/




































































































































































































































































Текст
С.М. Бебия
леей
С.М. Бебия
ПИХТОВЫЕ ЛЕСА КАВКАЗА
Монография
Издательство Московского государственного университета леса Москва - 2002
УДК 634.94
6Л2 Бебия С.М. Пихтовые леса Кавказа. Монография М.: МГУЛ, 2002. -270 с.: ил.
В монографии центральное место отведено лесообразовательным процессам пихтовых лесов Кавказа. Освещены генезис и закономерности флороценотической, эколого-географической, лесотипологической дифференциации и возрастной динамики одного из древних и сложных лесных сообществ с господством пихты кавказской, намечены пути и методы рационального их использования и охраны. Предназначена для научных работников и специалистов лесного хозяйства, преподавателей и студентов соответствующих вузов, а также для лиц, интересующихся проблемами охраны и рационального использования биоразнообразия, биологии вообще.
Табл. 43, ил. 29, библиогр. - 381 назв.
Рецензенты: доктор биологических наук, профессор, заслуженный лесовод
РФ Н.Г. Васильев (кафедра лесоводства и геологии Московской сельскохозяйственной академии);
доктор с.-х. наук, профессор И.П. Коваль (научно-исследовательский институт горного лесоводства и экологии леса)
Автор - Сергей Михайлович Бебия, доктор биологических наук
ISBN 5-8135-0105-3
©Бебия С.М., 2002
© Московский государственный университет леса, 2002
ПРЕДИСЛОВИЕ
I (роблома сохранения биологического разнообразия ставшая, одной Н1 дстуальнойших в современном мире, выдвинула перед исследователями рид первоочередных задач, среди которых - задача длительного и всестороннего изучения растительного покрова территорий, сохраняющих и в HHCTONiHeo время черты первобытности и девственности, занимает особое место
Пихтовые леса Кавказа, комплексному исследованию которых по-<жященп предлагаемая читателям книга видного исследователя горных ле-Киикпча» доктора биологических наук С.М. Бебия, в полной мере отвечает такому определению. Их изучение может дать весьма ценную информацию о происхождении, природе, эволюции, структуре и динамике девственных лесов Евразии. Без изучения истории лесов в прошлые эпохи нель-1и нонан» многие важные стороны биологии и экологии древесных пород, а щите жизнь лесных биогеоценозов. Большое значение исследованиям по истории формирования лесного покрова придавали известные отечественные ученые лесоводы Г.Ф. Морозов, В.Н. Сукачев, А.Б. Жуков, Б.П. Ко-ЮН'НИМОП
Рнбсггй С.М. Бебия, по сути дела, монотрафическое исследование НИМ1Ы кавказской и пихтовых лесов. Она написана по материалам собственным исследований, проводившихся автором в течение более 30 лет. Конечно, он не был «первопроходцем», но монографию с полным правом Мпаню назвать новой работой о пихтовых лесах Кавказа. В прежние временя зге работа могла быть «руководством к действию» для ведения хо-imHctmm и пихтовых лесах. Сейчас ситуация другая, но пихта кавказская фнйВ1 согни лот и надо надеяться, что для нее настанут лучшие времена, koi ли лесным хозяйством вновь будут заниматься всерьез.
Поражает и объем, и масштаб выполненных исследований. Автор нПстойтшН'НО знаком не только с пихтовыми лесами Кавказа. Он изучал ним IАрники европейской части России, Прибалтики, Германии, Карпат, I Hip, Урала, Тянь-Шаня, Сахалина, Курильских островов, Японии, Тайваня Ьстушюпно, появление настоящей книги - отрадное событие в лесово-лст ванной литературе.
I'M Бебия является подлинным новатором лесоводственной науки. IЖ неустанно ищет новые пути решения основных задач лесоведения и ле-VHHUMCTAA
Огромное количество фактов, черпаемых из живой природы, в сочетании е широкими теоретическими обобщениями, позволило С.М. Бебия HVMeiim а работе впервые все. аспекты лесообразовательного процесса ним 1оаых лесов Кавказа на уровне целостной лесной формации. Охарактеризованы генезис, история формирования и закономерности современного римнространсния лесов с господством пихты кавказской. Вывялены основ
ные закономерности возрастной структуры, строения и динамики древостоев и типологической дифференциации лесов.
Главное, что хотелось бы отметить - это проходящую через всю работу мысль о реликтовости, об уникальности, самобытности пихтовых лесов и необходимости бережного, рачительного отношения к ним, как к неповторимому явлению природы. В работе впервые приводится созологиче-ская оценка пихтовых лесов, редких видов растений и фитоценозов в них.
Автор использовал классическую лесоводственную методику, в ряде случаев разрабатывал собственные приемы оценки.
С большой полнотой и скрупулезностью анализируются различные точки зрения генезиса пихтовых лесов и их распространение на Кавказе, причем автор демонстрирует хорошее знание литературы как отечественной, так и зарубежной. О пихте кавказской автор говорит как о достаточно самостоятельном виде, сформировавшемся в Колхидском рефугиуме и убедительно обосновывает свою позицию.
Впервые имеющаяся информация об эколого-биологических и лесо-водственных^особенностях пихты кавказской дана с большой полнотой, причем многие сведения являются новыми. В ряде случаев автор вносит уточнения и исправления.
Большой интерес представляет для читателей раздел книги, в котором освещены основные закономерности строения и возрастной структуры пихтовых лесов. Автор подробно анализирует позиции разных исследователей в вопросе о том, какие леса следует считать "девственными". Это не схоластический вопрос, поскольку мы должны говорить на одном понятном языке. Для Кавказа С.М. Бебия считает возможным разделить все леса на две большие группы - естественные и искусственные. Первые он делит на девственные и хозяйственные, формирующиеся в результате целенаправленной деятельности человека.
Столь же тщательно и всесторонне изучены и типы пихтовых лесов. На Кавказе сложилось свое лесотипологическое направление, главными идеологами которого были А.Г. Долуханов и Л.Б. Махатадзе. Это направление восприняло идеи биогеоценологического подхода в понимании типа леса В.Н. Сукачева и концепцию динамизма лесного покрова Б.П. Колесникова. С.М. Бебия выступает как представитель Кавказского направления и имеет на это все основания. В начале раздела изложено теоретическое кредо автора, далее следует описание основных групп типов пихтовых лесов Кавказа. Такого рода классификация разработана впервые.
Глубокое и разностороннее изучение эколого-биологических и лесо-водственных особенностей пихты кавказской и пихтовых сообществ в целом позволило автору книги предложить хорошо продуманную систему мероприятий по оптимизации использования и охраны лесов этой формации.
Безусловно, идеи и основные выводы автора, будучи в некоторых частях спорными, могут быть очень плодотворными, поскольку они чрез-
вычвйно актуальны, способны вызвал» широкую дискуссию и привлечь внимвнио исследователей к вопросам эволюции девственных лесов.
Книгу завершает весьма обстоятельный список использованной ли-|«рюуры и резюме на английском языке. Большое количество оригинальным рисунков и таблиц также прядают ей особую ценность.
II звключении еще раз хочу подчеркнуть, что монография С.М. Бе-0|И это фундаментальное исследование крупной научной проблемы, имеющее при этом важное прикладное значение. Сведения, приводимые в мнит, помогут широкому кругу лесоводов, ботаников, природоохранных рвб(м пиков, а также студентам и преподавателям высших учебных заведений (шгсотвенно-научного и лесохозяйственного профилей получить современное представление о природе и значении пихтовых лесов Кавказа. О Ю1И1В можно сказать: "Все - о пихтовых лесах Кавказа". Для специалистов ня книга представляет непреходящую ценность.
Член-корреспондент РАН Л .П. Рысин
Рис. 1. Пихта кавказская (Нордманна) - Abies normanniana (Stev.) Spach
ВВЕДЕНИЕ
В практике ботанических и лесоводственных исследований изучение леве I пенных лесов отдельных регионов занимает особое место, поскольку они представляют огромный фактический материал для познания закономерностей формирования и динамики растительного покрова не только исследуемого региона, но гораздо больших территорий (Толмачев, 1948; (Чвнюкович, 1960; Walter, 1968; Leibundgut, 1982). В этом отношении обвинение происхождения и эволюции пихтовых лесов Кавказа, изучение ле-гообрпчовательного процесса в них, которым посвящена настоящая работа, предел являют большой научный интерес. Они также чрезвычайно важны дли решения проблем рационального использования пихтарников, для оп-1ММН1ЛЦИИ использования и охраны экосистемы всего Кавказа.
Развитие теории лесообразовательного процесса началось не так давно (Основоположником учения о лесообразовательном процессе считается I* 11 Колесников. Однако полного представления о лесообразовательном процессе еще не сформировалось. Лесообразовательный процесс понимает кпк процесс возникновения, развития, разрушения и смены лесных еообщес гв во времени в результате взаимовлияния биотических, абиотических факторов и антропогенного воздействия. В таком представлении поим । не лесообразовательного процесса базируется на учении о строении, возрастной структуре и динамике древостоев, разработанном Б.А. Ивашкевичем и Г. Лейбундгутом (Leibundgut), а также на теории типов леса и их /шнамихи, созданном Г.Ф. Морозовым, В.Н. Сукачевым, Б.П. Колесниковым. И С Мелеховым.
I Inine представление об особенностях и закономерностях лесообра-uiBAWJibiioro процесса пихтовых лесов Кавказа базируется на изучении их । вне шеи, современного распространения и природных условий нроичрпстания, исследований основных морфо^биоэкологических особен-шчп'сй к лесоводственных свойств пихты кавказской, процессов возобнов-менив лесов, их строения, возрастной структуры и развития, биогеоценоти-•нч'ких особенностей и динамики, а также реакции лесов на проведение хо-нШ тонных мероприятий.
Пихта является одной из важнейших древесных пород, слагающих вынншпродуктивные леса на значительных площадях горных систем се-ив|шо| о полушария. Эти леса выполняют весьма важные и разнообразные нн1|в тыс функции, имеют большое хозяйственное значение. Потребность в рАШосгороннем использовании этой породы в хозяйстве с каждым Годом НИ ЦНИИ ACT.
Чрезмерному антропогенному воздействию пихтовые леса подверга-4Hvk издревле. За последние два столетия доля пихтарников в лесном фонов многих регионов мира сильно сократилась. В Европе процесс уменыпе-
ния площади пихтовых лесов продолжает прогрессировать (Мауег, 1969). В отдельных странах отмирание пихты носит катастрофический характер, девственные пихтарники здесь сохранились лишь на нескольких сотнях, а то и десятках гектаров (Leibundgut, 1982). В Австрии, к примеру, по данным журнала Unsere Umwelt (1985), в 1983 году площадь пихтовых лесов, подверженных заболеваниям различной степени, составила 75 %, в 1984 году эта величина достигла 87 %.
Решение проблемы пихты - сохранения, восстановления и рационального использования лесов - стало одной из актуальных задач современного лесоводства и находится в центре внимания мировой лесо-водственной науки и практики. В частности, по линии Международного союза лесных исследовательских организаций (IUFRO) предложены важные методические рекомендации по изучению девственных лесов и разработке принципа, "сообразного природе" (NaturgemaBe) лесоводства (Leibundgut, 1982).
Сократились ц площади лесов Кавказа с господством пихты кавказской в результате их длительной нерациональной эксплуатации. Пихта кавказская - Abies nordmanniana (Stev.) Spach - (рис.1) - реликт третичного периода, одна из продуктивных, ценных, главных лесообразующих пород, наиболее мощное дерево Кавказа и всей Западной Евразии. Средний запас древесины на га составляет 700-800 куб. м, максимальный на отдельных участках - 2 тыс. куб. м (Орлов, 1951). Средний объем хлыста деревьев верхнего яруса составляет 8-10 куб. м. Эти данные свидетельствуют о больших потенциальных возможностях выращивания высокопроизводительных древостоев данной породы.
Важнейшей ценностью пихтарников, имеющих исключительно большое значение в условиях горной системы Кавказа, и особенно Колхидского рефугиума, следует считать их защитные и средообразующие функции. Произрастая в верхней горной зоне на значительных площадях (320 тыс. га) Кавказа, пихтовые леса выполняют первостепенную почвозащитную, водоохранную, климаторегулирующую роль. Образуя целые реликтовые ландшафты, эти леса являются хранителями уникального генофонда многообразия древнего живого мира, редких природных комплексов, источниками ценных видов растений для интродукции.
Добывание древесины в этих лесах не может быть основной целью, как это происходит в настоящее время. Ведение многоцелевого, рационального хозяйства в них, с учетом приоритетных отраслей, является важнейшей задачей. Вместе с тем еще с середины прошлого столетия на Кавказе широко использовалась пихтовая древесина для различных целей. Леса подвергались бессистемной рубке.
В последние десятилетия актуальность изучения пихтарников Кавказа возросла в связи с их интенсивным хозяйственным освоением. В них
проводятся рубки главного пользования. В частности, проводимые в этих лесах промышленно-выборочные и постепенные рубки с использованием нерациональной технологии лесосечных работ не отвечают природе пихтовых лесов, снижают их производительность. Нагрузка на пихтарники все больше возрастает. В них также осуществляется массовая заготовка разнообразного растительного сырья, чрезмерная рекреация, пастьба скота и г. д. В результате нерациональных рубок в Грузии и Абхазии, например, псе доступные площади пихтарников освоены, большая часть из них растроена, нуждается в научно обоснованном лесовосстановлении (Гулисашвили, 1967; Бебия, 1973). Здесь девственных лесов осталось очень мало. Решение проблемы пихты, в том числе и пихты кавказской, возможно лишь на основе глубоких знаний природы лесов, основанных на всестороннем исследовании биоэкологических, лесоводственных, фитоценотиче-ских свойств породы, процессов и событий в жизни леса. Однако в отношении пихты мы все еще не располагаем достаточно большим объемом всесторонней биологической научной информации, по сравнению, например, с елью. Информация в последние 40 лет, опубликованная в различных сводках (Малеев, 1936, 1940; Давыдов, 1940, 1956; Гроссгейм, 1948; Герасимов, 1948; Орлов, 1951; Каппер, 1954; Толмачев, 1954; Маценко, 1963, 1964; Долуханов, 1964, 1989; Фалалеев, 1964; Бицин, 1965; Махатадзе, 1967; Манько, 1967; Мауег, 1969, 1972; Liu, 1971; Гулисашвили и др., 1975; Ьобров, 1978; Krussmann, 1983; Крылов и др., 1986; Rushforth, 1987; Nakamura, 1992), указывает на определенную полноту и географию исследований пихты в разных регионах мира. Здесь мы приводим лишь важные и содержательные, с нашей точки зрения, работы о роде Abies Mill., а также in дельных его видах. И все же епде недостаточно данных для решения таких важных задач, как полной инвентаризации генофодца видов рода пих-। в, установления генезиса и истории их формирования; географии коренных лесных экосистем; экологической оптимизации использования лесных жосистем с практическим решением вопроса заповедования остатков дев-г| венных пихтарников. В настоящей работе, посвященной изучению пих-ювых лесов Кавказа, решаются некоторые из перечисленных выше задач.
Пихта кавказская и леса с ее господством также изучены недостаточно Исследования их осуществлялись, в основном, по отдельным районам мровла пихты, фрагментарно, не как целостная формация. Здесь пихтовые >юсм изучались в пределах единой комплексной формации темнохвойных »|всоп вместе с еловыми лесами (Малеев, 1936; Орлов, 1951; Гулисашвили, 1956; Долуханов, 1964; Махатадзе, 1967 и другие).
Недостаточно полная изученность пихтарников Кавказа как отдель-шИ1, самостоятельной формации, безусловно, не давала полного представ-1НН1ИМ об этих лесах и тормозила решение многих теоретических вопросов н практических задач. Сведения, имеющиеся в научной литературе, в раз
личных дендрологических сводках по вопросам биологии, морфологии, экологии и лесоводственных свойств пихты кавказской, зачастую противоречивы (предельные размеры, продолжительность жизни, теневыносливость, отношение к влажности воздуха и почв; строение, возрастная структура и развитие древостоев и т. д.). Не полностью были выяснены также происхождение и закономерности пространственного распространения пихтовых лесов в зависимости от основных экологических факторов среды. Как завершающая стадия не были получены полные данные о типологическом разнообразии лесов с господством пихты кавказской, их структуре и динамике, продуктивности, а также реакции лесов на различные хозяйственные мероприятия. Впервые эти вопросы в отношении пихты кавказской и лесов с ее господством на уровне единой целостной формации освещаются в данной работе по результатам наших многолетних (начиная с 1965 года) комплексных исследований на постоянных пробных площадях в пихтарниках Абхазии и целевых экспедициях, проведенных по всему ареалу ее на Кавказе, за исключением Сев. Анатолии. Использованы также работы В.П. Малеева (1936), В.А. Поварницина (1936); В.З. Гулисашвили (1940, 1964, 1972); А.Г. Долуханова (1964); Г.Н. Гигаури (1965); Л.Б. Махатадзе (1967), посвященные пихтовым (темнохвойным) лесам южного макросклона и М.В. Герасимова (1948); А.Я. Орлова (1951); П.Н. Ушатина (1962); Л.В. Бицина (1965); К.Ю. Голгофской (1967) - пихтарникам северного макросклона Кавказа.
Для сравнения и получения более широкой информации о пихте нами были осуществлены также рекогносцировочные обследования темнохвойных лесов Дальнего Востока, Европейского севера России, Литвы, Германии, горных систем Татр, Карпат, Урала, Тянь-Шаня, островов Сахалина, Кунашира, Шикотана, Хоккайдо, Тайваня. Все эти исследования послужили научной основой для разработки системы мероприятий на лесотипологической основе по оптимизации использования, воспроизводства и охраны пихтарников Кавказа с учетом их природных особенностей и региональной специфики. За время работы был собран большой фактический материал: заложено более 130 таксационных пробных площадей (38,5 га); на 4 постоянных пробных площадях учитывались результаты экспериментальных рубок, выполненных на площади около 30 га; осуществлено более 100 полных и около 1800 маршрутных лесотипологических описаний; обмерено около 40 тыс. учетных деревьев (из которых 120 распилены для анализа хода роста) и более 1 тыс. модельных экземпляров подроста и др.
Изучение пихтарников Кавказа велось маршрутным и маршрутнодетальным методами, при которых производилось большое количество биометрических, таксационных и других измерений изучаемых объектов. Особенности методики зависели от целей исследований, от характера объекта (древостой, лесосека, стационарная или временная пробная площадь и
। д.). Эффективному проведению исследований способствовали применение таких методических пособий как "Методические указания" IIII. Сукачева и С.В. Зонна (1961), принципы классификации растительно-ст на эколого-генетической основе, развитые в трудах Б.П. Колесникова и кавказских лесотипологов. В ходе работ были также использованы разработанные автором новая шкала лесоводственной классификации подроста (Ьебия, 1972), шкала оценки естественного возобновления (Бебия, 1997), метод классификации деревьев в разновозрастном лесу и оценка жизненного состояния древостоев (Бебия, 1999). В целом при изучении пихтарников был использован системный метод исследований. Все вопросы, освещаемые в работе, рассмотрены на уровне единой системы формации лесов с юсподством пихты кавказской. Это дало возможность более глубоко обобщить результаты исследований и выполнить работу как целостную и свершенную.
Автор выражает свою признательность академику Академии наук СССР А.Л.Тахтаджяну, член-корреспондентам Академии наук России JI II Рысину, В.Н. Павлову, академикам Академии наук Белоруссии Н И. Парфенову, Академии наук Азербайджана И.С. Сафарову, профессора В.А. Алексееву, Л.Б. Махатадзе, Н.Г. Васильеву, К.Ю. Голгофской, а также ныне покойным академикам АН Грузии В.З. Гулисашвили и А А. Колаковскому, докторам биологических наук А.Г. Долуханову и (' А. Дыренкову за ценные советы при выполнении данной работы.
Ценную информацию об итогах интродукции пихты кавказской в Европе прислали нам проф. Г. Томазиус и доктор П. Шмидт из Секции лесоводства Дрезденского Технического университета (Тарандт, Германия).
Большую помощь в осуществлении наших зарубежных экспедиционных исследований оказали проф. Г. Томазиус, академики Б. Керестеши из Института леса (Будапешт, Венгрия), Ф. Эрендорфер из института ботаники (Вена, Австрия), Ф. Бенчат из Словацкой АН (Братислава), доктора Ж Дебреци из Международного дендрологического института (Медфилд, (‘IIIА), И. Рац из Музея истории природы (Будапешт), профессора Дж. Янг, Ф 11ан доктор Я. Цэн из Тайваньского научно-исследовательского инсти-। yin леса (Тайпей, Тайвань), проф. М. Каджи из Токийского лесного уни-иорситета (Токио, Япония). Интересующую нас литературу прислали I Томазиус, Ф. Эрендорфер, М. Каджи, Я. Цэй, а также профессора Г. Лей-бунд|ут из Института лесоводства (Цюрих, Швейцария) и Г. Маер из Ин-г in гута лесоводства Университета земельных культур (Вена). Мы выражаем благодарность всем этим ученым. Мы также признательны Г.К. Солнцеву, док. с-х. наук, директору Института горного лесоводства и экологии икса и проф И.П. Ковалю, заместителю директора этого института, М.Д. Хе-।шурову, Генеральному директору санатория и дендропарка им. М II Фрунзе, лесоводам М.М. Бебия, А.З. Кобахия, и коллегам по работе
из Института ботаники ДНА канд. биолог, наук Т.А. Гуланян, Е.В. Лакоба, О.О. Васильевой, а также Е. Подлушани и И. Ретезар (Будапешт) за оказанную организационную и техническую помощь при сборе полевого фактического материала и оформлении работы.
I KI I КЗ ИС ПИХТОВЫХ ЛЕСОВ И ИХ РАСПРОСТРАНЕНИЕ НА КАВКАЗЕ
Происхождение и систематика рода пихты
Среди десяти родов, включенных в семейство сосновых, род пихта -.4 hit* Mill, является одним из древнейших. Возможно, этим объясняется । <> । факт, что вопросы происхождения рода пихта, времени и путей форми-ропания пихтовых лесов до сих пор остаются дискуссионными. Решение их HMCOI большое научное и практическое значение. Изучение истории формирования флоры пихтовых лесов позволяет объяснить основные проблемы систематики рода, миграции видов, изменения структур и состава лесных формаций (Крылов и др.,1968). В практическом отношении все это итн’обстпуст разработке и применению эффективных систем лесовод v । пенных мероприятий при использовании и воспроизводстве лесов.
По В.Г. Боброву (1978), палеоботанические и другие данные, отно-। ЯП1ИОСЯ к роду пихта, довольно значительны и с определенностью указываю! нп неогеновый возраст представителей этого рода. Однако исследования других ученых (Кеппен, 1885; Малеев, 1940, 1948; Szafer, 1954; Шоо, 19ХК, Маценко, 1964; Кондратюк, 1965;.Liu, 1971; Колаковский, 1974; Чочиева, 1985; Крылов и др., 1986; Axelrod, 1973, 1990 и других) указывают на более древний возраст появления как рода пихта, так и отдельных его представителей. Нами, на основании литературных источников, составлена гипотетическая схема возникновения одной из секций рода, куда болыпин-с гном систематиков включен вид пихта кавказская (рис. 2).
Несомненно, современное распространение видов пихт и пихтовых лесов представляется естественным продолжением • их геологического прошлого.
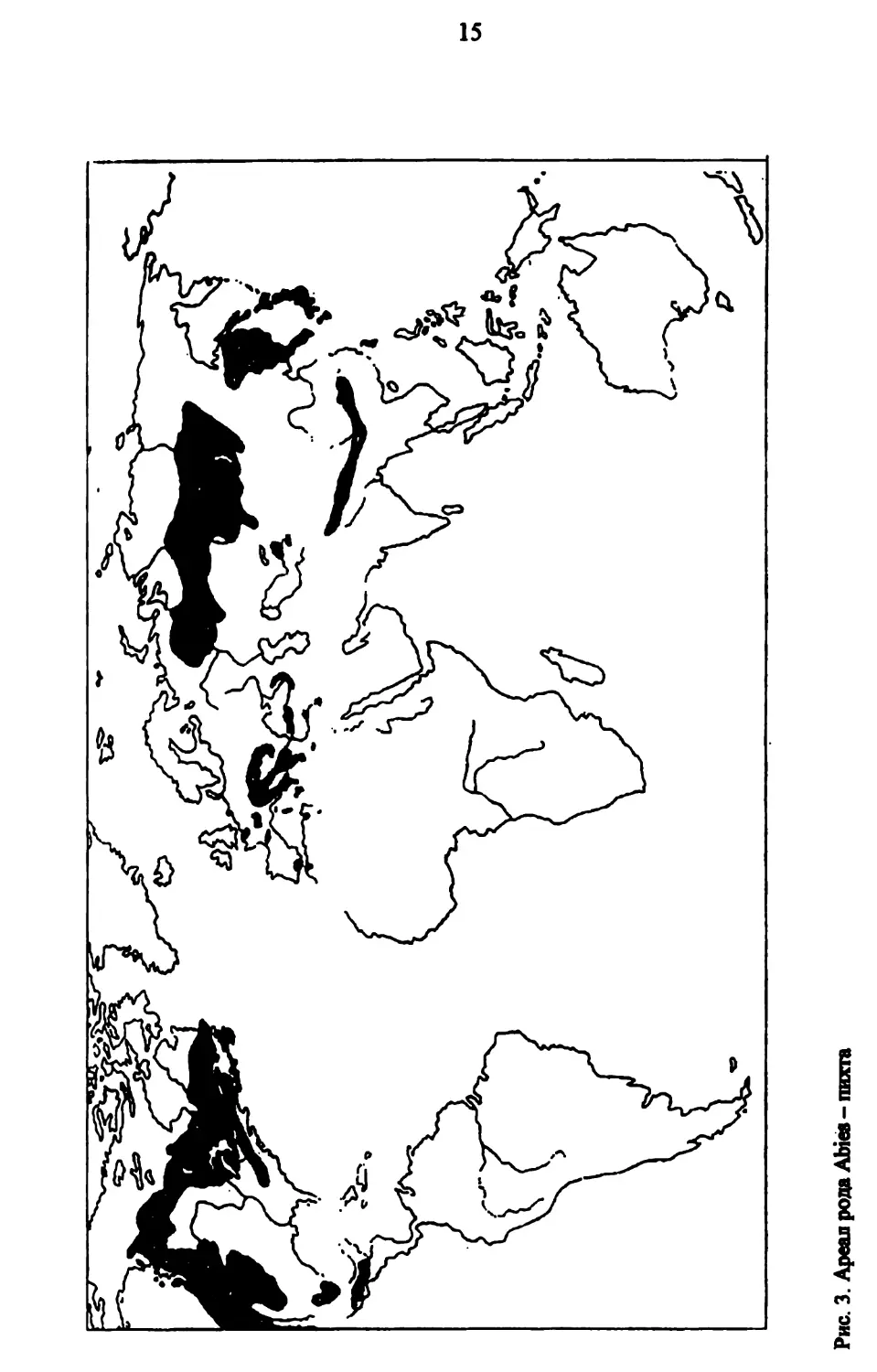
Основные ареалы современных видов пихт находятся в области вне-фопического Северного полушария (рис. 3). Именно эти области, в частности, Северной и Восточной Азии, Северной Африки, Центральной и Южной Европы и Северной Америки принято считать и областями древнего формирования родов “елоподобных”, в том числе и пихты, а также темпохвойной лесной формации - тайги (Толмачев, 1954; Юрцев, 1972).
Крайнюю точку зрения высказал по этому вопросу П.И. Дорофеев (1972), согласно которой родиной темнохвойных являются средние и высокие широты Голарктики, и автор полагал, что некоторые темнохвойные проникли в горы Южной Европы и Кавказа в конце олигоцена, немного позже - в горы Европы и еще позже в Китай и Гималаи. В плиоцене темно-хвойные определяли растительный ландшафт от Западного Приуралья до 11олыпи. Согласно этой гипотезе в указанное время здесь и сложилась таежная темнохвойная формация.
Четвертичный ( Голоцен 8 -г V) — hi — 0-0.1
0.1-3
Плейстоцен
3-10
Кайнозой Третичный Плиоцен ch 2. Nordmannianae Martz — 3. Albae Ma 4. Firmae Г
— 10-25
Миоцен
Олигоцен 25-40
40-60
Эоцен
60 -70
1 Палеоцен
70-80
Меловой Меловой Верхний о. сл 2
l.Abi< 80 -100
Средний
Млн. лет
Рис. 2. Гипотетическая схема возникновения представителей серии пихт (2, 3, 4) секции Abies Spach (1) и пихты Нордманна (5).
Следует отметить, что вопрос о роли миграции растений и автохтонного развития элементов флор и формирования различных флористических комплексов, является одним из узловых вопросов исторической географии растений, в частности, пихты. Объясняется это крайней скудностью и недостаточностью фактических данных по ископаемым пихтам в горных условиях.
Безусловно, одно из важнейших событий плиоцена - широкое распространение темнохвойной тайги. Существует несколько теоретических толкований по этому поводу, дискутируемых по сей день. Наиболее значительные из них: вычленение тайги из сложных хвойно-широколиственных лесов на месте (Васильев, 1958), представление о надвигании ее с Севера (Попов, 1957) или о выходе с гор на равнины (Толмачев, 1954). Из многих точек зрения, высказанных по поводу происхождения и формирования тайги, нам представляется наиболее логичной, и в нашем случае наиболее приемлемой, точка зрения А.И. Толмачева. В противоположность П.И. Дорофееву, А.И. Толмачев пришел к выводу об автохтонном горном проис-
Рис. 3. Ареал рода Abies - пихта
хождении темнохвойной тайги в районах умеренной Азии намного раньше, чем появились хвойные леса равнин Севера. Палеоботанические материалы последних лет и анализ палеографической обстановки земли позволили подтвердить не только верность данной гипотезы, но и установить горное происхождение рода пихты в районах древней Катазии, т.е. регионов Центрального Китая и Северной Индии, которые не испытывали разрушительных ледниковых периодов. Еще Ч. Дарвин (1937) и А. Гумбольдт (1936) придавали большое значение горным системам в вопросе флорогенеза, и в настоящее время можно считать доказанным, что горные системы являются ареной эволюции растительного мира (Комаров, 1921, 1961; Малеев, 1941; Вульф, 1944; Криштофович, 1946; Федоров, 1957; Крашенинников, 1974; Гребенщиков, 1972; Колаковский, 1974)..
Изучение характера и закономерностей распространения флоры и растительности горных регионов Северного полушария, в частности, особенностей распространения пихт, позволило А.А. Колаковскому (1974) выделить впервые средиземногорную фитогеографическую область с флористической эмблемой “флоры пЛхтьГ, куда и включены все пихты, в том числе и пихта кавказская. Это положение подтверждается и нашими исследованиями флористического состава пихтарников Кавказа, в результате которого выявлено, что элементы с горным ареалом составляют 80,5 % (табл. 1).
В настоящее время в Южной, Центральной и Восточной Азии произрастает почти половина - 28 видов пихт, в том числе в горах Юньнань и Сычуан - ряд пихт, входящих в наиболее древнюю секцию Bracteatae (Ма-ценко,1964). Время образования этой секции пихт относится к раннемеловой эпохе с возрастом в 120 млн. лет (Маценко, 1964; Крылов и др., 1986). В последующие геологические периоды характер горных образований Евразии, в том числе Кавказа, а также и Америки, в целом, был весьма благоприятен для установления между отдельными, горными областями флористических связей (Кеппен, 1885; Гроссгейм, 1936; Вульф, 1944). Возможность былого обмена элементами флоры между Гималаями и Кавказом через Переднюю Азию считается неоспоримой ботанико-географами, начиная с А. Энглера (Малеев, 1941; Горчаковский, 1954; Федоров, 1958; Матюшкин, 1982). Следовательно, пихты могли распространяться по горным системам всей Евразии и Америки. Так, уже в палеоген-олигоцене в горах формируются не только видовые секции пихт, но и сами виды (Криштофович, 1946;Сочава, 1946; Гребенщиков, 1972).
По С.В. Сябряй (1992), формирование пихтовых лесов в Альпах, Карпатах и в других горах Средней Европы относится к концу палеогена. Плиоценовые леса на юге Польши, состав которых установлен с большой полнотой исследованиями В. Шафера (Szafer, 1946-1947), в большой степени напоминали современные хвойно-широколиственные леса Восточной Азии.
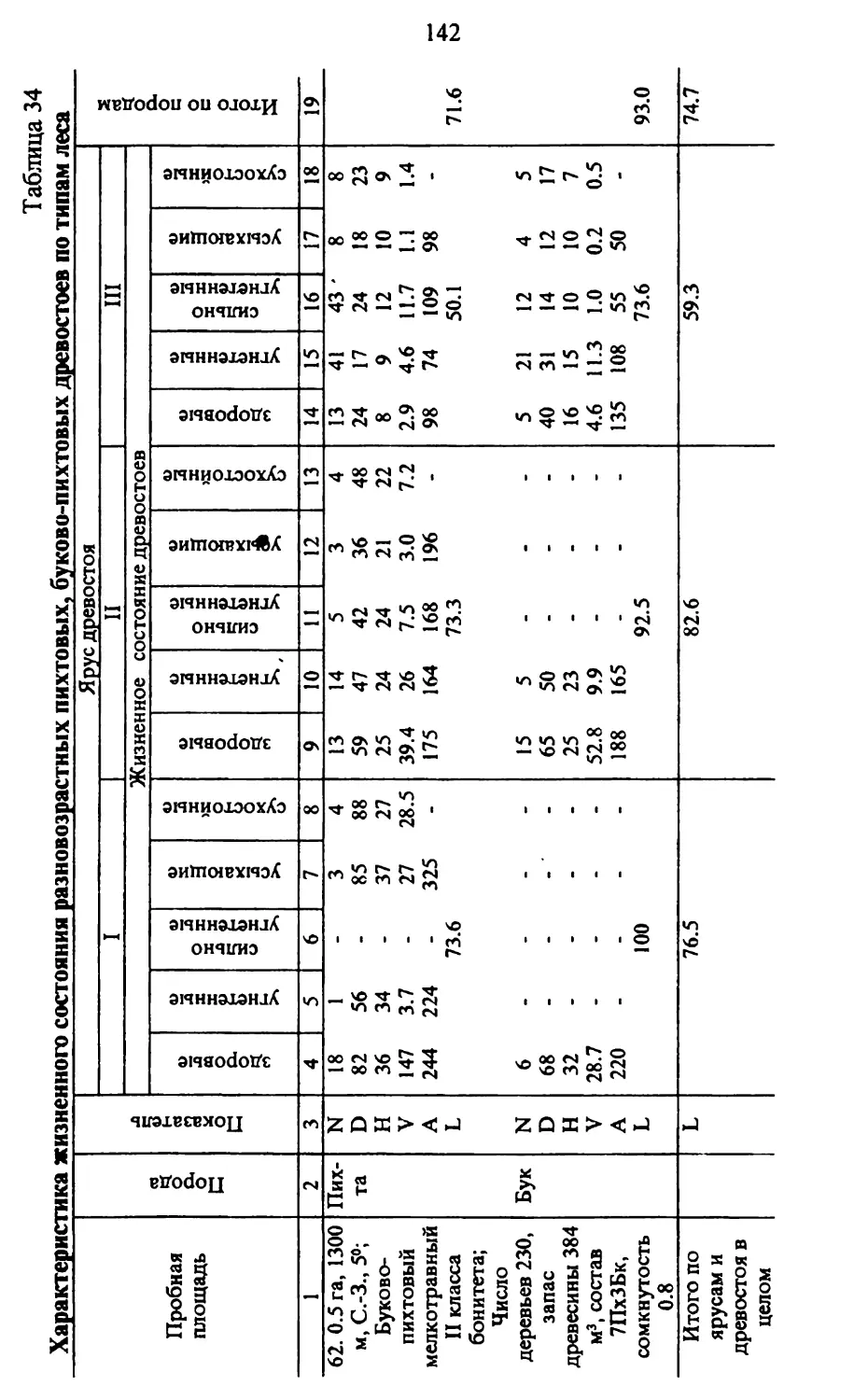
Таблица 1
1 Флористический состав пихтарников Кавказа
Наименование вида Географические элементы флоры
Реликты 3 I Г) 1 Кавказские эндемы Средиземногорные Средиземноморские Бореальные
2 3 4 5 6 7 8
Цргигсно - кустарниковые растения
1 Abies nordmanniana + + +
) Acer platanoides +
1 A. pseudonlatanus + +
'1 A. trautvetteri + + +
5 A. sosnowskyi + + +
Л Alnusbarbata + + +
/ Betula litwinowii + + +
н B. megretica + +
9 Buxus colchica + + +
К) Carpinus betulus + +
II C. caucasica ' + + +
12 C. orientalis 4- +
II Castanea sativa + +
14 Cerasus avium +
IS Corylus avellana + +
16 C. colchica + + +
17 Cotinus coggygria + + +
18 Daphne mezereum +
19 Daphne caucasica + +
20 D. pontica + + +
21 D. pseudosericea + + +
22 Euonymus earopaea +
21 E. latifolia +
24 E. verrucosa +
25 Fagus orientalis + + +
26 Fraxinus excelsior +
27 Ilex colchica + + +
28 Juniperus hemisphaerica + +
29 J. oblonga + +
10 laurocerasus officinalis + +
11 Lonicera caucasica + +
12 Philadelphus caucasicus + + +
13 Picea orientalis + + +
14 Pinus kochiana + + +
15 Populus tremula +
Продолжение табл. 1
1 2 3 4 5 6 7 8
36 Pyrus caucasica + + +
37 Quercus iberica + + +
38 Q. petraea + +
39 Q. pontica + . + +
40 Rhamnus imeretina + + +
41 Rhodedendran caucasicum + + +
42 Rh. luteum + +
43 Rh. ponticum + + +
44 Rh. ungemii + + +
45 Ribes cdpinum +
46 Ribes biebersteinii + +
47 Ruscus colchicus + + +
48 Salix caprea + +
49 S hastata +
50 Sambucus nigra +
51 Sorbus boissieri + +
52 S. graeca * +
53 S. migarica + +
54 S. subfusca + +
55 Staphylea colchica + + +
56 Swida koenigii + + +
57 Taxusbaccata + +
58 Tilia begoniifolia + + +
59 Ulmus glabra + + +
60 Vaccinium arctostaphylos +. + +
61 Viburnum opulus + +
62 V. orientate + + +
Итого видов - 62 43 22 14 56 1 8
%. - 100 69,4 35,5 22,6 90,3 1,6 12,9
Травянистые растения
1 Achillea biserrala + +
2 Aconitum nasutum + +
3 Actaea spicata + +
4 Aristolochia iberica + +
5 Aruncus vulgaris +
6 A. caucasicum + + +
7 Asperula caucasica + +
8 A. odorata +
9 Astrantia biebersteinii + +
10 A. maxima + +
11 Athyrium distentifolium +
12 A. filix-femina +
13 Atropa bella-donna + +
14 Blechnum spicant + +
15 Brachypodium pirmatum +
16 B. silvaticum +
17 Bupleurum polyphyllum + +
Продолжение табл. 1
1 2 3 4 5 6 7 8
IN (alamagrostis arundinacea +
|9 ('alamintha grandijlora + +
Л1 ('ampanula cordifolia + + +
Л (ardamine impatiens +
Л (pectinata + +
Л ('arex pendula + +
л (silvatica + +
л ('entaurea abbreviata +
я» ('ephalaria gigantea + +
л ('cphalanthera longifolia +
,'И Chaerophyllum aureum +
)*> ('.icerbita petiolata • + +
10 ('. pontica + +
Н C. prenanthoides + +
и Circaea alpina + + +
11 ('. lutetiana + + +
н ('irsium dealbatum + +
и ('. hypoleucum +
16 C. incanum +
1/ Colchicum speciosum + +
IM Convallaria transcaucasica + + +
19 Coronilla varia +
40 Corydalis caucasica + +
41 Dactylis glomerata + +
4/ Dentaria bulbifera + +
И D. quinquefolia + +
14 Digitalis schischkinii + +
41 Dryopteris carthusitoia + +
46 D. linneana + +
1/ D. filix-mas + + +
4N D. oreades '+ +
49 1). pseudomas +
10 Epilobium montanum +
М E. prionophyllum
1/ Epipogium aphyllum +
11 Euphorbia macroceras + +
14 Euphorbia sauamosa + +
и b'estuca drymeja + +
16 Eragaria vesca +
1/ (jade Ilia lactiflora + + +
1N Galium rotundifolium +
19 G. valantioides + +
60 Gentiana schistocalyx + + +
б| Geranium gracile + +
h} G. robertianum + +
С» 1 G. sylvaticum + +
64 Goodyera repens +
Продолжение табл. 1
1 2 3 4 5 6 7 8
65 Helleborus caucasicus + + +
66 Heracleum asperum +
67 H. ponticum + +
68 Impatiens noli-tangere +
69 Lapsana grandiflora +
70 Lathyrus aureus +
71 L. laxiflorus +
72 L. roseus + +
73 L. vemus +
74 Ligularia renifolia + +
75 Ligusticum alatum + +
76 L physospermifolium + +
77 Lilium monadelphum + + +
78 Listera cordata +
79 Lusula multiflora +
80 L. sylvatica + +
81 Matteuccia stnMopteris +
82 Melampyrum arvense + +
83 M. elatius +
84 Milium effusum +
85 M. schmidtianum + +
86 Mycelis muralis +
87 Myosotis amoena + +
88 Neottia nidus-avis +
89 Origanum vulgare +
90 Orthilia secunda +
91 Oxalis acetosella +
92 Pachyphragma macrophyllum + + +
93 Paris incompleta + + +
94 Petasites albus +
95 P.hibridus +
96 Platanthera bifolia + +
97 Poa nemoralis + +
98 Polygonatum multilorum +
99 Polygonatum ovatum + +
100 P. verticillatum +
101 Polypodium vulgare + +
102 Polystichum setiferum +
103 P. woronowii + +
104 Potentilla micrantha + +
105 Prenanthes purpurea +
106 Primula auriculata + +
107 Prunella vulgaris +
108 Pulmonaria mollissima +
109 Pyrethrum macrophyllum +
НО Pyrola media +
111 Ranunculus grandiflorus + +
Продолжение табл. 1
1 2 3 4 5 6 7 8
112 Rubus buschii + +
113 R. caucasicus + +
114 R. hirtus +
115 R. platyphyllos + +
116 Salvia glutinosa +
117 Sambucus ebulus +
НК Sanicula europaea + +
119 Saxifraga cymbalaria + +
120 Scrophularia chrysantha + +
121 S. nodosa +
122 Sedum stoloniferum + +
123 Senecio caucasicus + +
124 S. correvonianus + + +
125 S. platyphylloides + +
126 S.propinquus + +
127 Serratula quinquifolia +
128 Silene multifida + +
129 Solidago virgaurea +
ВО Stachys silvatica + +
HI Stellaria holostea +
112 Symphytum asperum + + +
133 S. grandiflorum + + +
134 Telekia speciosa + +
Н5 Trachystemon orientalis + + +
116 Trifolium ambiguum + +
137 T medium +
1 18 Urtica dioica 4-
139 Vaccinium myrtillus + +
140 Valeriana alliarifolia + +
141 V. colchica + + +
142 Veratrum lobelianum + + +
14.3 Veronica officinalis +
144 V. peduncularis + +
145 Vicia crocera + +
146 Viola alba +
147 V. reichenbachiana +
148 V. sieheana +
Итого - 148 видов 34 32 29 109 1 51
% - 100 23,0 21,6 19,7 73,7 0,7 34,5
В целом, видов - 210 77 54 43 169 2 59
%-100 36,7 25,7 20,5 80,5 1,0 28,1
Примечание. Названия растении приводятся по А.А. Колаковскому (Флора Абхазии, I-IV, 1980 - 1986) и С.К. Черепанову (Сосудистые растения России., 1995).
