/



































































































































































































































































Текст
БИБЛИОТЕКА
БИБЛИОТЕКА «ЗНАНИЕ»
А.Г. ТОМИЛИН
В мире
китов и
дельфинов
ИЗДАНИЕ 2-е, ПЕРЕРАБОТАННОЕ
И ДОПОЛНЕННОЕ
Издательство «Знание» Москва 1980
ББК 28.082
Т56
Томилин А. Г.
Т56 В мире китов и дельфинов.— 2-е изд.,
перераб. и доп.— М.: Знание, 1980.— 224 с.,
16 л. ил.— (Библиотека «Знание»).
65 к. Тир. 100 000 экз.
Книга известного специалиста по морским млекопитаю-
щим, доктора биологических на^к А. Г. Томилина посвящена
современным проблемам биологии китов и дельфинов. Автор
рассматривает удивительные приспособления этих млекопи-
тающих к водному образу жизни, сложное поведение, рефлек-
сы и инстинкты китообразных, их отношение к людям.
Сейчас дельфинами интересуются ученые разных специаль-
ностей — кораблестроители, гидроакустики, гидромеханики,
физиологи, медики. Автор рассказывает о перспективах ис-
пользования дельфинов в рыбном промысле, в исследовании
океана, о возможности одомашниванйя некоторых видов этого
семейства.
Книга рассчитана на широкий круг читателей.
21009—009
Т 073(02) —80 0 0000
ББК 28.082
57.026.2
© Издательство «Знание», 1974 г., доп.
Глава I
НАШЕСТВИЕ
СЕНСАЦИИ
«Дельфины — люди моря», «Мы сталкиваемся с не-
человеческим мышлением, может быть, более слож-
ным, чем наше», «Наши младшие братья по разуму»,
«Впервые интеллектуальное первенство человека ста-
вится под вопрос», «Люди или дельфины?», «Разум в
океане», «Дельфины любят блондинок» — такими оше-
ломляющими заголовками запестрели газеты и жур-
налы Запада в 1961 году. На рядового читателя обру-
шился поток «дельфиновых сенсаций».
Нельзя сказать, что до этого дельфины не привле-
кали к себе внимание человека. Культ дельфина был
широко распространен еще более 2000 лет назад. Древ-
ние сделали дельфина морским богом, его изображали
на монетах, складывали о нем легенды, в его честь на-
зывали города и населенные территории.
Время стерло былую славу дельфина, но во второй
половине XX века о нем вдруг заговорили с новой си-
лой. Однако в отличие от древних в современных сен-
сациях это животное возвели на пьедестал человека,
наделили человеческим мышлением и речью.
Начало сенсациям положил американский нейро-
физиолог Джон Лилли, тонкий экспериментатор, прово-
дивший опыты над дельфинами на Виргинских остро-
вах в хорошо оснащенной лаборатории на острове Сент-
Томас. Подводя итоги своих исследований, этот ученый
провозгласил, что первым животным на планете, ко-
торое установит с человеком сознательный контакт,
будет дельфин. По своим качествам дельфин будто бы
вплотную приближается к человеку и отличается от
всех остальных животных. Дельфины, по убеждению’
3
Лилли, обладают разумом и коллективной памятью.
Но поскольку они не имеют письменности, так как ли-
шены рук, жизненный опыт дельфинов передается
младшим сородичам и потомкам устно: «Примерно
так же, как передавались знания у примитивных че-
ловеческих племен — через народные сказания и ле-
генды — изустно от одного поколения другому, кото-
рое в свою очередь запоминало их и передавало даль-
ше. Способность к быстрому и прочному запоминанию,
необходимая при таком обучении, требует очень круп-
ного мозга. Наша письменность, книгопечатание и
другие способы хранения информации вне мозга в зна-
чительной степени освобождают нас от необходимости
запоминания. Дельфинам же приходится все хранить
в памяти, поскольку у них нет ни библиотек, ни кар-
тотек...»
Комментаторы Лилли шли дальше, заявляя, что в
недалеком будущем дельфины станут читать капита-
нам и штурманам лекции о кораблевождении, войдут
в разумный контакт с обитателями других миров на
особом языке «линкос» (лингвистика космоса), осно-
ванном на математической логике?! Согласно этой кон-
цепции, дельфины примут участие в прогрессе науки,
но не как подопытные животные, а как коллеги уче-
ных и носители знаний. Сам Лилли считал своего луч-
шего дельфина Эльвара способным экспериментиро-
вать над человеком; в одной из статей в научном жур-
нале «Саенс» ученый даже благодарил дельфина за
сотрудничество.
Американский ученый Форрест Вуд в книге «Мор-
ские млекопитающие и человек» пишет: «Наблюдения,
проведенные в океанариумах, не дают оснований счи-
тать, что дельфины общаются друг с другом на уровне,
сравнимом с уровнем полноценного языкового обще-
ния у людей».
Постепенно туман очеловечивания дельфинов рас-
сеялся, шум вокруг них, продолжавшийся несколько
лет, затих. Однако интерес ученых к дельфинам
отнюдь не ослабел, а, напротив, возрос. Сейчас дельфи-
нами интересуются люди самых разных специально-
стей: гидробионики, физиологи, гидроакустики, судо-
строители, экологи, океанологи, этологи (этология —
наука о поведении животных), конструкторы подводных
аппаратов, гидродинамики, психологи, медики, нейро-
4
анатомы, нейрохирурги — все они, каждый в своей
области, находят у этих животных предмет для своих
изысканий. Одних интересует моделирование кожи
дельфинов при создании обшивки быстроходных ко-
раблей, других — секреты быстрого и безвредного по-
гружения, третьих — тайны работы головного мозга,
четвертых — принципы устройства чувствительного,
портативного и устойчивого к помехам гидролока-
тора.
Взгляд на местоположение дельфинов на система-
тической лестнице к настоящему времени сложился до-
вольно противоречивый. Одни ученые считают, что эти
животные стоят впереди обезьян, в том числе и чело-
векообразных, а другие отводят им более скромное
место — рядом с собакой или между собакой и обезья-
ной.
Чтобы лучше познакомиться с китообразными, со-
вершим экскурсию в мир вторичноводных животных и,
не вдаваясь в фантастику, рассмотрим достижения на-
уки в изучении этих замечательных обитателей моря.
Глава II
ПОМОЩНИКИ
И ДРУЗЬЯ ЧЕЛОВЕКА
В МОРЕ
Мелкие китообразные с давних пор снискали себе
симпатии человека, оказывая ему услуги в рыбной
ловле и спасая утопающих. Древнеримский ученый,
автор «Естественной истории» (в 37 томах) Плиний
Старший, погибший в 79 году новой эры во время
извержения Везувия, описал, как на Средиземном мо-
ре дельфины способствовали успешному промыслу, ме-
шая косякам кефали выходить из мелководного зали-
ва Латера. Рыбаки никогда не обижали своих помощ-
ников и после удачного лова вознаграждали их частью
добычи.
О ночном рыболовстве с помощью дельфинов на ост-
рове Эвбея (Греция) поведал миру еще во II веке рим-
ский писатель и ритор Элиан Клавдий в своем 17-том-
ном произведении «О природе животных». В тихую
погоду рыбаки выходили на лов и в темноте зажигали
огни, на которые шли рыба и дельфины. Освещенную
рыбу, блокированную между лодками и дельфинами,
легко поражали острогой.
По описанию новозеландского журналиста Антони
Алперса, «совместное рыболовство» развивалось в ряде
мест и в нашем веке. В Неаполитанском заливе рыба-
ки опускали в воду электрофонари, свет которых соби-
рал массу мелких рачков, а те привлекали стаи сар-
дин, анчоусов и головоногих моллюсков. Вылов рыбы
сетями начинали тогда, когда появлялись дельфины,
которые сдерживали рыбные косяки и мешали им рас-
сеиваться.
Лов китов-горбачей с помощью хищных дельфи-
нов — косаток производили китобои Австралии в за-
6
ливе Туфолд: киты, возвращаясь с антарктических по-
лей нагула, заходили зимой в этот залив, возможно, в
поисках мест размножения. Здесь же появлялись ко-
сатки, запирая выход в открытое море. Сильно возбу-
жденные, косатки выпрыгивали из воды, привлекая
внимание людей. Тогда на лодках срочно выезжали
китобои и, пользуясь тем, что косатки прижимали ки-
та к берегу, после непродолжительного преследования
гарпунили его, а тушу оставляли на якоре, чтобы хищ-
ники могли полакомиться огромным языком горбача.
Особенно славилась косатка Старый Том: во время
охоты она зубами держалась за гарпунный линь, чем
затрудняла буксировку лодок раненым китом. Почти
50 лет Старый Том «служил» человеку и погиб в
1930 году, вероятно, от старости. За «заслуги» перед
людьми скелет косатки рыбаки выставили на главной
улице городка Иден.
Аналогичное поведение наблюдали у дельфинов в
реках Южной Америки и Юго-Восточной Азии. В со-
роковых годах «услугами» амазонской инии пользо-
вались жители поселка Мото-Гросс на реке Тапажос:
ночью при свете карбидных фонариков они били остро-
гой рыбу, которую иния загоняла на мелкие места и
не выпускала отсюда. Небольшой, с шаровидной го-
ловой иравадийский дельфин, загонявший рыбу в рас-
ставленные сети на реке Иравади в Бирме, был даже
предметом судебных споров между соседними поселка-
ми, поскольку каждый считал его своим.
А вот что рассказал корреспондент ТАСС Г. Тара-
нович из практики советского рыболовного траулера в
Северном море. Крупный дельфин, попавший в рыбо-
ловные сети, был отпущен на волю. После этого он
каждый день подплывал к траулеру в одном и том же
квадрате моря и подолгу резвился возле судна. Од-
нажды он появился вместе со стадом сородичей; по-
следние вскоре исчезли, а старый знакомый команды
продолжал крутиться вблизи корабля, высоко выпры-
гивая из воды. Причина возбуждения стала понятной,
как только включили эхолот: прибор указал на боль-
шой косяк рыбы, после чего последовал очень удачный
лов.
Этот дельфин ежедневно приплывал к судну, и за
экспедицию траулер выполнил три плана. Когда месяц
спустя судно вошло в район, где был отпущен дель-
фин, тот снова появился и в течение четырех экспеди-
ций наводил траулер на косяки рыб.
Контакты между дельфинами и человеком возника-
ли не только на почве пищевого подкрепления при «со-
вместном рыболовстве», но и когда животные прояв-
ляли сильно развитое любопытство, называемое ориен-
тировочным рефлексом. Примером тому служат «руч-
ные дельфины» : обитая в море, они перестают бояться
человека и приближаются к нему по своей воле. Такие
дельфины быстро становятся знаменитостями.
В 1955 году в местечке Опонони в Новой Зеландии
молодая самка афалины Опо-Джек почти каждый день
стала появляться среди купальщиков. Она вплотную
подплывала к ним, позволяла себя гладить (рис. 1 вкл.)
и даже сажать себе на спину детишек, совершала
прыжки из воды и играла в мяч, высоко подбрасывая
его в воздух. Так продолжалось почти год, пока люби-
мец публики не был обнаружен мертвым на берегу,
несмотря на специальный указ новозеландского прави-
тельства, запрещавший трогать дельфинов в бухте
Хакьянга Харбор. В память об Опо-Джеке жители го-
родка соорудили ему памятник в виде каменной ста-
туи дельфина. Подобные истории повторялись в раз-
ных частях мира. В 1965 году в заливе Ферт-оф-Форт
в Шотландии появился ручной дельфин-афалина Чар-
ли. Он давал «спортивные представления» близ ку-
рортного поселка Эли, соревнуясь в скорости с катера-
ми и водными лыжниками, и несколько месяцев дру-
жил с купальщицей-подростком Джейн Свенсон.
В августе 1968 года возле пляжа города Евпатории
целый месяц упорно держалась молодая ручная афа-
лина Альфа, которую отдыхающие кормили рыбой,
брали на руки, гладили, играли с ней. Она любила
подплывать к причалу и выпрашивать подачку у рыба-
ков, сидящих с удочками (рис. 2 вкл.). В том же году
в Австралии на пляже города Мельбурн в группу ба-
рахтающихся в воде детей заплыл дельфин и несколь-
ко часов играл с ними. Детишки цеплялись за спинной
плавник дельфина и плавали с ним у берега. Только
к вечеру удивительный зверь уплыл в океан.
В 1972 году объявилась ручная афалина Дональд
в водах Британии у острова Мэн. Она подплывала к
водолазам, любила общество одной спортсменки и в те-
чение пяти лет вступала в контакт с сотнями людей.
8
Водолаза Горация Добса она узнала год спустя после
знакомства и проявила к нему особое расположение.
Воспользовавшись этим, Добс сделал кинофильм для
Ньюйоркширского телевидения и опубликовал книгу
«Дружба с дельфином».
Ручные дельфины были хорошо известны у древних
греков и римлян на побережье Средиземного моря, о
чем упоминают Аристотель, Плутарх, Геродот, Оппиан
и другие ученые и писатели античного мира. Плиний
Старший описал несколько подобных случаев.
Мальчик Дионисий из древнегреческого города
Ясос близ Милета подружился с дельфином, который
возил его на себе в присутствии толпы, собиравшейся
на берегу. В том же городе другой мальчик — Хермес
катался на дельфине по морю, но во время шторма по-
гиб, и дельфин доставил его к берегу. Наиболее про-
должительной дружба с дельфином была у подростка
из поселка Байанум, расположенного на берегу лагуны
Лукрина (ныне Пеццуоли близ Везувия): животное ре-
гулярно, на протяжении нескольких лет перевозило
мальчика через лагуну, сообщавшуюся с морем, в шко-
лу и обратно домой. По зову школьника — «Сима, Си-
ма!»— дельфин подплывал к нему и брал пищу из
рук. Когда мальчик заболел и умер, дельфин еще дол-
гое время продолжал приплывать на место их бывших
встреч.
Ленинградский нумизмат А. Быков, исследуя ан-
тичные монеты из порта Таренто IV века до нашей
эры с изображением человека, оседлавшего дельфина,
пришел к выводу, что древние смотрели на дельфина
как на средство передвижения в море.
Много позже Наполеон мечтал форсировать Ла-
Манш для завоевания Англии с помощью дельфинов,
посадив на них своих воинов.
Среди древнегреческих легенд и преданий есть и
такая легенда, описанная Геродотом: певец Арион пос-
ле плавания в Грецию был пленен пиратами. Зло-
умышленники ограбили его и решили выбросить в мо-
ре, но перед смертью позволили ему сыграть на лире.
Когда певца бросили за борт, его спас дельфин, при-
влеченный звуками лиры. Он поднял Ариона на спину
и вынес его к. мысу Матапан на полуострове Пелопон-
нес в Средиземном море.
Подобные предания существуют и у полинезийцев:
9
оказавшихся в воде людей, чьи лодки потерпели кру-
шение, тоже спасали дельфины.
Сходство древнегреческих и полинезийских легенд
подтверждает реальность тех происшествий, на основе
которых они создавались. Между тем почти до наших
дней эти легенды считались вымыслом. Но неожидан-
но аналогичные случаи повторились в 1943 году во
Флориде и в 1966 году в Суэцком заливе. Первый слу-
чай был описан в американском журнале «Нэтчураль
Хистори»: жена адвоката купалась близ берега и по-
пала на глубокое место. Теряя сознание, она почув-
ствовала, как ее кто-то сильно толкнул. Оказавшись на
берегу, она хотела поблагодарить спасителя, но вблизи
лишь нырял дельфин. Подбежавший очевидец расска-
зал женщине, что ее вытолкнуло из воды животное.
Второй случай описал в «Известиях» корреспондент
Л. Коровин: 4 июня 1966 года каирский инженер Мах-
муд Вали выехал на рыбную ловлю и попал в шторм.
Мотор лодки заглох, и человека отнесло на десятки ки-
лометров от берега. Большая волна выбросила его в
бушующую стихию. Ухватившись за спасательный
матрас, потерпевший пролежал на нем более суток.
Еще ночью он почувствовал, как стайка дельфинов
стала толкать матрас. К вечеру странная группа при-
близилась к берегу, ее заметили с поста, и инженер
был поднят на подошедший катер. Эти реальные слу-
чаи заставили по-новому оценить склонность дельфи-
нов спасать человека.
Что же представляют собой дельфины и как они
живут?
Глава III
В Мировом океане обитает около полусотни разных
видов дельфинов. Вместе с китами они составляют от-
ряд китообразных. Лишь формой тела да тем, что жи-
вут в воде, китообразные напоминают рыб. Во всем же
остальном — это настоящие млекопитающие. Они теп-
локровны, дышат легкими, рождают живых детены-
шей и выкармливают их молоком.
Не менее 70 миллионов лет назад наземные предки
китообразных перешли жить в воду и в конце концов
полностью утратили связь с сушей. У них развилась
торпедообразная, легко обтекаемая форма тела, так
как при передвижении в новой среде сопротивление
увеличилось в 800 раз! Строение тела и образ жизни
неузнаваемо преобразились. Передние ноги преврати-
лись в уплощенные и жесткие грудные плавники: это
своеобразные рули высоты, поворота и торможения,
используемые для балансирования. Задние ноги исчез-
ли, как и поддерживающий их таз и крестцовый отдел
позвоночника. От таза сохранились лишь маленькие
косточки в мускулатуре по обе стороны гибкого позво-
ночника.
Главным органом передвижения стал хвост — мус-
кулистый, сжатый с боков стебель, спереди незаметно
переходящий в вытянутое туловище, а сзади заканчи-
вающийся широким треугольным хвостовым плавни-
ком.
Очень упругий хвостовой плавник расположен
горизонтально и на заднем крае поделен глубокой вы-
емкой на правую и левую лопасти. На спине большин-
ства китообразных имеется спинной плавник: он при-
11
дает устойчивость телу при движении в воде. Как и в
хвостовых лопастях, в нем нет скелета.
Упругая и гладкая кожа лишена волос, потовых и
сальных желез. Только отдельные волоски несколько
дней держатся на морде и подбородке новорожденных
дельфинов и потом выпадают, а у усатых китов сохра-
няются в течение всей жизни, выполняя роль вибрисс,
или осязательных щетинок.
Под кожей развился могучий пласт жира как за-
щита от охлаждения и запас энергии на случай голо-
да. Самый толстый слой подкожного жира залегает на
наиболее пассивных участках тела — на брюхе и меж-
ду грудными плавниками (у гренландского кита этот
слой достигает 0,5 м). На рабочих же органах, таких,
как хвостовой стебель, толщина жирового пласта быва-
ет в 2—3 раза меньше. Под слоем жира в задней тре-
ти тела залегают две молочные железы с одним сос-
ком каждая. Соски скрыты в продольных кожных кар-
манах, расположенных снизу по бокам мочеполовой
щели, и только у кормящих самок выступают наружу.
Окраска тела у одних видов имеет маскирующее,
а у других — сигнально-опознавательное значение.
Броские ярко-белые пятна на теле важны для сороди-
чей, чтобы не сталкиваться во время гона, игр, стре-
мительного плавания в стайках.
У многих китообразных окраска меняется с возра-
стом: одни, как белуха, родившись черными, затем
становятся серыми, голубыми и, наконец, белыми;
другие, как пятнистый дельфин, появляются на свет
однотонно-серыми или темными, а потом покрываются
пятнами; третьи, как бутылконосы или клюворылы, к
старости «седеют» с головы.
Кожные заболевания (дерматомикозы, парша, стри-
гучий лишай, рожистые воспаления), а также деятель-
ность эктопаразитов, травмы, порезы приводят к по-
светлению окраски, появлению пятен разной интенсив-
ности, которые со временем пигментируются вновь.
У ряда видов наблюдается сильная индивидуальная
изменчивость с момента родов, например, в форме, ве-
личине и количестве белых пятен на брюхе у южных
китов, горбачей, кашалотов. Иногда белые пятна оста-
ются после укусов мелких акул.
Как редкое явление у китообразных встречается
альбинизм и меланизм. В первом случае у родителей
12
с нормальной — не белой — окраской рождается аль-
бинос: чисто белый с красными глазами. В дальней-
шем этот потомок рождает нормально окрашенного де-
теныша, как о том свидетельствуют экспонаты — мор-
ская свинья-альбинос и ее созревший темный зародыш,
выставленные в музее Новороссийской биостанции. Во
втором случае у нечерных родителей рождается совер-
шенно черный детеныш — меланист, который впослед-
ствии рождает тоже меланиста. Такую черную самку
белокрылой морской свиньи поймали недавно японцы
у берегов префектуры Камаиши. Она была в паре с та-
ким же черным самцом, а в ней находился черный за-
родыш с белым кончиком морды.
Голова китообразных не обособлена от туловища,
и спина как бы продолжается до самого «носа». Нозд-
ри открываются на темени одним отверстием (у зуба-
тых китов) или двумя (у усатых китов). Это отверстие
называют дыхалом. Дыхало открывается особыми
мышцами лишь на момент короткого дыхательного
акта — слитного выдоха-вдоха, а все остальное время,
называемое дыхательной паузой, плотно закрыто. Ре-
гулируется дыхательный акт рефлексами вынырива-
ния.
Запирающее устройство можно грубо уподобить ре-
зиновой пробке, которая, лишь на миг растягиваясь,
пропускает воздух при выдохе-вдохе и тут же отводит-
ся назад, автоматически запирая ноздрю. Поэтому во-
да, если только животное не всасывает ее по своему
желанию, не может попасть в дыхательные пути
через дыхало. Это исключено и со стороны рта, посколь-
ку гортань устроена так, что воздухоносный путь от-
делен от пищевого: ни вода, ни пища изо рта не попа-
дают в дыхательное горло даже в момент дыхания.
Однако дельфинов можно обучить активно всасывать
воду в носовой канал через дыхало и выбрасывать ее
на один-два метра сильной струей или в виде распы-
ленного фонтана. Когда в воздухе прохладно, то при
выдохе над поверхностью моря взлетает конденсиро-
ванный пар — фонтан; иногда в нем заметны и распы-
ленные брызги случайно попавшей или засосанной в
ноздри воды.
По форме фонтанов китобои различают виды китов
на расстоянии в несколько километров (рис. 3 и
4 вкл.).
13
Легкие китообразных яесьма упруги и эластичны;
легочная ткань приспособлена к быстрому сжатию и
расширению. В легких сильно развиты хрящевые коль-
ца даже в мелких бронхах, а у дельфинов и в бронхио-
лах, запирающихся кольцевыми мышцами — сфинкте-
рами.
Китообразные могут долго находиться под водой и
глубоко нырять, экономно расходуя кислород. Как они
этого достигают, будет рассказано позже.
Жизнь в воде, с полным отрывом от суши, наложи-
ла глубокий отпечаток на способы регуляции тепла у
этих млекопитающих. Нормальная температура их те-
ла близка к 36—37°. Между тем им все время прихо-
дится плавать — то очень быстро, то медленно, нырять
на разную глубину в условиях меняющейся темпера-
туры среды. У многих китов перемена условий усугуб-
ляется еще сезонными миграциями из холодных морей
в теплые и обратно. Все это говорит о том, что кито-
образные должны обладать очень совершенными термо-
регуляторами. Оказалось, их роль выполняют плавни-
ки. Функция таких регуляторов тепла была открыта
советскими учеными в 1947 году.
Добычу, обычно только живую, китообразные за-
глатывают целиком, не пережевывая. В зависимости
от способа питания отряд делится на два подотряда:
зубатые киты (хваталыцики) и усатые киты (фильтро-
вальщики). Первые либо хватают добычу поштучно,
удерживая ее зубами, либо с помощью движения язы-
ком всасывают по нескольку рыб в момент открыва-
ния рта. Вторые ловят рачков или рыб сразу больши-
ми партиями в местах их массовых скоплений, проце-
живая пищу цедильным аппаратом — китовым усом
(рис. 5 вкл.). К зубатым китам относятся семейства
кашалотов, дельфинов, клюворылых и речных дель-
финов, всего около 70 видов, а к усатым — семейства
полосатиков, гладких и серых китов, всего 10 видов.
Китообразные очень прожорливы, желудок у них
многокамерный, состоит из 3—9 отделов. Он способен
растягиваться и вмещает у синих китов до 1,5 т, у фин-
валов — до 1, а у сейвалов — до 0,5 т рачков. Кал
дельфинов и китов всегда жидкий. Вместе с пищей ки-
тообразные неизбежно заглатывают соленую воду, но
от избытка соли освобождаются благодаря особому
строению почек и частому мочеиспусканию. Почки
14
крупные, составлены из множества мелких долек, хо-
рошо отделенных и собранных в гроздья. У дельфинов
в одной почке насчитывают сотни долек, а у усатых
китов — тысячи.
Китообразным требуются большие скопления кор-
ма, которые обусловливают и величину их стад. В за-
висимости от вида пищи китообразные предпочитают
те или иные зоны океана. Одни (планктоноеды — глад-
кие киты) питаются преимущественно в открытом мо-
ре близ поверхности воды на массовых скоплениях
мелких рачков. Другие (бентосоеды — серые киты) лю-
бят мелководья, где заглатывают придонных и донных
рачков; третьи (рыбоеды — большинство дельфинов)
охотятся за стайной рыбой как вдали, так и вблизи от
берегов, изредка посещают реки; четвертые (планкто-
но-рыбоеды — полосатики) кормятся в поверхностной
толще моря ракообразными, рыбой, реже головоногими
моллюсками. Хищники (косатки) пожирают рыб и
морских млекопитающих, нападая на них в любой зо-
не океана. Моллюскоеды (кашалоты, клюворылы, се-
рые дельфины и др.) поедают преимущественно голо-
воногих моллюсков, иногда на больших глубинах. Есть
и постоянные обитатели рек, питающиеся пресновод-
ными рыбами и различными беспозвоночными (речные
дельфины).
Массовые скопления корма образуются в летнее по-
лугодие в сравнительно холодных водах, в зонах пере-
мешивания теплых течений с холодными и особенно
близ кромки плавучего льда. Обилие корма и привле-
кает сюда китов нагуливать жир. Зимовать же они
уходят в теплые воды, где мало пищи, но легче сохра-
нять тепло, живя за счет накопленного летом жира.
Наиболее упитаны киты осенью, в конце нагула, а са-
мыми тощими бывают весной. Дельфины же наиболее
худосочны летом.
Некоторым китообразным свойственны возрастные
и половые группировки: ведь доступность корма не-
одинакова для всех особей. Поэтому, например у ка-
шалотов (рис. 6 вкл.), в группы подбираются животные
с равными физическими возможностями — скоростью
передвижения, способностью к нырянию в погоне за
пищей; по этой же причине самцы часто отделяются
от самок.
15
Усатые киты на полях нагула держатся в одиноч-
ку, парами или группами. Зоолог М. Н. Тарасевич из-
учала структуру китового стада в Аляскинском зали-
ве. Оказалось, что у финвалов группы образованы ли-
бо самцами (до восьми китов в каждой), либо самками
(до шести особей), или же теми и другими, но если пре-
обладают самцы, численность групп бывает в полтора-
два раза больше (до 16 голов), чем в случаях преобла-
дания самок. Видимо, у самцов тенденция к стайности
выражена резче, чем у самок.
Дельфины живут семьями, в которые входят потом-
ки нескольких поколений. Такие семьи, вероятно, и
представляют собой те поисковые группы, которые на-
блюдаются в районах концентрации корма. Группы
объединяются во временные, иногда очень многочис-
ленные стада, распадающиеся по мере рассеивания
скоплений корма. Иногда же (у гринд) семьи сходятся
в отдыхающие группы по нескольку десятков голов и
лежат на поверхности, выставив из воды морды и
спинные плавники. Временами семьи дельфинов во
главе с вожаком-самцом объединяются для совместных
действий против крупных акул и распадаются, когда
минует опасность.
Плодится большинство китообразных через два го-
да, но некоторые дельфины спариваются, еще не закон-
чив выкорма детенышей, и размножаются ежегодно.
Беременность у разных видов продолжается от 10 до
18 месяцев. Далеко мигрирующие виды (полосатики)
рожают главным образом зимой в относительно теп-
лых водах, а не совершающие больших миграций
(дельфины) — летом, но в обоих случаях при благопри-
ятных температурных условиях.
Единственный, хорошо развитый детеныш рождает-
ся очень крупным — от V4 до 7г длины тела матери
(этому способствует отсутствие таза и задних конечно-
стей). Изредка в одной самке находят несколько заро-
дышей. У синего кита однажды обнаружили семь, а
у финвала и сейвала — по шесть зародышей. Это сви-
детельство былого многоплодия предков китообразных
и последующего уменьшения плодовитости ради совер-
шенствования развития одного детеныша. Лишние
зародыши рассасываются еще в эмбриональном перио-
де, и только очень редко могут родиться двойни. Со-
ветские и японские ученые находили у горбатого
16
кита и морщинистозубого дельфина «сиамских близ-
нецов» — по два сросшихся зародыша.
Роды происходят под водой. Пуповина рвется у са-
мого брюха, где она менее прочная. Плод выходит
хвостом вперед. За лактационный период у усатых ки-
тов детеныши увеличиваются почти вдвое, а у зуба-
тых — на одну треть первоначального размера. Ко
времени перехода на самостоятельное питание у пер-
вых резко увеличивается китовый ус, а у вторых про-
резаются зубы.
Половая зрелость наступает в возрасте от 3 до
6 лет, но замедленный рост тела еще продолжается
долго. Когда скелет полностью окостеневает и все эпи-
физы позвоночника срастаются с телами позвонков,
тогда наступает физическая зрелость. Процесс окосте-
нения позвоночника начинается с обоих его концов,
притом с хвостового идет быстрее, чем с головного, и
заканчивается в грудном отделе.
Киты живут до 50, а дельфины до 30 лет. Возраст
китов определяют несколькими способами: подсчиты-
вают рубцы, остающиеся от редукции желтого тела на
поверхности яичников, или слои в ушных хитиноподоб-
ных пробках (рис. 1), устанавливают число «шагов»
на усовой пластине цедильного аппарата, либо коли-
чество концентрических слоев на поперечном сечении
в роговых трубочках пластины, куда заходят сосочки
пульпы с кровеносными сосудами, или же количество
слоев в плотной барабанной кости — булле. У зубатых
китов подсчитывают число дентиновых слоев на
поперечных и продольных срезах зубов.
Распространение большинства видов китообразных
очень широкое, чему способствует отсутствие резко вы-
раженных преград в океане. Тем не менее киты живут
локальными стадами и даже при очень далеких мигра-
циях, как правило, не переходят экватора. Местные
группировки китов на полях нагула ученые устанав-
ливают методом серологических реакций, т. е. с по-
мощью биологических и химических свойств сыворот-
ки крови, неодинаковых у разных локальных стад.
Есть холодолюбивые виды, живущие в полярных и
субполярных водах (белухи, нарвалы, гренландские
киты), теплолюбивые (полосатик Брайда), тропические
и субтропические (многие мелкие дельфины, карлико-
вые кашалоты) и виды с весьма широким ареалом,
17
включая космополитов (почти все полосатики, каша-
лоты, косатки и др.).
В результате приспособления к сезонным условиям
питания и размножения сформировалось несколько
биологических групп китообразных. Виды одной груп-
пы совершают строго регулярные миграции в пределах
Северного или Южного полушария; на зиму усатые
киты, часть клюворылых и кашалоты идут в низкие
широты для родов, а на лето следуют в умеренные и
холодные воды. Нагулять жир в Арктике и Антаркти-
ке легче, так как здесь планктонных организмов в
10—20 раз больше, чем в тропиках. Виды другой груп-
пы перемещаются тоже на значительные расстояния,
но менее правильно и с нарушением сезонных сроков
(мад&е косатки, гринды, отчасти сейвалы, нарвалы
и др.). Виды третьей группы ведут сравнительно осед-
лый образ жизни; их кочевки проходят в пределах не-
большой акватории (афалины, речные дельфины, серые
дельфины и др.), а у некоторых отмечена даже тен-
денция занимать определенный участок, например у
инии в реке Амазонке и ее притоках.
Наиболее загадочна первая группа. Ее изучение ве-
дут с помощью мечения: в кита стреляют из гладко-
ствольного ружья металлической цилиндрической мет-
кой, на которой ставится номер и адрес для возврата.
Кит плавает с меткой, пока его не убивают. Метку
возвращают владельцу, по ее номеру восстанавливают
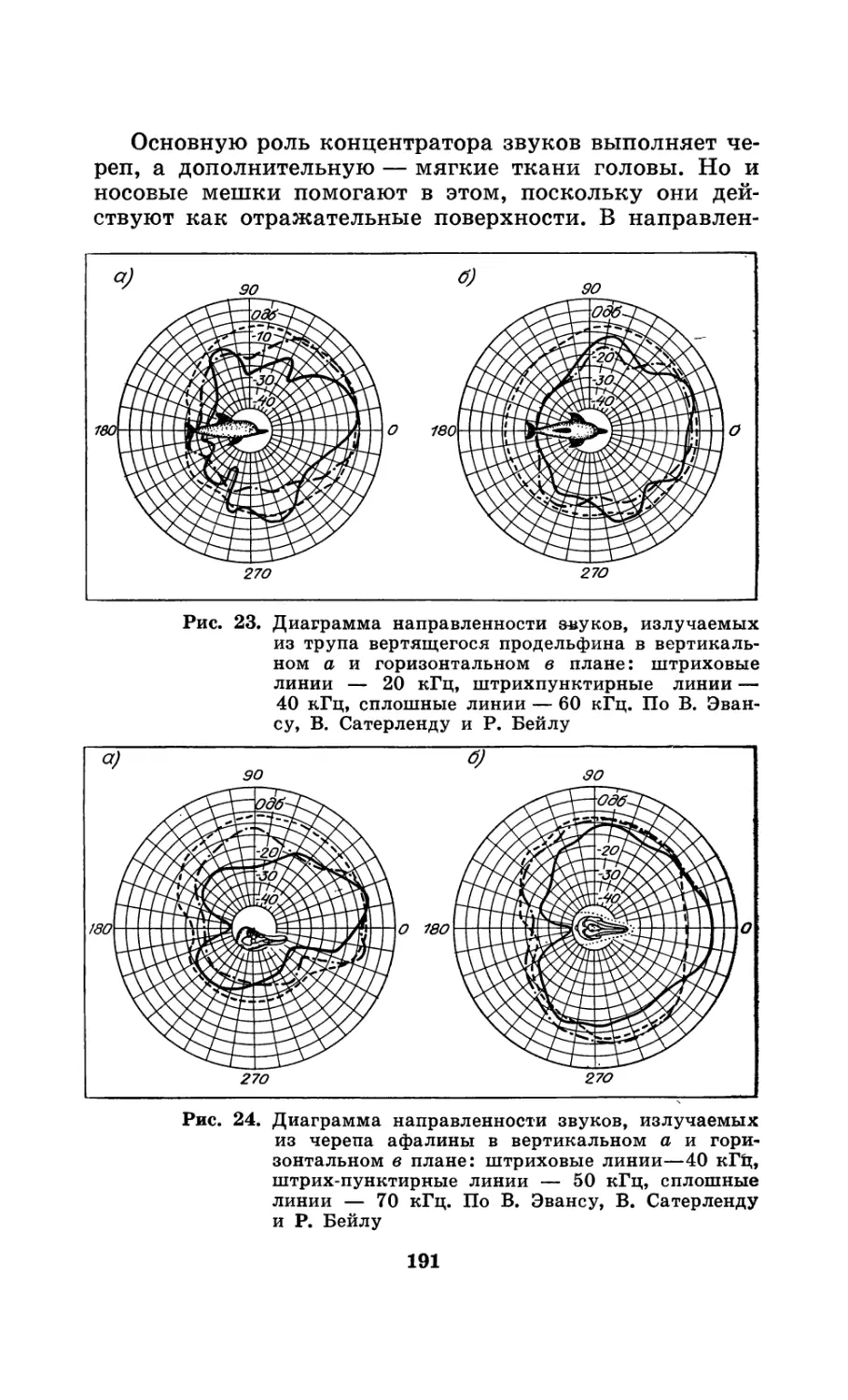
Рис. 1. Ушные пробки финвалов. По количеству слоев в них
определяют возраст усатых китов
18
место и дату мечения, а следовательно, и приблизи-
тельный путь животного. С 1932 по 1973 год в Миро-
вом океане пометили более 20 500 усатых китов и
кашалотов, а обратно получили 1798 меток (около
8,8%). Данные мечения показали, что киты мигриру-
ют лишь в пределах своих областей, придерживаясь
определенных направлений, а может быть, и путей.
При далеких странствиях они безошибочно, из года в
год, возвращаются в одни и те же районы и даже бух-
ты. Бывало, что меченого кита добывали много лет
спустя в том же сезоне и там же, где он был помечен,
а в другие сезоны убивали на расстоянии в тысячи
километров от места мечения. Например, в проливе
Кука в Новой Зеландии были взяты два горбача спус-
тя 10 и 18 лет, после того как были помечены в том же
самом проливе, а в Антарктике добыт финвал лишь в
350 км от того места, где он был помечен 30 лет назад.
Так были выявлены удивительные способности ки-
тообразных ориентироваться в океане ночью и днем, в
бурю и в штиль, на глубинах и у поверхности. Нет
сомнений, что киты, совершая тысячекилометровые
маршруты, точно выходят к цели — в места сезонных
концентраций пищи — с помощью совершенных орга-
нов чувств (анализаторов), о чем речь пойдет ниже.
В последнее время мечение проводят и на дельфи-
нах, миграции которых изучены еще крайне слабо.
В 1969—1974 годах калифорнийский биолог Вильям
Перрин с сотрудниками пометил в восточной части
Тихого океана 1727 пятнистых и 242 вертящихся
продельфина, которые случайно попались в тунцелов-
ные сети. У первого вида получено обратно 24 метки,
которые вернулись через разные сроки — от 1 до
502 дней; за это время метки были перенесены дель-
финами от пункта мечения на расстояние от 13 до
2415 км. У второго вида вернулись 5 меток через 38—
776 дней и переместились за это время на дистанцию
от 333 до 733 км.
Удачный опыт мечения проведен в Канадской Арк-
тике Дэвидом Сержантом: в устье реки Тюленьей (за-
падная часть Гудзонова залива) летом 1967 и 1968 го-
дов 700 гарпунных меток всадили в спины белух во
время их преследования и 118 дисковидных меток при-
крепили к краю спинного гребня белух, обсохших в
мелких водах во время отлива. Три гарпунные метки
19
были обнаружены в животных, пойманных сетями, из
них одна год спустя в том же районе, а две — спустя
5—7 недель в 300 и 800 км от места мечения.
Метки помогают раскрывать секреты китообразных,
глубоко спрятанные под поверхностью моря. Большую
роль в выяснении многих вопросов призваны сыграть
радиометки, аналогичные тем, какие применяли в Сан-
Диего доктор В. Эванс и другие ученые на сосунках
серых китов. Одного такого китенка (Гиги-2), жившего
в океанариуме, выпустили обратно в море с прикреп-
ленным к его спине радиопередатчиком. Связь с
животным поддерживали в течение 53 дней. Со следя-
щего судна и самолета установили наибольшую дыха-
тельную паузу китенка 16 мин 32 с и глубину погру-
жения 170 м. Потом молодой кит совершил миграцию
на север и через год был обнаружен у берегов Кали-
форнии в районе выпуска; здесь его опознали по бело-
му хвостовому плавнику.
Недавно американцы, биолог В. Шевилл и инженер
В. Уоткинс, успешно испытали радиометку весом 75 г,
которую бросали с вертолета в спину выныривающего
гладкого кита. Следящее судно с антенной в 15 м вос-
принимало радиосигналы метки в момент выхода ки-
та на поверхность с расстояния 6—7 км.
Хотя радиофицированный кит точно докладывает
о своем маршруте, метка не нашла широкого приме-
нения из-за сильного уменьшения численности китов.
На мигрирующих китах поселяется масса разных
организмов: это наружные кожные паразиты, кварти-
ранты, нахлебники и, возможно, симбионты. Все они
составляют на теле китов странствующее сообщество
и находятся в сложных взаимоотношениях друг с дру-
гом и со своим хозяином — китом. Такой комплекс
организмов, перемещающийся вместе с китом, называ-
ют странствующим биоценозом.
Из паразитов на наружном покрове китов, особен-
но серых и горбатых, поселяются и многочисленные
китовые вши — ракообразные из отряда бокоплавов,
питающихся кровью хозяина; и паразитические вес-
лоногие раки-пенеллы червеобразной формы, на 5—
7 см внедряющиеся в кожу кита передним концом те-
ла ; и миноги — из класса круглоротых, присасываю-
щиеся ротовой воронкой для питания тканями и
кровью китообразного. Кожа китов на полях нагула
обрастает зеленой пленкой, состоящей из множества
одноклеточных диатомовых водорослей, которые ис-
пользуют эпидермис как питательную среду и внедря-
ются в него протоплазматическими тяжами.
Из квартирантов («эпифитов») находят приют на
коже кита усоногие раки (коронулы (рис. 2), ксеноба-
лянусы), на известковых домиках которых в свою оче-
редь поселяются морские уточки из рода конходерма;
и рыбы-прилипалы — временные квартиранты, при-
крепляющиеся уплощенной головой-присоском к телу
гигантских млекопитающих, чтобы передвигаться
вместе с ними на тысячи километров. Одни кварти-
ранты из усоногих раков представляю^* одновременно
и паразитов, так как выделяют из перепончатого осно-
вания своего домика фермент, растворяющий белок
кожи китов для использования его в пищу, а другие —
и нахлебников, так как поедают частицы жидкого
кала кита и остатки его пищи, вымываемой изо рта.
Поэтому они поселяются обычно вокруг анального от-
верстия и на подбородке кита.
Настоящие нахлебники, или комменсалы, живут в
ротовой полости китов на клейкой поверхности усовых
пластин. Они не причиняют вреда хозяину и питают-
ся остатками его пищи. К нахлебникам относятся: и
маленькие, весьма многочисленные рачки-баленофилу-
сы, очищающие поверхность усов от липкого налета,
образованного остатками пищи; и крохотный круглый
червь одонтобиус, поселяющийся в деснах между усо-
Рис. 2. Раковины коронулы на теле горбатого кита
21
выми пластинами; и микроскопические одноклеточ-
ные организмы — жгутиковые гематофагусы. Эпизоди-
ческих, временных нахлебников представляют морские
птицы — чайки и трубконосые, подбирающие рачков
или рыб, выпадающих из пасти кормящихся китов во
время их выныривания на поверхность, или извлекаю-
щие добычу в водоворотах на месте ныряния китов.
Альбатросы иногда даже следят за подводным движе-
нием кита и летят в ту сторону, куда он перемещается.
В странствующем биоценозе есть и симбиотйческие
отношения. Например, некоторые птицы выступают
как взаимополезные сожители: они садятся на тело
кита и склевывают китовых вшей. Однако явление
симбиоза в странствующем биоценозе изучено очень
слабо.
Во время миграций кита численность его кварти-
рантов, нахлебников, эктопаразитов значительно меня-
ется, а некоторые из них временно выпадают из сооб-
щества в одних широтах и появляются в других. Так,
например, желтой пленкой, составленной из несколь-
ких десятков видов диатомовых водорослей, киты об-
растают на полях нагула в холодных широтах, притом
независимо от скорости своего хода. На пути в теплые
воды — к местам размножения — киты теряют эту
пленку, но, приходя на поля нагула, покрываются ею
вновь. Чем дольше они пребывают в холодных водах,
тем пленка становится толще, а желтая окраска интен-
сивнее. Китовые вши, облепляющие ссадины, ранки,
места порезов на теле хозяина, свободно перемещаются
по его коже и заражают других китов при контакте
с ними во время спаривания или кормления детены-
шей молоком, что бывает преимущественно в теплых
водах. Как правило, у каждого вида кита свои виды
китовых вшей.
Обычно на прибрежных китах-тихоходах (серых и
горбачах) обрастание представлено гораздо богаче, чем
на скороходах. Быстроходные полосатики обрастают
усоногими раками и китовыми вшами редко, значи-
тельно чаще — эктопаразитами пенеллами (из веслоно-
гих раков). По-видимому, сильные потоки воды, возни-
кающие при быстром ходе, не позволяют китовым
вшам и квартирантам в молодой стадии удерживаться
на коже своих стремительных хозяев. В виде редкого
исключения морские уточки ютятся на китовом усе и
22
на зубах китообразных. В свою очередь, на морских
уточках (конходерма ауритум) поселяются карликовые
морские уточки (конходерма виргатум, рис. 3). Они
прикрепляются также к телу паразита пенеллы и из-
редка на обнаженных костях поврежденных грудных
плавников китов.
В странствующих биоценозах можно различать
«ядро» и «временных попутчиков». «Ядро» составляет
сам кит со всеми его поселенцами или группа китов,
если миграция совершается локальным стадом, семьей
либо распыленным косяком. «Временные попутчики»
связаны с «ядром» непрочно и встречаются в самых
различных подвижных комбинациях в период нагула
на кормовых полях или во время дальних миграций.
«Попутчики» могут эпизодически выступать по отно-
шению к самим китам то как пищевые конкуренты
(дельфины, тюлени, пингвины, планктоноядные и хищ-
Рис. 3. Карликовые конходермы облепляют тело паразитическо-
го веслоногого рака-пенеллы, который прикрепляется к
телу кашалота
23
ные рыбы), то как симбионты и нахлебники (морские
птицы), то как промежуточные хозяева внутренних па-
разитов, то как враги (косатки, крупные акулы, опас-
ные для сосунков), либо как наружные паразиты (ми-
ноги), или временные квартиранты (рыбы-прилипалы).
На китовых кормовых полях встречаются многие
виды морских птиц — буревестники, альбатросы, моев-
ки, конюги, топорки, качурки и др. По птицам кито-
бои и рыбаки с далекого расстояния определяют скоп-
ление китов, рыб, кормового планктона и головоногих
моллюсков.
Жизнь миниатюрного странствующего сообщества
усложняется тем, что сами киты мигрируют не в оди-
ночку, а группами, семьями или рассеянным стадом.
Миграции китов и размещающиеся на них странст-
вующие биоценозы развились в ходе длительной эво-
люции. Обширные перемещения китов способствуют
более полному использованию кормовых источников
океана, притом в самое подходящее время — в сезон
наибольшей концентрации пищи. Эта сезонность исто-
рически обусловила строгую регулярность самих миг-
раций: киты научились точно и в сроки выходить в
области, максимально богатые кормами.
В настоящее время из китообразных эволюционный
расцвет переживает семейство дельфинов.
Наука очень мало знала о биологии китообразных
вообще. Лучше были изучены крупные китообразные,
которых промышляли с давних пор, а мелкими инте-
ресовались мало. Но положение за последние десятиле-
тия резко изменилось в связи с тем, что мелких кито-
образных стали содержать в неволе. В годы войны
дельфины не раз вызывали ложные тревоги военно-
морских звукометрических станций, которые принима-
ли голоса дельфиньих стад за звуки вражеских под-
водных лодок. Для предотвращения дорогостоящих
ошибок в такой ситуации возникла необходимость
срочно исследовать биогенные звуки моря, а в связи
с этим и поведение мелких китообразных.
Ведущую роль в изучении дельфинов сыграли боль-
шие океанариумы — искусственные бассейны с мор-
ской водой на берегу моря, предназначенные для дли-
тельного содержания морских млекопитающих.
Глава IV
ДЕЛЬФИНЫ
В ОКЕАНСКИХ
ЦИРКАХ
Попытки содержать китообразных в неволе пред-
принимались давно. Сто лет назад удачный опыт та-
кого рода был проведен с самцом белухи, пойманным
в заливе Св. Лаврентия. В искусственном бассейне го-
рода Бостона этот самец прожил два года: он сам под-
плывал за пищей, позволял себя гладить и, запряжен-
ный в тележку, для развлечения публики возил одну
даму вокруг бассейна.
Удачно содержал дельфинов Нью-Йоркский аква-
риум в 1912—1913 годах. Организатор этой затеи аме-
риканский зоолог Чарлз Таунсенд связался с дельфи-
новыми промыслами. На мысе Гаттерас уже сотню лет
добывали дельфинов ради коммерческих целей. От-
сюда и вывезли партию афалин, которых полюбила
нью-йоркская публика за их жизнерадостный нрав и
прыжки. Но животные прожили в аквариуме лишь
21 месяц и все погибли от кожной болезни.
В 1936—1937 годах в Туркмении гастролировал
передвижной цирк «Адыгей». В цирке показывали об-
ученных дельфинов, добытых в Черном море. Во время
аттракциона артистка плавала с животными и корми-
ла их с рук рыбой. Дельфины содержались в пресной
воде и лишь иногда для них удавалось привозить мор-
скую воду.
Об этом первом у нас публичном выступлении
дельфинов рассказал бывший артист цирка «Адыгей»
М. И. Старчевский.
Лишь в 1938 году в курортном городке Сент-Ога-
стин во Флориде открылся первый океанариум, назван-
ный «Морской студией».
25
Диаметр бассейна был 23 м и глубина 3,7 м. Вна-
чале надеялись привлекать публику причудливыми
тропическими рыбами. Но как только сюда поместили
дельфинов, они затмили славу разноцветных рыб, мас-
сивных морских черепах, лангустов, гигантских три-
дакн. Большой поток зрителей на водные представле-
ния с участием дельфинов убедил в исключительной
выгодности этих мероприятий.
Сюда был приглашен из Гамбурга дрессировщик
морских львов Адольф Фрон, благодаря которому этот
океанариум превратился в прибыльный дельфиний
цирк.
Выяснилось, что дельфины быстро привыкают к не-
воле, свободно принимают пищу из рук человека, лег-
ко обучаются различным трюкам. Используя классиче-
ский метод дрессировки с последующим вознагражде-
нием, А. Фрон отработал ряд номеров и с большим
успехом выступал перед публикой.
Пример с «Морской студией» оказался заразитель-
ным, и океанариумы с прирученными дельфинами ста-
ли открываться один за другим. Ныне сооружения для
содержания дельфинов — дельфинарии или океанариу-
мы имеются в 20 странах: в США более 13 (в том чис-
ле на Гавайских островах, близ Гонолулу), в Японии
21 (наибольший в г. Эносима), 15 в Англии (в Лондоне,
Брайтоне, Ланкастере, Морекамбе, Клитхорпсе, Винд-
зоре и др.), шесть в Австралии (в том числе в Сиднее
и Брисбене), пять в ФРГ (Дуйсбурге, Гамбурге и дру-
гих городах), восемь в Канаде, три в Индии. По два
океанариума функционируют в Южной Африке
(в Порт-Элизабете и Дурбане) и Голландии; по одному
имеют Новая Зеландия (Нейпир), Норвегия (Берген),
Дания (Стриб), Монако. В СССР также построен бас-
сейн для дельфинов (рис. 7 вкл.)
Наибольший опыт содержания дельфинов в неволе
накопили США (с 1938 г.) и Япония (с 1954 г.).
В США в неволе находилось 16 видов китообразных.
Известность получили океанариумы «Морская сту-
дия», «Морской аквариум» в г. Майами (построен
в 1955 г.; круглый бассейн диаметром 24 м и глуби-
ной 7 м); «Морской театр» в местечке Исламерада во
Флориде; Флоридский гальфариум в Форт Уэлтон Бич
на берегу Мексиканского залива.
26
Самый большой в мире калифорнийский океанари-
ум «Мэриленд» (рис. 8 вкл.) сооружен в 1954 году
близ Лос-Анджелеса в местечке Палос-Верде. В этом
океанариуме 14 бассейнов, из них три огромных — ем-
костью 2—2,5 тыс. куб. метров чистой морской воды.
В них и показывают морских млекопитающих — дель-
финов и ластоногих. На трибунах вокруг самого боль-
шого бассейна может разместиться около 3000 зрите-
лей. В океанариуме содержат девять видов дельфинов,
из них шесть видов мелких и средних (афалина, тихо-
океанский короткоголовый дельфин, вертящийся про-
дельфин, пятнистый продельфин и др.) и три вида
крупных (косатка, гринда и малая косатка). За день
дельфинов посещают около 18 тысяч зрителей. Для
своих морских обитателей океанариум покупает 165 г
свежемороженой рыбы в год. Фильтрационная система
пропускает в танки ежедневно 45 тыс. куб. метров чис-
той морской воды. Столько воды потребляет за сутки
современный город со 160-тысячным населением. Не-
сколько меньших дельфинариев имеется в американ-
ских городах Сан-Диего («Морской мир»), Чикаго,
Сиэтл, Нью-Йорк («Ниагара-Фолс»). В последнем со-
держали даже нарвалов и белух. Построены также
морские лаборатории с бассейнами для дельфинов в
Вудс-Холе, на островах Бимини и Сент-Томас.
В Сан-Франциско в «Голден Гейт-Парке» в большом
аквариуме отдельно от морских дельфинов содержат
пресноводных амазонских иний. Стеклянные стенки
позволяют посетителям наблюдать за подводными дви-
жениями дельфинов и слышать их голоса, усиливае-
мые через специальные приборы.
В Японии, по сообщению доктора Масаюки Нака-
дзима, содержали 14 видов китообразных в 21 бассей-
не разных городов страны. Восемь из этих бассей-
нов — вполне современные океанариумы. В Эносим-
ском дельфинарии с 1957 года побывали 118 особей
девяти видов китообразных, в том числе такие редкие,
как морщинистозубый дельфин, малайский продель-
фин, бесклювый дельфин, тропическая гринда. В дру-
гом известном японском океанариуме в г. Ито с
1958 года содержалось восемь видов китообразных,
среди них карликовая косатка, малайский и полоса-
тый продельфин. Построены и действуют аквариумы и
дельфинарии также в городах Симоносеки, Токусима,
27
Нагасаки, Хиросима, Осака, Нумазу (Мито), Киото,
Симода, Тоба, Сасебо, Яцу, Акаси, Абасири. Пока толь-
ко в одной Японии содержали бесперую морскую
свинью, оказавшуюся более способной к обучению, чем
близкая к ней обыкновенная морская свинья (рис. 9
вкл.).
Мировой опыт содержания китообразных в неволе
показывает, что лучше всех для этого приспособлена
афалина. Она хорошо контактирует с человеком и жи-
вотными (например, с обезьяной, рис. 10 вкл.), любо-
знательна, смела, но легко утомляется. Гринды, по оп-
ределению Карен Прайор, спокойны, иногда сонливы и
упрямы, медленно меняют приобретенные привычки,
крупные самцы неуклюжи в движениях и обычно
сердиты. Малые косатки оживленны, быстро возбуди-
мы, охотно сближаются с другими китообразными и
с людьми, но иногда и агрессивны. Морщинистозубые
дельфины более смелы и любопытны, чем афалины,
пригодны для выполнения сложных задач, но пред-
ставляют потенциальную опасность для пловца из-за
своего темперамента, злопамятности и обидчивости.
Длинномордые дельфины застенчивы, легко уязвимы в
присутствии других дельфинов; это прекрасные акро-
баты, свободно обучаются прыгать и переносить объек-
ты или нажимать на рычаг. Вертящиеся дельфины без
затруднения обучаются играть и плавать с человеком,
не причиняя ему вреда, легко копируют его движения,
что позволяет использовать их для подводного бале-
та. Сравнительно плохо переносят неволю морские
свиньи — они флегматичны, малоподвижны, слабо
контактируют с человеком.
В неволе жили даже усатые киты. В г. Мито в бас-
сейне длиной 70 м и глубиной 12 м, отгороженном от
моря металлической сетью, три раза содержали мало-
го полосатика сроком от двух недель до трех месяцев.
Кит питался здесь скумбрией и всегда плавал по кру-
гу, выныривая в одном и том же месте.
Первыми из усатых китов побывали в океанариу-
мах детеныши серых китов с кличками Гиги-1 и Ги-
ги-2. Гиги-1 — самка месячного возраста длиной 5 м
и весом 1,7 т — была загарпунена в 1965 году в лагу-
не Скаммона. Детеныша опутали сетью, подняли на
судно и через пять дней в купальном бассейне доста-
вили в г. Сан-Диего (Калифорния). На шестой день пос-
28
ле ранения из его тела извлекли гарпун; только тогда
китенок принял пищу — 15 л сливок, введенных ему
с помощью шланга, 15 кг головоногих моллюсков, 4 кг
пеламиды, витамины, минеральное масло и антибиоти-
ки. Кит хорошо переносил неволю, но погиб' через
45 дней от инфекции.
Другой — двухмесячный сосунок Гиги-2 (тоже сам-
ка) — в океанариуме того же города провел 12 меся-
цев и 13 марта 1972 года был выпущен обратно в род-
ную стихию. За это время он увеличился в длину на
231 см (с 584 до 815 см) и в весе на 4,4 т (с 1952 до
6350 кг).
