бывают удачными; иногда они
начинают садку сбоку, спереди, но, переступая задними ногами, в конце
концов принимают правильную позу. В среднем у одно-двухлетних самцов она
длится 22 с (от 8 до 120 с), у взрослых особей — 6 с. Две-три садки
могут идти через несколько минут одна после другой, затем косули более часа
отдыхают. После эйякуляции самец тяжело опускается на самку и почти всег-
234
да ложится на землю. Самка для лежек выбирает самые укромные места.
Партнер нередко теряет ее и энергично отыскивает по следам.
В естественных условиях самку в течке обычно преследует один взрослый
самец, очень редко два и в исключительных случаях три или четыре. Как
только она останавливается, рядом с ней остается доминирующий самец, который
при приближении остальных принимает позу угрозы и держит их на
расстоянии более 30 м. В этой ситуации четко видна иерархия: субдоминант
отгоняет третьего, тот — четвертого, самого молодого зверя, и все они
кружат возле тонной пары. После садки доминант ложится, а отошедшую
в этот момент самку могут покрыть второй и даже третий самец, и так за
несколько часов многократно (до 27 садок). При бегстве самки все самцы
устремляются за ней, следуя друг за другом согласно занимаемому
иерархическому положению.
В небольших вольерах ситуация многократного покрытия самок всеми содержащимися здесь
самцами представляется типичной. Например, в вольере под Москвой спаривание одной из них
проходило следующим образом. Два самца сибирской косули (трехлетний и однолетний),
содержавшиеся вместе, в первый день течки не смогли покрыть двухлетнюю самку даже после
продолжительной погони. Во второй день им удалось сделать это лишь после того, как она,
обессиленная долгим преследованием, обежав более 40 раз вокруг дерева, упала на тропу.
Первым спарился с ней, подняв ударами рогов, взрослый самец, и спустя две минуты —
молодой. Затем через три часа отдыха за последующее 5,5 часа до наступления темноты они
сделали 18 садок, из них молодой — 12, взрослый — 6, причем при приближении
последнего она всегда убегала и он покрывал ее после погони, а от однолетнего самца, которого
она боялась явно меньше, в 9 случаях из 12 не убегала. На следующее утро было
зарегистрировано еще 7 садок, и затем после продолжительного отдыха самка перестала
допускать партнеров к садкам.
При высокой численности самок вероятна и противоположная тенденция;
один самец может в один и тот же день поочередно покрывать
одновременно двух пришедших в охоту самок (Holmes, 1973).
Оплодотворенная самка при приближении самца ведет себя иначе, чем в
начале течки; она приседает и мочится, а самец долго, около минуты,
обнюхивает мочу на земле, "флемует" и начинает кормиться или, реже, вновь
начинает преследовать ее, но та при его приближении снова мочится, и он
после обнюхивания мочи прекращает погоню. Через час-два или более самец
снова начинает преследование этой самки и вновь наблюдается та же картина.
Однако партнер еще одни или двое суток продолжает держаться рядом с самкой.
Гон у косуль обычно длится 2—5 дней. В это время самец практически
перестает кормиться, теряет осторожность и не покидает самку даже при явной
опасности. Неоплодотворенные самки могут приходить в "охоту" еще раз с
промежутками от 9 до 84 дней, что говорит об отсутствии у них четкого
ритма течек (см. главу "Размножение").
Радиопрослеживанием установлено, что во время гона территориальная
система в целом не нарушается (Соколов, Данилкин, 1981; Sempere, 1982). Этому
во многом способствует то обстоятельство, что участок обитания самки часто
находится в пределах территории самца (см. главу "Структура популяций"),
и он обычно следует за ней, не предпринимая попыток изменить направление
ее передвижения. Однако в природе мы наблюдали в 4 случаях, как самцы
на границах своих территорий бросались наперерез самкам и вынуждали их по-
235
Рис. 84. Поведение косуль при спаривании
вернуть назад. В вольерах самец, преследующий самку в течке, также пытается
удержать ее в той части загона, где нет соперников.
Территориальность во время гона способствует уменьшению контактов и
возможных столкновений между самцами. Тем не менее "гонная" пара во время
быстрого бега нередко выходит за пределы территории самца, что приводит
к нарушениям границ. Соседние взрослые самцы в этом случае явно проявляют
исследовательское поведение и идут по их следу, но лишь в исключительных
случаях присоединяются к ним. Чаще же нарушение границ вызывает у них
агрессивную реакцию и усиленную маркировку участка.
Есть наблюдения, что самка в течке предпочитает определенного самца и
может спариваться с ним на протяжении нескольких лет. Замечено также, что
237
в конце гона увеличивается частота "рейдов" косуль за пределы своих обычных
участков, очевидно в поисках полового партнера. В этот период самцы
особенно легко реагируют на звуки, сходные с писком и часто становятся
добычей охотников, имитирующих этот сигнал (Kerchagl, 19S2; Kurt, 1968;
Bayern, 1975; Raesfeld et al., 1970; EUenberg, 1978; Stubbe, Passarge, 1979; и др.).
Половое поведение европейской и сибирской косуль, как показывают наши
сравнительные исследования и анализ литературы, идентично.
В связи с летними сроками гона, специфическим половым поведением и
наличием у самок косуль, единственных среди копытных, беременности с
уникальной эмбриональной диапаузой, возникает вопрос: каким образом в процессе
эволюции появились эти особенности и насколько они взаимосвязаны? В
главе "Размножение" на этот счет уже была предложена гипотеза, которую мы считаем
необходимым развить.
Выше достаточно убедительно показано, что у косуль эволюционная стратегия
была направлена в русло территориальности в летний период и именно к этому
моменту происходит максимальная активизация многих функций организма
самцов. Гон у их непосредственных предков, скорее всего, приходился на
осенние месяцы, о чем свидетельствуют осенний всплеск половой активности у
косуль и осенние сроки гона у ближайших родственников — представителей
семейства Cervidae. Преждевременное в связи с территориальностью достижение
самцами наивысшей половой потенции приводило, видимо, к преследованию
самок, обитающих на их территориях, еще до появления у них явных
признаков течки. Такие случаи наблюдаются в природе и в настоящее время
(Соколов, Данилкин, 1981).
Преждевременное, но энергичное и продолжительное преследование самок
возбужденными и агрессивными самцами (которых самки очень боятся и стремятся
убежать от них), вероятно, вызывало резкую активизацию функций их
организма. Очевидно, что, преследуя самок, самцы как бы стимулируют их к
спариванию3. Преждевременному спариванию во многом способствует также и то
обстоятельство, что после продолжительного преследования самец может
покрывать вконец обессиленную самку независимо от того, хочет она этого или нет.
Постепенно в процессе эволюции сдвиг сроков спаривания с осенних на летние
месяцы закрепился, обусловив и сдвиг начала течки, но и сейчас мы
наблюдаем характерное продолжительное преследование самцами самок перед
спариванием, необходимое для их стимуляции. Оплодотворенная преждевременно
яйцеклетка тем не менее не может развиваться вплоть до "нормальных"
зимних сроков беременности, чем и объясняется появление у косуль
эмбриональной диапаузы.
Предлагаемая гипотеза, на наш взгляд, отражает взаимосвязь и
эволюционного, и этологического, и физиологического аспектов этого уникального явления.
Не исключено, что сходным образом латентный период беременности мог
образоваться и у некоторых других млекопитающих.
'Стимуляция самок энергичными самцами, как известно, с успехом применяется в
звероводстве и иа животноводческих фермах. .
238
Ориентировочно-исследовательское
и оборонительное поведение
Под ориентировочно-исследовательским поведением обычно понимают
совокупность реакций, направленных на улучшение восприятия и поиск источника
раздражения, при этом увеличивается чувствительность животного к
стимуляции и возрастает его готовность к действию. С другой стороны,
ориентировочно-исследовательское поведение очень тесно связано с оборонительным
поведением, в большинстве случаев предшествует ему, и если раздражитель
очень силен, то ориентировочная реакция сменяется оборонительной. Во многих
случаях происходит конфликт между первым (любопытство) и вторым (страх), что
может вызвать смещенную реакцию (Павлов, 1938; Соколов Е., 1966; Шовен,
1972; Хайнд, 1975).
У косуль ориентировочно-исследовательское и оборонительное поведение
складывается из отдельных элементов (унитарных реакций; по: Крушинский, 1960):
ориентировочной позы, сигнальных прыжков, звуковых сигналов, реакции испуга,
сближения с другими особями, предупреждения об опасности "небеганием",
поиска и уточнения раздражителя, бегства, затаивания. В различных ситуациях
не все эти элементы могут присутствовать и могут идти в .разной
последовательности.
В периоды активности на ориентировочно-исследовательское поведение косули
тратят от 1/4 до 1/2 времени (в зависимости от ситуации, фактора
беспокойства, индивидуальных особенностей и др.). Даже в отсутствие раздражителя
они периодически поднимают голову, осматриваются и пронюхиваются. Однако
зрение у зверей слабое, и не составляет большого труда (при определенном
навыке и маскировке) из-под ветра подойти к пасущемуся животному на 20—50 м,
оставаясь неподвижным как только косуля принимает ориентировочную'позу.
Ориентировочная поза (см. рис. 85) — голова поднята высоко над
землей, мускулатура напряжена, волосы "зеркала" распушены, уши насторожены и
ориентированы по направлению к раздражителю, глаза широко раскрыты, ноздри
втягивают воздух — особенно четко проявляется у зверей, когда они чем-то
обеспокоены. Косуля может опускать голову, как бы кормясь, и вдруг вновь
резко вскидывать ее вверх, продолжая исследование. Отчетливо видно, как она
часто смачивает языком ноздри, возможно для улучшения восприятия запаха.
Ориентировочная поза нередко сопровождается уринацией и дефекацией.
Хронометраж поведения показывает, что ориентировочная поза принимается
в среднем через 36,2 с, но настороженность изменяется в разные дни, в
течение одного дня, заметно ослабевая в сумерки. Уровень "бдительности" у
косули сравним с таковым у оленя, но значительно выше, чем у лося и лани
(Pelosse, 1976).
При групповом образе жизни ориентировочная поза у одного зверя
немедленно вызывает настораживание соседних особей, сближение их и последующее
ориентировочно-исследовательское поведение всех. Если раздражитель себя "не
проявляет", косули успокаиваются или продолжают исследование. Одним из его
элементов являются звуковые сигналы.
Звуковые сигналы (лай, топание). Лаять на раздражитель может
одна или одновременно несколько косуль. При этом характерно для них
высокое поднимание и твердая постановка передних ног на землю (в отли-
239
V
Рис. 85. Ориентировочная поза
чие от гребущих движений передними ногами в других ситуациях) при
продвижении животного в направлении раздражителя либо от него. В качестве
примера приведем описание поведения группы сибирских косуль из 6 особей
(март, Южный Урал).
Группа лежит. Появляется раздражитель (человек на расстоянии 70 м от них,
направление ветра — от косуль). Самка, заметившая «движущийся предмет,
встает, принимает ориентировочную позу и передвигается на 2—3 м в
сторону от раздражителя, твердо ставя передние ноги на землю. Вслед за ней
встает вся группа и косули принимают ориентировочную позу. Лай самки —
37 раз подряд.
Раздражителя нет (человек стоит на месте), проходит 18 мин — все члены
группы поочередно ложатся. При наличии раздражителя (движение человека)
встает самец, на этот раз первым заметивший опасность, — ориентировочная
поза — встают еще две самки — ориентировочная поза — движение самца с
твердой постановкой передних ног на землю — сигнальный прыжок и бегство —
вся группа обращается в бегство без предварительной ориентировочной позы —
через 200 м звери останавливаются — ориентировочная поза у всех.
Эти звуковые сигналы как бы служат выражением эмоционального состояния
особи в ответ на раздражитель и выполняют функции предупреждения других
особей об опасности и, вероятно, запугивания врага.
Сигнальные прыжки почти всегда бывают после того, как косуля
заметила опасность. Пройдя или пробежав несколько метров в сторону от
раздражителя, зверь прыгает высоко вверх и приземляется, щелкая скакательными
суставами ног. Прыжок одной особи тотчас вызывает ориентировочную реакцию
других и бегство.
В высокой траве косули прыгают вверх через несколько десятков метров.
Однако летом, когда животные ведут одиночный образ жизни, основная функция
240
этих прыжков, скорее всего, не сигнальная: они служат больше для обзора
местности и выбора направления движения.
Поведение испуга выражается прежде всего внезапным "шараханьем"
зверей от реальной или мнимой опасности без предварительной ориентировочной
позы. Тем не менее и в этом случае, отбежав на 10—30 м, животные
принимают ориентировочную позу и только после этого спасаются бегством, если
опасность очевидна. При групповом образе жизни косули при испуге
сближаются и все принимают ориентировочную позу. Внезапно "шараханье"' от
мнимой опасности гораздо чаще бывает в закрытом биотопе (в лесу) и в больших
по количеству особей группах. Это, на наш взгляд, одна из причин, объясняющих,
почему в группах косуль, живущих в лесу, редко бывает более 10 особей.
Уточнение опасности обонянием. Слух и зрение позволяют косулям
выявить наличие раздражителя, тогда как уточнение информации об источнике
беспокойства достигается во многих случаях при помощи обоняния. В таких
ситуациях они стремятся зайти к источнику беспокойства под ветер, чтобы
уловить запах.
Следует заметить, что обонятельный орган является важнейшим из органов
чувств косуль. Обонятельная поверхность ноздрей у них превышает 90 см2 (у
человека 2,5 см2), число обонятельных клеток достигает 300 млн (у
человека примерно 30 млн) (Kolb, 1979). Курт (Kurt, 1965) подсчитал, что из
42 элементов социального поведения обонятельным восприятием вызывается 26,
акустическим — 13 и оптическим — только 3, что вполне подтверждает
предположение о прекрасном обонянии и об относительной слабости зрения
у этих животных.
Дальность обнаружения ими по запаху одного и того же объекта не всегда
бывает одинаковой: при ветреной погоде на открытой равнинной местности
животные обнаруживают человека за 200—400 м, а в лесу — за 100—200 м.
При тихой погоде это расстояние сокращается в 2—5 раз, а против ветра
к ним можно подойти, маскируясь, вплотную.
Предупреждение об опасности "набеганием". Зверь, заметивший
опасность, иногда уже после уточнения раздражителя с помощью обоняния, не
убегает прочь, а подбегает к другой или другим особям группы, находящимся
на лежке и не подозревающим об опасности. Обычно он пробегает рядом с
лежащими животными или, очень редко, перепрыгивает через них и вызывает тем
самым беспокойство или бегство последних. Такое поведение в большинстве
случаев проявляют телята в возрасте от 5 до 10 мес по отношению к своей
матери или более молодые звери по отношению к старшим, скорее всего
из-за того, что при опасности они стремятся; сблизиться с "вожаками".
Анализ ситуаций показывает также, что иногда косули предупреждают об
опасности своих партнеров сознательно. Вот один из примеров: спугнутый
с лежки взрослый самец после ориентировочной реакции зашел под ветер,
учуял человека, но не убежал прочь, а повернул назад в сторону
наблюдателя и пробежал рядом с другим двухлетним самцом, местонахождение
которого на лежке ему было, очевидно, известно. Остановившись от последнего
примерно в 20 м, он, повернув голову в сторону человека, выждал минуту,
затем вернулся, еще раз набежал на все еще лежащего самца и перескочил
через него, вызвав беспокойство, ориентировочную реакцию и следование
за собой.
16. Европейская и сибирская косули
241
Ршс.М.Следование за лидером
Затаивание. При явной опасности телята ложатся и прижимают голову
к земле (см. выше), а взрослые особи стоят неподвижно, наклонив голову,
или же, так же как и телята, ложатся. Последнее особенно характерно
для зверей, живущих в открытых степных биотопах.
Бегство и следование за лидером (рис. 86). В группе косуль
бегство одной особи вызывает немедленное бегство остальных. Примерно через
30—250 м они останавливаются, принимают ориентировочную позу, и, если
опасность установлена, бегство продолжается. Если животных не преследуют,
они убегают в лесу обычно не далее 300 м, в поле дистанция бегства
значительно больше.
В лесных и лесостепных стациях расстояние, с которого животные начинают
бегство от человека или транспорта, чаще всего находится в пределах 20—150 м.
Там, где велик пресс охоты, проводимой загонным способом, с подхода или
с автомобиля, эта дистанция значительно больше. В популяциях, видимо,
происходит своеобразный отбор особей с определенным оборонительным поведением:
выживают лишь животные, знакомые с подобной опасностью и научившиеся
ее избегать, а опыт старших усваивают молодые звери путем подражания
при групповом образе жизни.
Следует отметить, что наиболее реактивные особи у косуль — сеголетки.
Во многих случаях бегство при явной или мнимой опасности начинают именно
они. Взрослые животные обращаются в бегство почти всегда последними, но
сразу же становятся лидерами. Если явной опасности нет, вожаки, которыми
обычно являются самки (см. главу "Структура популяций"), успокаиваются и к ним
присоединяются молодые звери. Таким образом, взрослые особи на основании
своего индивидуального опыта показывают модель поведения, наилучшим образом
противодействуя всякому беспокойству без причины, что в конечном счете
необходимо для сохранения энергии и нормальной жизнедеятельности группы.
При опасности у косуль особенно отчетливо проявляется поведение типа
"руководство—следование": члены группы бегут за лидером цепочкой, чему в
значительной мере способствует "морфологическая поддержка" — распушенное
хвостовое "зеркало" (Северцов, 1951), служащее следующим сзади животным
ориентиром при быстром беге (см. рис. 86). Адаптивный механизм
"руководство—следование" особенно важен для молодых зверей, обеспечивая
необходимую связь с семейной группой и позволяя им выжить при опасности
благодаря опыту взрослых.
Активность
Активность косуль определяется многими обстоятельствами и может меняться
в течение года, сезона, суток, зависит от пола, возраста, степени беспокойства,
наличия корма, климатических и других факторов внешней среды. В то же
время в течение суток наблюдается определенная закономерность в периодичности
ее проявления: пастьба и передвижения чередуются с отдыхом и пережевыванием
пищи обычно от 6 до 12 раз в сутки, т.е. активность зверей имеет
полифазный характер, причем утренний и вечерний ее периоды наблюдаются
практически всегда и они наиболее продолжительны (рис. 87). Максимальная активность
как отдельных животных, так и групп преимущественно связана с восходом и
заходом солнца (Mottl, 1957; Bubenik, 1960, 1962; Lochmann, 1965; Eiberle, 1967;
243
Vincent et ak, 1979; Turner, 1980; Cederlund, 1981; Janeau et al., 1981; Sempere,
1982; Соколов, Данилкин, 1981; и др.).
Время активности с энергетической точки зрения определяется балансом
между расходом энергии и ее поступлением с пищей (Turner, 1978, 1979).
В связи с этим соотношение продолжительности фаз питания,
пережевывания пищи и отдыха весьма зависит от структуры и качества корма и
различается по сезонам года. Весной при мягком, легко перевариваемом корме
на жвачку они тратят меньше времени, чем на пастьбу, тогда как в другие
сезоны это соотношение противоположное. Потребность в отдыхе наиболее
высока зимой, что, очевидно, тоже связано с экономией энергии (Lochman, 1965).
По данным радиотелеметрии "зимний ритм" косуль замедлен и имеет около
6 пиков активности, тогда как "летний ритм" — 9, но паттерны суточной
активности у одной и той же особи могут в разные дни существенно отличаться
(Berg, 1978). Эти сведения совпадают с наблюдениями Элленберга (EHenberg,
1978) в вольерах. Зимой при снежном покрове он регистрировал от 4 до 8
периодов активности, летом — от 8 до 14; у самок в любой период года
отмечено меньшее их число (на 13%) по сравнению с самцами. Снижение
активности косуль зимой, как считает Элленберг, не только реакция на
ухудшившееся качество корма и его замедленную скорость переваривания, а имеется
к тому же наследственная физиологическая основа приспособленности к
уменьшенному количеству корна.
Семпере -(Sempere, 1982), напротив, нашел среднее число периодов суточной
активности относительно стабильным в течение всего года (6,7±0,2). В среднем
общее время Суточной активности при весеннем и осеннем равнодействии
практически равно общей длительности отдыха, несколько снижается (до 43,3%)
зимой и увеличивается весной (до 55,2%), что коррелирует с увеличением
продолжительности дня (см. рис. 87).
Ритм активности особей в группе косуль в значительной мере
синхронизирован, особенно в сумерках (Mertens, 1984). Социальным механизмом,
координирующим его, вероятно, служит поведение типа "руководство—следствие".
Синхронизация активности семьи происходит только с 2,5—3-месячного возраста
детей (см. рис. 63). Самцы пасутся и отдыхают дольше и тратят меньше
времени на ориентировочные реакции, чем самки. При увеличении размера
группы время пастьбы возрастает, продолжительность ориентировочных реакций
уменьшается, но число периодов пастьбы и отдыха не меняется. Длительность
отдыха и социальных контактов при разном количестве особей в группе
примерно одинакова (Btittner, 1980).
Косули полевой популяции с октября по апрель активны днем от 4 до 7 раз,
что составляет примерно 33,4% светлого времени суток, из него на питание
расходуется 76,6%, чистку стоя — 7,8% и на другие действия — 4,1%.
Остальное время они проводят на лежке, причем жвачка занимает почти треть его,
сон — 13%- и чистка лежа — 4,6%. В середине зимы число кормящихся
особей ниже (54%), чем весной (до 74%). Самцы зимой в течение суток
имеют на один период активности больше, чем самки, но общая
продолжительность активности у них примерно одинакова. Полевые косули днем
активнее лесных, они меньше времени тратят на жвачку и больше времени
пасутся, что связано с разно качественностью пищи на полях и в лесу (Zejda et al., 1985).
244
впн1гоэ BtfoxBt и вНохэоя Bwsda — р JBirBHJHaoHtfBd эиахэхЛэхо — f :чхэоняихяв — ^ txntfxo — /
(Z86I 'awduias
) кинваижэи'эойиои&'вй ииннв» ои (у) airaduB я и (ь-) эёванв а Ч1ГЛоол чхэонаиххв «вньохХэ 'Lt '3Hd
* -<
ШИН
<([
г , SIIIIHI
/Л ШПИНИ
' ' "пни и
m;ni;j
ОНЮШНШШН! С
IIIIIIII
ir
lllillllll.il
1111
&
III IIISSI
IISIIIIIII
г—1—II I I—I—I—I—I—I—I—I—|—I—г
В / 5 Г I п?? Id Bl 1.1 SI 5.7 // В
/чщ
i г i—i i i i i i—i i i i i i i—г
в /, Ч У. I ОМ 17. Ill 1.1 51 P.I Ч 6
В жаркие летние дни косули кормятся реже, чем в прохладные и дождливые,
в морозную погоду кормежки продолжительнее.
При повышении энергетических затрат взрослые самки сокращают свою
локомоторную активность, в то время как самцы увеличивают ее (Turner, 1978,
1979). Весной и летом подвижность животных наиболее высокая ночью и в
сумерках, зимой — в начале дня (Bideau et al., 1983). Средняя скорость.перемещения
у европейской косули, по данным Семпере, в территориальный период выше
(1,26±0,19 км/ч), чем в атерриториальныи (0,57±0,05 км/ч), и максимальна
в период гона — 6,2 км/ч. Аналогичная тенденция прослеживается и у сибирской
косули (Соколов, Данилкин, 1981). Быстрые передвижения в течение лета
позволяют самцам обеспечить наблюдение и регулярный контроль за границами
территорий. Шиз кие скорости движения зимой связаны с экономией энергии.
Визуальные и биотелеметрические наблюдения показывают, что косули при
мнимой опасности могут вставать с лежки и снова ложиться здесь же рядом.
Число периодов, в течение которых животные находятся на ногах (т.е. стоят или
передвигаются), в среднем за сутки достигает 22 (от 12 до 30) (Gonzales,
Angibault, 1987). Это число, видимо, тем больше, чем выше антропогенный
пресс и плотность населения других видов крупных животных. Там, где фактор
беспокойства очень велик, особенно при промысле, суточная периодичность
нарушается настолько, что косули становятся активными лишь в сумеречные и
ночные часы и вскакивают с лежки при любом подозрительном шорохе.
Соответственно у них может измениться нормальный ритм пищеварения со
всеми вытекающими отсюда негативными последствиями.
Глава десятая. Численность
Численность и плотность населения
Динамика численности косуль совпадает с общей тенденцией движения их
ареала, который при снижении поголовья заметно сокращался и при увеличении —
расширялся. В историческом плане эти изменения прослежены в главе "Ареал".
В послеледниковое время наивысшая численность зверей, скорее всего,
была во второй половине второго тысячелетия. По свидетельству очевидцев
леса в Европе "кишели" косулями, их добывали "неимоверное количество", а на
Урале, в Сибири и на Дальнем Востоке этих животных было "необыкновенное
множество" — во время миграций стада насчитывали сотни особей (Кириков,
1959, 1966, 1983; Пржевальский, 1870; Сабанеев, 1872, 1875; Туркин, Сатунин,
1902; Аверин, 1960 а,б). Еще во второй половине XIX в. лишь в Амуро-Уссу-.
рийском крае их поголовье, вероятно, достигало нескольких миллионов
(Бромлей, Кучеренко, 1983).
С XVIII в. из-за прямого истребления и изменения биотопов численность
косуль резко снижается в Западной Европе, в XIX в. — в Восточной Европе, в
начале XX в. — в Сибири и Казахстане и с середины его — на Дальнем Востоке, т.е.
по мере усиления хозяйственной деятельности человека в регионах. Во многом этому
процессу способствовали появление дальнобойного оружия и чрезмерный
промысел. Только в России в конце прошлого века добыча исчислялась сотнями
тысяч особей.
В такой же последовательности с запада на восток ареала благодаря охране
копытных и истреблению крупных хищников с XIX в. начинается увеличение
поголовья косуль. В годы войн на территории Европы оно снова резко сокращалось,
но быстро восстанавливалось. В Венгрии, например, с 10 тыс. особей в 1946 г.
возросло до 220 тыс., в Болгарии с 24 тыс. в 1925 г. — до 120 тыс., в Румынии
с .16 тыс. в 1950 г. — до 290 тыс. Примерно на порядок увеличилась
численность косуль и в других странах Европы, достигнув в настоящее время
максимума. В Восточной Европе, на Урале и в Сибири этот процесс начался с
30-х годов текущего столетия и привел в 70-е годы к полному восстановлению
былого ареала. Динамика численности животных на востоке европейской части
СССР в 60—70-е годы XX в. показана в табл. 12. Однако в 70-е годы рост поголовья
косуль в большинстве регионов СССР практически прекратился, а в Азии
их численность снова снизилась более чем на 100 тыс., особенно заметно (в 3—7 раз)
в Новосибирской и Кемеровской областях (с 11 до 2 тыс.), в Красноярском крае
(с 60 до 10 тыс.), в Туве (с 45 до 6 тыс.), в Читинской области (с 100 до 25 тыс.), в
Хабаровском крае (с 30 до 10 тыс.) (Метельский, 1976; Данилкин, 1982, 1985).
В начале 70-х годов в Амурской области поголовье достигало 100 тыс. особей
(Морозов, 1975), в 1981 г. — 35 тыс. В целом на юге Дальнего Востока оно
сократилось с 165—190 тыс. в 60-х годах до 58—67 тыс. в 1979 г. (Бромлей, Кучеренко,
247
Таблица 34. Плотность иаеслспия европейской и сибирской косуль в разных страпах
ко второй лоловинс 80-х годов XX в.*1
Страна
СССР
Польша
ГДР
Румыния
Чехословакия
Югославия
Венгрия
Болгария
ФРГ
Австрия
Франция
Дания
Швейцария
Нидерланды
Бельгия
Люксембург
Великобритания
Италия
Испания
Норвегия
Финляндия
Швеция
Албания
Греция
Турция
Сирия
Иран
Ирак
Итого
СССР
Монголия
Китай
Корея
Итого:
В целом для Capreolus
Численность,
тыс. особей*2
380
500
310
290
290
350
220
120
1700
1000
300
140
120
25
25
13
9007(400)
100
20
150
5
400
(10)
(5)
(100)
(5)
(5)
(5)
7488
315
100
(250)
(10)
675
8163
Площадь
ареала
Европейская
1916
313
108
237
128
256
93
111
248
84
551
43
41
41
30
3
173
58
89
302
53
450
(14)
<7)
(602)
(14)
(14)
(14)
5993
1 5
.тыс.км
лесов в ареале
косуля
478
84
30
64
45
87
14
33
74
37
32
5
10
3
6
_
20
14
40
72
. 32
256
(7)
(3)
(150)
—
—
—
1610
Сибирская косуля
4660
484
(2187)
(56)
7387
13380
41В скобках — очень приблизительные данные; прочерк
*2По литературным сведениям.
2012
44
(175)
(39)
2270
3860
Число особей на 1000 га
ареала
2,0
16,0
28,7
12,2
22,7
13,7
23,7
10,8
68,5
119,2
5,4
32,6
29,3
6,1
8,2
50,0
52,0
17,3
2,3
5,0
1,0
8,9
(7,1)
(7,1)
(1,7)
(3,6)
(3,6)
(3,6)
12,5
0,7
2,1
(1,1)
(1.8)
0,9
6,1
— отсутствие данных.
*3 Площади — по Географическому энциклопедическому словарю (1986), БСЭ (1970—
турным сведениям с корректировкой на ареал.
леса в ареале
8,0
59,2
102,6
45,2
64,7
40,2
157,1
36,0
228,5
271,0
22,7
269,2
116,5 '
75,8
41,7
—
450,0
71,4
5,0
20,8
1,5
15,6
(14,3)
(16,7)
(6,7)
—
—
—
46,5
1,6
22,7
(14,3)
(2,6)
3,0
'21,1
978) и литера-
248
ЧишЯшатт
Рис. 88. Соотношение площади ареала и численности европейской и сибирской косуль в Евразии и в
СССР во второй половине 80-х годов XX в.
1 — европейская косуля; 2 — сибирская косуля, 3 — доля косуль, обитающих в СССР. Европейская
косуля: 4 — СССР, 5 — страны Передней Азии, 6 — Западная Европа, 7 — РСФСР, 8 — Украина, 9 —
Белоруссия, 10 — Молдавия, // — Литва, Латвия, Эстония, 12 — республики Закавказья. Сибирская
косуля: 13 — страны Центральной и Восточной Азии (Монголия, Китай, Корея), 14 — Казахстан, 15 —
Средняя Азия
249
Таблица 35. Плотность населения европейской и сибирской косуль в регионах СССР во второй
половине 80-х годов XX в.
Республика
Численность,
тыс. особей*'
Площадь, тыс. км
ареала лесов в ареале*2
Число особей на 1000 га
ареала леса в ареале
Европейская косуля
РСФСР
Украина
Белоруссия
Молдавия
Литва
Латвия
Эстония
Грузия
Армения
Азербайджан
Итого
РСФСР
Казахстан
Узбекистан
Киргизия
Итого
В целом по СССР
63,6
144,2
22,8
9,8
40,0
44,3
38,4
11,8
4,1
3,1
382,1
291,2
14,8
0,5
7,3
313,8
695,9
816
600
207
34
65
64
45
44
13
28.
1916
Сибирская
3993
576
8
83
4660
6493*'
228
86
72
3
16
22
16
22
3
10
478
косуля
1950
47
8
7
2012
2470
0,8
2,4
1,1
2,9
6,1
7,0
8,5
2,7
3,2
1,1
2,0
0,7
0,3
0,6
0,9
0,7
1,1
2.8
16,8
3,2
32,7
25,0
20,0
24,0
5,4
13,7
3,1
8,0
1,5
3,1
0,6
10,4
1,6
2,8
*' По данным Управлений охотничьим хозяйством, Госкомитетов по охране природы республик и
Госагропрома СССР с корректировкой.
*2По Атласу СССР (1984) и Лесной энциклопедии, т. 1—2 (1985—1986) с корректировкой на площадь
ареала.
*'С учетом площади наложения ареалов европейской и сибирской косуль.
1983). В самые последние годы катастрофическое падение численности
приостановилось благодаря природоохранным мерам и запрету промысла, однако многие
популяции находятся на грани уничтожения.
В настоящее время население косуль распределено в Евразии следующим
образом. По нашим расчетам площадь ареала Capreolus составляет 13,4 млн км2, из них
европейская косуля занимает 6 млн, сибирская — 7,4 млн км2. Общая численность
животных оценивается в 8,2 млн. особей, но поголовье первой — 7,5 млн,
второй — около 700 тыс. (табл. 34).
В Западной Европе сосредоточено примерно 85% всего населения и 93% —
европейской косули (7 млн), причем половиной запасов последней (3,5 млн)
владеют три страны: ФРГ, Австрия и Англия (рис. 88).
Практически половина ареала косуль приходится на территорию бывшего
СССР (6,6 млн км2) и всего лишь 8,5% населения (700 тыс. особей). Ареал
европейской косули здесь составляет треть видового (1,9 млн км2), тогда как численность
250
Рис. 89. Динамика численности и добычи косуль тыс. особей
в СССР т
1 — общая численность; 2 — численность
европейской, 3 — сибирской косуль; 4 — добыто у п.,
всего
600
500
400
300
200
100 V
(около 400 тыс.) — 5%; ареал
сибирской косули (63%), но поголовье (около
300 тыс.) не достигает и.половины
видовой (табл. 35, рис. 88).
В России (табл. 35), где площадь,
занятая животными (4,7 млн. км2),
составляет 35% от площади всего
ареала косуль, насчитывается лишь 350 тыс.
особей, или 4,3% от всего поголовья.
Здесь сосредоточена половина
общесоюзного запаса этих копытных, в том
числе 93% (290 тыс.) поголовья сибирской и 17% (60 тыс.) европейской косуль.
Две трети ресурсов последней находятся на Украине (140 тыс.) и в Прибалтике
(120 тыс.).
Динамика численности косуль в б. СССР и по регионам страны показана на
рис. 89 и 90. Отчетливо видно, что в последние десятилетия поголовье животных
1971
1976
1981
1906
тыс. особей
400
1961
Рис. 90. Динамика численности и добычи косуль по регионам б. СССР
А — в РСФСР: / — общая численность; 2 — численность сибирской, 3 — европейской косуль,
4 — добыто всего; Б — поголовье на Украине (2), в Белоруссии (3), в Молдавии (4): 1 — общая
численность, 5 — добыто на Украине; В — в Прибалтике: / — общая численность, 2 — в Литве, 3 — в Латвии,
4 — в Эстонии, 5 — общая добыча; Г — в республиках Средней Азии: / — общая численность,
2 — в Казахстане, 3 — в Киргизии, 4 — в Узбекистане; Д — в республиках Закавказья: / — общая
численность, 2 — в Грузии, 3 — в Азербайджане, 4 — в Армении
251
180
160
140
120
100
во
60
40
20
-
-
-
-
-
-
-X/
' Г
1960-1965
1971
1976
1981
1986
210
180
150 "
120 -
SO
60 -
30
1936
тыс. особей
32
28
24
20
16
12
8
4
-ГуТ'ГТТ'ГГГТТ"] 1"1 I I 1"1 I 1 I ['"ГТТ I 1 I 1 I 1 р"ГТ I I I 1 1 I | I
1946 1956 1966 1976 1986
тыс. особей
32
2В
24
20
16
12
8
4
<w>-^-<f—o—o-^>—»-*j—■?—Р-Р-?-^
г* -•--•-
252
1976 1981 /386
Рис. 90 (окончание)
1976
1981
1986
r!
^*$3к
^^яС,-"^^
— * /A
• •■■.■;»
+ + з
\Ж*
1 III s
'ЯР
:::|^^±^-Угу
W^^^r
Рис. 91. Плотность населения косуль (особей) на 1000 га площади ареала
/ — менее 1; 2 — 1—5; 3 — 5—10; 4 — 10—20; 5 — 20—50; 6 — более 50
м
01
Рис.92.Плотность населения косуль (особей) на 1000 га леса в ареале
/ — менее 1; 2 — 1—5; 3 — 5—10; 4 — 10—20; 5 — 20—50; б — 50—100; 7 — более 100
Таблица 36. Плотность населения европейской и сибирской косуль в областях РСФСР во второй
половине 80-х годов XX в.
Район, область*1
1
Численность*2,
тыс. особей
2
Площадь*3, тыс. км2
ареала
3
лесов в ареале
4
Число особей на 1000 га
ареала
5
леса в ареале
6
+Калининградская обл.
Северо-Западный район
+Ленинградская обл.
+ Псковская обл.
Центральный район
+Брянская обл.
+Калининская обл.
+Калужсхая обл.
+Московская обл.
+Орловская обл.
+Рязанская обл.
+Смоленская обл.
+Тульская обл.
Центрально-Черноземный
район
+Белгородская обл.
+Воронежская обл.
+Курсгая обл.
+Липецкая обл.
+Тамбовская обл.
Поволжский район
++Волгоградская обл.
Куйбышевская обл.
Пензенская обл.
Саратовская обл.
Ульяновская обл.
Башкирия
Татария
Северо-Кавказский район
+Краснодарский край
++Ставропольский край
+Ростовская обл.
+Дагестан
+Кабардино-Балкария
+Северная Осетия
+Чечено-Ингушетия
Уральский район
Курганская обл.
Оренбургская обл.
Свердловская обл.
Челябинская обл.
10,0
4,1
0,7
3,4
19,7
4,7 ■
0,6
4,3
3,1
1,8
0,7
2,5
2,0
9,8
4,3
2,1
1,7
1,3
0,4
20,2
3,4
4,3
0,3
2,7
0,6
7,8
1,1
18,8
5,4
2,3
2,0
2,6
2,0
2,0
2,5
32,4
13,8
3,9
5,3
9,4
15,1
96,2
40,8
55,4
285,9
34,9
54,8
29,1
35,7
24,7
31,2
49,8
25,7
167,7
27,1
52.2
30,0
24,1
34,3
461,9
69,1
53,4
25,3
88,9
37,2
143,0
45,0
181,9
38,0
14,0
76,9
22,5
7,0
5,6
17,9
319,5
71,0
95,9
64,7
87,9
2,8
49,7
24,1
25,6
107,1
12,1
27,9
14,6
15,6
2,3
10,3
20,4
3,9
17,9
2,3
.6.0
2,9
2,4
4,3
109,5
6,3
7,7
10,1
6,5
10,7
59,8
8,4
44,4
19,8
6,6
5,1
4,9
2,0
2,0
4,0
94,0
16,0
7,0
45,0
26,0
6,6
0,4
0,2
0,6
0,7
1,3
0,1
1,5
0,9
0,7
0,2
0,5
0,8
0,6
1,6
0,4
0,6 "
0,5
0,1
0,4
0,5
0,8
0,1
0,3
0,2
0,5
0,2
1,0
1,4
1,6
0,3
1,2
2,8
3,6
1,4
1,0
1,9
0,4
0,8
1,1
35,7
0,8
0,3
1,3
1,8
3,9
0,2
2,9
2,0
7,9
0,7
1,2
5.1
5,5
18.7
3,У)
5,9
5,4
0,9
1,8
5,4
5,6
0,3
4,2
0,6
1,3
1.3
4,2
2,7
3,5
3,9
5,4
10,2
9,8
6,3
3,4
8,6
5,6
1,2
3,6
Таблиц* 36 (окончание)
1
2
3
4
5
6
Западно-Сибирский район
Алтайский край
Кемеровская обл.
Новосибирская обл.
Омская обл.
Томская обл.
Тюменская обл.
Восточно-Сибирский район
Красноярский край
Иркутская обл.
Читинская обл.
Бурятия
Тува
Дальневосточный район
Приморский край
Хабаровский край
Амурская обл.
Якутия
В целом по РСФСР
В том числе:
европейская косуля
сибирская косуля
22,0
11,4
2,3
3,8
1,5
1,2
1,8
126,4
10,5
40,0
38,4
28,0
9,5
91,4
14,5
22,6
49,0
5,3
354,8
63,6
291,2
592,1
170,2
49,2
168,8
115,3
39,4
49,2
1251,1
282,7
230,6
399,2
212,1
126,6
1354,2
140,6
175,8
331,9
705,9
4725,6
815,9
3992,8
218,8
54,2
30,9
44,7
45,0
24,4
19,6
831,9
171,2
191,3
260,0
142,1
67,3
681,9
110,0
130,9
230,0
418,0
2158,0
228,2
1950,1
0,4
0,7
0,5
0,2
'0,1
0,3
0,4
1,0
0,4
1,7
1,0
1,3
0,8
0,7
1,0
1,3
1,5
0,1
0,8
0,8
0,7
1,0
2,1
0,7
0,8
0,3
0,5
0,9
1,5
0,6
2,1
1,5
2,0
1,4
1,3
1,3
1,7
2,0
0,1
1,6
2,8
1,5
*' + — европейская косуля; ++ — европейская и сибирская вместе, без знака — сибирская косуля.
*2 По данным Главохоты РСФСР с уточнениями.
*3 Земельный фонд РСФСР (1985); с корректировкой на ареал.
в целом стабилизировалось на низком уровне, а в республиках Закавказья и
Прибалтийском регионе уменьшилось.
В настоящее время на каждые 1000 га площади ареала в среднем приходится
12 европейских косуль и 1 сибирская. Наивысшие показатели плотности населения:
в Австрии — 119, ФРГ — 68 и Великобритании — 52, наименьшие: в СССР,
Финляндии, Испании и Монголии — по 1—2. Вероятно, в странах Малой
Азии и Китае они примерно такого же низкого порядка (см. табл. 34 и рис. 91).
Показатели плотностей, рассчитанные на площадь ареала, хотя и наглядно
представляют распределение населения косуль в географическом аспекте, в
целом ниже реальных, поскольку животные распространены лишь на пригодной
для их жизни площади. К сожалению, сравнение по этому показателю пока
невозможно из-за отсутствия данных по большинству регионов. При низкой
численности зверей наиболее близки к истинным будут значения их плотности на
1000 га лесных угодий, при высокой — промежуточные между ними и
рассчитанными на общую площадь, так как животные занимают и полевые угодья.
В среднем на каждые 1000 га лесных угодий в ареале обитает 47 европейских
косуль и около трех сибирских. Наивысшие плотности их населения в Западной Ев-
256
Таблица 37. Плотность населения косуль в б. СССР на территориях с относительно слабым прессом
промысла или хищников
Регион (заповедник)
Число особей на 1000 га
угодий
Авторы
Калининградская обл.
Литва
Латвия
Белоруссия
(Беловежская Пуща)
Курская обл.
(Центрально-Черноземный)
Белгородская обл.
(Лес на Ворскле)
Южный Урал
(охраняемые угодья)
(Ильменский)
Казахстан
(Алма-Атинский)
Амурская обл.
(Хинганский)
28
32
40
23—46
31—66
50
33—70
18—33
25-36
36
Романов, Ромашин,
Блузма, 1975
Гарозс, 1987
Шостак, 1978
Гусев, 1984
Соколов и др., 1986
Соколов, Данилкин,
Дворников, 1984
Жиряков, 1980
Дарман, 1986
1982
1981
ропе (в среднем 72 особи): в Венгрии и Швейцарии — более 100, в ФРГ,
Австрии, Дании и Великобритании свыше 200 особей. Наиболее низкие
показатели в СССР, Финляндии, Испании и, вероятно, в странах Малой Азии
(табл. 34, рис. 92).
Плотность населения косуль в пределах ареала в значительной степени
является показателем успешности ведения охотничьего хозяйства и состояния
популяций. Очевидно, что поголовье зверей в Австрии, ФРГ и Великобритании
чрезмерно и здесь может происходить биологическая деградация популяций.
С другой стороны, на большей части территории России плотность почти
на порядок ниже возможной (табл. 36).
В то же время на большинстве территорий со слабым прессом браконьерского
промысла и там, где численность хищников сравнительно невелика, плотность
населения косуль превышает 20 особей на 1000 га угодий практически в любом
регионе (табл. 37).
Эти цифры наглядно отражают потенциальные возможности охотничьего
хозяйства. Расчеты показывают, что при повышении плотности населения косуль
хотя бы до 10 особей на 1000 га площади ареала, что в 2 раза ниже ныне
существующей в Западной Европе, их поголовье лишь в России может достигнуть 3 млн особей.
Скорее всего, примерно такая численность этих копытных была в России в XIX в.
17. Европейская и сибирская косули
257
Факторы, определяющие численность
населения популяций
Численность населения,популяций определяется, с одной стороны, экологической
емкостью среды обитания, с другой — факторами, влияющими на два
противонаправленных процесса — рождаемость и смертность. Если они сбалансированы,
поголовье (при неизменных условиях существования) может некоторое время
оставаться на относительно стабильном уровне. Однако на практике эта ситуация
встречается редко. Под воздействием промысла, хищников, погоды и других
факторов внешней среды происходит изменение численности.
Косули относятся к видам, движение численности которых не имеет частых
скачкообразных пиков, характерных для многих мелких млекопитающих.
Поголовье разреженной популяции косуль растет весьма медленно, но с каждым
годом абсолютный прирост увеличивается, и затем наступает период бурного
роста. Фаза пика численности непродолжительна. Истощаются кормовые ресурсы,
перенаселенная популяция находится на пределе экологических возможностей
и очень чувствительна к воздействию неблагоприятных факторов среды.
Первая же суровая многоснежная зима становится причиной массовой гибели
животных, что приводит к резкому снижению поголовья и сопровождается
иногда периодом депрессии. При новом подъеме численности прежний высокий
уровень плотности населения обычно уже недостижим из-за худших условий
питания, вызванных недавней перенаселенностью угодий.
Иллюстрацией может служить динамика численности европейской косули
в послевоенный период в Литве (рис. 93). Интенсивное уничтожение волка в
60—70-е годы и эффективные природоохранительные мероприятия
способствовали быстрому увеличению поголовья, но эксплуатация ресурсов началась с
опозданием и была недостаточно интенсивной, что привело к перенаселенности угодий
в некоторых районах республики, а позднее к массовому падежу животных
после суровых зим в 19.69/70 и 1975/76 гг.
В биотопах с хорошими кормовыми и защитными условиями численность
растет интенсивнее, чем в низкокачественных местообитаниях. К примеру,
на юго-востоке Литвы, где преобладают малокормные чистые сосновые леса,
средний годичный прирост популяции был бы меньше (13,8%), чем в
районах богатых смешанными и лиственными лесами (Блузма, 1975). В результате
на юго-востоке пик численности наступил на 3—7 лет позже (рис. 94).
Один из важнейших критериев динамики численности — скорость роста
населения популяций, представляющая баланс между уровнем воспроизводства
и смертности животных. Воспроизводство популяций косуль, как показано выше,
при нормальных условиях жизни и при умеренной плотности населения
относительно стабильно. Темп размножения падает лишь при чрезмерной плотности,
вероятно, превышающей 20—30 особей на 100 га. Следовательно, в большинстве
популяций скорость роста, как и динамика в целом, зависят главным образом от
факторов, вызывающих смертность (табл. 38), — чем ниже смертность, тем
быстрее увеличивается поголовье и тем выше становится уровень плотности
населения.
Факторы, влияющие на динамику численности косуль (естественные и
антропогенные, внешние и внутрипопуляционные, биотические и абиотические), всегда
взаимосвязаны и действуют комплексно. Тем не менее в конкретных ситуациях
258
Косуля
30 h тыс. особе и
Волк
тыс. особей -
1935 1945 1955 19Б5 1975 1985
Рис. 93. Динамика численности европейской косули (/) и волка (2) в Литве (1934—1987 гг.)
Столбиками показан уровень добычи косули
1955 I960 1365 1970
Рис. 94. Динамика численности европейской косули в отдельных районах Литвы в 1952—1974 гг.
Районы: / — восточные; 2 — северные; 3 — западные; 4 — южные; 5 — юго-восточные
методически оправдано выделение из них одного или нескольких, играющих
ключевую роль.
Биотопы. Емкость угодий ограничена и зависит от кормовых, защитных
и других свойств биотопа, удовлетворяющих жизненные потребности животных.
Изменение биотопов под воздействием естественных причин (пожаров) или
антропогенного преобразования (вырубки лесов, лесопосадок, распашки целинных
259
Таблица 38. Основные факторы смертности косуль в разных регионах
Регион
*
Антропогенные факторы
Всего
В том числе
браконьеры
транспорт
с/х машины
Швейцария (Lemann, Sagesser, 1986), «=12950
Польша (Pielowski, 1984), л = 81
Калининградская обл. (Романов, Ромашин, 1982),
« = 78
Белгородская обл. (наши данные), л =42
Украина (Карпенко, 1966), л = 64
Карпаты (Владышевский, 1968), л=99
Северо-Запад СССР (Верещагин, Русаков, 1979),
л=82
Ильменский заповедник (Филонов, 1974), л=1868;
(Дворников, 1982, 1984), л>122
Урал (наши данные), л= 164
Алма-Атинский заповедник (Жиряков, 1979), л=36
Забайкалье (Смирнов, 1978), л=46
Хинганский заповедник (Дарман, 1982а,б), л=69
82,5
35,8
57,5
52,3
12,5
2,4
32,6
11,3
34,1
52,2
24,6
6,2
37,0
45,2
12,5
32,6
7,8
29,3
24,6
13,5
7,4
6,8
1,2
3,5
3,0
5,5
1,2
12,3
7,1
1,8
степей и др.) заметно сказывается на динамике численности косуль, поскольку
существенно- меняется емкость угодий. Появление зарастающих гарей и лесосек,
посадка лесных культур повышают качество местообитаний, способствуя росту
поголовья. Напротив, распашка степей и мелиорация чаще вызывают обратный
процесс, особенно в глубокоснежных районах, где сельскохозяйственные поля
практически не пригодны для обитания. Интенсивная лесохозяйственная и
сельскохозяйственная (в ряде регионов) деятельность человека и "окультуривание"
лесных ландшафтов в целом благоприятно для косуль, что видно по значительным
цифрам их численности в наиболее урбанизированных биотопах в Западной и
Центральной Европе (табл. 34) и лишь в отдельных частях ареала сказывается
отрицательно на популяции (см. главы "Среда обитания" и "Хозяйственное
значение...").
Корма. Благодаря выработавшимся в процессе эволюции адаптациям к
зимним неблагоприятным условиям (резкое снижение обмена веществ и уменьшение
потребности в питании, миграции из глубокоснежных районов) в целом гибель
косуль в пределах естественного ареала от недостатка кормов проявляется лишь в
самые многоснежные и суровые зимы и именно в это время оказывает прямое
и довольно существенное влияние на динамику численности. В обычные годы
дефицит зимных кормов заметен лишь при высокой плотности населения косуль.
В Литве, например, при плотности 15—35 особей на 100 га наблюдается
полная деградация зимних пастбищ в лиственных лесах с примесью ели.
260
Причина гибели, %
Хищники
Всего
В том числе
волк
рысь
собаки
лисицы
Истощение t
болезни
Другие
причины
Неизвестна
0,9
9,9
19,6
26,1
42,1
72,7
45,1
57,8
80,7
58,5
55,6
45,6
53,6
—
—
—
4,7
10,9
36,4
23,2
50,4
—
48,2
30,5
8,7
33,3
—
—
—
—
—
19,2
14,6
7,0
78,9
8,5
11,1
36,9
15,9
0,9
9,9
6,8
16,7
10,9
—
4,9
0,4
1,8
1,8
—
—
2,9
—
—
4,7
20,3
7,1
2,4
—
—
—
—
—
6,0
19,7
4,1
—
18,7
—
31,7
3,6
—
—
5,6
2,2
2,1
3,7
1,4
7,1
—
—
4,9
1,7
•1,8
—
8,2
—
2,9
8,5
30,9
14,2
26,7
—
15,9
3,3
4,4
7,3
27,8
—
18,8
В смешанных елово-лиственных лесах и сосновых с примесью лиственных пород
переиспользование пастбищ происходит при плотности 6—8 голов на 100 га леса.
Несоответствие между кормовой базой угодий и количеством зверей кончается
массовым падежом (Падайга, 1965).
Не меньшую роль играет и качественный аспект питания. Из работ американских
экологов ( Smith et al., 1956; Dasmann, 1966; Ullrey et al., 1967) известно, что суточное
потребление протеинов дикими копытными должно составлять не менее
13%. Критический уровень — 7—8%, после чего от качественного, голодания может
погибнуть до 40% поголовья. Содержание протеина в основных зимних кормах
косуль находится ниже необходимой нормы, а перевариваемость их хуже, чем летом,
в 1,75 раза (Drozdz, 1979). Поэтому зимние кормовые ресурсы животных
определяются в основном не количеством, а питательностью (табл. 39) и
возможностью их переваривания (табл. 40).
По мнению Айсфельда (Eisfeld, 1974a, 1976), снабжение белком все же
не самое "узкое место" в питании косуль. Решающий фактор — содержание
усваиваемой в единицу времени энергии, т.е. энергетическая ценность корма.
При низкой энергетической ценности пищи или возрастающей потребности в
энергии достигается такое состояние, при котором из-за максимального заполнения
рубца потребление корма не может быть увеличено и возникает голод при
переполненном желудке. Эта ситуация усугубляется с ростом плотности
населения, поскольку из-за выборочного поедания ценных в кормовом отношении
261
растений или их частей уменьшается их количество и соответственно снижается
энергетическая ценность пищи. В конечном счете некачественное питание приводит к
гибели части популяции.
Обычно падеж животных наблюдается в самом конце зимы и начале весны.
Вероятно, он происходит от того, что как раз в это время из-за удлинившегося
светового периода через активацию желез внутренней секреции
интенсифицируется обмен веществ, однако повышенная потребность в энергии не может быть
удовлетворена из-за недостатка качественной пищи (Ellenberg, 1978).
При высокой плотности популяции животные могут испытывать энергетическое
голодание не только зимой, но и летом (Bobek, 1977). От качества, разнообразия
и обилия пищи в летний период зависят интенсивность размножения, рост и
развитие молодняка, физиологическое состояние самок. При недостатке
качественного корма появляются признаки аномального состояния популяции —
снижается плодовитость, сеголетки отстают в росте и развитии, создаются предпосылки
для повышенной зимней смертности.
Елленберг (Ellenberg, 1978) экспериментально установил, что при плохом
питании матерей в конце беременности и в первые дни после рождения
наблюдается значительная (до 75%) гибель новорожденных и мелких детенышей
преимущественно женского пола. Недостаточная пища перед гоном приводит к низкому
проценту овуляции у взрослых самок или полному ее отсутствию у молодых.
Плохо упитанная к началу гона мать рожает преимущественно детенышей
мужского пола, причем пределы сдвигов полового соотношения достигают 3:1.
Отсюда при' бедных пищевых ресурсах происходит повышенная смертность
особей, торможение скорости воспроизводства и оборота популяции и массовая
эмиграция молодых самцов. Напротив, при благоприятной пищевой ситуации
достигается высокая продуктивность популяции благодаря высокому уровню
плодовитости, рождению преимущественно (3:1) детенышей женского пола,
низкой смертности молодых особей.
В естественных условиях факты снижения воспроизводственного потенциала
популяции проявляются, если косули летом сильно повреждают 30—40% всех
побегов (Приедитис, 1985). А. Приедитис считает, что дефицит летних кормов
возместить нельзя, в то время как дефицит зимних кормов сравнительно
легко компенсируется правильной зимней подкормкой.
Этологическяя регуляция. Авторегуляторные процессы в популяции направлены
на приведение численности животных в соответствие с емкостью среды обитания,
как бы предохраняя ее от деградации, возможной при чрезмерном поголовье.
В основе их, помимо изменения уровня воспроизводства, лежат пространственно-
этологические отношения зверей, и прежде всего территориализм взрослых
самцов.
Главная функция территориальности у косуль и ее основное следствие —
рассредоточение особей в пространстве и регуляция численности популяции
непосредственно перед появлением нового поколения, для выращивания которого
нужен большой запас качественных кормов (Strandgaard, 1972). Изгнав молодых
особей, взрослый самец создает на своем участке для обитающих здесь беременных
и лактирующих самок зону слабой пищевой конкуренции, повышая тем самым
шансы на выживание нового, обычно своего же потомства (Ellenberg, 1978).
Территориальное поведение играет в данном случае роль важного гомеостатиче-
262
Таблица 39. Питательная ценность (содержание протеина, %) некоторых основных кормов косуль
(из Ларин и др., 1951; Юргенсон, 1973)
Растения
Лето
Зима
Сосна обыкновенная
, побеги
зеленая хвоя
Ель, зеленая хвоя
Пихта, хвоя
Береза, листья
побеги
Осина, листья
побеги
кора
Ива козья, листья
побеги
Дуб, листья
побеги
Иван-чай (кипрей)
Клевера
Люцерна
Вейник (разные виды)
13,0-18,8
8,9-10,5
12,7-18,5
17,8
17,4—18,9
12,4-15,5
14,0-17,5
14,9—17.7
16,5-24,7
18,8-27,3
7,9-8,9
6—9
4,6
4,6
6,7
6,6
4,75—7,4
4,3
4,0-8,1
3,1-6,3
6,2-8,5
5,3-6,6
14,4-15,2
(сено)
15,2-22,4
(сено)
6 (сено)
Таблица 40. Перевариваемость (%) любой комбинации используемых для корма растении,
предварительно отобранных человеком (из: Ellenberg, 1978)
Месяц
Максимум
Минимум
V
87
67
VI
72
65
VII
63
54
VIII
—
IX
60
59
X.
60
52
XI
45
41
XII
42
I
48
35
II
43
III
52
31
IV
44
37
ского механизма, поддерживающего уровень населения популяции в состоянии
устойчивого равновесия с утилизируемой ею средой по принципу лишения
части членов популяции тех преимуществ, которые дает обладание территорией
(Панов, 1983).
Выше показано, что в малочисленных популяциях косули распределены по
площади неравномерно, концентрируясь в наиболее благоприятных стациях,
где самцы конкурируют за территорию, а самки — за родовые участки.
Фактически этологический механизм регуляции численности населения "включается"
с момента занятия самцами территории, т.е. задолго до предельного насыщения
животными угодий и истощения ресурсов среды обитания. Плотность населения
косуль в лучших биотопах раньше, чем в худших, достигает определенной
величины, при которой стабилизируется. Этот порог, вероятно, определяется
"закодированными" для вида максимальными размерами территорий самцов
263
'''' I I I 1 L_
1973 1381 1983 1985 1987
Рис.95.Динамика численности сибирской косули в Хинганском заповеднике и на сопредельных
территориях (Амурская обл.) при действии разных факторов (по: Дарман, 1986; с дополнениями)
1 — при влиянии охотничьего промысла и хищников (Архаринский госпромхоз); 2 — только
хищников (Хинганский заповедник); i — при отсутствии охоты и хищников (филиал Хинганского
заповедника)
и участков обитания самок, коррелирующих с их энергетическими потребностями
при имеющейся массе тела.
Исследования пространственно-этологической структуры популяций
европейской и сибирской косуль (Соколов, Данилкин, 1981; Данилкин, Минаев, 1988)
показали, что в регионах с общей низкой численностью (3—5 особей на 1000 га
пригодных угодий) в летний период "пороговая" величина плотности населения
животных в лучших угодьях равняется 40—50 особям на 1000 га при размерах
территорий самцов, составляющих около 1(00 га. Этот уровень "социальной" плотности
благодаря эмиграции молодых особей сохраняется относительно неизменным
долгое время, видимо до момента полного заполнения соседних угодий. Затем
из-за всевозрастающей конкуренции в лучших стациях происходит уменьшение
размеров территорий и участков обитания, приводящее к достижению популяцией
нового более высокого уровня плотности населения.
Именно из-за явления "пороговых" величин, служащего адаптацией к
ресурсосбережению, рост неэксплуатируемой популяции в хороших по качеству стациях
вдруг прекращается (явно не достигнув пика), стабилизируется на уровне
гораздо меньшем, чем позволяет емкость угодий, или даже несколько снижается
(см. рис. 95). Незнание этой особенности динамики численности косуль нередко
приводит к неверным выводам при расчете целевой плотности населения в
охотничьих угодьях.
Авторегуляция осуществляется и при очень высокой плотности населения
(около 25 особей на 100 га), если существует возможность эмиграции молодых
особей на сопредельные участки, где, в частности, ведется интенсивный промысел
(Strandgaard, 1972).
Таким образом, у косуль этологический механизм регуляции (путем
рассредоточения особей) "работает" в основном на уровне внутрипопуляционных
группировок. Как ни парадоксально, он эффективен лишь при сравнительно невысокой
плотности населения популяции и в целом направлен не на ограничение роста
численности, а, напротив, способствует увеличению поголовья, так как
благоприятствует расселению особей и созданию лучших условий для выращивания
8
л
^
i$
м
*
'>
1
«d
50
40
30
20
10
264
нового потомства. При общей предельно высокой плотности этологическая
регуляция теряет значимость, поскольку на место эмигрировавших особей с соседних
участков приходят другие и заметного снижения численности популяций
не происходит. Аналогичная картина наблюдается в вольерах, где молодые особи
не имеют возможности покинуть загороженный участок.
На увеличение общей численности популяция реагирует постепенным
уменьшением участков обитания и территорий особей. Это, в свою очередь, приводит
к ухудшению условий питания и соответственно к снижению ее качества:
размеры тела и масса зверей через несколько поколений уменьшаются, снижается
плодовитость и становится более высоким процент яловости истощенных самок.
Они рожают чаще детенышей мужского пола, увеличивается смертность телят,
а оставшиеся в живых позднее включаются в размножение, т.е. происходит
торможение скорости воспроизводства и оборота популяции, которая "стареет"
и становится особенно чувствительной к неблагоприятным воздействиям.
Регуляция численности в перенаселенной популяции, таким образом,
осуществляется в большей степени не этологическими, а физиологическими
процессами, направленными на ограничение темда ее роста. Тем не менее при отсутствии
других лимитирующих факторов (в первую очередь хищников) авторегуляционные
процессы "срабатывают" со значительным опозданием и в целом оказываются
недостаточно эффективными для поддержания численности на уровне,
соответствующем емкости угодий. К тому же в переуплотненной популяции плодовитость
самок все-таки остается высокой (см. главу "Размножение"), что ведет к
дальнейшему ее росту, дальнейшему истощению пищевых ресурсов и при
неизменных условиях к гибели многих особей.
Этот вывод о неспособности популяций косуль самим решать проблему
перенаселения, на наш взгляд, весьма важен в связи с природоохранительными
тенденциями, поскольку служит основанием для хозяйственного способа
регулирования численности там, где крупные хищники по многим причинам нежелательны и
отсутствуют.
Сезонные миграции. Сезонные миграции популяций или отдельных особей
возникают в ответ на неблагоприятные изменения среды обитания и, казалось бы,
не связаны с регуляцией численности. Однако на путях миграций часто происходит
массовая гибель животных при переправах через реки, резко усиливается пресс
хищников, в экстремальных условиях погибают наиболее слабые особи (молодые и
старые звери и беременные самки), что существенно сказывается к тому же
на половом и возрастном составе популяции и ее воспроизводстве. Нередко
часть мигрантов остается жить в новых местах, так же способствуя снижению
численности исходной популяции (см. главу "Перемещения").