Безусловно, пихтовые леса представляют собой наиболее древнюю формацию, и в прошлом они были распространены значительно шире, что
подтверждается палеоботаническими данными, изоляцией ареалов большинства видов пихт, произрастающих на территории бывшего СССР. В конце неогена на северо-востоке Азии и на Аляске повсеместно господствовали темнохвойные леса (Юрцев, 1972; Хохряков, 1989). С середины эоцена, примерно 45 млн. лет назад, темнохвойные леса из пихт, ели, лиственницы образовывали самостоятельный высотный пояс на западе Северной Америки (Axelrod, 1990). Следовательно, уже в это время в верхнем горном поясе произрастали представители умеренной флоры в окружении равнинных тропических лесов, распространение растительности носило четко выраженный характер вертикальной поясности и пояс темнохвойных лесов из пихт и елей был представлен достаточно широко.
Таким образом, пихта - типично средиземногорный древний род, известный с палеогена, представленный в настоящее время видами более или менее обособленными, ограниченными ареалами. Только 2-3 вида встречаются с более широкими ареалами в северных широтах.
Систематика р<да пихт очень сложна. Изучение и критический пересмотр уже известных видов продолжаются и по сей день (Jain, 1975). К примеру, по сообщению журнала IDRI (№ 6, 1994), дендролог Ж. Дебреци в Мексике обнаружил и описал три новых вида пихт и две разновидности пихты гватемальской. *
Наиболее полная, на наш взгляд, флористическая обработка пихт Восточного полушария была осуществлена А.Е. Маценко (1964). Ею же проведен систематический обзор всех пихт мира (Маценко, 1963). Позже этому вопросу были посвящены и работы Д.П. Воробьева (1968), Т. Liu (1971), Е.Г. Боброва (1978), Г.В. Крылова с соавторами (1986), A. Faijon (1990). Уточнение видов пихт и их краткое описание приводятся также в ряде капитальных сводок последних лет по дендрологии (Bean, 1980; Krussmann, 1983; Rushforth, 1987; Mesterhazy, 1995 и др.). По данным этих авторов, в настоящее время в роде пихта насчитывается около 56 видов, из которых на Кавказе произрастает единственный - пихта кавказская, или Нордманна, Abies nordmanniana (Stev.) Spach.
В 1835 году А.Д. Нордманн, директор ботанического сада и профессор Ришельевского лицея в Одессе, проводя флористические исследования в Западном Закавказье, впервые описал видовые особенности пихты кавказской и указал на ее исключительно ценные качества. Позже название пихта кавказская было изменено по правилам ботанической номенклатуры на пихту Нордманна.
В систематическом отношении и по своему происхождению пихта кавказская наиболее тесно связана с группой средиземноморских видов (Долуханов, 1989). По А.Е. Маценко (1963), пихта кавказская относится в отдельный видовой ряд - Nordmannianae Matz, который, кроме этого вида, охватывает еще два анатолийских - пихту Борнмюллера и пихту троянскую.
11рсдставители видов пихты этой серии характеризуются клиновидными семенными чешуями, расширенными в верхней части, и линейно ок-pyi лыми кроющими чешуями, отогнутыми наружу (см. рис. 1). Общий аре-.»н них видов эвксинский, и обособление их в причерноморских регионах, ни мнению Е.Г. Боброва (1978), может быть отнесено к концу плиоцена.
I Io В.З. Гулисашвили (1951), пихта кавказская в Западной части Кавки ы появилась несколько раньше, в третичную эпоху в конце миоцена »iipMaicKoro века. По О.Г. Капперу (1954), ископаемые остатки пихты кав-кп и ной, найденные в долине р. Сунжи, относятся к периоду нижнего сарма hi В таком случае, формирование ее биоэкологических и ценотических • (»бснностей, надо полагать, происходило автохтонно в конце миоцена-н'1иоцена (рис. 2).
Интересно, что к секции Abies Spach относится видовой ряд Firmae \fdi -, который включает еще один совершенно обособленный японский пнд пихта крепкая - A. firma Siebold et Zucc. (см. рис 2). Этот факт свиде-кльегвует о довольно тесных генетических связях средиземноморских и тксинских видов с этим остаточным дальневосточным видом, об их геоло-। и песком прошлом и общем предке. Начало их разрыва может быть отне-< спо к олигоценовому времени (Бобров, 1978). В последующие геологиче-»к не периоды пихта кавказская формируется в достаточно самостоятельный и обособленный вид с четко выраженным ареалом в КолЯидскоКГре-фу। иумс. В настоящее время - это одна из ценных редЩсговых, эндемичных. лесообразующих древесных пород Кавказа, о которой еще Ф.К. Арнольд (1891) восторженно писал, что “ ..на/кавказе растет Abies nonhnanniana, составляющая в настоящее врешгпредмет удивления и вос-юргов лесоводов всей Европы”. Безусловно, <яК. Арнольд был абсолютно нрав.
Кавказ как рефугиум реликтовой дендрофлоры и позиция пихты в ней
В настоящее время можно считать доказанным, что основные лесообразующие породы современных лесов Кавказа, в том числе пихтовых, произрастали здесь в третичном периоде, во всяком случае они были широко представлены в плиоцене. Благодаря наличию на Кавказе рефугиаль-н|.IX регионов (Колхида, Талыш) они пережили периоды климатических репрессий и уже в это время преобладал тип лесной растительности с четко выраженной вертикальной поясностью (Сочава, 1949; Тумаджанов, 1955; Нойштадт, 1957; Колаковский, 1974; Колаковский, Шакрыл, 1976; Чочиева I *>К5). В частности, в это время, по крайней мере в Западной части Болыпо-। о Кавказа, существовали пояса широколиственных и хвойных лесов, в том числе пихтовых и буковых.
По А.А. Колаковскому (1961, 1974), растительный мир Колхиды еще в сармате был дифференцирован на три основных вертикально поясных фитоландшафта - низинных и приречных лесов, лесов нижнего горного пояса и лесов среднего и верхнего поясов. Эти фитоландшафты были представлены разнообразными лесными формациями. Леса среднего и верхнего горного поясов состояли из листопадных широколиственных, преимущественно буковых лесов, а также смешанных и хвойных лесов умеренного климата.
На существование растительных поясов на Кавказе в этом периоде указывают не только палеоботанические данные, но и тот факт, что многие реликты здесь не являются термофильными. Среди них много достаточно зимостойких видов, в том числе пихта кавказская, ель восточная, сосна Коха, бук восточный, рододендрон кавказский и другие представители горных, высокогорных и альпийских элементов. Это подтверждается также и значительной амплитудой их биологической приспособленности к многообразию экологических факторов, экотопов, от напряжения которых возможны существенные инверсии. К примеру, сосна Коха в Абхазии произрастает от 300 до 2000 м над уровнем моря, каштан посевной растет от берега моря до 1500 м. Они являются лесообразующими породами в средне-и верхнегорном поясе.
Неотектонические движения, создавшие здесь современный горный рельеф, оказали, несомненно, существенное воздействие на характер распространения флоры и растительности. Вертикальная смена ландшафтных поясов закономерно обуславливала и распространение растительности в горах в поясном порядке в строгой зависимости от климатических, главным образом, гидротермических условий (Докучаев, 1948; Быков, 1954; Станюкович, 1973). При этом, ширина пояса растительности по абсолютной высоте могла весьма различаться в зависимости от быстроты смен экологических условий по склону, а также от амплитуды экологической требовательности данной растительности. На небольшом расстоянии могли складываться переходные зоны с ценозами смешанного состава с элементами различных биоморф. Такая закономерность четко выражена при распространении растительного покрова современных горных систем Евразии и других регионов Земли (Лавренко, 1938; Мишич, 1974). Если учесть и тот факт, что горные регионы отличаются необычайным разнообразием экотопов, то становится ясной закономерность и объективность богатства флоры растительности Кавказа уже с третичного периода, вертикальный характер их распространения.
Такая растительность в эпоху орогенеза сохранилась в Колхиде и Та-лыше вплоть до начала эоцена, значительно дольше, чем в окружающих их областях (Альбов, 1896; Баранов, 1952; Гроссгейм, 1936; Сафаров, 1981). Такой точки зрения придерживается и И.А. Ильинская (1980). Ее исследо-
niiiiHN, на примере систематического состава ископаемых буковых, под-пи^рдили существование в нижнем палеогене на юге бывшего СССР веч-н'меленых субтропических лесов, слагающихся видами, близкими к современным видам субтропической флоры Южного Китая и более южных тер-|ц| ।орий Азии. По ее мнению, остатки этой флоры просуществовали в Колчине до начала плиоцена. Мы склонны разделить такую точку зрения. 1'ли пейс черты и закономерности развития растительного покрова Колхиды и |рс!ичном периоде, а также его необычное флористическое богатство и икальньгй характер распространения можно наблюдать и сегодня на <ндровах Юго-Восточной Азии, с достаточно высоким горным рельефом (Краснов, 1912; Вальтер, 1974). В частности, на острове Тайвань нам пред-< । ввилась возможность ознакомиться с такой растительностью во время жспсдиции в 1997 году (Бебия и др., 1999), на чем следует вкратце останови i вся.
По своему широтному положению Тайвань, подобно Южному Киппс ), располагается в зоне, переходной от умеренного к тропическому поя-। у Северный тропик пересекает остров почти посередине. Однако островные условия, а также горный, сложный топограф!|чес1аЭ"харакгер ландшафта, обусловили специфическое многообразцсГклиматических условий и вертикальный характер распространения естественного растительного покрова острова. Здесь, на территории, которая почти в три раза меньше территории Кавказа, естественная флора содержит около 4100 видов сосуди-СИ.1Х растений, в том числе 1069 древесных. Среди них многочисленные представители вечнозеленых лавровых и буковых, которые были широко представлены в колхидском рефугиуме в третичном периоде. Эндемы со-с гавляют порядка 43 % от общего числа видов флоры острова.
Достаточно высокие горы до 4000 м обусловили наличие на этом острове климатов от тропического до субарктического. До 500 м над уровнем моря климат тропический, обусловливающий существование фикусо-махилового растительного пояса. На отметках 500—1500 м климат субтропический, растительный пояс махило-кастанопсисовый. От 1500 до 2500 м климат теплоумеренный, растительный пояс вечнозеленых дубов. На высоте 2500—3100 м простирается пояс смешанных тсуго-еловых лесов. Пояс пихтовых лесов здесь приурочен к субальпийской зоне на высоте 3100— 1600 м с умеренно-холодным климатом. Выше 3600 м простирается альпийская зона с альпийской растительностью и субарктическим климатом (Hora of Taiwan, 1,1994).
Даже беглый анализ состава современной дендрофлоры острова и характера распространения его растительного покрова дает достаточное основание для выдвижения гипотезы о существовании близкого сходства с растительным покровом Колхиды в эоцене, а также указывает на общие закономерности вертикально-поясного распространения растительности.
Особенно это заметно на примере распространения пихтовых лесов, приуроченности их к верхней горной зоне в древней Колхиде и на острове Тайвань в настоящее время.
Естественно, что третичный, да и четвертичный периоды на Кавказе и прилегающих областях, были эпохой непрерывного изменения и эволюции растительного покрова, вызванного изменениями климатических условий. Однако в горных условиях эти изменения носили более сложный и дифференцированный характер, чем на равнинах. При трансгрессии уровня моря и соответствующем потеплении или континентализации климата в горных условиях существовала возможность инверсии не только отдельных представителей флоры, но и растительных поясов, способствовавших сохранению представителей субтропической, теплоумеренной флоры и растительных комплексов, в том числе пихтовых формаций на Кавказе, с последующей трансформацией элементов флор отдельных растительных поясов в разные пояса. Этим, отчасти, можно объяснить произрастание в Колхиде отдельных г^едставителей современной умеренной и альпийской флоры в нижнем поясе растительности с влажным теплым климатом (Populus tremula, Cerasus avium, Viburnum opulus, Coronilla varia, Oxalis ace-tosella, Stachys silvatica, Sanicula europaea\ а также встречаемость вечнозеленых и теплолюбивых листопадных элементов в верхне-горном лесном и субальпийском поясе (Ilex colchica, Laurocerasus officinalis, Staphylea col-chica, Kastanea sativd).
Нельзя не согласиться с мнением К.И. Чочиевой (1985), которая считает также, что с конца кимерийского времени в Колхиде происходило не становление новой флоры как таковой, а расширение ареала летнезеленых и, по всей вероятности, хвойных лесов, за счет сокращения площади распространения, а затем и полного распада и исчезновения формации вечнозеленых влажносубтропических лесов. Обосновывается ею такое предположение отсутствием в посткиммерийских флорах Колхиды какого либо рода, который не был бы представлен и во флорах предшествующих эпох геологической истории региона.
Ясно, что сложившиеся современные группы лесных комплексов, биогеоценозов на Кавказе сформировались автохтонно из третичных лесных флор. Однако современные реликтовые леса не тождественны третичным. Они беднее по флористическому составу. Вместе с тем, эти леса обогатились новыми, более молодыми видами. Но при этом они сохранили основные черты своего развития, ценотического состава и структуры. К примеру, основной эдификатор и соэдификаторы всех коренных типов леса пихтарников Кавказа представлены реликтами и эндемами такими, как пихта кавказская, ель восточная, бук восточный, падуб колхидский, черника кавказская и многими другими (см. табл. 1).
Отличительной особенностью пихтовых лесов Кавказа является относительное флористическое богатство их состава (табл. 2). Здесь произрастает более 62 видов древесно- кустарниковых и 148 травянистых растений.
В целом флора пихтарников состоит из. различных по происхождению видов, что особенно заметно в скально-лесных комплексах, на известняковых склонах и по верхней границе пихтовых лесов. Это объясняется разнообразием экотопов и стаций в указанных условиях.
Нами флорогенетическим анализом выявлено, что в дендрофлоре пихтарников реликты составляют до 70 %, колхидские эндемы - 36 %, общекавказские - 23 % и около 40 % являются евроазиатскими элементами (см. табл. 2). Примерно 90 % представителей дендрофлоры являются элементами средиземногорной флористической области и лишь около 13 % составляют более молодые бореальные элементы, которые в основе своей также можно считать выходцами из высокогорной флоры.
/ Таблица 2
Флорогенетическая характеристика пихтарников Кавказа
Географические элементы флоры ^сло видов
древесных травянистых всех растений
шт. % % шт. %
Реликты 43 69,4 34 23,0 77 36,7
Колхидские эндемы 22 35,5 32 22,6 54 25,7
Кавказские эндемы 14 22,6 Г9 19,7 43 20,5
Средиземногорные 56 90,3 /109 73,7 169 80,5
Средиземноморские 1 1.6 / 1 0.7 2 1,0
Бореальные 8 12,9 у ' 51 34,5 59 28,1
Итого . 62 148 100 210 100
Такое высокое содержание во флоре пихтовых лесов Кавказа реликтовых и эндемичных элементов, особенно среди древесных растений, несомненно, свидетельствует не только о древнем автохтонном формировании их основного флористического ядра, но и позволяет утверждать, что и нетронутые пихтовые ценозы в целом здесь сохранили черты реликговости.
С этой точки зрения весьма интересны представления В.Б. Сочава (1947). Согласно им утверждать однозначно, что фитоценоз, в составе которого представлены, хотя бы даже обильно, реликтовые виды, является древним фитоценотическим типом, на основании формальных флористических соображений, крайне рискованно. Свое представление по данному вопросу В.Б. Сочава обосновывает следующими положениями.
1. Влияние человека, нарушая соотношение между компонентами фитоценоза, сложившееся в процессе естественного отбора, а тем самым изменяя и экологическую обстановку, часто содействует распространению реликтов, которые при естественном положении вещей устраняются с территории.
2. Реликтовые растения, имеющиеся в незначительном обилии в естественных фитоценозах, иногда получают массовое распространение на вторичных местообитаниях. В силу этого происходит расширение ареала реликта (Dioscorea caucasica Lipsky).
3. У многих колхидских реликтов реликтовой и длительностабильной является лишь часть их ареала, а не вся область распространения на Кавказе.
4. Некоторые реликты окончательно порвали связь с коренными фитоценозами. Они блуждают по территории, поселяясь в местах, где обнаруживается подходящий субстрат или ослабляется роль того или иного эдификатора. К этому типу относится, например, диоскорея кавказская и толстостенка крупнолистная.
5. Реликты типа диоскореи-толстостенки составляют особый фито-ценотический тип, который хотя и представлен видами, утратившими конкурентные способности, но может существовать в соответствующей климатической зоне неопределенно долгое время, поскольку нарушение структуры коренных фитоценозов, ослабляющее конкуренцию, непрерывно сопровождает развитие {Растительности любого региона.
Безусловно, суждение по поводу того, что фитоценоз реликтовый или нет, возможно лишь в результате биогеоценотических и флорогенетических сопоставлений с учетом позиции реликтов в фитоценозе. Рассматривая пихтарники Кавказа с этой позиции, отметим следующее.
Как известно, в голоцене произошло становление человеческого общества, которое стало использовать ресурсы как живой, так и неживой природы и превратилось, по выражению В.И. Вернадского (1926), в “могучую геологическую силу”. Хозяйственная деятельность человека нарушила спонтанное развитие природы и вызвала разнообразные сукцессии биогео-ценотического покрова. По Г. Кларку (1953), например, распространение земледелия привело к быстрой деградации средиземноморских вечнозеленых лесов.
Несомненно, пихтовые леса Кавказа могут носить следы антропогенных воздействий (рубки, пастьба скота, рекреации и пр.). Однако эти воздействия начались здесь лишь с середины прошлого столетия. До этого пихтарники Кавказа были малодоступны для человека. Я.С. Медведев (1919) в отношении пихтарников Абхазии указывает, что это вековые, в полном смысле девственные леса, куда и поныне почти не проник топор человека. Проведенные исследования подтвердили, что существенных антропогенных трансформаций и влияния их на развитие и преобразование пихтовых лесов в историческом плане не наблюдается. Основные формы антропогенных трансформаций в пихтарниках выражаются, в основном, в появлении производных фитоценозов, которые практически в течение онтогенеза одного поколения основного эдификатора пихты, вновь восстанавливаются в исходные коренные. Такое положение обусловлено биоэко
логией пихты, природными условиями произрастания пихтарников и закономерностями лесообразовательного процесса в них, о которых более подробно пойдет речь в последующих главах.
Проведенные нами исследования показали, что позиция пихты кавказской во всех коренных типах леса достаточно устойчивая и встречаемость ее самая высокая - 100 % при фитоценотичном индексе равном 5400 (табл. 3). Устойчивыми являются позиции и большинства других реликтов девственных пихтовых лесов, которые играют существенную роль в их сложении. Встречаемость таких реликтов составляет 17-83 % с фитоцено-тическим индексом 34-1328. Каждый из них характерен и постоянен, как минимум, в двух типах леса. Лишь 13 видов из 84 (15,4 %), таких как Acer wsnowskyi, Castanea sativa, Quercus iberica и некоторые другие (табл. 3) можно отнести к факультативным.
Таблица 3
Реликтовые растения коренных типов леса пихтарников Кавказа
(данные 56 пробных площадей)
Наименование растений Обилие видов (балл) по типам леса Встречаемость, % Фитоценотичс-ский кодекс
I П Ш IV V VI vn IX X XI XU
1 2 3 4 5 6 1 8 / 9 10 11 12 13 14 15
Деревья
Ables nordmanniana 5 5 5 4 5 4 5 5 4 4 4 100 5400
Fagus oriental is 2 1 - 1 1 3 /\ 1 - 1 2 3 83 1328
Picea orientalis 1 1 - 1 1 1 1 1 1 67 536
Acer platanoides 1 - - - - /- 1 • 1 1 1 1 - 50 300
Acer pseudoplatanus - - - 1 1 1 1 • - - 50 300
litmus glabora - - 1 - 1 1 ! 1 - - 1 - 50 300
('arpinus betulus 1 1 - - - - - - - 1 - - 25 75
Quercus petrea 1 1 1 - 25 75
Tilla begoniifolia - 1 , - 1 1 - 25 75
Alnus barbata - - - - - • 1 1 - - - - 17 34
Hetula litwinovii 1 1 17 34
('arpinus caucasica 1 1 17 34
Fraxinus excelsior - - - - • - - - 1 - - 1 17 34
Pinus kochiana - 1 1 17 34
Acer trautvetteri • 2 8 16
Acer sosnowskyi 1 8 8
Castanea sativa - - - - - - - • - 1 • • 8 8
Pirus caucasica - 1 1 8 8
Quercus iberica - 1 1 8 8
Taxus baccata - 1 1 8 8
Кустарники
Vaccinium
arctostaphylos 1 - 2 - 1 1 2 - 5 2 2 2 75 1350
Ilex colchica 1 - - 3 1 • 1 - 3 2 2 3 67 1072
Ixnirocerasus offici- - 2 1 - 5 1 33 297
nalis
Hhododerubon ponticum - 1 5 1 - 25 175
Продолжение табл. 3
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Buxus colchica - - - - - - 1 1 - - 4 - 25 150
Quercus pontica - - - - - - - - - 1 2 3 25 150
Rhododentboncaucasica 1 1 - 3 25 125
Rhododendron ungemli - 1 4 - - 17 85
Corylus avelana - - - 1 - 1 - 1 - - - - 25 75
Phyladelphuscaucasicus 1 1 ' 1 25 75
Rhododendron luteum 1 - - - - - - - 1 - - 1 25 75
Viburnum orientalis - - - - - - - 1 1 - - 1 25 75
Cotinus coggigria 2 - - 2 - 17 68
Carpinus orientalis 1 - 1 - 17 34
Corylus colchica 1 1 - 17 34
Junlperus hemlspherica 1 - 1 17 34
Rhamnus imeretina - - - - - - - • - 1 - 1 - 17 34
Ruscus colchicus - - 1 1 - 17 34
Swida koenigii 1 - 1 - 17 34
Viburnum opulus - 1 - 1 - - 17 34
Daphne alboviana 2 8 16
Daphne pseudosericea Травы - ( - 1 8 8
Festuca drymeia 5 2 2 2 2 1 2 1 2 - 2 2 92 2116
Dryopteris felix-mas 2 1 2 - 1 1 5 2 1 1 - 2 83 1328
Calamintha grandiflora 2 2 1 1 2 - 1 1 1 - - 1 75 900
Paris incompleta 1 1 1 1 2 - 1 1 1 - - 1 75 750
Dentaria bulbifera 1 '1 2 - 1 1 1 1 1 - - - 67 603
Sanicula europaea 2 1 - 4 1 - 1 1 I - - - 58 638
Symphytum gyaubflorum 1 - - 2 1 3 1 1 - - - - 50 450
Circea lutetiana 1 - - 1 1 2 - 1 - - - 1 50 350
Valeriana alliarijblia 1 1 - - - - 1 i - - 1 2 50 350
Gentiana schistocalyx 1 1 - - 1 - - 1 - - - 1 42 210
Luzula silvatica 1 5 - - - - 2 1 - - - - 33 297
Trachystemon orientalis 1 - - - - 5 1 - 2 - - - 33 297
Circea alpina Pachyphragma - - • 1 • - 1 1 - - 2 33 165
macrophyllum 1 - - 1 - 2 1 - - - - - 33 165
Actaea spicata 1 1 - - - - 1 1 - - - - 33 132
ffelleborus Caucasians 1 • - - - - 1 1 - - 1 - 33 132
Stachys silvatica 1 - - - - - - - 1 - 1 1 33 132
Polypodium vulgare 1 2 3 - 25 150
Symphitum asperum - - - - - - 2 1 - - 2 25 125
Blechnum spicant - - - 1 - - - 2 1 - - 25 100
Atropa belladonna - - - 1 - - 1 1 - - - 25 75
Campanula cordifolia 1 1 1 25 75
Carex pendula 1 - - - - 1 - 1 - - - 25 75
Dentaria quinquifolia Convollaria 1 - 1 - - - 1 - - - - 25 75
transcaucasica - 1 3 - 17 68
Auruncus caucasicum - - - - - - 1 1 - - - 17 34
Godelia lactiflora - - - - - - - 1 - - 1 17 34
Lilium monadelphum - 2 8 16
Saxifraga cymbalaria - 2 - • 8 16
।ndlcKiie (XII); пихтарник ясменниковый la -1 классов бонитета (Ш), пихтарник под-пг( пиковый 1-П классов бонитета (IV), пихтарник ясменниковый П-Ш классов бони-। •» । л (V), пихтарник трахистемоновый I-П классов бонитета (VI), пихтарник кочедыжниковый la—I классов бонитета (VII), пихтарник страусоперовый I-П классов бонитета (VIII), пихтарник черничниковый I-П классов бонитета (IX), пихтарник рододендроновый П-Ш классов бонитета (X), пихтарник лавровишневый П-Ш классов бонитета (XI) Обилие вида для кустарниковых и травянистых растений определялось при учете н пощади покрытия, для деревьев - по запасу древесины на пробной площади. Применена шкала оценки обилия видов: проективное покрытие, запас до 20 % - 1 балл, 21-•1(| % 2 балла, 41-60 % - 3 балла, 61-80 % - 4 балла и 81-100 % - 5 баллов. Показате-*ш встречаемости и фитоценотического индекса вычислены по В.И. Василевичу (1969).
Встречаемость их составляет 8 % с фитоцевютическим индексом от 8 до 16. Однако и они не обнаруживают полного разрыва с коренными фито-ценозами, каждый из них является характерным и постоянным по крайней мере для одного типа леса. /
Как отмечалось выше, в период лешшковой депрессии с конца тре-1ичкого периода в Колхиде существовали достаточно благоприятные условия для сохранения не только тепдериеренных буковых, каштановых, а (пкже пихтовых лесов, но-и-етд^льных представителей тропических элементов, в частности нежного папоротника реликта - хименофеллюма -Hvmenophyllum timberidgense, произрастающего ныне в Аджарии. Его существование возможно лишь при исключительно постоянной влажности ущелий (Колаковский, 1961). Следовательно, надо полагать, что в Колхиде практически непрерывно с конца третичного периода существовал гумид-ный, достаточно теплый климат, благоприятствовавший сохранению этих элементов флоры. Можно с уверенностью утверждать, что именно своеобразные, характерные рефугиальные природные условия Колхиды обусловили сохранение здесь реликтовой черты развития современных спонтанных пихтарников.
Таким образом, результаты наших исследований (Бебия, 1987,1994), а *гакже данные палеографических и палеоботанических обобщений (Гули-сашвили, 1960; Колаковский, 1974 и другие) свидетельствуют об автохтонном горном происхождении пихтовых лесов Кавказа и подтверждают известную концепцию о горном происхождении тайги (Толмачев, 1954; Szafer, 1954; Маценко, 1964; Гребенщиков, 1990; Сябряй, 1992). Темнохвойные элементы в составе флоры Кавказа появились в олигоцене, одновременно с заселением образовавшихся горных систем Кавказа. В миоцен -
плиоцене в средней и верхней горной части Колхиды формируется выраженный пояс темнохвойных лесов с доминированием представителей родов Abies, Picea, Tsuga, близких к современным. В конце плиоцена здесь произрастали обширные леса из пихты кавказской с примесью ели восточной, бука восточного и других представителей ценотического состава современных пихтарников. Высокая степень преемственности ценотического состава между плиоценовыми и современными пихтарниками обусловлена существованием вертикальной растительной поясности и рефугиальных условий в Колхиде. Здесь во время максимального оледенения глетчеры опускались не ниже 800 м (Гулисашвили, 1960), и пихтарники могли сместиться на более низкие гипсометрические отметки, пережить суровый климат с наименьшими изменениями их видового состава и структурно -ценотического развития. Этому способствовал и ряд биоэкологических и ценотических особенностей пихты, который был выработан в процессе ее эволюции.
В настоящее время пихта кавказская является одной из важнейших древесных пород, образующих леса на значительной площади Западного Кавказа. Леса с ее преобладанием занимают здесь 320 тыс. га, или 6,9 % от всей покрытой лесом площади (Крылов и др., 1986). Однако в пихтарниках Кавказа, как показали исследования (Беленко, 1964; Бицин, 1965; Бебия, 1967-1983; Гулисашвили, 1970; Веселов, 1973), проводятся чрезмерно интенсивные рубки с применением бессистемной технологии лесосечных работ, которые не отвечают природе лесов и приводят к ослаблению позиции пихты, сокращению площади лесов с ее господством.
Вместе с тем реликтовые пихтарники Кавказа являются большой уникальностью не только в региональном, но и глобальном масштабе, что выдвигает требование и необходимость рационального их использования и охраны.
ПРИРОДНЫЕ УСЛОВИЯ И ЗАКОНОМЕРНОСТИ СОВРЕМЕННОГО РАСПРОСТРАНЕНИЯ ПИХТОВЫХ ЛЕСОВ КАВКАЗА
Природные условия произрастания пихтовых лесов
Природные условия произрастания пихтовых лесов освещены в трудах многих исследователей темнохвойных лесов Кавказа (Альбов, 1882; Малеев, 1940; Герасимов, 1948; Орлов, 1951; Гулисашвили, 1964; Махатад-зе, 1967; Голгофская, 1967; Веселов, 1973; Бебия, 1987 и др.). Общие положения по этим вопросам излагаются также в капитальных сводках по физической географии Кавказа, в частности, в работах Н.А. Гвоздецкого (1963), Б.П. Алисова (1969), Ф.Н. Милькова и Н.А. Гвоздецкого (1969), Ф.Н. Милькова (1977), в сборнике Альпы - Кавказ (1980), в климатических справочниках по Кавказу и других источниках. Однако цели и задачи наших исследований пихтовых лесов обусловили проведение специального анализа существующих представлений о природных условиях произрастания этих лесов для более правильного понимании современного состояния и тенденции их дальнейшего развития. /
Исследования вышеотмеченных авторов и результаты наших наблюдений показали, что природные условия произрастания пихтовых лесов сильно варьируют, в зависимости от морфологии гор, высоты над уровнем моря и других факторов средььОдаако широкое распространение их на Западном Кавказе укаЗываетгЗто развитие темнохвойных лесов из пихты и ели восточной самым тесным образом связано с определенным высотноклиматическом поясом. Этот пояс на южном макросклоне Кавказа простирается от 1000 до 2000 м над ур. моря (Долуханов, 1989), а на северном макросклоне от 1100 до 1950 м (Орлов, 1951). Результаты наших наблюдений о вертикально-поясном распространении пихтовых лесов несколько отличаются от этих показателей (рис. 4).
И на них мы остановимся более подробно в следующей главе. Ниже вкратце излагаются основные характерные особенности природных условий пояса пихтовых лесов.
Рельеф и геологическое строение
Рельеф в ареале пихты горный и сильно расчлененный. В орографическом отношении территория характеризуется, в основном, среднегорным рельефом с волнисто-увалистой, вогнуто-выпуклой, иногда и пенеплезиро-ванной, главным образом по северному макросклону, поверхностью водоразделов и довольно крупными, нередко с обрывистыми нижними частями,
2500
|-ПИХТА i -ЕЛЬ
5
77
0 БУК ^-КАШТАН
4 М
2000
^-ДУБ
Ф ГРАБ
V ТРАВЫ
КРИВОЛЕСЬЕ
500 >
Mr \Z Mr Mr Mr ЙГ МГ
Рис. 4. Растительные пояса Западного Кавказа:
1 - субтропических смешанных лесов; 2 - каштановых лесов; 3 - буковых лесов;
4 - пихтовых лесов; 5 - субальпийский; 6 - дубовых лесов; 7 - степной
склонами. Глубина расчленения, в зависимости от амплитуд неотектониче-ских поднятий, может достигать до 1700 м.
В геологическом строении здесь преобладают глинистые сланцы и песчаники юры. По крутым склонам в руслах рек, зачастую, обнажаются кристаллические породы триаса и палеозоя. Встречаются выходы массивных известняков триаса. Материнские породы - юрские глинистые и кристаллические сланцы, песчаники являются благоприятными для развития буково-пихтовых и пихтовых древостоев на бурых горно-лесных, преимущественно среднесуглинистых почвах различной мощности, степени ске-летности и увлажнения (Голгофская, 1967).
Для верхней части пояса пихтовых лесов характерен также высокогорный тип рельефа с резкими формами, обусловленными преобладанием । вердых кристаллических пород докембрийских сланцев и гранитов. Резко пересеченный рельеф высокогорья, с структурными обнажениями горных пород, каменистыми осыпями и скалами, обусловливает разорванность и фрагментарность лесной растительности. Особый интерес представляют массивы триасовых известняков, на которых развиваются пихтовые леса с богатой эндемичной флорой и эндемичными лесными биогеоценозами. На этих массивах широко распространены карстовые явления. Значительной интенсивности достигают карстовые процессы в районах Лагонакского плато и массивов Фишт и Оштен на Северном Кавказе и в районах Гагринского и Бзыбского хребтов на южном макросклоне. Здесь много котловин, воронок, пещер, колодцев и карстовых долин в верхнеюрских известняках (Гвоздецкий, 1963). Карстовые регионы бедны водными ресурсами, реками, и лесная растительность в них является важнейшим и основным регулирующим фактором гидрологического режтаа.
В целом гидрологическая сеть территорий пихтарников Кавказа богата и разнообразна. Представлена онашюгочисленными реками, озерами, источниками. Гидрологический ре^ейм в них во многом определяется состоянием лесов на водосборныхрйлонах.