В современных океанариумах дельфины разных ви-
дов демонстрируют перед многочисленными зрителями
различные трюки: метко бросают мяч в баскетболь-
ную корзину, высоко (до 5—6 м) выпрыгивают из
воды, делают стойку на голове, ходят на хвосте, вы-
ставившись всем телом из воды, буксируют пловцов,
звонят в колокол и многие другие.
Интересную программу представлений дельфинов
демонстрирует в трех бассейнах Калифорнийский оке-
анариум. В одном зритель видит кормление дельфинов
и их беспорядочные одиночные, парные и групповые
прыжки. Во втором дается настоящее представление
самых крупных дельфинов — косаток: два дрессиров-
щика верхом на спинах двух огромных, 5—6-метро-
вых хищников мчатся по кругу вдоль стенок бассей-
на, а третье животное без седока сопровождает их. За-
тем одна косатка почти целиком вертикально вылета-
ет из воды и падает в туче брызг; она подплывает к
борту бассейна и выставляет морду с широко раскры-
той пастью, в которую дрессировщик кладет свою го-
лову. Потом хищный кит выбрасывает из воды хвост и
размахивает им, как бы приветствуя публику. Нако-
нец, в третьем бассейне-танке выступают дрессирован-
ные афалины, гринды и полосатые дельфины. Живот-
ные совершают согласованные групповые прыжки
(рис. 11 вкл.), катают на лодке собаку, играют в мяч
и, глядя в ноты, «поют» в микрофон, прыгают через
горящий обруч или турники. Одна из афалин на брюхе
выползает из воды на наклонную сцену и издает гром-
кие сигналы в микрофон. Другая афалина, повернув-
шись брюхом вверх, постепенно выходит на стойку и,
опираясь только на хвостовые лопасти, шесть секунд
29
двигается задом наперед в вертикальном положении.
В нашей стране дельфинарий начал функциониро-
вать в городе Батуми 6 ноября 1974 года при Грузин-
ском отделении ВНИРО (рис. 12 и 13 вкл.). Располо-
жен он в 200 м от берега моря. Центральное место в
нем занимают три бассейна общей емкостью около
3500 куб. метров: большой в виде прямоугольника
(размером 36 X 14 м и глубиной 5 м) и два круглых
(диаметром по 10 м), соединенных с первым коротки-
ми шлюзами. Вдоль большой стороны бассейна напро-
тив шлюзов трибуна для зрителей (пять рядов,
520 мест). Между двумя бассейнами построена будка
для наблюдений. С крыши будки на высоте 3 м над
водой выдвигаются мостки, с них тренер дает рыбу
выпрыгивающим дельфинам. В бетонных стенках дель-
финария в два этажа врезаны 32 смотровых окна:
к верхним можно подойти по широкому коридору, под-
нявшись по лестнице, а к нижним — прямо с земли.
Мощные насосы заполняют бассейны морской водой
за 6 ч, а вся вода вытекает за 2 ч. Таким образом за
сутки вода в дельфинарии может обновляться 4 раза.
Для Батумского дельфинария афалин ловят у бере-
гов Северного Кавказа. Перевозка их требует особой
осторожности. Лучшие результаты достигнуты при
транспортировке их в ваннах с водой.
Дельфины в Батумском дельфинарии зимуют под
открытым небом, без подогрева воды (в декабре темпе-
ратура воды в бассейне не опускается ниже 15° С). На-
учный руководитель дельфинария обучает их простым
и сложным навыкам с помощью жестов. Общее на-
правление работы этого учреждения демонстрацион-
ное и научно-исследовательское.
В большом бассейне живет группа афалин, которые
резвятся, играют в мяч, подбрасывая его в воздух или
притапливая, таскают разные предметы и отбирают их
ДРУГ У Друга. Нередко животные подражают один дру-
гому, например, один из них начинает громко шлепать
на ходу своим хвостом, его тут же копирует один дель-
фин, затем второй, третий, четвертый, и каждый бьет
по 3—4 раза. В круглые бассейны отсаживают забо-
левших дельфинов или тех, с которыми отрабатывают
номера выступлений.
Животные демонстрируют перед публикой цирко-
вую программу: наперегонки плывут за цветными
30
кольцами, которые тренер бросает на середину бассей-
на, надевают их на клюв и спешат вернуть их челове-
ку. Нечто подобное проделывают с мячом: завладев
им, они помещают его на клюв, затем, тонко баланси-
руя, подплывают к баскетбольной корзине и резким
кивком головы точно закидывают мяч в цель. Потом
выбрасываются на помост, расположенный в четверти
метра над водой, либо достают с него кольцо, надева-
ют на шею и, сползая в воду, отдают его в руки дрес-
сировщика.
Шумный успех вызывает буксировка пловца, кото-
рый, заняв место между двумя дельфинами и ухватив-
шись за их спинные плавники, мчится на этой паре
через весь бассейн и возвращается к плотику (рис. 14
вкл.). Демонстрируется буксировка ярко-желтой лодки
со стоящим в ней человеком. Затем следуют синхрон-
ные прыжки трех дельфинов через подвешенные над
водой обручи.
В заключение программы необычные артисты
устремляются к высоко поднятому мостику, на кото-
ром стоит тренер и награждает рыбой тех, кто выпры-
гивает на высоту 3 м (рис. 15 вкл.).
С давних пор в цирке занимаются дрессировкой ди-
ких животных. Дельфины, поощряемые лакомством,
быстро обучаются отвечать на голос дрессировщика,
его свист, жесты, запоминают свою кличку и реагиру-
ют на нее, подплывают на зов и постепенно выполня-
ют все более и более усложняющиеся действия. Опыт-
ные дрессировщики безошибочно узнают настроение
дельфинов по едва заметным выражениям их глаз, рта,
положению тела. Сообразительность животных зависит
от индивидуальных особенностей, и некоторые дель-
фины тонко понимают, чего от них хотят.
Обучение начинается так: дельфину бросают рыбу
после каждого свистка, и когда свисток прочно ассо-
циируется с появлением пищи, дрессировщик обучает
животное другим реакциям на тот же сигнал, например
прыжкам из воды. Тренер выжидает, когда дельфин
начнет выпрыгивать по своему желанию, тогда пода-
ется свисток и бросается рыба. Так повторяется много
раз. В итоге прыжки связываются в мозгу дельфина
с наградой — рыбой. Способные «ученики» переходят
к освоению более сложных навыков — учатся бросать
мяч, прыгать через обручи.
31
Тренировкой достигается удивительная точность,
например, в прыжках двух афалин, которые по сигна-
лу мчатся навстречу друг другу и, сблизившись в цент-
ре одновременно, в доли секунды свечкой вылетают
вверх. Дельфины берут своим клювом рыбу из рук или
сигару изо рта дрессировщика на высоте 4 м. Развив
высшую скорость, они в нужный момент с большой
точностью, поражающей зрителей, вылетают из воды
вверх, успевая скорректировать направление в тот мо-
мент, пока хвост еще не оторвался от воды. На Гавай-
ских островах вблизи Гонолулу в особом бассейне,
напоминающем тропическую лагуну, демонстрируют
одновременно прыгающее стадо из шести малайских
продельфинов. Иногда такой ансамбль составляют
дельфины нескольких видов.
Обучить животное гораздо легче, если требуемое от
них действие совпадает с обычными его движениями.
Поэтому обучение начинается с использования естест-
венных способностей животного, с усиления какой-ли-
бо его привычки, с совершенствования того, что зало-
жено в его природе. Так, у дельфинов чаще всего раз-
вивают их склонность к прыжкам (рис. 16 вкл.). Для
отработки некоторых навыков нужно длительное вре-
мя. Несколько месяцев требуется афалине, чтобы ос-
воить прыжки через обруч. Вначале, вознаграждая ла-
комством, ее заставляют проплывать через обруч близ
поверхности воды, потом через обруч, едва поднятый
над водой. Потом высоту прыжка наращивают, подни-
мая обруч все выше и выше.
Дельфины (афалины, гринды, косатки) менее по-
слушны в период полового возбуждения.
При дрессировке нельзя допускать перенапряжения
животного (стресс-фактор). Это может сорвать обуче-
ние. Дрессировщик, отрабатывая какой-либо номер, от-
бирает наиболее подходящих для этого исполнителей.
Тренировку дельфинов в океанариумах стремятся на-
чинать в самом молодом возрасте, почти сразу после
их рождения. Но часто приходится иметь дело со
взрослыми особями, недавно пойманными в море.
В этом случае большую помощь дрессировщикам ока-
зывает подсадка к диким животным уже обученных
дельфинов, которым начинают подражать новички.
Рефлекс подражания у дельфинов развит превос-
ходно, и он определяет успех дрессировки, так как по-
32
зволяет случайную реакцию сделать достоянием всей
группы. Если один дельфин в стайке научился приги-
бать сеть и таким путем выходить из окружения, опыт
тут же перенимают другие. Дрессировщик Н. С. Ба-
рышников заснял на кинопленку, как афалина пере-
ползала из одного смежно расположенного бассейна з
другой через низкий барьер, возвышающийся на 5 см
над водой, и как вслед за ней то же самое делала дру-
гая. Подражание значительно сокращает сроки обуче-
ния дельфинов, а те, которые долго находились вместе
с дрессируемыми сородичами, могли выполнять неко-
торые действия даже без специальной тренировки.
В Калифорнийском океанариуме малая косатка об-
училась трюкам только на основе наблюдения за свои-
ми компаньонами-афалинами, с которыми работали
тренеры. В другом случае атлантические афалины, по-
мещенные в один бассейн с тропическими вертящими-
ся продельфинами, переняли у них манеру вращаться
по оси во время высокого прыжка в воздух. Поскольку
ареалы обоих видов не совпадают, животные не могли
обучиться этому приему на воле. Еще пример подра-
жания: во Флоридском дельфинарии одна афалина в
прыжке зубами схватывала мяч и таким способом под-
нимала флаг перед представлением. Этого дельфина
перевели в другое место и для выполнения того же но-
мера использовали вторую афалину. Однако из-за
ошибки в тренировке новый исполнитель стал подни-
мать флаг иначе — ударом по мячу мордой. Вскоре
этот дублер погиб, а его номер перешел к третьему
дельфину, жившему вместе с погибшим в одном танке.
Третий исполнитель перенял те же ошибки, которые
допустил его предшественник, и тоже стал поднимать
флаг с помощью удара по мячу.
Для океанариумов и лабораторий в настоящее вре-
мя отлавливают сотни дельфинов разных видов. Су-
ществует несколько способов лова. Из них очень по-
пулярным был отлов живых дельфинов с помощью
кошелькового невода (аломана). Несколько рыбацких
судов и лодок окружали стадо мелких китообразных
в море, потом расставляли полукругом невод, загоня-
ли туда животных и смыкали концы этой подковы.
При загоне стада ловцы на лодках стучали булыжни-
ками под водой: тем самым они мешали косяку дель-
финов уходить за пределы кольца, образованного лод-
2 9-1984
33
ками, и загоняли их в расставленный невод. Стук кам-
ней вызывал у дельфинов бегство в противоположную
сторону от устрашающего звука. Выход стада через
все более и более суживающиеся ворота также пред-
упреждали ударами булыжников. Жившие в морском
вольере мелкие дельфины — морские свиньи Машка
и Витя сильно вздрагивали в ответ на стук камней,
находясь в 15—20 м от источника звука; их дыха-
тельная пауза после первого удара возрастала в 2—3
раза против обычной. На последующие удары они
реагировали слабее. Столь же остро реагировали на
стук булыжников и дельфины-белобочки.
Постукивание камней применяют туземцы Соло-
моновых островов в Тихом океане на острове Малаита
при лове малайских продельфинов. Жители островов
используют зубы дельфинов как сырье для изготовле-
ния ожерелий и как денежную единицу в местной тор-
говле. Туземцы выезжают в море на быстроходных
пирогах, но без сетей и, выстроившись фронтом, гонят
животных к берегу. Стук камней вызывает панику в
стаде, и оно мчится прочь от лодок к суше. Очутив-
шись близ берега на мелководье, дельфины встают вер-
тикально головой вниз, как бы пытаясь зарыться ры-
лом в песок. Таких животных жители вытаскивают
из воды.
Групповой лов дельфинов — дорогостоящее пред-
приятие. В нем участвует много людей. Поэтому сейчас
за рубежом разрабатываются способы индивидуально-
го лова во время игр дельфинов возле судов. Для этого
чаще применяют небольшую сеть: с помощью шеста
ее ухитряются надевать на голову животного с быстро
идущего катера. Пользуются и газовой пушкой, из
которой выстреливают сеть в воду, когда животное
стремительно проносится возле носа судна и по инер-
ции заплывает в снасть.
Шансы на успех увеличиваются, когда дельфин
оседлывает корабельную волну и плывет, не работая
хвостом; с бака на него бросают специальную клеш-
ню-хапун, которая автоматически схватывает живот-
ное за суженную часть хвоста.
Для новозеландского и австралийского океанариу-
мов афалин ловят в мелких и узких заливах, куда они
заходят питаться. Выход из залива перегораживают
крупноячеистым неводом, а затем животных опуты-
34
вают маленькой сетью и с помощью более крупной вы-
таскивают на берег.
Оригинально ловит афалин сотрудник Флоридского
океанариума Мильтон Сантини: с катера, идущего на
полном ходу, он бросается с ручной сетью на спину
афалины; ухватившись за спинной плавник и прижи-
маясь к дельфину, он гладит и успокаивает его, а по-
том накидывает на него сеть.
Извлеченные из воды небольшие и средние дельфи-
ны обычно лежат спокойно, не сопротивляются, не
бьются, не наносят себе повреждений. Гринда борется
до тех пор, пока ее хвост находится в воде, но прекра-
щает борьбу, если он вытащен из воды. Тепловой удар
и шок часто мешают отлову и последующему исполь-
зованию дельфинов. Близ Сухуми был такой случай:
совершенно здоровый дельфин-белобочка, проплываю-
щий под мостками причала, был схвачен за спинной и
грудной плавники и вытащен из моря; он погиб через
четверть часа от эмоционального шока и теплового
Удара.
Из-за этих причин многие пойманные дельфины по-
гибают еще по дороге к бассейну. Поэтому нужны
строгие меры предосторожности как при лове, так и
при их транспортировке. Крупных животных типа
гринд или белух поднимают из воды на мягких носил-
ках (рис. 17 вкл.). Это предохраняет животных от
травм, которые впоследствии могут вызывать тяжелые
заболевания. Дельфинов среднего размера, таких, как
афалины, успешно перевозят в ваннах с морской во-
дой. При транспортировке без воды можно использо-
вать пенопластовые матрасы или висячие гамаки, пред-
охраняющие животных от ушибов. С помощью губки
или тряпки дельфинов обильно смачивают водой. Не
допускают облучения животных солнечными лучами,
а если это происходит, то место ожога срочно смазыва-
ют спермацетом или минеральным маслом. При соблю-
дении мер предосторожности дельфинов можно пере-
возить без воды двое, а в ящиках с водой — трое суток.
Опасно перевозить их в ваннах, окрашенных суриком,
так как может произойти свинцовое отравление.
Вошла в практику перевозка дельфинов самолета-
ми. Давление в кабине самолета не должно опускаться
ниже уровня, какой бывает в 1,5 км от земли вне са-
молета. В противном случае животные начинают бить-
2*
35
ся, тяжело дышать, обдирают кожу на голове и плав-
никах, стонут, мочатся, вздрагивают. Эти симптомы
исчезают, когда самолет снижается до соответствую-
щей высоты. Однажды после 7 ч полета и 9 ч езды на
автомашине из' Нью-Йорка в Амстердам были достав-
лены афалины, и в хорошем состоянии.
Во время длительной перевозки глаза дельфинов
нужно закрывать влажной тканью во избежание появ-
ления катаракты, а самих животных, если они лежат
на полу, переворачивать, чтобы не было пролежней и
омертвения кожи, что чаще бывает на груди между
плавниками.
Афалины могут жить в бассейнах различной глу-
бины, но не менее 0,5 м. Стенки водоема должны быть
гладкими, без выступов и углов, а вода — чистой и
морской. В пресной воде у морских дельфинов уже че-
рез 7—10 дней начинает сползать кожа, затем мутне-
ют глаза. В полупресных водах, где соленость пони-
жалась на 1% по сравнению с океанской, афалина ис-
пытывала недомогание, кожа ее огрубевала, но все это
проходило через 1—2 дня, если животное помещали
в морскую воду.
В неволе дельфины болеют различными нервными,
желудочными и многими другими болезнями, иногда
гибнут от стафилококковой инфекции, рожистых вос-
палений, инфарктов и инсультов, злокачественных
прыщей, опухолей и цирроза печени, абсцессов и вос-
паления легких. Признаком заболевания органов дыха-
ния служит выделяющаяся из дыхала черная зловон-
ная слизь. Повреждения скелета возникают вследствие
ушибов, ударов, травм, получаемых при транспорти-
ровке или в результате драк, вызываемых подсадкой
в одну и ту же группу нескольких взрослых самцов.
Случалось, что афалины гибли спустя месяцы пос-
ле доставки — от тромбоза сердца, пневмонии, легоч-
ных паразитов, пищевого отравления мороженой ры-
бой и других болезней. Тяжелые формы кожного забо-
левания афалин в Батумском дельфинарии, например,
сопровождались плотным смыканием век одного или
обоих глаз и в редких случаях S-образным искривле-
нием позвоночника в области хвостового стебля
(рис. 4). Больным дельфинам дают проглатывать ви-
тамины (ундевит, С и пангексавит), вкладываемые в
рыбу; вводят внутримышечно бициллин-3, бициллин-5,
36
стрептомицин и часами выдерживают заболевших в
ванне с раствором марганцевокислого калия.
В практике зарубежных океанариумов дельфинов,
вновь поступающих в танк (бассейн), иммунизируют:
вводят в кровь различные антибиотики, и повторяют
это через каждые полгода.
Большую опасность для дельфинов представляет их
склонность заглатывать инородные предметы. В Кали-
форнийском океанариуме по этой причине погибли во-
семь обученных дельфинов. Директор Эносимского
океанариума Масаюки Накадзима сообщил о судьбе
118 дельфинов девяти разных видов, живших здесь в
1957—1965 годах; 92 из них погибли от различных
болезней и в том числе 18 особей пяти видов пали из-
за проглоченных инородных тел. Заглатывались такие
предметы, как полиэтиленовые мешки, нейлоновые ве-
ревки, носки, резиновые мячи и трубки, пластмассо-
вые игрушки, куски бетона, гравий, тряпки, проволока,
ключи от двери, рыболовные крючья, перья, монеты,
гребни, пробки, консервные банки, металлические
крышки от бутылок, футляр от фотоаппарата н т. д.
Наибольший вес проглоченного предмета у афалины
длиной 279 см достигал 5 кг (стеклянная бутылка).
Заглатывание предметов не зависело от возраста жи-
вотного и длительности его пребывания в неволе.
В Калифорнийском океанариуме гринда проглотила
резиновую трубку размером 45 X Ю см, а афалина —
веретеновидный стержень величиной 30 X 4,5 см.
В случае заглатывания инородных предметов грин-
Рис. 4. Искривление хвостового стебля самца-афалины после зи-
мовки в неподогреваемом бассейне.
37
ды, малые косатки, афалины иногда сами подплывают
к человеку, принимают от него помощь и позволяют
засовывать руку глубоко в глотку (рис. 18 вкл.). Мел-
кие предметы, такие, как свисток, выходили из кишеч-
ника через сутки, после того как животным давали
минеральное масло.
В том же океанариуме калифорнийские зоологи Да-
вид и Мельба Колдуэллы описали странный случай.
Самка обыкновенного дельфина, сопровождаемая груп-
пой тихоокеанских дельфинов, таскала своего мертво-
рожденного детеныша, пока его не отбил огромный
самец гринды. Он носил маленький труп в зубах в те-
чение 30 мин, а затем его проглотил.
В море дельфины заглатывают инородные предметы
крайне редко, о чем свидетельствуют результаты на-
ших многочисленных вскрытий желудков дельфинов-
белобочек.
Поведение животных в природе, конечно, бывает
иным, чем в искусственных бассейнах: привычные ре-
акции в тисках узкого пространства в какой-то мере
искажаются даже в самых больших океанариумах.
Что меняется в этологии (поведении) китообразных и
как они живут в неволе?
Глава V
ПОВЕДЕНИЕ
УЗНИКОВ
Жизнь китового или дельфинового стада в море
очень сложна, и судить о ней можно лишь изучив
взаимоотношения особей внутри семейных и иных
группировок. Но пока практически сделать это возмож-
но только на дельфинах-узниках в условиях неволи.
Дельфины-новички в аквариуме свистят в течение
почти двух недель, притом ночью чаще, чем днем.
Привыкнув к новым условиям, они издаю!1 звук реже,
но всегда это делают'при возбуждении: в погоне за
рыбой или когда их тревожат. Встревоженные, они
плавают стайкой и в первое время издают громкий и
частый свист. Отделенный в такой момент от группы,
возбужденный дельфин стремится как можно скорее
соединиться с остальными. Когда возбуждение прохо-
дит, стайка распадается и свист прекращается. Силь-
нейший шум афалины поднимают при кормежке и ког-
да к ним пускают акул. Ко всякому новому предмету,
появляющемуся в океанариуме, необученные дельфи-
ны относятся настороженно в течение одного или не-
скольких дней.
Дельфины, обжившиеся в океанариумах, ведут себя
весьма разнообразно: они резвятся, играют, ловят до-
бычу и принимают пищу, спариваются и рождают де-
тенышей, воспитывают потомство, проявляют агрессию
по отношению к своим сородичам и редко — по отно-
шению к человеку. Различают несколько форм их по-
ведения : кормовое, игровое, половое, иерархическое,
агрессивное. Рассмотрим их подробнее.
Кормовое поведение связано с добыванием пи-
щи. Приучить дельфинов кормиться в бассейне иногда
39
удается с большим трудом. Многие афалины берут ры-
бу с первого дня неволи; такие особи лучше поддают-
ся дрессировке. Более робки дельфины-белобочки и
морские свиньи. Некоторые из дельфинов долго голо-
дают (например, косатка Моби-Долл в загоне Инглиш-
Бэй в бухте г. Ванкувер начала питаться лишь на 57-й
день), а иногда отказываются от пищи и погибают.
В морских вольерах и в океанариумах дельфины
предпочитают мертвую рыбу живой, на поимку кото-
рой они должны затрачивать усилия. Это упрощает
проблему их питания, поскольку мороженую рыбу до-
ставлять легче, чем живую, но в этом случае им необ-
ходимо дополнительно давать витамины. Животных
приучают к питанию разными сортами рыбы (сельдь,
треска, мойва, горбыль, кефаль, хамса, ставрида, сма-
рида и др.) и особенно теми, какими питаются их со-
родичи в море (рис. 19 вкл.).
Суточная норма питания дельфинов сильно варьи-
рует как в зависимости от особенностей индивидуумов,
так и в зависимости от свойств вида. Канадский зоо-
лог Дэвид Сержант, обобщивший опыт кормления
дельфинов в океанариумах, установил для разных ви-
дов китообразных нормы суточного рациона (в про-
центах от веса тела): для морской свиньи — 10,8%, бе-
локрылой морской свиньи — 11,3, тихоокеанского ко-
роткоголового дельфина — 7,9%. Для более крупных
животных цифры уменьшаются: у афалины — 4,2—
5%, белухи — 5,1—5,5, малой косатки — 4,7, у гринды
и косатки — 4 %. Однако могут быть значительные от-
клонения от этих норм. Прожорливым оказался моло-
дой самец морской свиньи, или азовки, Витя, весив-
ший 20 кг: он поедал за сутки 3 кг рыбы, что состав-
ляло 15% от веса его тела.
В-неволе при готовом питании у дельфинов притуп-
ляются охотничьи рефлексы и реакции лова добычи.
Азовки Машка и Витя, жившие то в искусственном
бассейне, то в морском вольере из капроновой сети,
получали пищу 2—3 раза в день, но даже в
голодном состоянии они не обращали внимания на
многочисленные стаи рыб, плававших в вольере. Пищу
азовкам давали по сигналу — после шлепка ладонью
по воде. Схватив рыбку из рук, азовка обязательно со-
вершала в вольере полный круг диаметром 3—6 м по
часовой стрелке и только после этого подплывала за
40
следующей рыбкой. Она не изменяла этому правилу,
даже если сигнал раздавался досрочно, т. е. когда она
только начала движение по кругу. В таких случаях
азовка никогда не бросалась к предложенной рыбе
( разу, а делала это, лишь завершив полный круг. По-
пытки сбить дельфинов с такого ритма были безуспеш-
ны. Движение по кругу, чаще по часовой стрелке,—
обычный способ движения китообразных в искусствен-
ном бассейне и особенно для тех, которые родились в
море. В неволе у афалин отработалась поза выпраши-
вания пищи: животные выставляют из воды голову с
широко открытым ртом и ждут подачки (рис. 20 вкл.).
В 1968 году мы с Д. А. Морозовым впервые уста-
новили еще никем не описанное явление, замеченное
на морской свинье и оказавшееся общим для всех
видов подотряда зубатых китов — всасывавание до-
бычи. Механизм засасывания пищи осуществляется
одновременно приоткрыванием рта и втягивающим
движением языка дельфина. Вначале думали, что пи-
щу всасывают лишь дельфины с широкой мордой
(обыкновенные и бесперые морские свиньи, белухи,
серые дельфины). Но с помощью тщательно- проведен-
ной кинофотосъемки мы обнаружили это явление так-
же у дельфинов с узкой длинной мордой. Взрослой
афалине Еве бросали на поверхность воды горстями
мелкую рыбу (хамсу) и процесс ее поедания снимали
на кинопленку. На кадрах было четко видно, как афа-
лина спокойно, без суетливых движений быстро втяги-
вала рыбок, едва приоткрывая рот. Рыбки поступали
в рот головой вперед с расстояния 5—10 см от краев
клюва зверя и притом иногда сразу по 2—3 штуки.
Прием всасывания добычи сокращает срок кормеж-
ки дельфина, избавляя его от резких движений мор-
дой; в скоплениях рыбных косяков это меньше распу-
гивает рыб, а также позволяет зверю брать добычу да-
же в узком пространстве, где скованы его охотничьи
навыки. Морские свиньи Витя и,Машка и дельфин-бе-
лобочка Люська могли засасывать мертвых рыб в ван-
не на расстоянии 10 см от рыла. Когда близ головы
афалины Евы на поверхность бассейна пускали живых
рыбок поодиночке, она бросалась преследовать добы-
чу и прибегала к комбинированным действиям — и
всасыванию и хватанию с резкими поворотами головы.
Ширина открывания пасти строго определялась раз-
41
мерами засасываемого корма. Засасывание добычи бы-
вает и в естественной обстановке.
Возможно, в связи со вкусовой функцией и функ-
цией засасывания добычи язык зубатых китов снаб-
жен очень крупными нервами.
Роль подвижного языка очевидна и при кормлении
детеныша-сосунка. Дельфиненок, обхватывая языком
материнский сосок, обеспечивает герметичность заку-
порки (ни одна капля молока не проникает в воду)
и очень цепкое прикрепление: даже при крутых и не-
ожиданных поворотах матери детеныш крепко дер-
жится за ее сосок.
Наблюдение над механизмом поедания пищи дель-
финами и анализ отснятых кинокадров дают основа-
ние заключить, что такой прием используют не только
зубатые киты — рыбоеды, но и моллюскоеды (каша-
лоты и клюворылые киты). Форма и устройство голо-
вы кашалота отлично приспособлены для выполнения
подобной функции. Незначительно отводя нижнюю че-
люсть и отодвигая язык, как поршень, назад, кашалот
засасывает в рот стремительно двигающихся скольз-
ких головоногих моллюсков. Этому способствуют по-
катые, сходящиеся вниз к краям рта «щеки», шерша-
вая поверхность ротовой полости, отсутствие верхних
зубов и длинное углубление, в которое входит нижняя
челюсть при закрытом рте. Большая длина нижней
челюсти и углубление для нее способствуют всасыва-
нию моллюсков сразу по нескольку штук за один при-
ем, быстрому наполнению огромного желудка, вме-
щающего сотни кальмаров по 20—30 см длиной.
Прием засасывания пищи позволяет кашалотам да-
же с изуродованными челюстями эффективно ловить
добычу и сохранять нормальную упитанность (рис. 21
вкл.). Предположение же о том, что кашалот при охо-
те на кальмаров плавает с широко раскрытой пастью
и привлекает их своими блестящими зубами, видимо,
лишено оснований.
Приемом всасывания пользуются и клюворылые
киты, у которых верхние зубы исчезают, а нижние со-
кращаются до 1—2 пар. Две расходящиеся складки
на их горле способствуют большой подвижности вса-
сывательного аппарата.
Фильтровальщики тоже могут засасывать добычу.
Представление о жесткости губ усатых китов, якобы
42
не способных к самостоятельному движению, оказа-
лось ложным. Детеныш серого кита Гиги-2, содержав-
шийся в океанариуме (мы о нем уже говорили), эф-
фективно подбирал головоногих моллюсков со дна бас-
сейна... с закрытым ртом! Для этого он раздвигал
небольшой участок плотно сомкнутых верхних и ниж-
них губ и, манипулируя языком, что приводило к уве-
личению полости рта, всасывал сильную струю, а вме-
сте с водой и лежащих на дне бассейна кальмаров.
Жидкость удалялась изо рта через щели между усо-
выми пластинами.
Некоторые дельфины во время охоты применяют
тактику блокирования или окружения добычи. В сен-
тябре 1969 года зоолог Д. А. Морозов с близкого рас-
< тояния с лодки пронаблюдал процесс охоты стай-
ки афалин за косяком рыбы. Косяк луфарей как
карусель кружился на одном месте, а в 100 м от них
полтора десятка дельфинов, растянувшись цепочкой,
плавали по дуге вперед-назад и не пускали рыб в от-
крытое море. От группы патрулирующих дельфинов
поочередно отделялась то одна, то другая пара и бы-
стро устремлялась в самую гущу кружащихся рыб.
Здесь оба зверя возбужденно прыгали, хватали круп-
ных луфарей, иногда подбрасывали их в воздух, а за-
тем возвращались к патрулирующей группе. Им на
смену в центр «рыбной карусели» бросалась другая
пара, потом третья и т. д. Охота продолжалась 20 мин.
Круговое движение рыбьей стаи вызывают дельфи-
ны: сначала они преследуют растянувшийся вдоль
берега рыбий косяк, уплотняют его, потом резким за-
ходом со стороны моря заставляют головную часть ко-
лонны сблизиться с хвостовой, в результате чего вытя-
нутая стая начинает двигаться по спирали, диаметр ко-
торой постепенно уменьшается. Когда передние рыбы
примкнут к самым задним, движение косяка становит-
ся круговым. Такой стоячий косяк позволяет дельфи-
нам длительное время кормиться на одном месте.
Дельфины умеют направлять рыбные косяки в нуж-
ную им сторону. Черноморские рыбаки не раз наблю-
дали, как афалины, двигаясь фронтом, загоняли кося-
ки рыб в Балаклавскую бухту через ее узкие ворота.
Дельфины способны выгонять одиночных рыб даже на
сушу. В небольшом бассейне мы выпускали вблизи
афалины Евы живых анчоусов. Дельфин, преследуя и
43
прижимая рыбок к бассейну, вынуждал их выбрасыва-
ться на выступающий над водой верхний край бетони-
рованной стенки водоема.
Много изобретательности проявляют афалины при
ловле добычи. Одна афалина на глазах рыбаков загна-
ла саргана в лощинку и вынудила его выброситься на
берег. Но, увлеченная охотой, она сама оказалась на
берегу; рыбаки сняли ее с камней и вытолкали в море.
Афалина любит подбрасывать пойманную крупную
рыбу (лобана или кефаль) высоко в воздух, чтобы схва-
тить ее на лету и перекусить на три части; середин-
ку рыбы она проглатывает сама, а голову и хвост ис-
пользуют чайки (так же перекусывают крупных рыб
и малые косатки). Привычку подбрасывания у афали-
ны, связанную с большой подвижностью головы, уси-
ливают тренировкой: кивком головы ее обучают вы-
брасывать из воды резиновые круги, забрасывать мяч
в баскетбольную сетку, подкидывать его в воздух.
Пищевые отношения у дельфинов типичны для жи-
вотных и далеки от гуманности, которую им приписы-
вают. Мельба и Давид Колдуэллы наблюдали во Фло-
ридском океанариуме отношение афалин к больному
сородичу. Он пытался брать пищу, но ее безжалостно
выхватывали прямо изо рта здоровые дельфины,
живущие в том же танке; когда один из них отходил
от больного, его место тут же занимал другой. Они так
поступали даже в тех случаях, когда пищу «вориш-
кам» давали в изобилии. «Обкрадываемого» больного
обычно изолируют от «жадных» сородичей и перево-
дят в другой танк, где он поправляется и приобретает
способность бороться за пищу в общем танке. Если
больному угрожает удушье и он подает сигнал бедст-
вия, тут начинает действовать инстинкт сохранения
вида, в силу которого сородичи спасают пострадавше-
го от гибели.
Родители обучают детенышей приемам ловли и по-
едания добычи. В водах Гавайских островов на глазах
зоологов малая косатка кормила малыша: держа во
рту рыбу-корифену, она подставляла ее детенышу, а
тот отрывал куски от рыбы и проглатывал их.
Игровое поведение наблюдается сразу же, как
только дельфины привыкнут к неволе и перестанут из-
давать тревожные свисты. Когда животные сыты, они
начинают играть. Естественная игривость — свойство
44
всех дельфинов, особенно в молодом возрасте. Обжив-
шиеся звери часами носятся по бассейну, забавляясь
подложенными для этого игрушками, проявляя боль-
шую изобретательность в играх.
В естественных условиях у дельфинов много време-
ни уходит на разыскивание и добывание корма. Био-
логический смысл игр в море состоит в тренировке,
необходимой для лова добычи и поддерживания охот-
ничьих привычек. В неволе при готовых кормах значе-
ние игр приобретает иной смысл, прежде всего как
компенсация недостаточности движений в замкнутом
пространстве. Игрой здесь заняты животные всех воз-
растов. Поводом для игры может стать любой пред-
мет, одушевленный и неодушевленный, появившийся
в океанариуме и привлекший внимание зверя: мяч, на-
тянутые веревки, резиновые игрушки. Дельфины ловят
за хвост и тут же отпускают рыб, а иногда превраща-
ют их в лохмотья, преследуют акул и скатов, иногда
отнимают предметы друг у друга. Афалины даже с
одним и тем же объектом играют по-разному. Один из
зрителей любил забавляться тем, что пускался впере-
гонки с афалиной: он бежал по борту четырехуголь-
ного водоема, а дельфин быстро плыл рядом, стремясь
обогнать человека. Во Флоридском бассейне дельфин
Флиппер часами играл с плавающей собачкой.
Животные ловко балансируют, пытаясь удерживать
па грудных плавниках или морде плавающие перья,
веточки, бумажки и т. п. Одна молодая афалина люби-
ла запускать свою морду под большую черепаху и,
поднимая ее, толкала на противоположную сторону
бассейна. Афалинам в бассейне нравится стремитель-
но мчаться на плывущего человека и внезапно оста-
навливаться в метре от него. Они любят ритмически
хлопать хвостом по поверхности воды; афалина Ева в
небольшом бассейне ловко ударяла по воде всей пе-
редней половиной тела, а хвостовую часть удерживала
вертикально в воде так, что изгибалась при ударе в ви-
де буквы Г.
Особую игру представляет у китообразных так на-
зываемый «лобтайлинг», описанный у кашалотов: ги-
ганты встают головой вниз, хвост высоко выставляют
на воздух и сильно бьют им по воде с огромным вспле-
ском и грохотом, слышным за милю. Возможно, пове-
дение Евы было аналогично лобтайлингу кашалотов.
45
Дельфинам нравится, если во время игр их повора-
чивают за хвост по оси тела. Тогда они, притапливая
ДРУГ друга, стараются поближе оказаться к забавляю-
щему их человеку.
В качестве забавы дельфины иногда пускают из
дыхала струи воды. В 1946 году мы установили, что,
если дельфинам заливать воду в дыхательные пути,
они выбрасывают ее в виде очистительных фонтанов,
при этом никакого кашля у них не возникает и попа-
дание жидкости в верхние дыхательные пути для них
совершенно безопасно. Некоторые дельфины сами ак-
тивно вбирают воду в носовой канал и выпускают ее
фонтаном. Это они делают в море, и этому же их об-
учают в неволе (рис. 22 вкл.). В американских океана-
риумах за подобные способности афалины получали
клички — Сквирт (Брызгалка), Сплеш (Плескун), Спрэй
(Разбрызгиватель). Спрэй, например, был обучен на
цирковых представлениях тушить такой струей импро-
визированный пожар или костер.
Активное всасывание воды дельфинами в природ-
ных условиях подтвердил голландский зоолог П. Ван
Бри, который нашел в носовых воздушных мешках
дельфина Гектора трех рачков-изопод, попавших из
окружающей среды.
Агрессивное поведение дельфинов наблюда-
лось в океанариумах и искусственных бассейнах по
отношению к сородичам, к особям других видов и к че-
ловеку. Проявляют агрессию обычно уже обжившиеся
дельфины, утратившие чувство страха к людям. Лишь
недавно отловленные животные робко лежат на полу
(без воды), тогда как пожившие в неволе могут, даже
в этих условиях ударить двостом. Чаще дельфины од-
ного вида нападают на представителей другого вида.
В океанариумах зарегистрированы случаи атаки афа-
линой серых, пятнистых и обыкновенных дельфинов,
гринд, карликового кашалота. Серые дельфины набра-
сывались на гринд, пятнистых и тихоокеанских корот-
коголовых дельфинов. Бывали случаи, когда дельфи-
ны в океанариумах забивали насмерть особей других
видов того же семейства.
Нередко дельфины атакуют и убивают своих есте-
ственных врагов — акул, опасных для сосунков. Даже
на теле взрослых афалин неоднократно замечали ха-
рактерные следы акульих зубов. Враждебное отноше-
46
ттие дельфинов к хищникам-рыбам остается и в неволе.
Однако связь между ними сложная и далеко не всегда
ясная. Рыбаки Флориды считают, что дельфины и аку-
лы относятся друг к другу как собаки и кошки. Афа-
лины нередко решительно избивают головой и хвостом
небольших акул, подсаженных к ним в один танк, но
сторонятся и опасаются крупных хищных рыб.
Французский океанолог Жак-Ив Кусто провел в тро-
пической Атлантике опыт: загарпунил дельфина и
пронаблюдал, как отнеслась к раненому группа акул.
Пока дельфин был жив, акулы боялись нападать, но
как только он погиб, одна из акул тотчас же набро-
силась на него и вспорола ему брюхо; не замедляя
скорости, она с хода отсекала огромные куски от тела
жертвы с такой легкостью, «словно это было сливочное
масло». На одном из исследовательских судов прона-
блюдали встречу акул и обыкновенных дельфинов: с
полсотни дельфинов-белобочек жались к бортам кораб-
ля, идущего со скоростью 3—4 узла, а вблизи нахо-
дились акулы. Создавалось впечатление, что дельфины
объединились не только для защиты своих 6—8 дете-
нышей, но и для обороны друг друга. Некоторые из
дельфинов были ранены.
Научный сотрудник В. Максимов наблюдал с тун-
це л овной базы «Солнечный луч», как на стайку отды-
хавших дельфинов напала большая акула, но жестоко
поплатилась за это. Дельфины, действуя согласованно,
выжимали хищницу из воды и заставляли ее выпрыги-
вать в воздух. За какую-то минуту тело акулы покры-
лось большим количеством ран, и она, окровавленная
и неподвижная, осталась лежать на поверхности.
Полагают, что дельфины в каждом случае способ-
ны оценить, представляют ли для них акулы угрозу.
Если хищники крупные и агрессивные, дельфины пы-
таются спасаться, но присутствие в дельфиньем стаде
новорожденных и раненых может вынудить млекопи-
тающих к яростной обороне
1 Зарубежный ученый Леонгард Шульц подсчитал, что за восемь
лет акулы в теплых морях напали на 1410 человек, из них
477 пловцов погибли. Сотрудник Калифорнийского океанариума
Давид Браун предложил помещать в одежду летчиков мини-
атюрный проигрыватель с записью сигналов бедствия дельфинов;
в случае аварии самолета над морем летчик, включив запись,
сможет привлечь дельфинов, которые отпугнут акул от его на-
дувной спасательной лодки.
47
Наиболее интересны сравнительно редкие случаи
нападения вообще-то миролюбивых дельфинов на тре-
неров. Атаковать может как здоровый дельфин, так
и больной, особенно когда человек досаждает зверю
или озлобляет его, неловко пытается помочь больному
животному. Проявление агрессивности наблюдалось
у карликовых косаток, гринд, афалин и других ви-
дов.
Карликовая косатка (самец длиной 216 см) в Гавай-
ском океанариуме бросалась на людей с типичной по-
зой угрозы («щелканье челюстями»). У гринды агрес-
сивность проявилась после 14 месяцев содержания ее
в Калифорнийском океанариуме без общения с други-
ми дельфинами; она подступила к водолазу с откры-
той пастью, сбила его взмахом хвоста и ударила так,
что тот потерял сознание. Когда эту гринду перевели
в большой бассейн, где жили ее сородичи, агрессив-
ность ее исчезла.
Знаменитая дрессированная афалина Таффи, уча-
ствовавшая в испытаниях морской лаборатории
«Силэб-2», неоднократно кусала руки своей дрессиров-
щице, обучавшей ее находить разные предметы на дне
моря, и в заключение сломала ей ребро.
Известен случай, когда две взрослые самки-афа-
лины нападали на человека, приближавшегося к дель-
финам с целью их отлова. Звери стремительно подпльь
вали к ловцу с раскрытой пастью и хвостом ударяли
по воде так, что сбивали его с ног. Некоторые иссле-
дователи замечали агрессивные реакции со стороны
афалины, когда тренер настойчиво пытался войти в
контакт с дельфином, не считаясь с утомлением жи-
вотного. Дельфин при этом мчался на человека с от-
крытой пастью, затем круто поворачивался и отшвы-
ривал тренера спинным плавником и хвостовым стеб-
лем.
Однажды специально вели наблюдения над агрес-
сивной афалиной-самцом (длиной 230 см, весом 220
кг), нападавшим на тренера во время занятий. Этот
дельфин несколько раз толкал дрессировщика рылом,
больно задевая грудным плавником, и даже ударил
его хвостовым стеблем по животу так, что человек
едва не потерял сознание. В других случаях этот
дельфин налетал на тренера с открытой пастью, угро-
жающе махал хвостом возле его лица и, подныривая,
48
ухитрялся наносить ему удар спинным плавником по
грудной клетке. Как-то тренеру бросили палку, и ею
он стукнул нападающего самца; в дополнение к этому
зверя оставили голодным. После такого урока агрес-
сивность животного исчезла. Физическим наказанием
и голодом погасили злобную реакцию дельфина. Счи-
тают, что раздраженность самца была вызвана изоля-
цией его от самок в начале гона. Возможно, было и
случайное подкрепление, когда во время нападения
дельфину удавалось схватывать выпускаемую из рук
человека рыбу.
Агрессивное поведение наблюдалось во многих
дельфинариях мира. Самка афалины по кличке Марья
(длиной 2,5 м) вскоре после пуска ее в бассейн силь-
но укусила ладонь сотрудницы биостанции, которая ко-
снулась язв на теле зверя при пересадке его из ван-
ны в бассейн. Еще до укуса Марья издавала дыхалом
короткие звуки, слышимые как «хрр-хрр-хрр».
Другой еще необученный крупный дельфин-афали-
на по кличке Федя (длиной 3 м), находясь в морском
вольере, ударил хвостом девушку в тот момент, когда
та, подплыв к сети и опершись о поплавки, выстави-
лась по пояс из воды. Девушка потеряла сознание, но
ее подхватили аквалангисты и доставили на берег.
Удары афалины испытал на себе дрессировщик
Н. С. Барышников: его избил крупный самец (рис. 23
вкл.), после того как дрессировщик удержал возбуж-
денное животное за спинной плавник и помешал ему
преследовать самку. Самец несколько раз толкал
Н. С. Барышникова спинным плавником, а затем на-
нес ему пять сильных ударов. Удар наносился кром<
кой хвостового стебля при круто согнутых вниз хвосто-
вых лопастях. Позже этот же дельфин избил еще од-
ного водолаза: у того из-под костюма торчал кусок
веревки, с помощью которой не раз отлавливали аг-
рессивное животное.
Чаще всего агрессивное поведение дельфина в от-
ношении человека заканчивается лишь демонстрацией
угрозы, но и при таких проступках бывают полезны
ответные действия, например лишение пищи и др.
Активно-принудительное общение с дельфином за-
метно сокращает срок адаптации его к человеку. Для
этого тренер часто спускается в бассейн и навязывает
себя животному в партнеры для игр. Эта назойливость
49
иногда вызывает недружелюбную реакцию дельфинов,
В такой ситуации одна афалина неоднократно опо-
рожняла кишечник вблизи тренера, как только тот
начинал ей надоедать.
Защитные реакции у дельфинов могут принимать
разные формы, помимо прямой агрессии. Суть их за-
ключается в том, чтобы помешать действиям человека,
к которым животное относится отрицательно. Иногда
подобная реакция выглядит как защита своего со-
родича. Однажды в небольшом бассейне снизили уро-
вень воды для того, чтобы поймать дельфинов для
лечения. Самец афалины Андрей атаковал врача, как
только он сделал укол (бициллин-3) самке Милке и
она издала вибрирующий свист. Врач был мгновенно
сбит с ног самцом, который, однако, не проявил агрес-
сии, когда такой же укол был сделан ему самому.
В океанариуме «Силайф-Парк» (о. Оаху) дрессиров-
щик не раз поглаживал ладонью афалину-мать, но ко-
гда он же погладил детеныша, мать ударила человека
хвостом по плечу.
Зоологи Д. Браун и Д. и М. Колдуэллы описыва-
ют, как малая косатка помогала обыкновенному дель-
фину, когда того отлавливали. Дельфин начинал сви-
стеть. В ответ на свист к ловцам подплывала малая
косатка и выталкивала дельфина из рук людей. Ко-
гда же того поймали вторично, малая косатка схвати-
ла ногу тренера и мягко сжимала ее до тех пор, пока
дельфина не отпустили.
Интересный способ воспрепятствовать отлову был
замечен в океанариуме «Мэриленд» и описан теми же
зоологами. Здесь четырем афалинам в течение двух
недель делали инъекцию — болезненный укол, воспри-
нимаемый животными весьма неохотно. Для этого их
каждый раз вылавливали в большом бассейне и пере-
водили в малый мелководный танк. Отлов производи-
ли с помощью сетчатого загона с раскрывающимися
и закрывающимися воротами. Когда появлялись люди
со шприцами и налаживали сетчатый загон, три дель-
фина пытались убегать от ловцов в дальний конец бас-
сейна, но всегда безуспешно. Четвертый же, самый со-
образительный, решил задачу иначе: он подпирал во-
рота своим массивным телом, а когда створки все же
открывали, он пытался закрывать их силой.
Из специфических форм поведения и оборонитель-
50
ных реакций, проявляемых дельфинами в неволе, об-
ращает на себя внимание и «пришпиливание» ко дну
бассейна.
Пришпиливание ко дну наблюдалось у афа-
лин в океанариумах «Морская студия» и «Мэриленд».
Афалины-самки применяли этот прием для наказания
непослушных детенышей, назойливых самцов. Но из-
вестны попытки так покарать и надоедливого трене-
ра. В «Морской студии» сосунок афалины неоднократ-
но нарушал обычный порядок и уплывал прочь от ма-
тери. В ответ на это самка, плывя над детенышем,
улучила момент и прижала его ко дну танка рылом и
так держала с полминуты.
Половая возбудимость, связанная с кожными раз-
дражениями, у афалины замечается с раннего возра-
ста, вскоре после рождения. Сосунок-самец обучается
половой игре с матерью, имея всего лишь несколько
недель от роду. Млечные железы, находящиеся по бо-
кам мочеполовой щели, привлекают внимание детены-
ша именно на этот участок тела. С другой стороны,
клюв кормящегося сосунка раздражает самку-мать.
Детеныш в такое время, как показала магнитофонная
запись, издает особый визг. В подобных случаях мать
урезонивала детеныша: кусая сосунка, она держала
его прижатым ко дну танка, где он не мог дышать ка-
кой-то период времени.
В Калифорнийском океанариуме видели двух са-
мок, толкавших лишь недавно родившихся детенышей
вниз и прижимавших их клювом ко дну бассейна.
Такие приемы эффективно заглушали возбуждение де-
теныша. Давид и Мельба Колдуэллы опубликовали
фотоснимок, на котором было показано, как две афа-
лины-самки прижали к жесткому дну водоема моло-
дого самца; это сопровождалось записанными на маг-
нитофонную ленту громкими криками наказанного.
В «Силайф-Парке» на Гавайских островах один во-
долаз нередко дразнил малую косатку, и однажды она
рылом прижала его ко дну водоема. К счастью, запа-
са воздуха хватило на те несколько минут, пока че-
ловек был пришпилен, и это не причинило ему вреда.
Аналогичный прием испытал на себе один назойли-
вый дрессировщик: он мешал плавать афалине-самцу,
удерживая его за рыло. Тогда дельфин, выбрав мо-
мент, на двухметровой глубине прижал ладонь трене-
51
ра ко дну бассейна и держал несколько секунд, пока
человек не вырвал руку. При подъеме же на поверх-
ность дельфин сопровождал этого человека и, когда
тот положил ему руку на клюв, снова сделал попыт-
ку утянуть тренера вниз.
Иерархическое поведение у дельфинов скла-
дывается в результате конкуренции из-за пищи, и оно,
видимо, чаще и резче проявляется в неволе, чем в
море. Первую попытку изучить иерархию (определен-
ный порядок подчиненности) в стаде дельфинов пред-
приняли американские зоологи М. Таволга и Ф. Эса-
пян в океанариуме «Морская студия». Обычно семейка
группируется из самца, двух или большего числа са-
мок, почти взрослых детенышей и молодняка теку-
щего сезона. В группе над всеми особями стада гос-
подствует сильнейший взрослый самец: он первым бе-
рет пищу, его все побаиваются и во всем уступают
ему. Непокорных доминирующий самец устрашает
особым жестом — широко раскрывает рот и щелкает
челюстями, а иногда кусает. Он проявляет агрессив-
ность, когда его на это провоцируют, а обычно быва-
ет миролюбив, чаще держится в одиночку и только в
брачный период плавает в сопровождении самок. Сле-
дующее место в доминировании занимают взрослые
самки. Среди них выделяется главная, наиболее актив-
ная самка, а другие, как правило, следуют за ней. На-
конец, самую покорную часть стада составляют дете-
ныши и молодые особи. Они подчиняются всем взрос-
лым членам стада.
У обыкновенных морских свиней при кормежках,
в вольере, по нашим наблюдениям, всегда доминиро-
вала более крупная самка Машка (длиной 144 см) над
молодым самцом Витей (длиной 120 см). Она оттесня-
ла его и хватала пищу первой.
О существовании у дельфинов иерархии в природ-
ной обстановке в какой-то мере свидетельствуют мно-
гочисленные, в том числе и давние, следы покусов на
теле вылавливаемых дельфинов, словно нанесенные
острыми зубьями гребенки. Дельфины пускают в ход
зубы для поддержания иерархии среди сородичей, но
еще чаще используют для этого удары хвостом и го-
ловой.
Иерархическое поведение не предотвращает драк.
Во время половой активности один доминирующий са-
52
мец сам был сильно покусан: на его хвостовом стеб-
ле, лопастях, возле гониталий и вокруг морды имелись
многочисленные следы зубов, свидетельствующие об
ответных действиях.
В период брачных игр доминирующие дельфины
жестоко преследуют соперников и сильно их кусают.
В такое время тело гонимых самцов бывает густо ис-
полосовано острыми зубами конкурентов. В Батумском
дельфинарии молодых афалин-самцов Боцмана и Мо-
ряка приходилось держать отдельно от стада, посколь-
ку к ним нетерпимо относился доминирующий самец
Персей. В декабре 1974 года нам пришлось быть сви-
детелями гибели одного отверженного самца (длиной
235 см), смерть которого, как показало вскрытие, на-
ступила от кровоизлияния в мозг в результате стрес-
сового состояния. Самец был отловлен у берегов Ана-
пы за полгода до своей гибели. До тех пор пока он не
проявлял интереса к самкам, он мирно уживался с
другими дельфинами то в большом бассейне Батум-
ского дельфинария, то в малом танке (для тюленей).
Однако в ноябре, когда начались брачные игры, меж-
ду крупными самцами возникли конфликты, и отно-
шение Персея к более слабому сопернику стало не-
выносимым. Последний отделился от группы, резко
снизил активность, полностью утратил аппетит. Все за-
кончилось инсультом в левое полушарие мозга. На
правой стороне трупа было насчитано 44 и на левой
32 сильнейших покуса. В каждом покусе было до де-
сяти глубоких параллельных борозд-ран от зубов мо-
гучего конкурента. Этот случай показывает, что нерв-
ная перегрузка, конфликты во время гона и упорное
преследование могут закончиться даже смертью угне-
тенного животного.
Половое поведение дельфинов связано с раз*
множением. Американские зоологи М. Таволга, Ф. Эса-
пян, Ф. Вуд, Г. Критцлер, А. Макбрайд и О. Хебб
вскрыли многие секреты размножения китообразных.
В океанариумах были установлены некоторые общие
черты поведения дельфинов во время гона, беремен-
ности, родов и воспитания потомства.
Способы ухаживания у разных видов дельфинов
более или менее однотипны и сводятся к контактным
действиям: при перекрестном или параллельном пла-
вании животные стараются коснуться друг друга на
53
быстром ходу, задеть партнера плавниками, прижать-
ся к его телу либо потереться мордой.
Поведение самца и самки перед спариванием (обыч-
но весной) напоминает игру в салки: животные го-
няются друг за другом, стараясь задеть тело партнера,
мягко покусывают челюстями или поглаживают один
другого головой (вроде того, как это бывает у копыт-
ных животных), а подчас высоко выпрыгивают из во-
ды. Самец нетерпимо относится к своим соперникам
и отгоняет их от самки, устрашающе открывая рот и
хлопая челюстями; перед самкой он изгибается в раз-
ных эффектных позах. В период гона и спаривания
самцы изредка визгливо взлаивают, но делают это го-
раздо чаще, если самка отплывает в сторону или при-
ближается к другому самцу.
Утробное развитие зародыша продолжается около
года. Во второй половине беременности самка уединя-
ется, утрачивает резкость движения и по мере прибли-
жения родов занимается своеобразной гимнастикой —
последовательно изгибает спину и хвостовой стебель.
На это она затрачивает десятки минут ежедневно.
Предродовая гимнастика не замечалась у афалин Ба-
тумского дельфинария, так как беременные самки до
самого дня родов выступали в демонстрационной про-
грамме. В начале родов из полового отверстия самки
показывается хвост дельфиненка; хвост может не-
сколько раз выставляться и прятаться вновь (рис. 24
и 25 вкл.). Выход плода продолжается от четверти ча-
са до двух часов и вызывает сильнейшее возбуждение
всего стада.
Пуповина рвется при первом же натяжении, из мес-
та разрыва лишь иногда выделяется несколько ка-
пель крови. Самки окружают рожающую афалину и ее
детеныша с боков, а иногда и снизу (такое же построе-
ние при родах у кашалотов наблюдалось с вертолета
в Атлантическом океане близ острова Тристан-да-
Кунья). Роль этих «тетушек-повитух» заключается в
том, чтобы, сопровождая роженицу, защищать малы-
ша от нападения акул или от атак взрослых дельфи-
нов-самцов, а при случае и оказать молодому помощь,
если он почему-либо не будет в состоянии двигаться
сам. Обычно же новорожденный, очутившись на сво-
боде, медленно плывет в окружении эскорта самостоя-
тельно, без всякой поддержки, по едва восходящей
54
линии к поверхности, чтобы через какое-то время про-
извести первый дыхательный акт при первом выныри-
вании. Этот акт осуществляется еще без всякого жиз-
ненного опыта, как безусловный рефлекс, при котором
раздражителем служит ощущение смены среды «во-
да— воздух». Хвостовые лопасти только что родивше-
гося детеныша еще свернуты в трубочки, а спинной
плавничок пригнут к телу (рис. 26 вкл.), но вскоре при
работе хвостом лопасти и плавничок распрямляются,
становятся очень упругими и в дальнейшем не опада-
ют. С этого времени малыш начинает активно двигать-
ся со взрослыми (причем работает хвостом вдвое ча-
ще, чем они), и у него стремительно нарастают по-
веденческие реакции.
