


























































































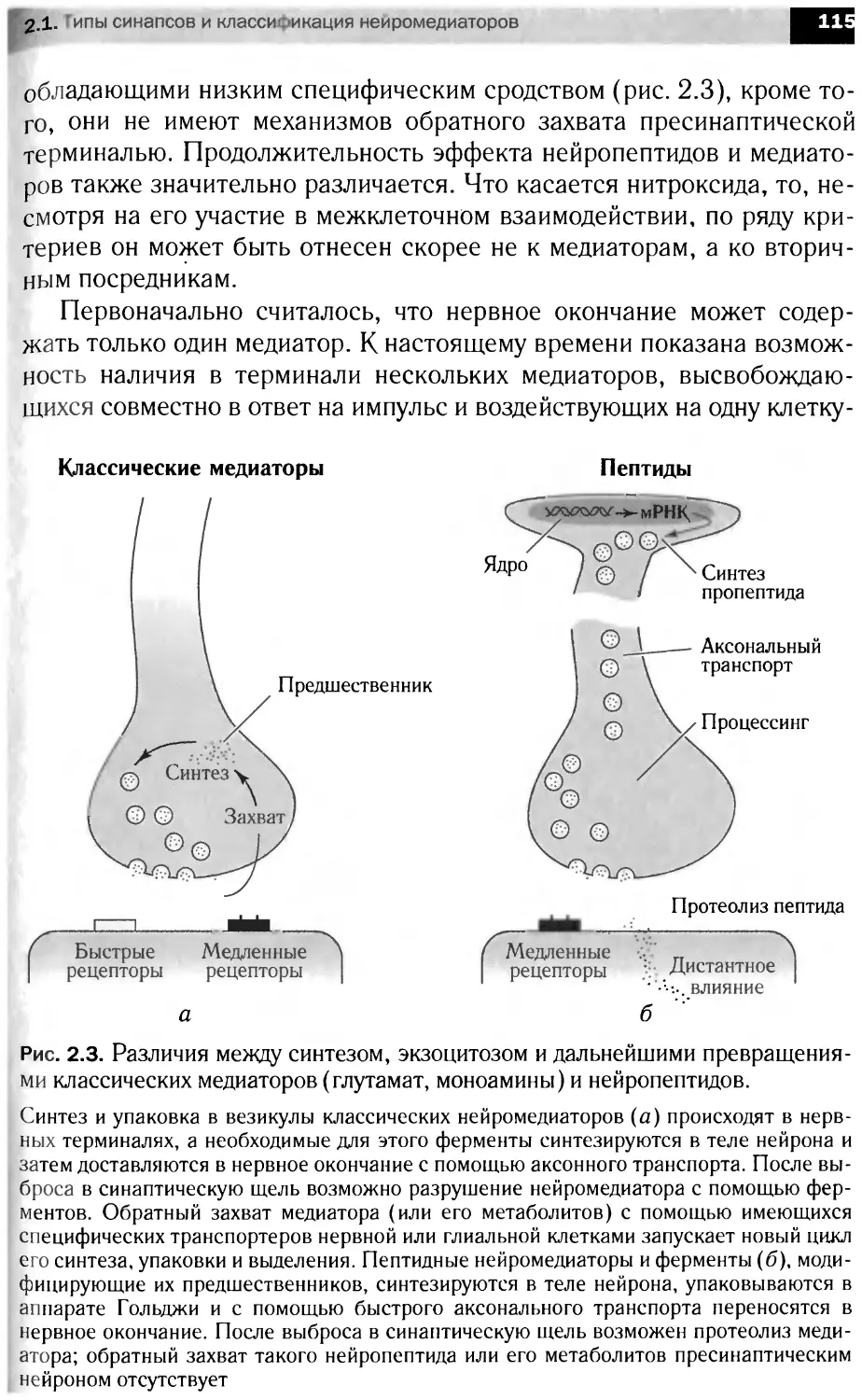


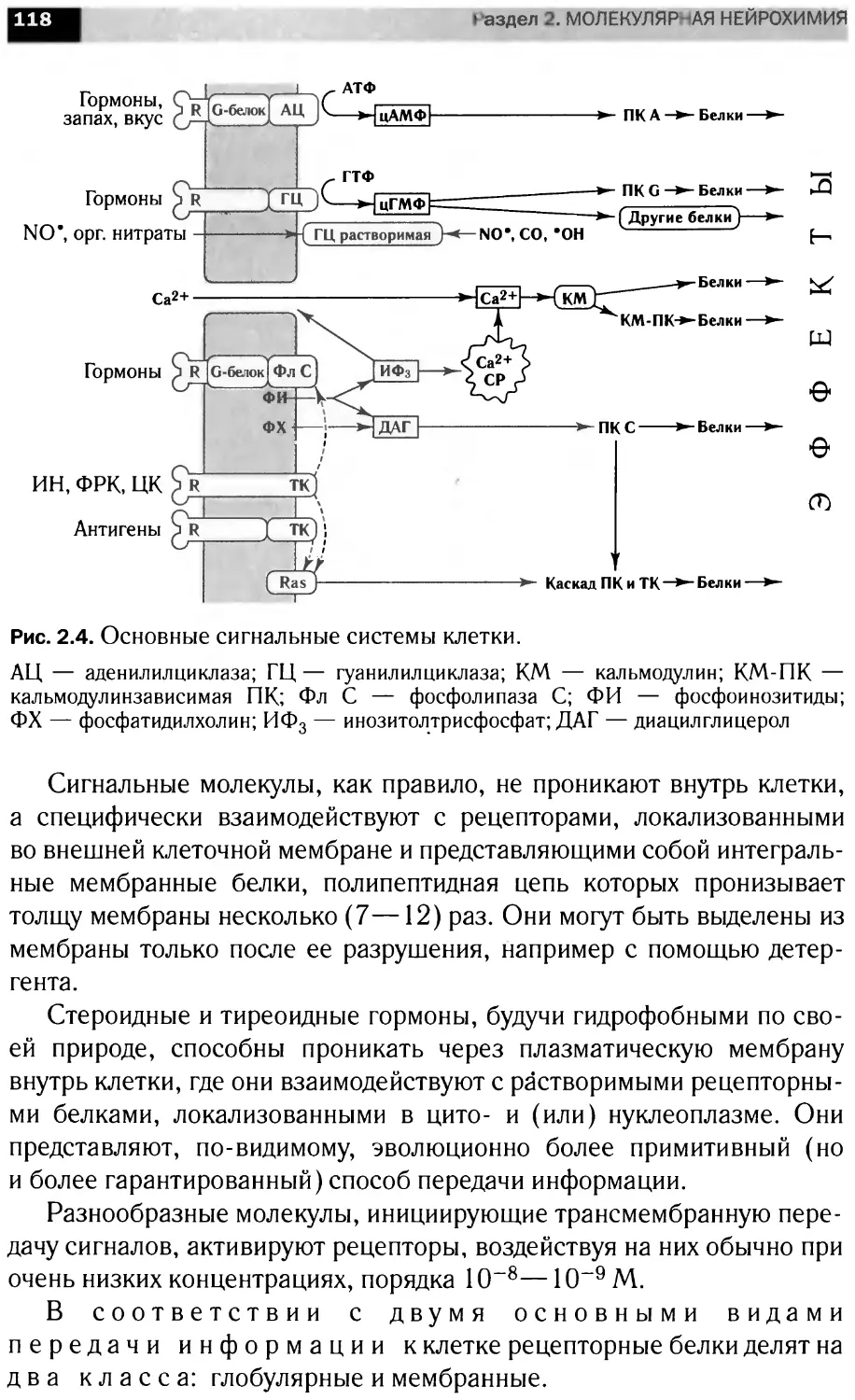





































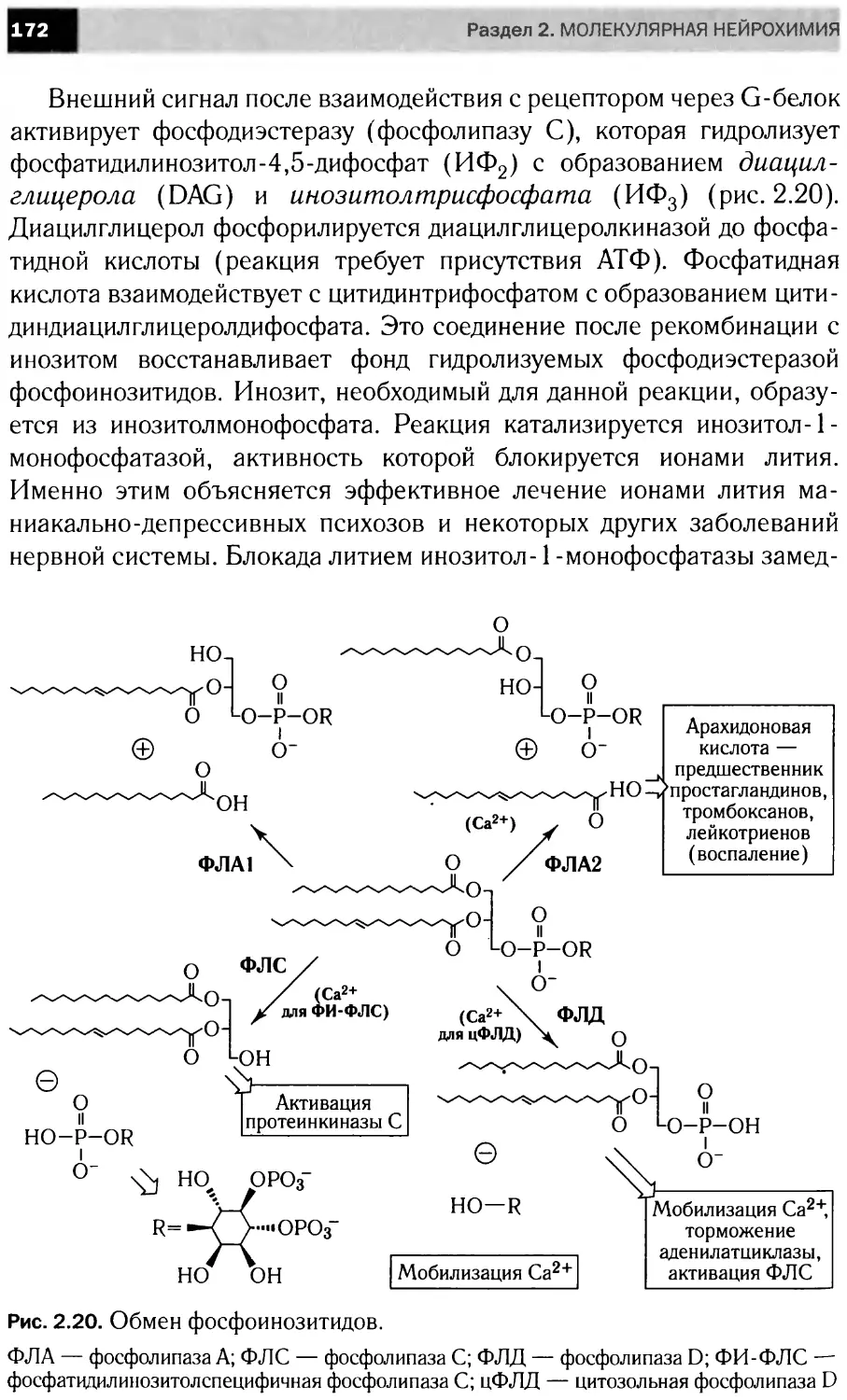















































































































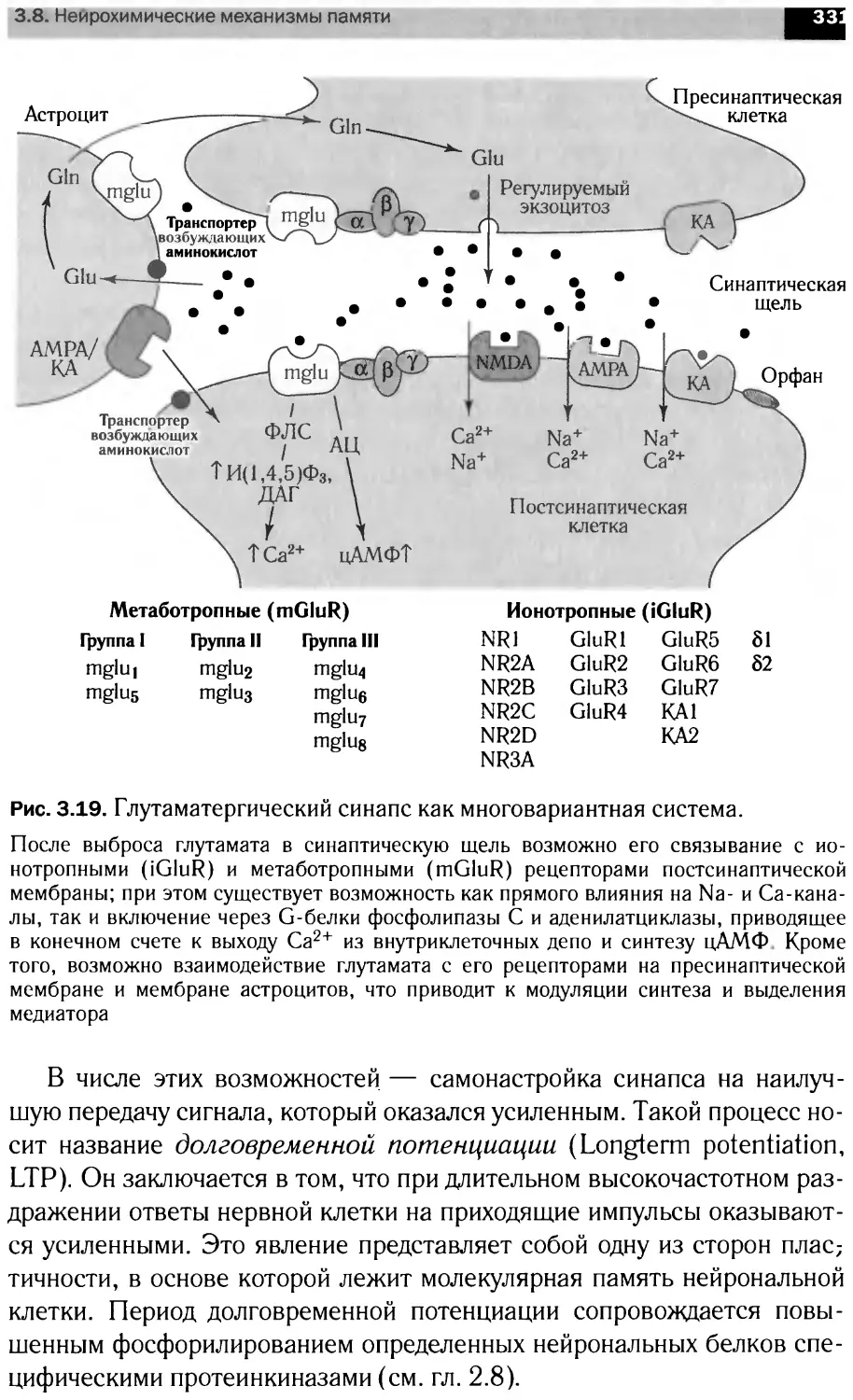

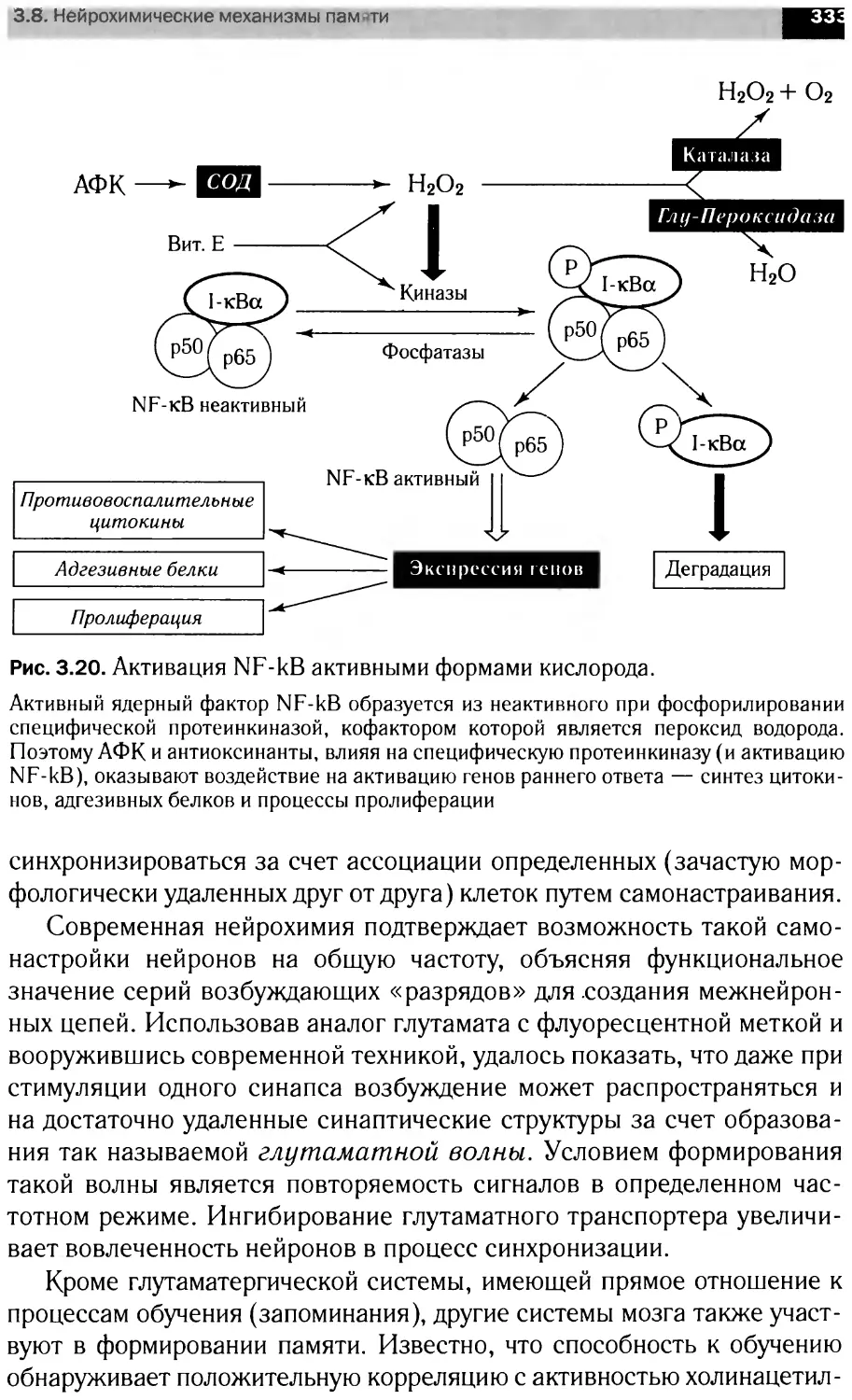



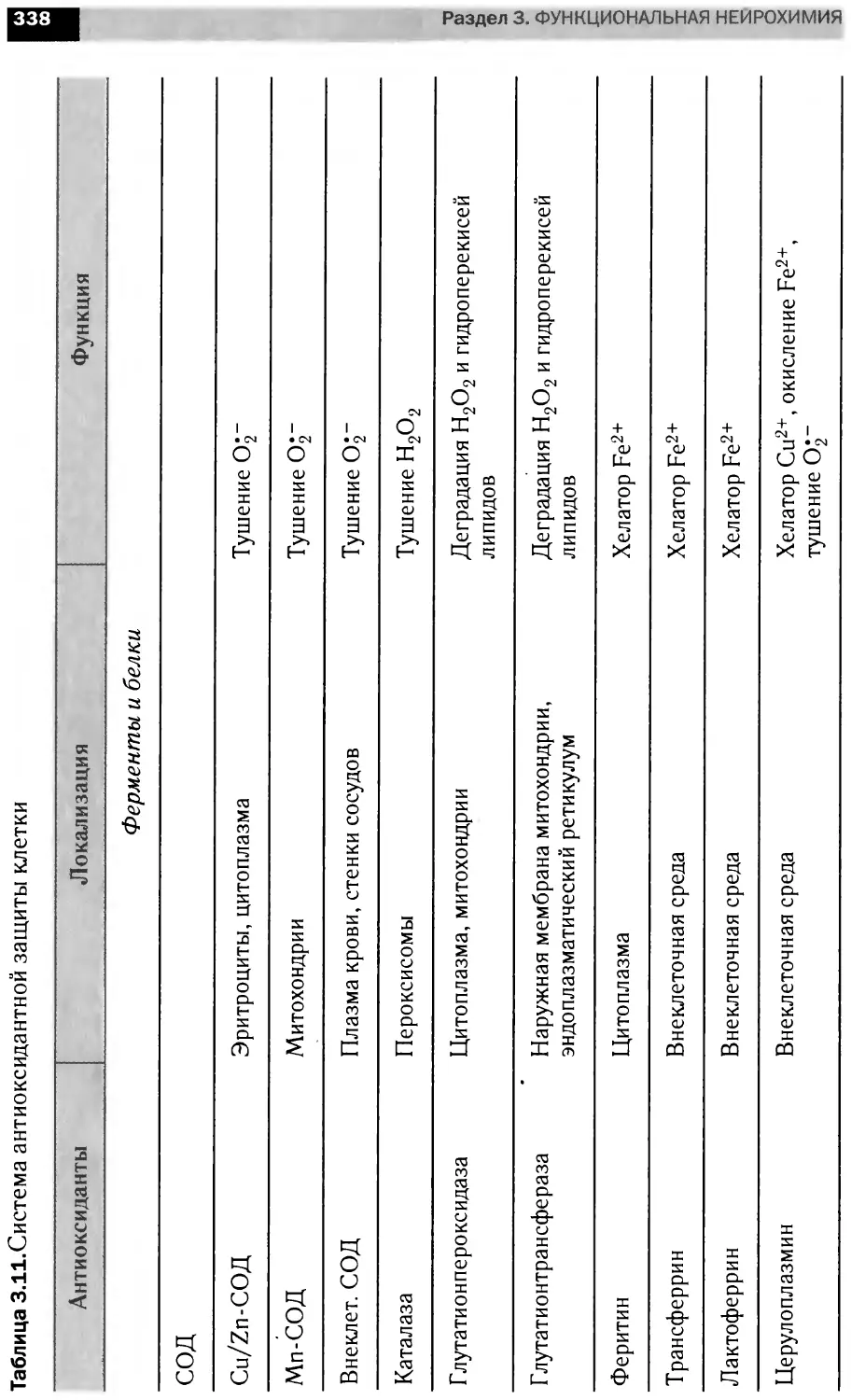






















































Автор: Болдырев А.А. Ещенко Н.Д. Илюха В.А. Кяйвяряйен Е.И.
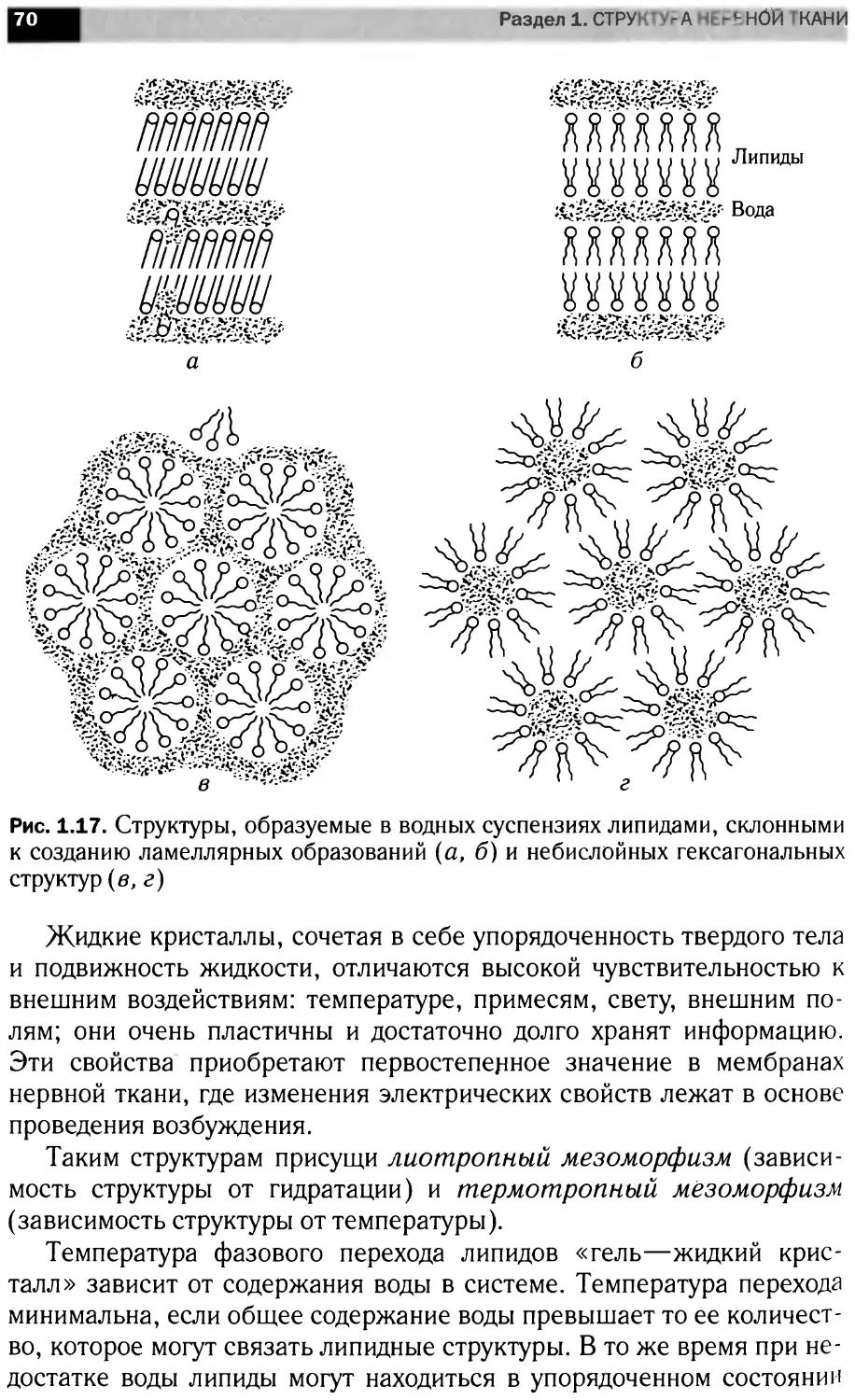
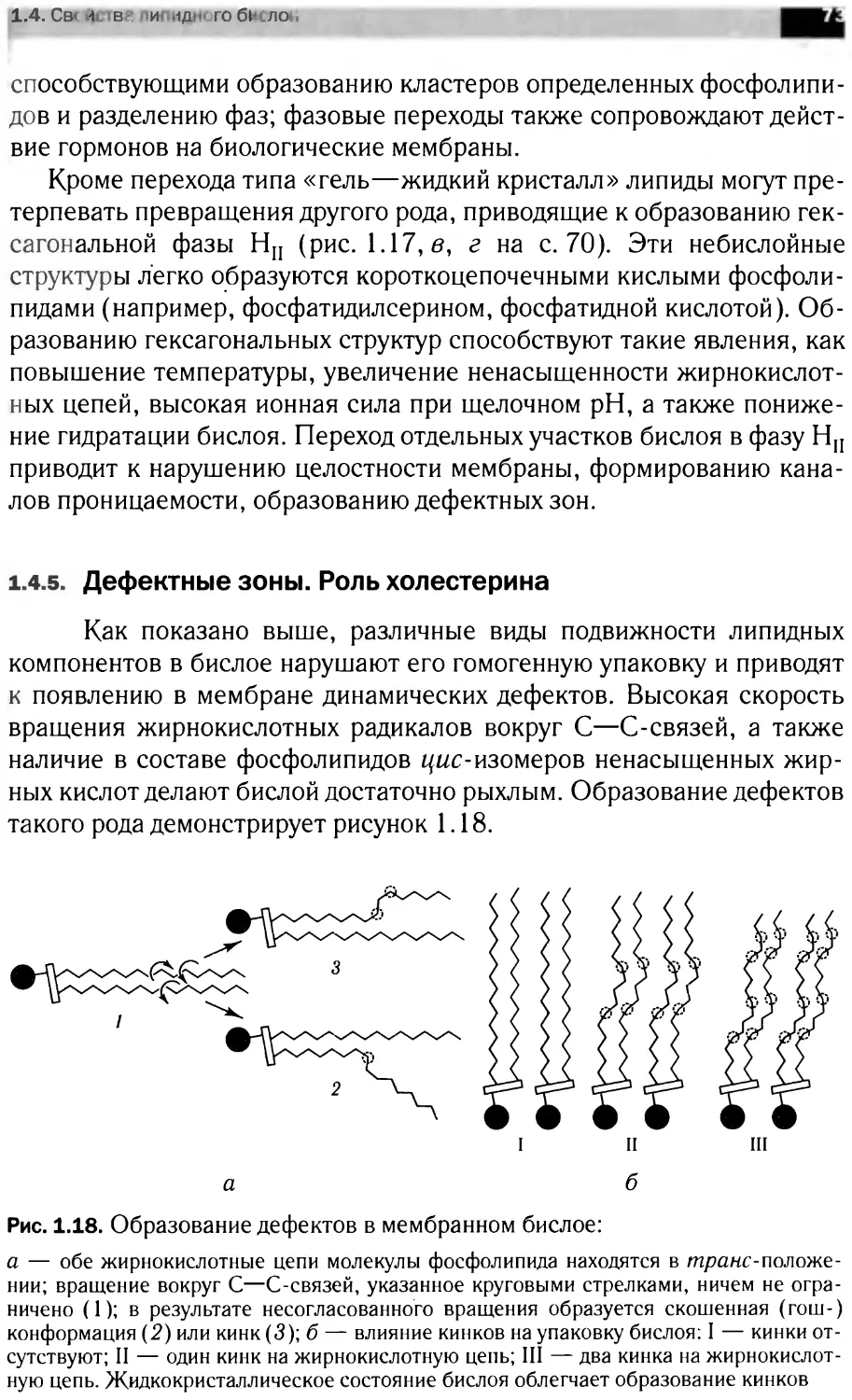
Теги: материальные основы жизни биохимия молекулярная биология биофизика общая биохимия химия эмбриология нейрохимия
ISBN: 978-5-358-03120-3
Год: 2010




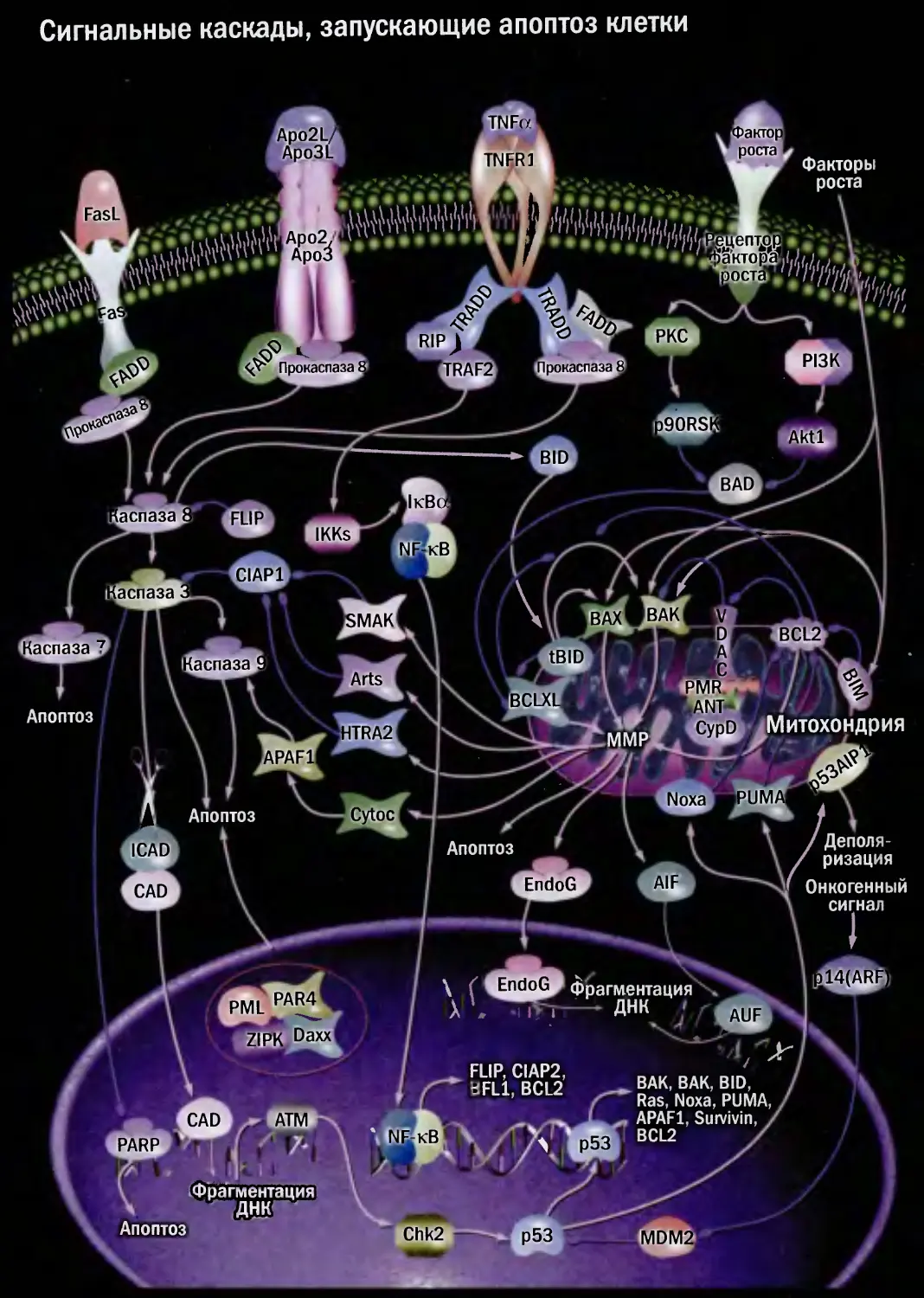
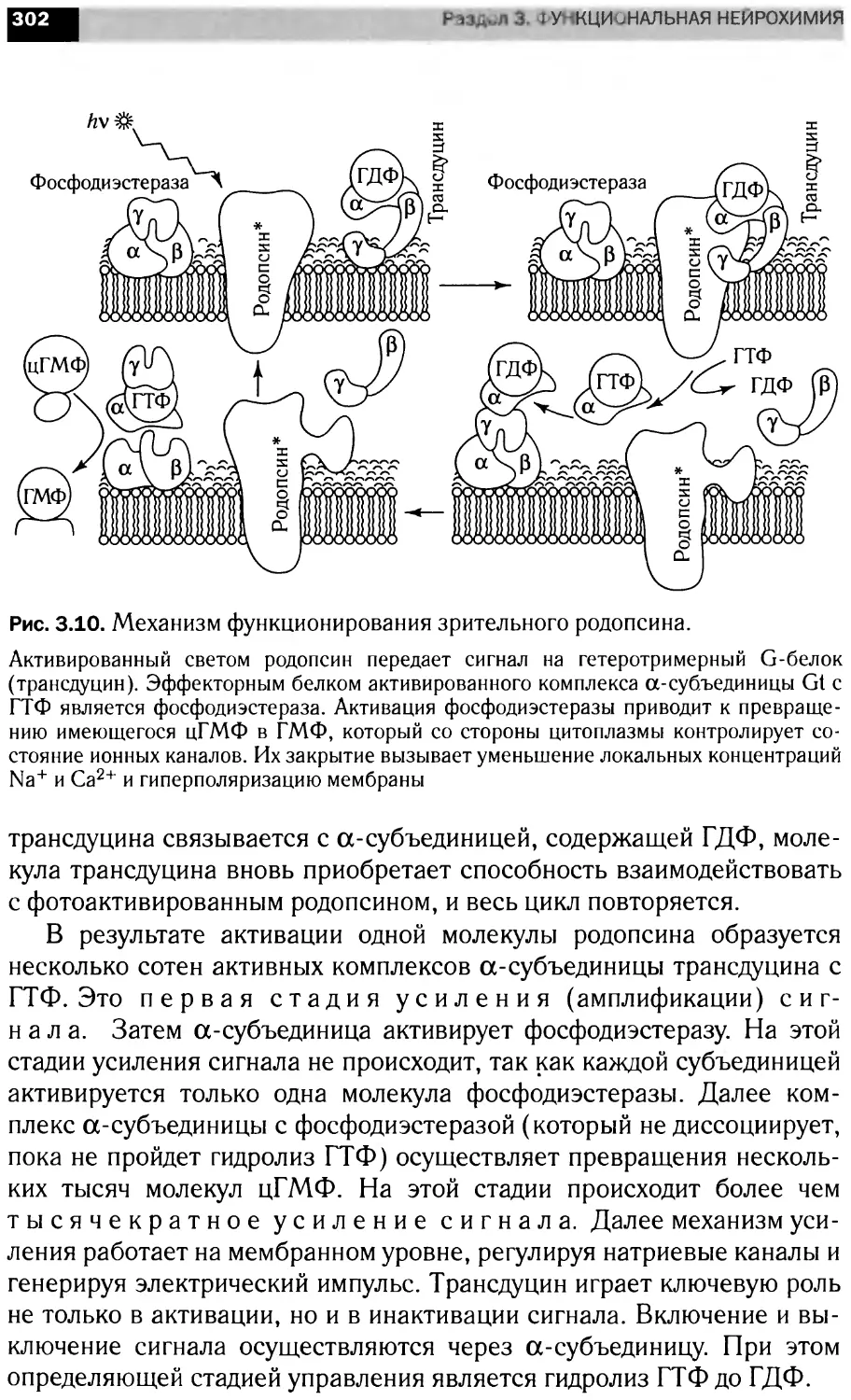
Сигнальные каскады, запускающие апоптоз клетки
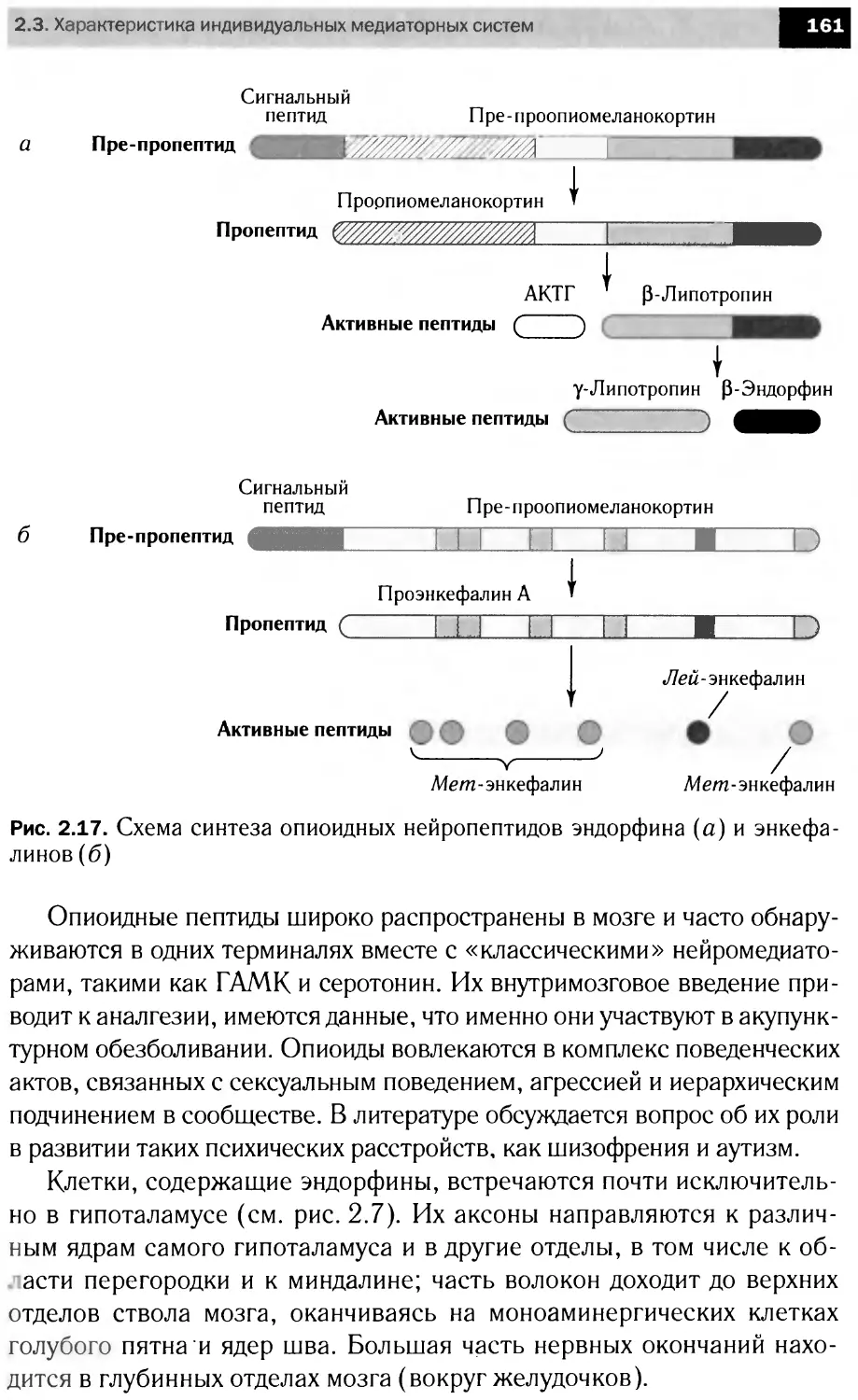
Апоптоз
Митохондрия
Апоптоз
Апоптоз
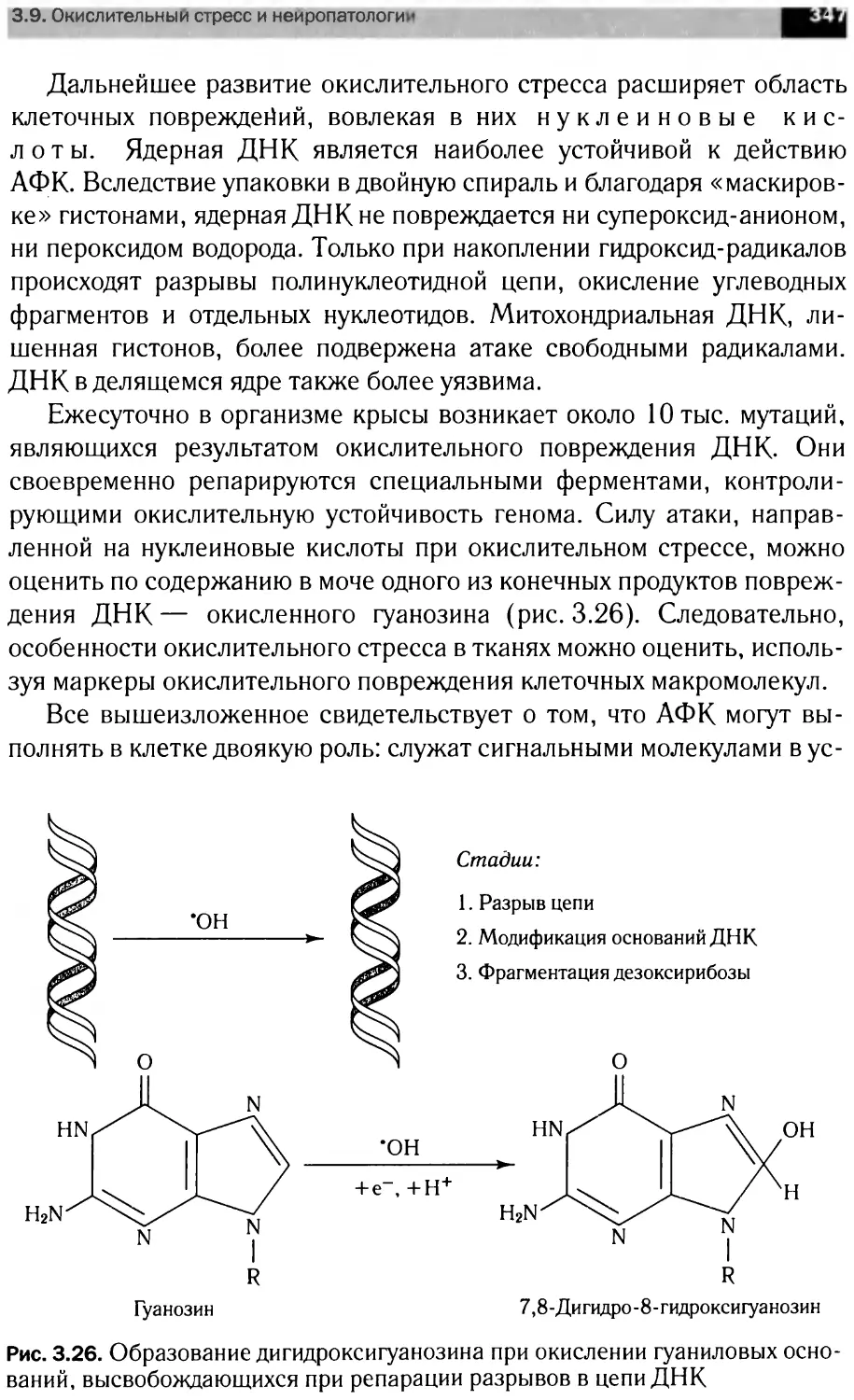
Фрагментация
FLIP, CIAP2,
BFL1, BCL2
Факторы
роста
ВАК, ВАК, BID,
Ras, Noxa, PUMA,
APAF1, Survivin, /
BCL2 /
/ Деполя-
' ризация
Онкогенный
сигнал
Апоптоз
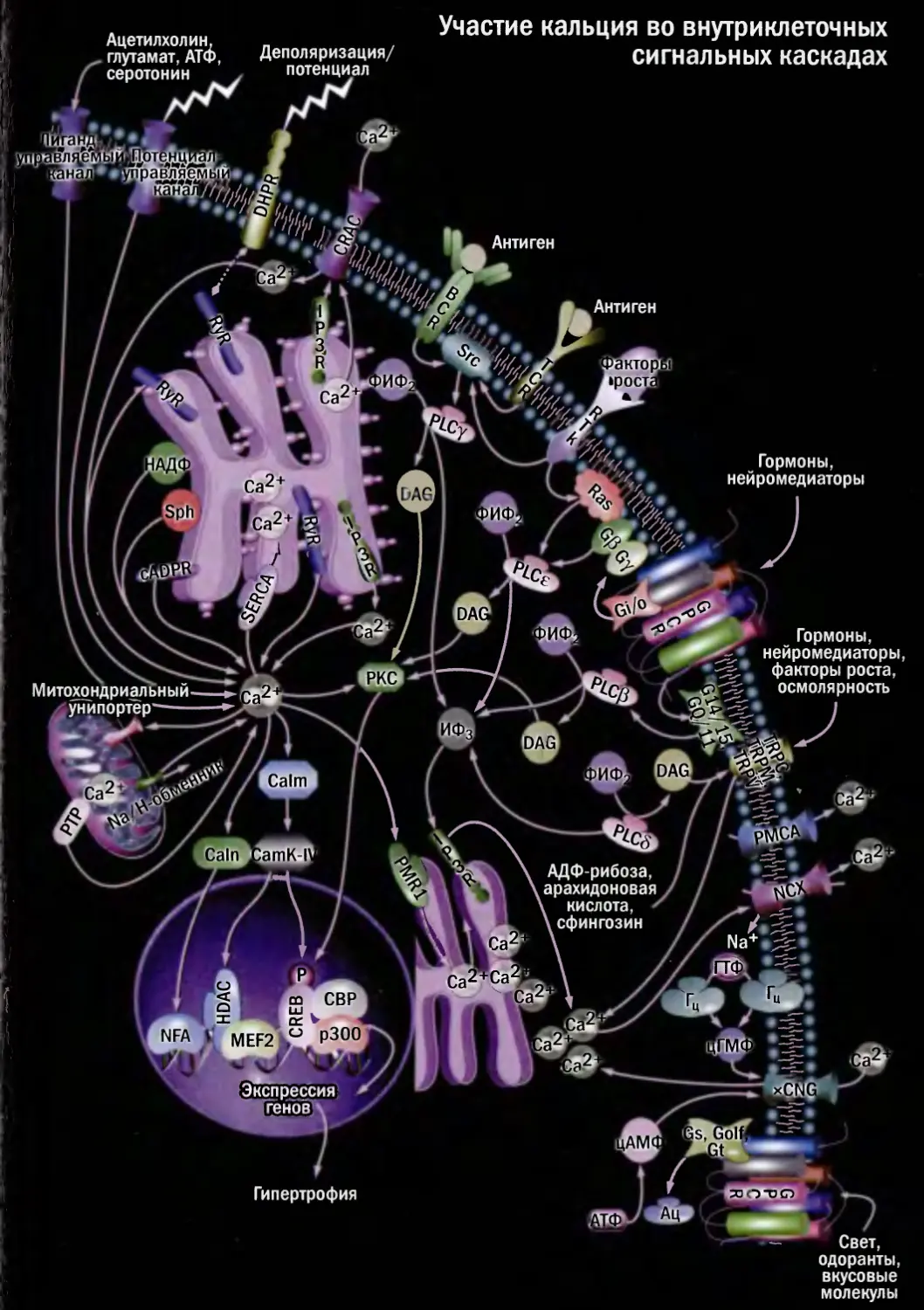
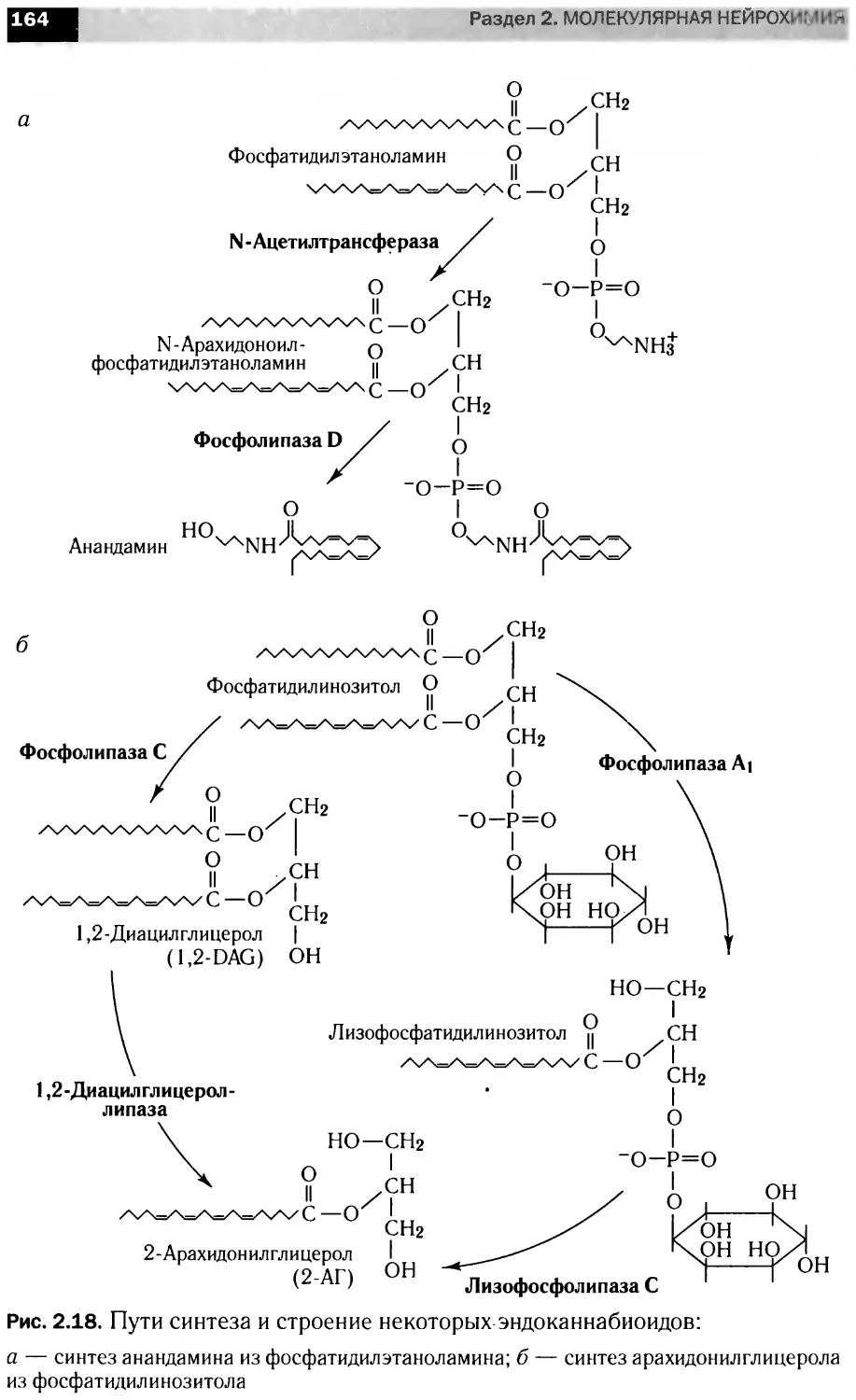
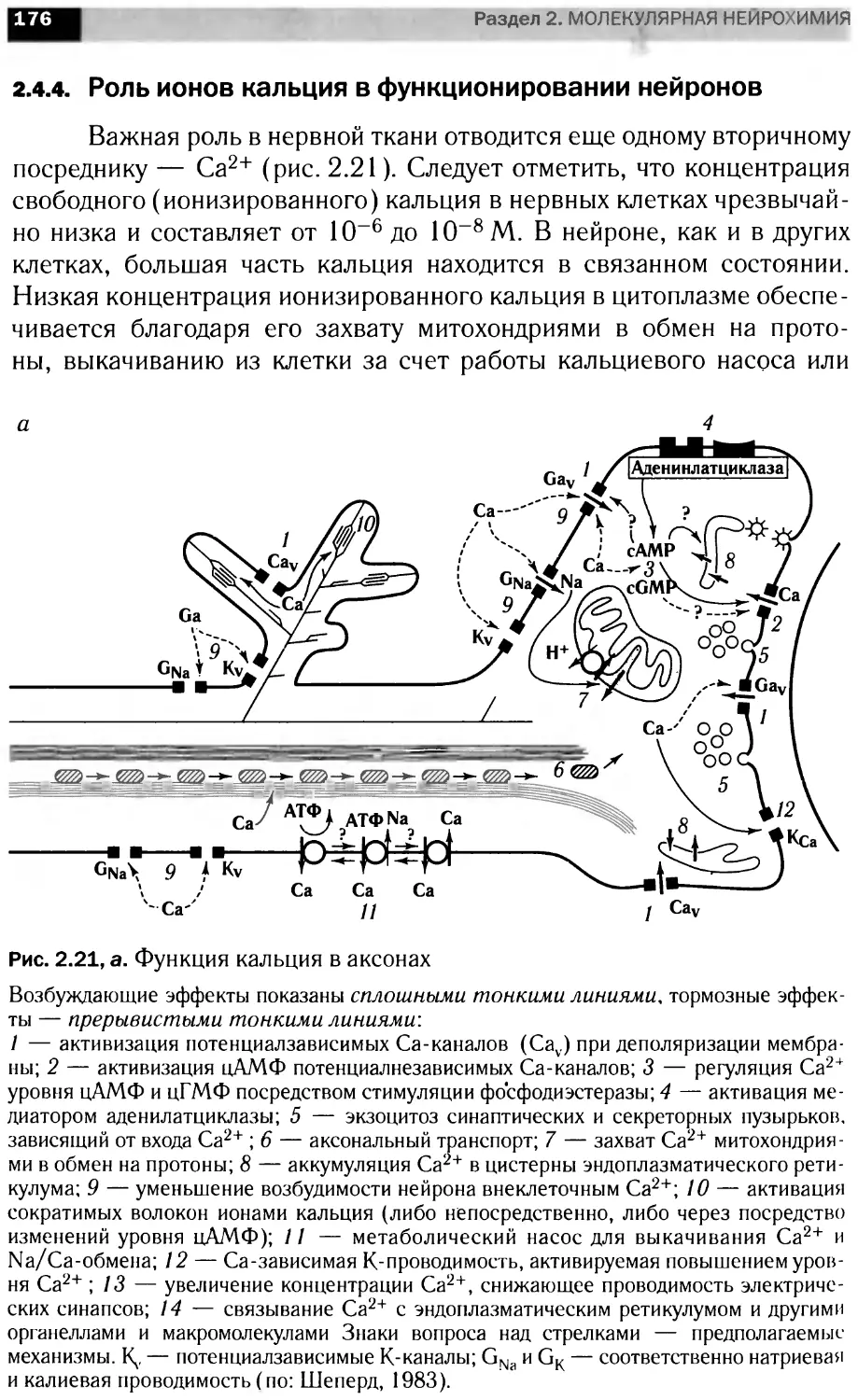
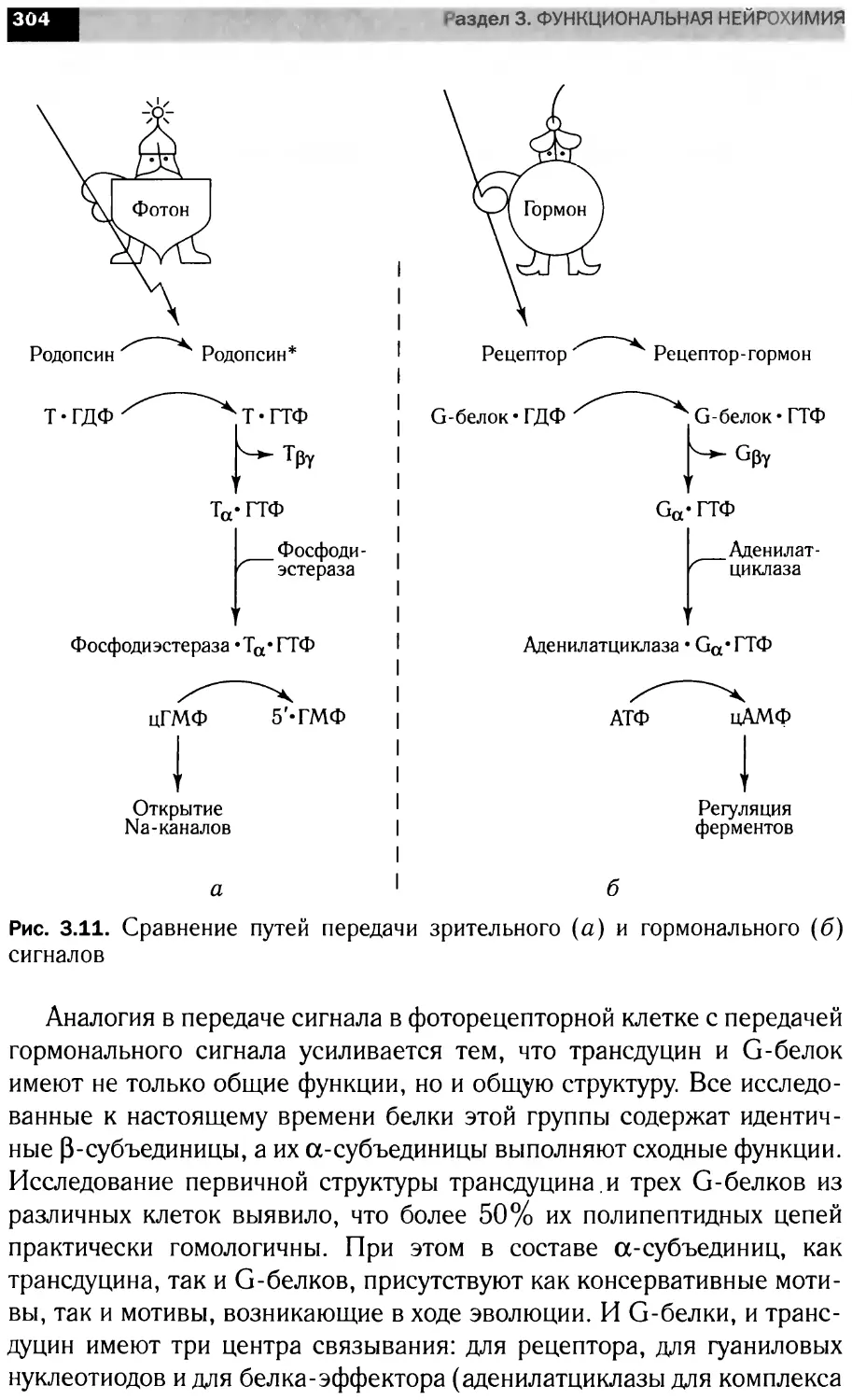
Участие кальция во внутриклеточных
сигнальных каскадах
ж®
Антиген
Антиген
.f
Гормоны,
нейромедиаторы
Ацетилхолин.
глутамат, АТ<Ь, Деполяризация/
серотонин потенциал
Na+.
Гипертрофия
Свет,
нейромедиаторы,
факторы роста,
осмолярность
Митохондриальный
унипортерт
кислота,
сфингозин
одоранты,
вкусовые
молекулы
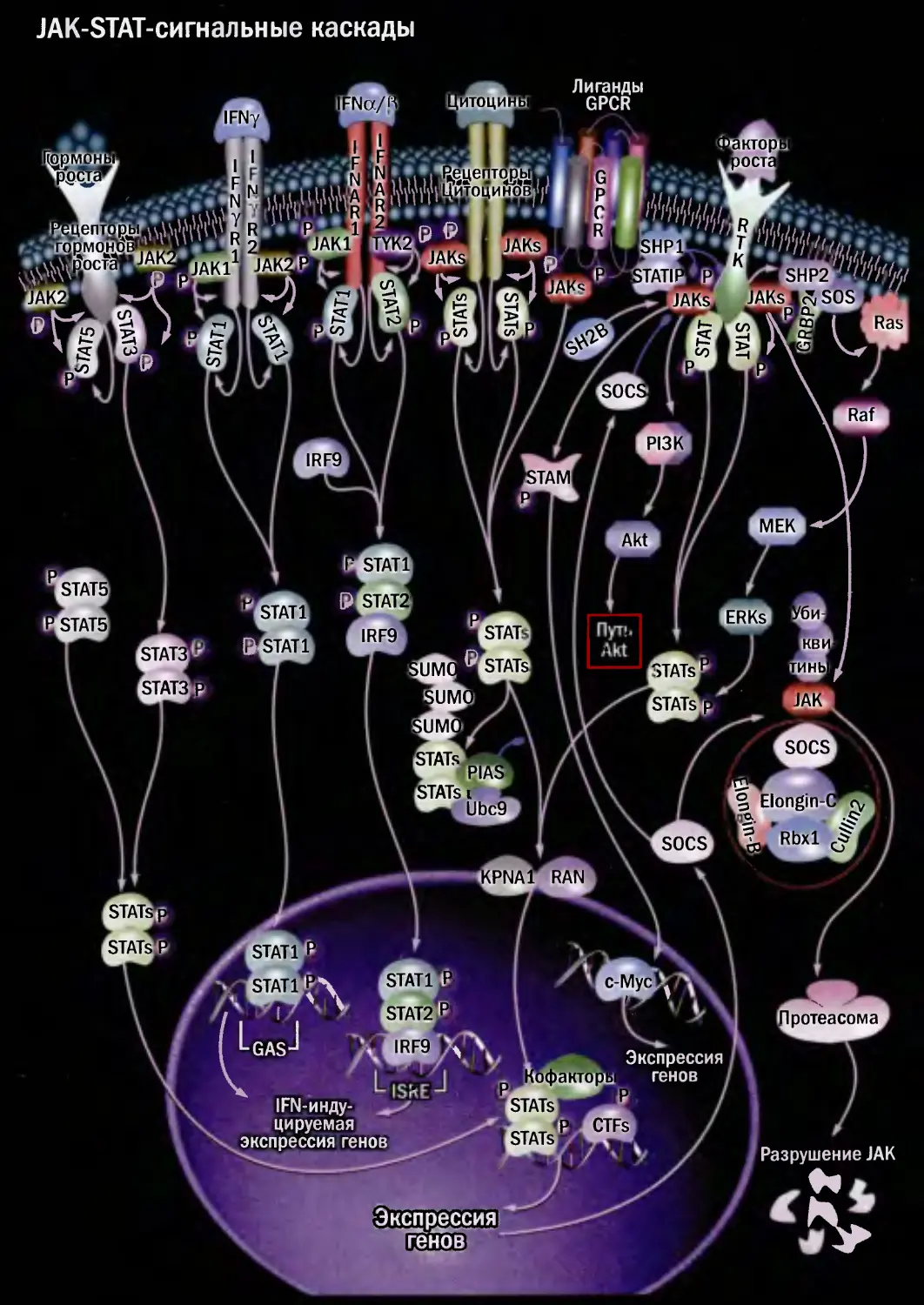
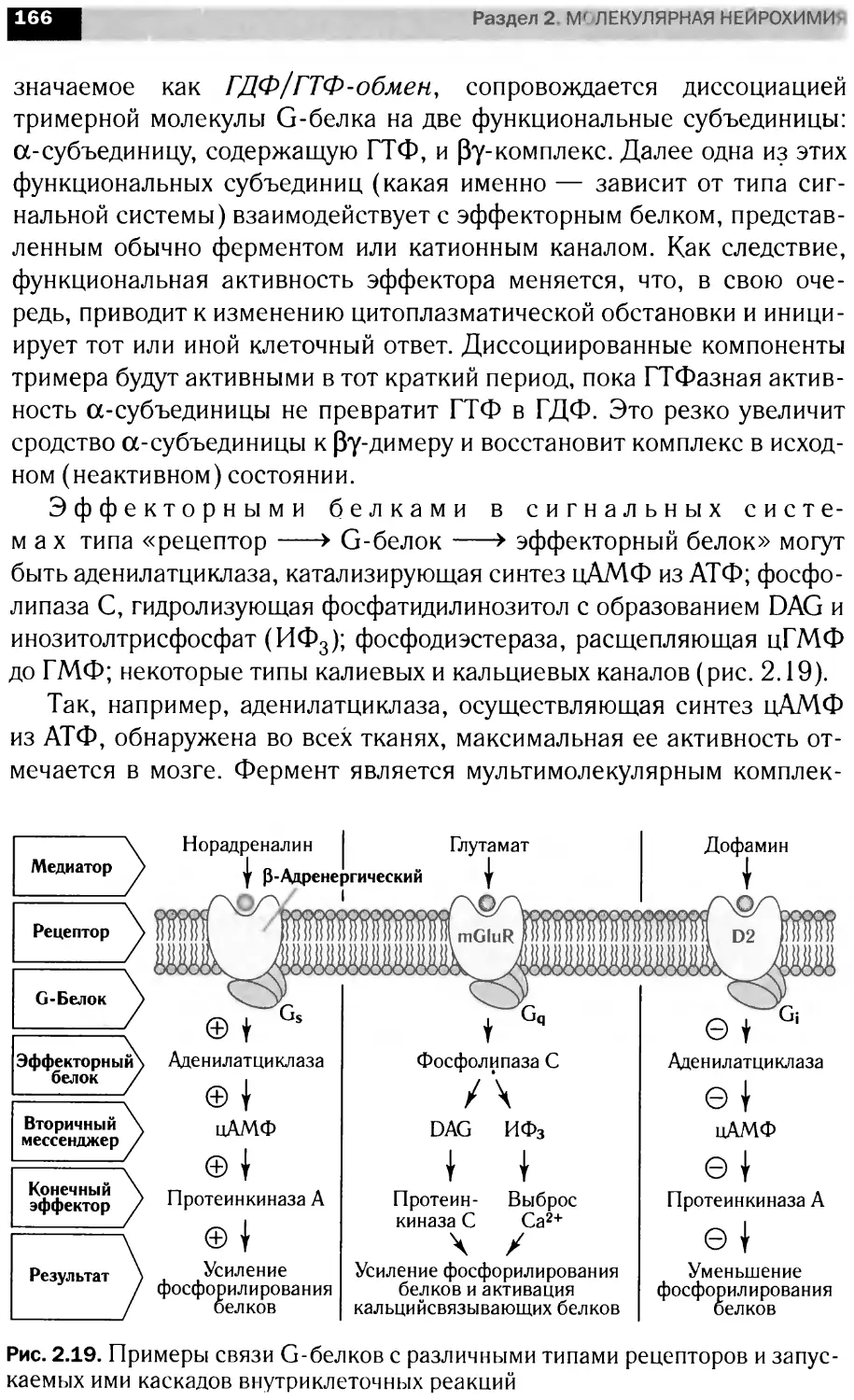
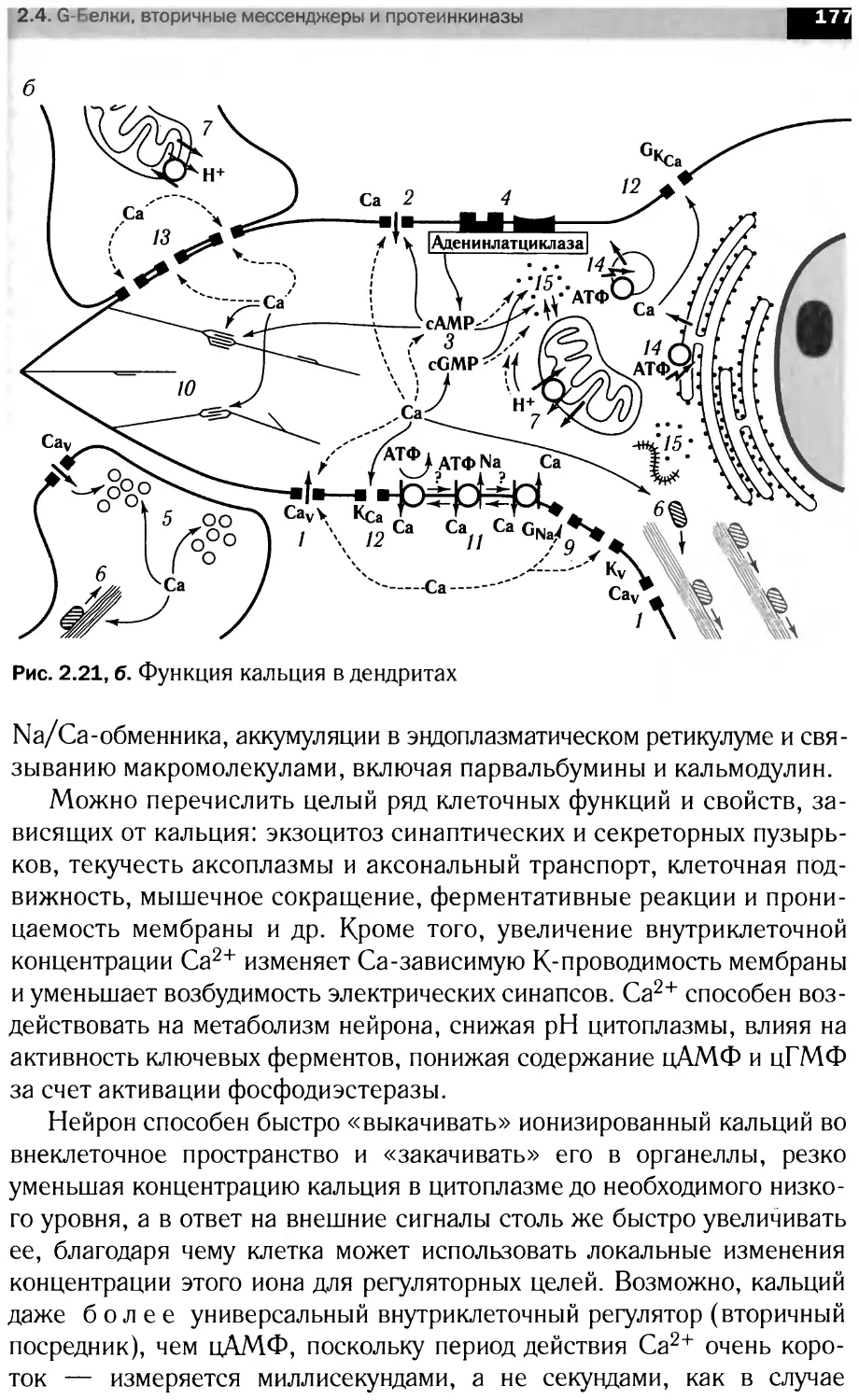
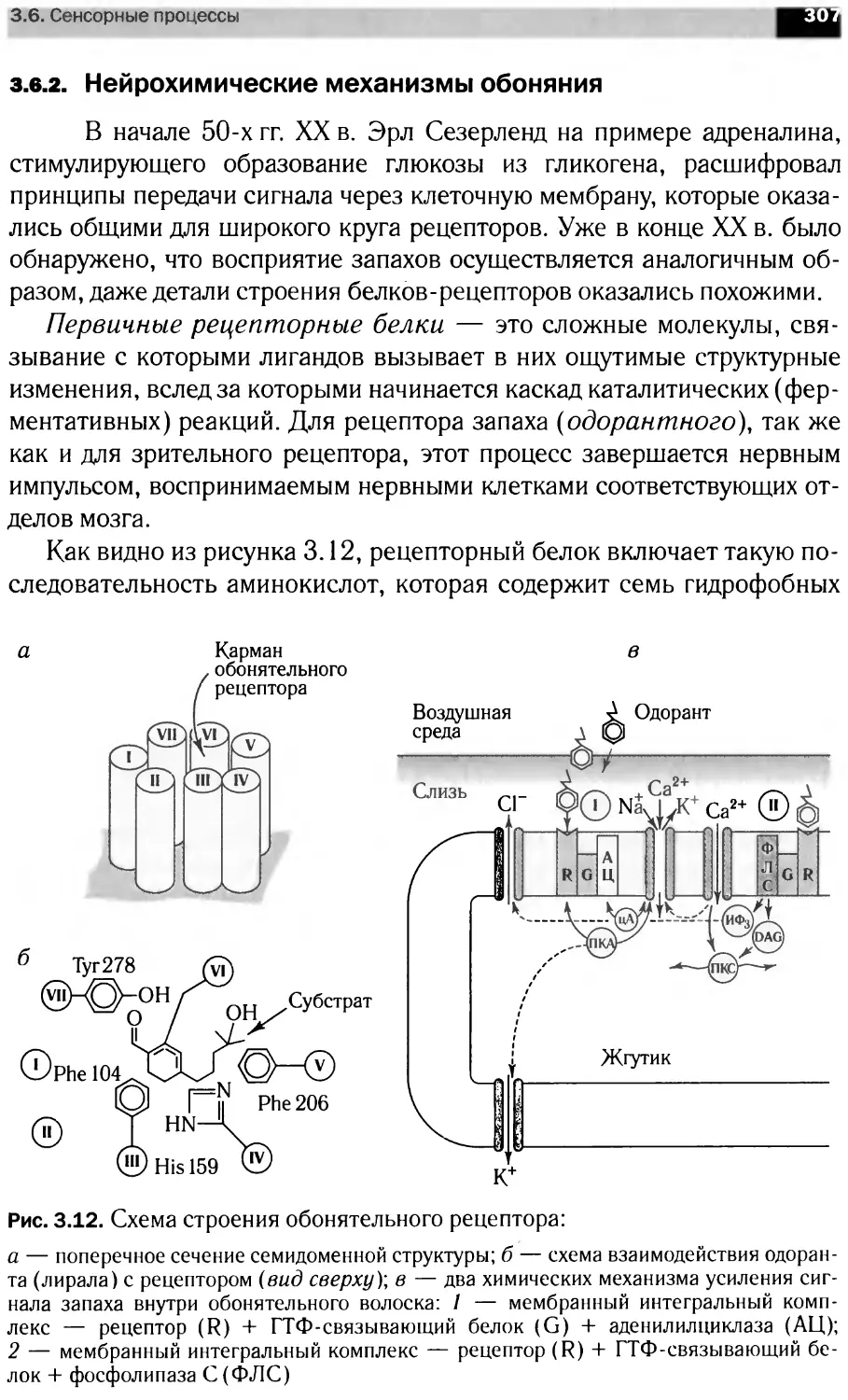
JAK-STAT-сигнальные каскады
1Т0ЦИ1
[ито циней
IAK1
SHP
□АКТ
SOC!
PI3K
IRF9
JAM
МЕК
STAT1]
STAT!
STAT1
STAT:
ERKs
STAT5]
STATsl
IRF9
ETstati
STATv
STATj
STATs
rSTAT3’
STATs1
SOCS
rSTAT<
Elongin
SOCS
:pna:
RAN
STATs
STATsl
STAT1
STAT1
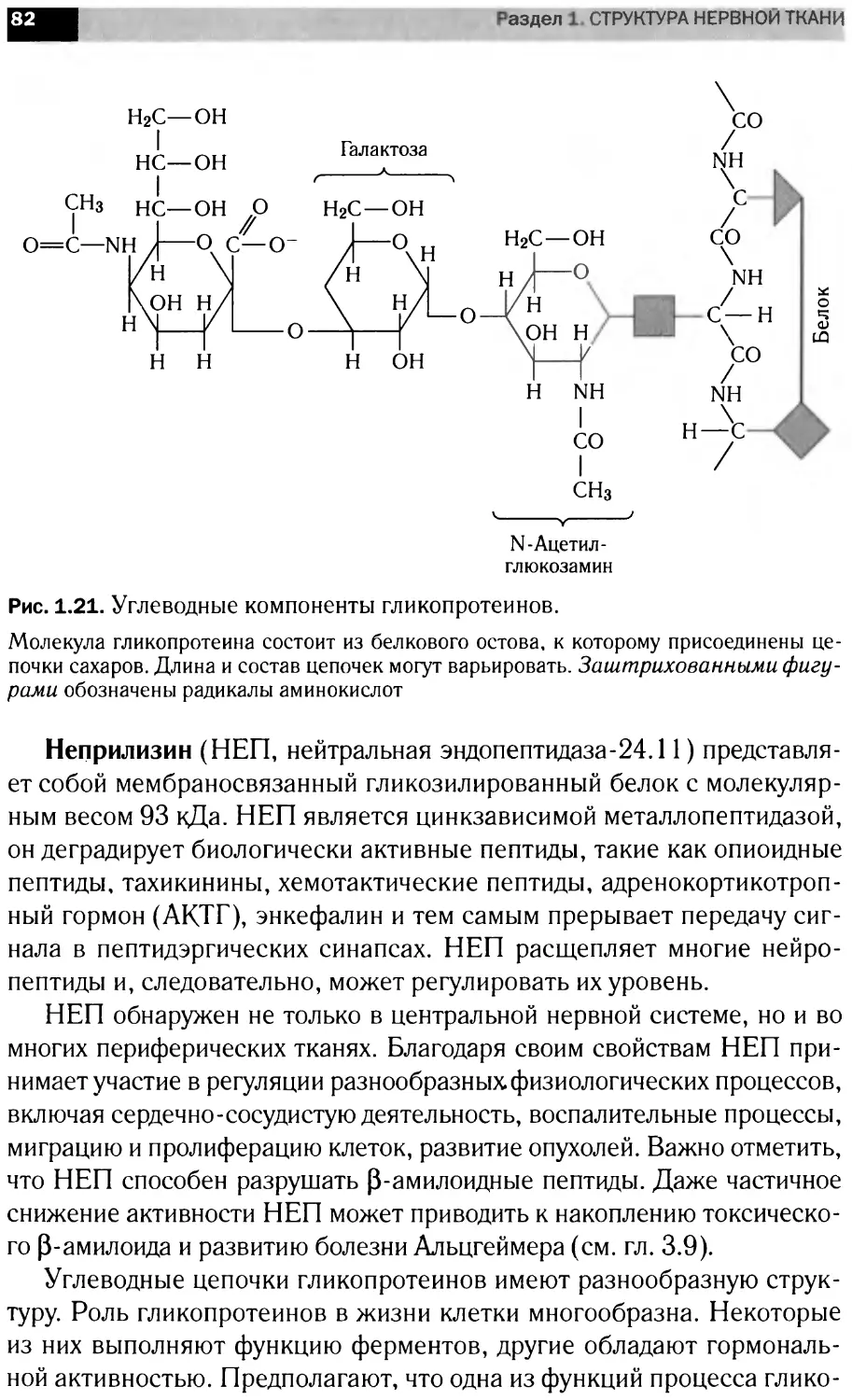
c-Mycl
STAT1
STAT2
1ротеасома
IRF9
STATs]
IKTO|
lOCTi
JATIPgl
r<JAKf
>TATs i Ml
A Ubc9
Гр’mohJ
[роста
КЪфактб|
STATsl^
SHP2
SOS
Разрушение JAK
[генов
цируемая
экспрессия генов
Экспрессия
генов ,
Лиганды
GPCR
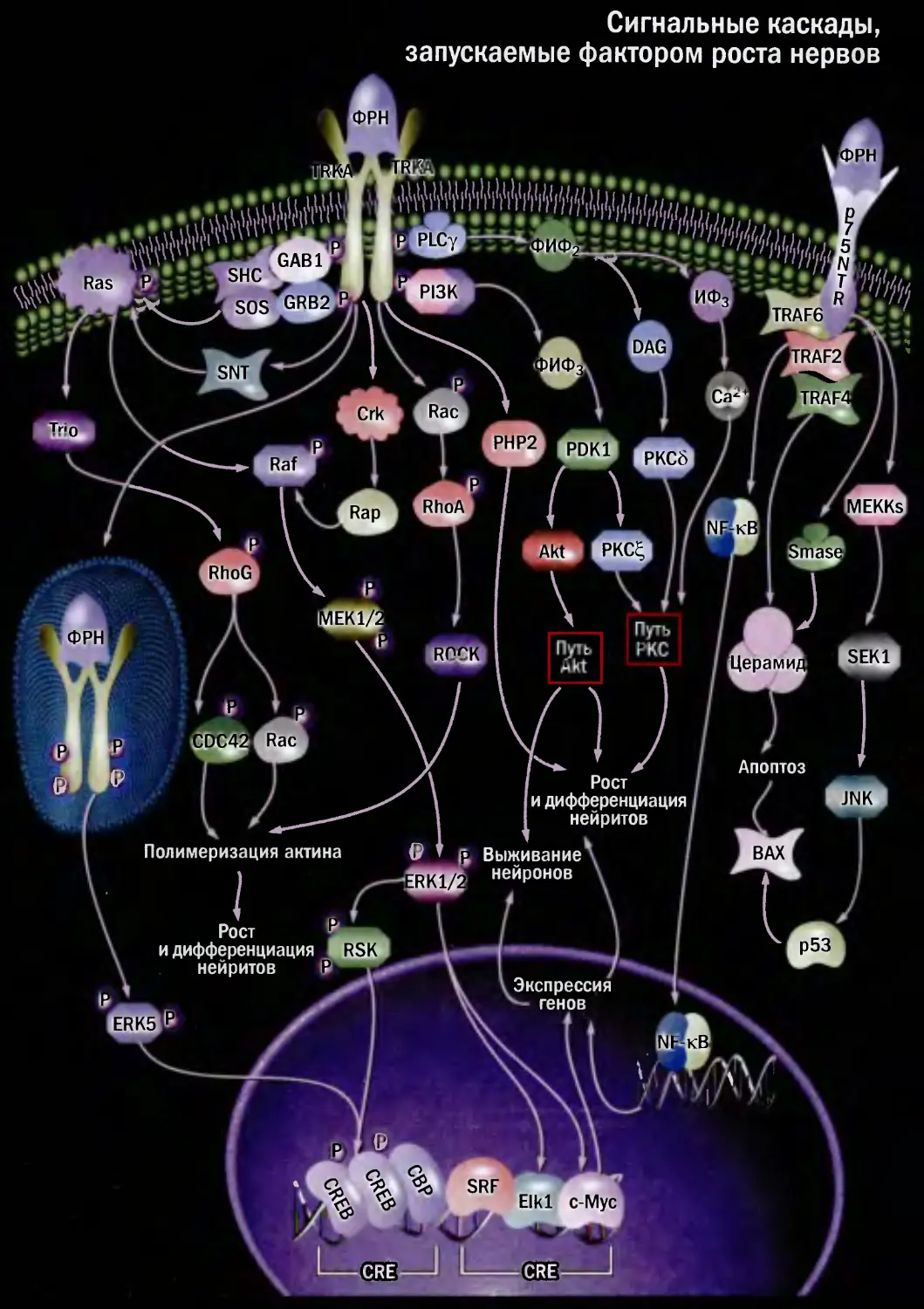
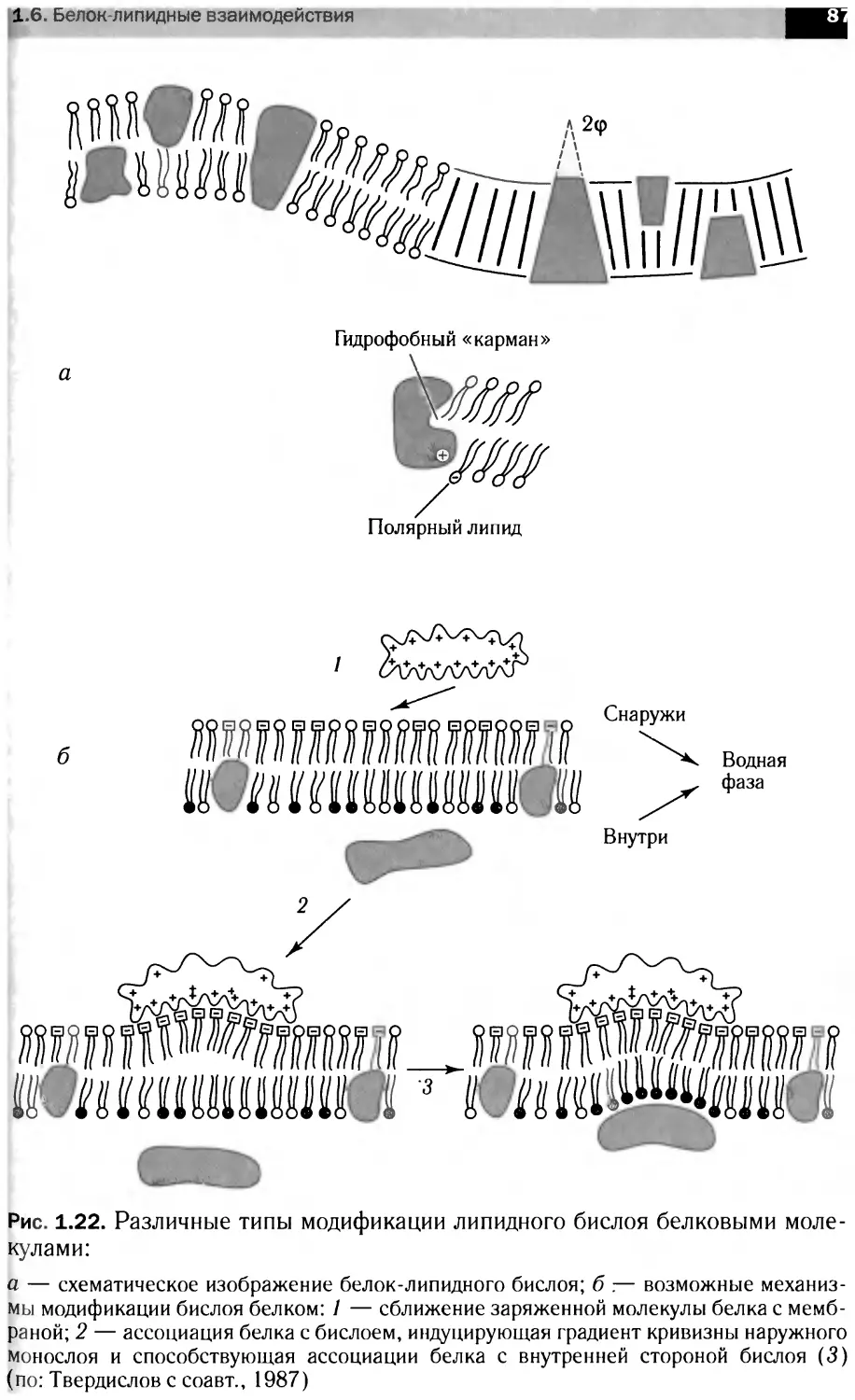
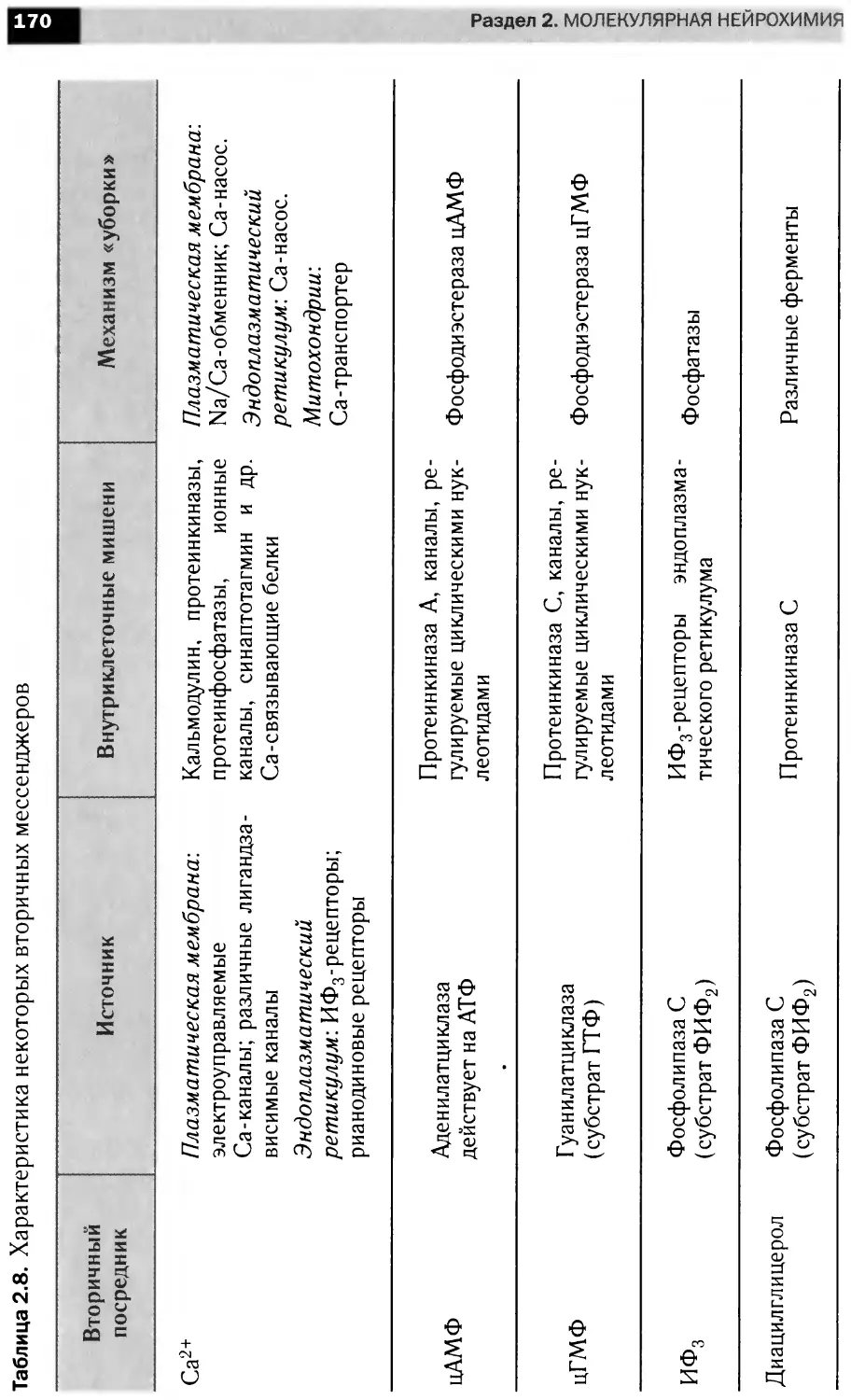
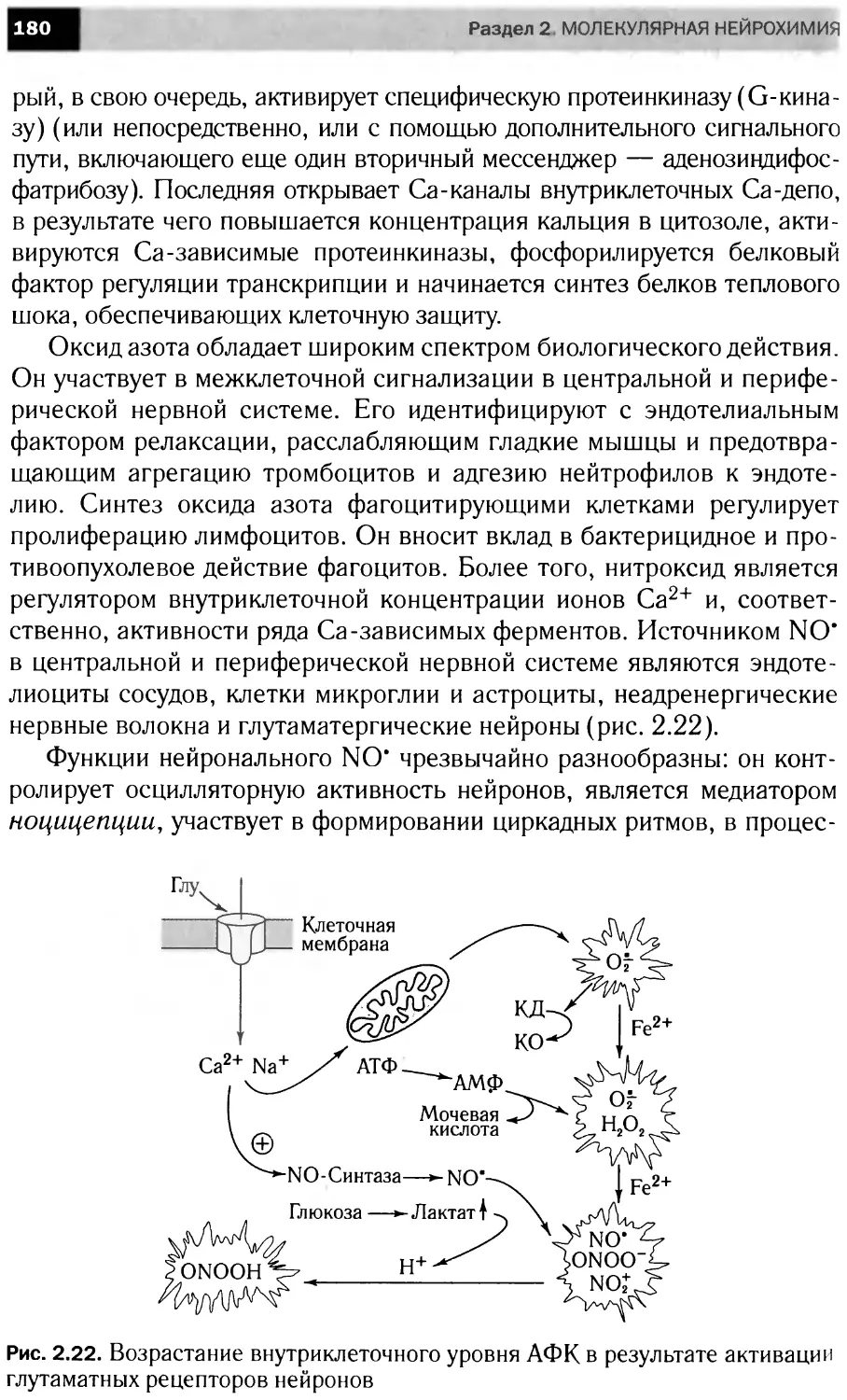
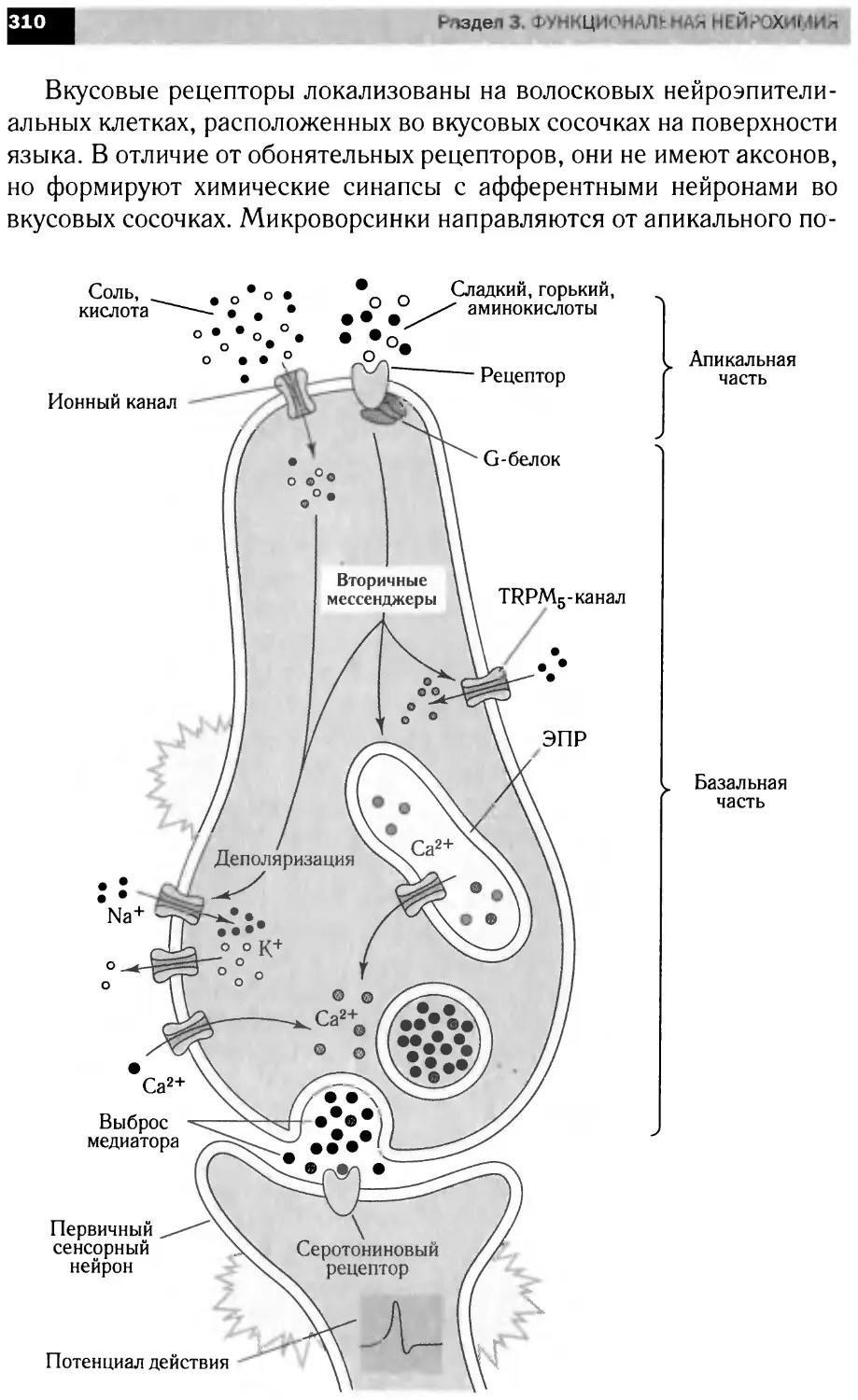
Сигнальные каскады,
запускаемые фактором роста нервов
1-КС
Апоптоз
Полимеризация актина
CRE
CRE
Экспрессия
генов
Рост
и дифференциация
нейритов
Выживание
нейронов
' Рост
и дифференциация
нейритов
Авторы:
А. А. Болдырев, Н. Д. Ещенко,
В. А. Илюха, Е. И. Кяйвяряйнен
Рецензенты:
кафедра эмбриологии Московского государственного
университета им. М. В. Ломоносова
(зав. кафедрой д-р биол. наук, проф. В. А. Голчченков)\
зам. директора Института высшей нервной деятельности
и нейрофизиологии РАН,
д-р биол. наук, проф. Н. В. Гуляева
(лаборатория функциональной биохимии нервной системы
ИВНДиНФ РАН)
Нейрохимия учеб, пособие для вузов / А. А. Болды-
Н45 рев, Н. Д. Ещенко, В. А. Илюха, Е. И. Кяйвяряйнен;
науч. ред. Ю. А. Владимиров. — М. : Дрофа, 2010. —
398, [2] с. : ил.
ISBN 978-5-358-03120-3
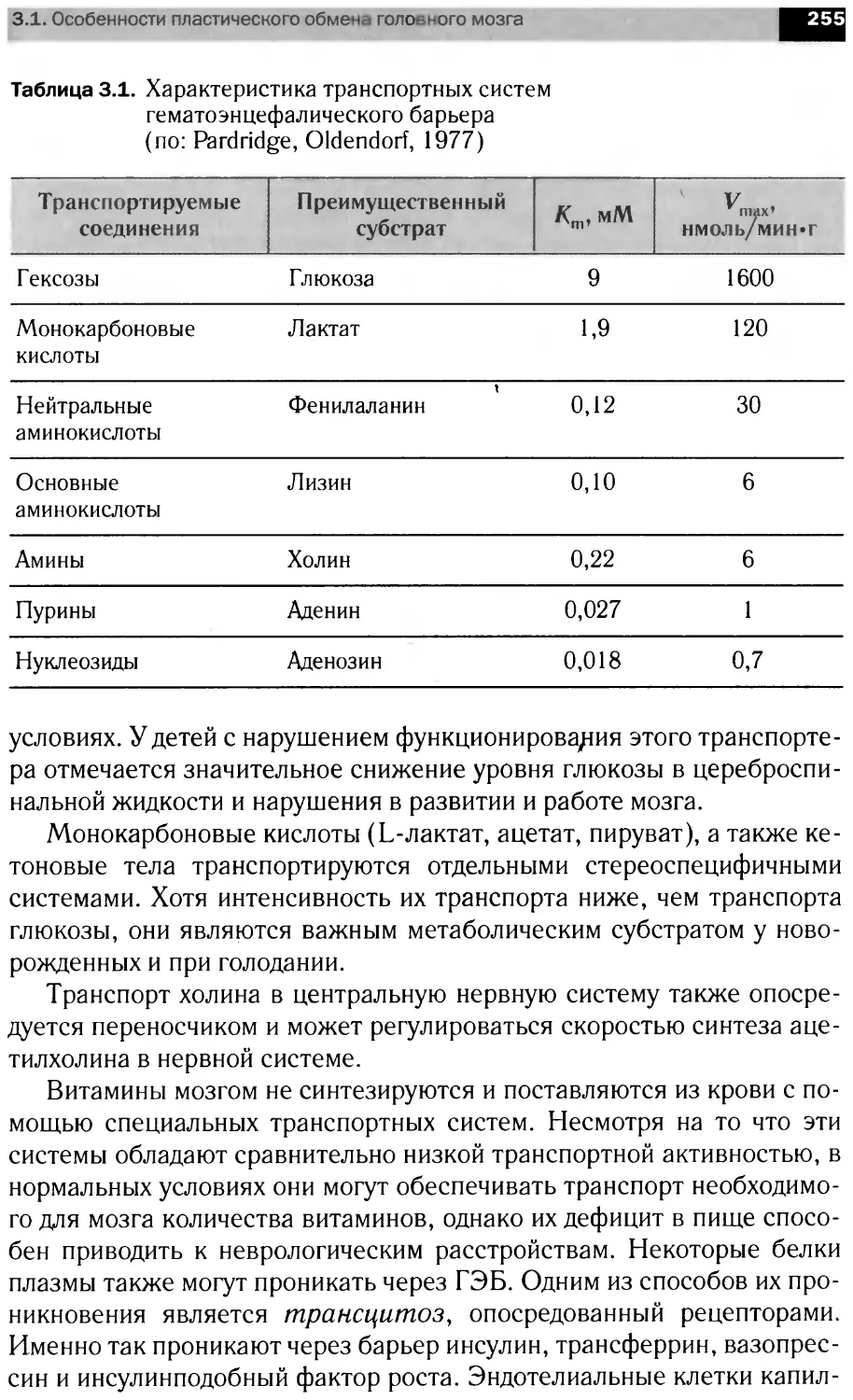
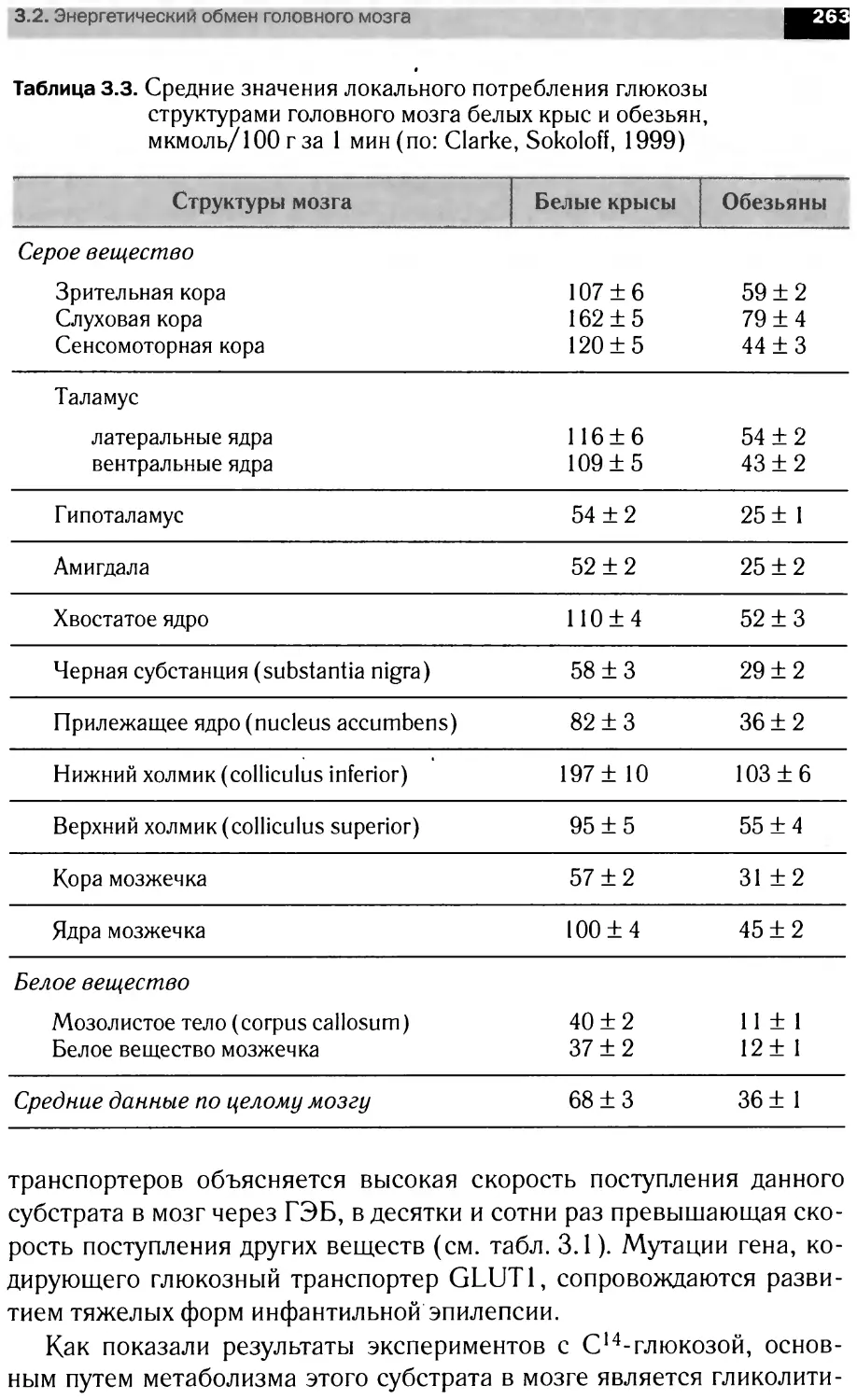
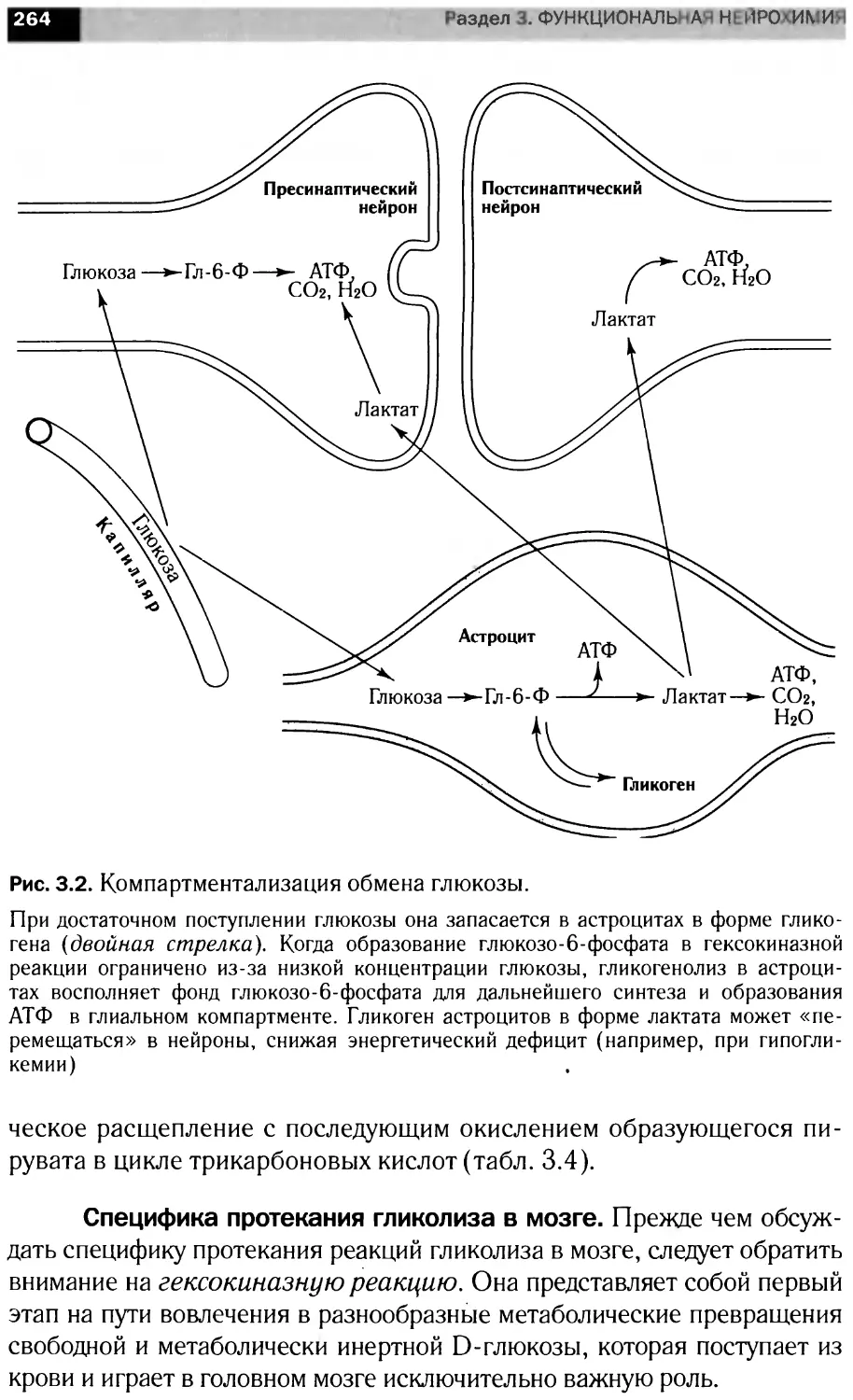
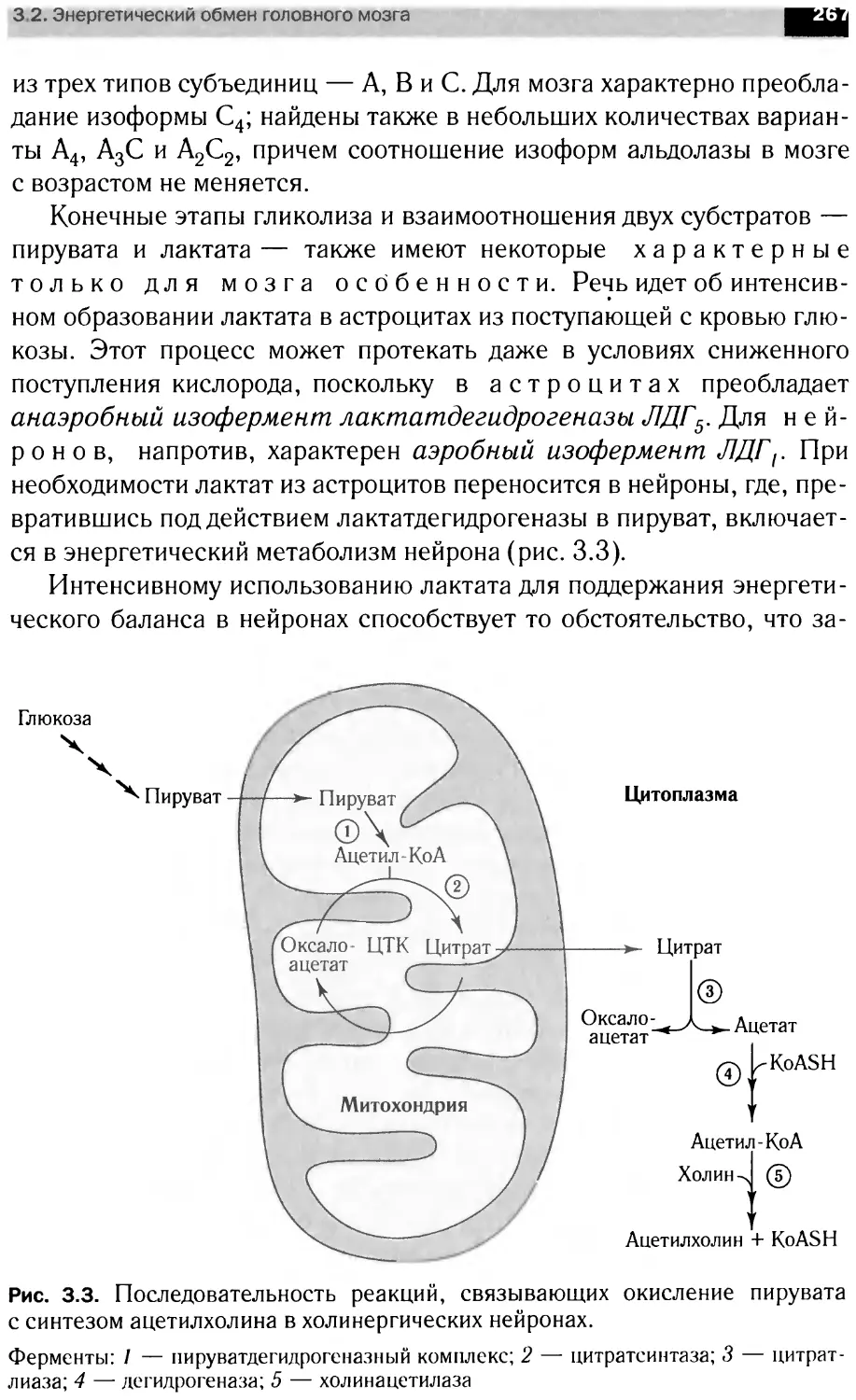
В книге рассмотрены вопросы мембранной организации нервной тка-
ни, особенности энергетического обмена мозга роль сигна ьных мол< кул
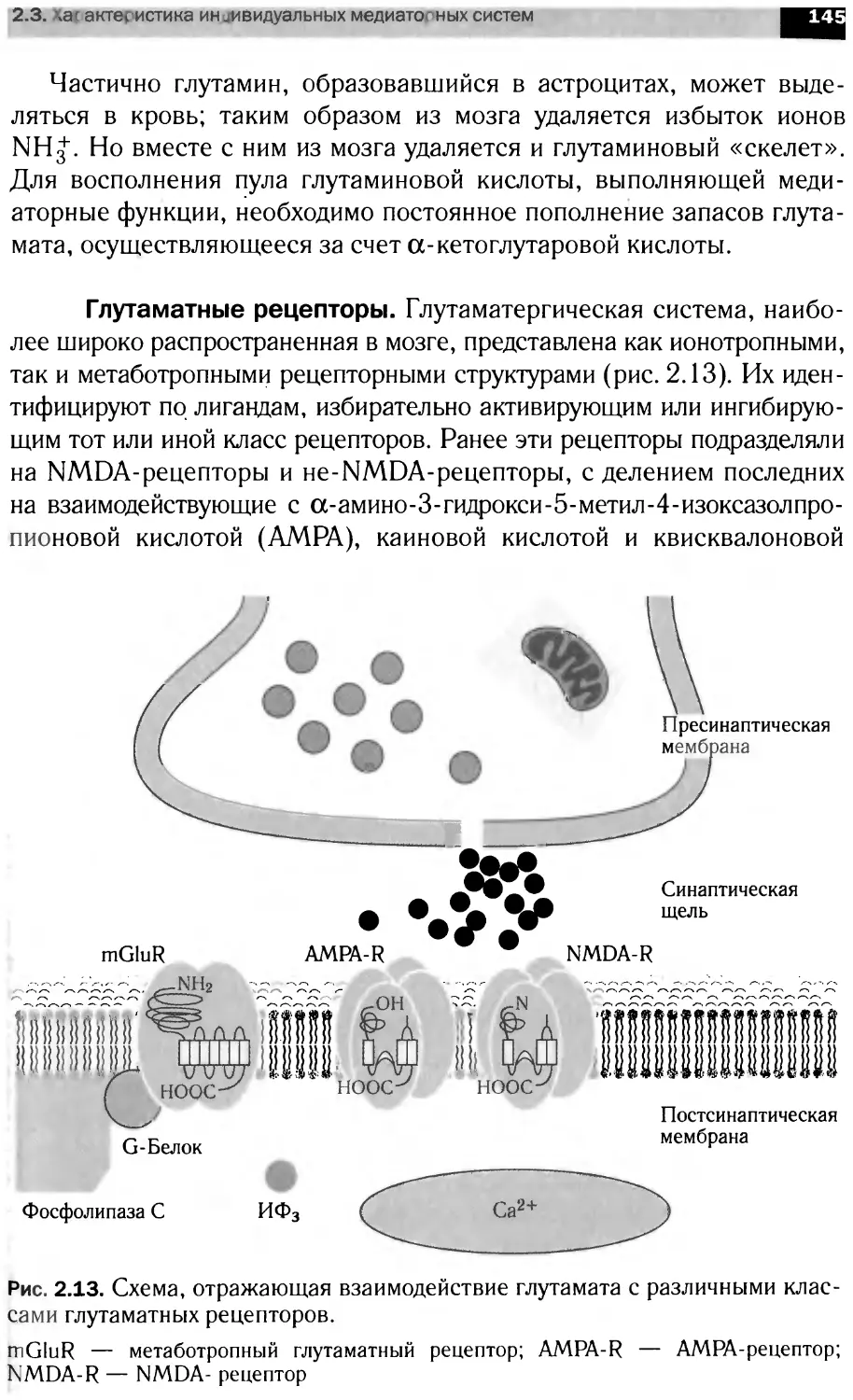
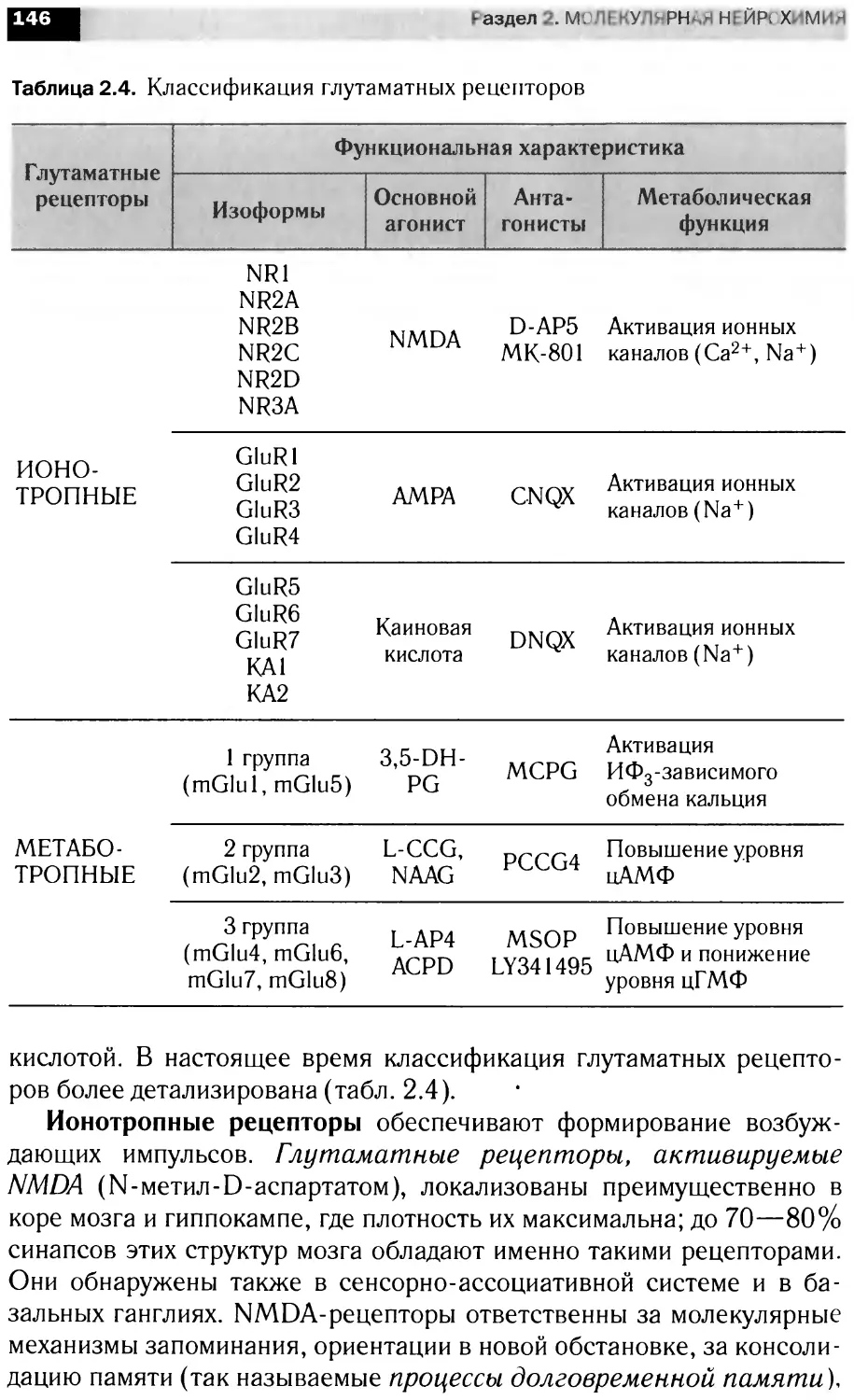
в анализе и передаче информации на уровне нейрональны клеток, бщие
принципы интеграции метаболизма мозга. Материал представлен в трех
разд< пах: <Структура н рвной ткани». Молекулярная нейрохимия» и
«Функциональная нейрохилия».
Для студентов вузов, специализирую щихся в области биохи-
мии, фар "акологии, физиологии и психологии.
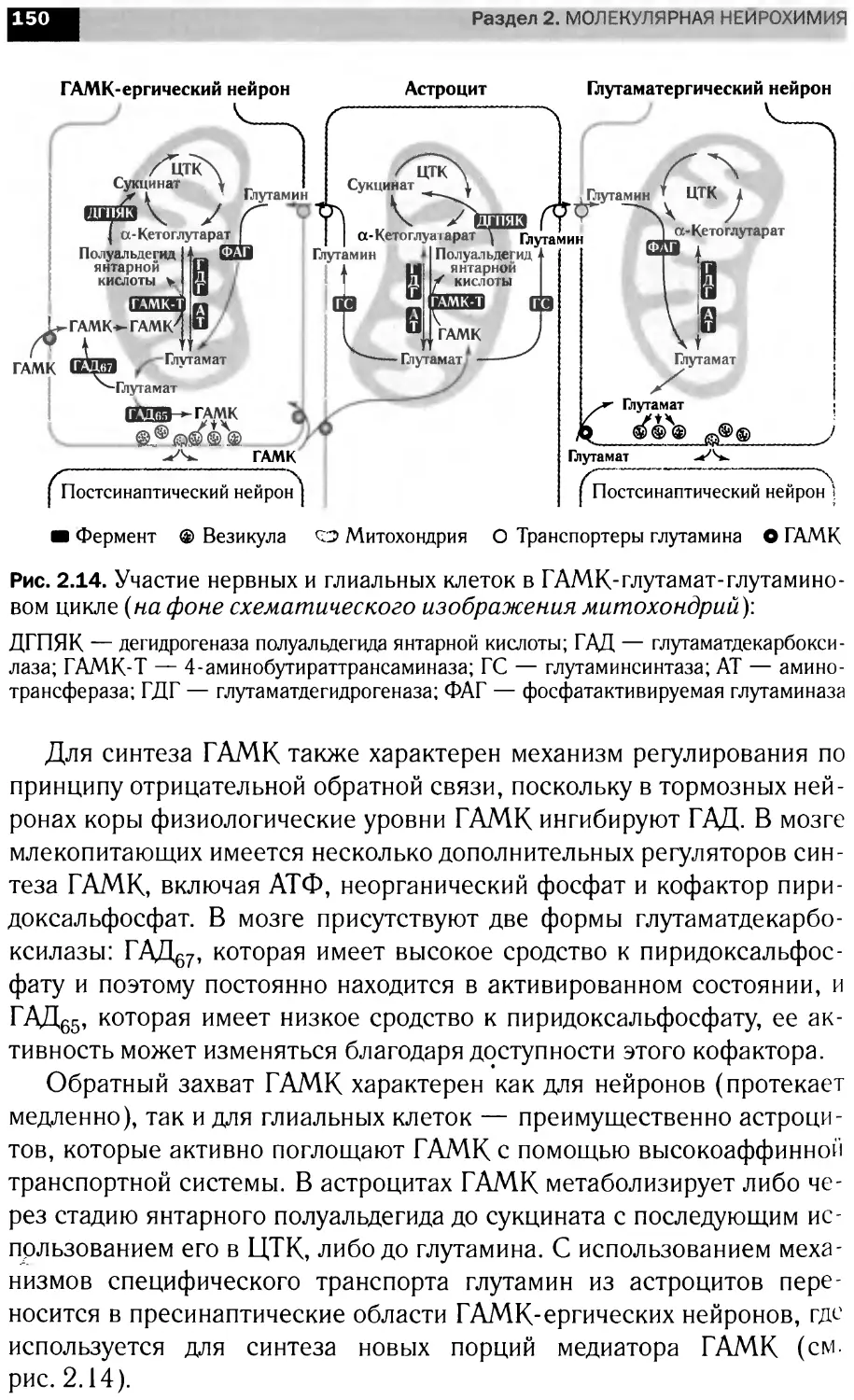
УДК 577.1(075.8)
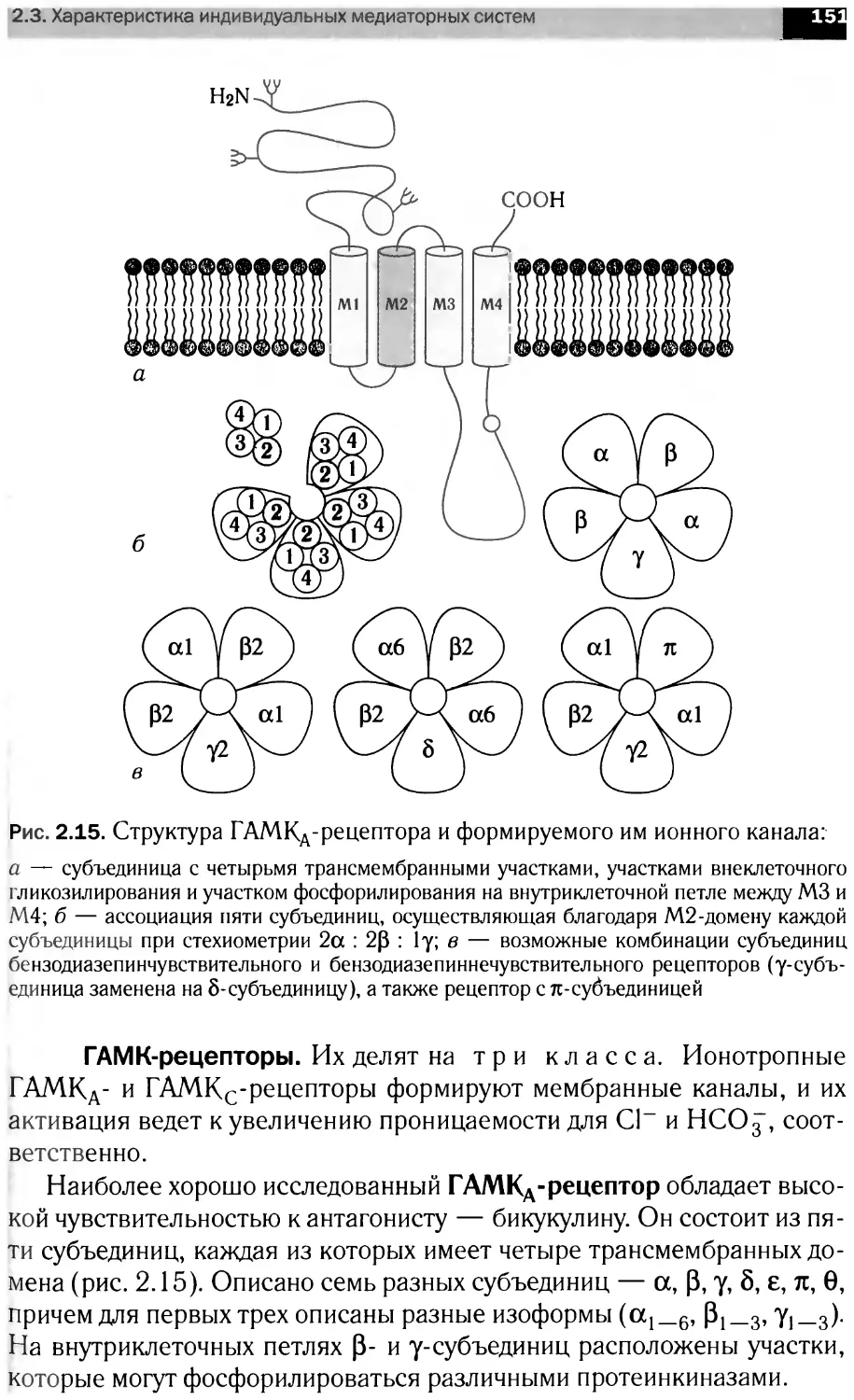
ББК 28.072я73
ISBN 978-5-358-03120-3
© ООО «Дрофа», 2010
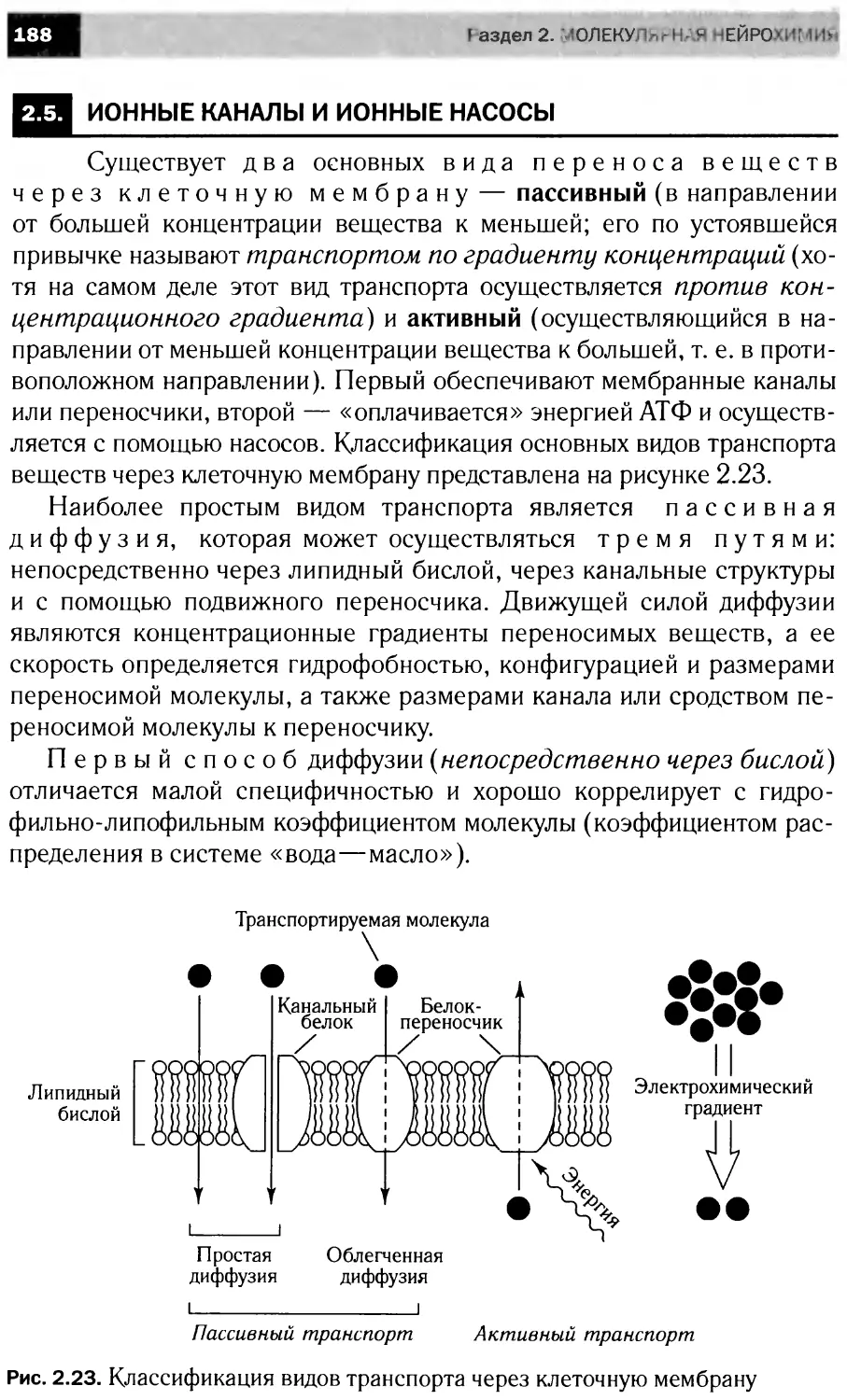
ПРЕДИСЛОВИЕ
Данное учебное пособие создавалось на основе «живого», по-
стоянно читаемого курса, что было непростой задачей, посколь-
ку огромная скорость накопления фактических данных в облас-
ти нейронаук стимулирует процессы формирования новых идей
и разрушения старых концепций.