Существенны в этих^яесах процессы речной эрозии и денудации в условиях избыточНипГ^влажнения. Однако лес препятствует развитию этих процессов. Роль леса здесь, как подчеркивал Г.Н. Высоцкий (1950), неоспорима. Ведущая роль леса в сокращении поверхностного стока воды и предотвращения эрозий почвы доказана многочисленными исследованиями ученых разных стран в различных климатических и почвенных условиях (Китридж, 1951; Ткачейко, 1955; Гулисашвили, 1956; Rubner, 1960; Молчанов, 1960; Беленко, 1968; Протопопов, 1975; Побединский, 1979; Смит, 1985). По нашим исследованиям (Бебия, 1973), в девственных пихтовых древостоях поверхностный сток воды и смыв почвы минимальный, эрозия почвы практически отсутствует.
В геологическом и историческом отношении формирование рельефа носит динамичный характер и это существенно влияет на динамику растительности.
Современное развитие рельефа в разных частях ареала пихты, в зависимости от неоднородности физико-географических процессов, происходит неодинаково. Например, в нижней части пояса пихтовых лесов развитие рельефа протекает менее интенсивно, чем в верхней, на южном склоне гор - более интенсивно, чем на северном. На известняковых массивах развитие рельефа протекает интенсивнее по сравнению с неизвестняковыми.
На характер и интенсивность развития рельефа существенное влияние может оказывать хозяйственная деятельность человека, например руб
ки леса, пастьба скота, лесные пожары и пр. Поэтому правильное ведение лесного хозяйства в пихтарниках имеет огромное значение для предотвращения негативных природных процессов, для сохранения устойчивой позиции пихты в коренных типах лесов.
Климат
Важнейшими природными факторами развития пихтовых лесов являются климатические, главным образом, гидротермические условия их произрастания (Гулисашвили, 1964; Franklin, 1981). Известно, что горные поднятия обусловливают вертикальную высотную зональность климата (Берг, 1938; Докучаев, 1948). На Кавказе климат природной зоны темнохвойных лесов из пихты и ели, по В.З. Гулисашвили (1964), может быть отнесен к типу достаточно влажного с коротким прохладным летом и холодной зимой или к умеренно холодному типу климата по Г. Майру (Маут, 1925) и Г. BanbTepy<(Walter, 1970).
Вместе с тем, климатические условия в области произрастания темнохвойных лесов Кавказа отличаются значительным разнообразием, резко меняясь с востока на запад. Меняются они на различных высотных уровнях, в зависимости от рельефа и морфологии гор (Орлов, 1951). Этим следует объяснить большое многообразие лесов по составу, строению, производительности, типам леса.
Однако, несмотря на различие климатических условий, влияющих на характер роста пихты и ели, на развитие лесов, климат природной зоны темнохвойных лесов Кавказа в целом характеризуется холодной зимой с продолжительным и устойчивым снежным покровом и морозами, достигающими -32°. Лето короткое, прохладное. Средняя температура января ниже нуля и колеблется от -3,2 до -7,4°. Средняя температура июля 12,8-16,8°. Продолжительность вегетационного периода три - четыре месяца. Количество осадков от 700 до 2500 мм в год. Распределение осадков по месяцам неравномерное, с минимумом в зимние месяцы. Высокой является также относительная влажность воздуха от 71 до 81 % (Гулисашвили, 1964). Для зоны характерны туманы. Некоторые исследователи считают туманы одной из важнейших характерных особенностей произрастания пихтовых лесов. Подчеркивается приуроченность зоны пихты к зоне туманов (Толмачев, 1954; Колаковский, 1961; Wang, 1968; Карпов, 1969; Спурр, Барнес, 1984; Nakamura, 1992). Однако, на наш взгляд, такое мнение справедливо лишь в отношении пихтарников, на произрастание которых оказывает существенное влияние океанический климат.
Почвы
Почва, наряду с климатом, - важнейший экологический фактор, определяющий существование и характер развития леса. Составляя неотъемлемую часть леса как биогеоценоза, она находится в постоянном взаимодействии с лесом на уровне лес - почва - лес. Почва оказывает влияние на состав растений лесного сообщества - от напочвенного покрова до древо-сгоя. От нее зависит возобновление леса, а также распространение древесных пород в пространстве и во времени в пределах их естественного ареала.
Влияя на рост и развитие леса, на состав и строение, на лесообразо-luiтельные процессы в целом, почва в значительной мере определяет также количественную и качественную тфо^Ктивность леса. Почва может быть использована в качестве одноГбиз ключевых диагностических признаков при выделении типов лер£
Почвы под лесами с преобладанием пихты кавказской изучены неодинаково. Опубликованные материалы по их характеристике указывают па неоднородность почвенного покрова в различных частях ареала, но не отражают полной, целостной картины. Вопросы географии, классификации и генезиса этих почв все еще дискутируются. Существующие сложности и противоречия в этих вопросах объясняются существенными различиями ведущих факторов почвообразования в ареале пихты. Эти факторы - сильная расчлененность территорий, разнообразие литологического состава горных пород, варьирование климатических условий, активность геомор-(|юлогических процессов, состав и характер растительного покрова и другие.
Наиболее полно изучены почвы темнохвойных лесов Северного Кавказа, в том числе почвы под пихтовыми лесами. Им посвящены работы С Л. Захарова (1914, 1948); Л.И. Прасолова (1929); С.А. Захарова, Н.А. Буша (1929); КЛ Богатырева (1946); МВ. Герасимова (1948); СВ. Зонна (1950); А.Я. Орлова (1951); К.Ю. Голгофской и др. (1967); В.И. Веселова (1973); Я.Г. Горчарука (1978).
Более или менее обобщенные сведения о почвах пихтарников южного макросклона находим в работах Л.И. Прасолова (1947); М.Н. Сабашвили (1948), В.М. Фридланда (1956); В.А. Ссорина и др. (1956); В.З. Гулисашвили (1964); Т.Ф. Урушадзе (1987).
Наши исследования почвенного покрова проводились при закладке пробных площадей для изучения лесообразовательных процессов в пихтарниках как на южном, так и на северном макросклоне Кавказа. На пробной площади закладывалось по одному почвенному разрезу в местах, характерных для пробы по типу леса. Описание почвенных разрезов производили по методике С.В. Зонна (1957), с некоторыми поправками. Всего заложено 18 почвенных разрезов.
Для выявления механических, физико-химических свойств почвы из почвенных разрезов брались образцы, которые затем направлялись в почвенную лабораторию Тбилисского института леса для проведения анализа почв. В данной работе физико-химические свойства почв нами охарактеризованы по сводкам аналитических данных Тбилисского института леса (Урушадзе, 1987), а также использованы материалы исследований А.Я. Орлова (1951) и других ученых по почвам пихтарников Северного Кавказа.
Совокупный анализ опубликованных работ, а также результаты наших исследований позволили несколько по-новому осветить основные закономерности формирования почвенного покрова в пихтарниках Кавказа и его роль в лесообразовательном процессе.
Известно, что под темнохвойными лесами формируются бурые лесные почвы. Эти почвы являются наиболее распространенными горными почвами Кавказа. Различные варианты этих почв имеют широкое распространение также под темнохвойными и буковыми лесами в горных системах Евразии, Дальнего Востока, Японии, Северной Америки (Tavernier and Smith, 1957; Розанов, 1961; Васильев и др.,1961; Иванов, 1967; Фирсова и др., 1972; Nagatsuka, 1975; Endo, 1977; Зонни др., 1978; Kaji, 1982; Jang, 1996).
Как показали исследования, под пихтовыми лесами Кавказа формируются, главным образом, бурые лесные кислые и бурые лесные псевдо-оподзоленные почвы.
Морфологическое строение этих почв можно рассмотреть на примере нескольких типичных разрезав, заложенных нами в наиболее распространенных типах леса в пихтарниках Абхазии.
Разрез 1 (7) заложен в пихтарнике овсяницевом типе леса 1 класса бонитета. Склон северо-восточной экспозиции, 20° крутизны, 1100 м над ур. моря. Состав древостоя 9Пх1Бк, ед. Кл, полнота 0,8, сомкнутость полога древостоя 0,8. Травяной покров: овсяница горная, ясменник, ежевика, папоротник. Проективное покрытие 0,6.
Ао - 0-3 см: Неразложившаяся подстилка из хвои, веточек. Переход четкий.
А1-3-12 см: Буровато - темно - коричневый тяжелый суглинок, свежий, мелкозернистый - порошистый, рыхловатый, много корней. Включение сланцев до 25 % по объему. Переход четкий.
В1 - 12-32 см: Буровато - светло - коричневый тяжелый суглинок, комковато-порошистый, несколько уплотненный, свежий. Включение сланцев до 35 %. Много корней. Переход постепенный.
В2 - 32-62 см: Буровато - светлая с желтоватым оттенком, тяжелосуглинистая, комковато-зернистая, среднеуплотненная, влажная. Встречаются корни. По ходам корней гумусовые затеки. Включение сланцев до 35 %. Переход нечеткий.
С^б2-95 скс^Р^и^^светло-желтьш оттенком тяжелый суглинок, / зернисто-комковатый, слабоуплотненный, Включение
/ сланцев 40-50% по объему. С глубины 100 см сплош-
ной пласт сланца.
Вскипание от НС1 не наблюдается. Почва - типичная бурая лесная кислая.
Разрез 2 (12) заложен в буково-пихтовом падубовом типе леса 1 класса бонитета. Склон 25°, юго-западной экспозиции, 1300 м над ур. моря. Состав древостоя 7ПхЗБк. Полнота 1,08, сомкнутость 0,9. Подлесок - падуб, реже черника, проективное покрытие 0,6, неравномерное. Травяной покров: овсяница горная, ежевика, папоротник, проективное покрытие 0,4.
Ао - 0-5 см: Неразложившаяся подстилка из хвои, веточек. Переход четкий.
А1 - 5-20 см: Буровато-светловатая, тяжело-суглинистая, зерни-сто- порошистая, свежая, слабоуплотненная. Много корней. Мелкоскелетная, скелет до 40 % объема. Переход постепенный.
Bi - 20-46 см: Буровато-светлая, глинистая, влажная, порошистая, среднеуплотненная. Много корней. Скелет составляет до 60 % объема. Переход постепенный.
В2 - 46-80 см: Буроватая, мелкоскелетная до 60 % по объему. Глинистая, порошистая, влажная, среднеуплотненная. Мало корней. Переход постепенный.
ВС - 80-100 см: Буроватая со слабым коричневым оттенком, глинистая, сильно мелкоскелетная, до 80 % по объему, влажная, среднеуплотненная. Переход постепенный.
С - 108-140 см: Сильноскелетная, содержание мелкозема незначительное 20-25 % по объему. Материнская порода -глинистый сланец в форме пластинчатых обломков.
Вскипание от НС1 не наблюдается. Почва - типичная бурая лесная кислая.
Разрез 3 (24) заложен в буково-пихтовом ясменниковом типе леса 1 класса бонитета. Склон южной экспозиции, крутизна 10°. Высота 1000 м над ур. моря. Состав древостоя 6Пх4Бк. Полнота 1,1, сомкнутость полога
древостоя 0,7. В покрове: ясменник, овсяница горная, кислица, единично папоротник, ежевика. Покрытие 0,6.
Ао - 0-2 см: Неразложившаяся подстилка из хвои, листьев, орешков бука, веточек. Переход четкий.
А1 - 2-10 см: Буровато-серая, суглинистая, зернистая, рыхловатая, свежая. Корней много. Переход четкий.
В1 - 10-35 см: Бурая, глина комковато-зернистая, уплотненная, свежая. Корней много. Включение сланцев до 25 % по объему. Переход четкий.
В2 - 35-60 см: Бурая с желтым оттенком, глинистая, уплотненная, свежая, комковато-зернистая. Наблюдаются гумусовые затеки. Включение сланцев до 30 % по объему. Переход постепенный.
С-60-105 см: Бурая, светловато-желтая. Тяжелосуглинистая, комковато-зернистая, плотная, свежая. Включение сланцев до 50 % по объему. Переход постепенный.
Вскипание tr НС1 не наблюдается. Почва - типичная бурая лесная кислая.
Из приведенных описаний почвенных разрезов, расположенных на гипсометрических отметках от 1000 до 1700 м, следует, что значительных различий в геоморфологии описанных почв не наблюдается. Характерным для всех разрезов является слабая выраженность гумусового горизонта, бурая окраска всего профиля, сильная скелетность, комковато-порошистая структура и среднеуплотненное сложение. Загруженность корней падает с глубиной 50-60 см. Ниже 80 см корни практически не встречаются. Такие почвы формируются, главным образом, в свежих типах леса - в пихтарнике ясменниковом, подлесниковом, падубовом, черничниковом, ежевичном. Характеризуясь умеренным увлажнением всего профиля и хорошей дрени-рованностью, бурые лесные почвы кислые, имеют наиболее оптимальные условия для формирования высокопродуктивных лесных биогеоценозов. На таких почвах произрастают высокопроизводительные древостои пихтарников с запасом более 700 куб. м на га.
По морфологическому строению к характеру формирования бурые лесные псевдопоодзоленные почвы несколько отличаются от вышеописанных бурых почв. Основное отличие заключается в светлой, белесоватобурой окраске верхнего горизонта почвы. Однако в нижних отметках разреза окраска становится несколько темнее. Дифференциация почвы по генетическим горизонтам слабо выражена.
Приводим описание морфологических особенностей бурой лесной псевдооподзоленной почвы на примере разреза 17 (76). Он заложен на Северном Кавказе в районе пастбища Абаго Кавказского биосферного запо
ведника, на высоте 1750 м над уровнем моря. Склон северо-западной экспозиции, 10° крутизны. Тип леса - пихтарник папоротниковый 1 класса бонитета. Состав древостоя 8Пх2Бк, ед. Кл. Полнота древостоя 0,8, сомкнутость полога 0,7. Напочвенный покров развит, состоит из папоротника мужского, трахистемона восточного, ясменника, герани Роберта, кочедыжника женского. Проективное покрытие 0,8.
Ао - 1-2 см: Лесная подстилка из опада хвои и веточек.
А - 2-20 см: Светло-бурый, белесоватый суглинок, слабовыражен-
ной комковато-порошистой, рыхлой структуры. Влажный. Включения - скелетные обломки песчаника. Переход постепенный.
В - 20-42 см: Несколько светлее, тяжелосуглинистая, влажная, слабо уплотненная, порошистая. Включения - обломки песчаника различной величины. Переход слабо выражен.
ВС - 42-76 см: Бурый тяжелый суглинок, с ржавыми пятнами, влажный, крупно-комковатый, уплотнен. Крупные камни песчаника. Переход выражен.
С - 76-100 см: Тускло-буроватый тяжелый суглинок. Влажный, уплотнен. Включение из крупных камней песчаника.
Вскипания от воздействия НС1 не наблюдается.
Обращает на себя внимание с глубины 30 см значительное количество конкреций полуторных окислов в виде пятен черного и ржавого цвета. Максимум корней наблюдается до глубины 45 см, ниже 70 см корни редки. По всему профилю встречаются скелетные обломки песчаника. Особенно их много с глубины 70 см.
К отличительным особенностям бурых лесов псевдооподзоленных почв следует отнести также приуроченность их к верхней части пояса пихтарников с высоты 1600 м над уровнем моря и сравнительно ограниченное распространение их в ареале пихты.
Развиваются бурые лесные псевдооподзоленные почвы, в основном, во влажных типах леса, в частности, пихтарнике папоротниковом. В этих почвах наблюдается постоянное увлажнение, зачастую переувлажнение верхних горизонтов А - В, которое способствует иногда поверхностному оглеению. Создаются несколько неблагоприятные условия для естественного возобновления и формирования высокопроизводительных древостоев пихты. Участие сопутствующей породы ели в составе древостоев заметно возрастает.
В целом в физико-химическом отношении бурые лесные почвы под пихтовыми лесами характеризуются сильнокислотной реакцией (PH составляет от 4,0 до 7,0) и ненасыщенностью почвенного поглощающего
комплекса (50-30 %). Почвы менее оглеены, высоко и глубоко гумусированы. Количество гумуса в горизонте А колеблется от 5 до 19 %, а на глубине 100 см до 5 %.
Распределение SiCh и Р2О3 по профилю указывает на некоторое их перераспределение.
Характеризуясь высокими благоприятными физико-химическими свойствами и достаточным увлажнением, бурые лесные почвы на отметках 1200-1500 м над уровнем моря способствуют формированию наиболее производительных пихтовых, буково-пихтовых древостоев 1 и 1а классов бонитета с запасом древесины до 1500 куб. м на га.
Основные закономерности широтного и высотного распространения пихтовых лесов
Большинство исследователей пихтарников Кавказа рассматривают их вместе с еловыми лесами, объединяя обе лесные формации в единую комплексную формацию темнохвойных лесов. И это не случайно. Экология этих лесообразователей во многом сходна. Развитие их связано с общей высотно-климатической зоной, поэтому ареалы распространения пихты кавказской и ели восточной несколько совпадают, но не полностью (рис. 5, A-В). Различаются и основные закономерности распространения пихтовых и еловых лесов.
В биогеоценотическом лесоводственно-экологическом отношении пихтовые леса существенно отличаются от еловых лесов. Они требуют дифференцированного подхода и проведения разных систем лесоводствен-ных мероприятий при решении практических задач. Поэтому пихтовые леса следует изучать отдельно от еловых лесов. Это важно и потому, что пихтовые леса, занимая третье место по своей площади и хозяйственной значимости после буковых и дубовых лесов на северном макросклоне Кавказа, и второе после буковых лесов на южном макросклоне, играют значительную ландшафтообразующую и экологическую роль в экосистеме всего Западного Кавказа.
В экологическом отношении пихта по сравнению с елью характеризуется большей мезофильностью и мезотрофностью, несколько большей тепл о любив остью. В силу этого ареал ели, по сравнению с пихтой, сдвинут к востоку (рис. 5, В).
Если сравнить крайние пункты встречаемости пихты и ели как видов, то они почти совпадают. Но если исходить из наличия лесов, то древостои с преобладанием пихты расположены в западной части, а ели в восточной - в Южной и Восточной Грузии, где количество осадков значительно меньше. Такая же закономерность распространения пихты и ели наблюдается и на северных склонах западной части Большого Кавказа (Гулисашви-
Рис. 5. Ареалы видов: А - Abies nordmanniana (Stev.) Spach, В - Picea orientalis (L.) Link, ли и др., 1975). Таким образом, границы пихтовых лесов на Кавказе проходят западнее границы еловых. Однако границы ареала пихты шире, чем границы лесов с ее господством.
На Северном Кавказе единичные деревья и группы пихты встречаются в бассейнах рек: Пшеха, М. Лаба и Б. Лаба. Восточнее реки Уруп площадь пихтовых лесов резко уменьшается (Орлов, 1951). Здесь пихта встречается преимущественно с елью в виде единичных примесей к еловым лесам (Растительность европейской части СССР, 1980). В верховьях бассейна реки Кубани пихта полностью выклинивается. Самое восточное местонахождение пихты по северному макросклону Кавказа, по В.З. Гулисашвили (1964), отмечено по реке Тютюнсу бассейна реки Терек.В Закавказье, по нашим наблюдениям, обширные леса с господством пихты произрастают от района Сочи (верховья реки Бзыч) через Абхазию до бассейна реки Рио-ни включительно. Далее, на востоке преобладают еловые леса. За пределами Колхиды по Большому Кавказу пихта встречается как примесь в еловых лесах, в глубине верховий ущелий, почти до бассейна реки Большой Лиах-ви. Последний пункт произрастания пихты отмечен в Чапарукском ущелье (Махатадзе, 1966).
Обширные леса с господством пихты произрастают по северным склонам Аджаро-Имеретинского хребта и по северным склонам Шавшет-ского хребта, по водоразделу которого проходит государственная граница Грузии с Турцией. Южная и юго-западная граница ареала пихты, как и ели, переходит на территорию Турции; в пределах Колхидского рефугиума пихта произрастает на Понтийском хребте (см. рис 5).
В филогенезе формирование и трансформация биоэкологических, лесоводственных, фитоценотических свойств пихты кавказской происходили в горных условиях. Не только пихта кавказская, но и весь род пихты характеризуется однотипной экологией, приуроченностью к горным образованиям, и все пихты, как правило, являются горными лесообразующими породами. Этим и объясняется, что оптимальные условия произрастания и господства пихты кавказской находятся в верхнелесном горном поясе от 1000 до 1800 м над уровнем моря (см. рис. 4), где количество осадков составляет не менее 750-900 мм при оптимуме 1800-2500 мм с летним максимумом. По нашим исследованиям, в Абхазии пояс пихтовых лесов простирается от 900 до 1800 м надбровием моря. Небольшие участки пихтарников здесь по ущельям рек спускаются и до 600 м. На Кавказе пихтовые леса не образуют сплошного непрерывного пояса. Местами они замещаются буковыми лесами. Пояса пихтовых и буковых лесов весьма близкие и сходные в био-экологическом отношении. По нашим исследованиям, обе формации, пихтовая и буковая, близки в ценотическом отношении. Общность их ценотического состава превышает 70 % (Бебия, 1987). Бук, как и пихта, является мезофильной, мезотрофной, горной лесообразующей породой. Обращает на себя внимание и то, что ареал бука на Кавказе включает полностью ареал пихты.
Однако все еще остаются неясными дискуссионными вопросы - почему не происходит расширение естественного ареала пихты кавказской в районы произрастания буковых лесов и, почему в пределах своего ареала она не везде образует сплошной пояс и местами замещается буком.
Объяснить эти явления низкой конкурентоспособностью пихты по отношению к буку и другим видам в поясе пихтовых лесов не всегда удается. По нашим многолетним наблюдениям, позиция пихты кавказской в пределах ареала лесов с ее господством достаточно устойчивая. Во всех типах лесного биогеоценоза смена ее сопутствующими видами елью, буком, грабом и другими, о чем будет изложено более подробно в разделе, посвященном возобновлению пихты, не наблюдается.
Объяснить такую загадку природы эдафическими факторами и влиянием подстилающей материнской породы на пихту оказалось также недостаточным. По А.Г. Долуханову (1940), характер распространения пихты кавказской можно объяснить экологическими факторами, в частности, характером увлажнения воздуха, конкретных условий местопроизрастания и
влиянием Черного моря. По его мнению, ближайшие к морю склоны, обращенные непосредственно к Колхидской низменности, заняты в основном буковыми лесами, и позиция пихты в этих условиях незначительна. С удалением от моря и ослаблением влияния морского климата в глубине ущелий, как бы дугой, окаймляя буковые леса, на тех же гипсометрических отметках произрастают, в основном, буково-пихтовые и пихтовые леса. Действительно, такая закономерность произрастания пихты на южном макросклоне Западного Кавказа наблюдается. Однако, по нашему мнению, не только сильным и прямым влиянием морского климата следует объяснять такую инверсию пихты в пределах ее ареала на Кавказе. К примеру, как объяснить, что высокобонитетные пихтовые леса произрастают и на южном склоне известнякового Гагринского хребта, расположенного в непосредственной близости от моря и находящегося под прямым и сильным влиянием морского климата. Немаловажную роль в размещении пихтовых лесов играют отличительные геоморфологические особенности того или иного хребта и связанные с этим специфические условия местопроизрастания. На северном макросклоне Кавказа размещение пихтовых лесов, к примеру, находится в непосредственной зависимости от влияния сухого климата степей и лесостепей Предкавказья, Кума-Манычской впадины (Орлов, 1951).
По В.З. Гулисашвили (1940, 1951), более широкому распространению пихты и ели на Кавказе препятствуют не только недостаток тепла и влаги, но и высокие летние температуры и теплые зимы. Это подтверждается также и нашими наблюдениями. К примеру, в условиях влажного и теплого климата, в Сухумском субтропическом дендропарке, расположенном на высоте 15 м над уровнем моря, пихта кавказская, высаженная на склоне восточной экспозиции, крутизной 30° на кислых дренированных почвах достигла предельного роста в возрасте 85 дет и погибла. Она имела высоту всего 17,5 м и диаметр 100 см. Дерево плодоносило и давало полнозернистые семена. Анализ хода роста ствола показал, что дерево до 60 лет росло интенсивно, достигнув диаметра 80 см. Текущий прирост по диаметру был выше среднегодичного и составлял 0,6-1,0 см. В последующие годы текущий прирост стал падать и к 80 годам он составлял 0,3-0,2 см. Дерево имело рыхлую и низкокачественную древесину. В естественных условиях произрастания пихта кавказская достаточно долговечна и своего предельного биологического возраста она достигает не ранее, чем к 250 годам.
Представляют интерес и результаты другого опыта. На экспериментальных участках Абхазской лесной опытной станции в г. Очамчира, расположенной на высоте 12 м над уровнем моря, была осуществлена посадка монокультуры пихты кавказской в количестве 150 саженцев, с размещением 2,5x2,5 м. До 20-летнего возраста деревья пихты росли нормально, их
средняя высота составила 7,2 м, с колебанием от 5 до 8,2 м. В возрасте 25 лет деревья стали высыхать. В возрасте 38 лет в монокультуре сохранилось лишь 7 экземпляров деревьев пихты плохого жизненного состояния. В то же время, по данным Бейна (Bean, 1980), в Англии самый старый интродуцированный экземпляр пихты кавказской, произрастающий в графстве Виклов во влажном умеренно-холодном климате, в возрасте 133 года имел в высоту 40,8 м, диаметр 104 см и продолжал расти нормально.
Несомненно, все эти данные указывают на отрицательное и ограничивающее влияние высоких летних температур и теплых зим на распространение пихты в более низкие поясные отметки, за пределами ее нижней границы.
Существенным фактором, ограничивающим распространение пихтовых лесов, также следует считать и весенние заморозки, которые повреждают всходы пихты сильнее по границам ареала, чем в оптимальных условиях ее распространения. Например, в верхнем пределе своего распространения пихта достигает субальпийского пояса до 2000 м, произрастая небольшими группами или единичными экземплярами в буково-березовом криволесье. Однак? здесь она часто повреждается морозом, имеет сильно сбежистый ствол с уродливой кроной. Характерно, что ограничивающим фактором распространения пихты белой вверх в горы является также недостаток тепла (Комендар, 1966; Leibundgut, 1984). По К. Рубнеру (Rubner, 1969); для нормального роста и развития пихты белой необходимо не менее трехмесячного безморозного вегетационного периода со средней температурой не ниже + 13°.
В заключение следует подчеркнуть, что характер высотного и широтного естественного ареала пихты определяется не столько влиянием на ее развитие отдельных ведущих факторов среды, сколько их комплексным воздействием на нее в конкретных условиях местопроизрастания.
Видовой состав пихтовых лесов и взаимоотношение пихты с сопутствующими породами
В целом, по всему своему ареалу, пихта растет вместе с буком восточным. На западе ареала она растет как примесь в буковых лесах, затем по направлению к востоку примесь пихты увеличивается, в центральной части ареала уже формируются чистые пихтовые леса. В восточной части она растет вместе с елью восточной, постепенно убывая в восточном направлении и участвуя как примесь в древостоях еловых и елово-буковых лесов. Кроме бука и ели, в пихтовых лесах сопутствующими породами являются также клены: остролистый, явор, Траутфеттера, ильм шершавый, липа кавказская, тис ягодный, сосна Коха, дуб грузинский, дуб скальный, береза Литвинова и другие (см. табл. 1). Все эти породы являются реликтами, многие из них - эндемами Кавказа и Колхиды.
Климатические факторы играют большую роль во взаимоотношениях пихты, бука и ели. Так, пихтовые леса приурочены к районам, характеризующимся большим количеством осадков и повышенной влажностью воздуха. В силу этого пихтарники распространены, в основном по бассейнам рек. В то же время, пихта, бук и ель, как уже отмечалось, обладают рядом общих биоэкологических черт. Длительное сосуществование их привело к созданию устойчивых смешанных буково-пихтовых, елово-пихтовых лесов, которые характеризуются многими общими экологическими, ценотиче-скими чертами и флористическим составом. Поэтому на Кавказе пихта образует как чистые, так и смешанные с буком и елью насаждения высокой производительности. При этом по флористическому составу пихтовые леса ближе к буковым, чем к еловым. К примеру, такие типологические эдифи-каторы, как рододендрон понтийский, лавровишня лекарственная, падуб колхидский, характерные для пихтовых, пихтово-буковых и буковых лесов, практически не встречаются в еловых лесах.
Важнейшим фактором, обусловливающим географию пихты на Кавказе и ее взаимоотношение с сопутствующими породами, являются ее био-экологические особенности, выработанные в процессе эволюции. Некоторые из этих особенностей необходимо здесь подчеркнуть. Так, пихта является холодоустойчивой породой, вследствие чего основные массивы лесов с ее господством размещаются в верхней горной зоне и достигают границы субальпийских лесов. С приближением к субальпийской границе в составе пихтовых лесов существенную роль начинают играть сопутствующие лиственные породы: бук, клен остролистый, явор, Траугфеттера, береза Литвинова, береза Медведева. Появляется сосна Коха.
На южном макросклоне, в частности в Рицинском национальном парке, по склонам юго-восточной экспозиции, на маломощных щебнистых почвах, на ограниченных площадях пихта образует иногда низкобонитет-ные смешанные с сосной Коха, дубом иберийским и осиной древостои или отдельные их группировки. В суховатых условиях местопроизрастания грабинник восточный, вяз шершавый, клен остролистный, а на свежих и влажных условиях - граб кавказский, ольха клейкая, лучше возобновляются, чем пихта, и формируют отдельные группировки из лиственных деревьев. Однако этот процесс временный и обратимый. Позже под пологом лиственных деревьев пихта начинает постепенно поселяться и возвращать свои позиции в древостое. Но на это требуется большой период времени, несколько десятков лет.
Пихта может произрастать на среднеувлажненных почвах, но избыточного увлажнения почв не переносит. На таких участках пихту может сменить ольха, граб, реже бук.
На глубоких свежих почвах пологих склонов пихта развивает толстый, но короткий стержневой корень и несколько глубоко залегающую
мощную горизонтальную корневую систему (рис 6, А-П). Корни проникают в почву до 1,3 м глубины. Но от всей массы корней 47 % приходится на глубину до 0,5 м, примерно 33 % - на глубину 0,5-0,9 м и лишь около 20 % корней распространено на глубину 0,9-1,3 м. В этих же условиях местопроизрастания бук развивает более поверхностную корневую систему, чем пихта (рис 6, A-I).
Глубина проникновения корней составляет 0,6 м. Примерно 77 % массы корней расположены на глубине до 0,3 м и остальная - 23 % - приходится на глубину 0,3-0,6 м. Из этих данных следует, что пихта и бук являются взаимодополняющими породами по отношению к почвенным условиям. На таких глубоких почвах сопроизрастание бука с пихтой в виде примесей вполне закономерно. Пихта, обладая относительно более глубокой корневой системой и более глубоким размещением биологически активных корней, чем у бука, способствует развитию смешанных буковопихтовых лесов. Древостои, осваивая разные горизонты почвы, способствуют высокой производительности такого леса, в котором сопроизрастание двух мощных эдификбгоров носит устойчивый характер. Кроме того, пихта в таких древостоях ветроустойчива. С этой точки зрения интересны также исследования В.И. Гниденко (1967) и Я.А. Сабана (1982). По их данным, темнохвойно-буковые древостои Карпат биологически более устойчивые и продуктивные, чем чистые буковые и чистые хвойные.
В более сухих условиях местопроизрастания в пихтарнике овсянице-вом, на более мощных, скелетированных почвах пихта развивает сравнительно глубокую, но поверхностную корневую систему (рис 6, Б - Ш-IV). Хотя корни проникают в почву до 1,1 м глубины, но основная биологически активная масса их, 87,7 %, распространена на глубине 0,5 м. В таких условиях местопроизрастания обычно формируются чистые пихтовые древостои, с единичной примесью в их составе дуба скального на Северном Кавказе и дуба иберийского на южном макросклоне Кавказа. На таких почвенных условиях пихта бывает ветровальной. Особенно она страдает вет-ровальностью в чрезмерно изреженных рубками древостоях. На таких участках позиция пихты ослаблена по сравнению с сопутствующими лиственными породами (дуб, ильм, клен). Формируется производный тип леса.
Все эти особенности пихты чрезвычайно важны, и необходимо учитывать их при решении практических задач в лесу.
A
Б
Рис. 6. Распределение корней (%) по почвенному профилю (м) пихты кавказской (светлый прямоугольник) и бука восточного (затемненный) в пихтарнике (ясменниково-кисличниковом) 1а класса бонитета (А) и пихтарнике овсяницевом II класса бонитета (Б); I - бук: диаметр 104 см, высота 40 м возраст 230 лет; П - пихта, диаметр 80 см, высота 43 м, возраст 240 лет; III -пихта, диаметр 32 см, высота 26 м, возраст 115 лет; IV - пихта: диаметр 90 см, высота 37 м, возраст 270 лет
Лесорастительное районирование области распространения темнохвойных лесов
Как отмечалось выше, природные условия в области произрастания темнохвойных лесов Кавказа отличаются значительным разнообразием, меняясь с востока на запад, на различных высотных уровнях, в зависимости от морфологии гор и других факторов. Отсюда и большое многообразие лесов по составу, строению, производительности, типам леса. В связи с этим изучение растительного покрова, в частности лесообразовательных процессов, в пихтарниках целесообразно осуществлять в рамках определенных лесорастительных условий. Это необходимо и потому, что природное лесорастительное районирование облегчает планирование основных лесохозяйственных мероприятий (рубки главного пользования, подбор ассортимента пород при лесовосстановительных работах и другие), а также рациональное использование и охрану лесов.
В нашем представлении лесорастительный район - это такая территория, которая бол?е или менее однородна в климатическом, почвенном и растительном отношении. Следовательно, в основе лесорастительного районирования лежит геоботаническое районирование, которое лучше всего отражает климат и почву.
Практически все исследователи лесов Кавказа районирование области распространения пихтовых лесов рассматривают в комплексе с темнохвойными (пихтовыми и еловыми) лесами.
Следуя данному принципу, мы, при составлении лесорастительного районирования темнохвойных лесов, старались, прежде всего, обратить особое внимание на характер распространения пихтовых лесов. В ареале темнохвойных лесов, в зависимости от определенных природных условий местопроизрастания (лесорастительный район), охарактеризована ценоти-ческая роль пихты и ели, их взаимоотношение по отдельным районам.
Наиболее удачное, на наш взгляд, лесорастительное районирование темнохвойных лесов Кавказа выполнено Л.Б. Махатадзе (1966), в котором область распространения темнохвойных лесов расчленена на одиннадцать лесорастительных районов. В нем были учтены районирования, составленные ранее Н.И. Кузнецовым (1909), Я.С. Медведевым (1915), Р.А. Еленев-ским (1940), А.А. Гроссгеймом (1948), А.Я. Орловым (1951), В.З. Гулисашвили (1956), А.А. Колаковским (1961), В.Н. Овсянниковым (1964). И все же, лесорастительное районирование Л.Б. Махатадзе нельзя считать достаточно полным. В него не включена, например, понтийская часть ареала (Сев.Анатолия) темнохвойных лесов за пределами административной границы Грузии, расположенная на территории Турции. Сев. Анатолия, безусловно, является составной частью Колхидской (Колаковский, 1961) или Эвксинской (Малеев, 1938) ботанико-географической провинции. В ней по
северным склонам Понтийского хребта, простирающегося на западе несколько дальше Трапезунда, широко представлены представители колхидской флоры, в том числе колхидских эндемов (Aquilegia colchica, Epimedium pubigerum, Quercus hartwissiana, Hedera colchica, Ilex colchica, Trachystemon orientalis, Ruscus colchicus и другие). Здесь достаточно хорошо развиты темнохвойные леса из пихты кавказской и ели восточной (Davis, J965). С нашей точки зрения, Сев. Анатолия должна быть выделена в отдельный геоботанический (лесорастительный) район темнохвойных лесов. В таком случае, область распространения темнохвойных лесов Кавказа составляет не одиннадцать лесорастительных районов, как у Л.Б. Махатад-зе, а двенадцать (рис. 7).
Ниже приводим описание всех 12 лесорастительных районов темнохвойных лесов Кавказа по результатам наших исследований, а также по данным А.Я. Орлова (1951), В.З. Гулисашвили (1956), Л.Б Махатадзе (1967), К.Ю. Голгофской (1967).
1. Западный Северо-Кавказский район. В него входят бассейн реки Белой с ее крупными притоками Пшеха, Киша и Сахрай. Характеризуется сравнительно более мягким и сглаженным рельефом по сравнению с двумя
Рис. 7. Схематическая карта лесорастительного районирования темнохвойных лесов Кавказа
расположенными восточнее районами (2, 3), что обусловливает образование более мощного и развитого почвенного покрова. Климат мягкий и влажный, сказывается влияние Черного Моря. В этом районе преобладают буковые и смешанные буково-пихтовые, в меньшей степени чистые пихтовые леса. Ель восточная встречается лишь единично в лесах и небольшими группировками по каменистым склонам. Пояс буково-пихтовых, пихтовых лесов здесь проходит на высоте 1500-1900 м над уровнем моря. На северном склоне Большого Кавказа этот район наиболее богат элементами колхидской флоры. Хорошо представлены типы леса с колхидским вечнозеленым (рододендрон понтийский, лавровишня, падуб) и листопадным (черника кавказская) подлеском, папоротниковые, особенно из страусника, и разнотравные. Реже распространены овсяницевые типы леса.
2. Средний Северо-Кавказский район объединяет территорию бассейнов рек Малой и Большой Лабы. Преобладают пихтовые и реже еловопихтовые леса. Представителей колхидской флоры здесь значительно меньше. Лавровипфя, рододендрон встречаются единично или небольшими участками, преимущественно в западной части района. Пояс пихтовых лесов проходит примерно на высоте 1300-1800 м, со значительным преобладанием чисто пихтовых древостоев. Ель встречается местами как незначительная примесь древостоев по более сухим и каменистым склонам. Распространены здесь в основном ежевично-папоротниковые типы леса. По выпуклости рельефа, крутосклонам произрастают овсяницевые типы леса. Из-за сложных орографических условий пояса буковых и пихтовых лесов четко не выражены и местами глубоко вклиниваются друг в друга.
3. Восточный Северо-Кавказский район объединяет бассейны рек Урупа, Б. Зеленчука, М. Зеленчука, а также верховья рек Теберды и Кубани. Район отличается наиболее засушливым и континентальным климатом, по сравнению с двумя предыдущими районами, и пихтовые леса здесь произрастают в значительно меньшей степени. К востоку площади темнохвойных лесов резко уменьшаются, они замещаются сосновыми и березовыми лесами. Темнохвойные леса больше всего развиваются по ущелью реки Теберда, где климат более ровный и влажный. В данном лесорастительном районе встречаются ожиковые типы леса с господством пихты. Слабо развиты папоротниковые и разнотравные типы леса. Преобладают сухие и свежие типы леса. Значительно в меньшей степени встречаются представители колхидской флоры, чем представители бореальной флоры.