Родившийся детеныш занимает определенное поло-
жение относительно матери и «тетки-повитухи»: он
плывет между двумя самками до первого выдоха-вдо-
ха, а потом все время находится в движении, подчиня-
ясь реакции следования. Такая реакция проявляется
с первых же часов, когда начинает функциониро-
вать пожизненно действующий механизм саморегуля-
ции упругости плавников. Неукоснительное местополо-
жение возле матери позволяет детенышам китообраз-
ных не терять родителя и не сбиваться при плавании
в многочисленном стаде, а также соблюдать оптималь-
ную индивидуальную дистанцию и не попадать под
удары хвоста сородичей.
Реакция следования родившегося детеныша рядом
с матерью представляет собой унаследованный реф-
лекс: пройдет не одна неделя, пока такая реакция на-
чнет ослабевать, и детеныш, усовершенствовав коорди-
нацию своих движений, получит наконец некоторую
независимость (рис. 27 вкл.). Спустя несколько часов
после родов детеныш ощупывает рылом • тело матери
(зубов у него еще нет). Выступающий из жесткого
кармана мягкий сосок он находит, руководствуясь, ве-
роятно, осязанием. Захватив кончиком рта правый
или левый сосок, детеныш получает сильную струю
молока, бьющую под давлением мышц млечной же-
лезы (рис. 28 вкл.). Вначале он кормится через каж-
дые 10—30 мин, а потом интервалы между кормежка-
ми увеличиваются. В первые две недели кормящая
самка поворачивается на бок, чтобы детенышу было
удобнее захватывать сосок (в такой момент кормящие
55
самки усатых китов нередко выставляют из воды груд-
ной плавник).
В неволе мать не позволяет малышу отплывать от
себя дальше нескольких метров и пресекает его попыт-
ки общаться с другими дельфинами, отгоняя их «щел-
каньем челюстей». Двухнедельный дельфиненок отлу-
чается от самки, но большую часть времени до самого
конца лактации все же проводит возле матери. Более
опытные самки-афалины предоставляют своему де-
тенышу большую свободу, чем первородящие: они не
бросаются на визг испуганного малыша, а сначала
оценивают обстановку, в которой был подан сигнал, и
лишь тогда предпринимают какое-либо действие.
В стаде дельфинов, где имеется одновременно две кор*
мящие самки, наблюдали, как оба сосунка остаются
под присмотром одной из матерей на то время, пока
другая кормится.
Детеныш питается очень жирным (до 54%) моло-
ком от четырех (мелкие дельфины) до 13 месяцев (ка-
шалоты), а в неволе даже до двух лет (афалины).
В дельфиновом молоке содержится жира в 13 раз и
белка в 4 раза больше, чем в молоке коровы. Поэтому
детеныш афалины растет очень быстро: через полгода
его первоначальная длина (1 —1,3 м) увеличивается до
1,8 м, через год — до 2,1 м, а через два года — до
2,5 м. Впервые малыши афалины берут рыбу в возра-
сте 3,5 месяца, а молодые гринды пробуют головоно-
гих моллюсков в 6 месяцев. Хотя в море лактацион-
ный период дельфинов, вероятно, короче, тем не менее
в сравнении со сроками других млекопитающих он
все же очень велик. Это и создает у зубатых китов бла-
гоприятные условия для обучения и перенимания опы-
та взрослых. Возможно, и у усатых китов молочный
период длится не полгода, как считали прежде, а вдвое
больше. В том уверяет южноафриканский зоолог Петер
Бест, извлекший молоко из желудка 15-метрового фин-
вала, имевшего возраст минимум около года.
В искусственных бассейнах приходилось неодно-
кратно наблюдать брачные игры между афалинами и
дельфинами-белобочками с излучением интенсивных
сигналов. Большинство исследователей относит случаи
межвидового и межродового спаривания китообраз-
ных к категории половых игр. Давид и Мельба Кол-
дуэллы пишут, что половая активность у дельфинор
56
«является как бы незапрещенной и свободно протекаю-
щей игрой».
В океанариумах привязанность между самцом и
самкой из разных родов оказывалась иногда более
сильной, чем привязанность между самцом и самкой
одного и того же вида. Этого в море, конечно, не быва-
ет. Гринда-самец Бимбо, проживший в Калифорний-
ском океанариуме три с половиной года вместе с сам-
кой тихоокеанского короткоголового дельфина Дэби,
явно предпочитал оставаться с ней, а не с самкой
гринды. Когда Дэби погибла, Бимбо, схватив ее за
хвост или грудной плавник, четыре часа носился с ней
по бассейну, широко раскрыв глаза, выталкивал ее
на поверхность и пресекал все попытки служителей
отобрать у него труп. Лишь когда погибшую, поддев
острогой, удалили из бассейна, крайнее возбуждение
самца улеглось, и он постепенно вошел в норму.
Половая игра наблюдалась между самцами как од-
ного и того же вида, так и разных видов. Это отмеча-
лось у всех трех видов черноморских дельфинов и у
молодых самцов-афалин Моряка и Боцмана в Батум-
ском дельфинарии. Гомосексуальная и мастурбацион-
ная активность отмечена у морских свиней, живущих
в бассейне города Стриб в Дании. Австралийский био-
лог Д. Э. Гаскин, установивший у кашалотов в водах
Новой Зеландии шесть типов группировок, обращает
внимание на уединяющиеся пары молодых самцов,
между которыми предполагается такая игра.
Объектами сексуальной игры у афалин могут стать
животные даже из других классов (черепахи, акулы).
Видели, как самец-афалина плавал на дне танка с уг-
рем, обвившимся вокруг копулятивного органа дельфи-
на. А однажды явственно наблюдали 6—7-летнюю
самку-белобочку, которая терлась гениталиями о склад-
ки брезента ванны и делала это так неистово,
что выплескивала воду.
Сложность и многообразие форм сексуального и иг-
рового поведения китообразных позволяют некоторым
ученым сближать этих животных по уровню развития
с обезьянами (шимпанзе).
Глава VI
АФАЛИНЫ
РАЗМНОЖАЮТСЯ
В НЕВОЛЕ
Афалина стала наиболее предпочитаемым дельфи-
ном для содержания в океанариумах не только благо*
даря своим способностям к обучению, но и вследствие
того, что она плодится в условиях неволи. Теперь уже
есть целые популяции афалин, которые никогда не ви-
дели моря.
В дельфинарии «Морская студия» афалина Пэджи
была поймана еще сосунком и в возрасте 15 лет весила
240 кг. В неволе она плодилась 4 раза, а самец того же
вида Хэппи был отцом 12 детенышей, рожденных
разными самками. Афалина Спрэй родилась в неволе
и здесь же родила первого детеныша в семилетием воз-
расте (у нее не оказалось молока, и детеныш погиб че*
рез две недели). К 1961 году в «Морской студии» уже
насчитывалось 16, а в японском океанариуме «Эноси-
ма» с 1957 по 1968 год — 25 случаев родов (3 — у се-
рого дельфина и 22 — у афалины).
В Калифорнийском и Флоридском океанариумах
США наблюдалось межродовое спаривание дельфинов.
Подобные случаи происходили между самкой обыкно-
венного дельфина и самцом афалины, самцом афали-
ны и самкой тихоокеанского короткоголового дельфи-
на, самцом тихоокеанской гринды и самкой малой ко-
сатки и, наконец, самцом амазонской инии и самкой
длинноклювого дельфина соталии. Животные в Зто
время издавали своеобразные звуки, а из дыхала вы-
рывался поток пузырьков воздуха. Гибриды от такого
спаривания не получались. Но в октябре 1971 года в
океанариуме «Силайф-Парк» впервые родился потомок
от скрещивания самца афалины с самкой морщинисто-
58
зубого дельфина, а в Японии (г. Вносима) 29 сентября
1978 года получен гибрид серого дельфина и афалины.
В Батумском дельфинарии с 1974 по 1978 год было
шесть случаев родов афалин, причем зачатие каждой
из них происходило здесь же в бассейне. Отцом всех
детенышей был крупный, доминирующий в группе са-
мец Персей. Длина новорожденных была 1—1,3 м, а
вес 10,5—15 кг. Дважды роды произошли ночью и
каждый раз сопровождались громким свистом дель-
финов ( в одном случае роды оказались преждевре-
менными). Заметим, что в стрессовой обстановке при
отлове дельфины могут абортировать как в море, так
и в океанариумах (рис. 29 вкл.).
Перед рождением дельфинят в поведении будущих
матерей не замечалось отклонений, так как все они до
последнего дня принимали участие в выступлениях пе-
ред публикой и даже исполняли все высотные прыж-
ки. Обремененные же потомством, они в корне меняли
свою этологию (поведение), подчиняясь новым обязан-
ностям воспитания молодняка. Так, например, когда
неожиданно для всех у «звезды» выступлений Васили-
сы появился детеныш, сразу было заброшено все
«искусство»: она отказалась выступать перед публи-
кой и склонила к тому же свою подругу — тетушку
новорожденного. Обе самки, к досаде зрителей, все
внимание обратили теперь на малыша, и никто не
знал, когда они вернутся на «подмостки» водной сце-
ны. Через пять недель малыш погиб. Мать целую ночь
выталкивала безжизненное тело дельфиненка, но... на-
прасно. После смерти дельфиненка Василиса за не-
сколько дней полностью восстановила свои навыки.
Послед в бассейне обнаруживали тетки-повитухи.
У одной и той же родящей самки тетка-повитуха в
разные сезоны могла меняться.
У первых двух новорожденных спинной плавник
выпрямлялся вскоре же после рождения, за несколько
десятков минут, у третьего (детеныша Василисы) —
только на второй день, а у преждевременно родившего-
ся так и не выпрямился до момента его гибели (к кон-
цу вторых суток). Вероятно, в море, где дельфины не
стеснены в движениях, значительной задержки в вы-
прямлении плавников не бывает.
Первые минуты жизни, когда формируется поведе-
ние,— наименее изученная область в биологии кито-
59
образных. Имеются лишь отдельные наблюдения в раз-
ных дельфинариях, еще не собранные воедино.
Детеныши в Батумском дельфинарии погибали че-
рез разные сроки. Первый в возрасте шести дней по-
платился жизнью из-за отсутствия молока у матери
Маши. Погибшего обнаружили в санитарный день и
хотели достать с помощью лодки. Однако Маша все
время носила его на роструме, не допускала к нему
людей, лязгала зубами и таранила лодку. Тогда в бас-
сейн спустились двое водолазов, но дельфины дружно
встали стеной, и снова усилия сотрудников дельфина-
рия остались тщетными. Дельфиненка удалось до-
стать, только спустив воду из бассейна.
Второй детеныш (у афалины Мамаши) умер от дер-
матомикоза в возрасте почти шести месяцев. За восемь
часов до смерти (17 мая 1976 г.) он плавал в малом
бассейне на поверхности воды, не заныривая и иногда
заваливаясь на бок. Мать, держась рядом с ним, пы-
талась помочь малышу, часто притапливала и вытал-
кивала его рылом, стремясь создать какую-то ритми-
ку в своих действиях. Когда малыш приближался к
сетке, отделявшей малый бассейн от большого, Мама-
ша резко отбрасывала его отсюда и не отпускала от
себя. Иногда детеныш клал рострум на спину матери,
которая тут же замирала. В поисках опоры ослабев-
ший детеныш неоднократно забрасывал свой хвост на
материнский. Потом он стал заваливаться на бок еще
чаще и по 20 секунд лежал без движения. За все это
время другие дельфины не оказывали погибающему
никакой помощи, но мать продолжала поддерживать
и выталкивать его на поверхность. За четверть часа до
кончины у детеныша последовали судороги, он начал
тонуть, и мать предприняла последние безуспешные
усилия по спасению дельфиненка. Уже погибшего, она
носила его на своей спине. При попытках людей из-
влечь труп самка таранила лодку и даже пыталась
забраться в нее, когда туда втащили дельфиненка.
Остальные дельфины, находившиеся за сетью, рвались
к месту событий в сильнейшем возбуждении.
Третий детеныш погиб в возрасте 37 дней от аб-
сцидирующей гельминтозной пневмонии, вызванной
легочным паразитом из круглых червей (Стенурус ова-
тус). Как показало вскрытие, легкие его были сильно
поражены глистами. Самка Василиса спасала (вытал-
60
кивала) погибшего детеныша до тех пор, пока труп не
вытащили из воды.
В четвертом случае причина гибели была, вероят-
но, связана с тем, что детеныш (второй у самки Мама-
ши) родился слабым, недоразвитым, имея лишь 85 см
в длину и 10,5 кг веса, вместо нормы 100 см и 16 кг.
Он прожил только двое суток. Абортирование было
вызвано, как предполагают, стрессом во время перено-
са беременной самки из большого в малый (круглый)
бассейн. Роды произошли спустя 11,5 месяцев после
гона, который был зарегистрирован и снят на кино-
пленку 13 июля 1976 года. В период гона дельфины
обычно не прекращают своих выступлений перед зри-
телями, а Персей даже совершал высотные прыжки
с выставленным эрегированным копулятивным орга-
ном. Гон продолжался три дня: он протекал периода-
ми по получасу и больше и возобновлялся несколько
раз в день. При этом животные (Персей и самки Ма-
маша, Маша и Инга) держались плотной группой, то
стремительно и беспорядочно носились в разных на-
правлениях по поверхности большого бассейна, то пла-
вали замедленно либо останавливались, раскинувшись
в разных позициях и ракурсах. Самец отдавал явное
предпочтение Мамаше, у которой наблюдалась поза
подставления: он мордой поглаживал голову избран-
ницы, мягко покусывая бока, «обнюхивал» ее генита-
лии, гонялся за ней и прыгал, а иногда в возбуждении
ударял хвостом по воде до 20 раз. Другие самки дер-
жались тут же, рядом — крест-накрест, одна над дру-
гой, касались друг друга, но при этом никакой борьбы
между «соперницами» не замечалось. Отношение мате-
ри к новорожденному в общих чертах было таким же,
как и в первых трех случаях.
Пятый детеныш (второй у самки Василисы) погиб в
середине лета 1978 года в возрасте десяти дней, имея
длину 115 см и вес 18 кг. Гибель малыша наступила
в результате травмы, возможно случайной, получен-
ной от удара Персея, когда тот проявил половую ак-
тивность к самке-матери.
Шестой детеныш в Батумском дельфинарии родил-
ся у самки Инги летом 1978 года и живет до сих пор.
Впервые наблюдали, как роль сопровождающей тетки-
повитухи выполнял... отец Персей: он заботливо отно-
сился к дельфиненку и играл с ним, а самка не отго-
61
няла его от малыша и даже принимала участие в сов*
местном с ним аттракционе. Вопреки этому самцы
афалин в других дельфинариях не вступали в контакт
с сосунками и не опекали их.
Дыхательные ритмы в первые дни и недели жизни
у всех малышей были синхронными с материнскими:
выдох-вдох у взрослого дельфина и детеныша совпадал
по времени. Видимо, мать приспосабливается к более
частому дыханию дельфиненка и укорачивает свою
естественную дыхательную паузу, которая обычно
вдвое больше, чем у сосунка. Тем не менее нами заре-
гистрировано, что детеныши-афалины уже в первые
дни могут находиться под водой более минуты.
С каждой неделей у детенышей все согласованнее
становятся движения и возрастает независимость пе-
редвижения. С третьей недели у них намечается игро-
вое поведение, они могут произвольно замедлять ско-
рость, играть с предметами, находящимися на поверх-
ности бассейна, поворачиваться на спину, плавать
автономно вдоль и поперек бассейна. Однако мать все-
таки держит их в поле зрения и в случае приближе-
ния какого-либо животного тотчас же оказывается ря-
дом с малышом. Однажды видели, как самка-мать,
перевернувшись на спину, подплыла под дельфиненка
и грудью подняла его над водой, поддерживая грудны-
ми плавниками и похлопывая ими тельце сосунка.
Возможно, это было наказание непослушного дете-
ныша.
По наблюдениям Я. И. Близнюка и К. А. Джинча-
радзе, в месячном возрасте сосунок совершает обороты
вокруг продольной оси тела, плавает на спине, произ-
водит резкие суетливые движения. К двум месяцам он
плавает в основном самостоятельно, у него развивает-
ся манипуляторная активность: берет в рот, выпуска-
ет и снова хватает рыбу, нередко следует с другими
самками, носит на роструме листья и т. д. В трехме-
сячном возрасте четко проявляется подражательная
способность, еще более улучшается координация в дви-
жениях: детеныш способен зависать в воде, внезапно
останавливаться, легко подбрасывать вверх и ловить
падающую рыбу и т. д. В четыре месяца его поведение
приближается к поведению взрослых: он может до
20—30 мин держаться отдельно от матери, приобре-
тает двигательные навыки, манипулирует предметами
62
при помощи хвостового и грудных плавников, ростру-
ма, подбрасывает вверх воду нижней челюстью, пла-
вает с открытым ртом и т. д.
Тонкие наблюдения провели Н. Р. Ванникова и
С. Д. Вронская, изучая взаимоотношения между роди-
телями и сосунками. Им удалось проследить, как сам-
ки-афалины мягко, с легким нажимом поглаживали
рострумом живот детенышей, видимо способствуя тем
самым их нормальной дефекации. Однажды явно боль-
ному детенышу со вздутым животом самка-мать про-
извела кончиком рыла серию резких толчков в приа-
нальную область дельфиненка, после чего у того вы-
шел плотный комок кала. Все это напоминает действие
наземных хищников, копытных и грызунов, которые
в подобных случаях делают массаж языком, вылизы-
вая околоанальную область сосунков.
Глава VII
ДЕЛЬФИНЫ
УСЛОЖНЯЮТ
НАВЫКИ
В Батумском дельфинарии удалось пронаблюдать
сложные формы поведения афалин.
Наблюдения над сложными формами поведения
дельфинов мы провели совместно с научным руководи-
телем Батумского дельфинария Я. И. Близнюком. Нас
интересовало, как у этих китообразных усложняется
навык на базе уже сложившихся ранее поведенческих
реакций и как в процессе обучения появляются новые
действия (рис. 30 вкл.).
В Батумском дельфинарии содержится группа из
восьми афалин. Животные были обучены приносить
бросаемые на поверхность воды разноцветные коль-
ца. Закрепленное действие — одеть на рострум лежа-
щее на поверхности воды кольцо, подплыть с ним к
человеку и подать его в руки — открыло животным
возможность проявлять собственную инициативу и
подносить самые разные предметы: подбираемые в
толще воды и на дне бассейна целлофановую пленку,
кусочки толя, капроновые шнурки и обрывки веревок,
мотки лески, ветки и листья деревьев, заносимые вет-
ром на поверхность водоема.
Особенно не просто было доставлять на роструме
кафельные плитки, которые отставали от пола бассей-
на: их дельфины находили на дне, на глубине 5 м, и
четко балансируя, на клюве подносили человеку. Так
делали Персей, самая умудренная опытом десятилет-
няя самка Василиса, самка Инга и другие.
Однажды после сильного ветра на поверхности бас-
сейна появилось много сорванных эвкалиптовых ли-
стьев, и дельфины занялись «уборкой». Они складыва-
64
ли листики на низко расположенный над водой по*
мост. Сначала это проделала одна афалина, а затем
к забаве подключились и другие. Так на плотике была
собрана небольшая кучка листьев.
В феврале 1976 года чистка бассейна не проводи-
лась и стенки его обросли водорослями. Один из дель-
финов сорвал пучок этих растений и принес человеку,
а через некоторое время то же самое стали делать
остальные афалины, кроме Персея.
После родов самки Маши (30 октября 1975 г.) ее
послед подобрала Василиса. Потаскав некоторое время
находку на своем клюве, она поднесла ее к борту во-
доема, а затем по жесту оказавшегося рядом дресси-
ровщика, резким кивком головы выбросила плаценту
к его ногам. Месяц спустя родила самка Мамаша. Ее
послед доставила к борту афалина Маша и рылом
подала скользкую ткань человеку, притом без его
команды.
Возможно, к подобным случаям относится и тот,
когда молодой самец, выпущенный в море, поймал ке-
фаль, подплыл с рыбой во рту к дрессировщику и по-
дал ее в руки. Не исключено, что такая реакция была
уже подготовлена в бассейне той своеобразной и часто
практикуемой игрой, при которой дельфины кивком
головы подбрасывали рыбу на 0,5—1 м выше уровня
воды (это нередко в естественных условиях наблюда-
ли рыбаки). Если во время таких игр рыбы не оказы-
валось, афалины отрыгивали ее из своего желудка, и
в тот же момент на поверхность поднимался пузырь
воздуха объемом до литра. Особенно любила так забав-
ляться Василиса: отрыгнутую рыбу после многократ-
ных подбрасываний она заглатывала и отрыгивала
бесконечное число раз, иногда кивком головы бросала
ее далеко в сторону, но тут же подхватывала и снова
подкидывала вверх, пока размочаленный кусок не рас-
падался на мелкие части.
Видимо, близким к подобным реакциям был и та-
кой случай: в одном из бассейнов афалина ловко бро-
сила изо рта медузу, которая попала в проходящего
мимо человека. Действие подкрепили, и дельфин стал
регулярно швырять медуз в находящихся поблизости
людей.
Интересно вели себя животные, когда дрессиров-
щик подвесил ведро с рыбой за сетку невысоко над во-
3 9-1984
65
дой: чуть приподнявшись из воды, они клювом накло-
няли ведро и доставали рыбу. Возможно, на основе
этого опыта один из дельфинов умудрился достать ван-
ночку с рыбой, стоявшую на помосте. Для этого он на-
давил на край ванночки кончиком рыла, опрокинул ее
и поживился добычей.
Любопытное наблюдение проведено в начале октяб-
ря 1976 года. Самку Мамашу отделили от группы и
посадили в коридор шлюза, соединяющего большой и
круглый бассейны; от остальных сородичей ее отде-
ляла капроновая сеть. Здесь Мамашу в изобилии кор-
мили рыбой. Однажды, когда она насытилась, а ей
продолжали подбрасывать ставридок, заметили, как
она стала подкармливать излишками своих сородичей:
для этого с рыбкой во рту приближалась к сетке и
сквозь ячею ловко выталкивала ее. Толпившиеся воз-
ле сетяной перегородки дельфины жадно подхватыва-
ли подачку. Это было проделано не менее 10 раз.
Особенно прославилась в дельфинарии как «непод-
ражаемый выдумщик» Василиса. В прошлом она, на-
пример, сама освоила искусство переползать из одно-
го бассейна в другой через каменный барьер высотой
около 20 см. Когда у Мамаши был еще жив детеныш
и его отсадили в коридор, мать проявила сильное бес-
покойство и старалась соединиться с ним. Однако это-
му мешал страх перед узким проходом, который нуж-
но было пройти, прежде чем попасть в коридор. На
помощь пришла Василиса: она зашла к детенышу и
вывела его из коридора в большой бассейн.
Василиса находила для своей забавы такие объек-
ты, которые обычно оставляли без внимания ее соро-
дичи. Например, она подставляла открытую пасть и
часами ловила холодные капли, падающие с сосулек,
висящих на крыше тренерской будки. Эта афалина
вносила совершенно новые оттенки в отрабатываемые
действия. Именно так она поступила с лодкой, кото-
рую обычно спускали в воду лишь на представлениях
(тогда Василиса надевала на рыло кольцо, привязан-
ное к лодке, и буксировала ее со стоящим «пассажи-
ром»). Однажды лодку почему-то не втащили на су-
шу. Василиса поднырнула под нее, пристроилась подо
дном и начала возить ее по бассейну, в заключение
подвезла к помосту и стала ожидать награды. Рыбку
ей дали, а лодку оттолкнули. Афалина бросилась к
66
лодке и снова подтащила ее на себе к помосту. Пять
раз повторялось одно и то же. Навык был закреплен.
Дельфины, когда им бросали на поверхность воды
мяч, любили носиться с ним и отнимать его друг у
друга. Мы наблюдали, как дельфин стремительно тол-
кал мяч, но в тот момент, когда его настигли сороди-
чи, на ходу швырнул предмет забавы назад, по возду-
ху и сам мгновенно повернулся, а все партнеры про*
скочили вперед. Это был прием «игрока», отлично ско-
ординировавшего свои движения, чтобы обмануть пре-
следователей.
У дельфинов манипуляции с мячом довольно мно-
гообразны, и в этом опять-таки превзошла всех Васи-
лиса. Она могла погрузиться с мячом во рту и так пла-
вать под водой, стремительно толкать его по поверх-
ности бассейна частыми и короткими ударами рыла
(другие обычно носили его на роструме, рис. 31 вкл.)
либо подбрасывать хвостом (рис. 32 вкл.). Когда Ва-
силису обучали закидывать в баскетбольную корзину
мяч, подаваемый ей тренером, она по собственной ини-
циативе усложняла свой навык: пока тренер был за-
нят другими дельфинами, Василиса, забросив мяч, ста-
ла вертикально выставляться из воды и дотягиваться
до корзины, затем мягким ударом выбивала из нее
мяч, подхватывала его на лету и снова забрасывала в
ту же корзину (до этого Василиса виртуозно умела го-
нять мяч по бассейну и схватывать его на лету, если
его бросал тренер). Ей понадобилось две недели, чтобы
она научилась подплывать с мячом во рту к подве-
шенной корзине и точным кивком головы забрасывать
его в цель. После того как этот навык был закреп-
лен, лишь два дня ушло на его усложнение. Подкреп-
ленные наградой, эти действия теперь демонстрируют-
ся перед публикой в качестве нового навыка, в основе
которого лежат поэтапно выработанные серии двига-
тельных актов, объединенных в общую цепь.
Еще одно усовершенствованное действие Васили-
сы — прицельное обливание водой из дыхала или рта
и оперирование фонтанами. Фонтанировать она научи-
лась несколько лет назад, когда жила вместе с груп-
пой дельфинов в другом бассейне. Здесь она старалась
привлечь внимание человека тем, что набирала воду в
рот и выпускала ее тонкой струйкой. Потом она ста-
ла вбирать жидкость в верхние дыхательные пути че-
3*
67
рез щель дыхала, чуть погрузив голову. Объем воды,
выпускаемой фонтаном, мог достигать полутора ста-
канов. Замечательно еще то, что струю фонтана Васи-
лиса могла разбить на несколько порций (3—5) и вы-
пускать их с небольшим интервалом. Если фонтан ока-
зывался недостаточным и за него не давали награды,
она тут же исправляла ошибку и пускала полную
струю на высоту 4 м. В совершенстве управляя фонта-
ном, Василиса стала пользоваться струей воды как
подсобным инструментом для разнообразных действий.
Заметили, как прицельной струей изо рта она однаж-
ды столкнула с помоста мяч в воду (рис. 5), а затем
время- от времени прибегала к такому приему, чтобы
доставать рыб или кольца с тренировочного помоста
либо борта бассейна. Фонтаном животное ухитрялось
обливать с головы до ног и своего тренера.
Дрессировщик Роин Иосава обучил эту афалину
точным фонтаном с расстояния двух метров тушить
огонь, разводимый в бетонной муфте на борту дельфи-
нария. Фонтаны она использовала также и для заба*
вы, притом без всякого подкрепления: например, пу-
скает струю вверх, а затем ловит ее ртом или ухитря-
ется подставлять под нее свой хвост либо грудной
плавник. Так она могла забавляться часами, а иногда
и ночь напролет, отделившись от остальной группы.
Другие дельфины — Боцман, Моряк, Мамаша —
Рис. 5. Афалина Василиса пытается смыть мяч с мостков стру-
ей воды изо рта
68
обрызгивали людей хвостом, при этом никакой связи
с цветом одежды посетителей не замечалось. Скорее
всего, обрызгивание у них, как и у Василисы, служит
способом привлечь к себе внимание человека, близко
стоящего у края бассейна и «не замечающего» дель-
финов. Не исключено, что таким способом проявля-
лось и легкое недовольство тренером: когда из двух
молодых самцов, отсаженных в круглый бассейн, на-
рочито внимание уделяли лишь одному —Моряку или
Боцману, то «обиженный» окачивал тренера водой.
При работе с кольцами или с любым новым объек-
том дельфины вначале проявляют настороженность
(пассивно-оборонительная реакция), при этом бдитель-
ность может обнаружиться по поводу едва заметного
изменения в этом объекте. Например, дельфины обу-
чены работать с резиновым кольцом; при его замене
на металлическое кольцо такой же формы и окраски
животное длительное время отказывалось надевать его
на рыло. Если к уже освоенному кольцу подвязывали
шнурок, снова следовал отказ от работы, и требова-
лось некоторое время, чтобы преодолеть пассивно-обо-
ронительную реакцию. Достигается это быстрее всего
на основе подражания, когда страх перед новым пре-
одолен каким-либо членом группы.
Как известно, подражание — лучший способ быст-
рого обучения дельфинов, которые часто прибегают к
копированию действий своих сородичей. Немало раз-
личных элементов поведения восприняли от Василисы
ее сообитатели. Так, Василиса стала первой надевать
на голову кольцо, лежавшее в полуметре от края по-
моста. Сначала она апортировала это кольцо, а когда
оно было «позабыто» на помосте, сделала попытку до-
стать его, навалившись на него своим рострумом. Это
подкрепили и в последующем выработали сложный на-
вык: высоко высунувшись из воды (значительно выше
края помоста), дельфин кладет рострум на кольцо и,
нажимая, чуть оттягивается назад; тогда кольцо за-
прокидывается на голову и животное сползает в воду,
а затем, выставляясь вертикально, мордой подает
кольцо в руки человека.
Интересным представляется навык — выбрасыва-
ние дельфинов на помост. Сложность этого действия
заключается в том, что для китообразных выход на
сушу в природной обстановке часто бывает смертель-
69
ным из-за теплового удара. У афалин удалось преодо-
леть эту естественную боязнь очутиться на суше. Пер-
вой выскакивать на помост стала опять же Василиса,
а за ней последовали Персей и Инга.
В настоящее время у дельфинов вырабатывается
ряд сложных навыков — стояние на хвосте, синхрон-
ные высотные прыжки (рис. 33 вкл.) и прыжки через
обручи в парном и тройном исполнении и т. д. Трене-
ры замечали, как при многократных повторениях од-
ного и того же задания афалины иногда отвлекаются
и превращают опытную ситуацию в игровую.
Наблюдения, проведенные в Батумском дельфина-
рии, показали:
1. У дельфинов в основе новых приобретаемых на-
выков лежат, как правило, уже имеющиеся закреп-
ленные реакции. На их основе животные легко услож-
няют свои поведенческие акты, присоединяя к ним эк-
страполяционные (с предусмотренным результатом)
элементы поведения, и используют навыки в иных по-
добных ситуациях. Условный же рефлекс действует
лишь в конкретной, данной обстановке. Таким обра-
зом, наблюдаемые сложные формы поведения соответ-
ствуют объективным элементам ситуации и упрочня-
ются по законам временных связей в определенной
последовательности цепи простых реакций.
2. У афалин высоко развит как ориентировочный
рефлекс, так и пассивно-оборонительная реакция. По
существу, весь начальный период обучения можно рас-
сматривать как подвижную систему взаимоотношений
двух реакций: пассивно-оборонительной и ориентиро-
вочного рефлекса. В дальнейшем с освоением обста-
новки на смену этим реакциям приходит пищедобыва-
тельный рефлекс.
3. Подвижная система поведенческих реакций по-
могает дельфинам тонко приспособляться к новой об-
становке при жизни в неволе, в условиях группового
существования и иерархического режима в группах.
В практике содержания дельфинов в неволе за по-
следние годы сделаны большие успехи в изучении
поведения гигантских представителей семейства дель-
финовых, овеянных зловещей славой убийц морских
зверей. Новые сведения о косатках изменили мнение
человека об этих страшных морских хищниках. Ко-
сатки стали настоящими фаворитами публики.
Глава VIII
ЛЮБИМЦЫ
ЗРИТЕЛЕЙ —
ТИРАНЫ КИТОВ
Мрачная слава косаток держалась веками. «Мор-
ской волк», «волк в китовой шкуре», «разбойник»,
«кит-убийца» — так называли этих животных во мно-
гих странах. Еще Карл Линней назвал косатку мучи-
телем и тираном китов. И действительно, весь ее об-
лик свидетельствует о хищничестве. Это самое круп-
ное животное из дельфинов: новорожденные достига-
ют 2,75 м, самки — 7, а самцы— 10 м. Крупнейший
самец был взят 3 июня 1927 года в Кандалакшском
заливе Белого моря в группе из 30 косаток. Его ске-
лет хранится в Зоологическом институте в Ленинграде.
Косаток легко распознают по огромному спинному
плавнику, который сзади виден как коса (рис. 6), у
взрослых самцов он по высоте достигает роста челове-
ка, у самок — вдвое ниже. Окраска на брюхе очень
характерна (рис. 34 и 35 вкл.). На висках притуплен-
ной головы по ярко-белому овальному пятну, позади
спинного плавника белое седло (только у самцов). Все
яркие отметины на теле косаток, видимо, помогают
им держать правильный строй в стаде, не наскакивая
друг на друга при быстром ходе (скорость их может
достигать 55 км/ч).
Другое значение отметин — разбивка контура тела
для маскировки.
Распространены косатки от окраинных морей Арк-
тики до Антарктики. Мигрируют нерегулярно. В уме-
ренных и умеренно холодных водах собираются в
местах скопления корма. Особенно много их бывает
возле работающих китобойных флотилий. Являясь кос-
мополитами, косатки отсутствуют в Черном море и в
71
то же время заходят в пресные устья больших рек,
поднимаются против течения на десятки километров
и живут там неделями. Одиночные косатки ныряют на
1—3, а в стадах — на 4—6 мин (максимум до 12
мин); на поверхности дышат от 2 до 6 раз через раз-
ные промежутки времени, в среднем через полминуты.
Вблизи берегов и на мелких местах они дышат менее
регулярно, чем на глубоких и вдали от берегов. За од-
ну дыхательную паузу звери могут проплыть сотни
метров.
Главная пища косатки — стайная рыба, головоно-
гие моллюски и морские млекопитающие (почти все
виды ластоногих, усатых китов и дельфиновых). Из
рыб она поедает сельдь, кету, чавычу, палтуса, мак-
рель, тунцов, пеламид, одноперых терпугов и т. д.
Больше других рыб любит треску, камбалу, лососевых,
а также акул. В ассортименте редких кормов отмеча-
лись пингвины, крохали, казарки, калан, электриче-
ский скат и даже морская черепаха. В связи с плото-
ядностью ее кишечник заметно укорочен: он лишь
в 8 раз длиннее тела (у афалины же — в 14—
16 раз).
Одесский зоолог В. И. Шевченко исследовал 49 же-
лудков косаток в Антарктике и 30 желудков в уме-
ренно теплой зоне океана. В первом случае было пять
пустых желудков, в большинстве встречались остатки
малых полосатиков, ластоногих, реже рыбы и кальма-
ры. Во втором случае пустых желудков было 11, а в
Рис. 6. Косатки, подплывая к шлюпке, проявляют любопытство.
Во время войны высокий спинной плавник самцов не
раз бывал причиной огневых залпов с военных судов
72
других находились остатки дельфинов, ластоногих и
зубатых китов средних размеров. Кроме того, отпечат-
ки зубов от укуса косаток обнаружены на теле 53%
осмотренных финвалов, 24% сейвалов, 6% малых по-
лосатиков и 65% кашалотов. Это свидетельствует о
том, что некоторым жертвам все же иногда удается
вырываться от косаток. На теле тюленей-крабоедов
и котиков-секачей изредка находят зажившие следы
крупных зубов, а у серых китов замечают откусанные
края грудных и хвостовых плавников.
Хищный нрав косаток в водах северо-западной
части Тихого океана подтверждают японские исследо-
ватели Масахару Нишиваки и Чикао Ханда: из 364
проанализированных ими желудков 64 % содержали
рыб и головоногих моллюсков, 29 — остатки китов и
дельфинов (преимущественно белокрылых морских
свиней) и 7% ластоногих. Желудки десяти косаток,
убитых между островом Кодиак и Сан-Мигуэлом в се-
веро-восточной части Тихого океана, изучил амери-
канский зоолог Д. Райс и нашел в них объедки трех
калифорнийских морских львов, четырех сивучей, се-
ми морских слонов, двух белокрылых морских свиней,
одного малого полосатика, двух акул, кальмара, пару
палтусов и рыб из рода ламприс.
И все же прожорливость косаток сильно преувели-
чивалась. Датский цетолог Д. Эшрихт однажды из-
влек из желудка взрослой косатки остатки 13 морских
свиней и 14 тюленей. Эти объедки комментаторами ра-
бот Эшрихта были приняты за целых животных, и
версия о невероятном обжорстве косаток была увеко-
вечена.
Инспектора, летавшие на малой скорости в районе
китобойного промысла в Антарктике, наблюдали,
как крупные полосатики успешно отгоняли косаток,
которые приближались к китовым детенышам. Воз-
можно, из-за отпора китов косатки редко нападают
на крупных полосатиков. Гораздо чаще от хищников
страдают мелкие китообразные и ластоногие.
Поведение этих хищников зависит от характера пи-
щи. Обычно они нападают стадом. В этом им помога-
ют высокий уровень координации действий между чле-
нами группы и совершенная коммуникация. Косатки,
преследуя косяки рыб, медленно плывут с частично
или полностью выставленным спинным плавником
73
под самой поверхностью воды, проплывая расстояние
до полукилометра. Они могут мирно пастись на рыбе
рядом с рыбоядными китами, не обращая на них
внимания и не всегда отваживаясь нападать на кита.
Для расправы с ним они пользуются особыми приема-
ми: атаку начинают (обычно с головы) взрослые сам-
цы, а самки и молодняк ожидают исхода борьбы в сто-
ронке. Сорок крепких и уплощенных зубов, сидящих
в расширенных челюстных костях, обеспечивают
страшный прикус косаток, когда те терзают кита и
топят его, не позволяя подняться на поверхность для
вдоха. Звери открывают пасть у кита, впиваются зуба-
ми в его горло, рвут массивный и жирный язык, ку-
сают плавники. При рывках добычи косатки отгреба-
ют с помощью грудных плавников назад. Это свойство
редкое среди китообразных.
В июне 1958 года близ Командорских островов с
катера «Беринг» наблюдали нападение стада косаток
на финвала: звери, заходя кругами, рвали тело кита
и через полчаса израненного и покусанного затащили
вглубь, откуда поднимались большие водовороты.
Нечто аналогичное наблюдал зоолог Д. Ханкок в
1965 году при нападении семи косаток на малого поло-
сатика в заливе Беркли (Британская Колумбия). Три
взрослых самца утопили кита за первые же полчаса
борьбы, а затем вся группа хищников выедала под во-
дой язык, куски сала и мясо жертвы, которая всплы-
ла из воды на следующий день.
В Калифорнийском заливе группа из 15—20 коса-
ток напала на сотню дельфинов. Двигаясь вблизи
жертв, хищники все больше и больше суживали круг,
а затем одна из хищниц ворвалась в центр и расчле-
нила стадо; тогда погибло много дельфинов, и вода
была красной от крови. Такую тактику окружения
стада косатки применяют к моржам, тюленям, плот-
ным косякам рыб. У мелких и тихоходных морских
свиней косатка может на ходу откусывать хвостовые
лопасти. В водах Гренландии от нее страдают белухи,
у тюленей она иногда отсекает лишь голову. Плохо
приходится тюленям и котикам, если те застигнуты
косаткой далеко от берега. Им остается только зата-
иться в воде. Но если вблизи берег, ластоногие спешно
выбираются на сушу. У берегов Калифорнии видели.
74
как косатка в течение 20 мин подбрасывала в воздух,
точно мячик, тело морского льва.
~ Иногда панику морских зверей, вызванную косат-
ками, используют охотники. В июле 1965 года в Ча-
унскую губу (Восточно-Сибирское море) зашло шесть
косаток. Кольчатые нерпы начали стремительно вы-
скакивать на сушу, и тогда местные охотники прямо
на берегу добыли 60 нерп.
В прежние годы косатки встречались близ котико-
вых залежек гораздо чаще. Караульщики лежбищ еще
в 1933 году наблюдали близ острова Беринга стада по
60 хищников. Звери патрулировали берег и уничтожа-
ли котиков, заходивших кормиться в воду. Теперь слу-
чаев нападения на котиков почти не бывает. Возможно,
что снижение интереса косаток к живым китам и тю-
леням было связано с возросшей деятельностью кито-
бойных флотилий, и питание отходами промысла смяг-
чило нрав хищников.
В опасных ситуациях косатки ведут себя миролю-
биво. Осенью 1955 года в Антарктике в Канале прин-
ца Густава льды отрезали от чистой воды 60 косаток.
Звери находились в сильно возбужденном состоянии,
на 1—2 м выставляли из воды головы и осматрива-
лись. Они не пытались нападать на многочисленных
тюленей-крабоедов и малых полосатиков, тоже отре-
занных от моря. Не трогали они и людей, подходив-
ших к краю льдин и палкой касавшихся хищников.
Косатки провели в этой полынье четыре месяца и вы-
брались на чистую воду, преодолев пояс сплошных
льдов шириной в 65 км.
За последние годы грозная репутация косаток зна-
чительно поколебалась, после того как были проведе-
ны два замечательных опыта.
Первый начался 16 июля 1964 года. В водах Бри-
танской Колумбии близ острова Сатурна загарпунили
самца-косатку 467 см длиной и весом 1090 кг. Ране-
ное животное доставили в бухту Ванкувер, оказали
ему медицинскую помощь (инъенкции пенициллина,
тиамина, витамина В12 и гидроксизина). Сначала его
поместили в док, а затем в загон, отгороженный от
залива проволочной сетью. Площадь загона была
14 X 23 м, а глубина 3—7 м.
Животное ошибочно приняли за самку и назвали
Моби-Долл.
75
В течение 57 дней косатка отказывалась принимать
пищу, а потом стала поедать по 45—90 кг рыбы в
день. За месяц ей скормили около 16 ц трески, лосо-
севых и терпуговых рыб. Головоногих моллюсков и
лошадиное мясо она поедала неохотно. В загоне ко-
сатка плавала по кругу против часовой стрелки со
скоростью 2—3 узла. В среднем она погружалась на
полторы минуты и выныривала на несколько секунд
для дыхания. Для кормежки ее подзывали к кормово-
му плотику, ударяя рыбой о поверхность воды, и дава-
ли 12—15 крупных рыб прямо из рук. Кормящий гла-
дил кита по голове, а потом, описывая круг над голо-
вой зверя, заставлял его поворачиваться брюхом
вверх. Так кит научился подставлять свое брюхо для
почесывания.
Когда кормежка внезапно прекращалась, животное
проявляло неудовольствие, вращая хвостом и хлопая
грудными плавниками. Перед кормежкой зверь воз-
бужденно выпрыгивал из воды. Через 86 дней инте-
ресный опыт приручения прервался из-за смерти жи-
вотного от кожных грибков. Смертельному исходу бо-
лезни способствовали ядовитые стоки реки Фрейзер,
питающей залив.
Еще более примечательным был второй случай
приручения 3,5-тонного самца длиной 655 см по клич-
ке Наму. Он был пойман в сети в июне 1965 года у
острова Наму (Британская Колумбия). Сюда немедлен-
но вылетел директор аквариума города Сиэтла Эдвард
Гриффин и купил у рыбаков живую косатку за 8000
долларов. Спешно построили клетку размерами 18 X
Х12Х5 м, в которой животное в течение месяца бук-
сировали на расстояние 830 км, не вынимая из воды.
С первого дня транспортировки Гриффин приступил к
укрощению хищника и добился больших успехов. Пер-
вый контакт человека и хищника состоялся еще во
время перевозки: Гриффин смело подплыл к зверю в
клетке и поскреб ему щеткой голову и подбородок. Это
повторялось при каждой встрече; позже, когда косатку
поместили в загон бухты города Сиэтла, она стала по-
зволять человеку садиться на себя, стремительно вози-
ла его по загону, подолгу резвилась с ним, то услуж-
ливо подставляя спину или брюхо, то внезапно сбрасы-
вая его в воду. В первое время Наму голодал, но через
месяц стал съедать до 1,5 ц красной рыбы в день.
76
В этих двух экспериментах были открыты весьма
ценные качества косаток: они хорошо уживаются в
неволе, легко обучаются и поддаются дрессировке.
Это очень пригодилось для организации зрелищных
мероприятий. В аквариумах Лос-Анджелеса, Майами,
Сан-Диего теперь наряду с другими видами дельфи-
нов показывают и гигантских косаток, иногда в од-
ном и том же танке с дельфинами, на которых они
нападают в море. Косатки выступают перед публикой
даже в небольших бассейнах, лишь втрое превышаю-
щих длину их тела. Эти звери оказались весьма по-
слушными. Они подчиняются воле человека, стреми-
тельно катают его по волнам бассейна, выпрыгивают
во весь рост, подают голос, берут рыбу изо рта тренера.
Эдвард Гриффин первый убедился, что косатка безо-
пасна для человека. Однажды она вместе с рыбой втя-
нула в пасть ногу тренера, но тут же вытолкнула ее
обратно.
Все рассказы о нападении косаток на лодки объяс-
няются любопытством (сильно развитым ориентировоч-
ным рефлексом), проявляемым хищниками, но кото-
рое внешне выглядит как атака. В связи с этим пока-
зателен случай с охотоведом А. М. Николаевым. Он и
его двое спутников в мае 1965 года шли на моторной
шлюпке в полусотне метров от берега в бухте Качаева
близ острова Уруп. Вдруг впереди в 150 м вынырнуло
4—6 косаток. Шлюпка резко повернула к берегу, а ко-
сатки устремились к шлюпке. В 20 м от суши мотор
был выключен, и лодка продолжала двигаться по
инерции. Когда до берега осталось 7 м, зверей отделя-
ли 25—30 м, а в 2—3 м от суши они настигли людей,
но удара по лодке не нанесли.
Подобные рассказы, чуть подправленные воображе-
нием «атакуемых», иногда преподносятся как факты
нападения косаток. На самом же деле достоверных
случаев нападения косаток на людей не существует.
Марокканские ныряльщики утверждают, что косатки
подплывают к человеку и, удовлетворив свое любо-
пытство, удаляются, не причинив ему вреда. Теперь
уже поймано более 30 косаток, которые поступили
для содержания в разные океанариумы.
Достопримечательна поимка сразу семи косаток на
основе использования инстинкта сохранения вида,
очень развитого у китообразных. Весной 1968 года в
77
Британской Колумбии в заливе. Гарден-Бэй, севернее
Ванкувера, на скалах застрял молодой самец весом
1,6 т. Своим сигналом бедствия он привлек еще шесть
косаток во главе с четырехтонным самцом. Все звери
были пойманы с помощью рыболовных снастей и по-
мещены недалеко от Ванкувера в бухте Пендер в воль-
ерах, сооруженных из сетей, подвешенных на колодах.
Ни одна из косаток не ушла в море и не сделала по-
пытки уйти прыжком через колоды или разорвать
сеть. Из пойманных косаток две попали в Калифор-
нийский и одна в Майамский океанариумы, одна — в
Ванкуверский аквариум, а три оставлены на китовой
станции в заливе Гарден-Бэй в распоряжение канад-
ского психолога Пауля Спонга для исследования их
органов чувств. Здесь зверей поместили в вольеры
(размерами 21X18X4,5 м), построенные из сетей, под-
вешенных на бревнах-поплавках. Косатки за короткое
время подружились с человеком. Они показали превос-
ходное зрение, близкое к остроте зрения кошек, а глу-
бина их сна приближалась к глубине сна человека и
наземных млекопитающих. Когда часть загородки бы-
ла нарушена, четырехтонный самец Скукум-Цецил
медленно выплыл в открытое море, а другие остались.
В дельфинарии «Си-Уорлд» (г. Сан-Диего) очень по-
пулярна косатка Шаму. Наездник надевает на нее пла-
стиковый обруч, держится за него во время высоких
прыжков и стремительной скачки, словно ковбой на
ретивом гигантском мустанге.
Самец по кличке Хуго, попавший в океанариум
Майами, миролюбиво живет в одном танке с обыкно-
венными дельфинами. Вначале (к трем годам) он имел
3,9 м длины и весил 680 кг, через два года вырос до
5,3 м и достиг 1,7 т веса, а к семи годам вырос
до 6 м и весил 2,1 т. Хуго работает за пищевое воз-
награждение и, если его не получает, ударом хвоста
окачивает людей каскадом брызг. Позже Хуго просла-
вился своим «носом»: в океанариуме он наскочил на
подводное окно и лишился кончика рыла. Но за четы-
ре месяца кончик рыла полностью восстановил перво-
начальную форму. Так была открыта еще одна инте-
ресная черта у китообразных — способность их тканей
к быстрой регенерации.
Еще недавно раздавались голоса — объявить
косатку «вне закона», как злейшего хищника, и назна-
78
чить премии за ее истребление. Однако вред, наноси-
мый косатками в море, явно преувеличивался. Дли-
тельные наблюдения биолога-охотоведа С. В. Марако-
ва на Командорских островах показали, что ныне
косатки здесь не причиняют серьезного ущерба ни ка-
лану, ни котикам. Эти хищники в первую очередь на-
падают на ослабевших и больных морских млекопи-
тающих либо на их неосторожных детенышей. Они вы-
полняют определенную функцию в естественном отбо-
ре: выступают в роли санитаров, выбраковывающих
ослабленных и старых животных, предотвращая у по-
следних появление хилого потомства. Учитывая эти
обстоятельства, «смертный приговор» для косаток явно
преждевремен. Косатки решительно не заслуживают
ни интенсивного преследования, ни жестокого отноше-
ния и в будущем еще могут сильно пригодиться чело-
веку. А то, что они иногда выедают язык у буксируе-
мых китов и таким способом расхищают запасы жи-
ра, от этого их можно отучить. Достаточно бывает
пушечного выстрела по любителям китовых языков —
и они надолго запоминают полученный урок и держат-
ся на большом расстоянии от судов, буксирующих
китов.
Перспективы же приручения косаток довольно за-
манчивы. Их уже сейчас обучают работать в откры-
том море. Форрест Вуд описывает, как две косатки,
пойманные в 1968 году в водах штата Вашингтон, бы-
ли обучены нырять по команде: Ахаб (самец длиной
5 м) — на глубину до 260 м за срок 7 мин 40 с, а Из-
маил (длиной 6 м) — до 152 м. Косаток, вероятно,
можно будет использовать, например, для лова круп-
ных акул, для охраны водолазов и аквалангистов от
нападения хищных рыб. Быстроходность косатки, от-
работанная в ходе эволюции (для преследования своих
жертв), ныне привлекает внимание многих исследова-
телей.
Гидродинамическое совершенство формы тела китов
и дельфинов, строение и демпферные свойства их кож-
ного покрова, способность ныряния на значительные
глубины, строение и функции гидролокатора и многие
другие особенности организации китообразных пред-
ставляют в настоящее время большой интерес для био-
ники.
79
Глава IX
СКОРОХОДЫ
МОРЕЙ
В мире существует почти полтора миллиона разных
видов животных, и все они отлично адаптированы к
окружающей среде. Это результат естественного отбо-
ра, в котором выживают наиболее приспособленные и
гибнут неприспособленные. Естественный отбор — не-
прерывно действующий механизм, который оттачива-
ет до совершенства любые признаки всякого вида, наи-
более соответствующие условиям жизни в данной сре-
де. Для водных млекопитающих важное преимущество
в борьбе за существование дает быстрота передвиже-
ния. Завоеванию дельфинами Мирового океана способ-
ствовали их быстроходность и отличная звуковая ори-
ентация, обеспечиваемая совершенно устроенным гид-
ролокатором. Изучение приспособлений быстро плава-
ющих водных животных может подсказать ценные
идеи для повышения скоростных качеств судов. Осо-
бенно достойны изучения дельфины (рис. 36 и 37 вкл.),
которые стремительно передвигаются и совершают ги-
гантские прыжки при сравнительно слабом расходе
энергии (рис. 7): ведь в течение многих тысячелетий
они приспособлялись догонять быстроходных рыб, что-
бы питаться Ч
Скорость дельфинов зависит, конечно, от хорошей
обтекаемости как всего тела, так и его отдельных
частей.
1 Остается загадкой, как в желудок тихоокеанского кашалота
попали три полных меча — по 80 см длиной — самой быстро-
ходной меч-рыбы, извлеченных при его разделке на китобазе
«Дальний Восток» в 1969 году.
80
Главный локомоторный орган китообразных —
хвост. Хвостовой стебель совершает удары вверх-вниз,
а лопасти работающего хвоста принимают разные
углы наклона к продольной оси стебля: когда стебель
идет вниз, лопасти поворачиваются вверх, и наоборот.
Вращательных движений при плавании хвост не про-
изводит. Частота и размах ударов хвоста и степень на-
клона хвостовых лопастей влияют на быстроту плава-
ния. Наблюдения показывают, что некоторые дель-
фины, плывущие возле носа быстроходных катеров, мо-
гут двигаться со скоростью до 65 км/ч. Однако в этих
случаях животные используют толкающую силу «осед-
ланной» корабельной волны, и такая скорость не свой-
ственна им при свободном движении в море. В экспе-
риментах, проведенных на Гавайских островах с ма-
лайскими продельфинами, специально выпускаемыми
для скоростного пробега, Ланг и Прайор установили
максимальную скорость не более 40,6 км/ч (11,3 м/с).
В 1937 году близкую цифру (36 км/ч, или 10 м/с) на-
шли советские исследователи В. Шулейкин, В. Лукья-
нова и И. Стась для дельфина-белобочки. У гринды
максимальная кратковременная скорость в Калифор-
нийском океанариуме была 40—48 км/ч, а у косаток,
по наблюдениям с судна «Монтерей», варьировала в
течение 20 мин от 38 до 55 км/ч. Даже самые бы-
строходные дельфины способны двигаться с большой
скоростью недолго (лишь несколько минут или даже
секунд).
Удивляло то, что, по расчетам гидродинамиков,
дельфин не может развивать скорость более 20 км/ч,
а в действительности он превышает эту цифру, напри-
мер, в 2 раза.
Каким образом обеспечивается высокая скорость
передвижения китообразных? Еще в 1936 году амери-
канец Джемс Грэй, изучавший движение водных жи-
вотных, заинтересовался необычайной легкостью пла-
вания дельфинов. Грэй буксировал по бассейну жест-
кую модель дельфина, замеряя сопротивление воды.
В результате он пришел к поразительному выводу:
дельфины развивают такую высокую скорость, кото-
рая по расчетам выходит за пределы их мускульной
силы. Получалось, что дельфины либо должны иметь
неестественно высокую мощность мышц, в 7 раз боль-
шую, чем у любого млекопитающего (что невозможно),
81
либо им удается каким-то способом уменьшать сопро-
тивление воды. Но каким? Это необъясненное в то
время явление — двигаться быстрее своих возможно-
стей— получило название «парадокс Грэя».
Следует заметить, что «парадокс Грэя» признают
не все исследователи, в том числе Форрест Вуд. Он от-
рицает «парадокс Грэя», так как дельфины не облада-
ют той быстроходностью, которую им приписывают.
Факты же обгона китообразными кораблей он объяс-
няет так: «...имея собственную скорость, например
около 15 узлов, и двигаясь почти под прямым углом
к кораблю, дельфин, умело скатываясь с носовой вол-
ны, может обогнать судно, плывущее со скоростью
30 узлов. Прекратив затем собственное движение от-
носительно корабля и оседлав его носовую волну, дель-
фин может двигаться вместе с кораблем, какова бы ни
была скорость последнего». Но не исключается и то,
что дельфин способен совершить рывок и самостоя-
тельно обогнать быстро движущееся судно.
Чтобы выяснить секрет «парадокса Грэя», обратили
внимание на мускулатуру. Главный движитель кито-
образных — могучий хвост — приобрел необычную си-
лу в связи с коренным преобразованием мускулатуры.
Эти изменения подробно изучил В. А. Родионов. Он за-
Рис. 7. Различные прыжки обыкновенных дельфинов: а — по-
логий; б — высотный (по вертикали); в — с плаванием
на спине; г — с поворотом вокруг оси тела. По Г. Пил-
лери
82
метил отчетливое упрощение мышечной системы кито-
образных, насчитав у дельфинов лишь 112 мышц
вместо 170 у наземных млекопитающих. Причины это-
го заключаются в глубоком адаптивном переустрой-
стве отряда: в перемещении локомоторной функции
на хвостовую часть тела, в редукции задних конечно-
стей и в специализации передних (грудных) плавни-
ков, в слиянии одних мышц и исчезновении других, в
изменении функции комплексов мышц соответственно
требованиям водной среды.
В мышечной системе позвоночника особенно выде-
ляется роль мощной двигательной мускулатуры, на-
правленной на обеспечение быстрого передвижения
китообразных в воде и достижение высокой маневрен-
ности. Упрощение мускулатуры позвоночника сопро-
вождается гипертрофическим развитием двух пар му-
скульных комплексов (длиннейшей и гипаксиальной
мышц), лежащих над и под позвоночным столбом и
выполняющих основную нагрузку при работе хвостово-
го движителя. Обе мышцы составляют почти 7б часть
веса всех мышц тела. При этих условиях огромная
масса мускулатуры позвоночного столба концентри-
рует свои усилия лишь на работе хвоста, на его ударах
вверх-вниз (поэтому вес мышц, осуществляющих боко-
вые изгибы хвоста, в 15 раз меньше веса мышц, дви-
гающих хвостом в вертикальной плоскости). Эффектив-
ность работы мышц возрастает за счет очень высоких
остистых отростков позвонков, к которым приклады-
вается усилие мышц при увеличенном плече рычага.