В книге сделана попытка объединить отечественную традицию
преподавания нейрохимии как науки, лежащей на стыке основных
биологических дисциплин (биохимии, молекулярной биологии,
физиологии), с современным взглядом на нейрохимию как само-
стоятельную составляющую естествознания, имеющую свой пред-
мет, специфические задачи и оригинальные методы исследования,
науку, являющуюся основой наших сегодняшних представлений о
молекулярных механизмах клеточной сигнализиции.
При написании пособия был использован предшествующий
опыт преподавания нейрохимии, в первую очередь академиком
Игорем Петровичем Ашмариным.
И. П. Ашмарин — автор получив-
шего широкую известность и до сих
пор единственного отечественного
учебника по данной дисциплине
(Нейрохимия. М., 1996 и его пере-
работанное издание: Биохимия моз-
га. СПб., 1999).
Построение учебного пособия в
целом соответствует структуре лек-
ционных курсов по общим вопро-
сам нейрохимии, читаемых в МГУ
им. М. В. Ломоносова и Санкт-Пе-
тербургском государственном уни-
верситете. В компоновке матери-
ала нашел отражение личный педа-
гогический опыт авторов.
И. П. Ашмарин
Основоположник отечествен-
ной нейрохимии
В книге три раздела: «Структура нервной ткани», «Молеку-
лярная нейрохимия» и «Функциональная нейрохимия». Пред-
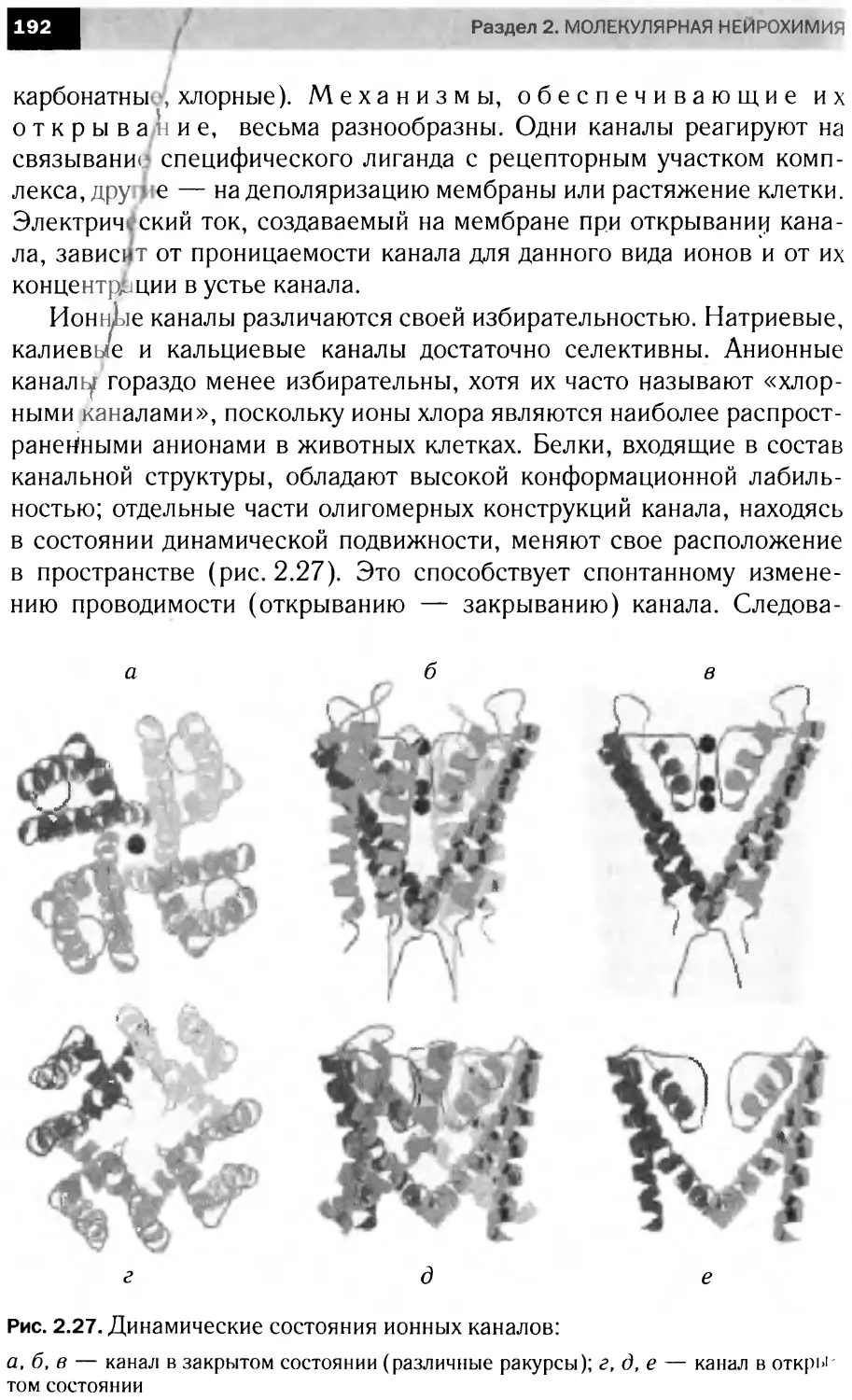
лагаемое разделение материала достаточно условно и служит
лишь для облегчения его усвоения.
Следует иметь в виду, что многие проблемы современной
нейрохимии не нашли отражения в данном пособии. Будучи свя-
занными рамками учебных часов, авторы вынуждены были
предпочесть широту изложения материала глубине анализа ча-
стных аспектов функционирования возбудимых структур. Заин-
тересованный читатель должен рассматривать эту книгу в каче-
стве введения в проблемы современной нейрохимии.
ОТ НАУЧНОГО РЕДАКТОРА
Биология во второй Головине XX в. характеризовалась такими
же кардинальными открытиями, как физика — в первой его по-
ловине. Разгадка строения двойной спирали ДНК стимулиро-
вала бурное развитие молекулярной биологии, биохимической
генетики, биотехнологии. Создание гибридомных технологий,
успехи в исследовании стволовых клеток, проникновение в
мембранные механизмы работы нервной системы способство-
вали тому, что к началу нового XXI в. нейрохимия оказалась
в числе наиболее востребованных дисциплин.
Интерес к нейронаукам частично вызван успехами в по-
знании механизмов работы нервной клетки (фактически, каж-
дая третья Нобелевская премия в последнее десятилетие была
присуждена за исследования, так или иначе имеющие отноше-
ние к наукам о мозге человека). Но главное — это огромная во-
стребованность нейронаук как фундамента для понимания мо-
лекулярных нарушений, лежащих в основе нейродегенератив-
ных заболеваний, как наследственных, так и вызванных
антропогенными факторами.
Последнее десятилетие XX в., по предложению Всемирной
организации здравоохранения, было названо «десятилетием
мозга», однако по его истечении вопреки ожиданиям общества
стала очевидной недостаточность наших знаний в этой области.
Подобная неоднозначность состояния нейрохимии как науки
привлекает внимание биологов разных специализаций.
Курс нейрохимии, читаемый на биологических факультетах
многих вузов страны, остро нуждается в учебной литературе.
Данное издание поможет решить эту проблему.
Специфика изложения материала в книге, ее структура
обусловлены опытом экспериментальной и педагогической ра-
боты авторов: имеет место явное превалирование вопросов мо-
лекулярной (мембранной) нейрохимии. Однако не исключено,
что, исходя из вышеназванных особенностей момента, это мо-
жет стать одним из достоинств данного учебного пособия.
В числе других — ясное и систематическое изложение матери-
ала, широта охвата нейрохимических проблем, разнообразие
иллюстративного материала.
Хочется пожелать, чтобы книга нашла своего заинтересо-
ванного читателя, для которого нейрохимия станет полем само-
стоятельной научной деятельности.
Академик РАМН, профессор
Ю. А. Владимиров
ВВЕДЕНИЕ
P. Е. Владимиров
Выдающийся российский био-
химик, известный работами
в области окислительного ме-
таболизма мозга
Конец XX в. ознаменовался дифференцировкой естественно-
научных дисциплин, существенным расширением задач и ме-
тодов научных исследований в области биологии. В результате
классические биологические науки (биохимия, физиология, гис-
тология) дали начало целому ряду новых научных направле-
ний, в том числе цитологии, молекулярной биологии, энзимо-
логии, биоэнергетике. Особое место среди них заняла нейро-
химия — наука о молекулярных превращениях, лежащих в
основе таких специфических проявлений живого, как возбу-
димость, способность анализировать поступающие к клетке
извне сигналы и специфическим образом на них реагировать.
Динамичное развитие нейрохимии привело к тому, что в на-
стоящий момент она рассматрива-
ется не только как самостоятель-
ная отрасль естествознания, но и
как методологическая основа для
совершенствования наших знаний в
эмбриологии, клеточной биологии,
экспериментальной и клинической
неврологии и других областях био-
логии и медицины. Курсы нейрохи-
мии на протяжении многих лет чи-
таются во всех крупных универси-
тетах нашей страны.
На страницах этой книги нель-
зя не вспомнить выдающихся оте-
чественных ученых, стоявших у исто-
ков нейрохимии, благодаря таланту
и упорству которых данная наука до-
стигла сегодняшних высот: И. П. Аш-
марина, Е. С. Лондона, Е. М. Креп-
А. В. Палладии
Основоположник украинской
школы нейрохимиков. Ос-
новные труды — по биохи-
мии нервной ткани и голов-
ного мозга
са, Г. Е. Владимирова, Н.Н. Деми-
на, М. И. Прохорову, Г. X. Бунатяна,
П. А. Кометиани, А. В. Палладина,
А. А. Галояна, Т. М. Турлаева. Сов-
ременная нейрохимия обязана им
актуальностью своей тематики. В то
же время нейрохимия, как, может
быть, никакая другая наука, раз-
вивается в тесном взаимодействии
ученых разных стран: Международ-
ное нейрохимическое общество —
одно из наиболее многочисленных
среди всех естественнонаучных со-
обществ в мире. На сегодняшний
день знание основ нейрохимии яв-
ляется непременным условием по-
лучения современного биологичес-
кого образования.
СТРУКТУРА НЕРВНОЙ ткани
НЕРВНАЯ ТКАНЬ
Нервная ткань — это взаимосвязанная система нервных клеток и
нейроглии, выполняющая специфические функции восприятия сигна-
лов, выработки ответного импульса и его передачи. Нервная ткань яв-
ляется основой строения нервной системы, регулирующей функции всех
тканей и органов, их интеграцию в организме и связь с окружающей
средой. Нервная система обеспечивает координацию всех жизненных
процессов в организме и его взаимодействие с внешней средой.
Анатомически нервную систему делят на центральную и перифери-
ческую. ^центральной нервной системе (ЦНС) относят головной и
спинной мозг, периферическая нервная система (ПНС) объединяет
периферические нервные узлы, стволы и нервные окончания.
В головном мозге различают серое и белое вещество. Большая
часть серого вещества располагается на поверхности мозга и в моз-
жечке, формируя их кору. Меньшая часть образует многочисленные
ядра ствола мозга.
Нервная ткань состоит в основном из нейрональных и глиальных
клеток. Нейроны (neuronia, нейроциты) — основные компоненты
нервной ткани. Нейроглия обеспечивает существование и функци-
онирование нервных клеток, осуществляя опорную, трофическую,
разграничительную, секреторную и защитную функции. Нейроны яв-
ляются высокоспециализированными клетками нервной системы, от-
ветственными за рецепцию, обработку импульсов, их проведение и
передачу на другие нейроны, мышечные или секреторные клетки.
Нейроны выделяют нейромедиаторы и иные вещества, передающие
информацию другим клеткам.
1.1.1. Нейрон
Морфологически и функционально самостоятельная единица.
С помощью отростков {аксона и дендритов) осуществляет контакты
с другими нейронами, образуя рефлекторные дуги — звенья, из ко-
торых построена нервная система.
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
В зависимости от функций в рефлекторной дуге различают аффе-
рентные (чувствительные), ассоциативные и эфферентные (эф-
фекторные) нейроны. Афферентные нейроны воспринимают импуль-
сы, эфферентные передают их на ткани рабочих органов, побуждая их
к действию, а ассоциативные нейроны обеспечивают межнейрональ-
ные связи. Рефлекторная дуга пред-
ставляет собой цепь нейронов, связан-
ных друг с другом синапсами и обес-
печивающих проведение нервного им-
пульса от рецептора чувствительного
нейрона до эфферентного окончания
в рабочем органе.
Нейроны отличаются большим раз-
нообразием форм и размеров. Диаметр
тел гранулярных клеток коры мозжеч-
ка — около 10 мкм, а гигантских пира-
мидных нейронов двигательной зоны ко-
ры большого мозга — 130— 150 мкм.
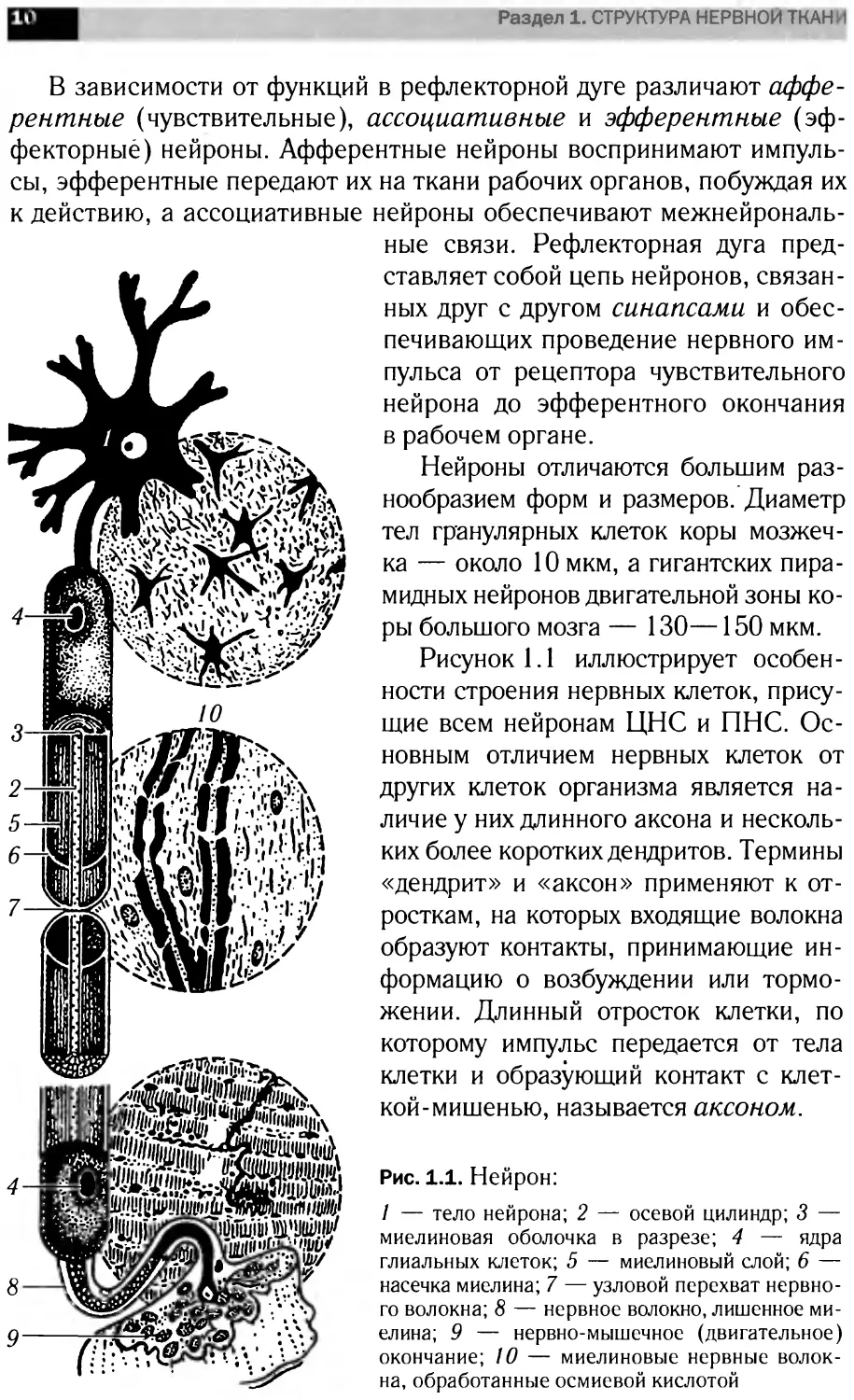
Рисунок 1.1 иллюстрирует особен-
ности строения нервных клеток, прису-
щие всем нейронам ЦНС и ПНС. Ос-
новным отличием нервных клеток от
других клеток организма является на-
личие у них длинного аксона и несколь-
ких более коротких дендритов. Термины
«дендрит» и «аксон» применяют к от-
росткам, на которых входящие волокна
образуют контакты, принимающие ин-
формацию о возбуждении или тормо-
жении. Длинный отросток клетки, по
которому импульс передается от тела
клетки и образующий контакт с клет-
кой-мишенью, называется аксоном.
Рис. 1.1. Нейрон:
/ — тело нейрона; 2 — осевой цилиндр; 3 —
миелиновая оболочка в разрезе; 4 — ядра
глиальных клеток; 5 — миелиновый слой; 6 —
насечка миелина; 7 — узловой перехват нервно-
го волокна; 8 — нервное волокно, лишенное ми-
елина; 9 — нервно-мышечное (двигательное)
окончание; 10 — миелиновые нервные волок-
на, обработанные осмиевой кислотой
1.1. Нервная ткань
По количеству отростков различают униполярные нейро-
ны, имеющие только аксон (у высших животных и человека обычно не
встречаются), биполярные нейроны, имеющие аксон и один дендрит, и
мулътиполярные нейроны, имеющие аксон и много дендритов (рис. 1.2).
Иногда среди биполярных нейронов встречается псевдоуниполярный, от
тела которого отходит один общий вырост — отросток, разделяющийся
затем на дендрит и аксон. Псевдоуниполярные нейроны присутствуют в
спинальных ганглиях, биполярные — в органах чувств. Большинство
нейронов мультиполярны. Их формы чрезвычайно разнообразны.
Аксон и его коллатерали разветвляются на несколько веточек, на-
зываемых телодендронами, последние заканчиваются терминальны-
ми утолщениями. Аксон содержит митохондрии, нейротубулы и нейро-
филаменты, а также агранулярный эндоплазматический ретикулум.
Трехмерная область, в которой ветвятся дендриты одного нейрона,
называется дендритным полем. Дендриты представляют собой истин-
ные выпячивания тела клетки. Они содержат те же органеллы, что и тело
клетки: хромафильную субстанцию (гранулярный эндоплазматический
ретикулум и полисомы), митохондрии, большое количество микротрубо-
чек (нейротубул) и нейрофиламентов. Благодаря дендритам рецепторная
поверхность нейрона увеличивается в 1000 и более раз. Так, дендриты
грушевидных нейронов (клеток Пуркинье) коры мозжечка увеличивают
площадь рецепторной поверхности с 250 до 27 000 мкм2; на поверхности
этих клеток обнаруживается до 200 000 синаптических окончаний.
Рис. 1.2. Типы нервных клеток:
а — униполярный нейрон; б — псевдоуниполярный нейрон; в — биполярный нейрон;
г — мультиполярный нейрон
12
г аздел 1. СТРУКТУРА НГЦНОЙ ТКАНИ
Не все нейроны соответствуют простому строению клетки, пока-
занному на рисунке 1.1. У некоторых нейронов отсутствуют аксоны.
Есть клетки, дендриты которых могут проводить импульсы и образо-
вывать связи с клетками-мишенями. Ганглиозная клетка сетчатки
глаза соответствует схеме стандартного нейрона с дендритами, телом
и аксоном, а у фоторецепторных клеток нет очевидных дендритов и ак-
сона, поскольку они активируются не другими нейронами, а внешними
стимулами (квантами света).
Тело нейрона содержит ядро и другие внутриклеточные ор-
ганеллы, общие для всех клеток. Подавляющее большинство нейронов
человека имеют одно ядро, расположенное чаще в центре, реже — экс-
центрично. Двухъядерные и тем более многоядерные нейроны встреча-
ются крайне редко. Исключение составляют нейроны некоторых гангли-
ев вегетативной нервной системы. Ядра нейронов имеют округлую фор-
му. В соответствии с высокой метаболической активностью нейронов
хроматин в их ядрах диспергирован. В ядре имеется одно, иногда два-три
крупных ядрышка. Усиление функциональной активности нейронов
обычно сопровождается увеличением объема (и количества) ядрышек.
Плазмалемма (плазматическая мембрана) нейрона обладает спо-
собностью генерировать и проводить импульс, ее структурными ком-
понентами являются белки, функционирующие как селективные ион-
ные каналы, а также рецепторные белки, обеспечивающие реакции
нейронов на специфические стимулы. В покоящемся нейроне транс-
мембранный потенциал составляет величину 60—80 мВ.
При окрашивании нервной ткани анилиновыми красителями в ци-
топлазме нейронов выявляется хромофилъная субстанция, обнару-
живаемая в виде базофильных зерен различных размеров и форм. Ба-
зофильные зерна локализуются в перикарионе и дендритах нейронов,
но никогда не обнаруживаются в аксонах и их конусовидных основани-
ях — аксональных холмиках. Их окраска объясняется высоким содержа-
нием рибонуклеотидов. Электронная микроскопия показала, что хромо-
фильная субстанция включает цистерны эндоплазматического ретикулу-
ма, свободные рибосомы и полисомы. Гранулярная эндоплазматическая
сеть синтезирует нейросекреторные и лизосомальные белки, а также ин-
тегральные белки плазматической мембраны. Свободные рибосомы и
полисомы синтезируют белки цитозоля (гиалоплазмы) и неинтегральные
мембранные белки.
Для поддержания целостности и выполнения специфических функций
нейронам требуются разнообразные белки. Для аксонов, не имеющих ор-
ганелл, синтезирующих белок, характерен постоянный ток цитоплазмы
от перикариона к терминалиям со скоростью 1 —3 мм в сутки (см. гл. 3.3).
1.1. Нервнаяткан.
Аппарат Гольджи в нейронах хорошо развит. При световой микро-
скопии он выявляется в виде различных по форме зернышек, извитых
нитей, колечек. Его ультраструктура обычна. Пузырьки, отпочковываю-
щиеся от аппарата Гольджи, транспортируют белки, синтезированные в
гранулярном эндоплазматическом ретикулуме, либо к плазматической
мембране (интегральные мембранные белки), либо к терминалям (ней-
ропептиды, нейросекрет), либо в лизосомы (лизосомальные гидролазы).
Митохондрии обеспечивают энергией разнообразные клеточные
функции, включая такие процессы, как транспорт ионов и синтез бел-
ков. Нейроны нуждаются в постоянном притоке глюкозы и кислорода
с кровью, и прекращение кровоснабжения головного мозга губитель-
но для нервных клеток (см. гл. 3.2).
Лизосомы участвуют в ферментативном расщеплении разнообраз-
ных компонентов клетки, в том числе белков-рецепторов.
Из элементов цитоскелета в цитоплазме нейронов присутст-
вуют нейрофиламенты (диаметром 12 нм) и нейротубулы (диа-
метром 24—27 нм). Пучки нейрофиламентов {нейрофибриллы) об-
разуют в теле нейрона сеть, в его отростках они располагаются парал-
лельно. Нейротубулы и нейрофиламенты участвуют в поддержании
формы нейрональных клеток, в росте отростков и осуществлении
аксонального транспорта.
Способность синтезировать и секретировать биологически актив-
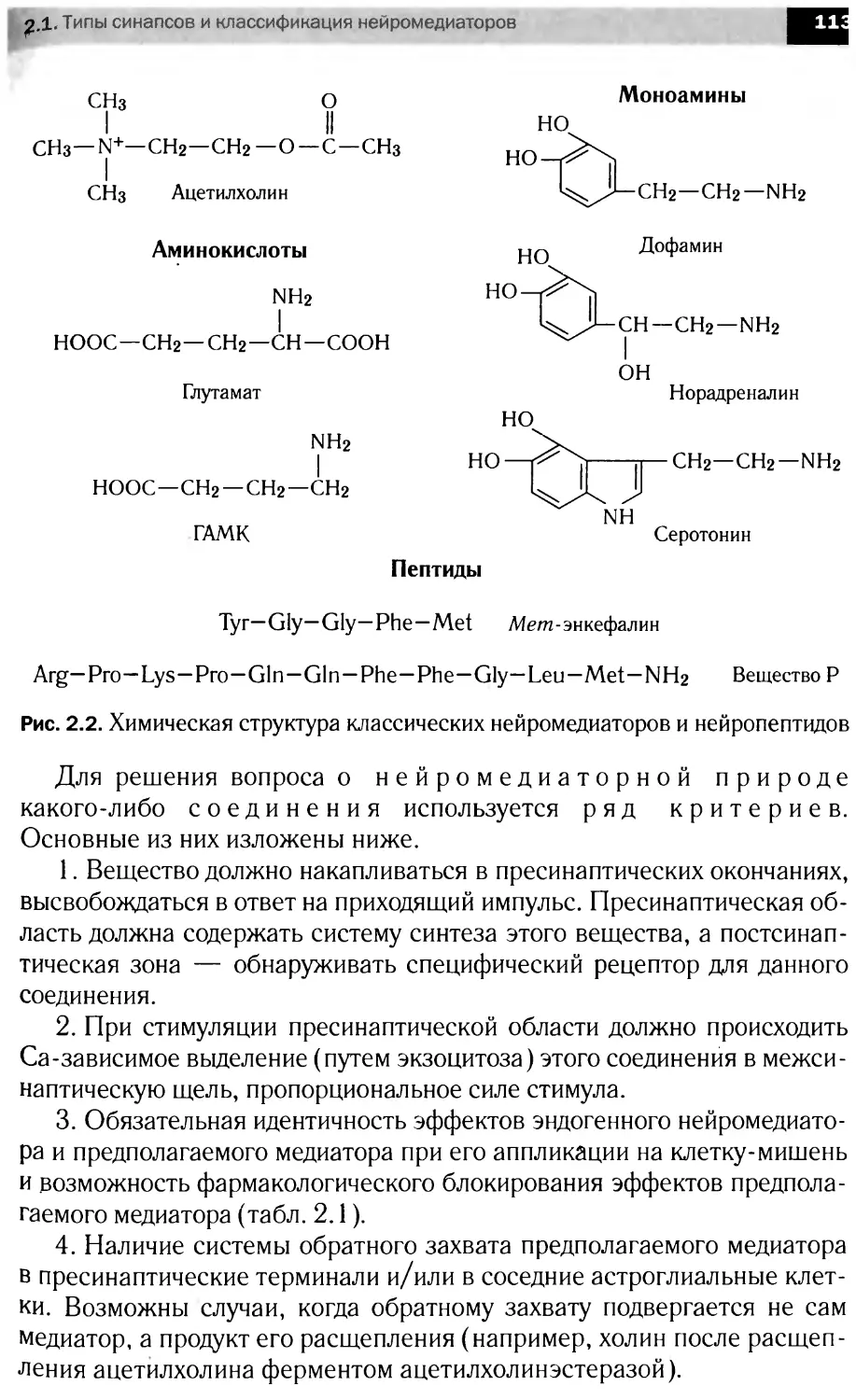
ные вещества, в частности медиаторы (ацетилхолин, норадреналин,
серотонин и др.), свойственна всем нейронам. Существуют нейроны,
специализирующиеся преимущественно на выполнении этой функ-
ции, например клетки нейросекреторных ядер гипоталамической об-
ласти головного мозга.
Секреторные нейроны имеют ряд специфических морфо-
логических признаков. Они крупные; хромофильная суб-
станция располагается преимущественно на периферии тела таких
нейронов. В цитоплазме самих нервных клеток и в аксонах находятся
различной величины гранулы нейросекрета, содержащие белки, а в
некоторых случаях — липиды и полисахариды. Гранулы нейросекрета
выводятся в кровь или в ликвор. Многие секреторные нейроны имеют
ядра неправильной формы, что свидетельствует об их высокой функ-
циональной активности. Секреторные гранулы содержат нейрорегу-
ляторы, обеспечивающие взаимодействие нервной и гуморальной сис-
тем организма.
Нейроны — высокоспециализированные клетки, су-
ществующие и функционирующие в строго определенной среде. Та-
кую среду им обеспечивает нейроглия, которая выполняет следующие
14
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
функции: опорную, трофическую, разграничительную, защитную,
секреторную, а также поддерживает постоянство среды вокруг нейро-
нов. Различают глиальные клетки центральной и периферической
нервной системы.
1.1.2. Глия
Глия центральной нервной системы. Клетки глии ЦНС делят-
ся на макроглию (глиоциты) и микроглию.
К макроглии относятся эпецдимоциты, астроциты и олигодендроциты.
Эпендимоциты выстилают стенки внутренней полости желудоч-
ков головного мозга и центральный канал спинного мозга. Это клетки
цилиндрической формы. Они образуют слой эпителиального типа.
Между соседними клетками имеются пояски сцепления, пространство
между которыми заполнено цереброспинальной жидкостью. Боль-
шинство эпендимоцитов имеют подвижные реснички, вызывающие
ток цереброспинальной жидкости. Цитоплазма эпендимоцитов содер-
жит многочисленные митохондрии, аппарат Гольджи, расположенный
над ядром, и слаборазвитый эндоплазматический ретикулум.
Астроциты — клетки с большим количеством отростков, бедные
органеллами. Они выполняют в основном опорную и разграничитель-
ную функции. Различают протоплазматические астроциты, локали-
зующиеся в сером веществе центральной нервной'системы, и волок-
нистые астроциты, присутствующие в белом веществе.
Протоплазматические астроциты характеризуются короткими, силь-
но ветвящимися отростками и светлым сферическим ядром. Астроци-
ты имеют 20—40 длинных, слабо ветвящихся отростков, в которых
много фибрилл, состоящих из филаментов диаметром 10 нм. Отрост-
ки астроцитов тянутся к базальным мембранам капилляров, к телам и
дендритам нейронов, окружая их синаптические контакты, а также к
мягкой мозговой оболочке, образуя пиоглиалъную мембрану, грани-
чащую с субарахноидальным пространством. Подходя к капилляру, от-
ростки астроцитов полностью окружают его, образуя расширенные
«ножки». Подобными отростками астроцитов окружено до 95% по-
верхности капилляров. Астроциты накапливают и передают вещества
от капилляров к нейронам, захватывают из внеклеточной среды избы-
ток ионов калия и других веществ, таких как нейромедиаторы, накап-
ливающихся там после интенсивной нейрональной активности.
Олигодендроциты имеют по сравнению с астроцитами более мел-
кие и более интенсивно окрашивающиеся ядра. Их отростки немного-
численны. Олигодендроциты присутствуют как в сером, так и в белом
1.1. Нервна 0 ткань
веществе. В сером веществе они локализуются вблизи перикарионов.
В миелинизированных нервных волокнах белого вещества их растяну-
тые плазматические мембраны образуют миелиновый слой, причем, в
противоположность нейроглии периферической нервной системы, один
олигодендроцит может участвовать в миелинизации нескольких аксо-
нов. Цитоплазма олигодендроцитов содержит много митохондрий, раз-
витый аппарат Гольджи, цистерны гранулярного эндоплазматического
ретикулума, многочисленные микротрубочки.
Микроглия представляет собой фагоцитирующие клетки, относя-
щиеся к системе мононуклеарных фагоцитов и происходящие из ство-
ловой кроветворной клетки. Функция микроглии — защита от ин-
фекции и повреждения, а также удаление продуктов разрушения нерв-
ной ткани. Клетки микроглии небольшие по размерам, имеют тела
продолговатой формы. Их короткие отростки несут на своей поверх-
ности вторичные и третичные ответвления, что создает впечатление
колючести клетки. В отличие от других типов нейроглии, имеющих
сферические ядра, ядра микроглии продолговатые с компактным хро-
матином. Описанная морфология характерна для типичной (ветвис-
той, покоящейся) микроглии ЦНС.
В развивающемся мозге млекопитающих обнаруживается и иная
форма микроглии — амебоидная микроглия, активно фагоцитирую-
щая в раннем постнатальном периоде, когда гематоэнцефалический
барьер еще не вполне развит и вещества из кровяного русла легко по-
падают в ЦНС. Созревая, амебоидные микроглиальные клетки пре-
вращаются в ветвистую микроглию. Ветвистая микроглия встреча-
ется как в сером, так и в белом веществе.
После травмы в поврежденной области мозга появляется реак-
тивная микроглия. Она не имеет ветвящихся отростков. В цито-
плазме клеток реактивной микроглии присутствуют плотные тельца,
липидные включения, лизосомы. Есть данные о том, что реактивная
микроглия формируется вследствие активации клеток ветвистой мик-
роглии при повреждении ЦНС.
Глия периферической нервной системы (периферическая ней-
роглия). Отличается от макроглии ЦНС — ее клетки происходят из
нервного гребня. К периферической нейроглии относятся шванновские
клетки (нейролеммоциты) и глиоциты ганглиев. Нейролеммоциты
формируют оболочки отростков нервных клеток в нервных волокнах пе-
риферической нервной системы. Глиоциты ганглиев окружают тела
нейронов в нервных узлах и участвуют в обмене веществ нейронов.
16
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
1.1.З. Нервные волокна
Отростки нервных клеток, покрытые глиальными клетками, на-
зывают нервными волокнами. Их делят на два класса: немиели-
низированные и миелинизированные. В составе нервных волокон ча-
ще всего (за исключением чувствительных нервов) находятся аксо-
ны, которые также называют осевым цилиндром нервного волокна.
В ЦНС оболочки отростков нейронов образуют отростки олигоденд-
роцитов, а в периферической нервной системе — шванновские гли-
альные клетки.
Немиелинизированные нервные волокна. Находятся преиму-
щественно в структурах вегетативной нервной системы. Нейролеммо-
циты оболочек немиелинизированных нервных волокон, располагаясь
плотно, образуют тяжи, в которых на определенном расстоянии друг
от друга видны овальные ядра. В нервных волокнах внутренних орга-
нов в таком тяже имеется, как правило, не один, а несколько (10—20)
осевых цилиндров, принадлежащих различным нейронам. Такие осе-
вые цилиндры могут, покидая одно волокно, переходить в соседнее.
Волокна, содержащие несколько осевых цилиндров, называют волок-
нами кабельного типа. Оболочки нейролеммоцитов очень тонкие,
поэтому под световым микроскопом они выявляются как однородный
тяж цитоплазмы, «одевающий» осевые цилиндры.
Миелинизированные нервные волокна. Встречаются как в
ЦНС, так и в периферической нервной системе. Они значительно тол-
ще немиелинизированных нервных волокон: диаметр поперечного се-
чения колеблется от 2 до 20 мкм. Состоят, так же как и немиелинизи-
рованные нервные волокна, из осевого цилиндра, «одетого» оболоч-
кой из нейролеммоцитов, но более сложной по строению.
В сформированном миелиновом волокне различают два слоя
оболочки: внутренний, более толстый миелиновый слой, и наруж-
ный, тонкий, состоящий из цитоплазмы, ядер нейролеммоцитов и ней-
ролеммы (плазматической мембраны).
В процессе формирования мозга аксон погружается в желобок на
поверхности нейролеммоцита. Когда края желобка смыкаются, обра-
зуется двойная складка плазмалеммы нейролеммоцита — мезаксон.
Мезаксон удлиняется, концентрически наслаивается на осевой ци-
линдр и образует вокруг него плотную слоистую зону — миелиновый
слой. Миелинизированная оболочка формируется в результате много-
кратного закручивания глиальной клетки вокруг аксона. Следователь-
1.1. Нервная ткань
17
но, миелин — это слой относительно богатых фосфолипидами плаз-
матических мембран глиальных клеток (олигодендроцитов в ЦНС
и шванновских клеток в ПНС).
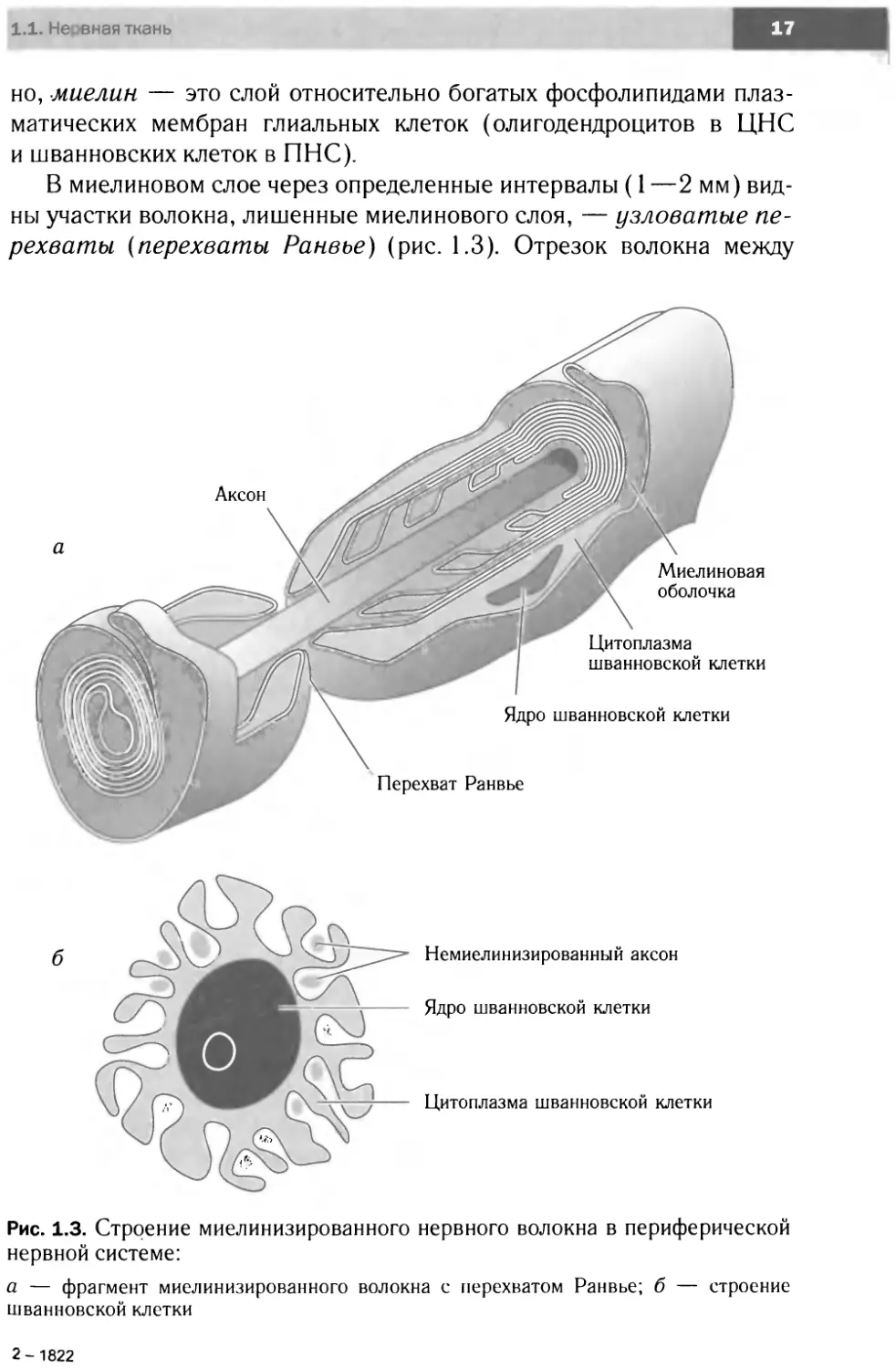
В миелиновом слое через определенные интервалы (1 —2 мм) вид-
ны участки волокна, лишенные миелинового слоя, — узловатые пе-
рехваты (перехваты Ранвье) (рис. 1.3). Отрезок волокна между
Немиелинизированный аксон
Ядро шванновской клетки
Цитоплазма шванновской клетки
Рис. 1.3. Строение миелинизированного нервного волокна в периферической
нервной системе:
а — фрагмент миелинизированного волокна с перехватом Ранвье; б — строение
шванновской клетки
2-1822
18
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
смежными перехватами — межузловой сегмент — формируется оди-
ночной шванновской клеткой (или отростком олигодендроцита). От-
сутствие миелинового слоя в области узловых перехватов объясняет-
ся тем, что в данном участке волокна заканчивается один нейролем-
моцит (или отросток олигодендроцита) и начинается другой. Осевой
цилиндр в этом месте частично покрыт интердигитирующими, т. е.
разобщенными, пальцевидными отростками нейролеммоцитов. В об-
ласти перехватов Ранвье аксональная мембрана напрямую контакти-
рует с внеклеточной средой. Наличие большого числа митохондрий в
области перехватов Ранвье свидетельствует о высокой метаболиче-
ской активности аксона в этих участках. Следует отметить, что ветвле-
ние аксона также происходит в области перехватов Ранвье.
Длина межузлового сегмента, так же как и толщина миелинового
слоя, зависит от толщины осевого цилиндра. В миелиновом слое пери-
одически встречаются узкие насечки, так называемые насечки Шмид-
та—Лантермана. Насечка миелина представляет собой участок
миелинового слоя, в котором завитки мезаксона лежат неплотно друг
к другу, образуя спиральный туннель (место расслоения миелина),
заполненный цитоплазмой нейролеммоцита.
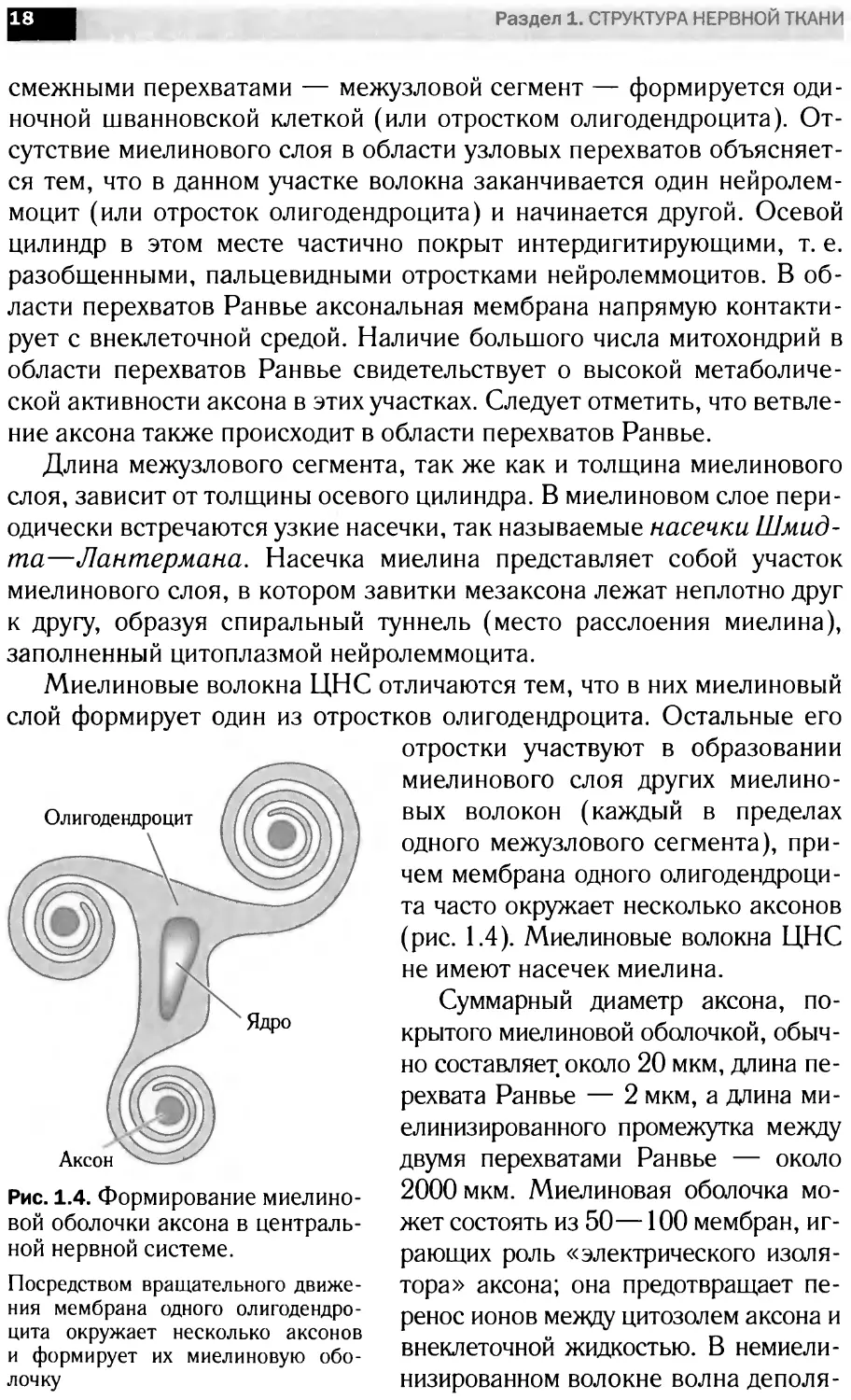
Миелиновые волокна ЦНС отличаются тем, что в них миелиновый
слой формирует один из отростков олигодендроцита. Остальные его
Рис. 1.4. Формирование миелино-
вой оболочки аксона в централь-
ной нервной системе.
Посредством вращательного движе-
ния мембрана одного олигодендро-
цита окружает несколько аксонов
и формирует их миелиновую обо-
лочку
отростки участвуют в образовании
миелинового слоя других миелино-
вых волокон (каждый в пределах
одного межузлового сегмента), при-
чем мембрана одного олигодендроци-
та часто окружает несколько аксонов
(рис. 1.4). Миелиновые волокна ЦНС
не имеют насечек миелина.
Суммарный диаметр аксона, по-
крытого миелиновой оболочкой, обыч-
но составляет около 20 мкм, длина пе-
рехвата Ранвье — 2 мкм, а длина ми-
елинизированного промежутка между
двумя перехватами Ранвье — около
2000 мкм. Миелиновая оболочка мо-
жет состоять из 50—100 мембран, иг-
рающих роль «электрического изоля-
тора» аксона; она предотвращает пе-
ренос ионов между цитозолем аксона и
внеклеточной жидкостью. В немиели-
низированном волокне волна деполя-
1.1. Нервная ткань
ризации мембраны распространяется вдоль всей аксолеммы, не пре-
рываясь, в миелиновом же волокне она выявляется только в области
перехватов Ранвье.
Скорость передачи импульса миелинизированными волокнами
больше, чем немиелинизированными. Тонкие волокна, бедные миели-
ном, и немиелинизированные волокна проводят нервный импульс со
скоростью 1—2 м/с, тогда как толстые миелинизированные волок-
на — со скоростью 5—120 м/с.
Миелин — электрический изолятор, однако его функция облегче-
ния проводимости в аксонах не имеет точной аналогии с электриче-
скими схемами. Когда мембрана в перехвате Ранвье возбуждена, ток
не может течь через оболочку высокого сопротивления и поэтому де-
поляризует мембрану в следующем перехвате. Активное возбуждение
аксональной мембраны перескакивает от одного перехвата к другому;
эту форму распространения импульса называют сальтаторной, или
прыгающей, проводимостью (от лат. saltatore — подскакивать).
Таким образом, миелин создает прерывистую изоляцию
нервного волокна.
1.1.4. Нервные окончания
Нервные волокна заканчиваются концевыми аппаратами —
нервными окончаниями. Различают три группы нервных оконча-
ний: концевые аппараты, образующие межнейрональные синапсы и
осуществляющие связь нейронов между собой; эффекторные оконча-
ния, передающие нервный импульс на ткани рабочего органа; рецеп-
торные (аффекторные) окончания.
Эффекторные нервные окончания бывают двух
типов — двигательные и секреторные. Двигательные нервные
окончания аксонов двигательных клеток вегетативной нервной сис-
темы осуществляют передачу нервного импульса на ткани рабочих
органов.
Двигательные окончания в поперечнополосатых мыш-
цах называются нервно-мышечными окончаниями. Плазмалеммы
ветвей аксона и мышечного волокна разделены синаптической щелью
шириной 50 нм, которая заполнена аморфным веществом, богатым
гликопротеидами. Мембрана мышечного волокна образует многочис-
ленные складки, формирующие синаптическую щель. Примембран-
ный слой, который характеризуется обилием митохондрий, скоплени-
ем круглых или слегка овальных ядер, образует постсинаптическую
часть синапса.
20
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
П. А. Кометиани
Основоположник школы гру-
зинских нейрохимиков, автор
первого национального учеб-
ника по биохимии, выдаю-
щийся ученый, известный
пионерскими исследованиями
в области биоэлектрической
активности клеток, биохимии
высшей нервной деятельнос-
ти и нейрохимических основ
памяти
Ветви нервного волокна в нервно-
мышечном синапсе характеризуются оби-
лием митохондрий и многочисленными
пресинаптическими пузырьками, содержа-
щими характерный для этого вида оконча-
ний медиатор — ацетилхолин. При воз-
буждении ацетилхолин поступает через
пресинаптическую мембрану в синапти-
ческую щель на холинорецепторы постси-
наптической (мышечной) мембраны, вы-
зывая ее возбуждение (см. гл. 2.3). Меж-
синаптическая область содержит фермент
ацетилхолинэстеразу, разрушающую ме-
диатор и ограничивающую этим срок его
действия. Нарушения в нервно-мышеч-
ных окончаниях вызывают различные
мышечные заболевания, например mias-
tenia gravis, характеризующееся прогрес-
сирующей мышечной слабостью. При
этом заболевании в крови циркулиру-
ют антитела против ацетилхолиновых ре-
цепторов сарколеммы (мембраны мышеч-
ного волокна). Эти антитела связывают-
ся с холинорецепторами постсинаптиче-
ской мембраны и инактивируют их, нарушая нервно-мышечное взаи-
модействие.
Двигательные нервные окончания в гладкомышечной
ткани менее дифференцированны. Они представляют собой утол-
щения нервного волокна, контактирующего с гладкомышечными
клетками. Сходное строение имеют секреторные нервные окон-
чания.
Рецепторные нервные окончания — рецепторы, ко-
торые воспринимают различные сигналы как из внешней среды, так и
от внутренних органов. Соответственно выделяют две большие
группы рецепторов: экстерорецепторы (реагирующие на
внешние сигналы) и интерорецепторы (реагирующие на внутренние
сигналы). К экстерорецепторам относят слуховые, зрительные, обо-
нятельные, вкусовые и осязательные рецепторы. К интерорецепторам
относят висцерорецепторы (сигнализирующие о состоянии внутрен-
них органов) и вестибулопроприорецепторы (рецепторы опорно-дви-
гательного аппарата).
1.1. Нервная ткань
Специализированные нейросенсорные клетки органов зрения яв-
ляются фоторецепторами, преобразующими внешнюю энергию
света в нервный импульс. Сетчатка глаза — сенсорная рецепторная
часть зрительного анализатора, в которой под воздействием света
происходят фотохимические превращения зрительных пигментов, фо-
тотрансдукция, изменение биоэлектрической активности нейронов и
передача информации о внешнем мире в подкорковые и корковые зри-
тельные центры (см. 3.6.1).
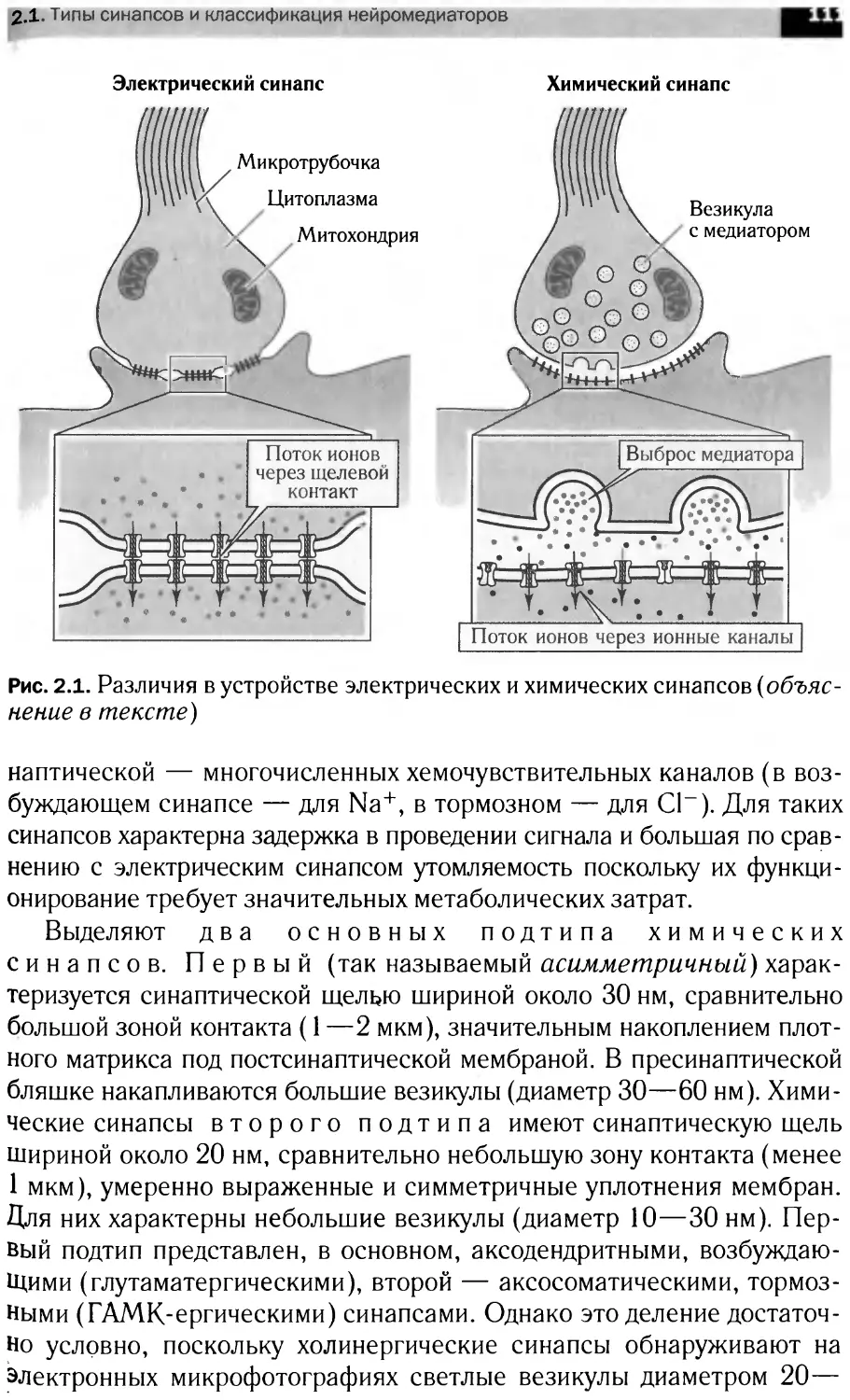
1.1.5. Синапсы
Синапсы — это структуры, предназначенные для передачи им-
пульса с одного нейрона на другой или на мышечные и железистые
структуры. Синапсы обеспечивают проведение импульса по цепи ней-
ронов, т. е. определяют направление проведения импульса. Только
импульс, достигающий нервных окончаний аксона, может с помощью
синапсов передать возбуждение на другой нейрон, мышечную или же-
лезистую клетку. В зависимости от способа передачи импульса синап-
сы бывают химическими или электрическими (см. гл. 2.1).
Нервное окончание аксона представляет собой пресинаптиче-
скую часть синапса, а область второго нейрона, с которой она
контактирует, — его постсинаптическую часть. В пресинаптиче-
ской части находятся синаптические пузырьки, многочисленные мито-
хондрии и отдельные нейрофиламенты. Форма и содержимое синапти-
ческих пузырьков связаны с функцией синапса. Так, в синапсах, в ко-
торых передача импульса совершается с помощью ацетилхолина
(холинергические синапсы), обнаруживаются округлые прозрачные
пузырьки диаметром 30—50 нм; в синапсах, в которых в качестве
нейромедиатора используется норадреналин (адренергические си-
напсы), синаптические пузырьки имеют диаметр 50—90 нм, в них
видна плотная сердцевина диаметром 15—25 нм.
Пресинаптическая мембрана — это мембрана клетки, передаю-
щей импульс (аксолемма). Межсинаптическая щель между пре- и
постсинаптическими мембранами имеет ширину 20—30 нм. Пре- и
постсинаптические мембраны находятся в тесном контакте, осуществ-
ляющемся с помощью филаментов, пересекающих синаптическую
щель. Постсинаптическая мембрана — это участок плазмалеммы
клетки, воспринимающий сигнал медиатора. Данная область содер-
жит рецепторные белки, способные воспринимать сигнал соответст-
вующего нейромедиатора.
22
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
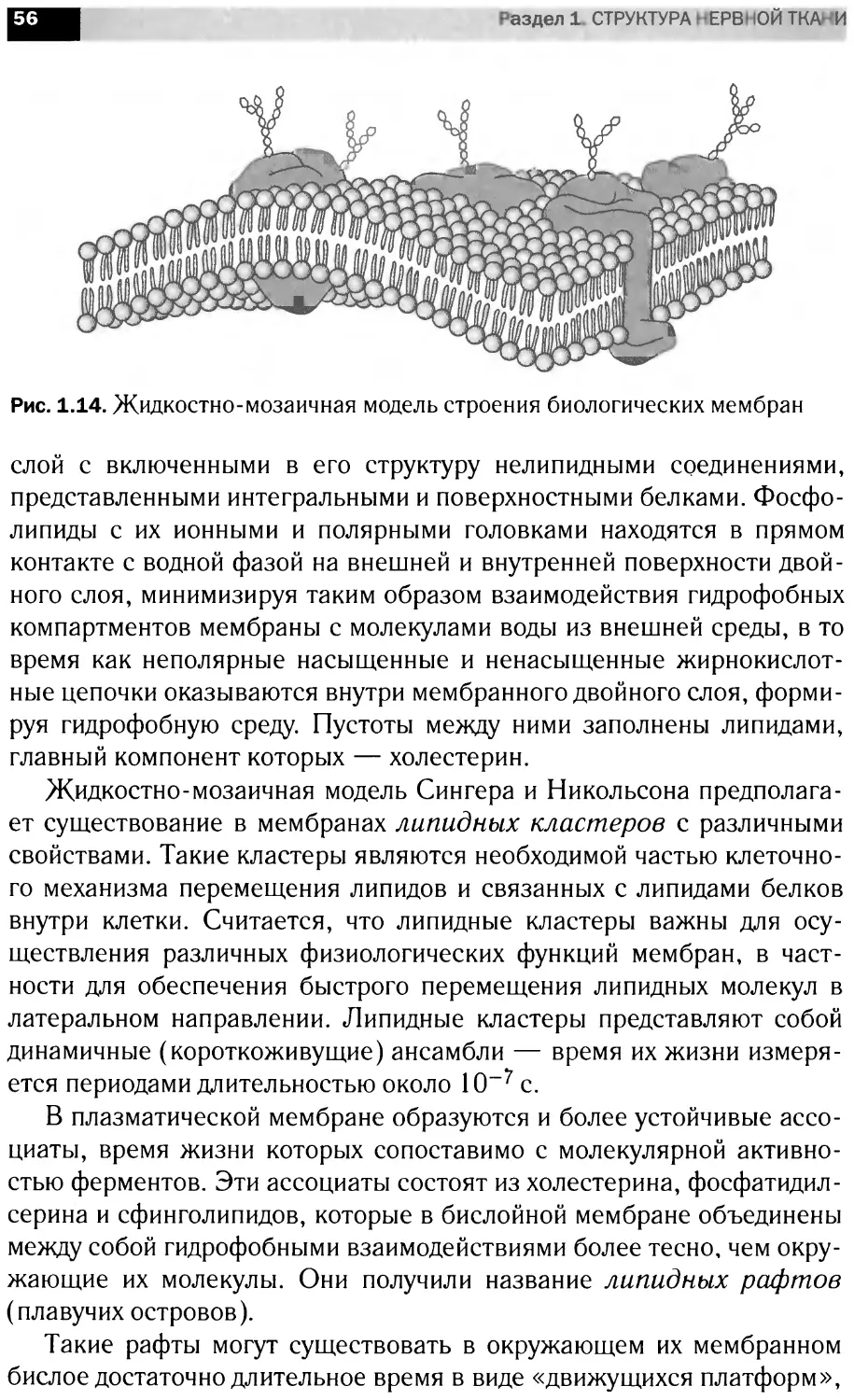
1.2.
ОСОБЕННОСТИ ЛИПИДНОГО И БЕЛКОВОГО СОСТАВА
НЕРВНОЙ ТКАНИ
Изучение липидов нервной системы составляет существенную
часть нейрохимических исследований. Существует несколько очевид-
ных свидетельств важной роли липидов головного мозга как структур-
ных компонентов, а также их участия в функциональной активности
мозга.
Среди всего многообразия органов мозг в наибольшей степени обо-
гащен липидами (приблизительно 50% сухой массы). Он содержит
уникальную структуру — миелиновую оболочку, которая по сравнению
с другими тканями и субклеточными компонентами имеет самую высо-
кую концентрацию липидов. Липидный состав нервной ткани практи-
чески постоянен и мало меняется даже под влиянием внешних факто-
ров (диета, гормоны, фармакологические вещества, стрессы), которые
существенно изменяют липидный состав висцеральных органов и плаз-
мы. Это — следствие защищенности ЦНС от различных внешних воз-
действий. Однако следует помнить, что в период развития нервной сис-
темы в липидном составе наблюдаются существенные изменения.
Существуют генетически детермированные метаболические болез-
ни, в механизм развития которых вовлекаются липиды мозга. Возмож-
ности идентификации аномальных липидов и поиска лежащих в осно-
ве их синтеза ферментативных дефектов дают стимул не только для
изучения таких патологических условий, но и для исследования химии
и метаболизма липидов мозга в целом.
В организме липиды выполняют две основные функции:
1)источника химической энергии, запасенной в триглицеридах;
2) структурного компонента клеточных мембран. Мозг содержит ми-
нимальные количества триглицеридов. Вся сложнейшая деятельность
нервной ткани опосредуется мембранами, в формировании и функци-
онировании которых липиды принимают непосредственное участие.
1.2.1. Липидный состав
Фосфолипиды, гликолипиды, стероиды. Липидный состав
мозга является уникальным не только из-за высокой общей концент-
рации липидов, но также из-за специфики. Почти все липиды голов-
ного мозга представлены тремя основными группами:
1)фосфолипидами; 2) гликолипидами; 3) стероидами, причем кон-
центрация фосфолипидов особенно высока. Распространение и свой-
ства фосфолипидов изучены наиболее детально.
1.2. Особенности липидного и белкового состава нервной ткани
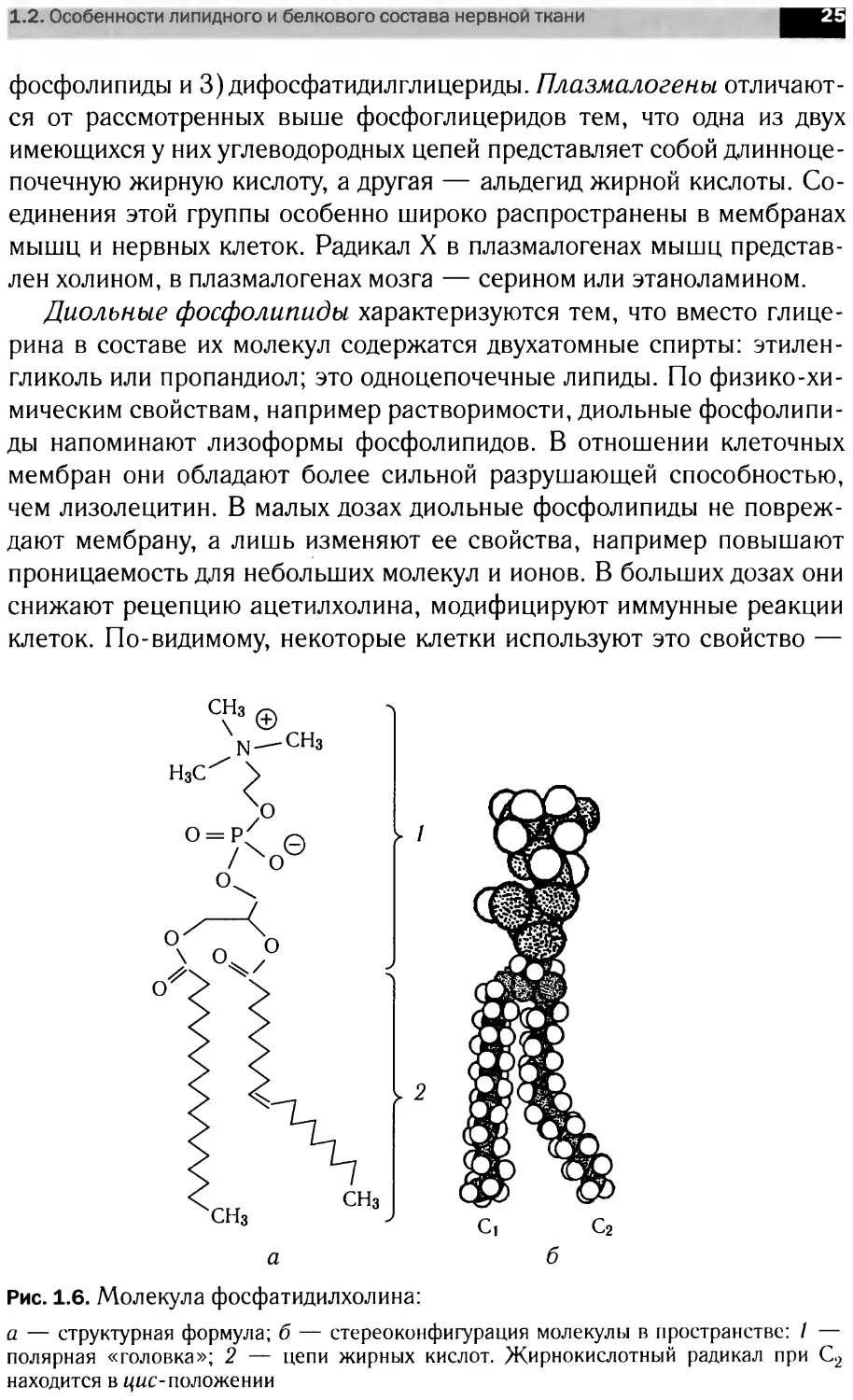
Фосфолипиды подразделяют на две
группы: глицерофосфолипиды (про-
изводные фосфатидной кислоты — фос-
фатидилхолин, фосфатидилэтаноламин,
фосфатидилсерин, фосфатидилинозит) и
сфингофосфолипиды (производные це-
рамида, сфингомиелины).
Глицерофосфолипиды представляют
собой производные фосфатидной кис-
лоты, к гидроксилу фосфорной кислоты
Ri R2
I I
ООН о
III II
Н—С>-С2-С3-О-Р-О-Х
III I
Н Н Н О-
Рис. 1.5. Схематическое стро-
ение молекулы фосфолипида.
X — полярная головка; R, и R2 —
жирнокислотные радикалы
которой сложноэфирной связью присоединен радикал X (рис. 1.5,
табл. 1.1); Rj и R2 — ацильные остатки жирных кислот, содержащие
от 12 до 18 атомов углерода (как правило, четное количество).
Таблица 1.1. Классификация фосфолипидов
по структуре полярных радикалов
Радикал 1,2-диацилфосфатидной кислоты Фосфолипид
-ОН Фосфатидная кислота
zch3 -och2-ch2-+n^ch3 хсн3 Фосфатидилхолин (лецитин)
-och2-ch2-nh+ Фосфатидилэтаноламин (кефалин)
-осн2-сн-соо~ 1 NH3+ Фосфатидилсерин
-о-сн-сн-соо- СН3 NH3 Фосфатидилтреонин
-ОСН2-СН-СН2 2 1 1 он он Фосфатидилглицерин
ОН ОН
ОН
Фосфатидилинозит
24
Раздел 1. СТРУКТУРА ЬЬРБ.ЧОЙ ТКАНИ
Таблица 1.2. Липидный состав некоторых биологических мембран,
% от суммарного количества липидов
(по: Болдырев и др., 2006)
Липид Миелин человека Эритроциты человека Митохондрии сердца быка £. coli
Фосфатидная кислота 0,5 1,5 0,0 0
Фосфатидилхолин 10.0 19,0 39,0 0
Фосфатидилэтаноламин 20,0 18,0 27,0 66
Фосфатидилглицерол 0,0 0,0 0,0 18
Фосфатидилинозит 1.0 1,0 7,0 0
Фосфатидилсерин 8,0 8,0 0,5 0
Кардиолипин 0,0 0,0 23,5 14
Сфингомиелин 8,5 17,5 0,0 0
Гликолипиды 26,0 10,0 0,0 0
Холестерин 26,0 25,0 3,0 0
Первая часть сложных слов, используемых для обозначения фос-
фолипидов, потерявших одну из двух ацильных цепей, — «лизо». Ли-
зофосфолипиды обнаруживаются в мембранах в небольших количест-
вах — появление этих веществ приводит к нарушению структуры би-
слоя и лизису клеток.
Характеристика липидного состава некоторых мембран животных
представлена в таблице 1.2. Как видно из таблицы, основными липи-
дами мембран животных клеток являются глицерофосфолипиды: фос-
фатидилхолин и фосфатидилэтаноламин. Структура фосфатидилхоли-
на представлена на рисунке 1.6.
Наиболее часто встречающиеся фосфолипиды построены по еди-
ному плану, их молекулы стерически хорошо соответствуют друг другу.
В то же время огромное разнообразие фосфолипидов обеспечивается
различием жирных кислот, входящих в состав их молекул. Так, су-
ществует несколько десятков природных видов фосфатидилхолина,
причем диолеилфосфатидилхолин сильно отличается по своим свойст-
вам от дипальмитоилфосфатидилхолина.
Существует несколько групп фосфолипидов, отличающихся стро-
ением от приведенных в таблице 1.2: 1) плазмалогены; 2)диольные
1.2. Особенности липидного и белкового состава нервной ткани
25]
фосфолипиды и 3) дифосфатидил глицериды. Плазмалогены отличают-
ся от рассмотренных выше фосфоглицеридов тем, что одна из двух
имеющихся у них углеводородных цепей представляет собой длинноце-
почечную жирную кислоту, а другая — альдегид жирной кислоты. Со-
единения этой группы особенно широко распространены в мембранах
мышц и нервных клеток. Радикал X в плазмалогенах мышц представ-
лен холином, в плазмалогенах мозга — серином или этаноламином.
Диольные фосфолипиды характеризуются тем, что вместо глице-
рина в составе их молекул содержатся двухатомные спирты: этилен-
гликоль или пропандиол; это одноцепочечные липиды. По физико-хи-
мическим свойствам, например растворимости, диольные фосфолипи-
ды напоминают лизоформы фосфолипидов. В отношении клеточных
мембран они обладают более сильной разрушающей способностью,
чем лизолецитин. В малых дозах диольные фосфолипиды не повреж-
дают мембрану, а лишь изменяют ее свойства, например повышают
проницаемость для небольших молекул и ионов. В больших дозах они
снижают рецепцию ацетилхолина, модифицируют иммунные реакции
клеток. По-видимому, некоторые клетки используют это свойство —
а
Ci С2
б
Рис. 1.6. Молекула фосфатидилхолина:
а — структурная формула; б — стереоконфигурация молекулы в пространстве: / —
полярная «головка»; 2 — цепи жирных кислот. Жирнокислотный радикал при С2
находится в цис-положении
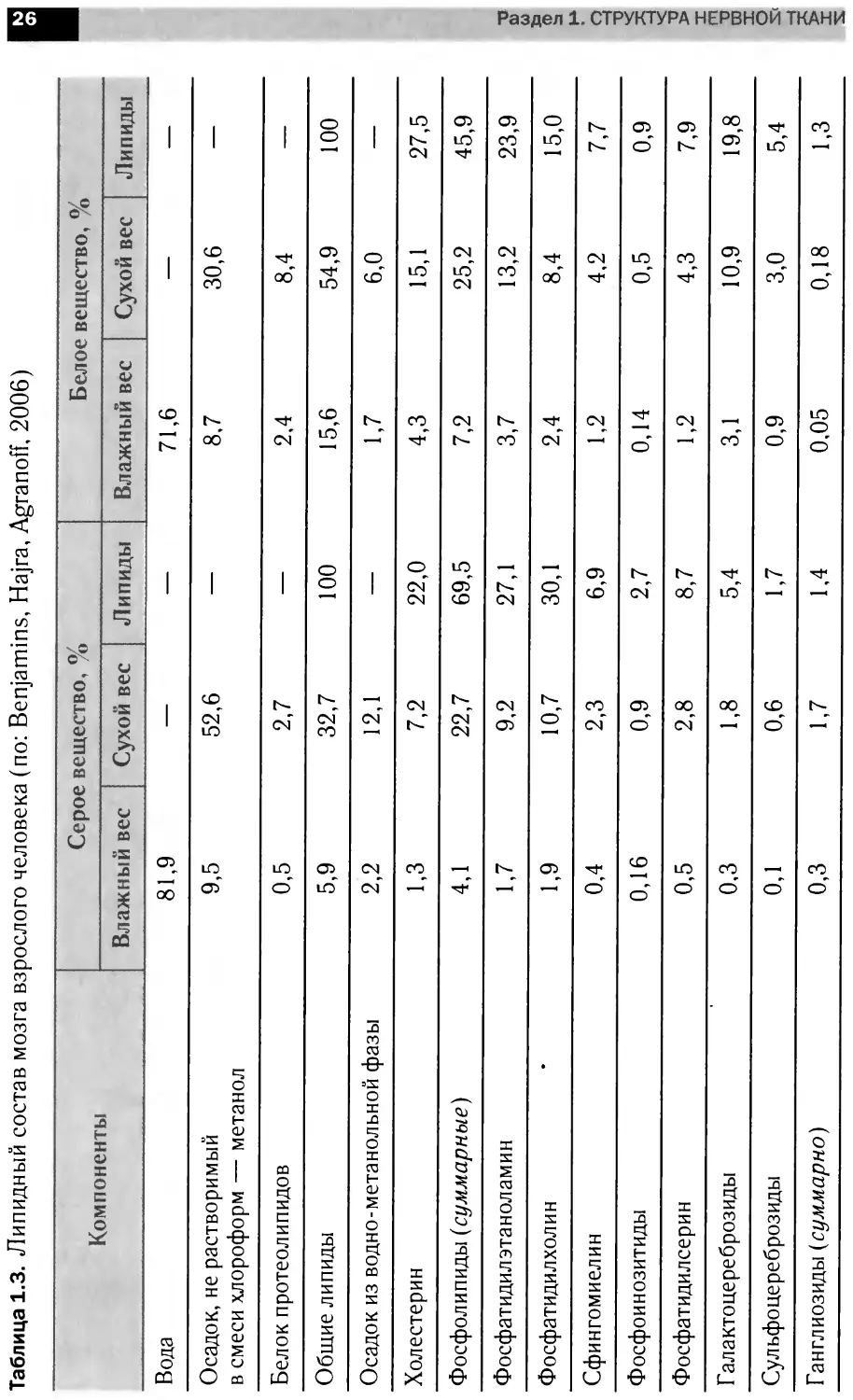
Таблица 1.3. Липидный состав мозга взрослого человека (по: Benjamins, Hajra, Agranoff, 2006)
Компоненты Серое вещество, % Белое вещество, %
Влажный вес Сухой вес Липиды Влажный вес Сухой вес Липиды
Вода 81,9 — — 71,6 — —
Осадок, не растворимый в смеси хлороформ — метанол 9,5 52,6 — 8,7 30,6 —
Белок протеолипидов 0,5 2,7 — 2,4 8,4 —
Общие липиды 5,9 32,7 100 15,6 54,9 100
Осадок из водно-метанольной фазы 2,2 12,1 — 1,7 6,0 —
Холестерин 1,3 7,2 22,0 4,3 15,1 27,5
Фосфолипиды (суммарные) 4,1 22,7 69,5 7,2 25,2 45,9
Фосфатидилэтаноламин 1,7 9,2 27,1 3,7 13,2 23,9
Фосфатидилхолин 1,9 10,7 30,1 2,4 8,4 15,0
Сфингомиелин 0,4 2,3 6,9 1,2 4,2 7,7
Фосфоинозитиды 0,16 0,9 2,7 0,14 0,5 0,9
Фосфатидилсерин 0,5 2,8 8,7 1,2 4,3 7,9
Г алактоцереброзиды 0,3 1,8 5,4 3,1 10,9 19,8
Сульфо цереброз иды 0,1 0,6 1,7 0,9 3,0 5,4
Ганглиозиды (суммарно) 0,3 1,7 1,4 0,05 0,18 1,3
Раздел 1. СТРУКТУРА НЕРВНОЙ
1.2. Особенности липидного и белкового состава нервной ткани
27|
Таблица 1.4. Содержание (%) индивидуальных фосфолипидов
в коре мозга крысы (по: Туманова, 1999)
Фосфолипиды Нейроны Нейроглия
Лизофосфатидилхолин 3,9 1,9
Фосфатидилхолин 46,1 46,9
Сфингомиелин 6,7 9,5
Фосфатидилсерин 9,1 7,1
Фосфатидилинозит 77 5,9
Фосфатидилэтаноламин 25,1 24,9
Фосфатидная кислота 1,8 3,6
начинают интенсивно синтезировать диольные липиды в период быст-
рого роста и прекращают их образование, когда клеточный рост за-
медляется. Возможно, это связано с тем, что в период роста клеток их
мембраны должны быть более лабильными.
Дифосфатидилглицериды образуются в результате соединения
двух молекул фосфатидилглицерида. Наиболее широко распростра-
ненным представителем этой группы фосфолипидов является кар-
диолипин — непременный компонент митохондриальных мембран,
выделенный первоначально из сердечной мышцы.
Значительная часть фосфолипидов в мозге представлена фосфати-
дилэтаноламинами (на долю которых приходится 25—30% суммарно-
го содержания) и фосфатидилхолинами (табл. 1.3). Состав фосфоли-
пидов обогащенных фракций нейронов и нейроглии коры мозга крысы
представлен в таблице 1.4.
Сфинголипиды. Как упоминалось выше, кроме глицерофосфоли-
пидов в группу фосфолипидов входят и сфинголипиды, которые можно
представить как производные церамида (жирнокислотного эфира нена-
сыщенного аминоспирта сфингозина) и монофосфорных эфиров спиртов.
В случае наиболее распространенного сфинголипида — сфингомиели-
на — таким эфиром является фосфорилхолин; один из его углеводород-
ных хвостов — длинная алифатическая цепь сфингозина, а другой —
этерифицированная жирная кислота (рис. 1.7). Основным блоком всех
сфинголипидов является сфингозин, который представлен длинноцепо-
чечным аминодиолом с одной ненасыщенной связью.
28
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
Олеиновая кислота
Рис. 1.7. Молекула сфингомиелина
(церамидфосфорилхолина).
Пунктиром обведена сфингозиновая
группировка
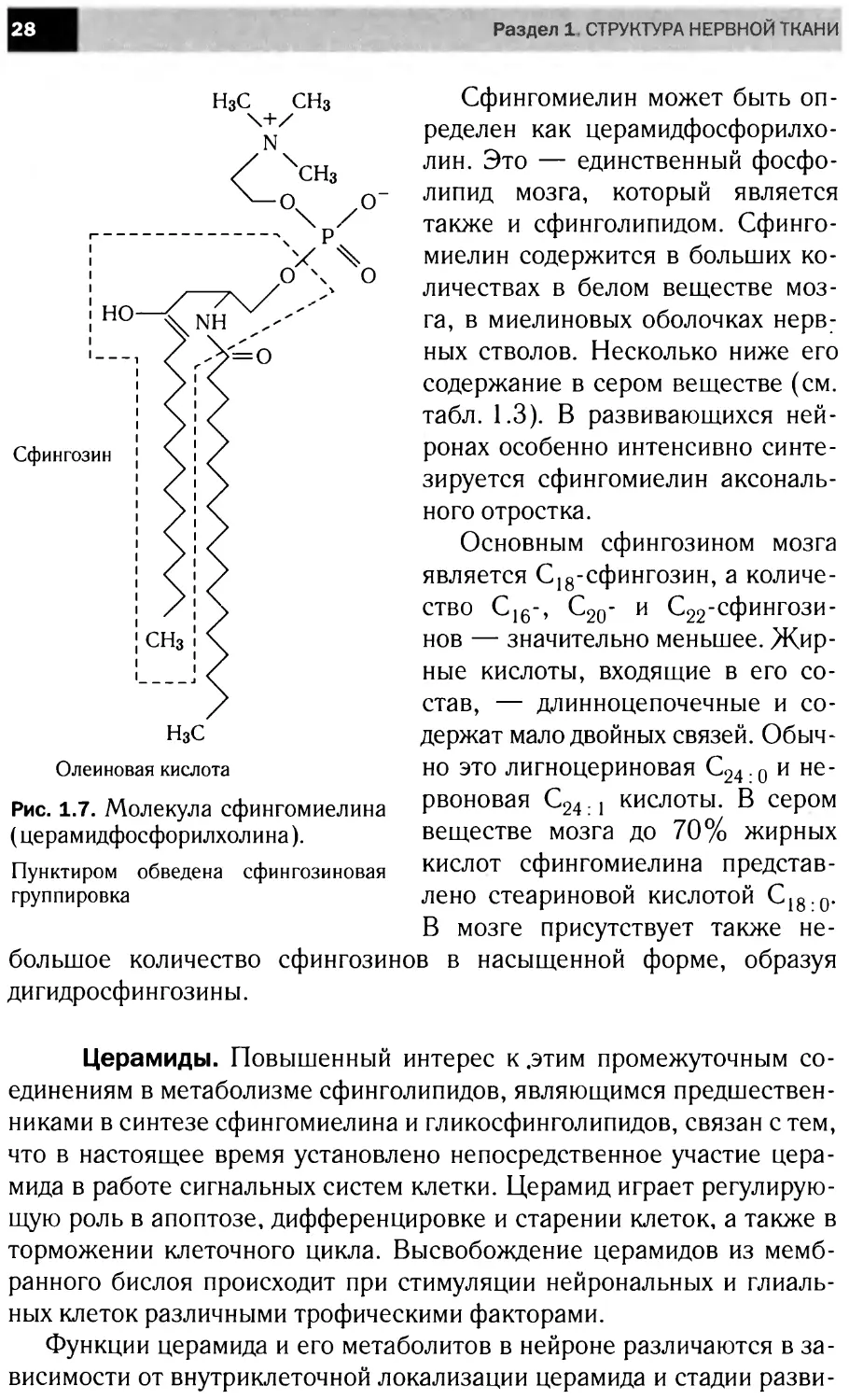
Сфингомиелин может быть оп-
ределен как церамидфосфорилхо-
лин. Это — единственный фосфо-
липид мозга, который является
также и сфинголипидом. Сфинго-
миелин содержится в больших ко-
личествах в белом веществе моз-
га, в миелиновых оболочках нерв-
ных стволов. Несколько ниже его
содержание в сером веществе (см.
табл. 1.3). В развивающихся ней-
ронах особенно интенсивно синте-
зируется сфингомиелин аксональ-
ного отростка.
Основным сфингозином мозга
является С18-сфингозин, а количе-
ство С16-, С20- и С22-сфингози-
нов — значительно меньшее. Жир-
ные кислоты, входящие в его со-
став, — длинноцепочечные и со-
держат мало двойных связей. Обыч-
но это лигноцериновая С24.0 и не-
рвоновая С241 кислоты. В сером
веществе мозга до 70% жирных
кислот сфингомиелина представ-
лено стеариновой кислотой С18,0.
В мозге присутствует также не-
большое количество сфингозинов в насыщенной форме, образуя
дигидросфингозины.
Церамиды. Повышенный интерес к .этим промежуточным со-
единениям в метаболизме сфинголипидов, являющимся предшествен-
никами в синтезе сфингомиелина и гликосфинголипидов, связан с тем,
что в настоящее время установлено непосредственное участие цера-
мида в работе сигнальных систем клетки. Церамид играет регулирую-
щую роль в апоптозе, дифференцировке и старении клеток, а также в
торможении клеточного цикла. Высвобождение церамидов из мемб-
ранного бислоя происходит при стимуляции нейрональных и глиаль-
ных клеток различными трофическими факторами.
Функции церамида и его метаболитов в нейроне различаются в за-
висимости от внутриклеточной локализации церамида и стадии разви-
1.2. Особенности липидного и белкового состава нервной ткани
тия нейрона. Рост и развитие нейронов связаны с изменением метабо-
лизма церамида. Синтез гликозилцерамида из церамида требуется для
роста аксонов в культуре нейронов гиппокампа, а образование цера-
мида из сфингомиелина с помощью сфингомиелиназы стимулирует
ранние стадии развития этих клеток, а именно — формирование
минорных нейрональных отростков и образование аксонов.
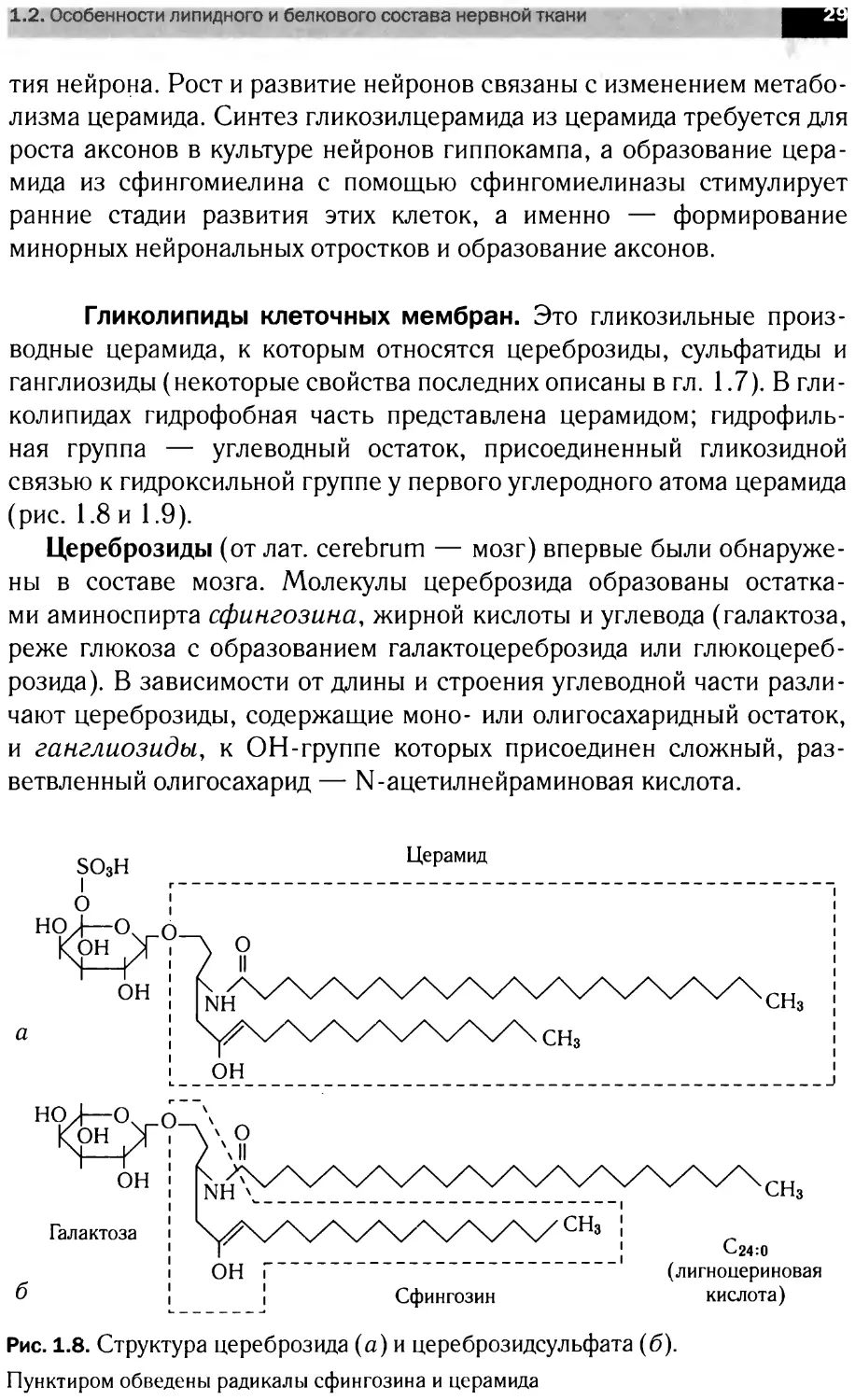
Гликолипиды клеточных мембран. Это гликозильные произ-
водные церамида, к которым относятся цереброзиды, сульфатиды и
ганглиозиды (некоторые свойства последних описаны в гл. 1.7). В гли-
колипидах гидрофобная часть представлена церамидом; гидрофиль-
ная группа — углеводный остаток, присоединенный гликозидной
связью к гидроксильной группе у первого углеродного атома церамида
(рис. 1.8 и 1.9).
Цереброзиды (от лат. cerebrum — мозг) впервые были обнаруже-
ны в составе мозга. Молекулы цереброзида образованы остатка-
ми аминоспирта сфингозина, жирной кислоты и углевода (галактоза,
реже глюкоза с образованием галактоцереброзида или глюкоцереб-
розида). В зависимости от длины и строения углеводной части разли-
чают цереброзиды, содержащие моно- или олигосахаридный остаток,
и ганглиозиды, к ОН-группе которых присоединен сложный, раз-
ветвленный олигосахарид — N-ацетилнейраминовая кислота.
Рис. 1.8. Структура цереброзида (а) и цереброзидсульфата (б).
Пунктиром обведены радикалы сфингозина и церамида
30
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
Длинные
алифатические цепи
Цереброзид (галактоцереброзид)
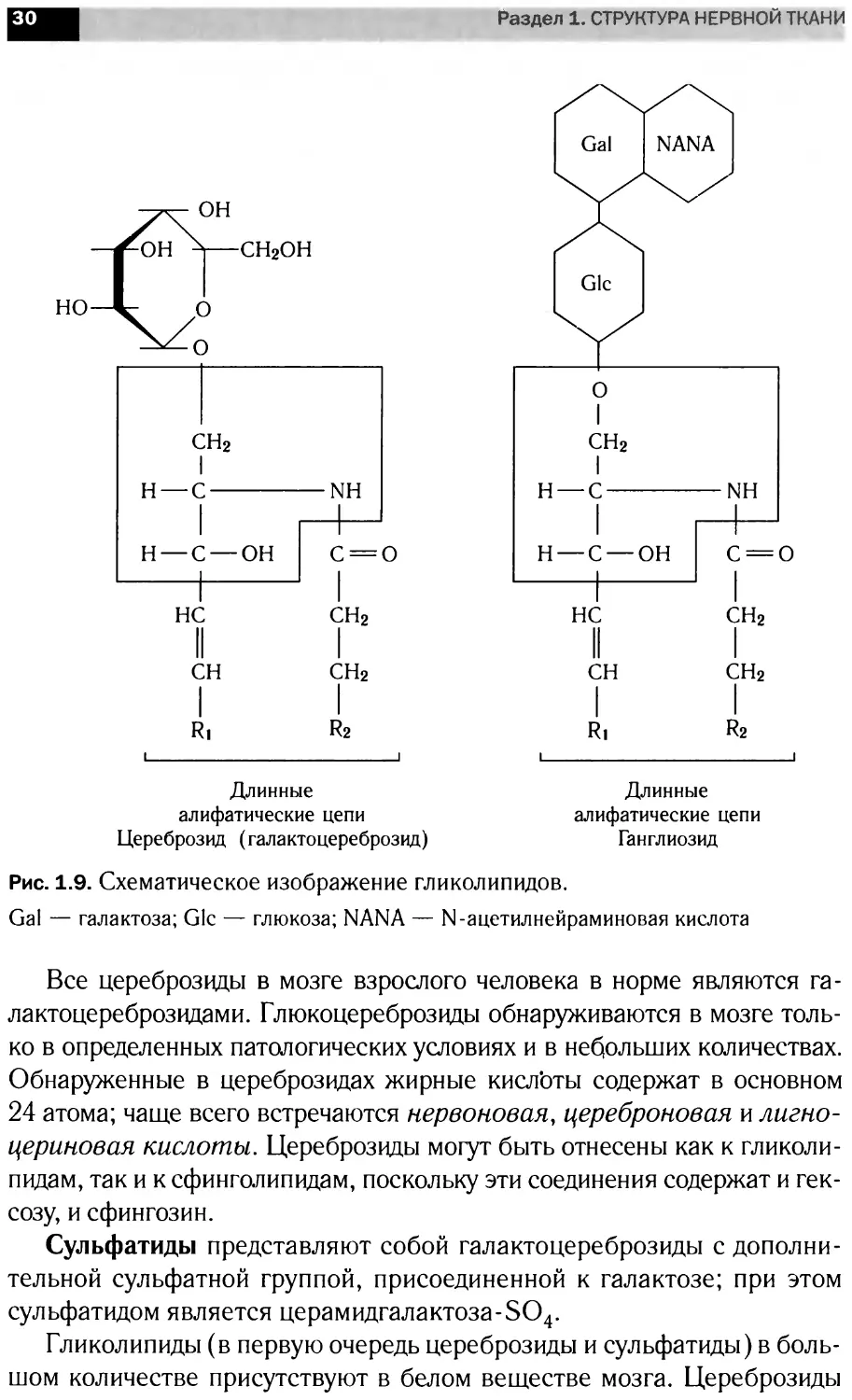
Рис. 1.9. Схематическое изображение гликолипидов.
Gal — галактоза; Glc — глюкоза; NANA — N-ацетилнейраминовая кислота
Длинные
алифатические цепи
Ганглиозид
Все цереброзиды в мозге взрослого человека в норме являются га-
лактоцереброзидами. Глюкоцереброзиды обнаруживаются в мозге толь-
ко в определенных патологических условиях и в небольших количествах.
Обнаруженные в цереброзидах жирные кислоты содержат в основном
24 атома; чаще всего встречаются нервоновая, цереброновая и лигно-
цериновая кислоты. Цереброзиды могут быть отнесены как к гликоли-
пидам, так и к сфинголипидам, поскольку эти соединения содержат и гек-
созу, и сфингозин.
Сульфатиды представляют собой галактоцереброзиды с дополни-
тельной сульфатной группой, присоединенной к галактозе; при этом
сульфатидом является церамидгалактоза-5О4.
Гликолипиды (в первую очередь цереброзиды и сульфатиды) в боль-
шом количестве присутствуют в белом веществе мозга. Цереброзиды
1.2. Особенности липидного и белкового состава нервной ткани
и сульфоцереброзиды рассматривают как специфические гликолипиды
мембран глиальных (прежде всего олигодендроглиальных и шваннов-
ских) клеток, поэтому особенно высоко содержание этих липидов в ми-
елине. Количество цереброзидов в астроцитах и особенно в мембранах
нейронов заметно ниже, чем в олигодендроцитах. Нарушение обмена
цереброзидов у человека приводит к психическим заболеваниям.
Ганглиозиды определяются как сфингогликолипиды, содержащие
сиаловую кислоту. Сиаловой кислотой называют N-ацилнейрамино-
вую кислоту, NANA (при этом сиаловая кислота в ганглиозидах мозга
всегда ацилирована). Ганглиозиды обнаружены во всех типах клеток
нервной системы, но наиболее богаты ими мембраны нейронов, преж-
де всего — мембраны синаптических окончаний.
Ганглиозиды не только являются структурным компонентом нейро-
нальных мембран, но и обладают регулирующим влиянием на процес-
сы пролиферации (см. гл. 1.7) и регенерации клеток, а также выражен-
ным нейротрофическим и нейритогенным эффектами; они способны
предотвращать дегенерацию культивируемых нейронов мозжечка, вы-
зываемую глутаматом или каинатом. Кроме того установлено, что ганг-
лиозиды ингибируют перекисное окисление липидов в изолированных
синаптосомах мозга крысы.
Известно, что ключевым моментом в реализации нейротоксического
(экзайтотоксического) действия возбуждающих аминокислот при
взаимодействии их с глутаматными рецепторами является гиперактива-
ция ионных каналов, сопровождающаяся избыточным поступлением
Са2+ в клетки и активацией Са2+-зависимых протеиназ и фосфолипаз,
что в итоге приводит к необратимому повреждению нейронов. Имеются
данные, указывающие на наличие в этих процессах свободнорадикаль-
ных реакций. Установлено, что нейрональная дегенерация, вызванная
каинатом, опосредуется супероксидными радикалами, генерируемыми
ксантиноксидазой. Защитный эффект мембранных ганглиозидов, по-ви-
димому, обусловлен образованием доменов, обогащенных этими липи-
дами, которые оказываются способными стабилизировать нейрональ-
ные мембраны и в конечном итоге защищать нейроны от окисления.
Природной функцией ганглиозидов в нейрональных мембранах яв-
ляется участие в дифференцировке нейрональной ткани. Ганглиозиды
лимфоцитов определяют видоспецифичность и регулируют межкле-
точные контакты.
Стероиды. Это спирты со стерановым скелетом, к которым от-
носятся как немембранные липиды (из них наиболее важны стероид-
ные гормоны), так и структурные компоненты мембран. В перечень
32
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
нм
3 Г
2
1
а
б
О
Рис. 1.10. Структурная формула холестерина (а) и его упаковка между двух
молекул фосфолипидов (б).
Звездочкой отмечен гидроксил, который формирует эфирную связь с жирной кислотой
при образовании эфиров холестерина: / — область полярных «головок»; 2 — область,
упорядочиваемая холестерином; 3 — область подвижных жирнокислотных цепей
мембранных компонентов стероидного ряда входят холестерин, си-
тостерин, тетрахименин. В тканях животных распространен хо-
лестерин. В растительных клетках холестерин не обнаружен, его за-
меняют фитостерины. У бактерий стероиды отсутствуют. Холестерин
и его эфиры — непременные составляющие плазматических мембран
клеток животных (рис. 1.10).
Молекула холестерина не содержит длинных жирнокислотных цепо-
чек, она состоит из четырех плоских колец, к одному из которых (шести-
членному) присоединена полярная гидроксильная группа (—ОН), а наи-
более отдаленное от него пятичленное кольцо соединено с разветвлен-
ной углеводородной цепочкой из восьми атомов углерода. Таким
образом, молекулы холестерина имеют полярную головку и вытянутую в
длину неполярную часть. Поэтому они хорошо встраиваются в бислой-
ные липидные структуры, особенно в области мембранных дефектов,
образующихся в результате фазового перехода липидов (рис. 1.11).
Особенно много холестерина содержится в наружных мембранах.
Например, в плазматической мембране клеток печени холестерин
составляет около 30% всех мембранных липидов.
Холестерин является единственным стеролом, представленным в
значительном количестве в мозге взрослого человека. У человека сред-
няя концентрация холестерина в ЦНС выше, чем в любой другой ткани
(свыше 20 мг/г). Несмотря на то что мозг составляет всего около 2%
1.2. Особенности липидного и белкового состава нервной ткани
Рис. 1.11. Встраивание холестерина в жидкокристаллический бислой:
а — кристаллическая упаковка липидов в бислое; б — разрыхление бислоя в резуль-
тате фазового перехода (сопровождается уменьшением толщины бислоя и возрастани-
ем его площади); в — встраивание холестерина в жидкокристаллический бислой, уве-
личивающее его толщину
веса тела, почти 25% общего количества холестерина, присутствую-
щего в организме, локализовано в этом органе, причем большая его
часть находится в миелине. Довольно высокое содержание холестерина
характерно и для плазматических мембран других клеток нервной тка-
ни. Например, в плотных миелиновых мембранах холестерин и фосфо-
липиды содержатся в соотношении 1 :0,76; в олигодендроглии —
1 : 2,2; в астроглии — 1 : 2,5; в нейронах — 1 : 3,5 (табл. 1.5).
Небольшие концентрации десмостерина и эфиров холестерина об-
наружены в ЦНС человека и некоторых животных на ранних стадиях
развития, а в ЦНС взрослых организмов все стероиды представлены
преимущественно неэстерифицированным холестерином. В ЦНС хо-
лестерин находится в основном в олигодендроглии и в плазматической
мембране астроцитов и нейронов.
Таблица 1.5. Состав и содержание липидов основных типов
нервных клеток и миелина мозга крысы,
мкмоль/мг сухой массы (по: Туманова, 1999)
Липиды Астроглия Нейрон Олиго- дендроглия Миелин
Холестерин 14,100 6,610 10,800 54,900
Цереброзиды 0,689 0,513 2,610 22,000
Сульфатиды 0,142 0,090 0,472 2,890
Общие фосфолипиды 35,600 22,400 23,400 41,800
Ганглиозиды 0,582 0,223 0,239 0,0453
Молярное соотношение'.
холестерин 1 1 1 1
цереброзиды 0,05 0,075 0,25 0,40
фосфолипиды 2,5 3,5 2,2 0,76
34
Раздел 1. СТРУКТУРА НЕРВНЕЙ ТКАНИ
Метаболизм холестерина в ЦНС существенно отличается от метабо-
лизма в других органах и тканях. У всех животных организмов наиболь-
ший рост и дифференцировка ЦНС наблюдаются в ранний постнатальный
период, и холестерин, расходуемый на формирование плазматических
мембран и компактного миелина, образуется исключительно в ходе очень
интенсивного синтеза de novo в мозге (главным образом в астроцитах).
В отличие от других тканей организма, где интенсивный синтез хо-
лестерина продолжается в течение всей жизни, в мозге человека и
животных скорость обновления этого липида значительно снижается с
возрастом. Так, в мозге взрослого человека скорость обновления хо-
лестерина составляет всего 0,03% в день (по сравнению с 0,7% вдень
для организма в целом).
В настоящее время нет доказательств поступления холестерина в
головной или спинной мозг из крови. У взрослых организмов скорость
его синтеза превышает потребность в нем, и экскреция холестерина из
ЦНС происходит с образованием 24-(C)-гидроксихолестерина. Уро-
вень холестерина в крови и интенсивность его обмена имеют большое
значение для развития ряда нейродегенеративных болезней, в частнос-
ти болезни Альцгеймера.
Сравнивая молярное содержание основных классов липидов в спе-
циализированных клетках мозга (см. табл. 1.5), можно увидеть, что
олигодендроглия и миелин обогащены цереброзидами, а в нейронах и
астроглии более высоко содержание фосфолипидов. Это лишний раз
подтверждает то, что плазматические мембраны нейронов и глиаль-
ных клеток отличны от миелина.
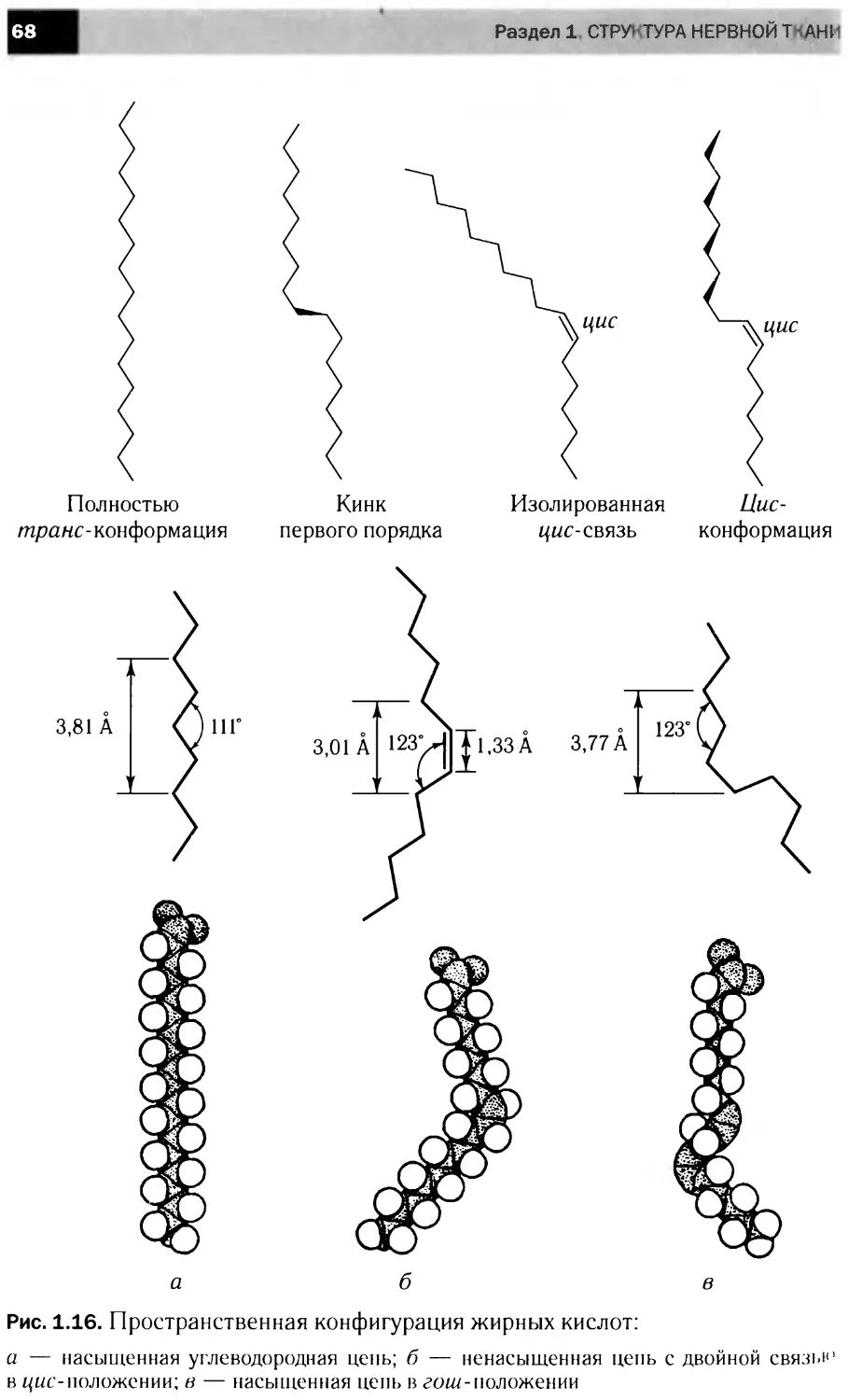
Жирные кислоты. В состав молекул как фосфо-, так и гликоли-
пидов входят ацильные радикалы различных жирных кислот (табл. 1.6).
Как правило, жирнокислотные радикалы имеют четное количество
атомов углерода — от 14 до 24; чаще всего встречаются жирные кис-
лоты с 16 или 18 атомами углерода. Жирные кислоты с нечетным чис-
лом атомов встречаются только в составе цереброзидов и ганглиози-
дов. Наиболее распространены кислоты С16, С18 и С20. Жирные кис-
лоты различаются как длиной цепи, так и степенью ненасыщенности,
которая может быть различной; чаще встречаются ненасыщенные
жирные кислоты 18 : 1, 18:2, 18 : 3 и 20 : 4. Первое число в соотно-
шении обозначает длину ацильной цепи, а второе указывает на число
содержащихся в ней двойных связей. Ненасыщенные кислоты мозга
могут содержать от одной (моноеновые) до шести (полиеновые) двой-
ных связей. Особенностью липидов мозга является относительно боль-
шое содержание длинноцепочечных полиеновых кислот — С20.4, С20.5,
1.2. Особенности липидного и белкового состава нервной ткани
Таблица 1.6. Некоторые свойства жирных кислот,
входящих в состав мембранных липидов
Соединение Тривиальное название Молекулярная масса, Да Температура плавления,°C
^12:0 Лауриловая 200,3 44,2
^14:0 Миристиновая 228,4 53,9
^16:0 Пальмитиновая 256,4 63,1
^17:0 Маргариновая 270,4 61,3
^18:0 Стеариновая 284,5 69,6
^20:0 Арахиновая 312,5 76,5
^22:0 Бегеновая 340,6 81,5
^24:0 Лигноцериновая 368,5 86,0
С|6: 1(9) Пальмитоолеиновая 254,4 -0,5
С|8: 1(9с) Олеиновая 282,5 13,5
С|8: l(9t) Элаидиновая 282,5 44,5
С|8: 1(7) Вакценовая 282,5 44,0
^24 : 1(9) Нервоновая 366,6 42,5
С|8:2(9, 12) Линолевая 280,5 -5,0
^18:3(9, 12, 15) Линоленовая 278,4 -10,0
^20:4(5,8, 11, 14) Арахидоновая 304,5 -49,5
О>2:5(7, 10, 13, 16, 19) Клупанодоновая 330,5 -45,0
^22:6(4, 7, 10, 13, 16, 19) Докозогексаеновая 328,5 -44,1
С22.6. Если ненасыщенной является лишь одна жирнокислотная цепь,
то она присоединена ко второму углеродному атому глицерина. Почти
все природные кислоты характеризуются цис-конфигурацией двойных
связей. Цепь в такой конфигурации имеет излом, что нарушает упа-
ковку липидных молекул в бислое (см. гл. 1.4).
Отдельные классы липидов мозга характеризуются своим набором
Жирных кислот. Имеет место также определенная специфичность жир-
з*
36
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
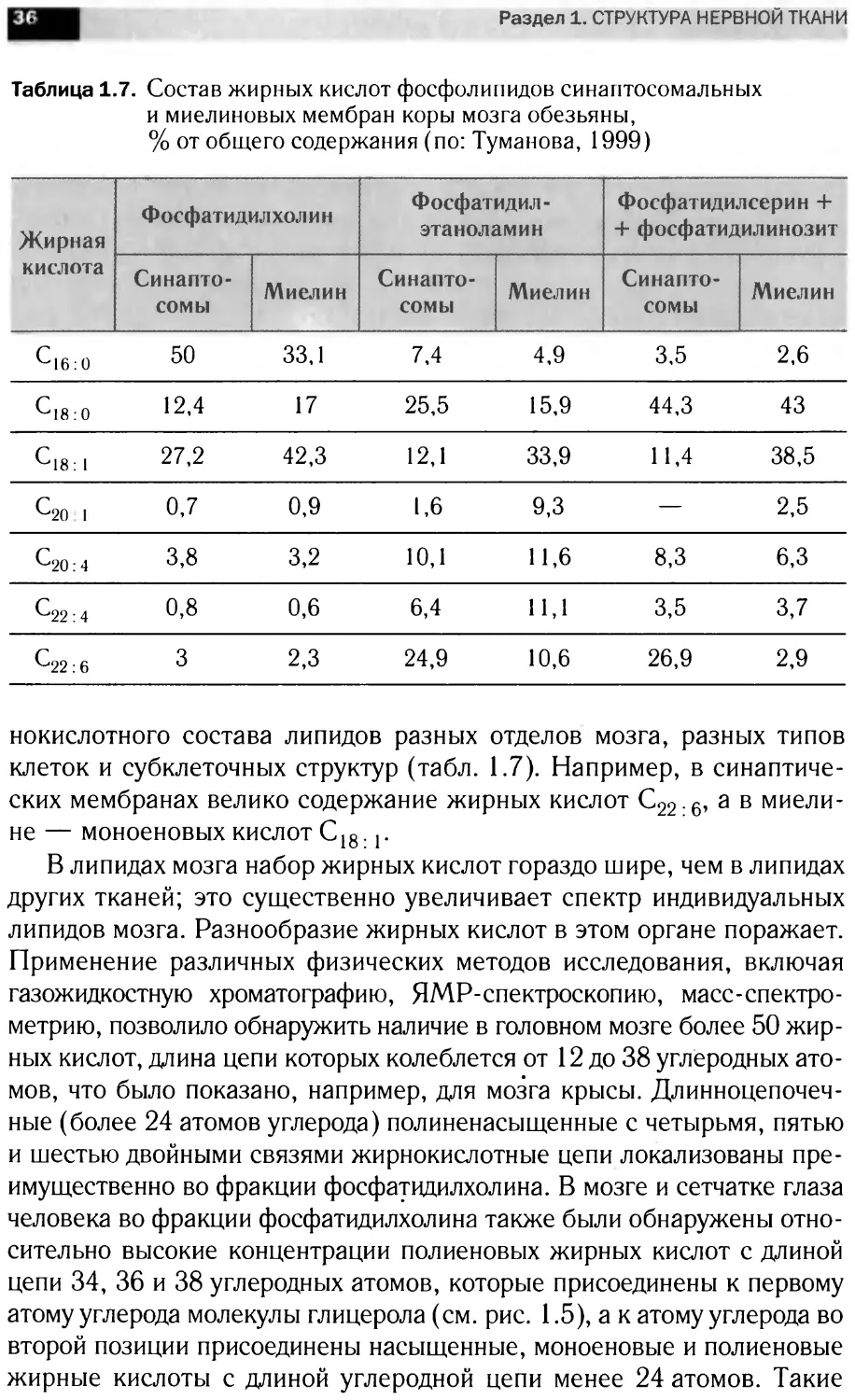
Таблица 1.7. Состав жирных кислот фосфолипидов синаптосомальных
и миелиновых мембран коры мозга обезьяны,
% от общего содержания (по: Туманова, 1999)
Жирная кислота Фосфатидилхолин Фосфатидил- этаноламин Фосфатидилсерин + + фосфатидилинозит
Синапто- сомы Миелин Синапто- сомы Миелин Синапто- сомы Миелин
6*16:0 50 33,1 7,4 4,9 3,5 2,6
6*18:0 12,4 17 25,5 15,9 44,3 43
С18: 1 27,2 42,3 12,1 33,9 11,4 38,5
6*20 1 0,7 0,9 1,6 9,3 — 2,5
6*20:4 3,8 3,2 10,1 11,6 8,3 6,3
6*22:4 0,8 0,6 6,4 11,1 3,5 3,7
6*22:6 3 2,3 24,9 10,6 26,9 2,9
нокислотного состава липидов разных отделов мозга, разных типов
клеток и субклеточных структур (табл. 1.7). Например, в синаптиче-
ских мембранах велико содержание жирных кислот С22.6, а в миели-
не — моноеновых кислот С18. г
В липидах мозга набор жирных кислот гораздо шире, чем в липидах
других тканей; это существенно увеличивает спектр индивидуальных
липидов мозга. Разнообразие жирных кислот в этом органе поражает.
Применение различных физических методов исследования, включая
газожидкостную хроматографию, ЯМР-спектроскопию, масс-спектро-
метрию, позволило обнаружить наличие в головном мозге более 50 жир-
ных кислот, длина цепи которых колеблется от 12 до 38 углеродных ато-
мов, что было показано, например, для мозга крысы. Длинноцепочеч-
ные (более 24 атомов углерода) полиненасыщенные с четырьмя, пятью
и шестью двойными связями жирнокислотные цепи локализованы пре-
имущественно во фракции фосфатидилхолина. В мозге и сетчатке глаза
человека во фракции фосфатидилхолина также были обнаружены отно-
сительно высокие концентрации полиеновых жирных кислот с длиной
цепи 34, 36 и 38 углеродных атомов, которые присоединены к первому
атому углерода молекулы глицерола (см. рис. 1.5), а к атому углерода во
второй позиции присоединены насыщенные, моноеновые и полиеновые
жирные кислоты с длиной углеродной цепи менее 24 атомов. Такие
1.2. ОСОбёННг. тг ъ гидн I g # 6efHиг«.СТль? И Тч?ии
3
необычные молекулярные формы фосфатидилхолина позволяют пред-
положить его физиологическую значимость для функционирования
мозга.
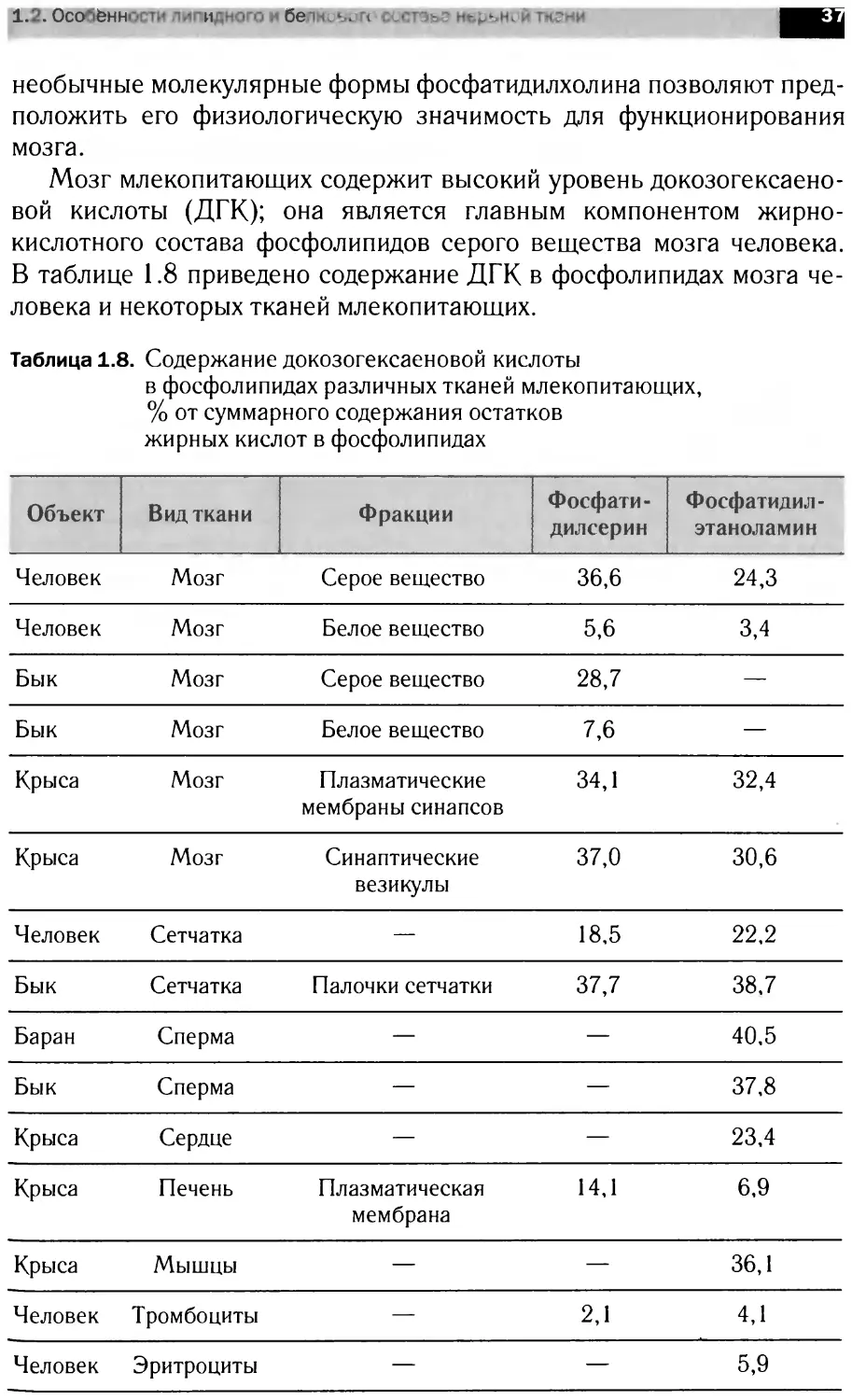
Мозг млекопитающих содержит высокий уровень докозогексаено-
вой кислоты (ДГК); она является главным компонентом жирно-
кислотного состава фосфолипидов серого вещества мозга человека.
В таблице 1.8 приведено содержание ДГК в фосфолипидах мозга че-
ловека и некоторых тканей млекопитающих.
Таблица 1.8. Содержание докозогексаеновой кислоты
в фосфолипидах различных тканей млекопитающих,
% от суммарного содержания остатков
жирных кислот в фосфолипидах
Объект Вид ткани Фракции Фосфати- дилсерин Фосфатидил- этаноламин
Человек Мозг Серое вещество 36,6 24,3
Человек Мозг Белое вещество 5,6 3,4
Бык Мозг Серое вещество 28,7 —
Бык Мозг Белое вещество 7,6 —
Крыса Мозг Плазматические мембраны синапсов 34,1 32,4
Крыса Мозг Синаптические везикулы 37,0 30,6
Человек Сетчатка — 18,5 22,2
Бык Сетчатка Палочки сетчатки 37,7 38,7
Баран Сперма — — 40,5
Бык Сперма — — 37,8
Крыса Сердце — — 23,4
Крыса Печень Плазматическая мембрана 14,1 6,9
Крыса Мышцы — — 36,1
Человек Тромбоциты — 2,1 4,1
Человек Эритроциты — — 5,9
38
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
Из таблицы видно, что значительное количество ДГК содержится в
клетках внутренних органов и мышц, а также в палочках сетчатки
глаз. Однако в нервной ткани содержание ДГК наиболее высоко. Воз-
можно, что высокий уровень докозогексаеновой кислоты в синаптиче-
ских мембранах необходим для активного транспорта ионов, так как
активность Na/K-АТФазы в них зависит от присутствия в составе фос-
фолипидов полиеновых кислот.
Изменения концентрации ДГК в мозге сопровождаются нарушения-,
ми познавательной и поведенческой активности животных (табл. 1.9).
Установлено, что при уменьшении содержания ДГК в мозге крыс на-
блюдается ухудшение зрительных и обонятельных функций при выпол-
нении ими обучающих тестов. Дополнительно к ухудшению выполнения
задач ассоциативного обучения у таких крыс обнаруживаются потеря
пространственной памяти и ослабление обоняния. Таким образом, мож-
но заключить, что уменьшение содержания ДГК в мозге затрагивает су-
щественные функции высшей нервной деятельности. В мозге имеются
регуляторные механизмы, поддерживающие степень ненасыщенности
и специфический жирнокислотный состав липидов.
Таблица 1.9. Влияние жирных кислот на биохимические
и физиологические показатели состояния мозга
Жирные кислоты (ЖК) Положение первой двойной связи Свойства
Линоленовая Эйкозогексаеновая («з)18:3 (п3)20:5 Незаменимые ЖК. Увеличива- ют число дендритов и плотность синапсов в развивающемся моз- ге. Находятся преимущественно в сером веществе. Крысы, дефи- цитные по (л3), плохо обучаются.
Докозогексаеновая (л3)22:6 При систематическом приеме алкоголя соотношение (л3)/(л6) в мозге снижается
Линолевая Арахидоновая (п6) 18:2 (п6)20:4 Равномерно распределены в тка- нях, количество не зависит от со- става пищи. Избыток вызывает замещение в мембранах докозо- гексаеновой на арахидоновую, диссоциацию от мембран белков RAS и RAF, которые активируют каспазу 3 и вызывают апоптоз
1.2. Особенности липидного и белкового состава нервной ткани
1.2.2. Биологические функции мембранных липидов
Клеточная мембрана может содержать более 100 различных ти-
пов липидных молекул. Почему их так много и почему каждая мембра-
на имеет уникальный липидный состав, пока неясно. Очевидно, что
соотношение липидов в различных мембранах мозга не является слу-
чайным. Становится все более очевидным то, что липиды активно уча-
ствуют в процессах, обеспечиваемых мембранами.
Рассмотрим факторы, определяющие специфику
липидного состава мембран нервной ткани.
1. Смесь липидов обязательно должна быть способной к образова-
нию стабильного бислоя, в котором функционируют белки.
2. Некоторые липиды способствуют стабилизации сильно искрив-
ленных участков мембраны, образованию контактов между мембранами
или связыванию определенных белков, поскольку форма подобных мо-
лекул соответствует нужной упаковке бислоя на тех или иных участках
мембраны. По форме липидных молекул можно определить роль от-
дельных липидов‘в бислое, например в стабилизации участков мембра-
ны с большой кривизной и упаковке молекул вокруг мембранных бел-
ков. Так, форма «конус», которая присуща таким липидам, как фосфа-
тидилэтаноламин (ненасыщенный), кардиолипин, фосфатидная кислота,
фосфатидилсерин (pH < 4,0), моногалактозилдиглицерид, способствует
образованию вогнутой формы мембранного бислоя; форма «переверну-
тый конус», свойственная лизофосфолипидам, приводит к образованию
выпуклых участков липидного бислоя мембран.
3. Ряд липидов — важные биорегуляторы. Некоторые мембранные
липиды служат предшественниками вторичных посредников при пере-
даче гормонального сигнала. Наиболее изучены в этом отношении про-
изводные фосфатидилинозитола в плазматических мембранах эукари-
отических клеток. Так, фосфатидилинозитол-4,5-бисфосфат (ФИФ2)
под действием фермента фосфолипазы С гидролизуется до диацилгли-
церола (DAG) — активатора протеинкиназы С и инозитол-1,4,5-трис-
фосфата (ИФ3) — регулятора кальциевого обмена в клетке (см. гл. 2.4).
DAG, ИФ3, протеинкиназа С и ионы кальция — участники фосфоино-
зитольной системы передачи сигнала, причем два первых образуются
из мембранных компонентов, третий является амфипатическим мемб-
ранным белком, а кальций — это ион, проникающий через специаль-
ные каналы на клеточной мембране и запускающий этот процесс.
4. Отдельные липиды необходимы для поддержания оптимальной
активности ряда ферментов. Они формируют среду для функциониро-
вания мембранных белков, способных принимать нативную конформа-
.о
Раздел 1. СТНУН7 EPBt И TH Н
цию лишь в гидрофобном окружении. Ферменты, лишенные специфи-
ческого липидного окружения, или не проявляют каталитической ак-
тивности, как ионные насосы (транспортные АТФазы), или проявляют
отличные от нативных ферментов свойства. Такова, например, мито-
хондриальная креатинкиназа — фермент, катализирующий образова-
ние креатинфосфата и принимающий участие в поддержании энергети-
ческого баланса в мозге. Для его нормальной активности требуется
взаимодействие с кардиолипином внутренней мембраны митохондрий.
5. Ганглиозиды, как полагают, играют важную роль в регуляции
роста и дифференцировки клеток (см. гл. 1.7), участвуя в процессах
клеточной адгезии.
6. Некоторые липиды выполняют «якорную» функцию, например к
молекуле фосфатидилинозитола через олигосахарид могут присоеди-
няться специфические белки наружной поверхности клетки и при этом
образуется фосфатидилинозитолгликан. Примером такого «заяко-
ренного» белка может служить ацетилхолинэстераза, катализирующая
гидролиз ацетилхолина в синаптической щели. Этот фермент фиксиру-
ется на постсинаптической мембране, ковалентно присоединяясь к
фосфатидилинозитолгликану. При участии фосфолипазы С (гидроли-
зующей мембранные липиды) может происходить модификация мемб-
раны и отделение белков от внешней поверхности клетки.
7. Липиды могут быть аллостерическими активаторами мембранных
ферментов (например, фосфатидилсерин активирует протеинкиназу С).
Фермент протеинкиназа С катализирует реакции фосфорилирования
белков. Семейство протеинкиназ С (ПК С) включает несколько изофер-
ментов — а-, р- и у-изоформы ПК С. Изоформа у-ПК С локализована
исключительно в нейронах головного и спинного мозга, в то время как а-
и p-ПК С присутствуют и в других тканях. В головном мозге протеин-
киназа С встречается преимущественно в мозжечке, гиппокампе и коре.
В неактивной форме протеинкиназа С находится в цитозоле. Однако пос-
ле стимуляции клетки (приводящей к повышению в ней концентрации
ионов кальция) фермент изменяет свою конформацию и приобрета-
ет способность ассоциироваться с плазматической мембраной. Функци-
онально активная протеинкиназа С представляет собой комплекс, содер-
жащий мономер фермента, молекулу диацилглицерола, один или более
ионов кальция и четыре молекулы фосфатидилсерина. Обнаружено, что
мутация гена ПК С вносит вклад в развитие болезни Паркинсона. Уста-
новлено также участие изоформы у-ПК С в развитии ишемического ин-
сульта. Одним из центральных событий в развитии ишемического ин-
сульта является высвобождение экзайтоксической аминокислоты глута-
мата, приводящее к смерти нейрона. Через рецепторы NMDA глутамат
1.2. Особенности липидного и белкового состава нервной ткани
41
вызывает приток ионов Са2+ и Na+ в клетку, а также генерацию свобод-
ных радикалов. Изоформа у-ПК С быстро активируется в течение ише-
мии, соответственно ингибирование ПК С снижает вызванную NMDA
нейротоксичность. Использование мышей с подавленным синтезом ПК С
позволяет предположить, что у-ПК С может играть отрицательную роль
в процессе мозговой ишемии; как показано с помощью гистологических
методов, мыши нокаута ПК С имеют значительно меньше инфарктов по
сравнению с животными дикого типа.