4. Туапсинско-Сочинский район - крайняя северо-западная часть Колхиды, начинается несколько западнее Туапсе и на юго-востоке доходит почти до Гагры. Район сравнительно обеднен элементами колхидской флоры. Вечнозеленый подлесок встречается в основном в юго-восточной части. Пояс пихтовых лесов от 900 до 1900 м над уровнем моря, характеризу-
стоя преобладанием пихтовых и буково-пихтовых лесов, елово-пихтовых лесов очень мало.
5. Абхазско-Сванетский район охватывает территорию Колхиды от реки Псоу на северо-западе до, примерно, бассейна реки Риони на юго-востоке. Здесь на значительной площади расположены известняки, которые изобилуют богатством колхидских реликтовых, эндемичных элементов флоры и растительности. Пояс пихтовых лесов, со значительным преобладанием чистых пихтовых и буково-пихтовых древостоев, проходит на высоте от 900 до 1900 м над уровнем моря. Здесь, восточнее реки Ингур встречаются также елово-пихтовые леса. Границы пояса буковых и пихтовых лесов часто заходят друг за друга. Хорошо развиты типы леса с вечнозеленым и листопадным колхидским подлеском (рододендрон понтийский, падуб, лавровишня, иглица, черника кавказская), а также папоротниковые, разнотравные, с субальпийским высокотравьем и менее - овсяницевые типы леса.
6. Рача-Имеретинский район занимает левобережье реки Риони от г. Кутаиси и выше, а также Малый Кавказ - восточную половину северных склонов Аджаро-Имеретинского хребта до горы Мепис-Цхаро на западе. Восточная граница района проходит по водоразделу Сурамского хребта. На северо-западе граничит с Абхазско-Сванетским районом, а на юго-западе с Гурийским. Пояс темнохвойных лесов на высоте 1500-1900 м представлен елово-пихтовыми и еловыми лесами. Чисто пихтовые и буково-пихтовые леса занимают сравнительно небольшие площади. Пояс темнохвойных лесов на Аджаро-Имеретинском хребте выражен хорошо, однако по южным склонам Главного Кавказского хребта он несколько оттеснен к верховьям ущелий. Местами пояса буковых и темнохвойных лесов заходят друг за друга. Темнохвойные леса произрастают в основном по глубоким ущельям и их верховьям. Район сильно обеднен элементами колхидской флоры. Це-нотическая позиция вечнозеленого подлеска здесь выражена слабее, чем в предыдущем районе, однако падуб развит хорошо и лучше, чем в соседних к западу более влажных районах.
7. Гурийский район, занимая территорию от северных и северо-западных склонов Аджаро-Имеретинского хребта до горы Мепис-Цхаро на востоке, является самым влажным районом распространения темнохвойных лесов, где количество осадков колеблется от 2500 до 2700 мм. в год. Пояс темнохвойных лесов на высоте 1500-1900 м представлен в основном пихтово-буковыми, пихтовыми и пихтово-еловыми лесами. Еловые леса занимают меньше площади и приурочены к каменистым и крутым склонам. Местами пояса темнохвойных и буковых лесов заходят друг за друга. Ценотическая позиция вечнозеленого подлеска (лавровишня, рододендрон понтийский, а также эндемичный вид для данного района - рододендрон
Унгерна) сильно выражена. Подлесок занимает обширные площади, образуя непроходимые заросли, препятствуя естественному возобновлению.
8. Аджарис-Цхальский район занимает бассейн реки Аджарис-Цхали. Осадков здесь меньше, чем в предыдущем районе, до 1400 мм. Пояс темнохвойных и темнохвойно-буковых лесов простирается от 1000 до 1850 м над уровнем моря. В составе темнохвойных лесов ель значительно преобладает над пихтой. Доминируют сухие и светлые типы леса, реже встречаются влажные типы леса. Ценотическая позиция вечнозеленого подлеска не существенна.
9. Месхетский район характеризуется сравнительно сухим и континентальным климатом, суровой зимой. Осадков выпадает не более 800 мм. По характеру растительности и развития темнохвойных лесов этот район сильно отличается от всех остальных. Представители колхидского элемента флоры здесь практически отсутствуют, сравнительно лучше представлен бореальный флористический элемент. Характерно также, что в этом районе почти полностью отсутствует бук. Хорошо развит пояс хвойных лесов из ели и сосны. К востоку сосновых лесов значительно больше. Небольшие участки пихтовых и пихтово-еловых лесов расположены вблизи Годерско-го перевала (урочище Бешуми) и выше курорта Абастумани.
10. Боржомский район охватывает склоны западной части Триалет-ских гор и расположен на стыке трех ботанико-географических районов Кавказа (по А.А. Гроссгейму, 1948): Колхиды, Юго-Западного и Центрального Закавказья. Для района характерно обилие бореальных элементов флоры. Сравнительно хорошо представлена и колхидская флора. Но ценотическая позиция вечнозеленого подлеска здесь не существенна и развит он лишь в глубине ущелий, в южной части района. Верхний предел пояса темнохвойных лесов проходит на высоте 1900 м. Пояс буковых лесов отсутствует. Еловые леса местами вклиниваются в дубовый пояс и спускаются до высоты 700 м (в Ахалдаба и Квишхети), достигая берега реки Куры. Площади чисто пихтовых и елово-пихтовых лесов здесь незначительны, приурочены к влажным ущельям бассейна реки Недзура и верховьям реки Гуджаретис-Цхали. Преобладают свежие типы леса с моховым покровом или мелкотравьем. Субальпийские хвойные и хвойно-лиственные типы леса с папоротниками и высокотравьем представлены хорошо.
11. Верхне-Карталинский район - самый восточный район области распространения темнохвойных лесов. Включает южные склоны Главного Кавказского хребта, а также северные склоны Триалетского хребта, входящего в систему гор Малого Кавказа. В этом районе площади темнохвойных лесов заметно убывают к востоку. На востоке темнохвойные леса представлены отдельными островками и группировками. Последние островки этих лесов находятся юго-западнее Тбилиси возле села Манглиси. Пояс темнохвойных лесов на высоте 1500-2000 м лучше всего выражен на Триа-
детском хребте в Верхне-Карталинском районе, где преобладают еловые, елово-буковые, реже елово-пихтовые леса. Небольшие участки чистых пихтовых лесов встречаются в верховьях рек Дзами, Тана и Тедзами. В темнохвойных лесах района преобладают мелкотравные и злаковые типы леса. Встречаются типы леса с блестящими мхами.
12. Северно-Анатолийский район занимает северные склоны Понтийского хребта, начиная от границ Аджарис-Цхальского и Месхетского районов на северо-востоке, заходит за Трабзон на юго-западе. В данном районе хорошо представлены элементы колхидской флоры, развит подлесок из вечнозеленых (лавровишня, рододендрон понтийский, падуб, иглица) и листопадных (черника кавказская) кустарников. Хорошо выражен пояс пихтовых лесов на высоте 1000-1750 м (Малеев, 1938; Davis, 1965). Площади буково-пихтовых и чисто пихтовых лесов значительные, большей частью по ущельям рек и их верховьям. В меньшей степени развиты еловопихтовые леса в восточной части, где они произрастают по каменистым склонам. Пояса пихтовых и буковых лесов зачастую заходят друг на друга.
ОСНОВНЫЕ ЧЕРТЫ МОРФОЛОГИИ, БИОЛОГИИ, экологии ПИХТЫ КАВКАЗСКОЙ И ЕЕ ЛЕСОВОДСТВЕННЫЕ
ОСОБЕННОСТИ
Морфология, биология и экология пихты
Пихта кавказская (абхазское название “апса”) - дерево первой величины, достигающее свыше 50 м высоты и более 1,5 м диаметра на уровне груди со стройным полнодревесным стволом цилиндрической формы. Основания стволов из-за сильно развитых корневых лап на горных склонах часто бывают утолщенными и ребристыми.
Отличается густой конусовидной величественной кроной (см. рис. 1). В свободном стоянии крона низко опущенная, в густых древостоях она составляет 1/3 - 1/5 часть высоты ствола. В период быстрого роста в высоту верхушка кроны острая, впоследствии становится овальной.
Молодые побеги блестящие, желтовато-зеленые, опушены, позднее становятся буровато-коричневыми и теряют опушение.
В раннем возрасте кора на стволе серая, гладкая. К возрасту 120 лет кора сплошь покрывается большими продольными трещинами и приобретает темно-бурый цвет (см. рис. 1). Хвоя длиной 2-4 см, шириной 2-2,5 мм, с тупой или округлой выемчатой верхушкой. Сверху она темнозеленая, блестящая, с продольными бороздками, снизу с двумя яркобелыми полосками из 10-15 рядов устьиц. Живет хвоя 9-12 лет. Расположена на побегах густо, неясно - гребенчато, так как побег покрыт сверху рядом настильно расположенных хвоинок. На боковых побегах зачастую хвоя имеется лишь на приростах последних трех лет. Почки крупные, красновато-коричневые, слабоопушенные, не смолистые. “Цветет” пихта в мае в период распускания хвои. Женские шишки длиною 12-20 см, 4-5 см толщины, молодые зеленого цвета, к осени буро-коричневые, смолистые, на ветках прямостоячие. Семенные чешуйки широкие, вверху покрыты нежным войлоком, цельнокрайние; кроющие - яйцевидной формы, у основания суженные, кверху расширенные. Семена крупные, блестящие, со светло-коричневыми крылышками. По нашим исследованиям, размеры семян пихты кавказской могут достигать длины 12,5, ширины 5,5 и толщины 3,7 мм. Созревают семена в сентябре. Опадают одновременно с раскрыванием шишки - в октябре-ноябре, оставляя на ветках крупные шероховатые стержни. Вес одной тысячи семян составляет до 80 г, всхожесть 57 % (Бебия, 1973). В специальной литературе указывается, что всхожесть семян пихты кавказской равна лишь 17-20 % при весе одной тысячи семян 64 г (Дендрофлора Кавказа, т. 1, 1959). Всхожесть семян сохраняется до двух лет.
У пихты иногда наблюдается смена главного побега при его повреждении или поломке. Взамен обломанной вершины из почек начинает раз-
виватъся новая, или очень редко ее замещает одна из боковых ветвей. Смена главного побега наблюдается также при повреждении верхушечных почек у деревьев молодого возраста. Двухвершинность у пихты наблюдается часто.
Смена главного побега у пихты происходит не всегда. Она зависит от возраста и жизненного состояния дерева. В спонтанном сомкнутом лесу у деревьев перестойной, сильно угнетенной, усыхающей категории смена главного побега не происходит.
Исследования показали, что количество стволов со сменой главного побега в пихтарнике овсяницевом I класса бонитета в среднем составило 3,4 % от общего числа деревьев толще 6 см и выше 4 м (табл. 4). Среди подроста высотой 0,5 - 4 м количество растений со сменой главного побега больше - 5,8 %.
Таблица 4
Встречаемость особей пихты со сменой главного побега _____в пихтарнике овсяницевом I класса бонитета__
Де ревья по я] русам Подрост высотой, м
I П Ш всего 0,5-1 1-2 2-3 3-4 всего
124 212 496 832 472 116 58 194 840
3 7 18 28 16 9 2 22 49
4,4 3,3 3,6 3,4 3,4 7,8 3,5 и,з 5,8
Примечание. Первый горизонтальный ряд - число деревьев и подроста на 1 га, второй ряд - число особей со сменой главного побега, третий ряд - то же в процентах. Пробная площадь заложена в Рицинском национальном парке на высоте 1250 м, склон юго-западной экспозиции, 40° крутизны. Состав древостоя 9Пх1Бк, сомкнутость 0,9.
У пихты хорошо выражен также отрицательный геотропизм. Этим отчасти можно объяснить отсутствие на Кавказе криволесья пихты в субальпийском поясе.
В целом для пихты кавказской характерна сравнительно низкая морфологическая пластичность, что особенно хорошо прослеживается в экстремальных для нее условиях обитания. Возможно, этим объясняется тот факт, что у пихты кавказской, в естественных условиях местопроизрастания, отмечены, официально признаны, лишь одна форма и одна ее разновидность. В культуре описано около 33 культиваров. Для сравнения у пихты белой, близкой в систематическом отношении и отнесенной к той же секции, что и пихта кавказская, выделены 3 формы, 9 разновидностей и 53 культивара (MesterMzy, 1995).
Сравнительно низкую морфологическую пластичность пихты кавказской можно рассматривать как проявление общебиологической закономерности, отражающей реликтовую черту вида, обусловленную произрастанием его в изолированном от других видов колхидском рефугиуме. Как известно, реликты - специализированные формы, характеризуются ограни
ченными ареалами, становятся редкими или вымирают, тогда как генерализованные формы дают начало новой производной группе, адаптированной к новым условиям существования. (Берг, 1922; Северцов, 1967; Завадский, 1968; Тахтаджян, 1970; Тимофеев-Рисовский и др., 1977; Гаузе, 1984; Грант, 1991).
В ареалогическом отношении пихта белая также характеризуется более широким распространением, чем пихта кавказская. Эти характерные черты пихты кавказской дают основание предполагать, что она представляет собой, по сравнению с пихтой белой, более старую ветвь в своей секции. Эти же черты свидетельствуют также о более или менее завершенном процессе формировании и обособлении ее как самостоятельного вида.
Пихта - порода относительно холодоустойчивая, выдерживает кратковременные морозы до -25°. Вследствие этого основные массивы ее размещаются в верхней горной зоне и достигают субальпийской границы лесной растительности. По своей морозоустойчивости она уступает пихте белой. К примеру, в условиях г. Москвы пихта кавказская, по данным М.С. АлександрЛой (1969), обмерзает до высоты снежного покрова, а пихта белая не обмерзает.
Пихта чувствительна к высоким летним температурам. Всходы ее страдают от заморозков и прямой солнечной радиации. В силу этого, по мнению большинства исследователей пихтовых лесов (Герасимов, 1948; Орлов, 1951; Долуханов, 1964 и др.), а также, по нашим исследованиям (Бебия, 1978), пихта кавказская не возобновляется семенным путем на лесосеках сплошных рубок, на полянах, в окнах большого диаметра, более 40 м. В табл. 5 приводятся данные наших наблюдений за выживаемостью всходов пихты вне полога леса на открытой площади. Наблюдения проводились в пихтарниках Кучба-Яштинского лесничества Гагринского лесхоза в квартале 18, на высоте 1100 м над уровнем моря. Тип леса - буковопихтовый I класса бонитета. Сомкнутость полога древостоя 0,7. Место закладки учетных площадок ровное. Наблюдения проводились весной и осенью в год после обильного урожая семян пихты. Учетные площадки, размерами 1 кв. м, закладывали на прогалине на расстоянии 5, 10, 20, 30 м в южном направлении от стены леса пихтового древостоя. Одна площадка (контрольная) была заложена под пологом леса. Наблюдения опыта проводились в трех повторностях. Предварительно перед опадением семян на площадках проведено глубокое рыхление поверхности почвы с удалением покрова.
Из табл. 5 видно, что из появившихся весной всходов пихты на самой удаленной от стены леса учетной площадке 5 к осени не сохранилось ни одного. Под пологом леса (площадка 1) сохранность всходов пихты к осени составила 58 %.
Таблица 5
Выживаемость всходов древесных пород на учетных площадках
№ учетных площадок Расстояние от стены леса, м Количество всходов
весной (25.05) осенью (25.09)
пихта, шт. лиственные породы, шт. пихта, шт./ % лиственные породы, шт./ %
1 — 57 31 33/58 19/61
2 5 38 47 8/21 22/57
3 10 46 34 3/7 22/65
4 20 23 32 — 9/28
5 30 2 18 - 4/22
Примечание. В составе лиственных пород доминирует граб кавказский, реже встреча* ются бук, ильм эллиптический.
Изучение влияния прямого солнечного освещения на всхожесть пихты было продолжено при выращивании сеянцев в холодном парнике (табл. 6). Часть посевов отенялись драночными щитами, другую часть оставляли без отенения и использовали как контрольную группу. Освещение в парниках измерялось объективным люксметром ОЛ - 3 с селеновым фотоэлементом К-20.
Таблица 6
Влияние отенения на выращивание однолетних сеянцев пихты
Варианты Освещен- Грунтовая Появилось Осталось осенью Высота
опыта ность, всхожесть всходов, здоровых всходов всходов
люкс/% % шт./пог. м шт./пог. м % мм
Посев без
отенения
(контроль) 142200/100 26 27 2 7,4 35
Посев с оте-
нением 48300/34 35 32 18 56,2 31
Выявлено, что грунтовая всхожесть семян на посевах с отенением на 9 % выше, чем в контроле, где осенний отпад сеянцев весьма значительный, составил 92,6 %, в то время как на посевах с отенением эта величина равна 43,8 %. Под щитом выход сеянцев пихты с одного погонного метра оказался в 8-9 раз выше, чем в контроле. Результаты этих опытов, безусловно, подтверждают достаточно высокую чувствительность всходов и сеянцев пихты к прямому солнечному освещению, что необходимо учитывать при выращивании пихты в первые два года в питомнике. Существенного влияния отенения сеянцев на третий год в нашем опыте не обнаружено.
Наступление фенологических фаз у пихты зависит от множества факторов: высоты над уровнем моря, почвенных условий, освещенности и других. Наблюдение за ростом побегов пихты на станционарных площадях наших опытных рубок в Кучба-Яштинском лесничестве Гагринского лесхоза в течение трех лет подряд показало, что продолжительность роста по
бегов пихты в среднем составила 63 дня. Они распускаются в конце мая, и завершается прирост по высоте к концу июля. Наблюдения за одним и тем же подростом пихты проводились на высоте 1100 м над ур. моря, в окне группово-выборочных рубок и под пологом леса в количестве 10 экземпляров в каждом варианте. Древостой - буково-пихтовый, мелкотравный I класса бонитета, состав 6Пх4Бк, сомкнутость 0,7.
Пихта кавказская - довольно влаголюбивая, мезофильная порода. Сухих ветров и вообще сухости не переносит. Требовательна к влажности воздуха. Эти факторы в значительной мере определяют высотный и широтный ареал ее распространения. Произрастает она на среднеувлажнен-ных почвах, но избыточного увлажнения почв не переносит. К богатству почвы требовательна и только на глубоких бурых лесных почвах характеризуется высокой производительностью. На глубоких почвах пологих склонов развивает толстый, но короткий стержневой корень и глубоко залегающую мощную корневую систему (см. рис. 6). Здесь она ветроустойчива. На сыром делювии около ручьев пихта кавказская развивает поверхностные корни и ввгровальна. Пихту кавказскую, по отношению к ветру, следует относить к породе переходной от ветроустойчивой к ветровальной. Нами отмечены в бассейнах рек Псоу (Аибгинское лесничество, кв. 21, 22), Геги (Черкесско-Полянское лесничество), Лашапса (Рицинский госзапо-ведник) участки ветровала пихты.
Пластичность корневой системы пихты проявляется и в ее способности произрастать, практически, в беспочвенных условиях. Она растет на крутых склонах с малоразвитыми почвами, на лавинных комплексных субстратах. Последняя особенность подтверждает не только ореофлорогенез и ореофитность пихты, но и устойчивость современной ее позиции в горных условиях. В подтверждение этой мысли мы сошлемся на высказывания В.Л. Комарова (1953). Он отмечал, что растения горных склонов часто имеют субстратом не почву, а непосредственно ту или другую породу или немного наноса, удержанного неровностями породы. Такое заключение подтверждается данными другого видного ученого Т. Накамура (Nakamura, 1992), проводившего исследования сукцессии и дифференциации лесных сообществ в зависимости от почвенно-грунтовых условий в субальпийской зоне на горе Фуджи. Он приводит описание двух типов пихтового леса: Abies veitchii - Dryopteris austriaca, Abies veitchii - Vaccinium axillare, которые успешно развивались на, практически, беспочвенном, чисто шлаковом и вулкано-лавовом субстрате.
Пихта кавказская - порода теневыносливая. Одной из ее отличительных особенностей является способность произрастать в широком диапазоне световых условий (Орлов, 1951; Долуханов, 1964; Бебия, 1972; Иванов, 1984). Впрочем, все виды пихт растут при полном солнечном освещении, а также при очень сильном затенении, в частности, под сомкнутым пологом
материнского древостоя, куда проникает не более 5-7 % дневного света. По исследованиям Л.А. Иванова и И.Л. Коссовича (1946), пихта белая при средней интенсивности света (30 % от полного солнечного освещения) отличается самой высокой фотосинтезирующей способностью среди хвойных после лиственницы, которой она уступает лишь при полном - 100 %-ном солнечном освещении. При освещении, равном 1 %, у пихты баланс между поглощением и выделением углекислоты остается положительным, равным 0,13, а у ели европейской отрицательным (-0,06). Известно также, что породы, сильно фотосинтезирующие, являются энергично растущими (Алексеев, 1975) Эго относится и к пихте кавказской, которая, например, в свободном стоянии в возрасте 30 лет может иметь высоту 6 м (рис. 8, IV).
Вместе с тем она способна расти в условиях сильного угнетения до столетнего возраста в виде угнетенного подроста с высотой 1 м (рис. 8, Ш). Однако при освещении такого угнетенного подроста до 50-60-летнего возраста он может оправиться и в дальнейшем расти нормально (рис. 8, II). В течение жизни пихта может претерпеть такое угнетение неоднократно, но с более коротким периодом угнетения (Бебия, 1972).
Высота
Рис. 8. Ход роста подроста пихты: I - в окне, II - (угнетенный), Ш - (усохший) под пологом древостоя, IV - в свободном стоянии
В обзоре, посвященном изменению светового режима растительных сообществ, В. Транквилини (Tranquilini, 1960) приводит шкалу относительного светолюбия подроста пихты белой, растущего в условиях умеренного климата 2,3-1,7 тыс. люкс.
Разумеется, теневыносливость пихты вовсе не означает, что она порода тенелюбивая. Для хорошего роста и развития ей необходимы нормальные световые условия.
В оптимальных условиях роста пихта нуждается в полном освещении. Однако подрост сильно угнетенной категории гибнет при внезапном полном освещении на лесосеке. К этому вопросу мы вернемся в дальнейшем при изложении классификации подроста по его жизненному состоянию.
Теневыносливость пихты нам представляется как один из способов вынужденной адаптации к произрастанию длительное время под материнским пологом древостоя, приобретенной в процессе борьбы за существование.
Пихта кавказская успешно возобновляется и хорошо произрастает под пологом материнского древостоя сомкнутостью 0,5-0,6. Однако специальные исследования показали, что наиболее интенсивный фотосинтез, лучшее развитие и максимальный прирост у нее при 60-100 % полной освещенности (Бебия, 1973). Хотя пихта в раннем возрасте и в угнетенном состоянии растет медленно, она является быстрорастущей. В благоприятных условиях с 10-летаего возраста прирост по высоте может достигать 60-70 см в год (рис. 8, IV).
О светолюбии пихты свидетельствует и характер разрастания кроны подроста в горизонтальном направлении в условиях затемнения. Высокая освещенность необходима ей также для нормального “цветения” и “плодоношения”. Этим объясняется тот факт, что в лесу семяносят деревья пихты, главным образом, верхнего яруса.
Пихта кавказская, по сравнению с елью восточной, характеризуется хорошим очищением ствола от сучьев. В зависимости от полноты и сомкнутости древостоя оно может составлять до 85 % общей высоты ствола с выходом деловой категории древесины 90-92 %. Интенсивный рост у пихты наблюдается до глубокой старости (Ушатин, 1962). При этом отдельные экземпляры, по данным М.В. Герасимова (1944), доживают до 800 лет. Однако, по нашим наблюдениям, предельный возраст пихты кавказской не превышает 500 лет (табл.7).
В Кучба-Яштинском лесничестве в квартале 18 нами было обнаружено пихтовое дерево высотой 62 м и диаметром ствола на высоте груди 196 см, возраст его составлял около 400 лет.
Таблица 7
Некоторые таксационные показатели старовозрастных экземпляров пихты кавказской
Район, высота над ур. моря, м Высота пня, м Диаметр на высоте пня, см Число годичных слоев на высоте пня Высота дерева, м
Черкесско-Полянское 0,9 123 328 41
лесничество, 1100 0,7 142 276 —
1,0 102 314 —
Кучба-Яштинское лесничество, 1100 1,3 196 361 62
Рицинский заповедник, 1200 0,8 158 248 —
Красная поляна, Пслух, 950 0,7 128 375 —
Кавказский биосферный 0,9 112 457 -
заповедник, 1100 0,7 78 341 —
То же, 1000 0,7 130 400 -
Некоторые особенности семяношения пихты кавказской
Изучение семяношения пихты кавказской имеет важное значение для выявления процесса формирования естественного возобновления. Возобновление лесов зависит от многих факторов и, прежде всего, от продуктивности семяношения деревьев, составляющих насаждения.
Вопросу семяношения пихты кавказской посвящено мало работ. Исследование по данному вопросу проводили А.И. Соснин (1939) и И.И. Хуторцев (1960) в лесах Северного Кавказа (Кавказский заповедник), Т.М. Джапаридзе и Т.Ф. Урушадзе (1973) в лесах Западной Грузии (Бор-жомский лесхоз). Сведения по этому вопросу приведены в работе В.З. Гулисашвили (1970). Нами было изучено семяношецие пихты в пихтарниках Абхазии (Бебия, 1973). В работах А.И. Данилова (1951), Т.П. Некрасовой (1962), А.И. Савченко и А.П. Бугкуте (1965) освещен вопрос семяношения пихты сибирской. Семяношение пихты белой изучали Я. Нетолицкий (1955), А.И. Швиденко (1980) и другие. Весьма интересные исследования этого процесса проводились у североамериканских видов пихт (Eis, 1973; Owens, 1977). Однако исследования цитируемых авторов не раскрывают все стороны вопроса семяношения пихты. Ниже приводим результаты наших исследований по этому вопросу.
Известно, что пихта кавказская образует, в основном, разновозрастные древостои (Давыдов, 1940; Бицин, 1965; Бебия, 1972 и др.). Считается, что насаждения этих лесов состоят из нескольких возрастных поколений или ярусов, чаще из трех. Наши исследования по этому вопросу более под
робно излагаются далее в разделе, посвященном возрастной структуре пихтарников.
В первое возрастное поколение входят деревья спелые и перестойные. Во второе - приспевающие и средневозрастные группы, а третье - молодые деревья. В силу этого семяношение деревьев в древостое одновозрастном и разновозрастном будет отличаться, так как дифференциация в процессе борьбы и конкуренции проходит в одновозрастных и разновозрастных древостоях по-разному (Гулисашвили, 1970; Бебия, 1973).
По исследованиям А.Н. Соболева и А.В. Фомичева, в одновозрастном сомкнутом древостое семяносят лишь деревья высших классов Крафта, или роста (Морозов, 1949). В разновозрастных древостоях семяношение деревьев обусловливается возрастной структурой, строением древостоя и характеризуется разными показателями семяношения.
Исследования семяношения пихты проводили в разновозрастных лесах Кучба-Яштинского лесничества Гудаутского лесхоза. Целью работы явилось изучение семяношения пихты в разновозрастных нетронутых древостоях и древостоях, пройденных добровольно-выборочными и группововыборочными рубками; определение количественных и качественных показателей семян, собранных с деревьев разных возрастных поколений. Семяношение изучали на пробных площадях 32, 34, 37, 38. Пробные площади 37 и 38 размером 1,6 га каждая заложены в наиболее распространенных типах лесов с нетронутым разновозрастным древостоем. Пробная площадь 32 - 1,0 га заложена на лесосеке добровольно-выборочных рубок трехлетней давности и 34 - 1,0 га на лесосеке группово-выборочных рубок трехлетней давности. Исследования семяношения проводились в годы хорошего урожая пихты. На пробных площадях проводили подробное лесово-дственно-таксационное описание древостоя. Изучали характер семяношения деревьев разных возрастных поколений. Учитывали деревья семяно-сящие и несемяносящие. Урожай деревьев оценивали глазомерно по методу В.Г. Каппера (1954). Данные об урожайности семян получили при учете на модельных деревьях. Рядом с пробными площадями в идентичных лесорастительных условиях были вырублены модельные деревья по возрастным поколениям. Для каждой модели учитывали количество всех шишек и собирали их в отдельности. В лабораторных условиях шишки взвешивали с точностью до 0,001 г, измеряли их длину и толщину с точностью до 0,1 мм. Извлечение семян из шишек и обескрыливание проводили ручным способом. Семена из шишек подсчитывались и взвешивались. Так как семена пихты в аппаратах плохо проращиваются, качество семян определяли взрезыванием. Данные учета обрабатывали методом вариационной статистики, определяли среднее значение показателей и его ошибку.
В результате учета были получены такие сведения: длина и толщина шишек, их вес, количество семян в шишках, общий вес семян, выход семян
и I шишек, средний вес 1000 шт. семян, процент полнозернистых и пустых семян. Были определены общая урожайность деревьев по возрастным поколениям и интенсивность семяношения их.
Результаты наших исследований показали, что в нетронутых разновозрастных пихтарниках семяносят, главным образом, деревья I и П возрастных поколений. Деревья Ш поколения семяносят единично. Так, из общего количества семяносящих деревьев на долю 1 и П поколений приходится 97,3-100 % деревьев на 1 га (табл. 8, проб. пл. 37, 38). На долю деревьев Ш поколения приходится всего 2,7 % (проб. пл. 38). В целом для древостоев количество семяносящих деревьев составляет 20,5-22,1 % от общего их количества на 1 га. В пределах возрастных поколений семяносят деревья: в I поколении - 91,5-93,5 %, во П - 33,0-35,8 % и в Ш поколении - 1 % от общего количества деревьев в поколении. Следует отметить, что деревья Ш поколения семяносят только в естественных окнах. Под сомкнутым пологом леса они не семяносят. Количественные показатели на проб. пл. 38 лучше, чем на проб. пл. 37, что объясняется более благоприятными условиями освещения, так как проба первая заложена в древостое меньшей сомкнутости полога и большей крутизны склона. Пробная площадь 37 заложена в древостое с сомкнутостью полога 0,8 на склоне 8-10 градусов юго-восточной экспозиции. Сомкнутость полога древостоя на проб. пл. 38 составляет 0,7, уклон местности 22 градуса, экспозиция южная (табл. 8).
Таблица 8
Семяношение деревьев пихты в разновозрастных древостоях
№ пробной площади, ее характеристика Возрастное поколение (ярус) Деревья пихты на 1 га
всего с шишками с шишками в % от общего количества в поколении
шт. % шт. %
37 1.0 га, юго-восточная экспозиция, 8-10°, 1300 м I 59 14,1 54 62,8 91,5
н.у.м., пихтарник ясменниковый, бонитет 1а, П 97 23,2 32 37,2 33,0
9Пх1Бк, ед. Кл., сомкнутость полога 0,8 Ш 269 62,7 - - -
Итого 419 100 86 100 20,5
38 1,0 га, южная экспоз.,22°,1300 м н.у.м., пихтар- I 46 13,7 43 58,1 93,5
ник ясменниковый, бонитет I, 8Пх2Бк, сомкну- II 81 24,2 29 39,2 35,8
тость полога 0,76 III 208 62,1 72 2,7 1,0
Итого 335 100 144 100 22,1
32 1,0 га, юго-восточная экс поз., 20°, 1200 м I 34 13,9 31 44,3 91,2
н.у.м., буково-пихтовый ясменниковый, бонитет п 50 20,5 36 51,4 72,0
I, бПх4Бк, сомкнут, полога 0,6 ш 160 65,6 3 4,3 1,9
Итого 244 100 70 100 28,7
34 1,0 га, юго-восточная экспозиция, 10-12°, 1200 м I 18 6,8 15 50.0 83,8
н.у.м., буково-пихтовый ясменниковый бонитет п 47 17,9 13 43,3 27,8
1а, 6Пх4Бк, сомкнутость полога 0,7 III 198 75,3 2 6,7 1,0
Итого 263 100 30 100 11,4
Освещение является важным фактором среды, определяющим в значительной степени семяношение деревьев в насаждении.
Совершенно иной характер носит семяношение деревьев в одновозрастных древостоях. Ссылаясь на слова Г.Ф. Морозова (1949), по исследованиям А.Н. Соболева и А.В. Фомичева, в семенной год в древостоях ели, в основном, семяносят деревья господствующего класса Крафта: I - 45 %, II - 29 %, Ш - 24 %. На долю IV класса деревьев приходится всего 2 %; деревья V класса вовсе не семяносят. Следовательно, на долю господствующих классов приходится 98 % семяношения. Следует подчеркнуть, что господствующие классы Крафта представлены деревьями одного яруса и возраста, но различающихся развитием и ростом.
Учет шишек на модельных деревьях по возрастным поколениям показал, что в нетронутом разновозрастном древостое урожай шишек на дереве пихты I поколения в среднем составил 83 ± 5 шт. Дерево П поколения се-мяносило 35 ± 3 шишки и дерево Ш поколения - 12 ± 1 шишка. Из рис. 9 видно, что если Лггенсивность семяношения дерева I поколения принять за 100 %, то для П поколения она составит 42,2 %, для Ш поколения - 14,4 %. По данным А.В. Фомичева, для одновозрастных лесов, если урожайность одного дерева I класса принять за 100 %, то урожайность дерева II класса составит 53,4 %, ПГ класса - 33,5 %, IV класса - менее 0,2 % и V - 0 %.
Изучение семяношения деревьев пихты на лесосеке добровольно -выборочных рубок трехлетней давности (проб. пл. 32) показало, что ряд показателей семяношения деревьев в этих условиях выше, чем в нетронутом лесу. Видимо, такое явление обусловлено тем, что после таких рубок на лесосеке создаются более благоприятные условия роста, развития и семяношения деревьев. Так, в семяношении II поколения на пробной площади 32 участвуют на 36-39 % больше деревьев, чем в нетронутых древостоях пробных площадей 37 и 38 (см. табл. 8). На пробной площади 32 во П поколении плодоносят 72 % от общего количества их в поколении. На пробных площадях 37 и 38 эта величина соответственно составила 33 и 36 %. Из этой таблицы видно, что доля семяносящих деревьев П поколения на пробной площади 32 составила 51 % от количества семяносящих во всех поколениях, что на 12-14 % больше, чем на пробных площадях 37 и 38. В целом процент семяносящих деревьев от общего количества их на 1 га на пробной площади 32 также выше на 7-9 %, чем на пробных площадях 37 и 38, где он соответственно составляет 29, 20 и 22 %.
Более благоприятные условия семяношения на пробной площади 32 обусловили и семяношение деревьев III поколения в количестве 1,9 % от их общего в поколении, что составило 4,3 % от количества семяносящих деревьев всех поколений. Тем не менее, деревья Ш поколения семяносят незначительно.
11 !;
-4:1— - --- 4k • : ' - Jk- >
11X3/? <*2,^^
Рис. 9. Интенсивность семяношения деревьев пихты I, II, и III возрастных поколений (ярусов)
Слабо семяносили деревья пихты III поколения и в окнах через три года после проведения группово-выборочных рубок. Так, на пробной площади 34 в Ш поколении семяносили деревья пихты до 1 % от общего количества в поколении. Они составили около 7 % от количества семянося-щих деревьев в поколении. Группово-выборочные рубки улучшили условия семяношения деревьев II поколения. Доля семяносящих деревьев II поколения на пробной площади 34 составила 43 %, что выше на 4-6 %, чем в нетронутых древостоях на пробных площадях 37 и 38 (см. табл. 8).
Исследования качественных показателей семян, собранных с модельных деревьев разных поколений, показали, что такие показатели, как длина, толщина и вес шишки, выход семян из одной шишки в граммах и штуках, вес 1000 шт. семян у деревьев I поколения превосходит таковой у деревьев II и Ш поколений (табл. 9). Выход полнозернистых семян у шишек с деревьев I поколения также на 6 % выше, чем у деревьев II поколения и на 34 %, чем у деревьев III поколения. Выход полнозернистых семян из шишек для деревьев I поколения составил 75 %, для П - 69 % и для III - 41 %.
Таблица 9
Урожай, количественные и качественные показатели шишек и семян ___________деревьев пихты по возрастным поколениям__________
Показатели Возрастное поколение деревьев
I П III
Число шишек, шт. 83 ±5 35±3 12 ±1
Длина шишки, мм 16,2 ± 0,4 15, ±03 13Д ±0,5
Толщина шишки, мм 4,7 ±0,2 4,5 ± 0,2 4,1 ±0,1
Вес шишки, г 101,0 ±3,7 91.5 ±1,6 67,8 ±2,5
Выход семян из одной шишки, г 36,5 ± 1,6 20,0 ±1,8 11,4 ±0,8
Выход семян из одной шишки, шт. 339 ± 14 280 ± 11 214 ± 8
Выход полнозернистых семян из одной шишки, шт. 255 192 89
Выход полнозернистых семян из одной шишки, % 75,3 68,6 41,4
Вес 1000 семян, г 108,4 ±2,13 73,6 ±2,58 51,9 ±2,56
Исследования показали, что на пробной площади 38 - 82 % урожая шишек на 1 га приходится на долю деревьев I поколения, 18 % - на долю деревьев II поколения (табл. 10). Урожай шишек Ш поколения составил менее 1 %.
Таблица 10
Урожайность древостоя пихты по возрастным поколениям __________:____на пробной площади 38_____________
Возрастное поколение Шишек Семян
шт. % всех ВТ. ч. полнозернистых
кг % шт. % шт. %
I 3569 81,6 130,26 87,6 1209891 84,3 910096 85,7
II 783 17,9 15,70 10,6 219240 15,2 150336 14,1
Ш 24 0,5 2,73 1,8 5136 0,5 2136 0,2
Итого 4376 100 148,69 100 1434267 100 1062567 100
Деревья I поколения дали 87 % урожая семян в кг, который по количеству семян составил 84 %. Эти показатели для деревьев П поколения соответственно составили 11% и 15 %, для деревьев Ш поколения - примерно 2 % и 1 %. В целом для древостоя на 1 га урожай составил: шишек -4376 шт., семян - 149 кг или 1433 тыс., в том числе полнозернистых -1062 тыс. семян (табл. 10). Более обобщенные данные по семяношению пихты приводятся на рис. 10.
Наши многолетние рекогносцировочные наблюдения показали, что в условиях Абхазии периодичность семяношения пихты выражена слабо и колеблется от 2 до 5 лет.
Таким образом, из вышеприведенных данных следует, что семяноше-ние деревьев в разновозрастных пихтарниках отличается от семяношения деревьев одновозрастных древостоев. В одновозрастных древостоях семя-
Девственный лес
ШИШЕК - 4376 кг/га
СЕМЯН - 149 кг/га
СЕМЯН - 1433000 шт/га
ПОЛНОЗЕРНИСТЫХ - 74 %
СЕМЯНОШЕНИЕ ПО ЯРУСАМ
Рис. 10. Семяношение в пихтовом лесу
носят деревья высшего класса Крафта, которые образовывают одновозрастное поколение, но различаются ростом и развитием. В разновозрастных-пихтарниках семяносят пихты I и П поколений, то есть первых и вторых ярусов. Деревья Ш поколения (Ш ярус) семяносят единично в естественных окнах. Количественные и качественные показатели семян деревьев I поколения превосходят таковые деревьев П и Ш поколений. Нетронутые разновозрастные пихтарники обсеменяются количественно и качественно вполне достаточно для беспрерывного естественного возобновления. Добровольно-выборочные и группово-выборочные рубки в разновозрастных пихтовых древостоях способствуют улучшению условий семяношения деревьев всех возрастных поколений.