Фактически повышенная смертность у косуль во время сезонных миграций на
большей части ареала является одним из основных и достаточно
эффективных способов регулирования численности на популяционном уровне,
направленном в отличие от этологических внутрипопуляционных механизмов на ее
снижение.
Погода. Климатические факторы не только определяют область
распространения косуль, но некоторые из них, в частности погода, очень существенно влияют
на динамику численности, вызывая ее резкие колебания. Гибель косуль в
суровые зимы равна или превышает потери от промысла, а при взаимодействии
еще и с хищниками представляет катастрофу для популяций. Особенно показа-
265
тельна в этом отношении ситуация, сложившаяся на Дальнем Востоке в необычно
многоснежную зиму 1972/73 г., когда поголовье бедствующих животных,
преследуемых больше, чем обычно, браконьерами и хищниками, сократилось в
3—4 раза (Кучеренко, Швец, 1977). Суровая зима 1940/41 г. вызвала массовую гибель
зверей (преимущественно сеголеток и беременных самок) на Урале. Сильно
возросшее поголовье волков и браконьерство обусловили продолжительный период
депрессии популяций в регионе (Кириков, 1952; Ушков, 1954; Филонов, 1974).
Эта же многоснежная зима и усилившееся хищничество волка привели к почти
полному исчезновению косуль в Северном Казахстане. Такая ситуация повторилась
зимой 1945/46 г. и в 1956—1960 гг. по всему Казахстану. В горной части
юго-востока республики наибольший урон принесли многоснежные зимы 1965/66
и 1975/76 гг. (Слудский и др., 1984).
На севере ареала европейской косули основная естественная причина
смертности — истощение. Доля этого фактора достигает на северо-западе СССР 32%
(Верещагин, Русаков, 1979), в Швеции —' 58,8% (Borg, 1970); в суровую зиму
1976/77 г. смертность здесь была почти на порядок выше, чем в обычные зимы,
особенно среди сеголеток (Cederlund, Lindstr6m, 1983).
В Польше в зимние сезоны 1969/70, 1976/77 и 1978/79 гг. потери составили треть
от осеннего поголовья. Очень велики они были в полевых популяциях (до 51%),
среди молодых (до 77—90%) и старых особей, причем смертность здесь возрастала с
увеличением плотности популяции (Fruzinski, Labudzki, 1982; Kaluzinski, 1982).
Редкие морозные зимы вызывали резкое снижение поголовья косуль и в ГДР,
особенно в многочисленных популяциях. Возрастной состав погибших животных
значительно отличался от возрастной структуры популяции (рис. 96, А,Б); среди
павших также преобладали молодые и старые особи и самки (Stubbe, Passarge,
1979; Briedermann, Ahrens, 1980; Stubbe, 1987).
Массовый падеж косуль от истощения, воспаления желудочно-кишечного
тракта и легких, зарегистрирован многоснежной зимой 1979 г. в Калининградской
области (Романов, Ромашин, 1982). В Беловежской Пуще глубокоснежной зимой
1969/70 г. поголовье сократилось втрое (Смоктунович, 1980). В Брянской области
падеж наблюдался зимой 1964 и 1967 гг., когда высота снежного покрова
достигла 70 см и сопровождалась продолжительной низкой (до —30°)
температурой воздуха; первыми погибали старые особи и молодые. В поисках
корма звери выходили на дороги и скапливались у стогов сена, где часто
становились жертвами бродячих собак и волков (Ватолин, 1975).
В Литве в суровые зимы 1969—1971 гг. перенаселенная популяция сократилась
в среднем на 34,4%, только в 60-е годы погибло около 17 тыс. особей.
Среди павших животных половину составили молодые звери, 34% — старые и
16% — средневозрастные (Блузма, 1975). В Латвии падеж достигал 12—20% от всего
поголовья (Приедитис, 1975). В Эстонии (Рандвеэр, 1985) резкое снижение
численности, происходящее после тяжелой зимовки, дополнялось значительным прессом
промысла и хищников (см. рис. 97). В 1939/40 г. здесь пало около 35—40%, в
1981/82 г. — 15—20% поголовья, в основном сеголетки (56—71%).
В Прибалтийских'республиках суровые зимы, пожалуй, самый важный фактор
динамики численности. Наибольшее значение имеют продолжительные низкие
температуры, высокий снежный покров, а также критические явления: сроки
наступления зимы и суровость ее середины, время наступления весны и состояние
погоды непосредственно во время рождения телят. Суровой зимой основным
266
3 5 7 9 11 13 15 17
Плотность j особей 1100 га
Рис. 96. Возрастной состав (%) косуль, погибших в суровую зиму (£), и зависимость уровня падежа от
плотности населения популяций (В) (из: Stubbe, Passarge, 1979)
А — возрастная структура идеальной популяции
лимитирующим фактором служит общее число дней, со снежным покровом и
снегопадами; умеренной зимой — температурный режим; мягкой — число
дней со снегопадами и со снежным покровом в апреле. Весной лимитирующим
оказывается температурный режим во время отела, дождливая и прохладная
погода в первые недели жизни детенышей (Кийли, 1985).
В.И. Падайга (1971), напротив, считает основными факторами, определяющими
зимний падеж косуль, истощение кормовой базы и паразитарные болезни,
напрямую связанные с плотностью населения копытных. Высота снежного покрова
лишь обостряет эту зависимость, поскольку при увеличении плотности населения
смертность возрастает более резко, чем при соответствующем увеличении высоты
снежного покрова. К такому же выводу пришел и Е.А. Смоктунович (1980),
исследовавший динамику численности копытных в Беловежской Пуще.
267
тыс. особей
/;7r 6Sr
1.0
*Й7
!••
0.1
50
35 -\
I
20
\ А Л
\ А / \ ' /
■ \/ V
- \ /
'1 ^-^
\
\
1
1
\
\
\
^У/
/ ••■' Г
1 1 1
\ * А -
Ж
VNfr
\
:
-J I'.'r'.'j 1 1
WO
140 Ц
I
woX
1
1955
то
1965
1970
1975
1900 Гиды
Рис.97. Динамика численности европейской косули в Эстонии и факторы, ее определяющие (по: Ранд-
веэр, 1985; с изменениями)
/ — численность, 2 — добыча косули; 3 — численность хищников; 4 — индекс характера зимы —
0,5(^4 + В), где А — средняя температура трех самых холодных месяцев (в % от средней температуры
за истекший период), В — средняя высота снежного покрова в декаду с максимальным снежным
покровом (в % от средней высоты покрова за изученный период)
Воздействие тяжелых условий зимовки выражается не только в гибели
истощенных животных, но и в снижении уровня воспроизводства и большей, чем
обычно, смертности детенышей (Borg, 1970; Ellenberg, 1978).
Болезни и паразиты. Косули подвержены инфекционным и паразитарным
заболеваниям, большинство из которых являются общими для многих видов
диких и домашних копытных. Паразитофауна и болезни косуль изучены
относительно хорошо (Ne£as, 1975; Stubbe, Passarge, 1979), однако их роль в динамике
численности недостаточно ясна.
Массовые заболевания нередко возникают как следствие недоедания в
результате чрезмерной численности диких копытных и деградации их пастбищ
(Горегляд, 1970). Степень зараженности некоторыми Паразитами обычно прямо
пропорциональна плотности населения зверей (Падайга, Марма, 1970; Падайга,
1971). Интенсивность инвазии зависит также от характера леса, почвы и
влажности местообитаний (Пужаускас, 1970). Гельминты, однако, сравнительно
редко становятся непосредственной причиной гибели хозяев; чаще их воздействие
носит косвенный характер, вызывая ослабление организма хозяина и популяции в
целом и приводя к массовой гибели зверей, особенно в суровые зимы (Рыков-
ский, 1970; Приедитис, 1970).
Действительно, доля животных, погибших в результате паразитозов, особенно
велика в Центральной Европе (где плотность населения косуль максимальна) и
может достигать 40% от общего числа павших (Siefke, 1966). В Чехословакии из
268
1285 исследованных в 1972—1981 гг. трупов косуль 46,3% отнесены к потерям
от паразитарных заболеваний, наиболее губительными из которых были
легочные гельминтозы (около 70%). Максимальная смертность отмечалась в осенне-
зимние месяцы (Sterba, Zamek, 1985). '
В Швеции болезни и паразиты были причиной гибели косуль в 17,7% случаев
(Borg, 1970). В Великобритании урон популяциям возникает в результате
легочного нематодоза (иногда погибает до 16% сеголеток) и трематодоза
печени (McDiarmid, 1974). В Болгарии зарегистрирована гибель от заразного
энцефаломиелита (Габрашанский, 1970). Бешенство также было причиной смерти
зверей в ряде районов Европы (Stubbe, Passarge, 1979).
В Крымском заповеднике их гибель вызывалась сибирской язвой (2,2%),
паразитами крови (6,6%), внутренними болезнями (5,4%), легочно-глистной
инвазией (1,6%), кишечными гельминтами (0,6%) (Рухлядев, 1941). В одном
из охотничьих хозяйств Волынской области зимой 1961/62 г. предположительно от
эпизоотии листериоза погибло около 25% популяции (Адамович, Ойцось,
1963). Систематический падеж косуль от клещевого токсикоза отмечен в
Ставропольском крае (Никольский, Позов, 1973).
С другой стороны, многие исследователи не отмечают заметного воздействия
болезней и паразитов на численность косуль на большей части ареала, где
плотность их населения сравнительно низка (см. табл. 38).
Конкуренты. Различия в составе рациона позволяют косулям и другим
копытным существовать в одном и том же биотопе и избегать при этом
жесткой пищевой конкуренции (Hearney, Jennings, 1983; Schr6der, Schr6der, 1985).
Однако в отдельные периоды и в определенных биотопах трофические ниши
зверей перекрываются и некоторые близкородственные животные (олени, лани и
лоси), а также живущие в сходных биотопах серны и зайцы могут являться
серьезными конкурентами за пищу (Владышевский, Ельский, 1974; Kalizinski,
Bresinski, 1976; Long, 1980; Смоктунович, 1983; Picard, 1985).
Косули остро нуждаются в наиболее питательном и легко перевариваемом
корме, тогда как другие жвачные обходятся более грубым кормом. Именно по
причине конкуренции из-за качественных кормов косули оказываются в менее
выгодном положении, чему способствуют еще и сравнительно небольшие размеры
тела и, следовательно, меньшая "кормовая зона" (Ellenberg, 1978). Конкуренция
возрастает по мере увеличения высоты снежного покрова, поскольку в этот период
в диете копытных основными становятся веточные корма (Дулькейт, 1964;
Roucher et al., 1985; Гордиюк, 1986). Благородный и пятнистый олени, а также лань
явно вытесняют косуль из лучших угодий и мешают использовать зимнюю
подкормку, отгоняя от кормушек. В некоторых районах определенное давление на
них оказывает лось (Киселев, 1976).
Численность разных видов диких копытных может увеличиваться до тех пор,
пока не наступает истощение кормовой базы. Рост популяции косуль начинает
замедляться при плотности населения благородных оленей около 20 особей на
1000 га, при 30 — практически прекращается, а при дальнейшем увеличении
поголовья оленей сопровождается резким снижением численности (Владышевский,
1968; Скриба, 1975; Шостак, 1978; Смоктунович, 1980, 1983).
Один из ярких примеров — изменение численности сибирской косули и марала
в заповеднике Богдо-Ула в Монголии. По сообщениям П.К. Козлова (1924), в
начале 30-х годов XX в. поголовье косуль в лесах заповедника было столь
269
велико, что не поддавалось никакому учету. До 50-х годов они были обычным
видом при сравнительно низкой численности марала, но в середине 70—80-х годов,
когда плотность населения оленей достигла 55—90 особей на 1000 га, косули
совсем исчезли с территории заповедника (Данилкин, Дуламцерен, 1981).
Пожалуй, еще более серьезную конкуренцию косулям составляет домашний скот,
который уничтожает их пищу во многих районах практически полностью
(Лавов, 1978; Николаев, 1982; Pereira, 1985). Отрадно, что переход
животноводства в Западной Европе на безвыпасное (стойловое) содержание скота
привел к резкому сокращению конкурентных отношений и уменьшению
беспокойства, увеличилась площадь пригодных стаций и все это способствовало
росту поголовья косуль.
Говоря о конкуренции за пищу, нельзя не отметить и положительную роль
других копытных. В зимнее время кабаны и домашние копытные (в азиатской части
ареала), раскапывая снег, предоставляют косулям место кормежки, и они охотно
пользуются этими покопками. Во многих многоснежных районах России
благополучие зимнего существования косуль сейчас тесным образом связано с оленем,
лосем и в первую очередь с кабаном — животными, прокладывающими тропы
в снегу, облегчая тем самым передвижение из одного биотопа в другой.
Хищники. На востоке ареала европейской косули и почти на всем ареале
сибирской они оказывают особенно существенное воздействие на численность
копытных. Волк и рысь играют в этом ведущую роль.
Хищничество волка многократно усиливается при многоснежной зиме и
образовании наста, затрудняющего передвижение жертв. На Дальнем Востоке
два-три волка за сутки давят по 12—14 косуль, в местах зимней концентрации
каждый хищник уничтожает за снежный период по 20—30, а за год до 35—45 особей.
Ориентировочно в Приморском крае ежегодно гибнет от них 2 тыс. косуль
и в Хабаровском крае — до 1,5 тыс. В лесных биоценозах уничтожается
20—25% популяции (Кучеренко, 1976; 1979; Кучеренко, Зубков, 1980; Кучеренко,
Швец, 1977; Бромлей, Кучеренко, 1983). В Северном Забайкалье волки губят
28,5—32,4% осеннего поголовья, причем добывают животных любого пола и
возраста. После истребления хищников на опытном участке площадью 4,5 тыс. км2
за два последующих сезона плотность населения косуль выросла с 18 до 27 особей
на 1000 га угодий (Лавов, 1974, 1982).
В Восточной Сибири они также в основном кормятся косулей, а в период
миграции, как и на Дальнем Востоке, следуют за стадами (Фетисов, 1953). В Якутии
особенно весом их пресс в многоснежные годы; после зимы 1957/58 г. поголовье
копытных в бассейнах рек Токко и Чары уменьшилось более чем в 10 раз (Тавров-
ский и др., 1971). На Урале в Ильменском заповеднике в период с 1936 по 1946 г.
волки уничтожали ежегодно от 10 до 29% популяции. Остатки косуль находили
в 95,9% исследованных экскрементов хищников (Аверин, 1949; Филонов, 1974).
В Жигулевском заповеднике за одну ночь 9 марта 1977 г. волки,зарезали 6 косуль
(взрослого самца и пять самок) (Белянин, 1979).
В Казахстане волк также наносит наибольший урон косулям при
глубокоснежье и настах, которые особенно часты в северной части республики. В Южном
Прибалхашье в разные годы их доля в питании хищника составляет от 2,9 до
98,2% (Слудский, 1953; Слудский и др., 1984), в Заилийском Алатау — 15%
(Жиряков, 1980).
Волки явно угнетающе действуют на популяцию косуль в Литве, что особенно
270
/
•/
у Ч
Рис.98. Изменение соотношения численности (/) и добычи (2) %
косуль и численности (3) и добычи (4) хищников (волка и рыси) МО
в Латвии (из: Гарозс, 1987) „-
ПВО
ярко проявилось в послевоенные годы (см. рис. 93). ^20
В Латвии их поголовье стало расти лишь в XVIII в., 380
когда начиная с 1825 г. стали выплачивать пре- ^0
мии за убитого хищника и их стало гораздо 300
меньше (Гарозс, 1987). В Белоруссии в Припят- 260
ском заповеднике эти копытные составляют 220
11,1% пищи волка (Литвинов и др., 1979). В Одес- МО - /
ской области стая волков из 4—5 особей за зиму НО у
истребляла не менее 60 косуль (Гурский, На- Ю0
заренко, 1969). ВО
Ущерб популяции косуль от рыси в ряде 20 h_
регионов превышает волчий. За год только в 1973 1975 1977 1979 1981 19ВЗ 198
Амурской области гибнет от нее до 6—10 тыс.
особей (Дымин, 1975а; Кучеренко, Шв^ц, 1977;
Бромлей, Кучеренко, 1983). Этот хищник при обилии и легкой доступности
жертвы давит их гораздо больше, чем требуется для питания, убивая почти
ежедневно, а иногда и до 2—3 в день (Дымин, Юдаков, 1967).
Косули являются главным компонентом их рациона в Забайкалье (Смирнов,
1978) и на Алтае (Дулькейт, 1953). Зимой 1969/70 г. на одном из участков
площадью 50 км2 в Южном Прибайкалье пять рысей сократили численность
косули на 80% (Лавов, 1970). В Приангарье рыси следуют за мигрирующими
стадами и их местообитания приурочены к зимовочным стациям этих копытных,
остатки которых отмечены в 100% экскрементов хищников (Ельский, Шишикин,
1979). На Урале этот хищник уничтожает от 5,3 до 45% поголовья (Киселев,
1974, 1979). В Грузии доля косули в числе жертв составляет 45% (Бараташвили,
1982), в Швейцарских Альпах — 54,5%. С помощью радиопрослеживания здесь
установлено, что взрослая рысь в среднем убивает одно копытное за 6,6 дня, а самка
с двумя детьми — 1 за 2,7 дня, т.е. годовая потребность одного хищника —
около 60 косуль (Brietenmoser, Haller, 1987).
Очень часто во многих районах наблюдается совместнее давление волка и
рыси, препятствующее быстрому восстановлению численности популяции косуль.
Это следует из данных, представленных В. Гарозсом (1979, 1987) по Латвии
(рис. 98) и Т. Рандвеэром (1985) по Эстонии (рис. 97).
На Северо-Западе СССР эти хищники становились причиной гибели косуль в
37,8% случаев (Верещагин, Русаков, 1979). В Карпатах в многоснежную зиму
1975/76 г. из 215 погибших косуль на их долю отнесено 155 (67,3%); волки были
причиной смерти в 22,7% случаев, рысь — в 44,6% (Керечун, 1979). На Дальнем-
Востоке из 625 зарегистрированных случаев смерти косуль от хищников доля
волка составляет 64,5%, рыси — 28,7%, тигра — 3% и харзы — 2,8% (Раков, 1979).
Хищничество волка и рыси нередко находится в определенном балансе (Бромлей,
Кучеренко, 1983; Матюшкин, 1985). При снижении численности волка и
ослаблении его влияния на популяцию жертвы возрастает хищническая роль рыси.
Например, в Ильменском заповеднике в предвоенные годы на долю
многочисленного в этот период волка приходилось 67,5% из числа всех погибших косуль,
271
Таблица 41. Сравнительная характеристика гибели косуль в Ильменском заповеднике
в 1936—1970 гг. (по: Филонов, 1974)
Причина гибели
Встречаемость погибших животных по периодам, %
1935—1941
1942—1946
1947—1951
1952—1956
1957—1961
1962—1966
1967-1970
Волк
Браконьеры
Рысь
Болезни и истощение
Травмы
Беркут
Собаки
Не установлена
67,5
6,5
3,0
13,7
0,9
4,4
0,0
4,0
47,0
51,3
0,7
0,0
0,3
0,1
0,0
0,6
59,6
15,0
0,0
0,0
13,4
—
2,4
9,6
50,0
13,4
13,1
6,1
6,0
—
0,9
10,5
42,0
9,6
13,6
0,0
9,4
—
2,2
23,2
18,7
20,7
34,5
5,2
5,2
—
2,8
12,9
0,0
10,0
84,0
1,0
1,0
—
0,0
4,0
а на долю рыси — только 3%, в послевоенные годы при резком снижении
численности волка гибель от него сократилась практически до нуля, зато
смертность от рыси увеличилась до 84% (Филонов, 1974, 1975) (табл. 41).
Аналогичная ситуация зарегистрирована в Хинганском заповеднике в Амурской
области (Дарман, 1982а,б).
Значительный ущерб поголовью косули, сравниваемый в некоторых случаях с
волчьим, наносят бродячие собаки. На Украине на их долю приходится 10,9%
(столько же давят и волки) от числа найденных погибшими (Карпенко, 1966), в
Жигулевском заповеднике — 22% (волки 18,5%) (Белянин, 1980а,б), в заповеднике
"Лес на Ворскле" Белгородской области и его окрестностях — 16,7% (см. табл. 38).
Известны случаи охоты собак, иногда стаями, в Казахстане (Слудский и др.,
1984; Плешак, 1980).
Еще более губительное воздействие оказывают гибриды волка и собаки. В
одном из районов Челябинской области за 8 лет с 1971 г. несколько их стай почти в
5 раз сократили численность популяции косуль, насчитывающей первоначально
около 500 особей (Соколов, Данилкин, 1981; Данилкин, 1982).
Лисица, очевидно, также может оказывать в ряде районов значительное
влияние на численность косуль и ее динамику, существенно снижая уровень
прироста (Raesfeld, 1956; Borg, 1962; Pielowski, 1970; Strandgaard, 1972; Wandeler,
1975; Stubbe, Passarge, 1979; Cederlund, Lindstrom, 1983). В Беловежской Пуще
периоды увеличения поголовья этих копытных совпали с годами интенсивного
истребления лисицы (Владышевский, 1968). В Крыму на их долю приходилось
более 50% случаев гибели косуль (Рухлядев, 1948), на Украине — до 20%
(Карпенко, 1966). В Казахстане остатки косулят часто находят возле нор
и в экскрементах лисиц. Наблюдались случаи преследования ими детенышей и
даже взрослых особей в многоснежные зимы (Слудский и др., 1984). Аналогичные
ситуации отмечены в Якутии (Тавровский и др., 1971) и Приангарье (Ельский,
Шишикин, 1979).
Вредоносная деятельность шакала, видимо, сравнима с таковой лисицы.
Гораздо меньший ущерб причиняют другие хищники: росомаха, харза, тигр,
бурый медведь; новорожденных иногда истребляют барсуки, енотовидные собаки,
куницы, беркуты, степные орлы, лесной кот, снежный барс (Фетисов, 1953;
272
Арабули, 1966; Айзин, 1969; Бромлей, Кучеренко, 1983; Слудский и др., 1984;
Жиряков, Джансынаев, 1986). Кабаны при высокой плотности населения могут
также причинять значительный урон, уничтожая косулят (Stubbe, Passarge, 1979).
Избирательна ли роль хищников в популяции косуль? Мнения на сей счет
противоречивы, особенно в отношении рыси (Малафеев, Кряжимский, 1984).
По данным Бубеника (Bubenik, 1966a), смертность самок от волка и рыси почти в
2 раза превышает гибель телят и самцов. По сведениям К.П. Филонова (1977),
доля самцов в пище волка (53%) почти в 2 раза больше, чем самок (26%) и
телят (21%), в то время как рысь в 90% случаев давит взрослых самок и
лишь в 10% — самцов. М.Н. Смирнов (1978), напротив, нашел, что в Западном
Забайкалье сеголетки чаще становятся жертвой рыси (50%), чем взрослые
и полувзрослые самки (35,7%) и самцы (14,3%). В Чехословакии соотношение
самец:самка:теленок составляет 1:1,8:0,9 (Hell, 1979). По нашим наблюдениям
(и =14) сеголетки гибнут от рыси реже (21%), чем взрослые самки (64,0%).
Половина из этих животных была задавлена хищником и нетронута, все особи не имели
признаков болезни, истощения и травм. Соотношение погибших от гибридов волка
и собаки (л=79) было иным: самцы 25,3, самки 39,3 и сеголетки 35,4%; явные
отклонения от нормы обнаружены лишь у 5 особей (раны и дефект черепа).
Бродячие собаки обычно давят более слабых молодых зверей, бегущих в группе
последними. Лисицы, шакалы и другие мелкие хищники в основном уничтожают
детенышей, невзирая на их пол и состояние.
Антропогенные факторы. Среди факторов непосредственно вызывающих
смертность косуль в настоящее время наиболее важное значение имеет промысел. Изъятие
части популяции, не превышающее ее восстановительных возможностей,
предотвращает перенаселение, сохраняет среду обитания от чрезмерной утилизации и
стимулирует процессы воспроизводства, позволяя достичь наибольшей
продуктивности. Чрезмерный промысел, напротив, губительно отражается на популяциях.
Размер добычи животных в пределах ареала существенно различается
(см. табл. 42). Наиболее интенсивно поголовье косуль эксплуатируется в странах
Западной Европы, где ежегодно отстреливается 20—40% особей в популяциях.
Наивысшие показатели достигнуты в ГДР (от 30 до 60%), ФРГ, Дании, Швейцарии
(около 40%), минимальны они (до 10%) в СССР, Норвегии, Испании, Болгарии
и некоторых странах Азии.
Однако в регионах, где развито браконьерство, официальная статистика не
отражает действительных размеров добычи.
Распространенность браконьерства иллюстрируют следующие данные. В
Свердловской области браконьеры уничтожают до 10% от общей численности
(Киселев, 1979). В Амуро-Уссурийском регионе на сезон охотникам-любителям
выдавалось до 2 тыс. лицензий на отстрел, а фактически животных по ним
добывали в 3—4 раза больше (Кучеренко, 1976). В Амурской области браконьерская
добыча превышает официальную в 5—6 раз (Морозов, 1975). В Центральной
Якутии в 1963 г. насчитывалось около 10 тыс. косуль, причем близ г. Якутска —
5,5 тыс; в 1970 г. из-за неумеренного браконьерского промысла осталось
только 800 особей (Попов и др., 1980). На Тянь-Шане от браконьеров
гибнет До 58,3% животных (Белоусов, 1978)1. М.Н. Смирнов (1978), изучавший
'Л.В.Жирнов (1958) приводит случай, когда одной из экспедиций за три месяца (апрель, май и
июнь) в Нарынкольском районе было добыто 120—125 косуль.
18. Европейская и сибирская косули
273
косулю Западного Забайкалья, пишет: "Практика работы охотнадзора показывает,
что незаконный отстрел косуль браконьерами в пределах региона носит еще
массовый характер. Он отмечается во всех районах и во все сезоны года.
Браконьеры хорошо вооружены, в их руках всевозможные виды транспорта.
Браконьерские солонцы есть почти в каждой пади, в ряде районов развит
петельный лов зверей, в последние годы все шире применяется охота из-под
фар, загонная охота с использованием автомобиля... Нелегальный отстрел
косули в Западном Забайкалье до сих пор превышал разрешенный не менее чем в
2 раза". О таких же фактах круглогодичного браконьерства сообщают и
другие исследователи (Шаргаев, 1972; Николаев, 1980). Суждения практиков
охотничьего хозяйства по этому вопросу еще более категоричны — во многих
районах, где охрана угодий слабая или ее нет, браконьеры уничтожают ежегодно от
трети до половины всех запасов или истребляют зверей полностью.
К сожалению, уровень браконьерства высок даже* в заповедниках, заказниках и
на других особо охраняемых территориях (табл. 38). Радиопрослеживание косуль
показало, что из 40 зверей, оснащенных радиопередатчиками в заповеднике
"Лес на Ворскле" в Белгородской области, браконьеры убили или ранили 8, т.е.
20% от числа маркированных (Соколов и др., 1986).
В последние годы широкое распространение получило браконьерство под
прикрытием лицензий. Анонимным опросом охотников Дальнего Востока
выявлено, что из 66 человек, получивших когда-либо лицензии, лишь один добывал по
одному зверю, остальные от 2 до 20, в среднем 3—8 (Данилкин, Дарман, 1987).
М.В. Попов и др. (1980) считают бесконтрольный лицензионный отстрел основной
причиной резкого сокращения ресурсов косуль в Якутии. По сути, здесь он
превратился в легальную форму браконьерства. При наличии одной лицензии
добывалось по нескольку десятков животных, широко применялось самое
современное воинское автоматическое нарезное оружие, велась стрельба с
автомашин из-под фар, стрельба на заячьих охотах дробью, уродовавшая животных.
Мигрирующие популяции подвергаются наибольшему угнетению, поскольку
во время массового хода зверей на их пути "традиционно" возникают десятки
и сотни браконьеров, среди них особенно опасны моторизованные и оснащенные
автоматическим оружием. В Амурской области, например, в 1983—1985 гг. было
вскрыто несколько случаев, когда браконьеры за одну ночь из-под фар
бронетранспортера, тракторов и других машин отстреливали от 6 до 17 особей; всего за 1984—
1986 гг. за незаконную охоту на косуль здесь задержано более тысячи человек.
Таким образом, действительные размеры добычи косуль в СССР во много раз
больше данных легального отстрела и не поддаются точному учету (Метельский,
1978; Лавов, 1978). По приблизительным расчетам здесь с учетом браконьерского
отстрела добывалось ежегодно не менее 25—30% (150—200 тыс. особей) от
общей численности (Данилкин, 1982, 1985а). Аналогичная ситуация с отстрелом
этих зверей и в Монголии (Данилкин, Дуламцэрэн, 1981).
Другой существенный антропогенный фактор смертности — транспорт.
В некоторых районах Швейцарии от него гибнет от 8 до 27% косуль (Eiberle,
1972). В ФРГ и Австрии на автомагистралях ежегодно погибает от 50 до ПО тыс.
особей, что составляет около 3—7% поголовья, и примерно столько же — под
ножами и колесами быстроходных сельскохозяйственных машин. В ряде
регионов Центральной Европы потери достигают 1—5 особей на 100 га (Stubbe,
Passarge, 1979; Beklova et al., 1982; Engl, 1982; Kaluzinski, 1982b).
274
В агроценозах имеет место смертность косуль в результате отравления мине-
ральными удобрениями и ядохимикатами, особенно при неправильном их
применении и хранении, однако этот фактор менее значим.
Гораздо более существенно косвенное влияние антропогенной деятельности:
уничтожение биотопов, беспокойство, вызванное присутствием людей, и
конкуренция со стороны домашних животных. Много косуль гибнет также во
время лесных пожаров (Stubbe, Passarge, 1979; Дарман, 1982а), особенно
весенних, которые нередко практикуются местным населением для улучшения
сенокосных угодий. В Казахстане они продолжаются по нескольку дней, а
иногда и недель, уничтожая все живое (Слудский и др., 1984).
Итак, факторы, влияющие на динамику численности популяций косуль,
хотя и носят компенсаторный характер (табл. 38), в большинстве случаев действуют
комплексно. На западе ареала, где хорошо развито охотничье хозяйство,
численность определяется в основном антропогенными факторами, из которых ведущую
роль играет промысел и потери от автотранспорта и сельскохозяйственных
машин. Среди естественной смертности значительна роль паразитарных
заболеваний, особенно при высокой плотности'населения зверей. Потери от хищников
здесь сведены до минимума, хотя в ряде районов лисица наносит существенный
урон детенышам косуль. Сравнительно редкие суровые зимы приводят к
повышенной гибели ослабленных зверей (преимущественно молодых и старых,
зараженных паразитами, больных), в целом компенсируя отсутствие отбора,
осуществляемого в других регионах хищниками, и время от времени сокращают численность
популяций. Очевидно также, что охотники пока не в состоянии взять на себя
полностью функции хищников в охотничьих угодьях в поддержании плотности
населения косуль на уровне, обеспечивающем здоровье популяций.
На севере ареала возрастает роль естественной гибели животных от истощения,
ограничивающее их численность, болезни здесь сравнительно редки, однако
заметно повышается хищничество, особенно в многоснежные годы.
На востоке ареала европейской косули и на всем ареале сибирской косули
(в частности, в России) основными факторами, определяющими в настоящее
время их численность, служат хищники и браконьерский промысел. Доля
официальной добычи здесь мала, однако вместе с браконьерским отстрелом в
большинстве регионов .эквивалентна или превышает потери популяции от других
причин. Гибель косуль от болезней и паразитов в разреженных популяциях
ничтожна (табл. 38). Значительную роль в регуляции населения многих
популяций играют сезонные миграции и время от времени суровые многоснежные зимы,
при которых повышается пресс хищников и браконьеров.
Из хищников .наибольшее влияние на динамику численности косуль оказывают
волк и рысь, замещающие один другого или оказывающие совместное давление на
популяции. При их высоком прессе доля косуль, погибших от болезней,
повсеместно мала. Говорит ли это обстоятельство в пользу "санитарной" функции
хищников? И да и нет! Нет сомнений, что в какой-то степени волк и рысь
способствуют оздоровлению популяций косуль, выбирая иногда наиболее
ослабленных и больных животных. Однако эти копытные —настолько легкая добыча для
хищников, что здоровые полноценные животные становятся их жертвой ничуть не
реже. При увеличении численности волка и рыси рост поголовья косуль
затормаживается, а затем численность жертвы снижается до минимума. При таком
губительном воздействии на популяцию "санитарная" роль хищников
проблематична.
Глава одиннадцатая. Хозяйственное значение,
рациональное использование ресурсов и охрана
Европейская и сибирская косули — наиболее многочисленные и повсеместно
распространенные виды среди диких копытных животных Евразии, что делает
их основным или одним из основных объектов охоты. Численность первых
достигает 7,5 млн особей, вторых .— около 1 млн (табл. 42). Ежегодная добыча
европейской косули находится в пределах 2 млн, сибирской — не менее
200 тыс. (по экспертной оценке с учетом браконьерского отстрела). В странах
Западной и Центральной Европы на 1000 га охотничьих угодий каждый год
отстреливается от 5 до 30 особей. При оптимальных плотностях населения
популяций и рациональном промысле мясная продуктивность с этой площади может
достигать 200—600 кг; в целом она исчисляется десятками тысяч тонн.
Промышленность ежегодно использует более 1,5 млн шкур косуль. Из них
вырабатывается замша, велюр, фото кожи, кожи с лицевым покрытием (для верха обуви и
подкладки), обладающие хорошими гигиеническими свойствами (Малюгин,
1981). Часто шкуры используются в домах как декоративное украшение.
Иногда из них шьют шапки, однако они быстро изнашиваются.
Неокостеневшие рога (панты) могут служить ценным сырьем для медицинской
промышленности. Биологическая активность препаратов, полученных из пантов
сибирской косули, практически равна таковой из пантов пятнистого оленя, что
открывает перспективы использования их в медицине (Размахнин и др.,
1976). Окостеневшие рога пригодны для производства ручек, пуговиц и разных
украшений.
Как объект промысловой охоты косули имели особенно большое значение
в Сибири и на Дальнем Востоке. По приблизительным расчетам в конце
прошлого века в России ежегодно добывалось до 500 тыс. этих зверей (Туркин,
Сатунин, 1902). Лишь в Горном Алтае добыча достигала порой 200 тыс. особей
(Собанский, 1988). Можно с уверенностью говорить, что сибирские косули во
многом способствовали освоению человеком новых районов в Азии, "заменяя"
здесь домашний скот (Черкасов, изд. 1962; Соловьев, 1922). Вот что пишут по
этому поводу Н.В. Туркин, К.А. Сатунин (1902) (см. также Ветлицын, 1902): "Охота
за косулями по многочисленности этих животных и по легкости и безопасности
самой охоты составляет в Сибири обширную отрасль зверопромышленности...
В крае не существует значительного скотоводства, могущего хотя
сколько-нибудь обеспечить рынки мясом. Скот привозят либо из Китая, либо
из Кореи. Трудность приобретения скота за границею и большой риск мясоторгов-
цев вследствие весьма частых падежей от эпизоотии создало в крае монополию и
ненормально высокие цены на мясо. Привозимая на базар зимой дичь регулирует это
зло и сбивает цены на мясо. Она дает возможность небогатым классам
иметь к столу дешевый продукт. Необходимо еще сказать, что и шкуры коз...
имеют широкое применение. В крае не существует овцеводства: по почвенным или
276
Таблица 42. Численность и добыча европейской я сибирской косуль в 80-х годах XX в.
Страна
Общая
численность, тыс. особей
Отстрел,
тыс. особей
Страна
Общая
численность, тыс. особей
Отстрел,
тыс особей
Европейская косуля
Европейская косуля
СССР
Польша
ГДР
Румыния
Чехословакия
Югославия
Венгрия
Болгария
ФРГ
Австрия
Швеция
Норвегия
Финляндия
Примечание.
350—400
500
310
290
290
350
220
120
1600—1700
1000
400
150
5
10—40(100)
115—128
100—188
30—35
90
60
41—80
7—14
550—687
130—237
80—125
11
2
Франция
Дания
Италия
Швейцария
Нидерланды
Бельгия
Испания
Люксембург
Великобритания
СССР
Монголия
Китай
Корея
В скобках дана экспертная оценка.
270—300
140
100
120
25
25
20
13 :
900(400)
Сибирская косуля
300—350
100—150
(200—300)
(Ю)
62—82
39—59
10
45
5—6,5
8—9
2
5
250
5—10(100)
(30)
(50)
(3)
Таблица 43. Оценка трофеев европейской косули на международной выставке в Болгарии
(г. Пловдив, 1981)
Страна
СССР
Польша
ГДР
Румыния
Чехословакия
Югославия
Золотые медали
59
194
111
376
186
57
Страна
Венгрия
Болгария
ФРГ
Австрия
Великобритания
Золотые медали
121
92
9
23
40
климатическим причинам овцы здесь не водятся. Зато шкуры диких коз с лихвой
вознаграждают жителей. Косулий мех и легок, и очень тепел. Поэтому косулья
доха не только заменяет бараний тулуп, но и предпочитается этому последнему,
особенно при дальних зимних поездках. Кроме дох из барловой козы делают
тулупчики. Гольды делают халаты и спальные мешки... Буряты и тунгусы
делают, сверх того, из козлин шубы, шаровары, унты и пр. Из дырявых
шкур или с которых уже облезла шерсть, выделывают замшу на наволочки,
летние шаровары и пр."
В настоящее время в большинстве европейских стран охота на косуль в
значительной мере утратила свой утилитарный промысловый характер, когда
зверей добывали ради мяса или шкуры. Сейчас все больше развивается спортивное
направление. Охотники к тому же имеют возможность добыть трофеи (рога),
277
ценность которых во многих случаях неизмеримо выше мясной продукции.
Лучшие трофеи являются национальным достоянием и демонстрируются на
международных выставках, а стоимость их достигает нескольких тысяч долларов.
Количество и качество классных трофеев в определенной степени
свидетельствует об уровне, достигнутом в управлении популяциями в отдельных странах
(табл. 43).
В ГДР с 1954 по 1983 г. добыто 219 самцов, рога которых оценены золотой
медалью. В Югославии за последние 10 лет такую оценку получили 109, а в
Венгрии за последние 20 лет — 2395 трофеев. В Швеции с 1973 по 1980 г. золотой
медалью оценен 171 трофей. Самцы с мощными рогами во всевозрастающей мере
добываются в Великобритании; только в 1983 г. получено 34 золотых, 37
серебряных и 26 бронзовых медалей. В России трофейное направление в
охотничьем хозяйстве развито слабо, поэтому на международных выставках выставляется
незначительное число классных рогов. В то же время разреженные популяции
здесь очень качественные и можно ожидать хороших трофеев.
Говоря о косулях как об охотничьих видах, нельзя не упомянуть о роли
охотников в их воспроизводстве: они рхраняют животных, подкармливают,
улучшают естественные биотопы, отстреливают крупных хищников, проводят
ветеринарные и другие мероприятия, направленные на сохранение и увеличение
поголовья. Именно благодаря охотникам численность этих зверей во многих
странах многократно возросла и всюду в Европе они стали фоновым видом.
Живя в антропогенном ландшафте вблизи жилья человека и не боясь его
(там, где зверей не преследуют), косули теперь являются неотъемлемым
компонентом природы, рекреационным животным, украшающим леса и поля. Однако
чрезмерная численность отрицательно сказывается на их качестве, ведет к
биологической деградации; они могут нанести также существенный вред лесному
хозяйству. Чтобы этого не случилось, необходимо управлять популяциями
и рационально использовать ресурсы косуль.
Управление популяциями
и рациональное использование ресурсов
Экология и поведение европейской и сибирской косуль в целом сходны (Соколов,
Данил кин, 1981). Этот вывод имеет принципиальное значение для охотничьего
хозяйства, позволяя применять одинаковые методы при управлении популяциями1.
Управлять популяциями косуль, т.е. регулировать их численность (плотность
населения) с учетом необходимости максимального для данных условий
использования биологической продуктивности можно и нужно путем
выборочного отстрела и воздействия на половой и возрастной состав.
Очень важно понимать, что при одной и той же численности животных, но
при разном половом и возрастном составе продуктивность популяции будет разная.
Не менее важно осознать, что в эксплуатируемой популяции необходимы
'Следуя Е.Е. Сыроечковскому (1986), мы предлагаем для практических целей смелее Использовать
условное понятие "популяции" для любой демографической группы животных, руководствуясь
географическими признаками, рамками административных границ и границ охотничьих хозяйств в
частности. Тем не менее и в этом случае следует стремиться к выделению природных популяций, —
длительное время существующих независимо друг от друга, достаточно многочисленных и со
своими биологическими параметрами групп особей (Яблоков, 1987).
278
такие нормы изъятия, которые стимулировали бы естественные механизмы
повышения плодовитости, т.е. популяция должна поддерживаться на уровне
максимальной продуктивности. Это основа рационального ведения охотничьего
хозяйства по косуле.
Плотность населения
При имеющихся в настоящее время охотхозяйственных условиях,
характеризующихся отсутствием во многих европейских регионах крупных хищников и
интенсивным использованием человеком сельскохозяйственных и лесных культур,
численность косуль должна быть приведена в соответствие с интересами лесного и
сельского хозяйства. Отсюда выводится понятие экономически допустимой
(или хозяйственной) плотности их населения, при которой вред для сельского и
лесного хозяйств не должен превышать допустимых пределов или же может быть
надежно предотвращен с помощью имеющихся средств (Ueckermann, 1956,
1959; Miiller, 1965; Падайга, 1965; Wagenknecht, 1971).
Хозяйственная плотность населения косули базируется на емкости угодий, т.е.
по существу на имеющихся условиях обитания, в первую очередь связанных с
кормодобыванием, и отражает взаимосвязь охотничьего, лесного и сельского
хозяйства.
Этому противостоит понятие биологически допустимой (биологической)
плотности населения дичи, при которой не возникает никаких отрицательных
явлений в развитии популяции. В качестве ее критерия берется конституция
и кондиция зверей и размножение.
Различают также понятие предельно допустимой плотности населения зверей,
значение которой по культурным и научным соображениям может быть выше
хозяйственной или биологической плотности, что имеет место в заповедниках,
заказниках, научных участках, в зонах пригородного отдыха и на других особо
охраняемых территориях.
Оптимальной плотностью населения косуль, по нашему мнению, следует
считать такую плотность, при которой не происходит биологической деградации
популяции, достигшей наивысшей продуктивности, а вред лесному и сельскому
хозяйствам не превышает допустимых пределов.
При установлении хозяйственной плотности населения учитывают следующие
аспекты: кормовую емкость биотопа, хозяйственные (экономические) задачи,
наличие других видов копытной дичи, интересы лесного и сельского хозяйства.
Ее значения особенно зависят от типа и качества угодий и значительно
колеблются даже в пределах одного и того же охотничьего хозяйства.
Ю.А. Дарман считает, например, что сибирская косуля в Амурской области
не наносит существенного вреда лесному хозяйству, если ежегодно потребляет
зимой от 30 до 60% осеннего запаса веточных кормов (табл. 44). Отсюда в
зависимости от типа угодий он выводит цифры хозяйственной плотности,
составляющей в среднем 40—80 особей на 1000 га.
Определение качества охотничьих угодий (бонитировка) заключается в
суммарной оценке условий существования вида. В отношении косули наиболее
известны методы, предложенные Еккерманом (Ueckermann, 1951), Мюллером
(Miiller, 1964) и Моттлем (Mottl, 1954). Практически в каждой европейской стране
разработаны свои шкалы оптимальной или допустимой численности (плотности
279
Таблица 44. Хозяйственная плотность населения сибирской косули в зоне дубово-черноберезовых
лесов и влажной лесостепи в Амурской области (по: Ю.А. Дарман, личное сообщение)
Тип угодий
Запас древесно-веточных
кормов (в сыром весе),
кг/га
Допустимое изъятие,
кг/га
Целесообразная
плотность, особей на
1000 га
Дубняки
Черноберезники
Белоберезники
Осинники
Липняки
Заросли лещины
Ерниковые заросли
В среднем по лесу
103,5
205,8
46,3
68,9
38,2
410,8
221,2
= 100
31-62
62-123
14-28
21-41
12-23
123-246
66—133
30—60%, осеннего
запаса
39-77
77—154
18-35
26-51
15—29
154—308
83—166
40—80
Таблица 45. Оптимальная численность европейской косули на 1000 га охотничьих угодий
Регион
Классы бонитета
I
II
III
IV
V
VI
Центральные области
ской части СССР
Украина, зоны:
равнинная
предгорная
горная
ГДР
евро
пей-
80
60
70
40
90-120
80—50
30
35
25
80—100
50—30
15
15
10
70-80
30-10
10
60—70
50—60
40-50
населения косуль) в угодьях разных бонитетов с учетом специфики охотничьего
хозяйства. Они отличаются как по количеству классов бонитета, так и по
средним цифрам плотностей, однако порядок величин при этом сходен.
В качестве примера приведем шкалу, рекомендованную для Центральных
областей европейской части СССР (Данилов и др., 1965), Украины (Рудышин
и др., 1979) и ГДР (Stubbe, Passarge, 1979) (табл. 45).
В современном охотничьем хозяйстве хозяйственная плотность населения
косуль колеблется в пределах от 30 до 120 особей на 1000 га пригодных угодий.
При особенно благоприятных условиях поголовье на той же площади может
достигать 200 и более особей. Эти цифры в основном и служат ориентиром для
практиков охотничьего хозяйства с определенной корректировкой, исходя из
возможностей и целей охотничьего и лесного хозяйства.
До достижения уровня плотности в регионе 10 особей на 1 тыс. га, звери
нуждаются в жесткой охране, поскольку кормовая емкость угодий в пределах
их естественного ареала позволяет значительно увеличить численность.
Умеренное хозяйственное использование ресурсов популяции возможно, если плотность
ее населения превышает 20 особей на 1 тыс. га пригодных угодий. Заметим, что
280
достижение такого уровня вполне реально в любом охотничьем хозяйстве в
пределах естественного ареала. Гораздо большую трудность представляет
поддержание численности животных на оптимальном уровне, т.е. собственно
регуляционные мероприятия, основной задачей которых является приведение плотности
населения животных в соответствие с имеющимися условиями окружающей среды.
Они необходимы, если плотность становится выше 50 особей на 1 тыс. га (см.
рис. 109).
Соотношение полов
После установления пределов хозяйственной плотности населения дичи
определяется целесообразное соотношение полов. От него во многом зависит размер
прироста.
Соотношение полов в естественной популяции косуль с увеличением возраста
неуклонно сдвигается под действием природных факторов в пользу самок (см. главу
"Структура популяций"). Это положение особо принимают во внимание при
оценке плотности населения дичи и прогнозировании размеров прироста
популяции. Чрезмерное возрастание численности может привести к резкому
увеличению потрав в лесном хозяйстве. Для противодействия этому в
переуплотненных популяциях планируют повышенный отстрел самок. В районах,
характеризующихся относительно высокими потерями косуль, но при достаточно высоком
уровне плотности, желательно половое соотношение 1:1. Очень высокие потери в
полевых угодьях, причиняемые уборочными машинами, неправильно
применяемыми средствами агрохимии, транспортом и др., могут быть скомпенсированы
снижением отстрела самок. В областях, где плотность населения косуль
находится ниже допустимых с хозяйственной или биологической точек зрения
пределов, необходимо полностью запрещать отстрел самок, и в первую очередь
половозрелых.
Проведение регуляционных мероприятий в популяциях (управление
популяциями) возможно лишь при выборочном отстреле.
Возраст и возрастные категории
Природные популяции косуль в основном состоят из особей не старше 10 лет, что
подтверждается всеми новейшими исследованиями. Популяции, управляемые
человеком, должны поддерживаться в том возрастном диапазоне, в котором
особи имеют максимальную массу тела и рогов и наивысшую воспроизводительную
способность.
Пределы возраста самцов определяются прежде всего временем кульминации
развития рогов. Известно, что в одной и той же популяции есть особи с
ускоренным и запаздывающим развитием — у одних рога бывают лучшего
качества в возрасте 3 лет, у других — в 6—7 лет или позднее, но у большинства
самцов все же они наиболее развиты в возрасте от 3 до 7 лет.
Пределы возраста самок определяются помимо конституции
воспроизводительной способностью, которая максимальна с 3-летнего возраста и начинает
снижаться с 7 лет (см. главу "Размножение").
Следовательно, для управляемой человеком популяции косуль желателен
возрастной диапазон от 3 до 7 лет. Поддержание его на этом уровне даст
возможность получения максимальной продуктивности и хороших трофеев.
281
В популяциях косуль особенно высок естественный отход молодых и старых
особей. В связи с этим рациональное использование ресурсов должно
базироваться на интенсивном отстреле молодняка дичи обоих полов, незначительном
изъятии из популяции особей среднего возраста и отстреле всех старых зверей.
В разных странах приняты различные пороги возрастных классов. Однако опыт
показывает, что в практике охотничьего хозяйства обычно достаточно различать
четыре возрастные категории: 1) молодые — до 1 года, 2) годовалые, 3)
средневозрастные — от 2 до 7 лет и 4) старые.
При оптимальном уровне плотности населения косуль наиболее рационально,
на наш взгляд, такое соотношение добычи: молодые (до 1 года) — 60—70%,
средневозрастные — 10—15% и старшего возраста — 20—25%. Годовалых самцов
отстреливают выборочно в зависимости от качества рогов (см. рис. 109).
Учет численности
Нельзя целенаправленно управлять популяциями косуль, не зная их численность.
Однако вряд ли имеется какой-либо метод учета численности этих животных,
который можно было бы назвать абсолютно надежным и точным. Все
применяемые на практике методы служат только для примерной оценки.
Сложность в определении численности косуль можно показать на примере
нескольких известных опытов. В одной из вольер площадью 75 га по учетным
данным находилось 12 особей; тотальный отстрел дал 25 (Ueckermann, 1951).
В Дании на участке леса площадью 340 га с оценочной численностью зверей в
70 особей добыто 312, т.е. в 3 раза больше, чем учтено. На другом участке
окруженного забором леса площадью 220 га, где насчитывали 125 животных,
отстреляли 161, но даже после этого там остались косули (Andersen, 1953; Strand-
gaard, 1972).
Ошибка при определении численности косуль, составляющая от 20 до 100%,
обычна. В отдельных случаях она может быть существенно более высокой, особенно
при методах относительного учета. Завышение численности при оценках происходит
гораздо реже, чем занижение, и это следует иметь в виду при планировании
отстрела.
Для оценки численности обычно применяют три группы методов:
1) визуальные наблюдения (сюда входит и метод прогона дичи);
2) учет следов или помета;
3) статистический расчет численности на основании добычи.
Относительно точна оценка численности по усовершенствованному способу
Петерсена, или Линкольн-индексу (Bailey, 1951). Предпосылкой его применения
служит мечение по меньшей мере 75% особей в популяции (Strandgaard, 1972,
1975). При меньшем их количестве должно быть большое число наблюдений.
Формула расчета численности такова:
JC + 1 '
где N — численность популяции; М — количество меченых особей; п — общее
число наблюдений; х — число встреч меченых особей.
К сожалению, при всех достоинствах этого метода он трудоемок и пригоден
только для научных исследований.
282
Хороший метод учета — круглогодичное наблюдение за популяцией косуль,
что дает возможность не только выяснить численность зверей в локальном
районе, но и позволяет установить соотношение полов и возрастных классов, т.е.
определить структуру популяций и тенденции ее развития. Однако он требует от
специалиста или охотника квалифицированной регистрации каждого
проведенного наблюдения, занимает много времени и пригоден в основном для научных
исследований. Наиболее эффективна для этой цели комбинация засад со скрадыва-
нием (Boisaubert, 1979).
Часто рекомендуемый метод учета численности населения зверей в контрольные
дни весной в самом начале периода вегетации может быть использован только
в хвойных лесах. В лиственном лесу в это время происходит концентрация
животных, вызванная появлением первоцветов, что приводит к завышению
численности.
Наилучшее время для подсчета численности косуль в лесах — конец зимы и
начало весны, но и в самом благоприятном случае в течение одного дня можно
видеть лишь около 50% косуль, обитающих в данном районе (Strandgaard,
1972; Pedroli et al., 1981).
В полях коллективные многократные засидки в основных биотопах позволяют
получить совершенно точные данные. Относительно быстро численность полевых
косуль определяют путем линейной таксации с автомобиля (Zejda, 1984, 1985).
Учет на маршрутах длиной 30 км предпочтительнее, чем на 10-километровых.
Метод учета численности через индекс плотности на километр дистанции
(Vincent, 1982) все же более пригоден для определения тенденции развития
численности популяции, чем для абсолютного учета. Подсчет дичи на трансектах
неопределенной ширины на открытых ландшафтах дает надежный результат
(Дарман, 1987).
Учет дичи с помощью прогона относительно точен, но трудоемок, требует
8—10 загонщиков и 6—8 наблюдателей. Иногда для прочесывания района
привлекают более 100 человек (Stangl, Margl, 1977).
Все методы оценки численности косуль путем прямых наблюдений за ними
трудоемки, но имеют то преимущество, что по их результатам к тому же можно
делать выводы о половой и возрастной структуре популяции, ее качестве и
приросте. Методы учета животных по следам их жизнедеятельности более
просты, однако они до сих пор недостаточно развиты.
Наиболее распространен маршрутный учет животных по следам на снегу
(Кузякин, 1979), но в отношении косуль он дает очень большие погрешности,
поскольку животные живут маленькими группами на очень небольшом участке,
активны днем и тяготеют к опушкам. Более точен "спиральный" метод подсчета
(Berge, 1969); при этом следы ищут независимо от дорог и просек по сужающимся
окружностям (спирали), начиная с внешних краев участка в направлении центра.
Лучшее время для подсчета следов — через 5—6 ч по окончании снегопада.
В конце зимы следует учитывать возрастание активности косуль. Значительное
влияние на количество следов может оказать структура леса, равно как и
эффект краевой зоны (Btittner, 1983).
Заключение об относительной численности дичи можно сделать исходя из
частоты встречаемости помета на контрольных площадях, которые перед
наступлением зимы должны быть очищены от всех остатков помета. Частота
дефекации животных варьирует в зависимости от времени года, питания, возраста,
283
пола, условий обитания, что может оказать заметное влияние на точность
учета. Например, в Польше найдено, что в вольере косуля оставляет в сутки
весной 14,9, летом 5,3, осенью 4,2 и зимой 3,75 кучек помета (Dzieciolowski,
1976). В Литве зимой их в среднем бывает 15,6 (Падайга, 1970; Падайга, Марма,
1979), а на Дальнем Востоке — 36,2 (Дарман, 1986).
Формула для определения плотности населения дичи в оседлых популяциях
таким способом следующая (Briedermann, 1982):
„ М-10000
где N — численность популяции; М — число дефекаций; п — количество контрольных
площадок; S — размер контрольной площадки, в м2; / — частота дефекаций
одной особи; / — период исследований в днях.
Все методы учета численности по количеству лежек, пропорциям тела или по
степени потравы растений не пригодны для практической работы.
В настоящее время в охотничьем хозяйстве широко распространено определение
"предшествующей" численности на основе многолетней статистики добычи.
Предпосылки для этого следующие: данные должны соответствовать фактическому
отстрелу; естественные потери считаются незначительными; прирост и соотношение
полов отвечают ситуации, сложившейся в действительности. Среднюю цифру
годовой добычи за последние 4 года считают эквивалентной приросту.
Множительный коэффициент при условном приросте самок дичи, составляющем
110%, — 1,86; при 100%-ном приросте расчетная численность определяется его
удвоенным значением при соотношении полов 1:1. Этот метод также не дает точных
данных, однако в отдельных регионах применение его возможно для принятия
директив в отношении снижения или увеличения размеров добычи.
Существенно более точный результат в отношении какой-либо популяции
получается, если, кроме того, принимается во внимание возраст и пол добытых
особей (Stubbe, 1966,1979). Тем не менее погрешность данного метода заключается в
неточном определении возраста и в неверной оценке уровня естественной
смертности косуль.
На азиатской части ареала в степной и лесостепной зонах наиболее
предпочтительны авиаучеты. Практически только так можно подсчитать численность
мигрирующих популяций. Учет с самолетов и вертолетов в целом оказывается более
экономичным, эффективным и точным, чем наземный. Для работ на борту
необходимы 3 человека, оптимальная высота полета 100—150 м (Егоров, Попов, 1964;
Конечных и др., 1964; Поле, 1966; Попов, 1970; Смирнов, 19706; Байкалов, Семенов,
1971; Шутеев, 1973; Николаев, 1982).
Кроме перечисленных выше, существует много других способов учета
численности (Berge, 1969), но все они страдают отсутствием точности.
Таким образом, для учета косуль пока нет достаточно надежного и точного
метода, но, несмотря на это, нельзя отказываться от оценки численности,
поскольку это основа хозяйственного использования. В зависимости от местных
условий предпочтение следует отдавать одному из методов или комбинации
нескольких. На наш взгляд, только круглогодичные наблюдения за локальной
популяцией смогут дать наиболее точные сведения как о численности, так и о
соотношении полов, структуре возрастных классов, приросте и качестве
популяции.
284
Планирование промысла
При планировании отстрела особое внимание обращают на следующие правила,
которые уже подчеркивались выше. В популяциях существует" тенденция сдвига
соотношения полов в пользу самок, что при высокой плотности населения
быстро приводит к чрезмерной численности животных. При избыточной
численности косуль необходим повышенный отстрел самок (более 50%). В тех районах, где
плотность населения косуль не достигла оптимального уровня, и в полевых
угодьях при высокой естественной смертности доля самок в добыче не должна
превышать 50%. В многочисленных популяциях планируется высокий уровень
отстрела молодняка, незначительный — особей среднего возраста и более
высокий — старых. При очень низкой плотности населения промысел вести не
следует.
Следовательно, плотность населения косуль, имеющаяся и
хозяйстве н н о-целесообразная представляет собой основу планирования и
рационального использования популяции в охотничьем хозяйстве.
В некоторых странах (в частности, России, Казахстане, Монголии)
планирование хозяйственного использования ресурсов косуль ведется исходя из
общей численности животных по региону без учета плотности их населения и
емкости угодий. На практике это приводит к тому, что любая численность
зверей (и 100 тыс., и 1 тыс., и 100 особей) считается промысловой и их повсеместно
добывают, невзирая на пол и возраст, даже если поголовье в пределах
региона в десятки раз ниже оптимального уровня (Данилкин, 1985). Причем
установленные нормы добычи часто остаются неизменными и тогда, когда
общая численность зверей в регионе сокращается в несколько раз.
Не везде и не всегда в планах добычи предусматриваются нормы отстрела по
полу и возрасту. Без этого отстрелу подвергаются обычно самые крупные
высокопродуктивные особи. Такая практика планирования хозяйственной
деятельности от достигнутого и без половозрастной дифференциации порочна и
оказывает губительное действие на популяции.
Нерациональные методы планирования часто усугубляются нерациональными
способами охоты. Косуль добывают в основном в снежный период при
коллективных охотах загонным способом, при котором охотник, как правило, стреляет
крупную косулю — лидера группы. А лидером групп у косуль в 80—90% случаев
являются половозрелые самки. Таким образом, нередко при очень низкой
численности, требующей безусловного восстановления, изымается репродуктивное
ядро популяции — половозрелые самки, тогда как оставшиеся без матери
телята нередко обречены на гибель, особенно в многоснежные зимы.
Такой способ "ведения охотничьего хозяйства" нельзя назвать иначе, как
полной бесхозяйственностью, и далеко не случайно, что там, где он имеет
место, плотность населения животных не превышает 0,01—2 особи на 1000 га
пригодных угодий.
На протяжении огромного ареала условия обитания косуль и охотничьи
традиции различны, тем не менее везде необходимо стремиться к наиболее
рациональным методам планирования, которые уже апробированы многолетней
практикой, по крайней мере в отношении оседлой европейской косули и оседлых
популяций сибирской. В то же время в отношении многих мигрирующих популяций
обычные способы планирования явно неприемлемы, поскольку мигрирующие
285
стада, перемещаясь из одного административного района в другой, могут
подвергаться неумеренному промыслу.
Знания о мигрирующих популяциях весьма скудны: еще ни разу ни одна из
них не была маркирована и прослежена в течение длительного времени. Поэтому
конкретные рекомендации по управлению такими популяциями дать весьма
затруднительно. Однако многие из них имеют весьма четкие экологические и
географические границы (см. главу "Перемещения"), а численность особей
поддается относительно точному предпромысловому учету с самолетов или вертолетов.
Известны и традиционные пути миграций.
В данном случае, вероятно, допустимо планировать промысел исходя из
общей численности локальной популяции, ежегодно устанавливая определенные
квоты добычи для каждого конкретного охотничьего хозяйства на пути ее
миграции, а также на местах зимовок и летнего обитания. Разумеется, и здесь должен быть
дифференцированный по полу и возрасту выборочный отстрел и именно в
таких популяциях процент изъятия молодняка должен быть максимальным при
максимально щадящем режиме для половозрелых самок.
В принципе при наличии разнообразных знаний о конкретной популяции
возможно построить имитационные модели управления, включающие экономические
критерии, т.е. смоделировать оптимальное охотничье хозяйство (Lomnicki,
1975; Law, 1979; Bobek, 1980). Существующие модели пока что далеки от
идеальных, но все же по мере развития науки и техники будущее за ними.
Сроки охоты
В разных странах приняты разные сроки охоты на косуль (табл. 46).
В большинстве европейских стран сроки добычи дифференцированы по полу
и возрасту и сезонам года. Самцов отстреливают преимущественно с мая—июня по
октябрь, самок и детенышей — с сентября—октября по декабрь—январь.
Отстрел самцов ведется в трофейных и селекционных целях в оптимальные для
этого сроки — когда у них есть рога и качество особей легко определить.
С сентября косули начинают вести групповой образ жизни и самок с подросшими
детенышами уже возможно оценить по качеству.
При массовом отстреле животных осенью оставшимся особям достанется
больше пищи зимой, что в Целом благоприятно скажется на качестве и
продуктивности популяции, на сохранении биотопов и к тому же будет способствовать
снижению вреда лесному хозяйству.
Дифференцированный выборочный отстрел требует высокого профессионального
уровня охотников, умения и навыков в определении пола и возраста зверей.
Определение пола и возраста
Определение пола, как правило, не составляет затруднений. Летом самцов
легко распознать по рогам, зимой по расположенному на пенисе длинному
пучку волос, хорошо различимому под брюхом. Труднее определить годовых
самцов с едва отрастающими рожками; здесь следует обращать внимание
на мошонку. Самки косуль в отличие от самцов летом безроги. Зимой же
их просто опознать по выдающемуся из вульвы пучку волос, который четко
выделяется желтым пятном на фоне белого зеркала (рис. 99).