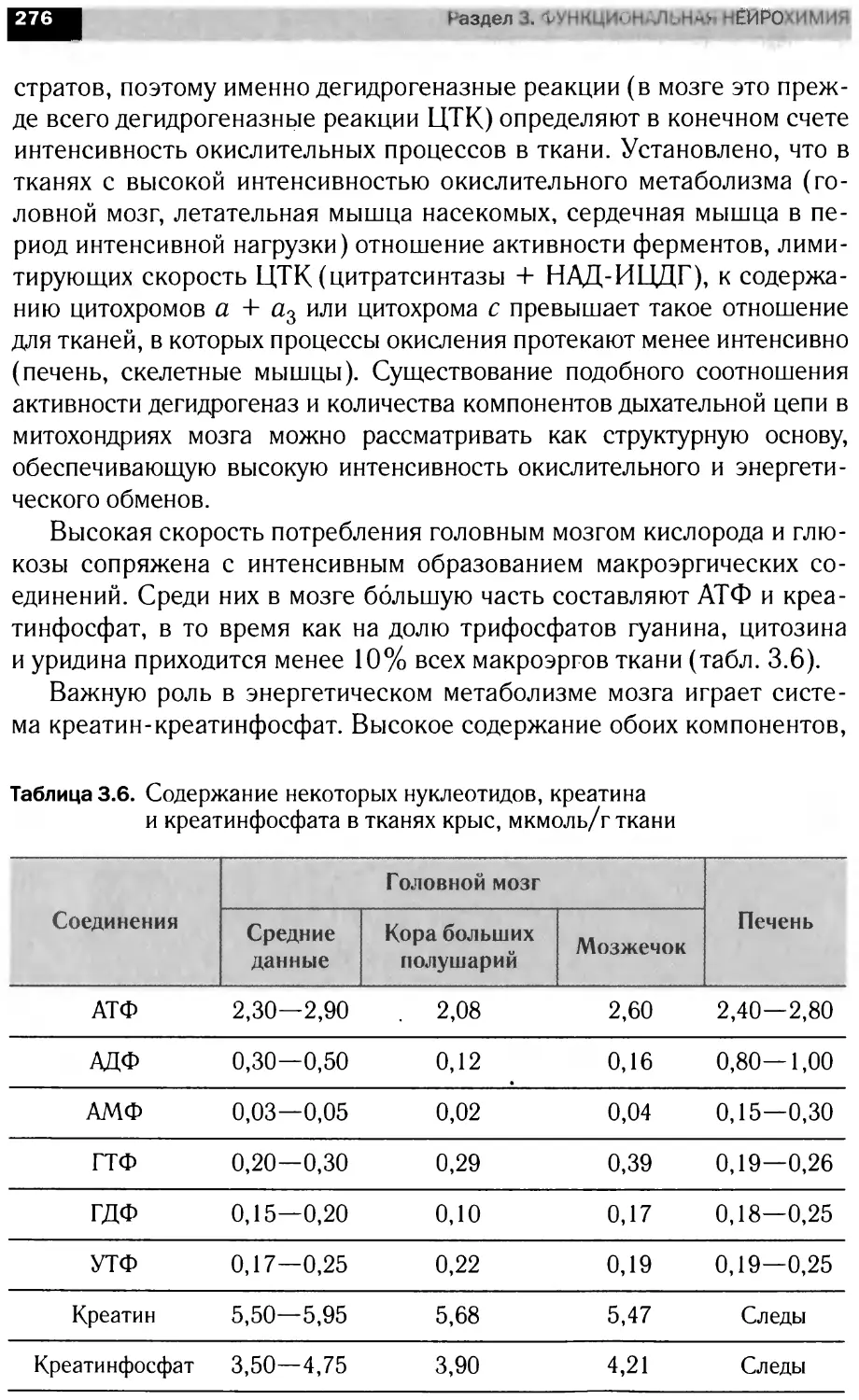
1.2.з. Миелин
Миелин — уникальное образование, организация которого по-
зволяет проводить по нервному волокну электрический импульс с ми-
нимальной затратой энергии (см. гл. 1.1). Миелиновая оболочка —
высокоорганизованная многослойная структура, состоящая из сильно
растянутых и модифицированных плазматических мембран шваннов-
ских (в ПНС) и олигодендроглиальных (в ЦНС) клеток (рис. 1.12).
Содержание воды в миелине составляет около 40%. Отличитель-
ная особенность миелина по сравнению с другими клетками заключа-
ется в том, что он содержит в среднем 70% липидов и 30% белка
(в расчете на сухой вес). Большинство биологических мембран имеет
более высокое значение отношения белков к липидам.
Рис. 1.12. Мезаксон и образование миелиновой оболочки на примере миелина
в аксонах периферической нервной системы:
/ — аксон; 2 — шванновская клетка; 3 — единичная мембрана; 4 — мезаксон; 5 —
щель; 6 — цитоплазма шванновской клетки; 7 — миелиновая оболочка; 8 — проме-
жуточный период; 9 — главный период
42
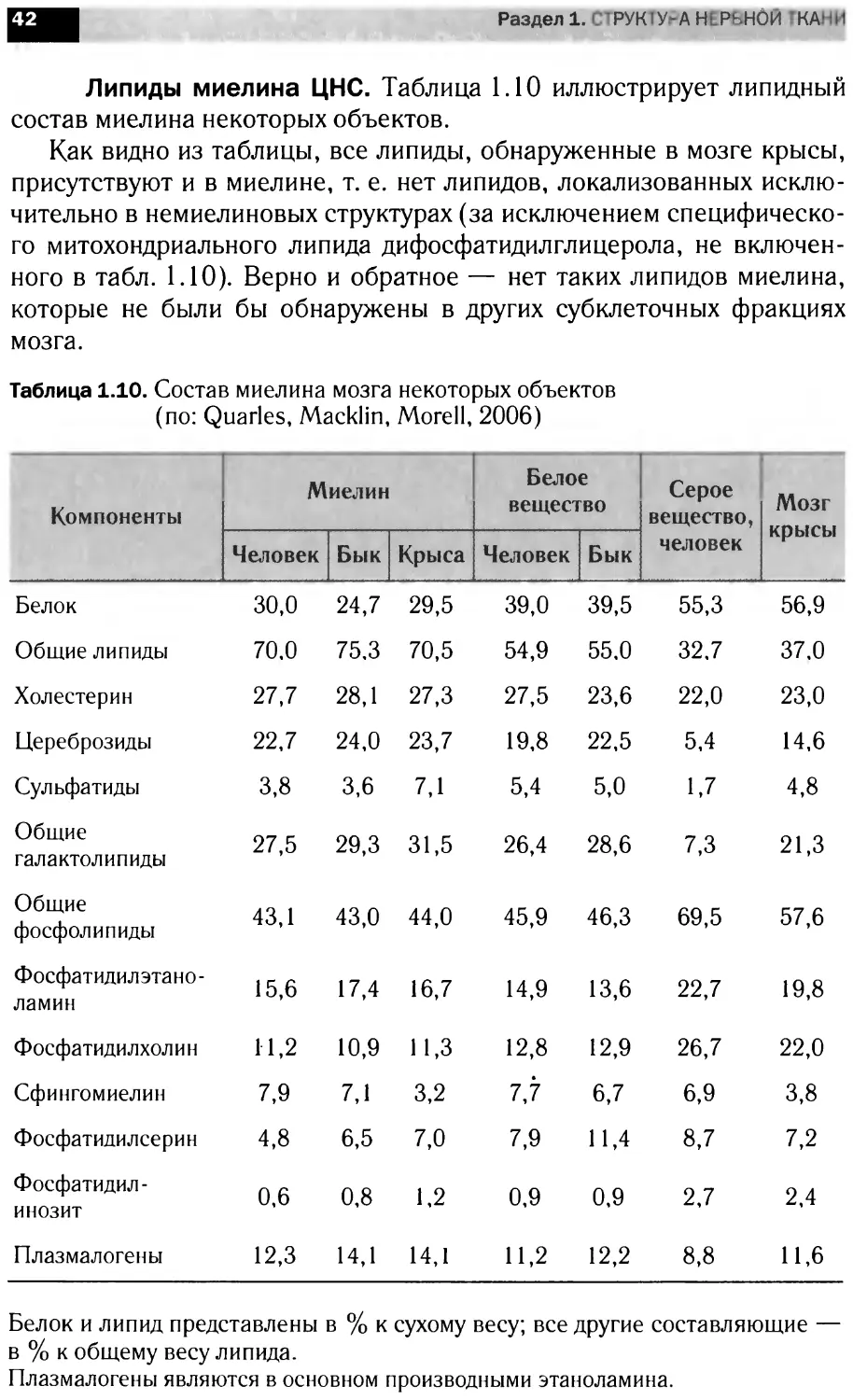
Раздел 1. тРУК1У-АНЕРЬН0й’КАНИ
Липиды миелина ЦНС. Таблица 1.10 иллюстрирует липидный
состав миелина некоторых объектов.
Как видно из таблицы, все липиды, обнаруженные в мозге крысы,
присутствуют и в миелине, т. е. нет липидов, локализованных исклю-
чительно в немиелиновых структурах (за исключением специфическо-
го митохондриального липида дифосфатидилглицерола, не включен-
ного в табл. 1.10). Верно и обратное — нет таких липидов миелина,
которые не были бы обнаружены в других субклеточных фракциях
мозга.
Таблица 1.10. Состав миелина мозга некоторых объектов
(по: Quarles, Macklin, Morell, 2006)
Компоненты Миелин Белое вещество Серое вещество, человек Мозг крысы
Человек Бык Крыса Человек Бык
Белок 30,0 24,7 29,5 39,0 39,5 55,3 56,9
Общие липиды 70.0 75.3 70,5 54,9 55,0 32,7 37,0
Холестерин 27,7 28,1 27,3 27,5 23,6 22,0 23,0
Цереброзиды 22,7 24,0 23,7 19,8 22,5 5,4 14,6
Сульфатиды 3,8 3,6 7,1 5,4 5,0 1,7 4,8
Общие галактолипиды 27,5 29,3 31,5 26,4 28,6 7,3 21,3
Общие фосфолипиды 43,1 43,0 44,0 45,9 46,3 69,5 57,6
Фосфатидил этано - ламин 15,6 17,4 16,7 14,9 13,6 22,7 19,8
Фосфатидил хол и н 11,2 10,9 1 1,3 12,8 12,9 26,7 22,0
Сфингомиелин 7,9 7,1 3,2 7,7 6,7 6,9 3,8
Фосфатидил се р и н 4,8 6,5 7,0 7,9 11,4 8,7 7,2
Фосфатидил- инозит 0,6 0,8 1,2 0,9 0,9 2,7 2,4
Плазмалогены 12,3 14,1 14,1 11,2 12,2 8,8 11,6
Белок и липид представлены в % к сухому весу; все другие составляющие —
в % к общему весу липида.
Плазмалогены являются в основном производными этаноламина.
1.2. Особенности липидного и шелкового состава нервной ткани
Цереброзид — наиболее типичный компонент миелина. За исклю-
чением самого раннего периода развития организма, концентрация
цереброзида в мозге прямо пропорциональна количеству в нем миели-
на. Только Vg общего содержания галактолипидов в миелине встреча-
ется в сульфатированной форме. Цереброзиды и сульфатиды играют
важную роль в обеспечении стабильности миелина.
Для миелина также характерен высокий уровень его главных липи-
дов — холестерина, общих галактолипидов и содержащего этано-
ламин плазмалогена. Установлено, что до 70% холестерина мозга
находится в миелине. Поскольку почти половина белого вещества
мозга может состоять из миелина, очевидно, что в мозге содержится
наибольшее количество холестерина по сравнению с другими органа-
ми. Высокая концентрация холестерина в мозге, особенно в миелине,
определяется основной функцией нейрональной ткани — генериро-
вать и проводить нервные импульсы. Большое содержание холестери-
на в миелине и своеобразие его структуры приводят к уменьшению
ионной утечки через мембрану нейрона (вследствие ее высокого со-
противления).
Фосфатидилхолин также является существенной составной частью
миелина, хотя сфингомиелин содержится в относительно незначи-
тельном количестве.
Как видно из таблицы 1.10, липидный состав как серого вещества,
так и белого вещества мозга отчетливо отличается от такового у ми-
елина. Состав миелина мозга всех изученных видов млекопитающих
почти одинаков; имеют место лишь незначительные различия (напри-
мер, миелин крысы имеет меньше сфингомиелина, чем миелин быка
или человека). Существуют некоторые вариации и в зависимости от
локализации миелина, например миелин, изолированный из спинного
мозга, имеет более высокое значение отношения липида к белку, чем
миелин из головного мозга.
В состав миелина входят также полифосфатидилинозиты, из ко-
торых трифосфоинозитид составляет от 4 до 6% общего фосфора ми-
елина, а дифосфоинозитид— от 1 до 1,5%. Минорные компоненты
миелина включают по крайней мере три эфира цереброзида и два ли-
пида на основе глицерина; в составе миелина также присутствуют не-
которые длинноцепочечные алканы. Миелин млекопитающих содер-
жит от 0,1 до 0,3% ганглиозидов. В миелине содержится больше
моносиалоганглиозида GMj по сравнению с тем, что обнаруживается
в мембранах мозга. Миелин многих организмов, в том числе и чело-
века, содержит уникальный ганглиозид сиалозилгалактозилцера-
мид GM4.
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНЙ
Липиды миелина ПНС. Липиды миелина периферической и
центральной нервной системы качественно подобны, но между ними
есть количественные различия. Миелин ПНС содержит меньше це-
реброзидов и сульфатидов и значительно больше сфингомиелина, чем
миелин ЦНС. Интересно отметить наличие ганглиозида GMH харак-
терного для миелина ПНС некоторых организмов. Различия в составе
липидов миелина центральной и периферической нервной системы не
столь существенны, как их различия по белковому составу.
Белки миелина ЦНС. Белковый состав миелина ЦНС более прост,
чем других мембран мозга, и представлен главным образом протеолипи-
дами и основными белками, которые составляют 60—80% от общего
количества. Гликопротеины присутствуют в гораздо меньших количест-
вах. Миелин центральной нервной системы содержит уникальные белки.
Белковый состав миелина мозга человека и крысы проиллюстриро-
ван на рисунке 1.13. Для миелина ЦНС человека характерно количест-
170KG, SAG
MAG
Т
CNPase
в г
170KG, SAG
MAG 4* — MAG
ST
CNPase
Po
MBP
MBP, P|
MBP
p2
MOG
PLP
MBP
DM-20
MBP
MBP
MBP
PMP-22
MBP, P]
MBP
MBP
P2
MOG
PLP
MBP
DM-20
MBP
MBP
MBP
Рис. 1.13. Электрофореграмма белков миелина в полиакриламидном геле:
а — миелин ПНС человека; б — миелин ЦНС человека; в — миелин ПНС крысы;
г — миелин ЦНС крысы. Указаны некоторые характерные белковые полосы
1.2. Особенности липидного м белковой? состава нервной тк ни
венное превалирование двух белков: положительно заряженного ка-
тионного белка миелина (myelin basic protein, МВР) и протеолипи-
да миелина (myelin proteolipid protein, PLP). Эти белки — главные
составные части миелина ЦНС всех млекопитающих.
Миелиновый протеолипид PLP (proteolipid protein), также извест-
ный как белок Фолча, имеет способность растворяться в органиче-
ских растворителях. Молекулярная масса PLP составляет приблизи-
тельно 30 кДа (Да — дальтон). Его аминокислотная последователь-
ность чрезвычайно консервативна, молекула формирует несколько
доменов. Молекула PLP включает три жирные кислоты, как правило,
пальмитиновую, олеиновую и стеариновую, соединенные с аминокис-
лотными радикалами эфирной связью.
Миелин ЦНС содержит несколько меньшие количества другого
протеолипида — DM-20, названного так по его молекулярной массе
(20 кДа). И анализ ДНК, и выяснение первичной структуры показали,
что DM-20 образуется в результате отщепления 35 аминокислотных
остатков от белка PLP. В процессе развития DM-20 появляется рань-
ше, чем PLP (в некоторых случаях даже до появления миелина); пред-
полагают, что в дополнение к структурной роли в образовании миели-
на он может участвовать в дифференцировке олигодендроцитов.
Вопреки представлениям о том, что PLP необходим для формиро-
вания компактного мультиламеллярного миелина, процесс образова-
ния миелина у мышей, «нокаутированных» по PLP/DM-20, происхо-
дит лишь с незначительными отклонениями. Однако у таких мышей
уменьшена продолжительность жизни и нарушена общая подвиж-
ность. Напротив, естественно происходящие мутации в PLP, в том
числе его повышенная экспрессия (normal PLP over-expression), име-
ют серьезные функциональные последствия. Следует отметить, что
существенные количества белков PLP и DM-20 представлены в ЦНС,
матричная РНК для PLP есть и в ПНС, и небольшое количество белка
там синтезируется, но не включается в миелин.
Катионный белок миелина (МВР) привлекает внимание исследо-
вателей вследствие его антигенной природы — при введении живот-
ным он вызывает аутоиммунную реакцию, так называемый экспери-
ментальный аллергический энцефаломиелит, который представляет
собой модель тяжелого нейродегенеративного заболевания — рассеян-
ного склероза.
Аминокислотная последовательность МВР у многих организмов
высоко консервативна. МВР расположен на цитоплазматической сто-
роне миелиновых мембран. Он имеет молекулярную массу 18,5 кДа
и лишен признаков третичной структуры. Этот основный белок обна-
руживает микрогетерогенность при электрофорезе в щелочных усло-
виях.
4G
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
Большинство исследованных млекопитающих содержали различ-
ные количества изоформ МБР, имеющих существенную общую часть
аминокислотной последовательности. Молекулярная масса МБР мы-
шей и крыс — 14 кДа. МБР с малой молекулярной массой имеет та-
кие же аминокислотные последовательности на N- и С-терминальных
частях молекулы, как и остальной МБР, но отличается редукцией око-
ло 40 аминокислотных остатков. Соотношение этих основных белков
изменяется в процессе развития: зрелые крысы и мыши имеют боль-
ше МБР с молекулярной массой 14кДа, чем МБР с молекулярной
массой 18 кДа. Две другие изоформы МБР, также обнаруживаемые во
многих организмах, имеют молекулярную массу 21,5 и 17 кДа, соот-
ветственно. Они образованы присоединением к основной структуре
полипептидной последовательности массой около 3 кДа.
При электрофоретическом разделении белков миелина выявляют-
ся белки с более высокой молекулярной массой. Их количество зави-
сит от вида организма. Например, мышь и крыса могут содержать та-
ких белков до 30% от общего количества. Содержание этих белков
также изменяется в зависимости от возраста животного: чем оно мо-
ложе, тем меньше в его мозге миелина, но тем больше в нем белков
с более высокой молекулярной массой.
Фермент 2\ 3'-циклический нуклеотид 3'-фосфодиэстераза (CNP)
составляет несколько процентов от общего содержания миелинового
белка в клетках ЦНС. Это гораздо больше, чем в других типах клеток.
Белок CNP — не главный компонент компактного миелина, он скон-
центрирован лишь в определенных участках миелиновой оболочки,
связанной с цитоплазмой олигодендроцита. Белок локализован в ци-
топлазме, но часть его связана с цитоскелетом мембраны — F-акти-
ном и тубулином. Биологическая функция CNP может заключаться в
регулировании структуры цитоскелета для ускорения процессов роста
и дифференциации в олигодендроцитах.
Миелинассоциированный гликопротеин (MAG) — минорный в
количественном отношении компонент очищенного миелина, имеет мо-
лекулярную массу 100 кДа (см. рис. 1.13), содержится в ЦНС в неболь-
шом количестве (менее 1 % от общего белка). MAG имеет единствен-
ный трансмембранный домен, который отделяет сильногликозилиро-
ванную внеклеточную часть молекулы, составленную из пяти подобных
иммуноглобулину доменов, от внутриклеточного домена. Его полная
структура подобна белку адгезии нейрональной клетки (NCAM).
MAG не присутствует в компактном, мультиламеллярном миелине,
но находится в периаксональных мембранах олигодендроцитов, обра-
зующих слои миелина. Напомним, что периаксональная мембрана
1.2. Особенности липидного и белкового состава нервной ткани
4?
олигодендроцита — наиболее близко расположена к плазматической
мембране аксона, но тем не менее эти две мембраны не сливаются, а
разделены экстраклеточной щелью. Подобная особенность локализа-
ции MAG, а также то, что этот белок относится к иммуноглобулиново-
му суперсемейству, подтверждает участие его в процессах адгезии и
передачи информации (сигналинга) между аксолеммой и миелинобра-
зующими олигодендроцитами в процессе миелинизации. Кроме того,
MAG — один из компонентов белого вещества ЦНС, который инги-
бирует рост нейритов в культуре ткани.
Из других гликопротеинов белого вещества и миелина следует отме-
тить минорный миелинолигодендроцитарный гликопротеин (Myelin-
oligodendrocytic glycoprotein, MOG). MOG является трансмембранным
белком, содержащим единственный иммуноглобулинподобный домен.
В отличие от MAG, который расположен во внутренних слоях миелина,
MOG локализован в его поверхностных слоях, в силу чего может уча-
ствовать в передаче внеклеточной информации к олигодендроциту.
Малые количества характерных белков мембран могут быть иден-
тифицированы в результате электрофореза на полиакриламидном ге-
ле (ПААГ) (например, тубулин). Электрофорез высокого разрешения
демонстрирует наличие других незначительных полос белка; они могут
быть связаны с присутствием ряда ферментов миелиновой оболочки.
Белки миелина ПНС. Миелин ПНС содержит как некоторые уни-
кальные белки, так и несколько общих с белками миелина ЦНС белков.
Ро — главный белок миелина ПНС, имеет молекулярную массу
30 кДа, составляет более половины белков миелина ПНС. Интересно
отметить, что хотя он отличается от PLP по аминокислотной последо-
вательности, путям посттрансляционной модификации и структуре,
тем не менее оба эти белка имеют одинаково важное значение для
формирования структуры миелина ЦНС и ПНС.
Содержание МБР в миелине ПНС составляет 5—18% от общего
количества белка, в отличие от ЦНС, где его доля достигает трети все-
го белка. Те же четыре формы белка МБР с молекулярными массами
21, 18,5, 17 и 14кДа, соответственно, обнаруженные в миелине
ЦНС, присутствуют и в ПНС. У взрослых грызунов МБР с молеку-
лярной массой 14 кДа (по классификации периферийных миелиновых
белков ему присвоено название «Рг») является самым значительным
компонентом всех катионных белков. В миелине ПНС присутствует и
МБР с молекулярной массой 18 кДа (в этом случае он носит название
«белок Р1»). Следует отметить, что важность семейства белков МВР
не так велика для миелиновой структуры ПНС, как для ЦНС.
43
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
Гликопротеины миелина ПНС. Компактный миелин ПНС со-
держит гликопротеин с молекулярной массой 22 кДа, названный пе-
риферийным миелиновым белком 22 (РМР-22), доля которого со-
ставляет менее 5% от общего содержания белков. РМР-22 имеет че-
тыре трансмембранных домена и один гликозилированный домен.
Этот белок не играет значительной структурной роли. Однако анома-
лии гена ртр-22 ответственны за некоторые наследственные невро-
патологии человека.
Несколько десятилетий назад считалось, что миелин создает инерт-
ную оболочку, которая не выполняет никаких биохимических функций.
Однако позже в миелине было обнаружено большое количество фермен-
тов, вовлекаемых в синтез и метаболизм компонентов миелина. Ряд фер-
ментов, присутствующих в миелине, включается в метаболизм фосфо-
инозитидов: фосфатидилинозитолкиназа, дифосфатидилинозитолкиназа,
соответствующие фосфатазы и диглицеридкиназы. Эти ферменты пред-
ставляют интерес вследствие высокой концентрации в миелине поли-
фосфоинозитидов и их быстрого обмена. Есть свидетельства присутствия
в миелине мускариновых холинергических рецепторов, G-белков, фос-
фолипаз С и D, протеинкиназы С.
В миелине ПНС обнаружена Na/K-АТФаза, осуществляющая транс-
порт одновалентных катионов, а также 5'-нуклеотидаза. Наличие этих
ферментов позволяет предположить, что миелин может принимать
активное участие в аксональном транспорте.
1.2.4. Белки нервной ткани
Многие белки нейронов и глиальных клеток близки или полно-
стью идентичны белкам из других органов и тканей человека и живот-
ных в силу общности базовых процессов жизнедеятельности. Тем не
менее к настоящему времени выделена и идентифицирована довольно
многочисленная группа нейроспецифических белков, которые экс-
прессируются исключительно (или преимущественно) в клетках нерв-
ной ткани и участвуют в выполнении функций, присущих только этой
ткани. Многие из таких белков представляют собой глико- или липо-
протеиды. Они участвуют в формировании рецепторных структур,
в процессах адгезии клеток при формировании нейронных ансамблей,
в перестройке межклеточных контактов в ходе онтогенеза, в работе
синаптического аппарата, в формировании долговременной памяти
и в других процессах.
Необходимо, однако, подчеркнуть, что поскольку нервная система
функционирует как единое целое, то невозможно в ряде случаев рас-
1.2. Особенности липидного и белкового состава Hei вной ткани
сматривать нейроспецифические белки отдельно от белков, присутст-
вующих в любой другой ткани.
Специфичность белков нервной ткани определяется по
нескольким критериям:
1) наличие белка исключительно или преимущественно в тканях
нервной системы, причем во втором случае количество белка в нерв-
ных клетках должно существенно превышать содержание его в других
тканях организма;
2) изучаемый белок должен участвовать в реализации специфиче-
ских функций нервной ткани (например, в процессах генерации и про-
ведения нервных импульсов, в работе синаптического аппарата, в ме-
ханизмах обучения и формирования долгосрочной памяти и др.);
3) наличие тесной корреляции между биологической активностью
белка и функциональным состоянием нервной системы.
Существование нейроспецифических белков обусловлено особен-
ностями экспрессии генов в мозге. Известно, что в нервной ткани ак-
тивно работают до 8—10% генов, что заметно выше, чем в других
тканях организма, где обычно функционирует не более 2—4% генов.
Именно разнообразие экспрессируемых уникальных генов определяет
синтез множества нейроспецифических белков.
Установлено, что в большинстве нервных клеток очень велико со-
держание РНК. Средняя величина соотношения РНК/ДНКв мозге мо-
жет достигать 50 и лишь в редких популяциях клеток она составляет
всего 3. Для сравнения напомним, что в клетках таких активно метабо-
лирующих органов, как печень, почки, поджелудочная железа, соотно-
шение РНК/ДНК составляет в среднем 2,0—4,5. Высокие значения
соотношения РНК/ДНК обеспечиваются главным образом за счет
большой концентрации рРНК, что указывает на наличие в мозге мощ-
ного, активно работающего рибосомального аппарата синтеза белка.
Огромное качественное разнообразие белков клеток нервной ткани
определяется многообразием мРНК. Несмотря на то что количество
мРНК составляет менее 5% от общей массы РНК клетки, в разных ней-
ронах (группах нейронов) или глиальных клетках могут экспрессировать-
ся различные гены, поэтому так велико число вариантов мРНК в мозге.
Использование методов молекулярного клонирования позволи-
ло разделить всю совокупность синтезирующихся в
мозге мРНКна четыре группы. Первая группа мРНК
(около 18%) экспрессируется во всех тканях с одинаковой интенсив-
ностью; вторая группа (около 26%) также экспрессируется
во всех тканях, но в разной степени. По-видимому, эти две группы
мРНК обеспечивают синтез общих для всех тканей белков, участвую-
50
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
щих в основных процессах жизнедеятельности организма. Третья
группа мРНК (около 30%) представляет собой нейроспецифичные
мРНК, не обнаруженные в других тканях. К четвертой группе
(около 26%) относятся особые нейроспецифичные мРНК, количество
которых в целом мозге крайне мало и которые обнаруживаются лишь
с помощью высокочувствительных специальных методических прие-
мов в отдельных структурных образованиях или даже в отдельных не-
больших популяциях нейронов. Таким образом, более половины син-
тезируемых в мозге мРНК являются нейроспецифичными,
обеспечивающими своеобразие белкового спектра данной ткани.
Определенный вклад в создание разнообразия нейроспецифичных
белков вносит альтернативный процессинг пре-мРНК, т. е. образова-
ние нескольких вариантов зрелых мРНК из одного первичного транс-
крипта в ходе альтернативного сплайсинга (соединение разных комби-
наций экзонов) или альтернативного З'-процессинга (использование
разных сигналов полиаденилирования). Наглядным примером роли
альтернативного сплайсинга служит образование целого семейства
метаботропных рецепторов глутамата (mGluR). К настоящему време-
ни описано восемь подклассов этих рецепторов и для mGluR 1,
mGluR4, mGluR5 и mGluR7 установлено наличие вариантов, образо-
вавшихся в результате альтернативного сплайсинга пре-мРНК. Эти
варианты рецепторов различаются относительно короткими участка-
ми (38 аминокислотных остатков), которые предшествуют четвертому
трансмембранному домену и представляют собой петлю белковой це-
пи рецептора, находящуюся в цитоплазме. Существование альтерна-
тивных вариантов каждого из подклассов метаботропных рецепторов
глутамата имеет значение с функциональной точки зрения, поскольку
они обладают разными кинетическими свойствами, разной чувстви-
тельностью к фармакологическим агонистам и антагонистам и лока-
лизованы в разных структурах ЦНС.
Другими примерами, подтверждающими значимость процесса аль-
тернативного сплайсинга для функционирования различных структур
нервной системы, могут быть четыре формы тирозингидроксилазы,
обнаруженные в ЦНС человека; три формы ацетилхолинэстеразы
в электрическом органе ската и др.
К настоящему времени с помощью различных биохимических и мо-
лекулярно-биологических методов выявлено более 200 нейроспеци-
фичных белков. К сожалению, информация о многих из них сводится
пока лишь к сообщениям о выделении и описаниям ряда их физико-
химических и антигенных свойств; данные же об участии этих белков
П
I
1.3. Современные представления об организации нейрональной мембраны
5:
1.3.
в той или иной специфической функции в мозге весьма скудные и про-
тиворечивые.
Описание всех известных нейроспецифичных белков не входит в на-
ши задачи; некоторые белки будут охарактеризованы в соответствую-
щих разделах, где рассматриваются те или иные функции нервной ткани
(в частности, выше были рассмотрены специфичные белки, входящие
в состав миёлина центральной и периферической нервной системы).
СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ
ОБ ОРГАНИЗАЦИИ НЕЙРОНАЛЬНОЙ МЕМБРАНЫ
Не вызывает сомнений тот факт, что основные принципы струк-
турной организации всех мембран животных клеток по сути одинако-
вы. Любая клетка имеет наружную мембрану — ее называют плаз-
матической. Она играет роль преграды, отделяющей живое содер-
жимое клетки от ее окружения. Но плазматическая мембрана — не
просто клеточная оболочка. Она регулирует поступление молекул и
ионов в клетку и выход их наружу. Кроме того, в состав плазматиче-
ской мембраны входят различные ферменты, природа которых зависит
от особенностей данной клетки. Плазматическая мембрана содержит
специализированные компоненты, участвующие в межклеточных кон-
тактах и взаимодействиях, в гормональном ответе и системах транс-
порта через мембрану как малых, так и больших молекул.
Вся сложнейшая деятельность нервной ткани опосредуется мемб-
ранами. В клетках нервной системы представлено несколько
типов высокоспециализированных мембран: сома-
тические мембраны мульти- и униполярных нейронов, мембраны денд-
ритов, аксонного холмика, миелинизированных и немиелинизированных
аксонов, по которым распространяется потенциал действия, мембраны
рыхлого и компактного миелина, мембраны синаптических пузырьков,
пре- и постсинаптические мембраны, мембраны клеток макро- и мик-
роглии. Возбудимость этих мембран колеблется в широких пределах от
высоковозбудимых (синаптические, мембраны аксонного холмика) до
относительно устойчивых мультимембранных структур миелина.
Мембраны нейронов, как и мембраны клеток других тканей, состо-
ят из организованных в бислой липидов и встроенных в липиды белко-
о
вых молекул. Толщина бислоя составляет 50—60 А (ангстрем).
В состав биологических мембран входят вещества трех классов,
обмен которых составляет основу метаболизма: белки, липиды и угле-
воды. Белки составляют 40—60%, остальное приходится на долю ли-
52
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
пидов. Основная масса углеводов является компонентом сложных ли-
пидов (гликолипидов) или сложных белков (гликопротеидов). Белко-
вый состав мембран чрезвычайно разнообразен, он в значительной
мере определяет свойства мембран и их функциональную активность.
Для мембран нервной ткани характерно наибольшее структурное
разнообразие липидов по сравнению с мембранами других орга-
нов. Это существенно отличает нервную ткань от других органов и
тканей (см. гл. 1.2). Именно специфические липиды в значительной
мере определяют сложность и своеобразие мембран и надмолекуляр-
ных образований (таких, как миелин) в нервной системе (см. гл. 1.3).
Для нервной ткани характерно наличие таких липидов, как ганглиози-
ды, галактоцереброзиды, полифосфоинозитиды, отсутствующих в дру-
гих тканях, либо обнаруживающихся в них в незначительных количе-
ствах. Эти липиды определяют функциональные особенности и струк-
туру гликокаликса нейронов (см. гл. 1.7).
Другой компонент мембран нейронов — углеводы — обнару-
живается в составе мембран лишь в соединении с белками (гликопро-
теинами и протеогликанами) и липидами (гликолипидами). В мембра-
нах гликозилировано около 10% всех белков и от 5 до 26% липидов
(в зависимости от объекта). В числе углеводных компонентов глюко-
за, галактоза, их амины, нейраминовая кислота, фукоза и манноза.
Углеводные компоненты мембранных структур в подавляющем
большинстве ориентированы во внеклеточную среду. Их функции свя-
заны с контролем за межклеточными взаимодействиями, поддержани-
ем иммунного статуса клетки, обеспечением стабильности белковых
молекул в мембране.
Очень важна роль углеводного компонента белковых молекул в
формировании специфических функций мембранных белков и липи-
дов. Многие белковые молекулы, особенно биологически активные
вещества (например, нейропептиды), синтезируются в виде крупных,
неактивных предшественников, которые затем расщепляются специ-
фическими протеазами с формированием «зрелых» биологически ак-
тивных продуктов. Деятельность протеаз контролируется уровнем
гликозилирования белков. Так, многие белки, синтезируемые вначале
как гликопротеины, в дальнейшем в результате процессинга теряют
олигосахаридную часть.
Биологические мембраны играют значительную роль в осуществ-
лении почти всех биологических функций клетки. Их молекулярная
организация обеспечивает инфраструктурную основу для всех специ-
альных функций, осуществляемых в клеточной мембране.
1.3. Современные представления об организации нейрональной мембраны
53
Несмотря на многообразие различных типов клеток, их мембраны
выполняют общие биологические функции. Прежде все-
го, они отграничивают живое содержимое от внешней среды, а
внутри клетки организуют компартменты с различными
свойствами. С их помощью происходит отделение содержимого ком-
партментов от цитоплазмы. В каждом компартменте мембраны обес-
печивают сохранение специфических физико-химических условий.
Поэтому по разные стороны мембраны такие условия среды, как кис-
лотность, концентрация растворенных веществ, электрический потен-
циал, как правило, неодинаковы. Для нервной ткани характерна от-
четливо выраженная компартментализация метаболизма — как в
разных отделах мозга, так и в различных субклеточных структурах
нейрона или в элементах системы нейрон—нейроглия. Наиболее от-
четливо это продемонстрировано на примере метаболизма аминокис-
лот и реакций энергетического обмена — существование цитоплаз-
матического и митохондриального метаболических компартментов,
отличающихся субстратами окисления, скоростью и направлением
протекания отдельных реакций (см. гл. 3.2). Важным примером ком-
партментализации метаболических процессов служат специфические
биохимические реакции, протекающие в синаптических окончаниях
и определяющие функционирование синапсов.
Мембраны не только разделяют клетку на отдельные компартмен-
ты, но и участвуют в формировании метаболичес-
ких сигналов, которые передаются через мембрану в клетки.
Это может осуществляться в виде физического переноса ионов или
молекул через мембрану или при помощи конформационных измене-
ний, индуцируемых в мембранных компонентах. Таким образом мемб-
раны «контролируют» проникновение в клетку и выход из нее метабо-
литов. Благодаря рецепторам они реагируют на внешние сигналы
и подвергают их трансформации (т. е. способны классифицировать и
избирательно модулировать сигналы — усиливать важные и снижать
до уровня шумов второстепенные), вычленяя и передавая внутрь клет-
ки существенную информацию.
Мембраны способны обеспечивать образование и
поддержание разности потенциалов, а также транс-
портировать мембранный потенциал вдоль по мембранным индукто-
рам, давая возможность использовать этот специфический вид энер-
гии в разных частях клетки.
С мембранами связано функционирование многих
клеточных ферментов. Мембраны оказывают большое
влияние на процессы, протекающие внутри клетки, изменяя их актив-
54
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
ность. Некоторые ферменты активны только тогда, когда они при-
креплены к мембране; другие, наоборот, в этом состоянии не проявля-
ют активности и начинают действовать лишь после отъединения и пере-
хода в цитоплазму. Поэтому важным свойством мембран является
способность создавать специальные условия для протекания реакций,
осуществляемых гидрофобными белками. Одновременно мембранные
липиды регулируют взаимодействие белков, погруженных в толшу мемб-
ран. Некоторые ферменты образуют своеобразные мембранные ансамб-
ли, осуществляющие цепь последовательных превращений именно бла-
годаря тому, что их компоненты объединены общностью локализации,
организованы мембраной. Благодаря этому обстоятельству повышается
суммарная эффективность каталитического процесса. Существуют фер-
менты, которые, действуя на мембраносвязанные субстраты, тем самым
участвуют в биогенезе мембран.
При том или ином участии мембран осуществляется большинство
жизненно важных функций организма, например репликация прока-
риотической ДНК, биосинтез белков и их секреция, биоэнергетиче-
ские превращения, а также функционирование систем гормонального
ответа. Важная функция мембран связана с координацией множества
химических реакций, протекающих в клетке: в мембранах различные
ферменты объединяются в единый конвейер, в котором каждый фер-
мент действует в строгом соответствии с остальными.
Мембраны участвуют во взаимодействии клеток
со средой. Это свойство лежит в основе обеспечения специфи-
ки межклеточных контактов, адгезии нейрональных клеток и т. д. (см.
гл. 1.7).
Большинство мембран кроме общих выполняют и специаль-
ные функции. Для того чтобы организм мог функционировать как
единое целое, между его клетками должна осуществляться взаимо-
связь — перенос разнообразных химических веществ и информации.
В передаче информации участвуют, например, гормоны, доставляе-
мые к клеткам кровью. В нервной системе передача информации осу-
ществляется прежде всего с помощью нервных импульсов. Мембраны
нервных клеток генерируют электрический потенциал.
Способность вырабатывать электрические сигналы — важнейшее
функциональное свойство нервных клеток, определяемое своеобрази-
ем молекулярной организации наружной мембраны нейрона, в кото-
рую встроены специфические белковые комплексы — так называе-
мые ионные каналы и ионные насосы. Особую роль в связи с этим
играет специфика строения нейрональных мембран (в том числе ми-
елина), способных к генерации и распространению электрического
1.3. Современные представления об организации нейрональной мембраны
55
импульса. Распространение по нерву импульса — процесс, ограни-
ченный плазматической мембраной, определяемый также специфи-
ческими изменениями мембранной структуры, скоординированными
во времени и в пространстве.
Некоторые клетки, например палочки сетчатки глаза, имеют высо-
коспециализированные мембраны, позволяющие выполнять уникаль-
ные функции преобразования энергии света в нервный импульс. Сен-
сорная система обеспечивает восприятие действующих на организм раз-
дражителей, преобразование их сигналов в нервный импульс, передача
которого по нервным путям в подкорковые и корковые центры ЦНС
обеспечивает анализ поступившей информации (см. гл. 3.7). В передаче
информации и формировании ответной реакции ЦНС принимают учас-
тие контактные зоны между нервными клетками — синапсы.
Мембраны синаптических структур обеспечивают осуществление
межнейрональных и периферических связей, а также анализ, отбор и
модуляцию сигнала с помощью химических и электрических механиз-
мов. На одном нейроне может быть от нескольких десятков до несколь-
ких тысяч синапсов. Именно благодаря синаптическим образованиям
происходит формирование временных и стабильных функциональных
ансамблей нейронов, осуществляются межнейронные, нейромышеч-
ные и нейросекреторные контакты.
Относительно немного известно о механизмах формирования си-
напсов в ЦНС. В течение всей жизни организма синапсы непрерывно
развиваются как структурно, так и функционально. Постоянное раз-
витие синапсов является важным условием обучения и формирования
памяти в нормальном мозге. Утрата нейроном такой способности яв-
ляется первопричиной различных нейродегенеративных и психологи-
ческих расстройств.
Для изучения функций мембран было предложено много моделей
молекулярной организации этой структуры. Среди них — динамиче-
ская жидкостно-мозаичная модель С. Сингера и Дж. Никольсона,
существенно усовершенствованная после ее первого описания в 1974 г.
и ставшая ныне широко признанной моделью, применимой к большин-
ству биологических мембран.
В рамках данной модели мембрана представляет собой фосфоли-
пидный бислой, в который погружены свободно диффундирующие бел-
ки (рис. 1.14). Созданию этой модели способствовало развитие совре-
менных подходов к изучению свойств мембранных белков, динамиче-
ских свойств мембран и их взаимосвязи с мембранными функциями.
Согласно жидкостно-мозаичной модели, клеточная мембрана со-
стоит, в основном, из молекул фосфолипидов, организованных в би-
56
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАнИ
Рис. 1.14. Жидкостно-мозаичная модель строения биологических мембран
слой с включенными в его структуру нелипидными соединениями,
представленными интегральными и поверхностными белками. Фосфо-
липиды с их ионными и полярными головками находятся в прямом
контакте с водной фазой на внешней и внутренней поверхности двой-
ного слоя, минимизируя таким образом взаимодействия гидрофобных
компартментов мембраны с молекулами воды из внешней среды, в то
время как неполярные насыщенные и ненасыщенные жирнокислот-
ные цепочки оказываются внутри мембранного двойного слоя, форми-
руя гидрофобную среду. Пустоты между ними заполнены липидами,
главный компонент которых — холестерин.
Жидкостно-мозаичная модель Сингера и Никольсона предполага-
ет существование в мембранах липидных кластеров с различными
свойствами. Такие кластеры являются необходимой частью клеточно-
го механизма перемещения липидов и связанных с липидами белков
внутри клетки. Считается, что липидные кластеры важны для осу-
ществления различных физиологических функций мембран, в част-
ности для обеспечения быстрого перемещения липидных молекул в
латеральном направлении. Липидные кластеры представляют собой
динамичные (короткоживущие) ансамбли — время их жизни измеря-
ется периодами длительностью около 10-7 с.
В плазматической мембране образуются и более устойчивые ассо-
циаты, время жизни которых сопоставимо с молекулярной активно-
стью ферментов. Эти ассоциаты состоят из холестерина, фосфатидил-
серина и сфинголипидов, которые в бислойной мембране объединены
между собой гидрофобными взаимодействиями более тесно, чем окру-
жающие их молекулы. Они получили название липидных рафтов
(плавучих островов).
Такие рафты могут существовать в окружающем их мембранном
бислое достаточно длительное время в виде «движущихся платформ»,
1.3. Современные представления об организации нейрональной мембраны
в которых концентрируются специфические белки. Липидный бислой
в рафтах более асимметричен, чем в других частях бислоя, при этом
его наружный слой обогащен сфинголипидами и гликосфинголипида-
ми, а внутренний — фосфатидилсерином и глицерофосфолипидами.
Сфинголипиды ассоциированы друг с другом в латеральном направле-
нии межзарядными взаимодействиями полярных голов, а с липидным
бислоем связаны насыщенными жирнокислотными цепями. Все пус-
тоты между ассоциатами сфинголипидных, глицеролипидных и фос-
фолипидных групп и их жирнокислотными цепями заполнены молеку-
лами холестерина. Внедрение холестерина «уплотняет» жирнокислот-
ные цепи липидов. Таким образом, при формировании липидных
рафтов холестерин служит своеобразным «наполнителем».
Синтез сфинголипидов и модификация их головок завершается, в
основном, в аппарате Гольджи. Холестерин синтезируется в эндоплаз-
матическом ретикулуме. Ассоциация холестерина с молекулами сфин-
голипидов происходит в аппарате Гольджи, откуда эти ассоциаты
транспортируются в плазматическую мембрану. Передвижение таких
ассоциатов к поверхности плазматической мембраны позволяет увели-
чить содержание в ней холестерина; встраивание доставленных компо-
нентов в плазматическую мембрану происходит в процессе эндоцитоза.
Мембранные рафты — относительно небольшие, весьма подвиж-
ные и довольно нестабильные структуры. Их размер и состав непостоян-
ны. Подвижность позволяет мембранным рафтам облегчать перенос
специфических белков от аппарата Гольджи (/rans-Golgi network —
TGN) к поверхности клетки, а также вдоль плазматической мембраны
или от плазматической мембраны к внутренним мембранам клетки. Раф-
о
ты имеют размеры около 5 А. Индивидуальные рафты в ответ на специ-
фическое физиологическое воздействие могут сливаться с образованием
больших структур. Объединение рафтов индуцируется церамидом.
Распределение липидных рафтов в клеточных мембранах зависит
от типа клеток. Поскольку почти 25% общего количества холестери-
на, присутствующего в организме человека, локализовано в мозге, и
его большая часть сосредоточена в миелине, то участие холестерина в
формировании липидных рафтов очень велико. В нейронах липидные
рафты чаще образуются в сомальных и аксональных мембранах, чем в
соматодендритах. Липидные рафты обнаружены также в постсинапти-
ческих участках нейронов.
В плазматической мембране рафты служат для прикрепления спе-
цифических белков, вовлекаемых во многие процессы, такие как пе-
редача сигнала (signal transduction), эндоцитоз, рецепция болезне-
58
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
творных микроорганизмов и адгезия клеток. Рафты заякоривают гло-
булярные интегральные белки на внешней и на внутренней стороне
бислоя. Рафты включаются в процесс передачи сигнала через такие
рецепторы, как рецептор эпидермального фактора роста, инсулино-
вый рецептор. В их действие вовлекаются тирозинкиназы, G-белки,
Ras, аденилатциклаза, изоферменты протеинкиназы С и др. Рафты
являются местами связывания многих внеклеточных лигандов.
Обнаружение мембранных рафтов, установление их свойств и
функций потребовало очередной модификации жидкостно-мозаичной
модели мембраны, и этот процесс будет продолжаться по мере совер-
шенствования наших знаний. Клеточная мембрана в современном
представлении все более отличается по своим свойствам от «класси-
ческого» липидного бислоя. Тем не менее жидкостно-мозаичная мо-
дель со всеми ее модификациями служит концептуальной основой для
объяснения многих мембранных феноменов. Сложность создания еди-
ной модели биологических мембран связана с огромным разнообрази-
ем их функций.
СВОЙСТВА ЛИПИДНОГО БИСЛОЯ
1.4.1. Фосфолипиды как структурная основа бислоя
Мембранные липиды построены по единому плану, их стерео-
конфигурация имеет общие черты. По этой причине, оказавшись в вод-
ном растворе, фосфолипиды ведут себя сходным образом: проявляют
стремление создать ансамбли из множества молекул. Тем не менее ли-
пиды плохо растворяются как в полярном растворителе — воде (меша-
ют неполярные «хвосты»), так и в неполярной среде — масле (меша-
ют полярные «головки»). Самое энергетически выгодное для них
расположение — мономолекулярный слой на поверхности раздела
между водой и маслом, в этом случае их «хвосты» погружены в масло.
Для того чтобы подчеркнуть различное отношение к воде и к маслу,
полярные «головки» называют гидрофильными, а «хвосты» — ли-
пофильными. Соответственно, липиды, молекулы которых содержат
как гидрофильную, так и липофильную группировки, относят к амфи-
фильным соединениям.
Строение липидных агрегатов в воде может быть чрезвычайно раз-
нообразным. Оно зависит не только от природы самого липида, но и от
его концентрации, температуры и присутствия других веществ. Боль-
шинство липидов плохо растворимы в водной фазе! Лишь при очень
I?
I
1.4. Свойства липидного бислоя
51
П. Квин
Известный исследователь
структурно-функциональных
свойств мембранных липи-
дов, один из первых ученых,
определивших функциональ-
ную роль олигомерной орга-
низации мембранных белков
(Великобритания)
большом разбавлении липиды находятся
в воде в виде отдельных молекул. Даже
при незначительном повышении концент-
рации липида его молекулы объединяют-
ся в замкнутые агрегаты, так называемые
мицеллы. Концентрацию, при которой
происходит это объединение, называют
критической концентрацией мицелло-
образования (ККМ). Для большинства
мембранных липидов она составляет ве-
личину, меньшую 1 %.
При увеличении концентрации липида
в воде и/или понижении температуры
среды количество мицелл растет. В раз-
бавленных водных растворах липиды об-
разуют преимущественно шарообразные
мицеллы — микроскопические капельки,
у которых полярные «головки» обраще-
ны наружу, а неполярные «хвосты» —
внутрь. В масле происходит аналогичный
процесс, но в данном случае мицеллы
оказываются «вывернутыми наизнанку». При увеличении концентра-
ции липидов в воде происходит дальнейшее изменение структуры
липидных ансамблей.
При относительно малом содержании воды фосфолипиды способ-
ны образовывать несколько типов жидкокристаллических структур:
ламеллярную (слоистую) структуру, состоящую из чередующихся
липидных бимолекулярных слоев, прослоенных водными промежутка-
ми, а также цилиндрические структуры, окруженные водной фазой.
Одна структура может переходить в другую при изменении концентра-
ции воды и/или ее температуры. Часто липиды одновременно образу-
ют агрегаты различных типов, которые могут переходить друг в друга.
Такое поведение называют мезоморфизмом. Если переход одних аг-
регатов в другие зависит от содержания воды, говорят о лиотропном
мезоморфизме. Когда он осуществляется под влиянием изменения тем-
пературы — о термотропном мезоморфизме. Такие фазовые перехо-
ды возможны и в биологических мембранах, они играют большую роль
в жизни клеток.
Если концентрация липидов в воде высока, то мицеллы сливаются
и образуют плоские бимолекулярные слои, являющиеся аналогами
структуры мембранного липидного бислоя.
60
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
Наличие у молекул липидов двух частей — полярной «головки» и
неполярных «хвостов» — имеет прямое отношение к их способности
самопроизвольно образовывать мембраны — из таких молекул проис-
ходит спонтанная самосборка мембранного бислоя. В бислойных струк-
турах полярные «головки» липидов обращены к воде, а гидрофобные
«хвосты» ориентированы внутрь бислоя. Как искусственные, так и ес-
тественные мембраны замкнуты сами на себя, образуя полые вакуоли,
пузырьки, везикулы, замкнутые мешочки или трубчатые образования.
Природные фосфолипиды — выраженные амфифилы. Величина ККМ
для них очень мала. Другими словами, уже при низких концентрациях
и комнатной температуре они образуют упорядоченные мицеллы, сли-
вающиеся при повышении концентрации фосфолипида в однослойные
или многослойные агрегаты, которые называют липосомами.
Бислойная структура стабилизируется гидрофобными взаимодейст-
виями в области ацильных цепей и полярными взаимодействиями на
границе раздела водной и липидной фаз. В основе полярных взаимо-
действий действуют ионные, диполь-дипольные, водородные и вандер-
ваальсовы связи. Все они являются слабыми. Важная роль слабых
взаимодействий в стабилизации бислоя объясняется их высокой плот-
ностью. Не менее важна роль и гидрофобных взаимодействий. Так как
неполярным «хвостам» липидов термодинамически невыгодно взаимо-
действовать с упорядоченной структурой воды, стабилизированной во-
дородными связями, липиды стремятся избежать взаимодействия с во-
дой и объединяются в агрегаты. Такое «притяжение» между молекула-
ми, вызываемое присутствием молекул воды, называют гидрофобным
взаимодействием. Гидрофобные силы определяют как способность
аполярных групп к тесному контакту в водных средах, которая обеспе-
чивает вытеснение воды из образуемых агрегатов.
В попытках отождествления свойств искусственных липидных агре-
гатов со свойствами живых клеток необходимо учитывать влияние со-
лей на липидные агрегаты. Липидная мицелла с суммарным отрица-
тельным зарядом будет не безразлична к солевому составу окружаю-
щей ее среды. На поверхности раздела между водой и липидом
создается разность потенциалов. Если к водной среде добавить хлорис-
тый натрий, то положительно заряженные ионы натрия (Na+) будут
связываться с поверхностью мембраны, а отрицательно заряженные
ионы хлора (С1“) — отталкиваться в водную фазу. Обычно примемб-
ранную область разделяют на две части — ближайшую к поверхности
липида, в которой ионы натрия стабилизированы (в ней концентрация
ионов выше — сольватация), и внешнюю, диффузную часть, в которой
ионы передвигаются более свободно.
1
I
1.4. свойства липидного оислоя
С
Как известно, в воде присутствуют (в небольшом количестве) по-
ложительно заряженные ионы водорода и отрицательно заряженные
гидроксильные группы. Ионы водорода притягиваются отрицательно
заряженной поверхностью мицеллы, а гидроксильные группы оттал-
киваются в водную фазу Поэтому величина кислотности среды вблизи
поверхности мицелл отличается от уровня кислотности водного рас-
твора. И по’ своим физическим свойствам вода вблизи поверхности
мицеллы заметно отличается от обыкновенной воды (например, она
не замерзает при О °C). Таким образом, у поверхности липидов в воде
существенно изменяются концентрация солей, физические свойства
и уровень кислотности.
Форма и размеры образуемых липидных ассо-
циатов зависят от многих факторов: от длины ациль-
ных цепей фосфолипидов; от жирнокислотного состава фосфолипи-
дов; от степени ненасыщенности ацильных цепей фосфолипидов; от
структуры полярной части молекулы.
Рассмотрим перечисленные факторы подробнее. Упаковка жирно-
кислотных цепей в мицеллах зависит от длины углеводородной цепи.
При малых значениях п (до 16) количество молекул, необходимое для
формирования мицеллы, таково, что в ее объеме жирнокислотные це-
пи располагаются достаточно свободно. Структурную стабильность
таких мицелл поддерживают полярные связи. При возрастании длины
жирнокислотной цепи плотность упаковки в мицеллах увеличивает-
ся быстрее, чем их размер, и внутреннее содержимое мицелл стано-
вится более компактным благодаря усилению связей между гидрофоб-
ными цепями. Гидрофобные взаимодействия зависят от степени кон-
такта между ацильными цепями — эффективность этих взаимодей-
ствий обратно пропорциональна подвижности цепей. Один из важных
факторов, регулирующих подвижность, — наличие в цепи двойных
связей.
Подавляющее большинство природных жирных кислот содержит
четное количество атомов углерода. Ранее это связывали с удобства-
ми превращения данных соединений в процессе окисления, поскольку
при этом образуются двууглеродные фрагменты ацетил-КоА. Позже
было замечено, что жирные кислоты с нечетным числом атомов угле-
рода в цепи также подвергаются окислительным превращениям (с об-
разованием трехуглеродного малонил-КоА на завершающей стадии
окислительного процесса). В то же время обращает на себя внимание
разная стабильность мицелл, образуемых жирными кислотами с чет-
ным и нечетным числом атомов углерода. Об этом свидетельствует и
величина энергии, которую необходимо сообщить структуре для пере-
62
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
Таблица 1.11. Зависимость температуры плавления
и энтальпии фазового перехода жирных кислот
от длины их углеродной цепи
Число атомов углерода в жирнокислотной цепи Температура плавления, °C Изменение энтальпии (Я0), ккал/моль
Счз -5,4 6,8
Си 5,9 10,8
С15 10,0 8,3
^16 18,2 12,8
С17 22,0 9,7
^18 28,2 14,8
^19 32,0 12,0
^20 36,7 16,8
вода ее из кристаллического состояния в жидкое. Из таблицы 1.11
видно, что изменение энтальпии для жирных кислот с четным количе-
ством атомов углерода заметно выше, следовательно, образуемые ими
ассоциаты структурно стабильнее.
Углеводородные цепи липидных молекул бывают двух
видов: насыщенные и ненасыщенные, причем у ненасыщенных це-
пей может быть одна или несколько двойных связей.
Если мономолекулярная пленка состоит из смеси насыщенных и нена-
сыщенных фосфолипидов, то в месте расположения двойных связей
упорядоченность бислоя нарушается, так как цепи не будут распола-
гаться строго параллельно друг другу. Гидрофобные взаимодействия
при этом ослабевают. Поэтому смеси липидов, содержащих как насы-
щенные, так и ненасыщенные цепи, при одной и той же температуре
являются более «жидкими», чем смеси, построенные из липидов, со-
держащих только насыщенные цепи.
Это свойство имеет важное значение для функции биологических
мембран в живой клетке. Для того чтобы клетка могла осуществлять
процессы жизнедеятельности, липиды, входящие в состав ее мембран,
должны обязательно содержать разупорядоченные участки. Только
при соблюдении этого условия возможны правильное функциониро-
вание мембранных белков и транспорт различных веществ через
1.4. Свойства липидного бислоя
мембрану. Например, было показано, что в мембранах бактерий при
замене ненасыщенных липидов на насыщенные скорость транспорта
веществ снижается в 20 раз.
В связи с вышеизложенным становится понятным столь чуткое ре-
агирование клеток на изменение температуры окружающей среды в
стремлении к обеспечению мембраны разупорядоченными участками.
Как только температура снижается, в бактериальных клетках проис-
ходит замена насыщенных липидов в мембранах на ненасыщенные.
Глубина таких изменений пропорциональна температурному сдвигу.
То же самое имеет место в клетках растений. Организм человека и
теплокровных животных обеспечивает достаточно «жидкое» состоя-
ние клеточных мембран, поддерживая постоянную температуру тела
(более высокую, чем в окружающей среде). У некоторых животных
эта способность выражена весьма наглядно. Например, в ногах пинг-
вина и северного оленя температура тканей падает по мере удаления
от корпуса тела, соответственно мембраны клеток в них содержат по-
степенно возрастающее количество ненасыщенных жирных кислот.
Способность липидов к самоорганизации (самосборке) зависит не
только от природы их углеводородных цепей, но и от природы и поляр-
ных «головок». Они несут либо отрицательный заряд, либо одновре-
менно отрицательный и положительный заряды, нейтрализующие друг
друга. Последний тип липидов в большинстве клеточных мембран явля-
ется преобладающим. И это не случайно. Липидам с отрицательно заря-
женными «головками» трудно объединяться в агрегаты, так как между
«головками» действуют электростатические силы отталкивания. В слу-
чае липидных молекул, несущих два заряда, их «головки» упакованы та-
ким образом, что отрицательный заряд одной молекулы ориентирован
на взаимодействие с положительным зарядом соседней молекулы. Это
позволяет полностью реализовать выгоду как межзарядных, так и гид-
рофобных взаимодействий.
1.4.2. Трансмембранная асимметрия липидов
Клеточные мембраны замкнуты, причем их внутренние и внеш-
ние поверхности различаются по липидному и белковому составу. Эту
особенность мембран называют трансмембранной асимметрией.
Данное свойство является характерной особенностью природных кле-
точных мембран, определяющей их роль в передаче сигналов в клетку.
Известно несколько механизмов, обеспечива-
ющих асимметричное распределение фосфолипи-
дов в мембране. Один из них связан с термодинамической веро-
64
Раздел 1.: Ta-v н '.> —* ънОй ТКАНИ
ятностью распределения липидов в соответствии со стерео-
конфигурацией их молекул. Асимметрия в распределении
липидов в бислое возникает прежде всего потому, что при замыкании
мембранного бислоя липиды с более объемными полярными «голов-
ками» за счет градиента кривизны мембраны стремятся остаться в на-
ружном монослое, так как в нем больше площадь поверхности, прихо-
дящаяся на полярную «головку». По этой причине основная часть
фосфатидилхолина, сфингомиелина, полифосфоинозитидов, холесте-
рина, цереброзидов и сульфатидов локализована в наружном слое, а
аминофосфолипиды (фосфатидилэтаноламин и фосфатидилсерин) на-
ходятся во внутреннем слое, обращенном в цитоплазму. Это соответ-
ственно приводит к различиям в заряде фосфолипидов и гидратации
их полярных «головок» по обе стороны бислоя.
Неодинакова и степень ненасыщенности жирнокислотных цепей в
составе монослоев: во внутреннем обнаруживается 2/3 всех двойных
связей в жирных кислотах липидов, а в наружном — только */3.
Второй механизм поддержания асимметрии бислоя реали-
зуется за счет различий в составе среды по обе его
стороны в условиях нативной клетки. С внеклеточной стороны
мембрану окружает среда с высоким содержанием натрия и кальция, а
со стороны цитоплазмы мембрана контактирует со средой, имеющей
повышенную концентрацию магния и калия. Различия ионного соста-
ва вне- и внутриклеточной среды также играют роль в формировании
и поддержании изгибов мембраны, т. е. «закрепляют» асимметрию
бислоя. Именно этот фактор обеспечивает создание градиента кри-
визны, складок, сморщиваний, облегчает отшнуровку частей биологи-
ческой мембраны в виде везикул, что имеет значение для реализации
межклеточных взаимодействий.
Асимметрия бислоя обеспечивается также ферментами ли-
пидного обмена, к которым прежде всего относятся фосфоли-
пазы, ферменты обмена холестерина и метилазы фосфатидилэтанола-
мина. Метилирование фосфатидилэтаноламина с превращением его в
фосфатидилхолин осуществляется в два этапа и происходит в раз-
ных слоях липидного матрикса. Образование монометилфосфатидил-
этаноламина под влиянием метилтрансферазы I осуществляется во
внутреннем слое*, где и локализован фермент. Монометилфосфатидил-
этаноламин переходит из внутреннего слоя во внешний, там под дей-
ствием метилтрансферазы II завершается его превращение в фосфа-
тидилхолин.
Этот транслокационный процесс меняет «жидкостность» мембра-
ны и рассматривается как фактор, стимулирующий важные процессы
1.4. Свойства липидного бислоя
в мембране: связывание рецепторов с лигандами, Са-индуцируемое
освобождение медиаторов из синаптических окончаний, активацию
мембранных АТФаз.
Асимметричность мембранного бислоя может поддерживаться
транспортом липидов: спонтанным, везикулярным или про-
исходящим с участием липидпереносящих белков. Низкомолекуляр-
ные липидпереносяшие белки обладают различной специфичностью к
разным липидным молекулам (от высокоспецифических, обеспечи-
вающих обмен одного или двух компонентов, до сравнительно мало-
специфических, связывающих и переносящих липиды разных клас-
сов), Ъни также вносят свой вклад в поддержание асимметрии мемб-
ран. Эти белки транспортируют липиды, образуя с ними устойчивые
гидрофильные комплексы.
1.4.3. Подвижность компонентов липидного бислоя
Структурная организация липидного бислоя мембраны не лиша-
ет его компоненты динамичности, которая возникает из-за передви-
жения отдельных липидных молекул в пределах мембранной структуры.
В изотропной жидкости молекулам присущи разные виды под-
вижности: вибрационные колебания, вра-
щение, трансляционные движения. В а н и-
зотропном бислое, напротив, молеку-
лярная подвижность его компонент упоря-
дочена. Различные типы подвижности про-
иллюстрированы рисунком 1.15.
Молекулы фосфолипидов способны к
нескольким видам подвиж-
н о с т и в бислое: изменение ориентации
полярных «головок»; латеральная под-
вижность; колебательная подвижность
ацильных цепей; образование кинков и их
перемещение вдоль ацильных цепей в по-
перечном направлении; ротационная под-
вижность (вращение вокруг длинной оси);
переход с одной стороны бислоя на дру-
гую (по типу «флип-флоп»); выход из
бислоя.
Рассмотрим некоторые виды подвиж-
ности молекул фосфолипидов подроб-
нее.
5
Рис. 1.15. Виды подвижнос-
ти липидных компонентов
в бислое:
/ — изменение ориентации по-
лярных «головок»; 2 — лате-
ральная диффузия в двумер-
ном пространстве бислоя; 3 —
быстрые колебания жирно-
кислотных цепей относительно
перпендикулярной оси; 4 —
образование кинков и их про-
движение вдоль ацильных це-
пей; 5 — вращательная подвиж-
ность; 6 — медленный обмен
между компонентами монослоев
мембраны («флип-флоп»)
5-1822
66
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
Латеральная подвижность. Способность липидов перемещать-
ся в плоскости мембраны показана многими экспериментами. Напри-
мер, измерено, что в мицеллах яичного лецитина при 25 °C молекула
преодолевает путь, равный 2,5 мкм, за 1 с. Таким образом, латеральная
диффузия в упорядоченной мембране позволяет компонентам бислоя
перемещаться с относительно высокой скоростью. Она делает возмож-
ным образование липидных кластеров. Латеральная диффузия имеет
место даже при температуре, структурирующей мембрану в кристалли-
ческом состоянии. Объяснить этот факт можно латеральной неоднород-
ностью мембраны, наличием в бислое дефектных зон, в которые могут
вытесняться молекулы из соседних упорядоченных областей.
Упорядоченность мембран, измеряемая с помощью ЭПР-спектро-
скопии, снижается при движении к метильному радикалу ацильной це-
пи, что свидетельствует об увеличении подвижности жирнокислотных
цепей в этом направлении.
Различные конфигурации молекул жирных кислот, возникающие
при поворотах (вращении) вокруг единичной С—С-связи, называют
ротамерами или конформерами, а изменение конформации молеку-
лы за счет таких поворотов носит название транс-гош-изомериза-
ции. Вращательная подвижность молекул фосфолипидов, измеренная
методом ЯМР-спектроскопии, показала, что подвижность гидрофоб-
ных сегментов цепи повышается в направлении от сложноэфирной
связи к метильной группе, т. е. к центру бислоя.
Трансбислойный переход («флип-флоп»), В искусственных ли-
пидных мембранах такие переходы осуществляются достаточно мед-
ленно, например полупериод перехода молекул холестерина с одной
стороны бислоя на другую в липосомах из фосфатидилхолина занима-
ет более 24 ч, однако в принципе такой переход возможен. Даже в
присутствии липидпереносящих белков перемещение из наружного
слоя во внутренний занимает более 4 ч, а в обратную сторону — бо-
лее 10 ч. Повышение температуры до 80 °C не ускоряет «флип-фло-
па», но он может быть существенно ускорен в присутствии окислен-
ных липидов, лизолецитина. Как правило, холестерин подвергается
более быстрому «флип-флопу», чем фосфолипиды.
Молекулы липидов не могут преодолеть липидный бислой в попереч-
ном направлении путем простого физического перехода с одной стороны
бислоя на другую, если в мембране нет особых ферментов — трансло-
каз. Поэтому довольно большую частоту миграции липидов в биологи-
ческих мембранах с одной стороны на другую связывали с активностью
этих ферментов. Однако вероятно, что гетерогенность липидного соста-
1.4. Свойства липидного бислоя
6
ва биологических мембран и образование короткоживущих внутренних
гексагональных мицелл увеличивает вероятность неферментативного
.<флип-флоп»-перехода в природных мембранах. Результаты, получен-
ные с помощью различных физических методов, указывают на возмож-
ность образования в природных мембранах гексагональной фазы (вы-
вернутых везикул), в которую вовлекаются молекулы липидов с одной
стороны бислоя, а возвращаться они могут в другую. Следует отметить,
что «флип-флоп» запускает функционально важные события в мембра-
не. Латеральная и вращательная диффузия липидов также оказывает
регулирующее влияние на активность мембранных белков.
Важной для функционального состояния мембранных белков явля-
ется и колебательная подвижность жирнокислотных «хвостов» отдель-
ных молекул фосфолипидов. Вблизи метильного конца она осуществ-
ляется независимо каждой молекулой, но с приближением к полярной
«головке» и возрастанием плотности упаковки (особенно начиная с 9-го
углеродного атома, ближе которого к поверхности бислоя обычно не
встречается цис-двойных связей) подвижность уменьшается.
По своей структурной конфигурации насыщенные жирные кислоты
сильно отличаются от ненасыщенных. Насыщенные жирные кислоты мо-
гут иметь множество различных конформаций вследствие высокой свобо-
ды вращения вокруг одиночных С—С-связей. Энергетически наиболее
выгодной является транс-конформация. Ненасыщенные жирные кисло-
ты имеют жесткую структуру, поскольку вращение вокруг двойных связей
невозможно. Они существуют либо в транс-, либо в цис-конфигурации.
(Различие между терминами конформация и конфигурация заключает-
ся в том, что переход между конформациями осуществляется за счет из-
менения взаимного расположения между частями молекул без разрыва и
образования новых ковалентных связей; переход же между разными кон-
фигурациями требует разрыва ковалентных связей и их замыкания в дру-
гом месте. Так, переход между D- и L-стереоконфигурациями аминокис-
лот невозможен без разрыва и замыкания ковалентных связей, а цис-
транс -переход между разными конформационными состояниями жир-
ных кислот легко осуществляется по принципу тепловых движений.)
Ненасыщенные жирные кислоты содержат двойные связи почти
всегда в цис-конфигурации (рис. 1.16), транс-ненасыщенные жир-
ные кислоты в природе почти не встречаются. Исключение составляет
лишь вакценовая кислота — конфигурационный аналог олеиновой
Кислоты. Ее температура плавления составляет 44 °C, в то время как
°леиновая кислота плавится при температуре 13,5 °C.
г Цис-конфигурация двойной связи обусловливает изгиб цепи под
Углом около 30°. По этой причине цис-ненасыщенные жирные кисло-
68
Раздел 1 СТРУКТУРА НЕРВНОЙ ТКАНИ
Полностью
транс- конформация
Цис-
конформация
Рис. 1.16. Пространственная конфигурация жирных кислот:
а — насыщенная углеводородная цепь; б — ненасыщенная цепь с двойной связы*’
в цис-положении; в — насыщенная цепь в гош-положении
1.4. Свойства липидного бислоя
ты с одной двойной связью вызывают локальные возмущения бислоя.
При этом длина такой цепи несколько уменьшается, а занимаемый ею
объем возрастает. В области локализации двойных цис-связей сосед-
ние участки цепи образуют изгибы (так называемая гош-форма).
Повышение температуры среды увеличивает тепловую подвиж-
ность жирнокислотных цепей и облегчает возникновение изгибов. Ес-
ли изгибы, соответствующие гош-конформации^ появляются на близ-
лежащих участках жирнокислотной цепи, эта область может прини-
мать вид петли или полости (кинк). В результате взаимопереходов
транс- и гош-конформаций (так называемого транс-гош-перехода)
кинки могут «скользить» вдоль цепи, обеспечивая перемещение их
содержимого поперек мембраны. Таким образом может осуществ-
ляться диффузия захваченной воды через гидрофобный бислой.
При повышении плотности упаковки бислоя конформационная под-
вижность С—С-связей снижается. В таком бислое подвижность цепей
ограничена согласованными колебаниями около точки прикрепления
жирнокислотных радикалов к полярной «головке» фосфолипида. В этой
ситуации в бислое наиболее предпочтительны две конформации цепи:
когда вся цепь находится в транс-конформации или при наличии струк-
туры типа «двойной гош», когда изгибы, возникающие на двух соседних
участках цепи вследствие образования гош-конформации, компенсиру-
ют друг друга, и вся цепь в целом укорачивается без существенного уве-
личения площади, приходящейся на одну молекулу.
1.4.4. Фазовые переходы мембранных липидов
Фазовое состояние, в котором в данный момент находятся липи-
ды бислоя, — гелеобразное или жидкокристаллическое — оказывает
влияние на все типы молекулярной подвижности липидных молекул.
Липиды обладают замечательным свойством — они способны к
фазовым переходам в физиологических условиях. При определенных
температурах, строго характерных для каждого вида липидов, в водной
среде липидные мицеллы (рис. 1.17, а, б), образуемые фосфолипида-
ми (ламеллярные, мицеллярные, гексагональные), могут находиться в
Упорядоченном (гелеобразном) или в разупорядочен-
н о м (мезофазном), так называемом жидкокристалличес-
ком, состоянии. Жидкие кристаллы — это анизотропные жидкости:
оптически они сходны с кристаллами, так как проявляют разные свой-
ства в различных направлениях, а механически подобны жидкости
(изотропны), текут в зависимости от вязкости.
70
Раздел 1. СТРУIг »А Hl гЪ НОИ I КАНИ
шшял
щшг~
Вода
Шш
шот
’ б
ши
‘Шт
иуяи
-. Л. К’Г- ;• - 'Л.
а
Рис. 1.17. Структуры, образуемые в водных суспензиях липидами, склонными
к созданию ламеллярных образований (а, б) и небислойных гексагональных
структур (в, г)
лВ'
тш'
Ж
Жидкие кристаллы, сочетая в себе упорядоченность твердого тела
и подвижность жидкости, отличаются высокой чувствительностью к
внешним воздействиям: температуре, примесям, свету, внешним по-
лям; они очень пластичны и достаточно долго хранят информацию.
Эти свойства приобретают первостепенное значение в мембранах
нервной ткани, где изменения электрических свойств лежат в основе
проведения возбуждения.
Таким структурам присущи лиотропный мезоморфизм (зависи-
мость структуры от гидратации) и термотропный мезоморфизм
(зависимость структуры от температуры).
Температура фазового перехода липидов «гель—жидкий крис-
талл» зависит от содержания воды в системе. Температура перехода
минимальна, если общее содержание воды превышает то ее количест-
во, которое могут связать липидные структуры. В то же время при не-
достатке воды липиды могут находиться в упорядоченном состоянии
I 1.4. Свойства липидного бислоя
и при температуре, выше критической. Таким образом, температура
фазовых переходов липидов, осуществляющихся по типу «гель —
жидкий кристалл», зависит от содержания воды в системе.
Примером существенного влияния воды на фазовые переходы би-
слоя является модификация мембраны под влиянием перекисного окис-
ления липидов (ПОЛ), способного снижать температуру фазового пере-
хода. Это явление объясняется увеличением содержания воды в бислое.
Дело в том, что гидратация бислоя зависит от присутствуя заряженных
фосфолипидов. Наиболее сильно она возрастает при индукции ПОЛ.
Образующиеся в качестве промежуточных продуктов гидроперекиси яв-
ляются высокогидрофильными, они эффективно разрушают бислой,
увеличивая доступность гидрофобных доменов мембраны для молекул
воды. Именно по этим причинам ПОЛ имеет такое «разрушительное»
влияние на клеточные мембраны.
Термотропные фазовые переходы липидов в мембране происходят в
сравнительно широком температурном интервале (в несколько граду-
сов). Это обусловлено тем, что в бислое более «жидкая» фаза возника-
ет в окружении другой, более упорядоченной. Сосуществование в ли-
пидном бислое двух фаз устанавливает между ними сложное равнове-
сие, приводя к снижению кооперативности фазового перехода. Обычно
кооперативные фазовые переходы липидов в мембране затрагивают не-
сколько сотен молекул. В нативной мембране постоянно находится
большое количество кооперативных единиц той или иной фазы. Гетеро-
генность мембран выражается, в частности, в наличии липидных рафтов.
На температуру фазового перехода большое влияние оказывают
природа липидных молекул, длина углеводородного скелета, наличие
цис- и транс-двойных связей, структура полярных групп.
При переходе липидов из гелеобразного в жидкокристаллическое
состояние одновременно происходит несколько событий: возрастает
вращательная подвижность липидов, увеличиваются колебательная
подвижность жирнокислотных радикалов относительно С—С-связей и
скорость латеральной диффузии. Все это связано с изменением геомет-
рических размеров бислоя из-за латерального расширения площади,
занимаемой каждой молекулой липида. Например, площадь, занимае-
мая 2С16-фосфатидилхолином, меняется от 0,49 до 0,58 нм2, среднее
расстояние между молекулами увеличивается от 0,49 до 0,52 нм, а тол-
щина мембраны уменьшается почти на 0,5 нм, т. е. латеральное расши-
рение слоя компенсируется его утончением. Гидрофобный объем
мембраны увеличивается примерно на 1,5%.
Жидкокристаллическое состояние доменов мембраны, увеличивая
вращательную подвижность жирнокислотных радикалов относительно
С—С-связей, способствует m/хшс-гош-изомеризации (ротамеризации).
72
раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
В мембранном бислое облегчается образование различных конформе-
ров жирных кислот. При повороте на ±120° относительно транс-кон-
формации появляются аош( + )- и гош( — )-конформации. Из-за неод-
новременное™ проявления этих свойств в ходе фазовых перестроек
бислоя более упорядоченные области мембраны сосуществуют с уже
расплавленными — наблюдается фазовое разделение. В данных усло-
виях для мембраны характерно появление разного рода структурных
дефектов, индуцирующих «сплывание» белковых молекул друг с дру-
гом, а также встраивание в бислой молекул холестерина, повышение
его проницаемости для ионов и т. д.
Наличие фазовых переходов, индуцируемых температурой, — объ-
ективная характеристика бислоя, состоящего из различных по свойст-
вам компонентов. Фазовые переходы липидов при постоянной темпе-
ратуре среды могут быть вызваны изменениями заряда их полярных
групп, возникающими при изменениях pH, ионной силы, концентрации
ионов. Доказано, что температура фазового перехода есть функция ве-
личины и плотности заряда липидных молекул. Любое увеличение за-
ряда полярных групп благоприятствует сохранению разупорядоченного
(жидкокристаллического) состояния ввиду латерального электростати-
ческого отталкивания, тогда как уменьшение заряда обусловливает пе-
реход в гелеобразное состояние.
Важным способом изменения поверхностного заряда липидов в
физиологических условиях является адсорбция катионов. Связывание
катионов заряженными липидами зависит от поверхностного потенци-
ала, значительно различающегося в разных фазовых состояниях, что
определяется молекулярной упаковкой липидных молекул.
Адсорбция или десорбция катионов на мембранной поверхности мо-
жет инициировать фазовые переходы липцдов. При определенных фи-
зиологических условиях структурные изменения липидов вызывают вы-
свобождение двухвалентных катионов с поверхности мембраны. Так, при
переходе «гель—жидкий кристалл» с липидной поверхности высвобож-
даются ионы кальция. Са2+ и Mg2+ стабилизируют организованную
структуру, повышая температуру фазового перехода, а одновалентные
катионы вызывают противоположный эффект. Другими словами, двухва-
лентные катионы благоприятствуют гелеобразному, а одновалентные —
жидкокристаллическому состоянию мембраны. Таким образом, поверх-
ность липидов может рассматриваться как резервуар катионов, объем
которого регулируется структурными изменениями мембраны.
Таким образом, фазовые переходы в мембране индуцируются не
только изменениями температуры. Они могут быть вызваны сдвигом
pH среды, электрическим потенциалом, двухвалентными катионами.
1.4.CBv4 iBi ИЛИД м ГОбГ ло<
способствующими образованию кластеров определенных фосфолипи-
дов и разделению фаз; фазовые переходы также сопровождают дейст-
вие гормонов на биологические мембраны.
Кроме перехода типа «гель—жидкий кристалл» липиды могут пре-
терпевать превращения другого рода, приводящие к образованию гек-
сагональной фазы Нп (рис. 1.17, в, г на с. 70). Эти небислойные
структуры легко образуются короткоцепочечными кислыми фосфоли-
пидами (например, фосфатидил серином, фосфатидной кислотой). Об-
разованию гексагональных структур способствуют такие явления, как
повышение температуры, увеличение ненасыщенности жирнокислот-
ных цепей, высокая ионная сила при щелочном pH, а также пониже-
ние гидратации бислоя. Переход отдельных участков бислоя в фазу Нп
приводит к нарушению целостности мембраны, формированию кана-
лов проницаемости, образованию дефектных зон.
1.4.5. Дефектные зоны. Роль холестерина
Как показано выше, различные виды подвижности липидных
компонентов в бислое нарушают его гомогенную упаковку и приводят
к появлению в мембране динамических дефектов. Высокая скорость
вращения жирнокислотных радикалов вокруг С—С-связей, а также
наличие в составе фосфолипидов £{ис-изомеров ненасыщенных жир-
ных кислот делают бислой достаточно рыхлым. Образование дефектов
такого рода демонстрирует рисунок 1.18.
Рис. 1.18. Образование дефектов в мембранном бислое:
а — обе жирнокислотные цепи молекулы фосфолипида находятся в транс-положе-
нии; вращение вокруг С—С-связей, указанное круговыми стрелками, ничем не огра-
ничено (1); в результате несогласованного вращения образуется скошенная (гош-)
конформация (2) или кинк (3); б — влияние кинков на упаковку бислоя: I — кинки от-
сутствуют; II — один кинк на жирнокислотную цепь; III — два кинка на жирнокислот-
ную цепь. Жидкокристаллическое состояние бислоя облегчает образование кинков
74
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
Рассмотрим насыщенные углеводородные цепи. Наиболее ста-
бильна транс-конформация (/), в которой цепь максимально вытя-
нута и не меняет своего направления, тогда как в гош-конформации
ее направление меняется (2) и образуется дефект упаковки. Последо-
вательность гош—транс—гош для трех смежных С—С-связей при-
водит к появлению в цепи излома (кинка), в результате чего участки
цепи выше и ниже излома оказываются значительно смещенными
друг относительно друга (3) — образуется дефект. Возможность фор-
мирования таких структур обеспечивает возникновение подвижных
дефектов, способных захватывать целые кластеры мембраны. Их кон-
центрация и подвижность зависят от температуры.
В случае ненасыщенных жирных кислот почти все двойные связи
в мембранных липидах находятся в ^uc-форме. Как и в случае гош-
формы, это приводит к изменению угла наклона цепи — образует-
ся дефект. Такие пространственные дефекты бислоя, индуцируемые
включением в него ненасыщенных жирных кислот, являются более
стабильными. В результате появления дефектов бислой становится
более рыхлым.
Рассмотрим вызванное дефектами изменение геометрии бислоя.
Растянутая углеводородная цепь жирной кислоты, содержащая 18 уг-
о
леродных атомов, имеет длину примерно 23—24 А, что возможно в
случае геля, т. е. в случае высокоструктурированного (упорядоченно-
го) состояния липидов. По этой причине можно было бы предполо-
жить, что бислой, составленный из фосфолипидов, жирные кислоты
которых содержат 18—20 углеродных радикалов, должен иметь тол-
о
щину более 47 А. Однако реальная толщина мембраны редко превы-
шает это значение (она составляет на 15—20% меньшую величину).
Это указывает на то, что жирнокислотные радикалы бислоя не рас-
прямлены полностью, а образуют рыхлые структуры. В результате
этого площадь мембраны оказывается несколько большей, чем следу-
ет из расчета теоретического пространства, приходящегося на одну
молекулу фосфолипида. Встраивание холестерина в бислой делает
возможным изменение формы мембран в результате значительной де-
формации обеих сторон липидного бислоя, так как в отличие от фос-
фолипидов холестерин может легко перемещаться из одного монослоя
в другой. При этом несколько возрастает и толщина бислоя.
Гетерогенность бислоя и образование в нем липидных кластеров
способствуют разделению (сегрегации) фаз в мембране. Латеральное
разделение липидных молекул в плоскости бислоя — важная особен-
ность мембраны. Особая сегрегирующая роль в мембране принадле-
1.4. Свойства липидного бислоя
жит холестерину. При низких концентрациях его в мембране происхо-
дит латеральное разделение фосфолипид-холестериновых комплексов
и свободных молекул фосфолипидов. При этом холестерин взаимодей-
ствует в первую очередь с теми молекулами фосфолипидов, которые
имеют низкую температуру фазового перехода. Благодаря этому в
бислое будут существовать как области только «жидкие» и только
«твердые»,’ так и области смешанные, где обе фазы сосуществуют.
Наличие разных по плотности областей в пределах одной мембраны
изменяет ее сжимаемость, что сказывается на глубине погружения
мембранных белков и на эффективности их работы.
Необходимо отметить, что кроме сегрегирующего холестерин ока-
зывает и другое важное воздействие на структуру и физические свой-
ства липидного бислоя. Встраивание холестерина в фосфолипидный
бислой вызывает как нарушение упаковки жирнокислотных цепей,
так и уменьшение их подвижности. Эти эффекты холестерина называ-
ют соответственно разжижающим и конденсирующим. При темпе-
ратуре, превышающей точку фазового перехода фосфолипида, холес-
терин уменьшает подвижность его углеводородных цепей. Так, при до-
бавлении холестерина к фосфатидилхолиновым мембранам площадь
молекулы фосфатидилхолина уменьшается с 0,96 до 0,56 нм2. Этот
уплотняющий эффект холестерина максимален в районе центрального
участка жирнокислотных радикалов и ослабевает в направлении кон-
цевых метальных групп. Но при температуре ниже точки фазового пе-
рехода фосфолипидов холестерин разжижает углеводородную область
бислоя.
Наличие в углеводородных цепях кинков, двойных связей и других
особенностей приводит к увеличению площади поперечного сечения
о
углеводородной цепи (минимальное ее значение составляет около 19 А2)
при ее полностью транс-конфигурации; это может иметь важные по-
следствия для упаковки липидов в бислое. В жидкокристаллической
фазе появление в цепи сош-конформеров увеличивает эффективное
о
поперечное сечение углеводородных цепей по меньшей мере до 50 А2.
Толщина бислоя уменьшается за счет наличия в цепях гош-конфор-
меров, приводящих к разупорядочиванию цепей, причем сами цепи
в целом растянуты и расположены перпендикулярно поверхности
бислоя.
В фазе геля насыщенные углеводородные цепи фосфолипидов на-
ходятся преимущественно в транс-конфигурации. Минимальная пло-
щадь поперечного сечения молекулы диацильного фосфолипида со-
о
ставляет около 38 А2. Примерно такую же площадь занимает полярная
76
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
головка фосфатидилэтаноламина, поэтому насыщенные фосфатидил-
этаноламины в фазе геля упаковываются таким образом, что их ацильные
цепи располагаются перпендикулярно плоскости бислоя (как в липидных
кристаллах).
На рисунках и схемах жирнокислотные цепи фосфолипидов в би-
слое, как правило, представляют скошенными относительно перпен-
дикулярной оси мембран (см. рис. 1.18). Угол наклона цепей опреде-
ляется природой полярной «головки» молекулы: наклон возникает в
том случае, когда объем гидратированной «головки» молекулы боль-
ше площади сечения ее «хвостовой» части. Например, в случае крис-
таллов фосфатидилхолина минимальная площадь, приходящаяся на
о
одну полярную «головку», составляет примерно 50 А2, поэтому ди-
о
пальмитоилфосфатидилхолин (38 А2) в фазе геля не может упаковы-
ваться так, как фосфатидилэтаноламин. В этом случае ацильные цепи
дипальмитоилфосфатидилхолина отклоняются от нормали к бислою
на 30°, благодаря чему их поперечное сечение увеличивается, чем и до-
стигается соответствие размерам полярной «головки». При этом угле-
водородные цепи сохраняют полностью транс-конформацию. В слу-
чае когда эти величины равны (например, для фосфатидилэтанолами-
на), скоса практически не наблюдается.
Наличие перечисленных особенностей обеспечивает определен-
ную рыхлость упаковки мембранного бислоя при нормальных услови-
ях, которая в литературе часто обозначается термином жидкост-
носпгь, смысл которого противоположен термину микровязкость.
1.4.6. Микровязкость мембран
Для мембран характерна высокая степень текучести бислоя,
обеспечивающая способность липидов и белков к латеральной диффу-
зии. Повышенная плотность упаковки (структуризация) бислоя увели-
чивает сопротивление диффузии молекул, транспортируемых через
мембрану, повышает ее микровязкость. Следовательно, скорость пере-
мещения молекул зависит от микровязкости мембран, которая, в свою
очередь, определяется относительным содержанием насыщенных и не-
насыщенных жирных кислот в составе липидов. Микровязкость мемб-
ран меньше, если в составе липидов преобладают ненасыщенные жир-
ные кислоты, и больше при высоком содержании насыщенных жирных
кислот. На этот параметр влияют также размеры углеводородных
«хвостов» липидов, с увеличением длины которых микровязкость би-
слоя уменьшается.
1.5. Мембранные белки
1.5.
МЕМБРАННЫЕ БЕЛКИ
Липиды — относительно низкомолекулярые соединения с молеку-
лярным весом менее 1000 Да (1 кДа), в то время как мембранные белки
имеют молекулярную массу, превышающую, как правило, десятки тысяч
дальтон. Клеточные мембраны содержат большое число различных бел-
ков, молекулярная масса которых колеблется от 10 до 240 кДа.
Последовательность аминокислотных радикалов в пептидной цепи,
как правило, содержит информацию, диктующую упаковку белковой
молекулы в пространстве. Поэтому уже в ходе синтеза белковой моле-
кулы происходит спонтанное комплектование вторичной и третичной
структур белка.
Функции белков определяются особенностями их структуры и упа-
ковки. Белковые глобулы являются примером третичной структуры —
на ее уровне формируется специфическая ферментативная, рецептор-
ная и другие виды активности белковой молекулы. При ассоциации
нескольких глобул в олигомерные ансамбли формируется четвертич-
ная структура, придающая белку способность тонкого регулирования
осуществляемых им функций.
Форма образующихся глобул в большой степени зависит от соотно-
шения полярных и аполярных аминокислотных радикалов в образую-
щемся белке. В водной фазе белковые молекулы стремятся принять
такую форму, чтобы гидрофобная сердцевина была «одета» заряжен-
ными или нейтральными группами гидрофильных аминокислот. При
«невыгодном» соотношении различных по природе аминокислот для
защиты от атаки молекулами воды обращенных к внешней среде гидро-
фобных радикалов белки вынуждены образовывать ассоциаты — гете-
ро- или гомоолигомеры. Другой способ их стабилизации — внедрение
в гидрофобные структуры клеточных мембран.
Спонтанное образование белковых глобул, происходящее уже в
процессе синтеза белковой молекулы, имеет место в случае относи-
тельно просто устроенных белков. Более сложные белковые структу-
ры принимают окончательную форму с помощью специальных бел-
ков-шаперонов в ходе так называемого процессинга, или созревания
(maturation). Но и в этом случае для большого количества белковых
молекул клеточная мембрана является областью, внедрение в кото-
рую обеспечивает белковой молекуле большую стабильность или воз-
можность осуществления специфических реакций.
В ряде случаев для решения вопроса о цитоплазматической или
мембранной локализации белка важным является не общее соотноше-
ние гидрофобных и гидрофильных аминокислотных радикалов в первич-
7й
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
ной структуре, а наличие участков первичной структуры, содержащих
протяженные гидрофобные цепи. Именно эти участки «нуждаются»
в маскировке гидрофобным окружением.
1.5.1. Локализация и подвижность белков в мембранном бислое
Мембраны содержат от 20 до 80% белка (по весу). При этом в
разных мембранах количество белков существенно различается. Так,
в мембранах митохондрий доля белков составляет около 75%, а в
плазматической мембране олигодендроцитов — около 25%. Так как
о
липидные молекулы имеют небольшой размер (около 5 А) и неболь-
шую молекулярную массу, их число в несколько десятков раз больше
числа белковых молекул. Поэтому белковые молекулы фактически
внедрены в липидный бислой мембраны.
Белки различаются расположением в мембране (рис. 1.19). Моле-
кулы интегральных белков могут глубоко проникать в липидный
бислой или даже пронизывать его (в этом случае их называют транс-
мембранными белками). Периферические белки прикрепляются к
мембране разными способами. Интегральные белки содержат транс-
мембранные домены, которые состоят преимущественно из неполяр-
ных аминокислотных радикалов и могут пересечь бислой один или
несколько раз. Трансмембранные домены обычно имеют структуру
Внутренняя поверхность мембраны
Рис. 1.19. Встраивание белков в липидный бислой.
Трансмембранные белки- 1 — гликофорин; 2 — адренорецептор; поверхностные
белки: 3 — белки, связанные с интегральными белками (сукцинатдегидрогеназа);
4 — белки, ассоциированные с полярными «головками» липидного бислоя (протеин-
киназа С); 5 — белки, «заякоренные» в мембрану с помощью короткого гидрофобного
концевого домена (цитохром 65); 6 — белки, «заякоренные» в мембрану за счет жир-
нокислотного радикала, ковалентно присоединенного к белковой молекуле (G-белок)
1.5. Мембранные белки
ос-спирали. Интегральные белки могут формировать гидрофильные
домены на одной или обеих поверхностях бислоя.
Периферические белки отличаются от интегральных меньшей глу-
биной проникновения в бислой и более слабыми белок-липидными
взаимодействиями (т. е. меньшей зависимостью от бислоя). Перифе-
рические белки могут обратимо менять свой статус, прикрепляясь к
мембране на определенное время (такие белки называют амфипати-
ческими). Прикрепляясь к мембране, они взаимодействуют либо с ин-
тегральными белками, либо с поверхностными участками липидного
бислоя, приобретая новые свойства.
Деление мембранных белков на периферические и интегральные
определяется их структурой, количеством гидрофобных аминокислот
и их расположением в первичной структуре, т. е. всеми теми свойст-
вами, которые обеспечивают взаимодействие белка с бислоем. Для
амфипатических белков существуют специальные сигналы, стимули-
рующие их ассоциацию с мембраной (часто таким сигналом является
фосфорилирование белков специфическими киназами, изменяющее
их третичную структуру и гидрофобность, точнее — лиотропность).
К таким белкам, например, относят протеинкиназу С.
Локализованный в мембране гидрофобный домен или «якорь» яв-
ляется еще одним характерным элементом структуры мембранных
белков. С помощью такой структуры происходит закрепление перифе-
рических белков в мембране. В качестве якоря может выступать жир-
нокислотный радикал (чаще всего — пальмитоил или миристоил),
ковалентно присоединяющийся к белковой молекуле.
Диаметр интегральных мембранных белков в среднем составляет
о о
80 А, но встречаются и более крупные белки — размером до 350 А.
Обычно эти белки обладают выра-
женной асимметрией (см. рис. 1.19).
Некоторые из трансмембранных
белков пронизывают мембрану один
раз (как гликофорин) — они называ-
ются битхтическими, другие имеют
несколько участков (доменов), последо-
вательно пересекающих бислой, — это
политопические белки (рис. 1.20).
Монотопические белки относятся к
периферическим белкам.
В качестве примера рассмотрим
несколько белков, по-разному свя-
занных с мембранами.
Рис. 1.20. Монотопическая (а),
битопическая (б) и политопи-
ческая (в) локализация белков
в мембране
80
Раздел ^ СТРУКТУРА НЕРВНОЙ ТКАНИ
Белок-предшественник амилоида (АРР) — трансмембранный (ин-
тегральный) белок, известный в связи с участием одного из продуктов
его протеолиза в развитии тяжелой патологии мозга — болезни Альц-
геймера. Этот белок состоит из 695—751 аминокислотного остатка,
экспрессируется в различных органах и тканях животных. Из извест-
ных трех изоформ этого белка в коре мозга присутствует в основном
более короткая форма АРР695, состоящая из 695 аминокислотных ос-
татков, которая фиксирована в мембранах нейронов так, что N-конце-
вой фрагмент из 625—630 остатков ориентирован во внеклеточное
пространство, а короткая (С-концевая) часть локализована в цито-
плазме клеток. По-видимому, АРР участвует в организации межней-
ронных контактов, особенно в области нервных окончаний. Экстра-
клеточные фрагменты белка АРР могут отщепляться протеиназами и,
как установлено, стимулируют развитие нейритов, участвуя в форми-
ровании следов памяти.
Среди многих регуляторных белков нейрональной мембраны осо-
бый интерес в связи с участием в развитии тяжелой патологии
мозга — болезни Альцгеймера — представляет так называемый бе-
лок р-АРР (P-amyloid precursor protein) — предшественник Р-амило-
идного пептида (P-А), образующего на поверхности нейронов больно-
го мозга структурные бляшки. Участие Р-АРР в возникновении и раз-
витии болезни Альцгеймера описано в 3.3.9.
АРР и продукты его обмена выполняют важные функции в жизне-
деятельности нейрональных клеток-. Эти белки обладают нейротрофи-
ческими и нейропротекторными свойствами, модулируют рост нерв-
ных окончаний и синаптогенез. Они участвуют в обеспечении возбуди-
мости нервных клеток и синаптической пластичности, а также в
процессах обучения и памяти. Р-АРР обладает многими характеристи-
ками, свойственными мембраносвязанным рецепторным молекулам,
так как может функционировать как рецептор, сопряженный с G-бел-
ками, опосредованно способствуя запуску каскада внутриклеточных
сигнальных реакций.
Цитохром 65 имеет единственный гидрофобный сегмент, с по-
мощью которого заякоривается на цитоплазматической поверхности
микросомальной мембраны. Это монотопический белок. Интересный
пример регулируемого взаимодействия с мембраной был обнаружен
при изучении аминокислотной последовательности интактной формы
микросомального цитохрома Ь5 (см. рис. 1.19). Этот белок участвует в
различных окислительно-восстановительных реакциях в качестве пе-
реносчика электронов. Его относительно короткий участок пептидной
цепи вблизи С-конца сплошь состоит из гидрофобных аминокислот.
| 1.5. Мембранные белки
81
Ес. 1И гидрофобный «якорь» удалить с помощью протеолиза, гемсвя-
зывающий домен теряет связь с мембраной и высвобождается в среду.
Мембранные белки битопического типа часто вовлекаются в переда-
чу сигналов через клеточную мембрану. Например, некоторые из рецеп-
торактивируемых цитоплазматических тирозинкиназ являются битопи-
ческими белками.
Белки ионных каналов, транспортные АТФазы (саркоплазматиче-
ская Са2+-АТФаза, см. гл. 2.5) и большинство рецепторных структур
(см. гл. 2.2) относятся к политопическим белкам.
Белки в бислое достаточно лабильны и могут совершать различные
виды движений; их подвижность и возможность ассоциации друг с дру-
гом определяются липидным окружением. Некоторые мембранные
белки легко перемещаются вдоль бислоя. Например, фосфолипаза А,
связываясь с клеточной мембраной (с цитоплазматической стороны),
может латерально перемещаться по поверхности бислоя и гидролизо-
вывать несколько тысяч фосфолипидов до тех пор, пока не отделится от
мембраны. Латеральная диффузия интегральных белков в мембране
ограничена их большими размерами, взаимодействием с другими мемб-
ранными белками, а также элементами цитоскелета или внеклеточного
матрикса. Однако она также имеет вполне измеримую величину.
Интегральные белки иммобилизуют окружающие их липидные моле-
кулы в так называемый аннулярный слой, в котором эти молекулы об-
ладают ограниченной подвижностью — гидрофобные белок-липидные
взаимодействия имеют ту же природу, что и гидрофобные взаимодейст-
вия между липидами, и зависят в основном от площади контактов. Таким
образ* >м, внедрение интегральных белков в мембрану изменяет свойства,
упаковку и подвижность мембранных липидов. Количество связанных
липидов зависит от насыщенности мембраны белками. Периферические
белки оказывают менее значительное воздействие на состояние и под-
вижность ацильных цепей фосфолипидов.
Важный белковый компонент мембраны — гликопротеины —
белковые молекулы, содержащие углеводные цепочки (рис. 1.21). Пе-
риферические белки или домены интегральных белков, расположенные
на наружной поверхности мембраны, почти всегда гликозилированы.
Молекулы клеточной адгезии нейронов (NCAMs) принадлежат к ши-
роко распространенному семейству гликопротеинов клеточной поверх-
ности, внеклеточные домены которых структурно напоминают домены
иммуноглобулинов. Некоторые трансмембранные гликопротеины осу-
ществляют взаимодействие между цитоскелетом и внеклеточным мат-
риксом. Интегрины являются основными представителями трансмемб-
ранных рецепторов с этими свойствами.
82
Раздел 1 СТРУКТУРА НЕРВНОЙ ТКАНИ
Н2С—ОН
I
НС—ОН
СНз
N-Ацетил -
глкжозамин
Рис. 1.21. Углеводные компоненты гликопротеинов.
Молекула гликопротеина состоит из белкового остова, к которому присоединены це-
почки сахаров. Длина и состав цепочек могут варьировать. Заштрихованными фигу-
рами обозначены радикалы аминокислот
Неприлизин (НЕП, нейтральная эндопептидаза-24.11) представля-
ет собой мембраносвязанный гликозилированный белок с молекуляр-
ным весом 93 кДа. НЕП является цинкзависимой металлопептидазой,
он деградирует биологически активные пептиды, такие как опиоидные
пептиды, тахикинины, хемотактические пептиды, адренокортикотроп-
ный гормон (АКТГ), энкефалин и тем самым прерывает передачу сиг-
нала в пептидэргических синапсах. НЕП расщепляет многие нейро-
пептиды и, следовательно, может регулировать их уровень.
НЕП обнаружен не только в центральной нервной системе, но и во
многих периферических тканях. Благодаря своим свойствам НЕП при-
нимает участие в регуляции разнообразных физиологических процессов,
включая сердечно-сосудистую деятельность, воспалительные процессы,
миграцию и пролиферацию клеток, развитие опухолей. Важно отметить,
что НЕП способен разрушать Р-амилоидные пептиды. Даже частичное
снижение активности НЕП может приводить к накоплению токсическо-
го [3-амилоида и развитию болезни Альцгеймера (см. гл. 3.9).
Углеводные цепочки гликопротеинов имеют разнообразную струк-
туру. Роль гликопротеинов в жизни клетки многообразна. Некоторые
из них выполняют функцию ферментов, другие обладают гормональ-
ной активностью. Предполагают, что одна из функций процесса глико-
к.5. Мембранные белки
зилирования — обеспечить возможность транспорта через клеточную
мембрану белкам, синтезированным в аппарате Гольджи. Гликопро-
теины, расположенные на поверхности клеток, ответственны за такие
важные процессы, как взаимное распознавание клеток — с их по-
мощью устанавливают межклеточные контакты в многоклеточном ор-
ганизме. Подобные контакты необходимы для адгезии клеток.
1.5.2. Функции мембранных белков
В отличие от липидов, мембранные белки трудно классифици-
ровать по структуре, целесообразнее подразделять основные виды
мембранных белков исходя из их функций. Как правило, именно белки
ответственны за функциональную активность мембран. К таким бел-
кам относятся разнообразные ферменты, транспортные белки, рецеп-
торы, канальные белки, белки, образующие водные поры (аквапори-
ны), а также различные структурные и регуляторные белки, которые
обеспечивают многообразные функции клеточных мембран.
По биологической роли мембранные белки можно разделить на
четыре группы: 1)белки-ферменты, обладающие каталитиче-
ской активностью; 2)рецепторные белки, специфически связываю-
щие те или иные вещества; 3) структурные белки; 4) белки, обес-
печивающие межклеточные взаимодействия.
Белки-ферменты наиболее распространены среди всех мембранных
белков. В их число входят как интегральные (мембранные АТФазы, вы-
полняющие транспортную функцию), так и периферические (ацетил-
холинэстераза, кислая и щелочная фосфатазы, РНКаза) белки.
Транспортные белки (см. гл. 2.5) создают устойчивые потоки опреде-
ленных веществ и ионов через мембраны. Транспорт ионов приводит к
возникновению трансмембранного потенциала во всех клетках, а так-
же к его изменениям в нервной и мышечной клетках. Последнее явле-
ние лежит в основе таких важнейших свойств этих тканей, как возбу-
димость и проводимость.
Рецепторными белками называют белки, специфически связываю-
щие те или иные лиганды, участвующие в передаче сигналов от одних
клеток к другим (см. гл. 2.2). Такая передача осуществляется различны-
ми способами. Часто рецепторы входят в состав более сложных мемб-
ранных комплексов, содержащих белки-исполнители. Например, в нерв-
ных и нервно-мышечных синапсах сигнальной молекулой (медиатором)
является определенное низкомолекулярное вещество, а плазматическая
мембрана содержит специальные рецепторные белки, соединенные с
ионными каналами, изменяющие свои свойства при связывании рецеп-
84
Раздел! СТРУКТУРА НЕРВНОЙ ТКдНИ
тора с лигандами. Эта реакция обеспечивает проницаемость мембраны
для различных ионов (натрия, калия, кальция, хлора) и формирует воз-
буждающий потенциал. Некоторые рецепторы (например, никотиновый
холинорецептор) сами являются ионными каналами (за счет включения
в рецепторный ансамбль дополнительных белковых субъединиц).
В ряде случаев рецепторный белок не является ионным каналом,
но связан с внутриклеточным сигнальным каскадом, активация кото-
рого происходит при связывании с рецептором лиганда, несущего ин-
формацию. В результате активации таких рецепторов (их называют
метаботропными, в противоположность ионотропным, регули-
рующим ионные потоки через мембрану) возникает каскад химических
реакций, управляющих клеточными функциями через изменения ме-
таболизма (отсюда и название этих рецепторов). Активация метабо-
тропных рецепторов лигандами (их можно считать первичными сиг-
нальными молекулами, или первичными мессенджерами) приводит к
выработке в цитоплазме активируемой клетки вторичных сигнальных
молекул (вторичных мессенджеров).
Структурные белки (см. гл. 1.7) придают клеткам и органеллам
определенную форму; обеспечивают те или иные механические свой-
ства (например, эластичность) плазматической мембране; осуществ-
ляют связь мембраны с цитоскелетом, а в случае ядерной мембраны —
с хромосомами.
Структурные мембранные белки, как правило, лишены фермента-
тивных свойств (возможно они просто пока мало изучены в химиче-
ском отношении). Их исследование затрудняется главным образом
двумя обстоятельствами. Во-первых, структурные белки «немы» —
не обладают известной ферментативной активностью. Во-вторых,
структурные белки имеют в составе своих молекул обширные гидро-
фобные участки. При очистке они образуют тесные ассоциаты друг
с другом или с липидами, что усложняет их изучение.
Нейроспецифический белок В-50 — один из основных фосфори-
лируемых структурных белков плазматических мембран синаптиче-
ских контактов. Методами иммунохимии установлено, что он локали-
зован преимущественно в пресинаптических мембранах. Молекуляр-
ная масса белка 48 кДа. Он является эндогенным субстратом
(диацилглицерол + Са2+)-зависимой протеинкиназы С. Активаторы
протеинкиназы С стимулируют процесс синаптической передачи в
срезах гиппокампа. Фосфорилирование белка В-50 приводит к увели-
чению времени возбужденного состояния синапса, что способствует
удержанию ионных каналов в активированном (открытом) состоянии
(в некоторых публикациях этот феномен называют состоянием «про-
1.5. Мембранные белки
85
торенности» синапса). Влияние фосфорилированного белка В-50 на
метаболизм фосфоинозитидов может быть одной из причин этого фе-
номена. Интересно, что в процессе старения организма интенсивность
фосфорилирования белка В-50 в мозге снижается, что, возможно,
и обусловливает снижение пластичности синапсов.
Еще одно доказательство роли процессов фосфорилирования бел-
ка В-50 в функционировании синапсов (на примере пептидергических
синапсов) получено в экспериментах in vitro, подтвердивших, что ней-
ропептид — фрагмент АКТГ1_24 — в 10 раз более эффективно тор-
мозит фосфорилирование В-50 в синаптических мембранах из сеп-
тальной области мозга, чем в мембранах целого мозга.
В группу мембранных белков также входят множество белков-
ферментов, образующих ионные каналы, — Na/K- и Са-АТФазы (см.
гл. 2.5), рецепторные белки (см. гл. 2.2), синапсины (см. гл. 3.4) и др.
Плазмин — сериновая протеиназа, в плазме крови действует в ос-
новном как тромболитический фермент, а также деградирует многие
компоненты внеклеточного матрикса. В мозге плазмин вовлекается в
осуществление многочисленных функций, таких как нейрональная
пластичность, обучение и память. Активация плазминогеновой систе-
мы наблюдается в мозге во время и в первые дни после инсульта. При
болезни Альцгеймера, напротив, происходит снижение уровня плаз-
мина в тканях мозга.
Входящий в состав плазматических мембран нейронов плазмин на-
ходится в ассоциации с богатыми холестерином рафтами, которые
считаются местом преимущественного образования амилоидного (3-А.
Это свидетельствует о наличии функциональной связи между плазми-
ном, холестерином и метаболизмом мозга.
Эндотелинконвертирующий фермент (endothelin-converting enzyme,
ЕСЕ-1) является еще одним амилоиддеградирующим ферментом, ко-
торый на 37% гомологичен НЕП по аминокислотной последователь-
ности. ЕСЕ-1 тоже является мембраносвязанной цинкзависимой ме-
таллопротеазой, также как и НЕП он способен расщеплять большое
число биологически активных веществ, включая брадикинин, нейро-
тензин, ангиотензин-1 и Б-цепь инсулина. В отличие от НЕП, ЕСЕ-1
существует в виде димеров, субъединицы которых соединены дисуль-
фидной связью.
Эти две металлопротеазы (ЕСЕ-1 и НЕП) различаются и по чувст-
вительности к ингибиторам. Для ингибирования НЕП требуются на-
Номолярные концентрации тиорфана и фосфорамидона, в то время
Как ЕСЕ-1 ингибируется микромолярными концентрациями фосфора-
мидона и не чувствителен к тиорфану.
86
Раздели — 'ЕР ОИТКА
1.6.
ЕСЕ-1 обнаружен во многих органах и тканях. Наиболее обогаще-
ны этим ферментом эндотелиальные клетки, он также экспрессирует-
ся в нервной ткани и мышцах. В гладкомышечных клетках ЕСЕ-1 на-
ходится в комплексе с а-актиновыми филаментами.
В настоящее время известны четыре изоформы ЕСЕ-1 че-
ловека (la, 1Ь, 1с и Id), которые не имеют существенных каталитиче-
ских отличий, но различаются по внутриклеточной локализации. Изо-
формы 1а, 1с и Id находятся на поверхности клетки, а ЕСЕ-16 явля-
ется внутриклеточной формой, локализованной в аппарате Гольджи.
Обнаружен еще один белок, подобный ЕСЕ-1, который локализо-
ван преимущественно в мозге и в незначительных количествах —
в эндотелиальных и гладкомышечных клетках. Его первичная структу-
ра на 59% идентична аминокислотной последовательности ЕСЕ-1. Он
обозначается как ЕСЕ-2 и отличается от ЕСЕ-1 более кислым рН-оп-
тимумом.
БЕЛОК-ЛИПИДНЫЕ ВЗАИМОДЕЙСТВИЯ
Анализ белок-липидных взаимодействий в мембране позволяет
выделить четыре основных типа контактов (рис. 1.22).
Наиболее распространенный вариант взаимодействия — это когда
внедряющийся в бислой белок вызывает локальное возрастание упо-
рядоченности части липидной массы и формирование аннулярного
слоя (рис. 1.22, а). С помощью метода ЭПР-спектроскопии показа-
но ограничение подвижности и характера упаковки углеводородных
цепей липидов под влиянием различных интегральных и перифери-
ческих белков (цитохром с-оксидаза, цитохром с, полилизин, родоп-
син и др.) вплоть до четвертого слоя липидов, окружающих молекулу
белка.
Второй этап взаимодействия — это прикрепление периферических
белков (не проникающих на всю глубину бислоя) к мембране, вызываю-
щее эластическую деформацию одной стороны бислоя (рис. 1.22,6).
Такое влияние белков на физико-химические параметры бислоя так-
же имеет определенные последствия. Именно им определяется облег-
чение взаимодействия мембранных рецепторов с инсулином и другими
гормонами.
Третий тип соответствует ситуации, когда в результате ярко выра-
женной гидрофобности белка могут иметь место как локальная дефор-
мация, так и резкое изменение градиента кривизны бислоя, что проис-
ходит в случае взаимодействия с мембраной цитохрома 65.
1.6. Белок-липидные взаимодействия
8
Полярный липид
Рис. 1.22. Различные типы модификации липидного бислоя белковыми моле-
кулами:
а — схематическое изображение белок-липидного бислоя; б — возможные механиз-
мы модификации бислоя белком: 1 — сближение заряженной молекулы белка с мемб-
раной; 2 — ассоциация белка с бислоем, индуцирующая градиент кривизны наружного
монослоя и способствующая ассоциации белка с внутренней стороной бислоя (3)
(по: Твердислов с соавт., 1987)
ьв
раздел 1. СТРУКТУРА T-JEPBHOii ТКАНИ
Наконец, сочетание гидрофильных и гидрофобных свойств белко-
вой молекулы может обеспечить не только проникновение белка че-
рез бислой, но и существенное давление на него, приводящее к изме-
нению упаковки и толщины бислоя — сжиманию одних частей и рас-
ширению других (рис. 1.22, а).
В основе белок-липидных взаимодействий лежат межмолекулярные
дисперсионные, электростатические и гидрофобные силы. Определен-
ную роль играют в этом и водородные связи. Гидрофобные силы являют-
ся доминирующими в связывании мембранных компонентов. Некоторые
мембранные белки увеличивают свою гидрофобность с помощью кова-
лентного присоединения липидных компонентов, чаще всего в этом ка-
честве выступают миристиновая С14.0- или пальмитиновая С16.0-жирные
кислоты или гликозилфосфатидилинозитол. Белки, связанные с жирными
кислотами, локализованы в основном на цитоплазматической поверхно-
сти плазматической мембраны, а белки, связанные с гликозилфосфа-
тидилинозитолом, — на наружной ее стороне.
В структуре многих мембранных белков, как правило, четко выяв-
ляются участки, ответственные за их биологическую активность.
Очень часто биологически активный участок состоит преимуществен-
но из полярных аминокислот, тогда как структурные домены постро-
ены главным образом с участием аминокислот с неполярными радика-
лами. Поэтому можно предположить, что полярная часть мембранно-
го белка контактирует с цитоплазмой и с полярными «головками»
липидов и обеспечивает функциональную активность белка, а непо-
лярная часть контактирует с углеводородными цепями липидных мо-
лекул и обеспечивает структурную устойчивость молекулы. Следова-
тельно, мембранные белки обычно взаимодействуют с мембраной с
помощью нековалентных взаимодействий двух ти-
пов: электростатических (на уровне полярных «головок» фосфо-
липидов) и гидрофобных (в толще бислоя).
Анализ аминокислотного состава некоторых мембранных белков
показал, что они содержат примерно столько же полярных аминокис-
лот, сколько и обычные водорастворимые белки. Причина их гидро-
фобности кроется не в аминокислотном составе, а в порядке чередова-
ния аминокислотных остатков — гидрофобные аминокислотные ради-
калы сконцентрированы в гидрофобные домены. Эти белки упакованы
в мембранный бислой таким образом, что их неполярные участки кон-
тактируют с гидрофобными участками бислоя.
Белки, функционирующие в пределах бислоя, обладают способно-
стью формировать в мембране межбелковые функциональные струк-
туры — олигомерные ассоциаты. Белок-белковые взаимодействия
1.6. Белок-липидные взаимодействия
проявляются в виде обратимой внутримембранной агрегации мемб-
ранных белков, часто сопровождающейся изменением функциональ-
ной активности системы. Циклы агрегация-дезагрегация белков в
клеточных мембранах — широко распространенное явление, прояв-
ляющееся при пиноцитозе, на ряде стадий клеточного цикла, при
взаимодействии и слиянии мембран и т. д.
Активность некоторых мембранных ферментов зависит от присутствия
определенных липидов. Например, для достижения максимальной актив-
ности Na/K-АТФазы требуются фосфатидилсерин, фосфатидилглицерол.
Эти ферменты проявляют специфичность по отношению к полярным «го-
ловкам» липидов, но их активность мало зависит от жирнокислотного со-
става. В противоположность этому функциональная активность, напри-
мер, родопсина коррелирует с длиной углеводородных цепей липидов.
Холестерин играет роль регулятора микровязкости липидной части
мембраны, изменяя подвижность жирнокислотных цепей фосфолипи-
дов, необходимых для нормального функционирования мембранных
компонентов. Содержание холестерина в клеточной мембране, вызы-
вая изменение микровязкости, влияет на конформацию мембранных
белков, таких как субъединицы рецепторов и ионных каналов, тем
самым регулируя их активность.
Примером модификации мембран липидного бислоя белками-фер-
ментами является действие фосфолипаз, которые играют значитель-
ную роль в функционировании мембранных структур мозга. Активация
фосфолипазы А2 постулирована как один из ключевых звеньев транс-
мембранной передачи химических сигналов. Фосфолипаза С принима-
ет участие в метаболизме фосфолипидов, который сопряжен с регуля-
цией транспорта ионов кальция внутрь нервной клетки. Фосфолипа-
за D участвует в метаболизме церамидов — важных составных компо-
нентов липидов мозга.
Активация фосфолипаз в определенных условиях физиологически
обусловлена. Функционирование этих ферментов обеспечивает не толь-
ко протекание необходимых метаболических процессов, но и одновре-
менную модификацию липидного бислоя мембран нейронов и глиальных
клеток. Обработка фосфолипазой приводит к изменению параметров
процесса возбуждения — длительности возбуждающего потенциала и
величины потенциала покоя. Воздействие фосфолипаз на мембраны си-
наптических окончаний мозга приводит к их деполяризации.
Функционирование биологических мембран реализуется за счет
поддержания на определенном уровне ряда их физико-химических ха-
рактеристик, что требует наличия биохимических механизмов стаби-
лизации биомембран.
90
раздел 1. Г’* ю • - • нг» t jPITKAHi
ЦИТОСКЕЛЕТ И ГЛИКОКАЛИКС
1.7.1. Белки цитоскелета
Структурные белки мембраны связаны со стороны цитоплазмы
с примембранными белками, создающими белковые компоненты ци-
тоскелета. Структура цитоскелета довольно лабильна, его перестрой-
ки происходят постоянно и с большой скоростью. Изменчива и связь
цитоскелета с мембранными белками.
Ориентация белков цитоскелета, а также относительное располо-
жение индивидуальных белков этой структуры достаточно хорошо ис-
следованы на примере эритроцитов. Установлено, что основные прин-
ципы организации цитоскелета являются общими для многих типов
клеток. В состав цитоскелета входят 12 различных белков с молеку-
лярной массой от 25 до 250 кДа (рис. 1.23). Функции ряда белков ус-
тановлены. Эти белки придают мембране устойчивость к действию
внешних факторов без утраты определенной подвижности ее компо-
нентов, необходимой для обеспечения транспортных функций мемб-
ран. Одновременно через белок анкирин (белок полосы 3) интеграль-
ные белки мембран связываются с молекулами спектрина — основ-
ного белка цитоскелета, образующего двумерную сеть, к которой
прикрепляется актин. Актин цитоскелета не существует в виде отдель-
а
Рис. 1.23. Цитоскелет клетки:
а — схема расположения белков в цитоскелете эритроцитов: / — спектрин; 2 — ан-
кирин; 3 — белок полосы 3,4 — белок полосы 4.1; 5 — белок полосы 4.9; 6 — олигомер
актина; 7 — белок полосы 6; 8 - гликофорин; 9 — липидный бислой; б — электрофо-
реграмма белков цитоскелета: 1, 2 — спектрины; 2.1 — анкирин; 5 — актин; 6 — не-
идентифицированный белок; 7 — тропомиозин; 8, 9, 10 — гликофорины; остальные
белки обозначены цифрами, соответствующими их положению на электрофореграмме
1,7. Цитоскелет и гликокаликс
9:
1
I
них глобул, а образует микрофиламенты — сократительный аппарат
цитоскелета.
С промежуточными филаментами цитоскелета связана также на-
ружная ядерная мембрана. Они фиксируют положение ядра в объеме
цитоплазмы. Митохондрии перемещаются в клетке также при участии
элементов цитоскелета. К цитоскелету примыкают микротрубоч-
ки (нейротубулы), образуемые при полимеризации глобулярного
белка тубулина.
Основная роль микротрубочек заключается в обеспечении при-
мембранного транспорта веществ, секреции, эндоцитоза. Микротру-
бочки и микрофиламенты цитоскелета обеспечивают противодействие
клетки изменению ее объема и придают эластичность мембране.
Цитоскелет придает мембране дополнительную прочность. Однако
роль структурных белков не ограничивается лишь функцией остова, на
котором крепятся липиды и ферменты. Известно, например, что струк-
турные белки взаимодействуют со многими низкомолекулярными со-
единениями, в том числе с АТФ, и могут влиять на работу мембранных
ферментов. Достаточно заменить одну аминокислоту в составе струк-
турного белка митохондрий, для того чтобы нарушить многоэтапный
процесс сборки ферментов митохондриальной дыхательной цепи. Бла-
годаря динамическим свойствам цитоскелета изменения в его структу-
ре позволяют клетке проявлять гибко-эластичные свойства, изменять
свою форму. В той области клетки, где спектриновая сеть цитоскелета
разрушается, могут образовываться впячивания или возникать отшну-
ровывающиеся части клетки.
Специфические структурные белки нервной ткани обеспечивают ее
общую динамичность и механическую подвижность синаптических кон-
тактов нейрональной клетки, участвуя в самосборке и распаде специ-
фических структур — нейротубул, нейрофиламентов и других преси-
наптических и постсинаптических образований, в переносе различных
соединений между разными областями нейрона, а также в поддержании
и модуляции пространственного положения частей нейрона.
Механические функции клеток требуют взаимодействий между ин-
тегральными мембранными белками и цитоскелетом. Эти функции
включают эндо- и экзоцитоз, формирование объединений клеток и ре-
гулирование их формы.
Спектрин-анкириновая сеть. В большинстве клеток основная
структурная связь плазматических белков и цитоскелета опосредова-
на взаимодействиями между анкирином и различными интегральными
Мембранными белками, ионными насосами и ионными каналами. Ан-
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
кирин соединяется с гетеродимером спектрина и таким образом обес-
печивает прикрепление цитоскелета к плазматической мембране.
Длинная фибриллярная молекула спектрина состоит из двух полипеп-
тидных цепей (220 и 240 кДа). Димеры спектрина формируют сеть,
располагающуюся параллельно плазматической мембране. Нейро-
ны содержат две изоформы спектрина — спектрин I (эритрин) и
спектрин 11 (фодрин). Спектрин II обнаружен во всех частях нейро-
нов, включая аксоны и синаптические окончания, тогда как спектрин I
встречается только в теле нейрональной клетки и в дендритах. Обра-
зуемая ими спектриновая сеть связывается с актиновыми нейрофила-
ментами. Эти ассоциаты, возможно, являются динамическими обра-
зованиями, но, может быть, они просто снижают подвижность белков
в плоскости мембран. Например, фосфорилирование анкирина может
изменить его сродство к спектрину.
Фодрин — нейроспецифический белок, который принимает учас-
тие в процессах синаптической передачи. Это структурный белок
постсинаптических мембран глутаматергических синапсов. Его моле-
кулярная масса составляет 230 кДа. Фодрин проявляет способность
блокировать активность рецепторов глутамата. Повышение концент-
рации ионов Са2+ вблизи постсинаптической мембраны активирует
мембранную Са-зависимую протеиназу кальпаин, расщепляющую
фодрин. Это возвращает активность глутаматным рецепторам, ранее
экранированным фодрином; повышение проводимости синапсов при
этом наблюдается в течение 3—6 суток.
Внутриклеточный мембранный транспорт — важный процесс для
эукариотических, в том числе нервных, клеток. Внутриклеточные
мембраны организованы в компартменты, и транспорт веществ между
компартментами требует высокой специфичности и регулирования.
Межкомпартментный транспорт обычно осуществляется с помощью
везикул, образующихся в компартменте, являющемся донором, кото-
рые затем транспортируются и присоединяются к акцепторному ком-
партменту. Белок клатрин впервые был выделен из мембран так
называемых окаймленных пузырьков, принимающих участие в эндо-
цитозе и быстром внутриклеточном транспорте веществ. Этот фиб-
риллярный белок (180 кДа) образует характерный многоячеистый чехол
из пяти- и шестиугольных пептидных цепей на поверхности окаймленных
пузырьков. Данная сеть формируется в результате нековалентных свя-
зей между «трехлопастными» молекулами клатрина, каждая из кото-
рых состоит из трех тяжелых и трех легких полипептидных цепей двух
типов. Легкие цепи клатрина активно связывают Са2+ в молярном
соотношении 1:1.
11.7. Цитоскелет и гликокаликс
Помимо клатрина в состав такого чехла входит так называемый
«белок сборки чехла» (50 кДа), который может подвергаться обрати-
мому фосфорилированию для регулирования процесса эндоцитоза.
При полимеризации клатрина на цитоплазматической стороне плаз-
матической мембраны в результате ее впячивания образуются пу-
зырьки с заключенными в них мембранными рецепторами. Везикулы,
одетые «клатриновым чехлом», также вовлекаются в экспорт белка.
Функция клатрина состоит в обеспечении некоторых форм эндоци-
тоза, в особенности процесса так называемой интернализации —
«втягивания» в цитоплазму клетки больших групп пептидных рецеп-
торов, связавшихся со своим лигандом. В последнее время эта форма
транспорта веществ в цитоплазму нервной клетки привлекает все
большее внимание. В данном процессе Rab-белки (являющиеся по
своей природе ГТФазами) играют центральную роль, регулируя обра-
зование везикул, их подвижность и слияние. Они принадлежат к се-
мейству G-белков. Большое количество этих белков было идентифи-
цировано в клетках млекопитающих; каждый из них, как оказалось,
регулирует определенный шаг в цепи молекулярных событий, связан-
ных с процессами образования, транспорта и слияния везикул.
НаЬ5-ГТФаза — регулятор ранних этапов эндоцитоза в клетке.
Этот белок играет важную роль в формировании везикул «клатрино-
вого чехла» в плазматической мембране, в их слиянии с первичными
эндосомами, а также в их слиянии между собой; он также важен для
нейрофиламентзависимой подвижности эндоцитозных везикул.
К актомиозиноподобным белкам ЦНС относится нейростенин. Он
состоит из двух белков — нейрина и стенина. Взаимодействуя меж-
ду собой, они образуют комплекс — нейростенин (47—50кДа). По
структуре и функциям он имеет много общего с мышечным актоми-
озином, хотя и не идентичен ему.
Нейростенин обладает АТФазной активностью и активируется
ионами Са2+ и Mg2+. Количество нейростенина в мозге составляет
около 1 —1,5% от общего белка; в синаптических образованиях его
содержание достигает 8—10%. Нейрин локализован преимуществен-
но в пресинаптических мембранах, а стенин — на наружной поверх-
ности мембран везикул. С образованием нейростенина в присутствии
АТФ и ионов Са2+ связывают контакт везикул с пресинаптическими
мембранами. Полагают, что сократительные белки мозга, в том числе
нейростенин, участвуют в раскрытии везикул и выходе нейромедиато-
ра в цитоплазму и синаптическую щель. В локальном изменении
микровязкости мембраны в области слияния с ней везикул (синапти-
ческих пузырьков или эндосом), называемом иногда «локальным
*4
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
плавлением», важную роль играют также синапсины и другие
Са-связывающие белки.
Некоторые периферические белки мембраны могут также рас-
сматриваться как белки цитоскелета — они ковалентно заякоривают-
ся к внешней поверхности мембраны через гликозилированные мемб-
ранные фосфоинозитиды. Примером таких белков являются щелочная
фосфатаза, 5'-нуклеотидаза, одна из форм ацетилхолинэстеразы и
гликопротеин NCAM. Белки цитоскелета спектрин и аннексины (се-
мейство белков цитоскелета, которые образуют Са-зависимые ассо-
циаты с мембранами клетки в области локализации фосфатидил сери-
на) также специфически концентрируются у клеточной мембраны в
местах локализации фосфатидил инозита и фосфатидилсерина, соот-
ветственно. Некоторые белки цитоскелета становятся мембраносвя-
занными в ответ на активацию кальцием. Это протеинкиназа С, фос-
фолипаза А2 и синаптотагмин.
Часто связывание с мембраной вызывает аллостерическое измене-
ние гидрофобности белков и модифицирует их свойства. Один из са-
мых известных случаев такого рода — Са-зависимое связывание
кальмодулина с другими белками.
1.7.2. Гликокаликс
Экстраклеточный матрикс, или так называемый гликокаликс,
влияет на многие макромолекулярные процессы, протекающие с учас-
тием клеточной мембраны: ионный обмен, проницаемость, эндо- и эк-
зоцитоз, межклеточные контакты, морфогенетическую и тканеспеци-
фическую агрегацию клеток.
Гликокаликс представляет собой внешний по отношению к клеточ-
ной мембране слой. Он состоит из гликопротеинов, протеогликанов и
глюкозаминогликанов и связывается с мембранными структурами с по-
мощью специальных белков, объединяя цитоскелет, мембрану и вне-
клеточный матрикс в подвижную структуру (рис. 1.24). Как показали
электронно-микроскопические исследования, гликокаликс имеет вид
о
рыхлого волокнистого слоя толщиной 30—49 А, покрывающего всю
поверхность клетки.
В состав гликокаликса входят полисахаридные цепочки мембранных
интегральных белков — гликопротеидов. Они содержат такие углево-
ды, как манноза, глюкоза, сиаловая кислота и др. Углеводные гетеро-
полимеры гликокаликса образуют ветвящиеся цепочки, между которы-
ми располагаются свободные гликолипиды и протеогликаны. Слой гл и-
L.7. Цитоскелет и гликокаликс
9£
’ис. 1.24. Взаимодействие цитоскелета с гликокаликсом:
— протеогликан; 2 — коллаген; 3 — фибронектин; 4 — молекулы актина; 5 — ин-
егральные белки мембраны, подобные белку полосы 3
юкаликса сильно обводнен, имеет желеобразную консистенцию, что
начительно снижает диффузию различных веществ из клетки. Здесь
ке могут накапливаться выделенные клеткой гидролитические фер-
менты, участвующие во внеклеточном расщеплении полимеров до мо-
юмерных молекул, которые затем транспортируются в клетку.
Гликокаликс содержит внешние компоненты рецепторов гормо-
юв, нейромедиаторов, ростовых факторов, нейропептидов, антиге-
юв. Он представляет собой комплексную динамичную систему, тесно
ссоциированную с мембраной: изменения свойств отдельных компо-
нентов мембраны приводит к глубоким изменениям матрикса.
Специфичность гликокаликса определяется первичной структурой
нлигосахаридной части гликопротеинов, гликолипидов, гликозамино-
гликанов; их конформацией и положением относительно плоскости
мембраны; площадью, занимаемой гликопротеинами и гликолипидами.
96
Раздел ]. СТРУКТУРА НЕРВНОЙ ТКАН.
Состав, структура и динамическая подвижность гликозилирован-
ной «шубы» гликокаликса влияют на функции деления, роста, диффе-
ренциацию и гибель клетки. Особая роль в этом принадлежит глико-
липидам и главным образом кислым гликолипидам — ганглиозидам.
Общая характеристика ганглиозидов. Ганглиозиды— спе-
цифические липиды гликокаликса нейрональных мембран. Кислые гли-
колипиды — ганглиозиды находятся в нервной ткани в высоких кон-
центрациях (табл. 1.12) и обнаруживаются преимущественно в возбу-
димых мембранах.
Ганглиозиды — общее название гликосфинголипидов, в составе
которых присутствует сиаловая кислота. Они содержат гидрофобную
церамидную часть и гидрофильную, богатую заряженными группами
олигосахаридную часть. На рисунке 1.25 приводится структура моно-
сиалоганглиозида головного мозга. Гидрофобная часть ганглиозидов
включает две длинные углеводородные цепочки — углеводородную
цепочку сфингозина и жирную кислоту, которая связана с аминогруп-
пой сфингозина пептидной связью. Обращает на себя внимание почти
полное отсутствие в ганглиозидах мозга гидроксикислот, кетокислот и
разветвленных жирных кислот. Состав сфингозиновых оснований
ганглиозидов головного мозга не отличается большим разнообразием.
Таблица 1.12. Содержание ганглиозидов в тканях человека,
нмоль липидосвязанной сиаловой кислоты
на 1 г свежей ткани (по: Ledeen, 1983)
Ткань Концентрация
Нервная ткань
Серое вещество головного мозга 2850—3530
Белое вещество головного мозга 900—1570
Серое вещество спинного мозга 751
Белое вещество спинного мозга 450
Сетчатка 366
Седалищный нерв 259
Надпочечники 407—757
Печень 214
Мышцы 52
Плазма и,з
Спинномозговая жидкость 0,841
^,7. Цитоскелет и гликокаликс
97
NH ОН
О=<
СН2ОН СН2ОН СН2ОН СН2ОН [
ОН NHCOCH3 он он
CH3COHN
соон
неон
сон
СН2ОН
ОН
Рис. 1.25. Структура моносиалоганглиозида
В ганглиозидах встречаются главным образом четыре основания:
сфингозин (4-сфингенин) и дигидросфингозин (сфинганин), содержа-
щие 18 атомов углерода, а также С20-сфингозин (4-эйкозасфенгенин)
и С20-дигидросфингозин (эйкозасфинганин).
Таким образом, углеводородная часть ганглиозидов мозга млеко-
питающих достаточно консервативна относительно длины, числа и
места двойных связей, а также присутствия метильных групп.