Несомненно, семяношение пихты кавказской - сложное биологическое явление в жизни леса. Поэтому оно зависит от множества факторов, таких как биоэкология самой пихты, условия местопроизрастания, индивидуальные особенности дерева, географические и связанные с ними климатические условия и множество других факторов. В силу этого, количественные и качественные показатели семяношения пихты могут изменяться в зависимости от влияния комплекса факторов на семяношение как отдельных деревьев, так и в целом древостоя. Результаты наших исследований не могут дать исчерпывающего ответа на все вопросы, связанные с семяно-шением пихты. Необходимы более глубокие и стационарные исследования
по этим вопросам. Однако закономерности семяношения, установленные нами для разновозрастных пихтарников, вполне объективны и могут служить практической основой для проектирования конкретных мероприятий по лесному хозяйству, лесным культурам и лесосеменному делу.
Естественное возобновление пихты под пологом леса
Наши исследования по семяношению пихты показали, что валовый урожай семян в разновозрастных пихтарниках вполне может обеспечить обсеменение лесных площадей. В силу этого, под пологом леса обычно имеется самосев и подрост, которые всегда способны заменить материнский полог. Ниже освещаются результаты наших исследований по естественному возобновлению пихты под пологом леса в девственных пихтарниках.
Изучение особенностей и закономерностей формирования возобновления под пологом леса имеет практическое и научно-теоретическое значение. Оно необхо^рмо для обоснования способов рубок и многих других лесохозяйственных, лесокультурных мероприятий.
Естественное возобновление пихты кавказской изучали ряд исследователей в Закавказье (Гулисашвили, 1948; Долуханов, 1964; Попов, 1955; Ссорин, 1955; Бахсолиани, 1968; Бебия, 1973; 1978) и на Северном Кавказе (Орлов, 1951; Солнцев, 1954; Иваненко, 1956; Голгофская, 1967; Веселов, 1973). Однако выводы этих исследователей по данному вопросу противоречивы, что следует объяснять различными физико-географическими условиями объектов исследования и неодинаковыми методическими подходами к изучению возобновления.
Известно, что ход естественного возобновления - прорастание семян, появление самосева и превращение его в подрост под пологом леса зависит, главным образом, от типа леса и сомкнутости материнского полога (Морозов, 1949). Поэтому основные закономерности формирования естественного возобновления пихты мы изучали по типам леса при различной сомкнутости полога с учетом экспозиции, крутизны склона и абсолютной высоты.
При изучении возобновительных процессов под пологом леса устанавливали количественное и качественное состояние подроста на единице площади, его возрастную структуру, состав и характер распределения по площади леса.
В процессе работы мы пользовались методикой А.В. Побединского (1966) с некоторыми дополнениями и уточнениями. Возобновление изучали на пробных площадях, заложенных для таксационной характеристики древостоев или при описании типов леса. Размер пробных площадей составлял 0,25-0,5-1,0 га. Закладывали их в наиболее распространенных, типичных группах типов леса.
Количественное и качественное состояние возобновления устанавливали при учете его на площадках размером 3x3 м, заложенных через одну по диагоналям пробной площади в количестве 40-46 площадок на 1 га. На учетных площадках проводили учет всходов и подроста с подразделением их по породам и возрастным группам: 1-2 года, 3-5 лет, 6-10 лет, 11 лет и старше. Визуально определяли равномерность пространственного распределения подроста по площади. К подросту относили молодые деревца диаметром на высоте груди до 6 см. Возраст мелкого подроста определяли по мутовкам, а крупного считали на срезе диаметра у основания. Подрост с 6-летнего возраста распределяли на благонадежный (б/н) и неблагонадежный (н/б). Качественное состояние подроста устанавливали визуально, исходя из ряда морфологических и дендрометрических признаков, таких как прирост по высоте главного и боковых побегов, форма кроны, протяженность и охвоение кроны и других. Метод классификации подроста пихты кавказской, разработанный нами, будет изложен ниже более подробно.
Оценка естественного возобновления вначале производилась по шкале Гулисашвили (1956). Позже нами была разработана и применена новая, более совершенная шкала возобновления, которая излагается отдельно ниже. Возобновление под пологом леса изучали всего на 48 пробных площадях, на которых были заложены 1020 учетных площадок.
Результаты наших исследований подтвердили зависимость хода естественного возобновления пихты под пологом леса в разновозрастных пихтарниках от сомкнутости материнского полога в пределах группы типов леса.
Исследования показали, что в овсяницевой группе типов благоприятные условия для естественного возобновления складываются при сомкнутости материнского полога, равной 0,7 (табл. 11). При такой сомкнутости полога во всех возрастных градациях возобновление протекает с преимуществом пихты (проб. пл. 20). Общее количество всходов составляет 6,50 тыс. и благонадежного подроста 8,26 тыс. шт. на 1 га, в том числе крупного подроста - 2,52 тыс. шт. Сомкнутость полога ниже 0,5 обусловливает худшие условия для возобновления. На пробной площади 28, заложенной в буково-пихтовом овсяницевом типе леса, сомкнутость полога равна 0,5. Общее количество всходов составляет 5,00 тыс., а подроста 3,20 тыс. шт./га, в том числе крупного - 1,00 тыс. шт. Незначительное количество подроста в возрасте 6-10 лет, равное 800 шт./га, указывает на неблагоприятные условия перерастания всходов в подрост.
Неудовлетворительное возобновление на пробной площади 48, где сомкнутость полога составляет 0,4. Низкая сомкнутость полога обусловила неблагоприятные условия не только для перехода всходов в подрост, но и для появления всходов. Так, общее количество всходов на пробной площади 48 составило всего 1,70 тыс. шт./га, а количество подроста - 1,90 тыс.
шт., в том числе крупного 0,70 тыс. шт./га (см. табл. 11). Высокая сомкнутость полога, превышающая 0,7, также не благоприятствует хорошему естественному возобновлению. На пробной площади 41 сомкнутость полога составляет 0,8. Возобновление здесь представлено в основном всходами (7,5 тыс. шт./га) и подростом 3-10 летнего возраста (2,12 тыс. шт./га). Количество же крупного подроста старше 11 лет не превышает 0,33 тыс. шт. Кроме того, на данной пробной площади наблюдается также наличие неблагонадежного подроста в количестве 0,80 тыс. шт./га. Неудовлетворительное возобновление на пробной площади 48, где сомкнутость полога составляет 0,4. Низкая сомкнутость полога обусловила неблагоприятные условия не только для перерастания всходов в подрост, но и для появления всходов. Так, общее количество всходов на пробной площади 48 составило всего 1,70 тыс. шт./га, а количество подроста - 1,90 тыс. шт., в том числе крупного 0,70 тыс. шт./га (см. табл. 11). Высокая сомкнутость полога, превышающая 0,7, также не благоприятствует хорошему естественному возобновлению. На пробной площади 41 сомкнутость полога составляет 0,8.
• Таблица 11
Динамика естественного возобновления в пихтовом и буково-пихтовом __________________типах леса овсяницевой группы_________________
№ пробной площади, ее характеристики Сонки угость полога Порода Количество всходов и подроста (тыс. шт./га) в возрасте (лет)
1-2 3-5 6-10 11 и старше Итого б/н подроста
б/н н/б б/н н/б
48
0,25 га, юго-западная экспозиция. 0,4 Пихта 1,50 0,60 0,40 — 0,40 — 1,40
уклон 25°, абсолютная высота Ель 0,20 0,10 — 0,10
1350 м, лесосека 8-летнего возраста, пихтарник овсяницево- Бук - - 0,10 - 0,30 - 0,40
ежевичный. бонитет П, состав древостоя 9П1БедЕ Итого 1,70 0,70 0,50 - 0,70 — 1,90
28
0,5 га, юго-восточная экспозиция. 0,5 Пихта 4,80 1,20 0,60 0,70 2,50
уклон 30*, абсолютная высота Бук 0,20 ОДО 0,20 0,20 — 0,60
1200 м, лесосека 8-летнего возраста, буково-пихтовый овсяни-цевый, бонитет П, состав древостоя 8П2Б Итого 5,00 1,40 0,80 - 0,90 - 3,10
20
0,25 га, южная экспозиция, уклон 0,7 Пихта 6,00 2,80 1,82 0,20 1,62 0,4 6,24
28*. абсолютная высота 1100 м. Бук 0,30 0,22 0,50 — 0,90 — 1,62
буково-пихтовый овсяницсвый, бонитет I, состав древостоя 7ПЗБ Липа 0,20 0,10 0,30 - - - 0,40
Итого 6,50 3,12 2,62 0,20 2,52 0,4 8,26
41
0,25 га, юго-восточная экспози- 0.8 Пихта 6,00 0,41 0,62 0,20 0,21 0,6 1,24
ция, уклон 28*, абсолютная высо- Ель 0,30 0,01 — 0,01 — 0,02
та 1100 м, пихтарник овсянице- Бук 1,00 1,10 0,10
вый, бонитет П, состав древостоя Липя 0,20 0,01 0,11
8П2Л + ЕБ Итого 7,50 1,52 0,62 0,20 0,33 0,6 2,47
Примечание, б/н - благонадежный, н/б - неблагонадежный подрост.
Возобновление здесь представлено в основном всходами (7,5 тыс. шт./га) и подростом 3-10 летнего возраста (2,12 тыс. шт./га). Количество же крупного подроста старше 11 лет не превышает 0,33 тыс. шт. Кроме того, на данной пробной площади наблюдается также наличие неблагонадежного подроста в количестве 0,80 тыс. шт./га.
Следует отметить, что овсяницевые группы типов леса приурочены в основном к юго-восточным, южным, юго-западным склонам. В силу этого при сомкнутости полога 0,5 и ниже наблюдается разрастание травостоя из овсяницы, а также ежевики, которые резко препятствуют появлению всходов и превращению их в подрост. В высокосомкнутых же насаждениях благоприятные условия складываются для самосева до трех лет. Недостаток освещения под пологом высокосомкнугого насаждения обусловливает плохой рост и развитие подроста старше трехлетнего возраста.
В разнотравной группе типов лучшее естественное возобновление наблюдается при сомкнутости полога 0,6. Так, на пробной площади 10 (табл. 12), где сомкнутость полога равна 0,6, во всех возрастных группах возобновление протекает за счет пихты с незначительной примесью бука. Общее количество всходов в этом случае составило 22,8 тыс. шт./га, подроста 9,08 тыс. шт., в том числе крупного 2,08 тыс. шт. Неплохое возобновление и на пробной площади 1, где сомкнутость полога равна 0,5. Однако количество крупного подроста в возрасте 11 лет и старше в этом случае меньше, чем при сомкнутости полога 0,6 (проб. пл. 10) и составляет 1,28 тыс. шт./га. Количество всходов равно 12,29 тыс. шт., всего подроста 7,68 тыс. шт./га. Значительно хуже и возобновление при сомкнутости полога ниже 0,5. Так, на пробной площади 50, заложенной в разнотравной группе типов леса, сомкнутость полога была снижена после рубки до 0,4, что привело к образованию на данной площади производного типа - ясменниково-ежевичного. В разнотравной группе типов леса при снижении полноты древостоя и сомкнутости полога ниже 0,5 наблюдается разрастание широкотравья и ежевики. При сомкнутости полога 0,4-0,3 степень покрытия их достигает 0,7-0,8. Развитый травяной покров и ежевика на пробной площади 50 обусловили ухудшение условий появления всходов, количество которых не превышает 2,32 тыс. шт./га (см. табл. 12). Отсутствие подроста пихты и незначительное количество ели (0,3 тыс. шт.) в 6-10 летней возрастной группе указывает, что всходы, достигнув возраста 3-5 лет, не выдерживают конкуренции травяного, ежевичного покрова и погибают. Количество же подроста в возрасте 11 лет и старше, равное 2,25 тыс. шт./га, появилось до снижения сомкнутости полога до 0,4, так как возраст его превышает возраст лесосеки.
Таблица 12
Динамика естественного возобновления в пихтовом, буково- пихтовом __________________типах леса разнотравной группы___________________
№ пробной площади, ее характеристики Сомкнутость полога Порода Количество всходов н подроста (тыс. штУга) в возрасте (лет)
1-2 3-5 6-10 11 и старше Итого б/н подроста
б/н н/б б/н Н/б
50
0,25 га, равнжма, абсолютная высота 0,4 Пихта 2,00 0,42 1.11 1.53
1000 и, лесосека 9-летиего возраста, буково-пихтовый ясмоюшково- Ель 0,12 0,10 0.30 - 0.54 - 0.94
ежевичный, бонитет 1, состав древо- Бук 0,20 - • - 0.60 - 0.60
стоя 8Пх2Бк ед. В Итого 2,32 0,52 0.30 - 2.25 - 3.07
1
0,5 га, юго-западная экспозиция. 0,5 Пихта 12.07 2.90 1.20 0.42 4.52
уклон 18е, абсолютная высота 1650 и, Бук 0.22 0.70 1.80 0.86 3.16
лесосека 9-летнего возраста, буково-пихтовый ежевнчно - ясментпювый, Итого 12.29 3.60 3.00 - 1.28 - 7.68
бонитет I, состав древостоя 7ПхЗБк
10
0,5 га, северо-западная экспозиция. 0,6 Пихта 12.20 3.20 2.80 2.02 0.21 8.02
уклон 16°, абсолютная высота 1600 м, буюово-пжхтовый ЯСМСНШХОВЫЙ, ф бонитет I. состав древостоя 7ПхЗБк Бук Итого 10.60 22.80 0.20 3.40 0.40 3.20 - 0.46 2.48 0.21 1.06 9.08
21
0,5 га, северо-западная экспозиция. 0,7 Пихта 10.80 2.20 1.20 0.84 0.26 4.24
уклон 25°, абсолютная высота 1400 м. Бук 5.80 0.80 0.80 - 0.41 - 2.01
состав древостоя 911x1 Бк Итого 16.60 3.00 2.00 - 1.25 0.26 6.25
22 *
0,25 га, северо-западная экспозиция. 0,8 Пихта 40.80 0.20 0.40 0.40 0.64 0.22 1.24
уклон 15°, абсолютная высота 1400 м. Бук 6.00 3.00 0.21 0.82 - 3.82
пвхтярпик ясмсннккомь кислячховый, бонитет 1, состав древостоя 10Пх+Бк Итого 46.80 0.20 3.40 0.61 1.46 0.22 5.06
Хорошее возобновление наблюдается также на пробной площади 21, в которой сомкнутость полога 0,7. Количество всходов в этом случае составило 16,60 тыс. шт./га. Общее количество подроста 6,75 тыс. шт. Возобновление во всех возрастных группах протекает за счет пихты. Наличие угнетенного подроста неблагонадежной категории в количестве 0,26 тыс. шт. указывает на ослабление оптимальных условий его роста.
В разнотравной группе типов сомкнутость полога 0,8 благоприятствует появлению всходов. Общее количество их составляет 46,8 тыс. шт./га. Однако высокая сомкнутость полога не вполне способствует условиям формирования подроста. Поэтому количество подроста пихты в этом случае составило всего 1,24 тыс. шт./га, а количество неблагонадежного подроста - 0,83 тыс. шт. Характерно, что в составе подроста преобладает бук, количество которого равно 3,82 тыс. шт./га. Кроме того, количество неблагонадежного пихтового подроста превышает количество неблагонадежного букового подроста и составляет соответственно 0,62 тыс. шт. и 0,21 тыс. шт./га. Следовательно, можно полагать, что при высокой сомкнутости полога буковый подрост возобновляется, растет и развивается сравнительно лучше, чем пихтовый.
Несколько иной характер естественного возобновления в папоротниковых группах типов леса (табл. 13). В целом папоротниковые пихтарники характеризуются слабым естественным возобновлением. Более или менее хорошее возобновление наблюдается при сомкнутости полога 0,5. Так, на пробной площади 9 сомкнутость полога равна 0,5. Подрост, общее количество которого равно 6,90 тыс. шт./га, представлен с преобладанием пихты и некоторым участием бука и клена. Однако количество всходов незначительно и равно 1,70 тыс. шт./га. Более благоприятные условия для появления всходов складываются в папоротниковых группах типов при сомкнутости полога 0,7 (проб. пл. 6) и 0,6 (проб. пл. 18). Количество всходов в первом случае составило 12,8 тыс. шт., во втором - 9,6 тыс. шт./га. Подрост лучше формируется на пробной площади 18, где сомкнутость полога равна 0,6, а количество подроста 4,2 тыс. шт./га. На пробной площади 6 при сомкнутости полога 0,7 общее количество подроста меньше и равно 1,80 тыс. шт./га.
Опускание сомкнутости полога ниже 0,5 обусловливает резкое ухудшение условий возобновления в папоротниковой группе типов лесов. Так, на пробной площади 5, где сомкнутость полога была снижена рубкой главного пользования до 0,3, на девятый год после рубки возобновление шло явно неудовлетворительно.
Таблица 13
Динамика естественного возобновления в пихтовом, буково-пихтовом ________________типах леса папоротниковой группы_______________
№ пробной площади, ее характеристики Сомкнутость полога Порода Количество всходов и подроста (тыс. шт./га) в возрасте (лет)
1-2 3-5 6-10 11 и старше Итого б/н подроста
б/н н/б б/н н/б
5
0,5 га, южная экспозиция, уклон 0,3 Пихта 1,20 2,40 - 2,40
10°, абсолютная высота 1400 м, Буж 0,20 0,50 0,20 0,10 - 0,80
лесосека 9-летнего возраста, Клен 0,20
буково-пихтовый кочедыжниковый, бонитет П, состав древостоя 7ПхЗБк+Кл 9 Итого 1,60 2,90 0,10 - 0,10 - 3,20
0,5 га, равнина, абсолютная вы- 0,5 Пихта 0,30 1,70 1,50 1,60 - 4,80
сота 1300 м, лесосека 7-летнего Бук 1,20 0,90 0,50 0,20 1,60
возраста, буково-пихтовый ко- Клен 0,20 0,50 0,50
чедыжниковый, бонитет I, состав древостоя 7ПхЗБк+Кл, Ил 18 Итого 1,70 3,10 2,00 - 1,80 - 6,90
0,25 га, юго-западная экспози- 0,6 Пихта 7.00 2,40 0,70 0,20 3,30
ция, уклон 5*, абсолютная высо- Бук 2,60 0,70 0,10 - 0,10 - 0,90
та 1200 м, буково-пихтовый кочедыжниковый, бонитет I, состав древостоя 7ПхЗБк 6 Итого 9,60 з.ю 0,80 - 0,30 - 4,20
0,25 га, западная экспозиция, 0,7 Пихта 6,80 0,40 0,20 0,40 0,2 1,00
уклон 8*, абсолютная высота Бук 6,00 - 0,20 - 0,40 - 0,60
1400 м, пихтарник страусоперо- Клен 0,20 0,20
вый, бонитет 1а, состав древостоя 9Пх1Бк + Кл Итого 12,80 0,60 0,40 - 0,80 0,2 1,80
Всходы пихты доживают только до 3-5 лет, затем гибнут. Количество всходов в таких условиях не превышает 1,6 тыс. шт./га, подроста - 3,2 тыс. шт. Наблюдается полное выпадение подроста пихты старше 6 лет. В группе типов леса с подлеском из черники кавказской наиболее хорошее возобновление наблюдается при сомкнутости полога 0,6 и 0,7. (табл. 14). Общее количество всходов при полноте 0,6 (проб. пл. 11) составило 18,4 тыс. шт., подроста - 9,42 тыс. шт., в том числе крупного 1,02 тыс. шт./га. Возобновление во всех возрастных группах представлено за счет пихты с некоторым участием бука.
На пробной площади 7 сомкнутость полога равна 0,7. Количество всходов 48,0 тыс. шт., подроста - 6,6 тыс. шт., в том числе крупного -2,0 тыс. шт./га.
В группе типов леса с подлеском из черники кавказской наиболее хорошее возобновление наблюдается при сомкнутости полога 0,6 и 0,7. (табл. 14). Общее количество всходов при полноте 0,6 (проб. пл. 11) составило 18,4 тыс. шт., подроста - 9,42 тыс. шт., в том числе крупного 1,02 тыс. шт./га. Возобновл&ие во всех возрастных группах представлено за счет пихты с некоторым участием бука. На пробной площади 7 сомкнутость полога равна 0,7. Количество всходов 48,0 тыс. шт., подроста - 6,6 тыс. шт., в том числе крупного - 2,0 тыс. шт./га.
Более высокая сомкнутость полога, хотя и обеспечивает появление всходов в значительном количестве, однако не благоприятствует формированию подроста пихты. Так, на пробной площади 8 при сомкнутости полога 0,8 количество всходов составило 40,4 тыс. шт., подроста - 6,2 тыс. шт./га. На пробной площади 5, где сомкнутость полога равна 0,9, количество всходов равно 16,0 тыс. шт., подроста всего 1,6 тыс. шт./га. Количество неблагонадежного подроста - 2,80 тыс. шт./га. В составе благонадежного подроста преобладает бук.
Сравнительно неплохие условия для появления всходов и развития подроста до пятилетнего возраста под пологом леса сомкнутостью 0,4. На пробной площади 43 общее количество всходов составило 16,2 тыс. шт., а подроста до пяти лет - 3,9 тыс. шт./га. Незначительное количество подроста возрастной группы 6-10 лет в количестве всего 0,50 тыс. шт. указывает на неблагоприятные условия формирования возобновления после снижения сомкнутости до 0,4.
Количество же подроста возраста 11 лет и старше - 2,4 тыс. шт. появилось до снижения сомкнутости полога, так как пробная площадь 43 заложена на лесосеке девятилетнего возраста. Снижение сомкнутости полога обусловило образование производного черничниково-ежевичного типа леса. После снижения сомкнутости полога до 0,4 черника и разросшаяся ежевика отрицательно влияют на ход возобновления.
Таблица 14
Динамика естественного возобновления в пихтовом, буково-пихтовом _____типах лесов с колхидским подлеском (черника кавказская)__
№ пробной площади, ее характеристики Сомкнутость полога Порода Количество всходов и подроста (тыс. шт./га) в возрасте
(Й
1-2 3-5 6-10 11 и старше Итого б/н подроста
б/н н/б б/н н/б
43
0.3 га, юго-восточная экспозиция. 0,4 Пихта 13,60 2,70 0,20 0,20 3,10
уклон 14°, абсолютная высота 1700 м, лесосека 9-ллнего возраста, пихтарник черничяиково-сжевичкый, бонн- Ель 0,30 - 0,10 - 0,10
Бук 2,30 1,20 0,30 - 1,10 - 2,60
тот II, состав древостоя 9Пх1 Бк ед Е Итого 16,20 3,90 0,50 - 2,40 - 5,80
11
0.25 га, западная экспозиция, уклон 0,6 Пихта 6,20 3,80 2,00 0,82 6,62
15*. абсолютная высота 1400 м, буково-пихтовый чериичниковый, бони- Бук 12,20 2,40 '0,20 - 0,20 - 2,80
тет П, состав древостоя бПх4Бк Итого 18,20 6,20 2,20 - 1,02 - 9,42
7
0,25 га, западная экспозиция, уклон 0,7 Пихта 22,30 3,00 1,00 0,20 1,00 0,20 5,00
20°, абсолютная высота 1500 м, буково-пихтовый черничяиковый, бонитет Бук 25,00 - 0,60 - 1,00 0,20 1,60
I, состав древостоя 8Пх2Бк Итого 48,00 3,00 1,60 0,20 2,00 0,40 6,60
8
0,25 га, западная экспозиция, уклон 0,8 Пихта 27,40 2,60 0,80 0,20 0,80 0,40 4,20
30°, абсолютная высота 1500 м, пихтарник черничяиковый, бонитет П, состав древостоя 9Пх 1 Бк Бук Итого 20,00 40,40 1,60 3,60 0,40 1,20 0,20 0,40 0,60 1,40 0,40 0,80 2,00 6,20
5
0,25 га, восточная экспозиция, уклон 0,9 Пихта 12,00 0,40 1,00 0,40
8*. абсолютная высота 1200 м. буково-пихтовый чсрничниковый, бонитет 1, состав древостоя 8Пх2Бк Бук Итого 4,00 16,00 0,40 0,40 0,40 0,40 1,00 1,00 0,40 0,80 0,80 1,80 1,60 1,60
В целом динамика естественного возобновления по группам типов леса отображена на рис. J1. На нем видно, что лучшим естественным возобновлением характеризуются разнотравные пихтарники. Удовлетворительно возобновляются пихтарники овсяницевые и с подлеском из черники кавказской. Папоротниковые пихтарники характеризуются слабым естественным возобновлением.
В пределах группы типов лучшее формирование возобновления наблюдается при сомкнутости полога 0,6-0,7 для овсяницевых групп типов, 0,5-0,6 - для разнотравных, 0,6-0,7 - в группе с подлеском из черники кавказской. В пихтарниках папоротниковой группы типов леса сравнительно лучшее возобновление при сомкнутости полога 0,6.
Для установления различия в росте нами изучена дендрометрическая характеристика пихтового подроста, произрастающего под пологом леса сомкнутостью 0,3, 0,6 и 0,9. Для этой цели в каждом варианте среди равномерно растущего пихтового подроста вырубали по 25 штук. У каждого подроста определяли возраст, измеряли высоту, диаметр у корневой шейки, прирост по диаметру за последние пять лет. С верхней части кроны брали по 100 штук хвои двухлетнего возраста и в лабораторных условиях взвешивали с точностью до 0,01 грамма в воздушно-сухом состоянии.
0,2 0,3 0,4 0,5 0,6 0,7 0,8
Сомкнутость полога
— -I.-------------П.------------ш.--------------IV
Рис. 11. Естественное возобновление пихты группы типов леса:
I - овсянициевые, II - разнотравные, Ш - папоротниковые, IV - черничнико-вые; возобновление: 1 *- очень слабое, 2 - слабое, 3 - удовлетворительное, 4 -хорошее
Взвешивание производили в трехкратной повторности. Установлено, что пихтовый подрост, произрастающий под пологом леса, сомкнутостью 0,6, характеризуется лучшим ростом, чем подрост под пологом сомкнутостью 0,9 (табл. 15). Разность в приросте по диаметру между ними составила 10 %, по высоте 50 % в пользу подроста, произрастающего под пологом сомкнутостью 0,6. Важным показателем роста подроста является вес хвои. Вес 100 шт. хвои в воздушно-сухом состоянии в первом случае на 33 % больше.
Лучшие дендрометрические показатели роста у подроста под пологом сомкнутостью 0,6 обусловлены, главным образом, более благоприятным режимом освещенности. По этой же причине под пологом леса сомкнутостью 0,3 подрост пихты растет и развивается еще лучше, чем под пологом сомкнутостью 0,6 и 0,9. У такого подроста прирост по диаметру на 29 %, прирост по высоте на 31 % и вес хвои на 10 % выше таковых у подроста под пологом сомкнутостью 0,6. Однако, как выше было отмечено, в первом случае наблюдается во всех группах типов леса неудовлетворительное количественное возобновление. Следовательно, сомкнутость полога, равную 0,6, можно считать оптимальной для естественного возобновления и фор-
Таблица 15
Биометрическая характеристика подроста пихты под пологом леса
Характеристика подроста
Сомкнутость полога возраст, лет высота, см диаметр у шейки корня, см прирост по диаметру за последние 5 лет, см/% годичный прирост по высоте за последние 5 лет, см/% вес 100 штук воздушно-сухой хвои, г/%
0,3 31 14517 3,010,2 2,6610,13/129 8,910,4/131 0,54 + 0,03/110
0,6 37 13015 2,710,1 2,0510,06/100 6,910,4/100 0,49 ±0,03/100
0,9 52 16018 3,610,2 1,8510,09/90 2,810,2/41 0,33 ± 0,02/67
мирования подроста пихты кавказской под пологом леса как количественно, так и качественно. Поэтому при проведении рубок главного пользования в пихтовых лесах можно допустить выборочные способы рубок с обязательным сохранением после рубки сомкнутости полога 0,6.
Следует отметить, что возобновление под пологом в пихтовых лесах зачастую носит куртинный характер, приуроченный к естественным окнам. В силу этого в пихтарниках на склонах до 25° возможно применение и группово-выборочных способов рубок. Известно, что группововыборочные рубки - возобновительные рубки (Ткаченко, 1955). Окна таких рубок должны обеспечить хорошее естественное возобновление главной породы. Однако появление всходов и формирование подроста во многом зависят от формы, размера окон, крутизны склона, их расположения и экспозиции.
Возобновление пихты в естественных окнах
Возобновление пихты под пологом леса часто носит групповой характер. В силу этого еще К. Гаером было обосновано соответствие группововыборочных рубок природе темнохвойных лесов (Мелехов, 1966). Эти рубки допускаются на склонах до 25°. Применение этих рубок в группах типов леса папоротниковых и с подлеском из колхидских элементов (падуб, лавровишня, черника, рододендрон) требуют дополнительных мероприятий по содействию естественному возобновлению. Поэтому возобновление в окнах изучали в наиболее распространенных - овсяницевой и разнотравной группах типов леса на склонах до 25° с учетом величины, формы окна, крутизны и экспозиции склона.
Окна, в которых изучали возобновление, возникли естественным путем в пихтовых, буково-пихтовых лесах в результате отмирания, вывала перестойных и фаутных деревьев, или ветровала, снеголома. В целом изучено в овсяницевой группе типов леса 11 окон, в разнотравной - 14 окон.
При изучении возобновления окна разделяли по величине площади и размерам диаметров на три группы: малые - площадью до 200 м2, диаметрами до 15 м; средние - площадью 200-500 м2, диаметрами 16-24 м и большие - площадью свыше 500 м2, диаметрами больше 25 м. Диаметры окон измеряли по направлению с севера на юг и с востока на запад. По форме окна различали округлые и эллипсоидные. По крутизне склона окна распределены на две группы: окна на пологих склонах до 15° и окна на склонах средней крутизны 16-25°. Учет возобновления в окнах вели по всей площади при разбивке их площадками 2x2 м. На площадках учитывали количество всходов и подроста по породам. У подроста измеряли высоту и прирост по высоте последнего года. Данные учета на площади окна при обработке материалов переводили на 1 кв. м.
Исследования показали, что в овсяницевых пихтарниках на склонах до 15° (табл. 16) наиболее благоприятные условия для естественного возобновления в о^сне среднего размера диаметрами 18x20 м (окно № 2). Количество всходов в таком окне составило 1,43 шт./кв. м, подроста 1,59 шт./кв. м. В возобновлении преобладает пихта, доля участия ее составляет 84%. Неплохое возобновление наблюдается в окне № 3 диаметром 20x22 м. Однако количество всходов и подроста в этом случае меньше, чем в окне № 2, и составляет соответственно 1,05 и 0,85 шт./кв. м. Такое явление можно объяснить тем, что в окнах с увеличением их размера повышается интенсивность света, который способствует разрастанию травяного покрова, препятствующего возобновлению. Такое положение хорошо прослеживается в окнах больших диаметров. Так, в окне № 4 диаметрами 24 х 26 количество всходов составляет 1,02 шт./кв. м, подроста - 0,64 шт./кв. м. В окне № 5 диаметрами 30x32 количество всходов и подроста еще меньше и составляет соответственно 0,42 и 0,31 шт./кв. м.
Значительно хуже происходит возобновление в окнах малых размеров на пологих склонах. В окне № 1 диаметры равны 10x12 м, количество всходов составило 0,88 шт./кв. м, подроста всего 0,21 шт./кв. м.
Следует отметить, что подрост пихты лучше растет в окнах больших размеров: Так, средний подрост последнего года по высоте у пихтового подроста в окне № 5 (большого размера) составляет 3,2 ± 0,1 см при средней высоте подроста 0,84 ± 0,03 м. В окне № 3 (среднего размера) при средней высоте подроста пихты 0,94 ± 0,02 м средний текущий прирост по высоте составил 2,4 ±0,1 см. Максимальный текущий прирост по высоте у подроста пихты в первом случае равен 16 см, во втором 14 см (см.табл. 16).
На склонах средней крутизны 16-25° в овсяницевой группе типов пихтарников наиболее хорошее возобновление наблюдается в окне № 7 диаметрами 16x17 м. Количество всходов составляет 2,58 шт./кв. м, подроста 1,46 шт./кв. м. Доля участия пихты в возобновлении достигает 79 %.
Таблица 16
Естественное возобновление пихты в окнах овсяницевой группы типов леса
Номера окон, диаметры по направлениям С-Юх В-3 в м, площадь в кв. м экспозиция уклон Порода Всходы шт./кв. м Подрост
шт./кв. м Высота, м Прирост по высоте последнего года, см Процент участия пород
макс. мин. средний
1 2 3 4 5 6 7 8 9
1
10x12,95 Пихта 0,50 0,18 0,6810,02 4,0 0,5 1,510,1 86
юго-восточная, Ель 0,02 0,01 1,00 3,0 0,2 - 5
4° Бук 0,36 0,02 1,62 8,0 2,0 - 9
Итого 0,88 0,21 - - - - 100
2
18x20,283, Пихта 1,21 1,34 0,4410,02 15,0 0,5 1,910,1 84
юго-восточная, Ель 0,21 0,10 0,36 6,0 0,2 - 6
10° Бук 0,01 0,15 0,72 22,0 2,0 - 10
Итого 1,43 1,59 - - - - 100
3
20x22,346, Пихта 0,82 0,61 0,9410,02 14,0 0,5 2,410,1 72
равнина Ель 0,02 0,12 0,72 4,0 0,2 - 14
Бук 0,21 0,12 1,02 24,0 2,0 - 14
4 Итого 1,05 0,85 - - - - 100
24x26,490, Пихта 0,78 0,53 0,49Ю,02 16,0 1,0 2,010,1 83
юго-восточная, Ель 0,12 0,10 0,47 15,0 0,5 - 15
0° Бук 0,11 0,01 0,32 24,0 3,0 - 2
Ильм 0,01 - - - - - -
5 Итого 1,02 0,64 - - - - 100
30x32,754, Пихта* 0,20 0,19 0,8410,03 16,0 0,8 3,210,1 61
юго-западная, Ель 0,18 0,11 0,70 8,0 1,0 - 36
5е Бух 0,04* 0,01 0,42 12,0 2,0 - 3
Итого 0,42 0,31 - , - - - 100
6
10x10,79, Пихта 0,90 0,46 0,9810,04 5,0 0,6 1,510,1 73
юго-восточная, Ель 0,15 0,07 1,00 3,0 0,2 - 11
25° Бук 0,23 0,10 1.18 14,0 2,0 - 16
Итого 1,28 0,63 - - - - 100
7
16x17,214, Пихта 1,83 1,16 0,8210,03 15,0 1,0 2.8Ю.1 79
южная, Бук 0,75 0,30 0,96 22,0 4,0 - 21
20° Итого 2,58 1,46 - - - - 100
8
14x26,286, Пихта 2,35 0,98 0,8510,03 14,0 0,8 2,210,1 83
южная, Ель 0,13 0,08 0,28 9,0 0,5 - 7
22° Бук 0,42 0,12 0,72 18,0 2,0 - 10
Итого 2,90 1,18 - - - - 100
9
30x12,283, Пихта 0,36 0,28 1,1610,03 16,0 1,0 2,710,1 64
юго-восточная, Ель 0,12 0,10 0,83 8,0 1,о - 22
18° Бук 0,08 0,06 1,30 20,0 4,0 - 14
Итого 0,56 0,44 - - - - 100
Продолжение табл. 16
1 2 3 4 5 6 7 8 9
10 20x21,330, Пихта 0,49 0,36 0,52±0,02 13,0 1,0 1,8±0,1 61
северо-западная. Бук 0,19 0,21 0,98 30,0 2,0 - 36
24е Клен 0,02 0,02 0,56 12,0 2,0 - 3
Итого 0,70 0,59 - - - 100
11 28x30,662, Пихта. 0,10 0,09 0,90±0,04 17,0 1,0 4,89±0,2 56
юго-западная, Ель 0,12 0,06 0,56 14,0 1,о - 38
20° Клен 0,01 0,01 0,84 12,0 з.о - 6
Итого 0,23 0,16 - - - - 100
Хорошее возобновление и в окне № 8 эллипсоидной формы, вытянутом поперек склона, диаметрами с севера на юг 14 м, с запада на восток 26 м. Количество всходов в этом окне равно 2,96 шт./кв. м, подроста - 1,18 шт./кв. м. Возобновление протекает за счет пихты, доля участия которой равна 83 %. В окне № 9 эллипсоидной формы, вытянутом вдоль склона, большим диаметром с севера на юг равным 30 м, наблюдается слабое возобновление. Количество всходов равно 0,56 шт./кв. м, подроста - 0,44 iirr./кв. м. С увеличением размера окон на склонах средней крутизны количество всходов и подроста уменьшается. В окне № 11 диаметрами 28x30 м количество всходов составляет всего 0,23 шт./кв. м, подроста 0,16 шт./кв. м. В окнах малых размеров на склонах средней крутизны наблюдается сравнительно лучшее возобновление (окно № 6), чем на пологих склонах (окно № 1).
Экспозиция играет существенную роль в процессе естественного возобновления. На пологих склонах восточной, юго-восточной и южной экспозиции лучшее возобновление отмечено в окне среднего размера диаметрами 18x20 м (окно № 2), а на юго-западной, северо-западной экспозициях можно отметить окно диаметрами 20x22 м (окно № 3) с хорошим возобновлением. На склонах средней крутизны восточной, юго-восточной и южной экспозиции лучшее возобновление наблюдается в окнах среднего размера диаметрами 16x17 м (окно № 7) округлой формы и диаметрами 14x26 м (окно № 8) эллипсоидной формы, вытянутых поперек склона. На юго-западной, северо-западной экспозициях лучшее возобновление в окнах среднего размера диаметрами 20x21 (окно № 10).
Как на пологих, так и на склонах средней крутизны лучший рост и развитие подроста наблюдаются в окнах больших и средних размеров. Однако, учитывая и количественную характеристику возобновления в овсяницевой группе типов леса, на пологих склонах оптимальными окнами для формирования возобновления следует считать окна средних размеров диаметрами 18-20 м и площадью 280-300 кв. м, на склонах средней крутизны окна средних размеров диаметрами 16-18 м округлой формы площадью 200-250 кв. м, а также эллипсоидной формы, вытянутые поперек склона, с
меныпим диаметром по направлению с севера на юг до 15 м, площадью 280-300 кв. м.