Наконец, важно и то обстоятельство, что китообразные
в воде находятся в состоянии, близком к невесомости;
поэтому в отличие от наземных обитателей им не при-
ходится тратить мускульную силу на преодоление соб-
ственного веса, и они расходуют ее целиком на дви-
жение как дополнительный источник энергии.
Однако мускульный аппарат далеко не исчерпывает
адаптаций дельфинов к скороходности. В XX гла-
ве мы расскажем об открытии важнейшей адаптации
китообразных к быстрому плаванию, заключающейся
в саморегуляции гидроупругости плавников. В данной
же главе ограничимся описанием приспособлений в
покровах.
О ламинаризации потока кожей китообразных сви-
детельствуют некоторые прямые наблюдения над дель-
83
финами, плавающими в условиях биолюминесценции
(свечения) моря. Еще в годы второй мировой войны
английский биолог Г. А. Стефен, служивший на фло-
те, наблюдал замечательный факт: ночью при тихой
погоде в светящемся море за стремительно плывущим
дельфином остаются лишь два огненных шнура, за
плывущим тюленем — широкое полыхающее поле.
Аналогичные наблюдения были проведены в 1967
году в водах Калифорнии с исследовательского судна-
катамарана «Си-Си» («Смотри море»), принадлежаще-
го американскому центру подводных исследований: из
прозрачного трехметрового шара, опускающегося под
днище катамарана, Вильям Эванс и Лари Мак-Кинли
ночью отлично видели, как у быстро плывущих корот-
коголовых дельфинов от кончиков спинного и обоих
грудных плавников протягивалась назад светящаяся
узкая полоска и три такие же узкие полоски тянулись
от двух уголков и серединной выемки хвостового плав-
ника. Таким образом, дельфин плывет, почти не на-
рушая спокойного состояния воды.
Гидродинамические свойства дельфина оказались
совершеннее, чем в конструкциях подводных тел,
созданных человеком. Этот факт заинтересовал зару*
бежных конструкторов. Гидродинамик инженер Макс
Крамер в США в 1958—1960 годах показал, что со-
противление воды, испытываемое дельфином при дви-
жении, в 10 раз меньше, чем сопротивление при дви-
жении модели того же размера и формы с обычной об-
шивкой. Стало очевидно, что быстроходность китооб-
разных зависит не только от формы тела и сильной
мускулатуры хвоста, но и от их «волшебной» кожи —
упругой, эластичной, несмачиваемой.
Обнаружено также, что в коже дельфина все вре-
мя отшелушивается тонкий наружный слой. Это позво-
ляет ему постоянно очищаться от водорослей и других
морских организмов, которые снижали бы его скорость.
У трупов китов, лежащих на воздухе, верхний слой эпи-
дермиса затвердевает и легко отделяется вроде целло-
фановой пленки.
Кожа дельфинов привлекла внимание судостроите-
лей, и американская фирма «Раббер компани» смоде-
лировала гладкую обшивку для судовых корпусов. За-
интересовались дельфиньей кожей и военно-морские
силы США, чтобы имитировать аналогичное каучуко-
84
вое покрытие и тем самым повысить скорость подвод-
ных лодок.
Какие же приспособления в коже позволяют дель-
фину быть «скороходом морей» и отличным прыгу-
ном?
Как мы говорили, некоторые ученые объясняли
«парадокс Грэя» особым строением мускулатуры дель-
фина. Однако существуют и другие гипотезы. Так, по
одной из них стремительно плывущий дельфин может
уменьшать вихри в потоке, обтекающем его тело, пу-
тем передачи тепла вдоль тела от одного участка ко-
жи к соседнему. По другой гипотезе, дельфин умень-
шает сопротивление воды при увеличении скорости пу-
тем гашения завихрений, возникающих в пограничном
слое жидкости.
При движении в воде всякое тело испытывает со-
противление. Сила сопротивления зависит от формы
тела и от строения потока около тела. Даже у хорошо
обтекаемых тел можно различать несколько областей/
У носа или головного конца в пограничном слое (вбли-
зи стенки тела) обтекание упорядоченное (ламинарное)
и сила трения минимальная. Затем на некотором рас-
стоянии от носа ламинарное обтекание нарушается и
становится турбулентным: в пограничном слое возни-
кают вихри (поперечные движения частиц жидкости),
и сопротивление на этом участке резко возрас-
тает.
Еще дальше — к корме или хвосту — может про-
изойти срыв обтекания, отделение потока с образова-
нием еще больших вихрей.
При конструировании обтекаемого тела основная
задача — не допустить раннего перехода ламинарно-
го течения в турбулентное и срыва потока. Турбулиза*
ция пограничного слоя всегда связана с первоначаль-
ным нарушением устойчивости течения. Гибкая кожа
при известных условиях будет задерживать возник-
новение нарушения устойчивости в пограничном
слое.
Американский инженер М. Крамер пытался найти
в коже дельфина ту структуру, которая способствует
уменьшению сопротивления воды. В коже дельфина
есть два основных слоя — эластичный наружный (эпи-
дермис) и лежащий под ним упругий внутренний (дер-
ма с высокими сосочками и жировым отложением). По
85
мнению ученого, наружный слой выгибается и пру-
жинит под давлением воды, подобно поролоновому
матрацу, который прогибается в определенных местах
под тяжестью тела.
В 1958 году М. Крамер в какой-то мере пытался
учесть строение кожи дельфина при создании им
искусственного покрытия ламинфло (от слов «лами-
нарфлоу», т. е. ламинарное течение), имитирующего
кожу дельфина. Модель, одетая в оболочку ламинфло
и испытанная в потоке воды при скорости 70 км/ч,
имела сопротивление жидкости на 60% меньшее, чем
контрольная модель.
Оболочка ламинфло была сделана из трех резино-
вых слоев общей толщиной 3,5 мм: гладкого верхне-
го (0,5 мм), среднего эластичного с гибкими столбико-
видными выступами (2,5 мм) и нижнего слоя (0,5 мм),
примыкающего к корпусу модели. В пространство
между выступами вводилась демпфирующая, то есть
погашающая колебания, кремнийорганическая жид-
кость, заключенная между нижним слоем и крышей
среднего слоя. Гладкий верхний слой имитировал эпи-
дермис кожи дельфинов, средний — дерму с ее колла-
геновой и жировой тканями, а нижний играл роль
опорной пластины. Позже Крамер предложил более
тонкое покрытие (2,5 мм) с удлиненными гибкими гре-
бешками вместо столбиков.
Однако искусственное покрытие ламинфло не мо-
жет до конца соперничать с кожей дельфина, в кото-
рой демпфирование осуществляется гораздо совершен-
нее. Главная причина гидродинамического совершенст-
ва дельфинов заключается в том, что они активно уп-
равляют своей кожей, к которой подходит большое ко-
личество нервов. Тонко улавливая силу сопротивления
трения и меняющееся давление в тех или иных участ-
ках кожи, дельфин может активно предотвратить на-
чальные фазы процесса турбулизации потока. От рецеп-
торов кожи плывущего дельфина поступает точная ин-
формация в головной мозг, и отсюда в виде биотоков
по двигательным нервам подается команда в тот от-
ветственный участок кожи и кожной мускулатуры, где
в данный момент ощущается возросшее сопротивле-
ние воды. Немедленно следует соответствующая двига-
тельная реакция кожной мускулатуры и поверхности
кожи,
86
Главная адаптация кожных покровов китообраз-
ных к скоростному плаванию — самонастройка демп-
фирования кожи. Она осуществляется сосудистой и
нервной системами в покровах, сложным аппаратом
мельчайших кожных сосочков, основание которых
составляют дермальные валики, отображающие направ-
ление водяных струй вокруг плывущего китообразного
(В. Е. Соколов и др.). Кожные валики ориентированы
вдоль тела. Демпфирование в основном выполняет
сосочковый слой, обильно снабженный кровеносными
сосудами и нервами. Каждый сосочек благодаря увели-
чению или уменьшению просвета кровеносных сосудов
на различных скоростях плавания обладает пере-
менной упругостью. В целом по всей коже это со-
здает условия для гашения турбулентных пульса-
ций в соответствии с той или иной скоростью пла-
вания. Такое регулирование переменного демпфирова-
ния совершается рефлекторно. О рефлекторной регу-
ляции свидетельствует также смена окраски кожи —
порозовение брюха дельфина в зависимости от ско-
рости его плавания на ограниченном расстоянии за-
плыва, от его динамической нагрузки. Мы отмечали
появление слабо-розового цвета у одной и той же
взрослой афалины при возрастании динамической на-
грузки и увеличении средней скорости хода животного
на 20%.
В настоящее время кораблестроители уже пытаются
заимствовать некоторые качества живых моделей —
китообразных, чтобы получить выигрыш в скорости
судов. Появляются корабли с китовидной формой и
дельфинообразными обводами носовой части. Такая
конструкция позволила заметно повысить скорость
хода судов при той же мощности двигателей. Известно,
что в судостроении в настоящее время применяются и
другие варианты судов рыбовидной формы. В част-
ности, имеется ряд проектов полупогруженных судов
со скрытым подводным корпусом и специальными над-
стройками, выступающими над водой. При определен-
ных условиях такие суда могут быть более выгодны-
ми. Таким образом, в судостроении применяются и
китовидные и рыбообразные формы судов. Японский
профессор Тако Инуи на модели корабля «Куренаи
Мару» показал, что китовидная форма корпуса судна
более пригодна для движения в море, чем острокиле-
87
вая, и кораблестроители построили океанское судно о
китовидным корпусом. Новое судно оказалось болев
экономичным: оно обеспечивало ту же скорость и гру-
зоподъемность, хотя мощность его двигателей была на
25 % меньше, чем у судов того же назначения.
Дельфины замечательно используют законы гидро-
динамики, когда «оседлывают» волны, т. е. пассивно
Скользят на волне возле носа идущего судна. Хвосто-
вые плавники при этом не работают, а тело движется
(иногда сотни метров) вместе с волной, возникающей у
носа корабля. Поскольку корабль следует вперед, он
создает гидродинамическое поле впереди носа, кото-
рое дельфины тонко обнаруживают своим кожно-му-
скульным чувством. Это подтверждается подводными
фотографиями ученых США Э. Феджера и Р. Бакку-
са: позы пристроившихся к кораблю дельфинов свиде-
тельствуют, что животные используют давление поля
в корабельной волне всем телом, а не только нижней
поверхностью хвостовых лопастей (рис. 38 вкл.).
Действие гидродинамического поля дельфины мо-
гут ощущать на расстоянии нескольких метров впере-
ди и в сторону от носа корабля, притом не только близ
поверхности, но и на глубине погружения носовой
части судна. Стайки этих юрких животных держатся
возле носа кораблей обычно от нескольких минут до
одного часа и выстраиваются временами в два или4 три
этажа. Пока не найдено нужное положение, дельфи-
ны крутятся в воде, стремительно ныряют, энергично
работают плавниками, а потом плывут какую-то часть
времени пассивно, затем снова чередуют активное по-
ведение с пассивным скольжением и т. д.
Для оседлывания корабельных волн дельфинам
нужна высокая скорость. Стайки быстроходных бело-
крылых морских свиней редко следуют возле судов,
идущих со скоростью 17 км/ч, и то лишь на 1—2 мин,
но очень часто и на более продолжительный срок при-
страиваются к носу судов, если те идут со скоростью
20—30 км/ч.
Недавно немецкий исследователь Генрих Гертель
установил, что пассивно движущийся дельфин, осед-
лавший корабельную волну, испытывает разное сопро-
тивление воды в зависимости от глубины его погруже-
ния: в 50 см от поверхности сопротивление воды
бывает максимальным, а на глубине 1,5—2 м стано-
88
вится минимальным. Если дельфин способен так тонко
улавливать разницу в сопротивлении воды на разной
глубине в пределах двухметрового слоя, то он, вероят-
но, широко использует эту способность и в жизни,
уменьшая сопротивление воды путем регулирования
глубины своего хода в поверхностной толще моря.
Явление «оседлывания волны» объясняет, почему
дельфины, двигаясь близ носа быстроходных судов, име-
ют большую скорость в сравнении с их естественной.
В 1961 году у китообразных было объяснено другое
интересное для гидробиоников явление, которое кали-
форнийские зоологи К. Норрис и Д. Прескотт назвали
«эшелонированным плаванием». Оказалось, что дете-
ныш, плывущий рядом с кормящей маткой, выбирает
наиболее выгодное положение относительно ее тела-
Это экономит силы молодого животного: сосунок, ис-
пользуя давление гидродинамического поля, создаю-
щегося вокруг стремительно идущей самки, может
плыть пассивно, как дельфины при оседлывании кора-
бельных волн, и не отстает от родителя, несмотря на
свои еще слабые удары хвоста.
Таким образом, с действием гидродинамического
поля дельфины знакомятся очень рано, плавая бок о
бок с родителями. Не затрачивая больших усилий, де-
теныш словно «едет на самке», а самка все время чув-
ствует «буксируемого» ею сосунка. Взрослый дельфин
может «буксировать» молодого, если тот придержи-
вается строго определенного положения: детеныш рас-
полагается рядом и параллельно боку тела матери,
притом так, чтобы конец морды детеныша не выдавал-
ся вперед за грудной плавник родителя.
Вначале сосунок редко отплывает от матери и
тут же снова пристраивается в излюбленном месте.
Иногда он занимает положение между спинными плав-
никами двух взрослых дельфинов. В океанариумах на-
блюдали, как детеныш, спасаясь от преследования
крупного самца афалины, пристраивался к материн-
скому боку, чтобы поддержать высокую скорость.
Эшелонированное плавание свойственно и китам.
Нам не раз приходилось видеть сосунков полосатиков
и всегда только очень близко возле родителя. При вы-
ныривании юркое тело детеныша появлялось на по-
верхности одновременно с массивным телом кита*
89
Случалось, что мелкие дельфины «оседлывали»
более крупных китообразных другого вида. В океана-
риумах замечали, как афалины, полосатые и обыкно-
венные дельфины пристраивались к плывущим взрос-
лым гриндам. Вероятно, по этой же причине в стадах
гринд в море изредка присутствуют небольшие дель-
фины другого вида. Поскольку такая буксировка за-
держивает скорость хода крупного животного, то по-
следнее нередко прогоняет от себя наездника.
Английский ученый Бриан утверждает, что стадо
косаток, выстроенное фронтом, плывет быстрее, чем
одиночки. Изучение всех этих закономерностей при
движении китообразных строем, шеренгой, тесным
стадом представляет определенный практический ин-
терес. Это могло бы иметь значение, например, при
разработке конструкций катамаранов (двухкорпусных
судов), при поисках более выгодного взаиморасполо-
жения судов и катеров при их групповом передвиже-
нии или наилучшего размещения лодок во время гонок,
соревнований и т. п.
Человек повторил форму китообразных в контурах
подводных кораблей. Дельфин облегчил задачу чело-
века в поисках такой формы и выдал ему один из се-
кретов своей быстроходности: люди построили под-
водную лодку с дельфинообразными обводами. Тако-
вы, например, формы таинственно погибшей в 1963 го-
ду американской атомной подводной лодки «Трешер»
и экспериментальной подводной лодки США «Доль-
фин», которые развивали скорость до 50 км/ч.
Мысли гидробиоников и океанологов связаны сей-
час с изучением загадочных океанских глубин. В ка-
ком направлении следует вести поиски, чтобы увели-
чить глубины погружения и продолжительность ны-
ряния человека без технического снаряжения? Не по-
может ли в решении этой проблемы изучение приспо-
соблений лучших ныряльщиков мира — китов и дель-
финов?
Глава X
ПОКОРИТЕЛИ
МОРСКИХ ГЛУБИН
В настоящее время свободно ныряющий человек
опускается лишь в самые верхние слои моря. Ловцы
губок, искатели жемчуга и кораллов в Австралии, Япо-
нии и других странах ныряют на 40—50 м и находят-
ся под водой до 4 мин.
На глубину 60 м у Багамских островов нырнул,
пробыв под водой 2 мин, француз Жак Майоль. Он
установил мировой рекорд погружения без акваланга.
Этот рекорд вскоре перекрыл на 13 м Роберт Крофт.
Но это ничтожно мало в сравнении с океанской безд-
ной. Человек завоевывает каждый метр морской тол-
щи ценой огромных усилий.
Гораздо большего в этом достигли морские млеко-
питающие, которых природа тренировала миллионы
лет, и они стали непревзойденными ныряльщи-
ками.
Из ластоногих глубже всех ныряет антарктический
тюлень Уэдделла. Англичанин доктор Геральд Куйман
прикреплял к его телу манометр и из 380 ныряний за-
регистрировал одно погружение в проливе Мак-Мёрдо
на глубину 600 м.
Максимальная скорость погружения при этом ока-
залась 104 м/мин.
Все китообразные — отличные пловцы. Регулируют
они свою плавучесть, выпуская под водой то большее,
то меньшее количество воздуха из дыхательных путей.
Но способности к нырянию у них весьма различны.
Усатые киты, питающиеся близ поверхности, обычно
глубоко не ныряют (не более чем на 100 м) и только
раненные гарпуном опускаются до 350—500 м.
91
Не глубоко, но очень легко ныряют дельфины, пи-
тающиеся пелагическими видами рыб. Исследователь
США Кеннет Норрис на Гавайских островах обучил
морщинистозубого дельфина Поно нырять по сигналу
50 раз за 105 мин на глубину до 30 м. На подъем с
такой глубины дельфин затрачивал лишь 18 с, тогда
как водолазу в мягком скафандре потребовалось бы
для этого не менее 10 ч, если он пробыл на такой
глубине 10 мин.
Значительно глубже погружаются особи видов
дельфинов, потребляющие придонную пищу. Обучен-
ная афалина, к которой прикреплялись манометр, осо-
бый звуковой сигнализатор и датчик глубин, в опы-
тах Сэма Риджуэя в 1966 году ныряла до 170 м. При
этом максимальная скорость погружения у нее дости-
гала 143 м/мин.
Ряд экспериментов провел центр подводных иссле-
дований США в Пойнт-Магу на афалине — знамени-
том самце Таффи (см. о нем в гл. XXII), обученном
нырять по команде. При погружении у него замечено
сдавливание грудной клетки, причем первые признаки
коллапса обнаруживались аппаратурой подводного те-
левидения уже на глубине 10 м. В рекордных погруже-
ниях Таффи достигал глубины 300 м за 4 мин 45 с.
Это точно зарегистрировал прибор (плунжер) и фото-
снимок животного. Таффи нырял на заданную глуби-
ну и после 4-минутной дыхательной паузы выдыхал
воздух под погруженную воронку воздухосборника.
После столь глубокого погружения состав выдыхаемо-
го воздуха приходил в норму только через 8—10 ды-
хательных актов. Дельфин в экспериментальных усло-
виях сделал 370 глубоких ныряний вдоль кабеля, а
однажды в течение только одного часа он сделал де-
вять погружений на 200 м и глубже.
Судя по характеру пищи, афалина в Гвинейском
заливе может нырять на 400—500 м, как это утвер-
ждает французский биолог П. Ранкурель. В наши дни
погружение на 500 м выполняет в Сан-Диего дрессиро-
ванная гринда: она находит опущенную на дно моря
ракету со звуковым сигнализатором, прикрепляет к
ней особое устройство с жидким газом; газ наполня-
ет мешок и поднимает ракету на поверхность моря.
Однажды, сообщает Форрест Вуд, она нырнула на глу-
бину 610 м, затратив на это 13 мин.
92
Из китообразных бесспорное место рекордсмена-
ныряльщика занимает кашалот, способный задержи-
вать дыхание до полутора часов. Между прочим,
экспедиция французского океанолога Ж.-И. Кусто в
Индийском океане с судна «Калипсо» установила впол-
не добродушное отношение этих исполинов к людям:
аквалангисты и кинооператоры, забрасываемые на
вспомогательных шлюпках в стадо кашалотов, бес-
страшно прыгали в воду, забирались китам на спину,
хватали их за плавники, метили и фотографировали
животных (то же самое они проделывали с горбачами,
финвалами и серыми китами в Тихом океане) (рис. 39
вкл.). Чтобы остановить кашалота, быстроходная лод-
ка «Зодиак» начинала кружить близ него, и это позво-
ляло аквалангистам вплотную подплывать к зверю.
Записи американских ученых Р. Баккуса и В. Ше-
вилла, проведенные с помощью эхолота, показали сво-
бодное погружение кашалотов в одном случае на 520,
а в другом — на 310 м.
Американский геолог Брус Хизен специально изу-
чал повреждения глубоководных телеграфных кабе-
лей, причиненные кашалотами. Четырнадцать раз из
глубин вместе с кабелем извлекали запутавшихся в
нем кашалотов. Чаще всего смертельные петли обви-
вали хвост и длинную нижнюю челюсть задохнувших-
ся китов. Пять из них погибли на глубине в один ки-
лометр (от 910 до 1128 м), семь — под столбом воды
от 120 до 855 м и два — на неустановленной глубине.
Некоторые исследователи • (доктор А. В. Яблоков)
считают, что кашалоты могут погружаться на 2,2 км.
В 1970 году американские ученые методом триангуля-
ции (т. е. методом засечки локационных щелчков ны-
ряющих кашалотов, произведенной из трех разных
пунктов на поверхности моря) определили максималь-
ную глубину погружения этих китов в 2,5 км. Кабели,
эхолоты и метод триангуляции засвидетельствовали,
что кашалот ныряет в царство вечной тьмы. Такие же
способности имеют и клюворылые киты (рис. 40 вкл.),
они тоже остаются под водой более часа, питаются
крупными глубоководными головоногими моллюсками
и снабжены спермацетовым органом.
Уже не раз в Антарктике на наших китобойных
судах в желудках кашалотов обнаруживали гигант-
ских кальмаров из рода архитеутис (рис. 41 вкл.), а
93
в 1971 году китобойная матка «Юрий Долгорукий»
привезла для музея кальмара, у которого длина туло-
вища была 1,5 м, щупальца—10 м. Специалисты,
например И. И. Акимушкин, утверждают, что охотни-
чий горизонт кашалота лежит на глубине 1—1,5 км.
Невольно возникают вопросы: какими приспособ-
лениями обладают эти удивительные водолазы? В чем
заключаются секреты их продолжительного ныряния?
Что позволяет некоторым китообразным погружаться
на чудовищную глубину и выдерживать колоссальное
давление в 100—200 атмосфер?
Такие необычайные способности морских млеко-
питающих определяются многими их морфологически-
ми, физиологическими и экологическими приспособ-
лениями.
Само по себе увеличивающееся давление при погру-
жении, видимо, не опасно для тканей китообразных,
поскольку их тело на 2/3 состоит из воды, а. вода
практически несжимаема. Выносливость ныряющего
кита к большому давлению объясняется уравновеши-
ванием давления внутри его тела (в полости, органах
и тканях) с внешним гидростатическим давлением.
Первостепенное значение при этом имеют следую-
щие физиологические приспособления.
Во-первых, у китообразных понижена чувствитель-
ность дыхательного центра к накоплению углекисло-
ты в крови, и дыхательный акт их возбуждается на-
ступающим недостатком кислорода.
У наземных же млекопитающих акт дыхания воз-
буждается накоплением углекислоты в крови, и они
совершают дыхательный акт еще при значительном
количестве кислорода в легочном воздухе.
Таким образом, китообразные гораздо полнее, чем
наземные млекопитающие, используют кислород в
крови и в легких и могут надолго задерживать дыха-
ние. Кроме того, очень упругая структура легких, при-
способленная к быстрому сжатию и расширению, обес-
печивает обновление воздуха в легких за одно дыха-
ние на 80—90%, а у человека лишь на 15%.
Во-вторых, у китообразных весьма высокое содер-
жание мышечного гемоглобина или миоглобина — ды-
хательного пигмента (например, в мышцах у дельфи-
на-белобочки его почти столько же, сколько гемогло-
бина в крови). Кит при дыхании на поверхности на-
94
сыщает кислородом и миоглобин и гемоглобин крови,
а следовательно, ныряет с увеличенным (до 40%) коли-
чеством кислорода, отдаваемого тканям во время ды-
хательной паузы. Миоглобин обеспечивает кислородом
главным образом работающие мышцы. Японские уче-
ные подсчитали, что в грамме мышц кашалота мйб-
глобина содержится в 8—9 раз больше, чем в грамйе
мышц быка.
В-третьих, во время ныряния у китообразных почти
вдвое замедляется частота ударов сердца \ уменьша-
ется пропускная способность сосудов мышц и перерас-
пределяется ток крови, чему способствует система
многочисленных сфинктеров в венах. Кровь начинает
двигаться медленнее, и ткани получают кислород в
уменьшенном количестве; это усугубляется действием
сфинктеров, задерживающих кровь в большой полой
вене. Благодаря такому перераспределению кровь при
нырянии снабжает кислородом в первую очередь голов-
ной и спинной мозг и сердечную мышцу, наиболее
чувствительные к кислородному голоданию. Кольце-
видные мускулы-сфинктеры, словно «экономный снаб-
женец» в энергетическом хозяйстве кита, перекрыва-
ют часть вен, несущих кровь из разных участков тела,
и переводят на голодный паек многие органы и мыш-
цы ныряющего кита, которые довольствуются лиШь
тем кислородом, который своевременно запасен мйо-
глобином.
В процессе ныряния возрастает кислородная задол-
женность в мышцах, где происходит бескислородный
гликолиз и накапливается конечный продукт расщеп-
ления углеводов — молочная кислота. Однако она по-
является в крови тогда, когда животное уже будет на
поверхности. Выключение из системы кровообращения
мускулатуры приводит к тому, что молочная кислота,
накапливающаяся при мускульной работе, не разно-
сится по телу, а остается в мышцах. Но как только
кит вынырнет и в мышце восстановится нормальное
кровообращение, молочная кислота сразу появляется
в крови.
1 Чем меньше размеры тела китообразных, тем чаще их пульс.
Кардиограммы показали частоту сердцебиения на поверхности
воды и на глубине: у афалины 110 и 50 ударов, у белухи —
30 и 16, а у раненого кита-полосатика — 30 и 15 ударов в
минуту.
95
В-четвертых, кислородная емкость крови у китооб-
разных на 1/4—1/3 выше, чем у человека.
В-пятых, среди механизмов, обеспечивающих бес-
перебойное снабжение мозга кислородом, важнейшее
значение имеет «чудесная сеть» — тонкое сплетение
артерио-венозных капилляров, служащее хранилищем
крбви, богатой кислородом. Эта сеть имеется на стен-
ках грудной полости, в шейной области, но особенно
сильно развита вокруг спинного и головного мозга.
Крупных артерий, по которым бы кровь поступала в
головной мозг, у дельфинов не обнаружено. Чудесная
сеть создает для мозга резервы кислорода и служит
кислородным депо. Емкость депо, возможно, увеличи-
вается и за счет жировой массы, окружающей чудес-
ную сеть: ведь кислород растворяется, например, в ка-
шалотовом жире и спермацете в 7 раз лучше, чем в
воде. Предполагают, что есть еще неизученные устрой-
ства, быстро переносящие кислород из крови в жиро-
вую массу, окружающую чудесную сеть.
Способ действия этой сети точно не установлен, но
полагают, что она может быстро наполняться кровью
и опорожняться, регулируя кровяное давление во вре-
мя погружения и выныривания, когда резко меняет-
ся внешнее давление среды.
Исследователи из штата Флорида Юджин Нейгел с
сотрудниками, изучая под лучами Рентгена кровооб-
ращение мозга у анестезированной афалины, устано-
вили еще одну важную роль чудесной сети: она га-
сит толчки крови, вызванные сокращением сердца, и
подает кровь в мозг равномерным потоком под посто-
янным давлением. Погашение пульса обусловлено бы-
стрым вбиранием большого количества крови в веноз-
ную часть чудесной сети и быстрого ее опорожнения
за счет мускульных стенок мелких артерий.
Среди морских млекопитающих чудесная сеть силь-
нее всего выражена у кашалотов и полностью отсутст-
вует у самых плохих ныряльщиков — сирен.
Все перечисленные приспособления определяют лег-
кость и продолжительность ныряния китообразных.
Одной из причин, мешающих водолазу опускаться
на глубины, является кессонная болезнь. Как извест-
но, с погружением на каждые 10 м давление воды воз-
растает на одну атмосферу, и уже на глубине, напри-
мер, 1000 м оно будет составлять 11 атмосфер. Чтобы
96
Рис. 1. Молодая афалина Оно-Джек подставляет свое тело для
поглаживания. Новая Зеландия. Фото С. Хилла
Рис. 2. Афалина Альфа на пляже Евпатории среди купальщи-
ков. Лето 1968 г.
Рис. 3. Фонтан южного кита с двумя широко расходящимися
струями
Рис. 4. Фонтан малого полосатика — с одной струей в виде опро-
кинутого конуса. Видны Две открытые ноздри.
Рис. 5. Цедильный аппарат для процеживания пищи у финвала.
Пластины расположены по обе стороны узкого неба. По Л. X.
Мэтьюсу
Рис. 6. Группа самок кашалотов с детенышами. Снято с само-
лета. Фото японской компании Суиса-Коку
Рис. 7. Дельфинарий на Черном море (СССР)
Рис. 8. Океанариум «Мэриленд» (Калифорния). Фото К. Брауна
Рис. 9. Бесперая морская сви-
нья в океанариуме г. Хироси-
ма
Рис. 10. Обезьяна шимпанзе
кормит афалину в Эносим-
ском океанариуме (Япония)
Рис. 11. Дружный прыжок короткоголовых тихоокеанских дель-
финов в Калифорнийском океанариуме
Рис. 12. Батумский дельфинарий. Тренер наедине с афалиной.
Фото автора
Рис. 13. Дельфины-афалины и очковый пингвин в Батумском
дельфинарии
□гйЖ
Рис. 16. Выпрыгнувшая афалина. Естественная склонность к
прыжкам усиливается дрессировкой
Рис. 17. Подъем гринды
из воды на мягких но-
силках. Фото К. Брауна
Рис. 18. Извлечение заглоченного предмета у гринды. Фото
К. Брауна
Рис. 19. Кормеж-
ка афалин в Ба-
тумском дельфи-
нарии
Рис. 20. Стайка афалин в океанариуме выпрашивает пищу
Рис. 21. Кашалот даже с искривленной нижней челюстью хорошо
упитан: дефект не мешает ему засасывать в рот моллюсков
Рис. 22. «Звезда» Батумского дельфинария Василиса дает высот-
ный фонтан. На переднем плане самец Персей
Рис. 23. Крупный самец-афалина (Зм), не раз проявлявший свою
агрессивность по отношению к тренерам
Рис. 24. Начало родов у афалины (хвост только показался). Фото
океанариума «Мэриленд»
Рис. 25. Более поздняя стадия родов у афалины (детеныш вышел
наполовину, виден пупочный канатик). Фото океанариума «Мэ-
риленд»
Рис. 27. Реакция
следования у де-
теныша афали-
ны: малыш стро-
го придерживает-
ся материнского
бока. Фото океа-
нариума «Мор-
ская студия». По
Э. Кэй
Рис. 26. Самка обыкновенного дельфина с новорожденным, у
которого еще не выпрямился спинной плавник. Фото Ф. Логэна
и Ф. Робсона
Рис. 28. Афалина-мать впрыскивает молоко в рот детенышу,
а «тетка-повитуха» на страже, чтобы защищать малыша. Фото
океанариума «Морская студия». По Э. Кэй
Рис. 29. Выкидыш афалины
Рис. 30. В Батумском дельфинарии. Обучение командам с по-
мощью жестов
Рис. 31. Афалина балансирует мячом на роструме
Рис. 32. Манипуля-
ция хвостом (афали-
на подбрасывает
мяч) в Батумском
дельфинарии
Рис. 33. Синхронный прыжок афалин в Батумском дельфинарии
Рис. 34. У косатки очень характерный рисунок на брюхе. В море
это можно видеть в момент ее мощного прыжка
Рис. 35. Момент выступления косатки в океанариуме «Си-Уорлд»
Рис. 36. Мчащееся стадо обыкновенных дельфинов. Фото В. Эван-
са
Рис. 37. Один из самых быстроходных китообразных — вертя-
щийся продельфин. Славится вращением тела в высотных прыж-
ках из воды; за прыжок он успевает сделать два с половиной
оборота. Фото Л. Джекобса
Рис. 38. Пятнистые продельфи-
ны оседлали корабельную вол-
ну. Фото сотрудников Флорид-
ского океанариума
Рис. 40. Один из лучших ныря-
льщиков — плосколобый бутыл-
конос (самец длиной 7 м). Фото
Ю. А. Михалева
Рис. 41. Гигантский кальмар
архитеутис (длина 9,5 м) из
Рис. 39. Помеченный легким желудка кашалота, добытого
гарпуном серый кит погружает- флотилией «Советская Россия»,
ся вблизи лодки. Фото экспеди- Фото Ю. А. Михалева
ции Жак-Ива Кусто
Рис. 42. Афалине закрывают глаза присосками для опыта по
слуховой ориентации. Фото океанариума «Мэриленд»
Рис. 43. Акустический глаз кашалота; видна пузырчатая ткань,
возможно выполняющая роль сетчатки. Фото В. А. Козака
Рис. 44. Финвал на палубе «Советской Украины». Фото В. И.
Шевченко
Рис. 45. Горбатый кит, круто вынырнувший на поверхность. Фото
В. И. Тройнина
Рис. 46. Квартет афалин в океанариуме. Фото Л. Джекобса
Рис. 47. Афалины:мать Василиса и «тетка» спасают новорожден-
ного. Батумский дельфинарий
Рис. 48. Самка обыкновенного дельфина выталкивает новорож-
денного детеныша в Нейпирском океанариуме. Фото Л. Логэна
и Ф. Робсона
Рис. 49. Детеныш афалины спасает ослабевшую мать; его дейст-
вия сводились к выталкиванию то хвостового стебля, то брюха,
то головы больной самки
Рис. 50. Гринды спасают больного сородича (в центре). Фото
М. и Д. Колдуэллов
Рис. 51. Быстро плывущее стадо кашалотов. Фото сотрудников
Японского китового института
Рис. 52. Обсохшее стадо гринд. По Ван Хил Дудоку
Рис. 53. Гибель стада кашалотов (46 самок и 13 самцов) на пля-
же Окита-Бич (Новая Зеландия, март 1970 года): вверху — послед-
ние минуты в воде; внизу — полное обсыхание. Фото Ф. Робсона и
П. Ван Бри
Рис. 54. Стадо белух в заливе Каннигем (Канадский архипелаг).
Видны самки с детенышами. Снимок с самолета. По Э. Митчел-
лу и Д. Сержанту
Рис. 55. Строй дельфинов у носа корабля. У всех одновременно
открыто дыхало. Фото В. Вольфа
Рис. 56. Кашалот, отдыхающий на поверхности моря. Снимок
с самолета. По М. Нисиваки
Рис. 57. Молодые афалины Моряк и Боцман во время дрессиров-
ки в Батумском дельфинарии
Рис. 58. Дельфина-афалину обучают апортировке спасательного
круга в Батумском дельфинарии
Рис. 59. Афалина Таффи — участник испытаний «Силэб II*.
Свободно плавает в море и возвращается по сигналу человека.
Фото Торштейна Телля
Рис. 60. Сейвалы на палубе китобойной матки
Рис. 61. Кашалоты у борта китобойной матки
уравновесить такое давление, водолазу подают по
шлангу воздух под напором, соответствующим глуби-
не погружения. На большей глубине водолаз вдыхает
больше воздуха и по весу. При увеличении давления
растворимость воздуха в крови возрастает, и мало-по-
малу газом насыщаются все ткани тела. Быстрый
подъем водолаза с глубины может оказаться для него
смертельным, так как с падением давления пузырьки
азота выделяются в кровь и могут закупорить кро-
веносные сосуды. Вот почему водолаза поднимают с
остановками в декомпрессионных камерах.
Киты и дельфины не подвержены кессонной болез-
ни даже при стремительном подъеме с глубин в сотни
метров. Разгадка проста: все китообразные, совершив
один — три дыхательных акта (дельфины) или целую
серию таковых (киты), погружаются в отличие от во-
долазов всегда только с одной порцией воздуха, при-
том воздуха нормального давления. Поэтому азот по-
падает в их кровь в безопасном количестве.
При известных условиях время, необходимое водо-
лазам для декомпрессии, можно сократить. В годы
Отечественной войны водолазов поднимали с глубины
180 м за 3/4 ч вместо 7 ч. Дело в том, что для дыха-
ния им подавали вместо воздуха газовую смесь гелия
с кислородом. Гелий, как и азот, тоже нейтральный
газ, но он попадает в кровь быстрее, меньше раство-
ряется в ней и скорее исчезает из нее. Поэтому такая
смесь безопаснее для человека.
Вскоре выявился иной путь покорения морских
глубин: замена тяжеловесного снаряжения водолаза
более легким. В 1943 году Жак-Ив Кусто и инженер
Эмиль Ганьян дали миру акваланг, или подводные
легкие. Этот аппарат позволяет любому тренированно-
му пловцу погружаться на глубину 40—50 м. Позже
Ж.-И. Кусто сам опускался с аквалангом на 100 и
140 м. Ныне, при дыхании смесью кислорода и гелия,
глубина ныряния человека с аквалангом в мягком
скафандре увеличилась до 330 м. Однако настоящее
освоение морских глубин невозможно без подводных
домов.
Чтобы использовать несметные богатства водной
толщи и недр, скрытых морем, по инициативе Жак-
Ива Кусто начато изучение глубин океана с помощью
подводных поселений.
6 9-1984 97
В сентябре 1962 года в Средиземном море близ
Марселя на глубине 11 м был установлен первый под-
водный дом «Преконтинент I», в котором два акванав-
та жили в течение недели. Они доказали возможность
продолжительного существования человека под водой.
Через год на глубине 11 м в Красном море на жест-
ком дне кораллового рифа Шааб-Руми была установ-
лена стальная пяти лучевая «морская звезда», состоя-
щая из пяти комнат, и недалеко от нее два дома иной
конструкции: один в виде цилиндра (на глубине 27 м),
другой — грибовидной формы. Группа людей пробыла
в «подводной деревне» «Преконтинент II» месяц.
В сентябре — октябре 1965 года шесть акванавтов
(в их числе сын Жак-Ива Кусто Филипп) три недели
вели исследования в шаре-лаборатории «Преконти-
нент III» (диаметром 5,7 м), установленном в Среди-
земном море близ мыса Феррато на глубине 110—
130 м. Они дышали газовой смесью гелия (98%)
и кислорода (2%) при давлении в 11 атмосфер, выплы-
вали с аквалангами из своего шара для проведения
опытов и наблюдений.
Эксперимент французских ученых («Преконти-
нент II») повторили американские исследователи, орга-
низовавшие подводные морские лаборатории в 1964 го-
ду «Силэб I» (близ Бермудских островов на глуби-
не 60 м) и в 1965 году «Силэб II» (в Калифорнии близ
местечка Ла-Холья, на глубине 64 м). В первой рабо-
тали 4 человека 11 дней, а во второй — 28 человек. Их
разделили на три группы, жившие под водой по 15 су-
ток каждая. Руководитель экспедиции «Силэб I» и
«Силэб II» Джордж Бонд полагает, что человек в бу-
дущем сможет организовать лабораторию на глубине
700—1000 м. Почти одновременно с работой «Си-
лэб II» на глубину 130 м была опущена на двое суток
«палатка» ученого Эдвина Линка с двумя акванавта-
ми — американцем Джоном Линдбергом и бельгийцем
Робертом Стенуи. На еще большей глубине (150 м) бы-
ло поселение «Преконтинент IV» в 1967 году в Аден-
ском заливе. В перспективе лежит штурм глубины бо-
лее 400 м.
С помощью специальных камер люди уже побыва-
ли на самых больших глубинах Мирового океана:
23 января 1960 года швейцарский пилот Жак Пиккар
и наблюдатель Дон Уолш в батискафе «Триест» достиг-
98
ли дна тихоокеанской котловины Челленджер в Мари-
анской впадине. Спускались они на глубину 10 910 м
пять часов, а поднимались три с половиной часа и
двадцать минут провели на дне. Здесь на металличе-
скую гондолу давило 200 000 т воды.
Сейчас стоит очень важная задача — расширить
возможности подводного плавания человека без спе-
циального снаряжения, в частности без громоздкого ак-
валанга. Проблема эта очень сложная, и успех будет
определяться тем, удастся ли в организме ныряль-
щика создать условия, свойственные морским млеко-
питающим. Подобную задачу можно решить лишь
путем тщательного изучения физиологических измене-
ний и адаптаций в организме ныряльщика, познания
физиологии китообразных, в первую очередь их дыха-
ния и кровообращения. Перспективными в этом, веро-
ятно, были бы поиски такой газовой смеси для дыха-
ния, которая практически не растворялась бы в крови
и устраняла вскипание газа в кровеносных сосудах.
Возможно, в решении проблемы свободного погру-
жения человека без специального снаряжения помогут
тончайшие пропускающие кислород в воде избиратель-
ные кремнийорганические пленки, подобные тем, ка-
кие испытывал на хомяках инженер США Уолтер
Рабб. Эта пленка не пропускает воду, но проницаема
для газов, в первую очередь для кислорода и углекис-
лого газа. Она может выполнять ту же роль, что и
жабры рыб: свободно впускать кислород из области
высокого парциального давления в область низкого и
в обратном направлении пропускать углекислоту.
В США ученые заняты идеей создания искусственных
жабр в виде портативного аппарата, который будет
подавать кислород в кровь ныряльщика без помощи
легких. Шланги такого аппарата должны подключать-
ся к аорте путем операции. Если удастся заполнять
легкие какой-либо стерильной пластической массой,
то человек с помощью искусственных жабр, по мне-
нию Жак-Ива Кусто, сможет погружаться на двухки-
лометровую глубину. Опыты на собаках, в легкие ко-
торых нагнетали воду, насыщенную кислородом, а по-
том удаляли ееу показали, что легочное дыхание после
подобных манипуляций может быть восстановлено у
некоторых наземных млекопитающих без пагубных
последствий.
6*
Глава XI
СУДЬБА
АНАЛИЗАТОРОВ
В связи с глубоким погружением китообразных воз-
никает вопрос: как они ориентируются и ловят добы-
чу в малопрозрачной водной толще, а иногда и в цар-
стве вечной тьмы, ночью и днем, как определяют путь
при далеких миграциях и точно выходят к цели, от-
даленной на тысячи километров? Каковы механиз-
мы этой изумительной подводной ориентации и нави-
гации?
Каждому ясно, что в этом им помогает высокораз-
витая нервная система. Деятельность мозга невозмож-
на без непрерывного притока информации из окружа-
ющей внешней и внутренней среды. Эта информация
поступает в организм через органы чувств (анализато-
ры), которые справедливо называют «окнами в мир».
Без органов чувств, доставляющих информацию о се-
зонных и суточных изменениях окружающей обста-
новки и о многих других внешних раздражителях, не
было бы у китообразных ни великолепной ориенти-
ровки в пространстве, ни правильных миграций, ни
случаев возвращения в одни и те же бухты после гран-
диозных странствий.
В новой среде понадобилось коренное переустрой-
ство всех анализаторов, полученных в наследство от
наземных предков. Одни анализаторы приобрели пер-
востепенное значение, другие — второстепенное, тре-
тьи — исчезли.
Редукции у китообразных подверглось обоня-
ние — ощущение запахов посредством органа, распо-
ложенного в носу. Сама по себе жизнь в воде еще не
ведет к исчезновению органа обоняния. Вспомним хо-
100
тя бы акул, которые идут на запах крови с далеких
расстояний, и у них сильно развиты обонятельные доли
переднего мозга. У рыб орган обоняния развивался и
приспосабливался к улавливанию запахов в воде, а у
наземных предков китообразных запахи проникали
через ноздри вместе с вдыхаемым атмосферным возду-
хом. С переходом в воду потомки этих предков утрати*
ли обоняние, так как молекулы пахучих веществ мог-
ли проникать в носовой канал лишь в момент очень
короткого вдоха, после длительной дыхательной пау-
зы, проводимой под водой с закрытыми ноздрями.
Воздух над океаном был чистым, и запахи в нем поте-
ряли всякое значение для живущих в воде китов.
Поэтому обонятельные луковицы мозга, обонятельные
нервы и обонятельный эпителий в решетчато-раковин-
ном отделе черепа либо полностью исчезли (зубатые
киты), либо сохранились лишь в зачаточном виде, при-
том лучше у зародышей (усатые киты). Растворен-
ные же в воде химические вещества, в том числе и
запахи пищи, кала и мочи сородичей, стали раздражи-
телями другого органа чувств — вкуса.
Вкус у китов считали слабо развитым или даже ис-
чезнувшим на основании того, что у них тонкий вку-
совой нерв и в их желудках встречаются несъедобные
предметы (булыжники, галька, куски Дерева и многое
другое). Но камни, по-видимому, заглатываются для
перетирания пищи и потому не могут свидетельство-
вать о слабом развитии вкуса. Другие инородные пред-
меты заглатываются случайно при процеживании це-
дильным аппаратом всего, что попадает в сферу
фильтрации, когда невозможно отделить съедобное от
несъедобного. Ученые, например Э. И. Черный и
А. В. Яблоков, полагают, что китообразные способны
тонко различать разную соленость воды и обнаружи-
вать своих сородичей по их «химическим следам» —
по моче и фекалиям. Опресненные устья рек посещают
многие морские свиньи, белухи, дельфины-соталии, ор-
целлы, малые полосатики, а серые киты находят полу-
пресные лагуны, чтобы очищать кожу от китовыд
вшей, погибающих от опресненной воды.
Далеко не все китообразные могут долго пребывать
в пресной воде. Некоторые океанские дельфины тяже-
ло переносят недостаточную соленость: они заболева-
ют прыщами, рожистыми воспалениями, грибковыми
101
поражениями кожи и погибают, как это было с косат-
кой Моби-Долл. Калифорнийские зоологи Сибеналер,
Мельба и Давид Колдуэллы описали, как в Мексикан-
ском заливе афалины по нескольку недель остаются в
опресненных заливчиках, кормясь сельдевыми рыба-
ми, которые здесь мечут икру. В это время на коже
дельфинов выступает сыпь. Соленость Черного моря
вдвое меньше солености океана. Не этим ли объясня-
ется отсутствие здесь китов и некоторых видов дель-'
финов, встречающихся в Средиземном море? Лишь три
вида дельфинов смогли приспособиться к жизни в
Черном море, обособившись в самостоятельные под-
виды: дельфин-белобочка, афалина и морская свинья.
Первый заселил зону открытого моря, вторая и
третья — прибрежную зону, причем морская свинья
заняла также мелководное и наиболее опресненное
Азовское море (11%о).
Черноморские дельфины, приспособившись к усло-
виям пониженной солености (18%о), вероятно, не выхо-
дят через проливы и не общаются со средиземномор-
скими популяциями, а средиземноморские китообраз-
ные, привыкшие к океанской солености (35%о), не
заходят в Черное море. Джон Лилли описал случай,
как одна афадина, находя^ в бассейне его лаборато-
рии на Виргинских островах, пришла в сильнейшее
возбуждение, когда вблизи проплывала стайка сороди-
чей. Ученый полагает, что дельфин узнал о близости
стайки по «вкусу» (химическому составу) той воды,
которая по шлюзу вливалась из моря в бассейн. Вкусо-
вые ощущения, по-видимому, воспринимаются валико-
видными сосочками, расположенными по краю языка.
Вообще орган вкуса у китообразных изучен еще очень
слабо. Однако за последние годы в этой области сдела-
ны интересные находки.
Советский морфолог доктор А. В. Яблоков в
1956 году нашел у корня языка белухи, а потом и у
других видов зубатых китов несколько ямок, покры-
тых призматическим эпителием. Исследователь связал
функцию ямок с хеморецепцией, специализированным
восприятием вкусовых ощущений. Московские биоло-
ги академик В. Е. Соколов и О. В. Волкова также счи-
тают, что ямки на языке дельфинов несут, сенсор-
новкусовую функцию, а обильные здесь слизистые
железы выносят из организма китообразных соли, по-
102
падающие в их пищеварительный тракт вместе с мор-
ской водой. Сотрудник Московского университета
В. Б. Кузнецов экспериментально показал разную
реакцию на разные вещества в растворе, которым про-
мывали ротовую полость подопытных дельфинов-бело-
бочек и морских свиней. Химические вещества в ма-
лых концентрациях, по мнению этого ученого, воспри-
нимаются хеморецепторами тройничного нерва.
Осязание китообразных, как показывают на-
блюдения над живыми особями, развито достаточно
хорошо. Кожа играет важнейшую роль в восприятии
раздражений, поступающих из внешнего мира. Она не
ороговевает сверху (роговой слой быстро слущивает-
ся), и это способствует сохранению высокой кожной
чувствительности. Кожные рецепторы (тактильные,
температурные, болевые) сигнализируют животному о
прикосновениях к телу, о температуре среды, о боле-
вых ощущениях, о меняющемся давлении воды. Вод-
ная среда с ее сравнительно стабильными окружаю-
щими условиями как будто мало способствует разви-
тию высокой кожной чувствительности китообразных.
Однако данные показывают другое.
Как показали гистологические исследования, в
кожном покрове китообразных (особенно на голове
дельфинов) расположены пучки нервных волокон с их
окончаниями, сложные клубочковые рецепторы и ин-
капсулированные чувствительные аппараты типа колб
Краузе и фатер-пачиниевых телец. Чувствительные
окончания этих нервных структур осуществляют, по
мнению ученых, термо-, баро- и механорецепцию
и служат рецепторами-датчиками, которые выполня-
ют прямую и обратную связь с мозгом. Возможно,
что очень высокие сосочки дермы, помимо выполне-
ния своей демпферной функции, предотвращающей
развитие вихревых потоков, играют также роль так-
тильных органов.
Редукция волосяного покрова, потовых и сальных
желез не притупила у китообразных кожной чувстви-
тельности. Смена среды, ощущаемая при вынырива-
нии, служит им сигналом для открывания дыхала и
дыхательного акта (поэтому к лобному выступу дель-
финов, выставляющемуся из воды первым, подходит
толстый чувствующий нерв). В ответ на легкое при-
косновение к коже дельфины открывают и закрывают
103
глаза. Хорошо иннервированная кожа китообразных
способна управлять пограничным слоем воды с по-
мощью тонких двигательных механизмов, тесно свя-
занных с тактильной чувствительностью наружного
покрова. Двигательные реакции кожи способствуют
сохранению обтекаемых потоков вокруг тела быстро
передвигающихся дельфинов. Высокая тактильная
чувствительность их кожи смогла развиться при жиз-
ни в условиях относительной невесомости погружен-
ного в воду животного.
Имеющиеся на морде усатых китов десятки волос-
ков, луковицы которых богато оплетены нервными
окончаниями, указывают на высокую кожную чувст-
вительность в этой части тела. Волоски — это своеоб-
разные вибрйссы: при прикосновении к ним они дей-
ствуют как рычаги, увеличивающие силу внешнего
раздражения на нервные окончания. Усатые киты, на-
талкиваясь в воде на крошечных рачков, легко опре-
деляют густоту скопления корма при любом освеще-
нии и часто кормятся ночью.
Зубатые киты, питающиеся одиночной и сравни-
тельно крупной добычей, в осязательных волосках не
нуждаются и утрачивают их еще в утробе матери или
вскоре после рождения. На месте выпадения волосков
остаются ямки специфического строения. Иннервация
их оказалась настолько богатой, что английские уче-
ные Элизабет Палмер и Г. Уэдделл рассматривают эти
ямки как спидометры — чувствительные приборы
дельфинов для измерения скорости движения и вос-
приятия низкочастотных колебаний. Лишь речные
дельфины с недостаточно развитым зрением, живущие
в мутной воде и роющиеся в илистом дне, сохраняют
волоски на клюве в течение всей жизни.
Китообразным во время миграций и при нырянии
приходится испытывать то тепло, то холод, и можно
полагать, что они тонко ощущают температуру среды,
о чем косвенно свидетельствует их превосходная ре-
гуляция тепла и тот факт, что многие киты избегает
тропических вод.
Кожу китообразных называют волшебной, так как
с кожным раздражением связано не только их осяза-
ние, но и исключительная быстроходность, теплорегу-
ляция, проявление инстинкта сохранения вида, пове-
104
дение во время сна и преследования судов, поощрение
во время дрессировки и многое другое.
Очень мало известно о проприоцептивной чувстви-
тельности китообразных (раздражение нервных окон-
чаний при сокращении, растягивании скелетных
мышц). Однако совершенство, с которым они двигают-
ся, позволяет думать, что у них хорошо развит про-
приоцептивный аппарат.
Зрительный анализатор — важнейший сре-
ди анализаторов у дневных наземных млекопитаю-
щих. С его помощью человек, например, получает
80—85% информации из внешней среды. Зрением вос-
принимается окружающий мир — величина, форма,
цвет, яркость, освещение и отдаленность предметов, от
которых отраженный свет поступает в глаз. Море, как
малопрозрачная среда, не благоприятствует сильному
развитию зрения водных млекопитающих. Свет здесь
быстро поглощается, и уже на глубине 500 м практи-
чески наступает полная темнота.
Глаз крупных китов весит около 1 кг, а у мелких
дельфинов достигает величины глаза собаки. Глазное
яблоко, спереди уплощенное, напоминает форму не-
сколько сдавленного шара. Щель глаза закрывается
веками, лишенными ресничек. Слезные железы отсут-
ствуют, а поверхность глаза смазывается прозрачным
и густым белковослизистым выделением гардеровых
желез. Оно предохраняет глаз от вредного действия
морской воды. Белковая оболочка (склера) толстая и
плотная (сосудистая — пятислойная, как у копытных).
Родятся китообразные с открытыми глазами. В воде
они видят лишь на коротком расстоянии. Раньше ду-
мали, что на воздухе они близоруки, так как их хру-
сталик шаровидный,, а ресничные мышцы исчезли. Од-
нако у них довольно острое зрение и хорошо развитый
зрительный нерв. В одном квадратном миллиметре сет-
чатки обнаружено у морской свиньи 200 000 палочек,
у белухи — 150 000, у бутылконоса — 143 000 и у
финвала — 62 000 палочек. Колбочки в сетчатке кито-
образных либо отсутствуют, либо крайне малочислен-
ны. По этой причине считают, что китообразные не
различают цветов.
Благодаря особой отражательной оболочке, содер-
жащей большое количество кристалликов гуанина,
глаза дельфинов могут отражать свет подобно зерка-
105
лу. Поэтому ночью глаза дельфинов светятся как у
кошек.
Предполагают, что аккомодация глаза китообраз-
ных достигается за счет «игры» радужины, которая
придает зрачку форму то узкой щели (при ярком све-
те — в воздухе), то круглого отверстия (при тусклом
освещении — в воде). В первом случае свет, пропускае-
мый через узкую, но высокую щель зрачка, проходит
словно через двояковогнутые очки, так как кривизна
и толщина роговицы бывает наименьшей в центре и
наибольшей у краев. Во втором случае свет, пропуска-
емый через круглое отверстие радужины, проходит че-
рез центральную (тонкую) часть роговицы и минует
ее толстую часть. Кроме того, роговица способна уси-
ливать свою кривизну под действием глазных мышц.
Все это обеспечивает разную преломляющую способ-
ность глаза, и китообразные могут хорошо видеть как
в воде, так и на воздухе.
Косатки, кашалоты, полосатики и серые киты
осматриваются, поднимая голову над водой; дельфины
в неволе следят за бросаемой рыбой и кидаются туда,
куда она должна упасть, а иногда схватывают ее да-
же в воздухе. Они точно хватают рыбу из рук дрес-
сировщика с высоты 5 м. Дельфины свободно пово-
рачивают глаз и мигают, если перед ними мелькает
какой-либо мелкий предмет. Зрение китообразных пре-
имущественно монокулярное и в меньшей мере стерео-
скопическое. Поскольку глаза их расположены по бо-
кам вытянутой головы, далеко от переднего кончика
морды, кашалоты, многие киты и часть дельфинов не
могут видеть предметов впереди своего рыла. Лишь у
дельфинов с маленькой мордой поле зрения левого
глаза может частично перекрываться полем зрения
правого глаза, и в этом случае их зрение будет частич-
но стереоскопическим.
Многие дельфины и даже косатки при разглядыва-
нии предметов в бассейне ложатся на бок и исследуют
их при помощи то одного, то другого глаза.
Капитан-китобой начала XIX века Вильям Скорсби
описал, как гренландские киты, проходя под килем
его корабля, поворачивались на бок, чтобы рассмот-
реть, что лежит над ними.