Разветвленная олигосахаридная часть присоединена P-связью к
ОН-группе первого атома углерода сфингозина. Большинство гангли-
озидов мозга имеют общую нейтральную углеводную часть, содержа-
щую первой в последовательности молекулу глюкозы, затем молекулу
галактозы (интернальная галактоза), ацетилированный галактозамин,
вторую молекулу галактозы (терминальная галактоза) и различное
число молекул N-ацилнейраминовой кислоты (NANA), прикреплен-
ных либо к интернальной, либо к терминальной галактозе.
Олигосахаридная часть является доминирующей в проявлении фи-
зических, химических и иммунологических свойств молекул ганглиози-
дов. Различия в строении олигосахаридной части порождают высокую
гетерогенность этих соединений. К настоящему времени в нервной тка-
ни м. юкопитающих выявлено свыше 50 различных ганглиозидов, при-
чем число их продолжает увеличиваться. Недавно описаны гекса- и
Декасиалоганглиозиды, имеющие соответственно шесть или десять
сиаловых кислот на церамидный остаток.
В силу большей, чем у фосфолипидов, гидрофобности ганглиозиды
Повышают жесткость липидного бислоя и тесно взаимодействуют с фос-
фолипидами и интегральными белками мембраны.
Увеличение числа углеводородных атомов и ненасыщенности
сфингозина, изменение природы жирной кислоты ганглиозидов вызы-
98
раздел 1. СТРуКТЧ’рд НЕРВНОЙ ТКАНИ
вают конформационные изменения в близлежащих белках. Церамид-
ная часть молекулы участвует в выборе определенного состава фос-
фолипидов, белков и холестерина в окружении индивидуальных ганг-
лиозидов.
Организация ганглиозидов в мембранах головного мозга.
Ганглиозиды обнаружены фактически в каждом типе клеток и
большинстве субклеточных образований ЦНС. Ганглиозиды локализо-
ваны главным образом в синаптических мембранах (которые составля-
ют примерно 6% сухой массы мозга), причем обнаружена корреляция
между накоплением ганглиозидов и интенсивностью синаптогенеза
(образования синапсов, ветвления аксонов) во время формирования
мозга. Использование специальных методов показало, что ганглиозиды
локализованы на наружной стороне пре- и постсинаптических терми-
налей, принимающих непосредственное участие в передаче нервного
импульса.
Молекулы ганглиозидов в мембране высокоподвижны, что, с одной
стороны, создает локальную неустойчивость мембраны, а с другой —
поддерживает ее целостность. Молекулы ганглиозидов не подвержены
«флип-флопу», но способны к латеральной диффузии с изменяющейся
в широких пределах скоростью.
Несмотря на большую подвижность, ганглиозиды не вносят ха-
отичности в распределение компонентов мембраны. Это достигается,
во-первых, образованием ковалентных связей между олигосахарид-
ными цепочками гликопротеинов и гликолипидов, образующих в
мембране устойчивые комплексы. Во-вторых, гликолипиды и глико-
протеины могут соединяться с периферическими гликозаминоглика-
нами, которые, как правило, свободно диффундируют в бислой и обра-
зуют с гликолипидами и гликопротеинами ионные и водородные связи,
создавая своеобразный латекс (рис. 1.26). В-третьих, ограничение ла-
терального движения гликолипидов достигается сосредоточением их в
определенных областях с повышенной вязкостью. В-четвертых, то-
пографию поверхности мембраны стабилизирует цитоскелет клетки.
Различные поливалентные лиганды гликопротеиновой природы
вовлекают цитоскелет в перераспределение мембранных гликолипи-
дов в определенные структурные ассоциаты. Степень их агрегации за-
висит от взаимодействия олигосахаридных структур с лектинами.
Как правило, плотные массы олигосахаридных цепочек гликопро-
теинов служат центром, вокруг которого увеличивается упаковка
ганглиозидов. Нековалентное кооперативное взаимодействие ганглио-
зидов приводит к тому, что в участках их скопления резко возрастает
г 7. Цитоскелет и гликокаликс
99
Гликозилированная
часть белка
..зжжжжжжж
Белковая молекула
Рис. 1.26. Схематическое изображение упаковки молекулы гликолипида
в мембране.
Гликозильные радикалы белка включаются в состав гликокаликса, белковая молекула
погружена в бислой
величина отношения ганглиозидов к фосфолипидам. В результате
микровязкость данных локусов мембраны увеличивается, ганглиозид-
ные кластеры из-за взаимного отталкивания отрицательно заряжен-
ных сиаловых кислот становятся нестабильными и мембранный по-
тенциал в этом локусе локально возрастает.
Участки, занятые заряженными молекулами ганглиозидов, имеют
повышенное сродство к экзогенным гидрофильным лигандам, а облас-
ти, свободные от ганглиозидов, осуществляют гидрофобное взаимо-
действие с лигандами. Взаимодействия такого рода вызывают разно-
образные структурные перестройки в мембране, которые оказывают
влияния на состояние клетки.
Участие ганглиозидов в дифференциации клеток. Ганглиози-
ды — важные элементы клеток, которые выполняют разнообразные
биологические функции, включая клеточное узнавание, адгезию и пе-
редачу сигналов. Ганглиозиды могут оказывать влияние на такие фи-
зиологические процессы, как эмбриогенез и дифференциация нейро-
нальных клеток и лейкоцитов. Участвуя в дифференциации клеток,
ганглиозиды вызывают их морфологические изменения, проявляя
нейритогенный эффект. Структура и количество ганглиозидов меня-
ются в процессе развития клеток по мере формирования нейрон-ней-
рональных взаимодействий. Многочисленные исследования показали,
100
раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
что уровень ганглиозидов во время клеточной дифференциации замет-
но повышается, что хорошо коррелирует с изменением активности гли-
козилтрансфераз, ответственных за биосинтез этих липидов. Актив-
ность гликозилтрансфераз может регулироваться на нескольких уров-
нях, включая транскрипционный и посттрансляционный контроль.
Таким образом, уровень и пространственное распределение ганглиози-
дов в нервной системе регулируются биосинтезом этих специфических
липидов в процессе развития организма.
Ганлиозиды участвуют в межклеточных взаимодействиях, регули-
руют активность рецепторных тирозинкиназ в плазматической мемб-
ране, также как и рецепторов эпидермального фактора роста, фактора
роста нерва, инсулина, и таким образом могут регулировать процессы
клеточной сигнализации. Воздействие ганглиозидов на рецепторы
фактора роста осуществляется в три этапа: вначале — модуляция за-
крепления лиганда ганглиозидами; далее — регулирование димериза-
ции рецептора и в завершение — вовлечение ганглиозидов в актива-
цию рецептора.
Ганглиозиды распределены по поверхности клетки неравномерно и
в комплексе с сфингомиелином и холестерином входят в состав специ-
фических рафтов, которые являются физиологической средой для раз-
мещения разнообразных рецепторных белков.
1.7.З. Белки гликокаликса
Значительная часть (до 90%) всех углеводов и их производных,
содержащихся в головном мозге в связанном виде, приходится на до-
лю гликопротеинов. Гликопротеины являются непосредственными
участниками межклеточных контактов, обеспечивая взаимное узнава-
ние определенных нейронов, они участвуют в синаптической передаче,
рецепторных реакциях, формировании и хранении памяти, входят в
состав сложных надмолекулярных образований синаптических мемб-
ран и других цитоструктурных элементов.
В этих белках углеводный компонент характеризуется более высо-
кой метаболической активностью по сравнению с пептидной частью
молекулы. Обращает на себя внимание тот факт, что гликопротеины,
содержащие гиалуроновую кислоту, хондроитинсульфат, гепаринсуль-
фат, сосредоточены в перикарионе нейрона, в аксоне и нейроглии
(астроцитах, олигодендроцитах), но отсутствуют в мембранах синап-
тических окончаний и митохондрий.
Ввиду большого разнообразия гликопротеинов до сих пор не раз-
работан единый принцип их классификации. Обычно гликопротеины
j.,7. цитоскелет и гликокаликс
101
делят на д в е основные группы по соотношению белков и
углеводов в составе их молекул. Первая группа гликопро-
теинов содержит от 5 до 40% углеводов и их производных. Белковая
часть этих гликопротеинов сходна с альбуминами и глобулинами.
Между пептидными и углеводными компонентами осуществляются не
только ковалентные, но и водородные, гидрофобные и вандерваальсо-
вы связи.
Вторая группа гликопротеинов содержит большее количество
углеводов — от 40 до 85%; в их состав иногда входят липидные ком-
поненты. В последнем случае образуются более сложные комплек-
сы — гликолипопротеины. В состав одного из гликолипопротеинов,
выделенных из серого вещества головного мозга человека (180 кДа),
входят 208 остатков галактозы, 26 остатков глюкозы, 36 — галакто-
замина, 150 — нейраминовой кислоты, 100 — лигноцериновой кис-
лоты, 100 — сфингозина. Пептидная часть состоит из 61 аминокис-
лотного остатка: 13 — глутамата, 10 — глицина, 10 — пролина, 8 —
серина, 6 — аланина; остальные аминокислоты содержатся в незна-
чительных количествах. Как видно, пептидная часть молекулы доволь-
но монотонна по составу.
Углеводный компонент гликопротеинов включает в первую оче-
редь N-ацетилнейраминовую кислоту и N-ацетилгалактозамин; эти
соединения играют специфическую роль, определяя, по-видимому,
своеобразие внешних участков пространственной структуры глико-
протеинов. Пептидная часть представляет собой стабильную основу
(каркас) молекулы, которая фиксирована непосредственно в мембра-
не, в то время как углеводный компонент расположен на ее поверхно-
сти. Прослеживается аналогия с молекулярной структурой ганглиози-
дов, у которых каркасом служит церамидная часть (жирнокислотный
эфир сфингозина), а углеводные компоненты и их производные (га-
лактозамин, N-ацетилнейраминовая кислота и др.) составляют наибо-
лее вариабельную и специфичную часть молекулы.
Первыми нейроспецифическими гликопротеинами, выделенными
из ткани мозга, были цитозольные (растворимые) гликопротеины,
однако в дальнейшем оказалось, что многие из них существуют
и в мембраносвязанной форме.
Особый интерес представляют поверхностные гликопротеины,
участвующие в клеточной адгезии. Довольно хорошо исследовано
шесть таких белков: D2 (139 кДа), NCAM, К4, BSP-2 (семейство бел-
ков 120—180 кДа), Ng-CAM и L-1. Первые четыре белка обеспечи-
вают гомотипическую адгезию между нейронами. Характерной осо-
102
Раздел 1. СТРукТУг АнEr t HLЯ ТнАНИ
бенностью является модификация их структуры в ходе онтогенеза,
которая затрагивает, в основном, углеводную часть молекулы.
И в эмбриональный период (во время интенсивной миграции нейро-
нов), и в течение постнатального развития (в стадии активного синап-
тогенеза) нейроспецифические белки клеточной адгезии представлены
в значительной мере полисиалогликопротеинами. В мозге взрослых
животных они модифицируются в олигосиало- или асиалогликопроте-
ины, состоящие из двух-трех полипептидных цепей. Предполагается,
что модуляция адгезии происходит именно благодаря изменению числа
остатков сиаловых кислот в полисиалогликрпротеинах.
Определенные трансмембранные гликопротеины могут способст-
вовать взаимодействию между цитоскелетом и внеклеточной матри-
цей, таковыми являются белки интегрины.
Кадгерины — Са-зависимые белки адгезии, которые в значитель-
ной степени ответственны за слипание клеток. Внутри клеток они ас-
социированы с актиновыми нейрофиламентами с помощью белков,
названных катенанами.
Молекулы адгезии нейрональных клеток (NCAM) принадлежат
широко распространенному семейству гликопротеинов клеточной по-
верхности, внеклеточные домены которых структурно подобны доме-
нам иммуноглобулинов (рис. 1.27). Два из них представляют собой
трансмембранные гликопротеиназы с идентичными внеклеточными
доменами и различными цитоплазматическими доменами. Третий не
относится к трансмембранным белкам и не участвует в процессах кле-
точной сигнализации, поскольку является внеклеточным гликопро-
теином, «заякоренным» в мембрану ковалентной связью с гликозил-
фосфатидилинозитолом.
По сравнению с белком NCAM, влияющим на межнейронные кон-
такты, белок Ng-CAM содержит меньше сиаловых кислот. Он лока-
лизован исключительно на поверхности плазматической мембраны
нейронов и появляется в онтогенезе позднее, чем гликопротеин
NCAM.
В постсинаптических уплотнениях и участках синаптических соеди-
нений обнаружен целый ряд других гликопротеинов: NSA-3, МВА-2,
Thy-I. Они могут служить субстратами для протеиназ и сиалидаз, под
действием которых происходят локальные модификации структуры
гликопротеинов в ответ на изменение функционального состояния си-
напса. Интересно, что аминокислотная последовательность гликопро-
теина Thy-1 обнаруживает гомологию с вариабельными доменами им-
муноглобулина. Роль этого гликопротеина на поверхности нейронов
остается невыясненной.
,Л. Цитоскелет и гликокаликс
DCC-R
Ll,Ng-CAM
COO- COO- COO- COO-
NHJ
UNC-5,
Netrin-R
NH3
FGR-R
NH3
Домены
тромбоспондина
I типа
coo- coo-
1 2 3 4 5 6 7 8
Eph А, В
: DPTP69D
NHJ
Cys
L
Cys
Домен П П
протеин- т
киназы
соо- соо-
Домены
тирозин-
фосфатазы
соо-
10 11
Рис. 1.27. Структура различных представителей суперсемейства иммуно-
глобулинов:
1—3 — белки только с иммуноглобулиновыми доменами во внеклеточном сегменте;
4—7 — белки с иммуноглобулиновыми и фибронектиновыми доменами во внеклеточ-
ном сегменте; 6 — белок с GPI-якорем; 8 — белок с иммуноглобулиновыми и тром-
боспондиновыми доменами во внеклеточном сегменте; 9 — белки-рецепторы с проте-
инкиназными доменами; 10 — белок-рецептор, содержащий области, обогащенные
цистеином; / / — белок-рецептор с протеинфосфатазными доменами. Члены надсе-
мейства иммуноглобулинов характеризуются многочисленными повторами дисульфид-
ных мостиков, гомологичными доменам, впервые описанным в постоянной части мо-
лекул иммуноглобулина. Многие из этих молекул клеточной адгезии также содержат
несколько доменов, сходных с повторами в фибронектине III типа (показано в виде
прямоугольников)
На плазматических мембранах мозга обнаружены также относи-
тельно низкомолекулярные гликопротеины, которые обладают спо-
собностью ингибировать клеточное деление и синтез белка в культуре
нормальных клеток мозга. По своей структуре это небольшие глико-
протеины, как содержащие фукозу (30 и 45 кДа), так и не содержащие
этот моносахарид (6—10 кДа).
Агрегация ганглиозидов и гликопротеинов на поверхности мембра-
ны важна для поддержания контактов между клетками, поскольку
конгломераты молекул обеспечивают более устойчивые контакты, чем
молекулы, дисперсно разбросанные по клеточной поверхности. Подоб-
ные агрегаты могут содержать различные рецепторы или несколько ко-
пий одного рецептора, составлять единый рецепторный комплекс из
гликолипидов и гликопротеинов. Зона мембраны, в которой происхо-
дит восприятие информации, передача ее в клетку, а также иницииру-
ется связь с клеточным ядром, представляет собой обширную систему
гетерогенных гликозилированных молекул. Эта область является свое-
104
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
образным «распределительным щитом» регуляторных сигналов, где
молекулы гликопротеинов и ганглиозидов могут играть роль триггеров
(регуляторов или трансдукторов), выполнять функции сигнальных мо-
лекул на стадии дифференцировки и участвовать в определении видо-
вой и тканевой специфичности.
Таким образом, можно заключить, что гликопротеины и гликоли-
пиды участвуют в осуществлении специфических функций нервной
ткани, в большей степени — в формировании контактов между раз-
личными нейронами.
1.8.
МЕМБРАННЫЕ МЕХАНИЗМЫ АДАПТАЦИИ
Исследования фазовых переходов в природных мембранах под-
водят к одному неожиданному заключению: при всем разнообразии
липидного состава клеточных мембран их свойства таковы, что усло-
вия проживания разнообразных объектов находятся вблизи темпера-
туры их фазовых переходов (или разделения фаз) (Шноль, 1978). Это
обстоятельство можно рассматривать в пользу утверждения о том, что
живые системы используют изменения упаковки мембранных липидов
Е. М. Крепе
Основоположник отечествен-
ной школы функциональной
нейрохимии, длительное вре-
мя возглавлявший исследова-
ния в этой области в качестве
директора Института эволю-
ционной физиологии и биохи-
мии им. Л. А. Орбели
как фактор контроля за функцией мемб-
ранных белков.
За счет чего происходит изменение
микровязкости липидов при изменении
условий среды? Ведь живые системы ис-
пытывают и смену температур (например,
холоднокровные животные при смене се-
зонов, гибернирующие в условиях зим-
ней спячки), и изменения солености и ос-
мотичности среды (проходные рыбы, ухо-
дящие для нереста из морей в пресные
реки), да и многие другие факторы ло-
кально воздействуют на упаковку липид-
ного бислоя, изменяя его микровязкость в
ходе клеточного цикла или экстремальных
воздействий.
Интересное наблюдение было сдела-
но Е. М. Крепсом (1981) при сопоставле-
нии липидного состава нервной ткани жи-
вых существ, намеренно далеко находя-
щихся друг от друга на эволюционной
лестнице (рис. 1.28). Оказалось, что со-
1.8. Мембранные механизмы а даптации
Та
Рис. 1.28. Фосфолипиды в эволюции животного мира (в % к суммарному фос-
фору фосфолипидов):
/ — фосфатидил инозит; 2 — фосфатидилхолин; 3 — сфингомиелин; 4 — фосфати-
дилсерин; 5 — фосфатидилэтаноламин; 6 — кардиолипин (по: Крепе, 1981)
отношение основных классов липидов в мозге разных видов, не объ-
единенных ни генетической, ни экологической общностью, порази-
тельно близко. Способностями к ориентации в пространстве, к обуче-
нию, абстрактному мышлению собака, бабочка и улитка разительно
отличаются друг от друга, но соотношение основных классов липидов
в их нервной ткани очень похоже. Действительно, надо признать, что
эволюция функций мозга осуществлялась не в результате совершен-
ствования липидного состава, а не затрагивая его. Уникальное соот-
ношение основных классов мембранных липидов было найдено приро-
дой на самой заре зарождения жизни и с тех пор сохранялось как неос-
поримое достижение эволюции.
А что же позволяло мозгу (а с ним и всему организму) наилучшим
образом приспосабливаться к изменяющимся условиям среды — при
смене сезонов, сред обитания, природных катаклизмах? Обширными
исследованиями Е. М. Крепса и его коллег (Институт эволюционной
физиологии и биохимии им. Л. А. Орбели в Ленинграде, теперь —
Санкт-Петербурге) было продемонстрировано, что тонкая адаптация
мембран к изменяющимся условиям жизни организмов осуществляет-
ся почти исключительно за счет изменений жирнокислотного состава
липидов. Следовательно, можно заключить, что клеточные мембраны
являются мишенью для адаптации.
106
Раздел 1.: Iгм; , г . г- г г «гнОЙ IКАНИ
Стимулом к развертыванию адаптационной программы могут слу-
жить сами факторы внешней среды (температура, гидростатическое
давление), хотя молекулярные механизмы активации соответствую-
щих ферментов пока не выяснены. По скорости осуществления меха-
низмы адаптации можно разделить на краткосрочные и долгосроч-
ные. Первые реализуются за счет изменения активности имеющихся
ферментов, осуществляющих обмен клеточных липидов (десатураз,
метилаз, ацилаз, фосфолипаз), вторые представляют собой экспрес-
сию адаптационных генов.
Определен ряд основных механизмов, используемых клеточными
мембранами для приспособления к увеличению уровня солености сре-
ды обитания, понижению температуры, возрастанию гидростатиче-
ского давления (важно для глубоководных и мигрирующих рыб) и т. д.
Во-первых, это активация мембранных десатураз и возрас-
тание ненасыщенности жирнокислотных цепей фосфолипидов
(главным образом кислых). Во-вторых, это активация фосфо-
липазы Л2 и замена в молекулах фосфолипидов коротких жирно-
кислотных цепей более длинными. В-третьих, это включение
в мембрану дополнительного количества холестерина. Все эти
процессы осуществляются достаточно быстро и обратимо, что обеспе-
чивает снижение микровязкости мембран. Перечисленные компенса-
торные механизмы относятся к так называемой фенотипической
адаптации и позволяют мембранным белкам осуществлять свою
функцию даже при тех изменениях среды, которые в обычных услови-
ях привели бы к резкому возрастанию микровязкости мембран и сде-
лали бы невозможным проявления жизнедеятельности.
Возрастные изменения липидного состава мембран также выража-
ются в изменении набора жирных кислот в молекулах липидов. Объ-
ективным показателем возрастных изменений в мозге крыс служит
значительное повышение доли ненасыщеных жирных кислот в составе
липидов. Так, в период от 10-годо 90-го дня постнатальной жизни ин-
декс ненасыщенности жирных кислот, входящих в состав фосфатидил-
холина и фосфатидилэтаноламина, возрастает в 1,5—2 раза, в соста-
ве сфингомиелина — более чем в 2 раза. Соответственно этому
уменьшается содержание короткоцепочечной насыщенной пальмити-
новой кислоты С16.0 и растет уровень докозогексаеновой С22.б и тет’
ракозотетраеновой С24.4. Интересно, что количество С18Ч (олеино-
вой кислоты) повышается при этом в фосфатидолхолине, но снижает-
ся в сфингомиелине (Гуляева с соавт., 2002). Остается неясным,
какие изменения носят адаптационный, а какие — нежелательный,
связанный со старением характер.
просы и задания
ВОПРОСЫ И ЗАДАНИЯ
1. Опишите особенности строения нейрональной клетки.
2. Каковы специфические морфологические признаки секреторных ней-
ронов?
3. Назовите функции, обеспечивающие взаимодействие нервной и гумо-
ральной систем организма.
4. Перечислите и охарактеризуйте типы клеток нейроглии.
5. Как происходит образование миелина в ЦНС и ПНС?
6. Какова структура и функции миелина в ЦНС и ПНС?
7. Назовите типы нервных окончаний аксонов и дайте их краткие харак-
теристики.
8. Опишите роль синапсов в проведении нервного импульса.
9. Каковы особенности липидного состава нейрональной мембраны?
10. Охарактеризуйте роль ганглиозидов и цереброзидов в регуляции функ-
ционального состояния нейрональных мембран.
11. Какова роль докозогексаеновой кислоты в функциональной активнос-
ти головного мозга?
12. Какова роль галактолипидов (цереброзидов и сульфатидов) и холесте-
рина в структуре миелина?
13. Охарактеризуйте миелинспецифичные белки.
14. Чем обусловлено качественное разнообразие белков нервной ткани?
15. Перечислите специфические функции нейрональных мембран.
16. Какие контакты образуют мембраны синаптических структур?
17. Опишите модель молекулярной организации биологических мембран
Сингера и Никольсона.
18. Охарактеризуйте свойства липидных рафтов в нейрональных мембра-
нах.
19. Чем обусловлено разнообразие строения липидных ассоциатов в воде?
20. Какие особенности структуры фосфолипидов приводят к спонтанному
образованию бислоя?
21. От каких параметров структуры липидных молекул зависят форма
и размеры липидных мицелл?
22. В чем причины трансмембранной асимметрии фосфолипидов в мемб-
ране?
23. Какими параметрами регулируется фазовый переход типа «гель—
жидкий кристалл»?
24. Назовите основные различия в свойствах мембранных и водораствори-
мых белков.
25. Благодаря каким факторам осуществляется стабилизация структуры
и функций мембранных белков?
26. Назовите виды подвижности мембранных белков, оцените их времен-
ные характеристики.
27. Охарактеризуйте способы взаимодействия белков с клеточной мемб-
раной.
Раздел 1. СТРУКТУРА НЕРВНОЙ ТКАНИ
28. Каково значение образования белковых олигомеров в мембране?
29. Какой тип взаимодействий между белками и липидами в мембранах
является доминирующим?
30. Опишите циклы агрегации—дезагрегации белков в мембране. Каково
их биологическое значение?
31. Охарактеризуйте взаимодействие белков и липидов в мембране на
примере процесса активации мембранных фосфолипаз.
32. Охарактеризуйте структурное и функциональное разнообразие белков
цитоскелета нейрональной клетки.
33. Какие липиды определяют функциональные особенности и структуру
гликокаликса нейронов?
34. Приведите примеры гликопротеинов гликокаликса, указав их локали-
зацию и клеточные функции.
35. В чем заключается биологическая роль ганглиозидов?
36. Охарактеризуйте роль агрегатов ганглиозидов и гликопротеинов в ре-
ализации специфических функций нервной ткани.
37. Каким образом мембранные липиды контролируют функции мембран-
ных белков?
38. Назовите краткосрочные и долгосрочные механизмы приспособления
клеточных мембран к изменяющимся условиям среды.
39. Каковы молекулярные механизмы адаптации клеточных мембран, про-
текающие с помощью изменений микровязкости липидного бислоя?
40. За счет чего изменяется индекс ненасыщенности мембранных липи-
дов?
РАЗДЕЛ 2
МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
2.1.
ТИПЫ СИНАПСОВ И КЛАССИФИКАЦИЯ НЕЙРОМЕДИАТОРОВ
2.1.1. Синапсы в нервной системе
Понятие «синапс» было введено в конце XIX в. Ч. Шеррингто-
ном, подразумевавшим под этим термином структуру, которая опосре-
дует передачу сигнала от окончания аксона к эффектору — нейрону,
мышечному волокну, секреторной клетке. В ходе изучения синапсов
морфологами, физиологами, биохимиками и фармакологами выяви-
лось их значительное разнообразие, при этом обнаружились общие
черты в строении и фукционировании; в итоге были выработаны прин-
ципы классификации синапсов.
Морфологический принцип классификации синапсов
учитывает то, какими частями двух клеток они образованы и как они
расположены на поверхности воспринимающего нейрона (на теле
клетки, на стволе или «шипике» дендрита, на самом аксоне). Соответ-
ственно различают синапсы аксо-аксональные, аксо-дендритные,
аксо-соматические. Однако эта классификация не объясняет ни
функциональной роли, ни механизма работы синапса.
Морфологически синапс представляет собой структуру из двух
демиелинизированных образований — утолщенного синаптичес-
кого окончания (синаптической бляшки) на конце акцона и участка
мембраны иннервируемой клетки, через синаптическую щель кон-
тактирующего с пресинаптической мембраной. Основной функ-
цией синапса является передача сигнала. В зависимости от способа
передачи сигнала выделяют химические, электрические и смешан-
ные синапсы. Они различаются по принципу работы.
Механизм проведения возбуждения в электрическом синапсе ана-
логичен механизму проведения возбуждения в нервном волокне — ПД
пресинаптических окончаний обеспечивает деполяризацию постсинап-
тической мембраны. Такая передача возбуждения возможна благодаря
особенностям строения синапсов этого типа — узкая (около 5 нм) си-
наптическая щель, большая площадь контакта мембран, наличие попе-
речных канальцев, соединяющих пресинаптическую и постсинаптическую
110
к аздел 2. ‘.’ОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Р. Н. Глебов
Известный российский ней-
рохимии, автор цикла работ
по функциональной нейрохи-
мии синапсов
мембраны и снижающих электрическое
сопротивление в области контакта. Наи-
более распространены электрические си-
напсы у беспозвоночных и низших позво-
ночных. У млекопитающих они обнаруже-
ны в мезенцефальном ядре тройничного
нерва между телами нейронов, в вестибу-
лярном ядре Дейтерса между телами клеток
и окончаниями аксона и между «шипика-
ми» дендритов в нижней оливе. Электриче-
ские синапсы образуются между нервны-
ми клетками, однотипными по структуре
и функциям.
Для электрической синаптической
передачи характерно отсутствие синап-
тической задержки, проведение сигнала в
обоих направлениях, независимость пе-
редачи сигнала от потенциала пресинап-
тической мембраны, устойчивость к изменениям концентрации Са2+ и
Mg2+, низкой температуре, некоторым фармакологическим воздейст-
виям, а также слабая утомляемость, поскольку передача сигнала не
требует значительных метаболических затрат. В большинстве таких
синапсов наблюдается «эффект выпрямления», когда сигнал в синап-
се передается только в одном направлении.
В отличие от электрических синапсов с прямой передачей
возбуждения, в значительно большем количестве в нервной сис-
теме позвоночных представлены химические синапсы (синапсы сне-
прямой передачей сигнала). В химическом синапсе нерв-
ный импульс вызывает высвобождение из пресинаптических оконча-
ний химического посредника — нейромедиатора, который диффунди-
рует через синаптическую щель (шириной в 10—50 нм) и вступает во
взаимодействие с белками-рецепторами постсинаптической мембра-
ны, в результате чего генерируется постсинаптический потенциал
(рис. 2.1). Химическая передача обеспечивает одностороннее прове-
дение сигнала и возможность его модуляции (усиление сигнала, а так-
же конвергенция многих сигналов на одной постсинаптической клет-
ке). Возможность модуляции в процессе передачи сигналов в синапсах
химического типа обеспечивает формирование на их основе сложных
физиологических функций (обучение, память и т. д.). К особенностям
ультраструктуры химического синапса можно отнести широкую синап-
тическую щель, наличие в синаптической бляшке везикул, заполнен-
ных медиатором, при помощи которого передается сигнал, а в постен -
2.1. Типы синапсов и классификация нейромедиаторов
Электрический синапс
Рис. 2.1. Различия в устройстве электрических и химических синапсов (объяс-
нение в тексте)
Химический синапс
наптической — многочисленных хемочувствительных каналов (в воз-
буждающем синапсе — для Na+, в тормозном — для С1“). Для таких
синапсов характерна задержка в проведении сигнала и большая по срав-
нению с электрическим синапсом утомляемость поскольку их функци-
онирование требует значительных метаболических затрат.
Выделяют два основных подтипа химических
синапсов. Первый (так называемый асимметричный) харак-
теризуется синаптической щелью шириной около 30 нм, сравнительно
большой зоной контакта (1 —2 мкм), значительным накоплением плот-
ного матрикса под постсинаптической мембраной. В пресинаптической
бляшке накапливаются большие везикулы (диаметр 30—60 нм). Хими-
ческие синапсы второго подтипа имеют синаптическую щель
Шириной около 20 нм, сравнительно небольшую зону контакта (менее
1 мкм), умеренно выраженные и симметричные уплотнения мембран.
Для них характерны небольшие везикулы (диаметр 10—30 нм). Пер-
вый подтип представлен, в основном, аксодендритными, возбуждаю-
щими (глутаматергическими), второй — аксосоматическими, тормоз-
ными (ГАМК-ергическими) синапсами. Однако это деление достаточ-
но условно, поскольку холинергические синапсы обнаруживают на
электронных микрофотографиях светлые везикулы диаметром 20—
112
кгздел 2. МОПЕКуЛяРНАя НЕЙРОХИМИЯ
40 нм, а моноаминергические (особенно с норадреналином) — боль-
шие плотные везикулы диаметром 50—90 нм.
Еще один принцип классификации синапсов — по веществу,
используемому в качестве медиатора (холинергиче-
ские, адренергические, пуринергические, пептидергические и т. д.).
Несмотря на то что в последние годы было показано, что в одном
окончании могут функционировать медиаторы различной природы, эта
классификация синапсов до сих пор остается широко употребляемой.
2.1.2. Медиаторы, нейромодуляторы, комедиаторы
Нейромедиатор (нейротрансмиттер, нейропередатчик) —
это вещество, которое синтезируется в нейроне, содержится в преси-
наптических окончаниях, высвобождается в синаптическую щель в от-
вет на нервный импульс и действует на специальные участки постси-
наптической клетки, вызывая изменения мембранного потенциала и
метаболизма клетки.
А. Лайта
Нейрохимик, широко извест-
ный своими работами по изу-
чению регуляторных функций
нейромедиаторов в мозге, в
частности гамма-аминомасля-
ной кислоты и глицина, а так-
же процессов повреждения и
восстановления в мозге; автор
и редактор периодического из-
дания «Handbook of neuro-
chemistry» (США)
До середины прошлого века к меди-
аторам относили только амины и амино-
кислоты, однако открытие нейромеди-
аторных свойств у пуриновых нуклеотидов,
производных липидов и нейропептидов
значительно расширило группу медиато-
ров. В конце прошлого века было показа-
но, что некоторые АФК также обладают
свойствами, сходными с медиаторами.
По химическому строению медиаторы
являются неоднородной группой (рис. 2.2).
В нее входят эфир холина (ацетилхолин);
группа моноаминов, включающая катехо-
ламины (дофамин, норадреналин и адре-
налин); индоЛы (серотонин) и имидазолы
(гистамин); кислые (глутамат и аспартат)
и основные (ГАМК и глицин) аминокис-
лоты; пурины (аденозин, АТФ) и пепти-
ды (энкефалины, эндорфины, вещество Р).
К этой же группе примыкают вещества,
которые не могут быть классифицирова-
ны как истинные нейромедиаторы — сте-
роиды, эйкозаноиды и ряд АФК, прежде
всего NO.
!,1. Типы синапсов и классификация нейромедиаторов
11
I II
СНз—N+—СН2—СН2—О—С—СНз
I
СНз Ацетилхолин
Аминокислоты
nh2
I
ноос—сн2—сн2—сн—соон
Глутамат
NH2
I
НООС—сн2 — сн2—сн2
ГАМК
Моноамины
Норадреналин
НО
Пептиды
Туг—Gly—Gly—Phe—Met Л4ет-энкефалин
Arg-Pro—Lys-Pro-GIn-GIn-Phe-Phe-Gly—Leu-Met-NH2 Вещество P
Рис. 2.2. Химическая структура классических нейромедиаторов и нейропептидов
Для решения вопроса о нейромедиаторной природе
какого-либо соединения используется ряд критериев.
Основные из них изложены ниже.
1. Вещество должно накапливаться в пресинаптических окончаниях,
высвобождаться в ответ на приходящий импульс. Пресинаптическая об-
ласть должна содержать систему синтеза этого вещества, а постсинап-
тическая зона — обнаруживать специфический рецептор для данного
соединения.
2. При стимуляции пресинаптической области должно происходить
Са-зависимое выделение (путем экзоцитоза) этого соединения в межси-
наптическую щель, пропорциональное силе стимула.
3. Обязательная идентичность эффектов эндогенного нейромедиато-
ра и предполагаемого медиатора при его аппликации на клетку-мишень
и возможность фармакологического блокирования эффектов предпола-
гаемого медиатора (табл. 2.1).
4. Наличие системы обратного захвата предполагаемого медиатора
в пресинаптические терминали и/или в соседние астроглиальные клет-
ки. Возможны случаи, когда обратному захвату подвергается не сам
медиатор, а продукт его расщепления (например, холин после расщеп-
ления ацетилхолина ферментом ацетилхолинэстеразой).
114
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Таблица 2.1. Влияние лекарственных препаратов на различные
этапы медиаторной функции в синаптической передаче
Этапы Модифицирующее влияние Результат воздействия
Добавка предшественника t
Синтез медиатора Блокада обратного захвата 1
Блокада ферментов синтеза 1
Накопление Торможение захвата в везикулы и
Торможение связывания в везикулах и
Стимуляция тормозных ауторецепторов 1
Выделение (экзоцитоз) Блокада ауто рецепторе в t
Нарушение механизмов экзоцитоза 1
Действие Эффекты агонистов на рецепторы t
на рецепторы Блокада постсинаптических рецепторов I
Блокада обратного захвата нейронами и/или глией t
Разрушение медиатора Торможение разрушения в нейронах t
Торможение разрушения в синаптической щели t
Применение различных методов тестирования медиаторной функ-
ции, в том числе и самых современных (иммуногистохимических, ре-
комбинантных ДНК и др.), затруднено из-за ограниченной доступности
большинства индивидуальных синапсов, а также из-за ограниченности
набора средств направленного фармакологического воздействия.
Попытка дать определение понятия «медиаторы» наталкивается на
ряд трудностей, поскольку в последние десятилетия значительно рас-
ширился список веществ, которые выполняют в нервной системе ту
же сигнальную функцию, что и классические медиаторы, но отличают-
ся от них по химической природе, путям синтеза, рецепторам. Прежде
всего, сказанное относится к обширной группе нейропептидов, а так-
же к АФК, и в первую очередь к оксиду азота (нитроксиду, NO), для
которого медиаторные свойства описаны достаточно хорошо. В отли-
чие от «классических» медиаторов, нейропептиды, как правило, име-
ют больший размер, синтезируются с невысокой скоростью, накапли-
ваются в небольших концентрациях и связываются с рецепторами,
2.1. Г ипы синапсов и классификация нейромедиаторов
115
обладающими низким специфическим сродством (рис. 2.3), кроме то-
го, они не имеют механизмов обратного захвата пресинаптической
терминалью. Продолжительность эффекта нейропептидов и медиато-
ров также значительно различается. Что касается нитроксида, то, не-
смотря на его участие в межклеточном взаимодействии, по ряду кри-
териев он может быть отнесен скорее не к медиаторам, а ко вторич-
ным посредникам.
Первоначально считалось, что нервное окончание может содер-
жать только один медиатор. К настоящему времени показана возмож-
ность наличия в терминали нескольких медиаторов, высвобождаю-
щихся совместно в ответ на импульс и воздействующих на одну клетку-
Пептиды
Рис. 2.3. Различия между синтезом, экзоцитозом и дальнейшими превращения-
ми классических медиаторов (глутамат, моноамины) и нейропептидов.
Синтез и упаковка в везикулы классических нейромедиаторов (а) происходят в нерв-
ных терминалях, а необходимые для этого ферменты синтезируются в теле нейрона и
затем доставляются в нервное окончание с помощью аксонного транспорта. После вы-
броса в синаптическую щель возможно разрушение нейромедиатора с помощью фер-
ментов. Обратный захват медиатора (или его метаболитов) с помощью имеющихся
специфических транспортеров нервной или глиальной клетками запускает новый цикл
его синтеза, упаковки и выделения. Пептидные нейромедиаторы и ферменты (б), моди-
фицирующие их предшественников, синтезируются в теле нейрона, упаковываются в
аппарате Гольджи и с помощью быстрого аксонального транспорта переносятся в
нервное окончание. После выброса в синаптическую щель возможен протеолиз меди-
атора; обратный захват такого нейропептида или его метаболитов пресинаптическим
нейроном отсутствует
116
Раздел 2 МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
мишень — сопутствующие (сосуществующие) медиаторы (коме-
диаторы, котрансмиттеры). При этом происходит накопление
разных медиаторов в одной пресинаптической области, но в разных
везикулах. Примером комедиаторов могут служить классические ме-
диаторы и нейропептиды, которые различаются местом синтеза и, как
правило, локализуются в одном окончании. Высвобождение комеди-
аторов происходит в ответ на серию возбуждающих потенциалов оп-
ределенной частоты.
В современной нейрохимии кроме нейромедиаторов выделяют ве-
щества, модулирующие их эффекты, — нейромодуляторы. Их дей-
ствие носит тонический характер и более продолжительно во времени,
чем действие медиаторов. Эти вещества могут иметь не только нейро-
нальное (синаптическое), но и глиальное происхождение и не обяза-
тельно опосредоваться нервными импульсами. В отличие от нейро-
медиатора модулятор действует не только на постсинаптическую
мембрану, но и на другие части нейрона, в том числе и внутрикле-
точно.
Различают пре- и постсинаптическую модуляцию (см. гл. 2.7).
Понятие «нейромодулятор» является более широким, чем понятие «ней-
ромедиатор». В ряде случаев медиатор может являться и модулято-
ром. Например, норадреналин, высвобождающийся из симпатического
нервного окончания, действует как нейромедиатор на al-рецепто-
ры, но как нейромодулятор — на а2-адренорецепторы; в последнем
случае он опосредует торможение последующей секреции норадрена-
лина.
Вещества, осуществляющие медиаторные функции, различаются
не только по химическому строению, но и по тому, в каких компарт-
ментах нервной клетки происходит их синтез. Классические низкомо-
лекулярные медиаторы синтезируются в аксонной терминали и вклю-
чаются в маленькие синаптические пузырьки (50 нм в диаметре) для
хранения и высвобождения. NO также синтезируется в терминали, но,
поскольку не может быть упакован в пузыръки, то сразу же диффунди-
рует из нервного окончания и оказывает воздействие на мишени. Пеп-
тидные нейромедиаторы синтезируются в центральной части нейрона
(перикарионе), упаковываются в большие везикулы с плотным цент-
ром (100—200 нм в диаметре) и транспортируются с помощью аксо-
нального тока к нервным окончаниям.
Ацетилхолин и катехоламины синтезируются из циркулирующих в
крови предшественников, тогда как аминокислотные медиаторы и
пептиды в конечном счете образуются из глюкозы. Как известно, ней-
роны (как и другие клетки организма высших животных и человека) не
2.2. Общая характеристика мембранных рецепторов
11
2.2.
могут синтезировать триптофан. Поэтому первым шагом, ведущим к
началу синтеза серотонина, является облегченный транспорт трипто-
фана из крови в мозг. Эта аминокислота, как и другие нейтральные
аминокислоты (фенилаланин, лейцин и метионин), транспортируется
из крови в мозг специальными переносчиками, относящимися к се-
мейству переносчиков монокарбоновых кислот. Таким образом, одним
из важных факторов, определяющих уровень серотонина в серотони-
нергических нейронах, является относительное по сравнению с други-
ми нейтральными аминокислотами количество триптофана в пище.
Например, добровольцы, которых кормили пищей с низким содержа-
нием белка в течение одного дня, а затем давали смесь аминокислот,
не содержавшую триптофана, демонстрировали агрессивное поведе-
ние и изменение цикла «сон—бодрствование», что связано со сниже-
нием уровня серотонина в мозге.
ОБЩАЯ ХАРАКТЕРИСТИКА МЕМБРАННЫХ РЕЦЕПТОРОВ
Важное свойство всех живых существ — способность восприни-
мать, перерабатывать и передавать информацию. Несмотря на разнооб-
разие систем обработки информации, функционирующих в животных и
растительных организмах, все они основаны на едином принципе. Про-
цесс получения информации, как правило, начинается с взаимодействия
сигнала (химического агента, кванта света, механического воздействия
и др.) с рецептором, представленным, как правило, мембранным бел-
ком. Следующий этап — передача информации в центр переработки
информации, находящийся внутри клетки. Этот процесс происходит с
помощью вторичных мессенджеров (посредников). В ответ на полу-
чение сигнала вторичного мессенджера в клетке происходит биохими-
ческая модификация специализированных молекул-эффекторов, через
которые и формируется ответ биологической системы (рис. 2.4).
Именно по такому принципу функционируют нервная, гормональ-
ная и иммунная системы животных, на такие же стадии могут быть
разложены и фотобиологические процессы, протекающие в организ-
ме как животных, так и растений. Общий принцип действия всех сис-
тем приема и передачи информации связан не только с химической
модификацией мембранных белков, но и с изменением концентрации
ионов внутри клетки, с изменением мембранного потенциала. В по-
следнее время выяснилось, что этот процесс играет важную роль не
только в функционировании нервной ткани, но и при переработке ин-
формации в тромбоцитах, лимфоцитах, тучных клетках.
118
раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
______। АТФ
с'М[а^(—>-^АМФ]------------»-
ПК А —>- Белки —►-
Рис. 2.4. Основные сигнальные системы клетки.
АЦ — аде нил ил цикл аза; ГЦ— гуанил ил циклаза; КМ — кальмодулин; КМ-ПК —
кальмодулинзависимая ПК; Фл С — фосфолипаза С; ФИ — фосфоинозитиды;
ФХ — фосфатидилхолин; ИФ3 — инозитолтрисфосфат; ДАГ — ди ацил глицерол
е
Сигнальные молекулы, как правило, не проникают внутрь клетки,
а специфически взаимодействуют с рецепторами, локализованными
во внешней клеточной мембране и представляющими собой интеграль-
ные мембранные белки, полипептидная цепь которых пронизывает
толщу мембраны несколько (7—12) раз. Они могут быть выделены из
мембраны только после ее разрушения, например с помощью детер-
гента.
Стероидные и тиреоидные гормоны, будучи гидрофобными по сво-
ей природе, способны проникать через плазматическую мембрану
внутрь клетки, где они взаимодействуют с растворимыми рецепторны-
ми белками, локализованными в цито- и (или) нуклеоплазме. Они
представляют, по-видимому, эволюционно более примитивный (но
и более гарантированный) способ передачи информации.
Разнообразные молекулы, инициирующие трансмембранную пере-
дачу сигналов, активируют рецепторы, воздействуя на них обычно при
очень низких концентрациях, порядка 10-8—10-9 М.
В соответствии с двумя основными видами
передачи информации к клетке рецепторные белки делят на
два класса: глобулярные и мембранные.
2.2. исщая характеристика меморанных рецепторов
119
Глобулярные белки представляют собой свободноплавающие в ци-
топлазме соединения, обладающие высоким сродством к определен-
ным лигандам. Появление в цитоплазме лигандов для этих рецепторов
приводит к изменению свойств последних и выполнению ими своих
функций. Например, цитоплазматическая протеинкиназа С, связывая
ионы кальция, изменяет свою конформацию, формирует гидрофобный
участок для «заякоривания» на мембране и начинает атаковать мемб-
ранные субстраты. Другим примером такого способа управления явля-
ется регуляция синтеза некоторых клеточных белков альдостероном:
появляясь в крови, этот гидрофобный гормон легко проникает через
клеточную мембрану, находит в цитоплазме свой рецептор и образует с
ним комплекс, способный проникать в ядро, где инициирует экспрес-
сию соответствующих генов.
Иной тип передачи информации осуществляют интегральные
белки, которые связывают сигнальные вещества на внешней стороне
мембраны и за счет изменения своей конформации генерируют новый
сигнал на внутренней стороне мембраны.
Мембранный рецептор, вне зависимости от природы связывающе-
гося с ним эффектора, имеет общий план строения: участок,
расположенный вне клетки, внутримембранный участок и внутрен-
ний участок, ориентированный в цитоплазму. Внешний и внутренний
участки рецептора являются вариабельными, его внутримембранная
часть более консервативна. N-конец рецептора (внешний) специфи-
чен к внешнему сигналу, тогда как внутренний С-конец — к ассоци-
ированному с рецептором внутриклеточному белку. Свойства последне-
го и определяют, с какой из внутриклеточных сигнальных систем будет
осуществляться взаимодействие и каким образом будет формироваться
физиологический ответ клетки. Сами сигнальные молекулы при этом
обычно не проникают внутрь клетки, а сигнал преобразуется в резуль-
тате модификации мембранных белков, которая приводит к образова-
нию (высвобождению) молекул-посредников — вторичных мессенд-
жеров.
В общем виде передача сигнала через мембрану
может быть сведена к трем основным стадиям:
1) взаимодействие сигнальной молекулы с рецептором;
2) конформационная перестройка рецепторной молекулы и изме-
нение функции специализированных мембранных белков-посред-
ников;
3) образование «вторичных мессенджеров» — сравнительно не-
больших молекул или ионов, диффузия которых в клетке к определен-
ным субклеточным структурам обеспечивает распространение сигнала.
12U
раздел 2. ‘/ОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Механизмы передачи информации в живых системах универсальны —
все громадное разнообразие сигналов, полученных различными рецеп-
торами, преобразуется по единому принципу за счет идентичности вто-
рой и третьей стадий передачи информации через мембрану (см.
рис. 2.4).
Различают три типа рецепторов. Рецепторы первого
типа являются белками, имеющими одну трансмембранную по-
липептидную цепь. Это аллостерические ферменты, активный центр
которых расположен на внутренней стороне мембраны. Многие из них
являются тирозиновыми протеинкиназами. К этому типу принадлежат
рецепторы инсулина, ростовых факторов и цитокинов. Связывание
сигнального вещества ведет к димеризации такого рецептора. При
этом происходит активация фермента и фосфорилирование остатков
тирозина в ряде белков. В первую очередь фосфорилируется молекула
рецептора по принципу аутофосфорилирования.
Рецепторы второго типа представляют собой олигомерные
мембранные белки, образующие лигандактивируемый ионный
канал. Связывание лиганда ведет к открыванию канала для ионов
Na+, К+, Са2+ или С1“. По данному механизму передают возбуждаю-
щий сигнал такие нейромедиаторы, как ацетилхолин (никотиновые
рецепторы), глутамат (ионотропные рецепторы, см. далее), у-амино-
масляная кислота (ГАМКд-рецептор).
Ансамбли рецепторов с ионными каналами — это интегральные
мембранные олигомерные белки, состоящие из нескольких субъеди-
ниц, полипептидная цепь которых многократно пересекает наружную
клеточную мембрану. Кроме канальных структур они одновременно
содержат рецепторные участки на внешней стороне мембраны, кото-
рые способны специфически связывать лиганды, изменяющие ионную
проводимость канала. Ионные каналы этих рецепторов могут нахо-
диться в трех основных состояниях: 1) закрытое состояние (в отсутствие
внешнего сигнала); 2) активированное (открытое) состояние (первич-
ный мессенджер связан с рецептором); 3) инактивированное (закры-
тое) состояние — может наступать, когда первичный мессенджер еще
связан с рецептором.
Эти рецепторы получили название быстрых ионотропных, по-
скольку время их действия составляет несколько десятков миллисе-
кунд (рис. 2.5).
В связи с тем что в состав рецептора может входить несколько
субъединиц, различающихся по структуре, существуют его многочис-
ленные изоформы, позволяющие варьировать основные свойства ре-
цепторов.
2.2. Общая характеристика мембранных рецепторов
Рис. 2.5. Взаимодействие быстрых (ионотропных) и медленных (метаботроп-
ных) рецепторов с ионными каналами.
Связывание медиатора с ионотропным рецептором приводит к быстрому открыванию
ионного канала. При связывании медиатора с метаботропным рецептором возможны
несколько путей влияния на ионный канал: непосредственно через G-белки, через вто-
ричные посредники, а также путем фосфорилирования, связанного с активностью про-
теинкиназ
Рецепторы третьего типа являются медленнодействую-
щими. Они сопряжены с ГТФ-связывающими белками (G-белки,
обладающие ГТФазной активностью). Полипептидная цепь этих бел-
ков включает семь трансмембранных сегментов (колонн). Такие ре-
цепторы передают сигнал с помощью ГТФ-связывающих белков на
белки-эффекторы, расположенные на соседних участках мембраны
или в цитоплазме. Функция рецепторов данного типа заключается в
тонкой модификации клеточного метаболизма, поэтому их называют
метаботропными. К этой группе относятся рецепторы катехолами-
нов, серотонина, ГАМКв-рецептор, мускариновый ацетилхолиновый
рецептор (jwXP), глутаматные метаботропные рецепторы, рецепторы
нейропептидов и ряд других. Эффекты, связанные с этими рецептора-
ми, развиваются в течение нескольких секунд или минут.
Рецепторы, ассоциированные с ферментами, по своей субъеди-
ничной структуре весьма разнообразны. Как правило, они представ-
лены мономерными белками, но в зависимости от присутствия пер-
вичного мессенджера могут объединяться в олигомерные мембран-
ные ансамбли. Практически у всех этих рецепторов полипептидная
122
► аздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
цепь пересекает клеточную мембрану всего один раз. Общим для них
является то, что участок для восприятия первичного сигнала лока-
лизован на стороне мембраны, обращенной во внеклеточное про-
странство.
По механизму взаимодействия с цитоплазма-
тическими мишенями рецепторы данного типа разделяют на
две группы.
Первая группа включает рецепторы-ферменты, каталитический
центр которых локализован с цитоплазматической стороны мембра-
ны; он активируется при действии на рецептор внешнего сигнала
(рис. 2.6). К рецепторам этой группы относится обширное семейство
тирозиновых протеинкиназ, способных как к аутофосфорилированию
(по тирозиновым остаткам), так и к фосфорилированию тирозиновых
остатков других белков-мишеней. К этой же группе относятся ре-
Рис. 2.6. Основные типы мембранных рецепторов:
а — рецепторы, сопряженные с G-белками; б — рецепторы — ионные каналы; в —
рецепторы, ассоциированные с ферментами. С — внешний сигнал; Рц — рецептор-
ный белок («*» при компоненте сигнальной системы означает, что он находится в со-
стоянии «включено»); а — а-субъединица G-белка, которая может находиться в свя-
занной с гуанозицдифосфатом (ГДФ) или гуанозинтрифосфатом (ГТФ) форме; ру —
функционирующий как единое целое комплекс Р- и у-субъединиц G-белка; Р — фос-
фатный остаток (остатки), ковалентно связанный с рецептором; Эф — эффекторы
2.2. Общая характеристика мембранных рецепторов
цепторы, обладающие протеинфосфатазной активностью, которые
дефосфорилируют фосфотирозиновые остатки белков-мишеней. Как
киназы, так и фосфатазы вовлекаются в регуляцию таких важнейших
событий, как деление и дифференцировка клеток, развитие иммунного
ответа.
Вторая группа рассматриваемых рецепторов собственной фер-
ментативной активностью не обладает. Однако в присутствии внешне-
го сигнала эти рецепторы приобретают способность связывать цито-
плазматические протеинтирозинкиназы, которые неактивны в сво-
бодном состоянии, но активируются в комплексе с рецептором и
осуществляют его фосфорилирование. Включение фосфатных остат-
ков в такой рецептор-«якорь» создает условия для связывания с ним
других белков-мишеней, которые также фосфорилируются, обеспечи-
вая тем самым передачу сигнала. В эту группу входят рецепторы, уча-
ствующие в развитии иммунного ответа: рецепторы антигенов и ре-
цепторы цитокинов, интерлейкинов.
Большинство белков-эффекторов осуществляют свои функ-
ции в результате фосфорилирования различными протеинкиназа-
ми. Многочисленное семейство протеинкиназ (ПК) подразделяется на
группы по м е х а н и з м у активации и по типу субстра-
та (см. 2.4.6). ПК А активируется цАМФ, ПКО — цГМФ, ПК С —
диацилглицеролом, Са2+-кальмодулинзависимые ПК — ионами каль-
ция.
Молекула протеинкиназы состоит из двух субъединиц: ре-
гуляторной и каталитической. Связывание активатора (напри-
мер, цАМФ, цГМФ) с регуляторной субъединицей приводит к отде-
лению каталитической субъединицы протеинкиназы (она становится
активной) и фосфорилированию соответствующего белка.
Классификация по типу субстрата зависит от того, какая амино-
кислота белка-мишени фосфорилируется данным ферментом — из-
вестны тирозинкиназы и серин-треониновые протеинкиназы. Многие
протеинкиназы являются растворимыми белками, другие относятся к
мембранным белкам, имеющим трансмембранные домены или «за-
якоренным» на мембране с помощью жирных кислот.
В каждой клетке обычно функционирует несколько разных рецеп-
торов к одному и тому же химическому сигналу (например, а- и Р-ад-
ренорецепторы). В дополнение к этому клетки обычно проявляют чув-
ствительность одновременно к нескольким регуляторным молеку-
лам (нейромедиаторам, гормонам, простагландинам, факторам роста).
Каждый из этих регуляторов рождает сигнал характерной продолжи-
тельности и амплитуды. На уровне исполнительных систем клетки мо-
жет происходить как потенцирование, так и взаимное тушение разных
124
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
2.3.
регуляторных сигналов. Глиальные клетки также имеют рецепторы,
однако связывание нейромедиатора с последними не ведет к генера-
ции потенциала действия.
Вещества, вызывающие тот же эффект, что и природные медиато-
ры, принято называть агонистами, а лиганды, которые связываются
с рецептором, препятствуя его взаимодействию с агонистом, хотя и не
вызывают сами биологического эффекта, — антагонистами.
Один и тот же медиатор может взаимодействовать с рецепторами
разных типов и подтипов, при этом достигается различный эффект как
по времени наступления эффекта, так и по его результату. Оказалось,
что многие заболевания нервной системы связаны не только с нару-
шением синтеза и выделения нейромедиаторов, но и с функциониро-
ванием рецепторного аппарата нервной клетки, и лечение некоторых
патологий стало возможным благодаря знанию строения и функци-
ональных особенностей рецепторов.
ХАРАКТЕРИСТИКА ИНДИВИДУАЛЬНЫХ
МЕДИАТОРНЫХ СИСТЕМ
Характеристику отдельных нейромедиаторов целесообразно де-
лать с учетом того, в каких структурах мозга обнаруживается их мак-
симальная концентрация. Именно такой подходе последующим элек-
трофизиологическим и фармакологическим анализом роли этих струк-
тур позволил выявить функцию большинства медиаторов в нервной
системе. Химические синапсы распределены в нервной ткани не слу-
чайным образом, а организованы в определенные группы (рис. 2.7).
Для того чтобы уверенно картировать какие-либо медиаторные пути в
головном мозге, необходимо иметь надежные специфические марке-
ры, с помощью которых можно визуализировать интересующие иссле-
дователя межклеточные взаимодействия.
В зависимости от изучаемой нейромедиаторной системы применяют
определенные методические подходы. В случае если сущест-
вует возможность получения окрашенного (флуоресцирующего) произ-
водного естественного нейромедиатора, для выявления групп клеток,
содержащих этот нейромедиатор, применяют соответствующий спект-
ральный подход (в последнее время — с использованием конфокально-
го микроскопа). С учетом того что в метаболизме ряда медиаторов при-
сутствует механизм обратного захвата, применяют медиатор, меченный
радиоактивными изотопами, и последующую авторадиографию. Еще
один способ, используемый как для локализации «классических» меди-
2.3. Характеристика индивидуальных медиаторных систем
аторов, так и для выявления нейропептидов и других модуляторов, —
применение специфических антител. В случае нейропептидов — к са-
мому нейропептиду, в случае нейромедиаторов — к синтезирующим
их ферментам или к специфическим рецепторам. Используются так-
же различные количественные способы оценки высвобождения ней-
ромедиаторов с последующим измерением электрического ответа си-
напса.
Система ацетилхолина. Система ацетилхолина (АХ) широко
представлена в разных отделах нервной системы (см. рис. 2.7). Основ-
ная локализация холинергических нейронов — медиальное ядро пере-
городки, диагональная связка, базальное гигантоклеточное ядро. Ак-
соны этих нейронов проецируются на гиппокамп, таламус, кору боль-
ших полушарий и ряд других структур. В спинном мозге АХ играет
роль нейромедиатора в синапсах, образуемых а-мотонейронами на
клетках Реншоу. В вегетативной нервной системе АХ — медиатор
всего парасимпатического отдела, а в преганглионарных нервных
окончаниях — симпатического отдела.
Выявление этого медиатора в нервных окон-
чаниях возможно по наличию в них фермента его синтеза (холин-
ацетилтрансферазы) или гидролизующего его фермента (ацетилхолин-
эстеразы). Первый способ является более надежным, так как ацетил-
холинэстераза широко распространена в различных тканях организма.
Для выявления рецепторов АХ применяются также радиоиммунологи-
ческие методы.
Холинергические системы участвуют в таких функциональных про-
явлениях мозга, как память, локомоторная функция, в частности меха-
низмы инициации подвижности, создание двигательных стереотипов,
регуляция уровня бодрствования (в этих процессах участвуют ретику-
лярная формация ствола мозга, базальные ганглии).
При паркинсонизме, наряду с нарушением дофаминергической
трансмиссии, отмечается гиперактивность некоторых холинергиче-
ских систем мозга. При поражении холинергических связей в перифе-
рической нервной системе, в частности в нервно-мышечных синапсах,
возникают симптомы «усталости» или «слабости» мышц. При неко-
торых неврологических заболеваниях, сопровождающихся резкими
неконтролируемыми движениями (хорея Гентингтона) и слабоумием
(болезнь Альцгеймера), наблюдается дегенерация пирамидных клеток
гиппокампа, к которым подходят аксоны холинергических нейронов из
других структур.
Ацетилхолин
Неокортекс
Черепномозговые
мотонейроны
Спинальные
мотонейроны
Пре ганглионарные
вегетативные нейроны
Дофамин Вещество Р НейропептидУ
Норадреналин
Серотонин
Глутамат/Аспартат
ГАМК
Рис. 2.7. Распределение нейронов, содержащих различные ме-
диаторы и нейропептиды, в головном мозге млекопитающих.
БШ — бледный шар; ВД — верхнее двухолмие; ИПЯ — интрапедун-
кулярные ядра; ВПП — вентральное поле покрышки; ГИ — груше-
видная извилина; ГК — гиппокамп; ГП — голубое пятно; ГТ — гипо-
таламус; ДЯ — дугообразное ядро; ЗР — задний рог спинного мозга;
ЛПП— латеральное поле покрышки; М — миндалина; ОБ — обоня-
тельный бугорок; ОЖС — околожелудочковое серое вещество; ОД —
обонятельная луковица; П — перегородка; NA —nucleus accumbens;
CM — спинной мозг; СМГ — спинномозговой ганглий; СНСМ — се-
ротонинергические нейроны спинного мозга; Стр — стриатум; Т — та-
ламус; У — уздечка; ЧС — черная субстанция; ЯМ — ядра мозжечка
Раздел 1. МОЛЕКУЛЯРНАЯ НЕИРОХИМИЯ
2.3. Характеристика индивидуальны, медиаторных систем
АХ является преимущественно возбуждающим нейромедиатором,
реже выполняет тормозные функции (например, в синапсах ответвле-
ний блуждающего нерва, иннервирующих миокард). Действие АХ мо-
жет быть длительным или кратковременным, возбуждающим или тор-
мозным, локальным или дистантным (см. далее).
Ацетилхолин синтезируется из холина и ацетил-КоА цитозольным
ферментом холинацетилтрансферазой. Основными источни-
ками холина для образования нейромедиатора
служат:
1) холин, образующийся при гидролизе нейромедиатора, диффун-
дировавшего от рецептора, и находящийся в синаптической щели
(система «обратного захвата» обеспечивает перенос молекул холина
в пресинаптическое окончание; около половины холина, поступающе-
го в холинергическую аксонную терминаль, является продуктом гид-
ролиза ранее высвобожденного ацетилхолина);
2) холин, поступающий из плазмы крови через гематоэнцефаличе-
ский барьер;
3) холин, образующийся при гидролизе фосфатидилхолина.
Перенос холина в пресинаптические окончания осуществляется
высокоаффинной (Км 1—5 мкМ) Na-зависимой транспортной систе-
мой, которая обнаружена только в холинергических нейронах. Во всех
других тканях в транспорте холина участвуют низкоаффинные (Км
10—100 мкМ) переносчики. Средняя концентрация холина в плазме
крови составляет ЮмкМ. Поступление холина в пресинаптические
окончания холинергических нейронов является этапом, лимитирую-
щим скорость синтеза нейромедиатора.
Поскольку реакция, катализируемая холинацетилтрансферазой,
является обратимой, уровень ацетилхолина контролируется соотно-
шением концентраций субстратов и продукта реакции.
Так, в условиях покоя накопление ацетилхолина ограничено одно-
временным гидролизом, который осуществляется внутриклеточной
ацетилхолинэстеразой, и ее фармакологическое ингибирование в нерв-
ном окончании вызывает повышение содержания медиатора. Таким об-
разом, уровень, до которого происходит накопление ацетилхолина, ус-
танавливается в результате динамического равновесия между его син-
тезом и распадом. Благодаря этому обеспечивается наличие постоянно
доступного необходимого количества медиатора. Следует отметить, что
в везикулах ацетилхолин защищен от разрушения. Кроме того, заклю-
чение ацетилхолина в везикулы приводит к снижению его концентра-
ции в цитоплазме, что также влияет на равновесие реакции, сдвигая
его в сторону синтеза медиатора.
128
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Холинорецепторы. Холинорецепторы (ХР) подразделяют на
никотиновые (нХР) и мускариновые (мХР). Как для первых, так и
для вторых агонистом является ацетилхолин, однако первый класс ре-
цепторов активируется также и никотином, а второй — мускарином
(соединением, содержащимся в мухоморе Amanita muscarina).
Успех в исследовании рецепторов во многом обеспечен примене-
нием природных токсинов, в первую очередь ядов пчел, пауков, змей и
некоторых земноводных. Эти яды содержат фосфолипазы, разрушаю-
щие мембранные фосфолипиды, что приводит также к нарушению пе-
редачи сигнала в клетке. Но наиболее простой путь для хищников па-
рализовать жертву — ингибировать ее «быстрые» рецепторы. В ре-
зультате такой «эволюции» животные различных классов (рыбы,
земноводные, змеи, насекомые) приобрели способность синтезиро-
вать яды — блокаторы «быстрых» рецепторов.
а-Бунгаротоксин из яда змей семейства эланидов (кобры, мамбы
и др.) способен селективно и цеобратимо связываться с нХР. Это его
свойство позволило идентифицировать рецепторный белок и очистить
его. Первичная структура ХР вначале была установлена для белка из
электрического органа ската (в этой ткани концентрация рецепторно-
го белка на два порядка превышает его концентрацию в других тканях,
что существенно облегчает процедуру выделения). Позднее на основе
исследования соответствующих генов первичная последовательность
холинорецептора была определена для тканей человека, теленка,
цыпленка и некоторых других животных.
Быстродействующие никотиновые холиноре-
цепторы делят на рецепторы мышечного и ганглионарного ти-
пов. Мышечные нХР, более чувствительные к бунгаротоксину и тубо-
курарину, локализованы преимущественно в местах контакта оконча-
ний аксонов со скелетными мышцами. Рецепторы ганглионарного
типа, более чувствительные к никотину и бензогексонию, сосредото-
чены в вегетативных ганглиях и хромаффинной ткани. В головном
мозге обнаружены оба типа нХР; максимальная плотность рецепто-
ров ганглионарного типа найдена в ядрах таламуса, а также в гипота-
ламусе и коре больших полушарий. Данный тип рецепторов может
быть локализован не только на пост-, но и на пресинаптических мемб-
ранах; в последнем случае они участвуют в модуляции активности до-
фамин-, серотонин-, глутамат- и пептидергических синапсов. нХР мы-
шечного типа найдены в коре, гипоталамусе, гиппокампе, в некоторых
зонах моста и продолговатого мозга.
н-Холинорецептор состоит из четырех видов субъеди-
ниц — а, р, у и 8; молярное соотношение этих субъединиц в белке —~
2-3. Характеристика индивидуальных медиаторных систем
2 : 1 : 1 : 1» а молекулярные массы составляют 40, 50, 60 и 65 кДа, со-
ответственно. Все субъединицы рецептора гликозилированы, на долю
углеводного компонента приходится около 20 кДа. Субъединицы, поли-
пептидные цепи которых четыре раза пронизывают липидный бислой,
взаимодействуют с белками тубулинового и актинового цитоскелета.
Первичная последовательность всех субъединиц холинорецептора
у различных классов животных.весьма консервативна: гомология пре-
вышает 50%, а для некоторых субъединиц (например, ос-субъединица
электрического ската и человека) — 80%. Из изолированных а-, р- и
8-субъединиц ХР электрического ската и у-субъединицы ХР теленка
удалось реконструировать холинорецептор, полностью обладающий
функциональной активностью.
Пять субъединиц холинорецептора в последовательности а—р—
а—у—8 образуют канал, проницаемый для ионов натрия и калия (в
меньшей степени — для кальция и некоторых органических катионов)
(рис. 2.8). ХР локализован на постсинаптической мембране клетки и
при связывании ацетилхолина изменяет свою конформацию таким об-
разом, что через устье, сформированное канальными субъединицами,
внутрь клетки устремляются ионы Na+. Происходит деполяризация
Рис. 2.8. Схематическое изображение никотинового холинорецептора:
а — общий вид; б — структура ионного канала
130
Раздел 2. МОЛЕЮ 1ЯРНАЯ НЕ ЙРОХИМИЯ
мембраны, что приводит к выходу К+ из клетки. Ток К+ возвращает
потенциал мембраны к исходной величине. В процессе перезарядки
мембраны, осуществляемой потенциалом действия, через этот же ка-
нал внутрь клетки могут входить ионы Са2+. Следовательно, канал не
является высокоизбирательным в отношении катионов. В то же время
это быстродействующая регуляторная система — потенциал дейст-
вия, вызываемый ацетилхолином, возникает и затухает в тканях мозга
за 1 —2 мс, благодаря чему синапс может проводить от аксона на ин-
нервируемую клетку до 500имп/с. В нервно-мышечном синапсе по-
тенциал действия длится значительно дольше (80— 120 мс).
Такое быстрое развитие потенциала действия оказывается воз-
можным благодаря высокой скорости обратимого связывания ацетил-
холина с рецептором, а также эффективному разрушению ацетилхо-
линэстеразой. Разумеется, не менее важен и механизм открывания
канала за счет конформационных переходов, происходящих в наносе-
кундном интервале. Бесперебойное функционирование холинергиче-
ского синапса требует большого запаса ацетилхолина, который синте-
зируется впрок, накапливаясь в везикулах пресинаптической зоны.
Кроме того, в клетках должны существовать достаточно высокие гра-
диенты ионов Na+ и К+ по обе стороны плазматической мембраны,
которые создаются и поддерживаются Na/K-насосом.
Быстродействие ацетилхолина (как и других нейромедиаторов) опре-
деляется также особенностями морфологического образования, находя-
щегося между аксоном и иннервируемой клеткой, — синапса. Благода-
ря тому что расстояние между пре- и постсинаптической мембранами
о
составляет всего 300—500 А, а холинорецепторы сконцентрированы в
виде кластеров вблизи участков секреции ацетилхолина, сразу же после
выброса нейромедиатор оказывается в месте его рецепции. Кроме того,
при раздражении аксона происходит выброс нескольких квантов аце-
тилхолина, так что молекулы этого нейромедиатора насыщают большое
количество ХР и вызывают формирование потенциала концевой плас-
тинки (ПКП). Этот сдвиг трансмембранного потенциала (ТМП) до по-
рогового уровня и вызывает развитие потенциала действия.
Холинергические рецепторы никотинового типа имеют довольно
низкое сродство к ацетилхолину. Полумаксимальное насыщение ре-
цепторов наблюдается в присутствии 10-4М ацетилхолина, поэтому,
как только ацетилхолинэстераза (локализованная в межсинаптиче-
ской области) начинает процесс гидролиза, снижение уровня АХ при-
водит к диссоциации этого нейромедиатора от рецептора и канал воз-
вращается в закрытое состояние.
2.3. Характеристика индивидуальных медиаторных систем
Как известно, сродство рецептора к лиганду (нейромедиатору, гор-
мону или др.) определяется соотношением скоростей диссоциации и
ассоциации лиганд-рецепторного комплекса. При константе диссоци-
ации 10-4—10-3 М скорость диссоциации ацетилхолина от рецептора
составляет доли миллисекунды, что очень важно для быстрого восста-
новления синаптической передачи.
По «канальному» механизму работают многие рецепторы, имею-
щие близкую молекулярную массу и, как правило, состоящие из не-
скольких субъединиц. В зависимости от селективности ионов, про-
пускаемых каналом, эти рецепторы вызывают деполяризацию или
гиперполяризацию мембраны. Вероятно, «ворота» каналов всех
этих рецепторов открываются по единому механизму, так как у неко-
торых субъединиц различных рецепторов найдены гомологичные
участки.
Совершенно иную, отличающуюся от холинергического рецептора
никотинового типа, структуру и другой механизм функционирования име-
ют так называемые мускариновые холинергические рецепторы (тиХР)
(рис. 2.9), которые локализованы преимущественно пресинаптически.
На эти рецепторы не действует никотин, но они легко активируются
мускарином, а также, разумеется, и ацетилхолином, константа сродст-
ва к которому имеет величину порядка 10-6 М.
Существуют по крайней мере пять изоформ мускариновых
рецепторов, причем все они близки по структуре (их полипептидная
цепь семь раз пронизывает мембрану) и сопряжены с G-белками, но
передают сигнал разным системам внутриклеточной сигнализации.
Так, например, мускариновые холинорецепторы могут стимулировать
фосфолипазу С, которая гидролизует фосфоинозитиды (jmXPj 3 5), или
ингибировать аденилатциклазу, а также активировать К-каналы плаз-
матической мембраны (?иХР2, ?иХР4).
Ответ тиХР растянут во времени, эффекты развиваются спустя
одну-две минуты после взаимодействия с лигандом и гасятся за несколь-
ко десятков минут.
Столь разительное отличие в скоростях проведения ацетилхоли-
нового сигнала между никотиновым и мускариновым -рецепторами
объясняется, прежде всего, разной кинетикой связывания лиганда с
соответствующими рецепторами (диссоциация ацетилхолина от мус-
каринового рецептора происходит весьма медленно), многоэтапным
каскадом проведения сигнала в случае мускариновой регуляторной
системы (необходимо последовательное взаимодействие рецептора с
соответствующим G-белком, а затем — G-белка с определенным
132
Раздел 1. МсЛыУлЯРнАч НЕЙРО *ИМИ^
ферментом или каналом), а также сравнительно медленно проте-
кающими реакциями образования (высвобождения) вторичных по-
средников, фосфорилирования (дефосфорилирования) белков-ми-
шеней.
Подтипы (изоформы) тиХР различаются как по локализации
в структурах мозга и в возбудимых клетках различных органов, так и
по вызываемым ими физиологическим эффектам.
Работа этих рецепторов связана с процессами психоэмоционального
восприятия и распознавания образов в центральной нервной системе,
с регуляцией секреторной функции слюнных желез и со многими дру-
гими реакциями организма.
Рис. 2.9. Процессы, происходящие при активации мускариновых холиноре-
цепторов
2.3. Характеристика индивидуальных медиаторных систем
Норадреналин (НА). Наряду с адреналином и дофамином, но-
радреналин относится к катехоламинам. Локализация норадренерги-
ческой системы — мост (голубое пятно, латеральная ретикулярная
формация моста), продолговатый мозг и ядро одиночного тракта (см.
рис. 2.7). Многочисленные (несколько сотен) нейроны голубого пятна
образуют диффузные проекции большой протяженности, достигая
практически всех отделов ЦНС — коры больших полушарий, лимби-
ческой системы, таламуса, гипоталамуса, спинного мозга.
Норадреналин является медиатором во всех постганглионарных
симпатических окончаниях (за исключением потовых желез). В ЦНС,
как правило, он выполняет функции тормозного медиатора (кора), ре-
же — возбуждающего (гипоталамус) медиатора. Норадреналин ней-
ронов голубого пятна способен диффузно выбрасываться в нейро-
пиль.
Нисходящие норадренергические пути спинного мозга вовлекают-
ся в регуляцию мышц сгибателей и сосудистого тонуса. Нарушение
обмена норадреналина связывают с возникновением ряда психоэмо-
циональных расстройств.
Воздействие на метаболизм НА фармакологическими средствами,
компенсирующими дефицит или избыток этого нейромедиатора, в ря-
де случаев способствует снижению симптоматики шизофрении, мани-
акально-депрессивных состояний.
Адреналин. Секретируется диффузно (в мозговом слое надпо-
чечников) и выполняет в первую очередь роль гормона. Тела нейро-
нов, содержащих фермент, осуществляющий метилирование норадре-
налина, обнаруживаются в нижних отделах моста и в продолговатом
мозге. Нисходящие пути достигают центрального серого вещества и
ядер гипоталамуса (в частности, паравентрикулярного и дорзомеди-
ального).
Нейромедиаторная роль адреналина сомнительна; нейромедиато-
ром адренергических нейронов является, очевидно, НА. Адреналин
способен играть роль модулятора, высвобождаясь диффузно.
Дофамин. Системы использования дофамина в качестве нейро-
медиатора сосредоточены в среднем мозге (черная субстанция, вент-
ральная покрышка), обонятельной луковице, гипоталамусе и пери-
вентрикулярной области продолговатого мозга. Локализация дофами-
нергических нейронов в ЦНС показана на рисунке 2.7.
Дофамин — первое нейроактивное вещество в цепи синтеза кате-
холаминов (рис. 2.10). В нейронах он выявляется при обработке сре-
134
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
зов мозга парами формальдегида или глиоксалевой кислотой; при
этом дофамин и близкие к нему моноамины проявляют характерную
флуоресценцию. В настоящее время этот способ выявления дофамина
дополнен иммуногистохимическими и другими биохимическими мето-
диками.
Имеются две основные группировки дофамино-
вых нейронов — ядра гипоталамуса и среднего мозга (дугооб-
разное ядро, черная субстанция и вентральная покрышка). Дофами-
новые нейроны черной субстанции дают проекции в хвостатое и чече-
вицеобразное ядра (которые вместе называются стриатум, или по-
лосатое тело) базальных ганглиев, образующих нигростриарный
путь. При болезни Паркинсона данные клетки дегенерируют, вслед-
ствие чего и происходит утрата дофаминергических синапсов в стри-
атуме. Именно это считают причиной таких типичных для паркин-
сонизма двигательных расстройств, как ограниченность движений и
тремор рук. Для лечения паркинсонизма применяют большие дозы
L-ДОФА — предшественника дофамина, что, к сожалению, чревато
существенными побочными эффектами. Фармакологические препара-
СООН
Тирозин РрСНг- CH-NH2
Тирозингидроксилаза
" соон
ДОФА H°^yCH2-CH-NH2
НО1^
ДОФА-декарбоксилаза
Рис. 2.10. Этапы синтеза норадреналина
2.3. Характеристика индивидуальных медиаторных систем
135
ты, содержащие метаболиты дофамина, проникающие в головной
мозг, способны смягчить симптомы болезни.
Образования лимбической системы, иннервируемые через мезо-
лимбические пути от покрышки среднего мозга, ответственны за такие
функции, как эмоции и агрессивное поведение; что касается лобных
отделов коры, то они играют важнейшую роль в осуществлении выс-
ших когнитивных функций. Есть прямые данные о наличии связи меж-
ду развитием шизофрении у человека с нарушениями как метаболизма
дофамина, так и функционирования дофаминергической медиаторной
системы.
В ряде отделов мозга существуют и «короткоаксонные» дофами-
нергические клетки. В гипоталамусе такие клетки посылают аксоны к
срединному возвышению и оказывают модулирующее влияние на сек-
рецию либеринов нейроэндокринными клетками. Дофаминергические
нейроны сетчатки, обонятельной луковицы и зрительных структур кры-
ши среднего мозга участвуют в организации местных нейронных сетей
(см. гл. 3.6). Кроме того, существует система дофаминергических кле-
ток вокруг четвертого желудочка, простирающаяся внутри ствола моз-
га до гипоталамуса.
Синтез катехоламинов (дофамина, норадреналина, адреналина) осу-
ществляется из общего для всех катехоламиновых медиаторов пред-
шественника — тирозина. На первой стадии аминокислота гидрокси-
лируется ферментом тирозингидроксилазой с образованием диоксифе-
нилаланина (ДОФА). Далее под действием фермента декарбоксилазы
ароматических L-аминокислот из ДОФА образуется дофамин, кото-
рый затем превращается в норадреналин под действием дофамин-р-
гидроксилазы (см. рис. 2.10). Первые две реакции происходят в цито-
плазме, а затем дофамин транспортируется в везикулы, где превраща-
ется в норадреналин дофамин-Р-гидроксилазой, связанной с мембра-
ной везикул. Основная часть норадреналина хранится внутри везикул;
некоторое его количество проникает обратно в цитоплазму, где под-
вергается распаду под действием моноаминоксидазы (МАО).
Дофаминергические нейроны содержат тирозингидроксилазу и де-
карбоксилазу ароматических L-аминокислот, но в них отсутствует до-
фамин-р-гидроксилаза. Другие нейроны, так же как и клетки надпо-
чечников, высвобождают адреналин, который образуется из норадре-
налина поддействием фенилэтаноламин-М-метилтрансферазы.
Как правило, такие многоступенчатые процессы регулируются за
счет способности конечного продукта к ингибированию первого фер-
мента. Тирозингидроксилаза ингибируется как норадреналином, так
136
г ♦ j,4._ n 2. МОпЕК1 ййН.ч-. ЕЙР< -HM.i-
и дофамином и адреналином, по мере накопления которых происходит
торможение их синтеза и установление состояния равновесия: ско-
рость синтеза уравнивается со скоростью распада и высвобождения
(см. рис. 2.10).
Доказательством того, что лимитирующий скорость процесса этап
контролируется по принципу отрицательной обратной связи, явились
эксперименты по изменению концентрации норадреналина в термина-
ли. Добавление медиатора в инкубационную среду приводило к уве-
личению его захвата терминалью и снижению скорости его синтеза,
а стимуляция, вызывающая выброс норадреналина, наоборот — уси-
ливала синтез медиатора.
Описанные выше регуляторные механизмы работают быстро, изме-
няя скорость синтеза в нервных окончаниях. Но существуют также ме-
ханизмы долговременной регуляции. Прекрасным примером такой ре-
гуляции является ответ симпатической нервной системы у животных,
подвергнутых продолжительному стрессу. При действии стресса на ор-
ганизм активируются симпатические нейроны, что ведет к трех-, четы-
рехкратному увеличению уровней тирозингидроксилазы и дофамин-0-
гидроксилазы в телах и терминалях симпатических нейронов, происхо-
дящему благодаря синтезу новых молекул ферментов. Другие фермен-
ты синтеза и деградации норадреналина, такие как декарбоксилаза аро-
матических L-аминокислот и МАО, не подвержены такому влиянию.
Повышенный уровень глюкокортикоидов при продолжительном стрес-
сорном воздействии индуцирует синтез катехол-М-метилтрансферазы,
что способствует ускорению образования адреналина.
Адренорецепторы. Присутствуют в нервной ткани в относи-
тельно небольшом количестве. В зависимости от связывания адрено-
тропных лигандов адренорецепторы подразделяют на два клас-
са — а и р, которые в свою очередь могут быть подразделены на
подклассы (рис. 2.11). Первоначально p-рецепторы были описа-
ны в различных периферических тканях: Pj *— в сердце, как усиливаю-
щие силу и частоту сокращений, Р2 — в бронхах, как вызывающие ре-
лаксацию гладких мышц.
Все адренорецепторы относятся к суперсемейству рецепторов,
связанных с G-белками, и обладают типичной 7-доменной трансмемб-
ранной структурой. 0Cj-Подкласс характеризуется высоким сродством
к антагонисту празозину и низким сродством к йохимбину. Их актива-
ция приводит к увеличению внутриклеточного содержания Са2+ через
включение фосфолипазы С и ИФ3/ОАС-зависимых путей регуляции.
Возможны также воздействия на дегидропиридинчувствительные и де-
2.3. V> . -.Pi . . .'НдИеР^уо HU* Мг.Д1о1срНЬ1>СИ -M
13
Адренорецепторы
alL a1A a1B alD a2A/D a2B a2C
Рис. 2.11. Классификация адренорецепторов
гидропиридиннечувствительные Са-каналы, фосфолипазу D, фосфоли-
пазу Д2, синтез арахидоновой кислоты и активацию протеинкиназы С,
а также активацию MAP-киназы, играющей важную роль в пролифе-
ративных процессах.
Для ос2-подкласса адренорецепторов характерно высокое сродство
к йохимбину. Их активация приводит к тормозным эффектам благода-
ря уменьшению синтеза цАМФ, усилению выхода К+ и уменьшению
выхода Са2+. Различные изоформы этих рецепторов отличаются срод-
ством к празолину: ос2А-изоформа (из тромбоцитов) обладает низ-
ким, а а2В-изоформа (из неонатальных легких) — высоким сродством
к празолину.
В отличие от ос2-адренорецепторов, p-адренорецепторы связаны с
Сделками и обладают высоким сродством к пропраналолу; они уси-
ливают синтез цАМФ. Исключением из этой группы является толь-
ко р3-изоформа, которая не обнаружена в мозге. Степень связывания
Р"рецепторов с различными катехоламинами сильно различается, в то
время как для ос-адренорецепторов подобные различия менее выра-
жены.
Для а2- и р2-адренорецепторов характерна пресинаптическая ло-
кализация, в то время как для остальных адренорецепторов — пре-
имущественно постсинаптическая. Адренорецепторы широко пред-
ставлены в периферических тканях. В мозге они обнаружены в коре
больших полушарий и в мозжечке (ос2А-подтип), в хвостатом ядре
(а2А- и а2В-подтипы) и в ряде других структур. Несмотря на то что чис-
ло адренорецепторов в мозге относительно невелико, они играют за-
метную роль в регуляции психоэмоционального состояния, и наруше-
ние их работы вносит определенный вклад в развитие ряда нейропато-
логий (шизофрения, депрессивные состояния и др.).
138
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИ*
Дофаминовые рецепторы. Активируются дофамином, являют-
ся метаботропными и осуществляют свои функции, изменяя актив-
ность аденилатциклазы. Выделяют несколько их подклассов, ко-
торые различаются по сродству к дофамину и его агонистам (бром-
криптину и апоморфину) и антагонистам (хлоропромазину, гало-
перидолу, клозапину), локализации в структурах мозга, направленнос-
ти влияний на аденилатциклазу и способности тормозить выброс про-
лактина (табл. 2.2).
Дофаминовые рецепторы участвуют в контроле двигательных
функций (стереотипии, вращение при разрушении нигро-стриарного
тракта и др.) и психических реакций. Они вовлечены также в работу
системы внутреннего подкрепления. Нарушение функции дофамино-
вых рецепторов играет заметную роль в патогенезе таких заболева-
ний, как паркинсонизм, шизофрения и др.
Таблица 2.2. Характеристика дофаминовых рецепторов
Класс D1 D2
Подкласс D1A DIB D2A D2B D2C
Классификация D1 D5 D2 D3 D4
Количество аминокислотных остатков (человек) 446 477 414/443 440 387
Сродство к дофамину Низкое Среднее Среднее Высокое Высокое
Активация АЦ t t l (!) (!)
Оборот ИФ3 (t) — t — —
Вход Са2+ (t) — 1 1
Выход К+ — — t — —
Распределение в мозге
Стриатум -1- -1-
Гиппокамп -1- -1- +
Гипоталамус -1- -1- -1-
Черная субстанция -I- -I-
2.3. Характеристика индивидуальных медиаторных систем
13
Серотонин (5-гидрокситриптамин, 5-НТ). Относится к моно-
аминам. Серотонинергические нейроны составляют ядра шва в рост-
ральной части моста мозга (см. рис. 2.7). Они дают проекции на лим-
бическую систему,.базальные ганглии, кору больших полушарий. Зо-
ны иннервации серотонинергических нейронов сходны с проекциями
норадренергических нейронов. В продолговатом мозге находятся се-
ротонинергические нейроны, аксоны которых образуют нисходящие
пути в ствол мозга и в спинной мозг. Кроме того, серотонин обнаружен
в нейронах гипоталамуса, черной субстанции, спинном мозге. Серото-
нин синтезируется и вне центральной нервной системы, особенно вы-
соко его содержание в тромбоцитах и кишечнике.
Серотонинергическая система играет важную роль в регуляции
двигательной активности, эмоционального, пищевого и полового по-
ведения, терморегуляции, участвует в контроле нейроэндокринных
систем. Серотонин может выполнять функцию не только нейромеди-
атора, но и нейромодулятора. Серотонинергическая система, так же
как и норадренергическая, по-видимому, оказывает генерализованное
влияние на цикл «сон—бодрствование», восприятие сенсорных и бо-
левых сигналов и высшие когнитивные функции. Этот нейромедиатор
(наряду с рядом других) обеспечивает функционирование системы
внутреннего подкрепления.
В опытах на головном мозге позвоночных были выявлены и воз-
буждающие, и тормозные эффекты серотонина. Как показали недав-
ние исследования, такие препараты, как LSD и трициклические анти-
депрессанты, воздействуют на серотонинергическую систему как на
пресинаптическом, так и на постсинаптическом уровне. Есть данные о
том, что у депрессивных больных содержание серотонина в головном
мозге снижено; лекарственные средства, способные понижать в си-
напсе уровень моноаминовых нейромедиаторов (дофамин, норадрена-
лин и серототин), вызывают депрессию, тогда как препараты, приме-
няемые для лечения депрессии, обычно повышают содержание этих
нейромедиаторов или усиливают их действие.
Серотонин синтезируется из триптофана. Первая (лимитирующая)
стадия этого процесса протекает с помощью фермента триптофангид-
роксилазы, который превращает триптофан в 5-гидрокситриптофан
(рис. 2.12). Последний декарбоксилируется с образованием серотони-
на при участии декарбоксилазы ароматических L-аминокислот, т. е.
того же самого фермента, который превращает ДОФА в дофамин. Вы-
свобождение серотонина из нейронов во время их стимуляции вызы-
вает повышение скорости превращения триптофана в 5-гидрокси-
триптофан. Предполагается, что это происходит благодаря изменению
140
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
NH
СООН
I
сн2—CH—nh2
Моноаминоксидаза А
Альдегиддегидрогеназа
5-Гидрокси индолуксусная
кислота
Рис. 2.12. Этапы синтеза и метаболизма серотонина
свойств триптофангидроксилазы, вызванному Са-зависимым фосфо-
рилированием, сходным с эффектами стимуляции тирозингидроксила-
зы. Как и в случае тирозингидроксилазы, для функционирования трип-
тофангидроксилазы необходим кофактор тетрагидробиоптерин, при-
сутствие которого будет определять возможность синтеза серотонина.
Если в середине прошлого века было известно лишь два класса ре-
цепторов серотонина (5-гидрокситриптамина, 5-НТ), то к настоя-
щему времени их уже четырнадцать (табл. 2.3). Подавляющее
большинство рецепторов серотонина — метаботропные рецепторы,
связанные с G-белками и имеющие характерную 7-доменную транс-
Таблица 2.3. Характеристика серотониновых рецепторов
Тип Действие Подтип Вторичный посредник1 Селективный/ преимущественный агонист Селективный антагонист
5-НТ1А Gi/o/z —► | цАМФ 8-OH-DPAT WAY 100635
5-HT1b/d Gi/o —► 1 цАМФ СР 93129 SB 224289
5-HTj Гиперполяризация 5-НТю N/A BRL 15572
5-НТ1Е Gi/o(?) — | цАМФ N/A N/A
5-HT1f LY 334370 N/A
5-НТ2А Gq/11 —► активация фосфолипазы С, 1 калиевой проводимости, | входа Са2+, активация фосфолипазы А2 и СР 94253 MDL 100907
5-НТ2 Деполяризация 5-НТ2В BW 273С86 SB 204741
5-НТ2С пути арахидоновой кислоты МК212 SB 242084
5-НТ3 Деполяризация Лигандуправляемый Na/K- канал 2-Метилсеротонин Ондасетрон
5-НТ4 Деполяризация Gs —> f цАМФ SDZ 216454 GR 113808
5-НТ5 Не известен Не известен N/A2 N/A
5-НТ6 Деполяризация Gs —* Т цАМФ N/A SB 271046
5-НТ7 Деполяризация N/A SB 258719
2.3. Характеристика индивидуальных медиаторных систем
1 Свойства G-белков см. в гл. 2.4.
2 N/A — селективный агонист или антагонист отсутствует.
142
Раздел 2. ML.nt КУЛЛРНл.я HI ЙРо,ИМИ>.
мембранную структуру. Они найдены как в разных структурах мозга
(кора больших полушарий, лимбическая система, базальные ганглии,
миндалевидный комплекс, черная субстанция, область вентральной
покрышки и др.), так и на периферии. Их локализация может быть как
пре-, так и постсинаптической. Исключением является подтип серото-
нинового рецептора 5-НТ3; он относится к быстродействующим
ионотропным рецепторам, канал которых пропускает ионы К+ и Na+.
Кроме того, некоторые из этих рецепторов регулируют выделение
других медиаторов (дофамина, норадреналина, пролактина и глутама-
та) и действуют как гетерорецепторы. В зависимости от типа, они мо-
гут использовать в качестве эффекторных ферментативных систем
аденилатциклазу или фосфолипазу.
Рецепторы серотонина участвуют в регуляции циркадных ритмов и
терморегуляции. Некоторые типы серотониновых рецепторов явля-
ются компонентами системы внутреннего подкрепления. Из поведен-
ческих и физиологических эффектов, связанных с этими рецепторами,
следует отметить состояния страха, паники, психозы, навязчивые со-
стояния, галлюциногенные эффекты.
Существует характерный двигательный синдром, регулируемый
5-НТ2-рецепторами, — синдром «отряхивания мокрой собаки» («wet
dog shakes»), его используют в качестве теста в исследованиях серо-
тонинергической системы.
Периферические эффекты активации рецепторов серотонина всех
классов весьма многообразны. Главные из них состоят в сокращении
сосудов, в реализации положительных хроно- и инотропных воздейст-
вий на сердце.
Гистамин. Гистаминергическая система весьма своеобразна по
своей локализации и функциям. Важным этапом исследования лока-
лизации элементов гистаминергической системы явилось открытие
специфического ингибитора синтеза гистамина — а-фторметилгиста-
мина. Нейроны, продуцирующие гистамин; сосредоточены в ограни-
ченной области мозга — туберомамиллярных ядрах заднего гипотала-
муса. Эта группа нейронов распространяет свои эфферентные волок-
на практически на все отделы мозга: повсеместно обнаруживаются
как постсинаптические, так и пресинаптические рецепторы гистами-
на. Столь широкая локализация рецепторов гистамина позволяет по-
нять разнообразные функции гистаминергической системы. Гистамин
стимулирует двигательную активность, половое поведение, участвует
в регуляции цикла «сон—бодрствование», подавляет восприятие бо-
ли. Усиливая жажду, он подавляет инстинкт добывания пищи. Гиста-
jp. ларактеристика индивидуальных медиаторных систем
мин входит в число факторов, которые участвуют в повышении арте-
риального давления, в терморегуляции (снижении температуры тела)
и (косвенно) в стимуляции энергетического обмена мозга, усиливая
кровообращение. Интересно, что гистамин реализует эти функции не
только в рамках классических синапсов, часть гистамина выделяется
так называемыми открытыми нервными окончаниями и способна к
распространению по межклеточным жидкостям, в том числе через
ликвор. Многие гистаминергические нейроны содержат и другие ме-
диаторы, такие как ГАМК, вещество Р или энкефалин.
Рецепторы гистамина. К настоящему времени выявлено и ис-
следовано три класса рецепторов гистамина: Hj и Н2 — постси-
наптические и Н3 — пресинаптические. Последние найдены не только
в гистаминергических, но и в терминалях аминергических, холинерги-
ческих и пептидергических нейронов.
Все три класса рецепторов связаны с G-белками и являются мета-
ботропными. Активация Hj-рецепторов приводит к образованию ИФ3
и DAG, а Н2 — к активации аденилатциклазы.
Ингибиторами Ht-рецепторов служат димедрол, фенкарал, супрас-
тин, получившие широкое применение в медицине. Хорошо изученным
ингибитором Н2-рецепторов является циметидин — известный проти-
воязвенный агент. Пресинаптические рецепторы гистамина класса Н3
являются ауторецепторами, подавляющими выход гистамина из нерв-
ных окончаний.
Гистаминовые рецепторы обнаруживаются во многих структурах
мозга: коре, гиппокампе, лимбических структурах и гипоталамусе.
Отмечается участие этих рецепторов в регуляции цикла «сон—бодр-
ствование».
Аминокислотные нейромедиаторы в соответствии с их функцией
делятся на две группы — возбуждающие (глутамат, аспартат)
и тормозные (ГАМК, глицин, таурин).
Глутаминовая кислота. Является главным возбуждающим ме-
диатором, выявляется во всех отделах ЦНС (см. рис. 2.7), очевидно,
благодаря тому, что служит не только нейромедиатором, но и пред-
шественником других аминокислот (см. гл. 3.1). Проекции глутама-
тергических нейронов находятся в коре больших полушарий, обоня-
тельной луковице, гиппокампе, черной субстанции, мозжечке, сетчат-
ке. Глутаматергические синапсы обнаружены в миндалине, стриатуме,
на клетках-зернах мозжечка. В спинном мозге глутамат выполняет
144
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
функцию нейромедиатора в первичных афферентных волокнах дор-
сальных корешков.
Выявление глутаматергических связей в головном мозге проводит-
ся в соответствии с идентифицированными местами высвобождения
нейромедиатора, а также с помощью моно- и поликлональных антител
к глутаматным рецепторам.
Нарушение глутаматергической передачи связано с целым рядом
патологических состояний нервной системы: эпилепсией, расстройст-
вами вестибулярной системы, ишемическим повреждением мозга.
Глутаминовая кислота практически не проникает через гематоэнце-
фалический барьер (ГЭБ), поэтому в качестве терапевтического ле-
карственного средства при хронической недостаточности аминокис-
лотного обмена, вегетососудистой дистонии и эпилепсии рассматри-
ваются некоторые ее аналоги, способные преодолевать ГЭБ.
Глутамат, являющийся основным возбуждающим медиатором
в мозге, может синтезироваться несколькими пу-
тями. В нейронах глутамат, использующийся как нейромедиатор,
первоначально синтезируется из углерода окисляющейся глюкозы че-
рез стадию образования кето глутарата в реакциях ЦТК.
После выброса в синаптическую щель и взаимодействия с рецеп-
торами пресинаптической мембраны диссоциировавший от рецепто-
ров нейромедиатор, а также избыточные количества
глутамата удаляются из синаптической щели преимущест-
венно двумя способами. Небольшие количества глутамата
подвергаются «обратному захвату» и относительно малоактивным
транспортером возвращаются в пресинаптическое окончание. Основ-
ная часть глутаминовой кислоты поглощается астроцитами, причем
в данном случае действует очень активный транспортер глутамата.
В астроцитах глутаминовая кислота аминируется глутаминсинтазой до
глутамина. Следует отметить, что глутаминсинтаза — очень активный
фермент, локализованный преимущественно в этом типе глиальных
клеток. О важности глутаминсинтазной реакции для мозга говорит тот
факт, что на долю этого фермента приходится до 0,2% суммарного
содержания белка в ткани.
Из астроцитов глутамин поступает в пресинаптические окончания
глутаматергических нейронов, где превращается в глутамат при помо-
щи активируемой фосфатом формы фермента глутаминазы. Специ-
фичный для мозга транспортер неорганического фосфата локализован
избирательно на терминалях глутаматергических нейронов и, таким
образом, может регулировать пополнение пула глутамата в пресинап-
тических окончаниях.
2.3. характеристика индивидуальных медиаторных систем
145|
Частично глутамин, образовавшийся в астроцитах, может выде-
ляться в кровь; таким образом из мозга удаляется избыток ионов
NH^-. Но вместе с ним из мозга удаляется и глутаминовый «скелет».
Для восполнения пула глутаминовой кислоты, выполняющей меди-
аторные функции, необходимо постоянное пополнение запасов глута-
мата, осуществляющееся за счет а-кетоглутаровой кислоты.
Глутаматные рецепторы. Глутаматергическая система, наибо-
лее широко распространенная в мозге, представлена как ионотропными,
так и метаботропными рецепторными структурами (рис. 2.13). Их иден-
тифицируют по лигандам, избирательно активирующим или ингибирую-
щим тот или иной класс рецепторов. Ранее эти рецепторы подразделяли
на NMDA-рецепторы и He-NMDA-рецепторы, с делением последних
на взаимодействующие с а-амино-3-гидрокси-5-метил-4-изоксазолпро-
пионовой кислотой (АМРА), каиновой кислотой и квисквалоновой
Рис. 2.13. Схема, отражающая взаимодействие глутамата с различными клас-
сами глутаматных рецепторов.
mGluR — метаботропный глутаматный рецептор; AMPA-R — АМРА-рецептор;
NMDA-R — NMDA- рецептор
146
► аздел 2. Mt ,’lf КУЛяРН^Я HF ЙРЧ ХИМИЯ
Таблица 2.4. Классификация глутаматных рецепторов
Глутаматные рецепторы Функциональная характеристика
Изоформы Основной агонист Анта- гонисты Метаболическая функция
NR1 NR2A NR2B NR2C NR2D NR3A NMDA D-AP5 MK-801 Активация ионных каналов (Са2+, Na+)
ИОНО- ТРОПНЫЕ GluRI GluR2 GluR3 GluR4 АМРА CNQX Активация ионных каналов (Na+)
GluR5 GluR6 GluR7 КА1 Каиновая кислота DNQX Активация ионных каналов (Na+)
КА2
1 группа (mGlui, mGlu5) 3,5-DH- PG MCPG Активация ИФ3-зависимого обмена кальция
МЕТАБО- ТРОПНЫЕ 2 группа (mGlu2, mGlu3) L-CCG, NAAG PCCG4 Повышение уровня цАМФ
3 группа (mGlu4, mGlu6, mGlu7, mGlu8) L-AP4 ACPD MSOP LY341495 Повышение уровня цАМФ и понижение уровня цГМФ
кислотой. В настоящее время классификация глутаматных рецепто-
ров более детализирована (табл. 2.4).
Ионотропные рецепторы обеспечивают формирование возбуж-
дающих импульсов. Глутаматные рецепторы, активируемые
NMDA (N-метил-О-аспартатом), локализованы преимущественно в
коре мозга и гиппокампе, где плотность их максимальна; до 70—80%
синапсов этих структур мозга обладают именно такими рецепторами.
Они обнаружены также в сенсорно-ассоциативной системе и в ба-
зальных ганглиях. NMDA-рецепторы ответственны за молекулярные
механизмы запоминания, ориентации в новой обстановке, за консоли-
дацию памяти (так называемые процессы долговременной памяти),
2.3. Характеристика индивидуальных медиаторных систем
147
принимают участие в формировании нейрональной пластичности,
в процессах интегративной деятельности мозга.
Большое внимание к NMDA-рецепторам обусловлено тем, что
благодаря этим рецепторам осуществляется медленная длительная
деполяризация нейронов. Именно с их функцией связаны долговре-
менная потенциация в гиппокампе, процессы ноцицепции, эпилепти-
формная активность, а также экзайтотоксические эффекты глутамата.
NMDA-рецептор имеет гетеромерную структуру. Он состоит из од-
ной NR1- и нескольких NR2-субъединиц, которые определяют функ-
циональное разнообразие его изоформ. NMDA-рецептор не является
специфичным каналом, поскольку может пропускать как ионы каль-
ция, так и ионы натрия.
Молекула NMDA-рецептора содержит пять различных участ-
ков, взаимодействие с которыми приводит к изменению его актив-
ности: участок связывания нейромедиатора; участок внутри канала,
который связывает фенциклидин и родственные соединения; модуля-
торный участок связывания полиаминов; стрихниннечувствительный
участок связывания глицина, являющегося коагонистом глутамата, а
также потенциалзависимый Mg-связывающий участок, позволяю-
щий блокировать канал, который восстанавливает свою работоспо-
собность только после деполяризации мембраны.
Следует обратить внимание на специфику регуляции NMDA-глута-
матных рецепторов, отличающую их от других известных ионотропных
рецепторов. Прежде всего, наряду с хемочувствительностью к лиганду
они обладают выраженной зависимостью от величины электрического
потенциала, что обусловлено потенциалзависимой блокадой канала
ионами Mg2+. Далее, NMDA-рецепторы весьма чувствительны к вели-
чине pH среды; частота открытия ионного канала снижается по мере
возрастания концентрации водородных ионов. Еще одна особенность
этих рецепторов — наличие коактиватора (глицина); связывание гли-
цина абсолютно необходимо для функционирования рецептора, по-
скольку в его отсутствие рецептор не активируется глутаматом.
Ионотропные АМРА и каинатные рецепторы (в большинстве
натриевые каналы) являются гомо- или гетеромерными пентамерами.
В большинстве областей ЦНС с активацией этих рецепторов связан
вход Na+ в клетку и быстрое формирование мембранного потенци-
ала (в виде деполяризации). Большая часть каналов каинатных и
АМРА-рецепторов не проницаемы для Са2+, в то же время вызывае-
мая ими деполяризация мембраны активирует потенциалзависимые
Са-каналы, что позволяет зарегистрировать Са-токи при активации
нейронов каинатом или АМРА.
148
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Интересно, что каинатные рецепторы Glu5R локализуются на пре-
синаптической мембране нейронов, выделяющих глутамат, и являют-
ся, таким образом, возбуждающими ауторецепторами, усиливающими
выброс медиатора в синаптическую щель. Наблюдение процесса фор-
мирования синаптических контактов обнаружило явление «переска-
кивания» каинатных рецепторов с постсинаптической зоны на преси-
наптическую мембрану, сопровождающее совершенствование синап-
тического проведения.
Метаботропные рецепторы глутамата, с характерной для этой
группы рецепторов 7-доменной мембранной структурой, имеют раз-
нообразный состав; к настоящему времени описано восемь типов этих
рецепторов. Для них характерна как пост-, так и пресинаптическая
локализация и экспрессия не только в нейрональных, но также и в
глиальных клетках. Эти рецепторы принимают участие в формиро-
вании механизмов памяти, восприятия боли, тревожных состояний,
а также в процессах нейродегенерации.
Следует добавить, что во многих случаях ионотропные (как NMDA,
так и не-NMDA) и метаботропные глутаматные рецепторы локализо-
ваны на мембранах вблизи друг от друга и действие их является согла-
сованным и взаимодополняющим.
Аспарагиновая кислота. Наиболее высокое содержание аспар-
тата найдено в среднем мозге. В спинном мозге аспартат содержится в
дорзальном и вентральном сером веществе. Предполагается нейроме-
диаторная роль аспартата в возбуждающих интернейронах, которые
регулируют различные спинномозговые рефлексы.
у-Аминомасляная кислота (ГАМК). Основным тормозным ме-
диатором головного мозга служит у-аминомасляная кислот. Эта спе-
цифическая аминокислота, в отличие от обычных аминокислот, не
входит в состав первичной структуры белков и синтезируется исклю-
чительно в головном и спинном мозге. ‘Не менее трети (у некоторых
видов до 50%) синапсов головного мозга используют в качестве тор-
мозного медиатора ГАМК. Области распространения ГАМК-ергиче-
ских нейронов показаны на рисунке 2.7 (с. 126). Интересно, что боль-
шая часть данных нейронов образует лишь внутренние связи в таких
отделах, как кора большого мозга, обонятельные луковицы, гиппо-
камп, мозжечок и сетчатка (исключение составляют лишь короткие
проекционные пути от стриатума к черной субстанции и от коры моз-
жечка к его ядрам). К областям, содержащим наиболее высокую кон-
центрацию ГАМК, относятся черная субстанция, бледный шар, гипо-
й
2.3. Характеристика индивидуальных медиаторных систем
14
таламус и мозжечок. Эта аминокислота содержится преимущественно
в сером веществе головного и спинного мозга. Высокие концентрации
ГАМК найдены также в горизонтальных клетках сетчатки; предпола-
гается, что ГАМК обеспечивает обратную связь и латеральное тормо-
жение в слое горизонтальных клеток (см. гл. 3.6).
Для выявления ГАМК в мозге используется широкий набор мето-
дов, в том числе иммуноцитохимическое выявление глутаматдекар-
боксилазы — фермента, участвующего в синтезе этого медиатора, а
также иммуногистохимическое выявление ГАМК-рецепторов и иссле-
дование распределения радиоактивной ГАМК.
ГАМК-ергическая система, как основная тормозная система моз-
га, участвует в реализации многих функций, в том числе в регуляции
моторной активности, формировании эмоционального поведения, в
осуществлении условных рефлексов, организации процессов обуче-
ния и памяти у млекопитающих. При этом она тесно взаимодействует
с другими нейромедиаторными системами мозга: дофаминергической,
холинергической, глутаматергической и др., модулируя их функции.
Существует большое количество данных о вовлечении системы
ГАМК в механизмы многих расстройств нервной системы. Например,
для хореи Гентингтона — наследственного неврологического заболе-
вания — характерен специфический дефицит ГАМК в мозге. При
этом заболевании возникают непроизвольные движения и происходит
постепенное разрушение полосатого тела. Нарушение работы
ГАМК-ергической системы имеет место при эпилепсии, тревожных
состояниях и ряде других нейропатологий.
Реакция синтеза ГАМК была впервые описана для мозга Е. де Ро-
бертасом в 1959 г. как часть специфического пути метаболизма глю-
козы, который позволяет миновать некоторые реакции цикла Кребса
(ГАМК-шунт). Благодаря этому а-кетоглутарат превращается в глу-
тамат, затем в ходе декарбоксилирования — в ГАМК. После выпол-
нения медиаторной функции ГАМК вначале превращается в янтарный
полуальдегид, а затем окисляется до сукцината, который может быть
далее окислен в ЦТК(рис. 2.14).
ГАМК синтезируется из глутамата ферментом глутаматдекарбо-
ксилазой (ГАД), обнаруженной только в ГАМК-ергических нейронах.
Таким образом, главная функция ГАМК-шунта заключается не в
окислении а-кето глутарата с образованием сукцинил - КоА (т. е. мета-
болита, способного участвовать в субстратном фосфорилировании для
получения макроэргов), а в синтезе основного тормозного медиатора
ГАМК (см. гл. 3.2).
150
Раздел 2. МОЛЕКУЛЯРНАЯ НЕИРОХИМИЯ
ГАМК-ергический нейрон
Астроцит
Глутаматергический нейрон
fПостсинаптический нейрон
^Постсинаптический нейрон )
Фермент @ Везикула Митохондрия О Транспортеры глутамина О ГАМК
Рис. 2.14. Участие нервных и глиальных клеток в ГАМК-глутамат-глутамино-
вом цикле (на фоне схематического изображения митохондрий}’.
ДГПЯК — дегидрогеназа полуальдегида янтарной кислоты; ГАД — глутаматдекарбокси-
лаза; ГАМК-Т — 4-аминобутираттрансаминаза; ГС — глутаминсинтаза; АТ — амино-
трансфераза; ГДГ — глутаматде гидро ген аза; ФАГ — фосфатактивируемая глутаминаза
Для синтеза ГАМК также характерен механизм регулирования по
принципу отрицательной обратной связи, поскольку в тормозных ней-
ронах коры физиологические уровни ГАМК ингибируют ГАД. В мозге
млекопитающих имеется несколько дополнительных регуляторов син-
теза ГАМК, включая АТФ, неорганический фосфат и кофактор пири-
доксальфосфат. В мозге присутствуют две формы глутаматдекарбо-
ксилазы: ГАДб7, которая имеет высокое сродство к пиридоксальфос-
фату и поэтому постоянно находится в активированном состоянии, и
ГАДб5, которая имеет низкое сродство к пиридоксальфосфату, ее ак-
тивность может изменяться благодаря доступности этого кофактора.
Обратный захват ГАМК характерен как для нейронов (протекает
медленно), так и для глиальных клеток — преимущественно астроци-
тов, которые активно поглощают ГАМК с помощью высокоаффинной
транспортной системы. В астроцитах ГАМК метаболизирует либо че-
рез стадию янтарного полуальдегида до сукцината с последующим ис-
пользованием его в ЦТК, либо до глутамина. С использованием меха-
низмов специфического транспорта глутамин из астроцитов пере-
носится в пресинаптические области ГАМК-ергических нейронов, где
используется для синтеза новых порций медиатора ГАМК (см.
рис. 2.14).
2.3. Характеристика индивидуальных медиаторных систем
151
Рис. 2.15. Структура ГАМКд-рецептора и формируемого им ионного канала:
а — субъединица с четырьмя трансмембранными участками, участками внеклеточного
гликозилирования и участком фосфорилирования на внутриклеточной петле между М3 и
М4; б — ассоциация пяти субъединиц, осуществляющая благодаря М2-домену каждой
субъединицы при стехиометрии 2ос : 2р : 1у; в — возможные комбинации субъединиц
бензодиазепинчувствительного и бензодиазепиннечувствительного рецепторов (у-субъ-
единица заменена на 8-субъединицу), а также рецептор с я-субъединицей
ГАМК-рецепторы. Их делят на три класса. Ионотропные
ГАМКд- и ГАМКс-рецепторы формируют мембранные каналы, и их
активация ведет к увеличению проницаемости для С1“ и НСО3, соот-
ветственно.
Наиболее хорошо исследованный ГАМКд-рецептор обладает высо-
кой чувствительностью к антагонисту — бикукулину. Он состоит из пя-
ти субъединиц, каждая из которых имеет четыре трансмембранных до-
мена (рис. 2.15). Описано семь разных субъединиц — а, 0, у, 5, е, я, 0,
причем для первых трех описаны разные изоформы (а1— б, 0j_3, Yi— 3).
На внутриклеточных петлях 0- и у-субъединиц расположены участки,
которые могут фосфорилироваться различными протеинкиназами.
152
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Работа рецептора модулируется целым рядом соединений, в том
числе фармакологическими препаратами (пикротоксином, анестети-
ками, бензодиазепинами, стероидами, барбитуратами), причем эти ве-
щества связываются с рецептором в участках, отличных от мест свя-
зывания с ним ГАМК, ее агонистов или антагонистов. Особое место
среди этих соединений занимают бензодиазепины в связи с их широ-
ким применением в качестве лекарственных препаратов снотворного,
противосудорожного, нейротропного действия. Детальные исследова-
ния механизмов взаимодействия бензодиазепинов с ГАМКд-рецеп-
торным комплексом привели к открытию эндогенных лигандов пеп-
тидной природы — эндозепинов. Присоединение этих регуляторных
пептидов к бензодиазепиновому сайту тормозит открытие С1-канала,
вызванное присоединением медиатора (ГАМК) к рецептору. Препара-
ты группы бензодиазепинов блокируют сайт связывания эндогенных
эндозепинов и удлиняют продолжительность существования открытых
ионных каналов, не влияя на их число и скорость транспорта хлора.
Барбитураты, в отличие от бензодиазепама, способны активировать
хлорные каналы и в отсутствие ГАМК.
ГАМКв-рецептор представляет собой связанный с G-белком ме-
таботропный рецептор; он влияет на активность аденилатциклазы,
подавляет выделение медиатора путем торможения Са-каналов или
гиперполяризирует постсинаптическую клетку в результате прямой
активации К-каналов (рис. 2.16). Эти рецепторы локализованы глав-
ным образом в периферической нервной системе. В ЦНС данный тип
рецепторов обнаружен на пресинаптических терминалях, выделяю-
Рис. 2.16. Структура и принцип функционирования ГАМКВ-рецептора.
Активированная Р/у-субъединица запускает открытие канала, а ос-субъединица оказы-
вает тормозящее действие на аденилатциклазу
2.3. Характеристика индивитальных 11- диаторных систем
is:
Ё1
I
щих различные медиаторы, в том числе и ГАМК. Антагонистом данно-
го типа рецепторов служит баклофен.
ГАМ!^-рецептор имеет сходную с ГАМКд-рецептором пентамерную
структуру, но в его состав входит специфическая субъединица р. Его от-
личительными чертами являются нечувствительность к бикукулину и
способность формировать гомомерный рецепторный комплекс только из
р-субъединиц, обладающий более высоким сродством к ГАМК, чем мно-
гие ГАМКд-рецепторы, состоящие из комбинации а, [3 и у-субъединиц.
Глицин. Выполняет нейромедиаторную роль в стволе головного
мозга и в спинном мозге млекопитающих, где опосредует постсинап-
тическое торможение мотонейронов, высвобождаясь из окончаний
клеток Реншоу. Глицин является нейромедиатором также в тормозных
интернейронах промежуточного мозга и ретикулярной формации про-
долговатого мозга. Для глутаматных рецепторов определенного клас-
са (см. NMDA-рецепторы) он является модулятором. В сетчатке наря-
ду с ГАМК глицин выполняет функции тормозного медиатора.
Локализация глицинергических нейронов устанавливается авторадио-
графически. У больных с некоторыми врожденными метаболическими
аномалиями, связанными с повышенным содержанием глицина в ткани
мозга и в крови, может развиваться гиперглицинемия, сопровождаю-
щаяся симптомами нарушения ряда психоэмоциональных функций.
Глициновые рецепторы. Эти рецепторы активируются рядом
аминокислот, такими как глицин, 0-аланин, таурин, и избирательно
блокируются стрихнином. Наибольшая их плотность обнаружена в
области ядер подъязычного и тройничного нервов, локализованных в
продолговатом мозге. Участки связывания стрихнина найдены также
в ретикулярных ядрах продолговатого мозга, моста, среднего мозга,
в передних и задних рогах серого вещества спинного мозга.
Строение глицинового рецептора аналогично таковому у ГАМКд-ре-
цептора, с той лишь разницей, что в состав пентамера входят четыре
а-субъединицы и одна [3-субъединица. На постсинаптической мембране
эти рецепторы образуют канал, обеспечивая проницаемость для хлора,
за счет чего при возбуждении способны формировать тормозный пост-
синаптический потенциал (ТПСП).
Находясь на пресинаптической мембране, глициновые рецепторы
способны вызывать ее деполяризацию, активируя Са-каналы и увели-
чивая выброс медиатора. В отличие от ГАМКд-рецепторов, глицино-
вый рецептор нечувствителен к бензодиазепаму, относительно нечув-
ствителен к барбитуратам и блокируется некоторыми стероидами.
154
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Пурины. Нейромедиаторами являются и некоторые пурины. Су-
ществует два основных типа пуринергической пе-
редачи сигнала. При первом типе передачи основным нейроме-
диатором служит аденозин, при втором — АТФ и его производное ди-
аденозинтетрафосфат. В качестве нейромедиаторов пуринергической
системы могут выступать также и другие производные пуринов и (как
установлено в последние годы) пиримидинов. В большинстве случаев
АТФ является котрансмиттером других медиаторов. Благодаря АТФ
осуществляется неадренергическая и нехолинергическая иннервация
гладкой мускулатуры кишечника. В периферических органах и тканях,
получающих симпатическую и парасимпатическую иннервацию. АТФ
используется как медиатор прямого действия. Частично АТФ выбра-
сывается из синапсов вместе с другими медиаторами, другая его часть
имеет цитоплазматическое происхождение.
АТФ служит медиатором во всех синапсах, образуемых метасимпа-
тическим отделом вегетативной нервной системы на гладких мышцах.
Аденозин играет роль эндогенного антиконвульсанта, а также вещест-
ва, регулирующего начальные стадии сна. Он оказывает главным об-
разом тормозное влияние на ряд возбуждающих синапсов. Аденозину
приписывается роль нейромодулятора метаботропных глутаматных
рецепторов.
Рецепторы пуринов. Их делят на два класса: Pj (преси-
наптические, активируемые аденозином) и Р2 (постсинаптические, ак-
тивируемые АТФ и диаденозинтетрафосфатом). Рецепторы аденозина
Pj-класса являются медленными метаботропными рецепторами. Сре-
ди них есть рецепторы, подавляющие системы синтеза цАМФ (AJ, и
рецепторы, активирующие синтез цАМФ (А2). Довольно специфиче-
скими ингибиторами Aj-рецепторов являются теофиллин и кофеин.
Агонистом Aj-рецепторов служит не только аденозин, но и АМФ, при-
чем эти соединения могут иметь как синаптическое, так и экстракле-
точное происхождение (т. е. образовываться в других клетках). Это
позволяет понять, почему истощение энергетических ресурсов мозга,
приводящее к накоплению АМФ и аденозина, активирует Aj-рецепто-
ры и оказывает успокаивающее (седативное) и противосудорожное
действие. Иными словами, один из физиологических эффектов данных
рецепторов — защита энергетических ресурсов мозга в экстремаль-
ных ситуациях. Такой же механизм лежит в основе участия аденозино-
вых рецепторов в регуляции цикла «сон—бодрствование».
Рецепторы Р2-класса в свою очередь подразделяются на две
группы— быстрые ионотропные (Р2х) и медленные метаботроп-
ные (P2Y) (к настоящему времени известно шесть изоформ первых
2.3. Характеристика индивидуальных медиаторных систем
15!
I
и семь изоформ вторых). АТФ, воздействуя на быстрые рецепторы,
вызывает гипотензию, стимулируя в то же время сократительную
деятельность сердца.
Следует отметить, что пуринергические рецепторы, локализован-
ные пресинаптически, способны модулировать работу других меди-
аторных систем, в первую очередь холин- и адренергической.
Нитроксид (NO). Образуется из аргинина под действием фер-
мента NO-синтазы. Аналогично классическим нейромедиаторам, в
пресинаптических терминалях из L-аргинина и молекулярного кисло-
рода с помощью фермента NO-синтазы образуется NO:
L-аргинин + НАДФН + О2------> цитруллин + NO + НАДФ+.
В настоящее время выделяют три изоформы NO-синтазы
(NOS). Их классификация основывается на анатомической локализа-
ции и механизмах регуляции.
По механизму активации NO-синтазы делят на консти-
тутивные и индуцибельную (iNOS). Две конститутивные,
обнаруживаемые главным образом в эндотелии и нейронах, получили
свое название по месту локализации — эндотелиальная (eNOS) и
нейрональная (nNOS). Эндотелиальная изоформа постоянно присутст-
вует в эндотелиальных клетках и обеспечивает кратковременный выброс
из них NO в ответ на активацию мембранных рецепторов и повышение
внутриклеточной концентрации Са2+. Нейрональная NO-синтаза отве-
чает за кальцийзависимое высвобождение NO из нервных клеток.
Индуцибельная NO-синтаза присуща макрофагам, гладко-
мышечным клеткам и печени. Она индуцируется действием на клетки
цитокинов и бактериальных эндотоксинов и обусловливает долговре-
менный синтез больших количеств NO. Конститутивные NO-синтазы
постоянно присутствуют в клетках, их активность находится в прямой
зависимости от уровня ионов Са2+, а регуляция осуществляется через
кальмодулин, являющийся кофактором этого фермента. Индуцибель-
ная форма не зависит от содержания кальция и транскрипционно ре-
гулируема. Глюкокортикоиды подавляют экспрессию только индуци-
бельной NO-синтазы.
Основными источниками NO в центральной и периферической
нервной системе являются эндотелиоциты сосудов, клетки микроглии
и астроциты, а также глутаматергическйе нейроны. В нейронах
NO-синтаза связана с ионотропным NMDA-рецептором глутамата.
Ионы кальция, входящие через активированный NMDA-рецептор,
связываются с кальмодулином, и кальций-кальмодулиновый комплекс
активирует NO-синтазу.
156
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
NO является водо- и жирорастворимой радикальной молекулой,
способной диффундировать из клетки, в которой образуется, в сосед-
ние клетки, где активирует фермент гуанилатциклазу. NO был впер-
вые описан как регулятор артериального давления, опосредующий
расширение сосудов при воздействии ацетилхолина.
В отличие от классических нейромедиаторов, NO не может накап-
ливаться в синаптических везикулах, освобождаться нервными оконча-
ниями и взаимодействовать с соответствующими рецепторами. Вместо
этого NO равномерно диффундирует от места синтеза в близлежащие
клетки, и его распространение лимитируется только временем жизни.
Это время достигает нескольких (до 10) секунд, вследствие чего его
сигнал может распространяться на относительно большие расстояния.
Такой тип сигнализации, промежуточный между прямой синаптиче-
ской передачей и высвобождением гормонов эндокринными органами в
кровеносное русло, называется паракринным. NO, благодаря своей
способности проникать через нейрональную мембрану, может коорди-
нировать активность ряда близлежащих клеток и, влияя на синаптиче-
скую пластичность, объединять нейроны
в локальные нейрональные сети. Следова-
тельно, в нервной системе NO выступает
как своеобразный вторичный мессенджер.
Выяснение сигнальных путей, в которые
вовлекается NO, привело к пониманию
механизма действия нитроглицерина, уже
давно используемого для временной ком-
пенсации сосудистых спазмов. Нитрогли-
церин является донором NO и особенно
эффективен для расширения коронарных
артерий. Его действие приводит к увеличе-
нию кровоснабжения сердца и устранению
боли, возникающей при ишемической бо-
лезни сердца (приступы стенокардии).
Отмечается важная роль NO в разви-
тии процесса повреждения нейронов при
избытке глутамата и гиперактивации глу-
таматных рецепторов (эксайтотоксичность
глутамата). Показано участие NO в ноци-
цепции. Многими исследователями отво-
дится существенная роль этому соединению
в долговременной потенциации, наблюдае-
мой в гиппокампе. Такая потенциация свя-
зана с активацией NMDA-рецепторов, ко-
С. Е. Северин
Основоположник отечествен-
ной биохимии, организатор
кафедры биохимии в МГУ
им. М. В. Ломоносова, руко-
водивший ею более 50 лет,
инициатор развития новых
направлений биохимической
науки — биоэнергетики, ней-
рохимии, мембранологии, из-
вестен оригинальными иссле-
дованиями биологической ро-
ли нейропептвда карнозина
2.3. Характер истина индивид сальных медиаторных систе*
15
торые регулируют состояние пресинаптической терминали. Блокада
NO-синтазы тормозит эту потенциацию.
Развитие ряда нейродегенеративных заболеваний сопряжено с на-
рушением баланса между синтезом NO и супероксида. Взаимодейст-
вие данных активных радикалов приводит к образованию пероксинит-
рита — высокоактивного окислителя. Следовательно, одновременное
накопление этих двух соединений является фактором окислительного
стресса (см. гл. 3.9).
Таким образом, хотя оксид азота не является классическим меди-
атором (поскольку не имеет мембранного рецептора), пути его обра-
зования в мозге аналогичны таковым для типичных медиаторов нерв-
ной системы.
Пептидные медиаторы. Нейропептцды составляют достаточно по-
лифункциональную группу регуляторов (табл. 2.5). Некоторые из них
удовлетворяют критериям нейромедиаторов. С известной долей осторож-
ности сюда можно отнести вещество Р, вазоактивный интестинальный по-
липептид (ВИП), соматостатин, нейропептидУ, люлиберин. Значительное
число нейропептцдов (опиоидные пептиды, кортикотропин и его фрагмен-
ты, кортиколиберин, тиролиберин, кокальцигенин, холецистокинин, бом-
безин, галанин, нейротензин, ангиотензин, атриопептцды, брадикинин,
вазопрессин, окситоцин и др.) обладают свойствами нейромодуляторов.
Таблица 2.5. Физиологическая роль некоторых нейропептидов
Пептид Участие в регуляции физиологических функций или состояний
Опиоиды Боль, тревога, эмоции, функционирование системы внут- реннего подкрепления
Тахикинины Воспаление, седативные эффекты
Холецистокинин Поиск пищи, модуляция функций дофаминергической сис- темы, восприятие боли
НейропептидУ Поиск пищи, эмоции, нейрональная возбудимость
Вазопрессин Обучение и память
Соматостатин Аналгезия
Нейротензин Терморегуляция, терморецепция, аналгезия, восприятие боли, модуляция функции дофаминергической системы
Галанин Сенсорная передача
158
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Многие пептиды, известные как гормоны, одновременно проявля-
ют свойства нейромедиаторов. Некоторые из них участвуют в форми-
ровании эмоций, другие, как вещество Р и опиоиды, вовлечены в про-
цесс восприятия боли; меланоцитстимулирующий гормон, адренокор-
тикотропин и p-эндорфин регулируют ответ на стрессорные реакции.
Среди нейропептидов можно выделить пептиды опиоидные, гипо-
физарные, а также гипоталамические «рилизинг-гормоны».
Вещество Р оказалось первым веществом пептидной природы, у
которого были обнаружены свойства нейромедиатора. Его название
связано с тем, что оно было выделено из ацетонового порошка (pow-
der) мозга и кишечника. Оно, в отличие от «классических» медиато-
ров, синтезируется не в синаптических окончаниях, а в перикарионе
первичных сенсорных нейронов спинномозговых ганглиев, депониру-
ется в оптически плотных пузырьках, перемещается к пресинаптиче-
скому окончанию посредством быстрого аксонального транспорта и
высвобождается из сенсорных нейронов под влиянием деполяризации
в присутствии Са2+. При аппликации на дорсальные рога спинного
мозга вещество Р вызывает мощное возбуждение сенсорных нейронов
второго порядка.
Этот пептид, состоящий из 11 аминокислотных остатков, обнару-
живается в высоких концентрациях в гиппокампе, неокортексе, а так-
же в афферентных С-волокнах периферических нервов, участвующих
в передаче информации о болевых и температурных сигналах. Веще-
ство Р является медиатором, опосредующим передачу сенсорной ин-
формации в спинном мозге, его выделение может блокироваться опи-
оидами из интернейронов спинного мозга.
Вещество Р найдено в нескольких специфических коротких проек-
ционных путях (см. рис. 2.7). Наличие его в нигростриальных волок-
нах позволило предположить, что оно служит в данном месте типич-
ным нейромедиатором, а значит, играет важную роль в функциях ба-
зальных ганглиев по организации движения. Вещество Р содержится
также в клетках спинномозговых ганглиев. Деполяризующее действие
вещества Р на мотонейроны в 200 раз сильнее, чем действие глутама-
та. Однако эффект его развивается медленно, тогда как входные сиг-
налы, поступающие по многим волокнам задних корешков, передают-
ся быстро и раздельно.
Вещество Р инициирует медленные возбуждающие постсинаптиче-
ские потенциалы, которые связаны с регуляцией тонуса гладких мышц.
Наряду с функцией возбуждающего нейромедиатора первичных сен-
сорных нейронов вещество Р способно оказывать модулирующее влия-
ние, в частности усиливая десенситизацию никотиновых холинорецеп-
2.3. Характер истина индивидуальна. медиаторнь.х систем
591
торов. В одних и тех же синаптических окончаниях вещество Р может
обнаруживаться вместе как с «классическими» нейромедиаторами (се-
ротонин, АХ, ГАМК, катехоламины), так и с другими пептидами (вазо-
активный интестинальный пептид, опиоидные пептиды, соматостатин,
нейротензин).
НейропептидУ состоит из 36 аминокислотных остатков. Он выяв-
ляется иммуноцитохимическими методами в центральной и перифери-
ческой нервной системе в значительных концентрациях (см. рис. 2.7).
Особенно высокое содержание нейропептида Y свойственно перифе-
рическим нервам сердечно-сосудистой системы и регулирующим ее
работу нервным центрам. При этом обнаруживается сосуществование
и взаимодействие нейропептида Y с норадреналином (характерно для
симпатических нервов гладких мышц кровеносных сосудов и других
органов). При введении в мозг этот нейропептид вызывает ряд биоло-
гических эффектов — увеличение потребления пищи, седативный
эффект, изменения в процессах запоминания и циркадных ритмах.
Присутствие и высвобождение вазоактивного интестинального
(поли)пептида (ВИП) зарегистрированы во многих отделах нервной
системы, прежде всего в коре больших полушарий и в вегетативной
нервной системе. Нейромедиаторная функция ВИП отмечена для
постганглионарных нейронов вегетативной нервной системы, которые
участвуют в расслаблении гладких мышц кровеносных сосудов, дыха-
тельных путей, кишечника. Кроме того, взаимодействуя с мускарино-
выми холинергическими рецепторами, ВИП способен проявлять ней-
ромодуляторную функцию. Так, обнаружено реципрокное взаимодей-
ствие этого пептида и АХ.
Соматостатин представляет собой пептид из 14 аминокислот. Его
название обусловлено тем, что, образуясь в гипоталамусе, этот нейро-
гормон тормозит секрецию гормона роста (соматотропина) клетками
гипофиза. Нейромедиаторные функции соматостатин выполняет в ве-
гетативных ганглиях и в клетках внутренних органов, а также в нейро-
нах спинномозговых ганглиев, гипоталамусе, миндалинах и коре боль-
шого мозга (см. рис. 2.7). Он обнаружен в норадренергических пост-
ганглионарных симпатических нейронах. Введение соматостатина в
желудочки мозга приводит к снижению двигательной активности. При
ионофоретическом подведении к отдельным нейронам это вещество,
подобно некоторым другим пептидам, оказывает медленное тормозное
действие. Соматостатин увеличивает амплитуду входящих К-токов в
нейронах энтеральных ганглиев. Обнаружено снижение уровня этого
пептида при болезни Альцгеймера.
160
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРО* ИМИЯ
Опиоидные нейропептиды (табл. 2.6) получили свое название
из-за способности связываться с теми же постсинаптическими рецеп-
торами, которые активируются опием, точнее его активным компо-
нентом морфином. Поиск эндогенных лигандов этих рецепторов при-
вел к открытию эндорфинов, обладающих, как и морфин, аналитиче-
скими свойствами. Обнаружено более 20 опиоидных пептидов,
которые делятся на три группы: эндорфины, энкефалины и ди-
норфины. Представители каждой из групп образуются в ходе процес-
синга из высокомолекулярных белков-предшественников: проопио-
меланокортина, продинорфина и проэнкефалина А (рис. 2.17).