Данные, характеризующие естественное возобновление в окнах в разнотравной группе типов пихтарников (табл. 17), свидетельствуют о том, что на пологих склонах до 15° в окнах среднего размера диаметрами 20x22 м (окно № 14) и 20x20 м (окно № 17) наблюдается наилучшее естественное возобновление, количество всходов в этих окнах достигает 1,04 и 2,33 шт./кв. м, подроста - 1,73 и 1,49 шт./кв. м. Доля пихты в подросте составила 84 и 74 %. Удовлетворительное возобновление и в окне № 15 диаметрами 28x31 м, где количество всходов равно 0,58 шт./кв. м, подроста - 0,72 шт./кв. м. В окне большого диаметра 35x37 м наблюдается неудовлетворительное возобновление. Количество всходов и подроста в таком окне не превышает 0,35 и 0,14 шт./кв. м. Необходимо отметить, что в окне такого размера, по нашим наблюдениям, в буково-пихтовом ясменниковом типе леса II класса бонитета (Кучба-Яштинское лесничество Гудаутского лесхоза кв. 18) происходит ветровал деревьев. Слабое возобновление также в окне № 12 малого размера диаметрами 11x12 м. Количество всходов в этом окне составило 0,82 шт./кв. м, однако количество подроста не превышает 0,23 шт./кв. м. В возобновлении значительное участие принимает буковый подрост. Доля участия пихты 65 %, бука - 35 %.
На склонах средней крутизны в разнотравной группе типов леса наилучшее возобновление наблюдается в окнах № 23, 22 и 20 (см. табл. 17). Окна № 23 и 22 - округлой формы, среднего размера. Диаметры их равны 17x18 и 20x20 м. Окно № 20 имеет эллипсоидную форму и вытянуто по горизонтали направлением меньшего диаметра с севера на юг. Диаметры его равны 16x24 м. Количество всходов в таких окнах составило соответственно, 1,72, 1,84, и 1,26 шт./кв. м. Количество подроста в них равно 1,42, 1,29 и 1,61 шт./кв. м. В окнах большего размера, налжлонах средней крутизны отмечено худшее возобновление. Так, в окне № 24 диаметрами 26x25 м всходы имеются в количестве 0,41 шт./кв. м, подрост 0,50 шт./кв. м. В окне № 25, где диаметр еще больше (30x32 м), хотя количество всходов значительно и составляет 0,98 шт./кв. м, количество подроста не превышает 0,34 шт./кв. м. В таких окнах, по нашим наблюдениям, происходит разрастание широкотравья, препятствующего возобновлению.
Слабое возобновление в окне № 21 диаметрами 28x14 м. Оно имеет эллипсоидную форму и вытянуто вдоль склона. Количество всходов в этом окне составляет 0,89 шт./кв. м, подроста - 0,69 шт./кв. м. Окно № 16 малого размера, диаметром 11x12 м. Количество всходов в нем достигает 1,88 шт./кв. м, но незначительное количество их перерастает в подрост, всего 0,57 шт./кв. м.
Естественное возобновление пихты в окнах
(мелкотравная группа типов леса)
Номера окон, дна-
метры по Всходы Подрост
направлениям Порода ШТ./КВ. пггУкв. Высота, Прирост по высоте Процент
С-ЮхВ-Звм, м м м последнего года, см участия
площадь в кв. м пород
экспозиция уклон макс. мин. средний
1 2 3 4 5 6 7 8 9
12
11x12,104, Пихта 0,43 0,15 0,4610,02 5,0 0,3 1,410,1 65
равнина Бук 0,39 0,08 1,20 12,0 2,0 - 35
Итого 0,82 0,23 - - - - 100
13
14x18,198, Пихта 1,20 0,48 0,7210,03 8,0 0,5 2,310,1 81
юго-восточная. Бук 0,44 о,п 1,04 10,0 2,0 - 19
8° Итого 1.64 0,59 - - - - 100
14
20x22,346, Пихта 0,85 1,45 1,2210,05 16,0 1,0 3,510,2 84
юго-западная. • Бук 0,18 0,28 1,28 24,0 4,0 - 16
6° Ильм 0,01 - - - - - -
Итого 1,04 1,73 - - - 100
15
28x31.683, Пихта 0,23 0,53 0,9410,03 20,0 2,2 5,110,2 74
южная, .Бук 0,24 0,18 1,22 22,0 6,0 - 25
10° Ильм 0,01 0,01 0,80 • - - 1
Итого 0,58 0,72 - - - 100
16
35x37,1077 Пихта 0,22 0,06 1,0810,04 19,0 2,0 4,810,2 43
юго-восточная, Бук 0,10 0,06 0,96 36,0 4,0 - 43
8° Ильм 0,03 0,02 1,38 - - - 14
Итого 0,35 0,14 - - - 100
17
20x20,314 Пихта 1,86 1,24 0,7610,03 16,0 1,0 2,510,1 83
восточная, Бук 0,47 0,25 1,00 26,0 2,0 - 17
10° Итого 2,33 1,49 - - - - 100
18
11x11,95 Пихта 1,24 0,42 1,0210,03 6,0 1,810 - 74
юго-восточная. Бук 0,63 0,14 1,32 14,0 ,1 - 24
22° Ильм 0,02 0,01 0,80 - 2,0 - 2
Итого 1,88 0,57 - - - - 100
19
9x12,85, Пихта 0,85 0,34 0,4810,02 5,0 0,5 1,310,1 74
южная. Бук 0,14 0,12 0,92 12,0 3,0 - 26
20° Итого 0,99 0,46 - - - - 100
20
16x24,301, Пихта 0,92 1,46 0,8810,03 12,0 0,5 З.ЗЮ.1 91
южная. Бук 0,33 0,15 1,04 16,0 2,0 - 9
18° Клен 0,01 - - 20,0 - - -
Итого 1,26 1,61 - - - - 100
21
28x14,308 Пихта 0,67 0,56 0,6210,02 12,0 0,5 2,110,1 80
юго-восточная. Бук 0,22 0,14 1,20 18,0 2,0 - ’ • 20
20° Итого 0,89 0,69 - - - - 100
Продолжение табл. 17
1 . 2 3 4 5 6 7 8 9
22 20x20,314 Бук 0,35 0,24 1,06 26,0 4,0 19
югозападная, Итого 1,84 U9 - - - - 100
24° 23 17x18.240 Пихта 1,29 1,28 0,64 ±0,02 14,0 1.0 2,7±0,1 90
юго-восточная. Бук 0,43 0,14 2,04 32.0 4.0 10
25° Итого 1,72 1.42 - - - - 100
24 26x25,510 Пихта 0,18 0,38 1,32±0,05 18,0 2,0 6,2±О2 76
южная. Бук 0,23 0,12 0,48 16,0 5.0 24
20° Итого 0,41 0,50 - - - - 100
25 30x32,754 Пихта 0,38 0,29 0,56±0,02 16,0 1.0 3,2±0,1 66
северозападная. Бук 0,58 0,15 1,02 22,0 5,0 - 34
18° Клен 0.02 - - - - - -
Итого 0,98 0,44 - - - - 100
В разнотравной группе типов леса на пологих склонах восточной, юго-восточной, южной экспозиции хорошее возобновление наблюдается в окнах среднего размера диаметрами 20x20 м (окно № 17). На склонах средней крутизны указанных румбов лучшее возобновление в окнах среднего размера диаметрами 17x18 м (окно № 23). На этих же склонах юго-западных, северо-западных румбов хорошо возобновляются окна диаметрами 20x20 м (окно № 22). Как на пологих, так и на склонах средней крутизны подрост пихты лучше всего растет в окнах среднего и большего диаметров. В таких окнах подрост отличается большим текущим приростом по высоте, чем bl окнах малых размеров. Так, например, в окне № 24 (большого размера) текущий прирост по высоте у подроста пихты равен 6,2 ± 0,03 см при высоте его *1,32 ± 0,05 м. В окне № 18 (малого размера) при высоте пихтового подроста 1,02 ± 0,03 м текущий прирост составил всего 1,8 ± 0,1 см. Подрост бука лучше всего растет в окнах среднего размера (окна № 17, 14,22, 23; табл. 17).
Таким образом, учитывая количественное состояние возобновления, рост подроста в разнотравной группе типов леса, оптимальные условия для возобновления пихты на пологих склонах складываются в окнах средних размеров диаметрами от 20 до 28 м. Площадь их составляет 300-600 кв. м, форма округлая. На склонах средней крутизны хорошее формирование возобновления можно ожидать в окнах средних размеров округлой формы диаметрами 18-20 м, площадью 250-300 кв. м. Оптимальны также окна эллипсоидной формы, вытянутые по горизонталям. Маленький диаметр таких окон по направлению с севера на юг не должен превышать 18 м.
Динамика естественного возобновления в окнах различного диаметра на пологих и средней крутизны склонах в пределах группы типов леса наглядно отражена на рис. 12.
Из этого рисунка видно, что на пологих и на склонах средней крутизны в окнах средних и больших размеров лучшее возобновление наблюдается в разнотравной группе типов леса (кривые 3,4). Окна малых размеров лучше возобновляются в овсяницевой группе типов леса (кривые 1,2).
Рис. 12. Возобновление в окнах различного диаметра:
1 - на пологих склонах до 15°, 2 - на склонах 18-25 в овсяницевой группе типов леса; 3 - на склонах до 15°, 4 - на склонах 18-25° в разнотравной группе типов леса
Следует отметить, что возобновление в окнах может носить разнообразный характер. Однако проведенные нами исследования дают возможность сделать достаточно обоснованные выводы по применению группововыборочных способов рубок в пихтарниках Кавказа.
Лесоводственная классификация подроста пихты
На основании изучения пихтового подроста различного качества под пологом высокополнотного насаждения и процесса восстановления его жизнеспособности на лесосеках предлагается метод классификации подроста по категориям жизнеспособности для горных пихтовых лесов Кавказа. Приводятся конкретные показатели морфологических признаков по категориям жизнеспособности.
Под пологом леса из пихты кавказской всегда имеется разновозрастный подрост разного качества. Важность сохранения его при лесозаготовках, особенно в горных условиях, в настоящее время не вызывает сомнений. Подрост - это ценный резерв для формирования будущего насаждения. По исследованиям А.В. Побединского (1967), Н.Е. Декатова и А.Н. Кузнецова (1968), В.П. Тимофеева (1968), с помощью сохраненного при лесозаготовках подроста можно сформировать биологически устойчивые и продуктивные насаждения, сократить трудовые затраты и ускорить выращивание леса на 20-35 лет. Кроме того, сохраненный подрост в горных условиях способствует обеспечению водоохранно-почвозащитных функций.
В силу различных экологических условий, главным образом разной освещенности, подрост пихты под пологом материнского насаждения обладает неодинаковой жизнеспособностью. В хорошо освещенных местах он растет и развивается лучше, под сильно сомкнутым материнским пологом пихтовый подрост угнетен, слабо развит и плохо растет. В процессе филогенеза у пихты выработались определенные биоэкологические особенности, в том числе способность произрастать под пологом при неблагоприятном световом режиме. М.В. Герасимов (1948), А.Я. Орлов (1951) и А.Г. Долуханов (1964) отмечали способность пихты кавказской длительное время произрастать под пологом. При изреживании и улучшении светового режима угнетенный пихтовый подрост начинает расти лучше. Чем сильнее был угнетен подрост, тем дольше он восстанавливает свою жизнеспособность и труднее переносит новые условия. При резком изменении условий произрастания, как отмечал Г.Ф. Морозов (1949), подрост может погибнуть.
В лесоводственной практике по качеству подрост принято подразделять на благонадежный и неблагонадежный (Гулисашвили, 1956; Победин-ский, 1967; Бебия, 1973). А.М. Савченко (1965), изучавший выживаемость подроста пихты сибирской на вырубках, классифицирует его на три кате
гории жизнеспособности: надежный, менее надежный и ненадежный. Д.И. Дерябин, А.Д. Букштынов (1970) при изучении лесоводственного значения хвойного подроста подразделяли его на жизнеспособный и сомнительный. В.Д. Касимов (1960) классифицирует еловый подрост под пологом леса на устойчивый, менее устойчивый и ненадежный.
Существует мнение, что пихтовый подрост, который способен длительное время произрастать в угнетенном состоянии, а в условиях достаточной освещенности восстанавливать жизнеспособность, всегда благонадежен. С таким мнением согласиться нельзя.
Улучшение или ухудшение роста подроста под пологом - это приспособление пихты к изменяющимся условиям окружающей среды. Однако в природе ни один живой организм, в том числе и растение, не располагает способностью к беспредельному приспособлению к значительным изменениям внешней среды. Поэтому при учете пихтового подроста под пологом леса необходимо классифицировать его по категориям жизнеспособности.
Ныне применяемые методы учета подроста хвойных пород под пологом леса разработаны в основном для равнинных и малопригодны для горных лесов. Закономерности приспособлений подроста в горных условиях к изменению внешней среды и пределы пластичности приспособления не выявлены. Не установлены конкретные изменения морфологических признаков, характеризующие неблагонадежный пихтовый подрост под пологом леса. Слабо изучена динамика восстановления жизнеспособности угнетенного подроста на лесосеках после рубки.
Цель наших опытов - изучить морфологические особенности роста и развития подроста пихты под пологом высокополнотного насаждения, установить морфологические признаки, характеризующие подрост по категориям жизнеспособности, проследить рост и развитие подроста различных категорий в процессе восстановления жизнедеятельности в окнах группово-выборочных рубок, разработать методику классификации подроста.
Исследования проведены на стационарных пробных площадях, заложенных в окнах группово-выборочных рубок, в 1967-1972 гг. в хвойных лесах Кучба-Яштинского лесничества Гудаутского лесхоза. В этих лесах преобладают разнотравные группы типов леса. Они характеризуются хорошим естественным возобновлением и высокой производительностью (бонитет I). Опыты по изучению жизнеспособности пихтового подроста и его выживаемости заложены в буково-пихтовом ясменниковом типе леса I класса бонитета. Высота над уровнем моря 1200 м. Экспозиция юго-восточная, уклон 10-15°. Почвы - мощные лесные буроземы. Древостой разновозрастный, состав 6Пх4Бк ед. Ил. Количество деревьев на 1 га 455, из них в первом ярусе 82. Средняя высота 39 м, средний диаметр 72 см. Во втором ярусе произрастает 92 дерева со средней высотой 25 м и диаметром 36 см. Количество деревьев на 1 га в третьем ярусе 281 шт. Их средняя вы
сота 12 м, средний диаметр 16 см. Общая сомкнутость древостоя 0,9, полнота 0,8, запас 752 куб. м/га. В напочвенном покрове, степень покрытия, которого не превышает 0,2, преобладают ясменник душистый, герань Роберта, реже встречаются овсяница горная, кислица, фиалка лесная, вороний глаз.
Естественное возобновление под пологом леса хорошее. Так, самосев до двух лет составляет 6,65 тыс. шт. на 1 га, подрост 3-5 летний -1,10 тыс. шт., 6-10 летний - 1,05 тыс. шт., а свыше 10 лет - 3,9 тыс. шт., в том числе пихты высотой 0,5 м и выше 3 тыс. шт. на 1 га.
Нами изучено 180 штук пихтового подроста в четырех окнах группово-выборочных рубок диаметрами 20x20 м и свыше 1600 ппук - под пологом высокополнотного леса. В окнах изучали один и тот же подрост до и после рубки. Возраст его колебался от 20 до 70 лет, высота - от 0,5 до 4 м (табл. 18).
Исследования показали, что различные экологические условия под пологом леса обусловили формирование разнокачественного подроста, отличающегося внешними морфологическими признаками. Это легко определяемые в полевых условиях внешние морфологические признаки были положены в основу классификации пихтового подроста. Следует отметить, что В.Д. Касимов (1960), З.Н. Науменко (1962), Д.А. Комиссаров и Л.П. Штейнвольф (1967), Д.И. Дерябин и А.Д. Букштынов (1970) также предлагают различные классификации елового подроста разных категорий жизнеспособности, основанные на морфологических признаках. Основными при оценке качества подроста хвойных пород Ю.А. Злобин (1970), Г.Г. Безумова (1977) считают морфологические критерии.
Наши исследования зависимости жизнеспособности пихтового подроста от морфологических признаков (средний годичный прирост в высоту и боковых побегов по длине, отношение прироста пр длине побегов второго порядка к приросту по высоте главного побега, процент протяженности кроны, отношение ширины кроны к ее протяженности, количество живых веток, форма кроны, характер и цвет коры, соотношение световой и теневой хвои) дали возможность, условно, разделить подрост на четыре категории жизнеспособности (см. табл. 18). При дифференцированном изучении подроста различного качества и выживаемости его в окнах группововыборочных рубок выявлены морфологические критерии, характеризующие благонадежный и неблагонадежный подрост.
К первой категории отнесен подрост, развивающийся в условиях слабого угнетения. У него значительный годичный прирост главного и боковых побегов, прирост главного побега превышает 3 см, боковых по длине -9-11 см. Протяженность кроны по стволу превышает 30 % общей высоты. Живых веток в подросте в среднем больше 30. Крона имеет пирамидальную форму, в ней преобладает световая хвоя. Охвоение густое, хвоя крупная, темно-зеленого цвета. Кора светло-коричневая, гладкая. Возраст 20-40 лет.
Таблица 18
Характеристика пихтового подроста по категориям жизнеспособности
Показатель Категории подроста
I П Ш IV
Высота, см 272/742 184/552 145/286 102/200
Возраст, лет 36/30 43/28 56/35 62/28
Численность живых веток 40/742 25/552 17/286 9/200
Протяженность кроны, см 120/32 45/25 22/15 14/14
то же, % 742 552 286 200
Максимальная ширина кроны, см 212/742 130/552 94/286 80/200
Отношение ширины кроны к ее протяженности 1,8/742 2,9/552 4,3/296 5,7/200
Прирост по высоте, см : до рубки (средний за пять лет) 3,86/42 1,82/52 0,98/46 0,20/40
через год после рубки 4,64/42 1,70/52 0,84/46 0,35/40
через два года после рубки 15,28/42 4,38/52 1,44/46 -/40
Прирост побегов второго порядка по длине, см : • до рубки 10,30/42 8,30/52 6,50/46 5,30/40
через год после рубки 10,40/42 7,70/52 6,40/46 4,60/40
через два года после рубки 14,50/42 8,80/52 5,60/46 4,0/40
Отношение прироста по длине побегов второго порядка к приросту по высоте того же года : до рубки 2,80/42 4,56/52 6,63/46 26,5/40
через год после рубки 2,24/42 4,53/52 7,61/46 23,0/40
через два года после рубки 0,94/42 2,01/52 3,89/46 -/40
Примечание. В числителе значение показателей, в знаменателе - численность обмеренного подроста.
Подрост второй категории развивается в условиях угнетения средней степени. Годичный прирост по высоте достигает 3 см, прирост боковых побегов 7-9 см, протяженность кроны - 20-30 % общей высоты. Форма кроны зонтико-пирамидальная. Количество живых веток достигает 30. Ох-воение средней густоты. Хвоя верхней части кроны световая, на нижних ветках - бледно-зеленая. Кора коричневая. Возраст подроста 30-50 лет.
Подрост третьей категории формируется в условиях достаточно сильного угнетения. Годичный прирост по высоте не превышает 2 см. Прирост боковых побегов доходит до 7 см. Протяженность кроны составляет 14-22 % от общей высоты, количество живых веток - до 20 штук; ветки тонкие, расположены горизонтально, поэтому крона имеет зонтикообразную форму. Охвоение слабое, хвоя бледная, мелкая, в нижней части кроны желтеющая, осыпающаяся. Кора серого или бурого цвета. Возраст такого подроста превышает 40 лет.
Подрост четвертой категории жизнеспособности сильно угнетен. Прирост по высоте главного побега не превышает 1 см, иногда отсутствует,
прирост боковых побегов достигает 6 см, протяженность кроны 15 %. Крона типично зонтикообразная, состоит в основном из теневой хвои. Ветки тонкие, расположены горизонтально. Нижние ветки сухие. Живых веток не более 10-11. Охвоение слабое, хвоя мелкая, бледная, зачастую желтеющая, осыпающаяся. Кора бурого цвета, морщинистая. Ствол неровный, его нижняя часть покрыта мхом. Возраст такого подроста свыше 50 лет.
При восстановлении жизнеспособности подроста в окнах группововыборочных рубок важным показателем является прирост по высоте. Эта величина изменяется по-разному у различных категорий (см. табл. 18). Например, подрост IV категории жизнеспособности имеет незначительный прирост по высоте - 0,2 см. Эта величина в 26,5 раза меньше величины прироста боковых ветвей в верхней половине кроны, прирост которых равен 5,3 см.
Такой характер развития кроны подроста обусловил формирование зонтиковидной формы, когда ширина кроны намного превышает протяженность по стволу. Величину отношения ширины кроны к ее протяженности назвали “зонтичностью” кроны. У подроста IV категории зонтич-ность высокая - 5,7. После группово-выборочных рубок рост и развитие подроста изменяются. В первый год после рубки прирост по высоте подроста в окне увеличивается на 75 % и достигает 0,35 м. Прирост по длине побегов второго порядка верхней половины кроны уменьшается на 13,2 %, зонтичность также уменьшается. Однако на второй год у подроста в окне прирост по высоте сокращается до минимума или исчезает полностью. Затем подрост усыхает даже в освещенных окнах. Такой подрост имеет в основном теневую хвою.
Известно, что пихта меняет свою хвою раз в 10-12 лет, а теневая хвоя сильно угнетенного прироста не может приспособиться в первые годы к условиям резкого освещения (Иванов, Коссович, 1932). Слабо развитая корневая система способствует физиологическому усыханию подроста. Характерно, что в IV категорию попал подрост, средняя высота которого составляет 102 см. У подроста с такой высотой условия роста и развития хуже, чем у более высокого, который находится в сравнительно лучших условиях светового режима и менее угнетен. Аналогичное положение отмечали Д.А. Комиссаров и Л.П. Штейнвольф (1967) при изучении интенсивности фотосинтеза подроста ели в разных экологических условиях. По данным А.М. Савченко (1965), освещенность под пологом леса на высоте 2 м в 2-2,5 раза больше, чем над поверхностью почвы.
У более жизнеспособного подроста увеличивается средняя высота. Так, у подроста III категории она равна 145 см. В процессе восстановления жизнеспособности у подроста этой категории в окнах в первый год ухудшается интенсивность роста. Например, прирост по высоте до рубки под пологом составлял 0,98 см, в первый год после рубки в окне прирост
уменьшился до 0,84 см, или на 14,3 %. Только через два года прирост по высоте сравнялся с приростом до рубки и превысил его на 47 % (1,44 см). Уменьшение через два года после рубки величины отношения прироста по длине побегов второго порядка к приросту по высоте примерно в два раза указывает на тенденцию к развитию кроны пирамидальной формы. Однако зонтичность кроны у подроста высокая - 4,3. Рост и развитие подроста III категории в первые годы после рубки в окнах ухудшаются. И только через два года он начинает заметно расти. Ухудшение роста угнетенного подроста при попадании его на простор и резком изменении условий освещенности отмечали также исследователи темнохвойных лесов Кавказа А.Я. Орлов, А.Г. Долуханов, В.И. Мирзашвили и другие.
Аналогично развивается подрост второй категории. Например, при средней высоте подроста 184 см прирост его по высоте до рубки составлял 1,82 см. Через год после рубки величина прироста уменьшилась на 6,6 %. Однако через два года она превысила первоначальный прирост на 140 % и составила 4,38 см. У подроста развивалась крона пирамидальной формы. Отношение прироста по длине боковых ветвей в верхней половине кроны к приросту по высоте уменьшилось больше, чем в два раза, на второй год после рубки. Зонтичность кроны менее выражена и составляет 2,9. Следовательно, подрост этой категории восстанавливает жизнеспособность в окнах группово-выборочных рубок быстрее, чем подрост III категории.
Совершенно по-иному восстанавливается жизнеспособность у подроста I категории. В первый же год после рубки его прирост по высоте увеличился на 26 % , а через два года - еще в 3,3 раза. Возрастает также прирост побегов второго порядка. Он достигает 10,3 см под пологом и 14,5 см в окнах на второй год после рубки. Отношение прироста по длине боковых ветвей верхней половины кроны к приросту по высоте сокращается с 2,8 до 0,94 - почти в три раза. Крона подроста развивается пирамидально. Надо отметить, что пирамидальная форма кроны характерна для хорошего роста. У такой кроны большая площадь боковой поверхности и много световой хвои. Это определяет высокую продуктивность фотосинтеза (Ричардс, 1961; Жилкин, 1965; Тооминг, 1967; Кайрюкштис, 1967; Blackman, Black, 1959; Kuroiva, 1960; Rees, 1962; larvis, 1964; Wareing, 1966; и др.).
Таким образом, слабо угнетенный подрост в окнах после группововыборочных рубок мало страдает, и хотя некоторые экологические условия изменяются, его жизнеспособность восстанавливается в течение вегетационного периода. Через два года после рубки такой подрост в окнах растет в 2-4 раза быстрее, чем под пологом леса.
С понижением степени надежности подроста уменьшается число живых веток второго порядка и протяженность кроны, что объясняется усыханием веток в результате увеличения степени освещенности. И.Г. Серебряков (1962) отмечает, что в неблагоприятных условиях произрастания
ветвей отмирает больше, чем образуется вновь. Результаты наших исследований подтверждают это положение. По мнению П. Крамера и Г. Козловского (1983), отмиранию ветвей предшествует длительный период задержки роста.
С этой точки зрения весьма интересны исследования Д.А. Комиссарова и Л.П. Штейнвольфа (1967), Л.П. Рысина (1970), Кайрюкштиса (1969), Т.Г. Кривицкого (1984), проследивших за динамикой роста угнетенного подроста. Снижение жизнеспособности подроста ведет к увеличению зон-тичности кроны. Развитие зонтикообразной кроны у подроста пихты при сильном угнетении объясняется тенденцией к увеличению развития поверхности кроны и усиления фотосинтеза.
Однако нельзя забывать что возможности приспособления подроста пихты к неблагоприятным условиям произрастания не беспредельны. Если у подроста под пологом леса годичный прирост по высоте сократился до 1 см, численность живых веток второго порядка - до 17, протяженность кроны уменьшилась до 15 %, зонтичность кроны возросла до 4 и отношение прироста по длине побегов второго порядка верхней половины кроны к приросту по высоте достигло 6-7, то такой подрост способен выжить в окнах группово-выборочных рубок диаметром 20 м.
Подрост же с дендрометрическими показателями, превысившими указанные, обречен на гибель. Это положение подтверждено нашими исследованиями. В окнах группово-выборочных рубок часть подроста IV категории после рубки погибла, остальная находится в стадии усыхания. Подрост III категории, показатели которого в связи с рубкой значительно ухудшились, спустя два года начал заметно оправляться.
Таким образом, изучение процесса восстановления жизнеспособности пихтового подроста позволило установить дендрометрические параметры, характеризующие в основном две категории подроста: способность выдерживать новые экологические условия, создаваемые после группововыборочных рубок в окнах и неспособность выдерживать эти условия. Поэтому в практике при учете пихтового подроста под пологом леса достаточно делить его на две категории: благонадежный и неблагонадежный (рис. 13). В наших опытах подрост I-Ш категории оказался благонадежным, IV категории - неблагонадежным. Деление подроста на благонадежный и неблагонадежный имеет важное хозяйственное значение. Только численность благонадежного подроста определяет характер и качество естественного возобновления. Благонадежный подрост служит резервом формирования будущего насаждения, и его необходимо сохранять при эксплуатации лесов.
f - благонадежный
Рис. 13. Подрост пихты кавказской
g - неблагонадежный
Предлагаемый нами метод распределения пихтового подроста на категории жизнеспособности применим на лесосеках добровольно-выборочных рубок, где экологические условия после рубки меняются меньше, чем в окнах группово-выборочных рубок.
Шкала оценки естественного возобновления в пихтарниках
Естественное возобновление леса - процесс динамичный. Его успешность определяется множеством факторов, среди которых основными являются тип леса, структура насаждений, биоэкологические особенности древесных пород и лесорастительные условия.
Решать вопросы лесовозобновления невозможно без глубокого знания эколого-биологических свойств важнейших лесообразующих пород. Отсюда следует, что характер возобновления формации различных видов будет существенно различаться. Поэтому изучение возобновления в лесу и методы его оценки надо проводить по формациям.
Принципы и методологические подходы к изучению характера естественного возобновления и оценки его успешности могут быть общими.
Однако единой оценки успешности возобновления леса, которая бы в достаточной степени количественно и качественно отражала и биологические, и хозяйственные аспекты его, не существует.
В настоящее время имеется ряд шкал оценки естественного возобновления, построенных на общих принципах (Нестеров, 1954; Гулисашвили, 1956 и др.). Все они являются числовыми, основанными на количественной характеристике всходов и подроста под пологом леса. Применение их в пихтовых лесах Кавказа, как показали исследования, не дало желаемых результатов. Специальная же методика изучения и оценки возобновления для пихтовых лесов Кавказа не была разработана.
А.Г. Долухановым (1956) была предложена несколько иная шкала применительно к буковым лесам. Она также основана на числовом методе, но отличается своей простотой и практичностью применения. В ней количественная характеристика возобновления бука различных возрастов выражена в легко сравнимых цифрах. По А.Г. Долуханову, число всходов одного возраста условно принимается эквивалентным какому-то другому числу подроста иного возраста. В частности, 1000 шт. всходов однолетнего возраста эквивалентны 400 шт. - 2 лет, 300 шт. - 4 лет, 100 шт. старше 10 лет. В дальнейшем эта идея использована при разработке местных шкал оценки возобновления лесов Северного Кавказа (Калуцкий, 1972; Беленко, 1964). Однако без специальных экспериментов такая условность перевода подроста пихты низших возрастных рангов в высшие может привести к необъективной оценке характера лесообразования.
В настоящее время согласно Правилам рубок главного пользования в горных лесах Северного Кавказа (1993) за нормативное количество подроста под пологом древостоев пихты, обеспечивающее естественное возобновление вырубок, принято не менее 8 тыс. шт./га мелкого подроста высотой 0,1-0,5 м, 5 тыс. - 0,6-1,5 м и 4 тыс. ^крупного подроста (1,5 м). Однако и эта шкала не дает в полной мере объективной оценки естественного возобновления. В связи с этим нами для пихтовых лесов была разработана новая, на наш взгляд, более совершенная шкала оценки возобновления.
Исследования проводили в пихтарниках Кавказа, произрастающих на южном и северном макросклонах. В ходе изучения лесообразовательного процесса выявили роль всех основных факторов, влияющих на характер и успешность возобновления. С этой целью в девственных лесах и на лесосеках выборочных рубок глазомерно в определенных границах одного фитоценоза закладывали пробные площади размером 0,25-1 га, по диагонали через 3 м - учетные площадки (3x3м). На стационарных пробных площадях возобновление изучали в течение нескольких лет на одних и тех же учетных площадках (10x10м), самосев по возрастным группам, подрост -по разработанной нами методике диагностики и оценки состояния с деле
нием его по категориям жизнеспособности (благонадежный, неблагонадежный) и с учетом показателя встречаемости.
При оценке успешности возобновления леса количественные показатели шкал для разных пород могут различаться. Однако для этого недостаточно только числовых показателей всходов и подроста на 1 га. Существенным моментом при оценке естественного возобновления является характер пространственного размещения самосева под пологом: По нашим исследованиям, в спонтанных пихтовых лесах возобновление под пологом носит неравномерный характер. На учетных площадках пробных площадей количество самосева колеблется от нуля до нескольких десятков и сотен экземпляров. Усредненный числовой показатель возобновления в пересчете на 1 га, если не брать во внимание пространственное размещение самосева, приводит к неправильной оценке возобновления, что подтверждается и исследованиями других авторов (Орлов, 1951; Гулисашвили, 1956; Голгофская, 1967 и др.). Следовательно, для получения более объективной характеристики возобновления и определения его успешности необходимо в шкалу включить коэффициент встречаемости подроста на пробной площади. Он представляет собой отношение числа площадок с наличием возобновления (хотя бы по одному экземпляру) к общему их числу.
В пихтовых лесах целесообразно закладывать учетные площадки размером 3x3 м через каждые 3 м по диагонали пробной площади, что позволит изучить возобновление на площади не менее 200 кв. м (из расчета на 1 га). Репрезентативность учета будет обеспечена статистически. Если численность возобновления на 1 га по местным шкалам достаточна, то при коэффициенте встречаемости 0,7 и больше можно считать, что возобновление покрывает площадь достаточно равномерно и каких-либо мероприятий по содействию ему не требуется. Индекс встречаемости менее 0,5 указывает на неравномерность размещения возобновления. На участке имеется прогалина шириной до 50 м. Здесь необходимо проведение мероприятий по содействию возобновления или частичные культуры.
Основным критерием оценки успешности возобновления принято считать наличие в момент учета под пологом леса достаточного количества благонадежного подроста. Однако, если под пологом высокосомкнутого древостоя по мере роста крупномерного подроста не наступят благоприятные экологические условия (освещенность, влагообеспеченность и т. д.), значительная часть благонадежных экземпляров пихты может перейти в категорию неблагонадежных. Такой подрост не войдет в нижний ярус древостоя. В результате нарушается динамика формирования древостоя, прерывается его развитие по времени. В данном случае возобновление нельзя считать достаточным. Непрерывность развития древостоев - важнейшая характерная биологическая особенность пихтарников Кавказа.
Для более полной и объективной характеристики состояния возобновления целесообразно использовать также процентное отношение де
ревьев нижнего яруса древостоя. Его легко можно установить по материалам перечета деревьев на пробной площади. Численность деревьев нижнего яруса, превышающая 20 % при достаточном количестве подроста на 1 га по местной шкале, указывает на успешный характер естественного возобновления фитоценоза.
Основываясь на биоэкологических особенностях пихты и материалах многолетних исследований лесообразовательных процессов в пихтарниках Кавказа, мы рекомендуем шкалу оценки успешности естественного возобновления (табл. 19). В общем количестве возобновления на долю пихты должно приходиться не менее 60 %.
Таблица 19 ________________Шкала оценки естественного возобновления______________
Оценка возобновления Кол-во благонадежных экз. самосева главных и сопутствующих пород, тыс. шт./га в возрасте, лет Коэфф, встречаемости самосева Кол-во деревьев нижнего яруса в древостое, %
до 5 5-10 старше 10
Хорошее более 10 более 3 более 1,5 более 0,7 более 30
Удовлетворит. 10-5 3-1,5 1,5-1,0 0,7 - 0,5 30-20
Слабое 5-3 1,5- 1,0 1,0-0,5 0,5 - 0,3 20-10
Очень слабое менее 3 менее 1 менее 0,5 менее 0,3 1 менее 10
Рекомендуемая шкала была широко использована при анализе экспериментальных данных, выявлении основных закономерностей естественного возобновления и решении других вопросов, касающихся лесообразова-телъного процесса в пихтарниках. Она оказалась достаточно эффективной.
В табл. 20 представлены данные, характеризующие естественное возобновление в пихтарниках по типам леса и его оценка по шкалам разных авторов. Как видно, в пихтарнике ясменниково-поддесниковым (пр. пл. I) наличие 3181 экземпляра крупного подроста на 1 та вполне достаточно для оценки возобновления по всем шкалам как хорошего. Однако подрост по каким-то причинам не перерастает в достаточном количестве в деревья нижнего яруса древостоя (59 экз./га, 26,1 %).
Это может повлиять на непрерывность развития. Поэтому возобновление в данном случае по нашей шкале оценивается как удовлетворительное. Такая закономерность хорошо подтверждается и данными, полученными на пр. пл. 14 и 52.
Пихтарники папоротниковые, как правило, характеризуются недостаточным естественным возобновлением. Этим и следует объяснить тот факт, что на пр. пл. 40 нижний ярус древостоя представлен сравнительно очень малым числом деревьев (32 экз./га, 9,2 %). Крупного подроста на данной пробной площади насчитывается всего 330 шт./га, что явно недостаточно для обеспечения непрерывности развития древостоя во времени. Возобновление на пробной площади оценивается как очень слабое.
Таблица 20
Характеристика возобновления под пологом леса в пихтарниках _______________________и его оценка_____________________
Параметры Пр. пл. 1 Пр. пл. 40 Пр. пл. 14 Пр. пл. 52
Тип леса (класс бонн- Пихтарник Пихтарник коче- Среднстравно- Овсяницевый
тега) ясмснниково- дыжниковый (I) овсяяицевый, буково- буково-пихтовый
подлссниковын (I) пихтовый (1-1а) лес
Общая сомкнутость крон Число стволов пихты в 0,8 0,7 0,8 0.8
нижнем ярусе древостоя, экз./га (%) Количество подроста, шт./га 59(26,1) 32 (9,2) 96 (28,5) 40(13,8)
старше 10 лет 3181 330 848 1775
5 -10 лет 727 - 2500 -
до 5 лет 3727 - 27250 -
Кол-во всходов шт./га 2181 - - 2500
Возобновление по
шкале: С.М. Бебия удовлетворитель- очень слабое удовлетворительное очень слабое
ное
B.3. Гулисашвили хорошее тоже хорошее слабое
В.Г. Нестерова тоже плохое слабое удовлетворитель-
* ное
Примечание. Пр. пл. 1 и 40 - по данным С.М. Бебия; пр. пл. 14 - К.Ю. Голгофской; пр. пл. 52 - А.Я. Орлова.
Таким образом, на основании изложенного можно сделать следующие выводы.
Изучение естественного возобновления в лесу и методы его оценки должны осуществляться по формациям. Специальная методика для этого применительно к пихтовым лесам Кавказа не разработана. Использование существующих общих и местных числовых шкал малоэффективно. Для определения успешности естественного возобновления недостаточно только числовых показателей всходов и подроста на 1 га.
В шкалу оценки также должны быть включены коэффициент встречаемости подроста и относительное число деревьев нижнего яруса древостоя (в процентах). Предложенная новая шкала оценки успешности возобновления для пихтовых лесов является достаточно объективной для применения ее на практике и в исследовательских целях. Она позволяет установить более точную количественную и качественную характеристики естественного возобновления в конкретных типах леса и наметить системы мероприятий по рациональному использованию и повышению производительности лесов.
ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ СТРОЕНИЯ И ВОЗРАСТНОЙ СТРУКТУРЫ ПИХТОВЫХ ЛЕСОВ
Большинство исследователей темнохвойных лесов в естественных условиях произрастания отмечают разновозрастный и сложный характер их строения и структуры. По мнению исследователей темнохвойных лесов Севера (Тюрин, 1916; Ивашкевич, 1929; Мелехов, 1947; Воропанов, 1950; Алексеев и Молчанов, 1954; Корчагин, 1956; Ипатов, 1969; Столяров, 1979; Дыренков, 1984 и др.), разновозрастность присуща самой природе леса, и естественные девственные леса всегда разновозрастны и разновысотны. К аналогичным выводам пришли исследователи темнохвойных лесов Сахалина (Толмачев, 1954; Агеенко, 1963), Восточной Сибири (Попов, 1957), Сибири (Колесников, 1956; Крылов, 1986), Горной Шории (Ушатин, 1962), Северного Сихоте-Алиня (Васильев, 1962; Розенберг, 1974; Манько, 1967), Карпат (Давыдов, 1956; Ходот, 1959; Молоткова, 1966; Швиденко, 1967), Тянь-Шаня (Ролдугин, 1989) и других регионов. Такого мнения придерживаются и исследователи темнохвойных лесов Европы Лейбундгут (Leibundgut, 1978, 1982), Майер (Mayer, 1969, 1975), Шмидт-Вогг (Schmidt-Vogt, 1977), Томазиус (Thomasius, 1973), Колш (Kolhs, 1974), Вискот (Vyskot, 1978), Шарваш (Sarvas, 1959) и др.