Определение возраста — одна из важнейших и сложнейших проблем при-
286
Таблица 46. Сроки добычи косуль
Страна
Сроки добычи (число, месяц)
самцов
самок и детенышей
Германия (б. ГДР)
Польша
Румыния
Чехословакия
Венгрия
Болгария
Югославия
Германия (б. ФРГ)
Австрия
Дания
Швеция
Люксембург
Нидерланды
Норвегия
Бельгия
Великобритания
Испания
Италия
Швейцария
Лихтенштейн
Монголия
Россия
Казахстан
Эстония
Литва
15.5—15.10
1.5—15.10 (годовалые)
11.5—30.9
15.5—31.10
16.5—30.9
15.5—15.10
1.6—2i.l0
16.5—31.12
16.5(1.6)—15.10
16.5(16.6)—15(31)Л0
16.5—15.7
1.10—31.12
15.8—31.12
1.6—15.7
1.6—30.9
1.10—23.12
10.10—30.11
1.5—20.10
5.6—25.7
11.9—1.11
1.5—20.10
1.5—31.12
1.6—15.10
1.9—15.1
15.7—15.8 (европейская)
25.8—25.9 (сибирская)
1.10—15.1
1.10—15.12
15.7—15.8
1.9—30.9
15.5—30.6
1.9—1.1
1.9—31.1
1.6—31.1 (годовалые)
1.9—31.3 (детеныши)
1.10—31.1
1.6—28.2
1.9—31.12
1.9—15.2
1.10—31.10
1.10—31.12
1.9—31.1
1.8(1.9)—15(31).12
1.10—31.12
1.10—31.12
15.10—30.11
1.1—31.3
1.10—23.12
10.10—30.11
21.10(1.11)—28.2
1.5—15.12
1.5—15.12
7.9—15.11
1.9—31.12
1.9—15.1
1.10—15.1
1.10—15.12
1.9—31.10
19—31.10
хозяйственном использовании косуль. На расстоянии едва ли возможно точно
определить возраст животного старше года у самок и двух лет у самцов. Задача
существенно облегчается тем, что в эксплуатируемой популяции имеется небольшое
количество старых зверей; большинство же особей — молодые и среднего
возраста.
Детеныши отличаются от взрослых косуль размерами тела вплоть до следующей
весны. Фигура их с возрастом претерпевает определенные изменения. Годовалые
особи обоих полов не имеют массивного туловища, так что ноги их кажутся
сравнительно длинными, а круп слегка приподнят сзади; после осенней линьки эти
отличия в значительной степени исчезают. Двухлетние самцы внешне выглядят
более крепкими, чем годовалые, однако все еще стройными. Туловище 4—5-летних
самцов, достигших максимального веса, кажется приземистым, ноги короткими.
287
Рис.99.Отличительные половые признаки у самцов (А) и самок (£) косуль зимой (рис. В.М. Гудкова)
Рис. 100. Возрастные отличия телосложения у косуль
А — молодые; Б — средневозрастные; В — старые особи (самцы и самки) (рис. В.М. Гудкова)
Такая фигура характерна все время, пока самец находится в высшей точке
своего развития. Престарелые самцы часто снова приобретают форму тела,
свойственную молодым особям (рис. 100).
У годовалых самок летом отсутствует вымя. В зимней шерсти уверенное
различение их и более взрослых самок уже вряд ли возможно. Старые самки
обычно угловаты, костисты и тощи, однако во многих случаях они неотличимы
от более молодых.
288
Рис. 101. Возрастная изменчивость окраски головы самцов
А — молодые; Б — средневозрастные; В — старые (рис. В.М. Гудкова)
Другими признаками при определении возраста косуль могут служить форма
головы и шеи и окраска морды. У годовалых особей голова узкая; постепенно
она становится более широкой, особенно у самцов, и поэтому кажется более
короткой. Шея у последних тонкая и длинная, при движении поставлена
вертикально. С годами она становится толще, мощнее и наклоняется ниже. Тем не менее
следует учитывать ситуацию, в которой находятся животные: молодые при
кормежке тоже наклоняют шею; обеспокоенные взрослые самцы, напротив,
держат шею вертикально.
Оценка возраста животных по окраске морды возможна только в случае
полностью законченной линьки, приблизительно с Июня по август. С конца
августа окраска волос в результате осенней линьки снова начинает меняться, что
может привести к неправильному определению. Морда годовалых особей
имеет одноцветную темную, подчас прчти черную окраску. Однако у развитых
самцов белое пятно на носу уже хорошо выражено, у двухлетних самцов
оно всегда четко отграничено, но с возрастом увеличивается в размерах,
белая окраска теряется и переходит в серую. У стареющих самцов из-за седых
волос сзетлеет лоб, седина простирается до глаз и постепенно вся голова
становится серой. Светло-серые кольца вокруг глаз ("очки") служат отличительным
признаком старых самцов (рис. 101).
Окраска морды, да и вообще окраска зверя, оценивается наблюдателем
по-разному в зависимости от погоды и степени освещенности. Кроме того,
окраска подвержена очень большим индивидуальным различиям. Поэтому данный
метод годится лишь для определения молодых и старых животных, но не для
точной оценки возраста косуль.
Рога также используются для уточнения возраста самцов. Отсутствие
отростков всегда свидетельствует о том, что эти рога первые, но у части годовалых
особей отростки есть. У взрослых самцов рога без отростков весьма редки,
основания и стержни рогов и у них всегда утолщены.
19. Европейская и сибирская косули 289
Довольно надежным показателем возраста служит высота оснований рогов,
которая в связи с ежегодным сбрасыванием рогов год от года уменьшается.
Самцы с основаниями рогов, "посаженными" непосредственно на череп и частично
закрытыми шерстью, являются старыми.
Многие охотники часто допускают ошибку, рассматривая в качестве критерия
возраста прежде всего венец рогов. Так называемые "макушковые", или "венечные",
отростки наблюдаются во всех возрастных классах, но среди годовалых
животных практически нет особей с отростками рогов, направленных назад;
они встречаются только в более старших возрастных классах.
Сроки формирования, очистки от кожи и сбрасывания рогов также в значительной
степени зависят от возраста. Взрослые самцы сбрасывают рога первыми и
примерно на 3 недели раньше молодых образуют новые и очищают их от
кожи. У некоторых старых особей европейской косули рога полностью
формируются уже в конце февраля, у самцов среднего возраста — середине марта, тогда
как у однолетних особей их развитие начинается лишь в марте (см. рис. 67).
На образование рогов большое влияние оказывает общее физическое состояние
особей. В одном и том же возрасте животные с особенно хорошим
физическим состоянием очищают рога на несколько недель раньше, в результате
чего создаетя видимость их более старого возраста. Условия зимовки могут
повлиять на сроки формирования рогов для всех особей.
Отличительным признаком при определении возраста косуль в какой-то
мере служит линька. Весной первыми обычно линяют годовалые особи. Самцы
среднего возраста меняют окраску лишь к середине июня. Самки весной линяют
несколько позже, чем самцы, что связано с беременностью и лактацией.
Осенняя линька происходит в той же последовательности. Сначала летнюю
окраску теряют молодые особи, затем особи среднего возраста и последними
старые. Период с середины сентября до середины или конца октября — наиболее
благоприятное время для определения возраста косуль по этому признаку.
Запаздывание линьки чаще всего обусловлено заболеваниями или вызвано
нарушениями обмена веществ. Такие животные подлежат отстрелу.
Уточнению возраста косуль во многом способствует их поведение. Годовалых
особей можно наблюдать вместе с матерью сравнительно долго, иногда
вплоть до родов. Этому возрастному классу свойственно игровое поведение,
любопытность и меньшая осторожность. По поведению двухлетних и более
взрослых самцов точный возраст установить невозможно, однако сделать
вывод о "более молодых" и "более старых" животных удается. С годами
животные становятся более осторожными и недоверчивыми и на места кормежки,
как правило, выходят последними. При столкновениях уступает более молодой
независимо от развитости рогов и физической силы, у одновозрастных особей
победителем оказывается владелец территории. Будучи побежденным, молодой
самец отбегает на короткое расстояние и затем долго лает, старый самец
совсем не кричит или лает несколько раз.
Определение качества
Качество особей, соответствующее тому или иному возрасту, и качество
популяции в целом — основные критерии при выборочном отстреле. Размеры тела и
рогов, физическое состояние особей и популяции — важнейшие характеристические
признаки.
290
Качество какой-либо популяции в целом может ухудшиться под влиянием
болезней, но чаще оно снижается при сильном поражении эндопаразитами.
В этом случае поголовье сокращают и проводят ветеринарные мероприятия.
Животных с явными признаками болезни (запачканный зад при поносе,
растрепанная шерсть на спине вне периода линьки, кашель при поражении
легочными паразитами, отощание, хромота, ненормальное поведение), при
явных отклонениях'от нормального развития (париковые рога) — отстреливают.
Сравнительным показателем качества популяции служит средняя масса тела
самок и сеголеток, определяемая с сентября по декабрь отдельно для каждой
возрастной группы. К тому же осенью представляется хорошая возможность
оценить самок по плодовитости и качеству потомства.
При существующей корреляции между массой тела и степенью развития
рогов отбор особей по массе тела скажется и на качестве рогов.
Особенно тщательно следует вести отбор годовалых особей. Во многих
районах можно добывать всех зверей с маленькими или уродливыми рогами без
отростков.
При ухудшающемся качестве популяции, например при снижении средней
массы тела и рогов, следует обратить особое внимание на численность популяции и
условия питания зверей.
В конечном итоге каждая отдельная особь должна характеризоваться в
аспекте ее качества по сравнению со средним значением величины для данной
популяции, а популяция — по сравнению со средними показателями других
популяций вида или подвида.
Выборочный отстрел
Численность косуль во многих странах Европы достигла таких пределов,
которые превышают или могут превысить естественную кормовую емкость
биотопов, что, в свою очередь, может вызывать ухудшение качества популяции или
массовую гибель животных от недостатка пищи. Возникает необходимость в
соответствующем ограничении численности и в пристальном контроле за качеством
популяций. Исторически эту роль выполняли крупные хищники. Теперь же,
при отсутствии волков и рысей в большинстве стран, охотник обязан взять
на себя их функции.
В целом при хозяйственном использовании многочисленных популяций имеет
силу принцип "массовый регуляционный отстрел предшествует выборочному
отстрелу". Выборочный отстрел эффективен только после сокращения слишком
высокой плотности дичи. При правильном применении он существенно влияет как
на развитие численности, так и на качество популяции.
Основные принципы выборочного отстрела следующие:
1) отстрел начинается с наступлением сезона охоты, поскольку дичь в это время
еще относительно доверчива;
2) селекционный отстрел самцов должен быть завершен до наступления гона;
3) в первую очередь добываются все больные и уродливые животные;
4) отстреливаются все слабые детеныши и старые животные;
5) отсреливаются семейства косуль с плохими качественными показателями.
Отбор самцов начинают с однолетнего возраста с учетом физического развития,
массы тела и длины рогов, что является предпосылкой получения хороших
291
трофеев; ежегодно их добывают до 50—60%. 2—7-летних самцов, напротив,
щадят и в целях селекции отстреливают лишь при явных аномалиях (многост-
вольность рогов, париковые или "бараньи" рога и т.п.).
Выборочный отстрел детенышей относительно несложен, поскольку осенью
и зимой семья держится группой и, таким образом, существуют хорошие
возможности для сравнения. Всегда отстреливают слабейшего детеныша, невзирая на
его пол, из двойни отстреливают одного, из тройни — двух. Чем раньше это
произойдет, тем больше пищи достанется оставшемуся в живых и благодаря чему
он лучше развивается.
Оценка самок должна производиться на основании качества их приплода.
При плохом качестве следует стремиться к отстрелу всей семейной группы.
Выявление старых самок наталкивается на большие затруднения, и на практике
нередко приводит к тому, что под выстрел вместо старых особей попадают хорошо
развитые молодые самки.
Современные способы охоты
В настоящее время косуль добывают в основном с помощью нарезного
оружия и снарядов, минимальная скорость которых при попадании в цель
составляет не менее 800 м/с. Применяют оружие калибров:.5,6X57; 5,6X61; 6,5X57 и
нарезное оружие калибров 7 или 8 мм. На близком расстоянии хорошая
действенность достигается и при стрельбе пулями из гладкоствольного оружия.
Применение картечи приводит к большому числу подранков и во многих странах
запрещено.
Существуют разные способы добычи косуль. Наибольшее распространение
получили загоны, засады, скрадывание, охота с манком во время гона и отстрел с
подъезда. Тот или иной способ охоты выбирается в зависимости от условий
местности, численности дичи, традиций, наклонностей и возможностей охотника.
Засада является, пожалуй, самым предпочтительным способом охоты на
косуль, приводящим к минимальному беспокойству зверей на участке. Стрельба
производится из укрытия, с лестницы или с вышки. Зверей можно длительное время
наблюдать, надежно определять их пол, возраст и сравнивать по качеству.
Из засады по большей части представляется возможность сделать
прицельный выстрел без спешки. Самцов добывают этим способом чаще, чем скрадыва-
нием.
Для засады выбирают места, вблизи которых дичь часто выходит на кормежку:
опушки леса, привлекательные полевые культуры, кормовые поляны в лесу,
богатый кормом древостой, где звери бывают чаще всего по утрам и вечерам.
Если нет охотничьих вышек, то охотник становится по возможности у
ствола дерева, чтобы не выделяться на фоне окружающих предметов. Засада на
земле требует осторожности, неподвижности и бесшумности.
При выборе места засады обязательно учитывают направление ветра и то
обстоятельство, что в ветреную погоду оно столь переменчиво, что засада
нередко становится бессмысленной.
Охота скрадом наиболее эффективна в смешанном ландшафте и в пересеченной
местности. В крупных лесных массивах, при наличии густого подлеска, в камышах
скрадывать их невозможно, равно как и на высокостебельных зерновых и
кукурузных полях. Этот способ имеет свои преимущества по сравнению с засадой,
292
поскольку позволяет обследовать большой участок охотничьих угодий и дает
возможность увидеть большее количество зверей.
Охота скрадом требует максимальной концентрации внимания охотника, умения
бесшумно передвигаться и быстроты реакции. Часто случается так, что на его
пути оказывается косуля, отстрелять которую нет надобности или нет возможности.
В таких случаях осторожно отходят назад или спугивают ее, но так, чтобы она
не начала лаять. Охота может производиться в течение всего дня, так как всегда
какие-нибудь животные активны: зимой она наиболее успешна в полуденное
время; летом наилучшее время для этого утром с 6 до 9 ч, днем с 12 до 13 и весь
вечер начиная с 16 ч. В период гона самцы активны в течение всего дня.
Эффективность охоты во многом зависит от точного знания местности и
поведения косуль. Следует учитывать, что потревоженный зверь часто для
видимости продолжает кормиться, наблюдая при этом за человеком, — голова
его опускается, а затем резко вскидывается вверх. При таком поведении животного
прекращают движение и затаиваются.
Очень добычливым и удобным способом охоты является охота "с подъезда" на
легкой конной повозке, вдвоем.
К полевым косулям можно подъехать на расстояние выстрела, если приближаться
широкими кругами, постепенно сужая их. Стрелок незаметно падает на землю и,
после того как повозка уйдет, делает выстрел. Если есть хорошая сеть дорог
для охоты "с подъезда" аналогично используют автомобиль. Здесь также
дичь нельзя отстреливать с транспорта. Если звери не подпускают автомашину
на дальность выстрела, всегда можно подозревать, что здесь ранее производилась
такая стрельба.
Охота на "манок" заключается в приманивании самцов имитацией звуков,
издаваемых самкой в состоянии течки. Можно имитировать также писк
детеныша для привлечения самки в надежде на то, что за ней последует и самец.
Очень важно при этом выбрать правильную тональность звуков. Прежде
пищащие звуки имитировались с помощью букового листа. Отсюда и возникло
название "листовая охота" или "листковое время". Лист туго растягивают
обеими руками и подносят к сжатому рту. Затем, дуя на него, заставляют
край листа вибрировать. Чтобы добиться правильного звучания, необходима
длительная тренировка. В принципе для изготовления манка пригодны листья и
других видов деревьев, у которых нет пильчатых и зубчатых краев.
Под "листковым временем" обычно подразумевают вторую половину периода
течки, т.е. время приблизительно с 1 по 15 августа в Европе, а в Сибири и на
Дальнем Востоке — позднее этих сроков. В начале гона самцы в первую очередь
покрывают самок, которые находятся на их территории. Теперь они могут
иногда широко бродить по всему участку в поисках других самок, еще
находящихся в состоянии течки, и легко реагируют на пищащие звуки. Охотнику
важно помнить, что нельзя применять "лист" слишком рано (до начала августа)
и нельзя устраивать непрерывных пищащих "концертов" по всему участку.
Слишком часто используемые звуки делают косуль недоверчивыми, и этот участок
может очень быстро стать непригодным для охоты с помощью манка.
Для "листковой" охоты наиболее подходящи жаркие или теплые безветренные
дни, при такой погоде писки хорошо слышимы. Время суток особой роли не
играет, при правильной тональности звучания манка и в надлежащей ситуации
самцы охотно идут на звук. Охота "на манок" нередко применяется в комбинации со
293
скрадыванием или с засадой, при этом с успехом используют не слишком
высокие охотничьи вышки. С помощью манка эффективнее охотиться вдвоем:
стрелок располагается на расстоянии 30—50 м впереди человека, издающего
звуки. Этот способ позволяет отстреливать даже очень осторожных старых
особей.
Подманивают косуль всегда против ветра. Количество пищащих звуков,
которые издаются с интервалом 3—4 с, может варьировать от 3 до 10. Через
20—30 с их повторяют 2—3 раза. Начать манить надо тихо, так как неизвестно,
на каком расстоянии находится зверь. На одном месте не следует манить
дольше получаса. Звуки, имитирующие крик страха и писк детенышей, желательно
применять только в самых исключительных случаях, они очень беспокоят
животных.
В России, где широко практикуется коллективная охота, наибольшее
распространение получили загонные способы добычи животных, при которых загонщики
выгоняют зверей на находящихся в засаде стрелков. При загонной охоте
косули передвигаются быстро, в большинстве случаев стрелки не могут правильно
определить их пол и возраст. Чаще всего под выстрел попадают половозрелые
самки — лидеры групп.
Этот способ по возможности не должен использоваться при низкой численности,
поскольку практически полностью исключает выборочную добычу зверей.
С другой стороны, при необходимости регуляции многочисленных популяций он
может дать неплохие результаты. К тому же комбинация засады с медленным
загоном имеет то преимущество, что позволяет увидеть и отстрелять тех животных,
которые в иных ситуациях наблюдаются очень редко.
Особые трудности представляет собой отстрел полевых косуль в районах, в
которых нет укрытий. Здесь хорошо зарекомендовала себя охватывающая большую
территорию коллективная засада, при которой видимых издалека зверей
направляют на стрелков 1 или 2 загонщика.
В некоторых странах распространена охота с гончими. Она весьма азартна,
интересна и добычлива. Собаки долго преследуют косулю, по-видимому,
оставляющую сильный запах желез, и при встрече предпочитают ее зайцу, часто
бросая след последнего. Однако этот способ имеет много отрицательных сторон
и во многих странах запрещен. В морозную погоду разгоряченные преследованием
звери простужаются и нередко погибают от пневмонии, а при недостатке
кормов не могут полностью восстановить затраченную энергию. Лай собак
вызывает сильное беспокойство животных на участке, и они иногда покидают свой
район обитания. К тому же выборочный отстрел зверей на гону не всегда
возможен. Напротив, применение собак для розыска подранков вполне
оправданно и необходимо. Для этой цели вполне пригодны специально натасканные гончие,
терьеры, легавые, спаниели и другие породы, работающие по кровяному следу.
Правильное поведение стрелка перед выстрелом и после него является
предпосылкой успешного завершения охоты. При любом ее способе везде и
всегда прежде всего важна проблема безопасности людей. Дороги и населенные
пункты должны быть исключены из сектора обстрела, окружающую местность
желательно внимательно осматривать, с соседними стрелками поддерживается
связь с помощью свистка или жестов. При морозе учитывают возможность
рикошета пули от замерзшей земли или стволов деревьев.
Стрелять косулю всегда предпочтительнее в лопатку или в шею. При выстреле
294
необходимо внимательно следить за реакцией дичи, это поможет сделать выводы
о месте попадания пули и направлении бегства зверя. Если после выстрела
зверь не упал в пределах прямой видимости, то его поиск следует начинать
не* ранее чем через 10—15 мин.
При необходимости отстрела нескольких особей стрельба ведется быстро,
прежде чем дичь оправится от первого испуга. Если отстреливают только одну
особь из группы, то после выстрела выжидают в укрытии 10—15 мин. Косули
обычно успокаиваются и начинают снова кормиться. Затем стадо побуждают
к бегству с помощью свистка или окриков и подходят к добытому зверю.
После выстрела при мгновенном падении дичи надо оставаться наготове с
перезаряженным ружьем, в таких случаях зверь нередко снова встает и убегает.
Когда косуля после выстрела только настораживается, то следует предполагать
промах; если убегает, то необходимо обозначить место, где она была при
выстреле и тщательно осмотреть его. По цвету крови, клочьям шерсти или
осколкам костей могут быть сделаны выводы о том, куда попала пуля.
Исторические и запрещенные способы охоты
В прошлые века особенно широко применялись безружейные способы
охоты.
Добыча с помощью ям. Этот способ был распространен на Урале и в Сибири.
По свидетельству Туркина и Сатунина (1902), только в Каслинской лесной даче
на Южном Урале насчитывали тысячи ям и редкий охотник имел их менее
сотни; некоторые местности были совершенно изрыты ими. У каждой ямы
ставилась лакомая *приманка из сена, березовых или осиновых веников.
Внутри ямы делался сруб из бревен, чтобы стенки не осыпались. Сверху клали
несколько тонких жердей, постом прутья, мох, землю и все это заравнивали.
Однако, несмотря на предосторожности, в новую яму звери попадали редко;
они начинали "работать" только на следующий год.
Во многих районах устраивалась система ям, соединенная изгородью из
жердей. Длина изгородей к югу от Красноярска достигала 40 км (Корнилов.
1856), засеки в 20 км длиной видел на Дальнем Востоке Н.М. Пржевальский
(1870), в Забайкалье общая длина городьбы превышала 2 тыс. верст и здесь
были выкопаны более 20 тыс. ям (Бионкур, 1901). На каждую сотню ям с мая по
декабрь в некоторых местностях ловили до 8 лосей, 5 изюбрей и 20 косуль
(Кирилов, 1902).
Отлов капканами. Многие зверопромышленники на Урале и в Сибири
считали отлов косуль капканами более надежным, эффективным и менее
трудоемким способом, полагая, что один капкан заменяет двадцать ям. О добычливости его
и масштабах круглогодичного применения говорит хотя бы такой пример
(Туркин, Сатунин, 1902): "Известный Воскресенский охотник Лев Новгородцев,
который, занимаясь только девять лет охотничьим промыслом, убил, а главным
образом поймал в капканы более 500 косуль; старик-охотник Канциферов из
Иткуля насчитывает более тысячи убитых им коз, но это число, по мнению
других охотников, ниже действительного".
Отлов с помощью изгородей с ловчими двориками. Эти сооружения,
простирающиеся на десятки километров, строились богатыми зверопромышленниками на
путях миграций косуль. Н.В. Туркин и К.А. Сатунин (1902) так описывают
295
устройство одного из них: "По козьей тропе, на протяжении около 40 верст
тянулось "крыло", или "загонная огорода" из протянутых горизонтально длинных
вышиною в 2 и 3 аршина шестов, или слег. По крылу были устроены в разных
местах "дворы", или загоны, обгороженные точно так же, как и самое крыло,
и в загонах, по углам, "кошцы" или маленькие, глухие дворики. На середине крыла
жили в избе два сторожа, да по концам огороды, также в избушках, по одному
сторожу. Во время козьего хода сторожа ежедневно обходили огороды и где
замечали во дворах или кошцах забежавших коз, тотчас давали знать товарищам и
палками забивали до смерти попавшихся зверей".
Отлов пастями. Принцип отлова пастями сходен с отловом ямами или ловчими
двориками с той лишь разницей, что на их место устанавливали "огородные"
пасти (рис. \02,А). Косули шли вдоль изгороди и пытались пролезть в ворота, где и
попадали в пасть.
Кроме огородной, примеиялась поедная пасть (рис. 102,£), которая
устанавливалась на месте жировок косуль. Звери прельщались лакомой приманкой,
задевали насторожку, и на них сверху падала тяжелая колода (Черкасов,
1962).
Добыча луками и самострелами. Специально сделанные луки устанавливались на
тропах косуль, на солонцах, около стогов сена (рис. 102, В). Иногда вместо луков
охотники настораживали старые винтовки и дробовики. Этот способ очень
опасен для людей.
Отлов петлями. До сего времени широко применяется браконьерами, которые
расставляют петли на тропах. Его нельзя назвать особенно уловистым, так как
у косуль хорошо развито чутье и они по большей части обходят петли стороной,
особенно летом. Однако там, где зверей много, этот браконьерский способ
может нанести большой ущерб охотничьему хозяйству и должен искореняться.
Гоньба по насту. Наиболее распространенный в прошлом и самый
губительный способ добычи, приводящий к поголовному истреблению, по сути,
беззащитных животных, неспособных к передвижению по глубокому снегу с настом.
Размах его применения и последствия хорошо иллюстрируют сведения
Л.П. Сабанеева (1875), который пишет, что на Южном Урале весной 1851 г.
только в одной Каслинской даче было добыто более тысячи косуль: "В зиму пришло
их чрезвычайно много, в марте очень долго держался отличный наст, и большая
часть их была переловлена. Обыкновенно после хорошего наста число коз
значительно уменьшается и в следующую зиму иногда не добывается и половины".
В горном Алтае жители с. Уймон весной 1882 г. зарезали до 300 косуль,
согнав их с предгорий в степь, где наст не выдерживал животных (Никольский,
1883).
Не в меньшей степени такой хищнический способ добычи был распространен
и в Восточной Сибири. А.А. Силантьев (1898), ссылаясь на Лаврентьева (1891),
приводит такие факты: "В конце 70-х годов прошлого столетия крестьяне
деревни Балаганки в 50-ти верстах от г. Балаганска, имеющий до 50 дворов,
добыли по насту по 30 косуль на двор, погубив, следовательно, до 1500 голов за
несколько дней. После такой бойни косуль здесь не было 3 года. Жители с. Мотское
Иркутского округа в 1875 г. за неделю по насту затравили 500 косуль,
большинство из которых составляли беременные самки".
Подобное мы находим и у А.А. Черкасова (1962): "В южной части Нерчинско-
го горного округа до 1850 года диких коз было множество повсюду, даже в
296
Рис. 102. Исторические орудия промысла косуль (из: Черкасов, 1962)
А — огородная, Б — поедная пасти; В — лук
безлесных местах; но в зиму этого года снега были чрезвычайно глубоки,
отчего весною, когда образовался наст, все промышленники от мала до велика
бросились за козами и душили их десятками, заганивая на лошадях с собаками.
Вот почему в этой части Забайкалья в продолжении почти десяти лет, а особенно
в первой половине десятилетия, коз было чрезвычайно мало, так что в некоторых
участках этого округа они были выведены почти совершенно".
Как видим, итог такого способа добычи везде одинаков — чрезмерный
промысел настолько угнетал популяцию, что для ее восстановления требовались
годы.
Облавы. Один из самых распространенных способов охоты в Восточной
Сибири, особенно у бурят. В облавах участвовали сотни всадников, сгонявших
всех зверей с округи в одно место, где их окружали плотным кольцом
и расстреливали из луков (Фетисов, 1953).
Отстрел на солонцах. В Сибири, на Дальнем Востоке и в Монголии охота
на солонцах практикуется до сих пор. Нередко браконьеры для приманивания
косуль специально делают искусственные солонцы и оборудуют рядом удобные
засидки. Такие "засады" нам нередко приходилось встречать во всех этих
районах. Добыча косуль осуществляется с весны до осени. В большинстве стран
относится к запрещенным способам.
-Отстрел на миграционных путях. Наиболее добычливый способ с применением
оружия (или без него), практиковавшийся на Урале, в Сибири и на Дальнем Востоке.
Наиболее широко применялся отстрел мигрирующих зверей в местах их
переправ через реки. Они обычно одни и те же на протяжении многих лет, хорошо
известны охотникам, и те в недалеком прошлом съезжались сюда на лошадях
за 200—300 верст (Николаев, 1890), как съезжаются и сейчас уже на автотранспорте.
Иногда на переправах устраивали загоны из жердей — "рукава"; на Иртыше в них
ловили сразу до 200 зверей (Деев, 1911). В прошлом столетии в период массовых
миграций шла заготовка мяса косуль для нужд армии (Шперк, 1885).
Добыча с транспорта "из-под фар". Наиболее распространенный современный
браконьерский способ добычи, особенно на азиатской части ареала косуль.
Звери не боятся автотранспорта и ночью подпускают его на расстояние
выстрела. Ослепленные светом мощной фары или фонаря, они какое-то время
стоят неподвижно, что позволяет сделать выстрел. Ночная неприцельная
стрельба дает много подранков и опасна для окружающих. Невозможность найти
в темноте подранка приводит к бессмысленной гибели большого числа животных.
Этот способ запрещен повсеместно.
Обработка добытой дичи и свойства мяса
Добытая дичь должна быть незамедлительно выпотрошена на месте.
Стрелок прежде выясняет ее возраст, для чего ощупывает указательным пальцем
коренные зубы нижней челюсти, исследуя степень их стертости. Затем добычу
переворачивают на спину, по нижней стороне шеи делают продольный разрез кожи
и раскрывают трахею и пищевод. Трахею вырезают как можно дальше у грудины.
Пищевод с помощью ножа освобождают от мускулатуры и связывают узлом,
иначе при вытягивании пищевода в грудную полость может вылиться содержимое
желудка.
У самца отделяют пенис и семенники от брюшной стенки, но не до конца,
298
поскольку при дальнейшем потрошении возникает опасность, что вследствие
давления содержимое мочевого пузыря прольется на мясо.
Брюшную полость вскрывают осторожно, чтобы не разрезать кишечник,
приблизительно на уровне первоначального положения семенников. В отверстие вводят
указательный и средний пальцы, приподнимают ими брюшную стенку и разрезают
ее до грудины, не повреждая кишок. Для этой цели на некоторых охотничьих
ножах имеются специальные лезвия с тупым концом.
Разрезав брюшную стенку и развернув ее, захватывают пищевод и осторожно
вытягивают его наружу. Затем берут руками желудок и кишечник и вытаскивают
их вместе с печенью из брюшной полости наружу и кладут на правую сторону,
не отрывая прямую кишку.
Вскрывают таз, делая разрез ножом точно между ногами вплоть до
находящегося между костями хрящевого шва. В большинстве случаев его тоже можно
разрезать ножом, но иногда для этого требуется пила. Затем разламывают руками
тазовые кости, захватывают прямую кишку и мочевой пузырь и вытягивают их
назад, отделив от брюшной стенки вместе с пенисом. В этой операции следует
следить за тем, чтобы не повредить инструментом мочевой пузырь.
После этого взрезают расположенные по обе стороны от позвоночника вены
перед тазом и отделяют диафрагму. Затем захватывают трахею выше сердца,
отделяют сердце в том месте, где оно прикреплено к грудине, и вынимают
внутренности. Печень и селезенку отделяют от кишечника и дают всем внутренностям
остыть.
Выпотрошенную тушу поднимают, чтобы стекла кровь, и если имеется
возможность, туловище подвешивают вертикально. Взятый с собой в рюкзак крюк
может сослужить хорошую службу.
В случае нормального выстрела никакая дополнительная работа с тушей не
требуется, но при попадании пули в кишечник, желудок или в грудную клетку с
прорывом пищевода их содержимое попадает в полость тела. Такого рода загрязнения
удаляются с помощью крови. Зачастую содержимое желудка застревает в зоне
ребер между шкурой и тканями, Эти места должны быть вырезаны. Неьзя
производить очистку загрязнений травой, листьями веток или дождевой водой.
В случае попадания пули в область брюшины рекомендуется после вскрытия
брюшной стенки вынуть диафрагму и все внутренности за один прием. Благодаря
этому туловище дичи будет чище.
Перед реализацией дичи у самцов отделяют голову. Для этой цели ее
запрокидывают далеко назад и снизу делают поперечный разрез приблизительно на
высоте гортани у основания черепа (между черепом и первым шейным
позвонком). Затем голову отгибают еще дальше назад, пока череп не выскочит из сустава.
Полное отделение завершается при помощи ножа.
Туша, предназначенная для собственного употребления, должна повисеть вниз
головой 2—3 дня в зависимости от погоды. При этом происходит улучшение
качества мяса. Пока туша находится в подвешенном состоянии, с нее удобно
снять шкуру.
Мясо косуль, как и других диких копытных, обладает ценными свойствами
(табл. 47). Оно характеризуется высоким содержанием мышечной ткани и
относительно слабыми жировыми отложениями. Красный и темно-красный цвет
обусловлен высоким содержанием гемоглобина, а также миоглобина и других
красящих пигментов в крови. Малое содержание соединительной ткани придает
299
Таблица 47. Свойства маса косуль (по: Жнтенко, 1970; с изменениями)
Мышцы
73,0—75,3
Влага
70,3—73,1
А
14,0
Цинк
13,7
Белок
19,5-21,5
В.
0,71
Морфологический состав, %
Соединительная ткань
5,2—8,1
Химический состав, %
Жир Зола
17,2-18,5 1,0-1,1
Содержание витаминов, мг %
В: В«
1,0 0,83
Жировая ткань
1,2—4,0
Другие вещества
0,9—1,2
В,2
6,5
Количественный состав микроэлементов, мг %
Медь
2,19
Марганец Кобальт
0,12 1,26
Молибден
0,068
Кости и хрящи
15,5—17,7
Калорийность, кал
118—155
РР
15,8
Железо
67,1
ему нежность и мягкость. Мясо легко усваивается, обладает несомненными
кулинарными достоинствами, высококалорийно. Основную массу белков составляют
полноценные, содержащие все незаменимые аминокислоты, что говорит о высоком
качестве мяса как белкового продукта.
Основная ценность мяса косули заключается в богатом содержании в нем
витаминов и микроэлементов. В 100 г содержится столько витаминов, что их
достаточно для покрытия суточной потребности взрослого человека. По своему составу
и качеству оно относится к диетическим и лечебным продуктам, которые
рекомендуются при витаминной и минеральной недостаточности или при нарушении
обмена веществ (Житенко, 1970).
Высококачественным пищевым продуктом является и жир косуль, температура
плавления которого находится в пределах 47—48°С. Он также хорошо усваивается
организмом и обладает многими ценными биологическими свойствами. Во
многих странах мясо косули ценится выше мяса других диких и домашних
копытных.
Качество мяса в значительной степени зависит от пола, возраста и упитанности
зверей, но еще большее влияние оказывают условия его получения, т.е. способ
добычи животных. В мышцах длительно преследуемых зверей резко сокращается
количество витаминов. Поэтому при охоте с гончими или загонным способом
ценность мяса ниже, чем при применении других современных способов добычи.
Несравненно ниже по качеству мясо" подранков, добытых через большой
промежуток времени после выстрела.
Обработ
■нка трофеев
С черепа снимают остатки шкуры, отделяют нижнюю челюсть, язык, удаляют все
мышцы и глаза. Мозг измельчают с помощью ложечки или проволочного крючка
и вымывают сильной струей воды через отверстие в основании черепа. Чем
тщательнее перед вываркой будет удален мозг, тем легче будет вычистить и
обезжирить череп.
зоо
Большинство охотников распиливают череп для того, чтобы рога с черепной
коробкой и носовой костью укрепить на деревянной дощечке. Лучше всего это
делать специальной пилой после удаления мышц головы. Сейчас все больше и
больше получает распространение обычай препарировать рога с черепом целиком
и вешать на стену без дощечки. Хорошо развитые рога с целым черепом
производят более сильное впечатление и представляют большую ценность для
науки.
После грубой препаровки череп погружают в холодную воду по меньшей мере
на 24 ч, пока не будет вымыта вся кровь. При этом полезно оставить его в воде
на несколько дней, чтобы начался процесс гниения мышц. Тогда при варке они
отделяются от костей значительно лучше. Если рога необходимо отпрепарировать
вместе с верхней частью черепа, то перед варкой мясистую нёбную оболочку
разрезают поперек, иначе она стягивает и сырывает кости.
Лучше всего варить череп в чистой воде без добавления каких-либо реагентов.
Этим избегается агрессивное воздействие их на кости и сохраняется цвет рогов.
Продолжительность времени выварки зависит от возраста косули. Череп
вынимают, когда мышцы начнут отслаиваться от костей. Кости очищают от крупных
мышц, воду, содержащую жир, заменяют чистой теплой водой и варят дальше,
пока не станет возможным легкое отделение всех мышц.
Череп препарируют, вклеивают выпавшие зубы, высушивают и отбеливают,
многократно протирая ватным тампоном, смоченным теплым 5%-ным раствором
перекиси водорода. Для этого пользуются резиновыми перчатками или
работают с помощью пинцета.
Отбеливание может осуществляться и другим способом. Порошкообразный
мел смешивают с 5%-ным раствором перекиси водорода для получения кашицы.
Череп заворачивают в вату, смазанную этой кашицей, и кладут в мелкую миску
с 5%-ным раствором перекиси водорода. Благодаря подсасывающему действию
ваты, кашица постоянно остается увлажненной. Упакованный таким образом череп
оставляют на 24 ч. Затем вату снимают, череп сушат и чистят щеткой.
При отбеливании следует следить за тем, чтобы не смачивались основания рогов
и зубы, так как в противном случае они потеряют свою естественную окраску.
Не следует производить отбеливание 30%-ным раствором перекиси водорода, как
рекомендуется во многих справочниках. Столь концентрированный раствор
оказывает агрессивное воздействие на кость. Кроме того, применение его
неэкономично и может сильно повредить здоровью человека.
Отпиленный череп укрепляют на подогнанной по размерам и форме
деревянной дощечке. При этом важным является то, чтобы кости черепа равномерно
прилегали к подставке. Трофеи небольшого размера могут быть наклеены на
деревянную подставку. Все рога, параметры которых лежат в пределах медальных,
должны прикрепляться шурупами, так как при трофейной оценке их вес и объем
должны измеряться без подставки. Шурупы закручиваются сквозь доску в
основания рогов.
Особой обработки требуют парикообразные и подобные им рога. Для защиты
от насекомых и загнивания они инъецируются раствором мышьяка с
формалином: на 4 части воды берется 1 часть формалина и 1 часть насыщенного раствора
мышьяка. Для предотвращения съеживания мягких парикообразных или подобных
им уродливых рогов их подвергают парафинированию.
301
/
Рис. 103. Надбавки и скидки при определении массы рогов
(из: Briederman u.a., 1969)
Находящаяся в настоящее время в употреблении международная формула оценки
трофеев косуль такова:
Баллы
1. Длина левого ствола Среднее значение в см X 0,5
Длина правого ствола
2. Масса сухих рогов, г X 0,1
3. Объем рогов, см X 0,3
4. Баллы за красоту
Цвет
Зернистость
Основание (розетки)
Размах рогов
Острия на концах
Прибавки за правильность формы
и качество (с обоснованием)
Скидки за изъяны (с обоснованием)
0—4
0-4
0-4
0—4
0—2
0—5
0—5
Окончательная сумма
Руководство по измерениям. 1. Длина стволов рогов измеряется от нижнего края
розетки вдоль внешней стороны ствола до острия. Измерительную ленту нельзя
вдавливать в угол между верхним краем розетки и стволом, ее следует проводить
по всем изгибам ствола. Для определения максимальной длины нужно в
соответствующих случаях измерить длину рога до конца среднего или заднего
отростка. Длина измеряется с точностью до 0,1 см.
2. Масса сухих рогов. Рога взвешивают на весах с точностью до 1 г. Обычно
принятый вес относится к накоротко спиленным рогам с черепной коробкой и
носовыми костями. В случае взвешивания рогов с полной верхней частью черепа,
включая зубы, следует вычесть 90 г. При какой-либо ином виде спиливания
302
соответственно устанавливают прибавки или скидки (рис. 103). Сухими считаются
рога через 6 недель п!рсле их добычи. Если после препарирования прошло меньше
времени, то их вес уменьшают на 10%.
3. Объем рогов наиболее точно определяют с помощью специальных
гидростатических весов. При помощи столика емкость с водой поднимают на такую
высоту, чтобы стволы рогов по нижний край розеток погрузились в воду. Состояние
равновесия весов изменяется, и для его восстановления на чашку весов кладут гири,
показывающие массу рогов — меру их объема. Рога не должны соприкасаться
со стенками емкости с водой, задняя часть черепа не должна погружаться в воду.
Для примерной оценки объема рогов существует простой метод. Заполненную
водой стеклянную емкость ставят на платформенные или настольные весы и
измеряют ее массу с точностью до 1 г. Затем в воду погружают рога по нижний
край розеток. Получающееся при этом увеличение массы соответствует объему
рогов.
На конференции CIC в Арле (1965 г.) был утвержден в качестве допустимого
приближенный способ VOLZ для рогов, оцениваемых ниже 130 баллов. При
применении его массу рогов умножают на 0,23, получая таким образом сумму баллов
по массе и объему. Однако эти приближенные значения можно применять лишь
для региональной оценки трофеев.
4. Баллы за красоту.
Цвет:
светлые или искусственно окрашенные 0
от желтоватых до светло-коричневых 1
коричневые 2
темно-коричневые, без блеска 3
темно-коричиевые, блестящие, почти черные 4
Это относится к цвету стволов, но не к светло окрашенным выступающим зернам.
Зернистость:
гладкие или почти гладкие стволы 0
слабая зернистость, на обоих стволах лишь несколько крупных 1
или мелких зерен
посредственная зернистость и многочисленные маленькие зерна, 2
которые не полностью покрывают бока стволов
хорошая зернистость, при которой все стороны стволов пол- 3
ностью покрыты небольшими зернами
очень хорошая, богатая зернистость, все стороны стволов 4
плотно покрыты крупными зернами
Розетки:
— узкие и низкие 0
—средних размеров, форма шнура, немного зерен 1
— хорошие веицеобразные, сравнительно высокие 2
— сильные, с широкими и высокими бугристыми венцами 3
— очень сильные, с очень широкими и сильно зернистыми буграми 4
Розетки оцениваются при учете их соотношения с другими частями рогов.
Размах измеряется в том месте, где внутренние стороны обоих стволов наиболее
удалены друг от друга. В случае прямых стволов это бывает по большей части
303
на концах средних отростков, у искривленных — на внутренних сторонах самого
сильного выгиба рогов. Место измерений должно быть обнаружено методом
проб.
Размах:
очень узкий (менее' 30% длины стволов) и очень широкий 0
(более 75%)
узкий (30—35%) 1
средний (35—40%) 2
хороший (40—45%) 3
очень хороший (45—74%) 4
Острия на концах:
тупые и слаборазвитые 0
тупые, посредственно развитые 1
хорошо развитые, острые, белые, отполированные 2
Прибавка за правильность формы и качество:
Полностью симметричным рогам с уравновешенными пропорциями дополнительно присуждается
5 баллов. Количество баллов уменьшается с ухудшением пропорциональности и симметричности.
При оценке рогов с неравномерными стволами и отростками аномально
широкого размаха или пористых может быть снято до 5 баллов.
Разрешается присуждать или снимать не только целые, но и половинные
значения баллов, четвертые доли баллов при этом исключены. Премируются трофеи
европейской и сибирской косуль по сумме баллов отдельно:
Премии Европейская косуля Сибирская косуля
1-й приз (золотая медаль) 130 баллов и выше 160 баллов и выше
2-й приз (серебряная медаль) 115—129,9 140—159,9
3-й приз (бронзовая медаль) 105—114,9 130—139,9
В последние годы появилась ничем не оправданная тенденция оценивать трофеи
европейской и сибирской косуль по разным критериям: у последних, например,
рекомендовано дополнительно измерять длину отростков и т.п. (Фандеев,
Никольская, 1983). И уже в совершенно гипертрофированном виде это проявилось на
IV Всесоюзной выставке охотничьих трофеев, где сибирская косуля оценивалась
отлично от европейской по схеме, близкой к оценке трофеев оленей. Это привело
к тому, что итоговая сумма баллов трофея-победителя равнялась 363,7 балла, т.е.
много выше мирового рекорда (Каталог охотничьих трофеев. М., 1985).
По нашему мнению, оценка трофеев сибирской косули по "оленьему" или
иному принципу наносит серьезный ущерб трофейному делу. Нельзя забывать,
что С. capreolus L. и С. pygargus Pall, являются близкородственными видами.
При оценке трофеев следует принять во внимание результаты исследований
систематики косуль (см. главу "Изменчивость и таксономия"). Северо кавказская
косуля, в частности, является европейской, а не сибирской, как считалось ранее,
и должна быть отнесена к европейской группе. Кроме того, приведенные в настоящей
монографии диагнозы и морфометрические характеристики европейской и
сибирской косуль помогут решать часто возникающие споры о видовой принадлежности
трофеев.
304
Отлов \
Отлов косуль производится в разнообразных целях (в интересах науки, для
переселения в другие районы, для содержания в зоопарках) и представляет собой
хорошую возможность разреживания слишком многочисленных популяций. Существует
несколько способов отлова. ,
Отлов ловушками. Метод основан на заманивании животных с помощью корма
в специальное устройство. В лесу наиболее пригодны стационарные ловушки
средних размеров и небольшие передвижные.
Стационарные ловушки представляют собой сооружения из жердей (рис. 104)
или мелкоячеистой проволочной сетки в виде овала или прямоугольной формы
длиной до 20 м и шириной около 2—2,5 м, снабженные на узких сторонах опадными
или откидными воротами. Боковые стены ловушки иногда изготавливают из
отдельных планок высотой 2 м, которые прибивают гвоздями с такой частотой, чтобы
звери не могли застревать ногами в промежутках. В противном случае переломы
ног неизбежны. Кроме того, сверху прибивают продольные планки для отражения
прыжков, с тем чтобы высота ловушки достигла 2,8—3,0 м. По углам ловушки
можно встроить небольшие ловчие ящики.
Ворота ловушки удерживаются в поднятом состоянии с помощью шнура,
который по роликам проводится к столбу, стоящему в центре ловушки. По этому
столбу шнур идет к спусковому устройству, устроенному на высоте 50 см. При
автоматическом режиме работы ловушки по обе стороны столба натягиваются
тонкие рыболовные лески, идущие от спускового рычага устройства к боковым
стенам. При прикосновении к леске срабатывает спусковое устройство, шнур
освобождается и ворота падают вниз.
Спусковое устройство может также приводиться в действие человеком с помощью
шнура из засады. Однако при этом звери отпугиваются, так как чувствуют запах
человека. Преимущество такого отлова заключается в том, что шнур можно
потянуть к себе в тот момент, когда в ловушке находится нужная добыча.
Передвижная ловушка малых размеров представляет собой крепкий темный
деревянный ящик высотой 1—1,5 м, шириной 0,6—0,8 м и длиной 1,5—2 м, который
может закрываться с помощью подающей дверцы. Он имеет открытое днище с
доской, устроенной в виде педали, к которой прикреплен брусок со шнуром,
идущим к падающей дверце. При прикосновении к педальной доске брусок
высвобождается и падающая дверца закрывает выход. Противоположная стенка ящика
снабжена отверстием, в которое вставляют емкость с кормом. В период
подкормки косули должны иметь доступ к корму как снаружи, так и изнутри, а после
привыкания к кормушке — только изнутри.
Установка ловушек должна производиться с большой тщательностью.
Обязательно проверяют свободу движения падающих ворот, исправное
функционирование шнура и роликов, регулируют спусковое устройство. Зимой на шнурах
оседает иней, утяжеляет и натягивает их, что может привести к произвольному
срабатыванию спускового устройства. Спусковое устройство педальной доски
регулярно очищают, так как на него падает корм из кормушки, после чего оно
не всегда действует. Следует учитывать, что приманка почти всегда привлекает
птиц, поэтому она должна устанавливаться таким образом, чтобы спусковые
лески не могли использоваться птицами в качестве насестов. Иначе возможны
частые ложные срабатывания ловушек.
20. Европейская и сибирская косули
305
] У Г П: «
•-■„ч!* ,>';
■;;--'. ;^:
..:■■!% ■'■%. .. ja
; JJ* ■ "
'■/'•
1
■ :.>■ *>
Рис. 104. Стационарная ловушка
A — вид снаружи; Б — вид изнутри
В качестве приманки в ловушку выкладывают кормовую и сахарную свеклу,
силос из топинамбура, концентрированный корм, снопы овса, сено из люцерны
или клевера, яблоки или силос из яблочных выжимок и другие наиболее
предпочитаемые ими корма. Собственно отлов начинают лишь после того, как будет
хорошо поедаться корм. В сезон отлова нельзя производить подкормку вне
ловушек; в противном случае шансы на успех существенно снижаются.
Настороженные ловушки следует проверять рано утром. При значительной
численности рекомендуется также и одна вечерняя проверка, приблизительно
через час после наступления темноты.
Для манипуляций с пойманными животными требуется 2 человека. В ловушке
малого размера один из них выдвигает дверцу, а другой вытаскивает за задние
ноги косулю наружу. В ловушке среднего размера их загоняют в ловчие ящики
или ловчий коридор или же оба ловца вместе надвигаются на животных,
схватывают в углах ловушки и поднимают за передние и задние ноги спиной вниз.
В этом положении звери весьма ограничены в движениях и могут быть вынесены
из ловушки с небольшими затратами усилий.
В темное время суток косуль отлавливают в ловушках с помощью мощных
фонарей, ослепляя животных светом. Здесь требуется быстрота действий и на
1—2 человека больше, чем при дневном отлове. Нельзя оставлять животных в
ловушке слишком долго, иначе возникает опасность излишнего их беспокойства,
вызываемого людьми и собаками.
Отлов сетями. В период, когда косули не берут подкормку или когда они по
каким-то причинам не идут в ловушки, их можно отлавливать сетями. Успех этой
работы во многом зависит от правильного выбора места отлова. Чем выше
плотность населения животных, тем больше шансов поймать их.
Перед началом работ тщательно обследуют основные места обитания косуль
и выясняют пути их передвижения по участку. Тропы зверей чаще идут в
определенном направлении, что учитывается при организации загонов; сети
ставят перпендикулярно тропам, и загонщики двигаются параллельно им или
по ним.
Необходимо учитывать, что при опасности косули бегут с открытых мест —
лугов, полей, мелких колков, опушек — в лесной массив, а выгнать их из леса
в открытые места весьма сложно или невозможно. По этой причине сети
устанавливают в глубине леса, а загон начинают с опушки.
Лучшее время для отлова косуль — август—декабрь (январь). Зимой ловить
их удобнее при высоком снеге: не надо ставить сплошную линию сетей,
перегораживаются лишь основные тропы, что существенно облегчает и ускоряет
работу. К тому же они в этот период живут группами, и часто удается поймать
за один загон нескольких особей.
В январе.—марте у самцов начинают интенсивно расти новые рога; они очень
хрупкие и легко ломаются при неосторожном обращении с животными при отлове
и транспортировке. Звери с поврежденными рогами могут погибнуть, если не
применять препараты, останавливающие кровотечение. Однако при наличии
квалифицированной ветеринарной помощи в этот период вполне можно совместить отлов
с заготовкой пантов самцов, что значительно повысит доходность работы.
В апреле (мае) рога у самцов становятся твердыми, и их уже можно ловить
без риска, но как раз в эти сроки нежелательно ловить самок, так как они
находятся на последних месяцах беременности.
307
До августа самки кормят телят молоком, и, пока у последних еще не развит
стадный инстинкт и нет необходимых жизненных навыков, позволяющих им
существовать без матери, не следует отлавливать (для расселения) самок. К тому же
летом эффективность отлова будет весьма низка, так как они ведут в это время
одиночный образ жизни и выгнать их за пределы участков обитания нелегко.
Подросших телят поймать почти невозможно, поскольку при опасности они
затаиваются. Тем не менее для мечения в научных целях все же можно ловить конкретных
особей в пределах их участков.
В августе—сентябре сеголетки начинают постоянно следовать за матерью и
не затаиваются при опасности, как раньше, а семьи нередко объединяются в
группы. Животные с этого времени меньше привязаны к своему участку, и их
легче направить в сети.
Необходимо учитывать, что в промерзшую землю очень трудно вбить колышки
при установке сети. Звери же, попавшие в сеть, начинают биться и могут
травмироваться о мерзлую землю. Косуль лучше ловить, когда нет мороза или при
достаточно глубоком снеге в безветренную погоду. При сильном ветре сети часто
падают на землю, если колышки установлены недостаточно прочно.
Ловить косуль следует днем. Рано утром и вечером животные обычно кормятся,
и, устанавливая сети, можно их спугнуть и свести на нет кропотливую работу.
Днем же они чаще находятся на лежке и не так пугливо реагируют на различные
раздражители. Тем не менее в тех местах, где звери не особенно боятся человека
и транспорта, можно, увидев кормящихся косуль, быстро поставить сети на
вероятном пути их движения и сделать загон.
В охотничьих хозяйствах и на научных участках, где необходим регулярный
отлов особей, целесообразнее установить долговременные линии и после загона
сети не сматывать, а лишь спускать на землю. На следующий день их можно
быстро поднять.
Сети для отлова косуль изготавливают из прочного капронового шнура
толщиной 3—4 мм. Оптимальный размер ячейки сети 15X15 см для отлова крупной
сибирской косули и 10X10 см — для европейской. При меньшем размере ячейки
в нее не всегда проскакивает голова, звери плохо запутываются и сеть оказывается
малоуловистой. При большом размере ячейки, например 20X20 см, сеть не
задерживает сеголетов и даже взрослых с&мок. Ширина (высота) сети должна быть в
пределах 2,2—2,7 м. Сети высотой менее 2 м малоуловисты, а 3-метровые тяжелы
•и с ними трудно работать.
Общая длина сети 0,5—2,0 км в зависимости от рельефа местности, наличия
людей и транспорта. Мелкие островки леса лучше окружать сетями полностью,
в больших массивах эффективнее располагать их в виде буквы Е.
Линию сетей составляют из отдельных звеньев, длина которых может быть
различной. С короткими звеньями (25—30 м) удобно работать в зарослях, где
невозможно применить транспортные средства. К тому же при попадании косули
в сеть падают на землю только одно-два соседних коротких звена и не нарушается
цельность всей остальной линии, что позволяет одновременно поймать большее
количество зверей. Однако очень короткие звенья (15 м) сильные самцы сибирской
косули, зацепившись рогами за край, утаскивают за собой на несколько сот метров,
и были случаи, когда они выпутывались или летом их не могли найти вместе с сетями.
Со звеньями сетей длиной более 30 м трудно работать без применения
транспорта из-за их тяжести. При работе на дорогах и просеках с заранее приготов-
308
Рис. 105. Линия сетей
ленной линией колышков при наличии транспорта оптимальная длина звена сети
50—60 м.
При изготовлении сети капроновая дель сажается на более толстый (6—8 мм)
шнур ("тетиву") только с одной стороны без "усадки", которая применяется при
изготовлении рыболовных сетей. "Тетива" должна быть с каждой стороны на 1 м
длиннее отрезка дели. Эти концы шнура служат при необходимости для
связывания собранной сети. Для маскировки сети окрашивают синтетическими
красителями в темный цвет.
Сеть навешивают на вбитые в кору деревьев на 2-метровой высоте небольшие
гвоздики без шляпок или на 2-метровые колышки диаметром 3—4 см, в верхний
конец которых также забивают гвозди (их острие должно выступать наружу
примерно на 2 см), либо подбирают деревце с сучком в верхней части. Колышек
нижним заостренным концом вертикально вгоняется в землю.
Сеть вешается на колышки с внешней стороны от линии загона (рис. 105). Нижний
край сети не следует значительно подворачивать в сторону загона, как это делается
при отлове зайцев: он должен находиться примерно на линии колышков. Смысл
такой установки в следующем. Косуля на бегу проскакивает головой или рогами
в ячейку сети, от удара ее верхний край свободно соскакивает с сучка или гвоздя,
падает и накрывает зверя. Поэтому нельзя сеть устанавливать с внутренней стороны
от линии загона, наматывать "тетиву" на колышки или ветви деревьев или
привязывать сеть к деревьям. В этом случае косуля не сможет сбросить верхний край
сети и не запутается в ней. При установке линии края звеньев не связывают,
а скрепляют травой или сухой веточкой.
309
Техника установки сети следующая. Один из ловцов разносит или развозит
колышки и устанавливает их через 3,5—4 м один от другого. Другой выкладывает
через определенные интервалы мешки, в которые аккуратно уложены звенья сети.
Еще три ловца разматывают сети и навешивают их на колышки. Быстрее всего
работать двумя такими группами, устанавливая сети от середины линии к ее краям.
При определенном навыке километр сети одна группа устанавливает примерно за
2—3 ч, при готовой линии колышков — за 1 ч.
Сети нельзя устанавливать посредине полян, просек и дорог. Их необходимо
располагать как можно ближе к деревьям и кустам со стороны загона; так
они будут менее заметны.
Косули, бегущие от загонщиков, реагируют на сеть с расстояния от 3 до 40 м и
часто бегут параллельно линии или поворачивают назад. Поэтому ее края надо
обязательно загибать в сторону загона. Лишь внезапно напуганные и быстро
бегущие звери не успевают вовремя среагировать и попадают в нее. На этой
особенности и основана тактика отлова их сетями.
Для отлова необходимы две группы людей — ловцы и загонщики. Количество
тех и других может быть различным в зависимости от длины сетей, рельефа
местности, наличия транспорта и т.д., но желательно, чтобы на километр линии
сетей приходилось не менее 8 ловцов и 10 загонщиков.
Задача загонщиков — выставить зверей к сети и не дать уйти им назад. Обычно
цепь загонщиков с выдвинутыми вперед флангами с небольшим шумом двигается
в сторону сети, соблюдая равнение, заданное направление и дистанцию. При
подходе к сетям шум усиливают. Зимой, при недостатке загонщиков, один или
два человека, хорошо распутывающие следы, идя за косулями, могут выставить
их к линии сетей. В загонах можно применять специально обученных незлобных
охотничьих собак.
Задача ловцов — напугать бегущих животных и затем удерживать их в сети.
Ловцы располагаются перед линией на расстоянии примерно 40—50 м от нее
и в 70—100 м друг от друга и ждут появления косуль. Как и на охоте, на номерах
надо замаскироваться, нельзя шуметь, передвигаться и курить. Бегущих от
загонщиков животных ловец пропускает мимо и, как только оказывается позади, с
криком бежит за ним, а затем удерживает в сети, не давая биться о землю и деревья,
что нередко приводит к серьезным травмам.
У косули сильные задние конечности и острые копыта, поэтому безопаснее
подходить к ней со спины и удерживать за ноги. Необходимо учитывать, что
рога у самцов очень шероховатые и острые, из-за чего работать лучше в
брезентовых рукавицах и в ватных куртках.
Пойманному зверю закрывают глаза (для чего нужно1 сделать специальный
темный рукав с резинками на концах), выпутывают из сети и связывают ноги
мягким ремнем. Если отлов происходит зимой, его во избежание простуды кладут
на какую-либо подстилку: мешок, ватник и т.п. Если загон продолжается, ловцы
устанавливают упавшую сеть на колышки и быстро встают на свои места.
Животных нельзя долго держать со связанными ногами. Их метят и выпускают,
а если необходимо передержать, сажают в транспортные ящики и доставляют к
месту назначения, причем на близкое расстояние перевозят и без ящиков. Для
успокоения им вводят транквилизаторы, снимающие нервное напряжение и
купирующие двигательное возбуждение.
Мы испытали также беззагонный способ отлова. Летом линией сетей перекры-
310
вали пути перехода зверей к водопою или к кормовым полянам и оставляли ее в
настороженном состоянии на сутки, проверяя через каждые 3—4 ч. При большой
плотности населения косуль удается ловить таким образом преимущественно
самцов, вероятно менее осторожных и более подвижных. Однако в этом случае
при неправильной установке сетей (если верхний край по каким-то причинам не
падает на землю) весьма нередки случаи гибели животных. Кроме того, ночью
в сети попадают кабаны. Поэтому рекомендовать данный способ можно лишь для
научных целей при тщательной установке сетей на ровной местности и частой их
проверке.
В целом отлов косуль сетями, вероятно, приводит к большей гибели животных,
чем отлов живоловушками. По крайней мере следует иметь в виду, что он
неизбежен и может доходить до 5—10% от общего числа пойманных животных.
Основная причина гибели — травмирование в сетях (преимущественно разрывы
мышц задних ног и повреждения сухожилий). Травмы эти выявляются только через
несколько часов после отлова; животные не встают или волочат зад, стирая при
передвижении кожу на задних ногах до кости. Таких зверей необходимо помещать
для лечения в тесные клетки или, если после выпуска они оказались в природе,
не тревожить.
Иммобилизацию (обездвиживание) косуль обычно применяют уже после отлова
их в сети или ловушки и в вольерах. Отлов этим способом в природе, как мы
убедились, недостаточно эффективен: требуется много времени на выслеживание
животного; цель относительно невелика, и на большом расстоянии нет гарантии
точного попадания; толщина мышц мала, и нередко снаряд попадает в брюшину
или в кость, что приводит к травмированию или гибели животного. Не удается
пока отработать достаточную эффективную методику обездвиживания с помощью
добавок препаратов в подкормку; у косуль очень хорошее чутье, и звери
отказываются от такого корма.
Для обездвиживания косуль из известных препаратов, вероятно, лучше всего
подходит комбинация ксилазина (ромпуна) и кетамин-гидрохлорида в соотношении
1:2 из расчета 3 мг/кг массы животного. Эта доза при слабом действии может
быть увеличена на 50%. После внутримышечной инъекции первые проявления
действия препарата наступают спустя 2—4 мин, а через 5—15 мин происходит
полное обездвиживание. Препарат действует около 45 мин, однако остаточные
явления и сонливость проявляются на- протяжении 5—8 ч (Eisfeld, 1974).
На опыте мы убедились, что применение других препаратов, таких, как аминазин,
дитилин (листенон), диплацин, гексенал, барбамил, этаминал натрий, сернилен
и ромпун в принципе возможно, но менее удобно из-за большого объема, или
недостаточно быстрого и эффективного действия, или побочных нежелательных
явлений, приводящих к гибели животных.
Лучше препараты применять в комбинациях. Для успокаивания отловленных
животных можно применять комбинацию вполне доступных для использования
барбитурата этаминала с аминазином в соотношении 30—35:15—20 мг/кг для
вольерных и 25:10—15 мг/кг для свободноживущих животных. При передозировке
внутримышечно вводят 0,5 мл 10%-ного раствора коразола (Блузма, 1975).
Применение обездвиживания при отлове, передержке, транспортировке и при
манипуляциях с животными позволяет сохранить им жизнь.
311
Мечение
Мечение отловленных косуль безусловно необходимо. Без него невозможно
получить достоверные данные о перемещении зверей, их экологии и поведении,
немеченая популяция "безлика". Результаты мечения особенно ценны для научных
обобщений, на которых, по сути, и базируется рациональное охотничье хозяйство.