Таблица 2.6. Некоторые опиоидные пептиды
Название Аминокислотная последовательность
а-Эндорфин Туг— Gly—Gly—Phe1—Met— Thr—Ser—Clu—Lys—Ser—Gin— Thr—Pro—Leu—Vai—Thr
а-Неоэндорфин Tyr— Gly—Gly—Phe—Leu—Arg— Lys—Tyr—Pro—Lys
Эндорфины р-Эндорфин Tyr—Gly—Gly—Phe—Met—Thr— Ser—Glu—Lys—Ser—Gin—Thr— Pro—Leu—Vai—Thr—Leu—Phe— Lys—Asn—Ala—He—Vai—Lys— Asn—Ala—His—Lys—Gly—Gin
у-Эндорфин Tyr—Gly—Gly—Phe—Met—Thr— Ser—Glu—Lys—Ser—Gin—Thr— Pro—Leu—Vai—Thr—Leu
Энкефалины Лей-энкефалин Tyr— Gly—Gly—Phe—Leu
Мет-энкефалин Tyr— Gly—Gly—Phe—Met
Динорфины Динорфин А Динорфин В Tyr— Gly—Gly—Phe—Leu—Arg— Arg— He—Arg— Pro—Lys—Leu— Lys—Trp—Asp—Asn—Gin Tyr— Gly—Gly—Phe—Leu—Arg— Arg—Gin—Phe—Lys—Vai—Vai— Thr
1 Гомологичные участки отмечены курсивом.
2.3. Характеристика индивидуальных медиаторных систем
161
а
Сигнальный
пептид
Пре-проопиомеланокортин
Прорпиомеланокортин
Пропептид ГЖЖМ
Активные пептиды
АКТГ р-Липотропин
у-Липотропин р-Эндорфин
Активные пептиды ( ) НЬ
Сигнальный
пептид Пре-проопиомеланокортин
Пре-пропептид
Проэнкефалин А
Пропептид ( |..[J I....I I I И ~~Р
Активные пептиды
Мет-энкефалин
Лей-энкефалин
Мет-энкефалин
Рис. 2.17. Схема синтеза опиоидных нейропептидов эндорфина (а) и энкефа-
линов (б)
Опиоидные пептиды широко распространены в мозге и часто обнару-
живаются в одних терминалях вместе с «классическими» нейромедиато-
рами, такими как ГАМК и серотонин. Их внутримозговое введение при-
водит к аналгезии, имеются данные, что именно они участвуют в акупунк-
турном обезболивании. Опиоиды вовлекаются в комплекс поведенческих
актов, связанных с сексуальным поведением, агрессией и иерархическим
подчинением в сообществе. В литературе обсуждается вопрос об их роли
в развитии таких психических расстройств, как шизофрения и аутизм.
Клетки, содержащие эндорфины, встречаются почти исключитель-
но в гипоталамусе (см. рис. 2.7). Их аксоны направляются к различ-
ным ядрам самого гипоталамуса и в другие отделы, в том числе к об-
асти перегородки и к миндалине; часть волокон доходит до верхних
отделов ствола мозга, оканчиваясь на моноаминергических клетках
голубого пятна и ядер шва. Большая часть нервных окончаний нахо-
дится в глубинных отделах мозга (вокруг желудочков).
162
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
После открытия эндорфинов возникло предположение, что они,
действуя подобно препаратам опия, играют роль внутреннего регулято-
ра, подавляющего болевые ощущения или их модулирующего. Изуче-
ние подобного механизма представляет большой интерес: в частности,
весьма заманчиво было бы найти естественный заменитель морфина,
не вызывающий болезненного привыкания и побочных эффектов. Кро-
ме того, высказывались различные идеи относительно физиологиче-
ской роли эндорфинов; им приписывали всевозможные функции — от
участия в терморегуляции до поддержания самосознания. Сейчас, од-
нако, становится ясно, что для выяснения конкретной роли эндорфинов
в различных формах поведения потребуются длительные исследования.
Опиатные рецепторы относятся к медленным метаботропным и
наиболее изученным среди рецепторов нейропептидов. Первыми ус-
тановленными лигандами этих рецепторов были экзогенные вещест-
ва — морфин, дигидроморфин, норморфин, леворфанол. Из алкало-
идов растительного происхождения наиболее известны налоксон, пен-
тазоцин и налтрексон. Два последних обладают свойствами частичных
антагонистов. Эндогенные пептиды — эндорфины, энкефалины и ди-
норфины — действуют аналогично морфину и относятся к агонистам.
Идентифицировано четыре класса опиоидных рецеп-
торов: мю-, дельта-, каппа- и ORL-l (Opioid Receptor-Like 1)
(табл. 2.7), между которыми, однако, существует меньше различий по
взаимодействию с агонистами и антагонистами, чем в других группах
рецепторов. Опиоидные рецепторы связаны с Gs- и Gj-белками, их
тормозное действие заключается в закрытии Са-каналов (к-рецептор)
или в открытии К-каналов (ц-, 3-, ORL-1 -рецепторы). Это приводит к
уменьшению выброса медиатора или угнетению нейрональной возбу-
димости в зависимости от того, где локализованы рецепторы.
Таблица 2.7. Характеристика опиатных рецепторов
Эффектор Рецепторы
Мю (ц) Дельта (8) Каппа (к) ORL-l
Эндогенный лиганд р-Эцдорфин, меторфамид, эндоморфин-1 и-II Мет-энкефалин, лей-энкефалин, Р-эндорфин Динорфин А1 —13, динорфин А1—8, динорфин В Ноцицептин, орфанин FQ
Агонист Морфин, DAGO DPDPE, SNC-80, DSTBULET U50488H, бремазоцин, пентазоцин
Антагонист Налоксон, P-FNA Налтриндол, налоксон Налоксон, nor-BNI Ноцицептин, (1 — 13)NH2
2.4. G-Белки, вторичные мессенджеры и протеинкиназы
163
Опиоидные рецепторы довольно широко представлены в разных отде-
лах ЦНС: в коре, стриатуме, таламусе (преимущественно ц-рецепторы); в
ядрах моста мозга, амигдале, обонятельной луковице, глубоких слоях ко-
ры (5-рецепторы); в гипоталамусе, срединном возвышении (к-рецепто-
ры). Кроме того, опиоидные рецепторы найдены в структурах перифери-
ческой нервной системы и в клетках ненейронального происхождения
(клетки иммунной системы, сердца, плаценты и др.).
Опиоидные рецепторы играют важную роль в системе внутреннего
подкрепления, в процессах восприятия боли, могут участвовать в мо-
дуляции работы других медиаторных систем.
Каннабиоиды. Еще древние цивилизации обнаружили, что раз-
личные препараты индийской конопли (Cannabis sativa) могут вызы-
вать у человека релаксацию, эйфорию и многие другие психоэмоци-
ональные эффекты. Обнаружены рецепторы в головном мозге к ос-
новному действующему началу получаемой из этого растения
марихуаны — Д9-тетрагидроканнабиолу и позднее выявлены эндоген-
ные лиганды этих рецепторов, которые оказались производными ара-
хидоновой кислоты, — анандамин и 2-арахидонилглицерол. К настоя-
щему времени установлены пути синтеза этих соединений в мозге
(рис. 2.18). Наиболее распространенные рецепторы типа СВИ распо-
ложенные в различных областях мозга, отвечают за специфические
эффекты каннабиоидов: с рецепторами неокортекса связаны измене-
ния перцепции, базальных ганглиев и мозжечка — психомоторного
контроля, гиппокампа — изменения кратковременной памяти. В пе-
риферических отделах нервной системы локализован преимуществен-
но другой тип каннабиоидных рецепторов — СВ2.
G-БЕЛКИ, ВТОРИЧНЫЕ МЕССЕНДЖЕРЫ И ПРОТЕИНКИНАЗЫ
2.4.
2.4.1. G-Белки и вторичные мессенджеры
Как уже отмечалось ранее (см. гл. 2.2), существует обширная
группа рецепторов, в которых передача сигнала не осуществляется
прямо на эффектор, а опосредуется через цепь химических превра-
щений. В настоящее время при классификации клеточных сигналь-
ных систем учитывается природа вторичных мессенджеров и регули-
руемых ими клеточных реакций. Исходя из этого, к числу основных
сигнальных систем относят: циклоаденилатную; Са-инозитолтрис-
фосфатзависимую; липоксигеназную; НАДФН-оксидазную (суперок-
164
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХи: 1ИЯ
а
О
СН2
Фосфатидилэтаноламин
СН
—° сн2
N - Ацетилтрансфераза
сн2
-О-Р=О
N-Арахидоноил-
фосфатидилэтаноламин
сн
°^NH3
сн2
О
О
Фосфолипаза D
О
-О-Р=О
НО
Анандамин '^NH
Рис. 2.18. Пути синтеза и строение некоторых эндоканнабиоидов:
а — синтез анандамина из фосфатидилэтаноламина; б — синтез арахидонилглицерола
из фосфатидилинозитола
|
2.4. G-Белки, вторичные мессенджеры и протеинкиназы
16!
сидсинтазную); NO-синтазную. В последнее время особое внимание
привлекает МАР-киназная система (Mitogen Activated Protein Ki-
nase), регулирующая клеточный цикл. В большинство осуществляе-
мых этими системами реакций в качестве промежуточного звена вов-
лекаются так называемые G-белки.
Рецепторы, сопряженные с G-белками, GPCR(G-Pro-
tein Coupled Receptors), передают сигнал от первичных мессенджеров к
внутриклеточным мишеням с помощью цепи «рецептор ----> G-белок
----> эффекторный белок». Первичными сигналами для этих рецепто-
ров служат разнообразные молекулы, среди которых низкомолекуляр-
ные гормоны и нейромедиаторы (например, адреналин, норадреналин,
ацетилхолин, серотонин, гистамин), опиоиды, гормоны пептидной и
белковой природы (адренокортикотропин, соматостатин, вазопрес-
син, ангиотензин, гонадотропин, эпидермальный фактор роста), неко-
торые нейропептиды (см. гл. 2.3). В этот же ряд попадает множество
химических сигналов, воспринимаемых обонятельными и вкусовыми
сенсорными клетками, а также свет, рецептором для которого служит
пигмент фоторецепторных клеток родопсин (см. гл. 3.6).
Один и тот же первичный сигнал может передавать информацию
через несколько разных GPCR, следовательно, если число внешних
сигналов для GPCR составляет несколько десятков, то самих рецепто-
ров известно более 200.
Следующий за рецептором компонент сигнального каскада пред-
ставлен G-б е л к о м. Идентифицировано около 20 различных G-бел-
ков, наиболее распространенными являются Gs и G{, которые соответ-
ственно стимулируют и ингибируют аденилатциклазу; Gq, активирую-
щий фосфолипазу С; G-белки сенсорных клеток: фоторецепторных —
Gt (трансдуцин), обонятельных — G0|f и вкусовых — Gg. G-Белки —
это гетеротримеры, которые состоят из субъединиц трех вадов: а, р и у.
В естественных условиях последние две субъединицы функционируют
как единый ру-комплекс.
Важнейшая характеристика G-белков — присутствие
на а-субъединице центра связывания гуаниловых нуклеотидов. Если с
G-белком связан ГТФ, это соответствует его активированному состо-
янию (другими словами, G-белок находится в положении «включе-
но»). Если в нуклеотидсвязывающем центре присутствует ГДФ, это
соответствует «выключенному» состоянию белка.
Центральное событие при передаче сигнала от рецептора, на кото-
рый подействовал первичный сигнал, к G-белку состоит в том, что ак-
тивированный рецептор катализирует процесс обмена ГДФ, связан-
ного с G-белком, на присутствующий в среде ГТФ. Это событие, обо-
166
Раздел 2 МОЛЕКУЛЯРНАЯ НЕЙРОХИМИ
значаемое как ГДФ/ГТФ-обмен, сопровождается диссоциацией
тримерной молекулы G-белка на две функциональные субъединицы:
а-субъединицу, содержащую ГТФ, и ру-комплекс. Далее одна из этих
функциональных субъединиц (какая именно — зависит от типа сиг-
нальной системы) взаимодействует с эффекторным белком, представ-
ленным обычно ферментом или катионным каналом. Как следствие,
функциональная активность эффектора меняется, что, в свою оче-
редь, приводит к изменению цитоплазматической обстановки и иници-
ирует тот или иной клеточный ответ. Диссоциированные компоненты
тримера будут активными в тот краткий период, пока ГТФазная актив-
ность а-субъединицы не превратит ГТФ в ГДФ. Это резко увеличит
сродство а-субъединицы к ру-димеру и восстановит комплекс в исход-
ном (неактивном) состоянии.
Эффекторными белками в сигнальных систе-
мах типа «рецептор---> G-белок--> эффекторный белок» могут
быть аденилатциклаза, катализирующая синтез цАМФ из АТФ; фосфо-
липаза С, гидролизующая фосфатидилинозитол с образованием DAG и
инозитолтрисфосфат (ИФ3); фосфодиэстераза, расщепляющая цГМФ
до ГМФ; некоторые типы калиевых и кальциевых каналов (рис. 2.19).
Так, например, аденилатциклаза, осуществляющая синтез цАМФ
из АТФ, обнаружена во всех тканях, максимальная ее активность от-
мечается в мозге. Фермент является мультимолекулярным комплек-
Глутамат
Дофамин
Норадреналин |
{ ^-Адренергический
Рецептор
mGluR
G-Белок
( © + s
Эффекторный\ Аденилатциклаза
белок / ।
-----------/ 1
Фосфолипаза С
© + Gi
Аденилатциклаза
Вторичный
мессенджер
цАМФ
DAG
ИФ3
цАМФ
Конечный
эффектор
[киназа А
Протеин-
киназа С
Выброс
Са2+
Протеинкиназа А
РечАлпьтят \ Усиление
/ фосфорилирования
/ белков
Усиление фосфорилирования
белков и активация
кальцийсвязывающих белков
Уменьшение
фосфорилирования
белков
Рис. 2.19. Примеры связи G-белков с различными типами рецепторов и запус-
каемых ими каскадов внутриклеточных реакций
2.4. G-Белки, вторичные мессенджеры и протеинкиназы
16
сом, состоящим из рецепторной и каталитической субъединиц, первая
из которых обращена к наружной поверхности клеточной мембраны, а
вторая «смотрит» в цитоплазму. Ответ клетки на действие сигнала за-
висит от концентрации рецепторов на поверхности мембраны и степе-
ни сопряжения рецепторов с аденилатциклазой. Сопрягающим ком-
понентом являются G-белки, обладающие как ГТФ-связывающей,
так и ГТФ-гидролизующей активностью.
Весьма важно, что при передаче сигнала в каскаде «рецептор->
----> G-белок -> эффекторный белок» исходный внешний сигнал
может многократно усиливаться. Это происходит благодаря тому, что
одна молекула рецептора за время пребывания в активированном со-
стоянии (R*) успевает перевести в активированную форму (G*) не-
сколько молекул G-белка. Например, в зрительном каскаде «родоп-
син -----------> Gt-> цГМФ-зависимая фосфодиэстераза» на каждую моле-
кулу R* может образовываться несколько сотен или даже тысяч молекул
G*, а это означает, что на первой стадии каскада («R*-> G*») коэф-
фициент усиления внешнего сигнала составляет 102—103. Хотя на
следующей стадии каскада («G* ---> эффекторный белок») каждая
молекула G* взаимодействует только с одной молекулой эффекторно-
го белка, сигнал в данном случае также усиливается, поскольку на
каждую молекулу G* и, соответственно, активированного эффектор-
ного белка в цитоплазме появляется (исчезает) большое число моле-
кул вторичного мессенджера. Так, на второй стадии зрительного кас-
када одна молекула активированной цГМФ-фосфодиэстеразы способ-
на расщепить в секунду до 3 тыс. молекул цГМФ, служащего в
фоторецепторных клетках вторичным мессенджером. Поскольку уси-
ление внешнего сигнала на обеих стадиях суммируется, в конечном
счете коэффициент усиления сигнала при его прохождении через кас-
кад может достигать достаточно высоких значений: в зрительных клет-
ках усиление составляет величину порядка 105—106.
Ясно, что прекращение действия внешнего стимула должно сопро-
вождаться «выключением» всех компонентов сигнальной системы. На
уровне рецепторов это достигается, во-первых, в результате диссоци-
ации первичного сигнала из комплекса с GPCR, во-вторых, путем фос-
форилирования рецепторов под действием специальных протеинкиназ и
последующего связывания с модифицированным рецептором специаль-
ного белка (например, Р-арестина). G-Белки обладают способностью
гидролизовывать связанный с ними ГТФ до ГДФ, что обеспечивает их
инактивацию. И, наконец, чтобы переход клетки к исходному состоянию
завершился, специальные механизмы восстанавливают исходный уро-
вень вторичного мессенджера в ее цитоплазме. Например, цАМФ, ци-
168
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
топлазматическая концентрация которого повышается при передаче
сигнала в каскаде «Р-адренорецептор -> С5-белок ---> аденилат-
циклаза», затем гидролизуется цАМФ-фосфодиэстеразой до АМФ,
который не обладает свойствами вторичного мессенджера.
G-Белки ответственны за трансмембранную передачу сигналов мно-
гих гормонов и нейромедиаторов к разнообразным мишеням клетки. Че-
тыре G-белка к настоящему времени очищены до гомогенного состояния
и биохимически охарактеризованы: Gt (трансдуцин), Gs, G{ и Go. Оказа-
лось, что каждый из них имеет уникальные мишени (эффекторные бел-
ки). Gt активирует цГМФ-специфичную фосфодиэстеразу в наружных
сегментах палочек сетчатки; Gs и Gf, соответственно, стимулируют и ин-
гибируют аденилатциклазу и присутствуют во всех клетках; Go представ-
лен в большом количестве в клетках мозга и, по-видимому, ингибирует
электрочувствительный Са-канал в нейронах.
Ряд существенных G-белков пока не очищен. Так, Gp, который ис-
пользует в качестве мишени фосфатидилинозитолспецифичную фос-
фолипазу С, инициирует быстрый распад фосфатидилинозитола в
плазматической мембране и образование нескольких вторичных по-
средников. Другой G-белок, Gk, по-видимому, открывает К-каналы в
миоцитах сердечной мышцы и других клетках; Ое-белки участвуют в
регуляции экзоцитоза. В некоторых случаях один G-белок внутри
клетки может реагировать на связывание лиганда с одним из несколь-
ких разных рецепторов. Такая ситуация имеет место, например, для
белка Gk из ганглионарных клеток Aplysia. G-Белки могут иметь
больше одной мишени. В качестве примера можно привести белок Gt,
который в наружных сегментах палочек сетчатки глаза быка активи-
рует как цГМФ-фосфодиэстеразу, так и фосфолипазу А2. По-видимо-
му, в этих процессах участвуют различные субъединицы Gt.
Все G-белки прочно связаны с плазматической мембраной, за иск-
лючением трансдуцина, который может от нее диссоциировать. Ни од-
на из субъединиц G-белка не является трансмембранным белком. Од-
нако по меньшей мере у-субъединица ацилирована и может удержи-
ваться на мембране с помощью ковалентно связанной жирной
кислоты. В качестве такого якоря для закрепления в мембране G-бел-
ков часто используется миристат или пальмитат.
Наиболее распространенными мишенями G-белков являются адени-
латциклаза (для Gs и Go) и фосфолипаза С, ответственная за гидролиз
фосфатидилинозитола (для Gp). Модуляция аденилатциклазы приводит
к изменению внутриклеточной концентрации цАМФ, который, как из-
вестно, влияет на множество внутриклеточных процессов. Одним из по-
следствий увеличения содержания цАМФ является, например, стиму-
2.4. G-Белки, вторичные мессенджеры и протеинкиназы
169
ляция цАМФ-зависимой протеинкиназы (протеинкиназа А), которая,
в свою очередь, фосфорилирует специфические белковые субстраты.
Клетки содержат также два типа Са-зависимых протеинкиназ, акти-
вируемых соответственно Са 4- кальмодулином и Са + диацилглицеро-
лом и фосфатидилсерином (протеинкиназа С). Активность обеих этих
киназ регулируется вторичными посредниками, образующимися в про-
цессе обмена фосфатидилинозитола, который во многих клетках ини-
циируется путем G-белокзависимой активации специфической фосфо-
липазы С.
Молекулярные машины, обеспечивающие передачу сигнала от ре-
цепторов к внутриклеточным мишеням, состоят, как правило, из не-
скольких белковых компонентов, совокупность которых обычно име-
нуют каскадом передачи сигнала, или сигнальным каскадом.
Помимо белковых посредников в передачу сигнала внутри клетки
вовлекаются относительно небольшие молекулы, служащие вторичны-
ми посредниками, или мессенджерами (от англ, messenger — посыль-
ный). В русскоязычной литературе данные соединения называют как
вторичными мессенджерами, так и вторыми посредниками. В лю-
бом случае в содержание данного понятия вкладывается мысль о том,
что это соединение — следующее в цепочке передачи сигнала, но не
в коей мере — что оно менее значимое, т. е. вторичное.
Для обозначения сигнальных молекул обычно используется термин
«мессенджер», а не «посредник». Дело в том, что в цитоплазме в пе-
редачу сигнала вовлечены как разнообразные белки, так и малые мо-
лекулы, причем функционально все они являются посредниками меж-
ду рецептором, на который подействовал внешний стимул, и клеточ-
ным ответом. Однако между ними есть и принципиальное
различие: белки образуют своеобразную молекулярную машину,
которая, с одной стороны, воспринимает внешний сигнал, а с другой —
обладает ферментативной или иной активностью, модулируемой этим
сигналом, в то время как малые молекулы действительно служат лишь
посыльными (мессенджерами) между различными белками, полифер-
ментными комплексами или даже клеточными структурами. Самый из-
вестный пример такого посыльного — это уже упоминавшийся выше
цАМФ, среди других наиболее важных вторичных мессенджеров следу-
ет упомянуть циклический гуанозин-3',5'-монофосфат (цГМФ), ино-
зит-1,4,5-трисфосфат (обозначаемый как ИФ3), диацилглицерол (ДАГ),
ионы кальция (табл. 2.8). Следует отметить, что изменение уровня вто-
ричных посредников, например ионов кальция, может быть обусловлено
не только взаимодействием медиаторов с рецепторами, но и являться ре-
зультатом изменения мембранного потенциала.
Таблица 2.8. Характеристика некоторых вторичных мессенджеров
Вторичный посредник Источник Внутриклеточные мишени Механизм «уборки»
Са2+ Плазматическая мембрана-. электроуправляемые Са-каналы; различные лигандза- висимые каналы Эндоплазматический ретикулум-. ИФ3-рецепторы; рианодиновые рецепторы Кальмодулин, протеинкиназы, протеинфосфатазы, ионные каналы, синаптотагмин и др. Са-связывающие белки Плазматическая мембрана: Na/Ca-обменник; Са-насос. Эндоплазматический ретикулум: Са-насос. Митохондрии: Са-транспортер
цАМФ Аденилатциклаза действует на АТФ Протеинкиназа А, каналы, ре- гулируемые циклическими нук- леотидами Фосфодиэстераза цАМФ
цГМФ Гуанилатциклаза (субстрат ГТФ) Протеинкиназа С, каналы, ре- гулируемые циклическими нук- леотидами Фосфодиэстераза цГМФ
ИФ3 Фосфолипаза С (субстрат ФИФ2) ИФ3-рецепторы эндоплазма- тического ретикулума Фосфатазы
Диацил глицерол Фосфолипаза С (субстрат ФИФ2) Протеинкиназа С Различные ферменты
Раздел 2. МОЛЕКУЛЯРНАЯ
2.4. G-Белки, вторичные мессенджеры и протеинкиназы
1711
Наиболее характерные свойства вторичного
мессенджера следующие:
1) относительно небольшая по сравнению с биополимерами моле-
кулярная масса (посыльный должен с высокой скоростью диффунди-
ровать в цитоплазме);
2) способность быстро (по сравнению со временем передачи сигна-
ла) расщепляться (или подвергаться связыванию, как в случае Са2+),
в противном случае сигнальная система останется во включенном со-
стоянии и после того, как действие внешнего сигнала прекратится.
Отсутствие одного из этих свойств у вторичных мессенджеров мо-
жет оказаться для клетки фатальным в прямом смысле. Так, напри-
мер, форболовые эфиры, которые представляют собой структурные
аналоги диацилглицерола, но, в отличие от него, в организме не рас-
щепляются, способствуют развитию злокачественных опухолей. Это
происходит потому, что форболовые эфиры вовлекаются в работу сиг-
нальных систем, которые регулируют клеточное деление с помощью
диацилглицерола как вторичного мессенджера. Имитируя действие
диацилглицерола и обеспечивая передачу пролиферативного сигнала,
они не подвергаются расщеплению. В результате сигнальная система
остается постоянно включенной, и пролиферация клеток перестает
быть контролируемой.
В прекращении сигнала цАМФ и цГМФ участвуют фосфодиэсте-
разы, гидролизующие данные нуклеотиды. Это преимущественно рас-
творимые ферменты. Высокий уровень фосфодиэстеразы цГМФ об-
наруживается в постсинаптических окончаниях нейронов.
Фосфодиэстеразы представлены большим количеством изоформ, ко-
торые различаются по молекулярной массе и сродству к субстратам.
Циклонуклеотидактивируемая фосфодиэстераза гидролизует как цАМФ,
так и цГМФ, при этом они взаимно ускоряют гидролиз друг друга, что
свидетельствует о положительной кооперативности их активных центров.
Кроме циклических нуклеотидов, Са2+ и процесса ограниченного проте-
олиза фосфодиэстеразу активируют также полианионы, фосфолипиды,
жирные кислоты. Ингибиторами или активаторами данного фермента
являются многочисленные фармакологические вещества.
2.4.2. Роль мембранных фосфоинозитидов в передаче сигнала
Для того чтобы представить механизм действия фосфоинозити-
дов в качестве вторичных мессенджеров, необходимо более подробно
рассмотреть цикл их метаболических превращений
в мембране.
172
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Внешний сигнал после взаимодействия с рецептором через G-белок
активирует фосфодиэстеразу (фосфолипазу С), которая гидролизует
фосфатидилинозитол-4,5-дифосфат (ИФ2) с образованием диацил-
глицерола (DAG) и инозитолтрисфосфата (ИФ3) (рис. 2.20).
Диацил глицерол фосфорилируется диацил глицерол киназой до фосфа-
тидной кислоты (реакция требует присутствия АТФ). Фосфатидная
кислота взаимодействует с цитидинтрифосфатом с образованием цити-
диндиацилглицеролдифосфата. Это соединение после рекомбинации с
инозитом восстанавливает фонд гидролизуемых фосфодиэстеразой
фосфоинозитидов. Инозит, необходимый для данной реакции, образу-
ется из инозитолмонофосфата. Реакция катализируется инозитол-1-
монофосфатазой, активность которой блокируется ионами лития.
Именно этим объясняется эффективное лечение ионами лития ма-
ниакально-депрессивных психозов и некоторых других заболеваний
нервной системы. Блокада литием инозитол-1 -монофосфатазы замед-
НО-.
Арахидоновая
кислота —
предшественник
►простагландинов,
тромбоксанов,
лейкотриенов
(воспаление)
О
ФЛС)
ЧЭН
Активация
протеинкиназы С
HO-P-OR
I
о- ч
чэ-p-or
I
\ °”
(Са2+ \ ФЛД
для цФЛД)
о
LO-P-OH
НО, ОРОз"
О"
R=
-•ОРОз”
но—R
но он
| Мобилизация Са2~*~~|
Мобилизация Са2+,
торможение
аденилатциклазы,
активация ФЛС
е
о
е
Рис. 2.20. Обмен фосфоинозитидов.
ФЛА — фосфолипаза А; ФЛС — фосфолипаза С; ФЛД — фосфолипаза D; ФИ-ФЛС —
фосфатидил инозитолспецифичная фосфолипаза С; цФЛД — цитозольная фосфолипаза D
2.4. G-E-fcflKM, ети^имг^ MvCCcH^bpU M npvTvHHHHH23U
ляет синтез инозита в организме и ослабляет зависящие от метабо-
лизма фосфоинозитидов процессы, протекающие в нейронах.
Инозитолтрисфосфат вызывает высвобождение кальция из мемб-
ран эндоплазматического ретикулума в результате связывания со спе-
циальным рецептором. Несмотря на то что процесс выброса кальция
из внутриклеточных депо под действием фосфоинозитидов обнаружен
в целом ряде клеток, механизм, с помощью которого этот ион откры-
вает внутриклеточные кальциевые каналы, окончательно не выяснен.
При передаче сигнала с помощью фосфоинозитидов происходит обра-
зование двух вторичных мессенджеров: инозитолтрисфосфата и ди-
ацилглицерола.
Диацилглицерол выполняет роль вторичного мессенджера, так как
инициирует механизмы, в результате которых активируются два типа
протеинкиназ: протеинкиназа С и цГМФ-зависимая протеинкиназа.
При этом фосфолипаза С играет ключевую роль в процессе переноса
сигналов с рецепторов, которые непосредственно не связаны с адени-
латциклазой, но регулируют скорость метаболизма фосфоинозитидов.
Вклад каждой из ветвей передачи сигнала с помощью фосфоинози-
тидов может быть исследован с помощью химических соединений,
имитирующих действие одного из двух вторичных мессенджеров, об-
разующихся в результате гидролиза фосфоинозитидов и, следователь-
но, активирующих эти пути регуляции по отдельности. Действие DAG
имитируется форболовыми эфирами, так как эти соединения вызыва-
ют активацию протеинкиназы С. Форболовые эфиры — это соедине-
ния растительной природы, их выделяют из семян восточноазиатского
растения Croton tigtium. Форболовые эфиры резко ускоряют иници-
ацию опухолей канцерогенами. Действие инозитолтрисфосфата ими-
тируется кальциевыми ионофорами, в том числе А 23187.
Существует синергизм между двумя путями передачи сигнала с по-
мощью фосфоинозитидов. Так, например, в тромбоцитах форболовый
эфир и кальциевый ионофор, действуя совместно, вызывают такую
сильную секрецию серотонина, которая не может быть вызвана ни од-
ним из этих соединений в отдельности. Форболовый эфир и кальцие-
вый ионофор могут инициировать деление некоторых клеток. Это по-
зволило предположить, что сигнальные пути, связанные с обменом
фосфоинозитидов, могут контролировать не только секрецию гормо-
нов и транспорт ионов, но и пролиферацию клеток. Появились экспе-
риментальные доказательства того, что форболовые эфиры способст-
вуют опухолевому росту через активацию «диацилглицеролзависимой
ветви» фосфоинозитидного сигнального пути.
174
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Диацил глицерол может подвергаться дальнейшей деградации под
действием диацилглицероллипазы до арахидоновой кислоты, которая
превращается в разнообразные биологически активные метаболиты,
в том числе эйкозаноиды и простагландины.
Суммируя сказанное выше, можно утверждать, что при функци-
онировании рассмотренной регуляторной системы образуются по
меньшей мере три вторичных посредника: инозитол-
трисфосфат, DAG и арахидоновая кислота (последнюю в связи с ее
участием в регуляции свертывания крови многие исследователи также
относят ко вторичным мессенджерам). Каждый из них выполняет спе-
цифические функции, включая увеличение содержания внутриклеточ-
ного Са2+ и активацию Са-зависимых протеинкиназ. Эта сигнальная
система распространена достаточно широко, хотя в деталях механизм
регуляции ею специфических клеточных функций пока неясен.
2.4.3. Метаболизм фосфоинозитидов и регуляция
проницаемости мембран для Са2+
Каков же механизм увеличения внутриклеточной концентрации
Са2+, вызываемой инозитолтрисфосфатом и диацилглицеролом, обра-
зующимися при гидролизе фосфатидилинозит-4,5-бисфосфата? Сле-
дует иметь в виду, что, во-первых, для работы фосфолипазы С необхо-
дим ГТФ, т. е. активация рецептора реализуется через G-белок, а
во-вторых, метаболизм фосфолипидов обеспечивает по крайней мере
два процесса: индукцию входа кальция в клетку из внешней среды по
градиенту концентрации и высвобождение ионов кальция в цитозоль
из внутриклеточных депо.
В модельных системах было показано, что фосфатидная кислота
способна транспортировать через искусственные мембраны двухва-
лентные катионы с эффективностью, близкой к действию кальциевого
ионофора А 23187. При этом ионы магния транспортировались значи-
тельно медленнее, чем ионы кальция, что говорит о селективности
фосфатидной кислоты в качестве кальциевого ионофора.
Относительно механизма ионофорного действия фосфатидной кис-
лоты существует ряд гипотез, ни одна из которых в настоящее время
не может быть признана полностью доказанной. Возможно, фосфа-
тидная кислота каким-то образом модифицирует структуру липидного
бислоя, образуя участки с небислойной упаковкой молекул. При этом
на границе бислоя и монослоя образуются каналы для ионов кальция.
Несмотря на то что имеется много фактов, указывающих на то, что
в физиологических условиях под действием ИФ3 происходит высво-
2.4. h-te ьи tniHMiij... мессенджеры и i^c . нниин. зь
бождение кальция из цистерн эндоплазматического ретикулума, меха-
низм открывания Са-каналов неясен. ИФ3 вызывает высвобождение
Са2+ в клетках поджелудочной железы, стимулирует мобилизацию за-
пасенного кальция в процессе индуцируемого адреналином поступле-
ния глюкозы из печени в кровь. В ооцитах лягушки поток ионов Са2+
увеличивается вдвое в ответ на стимуляцию ИФ3. Секреция корти-
кальных гранул, осуществляющаяся яйцеклеткой сразу же после оп-
лодотворения (в результате чего образуется так называемая оболочка
оплодотворения), также вызывается ионами кальция, концентрация
которых в яйцеклетке резко увеличивается в ответ на внутриклеточ-
ную инъекцию трифосфоинозитидов.
На долю фосфатидилинозитола приходится лишь 2—8% всех фос-
фолипидов, содержащихся в клеточных мембранах эукариот. Этот
фосфолипид часто называют миоинозитолом, поскольку впервые он
был выделен из мышечного гомогената. Небольшая часть фосфатиди-
линозитола фосфорилирована в положении 4 или в положениях 4 и 5.
От 1 до 10% фосфатидилинозитола, присутствующего в мембране, при-
ходится на долю фосфатидилинозитол-(4,5)-бисфосфата, обозначаемо-
го как ФИФ2 или ФИ(4,5)Ф2. Этот компонент является, вероятно, пер-
вой мишенью для фосфатидилинозитолспецифичной фосфолипазы С,
которая активируется во многих клетках G-белками.
Последующий гидролиз молекулы приводит к быстрому распаду
фосфатидилинозитола и кратковременному нарастанию продуктов
распада. Например, во время активации тромбоцитов за 90 с дегради-
рует половина общего фонда фосфатидилинозитола. Продукты распа-
да участвуют в качестве вторичных мессенджеров во многих клеточ-
ных процессах. Исследование этой системы в деталях еще не законче-
но, но самая общая ее схема представлена на рисунке 2.20 (с. 172).
Начальными продуктами гидролиза ФИФ2 являются диацилглице-
рол и инозитол-( 1,4,5)-трисфосфат. Диацилглицерол остается свя-
занным с мембраной, а ИФ3 высвобождается в цитоплазму. Обыч-
но жирнокислотные компоненты фосфатидилинозитола представлены
стеариновой кислотой в положении 1 и арахидоновой кислотой
в положении 2.
ФИ3 служит вторичным посредником, и его основной функцией,
по-видимому, является мобилизация Са2+, аккумулированного в эндо-
плазматическом ретикулуме. Возможно, этот компонент путем прямо-
го связывания открывает Са-специфичные каналы в эндоплазматиче-
ском ретикулуме, что приводит к выходу ионов кальция из ретикулума
в цитоплазму, где его уровень возрастает в несколько раз (от 0,1 мкМ
до 5—10 мкМ).
176
Раздел 2. МОЛЕКУЛЯРНАЯ НЕИРОХИМИЯ
2.4.4. Роль ионов кальция в функционировании нейронов
Важная роль в нервной ткани отводится еще одному вторичному
посреднику — Са2+ (рис. 2.21). Следует отметить, что концентрация
свободного (ионизированного) кальция в нервных клетках чрезвычай-
но низка и составляет от 10-6 до 10-8М. В нейроне, как и в других
клетках, большая часть кальция находится в связанном состоянии.
Низкая концентрация ионизированного кальция в цитоплазме обеспе-
чивается благодаря его захвату митохондриями в обмен на прото-
ны, выкачиванию из клетки за счет работы кальциевого насоса или
Рис. 2.21, а. Функция кальция в аксонах
Возбуждающие эффекты показаны сплошными тонкими линиями, тормозные эффек-
ты — прерывистыми тонкими линиями'.
/ — активизация потенциалзависимых Са-каналов (Cav) при деполяризации мембра-
ны; 2 — активизация цАМФ потенциалнезависимых Са-каналов; 3 — регуляция Са24
уровня цАМФ и цГМФ посредством стимуляции фосфодиэстеразы; 4 — активация ме-
диатором аденилатциклазы; 5 — экзоцитоз синаптических и секреторных пузырьков,
зависящий от входа Са2+ ; 6 — аксональный транспорт; 7 — захват Са2+ митохондрия-
ми в обмен на протоны; 8 — аккумуляция Са2+ в цистерны эндоплазматического рети-
кулума; 9 — уменьшение возбудимости нейрона внеклеточным Са2+; 10 — активация
сократимых волокон ионами кальция (либо непосредственно, либо через посредство
изменений уровня цАМФ); // — метаболический насос для выкачивания Са2+ и
Na/Ca-обмена; 12 — Са-зависимая К-проводимость, активируемая повышением уров-
ня Са2+ ; 13 — увеличение концентрации Са2+, снижающее проводимость электриче-
ских синапсов; 14 — связывание Са2+ с эндоплазматическим ретикулумом и другими
органеллами и макромолекулами Знаки вопроса над стрелками — предполагаемые
механизмы. — потенциалзависимые К-каналы; G№ и GK — соответственно натриевая
и калиевая проводимость (по: Шеперд, 1983).
2.4. G-Белки, вторичные мессенджеры и протеинкиназы
17
Рис. 2.21, б. Функция кальция в дендритах
Na/Ca-обменника, аккумуляции в эндоплазматическом ретикулуме и свя-
зыванию макромолекулами, включая парвальбумины и кальмодулин.
Можно перечислить целый ряд клеточных функций и свойств, за-
висящих от кальция: экзоцитоз синаптических и секреторных пузырь-
ков, текучесть аксоплазмы и аксональный транспорт, клеточная под-
вижность, мышечное сокращение, ферментативные реакции и прони-
цаемость мембраны и др. Кроме того, увеличение внутриклеточной
концентрации Са2+ изменяет Са-зависимую К-проводимость мембраны
и уменьшает возбудимость электрических синапсов. Са2+ способен воз-
действовать на метаболизм нейрона, снижая pH цитоплазмы, влияя на
активность ключевых ферментов, понижая содержание цАМФ и цГМФ
за счет активации фосфодиэстеразы.
Нейрон способен быстро «выкачивать» ионизированный кальций во
внеклеточное пространство и «закачивать» его в органеллы, резко
уменьшая концентрацию кальция в цитоплазме до необходимого низко-
го уровня, а в ответ на внешние сигналы столь же быстро увеличивать
ее, благодаря чему клетка может использовать локальные изменения
концентрации этого иона для регуляторных целей. Возможно, кальций
даже более универсальный внутриклеточный регулятор (вторичный
посредник), чем цАМФ, поскольку период действия Са2+ очень коро-
ток — измеряется миллисекундами, а не секундами, как в случае
178
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
с цАМФ. Механизмы, контролирующие доступность свободного каль-
ция, разнообразными способами связаны с путями синтеза медиаторов и
общим метаболизмом клетки. Благодаря этому Са2+ играет важную
роль в долговременных процессах, лежащих в основе пластичности си-
напсов, распознавания объектов и консолидации памяти (см. далее).
Одним из ферментов, регулируемых Са2+, является фосфолипаза С,
которая при низкой концентрации Са2+ использует в качестве субстрата
преимущественно ФИФ2, но при более высокой концентрации Са2+, по
крайней мере in vitro — нефосфорилированный фосфатидилинозитол.
Наиболее важным ферментом, активируемым Са2+, является про-
теинкиназа С. Этот фермент находится в цитозоле до тех пор, пока
в ближайшем его окружении не появляется диацилглицерол и Са2+.
В присутствии обоих агентов протеинкиназа С приобретает способ-
ность связываться с мембраной, выбирая для этого кластеры, обога-
щенные фосфатидилсерином. В данных условиях наблюдается ее вы-
раженная активация, причем субстратами фермента становятся мемб-
ранные белки ближайшего окружения.
Эффекты на протеинкиназу С, сходные с действием диацилглице-
рола, оказывают форболовые эфиры, в связи с чем ее часто рассмат-
ривают как внутриклеточную мишень для этих соединений. В активи-
рованном состоянии протеинкиназа С эффективно фосфорилирует
белки-мишени по серину и треонину.
2.4.5. Другие вторичные посредники
Наиболее важными вторичными мессенджерами являются цАМФ,
цГМФ, Са2+, ИФ3, DAG и активные формы кислорода и азота.
цГМФ образуется из ГТФ с помощью фермента гуанилатциклазы.
В отличие от аденилатциклазы, этот фермент не связан с мембранны-
ми рецепторами. Однако образование цГМФ частично сопровождает-
ся активацией трифосфоинозитидного цикла, поскольку некоторые
его компоненты, по-видимому, стимулируют гуанилатциклазу. цГМФ
активирует специальные G-киназы, вызывающие фосфорилирование
многочисленных белков; физиологическая роль некоторых из них пока
не установлена. По-видимому, цГМФ может играть роль вторичного
мессенджера в клетках нервной ткани.
цГМФ образуется с помощью двух разных форм гуанил ил цикла-
зы — мембраносвязанной и цитозольной. Мембранный фермент акти-
вируется натрийуретическим гормоном (G-белки в этом не принимают
участия), а растворимый (цитозольный)— радикалами NO*, СО* и
*ОН. Последние представляют собой новый класс неорганических ре-
гуляторов, проявляющих свойства то межклеточных (NO, Н2О2), то
14. ,-Б^ 1 . »ч- f । иь । .1
внутриклеточных регуляторов. цГМФ активирует ПК G, но, кроме то-
го, изменяет активность и других белков-мишеней, включая ионные
каналы (как в случае реализации светового сигнала).
Еще одним видом вторичных мессенджеров, чья регуляторная
функция установлена лишь недавно, являются активные формы кис-
лорода (АФК). Они образуются в ходе окислительного метаболизма в
результате постадийного восстановления молекулярного кислорода.
Ранее предполагалось, что в нормальных условиях восстановление
кислорода до конечного продукта (молекул воды) в митохондриальных
мембранах протекает таким образом, что «утечки» промежуточных
продуктов не наблюдается, и процесс приводит к образованию АТФ.
Однако на самом деле в процессе восстановления могут образовы-
ваться различные АФК. Было установлено, что избыток АФК вреден
для клетки. В то же время сами эти соединения в нормальных условиях
выступают в роли вторичных мессенджеров. В настоящее время к сво-
боднорадикальным соединениям кроме АФК относят также NO-ради-
кал (активная форма азота) и пероксинитрит, который фактически яв-
ляется активной (хотя и нерадикальной) формой азота.
В последние годы выявлен широкий спектр физиологических эффек-
тов АФК, к которым, прежде всего, относятся регуляция клеточной про-
лиферации и тонуса сосудов, индукция транскрипции определенных ге-
нов. Показано функционирование АФК в качестве вторичных внутри-
клеточных мессенджеров. Так, АФК непосредственно участвуют в
активации онкогенов, а также гена, кодирующего главную форму фак-
тора транскрипции АР-1 в ответ на ионизирующую радиацию. О2 и
Н2О2 активируют фактор транскрипции NFkB, который вызывает экс-
прессию генов, кодирующих ряд цитокинов и вирусов, в том числе ВИЧ,
а NO* подавляет активацию NFkB, увеличивая индукцию ингибитора
фактора транскрипции NFkB и стабилизируя его. В индукции синтеза
белков теплового шока, повышающих резистентность клеток к высоким
температурам, радиации, токсическому действию ионов тяжелых метал-
лов и лекарственных препаратов, основная роль отводится перекиси во-
дорода. NO* и СО* связываются с гемовой частью гуанилатциклазы и
обратимо изменяют синтез цГМФ, тем самым управляя работой этих
важных внутриклеточных регуляторов внутриклеточной коммуникации.
Одним из наиболее изученных факторов межклеточных сигнализа-
ций является оксид азота (нитроксид, NO’-радикал). Радикал NO* яв-
ляется важнейшим нейромедиатором, интермедиатом иммунной систе-
мы и регулятором сердечно-сосудистой системы человека. В животном
организме NO’ работает как вторичный мессенджер и в перифериче-
ской, и в центральной нервной системе.
В результате повышения концентрации NO’ активируется цитозоль-
ная гуанилатциклаза, катализирующая превращение ГТФ в цГМФ, кото-
180
Раздел 2 МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
рый, в свою очередь, активирует специфическую протеинкиназу (G-кина-
зу) (или непосредственно, или с помощью дополнительного сигнального
пути, включающего еще один вторичный мессенджер — аденозицдифос-
фатрибозу). Последняя открывает Са-каналы внутриклеточных Са-депо,
в результате чего повышается концентрация кальция в цитозоле, акти-
вируются Са-зависимые протеинкиназы, фосфорилируется белковый
фактор регуляции транскрипции и начинается синтез белков теплового
шока, обеспечивающих клеточную защиту.
Оксид азота обладает широким спектром биологического действия.
Он участвует в межклеточной сигнализации в центральной и перифе-
рической нервной системе. Его идентифицируют с эндотелиальным
фактором релаксации, расслабляющим гладкие мышцы и предотвра-
щающим агрегацию тромбоцитов и адгезию нейтрофилов к эндоте-
лию. Синтез оксида азота фагоцитирующими клетками регулирует
пролиферацию лимфоцитов. Он вносит вклад в бактерицидное и про-
тивоопухолевое действие фагоцитов. Более того, нитроксид является
регулятором внутриклеточной концентрации ионов Са2+ и, соответ-
ственно, активности ряда Са-зависимых ферментов. Источником NO*
в центральной и периферической нервной системе являются эндоте-
лиоциты сосудов, клетки микроглии и астроциты, неадренергические
нервные волокна и глутаматергические нейроны (рис. 2.22).
Функции нейронального NO* чрезвычайно разнообразны: он конт-
ролирует осцилляторную активность нейронов, является медиатором
ноцицепции, участвует в формировании циркадных ритмов, в процес-
Рис. 2.22. Возрастание внутриклеточного уровня АФК в результате активации
глутаматных рецепторов нейронов
2.4. G-Белки, вторичные мессенджеры и протеинкиназы
г
сах термогенеза, обоняния, принятия пищи и воды, снижает тревож-
ность, регулирует выход нейромедиаторов; играет существенную роль в
процессах потенциации и, соответственно, долговременной памяти.
Нейрональный NO’ является важнейшим вторичным мессенджером, с
помощью которого нервная система управляет тонусом сосудов, снаб-
жающих кровью все системы организма, причем действие оксида азота
может быть прямым и опосредованным (например, стимулируя высво-
бождение вазопрессина). Показано участие оксида азота, выделяюще-
гося в синоатриальном узле, в автономном контроле сердцебиения.
Снижение содержания цитоплазматического цГМФ при активации
клеточных рецепторов L-глутаматом и его производными является
NO-обусловленным процессом. В зависимости от уровня NO* он мо-
жет или ингибировать, или потенцировать высвобождение глутамата
и аспартата, тем самым защищая нейроны от экзайтотоксических
эффектов медиатора. Избыточная продукция или недостаточно быст-
рая нейтрализация глутаминовой и аспарагиновой аминокислот, вы-
полняющих роль возбуждающих медиаторов в нейронах, приводит к
нейротоксическим эффектам. В основе токсического действия лежит
стимуляция АФК, способная приводить к окислительному поврежде-
нию тканей. Кроме того, NO* защищает головной мозг от ишемиче-
ских и нейротоксических инсультов, контролирует осцилляторную ак-
тивность нейронов.
Нет сомнений в том, что существует гораздо больше сигналтранс-
дукторных систем, чем уже описано в настоящее время. В конце про-
шлого века были обнаружены сигнальные функции у церамида, фос-
фатидной кислоты, циклоаденозиндифосфатрибозы и 5'-АМФ. Сейчас
интенсивно изучаются связанные с ними сигналпередающие реакции.
2.4.6. Протеинкиназы и фосфопротеинфосфатазы
В настоящее время установлено, что часть вторичных посредни-
ков выполняет свою функцию в нейронах, модулируя свойства внутри-
клеточных белков за счет фосфорилирования. Фосфорилирование бы-
стро, специфично и обратимо меняет свойства и функции белка. Белки
фосфорилируются разнообразными протеинкиназами (ПК), а под дей-
ствием протеинфосфатаз (ПФ) фосфатная группа легко отщепляется.
Обе группы ферментов находятся под регуляторным влиянием раз-
личных внутриклеточных вторичных посредников. Как правило, вто-
ричные посредники активируют сериновые и треониновые киназы, в то
время как внеклеточные сигналы реализуются через тирозиновые про-
теинкиназы. Некоторые из них являются узкоспециализированными,
р для других субстратом может быть широкий спектр белков (табл. 2.9).
Таблица 2.9. Классификация протеинкиназ (по: Гусев, 2000)
Класс Подкласс Семейство
Протеинкиназы Циклонуклеотидза - цАМФ-зависимые киназы
A, G, С висимые киназы цГМФ-зависимые киназы
Фосфолипидзависи- мые киназы Киназы, зависящие от кальция
Киназы, не зависящие от кальция
Са-кальмоду- Истинные Са-каль- Многофункциональные
линзависимые модулинзависимые Са-кальмодулинзависимые
протеинкиназы протеинкиназы протеинкиназы
Киназа фосфорилазы
АМФ-зависимые
АМФ-зависимые киназы
киназы
00
IQ
Возможный
субъединичный
состав
Некоторые регуляторы
ферментативной
активности
R2C2 цАМФ, фосфорилирование
С2 цАМФ, фосфорилирование
С Диацил глицерол, фосфолипиды, Са2+
CnRn
Са2+, кальмодулин,
фосфорилирование
(CRR'R")4R"4 Са2+, кальмодулин,
фосфорилирование
Раздел 2. МОЛЕКУЛЯРНАЯ
CR'R'
АМФ, фосфорилирование
Протеинкиназы Циклинзависимые С, М, G киназы Несколько семейств циклинзависимых киназ
МАР-киназы Стрессактивируемые киназы, р42, р44 киназы
Киназы гликогенсинтазы Киназы гликогенсинтазы Казеинкиназа II
Тирозиновые протеинкиназы Растворимые тирози- Более 10 семейств (srk, brk, новые киназы tec, csk, fes, abl, jak и т. д.) Мембраносвязанные Многочисленные семейства тирозиновые киназы киназ—рецепторов гормонов и факторов роста Киназы Киназы МАР-киназ и киназы МАР-киназы киназ МАР-киназ
Казеинкиназы I
Несколько семейств
Окончание табл. 2.9
CR Фосфорилирование
С Фосфорилирование
С Фосфорилирование
C2R2 Самофосфорилирование, полиамины
С Фосфорилирование, регуляторные белки
C2R2 Гормоны, фосфорилирование
С Фосфорилирование
С Mg2+
2.4. G-Б.гЛги еторим ы._ м, ссендм еиЬ' и пр< тсиньиназы
£1
184
Раздел 2. МОЛЕКУЛЯРНАЯ НЕИРОХИМИЯ
Степень фосфорилирования определяется балансом между этими фер-
ментами и отражает в интегральном виде состояние сигнальных путей.
Субстратом ПК и ПФ являются разнообразные ферменты, рецепторы,
ионные каналы и структурные белки. Так, обратимое фосфорилирова-
ние характерно для нХР, рецепторов ГАМК, глицина и гутамина, а так-
же для 5-НТ3-серотониновых рецепторов. Фосфорилирование нейро-
специфических белков играет важную роль в формировании синапти-
ческих везикул и высвобождении медиатора (см. гл. 3.4), а также в
аксональном транспорте (см. гл. 3.3). ПК переносят фосфатную группу
от АТФ на серин, треонин или тирозин субстрата.
цАМФ-Зависимые протеинкиназы (ПК А). Являются первич-
ным эффектором для цАМФ. ПК А представляет собой тетра-
мерный комплекс из двух каталитических и двух регуляторных
(ингибиторных) субъединиц. цАМФ активирует ПК А, связываясь с ре-
гуляторной субъединицей, что вызывает ее диссоциацию и активацию
каталитической субъединицы. Такое снятие тормозного влияния регу-
ляторного белка является общим механизмом влияния вторичных по-
средников на функционирование ПК. Каталитическая субъединица ПК А
фосфорилирует сериновые и треониновые остатки различных бел-
ков-мишеней. Этот домен сходен с таковыми у других киназ, особенно-
стью является лишь то, что ПК А начинает фосфорилирование в ответ
на сигнал, передаваемый цАМФ. ПК А существует в двух изоформах,
относительное количество которых варьирует в разных тканях.
Протеинкиназы G (ПК G). Другим распространенным классом
протеинкиназ являются ферменты, стимулируемые с помощью цГМФ, —
протеинкиназы G. В тканях млекопитающих их содержание невелико
(1—2% от общей протеинкиназной активности). Наиболее высок уро-
вень протеинкиназ G в мозжечке, сердечной мышце и легких, эти же
ткани содержат и наибольшее количество цГМФ, не превышающее,
впрочем, 10% от содержания в них цАМФ.
Фермент состоит из двух субъединиц, обладающих катали-
тической активностью и способных связывать циклический нуклеотид.
Скорость, с которой ПК G фосфорилирует субстрат, значительно ни-
же, чем у ПК А, что может быть основой для субстратной специфич-
ности этих двух киназ.
(Са + кальмодулин)-зависимые протеинкиназы (ПК В). Ней-
роны в изобилии имеют ПК этого типа, являющиеся мультифункци-
ональными серин-треониновыми киназами. К настоящему времени
2.4. G-br KP, tk, . J.- Mr • НДЖсч 1 И нс еТеИ»*|-,ИН. jb'
Та
обнаружено три типа протеинкиназ В, которые в порядке убыва-
ния молекулярной массы обозначаются как протеинкиназы В I, II и III.
Основные их различия заключаются в субстратной специфичности.
Так, протеинкиназа В I фосфорилирует лишь несколько белков в
нервной ткани, в то же время для киназы В II только в головном мозге
имеется более 10 субстратов. ПК В II образована —14 а- и Р-субъ-
единицами, каждая из которых имеет в своем составе каталитический
и регуляторный домены, а также другие домены, позволяющие осу-
ществлять олигомеризацию фермента в необходимом участке клетки.
Са2+ и кальмодулин активируют эту ПК, «отодвигая» регуляторный
щмен от каталитического участка. ПК В II фосфорилируют большое
количество субстратов, включая белки ионных каналов, а также бел-
ки, участвующие во внутриклеточной передаче сигналов.
Как установлено, протеинкиназа В II составляет около 0,4% от об-
щего белка головного мозга млекопитающих, что свидетельствует о
важной регуляторной роли фермента в ЦНС. В отличие от «цитоплаз-
матической» ПК В I, она преимущественно связана с мембранной
фракцией и сконцентрирована в области постсинаптических уплотне-
ний в тех отделах нервной системы, которые связаны с обучением
(гиппокамп у позвоночных и сенсорные нейроны у беспозвоночных).
ПК В II обладает свойствами двухфазного переключателя со стабиль-
ностью, необходимой для кодирования долговременной памяти.
Киназа легких цепей миозина является представителем второй
группы (Са + кальмодулин)-зависимых протеинкиназ. Регуляция с ее
помощью легких цепей миозина осуществляется по принципу «все
или ничего». Так, в отсутствие кальмодулина (КМ) фермент практиче-
ски не активен, а при действии комплекса Са + КМ его активность
возрастает в 200—300 раз.
Протеинкиназы группы С (ПК С). Являются еще одной важной
группой серин-треониновых киназ. Семейство протеинкиназы С обна-
ружено в разных тканях млекопитающих, их локализация лишена
строгой тканевой и видовой специфичности. К ПК С относят мономер-
ные киназы, активаторами которых являются такие вторичные посред-
ники, как DAG и Са2+. Наибольший активирующий эффект на ПК С
оказывают диацилглицерол и в меньшей меуе фосфатидилинозитол,
фосфатидилсерин и фосфатидная кислота. Диацилглицерол уве-
личивает сродство ПК С к фосфолипидам, при этом она становится
чувствительной к физиологическим концентрациям Са2+ в клетке. DAG
вызывает перемещение ПК С из цитозоля в плазматическую мембрану,
где она связывается с мембранным фосфатидилсерином и Са2+. В ре-
186
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
зультате этого происходит разблокирование аутоторможения фермента
и активация фосфорилирования различных белков. ПК С способна
взаимодействовать не только с плазматической мембраной, но и с эле-
ментами цитоскелета, перинуклеарного и ядерного пространства, где
также может фосфорилировать белки. Продолжительная активации ПК С
может быть вызвана форболовыми эфирами и канцерогенными вещест-
вами, обладающими DAG-подобной активностью. В составе изофер-
ментов семейства ПК С обнаружена также Са-независимая фосфоли-
пидактивируемая форма.
Как отмечалось, протеинкиназы, как и сопряженные с ними G-бел-
ки, имеют различную преимущественную локализацию в тех или иных
клетках нервной ткани. Таким образом, разные типы клеток мозга
приспособлены к восприятию сигналов, активирующих синтез различ-
ных вторичных посредников: цАМФ, цГМФ, ИФ3, DAG и АФК.
Тирозиновые протеинкиназы (ТК). Представлены двумя
классами киназ, переносящих фосфатную группу на тирозиновый
остаток соответствующего субстрата. Рецептор ТК является транс-
мембранным белком с внеклеточным доменом, способным связываться
с различными лигандами (фактором роста, нейротрофическим факто-
ром или цитокинами), а внутриклеточный каталитический домен отве-
чает за фосфорилирование соответствующих субстратов. Нерецептор-
ные ТК являются цитоплазматическими или мембраносвязанными
ферментами, которые активируются внеклеточными сигналами опос-
редованно. Фосфорилирование тирозиновых остатков встречается ре-
же, чем фосфорилирование сериновых и треониновых, и чаще всего
служит для связывания сигнальной молекулы с фосфорилируемым
белком. ТК играют важную роль в процессах роста и дифференциации
нервных клеток.
Митогенактивируемые протеинкиназы (МАР-киназы, МАРК).
В отличие от других протеинкиназ, которые напрямую активируются
вторичными посредниками, могут активироваться и другими способа-
ми, например при фосфорилировании их другими ПК. Первоначально
МАРК были идентифицированы как участники контроля за клеточным
ростом, но к настоящему времени известно их участие и во многих
других процессах.
В нейронах МАРК находятся в неактивном состоянии, но активи-
руются при их фосфорилировании другими киназами. Они являются
частью киназного каскада, когда одна ПК фосфорилирует
и тем самым активирует следующую за ней в каскаде. Внеклеточный
2.4. G Белки, вторичные мессенджеры и протеинкиназы ВАШ
сигнал, являющийся пусковым для этого каскада, такой как внекле-
точный фактор роста, связываясь с рецептором ТК, на следующем
этапе активирует мономерный G-белок Ras. МАРК, будучи активиро-
ванной, способна фосфорилировать факторы транскрипции, регули-
рующие экспрессию генов. Среди широкого набора субстратов МАРК
можно назвать другие ферменты, включая протеинкиназы и белки ци-
тоскелета.
Протеинфосфатазы (ПФ). Хорошо изучены серин-треонино-
вые протеинфосфатазы (ПФ I и II), демонстрирующие меньшую суб-
стратную специфичность, чем ПК. Их ограниченная специфичность
может объясняться тем, что каталитические субъединицы этих фер-
ментов гомологичны, но каждый имеет специфичную регуляторную
субъединицу.
Среди серин-треониновых фосфатаз в клетках млекопитающих
преобладают ПФ 1, дефосфорилирующие широкий круг белков. Их
активность регулируется различными ингибиторными белками, экс-
прессируемыми в нейроне. ПФ ПА является мультисубъединичным
ферментом с широким кругом субстратов. Присутствующая в нейро-
нах в больших количествах ПФ ИВ (кальцинейрин) активируется
Са-кальмодулином. Несмотря на то что, как и ПК IB, она имеет тот же
активатор, субстраты их действия не всегда совпадают. ПФ ИВ состо-
ит из регуляторной и каталитической субъединиц. Первоначально для
ее активации Са-кальмодулин присоединяется к каталитической субъ-
единице и оттесняет тормозной регуляторный домен.
Фосфатазы / ингибируются белковыми ингибиторами, относитель-
но нечувствительны к ингибированию АТФ и предпочтительно дефос-
форилируют Р-субъединицу киназы фосфорилазы. Фосфатазы // не
ингибируются термостабильными белковыми ингибиторами, чувстви-
тельны к ингибированию АТФ и преимущественно дефосфорилируют
ос-субъединицу киназы фосфорилазы.
Все ПФ II делятся на подтипы: ИА, ИВ (кальцинейрин) и ИС. Отли-
чительной особенностью фосфатаз ИС является регулируемость Mg2+,
а фосфатаз ИВ — зависимость от Са2+ и кальмодулина.
Активность фосфопротеинфосфатаз регулируется по «внешнему»
(субстратному) и «внутреннему» (фосфатазному) механизмам. В пер-
вом случае различные метаболиты связываются с субстратом (фосфо-
протеином), изменяя скорость дефосфорилирования; во втором слу-
чае лиганды связываются с фосфатазой и таким образом изменяют
фосфатазную активность.
188
I аздел 2. :лОЛЕКУЛлгН * чЕЙРО.хИМИ»
2.5.
ИОННЫЕ КАНАЛЫ И ИОННЫЕ НАСОСЫ
Существует два основных вида переноса веществ
через клеточную мембрану — пассивный (в направлении
от большей концентрации вещества к меньшей; его по устоявшейся
привычке называют транспортом по градиенту концентраций (хо-
тя на самом деле этот вид транспорта осуществляется против кон-
центрационного градиента) и активный (осуществляющийся в на-
правлении от меньшей концентрации вещества к большей, т. е. в проти-
воположном направлении). Первый обеспечивают мембранные каналы
или переносчики, второй — «оплачивается» энергией АТФ и осуществ-
ляется с помощью насосов. Классификация основных видов транспорта
веществ через клеточную мембрану представлена на рисунке 2.23.
Наиболее простым видом транспорта является пассивная
диффузия, которая может осуществляться тремя путями:
непосредственно через липидный бислой, через канальные структуры
и с помощью подвижного переносчика. Движущей силой диффузии
являются концентрационные градиенты переносимых веществ, а ее
скорость определяется гидрофобностью, конфигурацией и размерами
переносимой молекулы, а также размерами канала или сродством пе-
реносимой молекулы к переносчику.
Первый способ диффузии (непосредственно через бислой)
отличается малой специфичностью и хорошо коррелирует с гидро-
фильно-липофильным коэффициентом молекулы (коэффициентом рас-
пределения в системе «вода—масло»).
Липидный
бислой
Транспортируемая молекула
Простая Облегченная
диффузия диффузия
I_____________________I
Пассивный транспорт Активный транспорт
Рис. 2.23. Классификация видов транспорта через клеточную мембрану
2.5, Ионные каналы и ионные насосы
Второй способ — диффузия через каналы ограничена соот-
ветствием размеров переносимого соединения и внутренней поры ка-
налов, а также временем их жизни в открытом состоянии и плотно-
стью расположения каналов на мембране.
Третий способ диффузии (с помощью переносчика), кроме
всех вышеуказанных факторов, ограничен временем, необходимым
для пересечения переносчиком клеточной мембраны, и, соответствен-
но, физико-химическими свойствами мембранного бислоя.
Специфический переносчик, осуществляющий процесс пассивной
диффузии, может использоваться и для активного транспорта, но для
«оплаты» этого вида работы он должен соединяться с преобразовате-
лем энергии (см. рис. 2.23). Поскольку чаще всего в клетке для этих
целей используется энергия АТФ, системы активного транспорта со-
единены с АТФазами или сами представляют собой АТФазы, вклю-
чающие участки связывания и переноса требуемых соединений.
Наиболее распространенным типом активного транспорта являет-
ся транспорт через клеточную мембрану ионов натрия и калия,
приводящий к асимметричному распределению этих ионов по обе сто-
роны клеточной мембраны (низкий уровень натрия и высокий — ка-
лия внутри клетки, табл. 2.10).
Имеющиеся в клетке ионные градиенты часто используются в каче-
стве движущей силы для транспорта метаболитов. Данный вид транс-
порта называют вторичноактивным транспортом (см. рис. 2.23),
поскольку при этом АТФ используется не непосредственно, а через
создание промежуточного вида энергии, в связи с чем транспорт, осу-
ществляемый за счет энергии АТФ, стали называть первичноактив-
ным транспортом.
Таблица 2.10. Содержание ионов в клетках,
во внеклеточной жидкости и в морской воде
Объект^^ —~~’ Ионы Na+ к+ Са2+ Mg2+ СГ
Крыса мышцы 27 101 1,5 11,0 16
плазма крови 145 6,2 3,1 1,6 116
Лягушка мышцы 24 85 2,5 11,3 10
плазма крови 104 2,5 8,5 1,2 74
Осьминог мышцы 81 101 3,7 12,7 93
жидкость тела 525 12,2 11,6 57,2 480
Морская вода 440 9,5 9,6 56 535
190
Раздел 2. МОЛЕКУЛЯРНАЯ НЕИРОХИМИ^
/ 2
транспортируемых молекул
Рис. 2.24. Различия между канальным ( /) и переносчиковым (2) механизмами
переноса ионов
Для разграничения транспорта веществ (ионов) с помощью каналов
или переносчиков пользуются различиями в концентрационной зависи-
мости этих процессов: при транспорте вещества через канал, как прави-
ло, не наблюдается насыщения, в то время как транспорт с помощью
переносчика лимитируется концентрацией его молекул и поэтому дости-
гает плато вне зависимости от дальнейшего увеличения концентрации
переносимых соединений (рис. 2.24). Кроме того, транспорт через канал
в меньшей степени зависит от температуры, чем транспорт с помощью
переносчика: для последнего существует критическая температура, со-
ответствующая температуре фазового перехода мембранного бислоя.
Транспортные процессы по способу переноса классифицируют как
унипорт, симпорт и антипорт (рис. 2.25). Классификация транс-
портных процессов облегчает понимание их механизмов и особенностей.
В настоящем изложении основное внимание будет уделено особен-
ностям систем, осуществляющих транспорт ионов (как пассивный, так
и активный), поскольку они наиболее хорошо изучены.
Ионные каналы. Ионные каналы образованы ассоциатами поли-
топических мембранных белков, при этом они являются гомо- или гете-
роолигомерами. Наиболее просто устроены ионные каналы микроорга-
Унипорт Симпорт Антипорт
।__________________________________I
Котранспорт
Рис. 2.25. Способы переноса веществ через мембрану
>. Ионные каналы и ионные насосы
ic. 2.26. Аламетициновый канал: а — встраивание молекул (/) в бислой (2);
— вид канала со стороны бислоя
1змов. Так, продуцентом аламетицина является Trichoderma viride.
'шметициновый канал образуется из переменного количества субъеди-
щ, поэтому его размеры могут изменяться (рис. 2.26). Канал, создавае-
.1Й антибиотиком амфотерицином, имеет постоянный размер пор. В его
ютав обязательно должны включаться молекулы холестерина, поэто-
этот канал формируется только в холестеринсодержащей мембране,
оответственно, данный антибиотик оказывает воздействие только на
микроорганизмы, наружные мембраны которых содержат холестерин.
Следует вспомнить, что представление об антибиотиках возник-
) после обнаружения способности некоторых продуцируемых микро-
>ганизмами соединений формировать каналы в мембранах тех или
тых патогенных микроорганизмов. В природе эти соединения (ихчас-
) называют каналоформерами) синтезируются микроорганизмами в
феделенный период их развития для осуществления с помощью кана-
IB кратковременной связи и обмена информацией, в том числе и гене-
неской, между одноклеточными организмами (каналы, соединяющие
ie клетки, называют коннексонами). (Поэтому правильнее называть
иродные каналоформеры не антибиотиками, а симбиотиками.)
Молекулы-каналоформеры имеют пептидную природу. Ионные ка-
мы, образуемые в мембранах животных клеток, формируются также
) принципу олигомеров, но образующие их субъединицы обладают
фаздо большей молекулярной массой. В результате эволюционного
>вершенствования эти ионные каналы приобрели две важные
г о б е н н о с т и: механизм контроля за открыванием — за-
быванием канальной поры и большую избирательность.
В зависимости от природы пропускаемых ионов, каналы возбудимых
1еток делятся на катионные (Na-, К-, Са-каналы) и анионные (би-
192
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
карбонатньг , хлорные). Механизмы, обеспечивающие их
отк р ы вал и е, весьма разнообразны. Одни каналы реагируют на
связывание специфического лиганда с рецепторным участком комп-
лекса, другие — на деполяризацию мембраны или растяжение клетки.
Электрический ток, создаваемый на мембране при открываний кана-
ла, зависят от проницаемости канала для данного вида ионов и от их
концентрации в устье канала.
Ионное каналы различаются своей избирательностью. Натриевые,
калиевые и кальциевые каналы достаточно селективны. Анионные
каналу гораздо менее избирательны, хотя их часто называют «хлор-
ными каналами», поскольку ионы хлора являются наиболее распрост-
раненными анионами в животных клетках. Белки, входящие в состав
канальной структуры, обладают высокой конформационной лабиль-
ностью; отдельные части олигомерных конструкций канала, находясь
в состоянии динамической подвижности, меняют свое расположение
в пространстве (рис. 2.27). Это способствует спонтанному измене-
нию проводимости (открыванию — закрыванию) канала. Следова-
Рис. 2.27. Динамические состояния ионных каналов:
а, б, в — канал в закрытом состоянии (различные ракурсы); г, д, е — канал в открь'
том состоянии
2.5. Ионные каналы и ионные насось
тельно, фактор, вызывающий его активацию (мембранный потенци-
ал или конформационный сигнал от связывания лиганд. ), только
увеличивает вероятность открытого состояния канала. Время, в те-
чение которого канал находится в открытом состоянии, является
специфической характеристикой каналов каждого вида. Активация
канала означает возрастание вероятности перехода в открытое со-
стояние, а не увеличение продолжительности открытого состояния
канала. \
Устранение активирующего стимула приводит к деактивации ка-
нала (при спонтанном открывании канала его деактивация такж^про-
исходит спонтанно). Появление активирующего стимула в период,
когда канал находится в открытом состоянии, вызывает блокирова-
ние работы канала (для потенциалзависимых каналов применяется
термин инактивация, для лигандактивируемых — десенситиза-
ция). Помимо этого возможно также блокирование открытого ка-
нала, вызываемое крупной молекулой (например, токсина) или спе-
цифическим агентом (например, ионами магния), связывание которо-
го с некоторыми типами каналов препятствует проникновению других
ионов.
Каналы, активируемые лигандами, содержат'соответствующие цент-
ры или с внешней, или с внутренней стороны мембраны. Типичным при-
мером первого варианта является никотиновый холинорецептор кон-
цевой пластинки мышечного волокна (нХР) (см. гл. 2.2), второго —
Са-активируемые калиевые каналы нервной клетки, реагирующие на
внутриклеточную концентрацию ионизированного кальция.
Важная информация о свойствах ионных каналов была получена в
результате разработки в конце XX в. метода регистрации активности
одиночных каналов, так называемого «пэтч-клампа», существенные
данные об их структуре — с помощью экспрессии в ооцитах Xenopus
ионных каналов инъекцией соответствующих мРНК.
Ооциты не экспрессируют собственных ХР, однако после микро-
инъекции мРНКони не только изготавливают соответствующие рецеп-
торные белки, но и обеспечивают их сборку в функционально активные
комплексы. Такая модель весьма удобна для выяснения функциональ-
ной роли разных частей канала с применением метода направленных
мутаций (Site-directed mutagenesis).
На рисунке 2.28 представлена структура потенциалзависимого Са-ка-
нала, а в таблице 2.11 систематизированы некоторые свойства наибо-
лее полно исследованных ионных каналов.
Характерной особенностью ионных каналов является большое
разнообразие их изоформ. Особенно много изоформ обнаружено у
194
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
а
Внешняя I И
IV
Внешняя
Внутренняя
Рис. 2.28. Структура потенциалзависимого Са-канала:
а — топология -субъединицы: четыре гомологичных домена (I—IV) формируют
шесть а-спиральных трансмембранных элементов (представлены в виде цилиндров
(1 —6)). Домен IV обычно является сенсором, домены I и II формируют ионную пору;
б — субъединичная структура канала, содержащего (Xj -, а2-, р-, у- и 8-субъединицы
нХР и К-каналов. Тот факт, что каждая субъединица канала кодирует-
ся особым геном, открывает простор для альтернативного сплайсинга.
Кроме того, обнаружено, что при синтезе некоторых рецепторов про-
исходит редактирование РНК, приводящее к дополнительным вари-
ациям изоформ рецепторных белков.
Идентифицировано четыре класса опиоидных ре-
цепторов: |1-,8-,к- hORL-1 (Opioid Receptor-Like 1)(см. табл. 2.7),
между которыми, однако, имеется меньше различий по взаимодейст-
вию с агонистами и антагонистами, чем в других группах рецепторов.
Опиоидные рецепторы связаны с Gs- и Gj-белками, их тормозное дей-
ствие заключается в закрытии Са-каналов (к-рецептор) или в откры-
тии К-каналов (ц, 5, ORL-1). В зависимости от того, где локализова-
ны рецепторы, это приводит к уменьшению выброса медиатора или уг-
нетению нейрональной возбудимости.
Таблица 2.11. Свойства ионных каналов
Тип канала Лиганд Функция Олигомерный состав
Никотиновый холинорецептор концевой пластинки АХ Создание Na-тока в концевой пластинке Пентамер состава а2Р6с (в эмбриональной ткани — а2р&у)
Хлорные тормозные каналы ГАМК-активируемые Глициновые 5-НТ-активируемые ГАМК Глицин Серотонин Обеспечение синаптического торможения, препятствующего деполяризации мембраны Создание Na- и К-тока Гексамер Пентамер
Na-канал Са-канал пд пд Na-ток Са-ток Содержат четыре субъедини- цы, состоящие из четырех вну- тримембранных доменов из шести сегментов, соединенных цитоплазматическими петлями
Потенциал- зависимые каналы К-канал пд К-ток Состоит из четырех субъеди- ниц, каждая похожа на одиноч- ный домен Са-канала
Cl-канал пд Стабилизация ТМП Субъединица содержит один- надцать трансмембранных сег- ментов, димеры образуют в мембране два независимых ка- нала
2,5. Ионные накалы и ио-ннасосы
Окончание табл. 2.11
Тип канала
Лиганд
Функция
Олигомерный состав
АТФ-активируемые каналы
(активируемые пуринергическими
рецепторами)
АТФ Регуляция сократимости глад- По-видимому, пентамер, субъ-
кой мускулатуры единица содержит два транс-
мембранных сегмента и петлю,
формирующую пору
Ионотропные Глутумат (агонис- Формирование электрического Пентамеры, содержащие субъ-
(три вида) ты NMDA, АМРА, потенциала единицы с тремя трансмембран-
каинат) ными сегментами и петлей, фор-
мирующей пору
Глугам атн ы е -------------------------------------------------------------------------------------------------
рецепторы
Метаботропные Глутамат Управление метаболизмом че- Гексамер
(три группы, (3-HPG, рез сопряженные G-белки
всего восемь видов) N-ацетиласпар-
тилглутамат,
ACPD и др.)
Каналы, активируемые
циклическими нуклеотидами
цАМФ/цГМФ
Раздел 2. МОЛЕКУЛЯРНАЯ
Формирование ионных токов в Вероятно, тетрамер с субъеди-
сетчатке и обонятельном эпи- ницей из шести трансмембран-
телии ных сегментов
£
2.5. Ионные каналы и ионные насосы
T91
Для глутаматных рецепторов GluRB, GluR5 и GluR6 показано, на-
пример, что они имеют во втором трансмембранном сегменте либо
глу, либо арг (присутствие аргинина устраняет Са-проницаемость и
уменьшает проводимость канала). Несмотря на то что ДНК всех трех
изоформ содержит кодон для глутамата (CAG), в мРНК может по-
явиться CGG, кодирующий арг. Это происходит вследствие редакти-
рования уже образованной мРНК. В GluR6 также найдено дополни-
тельное редактирование (А/G) в первом трансмембранном сегменте.
Ионные насосы (транспортные АТФазы). Первичноактивный
транспорт. Ионные насосы представляют собой интегральные транс-
мембранные олигомерные белки, превращающие энергию АТФ в энер-
гию ионных градиентов. Они содержат центры связывания и гидролиза
АТФ, ионные центры и обладают высокой конформационной лабильно-
стью — способностью принимать несколько устойчивых конформаций,
различающихся сродством к переносимым ионам и ориентацией цент-
ров связывания этих ионов.
Энергия АТФ используется для «оплаты» перехода из конформации
с высоким сродством к переносимым ионам (на этой стадии ионы свя-
зываются ионными центрами) в конформацию с низким сродством (на
этой стадии ионы высвобождаются). Возвращение в исходную кон-
формацию происходит спонтанно (релаксационная кинетика) (Блюмен-
фельд, 1977; 2002). Термин «транспорт-
ные АТФазы» является синонимом тер-
мина «ионные насосы».
Промежуточным продуктом гидролиза
АТФ является фосфофермент, образую-
щийся при переносе терминального фос-
фата АТФ на белок с образованием мак-
роэргической ацилфосфатной связи. По
этой причине транспортные АТФазы от-
носят к АТФазам Р-типа.
История исследования транспортных
АТФаз началась в 1957 г., когда В. Хас-
сельбах обнаружил в составе микросо-
мальной фракции скелетных мышц (со-
держащей пузырьки саркоплазматического
ретикулума) АТФазную активность, акти-
вирующуюся очень низкими концентрация-
ми ионов кальция. Кд5 для нее составляла
около 5 • 10-7 М. Одновременно два иссле-
дователя, Б. Марш и Дж. Бендалл, описали
В. Хассельбах
Первооткрыватель Са-насо-
са, исследовавший его мо-
лекулярные свойства и опи-
савший его работу как
мембранного преобразовате-
ля энергии(Германия)
198
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
c. Эбаси
Нобелевский лауреат по фи-
зиологии и медицине, автор
оригинальных работ в облас-
ти функциональной биохи-
мии, открывший тропонин С
и описавший его функцио-
нальные свойства (Япония)
свойства так называемого «расслабляю-
щего фактора», добавление суспензии ко-
торого к глицеринированным волокнам,
сокращенным вследствие внесения в сре-
ду ионов Са, вызывает их релаксацию.
Эти феномены было трудно связать друг с
другом, поскольку они были обнаружены
за несколько лет до сделанного С. Эбаси
открытия тропонина С, ответственного за
Са-регуляцию сокращения.
Тем не менее очень быстро расслаб-
ляющий фактор Бендалла—Марша был
идентифицирован с саркоплазматическим
ретикулумом, и содержащейся в его мемб-
ранах Са-АТФазе была приписана роль
системы, связывающей кальций и вызы-
вающей расслабление глицеринированных
волокон (Бендалл, 1970).
Одновременно датский ученый
Й. К. Скоу приступил к изучению фено-
мена утраты ионной асимметрии, проис-
ходящей при хранении клеток при пониженной температуре. К тому
времени благодаря исследованиям А. Ходжкина было уже известно,
что процессы возбуждения обеспечиваются неравновесным распреде-
лением ионов натрия и калия на мембране возбудимых клеток (удоб-
ным объектом исследования служил гигантский аксон кальмара, но
немало опытов было поставлено и на скелетных мышцах лягушки).
Кратковременное выдерживание препаратов на холоде не влияло на их
жизнеспособность, недлительное хранение в холодильнике (при +4 °C)
уменьшало выраженность ионных градиентов и снижало возбудимость
аксонов. В 1954 г. А. Шатцман установил, что утрату Na-градиента
можно вызвать, выдерживая клетки в присутствии какого-либо сер-
дечного гликозида — строфантидина или уабаина. Лечебный эф-
фект этих соединений, выделенных из сока строфанта или экстрагиро-
ванных из наперстянки, был давно известен: они снижали сердечную
недостаточность, но это ничего не объясняло.
В 1957 г. Й. Скоу поставил простой опыт, результат которого че-
рез 40 лет был отмечен Нобелевской премией. Измельченные до го-
могенного состояния нервы краба были подвергнуты воздействию
различных концентраций хлористого натрия и хлористого калия в при-
сутствии магния и АТФ. Соотношение Na+/K+ изменялось от 0/150
до 150/0 мМ, и пропорционально изменению этого коэффициента из-
2.5. Ионные каналы и ионные насосы
199
менялась способность гидролизовывать
АТФ. При этом измерялась АТФазная ак-
тивность. Максимум АТФазной активнос-
ти приходился на 130мМ Na+ и 20 мМ
К+. АТФаза, активируемая совместным
присутствием натрия и калия, ингибирова-
лась уабаином. В опубликованной статье
Скоу сообщил, что им найдена фермента-
тивная система, ответственная за асим-
метричное распределение ионов натрия и
калия на мембране возбудимых клеток; он
назвал ее Na/K-зависимой Mg-активи-
руемой АТФазой (Скоу, 1957). С этих пуб-
ликаций Хассельбаха и Скоу и началась
история исследования участия транспорт-
ных АТФаз в функционировании возбуди-
мых клеток.
Са-насос. Ионы кальция выполня-
ют во многих тканях функции вторичного
мессенджера, что объясняется наличием
в клетках большого количества белков,
Й. Скоу
Нобелевский лауреат в об-
ласти биоэнергетики, перво-
открыватель Na/K-АТФазы,
впервые описавший ее функ-
циональные свойства, в том
числе молекулярные свойст-
ва фосфофермента и спо-
собность АТФ регулировать
сродство фермента к перено-
симым ионам (Дания)
обладающих высоким сродством к этому
иону: кальмодулин, кальпаин (кальцийнейрин в нервных клетках),
тропонин С (в скелетной мускулатуре). Поскольку сродство этих бел-
ков к кальцию находится в области 5* 10“6 — 5* 10-7М, для осу-
ществления такой регуляции клетка должна иметь систему «уборки»
ионизированного кальция, способную снижать его уровень до величи-
ны, меньшей 10“7 М. С этой задачей вполне справляется Са-насос
саркоплазматического (в других тканях — эндоплазматического) ре-
тикулума. Легкость выделения фракции ретикулума из скелетных
мышц в виде замкнутых везикул, содержащих Са-АТФазу с активным
центром, обращенным наружу, стимулировало массовые исследова-
ния кинетических свойств и транспортной функции Са-насоса. В итоге
было выяснено, что АТФаза исходно обладает высоким сродством к
кальцию и в присутствии Mg-АТФ быстро и обратимо фосфорилиру-
ется. Фосфофермент обладает способностью к изменению конформа-
ции, вследствие чего участок связывания ионов кальция оказывается
доступным по другую сторону мембраны, где ионы диссоциируют от
молекулы фермента.
При экспериментальном определении эффективности транспорта
кальция поначалу получали широко варьирующие значения величины
200
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
ц+ Закисление среды снижает Квых, облегчая выход кальция из СР
Рис. 2.29. Экспериментальное определение Са-АТФмакс для Са-насоса сарко-
плазматического ретикулума (СР).
Стрелками отмечено изменение концентрации оксалата натрия, проникающего через
мембрану везикул, и ионов кальция; последняя может быть рассчитана из произведе-
ния растворимости оксалата кальция и концентрации оксалат-иона. Зависимость ко-
эффициента Са-АТФ от рассчитанной концентрации кальция внутри везикул [Са2+ ]вн
позволяет определить максимальную величину Са-АТФ и константу для выхода каль-
ция из ретикулума
Са-АТФ (от 0,5 до 1,5); причины этого были неизвестны. Использова-
ние Са-преципитирующих анионов (оксалата), проникающих через
мембрану везикул, позволило измерять скорость транспортного про-
цесса против неизменного внутривезикулярного градиента концентра-
ции кальция, а варьирование концентраций оксалата позволило из-
мерить зависимость Са-АТФ при разных концентрациях [Са2+]вн.
Оказалось, что измеряемая величина аккумуляции кальция является
результирующей между величинами активного входа и пассивного вы-
хода кальция из ретикулума. Получение частично «разобщенных» пре-
паратов Са-насоса (в тот период ингибиторы бактериальных протеаз
еще не использовались широко) объясняло сильные различия резуль-
татов измерения значений Са-АТФ у разных авторов. Расчет макси-
мальной стехиометрии процесса на основе зависимости транспорта
кальция от величины [Са2+]вн привел к величине, равной 2 (рис. 2.29).
Использование этого приема позволило В. Б. Ритову впервые рассчи-
тать константу сродства системы кальциевого транспорта для ионов
кальция с внутренней стороны мембраны ретикулума. Она оказалась
равной 5* 10-4М. Поскольку, согласно эксперименту В. Хассельбаха,
эта величина с внешней стороны мембраны составляет (1 —5) • 10“7 М,
можно было заключить, что соотношение констант сродства достаточно
для того, чтобы создать на мембране ретикулума 1000—5000-кратный
градиент ионов кальция (Болдырев, 1978). Следовательно, Са-насос
2 5. VL ннпе нгн л >
20
ЕЬСа2АТФ
ЕЬСа2АТФ
ЕЬСа2АТФ
E2----------Е2ФН-------------Е2Ф ------------Е2ФСа2
Рис. 2.30. Схема гидролиза АТФ Са-насосом (последовательные стадии
пронумерованы)
способен обеспечить запас кальция в цистернах, достаточный для ре-
гуляции мышечного сокращения.
Процесс транспорта кальция осуществляется как унипорт, однако
создаваемый на мембране ретикулума потенциал немедленно тушится
легко проникающими через мембрану ионами магния или протонами.
Кинетическая схема, описывающая цикл Са-АТФазы, представлена
на рисунке 2.30. Образование комплекса Михаэлиса (АТФ Е2Са2+)
может осуществляться любым из двух указанных на рисунке путей
(стадии 1—3 или 4—6), но первый путь наиболее вероятен из-за ис-
ходно высокого сродства фермента к ионам кальция. Конформации
фермента, с которой начинается описание цикла, присвоен индекс Е1,
и фосфорилирование переводит фермент в конформацию Е2, в кото-
рой ионы кальция оказываются доступными по другую сторону мемб-
раны, причем сродство ионных центров к кальцию в этом состоянии
резко падает, что делает ионные центры свободными от диссоциирую-
щих ионов. После этого наступает процесс дефосфорилирования фер-
мента, вследствие чего фермент становится способен релаксировать в
исходную конформацию; при этом он восстанавливает высокое срод-
ство к кальцию и процесс может повториться.
Успехи в исследовании первичной структуры фермента, а в после-
дующем и рентгеноструктурный анализ разных конформаций Са-
о
АТФазы (последнее достижение — с разрешением 2,6 А) позволили
установить молекулярные передвижки, сопровождающие Е1 —Е2-пе-
реход, и наполнить формальную схему (см. рис. 2.30) конкретным со-
держанием (рис. 2.31).
Общая характеристика Са-насоса должна была включать и иссле-
дование роли липидного окружения. Впечатляющие результаты по ре-
конструкции очищенной Са-АТФазы в липосомы, составленные из
различных амфифи 1ьных соединений, включая детергенты, были про-
ведены В. И. Мельгуновым. Оказалось, что восстановление активное-
202
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Рис. 2.31. Топография Са-насоса в мембране
ти прямо пропорционально гидрофильно-липофильному коэффициен-
ту, характеризующему свойства соединения, используемого для приго-
товления липосом (рис. 2.32) (Болдырев, Мельгунов, 1984).
Эти опыты положили конец дискуссиям о специфической потреб-
ности каждого мембранного белка в наборе аннулярных липидов, ок-
ружающих белковые молекулы в мембране. Действительно, скорость
обмена аннулярных липидов, происходящего за счет латеральной
диффузии липидных молекул в мембране, составляет величину около
10“4 с (см. раздел 1), в то время как скорость работы транспорт-
ных АТФаз достигает величин порядка 60—100 Гц. Это означает, что
за время одного цикла аннулярные липиды могут успеть обновиться
много раз. Как видно из рисунка 2.32, решающими для сохранения
активности Са-АТФазы являются физические характеристики гид-
рофобной среды, а не специфические свойства молекул. Некоторые
неприродные детергенты (например, Brij 52) обеспечивают более вы-
сокую активность реконструированной АТФазы, чем та, которую мож-
2.5. Ионные каналы и ионные .
20;
10
s
8
6
4
2
• Brij 52
• Span 80
«Brij 30
Span 20* ,Tween2I
0
• LubrolPX
• Nonidet P-40
BriJ56 ••Triton X-100
• Myij 58
• Tween 20
_Q • Brij 35
Brij 58® eMyrj59
, *Тп1оп X-30
5 10 15 20
Гидрофильно-липофильный коэффициент
Рис. 2.32. Реконструкция Са-АТФазы в детергенты.
Восстановление активности происходит пропорционально величине гидрофильно-ли-
пофильного коэффициента детергента, ГЛК. Названия детергентов приведены в соот-
ветствии с Международной классификацией (по: Болдырев, Мельгунов, 1978)
но измерить в нативной мембране. Аналогичные опыты с митохонд-
риальной АТФазой почти одновременно были проведены Э. Рэке-
ром(1979).
Однако при реконструкции интегральных белков в искусственные
мембраны реконструированная система отличается от нативной. В о -
первых, реконструкция молекул в липидный бислой происходит
случайным образом, в результате чего часть молекул бывает ориенти-
рована нужным образом относительно сторон мембраны, а другая их
часть (примерно половина) — инвертирована. Во-вторых, в ходе
очистки и реконструкции теряются многочисленные минорные компо-
ненты нативной мембраны, обеспечивающие регуляцию процесса тран-
спорта. Нарушаются пути, связанные с регуляторным фосфорилиро-
ванием протеинкиназами, экстрагируются такие белки-регуляторы,
какфосфоламбаН' фосфолемман и др. Таким образом, эксперименты
по реконструкции функций мембранных белков решили важную зада-
чу — доказали, что свойства мембранных систем определяются из-
вестными физическими законами. Однако этот подход быстро исчер-
пал себя, и ему на смену пришли методы направленного мутагенеза,
искусственной индукции химерных белков и исследования их функций
в составе живой клетки.
204
₽-эздч.л 2. МСПЕКУЛЯРНАЯ НЕЙРОХИМИЯ
В. И. Мельгунов
Талантливый ученый, иссле-
довавший молекулярные свой-
ства транспортных АТФаз в
мышцах; осуществил успеш-
ную реконструкцию Са-на-
соса в различные детер-
генты, показав корреляцию
между эффективностью ре-
конструкции мембранных бел-
ков и гидрофобно-гидрофиль-
ным коэффициентом окру-
жения
Тем не менее одну важную особен-
ность транспортных АТФаз оказалось
возможным изучать с помощью классиче-
ских биохимических подходов. Эта осо-
бенность касалась их способности спон-
танно образовывать в мембране олиго-
мерные ассоциаты, в которых свойства
индивидуальных протомеров подверга-
лись модификации за счет воздействия
соседних молекул. Подробнее роль олиго-
мерных комплексов мембранных белков
будет рассмотрена на примере гетерооли-
гомера Na/K-насоса (см. далее).
Следует подчеркнуть, что в качестве ин-
тегральных мембранных структур мембран-
ные белки являются объектом регуляции со
стороны липидного окружения, поскольку
белок-белковые гидрофобные связи прин-
ципиально не отличаются от липид-липид-
ных или белок-липидных. Соответственно,
изменения микровязкости (плотности упа-
ковки) липидного бислоя, вызванные на-
правленным изменением жирнокислотного
состава мембранных липидов, отражаются
на легкости образования белковых ассо-
циатов в мембране и «запрещают» или «разрешают» реализацию взаи-
модействий протомеров белка в составе мембранного олигомера.
В наружной клеточной мембране обнаружен и другой тип Са-насо-
са, имеющий некоторую гомологию с Са-насосом ретикулума, хотя и
экспрессирующийся на разных генах. Последнее обстоятельство еще
раз демонстрирует наличие в геноме клетки многочисленных повто-
ров, которые приобретают смысл при конструировании различных
белков, обладающих близкими функциями. Разница между двумя
формами Са-насосов заключается и в сродстве к кальцию, и в реали-
зации разных регуляторных механизмов. Их сравнение показывает,
что живые системы не смогли бы удовлетворительно решить пробле-
му кальциевого гомеостаза, обходясь одним видом Са-насоса.
Na/K-насос. Термин «Na/K-насос» является синонимом тер-
мина «Na/K-АТФаза». Этот белок входит в состав наружных плаз-
матических мембран всех клеток животных в качестве интегрально-
го компонента, обеспечивающего превращение энергии гидролиза АТФ
2.5. Ионные каналы и ионные насосы
20!
Рис. 2.33. Расположение полипептидных цепей каталитической а-субъедини-
цы Na/K-АТФазы в мембране.
Показано расположение полярной области, содержащей центр связывания АТФ и фос-
форилирования (Р) и 10 трансмембранных доменов (Ml—М10). Черным выделены
аминокислотные радикалы, участвующие в формировании ионного канала в мембране
в энергию градиентов одновалентных ионов натрия и калия в стехио-
метрии 1 АТФ : 3 иона натрия : 2 иона калия. Функциональная едини-
ца фермента (называемая протомером) представлена глобулой, со-
стоящей из двух пептидных цепей — а- и 0-субъединиц (рис. 2.33).
Большая субъединица (а) имеет молекулярную массу около 100 кДа и
несколько последовательностей, составленных гидрофобными амино-
кислотами, что позволяет ей «прошивать» плазматическую мембрану
10 раз, при этом ее и С- и N-концы обращены в цитоплазму. В цито-
плазму клетки обращены и протяженные гидрофильные петли между
2-м и 3-м и между 4-м и 5-м сегментами. На последней петле распо-
ложен активный центр фермента. Ионные центры также формируют-
ся на а-субъединице. В связывании ионов принимают участие карбок-
сильные группы дикарбоновых аминокислот, локализованных на 3-м
и 6-м трансмембранных сегментах.
Р-Субъединица представляет собой гликопротеин, белковая часть
которого соответствует молекулярной массе 35 кДа, а полная масса ко-
206
Г аздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Э. Рэкер
Автор пионерских работ в об-
ласти биоэнергетики, функ-
ционирования молекулярных
машин — мембранных пре-
образователей энергии в ми-
тохондриях (США)
А. Аскари
Исследователь транспортных
ферментов, определил функ-
циональное значение олиго-
мерной организации Na/K-
АТФазы(США)
леблется в зависимости от типа ткани
и других факторов в пределах 45—
55 кДа. Эта субъединица пронизывает
мембрану один раз, и ее N-конец обращен
в цитоплазму. C-Концевая часть содержит
четыре цистеиновых остатка, соединенных
двумя дисульфидными связями. Эта часть
пептидной цепи имеет участки, подвергае-
мые гликозилированию.
Как типичная АТФаза P-типа, Na/K-
АТФаза обладает способностью находить-
ся в двух основных конформациях, обла-
дающих высоким сродством к ионам нат-
рия (Е1) или калия (Е2). Исходной кон-
формацией считается Е1, примерно 70%
молекул находится именно в этой конфор-
мации в среде, где не присутствуют ионы
натрия и калия. Переход между конфор-
мациями можно индуцировать добавлени-
ем соответствующих катионов, хотя для
этого требуются значительно большие кон-
центрации ионов, чем имеются в клетке.
Е2—Е1-Переход существенно ускоряет-
ся при повышении pH среды. Но наилуч-
шим активатором такого перехода явля-
ется АТФ, взятый в физиологических кон-
центрациях (>0,5мМ). АТФ увеличивает
сродство фермента к ионам натрия и сни-
жает сродство к ионам калия. Таким об-
разом, по завершении гидролитического
цикла трансформация Е2 в Е1 усиливает-
ся при связывании ферментом АТФ, если
он присутствует в концентрациях свыше
0,5 мМ. Это свойство фермента опреде-
ляет наличие двух констант связывания
АТФ: натриевая и калиевая конформации
фермента различаются по сродству к суб-
страту. Указанные конформации различа-
ются также и по чувствительности к. спе-
цифическому ингибитору уабаину (рис. 2.34). Центр связывания уаба-
ина формируется на конформации Е2, но отсутствует в конформации
Е1. По этой причине уабаин, как и другие сердечные гликозиды, более
2.5. Ионные канал.। и и. -ны,- и .г^:;
20
Рис. 2.34. Две конформации Na/K-АТФазы:
калиевая — Е2К (слева) и натриевая — ElNa (справа). Латинскими буквами обо-
значены некоторые последовательности аминокислотных радикалов в пептидной цепи
эффективно связывается с работающими молекулами фермента и
практически не влияет на фермент в неактивном состоянии.
В самом начале исследований свойств Na/K-АТФазы Й. Скоу за-
интересовался фосфорилированным интермедиатом, возникающим
при инкубации фермента с АТФ-Р32 в присутствии ионов натрия. Уро-
вень интермедиата снижается при последующей инкубации суспензии
с ионами калия или с С14-меченным АДФ. В последнем случае образу-
ется АТФ, несущий двойную метку, — С14 и Р32. При добавлении
ионов магния дефосфорилирующий эффект калия сохраняется, но
АДФ теряет способность вызывать дефосфорилирование ЕР.
Интерпретация этих данных Скоу была следующей: две конформа-
ции фермента способны находиться в фосфорилированном состоянии,
но образование Е1Р из АТФ обратимо, и фосфат может возвратиться
в адениловую систему. Е1 способен частично превращаться в Е2, этот
переход ускоряется ионами магния. Образующийся Е2Р может дефос-
форил ироваться (в присутствии ионов калия) только с образованием
неорганического фосфата. Таким образом, натрий выступал как акти-
ватор стадии фосфорилирования, а калий — стадии дефосфорилиро-
вания, причем магний ускорял Е1—Е2-переход. Постулировалось,
что энергия АТФ расходуется на перевод молекулы фермента из
Na-конформации в К-конформацию. В первом случае фермент связы-
вает натрий (со стороны цитоплазмы), во втором — отдает натрий и
связывает калий (с внешней стороны клетки) за счет изменившегося
сродства к этим ионам. В соответствии с изменяющимся сродством
208
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
а
Е-С + АТФ
хон
Н,8ОН
р Н,8ОН
Е-С ХО"---------
ХО-Р=О
хо-
Е-С + Н,8О-Р-Р=О
хон хо
Е"< /О>8н.8он //О /О'8
О—Р=О--------Е-С + Н,8О-Р-Р=О
хо хон хо
Рис. 2.35. Доказательство образования фосфофермента в ходе гидролиза АТФ:
а — зависимость активности от соотношения Na/K; б — схема, поясняющая включение
«тяжелого» кислорода в неорганический фосфат, образующийся при гидролизе АТФ
АТФазы к ионам натрия и калия находится и кривая зависимости актив-
ности фермента от соотношения Na+/K+ — она напоминает, по выра-
жению Скоу, шляпу Наполеона (рис. 2.35).
Поначалу эти представления вызывали, разнообразную критику, свя-
занную с необычностью свойств данной транспортной системы. Главное
возражение выдвинул Р. Пост, обративший внимание на то, что фосфо-
фермент можно получить только в среде, не содержащей ионов калия —
в его присутствии ЕР быстро гидролизуется. Если при одновременном до-
бавлении к препарату фермента натрия и калия ЕР не удается зарегистри-
ровать, означает ли это, что ЕР не образуется и в процессе циклической
работы фермента?
Ответ на этот вопрос был основным для выбора кинетической схемы
работы Na/K-АТФазы. Он был получен в опытах с применением тяжелой
воды Н2О18. Используя ее как среду реакции, можно было рассчитывать,
что в продукте АТФазы неорганическом фосфате будет обнаруживаться по
2.5. Ионные каналы и ионные насосы
209
крайней мере один атом «тяжелого» кис-
лорода. Если в ходе АТФазной реакции об-
разуется фосфофермент, время жизни ко-
торого сравнимо со скоростью О18-обмена
(см. рис. 2.35), в неорганическом фосфате
должно обнаружиться более одного атома
О18, причем величина включения должна
быть прямо пропорциональна -времени су-
ществования ЕР (Пантелеева, 1975). При
достаточно длительной инкубации фермен-
та в реакционной смеси, содержащей все
компоненты реакции, полный гидролиз до-
бавленного АТФ позволял накопить экви-
валентное количество Фн и измерить с
помощью масс-спектрометра соотношение
018/Р Эти опыты показали, что при рабо-
те Na/K-АТФазы в одну фосфатную группу
включается достоверно более одного атома
«тяжелого» кислорода, что соответствует
образованию ЕР в ходе работы фермента
(Болдырев, Твердислов, 1978).
Более того, оказалось, что увеличение
концентрации ионов магния в среде инкуба-
ции, активирующее гидролиз АТФ, снижало
величину 018/Р, а отклонение соотношения
Na+/K+ от оптимального (130/20 мМ) уве-
личивало этот коэффициент. Другими сло-
вами, магний увеличивал число оборотов
фермента (уменьшая время жизни ЕР), а
неоптимальное соотношение ионов-акти-
Р. Пост
Нейрохимик, известный ос-
новополагающими исследова-
ниями молекулярных свойств
Na/K-АТФазы; описал два
конформационных состоя-
ния фермента, отличающих-
ся сродством к переносимым
ионам; вместе с Р. Олберсом
обосновал схему постадий-
ного гидролиза АТФ, пред-
сказал наличие окклюдиро-
ванного состояния фермента
в процессе переноса ионов
через мембрану (США)
ваторов Na+ и К+ действовало в противоположном направлении (Смирно-
ва с соавт., 1977). Интересно, что в присутствии уабаина, являющегося
необратимым неконкурентным ингибитором этого фермента, длитель-
ность времени, необходимого для полного гидролиза АТФ, возрастала, но
значение коэффициента не менялось, поскольку уабаин выключал из ра-
боты часть молекул АТФазы, а оставшиеся активными молекулы продол-
жали работать по той же схеме. В настоящее время общепринятая схема
гидролиза АТФ Na/K-АТФазой включает стадии образования и превраще-
ния фосфофермента.
Анализ кривых активации фермента при изменении соотношения Na/K
в присутствии разных концентраций АТФ или нуклеозидтрифосфатов раз-
личной природы привел Скоу к еще одному важному заключению. Как
видно из рисунка 2.36, Ко 5 для натрия при низкой концентрации АТФ со-
ставляет около 115 мМ и снижается пропорционально увеличению кон-
центрации субстрата. Другими словами, АТФ способен увеличивать срод-
ство АТФазы к натрию (влияние на сродство к калию противоположно, но
его трудно заметить на приведенном рисунке из-за исходно большего
210
Раздел 2 МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
а
Na+
К+
Рис. 2.36. Зависимость активности Na/K-АТФазы от соотношения Na/K при
использовании АТФ (а) или других нуклеотидов (б).
Концентрация субстратов (если она варьировала) указана около каждой кривой
сродства фермента к этому иону). Замена АТФ на другие субстраты изме-
няет профиль ионной кривой в таком же направлении, что и снижение
концентрации АТФ. Данные результаты позволили утверждать, что АТФ
является не только субстратом, но и модулятором Na/K-АТФазы, регули-
рующим ее сродство к переносимым катионам.
Действительно, при тех концентрациях АТФ, которые имеются в возбу-
димых клетках, за окончанием АТФазного цикла и образованием Е2-кон-
формации фермента должен быстро следовать Е2—Е1 - переход, индуцируе-
мый высокими концентрациями АТФ. Действительно, субстратная зависи-
мость Na/K-АТФазы свидетельствует о том, что с ростом концентрации
субстрата наблюдается отклонение кривой от типичной «михаэлисовской»
зависимости и появление промежуточного плато. Строгий кинетический
анализ этой сложной субстратной кривой показывает, что она является сум-
2.5. Ионные каналы и ионные насосы
211
мой кривых — похожей на гиперболу при низких концентрациях субстрата и
выступающей в качестве сигмоиды при концентрациях АТФ свыше 0,5 мМ.
Первая часть зависимости, регистрируемой при низких концентрациях
субстрата, характеризуется отрицательной кооперативностью по АТФ (коэф-
фициент Хилла < 1). Сигмоида, возникающая при повышении концентрации
субстрата, отражает положительную кооперативность фермента по АТФ, эта
часть кривой соответствует коэффициенту Хилла, существенно превышаю-
щему 2 и зависящему от условий инкубации (температура, присутствие де-
тергентов). Как отрицательная, так и положительная кооперативности могут
иметь различную природу, основывающуюся и на внутрибелковых, и на меж-
белковых взаимодействиях (Корниш-Боуден, 1983; Курганов, 1978).
Для прояснения этого вопроса было проведено сравнение свойств на-
тивного (связанного с мембраной) фермента и фермента, солюбилизиро-
ванного неионным детергентом С12Е8. Оказалось, что первая часть суб-
стратной кривой осталась без изменений, сохранив отрицательную коопе-
ративность по АТФ; вторая же часть кривой исчезла (рис. 2.37). Поскольку
обработка детергентом разрушает белковые ассоциаты, полученные дан-
ные свидетельствуют о том, что сигмоидальная кривая отражает взаимо-
действие между белковыми протомерами, а отрицательная кооператив-
ность присуща протомеру как таковому.
Согласно классическим представлениям, ферменты, включающие в
свой цикл стадию релаксации, должны соответствовать кинетике Рэбина,
проявляя отрицательную кооперативность по субстрату (Болдырев, Мель-
гунов, 1984). То, что обработка детергентом не устраняет отрицательной
кооперативности по АТФ, подтверждает релаксационный характер работы
Na/K-насоса как молекулярной машины. В то же время зависимость ко-
оперативности в работе фермента от факторов, влияющих на упаковку
мембранного бислоя (температура, обработка детергентами, концентрация
АТФ), указывает на способность Na/K-насоса образовывать олигомеры,
функциональная активность которых может контролироваться АТФ.
Интересно, что субстратная зависимость Na/K-АТФазы после свободно-
радикальной атаки приобретает вид, типичный для случая невзаимодейст-
вующих протомеров. Другими словами, окислительное повреждение приво-
дит к утрате способности мембранных белков взаимодействовать друг с дру-
гом. Аналогично этому и Na/K-АТФаза, выделенная из мозга животных,
перенесших экспериментальную ишемию, при гидролизе АТФ демонстрирует
субстратную зависимость, характерную для солюбилизированного фермента.
Насколько важна способность фермента реагировать повышением ак-
тивности на возрастание концентрации АТФ, можно видеть на рисунке 2.38.
Измерение субстратной зависимости Na/K-АТФазы мозга при разных
значениях pH обнаруживает важную закономерность: снижение pH среды
приводит к «вырождению» сложной кривой в кривую, подобную гипербо-
ле. Это действие pH обратимо, и при восстановлении нормальной кислот-
ности среды (до pH 7,4) кривая восстанавливает типичную форму с проме-
жуточным плато. Если вспомнить, что защелачивание среды ускоряет
Е2—Е1-переход (а закисление среды замедляет его), становится понят-
ным, что манипуляции pH являются способом регулировать скорость ли-
212
Раздел 2 МОЛЕКУЛг^нАл НЕЙ/'ОХИМИК
а
б
АТФ, мМ
Рис. 2.37. Субстратные кривые:
а — мембранного фермента: кривая / — зависимость от концентрации АТФ нативно-
го фермента; кривые 2 и 3 получены методом математического разложения из кривой
Г, б — солюбилизированного фермента: кривая / — то же, что и на рис. 2.37, а\ кри-
вая 2 получена после солюбилизации фермента неионным детергентом С12Е8 (кривые
нормализованы приведением максимальной активности к единице)
митирующей стадии реакции. В мозге, где нарушения кровоснабжения
особенно губительны и могут привести к ацидозу, одним из следствий аци-
доза будет торможение Na/K-насоса за счет замедления лимитирующей
стадии. Это, конечно, приведет к сбережению клеточных запасов энергии
(Na/K-насос затрачивает около 40% всей энергии нейрона), но поставит
под угрозу жизнеспособность и функциональную активность клетки. Дело
в том, что с помощью специальных механизмов Na/K-АТФаза вовлекает-
ся в процессы внутриклеточной сигнализации и, находясь в рабочем со-
стоянии, препятствует накоплению активных форм кислорода (АФК) в
нейрональной клетке. Подавление работы фермента немедленно приведет
к накоплению АФК и созданию условий для окислительного стресса.
2.5. Ионные каналы и ионные насосы
2i:
Рис. 2.38. Субстратные кривые для Na/K-АТФазы при различных значениях
кислотности среды (указаны у каждой кривой)
Действие АТФ на Na/K-насос следует считать высокоспецифичным,
поскольку другие нуклеозидтрифосфаты, также являющиеся макроер-
гическими соединениями, гидролизуемые ферментом и обеспечиваю-
щие активный транспорт натрия и калия, не проявляют положительной
кооперативности в координатах Хилла. Данные по кооперативности
Na/K-АТФазы сведены в таблицу 2.12. В ней для сравнения приведены
результаты расчета коэффициентов Хилла по натрию и калию, величи-
на которых не зависит от свойств липидного окружения фермента.
Таким образом, было постулировано, что АТФ является не только
субстратом и источником энергии для транспорта ионов, но и модуля -
Таблица 2.12. Коэффициенты Хилла для Na/K-АТФазы по субстратам
и ионам-активаторам, измеренные в разных условиях
Анализи- руемый лиганд Условия инкубации
В области низких концентраций субстрата В области физиологиче- ских концентраций субстрата Обработка детергентом
АТФ 0,8—0,9 3,4 1,1
ГТФ 1,0 1,0 —
УТФ 1,0 1,0 —
Na+ — 2,3 2,3
к+ — 1,2 1,3
214
газдел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
тором Na/K-насоса, способным выполнять еще какую-то функцию, не
присущую другим макроергическим субстратам. В соответствии с
этим графики Аррениуса, которые отражают наличие липидного конт-
роля за работой мембранного фермента, резко различаются для АТФ и
других нуклеотидов. Как иллюстрирует рисунок 2.39, на кривой тем-
пературной зависимости гидролиза АТФ Na/K-АТФазой в координатах
Аррениуса выявляется анаморфоза в той же температурной области,
при которой наблюдается фазовый переход мембранных липидов. Более
того, солюбилизация фермента и реконструкция в искусственное ли-
пидное окружение меняют характер температурной зависимости та-
ким образом, что происходит сдвиг температуры перехода в об-
ласть, характерную для липида, применяемого для реконструкции
(при реконструкции фермента в «жидкий» диолеилфосфатидилглице-
рин (DOPG) перегиба на графике Аррениуса не наблюдается вообще).
Согласно общепринятой точке зрения (Блюменфельд, 1977; Болдырев,
Твердислов, 1978), анаморфоза на графике Аррениуса отражает чувст-
вительность межбелковых взаимодействий к фазовым перестройкам
мембранных липидов — до фазового перехода микровязкость липидов
в большей степени ограничивает белок-белковые взаимодействия,
чем при более высоких температурах, когда липиды оказываются упа-
кованными более рыхло.
Простое сравнение графиков Аррениуса для Na/K-АТФазы, гидро-
лизующей АТФ или другие нуклеотиды (УТФ, ГТФ), обнаруживает су-
ществование механизма, по которому АТФ выполняет свою роль регу-
лятора фермента. Дело в том, что графики Аррениуса для других суб-
Рис. 2.39. Температурная зависимость Na/K-АТФазы (а) и микровязкости
мембранного окружения (б), измеренной с помощью мембранного зонда — про-
изводного холестерина (спектр приведен сбоку, DH — соответствует измеряе-
мому параметру порядка); в — графики Аррениуса для Na/K-АТФазы, реконст-
руированной в различные фосфолипиды (DOPG, DMPG, DPPG, DSPG, PS).
Видно, что характер температурной зависимости определяется свойствами липидного
окружения фермента
>2.5. Ионные каналы и ионные насосы
215
стратов имеют линейную форму. Следовательно, можно предположить,
что фермент, который при гидролизе АТФ выступает как олигомер, ис-
пользуя другие субстраты, теряет способность к межбелковым контак-
там. Другими словами, модифицирующее действие АТФ выражается в
его способности организовывать молекулы Na/K-насоса в олигомер-
ные ансамбли. Следует иметь в виду, что кинетическое поведение фер-
мента может только свидетельствовать о его олигомерной природе. Со
временем появились прямые доказательства того, что Na/K-АТФаза
функционирует как олигомер. Одним из наиболее прямых физических
методов, позволивших рассчитать молекулярные размеры функциональ-
ной единицы, является метод «флаш-фотолиза» (в англоязычной лите-
ратуре «Time-resolved phosphorescence decay»). Ковалентное мечение
препарата Na/K-АТФазы флуоресцентными метками (например, эози-
низотиоциатаном) позволяет измерить поляризацию флуоресценции
метки после ее кратковременного возбуждения вспышкой лазера. Ха-
рактер затухания сигнала показывает, сколькими экспоненциальными
компонентами описывается измеряемая кривая, а скорость затухания по-
зволяет рассчитать молекулярные размеры глобулы, к которой прикреп-
лена метка. Метод имеет ряд ограничений (одним из них является нали-
чие значительных различий между молекулярными размерами метящих-
ся глобул с тем, чтобы можно было ответить на вопрос о количестве
экспоненциальных компонент, которые вносят вклад в формирование
кривой затухания сигнала). Соответствует успеху наличие достаточно
быстродействующей системы измерения и возможности максимально
быстрого начала измерения после окончания лазерной вспышки.
Применение данного подхода к исследованию Са-насоса, а затем и
Na/K-насоса, позволило установить, что в мембранных препаратах
транспортныхАГФаз эозинизотиоцианатом метятся частицы трех
сортов: сигнал от самых малоподвижных частиц пропадает после
обработки препаратов детергентом. Этот сигнал принадлежит метке,
«севшей» на крупные агрегаты: сигнал от наиболее быстрых частиц
принадлежит метке, вращающейся вместе с цилиндром, соответствую-
щим размеру протомера Na/K-насоса. Вращательная подвижность
этой метки не зависит от присутствия лигандов. Средние по враща-
тельной подвижности частицы соответствуют по крайней мере диме-
рам, состоящим из двух протомеров (аР)2. Измерение вращательной
подвижности этих частиц зависит от условий инкубации (табл. 2.13).
Последнее обстоятельство позволяет предполагать, что в ходе гид-
ролитического цикла интенсивность взаимодействия протомеров мо-
жет изменяться таким образом, что на одних стадиях цикла взаимо-
действия ослабевают, и протомеры работают как индивидуальные
216
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Таблица 2.13. Вращательная подвижность Na/K-АТФазы
(по: Boldyrev, Quinn, 1994)
pH Е2-Форма Е1-Форма
Быстрая Медленная Быстрая Медленная
6,0 2,4 13,2 2,2 6,9
6,5 2,5 12,2 2,4 7,9
7,0 2,4 и.о 2,4 7,7
7,5 2,4 9,8 2,3 8,4
8,0 2,4 10,3 2,4 9,4
8,5 2,3 5,9 2,4 9,7
белки, на других же стадиях они могут объединяться в более высоко-
молекулярные ансамбли. Возможно, именно это обстоятельство объ-
ясняет различия в данных, полученных в разных лабораториях.
Представляет интерес проследить, как было доказано это предположение
и выяснен механизм модифицирующего влияния АТФ на мембранные преоб-
разователи энергии. При сравнении структуры нуклеотидов (рис. 2.40) легко
обнаружить, что АТФ (в отличие от других рассматриваемых соединений)
имеет группу, способную протонироваться при физиологических значени-
ях pH. Другими словами, присоединяясь к активному центру, АТФ может
акцептировать протон от какой-либо подходящей группы фермента.
Обнаружилось, что такой группой является имидазольное кольцо гисти-
дина, локализованного недалеко от N-конца а-субъединицы, — отщепле-
ние 13-членного фрагмента, содержащего гистидиновый остаток, лишает
фермент способности воспринимать регулирующее действие АТФ. Таким
образом, АТФ, в отличие от уже протонированных ГТФ, УТФ и ИТФ, по-ви-
димому, способен делать то же самое, что делает защелачивание среды.
Аденин, А
Гуанин, G Цитозин, С Урацил, U Тимин, Т
Рис. 2.40. Формулы нуклеотидов (пунктиром обозначена ионогенная группа)
2.5. Ионные каналы и ионные насосы
21
Исключением, казалось бы, является ЦТФ, также имеющий группу,
способную к обратимому протонированию в физиологическом интервале
pH. На самом деле ЦТФ только подтверждает сделанное заключение, по-
скольку он проявляет схожие с АТФ регуляторные способности.
Таким образом, можно утверждать, что присоединение АТФ к активно-
му центру фермента, находящегося в конформации Е2, оттягивает протон
от N 13-остатка гистидина и вызывает целую серию изменений в располо-
жении водородных и солевых связей в протомере, приводящих в итоге к
конформационному переходу Е2—Е1 (другими словами, к понижению
сродства к калию и возрастанию сродства к натрию).
В последнее время получены подтверждения этим представлениям.
При инкубации фермента в среде с ионами натрия или калия были получе-
ны двумерные кристаллы разной конфигурации. Использование их для
рентгеноструктурного анализа позволило выявить, что натриевая (Е1)
конформация кристаллизуется преимущественно в виде протомера, а ка-
лиевая (Е2) — преимущественно в виде димера.
Следовательно, индуцированный АТФ Е2—Е1-переход сопровожда-
ется изменением размеров олигомерного комплекса (или количеством
входящих в его состав протомеров). Вопрос о том, происходит ли это из-
менение олигомерного состояния фермента и с какой стадией АТФазного
цикла оно связано, был решен с помощью метода молекулярной мише-
ни. Этот метод был предложен в 1968 г. Дж. Кепнером и Р. Мейси для оп-
ределения молекулярной массы мембранных белков. Суть его заключает-
ся в том, что мембранные препараты любой степени очистки подвергают
облучению пучком нейтронов (обычно используется кобальтовая пушка) и
далее измеряется зависимость остаточной активности от дозы облучения.
Авторы метода вывели эмпирическую зависимость степени угнетения
функции от дозы радиации:
Мг = 6,4 • 105/D37,
где Мг — молекулярная масса белка, кДа; D — доза радиации, снижаю-
щая измеряемую активность до 37% от исходной величины.
При помощи этого уравнения можно получить довольно точное значе-
ние молекулярной массы белка, осуществляющего анализируемую функ-
цию. Авторы проверили этот метод на недавно очищенной Е0-АТФазе ми-
тохондрий. Позже его применяли для оценки размеров рианодинового ре-
цептора. Ценность метода убывала по мере разработки методов очистки и
выяснения первичной структуры мембранных белков. Применение этого
подхода для оценки молекулярных размеров Na/K-АТФазы позволило
выяснить, какова эффективная масса фермента, осуществляющего так
называемые «частные реакции» общего цикла.
Как видно из таблицы 2.14, молекулярный размер белка, выпол-
няющего Na-АТФазную реакцию, соответствует массе ос-субъедини-
цы. Такими же размерами характеризуется белок, осуществляющий
К-зависимую фосфатазную реакцию. Весь Na/K-АТФазный цикл
218
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРГ 'ХИМИЯ
Таблица 2.14. Молекулярные размеры Na/K-АТФазы
на разных стадиях цикла (по: Болдырев, 1995)
Частные реакции Условия измерения Молекулярная масса, кДа В
Na-АТФаза Na+,Mg2+,ATO 95
К-фосфатаза К+, Mg2+, р-нитрофенилфосфат 90
Na/K-АТФаза 100 мкМ АТФ 135
Na/K-АТФаза 3 мМ АТФ 360
Na/K-АТФаза ЗмМ ГТФ 125
в присутствии низких концентраций АТФ (соответствующих области
отрицательной кооперативности; см. рис. 2.37) осуществляется белком
большего размера, сообразного массе ccp-протомера. Если концент-
рацию АТФ увеличить до физиологических значений (3 мМ), размер
функциональной единицы фермента существенно возрастет. Следова-
тельно, в ходе гидролитического цикла размеры АТФазного ансамбля
изменяются таким образом, что Na-зависимая АТФаза, Е1Р—Е2Р-
переход и К-зависимая фосфатаза осуществляются каждой ос-субъ-
единицей независимо. В завершении цикла принимает участие также
р-субъединица, а при увеличении концентрации АТФ наступает крат-
ковременная ассоциация ccp-протомеров в более крупные Е2-ассо-
циаты, которые с большей легкостью способны превратиться в Е1;
в этом и заключается причина стимулирующего действия АТФ:
п(Е2)----> (Е2)п----> и(Е1).
Как видно из примеров, в присутствии высоких концентраций
АТФ-фермент работает с гораздо большей молекулярной активно-
стью. В чем же заключается выигрыш от образования короткоживу-
щих олигомеров в ходе осуществления гидролитического цикла? Этот
выигрыш иллюстрирует рисунок 2.41. Суммарная реакция гидролиза
АТФ является экзергонической. Но частные реакции цикла могут
быть как экзергоническими, так и эндергоническими, т. е. протекать
как с выделением, так и с поглощением тепла. «Заслуга» АТФ заклю-
чается в организации работы олигомеров со сдвигом по фазе (синфаз-
но): экзергоническая стадия одного протомера совпадает с эндергони-
ческой стадией другого, происходит «подпитка энергией» соседних
белковых структур. Ясно, что субстраты, не вызывающие образования
олигомеров, гидролизуются Na/K-АТФазой с меньшей скоростью.
2.5. Ионные каналы и ионные насосы
219
Рис. 2.41. Изменение олигомерного состояния фермента в ходе АТФазного
цикла (от мономерного через окта-, тетра- и димерные комплексы при пере-
ходе от Е2КкЕ1№)
Таким образом, Na/K-насос является системой, работа которой ре-
гулируется субстратом и концентрацией переносимых ею ионов. С по-
мощью этой регуляции Na/K-насос настраивается на текущие нужды
клетки. Однако имеются и дополнительные способы регуляции, осу-
ществляющие тонкую настройку его работы.
Уже давно замечено, что некоторые препараты фермента содержат
низкомолекулярный белок, очистка которого происходит параллельно
с очисткой а-субъедин и цы. Он был назван у-субъединицей, но до по-
следнего времени его роль в работе Na/K-насоса не была выявлена.
Оказалось, что у-субъединица входит в число белков, для которых ха-
рактерна последовательность аминокислот (F—X—Y—D), в честь
которой их стали называть FXYD-белками. Молекулярная масса
FXYD-белков составляет 8—15 кДа, они выполняют разнообразные
регуляторные функции. В настоящее время описано около 10 таких
мембранных белков, в их числе фосфоламбан, сарколипин, фосфоле-
ман и у-субъединица Na/K-АТФазы. В клеточной мембране последняя
взаимодействует только с Na/K-АТФазой, в результате чего сродство
фермента к ионам натрия и калия снижается. Вне комплекса с фер-
ментом у-субъединица нестабильна и быстро разрушается.
Еще одна группа природных регуляторов Na/K-АТФазы — сердеч-
ные гликозиды. Наиболее употребим уабаин — гликозид растительно-
го происхождения, обладающий способностью при связывании «вы-
ключать» молекулы Na/K-АТФазы. Исследователи широко исполь-
зовали высокое сродство уабаина к ферменту для идентификации при-
сутствия этого белка в препаратах и оценки специфической активности
220
(вздел 2. МОЛЕКУЛУ.- —.- НЕЙРОХИМИЯ
В. Шонер
Автор основополагающих
работ в области молекуляр-
ной нейрохимии, посвящен-
ных исследованию олигомер-
ной организации транспорт-
ных АТФаз и их регуляции
эндогенными уабаинподоб-
ными соединениями (Гер-
мания)
Na/K-АТФазы, не задумываясь о причине
такой чувствительности фермента живот-
ного происхождения к соединению расти-
тельной природы. Однако в дальнейшем
были накоплены данные о том, что в тка-
нях животных синтезируется эндогенное
уабаинподобное соединение, выполняю-
щее функции регулятора Na/K-АТФазы.
Стационарный уровень этого соединения
очень низок, и В. Шонеру (университет
г. Гиссен, Германия) понадобилось не-
сколько десятков килограммов надпочеч-
ников крупного рогатого скота, чтобы вы-
делить несколько миллиграммов препа-
рата для его идентификации.
Оказалось, что инкубация с уабаином
вызывает всплеск образования АФК в
миоцитах (Xie, Askari, 2002) и нейронах
(Болдырев, 2000). АФК, в свою очередь,
вызывают активацию факторов трансля-
ции и экспрессию определенных белков.
Таким образом, Na/K-насос оказывается
вовлеченным во внутриклеточные сигнальные процессы (рис. 2.42).
Этим, в частности, объясняется заметная гипертрофия сердечной
мышцы у пациентов, вынужденных регулярно принимать сердечные
гликозиды для компенсации кальциевой недостаточности. Подобная
врачебная рекомендация основана на наличии в ряде клеток тесной
связи между Na/K-АТФазой и Na/Ca-обменником, в том числе в ми-
оцитах. При ингибировании Na/K-АТФазы сердечными гликозидами
происходит накопление в миоцитах ионов Na, которые можно обменять
на Са2+ внешней среды с помощью Na/Ca-обменника. Дополнитель-
ным результатом приема гликозидов является повышение уровня АФК
внутри клеток и активация синтеза клеточных белков.
Обнаружение эндогенного уабаина и установление участия Na/K-
насоса во внутриклеточной передаче сигналов вновь обратили внимание
исследователей на тот факт, что разные ткани демонстрируют различ-
ную чувствительность к сердечным гликозидам — сердечная мышца и
нервные клетки характеризуются высоким к ним сродством, а почки —
гораздо более низким. Причиной тому служит экспрессия различных
изоформ фермента в разных тканях. Известно несколько изоформ как у
а-, так и у р-субъединиц, экспрессируемых в разных тканях, причем из-
бирательность их по отношению друг к другу не является абсолютной.
2.5. Ионные каналы и ионные насосы
Рис. 2.42. Целесообразность работы Na-насоса в олигомерном состоянии.
При работе олигомера со сдвигом по фазе эндергоническая стадия цикла на одном про-
томере совпадает с экзергонической стадией цикла на другом протомере
В почках и печени экспрессируется только al -изоформа, в клетках глии
и скелетных мышцах — al и а2, а в нейронах зрелого мозга обнаружи-
вается исключительно аЗ-изоформа (табл. 2.15).
Таблица 2.15. Распространение изоформ Na/K-АТФазы
в различных тканях
Ткани а-Изоформы р-Изоформы
al а2 аЗ а4 ₽1 ₽2 ₽3
Почки + +
Тимус + +
Легкие + + +
Печень + + +
С рдце + + +
Скелетная мышца + + + + +
Гипофиз + + +
Семенники + + +
Мозг + + + 4- +
222
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Биологический смысл экспрессии различных изоформ Na/K-насо-
са в разных тканях заключается в различиях их чувствительности к ре-
гуляторным факторам. Так, al - изоформа устойчива к окислению и од-
новременно обладает низким сродством к уабаину по сравнению с дру-
гими изоформами фермента, которые, напротив, легко окисляются
активными формами кислорода; они характеризуются высокой чуст-
вительностью к сердечным гликозидам. В миоцитах сердечной мышцы
экспрессируются две изоформы а-субъединицы — al и а2, причем в
ходе развития первая частично замещается второй. Поскольку эти
частицы обладают различным сродством к уабаину, по характеру кри-
вой ингибирования активности нарастающими концентрациями уаба-
ина можно рассчитать долю присутствия разных изоформ АТФазы в
исследуемых препаратах. Интересно, что после экспериментального
инфаркта в сердце крыс наблюдается замена чувствительной к окис-
лению изоформы al на более устойчивую к окислению изоформу а2,
т. е. происходит возврат к экспрессии изоформ, присущих онтогенети-
чески менее зрелым клеткам (рис. 2.43).
Такой же результат был получен на мозге: после ишемического эпи-
зода наблюдалось уменьшение доли аЗ-субъединицы и возрастание до-
ли al-субъединицы. Поскольку первая субъединица экспрессируется
преимущественно в нейронах, а вторая — в глии, наблюдаемые изме-
нения, вероятно, отражают перестройку метаболизма поврежденных
нейронов мозга, аналогичную той, что наблюдается в условиях окисли-
Рис. 2.43. Смена изоформного состава Na-насоса при дифференцировке ми-
оцитов сердца (а) и изменение формы кривой ингибирования Na-насоса
уабаином после экспериментальной ишемии сердца взрослой крысы (б).
В первом случае процесс дифференцировки сопровождается частичной заменой а2-субъ-
единицы на al-субъединицу Na-насоса, во втором случае окислительный стресс при-
водит к обратному процессу
2.5. Иоанне канали i* ионные насосы
223
тельного стресса в сердце. Смысл этих изменений становится понят-
ным, если учесть, что al -субъединица резистентна не только к уабаину,
но и к АФК. Одной из особенностей первичной структуры фермента яв-
ляется более «замаскированное» расположение SH-групп в структуре
белка (кроме того, al-субъединица имеет на одну SH-группу меньше,
чем а2-субъединица). Таким образом, можно заключить, что преимуще-
ственное замещение менее резистентной к окислительному поврежде-
нию изоформы на более к нему резистентную является естественным
ответом ткани на окислительный стресс. Однако возникает вопрос о
том, что теряет ткань, лишаясь АФК-чувствительной изоформы, ведь
одновременно она является и уабаинчувствительной.
Индукция окислительного стресса в нейронах за счет инкубации с
NMDA приводит к избыточной продукции АФК, которые атакуют кле-
точные макромолекулы. Одной из главных мишеней для АФК в нейро-
не является Na/K-АТФаза, причем ее уабаинчувствительная форма.
Таким образом, в нейроне а2-субъединица представляет собой ми-
шень для АФК. Одновременно в активном состоянии Na/K-АТФаза
препятствует росту свободных радикалов. Следовательно, Na/K-
АТФаза выступает в качестве фактора окислительной устойчивости
нейрональной клетки, ее протектора от окислительного стресса. В от-
сутствие подобного протектора нейроны не способны использовать
АФК в качестве сигнальных молекул (см. рис. 2.42).
Не так давно (2005) в лаборатории Дж. Лингрела в США получены
изящные доказательства участия уабаинчувствительной АТФазы в
осуществлении еще одной функции организма. Известно, что один из
существенных участков первичной структуры а-субъединицы, опреде-
ляющий сродство к уабаину, представлен двумя аминокислотными ос-
татками у N-конца — L111 и N122. С помощью генно-инженерных
подходов были получены мыши, содержащие замены в этих аминокис-
лотах: LI 11R и N122D. Они не отличались от нормальных животных
по длительности жизни и показателям сердечно-сосудистой системы в
нормальных условиях, хотя Na/K-АТФаза в их почках обладала высо-
ким сродством к уабаину. Введение этим мышам АКТГ, индуцирующе-
го выделение из надпочечников эндогенного уабаинподобного соеди-
нения, повышало кровяное давление таким же образом, как и у конт-
рольных животных. Однако линия мышей, у которых в дополнение к
вышеуказанной замене а2-субъединица утрачивала высокое сродство
к уабаину, теряла способность отвечать адекватным повышением кро-
вяного давления на введение АКТЕ Уабаинподобный фактор выделял-
ся в кровь, но не находил своей мишени. Это указывает на то, что тка-
невая специфичность чувствительности Na/K-АТФазы к уабаину име-
ет прямое отношение к процессу регуляции тонуса сосудов.
224
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Изложенные данные иллюстрируют участие Na/K-насоса не толь-
ко в создании асимметричного распределения ионов по обе стороны
клеточной мембраны, но и в процессе восприятия клеткой различных
сигналов и передаче их в ее внутреннее пространство.
Трансмембранные транспортеры и обменники. Вторичноак-
тивный транспорт. Как уже отмечалось, при вторичноактивном транс-
порте в качестве движущей силы используется ранее накопленный гра-
диент ионов натрия. Наиболее наглядным примером вторичноактивно-
го транспорта является Na-зависимый транспорт в клетку аминокислот
и сахаров. Этот весьма важный процесс снабжает метаболические
процессы клетки необходимыми субстратами. Для нейрохимических
процессов наиболее существенным является вторичноактивный транс-
порт ионов и медиаторов. Поэтому далее будет рассмотрен транспорт
глутамата, Na/Ca-обмен и вторичноактивный транспорт хлора.