В работах М.В. Давыдова (1940), М.В. Герасимова (1948), В.И. Мирзашвили (1959), А.И. Ильина (1961), П.Н. Ушатина (1962), Л.В. Бицина (1965), Г.Н. Гигаури (1965), В.М. Ломова (1971), С.М. Бебия (1973) и других авторов приведены основные положения и материалы, характеризующие возрастную структуру, строение и таксацию разновозрастных темнохвойных лесов Кавказа. Несмотря на такой обширный список работ, посвященных изучению данного вопроса, "до настоящего времени не выработаны обобщенные, исчерпывающие положения. Выводы исследователей зачастую противоречивы.
Строение, возрастная структура любых лесов определяются лесорастительными условиями, биологическими и экологическими особенностями пород, образующих эти леса, и зависят также от их происхождения, дальнейшего развития и степени воздействия человека. Закономерности строения и возрастной структуры пихтарников Кавказа имеют свои особенности. Изучение этого вопроса имеет важное значение для познания биоэкологии вида, при инвентаризации лесного фонда, проектировании рациональных методов лесохозяйственных мероприятий.
Классификация лесов по их происхождению и влиянию экзогенных факторов
Вопрос классификации лесов в зависимости от истории их происхождения, а также от степени нарушенности естественного хода их развития в пространстве и во времени в результате влияния экзогенных факторов давно интересует исследователей.
Характеризуя степень нарушенности лесов, ученые подразделяли все леса на девственные, первобытные, вторичные и пр. Однако среди исследователей по лесоводству и геоботанике все еще нет единого подхода к понятию “девственный лес”.
В лесоводственной литературе долгие годы было принято считать отличительной особенностью девственного леса разновозрастность, разно-высотность и развитие лесов без влияния хозяйственной деятельности человека. Исходя из этого^ любые разновозрастные леса естественного происхождения отождествлялись с понятием девственного, или первобытного леса (Ивашкевич, 1929; Воропанов, 1950; Толмачев, 1954; Алексеев, Молчанов, 1954; Смолоногов, 1968; Шавнин, 1968; Корчагин, 1976; Вальтер, 1962 и др.).
В.М. Толмачев (1954), считая закономерной разновозрастность девственных темнохвойных лесов, к девственному относит лес, не испытавший когда-либо разрушающего влияния огня.
Аналогичного мнения придерживается и М.Г. Попов (1957). По его мнению, из-за лесных пожаров в настоящее время в Восточной Сибири не сохранилось девственной тайги.
Однако следует подчеркнуть, что в любой тайге всегда имело и имеет место разрушающее действие пожаров и, как отмечает М.М. Орлов (1927), нельзя считать лес вторичным, если он возник и продолжает развиваться после воздействия стихийных или природных явлений (пожара, бурелома), по закономерностям, характерным для девственных лесов. По его мнению, девственный лес отличается от хозяйственного лишь отсутствием рубок.
Исследователи темнохвойных лесов Севера (Воропанов, 1950; Алексеев, Молчанов, 1954; Корчагин, 1956 и др.) также пришли к выводу, что разновозрастность и разновысотность всегда присущи природе девственных лесов. Однако они считают, что одновозрастные еловые леса Севера, возникшие в результате пожаров, через несколько поколений снова превращаются в разновозрастный и разновысотный лес при отсутствии вмешательства в них пожаров и человека.
По данным В.К. Захарова и В.Е. Ермакова (1959), девственные кедровые леса Забайкалья одновозрастны, и возникли они после пожаров или ветровалов.
В работах Вессели (Wessely, 1853), Димица (Dimitz,1905), Чермака (Tschermak, 1910) указывается (цитируем по В.З Гулисашвили, 1970), что девственные леса очень близки по своей форме к выборочному лесу. Основанием для такого утверждения они считали то, что условия роста деревьев и характер возобновления в насаждении в девственном лесу такие же, как и в выборочном и даже в группово-выборочном лесу.
Следует отметить, что по исследованиям А.Я. Орлова (1951), Л.А. Кайрюкштиса (1969), А.М. Савченко (1970), И.В. Веселова (1973), С.Г. Синицина в соавторстве (1979) и других, особенности естественного возобновления в девственных лесах в ряде случаев могут мало отличаться от возобновления на лесосеках постепенных, а иногда и условно сплошных рубок. Однако считать равнозначными понятия “девственный лес” и “выборочный (хозяйственный) лес” нельзя, так как в них характер дифференциации деревьев, основные закономерности формирования, динамики и продуктивности лесов различны. К примеру, запас спелых одновозрастных темнохвойных древостоев, в частности, лесных культур, может быть большим по сравнению с девственными разновозрастными древостоями при равном возрасте господствующего поколения (Ткаченко, 1929; Cooper, 1961). По другим данным, текущий прирост в среднем выше всего в девственных древостоях (Rubner, 1960).
По Б.П. Колесникову (1974), для тайги девственными считаются леса, не испытавшие даже косвенного, отдаленного влияния хозяйственной деятельности человека. Такие леса, по его мнению, исчезли уже с начала XX века с лица земли, с чем, по нашему мнению, согласиться трудно. Леса, не подвергшиеся хозяйственной деятельности человека, но испытавшие разрушительное влияние пожаров, он относит к первобытным. Современные леса Б.П. Колесников разделяет на природные, антропогенные и хозяйственные.
По мнению В.Б. Сочава (1930), к первобытным относятся разновозрастные леса, не подвергшиеся посторонним воздействиям извне, но*уже в течение времени, соответствующего максимальной долговечности в определенных условиях породы эдификатора, не обнаруживающие следов смены растительности как результата восстановительного процесса в нарушенной ассоциации.
Здесь под первобытным, надо полагать, имеется в виду девственный лес. Как отмечает М.В. Герасимов (1948), термин “первобытный” содержит скорее всего понятие доисторического леса, действительно исчезнувшего с лица земли.
Близкое определение девственного леса в понимании В.Б. Сочава дает Баселер (Beseler, 1932), согласно которому девственный лес - это такая растительность, которая в отношении состава, строения и роста выступает как климатически обусловленный конечный член сукцессионного ряда
(Rubner, 1960). Следовательно, девственный лес в таком определении представляет климаксовую стадию развития лесного сообщества. В понимании Клементса (Clements, 1928), климаксовым называется сообщество, в котором завершена сукцессия. Центральный вопрос концепции климакса -относительная стабильность сообщества (Уиттекер, 1980).
Однако в отношении лесного сообщества понятие стабильность зачастую является условным. Учитывая флуктуацию среды, трудно ожидать, что сообщество будет абсолютно и постоянно стабильным в пространстве и во времени. На этот момент обращают внимание также В.З Гулисашвили (1970), Г. Вальтер (1982). В связи с этим девственный лес, как и всякое природное явление, не может быть заключительной стадией развития сообщества. Наоборот, климаксовое состояние является одной из стадий развития девственного леса. Он представляет систему, развивающуюся в пространстве и во времени под воздействием присущих ей внутренних и внешних противоречий.
Такой подход к Пониманию природы лесов естественного происхождения позволил Б.А. Ивашкевичу (1929) обобщить основные закономерности строения древостоев и направления динамических процессов, протекающих в сложных лесах Дальнего Востока, разработать теорию строения и развития древостоев “девственных лесов”, построить стадийную схему (8 стадий) развития одного поколения кедра в составе приморских “девственных лесов”.
Последующие исследователи дальневосточных лесов В.А. Розенберг, Ю.И. Манько (1959), Н.Г. Васильев, Б.П. Колесников (1962), Г.Э. Курин-цова (1973), Ю.И. Манько (1967, 1987) и другие, развивая теорию “девственного” леса Б.А. Ивашкевича, выявили основные закономерности возрастно-восстановительных смен, связанных с развитием лесных эдифика-торов, в том числе различных видов пихт, и установили несколько стадий или фаз развития в процессе жизни одного поколения в девственном лесу.
Классифицируя леса по происхождению, И.С. Мелехов (1980) разделяет их на естественные и искусственные. Он отмечает, что естественные леса могут быть девственными, если они не тронуты человеком и стихийными бедствиями; стихийно-естественными, созданными природой, но подвергавшимися стихийным или антропогенным влияниям; направленноестественными, возникшими естественно, но регулируемыми человеком с использованием сил природы. Леса, созданные посевом или посадкой, он относит к искусственным.
Развивая идею разделения лесов на естественные (природные) и искусственные, Ю.И. Манько (1987) разработал оригинальную схему классификации лесов Дальнего Востока в зависимости от их происхождения и влияния экзогенных факторов. В ней, как и у большинства других исследо
вателей, основным отличительным признаком в понимании девственного леса является хозяйственная деятельность человека.
Различие точек зрения у исследователей относительно понятия “девственный лес” обусловлено, главным образом, различной целью и неодинаковым методом подхода к классификации лесов по происхождению и влиянию экзо- и эндогенных факторов, а также региональными особенностями природы лесов. Как известно, закономерности возобновления, возрастной структуры и строения, продуктивность леса зависят не только от лесорастительных условий, биоэкологических особенностей лесообразующих пород, но определяются также характером происхождения и дальнейшего развития леса, степенью воздействия человека, географией их размещения. К примеру, горные леса Кавказа, в том числе и пихтовые, испытывали влияние человека с незапамятных времен (охотники, пастухи, заготовители кровельной драни и др.), однако из-за сложности рельефа гор и отсутствия дорог это воздействие человека носило несистематический и неинтенсивный характер. Кроме того, основные лесообразующие породы в этих лесах достаточно долговечны, доживают до более чем 300-летнего возраста. Поэтому следы подобных давних воздействий в этих лесах даже в течение жизни одного поколения древостоя обнаружить практически невозможно. Леса, как правило, сохраняют основные черты своего развития.
В силу специфических климатических условий Кавказа, влажности воздуха и большого количества осадков в пихтовых лесах до 2000 мм в год, пожары бывают редко. Однако и леса, испытавшие в далеком прошлом пожары, в силу биоэкологических особенностей эдификатора породы в течение жизни одного поколения способны восстанавливать первоначальные особенности развития. Следовательно, в лесах, где не ведется систематическое хозяйство, отличить девственный от “недевстаенного” леса практически невозможно, и отрицать существование девственных лесов нельзя.
Учитывая особенности развития лесов Кавказа при их классификации по происхождению и влиянию экзо- и эндогенных факторов, считаем целесообразным разделить леса на естественные и искусственные. При этом естественные леса могут быть девственными и хозяйственными.
К девственным следует относить леса, не подвергавшиеся разрушительным экзогенным воздействиям, и те, которые после воздействия этих факторов восстановили присущие им особенности и закономерности своего первоначального развития.
Хозяйственные леса формируются в результате целенаправленной деятельности человека.
Искусственные леса - фитоценозы, созданные путем посева или посадки.
К сожалению, девственных лесов, в том числе и пихтовых, на Кавказе с каждым годом становится все меньше. Поэтому определение понятия девственного леса и классификация лесов по происхождению необходимы не только с целью познания природы лесов, но и для разработки важнейших практических мероприятий по их охране и организации рационального хозяйства в них.
Строение и возрастная структура пихтовых лесов
Для установления особенностей строения и возрастной структуры древостоев нами были проведены исследования в пихтарниках бассейнов рек Ингур, Кодор, Бзыб, Псоу, Мзымта - в Закавказье и рек Белой, Тебер-ды - на Северном Кавказе. Такое размещение работ позволило наиболее полно изучить пихтовые леса в различных условиях местопроизрастания на Западном Кавказе.
Исследования проводились в девственных, чистых пихтовых и смешанных буково-пихтовых насаждениях. Экспериментальным материалом послужили данные 38 пробных площадей, заложенных в насаждениях преобладающих типов леса (табл. 21). Кроме того, использованы данные шести пробных площадей'М.В. Герасимова (1948), заложенных в пихтарниках Кавказского и Тебердинского госзаповедников на Северном Кавказе (1 -КГЗ, 2 - КГЗ, 3 - КГЗ, 1 - ТГЗ, 2 - ТГЗ, 3 - ТГЗ).
Таблица 21
Распределение пробных площадей по типам леса и бонитетам
Группа типов леса N пробных площадей по бонитетам Итого, штук
Та I II Ш
Пихтарники овсяницевые 65 12, 36,41,64, 67, 1 -КГЗ, 3 - КГЗ, 1 - ТГЗ, 2 -ТГЗ З-ТГЗ - 11
Пихтарники разнотравные 2, 34 1, 3, 4, 9, 16, 22, 23, 24, 27, 29,31,32,33,35 14,21, 61,62 - 20
Пихтарники папоротниковые 6, 26 15, 25, 28, 2-КГЗ 18 - 7
Пихтарники с колхидским подлеском 66 5,7 8,11 19 6
Итого 6 29 8 1 44
Размеры пробных площадей составляли 0,25-1,0 га. В них проводили сплошной перечет деревьев по 4 - сантиметровым ступеням толщины.
Деревья толщиной менее 8 см, высотой до 5 м относили к подросту. Высоту каждого дерева отмечали глазомерно, у 15-20 деревьев измеряли высотомером. Параллельно на лесосеках свежих рубок у срубленных де-реьев измеряли высоту, возраст и диаметр на высоте груди. Кроме того, на
пробных площадях 21, 23, 24, 25, 26, 27, заложенных в пихтарниках Аиб-гинского лесничества Гагринского лесхоза, нами были вырублены сплошь деревья толщиной 8 см и выше полосой вдоль стороны пробной площади шириной 5 м и длиной 100-120 м. На этих пробных площадях вырублены 120 деревьев пихты и проанализированы методом сложного анализа.
Использованы данные срубленных деревьев при проведении опытных рубок на пробных площадях 32, 34. На 12 пробных площадях составлены вертикальные профили древостоев.
В целом проанализировано 1877 срубленных деревьев пихты. Для выяснения таксационной структуры и строения пихтарников материалы исследования были обработаны методом вариационной статистики, которая дала возможность получить объективные количественные показатели и подтвердить достоверность выводов. В качестве методической основы при статистической обработке материалов были использованы работы А.В. Тюрина (1961), В.К. Захарова и др. (1962), К.Е. Никитина (1966), Н.Л. Леонтьева (1966). При установлении строения и возрастной структуры пихтарников основное внимание было уделено анализу таксационных показателей древостоя - возрасту, диаметру и высоте, так как эти показатели являются исходными данными для определения остальных таксационных показателей (запас, бонитет и др.). Результаты наших исследований по освещаемым вопросам, глубокий анализ литературных данных послужили материалом для установления закономерностей строения и возрастной структуры пихтарников. Они позволили обосновать рекомендуемые методы их таксации и некоторые лесохозяйственные мероприятия.
Ниже излагаются результаты исследования.
Данные наших исследований свидетельствуют об исключительно сложном строении и возрастной структуре пихтовых и буково-пихтовых насаждений.
Характерной чертой их является значительный диапазон колебаний диаметров ствола во всех типах леса. По нашим данным, они составили от 80 до 148 см для пихты и 8-120 см для бука. В табл. 22 приведены данные статистических показателей варьирования диаметров пихты для 10 пробных площадей по бонитетам и наиболее распространенным типам леса.
Степень варьирования диаметра во всех типах леса и классах бонитетов значительна (см. табл. 22). Так, в овсяницевых пихтарниках коэффициент вариации диаметра ствола пихты составил 54,4 % (пробная площадь 41). В разнотравных группах типов пихтарников эта величина равна 42,7-63,5 % (пробные площади 2, 21, 22), в папоротниковых пихтарниках -41,6-72,1 % (пробные площади 6, 13, 18). В группе типов пихтарников с колхидским подлеском коэффициент вариации диаметров пихты составляет 50,1-60,9 % (пробные площади 5, 11, 19). Значения коэффициентов вариации диаметров пихты, полученные нами, близки к данным, вычисленным по мате
риалам пробных площадей М.В. Герасимова (пробная площадь 1 КГЗ, 2 КГЗ, 3 КГЗ, 1 ТГЗ, 2 ТГЗ, 3 ТГЗ), а также к данным Л.В. Бицина (1965) для пихтарников Северного Кавказа и равные 30-73 %.
Таблица 22
Варьирование диаметра в пихтовых и буково-пихтовых _______насаждениях по типам леса и бонитетам
N пробных площадей Тип леса, бонитет Состав древостоя Предельное варьирование диаметра, см • Статистические показатели варьирования
М±тм о V Р
1 2 3 4 5 6 7 8
41 Пихтарник овсяницевый, I 8Пх2Л+ ЕБк 8-80 30.8±1.3 16.7 54.4 4.1
2 Пихтарник ясмен-никовый, 1-а 9Пх1Бк 12-132 49.6±1.9 30.7 61.9 3.8
22 Пихтарник кислИ-циево-ясменниковый, I ЮПх 16-120 52 0±1.4 22.2 42.7 2.6
21 Пихтарник ясменниковый, П 9Пх1Бк 8-88 35.3±1.3 22.4 63.5 3.7
6 Пихтарник страусе-перо вы й, 1а | 9Пх1Бк 8-116 48.5±1.8 29.5 60.8 3.6
13 Пихтарник кочедыжниковый, 9Пх1Бк 12-120 67.3±2.2 28.0 41.6 3.2
5 Буково-пихтовый черничниковый, I 8Пх2Бк 8-108 54.3±2.0 27.4 50.1 3.7
11 Буково-пихтовый черничниковый, II 6Пх4Бк 16-120 52.9±2.2 27.2 51.4 4.1
19 Елово-пихтовый падубоволавровишневый, 4Пх4Е 1С1Лп 8-84 24.8±0.9 15.1 60.9 3.7
111
Следует отметить, что значение коэффициента вариации диаметров, полученное нами, для разновозрастных пихтовых и буково-пихтовых насаждений в 1,3-3,6 раза превышает коэффициент вариации диаметра одновозрастных нормальных древостоев. Последний, по данным А.В. Тюрина (1961), колеблется в пределах 20-32 %. Исследования показали, что для пихтовых и буково-пихтовых древостоев характерно неравномерное распределение числа стволов по ступеням толщины.
Кривые на рис. 14 характеризуют распределение стволов пихты (кривая 1, 3) и бука (кривая 2) по ступеням толщины на пробной площади 32.
Ступени толщины, см
----Пихта - - - Бук — - Пихта
Рис. 14. Распределение стволов пихты по ступеням толщины
Она заложена в буково-пихтовом ясменниковом типе леса I класса бонитета, состав древостоя 6Пх4Бк. Кривая 3 характеризует распределение стволов пихты по ступеням толщины на пробной площади 23, заложенной в пихтово-ясменниковом типе леса I класса бонитета. Состав древостоя 9Пх1Бк.
Как видно из рис. 14, кривые распределения стволов по ступеням толщины многовершинные. При этом, с увеличением ступеней толщины наблюдается тенденция к уменьшению числа стволов и не соблюдается концентрация их вокруг среднего диаметра, как это свойственно одновозрастным древостоям.
В таких одновозрастных насаждениях определение среднего диаметра древостоя, как отмечал М.М. Орлов (1916), не имеет никакого теоретического и практического смысла. Об этом указывал позже и Б.А. Ивашкевич (1929) в отношении разновозрастных кедровых лесов Дальнего Востока. Сложный характер строения пихтовых лесов подтверждается значительной разновысотностью деревьев в насаждениях. Данные, свидетельствующие об этом, приведены в табл. 23. Она составлена для пробной площади 34, которая заложена в буково-пихтовом ясменниковом типе леса I класса бонитета. Древостой состоит из деревьев высотой до 46 м. В связи с этим полог насаждения имеет вертикально-сомкнутую форму. Распределение стволов по ступеням высот также многовершинно (рис. 15). Коэффициент вариации высот составил 47.9 (V = 47,9, ст = 7,2, Р = 3,0).
Значительны также колебания и изменчивость возраста деревьев в пихтарниках. В пределах одного таксационного выдела встречаются деревья пихты в возрасте до 500 лет. Распределение стволов по классам возраста носит неравномерный характер. В пределах классов возраста наблюдается значительное колебание диаметров стволов пихты. Так, в V классе возраста наименьший диаметр равен 12 см, наибольший - 44 см. В X классе представлены деревья диаметром от 12 до 80 см (табл. 24). Колебание возраста в пределах ступеней толщины достигает от 120 до 240 лет (табл. 25).
Данные, характеризующие распределение стволов пихты по классам возраста, приведены в табл. 24. По этим данным, коэффициент вариации возраста для деревьев пихты диаметрами на высоте груди от 12 до 100 см составил 49.1 % (V = 49.1, ст = 13.3, Р = 2.6). Эта величина для пихтарников Северного Кавказа, по данным Л.В. Бицина (1965), достигает 60 %.
Таблица 23
Распределение числа стволов пихты по ступеням высоты и толщины на пробной площади 34 __
Ступени толщины см Ступени высот, м Ито-го Средневзвешенная вы- сота, м
8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46
12 16 20 24 28 32 36 40 44 48 52 56 60 64 68 72 76 80 84 88 30 2 9 3 21 10 2 13 11 5 5 23 8 1 2 1 5 7 1 1 4 2 1 1 3 7 1 6 1 1 1 4 6 1 1 V 2 2 1 1 2 2 3 Г 1 1 5 2 1 1 2 1 1 1 1 1 2 1 1 1 1 1 1 1 1 1 1 1 78 56 34 21 10 15 6 5 5 5 1 1 3 2 1 2 9,4 11,4 13,7 18,6 20,1 21,8 22.7 25.2 26,6 26,8 32,0 35,0 363 35,0 38,0 43.0
Итого количество деревьев 32 12 33 29 36 4 12 2 7 1 11 7 14 6 10 8 - 3 • 3 2 2 • 2 • 1 2 2 2 - 1 • • - • • 1 245 14.9
Таблица 24
Распределение деревьев пихты по классам возраста и ступеням толщины
Ступени Классы возраста Ито- Средне-
толщины IV V VI vn VIII IX X XI XII ХШ XIV XV XVI XVII XVIII XIX XX XXI го взвешенный
см возраст, лет
12 9 5 3 8 1 3 1 - 30 по
16 12 4 6 10 6 3 1 - 42 113
20 4 3 3 1 1 2 1 - 15 113
24 2 13 2 - 1 1 2 1 22 102
28 - 2 2 3 - 1 2 - 1 - 11 143
32 - 2 2 2 3 - 4 1 - 1 - - 15 155
36 - 1 4 - 1 1 1 - 8 148
40 - 1 1 4 7 2 - 5 4 - 1 ' 25 174
44 1 2 2 1 4 2 1 - 1 - 1 - - - - - - 15 169
48 - - 2 1 3 6 2 2 1 2 - 1 - - - - - - 20 182
52 2 1 2 2 5 3 1 1 - - 1 - - - - - 18 187
56 1 2 - 2 3 1 - 1 2 1 13 199
60 - 3 2 1 1 4 1 1 3 - 1 - - - - 17 207 О
64 - 1 - 2 1 2 3 2 2 1 - - 1 - - - 15 211
68 - - - 1 1 - 2 - 4 2 - 3 - 1 - - - 14 254
72 - - - 1 3 1 1 6 1 3 - - - 1 1 - - 18 220
76 - - - 2 2 3 - 2 1 - 2 1 1 1 - - 15 258
80 - 1 - 1 - 2 - 3 - 1 - 1 - - 9 268
84 - - - - 1 - - 2 - 1 - 2 - 1 - - - 7 259
88 - - - 2 - 1 - - - 1 1 1 1 7 316
92 1 1 - - 1 - 2 - 1 1 - 7 307
96 2 1 1 2 1 1 2 - 1 - 11 299
100 1 2 - 2 - - 1 - 6 317
Итого 27 32 31 38 34 33 31 36 16 19 16 11 10 8 8 5 4 1 360 -
коли-
чество
деревьев
Таблица 25
Зависимость возраста и высоты деревьев от ступени толщины
Ступени толщины, см Возраст, лет Высота, м
макс. мин. средний колебан. макс. мин. средняя колебан.
12 190 70 ПО 120 12 7 9.4 5
16 190 70 113 120 16 8 11.5 8
20 190 70 113 120 18 10 13.3 8
24 210 70 102 140 21 12 16.3 9
28 230 90 143 140 24 15 18.8 9
32 250 90 155 160 27 18 22.8 9
36 250 90 148 160 29 21 24.4 8
40 270 90 174 180 29 22 26.0 7
44 290 90 169 200 32 22 272 10
48 290 ПО 182 180 32 25 27.9 7
52 310 ПО 187 200 34 24 28.6 10
56 310 ПО 199 200 34 27 30.4 7
60 330 130 207 200 40 32 34.6 8
64 350 ПО 211 240 42 33 37.6 9
68 350 150 254 200 44 38 38.6 11
72 370 130 220 240 44 35 39.3 9
76 370 170 253 200 44 35 39.4 9
80 370 150 268 220 44 37 41.2 7
84 350 170 259 180 44 39 42.4 5
88 410 210 316 200 43 39 42.0 4
92 390 210 307 180 43 37 40.2 6
96 390 230 299 160 44 40 42.5 4
100 390 270 317 120 44 42 43.0 2
Исследования показали, <jto в разновозрастных пихтарниках наблюдается сложное соотношение основных таксационных показателей древостоя, прежде всего диаметра, высоты и возраста. Соотношение между ними можно широко использовать для определения таксационных показателей древостоя. Нами выявлено, что между диаметром и высотой деревьев нет прямой функциональной зависимости. С этой целью изучили зависимость между высотой и диаметром большого числа деревьев пихты в разнотравной группе типов леса I класса бонитета. Данные приведены в табл. 26. Колебание высот от 7 до 44 м и коэффициент вариации, равный 54,7% (V = 54.7, ст = 10.3, Р =2,4), еще раз свидетельствуют о высокой степени изменчивости высот пихтарников. Статистическая обработка данных показала высокую корреляционную связь между высотой и диаметром пихты. Коэффициент корреляции составил 0.958, корреляционное отношение 0.989. Эти величины для данных пробной площади 34 (см. табл. 23) при меньшем числе стволов, соответственно равны 0.944 и 0.963. Связь между диаметром и высотой довольно высокая, и зависимость между ними приближается к прямолинейной функции. Следует отметить, что увеличению диаметра
Таблица 26
Распределение числа стволов пихты по ступейям высот и толщины
Ступени толщины см Ступени высот, м Ито-го Средневзвешенная высота, м
7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44
12 16 20 24 28 32 36 40 44 48 52 56 60 64 68 72 76 80 84 88 92 96 100 20 8 5 22 5 34 19 2 25 21 7 3 50 13 1 9 6 2 8 18 2 3 4 6 1 1 3 10 1 2 7 2 1 6 5 1 2 4 2 1 1 1 3 1 1 7 3 1 6 2 2 1 1 5 3 2 2 1 2 5 1 2 1 1 5 2 4 1 1 1 2 2 3 1 1 3 3 1 2 1 1 2 1 2 1 1 1 1 1 1 1 1 1 1 2 1 1 1 1 3 1 1 1 1 1 1 1 2 1 2 1 1 1 3 2 1 1 1 1 1 1 1 2 2 1 1 2 1 2 1 1 2 1 1 2 1 1 1 3 1 1 1 1 1 2 2 1 112 121 56 37 18 27 17 17 13 13 11 5 9 12 .9 8 7 5 5 4 5 4 2 9,4 11,5 13,3 16,3 18,8 22,8 24,4 26,0 27,2 27,5 28,0 30,4 34,6 37,6 38,3 39,3 39,4 41,2 42,2 42,0 40,2 42,5 43,0
Итого количество деревьев 20 13 27 55 53 67 17 28 14 15 11 13 6 3 6 12 11 14 11 13 10 11 6 2 3 5 4 7 4 4 5 5 6 7 9 6 5 9 517 18,4
Пихтарники Рицинского национального парка
Побег пихты кавказской
с шишками
Оз. Рица, водоохраннопочвозащитные пихтарники
Для пихты кавказской характерно групповое естественное возобновление
Пихтарники разнотравные
Пихтарники папоротниковые
Пихтарники разнотравные
Пихтарники скально-лесных комплексов
Субальпийские пихтарники
Пихтарники с субальпийским высокотравьем
Рододендрон кавказский (Rhododendron caucasicum Pall.)
Береза Литвинова (Betula litwinowii Doluch.) в субальпийском криволесье
Плодоносящий побег падуба колхидского (Ilex colchica Pojark.)
Оз. Малая Рица, следы лавин
Леса с господством сосны Коха (Pinus kochiana Klotzsch.)
Подрост пихты под пологом древостоя бука восточного (Fagus orientalis Lipsky)
Поперечные срезы подроста пихты кавказской неблагонадежной и благонадежной категорий
Лесовозная дорога в пихтовом лесу
Пихтарник, пройденный промышленно-выборочной рубкой
Субальпийские пихтарники, расстроенные пастьбой скота
Сведение субальпийских пихтарников пасторальными факторами
Южный Приют, пихтарники с нарушенной возрастной структурой
Субальпийская растительность
Начало р. Клич, впадающая в р. Гвандра у Южного Приюта
Начало р. Березовой у водораздельного Гл. Кавказского хребта
Древостой пихты сахалинской (Ab. sachalinensis (Schmidt) Mast.) с примесью березы Максимовича (Betula maksimowiciana Regel) и подлеском сазы (Sasa kurilensis (Rupr.) Makino et Shibata). Хоккайдо, 700 м н. yp. м.
Древостой пихты Кавакама (Ab. kawakamii (Hayata) Т. Ito). Тайвань, 3100 м н. ур. м.
Субальпийские пихтарники. На переднем плане заросли кустарников можжевельника (Juniper us squamata var. morrisonicola (Hayata) Li et Keng) и рододендрона (Rhododendron pseudochrisanthum Hayata). Тайвань, 3600 м н. ур. м.
Сбор семян с плюсовых деревьев для банка семян. Тайвань
Пихтарники на пути из Южного Приюта к перевалу Клухор
ствола пихты соответствует тенденция увеличения высоты. Например, средняя высота деревьев для ступени толщины 12 см соответствует 9.4 м (см. табл. 25), а ступени толщины 28 см соответствуют деревья со средней высотой 18.8 м и т.д.
Зависимость между ступенями толщины и высотой деревьев пихты графически изображена на рисунке 16.
Характерно также, что между высотой и возрастом не наблюдается прямой связи. Так, средний возраст деревьев толщиной 68 см составляет 254 года, средняя высота их 38,3 м, в то же время деревьям толщиной 72 см соответствуют средний возраст 220 лет и средняя высота 39.9 м (см. табл. 25).
Результаты статистического изучения зависимости показали наличие корреляционной связи между диаметром и возрастом древостоя пихты. При этом степень связи между диаметром и высотой более высокая и устойчивая, чем между диаметром и возрастом. Коэффициент корреляции диаметра и возраста составил 0.767, корреляционное отношение равно 0.781.
Выше указано, что характеризуемые величины для диаметра и высоты соответственно равны 0.944-0.958 и 0.963-0.982.
Обращает на себя внимание и то, что в разновозрастных пихтарниках изменчивость возраста существеннее, чем изменчивость высот. Так, амплитуды колебания возраста в пределах ступеней толщины относительно их средних величин составляют от 0.38 до 1.37. Характеризуемые величины для высот менее значительны и выражаются цифрами 0.05-0.69 (табл. 27).
В целом анализ структуры ценопопуляции и фитоценотической структуры сообществ пихты кавказской указывает на абсолютноразновозрастный характер строения и структуры древостоев.
Рис. 16. Зависимость между диаметром и высотой деревьев пихты
Таблица 27
Пределы и амплитуды колебания возраста и высоты деревьев пихты относительно средних их величин в пределах ступеней толщины
Ступени толщины, см Возраст Высота
максимальный минимальный амплитуда максимальная минимальная амплитуда
12 1.73 0.64 1.09 1.38 0.74 0.64
16 1.68 0.62 1 06 1.39 0.70 0,69
20 1.68 0.62 1.06 1.35 0.75 0.60
24 2.06 0.69 1.37 1.28 0.74 0.54
28 1.61 0.62 0.99 1.22 0.80 0.42
32 1.61 0.58 1.03 1.18 0.79 0.39
36 1.70 0.61 1.09 1.18 0.84 0.34
40 1.55 0.51 1.04 1.22 0.74 0.37
44 1.71 0.53 1.18 1.17 0.81 0.36
48 1.59 • 0.60 0.99 1.11 0.89 0.22
52 1.59 0.59 1.06 1.18 0.83 0.35
56 1.05 0.55 0.50 1.11 0.88 0.23
60 1.60 0.64 0.96 1.15 0.92 0.23
64 1.66 0.52 1.14 1.11 0.87 0.23
68 1.37 0.59 0.78 1.14 0.86 0.28
72 1.68 0.60 1.08 1.12 0.89 0.23
76 1.46 0.67' 0.79 1.12 0.88 0.24
80 1.38 0.56 0.82 1.07 0.90 0.17
84 1.35 0.65 0.70 1.03 0.92 0.11
88 1.29 0.66 0.63 1.02 0.93 0.09
92 1.27 0.70 0.57 1.07 0.92 0.15
96 1.30 0.77 0.53 1.03 0.94 0.09
100 1.23 0.85 0.38 1.01 0.97 0.05
Проведенные исследования показывают, что в девственных пихтарниках по площади преобладают абсолютно-разновозрастные древостои, в которых имеются деревья всех возрастов, вплоть до предельного на Кавказе возраста пихты (450-500 лет). При этом численность деревьев по группам (с колебанием возраста 40 лет) падает плавно от младших к старшим (рис. 17). Последнее свидетельствует о непрерывном, более или менее равномерном процессе естественного возобновления.
Абсолютно-разновозрастные пихтовые и буково-пихтовые древостои развиваются только в девственных лесах или в лесах, где экзогенные воздействия были очень слабыми и не оставили заметных следов.
Они формируются на основе непрерывного естественного возобновления и развития молодых поколений пихты под пологом материнских. Для нормального развития таких древостоев обязательно наличие в них спелой и старовозрастной части, постепенное отмирание которой и смена ее новой и обусловливают дальнейшее развитие данных древостоев.
Рис. 17. Распределение числа стволов пихты по группам возраста (1 - 60-100 лет, 2 -101-140 лет, ..., 9 - 380-420 лет)
Встречаются здесь, но значительно реже, и относительно разновозрастные насаждения, а также древостои с переходным типом возрастной структуры с относительно недолговременным циклом развития.
Относительно-разновозрастные древостои принадлежат к коренным типам леса, в которых естественное возобновление в пространстве и во времени происходит неудовлетворительно или носит прерывистый характер, например, в папоротниковом типе леса. Древостой в нем характеризуется значительной амплитудой колебания возраста (61-300 лет), однако, основная масса стволов сосредоточена в двух-трех классах возраста.
Таким образом, исследования свидетельствуют о чрезвычайно сложной возрастной структуре пихтарников Кавказа.
Примечательно, что значительных различий в возрастной структуре и строении чисто пихтовых и смешанных буково-пихтовых лесов, хотя они относятся к разным типам леса, нами не обнаружено. Степень изменчивости таксационных признаков в них практически одинакова. Аналогичные выводы отмечали в своих исследованиях Л.В. Бицин (1965) и Г.Н. Гигаури (1965).
Онтогенез пихты кавказской и возрастная динамика древостоев
В девственном лесу непрерывно происходят трансформации, связанные с изменением возраста и возрастного состояния основных лесообразующих пород, ярусов древостоя, а также с процессами естественного возобновления. Изменения, происходящие в насаждении в связи с онтогене
зом эдификатора, а также в связи с естественным возобновлением древесной растительности, В.Н. Сукачев (1964а) рассматривал на уровне одной из форм динамики фитоценозов.
Б.П. Колесников (1968) смены, происходящие одновременно с возрастными изменениями преобладающего поколения основной лесообразующей породы, называл возрастными и относил их к категории “онтогенетических смен типов насаждений”, куда включал также и такие формы динамики лесов, как “смены поколений” и “восстановительные, или диг-рессионно-демутационные смены”. Б.П. Колесников вполне обоснованно возрастные смены и смены поколений считал как две разные формы динамики древостоев.
С нашей точки зрения, возрастная динамика - непрерывный процесс возникновения, роста и развития, а также отмирания отдельных деревьев, групп деревьев в древостое и смены их новыми.
Возрастные изменения, связанные с непрерывным процессом возникновения, ро?га и развития, а также отмирания отдельных возрастных поколений древесных пород и смены их новыми, следует относить к форме возрастной смены поколений, к типу возрастно-восстановительной динамики лесов. В одновозрастном лесу отделить онтогенетическое развитие в древостое от смены поколений не представляет большой сложности. В девственных разновозрастных лесах онтогенетические изменения происходят одновременно с динамикой поколений, и различить их очень сложно.
В девственных пихтарниках Кавказа, в связи с постоянным процессом естественного возобновления древесных пород, граница между отдельными поколениями насаждений выражена нечетко. Рост и развитие деревьев в разновозрастном лесу носят сугубо индивидуальный характер и зависят преимущественно от места, занимаемого ими в фитоценозе. В силу этого даже одновозрастные особи характеризуются неоднородными морфометрическими показателями.
По Н.П. Кренке (1940), каждая особь в древостое, в процессе роста и индивидуального развития последовательно проходит ряд этапов онтогенеза от эмбрионального до старения. В разновозрастном лесу периодизация возрастных этапов онтогенеза древесных растений и увязка стадии онтогенеза особей с их календарным возрастом довольно сложны. Однако в верхнем пологе одновозрастных и разновозрастных древостоев, как правило, доминируют особи в близком возрастном состоянии.
В одновозрастных лесах возрастные стадии, установленные по календарному возрасту древостоев, четко определяют возрастное состояние особей, формирующих основной полог древостоя. Сообразно этому принципу была разработана широко известная лесоводственная классификация древостоев по возрасту: молодняк, жердняк, средневозрастный, приспевающий, спелый и перестойный древостой. Этот принцип был использован
также для выделения стадий возрастной динамики и в девственном лесу. Но при этом, возрастным сменам подчинены выделяемые Б.П. Колесниковым (1968) смены поколений, и обе формы динамики древостоев, в данном случае, выступают как единый тип в классификациях форм динамики девственного леса.
Теоретические основы изучения возрастных смен в разновозрастных лесах были заложены Б.А. Ивашкевичем (1929). Исходя из положения о скачкообразном развитии девственного леса, он разработал универсальную схему развития разновозрастного леса, в которой для онтогенеза каждого возрастного поколения преобладающей породы выделил 8 стадий развития (классов возраста).