В большинстве случаев в руки охотников и специалистов попадают детеныши
косуль в возрасте до 2 недель. Они при опасности затаиваются, что облегчает
их отлов. Мечение телят и взрослых животных обычно производится различными
типами номерных алюминиевых сережек и пластиковыми кнопками в ухо. Однако
эти метки незаметны и малоинформатйвны. Гораздо лучший результат можно
получить, используя те же самые ушные кнопки, но подкладывая под них с обеих
сторон уха тонкие пластинки из цветного пластика разной формы, размера, цвета.
Чем больше их размер, тем заметнее они, однако пластинки более 7 см в диаметре
мешают животному и возрастает возможность зацепления их за кусты. Такая
метка позволяет уже многократно идентифицировать зверя на большом расстоянии.
Ухо для маркировки прокалывают специальными щипцами, предварительно
протерев режущий край и саму метку спиртом. Далеко не все ушные метки
сохраняются длительное время, поэтому желательно вставлять их в оба уха.
Взрослых зверей лучше метить ошейниками с яркими номерами. Ошейник
заметнее ушной метки, дольше сохраняется, и возможно длительное, иногда
в течение всей жизни, наблюдение за особью, что позволяет получить
уникальный материал. Наиболее подходят для этой цели мягкие кожаные или
капроновые ошейники с большими номерными пластинками, однако изготовление их
дорого и сложно.
При необходимости номер на ошейник можно сделать так: плотную
полиэтиленовую пленку нарезают на равные прямоугольники, обезжиривают спиртом
и несколько листов склеивают через материю горячим утюгом до получения
достаточно прочной основы, на которую таким же образом наносят слой цветной
(лучше черной) пленки. Затем на цветное основание приклеивают цифры,
вырезанные из полиэтилена другого цвета (лучше красного) или цветной бумаги,
которые сверху закрывают 2—3 слоями прозрачной пленки, проглаживая утюгом.
Номер пришивают капроновыми нитками сбоку ошейника с двух сторон.
Для долговременных научных исследований полезнее метить животных и
ошейниками и ушными метками, но эффективнее всего — радиопередатчиками
(рис. 106). Зверей с ушными кнопками регистрируют чаще всего дважды (при отлове
и отстреле); с номерными ошейниками — десятки раз; радиопередатчик дает
возможность непрерывного наблюдения за ними в течение 1—2 лет, после чего
особи, маркированные еще и номером на ошейнике, встречаются длительное время.
Снабженные радиопередатчиками животные могут быть очень полезны при
охоте на других особей: по пеленгу можно быстро отыскать группу и отстрелять
нужного зверя. Если же меченая косуля находится в одиночестве, то при
медленном преследовании она всегда приведет охотника или исследователя к другим
животным, находящимся на этом участке. Этим способом можно с большой
точностью проводить их учет.
Таким образом, внедрение снабженных радиопередатчиками особей в
популяцию безусловно будет всесторонне полезно как для научных исследований,
так и.для практических целей. Радиопередатчики позволяют длительное слежение
312
ж
Рис. 106.Оснащение косули радиопередатчиком
за мигрирующими популяциями с помощью самолетов или искусственных
спутников.
Очень важны при мечении точные записи, для чего на каждую особь
заполняют карточку с указанием места и даты мечения, пола, возраста, номера ушной
метки и ошейника, цвета метки. Эти сведения должны находиться в охотничьем
хозяйстве или заповеднике. На маркированного зверя заводится регистрационная
карточка, куда заносят все сообщения о местах встреч и составе групп, наличии
и качестве потомства и т.п.
Передержка, транспортировка и выпуск
При перевозке в другие места животных, помеченных на месте отлова, сразу же
доставляют на сборный пункт в специально оборудованные вольеры или сараи.
Здесь они содержатся все то время, пока идет их накопление. Этот период
передержки необходим и в ветеринарных целях. Однако многолетняя практика
переселения косуль показывает, что в момент передержки и транспортировки
происходит их максимальный отход (до 80% от числа пойманных), причины которого
до конца не были выяснены. Считалось, что животные погибают от стресса, что,
скорее всего, имеет место. Однако сейчас ясно и то, что одной из основных причин
гибели являются травмы, полученные ими как в сетях и ловушках, так и во время
передержки. Цри приближении к загону человека или собак косули очень пугаются
и прыгают на стены, ударяются головой, туловищем, ломают ноги, позвоночник.
Особенно губительным для них оказывается повторный отлов на базе передержки.
Поэтому в целях исключения гибели животных мы рекомендуем полностью
313
отказаться от передержки на месте отлова. После отлова, иммобилизации и
мечения их желательно сразу же помещать в темные транспортные клетки, в
которых можно передержать 1—2 дня и, собрав партию, перевозить в них к месту
назначения. Перевозить на большие расстояния (свыше 3 сут) лучше молодых или ручных
особей, отловленных в раннем возрасте и выращенных в неволе.
Правильная транспортировка имеет большое значение для сохранения животных.
Предварительно проведенной иммобилизацией можно снизить опасность их стрес-
сирования и травмирования. Однако при сильном морозе обездвиживать зверей
не следует, так как при этом сильно снижается интенсивность кровообращения,
что может привести к гибели от переохлаждения.
В любом случае лучше перевозить животных в темных закрытых ящиках,
оборудованных двумя выдвижными дверцами и снабженных сбоку
вентиляционными отверстиями. Если требуется транспортировка на большое расстояние, то
в ящик со стороны головы зверя кладут корм: сено, измельченные яблоки, свеклу,
брюкву и др. и ставят воду. Растущие рога самцов защищают повязками, а верх
ящика в этом случае затягивают плотной тканью.
Размеры транспортных ящиков различны в зависимости от вида и подвида
перевозимых косуль: для сибирских особей длина и высота 130—150X110—120 см,
ширина 50 см; для европейских — 115X85X45 см. Стенки ящиков, особенно дверцы,
не должны иметь щелей, выступающих острых углов, планок, сучков, гвоздей и т.п.,
иначе звери будут сильно травмироваться.
При транспортировке следует избегать переохлаждения или перегрева животных.
По прибытии на место назначения ящики открывают с той стороны, где находится
голова зверя, и дают им возможность выйти самостоятельно. Некоторые животные
выскакивают наружу тотчас, другие, в особенности подвергнутые иммобилизации,
выходят наружу лишь спустя продолжительное время.
На месте выпуска косуль желательно передержать до весны в большой вольере,
где они могут найти в изобилии корм и укрытия в виде густых зарослей. Весной
вместе с самками лучше содержать не более одного взрослого самца. Иначе
неизбежны драки между ними и сильное беспокойство беременных самок.
Соотношение полов от 1:2 до 1:5 в пользу самок при расселении более
оправданно и с биологической и экономической точки зрения, чем обычно
рекомендуемое 1:1.
Выпускать животных на волю лучше за 3—4 недели до отела, т.е. примерно
через 2 недели после таяния снега, оставив открытыми ворота загона, куда они
могут свободно заходить на кормежку. В этот момент самцы сразу же станут
занимать территории, а самки выбирать родовые участки. Так их можно удержать
вблизи вольеры. При выпусках в другое время года или без передержки нет
гарантии, что звери останутся в данном охотничьем хозяйстве. Однако
запаздывание с выпуском может привести к боям взрослых самцов за территорию в вольере,
что может кончиться гибелью не только их, но и отдельных самок.
Выращивание детенышей
Практически ежегодно многим охотникам, сотрудникам охотничьих хозяйств,
заповедников, а также любителям природы приходится выращивать детенышей
косули. Чаще всего их приносят люди, экологически неграмотные, считающие
затаившегося теленка отставшим от матери. В таких случаях его немедленно
314
Таблица 48. Рекомендуемый корм для выращивания детенышей косули (по: Drescher-Kaden, 1974)
Возраст, дни
Число
порций
Суточная
потребность, мл
Рецептура пойла
соотношение
коровьего и
сухого молока
концентрация
эаменителя
молока, %
Комбикорм, г
1
2
3
4
5—10
11 — 15
16—20
21-30
31-40
41—50
51-60
с 60
5—7
5—7
5—6
5—6
4—5
4-5
4-5
4
4
3
2
1—2
200-300
300—400
400—450
400—500
500—700
700—800
800—900
800—1000
800—1000
700—800
500—600
0—500
2:
2:
3:
1;
Примечание. СП — 15-го дня следует давать траву и листья деревьев по потребности.
10
10
10—15
15—20
20
20
20
15-20
10-15
10
10
10
—
—
- —
—
—
—
0—25
25—100
100-150
150-200
200—250
более 300
доставляют назад; мать непременно найдет детеныша и примет. При выращивании
диких животных необходимо ставить в известность органы охотнадзора, которые
принимают решение о дальнейшей их судьбе.
Часто телят косули приходится искусственно выращивать в зоопарках, зоосадах,
при разведении на специальных косульих фермах и в загонах. К настоящему
времени накоплен достаточно богатый опыт.
Содержать детенышей косуль лучше всего в небольших открытых вольерах
с убежищами от сквозняка и дождя, оборудованных деревянными полами, или
в хлевах с деревянным полом, подстилкой из соломы и древесных стружек.
Молодые животные очень чувствительны к переохлаждению, поэтому при
снижении окружающей температуры ниже +12°С, устанавливают дополнительный
источник обогрева (инфракрасный излучатель).
Детеныши в первые недели жизни в естественных условиях прибавляют в весе
приблизительно 100—150 г в день. К достижению таких показателей следует
стремиться и при их искусственном выкармливании.
Молоко косули имеет высокое содержание сухих веществ, жира и белков (см.
табл. 19). Козье молоко (3,4% белков и 4,1% жира) или овечье (5,4 и 6,2%) лучше
подходят для выращивания детенышей, чем коровье. Смеси коровьего молока
с овсяным отваром (1:1) или с сухим молоком в пропорциях, зависящих от возраста
детеныша, дают при выкармливании хорошие результаты (табл. 48).
С успехом применяют также заменители молока, предназначенные для
выращивания телят коров (примерно 17% жира и 23% белков в сухом веществе) и для
выращивания ягнят (26 и 22% соответственно). Концентрация пойла — 180—200 г
заменителя молока на 1 л теплой кипяченой (в первые 10 дней жизни) или же
очень чистой воды. Иногда для кормления косулят используют цельное коровье
молоко с обязательным добавлением воды (80 г на 1 л). Потребность молока на
315
одного детеныша в этом случае 1,0—1,3 л в день и от 73 до 115 л за период выпаивания
(Tschiderer, 1973).
Считается, что период выкармливания детенышей косуль только молоком или его
заменителем составляет приблизительно 10—20 дней. Однако и в это время
детеныши должны иметь доступ к траве и свежесрезанным побегам лиственных
деревьев, а позднее — к комбикорму с возможно более высоким содержанием
белков и минеральных веществ. Через 2 месяца после рождения кормить
детенышей молоком прекращают, но увеличивают порцию естественных кормов и
комбикормов.
Наибольшие сложности возникают на начальной стадии привыкания их к
человеку. В первые 2—4 дня из-за стресса детеныши могут отказываться от молока
и их приходится кормить принудительно. В первые 2—3 недели вместе с кормом
им дают какой-либо антибиотик, а в случае появления признаков энтерита поят
теплым крепким чаем или слабым отваром дубовой коры, сокращая эквивалентно
порцию молока или его заменителя. Молоко должно иметь температуру 38—40°,
слишком холодное приводит к кишечным заболеваниям.
Для кормления применяют бутылку с резиновой соской. Остатки корма
ликвидируют, емкость и соски ежедневно стерилизуют. Теленку в период выпаивания
каждый день предлагают свежую воду из бутылки, но она всегда должна
находиться в свободном доступе в вольере. При кормлении детенышу производят
массаж в области пупка и заднего прохода, что способствует регулярному
опорожнению кишечника.
Наипервейшей заповедью должна быть гигиена и чистота всего, что окружает
детеныша. Для общего укрепления организма можно добавлять в корм витамины А
и Д, глюкозу, кальциферол, смесь минеральных веществ, кормовой мел и кормовые
дрожжи. Наконец, при выращивании одиночных детенышей нельзя забывать о
ежедневной индивидуальной игре с ними.
Содержание в вольерах
Косуль трудно содержать в неволе, особенно на маленькой площади: звери
подвержены стрессу, инвазиям, есть сложности с кормлением, самцы очень
агрессивны, поэтому они живут не в каждом зоопарке.
Общие требования к разведению косуль в неволе следующие.
1. Животных содержат в просторных вольерах, где имеются естественные или
искусственные укрытия от непогоды и беспокойства.
2. При большом количестве животных вольеры разделяют на секции, жела-
тельнопо 500—1000 м2, в которых взрослые самцы в летний период содержатся
по одному с 1—2 самками и их детенышами.
3. Беременным и лактирующим самкам по возможности выделяют отдельный
загон.
4. Питание животных должно быть обильным, полноценным и разнообразным.
5. Регулярно проводят ветеринарные мероприятия, особенно направленные
на борьбу с гельминтами.
Соблюдение этих правил позволит сохранить косуль и добиться интенсивного
воспроизводства. При условии хорошего содержания каждая половозрелая самка
приносит не менее двух детенышей ежегодно. Меньший прирост говорит о плохих
условиях содержания.
Вольеры огораживают проволочной сеткой высотой не менее 2—2,2 м, заглубляя
316
ее в землю на 25—50 см. Ворота снабжают бетонными или крепкими деревянными
порогами для предотвращения проникновения бродячих собак внутрь. С внешней
стороны вольеры целесообразно установить ловушки с опадными дверцами для их
отлова. Внутри загона на расстоянии 1 м от сетки на высоте 60—80 см желательно
сделать барьер из жердей, препятствующий подходу косуль к сетке. Иначе
агрессивные самцы будут подолгу бодать сетку при виде людей или собак, что приводит
к разрушению ограды или к гибели зверя — зацепившись рогами за сетку он
иногда ломает шейные позвонки и погибает. .
Вольеру предпочтительнее делать близкой к форме круга или эллипса,
устанавливая столбы с внешней стороны; это предотвратит травмирование животных
при ударах о стены и даст им возможность двигаться по кругу во время гона или
беспокойства. В период гона кормушки и другие сооружения размещают в центре
вольеры, что способствует проявлению полового поведения и позволяет
многократные пробежки вокруг предметов.
Минимальный размер секции при содержании взрослого самца и 1—2 самок
с детенышами — около 100 м2. В таких маленьких боксах в качестве грунта
рекомендуется гравий с песком, а вдоль изгородей укладывают плиты. Здесь же
устраивают полуоткрытый хлев с дощатым полом площадью около 12 м2. Примерно
такую же площадь занимает песочница, предназначенная для отдыха зверей всухую
погоду. Кормление производится в хлеву из кормушки, снабженной подвижной
крышкой, которую при еде они должны отталкивать. Хлев может служить
ловушкой; это необходимо для взвешивания, ветеринарных целей или перевода
в другой загон. Загоны должны сообщаться через 2 двери, которые открывают
во время гона, что позволяет животным бегать по кругу (Eisfeld, 1974).
Практика показывает, что либо косуль в загонах нужно как можно меньше
беспокоить, либо необходим более тесный контакт с ними. В первом случае корм
лучше засыпать в кормушки, стоящие у сетки, извне; во втором — размещать
в центре вольеры. Кормушка должна быть долговременной, переносной, удобной
для животных, с устройствами, предохраняющими корм от попадания влаги и
растаскивания его птицами и грызунами.
Питание косуль в неволе должно быть разнообразным и соответствующим
чувствительности пищеварительной системы. Помимо концентрированных кормов
(комбикормов, пшеницы, овса или овсяных хлопьев), в зависимости от времени
года им выкладывают различные корнеплоды и фрукты (нарезанные морковь,
свеклу, яблоки, груши; зимой — сухофрукты), траву, ветви и листья деревьев
и кустарников, сено хорошего качества, желуди.
При кормежке только сухим кормом (отруби, зерновая сечка) у них развивается
энтерит, который ликвидируется добавлением в рацион побегов лиственных
деревьев и сена, заготовленного до цветения трав. Такой корм хорошо поедается,
если его закладывать в виде сечки по 5—10 см длиной. Зимой лучшим кормом
является силос, богатый белками и восполняющий их физиологическую
потребность в воде (Dissen, 1983).
Для профилактики кишечных заболеваний им систематически дают вместо
воды отвар чая и приблизительно 6 раз в год проводят дегельминтизацию:
рекомендуется препарат тиабендазол из расчета 250 мг/ кг массы тела или тетрамизол
(нилверм) в дозировке 20 мг/кг. Наилучшим образом зарекомендовал себя
и мебендазол; скармливание корма с примесью этого медикамента не представляет
проблем (Tschirch, 1984).
317
В вольерах обращают внимание и на социальную жизнь животных; взаимная
нетерпимость приводит к потерям. Агрессивные самки, толкая и гоняя других
самок, могут вызвать у них выкидыши; агрессивные самцы представляют
опасность друг для друга и для самок, особенно весной, и их необходимо
своевременно отделять от других зверей. Однако повторное сведение животных перед
течкой снова вызывает затруднения, они не скоро привыкают друг к другу. Поэтому
в вольерах лучше работать с ручными особями.
Соблюдение перечисленных выше рекомендаций позволит снизить смертность
косуль в загонах, тем не менее она неизбежна.
При работе с самцами следует соблюдать осторожность, так как летом многие
из них весьма агрессивны по отношению к человеку и могут нанести серьезные
травмы. В таких случаях рога у них спиливают.
Биотехнические мероприятия
При планировании биотехнических мероприятий принимают во внимание
следующие аспекты биологии косуль.
Значительную часть года косули живут поодиночке или же небольшими семьями
более или менее равномерно распределенными по всему биотопу, при этом звери
предпочитают краевые зоны лесов с густым подлеском.
Для питания им требуется большое количество легко перевариваемого и
калорийного корма, максимальное летом и осенью. Запасание энергии является
решающим для нормального развития эмбрионов, роста молодых животных и для
развития рогов.
В суровые зимы с высоким снежным покровом возникают большие потери
животных, если не будут приняты соответствующие меры.
Качество популяции во многом зависит от условий существования, и в первую
очередь от качества угодий. Практика охотничьего хозяйства, показывает, что
разумными мероприятиями можно (и необходимо) повысить кормовую емкость
биотопов, т.е. можно значительно улучшить условия обитания животных. Только
таким путем реально добиться успехов в деле качественного улучшения какой-
либо популяции косуль и сохранить ее численность в районах с высоким снежным
покровом.
Улучшение биотопа. Мероприятия по улучшению биотопов наиболее просты
и эффективны, если они проводятся в комбинации с лесохозяйственным
устройством. Развитию наземной растительности, необходимой для питания животных,
способствует прореживание древостоя и удобрение лесопосадок, в частности
известкование в еловых лесах и внесение богатых азотом минеральных
удобрений.
Закладка посадок, предназначенных исключительно для питания косуль,
необходимо только в редких случаях. В большинстве охотничьих угодий имеется
достаточное количество пригодных для кормежки деревьев, которые нуждаются
лишь в уходе. Поскольку звери поедают однолетние побеги, то все деревья и пневую
поросль, предназначаемые для кормежки, ежегодно подрезают до доступной
высоты.
Наибольшую ценность представляют плодоносящие деревья: дубы, буки,
каштаны, яблони, груши, дающие высококалорийные и богатые витаминами плоды,
что способствует накоплению жировых запасов и обеспечивает благополучную
318
зимовку. К сожалению, не все деревья плодоносят каждый год; на большой урожай
бука можно рассчитывать лишь один раз в семь лет, одиночно стоящие дубы
плодоносят значительно чаще, а красный дуб и конский каштан — почти каждый
год. Плодоношение увеличивается при обрезке крон. Посадка таких деревьев
в охотничьих угодьях очень желательна. Высаживают их саженцами,
недосягаемыми для объедания и повреждения рогами, а на огороженных лесопосадках
их сажают по краям или по углам (при этом отпадает необходимость
дополнительной защиты). Порубочные остатки лиственных деревьев на лесосеках
не должны сразу уничтожаться и могут служить ценным кормом для животных.
В еще большей степени это относится к фруктовым садам. Во многих районах
европейской части России и Украины именно ветви яблонь при их обрезке зимой
служат основным источником корма косуль и только наличие такого корма
позволяет им выжить при глубокоснежье.
В полевых районах все мероприятия по посадке лесных полос и кустарников
способствуют улучшению биотопа. В первые годы закладки насаждений плотность
населения зверей поддерживают на низком уровне, чтобы предотвратить потравы
и поврежденя деревьев рогами. При большой их численности сажают
преимущественно такие деревья, которые животные избегают поедать.
Закладка кормовых площадей. Во многих биотопах обеднение и ухудшение
естественной наземной растительности, нередко вызванное большой плотностью
населения копытной дичи, усугубляется применением гербицидов, выпасом скота
и недостаточным уходом за полянами. В противовес этому необходимо ис-
кусствено создавать кормовые площади, повышая тем самым кормовую емкость
биотопа.
Если для благородных оленей, ланей, муфлонов и кабанов достаточно
небольшого количества крупных хорошо ухоженных кормовых полян, то для косуль
по возможности закладывают много небольших (около 0,5 га) площадок,
распределенных более или менее равномерно по всему участку.
Большинство площадей для кормежки должны быть долговременными.
В качестве таковых могут служить любые свободные открытые участки:
заброшенные поля, поляны, всевозможные просеки. Закладка кормовой площади состоит
в одноразовой пропашке с последующим засевом смеси клевера с разнотравьем.
Впоследствии сюда ежегодно вносят удобрения, каждый второй или третий год
боронуют и снова высевают клевер. В зависимости от степени поедания и условий
роста травы здесь 1 или 2 раза в год проводят сенокос, оставляя сено в кормушках.
Долговременные площадки имеют большое значение только в том случае, если они
постоянно снабжают зверей свежим зеленым кормом.
В лесных районах с преобладанием хвойных пород целесообразна закладка
кормовых площадей из саженцев осины, ивы козьей, рябины и черемухи высотой
80—160 см, при одновременном высеве смеси из клевера, гречихи, люпина и
внесении удобрений. Такие кормовые площадки во избежание зарастания нуждаются
в постоянном уходе за ними (Kromschoder, 1982). На подкормочных площадках
в лесу никогда не будут лишними ежевика, малина, черника, а также яррвая
пшеница, овес, горох, вика (Meidel, 1982).
Неоценимое значение для косуль имеют луга, но ничто не делает их более
непригодными, как застойная влага и как ее следствие появление непоедаемых трав,
равно как и повышение опасности заражения печеночной двуусткой. К важнейшим
мероприятиям по улучшению лугов относится мелиорация: сырые и закисленные
319
прежде всего избавляют от избытка воды с помощью дренажа или очистки старых
канав. Обильное внесение удобрений — извести, фосфорной кислоты, калия, навоза,
компоста и др. — дает существенное улучшение качества травостоя. Еще лучше
произвести полную перепашку луга и засев его заново подходящей для данной
местности смесью семян с преобладанием клевера. Со временем клевер вытесняется
другими травами и луг теряет ценность для косуль, в этих случаях весной после
боронования его снова подсевают. Получающие хороший уход луга дают отличное
сено для кормления дичи.
Хорошим кормом для косуль летом является сладкий люпин, горошек мохнатый,
сераделла, гречиха, клевер и люцерна. Последняя особенно ценна на кормовых
площадях, поскольку при хорошем уходе может служить кормовым растением
8—10 лет; скашивая ее 2—3 раза в год, получают отличное сено; к зиме она образует
высокую стерню, что делает эти площади пригодными для зимнего питания.
Косули также охотно поедают стебли и клубни топинамбура и картофеля, но
к топинамбуру как к корму им нужно привыкнуть. В районах с высоким снежным
покровом желательно использовать высокостебельные растения: озимую рожь,
посеянную в июле—августе, подсолнечник, кукурузу, иван-чай и др. На Украине
рекомендованы сильфия пронзеннолистная, борщевник Сосновского и гречиха
Вейриха (Рудышин и др., 1979).
На Дальнем Востоке превосходным кормом для сибирской косули оказалась
соя; зимой звери концентрируются возле полей, на которых после уборки осталось
жнивье. Животные, обитающие на соевых полях, даже в исключительно
многоснежные зимы, как, например, в 1972/73 г., остаются высокоупитанными (Сапаев,
1976), тогда как в других районах в это же время отмечалась массовая гибель
от истощения (Кучеренко, Швец, 1977). Вполне уместно в данном регионе
использовать на кормовых площадях преимущественно это растение. Эксперименты
с соей желательны и в других районах.
В разумном сочетании-интересов охотничьего и сельского хозяйства кроются
огромные потенциальные возможности интенсификации производства мясной
продукции дичи и косуль в особенности. При хорошо наложенном сотрудничестве
с сельским хозяйством возникают дальнейшие возможности улучшения подкормки
животных. Вдоль опушек леса или же среди полей могут оставляться или
специально засеваться полосы сельскохозяйственных культур шириной до нескольких
десятков метров, служащие зимой в качестве кормовых площадок. С точки зрения
сельского хозяйства допустимо, а иногда и желательно оставлять поля с высокой
стерней, что предотвращает землю от ветровой эрозии и задерживает снег; для
косуль они становятся основным зимним пастбищем. К примеру, даже после уборки
урожая на 1 га соевого поля остается такой запас корма, которого хватает в течение
зимы 5—10 особям (Дарман, 1986). Равным образом ценным кормом их могут
снабдить свекольные и картофельные поля, если после уборки они не
перепахиваются, а ботва складируется в кучи.
Устройство солонцов и водоемов. В охотничьих угодьях на каждые 50—100 га
оборудуется один солонец с размещением соли в 3—4 местах, чтобы доступ к ней
имели одновременно несколько особей. Солонцы желательно устраивать на
подкормочных площадках или вблизи.
Наиболее пригодна природная каменная соль, которую, однако, достать трудно.
Как правило, охотникам приходится прибегать к имеющимся в продаже солевым
брикетам, используемым для домашних животных. Лучшей является соль с добав-
320
Рис. 107. Косуля на искусственном солонце
ками микроэлементов, но для каждой местности необходимы свои компоненты.
Консультации по этому вопросу можно получить у зоотехников.
Звери не должны получать прямого доступа к соли, поэтому ее обычно кладут
на высоте о коло 2 м на столб или ствол дерева, защитив сверху от дождя. Влажности
воздуха достаточно для того, чтобы соль растворялась и раствор медленно стекал
по стволу на землю. Спустя некоторое время животные начинают слизывать его
и поедать землю (рис. 107). В таком солонце куска соли или брикета хватает на
длительное время.
Оригинальный способ устройства долговременных и экономичных солонцов
предложен охотоведом К. А. Морозовым (1982). В стволе спиленной толстой осины
с помощью бензопилы выпиливается углубление длиной 210 см и диаметром —
30 см. В образовавшееся корыто, установленное на земле, вносится смесь, состоящая
из глины (50%) и водного солевого раствора. На дно кормушки сначала засыпается
15 кг глины, 9 кг поваренной соли, 6 кг монокальцийфосфата, затем снова столько
же глины и в той же последовательности остальные вещества. Все это
равномерно заливается раствором микроэлементов, рекомендованных зоотехниками
для данной местности в качестве минеральной подкормки для домашних животных.
В полученную смесь добавляют 10—15 л воды и все перемешивают лопатой. Чтобы
звери не опрокидывали корыто, к нему с торцов прибивают жерди.
Круглогодичное применение такого солонца из расчета 1 на 1 тыс. га охотничьих угодий
позволило значительно повысить качество и продуктивность местной популяции
косуль.
Солонцы не должны размещаться в посадках, так как в результате кон-
21. Европейская и сибирская косули „,
центрации животных происходят значительные потравы и повреждения деревьев
рогами. В холодное время года не рекомендуется возобновлять солонцы, так как
соль вызывает повышенную потребность в свободной воде, которая при замерзших
водопоях не может быть полностью удовлетворена.
В сухой летний период косули получ\ют достаточное количество воды при
поедании влажной зеленой растительности. Однако в засушливых районах
необходимо устраивать искусственные водопои, очищать естественные и сохранять их.
В воде особенно нуждаются самки в период лактации. Искусственные водоемы
выгодно размещать вблизи густого кустарника и кормового участка: луга, поля,
подкормочной площадки. Это гарантированно приведет к образованию в этом
месте родовых участков самок и к концентрации зверей. Зимой все мероприятия
в этом плане обычно сводятся к выкладыванию сочного корма: силоса и свеклы.
Искусственная подкормка. Искусственная подкормка косуль безусловно
необходима, особенно в районах с высоким снежным покровом, и является
первейшей обязанностью охотников. Она имеет целью поддержать хорошее
физическое состояние популяции, создать предпосылки для успешного развития
приплода и мощных рогов у самцов, снизить возможный ущерб древесной
растительности, а в глубокоснежных районах — сохранить популяцию. Животным
должен быть создан минимум условий для выживания, чтобы избежать
значительных потерь от бескормицы. Следует учесть также, что детеныши, получавшие
осенью и зимой недостаточное питание, уже не обладают способностью
накапливать достаточные энергетические запасы и остаются хилыми даже при хорошем
питании в более зрелом возрасте.
Пространственная структура популяции зимой очень часто определяется
условиями питания. С помощью соответствующей системы подкормки можно влиять
на распределение, т.е. управлять популяциями в пространственном аспекте,
привлекая их в нужные районы и отвлекая от важных лесопосадок.
Реакция косуль на подкормку неодинакова в разных районах. В одних они
берут ее лишь в действительно голодное время, в других преимущественно
кормятся на подкормочных площадках и лишь отдельные звери не посещают их.
В малоснежные зимы звери могут вовсе не брать подкормку, если хватает
естественного корма (Ulbricht, 1986). В районах с низким снежным покровом при
умеренной численности косуль подкормка, таким образом, необходима только
в многоснежные зимы. Для того чтобы при внезапном наступлении суровой зимы
животные быстрее нашли подкормочные площадки, их комбинируют с солонцами
и ежегодно выкладывают небольшое количество привлекательного корма.
Зимой потребность в питании у косуль сильно сокращена и составляет в сутки
всего лишь 300—900 г сухого корма (см. главу "Питание"), что соответствует 2 кг
кормовой свеклы, 0,25 кг концентрированного комбикорма и 1—1,5 кг грубого
корма или силоса. Для покрытия дневной потребности в энергии достаточны
следующие варианты (Missbach, 1974):
1 кг картофеля и 0,2 кг овса;
1 кг яблочных выжимок и 0,2 кг овса;
1,5 кг силоса из свекольной ботвы и 0,2 кг овса;
1,5 кг кормовой свеклы и 0,2 кг овса.
Совокупная потребность косуль в пище не должна покрываться только
подкормкой; большую часть корма в обычные годы они должны находить сами
в природе, иначе уже вымерли бы во многих областях своего ареала. Интенсивная
322
подкормка приводит к тому, что некоторые больные и слабые особи переживают
зиму и способствуют распространению болезней и паразитов. Следовательно,
при ведении интенсивного охотничьего хозяйства должен быть налажен отстрел
всех слабых особей. В любом случае учитывают и экономический аспект
подкормки.
Обычно одну подкормочную площадку с несколькими кормушками (из расчета 1
на 10 особей) сооружают на каждые 100—200 га охотничьих угодий. При этом
принимают во внимание биотопическое распределение животных, вопросы
предотвращения потрав, возможности хозяйства и сам процесс подкормки.
Подкормочные площадки по возможности размещают в тихих, защищенных от ветра,
солнечных и хорошо просматриваемых местах, доступных для подъезда на
автомобиле или лошади даже при плохих дорожных условиях. Лучше всего пригодны
для этого светлые строевые и старые леса без подроста. Кормушки иногда
целесообразно устанавливать в чаще леса, где на расстоянии 10 м от них удаляют
нижние ветви деревьев (Amann, 1981a).
Косули должны иметь возможность заметить приближающуюся опасность
с большого расстояния. С другой стороны, в открытых местах можно издалека
наблюдать за животными с помощью бинокля, не беспокоя их. В любом случае
следует избегать того, чтобы при переходах к подкормочной площадке звери
пересекали бы автомобильные дороги с интенсивным движением или
железнодорожные пути.
В горах с наступлением снегопада происходит миграция животных вниз, поэтому
выкладывакЗт подкормку именно здесь. При высоте снежного покрова более 20—
30 см на участке обитания косуль применяют плужный снегоочиститель, что
облегчает им передвижение; они экономят энергию и получают более легкий
доступ к естественной пище и к подкормочным площадкам. Подкормочную
площадку целесообразно расположить таким образом, чтобы очищенные от
глубокого снега пути звездообразно сходились к ней и вокруг нее было достаточно
места, чтобы трактор со снегоочистителем мог свободно развернуться.
Полевых косуль труднее привлечь к местам подкормки. Для этой цели
подкормочные площадки располагают у небольших групп деревьев или на
защищенной от ветра местности, где они бывают особенно часто. В полевых районах
по договоренности с сельскохозяйственными организациями иногда очищают
от снега часть посевов. Корм в полях богаче белком, но здесь может ощущаться
недостаток грубых балластных кормов, в результате чего животные могут страдать
ацидозом. Поэтому здесь желательна подкормка грубым концентрированным
кормом при обеспечении'зверей водой (Dittrich, 1981).
На одной подкормочной площадке размещают от 4 до 5 кормушек, чтобы все
члены стада кормились одновременно. Кормушки сооружают таким образом,
чтобы корм был защищен от осадков и не возникало бы загрязнения его
испражнениями животных, а все виды кормов можно было бы хранить в течение
длительного времени (Winter, 1980; Amann, 1981b).
Идеальным образом отвечает этим требованиям комплексная магазинная
кормушка, все детали которой делают из дерева и обрабатывают летом защитными
составами. Если это сделать осенью, то от них могут исходить неприятные запахи,
отпугивающие зверей. Перед емкостью с комбикормом приколачивают гвоздями
продольные планки на расстоянии 13—14 см и поперечные — через 30 см одна от
другой для предотвращения опустошения кормушек ланями и оленями. Для крупной
323
а'>::* •
.:■ ?<<*
A
■ *f
"... >t- ■
v
i
^
_ ■■■<■■■&§!%:■'■.
-*V
«4
гй:
'■'>■
,;■:■ "*'"''' *' ....«■. .. ** .
4h Л^£9^ ^^^К •■'
Рис. 108. Кормушка (Л) и кормовой
стол (Б) для косуль
сибирской косули это расстояние увеличивают на 5 см. При необходимости
магазинные кормушки дополняют земляными бункерами для хранения силоса или
козлами для сена. Силос лучше выкладывать на кормовые столы (их размер
120X160 см при высоте от 40 до 70 см) (рис. 108).
Остатки корма возле кормушек регулярно убирают, а сами кормушки ежегодно
перемещают на 10—20 м. Землю весной посыпают известью и при необходимости
перекапывают, что значительно снижает возможность заражения косуль
паразитами. Зараженность гельминтами может быть уменьшена и с помощью
добавления в корм противоглистных средств.
Успех подкормки во многом зависит от качества корма, чем раньше трава будет
скошена и высушена, тем выше в ней содержание белка и легкоусвояемых
калорийных веществ и тем лучшего она качества. Минимальные потери питательных
веществ возникают при искусственной сушке зеленого корма. Хорошее сено
ароматно, плохое — пахнет гнилью и скармливать его нельзя. Наилучшим
является клеверное (Авотиньш, 1976; Ulbricht, 1986) или люцерновое сено. Сено
первого укоса несколько грубовато, и животные едят его менее охотно,
прессованное не любят, а плохое не едят даже во время голода.
Особенно ценный и богатый белком корм для косуль — гранулы зеленой
люцерны. Однако гранулированный корм менее интенсивно обрабатывается слюной
и имеет меньшую усвояемость, в желудок попадает меньше буферных веществ.
Поэтому подкармливать животных только гранулированным кормом не следует.
Веники из побегов лиственных пород деревьев, кустарников и.стеблей трав
всегда охотно поедаются косулями. Наиболее предпочитаемы веники из осины,
ивы, березы, клена, тополя, рябины, каштана посевного, ясеня, липы, вяза, граба,
малины, ежевики, гороха, крапивы, иван-чая, топинамбура. Сушат их по
возможности под навесами, защищая от дождя и прямых солнечных лучей. Тем не менее
для подкормки целесообразнее использовать не веники, побеги или кору деревьев
(Авотиньш, 1976), а гранулированный корм, приготовленный из порубочных
остатков с добавлением небольшого количества зерна.
На практике широко применяется скармливание косулям корнеплодов,
особенно свеклы, картофеля и клубней топинамбура. Корнеплоды богаты
углеводами с небольшим содержанием белков и минеральных веществ и, кроме того,
содержат много влаги, что очень необходимо зверям зимой. Из них наиболее
ценен картофель, так как он содержит вдвое больше усвояемого белка, чем свекла,
и богат витамином С. Из всех разновидностей свеклы косули предпочитают
сахарную. Клубни топинамбура лучше всего оставлять в земле, звери сами
выкапывают их передними конечностями.
В районах развитого садоводства отличный корм — фруктовые выжимки,
яблоки и груши — падалицы, которые складывают в кучи на кормовых
площадках. Свежие фруктовые выжимки пересыпают солью, предназначенной для
скота (35—40 кг на 1—1,5 м3), и утрамбовывают доской. Добавка соли
предотвращает их гниение и замерзание и привлекает животных. Кучи огораживают и затем
корм порциями раскладывают по кормовым столам.
В настоящее время широко распространенным методом консервации сочного
корма является силосование. Силос для кормления дичи должен содержать до
75—80% воды. В дневном рационе, составляющем 2—2,5 кг на одну особь, воды
примерно 1,5—2,0 кг. Такая форма ее потребления с кормом более благоприятна
для животных, чем непосредственно водопой или поедание снега.
32S
В силос рекомендуется добавлять смесь минеральных веществ и 5—10%
комбикорма. Под комбикормом подразумевается высококалорийный корм с большим
содержанием питательных веществ. К нему относятся все семена зерновых с 8—12%
содержанием сырого белка. Существенно более богаты белком (20—45%)
стручковые плоды гороха, фасоли и вики, однако они легко вызывают вздутие живота,
и косули не должны их получать более 100 г в день на одну особь. Ни в коем случае
нельзя скармливать зверям протравленный семенной материал. Исключительную
ценность представляют отходы производства масел: жмых, экспеллер и шрот
после экстракции, а также естественные высококалорийные корма: плоды,
буковые орешки и желуди.
Существует много рецептов приготовления кормовых смесей для косуль.
В ФРГ, в частности, применяется такой состав: до 40% общего веса составляет
высушенная яблочная барда, 10% — соевые шроты, 10% — мука из льняного
жмыха, зерна овса (20%), каштаны, желуди (9%), кормовая поваренная соль (1%),
смесь минеральных(5%) и тонизирующих (5%) препаратов (Siebe, 1972).
Отходы лесного хозяйства в виде летних порубочных остатков осины (кора,
побеги, листья) являются очень перспективным сырьем для изготовления
комбикормов (осиновая кора — 20%, побеги с листьями — 79%, зерно ржи — 1%). Зимний
рацион из 981 г корма и 125 г свежих древесных побегов обеспечивает нормальное
физиологическое состояние животных и позволяет рационально использовать
отходы древесины (Приедитис, 1980; Приедитис, Таугинас, 1981).
Комбикорма следует давать косулям только в комбинации с грубым и сочным
кормом при обеспечении водой.
Влияние косуль на лесные культуры
и защита растений
Обитание косуль в антропогенном ландшафте приводит к тому, что они включают
в свой корм или обдирают рогами культивируемые человеком растения и тем
самым наносят определенный вред. Размер ущерба зависит чаще всего от
плотности населения животных и от наличия доступного корма. Вредоносная
деятельность зверей отчетливо проявляется при плотности, превышающей 60—100 особей
на 1000 га.
Наиболее значительный вред они приносят, обкусывая побеги, особенно
верхушечные. Очень уязвимы в этом отношении растения высотой до 120 см (в районах
обитания сибирской косули — до 150 см), самые существенные повреждения
наносятся на высоте от 40 до 80—100 см. При большей высоте предпочитаются
боковые побеги, обкусывание которых не имеет существенного значения для
лесовозобновления. Растения ниже 40 см зимой часто бывают покрыты снегом и
труднодоступны.
В разных регионах степень воздействия животных на лесные культуры
неодинакова. Хотя звери в общем предпочитают лиственные деревья, наибольший ущерб
причиняется хвойным лесопосадкам, что обусловлено незначительной
способностью хвойных к регенерации. Чем более редок какой-либо вид в древостое, тем
сильнее он подвергается угнетению. Наиболее страдают от косуль посадки бука,
сосен и пихт в возрасте 2—4 лет. Культивирование этих видов практически
невозможно без защиты даже при средней плотности населения зверей.
Зимой пресс косуль на древесно-кустарниковую растительность особенно ощутим
326
в регионах с высоким снежным покровом. В Амурской области, например, при
плотности 15—20 особей на 1000 га за зиму используется в среднем 25% побегов,
при плотности 40—50 — свыше 75% (Дарман, 1986). Высокий урожай плодовых
деревьев, дуба и др. и подкормка могут в значительной степени снизить ущерб,
причиняемый лесным насаждениям при питании.
Серьезный вред зимой может быть нанесен и фруктовым деревьям.
Предпочтение при питании отдается посадкам яблони и вишни. Объедание побегов
неблагоприятно сказывается на формировании кроны и нежелательно в плодовых
питомниках.
Местами охотно посещаются косулями и повреждаются плантации овощей,
цветов, виноградников (Haisch, 1982). Однако на обширных сельскохозяйственных
полях ущерб столь мал, что во многих случаях остается незамеченным.
Напротив, косули способствуют повышению урожайности и увеличению биомассы
растений, если плотность их населения не превышает 10 особей на 100 га. В
вегетационный период на полях, предназначенных на зеленый корм, воздействие
косуль на посевы дало прирост зеленой массы на 5%. Потери (9%) на полях люцерны
наблюдались лишь при очень высокой плотности дичи (150 особей на 100 га)
(Kaluzinski, Bresinski, 1976).
Ущерб, наносимый деревьям рогами, возникает независимо от условий питания.
Чистка рогов от "бархата" происходит за сравнительно короткое время и не может
иметь серьезного значения, но при маркировке территории (см. главу "Поведение")
самцы губят значительное количество деревьев в возрасте от 5 до 10 лет. В
разреженных насаждениях опасность таких повреждений выше, чем в густых. Деревья
редких видов, стоящие на опушках или отдельно, обдираются рогами чаще всего.
Лиственницы и лжетсуги вероятно имеют привлекающие для них запахи и
наиболее подвержены обдиранию рогами. С марта—апреля по май—июнь, когда
маркировка происходит особенно интенсивно, ущерб в таких посадках бывает столь
велик, что нередко требуется дополнительная посадка деревьев.
В открытых ландшафтах особенно сильно повреждаются рогами межевые
лесопосадки (Pielowski, 1983). Высокая плотность зверей на таких участках может
поставить под вопрос успех закладки лесополос.
Максимальный вред, наносимый косулями лесным и сельскохозяйственным
культурам, происходит все же в результате питания. Из этого следует исходить
при разработке мероприятий по его предотвращению.
Наиболее важной из таких мер является поддержание на оптимальном уровне
плотности населения зверей (Loudon, 1982; Eiberle et al., 1983), при котором ущерб
может возникнуть лишь местами и иметь несущественный или допустимый
характер. Во многом предотвратить его можно всеми мероприятиями по улучшению
условий питания косуль, начиная с повышения качества биотопов, закладки
долговременных кормовых полей и площадок и кончая правильным размещением
кормушек и солонцов, о чем говорилось выше.
Снизить вред можно также с помощью ряда лесоустроительных мероприятий.
Основное — увеличение густоты посадок; небольшое расстояние между
растениями в рядах легко компенсируется для многих видов деревьев увеличением
расстояния между рядами. Посадка лиственных видов растений рядом с плантацией
хвойных деревьев может в значительной мере отвлечь животных от последних.
Ограничение применения гербицидов во многих случаях послужит сохранению
ценного корма для дичи и тем самым будет способствовать уменьшению размеров
327
вреда. Это в особенности относится ко всем культивируемым площадям, которые
зарастают черникой, вереском или другими кормовыми растениями.
Механическую защиту площадей применяют лишь в случае редких или особо
ценных видов деревьев. Для защиты посадок при незначительной высоте снежного
покрова достаточна изгородь высотой 150 см. При высоком снежном покрове
требуется двухметровый забор, но и он при плотном снеге не спасает от косуль.
Наиболее удобно применять проволочную сетку высотой 120—150 см с двумя или
тремя проволоками, натянутыми сверху. Деревянные изгороди нуждаются в частом
контроле и ремонте и менее надежны. Все заборы должны иметь ворота и перелазы
для людей; если они будут отсутствовать, то перелезание через изгородь приведет
к ее повреждению. На участках, где обитают кабаны, сохранению изгороди
помогает устройство откидных дверок, используемых только кабанами.
Следует учитывать, что стоимость огораживания 1 га посадок на 45% выше,
чем стоимость защиты отдельных растений. Лишь при огораживании площадей
размером 2 га и больше стоимость сооружения квадратной формы соответствует
стоимости защиты отдельных растении. При других формах изгороди ее длина
увеличивается и соответственно возрастает и стоимость. На площадях размером
в 3 га и больше вряд ли удастся поддерживать изгороди в состоянии, полностью
исключающем проникновение косуль (Tabbush, 1979).
Для кратковременной защиты сельскохозяйственных и садовых культур
применяют электроизгородь; для этого достаточно от 2 до 4 проволок на высоте 30, 50,
80 и 100 см над поверхностью земли. При установке электроизгородей
руководствуются инструкцией по применению. К сожалению, достоинства данного метода
снижаются из-за частого травмирования косуль о проволоку.
Очень часто для защиты леса рекомендуют различные химические средства;
только в ФРГ их 29 наименований, в Австрии — 15 (Ueckermann, 1985).
Опыление и обрызгивание деревьев требует меньших затрат времени, но большего
расхода вещества, чем при нанесении препаратов щетками или кисточками. Однако
все они имеют относительно кратковременный эффект, но все-таки более
продолжительный, чем в отношении других видов копытной дичи (Карпенко, 1966;
Стенин, 1968; Tschiderer, 1978).
Кратковременный эффект имеют и средства оптического и акустического
действия — животные очень быстро привыкают к ним. Отпугивающее действие
соломенных чучел может быть усилено, если они будут использоваться охотниками
в качестве прикрытия при засаде.
Защита отдельных растений применяется в основном для предотвращения
повреждений, наносимых рогами. Наиболее известны пластмассовые манжеты, которые
обладают тем преимуществом, что не врастают в дерево. Обычно достаточно
вбить вокруг защищаемого ствола 3 кола. В молодых лесополосах в течение 5 лет
обспечивают максимально возможный отстрел самцов. В других местах
максимально уменьшают число одно-двухлетних самцов, которые провоцируют
увеличение маркировочной активности у взрослых особей.
В заключение хотелось бы заметить, что только при сотрудничестве между
охотничьим, лесным и сельским хозяйствами можно существенно снизить вред
растениям, причиняемый косулями. Вопросы регулирования численности дичи
при этом наиболее важны. Мероприятия, направленные непосредственно на защиту
растений, требуются только в случае посадки особо ценных культур.
328
Охрана и восстановление численности
Охрану животного мира не следует полностью отождествлять с его рациональным
использованием, однако система рационального природопользования и охраны
очень часто взаимосвязаны и взаимодействуют в едином комплексе (Сыроечков-
ский, 1986). Это положение полностью относится и к косулям. Именно на их
примере становится особенно ясным, что вопросы охраны можно решить только
при рассмотрении всего этого комплекса.
Популяции европейской или сибирской косули являются или охотничьими
или охраняемыми в зависимости от состояния их населения. На западе ареала
в Центральной и Западной Европе вопрос об охране практически не стоит.
Напротив, на востоке ареала европейской косули и почти на всем ареале сибирской,
где численность очень низка, а некоторые популяции находятся на грани
уничтожения, он имеет первостепенное значение.
Особенно остра проблема охраны в Сибири и на Дальнем Востоке. В последние
десятилетия численность сибирской косули сократилась в Бурятии, Туве, Якутии,
Красноярском и Хабаровскомтсраях, Кемеровской, Новосибирской, Омской,
Свердловской и Читинской областях в 2—6 раз.
Основные причины создавшейся ситуации известны: слабая охрана, массовое
браконьерство, нерациональный экстенсивный способ ведения охотничьего
хозяйства, обезличка охотничьих угодий, отсутствие комплексных биотехнических
мероприятий, обилие крупных хищников, отсутствие культуры охоты (Данилкин,
1982, 1985).
Исследуя проблемы охраны и восстановления численности косуль в СССР
и в Монголии, мы пришли к выводу, что решить их нельзя, не улучшив охрану
и не изменив коренным образом систему ведения охотничьего хозяйства.
Организация охраны и восстановление численности. Основной принцип этой
работы — популяционный. Популяции, достигшие уровня промысловой
плотности, вводятся в хозяйственный оборот, другие, подорванные из-за
перепромысла и обилия хищников или природных катаклизм, временно выводятся
из эксплуатации.
Региональные (в масштабе области, края, республики) меры охраны
применяются в зоне интенсивного охотничьего хозяйства лишь в том случае, когда
численность (плотность) в целом сокращается ниже минимального промыслового
уровня. Это необходимо для скорейшего заселения биотопов путем естественного
расселения особей из густонаселенных районов. В противном случае потребуется
дорогостоящее и трудоемкое искусственное расселение.
Каковы же минимальные уровни плотности населения косуль, при которых они
должны выводиться из хозяйственного использования? На основании анализа
населения этих животных в Евразии (с учетом экологических и социальных условий,
плодовидности и факторов смертности) считаем, что усиленная охрана зверей
во всем регионе требуется, если плотность ниже 10 особей на 1 тыс. га пригодных
угодий в пределах естественного ареала (за исключением северных районов
обитания), для популяции — 20 особей (рис. 109).
Первое, с чего должна начинаться организация охраны и восстановления
численности косуль, — определение имеющегося и оптимального уровней плотности их
населения в области, крае, автономной республике, районах и, главное, в охотничьих
хозяйствах. При плотности по региону ниже минимального уровня желателен
329
Рис. 109. Концепция стратегии управления популяциями косуль
I — запрет промысла, 2 — выборочный отстрел; 3 — регуляция численности (% — доля\от общего
числа особей, планируемых к отстрелу)
полный запрет охоты, для популяции эта мера обязательна. При более высокой
плотности сохраняют запрет на добычу самок при выборочном отстреле самцов
вплоть до достижения оптимального уровня, индивидуального для каждого
охотничьего хозяйства или отдельных его участков.
Не исключено, что региональный и популяционный принцип охраны могут
войти в противоречие, особенно на азиатской части ареала, где на огромных
пространствах при слабо развитом охотничьем хозяйстве еще долго не удастся
достичь минимального промыслового уровня плотности населения косуль. В этом
случае отдельным хорошо организованным охотничьим хозяйствам, имеющим
благодаря охране и биотехническим мероприятиям многочисленную популяцию,
невыгоден запрет охоты по всему региону. При решении этого вопроса приоритет
должен отдаваться популяционному подходу.
Любой запрет охоты на диких животных требует безусловного претворения
его в жизнь в комплексе с другими мерами, способствующими снижению
смертности: собственно усилением охраны, отстрелом хищников, биотехническими
мероприятиями и т.п. Без этого запрет охоты не даст эффекта. В отношении косуль
есть хорошие примеры быстрого восстановления поголовья там, где звери не
подвергались неумеренному преследованию человеком. Их численность в Румынии,
Венгрии, ^Болгарии и других европейских странах в послевоенные годы за 20 лет
увеличилась более чем на порядок, причем последнее десятилетие популяции уже
эксплуатировались. Позитивные примеры есть и в России. С начала 60-х годов
поголовье косуль возросло в Орловской, Курской, Тульской, Липецкой, Рос-
ззо
товской, Псковской, Саратовской, Волгоградской, Калужской, Куйбышевской
областях и Татарии в 20—250 раз. Вряд ли таких показателей можно было
достигнуть без полного запрета на добычу в совокупности с отстрелом хищников
и усилением охраны.
Очень показательны в этом отношении цифры изменения численности
сибирской косули в сходных биотопах в Амурской области (Дарман, 1986). В угодьях
Архаринского госпромхоза при значительном прессе промысла и наличии крупных
хищников плотность населения косуль на протяжении десятилетия держится на
стабильно низком уровне — 10 особей на 1000 га. Рядом, в Хинганском заповеднике,
где промысла нет, но есть хищники, она колеблется в пределах от 10 до 20 особей.
После организации филиала Хинганского заповедника на месте охотничьих
угодий, где запретили охоту и уничтожили хищников, за 5 лет поголовье
увеличилось в 5 раз (со 100*—150 до 800 особей, плотность населения соответственно
с 8—9 до 50 особей) (см. рис. 95).
Без резкого снижения браконьерства практически невозможно увеличить
поголовье косуль. Это совершенно ясно. Необходимы меры, исключающие
возможность проявления браконьерства, и в первую очередь ужесточение наказания
за незаконный отстрел. Суммы иска должны достигать того предела, при котором
возбуждается уголовное дело. При лицензионной системе срок действия лицензий
должен быть минимальным, а контроль за правильным их использованием —
всемерно усилен. Тем не менее в интенсивном охотничьем хозяйстве при высокой
плотности дичи лицензионная система становится тормозом и не нужна как таковая
при наличии планового лимита отстрела.
Сохранению и восстановлению численности косуль во многом будет
способствовать организация заповедников, заказников и других строго охраняемых
территорий в местах концентрации зверей и на путях миграций.
Охрана биотопов. Прямое отношение к охране косуль имеет проблема
сохранения естественных биотопов. В Сибири, в Казахстане и на Дальнем Востоке
в последние десятилетия произведена распашка целинных земель, затронувшая
миллионы гектаров их типичных стаций. Разумеется, это крупномасштабное
меропрятие не могло не сказаться отрицательно на состоянии популяций,
лишенных наиболее кормных зимний угодий. Мигрирующие популяции были вынуждены
резко менять веками сложившиеся- пути миграций и места зимовок на новые, что
привело к гибели многих, стад. Это обстоятельство усугубилось еще большим,
чем обычно, браконьерским прессом во вновь осваиваемых человеком районах.
Сотни тысяч гектаров пригодных для обитания биотопов поглотили в последние
десятилетия водохранилища новых гидроэлектростанций. К тому же после
образования водохранилищ мигрирующие популяции не сразу находили новые пути,
что приводило к гибели животных на обширных водных переправах (см. главу
"Перемещения"). С созданием Зейского водохранилища, например, резко
сократилась численность верхнезейской популяции (Ильяшенко, 1984).
Следовательно, охранные мероприятия непременно должны касаться биотопов.
В этом плане нельзя не упомянуть и некоторые другие факторы, так или иначе
влияющие на их качество. Применение гербицидов на больших площадях нередко
лишает косуль и других копытных естественных кормов, которые лесохозяйст-
венные и сельскохозяйственные службы считают малоценным или вредным.
В районах интенсивного разведения копытных необходим более жесткий контроль
за их применением или запрет.
331
Охрана от хищников. Во многих районах хищники наносят очень большой
ущерб популяциям косуль, иногда превышающий браконьерский. Подробно
этот вопрос был рассмотрен в главе "Численность". Здесь же отметим, что
высокая численность волка в местах обитания косуль говорит о плохом ведении
охотничьего хозяйства и вряд ли такое хозяйство добьется успехов в увеличении
поголовья копытных. Особенно велик урон от них будет при высоком снежном
покрове с образованием наста. Копытные в это время становятся настолько
легкой добычей, что за несколько дней волки могут погубить значительную
часть популяции и свести на нет все усилия по увеличению поголовья.
Рысь также нежелательна в районах интенсивного разведения косуль. Ее
вредоносная деятельность тоже усугубляется при высоком снежном покрове, и,
кроме того, она не боится сооружений и давит зверей на подкормочных
площадках и даже в вольерах. Поэтому каждый хищник, появившийся в местах
концентрации копытной дичи, должен быть незамедлительно отстрелян.
В глубокоснежный период серьезную опасность для косуль представляют
бродячие и охотничьи собаки, легко догоняющие их. Кроме того, долго
преследуемые собаками животные могут впоследствии погибнуть при сильном морозе
от простудных заболеваний. Всех бродячих беспородных собак в охотничьих
угодьях следует отстреливать, а охотничьих отлавливать. Пастьба домашнего
скота в сопровождении собак здесь должна быть запрещена, -поскольку собаки
постоянно находятся в охотничьих угодьях и в массе давят молодняк дичи.
Шакал и лисица опасны в основном для детенышей и для животных,
находящихся в бедственном положении. Разумеется, они также нежелательны в тех местах,
где обитает косуля.
Совершенно ясно, что при обилии крупных хищников, особенно волка и
рыси, практически бесполезны все мероприятия, направленные на
восстановление численности и рациональное использование ресурсов косули.
Тем не менее эта проблема весьма дискуссионна, особенно в отношении
волка (см.: Филонов, Калецкая, 1985): там, где человек по тем или иным
причинам не способен искусственно регулировать сильно возросшее поголовье
копытной дичи, и при снижении качества их популяций хищники могут быть
полезны. В заповедниках, где пресс промысла сведен до минимума или его нет,
крупные хищники просто необходимы.
Охрана в агроценозях. Косули как ни один другой вид копытной дичи
приспособились к жизни в культурных ландшафтах. Однако жизнь в агроценозах
чревата постоянной опасностью, особенно подстерегающей их во время
механизированной уборки культур. Такая особенность поведения зверей, как
затаивание, приводит к гибели большого количества животных под ножами или колесами
современного быстроходного уборочного транспорта. В ФРГ только в 1979 г.
погибли от сельхозмашин 60 тыс. косуль (Prax. Naturwiss.Biol., 1986. Vol. 35, N 5).
Для предотвращения массовой гибели дичи разработаны специальные
агротехнические приемы уборки урожая и обработки полей. В частности, такой
простой агротехнический прием, как обработка поля с центра в направлении
к его краям, позволяет многим зверям своевременно перемещаться на соседние
еще не убранные участки, и он должен неукоснительно соблюдаться.
Для выпугивания затаившихся детенышей сельскохозяйственные орудия
оборудуют специальными приспособлениями: впереди косилок крепят пружинящие
рейки, а сбоку, в сторону нескошенного участка, волочащиеся цепи с кон-
332
сервными банками и другим грузом. Неплохо зарекомендовало себя механическое
устройство, разработанное в ФРГ специально для этой цели. Оно представляет
собой металлическую штангу, в которой на расстоянии 12 см друг от друга крепят
гибкие стальные прутья, образующие гребенку перед косилкой. С помощью
этого приспособления на 2000 га сенокосных угодий удалось спасти 83 косуленка,
9 зайцев и 96 фазанов (Nowak, 1981).
Следует учитывать психологию работников сельского хозяйства. По
собственной инициативе он вряд ли станут применять менее удобные им
агротехнические приемы или дооборудовать сельхозорудия специальными
приспособлениями. Значит, необходим постоянный контроль и техническая помощь самих
охотников, а необходимые устройства желательно уже предусматривать в
технических проектах сельхозмашин.
Перед тем, как поле или луг будет скошен, необходимо организовать его
прочесывание и выпугивание дичи силами охотников с собаками и любителей
природы, школьников. Чучела, белые полотнища, бумажные мешки, шелестящие
предметы со световыми отражателями, вращающиеся прожекторы,
установленные вечером накануне косьбы, будут способствовать отпугиванию косуль.
Особая проблема охраны возникает при использовании в сельском хозяйстве
минеральных удобрений и средств защиты растений от сорняков или вредителей.
Для копытных опасны минеральные удобрения, оставленные на полях в открытых
кучах. При недостатке минеральных веществ животные поедают их и погибают
(Гайдук и др., 1986). Даже однократное распыление химикатов с самолета
может вызвать гибель косуль от отравления. Летом 1982 г. в Курской области
мы были свидетелями гибели помеченного радиопередатчиком самца сразу же
после расплыления минеральных удобрений. Это же наблюдается и при
обработке лесов гербицидами и инсектицидами. Необходимо всемерное
ограничение применения химических методов борьбы с вредными растениями и
насекомыми и переход на более безопасные и эффективные биологические и
агротехнические методы.
Охрана на транспортных магистралях. На современных скоростных
автострадах гибель косуль от столкновения- с транспортом необычайно велика.
Только в ФРГ ежегодные потери составляют около 60—100 тыс. косуль (Hartwig,
1986), в Австрии — более 50 тыс. (Osterr. Weidwerk, 1986. N 10). К сожалению,
предотвратить гибель на дорогах сложно, однако некоторые охранные и
профилактические меры возможны. Прежде всего на участках постоянных переходов
животных должны быть установлены предупреждающие знаки и ограничена
скорость движения автомобилей до 60 км/ч. Эта мера спасет не только косуль,
но и повысит безопасность водителей транспортных средств. На дорогах, где
случаи столкновений особенно часты, необходимо устанавливать
предохранительную изгородь высотой около 1,5 м или электроизгородь. Различные
отпугивающие средства — чучела, столбики с рефлекторами, зеркала и другие
приспособления не очень эффективны. Особое внимание обращают на
правильное размещение подкормочных полей и площадок. Они должны устраиваться
вдали от дороги и по обе стороны от нее, с тем чтобы звери не делали
переходов на кормежку через дорогу. Это же касается и размещения солонцов
и водопоев. Чем больше будет оборудовано подкормочных площадок по всему
лесу, тем меньше необходимость животных в перемещении и соответственно
меньше потерь дичи.
ззз
Подкормка как охранное мероприятие. В глубокоснежные годы сохранение
поголовья косуль практически целиком зависит от того, насколько они будут
обеспечены поддерживающим кормом. Подкормка и другие биотехнические
мероприятия выступают в данном случае в роли важнейшего фактора охраны
и способствуют восстановлению численности дичи. Только там, где забота
о животных имеет неформальный характер, где звери обеспечены кормами
естественными и выращенными человеком, где имеется много кормушек для
них, будет полностью сохранено все поголовье. Только тогда можно ожидать
существенного прироста популяции, удачной охоты и хороших трофеев.
Реакклиматизация и охрана генофонда. Попытки восстановления ареала
и повышения численности косуль путем реакклиматизации предпринимались
неоднократно (см. главу "Перемещения"). Работа велась и с целью создания
популяций улучшенного качества. Тем не менее до настоящего времени нет
достаточно точных научных экспериментов, которые подтвердили бы
целесообразность переселения косуль для "освежения крови" аборигенных популяций.
Практически во всех проведенных до сих пор опытах были допущены
принципиальные ошибки, отсутствовали основополагающие сравнительные
исследования как аборигенных популяций, так и популяций-интродуцентов, недостаточно
сокращалась численность первых, завозилось слишком малое число вторых.