Регуляция процессов возбуждения осуществляется при условии до-
статочно быстрого устранения после генерации возбуждения из межси-
наптической щели молекул медиатора. В холинергической системе это
осуществляется благодаря наличию специфической и высокоактивной
АХ-гидролазы — ацетилхолинэстеразы (А¥Э), ответственной за уст-
ранение данного медиатора. АХЭ локализована в материале межсинап-
тической щели, и выделение АХ при возбуждении активирует его взаи-
модействие одновременно с двумя белками — ХР и АХЭ. Затратный ме-
ханизм обмена АХ, по-видимому, оправдывает себя, так как расход
энергии окупается быстротой реакции и высокой регулируемостью дви-
гательной мускулатуры. В глутаматергической системе нервной ткани
проблема устранения медиатора решена другим способом: в мембранах,
существующих рядом с нейронами глиальных клеток, выполняющих
функцию питания и «охраны» нервных клеток, существует специальная
система обратного захвата (re-uptake) медиатора, которая функциони-
рует по принципу вторичноактивного (Na-з^висимого) транспорта. Час-
тично эта система представлена и в мембране нервной клетки.
Re-uptake глутамата в глиальную клетку осуществляется одновре-
менно со входом трех ионов натрия и одного протона и сопровождается
выходом одного иона калия. Аналогичная система функционирует для
вторичноактивного транспорта нейтральных и кислых аминокислот в
гепатоциты. Аналогично глутамату подвергаются обратному захвату из
синаптической щели и другие медиаторы, функционирующие в других
функциональных отделах мозга (норадреналин, серотонин, ГАМК, гли-
цин). Транспорт ГАМК, глицина и моноаминов сопровождается входом
двух ионов натрия и одного иона хлора. Во всех случаях система рабо-
22
2.5. Ионные каналы и ионные насосы
тает электрогенно, следовательно, она может регулироваться измене-
ниями мембранного потенциала. Это свойство может иметь и свои не-
гативные последствия: деполяризация клеточной мембраны при нару-
шениях мозгового кровообращения будет вызывать массированный
выброс медиатора, усугубляя его экзайтотоксическое действие.
Аккумуляция медиатора из цитоплазмы в синаптические пузырьки
также осуществляется системой вторичноактивного транспорта, но
имеет иную природу: вместо градиента ионов натрия используется гра-
диент протонов, создаваемый Н-АТФазой, переносящей протоны из
цитоплазмы в синаптические пузырьки. В случае отрицательно заря-
женного глутамата осуществляется перенос одной молекулы медиатора
и одного иона хлора в ответ на выход из пузырька одного протона. Для
нейтральных ГАМК и глицина имеет место обмен одной молекулы ме-
диатора на один протон. Для положительно заряженных АХ и моноами-
нов наблюдается обмен одной молекулы медиатора на два протона.
Na/Ca-обмен определяет функциональную связь между системами
первичноактивного транспорта этих ионов в возбудимых клетках. Из-
вестны две системы, осуществляющие этот обмен. Одна из них обме-
нивает три иона натрия на два иона кальция. Она функционирует в
сердечной мышце и нервных клетках. Сродство ее к кальцию сущест-
венно ниже, чем у Са-насосов, но плотность молекул переносчика на
мембране велика и не уступает системе первичноактивного транспор-
та этих ионов. Направление обмена ионов зависит от функционально-
го состояния клетки (потенциала на мембране). При недостатке вну-
триклеточного фонда кальция в сердечной мышце предсердие выделя-
ет специальный ингибитор Na/K-насоса (близкий по структуре
уабаину), который вызывает частичное его торможение, что позволя-
ет обменять часть внутриклеточного натрия на внеклеточный кальций.
Избыток кальция в цитоплазме также способствует его обмену на
внеклеточный натрий. В состоянии покоя (при потенциале покоя око-
ло 75 мВ) активность Na/Ca-обменника незначительна, он активиру-
ется при деполяризации мембраны. В палочках сетчатки, где величина
потенциала покоя существенно меньше, описанный Na/Ca-обменник
надежно работать не будет. В данном случае имеет место иная система
переноса: четыре иона натрия в обмен на один ион кальция и один ион
калия. Несмотря на то что конечная стехиометрия транспорта та же
самая, зависимость процесса от мембранного потенциала такова, что
при внеклеточной концентрации ионов кальция около 2 мМ внутри-
клеточный уровень кальция может поддерживаться на отметке 1 нМ
(обменник первого типа в этих условиях может понизить внутрикле-
точную концентрацию кальция только до 400 нМ).
226
р«мл?.г 2, Me |ЕнуЛънНАЯ НЕЙРОХИМИЯ
Концентрация хлора внутри нервной клетки должна поддерживаться
на строго определенном уровне, поскольку от этого фактора зависит так
называемая синаптическая депрессия (гиперполяризация мембраны со-
здается направленным внутрь транспортом ионов хлора). Хотя Cl-активи-
руемая АТФаза была найдена в культивируемых нейрональных культурах,
большая часть хлора переносится через мембрану с помощью трех
различных систем вторичн о-a ктивного транспор-
та. Это Cl-бикарбонатный обменник и (С1 + К)-котранспорт, выво-
дящие ионы хлора из клетки, а также (Na + К + С1)-котранспорт, за-
качивающий хлор внутрь клеток. Хотя все системы транспорта хлора
работают в электронейтральном режиме, первая активно участвует в
регуляции внутриклеточного pH за счет обратимой диссоциации би-
карбоната. Первая система ингибируется специфическими ядами
(SITS, DIDS). Вторая и третья системы не чувствительны к этим ин-
гибиторам, но подавляются фуросемидом и буметанидом (Постнов,
Орлов, 1987). Отличием системы второго типа является ее нечувст-
вительность к внеклеточной концентрации натрия.
Все переносчики, осуществляющие вторичноактивный транспорт,
представлены интегральными мембранными белками, имеющими вну-
тримембранные сегменты, 10—12 раз пересекающие мембранный
бислой. Для (К + С1)-котранспортера характерно наличие протяжен-
ной петли между 4-м и 5-м трансмембранными сегментами (как и для
Na/K-АТФазы), а для Na/Ca-обменника — аналогичной петли между
5-м и 6-м сегментами.
2.6.
МЕХАНИЗМЫ ОТБОРА И УСИЛЕНИЯ СИГНАЛОВ
НА КЛЕТОЧНОЙ МЕМБРАНЕ
Метаботропные рецепторы, а также рецепторы, участвующие в
восприятии света и запаха, обладающие семидоменной трансмемб-
ранной структурой и связанные с G-белками, называют молекуляр-
ными сигнальными машинами. Традиционно механизмы сигнальной
трансдукции с их участием рассматриваются только на примере гете-
ротримерных G-белков, а также их взаимодействия с семейством ци-
топлазматических рецепторных тирозинкиназ Src и p-аррестина (см.
гл. 2.2). Именно разнообразием G-белков обеспечивается многообра-
зие эффектов, возникающих в результате взаимодействия лиганда с
рецептором. В последние годы было открыто множество других путей,
определяющих рецепторную специфичность (рис. 2.44).
Примером такой рецепторной специфичности является инициация
полипролинового сайта (pPRO) С-терминального участка mGluR-pe-
2.6. Механизмы отбо »а и силения сигналов на клеточной мембране
227
Рис. 2.44. Участие метаботропных рецепторов в передаче информации в клет-
ку (объяснение в тексте}
цептора. Этот сигнальный мотив способен связываться с ЭНЗ-участ-
ком белков-мишеней. Для mGluR-рецептора такими мишенями явля-
ются вспомогательные гомерные белки (homer proteins), способные
формировать димеры при активации рецептора. Эти димеры служат
мостиком, связывающим mGluR-рецептор с Са2+-мобилизующим
ИФ3-рецептором, который содержит pPRO-мотив. Пока недостаточ-
но ясно, является ли такое взаимодействие важным для сигнальной
функции, однако показано, что нарушение нормального синтеза го-
мерных белков сопряжено с рядом нейродегенеративных нарушений.
Указанный pPRO-мотив также обнаружен в третьей внутриклеточ-
ной петле дофаминовых рецепторов D4, которые могут связываться
с ЭНЗ-содержащими адаптерными белками Grh2 и Nek, обычно
ассоциированными с тирозинкиназной рецепторной сигнализацией
и с MAP-киназным каскадом. Это свидетельствует о том, что этот
pPRO-домен также может участвовать в сигнальной функции.
Другой способ передачи сигнала для адренорецепторов связан с
С-концевым участком молекулы. Они имеют повторяющийся мотив,
способный связываться с PDZ-доменом белков. Прототипом рецеп-
торов этого класса является, например, родопсин. Он связывается с
PDZ-содержащими вспомогательными InaD-белками, которые взаи-
модействуют с различными мембранными эффекторными белками,
включая фосфолипазу Ср, протеинкиназу С и TRP-ионные каналы.
228
Раздел 2 МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Другим примером взаимодействия С-терминали с PDZ-доменом
является 0-адренергический рецептор. В этом случае происходит свя-
зывание рецептора с NHERF (фактор регуляции Na/H-обменного
белка, Na/H-exchanger regulatory factor), приводящее к его актива-
ции. Таким образом, рецептор не через G-белки, а напрямую влияет
на функцию эффектора. Показано, что норадреналин активирует об-
менник, а мутация С-терминального участка рецептора полностью
устраняет такое влияние медиатора.
Примером, иллюстрирующим физиологические механизмы регуля-
ции синтеза NO, являются серотониновые 2В-рецепторы, которые
взаимодействуют с так называемой конститутивной NO-синта-
зой. Присоединение серотонина к рецептору этого типа активирует
NO-синтазу, в результате чего генерируется NO, проявляющий свой-
ства вторичного мессенджера и влияющий в этом качестве на многие
внутриклеточные процессы. G-Белки не участвуют в этом каскаде ре-
акций, так как их нейтрализация с помощью антител не влияет на ге-
нерацию NO.
Для NO-синтазы существует и другая возможность регуляции ка-
талитической активности через рецепторы, связанные с G-белками
(GPCR). Показано, что эндотелиальная NO-синтаза способна связы-
ваться с 4-м внутримембранным доменом В2-рецептора брадикинина
(или ангиотензина). В обоих случаях связывание агониста с рецепто-
ром приводит к активации фермента. Это также независимый от G-
белка способ трансмембранной передачи сигнала.
Наконец, Asn—Pro—X—X—Туг-мотив в 7-м трансмембранном
регионе некоторых рецепторов класса А может участвовать в связы-
вании и активации малых G-белков, таких как ARF и Rho, что приво-
дит к активации фосфолипазы Д. В данном случае связывание лиган-
да с рецептором также приводит к активации фермента без участия
G-белка.
Следовательно, GPCR-рецепторы могут использовать для переда-
чи сигнала не только гетеротримерные G-белки, и поэтому название
мембранотропные рецепторы является более адекватным, хотя и
не отражает некоторых особенностей функционирования этих рецеп-
торов, например их способности напрямую взаимодействовать с ион-
ными каналами или ионотропными рецепторами.
Многообразие сигнальных путей, очевидно, обусловливает нали-
чие огромного разнообразия «рецепторной специфичности», наблю-
даемой в нервной системе, а также возможность выбора системой
адекватных способов реагирования на внешние сигналы.
2.7. Развитие н ; внои системы и о; ми звание нервны- сетей
2291
2.7.
РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ
И ФОРМИРОВАНИЕ НЕРВНЫХ СЕТЕЙ
В настоящее время уже известно, что нейрогенез и формирова-
ние нервных сетей происходят не только в период эмбрионального
развития, но продолжаются и после рождения. Многие функциональ-
ные свойства нейронов развиваются в течение всей жизни организма
и могут быть не только ответной реакцией на повреждение или старе-
ние клетки, но и адаптационным ответом мозга.
Эмбриональное развитие нервной системы у позвоночных начина-
ется на стадии гаструлы с образования нервной пластинки. Область
гаструльц из которой в дальнейшем формируется нервная система,
представляет собой полоску эктодермы, клетки которой находятся под
влиянием факторов роста, подавляющих дифференцировку. Нервная
пластинка, состоящая из продолговатых нейроэктодермальных кле-
ток, утолщается по краю, который приподнимается и образует нерв-
ные валики. После слияния валиков друг с другом по средней линии
образуется полая нервная трубка. Некоторые из клеток, расположен-
ные по краям нервных валиков, остаются между нервной трубкой
и вышележащей эктодермой и формируют нервный гребень.
Как нейроны, так и глиальные клетки проходят в своем развитии
ряд этапов — рождение, миграция, дифференцировка, созревание и
гибель. Пролиферация клеток-предшественников (для нейронов это
нейробласты) является началом данного процесса. В ходе последова-
тельного ряда митотических делений формируются специфические
клетки. Таким образом, моментом рождения нейрона является по-
следний митоз, пройденный клеткой в своем развитии. Для определе-
ния этого момента используют меченый тимидин, который интенсивно
поглощается клетками перед митозом.
Важным индуктивным сигналом развития нейрональных клеток яв-
ляется ретиноевая кислота (производное витамина А), принадлежащая
к той же группе регуляторных соединений, что и стероидные и тиреоид-
ные гормоны. Их цитоплазматические рецепторы после проникновения
в ядро выполняют функции транскрипционных факторов. Ретиноевая
кислота активирует уникальный класс транскрипционных факторов —
ретиноидные рецепторы, модулирующие экспрессию большого числа
генов-мишеней (рис. 2.45). Другая группа сигнальных веществ, конт-
ролирующих развитие нейронов и глиальных клеток, представляет со-
бой пептиды и включает семейства фактора роста фибробластов (FGF)
и фактора трансформации роста (TGF). В TGF-семействе одним из
важнейших является белок морфогенеза кости (BMP), участвующий в
230
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Фактор роста фибробластов (FGF)
Белок морфогенеза кости (BMP)
Рис. 2.45. Основные сигнальные пути
индукции развития нервных клеток
эмбриона позвоночных.
Представлены схемы взаимодействия ли-
гандов, рецепторов и внутриклеточных
сигнальных молекул в процессе передачи
сигнала в ядро эмбриональной нервной
клетки позвоночных. Каждый компонент
принимает участие в формировании ней-
рональной эктодермы, а также в диффе-
ренцировке нейронов и глии в мозге
2.7. Развитие нервной системы и формирование нервных сетей
процессах, связанных с нейрональной индукцией и дифференциацией, в
том числе и на более поздних стадиях формирования нейрональной сети.
Наконец, представители Wnt-семейства (у позвоночных гомологичны
продукту гена бескрылости дрозофилы wingless) могут модулировать
различные этапы нейрональной индукции и дифференциации, прежде
всего дифференциацию нервного гребня.
Все эти вещества секретируются различными эмбриональными
тканями, включая нотохорду, нейрональную пластинку и нейрональ-
ную эктодерму, и воспринимаются соответствующими рецепторами
близлежащих клеток. Эти вещества различаются по свойствам их ре-
цепторов (локализации, виду и механизмам взаимодействия), а также
включаемым ими сигнальным каскадам. Рецепторы пептидов групп
FGF и BMP обладают собственной протеинкиназной активностью.
Рецепторы FGF являются тирозинкиназами; сигнальная молекула
связывается с рецептором совместно с комплексом протеогликан —
гепаринсульфат. При этом активируется внутриклеточный киназный
домен рецептора, что, в свою очередь, ведет к активации RAS/MAP-
киназного пути. Эффектом этого взаимодействия может являться мо-
дификация цитоскелета и цитоплазматических компонентов, отвечаю-
щих за форму и подвижность клетки, а также экспрессия генов, в том
числе и отвечающих за клеточную пролиферацию.
BMP-рецепторы относятся к серин-треониновым киназам, которые
фосфорилируют группу цитоплазматических регуляторных SMAD-бел-
ков, что приводит к перемещению их в ядро, где они модулируют экспрес-
сию генов. Название SMAD-белков образовано от названий двух обнару-
женных первыми белков этого семейства: Sma (skeletal muscle actin)
у C. elegans и Mad (mothers against decapentaplegic) у Drosophila.
Некоторые сигналы в ходе нейрогенеза передаются более сложным
путем. Так, передача сигнала Shh зависит от кооперативного связывания
двух поверхностных рецепторов с последующей их интернализацией. Об-
разованный комплекс влияет на перенос в ядро факторов транскрипции
(в том числе и Glil) и последующую модуляцию экспрессии генов. Эф-
фекты, оказываемые Shh, зависят от его концентрации. Так, высокая
концентрация Shh на поверхности хорды приводит к формированию кле-
ток базальной пластинки из клеток нервной трубки, более низкая кон-
центрация вызывает превращение клеток в висцеральные мотонейроны,
а еще более низкая концентрация позволяет клеткам развиваться по их
основному пути и становиться соматическими мотонейронами.
Рецепторы Wnt-сигналов после связывания с молекулами лиганда
инициируют каскад реакций, приводящих к деградации цитоплазмати-
ческих белковых комплексов и препятствующих проникновению 0-ка-
232
Раздел 2 МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Молекулы
внеклеточного матрикса (МВМ)
Нейрональный белок
клеточной адгезии (NCAM)
т^ анс 1\ кция
Кадгерины
Эфрины (Eph)
Семафорины
сигнализация
Eph
2.7. Развитие нервной системы и формирование % с г-их г.етё^
2331
тенина из цитоплазмы в ядро. Снятие такого ингибирования приводит
к экспрессии соответствующих генов. Именно с этими сигналами свя-
зано формирование нейронального гребня, гранулярных клеток моз-
жечка и нейронов переднего мозга.
Общим этапом развития для всех нервных клеток является про-
цесс их миграции с мест образования в места окончательной локали-
зации. Клетки глии и нейроны формируются из нервной трубки и
нервного гребня. Нейробласты нервного гребня способны мигриро-
вать через периферические ткани и давать начало различным типам
нейронов периферической нервной системы. Это передвижение осу-
ществляется благодаря конусу роста — особой структуре, которой за-
канчивается формирующийся аксон. Конус роста с помощью амебоид-
ных движений продвигается между клетками и, прикрепляясь к окру-
жающим структурам, «тянет» за собой всю клетку. Мембрана конуса
роста способна генерировать кальциевые потенциалы действия.
Аксоны нервных клеток могут достигать более метра в длину, обра-
зуя синапсы на мембране определенных клеток в области, в которой
имеется большое количество других потенциальных клеток-мишеней.
В последние годы благодаря изучению условий, способствующих рос-
ту аксона, была обнаружена большая группа веществ, связанных с
клеточной адгезией и межклеточным узнаванием, в том числе прича-
стных к формированию движения конуса роста.
По своим свойствам все эти лиганды и их рецепторы подразделяют-
ся на несколько семейств: молекулы внеклеточного матрикса и их ин-
тегриновые рецепторы; Са-независимые молекулы клеточной адгезии
(САМ), Са-зависимые молекулы клеточной адгезии (кадгерины); эф-
рины. Молекулы внеклеточного матрикса являются первыми фактора-
ми, влияющими на рост аксона. К ним относятся ламинин, коллаген и
фибронектин, способные образовывать макромолекулярные комплек-
сы в межклеточном пространстве (рис. 2.46). Эти вещества, секрети-
Рис. 2.46. Некоторые семейства лигандов и их рецепторов, участвующие в уп-
равлении ростом аксона.
В управлении ростом аксона принимают участие различные классы молекул: молеку-
лы внеклеточного матрикса являются лигандами интегриновых рецепторов; гемо-
фильные молекулы Са-независимой клеточной адгезии (NCAM) выступают одновре-
менно в роли лигандов и рецепторов; молекулы Са-зависимой клеточной адгезии (кад-
герины) способны к гомофильному взаимодействию; нетрин/ Slit-семейство
лигандов влияет через два типа рецепторов: DCC («Deleted in Colorectal Cancer») и
Robo; семафорины являются первичными сигналами отталкивания и могут быть свя-
заны с клеточной поверхностью или секретироваться, их рецепторы плексин и нейро-
пилин обнаруживаются в конусе роста; эфрины могут быть трансмембранными или
мембраносвязанными сигналами для Eph-рецепторов, являющихся тирозинкиназами
234
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
руемые клетками, при полимеризации создают локальные внеклеточ-
ные структуры, с которыми специфически реагирует обширная группа
рецепторов, именуемых интегринами. Данные гликопротеины, имею-
щие большую массу, состоят из двух или более сходных субъединиц,
удерживаемых вместе благодаря дисульфидным мостикам.
Было идентифицировано большое количество изоформ а- и
0-субъединиц интегрина. Каждая ос0-комбинация приводит к формиро-
ванию рецепторов с различными свойствами. Интегрины обеспечивают
структурные связи между белками внеклеточного матрикса и внутри-
клеточным актиновым цитоскелетом, регулируя форму клетки и ее миг-
рацию, а также активируют внутриклеточные сигнальные пути, которые
управляют ростом клетки, пролиферацией и дифференцировкой. Сами
интегрины не обладают киназной активностью или какими-то сигналь-
ными возможностями. Тем не менее их взаимодействие с ламинином,
коллагеном и фибронектином приводит к активации сигнального каска-
да, связанного с цитоплазматическими киназами и другими растворимы-
ми сигнальными молекулами, что в конечном счете ведет к росту и удли-
нению аксона. Это взаимодействие вызывает изменение концентрации
таких внутриклеточных вторичных посредников, как ионы кальция и ИФ3,
и активацию дополнительных внутриклеточных киназных путей.
Многие процессы функционирования нейронов (их дифференци-
ация, выживание, рост, ветвление нейритов) и образования нервных
волокон находятся под контролем нейротрофических факторов. Ини-
циация роста нейрита, а также определение пути его роста зависят от
сигналов извне, которые могут поступать из внеклеточного матрикса,
расположенного на соседних клеточных поверхностях молекул (на-
пример, NCAM), или диффузных секретируемых факторов (NGF,
BDNF и др.). Молекулы, которые управляют конусом роста, могут
действовать как аттрактанты или репелленты, на короткой или длин-
ной дистанции. Например, в периферической нервной системе клетки
нервного гребня не мигрируют по путям предварительно спроециро-
ванного аксона или вдоль структур глиальных клеток, как это харак-
терно для ЦНС. Их миграция полностью управляется механизмами
привлечения и отталкивания при контакте с поверхностью клеток и
компонентами внеклеточного матрикса.
Поверхность клеток и молекулы адгезии, расположенные во вне-
клеточном матриксе, управляют движением конуса роста при помощи
притягивающих или отталкивающих механизмов, действующих на ко-
роткой дистанции. Нетрины играют роль хемоаттрактантов, а семафо-
рины — хеморепеллентов, действующих на большой дистанции, для
многих типов аксонов. Эфрины и семейство Eph-рецепторов, связан-
2.7. Развитие нервной системы и формирование нервных сетей
235
них с тирозинкиназой, при помощи хемореппелентных механизмов
управляют процессом поиска пути растущим аксонам, миграцией кле-
ток и образованием связей между клетками.
Роль хеморепеллентов играют и семафорины — большое семейст-
во секреторных и трансмембранных белков. Первоначально они были
идентифицированы как факторы, управляющие ростом аксона у куз-
нечиков, но затем было показано, что гомолог этого белка у позвоноч-
ных коллапсин-1 приводит к ретракции конуса роста.
Еще одним фактором, влияющим на рост нейронов и глиальных
клеток, служит нейрональный белок клеточной адгезии (N-CAM),
принадлежащий к иммуноглобулиновому суперсемейству белков кле-
точной адгезии. Он кодируется одним геном, состоящим из 26 экзо-
нов, в результате альтернативного сплайсинга которого может обра-
зовываться около 120 изоформ N-CAM. Обычно в нервной системе
обнаруживаются три основные изоформы этого белка: N-CAM 120,
N-CAM 140, N-CAM 180, имеющие идентичные внеклеточные участ-
ки и различающиеся трансмембранными и внутриклеточными домена-
ми. Они различаются также по своим функциям, времени появления и
локализации: N-CAM 140 преимущественно экспрессируется в кону-
сах роста незрелых нейронов, N-CAM 180— в постсинаптических
уплотнениях и местах контактов незрелых нейронов, a N-CAM 120 —
в основном на глиальных клетках.
Внеклеточные домены N-CAM могут взаимодействовать как друг с
другом (гомофильное взаимодействие), так и с другими молекулами, на-
ходящимися на клеточной поверхности или являющимися компонента-
ми внеклеточного матрикса (гетерофильное взаимодействие). В резуль-
тате гетерофильного взаимодействия N-CAM 140 и N-CAM 180 с ре-
цептором фактора роста фибробластов (FGFR) происходит активация
фосфолипазы Cy(PLCy) и диацилглицерол(ОАС)-липазы, ведущее к на-
коплению инозитолтрифосфата (ИФ3) и арахидоновой кислоты (АА).
Арахидоновая кислота, в свою очередь, вызывает увеличение концент-
рации Са2+ внутри клетки как за счет активации Са-каналов, так и бла-
годаря выходу Са2+ из внутриклеточных депо. Активированный FGFR
(epidermal growth factor receptor) стимулирует митогенактивируемую
протеинкиназу (МАРК) через так называемые малые G-белки Ras, Raf,
МЕК. В результате включения этих сигнальных каскадов происходит
рост нейритов.
Гомофильное связывание N-CAM 140 приводит к взаимодействию
с нерецепторными тирозинкиназами Fyn и FAK(фокальная адгезивная
киназа), что также в конечном итоге вызывает активацию МАРК. Об-
разование кластеров N-CAM при гомо- и гетерофильном взаимодей-
236
кэз,.,..п 2. МОЛЕКУЛЯРНАЯ HEHPt ХИМИЯ
ствии N-CAM на поверхности клетки приводит к аутофосфорилирова-
нию ассоциированных в этих же местах молекул нерецепторной тиро-
зинкиназы Fyn. Затем в рафтах клеточной мембраны образуется
комплекс N-CAM-Fyn-FAK, и FAK аутофосфорилируется. Далее ак-
тивируются обе формы MAPK(Erkl и Erk2), фосфорилируется CREB
(cAMP-responsive element binding protein) и образуется c-fos. N-CAM
может также активировать путь проведения сигнала, зависящий от
ПкВ и ПкС.
Экспрессия N-CAM и N-кадгерина в клетках приводит к их агрега-
ции в культуре клеток, вытягиванию аксонов в сторону клеточных суб-
стратов (но не субстратов внеклеточного матрикса), а также объеди-
нению отдельных растущих аксонов в пучки.
На процессы роста нейронов могут оказывать влияние многие хими-
ческие факторы, ряд из которых воздействует специфически, например
фактор роста нервов (ФРН), который был идентифицирован первым
(рис. 2.47). Этот белок имеет аминокислотную последовательность,
сходную с инсулиноподобным фактором роста 1 (ИФР1). Он секретиру-
ется структурами, являющимися производными нервного гребня, и иг-
рает существенную роль в росте, созревании и регенерации нейронов.
Его связывание с высокоафинными рецепторами приводит в пер-
вую очередь к усилению процесса фосфорилирования тирозина. Вы-
званное лигандом образование рецепторного димера способствует
сближению внутриклеточных доменов тирозинкиназы и аутофосфори-
лированию. При этом включаются сразу несколько внутриклеточных
сигнальных путей — фосфорилирование тирозиновых остатков SNT
белка (sr/c-associated neurotrophic factor-induced tyrosine-phosphory-
lated target) и активация фосфолипазы С, фосфоинозитол-3-киназы
и каскада Ras-MAPK.
Стадия дифференцировки заключается в формировании специфи-
ческих структур нейрона, его свойств и связей. По времени она может
совпадать с другими стадиями, а в некоторых случаях начинается даже
до начала этапа миграции нейрона к месту его будущей локализации.
В процессе дифференцировки нейроны приобретают характерные для
них физиологические свойства. Они возбудимы уже на ранних стадиях
развития, однако возбуждающий потенциал в этот период формирует-
ся за счет ионов Са2+ в теле клетки или конусе роста. Заметим, что у
мышечных клеток такой механизм генерации потенциала сохраняется
и при достижении ими зрелости. Наличие кальциевых потенциалов
может влиять на подвижность конусов роста, а также на внедрение
в мембрану рецепторных белков, чувствительных к медиатору. Для
большинства нервных клеток характерен постепенный переход внача-
2.7. Ра звити- '.^л г. 4 " ИС Те М *< И 'Р- j.i.iy. с .ни,, н. < t hl \С<_тей
23
Рис. 2.47. Сигнальные каскады, запускаемые при взаимодействии фактора
роста нервов с двумя TrkA-рецепторами.
S—РО4 — фосфорилирование серина; Т—РО4 — фосфорилирование треонина; Y—РО4 —
фосфорилирование тирозина. ФРН — фактор роста нервов; SNT — связанная с sue и
терозинфосфорилируемая в ответ на нейтрофические факторы мишень (swc-associated
neurotrophic factor-induced tyrosine-phosphorylated target); Raf — рецептор тирозинкина;
Ras — Ras-подобная малая ГТФаза; She—8Н2-доменсодержащий адапторный белок
(Src homology 2 domain-containing transforming protein); RSK — киназа 40S-рибосомных
белков; МЕК — киназа MAP-киназ или киназа ER-киназ; SOS — белок-фактор обмена
нуклеотида; GRb2 — growth factor receptor bound protein; She, Grb-2, SOS — белки
MAP-киназного сигнального пути. При связывании димера фактора роста нервов с двумя
ТгкА-рецепторами происходит фосфорилирование внутриклеточным доменом тирозинки-
назы каждого из ТгкА-рецепторов остатков тирозина другого ТгкА-рецептора. При этом
запускаются четыре внутриклеточных сигнальных каскада, приводящих к росту и диффе-
ренцировке клеток. Активация фосфоинозитол-3-киназы (1Р3-киназы) способствует удли-
нению отростков и их выживанию. Фосфорилирование тирозина SNT приводит к диффе-
ренцировке. Активация фосфолипазы C-y(PLC-y) стимулирует MAP-киназы, которые
индуцируют экспрессию генов, дифференцировку и рост как непосредственно, так и через
фосфорилирование RSK. МАР-киназа также активируется через сигнальный каскад, ко-
торый включает связывание белков SHC, Grb-2, SOS и Ras, а также киназ Raf и МЕК
238
Раздел 2. МОЛЕКУЛЯРНАЯ НЕИРОХИМИЯ
ле на совместное использование для генерации потенциала ионов
Са2+ и Na+, а позже на использование для этих целей исключительно
ионов Na+.
Насколько сильно влияет на функциональные особенности нейро-
нов их окружение, демонстрируют эксперименты с выращиванием
нейронов симпатических ганглиев в культуре клеток. Так, если клетки
иной природы в культуре отсутствуют, в ней экспрессируются типич-
ные адренергические нейроны. Если же в культуре имеются еще и мио-
циты, то экспрессируются холинергические нейроны. Таким образом,
окружающая среда в значительной степени изменяет равновесие меж-
ду адренергическими и холинергическими свойствами одной и той же
клетки.
Определяется ли фенотип клеток нервного гребня до того, как
клетки начинают миграцию, или он формируется после достижения
ими своего окончательного месторасположения на периферии? Клет-
ки надпочечника эмбриона млекопитающих, выращиваемые в культу-
ре клеток, образуют либо хромаффинные клетки, либо клетки мозго-
вого вещества надпочечников, либо адренергические симпатические
нейроны. Глюкокортикоиды (синтезируемые клетками коры надпочеч-
ника) вызывают экспрессию ферментов, специфических для хромаф-
финных клеток. И наоборот, два белка, присутствующие в местах об-
разования симпатических ганглиев, — основной фактор роста фиб-
робластов (basic fibroblast growth factor, bFGF) и фактор роста нерва
(NGF) — в культуре клеток вызывают дифференцировку клеток-
предшественниц в симпатические нейроны.
Важным этапом в развитии нейронов является формирование си-
наптических контактов. Для этого нейрон должен в определенное
время прекратить миграцию в соответствующую область, занять оп-
ределенное место относительно соседних клеток, сформировать оп-
ределенным образом ориентированные дендриты и аксон, который
должен направить свои окончания к определенным областям и
сформировать связи с определенными клетками (телами или денд-
ритами). Было высказано предположение о наличии химического
сродства между аксонами и нейронами-мишенями, благодаря чему
обеспечивается восстановление связей при регенерации и нормаль-
ном развитии. Подтверждением этого предположения явились экспе-
рименты, в которых при перерезке одного из зрительных нервов у ам-
фибий и повороте глазного яблока на 180° регенерация зрительного
нерва приводила к восстановлению связей аксонов с прежними клет-
ками-мишенями.
2.7. Развитие нервной истомы и н ормирование нервных сетей
239
Временные характеристики формирования синаптического контак-
та детально изучены для нервно-мышечных синапсов. На ранних ста-
диях развития рецепторы АХ распределены диффузно вдоль поверхно-
сти неиннервированных мышечных волокон. Их плотность составляет
около нескольких сотен рецепторов на квадратный миллиметр. По ме-
ре приближения конуса роста мотонейрона к мышечному волокну в
клетке-мишени возникают деполяризующие потенциалы в ответ на
выделение АХ из конуса роста. После возникновения контакта частота
спонтанного выделения квантов АХ и величина синаптического потен-
циала быстро возрастают и в течение нескольких минут формируется
функциональная синаптическая связь.
Через несколько часов после первоначального контакта происхо-
дит накопление рецепторов АХ непосредственно под терминалью ак-
сона, а через один-два дня плотность рецепторов в этой области со-
ставляет уже несколько тысяч на квадратный миллиметр, и в областях
синапсов начинает накапливаться ацетилхолинэстераза. В течение не-
скольких последующих недель развития у многих видов у-субъединица
рецептора АХ заменяется на 8-субъединицу, что приводит к формиро-
ванию этих рецепторов, при этом плотность рецепторов под терми-
налью аксона постепенно увеличивается, достигая уровня, равного
примерно 104 на 1 мкм2. Вне синаптической зоны она составляет ме-
нее чем 10 рецепторов на 1 мкм2. При этом время полужизни рецеп-
торов увеличивается с одного до десяти дней.
На этапе созревания нейроны и нейрональные сети приобретают
свою окончательную форму. Трудность определения наступления дан-
ной стадии для нейрональных сетей заключается в том, что не все ней-
роны, составляющие сеть, заканчивают миграцию и дифференцировку
одновременно.
Одним из критериев созревания является миелинизация аксонов.
Начинаясь на заключительных этапах эмбриогенеза или в начале
постнатального развития (в зависимости от области мозга), она про-
должается достаточно долго.
Одной из важных особенностей процесса развития нервных клеток
является их запрограммированная гибель — апоптоз (Скулачев,
1996). Сигналами к его началу могут служить некоторые активные
формы кислорода (АФК) (см. гл. 3.9).
Апоптоз делает безвредной для организма утрату большого количе-
ства нейронов в процессе онтогенетического развития мозга. Это явле-
ние характерно не только для стареющих клеток, оно имеет место и на
ранних этапах эмбрионального развития. Доля погибших нейронов в от-
дельных областях мозга достигает 75%. Нарастание этого явления сов-
240
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
падает по времени с моментом установления нейронами связи с клетка-
ми-мишенями. Доля гибели клеток может зависеть от размеров мише-
ни. При этом нейроны, не установившие контактов с соседними
клетками, не получают сигналов от иннервируемых клеток и погибают.
На основе этого принципа происходит эффективный отбор нейронов
для формирования межнейрональных сетей и устранение избыточных и
неэффективных нейронов. Было обнаружено, что нейротрофические
белки, вырабатываемые в мышце (инсулиноподобный фактор роста,
нейротрофический фактор ресничек, фактор холинергической диффе-
ренцировки и др.), могут управлять развитием мотонейронов. При вве-
дении их эмбриону эти белки предупреждают гибель мотонейронов.
Главной причиной необратимости изменений структур нервной
системы после повреждений является то, что после завершения про-
цессов дифференцировки нейроны практически теряют способность к
делению: новые клетки не возникают или возникают в крайне незна-
чительном количестве. Исключения из этого правила очень редки, на-
пример обонятельные рецепторные клетки. Но вместе с тем у каждого
нейрона сохраняется способность к формированию новых отростков и
новых синаптических контактов. Поэтому нейроны могут формиро-
вать новые нейриты (отростки), восстанавливая утраченные связи.
При этом восстанавливаются и специфические синаптические связи,
как это, например, происходит в случае нейронов зрительного тракта.
Исследования на клеточном уровне позволили установить, что свойст-
ва клетки зависят от вида ее деятельности (уровня функционирова-
ния), с их помощью были выявлены некоторые молекулярные меха-
низмы такой зависимости.
Представляет интерес ряд закономерностей формирования новых
синаптических связей при перерезке нейрональных путей. Так, уста-
новлено, что если нервные клетки иннервируются различными путя-
ми, то перерезка одного из них приводит к тому, что синаптические
участки замещаются терминалями нейронов другого пути. Происходит
своеобразное разрастание нервных окончаний, подобное тому, кото-
рое имеет место в период развития нейронов, когда формируются ак-
соны, способные прорастать в направлении освободившихся мест.
Так, для голубого пятна, посылающего норадренергические волокна в
передний мозг и мозжечок, потеря ветвей, идущих в одну из этих
структур, приводит к разрастанию ветвей во второй структуре. Таким
образом клетка пытается восстановить генетически запрограммиро-
ванное количество соединений, хотя они и образуются в несоответст-
вующих местах. Очевидно, что конкуренция за синаптические контак-
ты сохраняется и у зрелых нейронов.
2.8. Механизмы синаптической пластичности
241
2.8.
МЕХАНИЗМЫ СИНАПТИЧЕСКОЙ ПЛАСТИЧНОСТИ
Способность синапсов к функциональным и морфологическим
перестройкам в процессе функционирования, их пластичность являет-
ся важным свойством всех химических синапсов центральной и пери-
ферической нервной системы. Выделяют кратковременные формы
синаптической пластичности, длящиеся от нескольких секунд до
нескольких минут, и долговременные формы, которые осуществля-
ются в течение часов, месяцев и лет. Последние формируются на ос-
нове кратковременных форм синаптической пластичности и лежат
в основе таких процессов, как обучение и память.
Синаптическая пластичность может быть связана с изменениями в
пресинаптическом или постсинаптическом нейронах. Постсинаптические
механизмы кратковременной пластичности могут быть обусловлены по-
вышением (потенциацией), либо снижением (десенситизацией) чувстви-
тельности постсинаптических рецепторов к медиатору. Долговременная
синаптическая пластичность может проявляться в виде долговременной
потенциации <ДВП), либо долговременной депрессии (ДВД). Эти про-
цессы приводят к различным перестройкам в клетке, в том числе и мор-
фологическим. Структурные изменения заключаются в росте и ветвле-
нии пресинаптических нервных окончаний, изменении количества рецеп-
торов на постсинаптической мембране, образовании новых синапсов.
Ранее (см. гл. 2.2) уже обсуждались механизмы пресинаптической
модуляции синаптической передачи. Как
уже неоднократно отмечалось, постси-
наптическая мембрана не является струк-
турой, пассивно воспринимающей сигнал.
Модуляция может происходить и на уров-
не структур постсинаптической мембра-
ны. Способность рецепторов адекватно
реагировать на сигналы обеспечивается
несколькими механизмами (рис. 2.48).
Если концентрация сигнальных молекул
долгое время остается высокой, рецепто-
ры будут находиться в активированном
состоянии (связанном с медиатором), при
котором они не способны воспринимать
новые сигналы от пресинаптической клет-
ки. Именно поэтому по истечении опреде-
ленного времени рецепторы должны инак-
тивироваться.
Н. Н. Демин
Один из основоположников
функциональной нейрохимии,
автор оригинальных работ по
биохимии памяти
242
Раздел 2. МОЛЕКУЛЯРН Я НЕЙРОХИМИЯ
Обратный захват
медиатора
Удаление медиатора
Разрушение
медиатора
Пресинаптическая
мембрана
Медиатор
Постсинаптическая
мембрана
Инактивация рецептора
Эндоцитоз рецептора
Негативная регуляция
рецептора
Рис. 2.48. Разные способы изменения активности рецепторов
Эффективным способом очистки синаптической щели от нейромеди-
аторов является их захват пресинаптическим окончанием и глиальными
клетками с использованием специфических транспортеров или разруше-
ние нейромедиаторов с помощью специфических ферментов (см. гл. 3.4).
В каждой клетке функционирует особая структура, регулирующая
ее чувствительность, и, прежде всего, чувствительность клеточных
рецепторов к сигналу. Обычно уровень сигналов, действующих через
систему трансмембранной сигнализации, связанную с G-белками, по-
вышается на короткое время, которого достаточно, чтобы произошло
образование нужного количества вторичных мессенджеров (цАМФ-
Са2+, ДАГ), вызывающих активацию соответствующих протеинкиназ
и последующее за этим фосфорилирование белков-мишеней. Если же
2.8. Механизмы синаптической пластичности
243
уровень сигнала сохраняется повышенным в течение более длитель-
ного времени (десятков минут или даже часов), происходит десен-
сибилизация соответствующего рецептора.
Мембранные протеинкиназы, присутствующие в плазматической
мембране практически всех клеток, фосфорилируют рецептор, десенси-
билизируя его к присутствию лиганда. Такие киназы могут фосфорилиро-
вать только гормон-рецепторный комплекс, поэтому чем дольше гормон
связан с рецептором, тем больше вероятность того, что рецептор будет
узнан и помечен киназой. Если подобное фосфорилирование не в состоя-
нии погасить сигнал, то спустя 15—30 мин происходит фосфорилирова-
ние рецептора второй протеинкиназой, которая активируется соответст-
вующим вторичным посредником (например, в случае Р-адренергических
рецепторов, активирующих аденилатциклазу, — цАМФ-зависимой про-
теинкиназой; в случае al-адренергических или Ml - и МЗ-холинергиче-
ских рецепторов, активирующих фосфолипазу С, — протеинкиназой С).
Вторичное фосфорилирование рецепторов нарушает их связь с G-белка-
ми, вследствие чего влияние медиаторов ослабевает. Если высокий уро-
вень гормона сохраняется в течение нескольких часов и перечисленные
выше механизмы десенсибилизации не в состоянии погасить регулятор-
ный сигнал, происходит эндоцитоз гормон-рецепторных комплексов, и
внутри клетки появляются рецепторосомы, изолирующие рецепторные
молекулы от сигнальных путей (рис. 2.49). Они могут вновь встроиться в
плазматическую мембрану, если со временем уровень гормона понизит-
ся. Если этого не происходит, они сливаются с лизосомами, после чего
рецепторы разрушаются. Очевидно, что восстановление чувствительнос-
ти клетки к данному гормону потребует нового синтеза рецепторов.
Так, например, полная десенсибилизация фоторецептора осуществ-
ляется специфической киназой, которая фосфорилирует его в активи-
рованном состоянии по сериновым и треониновым остаткам. Происхо-
дит этот процесс достаточно медленно, поскольку необходимо перенес-
ти семь — девять фосфатов на активированный рецептор. Но его
можно ускорить с помощью арестина, который присоединяет к рецеп-
тору 1—2-фосфатные группировки и предотвращает дальнейшую ак-
тивацию G-белка.
Десенсибилизация путем фосфорилирования активированных ре-
цепторов специфическими киназами — довольно часто используемый
механизм. Так, например, 0-адренергические рецепторы могут фос-
форилироваться специфической киназой, цАМФ-зависимой киназой
и протеинкиназой С. Фосфорилирование любой из них вызывает де-
сенсибилизацию рецептора, однако степень ее различается. Десенси-
билизация с помощью специфической, но не цАМФ-зависимой кина-
зы усиливается 0-аррестином, так же как и в случае фоторецепторов.
244
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
Рецептор нейрокинина 1
Фосфорилирование
и инактивация
Агон и сти иду ци рованный
эндоцитоз
Повторное
встраивание
рецептора
в мембрану
Рис. 2.49. «Эндоцитоз — экзоцитоз» рецептора нейрокинина 1 в клетке
Ионотропные рецепторы также инактивируются при длительном
воздействии на них активных лигандов. В этом случае и для их десенси-
билизации применяется фосфорилирование рецептора. Никотиновый
ацетилхолиновый рецептор фосфорилируется по у- и 8-субъединицам
цАМФ-зависимой киназой, по 8- и ос-субъединицам — протеинкиназой
С, а по Р", у- и 8-субъединицам — тирозинкиназой. При длитель-
ном воздействии ацетилхолина благодаря фосфорилированию происхо-
дит 7—8-кратное снижение чувствительности к нему рецептора. Подоб-
ным образом ГАМК-рецепторы, фосфорилируемые цАМФ-зависимой
протеинкиназой, снижают амплитуду рецепторного ответа. Регуляция де-
сенсибилизации ацетилхолинового и ГАМК-рецепторов с использовани-
ем активации протеинкиназ таким вторичным мессенджером, как цАМФ,
свидетельствует о том, что лиганды, регулирующие уровень внутрикле-
точных вторичных посредников, могут оказывать длительное влияние
и на возбудимость рецепторов, связанных с ионными каналами.
Вместе с тем все указанные способы адаптации постсинаптической
мембраны связаны с изменениями на функциональном уровне, они нс
вызывают изменения количества рецепторов. Способы адаптации пост-
синаптической мембраны, воздействующие на число рецепторов на по-
верхности мембраны, — это эндоцитоз (интернализация) рецепторов,
а также изменение уровня синтеза и деградации рецепторов.
2.8. Механизмы синаптической пластичности
245
Так называемая down-регуляция количества рецепторов в мембра-
не связана с изменением скорости двух взаимно сбалансированных
процессов — синтеза и деградации рецепторов. Долговременный эф-
фект медиатора может приводить к повторяющейся интернализации и
возврату рецептора в плазматическую мембрану. При этом часть ре-
цепторов утрачивается во время каждого подобного цикла из-за дег-
радации в лизосомах. В конечном счете это приводит к уменьшению
количества рецепторов. Мутации мускариновых и адренергических
рецепторов, при которых они не влияют на их десенсибилизацию, что
является подтверждением того, что down-регуляция и интернализация
рецепторов — это независимые процессы и, возможно, включаются с
помощью различных сигналов. Аналогичный результат — снижение
количества рецепторов в постсинаптической мембране — достигается
и уменьшением скорости их синтеза.
Следует иметь в виду, что при нормальной работе мозга синапсы
активируются не отдельными потенциалами действия, а их непрерыв-
ными потоками разной частоты и регулярности. Например, при стиму-
ляции пресинаптического волокна коротким залпом стимулов резуль-
тирующие постсинаптические потенциалы могут либо увеличиваться
(синаптическая фасилитация), либо уменьшаться (синаптиче-
ская депрессия).
Во многих синапсах повторная активация может вызвать не только
кратковременные, но и долговременные изменения синаптической
эффективности, продолжающиеся часы и даже дни. Два таких яв-
ления — долговременная потенциация (ДВП) и долговременная
депрессия (ДВД) — опосредуются работой вторичных мессенджеров.
При ДВП происходит быстрое и значительное увеличение концентра-
ции кальция в постсинаптической клетке, в результате которого за-
пускаются каскады систем вторичных мессенджеров; их активность
приводит к появлению дополнительных рецепторов в постсинаптиче-
ской мембране и увеличению чувствительности рецепторов. ДВД по-
является в ответ на сравнительно меньшее увеличение концентрации
кальция в постсинаптической клетке и сопровождается снижением
количества и чувствительности постсинаптических рецепторов.
Было отмечено, что при одновременной стимуляции получающих
иннервацию через два входа нейронов гиппокампа возникает ДВП.
При этом высокочастотная стимуляция одного из входов вызывала
обычные ПД, а низкочастотная стимуляция второго приводила лишь к
незначительной деполяризации постсинаптической мембраны. В пи-
рамидных клетках гиппокампа индукция ДВП происходит с участием
кальция, входящего через глутаматные NMDA-рецепторы. Эти рецеп-
246
Раздел 2. МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
торы (см. гл. 2.3) образуют необычные ка-
Д. Карпентер
Автор цикла электрофизио-
логических экспериментов
по биохимии нейромедиато-
ров, один из первых исследо-
вателей роли NMDA-рецеп-
торов в пластичности нерв-
ной системы (США)
тионные каналы — при нормальном по-
тенциале покоя клетки они заблокирова-
ны ионами магния. Канал активируется
при высвобождении ионов магния, вы-
званном деполяризацией мембраны. Боль-
шая часть глутаматчувствительных кле-
ток имеет в постсинаптической мембране
NMDA- и АМРА-рецепторы, которые ак-
тивируются при выделении глутамата из
пресинаптических терминалей. NMDA-
рецепторы обладают относительно высо-
кой кальциевой проводимостью, однако
возможность входа кальция определяется
величиной деполяризации, которая долж-
на быть достаточной для снятия блоки-
рующего действия магния. Именно этим
можно объяснить тот факт, что в экспери-
менте по активации ДВП стимуляция
только одного (слабого) входа не вызыва-
ла синаптической деполяризации, доста-
точной для снятия магниевого блока и входа кальция. Однако, когда эта
стимуляция сопровождалась деполяризацией, вызванной активацией
другого входа, NMDA-рецепторы оказывались разблокированными,
что приводило к входу кальция и возникновению ДВП.
Повышенная концентрация внутриклеточного кальция может ак-
тивировать целый ряд внутриклеточных биохимических путей
(рис. 2.50). Наиболее важны для индукции ДВП активация СаМК II и
цАМФ-зависимой протеинкиназы. СаМК II обнаружена в больших
количествах в постсинаптических уплотнениях шипиков дендритов, и
внутриклеточная инъекция блокаторов СаМК II предотвращает ДВП.
Кроме того, индукция ДВП нарушена у. мышей с генетическими на-
рушениями синтеза одной из субъединиц СаМКИ. Блокада цАМФ-
зависимой протеинкиназы также существенно уменьшает ДВП. При
достаточной деполяризации синапса ритмическая синаптическая ак-
тивация приводит к входу кальция, который связывается с кальмоду-
лином. Комплекс Са-кальмодулин активирует СаМК II, которая авто-
фюсфорилируется, превращаясь в форму, сохраняющую активность
спустя долгое время после возвращения концентрации кальция к на-
чальному уровню. СаМК II оказывает двойной эффект на синаптиче-
скую передачу: фосфорилирует АМРА-рецепторы в мембране, увели-
чивая их проводимость для ионов, и облегчает мобилизацию резерв-
2.8. Механизмы синаптической пластичности
24
Рис. 2.50. Механизм возникновения и поддержания ДВП в нейронах гип-
покампа.
Каналы NMDA-рецепторов при наличии в синаптической щели глутамата открывают-
ся только в случае деполяризации постсинаптической клетки. Ионы Са2+, проникаю-
щие в клетку через эти каналы, активируют протеинкиназы, которые за счет фосфори-
лирования субстратов способствуют встраиванию новых АМРА-рецепторов в постси-
наптическую мембрану. Увеличение количества рецепторов ведет к повышению
чувствительности к глутамату, выделяемому в синаптическую щель. ИКС — протеин-
киназа С; СаМКП — Са-кальмодулинзависимая ПК II типа
ных АМРА-рецепторов из цитоплазмы в плазматическую мембрану,
в результате чего большее число постсинаптических участков стано-
вится доступным для активации квантами глутамата, высвобождаемо-
го из терминали.
В случае когда несколько пресинаптических окончаний контактиру-
ют с постсинаптической мембраной дендритного шипика, содержащего
небольшое количество работающих АМРА-рецепторов или совсем не
содержащего их, выброс глутамата вызовет очень незначительный от-
вет или вообще не вызовет никакого ответа — такой синапс будет
«молчащим». Примерно половина дендритных шипиков содержит в ос-
новном NMDA-рецепторы, которые не дают ответа на глутамат в усло-
виях потенциала покоя. Однако медиатор повлияет на функциональное
состояние NMDA-рецепторов, что вызовет индукцию ДВП, и новые
глутаматные АМРА-рецепторы будут включены в постсинаптическую
мембрану «молчащего» синапса. Эти рецепторы начнут «отвечать» на
кванты медиатора, выбрасываемые из пресинаптической терминали,
и, соответствейно, увеличится ответ постсинаптического нейрона.
248
Раздел 2. МОЛЕКУЛЯРНАЯ НЕИРОХИМИЯ
В настоящее время существуют доказательства того, что при ДВП
увеличивается количество АМРА-рецепторов. Показано, что субъеди-
ницы АМРА-рецепторов доставляются в шипики дендритов после рит-
мической стимуляции, сопровождающейся активацией NMDA-рецеп-
торов. После стимуляции нейронов меченые рецепторы быстро появ-
ляются и в тех дендритах, где их раньше не было. Эти данные
позволяют предположить, что многие дендритные шипики являются
«молчащими» и получают полный набор АМРА-рецепторов после
ритмической стимуляции.
Достаточно убедительными являются данные о существовании
корреляции между способностью к обучению и наличием ДВП в ней-
ронах срезов гиппокампа. Так, оба этих параметра могут быть блоки-
рованы антагонистами NMDA-рецепторов или метаботропных глута-
матных рецепторов, либо блокаторами кальций-кальмодулинзависи-
мой протеинкиназы.
Низкочастотная стимуляция (один импульс в секунду) клеток гиппо-
кампа ведет не к быстрому, как при ДВП, а к медленному и менее зна-
чительному нарастанию концентрации кальция в постсинаптическом
нейроне (рис. 2.51). При этом происходит активация протеинфосфатаз
Рис. 2.51. Механизм возникновения и поддержания ДВД в нейронах гип-
покампа.
Низкоамплитудное увеличение концентрации Са2+ в постсинаптических нейронах поля
СА1 гиппокампа приводит к активации протеинфосфатаз, что вызывает интернализацию
АМРА-рецепторов постсинаптической мембраны. В результате этих процессов происхо-
дит снижение чувствительности к выделяемому в синаптическую щель,глутамату
2.8. Мех. HU3Mi синрптичсгк^ й пл. Тм :ти
24
и, как результат, интернализация постсинаптических АМРА-рецепто-
ров с соответствующим снижением чувствительности к глутамату.
Несколько иной механизм ДВД наблюдается в клетках Пуркинье
мозжечка. Эти клетки получают сигналы с лазающих и параллельных
волокон, причем каждый из них заканчивается на своем дендритном
шипике. Выброс глутамата параллельными волокнами приводит к ак-
тивации АМРА- и метаботропных глутаматных рецепторов. В резуль-
тате активации последних образуются такие вторичные мессенджеры,
как DAG и ФИФ3, которые могут взаимодействовать с Са2+, входя-
щим через потенциалчувствительные Са-каналы в случае поступле-
ния сигнала по лазающим волокнам. Это ведет к активации ПКС, ко-
торая запускает клатринзависимую интернализацию АМРА-рецепто-
ров в синапсах, контактирующих с параллельными волокнами. При
отсутствии активации со стороны лазающих волокон ДВД не наблю-
дается.
Чувствительность к медиатору в случае долговременной потенци-
ации обусловлена изменением количества уже имеющихся рецепто-
ров в мембране. Однако при ДВП наблюдаются как относительно бы-
стрые изменения, связанные со встраиванием в мембрану уже имею-
щихся рецепторов, так и более поздние компоненты возбуждения,
связанные с регуляцией синтеза новых рецепторов и структурными
изменениями постсинаптической мембраны (образование новых ши-
пиков на дендритах). Такие изменения вызваны тем, что активация
протеинкиназ приводит не только к встраиванию новых АМРА-рецеп-
торов в мембрану, но и, в свою очередь, к активации аденилатциклазы
и ПКА, которая оказывает влияние на процессы, протекающие в ядре
клетки. В результате происходит синтез новых рецепторов и формиро-
вание новых синапсов.
250
Раздел 2. Л Ей РОХИ. ИЯ
ВОПРОСЫ И ЗАДАНИЯ
1. Назовите принципы классификации синапсов и их основные типы
2. Охарактеризуйте свойства электрических и химических синапсов '
3’ тор»? РЗЗНИЦа Ме>МУ ТерМИНЭМИ <<нейРомедиатор» и «нейромодуля-
4. Перечислите основные критерии, позволяющие отнести соединение
к классу нейромедиаторов. мнение
5. Каковы основные этапы передачи информации в клетку обусловлен
ные рецепторами? -Усилен-
6. Каковы различия между интегральными и амфипатическими оепеп
торными белками? рсцеп-
7. Охарактеризуйте типы мембранных рецепторов. Каковы особенности
их строения и функционирования? синими
8. Охарактеризуйте систему обмена ацетилхолина: локализация в мозге
этапы синтеза фермента. тияе«
9. Какие типы холинорецепторов имеются в нервной ткани’ Каковы
принципы их классификации, структура и свойства? '
10. Каковы особенности локализации и синтеза катехоламинов в мозге’
11. Какие адрено- и дофаминовые рецепторы обнаружены в мозге’ Как
их классифицируют? Каковы их структура и свойства? Что происходит
с ними при различных нейродегенерациях? р^ислодит
12. Охарактеризуйте серотонинергическую систему и серотониновые пе
цепторы. е
13. Какие свойства глутаминовой кислоты определяют ее функциониоовв
ние в качестве нейромедиатора? Назовите классы глутаматных рецеп
торов. Какими свойствами они обладают? р^цен-
14. Каковы особенности ГАМК-ергической системы мозга?
15. Опишите свойства NO как нейромодулятора. Может ли это соелине
ние функционировать в качестве нейромедиатора? ^дине-
16. Назовите изоформы фермента синтеза NO. Каковы особенности каж
дои из них? тлликаж-
17. Каков механизм синтеза NO?
18. Назовите особенности синтеза, распределения в мозге и оеиеппии
пептидных нейромедиаторов. рецепции
19. Каковы основные принципы функционирования механизмов пеоедачп
информации в клетку? 1СР<=дачи
20. Каковы механизмы, этапы передачи сигнала в клетку через сопояжец
ные с G-белками рецепторы? н впряжен-
21. Перечислите типы вторичных мессенджеров. Каким образом они
функционируют? рсиим они
22. Каков механизм трансмембранной передачи сигнала с участием мемя
ранных фосфоинозитидов? J и см мемо-
23. Охарактеризуйте роль ионов кальция в качестве вторичного поспелей
ка в функционировании нервных клеток. ^средни-
вопросы и задания
2511
24. Какую роль играют циклические нуклеотиды, АФК и NO в процессах
клеточной сигнализации?
25. В чем заключается роль протеинкиназ и протеинфосфатаз в механиз-
мах клеточной сигнализации? Назовите основные классы этих фер-
ментов и дайте их краткую характеристику.
26. С помощью каких соединений можно имитировать трансмембранную
передачу сигнала?
27. Какова классификация транспортных процессов в клетке?
28. В чем различия между первичноактивным и вторичноактивным транс-
портом ионов через мембрану?
29. Охарактеризуйте ионные каналы. Каковы их структура, функции,
классификация?
30. Какова современная классификация транспортных АТФаз?
31. Опишите рабочий цикл транспортных АТФаз.
32. Как функционирует Na/K-АТФаза?
33. Какие изоформы Na/K-АТФазы известны? Каковы их функциональ-
ные особенности и локализация в тканях?
34. Какие трансмембранные транспортеры и обменники имеются в нерв-
ных клетках?
35. Охарактеризуйте известные механизмы отбора и усиления сигналов на
клеточной мембране.
36. Каковы основные этапы развития нейрона?
37. Какие сигнальные вещества контролируют развитие нейронов и гли-
альных клеток? Опишите механизмы их влияния на клетку.
38. Охарактеризуйте вещества, связанные с клеточной адгезией и меж-
клеточным узнаванием.
39. Какова роль апоптоза в развитии нервной системы?
40. Что такое «синаптическая пластичность»?
41. Перечислите механизмы модуляции синаптической передачи.
42. Что такое «долговременная потенциация» и «долговременная депрес-
сия»?
43. Что такое «синаптическая десенситизация»?
44. Каков механизм интернализации мембранных рецепторов?
ФУНКЦИОНАЛЬНАЯ НЕИРОХИМИЯ
3.1.
ОСОБЕННОСТИ ПЛАСТИЧЕСКОГО ОБМЕНА ГОЛОВНОГО МОЗГА
З.1.1. Гематоэнцефалический барьер (ГЭБ)
Метаболизм мозга, как никакого другого органа, зависит от ве-
ществ, поступающих с кровотоком. Многочисленные кровеносные со-
суды, обеспечивающие работу нервной системы, отличаются тем, что
процесс проникновейия веществ через их стенки является избира-
тельным. Эндотелиальные клетки капилляров головного мозга соеди-
нены между собой непрерывными плотными контактами, поэтому ве-
щества могут проходить только через сами клетки, но не между ними.
К наружной поверхности капилляров прилегают клетки глии — вто-
рого компонента ГЭБ. В сосудистых сплетениях желудочков мозга
анатомической основой барьера являются эпителиальные клетки,
также плотно соединенные между собой. В настоящее время гемато-
энцефалический барьер рассматривается не как анатомо-морфологи-
ческое, а как функциональное образование, способное избирательно
пропускать, а в ряде случаев и доставлять к нервным клеткам с по-
мощью активных механизмов транспорта различные молекулы. Таким
образом, барьер выполняет регуляторную и защитную функции
В головном мозге есть структуры, в которых ГЭБ ослаблен. Это,
прежде всего, гипоталамус, а также ряд образований на дне 3-го и 4-го
желудочков — самое заднее поле (area postrema), субфорникальный и
субкомиссуральный органы, а также шишковидное тело. Целостность
ГЭБ нарушается при ишемических и воспалительных поражениях мозга.
Гематоэнцефалический барьер исключительно важен для обеспече-
ния гомеостаза головного мозга, однако многие вопросы, касающиеся
его формирования, все еще окончательно не выяснены. Но уже сейчас
совершенно ясно, что ГЭБ представляет собой максимально выражен-
ный по дифференцированности, сложности и плотности гистогематиче-
ский барьер. Основная структурная и функциональная его единица —
эндотелиальные клетки капилляров мозга. ГЭБ считается окончательно
сформировавшимся, когда свойства этих клеток будут удовлетворять
двум условиям. Во-первых, скорость жидкофазного эндоци-
3.1. Особенности пластического обмена головного мозга
тоза (пиноцитоза) в них должна быть
крайне низкой. Во-вторых, между
клетками должны формироваться специ-
фические плотные контакты, для кото-
рых характерно очень высокое электри-
ческое сопротивление. Оно достигает ве-
личин 1000—3000 Ом/см2 для капилля-
ров мягкой мозговой оболочки и от 2000
до 8000 Ом/см2 для интрапаренхималь-
ных мозговых капилляров. Для сравне-
ния: средняя величина трансэндотелиаль-
ного электрического сопротивления ка-
пилляров скелетной мышцы составляет
всего 20 Ом/см2.
Проницаемость гематоэнцефалическо-
го барьера для большинства веществ в
значительной степени определяется их
свойствами, а также способностью нейро-
нов синтезировать эти вещества самостоя-
тельно. К веществам, которые могут пре-
одолевать этот барьер, относятся, прежде
М. И. Прохорова
Ученица и соратница
Е. С. Лондона, автор клас-
сических исследований энер-
гетического метаболизма
мозга, создатель школы
нейрохимиков в Ленинград-
ском (Санкт-Петербург-
ском) университете
всего, кислород и углекислый газ, а также различные ионы металлов,
глюкоза, незаменимые аминокислоты и жирные кислоты, необходимые
для нормального функционирования мозга (рис. 3.1; табл. 3.1). Транс-
порт глюкозы и витаминов осуществляется с использованием перенос-
чиков. Вместе с тем D- и L-глюкоза обладают различной скоростью
проникновения через барьер — у первой она более чем в 100 раз выше.
Глюкоза играет главную роль как в энергетическом обмене мозга, так
и в синтезе ряда аминокислот и белков.
Ведущим фактором, определяющим функционирование гематоэн-
цефалического барьера, является уровень метаболизма нервных
клеток.
Обеспечение нейронов необходимыми веществами осуществляет-
ся не только с помощью подходящих к ним кровеносных капилляров,
но и благодаря отросткам мягкой и паутинной оболочек, по которым
циркулирует цереброспинальная жидкость. Цереброспинальная жид-
кость находится в полости черепа, в желудочках мозга и пространст-
вах между оболочками мозга. У человека ее объем составляет около
100—150 мл. Благодаря цереброспинальной жидкости поддержива-
ется осмотическое равновесие нервных клеток и удаляются продукты
метаболизма, токсичные для нервной ткани.
254
раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
Кровоток
Холин Ионы Глюкоза Триптофан Тирозий
v | Мембрана у нервной клетки и
Холин ма+ Глюкоза Триптофан Тирозин
Рис. 3.1. Пути обмена медиаторов и
в обмене веществ (по: Шеперд, 1987)
роль гематоэнцефалического барьера
Прохождение веществ через ГЭБ зависит не только от проница-
емости для них сосудистой стенки (молекулярной массы, заряда и ли-
пофильности вещества), но также и от наличия или отсутствия систе-
мы активного транспорта.
Стереоспецифичным инсулиннезависимым транспортером глюко-
зы (GLUT-1), обеспечивающим перенос этого вещества через ГЭБ,
богаты эндотелиальные клетки капилляров мозга. Активность данного
транспортера может обеспечить доставку глюкозы в количестве, в
2—3 раза превышающем то, которое требуется мозгу в нормальных
3.1. Особенности пластического обмене головного мозга
255
Таблица 3.1. Характеристика транспортных систем
гематоэнцефалического барьера
(по: Pardridge, Oldendorf, 1977)
Транспортируемые соединения Преи муществен ны й субстрат Кт, мМ V max* нмоль/мин*г
Гексозы Глюкоза 9 1600
Монокарбоновые кислоты Лактат 1,9 120
Нейтральные аминокислоты Фенилаланин 0,12 30
Основные аминокислоты Лизин 0,10 6
Амины Холин 0,22 6
Пурины Аденин 0,027 1
Нуклеозиды Аденозин 0,018 0,7
условиях. У детей с нарушением функционирования этого транспорте-
ра отмечается значительное снижение уровня глюкозы в цереброспи-
нальной жидкости и нарушения в развитии и работе мозга.
Монокарбоновые кислоты (L-лактат, ацетат, пируват), а также ке-
тоновые тела транспортируются отдельными стереоспецифичными
системами. Хотя интенсивность их транспорта ниже, чем транспорта
глюкозы, они являются важным метаболическим субстратом у ново-
рожденных и при голодании.
Транспорт холина в центральную нервную систему также опосре-
дуется переносчиком и может регулироваться скоростью синтеза аце-
тилхолина в нервной системе.
Витамины мозгом не синтезируются и поставляются из крови с по-
мощью специальных транспортных систем. Несмотря на то что эти
системы обладают сравнительно низкой транспортной активностью, в
нормальных условиях они могут обеспечивать транспорт необходимо-
го для мозга количества витаминов, однако их дефицит в пище спосо-
бен приводить к неврологическим расстройствам. Некоторые белки
плазмы также могут проникать через ГЭБ. Одним из способов их про-
никновения является трансцитоз, опосредованный рецепторами.
Именно так проникают через барьер инсулин, трансферрин, вазопрес-
син и инсулинподобный фактор роста. Эндотелиальные клетки капил-
256
Иазде, । 3. 4. чКЦИОНдДЫкЯ НЕЙР< х МИЯ
ляров мозга имеют специфические рецепторы к этим белкам и спо-
собны осуществлять эндоцитоз белок-рецепторного комплекса (см.
рис. 2.49). Важно, что в результате последующих событий комплекс
распадается, интактный белок может выделяться на противополож-
ной стороне клетки, а рецептор вновь встраиваться в мембрану. Для
поликатионных белков и лектинов способом проникновения через
ГЭБ также является трансцитоз, однако он не связан с работой специ-
фических рецепторов.
Многие нейромедиаторы, присутствующие в крови, не способны
проникать через ГЭБ. Так, дофамин не обладает этой способностью, в
то время как L-ДОФА проникает через ГЭБ с помощью системы
транспорта нейтральных аминокислот. Кроме того, клетки капилляров
содержат ферменты, метаболизирующие нейромедиаторы (холинесте-
раза, ГАМК-трансаминаза, аминопептидазы и др.), лекарственные и
токсические вещества, что обеспечивает защиту мозга не только от
циркулирующих в крови нейромедиаторов, но и от токсинов.
В работе ГЭБ участвуют также белки-переносчики, осуществляю-
щие транспорт веществ из эндотелиальных клеток капилляров голов-
ного мозга в кровь, препятствуя их проникновению в мозг, например
Р-гликопротеид.
В ходе онтогенеза скорость транспорта различных веществ через
ГЭБ существенно изменяется. Так, скорость транспорта р-гидрокси-
бутирата, триптофана, аденина, холина, а также глюкозы у новорож-
денных существенно выше, чем у взрослых. Это отражает относитель-
но более высокую потребность развивающегося мозга в энергии
и макромолекулярных субстратах.
З.1.2. Особенности обмена липидов
Нервная система является второй после адипозной ткани по
концентрации в ней липидов и поэтому очень чувствительна к их пос-
туплению в организм с пищей. Особенно важно достаточное поступ-
ление незаменимых липидов в периоды дифференциации и созревания
нервной ткани. Для работы мозга необходимы линолевая (пЗ) и лино-
леновая (пб) полиненасыщенные жирные кислоты, которые не синте-
зируются в организме человека и животных, но являются предшест-
венниками в синтезе остальных полиненасыщенных жирных кислот.
Наблюдается прямая зависимость между поступлением этих кислот с
пищей и их содержанием, а также содержанием их производных в моз-
ге. Снижение концентрации этих кислот приводит к изменению мик-
3.1. Особенности пластического обмена головного мозга
ровязкости мембраны и в значительной мере изменяет активность
мембраносвязанных ферментов.
Одним из важных механизмов локальной регуляции свойств липид-
ного бислоя в нейрональных клетках, также влияющим на микровяз-
кость мембраны, являются реакции деацилирования—реацилирова-
ния липидов с заменой одной из жирных кислот. Такая замена происхо-
дит по второму положению глицеринового остатка. По такому же
механизму происходит преобразование диацилглицерола в моноацил-
глицерол. Показана возможность влияния на этот процесс медиаторов.
Так, норадреналин через аденилатциклазную систему (см. гл. 2.3) спо-
собен активировать фосфолипазу А2, что приводит к деацилированию
диацилглицеролов.
Синтез жирных кислот. Скорость синтеза жирных кислот
de novo определяется функциональным состоянием нервной ткани и
довольно сильно колеблется в зависимости от периода онтогенеза.
Так, у крыс липиды и свободные жирные кислоты наиболее интенсив-
но накапливаются в период с 10-го по 20-й день постнатального раз-
вития. Возможны как митохондриальный, так и немитохондриальный
пути синтеза жирных кислот, связанные с восстановительными реак-
циями и использованием для этих целей НАДФН. В ходе постнаталь-
ного онтогенеза наблюдается прямая зависимость между активностью
синтазы жирных кислот и концентрацией восстановленного НАДФН.
Максимальное количество НАДФ в мозге также отмечается в период
усиленного липогенеза. Суммарное содержание восстановленной и
окисленной форм НАДФ в мозге значительно меньше, чем в печени,
а с учетом его интенсивного использования в различного рода синте-
зах (жирных кислот, холестерина, сфингозина) для поддержания кон-
центрации на физиологическом уровне интенсивность пополнения
восстановленного кофермента должна быть очень высока. Это проис-
ходит в основном за счет дегидрогеназ, участвующих в окислении глю-
козо-6-фосфата, 6-фосфоглюконата, изоцитрата и малата. С возрас-
том в цитоплазме нейронов основную роль в пополнении запасов
НАДФН начинают играть дегидрогеназы пентозофосфатного пути
(до 70%), а в митохондриях — изоцитратдегидрогеназа.
Пути синтеза и метаболизма липидов в мозге совпадают с таковы-
ми, выявленными в других органах, однако имеются и некоторые осо-
бенности. При наследственных нарушениях метаболизма липидов (ли-
пидозах) у больных наблюдаются в той или иной степени выраженные
расстройства функций мозга.
258
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
Гидрофобные цепи жирных кислот и изопреноидов синтезируются
из ацетил-КоА. Биосинтез жирных кислот происходит путем удлине-
ния присоединением С2-звена в ходе циклического повторения четы-
рех последовательных реакций (конденсации, восстановления, дегид-
ратации и гидрогенизации), катализируемых мультифункциональной
синтазой жирных кислот. Этот цикл повторяется до тех пор, пока дли-
на цепи не достигнет определенного размера (>С12). Превращение
насыщенных жирных кислот в ненасыщенные происходит в эндоплаз-
матическом ретикулуме при участии десатураз. Таким путем в мозге
синтезируется олеиновая кислота из стеариновой. Однако полинена-
сыщенные жирные кислоты подобным образом синтезироваться не
могут: так, из олеиновой не может синтезироваться линоленовая кис-
лота. Поэтому соЗ(и2)- и со6(и6)-жирные кислоты могут быть получе-
ны преимущественно с пищей, в основном растительной. Основные
компоненты фосфолипидов мозга — полиненасыщенные жирные кис-
лоты арахидоновая (20 : 4, соб) и декозогексаеновая (22 : 6, соЗ) — фор-
мируются путем удлинения цепи и десатурации (образования двойных
связей) более коротких жирных кислот.
Ацетил-КоА является предшественником и для синтеза всех изо-
преноидов, в том числе и холестерина. При синтезе холестерина из
трех молекул ацетил-КоА образуется мевалонат (Сб), который затем
превращается в изопентенилдифосфат («активный изопрен»), поли-
меризующийся далее с образованием сквалена (С30). Циклизация
сквалена с отщеплением трех атомов углерода приводит к синтезу хо-
лестерина. Обмен холестерина в мозге идет очень медленно, и с воз-
растом его содержание постепенно увеличивается.
Синтез фосфолипидов начинается с sn-3-глицерофосфата, который
образуется из дигидроксиацетон-3-фосфата или в результате фосфори-
лирования глицерина. Этерификация его по С-1 длинноцепочечной
жирной кислотой приводит к образованию лизофосфатидов, а при по-
следующей этерификации по С-2 ненасыщенной жирной кислотой —
фосфатидов, которые являются ключевыми промежуточными продук-
тами синтеза жиров, фосфо- и гликолипидов. Из фосфатидных кислот
через промежуточное образование диацилглицерина идет синтез фос-
фатидилхолина (лецитина). При этом фосфохолиновая группа из хо-
линцитидиндифосфата переносится на диацилглицерин. Аналогичным
образом происходит синтез фосфатидилэтаноламина. Для трех указан-
ных липидов характерны взаимопревращения: фосфатидилсерин при де-
карбоксилировании превращается в фосфатидилэтаноламин, который в
результате метилирования S-аденозилметионином образует фосфати-
дилхолин. Фосфатидная кислота при взаимодействии с цитидин-5'-мо-
3.1. Особенности пластического обмена головного мозга
259]
нофосфатом образует цутидиндифосфатдиацилглицерин, который, ре-
агируя с инозитолом, образует фосфатидилинозит. Последний переходит
в плазматическую мембрану, где может быть фосфорилирован АТФ с
образованием фосфатидил инозит-4-фосфата и фосфатидил инозит-4,5-
дифосфата, являющегося субстратом ФЛС (см. гл. 2.4). Большинство
ферментов, участвующих в синтезе глицеролипидов, связаны с мембра-
нами эндоплазматического ретикулума или митохондрий (принимающие
участие в синтезе кардиолипина).
Сфинганин образуется при конденсации декарбоксилированного
серина с пальмитоил-КоА и последующем восстановлении с использо-
ванием НАДФН. После ацетилирования и дегидрогенизации он пре-
вращается в церамид, являющийся предшественником всех сфинголи-
пидов. При взаимодействии церамида с фосфатидилхолином образу-
ются сфингомиелин и ДАТ. Гликозилирование церамина приводит к
синтезу ганглиозидов, а присоединение галактозы — к образованию
сульфатидов или цереброзидов.
Метаболизм жирных кислот. P-Окисление жирных кислот
происходит путем последовательного отщепления двууглеродных фраг-
ментов, причем каждый этап катализируется отдельным ферментом.
Этот процесс идет в митохондриях и пероксисомах. Имеются некото-
рые отличия метаболизма кислот в каждом из компартментов. В мито-
хондриях отщепление водорода осуществляется с помощью ФАД-со-
держащего фермента и связано с окислительным фосфорилированием
и синтезом АТФ. В пероксисомах же этот этап катализируется флавин-
содержащей оксидазой, которая использует молекулярный кислород с
образованием в качестве продукта реакции пероксида водорода, кото-
рый затем разлагается каталазой. Пероксисомальному окислению под-
вергаются жирные кислоты с длинной (>С22) цепью, содержание кото-
рых в мозге превышает таковое в других тканях. При ряде генетических
нарушений (цереброгепаторенальный синдром Зельвегера и адрено-
лейкодистрофия) нарушения в процессе пероксисомального окисления
приводят к накоплению жирных кислот с очень длинной цепью, что
наиболее характерно для нервной ткани.
В дополнение к классическому Р-окислению в мозге наблюдается
также ос-окисление жирных кислот, особенно важное для галактоцереб-
розидов. Сравнительно высокая интенсивность этого процесса в мозге
может быть связана с наличием в нем большого количества жирных
кислот с нечетным числом атомов углерода и 2-гидроксижирных в га-
лактоцереброзидах. Генетическое заболевание, обусловливающее на-
рушение этого пути окисления (болезнь Рефсума), приводит к тому, что
260
Иазм и 3. ФУНКЦИОНАЛЬНАЯ НЕЙР0ХИМИ>
в нервной ткани накапливаются жирные кислоты с разветвленной
цепью, поступающие в организм в основном с пищей.
Особо следует остановиться на метаболизме арахидоновой кис-
лоты, образующейся из мембранных фосфолипидов под действием
фосфолипаз А2, С и D, которые активируются при стимуляции опре-
деленных рецепторов (см. гл. 2.4). Три возможных пути метаболизма
арахидоновой кислоты (циклооксигеназный с образованием простаг-
ландинов и тромбоксанов; липоксигеназный с образованием лейко-
триенов и других эйкозаноидов; с участием цитохрома Р450) приводят к
синтезу биологически активных веществ, которые работают в синапсе
как нейромодуляторы, участвуя в механизмах долговременной потен-
циации. Возможный путь метаболизма арахидоновой кислоты с учас-
тием цитохрома Р450 в головном мозге выражен слабо, но может быть
индуцирован различными факторами.
Фосфоглицериды гидролизуются специфическими фосфолипазами
(см. гл. 2.4), которые прямо или опосредованно регулируются клеточ-
ными рецепторами, а продукты их метаболизма являются вторичными
посредниками в сигналтрансдукторных системах.
Сфинголипиды метаболизируются присутствующими в лизосомах
специфическими гидролазами, которые стимулируются некаталитиче-
скими лизосомальными белками. Наследственные нарушения в работе
гидролаз или белков-регуляторов приводят к накоплению интермедиа-
тов обмена в лизосомах и к связанным с этим заболеваниям. Группа по-
добных заболеваний более обширна, чем для других классов липидов:
лимфогрануломатоз Фарбера, болезнь Нимана-Пика, Тей-Сакса, Гоше
(гликозилцерамидный липидоз), Крабе (галактозилцерамидный липи-
доз), метахроматическая лейкодистрофия (сульфатидный липидоз).
ЭНЕРГЕТИЧЕСКИЙ ОБМЕН ГОЛОВНОГО МОЗГА
Высокая интенсивность энергетического обмена — характер-
ная черта головного мозга, во многом определяющая его функци-
ональную активность. Первые данные, указывающие на очень высо-
кий уровень протекания окислительных процессов в мозге, были по-
лучены при определении скорости потребления кислорода (табл. 3.2).
Установлено, что по этому показателю мозг опережает такие крупные
и активно работающие органы, как сердце, почки, печень. Так, ско-
рость потребления кислорода корой больших полушарий головного
мозга в 1,7 раза выше, чем в сердце, в 2,2—2,3 раза выше, чем в поч-
ках, и почти в 3 раза превышает потребление кислорода печенью.
3.2.
3.2. Энергетический гбмен ЛОЯСфИСГи мезгз
261
Таблица 3.2. Средние данные по потреблению кислорода
и глюкозы головным мозгом, мкмоль/г« мин
Показатели Мозг человека Мозг крысы
Потребление кислорода 1,5—1,7 3,8—4,6
Потребление глюкозы 0,25—0,30 0,6—0,8
Дыхательный коэффициент 0,97—0,98 0,97
Масса головного мозга взрослого человека составляет около 2%
массы тела, но его доля потребления поступающего в организм кисло-
рода достигает 20—25%. Приведенные данные относятся к головно-
му мозгу в целом. Однако необходимо подчеркнуть, что с помощью
современных методов исследования (например, позитронно-эмисси-
онной томографии и др.) удалось установить наличие существенных
различий в потреблении кислорода отдельными структурами. Наибо-
лее активно процессы окисления протекают в коре больших полуша-
рий (особенно в слуховой и зрительной зонах), в таламусе, некоторых
подкорковых ядрах (скорлупа, внутреннее коленчатое тело и др.). Бо-
лее низкие значения потребления кислорода характерны для таких
структур, как амигдала, черная субстанция, гипоталамус, кора моз-
жечка, продолговатого мозга, а также для белого вещества.
Весьма заметны различия в потреблении кислорода между основ-
ными типами клеток мозга: по данным многих исследователей, на долю
глиальных клеток (суммарно) приходится от 17 до 30% общего количе-
ства поглощенного мозгом кислорода. Считается, что в среднем по-
требление кислорода нейронами протекает в 4—5 раз активнее, чем
глиальными клетками. Установлены также различия в интенсивности
протекания окислительных процессов в отдельных компартментах од-
ного и того же нейрона: больше кислорода потребляют структуры, ло-
кализованные в области перикариона и в синаптических окончаниях.
Высокая интенсивность потребления кислорода головным мозгом
объясняет, почему этот орган так чувствителен к гипоксии. Прекра-
щение снабжения мозга кислородом всего на 5—7 мин (не более
10 мин) приводит к необратимым и драматическим последствиям, в то
время как многие другие органы выдерживают без потери функци-
ональной активности значительно более длительные эпизоды гипо-
ксии. Известно также, что в патогенезе многих неврологических забо-
леваний лежат нарушения кровоснабжения мозга, т. е. нарушения
в бесперебойной доставке кислорода, а также субстратов окисления.
262
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
Е. С. Лондон
Крупнейший российский фи-
зиолог, радиобиолог, биохи-
мик, основоположник отече-
ственной функциональной
биохимии; впервые получил
количественные данные о
потреблении глюкозы тканя-
ми интактного мозга
Основным субстратом оки-
сления в мозге взрослых животных
и человека служит глюкоза; на это ука-
зывает величина дыхательного коэффи-
циента (см. табл. 3.2). Собственные запа-
сы этого вещества в мозге невелики. По
данным разных исследователей, они колеб-
лются в пределах 2,5—4,0мкмоль*г-1.
Отсюда становится понятной потребность
мозга в постоянном поступлении глюкозы
из крови.
Ни один орган не потребляет глюкозу
с такой скоростью и в таких количествах
как головной мозг, и ни для одной ткани
организма не отмечено такой острой по-
требности в этом субстрате окисления для
обеспечения функциональной активнос-
ти. Головной мозг взрослого человека и
животного использует около 70% глюко-
зы, выделяющейся из печени в общий
кровоток.
Средние данные, характеризующие потребление глюкозы мозгом в
целом, приведены в таблице 3.2. С помощью позитронно-эмиссион-
ной томографии, а также благодаря использованию содержащей ра-
диоактивную метку дезоксиглюкозы в настоящее время установлены
существенные различия в потреблении глюкозы отдельными структу-
рами мозга и отмечен четкий параллелизм между локальным потреб-
лением глюкозы и кислорода. Для упоминавшихся выше структур моз-
га, которые отличаются высоким потреблением кислорода, характер-
но и высокое потребление глюкозы (табл. 3.3).
Интенсивное поступление глюкозы из’ капилляров в клетки моз-
га обеспечивается работой специальных глюкозных переносчиков.
В первую очередь следует упомянуть активно экспрессирующийся в
эндотелиальных клетках, в нейронах и астроцитах очень активный
транспортер GLUT1, работающий в широких пределах концентрации
глюкозы в крови (от 2,75 до 16,50 мкмоль/мл). Количество этого
мембранного белка существенно увеличивается с возрастом парал-
лельно усилению функциональной активности мозга. Установлено,
что около 95% поступления в мозг глюкозы обеспечивается работой
ее транспортеров (рис. 3.2). Именно активной работой глюкозных
3.2. Энергетический обмен головного мозга
Таблица 3.3. Средние значения локального потребления глюкозы
структурами головного мозга белых крыс и обезьян,
мкмоль/100 г за 1 мин (по: Clarke, Sokoloff, 1999)
Структуры мозга Белые крысы Обезьяны
Серое вещество
Зрительная кора 107 ±6 59 ±2
Слуховая кора 162 ±5 79 ±4
Сенсомоторная кора 120 ±5 44 ±3
Таламус
латеральные ядра 116 ± 6 54 ±2
вентральные ядра 109 ±5 43 ±2
Гипоталамус 54 ±2 25 ± 1
Амигдала 52 ±2 25 ±2
Хвостатое ядро 110 ± 4 52 ±3
Черная субстанция (substantia nigra) 58 ±3 29 ±2
Прилежащее ядро (nucleus accumbens) 82 ±3 36 ±2
Нижний холмик (colliculus inferior) 197 ± 10 103 ±6
Верхний холмик (colliculus superior) 95 ±5 55 ±4
Кора мозжечка 57 ±2 31 ±2
Ядра мозжечка 100 ±4 45 ±2
Белое вещество
Мозолистое тело (corpus callosum) 40 ±2 И ± 1
Белое вещество мозжечка 37 ±2 12± 1
Средние данные по целому мозгу 68 ±3 36 ± 1
транспортеров объясняется высокая скорость поступления данного
субстрата в мозг через ГЭБ, в десятки и сотни раз превышающая ско-
рость поступления других веществ (см. табл. 3.1). Мутации гена, ко-
дирующего глюкозный транспортер GLUT1, сопровождаются разви-
тием тяжелых форм инфантильной эпилепсии.
Как показали результаты экспериментов с С14-глюкозой, основ-
ным путем метаболизма этого субстрата в мозге является гликолити-
264
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
Рис. 3.2. Компартментализация обмена глюкозы.
При достаточном поступлении глюкозы она запасается в астроцитах в форме глико-
гена (двойная стрелка). Когда образование глюкозо-6-фосфата в гексокиназной
реакции ограничено из-за низкой концентрации глюкозы, гликогенолиз в астроци-
тах восполняет фонд глюкозо-6-фосфата для дальнейшего синтеза и образования
АТФ в глиальном компартменте. Гликоген астроцитов в форме лактата может «пе-
ремещаться» в нейроны, снижая энергетический дефицит (например, при гипогли-
кемии)
ческое расщепление с последующим окислением образующегося пи-
рувата в цикле трикарбоновых кислот (табл. 3.4).
Специфика протекания гликолиза в мозге. Прежде чем обсуж-
дать специфику протекания реакций гликолиза в мозге, следует обратить
внимание на гексокиназную реакцию. Она представляет собой первый
этап на пути вовлечения в разнообразные метаболические превращения
свободной и метаболически инертной D-глюкозы, которая поступает из
крови и играет в головном мозге исключительно важную роль.
3.2. Энергетическим обмен головного мозга
265
Таблица 3.4. Интенсивность использования
глюкозо-6-фосфата в различных метаболических путях
в головном мозге и печени взрослых крыс
* Метаболический путь Доля глюкозо-6-фосфата, вовлекаемого в реакции, °о
Мозг Печень
Окисление до СО2 и Н2О в ходе аэробного гликолиза и ЦТК 80—90 Около 20
Синтез гликогена 5—7 20—25
Пентозофосфатный путь 2—3 5—15
Глюкозо-6-фосфатазная реакция Следы До 50
Другие реакции Менее 5 5—10
Катализируемое гексокиназой образование глюкозо-6-фосфата
является основным источником пула метаболитов гликолиза в мозге,
в отличие от других тканей, где этот пул может пополняться в резуль-
тате интенсивного расщепления гликогена (мышечная ткань, печень)
или за счет фосфорилированных триоз, образующихся в ходе метабо-
лизма фосфолипидов. Поэтому неудивительно, что активность гексо-
киназы в мозге, поданным многих исследователей, заметно выше, чем
в печени, почках, мышечной и других тканях. Тем не менее фосфори-
лирование глюкозы под действием гексокиназы относится к наиболее
медленным этапам и может лимитировать скорость гликолиза в мозге
при некоторых экстремальных или патологических состояниях.
Интересным и важным с точки зрения регуляции скорости гексо-
киназной реакции в мозге является вопрос о локализации фермента.
До 70% гексокиназы связано с внешней мембраной митохондрий, что,
по-видимому, способствует более эффективному и быстрому испольт
зованию образующегося в митохондриях АТФ для фосфорилирования
глюкозы. В других тканях доля связанного с митохондриальной мемб-
раной фермента заметно ниже (20—40%). Наиболее активно связы-
вается с мембраной доминирующая в мозге изоформа ГКТ Процесс
связывания фермента с мембраной является обратимым; соотноше-
ние связанной и солюбилизированной форм гексокиназы контролиру-
ется соотношением АТФ и АДФ, концентрацией продукта реакции и
неорганического фосфата, а также уровнем свободных жирных кис-
лот. Таким образом, в мозге в большей степени, чем в других тканях,
266
раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
скорость вовлечения глюкозы в метаболизм зависит от скорости гек-
сокиназной реакции.
Еще одним важным этапом гликолиза в мозге, лимитирующим ско-
рость перемещения потока метаболитов по гликолитической цепи, как
и в других тканях, служит фосфофруктокиназная реакция. В физио-
логических условиях в регуляции активности фосфофруктокиназы
наиболее важна роль аллостерических эффекторов АТФ и цитрата,
подавляющих активность фермента, и АДФ, обладающего, напротив,
стимулирующим эффектом. Дополнительными регуляторными факто-
рами служат АМФ, неорганический фосфат и продукт реакции фрук-
тозо-1,6-бисфосфат. Характерным для мозга является преобладание
В-изоформы фермента, наиболее чувствительной к ингибированию
АТФ. Кроме того, данная изоформа, в отличие от М-изофермента
(преобладающего в других тканях), сильно активируется фруктозо-
фосфатом. Определенную роль в регуляции активности фосфофрукто-
киназы в мозге играет и кальмодулинзависимое фосфорилирование
фермента, в котором участвует нейроспецифический белок S-100.
Таким образом, в головном мозге соотношение АТФ/АДФ (с по-
мощью различных механизмов) может одновременно и однонаправлен-
но контролировать две принципиально важные стадии метаболизма
глюкозы — гексокиназную и фосфофруктокиназную реакции. Такой
«двойной контроль» над активностью этих ферментов со стороны ко-
нечных продуктов энергетического обмена является характерной осо-
бенностью функционирования головного мозга.
Реакции гликолиза, включающие расщепление фруктозо-1,6-бис-
фосфата и последующие превращения фосфотриоз в физиологических
условиях обычно не лимитируют общую скорость гликолиза в мозге,
поскольку активность всех катализирующих их ферментов существен-
но выше, чем активность гексокиназы и фосфофруктокиназы. Рас-
сматривая эти этапы гликолиза в нервной ткани, можно отметить
специфичную локализацию изоферментов, енолазы. Напомним, что
фермент представляет собой димер, в состав которого входят субъеди-
ницы двух типов — а и у. В глиальных клетках (астроциты, микро-
глия) обнаружен лишь гомодимер оса, идентичный енолазе печени и
ряда других тканей, в то время как изоформа уу встречается только
в нейронах, и она идентична нейроспецифическому кислому белку
14—3—2. Гибридная форма ау, имеющая низкую активность, пред-
ставлена в клетках мозга в незначительном количестве.
Еще одним ферментом гликолиза, изоферментный спектр которого
в мозге отличается от такового в других тканях, является альдолаза.
Известно, что этот энзим существует в форме тетрамера, состоящего
3.2. Энергетический обмен головного мозга
из трех типов субъединиц — А, В и С. Для мозга характерно преобла-
дание изоформы С4; найдены также в небольших количествах вариан-
ты А4, А3С и А2С2, причем соотношение изоформ альдолазы в мозге
с возрастом не меняется.
Конечные этапы гликолиза и взаимоотношения двух субстратов —
пирувата и лактата — также имеют некоторые характерные
только для мозга особенности. Речь идет об интенсив-
ном образовании лактата в астроцитах из поступающей с кровью глю-
козы. Этот процесс может протекать даже в условиях сниженного
поступления кислорода, поскольку в астроцитах преобладает
анаэробный изофермент лактатдегидрогеназы ЛДГ5. Для ней-
ронов, напротив, характерен аэробный изофермент ЛДГ^ При
необходимости лактат из астроцитов переносится в нейроны, где, пре-
вратившись под действием лактатдегидрогеназы в пируват, включает-
ся в энергетический метаболизм нейрона (рис. 3.3).
Интенсивному использованию лактата для поддержания энергети-
ческого баланса в нейронах способствует то обстоятельство, что за-
Рис. 3.3. Последовательность реакций, связывающих окисление пирувата
с синтезом ацетилхолина в холинергических нейронах.
Ферменты: / — пируватдегидрогеназный комплекс; 2 — цитратсинтаза; 3 — цитрат-
лиаза; 4 — дегидрогеназа; 5 — холинацетилаза
268
г аздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРСХЙ ИИЯ
метная доля лактатдегидрогеназы (до 10% общей активности фермен-
та) связана с наружной мембраной митохондрий. Для сравнения: в
большинстве других тканей доля фермента, связанного с митохонд-
риями, гораздо ниже и составляет обычно менее 1 %. Поэтому вполне
обосновано предположение о том, что такая особенность в локализа-
ции лактатдегидрогеназы в мозге способствует более полному исполь-
зованию конечных продуктов гликолиза (лактата и пирувата) в окис-
лительных реакциях, протекающих в митохондриях.
Особенности метаболизма пирувата. Пировиноградная кис-
лота, образующаяся в цитоплазме в ходе аэробного гликолиза, пере-
носится в митохондрии активной протонзависимой транспортной сис-
темой. Эксперименты с С14-пируватом продемонстрировали высокую
скорость утилизации этого субстрата в головном мозге: для белых
крыс этот показатель составляет 1,3—1,4 мкмоль* мин-1 в расчете
на 1 г ткани, в то время как для печени тех же животных — всего
0,35—0,40 мкмоль • мин-1 в расчете на 1 г ткани.
Основным путем метаболизма пирувата в ми-
тохондриях мозга является вовлечение его в реакции ЦТК,
т. е. использование для поддержания энергетического баланса в орга-
низме. Сопоставление активности ферментов, катализирующих раз-
ные реакции с использованием пирувата, а также результаты опытов с
С14-субстратом показали, что 85—90% пировиноградной кислоты
подвергается окислительному декарбоксилированию в ходе пируват-
дегидрогеназной (ПДГ) реакции. В отличие от мозга, в большинстве
тканей животных и человека (печень, сердечная и скелетная мышцы,
почки и др.) в данной реакции используется лишь 15—25% пирувата,
но активно функционирует другой путь ввода этого метаболита в ЦТК,
а именно — пируваткарбоксилазная (ПК) реакция, приводящая к
образованию важного субстрата ЦТК щавелевоуксусной кислоты.
Необходимо подчеркнуть, что по данным многих исследователей,
доминирование дегидрогеназной реакции над карбоксилазной пред-
ставляет собой особенность метаболизма пирувата, характерную толь-
ко для мозга. Такое соотношение ПДГ- и ПК-реакций в мозге имеет
место не только в физиологических условиях, но и при функциональ-
ных отклонениях. Напротив, в других тканях изменение функциональ-
ного состояния или различные воздействия обычно сопровождаются
значительной активацией реакции карбоксилирования пирувата. На-
пример, в митохондриях печени при голодании или в условиях интен-
сивного глюконеогенеза, в сердце при интенсивной нагрузке скорость
реакции карбоксилирования пирувата существенно возрастает и мо-
3 2. ihj-o ew । 4И trf.leh । Et.n.iMGjr.
жет в 5—10 и более раз превысить скорость окислительного декар-
боксилирования субстрата в ПДГ-реакции.
Особую важность ПДГ-реакции для метаболизма головного мозга
подтверждает хорошо известный факт более высокой чувствительнос-
ти нервной ткани (по сравнению с другими тканями организма) к не-
достатку тиамина. Нарушение образования тиаминпирофосфата (ко-
фермента декарбоксилазного компонента ПДГ-комплекса) при Вj-ави-
таминозе приводит к нарушениям, наиболее ярко проявляющимся в
центральной и периферической нервной системе — полиневритам и др.
(болезнь бери-бери).
Еще одним обстоятельством, указывающим на большое значение
ПДГ-реакции для мозга, является то, что она представляет собой на-
чальный этап биосинтеза одного из ведущих нейромедиаторов — аце-
тилхолина (рис. 3.4). Установлено, что замедление окислительного де-
карбоксилирования пирувата в ПДГ-реакции всего на 10% резко тормо-
зит образование ацетилхолина и приводит к нарушению холинергической
трансмиссии.
Основным механизмом регуляции активности сложного мультифер-
ментного ПДГ-комплекса служит процесс фосфорилирования/дефосфо-
Рис. 3.4. Схематическое изображение двух метаболических компартментов:
«большой» (энергетический) в нейронах и «малый» (синтетический) в гли-
альных клетках (преимущественно в астроцитах)
270
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
рилирования декарбоксилазного компонента комплекса (пируватдекар-
боксилазы), который катализирует наиболее медленную стадию процесса
окислительного декарбоксилирования пирувата. Реакция фосфорилиро-
вания осуществляется специфичной калъмодулинзависимой ПДГ-ки-
назой, которая переводит фермент в неактивное состояние. Активная де-
фосфорилированная форма фермента образуется под действием фосфа-
тазы, не обладающей выраженной специфичностью. Скорость реакции
фосфорилирования, т. е. образования неактивной формы фермента, за-
висит от концентраций АТФ (субстрат реакции) и АДФ (конкурирует
с АТФ); реакция протекает в присутствии ионов Mg2+.
Определенную роль в контроле скорости окислительного декарбо-
ксилирования пирувата в физиологических условиях может играть изме-
нение концентрации конечного продукта реакции — ацетил-КоА. Одна-
ко для мозга значение этого регуляторного фактора невелико, в отличие
от других тканей, где активно протекает окисление свободных жирных
кислот и, возможно, накопление ацетил-КоА в количествах, сопостави-
мых с величиной/^, равной для ПДГ-комплекса (3—5)* 10-5М. Кроме
того, следует отметить, что возрастание концентрации ацетил-КоА име-
ет еще два следствия. Во-первых, ацетил-КоА служит положительным
эффектором ПДГ-киназы, т. е. способствует переходу ПДГ-комплекса в
неактивную фосфорилированную форму. Во-вторых, ацетил-КоА, явля-
ясь аллостерическим эффектором пируваткарбоксилазы, может значи-
тельно ускорять реакцию карбоксилирования пирувата до щавелевоук-
сусной кислоты.
Таким образом, в митохондриях мозга доминиру-
ющий путь метаболизма пирувата — пируватдегидро-
геназная реакция — контролируется главным образом изменением
соотношения АТФ/АДФ. Наряду с этим, в мозге, как и в других тканях
с высокой интенсивностью окислительных процессов, скорость
ПДГ-реакции зависит от соотношения НАД+/НАДН.
Расчеты, проведенные на основании опытов с 2С14-пируватом, по-
казали, что скорость использования для синтеза лимонной кислоты
образующегося в ПДГ-реакции ацетил-КоА в головном мозге в 3—
5 раз выше, чем в печени, сердце, почках. Максимальная активность
ПДГ-комплекса (около 1,8 мкмоль • мин-1 в расчете на 1 г ткани) лишь
незначительно превышает скорость потока пирувата в мозге взрослых
интактных крыс (1,3— 1,4 мкмоль • мин-1 на 1 г ткани), что свидетель-
ствует об эффективной регуляции работы ферментного комплекса. Ус-
тановлено, что в головном мозге крыс доля активной дефосфорилиро-
ванной формы ПДГ-комплекса составляет около 70% от общего со-
держания фермента, что гораздо выше, чем в других тканях: в сердце
3.2. Энергетическим обмен головного мозга
40—50%, в печени, жировой ткани — около 20%. Регуляция степени
фосфорилирования ПДГ-комплекса, т. е. скорости окислительного де-
карбоксилирования пирувата, играет существенную роль в модуляции
пластичности синапсов, поскольку обнаружена корреляция между со-
отношением фосфорилированная/дефосфорилированная форма фермен-
та и эффективностью процессов обучения.
Из изложенного выше ясно, что ПДГ-реакция служит основным
источником ацетил-КоА для дальнейшего окисления в цикле трикар-
боновых кислот.
Образование ацетил-КоА в ходе окисления свободных жирных
кислот и кетоновых тел протекает в головном мозге взрослых живот-
ных значительно медленнее, чем в процессе ПДГ-реакции. Это связа-
но с малой проницаемостью ГЭБ для жирных кислот и кетоновых
тел, а также с низкой активностью ферментов, участвующих в окис-
лении данных субстратов (прежде всего, с низкой активностью
ацил - Ко А- си нтазы).
Напротив, в мозге растущих животных, где активность этих фер-
ментов высока, а структуры ГЭБ еще не сформированы окончательно,
окисление свободных жирных кислот, и особенно кетоновых тел, идет
весьма интенсивно. Однако образующийся при этом ацетил-КоА рас-
ходуется не только на окисление в ЦТК, но главным образом исполь-
зуется для биосинтеза специфических липидов мозга. Интенсивное
окисление кетоновых тел характерно именно для периода активной
миелинизации, роста аксонов и дендритов.
Весьма незначительно образование ацетил-КоА в ходе метаболиз-
ма таких аминокислот, как тирозин, фенилаланин, лизин, лейцин, что
обусловлено очень низкой (по сравнению с глюкозой) проницаемо-
стью для них ГЭБ (см. табл. 3.1).
Окисление аминокислот, свободных жирных кислот и кетоновых тел в
зрелом головном мозге протекает в основном в «малом» (глутаминсинте-
зирующем) компартменте, морфологически приуроченном к глиальным
клеткам, преимущественно астроцитам (см. рис. 3.4); в этом особенно
ярко проявляется анаболическая функция ЦТК. Напротив, катаболиче-
ская роль ЦТК наиболее выражена в «большом» (энергетическом) ком-
партменте, который сосредоточен главным образом в нейронах.
Скорость метаболического потока, определенная
для мозга крыс (в расчете на 1 г ткани): «большой» (энергетический)
компартмент — 1,25—1,35 мкмоль ацетил-КоА за 1 мин; «малый»
(глутаминсинтезирующий) компартмент — 0,28—0,30 мкмоль аце-
тил-КоАза 1 мин.
272
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕИ^' • j 1ИЯ
Обмен метаболитов между нейрональным и глиальным компарт-
ментами происходит относительно медленно; скорость метаболиче-
ского потока составляет в среднем 0,14—0,15 мкмоль субстрата за
1 мин в расчете на 1 г ткани. Следует обратить внимание на то обсто-
ятельство, что транспорт глутамата, глутамина и ГАМК между нейро-
нами и окружающими их астроцитами играет важную роль в обеспече-
нии нормального функционирования глутамат- и ГАМК-ергических
нейронов (см. гл. 2.3).
Обсуждая специфику функционирования и регуляции скорости ЦТК
как единой метаболической системы, прежде всего следует обратить
внимание на наиболее медленные этапы синтеза и окисления трикарбо-
новых кислот. Образование лимонной кислоты, необратимое в физио-
логических условиях, катализируется цитратсинтазой, отрицательным
аллостерическим модулятором которой служит АТФ (табл. 3.5).
Приведенные в таблице 3.5 данные свидетельствуют о том, что в
физиологических условиях скорость синтеза лимонной кислоты конт-
ролируется уровнем АТФ (точнее, его долей в общем пуле адениновых
нуклеотидов в митохондриях) и концентрацией щавелевоуксусной
кислоты. Изменение концентрации другого субстрата (ацетил-КоА) в
данном случае роли не играет, поскольку фермент работает в условиях
полного насыщения ацетил-КоА.
В головном мозге взрослых животных и человека значительная
часть изоцитрата (до 65—70%) окисляется НАД-изоцитратдегидроге-
назой (ИЦДГ), в то время как в других тканях (печень, скелетные и
сердечная мышцы, почки и др.) в этой реакции используется лишь око-
ло 10% субстрата, а основной путь окисления изолимонной кислоты в
митохондриях, как и в цитоплазме, обеспечивается НАДФ-ИЦДГ. Сле-
дует отметить, что явное преобладание НАД-зависимого пути окисле-
ния изоцитрата характерно лишь для мозга взрослых животных. В моз-
ге растущих животных, особенно в период активной миелинизации, ос-
новная часть изолимонной кислоты окисляется в НАДФ-зависимой
реакции, которая служит одним из источников восстановительных эк-
вивалентов (НАДФН) для процессов липогенеза.
Регуляция скорости окисления изолимонной кислоты осуществля-
ется главным образом в результате изменения активности НАД-зави-
симой ИЦДГ, работа которой контролируется соотношением высоко-
и низкоэнергетических компонентов адениннуклеотидного пула. Эф-
фективность адениннуклеотидного контроля в определенной степени
зависит от внутримитохондриальной концентрации ионов Са2+. Со-
поставив механизмы регуляции НАД-ИЦДГ и цитратсинтазы, можно
сделать заключение о существовании специфической для мозга сопря-
Таблица 3.5. Основные регуляторные факторы, контролирующие скорость синтеза
и окисления трикарбоновых кислот в ЦТК
Регуляторные факторы Цитратсинтаза Изоцитратдегидрогеназа
НАД-зависимая НАДФ-зависимая
Аллостерические эффекторы АТФ — отрицательный эффектор АМФ, АДФ — положительные эффекторы; АТФ — отрицательный эффектор Отсутствуют
3.2. Энергетический обмен головного мозга
Субстратная 1. Оксалоацетат Изоцитрат активация Км = (1,6-4,0)- 10-6М. Км = (1,0—1,5)-Ю-М. Концентрация в митохондриях Концентрация в митохондриях — (2,0-5,0)-10-М. (1,о-3,0)- 10-М 2. Ацетил-КоА Км = (1,3—1,6)-10-М. Концентрация в митохондриях — (2,0—5,0)- 10—М Изоцитрат ^ = (2,0—2,6)- 10"6М. Концентрация в митохондриях (1,0—3,0)- 10-4М
*4
О)
274
Раздел 3. ФУНКЦИОНАЛЬНА и НЕЙРО* и МИ«
женной (одновременной и однонаправленной) регуляции скорости
этих двух наиболее медленных этапов ЦТК. Снижение доли АТФ (от-
рицательный эффектор цитратсинтазы) и возрастание уровня низко-
энергетических компонентов адениннуклеотидного пула — АДФ и
АМФ (положительные эффекторы НАДФ-ИЦДГ) приводят к одно-
временному ускорению обеих реакций. Более того, усиленное образо-
вание лимонной кислоты в цитратсинтазной реакции и сопровождаю-
щее его накопление изоцитрата в силу различий в величинах двух
дегидрогеназ (см. табл. 3.5) будет активировать только НАД-ИЦДГ.
Приведенные данные позволяют рассматривать цитратсинтазу и
НАД-ИЦДГ в митохондриях мозга как единый функциональный комп-
лекс, подобный комплексу гексокиназы и фосфофруктокиназы.
Такая сопряженная регуляция начальных этапов ЦТК и прямая за-
висимость их скорости от соотношения компонентов адениннуклеотид-
ной системы указывают на существование в мозге взрослого организма
(в отличие от других тканей) более выраженной корреляции между ин-
тенсивностью энергетического обмена и скоростью метаболического
потока в ЦТК.
Рассматривая начальные этапы ЦТК, следует отметить, что синтез
цитрата, наряду с ПДГ-реакцией, играет важную роль в мозге как
один из этапов синтеза нейромедиатора ацетилхолина (см. рис. 3.3).
Замедление цитратсинтазной реакции приводит к нарушению холин-
ергической трансмиссии.
Еще одним важным для метаболизма мозга этапом ЦТК являются
реакции образования и окисления а-кето глутарата. Кетоглутаратде-
гидрогеназный комплекс, окисляющий этот субстрат, по многим свой-
ствам, в том числе и по механизмам регуляции скорости реакции, схо-
ден с рассмотренным выше ПДГ-комплексом.
Значимость этого этапа ЦТК в мозге опре-
деляется, во-первых, тем, что он играет большую роль в под-
держании суммарной концентрации метаболитов цикла на стационар-
ном уровне. Это обеспечивается одновременным участием а-кетоглу-
тарата как в ЦТК, так и в глутаматдегидрогеназной и трансаминазных
реакциях. Перенос а-кетоглутарата из митохондриального в цито-
плазматический компартмент осуществляется транслоказой дикарбо-
новых кислот в обмен на малат. Окисление малата в митохондриях
НАД-малатдегидрогеназой повышает интрамитохондриальный уровень
оксалоацетата, что приводит к активации цитратсинтазы, т. е. к ускоре-
нию наиболее медленной реакции ЦТК. Во-вторых, взаимопрев-
ращения а-кетоглутарата и глутамата обеспечивают принципиально
важную для мозга возможность образования главного тормозного
3.2. Эш ; гкгич-г.скии O^IVг i- * л» *• и го мс /н-
медиатора — ГАМК в ходе глутаматдекарбоксилазной реакции. Наи-
более интенсивно эти реакции протекают в ГАМК-ергических нейро-
нах, где через ГАМК-шунт метаболирует около 10% а-кетоглутарата.
ГАМК-шунт представляет собой альтернативный вариант ЦТК,
характерный исключительно для нервной ткани. Последовательность
реакций ГАМК-шунта следующая:
1) превращение а-кетоглутарата в глутамат под действием глута-
матдегидрогеназы;
2) декарбоксилирование глутамата с образованием ГАМК;
3) образование янтарного полуальдегида из ГАМК, катализируе-
мое ГАМК-трансаминазой;
4) окисление янтарного полуальдегида специфической дегидроге-
назой до сукцината, который далее окисляется в ЦТК.
Таким образом, ГАМК-шунт отличается от универсальной после-
довательности реакций ЦТК тем, что в нем отсутствует стадия образо-
вания сукцинил-КоА, связанная с субстратным фосфорилированием и
образованием макроэрга, но появляется возможность синтезировать
нейромедиатор ГАМК- Рассмотренные взаимоотношения между ЦТК
и метаболизмом глутамата и ГАМК наиболее ярко проявляются, соот-
ветственно, в глутамат- и ГАМК-ергический нейронах и окружающих
их астроцитах (см. рис. 2.14).
Активность ферментов, катализирующих конечные стадии ЦТК, от
сукцината до оксалоацетата, в мозге, как и в других тканях, сущест-
венно превышает активность цитратсинтазы и НАД-ИЦДГ и, следо-
вательно, в физиологических условиях не лимитирует скорости потока
метаболитов в цикле.
Заканчивая рассмотрение ЦТК, следует подчеркнуть, что для мозга
характерна резко выраженная компартментализация реакций ЦТК.
К настоящему времени получены убедительные экспериментальные
доказательства гетерогенности митохондрий, выделенных из нейро-
нов, различных типов глиальных клеток или синаптических оконча-
ний. Более того, даже в пределах одной клетки (нейрона или астроци-
та) удалось установить наличие субпопуляций митохондрий, заметно
различающихся по активности ферментов ЦТК, что отражает наличие
компартментализации на субклеточном уровне. Существование по-
добных микродоменов внутри нейронов или астроцитов указывает на
их функциональную метаболическую специализацию.
Дыхательная цепь митохондрий мозга организована и функциони-
рует так же, как и в других тканях. Необходимо отметить, что скорость
окислительно-восстановительных превращений компонентов дыхатель-
ной цепи значительно превышает скорость реакций окисления суб-
276
►аздел 3. Ф -' • ЁИРО
стратов, поэтому именно дегидрогеназные реакции (в мозге это преж-
де всего дегидрогеназные реакции ЦТК) определяют в конечном счете
интенсивность окислительных процессов в ткани. Установлено, что в
тканях с высокой интенсивностью окислительного метаболизма (го-
ловной мозг, летательная мышца насекомых, сердечная мышца в пе-
риод интенсивной нагрузки) отношение активности ферментов, лими-
тирующих скорость ЦТК (цитратсинтазы + НАД-ИЦДГ), к содержа-
нию цитохромов а -Г а3 или цитохрома с превышает такое отношение
для тканей, в которых процессы окисления протекают менее интенсивно
(печень, скелетные мышцы). Существование подобного соотношения
активности дегидрогеназ и количества компонентов дыхательной цепи в
митохондриях мозга можно рассматривать как структурную основу,
обеспечивающую высокую интенсивность окислительного и энергети-
ческого обменов.
Высокая скорость потребления головным мозгом кислорода и глю-
козы сопряжена с интенсивным образованием макроэргических со-
единений. Среди них в мозге большую часть составляют АТФ и креа-
тинфосфат, в то время как на долю трифосфатов гуанина, цитозина
и уридина приходится менее 10% всех макроэргов ткани (табл. 3.6).
Важную роль в энергетическом метаболизме мозга играет систе-
ма креатин-креатинфосфат. Высокое содержание обоих компонентов,
Таблица 3.6. Содержание некоторых нуклеотидов, креатина
и креатинфосфата в тканях крыс, мкмоль/гткани
Соединения Головной мозг Печень
Средние данные Кора больших полушарий Мозжечок
АТФ 2,30—2,90 2,08 2,60 2,40—2,80
АДФ 0,30—0,50 0,12 0,16 0,80—1,00
АМФ 0,03—0,05 0,02 0,04 0,15—0,30
ГТФ 0,20—0,30 0,29 0,39 0,19—0,26
ГДФ 0,15—0,20 0,10 0,17 0,18—0,25
УТФ 0,17—0,25 0,22 0,19 0,19—0,25
Креатин 5,50—5,95 5,68 5,47 Следы
Креатинфосфат 3,50—4,75 3,90 4,21 Следы
3.2. Энергетический обмен головного мозга
а также значительная активность креатинкиназы позволяют рассмат-
ривать ее как мощную систему стабилизации уровня АТФ. В головном
мозге до 25—30% активности креатинкиназы сосредоточено в мито-
хондриях, где фермент локализован на внешней мембране. Равнове-
сие катализируемой им реакции сдвинуто в сторону образования кре-
атинфосфата. в отличие от цитоплазматического изофермента.
Вместе с АТФ/АДФ-транслоказой, находящейся на внутренней мемб-
ране митохондрий, креатинкиназа участвует в трансформации макроэрги-
ческих соединений и переносе их из одного клеточного компартмента в
другой. Последняя функция системы креатин-креатинфосфат приобрета-
ет особое значение в нейронах в связи с выраженной гетерогенностью
митохондриальных популяций в этих клетках. Другие возможные аспек-
ты работы системы креатин-креатинфосфат в мозге окончательно не ус-
тановлены. Однако следует отметить, что у мышей с «нокаутом» кре-
атинкиназы обнаружены нарушения пространственного обучения (ори-
ентации в пространстве) при сохранении двигательных навыков.
Стационарная концентрация АТФ в мозге (в среднем 2,3—
2,9 мкмоль • г-1 ткани) существенно не отличается от таковой в других
тканях, несмотря на наличие значительных различий в интенсивности
окислительного и энергетического метаболизма. Однако разница от-
четливо выявляется, если сопоставить данные о скорости образования
и утилизации макроэргических соединений (величина энергетического
потока, energy flux). Определенная с помощью различных методиче-
ских приемов, эта величина составляет в среднем для мозга человека
21, а для мозга крыс — 24—29 мкмоль «макроэргического» Р/г ткани
за 1 мин. В сером веществе мозга данный показатель заметно выше,
чем в белом веществе, и составляет в среднем 33—35 мкмоль «макро-
эргического» Р/г ткани за 1 мин (мозга крыс). Для сравнения можно
привести данные о величинах энергетического потока в других тканях
экспериментальных животных (крыс): сердце — 15,6 (в покое) и 21,4
(при интенсивной нагрузке); почки — 12,2; печень — 9,2 мкмоль «мак-
роэргического» Р/г ткани за 1 мин.
Высокая интенсивность образования и использования макроэрги-
ческих соединений в мозге подтверждается и результатами определе-
ния скорости обновления терминальной фосфатной группы АТФ в
экспериментах с 32Р. Величина tQ 5, установленная в экспериментах с
32Р, составляет в среднем около 3 с, причем для отдельных структур
мозга она может быть еще меньше.
К настоящему времени накоплены многочисленные эксперимен-
тальные данные, неоспоримо доказывающие наличие тесной связи меж-
ду интенсивностью энергетического обмена мозга и его функциональной
27а
г аздел i дИмНАЛСНАЯ НЁЙРОхГ М t Я
активностью. Пионерские работы Г. Мак-Ильвейна, Б. Съешио и дру-
гих нейрохимиков, выполненные еще в середине прошлого века, пока-
зали, что нарушения снабжения мозга кислородом и глюкозой, длящие-
ся даже всего лишь несколько минут, приводят к резким нарушениям
деятельности ЦНС и могут послужить причиной трагического исхода.
Большой вклад в изучение взаимосвязи между энергетическим мета-
болизмом и функциональной активностью уже не целого мозга, а его от-
дельных структур внесен Л. Соколовым с сотрудниками. Эти исследова-
тели впервые применили в своих работах 2-дезоксиглюкозу, предвари-
тельно установив, что она проникает через ГЭБ и используется в
гексокиназной реакции со скоростью, прямо пропорциональной скорос-
ти использования обычной D-глюкозы, дальнейших же метаболических
превращений не претерпевает. Использование радиоактивной (14С или
3Н) 2-дезоксиглюкозы с последующей авторадиографией срезов мозга
показало снижение на 35—60% потребления глюкозы структурами
зрительной системы (зрительная кора, латеральное коленчатое тело,
верхний колликул и др.) после односторонней зрительной депривации.
Контролем в данных экспериментах служили аналогичные структуры
контралатерального полушария того же животного, т. е. структуры, со-
ответствующие нормально функционирующему глазу.
В настоящее время для прижизненного изучения энергетического
метаболизма отдельных структур мозга интактного животного или че-
ловека используют методы позитронно-эмиссионной томографии,
ядерно-магнитного резонанса и др. Сопоставление результатов таких
исследований с данными, полученными при изучении различных сто-
рон метаболизма мозга in vitro, позволяет дать общую оценку энерге-
тических затрат на выполнение тех или иных функций.
Около 25% богатых энергией соединений, образующихся в мозге,
расходуется на поддержание общего метаболизма, т. е. на реакции
биосинтеза различных макромолекул, необходимых для постоянного
обновления структурных элементов клеток. Подобные энергетические
расходы имеют место и в других тканях. Следует лишь подчеркнуть,
что для мозга характерна более высокая по сравнению с другими тка-
нями интенсивность обновления фосфолипидов и встраивания их в
мембраны, а также высокая скорость обновления ряда нейроспецифи-
ческих белков, прежде всего белков синаптических окончаний.
Основная часть образующихся в мозге макроэргических соедине-
ний (около 75%) расходуется на выполнение функций, специфичных
только для нервной ткани. Представление о совокупности таких био-
химических и биофизических процессов дают материалы, приведен-
ные в таблице 3.7. Однако необходимо отметить, что конкретные де-
3.2. Энергетическим обмен головного мозга
Таблица 3.7. Основные энергозависимые процессы,
лежащие в основе специфических функций мозга
Функции Биохимические реакции Энерго- затраты, °О
Восстановление градиента ионов натрия и калия после проведения импульса Na/K-АТФазная реакция До 40
Функционирование синапти- ческих структур Синтез специфических белков, ли- по- и гликопротеидных комплексов синаптических структур; синтез и метаболизм медиаторов; хранение, транспорт, выделение, обратный захват медиаторов; функциониро- вание рецепторов 30—35
Хранение и переработка ин- формации (нейрологическая память) Синтез специфических белков, ней- ропептидов, нуклеиновых кислот, липо- и гликопротеидных комплек- сов Около 20
Аксональный и ретроград- ный ток Гидролиз АТФ при функционирова- нии сократительных белков се- мейств кинезинов, динеинов, ней- ромиозинов 2—3
Поддержание определенной пространственной ориента- ции структурных единиц ней- ронов (аксон, дендриты) Фосфорилирование структурных и сократительных белков 1—2
Примечание. Энергозатраты на выполнение отдельных функций выражены в про-
центах от суммарных (принятых за 100%) затрат макроэргических соединений,
расходуемых на все специфические для мозга функции.
тали биохимических реакций, лежащих в основе некоторых сторон
функциональной активности мозга, еще полностью не ясны. В первую
очередь это относится к такой сложнейшей функции нервной ткани,
как хранение и переработка информации.
Изложенные в данном разделе результаты исследований свиде-
тельствуют о том, насколько велика зависимость функциональной ак-
тивности нервной ткани от интенсивности энергетического обмена,
они объясняют, почему все структуры нервной системы, и прежде все-
го головной мозг, очень чувствительны к недостатку кислорода. Имен-
280
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
3.3.
но гипоксия — один из наиболее губительных факторов, быстро (в те-
чение нескольких минут) приводящий к резким, подчас необратимым
нарушениям функций структур мозга. Фатальной является ситуация,
когда в результате нарушения кровоснабжения мозга одновременно
снижается (или вовсе прекращается) поступление в мозг не только
кислорода, но и основного окисляемого субстрата — глюкозы.
АКСОНАЛЬНЫЙ ТРАНСПОРТ
Кроме нейромедиаторов в синаптических окончаниях нейронов
обнаруживаются и соединения (в основном белки), которые синтези-
руются в теле нейрона. Закономерным является вопрос о способах их
транспортировки по аксону. Первые доказательства наличия такого
транспорта были получены благодаря простому методическому приему:
на периферические нервы накладывали лигатуру, благодаря которой
аксональный транспорт прекращался и происходило набухание аксона
выше наложения лигатуры. Снятие лигатуры приводило к исчезнове-
нию утолщения — накопленные вещества начинали перемещаться
вдоль аксона. Транспорт веществ вдоль аксона получил название ак-
соплазматический ток. Он происходит со скоростью 1 —2 мм в день.
В дальнейшем для исследования этого процесса стали применять ло-
кальное охлаждение аксона, позволяющее обратимо замедлять аксо-
плазматический ток веществ.
Различные компоненты нервной клетки продвигаются по аксону с
различной скоростью (табл. 3.8). Наиболее медленно транспортируют-
ся структурные белки (тубулин) и нейрофиламенты, в то время как ми-
тохондрии и пузырьки, содержащие медиаторы, могут продвигаться со
скоростью до 400 мм в день. Для столь быстрого продвижения по аксо-
ну недостаточно объемного тока аксоплазмы, необходимы механизмы,
значительно ускоряющие продвижение этих частиц. Такое ускоренное
передвижение было названо аксональным транспортом. Белки и
органеллы движутся по аксонам как от тела нейрона к аксональным
окончаниям (антероградный транспорт), так и в обратном направ-
лении (ретроградный транспорт). Именно ретроградный транспорт
необходим для перемещения фактора роста нервов в тело клетки.
Интересно, что таким же способом в нейроны проникают и некоторые
вирусы. Способность активно переносить вещества по аксону использу-
ется для картирования синаптических контактов с использованием спе-
цифических маркеров, наиболее известный из которых — пероксидаза
хрена. Применение пероксидазы хрена в сочетании с флуоресцентными
3.3. Аксональный транспорт
2811
Таблица 3.8. Некоторые характеристики аксонального транспорта
(по: Lasek, Brady, 1982)
Тип аксонального транспорта Ско- рость, мм/день Т ранспорти руем ы й материал
быстрый, I 200—400 Гликопротеиды, гликоли- пиды, белки, медиаторы, ферменты, Са2+
промежуточный
11 50 Митохондриальные белки
ПРЯМОЙ: III 15 Миозиноподобные белки
IV 2—4 Актин, клатрин окаймлен- ных пузырьков, кальмоду- лин, ферменты
медленный, V 0,2—1,0 Белки нейрофиламентов, тубулин, т-белки аксо- плазмы
ОБРАТНЫЙ (ретроградный): быстрый 100—200 Нейроростовые факторы, лизосомальные фермен- ты, трофические вещест- ва, катаболиты
метками позволяет выявить ход отдельных аксонов в нервной системе,
их ветвление и локализацию их тел.
Установлено, что движение органелл осуществляется путем их
прикрепления к микротрубочкам (диаметром 25 нм) и последующего
постепенного перемещения вдоль них с использованием энергии АТФ.
Для микротрубочек характерна полярность, которая важна для рабо-
ты двух белков — кинезина и динеина, осуществляющих соответ-
ственно антероградный и ретроградный транспорт. Присоединение к
каждому из этих белков осуществляется благодаря специфическим
рецепторам на поверхности органелл или везикул.
Каждый из транспортируемых компонентов характеризуется своей
скоростью перемещения. Она в значительной степени зависит от раз-
меров транспортируемых частиц. Различают быстрый и медленный
аксональный (не путать с аксональным током!) транспорт. При
этом в ретроградном направлении не наблюдается медленного аксо-
нального транспорта. Из транспортируемых белков быстро перено-
сятся в основном белки, ассоциированные с мембраной, медленно —
282
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
растворимые белки. Ферменты, регулирующие синтез большинства
классических медиаторов, переносятся с высокой скоростью, скорость
же транспорта митохондрий не очень высока (50—100 мм вдень). Ес-
ли в антероградном направлении наблюдается перемещение функци-
онально активных органелл, то ретроградный транспорт обеспечивает
доставку в тело нейрона поврежденных и функционально неактивных
митохондрий. Компоненты цитоскелета транспортируются в виде со-
бранных полимеров, а не отдельных мономеров.
Рис. 3.5. Сравнение скорости транспорта метаболитов и проведения импуль-
сов в нервных волокнах с другими явлениями.
Точкой отмечен средний уровень скорости указанных процессов (но: Шеперд, 1987)
3.4. Межсинаптические контакты и принципы функционирования синапса ИЩ|
Необходимо учитывать, что такой вид транспорта осуществляется
как в аксонах, так и в дендритах. По сравнению с другими процессами,
происходящими в нервной клетке (рис. 3.5), транспортные процессы
являются очень медленными, и передвижение даже по самым корот-
ким аксонам (длиной порядка 1 мм и меньше) занимает много минут, а
по длинным — несколько часов. Очевидно, именно по этой причине
синтез нейромедиаторов осуществляется в непосредственной близос-
ти от места их использования, туда транспортируются ферменты, не-
обходимые для этого синтеза. Кроме того, с помощью аксонального
транспорта в терминали доставляются все белки, обеспечивающие
работу синапсов, — компоненты синаптических (пре- и постсинапти-
ческих) мембран, синаптических везикул и т. д.
3.4.
МЕЖСИНАПТИЧЕСКИЕ КОНТАКТЫ
И ПРИНЦИПЫ ФУНКЦИОНИРОВАНИЯ СИНАПСА
З.4.1. Этапы синаптической передачи
К настоящему моменту достаточно полно охарактеризована по-
следовательность нейрохимических процессов, происходящих в пери-
од подготовки и осуществления синаптической передачи. Сам процесс
химической передачи можно условно разделить на ряд этапов: синтез
медиатора и его накопление в нервных окончаниях (см. гл. 2.1; 3.1);
процесс высвобождения под действием нервного импульса; диффузия
в синаптической щели; взаимодействие с рецепторами (см. гл. 2.2);
изменения в постсинаптической мембране, приводящие к генерации
постсинаптических потенциалов и включению каскадов химических
реакций в нейроне (см. гл. 2.3; 2.5), а также включение механизмов
удаления различных медиаторов из синаптической щели.
Процесс синаптической передачи запускается нервным импульсом
(рис. 3.6). Синапс, образуемый окончанием аксона, в норме активирует-
ся импульсом, приходящим к этому окончанию, но синапс, образуемый
дендритом, может активироваться градуальными деполяризационными
синаптическими потенциалами внутри этого дендрита. Изменения мемб-
ранного потенциала приводят к открытию потенциалуправляемых каль-
циевых каналов пресинаптической мембраны. В связи с наличием кон-
центрационного градиента их открытие приводит к быстрому входу Са2+
в пресинаптическую терминаль и вызывает резкое увеличение его кон-
центрации в цитоплазме синаптического окончания. Нарастание кон-
центрации [Са2+]; является «триггером» каскада реакций, приводящих
284
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
Приход потенциала
действия в терминаль
Синтез
и накопление
медиатора
Формирование везикул
из плазматической
мембраны
Деполяризация мембраны,
открытие управление Са-каналов
Вход Са2+
через открытые каналы
Опосредованное
Са2+-слияние
везикул
с пресинаптической
мембраной
Экзоцитоз медиатора
и его диффузия
в синаптической щели
Возникновение возбуждающих
или тормозных постсинаптических
потенциалов, изменение
возбудимости постсинаптического
нейрона
Связывание медиатора
с рецепторами
постсинаптической
мембраны
Рис. 3.6. Последовательность событий, происходящих в синапсе (а), и ско-
рость рециркуляции синаптических везикул (б)
3.4. Межсинаптические контакты и принципы функционирования синапса
285
к передвижению синаптических пузырьков к пресинаптической мемб-
ране, слиянию мембран везикул с синаптической мембраной и в ко-
нечном счете к выбросу медиатора в синаптическую щель путем экзо-
цитоза.
Следует отметить, что если для высвобождения классических нейро-
медиаторов достаточно одного импульса, то для высвобождения нейро-
пептидов требуется целая серия импульсов. Для высвобождения нейро-
пептидов необходимо не локальное повышение концентрации Са2+
вблизи мембраны, а более значительное общее повышение его кон-
центрации в пресинаптическом окончании, что достигается при более
высокочастотной стимуляции. После попадания в синаптическую щель
медиатор диффундирует к постсинаптической мембране, где связывает-
ся с рецепторами. Связывание медиатора с рецептором вызывает изме-
нение состояния каналов на постсинаптической мембране — открытие
или закрытие в зависимости от медиатора и изменение потока ионов
внутрь или из постсинаптической клетки, в результате чего изменяется
мембранный потенциал постсинаптического нейрона (снижается или
повышается). При достижении критического порога деполяризации та-
кое изменение может привести к генерации потенциала действия в
постсинаптическом нейроне. На этом процесс передачи информации в
синапсе не заканчивается, поскольку для его нормального функциони-
рования необходимо очистить синаптическую щель от медиатора, а так-
же синтезировать и накопить его новые порции в везикулах.
Последовательные этапы процесса выделения медиатора требуют
времени. В нервно-мышечном соединении минимальная синаптиче-
ская задержка между началом пресинаптической деполяризации и
постсинаптической реакцией составляет около 0,5 мс. Относительно
длительное время синаптической задержки в химическом синапсе за-
трачивается на вход Са2+ в синаптическое окончание, экзоцитоз, диф-
фузию медиатора. Только !/10 ее часть (50 мкс) можно отнести на счет
времени диффузии через синаптическую щель. Синаптическую задерж-
ку часто используют при анализе синаптических сетей.
3.4.2. Квантовое выделение медиатора
Выделение молекул медиатора квантами было обнаружено в не-
скольких типах синапсов, и в настоящее время считается, что такой ме-
ханизм представлен во всех химических синапсах. Первоначально дока-
зательство существования такого выделения было получено Б. Кацем в
середине прошлого века. Экспериментируя с нейромышечным препа-
ратом (удобной моделью, поскольку синапсы между нейроном и мы-
286
ег ЗД. I 3 4 > НКЦИоН ЪНАь HEHF4 ХИМЛЯ
шечной клеткой имеют достаточно большие размеры и локализуются
на периферии), он изучал потенциалы концевой пластинки (ПКП), од-
ной из разновидностей ПД (см. 3.5.2). Стимуляция нерва приводила к
деполяризации постсинаптического мышечного волокна и формиро-
ванию ПКП. Однако было обнаружено, что наряду с такими потенциа-
лами (40—50 мВ) возникают и потенциалы с более низкой амплиту-
дой (менее 1 мВ), названные миниатюрными потенциалами кон-
цевой пластинки (МПКП).