Ученые США Уинтроп Келлог и Чарлз Райс в не-
большом бассейне во Флориде испытывали на 8-летней
106
ручной афалине Пэдди ее способность к распознава-
нию различных фигур с помощью зрения. Фигуры, вы-
ставляемые на мишень, состояли из. светящихся
кружков, треугольников, квадратиков, крестиков, ром-
биков и т. д. Когда дельфин обучился распознавать
эти фигуры, ученые комбинировали их в пары и пока-
зывали ему под водой и на воздухе. В опытах под во-
дой из 25 пар фигур-раздражителей дельфин успешно
различал 21 пару (84%). Результат был отрицатель-
ный, если фигуры помещались на воздухе, а рассмат-
ривались из-под воды. Распознаванию мешала рябь на
поверхности воды, искажавшая изображение фигур.
Острота зрения у Пэдди оказалась наилучшей на рас-
стоянии 1,8 м от мишени.
Насколько велика роль зрения у дельфинов при ло-
ве добычи и распознавании препятствий?
Зрение особенно важно в тех случаях, когда за-
труднено «акустическое разглядывание». Тонкое диф-
ференцирование жестов-команд в Батумском дельфи-
нарии афалины осуществляют при помощи зритель-
ного анализатора.
Биологи — француз Рене Бюснель,. датчанин
А. Дзидзик и норвежец С. Андерсен — провели серию
опытов над морской свиньей в бассейне диаметром
54 м. В бассейн вертикально в шахматном порядке по-
гружали 30' сетей из проволоки толщиной от 0,2 до
4 мм. Сюда выпускали морскую свинью и за 20 мин
свободного плавания подсчитывали, сколько раз
она натыкалась на сети и сколько раз их обходила.
В одной серии опытов она плавала с открытыми глаза-
ми, а в другой — с глазами, закрытыми резиновыми
колпачками. Подсчет показал: чем тоньше была сеть,
тем больше было сталкиваний. Сети из проволоки диа-
метром 0,2 мм «зрячая» морская свинья избегала в
77% случаев, а «ослепленная» —в 46%. Сети из ней-
лоновой лески толщиной 1,5 мм «зрячее» животное из-
бегало в 95%, а «ослепленное» — в 72%. Ясно, что
зрение помогает «зрячим» особям избегать препят-
ствия. Но с помощью чего делают то же самое «ослеп-
ленные» дельфины?
Хорошо известно, что некоторые и зубатые и уса-
тые киты кормятся по ночам, когда зрение не помога-
ет. У речного дельфина, живущего в мутной реке Ганг,
зрение редуцировалось из-за потери хрусталика. Силь-
107
но ослаблено оно и у китайского речного дельфина-ли-
потес. Во всех этих случаях китообразные при лове до-
бычи обходятся без помощи глаз.
Какой же орган заменяет им зрение? Натуралисты
давно заметили, что, если среда не позволяет млеко-
питающим использовать зрение, оно компенсируется
другим органом чувств. Таковым у китообразных ока-
зался слух. Мир дельфинов и китов — это мир зву-
ков. Именно слух стал в йх жизни самым важным
анализатором, совершенство которого выдвинуло ки-
тообразных по уровню развития на первое место среди
животных Мирового океана.
Море оказалось исключительно благоприятной сре-
дой для развития тонкого слуха у водных млекопи-
тающих: ведь звуки в воде распространяются почти в
5 раз быстрее, чем в воздухе, и на гораздо большее
расстояние. Вместе со слухом у зубатых китообраз-
ных развился очень совершенный гидролокатор, позво-
ляющий ориентироваться в окружающей среде при по-
мощи звуковых лучей. Для исследования водного про-
странства звуковыми лучами китообразные вначале
издают звуки, а потом улавливают эхо посланных зву-
ковых волн от окрестных предметов. По эху кито-
образные отлично ориентируются в воде независимо
от освещенности и глубины и получают весьма полную
информацию об окружающем — о пище, сородичах,
близости берега и о многом другом. Ориентация по
отраженным звукам и называется эхолокацией. Эхо-
локация — одно из средств обнаружения препятствия
и определения его положения в пространстве с по-
мощью посланного и отраженного звукового сигнала.
Этим средством, как теперь установлено, владеют
многие млекопитающие и даже птицы, например стри-
жи-саланганы, кроншнепы, живущие в пещерах аме-
риканские козодои. Эхолокацией пользуются некото-
рые ластоногие (например, тюлени), но лучше всего
она развита у рукокрылых и зубатых китов.
В гидролокаторе китообразных имеются как бы два
тесно связанных устройства: передающий ме-
ханизм и приемный механизм. Передающее уст-
ройство включает три пары воздушных мешков, тесно
связанных с носовым каналом, гортань, лобно-носовую
подушку или «жировую линзу» и рефлектор, образо-
ванный вогнутой передней поверхностью черепа. При-
108
емное устройство, к которому возвращаются отражен-
ные звуки, включает органы слуха, нижнюю челюсть
и, возможно, механорецепторы головы.
Передающий механизм эхолокационного
аппарата очень сложен. Главную роль в нем играют
воздушные мешки, расположенные в мягких тканях
головы над костными ноздрями. У разных видов дель-
финов воздушные мешки устроены по-разному. У боль-
шинства видов их три пары (рис. 8 и 9). Первая верхняя
пара — вестибулярные мешки — открывается в носо-
вой канал на 1 см ниже наружной щели дыхала.
К ним прикрепляется несколько слоев радиально рас-
положенных мышц, способных при сокращении изго-
нять воздух из этих резервуаров. Вторая пара — труб-
чатые мешки — залегает ниже первой пары в виде
двух подкововидных трубок вокруг ноздри. Они откры-
ваются с задне-боковой стороны носового канала, ни-
же основной мускульной пробки. По-видимому, воз-
дух, перекрытый пробкой, может циркулировать в них
без выхода наружу. Третья пара — нижние мешки —
залегает на межчелюстных костях и открывается как
раз впереди наружного края костных ноздрей, у само-
го основания мягкой части носового канала. Эта пара
связана со второй соединительными проходами. Все
мешки работают с помощью сложной системы носо-че-
люстных мышц (рис. 10). Выяснение роли каждого
мешка в механизме звукообразования — сложная за-
дача, требующая специальных экспериментов и осо-
бой методики.
Верхние мешки при издавании звуков могут играть
роль вместилищ, из которых воздух под давлением
мышц или вырывается наружу через узкую щель едва
приоткрывающегося клапана дыхала, или поступает в
нижележащие мешки.
В первом случае в воде видно, как из дыхала выле-
тает струйка мелких воздушных пузырьков и края
сомкнутой щели дыхала слабо шевелятся. Когда стя-
гивают кошельковый невод с окруженными дельфина-
ми, вода словно вскипает от таких пузырьков.
Во втором случае пузырьки в воде не видны, так
как воздух перегоняется в более глубокие мешки (во
вторую и третью пары), что регулируется мускулатурой
дыхала, мускульной пробкой и ее выступом в виде гу-
бы. Этот выступ американские ученые В. Эванс и
109
Д. Прескотт назвали «дребезжащим органом», так
как он, открывая и закрывая вход в парные трубча-
тые мешки, обусловливает щелканья и взрывы щел-
каний. По гипотезе Кеннета Норриса, мешки работают
так (рис. 11). В момент дыхательного акта мускульная
пробка носового канала втягивается, носовой проход
и дыхало открываются, а верхние мешки спадают.
В конце дыхательного акта дыхало плотно закрывает-
ся, верхние мешки наполняются воздухом, и носовой
канал блокируется мускульной пробкой так, что выше
ее оказываются верхние и средние, а ниже ее — ниж-
ние мешки. Ряд щелканий производится, как только
воздух из верхних мешков наполняет средние и отсю-
да начинает поступать через соединительные проходы
в нижние мешки. К концу ряда щелканий запас воз-
духа в верхних мешках истощается, и мускульная
Рис. 8. Схема расположения воздушных мешков у обыкновен-
ного дельфина: а, б — продольный разрез головы; в —
проекция сверху: 1—щель дыхала; 2 — верхний ме-
шок; 3 — его отверстие в носовой канал; 4 — мускуль-
ная пробка, запирающая носовой канал; 5 — трубчатый
мешок; 6 — соединительный проход с расширением;
7 — нижний (надмежчелюстной) мешок; 8 — его отвер-
стие в носовой канал; 9 — кости черепа; 10 — носовой
канал. По Е. В. Романенко, А. Г. Томилину и Б. А. Ар-
теменко
110
пробка готова открыться, чтобы эти мешки могли
вновь заполниться для следующей серии щелканий.
Дыхательный акт при этом не обязателен, так как
верхние мешки могут перезаполняться воздухом из
легких под давлением диафрагмы и мышц грудной
клетки.
Очевидно, функции мешков в звукообразовании
дифференцированы. В 1962 году Д. Лилли и Э. Миллер
наблюдали, как одна афалина щелкала левыми воз-
душными мешками, а свистела правыми, но могла де-
лать и то и другое одновременно. Левая и правая ча-
сти звукосигнального аппарата могут действовать не-
зависимо : правая — издавать свист в воздухе или под
водой, а левая — щелкать. Механизм звучания
эти ученые связывают не только с воздухоносными
мешками, но и с пневматическими синусами слуховой
области: произведенный щелчок возбуждает в системе
воздушных синусов и мешков резонирующие колеба-
Рис. 9. Схема продольного разреза головы дельфина: А — че-
реп; Б — жировая подушка; В — верхняя челюсть; ? —
носовой канал: 1 — носовое отверстие; 2 — верхний воз-
душный мешок; 3 — трубчатый мешок; 4 — соедини-
тельный проход с расширением; 5 — нижний (надмеж-
челюстной) мешок
111
Рис. 10. Поверхностные слои мышц дыхала дельфина-бе-
лобочки: 1— передненаружный мускул; 2 —
комиссура дыхала; 3— промежуточный мускул;
4 — ткань жировой подушки; 5 — задненаруж-
ный мускул; 6 — верхняя пара воздушных меш-
ков. По Р. Харрисону
112
Познакомимся с прием н ы м механизмом
гидролокатора китообразных.
Их отличный слух не снижается ни отсутствием
наружного уха, ни суженным, заросшим слуховым
проходом. Дельфины воспринимают не только звуки,
но и инфра- и ультразвуки, лежащие далеко за пре-
делами человеческого слуха.
Они прекрасно разбираются в многоголосом хаосе
моря и очень точно определяют, с какого направления
поступает звук, (в воде это сделать не может ни одно
наземное млекопитающее). Вибрация черепа под воз-
действием звуковых волн мешала бы этому, если бы
уши китообразных не были изолированы от костей
черепа.
Среднее и внутреннее ухо помещаются в двух очень
плотных костных образованиях: первое в барабанной,
а второе — в околоушной кости (рис. 12).
ОбЪ кости жестко связаны и подвешейы на корот-
кой сухожильной связке одна над другой в обширной
барабанной полости черепа и со всех сторон окружены
воздушными камерами, заполненными пеной из жиро-
вой эмульсии. Пена, содержащая миллионы воздуш-
ных пузырьков, поглощает все звуковые колебания,
которые проводят череп, мускулатура и сало. Остается
единственный путь для звука попасть во внутреннее
ухо — через наружный слуховой проход и цепь косто-
чек среднего уха. Но в 1964 году К. Норрис обнаружил
и другой путь передачи звука — через нижнюю че-
люсть. В 1968 году значение этого волновода экспери-
ментально подтвердили электрофизиологи США и Япо-
нии — Т. Буллок, А. Гринелл, Е. Икезоно, К. Камеда,
Ю. Катцуки, М. Номото, О. Сато, Н. Суга и К. Янаги-
сава. В это же время Дж. Маккормик, Е. Уивер, Дж.
Пейлин и С. Риджуэй в Пойнт-Магу изучили у анестези-
рованных афалин изменения электрических потенци-
алов улитки внутреннего уха в ответ на внешние зву-
ковые раздражения. В этих экспериментах звук
наилучшим образом проводила именно нижняя че-
люсть, которая задним концом, имеющим очень тонкие
наружные костные стенки, вплотную подходит к уш-
ной области дельфинов. Этот участок К. Норрис и рас-
сматривает как «акустическое окно» для прохождения
звука. При этом ход акустических лучей во время
114
эхолокации таков (рис. 13): локационный пучок, про-
изведенный воздушными мешками, а затем отражен-
ный от костной стенки черепа и преломленный жиро-
вой подушкой (акустической линзой), встречает на
своем пути, например, рыбу. Отраженные от рыбы
акустические лучи возвращаются в ухо дельфина:
сначала через кожу к самой тонкой части нижней
челюсти — костной мембране (0,3 мм толщиной), от-
сюда к внутричелюстному жировому телу и, наконец,
в ухо. При всем том имеет значение угол, под которым
отраженные звуковые волны падают на нижнюю че-
люсть, и в зависимости от этого они проводятся с раз-
Рис. 12. Ухо китообразных: а — усатых китов; б — зубатых ки-
тов. 1 — наружное ушное отверстие; 2 — слой кожного
сала; 3 — слуховой проход и его зарастающая часть;
4 — соединительная ткань; 5 — барабанная перепонка;
6 — ушные косточки (молоточек, наковальня, стремеч-
ко}; 7 —^барабанная полость; 8 — барабанная кость (бул-
ла); 9— каменистая кость; 10 — пространство, запол-
ненное пеной; 11—кости черепа; 12 — слуховой нерв;
13—ушная пробка; 14 — улитка. По Рейсенбаху де Ха-
ан и Пурвесу
115
личным эффектом. Коэффициент звуковой передачи
будет высоким, если угол падения звуковой волны к
нижней челюсти будет не менее 30°, точнее, от 30 до
90°.
В связи с этим , и рассматривается привычка дель-
фина покачивать («сканировать») головой при при-
ближении его к добыче.
Сильное развитие нервно-рецепторного аппарата в
нижней челюсти дельфинов может служить дополни-
тельным аргументом к концепции Норриса о поступ-
лении акустической информации через нижнюю че-
люсть.
Дельфины, слуховые отверстия которых закры-
вались присосками из латекса, продолжали спокойно
пользоваться гидролокацией. В опытах американских
и японских исследователей восприятие звуков через
нижнюю челюсть оказалось в 6 раз выше, чем через
слуховой проход.
Ушные проходы дельфинов — тонкие, нитевидные,
чуть изогнутые каналы — открываются на обеих сто-
ронах головы позади и чуть ниже уровня глаза. На
середине пути канал зарастает, но вновь появляется,
Рис. 13. Схема эхолокации дельфина (ультразвуковой прожектор
и акустическая линза в голове дельфина): 1 — воздуш-
ные мешки, излучающие звуки; 2 — жир9вая подушка;
3 — череп; 4 — нижняя челюсть, проводящая звуки в
ухо; 5 — барабанная кость и внутреннее ухо; 6 — пред-
мет лоцирования (рыба); 7 — излученные звуковые вол-
ны; 8 — отраженные звуковые волны. По К. Норрису,
с изменениями
116
входя в полость среднего уха. Барабанная перепонка
напоминает сложенный зонтик, вершиной упирающий-
ся в укороченную рукоятку молоточка. Молоточек тон-
ким концом связан с барабанной костью, толстым — с
наковальней. Все три слуховые косточки (молоточек,
наковальня и стремечко) плотные, миниатюрные, спе-
циализированные для передачи звуков и ультразву-
ков. Удельный вес их почти в полтора раза больше,
чем у человека. Амплитуда движения стремечка, при-
мыкающего к овальному окошечку, увеличивается по
сравнению с амплитудой движения барабанной пере-
понки почти в 30 раз, но во столько же раз теряет в
силе. Это важное приспособление к передаче звука в
воде, где амплитуда той же самой звуковой волны по
сравнению с воздушной средой уменьшена почти в
60 раз, но во столько же'раз усилено ее давление. Ба-
рабанная кость (булла) — плотная и полая внутри ка-
мера, у усатых китов — яйцевидная, а у дельфинов —
полуцилиндрическая. С помощью мышц эта кость мо-
жет совершать небольшие перемещения относительно
черепа, обеспечивая стереофонический или объемный
прием отраженных локационных щелчков.
Строение внутреннего уха сложнее. Жесткоподве-
шенная околоушная кость массивная и плотная, с ми-
нимумом пустот. Улитка и сильно уменьшенный ве-
стибулярный аппарат занимают в этой кости очень
скромное место. В улитке развиты вторичная спираль-
ная пластина и относительно маленький, жесткопбд-
вешенный кортиев орган. Слуховой нерв достигает
исключительно мощного развития. У афалины, напри-
мер, улитка почти такой же величины, как у челове-
ка, но слуховой нерв много толще: он достигает в се-
чении 5 мм.
У китообразных, по мнению голландского ученого
Рейсенбаха де Хаана, выполнены все условия для раз-
вития оптимального подводного слуха и восприятия
ультразвуковых частот. Советский исследователь
В. П. Зворыкин показал, что у дельфинов гораздо
сильнее, чем у человека, развиты подкорковые слухо-
вые центры, что, вероятно, связано со способностью
зубатых китов воспринимать ультразвуки. Височ-
ная же доля коры больших полушарий лишь по глу-
бине и количеству извилин превосходит височную до-
лю мозга человека, но по микроструктуре такого пре-
117
восходства нет. Видимо, справедливо мнение, что эхо-
локационная деятельность у дельфинов протекает
брлыпе на уровне подкорки, чем на уровне слуховой
коры мозга.
В океанариумах методом условных рефлексов уста-
новили частоты, на какие отвечают дельфины. Диапа-
зон воспринимаемых частот оказался от нескольких
десятков герц до 196 кГц. Это в 10 раз выше верхнего
предела слышимости человеческого уха. Американ-
ские биологи Барбара Лоренс и Вильям Шевилл обу-
чили одну афалину подплывать к кормовой площадке
в ответ на звуковой сигнал и каждый раз награждали
ее рыбой. Скормив ей 1200 рыб, они узнали, что афа-
лина отвечает на звуки от 150 Гц до 153 кГц. Однако
подопытное животное отвечало на 100 % сигналов
только в пределе частот от 150 Гц до 120 кГц: с повы-
шением частоты процент ответов резко снижался.
Смещение чувствительности слуха в сторону высо-
ких частот связано с тем, что в спектре звуков, издава-
емых дельфинами, имеются высокочастотные сигна-
лы. А для наземных млекопитающих известно: какую
частоту они излучают, такую и воспринимают.
Дельфины способны воспринимать также инфразву-
ки, например «голос шторма»: они обычно уходят из
прибрежной зоны перед надвигающейся бурей, угро-
жающей выбросить их на отмель. Очевидно, ими улав-
ливаются шум волн, омывающих их тело, всплески на
поверхности моря, гул, издаваемый косяками рыб, ра-
кообразными, водными млекопитающими, а также от-
раженные волны разных частот (эхо).
Способность воспринимать звуки в очень широких
пределах частот позволяет китообразным получать
исключительно полную информацию из окружающей
среды, что вполне компенсирует недостаточное разви-
тие их зрительного и вкусового анализаторов.
В соответствии с высоким развитием слуха у кито-
образных сформировались превосходный голосовой
аппарат и весьма богатая звуковая сигнализация. Се-
верный дельфин — белуха даже получил название
«морской канарейки» за свою способность хрюкать,
стонать, свистеть, издавать звуки, напоминающие плач
ребенка, женский пронзительный крик, шум толпы,
игру на флейте, щебетание птиц.
Точное определение источников звуков, в том числе
118
и биогенных, приобрело особое значение. Слух и голос
китообразных стали предметом тщательных экспери-
ментальных исследований, и в этой области были сде-
ланы важные открытия.
Каждый вид китообразных пользуется нескольки-
ми типами звуков. Для чего и при каких обстоятель-
ствах используют свой голос дельфины и киты? Како-
во при этом их поведение и ответные реакции сороди-
чей на эти сигналы?
Чтобы разобраться во множестве сигналов китооб-
разных, необходимо классифицировать издаваемые
ими звуки в зависимости от физической харак-
теристики и от биологического значения сиг-
налов для самих китообразных.
По физическим свойствам сигналы различают
непрерывные и прерывные, состоящие из серии им-
пульсов или щелчков. Джон Лилли и Элис Миллер
все звучания дельфинов по физическим свойствам
объединили в три класса: I — эхолокационные ульт-
развуковые щелкания (до 170 кГц), II — непрерывные
звуки — свисты частотой от 4 до 18—20 кГц и III —
звуки сложного спектра — комплексные волны высо-
кой амплитуды, называемые по внешней аналогии с
сигналами других животных кряканьем, мяуканьем,
лаем, воем, жужжаньем, ревом и т. д. Эти ученые от-
крыли, что один и тот же дельфин способен издавать
акустические колебания двух или даже всех трех клас-
сов одновременно. Это значительно увеличивает воз-
можные комбинации сигналов и сильно усложняет их
звучание.
По биологическому значению акустические
сигналы зубатых китов можно ус л о в н о подразде-
лить на две группы: одни издаются преимущественно
для ориентации, навигации, рекогносцировки, для ра-
зыскивания пищи (эхолокационные сигналы
первого класса), а другие — для связи или общения со
своими сородичами (коммуникационные сиг-
налы второго класса). Акустические же сигналы
третьего класса, по-видимому, используются и как
коммуникационные, и как эхолокационные. Рассмот-
рим вначале эхолокацию как способ ориентации
зубатых китов.
Глава XII
«АКУСТИЧЕСКИЕ
ГЛАЗА»
ДЕЛЬФИНОВ
Впервые явление ориентации по звуку было обна-
ружено у летучих мышей итальянцем Лаццаро Спал-
ланцани в 1793 году, а эхолокация у китообразных
открыта лишь в пятидесятых годах XX века.
Насколько точен эхолокационный аппарат дельфи-
нов и какова роль эхолокации в жизни китообразных?
По эхолокационным сигналам дельфины могут с боль-
шой точностью ориентироваться в пространстве, опреде-
лять глубину дна и близость берега, разный характер
грунта, величину и качество погруженных предметов,
находить вещи в мутной воде.
Действие гидролокатора у дельфинов хорошо изу-
чено в океанариумах.
На каком расстоянии дельфины могут обнаружи-
вать пищу с помощью своего гидролокатора? Ответ
получили В. Шевилл и Б. Лоренс в следующем опыте:
от лодки перпендикулярно ее борту протягивали раз-
делительную сеть длиной в несколько метров, и в тем-
ноте рыбу бесшумно опускали в воду то с носа, то с
кормы. Подопытный самец должен был заблаговре-
менно решать, с какой стороны сети ему плыть, чтобы
найти ожидаемую рыбу. Эту задачу он правильно ре-
шал с расстояния до 4,5 м. Дельфин оказался способ-
ным не только обнаруживать отдельные эха своих
скрипов от рыбки длиной 15 см, но мог отличать имен-
но это эхо от всех прочих звуков, отраженных от бере-
га, дна, камней, лодки и других предметов в пруду.
Точность исследования В. Шевилла и Б. Лоренса была
подтверждена физиком Джоном Скоттом специальной
проверкой на афалине Сальти. Этот дельфин слышал
120
звуки в диапазоне от 75 Гц до 150 кГц и наиболее чет-
ко воспринимал частоты от 20 до 100 кГц.
Профессор Флоридского университета психолог
Уинтроп Келлог с сотрудниками в морской лаборато-
рии близ гавани Аллигатор несколько лет изучал эхо-
локацию афалин. Опыты он проводил над двумя обу-
ченными дельфинами Альбертом и Бетти в бассейне
площадью 350 м2 и глубиной до 2 м. Илистое дно и
стенки бассейна хорошо поглощали звуки, не давая
эха. Дельфины, плавая, взмучивали воду так, что ви-
димость во время опыта не превышала 35—85 см.
Чтобы дельфины не могли видеть действий человека
ни в воде, ни в воздухе, эксперименты проводились
ночью. Звуки записывали специальной аппаратурой.
Предметы погружали в воду за фанерным экраном. Ес-
ли в водоеме было спокойно, дельфины лишь изредка
издавали скрипы. При холостом шуме (например, ког-
да капали жидкость на поверхность воды) животные
издавали короткий скрип и замолкали. Если в воду
бросали погружающийся несъедобный предмет, то
вслед за первым издавалось еще несколько скрипов
с промежутками в 1—2 с. Но когда кидали рыбу, сле-
довал целый залп скрипов, частота импульсов возра-
стала до нескольких сотен в секунду, и дельфин без-
ошибочно направлялся к цели. При этом он покачивал
головой из стороны в сторону на 10°, как бы прицели-
ваясь на объект посредством отраженного звука, и уве-
личивал частоту излучаемых щелчков по мере прибли-
жения к рыбе.
Темнота не мешала ему различать рыбу по разме-
рам. Крупной кефали (30 см) Альберт и Бетти явно
предпочитали вдвое меньшего пятнистого горбыля.
Если погружали обеих рыб вместе, афалины устрем-
лялись к предпочитаемой рыбе; в первых 16 испыта-
ниях самец ошибся только 4 раза, а в 140 последую-
щих — ни разу! Когда горбыля подвешивали позади
стеклянного экрана (он был виден, но недоступен для
эхолокации), а кефаль — впереди экрана (она была до-
ступна для ультразвукового распознавания), дельфин
не пытался ловить горбыля и довольствовался лишь
кефалью.
В последующих опытах в Калифорнийском океана-
риуме экспериментаторы стали «ослеплять» подопыт-
ных дельфинов, надевая им наглазники (рис. 42 вкл.),
121
и обучили их принимать пищу по сигналу. Оказалось,
что дельфины с выключенным зрением могут свобод-
но плавать в лабиринте из подвешенных металличе-
ских стержней, не касаясь их, и точно различать за-
граждения из тонкой проволоки и нейлоновых нитей.
Такие способности помогают дельфинам отличать раз-
ные виды корма (рыб, моллюсков, рачков) и выбирать
предпочитаемую пищу в условиях любой освещенно-
сти среды, безопасно плавать в щелях между выступа-
ми дна.
В бассейне лаборатории Пойнт-Магу Кеннет Норрис
и физиолог Рональд Тёрнер обучили взрослую самку
афалины Элис с надетыми наглазниками распознавать
два металлических шарика размером 3,7 и 6,2 см. Ша-
рики располагались в 60—120 см от животного и мог-
ли взаимно переставляться. Если самка выбирала
большой шарик (6,2 см), она получала в награду ры-
бу. Затем меньший шарик стали заменять все более и
более крупным, уменьшая разницу с большим шари-
ком. Когда меньший шарик взяли диаметром 5 см,
дельфин продолжал безошибочно выбирать большой
шарик (6,2 см). Когда разница в диаметрах шариков
уменьшилась до 0,8 см, афалина все же в большинстве
случаев делала правильный выбор. Этот опыт про-
демонстрировал поразительную точность распознава-
ния величины лоцируемых объектов. В других опы-
тах К. Норриса и его коллег «ослепленная» афалина
с расстояния 6—7 м отличала капсулу из желатина от
куска рыбы такого же размера.
Акустические лучи, проникая внутрь предметов, в
виде эха несут информацию о размере, форме и мате-
риале этих предметов.
Эхолокационные импульсы, похожие на разного
рода скрипы, животные посылают направленно с по-
мощью координированной работы воздушных мешков,
носового канала, лобно-носовой (жировой) подушки и
сложной системы мышц, расположенных в выпуклой
части головы. Лобно-носовая подушка, лежащая на
челюстных и межчелюстных костях, и вогнутая пе-
редняя поверхность черепа действуют как звуковая
линза и рефлектор: они концентрируют (фокусируют)
сигналы, излученные гидролокатором, и в виде звуко-
вого пучка направляют на лоцируемый объект. Дока-
зательства действия такого ультразвукового прожек-
122
тора получены с помощью экспериментов как в нашей
стране, так и за рубежом»
Особенно высока разрешающая способность к лока-
ции у дельфинов в горизонтальном плане и хуже —
в вертикальном. Они могут, находясь на поверхности
воды, лоцировать объекты и на глубине. Это для них
очень важно: ведь им приходится постоянно то выны-
ривать (чтобы дышать), то погружаться вглубь. Но
куски рыбы, находящиеся под нижней челюстью, они
лоцировать не могут. Вероятно, потому, что нижняя
челюсть служит экраном, отражающим звуковые вол-
ны, издаваемые системой воздушных мешков.
Возможно имеет значение и то, что лобно-носовая
подушка расположена наклонно к продольной оси
черепа.
В 1967 году американские исследователи Эванс и
Пауэлл доказали способность ослепленного дельфина
различать как одинаковые по площади пластины —
медную, алюминиевую и латунную, так и разные по
толщине (2,2 и 3,2 мм) медные пластины. Советские
исследователи также экспериментально показали, что
верхний порог слуха афалины достигает 180 кГц, и
это животное в условиях опыта различает одинаковые
по внешнему виду, но сделанные из разного материа-
ла предметы.
Оказалось, что дельфины могут различать геомет-
рические фигуры, помещенные внутри холщовых ме-
шочков, или даже положения поршня внутри метал-
лического цилиндра. Этого нельзя достичь с помощью
зрения, так как свет отражается от поверхности пред-
метов; ультразвуковые же лучи с их большей длиной
волны по сравнению с оптическими проникают внутрь
предметов. Это позволяет дельфинам с помощью аку-
стических волн исследовать не только размер и объем,
но и качество (материал) предметов и отлично «ви-
деть» ушами при выключенном зрении.
За последние грды появились интересные гипотезы
о действии приемной системы дельфина по голографи-
ческому принципу (Джон Дреер) либо по принципу
акустического видения (рис. 43 вкл.) При разработке
этих гипотез обращалось внимание на структуры над-
черепной части головы дельфина — на лобно-жировую
подушку, способную фокусировать звуки, на характер
взаимного расположения воздушных мешков, жировой
123
подушки и черепа, на гистологию эпителия воздушных
мешков. Эти гипотезы исходят из признания, что в
звуковидении дельфинов имеют значение кожные ре-
цепторы на голове и в надчерепных тканях, якобы
способные воспринимать акустическую информацию.
В 1969 году американский исследователь Джон
Дреер предположил, что дельфины воспринимают звук
системой рецепторов, распределенных на лобном
выступе головы.
В естественных условиях локационный аппарат ис-
пользуется китообразными на каждом шагу. Дельфи-
ны применяют его, например, для разведки препятст-
вий на их пути.
Калифорнийские исследователи Вильям Эванс и
Джон Дреер с судна «Си-Квест» наблюдали поведение
стайки афалин в лагуне Скаммона. На пути плывущей
группы из пяти афалин оказался звуковой барьер из
вертикальных алюминиевых труб. Животные, эхоло-
цируя, заметили барьер в 360 м от себя и замедлили
ход. Затем стайка повернула к берегу, а от нее отде-
лился разведчик. Он, лоцируя, подплыл к препятствию
и вернулся к группе; тогда вся стайка, издавая эхо-
сигналы, направилась к преграде, проплыла между
алюминиевыми трубами и продолжила путь в перво-
начальном направлении (рис. 14).
Рис. 14. Стайка афалин в лагуне Скаммона преодолева-
ет препятствие: движение с эхолокационными
щелканиями Э и свистами С: 1—прибрежная
отмель; 2 пролив; 3 — барьер; 4 — путь раз-
ведчика. По Д. Дрееру и В. Эвансу
124
Подобное наблюдение повторили Давид и Мельба
Колдуэллы и Сибеналер во время лова атлантической
афалины в узком заливчике во Флориде. Как только
одна стайка дельфинов вошла в залив, выход из него
перекрыли сетями. Тогда от стайки отделился вожак,
разведал преграду, а затем в сопровождении крупной
самки атаковал сеть. Через разрыв сети и через погру-
женные в воду края вся стайка ушла в море.
На основании опытов в океанариумах и наблюде-
ний в природе был сделан общий вывод, что эхолока-
ция у китообразных — главный способ распознавания
объектов, погруженных в воду. Скрипучими сериями
звуков высокой частоты они «ощупывают» окружаю-
щую обстановку и получают необходимую информа-
цию.
В условиях хорошей освещенности дельфины ори-
ентируются и с помощью зрения. Исследователи
Р. Бюснель и А. Дзидзик наблюдали в Средиземном
море, что белобочки в ясную и тихую погоду осуще-
ствляют «молчаливую навигацию» и пользуются сво-
им гидролокатором гораздо реже, чем при недостаю-
щем свете.
Эхолокация разъясняет многие таинственные явле-
ния в жизни мелких китообразных. Например, почему
они в мутных водах и ночью не ловятся в мелкоячеи-
стые сети, но попадают в крупноячеистые, «прозрач-
ные» для отраженных звуков, не касаются мин, когда
резвятся на минных полях; как речные дельфины —
инии и соталии, не повреждая себя, свободно плавают
в сумеречных лесах, заливаемых во время половодья
реки Амазонки, а обитатель мутного Ганга — слепой
дельфин платаниста, возможно, обходится вообще без
помощи зрения?
Принцип действия эхолокационного аппарата дель-
финов ныне широко применяется в технике в виде со-
наров. Термин «сонар» происходит от начальных букв
трех английских слов: sound — звук, navigation —
навигация и range — дальность. Звукоизлучающие
приборы посылают импульсы в воду, а их эхо достав-
ляет информацию о направлении и расстоянии до не-
видимых препятствий, опасных для навигации (айс-
берги, мели, рифы и т. п.). На этом же принципе по-
строена работа эхолотов при измерении глубин и по-
исках косяков рыб.
125
Такие приборы — верные помощники человека в
службе безопасности кораблевождения и в рыболовст-
ве. Однако эффективность природных локаторов ока-
залась гораздо выше технических характеристик суще-
ствующих сонаров и эхолотов. Гидролокатор дельфи-
на выгодно отличается от применяемых в морской
практике гидролокационных аппаратов более высокой
устойчивостью к помехам, большей шириной спектра
излучаемых колебаний, богатством модуляций по ин-
тенсивности и частоте повторения. Гидролокатор дель-
фина является ныне важнейшим прототипом для мо-
делирования в технике.
Естественные свойства дельфинового сонара объеди-
няют оба основных принципа современной техниче-
ской гидроакустики: активный поиск, когда по-
сылается пакет волн и улавливается эхо от него, и
пассивное подслушивание шумов и звуков,
присущих самим лоцируемым объектам.
Гидробиоников привлекает феноменальная точ-
ность локации дельфинов и замечательная портатив-
ность их локатора, превосходящая все образцы аппа-
ратов того же назначения, созданных человеком.
Рыбопоисковый эхолот «Дельфин» весит столько, сколь-
ко и афалина, тогда как гидролокатор афалины со-
ставляет ничтожную часть ее веса.
Удивление вызывает способность дельфинов выде-
лять из хаоса звуков только необходимые. В бассейн,
где жили афалины, с целью их дезориентации излу-
чались их же гидролокационные сигналы, записанные
ранее. Однако эти сигналы не могли сбить животных
и помешать точности их гидролокации.
В голове дельфинов имеется сложный вычислитель-
ный механизм, с помощью которого нужные эхо-сигна-
лы, отраженные от цели, отделяются от ненужных, от-
раженных от посторонних предметов, дна и поверхно-
сти. Столь замечательный сонар позволяет дельфи-
нам «видеть» ушами, «слушать» челюстями, а «раз-
говаривать» ноздрей и макушкой.
О чем же могут «разговаривать» дельфины?
Глава ХШ
ПОЮЩИЕ КИТЫ
и
«МОРСКИЕ ПОПУГАИ»
В 1951 году появились первые записи голоса афа-
лины, сделанные во Флоридском океанариуме Ф. Ву-
дом. К настоящему времени записан голос почти од-
ной трети всех видов отряда китообразных.
Интенсивные исследования открывают у китообраз-
ных все новые и новые сигналы нелокационного харак-
ра, т. е. коммуникационные сигналы, с помощью ко-
торых особи общаются друг с другом. Даже сравни-
тельно молчаливые усатые киты, как оказалось, вла-
деют различными сигналами, возможно, для связи
между собой. Еще недавно считали, что они издают
лишь звучные стоны низкой частоты от 20 до 1000 Гц;
такие стоны слышали у горбачей, сейвалов, финвалов
(рис. 44 вкл.), серых, бискайских, австралийских и
гренландских китов.
У австралийского гладкого кита биоакустики центра
подводных исследований США в водах Аргентины с
судна «Геро» в 1971 году записали более 200 низко-
частотных подводных звуков. Наиболее обычными сре-
ди них были сигналы, звучащие как отрыжка, часто-
той от 30 до 2200 Гц и средней продолжительностью
1,4 с. Были и специфические ревущие сигналы часто-
той до 1250 Гц и продолжительностью от 0,6 до 4,1 с.
Сенсацию произвела запись сотрудника Родайленд-
ского университета Пауля Перкинса: ему удалось
записать сигналы пары финвалов (крупных полосати-
ков), которые подошли к судну на 25—200 м. За 4 мин
микрофон зарегистрировал щебетанье и свисты часто-
той от 1,5 до 5 кГц, продолжительностью от 50 до
600 мс.
. 127
Стало ясно, что голосовые возможности китов явно
недооценивались.
Усатые киты издают звуки, используя для этого
мощный горловой мешок, соединенный с гортанью.
Однако механизм подачи звуков остается совершенно
не изученным. Калифорнийский исследователь Поул-
тер Томас в 1968 году записал голос серых китов в ла-
гуне Скаммона, где они выкармливают своих детены-
шей. Гидрофоны устанавливали в 1,5 м выше дна, на
середине входа в лагуну. Самки с детенышами, прохо-
дя мимо аппаратуры, проявляли высокую звуковую
активность. Они издавали скрежет продолжитель-
ностью в несколько секунд, серии импульсных звуков,
громкие сигналы, а во время тревоги, когда к китам
приближался вертолет,— звуки китайского гонга. Ча-
стота скрежета и звуков «гонга» достигала 12 кГц.
У горбатого кита Ховард Винн и Пауль Перкинс
записали сигналы частотой до 3 кГц, среди них рев,
продолжительный рев, крик, щебет, стон и храп. Та-
кие усложненные призывы имеют значение для инди-
видуальной и групповой рекогносцировки, организа-
ции индивидуального участка, для нахождения полов
в брачный период, поддержки контакта с мигрирую-
щими группами. Поддержка связи между сородича-
ми, как показывают наблюдения, возможна на рас-
стоянии до 10 миль.
Совершенно особые звуковые сигналы были откры-
ты Роджером Пайном и Скоттом Макуэем в 1971 году
на основе анализа записи голоса горбачей в море. Ока-
залось, что эти киты (рис. 45 вкл.) производят серию
прекрасных варьирующих звуков продолжительно-
стью от 7 до 30 мин, которые затем повторяют. По
признаку мелодичности такие звуки названы, как у
птиц, «пением». В звучании поющих горбачей замече-
ны четко выраженные индивидуальные черты. Даже
динамитные взрывы в океане, совпавшие с работой
записывающих приборов, не могли помешать вокаль-
ным занятиям этих животных. В отличие от песен
птиц, длящихся лишь несколько секунд, пение горба-
чей продолжается непрерывно до получаса и притом
без пауз между исполняемыми партиями (рис. 15).
Французские гидробиологи Филипп Кусто и Эжен
Лагорио в районе Бермудских островов ночью запи-
сали настоящий концерт, в котором участвовало около
128
сотни горбачей. Стадо издавало до тысячи разных
звучаний! Тут были и трели, и скрипы, и вой, и жуж-
жание, и рев оленя, и что-то вроде писка мышей. «Хо-
ровой ансамбль» горбатых китов напоминал звуки
группы школьников, вслух твердящих урок. «Кон-
церт» начался с того, что запел один кит, к нему при-
соединился второй, потом третий и, наконец, все про-
странство заполнилось звуками.
Какое значение имеет пение для горбачей — остает-
ся тайной. Роджер Пайн, X. Е. Винн и Л. К. Винн
считают, что эти песни специфичны для вида и испол-
няются лишь молодыми самцами в брачный период и
только в районах размножения — в водах Вест-Индии,
Гавайских и Бермудских островов. Песни, видимо, по-
могают поддерживать контакт между разбросанными
особями и передавать акустическую информацию, от-
ражают половую готовность самцов и служат средст-
вом распознавания полов.
Механизм подачи звука песен, кажется, связан с
вытеснением воздуха из одной полости головы кита в
другую. Возможно, что еще ранее записанные у фин-
валов и южных китов продолжительные флейтовые
звуки от низкого до высокого тона тоже относятся к
категории загадочного пения. Ясно одно: у усатых
китов комплекс звуков довольно разнообразен, и изуче-
ние их сулит еще много сюрпризов.
Все еще остается недоказанным, свойственна ли
усатым китам эхолокация. Сведения об этом противо-
речивы. С одной стороны, 22 мая 1969 года американ-
ские исследователи Петер Бемиш и Эдвард Митчелл с
борта норвежского зверобойного судна «Полярная
звезда» (в 100—800 м от животного) записали у си-
него кита ультразвуковые сигналы частотой 21 и
31 кГц; они могли бы быть использованы для эхоло-
кации. А с другой стороны, тот же Петер Бемиш про-
вел эксперимент с молодой самкой горбача, давший
отрицательный результат. Эта самка 2 июля 1976 го-
да попала в рыболовные сети в водах Ньюфаундленда
и была помещена на 29 дней в вольер из нейлоновых
сетей близ берега. Киту надели наглазники и предо-
ставили условия свободного плавания среди лабирин-
та из подвешенных алюминиевых стержней. Во время
опыта велась акустическая запись. Однако никаких
данных, указывающих, что кит пользовался сонаром,
7 9-1984
129
не было получено. Если усатые киты и имеют эхоло-
кацию, то гораздо менее развитую, чем дельфины.
Какио же коммуникационные и эмоциональные
сигналы производят самые «говорливые» из китообраз-
ных — дельфины? Среди их не локационных сигналов,
частотный диапазон которых уже, чем у локационных
секунды
О 5 10 15 20 25 30 35 40 45 50 55 60
Рис. 15. Звукозапись непрерывного «пения» двух горбатых ки-
тов продолжительностью по 10 минут. По Р. Пайну и
С. Макуэю
130
(у афалины не выше 50 кГц), можно различать свисты
и сигналы сложной формы, слышимые как лай, мяу-
канье, вой, визг, крики, кряки, птичьи трели и щебет,
взрывы, рев и т. д. Некоторые из этих звуков соответ-
ствуют сигналам питания, беспокойства, страха, спа-
ривания, бедствия и т. д. При этом были замечены ви-
довые и индивидуальные отличия в сигналах, по кото-
рым особей удавалось узнавать персонально, но для
этого ленту с записью голосов приходилось пропускать
с замедленной скоростью.
В 1964 году Д. Дреер и В. Эванс установили у
взрослой атлантической афалины 17 разнотипных сви-
стовых сигналов, у тихоокеанской афалины — 16, а у
тихоокеанского дельфина-белобочки — 19. В совокуп-
ности у них отметили 32 разнотипных сигнала, и из
них лишь восемь были общими для всех трех видов.
Детеныш атлантической афалины оказался «болт-
ливее» взрослых: он пользовался своим голосом гораз-
до чаще, но подавал лишь шесть сигналов. Видимо,
многообразие сигналов у дельфинов с возрастом уве-
личивается.
В Нью-Йоркском аквариуме с помощью акустиче-
ской аппаратуры М. Фиш и В. Маубрэй записали у мо-
лодого самца и двух взрослых самок-белух из Бри-
стольского залива девять разных типов звуков. Кри-
сталлический гидрофон, помещенный на глубине 1,5 м,
зарегистрировал щелканье и скрипы, обычный и моду-
лирующий свист, визг, рев (у самца, преследуемого
самками), пронзительный птичий крик, трели и гром-
кие удары, производимые после визга.
В 1970 году биоакустики США В. Шевилл, В. Уот-
кинс и К. Рэй записали и проанализировали голос ро-
дича белухи — ныне редчайшего полярного дельфи-
на — нарвала Ч Нарвал издавал звуки очень регуляр-
но и в отличие от других дельфинов не увеличивал
частоту щелканий по мере приближения к мишени.
Позже в сигналах этих животных были отмечены свои
«свистовые автографы».
Год спустя японские ученые К. Мизюэ, М. Ниси-
ваки и А. Такемура воспроизвели на пленке голос сле-
пого гангского дельфина платанисты: на 87% он со-
1 Впервые группу из четырех нарвалов удалось содержать в
Ванкуверском аквариуме в Канаде. Животные прожили в не-
воле четыре месяца и все погибли от гельминтоза легких.
7*
131
стоял из эхолокационных сигналов («скрипы весла») и
на 5% — из коммуникационных, напоминавших мяу-
канье кошки.
Среди зубатых китов менее всего одаренным счи-
тался кашалот. Ученые В. Шевилл и Л. Ворсингтон
впервые записали звуки небольшой группы кашалотов
в море. Эти звуки были очень однообразны, как стук
печатающей машинки. Однако сложное устройство
ноздрей кашалота указывало, что его «репертуар»
должен быть гораздо богаче. Это и подтвердили амери-
канские ученые П. Перкинс, М. Фиш и В. Маубрэй с
судна «Тридент» в теплых водах Атлантики: оказа-
лось, что в стаде вожак, самки и детеныши обща-
лись между собой восемью разными типами сигналов.
Среди коммуникационных сигналов были варьирую-
щие свисты частотой от 0,7 до 16,5 кГц, щебетанье
или чириканье, сложное и простое гудение, звуки, на-
поминающие пронзительные крики, отрывистый лай,
скрежет напильника и хрипы.
Стадный вид — гринда — в неволе, кроме щелка-
ний, скрипов и свистов, издает на поверхности воды
звук продолжительной отрыжки; в это время у нее
заметно шевелятся края дыхала. Отдыхая, она чмо-
кает плотно сжатыми губами дыхала, а когда пытает-
ся поднять морду из воды, щебечет и хнычет.
Французские ученые Р. Бюснель и А. Дзидзик
наблюдали стайку из 11 гринд в Средиземном море со
специального судна «Калипсо». Из стайки они загар-
пунили одну гринду и в течение 41 мин записали все
акустические сигналы, изданные ею и ее сородичами.
Среди сигналов, проанализированных с помощью
электроспектрографа и катодно-лучевого осциллогра-
фа, было установлено пять разных свистовых типов.
Среди них преобладали свисты продолжительностью
0,4—0,9 с и частотой 4—8 кГц. Были также продол-
жительные (1,4—2,7 с) модулирующие свисты, начи-
навшиеся при частоте 8,8 кГц, а затем понижавшиеся,
или свисты длительностью 0,7—0,9 с с частотой в на-
чале и конце 3 кГц, а в середине 4 кГц.
Крупнейшие из дельфинов — косатки тоже имеют
разнообразный «вокальный репертуар», в котором пре-
обладают низкочастотные крики (0,5—2 кГц) — ряды
быстрых импульсов с большим количеством гармоник,
132
взрывные щелканья (по 10—15 импульсов в каждом
взрыве) и щелканья с замедленным повторением. Это
установили В. Шевилл и В. Уоткинс (рис. 16) на ко-
сатке Моби-Долл в загоне Ванкуверской бухты. Аку-
стические излучения, видимо, помогают косаткам
обнаруживать трещины во льдах Антарктики и согла-
сованно двигаться на расстоянии в несколько километ-
ров друг от друга (наблюдения К. Норриса и Д. Пре-
скотта). Американский биоакустик Вильям Каммингс
наблюдал, как крик косаток, воспроизведенный под
водой, отгоняет гринд и морских львов от нерестилищ
головоногих моллюсков, а белух, зашедших в реки, за-
ставляет поворачивать назад.
Даже у весьма примитивных китообразных — реч-
ных амазонских иний исследователи Мельба и Давид
Колдуэллы и В. Эванс в аквариумах США записали
двенадцать разных типов коммуникационных звуков.
Рис. 16. Крик (а) и серия эхолокационных щелканий (б) косатки
Моби-Долл в загоне Ванкуверской бухты. По В. Шевиллу
и В. Уоткинсу
133
Запись суммарно длилась 11 ч у восьми животных,
живущих парами в четырех разных танках. Среди зву-
ков были крики, писк, лай, хныканье, треск, скрежет,
а также и эхолокационные скрипы, но менее выражен-
ные, чем у других видов дельфинов.
Итак, коммуникационные сигналы китообразных
звучат по-разному. Но что скрывается за каждым си-
гналом, какому поведению животного соответствуют
эти звучания? В какой ситуации они издаются и ка-
кие вызывают ответные действия у сородичей? Эти
вопросы еще только начинают проясняться. Ответы
животных на сигналы в неволе и в море не всегда сов-
падают. Тем не менее хотя бы предположительно рас-
смотрим значение некоторых сигналов, соответствую-
щих тому или другому поведению дельфинов.
В ситуации беспокойства, возбуждения и любопыт-
ства обычными из коммуникационных сигналов были
свисты. Такие сигналы издают вновь вводимые в
океанариум дельфины. Свист в океанариуме чаще все-
го возникал, когда там появлялись «странные» новые
предметы или животные (например, акулы), в отноше-
нии которых у дельфинов проявлялся ориентировоч-
ный (исследовательский) рефлекс. Возбужденные и
чем-нибудь встревоженные дельфины сбивались в стай-
ку и начинали быстро плавать вдоль стенок бассейна,
громко и часто пересвистываясь между собой. С тече-
нием времени ориентировочный рефлекс затормажи-
вался, стайка распадалась и свист прекращался.
Таким образом, свист (писк) с частотами от 7 до
18 кГц выдает возбужденное состояние дельфинов и
служит сигналом для образования стайки и
поддержания стайности. Детеныши начинают пользо-
ваться таким сигналом, как только рождаются и реа-
гируют на свистящий призыв самки.
Лай представляет собой короткие звуки длитель-
ностью 50—350 мс с частотой от сотен герц до 10 кГц.
В момент лая замечается движение краев клапана ды-
хала. Дельфины лают при эмоциональных ситуациях
во время сильного возбуждения, например, в период
кормления, при появлении в бассейне человека, а так-
же в брачный период при половых играх и во время
спаривания. В сезон гона визгливым или хриплым
лаем самец призывает самок.
134
Если лай афалин несколько удлинить во времени,
образуется что-то вроде мяуканья. Не известно, какие
звуки дельфины издают в море на местах концентра-
ции пищи, но в неволе во время кормежки они мяука-
ют и скрежещут.
Удлиненное мяуканье (от долей секунды до несколь-
ких секунд) дает так называемый вой частотой (у
афалин) от 1—2 до 80 кГц. Е. В. Шишкова записала
вой продолжительностью 0,5 с и частотой 60—5000 Гц
во время питания черноморских дельфинов. В опытах
А. А. Титова дельфины выли во время игры в мяч,
при локации рыбы с близкого расстояния и во время
поддразнивания. Он считает, что это сигналы просьбы
пищи и удовольствия.
Совсем другое значение имеет сигнал угрозы или
доминирования, слышимый как хлопок в ла-
доши и сопровождаемый быстрым открыванием и
закрыванием рта (жест укуса). Этот сигнал был описан
как «хлопанье челюстями». Крупный самец, возглав-
ляющий стадо, такими хлопками запугивает более
слабых сородичей. Самка подобными звуком и жестом
выражает недовольство, когда к ее детенышу или к
ней самой приближается агрессивно настроенный са-
мец. Молодые особи пользуются таким сигналом редко
и робко, а со временем — смелее и чаще. Поскольку
в ответ на «хлопанье челюстями» сородичи бросаются
в сторону, этот звук сочли как сигнал угрозы и
предупреждения. Когда животное в танке одно,
оно не пользуется подобным сигналом; не замечали
его и в группе у слабейших особей.
До 1962 года думали, что «хлопанье» дельфины
производят ударом нижней челюсти о верхнюю. Одна-
ко Джон Лилли заметил, что жест укуса сопровожда-
ется 3—4 громкими короткими щелканьями, создаю-
щими ложное впечатление челюстного хлопка, по-
скольку в воде хлопнуть челюстями невозможно. Хло-
пок производится голосовым аппаратом как сигнал
доминирования и чаще во время кормежки в стайках
в неволе. Доминирующая самка морской свиньи по-
добным сигналом вынуждала хилого самца бросать
добычу и подбирала ее сама. Она подчеркивала гос-
подство в группе и своими действиями: быстро при-
ближалась к более слабым сородичам и толкала их
135
рылом. Этот сигнал у нее отмечен как два-три после-
довательных крика частотой 2 кГц (рис. 17).
Интересен «пронзительный птичий крик», состоя-
щий из серии щелканий, повторяющихся от 220 до
400 раз в секунду. Он слышится в воздухе при от-
крытом, а в воде при закрытом дыхале.
У афалины на небольшой биостанции записаны
крики длительностью от долей секунды до 2 с со
спектром частот 1—49 кГц. Крики издавались при
отлове животных в танке, брачном поведении, играх
в мяч. Такие звуки могут выражать боль, голод, при-
зыв особи другого пола и т. д. На той же станции
у афалины зарегистрированы «взрывы»; они чаще все-
го отмечались в «переговорах» изолированных живот-
ных, соединенных каналом электроакустической свя-
зи. Длительность «взрывов» была 30—180 мс, а спектр
охватываемых частот 10—30 кГц. При замедлении
а ё
кГц кГц
2 Ц Время 0,1с
2 4 6 8 10 12 14- 16 Время 0,1с
Рис. 17. Сонограммы обыкновенной морской свиньи, записанные
Р. Бюснелем и А. Дзидзиком в танке, где жили три
самки и два самца: а — сигнал доминирования (изда-
вался самкой во время кормежки, как только самец
приближался к пище); б — сигнал боязни; в — сигнал
в момент начала спаривания
136
воспроизведения звукозаписи в 16 раз эти сигналы
воспринимаются на слух как серия стуков.
Американскими учеными у афалины открыт сиг-
нал тревоги или страха. Животные его изда-
ют при испуге, он слышится в воде через гидрофон
как резкий треск, легко перекрывающий по силе эхо-
локационные скрипы и «лай» дельфинов. На ухо че-
ловека, слушающего через гидрофон, этот звук дей-
ствует как удар. Механизм, с помощью которого
создается столь энергичный звук, не установлен. Реак-
цию испуга супруги Колдуэллы вызывали у афалин
вскоре после их поимки несколькими приемами:
ночью — ярким пучком света, который внезапно на-
правляли из фонарика в глаза выныривающего жи-
вотного, . а днем — моделью дельфина, которую очень
близко подводили к живым афалинам.
Во всех этих случаях наблюдалось вздрагивание
дельфинов.
Французские ученые Р. Бюснель и А. Дзидзик за-
писали слышимый в гидрофон резкий сигнал страха
у обыкновенного дельфина в Средиземном море во
время попыток поймать его за хвост хапуном или за-
гарпунить близ носа корабля (рис. 18). Сигнал вклю-
чал четыре-пять очень коротких свистов (0,12 с), сле-
дуемых друг за другом через 0,2 с. Частоты в начале
и в конце свиста были 14 кГц, а в середине — около
10 кГц. После подачи сигнала страха стайка обыкно-
венных дельфинов бросалась врассыпную. Ученые
предполагают, что сигналом страха можно отпугивать
стада обыкновенных дельфинов от рыболовных сетей.
Похоже, что этот же сигнал действует у обыкновен-
ных дельфинов в Черном море во время их загона в се-
ти с помощью телефона или во время промысла ма-
лайских продельфинов на Соломоновых островах (см.
IV главу).
Мельба и Давид Колдуэллы заметили, что резкие
ударные звуки, возможно напоминающие сигнал стра-
ха, действуют на дельфинов в неволе столь сильно, что
вызывают у них язву желудка. Когда в дельфинарии
проводили водопроводные трубы, из-за чего в течение
нескольких недель раздавался металлический звук,
шесть дельфинов заболели язвой 12-перстной кишки и
вскоре один за другим погибли.
137
Существует у дельфинов и сигнал боли, напо-
минающий громкий, отрывистый визг поросенка. Мы
слышали такой сигнал у дельфина-белобочки на Чер-
ном море при следующих обстоятельствах. Крупного
самца случайно положили хвостом на кучу соли на
палубе сейнера близ других дельфинов. Когда кри-
сталлики соли вонзились в кожу, самец начал изда-
вать короткие визги через каждую секунду. Визг слы-
шался четко с расстояния 200 м, несмотря на шум
мотора при ходе судна. Иногда визг прекращался ми-
нут на пять и возобновлялся с новой силой. Звук уси-
ливался, если дотрагивались ладонью до тела дельфи-
на, и прекратился лишь после того, как с животного
смыли соль.
Рис. 18. Сонограммы обыкновенного дельфина, записанные в Сре-
диземном море: а — свист и эхолокационные щелканья,
изданные одновременно одним и тем же дельфином;
б — сигнал испуга, произведенный в момент, когда дель-
фина пытались поймать за хвост хапуном. По Р. Бюс-
нелю и А. Дзидзику
138
К сигналам боли, видимо, относится также «свиня-
чий визг» у обсохших ремнезубов и клюворылых ки-
тов, оказавшихся на берегу в особо неблагоприятных
условиях — под палящими лучами солнца, на камени-
стом грунте и т. д. Сигнал боли — это редкий сигнал,
так как его издают далеко не все загарпуненные кито-
образные.