Хотелось бы подчеркнуть, что реакклиматизация сама по себе не приводит
к увеличению поголовья дичи, если не будут созданы необходимые условия
для нормальной жизни животных. А если они будут созданы, то вряд ли
понадобится искусственное расселение, поскольку естественным путем косули
способны довольно быстро занять пустующую экологическую нишу и быстро
увеличить численность. К тому же следует учитывать и негативные стороны
реакклиматизации: сложность отлова, перевозки и передержки; дороговизну
этого мероприятия; массовую гибель (иногда до 80%) отловленных животных,
особенно при перевозках на дальние расстояния; непредвиденные последствия
при завозе особей из других районов (новый генофонд, паразиты, болезни)
и др. Присовокупив к этому отрицательный опыт расселения косуль в СССР,
в целом не давший ожидаемого результата, мы приходим к выводу о
нецелесообразности проведения реакклиматизационных работ в широких масштабах,
тем более в настоящее время, когда животные снова распространены по всему
историческому ареалу.
Таким образом, работа по реакклиматизации косуль оправдана лишь в
ограниченных масштабах и в основном после стихийных бедствий, повлекших за
собой массовую гибель.
Основные научные предпосылки искусственного расселения были
сформулированы на совещаниях по вопросам акклиматизации животных в 1963 и
1973 гг.
1. Косуль можно выпускать только в тех районах, где высота снежного
покрова зимой не превышает 30—40 см.
2. Материалом для расселения могут служить особи из популяций, не
склонных к зимним миграциям (в особенности это касается сибирской косули).
3. Выпуски следует производить в пределах их ареала, в районы с
высокоорганизованным охотничьим хозяйством, в которых гарантируются хорошая
охрана, отсутствие крупных хищников и бродячих собак и интенсивные
биотехнические мероприятия, в первую очередь подкормка.
334
В свете последних исследований систематики и биологии косуль мы считали
бы необходимым к этим положениям добавить следующие.
4. Нельзя смешивать разные виды и подвиды косуль между собой.
5. Внутриобластное расселение более целесообразно и экономично.
Сибирскую косулю совершенно недопустимо интродуцировать в ареал
европейской. Видообразованию, как известно, предшествует длительный период развития
популяций в своебразных условиях среды. Подвиды являются формами, наиболее
приспособленными к специфическим "местным" условиям, поэтому переселение
животных в другие районы и смешение их с аборигенными популяциями других
подвидов может нарушить уникальный генофонд последних, также тысячелетиями
приспосабливаемый к местным условиям. Поэтому следует воздерживаться,
например, от завоза относительно мелких косуль тянь-шаньского подвида из
Забайкалья, с Дальнего Востока и Средней Азии на Урал и Поволжье, где
обитает крупный сибирский подвил, приспособленный к глубокоснежью.
Расселять животных в пределах их подвйдовых границ, относительно хорошо
установленных к настоящему времени (см. главу "Изменчивость и таксономия"),
видимо, можно. К сожалению, в генетическом отношении популяции косуль
изучены крайне слабо, поэтому дать какие-либо практические рекомендации,
направленные на охрану их генофонда, пока что не представляется возможным.
Внутриобластное расселение косуль, т.е. переселение косуль из близлежащих
популяций, таким образом, наиболее приемлемо для охотничьего хозяйства.
Благодаря близкому расстоянию сокращается время перевозки, не нужна
передержка и карантинные мероприятия, резко1 сокращается отход, упрощается
организационная работа, резко снижаются расходы и, наконец, выпущенные
животные попадают в относительно привычную для себя среду обитания
(в сходный биотоп и климат) и не вносят чуждый генофонд.
Многолетний опыт показывает, что наименьший отход животных происходит
тогда, когда искусственно расселяют молодых особей, выращенных человеком.
Поэтому в тех районах, где требуется восстановление численности, вполне
будет оправданна организация специальных ферм по выращиванию косуль.
Пропаганда охранных мероприятий. Во многих цивилизованных странах
приняты законы об охране и рациональном использовании животного мира.
Однако любой даже самый хороший закон будет мертв и забыт, если не будет
ежедневной просветительской природоохранной деятельности, если не осознана
крайняя необходимость в экологическом обучении населения, если закон не
подкреплен соответствующими нормативными подзаконными актами и не
осуществляется контроль за его неукоснительным соблюдением.
Охотничья культура и этика создаются веками и передаются
подрастающему поколению как национальное достояние. Не случайно в передовых
в охотничьем отношении странах практически исключено браконьерство,
а охотник является первейшим защитником природы. Культуре охоты и
охране природы надо учиться. К сожалению, в отдельных странах владельцами
оружия нередко становятся случайные люди, которые могут причинить
и причиняют природе немалый вред. Такие люди не должны и не могут быть
членами охотничьих обществ.
Воспитание культурного знающего охотника, таким образом, является
задачей первостепенной важности. Охотник на косулю должен хорошо знать биологию
и повадки зверей, основы управления популяциями, технику добычи и обра-
335
ботки, правила и сроки охоты, охранные и биотехнические мероприятия и
применять свои знания на практике. Разумеется, для этих целей необходима
специальная литература и учеба в поле под руководством специалистов.
Природоохранная пропаганда особенно необходима в местах выпуска косуль
и в тех районах, где требуется восстановление численности зверей. Местная
печать, радио и телевидение могут оказать в этом деле очень важную помощь.
В заключение хотелось бы подчеркнуть следующее. Европейская и сибирская
косули — самые перспективные охотничьи виды в агроценозах, и мы с
оптимизмом смотрим на их будущее. На востоке ареала европейской косули и на всем
ареале сибирской существуют огромные потенциальные возможности для
увеличения их поголовья. Только в России емкость угодий позволяет обитание не менее
4 млн особей, что на порядок больше, чем имеется здесь сейчас. Далеко не
исчерпаны возможности оптимизации промысла и в других странах. Авторы надеются,
что их опыт, а также труд сотен других исследователей, обобщенный здесь,
будет весьма полезен многим зоологам, охотоведам и охотникам, которым
мы желаем успехов в дальнейшем изучении биологии этих видов и удачной
охоты.
Приложение
Измерения тела, черепа и рогов1
А. Измерения тела (рис. 110)
1. Длина тела — от переднего края верхней губы до конца хвоста (без волос) —
измеряется лентой или тесьмой по дорзальной стороне по изгибам тела.
2. Высота в холке — от уровня холки по остистым отросткам позвонков по прямой
линии до конца копыта (обе передние конечности выпрямлены, соединены
вместе и ориентированы перпендикулярно позвоночному столбу).
3. Обхват туловища — мерной лентой позади передних конечностей вокруг
туловища.
Рис. 110. Схема измерения тела косули
/ — длина тела; 2 — высота в холке; 3 — обхват туловища; 4 — длина туловища; 5 — длина ступни;
6 — длина уха
' Методики измерений приняты на Координационном совещании специалистов по косуле
(Москва—Вильнюс, 28 мая — 10 июня 1979 г.). Измерения тела проводятся с точностью до 1 см,
черепа и рогов — до 1 мм.
22. Европейская и сибирская косули 337
Рис. Ш. Схема измерения черепа и рогов косули
/ — ма ксимальная длина черепа; 2 — кондилобазальная длина; 3 — основная длина; 4 — максимальная
ширина; 5 — скуловая ширина; б — межглазничная ширина; 7 — длина лицевой части; 8 —
максимальная длина носовых костей; 9 — длина верхнего ряда зубов; 10 — максимальная ширина мозговой
капсулы; // — длина нижней челюсти; 12 — длина нижнего ряда зубов; 13 — длина диастемы нижней
челюсти; 14 — расстояние между внутренними сторонами осевых стержней рогов; 15 — расстояние
между внешними сторонами осевых стержней рогов; 16 — максимальная длина рогов; 17 — максимальное
расстояние между рогами
4. Длина туловища — от переднего выступа плечевого сустава до заднего
края бедренного сустава (по прямой линии, спина животного выпрямлена,
конечности ориентированы перпендикулярно к позвоночному столбу).
5. Длина ступни — от пяточного бугра до кончика копыта по тыльной (плантар-
ной) стороне.
6. Длина уха — от нижнего края ушной вырезки до кончика уха без волос.
Б. Измерения черепа и рогов (рис. 111)
1. Максимальная длина черепа — от наиболее выступающих точек сзади и спереди.
2. Кондилобазальная длина — от наиболее выступающей назад точки
затылочных мыщелков до наиболее выступающей вперед точки межчелюстных
костей.
3. Основная длина — от нижнего края затылочного отверстия до наиболее
выступающей вперед точки межчелюстных костей.
4. Максимальная ширина — в наиболее широком месте на уровне глазниц.
5. Скуловая ширина — в наиболее широком месте на уровне скуловых дуг.
6. Межглазничная ширина, минимальная.
7. Длина лицевой части — от наиболее выступающей вперед точки
межчелюстных костей до переднего края глазницы.
338
8. Максимальная длина носовых костей — от наиболее выступающих вперед
и назад их концов.
9. Длина верхнего ряда зубов — по альвеолам.
10. Максимальная ширина мозговой капсулы — в наиболее широком месте
параллельно скуловым дугам.
11. Длина нижней челюсти — от заднего края сочленовного отростка по прямой
до переднего края зубной кости, без зубов.
12. Длина нижнего ряда зубов — по альвеолам.
13. Длина диастемы нижней челюсти — по альвеолам.
14. Расстояние между внутренними сторонами осевых стержней рогов — иа
уровне вершин лобных костей.
15. Расстояние между внешними сторонами осевых стержней рогов — на уровне
вершии лобных костей.
16. Максимальная длина рогов — среднее от измерений обоих рогов от нижнего края
розетки до его максимально удаленного конца по внешней стороне по
изгибам рога (измеряется мерной лентой).
17. Максимальное расстояние между рогами — от внутренних сторон рогов.
Пища европейской и сибирской косуль
Виды растений
1
Питание по сезонам года
зима
2
весна
3
лето
4
осень
5
Авторы
6
Abies alba
A. nephrolepis
A. nordmanniana
A. sibirica
A. sp.
Acer campestre
A. hyrcanum
A. laetum
A. negundo
A. platanoides
A. pseudoplatanus
A. tataricum
A. tegmentosum
*A sp.
Alnus barbata
A. fruticosa
A. glutinosa
A. incana
A. pisbescens
*A sp.
Amelanchier ovaiis
Amorpha fruticosa
Andromeda polifolia
Древесные породы [деревья, кустарники
Р
+
Р
PL
PL
+
LP
LS
LP
LP
LP
+
P
+
LPK
■+
LPS
+
Arctostaphylos uva-ursi +
Berberis amurensis
B. vulgaris
*Betula alba
B. costata
B. davurica
B. exilis
B. fruticosa
PS
LP
LP
LP
P
+
P
+
+
+
+
L
LP
+
LPK
+
LPS
+
+
PS
P
LP
LP
LP
P
+
L
LP
L
LP
+
L
L
P
+
L
P
P
LP
LP
, полукустарники]
P
+
PL
+
LP
LS
L
L
L
+
+
L
LP
+
LPS
+
PSL
LP
LP
К
4, 9, 12, 18, 38, 39, 40, 58
(53)
41
(45)
(47)
7,9, 11,28,38,40,41,55,59
(54)
41
7, 28, 31, 59
1, 7, 9, 13, 23,24,28,38,40,41
55, 56, 58, 59
9, 11, 12, 18, 19, 26, 28, 31, 33
39,40,41, 58
7,59
(52)
27,28,31,32,(53)
41
(51)
9, 13, 18, 24
9, 12, 22, 40, 58
(53)
12, 14,27, 32, (46), (51)
9
28, 29, 32, 40
13,23
23, 38, 39
(52)
9
39, (48)
(52)
(52)
(51)
(48), (52)
340
Таблица (продолжение)
1 2
*В. humilis
В, litwmovii
В. mandshurica
В. middendorfii
В. папа
В. penduh
*В. pubescens
В. raddeana
В. rotundifolia
*В. verrucosa
•В. sp.
Calluna vulgaris
•Caragana arborescens
С. microphylla
С. stenophylla
Carpinus betulus
С caucasica
С crientalis
С duinensis
С orientalis
Caulophyllum robustum
Castanea sativa
Cerasus fruticosa
С sp.
Chamaecyparis Laasoniana
Chamaepericlymenum canadense
С sibirica
Clematis vitalba
Cornus mas
С sanguinea
*C. tatarica
Coronilla emerus
С varia
•Corylus avellana
C. holophylla
С heterophylla
С mandshurica
LP
LP
LPS
LP
+
+
LP
LPS
LPS
LPS
P
+
+
+
+
+
P
LP
+
+
LPS
P
+
LS
S
3
LP
LP
+
LP
LPK
L
+
+
LP
LPS
LPS
LPS
LPK
LP
+
P
P
PS
+
LP
+
L
LP
4
LP
L
LP
LP
L
+
+
LPK
LPS
LPS
LP
LP
+
P
P
L
+
LP
+
LP
L
5
LP
PL
LPS
LP
+
+
+
LPK
LPS
LPS
LP
+
+
+
+
LP
P
L
L
+
+
+
+
+
+
LS
LS
6
1,(43), (45), (48), (50)
41
(52)
(51)
23
1,9, 12, 13, 18,22,24,28,30
1, 7, 13, 22, 24, 30, (43), (45),
(50), 56
41
(45)
(37), (40, (43), (45), (46), (47),
(49), (50), 55, 58, 59
23, 24, 26, 27, (49), (53), 58
1,9, 11, 13, 17,21,23,24,25,36
7, 13, 32, (45), (46), (47)
(48)
(48)
1,9, 11, 12, 13, 17, 18, 19,24,27,
30, 33, 38, 40, 55, 58
41
41
38,40
41,55
(52)
11,38
59
(43)
11
(52)
(51)
9,41
41, 38, 40, 55
9, 11,28,31.32,33,40
(53), (54), 58
9
39
1, 7, 9, 11, 12, 13, 18,24,26,30,
40, 41, (54), 56, 58, 59
(53)
(52)
(52)
341
Таблица (продолжение)
1
Cotinus coggygria
Crataegus curvisepala
С. kytrostyla
С. monogyna
С. oxyacantha
С. pentagyna
С. sanguines
Cytisus albus
С austriacus
С leucotrichus
C. nigricans
С ruthenicus
*C. sp.
Daphne laureola
*D. mezereum
•Deutzia amurensis
Elaeagnus argentea
Eleutherococcus senticosus
Empetrum nigrum
Erica cinerea
E. tetralix
Eunonymus europaea
E. latifolia
E. pauciflora
E. sacrosancta
E. verrucosa
E. sp.
Fagus orientalis
F. sylvatica
F. taurica
Frangula alnus
Fraxinus americana
.F. excelsior
F. mandshurica
F. ornus
F. oxycarpa
F. oxyphylla
Genista germanica
G. sagittalis
G. tinctoria
Gleditschia triacanthos
Grossularia reclinata
Hedera colchica
H. helix
2
+
P
+
P
+
P
+
+
+
+
L
L
LS
+
+
+
ррг
p
p
LS
+
+
PS
L
+
LPS
+
+
L
+
+
LS
+
L
3
+
+
+
L
+
+
LP-
+
+
+
+
L
L
+
P
+
LP
LP
L
+
LK
PL
L
4
+
P
+
L
L
+
4-
P
+
+
P
L
L
+
P
+
LP
L
L
+
LK
+
5
+
+
+
+
+
+
LPS
+
+
+
L
L
+
+
P
L
LPS
L
+
+
+
+
6
7
59
7
9, 11, 18,26,38,40,41
6, 9, 41
40,41
(50)
40
40
40
40
1
12, 13, 31, 38, 39, 41, (43), (44),
(51), (53)
9
9, (49)
(52), 58
7
(53)
23
25
21,25
7, 11, 18,24,40,59
41
(52)
(52)
7, 28, 58
(43), (53)
41
9, 11, 12, 14, 17, 18, 19, 26, 27
38, 39, 40, 58
(54)
7, 9, 13, 24, 28, 30, 58, 59
7
1, 7, 9, 11, 12, 18, 22, 24, 26,28
31,32,33,40,56,58
(53)
40
56
55
9
39
1,9,24
29
59
41
9, 11,24,25,41,55
342
Таблица (продолжение)
1
Hyssopus officinalis
Ilex aquifolium
Jasminum sp.
Juglans regia
Juniperus communis
J, sabina
*J. sp.
Larix dahurica
L. decidua
L. sibirica
*L. sp.
Ledum palustre
Ligustrum vulgare
Lonicera altmannii
L. caucasica
*L. caerulea
L. nigra
L. periclymenum
L. tatarica
*L. xylosteum
L. sp.
Malus domestica
M. mandshurica
M. orientalis
M, silvestris
M. sp.
Mespilus oxyacantha
Morus alba
Myrica gale
Nothofagus obliqua
N. procera
Ononis repens
Oxycoccus quadripetalus
Padus asiatica
P. racemosa
Phellodendron amurense
Picea contorta
*P. excelsa
P. falcata
P. laricio
P. nigra
P. obovata
P. omorica
P. pence
P. schrenkiana
P. sosnovskyi
2
P
+
LP
LPS
P
LSP
LP
+
LPS
P+
P
+
+
LPS
LP
+
PS
S
L
+
+
LP
+
+ ■
LP
LP
LP
L
LP
LP
+
LP
+
3
P
L
L
L
+
+
LPS
L
L
+
+
+
+
+
LP
+
+
L
+
L
LP
LP
L
LP
4
P
+
+
+
LK
L
+
+
5
P
+
+
+
LK
P
+
+
LPKS LP
L
L
LP
+
+
+
+
+
LPS
+
+
+
LP
+
LP
LP
LP
L
+
+
L
+
. +
LPSK
+
LPS
S
L
+
+
KP
+
L
LP
LP
L
+
6
20
9, 11,25
(53)
9
13, 23, 39
(54)
(46), 58
(48), (52)
13, 18
(48), (50)
11,(46), (47)
(49), (51), (52)
5,9, 17,28,31,40,55
(44)
41
(45), (50), 59
9
9, 11
7
9, 30, (49), 58
(47), (53), (54), (60)
7, 13, 22, 27
(52)
41
7, 9, 11, 13,23,40, 58,59
6, 17, 27
55
28
23
11
11
9
13, 23
(52)
7
(52), (53)
(44), (46), (47)
9, 11, 12, 13, 18, 19, 210 23, 24
26, 27, 30, 38, 39, (43), (49), (54)
56, 58
11,21
55
11,25
(50)
11
39
(44), (46), (47)
41
343
Таблица (продолжение)
P. sibirica
*Р. siNtstris
P. sp.
Pirus communis
P. malus
P. piraster
Populus alba
P. davidiana
P. marilandica
*P. nigra
*P. tremula
P. suaveolens
*P. sp.
Pru:ms avium
P. divaricate
P, domestica
P. fruticosa
P. mahaleb
♦P. padus
P. racemosa
P. spinosa
P. sp.
Pseuriotsuga menziesii
P. taxifolia
Pyrus sp.
Quercus boreaiis
Q. cerris
Q. conferta
Q. dalechampii
Q. frainetto
Q. iberica
Q. hartwissiana
Q. macranthera
Q. mongolica
Q. pedunculate
Q. pedunculiflora
Q. petraea
Q. pontica
*Q. robur
Q. rubra ■■>
Q. sessiliflora
p
LP
L
PS
+
P
+
+
LP
LP
+
+
PL
L
+
P
P
LP
+
■H
+
+
+
+
+
+
+
+
P
+
LS
L
LP
L
P+
LP
+
LP
L
LP
+
LP
+
P
P
+
+
+
LPS
LP
LP
P+
LP
LPS
L
P
+
P
LP
P
+
+
+
P
LP
+
LP
PS
L
LP
LPK
+
LPKS
L
P
S
+
+
+
+
+
+
LS
LS
P
+
LPS
(45)
1, 7, 9, 11, 12, 13, 16, 17. "8,19,
21, 23, 24,26,27,28,29,32,(37),
38, 39, (43), (46), (48), (49), (50),
(52), 55, 56, 58
(51), (60)
6, 7, 9, 13, 17, 27, 59
38,55
40
40
(52)
40
9, (37), 40, 42, (54)
1, 7, 12, 13, 18,22,23,24,27,30,
31,(37), 38, 39, 40, 41,(43), (45),
(48), (49), (50), (51), (53), 56,58,59
(51)
11, 18,27,28,29,32, (46), (47), (53)
9, 16
41
6
(50)
28
(47), (50), (53), 56
9, 18, 24, 31
7,9, 13, 16, 18,40,41, 55,59
6, 11,27,29, 32
9,21
11, 18
58
9
38,40
38
40
40
41
41
41
(52)
55,56
40
9,40
41
1,7,8,9, 11,14,18,22,24,26,30,
40, (54), 59
11,58
39,55
344
Таблица (продолжение)
1
Q. sp.
Rhamnus cathartica
R. frangula
Rhus cotinus
R. typhina
Ribes alpinum
R. grossularia
R. hispidulum
R. mand&huricum
*R. nigrum
R. rubrum
R. vulgare
R. sp.
Robinia pseudoatacia
Rosa acicularis
R. arvensis
*R. canina
R. cinnamomea
R. davurica
R. eglanteria
*R. pendulina
R. plafyacantha '
R. rugosa
*R. sp.
Rubus buschii
R. caesius
R. caucasicus
R. chamaemorus
R. fruticosus
R. hirtus
*R. idalus
R. saxatilis
R. serpens
*R sp.
Salix adenophyla
S. alba
S. americana
S. aquatica
S. arbuscula
S. atrocinerea
S. aurita
*S. caprea
2
LP
LP
L
LP
+
+
L
LP
LPS
LPS
+
S
LP
LP
+
S
L
L
L
L
+
L
+
L
LP
+
+
+
+
3
LP
LP
L
P
PL
P
L
P
LPS
LP
+
S
LP
LP
+
S
L
L
+
+
LK
LPS
+ '
+
+
+
+
4
LP
LP
L
LP
P
LP
L ,
L
P
LP
+
LS
LPK
LP
+
L
S
+
L
L
LPKS
LP
LK
LP
LP
+
+
+
+
LP
5
LPS
LP
L
+
LS
L
L
P
PL
LP
LP
+
S
LPKS
KP
+
+
LS
S
LPS
+
L
S
L
+
LPS
+
+
+
+
6
5, 12, 13, 17,25,27,28,31,38
9, 13, 18, 31,59
56
(54)
18
41
9
59
(52)
1,24, (48), (50), (53)
(48), (49), (51), (53)
9
(46), (47), (53)
5, 7, 9, 16, 18, 27, 28, 32, 40
(48), (52)
9
5,6, 7, 9, 23, 31, (37), 41, (49), 55,
58,59
(50)
(52)
13
9, 39, (60)
(44)
6
11, 13, 25, 26, 32, 38, (43), (47),
(46), (51), (60)
41
1,7,9,56,58,59
41
23
6,9, 11,24,25,26,31, 33
41, 38, 39, 59
1,7,9, 12,13,23,30,33,34,(37),
38, 39, (43), (46), (53), (54), 55,
56,59
1,2, 13
41
5, 12,13, 15, 16, 17, 19, 21, 24,
28, 29, 32, 38, (60)
18
9, 13,40,42
8, 14
8, 14
(45)
26
9, 13
1, 8,9, 11, 12, 13, 14, 19,24,(37),
40, 41, 39, (49), (50), (52), (53),
56, 58, 59
345
Таблица (продолжение)
S. chlorostaehya
*S. cinerea
S. cinerascens
S. fragilis
S. glauca
S. gmelinii
S. krylovii
S. linearifolia
S. livida
*S. myrsinifolia
S. myrtilloides
S. nastata
*S. pentandra
S. pseudocapr
S. pulchra
S. purpurea
S. rossica
S. schwerinii
S. sibirica
S. smithiana
*S. triandra
S. viminalis
S. xerophila
*S. sp.
Sambucus nigra
*S. racemosa
S. sibirica
S.'sp.
Sarothamnus scoparius
Schizandre chinensis
Sorbaria sorbifolia
Sorbus aria
*S. aucuparia
S. caucasigena
S. torminalis
S. tianschanica
S. sibirica
S. sp.
Spiraea chamaedryfolia
S. hypericifolia
S. media
Symphoricarpos albus
Syringa amurensis
Taxus baccata
Thelycrania sanguinea
Thuja occidentalis
Th. plicata
346
PL
+
PI,
P
+
PL
+
+
LP
+
+
P
+
+
+
+
P
P
PLS
LP
P
+
P
LPS
LP
LP
PS
+
L
P
+
PL
LPS
LP
+
LP
PLS
+
PLS
+
PLS
+
SP
LP
+
LP
P
L
+
LP
+
+
P
P
LPS
LP
+
+
LK
LPK
LP
LP
+
+
+
+
L
L
PS
+
PLK
LP
LP
+
LP
PLS
P
PLK
+
PLS
LP
SP
LP
+
LP
P
L
+
LP
PL
+
P
P
PLK
LP
LPKS
L
LK
+
LPK
LP
+
LP
+
+
L
L
L
L
PLKS
LP
LP
+
LP
LP
+
PL
PL
LP
+
LP
+
PL
P
+
+
L
PLS
P
P
KPL
LP
+
+
LP
+
LP
LPS
S
+
+
LS
+
PL
L
LPS
LP
LP
(48)
9, 18, 24, 40, (43), (50)
(48)
40, 59 .
(45) .
(48), (51), (53)
(52)
(43)
(37), (48), (50)
9, (50)
(52)
3, 10, 18
13, (50), 56
(44)
(51), (53)
8, 9, 14, 40
(51), (53)
(52)
(45)
8, 14
9, 18, (51), (53)
18
(48), (51), (52), (53)
1,7, 12, 13, 16, 18,22,23,27,
(46), (47), (49), (45), 55, (60)
9, 12, 13, 18, 19, 31, 40, 55,
1, 9, 12, 38, 39, (49), 56, 58,
(52)
16, 28, 32
9
(53)
(51), (53)
9, 11
1, 6, 9, 11, 12, 13, 17, 18,23,
30, (45), (49), (50), (54), 56, 58
41
9, 40,41, 55
(44)
(51), (53)
(43), (46), (47), (60)
(43), (45)
(50)
(51)
28
(53)
9, 11,41
7
11
1!
30,
59
59
24,
,59
Таблица (продолжение)
Tilia amurensis
*Т. cordata
•Т. platyphyllos
Т. taquetii
*Т. tomentosa
Т. sp.
Tsuga heterophylla
Ulex europaea
Ulmus campestris ,
U. carpinifolia
U.elliptica
U. foliacea
U. glabra
U. laevis
U. montana
U. pumila
U. scabra
U, sp.
Viburnum lantana
•V. opulus
V. orientale
V. sargentii
V. sp.
Viscum album
Vitis amurensis
V. vinifera
Zinnia sp.
Achillea asiatica
A. biserrata
*A. millefolium
A. ptarmica
*A. sp.
Aconitum boreale
A. cxcelsum
A. kusnezoffii
*A. lycoctonum
Acter alpinus
Actaea spicata
Acroptilon sp.
Adenophora liliifolia
A, sp.
Adonis amurensis
Adoxa moschatellina
•Aego podium podagraria
Aethusa cynapium
p
LP
LP
L
+
LP
+
+
+
+
+
P
+
P
+
LP
LP
+
PL
P
L
PS
ольные
LP
+
L
L
PL
LP
LP
L
L
LP
LP
LP
+
LP
+
+
P
+
+
+
L
PS
LP
LP
LP
+
L
+
P
+
+
+
L
L
LP
+
LP
+
+
+
P
PS
+
+
L
L
S
травянистые растения
LP
+
+
PK
PL
L
+
+
LP
LK
L
+
+
+
PK
L
PL
+
L
+
+
LP
+
LKS
LPS
+
+
+
PK
L
PLKS
+
LP
+
+
KLS
LKS
(52)
1,7,9, 13, 18,24,30,40,41,(54)
55, 56, 59
9, 40, (50)
(52)
28,(53)
40
11
21,25
38,55
7,30
41
40, 58, 59
55,56
7, 18, 56, 58, 59
38
(52)
7, 9, 24, 59
11, 28
9
9, 28,40,41,(50), 58
41
(52), (53)
59
7, 9, 24, 38,39,41, 56
(52), (53)
6, 35
(53)
(48). (51), (52)
41
7, 9, 15,(37)
9
(43), (53), 59, (60)
(50), (53)
(48)
(48)
9. (43)
(48)
9
59
(37), (50)
59
(52). (53)
13
2.9,7, 13. 24. 27. (43), (50), (54), 59
28
347
Таблица (продолжение)
1
•Agrimonia eupatoria
A. pilosa
A. sp.
Ajuga orientalis
*A. reptans
Alchimila micans
A. pratensis
*A. vulgaris
*A. sp.
Alectorolophus sp.
Althaea rosea
Alyssum sp.
Amaranthus retroflexus
A. sp.
.".npgpliis arvensi>
AnchuMi officinalis
Anemone dichotoma
A. nareisiflora
A. nemorosa
A. ranunculoides
A. sylvestris
A. sp.
Anethum graveolens
♦Angelica silvestris
*A. sp.
Anthemis cotula
A. tinctoria
A. sp.
Anthriscus silvestris
Aphanes arvensis
Apocynum sp.
Aquilegia glandulosa
A. vulgaris
A. sp.
Archangelica duccerens
Arctium nemorosum
A. sp.
Arctostaphylos sp.
Armoracia rusticana
Artemisia anethifolia
A. campestris
A. frigida
A. glauca
A. integrifolia
A. lanciniata
A. leporina
A. mongolica
A. sericea
2
+
+
+
LP
LP
LS
S
LS
LP
4-
LS
L
+
SLP
PSL
PSL
+
PSL
PSL
PL
3
+
+
+
+
+
+
+
+
LP
LS
LP
L
L.
LP
+
+
L
+
+
LK
+
+
KL
+
LKP
LP
SLP
+
PL
PL
+
L
PL
PL
VI
t
+
LP
+
+
L +
+
LPK
LP
+
LKS
S
LP
L
L
LP
+
+
LKS
LKS
LK
LK
LK
+
+
К
LPK
К
.-i_
SLP
PL
L
PL
+
L
PL
PL
5
+
KP
4.
+
+
+
LPK
P
S
LKS
KP
S
+
+
+
LKS
LS
LK
+
К
LPK
+
SLP
PLK
PLSK
+
PLS
PLK
PL
6
9, 13,(50)
(52)
(37). (43)
55
9. 13, 24. (54)
13
12
9. 23, (37), (39), (44)
12, 13. 30, (50), (54)
(46)
20
59
40
28, 31. 32
31.32
29
(52), (53)
(44)
9, 23, 24
(54)
(37)
(43), (46), (47). (53), (604
20
9, 13,(43), (45), .(46). (50)
(52). 59
27, 59
22
31
7,9, 13. 23. 41
27
(43)
(45), (46)
9
(43), (46), (51), (52)
(45), (46)
7
30,41
(51)
27
(48)
22
(48), (53)
(37), (43), (47)
(52)
(48), (52), (53)
(51)
(48)
(50), (51), (53)
348
Таблица (продолжение)
1
A. silversiana
A. stolonifera
A. tanacetifolia
A. tilesii
*А. vulgaris
A. sp.
Aruncus silvaticus
*А. vulgaris
Asarum europaeum
Asperula odorata
Aster incisus
Astragalus adsurgens
A. aksuensis
*A. caucasicus
A. frutieosus
A. glycyphyllus
A. melilotoides
A. onobrychus
A. uliginosus
A. sp.
Atriplex hastata
A. nitens
*A. patula
A. sibirica
A. sp.
Atropa belladonna
♦Barbarea orthoceras
*B. sp.
Bellis perennis
Berferos incana
Beta vulgaris
*Betonica officinalis
•Bidens tripartita
Brassica campestris
B. napus
B. oleracea
Bunias sp.
Bupleurum aureum
B. scorzonerifolium
Cacalia hastata
С sp.
Calamintha clinopodium
С montana
*Caltha palustris
С sp.
*Camelina microcai pa
Campanula bononiensis
C. cervicaria
С glomerata
2
PLS
PSL
+
L
P
L
+
+
LP
LP
LP
L
LP
LP
S
LP
LS
PL
+
+
LP6
+
L
L
L
L
L
PL
LP
LP
L
• s
+
L
3
PL
L
PL
+
L .
L
+
К
LP
LPK
К
LPK
L
LPK
L
SPL
+
L
L
LPc
+
+
L
L
L
L
PL
L
LP
LK
S
+
LK
4 |
1
PL
L
PL
+
+
L
+
К
LP
LPK
LPSK
L
LSPK
LS
PLK
LKS
S
LKS
L
+
L
L
+
+'
L
LP
PLK
L
LP
+
+
LK
S
+P
LK
+
5
PLK
PLK
+
LK
LP
L
+
LP
LPK
L
LSK
L
LPK
L
LPK
+
+
LP
LS
+
PK
LP6
LP6
' +
+ '■
L
L
L
LP
L
L
LP
+
+
L
S
+
LKP
+
6
(48), (51), (52), (53)
(52)
(48)
(51)
7,20,29,31,(37)
28, 30, 32, 59
(51), (53)
9, (46)
7, 22,24, 30
1, 7, 9, 24, 56, 59
(52)
(48)
(44)
41,(43)
(48)
1, 7, 9, 13, 55, 56
(48)
(45), (46)
(47)
(51)
31
7,31
15, 31, (37), (43)
(48)
28
9
(48), 59
22,(51)
9, 13
29, 40, 59
5, 7, 9, 17, 27, 28, 30, 31, 32, 59
9, 30, (43), (50), 59
(43), 59
30,41, 59
5, 9, 27
9, 11, 13,20
59
(50)
(48)
(50)
(46)
9,30
9
2, 9, 24, 30, 32, 39, (46), (48)
(47), (53)
27, 28, 31, 32,(46)
(37), (46), (48), (50),
(50)
9
349
Таблица (продолжение)
1
С. latifolia
С. patula
С. persicifolia
С. punctata
С. rapunculoides
С. rapunculus
С. rotundifolia
С. trachelium
Cannabis ruderalis
*Capsella bursa—pastoris
С. !>р.
Cardamine amara
С tlexuosa
С pratensis
C. tenera
Carlina vulgaris
Carpesium cernuum
*Carum buriaticum
*C. carvi
Centaurea cyanus
C. iberica
С jacea
С montana
С pseudophrygia
C. ruther.ica
C. scabiosa
С sp.
Centaurium sp.
Centiana sp.
Cerastium arvense
*C. caespitosum
C. semidecandrum
*C. sp.
Cerasus silvestris
Chaerophyllum aureum
С bulbosum
С hirsutum
С prescottii
*Chamaenerium angustifolium
Chelidonium majus
*Chenopodium album
Ch. polyspermum
Ch. rubrutn
Ch. vulgaria
Ch. sp.
Chrysanthemum corymbosum
*Ch. leucanthemum
Ch. segetum
T
', 2
i
P
+
+
LS
+
+
+
LP
LP
+
+
+
+
+
+ LP
+
L
L
+
+
PLS
LS
| 3
i
+
+
P
+
+
LS
+
+
L
LP
LPK
+
PL
+
+
+
+
+ '
L
LK
L
+
+
+
+
L
+
LKS
+
+
4
L
+
+
LP
L+
+
+
+
LS
+
LPK
L+PK
L
К
+
L
+
+
+
+
+
L
LK
PS
+
+
+
L
+
LKS
+
LK
5
+
+
LP
+
+
+
+
+
S
+
+
LPK
+PK
К
+
L
+
+
+
+
+
LPK
LK
PS
+
+
+
PSL
+
LKS
+
К
! 6
i
41
9, 13
9,24
(52)
7,41, 59
9
9
7, 9, 59
(37)
27,28, 31,32, 40, 41,(46), (52)
(51)
24
9
9
55
13
41
(48), 59
20, (37), 41, (46), (48)
13
41
1,9. 13, 56
9
59
(43)
(37). (51)
29. 30
(43)
(43)
40
9.(48)
27
13. (46), (51)
41
41. 55
41
9
(50)
9, 13. 17, 21,23, 24, (37), 39, (51).
(52). (53), (54), 59
7,59
27, (37), (43), (48)
27
(48)
7
28, 30, 31, 32
9
7, 9, 23, (45), (46)
27
350
Таблица (продолжение)
1 [ 2
Ch. vulgare
•Chrvsosplenium alternifolium
Ch. oppositifolium
*Cichorium intybus
Circaea intermedia
С lutetiana
•Cirsium arvense
C. heterofillum
C. incanum
С lanceolatum
С oleraceum
С palustre
C. rivulare
С sp.
Clematis sp,
Comarum palustre
ConioseMnum sp.
Consolida segetum
♦Convolvulus arvensis
С sp.
Coronilla sp.
Coriandrum sativum
Corydalis ambiqua
C. caucasica
С solida
Cotoneaster meianocarpa
*C. pyracantha
Crepis biennis
С capillaris
*C. paludosa
С praemorsa
C. sibirica
С tenuifolia
*C. sp.
Cynanchum acutum
C. scandens
С vincetoxicum
Cytisus caucasicus
С ruthenicus
С sp.
Daucus carota
Dasiphora fruticosa
•Delphinium elatum
D. grandiflorum
Dentaria bulbifera
D. quinquefolia
•Descurainia sophia
Dianthus versicolor
Digitalis sp.
*
L
L
Pi
+
+
+
L
LP
+
+
LS
LP
LP
+
+
PLS
P
LP
P
PL
+
+
I,P
LP
PL
+
L
3
+
L
L
+
LP
L
L
L
LP
L
a.
+
S
L
L
+
P
P
К
+
LPK '
LP
PLK
+
P
I
L
L
P
PL
L
4
+
+
+
+
+
KL
L
+
L
L
LP
L
+
L+
S
L
LP
+
L
P
К
+
+
LPK
L
+
+
LPK
LPK
+
L
PL
+
LK
+
5
~~""———-
+
+
P6
+
+
+
+
L
L
+
+
+
S
LP '
LP
SP
SP
К
+
+
+
PLK
+
+
LPKS
LPK
+
L
+
+
L
6
7
9, 22,30, (46), (53)
9
7, 17, (43), 59
9
9
13, 30,(37)
(45), (46)
41
(37)
7, 9, 13, 30
13
2
28, 31, 32, 59
59
1, 23
(43)
30
15,(37), 41,(48), (52)
28, 31, 32, 59
59
20
(52)
41
40
(48), (50), (52)
41, (46), (47)
9
9
9, 12, 30, (54)
(50)
(43), (44). (50)
(48)
27, (47), 59
7,59
9, 50
41
41
(37), (50)
59
7. 13, 20, 30
(48)
(50), 59
(48)
40.
41
31,(37)
(48)
39
351
Таблица (продолжение)
Diospyros lotus
Doronicum altaicum
Dorycnium herbaceum
Dracocephalum Ruyschiana
Dryopteris filixmas
D. sp.
Echium vulgare
Empetrum sp.
*Epilobium angustifolium
E. hirsutum
*E. montanum
E. obscurum
E. palustre
E. parviflorum
E. roseum
*E. sp.
Erigeron canadensis
Erodium sp.
Erophila verna
Eupatorium cannabinum
•Euphorbia discolor
E. dulcis
E. villosa
E. sp.
Eurotia sp.
F-agopyrum convolvulus
f. dumetorum
*F. sagittatum
fF. sp! \
Falcaria vulgaris
Ficaria verna
•Filipendula hexapetala
F. palmala
*F. ulmaria
*F.sp.
1-oeniculum vulgare
Fragaria moschata
F. orientalis
*F. vesca
F. viridis
»F. sp.
Galega orientalis
Galeopsis ladanum
G. tetrahit
G. sp. . '
Galium aparine
*G. boreale
G. hybridum
352
+
LP
SLP
LP
+
+
P
S
+
+
s
L
L
+
+
LP
L
+
S
S
+
L
P
PL
L
PSL
+
+
+
LPK
PL
+
+
LKS
LK
+
LP
+
+
+
LP
L
+
+
+
S
L
P
KL
+
L
L
+
LSP
L+
+
+
+
+
+
+
+
LPK
LP
+
+
+
LS
+
LK
L
L
+
LP
L
+
+
KL
L
+
+
L
LK
+
PL
P
LP
LSP
LK
+
+
+
+
+
+
LPK
13
LP
+
L
+
S
+
+
LPK
L
+
+
IP
L
+
KS
+
+
PL
KP
41
(45). (46)
40
(50)
41
(53)
30
(51)
42, (44), (45), (46), (48). (50)
24
9, 30, (37), 38, 39
9
9, 13
9
7
13, (43), (60)
40, 59
(46)
30.41
/(52), 59
9
9
22, 27
(43)
13, 28, 30, 31
13
7, (37)
32. (46)
31
7, 9, 22, 24, 30
13.(37)
(52)
2, 9, 13', 22, 23, 24, 27, 34. (37)
(50), (54)
26.(53)
20
12
(52)
7, 9 13, 19.22,23, 30, (37). 38, 4
(37)
12. 17,(47), 59
41
30
7, 9, 30, 31
13, 32
9, 30
(48), (50), (52), 56
56
Таблица (продолжение)
1
G. mollugo
G. palustre
G. pumilum
G. rotundifolium
G. rubioides
G. schultesii
G. silvaticum
*G. verum
G. vernum
*G. sp.
Genista tinctoria
Gentiana pneumonanthe
Geranium albiflorum
G: columbinum
G. collinum
G. dahurlcum
G. dissectum
G. macrorrhirum
ti. molle
G. palustre
*G. pratense
G. Robertianum
G. sanguineum
*.G silvaticum
G. Wlassovianum
*G. sp.
*Geum rivale
G. strictum
•G. urbanum
*G. sp.
Geutianaceae triflora
•Glechoma hederacea
Glycine sp.
Glycyrrhiza sp.
Gnaphalium silvaticum
Goniolimon sp.
Gypsophila muralis
Hedysarum alpinum
H. sibiricum
H. sp.
Helianthemum nummularium
Helianthus tuberosum
*H. sp.
Helleborus foetidus
Hepatica nobilis
Heracleum dissectum
H. flagellare
H. lachenalii
2
+
+
LS
+
+
P
+
+
L
L
L
LS
LS
*t
L
L
LPS
+
P6
L
+
+
+
3
Г 1
+
L
+
+
PL
+
+
+
LS
L
+
LP
+
+
+
+
LP
L
LK
+
LS
L
+
+
+
+
+
LP
LPKS
L
L
L
L
4
+
L
PK
+
+
+
KLS
+
LK
+
+
LSP
+
+
+
+
+
+
+
LPS
+
LK
+
LKS
L
+
+
+
+
+
LP
LPKS
L
+
+
5
+
L
+
+
+
KLS
+
+
+
+
PL
+
+
+
' +
+
LP
LK
+
LK
LS
PL-
L
L
+
+
+
LPKS
+
P6
+
+
+
6
9, 13
1
21
9
56,59
13
9
7, (48), (50), (52)
9, 13
13, 27, 28, 31(32,)(46)
(37), (50), (59) -"
7
(45)
9
(43), (44)
(52)
9,39
39
9
12, 13
13, (37), (48)
9,39
9, 13
7, (54)
(52)
13,32,(51)
9, 13, 23, 24, 30, (54), 59
(47)
7,9. 13, 22, 30, 31,32, (43), (50)
28, 30, 39, (60)
(52)
7, 9, 13, 28, 30, (37), 40, 59
(52)
(43)
9
(43)
27,59
(50)
(48)
(43), (44)
13
17
17,32,(52)
9
13, 22, 24, 30
(43), (45), (46), (47)
13
13
23. Европейская и сибирская косули
353
Таблица (продолжение)
Н.
Н.
Н. moellendorffii
Н. murorum
Н. pilosella
Н. sibiricum
Н. sosnowskyi
Н. sphondylium
»Н. sp.
•Humulus lupulus
Hylomecon vernalis
Hypericum acutum
H. androsaemum
H. centralis
H. desetangsii
*H. hirsutum
maculatum
montanum
H. perforatum
H. polugonifolium
H. pulchrum
H. sp.
Hypochoeris radicata'
Impatiens noli-tangere
I. parviflora
Inula hirta
I. salicina
Isopyrum thalictroides
Knautia arvensis
K. silvatica
Kochia sp.
Lactuca altissima
L. muralis
L. serriola
»L. sp.
Lamium album
L. amplexicaule
L. galeobdolon
L. maculatum
L. purpureum
L. sp.
Lampsana communis
Laserpitium hispidum
L. prutenicum
Lathyrus flolovii
L. gmelini
L. humilis
L. inermis
L. komarovii
*L. montanus
L. paluster
354
+
+
L
L
+
LP
+
+
+
+
+
+
+
+
+
+
+
+
+
L
LP
P
LPS
+
-
LP
К
LK
+
+
L
К
+
+
+
P
+
+
+
+
+
+
+
+
+
+
+
+
+
KL
L
L
LPK
PK
LP
+
L
LP
К
+'
LK
+
+
LKS
+
+
L+
+
+
К
+
•f
+
+
К
PK
+
+
+
+
+
L+
+
+
LK
+
+
KL
К
L
LP
LPK
P
LP
+
L
LP
К
+
LK
+
LS
+
+
+
L+
+
+
+
+
+
+
+
К
+
+
+
+
+
+
+
+
+
+
+
L
+
+
LPSK
LPK
+
-
(52)
9
7,9
(37), (43), (50), (51), (53)
41
9, 16, 42
13, 23, (43), (47), 59
13,(50)
(52)
9
41
1
9
9, (45), (46), (50), 56
9
9
1,9,13,30,39
41
9
12,59
13,30
9,24
9
1,59
56
13,24
9, 13, 59
9
(43)
7
39,56
7,41
(43), 59
13,27
40
9. 13, 24, 30
7, 9, 31
7, 13, 27, 40
27,59
7, 9, 59
56
1
(45)
(45), (50)
(48)
56
(52)
9, (43)
(37), (45), (46), (47)
Таблица (продолжение)
*L. pisiformis
*L. pratensis
L. silvestris
*L. tuberosus
*L. vernus
*L. sp.
Lavatera sp.
"Leontodon autumnalis
L. hispidus
Leonurus sp.
Lepidium campestre
L. sp.
Lespedera bicolor
Leucanthemum sp.
Leuzea carthamoides
Levisticum officinale
•Libanotis sibirica
L. seseloides
Ligularia sp.
Linaria sp.
Lonicera caerulla
Lotus corniculatus
L. uliginosus
Lupinus polyphyllus
Lychnis flos-cuculi
Lycopus europaeus
Lysimachia nemorum
L. nummularia
*L. vulgaris
♦Lythrum salicaria
Maackia amurensis
Malachium aquaticum
Malva neglecta
Matricaria chamoilla
M. maritima
M. sp.
♦Medicago falcata
M. glutinosa
M, lupulina
M. sativa
*M. sp. -^
Melampyrum pratense
M. silvaticum
M. sp.
•Melandrium album
M. sp.
•Melilotus albus
+
+
L
P
L
LP
+
L
+
+
+
P
S
KS
KS
+
LS
+
L
LS
+
+
+
+
+
+
+
L
+
PL
+
+
LS
+
-L
+
LK
+
+
PL
+
KS
+ .
+
+
L
-i-
L
LS
+
+
+
К
LK
+
PL
+
LK
+
+
PL
PL
L
+
LP
+
+
+
LK
+
+
+
+
PL
S
+
KS
+
L
+
LKS
+
+
LKS
+
+
+
К
+
L
+
LK
+
P
PL
L
+
LP
+
+
+
LK
+
+
+
+
+
L
+
S
LKS
+
+
LKS
+
+
LK
LKS
+
(37), (50), 59
9, 13, 30, 39, (48), (50), 59
1,24
31,(55)
13, 24, (54)
21, (47), (51), (53)
59
9, 30, (37)
9
(43)
27
32, 59
(52), (53)
59
(45), (46)
20
(37), (43), (50), 59
(52)
(44)
(46)
(43), (48)
9, 13, 30
9
20
9, 13
9, 40, 55
9,59
7,9
9, (50)
7, 9, (52)
(52)
30
28,31
20
22, 2"? 30
31, 32
7, 30, 40, (48)
41
9, 13, 30
5, 7, 9, 13, 16, 17, 27, 28, 31
38, 40, 41
41,(43), (47), 59
9, 13, 23, 30
39
12,23
9,29,31,(50)
31, 32, 59
9, 13, (48)
32,
355
Таблица (продолжение)
М. altissimus
М. dentatus
М. officinalis
М. melissophyllum
Mentha sp.
•Menyanthes trifoliata
Mercurialis perennis
Moehringia trinervia
Mycelis muralis
Myosotis arvensis
M. caespitosa
M. palustris
Nepeta sp.
Nitraria sp.
Onobrychis viciifolia
Origanum vulgare
Orobus vernus
*0. sp.
Oxalis acetosella
Oxyria sp.
Oxytropis altaica
O. caespitosa
O. glabra
O. myriophylla
O. uralensis
Paeonia lactiflora
Papaver rhoeas
P. sp.
Pastinaca pratensis
P. sativa
Pedicularis comosa
P. lasiostachys
Petasites sp.
Petroselimum crispum
P. hortense
Peucedanum morissonu
Phaseolus vulgaris
Phlomis tuberosa
Phyteum^ spicatum
Picris hieracioides
Pimpinella major
•P. saxifrage
*Pirola rotundifolia
P. sp.
*Pisum sativum
Plantago lanceolata
•P. major
L
+
+
+
L
L
LP
+
+
LP
LP
LP
+
s
S
+
LP
+
+
L
+
L
+
+
LKS
+
PL
PL
+
+
L
L
+
LP
+
+
LPK
LPK
LPK
LP
S
+
L
+
LP
+
+
LK
L
+
+
L
+
LK
+
+
LKSP
+
+
PL
L
+
+
LK
+
К
LP
+
LK
L
+
SL
LPKS
SL
LPKS
LPKS
KL
LKS
+
+
L
+
LP
+
+
+
L
LK
+
+
+
+
LK
+
+
LKP
+
+
+
+
+
+
К
P
P
+
К
L
+
+S
LPKS
s
LPKS
LPKS
L
S
+
+
L
+
LP
+
+
+
LK
+
+
+
+
LK
+
9
(48)
9
9
59
1,23,42, (43), (45), (46), (48), (50)
(52), (53), (54)
9, 12, 55
7,9
1,9, 13,24,30
9
55
9
59
(43)
9
1,41,59
(50)
(44), (46), 50, 59
9, 12, 13, 24, 30, 39
(51)
(45)
(48)
(45)
(48)
(48)
(52)
28, 31
32
(37)
9
(50), (60)
(43), (47)
(51), (53)
20
29
(43)
9
(37), (43), (48), (50)
9
9, 29
9
9, 13, 30,(37)
9, (48)
22
7, 9, 16, (46), 59
9, 12, 13, 27, 56, 59
13, (37), (48), (54)
356
Таблица (продолжение)
1
*Р. media
•P. sp.
Pleurospermum uralense
Poligonum divaricatum
Polemonium liniflorum
P.sp.
Polygala comosa
P. sibirica
Polygonum alpinum
*P. aviculare
P. bistorta
P. convolvulus
P. lapathifoiium
P. mite
P. paradoxum
P. persicaria
P. scabrum
P. viviparum
P. sp.
Portulaca oleracea
•Porentilla alba
P. anserina
P. erecta
P. intermedia
P. multifida
P. nivea
P. recta
P. reptans
P. sterilis
P. tanacetifolia
•P. sp.
Prenanthes purperea
Primula acaulis
P. elatior
P. macrocalyx
P. pallasii
P. patens
Prunella vulgaris
Pulmonaria angustifolia
P. mollissima
P. officinalis
P. rubra
Pulsatilla cernua
P. flavescens
P. patens
P. turczaninovii
P. sp.
Pyrethrum pulchellum
2
s
L
+
+
+
+
LS
+
+
+
L
P
+
+
LP
L
LP
+
+
SL
LPS
LPS
+
PL
3
+
s
+
LP
LP
LP
+
L
LP
+
+
S
+
L
+
К
+
L
+
PL
+
+
L
K+
L
LK
L
L
+
+
+
+
PL
+
LPSK
LPSK
+
PL
4
К
К
+
LK
LP
LP
+
LP
+
PL
LP
+
К
+
LP
LKS
+
LK
+
LP
LP
P
L+
+
LP
+
+
LK
LK
LS
+
+
+
К
+
LPSK
LPSK
+
5
К
KS
+
L
L
+
+
+
+
+
+
LS
+
К
+
LK
L
+
+
LKS
+
L
L
+
LS
+
+
+
PL
LPSK
LPSK
+
PLKS
6
7, 9, 30, (37), (48), (52), 55
23,28,31,32,39,(46), 59
(43), (47), (50)
(48)
(52)
(51)
13
(52)
(50), (51)
13, 15,(37)
(45), (50), (54)
59
30
30
(43)
15, 22, 30
(37)
(45), (46), (51), (53)
12,27,28,31,32, 34
27, 39, 40
1,(60)
9, 13
1, 13, 23, 30, 31
7
(48)
(51)
56
9, 13,41
9
(48)
27, 30, 39, (44), (51)
9,39
55
9, 12
(50)
(45)
(52)
9, 30
13, 42, 59
(43), (46), (50)
7,24
39
(46), (52)
(47), (51)
(48), (50)
(48)
(53)
(48)
357
Таблица (продолжение)
J
Pyrola sp.
♦Ranunculus acer
R. aconitifolius
R. arvensis
R. auricomus
R. bulbosus
*R. cassubicus
R. hicaria
R. japonicus
R. lanuginosus
R. montanus
R. nemorosus
R. poliantemum
*R. polyanthemos
*R. re pens
*R. sp.
Raphanus raphanistrum
R. sativus
Rhinanthus alectorolophus
R. alpinus
R. major
R. minor
Rhododendron dauricum
R. sp.
Roripa sp.
Rubus arcticus
R. caesius
R. idaeus
R. sachalinensis
R. saxatilis
•Rumex acetosa
R. acetosella
R. conglomeratus
*R. confertus
R. obtusifolius
R. orifolius
R. sanguineus
R. thyrsiflorus •
*R,sp.
Sagittatum sp.
Salsola sp.
Salvia glutinosa
S. pratensis
S. verticillata
•Sambucus ebulus
2
L
L
+
+
LP
L
L
L
L
LPS
+
PL
PL
+
L
+
+
LS
P
P
1
L
LK
LK
LK
+
+
+
LP
+
LK
L
+
L
+
L
+
+
KP
К
+
L
+
PL
LP
LK
LK
+
+
S
LK
+
К
P
4
LK
LK
L
LK
LP
LK
LK
L
+
L
LS
L
+
+
+
+
LP
+
LK
L+
LS
PLS
L
LK
LK
+
+
KS
L
S
+
LS
LK
+
L
К
К
+
5
L
L
LP
LK
LK
L
+
L
+
+
LPKS
LP
S
(52)
L
+
LKS
+
+
KS
s
+
+
+
к
+
6
(46), (51)
1,9,12,23,30, (45), (46), (47), (54),
56,59
9
41
7, 9, 22
13
(54), 59
40
(52)
9, 24, 30
39
9, 12
(45), (46)
7, (50), 59
7,9, 13, 24, 30, (43), (50), (54), 55
12,23,27, 30,31,(53)
22
28
9
12
12, 13
9,23
(48), (51), (52)
(46)
59
(52)
(43), (44)
(45), (46), (50)
(50)
7, 9, 13, 23, 30, (37), (45), (46),
(54), 59
9, 13, 23, 30
26
7, (37), (43), 59
9
41
9
30
12, 13, 21, 23, 24, 27, 28, 30, 31,
32, (44), (51), (52), (60)
(46)
(43)
41,59
9
56
9, 38, (46)
358
Таблица (продолжение)
1
S. racemosa
Sanguisorba minor
*S. officinalis
S. parviflora
Sanicula europaea
Saussurea amurensis
S. frolowii
S. latifolia
S. salsa
S. triangulata
Saxifraga sp.
♦Scabiosa columbaria
S. Fischeri
Scorzonera humilis
Scrophularia nodosa
Scutillaria altissima
S. galericulata
S. sp.
Sedum acte
S. Eversii
Selium carvifolia
Senecio aquaticus
S. jacobaea
S. silvaticus
S. sulphureus
S. vernalis
*S. vulgaris
*S. sp.
Serratula coronata
S. isophylla
*S. tinctoria
Sherardia arvensis
Silaum selinoides
Silaus sp.
Silene cucubalus
S. inflata
*S. nutans
S. venosa
*S. sp.
Sinapis alba
*S. arvensis
S. sp.
Sisymbrium officinale
Solanum dulcamara
S. lycopersicum
S. nigrum
S. tuberosum
2
L
PL
PL
L
LP
LP
P
L
+
P
P
+
+
PL
+
L
+
L
LP
LS
S
Pi
3 ■
+
L
L
L
LP
PL
PL
PL
+
SL
+
+
P
PL
+
L
+
+
+
+
+
LP
+
+
+
L
+
LP
+
S
рг
4
L
+
L+P
PL
К
LP
LK
LK
PL
PL
+
+
SL
+
L
LP
+
L
+
+
+
+
+
+
P
L
+
+
LK
KL
+
L
+
LS
+
+
L
+
L
5
L
+
P+S
PLS
LK
LP
+
+
L
PL
+
+
L
+
L
L
+
+
+
+
+
+
+
PL
PL
+
+
LK
LK
LP
LKP
+ '
+
+
L
+
LP6
6
(48)
9
9, (37), (43), (45), (46), (47), (48),
(50), (51), (52), (53), 59
(51), (52)
9, 55
(52)
(45)
(45)
(50), (51)
(52)
39
9, (46)
(48)
13
7, 9, 59
56
1,30
(52)
7
(44)
9
9
26,30
9
9
13
9, (37)
13, 27, (46), (47)
(48)
(50)
9, (50), 59
27
9
(47)
7,9
56
7, 13, (37)
(48)
27, (46)
27,2»
27, 32. (37)
31
(37)
9
9
9
5, 9. 15, 17, 27
359
Таблица (продолжение)
Г
1
Soldanella alpina
Solidago pacifica
*S. virgaurea
♦Sonchus arvensis
*S. oleraceus
*S. sp.
Spergula sp.
Spiraea aquilegifolia
Sp. chamaedryfolia
Sp. media
*Sp. salicifolia
Stachys palustris
St. silvatica
Stellaria alsine
•St. graminea
St. holostea
St. media
St. nemorum
*St..sp.
Stenactis annuus
Suaeda linifolia
♦Succisa pratensis
Symphytum grandiflora
S. officinale
S. tauricum
Tanacetum sibiricum
*T. vulgare
♦Taraxacum officinale
Teucrium chamaedrys
T. scorodonia
T. sp.
Thalictrum amurense
*Th. aquilegiifolium
T. minus
T. petalotdeum
T. simplex
T. squarrosum
T. sp.
Thelycrania alba
♦Thlaspi arvense
Thymus 'glabrescens
T. serpyllum
Torilis arvensis
T. japonica
Tragopogon orientalis
Tralictrum aquilegifolium
2
LP
+
+
LS
LP
+
LP
LP
+
L
+
L
LS
P
LP
+
LS
LP
LP
LP
LP
+
LP
L
S
+
LS
3
LP
+
+
L
+
L
L
L
L
+
+
L
L
LS
+
+
P
+
LKS
LP^
L+PK
LK
LP
+
LP
S
+
+
+
4
LP
К
+
+
LKS
S
L
+
L
L
L
L
L
+
+
LS
L
+
+
+
LS
+
К
L+PK
LP
К
+
LP
+
L
+
+
+
5
LP
+
+
+
L
+
L
L
+
L
+
+
L
L
SP
+
+
LPKS
+
LK
+
LP
L+PK
L
LP
+
L
SLPK
+
+
6
39
(52)
2, 7, 9, 13, (37), (50)
7, 13, 30, (37), (43), (50)
9. (50)
28, 31,(47), (53), 59
22
(48)
(43)
(48)
(48), (52), 59
15,56
I, 7, 9, 24
9
9, 13, 27, (37), (50)
7, 13, 24, 27
9, 13, 16,31,39,40
9,30
13, 22, 27, 28, 32, (46), 59
41
(43)
9, 13, 23, (50)
41
40
56
(48)
(37), 59
9, 12, 13, 15, 20, 23, 24, 26, 27,
28, 29, 30, 31, 32, (37), 40, (46),
(47), (54), (59)
55
9
39
(52)
9, 13, (47), (46), (54)
(48), (50)
(52)
(37)
(48)
(51), (52), (53)
(48)
28, 30, 32, (46), 59
29
13
41
7,9
(37), (43)
59
360
Таблица (продолжение)
*Т. pratensis
T.sp.
Trientalis europaea
Trifolium alpestre
*T. arvense
T. humens
T. hybridum
T. lupinaster
*T. medium
*T. montanum
*T. pratense
*T. re pens
T. spadiceum
Tripolium sibiricae
•T. sp.
Trollius asiaticus
T. chinensis
T. europaeus
*T. sp.
Tussilago farfara
Unula hirta
*Urtica dioica
*U. urens
»U. sp.
•Vaccinium arctostaphylos
V. myrtyllus
V. uliginosum
V. vitis idaea
Valeriana dioica
V. officinalis
V. transjenisensis
Verbascum thapsus
V. sp.
Verbena officinalis
Veronica agrestis
V. anagallis
V. beccabunga
V. chamaedrys
V. hederifolia
V. incana
V. longifolia
V. montana
V. officinalis
V. persica
V. sibirica
)
L
+
L
L
+
P
LP
LP
+
+
+
LS
L
LPS
LPS
LPS
+
+
+
+
+
+
L
L
+
+
L+
+
L
+
LP
+
+
L
LK
LK
P
LP
L
L
+
+
+
LS
LP
LP
LP
+
LP
+
+
+
+
+
L
L
+
+
+
LP
L+
+
L+
L
+
LKP
+
L+
LK
LK
LK
L
К
LP
+
+
+
LK
LK
LK
+
+
LP
+
+
+
+
L
LP
+
+
+
LP
+
+
+
LKP
+
+
LK
LK
LK
LP
LP
LPSK
+
+
+
LKS
LKS
LKS
+
+
+
+
+
+
+
+
L
LP
+
+
+
9, 41. 42, (51), (59)
59
23
13,59
(37), 41
41
(37)
(50), (52)
7, 9, (43), (50), 56, 59
9, 41, (50), 59
9, 11, 13, 16,22,27,30,34,(37),
40, 42, (48), (52)
9, 13, 16, 22, 26, 27, 30, (37), 40,
41, (48), (52)
41
(46)
1, 12, 13, 17, 21,23, 39, (47), (51),
(53)
(43), (45), (46), (48)
(52)
(50), (54)
(50), 59
15,30
(37)
7, 9, 12, 13, 24, 30, (37). 39, 39,
(50)
7, (37), (54)
28, 32, (43), 59
41, (53)
(48)
(48), (52)
(48), (50)
9
9,34
(52)
(37) -
' 29, 59
9
27
9
9
7, 9, 13, 30, 31, 59
27,40
(48)
59
9
9, 13
9, 27, 40
W
Таблица (продолжение)
1
V. spuria
V. teucrium
*V. jp.