Оба типа потенциалов оказались чувствительными к кураре —
блокатору ацетилхолиновых рецепторов. В результате постепенного
снижения концентрации Са2+ в среде было установлено, что амплиту-
да ПКП постепенно приближается к таковой у МПКП. Был сделан
вывод, что ПКП возникает в результате одновременного синхронного
высвобождения квантов медиатора. Одна везикула в мотонейроне
диаметром ~50 нм содержит около 10 000 молекул АХ. Выделение од-
ного кванта приводит к формированию МПКП.
3.4.3. Превращения синаптических везикул
в синаптическом окончании
Все этапы круговорота синаптических пузырьков (везикул)
контролируются белками, встроенными как в их мембрану, так и в
пресинаптическую мембрану, а также цитоплазматическими белками.
Среди них можно выделить следующие белки: Са-связывающие;
SNARE-ассоциированные; вовлекаемые в эндоцитоз; формирующие
каналы, транспортеры или рецепторы; ГТФ-связывающие белки. Все
они участвуют в одной или нескольких стадиях рециркуляции синапти-
ческих пузырьков. Так, два из них играют важную роль в слиянии ве-
зикул с аппаратом Гольджи, а также в подготовке везикул к слиянию
с пресинаптической мембраной — цитоплазматическая АТФаза NSF
(NEM-sensitive fusion protein), а также SNAP (soluble NSF-attach-
ment proteins). Работа этих белков контролируется SNARE-белками
(SNAP receptors). Один из представителей этой группы (синаптобре-
вин) находится в везикулярной мембране, в то время как два других
(синтаксин и SNAP-25) — в плазматической. Благодаря этим белкам
формируется комплекс, обеспечивающий контакт двух мембран.
Способность белков плазматической мембраны синтаксина и
SNAP-25 к связыванию кальциевых каналов и SNARE может играть
роль в колокализации кальциевых каналов и мест выброса медиатора.
Такие мембранные белки, как снапин, синтафилин и томозин, а также
цитоплазматический белок комплексин, связываясь с ними, регулиру-
3.ские нгакты . принципы фу акционирования синапса
287]
ют сборку или разрушение этого комплекса. SNARE-Белки не спо-
собны связывать кальций и, соответственно, реагировать на повыше-
ние его уровня в цитоплазме. Белком-сенсором, который регулирует
Са-зависимое выделение медиатора из везикул, является синаптотаг-
мин, присутствующий в везикулярной мембране. Кальциевый канал и
синаптотагмин также могут связываться друг с другом таким образом,
что синаптотагмин оказывается возле открытых каналов на участке
повышенной концентрации кальция. Связываясь с кальцием в опре-
деленном интервале концентраций, он изменяет степень своего взаи-
модействия с белками синаптических пузырьков и плазматической
мембраны. У мутантных животных с отсутствием гена, кодирующего
синаптотагмин, хотя и наблюдается повышенное спонтанное высво-
бождение медиатора, стимуляция аксона не оказывает эффекта или
вызывает лишь незначительное его высвобождение. Каким образом
происходят взаимодействие синаптотагмина с другими белками и его
участие в процессе слияния мембран, пока до конца не выяснено.
Накопление медиаторов в синаптических пузырьках осуществляется
специфическими транспортными белками. Обнаружено и описано ч е-
тыре везикулярных транспортера: транспортер всех био-
генных моноаминов (в английской транскрипции VMAT), переносчик
ГАМК и глицина, транспортер ацетилхолина (сокращенно называемый
VAChT) и транспортер глутамата. Каждый из них представляет собой ин-
тегральный мембранный белок, имеющий 12 трансмембранных доменов.
Мембраны синаптических везикул содержат также специальную АТФазу,
которая перекачивает протоны внутрь пузырьков, создавая в них поло-
жительный заряд и более кислую реакцию среды по сравнению с цито-
плазмой. Для того чтобы перемещать молекулы медиатора внутрь пу-
зырьков против градиента концентрации, везикулярные транспортеры
медиаторов используют энергию электрохимического и pH-градиентов.
Наиболее исследованным путем круговорота везикул является
путь классического эндоцитоза, при котором мембрана пузырька пол-
ностью сливается с мембраной в процессе экзоцитоза, и компоненты
везикулярной мембраны возвращаются обратно путем эндоцитоза в
виде фрагментов, покрытых клатрином (см. рис. 3.6). В формирова-
нии везикул из плазматической мембраны путем эндоцитоза важную
роль играет клатрин, молекулы которого благодаря своей структуре,
присоединяясь к мембране, формируют специфическую структуру ве-
зикулы. В этом процессе также участвуют вспомогательные цитоплаз-
матические белки (амфифизин, АР2, АР180). Конечный этап превра-
щения впячивания в полноценную везикулу и разрыва двух мембран
происходит с участием динамина и кальцийзависимой цитоплазмати-
288
раздел X ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
ческой ГТФазы, которая отщепляет покрытые клатрином пузырьки от
плазматической мембраны. Удаление клатриновой оболочки осу-
ществляется при участии специфической АТФазы (Hsp70) и цито-
плазматических белков ауксилина и синаптоянина. Вновь образован-
ные везикулы обратимо связываются с цитоскелетом с помощью си-
напсина. Фосфорилирование синапсина протеинкиназами приводит к
его отделению от везикулярной мембраны. Таким образом, в нормаль-
ных условиях активность эндогенных киназ и фосфатаз, регулируя
фосфорилирование синапсинов, может модулировать количество ме-
диатора, высвобождаемого из аксонной терминали.
Существует также гипотеза о том, что везикулы отсоединяются
сразу же после высвобождения их содержимого без слияния с плазма-
тической мембраной. Такая схема экзоцитоза может иметь место в си-
напсах, где необходимо выделение медиатора из небольшого числа пу-
зырьков, но с большой частотой.
Изучение аксонных терминалей моторных и симпатических нейро-
нов млекопитающих показало, что высвобождаются преимуществен-
но вновь синтезированные молекулы медиатора. Такие результаты да-
ют возможность предположить, что существует субпопуляция пузырь-
ков, которые подвергаются быстрому круговороту, в то время как
большинство везикул находится в резерве.
3.4.4. Удаление медиаторов из синаптической щели
Заключительным этапом передачи сигнала в химическом синап-
се является удаление медиатора из синаптической щели. В зависимос-
ти от природы медиатора механизмы этого процесса могут несколько
отличаться, однако в большинстве случаев происходит распад нейро-
медиатора под действием соответствующих ферментов и захват самого
медиатора или продуктов его распада глиальными клетками (астроци-
тами) или нервными окончаниями с дальнейшим использованием для
синтеза новых молекул медиатора.
Наиболее простой способ удаления медиатора — его
распад. Именно так из синаптической щели удаляется АХ, который
под действием ацетилхолинэстеразы (АХЭ) гидролизуется до холина и
ацетата. Значительная часть холина транспортируется обратно в нерв-
ное окончание, где используется для синтеза АХ. Для сравнения следу-
ет сказать, что если в постсинаптической мембране насчитывается око-
ло 104 рецепторов ацетилхолина на 1 мкм2, то концентрация каталити-
ческих субъединиц фермента составляет около 2600. Для того чтобы
связаться с рецептором, молекулы АХ должны пересечь пространство,
3.4. Межсинаптические контакты и принц ипу Функ! |Иони^ ования синапса
289
«заполненное» АХЭ. Поэтому доля выброшенных из нервного оконча-
ния молекул АХ, которые взаимодействуют, прежде всего, с постсинап-
тическими рецепторами, определяется отношением числа рецепторов к
числу молекул АХЭ. Приблизительно 20% молекул АХ связываются с
ферментом и 80% — с рецепторами. Связывание АХ с АХЭ и рецепто-
рами, а также его расщепление происходят очень быстро — в течение
0,1 мс. Концентрация АХ остается низкой из-за того, что фермент
может расщеплять АХ значительно быстрее (10 молекул/мс), чем он
высвобождается рецептором после закрытия ионных каналов.
Аналогично путем гидролиза прерывается действие АТФ — вне-
клеточная АТФ-дифосфогидролаза последовательно превращает его в
АДФ и АМФ, который, в свою очередь, превращается в аденозин под
действием фермента экто-5'-нуклеотидазы. Образующийся аденозин
подвергается действию аденозиндезаминазы, расщепляющей его до
инозина. Нейропептиды под влиянием пептидаз также расщепляются
до аминокислот, которые затем поступают в клетки.
Другим важным способом, позволяющим удалять медиатор
из синаптической щели, является его обратный захват (up-
take ) пресинаптическими нервными окончаниями
и/или глиальными клетками (астроцитами). Такой способ
характерен для дофамина, норадреналина, серотонина, глутамата, гли-
цина и ГАМК. Норадреналин, дофамин и серотонин после обратного
захвата в синаптические окончания могут быть опять упакованы в ве-
зикулы или окислены МАО. В случае ГАМК и глутамата наблюдается
их слабый захват в синаптические окончания, но активная переработка
в астроцитах с последующим транспортом в синаптические окончания
глутамина, из которого могут быть получены новые молекулы глутама-
та или ГАМК(см. рис. 2.14). Те медиаторы, которые транспортируются
в пресинаптические нервные окончания, могут быть вновь упакованы
в везикулы и использованы для синаптической передачи.
К настоящему времени известно два семейства белков, осуществ-
ляющих транспорт медиаторов через мембрану. Процесс захвата при
помощи специальных белковых транспортеров требует энергетических
затрат. Транспортеры норадреналина, дофамина, серотонина, ГАМК и
глицина состоят из 12 трансмембранных доменов и являются натрий- и
хлорзависимыми. Глутамат и аспартат захватываются с помощью бел-
ков, являющихся натрийзависимыми и имеющих 10 трансмембранных
доменов. Для транспорта используется энергия натриевого электрохи-
мического градиента, но стехиометрические параметры транспорта раз-
личаются. Например, с каждой молекулой глутамата внутрь клетки по-
ступает два-три иона натрия и один ион калия выводится наружу.
290
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
Транспортеры, переносящие медиаторы в нервные окончания (за
исключением серотонинового), отличаются от переносящих их в синап-
тические пузырьки. Соответственно, для каждого из них существуют
фармакологические специфические вещества-блокаторы. Так, кокаин
блокирует захват норадреналина в нервную терминаль, а резерпин —
в везикулы.
ЭЛЕКТРИЧЕСКИЕ И ХИМИЧЕСКИЕ МЕХАНИЗМЫ
ПЕРЕДАЧИ ИНФОРМАЦИИ
3.5.
3.5.1. Потенциал покоя
Еще в начале прошлого века было установлено, что между
внешней и внутренней поверхностями мембраны клетки устанавлива-
ется разность электрических потенциалов, связанная с неравномерным
распределением ионов на внешней и внутренней сторонах клеточной
мембраны. В состоянии покоя на мембране нейронов также поддержи-
вается постоянный электрический потенциал, при этом внутриклеточ-
ная среда заряжена отрицательно по отношению к внеклеточной.
Поскольку внутри клетки больше ионов калия и меньше ионов нат-
рия и хлора, чем снаружи, то каждый из этих ионов стремится проник-
нуть сквозь мембрану в направлении своего электрохимического гра-
диента. В состоянии покоя ионы натрия непрерывно перемещаются
внутрь клетки, снижая таким образом отрицательный заряд на мемб-
ране, а калий выходит из клетки. Если бы не механизмы компенсации,
то такое перемещение натрия и калия привело бы к изменению их вну-
триклеточных концентраций. Этого не происходит благодаря работе
Na/K-насоса (см. гл. 2.5).
В связи с наличием градиента концентрации скорость диффузии
ионов калия из клетки выше, чем в клетку. При этом оставшиеся в клет-
ке анионы, для которых мембрана непроницаема, создают суммарный
отрицательный заряд. Процесс диффузии продолжается до тех пор, пока
возникший на внутренней стороне мембраны отрицательный заряд не
начинает сдерживать диффундирующие из клетки, положительно заря-
женные ионы калия. В результате устанавливается динамическое рав-
новесие ме>кду количеством ионов калия, проходящих через мембрану в
обоих направлениях. Каждому значению градиента концентрации, т. е.
соотношению [К+]нар/[К+]вн, соответствует равновесный электростати-
ческий потенциал, определяемый уравнением Нернста:
Ек+ = RT\n Fх In ([К+]нар/[К+]В11).
3.5. Электрические и химические механизмы передачи информации ЙЙ&
Подставив соответствующие константы и перейдя к десятичным
логарифмам, получим:
Ек+ = ~ 58 lg([K+]Hap/[K+]BH).
Аналогичным образом можно рассчитать равновесный потенциал
для других ионов.
Для исследования электрических явлений в нервной ткани была
необходима экспериментальная модель. Основным требованием к та-
кому объекту были размеры, которые позволяли бы вводить микро-
электроды для регистрации электрических явлений, а также произво-
дить манипуляции с изменением внутриклеточной среды. Этим усло-
виям соответствовали гигантские аксоны кальмара, диаметр которых
достигает 0,5 мм. Учитывая, что для гигантского аксона кальмара
концентрации калия соотносятся как 1 : 20, из уравнения Нернста по-
лучим значение потенциала, равное —75 мВ; однако микроэлектрод,
введенный в клетку, регистрирует по отношению к внешней среде
мембранный потенциал, равный —70 мВ. Различия в значениях вы-
званы тем, что в действительности этот потенциал возникает не толь-
ко благодаря ионам калия.
Мембрана в определенной степени проницаема и для ионов натрия,
а поскольку для концентрации Na+ характерен противоположный гра-
диент по сравнению с градиентом концентрации К+, их поток, направ-
ленный в клетку, уменьшает измеряемый потенциал мембраны. По-
скольку ионы С1“ также могут диффундировать через мембрану, для
полного описания мембранного потенциала необходимо принимать во
внимание изменение концентраций трех ионов. Для этих ионов прони-
цаемость клеточной мембраны различается, в уравнение Нернста сле-
дует подставлять не их концентрации внутри и снаружи клетки, а эти
значения с учетом коэффициентов проницаемости (PCi, PNa, Рк). Необ-
ходимо также учитывать и тот факт, что ионы С1_ имеют отрицательный
заряд. Тогда мембранный потенциал Ет будет определяться формулой
PK[K+]„ap + PNa[Na+]„ap + Pcl[Cl-]n„
m g W+ ]вн + /WNa+L + ^С1[С1-1иар •
Предложенное Гольдманом данное уравнение сводится к уравнению
Нернста при условии, что рассматривается только один ион. Мембран-
ный потенциал нейрона практически эквивалентен К+-потенциалу, что
связано с низкой способностью Na+ проникать через мембрану в со-
стоянии покоя. Ионы хлора, хотя и проникают через мембрану по ка-
лиевому потенциалу, обладают отрицательным зарядом, а значит, не
участвуют в создании суммарного мембранного потенциала.
292
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
Постоянный отрицательный потенциал называют мембранным по-
тенциалом покоя. На его основе формируются все остальные элект-
рические потенциалы нервной клетки. Уменьшение степени поляриза-
ции мембраны (смещение мембранного потенциала к нулю и далее в
сторону положительных значений) называется деполяризацией, а уве-
личение степени поляризации мембраны — гиперполяризацией.
3.5.2. Локальный потенциал и потенциал действия
Рассмотренные выше электрические свойства нервной клетки
характерны для состояния покоя. Внешние стимулы, как электриче-
ские, так и химические, способны приводить к временному отклонению
мембранного потенциала от значений потенциала покоя — возбуждать
мембрану. При этом меняется ее ионная проводимость. Возбуждение
распространяется от стимулированного участка к близлежащим облас-
тям, в которых наблюдается изменение ионной проводимости, а следо-
вательно, и мембранного потенциала. Такое распространение (генера-
ция) возбуждения называют импульсом. В зависимости от того, рас-
пространяется ли импульс от участка возбуждения неизменным или
быстро уменьшается по мере удаления от него, различают потенциал
действия и локальный (или электротонический) потенциал.
При использовании гигантского аксона кальмара локальный по-
тенциал можно создать введением положительных зарядов, которые
вызовут деполяризацию мембраны. От места возникновения импульс
распространится на расстояние, определяемое внутренним сопротив-
лением аксона. Как у любого проводника, его сопротивление обратно
пропорционально диаметру, т. е. тонкие аксоны хуже проводят ло-
кальный потенциал, чем аксоны с большим диаметром. Передача ло-
кального потенциала зависит и от сопротивления мембраны аксона.
Поскольку мембрана — не очень хороший изолятор, часть введенных
положительных зарядов (например, ионов калия) вытекает наружу,
препятствуя деполяризации мембраны. G увеличением расстояния от
места введения импульс ослабевает.
Сравнение скорости проведения импульса в физических проводни-
ках и в нервных волокнах показало, что в последних она значительно
ниже: в крупных нервных волокнах лягушки составляет около 40 м/с.
Этот факт имеет первостепенное значение, так как из него следует, что
передача нервного импульса — это активный биологический про-
цесс, а не просто физическое проведение электричества по проводнику.
Возбудимостью и способностью генерировать импульсы обладают не
только нервные клетки. Как уже отмечалось, свойством возбудимости
3.5. Электрические и химические механизмы передачи информации
обладает скелетная мышца, а также мышца сердца. К генерации им-
пульсов способны самые различные клетки животных и даже растений.
Деполяризовать мембрану можно с помощью электрического тока.
При этом, если мембрана была деполяризована кратковременно и не-
значительно, наблюдается слабый эффект, и после выключения депо-
ляризующего тока мембранный потенциал быстро возвращается к ис-
ходному состоянию. При более интенсивной деполяризации измене-
ние потенциала до —40 мВ приводит к необратимому эффекту и
мембранный потенциал не возвращается к потенциалу покоя, а на-
блюдается спонтанно усиливающаяся деполяризация до тех пор, пока
знак потенциала не изменяется на противоположный. При достиже-
нии значения потенциала, равного приблизительно +40 мВ, происхо-
дит реполяризация. При этом мембранный потенциал на некоторое
время становится более электроотрицательным, чем потенциал по-
коя, и лишь затем медленно возвращается к начальному уровню.
В середине прошлого века английские физиологи А. Ходжкин и
А. Хаксли исследовали потенциал действия и заложили основы совре-
менного понимания механизмов его возникновения. Они показали,
что первоначально падение потенциала (деполяризация мембраны)
обусловлено «утечкой» ионов натрия. По достижении порогового зна-
чения ионные каналы в мембране открываются и пропускают ионы
натрия. Последующая реполяризация происходит благодаря откры-
тию специальных калиевых каналов и устремлению потока ионов ка-
лия в обратном направлении, т. е. изнутри наружу; одновременно про-
исходит инактивация (закрывание) натриевых каналов. При реполя-
ризации значения мембранного потенциала больше, чем при
потенциале покоя. Это различие медленно исчезает в результате за-
крывания калиевых каналов и восстановления натриевого потенциала
покоя. Инактивация каналов подразумевает, что следующий потенци-
ал действия не может реализоваться немедленно. Период покоя между
двумя потенциалами действия называют рефрактерным периодом.
Для выяснения механизмов возникновения потенциала действия
Ходжкин и Хаксли измеряли мембранные токи при пространственной
фиксации потенциала (импульс не распространяется по аксону, а удержи-
вается на необходимом уровне на определенном участке). Фиксировались
также изменения мембранных токов при уменьшении концентрации ионов
в омывающий аксон кальмара среде. Важным выводом явилось то, что
ионы натрия во внеклеточной среде необходимы для возникновения ПД и
при их отсутствии генерация ПД невозможна. Удаление ионов натрия из
среды приводило к тому, что наблюдался лишь более поздний выходящий
компонент тока, который увеличивался по мере уменьшения мембранного
потенциала. Выраженность этого компонента усиливалась с увеличением
294
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
деполяризации мембраны, т. е. по мере возрастания разницы между мемб-
ранным потенциалом и равновесным калиевым потенциалом. В то же вре-
мя отсутствие ионов калия во внутренней среде изменяет структуру ПД.
Вычитание позднего компонента из суммарной кривой ПД позволило по-
лучить кривую раннего компонента, который выявлялся при небольшом
уровне деполяризации. Максимальные его значения наблюдались при
мембранном потенциале, близком к 0, а при потенциале около +55 мВ он
менял свое направление. Эти выводы были подтверждены в дальнейших
экспериментах с использованием ядов, избирательно блокирующих на-
триевые и калиевые каналы.
На рисунке 3.7 представлена схема, иллюстрирующая изменение на-
триевой и калиевой проводимости и процесса деполяризации мембраны
(рис. 3.7, а), а также динамика вышеописанных процессов (рис. 3.7, б).
Для натриевой проводимости и степени деполяризации мембраны сущест-
вует положительная обратная связь, а для калиевой проводимости — от-
рицательная обратная связь. При этом скорость в этих двух циклах разли-
чается. При деполяризации натриевая проводимость быстро возрастает,
что увеличивает исходную деполяризацию мембраны, и в результате на-
триевая проводимость еще больше повышается. Благодаря этому свойству
потенциал действия обладает порогом и вызывает короткую рефрактер-
ность(т. е. невосприимчивость нервного волокна к новому раздражению), а
также характеризуется постоянством амплитуды при распространении на
б
Вход Na+
Усиление
деполяризации
мембраны х
а
Быстрый цикл,
положительная
связь
Открытие
Na-каналов
Мембранный
потенциал
Открытие
К-каналов
Медленный цикл,
отрицательная
г связь
Гиперполяризация
мембраны
Выход К+
Рис. 3.7. Деполяризация мембраны и входящий ток Na+ :
а — взаимосвязь между деполяризацией мембраны, увеличением натриевой проница-
емости и входящим током Na+, а также замедленным открытием калиевых каналов,
увеличением тока К+ и гиперполяризацией мембраны; б — кривые, характеризующие
изменение количества открытых Na- и К-каналов в зависимости от мембранного по-
тенциала.
3.5. Электрические и химические механизмы передачи информации
295
большие расстояния. Деполяризация же мембраны включает другой, более
медленный процесс — нарастающее увеличение калиевой проводимости,
причем в данном случае обнаруживается корреляция по типу отрицатель-
ной обратной связи. Именно эти нелинейные, регенеративные взаимоотно-
шения, благодаря которым натриевая и калиевая проводимости последова-
тельно (а не одновременно) увеличиваются, обусловливают различия меха-
низмов потенциала действия и синаптического потенциала.
3.5.3. Проведение потенциала действия
Распространение потенциала действия вдоль аксона обеспечи-
вается благодаря потенциалзависимым натриевым и калиевым кана-
лам (см. гл. 2.5). При потенциале действия деполяризация вызывает
открывание ионных каналов и, следовательно, вход Na+ в соседние
области мембраны (рис. 3.8). Это приводит к дальнейшей деполяриза-
ции мембраны и открыванию ионных каналов в близлежащих мемб-
ранных зонах.
Возникший сигнал распространяется в двух направлениях от точки
возникновения — к клетке и к окончанию аксона. Расстояние, на ко-
торое распространится этот ток по волокну, определяется соотноше-
нием сопротивления цитоплазмы и мембраны. Повышение сопротив-
ления мембраны, равно как и снижение сопротивления цитоплазмы,
приводит к тому, что ток будет распространяться на более длинные
Рис. 3.8. Проведение нервного импульса в немиелинизированном (а) и миели-
низированном (б) нервном волокне
296
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
расстояния. Скорость распространения возбуждения по волокну зави-
сит от его диаметра: при прочих равных условиях она тем выше, чем
больше диаметр волокна.
Подобное распространение нервного импульса от одного участка
мембраны к другому характерно для немиелинизированных нервных во-
локон. Таким образом, одним из способов увеличения скорости проведе-
ния возбуждения является увеличение диаметра волокна. Именно поэто-
му у некоторых беспозвоночных животных отдельные волокна бывают
очень крупными (например, гигантский аксон кальмара). У позвоночных
же немиелинизированные волокна принадлежат к наиболее мелким —
их диаметр колеблется от нескольких микронов до 0,2 мкм. Та же
задача — ускорение проведения сигнала — у них решается благодаря
использованию другой, более эффективной стратегии — передаче нерв-
ного импульса по миелинизированным волокнам.
Миелинизированные волокна (см. гл. 1.1) покрыты многослойной,
периодически прерывающейся оболочкой. Участки аксона, в которых
миелиновая оболочка прерывается, называются перехватами Ранвье.
Эти участки аксона имеют чрезвычайно высокую плотность натриевых
каналов: в них она достигает 12 000 на 1 мкм2. Ни в каких других отде-
лах нервной системы столь высокой плотности каналов не обнаружено.
Нервные импульсы в миелинизированных волокнах генерируются не на
всем протяжении, а только в перехватах Ранвье. Поскольку в участках,
покрытых слоями миелина, потенциалзависимых каналов мало, между
перехватами возможно лишь электротоническое распространение нерв-
ного импульса.
Благодаря наличию перехватов Ранвье нервный импульс как бы «пе-
рескакивает» от одного перехвата к другому — подобное проведение
называют сальтаторным. Такой тип проведения импульса весьма эф-
фективен, так как благодаря ему достигается максимальная скорость
проведения при минимальной площади активной мембраны (в районе
перехвата). При этом отпадает необходимость в увеличении диаметра
волокна и на проведение импульса затрачивается меньше энергии.
3.5.4. Постсинаптические потенциалы
и интеграция синаптических влияний
В синапсах и сенсорных нервных окончаниях наблюдаются
локальные потенциалы. В сенсорных нервных окончаниях
их называют рецепторными, или генераторными, потенциалами.
В отличие от ПД, который возникает по принципу «все или ничего»,
локальные потенциалы могут суммироваться, т. е. они способны увели-
3.5. Электрические и химические механизмы передачи информации
29
чиваться при последующих возбуждениях. В зависимости от того, при-
ближают ли синаптические потенциалы мембранный потенциал к по-
роговому значению (способствуют генерации ПД) или наоборот, раз-
личают возбуждающие постсинаптические потенциалы (ВПСП) и
тормозные постсинаптические потенциалы (ТПСП).
ВПСП в постсинаптической мембране возникают в ответ на воздей-
ствие такого медиатора, как ацетилхолин, вызывающего деполяриза-
цию мембраны. В случаях некоторых нейромышечных синапсов дан-
ный потенциал называют потенциалом концевой пластинки (ПКП).
Подобно локальному потенциалу, величина ВПСП быстро падает с
увеличением расстояния от места его возникновения и амплитуда за-
висит от концентрации медиатора, т. е. если концентрация ацетилхо-
лина увеличивается, деполяризация также усиливается. При достиже-
нии определенной величины деполяризации возникает ПД. Деполяри-
зация, вызываемая ацетилхолином, возникает в результате открытия
ионных каналов. Важно отметить, что эти каналы принципиально
отличаются от электровозбудимых каналов, обус-
ловливающих ПД, — они открываются под влиянием молекулы
медиатора. Эти каналы отличаются по своей ионной специфичности.
Так, ПД генерируется двумя различными потенциалзависимыми ион-
ными каналами (раздельно натриевыми и калиевыми), а активируе-
мый ацетилхолином ионный канал, формируемый рецептором АХ,
практически одинаково проницаем для ионов натрия и калия.
При ТПСП происходит гиперполяризация постсинаптической
мембраны — мембранный потенциал смещается в сторону более от-
рицательных значений. Такой результат может быть достигнут либо
закрыванием катионных каналов, либо открыванием анионных кана-
лов. Подобный процесс называется тормозным, так как он подавляет
возбудимость мембраны и таким образом ингибирует или тормозит де-
поляризующий ответ Na- и К-каналов и, соответственно, способность
генерировать ПД.
Идея о том, что управление мембранным потенциалом в различных
частях нейрона осуществляется главным образом путем взаимодейст-
вия между возбуждающими и тормозными синаптическими влияния-
ми, была впервые высказана Ч. Шеррингтоном. Позже такое взаи-
модействие стали называть интеграцией синаптических влияний.
Для понимания некоторых принципов этой интеграции удобно рас-
смотреть взаимодействие между одиночными ВПСП и ТПСП. Если
рядом с возбуждающим синапсом на нейроне расположен тормозный,
их активация приводит к генерации соответственно ВПСП и ТПСП.
298
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕИРОХИМИЯ
Если оба синапса возбуждаются одновременно, то под влиянием
ТПСП величина ВПСП уменьшается — мембранный потенциал в
меньшей степени приближается к пороговому уровню, чем при оди-
ночном ВПСП (рис. 3.9, а).
Широко распространенная точка зрения, согласно которой интег-
рация синаптических влияний заключается в простом алгебраическом
сложении противоположных синаптических потенциалов, не в полной
мере характеризует сложные взаимодействия между возбуждающими
и тормозными влияниями. Их взаимодействие в значительной степени
зависит также и от потенциала покоя мембраны. При любом значении
потенциала покоя ТПСП вызывает такие изменения ионных потоков,
при которых величина ВПСП уменьшается; при этом полярность
ТПСП может изменяться в зависимости от соотношения между вели-
чинами мембранного потенциала, с одной стороны, и £к и £С] — с дру-
гой. Так, если потенциал покоя равен равновесному потенциалу для
ТПСП (рис. 3.9, б), последний не регистрируется, а величина возни-
кающего одновременно с ним ВПСП снижается, что обусловлено
шунтирующим влиянием каналов, ответственных за тормозный потен-
циал. Если же потенциал покоя еще выше (рис. 3.9, в), то ТПСП пре-
вращается в деполяризующий потенциал — мембранный потенциал
-65
-70
-75
-80
а
мВ
ТПСП
Рис. 3.9. Интеграция ВПСП и ТПСП при различных уровнях мембранного
потенциала:
а — в — различные уровни мембранного потенциала клетки. ВПСП обусловлен сум-
марным входящим током положительных зарядов (выход К+ и вход Na+), в результате
чего мембранный потенциал смещается по направлению к равновесному потенциалу;
ТПСП вызван суммарным выходящим током положительных зарядов (выход К+ или
вход С1~), что приводит к удержанию мембранного потенциала на уровне покоя или не-
которой гиперполяризации мембраны. ТПСП вызывает изменения ионных проница-
емостей, при которых величина ВПСП уменьшается (пунктирная линия). Поляр-
ность ТПСП может изменяться в зависимости от мембранного потенциала и проводи-
мости К+ и С1~. Прерывистой линией на рисунке показано результирующее изменение
потенциала, характеристики которого определяются интеграцией обоих синаптических
потенциалов
3.6. Сенсорные процессы
299
стремится к равновесному потенциалу для ТПСП. Однако даже в этом
случае амплитуда ВПСП уменьшается, так как увеличивается прони-
цаемость каналов тормозного потенциала. Таким образом, тормозное
действие состоит не столько в гиперполяризации мембраны, сколько в
увеличении проницаемости для ионов, равновесный потенциал кото-
рых достаточно высок; при этом мембранный потенциал приближает-
ся к этому равновесному потенциалу.
Использование в нервной системе электрических сигналов для пе-
редачи информации не решало бы при отсутствии химических синап-
сов одну из важнейших ее задач — оценки и переработки этой инфор-
мации. Синапсы позволяют нейронам производить кодирование-деко-
дирование поступающей и отправляемой ими информации. На уровне
синапса осуществляется оценка информации и, в случае необходимос-
ти, значительное усиление (амплификация) сигнала с использованием
соответствующих химических каскадов (см. 2.4.1).
СЕНСОРНЫЕ ПРОЦЕССЫ
3.6.
з.в.1. Передача сигнала в фоторецепторных клетках сетчатки
Фоторецепторные молекулы воспринимают в качестве сигнала
квант света. Принципы передачи сигнала через мембрану у фоторецеп-
торов и у нейрональных клеток достаточно сходны. Зрительный родоп-
син локализован у большинства позвоночных в специализированных
фоторецепторных клетках двух типов — палочках и колбочках, высти-
лающих внутреннюю сторону сетчатки глаза. Палочки функционируют
в условиях слабого освещения. Они очень чувствительны к световым
сигналам, и при сильном освещении десенсибилизируются. При ярком
свете зрительный процесс обеспечивается колбочками. По-видимому,
именно колбочки отвечают за объемное изображение, реагируют на
перемещение предметов, передают цветовую гамму изображения.
В колбочках содержатся пигменты, поглощающие в различных об-
ластях спектра. В сетчатке человека различают три вида колбочек, по-
глощающих свет в коротковолновой, средней и длинноволновой облас-
тях видимого спектра. Именно эти различия в свойствах пигментов
колбочек лежат в основе цветного зрения. При слабом освещении кол-
бочки не принимают сигнала, и глаз не воспринимает цвета объектов.
Рецепторная мембрана палочек состоит из замкнутых дисков, не
соприкасающихся с цитоплазматической мембраной, в то время как в
колбочках она образует систему складок. Таким образом, в палочках
300
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕИРОХИМИЯ
для передачи сигнала от диска к плазматической мембране необходим
гидрофильный посредник.
Как палочки, так и колбочки условно делят на два сегмента:
наружный и внутренний. Наружный сегмент содержит фоторецептор-
ные мембраны, а внутренний специализируется на генерации энергии
и содержит аппарат, синтезирующий молекулы вторичных посредников,
участвующих в передаче сигнала. Из внутреннего сегмента формируется
синаптическое окончание, осуществляющее передачу сигнала к нерв-
ным волокнам. Наружный сегмент легко отделяется от внутреннего при
гомогенизации, что позволяет с помощью дифференциального центри-
фугирования выделять фоторецепторные мембраны в чистом виде.
Зрительный родопсин (как и бактериородопсин) содержит в активном
центре 11-ф/с-ретиналь. Белковую часть родопсина без ретиналя назы-
вают опсином, она функционирует как фермент. Первичные структуры
зрительного родопсина и бактериородопсина различны. Полипептидная
цепь родопсина состоит из 348 аминокислот. К остаткам аспарагина мо-
лекулы белка в положении 2 и 15 присоединены две олигосахаридные це-
пи. Как и у бактериородопсина, молекула зрительного родопсина пере-
секает мембрану семь раз, используя участки первичной структуры, со-
стоящие из гидрофобных аминокислот. Между собою они соединены
короткими цепями гидрофильных аминокислотных последовательнос-
тей. По-видимому, упаковка молекулы ретиналя в 7 а-спиральных ко-
лонн является оптима в качестве первичного акцептора многих внеш-
них сигналов.
Широкий разброс максимумов спектров поглощения зрительных
пигментов (от 360 до 620 нм) обусловливается природой аминокислот-
ных радикалов опсина и их расположением относительно 11 рети-
наля в хромофорном центре (Каламкаров, Островский, 2002). В па-
лочках сетчатки человека содержится родопсин с максимумом спектра
поглощения 500 нм, а в трех типах колбочек (сине-, зелено- и красно-
чувствительных) — с максимумами 420, 531 и 558 нм, соответственно.
Пигменты из колбочек сетчатки глаза человека, условно названные в
соответствии с областями поглощаемого ими света — красный, го-
лубой и зеленый, обнаруживают в своей структуре чередование гидро-
фобных и гидрофильных участков; при этом красный и зеленый пиг-
менты обладают высокой степенью гомологии с родопсином из пало-
чек, а голубой значительно от него отличается.
Бактериородопсин и родопсин из палочек сетчатки практически не
имеют гомологичных участков, однако они также формируют в мембра-
не структуры, состоящие в каждом случае из семи гидрофобных колонн.
В основе функционирования родопсина лежит светозависимая изо-
меризация ретиналя. При поглощении кванта света 11-^щ?-ретиналь
3.6. Сенсорные процессы
301
родопсина переходит полностью в форму. Эта изомеризация
запускает каскад реакций, сопровождающихся изменением спектра
поглощения молекулы родопсина. В результате поглощения родопси-
ном фотона происходит образование опсина и свободного ретиналя.
Другой результат этого процесса — гиперполяризация мембраны па-
лочек. В норме на мембране палочек регистрируется так называемый
«темновой ток», обеспечиваемый входом в клетку натрия и выходом
калия за счет работы Na/K-АТФазы (см. гл. 2.5). Трансмембранный
потенциал при этом составляет 20 мВ. Вспышка света вызывает ги-
перполяризацию мембраны до 70 мВ, причем чем интенсивнее свет,
тем сильнее выражена гиперполяризация мембраны.
Измерение мембранного потенциала индивидуальной клетки с по-
мощью микроэлектродной техники показало, что гиперполяризация
мембраны палочки является необходимым и достаточным условием
передачи светового сигнала через синапсы к зрительным нейронам.
Таким образом, поглощение кванта света ретиналем родопсина,
локализованным в фоторецепторном диске, должно привести к гипер-
поляризации плазматической мембраны палочек, непосредственно не
связанной с мембраной диска. Когда обнаружилось, что в передаче
сигнала участвует цГМФ, было сделано предположение, что его сиг-
нал воспринимается специальным белком-посредником. Этот белок был
назван трансдуцином, так как участвует в трансдукции (преобразо-
вании) светового сигнала в электрический. Оказалось, что трансдуцин
состоит из трех субъединиц — а, Р и у, способных к обратимой диссо-
циации.
Активированная светом молекула родопсина — метародопсин-11 —
образует специфический комплекс с трансдуцином, с а-субъединицей
которого связан ГДФ (рис. 3.10). Взаимодействие родопсина с трансду-
цином катализирует обмен ГДФ на ГТФ на а-субъединице. После этого
комплекс трансдуцина с родопсином диссоциирует, и практически одно-
временно происходит диссоциация трансдуцина на а-субъединицу, свя-
занную с ГТФ, и комплекс, состоящий из Р- и у-субъединиц. а-Субъеди-
ница трансдуцина в ГТФ-связанной форме активирует цГМФ-зависимую
фосфодиэстеразу. В инактивированном состоянии фермент состоит из
трех субъединиц — а, Р и у, причем последняя ингибирует фосфодиэсте-
разную активность. Активация фермента осуществляется за счет свя-
зывания содержащей ГТФ ос-субъединицы с у-субъединицей фосфодиэс-
теразы. При этом у-субъединица отделяется, а свободные а- и Р-субъ-
единицы осуществляют гидролиз ГТФ. Этот процесс протекает с очень
большой скоростью (до 4000 молекул в секунду).
Активирующее влияние трансдуцина на фосфодиэстеразу прекра-
щается после гидролиза ГТФ, затем комплекс Р- и у-субъединиц
302
3 ФУНКЦИОНАЛЬНАЯ НЕИРОХИМИЯ
Рис. 3.10. Механизм функционирования зрительного родопсина.
Активированный светом родопсин передает сигнал на гетеротримерный G-белок
(трансдуцин). Эффекторным белком активированного комплекса а-субъединицы Gt с
ГТФ является фосфодиэстераза. Активация фосфоди эсте разы приводит к превраще-
нию имеющегося цГМФ в ГМФ, который со стороны цитоплазмы контролирует со-
стояние ионных каналов. Их закрытие вызывает уменьшение локальных концентраций
Na+ и Са2+ и гиперполяризацию мембраны
трансдуцина связывается с а-субъединицей, содержащей ГДФ, моле-
кула трансдуцина вновь приобретает способность взаимодействовать
с фотоактивированным родопсином, и весь цикл повторяется.
В результате активации одной молекулы родопсина образуется
несколько сотен активных комплексов а-субъединицы трансдуцина с
ГТФ. Это первая стадия усиления (амплификации) сиг-
нала. Затем а-субъединица активирует фосфодиэстеразу. На этой
стадии усиления сигнала не происходит, так как каждой субъединицей
активируется только одна молекула фосфодиэстеразы. Далее ком-
плекс а-субъединицы с фосфодиэстеразой (который не диссоциирует,
пока не пройдет гидролиз ГТФ) осуществляет превращения несколь-
ких тысяч молекул цГМФ. На этой стадии происходит более чем
тысячекратное усиление сигнала. Далее механизм уси-
ления работает на мембранном уровне, регулируя натриевые каналы и
генерируя электрический импульс. Трансдуцин играет ключевую роль
не только в активации, но и в инактивации сигнала. Включение и вы-
ключение сигнала осуществляются через а-субъединицу. При этом
определяющей стадией управления является гидролиз ГТФ до ГДФ.
3.0. nj.vUcCCti
30'
Родопсин инактивируется с помощью специальной протеинкиназы.
Этот фермент присоединяет фосфатные группы к нескольким амино-
кислотам на одном конце полипептидной цепи опсина. Затем родопсин
образует комплекс с белком, называемым арестином, который бло-
кирует связывание трансдуцина и возвращает систему в исходное
«темновое» состояние.
Несмотря на то что механизм передачи сигнала от фоторецептор-
ного диска к плазматической мембране изучен достаточно подробно,
ряд вопросов остается невыясненным. Во-первых, не вполне понятна
роль ионов кальция. В некоторых работах было показано, что светоза-
висимое увеличение концентрации внутриклеточного кальция приво-
дит к гиперполяризации мембраны, исчезающей после удаления Са2+.
При этом хелатирующие Са2+ агенты снижают чувствительность фо-
торецепторной клетки к свету.
В последнее время активно обсуждается роль фосфоинозитидных
мессенджеров в передаче сигнала в фоторецепторной клетке. Показа-
но, что однократное освещение приводит к активации фосфоинозитид-
ного цикла. Освещение палочек активирует фосфолипазу А2; этот
процесс зависит от трансдуцина и ингибируется коклюшным токси-
ном. По-видимому, фосфоинозитидный цикл также является частью
механизма передачи сигнала в фоторецепторной клетке, однако меха-
низмы этого участия еще предстоит узнать.
Функционирование родопсина в фоторецепторных дисках сущест-
венно зависит от липидного окружения. В фоторецепторной мембране
низко содержание холестерина, а основные фосфолипиды, входящие
в ее состав (фосфатидилхолин — 40%, фосфатидилэтаноламин — 38%,
фосфатидилсерин — 13%), содержат большое количество полинена-
сыщенных жирных кислот (до 90%). Такой состав мембраны, по-види-
мому, обеспечивает высокую степень ее жидкостности, необходимую
для функционирования родопсина. В то же время большое количество
полиненасыщенных жирных кислот делает фосфолипиды сетчатки уяз-
вимыми для окислительного повреждения (см. гл. 3.9).
Механизмы работы фоторецепторов и метаботропных рецепторов
нейромедиаторов, сопряженных с G-белками, аналогичны (рис. 3.11).
Связывание гормона с рецептором приводит к активации G-белка, а воз-
буждение родопсина квантом света — к активации трансдуцина. Как ак-
тивация G-белка, так и активация трансдуцина включает связывание ГТФ
а-субъединицей. G-Белок активирует аденилатциклазу (АЦ), а трансду-
цин — фосфодиэстеразу (ФДЭ). Оба фермента осуществляют свои функ-
ции через циклические нуклеотиды: цАМФ участвует в регуляции фер-
ментов — эффекторов медиаторов, а цГМФ индуцирует открывание на-
триевого канала в плазматической мембране фоторецепторной клетки.
304
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
Рецептор-гормон
G-белок’ ГДФ
Т-ГДФ^ ^Т-ГТФ
Tfjy
Та-ГТФ
Фосфоди-
' эстераза
Фосфодиэстераза ’Та’ГТФ
g₽y
Аденилатциклаза • Ga* ГТФ
Аденилат-
циклаза
G-белок • ГТФ
ГТФ
цГМФ 5'-ГМФ
АТФ
цАМФ
Открытие
Na-каналов
Регуляция
ферментов
б
а
Рис. 3.11. Сравнение путей передачи зрительного (а) и гормонального (б)
сигналов
Аналогия в передаче сигнала в фоторецепторной клетке с передачей
гормонального сигнала усиливается тем, что трансдуцин и G-белок
имеют не только общие функции, но и общую структуру. Все исследо-
ванные к настоящему времени белки этой группы содержат идентич-
ные 0-субъединицы, а их a-субъединицы выполняют сходные функции.
Исследование первичной структуры трансдуцина.и трех G-белков из
различных клеток выявило, что более 50% их полипептидных цепей
практически гомологичны. При этом в составе a-субъединиц, как
трансдуцина, так и G-белков, присутствуют как консервативные моти-
вы, так и мотивы, возникающие в ходе эволюции. И G-белки, и транс-
дуцин имеют три центра связывания: для рецептора, для гуаниловых
нуклеотиодов и для белка-эффектора (аденилатциклазы для комплекса
3.6. Сенсооные процессы
медиатор—рецептор и фосфодиэстеразы в случае трансдуцина). Наи-
более консервативными последовательностями аминокислот обладают
центры связывания гуаниловых нуклеотидов.
Интересно, что участки связывания гуаниловых нуклеотидов в G-бел-
ках и трансдуцине оказались гомологичными областям связывания
ГТФ в белке совершенно другого класса — в так называемом факторе
элонгации. Этот фактор участвует в синтезе белка, образуя комплекс
цГМФ с молекулами аминоацил-тРНК и обеспечивая доставку амино-
кислот к месту удлинения полипептидной цепи. Цикл функционирова-
ния этого белка похож на цикл G-белков и трансдуцина — в основе
его лежит механизм расщепления связанного в активном центре
цГМФ. Можно предположить, что фактор элонгации является эволю-
ционным предшественником трансдуцина и G-белков, что можно рас-
сматривать как проявление единства путей биохимической эволюции.
Слой палочек и колбочек граничит со слоем, в котором размещены
нейроны трех различных типов — биполярные, гори-
зонтальные и амакриновые клетки. Биполярные клетки одним из от-
ростков образуют связи с рецепторными клетками, а другим — с ганг-
лиозными, своими аксонами формирующими зрительный нерв. Гори-
зонтальные клетки соединяют рецепторы с биполярными клетками с
помощью длинных тяжей, идущих параллельно слоям сетчатки. Подоб-
ным же образом амакриновые клетки соединяют биполярные клетки
с ганглиозными. Таким путем информация от фоторецепторов может
попадать непосредственно к биполярным, а затем — к ганглиозным
клеткам. Передача информации между рецепторными и биполярными
клетками может осуществляться и другим путем, который реализуется
через горизонтальные клетки.
Амакриновые клетки не имеют аксонов и классифицируются по
типам ветвлений их дендритных «деревьев». Насчитывается несколь-
ко десятков подтипов амакриновых клеток, образующих чрезвычайно
сложную, соединенную щелевыми контактами сеть.
Биполярные клетки не генерируют импульсы. Эти клетки образу-
ют синапсы с группой фоторецепторов, а также получают сигналы от
горизонтальных клеток, т. е. на них воздействует свет, падающий на
небольшой участок сетчатки, который определяется как рецептив-
ное поле (РП).
Различают два типа биполярных клеток: генери-
рующие «ON-ответ» при освещении в центре РП и генерирующие
«OFF-ответ» при освещении периферии РП. Если же биполярная
клетка в центре РП вызывает «OFF-ответ» (т. е. отвечает на выклю-
чение света), то при освещении периферии генерируется «ON-ответ».
306
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
Считается, что за такие антагонистические взаимоотношения отвеча-
ют главным образом горизонтальные клетки.
В темноте «ножки» колбочек выделяют глутамат на горизонталь-
ные клетки, поддерживая их в деполяризованном состоянии. Деполя-
ризованные горизонтальные клетки выделяют тормозный медиатор на
соседние ножки колбочек, что несколько уменьшает их деполяриза-
цию по сравнению с тем уровнем, который у них был в темноте. Когда
свет попадает на периферию РП, выделение глутамата из перифери-
ческих колбочек уменьшается, горизонтальные клетки гиперполяри-
зуются, выделение глутамата на основание колбочек в центре РП сни-
жается, вследствие чего они деполяризуются.
При использовании селективных рецепторных антагонистов выявлена
функциональная роль эндогенных трансмиттеров, таких как глутамат, яв-
ляющийся главным в передаче возбуждения в цепи фоторецепторов, би-
полярных и ганглиозных клеток; аспартат, специфичный для палочек, и
ацетилхолин, известный как трансмиттер холинергических амакриновых
клеток. Определена функциональная роль ГАМК и глицинергических пу-
тей, которые являются зрительными ингибиторами соответственно on- и
off-ганглиозных клеток. Тормозный нейромедиатор глицин обнаружива-
ется в биполярных клетках. Из множества нейропептцдов, обнаруженных
в сетчатке разных животных, в сетчатке человека выявлены только ней-
ропептид Y, соматостатин и субстанция Р, которые, наряду с регулярны-
ми нейротрансмиттерами и нейрорегуляторами, локализуются в различ-
ных амакриновых и ганглиозных клетках. Амакриновые клетки наряду с
нейропептидами содержат также ГАМК, глицин, ацетилхолин, дофа-
мин и серотонин. Имеются морфологические доказательства того, что
ацетилхолин и ГАМК содержатся в амакриновых клетках одного типа.
Нервные импульсы, возникающие в сетчатке, поступают по волокнам
зрительного нерва в расположенную в задней части затылочных долей
зрительную область коры. В мозге человека аксоны от левых половин
сетчатки обоих глаз направляются клевой половине зрительной коры, а
аксоны от правых половин сетчатки обоих глаз — к правой стороне зри-
тельной коры. Аксоны, идущие от носовых половин обеих сетчаток, пе-
ресекаются, образуя зрительный перекрест, или хиазму. Волокна, иду-
щие от височных (темпоральных) половин каждой сетчатки, продол-
жают идти с той же стороны (ипсилатерально), объединяясь вместе с
перекрещенным пучком аксонов из контрлатерального зрительного не-
рва. Они образуют зрительный тракт, который приводит к латеральным
коленчатым телам, верхним бугоркам четверохолмия, ядрам вспомога-
тельного зрительного тракта и претектальной области ствола мозга. Ак-
соны клеток латерального коленчатого тела образуют зрительную ра-
диацию и оканчиваются в основном в первичной зрительной коре.
3.6. Сенсорные процессы
3.6.2. Нейрохимические механизмы обоняния
В начале 50-х гг. XX в. Эрл Сезерленд на примере адреналина,
стимулирующего образование глюкозы из гликогена, расшифровал
принципы передачи сигнала через клеточную мембрану, которые оказа-
лись общими для широкого круга рецепторов. Уже в конце XX в. было
обнаружено, что восприятие запахов осуществляется аналогичным об-
разом, даже детали строения белков-рецепторов оказались похожими.
Первичные рецепторные белки — это сложные молекулы, свя-
зывание с которыми лигандов вызывает в них ощутимые структурные
изменения, вслед за которыми начинается каскад каталитических (фер-
ментативных) реакций. Для рецептора запаха (одорантного), так же
как и для зрительного рецептора, этот процесс завершается нервным
импульсом, воспринимаемым нервными клетками соответствующих от-
делов мозга.
Как видно из рисунка 3.12, рецепторный белок включает такую по-
следовательность аминокислот, которая содержит семь гидрофобных
в
Рис. 3.12. Схема строения обонятельного рецептора:
а — поперечное сечение семидоменной структуры; б — схема взаимодействия одоран-
та (лирала) с рецептором (вид сверху); в — два химических механизма усиления сиг-
нала запаха внутри обонятельного волоска: / — мембранный интегральный комп-
лекс — рецептор (R) + ГТФ-связывающий белок (G) + аденилилциклаза (АЦ);
2 — мембранный интегральный комплекс — рецептор (R) + ГТФ-связывающий бе-
лок 4- фосфолипаза С (ФЛС)
308
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
сегментов, содержащих от 20 до 28 остатков в каждом, что достаточно
о
для пересечения мембраны толщиной 30 А. Эти полипептидные уча-
стки свернуты в а-спираль. Таким образом, тело рецепторного белка
представляет собой компактную структуру из семи сегментов, пересе-
кающих мембрану. Такая структура интегральных белков характер-
на для опсина в сетчатке глаза, рецепторов серотонина, адреналина
и гистамина.
Для реконструкции структуры мембранных рецепторов пока недо-
статочно рентгеноструктурных данных. Поэтому в таких схемах в на-
стоящее время широко используются аналоговые компьютерные моде-
ли. Согласно этим моделям, обонятельный рецептор образован семью
гидрофобными доменами. Лигандсвязывающие аминокислотные остат-
ки формируют «карман», отстоящий от поверхности клетки на рас-
о
стояние в 12 А. Карман изображают в виде розетки, построенной одно-
типно для разных рецепторных систем. На рисунке 3.12 представле-
на схема взаимодействия молекулы лирала (синтетического одоранта)
с обонятельным рецептором крысы.
Связывание одоранта с рецептором приводит к включению одного
из двух сигнальных каскадов, открытию ионных каналов и генерации
рецепторного потенциала. Специфический для обонятельных клеток
G-белок (Goif) может активировать аденилатциклазу, что приводит к
увеличению концентрации цАМФ, мишенью которой являются кати-
онселективные каналы. Их открытие приводит ко входу Na+ и Са2+
внутрь клетки и деполяризации мембраны.
Увеличение концентрации внутриклеточного кальция вызывает от-
крытие Са-управляемых С1-каналов, что ведет к еще большей деполя-
ризации и генерации рецепторного потенциала. Тушение сигнала про-
исходит из-за снижения концентрации цАМФ, благодаря специфиче-
ским фосфодиэстеразам, а также в результате того, что Са2+ в
комплексе с кальмодулином связывается с ионными каналами и сни-
жает их чувствительность к цАМФ.
Другой путь тушения сигнала связан с активацией фосфолипазы С
и протеинкиназы С. В результате фосфорилирования мембранных
белков открываются катионные каналы и, как следствие, мгновенно
меняется трансмембранный потенциал, в результате чего также гене-
рируется потенциал действия. Таким образом, фосфорилирование
белков протеинкиназами и дефосфорилирование их соответствующи-
ми фосфатазами оказалось универсальным механизмом мгновенного
ответа клетки на внешнее воздействие.
30
3.6. Сенсорные п,- ^цессы
Аксоны, направляющиеся в обонятельную луковицу, объединены в
пучки. Слизистая оболочка носа, кроме того, содержит свободные
окончания тройничного нерва, часть которых тоже способна реагиро-
вать на запахи. В области глотки обонятельные стимулы могут возбуж-
дать волокна языкоглоточного (IX) и блуждающего (X) черепномозго-
вых нервов. Их роль в восприятии запахов не связана с обонятельным
нервом и сохраняется при нарушении функции обонятельного эпите-
лия при болезнях и травмах.
Гистологически обонятельная луковица подразделяется на не-
сколько слоев, характеризующихся клетками специфической формы,
снабженными отростками определенного типа с типичными видами
связи между ними.
На митральных клетках происходит конвергенция информации. В клу-
бочковом (гломерулярном) слое примерно 1000 обонятельных клеток
оканчиваются на первичных дендритах одной митральной клетки. Эти
дендриты также образуют реципрокные дендродендритные синапсы с
перигломерулярными клетками. Контакты между митральными и пери-
гломерулярными клетками являются возбулдающими, а противопо-
ложно направленные — тормозными. Аксоны перигломерулярных кле-
ток оканчиваются на дендритах митральных клеток соседнего клубочка.
Клетки-зерна также образуют реципрокные дендродендритные си-
напсы с митральными клетками; эти контакты влияют на генерацию
импульсов митральными клетками. Синапсы на митральных клетках
также являются тормозными. Кроме того, клетки-зерна образуют
контакты с коллатералями митральных клеток. Аксоны митральных
клеток формируют латеральный обонятельный тракт, направляющий-
ся в кору головного мозга. Синапсы с нейронами высших порядков
обеспечивают связь с гиппокампом и (через миндалину) с вегетатив-
ными ядрами гипоталамуса. Нейроны, отвечающие на обонятельные
стимулы, обнаружены также в орбитофронтальной коре и ретикуляр-
ной формации среднего мозга.
3.6.3. Механизмы работы вкусовых рецепторов
Механизмы восприятия вкуса и запаха во многом аналогичны, так
как оба ощущения активируются химическими стимулами, приходящими
из внешнего мира. Действительно, вкусовые стимулы, как правило, дей-
ствуют на рецепторы, связанные с G-белками, способами, весьма схо-
жими с теми, которые были описаны выше для обоняния. В то же время
некоторые вкусовые стимулы (в основном, соли и кислоты) действуют
непосредственно на мембранную проводимость рецепторных клеток.
310
- »здел 3. 4Л‘НКЦ;!
Вкусовые рецепторы локализованы на волосковых нейроэпители-
альных клетках, расположенных во вкусовых сосочках на поверхности
языка. В отличие от обонятельных рецепторов, они не имеют аксонов,
но формируют химические синапсы с афферентными нейронами во
вкусовых сосочках. Микроворсинки направляются от апикального по-
Рецептор
G-белок
Выброс
медиатора
Сладкий,горький,
аминокислоты
Соль,
кислота
Первичный
сенсорный
нейрон
TRPM5-канал
Са2+
Na+
Са2+
Потенциал действия
Серотониновый
рецептор
Ионный канал
Са2+
Апикальная
часть
Вторичные
мессенджеры
ЭПР
Базальная
часть
3.6. Сенсорные процессы
31В
люса вкусовой клетки в открытую пору вкусового сосочка, где вступа-
ют в контакт со вкусовыми стимулами (веществами, растворенными
в слюне на поверхности языка).
Начальные стадии хемосенсорного восприятия проходят во вкусовых
клетках, которые имеют рецепторы на апикальной части, расположен-
ной возле отверстия вкусовой поры (рис. 3.13). Подобно обонятельным
рецепторным клеткам, вкусовые клетки отмирают через ка>вдые две не-
дели и происходит регенерация новых клеток из базальных. Для каждого
из пяти воспринимаемых вкусов имеются отдельные типы рецепторов.
Вкус соли или кислоты создается непосредственным воздействием
ионов натрия или протонов на специфические каналы — амилоридчув-
ствительные Na-каналы, воспринимающие соленое, и Н-чувстви-
телъные каналы, воспринимающие кислое (рис. 3.14). Проникнове-
ние соответствующих зарядов внутрь вкусовой клетки ведет к деполяриза-
ции ее мембраны. Эта начальная деполяризация активирует потенциалуп-
равляемые Na- и Са-каналы в базолатеральной части вкусовой клетки,
что приводит к выбросу нейромедиатора на базальном участке вкусовой
клетки и генерации потенциала действия в ганглионарной клетке.
У человека и других млекопитающих рецепторы, воспринимающие
вкус сладкого и аминокислот, состоят из семи трансмемб-
ранных доменов и связаны с G-белком. Восприятие сладкого осуществ-
ляется благодаря паре рецепторов Т1R3 и Т1R2, а аминокислот — Т1R3
и T1R1. Рецепторы Т1R2 и T1R1 выявляются в различных частях рецеп-
торной клетки. При связывании с сахарами или другими стимулами слад-
кого T1R2/T1R3-рецептор инициирует каскад реакций, опосредуемый
G-белком, который ведет к активации фосфолипазы С (изоформа PLCp2)
и, соответственно, увеличению концентрации 1Р3 и открытию так назы-
ваемых TRP-Ca-каналов (специфических ТРРМ5-каналов), благодаря
работе которых: происходит деполяризация вкусовой клетки из-за увели-
чения внутриклеточной концентрации Са2+. T1R1/T1R3-рецептор при-
способлен для восприятия двадцати L-аминокислот, входящих в состав
белков, но не может воспринимать D-аминокислоты. Трансдукция
Рис. 3.13. Механизмы влияния вкусовых стимулов на различные рецепторы
и усиления вкусового сигнала.
Во вкусовой клетке катионселективные протонные каналы, связанные с восприятием
кислого и соленого, а также рецепторы сладкого, горького вкуса и аминокислот, связан-
ные с G-белками, локализуются в апикальной части. Расположенные в базальной части
клетки потенциалрегулируемые Na-, К- и Са-каналы обеспечивают выделение нейроме-
диаторов и передачу сигнала на первичный сенсорный нейрон. Эндоплазматический ре-
тикулум способен модулировать уровень внутриклеточного Са2+ и регулировать выде-
ление медиатора. ТКРМ5-каналы обеспечивают облегчение возникновения деполяриза-
ции мембраны при поступлении сигнала от связанных с G-белками вкусовых рецепторов
312
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
Соленый
Кислый
Амилоидчувствительный
Na-канал
Н-чувствительный
катионный канал
О1
О О
Рис. 3.14. Механизм вкусовой рецепции.
Соленый и кислый вкус воспринимается за счет катионселективных амилоридчувстви-
тельного Na+ и Н+-чувствительного протонных каналов, соответственно. В обоих слу-
чаях вход катионов через канал приводит к деполяризации клетки. Сладкий, горький
вкус и вкус аминокислот воспринимаются рецепторами, связанными с G-белками.
Воспринимаемые различными рецепторами сигналы во всех трех случаях передаются
через ФЛСр2 и ИФ3 на ТИРМ5-кальциевый канал, что ведет к его активации, входу
Са2+ и деполяризации клетки
3.6. Сенсорные процессы
3131
аминокислотного сигнала через этот рецептор осуществляется с ис-
пользованием того же сигнального каскада, что и для сахаров.
Другое семейство связанных с G-белками рецепторов, известное
как T2R, отвечает за восприятие горького вкуса. Существует
около 30 подтипов этих рецепторов, кодируемых 30 различными генами.
Эти рецепторы отсутствуют в тех клетках, в которых имеются T1R1-,
T1R2- или Т1 RS-рецепторы. Таким образом, рецепторы горького явля-
ются рецепторами особого класса. Передача сигнала о горьком вкусе
имеет сходный со сладким и вкусом аминокислот механизм передачи сиг-
нала, включающий специфический для вкусовых клеток G-белок густ-
дуцин (gustducin). Структурно этот белок на 90% гомологичен трансду-
цину — G-белку фоторецепторов. Такой же уровень сходства наблюда-
ется между трансдуцинами, функционирующими в палочках и колбочках.
У а-трансдуцина и а-густдуцина оказались идентичными последователь-
ности из 38 С-терминальных аминокислот.
Во многих продуктах, включая мясо, сыр и некоторые овощи,
встречается свободный глутамат. В виде глутамата натрия он исполь-
зуется в качестве пищевой приправы. Вкус глутамата переда-
ется сопряженным с G-белком метаботропным глутаматным рецепто-
ром, который специфически экспрессируется во вкусовых сосочках.
Методом условной вкусовой аверсии было показано, что как глутамат
натрия, так и специфичный агонист mGluR4-penenTopoB (метаботроп-
ных глутаматных рецепторов 4-го типа) L-AP4 вызывают сходные вку-
совые ощущения у крыс.
«Жгучий» вкус ряда продуктов — еще один пример много-
функциональности молекулярных рецепторов. Вкус перца восприни-
мается не самими вкусовыми клетками, а болевыми волокнами в язы-
ке, которые активируются капсаициновыми соединениями. Рецептор
капсаицина клонирован, и доказано, что это кальцийселективный ка-
тионный канал. Он образован волокнами небольшого размера (С-во-
локна), идущими от клеток спинальных ганглиев и сигнализирующими
о боли. Таким образом, природа снабдила перцы химической нацелен-
ностью на данный рецептор, возможно, чтобы отпугивать травоядных
путем активации болевых волокон.
Вкусовые клетки при стимуляции способны генерировать рецеп-
торный потенциал. При помощи синаптической передачи это возбуж-
дение передается афферентным волокнам черепномозговых нервов,
по которым поступает в виде импульсов в мозг. Барабанная струна —
ветвь лицевого нерва (VII) иннервирует переднюю и боковые части
языка, а языкоглоточный нерв (IX)— его заднюю часть. Вкусовые
почки надгортанника и пищевода иннервируются верхней гортанной
314
раздел 3. ФУНКЦИОНАЛ НАЯ НЕИРОХИМИм
3.7.
ветвью блуждающего (X) нерва. Разветвляясь, каждое волокно полу-
чает сигналы от рецепторов разных вкусовых почек. Амплитуда рецеп-
торного потенциала увеличивается вместе с концентрацией стимули-
рующего вещества. Деполяризация рецепторных клеток оказывает
возбуждающее, а гиперполяризация — тормозное действие на аффе-
рентные волокна. Волокна IX пары черепномозговых нервов особенно
сильно реагируют на вещества с горьким вкусом, а VII пары — силь-
нее на действие соленого, сладкого и кислого, причем каждое волокно
в большей степени реагирует на один определенный стимул.
Вкусовые волокна этих черепномозговых нервов оканчиваются в
пределах или вблизи от ядра одиночного пути продолговатого мозга,
связанного с вентральным постеромедиальным ядром таламуса. Аксо-
ны нейронов третьего порядка оканчиваются в постцентральной изви-
лине коры головного мозга. Ряд корковых клеток реагирует только на
вещества с одним вкусовым качеством, другие — также и на темпера-
турные и механические стимулы.
РЕГУЛЯЦИЯ ДВИГАТЕЛЬНОЙ АКТИВНОСТИ
Одна из важных функций нервной системы — организация про-
извольных движений и контроль за ними. Мышечные волокна, осу-
ществляющие согласованное перемещение тела в пространстве и дру-
гие виды произвольных движений, иннервируются специфическими
афферентными волокнами. Решение о характере движения принима-
ется на уровне ЦНС, вовлекает в сферу своего действия спинной мозг
и распространяется по мотонейрону к соответствующему эффекторно-
му органу (скелетной мышце). Существуют также сенсорные рецепто-
ры, сообщающие информацию об успешности сократительной реакции
в мотонейрон и в высшие отделы нервной системы, чтобы обеспечить
эффективную обратную связь в двигательной системе.
В настоящее время в общем виде можно сформулировать три
основных принципа организации двигательной
системы:
1) контроль организован иерархически;
2) основные двигательные команды контролируются наличием обрат-
ной связи, проявляющейся на каждом уровне двигательной системы;
3) двигательные команды от высших отделов ЦНС обеспечивают
правильное вовлечение в работу проксимальных и дистальных мышц.
Проксимальная мускулатура конечностей и туловища обеспечивает
осанку и осуществление ритмических движений (например, при дыха-
3.7. Регуляция двигательной активности
315
нии), в то время как дистальная группа мышц вовлечена в сложные
произвольные движения, в том числе и в те, которые основаны на обу-
чении (примером является виртуозная техника руки пианиста).
На более низком уровне организации движения — в спинном моз-
ге сенсорные нейроны образуют синапсы с мотонейронами, замыкая
цепи, обеспечивающие простые рефлексы, для реализации которых
нет необходимости вовлекать ЦНС. Вставочные нейроны спинного
мозга и ствола создают генераторы центрального ритма для координа-
ции множественных моторных единиц. «Надзирающий» контроль за
ними осуществляют нейроны двигательной коры, которые способны
к реализации сложных двигательных команд.
Единицей двигательной системы является одиночный
а-мотонейрон, иннервирующий группу мышечных волокон. Число
мышечных волокон составляет от нескольких единиц до нескольких ты-
сяч (например, в проксимальных мышцах конечностей). Возбуждение
мотонейрона приводит к одновременному сокращению всех составляю-
щих двигательную единицу мышечных волокон. Точность движений
обеспечивается изменением числа и времени вовлечения в сокращение
отдельных двигательных единиц. Так, сокращение камбаловидной мыш-
цы регулируется ста а-мотонейронами, в то время как вся мышца со-
держит около 25 000 мышечных волокон. Одиночное сокращение этой
мышцы складывается из ста «элементар-
ных» процессов. Тетаническое сокращение,
осуществляемое в результате суммирова-
ния следующих друг за другом возбуждаю-
щих импульсов, будет сложным резуль-
татом суперпозиции в активности а-мо-
тонейронов, каждый из которых может
генерировать собственную частоту потен-
циалов действия (ПД) и отличный от ос-
тальных единиц характер суммации.
Произвольная мускулатура иннерви-
руется нейронами с синапсами, имеющи-
ми химический (а не электрический, кото-
рый имеет место в некоторых синапсах
ЦНС) механизм передачи. Считается, что
химическая природа передачи возбужде-
ния установлена О. Леви в его получив-
ших широкую известность опытах, при-
несших ему впоследствии Нобелевскую
премию (Loewi, 1921).
О. Леви
Лауреат Нобелевской пре-
мии по физиологии и медици-
не, обосновавший концеп-
цию химической природы пе-
редачи возбуждения в сердце
(Австрия)
316
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕИРОХИМИЯ
Стимуляция
Блуждающий
Второе сердце
Стимуляция блуждающего
нерва первого сердца
Второе сердце
Рис. 3.15. Схема опыта Отто Леви:
а — схема установки; б — запись сократительной работы сердца. После стимуляции
блуждающего нерва у автоматически работающего сердца с сохраненной иннервацией
окружающий раствор приобретает способность тормозить сокращения другого сердца
Благодаря тому, что столетие назад стиль изложения научных фак-
тов существенно отличался от сухого информационного языка совре-
менной науки, мы имеем возможность оценить обстоятельства цент-
рального эксперимента, поставленного О. Леви. Предметом его ис-
следования был характер тормозящего действия блуждающего нерва
на ритмическую работу сердечной мышцы. Известный феномен тор-
можения сердечных сокращений в результате раздражения n. vagus
еще не имел своего объяснения, хотя был известен факт остановки
сердца от удара в солнечное сплетение.
О. Леви отмечал, что идея решающего эксперимента пришла к не-
му во сне, спросонок он записал план опыта, но поутру не смог восста-
новить смысла записанного. К счастью, сон повторился, и экспери-
ментатор не стал дожидаться утра, когда критический настрой ума мог
бы помешать потратить время на «такую’бессмыслицу». Суть опыта
заключалась в том, чтобы перфузат, полученный от сердца, испытав-
шего тормозящее действие блуждающего нерва, использовать для по-
давления сокращений другого сердца, не испытавшего вагусного конт-
роля. Леви писал1: «По серьезному размышлению я бы никогда не
поставил этого эксперимента. То, что блуждающий нерв может вы-
свобождать тормозящее вещество, было очень маловероятно. Еще
менее вероятным было то, что химическое вещество, которое предпо-
1 Цитировано по 3. Бак, 1977.
3.7. Регуляция двигательном активности
31
ложительно должно было действовать на
коротком расстоянии между нервным
окончанием и мышцей, освобождалось бы
в таком количестве, что сохраняло бы эф-
фективность в перфузирующем раство-
ре». Однако счастливое стечение обсто-
ятельств позволило О. Леви продемонст-
рировать, что перфузат из сердца лягушки
с сохраненной симпатической иннерваци-
ей, остановленного раздражением вагус-
ного нерва, способен вызывать остановку
сердца второй лягушки (рис. 3.15).
Опыты О. Леви послужили основани-
ем для утверждения, что возбуждение не-
рва вызывает выделение какого-то хими-
ческого агента, который и осуществляет
передачу информации на эффекторный ор-
ган. Позже этот химический агент был
идентифицирован с ацетилхолином, и ре-
зультаты, полученные О. Леви, были под-
А. Ф. Самойлов
Отечественный физиолог,
внесший вклад в функцио-
нальную характеристику нерв-
но-мышечного аппарата, ав-
тор блестящего эксперимен-
та, доказавшего химическую
природу передачи возбужде-
ния в синапсах
тверждены независимыми исследователя-
ми. На самом деле до сих пор приходится сомневаться в том, был ли
продемонстрирован описанный О. Леви эффект или он был следствием
сильного настроя на положительный результат. Дело в том, что в пер-
фузат изолированного сердца легко вымывается активная холинэстера-
за, которая препятствует накоплению ацетилхолина и делает «вагусный
эффект» перфузата маловероятным. Фактически опыт Отто Леви до
сих пор является классической демонстрацией химической передачи
возбуждения, но он эффективен при одном условии — демонстрация
должна происходить в присутствии ингибиторов холинэстеразы. Пер-
вое соединение с таким эффектом было описано только после получе-
ния О. Леви Нобелевской премии.
Интересно, что после положительного решения Нобелевского коми-
тета у О. Леви были сильные сомнения по поводу того, стоит ли ему согла-
шаться с этим решением — настолько смущало его самого отсутствие
четкости в воспроизведении описанного феномена. Под давлением друзей
он соглашается приехать на церемонию вручения, но обусловливает свое
согласие предварительной демонстрацией своего феномена. По свидетель-
ству присутствующих, результаты демонстрации удовлетворили только са-
мого номинанта (Бак, 1977). Но в 1936 г. премия все-таки была вручена,
и это подтвердило справедливость идеи химического механизма передачи
возбуждения.
318
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
Парадоксом в этой истории является то, что в тот же период, когда
О. Леви проводил описанные эксперименты, уже существовали неоспо-
римые доказательства этой идеи, причем они были получены в результате
безукоризненного эксперимента. Его провел русский физиолог А. Ф. Са-
мойлов, работавший в тот период в Казани. Самойлов измерял так на-
зываемый период задержки, характеризующий передачу возбуждения
через синапс (Самойлов, 1946). Собственноручно сконструированный Са-
мойловым кимограф, соединенный с пружиной, раскручивающей барабан
в момент подачи возбуждения на нервное волокно, позволял измерять за-
держку между подачей стимула и началом сокращения. Время задержки
экспериментатор измерял при разных температурах. В результате он об-
наружил, что с изменением температуры на 10 °C время задержки изме-
няется вдвое, т. е. так же, как зависит от температуры химическая реак-
ция. Температурный коэффициент задержки при передаче возбуждения по
нерву составлял 1,54. Самойлов сделал вывод о том, что в основе переда-
чи информации с нерва на мышцу лежит химический процесс, в то время
как передача возбуждения по нерву имеет физическую природу. Единст-
венным «недостатком» этой методически безупречной работы явилось то,
что она была опубликована в «Трудах Казанского университета», что сде-
лало ее неизвестной для мировой науки...
Так или иначе, но химическая природа передачи возбуждения в мы-
шечном синапсе была установлена. ПД, измеряемый при распростране-
нии возбуждения от синапса вдоль мышечного волокна, мобилизовывал
ионы кальция, высвобождаемые из саркоплазматического ретикулума,
инициируя образование актомиозина и сократительную реакцию. Изме-
рение ионных токов через мышечную мембрану показало, что форма
ПД соответствует сумме двух кривых, описывающих натриевый и ка-
лиевый токи через сарколемму, следовательно, перемещение через
мембрану мышечной клетки только этих двух ионов достаточно для со-
здания условий для ее возбуждения. Для сердечной мышцы было обна-
ружено существенное отличие — увеличение времени ПД за счет нали-
чия стадии «плато», вызванного входом ионов кальция из внешней сре-
ды, удлиняющим период деполяризации (рис. 3.16).
Если обратиться к таблице, описывающей физико-химические свой-
ства ионов натрия и калия (см. гл. 2.5), можно увидеть, что в покоящейся
клетке трансмембранный потенциал далек от равновесного потенциала
для ионов натрия. Открытие ионных каналов при деполяризации мемб-
раны и создание условий для выявления натриевого тока (направленного
внутрь волокна) вызывает деполяризацию мембраны, и в силу изменений
мембранного потенциала открывает возможность для входа калия в
клетку. Это создает вторую фазу ПД — реполяризацию мембраны (воз-
вращение ТМП к исходному состоянию). Запуском для активации по-
тенциалзависимых Na-каналов сарколеммы является сдвиг ТМП на
3.7. Регуляиия двигательной активности
319
10 мс
Рис. 3.16. ПД скелетной (а) и сердечной (б) мышц
1 мВ[।
10 мс
величину, превышающую порог возбудимости, позволяющий открыть
потенциалзависимые Na-каналы. Таким образом, необходимым фак-
тором для осуществления процесса возбуждения является создание
потенциала концевой пластинки (ПКП), выполняющего эту функцию.
Данная задача выполняется специфическим белком — никотино-
вым рецептором ацетилхолина, представляющим собой олигомерный
комплекс, который включает в свой состав ионный канал. Фактически
ионный канал управляется рецепторным аппаратом, который подает
сигнал о его открывании, когда с ним соединяется молекула лиганда —
ацетилхолина.
Из двух известных классов ацетилхолинового рецептора (никотино-
вые и мускариновые, см. гл. 2.3) в скелетной мускулатуре функциони-
рует н-ХР, представляющий собой сложный комплекс интегральных
белков, пронизывающих мембрану, состоящих из пяти различных
субъединиц (двух а и по одной — р, 8 и у), расположенных радиально с
углом около 72°. ос-Субъединицы содержат центры связывания ацетил-
холина. Внемембранная часть рецепторного комплекса выступает над
поверхностью мембраны на 5 нм. Ее диаметр в этой области составляет
около 8,5 нм (линейная длина молекулы ацетилхолина — 11 нм). Диа-
метр центральной поры, открываемой при активации холинорецептора,
достигает 0,7 нм, что в несколько раз больше диаметра переносимых че-
рез канал ионов. Современные представления об упаковке ХР в мемб-
ране систематизированы и представлены на рисунке 3.17.
Согласно современным представлениям, связывание АХ с ос-субъ-
единицами рецептора вызывает конформационные перестройки канал-
рецепторного комплекса, вследствие чего белковые цепи, формирую-
щие канал в мембране, расходятся и открывается возможность для
входа ионов натрия внутрь клетки (см. рис. 3.17).
320
Раздел 3 ФУНКЦИОНАЛЬНА^ НЕЙРО) и МИ
Рис. 3.17. Структура ХР: а — общий вид; б — вид сверху в закрытом и откры-
том состояниях
Высвобождение АХ в синаптическую щель осу-
ществляется двумя способами — в результате спонтан-
ного изливания содержимого одного или нескольких синаптических
пузырьков или вследствие синхронного выброса медиатора из не-
скольких десятков (сотен) пузырьков, происходящего в результате де-
поляризации пресинаптической мембраны. Наблюдающаяся в первом
случае активация затрагивает ограниченное количество рецепторов и
создает малоамплитудное возбуждение (так называемые миниатюр-
ные потенциалы концевой пластинки, МПКП). Амплитуда МПКП,
вызываемая высвобождением содержимого одной везикулы (выделе-
нием одного кванта АХ), составляет около 1 мВ. Установлено, что
квант состоит из 7000 (±10%) молекул медиатора. Поскольку воз-
буждение одной молекулы ХР вызывает деполяризацию мышечного
волокна примерно на 1 мкВ, в генерации каждого МПКП участвует
около 1000 молекул рецептора. Для открывания канала нужно в
2 раза большее количество молекул медиатора (две молекулы АХ на
3.7. Регуляция двигательной активности
321
одну молекулу ХР), в то же время экспериментально показано, что из
7000 молекул каждого кванта для взаимодействия с рецептором ис-
пользуется около 2600. Таким образом, при квантовом выделении АХ
эффективность его действия составляет около 30%. Остальная часть
молекул медиатора рассеивается в межсинаптическом пространстве и
подвергается гидролизу АХЭ. Аналогичная зависимость была получе-
на для глицинергических синапсов головного мозга миноги. Данные,
полученные для других синапсов нервной ткани, свидетельствуют о
том, что в реализации одной МПКП участвует от 15—65 молекул ХР.
При синхронизованном выбросе медиатора из десятков—сотен си-
наптических везикул наблюдается формирование потенциала, дости-
гающего порогового уровня, приводящего к возникновению ПД. Для
генерации ПД трансмембранный потенциал должен измениться на
10—12 мВ (на величину потенциала концевой пластинки, ПКП). Ес-
ли приход возбуждения в пресинаптическую часть синапса вызовет
выделение достаточного для этого количества медиатора, то в постси-
наптической области происходит формирование ПКП и возникает ПД.
Время жизни АХ в синаптической щели ограничено активностью
ацетилхолинэстеразы (АХЭ), которая гидролизует АХ, что приводит к
диссоциации комплекса АХ с ХР и закрытию канала. Увеличение час-
тоты возбуждения пресинаптической зоны будет приводить к суммации
возбуждающих потенциалов, продлевающей длительность и эффек-
тивность сокращения мышцы. В таком случае возникает тетаническое
сокращение, когда каждый следующий импульс вызывает дополни-
тельное сокращение не успевшей расслабиться мышцы.
Дальнейшее повышение частоты раздражений приведет к сниже-
нию эффективности сокращений за счет их попадания на период отно-
сительной невозбудимости (рефрактерный период). Наличие реф-
рактерного периода объясняется тем, что формирование ПД приводит
ко входу ионов натрия и выходу ионов калия из мышечных волокон.
Возрастание соотношения Na/K активирует Na/K-насос, который,
обеспечивая восстановление ионных градиентов, работает «против»
направления пассивных ионопотоков. Вклад Na/K-насоса в величину
ТМП можно определить, измеряя ТМП в присутствии специфического
ингибитора насоса уабаина. Аппликация уабаина вызывает деполяри-
зацию мембраны на 10—12 мВ, т. е. на величину, сравнимую с ПКП.
Это и объясняет наличие рефрактерности, появляющейся при слиш-
ком быстром следовании возбуждающих импульсов друг за другом.
Рефрактерный период является относительно кратковременным —
восстановление ионных градиентов осуществляется быстро. Но имеет
место и период абсолютной рефрактерности, когда дальнейшее
322
Раздел 3. ФУ кЦИсНл Пк НхЯ НГЙРОХИМИу
а
О
II
(CH3)3N+—СН2—СН2—О—С—СНз
Эфирный
сайт
Анионный
сайт
б
О
II
CH3-C-O-CH2-CH2-N+-(CH3)3
Анионный
сайт
CH3-C-O-CH2-CH2-N+-(CH3)3
Эфирный
сайт
Рис. 3.18. Два способа взаимодействия АХ с АХЭ: а — эффективный гидролиз
молекулы медиатора; б — неэффективное взаимодействие
увеличение частоты раздражений делает -мышцу нечувствительной к
раздражению, пришедшему в период реполяризации, вызванной выхо-
дом К+. Это явление было впервые описано в начале прошлого века
Н. Е. Введенским и носит в специальной литературе название «пес-
симум Введенского». Оно представляет собой природный механизм,
защищающий мышцу от повреждения.
Известен еще один способ защиты мышцы от чрезмерной нагрузки.
Он обусловливается избытком ацетилхолина, который не может быть
гидролизован АХЭ вследствие субстратного торможения. Быстрое
ферментативное разрушение АХ осуществляется за счет хорошего сте-
3.7. Регуляция двигательной активности
3231
рического соответствия его молекулы структуре активного центра.
Субстрат располагается в нем таким образом, что триметильный конец
молекулы попадает в гидрофобную область, а противоположный заря-
женный конец ориентируется у соответствующего заряда другой груп-
пы. Расстояние между центрами фиксации несколько превышает то,
которое требуется для длины ковалентной связи. Это облегчает водную
атаку, направленную на эфирную связь, и приводит к ее быстрому раз-
рыву. При большом избытке субстрата фермент-субстратный комп-
лекс образуется «неправильным» образом — к активному центру АХЭ
присоединяются две молекулы АХ. При такой оккупации фермента гид-
ролиз медиатора становится невозможным (рис. 3.18). В этих условиях
в синаптической щели накапливается АХ, что вызывает «перегрузку»
рецепторов, ответственных за десенситизацию. Десенситизация явля-
ется «внутренним» свойством нХР, который настроен на кратковре-
менное изменение конформации и транзиторное открывание канала.
Замедление диссоциации медиатора от рецептора увеличивает время
его жизни в оккупированном лигандом состоянии, но не может удер-
жать канал открытым. Скорость выхода рецепторов из десенситизиро-
ванного состояния регулируется специальными протеинкиназами.
Систематические исследования ХР из разных источников показа-
ли, что существует более 10 изоформ, специфичных для разных объ-
ектов (Турпаев, 1964). Они образуются
в результате альтернативного сплайсин-
га. Поскольку субъединицы ХР кодируют-
ся различными экзонами гена(ов) и обра-
зование конечной формы мРНК включает
их различные альтернативные комбина-
ции, возможно образование различных
изоформ. Специальный механизм опреде-
ляет, какая из мРНК будет использована
для синтеза.
Этот механизм обусловливает отличия,
выявляемые в структуре нХР эмбри-
ональной и взрослой форм ХР. В онтоге-
незе мышц происходит замена у-субъ-
единицы е-субъединицей, имеющей нес-
колько отличный аминокислотный состав.
Вследствие этого длительность сущест-
вования рецептора в возбужденном сос-
тоянии уменьшается, а проводимость воз-
растает.
Т. М. Турпаев
Блестящий ученый, извест-
ный своими работами в облас-
ти биологии развития, автор
первой монографии о молеку-
лярных свойствах холиноре-
цептора
324
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕИРОХИМИЯ
3.8.
В состав нХР электрического органа ската, представляющего со-
бой редуцированные мышечные элементы (электроциты), также вхо-
дит четыре вида субъединиц (а, 0, у, 5). Нейрональный нХР является
пентамером состава а203.
НЕЙРОХИМИЧЕСКИЕ МЕХАНИЗМЫ ПАМЯТИ
Эта область нейрохимии наиболее трудна для исследования. Хо-
тя молекулярные механизмы функционирования одиночной нервной
клетки изучены во многих своих проявлениях и сформулированы
принципы организации межнейрональных связей, все еще неясно, ка-
ким образом молекулярные свойства нейронов обеспечивают хране-
ние, воспроизводство и анализ информации. То обстоятельство, что
приобретенные знания (как и нравственные принципы) не передаются
по наследству, и новым поколениям приходится обучаться им заново,
позволяет считать, что обучение представляет собой процесс создания
новых межнейронных связей и запоминание информации обеспечива-
ется способностью мозга по необходимости воспроизводить эти связи
(активировать их). Однако современная нейрохимия еще не в состоя-
нии представить непротиворечивую теорию, описывающую то, каким
образом анализ факторов внешнего мира осуществляется в живом
мозге. Можно лишь очертить те проблемы, над которыми интенсивно
работают ученые разных областей нейробиологии.
Практически все виды животных в той или иной степени способны
анализировать изменения, происходящие во внешней среде, и адек-
ватно на них реагировать. При этом повторная реакция организма на
внешнее воздействие зачастую бывает иной, чем при первом столкно-
вении. Это наблюдение показывает, что живым системам присуща
способность к обучению. Они обладают памятью, сохраняющей лич-
ный опыт животного, который формирует поведенческие реакции
и может отличаться от опыта других индивидуумов.
Биологическая память разнообразна. Она присуща не только клет-
кам мозга. Память иммунной системы, например, надолго (часто на
всю жизнь) сохраняет информацию о когда-то попавшем в организм
чужеродном антигене. При повторной встрече иммунная система за-
пускает реакцию образования антител, позволяющую быстро и эф-
фективно победить инфекцию. Однако иммунная система «знает», как
реагировать на известный фактор, а встречаясь с неизвестным аген-
том, должна вырабатывать стратегию поведения заново. Нервная сис-
тема, в отличие от иммунной, может обучаться создавать стратегию
3.8. Нейрохимические механизмы памяти
325
поведения в новых обстоятельствах, основываясь на «жизненном
опыте», который позволяет выработать эффективную реакцию на не-
известный раздражитель.
Основными вопросами, на которые следует получить ответ при
изучении молекулярных механизмов памяти, являются следующие:
какие метаболические изменения происходят в нейронах при встрече с
внешним раздражителем, позволяющие сохранять полученную ин-
формацию в течение определенного (иногда длительного) времени; в
каком виде хранится полученная информация; каким образом она ана-
лизируется?
В процессе активного обучения, происходящего в раннем возрасте,
наблюдаются изменения в структуре нейронов, увеличивается плот-
ность синаптических контактов, возрастает соотношение глиальных и
нервных клеток. Трудно разграничить процесс созревания мозга и
структурные изменения, являющиеся молекулярными носителями па-
мяти. Однако ясно, что для полноценного развития интеллекта необ-
ходимо решать задачи, предъявляемые внешней средой (вспомните
феномен Маугли или проблемы адаптации к жизни в природе живот-
ных, выращенных в неволе).
В последней четверти XX в. были предприняты попытки детально
изучить морфологические особенности мозга А. Эйнштейна. Однако
результат был скорее разочаровывающим — никаких особенностей,
отличающих его от среднестатистического мозга современного чело-
века, не выявилось. Единственное исключение заключалось в некото-
ром (незначительном) превышении соотношения глиальных и нерв-
ных клеток. Значит ли это, что молекулярные процессы памяти не ос-
тавляют видимых следов в нервных клетках?
С другой стороны, довольно давно было выяснено, что ингибиторы
синтеза ДНК не влияют на память, в то же время ингибиторы транс-
крипции и трансляции ухудшают процессы запоминания. Значит ли
это, что определенные белки в нейронах мозга являются носителями
памяти?
Организация мозга такова, что основные функции, связанные с
восприятием внешних сигналов и с реакциями на них (например, с
двигательной реакцией), локализованы в определенных отделах коры
головного мозга. Тогда выработка приобретенных реакций (условных
рефлексов) должна представлять собой «замыкание связей» между
соответствующими центрами коры. Экспериментальное повреждение
этого центра должно разрушать память о данном рефлексе.
Однако экспериментальная нейрофизиология накопила множество
свидетельств того, что память о приобретенных навыках распределена
326
Раздел 3. ' iMchk ii <А.ъ HI ЙРЦХИРИЯ
по разным отделам мозга, а не сконцентрирована только в области,
отвечающей за рассматриваемую функцию. Опыты с частичным нару-
шением коры у крыс, обученных ориентироваться в лабиринте, по-
казали, что время, требующееся для восстановления нарушенного на-
выка, пропорционально объему разрушения и не зависит от его лока-
лизации.
Вероятно, выработка поведения в лабиринте включает анализ це-
лого набора факторов (обонятельных, вкусовых, зрительных), и участ-
ки мозга, ответственные за этот анализ, могут располагаться в раз-
личных областях мозга. Таким образом, хотя за каждый компонент по-
веденческой реакции отвечает определенный участок мозга, общая
реакция осуществляется при их взаимодействии. Тем не менее в мозге
обнаружены отделы, функция которых имеет непосредственное отно-
шение к процессам памяти. Это гиппокамп и миндалевидный комп-
лекс, а также ядра средней линии таламуса.
Совокупность изменений в ЦНС, связанных с фиксацией инфор-
мации (образа, типа поведения и т. д.), нейробиологи называют эн-
граммой. Современные представления о молекулярных механизмах
памяти говорят о том, что участие отдельных структур мозга в процес-
се запоминания и хранения информации заключается не в хранении
специфических энграмм, а в регуляции создания и функционирования
нейронных сетей, осуществляющих запечатление, фиксацию и восп-
роизведение информации.
В целом данные, накопленные при изучении поведенческих реф-
лексов и электрической активности мозга, свидетельствуют о том, что
и поведенческие, и эмоциональные проявления жизнедеятельности
не локализованы в определенной группе нейронов головного мозга, а
выражаются в изменении взаимодействий большого числа нервных
клеток, отражающих функционирование всего мозга как интегральной
системы.
Для описания протекания процесса запоминания новой информа-
ции во времени часто применяют термины краткосрочная память и
долговременная память. В краткосрочной памяти информация мо-
жет сохраняться от долей секунды до десятков минут, тогда как в дол-
говременной памяти информация иногда содержится всю жизнь. Для
превращения первого вида памяти во второй необходим так называе-
мый процесс консолидации. Иногда его выделяют в отдельную ста-
дию промежуточной памяти (табл. 3.9). Однако все эти термины, ве-
роятно, отражающие очевидные процессы, пока еще не наполнены
реальными биохимическими данными.
3.8. Нейрохимические механизмы памяти
Таблица 3.9. Виды памяти и их модуляция (по: Ашмарин, 1999)
Виды памяти Ингибиторы, воздействия
Кратковременная память Электрошок, л*-холинолитики (атропин, скопо- ламин), галанин, LiCl (введение в специфиче- ские отделы мозга)
Промежуточная память (консолидация) Ингибиторы энергетического обмена, уабаин, гипоксия, ингибиторы синтеза РНК и бел- ков (анизомицин, циклогексимид, пуромицин, актиномицин D, РНКаза), антитела к нейро- специфическим белкам (вазопрессину, белку S-100), 2-амино-5-фосфорновалериановая кис- лота (D-APV)
Долговременная (пожизненная) память Ингибиторы, необратимо нарушающие ее, неиз- вестны. Частично подавляется атропином, ди- изопропилфторфосфатом, скополамином
Кратковременная память. Кратковременная память, осуществ-
ляющая анализ информации, поступающей от различных органов
чувств, и ее обработку, реализуется при участии синаптических контак-
тов. Это кажется очевидным, поскольку время, в течение которого осу-
ществляются эти процессы, несоизмеримо со временем синтеза новых
макромолекул. Это подтверждается и возможностью ингибирования
кратковременной памяти синаптическими ингибиторами, и ее нечувст-
вительностью к ингибиторам синтеза белка и РНК (см. табл. 3.9).
Процесс консолидации занимает более длительное время и не вме-
щается в строго определенный интервал (продолжаясь от нескольких
минут до нескольких суток). Вероятно, на длительность этого периода
влияет как качество информации, так и состояние мозга. Информа-
ция, которую мозг сочтет несущественной, не подвергается консоли-
дации и исчезает из памяти. Остается загадкой, как решается вопрос о
ценности информации и каковы реальные нейрохимические механиз-
мы процесса консолидации. Сама длительность процесса консолида-
ции позволяет считать, что он является постоянным состоянием моз-
га, непрерывно осуществляющим «мыслительный процесс». Разно-
образный характер информации, поступающей в мозг для анализа, и
широкий спектр разных по механизму действия ингибиторов процесса
консолидации позволяют предполагать, что на этом этапе во взаимо-
действие вовлекаются разнообразные нейрохимические механизмы.
Применение соединений, указанных в таблице 3.9 в качестве инги-
биторов процесса консолидации, вызывает у подопытных животных
328
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
амнезию (потерю памяти) — неспособность воспроизвести вырабо-
танный поведенческий навык или предъявить для использования по-
лученную информацию.
Интересно, что некоторые ингибиторы проявляют свое действие
после предъявления запоминаемой информации (ретроградная ам-
незия), а другие — при применении в предшествующий этому период
(антероградная амнезия). Широко известны эксперименты по обу-
чению цыплят отличать зерно от несъедобных, но похожих по размеру
объектов. Введение в мозг цыплят ингибитора белкового синтеза
циклогексимида не мешало процессу обучения, но полностью пред-
отвращало закрепление навыка. Напротив, введение ингибиторов
Na-насоса (Na/K-АТФазы) уабаина полностью тормозило процесс
обучения, не оказывая влияния на уже сформированные навыки. Это
означает, что Na-насос вовлечен в формирование кратковременной
памяти, но не участвует в процессах консолидации. Более того, ре-
зультаты опытов с циклогексимидом указывают на то, что синтез но-
вых молекул белка необходим для осуществления процессов консоли-
дации, но не нужен для формирования кратковременной памяти.
Следовательно, обучение в период формирования кратковремен-
ной памяти включает в себя активацию определенных нейронов, а
консолидация — создание долговременных межнейронных сетей, для
закрепления взаимодействия в которых необходим синтез специаль-
ных белков. Не следует ожидать, что эти белки будут носителями
конкретной информации, их образование может быть «всего лишь»
побудительным фактором для активации межнейронных связей. Ка-
ким образом консолидация приводит к формированию долговремен-
ной памяти, которую нельзя нарушить, но можно воспроизвести по
первому требованию, остается неясным.
В то же время понятно, что за созданием стойкого навыка стоит спо-
собность популяции нейронов формировать сеть, в которой передача
сигнала становится наиболее вероятной, и эта способность мозга может
сохраняться длительное время. Наличие одной такой межнейронной се-
ти не мешает Нейронам вовлекаться в аналогичные иные сети. Поэтому
ясно, что аналитические способности мозга весьма велики, если не без-
граничны. Понятно также, что реализация этих способностей зависит от
интенсивности обучения, особенно в период созревания мозга в онтоге-
незе. С возрастом способность к обучению падает.
Обучаемость тесно связана со способностью к пластичности —
способности синаптических контактов к функциональным перестрой-
кам, происходящим в процессе функционирования, направленным на
синхронизацию нейрональной активности и создание межнейронных
3.8. Нейрохимические механизмы памяти
сетей. Проявление пластичности сопровождается синтезом специфи-
ческих белков, выполняющих известные (например, рецепторные) или
неизвестные функции. Одним из участников реализации этой програм-
мы является белок S-100, относящийся к аннексинам и обнаруживае-
мый в мозге в особенно больших количествах (он получил свое назва-
ние от способности оставаться растворимым при 100-процентном на-
сыщении сульфатом аммония при нейтральных значениях pH). Его
содержание в мозге на несколько порядков больше, чем в других тка-
нях. Он аккумулируется преимущественно в клетках глии и обнаружи-
вается вблизи синаптических контактов. Содержание белка S-100 в
мозге начинает увеличиваться через 1 ч после обучения и достигает
максимума через 3—6 ч, оставаясь на высоком уровне в течение не-
скольких суток. Введение антител к этому белку в желудочки мозга
крыс нарушает обучаемость животных. Все это позволяет считать бе-
лок S-100 участником создания межнейронных сетей.
Молекулярные механизмы пластичности нервной системы.
Пластичность нервной системы определяется как способность
нейронов воспринимать сигналы внешней среды, изменяющие жест-
кий детерминизм генома. Пластичность подразумевает возможность
изменения функциональной программы взаимодействия нейронов в от-
вет на изменения внешней среды (Хухо, 1989).
Молекулярные механизмы пластичности многообразны. Рассмот-
рим основные из них на примере глутаматергической системы. Как
было изложено ранее (см. гл. 2.3), в глутаматергическом синапсе од-
новременно обнаруживаются различные по свойствам рецепторы —
как ионотропные, так и метаботропные. Высвобождение глутамата в
синаптическую щель в процессе возбуждения приводит к активации
каинатных и АМРА-активируемых ионотропных рецепторов, вызы-
вающих деполяризацию постсинаптической мембраны. При величине
трансмембранного потенциала, соответствующей величине потенци-
ала покоя, NMDA-рецепторы не активируются глутаматом вследствие
того, что их ионные каналы блокированы Mg2+. По этой причине
NMDA-рецепторы не имеют шанса для первоочередной активации.
Однако при начинающейся деполяризации синаптической мембраны
ионы магния удаляются из участка связывания, что резко повышает
сродство рецептора к глутамату.
Активация NMDA-рецепторов вызывает вход кальция в постсинапти-
ческую зону через ионный канал, принадлежащий молекуле NMDA-ре-
цептора. Вход кальция наблюдается также и через потенциалзависимые
Са-каналы, активируемые вследствие работы каинатных и АМРА-глута-
330
раздел з. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЙ
матных рецепторов. В результате совокупности этих процессов в при-
мембранных областях постсинаптической зоны увеличивается содержа-
ние ионов кальция. Этот сигнал слишком слаб, чтобы изменить актив-
ность многочисленных ферментов, чувствительных к ионам кальция, но
достаточно значителен, чтобы активировать примембранную фосфо-
липазу С, субстратом которой является фосфоинозит, и вызвать на-
копление инозитолфосфатов и активацию инозитол-3-фосфатзависи-
мого высвобождения кальция из эндоплазматического ретикулума.
Таким образом, активация ионотропных рецепторов не только вы-
зывает в постсинаптической зоне деполяризацию мембраны, но и со-
здает условия для существенного повышения концентрации ионизиро-
ванного кальция. Тем временем глутамат активирует в синаптической
области и метаботропные рецепторы. В результате возникает воз-
можность приводить в действие соответствующие G-белки, «привя-
занные» к различным эффекторным системам. Могут активироваться
киназы, фосфорилирующие разнообразные мишени, в том числе и
ионотропные рецепторы, что модифицирует активность канальных
структур этих образований (см. рис. 2.23).
Более того, на пресинаптической мембране также локализованы
глутаматные рецепторы, которые тоже имеют шанс взаимодейство-
вать с глутаматом. Метаботропные рецепторы этой области синапса
связаны с активацией системы удаления глутамата из синаптической
щели, работающей по принципу обратного захвата глутамата. Этот
процесс зависит от активности Na-насоса, поскольку является вто-
ричноактивным транспортом (см. рис. 2.14 и гл. 2.5).
Активация имеющихся на пресинаптической мембране NMDA-pe-
цепторов также вызывает повышение уровня ионизированного каль-
ция в пресинаптической области синаптического окончания. Накопле-
ние ионов кальция синхронизует слияние синаптических везикул с
мембраной, ускоряя высвобождение медиатора в синаптическую щель.
Когда в синапс приходит серия возбуждающих импульсов и общая
концентрация свободных ионов кальция оказывается стойко повы-
шенной, может наблюдаться активация Са-зависимой протеиназы
кальпаина, который расщепляет один из структурных белков фодрин,
маскирующий глутаматные рецепторы и предотвращающий их взаи-
модействие с глутаматом. Таким образом, высвобождение медиатора
в синаптическую щель при возбуждении предоставляет разнообраз-
ные возможности, реализация которых может привести к усилению
или торможению сигнала, или к его выбраковке: синапс работает по
многовариантному принципу, и реализуемый в каждый момент путь
зависит от множества разнообразных факторов (рис. 3.19).
3.8. Нейрохимические механизмы памяти
Метаботропные (mGluR)
Группа I Группа II Группа III
mglui mglii2 mglu4
mglu5 mglu3 mglu6
mglu7
mglu8
Ионотропные (iGluR)
NR1 GluRl GluR5 51
NR2A GluR2 GluR6 52
NR2B GluR3 GluR7
NR2C GluR4 KAI
NR2D KA2
NR3A
Рис. 3.19. Глутаматергический синапс как многовариантная система.
После выброса глутамата в синаптическую щель возможно его связывание с ио-
нотропными (iGluR) и метаботропными (mGluR) рецепторами постсинаптической
мембраны; при этом существует возможность как прямого влияния на Na- и Са-кана-
лы, так и включение через G-белки фосфолипазы С и аденилатциклазы, приводящее
в конечном счете к выходу Са2+ из внутриклеточных депо и синтезу цАМФ Кроме
того, возможно взаимодействие глутамата с его рецепторами на пресинаптической
мембране и мембране астроцитов, что приводит к модуляции синтеза и выделения
медиатора
В числе этих возможностей — самонастройка синапса на наилуч-
шую передачу сигнала, который оказался усиленным. Такой процесс но-
сит название долговременной потенциации (Longterm potentiation,
LTP). Он заключается в том, что при длительном высокочастотном раз-
дражении ответы нервной клетки на приходящие импульсы оказывают-
ся усиленными. Это явление представляет собой одну из сторон плас;
тичности, в основе которой лежит молекулярная память нейрональной
клетки. Период долговременной потенциации сопровождается повы-
шенным фосфорилированием определенных нейрональных белков спе-
цифическими протеинкиназами (см. гл. 2.8).
332
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕИРОХИМИЯ
Одним из результатов увеличения уровня ионов кальция в клетке яв-
ляется активация Са-зависимых ферментов (кальпаина, фосфолипаз,
Са-кальмодулинзависимых протеинкиназ). Некоторые из этих фермен-
тов имеют отношение к образованию активных форм кислорода и азота
(НАДФН-оксидаза, NO-синтаза и др.). В итоге в активированном нейро-
не можно зарегистрировать накопление свободных радикалов, которые
рассматриваются как вторичные посредники регуляции метаболизма.
Важным, но не единственным результатом накопления свободных
радикалов в нейрональной клетке является активация так называемых
генов раннего ответа. Этот процесс является наиболее ранним и
скоропреходящим ответом клеточного ядра на сигнал свободных ради-
калов, активация этих генов наступает через 5—10 мин и продолжа-
ется несколько часов. К числу этих генов относят группы c-fos, c-jun,
c-junB, zif/268 и др. Они кодируют несколько обширных семейств
специфических белков-регуляторов транскрипции.
Активация генов немедленного ответа происходит при участии
ядерного фактора NF-кВ, который должен для реализации своего дей-
ствия проникнуть в ядро через ядерную мембрану. Его проникновению
препятствует то обстоятельство, что этот фактор, представляющий
собою димер из двух белков (р50 и р65), в цитоплазме находится в
комплексе с белковым ингибитором и не способен проникать в ядро
(рис. 3.20). Ингибиторный белок является субстратом для фосфори-
лирования специфической протеинкиназой, после чего диссоциирует
от комплекса, что открывает дорогу для NF-кВ в ядро. Активирующим
кофактором протеинкиназы является пероксид водорода, поэтому
волна свободных радикалов, захватывая клетку, вызывает ряд описан-
ных выше процессов, приводящих к активации генов раннего ответа.
Активация c-fos может также вызывать синтез нейротрофинов и об-
разование нейритов и новых синапсов. Долговременная потенциация,
вызываемая высокочастотной стимуляцией гиппокампа, приводит к
активации zif/268, кодирующего Zn-чувствительный ДНК-связываю-
щий белок. Антагонисты NMDA-рецепторов блокируют долговремен-
ную потенциацию и актвиацию zif/268.
Одним из первых, кто предпринял в 1949 г. попытку разобраться в
механизме анализа информации в мозге и выработать стратегию пове-
дения, был С. О. Хебб. Он высказал мысль о том, что для выполнения
этих задач в мозге должна образовываться функциональная ассоци-
ация нейронов — локальная межнейронная сеть. Уточнил и углубил
эти представления М. Розенблат (1961), сформулировавший гипотезу
«Unsupervised correlation base learning». Согласно развиваемым им
представлениям, в случае генерации серии разрядов нейроны могут
3.8. Нейрохимические механизмы памяти
Н2О2 + Ог
Рис. 3.20. Активация NF-kB активными формами кислорода.
Активный ядерный фактор NF-kB образуется из неактивного при фосфорилировании
специфической протеинкиназой, кофактором которой является пероксид водорода.
Поэтому АФК и антиоксинанты, влияя на специфическую протеинкиназу (и активацию
NF-kB), оказывают воздействие на активацию генов раннего ответа — синтез цитоки-
нов, адгезивных белков и процессы пролиферации
синхронизироваться за счет ассоциации определенных (зачастую мор-
фологически удаленных друг от друга) клеток путем самонастраивания.
Современная нейрохимия подтверждает возможность такой само-
настройки нейронов на общую частоту, объясняя функциональное
значение серий возбуждающих «разрядов» для .создания межнейрон-
ных цепей. Использовав аналог глутамата с флуоресцентной меткой и
вооружившись современной техникой, удалось показать, что даже при
стимуляции одного синапса возбуждение может распространяться и
на достаточно удаленные синаптические структуры за счет образова-
ния так называемой глутаматной волны. Условием формирования
такой волны является повторяемость сигналов в определенном час-
тотном режиме. Ингибирование глутаматного транспортера увеличи-
вает вовлеченность нейронов в процесс синхронизации.
Кроме глутаматергической системы, имеющей прямое отношение к
процессам обучения (запоминания), другие системы мозга также участ-
вуют в формировании памяти. Известно, что способность к обучению
обнаруживает положительную корреляцию с активностью холинацетил-
334
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
Т. Т. Березов
Создатель современной шко-
лы отечественной нейрохи-
мии, автор учебника «Меди-
цинская биохимия», основа-
тель кафедры медицинской
биохимии в Российском уни-
верситете дружбы народов
трансферазы и отрицательную — с фер-
ментом, гидролизующим этот медиатор —
ацетилхолинэстеразой. Ингибиторы холи-
нацетилтрансферазы нарушают процесс
обучения, а ингибиторы холинэстеразы
способствуют выработке оборонительных
рефлексов.
В формировании памяти принимают
участие и биогенные амины — норадре-
налин и серотонин. При выработке услов-
ных рефлексов с отрицательным (элек-
троболевым) подкреплением происходит
активация норадренергической системы,
а при положительном (пищевом) под-
креплении скорость метаболизма норад-
реналина снижается. Серотонин, напро-
тив, облегчает выработку навыков в ус-
ловиях положительного подкрепления и
отрицательно воздействует на формиро-
вание оборонительной реакции. Таким
образом, в процессе консолидации памя-
ти серотонинергическая и норадреналинергическая системы являются
своего рода антагонистами, и нарушения, вызываемые избыточным
накоплением серотонина, по-видимому, могут быть компенсированы
активацией норадренергической системы.
Участие дофамина в регуляции процессов памяти имеет многофактор-
ную природу. С одной стороны, выявлено, что он может стимулировать
выработку условных рефлексов с отрицательным подкреплением. С дру-
гой стороны, он снижает фосфорилирование нейрональных белков (на-
пример, белка В-50) и индуцирует обмен фосфоинозитидов. Можно пола-
гать, что дофаминергическая система участвует в консолидации памяти.
Нейропептиды, выделяющиеся в синапс.е в процессе возбуждения,
также вовлекаются в процессы формирования памяти. Вазоактив-
ный интестинальный пептид увеличивает сродство холинорецеп-
торов к медиатору в несколько тысяч раз, способствуя функциониро-
ванию холинергической системы. Гормон вазопрессин, выделяющий-
ся из задней доли гипофиза, синтезируемый в супраоптических ядрах
гипоталамуса, аксональным током переносится в заднюю долю гипо-
физа, где хранится в синаптических везикулах, и оттуда выделяется в
кровь. Этот гормон, а также гипофизарный адренокортикотроп-
ный гормон (АКТГ) постоянно функционируют в мозге в качестве ре-
гуляторов процессов памяти. Следует подчеркнуть, что этот эффект
3.8. Нейрохи ми । с к •• -- м^х ? н и эм г» г м яти
33
отличается от их гормональной активности — фрагменты данных со-
единений, лишенные этой активности, оказывают такое же воздейст-
вие на процесс обучения, как и целые молекулы.
Непептидные стимуляторы памяти практически неизвестны. Иск-
лючение составляют оротат и широко применяющийся в клинике
пирацетам. Последний представляет собой химический аналог гам-
ма-аминомасляной кислоты и относится к группе так называемых
ноотропных препаратов, одним из эффектов которых является уси-
ление мозгового кровообращения.
С изучением роли оротата в механизмах закрепления памяти связана
интрига, волновавшая умы нейрохимиков во второй половине XX в. Исто-
рия началась с опытов Дж. Мак-Коннела (J. McConnel, 1925—1990) по
выработке у примитивных плоских червей планарий условного рефлекса на
свет. После создания устойчивого рефлекса он разрезал планарий поперек
на две части и проверял обучаемость тому же рефлексу у животных, регене-
рировавших из обеих половинок. Сюрпризом было то, что не только особи,
получившиеся из головной части, обладали повышенной обучаемостью, но и
те, что регенерировали из хвоста, обучались гораздо быстрее, чем контроль-
ные особи. Для обучения тех и других требовалось в 3 раза меньше времени,
чем для особей, регенерируемых от контрольных животных. Мак-Коннел
сделал вывод о том, что приобретенная реакция кодируется веществом, ко-
торое накапливается как в головной, так и в хвостовой части тела планарий.
Воспроизведение результатов Мак-Коннела на других объектах встре-
тило ряд трудностей, вследствие чего ученого объявили шарлатаном, а
статьи его перестали принимать для публикации во всех научных журна-
лах. Разгневанный автор основал собственный журнал, где публиковал не
только результаты последующих экспериментов, но также и шаржи на
своих рецензентов и пространные описания экспериментов, проводимых
им в ответ на критические замечания. Благодаря такой уверенности Мак-
Коннела в своей правоте современная наука имеет возможность возвра-
титься к анализу этих оригинальных научных данных.
Обращает на себя внимание тот факт, что в тканях «обученных» планарий
обнаруживается повышенное содержание оротовой кислоты, которая являет-
ся метаболитом, необходимым для синтеза РНК- Результаты, полученные
Мак-Коннелом, можно трактовать следующим образом: условия для более
быстрого обучения создает повышенное содержание оротата у «обученных»
планарий. Когда исследуют обучаемость регенерировавших планарий, сталки-
ваются не с переносом памяти, а с переносом навыка к ее формированию.
С другой стороны, оказалось, что когда регенерация планарий осу-
ществляется в присутствии РНКазы, повышенную обучаемость демонст-
рируют только особи, полученные от головного фрагмента. Независимые
эксперименты, проведенные в конце XX в. Г. Унгара, позволили выделить
из мозга животных, обладающих рефлексом избегания темноты, 15-член-
ный пептид, получивший название скотофобин (индуктор боязни темно-
ты). По-видимому, и РНК, и некоторые специфические белки способны
создавать условия для запуска функциональных связей (межнейронных
сетей), аналогичных тем, которые были активированы у исходной особи.
336
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕИРОХИМИЯ
В 2005 г. исполнилось 80 лет со дня рождения Мак-Коннела, опыты
которого положили начало исследованию молекулярных носителей памя-
ти. На рубеже XX и XXI вв. появились новые методы геномики и протео-
мики, использование которых позволило выявить вовлечение низкомоле-
кулярных фрагментов транспортных РНК в процессы консолидации.
Новые факты дают возможность пересмотреть и концепцию о не-
участии ДНК в механизмах долговременной памяти. Обнаружение в
ткани мозга РНК-зависимой ДНК-полимеразы и наличие положитель-
ной корреляции ее активности с обучаемостью указывают на возмож-
ность участия ДНК в процессах формирования памяти. Обнаружилось,
что выработка пищевых условных рефлексов резко активирует опреде-
ленные участки (гены, ответственные за синтез специфических белков)
ДНК в неокортексе. При этом отмечается, что активация ДНК затра-
гивает главным образом участки, редко повторяемые в геноме, и на-
блюдается не только в ядерной, но и в митохондриальной ДНК, причем
в последней — в большей степени. Факторы, подавляющие память,
одновременно подавляют и эти синтетические процессы.
Таблица 3.10. Некоторые стимуляторы запоминания
(по: Ашмарин, Стукалов, 1996)
Специфичность действия Стимуляторы
Классы соединений Примеры веществ
Относительно специфичные агенты Регуляторные пептиды Вазопрессин и его аналоги, дипептид pEDA, АКТГ и его аналоги
Непептидные соединения Пирацетам, ганглиозиды
Регуляторы метаболизма РНК Оротат, низкомолекулярная РНК
Агенты широкого спектра действия Нейростимуляторы Антидепрессанты Фенилалкиламины (фенамин), фенилалкилоиднонимины (сиднокарб) Дигидрохлорид 2-(4-метил- 1-пиперазинил)-10-метил- 3,4-диазафеноксазина (азафен)
Модуляторы холинергической системы Холиномиметики, ингибиторы ацетилхолинэстеразы
3.9. Окислительный стресс и нейропатологии
337
3.9.
В таблице 3.10 приведены примеры соединений, стимулирующих
запоминание.
Возможно, что исследование участия ДНК в процессах формиро-
вания памяти позволит дать обоснованный ответ на вопрос, существу-
ют ли условия, при которых сформированные навыки или возникшие
впечатления могут быть наследуемы. Возможно, что генетическая па-
мять о давних событиях, пережитых предками, лежит в основе некото-
рых необъяснимых пока явлений психики.
Согласно остроумному, хотя и бездоказательному мнению, полеты во
сне, сопровождающие окончательное формирование зрелого мозга, пере-
живаемые каждым из нас в молодости, отражают ощущение полета, пере-
житого нашими далекими предками в пору, когда они ночевали на деревь-
ях. Недаром полеты во сне никогда не заканчиваются падением — ведь те
далекие предки, которые при падении не успевали схватиться за ветки, хо-
тя и переживали это ощущение перед смертью, но не давали потомства...
ОКИСЛИТЕЛЬНЫЙ СТРЕСС И НЕЙРОПАТОЛОГИИ
Как было рассмотрено ранее, свободные радикалы, в число ко-
торых входят активные формы кислорода и азота, выполняют функции
вторичных посредников, обеспечивая как циклическими нуклеотида-
ми, так и ионами кальция и другими сигнальными молекулами адап-
тацию клеточного метаболизма к изменяющимся условиям среды.
Свободнорадикальные соединения являются высокореакционноспо-
собными и поэтому короткоживущими (их характеристика была пред-
ставлена в разд. 2).
Стационарный уровень свободнорадикальных соединений в клетке
очень низок, поскольку регулируется специальной системой анти-
оксидантной защиты. В состав этой системы входят как белки-
ферменты, так и низкомолекулярные соединения — «тушители»
свободных радикалов, а также белковые и небелковые факторы,
ограничивающие образование активных форм кислорода (АФК)
(табл. 3.11).
Из таблицы 3.11 следует, что в состав небелковых анти-
оксидантов входят соединения растительного происхождения, не
синтезируемые в организме многих животных. Например, аскорбино-
вая кислота является витамином для человекообразных обезьян и че-
ловека, в то время как мартышки «умеют» синтезировать аскорбат.
Значит ли это, что при выборе «подходящего претендента» на роль
развитого интеллекта природе пришлось поступиться антиоксидант-
ной защитой мозга для обеспечения сигнальных функций АФК?
Таблица З.Н.Система антиоксидантной защиты клетки
Антиоксиданты Локализация
Ферменты и белки
сод
Cu/Zn-СОД Эритроциты, цитоплазма
Мп-СОД Митохондрии
Внеклет. СОД Плазма крови, стенки сосудов
Каталаза Пероксисомы
Глутатионпероксидаза Цитоплазма, митохондрии
Глутатионтрансфераза Наружная мембрана митохондрии, эндоплазматический ретикулум
Феритин Цитоплазма
Трансферрин Внеклеточная среда
Лактоферрин Внеклеточная среда
Церулоплазмин
Внеклеточная среда
Функция
Са)
Са)
00
Тушение О2
Тушение О2
Тушение О2
Тушение Н2О2
Деградация Н2О2 и гидроперекисей
липидов
Деградация Н2О2 и гидроперекисей
липидов
Хелатор Fe2+
Хелатор Fe2+
Хелатор Fe2+
Хелатор Си2+, окисление Fe2+,
тушение О2“
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕИРОХИМИЯ
Окончание табл. 3.11
Альбумин Внеклеточная среда Хелатор Си2+, тушитель ’ОН, LOO’, НОС1
Витамин Е Низкомолекулярные соединения Биомембраны Тушение ОН’, LOO’, HOCI и т. п.
Убихинон Биомембраны Тушение ОН’, LOO’, HOCI и т. п.
Каротиноиды Биомембраны Тушение ОН’, LOO’, HOCI, *О2
Витамин С Цитоплазма Тушение ОН’, О2“
Карнозин Цитоплазма Тушение ОН’, О2“, нейтрализация гипохлорита
Таурин Цитоплазма Нейтрализация гипохлорита
Глутатион Цитоплазма, митохондрии Тушение ОН’, О2“
Мочевая кислота Кровь Предотвращение перекисного окисления липидов
Билирубин Кровь Предотвращение перекисного окисления липидов
3.9. Окислительный стресс и нейропатологии
340
»аздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
Г. X. Бунатян
Один из основоположников
Содержание в тканях а-токофе-
рола зависит от характера питания чело-
века, и при недостатке его в пище наблю-
даются Е-авитаминозы. Не образуются в
организме животных и флавоноиды, яв-
ляющиеся активными «тушителями» сво-
бодных радикалов. С другой стороны, мно-
гие метаболиты способны выполнять анти-
радикальные функции, например мочевая
кислота, маннитол, глицерин. В услови-
ях сбалансированного метаболизма клетки
стационарный уровень АФК очень низок
(Ланкин с соавт., 2001; Меньшикова
с соавт., 2006).
отечественной нейрохимии,
автор приоритетных работ в
области биохимии мозга;
впервые установил участие
аскорбиновой кислоты в об-
мене фосфолипидов
Окислительный стресс. Дисбаланс
между образованием и нейтрализацией
АФК наблюдается при смене функци-
онального состояния клетки. Как было от-
мечено выше, активация нейронов приво-
дит к кратковременному росту АФК, что
указывает на их возможную сигнальную функцию. Благодаря этой сиг-
нализации, клетки приспосабливаются к изменяющимся условиям
функционирования. «Вспышка» АФК является кратковременной и ту-
шится за счет окислительно-восстановительного потенциала клетки.
Однако известны многочисленные отклонения от сбалансирован-
ного метаболизма, вызванные различными патологиями, а также воз-
растными изменениями или наличием патогенных факторов среды,
которые связаны с нарушением контроля за стационарным уровнем
АФК. Это могут быть генетические или возрастные дефекты антиок-
сидантных ферментов (часто СОД), загрязнение воздуха, воды и пи-
щевых продуктов тяжелыми металлами, профессиональные заболева-
ния (асбестовая пыль, содержащая паракват), провоцирующие об-
разование свободных радикалов, а также метаболические отклонения,
приводящие к избыточной продукции АФК (дефекты митохондриаль-
ной дыхательной цепи, нарушения обмена биогенных аминов и т. д.)
(рис. 3.21).
В этих условиях в тканях наблюдается стойкое возрастание уровня
свободных радикалов, не компенсируемое антиоксидантной системой;
это состояние носит название окислительного стресса. Содержание
свободнорадикальных соединений при окислительном стрессе резко
возрастает (табл. 3.12).
3.9. Окислительный стресс и нейропатологии
341
Метаболические
Рис. 3.21. Дисбаланс АФК при окислительном стрессе
Перепроизводство АФК при окислительном стрессе чревато окис-
лительной модификацией клеточных структур. В первую очередь по-
вреждаются клеточные белки. АФК способны модифицировать разно-
образные аминокислотные остатки, при этом кислородные радикалы
проявляют различные свойства (табл. 3.13).
В результате окислительной модификации некоторые белки-
ферменты теряют свои свойства, другие утрачивают чувствитель-
ность к регуляторным сигналам, например при нитрозилировании ра-
дикалов тирозина белковые молекулы перестают фосфорилироваться
тирозиновыми киназами.
Таблица 3.12. Содержание некоторых свободных радикалов
в норме и при окислительном стрессе, мкМ
Свободнорадикальные соединения и их метаболиты В норме При окислительном стрессе
Супероксид-анион кислорода 10 10
Пероксид водорода Отсутствует 100—120
Гидроксид-радикал < 0,015 нМ До 75
NO-радикал 5—10 <5
Пероксинитрит Отсутствует 100 и выше
Пероксильный радикал Отсутствует 100—120
342
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИИ
Таблица 3.13. Окислительная модификация
аминокислотных радикалов белков
Модифицируе- мая группа Окислитель Модификация Дополнительные замечания
Цистеин Супероксид- анион, ОН*, ОСП Сульфоновые, дисульфидные группы S—S-связи
Метионин ОН*, HOCI-, Н2О2, 'О2 Сульфоксиды Легко подвергаются дальнейшему окислению
Гистидин Супероксид- анион, *О2 2-Охо-гистидин Вызывает поперечные сшивки белков
Пролин, аргинин Различные АФК Образование полуальдегидов Активация поперечных сшивок
Триптофан Пероксинитрит ОН* Образование 6-нитротрипто- фана Образование флуоресцентных продуктов
Фенилаланин ОН* Образование битирозиновых радикалов
Тирозин АФК. ОСГ, NO Нитрование, хлоринирование или образование битирозиновых сшивок Ингибирование передачи клеточного сигнала через тирозинкиназы блокированием фосфорилирования тирозина
Концевые NH2-rpynnbi белков Гипохлорит Образование белковых карбонилов Легко образуют поперечные сшивки
При атаковывании белков АФК наиболее легко происходит
окисление Е-аминогрупп лизина, в результате чего образуются карбо-
нильные производные белков. Карбонильные группы атакуются
аминогруппами липидов или сахаров с образованием шиффовых ос-
нований. Их последующая перегруппировка приводит к образованию
так называемых продуктов Амадори (рис. 3.22); далее происходит
формирование прочных ковалентных сшивок, ограничивающих кон-
формационную лабильность белковых молекул.
3.9. Окислительный стресс и нейропатологии
343
Продукты Амадори
Кетоальдегиды
Рис. 3.22. Взаимодействие окисленных белков с углеводами и липидами при
окислительном стрессе
Рис. 3.23. Перекисное окисление липидов
344
раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
Аналогичные продукты образуются и при атаковывании амино-
групп белка молекулами сахаров, активированными свободными ра-
дикалами. Далеко зашедший процесс свободнорадикальной атаки
белков и углеводов является практически необратимым.
Провоцирующим фактором окислительного стресса является повы-
шенная концентрация глюкозы в крови, что имеет место при диабете.
Поэтому сопутствующими проявлениями диабета являются возникно-
вение незаживающих кожных ран, развитие катаракты хрусталика.
Липиды подвергаются окислительной модификации труднее,
чем белки. В первую очередь окисляются ненасыщенные жирнокис-
лотные цепи мембранных фосфолипидов, содержание которых осо-
бенно велико в клетках нервной системы. Инициация окисления ли-
пидов может быть осуществлена даже наименее реакционноспособ-
ным супероксид-анионом кислорода, в результате чего в мембране
происходит образование липидных гидроперекисей (поэтому процесс
и называют перекисным окислением липидов. ПОЛ) (рис. 3.23).
При образовании гидроперекиси в мембранном бислое возникает локаль-
ное «возмущение» упаковки, вызывающее активацию фосфолипазы и удале-
ние ацильной цепи, несущей перекисную группу. В области образования ли-
зофосфолипщюв образуется дефектная зона, которая может быть репариро-
LH
Липид
LH’ х
Липидный
радикал
О2
Свободно- w Окисленный Окисленная Репарированная
радикальная бислой . жирная кислота, мембрана
атака бислоя с нарушенной подвергающаяся
структурой восстановлению
Глу-Пер
LOO* х/Ч/Ч/Х
Пероксиль- I
ный 00*
радикал
Рис. 3.24. Репарация мембраны, поврежденной перекисным окислением липидов
3.9. Окислительный стресс и нейропап логии
вана с помощью ферментов (рис. 3.24). В случае когда клетка в состоянии
противостоять окислительной атаке на липиды, репарирующие системы ус-
певают починить образующиеся дефектные зоны. Поскольку в первую оче-
редь окисляются полиненасыщенные жирнокислотные радикалы, а при репа-
рации к образованным молекулам лизофосфолипидов можно присоединять
различные жирнокислотные радикалы, инициацию ПОЛ можно рассматри-
вать как способ изменения жирнокислотного состава мембранных липидов.
Более того, инициация ПОЛ приводит к высвобождению из мембран арахи-
доновой кислоты, являющейся незаменимым субстратом для образования
клеточных регуляторов (простагландинов, лейкотриенов, тромбоксанов).
Однако если процесс ПОЛ выходит из-под контроля, образуются бо-
лее стабильные продукты, в числе которых — малоновый диальдегид
(РАДА), а также короткоцепочечные летучие углеводороды (Каган, 1998).
Как видно из рисунка 3.25, различные молекулярные продукты могут яв-
ляться маркерами окисления липидов.
он
Восстановление
RCH2OH -<---------------
Гидроксиноненол
а-Токоферол
4 - Гвдрокси - 2 - ноненал ь (HNE)
Окисление
----------RCOOH
4-Гидрокси-
2-ноненоевая
кислота
GSH
Н3С(СН2)4СН-СН-СН2СНО
ОН SG
+Тиомочевина
-NH2
Аддукты
с белками,
липидами
и нуклеиновыми
кислотами
МДА
Циклизация
+ТБК
Биомаркер продукции
HNE
Рис. 3.25. Маркеры окисления липидов
346
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
В. Каган и А. Шведова-Каган
Ученые, плодотворно работающие в области функциональной нейрохимии, авто-
ры препарата «эмоксипин», применяющегося для коррекции окислительного
стресса у пациентов с нейропатологиями
Наиболее широко используется спектрофотометрическое определение
МДА по реакции с тиобарбитуровой кислотой (ТБК). Стационарный уро-
вень МДА в тканях очень низок, при индукции ПОЛ он повышается. Од-
нако этот тест является наименее эффективным. Во-первых, МДА явля-
ется конечным продуктом окислительного процесса; в определенных усло-
виях он может и не накапливаться, а его промежуточные продукты будут
нарастать. Во-вторых, с ТБК вступают в реакцию и другие соединения
(предшественники МДА), которые внесут свой вклад в окрашивание
опытных образцов. Поэтому оценить долю окраски, обеспечиваемую
только уровнем МДА, невозможно. При поверхностном анализе сам факт
усиления окраски проб после инкубации с ТБК может удовлетворительно
отражать степень индукции процесса ПОЛ или его подавления. Выража-
ясь более строго, этот тест отражает тканевой уровень ТБК-реактивных
продуктов (ТБК-РП). В большей степени характер окисления липидов
можно оценить при одновременном измерении нескольких продуктов (ди-
еновых конъюгатов, гидроксиноненалей) (см. рис. 3.24 и 3.25).
Тем не менее нарастание уровня МДА свидетельствует о том, что кле-
точные компоненты в этих условиях могут атаковываться данным альдеги-
дом, который будет индуцировать образование беспорядочных попереч-
ных сшивок между различными группами белков и липидов, способными
взаимодействовать с альдегидными группировками. Таким образом, выра-
женная активация ПОЛ, с одной стороны, нарушает целостность клеточ-
ной мембраны, а с другой — вызывает инактивацию мембранных белков,
снижая жизнеспособность клетки. Неудивительно, что эффективность
ПОЛ используется в клинической биохимии в качестве показателя разви-
тия окислительного стресса и патологического состояния тканей. Актива-
ция ПОЛ сопровождает развитие многих патологий.
3.9. Окислительным стресс и неиропатологии
Дальнейшее развитие окислительного стресса расширяет область
клеточных повреждений, вовлекая в них нуклеиновые кис-
лоты. Ядерная ДНК является наиболее устойчивой к действию
АФК. Вследствие упаковки в двойную спираль и благодаря «маскиров-
ке» гистонами, ядерная ДНК не повреждается ни супероксид-анионом,
ни пероксидом водорода. Только при накоплении гидроксид-радикалов
происходят разрывы полинуклеотидной цепи, окисление углеводных
фрагментов и отдельных нуклеотидов. Митохондриальная ДНК, ли-
шенная гистонов, более подвержена атаке свободными радикалами.
ДНК в делящемся ядре также более уязвима.
Ежесуточно в организме крысы возникает около 10 тыс. мутаций,
являющихся результатом окислительного повреждения ДНК. Они
своевременно репарируются специальными ферментами, контроли-
рующими окислительную устойчивость генома. Силу атаки, направ-
ленной на нуклеиновые кислоты при окислительном стрессе, можно
оценить по содержанию в моче одного из конечных продуктов повреж-
дения ДНК— окисленного гуанозина (рис. 3.26). Следовательно,
особенности окислительного стресса в тканях можно оценить, исполь-
зуя маркеры окислительного повреждения клеточных макромолекул.
Все вышеизложенное свидетельствует о том, что АФК могут вы-
полнять в клетке двоякую роль: служат сигнальными молекулами в ус-
Рис. 3.26. Образование дигидроксигуанозина при окислении гуаниловых осно-
ваний, высвобождающихся при репарации разрывов в цепи ДНК
348
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
Рис. 3.27. Двойственная роль свободных радикалов в клетке
ловиях нормального метаболизма клетки и являются фактором по-
вреждения, вызывающим необратимые нарушения клеточных макро-
молекул (рис. 3.27). Четко разграничить эти функции довольно трудно
(рис. 3.28), поскольку кроме клеточного уровня содержания АФК не-
обходимо учитывать и место их образования в клетке, а также эффек-
тивность ее системы антиоксидантной защиты.
Тем не менее одно из проявлений окислительного стресса имеет за-
ведомо положительное действие. Это способность мобилизовывать за-
щитные свойства организма и подготавливать его к перенесению чрез-
мерных нагрузок. Жизненные наблюдения отразили эту закономер-
ность в пословице: «Малые неприятности охраняют от больших бед».
Научное обоснование адаптационного значения окислительного стрес-
са в нашей стране было положено экспериментами Ф. 3. Меерсона
(1984—1993). Им было показано, что слабые индукторы окислитель-
ного стресса способны улучшать переносимость последующих испыта-
ний. Например, неизбегаемое электроболевое раздражение в течение
30 мин почти вдвое снижало смертность от последующего эксперимен-
тального инфаркта миокарда у лабораторных крыс. При этом метабо-
лические сдвиги, вызываемые инфарктом миокарда, были выражены
не до такой опасной степени. Ф. 3. Меерсон назвал это адаптацион-
ным проявлением окислительного стресса.
3.9. Окислительный стресс и неиропатологии
349
Рис. 3.28. Гравюра М. Эшера «Небо и вода»
К сожалению, продолжения эта работа не имела и спустя несколько
лет аналогичные данные были получены в отношении химического
стрессорного агента 3-нитропропионовой кислоты И. Кригельштайном,
который в 1997 г. назвал этот феномен химическим прекондициони-
рованием (chemical preconditioning). Термин, возможно, несколько
неуклюжий, но он прижился, поскольку элегантные опыты Меерсона
на тот период все еще оставались неизвестными. В настоящее время
описан целый ряд моделей прекондиционирования и показано, что в
его основе лежит активация некоторых генов раннего ответа, приво-
дящая к синтезу белков теплового шока (heat shock proteins).