Среди акустических сигналов китообразных выде-
ляется сигнал бедствия — чудодейственный сиг-
нал, при котором забывается собственная безопасность,
и животные, бросая все, мчатся к месту, откуда пода-
ется призыв о помощи. У разных видов кито-
образных этот сигнал звучит по-разному и особенно
действует в тех случаях, когда сородичу грозит опас-
ность задохнуться под водой. У афалины он слышится
в гидрофон как двуколенный продолжительный свист,
повторяющийся через разные интервалы до тех пор,
пока не подоспеет помощь. В первой части сигнала
свист постепенно нарастает в силе и частоте от 3 до
8—16 кГц, а во второй постепенно ослабевает от мак-
симума до минимума (рис. 19).
Сигнал бедствия у афалины Джон Лилли и Элис
Миллер вызывали двумя способами. В одном случае
они ввели афалине 27 мл раствора нембутала и отчет-
ливо слышали в гидрофон, как она издала сигнал бед-
ствия в тот момент, когда ее выпустили в бассейн с
водой. На сигнал сразу ответили два дельфина: они
поднырнули под нее и стали выталкивать ее к поверх-
ности для дыхания.
В другом случае тот же сигнал Джон Лилли вы-
зывал у афалины с помощью электростимуляции моз-
га, раздражая слабым током определенный участок
коры больших полушарий.
Реакцию на сигнал бедствия проследили сотрудни-
ки Флоридского океанариума на беременной афалине
Пэджи во время трудных и затянувшихся родов, вы-
званных тем, что плод ее выходил не хвостом, а голо-
вой вперед. Самка эта была сильно измучена, она бес-
сильно опускалась на дно, и оставаясь там неподвиж-
ной 2—3 мин, издавала тонкий продолжительный
свист. В ответ на это к ней бросались две самки и клю-
вами поднимали ее на поверхность.
О существовании у китообразных сигнала бедствия
догадывались гораздо раньше, чем его услышали и за-
139
писали. На чем основывается помощь сородичей тем
китообразным, которые терпят бедствие и подают свое-
образные сигналы SOS? Этот вопрос мы рассмотрим
в связи с инстинктом сохранения вида (XVI глава).
Установив обилие коммуникационно-эмоциональ-
ных сигналов, ученые начали изучать у дельфинов об-
мен сигналами. В заливе Назарет на острове Сент-То-
мас (Виргинские острова) нейрофизиологи Джон Лилли
и Элис Миллер разными приемами стимулировали из-
лучение сигналов у афалины. Они записывали звуки
этих животных при разных условиях: у изолирован-
ной особи; у общающихся между собой дельфинов,
расположенных то близко, то далеко один от другого;
Рис. 19. Сигнал бедствия у афалины: а, б, г — взрослых, в —
молодой. По Дж. Лилли
140
у свободно плавающих одиночек или стаек при вмеша-
тельстве или без вмешательства человека.
У изолированной афалины, лежавшей в узкой ван-
не, наиболее часто слышались свисты и неультразву-
ковые щелканья; у свободно плавающей одиночки —
те же звуки плюс эхолокационные щелканья. Когда
в опыте участвовали два животных, не видевших, но
слышавших один другого, они интенсивно свистели и
щелкали. Если же оба дельфина плавали, находясь
вместе, они поддерживали между собой связь — сви-
стели, щелкали, кричали, крякали и эхолоцировали.
Попав в океанариум, дельфины производят свои ес-
тественные сигналы в воде или в воздухе. Новички на-
чинают со слабых свистов и щелканий, но со временем
увеличивают громкость до такой степени, что у стоя-
щего вблизи человека появляется неприятное ощуще-
ние. Щелканья напоминают громкие удары в ладоши,
а свисты — самый сильный человеческий свист, но с
большей частотой.
Афалины в условиях опыта обменивались звуками,
поочередно исполняя продолжительные «монологи»,
«соло» и «дуэты». В «обменный репертуар» входили
свисты, свисты и щелканья, разреженные щелканья,
пронзительный крик, свисты и кряканье и т. д.
Обученные афалины обменивались звуками более
свободно. Когда два дельфина плывут вместе, они об-
мениваются довольно сложным комплексом звуков.
Удачную запись «дуэта» сделали у наиболее способ-
ных афалин — самца Эльвара и самки Тольвы. Было
замечено, что при обмене дельфины соблюдали опреде-
ленный порядок: когда один свистел или щелкал, дру-
гой молчал, и наоборот. Т. Лэнг и X. Смит продолжили
«телефонные» опыты над изолированными дельфина-
ми: через двухканальную электроакустическую связь
животные могли слышать друг друга и «переговари-
ваться» (рис. 20). Сигналы обоих «собеседников» (сам-
ца и самки) вначале записывались, а потом звуки од-
ного воспроизводились другому, вызывая у него опре-
деленную реакцию.
Большую помощь в изучении проблемы звуковой
сигнализации оказал ученым метод электрической сти-
муляции мозга, впервые примененный к дельфинам
Джоном Лилли. В разные участки больших полуша-
рий мозга живой афалины под местным наркозом
141
вживляли, пробивая кожу и череп, короткие и тонкие
трубочки из нержавеющей стали и через них в ткань
мозга вставляли электроды на разную глубину. Такой
операцией с помощью слабых токов пытались найти,
где, в каких участках мозга располагаются слуховой,
зрительный, моторный и другие центры, чтобы соста-
вить карту коры. Для этого электроды миллиметр за
миллиметром перемещали из одного участка коры
мозга в другой. Обезьяны при таких операциях кри-
чат, бьются, рыдают от боли, злобно рычат, а дельфин
спокойно молчит. Его молчание нарушалось внезапно,
когда погружающийся электрод вызывал изменение
электрического тока, за чем следовали сильные удары
хвоста и целый каскад звуков.
Электрическое раздражение мозга для дельфина
оказалось приятным, особенно в моменты включения и
выключения тока. Был открыт новый центр — «центр
удовольствия». Это проверили так: подопытного жи-
вотного научили замыкать и размыкать ток в собствен-
ном мозгу. Для этого над его рылом установили ком-
мутатор в виде прутика: теперь было достаточно лег-
кого движения головы вверх или вниз, как прутик раз-
мыкал или замыкал цепь. Дельфин стал очень часто
включать и размыкать ток. Так он научился раздра-
жать свой мозг, притом гораздо быстрее обезьяны.
Рис. 20. Схема опыта X. Смита и Т. Лэнга: изолированные афа-
лины в разных танках общались друг с другом с по-
мощью двухканальной электроакустической связи
142
Необученной афалине для этого понадобилось 20 испы-
таний, а обезьяне — сотни. При самораздражении моз^
га дельфин издавал разнообразные звуки. Чтобы их
слышать и записывать, над ноздрей животного укре-
пили микрофон с усилителем. Стереофонический ап-
парат с двумя лентами позволял экспериментатору за-
писывать на одной ленте звуки афалины, а на другой
под диктовку собственные наблюдения. Методом поощ-
рений (приятным раздражением мозга) от афалин до-
бивались излучения тех звуков из нескольких возмож-
ных, которые были желательны экспериментатору.
Таким способом одного дельфина Лилли заставил от-
казаться от ультразвуков и «петь» только на слыши-
мых частотах. При этом «певец» был настолько спо-
собным, что ему достаточно было двух-трех поощре-
ний, чтобы он выполнял требуемое от него дейст-
вие.
Некоторые звуки афалины издают лишь во время
контакта с человеком. Через несколько недель обуче-
ния в воздухе слышатся уже более сложные и про-
должительные звуки. Если дрессировщик изо дня в
день громко разговаривает вблизи подопытного дель-
фина, у последнего издаваемые звуки постепенно при-
обретают сходство с голосом человека.
Однажды, прослушивая ленты с голосом животно-
го, Джон Лилли, вперемежку с щелкающими й лаю-
щими звуками, услышал... взрывы смеха: оказалось,
что дельфин скопировал незадолго перед этим раздав-
шийся смех ученого, что и записал прибор. Последую-
щие опыты ученого подтвердили, что афалина может в
какой-то степени подражать голосу человека, а после
известной тренировки способна воспроизводить даже
простенькие фразы человеческой речи. Электрическая
стимуляция мозга сильно ускоряет обучение дельфи-
нов, так как раздражение «центра удовольствия» —
лучшее, чем даже пища, поощрение для животного.
Но афалина подражала голосу человека и без электри-
ческой стимуляции.
Сравнение виброграмм с записью голосов дельфи-
нов и человека подтвердило звукоподражание афали-
ны. Человек произносил слова в пределах колебаний
от 200 до 3000 Гц, а дельфин воспроизводил их в ча-
стотах от 1000 до 8000 Гц. Если магнитофонную ленту
с голосом дельфина, имитировавшим слова человека,
143
пропускали со скоростью, замедленной в 2—4 раза, то
достигали известного сходства с голосом человека.
Самые удачные наблюдения Лилли и Мюллер про-
вели над дельфином по кличке Эльвар. Вначале, с ию-
ля 1960 года, Эльвар жил один и общался лишь с тре-
нером. Наблюдатель говорил громко, чтобы звуки про-
никали под воду. В течение 15 месяцев Эльвар на-
учился воспроизводить звуки, подобные человеческой
речи и в воде и на воздухе, включая гласные, взрыв-
ные и свистящие. Частоты этих звуков у афалины бы-
ли выше, чем у взрослого мужчины или женщины, и
скорее напоминали голос ребенка: вначале его бол-
товню, а потом и отдельные слова.
Лежа в ванне, этот дельфин имел привычку наби-
рать в ноздрю воду и выбрызгивать ее вверх, а иногда
и в экспериментатора. Однажды Элис Миллер, обрыз-
ганная водой, крикнула ему «стоп ит» («прекрати
это»), и через несколько секунд в воздухе разда-
лось— «стоп ит». В другой раз человек сказал Эльва-
ру «бай-бай», а в ответ услышал (и записал на ленте)
звук «ба-баий».
После Элис Миллер дельфина стал обучать Джон
Лилли» Эльвар «учел» более грубый голос нового дрес-
сировщика и понизил нижние частоты в своих «сло-
вах» с 1000 до 450, а затем и до 200 Гц. Он был обу-
чен произносить свое имя и слова more (больше), speak
(говори), louder (громче), up (вверх) и фразы squirt
water (брызни воду), more, Elvar (больше, Эльвар) и
ДР.
Подражательные или «гомйнидные» звуки дельфи-
ны воспроизводят лишь после упорной тренировки.
Способность дельфинов к звукоподражанию остава-
лась незамеченной в шумных океанариумах. Для этого
открытия понадобилась хорошо оснащенная лабора-
тория, где экспериментировали с изолированными жи-
вотными в стеклянных ваннах, обложенных пенопла-
стом. Здесь животные могли издавать звуки не только
в воде, но и на воздухе, выставив дыхало.
Теперь голос дельфинов демонстрируют перед пуб-
ликой. В Калифорнийском океанариуме сразу четыре
дельфина, высунув головы из воды, производят серию
звуков, в шутку названных «Гимном Мэриленда»
(рис. 46 вкл.).
144
В дельфинариях между смежными отсеками
и загонами для афалин можно услышать множество
дельфиньих голосов (человек испытывает ощущение,
какое бывает возле клетки со множеством голосистых
птиц, например, волнистых попугайчиков). Его оглу-
шают самые различные визги, писки, свисты, крики,
хрипы и многие другие звучания. Звуковая активность
усиливается, когда с животными проводятся разного
рода эксперименты. Подопытные афалины выставля-
ются «по пояс» вертикально из воды и выдают такие
рулады, которые отчетливо слышатся за 200 м. В по-
добной многоголосице, конечно, невозможно разли-
чить подражательные звуки, существование которых
доказывал Дж. Лилли.
Способность к звукоподражанию и «гоминидные»
звуки натолкнули некоторых ученых, и прежде всего
Джона Лилли, на мысль о существовании у дельфинов
настоящего языка, как системы обмена абстрактной
информацией, аналогичной человеческой речи. Нацио-
нальное управление США по аэронавтике и изучению
космоса выделило Лилли 80 тысяч долларов на обуче-
ние афалины человеческому языку. Однако результа-
ты оказались разочаровывающими. Дельфин смог по-
вторить за человеком 12 бессмысленных слогов или в
лучшем случае имитировал лишь отдельные слова,
как это делают попугаи.
Глава XIV
ОБЛАДАЮТ ЛИ
ДЕЛЬФИНЫ РЕЧЬЮ?
Уровень развития животных определяется многими
признаками и прежде всего характером организации
нервной системы. Бесспорно, дельфины, как и весь
отряд китообразных, обладающие крупным, сложно
устроенным мозгом, стоят на высокой ступени разви-
тия. Выделяется прежде всего шестислойная кора боль-
ших полушарий, в которой, например у афалины,
насчитывают до 30 млрд, нервных клеток. Относитель-
ный вес ее мозга достигает 1,25% веса тела. Поверх-
ность мозга сильно увеличивается благодаря большо-
му количеству извилин. Площадь коры, скрытая
внутри извилин, составляет у дельфинов 75—85% всей
площади коры, что выше, чем у человека (64—67%).
Значительно развиты также промежуточный и сред-
ний отделы мозга и мозжечок.
Сразу же возникают вопросы: что может такой
мозг, для чего дельфинам столь сложная организация
нервной системы и правомерно ли ей приписывать
функции, специфические для человеческого сознания,
в частности речь?
Американский нейрофизиолог Джон Лилли отве-
чал на эти вопросы очень просто: мозг дельфина стоит
на одном (или даже более высоком) уровне с мозгом
человека и потенциально способен на то же самое, на
что и человеческий мозг. Такой орган, по мнению Лил-
ли, обеспечивает дельфинам речевое (словесное) обще-
ние друг с другом и в будущем позволит осмысленно
разговаривать с человеком, в результате чего наука
сделает грандиозный скачок вперед. Лилли исходил из
того, что есть критический размер мозга (1 кг), ниже
146
которого язык невозможен. Поскольку средний вес
мозга у афалины (1700 г) больше, чем у человека
(1400 г), то ум обоих следует поставить по крайней
мере в один ряд. Трудность же словесного общения
человека и дельфина временная, и объясняется она
тем, что первый слышит лишь небольшую часть сиг-
налов второго: ведь диапазон восприятия частот у
дельфинов в 10 раз выше, чем у людей.
Большинство биологов разных стран (Рене Бюснель,
Вильям Таволга, Грэгори Бэтсон, Форрест Вуд, Давид
и Мельба Колдуэллы и многие другие) отрицают су-
ществование настоящего языка у дельфинов. Сам же
Лилли не смог доказать правоту выдвинутой им ги-
потезы и в 1967 году закрыл свою лабораторию.
В поддержку языковой гипотезы Джона Лилли при-
водились следующие соображения:
сложный мозг дельфинов внешне сходен с челове-
ческим (вес, шаровидная форма, обилие извилин, соот-
ношение веса больших полушарий и мозжечка, коли-
чество нервных волокон в 1 см3);
дельфины очень понятливы и легко обучаются;
акустические сигналы, издаваемые сложным зву-
косигнальным аппаратом в верхней части дыхательно-
го пути, весьма многообразны (В. Эванс и Д. Дреер у
трех видов дельфинов выявили 32 разных свистовых
контура);
в свистах существуют индивидуальные различия
(«акустические автографы»), по которым можно рас-
познавать дельфинов персонально;
некоторые афалины способны подражать словам че-
ловека (о том свидетельствуют сонограммы, записан-
ные Джоном Лилли);
дельфины, посаженные в два отдельных бассейна,
между которыми работает электронная связь, проявля-
ют интерес к голосам сородичей (Томас Лэнг и X. Смит
наблюдали пересвистывание афалин, находившихся в
разных танках, но слышавших друг друга; по свиде-
тельству прессы, был осуществлен даже радиообмен
сигналами между дельфинами, находящимися во Фло-
риде и на Гавайских островах, с расстояния 8000 км);
в экспериментах со звукообменом подопытные дель-
фины не нарушают молчания, пока «говорит собесед-
ник» ;
и наконец, некоторые дельфиньи свисты повто-
147
ряются при воссоздании тех условии окружения, в ко-
торых эти сигналы производились раньше.
Все перечисленные доводы, хотя и свидетельствуют
о высоком развитии ума дельфинов, и о сложности
внутривидовой коммуникации, однако не доказывают
наличия у них языка и понятийного мышления с по-
мощью абстрактных категорий.
В самом деле, большой размер мозга с огромным
числом нейронов и множество извилин в нем не обяза-
тельно могли развиваться у дельфинов в связи с речью.
Вес мозга относительно веса тела сам по себе еще ни о
чем не говорит. У человека мозг составляет 7з4 веса
тела. Но есть обезьяны в Южной Америке, относи-
тельный вес мозга которых выше, чем у человека
(у капуцина — 1/18» а у черной коаты — до 715 веса
тела). Однако никто не считает этих низкоорганизо-
ванных широконосых обезьян «умнее» человека. С дру-
гой стороны, имеются резкие индивидуальные откло-
нения в весе этого органа от средней нормы: напри-
мер, мозг Анатоля Франса был на 400 г меньше, а у
И. С. Тургенева на 600 г больше нормы, принятой для
человека.
Далее, легко поддаются дрессировке и очень понят-
ливы не только дельфины, но и собаки, обезьяны, мор-
ские львы, лошади и другие животные.
Звукосигнальный орган разной сложности имеют
многие животные, начиная от насекомых (саранчовые)
и кончая птицами и млекопитающими.
Голосовая сигнализация дельфинов не является
уникальной в животном мире ни по богатству сигна-
лов, ни по характеру их использования. Многообразие
звуков, в сравнении с богатством звуков других живот-
ных, несколько преувеличивается. Немецкий ученый
Темброк, например, установил для лисицы 36 разных
сигналов. Еще больше их обнаружено у гамадрилов и
человекообразных обезьян. Во всех этих случаях сиг-
налы отвечают тем или иным отношениям между со-
родичами и повторяются стереотипно в соответствии с
обстановкой.
Индивидуальные оттенки в звуковой сигнализации
свойственны почти всякому животному, обладающему
голосом, а не только дельфинам; кто не знает, что хо-
зяин может по лаю определить свою собаку из стаи
в десяток голов?
148
Интерес к голосам сородичей и обмен сигналами
проявляются у многих животных, например в семьях
обезьян, в стайках синичек й т. п. Спровоцировать
ответ на искусственно подаваемые сигналы удается у
многих млекопитающих и птиц. На этом основана охо-
та с манком на рябчика и уток, с рогом на оленей
и т. д.
Дельфины могут в какой-то мере имитировать сло-
ва человека, но то же самое делают (только гораздо
чище, членораздельнее) и другие животные — попу-
гаи, скворцы, сороки, майны, не вкладывая в эти сло-
ва смысла, абстрактных понятий.
Со свистом дельфина иногда сравнивают свистовой
язык жителей некоторых деревушек в Пиренеях и на
Канарских островах. Но нельзя забывать, что свисто-
вой язык человека — это те же закодированные слова,
сигналы сигналов. Французский физиолог Рене Бюс-
нель специально изучал язык этих горных народов
и пришел к выводу, что по физическим свойствам
возможна аналогия между дельфиньими свистами и
свистовым языком человека, но она не доказывает су-
ществования языка у дельфинов. И методы, анализи-
рующие свистовой язык человека, не могут быть при-
менимы к акустическим сигналам животных, в том
числе и к свистам дельфинов.
Ныне часто ссылаются на опыты калифорнийского
психолога Джарвиса Бастиана, который в морской ла-
боратории военно-морского флота США в Пойнт-Магу
якобы пытался экспериментально проверить существо-
вание языка у афалин. Суть эксперимента была в том,
что афалину-самца Бэс обучили подражать действиям
самки Дорис — нажимать на один, другой или третий
контакты с сигнальными лампами. Когда обоих жи-
вотных в бассейне разъединяли непрозрачной перего-
родкой, чем исключали зрительный анализатор, то по-
дражание в нажимании на тот или иной контакт было
точным. Когда же перегородку делали еще и звуконе-
проницаемой (исключали слуховой анализатор), подра-
жание стало ошибочным. Но стоило в перегородке сде-
лать небольшое отверстие, через которое проходил звук,
как подражание вновь стало правильным.
Комментаторы опытов Д. Бастиана объяснили точ-
ность подражания тем, что Дорис будто бы передава-
ла информацию для Бэс с помощью языковых сигна-
149
лов. Однако так толковать этот опыт не решился даже
сам Бастиан, так как афалина Бэс могла следить за
местоположением и действиями Дорис с помощью свое-
го гидролокатора.
Как считает Форрест Вуд (а он проанализировал
эксперименты Бастиана), в этих опытах не было полу-
чено доказательств того, что Дорис понимала связь
между своими голосовыми сигналами и поведением
самца Бэса. Наоборот, была доказана полная непред-
умышленность действий Дорис. Вот почему ссылка на
опыты Бастиана неправомочна.
Советский нейрофизиолог член-корреспондент Ака-
демии наук СССР Н. П. Бехтерева, изучая психические
заболевания и нейрофизиологические процессы, при-
шла к выводу, что в ответ на слова, которые слышит
человек, в его мозгу возникают два кода — акустиче-
ский и смысловой (семантический). Первый связан со
звуковой характеристикой слова и свойствен также
животным, например в тех случаях, когда те реаги-
руют на свою кличку или исполняют команды с голо-
са дрессировщика. Второй связан со смысловой харак-
теристикой слов и присущ только человеку. Именно в
этом принципиальное отличие животных (в том числе
и дельфинов с их сильно развитым акустическим ко-
дом) от человека.
Язык, речь, слово свойственны только людям. Даже
самую сложную сигнализацию животных нельзя ото-
ждествлять с речью. С помощью языка могут быть на-
званы и классифицированы вещи, выражены отвлечен-
ные идеи, обсуждены и сделаны умозаключения,
обговорены любая концепция и ситуация. Язык рас-
сматривает события прошлого и будущего, называет
то, что отсутствует в данный момент. Язык дал нача-
ло цивилизации.
Животные не могут пользоваться словом сознатель-
но, как отвлеченным понятием, хотя некоторые из них
и могут скопировать (за награду) звук произносимого
слова. Так делают живущие в неволе попугаи, сквор-
цы, обезьяны или дельфины (в опытах Лилли), а у
Владимира Дурова жила собака, четко произносившая
слово «мама»; выговаривали слова «мама» и «папа»
также обезьяны. Однако еще ни разу даже самые ум-
ные животные, будь то пернатые или млекопитающие
150
(включая китообразных), не показали понимания ими-
тируемых ими слов.
Таким образом, доказательств того, что дельфины
пользуются языком, словесной (абстрактной) информа-
цией, до сих пор нет. За поведением дельфина не стоят
мыслительные процессы, аналогичные нашим. Ни по-
ведение, ни структурно-функциональная организация
коры не подтверждают, что у дельфинов имеются свой-
ства человеческой психики. Научный сотрудник ин-
ститута мозга В. С. Кесарев, сравнивая микрострук-
туры мозга человека и дельфина, нашел в них суще-
ственные отличия. Шестислойное строение коры —
общий признак мозга всех млекопитающих. Но орга-
низация нейронов в колонки — особенность мозга че-
ловека: оказывается, нейроны в виде зерен (2-й слой
коры), пирамид (3-й слой), треугольных клеток (5-й
слой) и веретен (6-й слой) располагаются друг под
другом, образуя вертикальные колонки, которые мо-
гут работать и самостоятельно, и в комплексе с сосед-
ними. В слуховом анализаторе степень такой упорядо-
ченности нейронов гораздо выше, чем в зрительном,
что, видимо, обусловлено восприятием речи. Все эти
сложные «архитектурные сооружения» человеческого
мозга локализуются в новой коре — наиболее позднем
эволюционном образовании,— занимающей 96% пло-
щади всей коры (древняя кора составляет лишь 0,6%).
У дельфинов же подобные колонки не обнаружены,
а межуточная кора (2-й, 3-й и 4-й слои), хотя и силь-
но разрослась, усложнилась и дифференцировалась, но
не дошла до стадии новой коры, характерной для че-
ловека.
Заключение об отсутствии речи у китообразных не
опровергают ни экстраполяционные рефлексы, уста-
новленные профессором МГУ физиологом Л. В. Кру-
шинским, ни его опыты по сложным формам поведе-
ния дельфинов, оперирующих эмпирической мерно-
стью фигур и воспринимающих различие плоских и
объемных образов.
Но если у дельфинов нет языка и понятийного
мышления, то чем можно объяснить развитие их ог-
ромного головного мозга? Причины и обстоятельства
для этого у них совсем другие, чем у человека: мозг
дельфинов обслуживает потребности жизни в водной
среде и определяет всю тонкость приспособительной
151
деятельности в этих условиях. В воде у китообразных
наиважнейшим органом чувств оказалось не зрение,
как у наземных млекопитающих, а слух в сочетании
с эхолокационным аппаратом.
Ведущую роль в развитии крупного мозга дельфи-
нов сыграла, видимо, эхолокация как важнейший спо-
соб ориентации зубатых китов в океане и главный
путь получения информации об окружающем. В есте-
ственных условиях эхолокационный аппарат исполь-
зуется ими на каждом шагу. Время между произведен-
ным щелчком-сигналом и возвратом его эха указывает
животным расстояние до любого объекта на их пути.
В связи с этим отрабатывается тончайшее управление
движениями своего тела. Для переработки поступаю-
щих эхо-сигналов потребовался высокоразвитый голов-
ной мозг. Не случайно в мозге дельфинов обнаруже-
ны некоторые преимущества перед мозгом человека
именно в области слуховой системы.
В мозге дельфинов сильно развиты подкорковые
образования, с которыми связана работа эхолока-
ционного аппарата и обработка звуковой информации.
Способность пользоваться ультразвуком у дельфинов
связана с отделами, лежащими в глубоких слоях мозга.
Советский ученый В. П. Зворыкин нашел у этих жи-
вотных черты превосходства в слуховой подкорке, раз-
витой сильнее, чем у людей; оказалось, например, что
по объему верхняя олива мозга у дельфина в 150 раз
больше, чем у человека.
Сильному развитию мозга дельфинов благоприят-
ствовали также:
большая скорость плавания и быстрая смена внеш-
них условий при нырянии (и то и другое устраняло
сенсорную недостаточность; относительное укрупнение
мозга, или цефализация, интенсивнее у скоростных ви-
дов);
выдвижение на первое место из органов чувств слу-
хового анализатора, воспринимающего в очень широ-
ком диапазоне акустические колебания (от десятков
герц до 170 кГц), что расширяло поток поступающей
информации;
работа локатора на высоких частотах, позволявшая
передавать больший объем информации и за более ко-
роткий срок, чем при работе на низких частотах;
отличная звукопроводимость моря и высокая (в 4,5
152
раза большая, чем в воздухе) скорость распростране-
ния звука в воде, вызывавшая необходимость мгновен-
ного ответа животного (поэтому дельфин на звуко-
вые раздражители отвечает гораздо быстрее чело-
века) ;
стадный и семейный образ жизни и совместная охо-
та за рыбой (на основе общения у дельфинов формиро-
вались различные коммуникационные сигналы — ра-
зыскивания и преследования добычи, страха, угрозы,
доминирования, бедствия и др.; эти сигналы компен-
сировали невозможность использования в воде мими-
ки и жестов);
продолжительное совместное пребывание детены-
шей и родителей, обогащавшее индивидуальным опы-
том молодых особей.
Все это и обусловило в ходе эволюции сильное раз-
витие центральной нервной системы дельфинов, при-
чем эхолокация, используемая «на все случаи жизни»,
явилась, вероятно, главным фактором в формирова-
нии крупного головного мозга зубатых китов *.
Это соображение можно подкрепить сопоставле-
нием кашалота и усатых китов. У последних эхолока-
ция не доказана, и мозг их, по общему признанию
специалистов, развит слабее, чем у зубатых китов. Ка-
шалоту чаще, чем другим зубатым китам, приходится
пользоваться эхолокацией, так как питается он в зоне
вечного мрака, на глубине до 1—2 км. Вероятно, по-
этому его мозг занял рекордное место среди живых
существ планеты как по абсолютному весу (9,2 кг у
самца длиной 14,9 м), так и по числу извилин на по-
верхности коры больших полушарий. Это не означает,
конечно, что кашалот — умнейшее животное в отряде,
но он, видимо, обладает крайне специфической ориен-
тацией и звуковидением, в основе которого лежит эхо-
локация (см. XII гл.).
Все это, вместе взятое, и привело к развитию ги-
гантской, нелепой по форме головы кашалота с ре-
кордной массой головного мозга.
1 Не противоречит ли этому положению тот факт, что рукокры-
лые и ластоногие, тоже пользующиеся эхолокацией, имеют
небольшой мозг? Однако рукокрылые живут в воздушной сре-
де, где эхолокация не может быть столь эффективной, как в
воде, а ластоногие имеют эхолокацию лишь в зачаточном со-
стоянии.
153
В заключение главы отметим, что надежды научить
афалину сознательно пользоваться человеческой ре-
чью наивны и беспочвенны, ибо нельзя наследственно
обусловленную и отработанную за миллионы лет при-
роду дельфина в короткий срок, с помощью даже са-
мой изощренной дрессировки переделать на человече-
ский лад, забывая при этом роль эволюции и силу на-
следственной информации.
Общение дельфинов с человеком в неволе ликви-
дирует их умственную вялость, ставит их в новые, не-
обычные ситуации, тренирует их мозг. При этом жи-
вотные проявляют такие реакции, которые далеко не
всегда укладываются в рамки условных рефлексов и
могут быть отнесены к элементарной рассудочной дея-
тельности.
От проблесков мышления животного до человече-
ского сознания — большая дистанция, однако путь к
понятийному мышлению проходил через эти ступень-
ки, через усложняющиеся формы поведения и элемен-
тарную рассудочную деятельность. И хотя сигнализа-
ция животных различной сложности была когда-то на-
чальным этапом развития речи человека, даже самые
изысканные формы сигнализации в мире животных
находятся на крайне низком уровне в сравнении с
уровнем языка разумного существа.
Глава XV
ЗАЧАТКИ
РАССУДОЧНОЙ
ДЕЯТЕЛЬНОСТИ
У ДЕЛЬФИНОВ
УЖИВАЮТСЯ
С ИНСТИНКТАМИ
Дельфины с их сильно развитым мозгом обладают
большими психофизиологическими возможностями.
Они великолепно ориентируются в море, многосторон-
не используют сонар и звуковую сигнализацию, спо-
собны к быстрому обучению. Такие формы поведения
высокоорганизованных животных, когда совершаются
как бы предусмотрительные поступки и проявляются
проблески сознания, Л. В. Крушинский назвал экстра-
поляционным поведением, элементарной рассудочной
деятельностью. Это следующая ступенька после услов-
ных рефлексов на пути к мышлению, когда развива-
ется в какой-то мере способность решать новые задачи
в незнакомых ситуациях.
Приведем несколько примеров. Во Флоридском
океанариуме резвятся афалины, неповоротливая мор-
ская черепаха выплывает на середину танка и мешает
игре. Тогда дельфины подплывают к черепахе и, дей-
ствуя клювом, удаляют ее из центра водного зеркала,
после чего продолжают игру. В том же океанариуме
самка афалины Элджи извлекала головоногих моллю-
сков, которые прятались в трещинах и пещерках на
дне бассейна: она стремительно проплывала над этим
местом и создавала хвостом такие вихри, что моллюс-
ка выбрасывало в незащищенную зону.
Биологи США Давид Браун и Кеннет Норрис в Ка-
лифорнийском океанариуме были свидетелями того,
как две афалины тщетно пытались вытащить мурену,
спрятавшуюся в небольшой расщелине скалистого дна
танка. Вскоре один из дельфинов, отплыв в сторону,
убил недавно впущенного в бассейн морского ерша-
155
скорпену, обладающего ядовитыми колючками, вер-
нулся с этой рыбой и сунул ее в убежище мурены.
Уколотая мурена выплыла из расщелины и была
поймана афалиной. Этот пример приводят (обычно
журналисты), как свидетельство «почти человеческой
разумности» дельфина. Но ершом-скорпеной афали-
на питается в море, и нельзя быть уверенным, что
она не повторила приемы, уже отработанные у нее
природой.
Примечательный случай описала в гавайском «Си-
лайф-Парке» в 1973 году биолог Карен Прайор: моло-
дые дельфины — черная косатка и афалина предпо-
читали находиться вместе в одном и том же бассей-
не, но их разъединили, поместив в двух смежных
водоемах, отделенных перегородкой. Животные стали
перепрыгивать ее. Тогда перегородку усилили более
высокой преградой. В ответ на это малая косатка
ночью сдвинула препятствие, и общение продолжа-
лось с помощью прыжков.
Аналогично этому поступала другая афалина: она
хитроумно избегала болезненного укола, мешая ее от-
лову — закрывала ворота загона (см. главу V). Ряд
примеров сложного поведения дельфинов приведен из
практики Батумского дельфинария в VII главе.
Очевидно, у афалины имеются проблески сознания
и элементы рассудочной деятельности, так же как и у
других видов млекопитающих и птиц. Поэтому описан-
ные случаи с дельфинами нельзя рассматривать как
что-то уникальное в мире животных, хотя эти кито-
образные довольно сообразительны и, вероятно, зани-
мают место где-то между собакой и обезьяной.
Несмотря на свой гигантский мозг и зачатки рас-
судочной деятельности, дельфины совершают и явно
неразумные поступки. Они зачастую не могут решать,
казалось бы, весьма простые задачи. Например, в
океанариумах дельфины гибнут от заглатывания по-
сторонних предметов — кусочков стекла, резины, вере-
вок, тряпок и т. д. Когда дельфинов держат на привя-
зи, они не могут выбрать для себя наиболее безвредные
формы поведения и запутываются на длинных повод-
ках так же, как домашние животные. Особенно опасно
привязывать дельфинов за столбы в воде: наматывая
бечевку вокруг столба и укорачивая ее, животные по-
гибают, лишив себя возможности выплывать для ды-
156
хания на поверхность. Чтобы распутать веревку, им
достаточно было бы сделать несколько кругов в обрат-
ном направлении, но сообразить этого они не могут.
Особенно странно дельфины ведут себя при отлове.
Обладая стремительным ходом и великолепным лока-
тором, они позволяют ловцам окружать и загонять се-
бя в сети, а иногда и на берег. На Фарерских островах
местные жители без всяких снастей выгоняют на мель
стада гринд в сотни голов, а туземцы острова Малаи-
та — малайских продельфинов.
Не лучше ведут себя и черноморские дельфины при
аломанном лове. Автор этой книги сам участвовал в
таком промысле и убедился, насколько малоразумны
действия дельфинов в это время: обметанные кошель-
ковым неводом, дельфины стремительно плавают в
нем, как в огромной чаше, и... не догадываются пере-
прыгнуть верхний край сети, чтобы вырваться на во-
лю. Они остаются в сетяной загородке дни, недели,
месяцы. У нас на Черном море в вольерах из капроно-
вой сети дельфины обычно живут целое лето, и не бы-
ло случая, чтобы они уходили с помощью прыжка
(разумеется, те, которые обучены прыгать через заго-
родку, как Флиппер, в расчет приниматься не могут).
В чем же причина такого нецелесообразного пове-
дения китообразных? Видимо, дело в том, что зачатки
рассудочной деятельности далеко не всегда позволяют
дельфинам решать задачи, связанные с преодолением
инстинктов, штампов в поведении, которые у кито-
образных сильно развиты в условиях водной жизни и
им необходимы. Дельфины привыкли находить выход
из сети и обходить препятствия под водой с помощью
своего сонара, а если тот не показывает разрыва в се-
ти, задача для необученных животных остается не-
решенной. Чтобы выйти из окружения с помощью
прыжка в воздух, нужно абстрактное мышление, кото-
рое у дельфинов оказалось недостаточным.
Глава XVI
РЕФЛЕКСЫ
ВЫНЫРИВАНИЯ
И ИНСТИНКТ
СОХРАНЕНИЯ ВИДА
В характере действий дельфинов наблюдаются две
странности: они спасают людей и в то же время сами
нелепым образом погибают в массовом количестве на
берегах (это явление получило название «массовое са-
моубийство»).
Первую странность нередко приводят как доказа-
тельство высокой разумности и абстрактного мышле-
ния дельфинов: раз дельфины спасают человека, вы-
талкивая его из воды,— значит, они будто бы пони-
мают, что для людей, дышащих легкими, необходимы
воздух и суша.
Но невольно возникает вопрос: как такие «смыш-
леные» животные, спасающие тонущего человека, до-
пускают собственную гибель и гибель целого стада со-
родичей на берегах?
Чтобы объяснить обе странности в поведении, не-
обходимо познакомиться с инстинктом сохранения ви-
да у китообразных.
В основе этого инстинкта лежит рефлекс вы-
ныривания, который был установлен нами в 1946
году следующим образом. Мы опустили живого дель-
фина в воду и, взяв за плавники, стали его периоди-
чески выставлять на воздух и погружать в воду. При
каждом выставлении дельфин производил выдох-вдох.
Оказалось, что выставление из воды у китообразных
служит безусловным раздражителем для открывания
дыхала и осуществления дыхательного акта.
Китообразные открывают дыхало, совершая корот-
кий выдох-вдох каждый раз, когда их голова выстав-
ляется из воды. В естественной среде дыхало нормаль-
158
но открывается, если рецепторы кожи ощущают в мо-
мент выныривания смену среды вода — воздух (реф-
лекторная регуляция дыхания).
Разумеется, если выставлять дельфинов из воды
очень часто, они будут отвечать дыхательным актом
лишь первые несколько раз, а потом наступит тормо-
жение, вызванное искусственным учащением дыха-
ния. Но минует какой-то срок, превышающий есте-
ственную дыхательную паузу, и животное опять нач-
нет реагировать подобным же образом при новых
выставлениях его из воды. Этот рефлекс выныривания
у диких дельфинов в естественной среде их обитания
проявляется очень четко, но в неволе (в суженных
пространствах) — как большое исключение. Например,
в морских вольерах, а тем более в искусственных бас-
сейнах дельфины выныривают иногда без дыхательно-
го акта, а в океанариумах могут даже часами лежать
на поверхности, медленно покачиваясь, и дышать с
разными интервалами, не заныривая. Исключения в
проявлении рефлекса выныривания тем значительнее,
чем дольше дельфин пробыл в неволе и чем меньше
искусственный водоем (с уменьшением глубины и пло-
щади бассейна живущие в неволе дельфины привыка-
ют к человеку быстрее).
Кроме описанного, мы установили и другой реф-
лекс выныривания: всякий раз почти одновременно
с выдохом дельфины совершают своим хвостом ры-
вок, чем обеспечивают к моменту вдоха оптимальное
поднятие тела из воды для совершения дыхательного
акта. Оба рефлекса исключают попадание воды в лег-
кие и позволяют китообразным безопасно дышать в
любую погоду как в бодрствующем состоянии, так и
во время сна.
В связи с рефлексами выныривания становится по-
нятно, почему на дельфинов не действуют наркотики
в обычных дозах, а в увеличенных вызывают гибель:
снотворные вещества приводят к потере чувствитель-
ности кожи, а во время сна это нарушает правиль-
ность дыхательного акта, что в условиях водной среды
для легочнодышащего животного равносильно смерти.
Рефлексы выныривания не мешают китообразным
в известных пределах управлять своим дыханием: на-
пример, паузой расчленять слитный дыхательный акт
па выдох и вдох, выдыхать воздушные пузыри под
159
водой и тем самым регулировать свою плавучесть.
Безусловные рефлексы выныривания имеют еще
одно важнейшее значение: они позволяют членам ста-
да спасать погибающего сородича, которому угрожает
удушье в воде, выталкивая его на поверхность. Вы-
ставление же пострадавшего из воды автоматически
вызывает у него дыхательный акт. Поэтому самой цен-
ной помощью для погибающего будет выталкивание
его из воды, чем стимулируется дыхание. В этом и
заключается у китообразных инстинкт сохра-
нения вида. Но чтобы инстинкт проявился, гибну-
щее животное должно подать сигнал бедствия. Услы-
шав такой сигнал, сородичи немедленно бросаются
на помощь и начинают выталкивать потерпевшее жи-
вотное из воды. При этом инстинкт сохранения вида
подавляет инстинкт самосохранения особи: помогаю-
щие оказывают помощь даже при смертельной опас-
ности для себя (например, во время охоты китобой-
цев). Таким образом природа заботится сохранить вид
в целом, а не отдельную особь. Взаимопомощь у кито-
образных — такое запрограммированное в их мозгу
поведение, над которым им не приходится «долго ду-
мать» : оно записано языком унаследованных рефлек-
сов и отшлифовано личным опытом.
В условиях водного образа жизни, когда существует
постоянная угроза задохнуться в воде, больные или
ослабевшие китообразные могут получить помощь
только от своих сородичей, которые в этом случае
вытолкнут их на поверхность. Поэтому в поведении
китообразных на первое место выдвигается взаимопо-
мощь, а агрессивность и пристрастие к индивидуаль-
ному участку, столь сильно развитые у ластоногих,
здесь исчезают.
Приведем несколько примеров. Американские зоо-
логи Д. Сибеналер и Д. Колдуэлл в 1956 году видели в
море, как афалины спасли одного дельфина, оглушен-
ного взрывом динамита: два здоровых животных под-
плыли к пострадавшему, поместили его тело на свои
грудные плавники и раскачивали то вверх, то вниз,
совсем как в наших опытах в 1946 году (см. выше).
Однажды научные работники Давид Браун и Кен-
нет Норрис поместили в Калифорнийский океанариум
самку обыкновенного дельфина. После поимки в море
она едва держалась на поверхности воды, и инъекция
160
адреналина ей не помогала. Для плавучести к ней при-
крепили четыре баллона — по два с каждой стороны.
Однако больную спасло другое: в танк подсадили сам-
ца, который и оказал «неотложную помощь» — заплы-
вал под самку и выталкивал ее к поверхности воды.
Сородичи выталкивают пострадавшего дельфина
не только рылом и плавниками, но также и брюхом,
перевернувшись спиной вниз, а иногда зацепив своей
хвостовой лопастью хвостовой стебель партнера или
схватив зубами за плавники (рис. 21).
Рис. 21. Разные способы оказания помощи у дельфинов (но на-
блюдениям Д. и М. Колдуэллов, Д. Брауна, Д. Сибе-
налера и Г. Пиллери): а, б, в, г, д — помощь гринды
мертвой самке полосатого дельфина путем выталкива-
ния за хвост, за гарпун, за грудной плавник, за хво-
стовой стебель и за спинной плавник; е — помощь осла-
бевшему сородичу путем закрючивания за хвост; ж —
поддержка пострадавшего с боков; з — помощь ране-
ному обыкновенному дельфину; и — помощь ослабевшей
амазонской инии
8 9-1984
1G1
Подобных методов у них много. 30 июня 1963 года
в мелководной лагуне на Черном море мы наблю-
дали, как оказывали помощь обыкновенные дельфи-
ны. Здесь содержали трех самок. Одна из них была в
тяжелом состоянии и плохо плавала. Когда она под-
плыла слишком близко к берегу, люди толкнули ее к
центру лагуны. Тут же заметили, как к ней с расстоя-
ния 50 м бросилась наиболее активная самка, она два-
жды подталкивала больную так, что та выставлялась
из воды на 10—15 см. Затем возле них очутилась
третья самка, и все трое стали плавать вместе.
Так же дельфины оказывают помощь детенышам и
новорожденным (рис. 47 вкл.). На фотоснимке, сделан-
ном в Нейпирском океанариуме (Новая Зеландия), за-
печатлен момент, когда самка обыкновенного дельфи-
на Бренда, сопровождавшая только что родившую сам-
ку Бэсс (рис. 48 вкл.), вытолкнула ее детеныша спустя
несколько минут после родов. Весь процесс появления
детеныша на свет (от первого усилия матери до пер-
вого вдоха дельфиненка) занял 14 мин. Нам известен
случай, когда в роли спасающего выступил годова-
лый детеныш, выталкивавший из воды больную афа-
лину-мать (рис. 49 вкл.).
Инстинкт сохранения вида развит не только у зу-
батых, но и у усатых китов, однако наблюдать его
удается очень редко. Если загарпунен детеныш горба-
ча или серого кита, то самка, несмотря на шум, вы-
стрелы гарпунной пушки и даже свое ранение, не по-
кидает сосунка, пока он еще подает признаки жизни.
К. Норрис и Дж. Прескотт рассказали, как в водах
Калифорнии в январе 1958 года косатка напала на
пятерых серых китов. Хищник несколько раз прибли-
жался к китам. Вскоре можно было видеть, как четве-
ро китов длиной 9—11 м поддерживали своими плав-
никами более крупного пятого, раненного косаткой.
Они «раскачивали» его близ поверхности и подталки-
вали снизу вверх. Когда косатка удалилась и опас-
ность миновала, водолаз приблизился к группе и за-
метил, что два кита поддерживали переднюю часть
тела пострадавшего, третий располагался снизу его, а
четвертый был скрыт остальными животными.
Особенно бурно реакция помощи проявляется у ка-
шалотов в косяках, состоящих из самок с детеныша-
ми, и в гаремах.
162
Японским исследователям Масахару Нисиваки и
Такашиме с самолета удалось снять момент сутолоки
кашалотов вокруг раненого, которому оказывалась по-
мощь. Животные при этом располагались, как лепе-
стки в ромашке, и выталкивали раненое животное,
помещенное в центре. Интересно, что сходные вытал-
кивающие движения кашалоты делали во время игры
с бревном на поверхности моря. Спасательные дей-
ствия у гринд удалось пронаблюдать в неволе: три
самки в океанариуме мягко выталкивали своими мор-
дами четвертую, лежащую на боку самку (рис. 50 вкл).
Через час больная выправилась, приняв нормальное
положение.
Гринды и афалины очень упорно проявляют ре-
акцию «выталкивания» даже по отношению к уже
погибшим сородичам, детенышам, а иногда лишь к
какой-то части их тела и притом даже полуразложив-
шейся. Как-то в прибрежных водах Флориды встре-
тили двух афалин, занятых выталкиванием головы
сосунка, откушенной от туловища крупной акулой.
В другом случае у берегов Эверглейдс (Национальный
парк в США) наблюдали взрослую афалину, выталки-
вающую из воды дельфиненка около метра длиной,
погибшего три дня назад от покусов акулы.
С борта судна «Жеронимо», отлавливавшего дель-
финов для Калифорнийского океанариума, убили мо-
лодую гринду, которая утонула. Но сородичи вынесли
ее труп на поверхность: он лежал поперек, на голове
взрослой гринды, которая выталкивала его из воды
при выныривании. Спасательная привычка у дельфи-
нов обнаружена не только по отношению к сородичам,
но и к особям других видов и родов (межвидовая и
межродовая помощь).
Когда в Калифорнийском океанариуме крупный са-
мец тихоокеанского белобокого дельфина погиб от шо-
ка и пошел ко дну, взрослая, афалина-самка (из дру-
гого рода!) нырнула на дно, просунула свою морду под
тело погибшего и дважды с силой толкнула его вверх.
Межродовая помощь была оказана двумя тихооке-
анскими белобокими дельфинами самцу белокрылой
морской свиньи. Пойманный в море и помещенный в
океанариум, этот самец беспомощно погрузился на
дно, но к нему с обеих сторон пристроились два взрос-
лых дельфина, подсунули свои морды под его плавни-
8*
163
ки и дружно выталкивали к поверхности. Этим они
были заняты и на следующий день.
Как видно, реакцию помощи (выталкивания из во-
ды) у дельфинов вызывает не только сигнал бедствия:
иногда для этого бывает достаточно одного вида непо-
движного тела больного сородича. Но если поступает
одновременно и сигнал бедствия, и зрительное раздра-
жение, такая реакция резко усиливается.
Иногда раздражителем может стать предмет, лишь
отдаленно напоминающий безжизненное тело сороди-
ча. Например, в Суджукской лагуне одна афалина в
течение нескольких часов поддавала своей головой
плававший кусок мяса весом в 5—7 кг. Эта забава бы-
ла прервана подплывшей лодкой. Бывали случай, ког-
да дельфины толкали перед собой мертвых черепах,
обрубки дерева, матрац и т. п.
Давид и Мельба Колдуэллы в океанариуме пыта-
лись искусственно вызвать у малообученных афали-
ны, гринды и полосатого дельфина «выталкивательное
поведение», вводя в танк мертвого детеныша или его
резиновую модель. Но опыты не увенчались успехом:
дельфины пугались и подальше отходили от этих объ-
ектов.
Теперь, когда мы познакомились с инстинктом со-
хранения вида, нам не трудно будет объяснить и слу-
чаи спасания дельфинами человека, и странное явле-
ние среди китообразных — «массовое самоубийство»
(человек далеко не всегда может заставить дельфинов
проявить такую привычку).
В легендах древних греков и римлян, полинезийцев
и моарисов восхваляется склонность дельфинов спа-
сать тонущих людей. Человек уже давно знал, что
дельфины оказывают помощь утопающим при кораб-
лекрушениях и разгоняют акул, собирающихся вбли-
зи человека. Мы в начале книги приводили подобные
факты. Аналогичный случай произошел и во время
второй мировой войны: четверо американских летчи-
ков, сбитых в воздухе над Тихим океаном, спаслись на
надувном резиновом плоту. В этом им помогли дель-
фины, которые подталкивали плот к острову.
Нечто подобное, по сообщению печати, недавно про-
изошло с супругами Рэмбо из Флориды. Они отправи-
лись на морскую прогулку в корабельной лодке, но
попали в бедственное йоложение, так как перестал ра-
164
ботать мотор. Лодку отнесло далеко в море. На второй
день их окружила стая акул, но вскоре появились
дельфины и прогнали хищных рыб. Только на пятый
день изменившийся ветер подогнал лодку совсем близ-
ко к берегу, и дельфины покинули ее.
Летом 1959 года в Карибском море потерпело ава-
рию от взрыва судно «Рио Атаро». Люди, оказавшие-
ся за бортом, привлекли акул. Но вскоре большая стая
дельфинов набросилась на хищников и отогнала их.
Все эти случаи толкуются в широкой печати как
благородные и сознательные поступки дельфинов, про-
диктованные их «гуманностью» и дружелюбием к че-
ловеку.
Что касается отношения дельфинов к акулам, то
оно легко объяснимо: ведь эти хищные рыбы, загла-
тывающие дельфинят,— естественные враги мелких
китообразных. И не удивительна антипатия к ним,
наблюдавшаяся и в океанариумах: когда в бассейн
впускали тигровых акул, афалины их- убивали. (Сов-
местное существование в пределах одного танка в кон-
це концов примиряло рыб и млекопитающих. В океа-
нариумах китообразные выталкивают из воды даже
этих смертельных врагов, притом очень настойчиво.
В Калифорнийском океанариуме девять суток подряд
афалины «спасали» акулу длиной 150 см. В море же
эти отношения остаются резко враждебными.)
Но чем объяснить помощь дельфинов гибнущим
людям в воде?
Как мы говорили, склонность дельфинов к вы-
талкивательным действиям проявляется не только по
отношению к своим сородичам, но и к животным дру-
гого вида и даже к самым различным неживым объек-
там.
Ясно, что если таким внешним раздражителем ока-
жется тело не акулы или сородича, а тонущего чело-
века, то и в этом случае последуют те же выталкива-
тельные действия, которые могут и впрямь спасти че-
ловека. Но в основе этих действий лежит инстинкт, а
не разум. Только этим можно объяснить бессмыслен-
ность действий дельфинов:' «спасание» давно мертвых
сородичей или своих исконных врагов — акул.
Чтобы предупредить ложное толкование спасатель-
ных действий дельфинов, разберем один аналогичный
пример «спасания» человека медведем. В заметке
165
«Похороненный медведем» газета «Вечерняя Москва»
рассказала о югославском крестьянине Дана Конста-
новиче: застигнутый грозой в лесу, он спрятался под
дубом, и при вспышке молнии увидел вблизи хищ-
ника. Следующий удар молнии пришелся в этот дуб,
и человек потерял сознание. Он пришел в себя лишь
на седьмой день в 200 м от дуба, засыпанный землей.
Медведь поступил с человеком как с добычей, зарыв
ее в землю, что инстинктивно делают многие хищники.
Но, по широко распространенному мнению, зарыва-
ние в землю — лучшая помощь для пораженных мол-
нией. Поэтому описанный факт легко истолковать в
духе антропоморфизма: медведь знал, как оказать по-
мощь человеку, пораженному молнией.
На подобное ложное умозаключение и наталки-
вают поклонники «разума» дельфина: дельфин якобы
понимает, что воздух необходим для человека, и по-
тому выталкивает тонущего из воды.
Некоторые пытаются доказать разумность спаса-
тельных действий дельфинов тем, что они выталки-
вают человека к берегу, а не в открытое море, и ссы-
лаются на пример с Махмудом Вали. Однако стоит
взглянуть на карту, как все разъясняется: Суэцкий
залив имеет вид реки длиной 400 и шириной 30 км.
Поэтому в какую бы сторону животные ни толкали
матрац с человеком, он все равно рано или поздно
столкнулся бы с берегом. Помог, вероятно, в этом и
ветер.
Возможно, дельфины в подобных ситуациях тол-
кают обессиленных людей в сторону открытого моря
столь же часто, как и в сторону берега.
Рассмотрим другую особенность в поведении ки-
тообразных, так называемые «самоубийства», которые
в действительности объясняются инстинктом сохране-
ния вида.
Жители приустьевых районов бывают свидетелями
заходов китообразных в реки. Для усатых китов та-
кие визиты заканчиваются трагически. Так, напри-
мер, в 1950 году погиб молодой финвал в реке Енисей
в 400 км от устья: зверь изрезал свое брюхо о камени-
стые выступы дна.
Более безопасны заходы дельфинов, которые умеют
выходить обратно в море. В начале лета 1966 года
жители Западной Германии на протяжении месяца
166
были очевидцами своеобразного поединка белухи и
человека. Белый кит около 6 м длиной случайно за-
плыл из Арктики в Северное море и отсюда проник
в реку Рейн, поднялся против течения до Бонна (более
350 км от устья), а затем, избежав все ловушки, вер-
нулся в море. Многочисленные попытки поймать его
живым для Дуисбургского дельфинария не имели
успеха: белуха, несмотря на грязные воды Рейна, с
помощью гидролокатора точно распознавала опасность
и ловко обходила расставленные на ее пути сети;
снотворные же пули на нее не действовали.
В прошлом зарегистрированы заплывы белух в ре-
ки Хатанга на 700 км, Юкон (Аляска) на 1100 км,
Аргунь на 2000 км от устья. Заходят в реки и дель-
фины Черного моря. Так, весной 1978 года интересный
визит в устье реки Маныч (приток Дона) нанесла стай-
ка афалин. Животные задержались близ станицы Ма-
нычской, здесь резвились, играли, чувствовали себя
«как дома». Услышав с берега музыку из включенного
транзистора, «путешественники» подплыли совсем
близко, затем один из них поднялся на хвосте, раски-
нув грудные плавники, и в такт музыке стал «тан-
цевать» (известно, что к мелодичной музыке дельфи-
ны неравнодушны).
Искусно владея локатором, дельфины обычно за-
канчивают вояж и возвращаются в родную стихию без
особого ущерба для себя. Гораздо драматичнее об-
стоит дело при обсыханиях китообразных. Это зага-
дочное явление с древности интригует человека; киты
или дельфины в одиночку либо группами близко под-
ходят к морскому берегу, а затем, будучи вполне здо-
ровыми, оказываются на суше. Здесь они барахтают-
ся, бьют хвостом, впадают в панику и погибают через
разное время от теплового удара (при вскрытии обна-
руживают переполненные кровью сосуды мозга) или же
от удушья, если в их легкие попадает много воды.
«Нахлебаться» водой животные успевают, когда при-
бой кидает их тело по отмели, и они, перевернутые
на бок, должны дышать при погруженном дыхале.
Сильные удары хвостом лишь вгоняют их тело в песча-
ный грунт. Попав в беду, животные издают громкие
крики, а кашалоты голосят так, что их вопли слы-
шатся за несколько километров.
Из-за «просчета» на берегу могут оказаться не
167
только одиночки, но и группы в десятки или даже сот-
ни голов. Молва о подобных бедствиях быстро разно-
сится по всему миру.
Весьма впечатляющи такие катастрофы со стадами
кашалотов (рис. 51 вкл.). Во Франции животные
вошли в гавань Одьерн 13 марта 1784 года во время
прилива, при сильном юго-западном ветре. На песча-
ном берегу оказалось 32 кашалота, в большинстве сам-
ки. Звери подавали признаки жизни несколько дней,
и их рев слышался более чем за четыре километра.
Нечто подобное случалось с кашалотами на бере-
гах Тасмании (в 1911 году на песчаный пляж острова
Перкинс выбросилось 36 самцов и одна самка), Австра-
лии (3 февраля 1972 года на пляже Гуннамата на по-
лукилометровом участке обсохло 36 китов), Средйзем-
ного моря, Голландии, Мексики, Калифорнии, а также
в устьях рек Темзы (погибла группа из 12 самцов),
Эльбы (стадо в 17 голов), Луары (5 кашалотов) и т. д.
Наиболее крупное обсыхание кашалотов произошло
18 марта 1970 года на берегу Новой Зеландии в 3 ми-
лях от г. Гисборна на пляже Окита-Бич: во время
шторма за два часа на песчаный берег протяженностью
в несколько сотен метров было выброшено 46 самок
и 13 самцов, (рис. 53 вкл.). В обсохшем стаде отсут-
ствовали вполне взрослые самцы; в числе самок 10 бы-
ло неполовозрелых, 36 половозрелых и из них две с
новорожденными детенышами длиной 2,4 м (самка)
и 4,6 м (самец).