Vicia amoena
•V. cracca
V. dumetorum
V. hirsuta
V. pseudorobus
V. unijuga
*V. sativa
*V. sepium
V. sibirica
*V. silvatica
V. temrifolia
V. venosa
V. villosa
*V. sp.
Viola hirta
V. mirabilis
V. odorata
V. Riviniana
V, silvatica
V. silvestris
V. suavis
V. tricolor
*V. sp.
Viscaria viscosa
V. vulgaris
Viscum coloratum
Vogelia paniculata
2
PL
+
LP
LPS
+
LP
LP
+
+
SP
LP
LPS
+
+
+
+
+
LS
LS
3
L
+
L
LPK
+
+
LPK
LPK
+
К
+
L
LP
+
+
+
+
+
+
L
L
+
LP
+
4
+
S
LPK
LK
L
LPS
LPS
L+
LK
L
LK
L
LPS
+
+
+
+
+
+
LS
+
+
LP
5
+
+
LK
LPSK
LPK
+
SK
+
SK
LKP
LKP
+
+
+
+
+
+
L
Однодольные травянистые растения
♦Agropyrum canumun
A. cristatum
A. intermedium
*A. re pens
A. tenerum
*A. sp.
•Agrostis alba
A. canina
A. capillaris
A. mongolica
A. setacea '
A. stolonifera
A. vulgaris
A. sp.
Allium lineare
A. oleraceum
A. ursinum
362
+
+
LS
+
LP
+
+
+
L
+
LP
LP
L
, +
LP
+
+
PL
L
L
+
LS
+
L
L
LPS
+
+
PL
L
+
LP«
+
S
+
+
+
PK
6
(50)
(37), (50)
29, 31,32, 39, (46), (60)
(48), (52)
1, 9, 15, 30, (37), (43), (45), (48),
(51), (52), (53), 56, 59
13
41
(48)
(48), (52)
27, (37), 41, (43)
1,9, 41, (45), (46), (54)
(44)
1,(45)
(50)
(52)
39
13, 27, 30, 41,(47), (53)
9,59
7
40
9,23
9,40
13,24
7
30,31
13, 23, 27, 28,32, (46), (52), 59
(50)
7
(52)
27
(50), 59
(48)
40
9, 13, 16, 17, 22, 31,(37), (52)
(48)
17, 28, (46)
41,(43)
7
41
(48)
25
30,40
13
13, 21, 36
(46)
7
9
Таблица (продолжение)
1
A. schoenoprasum
A. strictum
A. victoriale
A. sp.
Alopecurus pratensis
A. ventricosus
Andropogon ischaemum
Anthericum ramosum
Anthoxanthum odoratum
Apera spica-venti
Arrhenatherum clatius
Arum maculatum
Asparagus officinalis
A, sp.
Atropis distans
A. fatua
•Avena sativa
A. sp.
Avenastrum pubescens
•Brachyposium pinnatum
*B. silvaticum
Briza media
Bromus arvensis
B. erectus
B. hordeaceus
B. inermis
B. mollis
B. ramosus
B. sterilis
B. sp.
♦Calamagrostis arundinacea
С brachytricha
С canescens
С epiglios
C. langsdorffii
С Obtusata
*C. sp.
Carex acutiformis
С artrata
C. brizoides
С caespitosa
С capillaris
С digitata
С flava
С glacilis
С glauca
С Hostiana
C. hasiocarpa
2
3
SL
^LP
+
+
LK
S
L
S
+
L
S
L
+
L
+
L
+
L
- +
+
+
+
+
+
+
+
+
+
+
L
+
К
L
LK
LK
LK
+
+
+
L
+
LK
LK
LK
+
LK
К
LP
+
PL
PL
L
L
+
PL
+
+
+
L
+
+
4
SL
LP
L
К
L
LK
LK
LK
+
+
KS
s
LK
LK
LK
+
LK
L
S
К
+
L
+
L
L
+
+
+
PLSK
5
SK+
+
+
+
К
L
LK
LK
+
LKS
S
LK
+
LK
+
LK
L
+
L
+
+
+
+
L
+
+
+
+
+
+
6
(45), (46), (48)
(50)
(46)
(44), (45), (51), (52), (53)
9,40
(37), (43)
9,40
59
1,9
28
9,59
9
29
27
(37)
(37), (47)
7,9,12,13,17, (37), (46), (47), (48),
(52>
11,28,31
9, 13, 30
9, (50), 59
9, 13, 25, 40, 41, (47)
9
40
9
40
(37), (43)
9, 13
9,55
31,40
1,29
13, 24, (50)
(52)
13
(37), (43), (48), (50), (52)
(45), (48), (52), (53)
(45), (46)
1,7, 13, (47), (51), 59
9
(45), (50)
9
(48), (50)
22
9, 13
9
(45)
9
9
(45), (46)
363
Таблица (продолжение)
1
С. leporina
С. limosa
С. Meyeriana
С. montana
С. muricata
С. pallescens
С. panicea
С. pediformis
С. pendula
С. pilosa
С. pilulifera
С. pseudocyperus
С. remota
С. rostrata
С. Schmidtii
С. silvatica
С. strigosa
С. tomentosa
С. umbrosa
С. verna
♦С. vesicaria
С. wiluica
»С. sp.
Cephalanthera alba
С. longifolia
С. rubra
Colchicum speciosum
•Convallaria majalis
Cynodon dactylon
Cynosurus cristatus »
Cypripedium ventricosum
*Dactylis glomerata
•Deschampsia caespitosa
D. flexuosa
D. sp.
Digitaria sanguinalis
Digraphis arundinacea
Diplachne squarrosa
Echinochloa crus-galli
Echinops dahuricus
Eleocharis pauciflora ~
Elymus sibiricus
Epipactis atropurpurea
E. latifolia
E. sessilifolia
Eragrostis minor
2
L
L
L
+
+
+
L
L
L
L
L
L
L
L
+
L
L
+
L
L
L
+
+
LP
S
PL
L
3
l Г
L
PLSK
LPS
L
L
+
+
L
L
L
L
L
PLS
PLS
L
L
L
L
+
LP
LP
KPL
К
К
К
L
LK
L
LK
L
+
+
LPS
S
L
LPS
4
PL
PL
S
S
L
PL
PL
S
s
L
L
К
К
К
LK
L
LK
L
+
L
LPS
S
L
L
К
К
К
5
L
L
+
+
+
LS
L
LS
LS
L
+
+
+
К
К
к
LK
LP
LK
L
+
PL
L
К
К
К
6
9
(48), (51), (53)
(48)
9
9
9
9
(45)
9,41
7, 9, 40
9
55
9
(48)
(48)
9,41, 55
9
9
9
9
(50), (51), (53), 55
(51), (53)
1, 13, 16, 21,23, 24, 25, (37), 41
(46), (50), (52), (53), 59
9
9
9
41
2, 13, 39, (54)
40
9, 13
(52)
1,7,9, 13, 21,24, 27, 29, (37), 40
41, (43), 55, 56, 59
7, 9, 13, 25, 27, 30, (37), (50)
13,23
21
40
(50)
(48) ■
27, 28, 31, 32t 40
(48)
(48)
(51)
9
9
9
40
364
Таблица (продолжение)
1
♦Eriophorum vaginatum
Е. sp.
Festuca arundinacea
F. giganta
F. lenensis
*F. ovina
*F. pratensis
*F. rubra
*F. silvatica
*F. sulcata
F. sapina
F. valesiaca
»F. sp.
Funees bufonius
Galla palustris
*Glyceria fluitans
G. plicata
Habenaria linearifolia
Heleocharis palustris
Hemerocallis minor
Hierochloe dahuricum
Holcus lanatus
H. mollis
H. sp.
Hordeum brevisubulatum
H. distichum
*H. vulgare
H. sp.
Iris ensata
Juncus conglomeratus
J. effusus
•J. sp.
•Koeleria glauca
K. gracilis
♦Lilium martagon
L. puechellum
L. tenuifolium
Lolium multiflorum
L. perenne
Luzula albida
L. campestris
L. pilosa
L. silvatica
L. sp.
♦Majanthemum bifolium
•Melica nutans
Mentha sp.
Milium effusum
•Molinia coerulea
2
+
+
L
L
L
L
L
+
+
+
+
+
L
L
L
+
+
LS
L
S
L
L
PL
+
LP
L
L
L
L
L
L
L
+
LP
L
+
3
+
+
L
PL
LK
LK
LK
+
+
+
+
+
L
LP
+
+
LP
+
LP
LK
+
+
LS
L
L
+
+
LP
К
P
L
LK
LK
L
L
L
LK
+
L
К
LKS
+
1 4
LS
L
LK
LK
LK
L
L
LK
L
LP
LP
SLP
LK
LK
S
+
+
P+
+
К
PK
L
LK
LKS
KS
К
L
К
LKS
+
5
+
L
S
LK
LK
LK
+
+
■+
+
LS
LS
LK
L
+
L
L
S
L
P+
+
К
P
L
LK
LK
LKS
L
LK
+
L
LKS
+
6
23, 36, (50), (51), (52)
(52), (53)
9
7, 9, 41
(48)
9, 13, 27, (37)
9, 13, 22, 27, 30, (37), 41
9, 13, 27, 30, (37), 40
40, (50)
40, 41, (43)
(45)
40
1,2, 13, 21,36, 41,(43), (47)
(50)
(52)
9, (55)
9
(52)
30,40
(52)
(41)
9,27
9
21
(48)
5, 12, 16, 22, 32
9, (37)
11,28,30,31
(48), (53)
13
9,40
21, 22, 23, 30, 36, (46), 59
13, (43), (46)
(48)
9, (46), (51), (53), 59
(52)
(48)
9, 13
9, 31,40
9, 39, 40
9
9,23
9,40
13, 19
12, 13, 23, 24, (46), (48), (53)
1,13, (54), 59
41
1,9,24,55,59
1, 25, 30, (50) зет
Таблица (продолжение)
1
Nardus stricta
Orchis maculata
Paris incompleta
P. quadrifolia
*Phleum sp.
♦Phleum pratense
Phragmites communis
Platanthera bifolia
Poa alpina
*P. annua
P. botryoides
P. compressa
P. nemoralis
♦P. pratensis
P. trivialis
P. sibirica
P. stepposa
•P. sp.
Polygonatum humile
P. Multiflorum
*P. odoratum
*P. officinale
Puccinella tenuiflora
Ruscus hypophyllum
Sagittaria sp.
Scilla bifolia
S. nonscripta
S. sibirica
Scirpus silvaticus
♦Secale'cereale
S. viridis
S. sp.
Smilax excelsa
Stipa joannis
Triglochin maritima
Trisetum flavescens
•Triticum vulgare
Tulipa behmiana
Typha Laxmannii
♦Veratrum iobelianum
*V. nigrum
Zea mays
Z. sp.
Athyrium filix-femina
Cystopteris fragilis
366
2
+
L
+
+
+
+
+
L
L
LP
L
PL
PL
PL
LP
LP
L
S
+
1
L
LS
LS
3
+
+
L
+
LK
К
+
+
LK
LK
LK
+
+
L
L
L
LP
LPK
К
+
LP
L
L
LK
LS
+
+
PL
PL
LKS
4
+
L
+
LK
К
L
+
+
L+
LK
LK
LK
+
S
L
L
L
LP
LPSK
+
LP
LKS
LK
LK
KS
+
+
L
L
LKS
5
+
LK
+
+
LK
LK
LK
+
L
L
L
L
LP
LS
+
LP
LK
L
+
+
KS
Папоротники, хвощи и плауны
L
L
+
6
21
9
41
9
(43), 59
1,9, 13, 22, 27, (37), 41
7,42
9
41
13,40,(54^
(48)
13
7, 9, 13, 29, 30, 41
1,7,9,13,22,27, (37), 40,41, (44),
(48), (51)
9,41
(48)
(43)
13, 17,21,28,30,31,36,41,(50),
(52), 59
(52), (53)
7,59
9, (52)
(37), 40, (46), (50), 59
(48)
41
(53)
55
11
7
(50)
5,9,11,16,17,23,27,30, (46), (47)
40
28
41
(37)
(48)
9
5, 7, 9, 11, 16, 17,27,28,29,30,
31,32, (37), (47), (48), (52)
(43)
(43)
(45), (46), (53), 59
(45), (46), (47), 59
5, 11, 17,27,28,29,31, 32,40
(52)
7, 9, 12
13
Таблица (продолжение)
■
1
Dryopteris austriaca
D. carthusiana
D. disjuncta
D. filix-mas
D. oreopteris
D. spinulosa
D. thelypteris
D. sp.
Equisetum arvense
E. hiemale
E. palustre
E. scirpoides
*E. silvaticum
E. variegatum
E. sp.
Lycopodium annotinum
Polystichum lobatum
Pteridium aquilinum
P. sp.
Pteris aquilina
Struthiopteris spicant
Anomodon longifolium
Brachythecium salebrosum
Dicranium scoparium
Eurhynchium swartzii
Hylocomium splendens
H. sp.
Mnium hornum
M. sp.
Muscia sp.
Plagiothecium undulatum
Polytrichum sp.
Sphagnum sp.
*Cladonia furcata
*C. sp.
Evernia prunastri
Parmelia physodes
Physcia pulverulenta
*Usnea barbata
U. parmelia
♦Armillaria mellea
Boletus luteus
B. scabe/
•B. sp.
2
L
+
+
L
+
L
+
+
+
L
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
3
L
L
+
+
+
+
+
+
+
L
-:-
LP
L
Мхи
+
+
+
+
+
+
+
+
+
+
4
L
+
L
+
+
+
PL
+
+
L
+
L
L
+
+
+
+
Лишайники
+
Грибы
+
+
+
+
ГЛ
5
L
+
L
+
L
+
L
+
+
+
+
+
+
+
+
+
+
1 — i —
6
9, 12, 13
23
9
9,21,41
9
22
9
21, 26, 30
(51)
(46), (48)
(48)
(51)
13, (46), (48)
(51)
23, 28, 30, 59
9
9,39
9, 13, 21, 23, 39
26,59
(45), (46)
9,21
7
7
21
7
21
13
26
13
11,23,25,27
21
21
21
39, (46), (51), (53)
23, (46), (49), (51), (53)
7
23
7
1,38, 39, (46), (47), (51>
(46)
30, (37), (51)
(48), (51)
(37), (51), (52)
7, (43)
367
Таблиц* (окончание)
1
Fomitopsis pinicola
Hypholoma capnoidea
Lactarius deliciosus
L. forminosus
L. piperatus
L. vellereus
Polypodium vulgare
Polyporus brumalis
P. sp.
•Russula sp.
Tricholoma sp.
Ustilago major
*Fungi sp.
+
+
4
+
+
+
22
30
+ 4 (37), (45), (46), (48), (51), (60)
+ +4- (48), (60)
'+ 4- (43)
+ 4-4- (46), (48), (51), (52), (60)
39,41
4- 30
4- 30
+ (37), 41, (51), (52)
+ 30
+ 31
4- 4- 4- 4- 11,12,17,19.23,25,27,28,31,
39, 42, (53), (60)
Примечание. Полужирным даны виды растений, поедаемые сибирской косулей; не обозначены —
европейской; обозначены звездочкой — общие для них виды.
К — цветы, соцветия; L — листья; Р — побеги; Рс — подземные части; S — семена, плоды; 4- — растения
(без указания конкретной части), поедаемые в данный период года.
Номера без скобок обозначают авторов — исследователей питания европейской косули, в скобках —
сибирской.
Авторы и районы исследований
1 — Северцов, Саблина, 1953, СССР (Беловежская Пуща) [1]'; 2 — Саблина, 1955ч СССР
(Беловежская Пуща) [1]; 3 — Bukiewicz, 1960 (по; Szmidt, 1975), Польша [2]; 4 — Wagner,
1961, ЧСФР [1]; 5 — FiSer, 1962 (по: Netas, 1975),ЧСФР [3]; 6 — Mdhring, 1963, ФРГ [2];
7 — Новиков, Тимофеева, 1964, СССР (Белгородская область) [1, 2]; 8 — Ueckermann, 1964,
ФРГ [2]; 9 — KlOtzli, 1965, Швейцария [1]; 10 — Skotnicki, 1967 (по: Szmidt, 1975), Польша
[2]; 11 — Prior, 1968, Великобритания [1]; 12 — Fiser, Lochman, 1969, ЧСФР [3]; 13 — Siuda et al.,
1969, Польша [3]; 14 — Wagenknecht, 1969 (no: Szmidt, 1975), ГДР [2]; 15 — Holzhausen, 1970,
ФРГ [1, 2]; 16 — Knorr, 1973 (no: Stubbe, Passarge, 1979), ГДР [З]; 17 — Necas, 1975,
ЧСФР [1, 2, 3]; 18 — Szmidt, 1975; Польша [2]; 19 — Григоров, 1976, Болгария [3]; 20 — Stubbe,
Zdrner, 1977, ГДР [1, 2]; 21 — Henry, 1978a, Великобритания [3]; 22 — Балейшис, Прусайте,
1980, СССР (Литва) [3]; 23 — Cederlund et al., 1980. Швеция [3]; 24 — Gebczyriska, 1980,
Польша [3]; 25 — Jackson, 1980, Великобритания [3]; 26 — Hosey, 1981, Великобритания
[3]; 27 — Kaluzinski, 1982, Польша [3]; 28 — HoliSova et al., 1982, ЧСФР [3]; 29 — Matrai et al., 1983 (личное
сообщение) [3]; 30— Прусайте и др., 1983, СССР (Литва) [3]; 31 — HoliSova et al., 1984, ЧСФР [3]; 32 —
HoliSova et al., 1986a, ЧСФР [3,4]; 33 — Casanova, 1981, Италия[1]; 34 — Musi, Perco, 1975, Италия[1]; 35 —
Haisch, 1982, ФРГ; 36— Byron, 1981, Великобритания; (37) — Соколов, Данилкин, 1981, СССР (Южный
Урал) [1,3]; 38 —Петров и др., 1968; Болгария [1,3]; 39 —Колев, 1966, Болгария [1,3]; 40— AlmaSan et
al., 1969, Румыния [1,3]; 41 — Арабули, 19636, 1972,1977, СССР (Грузия) [1,3]; 42 — Верещагин, Русаков,
1979, СССР (Северо-Запад РСФСР) [1]; (43)— Слудский и др., 1984, СССР (Казахстан) [1,3]; (44) —
Антипин В.М., 1941, СССР (Казахстан) [3]; (45)— Пастернак, 1955, СССР (Алтай) [1]; (46)— Фетисов,
1953, СССР (Восточная Сибирь) [1,3];(47)— Зырянов, 1975, СССР (Восточная Сибирь) [1]; (48) —
Смирнов, 1978, СССР (Забайкалье) [1,3]; (49)— Владышевский, Ельский, 1974, СССР (Приангарье) [1];
(50)— Аверин, 1949, СССР (Урал) [1,3]; (51)— Егоров, 1965, СССР (Якутия) [1,3]; (52) —Дарман, 1987
(личное сообщение), СССР (Амурская область) [1,3]; (53)— Бромлей, Кучеренко, 1983, СССР (юг
Дальнего Востока) [1,3]; (54) — Габузов, 1960, СССР (Московская область) [1]; 55 — Даль, 1930, СССР
(Крым) [1,3]; 56— Лавров, 1929, СССР (Крым) [1]; (57)— Куражсковский, 1952, СССР (Северный
Казахстан) [I]; 58— Владышевский, Ельский, 1974, СССР (Карпаты) [1]; 59— Гусев, 1987 (личное
сообщение), СССР (Курская обл.) [1,3]; (60)— Айзин, 1969, СССР (Киргизия) [1,3].
' Методы сбора данных: [1] — полевые наблюдения, [2] — эксперименты, [3] — анализ
содержимого рубца, [4] — анализ помета.
Литература
Аверин Ю.В. Экология козули (Capreolus
capreolus pygargus Pall.) в Ильменском
заповеднике // Тр. Ильменского заповедника. 1949.
Вып. 4. С. 9—62.
Аверин Ю.В. Главнейшие изменения в
видовом составе фауны млекопитающих и птиц
Молдавии в XVII—XX столетиях // Бюл. МОИП.
Отд. биол. 1960а. Т. 65, вып. 2. С. 5—12.
Аверин Ю.В. Некоторые изменения в
составе охотничье-промысловой фауны Молдавии за
последние столетия // Охрана природы
Молдавии. 19606. Вып. 1. С. 125—132.
Авотииьш А. Корма для косули // Охота
и охотничье хоз-во. 1976. N 6. С. 36—37.
Адамович В., Ойцось И. Причины гибели
косуль в Волынской области // Там же. 1963.
N 4. С. 27—28.
Адольф Т.А. Некоторые данные по биологии
сайгака в Астраханских степях // Учен. зап.
Моск. гор. пед. ин-та им. В.П. Потемкина.
1954. Т. 28, вып. 2, С. 247—256.
Айзин Б.М. Копытные // Охотничье-промыс-
ловые звери Киргизии. Фрунзе, 1969. С. 78—111.
Алексеева Л.И. Териофауна раннего антропо-
гена Восточной Европы. М., 1977. 216 с. (Тр. геол.
ин-та; Вып. 300).
Алексеева Э.В. Млекопитающие плейстоцена
юго-востока Западной Сибири. М.: Наука. 1980.
137 с.
Алиев'С.Д. Фауна Азыхской пещеры — объект
охоты первобытного человека // 2-й съезд
Всесоюз. териол. о-ва: Тез. докл. М., 1978. С. 3—4.
Ащпипип В.М. Млекопитающие Казахстана:
Копытные. Алма-Ата: Казгосиздат, 1941. Т. 3. 107 с.
Арабу ли А.Б: Кочевки европейской косули
(Capreoius capreolus capreolus Linne) в Кахетии
и вызывающие их причины // Зоол. журн.
1963а. Т. 42, вып. 7. С. 1113—1115.
Арабули А.Б. О питании европейской косули
(Capreolus capreolus capreolus L.) на Цив-Гомбор-
ском хребте ,,'Сообщ. АН ГССР. 19636. Т.30,
N 4. С. 467-474.
Арабули А. Б. Косуля в Восточной Грузии:
Автореф. дис. ... канд. биол. наук. Тбилиси,
1966. 22 с.
Арабули А.Б. К изучению экологии косули
(Capreolus capreolus L.) на Кахетинском Кавка-
сиони // Тр. Ин-та зоологии АН ГССР. 1967.
С. 166—199. (Материалы к фауне Грузии; Вып. 2).
Арабули А.Б. Дикая свинья, косуля и олень в
Абхазии. Тбилиси, 1972. 99 с. На груз. яз.
Арабули А.Б. Кабан, косуля и олень на Малом
Кавказе. Тбилиси, 1977. 82 с. На груз. яз.
Бадер О.Н. Некоторые вопросы
палеогеографии Урала и Северо-Восточной Европы в свете
археологических данных // Материалы по
четвертичному периоду СССР. М.; Л.: Изд-во
АН СССР, 1950. Вып. 2. С. 140—149.
Бажанов B.C., Костеико Н.Н. Атлас
руководящих форм млекопитающих антропогена Ка-
захтана. Алма-Ата: Изд-во АН КазССР.1962. 111с.
Байкалов А.Ф., Семенов Г.К. Авиаучет
численности косули в Хакассии // Проблемы
охотничьего хозяйства Красноярского края.
Красноярск, 1971. С. 104—108.
Балейшис P.M.. Прусайте Я.А. Питание
европейской косули в небольшом лиственном лесу
Северной Литвы в 1975—1976 гг. ,/ Тр.
АН ЛитССР. Сер. В. 1980. Т. 1(89). С. 85-91.
Банников А.Г. Млекопитающие Монгольской
Народной Республики. М.: Изд-во АН СССР,
1954. 669 с.
Барабаш-Никифоров И.И. Звери
юго-восточной части Черноземного центра. Воронеж, 1957.
370 с.
Барабаш-Никифоров И. И.. Формозов А.Н. ,
Териология. М.: Высш. шк., 1963. Т. 1. 394 с.
Баранчеев Л.М. Массовые непериодические
миграции косуль в Амурской области ,,
Миграции животных. М., 1962. Вып. 3. С. 26—36.
Бараташвили Т. Рысь в Боржомском
заповеднике // Охота и охотничье хоз-во. 1982.
N 2. С. 12-13.
Барышников Г.Ф., Гарутт В.Е. и др. Каталог-
млекопитающих СССР. Л.: Наука, 1981. 455 с.
Баскин Л.М. Поведение копытных. М,: Наука,
1976. 295 с. '
Белое С.Н., Николаев В.В.. Унжаков В.В.,
Юдин Б.С. Движение численности копытных в
Новосибирской области и определяющие ее
факторы II Копытные фауны СССР. М.: Наука,
1980. С. 67—69.
Белоусов Н.И. Некоторые аспекты экологии
косули в условиях высокогорий II Проблемы
24. Европейская и сибирская косули
369
экологии позвоночных Сибири. Кемерово, 1978.
С. 164—171.
Белых В. И. Состав и распределение охот-
промысловой фауны млекопитающих Якутии //
Промысловая фауна и охотничье хозяйство
Якутии. Якутск. 1963. Вып. 1. С. 5-19.
Белышев Б.Ф. Распространение косули в
Западной Сибири // Охотник Сибири. 1934. N 11/12.
С. 27.
Белянии В.Н. Размещение и численность
косули в Жигулевском заповеднике // Копытные
фауны СССР. М.: Наука, 1975. С. 63.
Белянии В.Н. Волки в Жигулях //
Экологические основы охраны и рационального
использования хищных млекопитающих. М.: Наука,
1979. С. 86-87.
Белянии В.Н. Материалы по смертности
копытных в Жигулевском заповеднике // Копытные
фауны СССР. М.: Наука, 1980а. С. 122—123.
Белянии В.Н. Размещение и численность косули
(Capreolus capreolus pygargus Pall.) в
Жигулевском заповеднике // Физиологическая и попу-
ляционная экология животных. Саратов: Изд-во
Сарат. Ун-та, 19806. Вып. 6(8). С. 186—189.
Бибикова В.Н. Из истории голоценовой фауны
позвоночных в Восточной Европе // Природная
обстановка и фауна прошлого. Киев: Изд-во
АН УССР, 1963. Вып. 1. С. 119—146.
Бибикова В.И. О смене некоторых
компонентов фауны копытных на Украине в голоцене //
Бюл. МОИП. Отд. биол. 1975. Т. 80, N 6. С. 67—72.
Бионкур А.А. О состоянии ружейного дела в
России // Природа и охота. М., 1901. Кн. 8.
С. 10-16.
Бируля А.А. Предварительное сообщение о
млекопитающих из "кухонных отбросов" стоянки
каменного века на Верхнеленской юре близ
Иркутска // Докл. АН СССР. 1929. N 4. С. 91-93.
Блуэма П.П. Половые различия и возрастные
изменения веса и размеров тела Capreulus capreolus
(Cervidae) в Литве // 1-й Междунар. конгр.
по млекопитающим. М.: ВИНИТИ, 1974. Т." 1.
С. 72- 73.
Блузма П. П. Косуля в Литве (эколого-морфо-
логическая характеристика): Автореф. дис... канд.
биол. наук. М., 1975. 30 с.
Браунер А.А. К какому виду принадлежат
косули Южной России и Крыма? // Зап. Крым,
о-ва естествоиспытателей. Симферополь, 1915.
Т. 5. С. 112-114.
Браунер А.А. Сельскохозяйственная зоология.
Киев: Укриздат, 1923. 436 с.
Бромлеи Г.Ф.. Кучеренко СП. Копытные юга
Дальнего Востока. М.: Наука, 1983. 305 с.
Бромлеи Г.Ф. Отряд копытные ,,
Млекопитающие Зсйского заповедника. Владивосток,
1984. С. 107- 117.
Бурчак-Абрамович Н.Н.. Бурчак-Абрамо-
вич Д.Н. Отложения шюйсюисновых асфалмов
370
(битумов, киров) — ценные хранилища
палеонтологических объектов (млекопитающих, птиц,
беспозвоночных, растений и др.) , / 4-й съезд
Всесоюз. териол. о-ва: Тез. докл. М., 1986.
Т. 1. С. 11.
Вангенгейм Э.А. Палеонтологическое
обоснование стратиграфии антропогеновых отложений
севера Восточной Сибири // Тр. геол. ин-та. 1961.
Вып. 48. 176 с.
Вайгенгейм Э.А. Палеонтологическое
обоснование стратиграфии антропогена Северной Азии.
М.: Наука. 1977. 171 с.
Вангенгейм Э.А. Флеров К. К. Систематический
обзор основных отрядов млекопитающих. Отряд
Aniodactyla. Парнопалые // Стратиграфия СССР:
Четвертичный период. М., 1982. П/т 1. С. 326—
329.
Ватолин Б.А. Размещение, численность и
влияние экологических факторов на популяцию
косули в ландшафтах Брянской области //
Копытные фауны СССР. М.: Наука, 1975. С. 69—70.
Векуа А.К. Квабебская фауна акчагыльских
позвоночных. М.: Наука, 1972. 353 с.
Веиюков П.Н. Нижнеплиоценовая фауна
млекопитающих бессарабских песков / / Зап. Минерал.
о-ва. 1901. Т. 39, вып. 1. 33 с.
Веиюков П. Н. Фауна млекопитающих балтских
песков Подольской губ. / / Материалы но геологии
России. 1902. Т. 21. С. 123-133.
Верещагин Н.К. Байкальский як из
плейстоценовой фауны Восточной Сибири // Докл.
АН СССР. 1954. Т. 99, вып. 3. С. 455-458.
Верещагин Н.К. О прежнем распространении
некоторых копытных в районе смыкания Евро-
псйско-Казахстанских и Центральноазиагских
степей ,/ Зоол. журн. 1956. Т. 35, вып. 10, С. 1541-
1553.
Верещагин Н.К.Млекопитающие Кавказа. М.;
Л.: Изд-во АН СССР, 1959. 703 с.
Верещагин Н.К. Рецензия на книгу СВ. Ки-
рикова "Изменения животного мира в
природных зонах СССР (X1I1- XIX вв.): Степная зона и
лесостепь" , / Зоол. журн. 1960. Т. 39, вып. 7.
С. 1114 1116.
Верещагин Н.К. Охоты первобытного
человека и вымирание плейстоценовых
млекопитающих в СССР ,| Тр. Ин-та АН СССР
зоологии. 1971. Т. 49. С. 200-232.
Верещагин Н.К.. Барышников Г.Ф. Ареалы
копы i пых фауны СССР в антропогене , , Там же.
1980а. Т. 93. С. 3 20.
Верещагин Н.К.. Барышников Г.Ф.
Млекопитающие предгорного Северного Крыма в эпоху
палеолита (по кухонным остаткам из пещер
Чокурча. Староселье и Мамат-Коба) Там же.
19806. Т. 93. С. 26 49.
Верещагин Н.К..Батыров Б. Фрагмент
истории 1сриофауны Средней Азии Бюл. МОИП.
Out. биол. 1967. Т. 72. вып. 4. С. 104 115.
Верещагин U.K.. Оводов Н.Д. Исюрия фауны
Приморья Природа. 1968. N 9. С. 42 49.
Верещагин U.K.. Русаков О.С. Копытные
Северо-Запада СССР. Л.: Наука. 1979. 308 с.
Вершинин А.А. Расселение охотничьих
живо iпых в Подмосковье Вопросы повышения
продуктивности охотничьих угодий. М.. 1969.
С. 50 99.
Вет.шцин П. И. О "ходовой" коче в
Амурской области Природа и охота. 1902. N 11.
С. I 8.
Вие.юбокова И.А. О местонахожениях эоплей-
сгоиеновых млекопитающих в Павлодарском
Прииртышье Геология и геофизика.
Новосибирск. 1973. N 5. С. 123 125.
Вие.юбокова II. А. Палеон гологическос
обоснование стратиграфии эонлейс тисковых отложений
юга Западной Сибири: (По фауне
млекопитающих): Автореф. дис... канд. геол.-минерал, наук.
Новосибирск. 1974. 19 с.
Вш.юбокова II.А. К вопросу о ранней
эволюции меганерин Палсонюл. жури. 1981. N 4.
С. 105 117.
Вислобокова И.А. Ископаемые олени
Монголии // Тр. Советско-Монгольской палеонтол.
экспедиции. М.: Наука, 1983. Вып. 23. 76 с.
Владышевекий Д. В. О факторах численности
европейской косули // Зоол. журн. 1968. Т. 47,
вып. 3. С. 438—443.
Владышевекий Д.В., Ельский Г.М. Некоторые
закономерности зимнего питания оленя и
косули // Экология популяций лесных животных
Сибири. Новосибирск, 1974. С. 87—102.
Габрашаиекий П. Заразный энцефаломиелит у
косуль // Тр. IX Международ, конгр.
биологов-охотоведов. М., 1970. С. 640—642.
Габузов О.С. О питании косуль // Зоо. журн.
1960. Т. 39, вып. 12. С. 1892—1894.
Гайдук В.Е., Кузьмич А.И.. Блоцкая Е.С.
Влияние антропогенных факторов на охотничье-
промысловых зверей в Брестской области //
Антропогенное воздействие на популяции
животных. Волгоград, 1986. С. 124—134.
Гарозс В. Косули: почему их мало в наших
лесах // Наука и техника. 1987. N 1. С. 14—15.
Гаросс В.Я. Влияние рысей и волков на
латвийскую популяцию косули // Экологические
основы охраны и рационального использования
хищных млекопитающих. М.: Наука, 1979. С. 257—
261.
Гатих B.C. Распространение и численность
диких копытных на территории Припятского
заповедника // Копытные фауны СССР. М.: Наука,
1975.С. 73—74.
Гептнер В.Г., Насимович А.А., Банников А.Г.
Млекопитающие Советского Союза:
Парнокопытные и непарнокопытные. М.: Высш. шк„ 1961.
Т. 1. 776 с.
Гептнер В.Г., Цалкин В.И. Олени СССР
(систематика и зоогеография). М.: Моск. о-во
испытателей природы, 1947. 176 с.
Герасимов М.М. Поздние палеолитические
стоянки р. Белой // Бюл. комиссии по изуч.
1етвертичного периода. 1940. N 67. С. 100—101.
Гладков Н.А., Никольский Г.В. Материалы к
познанию фауны млекопитающих среднего и
нижнего течения реки Амударьи // Тр. Гос. зоол.
музея МГУ. 1935. Т. 2. С. 3—16.
Горбачев С.Н. Млекопитающие Орловской
губернии по новым данным 1910—-1914 гг. //
Материалы к познанию природы Орловской
губернии. 1915. N 21. С. 1—13.
Гордиюк Н.М. Взаимоотношения копытных в
Башкирском заповеднике // Роль крупных
хищников и копытных в биоценозах заповедников.
М., 1986. С. 70—81.
Горегляд Х.С. Основы борьбы с болезнями
диких животных // Тр. IX Междунар. конгр.
биологов-охотоведов. М., 1970. С. 575—579.
Граевская Б.М.. Золотарева Н.Н., Дания-
кин А.А., Елфимова С.С. Сравнительное
исследование метаболизма европейской (Capreolus
capreolus L.) и сибирской (Capreolus pygargus Pall.)
косуль // Копытные фауны СССР. М.; 1980.
С. 332-333.
Граевская Б.М., Золотарева Н.Н.,
Елфимова С.С. Биохимическая характеристика видового
статуса европейской и сибирской косуль //
Докл. АН СССР. 1988. Т. 303, N 5. С. 1259—
1261.
Грачев Ю.А., Смирнова Э.Д. Вертикальное
распространение и кочевки копытных в
Таласском Алатау // Копытные фауны СССР. М.:
Наука, 1980. С. 82—83.
Гречушкин B.C., Харченко В.И., МолоданГ.Н.,
Мглинец СМ. Современное состояние популяций
копытных Донецкой области / / Копытные фауны
СССР. М.: Наука, 1975. С. 78—79.
ГригорОв Г. Р. Проучвания върху храненето на
сърните (Capreolus capreolus L.) през зимния
период в длс "Лъг-ьт", Габровски окръг // Горско-
стоп. наука. 1976. Год. 13, N 6. С. 67—75.
Громов В. И. Палеонтологическое и
археологическое обоснование стратиграфии
континентальных отложений четвертичного периода на
территории СССР (млекопитающие, палеолит) // Тр.
Ин-та геол. наук. Сер. геол. 1948. Вып. 64. N 17.
521с.
Громов B.C. Морфологическая изменчивость,
поведение и систематика косули: Автореф. дис. ...
канд. биол. наук. М., 1986а. 27 с.
Громов B.C. О различных в окраске тела у
европейской (Capreolus capreolus) и сибирской
(С. pygargus) косуль // Зоол. журн. 19866. Т. 65,
вып. 11. С. 1693—1703.
Громов B.C. Возрастные и сезонные
потребности в кормах и энергии у европейских (Capreolus
capreolus) и сибирских (С. pygargus) косуль //
371
Актуальне проблемы морфологии и экологии
высших позвоночных. М., 1988а. Ч. 2. С. 495—
508.
Громов B.C. Закономерности роста
европейских (Capreolus capreolus) и сибирских (С. pygargus)
косуль // Зоол. жури. 19886. Т. 67, вып. 9.
С. 1381-1392.
Громов B.C., Данилкин А.А. Взаимоотношения
самки с детенышами у сибирской косули //
Сигнализация и экология млекопитающих и птиц.
М.: Наука, 1984. С. 93—107.
Громов B.C., Скулкин В.С. Географическая
изменчивость неметрических признаков черепа
косули (Capreolus capreolus) // Зоол. журн. 1986.
Т. 65, вып. 6. С. 911—923.
Громова В.И. Остатки млекопитающих из
раннеславянских городищ вблизи г. Воронежа //
Материалы и исследования по археологии СССР.
1948. Вып. 8.
Громова В.И. Краткий обзор млекопитающих
Европы. М.: Наука, 1965. 144 с.
Гурский И. Г. Кабан, косуля, лось и
благородный олень в северо-западном
Причерноморье // Копытные фауны СССР. М.: Наука,
1975. С. 79—80.
Гурский И.Г., Назаренко Л.Ф. Воздействие
волка на численность косули в Причерноморской
степи // Производительность и продуктивность
охотничьих угодий СССР. Киров, 1969. Ч. 2.
С. 168—169.
Гусев А.А. Трофодинамическая характеристика
популяций копытных Центрально-Черноземного
заповедника / / Растительноядные животные в
биогеоценозах суши: Материалы Всесоюз. совещ.
Валдай, 3—6 июня 1984. М., 1984а. С. 129—131.
Гусев А.А. Роль диких копытных в
функционировании биогеоценозов
Центрально-Черноземного заповедника: Автореф. дис. ... канд. биол.
наук. М., 19846, 19 с.
Давид А.И. Формирование териофауны
Молдавии в антропогене. Кишинев: Штиинца, 1982.
150 с.
Даль С.К. Материалы по систематике и
биологии крымской косули // Зап. Крым, о-ва
естествоиспытателей. 1930. Т. 12. С. 64—127.
Данилкин А.А. Звуковая сигнализация
сибирских косуль // Поведение млекопитающих. М.:
Наука, 1977. С. 239—243.
Данилкин А.А. Внутрипопуляционная
структура и поведение сибирской косули: Автореф. дис....
канд. биол. наук. М., 1978. 24 с.
Данилкин А.А. Ресурсы косули в СССР и их
рациональное использование // Промысловая
териология М.: Наука,1982. С. 108—115.
Данилкин А.А. Косули в России: меры охраны
и воспроизводства // Охота и охотничье хоз-во.
1985а, N З.С. 16-18.
Данилкин А.А. Кариотипы косуль (Capreolus
Gray) Евразии. Гипотеза о видообразовании //
372
Докл. АН СССР. 19856. Т. 285, N 6. С. 1513—
1516.
Данилкин А.А. Современные ареалы
европейской (Capreolus capreolus L.) и сибирской (С.
pygargus Pall.) косуль // Там же. 1985в. Т. 283,
N 3. С. 749—752.
Данилкин А.А. Европейская и сибирская
косули: подвиды или виды / / Охота и охотничье
хоз-во. 1986а, N 7. С. 16—18.
Данилкин А.А.Гибридизация европейской и
сибирской косуль // Там же. 19866, N 9. С. 16—
18.
Данилкин А.А. Современные представления о
систематике косуль (Capreolus Gray) и их
распространении // 4-й съезд Всесоюз. териол. о-ва:
Тез. докл. М., 1986в. Т.1. С. 53—54.
Данилкин А.А., Баскевич М.И. Кариотипы
косуль (Capreolus pygargus) Дальнего Востока //
Зоол. журн. 1987. Т. 66, вып. 2. С. 314—317.
Данилкин А.А., Дарман Ю.А. От дискуссий —
к действиям // Охота и охотничье хоз-во.
1987. N 4. С. 6—7.
Данилкин А.А., Дуламцэрэн С. Косуля в
Монголии // Там же. 1981. N 3. С. 44—45.
Данилкин А.А., Марков Г.Г. О
систематическом статусе косуль (Capreolus Gray)
Кавказа // Докл. АН СССР. 1985. Т. 283, N 1. С. 231 —
235.
Данилкин А.А., Марков Г.Г. К таксономии
косуль (Capreolus pygargus Pall.) Дальнего
Востока, Восточной Сибири и Тянь-Шаня // Изв.
АН СССР. Сер. биол. 1987. Вып. 2. С. 315—318.
Данилкин А.А., Марков ГГ., Герасимов С,
Николов X. Краниометрическое исследование
сибирской косули (Capreolus pygargus Pall.) //
Докл. АН СССР. 1985. Т. 281, N 2. С. 503—
506.
Данилкин А.А., Минаев А.Н. Пространственно-
этологическая структура популяции европейской
косули (Capreolus capreolus) в заповеднике "Лес
на Ворскле" // Актуальные проблемы морфоло-
нии и экологии высших позвоночных. М., 1988.
Ч. 2. С. 509—538.
Данилкин А.А., Чистова Т.Ю., Яценко В.Н.,
Чудиновская Г.А. О хромосомном полиморфизме
косуль // Популяционная изменчивость вида
и проблемы охраны генофонда млекопитающих.
М., 1983. С. 37.
Данилов Д.Н., Русанов Я.С, Рыковский А.С.
и др. Методическое руководство по
внутрихозяйственному устройству охотничьих хозяйств
Росохотрыболовсоюза. М.: Лесн. пром-сть, 1965.
138 с.
Данилов П. И. Появление кабана и косули в
Карелии // Вопр. экологии животных.
Петрозаводск,. 1974. С. 158—160.
Дарвин Ч. Происхождение видов. М., Л.:
Сельсхозгиз, 1935 (1859). 631 с.
Дарман Ю.А. Анализ смертности косули в
Хинганском заповеднике // Млекопитающие
СССР. 3-й съезд Всесоюз. териол. о-ва: Тез. докл.
М., 1982а. Т.1. С. 182—183. •
Дар май Ю.А. Крупные хищники в
Хинганском заповеднике //' Охрана хищных
млекопитающих Дальнего Востока. Владивосток, 19826.
С. 4-7.
Дар.чан Ю.А. Биотопическое распределение
косули Хинганского заповедника // Биология и
хозяйственное использование промысловых
зверей и птиц Сибири и Дальнего Востока. Иркутск,
1983. С. 43—51.
Дар май Ю.А. Биология косули Хинганского
заповедника: Автореф. дис.... канд. биол. наук. М.,
1986. 20 с.
Дарман Ю.А. Учет косули на трансектах
неопределенной ширины // Организация и методика
учета промысловых и редких видов
млекопитающих Дальнего Востока. Владивосток, 1987. С. 20—
23.
Дворников М.Г. Факторы, определяющие
смертность косули Ильменского заповедника //
Вопр. охотоведения. Пермь, 1982. С. 18—22.
Дворников М.Г. Экология и биогеоценоти-
ческая роль копытных в Ильменском
государственном заповеднике им. В.И. Ленина: Автореф.
дис... канд. биол. наук. Свердловск, 1984. 25 с.
Деев П. Естественноисторическое,
географическое и хозяйственно-экологическое описание
лесных дач Устькаменогорского, Бухтарминского
лесничеств Семипалатинской области //Из
Казенных лесов Акмолинской и Семипалатинской
областей. Омск, 1911. Вып. 1. С. 1—23.
Дементьев Г.П. Козуля. М.; Л.: КОИЗ, 1933.
52 с.
Демидов Г. Хроника // Сов. охотник. 1940, N 6.
С. 49.
Денисов Г.В. Итоги расселения копытных
животных в РСФСР в девятой пятилетке // Тр.
ВСХИЗО. М., 1978. Вып. 149. С. 24—49.
Динесман Л.Г., Князев А.В., Болд Г. К истории
охоты и скотоводства в Монголии // Бюл. МОИП.
Отд. биол. 1986. Т. 91, вып. 1. С. 31—37.
Динник Н.Я. Звери Кавказа. Ч. I.
Китообразные и копытные // Зап. Кавказ, отд. имп.
Рус. геогр. о-ва. 1910. Кн. 27, вып, I, 247 с.
Дмитриев В.В. Копытные звери Алтайского
заповедника и прилегающих мест (Восточный
Алтай и Западные Саяны) // Тр. Алтай,
заповедника. М., 1938. Вып. 1. С. 226—229.
Дмитриев В.В., Золотовский М.В., Фолита-
рек С. С. и др. Алтайский государственный
заповедник. М., 1937. 103 с.
Доппельмаир ГГ. Соболиный промысел на .
северо-восточном побережье Байкала.Улан-Удэ,
1926. 272 с.
Драгоев П. Проучвания върху биологията на
сърната (Capreolus capreolus capreolus L.) //
Горскостоп. наука. 1965. Год. 2, N 6. С. 501 —
513.
Дулицкий А.И., Кормилицин А.А:, Кормили-
цина В.В. Изучение копытных Крыма //
Копытные фауны СССР. М.: Наука, 1975. С. 88.
Дулькейт Г.Д. Значение рыси и росомахи как
хищников в природном комплексе алтайской
тайги // Преобразование фауны позвоночных
нашей страны. М.: МОИП, 1963. С. 147—152.
Дулькейт Г.Д. Охотничья фауна, вопросы и
методы оценки производительности охотничьих
■угодий Алтайско-Саянской горной тайги //
Тр. гос. заповедника "Столбы". 1964. Вып. 4. 352 с.
Дуров В.В. Динамика численности и сезонное
размещение косули в Кавказском заповеднике //
Копытные фауны СССР. М.: Наука, 1980. С. 147—
148.
Дымин В.А. Воздействие хищников на диких
копытных Верхнего Приамурья // Копытные
фауны СССР. М.: Наука, 1975а. С. 194—195.
Дымин В.А. Сезонные миграции косули в Верх-
.лем Приамурье // Копытные фауны СССР.
М.: Наука, 19756. С. 92—93.
Дымин В.А., Юдаков А.Г. Воздействие рыси
на промысловую фауну Верхнего Приамурья //
Охрана, рациональное использование И
воспроизводство естественных ресурсов Приамурья.
Хабаровск, 1967. С. 164—166.
Егоров О.В. Дикие копытные Якутии. М.:
Наука, 1965. 259 с.
Егоров О. В., Попов М.В. Аэровизуальный учет
диких копытных на территории Лено-Вилюйского
междуречья // Позвоночные животные Якутии.
Якутск, 1964. С. 15—26.
Елпатьевский П.В., Паничев A.M.
Геохимические особенности зверовых солонцов Сихотэ-
Алиня // Бюл. МОИП. Отд. биол. 1980. Т. 85,
вып. 6. С. 12—23.
Ельский Г.М. Экологическая избирательность
угодий косулей при многоснежности / / Копытные
фауны СССР. М.: Наука, 1975. С. 169—170.
Ельский Г.М. Поведение мигрирующих гр;:ш
косули в подтаежной зоне Красноярского
Приангарья // Групповое поведение животных. М.:
Наука, 1976. С. 108—110.
Ельский Г.М. Экологические особенности
зимнего питания косули в сосновых лесах
Красноярского Приангарья // Экология питания лесных
животных. Новосибирск, 1978. С. 58—70.
Ельский Г.М. Типы зимних угодий изюбря
и кЬсули на Хамар-Дабане // Фауна лесов
бассейна оз. Байкал. Новосибирск, 1979. С. 146—
154.
Ельский Г.М. Косуля Красноярского
Приангарья // Охота и охотничье хоз-во. 1983. .N 3.
С. 22—23.
Ельский Г.М., Шишикин
А.С.Взаимоотношения хищных и растительноядных млекопитающих
373
в местах зимних коцентраций в Приангарье / /
Экологические основы охраны и рационального
использования хищных млекопитающих. М.:
Наука, 1979. С. 265—266.
Емельянов А., Рыболов В. Косуля в
Ставрополье // Охота и охотничье хоз-во. 1968. N 8.
С. 13.
Ермолова Н.М. О фауне млекопитающих эпохи
палеолита и неолита Прибайкалья // Материалы
по этнографии. Л.: Всесоюэ. геогр. о-во, 1963.
Вып. 3. С. 27—64.
Ермолова Н.М. Териофауна долины Ангары
в позднем антропогене. Новосибирск: Наука,
1978. 220 с.
Жирное Л. В. К биологии косули
Центрального Тянь-Шаня // Учен. зап. Моск. гор. пед.
ин-та им. В.П. Потемкина. 1958. Т. 84, вып. 7.
С. 137—149.
Жиряков В.А. Влияние крупных хищников на
популяции диких млекопитающих в
Алма-Атинском заповеднике // Экологические основы
охраны и рационального использования хищных
млекопитающих. М.: Наука, 1979. С. 37—39.
Жиряков В.А. К экологии косули в Заилий-
ском Алатау // Копытные фауны СССР. М.:
Наука, 1980. С. 155—156.
Жиряков В.А., Джансынаев А.Д. Снежный
барс в Алма-Атинском заповеднике // Редкие
животные Казахстана: Материалы по 2-му изд.
Красной книги КазССР. Алма-Ата, 1986. С. 51—54.
Житенко П. В. Свойства мяса диких
животных / / Тр. IX Международ, конгр. биологов-
охотоведов. М., 1970. С. 932—938.
Закиров А. Хищные и парнокопытные //
Позвоночные животные Ферганской долины.
Ташкент: Фан, 1974. С. 185—201.
Залесский П.М. Прошлое и современное
распространение копытных в Западно-Сибирском
крае // Охотник Сибири. 1934. N 7/8. С. 28.
Зарудный Н.А. Краткий очерк охотничьего
промысла с Сырдарьинской области //
Туркестан, сел. хоз-во. 1915. N 8. С. 756—778.
Зверев М.Д. Млекопитающие Новосибирского
района // Тр. Новосибирского зоосада. Т. 1. Звери
и птицы Западной Сибири. Новосибирск, 1937.
С. 7—30.
Злобин Б.Д. Редкие виды копытных Кировской
области // Копытные фауны СССР. М.: Наука.
1975. С. 264—265.
Зырянов А.Н. Дикие копытные животные
заповедника "Столбы" и прилежащих районов //
Вопр. экологии. Красноярск, 1975. С. 224—338.
(Тр. гос. заповедника "Столбы"; Вып. 10).
Иванова И. К. О следах деятельности ископа-
мых гоминид в отложениях эоплейстоцена и
раннего плейстоцена Европы // Геология и фауна
нижнего и среднего плейстоцена Европы. М.:
Наука, 1972. С. 220—240.
Ильяшенко В.Ю. Влияние Зейского
водохранилища на наземных позвоночных животных
горнотаежных экосистем: Автореф.дис. ... канд.биол.
наук. М., 1984. 18 с.
Ишунин Г.И. Млекопитающие (хищные и
копытные) // Фауна Узбекской ССР. Ташкент:
Изд-во АН УэССР, 1961. Т. 3. 231 с.
Калниньш А.И. Охота и охотничье хозяйство
■ Латвийской ССР. Рига: Латвгосиздат, 1950.
540 с.
Кальке Г.Д. Семейство Cervidae Gray, 1821 //
Плейстоцен Тирасполя. Кишинев: Штиинца, 1971.
С. 137—156.
Капитонов В. И. Влияние летней засухи 1974
и 1975 гг. на копытных Казахского нагорья //
Копытные фауны СССР. М.: Наука, 1980. С. 163—
165.
Карпенко А. В. Биоэкологическое обоснование
системы мероприятий защиты леса от вредного
влияния косули в лесах левобережной лесостепи
УССР: Автореф. дис.... канд. биол. наук. Харьков,
1977. 21 с.
Каспаров А.К. Остатки млекопитающих из
позднепалеолитического поселения Сухотино-4 в
Забайкалье // 4-й съезд Всесоюз. териол. о-ва:
Тез. докл. М., 1986. Т. 1. С. 15.
Кашкаров Д.Н. Животные Туркестана.
Ташкент: УЗГИЗ, 1931. 448 с.
Керечун С Ф. Влияние хищников на поголовье
копытных в зоне Карпат / / Экологические основы
охраны и рационального использования хищных
млекопитающих. М.: Наука, 1979. С. 43—44.
Кийли Я. Анализ факторов динамики
численности косули // Изв. АН ЭССР. Биология.
1985. Т. 34, N 3. С. 216—225.
Кириков СВ. Птицы и млекопитающие в
условиях ландшафтов южной оконечности Урала.
М.: Изд-во АН СССР, 1952. 412 с.
Кириков СВ. Изменения животного мира в
природных зонах СССР (XIII—XIX вв.); степная
зона и лесостепь. М.: Изд-во АН СССР, 1959.
175 с.
Кириков СВ. Исторические акты как
основные источники для оценки запасов и определения
ареала охотничье-промысловых зверей и птиц
в XVI—XVIII — начале XIX в. // Ресурсы
фауны промысловых зверей в СССР и их учет.
М.: Изд-во АН СССР, 1963. С. 52—63.
Кириков СВ. Промысловые животные, среда и
человек. М.: Наука, 1966. 346 с.
Кириков СВ. Человек и природа степной
зоны. М.: Наука, 1983. С. 52—63.
Кирилов Н. Охотничье хозяйство в
Забайкальской области // Природа и охота. 1902.
Янв. С. 32—40.
Киселев А.А. Косуля и рысь в Свердловской
области // Охота и охотничье хоз-во. 1974.
N 12. С. 16—17.
Киселев А.А. К морфологической
характеристике косули двух популяций на Урале / / Тр. Ин-та
374
экологии растений и животных УНЦ АН СССР.
1975. Вып. 96. С. 112—115.
Киселев А.А. Распространение и численность
косули на Среднем Урале // Охотоведение
(использование и охрана лесных копытных). М.: Лесн.
пром-сть, 1976. С. 63—70.
Киселев А.А. Косуля на Урале: Автореф.
дис... канд. биол. наук. Свердловск, 1979. 23 с.
Кожамкулова Б.С. Зоогеографический анализ
состава и распределения антропогенной фауны
Казахстана в пространстве и времени //
Мезозойская фауна и флора Северо-Западного
Казахстана. Алма-Ата, 1977. С. 89—104. (Материалы
по истории фауны и флоры Казахстана; Т 7).
Кожамкулова Б.С. Позднекайнозойские
копытные Казахстана. Алма-Ата: Наука, 1981. 145 с.
Козлов П.К. Монгольский заповедник Богда-
Ула // Изв. Рус. геогр. о-ва. 1924. Т. 56, вып. 1.
С. 169—173.
Колее И. В. Проучвания върху храненето на
сърната в Чепинския район // Горскостоп. наука.
1965. Год. 3, N 1. С. 71-81.
Конечных Г.К., Аникин Р.К, Девяткин А. А.
Опыт авиаучета лося и косули в Центральной
Якутии // Позвоночные животные Якутии.
Якутск, 1964. С. 27—31.
Корелов М.Н. Фауна позвоночных Бостандык-
ского района / / Природа и хозяйственные условия
горной части Бостандыка. Алма-Ата, 1956. С. 259—
325.
Коржуев П.А., Тимошевская Г.В., Кадыко-
ва И.А., Спасская Г.Х. Рога как форма
адаптации дагестанского тура к высокогорным
условиям // Экология. 1978. N 2. С. 100—102.
Корнеев О. П. Визначник звирив УРСР. Кшв:
Рад. шк., 1952. 216 с.
Корнилов И. П. Олени Восточной Сибири.
Козуля // Вестн. естеств. наук. 1856. Т. 3, N 16.
С. 494—504.
Короткевич Е.Л. Новый вид косули из мэоти-
са Украины // Палеонтол. журн. 1965. N 4.
С. 60—67.
Короткевич ЕЛ. Фауна крупных
млекопитающих из плиоценовых отложений долины р. Кучур-
ган // Место и значение ископаемых
Молдавии в кайнозое СССР. Кишинев: Изд-во АН МССР,
1967. С. 77—84.
Короткевич Е.Л. Поздненеогеновые олени
Северного Причерноморья. Киев: Наук, думка,
1970. 196 с.
Короткевич Е.Л. Новый представитель рода
Procapreolus на территории Северного
Причерноморья // Вестн. зоол. 1974. N 6. С. 68—77.
Короткевич ЕЛ. Фоновые группы поздне-
миоценовых жвачных и история формирования
гиппарионовой фауны Восточной Европы:
Автореф. дис... канд. биол. наук. Киев, 1984. 47 с.
Короткевич О.Л. Hoei даю про
систематику i фшогешю викопных козуль роду
Procapreolus // Доп. АН УРСР. 1963. N 10. С. 1390—
1393.
Короткевич О.Л. Hoei энахщки викопно!'
козул1 роду Procapreolus на твдш СРСР //
Доп. АН УРСР. 1964. N3. С. 382—386.
Корш П. Дикие козы в Омской области //
Сов. охотник. 1941. N 4. С. 33.
Коеинцев П.А. Косуля из археологических
памятников Урала и Западной Сибири (эпоха
бронзы — средневековье) / /
Териология на Урале. Свердловск, 1981. С. 39—43.
Коеинцев П.А. Костные остатки диких
млекопитающих из археологических памятников
лесной полосы Урала и Западной Сибири (эпоха
бронзы — средневековье) // 3-й съезд Всесоюз.
териол. о-ва: Тез. докл. М., 1982. Т. 1. С. 42—43.
Коеинцев П.А. Териофауна Урала и Западной
Сибири в голоцене // 4-й съезд Всесоюз. териол.
о-ва: Тез. докл. М., 1986. Т. 1. С. 16—17.
Коеинцев П.А., Ражее Д.И. Новые данные
по териофауне Южного Урала и Зауралья в
голоцене // Млекопитающие в системе
природопользования на Урале. Свердловск, 1985. С. 17—18.
Крушинский Л. В. Формирование поведения в
норме и патологии. М.: Изд-во МГУ, 1960.
263 с.
Кузнецов Б.А. Зверовые промыслы Восточного
Забайкалья // Тр. по лесн. опыт, делу Центр,
лесн. опыт, станции. М., 1929. Вып. 6. С. 83—
106. ч
Кузнецов Б.А. Млекопитающие Казахстана.
М.: МОИП, 1948. 226 с.
Кузьмина Е.И. Формирование териофауны
Северного Урала в позднем антропогене // Тр.
Зоол. ин-та АН СССР. 1971. Т. 49. С. 44—
122.
Кузякин В.А. Охотничья таксация. М.: Лесн.
пром-сть, 1979. 199 с.
Кузякин В.А, Состояние ресурсов диких
копытных // Итоги науки и техники. 1985. С. 5—65.
(Сер. "Зоология позвоночных";Т. 13).
Куражсковский Ю.Н. К изучению экологии
косули в лесных насаждениях засушливых
областей // Охрана природы. 1952. N 15. С. 122—
126.
Кучеренко СП. Копытные млекопитающие
Амуро-Уссурийского района // Животный мир и
охотничье хозяйство Дальнего Востока.
Владивосток, 1976. С. 97—125.
Кучеренко СП. Волк юга Дальнего Востока / /
Экологические основы охраны и рационального
использования хищных млекопитающих. М.:
Наука, 1979. С. 117—118.
Кучеренко С, Зубков Ю. Волк юга Дальнего
Востока // Охота и охотничье хоз-во. 1980.
N 1. С. 20—23.
Кучеренко С, Швец В. Косуля
Амуро-Уссурийского края // Там же. 1977. N 3. С. 22—23.
Лавов М.А. Популяция косули на Витим-
375
ском плоскогорье // Вопр. зоологии. Томск:
Изд-во Том. ун-та, 1966. С. 206—207.
Лавов М.А. Косуля // Охота и охотничье
хоэ-во. 1970. N 8. С. 16—19.
Лавов М.А- Структура популяции косули
на Витимском плоскогорье // Бюл. МОИП.
Отд. биол. 1971. Т. 76, вып. 2. С. 56—59.
Лавов М.А. Структура и воспроизводство
популяции косули на Витимском плоскогорье //
Сб. науч.-техн. информ. ВНИИОЗ "Охота,
пушнина, дичь". Киров, 1974. Вып. 43. С. 8—12.
Лавов М.А. Распространение и примерная
численность копытных зверей в Красноярском крае / /
Вопр. экологии. Красноярск, 1975. С. 339—347.
(Тр. гос. заповедника "Столбы"; Вып. 10).
Лавов М.А. Косуля // Крупные хищники и
копытные звери. М., 1978. С. 190—220.
Лавов М.А. Волки — мифы и
действительность Ц Охота и охотничье хоз-во. 1982. N 7.
С. 20—21.
Лавров Н.П. Распространение косули в
Сибири // Охотник. 1927. N 5. С. 14—16.
Лавров Н.П. Географическое распределение,
биология и хозяйственное значение косули в
СССР // Тр. по лесн. опыт, делу Центр, лесн.
опыт, станции. 1929. Вып. 6. С. 49—82.
Лапсин Г.М., Соколов Г.А. Экология и
хозяйственное значение косули в отрогах
Кузнецкого Алатау // Проблемы охотничьего хоз-ва
Красноярского края. Красноярск, 1971. С. 65—
70.
Лаптев И.П. О распространении некоторых
млекопитающих в Западной Сибири // Заметки
по фауне и флоре Сибири. Томск, 1955. Вып. 18.
с. 3—6.
Лаптев И.П. Млекопитающие таежной зоны
Западной Сибири. Томск: Изд-во Том. ун-та, 1958.
'.85 с.
Ларин И.В., Агабабян Ш.М-. Работное Т.А.
и др. Кормовые растения сенокосов и пастбищ
СССР. М.; Л.: Гос. изд-во с.-х. лит., 1952.
Т. 2. 948 с.
Литвинов В.Ф., Ильясевич В.А., Лунин В.Ф.
Роль хищников в гибели копытных //
Экологические основы- охраны и рационального
использования хищных млекопитающих. М.: Наука,
1979. С. 44—46.
Лобачев B.C. Косуля в пустынях Приаралья //
Охота и охотничье хоз-во. 1972. N 8. С. 11.
Лобачев B.C. Численность копытных в пойме
р. Урал // Копытные фауны СССР. М.: Наука,
1980. С. 94—95.
Маак Р.К. Вилюйский округ Якутской области.
СПб., 1886. Ч. 2. 368 с.
Майр Э. Зоологический вид и эволюция. М.:
Мир, 1968. 513 с.
Майр Э. Принципы зоологической
систематики. М.: Мир, 1971. 454 с.