Тот или иной фактор, индуцирующий прекондиционирование, вы-
зывает увеличение стационарного уровня АФК, которые являются
стимуляторами протеинкиназы, фосфорилирующей комплекс NF-kB
с его ингибитором и освобождающей этот фактор транскрипции от ин-
гибиторного контроля (см. рис. 3.25). Дополнительным фактором, оп-
ределяющим, какие гены надо активировать, является семейство
Янус-киназ, которые также активируются АФК и в активном состоя-
нии фосфорилируют группу белковых факторов, инициирующих
транскрипцию. Эти белковые факторы участвуют в передаче сигнала
350
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
от клеточной мембраны к ядру; они носят название STAT-белков (Sig-
nal Transduction and Activation Translation Proteins). Фосфорилирова-
ние этих белков Янус-киназой происходит по тирозиновому радикалу
(данная киназа относится к семейству тирозиновых киназ), вследствие
чего они димеризуются и приобретают способность транспортиро-
ваться через ядерную мембрану. В ядре совместно с NF-кВ они обес-
печивают экспрессию различных белков теплового шока. На рис. 3.29
показана последовательность этих процессов и указаны STAT-белки,
обнаруженные в сердце крысы, успешно перенесшей эксперимен-
тальный инфаркт миокарда (в случае смертельного исхода после опе-
рации в сердце животного обнаруживаются другие STAT-белки).
Рис. 3.29. Роль СТАТ-белков в передаче сигнала от АФК к ядру.
При действии повреждающего фактора (L) и возникновении в клетке свободноради-
кальных повреждений создаются условия для активации G-белков, что вызывает генера-
цию АФК. Основной мишенью Янус-киназ в клетке являются белки, регулирующие экс-
прессию генов — СТАТ-белки (Signal Transduction Activating Translation Proteins).
Янус-киназы относятся к тирозиновым киназам, фосфорилирующим белки по тирозино-
вым остаткам. Фосфорилирование ими СТАТ-белков приводит к димеризациии послед-
них и облегчает проникновение через ядерную мембрану, где они и создают условия для
образования белков теплового шока. Дефосфорилирование СТАТ-белков клеточными
фосфатазами препятствует их проникновению в ядро и проявлению эффекта Янус-киназ.
Низкие концентрации АФК(или супероксид-ан ион кислорода) активируют Янус-киназы,
а более высокие концентрации (гидроксид-радикал) ингибируют фосфатазы. Образова-
ние пероксинитрита ведет к противоположному влиянию — модифиции тирозиновых ос-
татков СТАТ-белков и потере их чувствительности к Янус-киназам
3.9. Онй1 1,’ieni. tik'iTpeocM иропат-.логн.’
Одновременно с активацией Янус-киназ АФК (в основном, гидро-
ксид-радикал) ингибируют протеинфосфатазы, которые могут ограни-
чивать димеризацию STAT-белков и их способность проникать в ядро.
Но дальнейшее развитие окислительного стресса приводит к накопле-
нию пероксинитрита, который, как отмечалось ранее, способен нитро-
зилировать тирозиновые радикалы белков, тем самым делая их нечув-
ствительными к тирозиновым киназам (см. рис. 3.29). В итоге клетки
могут как получить шанс адаптироваться к стрессу, так и не получить
его. Следовательно, АФК обеспечивают многовариантность функци-
онирования живых систем.
Устоявшейся догмой клинической биохимии является представле-
ние о значительном терапевтическом эффекте антиоксидантов, приме-
нение которых широко рекомендуется при ряде патологий. Изложен-
ный материал свидетельствует о том, что антиоксидантная терапия не
является панацеей: с одной стороны, перегрузка антиоксидантами на-
рушает сигнальную функцию АФК, а с другой — свободнорадикальная
атака поврежденных клеток является способом вызвать клеточную
смерть и избавиться от недоброкачественных структур.
Нейродегенеративные заболевания. Многие патологии нерв-
ной системы объединяет одно важное обстоятельство — они протекают
на фоне повышенного уровня АФК, вызванного их усиленным образова-
нием или дефектами антиоксидантной системы. Таким образом, заболе-
вание протекает в условиях постоянного окислительного стресса. В боль-
шинстве случаев стрессирующее действие АФК усугубляет развитие за-
болевания, поскольку приводит к массовой гибели нейронов.
Рассмотрим некоторые из неврологических заболеваний.
Типичным примером избирательной гибели нейронов головного
мозга, как непосредственной причины заболевания, является генети-
чески детерминированное нарушение обмена дофамина в стриатуме,
хвостатом ядре и особенно в черной субстанции, приводящее к болез-
ни Паркинсона, выражающейся в нарушениях стереотипной двига-
тельной активности, ее координации и инициации. Впервые эта бо-
лезнь была описана Д. Паркинсоном в 1817 г. Развитие болезни при-
водит к гибели дофаминергических нейронов в указанных областях
мозга, причем вследствие взаимозаменяемости аналогичных нейронов
симптомы болезни проявляются лишь тогда, когда имеет место потеря
более половины нейрональных клеток.
Гибель дофаминергических нейронов осуществляется по механиз-
му апоптоза, стимулированного АФК, причем одним из источников
АФК является реакция, катализируемая высокоактивной моноамино-
352
раздел 3. ФУНКЦИОНАЛЬНАЯ НЕИРОХИМИЯ
оксидазой В (МАО В), которая разрушает биогенные амины, превра-
щая их в семиальдегид и супероксид-анион кислорода (а затем —
в пероксид водорода).
Установлено, что в развитие паркинсонизма вовлекается ряд ге-
нов: ген ос-синуклеина на 4-й хромосоме, паркин на 6-й и PARK3 —
на 2-й хромосомах.
Стандартное лечение паркинсонизма основано на стремлении ком-
пенсировать недостаток дофамина в мозге за счет введения в организм
его метаболического предшественника, проникающего через гемато-
энцефалический барьер — диоксифенилаланина (ДОФА). Вначале ле-
чение протекает достаточно результативно, в последующем требуются
все возрастающие дозы ДОФА. Они индуцируют в мозге экспрессию
МАО В, одновременно вызывая гиперактивность в других дофаминер-
гических областях мозга. Побочные эффекты от длительного лечения
выражаются в симптомах, типичных для шизофрении, одним из биохи-
мических проявлений которой является высокая активность дофами-
нергической системы в мозге.
Близкой моделью для изучения паркинсонизма на лабораторных
животных явилось использование нейротоксина 1 -метил-4-фенилтет-
рагидропиридина (МРТР), который под влиянием МАО В легко пре-
вращается в МРР-радикал, обладающий высоким сродством к дофа-
миновому транспортеру. Таким образом, введение животным МРТР
приводит к избирательному накоплению МРР-радикала в дофаминер-
гических структурах мозга и вызывает в них стабильный окислитель-
ный стресс, приводящий к быстрой гибели клеток.
Наиболее выраженные симптомы паркинсонизма под влиянием
введения МРТР проявляют мыши линии SAM (Senescence Accelera-
ted Mice), у которых накопление старческих признаков происходит в
ускоренном темпе. Оба фактора — ускоренное старение и действие
нейротоксина — взаимно усиливают друг друга. Использование этой
модели позволяет оценить эффективность лекарственных препаратов
при лечении паркинсонизма. Один из них, йа который делали ставку
фармакологи во второй половине XX в., — селегилин — является из-
бирательным ингибитором МАО В. Его применение позволяет сни-
жать используемую дозу ДОФА, что важно при длительном приеме.
Противоположным по нейрохимическому механизму веществом, ока-
завшимся удачным дополнением к стандартному протоколу терапии
паркинсонизма, явился нейропептид карнозин. Отсутствие токсиче-
ских эффектов даже при длительном применении, способность пони-
жать внутриклеточный уровень АФК и защищать активность суперок-
сиддисмутазы от свободнорадикальной атаки, а также, возможно,
3.9. Окислительный стресс и неиропатологии
и другие нейротрофические эффекты карнозина позволяют считать
этот природный нейропептид эффективным соединением, позволяю-
щим снимать побочные эффекты длительного приема ДОФА.
В самом конце XX в. для радикального лечения паркинсонизма бы-
ло предложено осуществлять пересадку в мозг пациента клеток мезен-
цефалона от человеческого плода. Особенности иммунологического
статуса мозга позволяют проводить пересадку чужеродных клеток без
боязни их отторжения. До настоящего времени уже осуществлено оп-
ределенное количество таких операций; наблюдение за подобными па-
циентами внушает оптимизм. Ведутся работы по культивированию кле-
ток мезенцефалона, которые позволят избегать использования абор-
тивного материала.
Как было описано выше, для глутаматергической передачи возбужде-
ния характерна высокая экзайтотоксичность — избыточная актива-
ция NMDA-рецепторов при многих нарушениях становится токсичной
для клеток, поскольку при этом происходит существенное увеличение
концентрации ионизированного кальция внутри нейрона. Нарушения
глутаматергической системы имеют место при шизофрении и, в допол-
нение к активации дофаминовой системы, вносят свой вклад при разви-
тии этой болезни в создание окислительного стресса в мозге.
Глутаматные рецепторы вовлекаются и в процесс развития судо-
рожных состояний (эпилепсия). В этих условиях повышенная возбу-
димость каинатных рецепторов вызывает стойкое возбуждение боль-
шого количества глутаматергических нейронов, охватывая обширные
зоны мозга. Провоцирующим фактором служит одновременное ослаб-
ление ГАМК-ергической системы, выполняющей в мозге функции
торможения. Барбитураты, применяющиеся для ослабления эпилеп-
тических припадков, действуют противоположным образом на два
класса рецепторов: подавляют активность ионотропных глутаматных
рецепторов и стимулируют тормозную активность ГАМКд-рецепто-
ров. Окислительный стресс, по-видимому, является вторичным фак-
тором, осложняющим течение болезни.
В основе эпилепсии (которой страдает 1—2% человеческой попу-
ляции) лежат многочисленные неврологические нарушения, затраги-
вающие разные отделы головного мозга. До недавнего времени была
известна лишь одна линия мышей с идентифицированным геном, вы-
зывающим фенотип кортикальной эпилепсии. За последние 15 лет вы-
явлено более 30 единичных мутаций, связанных с проявлениями этого
заболевания. Сложности проявления и взаимодействия дефектных ге-
нов обусловливают индивидуальный подбор химических препаратов
для выработки стратегии купирования эпилептических припадков.
354
газде л 3. 4 '• 11 н.ЦИ< 1iЛ/11 М-./* 11• , i - < к и М ИЯ
Болезнь Альцгеймера сопровождается прогрессирующей деге-
нерацией нейрональных клеток. Развитие болезни затрагивает м-хо-
линергические структуры коры и гиппокампа, сопровождается сниже-
нием активности холинацетилтрансферазы. Морфологические изме-
нения холинергических нейронов выражаются в накоплении так
называемых сенильных бляшек и нейрофибриллярных образований
в области нервных окончаний. Природа этих структур связана с нару-
шениями обмена мембранного белка, состоящего из 695—770 амино-
кислотных остатков и являющегося предшественником Р-амилоида,
Р-АРР (Р-Amyloid Precursor Protein). В нормальных условиях этот бе-
лок принимает участие в формировании межнейронных контактов и
вовлекается в процессы консолидаций памяти. При болезни Альцгей-
мера его обмен нарушается, в результате чего при протеолитическом
расщеплении из его средней части образуется потенциально токсич-
ный 42-членный пептид, получавший название Р-амилоида (P-А). Он
инициирует образование нейрофибриллярных структур, нарушающих
нормальную работу межсинаптических контактов. Нормальное функ-
ционирование предшественника Р-амилоида также нарушается, у па-
циентов наблюдаются прогрессирующие глубокие изменения процес-
сов запоминания.
Р-АРР представляет собой трансмембранный белок, его N-концевой
фрагмент находится во внеклеточном пространстве или внутри везику-
лярных клеточных органелл, в то время как короткая С-концевая часть
молекулы обращена в цитоплазму. В настоящее время известны три изо-
формы этого белка (р-АРР770, р-АРР751 и Р-АРР695), отличающиеся сте-
пенью гликозилирования. Они экспрессируются в различных клетках и
тканях животных разного уровня эволюционного развития. В головном
мозге присутствует в основном более короткая форма Р-АРРб95. В коре
головного мозга человека содержание этих форм соотносится следую-
щим образом: Р-АРР770: Р-АРР751: Р-АРРб95 = 1:10:20.
Существует семейство родственных Р-АРР белков, называемых
Р-АРР-подобными белками (APLP). Описаны две изоформы отличаю-
щихся друг от друга первичной структурой белков APLP1 и APLP2. Сле-
дует отметить, что у грызунов (мышей и крыс), в мозге которых отсутст-
вуют типичные для болезни Альцгеймера агрегаты P-А, аминокислотная
последовательность р-АРР на участке, соответствующем Р-А-пептиду,
отличается от таковой у человека по трем аминокислотам.
Замена в молекуле P-А крыс и мышей гистидина на аргинин лиша-
ет р-А способности связывать ионы цинка, необходимые для иници-
ации агрегации молекул пептида и образования фибрилл.
3.9. Окислительный стресс и нейропатологи*
355
Направленный протеолиз под действием специфических фермен-
тов, расщепляющих Р-АРР, приводит к образованию ряда раствори-
мых биологически активных продуктов. Выделяют три группы фер-
ментов (а-, Р- и у-секретазы), расщепляющих Р-АРР с образовани-
ем фрагментов разной длины. Крупные растворимые фрагменты
sAPPoc и sAPPp, получаемые в результате действия а- или Р-секретаз,
играют важную роль в процессах пролиферации нервных клеток и
развитии нервной ткани. sAPPa, образующийся в результате активации
a-секретазы, обладает не только нейропротекторными свойствами, но
также и свойствами эпидермального фактора роста, кроме того, он спо-
собствует обучению и формированию памяти. sAPPp также обладает
нейропротекторными свойствами и служит субстратом для у-секрета-
зы, под влиянием которой и образуется 42-членный P-А и короткий
цитоплазматический фрагмент, играющий регуляторную роль.
Таким образом, метаболизм Р-АРР может осуществляться как по
неамилоидогенному пути (под действием a-секретазы), так и по ами-
лоидогенному (токсическому) пути в результате действия Р- и у-секре-
таз. Поскольку a-секретаза расщепляет Р-АРР внутри последова-
тельности P-А, этот путь предотвращает образование Р-А-пептида.
Следовательно, повышение активности a-секретазы может явиться
одним из возможных путей профилактики болезни Альцгеймера.
Представителем семейства трансмембранных аспартатных проте-
аз, инициирующим образование P-А, является мемапсин (обозначае-
мый в англоязычной литературе P-site APP-cleaving enzyme, ВАСЕ1).
Фермент, гомологичный ВАСЕ1, получивший название ВАСЕ2, также
обладает Р-секретазной активностью по отношению к Р-АРР Дейст-
вие BASE1 приводит к образованию более короткого варианта Р-А,
также обладающего амилоидогенными свойствами, и более длинного
фрагмента Р-АРР, представляющего собой растворимый sAPPp. Со-
здание ингибиторов ВАСЕ1 в терапевтических целях следует прово-
дить с определенной осторожностью, поскольку существуют и другие
эндогенные субстраты этого фермента.
Известны еще два белка — пресенилины 1 и 2 (PS1 и PS2), при-
нимающие участие в амилоидогенном процессинге Р-АРР. Вовлечение
дефектного гена пресенилина 1, локализованного на 14-й хромосоме,
в развитие болезни Альцгеймера было показано Е. И. Рогаевым в
1999 г. (Институт молекулярной генетики, Москва). Пресенилины
также являются интегральными мембранными белками со свойствами
классических аспартатных протеиназ, таких как пепсин, ренин и ка-
тепсин Д. Пресенилины входят в состав мультипротеинового комп-
лекса, проявляющего у-секретазную активность.
356
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕИРОХИМИ>
В расщеплении молекулы Р-АРР в клетках мозга больных, пора-
женных болезнью Альцгеймера, помимо секретаз могут принимать
участие кальцийзависимые аспарагиновые-сериновые протеиназы,
каспазы, работа которых приводит к образованию токсичных продук-
тов. Их субстратом являются модифицированные фосфорилировани-
ем молекулы Р-АРР.
Контроль концентрации амилоидного пептида внутри клеток, в мес-
тах синапсов, вдоль нервных окончаний и во внеклеточной среде осу-
ществляется ферментами, участвующими в метаболизме P-А. Исследо-
вание свойств и функций ферментов, связанных с деградацией амилоид-
ного пептида, важно для понимания их роли в-процессах развития мозга
и возникновения данной нейропатологии. Существуют эндогенные био-
химические пути деградации амилоида P-А в ткани мозга, а именно его
гидролиз в физиологических условиях протеазами мозга, такими как не-
прилизин (НЕП), эндотелинконвертирующий фермент (ЕСЕ-1), инсу-
линдеградирующий фермент (ИДФ; инсулизин) и плазмин, которые
способны вызвать деградацию P-А как in vitro, так и in vivo. Это обес-
печивает мозгу естественную защиту против накопления P-А. При пато-
логических условиях снижение экспрессии или активности амилоидде-
градирующих ферментов может приводить к накоплению р-АРР и раз-
витию болезни Альцгеймера.
Токсический эффект самого р-амилоида не связан с ростом кон-
центрации АФК в нейрональных клетках, но процесс нейродегенера-
ции сопровождается развитием окислительного стресса в нейронах.
Возможно, что окислительный стресс и возникает при взаимодействии
накапливающегося Р-амилоида с факторами, индуцирующими заболе-
вание. Об этом свидетельствуют данные о возможном участии алюми-
ния в развитии болезни Альцгеймера. Эпидемиологические обследова-
ния показали, что частота встречаемости этого заболевания коррели-
рует с накоплением алюминия в почве и продуктах. Позже выяснилось,
что в мозге в области формирования сенильных бляшек также наблю-
дается накопление алюминия. Появились предположения, что алюми-
ний может провоцировать «неправильный» метаболизм белка-пред-
шественника Р-амилоида.
Нейрохимические эксперименты, однако, вначале не обнаружили
токсичного воздействия алюминия на метаболизм нейронов — небла-
гоприятный эффект вызывали только высокие дозы этого металла, су-
щественно превышающие предельно допустимые концентрации алю-
миния в воде и пищевых продуктах. В связи с этим в ряде стран мира, в
том числе в США, стали использовать алюминиевые коагуляты для
очистки питьевой воды. Однако последние исследования требуют пере-
3.9. Окислительный стресс и нейропатологии
357
смотра существующих представлений. Дело в том, что хотя алюминий
сам по себе действительно достаточно инертен и вызывает клеточную
смерть только при концентрациях выше 100 мкМ, в сочетании с факто-
рами индукции окислительного стресса он способен вызвать резкое
увеличение вероятности некроза клеток. Так, было обнаружено (Туне-
ва, 2005), что алюминий значительно усиливает экзайтотоксическое
действие NMDA, причем этот эффект развивается за короткий период
времени. Вероятно, действие алюминия на активные нейроны будет го-
раздо более токсическим, чем на покоящиеся. Поскольку нейроны в
мозге характеризуются постоянно высоким уровнем активности, ток-
сичность алюминия в условиях in vivo будет гораздо более выражен-
ной, чем обнаруживаемая в опытах in vitro.
Полиглутаминовые болезни. К числу наследственных заболе-
ваний, обращающих на себя внимание нейрохимиков, относят и семей-
ство аутосомальных нейродегенеративных расстройств, вызываемых
многократным повтором в ДНК кодона CAG, кодирующего глутамин в
полипептидных цепях белков. По этой причине подобные заболевания
называют полиглутаминовыми (поли-Q — от сокращенного наиме-
нования глутамина). При таком дефекте ДНК возникают многократные
CAG-повторы, приводящие к появлению в кодируемом белке протя-
женных полиглутаминовых участков.
На сегодняшний день описано девять таких заболеваний, включая
болезнь Хантингтона (HD, вызывается дефектом гена хантингтина
(Htt), расположенного на коротком плече 4-й хромосомы), спинобулъ-
барную мышечную атрофию и некоторые спиноцеребралъные
атаксии. Развитие неврологической симптоматики при этих заболева-
ниях характеризуется присутствием ядерных (а в некоторых случаях и
неядерных) нерастворимых агрегатов, обладающих положительной им-
мунореактивностью на мутантный белок и на убиквитин. Структура ин-
термедиатов, участвующих в образовании этих агрегатов, стадии, на ко-
торых данный процесс начинает наносить ущерб клеткам, и взаимоза-
висимость процесса агрегации, нейрональной дисфункции и смерти ней-
рональных клеток до сих пор не выяснены. Исследования, проведенные
на пациентах с полиглутаминовыми заболеваниями, а также на транс-
генных мышах, показали, что в большинстве случаев ядерные включе-
ния, образуемые дефектными белками, являются общей патологической
причиной этого вида заболеваний.
В настоящее время исследователи предпринимают значительные
усилия по поиску молекул, способных подавлять полиглутаминовую
агрегацию и цитотоксичность в терапевтических целях.
358
Раздел 3. ФУНКЦИОНАЛЬНАЯ НЕИРОХИМИЯ
На ранних стадиях заболевания накопление клеточных дефектов
предотвращается клеточными шаперонами — белками теплового
шока (Heat shock proteins, Hsp). Некоторые шапероны, например
Hsp70 и Hsp40, способны связываться с ядерными включениями, об-
разуемыми в ткани мозга дефектными белками. В различных клеточ-
ных моделях полиглутаминовых заболеваний обнаружено, что повы-
шенная экспрессия, как Hsp70, так и Hsp40, уменьшает образование
ядерных агрегатов и вызываемую ими цитотоксичность, а также сни-
жает клеточную смерть. Hsp27 тоже способен к подавлению клеточ-
ной смерти, опосредованной накоплением нейронального белка хан-
тингтина, содержащего 74 полиглутаминовых повтора. Однако в ряде
случаев оказалось, что подавление нейродегенерации не обязательно
сопровождается подавлением агрегации хантингтина.
Другой белок теплового шока — шаперонин TRiC также может
непосредственно взаимодействовать с полиглутаминовыми участками
хантингтина и ингибировать его агрегацию. Снижение уровня этого
шаперонина повышает полиглутаминовую агрегацию в клетках дрож-
жей и млекопитающих, а его повышенная экспрессия подавляет как
агрегацию Htt, так и смерть нейрональных клеток. Хорошим терапев-
тическим агентом против полиглутаминовой агрегации белков и кле-
точной смерти может служить дисахарид трегалоза — как в клеточных
моделях in vitro, так и в опытах in vivo на «мышиной модели» HD.
Молекулярный механизм снижения цитотоксичности полиглутами-
новых заболеваний за счет повышенной экспрессии шаперонов не впол-
не ясен. В недавних исследованиях систему шаперонов Hsp70/Hsp40
использовали не только для солюбилизации полиглутаминовых агрега-
тов, но и для усиленной деградации этих белков при участии 265-проте-
асом. Солюбилизация агрегатов сопровождалась существенным сни-
жением периода полужизни полиглутаминовых белков. Это указыва-
ет на то, что шапероны могут сдвигать равновесие между аморфным
(растворяемым в детергенте) и фибриллярным (устойчивым к дейст-
вию детергента) состояниями полиглутаминбвых агрегатов, в резуль-
тате чего протеолитическая система клетки может более эффективно
метаболизировать полиглутаминовые белки. Протяженные полиглу-
таминовые участки вовлекаются в убиквитинзависимый путь деграда-
ции белков и приводить к нейрональным повреждениям.
Дальнейшие исследования этих заболеваний необходимы для рас-
шифровки молекулярных механизмов патогенеза дегенерации нерв-
ных клеток и разработки будущих терапевтических подходов. По-ви-
димому, молекулярные шапероны могут явиться перспективными суп-
прессорами полиглутаминовой токсичности человека.
3.9. Окислительный стресс и неиропатологии
Ишемия головного мозга. Одними из наиболее часто встре-
чающихся возрастных заболеваний считают ишемические повреж-
дения головного мозга, являющиеся результатом острого наруше-
ния кровообращения (Верещагин с соавт., 1997; Верещагин, Сусли-
на, 2003). Они возникают как результат неспособности сосудов к
быстрой адаптации при повышенных физических нагрузках. Психиче-
ский стресс также является провоцирующим фактором.
Острые нарушения кровообращения приводят к появлению трех
важных факторов, вызывающих ишемическое повреждение ткани, —
гипоксии, гипогликемии и ацидоза. Эти факторы являются взаимо-
связанными, поскольку закупорка сосуда приводит к нарушению снаб-
жения глюкозой и кислородом, в результате энергетический метабо-
лизм ограничивается только анаэробными процессами, вызывающими
накопление кислых продуктов, устранение которых из тканей затрудне-
но (Гусев, Скворцова, 2001). Нарушения обмена в зоне повреждения
носят системный характер и, сохраняясь длительное время, приводят к
необратимым изменениям (рис. 3.30). В этих условиях в области по-
вреждения индуцируется массовая клеточная смерть, протекающая как
по пути апоптоза, так и по пути некроза. Смерть нейрональных клеток
является особенно критичной для мозга, поскольку влияет на его функ-
циональные свойства. Характер функциональных нарушений опреде-
ляется областью, в которой возникает повреждение.
Эпизод острого нарушения мозгового кровообращения, приводя-
щий к созданию ишемического очага в мозге, может и не вызвать губи-
тельных последствий, если будут использованы резервные механизмы
Мозговой кровоток,
мл/100 г в мин
ВИ Аноксическая
деполяризация
мембран,смерть
клетки
ВИ Энергетический
дефицит, глутаматная
«эксайтотоксичность»,
увеличение содержания
внутриклеточного Са2+
Г,- 1 Лактат-ацидоз,
цитотоксические отеки
Время от начала ишемии
до необратимого повреждения
Рис. 3.30. Нарушения мозгового кровотока (а) и повреждение ткани мозга (б)
при инсульте головного мозга (по: Гусев, Скворцова, 2001)
Снижение белкового
синтеза, селективная
экспрессия генов
360
газдел 3. ФУНКЦИОНАЛЬНАЯ НЕИРОХИМИЯ
кровоснабжения. Природные механизмы защиты связаны с индукцией
АФК и особенно оксида азота, поскольку NO является природным
фактором релаксации гладкомышечных стенок сосудов. В результате
этого включаются пути кровоснабжения по коллатералям, и последст-
вия гипоксии и другие опасные факторы ишемии устраняются. При де-
фиците природных механизмов регуляции эффективность медикамен-
тозных вмешательств зависит от времени начала лечения — терапев-
тическое окно для успешной борьбы с инсультом составляет 60—
180 мин. С удлинением этого периода результативность лечения суще-
ственно снижается.
Нарушения мозгового кровообращения, даже преходящие, вызы-
вают глубокие изменения метаболизма нервных и глиальных клеток,
выражающиеся в нарушениях высвобождения и обратного захвата
медиатора, неуправляемом возрастании концентрации ионизирован-
ного кальция в клетках, лавинообразной активации целого ряда фер-
ментов, разрушающих макромолекулы клеток.
Для глутаматергической системы ишемические повреждения реали-
зуются по описанному выше экзайтотоксическому механизму, приводя к
активации обоих типов клеточной смерти — и некроза, и апоптоза. Вы-
бор в пользу одного из них зависит от обширности зоны инсульта и, со-
ответственно, от выраженности повреждений. Как правило, в начале
патологического процесса имеет место некротическая гибель клеток,
апоптоз же проявляется в качестве отсроченной гибели. Повреждения
затрагивают как нейрональные, так и глиальные клетки, и нервная
ткань лишается защиты со стороны астроцитов.
Рано или поздно на смену гипоксии приходит реперфузия — кро-
воснабжение восстанавливается по коллатералям, активированным
или за счет собственных резервов организма, или вследствие терапев-
тических воздействий. При этом, чем раньше начинается лечение, тем
оно оказывается более эффективным. Стадия реперфузии знаменует-
ся всплеском продукции свободных радикалов, поэтому на этой стадии
антиоксидантная терапия особенно целесообразна.
На практике лекарственная терапия инсульта не слишком
успешна — в непосредственной области поражения и в окружающих
ее областях (в так называемой пенумбре) требования к лекарствен-
ной терапии различаются, а доступ лекарственных препаратов к пора-
женным тканям мозга затруднен. В целом заболевание характеризует-
ся серьезной инвалидизацией, и случаи полноценного восстановления
относительно редки.
Тем не менее введение в практику лечения ишемических поврежде-
ний мозга методов визуализации пораженной области (компьютерная то-
3.9. Окислительный стресс и нейропатологии
361
мография разных видов) позволяет определить размер поврежденных
участков мозга и контролировать изменения их состояния в ходе лечения.
Поданным компьютерной томографии, подтвержденным анализом
неврологической симптоматики, определенную эффективность демон-
стрируют лекарственные препараты, купирующие экзайтотоксич-
ность, — антагонисты АМРА- и NMDA-рецепторов, вещества, подав-
ляющие кальциевый ток внутрь нейронов, соединения, способные
нейтрализовывать свободные радикалы. Однако выбор лекарственной
терапии основывается на свойствах препаратов, оцениваемых in vitro.
Нет указаний на то, что метаболиты, образующиеся при введении этих
препаратов в кровь, не будут проявлять дополнительного (или проти-
воположного) эффекта. Действительно, очень часто вещества, реко-
мендуемые для восстановления кровоснабжения на основе предвари-
тельного тестирования, при клинических испытаниях не проявляют
прогнозируемой активности.
Накопление старческих признаков не ограничивается возрастанием
частоты проявления ишемических повреждений тканей. Имеет место
увеличение вероятности и других заболеваний, в том числе нейродеге-
неративных. Само физиологическое старение во многом является ре-
зультатом накопления различных ошибок метаболизма вследствие уси-
ливающегося дисбаланса в обмене АФК и все в большей степени прояв-
ляющегося окислительного стресса (Анисимов, 2004). Окислительный
стресс является одним из факторов накопления возрастных изменений
метаболизма, в том числе и в мозге. Он проявляется в ограничениях
процессов обучения, консолидации (мало затрагивая долговременную
память), в адаптационных ограничениях. Казалось бы, процесс старения
должен хорошо если не «лечиться», то «защищаться» антиоксидантной
терапией. К сожалению, высказанные ранее положения о двойственной
роли свободных радикалов в метаболизме клетки делают антиоксиданты
ограниченным по эффективности лекарством. В то же время энтузиасты
избирательной защиты клеток от АФК не теряют надежды найти такие
регуляторы метаболизма, которые бы оберегали ткани от нежелатель-
ных свободнорадикальных повреждений, увеличивая при этом биологи-
ческие пределы жизнедеятельности (Скулачев, 2006).
Раздел 3. ФУНКЦИОНАЛЬНА •, НЕЙР’ ХИМИЯ
ВОПРОСЫ И ЗАДАНИЯ
1. Что представляет собой гематоэнцефалический барьер, как происхо-
дит снабжение мозга метаболитами?
2. Почему D- и L-глюкоза проникает через ГЭБ с разной скоростью?
3. Каковы особенности обмена липидов в тканях мозга?
4. Каковы особенности обмена жирных кислот в мозге?
5. В чем заключается биологическое значение фосфоинозитидного об-
мена?
6. Каковы особенности энергетического обмена мозга?
7. Перечислите и охарактеризуйте пути образования ГАМК в мозге.
8. Что такае ГАМК-шунт?
9. Чем объяснить чувствительность мозга к недостатку кровоснабжения?
10. Дайте определение понятия «аксоплазматический ток». Каково его функ-
циональное значение?
11. Какова роль ионов кальция в функционировании синаптических кон-
тактов?
12. Что такое синаптическая задержка?
13. Назовите общие и отличительные черты электрических и химических
синапсов.
14. Что представляют собой синаптические везикулы и как они участвуют
в передаче сигнала через синапс?
15. Охарактеризуйте процесс квантового выделения медиатора.
16. Каковы свойства потенциала покоя и потенциала действия?
17. Каковы особенности передачи сигнала в фоторецепторных клетках?
18. В чем состоит биологическая роль родопсина?
19. К каким белкам относится трансдуцин и какова его роль в передаче
сигнала в фоторецепторной клетке?
20. Как происходит передача обонятельного сигнала?
21. Опишите свойства вкусовых рецепторов.
22. Каковы принципы организации двигательных систем?
23. Что такое произвольная мускулатура?
24. Охарактеризуйте молекулярные свойства мускариновых и никотино-
вых холинорецепторов.
25. Какова роль Na/K-насоса в создании трансмембранного потенциала?
26. Каков механизм участия Na/K-насоса в клеточной сигнализации?
27. Охарактеризуйте различные виды памяти.
28. Дайте определение понятия «энграмма».
29. Что такое консолидация памяти?
30. Охарактеризуйте феномен пластичности нервной системы.
31. Назовите регуляторные функции основных нейропептидов.
32. Что такое окислительный стресс?
33. Каков механизм действия клеточной системы антиоксидантной за-
щиты?
34. Перечислите основные маркеры окислительного стресса.
Вопросы и задания
363
35. В чем заключается двойственная роль свободных радикалов в функции
возбудимой клетки?
36. В чем заключается адаптационное значение окислительного стресса?
37. Что такое прекондиционирование, каковы его молекулярные основы?
38. Как связаны понятия «окислительный стресс» и «нейродегенеративные
заболевания»?
39. Охарактеризуйте свойства белков теплового шока.
40. Каковы природные механизмы защиты мозга от окислительного стресса?
ЛИТЕРАТУРА
Авдонин П. В., Ткачук В. А. Рецепторы и внутриклеточный кальций. М., 1994.
Альберте А., БрейД.Ю., Льюис Р. и др. Молекулярная биология клетки/
пер. с англ. М., 1994.
Анисимов В. Н. Приоритеты фундаментальных исследований в геронтологии:
роль русских ученых// Успехи геронтологии. 2003. Т. 12.
Антонов В. Ф., Смирнова Е. Ю., Шевченко Е. В. Липидные мембраны при
фазовых превращениях. М., 1992.
Ашмарин И. П. Загадки и откровения биохимии памяти. Л., 1975.
Ашмарин И. П., Каменская М. А. Нейропептиды в синаптической передаче //
Итоги науки и техники, физиология человека и животных. М., 1988.
Бак 3. Химическая передача нервного импульса. М., 1977.
Бендалл Дж. Мышцы, молекулы и движение. М., 1970.
Березин В. А. Характеристики мембранных и нейроспецифических белков //
Нейрохимия. 1984. Т.З.
Березин В. А., Гайдар Л. И. Специфические гликопротеины нервной ткани //
Нейрохимия. 1987. Т. 6.
Биологические мембраны. Двенадцать очерков о структуре, свойствах и функ-
циях мембран / под ред. Д. Парсона. М., 1978.
Биологические мембраны. М., 1992.
Биофизика / под ред. В. Ф. Антонова. М.,1996.
Биохимия мозга / под ред. И. П. Ашмарина. М., 1999.
Блум Ф., ЛейзерсонА., ХофстедтерЛ. Мозг, разум и поведение/пер. с англ.
М„ 1988.
Блюменфельд Л. А. Проблемы биологической физики. М., 1977.
Блюменфельд Л. А. Решаемые и нерешаемые проблемы биологической фи-
зики. М., 2002.
Болдырев А. А. Биохимические механизмы электромеханического сопряже-
ния. М., 1977.
Литература
365
Болдырев А. А. Биологические мембраны и транспорт ионов. М., 1985.
Болдырев А. А. Введение в биохимию мембран. М., 1986.
Болдырев А. А. Олигомерные ансамбли транспортных АТФаз в мембранном
бислое // Журнал эвол. биохим. физиол. 1995. Т. 31.
Болдырев А. А. Парадоксы окислительного метаболизма мозга// Биохимия.
1995. Т. 60.
Болдырев А. А. Карнозин: биологическая роль и возможности применения
в медицине. М., 1998.
Болдырев А. А. Карнозин и защита тканей от окислительного стресса. М., 1999.
Болдырев А. А. Дискриминация между апоптозом и некрозом нейронов под
влиянием окислительного стресса // Биохимия. 2000. Т. 65.
Болдырев А. А. Роль активных форм кислорода в функции нейронов//Усп.
физиол. наук. 2003. Т. 34.
Болдырев А. А., Куклей М.Л. Свободные радикалы в нормальном и ишеми-
ческом мозге // Нейрохимия. 1996. Т. 13.
Болдырев А. А., Мельгунов В. И. Транспортные АТФазы. М., 1984.
Болдырев А. А., Твердислов В. А. Молекулярная организация и механизм
функционирования Na-насоса. М., 1978.
Болдырев А. А., Кярвяряйнен Е. И., Илюха В. А. Биомембранология. Петро-
заводск; М., 2006.
Бородкин Ю. С., Шабанов П.Д. Нейрохимические механизмы извлечения
следов памяти. Л., 1986.
Бредбери М. Концепция гематоэнцефалического барьера/пер. с англ. М.,
1983.
Вартанян Г. А., Пирогов А. А. Механизмы памяти центральной нервной
системы / под ред. Г. А. Вартаняна. Л., 1988.
Введение в биомембранологию / под ред. А. А. Болдырева. М., 1990.
Верещагин Н. В., Моргунов В. А., Булевская Т. С. Патология головного моз-
га при атеросклерозе и артериальной гипертонии. М., 1997.
Верещагин Н. В., Суслина 3. А. Инсульт в глазах медицины и общества//
Вестник РАМН. 2003. Т. И.
Владимиров Ю. А., Азизова О. А., Деев А. И. и др. Свободные радикалы
в живых системах// Итоги науки и техники. Биофизика. 1992. Т 29.
Владимиров Ю. А., Арчаков А. И. Перекисное окисление липидов в биологи-
ческих мембранах. М., 1972.
Владимиров Ю. А., Битов В. Б. Механизмы работы Са-транспортной АТФа-
зы в мембранах саркоплазматического ретикулума / Биомембраны: струк-
тура, функции, медицинские аспекты. Рига, 1981.
366
Литература
Возбуждающие аминокислоты как трансмиттеры / под ред. К. С. Раев-
ского // Итоги науки и техники. ВИНИТИ. (Физиология человека и жи-
вотных). М., 1988.
ГеннисР. Биомембраны, молекулярная структура и функции/ пер. с англ.
М., 1999.
Глебов Р. Н., Крыжановский Г. Н. Аксональный ток веществ при различных
физиологических и патологических состояниях нервной системы // Усп.
совр. биол. 1976. Т. 82.
Глебов Р. Н., Крыжановский Г Н. Функциональная биохимия синапсов. М.,
1978.
Гомазков О. А. Нейротрофическая регуляция и стволовые клетки мозга. М.,
2006.
Гуляева Н. В. Неапоптозная функция каспазы-3 в нервной ткани//Биохи-
мия. 2003. Т. 68.
Гусев Е. И., Скворцова В. И. Ишемия головного мозга. М., 2001.
Дамбинова С. Л. Нейрорецепторы глутамата. Л., 1989.
Дюв К де. Путешествие в мир живой клетки. М., 1987.
Ещенко Н.Д. Биохимия психических и нервных болезней. СПб., 2004.
Ещенко Н. Д., Маслова М. Н. Практикум по свободнорадикальному окисле-
нию. СПб., 2005.
Заварзин А. А., Харазова А.Д., Молитвин М. И. Биология клетки: общая
цитология. Л., 1993.
Зенков Н. К, Ланкин В. 3., Менъщикова Е. Б. Окислительный стресс. Био-
химический и патофизиологический аспекты. М., 2000.
Зефиров Л. Н., Рахманкулова Г М. Медиаторы. Обмен, физиологическая
роль и фармакология. Казань, 1975.
Ивков В. Г., Берестовский Г. Н. Динамическая структура липидного бислоя.
М., 1981.
Ивков В. Г., Берестовский Г. Н. Липидный бислой биологических мембран.
М., 1982.
ИржакЛ. И. Очерки физиологии: В 2 ч. Сыктывкар, 2005. Ч. I; 2006. Ч. II.
Каламкаров Г. М., Островский М. А. Молекулярные механизмы зрительной
рецепции. М., 2002.
Климов П. К Физиологическое значение пептидов мозга для деятельности
пищеварительной системы. Л., 1986.
Конев С. В. Структурная лабильность биологических мембран и регулятор-
ные процессы. Минск, 1987.
Корниш-Боуден Э. Основы математики для биохимиков. М., 1983.
Литература
367
КостюкП. Г. Кальций и клеточная возбудимость. М., 1986.
КотыкА., Яначек К. Мембранный транспорт. М., 1980.
Крепе Е. М. Липиды биологических мембран. Л., 1981.
Курганов Б. И. Аллостерические ферменты. М., 1978.
Лайтфут Э. Явления переноса в живых системах. М., 1977.
Ланкин В. 3., Тихазе А. К, Беленков Ю. Н. Свободнорадикальные процессы
в норме и при патологических состояниях. М., 2001.
Левицкий Д. О. Кальций и биологические мембраны. М., 1990.
Мак-Ильвейн Г. Биохимия и центральная нервная система. М., 1962.
Маркин В. С., Чизмаджев Ю. А. Индуцированный ионный транспорт. М.,
1974.
Меерсон Ф. 3. Адаптационная медицина: Механизмы и защитные эффекты
адаптации. М., 1993.
Мембраны'. Ионные каналы. М., 1981.
Меныцикова Е. Б., Ланкин В. 3., Зенков Н. К и др. Окислительный стресс.
М.,2006.
Механизмы памяти: Руководство по физиологии. Л., 1988.
Михельсон М. Я., Зеймаль Э. В. Ацетилхолин. Л., 1970.
Нейрохимия / под ред. И. П. Ашмарина. М., 2001.
Николлс Дж., Мартин Р., Валлас Б., ФуксП. От нейрона к мозгу/пер.
с англ. М., 2003.
Николс Дж. Биоэнергетика: Введение в хемиосмотическую теорию. М., 1985.
Овчинников Ю. А., Иванов В. Т., Шкроб А. М. Мембраноактивные комплек-
соны. М., 1974.
Орлов С. Н. Кальмодулин: Общие проблемы физико-химической биологии/
Итоги науки и техники. ВИНИТИ. (Физиология человека и животных).
М., 1987.
Островский М. А. Молекулярные механизмы зрительной рецепции. М., 2002.
Палладии А. В., Белик Я. В., Полякова Н. М. Белки головного мозга и их об-
мен. Киев, 1972.
Пальцев М. А. Введение в молекулярную медицину. М., 2004.
Пальцев М. А., Иванов А. А., Северин С. Е. Межклеточные взаимодействия.
М., 2003.
Пантелеева Н. С. Миозин: О18-обмен и фосфорилирование. Л., 1975.
Певзнер Л. 3. Функциональная биохимия нейроглии. Л., 1972.
Пермяков Е. А. Кальцийсвязывающие белки. М., 1993.
368
Литература
Постнов Ю. В., Орлов С. Н. Первичная гипертензия как патология клеточ-
ных мембран. М., 1987.
Раевский К. С., Георгиев В. П. Медиаторные аминокислоты. М., 1986.
Регистрация одиночных каналов / под ред. Б. Сакмана, Э. Неера. М., 1987.
Розен В. Б. Основы эндокринологии. М., 1994.
Рубин А. Б. Биофизика. Биофизика клеточных процессов. М., 1987.
РэкерЭ. Биоэнергетические механизмы: новые взгляды. М., 1979.
Самойлов А. Ф. Избранные статьи и речи. Л., 1946.
Сергеев П. В., Шимановский Н. Л. Рецепторы физиологически активных
веществ. М., 1987.
Скулачев В. П. Биоэнергетика. Мембранные преобразователи энергии. М.,
1989.
Скулачев В. П. Энергетика биологических мембран. М., 1989.
Скулачев В. П. Феноптоз: запрограммированная смерть организма // Био-
химия. 1999. Т. 64.
Смагин В. Г., Виноградов В. А., Булгаков С. А. Лиганды опиатных рецепто-
ров: гастроэнтерологические аспекты. М., 1983.
Смирнов А. Н. Элементы эндокринной регуляции. М., 2006.
Современное естествознание: Энциклопедия. Т. 2. Общая биология. М., 2000.
Современное естествознание: Энциклопедия. Т. 8. Молекулярные основы
биологических процессов. М., 2000.
Соросовский образовательный журнал <http://journal.issep.ru>.
Сытинский И. А. ГАМК в деятельности нервной системы. Л., 1972.
Титов С. А., Крутько В. Н. Современные концепции механизмов старения//
Физиология человека. 1996. Т. 22.
Ткачук В. А. Основы молекулярной эндокринологии. М., 1989.
Турпаев Т. М. Медиаторная функция ацетилхолина и природа холинорецеп-
тора. М., 1962.
Физиология человека: В 3 т. / пер. с англ.; под ред. Р. Шмидта, Г. Тевса. М.,
1986.
Ходжкин А. Нервный импульс. М., 1965.
Хочачка П., Сомеро Дж. Биохимическая адаптация. М., 1988.
Хухо Ф. Нейрохимия. Основы и принципы / пер. с англ. М., 1990.
Ченцов Ю. С. Введение в клеточную биологию. М., 2004.
Чухрай Е. С., Полторак О. М., Кунаев Ф.Д. и др. Каталитические механиз-
мы обонятельной функции // Вестник МГУ. Сер. 2. Химия. 1988. Т. 29.
Литература
369
Шалабодов А. Д., Гусева Н. В. Основы мембранного транспорта/ под ред.
А. А. Болдырева. Тюмень, 2001.
Шеперд Г. М. Нейробиология / пер. с англ. М., 1987. Т. 1 —2.
Шеррингтон Ч. Интегративная деятельность нервной системы / пер. с англ.
М., 1969.
Armstrong С. М., Hille В. Voltage-gated ion channels and electrical excitability//
Neuron. 1998. Vol. 20.
Ashcroft F. M. Ion channels and disease. Boston, 2000.
Augustine G. J. How does calcium trigger neurotransmitter release? // Current
Opinion Neurobiol. 2001. Vol. 11.
Basic Neurochemistry / Ed. C. J. Siegel, B. W. Agranoff, R. W. Albers, P. R. Mo-
linoff. N.Y., 1993.
Bennett M. V. L. Electrical synapses, a personal perspective (or history)// Brain
Res. Rev. 2000. Vol. 32.
Boldyrev A., Bulygina E., Yuneva M., Schoner W. Na/K-ATPase regulates intra-
cellular ROS level in cerebellum neurons//Ann. N. Y. Acad. Sci. 2003.
Vol. 986.
Boldyrev A. A., Quinn P. E1/E2 type cation transport ATPases: evidence for
transient associations between protomers // Int. J. Biochem., 1994. Vol. 26.
BradfordH. F. Chemical Neurobiology / Ed. by W. H. Freeman. N. Y, 1986.
BrungerA. T. Structure of proteins involved in synaptic vehicle fusion in
neurons // Annu. Rev. Biophys. Biomol. Struct. 2001. Vol. 30.
Cell Organelles / Ed. R. G. Herrmann. Wien; N. Y., 1992.
ChangeuxJ.-P. Chemical signaling in the brain // Sci. Am. 1993. Vol. 269.
Chapman D. Phase transition and fluidity characteristics of lipids and cell
membranes//Quart. Rev. Biophys. 1975. Vol. 8.
Garthwaite J. Neural nitric oxide signaling// Trends Neurosci. 1995. Vol. 18.
GrewalS.S., YorkR.D., Stork P. J. S. Extracellular-signal-regulated kinase
signallingin neurons//Current Opinion in Neurobiol. 1999. Vol. 9.
HallZ. An introduction to molecular neurobiology. Sunderland, 1992.
Hanks S. K., Hunter T. The eukaryotic protein kinase superfamily: kinase (cata-
lytic) domain structure and classification // FASEB J. 1995. Vol. 9.
HeleniusA., SimensK. Solubilization of membranes by detergents// Biochem.
Biophys. Acta. 1975. Vol. 415.
Hodgkin A. L. Chance and design. Cambridge, 1992.
Hodgkin A. L. The conduction of the nervous impulse. Springfield, 1967.
Introduction to cellular signal transduction / Ed. A. Sitaramaya. Boston, 1999.
370
Литература
Junge D. Nerve and muscle excitation (3rd Edition)/ Ed. by M. A. Sunderland,
N. Y„ 1992.
Kagan V., Quinn P. Toward oxidative lipidomics of cell signaling// Antiox. Redox
Sign. 2004. Vol. 6.
Katz B. The release of neural transmitter substances. Liverpool, 1969.
Linas R. The squid giants synapse: a model for chemical synaptic transmission.
Oxford, 1999.
McIlwain H., Bacheland H. S. Biochemistry and central nervous system. L., 1971.
Nature'. Neurological disorders (Special Issue), 1999. Vol. 399 (SuppL).
NeherE. Nobel lecture: Ion channels for communication between and within
cells//Neuron. 1992. Vol. 8.
Neuroscience / Ed. by D. Purves et al. N. Y., 2004.
Neurotransmitters, drugs and brain function / Ed. by R. Webster. N. Y., 2001.
Nicholls D. G. Proteins, transmitters and synapses. Oxford, 1994.
Nowycky M. C., Fox A. P., Tsien R. W. Three types of neuronal calcium channel
with different calcium agonist sensitivity// Nature. 1985. Vol. 316.
Rothman J. E. Mechanisms of intracellular protein transport//Nature. 1994.
Vol. 372.
Singer M. S., Sheperd G. M. Molecular modeling interaction in the OR5 olfacto-
ry receptor// Neuroreport. 1994. Vol. 5.
SkouJ. The influence of some cations on an adenosine triphosphatase from pe-
ripheral nerves. Biochem. Biophys. Acta. 1957. Vol. 23.
SkouJ. Overview: The Na, K-pump// Methods EnzymoL 1988. Vol. 156.
Skulachev V. P. Bioenergetic aspects of apoptosis, necrosis and mitoptosis //
Apoptosis. 2006. Vol. 11.
SudhofT. C. The synaptic vesicle cycle: a cascade of protein-protein interac-
tions I/ Nature. 1995. Vol. 375.
Tucker W. C., Chapman E.R. Role of synaptotagmin in Ca2+ triggered
exocytosis // Biochem. J. 2002. Vol. 366.
XieZ., Askari A. Na/K-ATPase as a signal transducer// Eur. J. Biochem. 2002.
Vol. 269.
СОКРАЩЕНИЯ, ИСПОЛЬЗУЕМЫЕ В КНИГЕ
АДФ — аденозин-5'-дифосфат
АКТГ — адренокортикотропный гормон
АМФ — аденозин-5'-монофосфат
АТФ — аденозин-5'-трифосфат
АФК — активные формы кислорода
АРРр — р-амилоид
АХ — ацетилхолин
АХЭ — ацетилхолин эсте раза
АЦ — аденилатциклаза
ВИП — вазоактивный интестинальный (поли)пептид
ВПСП — возбуждающий постсинаптический потенциал
ГАД — глутаматдекарбоксилаза
ГАМК — у-аминомасляная кислота
ГМФ — гуанозин-5'-монофосфат
ГДФ — гуанозин-5'-дифосфат
ГТФ — гуанозин-5'-трифосфат
ГЭБ — гемоэнцефалический барьер
ДВД — долговременная депрессия
ДВП — долговременная потенциация
ДГК — докозогексаеновая кислота
L-ДОФА — L-дигидроксифенилаланин
ИДФ — инсулинде градирующий фермент, инсулизин
ИФ2 — инозитдифосфат
ИФ3 — инозит-1,4,5-трисфосфат
ИФР1 — инсулиноподобный фактор роста 1
ИЦДГ — изоцитратдегидрогеназа
КМ — кальмодулин
ЛДГ — лактатдегидрогеназа
МАО — моноаминоксидаза
МДА — малоновый диальдегид
МПКП — миниатюрный потенциал концевой пластинки
мРНК — матричная рибонуклеиновая кислота
>иХР — мускариновые холинорецепторы
НА — норадреналин
НАД — никотинамидадениндинуклеотид, окисленная форма
НАДН — никотинамидадениндинуклеотид, восстановленная форма
НАДФ — никотинамидадениндинуклеотидфосфат, окисленная форма
372
сокращения, используемые в книге
НАДФН — ни коти на мидаден и нди нуклеотидфосфат, восстановленная форма
НЕП — нейтральная эндопептидаза, неприлизин
нХР
ПААГ
ПД
ПДГ
ПК
Пк
ПКП
ПНС
ПОЛ
ПФ
РНК
РП
ТБК
— никотиновые холинорецепторы
— полиакриламидный гель
— потенциал действия
— пируватдегидрогеназа
— протеинкиназа
— пируваткарбоксилаза
— потенциал концевой пластинки
— периферическая нервная система
— перекисное окисление липидов
— протеинфосфатаза
— рибонуклеиновая кислота
— рецептивное поле
— тиобарбитуровая кислота
ТБКРП — ТБК-реактивные продукты
ТМП — трансмембранный потенциал
ТПСП — тормозный постсинаптический потенциал
тРНК — транспортная рибонуклеиновая кислота
УТФ — уридин-5'-трифосфат
ФДЭ — фосфоди эсте раза
ФИФ2 или
ФИ(4,5)Ф2— фосфатидилинозитол-4,5-бисфосфат
ФРН — фактор роста нерва
ХР — холинорецептор
цАМФ — циклический аденозин-3'-5'-монофосфат
цГМФ — циклический гуанозин-3',5'-монофосфат
ЦНС — центральная нервная система
АА — arachidonic acid (арахидоновая кислота)
AIF — apoptosis inducing factor (индуцируемый апоптозом фактор)
Akt — v-Akt murine thymoma viral oncogene homolog (гомолог онкогена
вируса мышиной тимомы)
АМРА — a-amino-3-hydroxil-5-methil-4-isoxasole-propionate
(а-амино-3-гидрокси-5-метил-4-изоксазолпропионовая кислота)
ANT — adenine nucleotide transporter (транспортер адениновых нуклеотидов)
APAF1 — apoptotic protease activating factor-1 (фактор, активируемый апоп-
тическими протеазами)
р-АРР — p-Amyloid Precursor Erotein (белок-предшественник р-амилоида)
APV — 2-amino-5-phosphonovalerate (2-амино-5-фосфоновалериановая кис-
лота)
ATM — ataxia telangiectasia mutated protein (мутировавший белок атак-
сической телеангиэктазии)
ВАСЕ1 — p-site APP-cleaving enzyme (мемапсин)
BAD — Bcl2-antagonist of cell death (Вс12-антагонист клеточной гибели)
BAK1 — Bcl2-antagonist killer-1 (киллер Вс12-антагонистов, проапоптиче-
ский белок)
Сокращения, используемые в книге
ВАХ — Bcl2-associated X-protein (Вс12-ассоциированный проапоптический
Х-белок)
Вс12 — В-cell leukemia-2 (фактор лейкемии В-клеток)
BciXL — Вс12- related protein long isoform (длинная изоформа Вс12-связанно-
го белка)
BCR — В-cell receptors (рецепторы В-клеток)
BDNF — brain-derived neurotrophic factor (нейротрофический фактор мозга)
bFGF — basic fibroblast growth factor (основной фактор роста фибробластов)
BID, BIM — Вс12 interacting proteins (белки, взаимодействующие с Вс12)
BMP — bone morphogenic protein (белок морфогенеза кости)
CAD — caspase-activated DNAse (активируемая каспазами ДНКаза)
cADPR — cyclic ADP-ribose (циклическая АДФ-рибоза)
Calm — calmodulin (кальмодулин)
CalmKIl,
CalmKIV — calm kinase-II and IV (кальмодулинзависимые киназы)
Caln — calcineurin (кальцинейрин)
CB — cannabinoid (каннабиноиды)
СВР — CREB binding protein (CREB-ассоциированные белки)
CDC42 — cell division cycle-42 (регулятор клеточного деления)
Chk2 — checkpoints factor-2 (контролирующий фактор)
CIAP1 — cellular inhibitor of apoptosis-1 (клеточный ингибитор апоптоза)
CNG — cyclic nucleotide gated channels (каналы, активируемые цикличе-
скими нуклеотидами)
CNP — 2':3'-Cyclic nucleotide 3'-phosphodiesterase (2':3'-циклический нук-
леотид З'-фосфодиэстераза)
CRAC — Ca-release-activated Ca channel (Са-каналы, активируемые высво-
бождением Ca2+)
CRE — cAMP-responsive element (элементы, активируемые цАМФ)
CREB — cAMP-responsive element binding protein (белки, связанные c CRE)
Crk — v-Crk avian sarcoma virus Ct 10 oncogene homolog (гомолог вируса
птичьей саркомы)
CTF — cardiotrophin (кардиотрофин)
CypD — cyclophilin-D (циклофилин D)
DAG — diacylglycerol (диацилглицерол)
Daxx — death domain (домены гибели)
DHPR — dihydropyridine receptors (дигидропиридиновые рецепторы)
DOPA — dihydroxyphenilalanine (дигидроксифенилаланин)
DO PG — (диолеил фосфатидил глицерин)
EAE — experimental allergic encephalomyelities (экспериментальный аллер-
гический энцефаломиелит)
ЕСЕ-1 — endothelin-converting enzyme (эндотелинконвертирующий фермент)
EndoG — endonuclease-G (эндонуклеаза G)
ERK — extracellular signal-regulated kinase (регулируемая экстраклеточны-
ми сигналами киназа)
FADD — Fas-associated via death domain (Fas-ассоциированные домены гибели)
FAK — focal adhesion kinases (фокальная адгезивная киназа)
FGF — fibroblast growth factor (фактор роста фибробластов)
FGFR — epidermal growth factor receptor (рецептор фактора роста фибро-
бластов)
374
Сокращения, используемые в книге
FLIP GAB — FLICE inhibitory protein (ингибиторный белок) — GRB-associated binding protein (GRB-ассоциированный связующий белок)
GAS GluR GLUT GPCR GRB — growth arrest-specific protein (ростзамедляющий специфический белок) — glutamate receptor (глутаматный рецептор) — glucose transporter (переносчик глюкозы) — G-protein coupled receptor (рецептор, сопряженный с G-белком) — growth factor receptor-bound protein (белок, связующий с рецепто- ром фактора роста)
HDAC 5-HT ICAD — histone deacetylases (деацетилазы гистонов) — 5-hydroxytryptamine, serotonin (серотонин) — inhibitor of caspase-activated DNase (ингибитор активируемых кас- пазой ДНКаз)
IFN IFNR IKK IP3 IP3R — interferons (интерфероны) — interferon receptors (рецепторы интерферонов) — I-KB-kinase (киназа I-кВ) — inositol-1,4,5-triphosphate (инозит-1,4,5-трисфосфата) — inositol-1,4,5-triphosphate receptor (рецептор инозит-1,4,5-трисфос- фата)
IRF ISRE — interferon regulatory factor(интерфероновый регуляторный фактор) — Ifn-stimulated response element (стимулируемый интерфероном эле- мент ответа)
JAK JNK KPN Al LTD LTP MAG — Janus tyrosine kinase (Janus-тирозинкиназы) — Jun N-terminal kinase (Jun N-концевая киназа) — kaiyopherin (importin) alpha 1 (импортин) — long-term depression (долговременная депрессия) — long-term potentiation (долговременная потенциация) — myelin-associated glycoprotein (миелинассоциированный гликопро- теин)
МАРК MBP MDM2 — mitogen activated protein kinase (киназа митогенактивируемого белка) — myelin basic protein (катионный белок миелина) — mouse double minute-2 homolog (онкоген, экспрессируемый на го- мологичных мышиным «double minute» хромосомах)
MEF2 MEK MEKKs MMP — myocyte enhancing factor-2 (фактор усиления миоцитов) — MAP kinase kinase (киназа МАР-киназы) — MAP/ERK kinase kinases (киназы киназ MAP/ERK) — mitochondrial membrane permeabilization (нарушение проница- емости митохондриальной мембраны)
MOG — Myelin-oligodendrocytic glycoprotein (миелин-олигодендроцитарный гликопротеин)
MPP MPTP — 1 -methyl-4-phenylpyridinium (метилфенилпиридин) — 1 -methyl-4-phenyl-1,2,3,6-tetrahydropyridine (метилфенилтетрагид- ропиридин )
NANA NCAM NCX NEM NFAT — N-acetyl-neuraminic acid (N-ацетилнейраминовая кислота) — neural cell adhesion molecules (молекулы клеточной адгезии нейронов) — Na/Ca exchanger(Na/Ca-обменник) — N-ethylmaleimide (N-этилмалеимид) — nuclear factor of activated T-cells (ядерный фактор активированных T-клеток)
Сокращения, используемые в книге
NF-kB — nuclear factor-kappaB (ядерный фактор)
Ng-CAM — neuroglial cell adhesion molecules (молекулы клеточной адгезии
нейроглии )
NGF — nerve growth factor (фактор роста нерва)
NHERF — Na/H-exchanger regulatory factor (фактор регуляции Na/H-обмен-
ного белка)
NMDA — N-methil D-aspartic acid (N-метил-О-аспартат)
NOS — nitric oxide synthase (NO-синтаза)
NSF — NEM-sensitive fusion protein (N-этилмалеимидчувствительный белок)
p53AIPl — p53-regulated apoptosis-inducing protein-1 (р53-регулируемый
апоптозом индуцируемый белок)
p75(NTR) — р75 neurotrophic receptor (нейротрофический рецептор)
p90RSKs — ribosomal-S6 kinases (рибосомальные киназы)
PARP — prostate apoptosis response protein-4 (индуцируемый апоптозом белок)
PDK — phosphoinositol dependent kinase (фосфоинозитолзависимая киназа)
PI3K — phosphatidylinositide-3 kinase (фосфатидилинозитид-3-киназа)
PIAS — protein inhibitors of activated STATs (белковые ингибиторы активи-
рованных STAT-белков)
PK — protein kinase (протеинкиназа)
PLC — phospholipase (фосфолипаза С)
PLP — proteolipid protein (миелиновый протеолипид)
РМСА — plasma membrane Ca2+-ATP^se (Са-АТФаза плазматической мембра-
ны)
РМР-22 — peripheral myelin protein-22 (миелиновый периферийный белок 22)
PMR1 — P-type Ca2+-transport ATPase (Са-АТФаза Р-типа)
PODs — promyelocytic leukemia oncogenic domains (онкогенные домены
промиелоцитарной лейкемии)
PS — presenilin (пресенилины)
РТР — permeability transition роге (временная пора, обеспечивающая про-
ницаемость)
PUMA — p53-upregulated modulator of apoptosis (р53-регулируемый модулятор
апоптоза)
Rac — Ras-related C3 botulinum toxin substrate (ботулиночувствительный
субстрат, родственный Ras )
Raf — киназа Ser/Thr
RAN — регуляторная СТРаза
Rap — regeneration-associated protein (белок, связанный с регенерацией)
Ras — rat sarcoma genes (гены саркомы крыс)
Rbx — ring subunit of ubiquitin ligases (кольцевая субъединица убикви-
тинлигазы)
Rho — ras-related homologue (гомолог, родственный Ras)
RIP1 — receptor-interacting protein-1 (белок взаимодействия с рецептором)
ROCK — Rho-associated coiled-coil-containing protein kinase (Rho-ассоцииро-
ванная биспиралисодержащая протеинкиназа)
RSK — ribosomal S6 kinase (рибосомальная SB-киназа)
RTK — receptor tyrosine kinases (рецепторные тирозинкиназы)
RyR — ryanodine receptor channels (каналы рианидинового рецептора)
SAM — Senescence Accelerated Mice (ускоренностареющие мыши)
SEK — SAPK/ERK Kinase (киназа SAPK/ERK)
376
Сокращения, используемые в книге
SERCA — sarco- and endoplasmic reticulum Ca2+ ATPase (Са-АТФаза сарко- и эндоплазматического ретикулума)
SH2B SHC Shh SHP SMAC — SH2B adaptor protein ($Н2В-адапторный белок) — SH2 containing protein (SH2-содержащий белок) — sonic hedgehog («звучащий ежик») — фосфатаза — second mitochondria-derived activator of caspase (вторичный актива- тор каспазы митохондриального происхождения)
SMAD Smase SNAP SNARE SNT — Sma (skeletal muscle actin) 4- Mad (mothers against decapentaplegic) — sphingomyelinase (сфингомиелиназа) — soluble NSF-attachment proteins — SNAP-receptor (SNAP-рецептор) — suc-associated neurotrophic factor-induced tyrosine-phosphorylated target
SOCS — suppressors of cytokine signaling (супрессор цитокиновой сигнали- зации)
SRE SRF STAM — serum response element (сывороточный транскрипционный элемент) — serum response factor (сывороточный транскрипционный фактор) — signal-transducing adapter molecules (адаптерные молекулы пере- дачи сигнала)
STAT — signal transducer and activator of transcription (переносчики сигнала и активаторы транскрипции)
STATIP SUMO tBID TCRs TGF TK TNFR TRADD — STAT-interacting protein (белок взаимодействия со STAT) — убиквитинподобный белок — truncated BID (укороченный проапоптический белок BID) — T-cell receptors (рецепторы Т-клеток) — transforming growth factor (фактор трансформации роста) — tyrosine kinases (тирозиновые протеинкиназы) — tumor necrosis factor receptor (рецептор фактора некроза опухоли) — tumor necrosis factor receptor-1 -associated death domain (домен ги- бели, ассоциированный с рецептором фактора некроза опухоли)
TRAF — TNF receptor associated factor (фактор, ассоциированный с рецеп- тором фактора некроза опухоли)
Trio — triple functional domain-PTPRF interacting (тройное функциональ- ное взаимодействие с доменом PTPRF)
TRKA TRP TRP TYK2 — tyrosine kinase receptor-А (рецептор, связанный с тирозинкиназой) — taste receptor (вкусовые рецепторы) ’ — transient receptor potential (кратковременный рецепторный потенциал) — non-receptor protein tyrosine kinase-2 (нерецепторная тирозиновая протеинкиназа)
Ubc9 VAChT — фермент убиквитинового пути — vesicular acetylcholine transporter (везикулярный транспортер аце- тилхолина)
VDAC — voltage-dependent anion channel (потенциалзависимый анионный канал)
VMAT — vesicular monoamine transporter (везикулярный транспортер био- генных моноаминов)
Wnt — wingless gene protein
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Авитаминоз Bj 269
Авитаминоз Е 340
Адамализины (цинкзависимые метал-
лопротеиназы) 82, 85
Аденилатциклаза 58, 132, 138, 142,
143, 152, 165, 166, 167, 168, 170,
176, 178, 243, 249, 257, 303, 304
S-Аденозил метионин 258
Аденозиндезаминаза 289
Аденозиндифосфатрибоза 181
Адреналин
— метаболизм 135
— пути 136
— распределение в мозге 133
— рецепторы 135, 165, 307
---агонисты 137
--- антагонисты 137
---структура 136—137
---типы 137
— синтез 135, 136
— функции 136
Адренокортикотропный гормон (АКТГ)
82, 165, 334
Аксон
— гигантский, кальмара 198, 291,292,
293, 296
— конус роста 233, 234, 235
— миелинизированный 17, 18, 19, 41,
51,239, 271
— немиелинизированный 18, 51
—формирование синапсов 21, 98,
109, 233, 238, 283
— функция 130, 279
Аксональный транспорт 48, 115, 158,
176, 177, 184, 280, 281,283
— быстрый 281
— медленный 281
— обратный (ретроградный) 279, 280
— промежуточный 281
— прямой (антероградный) 280
Аксональные холмики 12,51
Аксоплазматический ток 280
Актин 90, 96, 129, 234,281
Алкоголизм 38
Аллергический энцефалит 45
Альбумин 101,338
Альдолаза 266, 267
Альдегидде гидро ген аза 140
Альтернативный сплайсинг 50, 194,
235
Алюминий 356, 357
Амигдала 163, 261, 263
р-Амилоид 80, 82, 85, 354, 355, 356
Амилоидде градирующий фермент 85
АМРА(а-амино-3-гидрокси-5-метил-
4-изоксазол-пропионовая кисло-
та)-рецепторы 145, 146, 247, 249,
329, 361
— долговременная потенциация 241,
245, 246, 247, 248, 249, 332
— долговременная депрессия 241,245,
248, 249
— зрительная рецепция 306, 307
— изменения 246, 247
— клатринзависимая интернализация
249
— субъединицы 248
— структура 147
4-Аминобутират трансаминаза 150
Аминокислота
— аланин 101, 153
— аргинин 155, 197, 342, 354
— ароматические 135, 139
— аспартат 112, 143, 146, 148, 181,
289, 306
— аспарагиновая 181,356
378
Предметный указатель
— возбуждающие см. Глутамат; Ас-
партат
— ГАМК см. у-Аминомасляная кис-
лота
— гистидин 216, 217, 342, 354
— глицин 101, 112, 143, 147, 153,
184, 195, 224, 225, 287, 289, 306
— глутамин 31, 40, 50, 101, 112,
1 15, 120, 143, 144, 145, 147, 148,
149, 181, 196, 197, 224,225, 247,
248, 249, 272, 274, 275, 287, 289,
306, 313, 329, 330, 331,333, 353
— глутаминовая 144, 145, 181
— дикарбоновая 205
— источник энергии 22, 213
— компартментизация метаболизма
53
— лейцин 117, 271
— лизин 255, 271,342
— метионин 1 17, 342
— пролин 101,226, 342
— серин 25, 101, 123, 178, 181, 184,
185, 187, 237,243
— таурин 143, 153, 339
— тирозин 135, 184, 236, 237, 341,
342
— тормозные см. ГАМК; Глицин; Та-
урин
— треонин 176, 184, 237
— триптофан 116, 117, 139, 256, 342
— у-аминомасляная (ГАМК) 112,
148,335
— фенилаланин 117, 255, 271,342
— цистеин 103, 206, 342
— энергетические функции 140
L-Аминокислоты 135, 136, 139,309
D-Аминокислоты 309
Амнезия 328
— антероградная 328
— ретроградная 328
Амфифизин 287
Аналгезия 157
Анандамин 163, 164
Ангиотензин 85, 157, 165, 228
Анестетики 161
Анкирин 90—92
Аннексины 94, 329
Аннулярный слой 81,86
Антибиотики 191
Антидепрессанты 336
Антипорт 190
Антитела 20, 124, 144, 228, 324, 327,
329
Апоморфин 138
Апоптоз 28, 38, 239, 351,359, 360
Аппарат Гольджи 13, 14, 15, 57, 83,
86, 115, 286
2-Арахидон ил глицерол 163, 164
Арахидоновая кислота 35, 38, 137,
148, 174, 175, 235,258,260
Аргинин 155, 197, 342, 354
L-Аргинин 155
Арестин 167, 243, 302
Асимметричность мембранного би-
слоя 65
Астроциты 14,31—34, 100, 144, 145,
150, 155, 180, 262, 264, 266, 267,
269, 271,272, 275, 289, 331,360
АТФ-дифосфогидролаза 289
Ауксилин 288
N-Ацетилгалактозамин 101
N-Ацетилнейраминовая кислота
(NANA) 29—31,97, 101
Ацетил-КоА 61, 126, 149, 258, 259,
270,271,272, 273, 275
Ацетилхолин
— выделение 317
— метаболизм 40, 113, 127, 288
— преганглионарные нейроны 126
— рецепторы 20, 121, 129, 130, 131,
132, 165, 244, 286,319
-----классификация 127—133
-----функция 131
— синтез 13, 116, 126, 130, 267, 269,
274
— содержание 21,25, 124, 127, 130,
306,317,322
— структура 319
— функция 20, 120, 127, 130, 297,
306
Ацетилхолинэстераза (АХЭ) 20, 224,
239
— ингибиторы 336
— структура 40, 50, 83, 94
Ацилазы 106
Аэробное окисление 265
Базальное гигантоклеточное ядро 126
Базальные ганглии 126, 139, 142
Баклофен 153
Бактериородопсин 300
Барбитураты 152, 153, 353
Предметный указател -
379
Белок
— адгезии нейрональной клетки
(NCAM) 46, 54,81, 102, 180,
233, 235
— актин 90, 92, 95, 102, 128, 236,
281
— анкирин 90, 91,92
— вторичная структура 77
— гистоны 347
— гликолипопротеины 101
— гликопротеины 44, 46, 47, 48, 52,
81, 82, 94, 95, 98, 100, 101, 102,
103, 104, 108, 205, 234,364
— ГТФ-связывающие 286, 307
— кальмодулин 94, 118, 155, 169, 170,
177, 182, 185, 187, 199,246,308
— кальпаин 92, 199, 330, 332
— кальцинейрин 187
— катионный белок миелина 45
— кинезин 279, 281
— клатрин 92, 93, 281,287, 288
— коллаген 95, 233, 234
— комплексин 286
— концевые \Н2-группы 342
— белки мембранные
-----амфипатические 39
-----белки-ферменты 82, 83, 84, 120,
169
-----белки-эффекторы 119, 120, 123,
142, 166, 167, 168, 227, 302, 303,
304
-----белок р-АРР 80, 354, 355, 356
-----битопические 79, 81
-----интегральные 12, 56, 58, 78,
79, 81, 83, 91, 94, 97, 118, 119,
120, 184, 197, 203, 204, 226, 287,
308,319, 355
-----лиотропность 79
-----монотопические 79, 80
-----неинтегральные 79, 80
-----периферические 78, 79, 83, 86,
94,98
-----политопические 79, 81
-----рецепторные 12, 21, 80, 83,
84, 85, 100, 118, 1 19, 128, 194,
236, 307, 308
-----структурные 83, 84, 90, 91, 92,
184,280, 330
— метаболизм 51,356
— миелинассоциированный глико-
протеин 46
— миозин 185
— модуляторы мембран 116
— морфогенеза кости (BMP) 229,
231
— нейростенин 93
— нейротрофины 332
— нейротубулин 46, 47, 128, 281
— полисиалогликопротеины 102
— протеолипиды 26, 45
— р-амилоид (р-АРР) 80, 82, 85, 354,
356
— сборки чехла 93
— сиалогликопротеин GP-351 102,
103
— синапсины 85, 94, 288
— синаптобревин 286
— синаптотагмин 94, 170, 287
— синтаксин 286
— синтафилин 286
— снапин 286
— сократительные 93, 279
— томозин 286
— третичная структура 45, 77, 79
— тубулин 46, 47, 91, 128, 280, 281
— фодрин 92, 330
— цитоскелетные 46, 81, 84, 90—94
— р-АРР 78,354—356
— В-51 44
— BSP 101
— Са2+-связывающие 94, 170, 198,
286
— D3 138
— DM20 45
— G-белки 165, 168, 307, 308, 313
— К5 101
— L-2 101
— МАР-3 183, 237
— МВА-3 42, 102
— N-CAM 46, 81,94, 101, 102, 233,
234, 235, 236
— Ng-CAM 101, 102
— NSA-4 102
— NSF 286
-Р047
— PLP45
— РМР-22 48
— S-100 327, 329
— SNAP 286
— SNARE-ассоциированные 286
— Thy-2 102
380
Предметный указатель
Белки-транспортеры (переносчики)
— ацетилхолина (VAChT) 287
— биогенных моноаминов (VMAT) 287
— везикулярные 287
— вещества-блокаторы 128, 290
— ГАМК 287, 289, 290
— глицина 287, 289, 290
— глутамата 144, 287, 333
— глутамина 144
— глюкозы 254, 262, 263
— дофамина 289, 290
— норадреналина 289, 290
— серотонина 289
Белки теплового шока 179, 180, 349,
350,358
Белки-предшественники 52, 80, 160,
354
Белковые ассоциаты 77, 84, 88, 190,
204,211,302
— гетероолигомеры 92, 147, 165, 204
— гомоолигомеры 77
Белое вещество 9, 26, 37, 42, 96, 263
Белок-белковые взаимодействия 211,
214, 215, 219, 226, 228, 234, 287,
301,358
Бензогексоний 128
Бензодиазепам 152, 153
Бензодиазепины 151, 152
Бета-амилоид см. Р-Амилоид
Билирубин 339
Бледный шар 125, 148
Блок открытого канала 193
Болезнь
— Альцгеймера 34, 80, 82, 85, 126,
159, 354, 355, 356
— бери-бери 269
— Паркинсона 40, 126, 134, 138, 351,
352, 353
Брадикинин 85, 157
Бромкриптин 138
а-Бунгаротоксин 128
Вазоактивный интестинальный (по-
липептид (ВИП) 157, 159
Вазопрессин. 157, 181,327, 334, 336
Вакценовая кислота 67
Вегетативная нервная система 12, 16,
19, 126, 128, 154, 159
— парасимпатическая 126, 154
— симпатическая 126, 136, 154, 159
Везикулы 37, 60, 64, 67, 92, 93, 110—
112, 114—116, 127, 130, 135,
156, 184, 199,200, 283,285—290,
320, 321,330, 334
Везикулярные транспортеры 287
Вентральная покрышка 125, 134, 142
Вентральные ядра 263, 314
Вентральное постеромедиальное ядро
таламуса 314
Верхний холмик (colliculus superior)
263
Вещество Р 112, 143, 157, 158. 159
Витамин
— Е339
— С 337, 339
Вкусовые
— рецепторы 20, 310, 311
— стимулы 309, 311
— сосочки 310,311,313
Внутреннее подкрепление (внутреннее
вознаграждение) 138, 139, 142,
163
Водородные связи 88, 98
Возбуждающие постсинаптические По-
тенциалы (ВПСП) 297, 298,299
Вторичные мессенджеры (посредни-
ки) 117, 119, 120, 169
— Са2+ 169, 170, 171, 173, 174, 175,
242
— NO 156, 179, 180, 181,228
— АФК 178, 179
— диацилглицерол (DAG) 169, 170,
173, 174, 175, 242, 249
— инозитолфосфаты 173, 174, 175,
199, 303
— ИФ3169, 170, 173,249
— цАМФ 168, 170, 171. 178, 242, 244
цГМФ 169, 170, 171, 178
ГДД65 150
ГАД67150
G-белки 165
— активация 165, 167
— связывание 165
— типы 165
— структура 307
Галактолипиды 42, 43
Галактоцереброзиды 26, 29, 30, 52
Галанин 157, 327
Галлюциногенные эффекты 142
ГАМК-шунт 149,275
Предметный указатель
38:
Гамма-аминомасляная кислота
(ГАМК), см. также Медиаторы
112, 148, 335
— метаболизм 149, 150, 275
— пути 150, 224, 272, 287, 289
— распределение в мозге 148, 149
— рецепторы 120, 121, 149, 151, 152,
153, 184,244
--- агонисты 152
---антагонисты 152, 153
---структура 120, 150, 151, 152,
153
---типы 120, 121
---функции 121
— синтез 148, 149, 150, 275
— фоторецепция 306
— функции 143, 149, 306
— эпилепсии терапия 144, 149, 263,
353
Ганглиозиды
— ацетилирование 97
— в онтогенезе 99, 100
— иммунные свойства 97
— лечебное действие 336
— локализация 31,98
— межклеточное гликозилирование
100, 103,259
— нейритогенез 31,99
— номенклатура 97
— содержание 26, 33, 43, 96
— электрогенность 99
Гексокиназа см. Гексокиназная реак-
ция
Гексокиназная реакция 264, 265, 266,
274, 278
Гематоэнцефалический барьер (ГЭБ)
144, 252, 254, 255, 256, 263, 271,
278
Гены
— c-fos 332
— c-jun 332
— c-jun В 332
— zz//268 332
— pmp-22 48
— в ЦНС позвоночных 336
— экспрессия 49, 187, 204, 231,233,
238
Гепарин 100, 231
Гетерорецептор 142
Гиалуроновая кислота 100
Гидроксид-ради кал 341,347, 350
Гидроксилаза 135
Гидроксиноненали 345—346
5-Гидрокситриптамин (5-НТ) см. Серо-
тонин
5-Гидрокситриптофан 139
Гидроперекиси липидов 71,344
Гидрофобные
— связи 101,204
— взаимодействия 89
Гиперполяризация 141,226, 292,
294, 297, 298, 299, 301,302, 303,
314
Гипоксия 290, 327
Гипоталамус 128, 133, 134, 135, 138,
139, 143, 159, 161, 163, 252, 263,
309, 334
Гипофиз 159, 221,334
Гиппокамп
— долговременная потенциация 147,
156, 245, 248, 332
— долговременная депрессия 241,245,
248
— обучение 185, 248
— память 163
Гистамин
— метаболизм 142
— пути 142
— распределение в мозге 143
— рецепторы 142, 143, 165, 302
--- агонисты 143
--- антагонисты 143
---структура 307
---типы 143
---функции 142, 143
— структура 112
— синтез 142
— функции 142, 143, 165
Гистидин 216, 217, 342, 354
Гистоны 347
Гликоген 264, 265, 307
Гликозаминогликан 98
Гликозилтрансфераза 100
Гликокаликс 52, 94, 95, 96, 99
Гликолиз 264—268
Гликолипиды 22, 24, 30, 31,52, 94—
96, 98,99, 103, 258
Гликолипопротеины 101
Гликосфинголипиды 28, 57, 96
Глицерофосфолипиды 23, 24
Глицин 101, 112, 143, 147, 153, 184,
195, 224,225, 287, 289, 306
382
Предметный , к. гз )Тель
Глия
— функция 9, 14, 15
— гликолиз 262—268
— пентозофосфатный путь 265
Глутамат
—долговременная потенциация 245,
247
— метаболизм 115, 149, 150, 274, 275
— «молчащие» синапсы 247
— пути 141
— распределение в мозге 143
— рецепторы 31, 50, 92, 120, 121,
143—148, 150, 155J81, 196, 197,
249, 272,313, 329,330
---- агонисты 146
---- антагонисты 146
----структура 120, 121, 145, 197
----типы 50, 120, 121, 145, 146, 148
----функции 50, 92, 155, 196, 249,
313, 329
— синтез 144, 150
— функции 31,50, 120, 143, 181,272,
330
— экзайтоксичность 147, 156, 181,
353, 360
Глутамат-глутаминовый цикл 150
Глутаматдегидрогеназа 150,274
Глутаматдекарбоксилаза (ГАД) 149,
150
Глутаминаза 144, 150
Глутаминовый цикл 150
Глутаминсинтаза 144
Глутатион 339
Глутатионпероксидаза 338
Глутатионтрансфераза 338
Глюкоза
— гипогликемия 278, 280
— переносчики через ГЭБ 253, 255,
256
— потребление 13, 175, 254, 261,262,
263, 278
— содержание 254
— синтез медиаторов 144, 253
D-глюкоза 278
Глюкозо-6-фосфат 257, 264, 265
Глюконеогенез 269
Глюкоцереброзиды 30
Голубое пятно 125, 133
Гонадотропин (люлиберин) 157, 165
Гормоны 22, 31, 54, 73, 82, 86, 118,
119, 123, 133, 156, 158, 159, 165,
168, 178, 183, 243, 303, 334
Горизонтальные клетки 149, 305, 306
Гуанилатциклаза 156, 170, 178, 179
Гуанозин окисленный 347
Густдуцин (gustducin) 313
Деацилирование-реацилирование
липидов 257
Дегидрогеназа полуальдегида янтарной
кислоты 150, 275
Дезоксиглюкоза 262, 278
Декарбоксилаза ароматических ами-
нокислот 135, 136, 139
Дендрит 9, 10, 11, 12, 14,38, 92, 109,
НО, 177, 238, 246—249, 271,
279, 283, 309
Дендритное поле 11
Деполяризация 89, 292
— выделение нейромедиатора 153,
225, 329
— ионный ток 129, 130, 147, 153,
195, 246, 293, 308,318
— мембрана 109, 129, 147, 153, 176,
192, 225, 245, 246, 294, 308, 318,
329
— проводимость 109, 130, 192, 225,
292, 294, 295
Десатуразы 106, 258
Десенситизация 158, 241,323
Десмостерин 33
Детергент 118, 201—203, 211, 212,
213,215, 358
Диагональная связка 126
Диаденозинтетрафосфат 154
Диазепам 152—153
Диацилглицерол (ДАГ) 39, 40, 84,
118, 169, Г70, 171, 172, 173, 174,
175, 178, 182, 185, 235, 257
Дигидросфингозин (сфинганин) 97, 259
С20-дигидросфингозин (эйкозасфин-
ганин) 97
Диеновые конъюгаты 346
Дикарбоновые аминокислоты 205
Динамин 285
Динеин 279
Динорфины
— динорфин А 160, 162
— динорфин В 160, 162
Предметный указатель
383
Диоксифенилаланин (ДОФА) 135, 139,
256, 352,353
Диольные фосфолипиды 24, 25, 27
Дифосфатидил глицериды 25, 27
Дифференциация клеток 28, 31, 40,
45, 46, 99, 100, 104, 122, 186,
222, 229, 232, 236, 237, 238
ДНК 3, 45, 54, 114, 323, 332, 336,
337, 347
— репарация 347
— содержание 49
— тринуклеотидные повторы 199,
357
ДНК-полимераза 336
Долговременная депрессия (ДВД) 241,
245, 248, 249,251
Долговременная потенциация (ДВП)
241,245
— АМРА-рецепторы 246—249
— молекулярный механизм 245—247,
249
— свойства 245
— функция 246, 248
ДОФА-декарбоксилаза 134
Дофамин
— метаболизм 134
— пути 134
— распределение в мозге 134
— рецепторы 138
---- агонисты 138
----антагонисты 138
----типы 138
----функции 138
— структура 112, 133
— синтез 134—136
— функции 126, 135
Дофамин-Р-гидроксилаза 134
Дугообразное ядро 125, 134
Дыхательная цепь митохондрий 273,
274,340
Дыхательный коэффициент 261,262
Енолаза 266
Желудочки мозга 14, 125, 135, 159,
161,252, 253, 329
Жирные кислоты
— арахидоновая 35, 38, 137, 141,
174, 175, 235, 258, 262
— бегеновая 35
— Р-окисление 259
— вакценовая 35, 67
— длинноцепочечные 27, 28, 34, 36,
41,258
— докозогексаеновая 35, 37, 38, 106
— источник энергии 22
— клупанодоновая 35
— лауриловая 35
— лигноцериновая 30, 35, 101
— линолевая 35, 38, 256
— линоленовая 35, 38, 258
— маргариновая 35
— миристиновая 35, 88
— моноеновая 36
— нервоновая 30, 35
— олеиновая 35, 45, 67, 106, 258
— пальмитиновая 35, 45, 88
— пальмитоолеиновая 35
— пероксисомальное окисление 259
— полиеновые 34, 36
— синтез 257—259
— стеариновая 28, 35, 45, 175, 258
— цереброновая 30
— эйкозогексаеновая 38
— элаидиновая 35
«Звучащий ежик» (Shh) 230, 231
Зрительная кора 263, 278, 306
— радиация 305, 306, 238
Зрительный нерв 306
Изопреноиды 258
Изоцитратсинтазные реакции 274
Иммуногистохимические методы 114,
134,149
Импульс 7, 9, 10, 12, 19, 21, 41, 43,
49, 54,55, 98, НО, 112, ИЗ, 116,
146, 279, 283, 285, 292, 293, 295,
296, 302, 306, 307, 309, 315, 331
Инозитол-1,4,5-трисфосфат (ИФ3)
3, 9, 118, 137, 138, 141, 166, 169,
170,175
Инсулинде градирующий фермент
(ИДФ; инсулизин) 356
Инсульт 85, 181,359, 360
Интегративная функция 147
Интегриновые рецепторы 232—234
Интегрины 232, 234
Интерорецепторы 20
Инфаркт 41,222, 348, 350
Ионные каналы см. Каналы ионные
.Предметный указатель
Ионные насосы (транспортные
АТФазы) 40, 54, 197
Ишемия 40, 41, 144, 154, 181, 211,
222, 252, 359, 360
Йохимбин 136, 137
Кадгерины 232—233, 236
Каинатные рецепторы 148, 329
— функция 147, 351
— структура 149
Каиновая кислота 145, 146
Калиевые каналы см. также Каналы
ионные
— закрывание 293
— кальцийактивируемые 193
— кристаллография 217
— мускариновые рецепторы 132
— потенциалактивируемые 176
Кальпаин 92, 199, 330, 332
Кальциевый ионофор 173, 174
Кальциевый канал см. также Каналы
ионные
— зрительная рецепция 302
— потенциалзависимые 176, 177
— топология 202
— трансдукция сигнала 170, 235
Кальция ионы
— активация калиевых каналов 174
— долговременная потенциация 147,
156, 331,332
— долговременная депрессия 241,245,
248, 249
— выделение нейромедиатора 310—
311
— вторичный мессенджер 176—178
— NMDA-рецепторами связывание
147
Са-зависимые молекулы клеточной ад-
гезии (кадгерины) 232—233,276
Са-независимые молекулы клеточной
адгезии (САМ) 232—233
Каналы ионные
— анионные 191
— ацетилхолином активируемые 297
— вкусовых рецепторов функция
310—313
— десенситизация 158, 193, 323
— избирательность 129, 191, 192
— инактивация 193
— ионный градиент 197, 283, 293
— калиевые (К+) см. Калиевые
каналы
— кальциевые (Са2+) см. Кальцие-
вые каналы
— катионные 191
— катионселективные 311
— лигандуправляемые см. также
Ионотропные рецепторы
— молекулярная структура 129, 151,
152, 194,320
— натриевые (Na+) см. Натриевые
каналы
---амилорид-чувствительные
311,312
— пора 320
— потенциал действия 129, 292—296
— потенциалзависимые 147, 176, 177,
193, 194
— селективность 308, 310, 311
— хлорные (С1~) 111, 151, 152, 192,
195
---Са-управляемые 308
— топология 194
— Н-чувствительные 311
— TRPM5 310—311
Каннабиноиды 163—164
Кардиолипин 24, 39, 40, 105, 254
Карнозин 156,339, 352, 355
Каротиноиды 339
Каскад
— МАР-киназный 165, 183, 225, 231,
237
Каспазы (Са-зависимые аспарагино-
вые-сериновые протеиназы) 38,
256
Каталаза 259, 336, 348
Катенины 102
Катехоламины см. Дофамин; Нора-
дреналин; Адреналин; Медиаторы
Катионный белок миелина 46
— изоформы 46
Квант (порция) медиатора 285—286
Квискваловая кислота 145
а-Кетоглутарат 145, 149, 150, 274, 275
а-Кетоглутаратдегидрогеназа 150
Кетоновые тела 255, 271
Кимограф 318
Janus-Киназы 351,349, 350
Кинезин 279, 281
Кинины 80, 85, 157, 244
Кинк 65, 69, 73—75
Предметный указатель
385
Клатрин 92, 93, 281,287, 288
Клатринзависимая интернализация
рецепторов 93, 249
Клетки
— амакриновые 305
— биполярные 305
— ганглиозные (ганглионарные) 12
— горизонтальные 149, 305, 306
— зерна 309, 143
— митральные 309
— перигломерулярные 309
— Пуркинье 11,249
— Реншоу 126, 153
— адгезия 40, 48, 52, 58, 83, 101,
103, 232,235
Клозапин 138
Колбочки 299
Коллаген 95, 233, 234
Коллапсин-1 235
Компартментализация метаболизма
264, 275
Коннексоны 191
Конус роста 233
— семафорины 232—233
— структура 233
Кора мозжечка 10, 11,263
Кортикотропины 332
Коэнзим А (КоА) 61,258, 270—272
Креатинкиназа 40, 277
Креатинфосфат 40, 276, 277
Критическая концентрация мицелло-
образования (ККМ) 59
Кураре 286
Лактатдегидрогеназа (ЛДГ) 267
Лактоферрин 338
Ламеллярная (слоистая) структура 43,
59, 69, 70
Ламинин 232—234
Латеральная ретикулярная формация
моста 133
Латеральное коленчатое тело 278, 306
Лей-энкефалин см. Энкефалины
Лигандуправляемые ионные каналы
см. Каналы ионные
Лизосомы 13, 15, 243, 245, 260
Лизофосфатидилхолин 27
Лизофосфолипиды 39, 345
Лимбическая система 135, 142, 143
Лимонная кислота 270, 272, 274
Лиотропный мезоморфизм 59, 70
Липиды
— аллостерические активаторы 40
— астроглии 33, 34
— белого вещества 43, 47
— в онтогенезе 257
— дефектные зоны 66. 73, 342, 345
— жидко-кристаллическое состояние
59, 62, 69,71—73, 75
— кластеры 56, 66, 73, 74, 99, 178
— миелина 22, 24, 28, 33, 34, 36,
41—44
— нейронов 34
— олигодендроглии 31,33
— рафты 56—58, 71,85, 100
— серого вещества 37, 43, 103
— трансмембранная асимметрия
63—65
— фазовые переходы 69—73
— якорная функция 58, 78—81, 103,
119, 123, 168
Липидозы 260
Липосомы 60, 66, 201,202
Локальный (электротонический) по-
тенциал 292
Локомоторная функция 126
Люлиберин см. Гонадотропин
Магния ионы 174, 193, 201,207, 209,
246, 329
Макроглия 14, 15
Макроергические соединения
— АТФ 214
— ГТФ 214
— креатинфосфат 40, 276, 277
— фонд 264
Малоновый диальдегид (МДА) 345,
346
Малые G-белки, см. Ras, Raf, МЕК
МАО см. Моноаминоксидаза
Мевалонат 258
Медиальное ядро перегородки 124
Медиаторы
— адреналин 112, 133, 135, 136, 165,
175, 307
— аспартат 112, 143, 146, 181, 289,
306
— ацетилхолин 13, 20,21, 112, 116,
120, 122—133, 156, 165, 244,
267, 269, 274, 297, 306, 317, 319
— вазоактивный интестинальный пеп-
тид(ВИП)157, 159
— вещество Р 112, 143, 157— 159
Яредметныи указатель
— влияние на ауторецепторы 114,
143, 148
— выделение (экзоцитоз) 91, 113—
115, 168, 176, 177, 285, 288, 329
— ГАМК 112, 143, 146—153, 159,
161, 184, 224, 225, 272, 275, 285,
287, 306, 353
— гистамин 112, 142—143, 165, 308
— глицин 112, 143, 153, 184, 195,
224, 225, 289, 306
— глутамат 29, 40, 112, 115, 120,
143—148, 150, 156, 181, 196,
224, 225, 246—249, 274, 275,
285,289, 306, 329, 330, 331
— деградация 136
— дофамин 112, 126, 133—138
— катехоламины 116, 133, 135, 137,
159
— кванты 130, 239, 247, 285—286,
320, 321
— критерии 113
— классификация 112
— люлиберин 157
— моноамины 112, 115, 134, 139, 224
— накопление 225, 287
— нейропептидУ 157, 159, 306
— нитроксид 112, 113, 177, 178
— норадреналин 13, 21, 112, 116,
134, 135, 136, 142, 159, 165, 224,
228, 257, 290
— пептидные 52, 95, 112, 114, 116,
124, 163, 334,352
— предшественник 161
— пурины 112, 154, 196
— разрушения в нейронах 31
— разрушения в синаптической щели
40, 289, 320, 321,334
— серотонин (5-НТ) 12, 112, 116,
117, 121, 139—142, 161, 165,
173, 195, 224, 228, 289, 306, 334
—синаптические везикулы 93, 116,
285, 287
— синтез 116, 134—136, 274—275
— системы обратного захвата 113—
115, 124, 126, 144, 150, 224,279,
289, 290, 330, 360
— соматостатин 157, 159, 165, 306
— сопутствующие (котрансмиттеры,
сомедиаторы) 116
— таурин 143, 153, 339
— упаковка 115, 116, 289
Мезаксон 16, 18, 41
Мезенцефалон 353
Меланокортины 160
Мембрана
— везикулярная 286—288
— плазматическая 12, 13, 15, 32—
34, 39—41,45, 47, 51,55, 57, 78,
83—85, 89, 92, 93, 100, 102, 103,
118, 130, 168, 185, 186, 243, 245,
247, 259, 287, 288, 299, 300, 301,
303
— постсинаптическая 20, 21, 40, 51,
92, 109—111, 116, 129, 130, 153,
241,244—249, 283, 285, 288,297,
329
— пресинаптическая 20, 21, 84, 93,
109, НО, 128, 144, 153, 283, 285,
286, 320, 330, 331
— бислой 26, 39, 51,56, 59, 60, 65, 72,
76, 78, 88,99, 189,211,226, 344
— диффузия 188—189
— транспорт 188—192
— фазовые переходы 59, 71,72, 73
— ферменты 78
Мембранный потенциал
— амплитуда 123, 159, 244, 295, 297,
299,314,320
— возникновение 297, 298, 301
— запись 291
— ионный ток 159
— Na/K-насос 290, 293
— нервная клетка 291
— проницаемость 290, 291
Метародопсин-II 301
Метилазы 64, 103, 136
Метилтрансфераза 64
Метилфенилтетрагидропиридин
(МРТР)352
Метионин 117,342
Метод фиксации потенциала 293
Mez??-энкефалин 160, 162
Механизмы адаптации
— долгосрочные 106, 325
— краткосрочные 105, 106, 244, 359
Миелин 17, 18, 19, 24, 33, 34, 36,
41—46
Миелинассоциированный гликопро-
теин (MAG) 46
Миелинизация 15, 47, 239, 271,272
Миелинолигодендроцитарный гли-
копротеин (MOG) 47
Предметный указатель
387
Микровязкость мембран 76, 89, 99,
104, 106, 204,214
Микроглия 15, 266
Микротрубочки (нейротубулы) 11, 13,
91
Миндалевидный комплекс 142, 328
Миндалина 125, 143, 159, 161,309
Миниатюрные потенциалы концевой
пластинки (МПКП) 286, 320, 321
Миорелаксанты
— налоксон 162, 164
— налтрексон 162
Мицеллы 59—61,66, 67, 69
Модель мембраны 56, 58
— жидкостно-мозаичная Сингера и
Никольсона 55, 56, 58
Модуляция
— ауторегуляция 142
— постсинаптическая 241
— пресинаптическая 137, 148
Мозолистое тело (corpus callosum) 263
Молекулы-эффекторы 120, 122
Моноамины 112, 115, 134, 139, 224
Моноаминооксидаза (МАО) 135, 136,
289, 351—352
Моносиалоганглиозид 43, 97
Морфин 160, 162
Мотонейрон 126, 153, 198, 231, 239,
240, 286,314,315
Мочевая кислота 340
Мышечные волокна 19, 109, 193,
239, 296,314,315,318, 320, 321
Мягкая мозговая оболочка 14, 253
Навязчивые состояния 142
НАД-изоцитратдегидрогеназа 272,
274—276
Налоксон 162, 164
Насечки Шмидта-Лантермана 18
Натриевые каналы см. также Ион-
ные каналы
— вкусовые рецепторы 310—312
— влияние токсинов 127—128, 193
— гены 194,219, 221,222
— потенциал активируемые 176
— топология 205, 207
— фоторецепторы 301,302, 304
Na/K-насос (Na/K-АТФаза)
— Na-конформация 129, 206, 207
— график Аррениуса 214
— Е2—Е1 переход 206
— изоформы 220—224
— кинетическая схема работы 208—
211
— К-конформация 206, 207
— коэффициент Хилла 211
— олигомер 211,219, 221
— отрицательная кооперативность 211
— функциональная единица 218
Na/Ca-обмен 170, 176, 177, 220, 224,
225, 226
(Na + К + С1)-котранспорт 226
Насос электрогенный 225
Нейробласты 229, 233
Нейрогенез 229, 231
Нейроглия 9, 13, 15, 27, 53, 100
— макроглия (глиоциты) 14, 15
----астроциты 14, 31, 33, 34, 100,
144, 145, 150, 155, 180, 262, 264,
266, 267, 269, 271,272, 275, 287,
329, 358
----олигодендроциты 14, 15, 17,
18,31,45—47, 78, 100
----эпендимоциты 14
— микроглия 14, 15, 155, 180, 266
Нейрогормоны 133, 158, 159, 165, 334
Нейродегенеративные заболевания
3, 34,45, 55, 157, 227, 357
Нейромедиатор (нейротрансмиттер,
нейропередатчик), см. Медиаторы
Нейромедиаторные пути
— ацетилхолиновые 124, 125, 126
— ГАМК 125, 148
— глициновые 153
— глутаматные 143— 144
— дофаминовые 125, 134, 135
— катехоламиновые 125, 133—135
— моноаминовые 142
— норадреналиновые 133
— серотониновые 125, 139, 142
Нейромодуляторы 116, 139, 154, 157,
260
Нейрональный белок клеточной адге-
зии (N-CAM) 46, 81, 94, 101,
233,234
Нейроны (нейроциты)
— апоптоз 238, 351,359, 360
— ассоциативные 10
— афферентные (чувствительные) 10,
310
— биполярные 11,305
— миграция 229, 234
388
Предметный указател-
— моторные см. Мотонейрон
— мультиполярные 11
— разнообразие 10—12
— рецептивное поле 305, 306
— рост и развитие 17, 34, 234, 235,
237
— пирамидные 126
— псевдоуниполярные 11
— симпатические 136, 238
— униполярные 11
— эфферентные (эффекторные) 10
— элементы цитоскелета 46, 81, 84,
90—94
НейропептидУ 157, 159, 306
Нейропептиды 52, 95, 112, 114, 116,
124, 163, 334,352
— метаболизм 161,289
— предшественники 161
— распределение в мозге 125
— рецепторы 115, 121, 162
— синтез 1 15, 161
— структура 113, 160
— транспорт 115
— функции 157
Нейропилины 231 —232
Нейроспецифические
— белки 48, 49, 51, 84, 92, 102, 266,
278, 327
---антигенные свойства 45
— мРНК49, 50
Нейростенин 93
Нейротензин 157, 159
Нейротрофические белки 234, 240
Нейротубулы 11, 13, 91
Нейрофибриллы 13,354
Нейрофиламенты 11, 13, 21, 91, 93,
280, 281
Нековалентные взаимодействия
— гидрофобные 56, 60, 78, 81,88, 89,
101,204
— электростатические 63, 88
Некроз 357, 359, 360
Неокортекс 158, 163, 336
а-Неоэндорфин см. Эндорфины
Неприлизин 82
Неприродные детергенты 118, 201 —
203,211,212,215
Нерв блуждающий (X) 126, 309, 314,
316
— лицевой (VII) 313
— языкоглоточный (IX) 309, 313
Нервная пластинка 229, 231
Нервная система
— периферическая 9, 17, 41, 44, 45,
47, 48
— центральная 9
Нервная трубка 229, 231,233
Нервный гребень 15, 229, 231, 233,
234, 236, 238
Нервные волокна 10, 15, 16, 105,
180, 240, 282,292, 294,295
— миелинизированные 10, 15—19,
294
— немиелинизированные 16, 18, 19,
294
Нервные окончания 9, 10, 11, 19,21,
80
— двигательные 10, 19, 20
— рецепторные 19
— секреторные 9, 13, 19, 20, 109
Нервный импульс 19, 21, 43, 51, 54,
55, 98, 110, 112, 116, 283, 292,
295, 296, 306, 307
Нетрины 232—234
Нигростриатный путь 134, 138, 158
Нижняя олива 110
Нижний холмик (colliculus inferior) 263
Никотин 131
Нитроксид 112, 113, 177, 178
NMDA (1М-метил-Б-аспартат) рецеп-
торы 38, 145, 155, 156, 196, 223,
245—248, 329, 330, 332, 351,357
— структура 146, 147
— субъединицы 146
— функции 146, 147
NO-радикал 112, 156, 178—181,341,
342
Норадреналин, см. также Катехо-
ламины 13,81, 112, 159
— метаболизм 133, 135
— пути 125
— распределение в мозге 125, 133
— рецепторы 116, 142, 165
— синтез 13, 133—135
— структура 113
— транспортер 224, 289, 290
— функции 133, 257
NO-синтаза (NOS) 155, 157, 332
— индуцибельная (iNOS) 155
— конститутивная 155, 228
-----нейрональная (nNOS) 155
-----эндотелиальная (eNOS) 155,
228
Предметный указатель
389
Норморфин 162
Нуклеиновые кислоты
— содержание 47
-----в глин 47
-----в нейронах 47, 48
5-Нуклеотидаза 48, 94
Нуклеотиды
— структура 210,216
Нуклеотиды циклические 112, 170,
171, 184, 196, 303, 337
— вторичные посредники 169, 170,
178, 186
— ионные каналы 170, 196, 303
— метаболиз 166, 168, 170, 171, 177
— сигнал-трансдукторные системы
123, 166, 167, 168, 170, 179, 182,
184,301—303
— синтез 137, 154, 166, 170, 176,
178, 179,308
— цАМФ 170, 196
— цГМФ 123, 169, 170, 177, 196, 305
Обонятельная луковица 125, 134, 135,
143, 148, 169, 309
Обратный захват (re-uptake) меди-
атора 113—115, 124, 126, 144,
150, 224, 279, 289, 290, 330, 360
Обучаемость 328, 329, 335, 336
Обучение 36, 47, 65, 80, 85, 110,
157, 185, 241,248, 271,277, 324,
325, 328, 329, 333—335, 361
Одоранты 307
— восприятие 307
— трансдукция сигнала 307—309
Окислительное
декарбоксилирование 268—271
Окислительный стресс 157, 212, 222,
223, 340—344, 346—348, 351 —
353,356, 357, 361
Оксалоацетат 273—275
Оксид азота см. Нитроксид (NO)
Олигодендроциты 14, 15, 17, 18, 31,
45—47, 78, 100
On-/off-клетки 305, 306
Опиатные рецепторы 162
— агонисты 162
— антагонисты 162
— типы 162
-----дельта (8) 162, 163
-----каппа(к)162, 163
-----мю (ц) 162, 163
---ORL1 162
— функции 162
Опиоидные нейропептиды см. Эндор-
фины; Энкефалины; Динорфины
Опсин 300—302, 308
Орбитофронтальная кора 309
Оротат 335, 336
Оротовая кислота 335
Основной фактор роста фиброблас-
тов (bFGF) 238
Отрицательная обратная связь 136,
150, 294
Отрицательное (электроболевое) под-
крепление 334
Палочки 37, 38, 55, 168, 225, 299—
301,303, 305, 306,313
Память 20, 55, 80, 85, 100, НО, 126,
150, 151, 157, 241,327, 333—335,
359
— биологическая 324
— генетическая 337
— долговременная (долгосрочная) 48,
146, 181, 183, 185, 326, 327, 328,
336,361
— иммунологическая 324
— краткосрочная 163, 237, 326, 328
— локализация 326
— нейрологическая 279
— перенос 335
— пожизненная долговременная
(ДПП)327
— промежуточная 326, 327
— пространственная 38
Паника 142
Паракват 340
Паракринная сигнализация 156
Паркинсонизм 38, 126, 134, 135, 138,
351—353
Пассивная диффузия 188, 189
Патология
— ганглиозидозы 100, 104
— гликопротеины 48
— глюкоцереброзидозы 30
— состава миелина 45
Паутинная оболочка 253
ПДГ-киназа 270
Пептидазы 289
Передача сигнала
— стадии 119, 167,302,311
Перекисное окисление липидов (ПОЛ)
71,343—346
390
Предметный указатель
Перекись водорода 179, 338
Переносчик 117, 127, 186, 189, 190,
225, 226, 255, 256, 287
— подвижный 188
Перехваты Ранвье 17—19, 296
Перикарион 12, 15, 100, 116, 158,
261
Периферийный миелиновый белок 22
(РМР-22)48
Периферическая нервная система
(ПНС) 9, 17,41,44,45, 47, 48
Пероксидаза хрена 280
Пероксинитрит 157, 179, 341, 342,
350, 351
Пероксильный радикал 341
Пикротоксинин 152
Пиноцитоз 89, 253
Пиоглиальная мембрана 14
Пирацетам 335, 336
Пиридоксальфосфат 150
Пировиноградная кислота 268
Пируватдегидрогеназа (ПДГ) 267—
271,273
Пируваткарбоксилаза 270
Плазмалемма (плазматическая мемб-
рана) 12, 13, 15, 32—34,39
Плазмалогены 24, 25, 42, 43
Плазмин 85, 356
Пластичность нейронов 85, 147
Подвижность липидов 57, 61, 65—
67, 69,71,73, 75,81,86
— латеральная 54, 59, 65, 66, 71,98,
102
— трансбислойный переход («флип-
флоп») 65—67, 98
Позитронно-эмиссионная томография
261,262, 278
Полилизин 86
Полиненасыщенные жирные кислоты
см. Жирные кислоты
Полипролиновый сайт (pPRO) 226,
227
Полисомы 11,12
Постцентральная извилина коры го-
ловного мозга 314
Потенциал
— генераторный 294
— действия 51, 123, 130, 285, 292
-----все или ничего принцип 296—
297
-----временной ход 129
----ионные каналы 129, 293—294
----кальциевый 233, 236
----мембранная проницаемость 292
----проведение 292, 295—296
----сальтаторное проведение 19, 296
----фазы 294
----функция 129
— концевой пластинки (ПКП) 130,
286
— локальный 292
— мембранный см. Мембранный по-
тенциал
— покоя 12, 89, 247, 290—292
----проводимость 391
----ионная природа 225, 290
----гигантского аксона кальмара 291
----проницаемость мембраны 247,
291
— постсинаптический вызванный 110,
283
— рецепторный 296
— синаптический 296—299
— электротоническое распростране-
ние 292
Потенциация 181
— длительная (долговременная) 147,
156, 331,332
— кратковременная (посттетаничес-
кая) 241
Потребление кислорода 260
— головной мозг 260, 261,274
— дыхательная цепь митохондрий 340
— мозжечок 261
— продолговатый мозг 261
— промежуточный мозг 261
— средний мозг 261
Празозин 136
Пресенилины 355
Пресинаптическая модуляция 116,
241
Претектальная область ствола мозга
306
Прилежащее ядро (nucleus accumbens)
263
Природные каналоформеры 191
Проди норфи н 160
Продолговатый мозг 128, 133, 134,
139, 153, 261,314
Продукты Амадори 342
Пролин 101,342
Проопиомеланокортин 160
Предметный указатель
391
Простагландины 123, 174, 345
Протеинкиназа А
— изоформы 184
— механизм действия 169
— структура 182, 184
— фосфорилирование 182, 244
Протеинкиназа В
— изоформы 184—185
— ингибиторы 185
—локализация 185
— структура 122
— фосфорилирование 183, 244
Протеинкиназа С 58
— изоформы 40
— активаторы 39, 40, 84, 169, 170,
185
— активация 119, 123, 178, 185, 227,
243
— локализация 40, 48, 94
Протеинкиназа G 123
— изоформы 184
—локализация 180
— специфический субстрат 184
— структура 182
— участие в хранении информации 246
— фосфорилирование 183
Протеинкиназы
— митоген активируемые (МАР-ки-
назы) 183, 186—187, 235
— серин/треониновые 123
— тирозинкиназы (ТК) 120, 122, 183,
186, 244
— (Са+ кальмодулин )-зависимые
(ПК В) 123, 169, 184,332
Протеинфосфатазы 103, 122, 170, 181,
187, 248, 351
Протеолиз 80, 81, 115, 355
Протеолипид миелина (белок Фолча)
45
Процессинг
— белков 77, 355
— гликопротеинов 52
— пре-дгРНК 50
Проэнкефалин А см. Энкефалины
Психозы 142, 172
ПФ2В (кальцинейрин) 187
Рассеянный склероз 45
Регуляторные пептиды см. Нейропеп-
тиды
Резерпин 290
Репарация 344, 347
Реполяризация 293, 318, 322
Ретикулярная формация среднего мозга
309
11 -Z/wc-ретиналь 300, 301
Ретиноевая кислота 229
Ретиноидные рецепторов 229
Рефлекторная дуга 9, 10
Рефрактерный период 293, 321
Рецептивное поле (РП) 305, 306
Рецепторы
— агонисты 50, 114, 123, 127, 138,
141, 146, 152—154, 162, 194, 228
— аденозиновые 154
— адренергические 132, 136—137,
228, 243, 245
— активация 84, 131, 137, 142, 143,
147, 151, 155, 156, 174, 180, 183,
227, 250, 251,312,318,319, 329
— АМРА 145—147, 196, 246—249,
328, 361
— антагонисты 50, 123, 138, 141,
151 — 153, 162, 194, 248, 305,
306, 332, 334,361
— ассоциированные с ферментами
121,122
— ауторецепторы 114, 142, 148
— аутофосфорилирование 120
— ацетилхолиновые см. Холиноре-
цепторы
— вкусовые 310—314
-----T1R1 311,312
-----T1R2 311,312
-----T1R3 311,312
-----T2R 311,312
-----капсаицина 313
— ГАМК 120, 149, 151 — 153, 244,
353
— гетерорецепторы 142
— гистамина 142, 143, 165, 302
— глицина 153
— глутамата 31, 50, 92, 120, 121,
143—148, 150, 155, 181, 196, 197,
249, 272,313, 329, 330
— деградация 242, 245
— дофамина 123, 138, 141, 157, 225
— запаха (одорантные) 307—309
— инактивация 242
— ионотропные 84
— каината 147—149, 329, 351
— квисквалата 145
392
Предметный указатель
— кластеры 130, 235
— метаботропные 50, 84, 121, 138,
140, 145, 148, 154, 162, 196, 226,
227, 248, 249, 303, 313, 329—331
— мускариновые см. Холинорецеп-
торы
— не NMDA 145, 148
— NMDA 40, 41, 145—148, 153, 155,
156, 196, 223, 245—248, 329—
330, 353, 357, 361
— нейропептидов 115, 121, 162
— никотоновые см. Холинорецеп-
торы
— N-конец 119
— норадреналина 116, 142, 165
— опиатов 162—163
— пресинаптические 142, 143
— пуринов 154
— серотонина (5-НТ) 121, 139—142,
165, 184, 226, 307, 308, 334
— синтез 194, 243—245, 249
— С-конец 119
— сопряженные с G-белками 122,
165—168, 196, 303,315
---а-субъедин и ца 166
---Ру-комплекс 166
— сродство к лиганду 115, 130, 138
— типы 118—121
— тормозные 114, 137, 194, 195
— ферменты 121 —123
— «якорь» 122—123
— Wnt-сигналов 231 —232
Рецепторные белки 12, 48, 83, 116—
123
— глобулярные (цитоплазматические)
118—119
— интегральные 118—119
— мембранные 83, 119—120
— тирозинкиназы 123
Рибосомы 12
РНК
— нуклеотидные последовательности
197
— проблема переноса памяти 327,
335, 336
— синтез 49—50
— содержание 45, 49
Родопсин 86, 165, 227, 298—303
Рост клеток 27, 34, 234, 235, 237
Рота меры 66
Сальтаторная (прыгающая) проводи-
мость 19, 296
Самосборка мембранного бислоя 60,
63
Саркоплазматический ретикулум
197—201,318
Секретаза 355—356
Семафорины 233—235
Сенильные бляшки 354, 356
Сенсомоторная кора 263
Сердечная мышца 27, 168, 184, 220,
222, 225. 268. 272. 276. 316, 318.
319
Серое вещество 9, 26, 37, 42, 96,
125, 263
Серотонин (5-гидрокситриптамин,
5-НТ) 139
— метаболизм 140
— пути 139
— распределение в мозге 139
— рецепторы 121, 140—142, 165,
184, 226
---агонисты 141
---антагонисты 141
---структура 11, 140, 142, 307—
308
---типы 141
---функции 141
— структура 112
— синтез 11, 116, 139—140
— функции 142, 334
Сетчатка 21,96, 305
— амакриновые клетки 305
— биполярные клетки 305
— ганглионарные клетки 12
— колбочки 299
— палочки 37, 38, 55, 168, 225
— структура 305—306
— слои 305 ,
— функция 306
Сиаловая кислота см. N-Ацилнейра-
миновая кислота (NANA)
Сигнальные системы
— NO-синтазная 118
— липоксигеназная 163
— МАР-киназная 162
— НАДФН-оксидазная (супероксид-
синтазная) 162—163
— Са-инозитолтрисфосфатзависи-
мая 162
— циклоаденилатная 162
Предметный указател.
Симпорт 190
Синапс 10
— аксо-аксональный 109
— аксо-дендритный 109
— аксо-соматический 109
— асимметричный 111
— дентрито-дендритный 109
— дентрито-аксональный 109
— механизмы долговременной потен -
циации 241,245—248
— непрямой передачи сигнала 110
— нервно-мышечные 19, 283
— пластичность 241—249
— синаптосомы 31,36
— смешанный 109
— сомато-аксональный 109
— формирование 55, 238
— химический 21, 109—111
— электрический 21, 109—111
Синапсины 85, 94, 288
Синаптическая
— бляшка 109— 111
— депрессия 226, 245
— задержка 111,285
— фасилитация 245
— щель 21, 111
Синаптические пузырьки (везикулы)
83, 184
— выделение медиатора 93, 116, 285,
287
— оборот 93, 286—288
— рециркуляция 284
— типы 111 —112, 1 16
— экзоцитоз 284, 287
Синаптобревин 286
Синаптотагмин 94, 287
Синаптоянин 288
Синдром
— Зельвегера 259
— отряхивания мокрой собаки 142
— Паркинсона 40
Синтаксин 286
Скотофобин 225
Слуховая кора 261,263
СОД 338, 340
— внеклеточная 338
— Cu/Zn- 338
— Mg- 338
Соматостатин 157, 159, 165, 306
Сон—бодрствование цикл 117, 139,
142, 154
Соотношение РНК/ДНК49
Спектрин 90, 91,94
— изоформы 92
— строение 92
— функция 92
Спектрин-анкириновая сеть 90
Спинномозговая жидкость 96
Сплайсинг 50, 194, 235
СТАТ-Белки 350
Стенин 93
Стереотипии 138
Стероидные гормоны 118, 229
Стероиды 32
Страх 140
Стресс 22, 136, 157, 158, 212, 222,
223, 340—351,353, 356, 359, 361
Стриатум (полосатое тело) 125, 134,
138, 143, 148, 149, 163, 349
Стрихнин 147, 153
Строфантидин 198
Структуры
— канальные 188
Субарахноидальное пространство 14
Субъединицы 185
— каталитические 123, 167, 184—
187, 205
— регуляторные (ингибиторные)
123, 184, 185, 187
Сукцинатдегидрогеназа 78, 275
Сульфатиды 29, 30, 33, 42—44, 64,
259, 260
Сульфоцереброзиды 26, 31
Супероксид анион-радикал 31, 157,
341,342, 344, 347, 350, 352
Сфинганин 97, 259
4-Сфингенин 97
Сфингозин 25, 29, 97, 257
С20-Сфингозин (4-эйкозасфенгенин)
28, 97, 101
Сфингомиелин 23, 24, 26—29, 42—
44, 56, 57, 64, 96, 100, 105, 106,
259
Сфингофосфолипиды 23, 25
Таурин 143, 153, 339
Тахикинины 82, 157
ТБ К-реактивных продуктов (ТБК-РП)
346
Теофиллин 154
Терминаль
— пузырьки (везикулы) 115, 116, 290
— экзоцитоз 115
Предметный указатель
Терморегуляции 139, 142, 143, 157,
162
Терморецепция 157
Термотропный мезоморфизм 59, 70
Тетрагидробиоптерин 140
Д9-Тетрагидроканнабиол 163
Тетрахименин 32
Тиамин 269
Тиобарбитуровая кислота (ТБК) 346
Тиорфан 85
Тип аксонального транспорта 280—
283
Тирозин 120, 135, 184, 237, 238, 271
Тирозингидроксилаза 50, 135, 136, 140
Тирозиновых радикалов нитрозили-
рование 241,351
Тиролиберин см. Нейропептиды
ос-Токоферола см. Витамин Е
Токсины
— а-бунгаротоксин 128
— 1-метил-4-
фенилтетрагидропиридин 352
— пикротоксин 152
Томография позитронно - эмиссионная
261,262, 278
Тормозные постсинаптические потен-
циалы (ТПСП) 153, 297—299
Трансаминаза 274
Трансаминаза ГАМК 256, 275
Транс-гош изомеризация 66, 71
Транс-гош-переход 69
Трансдуцин 165, 168, 301—305, 313
Трансмембранные транспортеры и об-
менники 81, 224—226
Транспорт 255, 279
— аденин 255, 256
— аденозин 255
— аксональный 280—283
— амины 224, 255, 287
— вторично-активный 188, 189, 328
— гексозы 224, 255
— глюкоза 224, 254, 255, 262—263
— лактат 255
— лизин 255
— монокарбоновые кислоты 117, 224,
255
— натрий-зависимый 127, 224
— нейтральные аминокислоты 117,
224,256
— нуклеозиды 259
— основные аминокислоты 255
— первично-активный 188, 189
— пурины 255
— фенилаланин 117
— хлорзависимый 289
— холин 65, 127, 255, 256, 268
— GLUT1 254,262, 263
Трансферрин 255, 256, 338, 342
Триптофан 116, 117, 139
Триптофангидроксилаза 139, 140
Трициклические нейролептики 139
Тромбоксан 260, 345
Туберомамиллярные ядра 142
Тубулин 46, 47, 91, 128, 280, 281
Уабаин 198, 199, 206, 209, 219, 220,
222, 223,225, 321,327, 328
Убихинон 339
Унипорт 190, 201
Уравнение
— Гольдмана 291
— Нернста 290, 291
Усиление (амплификация) сигнала 299,
302
Фактор
— NF-kB 179, 332, 349, 350
— регуляции Na/H-обменного белка
228
— роста нервов NGF 236, 237, 280
— роста фибробластов (FGF) 229, 231
— трансформации роста (TGF) 229
— элонгации 305
Фенамин 336
Фенилаланин 117, 255, 271,342
Фенилалкиламины 331
Фенилалкилоиднонимины 336
Фенилэтаноламин-М-метилтрансфе-
разы 133
Фенотипическая адаптация 106
Феритин 338
Фибронектин 95, 103, 233, 234
Фодрин 92, 330
Форболовые эфиры 171, 173, 178, 186
Формальдегид 124
Формирование функциональной си-
наптической связи 238—240
Фосфатактивируемая глутаминаза
144,150
Фосфатидилглицерин 23—25, 27, 42,
89
Предметный указатель
Фосфатидилинозит 24, 27, 36, 39, 40,
94, 105, 164, 166, 168, 169, 172,
175,178,185,259
Фосфатидилинозитол-4,5-бисфосфат
(ФИФ2)39, 174, 175, 178, 259
Фосфатидилинозитолгликан 40, 48
Фосфатидилсерин 23, 24, 26, 27, 36,
39, 40, 42, 57, 64, 73, 89, 94, 105,
169, 178, 185, 256, 303
Фосфатидилтреонин 23
Фосфатидилхолин (лецитин) 23—
27, 36, 37, 42, 43, 64, 66, 71, 75,
76, 105, 116, 127, 259, 298, 303
Фосфатидил этанола мин (кефалин) 23,
24, 26, 27, 64, 76, 105, 106, 164,
258, 303
Фосфатидная кислота 23, 24, 27, 39,
73, 172, 174, 185, 258
6-Фосфоглюконатдегидрогеназа 297
Фосфодиэстераза 46, 166, 167, 170—
172, 176, 177, 302, 304
— изоформы 171
— ингибиторы 171
— структура 299
— фосфорилирование 118, 172—175,
303, 334
Фосфоинозитиды 26, 43, 48, 52, 64,
85, 94, 118, 172—175, 303, 334
Фосфоинозитолы 37
Фосфоламбан 203, 219
Фосфолемман 203
Фосфолипаза 31,81, 106, 127, 332
— А1 172
— А2 89, 137, 141, 168, 172, 257,
260, 303
— С 39, 40, 118, 132, 137, 141, 163,
170, 172—175, 178. 234, 237,
260, 329
— Д 172,228,260
— функция 89
— механизм действия 81
— субстратная специфичность 89
Фосфолипиды 22—23
Фосфопротеинфосфатаза
— активаторы 187
— ингибиторы 187
— локализация 187
— структура 187
Фосфорамидон 85
Фосфофруктокиназа 266, 274
Фосфофруктокиназная реакция 266
Фруктозо-1,6-бисфосфат 266
ос-Фторметилгистамин 140
Фукоза 52, 103
Хвостатое ядро 134, 137, 263, 351
Хемоаттрактанты 234
Хеморепелленты 234
Хемосенсорное восприятие 311
Химическое прекондиционирование
(chemical preconditioning) 349
С1-бикарбонатный обменник 226
(С1 + К)-котранспорт 226
Хлоропромазин 138
Холестерин 22, 26, 32—34, 42, 43
— сегрегирующая роль
Холецистокинины см. Нейропептиды
Холин ИЗ, 126, 127, 255
Холинацетилаза 267
Холинацетилтрансфераза 124, 125, 354
Холиномиметики 336
Холинорецепторы 125, 156, 332
— мускариновые (jwXP) 125, 131 —
133
— мышечные 20, 128
— никотиновые (нХР) 84, 125,
128—130, 193, 195,319
— субъединицы 128—130,319
Хондроитин 100
Хорея Гентингтона 126, 149
Хроматин 12
Хромофильная субстанция 12
Центральная нервная система (ЦНС) 9
Церамид 27—29
Цереброзиды 29—30
Цереброспинальная жидкость 253
Церулоплазмин 38
Цикл трикарбоновых кислот (ЦТК)
264, 273
2': З'-Циклический нуклеотид
З'-фосфодиэстераза (CNP) 46
Циклогексимид 327, 328
Циркадные ритмы 142, 159, 180
Цис-конфигурация 23, 35, 67—68
Цистеин 103, 206, 342
Цитозоль (гиалоплазма) 12, 18, 40
Цитоскелет 46, 81,84, 90—94
Цитохром
— Ь5 78, 80, 85
-Р450 2 60
— а + а3 276
— с 86, 276
396
Предмет»* ь и указатель
Цитохром с-оксидаза 86
Цитратсинтазная реакция 267, 272—
274
Цитруллин 155
Черная субстанция (substantia nigra)
125, 134, 138, 139, 142, 143, 148,
261,263, 351
Шапероны 75, 356, 358
Шванновские клетки (нейролеммо-
циты) 15—18, 41
Шизофрения 133, 135, 137, 138, 161,
352, 353
Шиффовы основания 342
Шмидта—Лантермана насечки 18
Шавелевоуксусная кислота 168, 272
Эйкозасфинганин 97
4-Эйкозасфингенин 97
Экзайтотоксичность 31, 147, 181,225,
353, 357, 361
Экзон 50, 323
Экспериментальный аллергический эн-
цефаломиелит 45
Экспрессия генов в мозге 231
Экстерорецепторы 20
Экстра клеточный матрикс см. Гли-
кокаликс
Экто-5'-нуклеотидазы 289
Эмоции 135, 157, 158
Энграмма 326
— перенос 329
Эндоплазматический ретикулум 11,
13—15, 57, 168, 170, 173
Эндопротеазы 52
Эндорфины 112, 160—162, 175, 177,
258, 259, 330, 338
— а-Неоэндорфин 160
— а-Эндорфин 160
— Р-Эндорфин 158, 160
— у-Эндорфин 160
Эндотелиальные клетки капилляров
86, 155, 252, 254—256
Эцдотелинконвертирующий фермент
(ЕСЕ-1)83, 356
Эндотелины см. Нейропептиды
Эндотелиоциты сосудов 153
Эндоцитоз (интернализация) рецеп-
торов 93, 243, 244, 256, 286
Энергетический обмен 260—280
Энергозависимые процессы 279
Энкефалины 82, 112, 143, 160, 162
— лей-энкефалин 160, 162
— juem-энкефалин 160, 162
Эпидермальный фактор роста 165
Эпилепсия 353
Эфрины 233, 234
Эффект выпрямления 110
Ядерно-магнитный резонанс (ЯМР)
278
Ядра мозжечка 125, 263
Ядра одиночного пути продолговатого
мозга 314
Ядро Дейтерса 110
ЯМР-спектроскопия 36, 66
АР 180 287
АР2 287
Р-АРР (Р-Amyloid Precursor Protein)
78, 354—356
В-50 84,85, 332
BSP-2 101
D2 101
DM-20 45
Hsp 70 288
K4 101
L-l 101
MBA-2 102
NCAM 46, 81,94, 101, 107, 233, 234
Ng-CAM 101, 102
NSA-3 102
КаЬ5-ГТФаза 93
Rab-белки 93
RAS/MAP-киназный путь 231,235—
237
SAM (Senescence Accelerated Mice)
352
SMAD-белки 229
SNAP-25 286
STAT-белки 350, 351
Thy-1 102
Wnt-семейства 231
СОДЕРЖАНИЕ
Предисловие.......................................................... 3
От научного редактора.............................................. 5
Введение 7
Раздел 1
СТРУКТУРА НЕРВНОЙ ТКАНИ
1.1. Нервная ткань ........................................... 9
1.1.1. Нейрон............................................. 9
1.1.2. Глия.............................................. 14
1.1.3. Нервные волокна.................................. 16
1.1.4. Нервные окончания................................. 19
1.1.5. Синапсы........................................... 21
1.2. Особенности липидного и белкового состава
нервной ткани................................................ 22
1.2.1. Липидный состав .................................. 22
1.2.2. Биологические функции мембранных липидов ......... 39
1.2.3. Миелин ........................................... 41
1.2.4. Белки нервной ткани............................... 48
1.3. Современные представления об организации
нейрональной мембраны........................................ 51
1.4. Свойства липидного бислоя............................... 58
1.4.1. Фосфолипиды как структурная основа бислоя ........ 58
1.4.2. Трансмембранная асимметрия липидов................ 63
1.4.3. Подвижность компонентов липидного бислоя.......... 65
1.4.4. Фазовые переходы мембранных липидов .............. 69
1.4.5. Дефектные зоны. Роль холестерина ................. 73
1.4.6. Микровязкость мембран............................. 76
1.5. Мембранные белки ....................................... 77
1.5.1. Локализация и подвижность белков
в мембранном бислое...................................... 78
1.5.2. Функции мембранных белков ........................ 83
398
Содержание
1.6. Белок-липидные взаимодействия ........................... 86
1.7. Цитоскелет и гликокаликс................................. 90
1.7.1. Белки цитоскелета.................................. 90
1.7.2. Гликокаликс........................................ 94
1.7.3. Белки гликокаликса................................. юо
1.8. Мембранные механизмы адаптации.......................... 104
Вопросы и задания ...................................... 107
Раздел 2
МОЛЕКУЛЯРНАЯ НЕЙРОХИМИЯ
2.1. Типы синапсов и классификация нейромедиаторов ......... 109
2.1.1. Синапсы в нервной системе ....................... 109
2.1.2. Медиаторы, нейромодуляторы, комедиаторы.......... 112
2.2. Общая характеристика мембранных рецепторов............. 117
2.3. Характеристика индивидуальных медиаторных систем ...... 124
2.4. G-Белки, вторичные мессенджеры и протеинкиназы......... 163
2.4.1. G-Белки и вторичные мессенджеры ................. 163
2.4.2. Роль мембранных фосфоинозитидов в передаче
сигнала ................................................ 171
2.4.3. Метаболизм фосфоинозитидов и регуляция
проницаемости мембран для Са2+ ......................... 174
2.4.4. Роль ионов кальция в функционировании нейронов .. 176
2.4.5. Другие вторичные посредники...................... 178
2.4.6. Протеинкиназы и фосфопротеинфосфатазы ........... 181
2.5. Ионные каналы и ионные насосы ......................... 188
2.6. Механизмы отбора и усиления сигналов
на клеточной мембране....................................... 226
2.7. Развитие нервной системы и формирование
нервных сетей............................................... 229
2.8. Механизмы синаптической пластичности................... 241
Вопросы и задания ....................................... 250
Раздел 3
ФУНКЦИОНАЛЬНАЯ НЕЙРОХИМИЯ
3.1. Особенности пластического обмена головного мозга .......... 252
3.1.1. Гематоэнцефалический барьер (ГЭБ)................ 252
3.1.2. Особенности обмена липидов ...................... 256
Со/^рние
3.2. Энергетический обмен головного мозга..................... 260
3.3. Аксональный транспорт ................................... 280
3.4. Межсинаптические контакты и принципы
функционирования синапса.................................... 283
3.4.1. Этапы синаптической передачи ..................... 283
3.4.2. Квантовое выделение медиатора .................... 285
3.4.3. Превращения синаптических везикул
в синаптическом окончании ............................... 286
3.4.4. Удаление медиаторов из синаптической щели......... 288
3.5. Электрические и химические механизмы передачи
информации.................................................. 290
3.5.1. Потенциал покоя .................................. 290
3.5.2. Локальный потенциал и потенциал действия.......... 292
3.5.3. Проведение потенциала действия .............. 295
3.5.4. Постсинаптические потенциалы и интеграция
синаптических влияний ..................................; 296
з.б. Сенсорные процессы......................................£ 299
3.6.1. Передача сигнала в фоторецепторных клетках л
сетчатки...........................................® 299
3.6.2. Нейрохимические механизмы обоняния ..............« 307
з.б.з. Механизмы работы вкусовых рецепторов.............ж 309
3.7. Регуляция двигательной активности ......................ж 314
3.8. Нейрохимические механизмы памяти ......................, 324
3.9. Окислительный стресс и нейропатологии .................j 337
Вопросы и задания .....................................I 362
Литература ......................................................1 364
Сокращения, используемые в книге ................................Ж 371
Предметный указатель...........................................ж.. 377