Известны случаи массовых обсыханий и с другими
видами. В газетах было напечатано сообщение из
Нью-Йорка о «самоубийстве» 150 китов-«пигмеев» (ма-
лых косаток), которые 11 января 1970 года выброси-
лись на песчаный берег Флориды близ г. Форт-Пирс.
Некоторых животных береговая охрана пыталась спа-
сти и отвозила в море, но они упорно возвращались на
прежнее место и снова выбрасывались на песок. Опе-
рация по спасению животных продолжалась с полу-
дня до ночи, но была безуспешной.
Год спустя (10 января 1971 г.) 29 китов, вероятно,
того же вида выбросились на берег необитаемого
островка Сан-Клементе неподалеку от Лос-Анджелеса.
Известно также, что 1 декабря 1966 года «совершило
самоубийство» стадо из 102 китов на берегу филип-
пинского острова Куйо (здесь, в том же самом месте
168
сорок лет назад погибло аналогичным образом боль-
шое стадо китов).
Чем можно объяснить столь странное поведение?
Толкования давались разные. В древности Плутарх
связывал проблему обсыхания с самоубийством живот-
ных. Этой причиной некоторые газеты нередко и сей-
час объясняют массовую гибель китов.
Много позднее после Плутарха описанное явление
связывали то с помешательством вожака, за которым
следовало стадо, то с болезнями, то с питанием на
мелководьях близ песчаных берегов в период шторма
или в ночное время, то с голодовками, вызываемыми
якобы непогодой и приводившими к истощению и обес-
силиванию животных, то с сильными ветрами, подго-
няющими пищу китообразных в опасную зону к бере-
гам, а иногда и с совершенно фантастическими причи-
нами.
В 1937 году мы изучали обстоятельства гибели ки-
тов на берегах Камчатки и установили, что обсыха-
нию способствуют разность колебания уровня воды
(значительные приливы — отливы, штормы, сильное
волнение) и легко размывающийся рельеф дна с барье-
рами, подводными песчаными косами, банками и т. п.
Стоит только киту, подошедшему с высоким валом
прибоя, коснуться отлогого дна, как последующие мел-
кие валы, нанося ил и песок, создают барьер, преодо-
леть который кит уже не в состоянии. Такие места в
Охотском море — устья рек Уды, Алы — петербург-
ский академик А. Ф. Миддендорф называл китовыми
ловушками. Опасные для китов места есть на берегах
почти всех материков.
Но все эти обстоятельства не отвечают на главный
вопрос: что нарушает ориентировку обладающих пре-
восходным локатором китообразных перед обсыханием
и какова основная причина катастроф, случающихся
с ними?
В поисках ответа на эти вопросы голландский ис-
следователь Ван Хил Дудок в 1962 году проанализи-
ровал 133 случая одиночных и стадных обсыханий
26 видов китообразных. Он выяснил типичную обста-
новку, в какой происходят обсыхания и когда подво-
дит эхолокационный аппарат.
Места, где наблюдались обсыхания, разбросаны в
различных частях земного шара. Это обычно низмен-
169
ные берега, подводные песчаные отмели, пляжи, га-
лечники или участки илистых наносов, мысы, высту-
пающие далеко в море. Обсыхание не обязательно (хо-
тя и чаще всего) происходит на песчаном пологом
грунте, но всегда в тех случаях, когда возникают
затруднения в навигации животных в связи с помеха-
ми для эхолокации в водной среде.
Обсыхания бывают чаще всего во время сильных
ветров и штормов: тогда поднимается со дна масса
воздушных пузырьков и частиц ила и песка. Они ухуд-
шают условия для эхолокации, так как мешают вос-
принимать отраженные сигналы. Немаловажная при-
чина «ослепления» гидролокатора — пологое песчаное
дно: падая на него, эхо-сигнал затухает, не возвраща-
ясь к животному (такое явление затухания называет-
ся акустическим клином). У китообразного нарушает-
ся ориентировка, и оно плывет навстречу опасному
месту. Еще одно обстоятельство имеет значение: когда
зубатые киты излучают низкочастотные щелканья, они
получают информационное эхо с дальней дистанции.
Приближаясь же к лоцируемому объекту — к добыче,
они излучают все более высокие частоты и концент-
рируют свое внимание на высокочастотном эхе; в этих
условиях легко пропускается информация о близком
расположении рокового берега, которую несут низкие
частоты. Поэтому во время кормежки опасность об-
сыхания резко возрастает.
Причиной осечки локатора могут быть и паразиты,
сильно понижающие восприимчивость слуха.
Наконец, возможно, что китообразные, будучи по-
томками наземных предков, бессознательно проявляют
свое влечение к спасительному берегу. В результате
возникает губительная конфликтная ситуация: с од-
ной стороны, суша дает необходимый кислород, а с
другой — приводит к тепловому удару при обсыха-
нии. У ослабленных одиночных дельфинов иногда про-
является неодолимая склонность приблизиться к са-
мому берегу на такую глубину, где можно было бы
выставиться из воды, опираясь на дно. Так, например,
в 1947 году мы замеряли температуру у больного
дельфина-белобочки в море возле самого берега на глу-
бине в полметра. При многократных попытках повер-
нуть животное головой в сторону моря, оно резко со-
противлялось этому и каждый раз поворачивалось к
170
берегу. Подобное же поведение заметил у погибающих
афалин Е. В. Романенко: дельфины подплывали уми-
рать на мелководье, упирались грудными плавниками
в дно и до самой агонии стояли на месте. Если их
отталкивали от берега, они делали круг на глубине и
вновь возвращались к берегу. Вероятно, такую реак-
цию у дельфинов следует истолковать как «зов пред-
ков», как поведение, обусловленное генетической па-
мятью о далеких временах жизни на суше.
Мотивы одиночных и групповых обсыханий неоди-
наковы. Случаи со стадами более сложны, чем с оди-
ночками, и вызываются они физиологическими или
зоопсихологическими причинами, физические же пово-
ды служат лишь начальным звеном в драме группо-
вого обсыхания.
Важнейшей причиной массового обсыхания явля-
ются уже знакомые нам инстинкт сохранения вида и
сигнал бедствия. Рассмотрим несколько случаев мас-
совой гибели китообразных на берегах.
Вот пример обсыхания семи обыкновенных дель-
финов в Новой Зеландии. Беда разразилась на скалах
острова Грейт-Барьер в заливе Хаураки, возле по-
селка Трайфена. Когда заметили животных, часть их
уже лежала на камнях, заливаемых водой, а осталь-
ные еще свободно плавали. Громкие сигналы разда-
вались даже в воздухе. Чтобы спасти животных, ку-
рортники, оказавшиеся поблизости, отвезли двух зве-
рёй подальше от берега и выпустили на глубоком
месте. Они надеялись, что плавающие у берега дельфи-
ны, услышав сигналы освобожденных, приплывут к
ним из опасного места сами, а остальные будут потом
перевезены людьми. Однако двое отвезенных тут же
вернулись назад к тем, что находились на мели. Все
попытки отогнать их от гибельного места не увенча-
лись успехом. Сигнал бедствия и инстинкт сохранения
вида не позволили животным покинуть своих сороди-
чей в беде. После беспокойной ночи и дня в живых
остались только два дельфина.
Эти примеры подсказывают, как происходит мас-
совое обсыхание и у других видов китообразных.
Показательно обсыхание малых косаток в 1934 го-
ду на острове Шри Ланка (Цейлон) в лагуне Мутур,
связанной с морем и поросшей мангровыми деревья-
ми. Группа из 97 животных зашла на мелкое место,
171
где было илистое дно и глубина иногда не превы-
шала метра. Здесь малые косатки проплавали несколь-
ко суток и погибли одна за другой. Почему же они
не смогли выйти в море? Очевидно, их локатор не мог
работать точно из-за мягкого и легко взмучиваемого
дна, поглощавшего эхо-сигналы. Поэтому несколько
животных оказались на мели, и последовавшие за этим
сигналы бедствия помешали остальным разыскать вы-
ход на чистую воду. Так катастрофа распространилась
на всю группу.
Стада малых косаток обсыхали во многих местах,
в том числе на берегах Шотландии — стадо в 127 го-
лов, на Чатэмских островах — несколько сотен сразу,
в Южной Африке близ Мамре — 200 особей, на ост-
рове Веланай близ Шри Ланки — 167 голов и т. д.
Самое массовое обсыхание крупных дельфинов про-
изошло в Аргентине 10 октября 1946 года: на песча-
ный пляж курортного городка Мар-дель-Плата выбро-
силось 835 малых косаток. Обсыхание развивалось
типичным образом и коснулось вначале немногих оди-
ночек, а затем охватило все стадо. Большинство груп-
пы составляли самки, некоторые с новорожденными
детенышами. Лишь немногие животные дожили до
конца следующего дня. Местные власти в санитар-
ных целях отбуксировали туши погибших далеко в
море.
Сходным образом обсыхают и стада гринд (рис. 52
вкл.). 14 марта 1955 года в район острова Уэстрей
(Оркнейские острова) зашло стадо гринд в мелкий
пролив глубиной около 5 м и почти пять суток пропла-
вало между двумя островами. С севера и с востока
можно было выйти к морю, но животные не сделали
этого, и в результате две группы — 16 и 37 голов —
очутились на отлогом каменистом берегу в 50 м одна
от другой. На протяжении километра вдоль линии бе-
рега погибло еще несколько одиночек.
Драма развивалась так. Во время непогоды в сре-
ду животные проникли в пролив с севера и оказались
с трех сторон окруженными островами. Очевидцы за-
метили в стаде кровоточащих животных, вероятно ра-
ненных незадолго до этого охотниками. Может быть, в
числе подранков был и тот самец, который в пятницу
осел на рифах со следами порезов. Сигналы бедствия
обсохшего самца, а возможно, и других ослабевших
172
подранков могли быть причиной задержки группы в
неблагополучном проливе. Вслед за тем на берегу за-
стряли еще несколько одиночек. Гринды возбужденно
плавали в опасном районе, отказываясь покинуть по-
павших в беду одиночек. Катастрофа разразилась в
ночь на воскресенье, во время поднявшегося шторма.
Среди животных возникла паника, сопровождавшаяся
пронзительным визгом и ужасающим шумом. Погибло
66 гринд.
Анализ всех этих случаев раскрывает главную
причину групповых обсыханий китообразных. Что же
происходит? Допустим, что одна особь из-за помех в
эхолокации попала на отмель и ей угрожает удушье.
В силу выработанных стадным образом жизни средств
общения — с помощью сигнализации она начинает по-
давать сигнал бедствия. Приняв этот сигнал, другие
животные устремляются на помощь в соответствии с
программой инстинкта сохранения вида, отработанной
естественным отбором. Но, подплыв на выручку, со-
родичи сами попадают под пагубное действие небла-
гоприятных условий района бедствия. В необычной,
тяжелейшей обстановке «спасатели», подгоняемые
инстинктом, с каждым метром приближаются к собст-
венной гибели. В результате бедствие одной особи, как
по цепной реакции, превращается в бедствие всей
группы: стремясь помочь первым пострадавшим, ста-
до в конце концов разделяет их печальную участь и
при соответствующих условиях целиком погибает на
берегу. Таким образом, «массовое самоубийство» ки-
тообразных объясняется «осечкой» гидролокатора и
проявлением инстинкта сохранения вида.
Но бывают среди китообразных «самоубийства» и
другого рода. Они наблюдались только в неволе, в
океанариумах у афалин, дельфинов-белобочек, гринд,,
клюворылых китов. Причины этого явления ничего
общего не имеют с инстинктом сохранения вида и сиг-
налами бедствия. Приведем несколько примеров.
В феврале 1956 года в овальный танк Калифорний-
ского океанариума поместили молодого настоящего
клюворыла длиной 340 см. Зверь вначале медленно
плавал по кругу вдоль стенок водоема, но потом начал
метаться и с большой скоростью наскочил на дальнюю
жесткую стенку, о которую сломал челюсть, и погиб.
Трагический случай имел место в сентябре 1969 года
173
в открытом бассейне биологической станции, куда при-
везли двух дельфинов-белобочек — Адама и Еву. Через
некоторое время Адама нашли мертвым, Ева на глазах
сотрудника станции В. М. Лекомцева «покончила с со-
бой» : она с налета дважды ударилась о каменные стен-
ки бассейна, разбила себе морду и через двадцать ми-
нут погибла.
Случай с афалиной Лэди наблюдали В. М. Белько-
вич и Н. Л. Крушинская в 1969 году. Сильно больной,
державшийся отдельно от группы сородичей, этот дель-
фин трижды (с интервалом в 15 и 60 мин) бросался на
стенку бассейна, причем в третий раз — со смертель-
ным исходом (смерть наступила через четверть часа
после удара).
Во всех описанных случаях причина бросков жи-
вотных на стенки бассейнов была, по-видимому, одной
и той же: состояние агонии. На промысле нам прихо-
дилось много раз видеть агонизирующих дельфинов на
палубе сейнера. Чаще всего агония сопровождалась
сильнейшими непроизвольными ударами хвоста. По-
добные взмахи хвоста в таком состоянии животных,
очевидно, и были причиной неуправляемых стреми-
тельных рывков дельфинов, приводивших к роковым
ударам о цементные стенки бассейнов на большой ско-
рости. Трактовка же подобного поведения' как предна-
меренное «самоубийство» в тяжелой ситуации не име-
ет ничего общего с действительной причиной — кон-
вульсивной работой движителя, ускоряющей гибель
обреченного животного.
Китообразным угрожают несчастья и другого по-
рядка — например, льды.
Глава XVII
БЕДСТВИЯ
СРЕДИ ЛЬДОВ
Морские млекопитающие заселили Мировой океан,
включая самые трудные для жизни полярные аквато-
рии, окутанные холодом и покрытые льдами. Море с
ледовым панцирем — смерть для животного, если оно
не может выставиться на поверхность для дыхания.
Спасение для них в том, чтобы пробить лед, сделать
продухи, поддержать полыньи, найти незамерзающие
«окна» в воде с помощью эхолокации и «голоса льдов»
либо своевременно покинуть опасные места. Массив-
ные гренландские киты спиной разламывают лед тол-
щиной до 20 см, а белухи — до 3—5 см. Редукция их
спинного плавника — несомненное приспособление к
хотя бы временному пребыванию среди льдов.
Дальше всех по направлению к Северному полю-
су заходят нарвалы (единороги). При интенсивном
образовании льда они, как и белухи, а иногда и вместе
с ними, надолго остаются в полыньях, не давая им за-
мерзать. Но даже эти «полярники» иногда испытыва-
ют трудности при смыкании льдов. Тогда самцы ору-
дуют бивнем. Спиральный орнамент на поверхности
бивня придает ему особую прочность для пробивания
льдов. Легко представить себе силу пробойного удара
двух-трехметрового костного стержня, если нарвал ве-
сом в 1—2 т, развив скорость до 30 км/ч, налетает
на льдину снизу. Вероятно, поэтому в одной коллек-
ции из 314 бивней, собранной эскимосами, 107 были
сломаны, при этом в зубном канале четырех бивней
оказалась костная пробка, образовавшаяся после по-
ломки. Через пробитые продухи дышит все стадо (сам-
ки бивней не имеют). У эскимосов такие «окна» во
175
льду называются «савссад», и когда в них находят
нарвалов, их без труда добывают прямо со льда.
В полыньях близ Гренландии местные жители по фон-
танам и далеко слышному реву иногда обнаруживают
затертых белух.
У белух кожа покрыта особым упругим слоем раз-
росшегося эпидермиса толщиной до 1 см, который пре-
дохраняет их от ран при разламывании льда. Однаж-
ды в Кандалакшской губе Белого моря торосами зажа-
ло стадо белух. Льды сели на отмели и не дали
животным уйти под водой, заставив их зимовать в по-
лынье. Белухи все время поддерживали продух и спи-
ной проламывали молодой лед. Однако этот природ*
ный опыт не был доведен до конца, так как зверей
перебили.
Ледовый плен — явление нередкое в холодных во-
дах. Кромка плавучих льдов привлекает многих уса-
тых китов, которые летом здесь нагуливают жир, ис-
пользуя огромные скопления ракообразных. Глубоко в
плавучие льды Антарктики проникают синие киты и
малые полосатики, а следом за ними и косатки. Не-
сколько лет назад в одной полынье зимовали 60 ко-
саток, которые были соблазнены расположенной по
соседству залежкой тюленей-крабоедов. Во льдах Ан-
тарктики малые полосатики собираются группами и
иногда попадают в труднопреодолимое широкое ледо-
вое кольцо. Тогда они поддерживают полынью, часто
выныривая либо плавая с вертикально выставленной
мордой выше уровня глаз и нередко к себе подпускают
людей. В Англии даже образовалось общество «Похло-
пай кита», в которое входят счастливцы, потрогавшие
живого гиганта. Принимая вертикальные позы, малые
полосатики, видимо, удовлетворяют свое любопытство,
снижают возможность травм и порезов о края льдин и
находят наиболее удобный способ выныривания в зам-
кнутом пространстве. В трудных ситуациях эти киты
пользуются проходами, оставляемыми во льдах кораб-
лями и движущимися айсбергами.
В борьбе со льдами обессиливают многие (особенно
мелкие) дельфины в северных водах на Балтике, в мо-
рях Дальнего Востока и даже в Азовском море.
Ледяной покров представляет, например, для мор-
ской свиньи, серьезную угрозу. Поэтому у восточных
176
берегов Гренландии она бывает лишь в годы с мини-
мальным количеством льдов. Массовая гибель этого
дельфина в суровую зиму отмечена в Остерсионе (Бал-
тийское море), когда, застигнутые быстрым образова-
нием льда, многие свиньи задохнулись; трупы их бы-
ли выброшены на острове Фаре, близ острова Готланд.
По той же причине около острова Борнхольм в течение
1929 года погибло так много свиней, что небольшие
суда в течение шести недель выловили несколько со-
тен мертвых дельфинов. Обычно не замерзающие в
этом районе воды привлекают на зиму животных из
смежных, замерзающих частей Балтики.
Вот что произошло в Азовском море, по описанию
корреспондентов АПН Л. Гранкова и Л. Лучкина:
«...Морской буксир «Калгуев», следовавший из
Жданова в Керчь, попал в полосу сплошного льда.
Пришлось ждать ледокол. Неожиданно в образовав-
шейся у борта судна полынье моряки увидели окро-
вавленного дельфина (морскую свинью.— А. Т.). Чле-
ны экипажа советского судна немедленно подняли на
борт пострадавшего дельфина и поместили его в бас-
сейн из брезента, куда поступала свежая морская во-
да. Прошло несколько минут, и у борта судна появил-
ся второй обессиленный дельфин. Животное само под-
плыло к спустившемуся на лед матросу и дало взять
себя на руки. Оба дельфина были сильно изранены,
они провели длительное время подо льдом в поисках
полыньи. Более трех часов пробыли дельфины на бор-
ту «Калгуева». Их раны были промыты и обработаны
стрептоцидом. А когда судно вышло из льдов на чи-
стую воду, моряки расстались со своими гостями».
О другом случае недавно поведал корреспондент
«Правды» В. Рябчиков: «Ночью в районе острова Иту-
руп подул сильный ветер и погнал льды на берег. На
рассвете сторож Курильского рыбзавода увидел, как
на льдинах барахтаются какие-то черные фигуры. Ока-
залось, что это были дельфины. Целое стадо, застиг-
нутое врасплох. Льды прижали животных ко дну у бе-
рега, и они никак не могли выбраться из западни.
На помощь дельфинам поспешили рыбаки местно-
го отделения колхоза «Заветы Ильича». Они взяли
трактор, лошадь, двое саней и двинулись в путь.
С трудом добравшись до места, люди принялись ше-
стами расталкивать льдины, освобождая дельфинов
177
из плена. Это было не так-то легко. Льдины оказались
тонкими и неустойчивыми. Всего из ледовых объя-
тий удалось вызволить 15 дельфинов. Вначале их вре-
менно поместили под пирсом, где была чистая вода, а
затем, как только позволили обстоятельства, привезли
в бухту Оля и выпустили в родную стихию».
Китообразные, попадающие в ледовый плен, силь-
но худеют, теряют в весе. Канадский исследователь
Фриман Мильтон зимой 1966/67 года наблюдал за ста-
дом белух в 150—200 голов, запертых тяжелыми льда-
ми в проливе Джонса в Баффиновом заливе. Полоса
сплошного льда без разводий и трещин, отделявшая бе-
лух от свободной воды, достигала в ширину 46—55 км.
Площадь полыньи постепенно уменьшалась, и на теле
узников в январе проступили следы истощения жиро-
вых ресурсов. Это хорошо было заметно в грудном от-
деле, где появились складки кожи. Однако хвостовые
лопасти оставались гладкими и упругими. Причина
этого заключалась, видимо, в нагнетанйи значительно-
го количества крови в лопасти хвоста в связи с его
локомоторной и терморегуляторной функциями.
Как видно, льды представляют неодинаковую опас-
ность для разных видов китообразных: наибольшую —
для заходящих гостей, не приспособленных пережи-
вать ледяные ловушки, наименьшую — для постоян-
ных обитателей северных морей — нарвалов, белух,
гренландских китов. Виды этой группы даже рожда-
ют детенышей в полыньях. Ледовый плен хотя и труд-
ное для них событие, но все же редко имеет губитель-
ные последствия. Льды для этих животных столь же
естественная среда, как и открытое море. Есть много
доказательств, что представители подсемейства белу-
ховых (рис. 54 вкл.) регулярно зимуют во льдах арк-
тических морей, и, вероятно, справедливо утверждение
А. В. Яблокова о массовых зимовках белух в районе
Великой Сибирской полыньи — к северу от Новоси-
бирских островов, Новой и Северной Земли.
Глава XVIII
СТРАННЫЕ
ПОПУТЧИКИ
ИЛИ БЕЗДУМНЫЕ
ЛОЦМАНЫ
С давних пор привлекает к себе внимание одна
странная малоизученная привычка китообразных —
следовать за идущими судами. Такое движение длится
или несколько минут, редко до получаса, или весьма
продолжительное время.
Впервом случае подобное поведение наблюда-
ется у мелких китообразных — стремительных дель-
финов: они с ходу пристраиваются к идущему судну,
резвятся чаще всего возле его носа, играют, иногда
двигаются строем и столь же внезапно покидают ко-
рабль (рис. 55 вкл.). Как мы уже видели, дельфины
очень любопытны и любят оседлывать корабельные
волны. х
Настоящую сенсацию вызвал своим поведением се-
рый дельфин в водах Новой Зеландии. В течение
24 лет (с 1888 по 1912 г.) его регулярно наблюдали
близ пароходов, курсировавших в проливе Кука между
портами Веллингтон и Нельсон. Он встречал и сопро-
вождал корабли в любое время суток, но в строго огра-
ниченном участке маршрута — в заливе Адмиралти,
на отрезке в 11 км, затрачивая на это в среднем по
20 мин. Как только судно, идущее в порт Нельсон,
достигало входа в пролив Пелорус, дельфин прекра-
щал кормежку, крупными прыжками устремлялся к
кораблю и сопровождал его до острова Дюрвиля. Встреч-
ные суда, следующие в порт Веллингтон, дельфин
экскортировал от острова Дюрвиля до пролива Пе-
лорус.
Не страшась шума и криков восторженных пасса-
жиров, дельфин начинал резвиться возле носа и бор-
179
тов корабля, оседлывая идущие от него волны. Если
одновременно шли два судна, для преследования выби-
ралось более быстроходное. Животное, казалось, без
всякого напряжения стремительно плыло возле судна,
делавшего 28 км/ч. Пелорус-Джек (дельфина назвали
по имени пролива) стал очень популярным среди жи-
телей острова, моряков и многочисленных туристов,
приезжавших сюда из разных концов земного шара.
Чтобы сохранить это животное, губернатор Новой Зе-
ландии 26 сентября 1904 года издал специальный
указ, запрещающий под угрозой штрафа «покушаться
на рыбоподобное млекопитающее — серого дельфина,
сопровождающего корабли в проливе Кука». Дельфин
достигал почти 4 м длины (измерение сделано по дли-
не лодки, когда он двигался рядом). Его серая окраска
с возрастом светлела.* Знаменитого дельфина убили
20 апреля 1912 года норвежские китобои. На набереж-
ной города Веллингтона люди поставили ему па-
мятник.
Чтобы восстановить черты поведения Пелорус-Дже-
ка, журналист Антони Алперс обратился к докумен-
там и газетам того времени, опрашивал местных жи-
телей и переписывался с очевидцами. В их письмах
отмечалась интересная деталь: Пелорус-Джек, двига-
ясь рядом с кораблями, терся своим телом об их борта
и почесывался о киль. Возможно, это и было причи-
ной его регулярного подхода к судам.
Проанализировав и ряд других наблюдений, можно
предположить, что китообразных привлекает к движу-
щимся судам возможность соприкасаться своим телом
с их скользким корпусом. Приятное кожное ощуще-
ние, получаемое при этом, вероятно, удерживает жи-
вотных возле идущего корабля. Воздействие на кож-
ные покровы — сильнейший стимул-раздражитель.
Как известно, в океанариумах дельфины положитель-
но реагируют на поглаживание их тела (для почесыва-
ния на дне океанариумов им устанавливают даже спе-
циальные щетки). Дрессировщики, применяющие для
подкрепления условных рефлексов поглаживание и по-
чесывание кожи дельфинов, быстро получают хоро-
шие результаты.
Аналогичные кожные ощущения, возможно, игра-
ют решающую роль при выработке условных рефлек-
сов и у китообразных, сопровождающих движущиеся
180
суда. При движении рядом с кораблями такие живот-
ные часто подкрепляют рефлекс, потирая свое тело о
борта судов и испытывая щекочущее действие водных
струй, возникающих при ходе судна. Эти же причины,
видимо, объясняют и «лоцманское» поведение серого
дельфина Пелорус-Джека. Кстати, в то время на Новой
Зеландии Пелорус-Джека никто не рассматривал как
лоцмана, а качество «проводить корабли через опас-
ный пролив со знанием дела и спасать от гибели ты-
сячи людей» ему приписали лишь недавно.
Итак, главная причина кратковременного движения
мелких китообразных с судами — кожные раздраже-
ния и сильно развитый ориентировочный рефлекс (про-
явление любопытства), а в некоторых случаях и склон-
ность к оседлыванию корабельной волны (см. IX гла-
ву).
Остается добавить, что жившие на воле дельфины
Опо-Джек и Чарли стали ручными также под влия-
нием аналогичных причин. Поощрение подплывающе-
го к купальщикам дельфина путем поглаживания его
тела могло постепенно усиливать эту привычку, пока
он не стал ручным.
Во втором случае наблюдаются факты уди-
вительно стойкого и загадочного сопровождения ко-
раблей крупными китообразными. Например, группа
горбачей сопровождала один пароход от мыса Горн
до Ливерпуля, а одиночка-горбач шел за судном «Чел-
ленджер» несколько дней. Еще один горбач упорно
двигался с кораблем от острова Св. Павла до берегов
Бразилии (свыше 1000 км). Рекорд побил крупнейший
из китов: в ноябре 1851 года парусный корабль «Пли-
мут» проходил мимо нескольких синих китов. Замети-
ли, как один из них отделился от сородичей и при-
строился к кораблю, а затем сопровождал его в тече-
ние 24 суток. Члены экипажа, опасаясь повреждения
судна огромным животным, старались отогнать его:
они выкачивали на него затхлую воду из трюма, бро-
сали в него бутылки, поленья, горящий уголь... но кит
не отставал, нырял под корабль и держался так близ-
ко от бортов, что брызги его фонтанов залетали в от-
крытые иллюминаторы. Неоднократно менявшаяся по-
года не могла приостановить движение назойливого
попутчика, и лишь когда корабль стал подходить к
берегу, кит исчез.
181
Длительное и упорное следование китов за кораб*
лями мы рассматриваем как своеобразный этологиче-
ский рецидив давно минувшего «младенчества», ког-
да после рождения у детеныша проявлялась реакция
следования с матерью (см. V главу). Тогда детеныш
испытывал положительные ощущения от «оседлыва-
ния волны» и «эшелонированного плавания» (глава
IX), что позволяло ему плыть рядом с родителем, не
затрачивая усилий. Полученный в детстве такой экс-
тероцептивный эффект (комплекс кожно-мускульных,
зрительных и других ощущений) запечатлялся, а мно-
го позже, уже в зрелом возрасте, проявлялся (вероят-
но, при стечении напоминающих обстоятельств) в ви-
де реакции следования за кораблями. Иногда это вы-
глядело как многодневное неодолимое движение кита
и судна.
Как видим, реакция следования бывает довольно
продолжительной. В связи с этим возникает вопрос:
есть ли у китообразных сон?
Глава XIX
СПЯТ ли киты?
Природа мудро решила проблему сна для разных
групп животного мира. Оказывается, спят даже пти-
цы на лету. Например, аисты дремлют в полете. Когда
утомится один, он перелетает в центр стаи и закрыва-
ет глаза на десять минут, не теряя направления и вы-
соты полета. Отдохнувший, он уступает место другому
аисту, а сам перемещается в хвост или голову стаи.
Это выяснили орнитологи с помощью датчиков, при-
крепляемых к летящим птицам. Информацию о рабо-
те сердца, крыльев и других органов датчики переда-
вали на скользивший неподалеку от летящих птиц
планер.
А спят ли киты? Когда, как и насколько глубоко,
утрачивают ли при этом чуткость и сознание, пробуж-
даются ли при каждом дыхательном акте и действу-
ют ли на них снотворные? Могут ли они почивать на
ходу в такой опасной для легочного дыхания среде,
как вода?
Наблюдения в море над спящими китообразными
очень редки и отрывочны (рис. 56 вкл.). На основе
этих наблюдений создавались противоречивые гипоте-
зы. Например, одни полагали, что китообразные вооб-
ще не спят, другие считали, что спят только на по-
верхности воды, а третьи утверждали, что сон бывает
лишь под водой.
В настоящее время кое-что начинает проясняться в
результате наблюдений и экспериментов в океанариу-
мах, согласующихся с представлением, что сон — это
особым образом организованная деятельность мозга.
Ночное поведение афалины, по наблюдениям
183
В. Ф. Флейнигена, состоит из периодов ясного бодр-
ствования, стереотипического плавания по кругу с ко-
ротким (на 20—30 с) закрыванием глаз и неподвижно-
го зависания с признаками сна.
Спит дельфин на поверхности или близ поверхности
воды (на глубине не более полуметра), с опущенным
вниз расслабленным хвостовым стеблем. Через каждые
0,5—2 мин слабый удар хвоста медленно доставляет
дельфина к поверхности (поскольку за время дыха-
тельной паузы он успевает чуть погрузиться), и тут
же в силу рефлекса выныривания происходит дыха-
тельный акт. Оба глаза, а иногда только один, быва-
ют закрытыми. Аналогичным образом спят у поверх-
ности и другие виды китообразных, различны лишь
продолжительность и глубина опускания между акта-
ми дыхания. Некоторые дельфины в неволе спят семь-
ями, касаясь друг друга грудными плавниками. Дете-
ныш дремлет возле хвостового стебля или спинного
плавника матери, они поднимаются для дыхания одно-
временно. Частота дыхания во время сна более или
менее равномерна, но в 2—3 раза реже, чем у бодрст-
вующих особей.
Это сон приповерхностный, пассивный. В 1967 году
биолог США Д. Маккормик описал у афалины и ко-
роткоголового дельфина другой тип сна — придонный,
активный: дельфин погружается на дно бассейна, где
выдерживает удлиненную дыхательную паузу (у афа-
лины около 4 мин), затем поднимается на поверхность,
быстро дышит несколько раз (афалина до 10—20 раз)
и снова надолго опускается ко дну.
Таким образом, у спящих дельфинов могут быть
две фазы сна:
1) глубокий (приповерхностный) сон, при котором
исчезает реакция на изменение обстановки, пропада-
ет брадикардия (не замечается урежения пульса во
время дыхательных пауз) и смыкаются веки; синх-
ронные движения хвостового плавника, совпадающие
с дыхательным актом, удерживают животное на по-
верхности воды;
2) легкий (придонный) сон — в виде дремоты, с
одним или обоими открытыми глазами; животные дол-
го лежат у дна, а вынырнув, совершают несколько ды-
хательных актов и снова залегают на дне. Физиолог
США Сэм Риджуэй наблюдал придонный сон в усло-
184
виях недостаточной солености воды. Но, видимо, он
бывает также при низкой (отрицательной) температуре
воздуха и охлажденной поверхности воды, как это бы-
ло в случае с афалинами Милкой и Андреем во время
зимовки в открытом бассейне биостанции.
Тот факт, что дельфины спят с одним открытым
глазом, дал повод Д. Лилли предположить, что у ки-
тообразных спит лишь одно полушарие мозга, а дру-
гое бодрствует. Экспериментально доказать поочеред-
ное развитие сна в двух полушариях и раскрыть свое-
образную регуляцию бодрствования и сна у этих жи-
вотных удалось советским ученым А. Я. Супину и
Л. М. Мухаметову. В 1975 году они провели электро-
физиологическое исследование мозга на семи свободно
плававших в маленьком бассейне афалинах, которым
были вживлены электроды для регистрации физиоло-
гических показателей деятельности мозга (электро-
энцефалограммы), глазодвигательных мышц (электро-
миограммы), сердца (электрокардиограммы) и легких
(пневмограммы). Эти данные на подопытных живот-
ных выявили стадии бодрствования и «медленного» сна
(глубокого, легкого и дремоты), но не удалось конста-
тировать быстрого (парадоксального) сна, сопровожда-
ющегося сновидениями. Подтвердились также ранее вы-
сказанные в литературе мнения, что дыхательный акт
дельфина не требует пробуждения.
На основании фактов многодневного следования ки<
тов за кораблями можно думать, что китообразные
спят на ходу и без сновидений (т. е. без парадоксаль-
ного сна), иначе они теряли бы судно. Бодрствование
же одного полушария во время сна позволяет нервной
системе нести дозорную службу, а следовательно, и
обеспечивать безошибочное и долговременное движе-
ние китов за кораблями. О том же, видимо, свидетель-
ствует и реакция неотрывного следования новорож-
денного за матерью в течение первых дней.
У большинства спящих наземных млекопитающих
сторожевую функцию несет слух, а у китообразных к
этому подключается и зрение. В опытах А. Я. Супина
и Л. М. Мухаметова, плавающая с одним открытым
глазом афалина пробуждалась, если в поле ее зрения
появлялся новый предмет.
В силу рефлексов выныривания и сторожевой дея-
тельности мозга дельфины трудно поддаются наркозу*
185
Еще недавно думали, что усыпить их вообще невоз-
можно. Но американские исследователи С. Риджуэй и
Дж. Маккормик разработали методику анестезирова-
ния дельфинов для полостных операций и электрофи-
зиологических экспериментов над мозгом. Дельфина
закрепляют в особый станок, вставляют ему в дыхало
и трахею дыхательную трубку, а затем дают вдыхать
под небольшим давлением закись азота (N2O). При
анестезии у дельфинов снижается артериальное давле-
ние— со 120—130 мм ртутного столба до 115 мм —
и исчезают хорошо заметные рефлексы: мигание век
при дотрагивании до роговицы, следящее движение
глаз за движением пальцев близ морды, подергивание
хвостом перед дыхательным актом. Анестезия быстро
проходила, если дельфинам давали вдыхать атмосфер-
ный воздух с кислородом.
Разработанная методика усыпления дельфина дала
многое для науки и способствовала превращению его
в новое лабораторное животное.
Глава XX
ДЕЛЬФИНЫ И НАУКА
За последние два десятилетия дельфины стали
объектом пристального изучения.
Как новое лабораторное животное, дельфин очень
перспективен: на нем можно изучать важные вопро-
сы физиологии, имеющие прямое отношение к меди-
цине, технике, гидродинамике, бионике и т. д. По сво-
ему миролюбивому характеру, высокому уровню ор-
ганизации, исключительной терпеливости к разного ро-
да опытам (в том числе к электростимуляции мозга),
по замечательным акустическим, эхолокационным, ги-
дродинамическим свойствам и многим другим ценным
качествам дельфин выдвигается на одно из первых
мест в списке экспериментальных животных. Есть уве-
ренность, что дельфин сыграет не меньшую роль в
будущих исследованиях, чем в свое время сыграли
«мученицы'науки» — лягушка и собака. На дельфинах
сделано много открытий в разных областях знаний.
В настоящее время на этих животных активно раз-
рабатываются проблемы гидробионики, среди которых
выделяются два главных направления: 1) гидродина-
мическое, включая изучение кожи с целью создания
обшивок для скоростных судов; 2) эхолокационное.
Первое, гидродинамическое, направление,
прежде всего строение и свойства кожного покрова,
частично рассматривалось в IX главе.
Как мы уже говорили, в ходе эволюции у китооб-
разных, передвигающихся в плотной водной среде, об-
разовались легко обтекаемая форма тела, упругий кож-
ный покров, способный задерживать появление турбу-
лентных пульсаций в пограничном слое воды, сформи-
187
ровался своеобразный локомоторный орган — хвосто-
вой плавник — эффективный машущий движитель»
приводимый в действие сильной мускулатурой.
Однако проблема передвижения китообразных с вы-
сокой скоростью была решена природой только тогда,
когда появились два важнейших приспособления —
«саморегуляция гидроупругости плавников» и «само-
настройка кожи к быстрому плаванию путем демпфи-
рования». Саморегуляция упругости плавников — не-
известное ранее явление — была открыта советскими
учеными С. В. Першиным, А. С. Соколовым и
А. Г. Томилиным в 1968 году на основе разносторон-
них исследований, проведенных на пяти видах дель-
финов и трех видах китов. Исследователи установили,
что в зависимости от скорости плавания у китообраз-
ных происходит автоматическое (рефлекторное) регу-
лирование упругости плавников (особенно хвостового)
при помощи специфических комплексных артерио-ве-
нозных сосудов, общего распределительного узла кро-
веносной системы и особой структуры покровных тка-
ней хвостового плавника, включающих покрытие из
сухожильных тяжей.
Хотя комплексные артерио-венозные сосуды в плав-
никах дельфинов были обнаружены одним из соавто-
ров открытия еще около 30 лет назад, физиологиче-
ское назначение этих сосудов было установлено лишь
в последние годы. Саморегулирование гидроупругости
плавников позволяет китообразным двигаться с огром-
ной скоростью и догонять юрких, скоростных рыб и
головоногих моллюсков, обеспечивает высокую манев-
ренность, возможность совершать высокие прыжки,
внезапные рывки, мгновенные остановки при стреми-
тельном ходе и т. п. Экстремальный режим плавания
китообразных создал необходимость изменения и ре-
гулирования упругих свойств плавников и прежде все-
го хвостового плавника. В плавниках китообразных
происходят синхронно два явления — и терморегуля-
ция, и саморегуляция упругости. С одной стороны,
чем быстрее китообразное плывет и энергичнее рабо-
тает хвостом, тем выше жесткость плавников, возра-
стающая за счет притока крови к ним. С другой сто-
роны, плавники нагреваются тем интенсивнее, чем
больше притекает к ним крови, и тем самым эффек-
тивнее отдают в наружную среду то избыточное тепло,
188
которое возникает при усиленной работе хвоста
(рис. 22).
Другой секрет китообразных заключается в реф-
лекторно регулируемом свойстве самонастройки их ко-
жи — гасить турбулентные пульсации, возникающие в
пограничном слое, и тем самым снижать гидродинами-
ческое сопротивление.
Дельфин при движении тонко использует в раз-
личных сочетаниях средства уменьшения гидродина-
мического сопротивления воды и управления погра-
ничным слоем на поверхности своего тела. Он может
улавливать гидродинамическое давление поля движу-
щихся судов.
Второе направление гидробионических исследова-
ний китообразных — эхолокационное. Дельфи-
ны отлично ориентируются в водной среде с помощью
превосходно развитого у них гидролокатора. В звуко-
вой ориентации немаловажное значение имеет направ-
ленность посылаемых ими звуков.
В 1957 году зоологи США Кеннет Норрис и В. Мак-
Фарлан, описывая новый вид морской свиньи из Ка-
лифорнии, заметили, что лобная поверхность ее чере-
па подобна форме параболического зеркала, а мягкие
части над челюстными костями напоминают линзу.
Возникла мысль, что голова морской свиньи может
концентрировать звуки, излученные воздушными меш-
ками, как рефлектор и фокусирующий аппарат. Так
зародилась гипотеза звукового прожектора и акусти-
ческой линзы дельфинов, красиво объясняющая точ-
ность, прицельность и дальность эхолокации. В даль-
нейшей разработке этой гипотезы принимали участие
ученые разных стран.
Как мы говорили, эхолоцирующий дельфин, при-
ближаясь к добыче, покачивает головой, как бы наце-
ливаясь на рыбу своим звуковым лучом, т. е. звуковые
волны дельфины посылают направленно.
Испытания показали, что с увеличением частоты
акустического излучения направленность звуков, обу-
словливаемая вогнутой передней поверхностью мозго-
вой части черепа и мягкими тканями головы, четко
возрастает, и звуковое поле суживается (рис. 23, 24).
Такую закономерность на продельфине и афалине
продемонстрировали, например, американские иссле-
дователи Эванс, Сатерленд и Бейл.
189
Рис. 22. Хвостовой плавник дельфина — это и локомоторный ор-
ган с переменной упругостью и теплорегулятор:
А — поперечный разрез через хвостовую лопасть
дельфина: 1 — кожа; 2 — слой сухожильных тканей;
3 — гиподерма с коллагеновыми волокнами; 4 — оди-
ночная вена; 5 — магистральный комплексный сосуд;
Б — комплексный сосуд: 6 — центральная толсто-
стенная артерия; 7 — тонкостенные вены, окружающие
артерию;
В — хвостовая лопасть с магистральным типом ветв-
ления сосудистой сети: 5 — магистральный комплексный
сосуд; 8 — распределительный узел потока крови; 9 —
последние хвостовые позвонки; 10 — выемка хвостового
плавника
190
Основную роль концентратора звуков выполняет че-
реп, а дополнительную — мягкие ткани головы. Но и
носовые мешки помогают в этом, поскольку они дей-
ствуют как отражательные поверхности. В направлен-
ные. 23. Диаграмма направленности звуков, излучаемых
из трупа вертящегося продельфина в вертикаль-
ном а и горизонтальном в плане: штриховые
линии — 20 кГц, штрихпунктирные линии —
40 кГц, сплошные линии — 60 кГц. По В. Эван-
су, В. Сатерленду и Р. Бейлу
Рис. 24. Диаграмма направленности звуков, излучаемых
из черепа афалины в вертикальном а и гори-
зонтальном в плане: штриховые линии—40 кГц,
штрих-пунктирные линии — 50 кГц, сплошные
линии — 70 кГц. По В. Эвансу, В. Сатерленду
и Р. Бейлу
191
ности сигналов, видимо, и таится секрет «ультразвуко-
вого разглядывания» дельфинами предметов на раз-
ных расстояниях. В связи со сменой звукового поля
дельфины могут даже менять форму жировой подуш-
ки. Ученому США Рэю в Нью-Йоркском аквариуме
удалось сфотографировать белуху в момент фокуси-
ровки звука: жировая подушка на ее голове была
нормально-округлой или заостренной формы (рис. 25).
У зубатых китов формирование эхолокационного
аппарата и новая роль черепа, как акустического ре-
флектора, отразились на неравномерном развитии ко-
стей на левой и правой стороне головы: асимметрия,
по мнению биологов Кеннета Норриса и Форреста Ву-
да, резче всего затронула область ноздрей и тот квад-
рант черепа, где генерируются звуки. Носовые про-
ходы, как это хорошо видно на черепе карликового ка-
шалота, специализируются: один — как воздухонос-
ный путь, а другой для выполнения звукосигнальной
функции. Так была объяснена асимметрия черепа зу-
батых китов, долго остававшаяся тайной для биоло-
гов мира.
Зубатые киты — это адаптивно измененные лока-
торщики, чей череп с мягкими частями, звукосигналь-
ный аппарат и орган слуха приспособились для гене-
рации звуков, их посылки, восприятия эха и анализа.
Все это наложило глубокий отпечаток на особенности
биологии, анатомии и физиологии группы.
В последние годы в США изучение эхолокации и
Рис. 25. Изменение формы лобно-носовой (жировой) подушки бе-
лухи: до (А) и в момент (Б) фокусировки. По К. Рэю
192
сигнализации дельфинов начали проводить прямо в
море на предварительно выдрессированных живот-
ных, послушно возвращающихся по команде экспери-
ментатора. Это имеет большие преимущества,, так как
в океанариумах и небольших водоемах стенки отра-
жают звуки и мешают точности замеров. Только рабо-
та в естественной обстановке позволит окончательно
выяснить дальность и точность локации дельфинов, а
также влияние глубины погружения на работу их со-
нара.
Недавно канадские исследователи К. Левенсон и
В. Лиапли предложили пользоваться акустическими
буями для выяснения распространения и относитель-
ной численности китообразных в какой-либо заданной
акватории. Этим ученым удалось получить данные о
горбачах Карибского моря с помощью сонарных бу-
ев, расставленных на морской поверхности с самоле-
та, которые записывали голоса китов.
Глава XXI
СИГНАЛЫ
ДАЛЬНЕГО ДЕЙСТВИЯ
И ЗВУКОВЫЕ МАЯКИ
В ОКЕАНЕ
До сих пор неясно — владеют ли усатые киты эхо-
локацией, а между тем именно они совершают огром-
ные сезонные миграции, осуществление которых воз-
можно лишь при отличной ориентации в океане. Голос
усатых китов беднее, чем у дельфинов.
Для лова малоподвижных рачков, скученных на
большом пространстве, усатым китам не требуется
столь точный «нащупывающий аппарат», какой ну-
жен зубатым китам для отыскания очень подвижной
и одиночной добычи. Осязательные волоски усатых
китов доставляют им информацию о пище на ближнем
расстоянии, однако это не доказывает, что они лише-
ны эхолокации вообще (см. XIII главу). Если только
они обладают эхолокацией, то пользование звуками
низких и средних частот дает им некоторое преимуще-
ство перед зубатыми китами в том, что позволяет по-
сылать сигналы на большие расстояния. Как известно,
дальность распространения звуков средних частот под
водой определяется в 15—20 км, а ультразвуков —
лишь в 3—5 км. Вероятно, усатым китам для даль-
ней локации низкочастотные звуки подходят боль-
ше, чем высокочастотные. Может быть, не случайно
стоны горбачей ассоциируются с миграциями, когда
особенно важна дальняя локация.
Мигрируют киты на 5—10 тыс. км. Как они ориен-
тируются в океане на таком громадном пути, возвра-
щаясь из года в год в одни и те же места? Это пока не
изучено. Очевидно, есть какие-то внешние раздражи-
тели, которые помогают усатым китам выходить к
цели.
194
Но что это за раздражители и как информация о
них поступает к мигрантам? Можно думать, что глав-
ным анализатором, улавливающим информацию для
правильного движения к цели, служит орган слуха.
Характер и рельеф дна, впадины, подводные хребты,
близость берега и другие особенности среды, вероят-
но, могут иметь значение ориентиров на далеком пу-
ти животных. На эти ориентиры у мигрирующих ки-
тов вырабатываются соответствующие рефлексы, игра-
ющие решающую роль при выборе нужного направле-
ния. Это и приводит китов каждый год в одни и
те же области, районы и даже бухты. Высокоразвитая
нервная система китообразных и их способность бы-
стро вырабатывать условные рефлексы вполне согла-
суются с таким представлением об ориентировке во
время миграций.
В акустике есть область, которая, возможно, свя-
зана с дальней ориентацией китов. Это подводные
звуковые каналы в океане, обнаруженные нашими
учеными Л. М, Бреховских и Л. Д. Розенбергом в
1946 году. По таким каналам-волноводам звуки, осо-
бенно низкочастотные и инфразвуки, проходят, не уга-
сая, тысячи километров. Например, взрыв динамита,
произведенный в водах Австралии, был услышан че-
рез несколько часов у Бермудских островов. В канале
создаются особо благоприятные условия для распро-
странения звуковых волн, отражающихся и от верхне-
го и от нижнего слоев воды разной плотности. Под-
водный звуковой канал образуется вследствие того, что
с увеличением глубины в море понижается темпера-
тура и повышается давление; при понижении темпе-
ратуры скорость звука уменьшается, а с увеличением
давления — возрастает. В результате этого на какой-
то глубине в определенной зоне звук проходит с мини-
мальной скоростью. Здесь и возникает подводный
канал, характеризующийся - слабым поглощением и
сверхдальним распространением звука. Иногда он под-
нимается близко к поверхности океана.
Подводные звуковые каналы позволяют мигрирую-
щим китообразным использовать звуковые маяки даль-
него действия. Такими маяками могут быть, например,
грохот прибоя, постоянного близ океанических остро-
вов и выступающих мысов, или шум ревущих сороко-
вых широт с их непрестанными штормами.
9*
195
По звуковым каналам, как по гигантским перего-
ворным трубам, возможна и сверхдальняя сигнализа-
ция китов, и отыскивание в период гона своих редких
сородичей, разбросанных на большом расстоянии друг
от друга. Не с таким ли сигналом был связан случай,
описанный еще Вильямом Скорсби, когда один грен-
ландский кит за 36 км почуял другого кита, которого
китобои ранили ручным гарпуном (без выстрела)?
В связи с возможным значением подводных звуко-
вых каналов для дальней сигнализации обращают на
себя внимание низкочастотные сигналы у настоящих
полосатиков, открытые учеными США В. Шевиллом,
В. Уоткинсом и Р. Баккусом. Это ритмично повторяю-
щиеся и сильные звуки частотой около 20 Гц. Акусти-
ки слышали их в течение 20 последних лет, но не зна-
ли их происхождения.
Недавно в Норвежском море исследователи Д. Уэ-
стон и Р. Блэйк слышали в гидрофон пять разных
типов низкочастотных звуков крупных полосатиков.
Одни звуки были похожи на хрюканье частотой 70 Гц
и продолжительностью 2 с, другие напоминали регу-
лярно повторяемые стоны общей продолжительностью
23 с и частотой 20 Гц; остальные занимали проме-
жуточное положение. Как издают такие сигналы уса-
тые киты и с какой целью — еще не выяснено, но,
учитывая дальность распространения низкочастотных
звуков, особенно по подводным каналам, можно ду-
мать об их важной роли в жизни китов.
Изучение проблемы звуковой сигнализации и рас-
шифровка биологического значения акустических сиг-
налов дельфинов обещают быть особенно успешными,
если при этом будут использованы методы звуковиде-
ния и новые приборы-скептроны, преобразующие зву-
ковые сигналы в световую картину, соответствующую
частотным спектрам сигналов. При этих условиях уве-
личиваются возможности расшифровки связи поведе-
ния особей с теми или иными звуковыми сигналами
и вскрытия закономерностей поведения стада.
Изучение эхолокации дельфинов должно привести
к реальной оценке возможностей использования аку-
стических свойств дельфинов в будущей практике че-
ловека, в его морских исследованиях.
Глава XXII
«ЗВЕЗДЫ МОРЯ»
СЛУЖАТ ЧЕЛОВЕКУ
В настоящее время успешно проводятся опыты с
дрессированными дельфинами не только в океанариу-
мах, но и непосредственно в море, в естественной об-
становке. Выработка у подопытных животных привыч*
ки возвращаться из родной стихии на базу открыла
широкие возможности использования мелких китооб-
разных в области морской биотелеметрии как носите-
лей современных датчиков и приемников для сбора
информации о микроклимате при исследовании океа-
на. Обученные и свободно плавающие в море дель-
фины подчиняются приказам человека, подаваемым с
помощью подводной локационной техники. При этом
бывают сюрпризы даже с отлично вышколенными жи-
вотными.
Форрест Вуд описал, как из загона через дыру в се-
ти ушли на волю две обученные афалины — знамени-
тый Таффи и Пег. Для их поисков был привлечен вер-
толет и несколько быстроходных спасательных кате-
ров. Таффи обнаружили на следующий день в 55 км
от загона. К дельфину с вертолета прыгнул тренер с
мешком рыбы. Беглец подплыл к человеку и стал жад-
но поедать рыбу; вскоре прибыл катер со снастью,
зверя заманили в ловушку и подняли на борт. Другой
блуждающий дельфин Пег был найден на третий день
после побега. С помощью призывных сигналов его при-
влекли к катеру, а затем тренер, бросая с борта го-
лодному зверю рыб, привел его в загон, преодолев рас-
стояние в 11 км.
Безвозвратный уход в море был и у нас. Заведую-
щий отделом морских млекопитающих Батумского
197
дельфинария Я. И. Близнюк с сотрудниками подгото-
вил для работы в родной стихии двух молодых сам-
цов-афалин — Моряка и Боцмана (рис. 57 и 58 вкл.).
В конце октября 1976 года обоих привезли в район
Геленджика и поместили в плавучую клеть в 50 м от
берега, где с целью адаптации намеревались передер-
жать их несколько дней перед выпуском. Но внезапно
разразившийся ураган разрушил плавучую клеть, и
дельфины, пробыв возле нее с полчаса, исчезли в бу-
шующем море. Показались они через четыре месяца
вблизи Геленджика (плавали между удочками рыба-
ков и выставлялись из воды в позе «просителя»), а
еще через полгода были встречены туристами возле
Сухуми: пассажиры с туристического поезда наблю-
дали, как вблизи берега играли в мяч два убежавших
дельфина.
Среди дельфинов, уже служащих человеку, имеют-
ся настоящие «звезды моря» — знаменитые афалины
Кейки и Таффи.
Зоолог Норрис по заданию центра подводных ис-
следований военно-морского флота США ведет обшир-
ную работу с дельфинами в океанологическом инсти-
туте на Гавайских островах (о. Оаху). В 1964 году он
обучил молодого самца-афалину по кличке Кейки
(«ребенок») плавать с особой сбруей, в которую встав-
лялись приборы, по команде развивать высокую ско-
рость и возвращаться в открытом море с расстоя-
ния около километра. Кейки много раз выпускали
в родную стихию для скоростных пробегов и по особо-
му сигналу он поспешно возвращался к катеру или в
свой плавучий загон из проволочной сетки. За несколь-
ко месяцев до опытов этот самец был выловлен в мо-
ре из небольшой стайки и прошел 70-дневную трени-
ровку в мелководной лагуне. Кейки очень любил, ко-
гда пловцы, в обществе которых он проводил время,
поглаживали его тело. Дельфины, обученные в районе
Гавайских островов, возвращались к человеку даже по-
сле нескольких дней свободного плавания в море. Но
однажды Кейки, выпущенный в море для очередного
эксперимента, уплыл и не вернулся на базу.
В конце лета 1965 года были проведены знамени-
тые опыты в морской лаборатории «Силэб II» с афа-
линой Таффи. Этот дельфин за короткое время при-
обрел почти мировую известность в свои десять лет
198
(рис. 59 вкл.). До семилетнего возраста он жил в про-
сторах Атлантического океана и часто подвергался на-
падениям акул, о чем свидетельствовали многочислен-
ные шрамы на его коже. Его поймали и сначала поме-
стили в аквариум «Морская жизнь» в Галфпорте (штат
Миссисипи), а через полтора года перевели в новый
бассейн в Пойнт-Магу. Животное в это время имело
длину 210 см и весило 122 кг.
В начале обучения Таффи часто нападал на дрес-
сировщиков, за что и получил кличку Таффи, что
значит «драчун». За полтора года до испытаний дель-
фина стали тщательно готовить к той роли, которая
выдвинула его в число звезд моря. Дрессировщик Уол-
лес Росс перенес обучение из. дельфинария в лагуну
и четыре месяца обучал дельфина носить сбрую, к ко-
торой прикреплялись различные инструменты, датчи-
ки, донесения, спасательные тросы, тренировал его с
надетыми наглазниками проплывать через обруч и
отыскивать дорогу по щелчкам зуммера. После пре-
бывания в лагуне дельфина восемь месяцев обучали
в море. Выпускаемый в родную стихию, Таффи стре-
мительно плавал, погружался на глубину до 90 м и
по сигналу человека послушно возвращался из откры-
того моря с расстояния более полкилометра. Наконец
наступил день проверки способностей Таффи.
У берегов Калифорнии начались полуторамесячные
подводные испытания по программе военно-морского
флота США под девизом «Человек в море». Подводная
лаборатория представляла собой цилиндр длиной 17 м
и диаметром 3,6 м. В помещение вел люк, через кото-
рый акванавты ежедневно выходили в море для раз-
ного рода работ. Над лабораторией бросило якорь суд-
но «Бэркон». Роль связного между поселенцами на
дне и людьми на корабле отлично выполнял Таффи.