Майр Э. Популяции, виды и эволюция. М.:
Мир, 1974. 460 с.
Майр Э„ Линсли Э., Юзингер Р. Методы и
принципы зоологической систематики. М.: Изд-во
иностр. лит., 1956. 352 с.
Малафеев Ю.М., Кряжский Ф.В. Питание
рыси и ее взаимоотношения с копытными на
Среднем Урале // Бюл. МОИП. Отд. биол. 1984. Т. 89,
вып. 4. С. 70—81.
Малюгин Ю.И. Исследования строения и
свойств шкур косуль и их рациональное
использование: Автореф. дис... канд. техн. наук. М.,
1981. 17 с.
Марвин М.Я. Млекопитающие Карелии.
Петрозаводск: Гос. изд-во Карельской АССР, 1959.
238 с.
Марков Г. Г. Сравнителен популационно-мор-
фологичен и генетичен анализ на род сърни
(Capreolus Gray, 1821): Автореф. дис.... канд. биол.
наук. София, 1985. 30 с.
Марков Г.Г., Данилкин А.А.. Герасимов С,
Николов Х.М. Сравнительный краниометрический
анализ европейской косули (Capreolus capreolus
L.) // Докл. АН СССР. 1985. Т. 282, N 2.
С. 489—493.
Марков Г. Г., Добриянов Д. Кариотаксономично
иэследване на сърната (С. capreolus L., 1758)
в България // ActaZool. Bulg. 1985. Vol. 28.
p. 241—244.
Марков Г.Г., Кехаиов И„ Данилкин А.А.,
Пешева П., Братанова Т. Сравнительное
иммунологическое исследование европейской
(Capreolus capreolus L.) и сибирской (С. pygargus Pall.)
косуль // Докл. АН СССР. 1985. Т. 281, N 5.
С. 1277—1281.
Марков К.К., Величко А.А. Четвертичный
период. М.: Недра, 1967. Т. 3. 440 с.
Марков К. К., Лизуков Г. И., Николаев В. В.
Четвертичный период. М.: Изд-во МГУ, 1965.
Т. 2. 435 с.
Маркович-Марьянович Е. Данные о
стратиграфии и фауне нижнего и среднего плейстоцена
Югославии / / Геология и фауна нижнего и
среднего плейстоцена Европы. М.: Наука, 1972. С. 153—
161.
Матюшкин Е.Н. Взаимоотношения с другими
хищными млекопитающими // Волк. М.: Наука,
1985. С. 355—370.
Мейер М.Н. Метод гибридизации в
систематике животных // Зоол. журн. 1986. Т. 65.
вып. 11. С. 1605—1613.
Мельников В.К. Сроки осеннего перемещения
косули, марала и медведя в бассейне Малого
Абакана (Западные Саяны) / / Фауна Сибири и ее
хозяйственное использование. Иркутск, 1978.
С. 3—5.
Метельский А.П. Факторы, определяющие
размещение косули в горных ландшафтах Сибири
376
и Дальнего Востока // Копытные фауны СССР.
М.: Наука, 1975. С. 114—115.
Метелъекий А. П. Распространение и
численность косули в Сибири и на Дальнем Востоке //
Охотоведение: (Использование и охрана лесных
копытных). M.s Лесн. пром-сть, 1976. С. 23—
63.
Мигулин О. Щкидни та корисни звири УкраМ.
XapbKie: Рад. селянин, 1927. 166 ic.
Мигулин О. ВизначникзвиривУкра'1'Hi. XapbKie:
Державне вид-во Укра'(ш, 1929. 95 с.
Миддендорф А. Путешествие на север и восток
Сибири: (Сибирская фауна) //СПб., 1869. Ч. 2,
отд. 5. 310 с.
Морозов К.А. Численность популяции косули
Верхнего Приамурья // Копытные фауны СССР.
М: Наука, 1975. С. 56—57.
Морозов К.А. Опыт минеральной подкормки
диких копытных в Амурской области //
Повышение продуктивности охотничьих угодий. М.,
1982. С. 43—49.
Наеимович А.А. К познанию минерального
питания диких животных Кавказского
заповедника // Тр. Кавказ, гос. заповедника. 1938. Вып. 1.
С. 103—150.
Наеимович А.А. Зима в жизни копытных
западного Кавказа // Вопр. экологии и
биоценологии. Л., 1939. Вып. 7. С. 3—91.
Наеимович А.А. Роль режима снежного
покрова в жизни копытных животных на
территории СССР. М.: Изд-во АН СССР, 1955. 401 с.
Наеимович А.А., Шубникова О.Н. Охотничьи
животные — использование, воспроизводство и*ох-
рана // Природные ресурсы Русской равнины в
прошлом, настоящем и будущем. М.: Наука,
1976. С. 202—217.
Наумов Н.П. Экология животных. М.: Высш.
шк., 1963. 618 с.
Наумов Н.П. Структура популяций и
динамика численности наземных позвоночных // Зоол.
журн. 1967. Т. 46, вып. 10. С. 1470—1476.
Наумов Н.П. Пространственные структуры
вида млекопитающих // Там же. 1971. Т. 50,
вып. 7. С. 965—980.
Наумов Н.П. Этологическая структура
популяций наземных позвоночных // Поведение
животных. М.: Наука, 1972. С. 37—39.
Наумов Н.П. Биологические (сигнальные) поля
и их значение в жизни млекопитающих / / Успехи
современной териологии. М.: Наука, 1977. С. 93—
108.
Никифоров М. Н. Запасы диких копытных в
Тувинской АССР и их использование //
Производительность и продуктивность охотничьих
угодий СССР. Киров, 1969. Ч. 2. С. 28—31.
Никифорова К. В. Кайнозой Голодной степи
Центрального Казахстана // Тр. Геол. ин-та.
1960. Вып. 45. 255 с.
Николаев В. Обзор 1889 г. // Природа и охота.
1890. Янв. С. 105—110.
Николаев В.В. О косуле сибирской (Capreolus
capreolus L.) в средней части Витимского
плоскогорья / / Фауна и экология позвоночных Сибири.
Новосибирск, 1980. С. 158—160.
Николаев В.В. Размещение и численность
косули в лесостепи Западной Сибири //
Размещение и численность позвоночных Сибири.
Новосибирск: Наука, 1962. С. 254—258.
Никольский А. А. Звуковой, предупреждающий
об опасности сигнал сурков (Marmota) как
видовой признак // Зоол. журн. 1976. Т. 55, вып. 8.
С:\ 214—1224.
Никольский A.M. Путешестви&чВ Алтайские
горы летом 1882 года // Тр. СПб. о-ва
естествоиспытателей. 1883. Т. 14, вып. 1. С. 150—218.
Никольский С, Позов С. Клещевой токсикоз
косуль // Охота и охотничье хоз-во. 1973.
N 10. С. 19.
Новиков Г.А., Тимофеева Е.К. Питание и ле-
сохозяйственное значение косули в лесостепных
дубравах // Бюл. МОИП. Отд. биол. 1964. Т. 69,
Вып. 2. С. 39—53.
Новиков Г.А., Тимофеева Е.К. Об экологии
косули в лесостепных дубравах // Зоол. журн. 1965.
Т. 44, вып. 3. С. 442—451.
Окладников А.П. Ленские древности. М.; Л.:
Изд-во АН СССР, 1950. 195 с.
Оцхели Т.А. К изучению размножения
косули // Сообщ. АН ГССР. 1964. Т. 34, вып. 3.
С. 677-682.
Паавер К.Л. Формирование териофауны и
изменчивость млекопитающих Прибалтики в
голоцене. Тарту, 1965. 494 с.
Павлинов И.Я., Россолимо О.Л. Систематика
млекопитающих СССР. М.: Изд-во МГУ, 1987.
282 с.
Павлов И.П. Двадцатилетний опыт
объективного изучения высшей нервной деятельности
(поведения) животных. М.; Л.: Биомедгиз, 1938. 771 с.
Павлов М.П., Корсакова И.Б., Лавров Н.П.
Акклиматизация охотничье-промысловых зверей
птиц в СССР. Киров, 1974. 458 с.
Падайга В. И. Значение косули в лесном
хозяйстве и система мероприятий по защите от
нее лесовозобновлений в Литовской ССР: Авто-
реф. дис. ... канд. биол. наук. Таллинн, 1965. 25 с.
Падайга В. И. Опыт учета численности
европейской косули по зимним экскрементам //
Тр. IX Междунар. конгр. биологов-охотоведов.
М, 1970. С. 350—352.
Падайга В.И. Факторы, определяющие зимние
падежи косуль (Capreolus capreolus) // Зоол. журн.
1971. Т. 50, вып. 10. С. 1546-1552.
Падайга В.И. Основные факторы среды,
определяющие плотность населения диких копытных
животных в лесных охотничьих угодьях / / Охот-
377
ничье хоз-во в интенсивном комплексном лесном
хоз-ве. Каунас: Гирионис, 1975. С. 37—39.
ПадаОга В. И. Экологические основы
управления численностью оленей в Литовской ССР:
Автореф. дис... д-ра. биол. наук. Тарту, 1984.
33 с.
Падайга В. И., Марма Б. Б. Зависимость
экстенсивности инвазии косуль (Capreolus capreolus L.)
некоторыми паразитами от плотности населения
и условий обитания // Тр. IX Междунар. конгр.
биологов-охотоведов. М., 1970. С. 667—672.
Падайга В.И., Марма Б.Б. О методике оценки
численности европейской косули по количеству
дефекаций // Экология. 1979. N 4. С. 101 — 103.
Паллас П.С. Путешествие по разным местам
Российского государства. СПб., 1976. Кн. 2. 571 с.
Паллас П.С. Путешествие по разным
провинциям Российской империи. СПб., 1809. Ч. 1. 657 с.
Панииев A.M. Природные минеральные ионо-
обменники — регуляторы ионного равновесия в
организме животных-литофагов // Докл. АН СССР.
1987. Т. 292, N 4. С. 1016-1019.
Панов Е.Н. Демонстративное поведение
животных // Природа. 1969. N 1. С. 51—59.
Панов Е.Н. Поведение животных и этологи-
ческая структура популяций. М.: Наука, 1983.423 с.
Пастернак Ф.А. Монография косули: Автореф.
дис. ... канд. биол. наук. М., 1952. 9 с.
Пастернак Ф.А. Материалы к систематике и
биологии косули / / Учен. зап. Моск. гор. пед. ин-та
им. В.П. Потемкина. 1955. Т. 38, вып. 3. С. 29—140.
Петров П., Драгоев П., Колее И. Сърната
в Българии. София: Земиздат, 1968. 236 с.
Пидопличко И.Г. О ледниковом периоде.
Киев: Изд-во АН УССР, 1951а. Вып. 2. 264 с.
Пидопличко И.Г. Новая форма мунтжака из
плиоценовых отложений юга УССР // Докл.
АН СССР. 19516. Т. 34, N 6. С. 647—679.
Шдопличко 1.Г. Матер1али до вивчення ми-
нулих фаун УРСР. Ки1в, 1956. Внп. 2. 189 с.
Плешак Т. В. Косуля в Заилийском Алатау //
Копытные фауны СССР. М.: Наука, 1980. С. 195—
196.
Побединский Г. Восстановление оленя и
косули // Охота и охотничье хоз-во. 1987. N 6.
С. 6—7.
Подаревский В. Б. Проблемы охотхозяйствен-
ной акклиматизации в Восточной Сибири.
Иркутск: Вост.-сиб. кн. изд-во, 1936. 118 с.
Поле В. Б. Авиавизуальный учет косули и лося
в Северном Казахстане // Тр. Ин-та зоологии
АН КазССР. 1966. Т. 26. С. 186—192.
Поле В.Б. Размножение косули в Казахстане / /
Там же. 1973. Т. 34. С. 135—144.
Поле В.Б. Систематическое положение косули
(Capreolus capreolus) Казахстана // Зоол. журн.
1974. Т. 53, вып. 12. С. 1833—1839.
Попов М.В. Учет численности косули с
самолета // Методы учета численности
промысловых животных Якутии. Якутск, 1970. С. 38—45.
Попов М.В., Соломонов Н.Г., Мордосов И.И.,
Лабутин Ю.В. Биология охотничье-промысловых
зверей Якутии. Новосибирск: Наука, 1980. 158 с.
Пржевальский Н.М. Путешествие в
Уссурийском крае в 1868—1869 годах. СПб., 1870.
297 с.
Приедитис А.А. Влияние зараженности
гельминтами ' на зимний отход косули (Capreolus
capreolus) // Тр. IX Междунар. конгр.
биологов-охотоведов. М., 1970. С. 709—713.
Приедитис А.А. Факторы, определяющие
зимнюю смертность косуль // Охотничье хоз-во
в интенсивном комплексном лесном хоз-ве.
Каунас: Гирионис, 1975. С. 62—64.
Приедитис А.А. Зимние корма косули //
Охота и охотничье хоз-во. 1980. N 1. С. 17.
Приедитис А.А., Оценка состояния
популяции косули в Латвийской ССР // Управление
популяциями диких копытных животных. М.,
1985, С. 79—94.
Приедитис А., Таугинас И. Кормовая
эффективность гранул из летних порубочных отходов
осины в зимнем рационе косули / / Тр. АН ЛитССР.
Геп. В. 1981. Т. 3(75). С. 149—158.
Простаков Н.И., Обтемперанский СИ.
Условия обитания и стациальное распределение
косуль в островных лесах севера Черноземного
Центра // Изучение и охрана природы малых
заповедных территорий. Воронеж. Изд-во
Воронеж, ун-та, 1986. С. 160—164.
^Прусайте Я.А., Балейшис P.M. Плодовитость
косули в условиях Северной Литвы //
Охотничье хоз-во в интенсивном комплексном лесном
хоз-ве. Каунас: Гирионис, 1975. С. 129—130.
Прусайте Я.А., Балейшис P.M., Блузма П.П.
Состав кормов европейской косули в
зависимости от лесистости обитаемой территории / / Тр.
АН ЛитССР. Сер. В. 1983. Т. 4(84). С. 84—98.
Прусайте Я.А., Блажис А.С, Балейшис P.M.
Интенсивность размножения и прирост популяции
Capreolus capreolus L. (Cervidae) в Литовской
ССР // I Междунар. конгр. по млекопитающим.
М.: ВИНИТИ, 1974. Т. 2. С. 144.
Прусайте Я.А., Блажис А.С, Балейшис P.M.
Интенсивность размножения и плодовитость
европейской косули в Северной Литве // Тр. АН
ЛитССР. Сер. В. 1977. Т. 3(79). С. 105-110.
Прусайте Я.А., Блажис А.С., Мицкус А.В.,
Блузма П. П. Динамика численности и структура
неэксплуатируемой популяции косули / / Там же.
1973. Т. 2(62). С. 115—125.
Прядко Э.И. О систематическом статусе
рода косуль (Capreolus) в свете гельминтологических
данных / / Зоол. журн. 1972. Т. 51, Вып. 7. С. 1092—
1093.
Пужаускас Р. Зависимость зараженности ко-
378
суль (Capreolus capreolus) в Литве гельминтами
от экологических факторов // Тр. IX Между-
нар. конгр. биологов-охотоведов. М., 1970.
С. 685—687.
Равский Э.И., Александрова Л.П., Ванген-
гейм Э.А. и др. Антропогеновые отложения юга
Восточной Сибири // Тр. Геол. ин-та. 1964.
Вып. 10S. 280 с.
Разевиг В.А. Косуля Ферганская // Семья
охотников. 1909. N 8. С. 160.
Размахнин В., Киселев А., Киданов Н. Панты
сибирской косули // Охота и охотничье хоэ-во.
1976. N 3. С. 27.
Раков Н.В. Распределение и численность косули
в Приамурье // Охотничье-промысловые звери.
М.: Россельхозиздат, 1965. С. 93—107.
Раков Н.В. О роли хищников в ограничении
численности копытных в Приамурье и
Приморье // Экологические основы охраны и
рационального использования хищных млекопитающих.
М.: Наука, 1979. С. 58—59.
Рандвезр Т.Э. Косуля в Эстонии //
Управление популяциями диких копытных животных.
М., 198S, С. 94—111.
Реймерс Н.Ф. Азбука природы. М.: Знание,
1980. 207 с.
Рихтер Г.Д. Роль снежного покрова в физико-
географическом процессе // Тр. ин-та географии
АН СССР. М.; Л., 1948. Вып. 40. 169 с.
Романов Ю.М., Ромашин А.В. Материалы по
экологии европейской косули в Калининградской
области // Вестн. зоологии 1982. N 3. С. 48—52.
Рубис Л. В. Питание косули (Capreolus
capreolus L.) в осенне-зимний период // Вестн.
АН БССР. Сер. биол. 1982. Вып. 3. С. 109—
111.
Рудышин М.П., Мурский Г.Н., Борух М.И.
Методические рекомендации по улучшению
кормовой базы охотничьих угодий. Львов, 1979.
С. 40.
Русаков О.С. Динамика ареала и численность
косули на Северо-Западе СССР // Фауна и
экология птиц и млекопитающих таежного Северо-
Запада СССР. Петрозаводск, 1978. С. 175—181.
Рухлядев Д. П. Паразитофауна, заболевания и
причина отхода диких млекопитающих животных
Крымского заповедника // Научно-методические
записки. 1941. Вып. 8. С. 78—79.
Рухлядев Д. П. Паразиты и параэитозы
диких копытных и хищных животных
горно-лесного Крыма / / Паразитофауна и заболевания
диких животных. М., 1948. С. 3—102.
Рыковский А.С. К вопросу о месте и роли
гельминтов в динамике биоценозов / / Тр. IX Меж-
дунар. конгр. биологов-охотоведов. М., 1970.
С. 592—597.
Сабанеев Л.П. Звериный промысел в
Уральских горах // Беседа. 1872. Кн. 6.
Сабанеев Л.П. Козуля и козлиный промысел
в Уральских горах // Природа. 1875. Кн. 4.
С. 1—21.
Саблина Т.Е. Копытные Беловежской пущи //
Тр. ИМЖ им. А.Н. Северцова. 1955. Т. 15.
191 с.
Саблина Т.Е. Адаптивные особенности питания
некоторых видов копытных и воздействие этих
видов на смену растительности // Сообщ. Ин-та
леса. 1959. Вып. 13. С. 32—43.
Савинов В. Охотничьи угодья Алтайского края.
Барнаул: Алт. кн. изд-во, 1961. 25 с.
Савинов В. Звери и птицы Алтая. Барнаул:
Алт. кн. изд-во, 1967. 187 с.
Савинов Е.Ф. Размножение и рост сибирского
козерога в Джунгарском Алатау (Казахстан) //
Тр. Ин-та зоологии АН КазССР.1962. Т. 17.
С. 167—182.
Сапаев В.М. Млекопитающие
сельскохозяйственного ландшафта Юга Приамурья //
Животный мир и охотничье хозяйство Дальнего Востока.
Владивосток, 1976. С. 126—132.
Сапоженков Ю.Ф. Распределение копытных в
Костромской области // Природное и
сельскохозяйственное районирование СССР: Материалы
7-й Всесоюз. науч. конф. по природ, и экон.-геогр.
районированию СССР для сел. хоз-ва. М., 1981.
С. 115.
Северцов С. Охотничья фауна Прибалтики //
Сов. охотник. 1941. N 1. С. 19.
Северцов С.А. Динамика населения и
приспособительной эволюции животных. М.; Л., 1941.
316 с.
Северцов С.А. О конгруэнциях и понятии
целостности вида // Проблемы экологии
животных. М.: Изд-во АН СССР, 1951а. Т. l.'C. 30—57.
Северцов С.А. Эволюция рогов некоторых
парнокопытных как турнирного оружия в боях
за самку // Проблемы экологии животных. М.:
Изд-во АН СССР, 19516. Т. 1. С. 58—59.
Северцов С.А., Саблина Т.Е. Олень, косуля
и кабан в заповеднике "Беловежская Пуща" //
Тр. Ин-та морфологии животных АН СССР.
1953. Вып. 9. С. 140—205.
Сержанин И. И. Млекопитающие Белорусской
ССР. Минск: Изд-во АН БССР, 1955. 311 с.
Силантьев А.А. Обзор промысловых охот в
России. СПб., 1898. 619с.
Скалон В.Н. О расширении козулей своего
ареала в Сибири // Природа. 1946. N 6.С. 73.
Скалон В.Н. Распространение и обзор диких
копытных в бассейне Олекмы // Биол. сб.
Иркутск, 1960. С. 150—158.
Скриба Г. В. Взаимоотношения европейских
оленей с лосями и косулями // Охотничье
хозяйство в интенсивном комплексном лесном
хозяйстве. Каунас: Гирионис, 1975. С. 101 — 102.
Словцо* И.'я. Позвоночные Тюменского округа
и их распространение в Тобольской губернии.
М., 1892. 78 с.
379
Слудский А.А. Выселение "таежных" зверей в
лесостепи и степи Западной Сибири и
Казахстана // Бюл. МОИП. Отд. биол. 1953. Т. 53,
вып. 2. С. 14—32.
Слудский А.А. Размножение джейрана //
Тр. Ин-та зоологии АН КазССР. 1956. Т. 6.
С. 78—108.
Слудский А.А., Байдавлетов Р.Ж. и др.
Млекопитающие Казахстана. Алма-Ата: Наука, 1984.
Т. 3, ч. 4. 231 с.
Смирнов М.Н. К вопросу территориальной
дифференциации населения косуль и связи с
условиями обитания в Западном Забайкалье и
Восточном Саяне // Изв. Иркут. с.-х. ин-та. 1970а.
Т. 26, вып. 3. С. 91—104.
Смирнов М.Н. О применении вертолета для
количественного учета диких копытных в горном
Забайкалье // Там же. 19706. Вып. 26. С. 105—
119.
Смирнов М.Н. Косуля в Западном Забайкалье:
Автореф. дис. ... канд. биол. наук. Иркутск,
1974. 29 с.
Смирнов М. Н. Прежнее и современное
распространение косули в Бурятии // Зоол. исслед.
в Забайкалье: Тр. Бурят, ин-та естеств. наук.
Улан-Удэ, 1975а: Вып. 13. С. 218—230.
Смирнов М.Н. О вокальной деятельности
косуль Западного Забайкалья // Там же. 19756.
С. 214—217.
Смирнов М.Н. Очерк поведения косули
(Capreolus capreolus pygafgus Pall.) в Западном
Забайкалье // Поведение млекопитающих. М.:
Наука, 1977. С. 221—243.
Смирнов М.Н. Косуля в Западном
Забайкалье. Новосибирск: Наука, 1978. 189 с.
Смирнов М.Н. Дикие копытные Тувинской
АССР // Копытные фауны СССР. М.: Наука,
1980. С. 104—105.
Смирнов М.Н. Распространение, ресурсы и
перспективы использования диких копытных
Тувы // Ресурсы охотничье-промыслового хоз-ва
и прогноз их использования. М., 1985. С. 141 —
152.
Смирнов Н.Г. Динамика териофауны и
изменчивость некоторых млекопитающих Среднего,
Южного Урала и Зауралья в голоцене: Автореф.
дис. ... канд. биол. наук. Свердловск, 1976а. 20 с.
Смирнов Н.Г. Некоторые особенности голоце-
нового этапа развития териофауны Урала //
История биогеоценозов СССР в голоцене. М.:
Наука, 19766. С. 213—221.
Смоктунович Е.А. Некоторые факторы,
определяющие динамику численности европейской
косули Беловежской Пущи // Заповедники
Белоруссии. 1980. Вып. 4. С. 132—139.
Смоктунович Е.А. Олень и косула в местах
совместного обитания // Охота и охотничье
хоз-во. 1983. N 10. С. 19.
Собанский Г. Г. Расширение ареалов и рост
380
численности копытных на Алтае // Бюл. МОИП.
ОтД. биол. 1986. Т. 91, вып. 1.С. 25—27.
Собанский ГГ. Миграции косули (Capreolus
capreolus pygargus Pall.) на- Алтае // Фауна,
таксономия, экология млекопитающих и птиц
(Фауна Сибири). Новосибирск: Наука, 1987.
С. 121-128.
Собанский Г. Г. Промысловые звери Горного
Алтая. Новосибирск: Наука, 1988. 159 с.
Соколов В.Е. Систематика млекопитающих. М.:
Высш. шк., 1979. 528 с.
Соколов В.Е., Громов B.C. Морфометрические
исследования черепа разных подвидов косули
(Capreolus capreolus) // Зоол. журн. 1985а. Т. 64,
вып. 5. С. 738—749.
Соколов В.Е„ Громов fi.C. Опыты по
гибридизации европейской и азиатской косуль // Докл.
АН СССР. 19856. Т. 285, N 4. С. 1022—1024.
Соколов В.Е., Громов B.C. Новая
таксономия современных косуль (Capreolus Gray, 1821) //
Изв. АН СССР. Сер. биол. 1988. N 4. С. 485—
493.
Соколов В.Е, Громов B.C., Бененсон И.Е.
Сравнительное исследование краниологических
признаков разных географических форм косули
(Capreolus capreolus L.) // Там же. 1986. N 1.
С. 85—95.
Соколов В.Е, Громов B.C., Данилкин А.А.
Онтогенез поведения сибирской косули (Capreolus
capreolus pygargus) // Зоол. журн. 1985. Т. 64,
вып. 6. С. 915—926.
Соколов В.Е, Громов B.C., Рутовская М.В.
Звуковая сигнализация у европейской (Capreolus
capreolus) и сибирской (С. pygargus Pall.)
косуль // Зоол. журн. 1987. N 3. С. 430—443.
Соколов В.Е., Данилкин А.А. Мечение
территории самцами сибирской косули (Capreolus
capreolus pygargus) на Южном Урале // Зоол.
журн. 1977. Т. 56, вып. 10. С. 1543—1556.
Соколов В.Е., Данилкин А.А. Сибирская
косуля. М.: Наука, 1981. 144 с.
Соколов В.Е„ Данилкин А.А.,Дуламцэрзн С.
Современное распространение и численность
лесных копытных в Монголии // Зоологические
исследования в МНР. М.: Наука, 1982.С. 37—56.
Соколов В.Е, Данилкин А.А., Марков ГГ.
Краниометрическое исследование сибирского
подвида сибирской косули (Capreolus pygargus
pygargus Pall.) // Изв. АН СССР. Сер. биол. 1986.
Вып. 3. С. 445—448.
Соколов В.Е., Данилкин А.А., Минаев А.Н.
Участки обитания европейской косули (Capreolus
capreolus L.) на востоке ареала // Докл. АН СССР.
1986а. Т. 29, вып. 5. С. 1267—1271.
Соколов В.Е, Данилкин А.А., Романенко ЕВ.
Радиопрослеживание самцов европейской косули
в Центрально-Черноземном заповеднике //
Млекопитающие. 3-й съезд Всесоюэ. териол. о-ва: Тез.
докл. М., 1982. Т. 1. С. 300—301.
Соколов В.Е., Марков Г.Г., Данилкин А.А.
Морфометрическая характеристика кариотипов
европейской (Capreolus capreolus L.) и сибирской
(С. pygargus Pall.) косуль // Изв. АН СССР.
Сер. биол. 1986. Вып. 5. С. 780—784.
Соколов В.Е., Марков Г.Г., Данилкин А.А.
и др. О видовом статусе европейской (Capreolus
capreolus L.) и сибирской (С. pygargus Pall.)
косуль (краниометрическое исследование) / / Докл.
АН СССР. 1985а. Т. 280, N 6. С. 1505—1509.
Соколов В.Е., Марков Г.Г., Данилкин А.А.
и др. Сравнительное краниометрическое
исследование развития европейской (Capreolus
capreolus L.) и сибирской (С. pygargus Pall.)
косуль // Там же. 19856. Т. 282, N 1. С. 243—248.
Соколов В.Е., Орлов В.Н., ЧуЬиновская Г.А.,
Данилкин А.А. Хромосомные различия двух
подвидов косули (Capreolus capreolus capreolus L.
и Capreolus capreolus pygargus Pall. / / Зоол. журн.
\Ш. Т. 57, вып. 7. С. 1109—1112.
Соколов В.е., Орлов В.Н., ЧуЬиновская Г.А.,
Данилкин А.А. О систематическом статусе
европейской и сибирской косуль // Копытные фауны
СССР. М.: Наука, 1980. С. 105-106.
Соколов В.Е., ЧуЬиновская Г.А.,
Данилкин А.А.. Орлов В.И. Хромосомный набор
косули Capreolus capreolus L. // Систематика и
цитогенетика млекопитающих: Материалы
Всесоюэ.симпоз. М.: Наука, 1975. С. 45—46.
Соколов В. Е., Шур хал А. В., Данилкин А.А.
и др. Сравнительный анализ электрофоретических
спектров белков крови и мышечной ткани
европейской (Capreolus capreolus L.) и сибирской
(С. pygargus Pall.) косуль // Докл. АН СССР.
1986. Т. 288, N 5. С. 1274—1276.
Соколов Е.Н. Нейронный механизм-
ориентировочного рефлекса / / 18-й Междунар. конгр.
психологов. V симпоз. М.: Наука, 1966. С. 31—33.
Соколов И.И. Копытные звери (отряды Peris-
sodactyla и Artiodactyla). Фауна СССР.
Млекопитающие. М.; Л.: Изд-во АН СССР, 1959. Т. 1,
вып. 3. 640 с.
Соколова СМ., Данилкин А.А. О питании
сибирской косули // Зоол. журн. 1977. Т. 56,
вып. 12. С. 1870—1873.
Соловьев Д. К. Промыслы копытных зверей .и
лесной дичи в России // Ежегодник Всерос.
союза охотников. 1922. С. 53—73.
Стекленев Е.П. Морфогенетическая
характеристика спермиев представителей семейства
оленьих в связи с их гибридизацией // Тр.
2-го Всесоюз. совещ. по млекопитающим. М.:
Изд-во МГУ, 1975. С. 55—62.
Стекленев ЕП. Особенности размножения
европейской косули на юге Украины //
Копытные фауны СССР. М.: Наука, 1980. С. 202—
203.
Стекленев ЕМ. Межвидовая гибридизация
благородного (Cervus elaphus L.) и пятнистого
оленя (Cervus nippon hortulorumTemm.) //
Цитология и генетика. 1986. Т. 20, N 2. С. 138—
142.
Стенин Г. Результаты от изпозуването на
химически средства за опозвоне на горските
культури от дичови повреди // Горскостоп.
1968. Т. 4, вып. 8. С. 50—54.
Субботин A.M. О миграциях косули в
центральных районах Красноярского края // Тр.
Киров. с.-х. ин-та (1971). 1973. Вып. 28. С. 72—77.
Сыроечковский ЕЕ. Северный олень. М.: Аг-
ропромиздат, 1986. 256 с.
Тавровский В.А., Егоров О.В., Кривошеее В.Г.
и др. Млекопитающие Якутии. М.: Наука, 1971.
650 с.
Татаринов К.А. Фауна неогеновых и антропо-
геновых позвоночных Подолии и Прикарпатья,
ее история и современное состояние: Автореф.
дис. ... д-ра биол. наук. Киев. 1970. 56 с.
Тимофеева ЕЕ. Косуля. Л.: Изд-во ЛГУ,
1985. 224 с.
Тимофеев-Ресовский Н.В., Воронцов Н.Н.. Яб-
локов А.В. Краткий очерк теории эволюции.
М.: Наука, 1977. 301 с.
Тимченко Н.Г. К истории охоты и
животноводства в Киевской Руси (позднее Поднеп-
ровье). Киев: Наук, думка, 1972. 204 с.
Тинберген Н. Поведение животных. М.: Мир,
1969. 93 с.
Тинберген Н. Осы, птицы, люди. М.: Мир,
1970. 334 с.
Тинберген Н. Мир серебристой чайки. М.:
Мир, 1974, 272 с.
Толстое СП. Древний Хорезм: Опыт исто-
рико-археологического исследования. М.: Изд-во
МГУ, 1948. 352 с.
Троицкий- В.Н. Охотничий промысел Чуно-
Ангарского района б. Канского округа // Изв.
Сиб. краевой науч. охотничье-промысловой
станции. Новосибирск: Сиб. охотпромстанции, 1930.
Вып. 1. С. 3—85.
Туркин Н.В., Сатунин К.А. Звери России.
М.: Туркина, 1902. 506 с.
Успенский Г.А., Лозан М.Н. Косуля в
Молдавии // Охрана природы Молдавии. Кишинев,
1965. Вып. 3. С. 150—166.
Устинов С. К. Кабарожья струя // Охота
и охотничье хоз-во. 1967, N 3. С. 9.
Устинов С.К, Лобанов П.Н. К эколого-
морфологической характеристике косули бассейна
реки Кулинги // Экология. 1983. N 2. С. 81—82.
Ушков С.Л. Промысловая фауна Ильменского
госзаповедника // Тр. Ильмен. заповедника. 1947.
Вып. 3, ч. 1. С. 87—159.
Ушков С.Л. Переходы косули на Южном
Урале // Бюл. МОИП. Отд. биол. 1954. Т. 59,
вып. 5. С. 9—12.
381
Фадеев Е.В. О выпусках косуль в охотничьих
угодиях России // Вести. МГУ. Биология. 1969а.
N 3. С. 16—22.
Фадеев Е.В. Результаты искусственного
расселения некоторых видов диких копытных
животных в охотничьих хозяйствах России / / Бюл.
МОИП. Отд. биол. 19696. Т. 74, вып. 1. С. 37—46.
Фадеев Е. В. Косуля европейской части СССР / /
Охота и охотничье хоз-во. 1981. N 5. С. 18—20.
Фандеев А.А., Никольская В.П. Охотничье-
промысловые звери и трофеи. М.: Россельхоэ-
издат. 1978. 173 с; 1983, 238 с.
Федосенко А. К. Поведение маралов (Cervus
elaphus sibiricus Sev.) во время гона в Джунгарском
Алатау // Поведение млекопитающих. М.: Наука,
1977. С. 124—134.
Фетисов А.С. Сезонные корма сибирской
косули в Иркутской области. Иркутск: Обл. изд-во,
1947. 15 с.
Фетисов А.С. Косуля в Восточной Сибири.
Иркутск: Обл. изд-во, 1953. 73 с.
Филонов К. П. Особенности населения
сибирской косули на Южном Урале // Охотоведение.
М.: Лесн. пром-сть, 1974. С. 26—40.
Филонов К. П. Экологическая эамещаемость
факторов смертности в популяциях копытных //
Копытные фауны СССР. М.: Наука, 1975. С. 206—
207.
Филонов К.П. Динамика численности
копытных животных и заповедность. Охотоведение.
М.: Лесн. пром-сть, 1977. 229 с.
Филонов К.П., Калецкая М.М. Воздействие
волка на диких копытных животных // Волк. М.:
Наука, 1985. С. 336—354.
Флеров К. К. О диагностических признаках рода
Capreolus Frisch. (сем. Cervidae) // Докл.
АН СССР. Сер. А., 1928. N 23. С. 479—484.
Флеров К.К. Некоторые новые данные о
косулях (Capreolus Frisch.) Восточной Азии / / Там же,
1929. N 18. С. 429—432.
Флеров К. К. Копытные звери (Ungulata)
Таджикистана // Звери Таджикистана. М.; Изд-во
АН СССР, 1935а. С. 93—130. (Тр. Таджик,
базы; Т. 1).
Флеров К.К. Копытные (Ungulata) арктических
стран // Звери арктики. Л.: Изд-во Главсев-
морпути, 19356. С. 105—264.
Флеров К. К.Морфология и экология олене-
образных в процессе их эволюции // Материалы
по четвертичному периоду. М.; Л.: Изд-во АН
СССР, 1950. Вып. 2. С. 50—69.
Флеров К. К. Кабарги и олени. Фауна СССР.
Млекопитающие. М.; Л.: Изд-во АН СССР,
1952. Т. 1, вып. 2. 256 с.
Формозов А.Н. Снежный покров в жизни
млекопитающих и птиц СССР. М.: МОИП, 1946.
141 с.
Хавесон Я. И. О значении скрещиваемости
форм, и плодовитости гибридов для определения
382
ранга систематической категории // Науч. конф.
по итогам науч-исслед. работы за 1958—1959 гг.
Вологда, 1960. С. 73—75.
Хайнд Р. Поведение животных. М.: Мир,
1975. 885 с.
Холодова М.В. Сезонные изменения в
потребности кормов у некоторых копытных // 4-й съезд
Всесоюз. териол. о-ва: Тез. докл. М., 1986. Т. 1.
С. 367-368.
Хороших П.П. Пещеры Забайкалья // Тр.
Кяхт. краевед, музея. Улан-Удэ, 1949. Т. 16,
вып. 1. С. 128—132.
Цалкин В. И. Материалы для истории
скотоводства и охоты в древней Руси // Материалы
и исследования по археологии СССР. 1956. N 51.
183 с.
Цалкин В.И. Домашние и дикие животные
Северного Причерноморья в эпоху раннего железа / /
Там же. 1960. N 53. С. 7—109.
Цалкин /{.//.Млекопитающие древней
Молдавии // Бюл. МОИП. Отд. биол. 1962а. Т. 67,
вып. 5, С. 36—50.
Цалкин В. И. К истории животноводства и охоты
в Восточной Сибири // Материалы и
исследования по археологии СССР. 19626. N 107.129 с.
Цалкин В. И. К истории млекопитающих
восточноевропейской лесостепи // Бюл. МОИП.
Отд. биол. 1963. Т. 68, вып. 2. С. 43—62.
Цалкин В. И. Древнее животноводство Уплемен.
Восточной Европы и Средней Азии. М.: Наука,
1966. 157 с.
Цаплюк О.Э. Сезонные особенности гисто-
структуры гонад косули // Копытные фауны
СССР. М.: Наука, 1975. С. 313—314.
Цаплюк О.Э. Возрастные и сезонные
особенности биологии размножения косули (Capreolus
capreolus L.) в Казахстане // Зоол. журн. 1977.
Т. 56, вып. 4. С. 611—618.
Царев Ю.С. Распространение и численность
лося, косули и северного оленя //Биологическое
районирование Новосибирской области.
Новосибирск, 1969. С. 182—190.
Чегорка П.Т. Фенотипические критерии
диагностики благородных и пятнистых оленей и
гибридов между ними, в смешанных популяциях //
Проблемы охраны генофонда и управления
экосистемами в заповедниках лесной зоны:
(Всесоюз. совещ. Березин. заповедник). М., 1986.
Ч. 2. С. 222—225.
Чельцов-Бебутов A.M. К зоогеографической
характеристике песчаных полупустынь Тургайской
области // Теоретические и прикладные аспекты
охраны природы и охотоведения: Сб. науч. тр.
Моск. вет. академии. М., 1976. Т. 84. С. 44—53.
Черкасов А.А. Записки охотника-натуралиста.
М.: Изд-во АН СССР, 1962. 504 с.
Шарапов Ш. Обзор местонахождений плиоцен-
нижнеплейстоценовых наземных млекопитающих
Таджикистана // Тр. Ин-та зоологии и парази-
тологии АН ТаджССР. 1972а. С. 216—231. (Вопр.
зоологии Таджикистана).
Шарапов Ш. Остатки млекопитающих из
неолитической стоянки Сай-Сайед (Южный
Таджикистан) // Там же. 19726. С. 216—231. (Вопр.
зоологии Таджикистана).
Шаргаев М.А. Об ареале сибирской косули
на Обь-Иртышском севере // IV межвуз. зооге-
ографическая конф. Одесса, 1966. С. 310—311.
Шаргаев М.А. Изменения численности и
размещения копытных Витимского плоскогорья под
воздействием антропогенного фактора //
Зоологические проблемы Сибири. Новосибирск: Наука,
1972. С. 496—497.
Шарле мань М. Зоогеография УРСР. Ки1в:
Вид-во УРСР, 1937. 234 с.
Шарфф Р.Ф. Европейоая животныя. М.:
Природа, 1918. 233 с.
Шахмарданов З.А., Рамазанов Х.М. Ареалы
копытных Дагестана // Копытные фауны СССР.
М.: Наука, 1980. С. 113.
Шварц С.С. Популяционная структура вида //
Зоол. журн. 1967. Т. 46,' вып. 10. С. U56—
1469.
Шварц С.С. Экологические закономерности
эволюции. М.: Наука, 1980. 278 с.
Швец В.Г. Сокращение численности косули в
Хабаровском Приамурье // Копытные фауны
СССР. М.: Наука, 1975. С. 142—143.
Швец В.Г. Влияние хозяйственной деятельности
человека на численность копытных в Приамурье / /
Копытные фауны СССР. М.: Наука, 1980. С. 54—
57.
Шилов И.А. Соотношение пространственной
и этологической структуры популяций
позвоночных животных // Поведение животных. М.:
Наука. 1972. С. 12—14.
Шилов И. А. Эколого-физиологические основы
популяционных отношений у животных. М.:
Изд-во МГУ, 1977. 261 с.
Шовен Р. Поведение животных. М.: Мир,
1972. 487 с.
Шостак СВ. Численные соотношения
европейского благородного оленя с другими
копытными // Заповедники Белоруссии. 1978. Вып. 2.
С. 130-138.
Шперк Ф. Россия Дальнего Востока // Зап.
имп. Рус. геогр. о-ва по общей географии. СПб.,
1985. Т. 14. С. 503.
Штуббе Г., Ерухголъц 3. Опыты по
гибридизации европейской и сибирской косуль (Capreolus
capreolus capreolus L. (1758) X Capreolus capreolus
pygargus Pall. (1771) // Зоол. журн. 1979. Т. 58,
вып. 9. С. 1398—1403.
Шурупов И. И. Роль "Зоообъединения" в
расселении охотничьих животных // Обогащение
фауны и разведение охотничьих животных. Киров,
1982. С. 116.
Шурхал А. В., Данилкин А. А., Марков Г.Г. и др.
Генетическая изменчивость косули по
совокупности биохимических маркеров генов // Фене-
тика популяций: Материалы 3-го Всесоюз. совещ.,
Саратов. М., 1985. С. 273—274.
Шутеев М.М. Опыт авиаучета косули в
Омской области // Тр. Омск. гос. заповедника.
1973. Вып. 9. С. 101 — 103.
Шухов И. Посадка косули в лесостепной
полосе Западной Сибири // Охотник и рыбак
Сибири. 1929. N 1. С. 20—21.
Щетинин В. И. Местная группировка косули
Зейского заповедника / / Копытные фауны СССР.
М.: Наука, 1975. С. 147.
Юргенсон П. Б. Охотничьи звери и птицы.
М.: Лесн. пром-сть. 1968. 308 с.
Юргенсон П.Б. Состояние и перспективы
охотничьих ресурсов // Тр. Завидов. заповед.
хоз-ва. 1969. Вып. 1. С. 79—208.
Юргенсон П.Б. Биологические основы
охотничьего хозяйства в лесах. М.: Лесн. пром-сть,
1973. 173 с.
Яблоков А.В. Популяционная биология. М.:
Высш. шк., 1987. 303 с.
Янсон Я. Морфологическая характеристика
местной популяции косуль (Capreolus capreolus
capreolus L.) // Тр. Латв. с.-х. акад. 1975.
Вып. 83. С. 52—55.
Янушевич А.И., Айзин Б.М., Кадыралиев и др.
Млекопитающие .Киргизии. Фрунзе: Илим, 1972.
464 с.
Янушевич А., Благовещенский И.
Промысловые звери и птицы Западной Сибири. Новосибирск,
1952. 287 с.
Aitken R.J. Delayed implantation in roe deer
(Capreolus capreolus) // J. Reprod. and Fert. 1974.
Vol. 39, N 1. P. 225—233.
Aitken R.J. Uterine secretion of fructose in the roe
deer // Ibid. 1976. Vol. 46, N 2. P. 439—
440.
Aitken R.J. Aspects of delayed implantation
in the roe deer (Capreolus capreolus) // Ibid. 1981.
Vol. 29, V 1. P. 83—95.
Aitken R.J., Burton J., Hawkins J.. Kerr-Wilson R..
Short R.V., Steven D.H. Histological and ultra-
structural changes in the blastocyst and reproductive
tract of the roe deer (Capreolus capreolus) during
delayed implantation // Ibid. 1973. Vol. 34,
N 3. P. 481—493.
Allen CM. Pigs and deers from the Asiatic
expedition // Amer. Mus. Novit. 1930. N 430.
P. 1—19.
Allen CM. The mammals of China and
Mongolia // Natural history of Central Asia. N.Y., 1940.
Vol. 11. Pt 2. P. 621—1350.
Almasan H., Grapini V., Manolache L., Hie E.
Contributii la cunoasterea compozitiei specifice
si dinamicii hranei la caprior (Capreolus
capreolus L.) // Cercetari d'e ecolugie animaUE. Bucuresti,
1969. P. 415—424.
383
Amann R. Grundsatzliches zur Rehwildfutterung //
Niedersach. JSger. 1981a. Bd. 26, N 22. S. 1130.
Amann R. Wissen um Reviereinrichtungen:
Der Saftfutter-Behalter fur Rehwild // Ibid. 1981b.
Bd. 26, N 22. S. 1131 — 1132.
Amoroso E.A., Finn C.A. Ovarian activity during
gestation, ovum transport and implantation //Theovary.
N.Y.; L., 1962. N 1. P. 451—537.
Amrud J., Nes N. The chromosomes of the
roe (Capreolus capreolus) // Hereditas. 1966.
Vol. 56, N 2. P. 217—220.
Andersen J. Analysis of a Danish roe-deer
population based upon the extermination of the
total stock // Dan. Rev. Game Biol. 1953.
Vol. 2. P. 127—155.
Aylmer F. What do the does do? // Deer.
1977. Vol. 4, N 4. P. 206—207.
Baccus R., Ryman N.. Smith M.H., Reuierwall C,
Cameron D. Genetic variability and differentiation
of large grazing mammals //J. Mammal. 1983. Vol. 64,
N 1. P. 109—120.
Bailey N.T.J. On estimating the size of mobile
populations from recapture data // Biometrica.
1951. Vol. 38. P. 293—306.
Baldwin R.L. Digestion and metabolism of
ruminants // BioScience. 1984. Vol. 34, N 4. P. 244—249.
Barclay E.N. Notes on the Roe-deer // Ann.
Mag. Nat. Hist. 1933. Vol. 10, N 12. P. 66—80.
Barth D., Gimenez Т., Hoffmann B.,Karg H.
Testosteronkonzentrationen im peripherien Blut beim
Rehbock (Capreolus capreolus): Seasonale und
rhythmische Veranderungen sowie Beziehungen zu
einigen Verhaltensweisen und anderen saisonabhan-
gigen Parametern // Ztschr. Jagdwiss. 1976. Bd. 22,
N 3. S. 134—148.
Bartos L., Hydnek J., Zirovnicky' J. Hibridization
between red and sika deer // Zool. Anz. 1981.
Bd. 207, N 5/6. S. 260—270.
Bayern A.. Bayern J. Ober Rehe in einem
steirischen Gebirgsrevier. Hamburg: Bauer, 1975.
207 S.
Becker-Dillingen J. Die Ernahrung des Waldes in
der freien Wildbahn. Donauwdrth, 1945. 251 S.
Beklovd M., Koubek P.. Pikula J., Zejda J. Game
losses during the harvest of perennial fodder plants //
Folia zool. 1982. Vol. 31, N 1. P. 37—54.
Berg F.-C. Zum Raum-Zeit-System des Rehes //
Allg. Forst. 1978. Bd. 33, N 3. S. 48—50.
Berge F. Takseringsmetoder for radyr, Capreolus
capreolus (L.) // Medd. Statviltundersok. Ser. 1.
1969. N 30. 45 S.
Bettmann H. Winterbrunft beim Rehwild // Ztschr.
Jagdwiss. 1961. Bd. 7. N 4. S. 169.
Bideau E., Vincent J.P., Quere J.P.. Angibault J.M.
Occupation de 1'espace chez le chevreuil (Capreolus
capreolus L., 1758). 1. Cas des mslles // Acta
Oecol. Oecol. Applic. 1983a. Vol. 4, N 2. P. 163—184.
Bideau E., Vincent J.P., Quere J.P.. Angibault J.M.
Note sur 1'evolution de I'association mere-jeune chez le
384
chevreuil (Capreolus capreolus L., 1758) etudiee par la
technique du radio-tracking // Mammalia. 1983b.
Vol. 47, N 4. P. 477—482.
BischoffT.L. W. Entwicklungsgeschichte des Rehes.
GiePen: Ricker, 1854. 36 S.
Blankenhorn H.J. Kitz-Markierungs-Aktion,
1970—1971 // JSger. 1975. N 9. S. 42—45.
Blant M. Dinamique de population, condition et
constitution du chevreuil (Capreolus capreolus L.,
1758) dans les cantons de Neuchatel et Vaud
(ouest de la Suisse). Neuchatel: Press univ., 1987.
160 p.
Blant M., Chappuis F., Maire J.A., Mermod C,
Pedroli J.C. La biologie du chevreuil Capreolus
capreolus (L.) dans le canton de Neuchatel (Suisse).
Neuchatel: Press, univ., 1982. 105 p.
Blauvelt H. Dynamics of the mother-newborn
relationship in goats // Group Proces. 1955.
Vol. 1. P. 211—258.
Bobek B. Summer food as the factor limiting roe
deer population size // Nature. 1977. Vol. 268, N 5615.
P. 47—49.
Bobek B. A model for optimization of roe deer
management in Central Europe // J. Wildlife
Manag. 1980. Vol. 44, N 4. P. 837—848.
Bobek В., Drotdt A., Grodtinski W.. Weiner J.
Studies on productivity of the roe deer population
in Poland // XI Intern. Congr. Game Biol. Stockholm,
1974. P. 115—123.
Boessneck J. Zur Grope des mitteleuropaischen
Rehes (Capreolus capreolus L.) in alluvial-vor-
geschichtlicher und frliher historischer Zeit // Ztschr.
Saugetierk. 1956. Bd. 21, N 3/4. S. 121 — 131.
Boisaubert B. Beitrag zur Untersuchung der
Entwicklung einer Methode zur Bestandsaufnahme
von Capreolus capreolus, die in Waldgebieten leben //
Bull. Mensuel. 1979. Vol. 40. P. 193—225.
Boisaubert B. Aussetzung von Rehwild — Vorlaufi-
ge Ergebnisse // Ibid. 1980. Vol. 41. P. 11—18.
Borg K. On mortality and reproduction of roe
deer in Sweden during the period 1948—1969 //
Viltrevy. 1970. Vol. 7, N 2. P. 121—149.
BorgK. Predationon'roe deer in Sweden//J.
Wildlife Manag. 1962. Vol. 26, N 2. P. 133—136.
Borowski S., Kossak S. The food habits of deer
in the Bialowieza Primeval Forest // Acta theriol.
1975. Vol. 20, N 32. P. 463—506.
Boule M. Les gisements des Mammiferes fossiles de
la Montagne de Perrier (Puy-de-Dome) // С. г. Somm.
Soc. geol. P., 1905. P. 32—33.
Bramley P.S. Territoriality and reproductive
behaviour of roe deer // J. Reprod. and Fert. Suppl.
1970. N 11. P. 43—71.
Bramley P.S. Management of roe deer (Capreolus
capreolus) in small deciduous woods in England //
Forestry. 1972. Vol. 45, N 2. P. 211—221.
Bravard A. Monographie de la Montagne de
Perrier, pres d'Istoire (Puy-de-Dome) et de deux
especes fossiles du genre Felis, decouvertes dans
I'une de ses couches d'alluvion. P.: Dufour et Docagne,
1928. 146 p.
Bresinski W. Grouping tendencies in roe deer
under agrecenosis conditions // Acta theriol. 1982.
Vol. 27, N 29. P. 427—447.
Briedermann L. Was ast unser Rehwild? // Unsere
Jagd. 1974. Bd. 24, N 4. S. 110—111.
Briedermann L. Der Wildbestand — die grosse
Unbekannte. В.: DLW, 1982. 212 S.
Briedermann L., Ahrens M. Bestande und Verbrei-
tung des Schalenwildes in der DDR // Unsere Jagd.
1980. Bd. 30, N 1. S. 4—6.
Briedermann £., Mehlitz S., Richter H, Trophaen-
schau und Trophaenbewertung beim Schalenwild.
В.: VEB Dt. Landwirtsch., 1969. 156 S.
Breilenmoser U., Haller H. Zur NahrungsSkologie
des Luchses Lynx lynx in den schweizerischen
Nordalpen // Ztschr. Saugetierk. 1987. Bd.-52, N 3.
S. 268—291.
Broom D.M., Johnson E. Responsiveness of
hand-reared roe deer to odours from skin glands // J.
Natur. Hist. 1980. Vol. 14, N 1. P. 41—47.
Bubenik A.B. Grundlagen der Wildernahrung.
В.: Bauernverl., 1959. 229 S.
Bubenik A.B. Le rythme nycthemeral et le regime
journalier des ongules sauvages; problemes theoriques;
rythme d'activite du chevreuil // Mammalia. 1960.
Vol. 24. P. 1—66.
Bubenik A.B. Das Verhalten des Rehes am Tage
und in der Nacht im Laufe des Jagres // Pirsch.
1962. Bd. 24/25. S. 944—947, 988—1000.
Bubenik A.B. Beitrag zur Geburtskunde und zu
den Mutter—Kind—Beziehungen des Rehes
(Capreolus capreolus L.) und Rotwildes (Cervus elaphus L.) /7
Ztschr. Saugetierk. 1965. Bd. 30, N 2. P. 65—128.
Bubenik A.B. Vliv rysa (Lynx lynx L.) a vlka
(Canis lupus L.) na strukture populaci srnEi (Capreolus
capreolus L.) a jeleni zvefe (Cervus elaphus L.) //
Lynx. 1966a. N 6. P. 7—10.
Bubenik A.B. Das Geweih: Entwicklung, Aufbau,
und Ausformung der Geweihe und Gehorne und ihre
Bedeutung fur das Wild und fur die Jagd. Hamburg;
В.: Рагеу, 1966b. 214 S.
Bubenik A.B. The significance of the antlers in
the social life of the Cervidae // Deer. 1968.
N. 1. P. 208—214.
Bubenik A.B. RehwildHege und Rehwildbiologie.
Emahrung und Beifutterung // Dt. Jager. 1970.
Bd. 38, N 15. S. 389—394.
Bump G. Acclimatization of game animals in the
United States // IX Intern. Congr. Game Biol.
Moscow, 1970. P. 136—141.
Burl W.H. Territoriality and home range concepts
as applied to mammals // J. Mammal. 1943. Vol. 24.
P. 346—352.
Banner K. Untersuchungen zum Einflup von Grdpe
und Zusammensetzung des Wintersprungs beim
Rehwild auf die Zeitdauer verschiedener Verhalten-
sweisen // Ztschr. Jagdwiss. 1980. Bd. 26, N 4.
S. 181—194.
Banner K. Winterliche Fahrtenzahlungen beim
Rehwild in Anhangigkeit von Witterung, Waldstruktur
und Jagddruck // Ibid. 1983. Bd. 29, N 2. S. 82—95.
Byron H.A.M. Distribution patterns of roe deer
(Capreolus capreolus) related to the availability of food
and cover//J. Zool. 1981. Vol. 194, N 2. P. 271—275.
Cadariu M„ Popovici N., Cismas M.A. Schimbari
ciclice sezoniere in hipofiza capriorului (Capreolus
capreolus L.) mascul // Stud. Univ. Babes- Bolyai. Ser.
biol. 1977. Vol. 22, N 2. P. 48—55.
Casanova P. II caprielo (Capreolus capreolus L.):
un tipice abitatere della macchia // Ital. forest
mont. 1981. Vol. 36, N 5. P. 251—256.
Cederlund G. Daily and seasonal activity pattern
of roe deer in a boreal habitat // Viltrevy.
1981. Vol. 11, N 1. P. 1—41.
Cederlund G. Mobility response of roe deer
(Capreolus capreolus) to snow depth in a boreal
habitat // Ibid. 1982. Vol. 12, N 2. P. 39—68.
Cederlund G. Home range dynamics and habitat
selection by roe deer in a Boreal area in Central
Sweden // Acta theriol. 1983. Vol. 28, N 30.
P. 443—460.
Cederlund G, Lindstrom E. Effects of severe
winters and fox predation on roe deer mortality //
Ibid. 1983. Vol. 28, N 7. P. 129—145.
Cederlund G., Ljungquist H., Markgren G., Stal-
felt F. Foods of moose and roe-deer at Grims6 in
central Sweden. Results of rumen content analyses //
Viltrevy. 1980. Vol. 11, N 4. P. 169—247.
Chaplin R.E.. Chapmann D.S.. Prior R. An
examination of the uterus and ovaries of some female
roe deer (Capreolus capreolus L.) from Wildshire
and Dorset, England // J. Zool. London. 1966.
Vol. 48. P. 570—574.
Chapmann D.I. Antlers—bones of contention //
Mammal. Rev. 1975. Vol. 5, N 5. P. 121 — 172.
Chapmann D.I., Chapmann N.G. Further
observations on the incidence of twins in Roe deer,
Capreolus capreolus // J. Zool. 1971. Vol. 165.
P. 505—509.
Corbet G.B. The mammals of the Palaearctic
region. A taxonomic review. L.; Ithaca, 1978.
314 p.
Cotta V. Asupra capriorului Siberian (Capreolus
capreolus pygargus Pallas) in Romania // Rev.
padurilor. 1969. Vol. 84, N 7. P. 345—348.
Cotta V.. Almasan H.A.. Babulia Т., Scutaru V.
RSspindirea si densitatea de effectiv a capriorului
in R.P.R. Caile pentru obtineria unui efectiv rational //
Stud, si cere. Inst. cere, forest. A. 1961. Vol. 22.
P. 5—25.
Croizet J.В., Joberl A. Recherches sur les osse-
mens fossiles du departement du Puy-de-D6W. P.,
1828. T. 1.226 p.
Czyzewska T. Nowy gatunek jelenia rodzaju Cer-
25. Европейская и сибирская косули
385
vocerus Khomenko z pliocenskiej brekcji kostnej z
Werow // Acta palaeontol. Pol. 1960. Vol. 5,
N 3. P. 283—318.
Czyzewska T. Deers from Weze and their
relationship with the Pliocene and recent eurasistic Cervidae //
Ibid. 1968. Vol. 13, N 4. P. 537—593.
Dasmann B. The wildlife biology. N.Y., 1966.
P. 61—65.
Dathe H. Zum Mutter-Kind-Verhaltnis bei Cervi-
den // Beitr. Jagdund Wildforsch. 1966. Bd. 5.
S. 83—88.
Dawkins B. Contribution to the history of the
deer of the European Miocene and Pliocene Strata //
Quart. J. Geol. Soc. London. 1878. Vol. 34.
P. 402—420.
Deperet Ch. Nouvelles etudes sur les Ruminants
pliocenes et quaternaires d'auvergne // Soc. geol.
Bull. P., 1884. Vol. 12. P. 247—284.
Dissert J. Untersiichungen fiber die Verdaulichkeit
von Rohnahrstoffen verschiedener .Futterrationen
an Rehwild (Capreolus с capreolus L.) und Ziegen
(Capra a. hircus L.) sowie Beobachtungen iiber
das Asungsverhalten von Gehege-Rehen: Diss. Dokt.
Landwirt.- Bonn, 1983. 100 S.
Dittrich G. Hinweise zur Winterfiitterung des
Rehwildes in Wald und Feldgebieten // Unsere
Jagd. 1981. Bd. 31, NILS. 332—333.
Drescher-Kaden V. Untersuchungen iiber Ver-
dauungstrakt im Pansen von Gamswild post
mortem // Tagungsber. I. Intern. Gamswild-Treffen.
Oberammergan, 1974. S. 26—33.
Drescher-Kaden" U. Untersuchungen am Ver-
dauungstrakt von Reh, Damhirsch und Mufflon.
Mitt. 1. Gewichtserhebungen und Kapazitatsmessun-
gen am Verdauungstrakt, insbesondere am Pansen —
Hauben — Raum von Reh, Damhirsch und Mufflon //
Ztschr. Jagdwiss. 1976. Bd. 22. S. 184—190.
Drescher-Kaden U., SeifelnasrE.A. Untersuchungen
am Verdauungstrakt von Reh, Damhirsch und
Mufflon. Mitt. 2. Rohnahrstoffe im Panseninhalt von
Reh, Damhirsch und Mufflon // Ibid. 1977.
Bd. 23. S. 6—11.
Droidt A. Seasonal intake and digestibility of
natural foods by roe deer // Acta theriol. 1979. Vol. 24,
N 13. P. 137—170.
Droidt A., Osiecki A. Intake and digestibility of
nature foods in roe deer // Ibid. 1973. Vol. 18, N 3.
P. 81—91.
Dzieciolowski R. Roe deer census by pellet-group
counts // Ibid. 1976. Vol. 21, N 24—31. P. 351—358.
Eberle J. Altersstruktur des Rehwildes in der
Blattzeit bestimmt // Unsere Jagd. 1986. Bd. 36, N 10.
S. 300.
Eiberle K. Beobachtungen iiber das Verhalten des
Rehwildes // Schweiz. Ztschr. Forstwiss. 1962.
Bd. 113, N 11. S. 660—668.
Eiberle K. Wildverteilungsstudien am Rehwild //
Ibid. 1967. Bd. 118, N 2. S. 45—56.
386
Eiberle K. Rehwildhege und StraPenverkehr //
Ibid. 1972. Bd. 123, N 4. S. 201—210.
Eiberle K. Rechnerische Moglichkeiten der Bestan-
deskontrolle beim Rehwild // Ibid. 1976. Bd. 127, N 5.
S. 312—326.
Eiberle K., Wenger C.-A. Zur Bedeutung der forstli-
chen Betriebsart fur das Reh // Ibid. 1983. Bd. 134,
N 3. S. 191—206.
Eisfeld D. Der Eiweis- und Energiebedarf des
Rehes, diskutiert anhand von Laborversuchen //
Verh. Ges. Okol., Erlangen, 1974a. S. 129—139.
Eisfeld D. Haltung von Rehen (Capreolus
capreolus L.) zu Versuchszwecken // Ztschr. Saugetierk.
1974b. Bd. 39, N 3. P. 190—199.
Eisfeld D. Ernahrungsphysiologie als Basis fur
die 6kologische Beurteilung von Rehpopulationen //
Rev. Suisse Zool. 1976. Bd. 83. S. 914—928.
Eisfeld D, Anspriiche von Rehen an die Qualitat
ihrer Nahrung // XVII Intern. Congr. Game Biol.
Brussels, 1985. Pt. 2. P. 1027—1034.
Ellenberg H. Wilddichte, Ernahrung und Vermeh-
rung beim Reh // Verh. Ges. Okol., Erlangen,
1974a. S. 59—76.
Ellenberg H. Die Korpergrdpe des Rehes als
Bioindikator // Ibid. 1974b. S. 141 — 154.
Ellenberg H. Zur Populationsdkologie des Rehes
(Capreolus capreolus L., Cervidae) in Mitteleuropa.
Miinchen, 1978. 211 S. (Spixiana; Suppl. 2).
Ellerman J.R., Morrison-Scott T.C.S. Check-list of
Palaearctic and Indian mammals 1758 to 1946.
L., 1951. 1227 p.
Engl D. Rehwildmarkierung // Osterr. Weidwerk.
1982. N 5. S> 210—212.
Espmark Y. Mother-young relations and
development of behaviour in roe deer (Capreolus
capreolus L.) // Viltrevy. 1969. Vol. 6, N 6. P. 460—
540.
Espmark У. Social behaviour of roe deer at winter
feeding stations // Appl. Anim. Ethol. 1974. Vol. 1,
N 1. P. 35—47.
Essen L. Das Rehwild in Schweden // Beitr.
Jagdund Wildforsch. 1966. Bd. 5. S. 143—148.
Esser W. Beitrag zur Untersuchung der Asung des
Rehwildes // Ztschr. Jagdwiss. 1958. Bd. 4, N 1.
S. 1—41.
Feng Z; Cai C, Zheng Ch. A checklist of the
mammals of Xizang (Tibet) // Acta theriol. sin.
1983. Vol. 4, N 4. P. 341—358.
Filler Z, Lochman J. Studium pfirozene potravy
jeleni a srnJi zvJFe v oblasti KrkonoS // Opera
Concontica. 1969. Vol. 6. P. 139—161.
Fruzinski В.. KaluzinskiJ., BaksalaryJ. Weight and
body measurements of forest and field roe deer //
Acta theriol. 1982. Vol. 27v N 33. P. 479—488.
Fruzinski В., Labudzki L Demographic processes in
a forest roe deer population // Ibid. 1982a.
Vol. 27, N 25. P. 365—375.
Fruzinski В., Labudzki L Sex and age structure of a
forest roe deer population under hunting pressure //
Ibid. 1982b. Vol. 27, N 26. P. 377—384.
Fruzinski В.. Labudzki L, Wlazelko M. Habitat,
density and spatial structure of the forest roe
deer population // Ibid. 1983. Vol. 28, N 16.
P. 243—258.
Gaal I. Unterpliozane Saugetierreste aus Hatvan in
Ungarn // Geol. hung. ser. palaeontol. 1943. Bd. 20.
S. 65—115.
Galamb С Tusnadi G. Исследование
структуры популяции косули (Capreolus c.L.) на основе
квантативных характеристик // Изв. фак. с.-х.
наук в г. Кестхей (Венгрия). 1973. Т. 15, N 9.
С. I—31. На венг. яз.
GebczyAska Z. Food of the roe deer and red deer
in theer in the Bialowieza Primeval Forest //
Acta theriol. 1980. Vol. 25, N 40. P. 487—500.
Geiger G., KrSmer A. Rangordnung von Rehwild
(Capreolus capreolus L.) und der Winterfutterung in
einem schweizerischen Jagdrevier // Ztschr. Jagdwiss.
1974a. Bd. 20, N 1. S. 53—56.
Geiger G., Kra'mer A. Rank-order of roe deer at
artificial winter feeding sites in a Swiss hunting
district // XI Intern. Congr. Game Biol. Stockholm,
1974b. P. 107—113.
Geist V. Horn-like structures as rank symbols,
guards and weapons // Nature. 1968. Vol. 200, N 5169.
P. 813—814.
Gelder R.G. Mammalian hybrids and genetic
limits // Amer. Mus. Novit. 1977. Vol. 2635. P. 1—25.
Georgii B. Corpora li'tea and adrenal weight in
roe deer // XI Intern. Congr. Game Biol. Stockholm,
1974. P. 125—132.
Gervais P. Zoologie et palaeontologie Franchise.
P., 1859. 544 p.
Gonzalez R., Angibault J.M. Etude du rythmed'ac-
tivite du chevreuil parbiotelemetrie: aspect methogolo-
gique; Premiers resultats en milieu forestier,// Bull.
Sfeca. 1987. Vol. 2, N 1. P. 159—164.
Gosso.w H. Waldstrukturen und Wildstandsent-
wicklung // Wald und Wild Seminar. Bonn.,
1977. S. 1—27.
Graczyk R. Charakter powiazart ekologicznych
populacji sarn (Capreolus capreolus Linnaeus, 1758) z
ekosystemami lesnym i polnym // Rocz. AR Pozn.
1978. Wol. 100. S. 23—29.
Graczyk R., Bereszynski A. Obserwacje nad twor-
zeniem sie, organizacja i liczebnoscia gesienno-
zimowych skupien sarn (Capreolus capreolus L.)
w ekosystemie polnym // Ibid. 1978. Wol. 100.
S. 31—38.
Gray J. On the natural arrangement of vertebrate
animals // London. Med. Reposit. 1821. Vol. 15.
P. 296—320.
Giinther G. Ein entschlossener Versuch. Sichere
Ermittlung vorhandener Wildbestande // Pirsch.
1978. Bd. 30, N 23. S. 1563—1567.
Guslavsson I. Chromosome studies in five species of
deer representing the four genera Alces, Capreolus,
Cervus and Dama // Mammal. Chromosomes
Newslett. 1965. Vol. 18. P. 149.
Guslavsson I.. Sundl CO. Karyotypes in five
species of deer (Alces alces L., Capreolus capreolus L.,
Cervus elaphus L., Cervus nippon nippon Temm. and
Dama dama L.) // Hereditas. 1968. Vol. 60,
N 1/2. P. 233—248.
HafferJ. Superspecies and species limits in
vertebrates // J. Zool. Syst. and Evol. 1986. Vol. 24,
N 3. P. 169—190.
Haisch E. Rehwildschaden im Weinbergen //
Wild und Hund. 1982. Bd. 85, N 14. S. 10.
Hallenorlh Th. Klassifikation der lebenden Sauge-
tiere: Artiodactyla // Handbuch der Zoologie.
В., 1963. Bd. 8. 167 S.
Hamilton W.J., HarrisonR.J.. YoungB.A. Aspects of
placentation in certain Cervidae // J. Anat. 1960.
Vol. 94. P. 1—33.
Hanley T.A. The nutritional basis for food
selection by ungulates // J. Range Manag. 1982. Vol. 35,
N 2. P. 146—151.
HanuS V., Filler Z. Telesny vjroi srnli zvere //
Pr. vyzk. Ostavi Lesn. 1981. Sv. 58. S. 191—212.
Harrington R. Evolution and distribution of the
Cervidae. Biology of deer production // Bull. Roy.
Soc. N.Z. 1985a. Vol. 22. P. 3—11.
Harrington R. Hybridisation in deer — its
detection and uses // Ibid. 1985b. Vol. 22. P. 62.
Harrison D.L. The mammals of Arabia. L.,
1968. Vol. 2. 381 p.
Hartwig D. Gefahrdung des Strassenverkehrs
durch Wild // Wild und Hund. 1986. Bd. 89, N 6.
S. 12—16.
Hearney A.W.. Jennings T.J. Annual foods of the
red deer (Cervus elaphus) and the roe deer (Caprelous
capreolus) in the east of England // J. Zooi.
1983. Vol 201, N 4. P. 565—570.
Hein J. Untersuchungen uber Wege zur Verbesee-
rung des Qualitat eines Rehwildbestandes: Diss.
G6ttingen, 1966.
Heintz E. Les cervides villafranchiens de France et
d'Espagne // Mem. Mus. Nat. hist, natur. N.S. Ser.
С 1970. Vol. 22, N 1. 303 p.
Hell P. SrnJia zver., Br.: Priroda, 1979. 310 s.
Hell P., HerzJ. Prispevok k taxonomii srnca
h6rneho europskeho (Capreolus с capreolus L.,
1758) na Slovensku // Bioldgia (CSSR). 1968. Sv. 23,
N 8. S. 623—644.
Hell P.. Herz J. RozSirenie, hustota osidlenia
a produkcia srnca horneho europskeho (Capreolus
с capreolus L., 1758) na Slovensku // Acta zoo-
techn. nitra. 1970. Vol. 20. P. 227—241.
Helle P. Food composition and feeding habits
of the roe deer in winter in central Finland //
Acta theriol. 1980. Vol. 25, N 3. P. 395—402.
Hennig R. Ober des Revierverhalten der Rehbo-
cke // Ztschr. Jagdwiss. 1962a. Bd. 8, N 2.
S. 61—81.
Hennig R. Ober einige Verhaltensweisen des
387
Rehwildes (Capreolus capreolus) in freier Wildbahn //
Ztschr. Tierpsychol. 1962b. Bd. 19, N 2. S. 223—
229.
Henry B.A.M. A comparison of the winter diet
of rpe deer and sheep // J. Zool. 1978a. Vol. 185,
N 2. P. 270—273.
Henry B.A.M. Diet of the roe deer in an
English conifer forest // J. Wildlife Manag. 1978b.
Vol. 42, N 4. P. 937—940. '
Herz J. Zhodnotenie polovnickeho obhospodaro-
vania zveri srnSej v okrese Trnava // Folia venatoria.
1971. Vol. 1. P. 201—213.
Herzog A., Hdhn H. Darstellung der Chromoso-
men aus Knochenmarkzellen beim Reh (Capreolus
capreolus) und Rottier (Cervus elaphus) // Ztschr.
Jagdwiss. 1967. Bd. 13. S. 118—121.
Hoffman В., Barth D„ Karg H. Progesterone
and estrogen levels in peripheral plasma of the pregnant
and non pregnant roe deer (Capreolus capreolus) //
Biol. Reprod. 1978. Vol. 19. P. 931—936.
Hofmann R.R. Die Verdauungsorgane des Rehes und
ihre Anpassung an die besondere ErnShrungsweise //
Wildbiolog. Inform. Jager. Stuttgart, 1978a. S. 103
— 112.
Hofmann R.R. Die ErnShrung des Rehwildes im
Jahresablauf nach Modell Weichselboden // Ibid.
1978b. S. 121 — 136.
Hofmann R.R., Geiger G., Kcrrig R. Vergleichend-
anatomische Untersuchungen an der Vormagen-
schleimhaut von Rehwild (Capreolus capreolus) und
Rotwild (Cervus elaphus) // Ztschr. Saugetierk.
1976. Bd. 41, N 3. S. 167—193.
HoliXovd V., Koienu I.. Obrtel R. The summer
diet of field roe bucks (Capreolus capreolus) in
Southern Moravia // Folia zool. 1984. Vol. 33,
N 3. P. 193—208.
HoliSovd V.. Obrtel R., Koienu 1. The winter
diet of roe deer (Capreolus capreolus) in the Southern
Moravian agricultural landscape // Ibid. 1982.
Vol. 31, N 3. P. 209—225.
НоШочй V., Obrtel R., Koienu I. Fragmentation
of food in roe deer (Capreolus capreolus) // Ibid.
,J985. Vol. 34, N 2. P. 101 — 109.
/ *~" HoliSovd V„ Obrtel R., Koienu I. Rumen content
i' vs. faecal analysis to estimate roe deer diets //
Llbid. 1986a. Vol. 35, N,1. P. 21—32.
Holtfovd V., Obrtel R., Koiena I. Seasonal
variation in the diet of field roe deer (Capreolus
capreolus) in Southern Maravia // Ibid. 1986b.
Vol. 35, N 2. P. 97—115.
Holler W., PrilkoferK. Graviditat und Geburt beim
Reh (Capreolus capreolus L.) // Physiol. Pathol.
Therapiemdglichk. 1980. Bd. 12. S. 368—374.
Holmes F. Following the roe. A natural history of
the roe deer. Edinburgh, 1973. 110 p.
Holzhausen O. Asungsbedingungen des Rehwildes
(Revier Ahnsen) // Fachwiss. Arbeit fur Biol.
Luneburg, 1970. 80 S.
HonackiJ.H., KinmanK.E.. KoepplJ.W. Mammals
388
species of the world. Lawrense (Kan.), 1982.
694 p.
Honak B. Mezi Kamerou a puSkou. Brno:
Naklad. Blok, 1970. 79 s.
Hosey G.R. Annual foods of the roe deer
(Capreolus capreolus) in the south of England // J.
Zool. 1981. Vol. 194, N 2. P. 276—278.
Hromas J. Relations between the trophy quality and
living conditions of roe deer in the CSSR //
Lesnictvi. 1983. Vol. 29, N 7. P. 591—610.
Huang W. Note on holocene gazo site of
Changdu, Xizang // Vertebrata Pal-Asiat. 1980.
Vol. 18, N 2. P. 163—168.
Hiibner F. Das Rehwild—Biologie // Weidwerk der
Welt. 1938. S. 197—200.
Jackson J.E. Feeding habits of roe deer //
Mammal. Rev. 1974. Vol. 4. P. 93—101.
Jackson J.E. The annual diet of the roe deer
(Capreolus capreolus) in the New Forest, Hampshire,
as determined by rumen content analysis // J. Zool.
1980. Vol. 192, N 1. P. 71—84.
Jacobi J. Kraniometrische Untersuchungen und
Stammesgeschichtliche Ableitung der Rehe (Capreolus
H. Smith) // Jena. Ztschr. Naturwiss. 1932. Bd. 67.
S. 326—345.
Jaezewski Z The induction of antler growth in
female Red deer // Bull. Acad. Pol. sci. Ser. sci. biol.
1976. Vol. 24, N 1. P. 61—65.
Jaezewski Z„ Doboszynska Т., Krzywinski A.
The induction of antler growth by amputation of
the pedicle in red deer (Cervus elaphus L.) males
castrated before puberty // Folia biol. (PRL).
1976. Vol. 24, N 3. P. 299—307.
Janeau G., Quer'e J.P., Spitz F., Vincent J.P.
Etude par radiotracking des variations saisonnieres
de l'etendue du domaine vital de cing chevreuils
(Capreolus capreolus L.) // Biol. Behav. 1981.
Vol. 6, N4. P. 291—304.
Jewell P.A. The concept of home range in
mammals // Symp. Zool. Soc. London. 1966.
vol. 18. P. 85—109.
Jiang P. Late pleistocene cave deposits of Antu,
Kirin // Vertebrata Pal-Asiat. 1975. Vol. 13,
N 3. P. 197—198.
Juon P. Ober neuere Erkenntnisse zur Frage der
Rehwildernahrung // Schweiz. Ztschr. Forstwiss.
1963. Bd. 114, N 3. S. 98—117.
Kahlke H.D. Grossaugetiere im Eiszeitalter.
Leipzig; Jena: Urania—Verl., 1955. 88 S.
Kahlke H.D. Cervidenreste aus den altpleistozanen
Tonen von Voigtstedt bei Sangerhausen. T. 1.
Die Schadel, Geweihe und Geh6rne // Abh. Dt. Akad.
Wiss. Berlin. Kl. Chem., Geol. Biol. 1958. Jg. 1958. N 9.
P. 1—51.
Kahlke H.D. Die Rhinocerotiden-Reste aus cten
Kiesen von Sussenborn bei Weimar. Das PleistozSn
von Sussenborn // Pal. Abh. PalaozooS. 1969.
Bd. 3, N 3/4. S. 667—709.
Kaluzinski J. Badania biometryczne i obserwacje
biologiczne sarny (Capreolus capreolus L.) populacji
polnej // Rocz. AR Pozn. 1978. Wol. 100. S. 73—
81.
Kaluzinski J. Dynamics and structure of a field roe
deer population // Acta theriol. 1982a. Vol. 27,
N 27. P. 385—408.
Kaluzinski J. Roe deer mortality due to
mechanization of work in agrocenoses // Ibid. 1982b. Vol. 27,
N 30. P. 449—455.
Kaluzinski J. Composition of the food of the roe
deer living in fields and the effects of their feeding
on plant production // Ibid. 1982c. Vol. 27,
N 30. P. 457—470.
Kaluzinski J., Bresinski W, The effect of the
european hare and roe deer populations on the
yields of cultivated plants // Ecology und management
of European hare populations. Warsaw, 1976. P. 247—
253.
Kampmann H. Untersuchungen fiber die Auswir-
kungen des Seitentriebverbisses durch Rehwiid auf das
Hohenwachstum von Fichtenpflanzen // Ztschr.
Jagdwiss. 1983. Bd. 29, N 4. S. 235—243.
Keibel F. Zur Entwicklungsgeschichte des Rehes //
Anat. Anz. 1899. Bd. 16. P. 64—65.
Kerschagl W. Rehwildkunde. Wien: Hubertusver-
lag, 1952. 272 S.
Kioroglanidis J. Nahrungswahl und Oberlegungen
zur Nahrungsnische bei Reh (Capreolus capreolus),
Rothirsch (Cervus elaphus) und Gemse (Rupicapra
rupicapra) im Nationalpark Berchtesgaden: Diss.
Ludwig-Maxmilian-Univ. Munich, 1981. 175 S.
KWtzli F. Qualitat und Quantitat der Rehasung.
Bern: Huber, 1965. 186 S.
Knorr #., Briedermann L. Die Ernahrung des
Feldrehes // Jagd-informationen, Feldrehbewirt-
schaftung 1 (5 Jahrgang). Inst. Forstwissenschaften.
Eberswalde, 1976. S. 40—49.
Kolb A. Das Scheinasen des Rehes // Ztschr.
Jagdwiss. 1979. Bd. 25, N 4. P. 201—207.
Komdrek J. Lesnicka zoologie. Pr., 1954. 287 s.
Kbnig R., Hofmann R.R., Geiger G. Differentiall-
rhorphologische Untersuchungen der resorbierenden
Schleimhautoberfleche des Pansens beim Rehwiid
(Capreolus capreolus) im Sommer und Winter //
Ztschr. Jagdwiss. 1976. Bd. 22. S. 191т-196.
Kossak S. The complex character of the food
preferences of Cervidae and phytocenosis structure //
Acta theriol. 1976. Vol. 21, N 24/31. P. 359—375.
Kossak S. Hand-rearing and care of a group of
roe deer // Ibid. 1981a. Vol. 26, N 11. P. 207—219.
Kossak S. Development of food habits in
roe-deer // Ibid. 1981b. Vol. 26, N 33. P. 483—
494.
Kassak S. Trophic relations of roe deer in a
fresh decidious forest // Ibid. 1983. Vol. 28,
N 6. P. 83—128.
Kromschbder D. VerbissflSchen fur Schalen-
wild // Wild und Hund. 1982. Bd. 85, N 1. S. 42—45.
Kuen #., Bubenik A.B. Botanische Pansenanalysen
bei Rotwild (Cervus elaphus hippelaphus), Rehwiid
(Capreolus capreolus) und Gamswild (Rupicapra
rupicapra) // Alpine Umweltprobleme. Ergebnisse des
Forschungsprojekts Achenkirch. В., 1980. T.V. Beitr.
Umweltgestaltung. A. Bd. 67. S. 41—60.
Kumerloeve H. Die SSugetiere (Mammalia) der
TUrkei. Die SSugetiere (Mammalia) Syriens und
des Libanon // Verfiff. Zool. Staatssamml. MUnchen,
1975. Bd. 18. S. 69—225.
Kurt F. Feldbeobachtungen und Versuche fiber
das Revierverhalten der Rehbdcke (Capreolus
capreolus L.) // Rev. Suisse Zool. 1966. Bd. 73, N 3.
S. 408—421.
Kurt F. Zusammenhange zwischen Verhalten und
Fortpflanzungsleistung beim Reh (Capreolus
capreolus) // Ztschr. Jagdwiss. 1968a. Bd. 14, N 3. S. 97—
106.
Kurt F. Das Sozialverhalten des Rehes (Capreolus
capreolus L.). Hamburg; В.: Parey, 1968b 102 S.
Kurt F. Rehwiid. MUnchen: BLV Verl., 1970.
174 S.
Kurten B. Pleistocene mammals in Europe //
Striae. 1986. Vol. 24. P. 47—49.
Law R. Harvest optimization in populations with
age distributions // Amer. Natur. 1979. Vol. 114,
N 2. P. 250—259.
Lehmann E.R. Chevreuils d'Asia et d'Europe.
Contribution a I'etude du genre Capreolus //
Mammalia. Ю58. Vol. 22, N 2. P. 262—270.
Lehmann E. Warum brunftet das Reh im
Hochsommer? // Dt. Jager. 1960a. N 10. S. 87.
Lehmann E. Das Problem der Grossenabnahme
(Deminutions—Tedenz) beim Reh // Ztschr.
Jagdwiss. 1960b. Bd. 6, N 2. S. 41—51.
Lehmann E. Die Farbe des Haarkleides (Decke)
beim Europaisch-Vorderasiatischen Reh als taxo-
nomisches Hilfsmittel // Ibid. 1966. Bd. 12, N 1.
S. 5—11.
Lehmann E. Das Sommerkleid des Rehs im
Klimagefalle // Umschau. 1971. N 12. S. 428—429.
Lehmann E. Einige Bemerkungen zum Sibirischen
Reh (Capreolus capreolus (pygargus) caucasicus
Dinnik, 1910) in Mitteleuropa // Ztschr. Jagdwiss.
1976. Bd. 22, N 2. S. 75—84.
Lehmann E„ Sagesser H. Capreolus capreolus
Linnaeus, 1758 — Reh // Handbuch der Saugetiere
Europas. Wiesbaden, 1986. Bd. 2. S. 233—264.
Leibundgut H. Contribution au theme «Foret et
gibier» // Schweiz. Ztschr. Forstwiss. 1976. Bd. 127,
N 4. S. 227—236.
Lent P. Mother-infant relationships in ungulates //
JVCN Publ. N.S. 1971. Vol. 1, N 24. P. 14—
53.
Li Ch., Wu W., Qiu 2. Chinese neogene:
subdivision and correlation // Vertebrata Pal-Asiat.
1984. Vol. 22, N 3. P. 164—178.
Liepmann H. Rohe gegen Solanin unempfindlich? //
Dt. Jager-Ztg. 1972. Bd. 90, N 12. S. 488.
Lincoln G.A. The role of antlers in the behaviour of
389
red deer // J. Exp. Zool. 1972. Vol. 182, N 2.
P. 233—250.
Lincoln G.A., Guinnes F.E. Effect of altered
photoperiod on delayed implantation and moulting in
roe deer // J. Reprod. and Fert. 1972. Vol. 31,
N 3. P 455—457.
Lochman J. Spotfeba iivin u srnSi zvife //
Sb. CSAZV. Lesnictvi. 1961. Sv. 6. S. 551.
Lochman J. Pastevni rytmus s denni reiim jeleni a
srnii zvere /7 Pr. vyzk. listavu lesn. CSSR. 1965.
Sv. 30. S. 103—140.
Lochman J.. Barth A. Otazka Skod zpusobovanych
zveTFi s hlediska fysiologie vySivy sparkate zvife //
Zpravy les. vyzkumu. Zbraslav—Strnady, 1965. Sv. 11,
N 5/6. S. 16—17.
Lochman J., SeMkova E., Melicharova A. Vyuiivani
krmiv sparkatou zveTFi. III. Vysledky zjiStovani
koeficientu stravitelnosti a delky doby pruchodu
potravy zaZivacim traktem // Prace VULHM,
1974. Sv. 45. S. 39—60.
Lockie J.D. Deer and their habitat. Forestry
Suppl. Oxford, 1967. P. 21-J-27.
Lomnicki A. Modeling and optimization of roe
deer productivity // Pol. Ecol. Stud. (PPL). 1975.
Vol. 1, N 2. P. 137—142.
Long M. Fair play for roe deer // Scot. Forest.
1980. Vol. 34, N 2. P. 93—97.
Lorenz K. On aggression. N.Y., 1967. 306 p.
Loudon A.S.J. The control of roe deer populations:
A problem in forest management // Forestry.
1978. Vol. 51, N 1. P. 73—83.
Loudon A. Too many deer for the trees? //
New Sci. 1982. Vol. 93, N 1297. P. 708—711.
Macarovici N. Restes de Mammiferes fossiles de la
Bessarabie meridionale // Ann. sci. Univ. Jassy.
1936. Vol. 22, N 1/4. P. 349—367.
Manolache L., Babulia T. CercetJri asupra hranei
suplimentare la caprior si lepure. Buc, 1969.
80 p.
Maublanc M.L., Bideau £., Vincent J.P. Elexibilite
de lorganisation sociale du chevereuil en fonction des
caracteristiques de I'environnement // Rev. ecol. 1987.
Vol. 42, N 2. P. 109—133.
Mayer H. Waldstruktur—Wildstand—Wildscha-
den—Jagdtechnik // Wald und Wild Seminar. Bonn,
1977. S. 28—34.
McDiarmid A. Mortality in deer // Mammal.
Rev. 1974. Vol. 4, N 3. P. 75—78.
Meidel E. Rehwildhege und Waldschutz // Wild
und Hund. 1982. Bd. 85, N 13. S. 4—6.
Meikle I. Return of the roe // Wildlife.
1975. Vol. 17, N 1. P. 4—7.
Melichar J. Das Abasen der Waldvegetation durch
das Rot- und Rehwild // Ztschr. Jagdwiss. 1960.
Bd. 6. S. 78.
Melichar J. Vyiivne hodnoty v potrave dfevin pro
parohatou zveT? // Myslivost. 1961. Sv. 10.
S. 151 — 152.
Merlens С Zur Verhaltenssynchronisation bei
390
gruppenweise gehaltenen Rehen (Capreolus
capreolus) // Behaviour. 1984. Vol. 89, N 3/4. P. 220—240.
Meyer P. Territoriumsmarkierung beim Ren
und Morphologie des sogenannten Stirnorgans:
Inang.—Diss. Honnover, 1968. 54 S.
Meyer-Brenken H. Schwarzes Rehwild // Wild und
Hund. 1986. Bd. 89, N 9. S. 40—45.
Missbach K. Die Winterfiltterung des Rehwildes //
Wildbiologie und Wildbewirtschaftung. 1984. Bd. 3.
S. 515—525.
Mbhring G. Zur Beerenahrung des Rehes
(Capreolus capreolus L.) // Waldhygiene. 1963. Bd. 5.
S. 68—69.
Mold S. Bonitace honbiSte se srnji zvefi //
Pr. vjzk. listavu lesn. CSSR. 1954. Sv. 7. S. 97—129.
Mottl S. Potrava srnfi zvere (Capreolus capreolus
capreolus L.) // Biologia. 1957a. Vol. 12, N 1.
S. 29—43.
Mottl S. Die Jagdwirtschaftlich erforderliche
Mindestgrope von Rehwildrevieren im Wald //
Ztschr. Jagwiss. 1957b. Bd. 3. S. 64—69.
Mottl S. Zur Frage der Wilddichte und der
Qualitat des Rehwildes // Beitr. Jagd- und Wildforsch.
1962. Bd. 11. S. 35—40.
Mottl S.. Janda M. NJktere vysledky studia о
vlivu prostfedi na srn£i zve? // Sb. Cs. akad.
zemedSls. ved. r. les. 1955. Sv. 28, N 4. S. 613—628.
Miiller H.J. Vorschlag zur Bestimmung der
Wilddichte nach den Asungsverhaltnissen der Jagdgebie-
te // Unsere Jagd. 1964. Bd. 14, N 25/28.
S. 51—56.
Miiller H.J. Untersuchungen zur Beurteilung der
wirtschaftlich tragbaren Schalenwilddichte im Wald.e
nach Wildschaden und Standort // Arch. Forstwiss.
1965. Bd. 14. S. 533—561.
Miiller K. Okologische Untersuchungen der Liege-
zonen beim Reh (Capreolus capreolus) // Rev.
Suisse Zool. 1973. Bd. 80, N 3. S. 667—670.
Miiri H. Beobachtungen und Experimente zum
Futterlernverhalten des Rehs (Capreolus capreolus) //
Ztschr. Saugetierk. 1978. Bd. 43, N 3. S. 171 — 186.
Miiri H. Futter-Tradition beim Reh. Individuelle
und soziale Mechanismen bei der Annahme und
der Vermeidung unbekannter Futtermittel: Diss.
Univ. Zurich, 1983. 98 S.
Musi /•'., Perco F. Alcune osservacioni sulla alimen-
tazione e sul comportamento des capriolo (Capreolus
capreolus L.) nei cansiglio orientale // Monti e
boschi. 1975. Vol. 26, N 5/6. P. 49—55.
Myrbirget S. Merkingsresultate XVI. Returns of
game species marked by the Norwegian Hunting and
Fishing Association, 1947—1962 // Medd. States
Viltundersokelser. Ser. 2. 1965. N 21. P. 1—24.
Myrberget S. Merking av radyr i Norge //
Fauna (Norge). 1973. Bd. 26, N 2. S. 97—101.
Myrberget S., Milton R. Capreolus capreolus
with 5 embryos // Fauna Norsk zool. foren.
tidsk. 1971. N 1. P. 67.
Nagy J.G., Regelin W.L. Comparison of digestive
organ size of three deer species // J. Wildlife
Manag. 1975. Vol. 39, N 3. P. 621—624.
Ne?as J. Pfispevek к paznani poietnosti, skladby
a organizace tlup srnJi zvere // Pr. vyzk. listavu
lesn. 1960. Sv. 20. S. 111 — 142.
Nefas J. Srnci zver. Pr.: SZN, 1963. 283 s.;
1975. 302 s.
Nei M. Genetic destance between populations //
Amer. Natur. 1972. Vol. 106. P. 283—292.
Neubert E. Beschlagenes Rehkitz // Unsere Jagd.
1970. Bd. 20, N 7. S. 212.
Nikolandic D. EkoloSke karakteristike populacije
srna na Belju // Jelen. 1968. Sv. 7. S. 73—95.
Nobis G. Zur Fauna der Fruhneolithischen Siedlung
Ovcarovo gorata, Bez. Targoviste (No—Bulgarien)//
Bonn. Zool. Beitr. 1966. Bd. 37, N 1. S. 1—22.
Novdkovu E.. Hanzl R. Pfispevek к urEeni
potencialu krajiny pro chov njkterych druhu zvefe //
Quaest. geobiol. 1974. Sv. 13. S. 5—82.
Nowak W. Methoden der Jungwildrettung //
Wild und Hund. 1981. Bd. 84, N 3. S. 150—153.
Nudds T.D. Forage "preference": theoretical
consideration of diet selection by deer // J. Wildlife
Manag. 1980. Vol. 44, N 3. P. 735—739.
Obrtel R., Holtfovd K, KoZenu I. The winter diet of
sike deer (Cervus nippon) in the Bouzovsko area //
Folia zool. 1985. Vol. 34, N 1. P. 1—22.
Oszlunyi J. Wood, bark, needles, leaves and
roots energy values of Pinus silvestris L., Picea excelsa
Link, and Fagus silvatica L. // Ekologia (CSSR).
1982. Vol. 1, N 3. P. 289—296.
Padaiga V. Okologie und Prinzipien der wirt-
schaftlichen Nutzung des Rehwildes (Capreolus
capreolus) in den Waldern des stldlichen Teils der
baltischen Sowjetrepubliken // Beitr. Jagd- und
Wildforsch. 1975. Bd. 9. S. 276—287.
Papageorgiou N.. Neophytou C, Spais A., Vavale-
kas C. Food preferences and protein and energy
requirements for maintenance of roe deer // J.
Wildlife Manag. 1980. Vol. 45, N 3. P. 728—732.
Pavlov M. Nouweaux mammiferes tertiaires
trouves en Russie // Bull. Soc. Imp. natur. Moscou,
1896. N 2. P. 12.
Pedroli J.C., Blant M., Chappuis F., M'aire J.A. La
biologie du chevreuil Capreolus capreolus (L.)
dans le canton de NeuchStel (Suisse). I. Recensement de
la population; comparaison de deux methodes //
Mammalia. 1981. Vol. 45, N 4. P. 467—472.
Pelosse J. Etude sur le comportement diattention
chez une population de chevreuils (Capreolus
capreolus) // Zool. Gart. 1976. Vol. 46, N 6. P. 441—451.
Pereira M.S.R. Effect of human and wolf
(Canis lupus) presence on a roe deer (Capreolus
capreolus) population in Northeastern Portugal (Sierra da
Nogueira) // XVII Congr. Intern. Union Game Biol.
Brussels, 1985. Pt. 2. P. 671—678.
Perzanowski K. The effect of winter food
composition on roe deer energy budget // Acta theriol.
1978. Vol. 23, N 31. P. 451—468.
Petrusewicz K. Dynamics, organization and
ecological structure of population // Ecol. Pol. Ser. A. T.
1966. Vol. 14, N 25. P. 412—436.
Picard J.F., Caburel A., Oieffe P. Etude du
regime alimentaire automnal et hivarnal du Cerf
(Cervus elaphus L.) et du Chevreuil (Capreolus
capreolus L.) par l'analyse des contenus stomacaus //
XVII Congr. Intern. Union Game Biol. Brussels,
1985. Pt. 1. P. 439—446.
Peilowski Z. Sarna. W-wa: PWRiL, 1970. 220 s.
Pielowski Z. Das Feldreh — eine Bereicherung
der Wildtierfauna unserer Feldreviere // Wildbiologi-
sche Informationen flir den Jager. Stuttgart, 1983.
Bd. 4. S. 29—35.
Pielowski Z. Some aspects of populations
structure and longevity of field roe deer // Acta theriol.
1984. Vol. 29, N 2. P. 17—33.
Pielowski Z, Bresinski W. Population
characteristics of roe deer inhabiting a small forest //
Ibid. 1982. Vol. 27, N 28. P. 409—425.
Pikula J., Koubek P., Kratochvil Z, Kux Z.
Age composition of roe deer population in
Czechoslovakia // Acta Sci. natur. 1985a. Vol. 19, N 5.
P. 1—46.
Pikula J., Koubek P., Kratochvil Z, Kux Z.
Reproduction of roe deer population in
Czechoslovakia // Ibid. 1985b. Vol. 19, N 6. P. 1—47.
Pomel A. Catalogue methodique et descriptif des
Vertebres fossiles decouverts dans le bassin hydro-
graphique sJperieur de la Loire, et surtout dans la
vallee de son affluent principal, l'Allier // Auvergne,
Ann. sci. P., 1853. T. XXVI. P. 81—229.
Pond K.R.. Ellis W.C.. Akin A.E. Ingestive
mastication and fragmentation of forages // J. Anim.
Sci. 1984. Vol. 58, N 6. P. 1567—1574.
Popovici N.. Crisan A., Gotea J. Modificari
sezoniere ale structurii ovarului la caprioara
(Capreolus capreolus L.) // Stud. Univ. Babes-Bolyai.
Ser. Biol. 1978. Vol. 23, N 2. P. 22—32.
Prieditis A. Influence of dry food and needles on
body weight and consumption of food substances in
roe deer, Capreolus capreolus L. // Acta zool.
fenn. 1984. Vol. 171. P. 213—216.
Prior R. The roe deer of Cranborne Chase:
An ecological survey. L.: Oxford Unid. press,
1968. 119 p.
Prior R. Roe stalking. Dorset: Bourne press,
1987. 130 p.
Pulliainen E. Occurence and spread of the roe deer
(Capreolus capreolus L.) in eastern Fennoscandia
since 1970 // Mem. Soc. fauna et flora fenn. 1980.
Vol. 56, N 1. P. 28—32.
RaesfeldF. Das Rehwild. Hamburg; В.: Parey, 1956.
328 S. (1965; 1970; 1978).
Ralcli/fe P.R., Rowe J.J. A biological basis for
managing red and roe deer in British commercial
forests // XVII Congr. Intern. Union Game Biol.
Brussels, 1985. Pt. 2. P. 917—925.
ReichholfJ. Jahreszeit — und Biotopabhangigkeit
391
der Rudelbildung beim Rehwild (Capreolus
capreolus L.) // Spixiana. Suppl. 1980. Vol. 3, N 2.
S. 193—208.
Reimoser F. Rehwildbejagung in einem dec-
kungsreichen Gebirgsrevier bei waldbaulicher Bet-
riebsumstellung // Zentr.-Bl. ges. Forstwiss. 1982.
Bd. 99, N 3. S. 167—170.
Reimoser F. Rehwildmarkierung in Niederoster-
reich // Osterr. Weidwerk. 1984. N 5. S. 6.
Reimoser /■'., Zand/ J. Zwischenergebnisse der
n—6. Rehwildmarkierung//Ibid. 1986. N 5. S. 6—10.
Reimoser /•"., Zand/ J. Rehwildmarkierung //
Ibid. 1987. N 5. S. 4—5.
Rieck W. Die Setzzeit bei Ren-, Rot- und Damwild
in Mitteleuropa // Ztschr. Jagdwiss. 1955. Bd. 2,
N 1. S. 6*—75.
Robin K. Raumliche Verschiebungen von markier-
ten Rehen (Capreolus capreolus L.) in einem
voralpinen Gebiet der Ostschweiz // Ibid. 1975.
Bd. 21, N 3. S. 145—163.
Roucher /■'., Turcheim В., Turcheim E. Roe deer and
trees joint management on an overcrowded woodland
estate in Northern Vosges, France // XVII Congr.
Intern. Union Game Biol. Brussels, 1985. Pt 1.
P. 415—420/
Riiedi K. Wildschadenbekampfung in Wald durch
Verbesserung der Asungverhaltnisse // Schweiz.
Jagd. Ztg. 1956. S. 14—18.
Rusterhotz M., Turner D.C. Versuche uber die
"NShrstoffweisheit" beim Ren (Caprfeolus
capreolus) // Rev. Suisse Zool. 1978. Bd. 85, N 4. S. 718—730.
Sagesser H. Ober den Einflup des Standortes auf
das Gewicht des Rehwildes (Capreolus с capreolus
/Linne 1758/) // Ztschr. Jagdwiss. 1966a. Bd. 12, N 2.
S. 54—62.
Sagesser H. Ober den Einfluss der Hohe auf
einige biologische Erscheinungen beim Reh
(Capreolus с capreolus) und bei der Gemse (Rupicapra
r. rupicapra) // Rev. Suisse Zool. 1966b. Bd. 73, N 2.
S. 422—433.
Sagesser H. Analyse der Setzzeiten (1965—1967)
beim Reh im schweizerischen Mittelland // Beitr.
Jagd- und Wildforsch. 1968. Bd. 6. S. 35—46.
Sagesser H., Kurt F. Ober die Setzzeit 1965
beim Reg (Capreolus с capreolus /L./) // Mitt.
Natur. Ges. Bern, 1966. Bd. 23. S. 21—38.
Savage D.E., Russell D.F.. Mammalian Palaeofau-
na of the World. L.: Addison-Wisley, 1983. 432 p.
Schafer E. Hegen und Ansprechen von Rehwild.
Munchen: BLV, 1974. 189 S.
Schaffer ./. Die Hautdriisenorgane der Saugetiere
mit besonderer Beriicksichtigung ihres histologishen
Aufbaues und Bemerkungen uber die Proktodaaldrii-
sen. В.; Wien: Urban und Schwarzenberg, 1940.
464 S.
Schams /)., Barlh D. Annual profiles of
reproductive hormones in peripheral plasma of the male roe
deer (Capreolus capreolus) // J. Reprod. and Fert.
1982. Vol. 66, N 2. P. 463—468.
392
Sehlosser M. Die fossilen Saugethiere Chinas,
nebst einer Odontographie der recenten Antilopen //
Abh. Bayer Akad. Wiss. 1903. Bd. 2, N 22.
S. 1—221.
Sehlosser M. Tertiary vertebrates from
Mongolia // Palaeontol. sinica. Ser. С Peking. 1924.
Vol. 1, N 1. 119 p.
Schroder J., Schroder W. Niche breadth and overlap
in red deer (Cervus elaphus L.), roe deer (Capreolus
capreolus) and chamois (Rupicapra rupicapra) //
III Intern, theriological Congr. Abstr. Pap. Helsinki,
1982. P. 222.
Schumacher S. Jagd und Biologie. В., 1939.
136 S.
Sedlaczek E. Zehn Jahre Hege im Rehwildrevier//
Pirsch. 1964. N 5. S. 148—155.
Seibe A. Futtermittel fur Rehe, Hirsche, Hasen od.
dgl.: Pat BRD, Kl. 53 d 4/04, (A 23 к 1/14),
N 1692488.
Sempere A. Utilisation et evolution du domaine
vital chez le chevreuil male europeen determiners
par radiotracking // Biol. Behav. 1979. Vol. 4,
N 1. P. 75—87.
Sempere A. Fonction de reproduction et caracteres
sexuels secondaires chez le chevreuil (Capreolus
capreolus L.): variations saisonnieres et incidences sur l'uti-
lisation du budget temps—espace. Frangois—Rabelais
(Tours), 1982. 306 p.
Shen S., Abies E.D., Qian-zhu X. The Chinise view
of wildlife // Oryx. 1982. Vol. 16, N 4. P. 340—347.
Short R.V., Hay M.F. Delayed implantation in
the roe deer Capreolus capreolus // Сотр. Biol.
Reprod. Mammals: Symp. Zool. Soc. L., 1966.
Vol. 15. P. 173—194.
Short R.V., Mann T. Androgenic activity in a
seasonally "breeding animal, the roe buck (Capreolus
capreolus) // J. Endocrinol. 1965. Vol. 31, N 19/20.
P. 1—2.
Short R.V.. Mann T. The sexual cycle of a
seasonally breeding animal, the roe buck (Capreolus
capreolus) // J. Reprod. and Fert. 1966. Vol. 12, N 2.
P. 337—351.
Siefke A. Die jagdwirtschaftliche Bedeutung para-
sitischer Wiirmer beim Reg- und Damwild //
Beitr. Jagd- und Wildforsch. 1966. Bd. 4. S. 135—
145.
Simpson G. The principles of classification and
a classification of Mammals. N.Y., 1945. 350 p.
(Bull. Amer. Mus. Natur. Hist. Vol. 85).
Siuda A.. Zurowski W.. Siuda H. The food of the
roe deer // Acta theriol. 1969. Vol. 14, N 18.
P. 247—262.
Smith /■.. BeesonK.. Price W. Chemical composition
of herbage browsed by deer in two wildlife
management areas // J. Wildlife Manag. 1956.
Vol. 20, N 4. P. 359—367.
Sokolov V.E.. Danilkin A.A. Einige Aspekte der
Systematik, Okologie und des Verhaltens des Sibiri-
schen Rehs (Capreolus capreolus pygargus, Pallas,
1771) // Beitrage Jagd- und Wildforsch. 1980.
Bd. 11. S. 276—288.
Southern H.N. The handbook of British
mammals. Oxford, 1964. 465 p.
Stangl W.G., Margl H. Rehwildstandserhebungen
in einem Mittelwaldbetrieb im Weinviertel // Mitt.
Forstl. Bundes-Versuch. Wien, 1977. N 122. S. 97—
113.
Slang/ W.G., Wenter W. Versuch einer Darstel-
lung der Wildstandsentwicklung auf Grund von
Abschussmeldungen bei Rehwild // Ibid. 1977.
N 122. S. 63—83.
Stark D. Territoriumsmarkierung bei Rehbocken
(Capreolus capreolus) // Saugetierk. Mitt. 1966.
Bd. 14, N 4. S. 325—326.
Sterba F., Zamek L. Parazitarni pfiJiny ztrat
srnfi zvefe v letech 1972—1981 // Veterinarstvi.
1985. Sv. 35, N 3. S. 131—135.
Stieve H. Anatomisch-biologische Untersuchungen
iiber die Fortpflanzungsfahigkeit des europaischen
Rehes // Ztschr. Mikrosk. und Anat. Forsch.
1950. Bd. 55. S. 427—530.
Strandgaard H. The roe deer (Capreolus capreolus)
population at Kalo and the factors regulating
its size // Dan. Rev. Game Biol. 1972a. Vol. 7, N 1.
P. 1—205.
Strandgaard H. An investigation of Carpora
lutea, embrionic development, and time of birth of
roe deer (Capreolus capreolus) in Denmark //
Ibid. 1972b. Vol. 6, N 7. P. 1—22.
Strandgaard H. Rehbestand und Regulation der
Rehdichte auf Kalo (Ost.-Jutland) // Allg. Forstztschr.
1975. Bd. 30. S. 1129—1131.
Strandgaard H. Untersuchungen iiber danisches
Rehwild in seiner Beziehung zur Umwelt // Beitr.
Jagd- und Wildforsch. 1977. Bd. 10. S. 234—243.
Strandgaard H. Densite population et croissance
des animaux chez le chevreuil: perspectives nouvelles //
Bull. Mens. Off. Nation. Chasse. No Sp. Scien. Tech.
Nov. 1978. P. 149—159.
Strothman A., Oslage H.-J. Untersuchungen zur
Jahresrhytmik der Nahrstoff- und Energieaufnahme
von Rehwild // Kleintierpraxis. 1986. Bd. 31,
N 3. S. 153—156.
Stubbe С Untersuchungen iiber einige die Qualitat
des Rehwildes beeinflussende Faktoren // Beitr.
Jagd- und Wildforsch. 1966. Bd. 5. S. 155—172.
Stubbe С Die Bestandesermittlung bei Rehwild im
Wald // Unsere Jagd. 1979. Bd. 29, N 2. S. 44—45.
Stubbe С Das Rehwild in unserer Kultur-
landschaft // Ibid. 1987. Bd. 37, N 5. S. 136—137.
Stubbe C, Smirnow M.N. Korperwachstum und
Korpergrbpe des Sibirischen Rehwildes (Capreolus
с pygargus Pallas, 1771) und Unterschiede zum
Europaischen Ren (Capreolus с capreolus L.) /7
Zool. Garten. N.F. 1972. Bd. 42, N 3/4. S. 166—175.
Stubbe C, Passarge H. Rehwild. В.: VEB Dt.
Landwirtsch.-Verl., 1979. 432 S.
Stubbe С Stubbe M„ Stubbe I. Zur Reproduktion
der Rehwildpopulation — Capreolus с capreolus
(L., 1758) — des Wildforschungsgebietes Hakel //
Hercynia. 1982. Bd. 19, N 1. S. 97—109.
Stubbe C. Stubbe W. Ergebnisse der Rehwildmar-
kierung im Wildforschungsgebietes Hakel // Unsere
Jagd. 1985. Bd. 35, N 6. S. P2—173; N 12.
S. 355.
Stubbe C, Zbrner H. Zum Bevorzugungsgrad
einiger GemUse-, GewUrz- und Arzneipflanzen durch
Rehwild // Beitr. Jagd- und Wildforsch. 1977.
Bd. 10. S. 222—226.
Stubbe #., Bruchholz S. Cber Bastardierungsver-
suche zwischen Europaischen und Sibirischen Rehen
(Capreolus capreolus L., 1758 x Capreolus
capreolus pygargus Pallas, 1771) // Ibid. 1980. Bd. 11.
S. 289—303.
SysaP.S., KaluzinskiJ. Possibility of freemartinism
in roe deer // Acta theriol. 1984. Vol. 29, N 1/10.
P. 133—137.
Szaniawski A. Proportions of the alimentary
tract in deer // Ibid. 1973. Vol. 18, N 18.
p. 343—347.
Szederjet A., Szederjei M. Geheimnis der Wel-
trekordes — das Reh. Budapest: Terra, 1971.
403 S.
Szmidt A. Food preference of roe deer in
relation to principal species of forest trees and
shrubs // Acta theriol. 1975. Vol. 20, N 20.
P. 255—266.
Tabbush P. Roe deer control and restocking in the
border forests // Scot. Forest. 1979. Vol. 33,
N 4. P. 290—294.
Thenius E. Ober die Entwicklung des Hornzapfens
von Miotragocerus // Anz. Osterr. Akad. Wiss.
Math, naturwiss. Kl. 1948. Bd. 157. S. 203—221.
Thenius E. Die Grossaugetiere des Pleistozans von
Mitteleuropa // Ztschr. Saugetirk. 1962. Bd. 27.
S. 65—83.
Tschiderer K. Aufzucht-, FUtterungs- und Hege-
versuche beim Rehwild (Capreolus capreolus L.) //
Ztschr. Jagdwiss. 1973. Bd. 19, N 4. S. 198—204.
Tschiderer K. "Neydhartiger Moor" im Einsatz
gegen Verbiss und Schalschaden bei Reh- und
Rotwild // Bodenkultur. 1978. Bd. 29, N 3.
S. 277—287.
Tschirch W. Zwolf Jahre praktische Parasiten-
bekampfung bei Europaischem und Sibirischem
Rehwild im Wildgatter Rothenburg // Beitr. Jagd- und
Wildforsch. 1984. Bd. 13. S. 286—289.
Turcek E.J. Effect of introductions on two
game populations in Czechoslowakia // Wildlife
Manag. 1951. Vol. 15, N 1. P. 113—114.
Turcek F.J. Untersuchungen Uber das Fegen
der Rehbtjcke an Geholzen // Ztschr. angew. Zool.
1962. Bd. 49, N 4. S. 429—438.
Turner D.C. Aktivitatsmuster freilebender Rene
im Verlauf des Fruhjahrs: optimale Ausnutzung der
393
Tageszeit // Kev. Suisse Zool. 1978. Bd. 85,
N 4. S. 710—712.
Turner D.C. An analysis of time — budgeting
by roe deer (Capreolus capreolus) in an agricultural
area // Behaviour. 1979. Vol. 71, N 3/4. P. 246—279.
Turner D.C. A multi-variate analysis of roe
deer (Capreolus capreolus) population activity // Rev.
Suisse Zool. 1980. Vol. 87, N 4. P. 991 — 1002.
Ueckermann D. RehwildbestandsermitUung anhand
des Verzehrs an Winterfiitterungen // Ztschr.
Jagdwiss. 1986. Bd. 32, N 2. S. 105—122.
Ueckermann E. Die Einwirkung des Standortes auf
Korpergewicht und Gehornbildung des Waldrehes:
Diss. Hann. Munchen, 1951. 150 S.
Ueckermann E. Rehwild und Standort // Anblick.
1952. Bd. 7. S. 32.
Ueckermann E. Das Rehwild auf Fehmarn //
Wild und Hund. 1956. Bd. 48. S. 376—377.
Ueckermann E. Wildstandsbewirtschaftung und
Wildschadensverhutung beim Rehwild. Neuwied,
1957. HIS.
Ueckermann E. D^er Rehwildabschup. Hamburg;
В.: Parey, 1964. 109 S.
Ueckermann E. Forstwirtschaft und Wild //
XVII Congr. Intern. Union Game Biol. Brussels, 1985.
Pt. 1. P. 371—383.
Ulbricht С Natiirliche Asung bevorzugt. Beo-
bachtungen bei der Rehwildfiitterung // Unsere
Jagd. 1986. Bd. 36, N 10. S. 301.
Ullrey D., Jouall W., Johnson #., Fay L. Protein
regirement of white-tailed deer fawn // J. Wildlife
Manag. 1967. Vol. 31, N 4. P. 679—685.
Velich А. О "masozrave" srn? // Stra? myslivosti.
Trebic. 1925. Sv. 3, N 10. S. 131 — 133.
Vincent J.P. SchStzung der Rehwildpopulation
im Walde. Vorstellung einer einfachen Methode //
Ztschr. Jagdwiss. 1982. Bd. 28, N 1. S. 58—63.
Vincent J.P.. Bideau E. Influence de la densite
sur 1'occupation de Г espace chez le chevreuil
forestier // Bull. SFECA. 1987. Vol. 2, N 1.
P. 165—168.
Vincent J.P., Bideau E.. Quere J.P., Angibault J.M.
Occupation de I'espace chez le chevreuil (Capreolus
capreolus L.). II. Cas des femelles // Acta oecol.
Oecol. appl. 1983. Vol. 4, N 4. P. 379—389.
Vincent J.P., Janeau C. Quere J.P., Spitz /■'.
Note sur la repartition et le rythme d'activite
du chevreuil (Capreolus capreolus) en foret ouverte//
Ann. Zool. Ecol. Anim. 1979. Vol. 11, N 2.
P. 145—157.
Viret J. Artiodactyla // Treite de paieontologie.
P.: Masson, 1961. T. 6. Vol. 1. P. 887—1027.
Vos A.. Brox P., Geist V. A review of social
behaviour of the North American Cervids during the
reproductive period // Amer. Midland Natur.
1967. Vol. 77, N 2. P. 390—417.
Voser-Huber MX., Nievergelt B. Das Futter-
wahlverhalten des Rehes in einem voralpinen Revier //
Ztschr. Jagdwiss. 1975. Bd. 21, N 4. S 197—215.
394
Wagenknecht E. Reifealter, Zielalter, Ermealter
beim Schalenwild // Beitr. Jagd- und Wildforsch.
1971. Bd. 7. S. 77—95.
Wagner H.A.- Waldbau und Wild // Anblick.
1961. Bd. 16. S. 237—240.
Walther /•'. Zum Kampf- und Paarungsverhalten
einiger Antilopen // Ztschr. Tierpsychol. 1958.
Bd. 15. N 3. S. 340—380.
Walther /■'. Einige Verhaltensbeobachtungen am
Dibatag (Ammodoreas clarkei. Thomas, 1891) //
Zool. Garten. N.F. 1963. Vol. 27. P. 233.
Walther I'. Verhalten der Gasellen. Wittenberg;
Zutherstadt, 1968. 144 S. (Die Neue Brehm Bucherei;
Bd. 373).
Walther I'.R. Territorial behaviour in certain
horned ungulates, with special reference to the
examples of Thomson's and Grant's gazelles // Zool.
Afr. 1972. Vol. 7, N 1. P. 303—307.
Wandeler A.I. Die Fortpflanzungsleistung des Rehs
(Capreolus capreolus L.) im Berner Mittelland // Jb.
Natur. Hist. Mus. Bern. 1972—1974. 1975. Bd. 5.
S. 245—301.
Wandeler A., Huber W. Gewichtswachstum und
jahreszeitliche Gewichtsschwankungen bei Reh und
Gemse // Rev. Suisse Zool. 1969. Bd. 76, N 3.
S. 686—694. ,
Weiner J. Model of the energy budget of an
adult roe deer // Pol. Ecol. Stud. 1975. Vol. 1,
N 2. P. 103—119.
Winter W. Ein praktischer Kraftfiitterautomat //
Pirsch. 1980. Bd. 33. N 19. S. 1319.
Wishart W. Hybrids of white-tailed and mule
deer in Alberta // J. Mammal. 1980. Vol. 61, N 4.
P. 716—720.-
Wurster D.. Benirschke K. The chromosomes of
twenty three species.of the Cervoidea and Bovoidea //
Mammal. Chromosomes Newslett. 1967. Vol. 8.
P. 226—228.
Wynne-Edwards V.C. Animal dispersion in
relation to social behaviour. Edinburgh; L., 1962. 653 ji.
Wynne-Edwards V.C. Population control in
animals // Sci. Amer. 1964. Vol. 211, N 2. P. 68—74.
Young C.C. On the fossil vertebrate remains
from localities 2,7 and 8 at Choukoutien //
Palaeontol. sin. Ser. С 1932. Vol. 7. N 3. P. 1 —
24.
Zdanskv O. Fossile Hirsche Chinas // Ibid.
1925. Vol.'2, N 3. P. 1—90.
Zejda J. Field groupings of roe deer (Capreolus
capreolus) in a lowland region // Folia zool.
1978. Vol. 27, N 2. P. 111 — 122.
Zejda J. Road strip transects for estimating
field roe deer density // Ibid. 1984. Vol. 33,
N 2. P. 109—124.
Zejda J. Field transects for roe deer census //
Ibid. 1985. Vol. 34, N 3. P. 209—215.
Zejda J.. Bauerovd Z. Home ranges of field roe
deer // Acta sci. natur. 1985. Vol. 19, N 1. P. 1—43.
Zejda J., Homolka M. Habitat selection and popu-
latiori density of field roe deer (Capreolus capreolus)
outside the growing season // Folia zool. 1980.
Vol. 29, N 2. P. 107—115.
Zejda J., Rebifkovd M., Homolka M. Study of
behaviour in field roe deer (Capreolus capreolus) //
Acta sci. natur. 1985. Vol. 19, N 12: P. 1—37.
Zernahle K. Zytogenetische Untersuchungen am
Europaischen Rehwild (Capreolus с capreolus L.,
1758), Sibirischen Rehwild (Capreolus с pygargus
Pallas, 1771) und deren Bastarden // Beitr. Jagd-
und Wildforsch. 1980. Bd. 11. S. 304—309.
Zhang T. Characteristics of mammal fauna and
ecogeography in Beijing area // Acta theriol. sin.
1984. Vol. 4, N i. P. 187—195.
Zfiuge Y. On the geographical distribution and
the mammalian fauna of Zhejiang province //
Ibid 1982. Vol. 4, N 2. P. 157—166.
Ziegler E. Beobachtungen Uber die Brunst und
den Embryo der Rene. Hannover, 1843. 87 S.
Zima J., Havrdnkovd J. Charakter prouzkovani
chromozomu v karyotypu srnce obecneho // Vet. med.
(CSSR). 1987. Ro6. 32 (60), N 1. S. 53—
58.
Zivkovic S., Isakovic J. Analiza kariotipa divljaci
kao metod genetiEkogo testiranija pri jihovom
unoSenju u loviSta // Simp, lovstvu. Beograd,
1972. S. 104—112.
Оглавление
От редактора S
Предисловие 6
Глава первая, Филогения, эволюция, систематика QE.JI. Короткевич,\А.А. Данилкин) 8
Глава вторы. Изменчивость и таксономия 22
История исследований (А.А. Данилкин) 22
Морфометрический анализ (А.А. Данилкин, Г.Г. Марков, К. Штуббе, А.Ю. Стручков) 25
Окраска как таксономический признак (А.А. Данилкин) 43
Цитогенетическая изменчивость (А.А. Данилкин) 47
Биохимический и иммунологический анализ (Г.Г. Марков, А.В. Шурхал, А.А. Данилкин) SO
Гибридологический анализ (А.А. Данилкин) 55
Таксономия v Capreolus в свете современных исследований (В.Е. Соколов, А.А. Данилкин,
Г.Г. Марков) 60
Глава третья. Ареал (А.А. Данилкин) 64
История ареала рода Capreolus 64
Ареал в XX'веке 68
Видовые ареалы европейской и сибирской косуль 76
Глава четвертая. Среда обитания (Я. Зейда, А.А. Данилкин) 86
Основные биотопы 86
Факторы, влияющие на биотопическое размещение 94
Приспособленность популяций к определенным биотопам и их изменениям 97
Глава пятая. Перемещения (А.А. Данилкин) 101
Оседлость и миграции 101
Естественное и искусственное расселение 119
Глава шестая. Питание (В. Голишова, Р. Обртел, И. Кожена, А. Данилкин) 124
Общая характеристика питания 124
Выбор корма и избирательность питания 126
Основные и второстепенные корма 127
Видовые, региональные и биотопические аспекты питания 129
Сезонные особенности питания 131
Количество потребляемой пищи 135
Минеральное питание и потребление воды 137
Глава седьмая. Размножение (К. Штуббе, А.А. Данилкин) 140
Половая зрелость 140
Спаривание 142
Беременность 146
Отел .'.. 149
Воспроизводство 153
Глава восьмая. Структура популяций (А.А. Данилкин) 160
Половая и возрастная структура 160
Социальная организация 165
Пространственно-этологическая структура 168
Глава девятая. Поведение (А.А. Данилкин) 185
Онтогенез и материнское поведение 185
Агрессивное поведение 195
396
Маркировочное поведение и химическая коммуникация 214
Звуковая сигнализация 226
Половое поведение 232
Ориентировочно-исследовательское 'и оборонительное поведение 239
Активность 243
Глава десятая. Численность (А.А. Данилкин. П.П. Блузма) 247
Численность и плотность населения 247
Факторы, определяющие численность населения популяций 258
Глава одиннадцатая. Хозяйственное значение, рациональное использование ресурсов и охрана
(К. Штуббе, А. Данилкин) 276
Управление популяциями и рациональное использование ресурсов 278
Охрана и восстановление численности .... 329
Приложение 337
Литература 369
Contents
From Editor 5
Foreword 6
Chapter one. Phylogeny, evolution and systematics (E.L. Korolkevich, A.A. Danilkin) 8
Chapter two. Variability and taxonomy 22
History of research (A.A. Danilkin) 22
Morphometric analysis (A.A. Danilkin, G.G. Markov, C. Stubbe, A. Yu. Struchkov) 25
Coloration as a taxonomic character (A.A. Danilkin) 43
Cytogenetics variability (A.A. Danilkin) 47
Biochemical and immunological analysis (G.G. Markov, A.V. Shurkhal, A.A.
Danilkin) 50
Hybridological analysis (A.A. Danilkin) ч ■. 55
Taxonomy of Capreolus in the light of modern research (V.E. Sokolov, A.A. Danilkin,
G.G. Markov) 60
Chapter three. Range (A.A. Danilkin) 64
History of the range of the genus Gapreolus 64
Range in the 20th century 68
Species ranges of the European and Siberian roe deer 76
Chapter four. Environment (J. Zejda, A.A. Danilkin) , 86
Main habitats 86
Factors affecting habitat distribution 94
Adaptation of populations to definite hahitats and their changes 97
Chapter five. Relocations (A.A. Danilkin) 101
Sedentary mode of life and migrations 101
Natural and artificial dispersal 119
Chapter six. Feeding (V. HolUSovd, R. Obrtel, I. Kofend, A. Danilkin) 124
General characterization of feeding 124
Choice of forage and selectiveness of feeding 126
Main and secondary forages 127
Species, regional and habitat aspects of feeding 129
Seasonal properties of feeding 131
The amount of food consumed 135
Mineral nutrition and water consumption 137
Chapter seven. Breeding (C. Stubbe, A.A. Danilkin) 140
Sexual maturity 140
Mating 142
Gestation 146
Parturition 149
Regeneration 153
Chapter eight. Populations structure (A.A. Danilkin) 160
Sex and age structure 160
Social organization 165
Spatial and ethological structure 168
Chapter nine. Behaviour (A.A. Danilkin) 185
Ontogenesis and maternal behaviour 185
398
Aggressive behaviour 195
Marking behaviour and chemical communications 214
Acoustical signalling 226
Sexual behaviour 232
Orientation-exploratory and defensive behaviour 239
Activity 243
Chapter ten. Numbers (A.A. Danilkin, P.P. Blusmd) 247
Numbers and population density 247
Number factors 258
Chapter eleven. Economic importance, rational use of resources and conservation (C. Stubbe,
A. Danilkin) 276
Population management and rational use of resources 278
Conservation and restocking of populations 329
Supplement 337
References 369
Научное издание
Европейская
и сибирская
косули
Систематика, экология,
поведение,
рациональное использование
и охрана
Утверждено к печати
Институтом эволюционной морфологии
и экологии животных
им. А.Н. Северцова
Российской академии наук
Редактор издательства Г.М. Орлова
Художественный редактор Н.Н. Михайлова
Технический редактор Г.П. Каренина
Корректор Н.Л. Голубцова
Набор выполнен в издательстве
на электронной фотонаборной системе
Подписано к печати 10.09.92
Формат 70x90 1/16. Бумага этикеточная
Гарнитура Тайме. Печать офсетная
Усл.печ.л. 29,3. Усл.кр.-отт. 32,8. Уч.-изд.л. 33,7
Тираж 650 экз. Тип. зак.3234.
Ордена Трудового Красного Знамени
издательство "Наука"
117864 ГСП-7, Москва В-485,
Профсоюзная ул., д. 9 0
Ордена Трудового Красного Знамени
1-я типография издательства "Наука"
199034, Санкт-Петербург В-34, 9-я линия, 12