Он же должен был защищать пловцов от хищных рыб,
но людей не потревожила ни одна акула. Дельфина
ежедневно привозили на вертолете из плавучего мор-
ского загона и выпускали в море. Пять дней Таффи ра-
ботал с акванавтами. В первый день он пугался, боял-
ся шума, необычной обстановки на дне. В ответ на
поданный сигнал бедствия он приблизился к «потер-
певшему» акванавту, чтобы оказать ему помощь, но
раньше чем тот схватил спасательный трос, ринулся к
поверхности. Вернувшись через несколько минут, он
199
все-таки держался на расстоянии от людей, и пловцы
не смогли получить инструменты, прикрепленные к
его сбруе.
В последующие дни служба Таффи наладилась:
дельфин, курсируя между глубоководной лабораторией
и судном, 35 раз передал в водонепроницаемом фут-
ляре письма и пакеты, доставлял акванавтам разного
рода инструменты. (По решению окружного отделения
профсоюза почтальонов города Сан-Луис (США), Таф-
фи был избран в почетные члены общества связистов,
как доставлявший почту жителям подводного дома
«Силэб II».) Он делал по 20 рейсов в день туда и об-
ратно, преодолевая путь в один конец за 45 с. Испыта-
ния проводились в очень мутной воде, где дальность
видимости не превышала 3 м, а иногда была и мень-
ше. Таффи был обучен оказывать помощь заблудив-
шимся акванавтам. Он был способен различать людей
индивидуально; узнавал «своих» даже тогда, когда
внешне их преображали скафандры, маски и аквалан-
ги. Услышав сигнальный свисток пловца, дельфин
бросался с поверхности вглубь, к подводной лаборатО'
рии, находил катушку с намотанным тросом, хватал
зубами его конец и подплывал к сигналящему челове-
ку, чтобы тот смог взять трос в руки. Таким образом
он 3 раза спасал пловцов, «заблудившихся» в десят-
ках метров от подводной станции.
Сходный опыт проводили на двух акванавтах, раз-
деленных расстоянием в 30 м: один держал катушку
с тросом, а другой имитировал заблудившегося пловца.
По сигналу Таффи находил конец троса у первого ак-
ванавта и передавал его в руки второго. Дельфин все
это проделывал за вознаграждение — куски мороже-
ной рыбы. Когда же один из акванавтов не дал на-
грады за выполненное действие, дельфин ударил об-
манщика хвостом.
Дальнейшую службу Таффи продолжил на одном
из ракетных полигонов США в Пойнт-Магу, где за че-
тыре месяца сэкономил военно-морскому ведомству
США 70 000 долларов: он успешно помогал искать на
морском дне дорогие телемеханические устройства
расцепки (спусковые подушки стоимостью в 4700 дол-
ларов), которые падали в море во время запуска ра-
кет. Такие устройства обычно погружаются в ил, и их
поиск бывает безрезультатным. Прикрепление же к
200
устройствам ультразвукового сигнализатора (автомати-
ческого зуммера) позволило Таффи находить их по
ультразвуку на большом расстоянии. Дельфин отлич-
но ориентировался по акустическому маяку, и когда
находил нужный объект, он сбрасывал надетый на
его рыло обруч, состоявший из двух половинок — тя-
желой и легкой. Тяжелая половинка оставалась у за-
тонувшего предмета, а легкая поднималась на поверх-
ность и служила буем, который легко находили лю-
ди. Аквалангисты доделывали все остальное.
В 1968/69 году Таффи участвовал в испытаниях
подводной лаборатории «Силэб III», которые закон-
чились преждевременно вследствие гибели акванавта
и технических неисправностей. Погиб этот дельфин в
1973 году в возрасте 17—18 лет от бактерий — раз-
рушителей мышечной ткани.
В~ последние годы начаты наблюдения за поведени-
ем дельфиньих косяков в открытом море с помощью
судна, снабженного кабиной с подводными окнами.
Интересный опыт в этом плане был проведен лабора-
торией подводных исследований в Сан-Диего, США (со-
общение «Транс Марине инженир», 1969, 81, № 6,
стр. 10—11): на спинном плавнике отловленного в мо-
ре дельфина были укреплены радиопередатчик (с ра-
диусом действия 10 миль) и источник питания, рассчи-
танный на 30 суток. Выпущенный в море, этот зверь
лишь вначале вел себя неспокойно, но уже через шесть
часов присоединился к стайке сородичей. Специально
оборудованный катер с радиопеленгатором следовал за
этим дельфином 14 суток в 2—4 милях от него. Пред-
полагают, что в дальнейшем с помощью датчиков
можно будет собирать пробы о физических свойствах
океана и данные о физиологическом состоянии живот-
ного; наблюдения будут поступать на береговую стан-
цию при каждом всплытии дельфина на поверхность.
Приобрели большую известность дельфины из Фло-
риды — афалины Долли и Дина, сонар которых испы-
тывали в бухте Мидл-Байт на острове Андрос (Багам-
ские острова). Этих животных, когда они уплыли в
море, с помощью батарейного сигнализатора подзывал
выходящий за ними катер и приводил за собой в мор-
ской загон.
Учитывая эхолокационные способности дельфинов,
их легкую обучаемость и положительный опыт с афа-
201
линами Кейки, Таффи, Долли, Дина и другими, за ру-
бежом разрабатываются военные аспекты использова-
ния мелких китообразных — для поисков подводных
ракетных установок, минных полей, затонувших судов,
ракет и торпед, для гидроакустического патрулирова-
ния участков моря, обнаружения подводных лодок,
бойцов-подводников противника, нарушителей морских
границ, для доставки шпионского оборудования в ино-
странные порты, для подводной связи и оказания по-
мощи при подводных авариях и т. д.
По словам вашингтонской газеты «Нэшеналь
джельбрехен», дельфин стал объектом беспрецедентно-
го изучения. Дельфинами занимаются флот, военно-
воздушные силы, Национальное управление по аэро-
навтике. Лаборатории военно-морского флота США
тренируют дельфинов для выполнения военных задач.
Например, их обучают плыть рядом с подводными лод-
ками, чтобы обеспечивать ложную акустическую ми-
шень. Под голоса дельфинов маскируются подводные
лодки. Сообщалось о том, что американские специали-
сты научили гринду поднимать торпеды с глубины
300 м. Этот процесс они засняли на пленку и продемон-
стрировали журналистам. Зрители видели, как гринда,
держа в зубах особое портативное грузоподъемное уст-
ройство, прикрепляет его к смертоносному снаряду.
Газовый генератор этого устройства наполняет газом
баллон, и торпеда поднимается со дна на поверхность
моря. Полагают, что таким способом гринда сможет
поднимать грузы весом до полутонны с глубины в пол-
километра.
Пентагон хочет поставить океанских животных —
дельфинов на службу армии. Как говорилось в сооб-
щении со ссылкой на сайгонского корреспондента
«Нью-Йорк дейли ньюс», американские военно-мор-
ские соединения в Южном Вьетнаме «используют бу-
рых дельфинов (афалин.— А. Т.) для подводной боевой
разведки». Эти животные прошли дрессировку в США.
По словам корреспондента, они «составляют часть
секретной военно-морской операции, имеющей целью
оценить полезность использования бурых дельфинов
на войне».
По сообщению исследователя Фарука Хуссейна, во
Вьетнаме афалина была использована как убийца с
ножом, прикрепленным к морде. Такой вооруженный
202
дельфин, обнаружив подводного лазутчика, стреми-
тельно наплывал на него и прокалывал ножом.
В печати появилась информация со ссылкой на
телевизионную компанию Си-би-эс: «Пентагон выде-
лил 30 миллионов долларов на совершенно секретную
программу подготовки дельфинов, моржей и морских
львов в военных целях... Основная роль, к которой
готовят морских животных дрессировщики Пентаго-
на,— быть живыми торпедами».
Между тем, имеются широкие возможности исполь-
зования дельфинов в самых различных областях
деятельности человека, например в области бионики,
медицины и физиологии, рыбного хозяйства, искусства
тренировки и обучения животных.
Дельфин — достойнейший объект для моделирова-
ния в технике, и уже достигнуты кое-какие практиче-
ские результаты. В США построили механического
дельфина «Порпез» с целью регистрации температуры
воды и передачи данных на базовый корабль. Снаряд
длиной 4 м и диаметром 0,5 м действует автономно и
может работать на глубине 170 м.
По принципу дельфиньих грудных плавников ин-
женером С. Роговым были сконструированы приспо-
собления у быстроходного маневренного глиссера («Из-
вестия», 1971, 4 декабря). При быстром ходе на бортах
кормовой части глиссера в сторону поворота штурва-
ла выдвигаются стреловидные крылья-накладки, кото-
рые действуют как грудные плавники дельфинов (рули
поворотов) и позволяют совершать крутые повороты.
Принцип эхолокации дельфинов заимствован при
создании миниатюрного ультразвукового аппарата для
ориентации аквалангистов при плавании в толще воды.
По типу строения гидролокатора дельфина американ-
ским ученым Л. Кэем разрабатывается портативный
эхолокатор, который позволит слепым людям сво-
бодно передвигаться по улицам и переходить пере-
крестки.
Ученому США Р. Пелту удалось снизить на 35%
потери на преодоление сопротивления трения при пере-
гоне жидкости по трубам путем имитации под дель-
финью кожу внутренней выстилки труб (выстилка бы-
ла сделана из уретановой смолы на полиэфирной ос-
нове). Такие трубы могут дать огромную экономию
203
при перекачке различных жидкостей (спирт, вода, па-
тока, жидкие газы) на большие расстояния.
По сообщению корреспондента ТАСС М. Рыбакова
от 21 апреля 1971 года, на промысле охотской сель-
ди успешно использовали голоса дельфинов. Рыбу
удерживали в неводе, имитируя звуки дельфинов с
помощью «пузырьковой завесы»: косяк сельди охва-
тывали шлангом со многими мелкими дырками и по-
давали в него воздух. Поднимающиеся вверх пузырьки
газа, булькая, напоминали сигналы дельфинов, поче-
му сельдь и не пыталась уйти из невода.
За рубежом разрабатывается программа использо-
вания дельфинов в области медицины и для разреше-
ния ряда физиологических задач. Все глубокие при-
способления дельфинов в дыхании, кровообращении,
водно-солевом обмене, урежение пульса и перераспре-
деление тока крови во время ныряния, бесперебойное
снабжение кислородом сердечной мышцы и мозга,
функциональные особенности сильно развитой чудес-
ной сети представляют большой интерес и для физио-
логов, и для медиков.
Врачи в США намереваются изучать проблемы рев-
матизма и сердечно-сосудистых заболеваний на дель-
финах. Их грудная клетка просматривается Х-лучами
при обнаружении пневмонии, снимаются кардиограм-
мы. Специалисты намерены искусственно вызывать у
дельфинов пороки сердца, изучать влияние физиче-
ских перегрузок на состояние сердечно-сосудистой си-
стемы обучаемых афалин, влияние той или иной дие-
ты на разного рода заболевания. Ставятся, например,
вопросы, как будет влиять чисто рыбная диета на этих
животных, как нервные раздражения (стук и т. д.)
стимулируют, например, язвенные заболевания.
Методика анестезирования этих животных, разра-
ботанная американскими учеными (см. главу XIX), от-
крыла путь для хирургического исследования живых
дельфинов, для полостных операций, электрофизиоло-
гических экспериментов над мозгом и т. д.
Явление поглощения пульса чудесной сетью, осо-
бенно в кровеносных сосудах головного мозга дельфи-
нов, интересует медиков и конструкторов для совер-
шенствования механического аппарата «сердце — лег-
кие», в котором ток крови также не имеет пульсаций.
Большой интерес для физиологии и медицины пред-
204
ставляет проблема изучения у китообразных водно-со-
левого обмена и внутреннего ионного регулирования.
До последнего времени считалось, что дельфины во-
ду не пьют. Но проведенные в 1970 году эксперименты
(Н. Тельфер, Л. Корнелл, Д. Прескотт) показали иное.
Дельфина-афалину содержали в ванне с растворенным
радиоактивным калием; оказалось, что калий прони-
кает в организм дельфина, но не через кожу, а через
рот с заглатываемой водой и затем выходит наружу
вместе с мочой. Как в этом случае организм дель-
фина справляется с избытком поступающей соли?
Прекращается ли у дельфинов, как у ластоногих, по-
чечное кровообращение и фильтрация почечных клу-
бочков на время погружения или имеются другие осо-
бенности в крайне своеобразных многодольчатых поч-
ках? Нет ли у них какого-либо опреснительного меха-
низма? Что им помогает уничтожить лишнюю соль?
Изучение этих закономерностей, возможно, поможет
изыскать пути безвредного потребления морской воды
для питья при кораблекрушениях или при вынужден-
ном длительном пребывании на море. Ведь в конце
концов почки регулируют и поддерживают в жидкой
среде организма тот состав солей, который содержится
и в море, откуда вышла жизнь. Весьма вероятно, что
именно с поступлением большого количества хлористо-
го натрия у китообразных интенсивно выделяется моча
и происходит частое мочеиспускание из миниатюрного
мочевого пузыря.
Осмотические изменения мочи дельфинов указыва-
ют, что у них имеется и значительное продуцирова-
ние метаболической воды.
Тщательное изучение физиологии дельфина помо-
жет раскрыть среди тайн этого животного и секреты
быстрого, глубокого погружения, которые будут ис-
пользованы для практических целей. Дельфин будет
тесно связан с перспективами всестороннего изучения
океана как будущей житницы человека.
При бурном развитии океанографии и создании но-
вой подводной техники возникает необходимость вести
морское хозяйство на научных основах. В Японии в
бухтах и заливах пытаются разводить ценные породы
рыб на «подводных пастбищах». В связи с этим воз-
никла идея воспитания «подводных пастухов». Про-
фессор океанографического института Токийского уни-
205
верситета Т. Куроки предложил программу «курса обу-
чения» афалин: в течение 12 лет дельфинов будут
обучать по команде человека изменять направление
движения рыбных косяков. Уже в настоящее время
то, что выполняют одиночные тренированные афали-
ны, позволяет представить себе их службу в области
рыбного хозяйства будущего.
Советский исследователь Е. В. Шишкова предлагает
использовать для обнаружения косяков рыбы подслу-
шивание и пеленгование громких дельфиньих сигна-
лов, издаваемых при кормежке. С помощью шумопе-
ленгаторов такие звуки можно улавливать с расстоя-
ния в несколько миль. Если пеленг на шумы не
смещается, значит, дельфины держатся в одном рай-
оне и, весьма вероятно, питаются рыбой. Тогда это
место следует проверить.
В Южноафриканском океанариуме в Порт-Элизабе-
те пытаются обучить афалин загонять рыбу в сети,
чтобы в дальнейшем применять их способности в мо-
ре. Однако натурный эксперимент еще не состоялся.
Тем не менее в Африке на побережье Тимирисского
залива (Мавритания) жители племени имрагенов с
давних пор ловят рыбу с помощью дельфинов. Когда
с дозорных вышек рыбаки заметят косяки рыб, они
сильными плещущими ударами по воде привлекают
дельфинов, которые, направляясь к берегу, способст-
вуют заходу рыбы в расставленные сети.
Известно, что калифорнийские и мексиканские ры-
баки разыскивают в океане дельфинов, так как легче
находят стаи тунцов, если ориентируются на морских
млекопитающих.
Очень интересные опыты проведены Вильямом
Эвансом в центре подводных исследований в Сан-Дие-
го в 1971 году. Он установил, что обыкновенные дель-
фины питаются главным образом ночью на скоплени-
ях рыб, на глубине 100—130 м, куда они ныряют,
затрачивая в среднем по 3—4 мин. Утром стадо дель-
финов разбивается на мелкие рассеивающиеся группы,
которые днем ныряют мало и остаются близ поверх-
ности. Ученый считает возможным увеличить эффек-
тивность рыбного промысла за счет сокращения вре-
мени поисков рыбных косяков; для этого суда должны
радиометкой пометить какого-либо дельфина и следо-
вать за ним до ночи, пока тот не выведет на крупные
206
скопления рыб. Проверка с эхолотом подтвердила це-
лесообразность такого использования «радиофициро-
ванного» дельфина: тот действительно ночью выводил
следящее судно на огромные глубинные косяки
рыб.
Новая методика тренировок дельфинов исключает
наказание, а допускает только ласку. Использование
в разных сочетаниях подкрепления пищевыми, игро-
выми, кожными и половыми раздражителями может
значительно повысить результативность их дресси-
ровки.
Другой путь повышения эффективности их обуче-
ния — тренировка с помощью показа, использование
склонности дельфинов к подражанию. Подражание у
высокоорганизованных животных имеет важнейшее
значение в выработке поведенческих реакций. Подса-
живание уже обученных дельфинов к необученным
сильно сокращает время их дрессировки. Теперь в не-
которых океанариумах обучение дельфинов начинают
с момента их рождения и последовательно проводят
тренировку в несколько стадий.
Успехи в методике тренировки дельфинов обещают
значительно расширить возможность практического ис-
пользования их и ставят вопрос об одомашнива-
нии этих млекопитающих.
Глава XXIII
БУДУЩЕЕ
ДЕЛЬФИНОВ
Накопившийся опыт работы с мелкими китообраз-
ными выявил реальную возможность одомашнить не-
которые виды дельфинов и прежде всего афалину с
целью широкого использования их в море. Будущие
обязанности первых морских домашних животных по
своему многообразию могут быть подобны тем, ка-
кие выполняет собака, одомашненная почти 20 тысяч
лет назад. Люди используют собаку для выполнения
самых различных действий: сторожевой и розыскной
службы, охоты, охраны и защиты, переноски и пере-
возки грузов, связи, как подопытных животных, по-
мощников геологов при разведке полезных ископаемых,
имеющих запах, для разыскивания повреждений в га-
зовой сети и т. д. Хорошо обученная собака пасет
скот,’ ищет мины, спасает утопающих, вывозит ране-
ных с поля боя, водит слепых...
Дельфина можно будет широко использовать в ка-
честве связиста при различные подводных работах,
как помощника океанологов при изучении океана и
сборщика необходимой информации.
Дельфина можно научить разведывать морское дно,
доставляя пробы грунта, перевозить грузы, спасать
утопающих, подавая им пояса или выталкивая из во-
ды, защищать человека от акул, быть буксировщиком
и поводырем пловцов и акванавтов в трудных для них
ситуациях. Он будет нести поисковую службу в море,
отыскивая различные объекты, интересующие челове-
ка, начиная от раковин с жемчугом и кончая зато-
нувшими судами. Обученные и одомашненные дельфи-
ны будут очень полезны в области рыбного хозяйст-
208
ва, как разведчики скоплений рыбных косяков, как
наводчики рыболовных траулеров и загонщики рыб-
ных стай. Лучшей ищейки и загонщика рыбных стай,
чем дельфины, трудно найти: ведь в данном случае
будут использованы его прирожденные рыбопоисковые
способности. Недаром так высоко ценят этого случай-
ного помощника рыболовства в некоторых районах
мира.
Насколько реальны все эти перспективы?
Испытания в водах Гавайских островов показыва-
ют, что дельфины очень послушны, исполнительны,
возвращаются по команде человека со значительных
расстояний и даже через несколько 'дней свободного
плавания в море.
У дельфинов, особенно афалин, есть все задатки,
благоприятствующие процессу одомашнивания. Они
легко приобретают навыки, быстро вырабатывают
условные рефлексы, стойко их сохраняя, положительно
относятся к ласке, миролюбивы, предпочитают опреде-
ленные виды пищи, точно подражают своим сороди-
чам. Все это расширяет возможности их поощрения и
закрепления у них рефлексов.
Очень важно, чтобы одомашниваемые дельфины бы-
ли оседлыми и держались в одной и той же мест-
ности.
Всем перечисленным условиям удовлетворяет афа-
лина. По поврежденному спинному плавнику рыбаки
опознавали афалину в одном и том же месте в тече-
ние трех лет. В другом случае афалина-альбинос ме-
сяцами держалась в проливе Св. Елены у берегов Юж-
ной Каролины, пока не была отловлена для дельфина-
рия.
Выпуски афалин из океанариумов в море подтвер-
дили их оседлость. Так, афалину Присциллу из Сент-
Огастинского океанариума выпустили в море, посколь-
ку она не проявила способности к обучению, а через
полтора года отловили в том же месте. Присцилла, по-
пав в бассейн, почувствовала себя «как дома», даже
сохранила привычку, выработанную ранее,— подолгу
лежать на дне бассейна.
История с Опо-Джеком и Чарли — молодыми афа-
линами — укрепляет уверенность в возможности одо-
машнивания этого вида: оба дельфина в различных
частях мира за короткое время превратились в совер-
209
шенно ручных животных и регулярно в течение мно-
гих месяцев посещали места с большим скоплением
людей.
Трудоемкий процесс одомашнивания афалин будет
особенно эффективным при содружестве разных спе-
циалистов — экологов, физиологов, дрессировщиков,
зоотехников, акустиков, ветеринаров.
Родившиеся в неволе и обученные афалины (а они
размножаются в океанариумах с 1940 г.) могут быть
основным костяком в создании стада домашних дель-
финов.
В связи с этой проблемой может возникнуть и за-
дача изучения межвидового и межродового спарива-
ния дельфинов. Известно, что английский зоолог
Ф. Фрейзер на берегу Ирландии встретил помесь ме-
жду афалиной и серым дельфином. В главе VI уже
приводились примеры спаривания дельфинов в неволе,
принадлежащих к разным видам и даже родам, а так-
же случаи получения межродовых гибридов. В лите-
ратуре описаны крупные киты с чертами двух разных
видов, например финвала и синего кита, сейвала и
финвала. Проблема получения межвидовых и межро-
довых гибридов изучена крайне слабо. В спаривании
разных видов дельфинов большую стимулирующую
роль играют, видимо, кожные раздражения, столь ча-
стые при жизни в узком замкнутом пространстве.
Изучение подобных фактов, вероятно, поставит перед
исследователями вопрос о возможности гибридизации
дельфинов, позволит наметить пути преодоления это-
логической и механической изоляции и, может быть,
приведет к гибридогенному формированию новых ви-
дов дельфинов.
Одомашненные дельфины, свободно плавая в море,
будут привлекать к себе диких, еще необученных со-
родичей. Так будет пополняться, помимо естественно-
го размножения, домашнее стадо дельфинов. В океане
появится много наших друзей, умных и исполнитель-
ных, готовых служить человеку не менее усердно, мно-
гообразно и преданно, чем старый четвероногий друг
людей — собака.
Разводить в неволе, возможно, будут и некоторые
виды китов. Так, привлекает к себе внимание проект
американского профессора Жиффора Пиншота — раз-
водить китов в небольших атоллах. Лагуны этих коль-
210
цеобразных коралловых островов, сообщающиеся с мо-
рем, были бы очень удобны для культивирования и
изучения китов. Проблему с питанием китов в атол-
лах профессор рекомендует решить путем «удобрения»
лагун. Для этого ветровой двигатель будет выкачивать
глубинную воду, насыщенную нитратами и фосфата-
ми, на поверхность. Перемешивание воды в лагуне
должно вызвать сильное развитие фитопланктона, а
вслед за ним и зоопланктона; все это привлечет стай-
ных рыб и обеспечит питание кита. Кит, если у него
не пропадет аппетит в «тропической квартире», упо-
добится огромному механизму, перерабатывающему
планктон в жир.
Доставить кита в атоллы помогут его же миграция
и радиометка: в Антарктике кита пометят радиомет-
кой, а та даст возможность следовать за ним до теп-
лых вод, где он и будет обметан с помощью техники
(сети и суда). Оставшийся небольшой путь до атолла
кит проделает в огромной клетке, аналогичной той,
в какой Тэд Гриффин доставил косатку Наму в бухту
г. Сиэтла...
Однако осуществлению этих планов может поме-
шать... сам человек.
Глава XXIV
МИР ДЕЛЬФИНОВ
И китов
В ОПАСНОСТИ
Разные виды животных, страдая от врагов, сурово-
го климата, болезней, паразитов и других неблагопри-
ятных факторов, стараются всеми средствами удер-
жаться на арене жизни. В ходе естественного отбора
они успешно приспосабливаются, меняя поведение, ин-
тенсивность воспроизведения. Однако эти же виды ста-
новятся совершенно бессильными перед натиском чело-
века, который изобретает хитроумные способы лова
животных. Он вносит настоящее опустошение в фау-
ну и флору, воздействуя на живую природу техниче-
ским прогрессом, разрушая экосистемы, изменяя лик
планеты и загрязняя окружающую среду. Настало
время, когда во имя сохранения человечества Власте-
лин Земли сам должен предпринять неотложные меры
по очищению среды, по защите ценных видов живот-
ных и растений — от кораллов до слонов, от водоросли
до баобаба, от индивидуума до целого биогеоценоза.
В бедственном положении сейчас оказались многие
морские млекопитающие, в первую очередь киты. Со-
вершенствование техники китобойного промысла пре-
допределило печальную судьбу неповторимых гигантов
океана. Тихоходные гладкие киты в основном были
выбиты в середине XIX века с помощью ручных гар-
пунов. В 1864 году норвежец Свенд Фойн изобрел гар-
пунную пушку, которая позволила легко справляться
с крупными и тонущими после убоя китами-полосати-
ками, ранее недосягаемыми для лодочного — ручного
промысла. Изобретение в 1904 году флотингов — пла-
вучих китобойных маток-фабрик разрешило проблему
охоты за китами вдали от берегов, а изобретение сли-
212
па (наклонного туннеля в корме матки, через который
киты втаскиваются на палубу) сняло последние огра-
ничения для разделки китов в самых отдаленных ча-
стях Мирового океана.
Промысел на китов стал пелагическим. Ввод в дей-
ствие дизельэлектроходных китобойцев, поисковых
вертолетов, совершенных многотоннажных маток, спо-
собных переработать до 60—100 китов в сутки, создал
реальную угрозу выбоя усатых китов на земном ша-
ре. Человек добрался до последнего и самого важного
резервата полосатиков — далекой Антарктики. Теперь
промысел охватил, особенно в прибрежных водах Нор-
вегии, Исландии, Японии, даже мелких китообраз-
ных — малых полосатиков, косаток, гринд, бутылко-
носов, плавунов, на которых раньше не обращали вни-
мания.
В условиях сильнейшей интенсификации китобой-
ного промысла проявляются внутрипопуляционные ме-
ханизмы ускоренного пополнения стада китов, в свя-
зи с чем: 1) учащаются случаи зачатия самок еще в
лактационный период, когда продолжается выкорм
детенышей; 2) сокращаются сроки полового созрева-
ния и 3) увеличивается число зародышей-двоен. Одна-
ко все это не может компенсировать слишком боль-
шие потери в популяции китов, численность которых
неуклонно падает.
За последние 100 лет в Мировом океане добыто
более 2,1 млн. китов, из них 18% —синих, 40 —
финвалов, 22 — кашалотов, 9 — сейвалов, 8,5 — горба-
чей и 2,5% — прочих. Это приблизительно столько же,
сколько насчитывалось в океане китов к началу меха-
низированного промысла. Охота на них в особо круп-
ных масштабах велась до начала второй мировой вой-
ны. Выделялись два рекордных года: сезон 1930/31,
когда в Антарктике было убито почти 30 тыс. синих
китов (это в 3 раза больше, чем их осталось сейчас на
планете), и сезон 1937/38, когда было добыто 55 тыс.
китов, из них только в Антарктике — 28 тыс. фин-
валов.
Для регулирования и ограничения китобойного про-
мысла в 1946 году была создана Международная
китобойная комиссия, включавшая представителей
20 стран. По принятым правилам Международного со-
глашения (Конвенции) запрещена охота на гладких,
213
серых, горбатых, синих китов и (с 1976 г.) на финва-
лов. На каждый год устанавливаются нормы или
квоты выбоя и разрешается промысел лишь в опре-
деленные сроки и в строго ограниченных акваториях
Мирового океана. Запрещается бой кормящих самок
с детенышами, неполовозрелых китов, не достигших
достаточной длины.
Ежегодная квота выбоя китов по мере сокращения
их запасов уменьшилась: с 16 тыс. условных китовых
единиц в 1948/49 году до 2300 единиц в 1971/72 году.
С 1972/73 года введена видовая квота, разрешающая
добывать в Антарктике 5 тыс. сейвалов (рис. 60 вкл.),
1950 финвалов, 5 тыс. малых полосатиков и 13 тыс.
кашалотов (рис. 61 вкл.), а в северной части Тихого
океана — 3 тыс. сейвалов, 650 финвалов и 10 тыс. ка-
шалотов в год. Хотя биологическая информация свиде-
тельствовала о перепромысле китов, Международная
китобойная комиссия была бессильна приостановить
падение численности китов, так как это затрагивало
национальные квоты. Мешало и то, что квоты устанав-
ливались на основе учета общей продукции, а не экс-
плуатации отдельных видов. В 1972 году этот недочет
был устранен и введена система международных на-
блюдений, но уже после того, как были в корне подо-
рваны запасы синих и горбатых китов и значительно
сокращена численность финвалов в Антарктике.
Несмотря на все эти меры, китобойный промысел
угасает.
Некогда огромная популяция усатых китов, насчи-
тывавшая в Мировом океане 1 млн. 445 тыс. голов,
ныне сократилась до 577 тыс., т. е. на 60%, а если из
расчетов исключить слабо затронутого промыслом
малого полосатика, то популяция уменьшилась
на 77%.
Американский ученый Виктор Шеффер объединил
данные международной китобойной статистики и от-
дельных исследователей в итоговую таблицу. В ней он
привел для разных видов китов следующую началь-
ную (девственную) и современную (на 1974 г.) числен-
ность популяции, а также процент оставшихся в Миро-
вом океане запасов от исходных: кашалотов было
922 тыс., осталось 641 тыс. (или 69%); финвалов — со-
ответственно 448 тыс. и 101 тыс. (22%); синих ки-
тов— 215 тыс. и 13 тыс. (6%); сейвалов — 200 тыс.
214
и 76 тыс. (38%); горбачей — 50 тыс. и 7 тыс. (14%);
малых полосатиков — 361 тыс. и 325 тыс. (90%); по-
лосатиков Бранда—100 тыс. и 40 тыс. (40%); юж-
ных китов — 50 тыс. и 2 тыс. (4%) и гренландских ки-
тов— 10 тыс. и 2 тыс. (20%). Как видно, сильнее все-
го пострадали гладкие, синий и горбатый киты.
В 1972 году английский исследователь Н. А. Ма-
кинтош подсчитал, что в Антарктике запасы финва-
лов уменьшились в 5 раз, синих китов — в 25, горба-
чей — в 30 раз.
Все это свидетельствует о том, что действенной ме-
рой спасения китов был бы общий запрет китобойного
промысла.
Ученые подсчитали, что для восстановления ан-
тарктических стад синих китов до 150 тыс. голов по-
требуется 50 лет, а для роста стада горбатых китов до
27 тыс. голов — 60 лет. То обстоятельство, что киты
живут локальными стадами, способствует их быстро-
му истреблению. Например, у берегов Габона (Афри-
ка) крупные полосатики исчезли после двухлетнего ин-
тенсивного промысла в 1949—1951 годах.
Очень поучительна в этом плане история с тремя
популяциями серых китов. Серый кит — превосход-
ный пример как благотворного влияния своевременно-
го запрета промысла на восстановление численности
редкого животного, так и опустошительного и непопра-
вимого воздействия на запасы чрезмерного промысла
и запоздалых мер. До войны (в 1937—1940 гг.) в бе-
рингово-чукотском стаде насчитывалось лишь несколь-
ко сотен серых китов, но запрещение охоты в 1947 го-
ду предотвратило их гибель, и через 20 лет числен-
ность выросла до 11 тыс. При сохранении полного за-
прета промысла это стадо, вероятно, достигнет своей
первоначальной величины (30 тыс.), которую оно име-
ло в первой половине XIX века. Другое, охотско-ко-
рейское, стадо, мигрировавшее между Охотским морем
и Корейским проливом, угасло после 1933 года, хотя
еще в 1910 году насчитывало около 1500 китов. Запо-
здалый запрет промысла уже не мог спасти эту по-
пуляцию.
Наконец, в Северной Атлантике несколько веков на-
зад существовала популяция серого кита, которого
здесь называли «скрэгвал». Она размножалась в мел-
ководных заливах Балтийского моря. Преследование
215
человека, а также обмеление балтийских заливов по-
губили и эту популяцию, последние остатки которой
дожили до XVII века.
Китообразные представляют важное звено в цепях
питания Мирового океана, создавая стабильность био-
логического круговорота веществ в экологической си-
стеме. Никто не знает, чем может закончиться устра-
нение из пищевой цепи океана усатых китов. Может
быть, это внесет нарушения в биологический кругово-
рот веществ, оказав влияние на массы фитопланктона
через промежуточные звенья — ракообразных, моллю-
сков и рыб. Фитопланктон, как известно, поддержива-
ет равновесие между содержанием углекислого газа и
кислорода в атмосфере Земли.
О возможности такого воздействия свидетельствуют
как приблизительные расчеты роли китов в биогеоце-
нозах океана, так и совпадения основных районов ки-
тобойного промысла с главными областями скопления
океанского фитопланктона.
Исключение китов из цепи питания будет неизбеж-
но вызывать резкие колебания численности то одних,
то других членов биоценоза. Следствием сокраще-
ния запаса китов и освобождения ранее занимаемой
ими ниши (ракообразных), вероятно, явилась вспышка
численности тюленей-крабоедов, достигших 6 млн., а
по другим данным 15 млн. голов. Этот тюлень, по
оценке зоологов, стал самым многочисленным ластоно-
гим Южного полушария.
Борьба человека за сохранение китов вписывается
в общую проблему борьбы за чистоту океана. Извест-
ный французский океанолог Жак-Ив Кусто в статье
«Океан на пути к смерти» (1970 г.) предостерегает:
«Океан умирает. Он болен по вине человека. За по-
следние полвека в море исчезло около 1000 видов жи-
вотных...» В другом месте он пишет: «Мы не можем
позволить навсегда отравить море в то самое время,
когда только-только начали постигать, что оно может
дать науке, философии, искусству, начали учиться
жить в его лоне».
Морские млекопитающие — яркие индикаторы за-
грязнения океана, так как они долго живут и в тече-
ние длительного времени накопляют в своих тканях
стронций, ядохимикаты, ртуть и т. д.
Мелкие китообразные довольно чувствительны к
216
ядовитым веществам в окружающей среде. В бассейне
одной биостанции погибли две афалины, до этого вы-
держанные в морской воде в ванне, покрашенной крас-
ным суриком. Ртутные соединения проникают в орга-
низм дельфинов, видимо, вместе с заглатываемой
водой и пищей. Концентрация ртути в разных тканях
взрослых афалин, установленная методом спектрофо-
тометрии, оказалась наибольшей в почках и печени и,
по исследованиям Р. Стикнэя и Уиндона Херберта, в
36—40 раз превышала концентрацию в коже и под-
кожном сале. Возможно, одной из причин слабого вос-
становления численности морских млекопитающих в
районах, где отсутствует промысел на них, является
недостаточная чистота воды. Сейчас много мелких ки-
тообразных гибнет в океане и по другим причинам.
Большие потери наносят рыболовные снасти. Ежегод-
но, например, в сети японских рыбаков попадает более
10 тыс. белокрылых морских свиней, а в тунцеловных
сетях тропической и субтропической зон океана поги-
бает 200—300 тыс. полосатых, вертящихся и пятнис-
тых дельфинов. Бережного и разумного отношения
требуют к себе и мелкие китообразные.
В СССР, как и в других странах, заведена «Красная
книга» со списком животных, существование которых
поставлено под угрозу. Это поможет сохранить для на-
шего Отечества ценные виды млекопитающих и птиц.
В этот список внесены из околоводных животных ка-
лан, бобр, выхухоль, белый медведь; из полуводных —
котик, морж; из водных — все гладкие киты, синий,
серый и горбатый киты,
Социалистическая система дает огромные преиму-
щества в деле охраны среды и в рациональном исполь-
зовании природных ресурсов, в создании наилучшей
формы взаимодействия с природой. В СССР, где уде-
ляется исключительно большое внимание вопросам
борьбы с загрязнением окружающей среды, охрана
природы является важнейшей государственной зада-
чей, делом всего народа. Это подтверждают правитель-
ственные постановления, законодательные акты и кон-
кретные мероприятия. В 1971 году вступили в дейст-
вие «Основы водного законодательства Союза ССР и
союзных республик». В этом законе содержатся обяза-
тельные для всех учреждений и граждан Советской
страны общие положения о порядке использования и
217
охраны рек, озер, морей и водохранилищ. В сфере ми-
ровой юриспруденции нет такого закона, поскольку не
в интересах частного предпринимательства затрачи-
вать крупные средства на строительство очистных со-
оружений, на мероприятия по охране природы.
Наша страна приняла существенные меры также по
защите китообразных. Введение запрета добычи мел-
ких китообразных в СССР в 1966 году встретило ши-
рокое одобрение со стороны ученых многих стран.
Хочется верить, что дельфины будут ограждены от
бессмысленного истребления, если примеру СССР по-
следуют другие государства, как это сделали Болгария
и Румыния.
Китообразные и ластоногие не только ценные для
человека животные, но и влиятельные члены биогео-
ценоза, своеобразные аккумуляторы в океане, перера-
батывающие органические вещества в полезную для
людей продукцию — жир и мясо. Этих животных нуж-
но во что бы то ни стало сохранить. Вероятно, недале-
ко то время, когда будут заниматься организацией
океанского продуктивного хозяйства, и в нем достой-
ное место займут морские млекопитающие.
Приложение
СИСТЕМАТИЧЕСКИЙ СПИСОК ВИДОВ ОТРЯДА
КИТООБРАЗНЫХ
39 родов и 77 видов
А. Подотряд УСАТЫЕ КИТЫ — Mysticeti (3 семейства, 6
родов и 10 видов). Во рту цедильный аппарат или китовый
ус.
I. Семейство ГЛАДКИЕ КИТЫ — Balaenidae. Ныне очень ред-
кие киты. Брюхо гладкое без продольных полос. Пластины
китового уса (по 230—400 пластин слева и справа) узкие,
высокие с волосовидной бахромой. Микропланктонофаги.
В семействе 3 рода и 3 крайне редких вида.
1) Греиладский кит — Balaena mysticetus L. Длина тела до
21 м. Пластины китовых усов до 3—4,5 м высоты, темные.
Живет в морях Арктики и Беринговом и Охотском морях
среди льдов.
2) Южный кит — Eubalaena glacialis Muller. Длина тела 17—
21 м: в Северной Атлантике подвид бискайский кит и в
Южном полушарии австралийский кит до 17 м, а в Северной
Пацифике японский кит до 21 м. На морде сверху большой
роговой нарост. Пластины китового уса до 2,6 м, темные.
Обитает в умеренных водах Мирового океана.
3) Карликовый кит — Caperea marginata Gray. Длина тела
до 6,4 м. Пластины усов до 70 см высоты, желтовато-белые.
Умеренные воды Южного полушария (близ Новой Зеландии,
Австралии, Ю. Африки и Ю. Америки).
II. Семейство ПОЛОСАТИКИ — Balaenopteridae. Стройные киты
с многочисленными продольными полосами на брюхе. Пла-
стины китовых усов низкие, не выше 1 м, по 260—470 штук
справа и слева, со щетиновидной бахромой. В семействе 2
рода и 6 видов.
4) Синий кит — Balaenoptera musculus L. Размер тела до
33 м. Пластины китового уса черные. Планктонофаг. Космо-
полит. Сохранилось в океане не более 13 тысяч голов.
-5) Финвал — Balaenoptera physalus L. Размер тела до 23 —
27 м. Китовый ус темно-серый с голубоватым оттенком, вы-
сотой до 90 см. Брюхо фарфорово-белое. Космополит.
6) Сейвал — Balaenoptera borealis Lesson. Длина до 18 м.
Пластины уса черные с тонкой волосовидной бахромой пе-
пельного цвета, до 80 см высоты. Космополит. Ныне основ-
ной объект китобойного промысла.
7) Полосатик Брайда — Balaenoptera edeni Anderson. Длина
тела до 15,5 м. Пластины китового уса темные со щетино-
видной бахромой. Теплые воды Мирового океана, ограничен-
ные изотермой в 20° С.
8) Малый полосатик — Balaenoptera acutorostrata Lacepede.
Длина до 10 м. Китовый ус белый, высотой до 25 см. Космо-
полит.
219
9) Горбатый кит — Megaptera novaeangliae Borowski. Длина
до 18 м. Китовый ус высотой до 1 м, пепельно-черного цвета.
Космополит.
III. Семейство СЕРЫЕ КИТЫ — Eschrichtiidae. Усовых пластин
по 130—180 справа и слева, белого цвета, с толстой бахро-
мой. На брюхе 2—4 глубокие борозды, расходящиеся назад.
Один вид.
10) Серый кит — Eschrichtius gibbosus Erxleben. Длина до
15 м. Живет в прибрежных водах Северной Пацифики.
Б. Подотряд ЗУБАТЫЕ КИТЫ — Odontoceti. Во рту от 1 до
240 зубов одновершинного типа (4 семейства, 33 рода и 68
видов).
IV. Семейство РЕЧНЫЕ ДЕЛЬФИНЫ — Platanistidae. Древней-
шие и наиболее примитивные зубатые китообразные. Рыло
узкое, сильно вытянуто, с длинным нижне-челюстным сим-
физом. Зубы с расширенными корнями и гребнями на корон-
ках. Обитают в пресных реках и озерах. В семействе 4 рода
и 5 очень малочисленных видов.
11) Гаагский дельфин — Platanista gangetica Lebeck. Длина
тела до 2,5 м. Зубов по 29—33 пары вверху и внизу. Хруста-
лик глаза утрачен. Обитает в реках Ганге, Брахмапутре и их
притоках.
12) Индский дельфин — Platanista indi Blyth. Длина до 2 м.
зубов по 26 пар вверху и внизу. Обитает в реке Инд и при-
токах.
13) Иния — Inia geoffrensis Blainville. До 2,5 м. Зубов по
33—34 пары вверху и внизу. Бассейны рек Амазонки, Ори-
ноко, Токантинса.
14) Китайский озерный дельфин — Lipotes vexillifer Miller.
Зубов по 33—36 пар вверху в внизу. Озеро Дунтинху и реке
Янцзы.
15) Лаплатский дельфин — Pontoporia blainvillei Gervais,
Orbigny. До 1,7 м. Зубов по 50—60 пар вверху и внизу. Жи-
вет в реке Ла-Плата и прибрежных водах Бразилии, Уругвая
и Аргентины.
• Семейство КАШАЛОТОВЫЕ — Physeteridae. Зубы только в
нижних челюстях. Рот открывается снизу головы. В семей-
стве 2 рода и 3 вида.
16) Кашалот — Physeter catodon L. Длина самцов до 20, са-
мок до 15 м. Зубов до 30 пар. Самцы космополиты, самки —
в теплых водах между 40° с. ш. и 40° ю. ш.
17) Карликовый кашалот — Kogia breviceps Blainville. Дли-
на тела до 3,4 м. Зубов 12—16 пар, только нижних, верхние
отсутствуют. Теплый пояс Мирового океана.
18) Малый карликовый кашалот — Kogia simus Owen. Длина
тела до 2,7 м. Зубов 8—11 пар внизу и 0—3 пары вверху.
Теплый пояс Мирового океана.
VI. Семейство КЛЮВОРЫЛЫЕ КИТЫ — Ziphiidae. Под горлом
2—4 борозды. Зубов 1—2 пары лишь в нижней челюсти
(исключение — тасманов клюворыл). На хвостовом плавнике
нет выемки. Глубоко и надолго ныряют. Питаются главным
образом головоногими моллюсками. В семействе 5 родов и
16 видов.
19) Северный плавун — Berardius bairdi Steineger. Самки до
220
12,5 м, самцы до 11 м. Обитает в Северной Пацифике от
Японии и Калифорнии до Берингова моря.
20) Южный плавун — Berardius arnouxi Duvernoy. До 11 м.
Живет в южных частях Тихого, Индийского и Атлантичес-
кого океанов.
21) Высоколобый бутылконос — Hyperoodon ampullatus For-
ster. Самцы до 9,4 м, самки до 8,7 м. Северная Атлантика.
22) Плосколобый бутылконос — Hyperoodon planifrons Flo-
wer. До 8 м. Южные части Тихого, Индийского и Атланти-
ческого океанов.
23) Настоящий клюворыл — Ziphius cavirostris Cuvier. До
8 м. Умеренный и теплый пояс Мирового океана.
24) Тасманов клюворыл — Tasmacetus shepherdi Oliver. До
9 м. Зубов до 19 пар вверху и 27 пар внизу. Воды Новой
Зеландии.
25) Антильский ремнезуб — Mesoplodon europaeus Gervais.
До 6,7 м. Умеренный и теплый пояс Атлантического океана.
- 26) Ремнезуб Тру — Mesoplodon mirus True. До 5 м. Умерен-
ные воды Северной и Южной Атлантики.
27) Атлантический ремнезуб — Mesoplodon bidens Sowerby.
До 6 м. Северная Атлантика.
28) Командорский ремнезуб — Mesoplodon steinegeri True.
До 6 м. Северная Пацифика.
29) Ремнезуб Карлхубса — Mesoplodon carlhubbsi Moore. До
6 м. Умеренная зона Северной Пацифики.
30) Японский ремнезуб — Mesoplodon ginkgodens Nishiwaki,
Kamiya. До 5 м. Воды Японии и Калифорнии.
31) Новозеландский ремнезуб — Mesoplodon bowdoini An-
drews. До 5 м. Воды Новой Зеландии.
32) Ремнезуб Лейрда — Mesoplodon layardi Gray. До 6 м.
Воды Ю. Африки, Новой Зеландии, Австралии, Фолкленд-
ских островов.
33) Ремнезуб Грэя — Mesoplodon grayi Haast. До 4,5 м. Воды
Новой Зеландии, Австралии, Патагонии, Южной Африки,
Фолклендских островов.
34) Ремнезуб Бленвиля — Mesoplodon densirostris Blainville.
До 4,4 м. Теплый пояс Мирового океана.
VH. Семейство ДЕЛЬФИНОВЫЕ — Delphinidae. Под горлом нет
борозд. Зубы и в верхних и в нижних челюстях (исключе-
ние — нарвал). В семействе 21 род и 44 вида.
35) Афалина — Tursiops truncatus Montagu. Длина до 3,6 м
в океане и до 3,3 м в Черном море. Зубов по 19—24 пары
вверху и внизу. Умеренные и теплые воды Мирового океана.
36) Обыкновенный дельфин — Delphinus delphis L. До 2,1 м
в Черном море и до 2,6 м в океане. Зубов 200—220. Обитает
в умеренных и теплых водах Мирового океана.
37) Тропический дельфин — Delphinus tropicalis Van Bree.
Дл. до 2 м. Зубов 240—258. Обитает в тропическом поясе
Мирового океана (побережье Сомали, Малабара, Пакистана,
Южно-Китайского моря).
38) Полосатый продельфин — Stenella coeruleoalbus Meyen.
До 2,5 м. Зубов по 44—50 пар вверху и внизу. Тропические,
субтропические и умеренные воды Мирового океана.
39) Малайский продельфин — Stenella dubia Cuvier. До 2,1 м.
221
Зубов по 38—45 пар вверху и внизу. Тропические части Ти-
хого и Индийского океанов.
40) Пятнистый продельфин — Stenella plagiodon Соре. До
2,15 м, Зубов 180—208 штук. Теплые воды Атлантики.
41) Уздечковый продельфин — Stenella frontalis Cuvier. До
1,8 м. Зубов 148—152. Теплые воды Атлантики, Индийского
океана и Средиземное море.
42) Вертящийся продельфин — Stenella longirostris Gray. До
2,1 м. Зубов до 204 штук. Тропические и субтропические во-
ды Тихого и Индийского океанов.
43) Амазонский дельфин — Sotalia fluviatilis Gervais, Deville.
До 1,6 м. Зубов до 122 штук. Амазонка и ее притоки.
44) Гвианский дельфин — Sotalia guianensis Van Beneden. До
1,6 м. Зубов до 122 штук. Прибрежные воды и устья рек
Гвианы и Венесуэлы.
45) Китайский белый дельфин — Sotalia chinensis Osbeck. До
2 м. Зубов до 128 штук. Реки Южного Китая.
46) Зондский белый дельфин — Sotalia borneensis Lydekker.
Зубов до 148—-156 штук. Живет в Сиамском заливе, Южно-
Китайском море.
47) Крапчатый дельфин — Sotalia lentiginosa Owen. До 2,4 м.
Зубов 144. Индийский океан от Шри Ланки до Южной Аф-
рики.
48) Свинцовый дельфин — Sotalia plumbea Cuvier. До 2,4 м.
Зубов до 148 штук. Индийский океан.
49) Западноафриканский дельфин — Sotalia teuszi Kiikenthal.
До 2,5 м. Зубов 120. Бухты и реки Западной Африки (от Се-
негала до Камеруна).
50) Морщинистозубый дельфин — Steno bredanensis Lesson.
До 2,4 м. Зубов 104. Умеренный и тропический пояс Миро-
вого океана.
51) Северный китовидный дельфин — Lissodelphis borealis
Peale. До 2,4 м. Зубов до 188 штук. Северная Пацифика.
52) Южный китовидный дельфин — Lissodelphis peroni Lace-
pede. До 2,3 м. Зубов 180. Умеренный пояс Южного полу-
шария.
53) Тихоокеанский белобокий дельфин — Lagenorhynchus
obliquidens Gill. До 2,3 м. Зубов 120. Северная Пацифика.
54) Атлантический белобокий дельфин — Lagenorhynchus
acutus Gray. До 2,7 м. Зубов до 156 штук. Северная Атлан-
тика.
55) Беломордый дельфин — Lagenorhynchus albirostris Gray.
До 3,0 м. Зубов 108. Северная Атлантика.
56) Крестовидный дельфин — Lagenorhynchus cruciger Quoy,
Gaimard. До 1,8 м. Зубов 112. Умеренный пояс Южного по-
лушария.
57) Тусклый дельфин — Lagenorhynchus obscurus Gray. До
1,8 м. Зубов 124. Умеренный пояс Южного полушария и
Северная Антарктика.
58) Южный белобокий дельфин — Lagenorhynchus australis
Peale. До 2,2 м. Зубов 120. Южные воды Южной Америки.
59) Широкомордый дельфин — Peponocephala electra Gray.
До 2,6 м. Зубов около 100. Встречается от Антарктики до
Японии.
222
60) Саравакский дельфин — Lagenodelphis hosei Fraser. До
2,4 м. Зубов 176. Воды Саравака, остров Борнео.
61) Серый дельфин — Grampus griseus Cuvier. До 3,7 м. Зу-
бов 2—7 пар только в нижней челюсти. Теплый и умеренный
пояса Мирового океана.
62) Обыкновенная гринда — Globicephala melaena Trail. До
6,5 м. Зубов до 48. Живет от умеренных вод Северного полу-
шария до Антарктики.
63) Косатка — Orcinus orca L. До 10 м. Зубов до 40. Космо-
полит.
64) Малая косатка — Pseudorca crassidens Owen. До 6 м. Зу-
бов около 40 штук. Умеренные и теплые воды Мирового
океана.
65) Карликовая косатка — Feresa attenuata Gray. До 2,4 м.
Зубов до 50 штук. Тропики и субтропики Тихого океана и
Атлантики.
66) Иравадийский дельфин — Orcaella brevirostris Owen. До
2,2 м. Зубов 48—52. Моря и устья рек Юго-Восточной Азии
от Мадраса до Бангкока.
67) Дельфин Коммерсона — Cephalorhynchus commersoni
Lacepcde. До 1,58 м. Зубов до 120 штук. Южная Атлантика
(пролив Магеллана, Фолклендские острова, остров Кергелен).
68) Дельфин Хевисайда — Cephalorhynchus heavisidei Van
Beneden. Длина тела около 1,2 м. Зубов 120. Воды Южной
Африки.
69) Дельфин Гектора — Cephalorhynchus hectori Van Bene-
den. До 1,8 м. Зубов до 128. От Южно-Китайского моря до
Новой Зеландии.
70) Чилийский дельфин — Cephalorhynchus eutropia Gray.
Около 1,35. м. Зубов до 128. Воды Чили.
71) Обыкновенная морская свинья — Phocoena phocoena L.
До 1,8 м. Зубов до 110. Северная Атлантика и Северная Па-
цифика.
72) Калифорнийская морская свинья — Phocoena sinus Nor-
ris, McFarland. До 1,6 м. Калифорнийский залив.
73) Очковая морская свинья — Phocoena dioptrica Lahille.
До 2 м. Зубов 84. Моря, омывающие Перу, Чили, Аргентину,
Южные Фолклендские острова, Южную Георгию.
74) Черная морская свинья — Phocoena spinipinnis Burmeis-
ter. До 1,8 м. Зубов 68. Воды Перу, Аргентины и Чили.
75) Бесперая морская свинья — Neophocoena phocoenoides
Cuvier. До 1,6 м. Зубов 80. Прибрежные воды Тихого и
Индийского океанов от Японии до мыса Доброй Надежды.
76) Белокрылая морская свинья — Phocoenoides dalli True.
До 2 м. Зубов до 104. Северная Пацифика.
77) Белуха — Delphinapterus leucas Pallas. До 6 м. Зубов 40.
Арктика и прилежащие моря.
«5 78) Нарвал — Monodon monoceros L. До 6 м. Зубов 1—2
(только верхние). Арктика.
ОГЛАВЛЕНИЕ
Глава I. НАШЕСТВИЕ СЕНСАЦИИ..................... 3
Глава II. ПОМОЩНИКИ И ДРУЗЬЯ ЧЕЛОВЕКА В
МОРЕ...................................... 6
Глава III. С СУШИ В ВОДУ...................... 11
Глава IV. ДЕЛЬФИНЫ В ОКЕАНСКИХ ЦИРКАХ . . 25
Глава V. ПОВЕДЕНИЕ УЗНИКОВ.................. 39
Глава VI. АФАЛИНЫ РАЗМНОЖАЮТСЯ В НЕВОЛЕ 58
Глава VII. ДЕЛЬФИНЫ УСЛОЖНЯЮТ НАВЫКИ . . 64
Глава VIII. ЛЮБИМЦЫ ЗРИТЕЛЕЙ — ТИРАНЫ КИТОВ 71
Глава IX. СКОРОХОДЫ МОРЕЙ...................... 80
Глава X. ПОКОРИТЕЛИ МОРСКИХ ГЛУБИН ... 91
Глава XI. СУДЬБА АНАЛИЗАТОРОВ..................103
Глава XII. «АКУСТИЧЕСКИЕ ГЛАЗА» ДЕЛЬФИНОВ 120
Глава XIII. ПОЮЩИЕ КИТЫ И «МОРСКИЕ ПОПУГАИ» 127
Глава XIV. ОБЛАДАЮТ ЛИ ДЕЛЬФИНЫ РЕЧЬЮ? . . 146
Глава XV. ЗАЧАТКИ РАССУДОЧНОЙ ДЕЯТЕЛЬНОСТИ
У ДЕЛЬФИНОВ УЖИВАЮТСЯ С ИН-
СТИНКТАМИ ........................... . . 155
Глава XVI. РЕФЛЕКСЫ ВЫНЫРИВАНИЯ И ИНСТИНКТ
СОХРАНЕНИЯ ВИДА..........................158
Глава XVII. БЕДСТВИЯ СРЕДИ ЛЬДОВ...............175
Глава XVIII. СТРАННЫЕ ПОПУТЧИКИ ИЛИ БЕЗДУМ-
НЫЕ ЛОЦМАНЫ..............................179
Глава XIX. СПЯТ ЛИ КИТЫ?.......................183
Глава XX. ДЕЛЬФИНЫ И НАУКА.....................187
Глава XXI. СИГНАЛЫ ДАЛЬНЕГО ДЕЙСТВИЯ И ЗВУ-
КОВЫЕ МАЯКИ В ОКЕАНЕ................... 194
Глава XXII. «ЗВЕЗДЫ МОРЯ-» СЛУЖАТ ЧЕЛОВЕКУ . . 197
Глава XXIII. БУДУЩЕЕ ДЕЛЬФИНОВ ................208
Глава XXIV. МИР ДЕЛЬФИНОВ И КИТОВ В ОПАСНОСТИ 212
Авенир Григорьевич Томилин
В МИРЕ КИТОВ И ДЕЛЬФИНОВ
Издание второе, переработанное и дополненное
Зав. редакцией М. Новиков. Редактор В. Климачева
Мл. редактор М. Вержбицкая. Художник В. Пантелеев
Худож. редактор М. Гусева. Техн, редактор С. Птицына
Корректор В. Калинина
ИБ № 2520
Сдано в набор 31.07.79. Подписано к печати 22.07.80. А 11700. Формат
бумаги 84X108732. Бумага книжно-журнальная. Гарнитура школьная.
Печать высокая. Усл. печ. л. 11,76+1,68 вкл. Уч.-изд. л. 11,69+2,16 вкл.
Тираж 100 000 экз. Заказ № 9—1984. Цена 65 коп.
Издательство «Знание». 101835, ГСП, Москва, Центр, проезд Серова, д. 4.
Индекс заказа 807707.
Головное предприятие республиканского производственного объединения
«Полиграфкнига» Госкомиздата УССР, 252057, Киев-57, Довженко, 3.