Идея Б.А. Ивашкевича получила дальнейшее развитие в трудах многочисленных исследователей - в частности, в трудах Б.П. Колесникова (1956), В.А. Розенберга (1972, 1978), Ю.И. Манько (1967, 1987), И.В. Семечкина (1970), Н.М. Глазова (1979), Э.Н. Фалалеева (1976), Д.П. Столярова, В.К. Кузнецова (1979), С.А. Дыренкова (1984) и других. Многие из них возрастную динамику разновозрастных лесов рассматривали на примере онтогенеза одного поколения преобладающей древесной породы, начиная с его появления и до отмирания.
Совершенно ясно, что для разных пород количество стадий развития будет различно. Каждая из них отражает отдельный этап онтогенеза и положение особей этого поколения в древостое.
Для лесов Западной Европы классификация стадий или фаз развития девственного леса была разработана классиком европейского лесоводства Гансом Лейбундгугом (Leibundgut, 1959) и его школой. Позже классификация Лейбундгута стала популярной и использовалась исследователями во многих странах мира. Она нашла развитие при освещении строения и динамики древостоев, связанных с особенностями лесообразовательного процесса в разных лесных формациях и регионах (Фалалеев, 1964; Мауег, 1969; Комин и др., 1970; Kolhs, 1974; Дыренков, 1976, 1984; Манько, 1986 и другие).
Учитывая широкую популярность этой классификации, а также стремление отдельных исследователей придать ей универсальный характер, считаем целесообразным остановиться на ней несколько подробнее.
По Г. Лейбундгуту (1959, 1982) девственный лес в своем жизненном процессе проходит следующие фазы (стадии) развития.
1. Оптимальная фаза (Optimalphase) - более или менее сомкнутые крупномерные древостои с высоким древесным запасом и в среднем с высокой жизненностью деревьев. В ранней оптимальной фазе верхняя часть полога сомкнута неполностью, в нее еще могут войти деревья из средней части полога. В поздней оптимальной фазе верхний полог плохо сомкнут.
2. Фаза зрелости (Alterphasa) - крупномерный древостой с отпадом отдельных деревьев и группы деревьев, имеющих высокий возраст. Ранней фазе зрелости соответствует максимальный для данного местообитания запас древостоя, поздней фазе зрелости - не менее половины максимального запаса. В этой фазе в верхнем пологе образуются окна, связанные с отпадом отдельных деревьев или их групп, но окна еще небольшие.
3. Фаза распада (Zerfallsphase) - крупномерный древостой с прогрессирующим изреживанием, при котором запас снижается ниже половины возможного максимального. Окна в пологе увеличиваются, возрастают прогалины, поляны.
4. Фаза возобновления (Verjungungsphase) - изреживание древостоя замедляется, в окнах и полянах имеется обильный подрост.
5. Фаза выборочного леса (Plenterwaldphase) - естественные древостои по строению становятся подобными выборочному лесу, сформирован ступенчатый вертикально сомкнутый полог.
6. Фаза молодого леса (Jungwaldphase) - более или менее однородный по сложению полога молодой древостой, образовавшийся из подроста, занявшего всю площадь после быстрого распада материнского древостоя; это молодняки, чащи и жердняки.
7. Фаза однородного толстомерного средневозрастного и приспевающего древостоя (Phase des gleichformigen starken Stangenholzes und Baumholzes), возникающего при дружном естественном возобновлении на большой площади; похожа на оптимальную, но не тождественна ей, запас древостоя еще незначителен, и деревья в верхней части полога характеризуются интенсивным ростом.
Нетрудно убедиться в том, что в основе данной классификации развития девственного леса лежит идея Б.А. Ивашкевича о возрастновосстановительных сменах поколений в древостоях, в которой нашли отражение, прежде всего, принципиальные аспекты циклического процесса развития древостоев. Такую схему возрастно-восстановительной динамики темнохвойных лесов мы наблюдали во многих регионах, в частности, на экспериментальных участках Верхне-Уссурийского стационара (ВУС 1) биолого-почвенного института ДВНЦ РАН. Схема динамики древостоев в этих лесах включает 6 стадий, каждая из которых длится 20 лет, и общая продолжительность цикла составляет около 120 лет (Розенберг, 1978; Манько, 1987).
Близкие схемы возрастной динамики древостоев рассматриваются для темнохвойных лесов Австрии (Мауег, 1969; Zukrigl, 1983), Европейского Севера России (Дыренков, 1984), Урала (Смолоногов, 1968; Комин и др., 1970), Сибири (Фалалеев, 1964) и других регионов. Однако рассмотренные схемы естественно не могут охватить всего многообразия возрас-
гной динамики лесов, особенно зависимости ее от условий местопроизрастания и структуры ценопопуляции древесных пород.
Общий подход в представлениях о динамике девственного леса, связанной с особенностями лесообразовательного процесса и различными типами возрастной структуры и строения древостоев в разных лесных формациях, может не всегда отражать истинное положение вещей. Об этом в определенной мере свидетельствуют полученные к настоящему времени материалы многих исследователей возрастной структуры и строения древостоев пихты кавказской (Ушатин, 1962; Долуханов, 1964; Бицин, 1965; Бебия и др., 1972; Веселов, 1973; Гигаури, 1973).
Как уже отмечалось выше, для девственных лесов характерны, в основном, два типа развития древостоев: когда отпад старых поколений происходит подеревно или мелкими группами, их место занимают новые более молодые (возрастные смены), и, когда отпад имеет явно выраженный групповой характер, т. е. происходит отмирание отдельных возрастных поколений древесных пород и смена их новыми (возрастновосстановительные или дигрессивно-демутационные смены).
Литературные данные и наши фактические материалы позволяют утверждать, что для пихтарников Кавказа характерен, главным образом, первый вариант развития древостоев. Хорошей иллюстрацией этой мысли могут служить сведения, приводимые в табл. 28. Из них видно, что во всех типах леса доля отмирающих и сухостойных деревьев среди спелой и старовозрастной части древостоя, высотой более 30 м, практически, незначительна и колеблется от 5.1 до 22.2%. Основная масса отпада сосредоточена в молодой части древостоя, что можно объяснить следствием естественного самоизреживания древостоя. Характер отпада в верхней части полога четко указывает на отсутствие признаков распада и смены ее новым.
По данным табл. 29, в девственных разновозрастных древостоях пихты кавказской общий отпад за 10-летний период колеблется от 46 до 107 м3/ га. Эти величины по отношению к запасу 1-го яруса (спелой и старовозрастной части) древостоя составили не более 11.2%.
Безусловно, эти данные подтверждают отсутствие в древостоях пихты отмирающего поколения из спелой и старовозрастной части древостоя и смены его новым более молодым, отсутствие выраженных возрастновосстановительных смен поколений в древостоях.
В настоящее время в литературе часто стали использовать профильные диаграммы, предложенные в свое время П.В. Ричардсом (1952) для изучения строения и структуры надземных частей фитоценозов, в том числе и древесных.
Таблица 28
Характеристика отпада в девственных насаждениях пихты кавказской
Ступени высоты, м Тип леса, класс бонитета
Пихтарник ясменниковый,П Пихтарник падубовый, 1а Пихтарник овся-ницевый, П Пихтарник папоротниковый, I
А Б В А Б В А Б В А Б В
6 22 6 7.1 32 4 11.1 124 68 29.3 84 32 66.7
8 12 4 4.8' 26 4 11.1 120 48 20.7 32 4 8.3
10 22 8 9.5 34 8 22.2 116 32 13.8 16 6 12.5
12 28 8 9.5 26 4 11.1 100 36 15.5 48 -
14 38 4 4.8 16 2 5.6 80 12 5.2 16 -
16 38 12 14.3 14 - 28 12 5.2 6 -
18 50 8 9.5 14 2 5.5 56 8 3.5 38 -
20 50 6 7.1 12 - 36 - 16 2 4.2
22 34 6 7.1 4 - 72 - 8 -
24 34 4 4.8 8 2 5.6 40 - 6 -
26 24 6 7.2 4 2 5.6 32 4 1.7 20 -
28 30 8» 9.5 14 - 44 - 12 -
30 16 4 4.8 8 - 48 - 22 2 4.1
32 14 - 14 - 52 8 3.4 20 -
34 6 - 14 2 5.6 48 - 10 -
36 4 - 6 4 11.1 24 4 1.7 14 -
38 2 - 6 2 5.5 14 -
40 8 10
42 6 - 14 2 4.2
44 4 - 14 -
46 - -
48 4 -
Итого 424 84 100 284 36 100 1020 232 100 420 48 100
Примечание. А - общее число деревьев на га; Б - число отмирающих и сухостойных деревьев на га; В - то же в %
Таблица 29
Характеристика отпада по данным опытных таблиц хода роста девственных пихтовых насаждений Кавказа (по Л.В. Бицину, 1965)
Бонитет, группа типов леса Средний возраст 1-го яруса (основная часть насаждения), лет Запас 1-го яруса, м3/га Общий отпад за 10-летний период
м3/га в % к запасу 1-го яруса
1а; овсяницевые, папоротниково- 140 959 107 11.2
разнотравные 270 1383 93 6.7
I; овсяницевые, папоротниково- 140 782 86 11.0
разнотравные, папоротниковые 270 1159 78 6.7
П; ожиковые, рододендроновые 140 626 64 10.2
270 980 64 6.5
Ш; мертвопокровные, вейниковые 140 • 490 46 9.4
270 791 50 6.3
Несмотря на то, что упомянутые диаграммы отличаются достаточно хорошей наглядностью (рис. 18), однако, как справедливо отмечает В.В. Плотников (1979), они в строгом смысле профилями древостоев не являются. Такие диаграммы, как правило, не дают в полной мере количественную морфометрическую характеристику строения и структуры древостоев, недостаточно полно отражают характеристику их динамики. В связи с этим, в наших исследованиях был использован новый способ количественного выражения представлений о вертикальной структуре фитоценозов, предложенный В.В. Плотниковым (1979), но несколько дополненный нами. Речь идет о способе обработки и обобщения данных пробных площадей или профильных трансектов со сплошным обмером высоты деревьев, протяженности крон и других показателей, связанных с дифференциацией сообществ по вертикали. Если пересечь вертикальную толщу древостоя системой горизонтальных плоскостей через определенный интервал, то ситуация в каждой из них будет одинакова, отражая изменения в профиле древостоя. В качестве основной характеристики ситуации в плоскости можно избрать численность особей, пересекающих данную плоскость. При этом высокое дерево будет учтено в большом количестве плоскостей, чем дерево меньшей высоты. В целом же, благодаря тому, что каждое дерево имеет определенную высоту, общее число наблюдений значительно превысит численность деревьев, достигших высоты наблюдаемой плоскости. Представляется возможным выразить вертикальную структуру древостоя в форме распределения условных дискретных количественных величин, именуемых
Рис. 18. Строение елово-пихтового древостоя I класса бонитета (Piceto-abietum vaccin-ioso-ilexosum)
В.В. Плотниковым “коэффициентами заполнения”. В данном случае коэффициент заполнения представляет собой численность особей в системе горизонтальных плоскостей вертикального профиля древостоя. Основой для анализа и синтетической характеристики древостоя будет распределение коэффициентов заполнения по разрядам высоты, выраженное в относительных величинах. На практике распределение коэффициентов заполнения можно легко получить, пользуясь ведомостью обмера деревьев при закладке профильных трансект в лесу и построения профильных диаграмм. В развитие представлений о вертикальной структуре и динамике девственных древостоев, из рис. 19-22 профильных диаграмм следует, что для древостоев пихты кавказской характерен коэффициент заполнения, величина которого равномерно снижается сверху вниз. Кривые распределения коэффициентов заполнения по разрядам высоты имеют вогнутую форму, иногда близкую к прямолинейной, что указывает на непрерывный характер вертикального строения и динамики древостоев. Только в папоротниковом типе леса кривая распределения коэффициента заполнения по разрядам высоты имеет заметно выпукло-вогнутую форму (рис. 23), что характеризует полог древостоя как относительно неравномерно сомкнутый.
Для сравнения на рис. 24 приводятся профильные диаграммы четырех фаз развития древостоев девственного пихтово-елового леса Шлосберг-Линца (Австрия) из работы Kolhs (1974). Кривые распределения коэффициента заполнения по разрядам высоты (см. рис. 24), составленные нами по этим профильным диаграммам, четко указывают на явно выраженный, неравномерный, скачкообразный характер вертикального строения полога и динамики древостоев. Кривые имеют вогнуто-выпуклую, выпуклую форму. Перегибы в них, отмеченные на рисунках вертикальной ломаной линией, определяют наличие разрывов в вертикальном строении полога древостоев, что характерно в основном для древостоев, которые развиваются по линии возрастно- восстановительных смен.
Интересно, что леса с господством пихты сахалинской на острове Хоккайдо, по нашим наблюдениям, также развиваются по линии близкой к возрастно-восстановительным сменам. Это наглядно отображено на рис. 25, из которого видно, что кривая распределения коэффициента заполнения по разрядам высот имеет выпукло-вогнутую форму.
И, наконец, на рис. 26 приводим изображение вертикального профиля древостоя ельника кисличника, составленного нами на стационарных лесотипологических пробных площадях Литовского НИИЛХ. Из рисунка видно, что кривая распределения коэффициента заполнения по разрядам высот имеет четко выраженную выпуклую форму, которая типична лишь для древостоев, развивающихся по типу возрастно-восстановительных смен поколений. Форма кривой на рис. 26 характерна для второй фазы развития древостоя (Alterphase) по Г. Лейбундгуту (1982).
Высота, м
Коэффициент заполнения, экз.
Коэффициент заполнения, экз.
Экз. Высота,
Коэффициент заполнения, экз.
Высота, м
Коэффициент заполнения, экз.
Экз. Высота
Коэффициент заполнения, экз.
Фазы развития:
Спелая
Старая
Конечная
Возобновительная
О
ю QO
Рис. 24. Диаграмма вертикальных профилей 4-х фаз развития пихтово-елового древостоя из Abies alba и Picea abies (Шлосберг-Линц, Австрия; по J. Kolhs, 1974)
Рис. 25. Диаграмма вертикального профиля елово-пихтового древостоя с подлеском сазы курильской. Породы: 1 - Ab. sachalinensis, 2 - Picea jezoensis, 3 - Betula ermanii, 4 - Ulmus laciniata, 5 - Tilia japonica, 6 - Acer mono, 7 - Quercus mongo-lica var. Grosseserrata, 8 - Acer japonicum
ШШПИффЮЯ
<---------------------1-----------------------1----------------------1 ----- .1______________________________I
0 10 20 30 40 50 м
Рис. 26. Диаграмма вертикального профиля древостоя ельника кисличного второй фазы развития (Alterphase - фаза зрелости по Leibundgut, 1982). Породы: ель -Picea abies, береза - Betula verrucosa
Итак, проведенные исследования позволяют сделать важные для теории и практики выводы о том, что древостои девственных пихтарников Кавказа развиваются по линии возрастной динамики с непрерывным процессом возникновения, роста и развития, а также отмирания отдельных деревьев, групп деревьев в древостое и смены их новыми. Возрастновосстановительный тип динамики путем смены отдельного отмирающего поколения древостоя новым, более молодым, для этих древостоев не характерен.
Эти выводы чрезвычайно важны и должны быть учтены при разработке и осуществлении различных лесоводственных мероприятий в лесу, прежде всего, при осуществлении рубок.
Эти выводы важны и потому, что они основательно опровергают выдвигаемый вновь и вновь отдельными “лесоводами” по конъюнктурным соображениям, порочный тезис о “перестойном” состоянии разновозрастных пихтовых насаждений Кавказа. В действительности же, на протяжении “обозримых отрезков времени эти леса не стареют” и не “молодеют”.
С нашей точки зрения, постоянный процесс естественного возобновления и сложная возрастная структура являются важнейшими факторами основополагающих приспособлений на уровне сообществ, которые обеспечивают пихтовым лесам Кавказа длительное процветание.
Ярусное строение пихтовых древостоев
Большинство исследователей разновозрастных темнохвойных лесов Кавказа, а также других регионов едины во мнении, что сложные соотношения элементов структуры разновозрастных насаждений исключают возможность таксации их как одновозрастных, нормальных древостоев (Давыдов, 1940; Leibundgut,1945; Ушатин, 1962; Фалалеев, 1964; Бицин, 1965; Бебия, 1973, 1989; Гулисашвили, 1970; Недялков, 1979 и другие). Усредненная таксационная характеристика таких древостоев затушевывает фактическое соотношение элементов структуры древостоя, что может привести к ошибочным выводам биологического и практического характера при их инвентаризации. Для этой цели следует считать целесообразным расчленение сложных по строению и возрастной структуре древостоев на отдельные составные части или ярусы. Расчленение разновозрастных насаждений на ярусы отвечает, прежде всего, интересам проектирования лесохозяйственных мероприятий, так как ярусное расчленение древостоев основано на размерности деревьев.
В основу расчленения древостоя на ярусы положены высота деревьев, закономерность распределения числа стволов по ступеням высот (см. табл. 26), соотношение между высотой и диаметром (см. рис. 16), а также закономерности строения нормальных насаждений. Наличие ярусов опре
делилось с помощью графиков распределения числа стволов по ступеням высот (см. рис. 15). Ступени высот с наименьшим количеством стволов принимали за границы ярусов. Такую методику применяли еще раньше М.В. Герасимов (1948), Г.Н. Гигаури (1965), В.М. Ломов (1971) при исследовании разновозрастных лесов Кавказа. Исследования показали, что разновозрастные пихтарники состоят, главным образом, из трех ярусов (см. рис. 18).
Установлено, что выделенные ярусы характеризуются закономерностями строения одинаковыми (I-П ярусы) или близкими (III ярус) с одновозрастными насаждениями. Колебание высот деревьев для первого яруса составляет 0.82-1.17 от среднего значения, для второго - 0.81-1.14 и для третьего яруса - 0.58-1.67. Характеризуемая величина для нормальных древостоев равна 0.8-1.2. В пределах ярусов наблюдается концентрирование числа стволов вокруг среднего диаметра (рис. 27). Коэффициент вариации диаметра для первого яруса составляет 19 %, для второго яруса - 23% и для третьего яр$са - 27%. Следовательно, выделенные ярусы являются вполне выраженными элементами насаждения.
Выше было установлено, что между диаметром и возрастом насаждения наблюдается корреляционная связь. Поэтому при установлении возраста выделенных ярусов в древостое руководствовались этой зависимостью.
Число стволов
Рис. 27. Распределение стволов пихты по ступеням толщины в пределах ярусов: 1 - третий ярус, 2 - второй ярус, 3 - первый ярус
Исследования показали, что возраст деревьев первого яруса превышает 181 год и состоит в основном из спелых и старовозрастных групп. Второй ярус представлен деревьями приспевающих и средневозрастных групп, возраст которых колеблется от 121 до 180 лет. Третий ярус состоит из молодых деревьев в возрасте до 120 лет. В табл. 30 приводятся основные параметры, характеризующие выделенные ярусы для разновозрастных пихтарников.
Таблица 30
Распределение разновозрастных пихтовых древостоев по ярусам
Ярусы, возрастные группы Показатели по бонитетам Возраст, лет
I а-1 П-Ш
Н, м D, см Н, м D, см
I, 31 и 56 и 28 и 52 и 181 и
спелые и перестойные выше выше выше выше выше
П> 21-30 32-52 19-27 28-48 121-180
приспевающие и средневозрастные
ш, 7-20 8-28 6-18 8-24 41-120
молодые
Производству рекомендуется проводить таксацию пихтарников по ярусам и устанавливать таксационные показатели для каждого яруса в отдельности. Для примера в табл. 31 приводится таксационная характеристика разновозрастного пихтового древостоя для насаждений пробной площади 31, заложенной в типе леса пихтарнике ясменниковом I класса бонитета.
Таблица 31
Характеристика древостоя на пробной площади 31
Таксационные показатели * •Ярус Всего древостоя
I II III
Состав ЮПх 7ПхЗБк' 4Пх5Бк 8Пх2Бк
Полнота 0.65 0.29 0.08 1.02
Сомкнутость - - - 0.9
Запас, м3/га 755 252 31 1038
Число стволов, шт/га 120 130 190 440
Площадь сечения, м2/га 44.35 20.40 4.94 69.69
Средний диаметр, см 69 45 18 -
Средняя высота, м 37 27 13 -
Возраст, лет 230 170 105 -
Из таблицы 32 видно, что в древостое преобладает запас I яруса, который составляет 755 м3, или 73% от всего запаса древостоя. Однако по количеству в насаждении доминируют деревья П и Ш ярусов (130+190) или 72,7 %.
Таксация разновозрастных насаждений по отдельным ярусам имеет важное значение для организации правильного пользования лесом. Такой
метод таксации также значительно сокращает время на полевые и камеральные лесотаксационные работы. При этом точность сортиментов не снижается (Бицин, 1965).
Проведенные исследования показали, что в девственных пихтовых древостоях количество деревьев первого яруса не превышает 30-40 % от общего числа всех деревьев на га. Превалирующее количество - 60-70 % деревьев составляют приспевающие, средневозрастные (П ярус) и молодые (П1 ярус) возрастные группы. Наличие деревьев всех возрастных групп, постоянный процесс естественного возобновления под пологом леса обеспечивают непрерывность развития разновозрастных насаждении. В силу этого пихта кавказская без воздействия внешних факторов среды не образует самостоятельных, обособленных насаждений молодого возраста с последующим прохождением нескольких фаз или циклов развития до стадии перестойного, отмечаемых для насаждении другого типа возрастной структуры лесов из пихф белой, ели европейской (Mayer, 1969; Дыренков, 1971; Kolsh, 1974; Leibundgut, 1982). В этом заключается отличительная особенность пихтарников Кавказа от темнохвойных лесов других регионов.
Ведение хозяйства в этих лесах следует осуществлять с учетом биоэкологических и лесоводственных особенностей вида. По данным наших экспериментальных исследований (Бебия, 1973, 1983), установлено, что природе пихтовых лесов, имеющих сложную форму строения и возрастную структуру, наиболее отвечает выборочная форма хозяйства, в частности, комплексно-выборочные и добровольно-выборочные рубки, при ведении которых в первом ярусе проводятся рубки главного пользования, во втором и третьем ярусах - рубки ухода. Эти рубки будут способствовать улучшению санитарного состояния лесов, повышению продуктивности, обеспечат естественное их воспроизводство и сохранение защитных функций.
Дифференциация деревьев в лесу, их классификация
В данном разделе изложены закономерности дифференциации деревьев в одновозрастных и разновозрастных лесах и метод их классификации. Приведены морфолого-биологические и морфометрические признаки жизненного состояния деревьев. Предложены способы расчета показателей и шкала состояния древостоев по ярусам.
Изучение процессов дифференциации деревьев по характеру роста и развития, морфологическим особенностям строения вегетативных органов служит фактической основой для оценки влияния борьбы за существование на формирование и развитие лесных сообществ. Вопросу дифференциации деревьев в лесу в процессе борьбы за существование уделяли большое внимание Г.Ф. Морозов (1949) и В.Н. Сукачев (1953). Ими были освещены основные особенности этого явления. Значительно меньше известно о за-
писимости этого процесса от типа леса, динамического статуса сообщества и условий среды в конкретных экологических ситуациях, поэтому исследования в этом направлении необходимо продолжить.
Основной целью наших исследований по данному вопросу явилось изучение особенностей лесных деревьев, закономерностей их роста, развития и дифференциации в лесу в зависимости от строения и возрастной структуры древостоев, а также разработка новой классификации деревьев и метода определения жизненного состояния древостоев.
Сведения о дифференциации деревьев и созданная на их основе классификация имеют не только теоретическое, но и большое практическое значение. С учетом классификации деревьев в разновозрастном лесу должны проводиться санитарные рубки, рубки промежуточного и главного пользования. При осуществлении выборочных рубок она может помочь в определении первоочередности назначения деревьев в рубку. Регулируя количество и качественное состояние деревьев в пределах яруса и между ярусами в разновозрастных лесах, можно создать оптимальные условия роста и развития деревьев и сократить таким образом срок выращивания высокопроизводительных древостоев.
Для решения поставленных задач в наиболее распространенных типах леса закладывали пробные площади размерами 0.25-0.5 га. В них таксационную характеристику древостоев устанавливали отдельно по каждому ярусу. Подрост /II/ до 4 м высоты учитывали сплошь с разделением его условно на две категории жизненного состояния - благонадежный (f) и неблагонадежный (g) /рис. 28). На пробных площадях морфологические и морфометрические признаки деревьев устанавливали для каждого в отдельности.
При изучении лесов мы применили классификацию деревьев, разработанную специально для разновозрастных древостоев (см. рис. 28). В основе ее - методика диагностики жизненного состояния деревьев и древостоев В.А. Алексеева (1989), но измененная и дополненная нами. В частности, методика оценки жизненного состояния древостоев разработана с учетом их строения и возрастной структуры, биоэкологии пород по типам леса. Данная классификация, по сравнению с ранее предложенными, позволяет доступными в полевых условиях способами более точно определить основные диагностические признаки жизненного состояния деревьев и рассчитать жизненное состояние древостоя - основного компонента лесных биогеоценозов.
Термин “жизненное состояние” или “жизненность” (дерева, древостоя), как отмечает В.А. Алексеев, более точно отражает цель и смысл осуществляемых оценок, чем термин “жизнеспособность”, “биологическая устойчивость” или “выносливость”, применяемые часто в биологической литературе (Спурр, Баране, 1984). Понятие “жизненность”, “жизненное со-
Рис. 28. Классификация деревьев и подроста по их жизненному состоянию в разновозрастном лесу: дерево, а -- здоровое, b - угнетенное, с - сильно угнетенное, d - усыхающее, е - сухостойное, подрост (П): f- благонадежный, g - неблагонадежный. Ярус: I - верхний, II - средний, Ш - нижний
стояние” в данном случае нами рассматривается как комплексный биологический параметр на уровне особей, группы особей (яруса или поколения), сообществ (древостоя). Отметим также, что понятие “жизненность” и “жизнеспособность” не синонимы, и между ними знак равенства ставить нельзя. Жизненность деревьев - оценка их жизненного состояния в момент наблюдения, выраженная морфометрическими признаками роста и развития, обусловленного конкуренцией и взаимодействием между деревьями в процессе развития сообществ и влиянием среды, мера устойчивости к неблагоприятным условиям. Жизнеспособность - оценка жизненного состояния особей, мера потенциальной устойчивости к неблагоприятным условиям и способность к онтогенетической адаптации. Потенциальный уровень жизнеспособности растений определяется, главным образом, их генотипом.
Закономерности дифференциации деревьев в разновозрастном лесу
В биологической науке, еще со времен Е. Уоллеса и Ч. Дарвина принято считать, что объектом конкуренции обычно служит какой-нибудь ресурс, запасы которого в данной среде ограничены (пища, место для прорастания семян и пр.). Исходя из этой концепции, большинство исследователей зачастую описывают жестокую борьбу между особями, объясняя ее ограничением ресурсов.
Исследования последних лет по данному вопросу (Simpson, 1964; Шмальгаузен, 1969; Гулисашвили, 1974; Тимофеев-Рисовский, 1974; Галл, 1976; Солбриг и др., 1982; Спурр и др., 1984 и другие) убеждают нас в том, что конкуренция -это скорее всего пассивный процесс, в котором наиболее эффективно функционирующая особь получает основную массу данного ресурса. Результат конкуренции состоит в том, что жизненное состояние одной из участвующих во взаимодействии особей возрастает, а жизненность другой - снижается.
В зависимости от сочетания факторов окружающей среды и генетических факторов одни деревья в лесу растут интенсивно, с хорошо развитой кроной, другие растут медленно, их кроны сравнительно менее развиты. Некоторые деревья сильно отстают в росте, заметно угнетены и со временем отмирают. Поэтому дифференциация деревьев в лесу биологически закономерна (Хильми, 1957), и классификация их должна быть осуществлена с учетом роста, развития и жизненного состояния деревьев.
Основные закономерности дифференциации деревьев и их классификация в одновозрастном лесу достаточно хорошо освещены и сформулированы в лесоводственной литературе (Мелехов, 1980). В настоящее время существует свыше 100 таких классификаций.
Признаки, по которым различаются деревья в разновозрастном многопородном лесу, чрезвычайно многообразны. В таком лесу подрост по
стоянно пополняет нижний ярус древостоя. В строении и возрастной структуре древостоя происходят постоянные изменения, что обусловливает произрастание в древостое разнообразных деревьев по возрасту, размерам, формам роста и развития. Если на ранних этапах рост деревьев подавляется и происходит их дифференциация, то в последующем на формирование этих деревьев совокупно воздействуют условия всей лесной среды (Мелехов, 1980). Усложняется связь между деревьями, усиливаются конкурентные взаимоотношения между ними. В силу этого дифференциация деревьев и процесс самоизреживания в таком разновозрастном лесу более сложный биологический процесс, чем в одновозрастном лесу. Поэтому классификация деревьев должна раскрывать биологическую сущность и характер дифференциации деревьев, отражать динамику формирования древостоев. С учетом этих представлений разработана излагаемая ниже шкала категорий жизненного состояния деревьев по их морфометрическим и морфологическим показателям. Она приемлема для классификации деревьев хвойных пород в разновофастных и одновозрастных лесах.
Классификация деревьев по их жизненному состоянию
а. Здоровое дерево не имеет внешних признаков угнетения, повреждения кроны и ствола. Крона хорошо развитая, симметричная, густая. Мертвые и отмирающие ветви единичны, приурочены к нижней части кроны. Вершина заостренная у интенсивно растущих деревьев или притупленная у старовозрастных спелых и перестойный деревьев. Хвоя зеленого и темно-зеленого цвета. Деревья среднего и верхнего яруса семеносят, в нижнем ярусе - редко. Выделить такое дерево в древостое не представляет трудности.
Ь. Угнетенное дерево - внешне заметно угнетено. Крона устремлена вверх, но зачастую несколько асимметрична, сдавлена с одной стороны. Вершина слегка заостренная, что указывает на некоторую общую подавленность в росте. Число мертвых и усыхающих ветвей в верхней части кроны может достигать до 30%. Густота кроны может быть снижена на 30% за счет преждевременного опадения хвои или в результате изрежива-ния скелетной части кроны, приводящих к ослаблению жизнеспособности дерева до 30%. Деревья верхнего яруса семеносят, среднего яруса - реже.
с. Сильно угнетенное дерево - угнетение внешне ярко выражено, крона, ствол или корневые лапы повреждены, жизненность дерева снижена и достигает до 60%. Крона зачастую сдавлена по меньшей мере с двух и более сторон. У деревьев второго и третьего ярусов она бывает зонтикообразной. Вершина притупленная, иногда - усыхающая. Ветвление по главному стволу близко друг к другу, что указывает на слабую интенсивность роста дерева. Густота охвоения кроны может быть до 60 %
за счет преждевременного опадения хвои или изреживания скелетной части кроны. Деревья верхнего яруса иногда могут семеносить.
d. Усыхающее дерево имеет разрушенную или разрушающуюся крону, густота ее менее 20%. В кроне более 70% ветвей сухие или усыхающие. Комлевая и средняя части ствола могут быть заселены стволовыми вредителями, имеется стволовая, корневая гниль. Жизненное состояние таких деревьев редко превышает 10%. Семеношение отсутствует.
е. Сухостойное - дерево усохшее, но продолжающее стоять в древостое.
Для оценки жизненного состояния деревьев мы использовали основные и дополнительные признаки, которые легко определить в полевых условиях при учете деревьев на пробных площадях. Применение нами приведенной выше шкалы классификации жизненного состояния деревьев при изучении лесообразовательных процессов в пихтовых лесах Кавказа подтвердило ее эффективность в практических и исследовательских целях.
Важнейшим показателем жизненного состояния деревьев в лесу может служить протяженность кроны по стволу. Данные наших исследований в разновозрастных пихтарниках по шести пробным площадям, с сомкнутостью полога древостоев равной 0.7-0.9, приведены в табл. 32. Так, протяженность кроны по стволу для здоровых деревьев составляет в среднем 55.511.78% от его высоты. Протяженность кроны у угнетенных и сильно угнетенных деревьев 41.3±2.18 и 35.811.8%. У усыхающих деревьев крона слабо выражена. Протяженность ее, хотя и составила в среднем 33,8±4.71%, но величина этого показателя от 11 до 58%, с коэффициентом вариации 48.2%. Однако отличить такое дерево в древостое не сложно.
Жизненное состояние дерева в значительной степени зависит от его высоты - более высокие деревья находятся в лучших условиях, чем те деревца, которых они перегнали в росте и начинают угнетать. К примеру, в пихтовом древостое среди низкорослых деревьев 6 пределах ступени высоты 4 м здоровые деревья составляют всего 5.1% от общего числа стволов этой ступени высоты.
Таблица 32
Протяженность кроны по стволу деревьев (%) разных категорий жизненного состояния в разновозрастных пихтарниках _______
Деревья Число деревьев x±s, max-min о V,% Р,%
Здоровые 278 55.311.78 69-38 7.74 14.1 3.2
Угнетенные 317 41.312.18 54-18 8.42 20.5 5.3
Сильно угнетенные 360 35.811.80 53-24 6.23 17.4 5.0
Усыхающие 125 33.814.71 58-11 16.31 48.2 13.9
Примечание. X - среднеарифметический показатель; Sx - ошибка среднеарифметического; о - среднеквадратическое отклонение; V - коэффициент вариации; Р - точность наблюдений
Деревьев угнетенной, сильно угнетенной, усыхающей и сухостойной категории значительно больше: соответственно 23.1, 34. 6, 21.8 и 15.4%. Однако для ступени высоты 24 м здоровых деревьев 39.1%, а угнетенных - 45.7, сильно угнетенных - 8.7, усыхающих - 2.2, сухостойных - 4.3% (табл. 33).
Таблица 33
Распределение деревьев разных категорий жизненного состояния по ступеням высоты в разновозрастных пихтарниках___________
Ступень высоты, м Всего деревьев Жизненное состояние деревьев
здоровые угнетенные сильно угнетенные усыхающие сухостойные
шт. шт. % шт. % шт. % шт. % шт. %
1 2 3 4 5 6 7 8 9 10 11 12
4 156 8 5.1 36 23.1 54 34.6 34 21.8 24 15.4
6 135 5 3.7 24 17.8 63 46.7 28 20.7 15 11.1
8 103 4 3.9 17 16.5 55 53.4 17 16.5 10 9.7
10 92 2 • 2.3 17 18.5 48 52.2 14 15.2 11 11.8
12 76 1 1.3 22 29.0 39 51.3 5 6.6 9 11.8
14 64 3 4.7 23 35.9 28 43.8 3 4.7 7 10.9
16 59 1 1.7 26 44.1 23 39.0 3 5.1 6 10.1
18 71 15 21.1 27 38.0 28 39.5 - - 1 1.4
20 69 12 17.4 43 62.3 9 13.0 2 2.9 3 4.4
22 45 17 37.9 22 48.9 2 4.4 2 4.4 2 4.4
24 46 18 39.1 21 45.7 4 8.7 1 2.2 2 4.3
26 42 19 45.2 15 35.7 3 7.2 4 9.5 1 2.4
28 51 32 62.7 15 29.4 3 5.9 - - 1 2.0
30 33 27 81.8 2 6.0 - - 2 6.1 2 6.1
32 48 41 85.4 4 8.3 1 2.1 - - 2 4.2
34 33 26 78.8 3 9.1 - - 3 9.1 1 3.0
36 18 12 66.7 - - - - 4 22.2 2 11.1
38 18 15 83.3 - - - - 2 11.1 1 5.6
40 7 5 71.4 - - - - 1 14.3 1 14.3
42 4 3 75.0 - - - - - - 1 25.0
44 6 6 100
46 4 4 100 - - - - - - - -
48 2 2 100
Всего 1182 278 23.5 317 26.8 360 30.5 125 10.6 102 8.6
Примечание. Приведены данные для шести пробных площадей
Таким образом, в разновозрастных древостоях с возрастанием ступени высоты доля здоровых деревьев увеличивается, достигая максимума в наивысших ступенях высоты. Доля деревьев остальных категории жизненности, в зависимости от степени их угнетения, существенно уменьшается с увеличением высоты деревьев, что может быть объяснено заметным ослаблением конкуренции между деревьями верхнего яруса древостоя. В целом же для древостоя соотношение деревьев разной категории жизненности
иное (см. табл. 33). Здоровые деревья составляют 23.5% от числа всех деревьев в древостое. Доля угнетенных и сильно угнетенных деревьев несколько выше: соответственно 26.8 и 30.5%. Усыхающих и сухостойных -10.6 и 8.6%. Однако эти процессы необходимо рассматривать с учетом типа леса, структуры и возрастного строения древостоя (табл. 34).
Выше нашими исследованиями было установлено, что пихтарники Кавказа отличаются разновозрастным и сложным характером строения древостоев. В практических целях в таких древостоях, как отмечалось выше, целесообразно условно выделять три яруса, в которых конкурентные отношения существуют как между деревьями внутри одного, так и между деревьями разных ярусов. Однако в зависимости от типа леса характер таких отношений функционально и биологически существенно различается в пространстве и во времени.
Известно, что для теневыносливых и мезофильных пород, какой является, например пихта кавказская, характерно неравномерное, зачастую групповое размещение деревьев в биоценозе (Гулисашвили, 1970; Бебия, 1987). В таких биогруппах одни деревья начинают отставать в росте и развитии, что может быть результатом старения, конкурентной борьбы, а также обусловлено генетическими факторами (Сукачев, 1953; Синнот, 1963; Райт, 1978). Другие деревья, занимающие господствующее положение в таких биогруппах, разрастаются, их кроны начинают отенять, тем самым заметно угнетать отстающие в росте деревья. Среди деревьев I яруса конкурентные отношения выражены слабо. В зависимости от типа леса возраст деревьев I яруса составляет 208 лет и выше (табл. 34). В таком возрасте среди деревьев I яруса конкурентные отношения, как видно, снижены до минимума, и процесс дифференциации деревьев заметно затухает. Поэтому Деревья I яруса преимущественно здоровые, угнетенные и сильно угнетенные в этом ярусе встречаются гораздо реже или полностью отсутствуют. Полного затухания конкурентных отношений не происходит. В отдельных типах леса в составе I яруса произрастают деревья угнетенной, реже и сильно угнетенной категории жизненности (см. табл. 34, пробные площади 61, 66).
Наличие усыхающих и сухостойных деревьев в составе деревьев I яруса обусловлено не столько конкурентными факторами, сколько достижением ими в конкретных условиях местопроизрастания предела своего биологического возраста. В зависимости от типа леса возраст усыхающих деревьев в I ярусе составил 239-340 лет.
В девственном лесу в процессе самоизреживания старовозрастные деревья отмирают. В лесу образуются естественные “окна”. С течением времени рост и развитие жизнеспособного подроста и деревьев II и Ш ярусов в таких окнах начинают ускоряться. Некоторые из них переходят из категории сильной степени угнетения в слабую.
Таблица 34
Характеристика жизненного состояния разновозрастных пихтовых, буково-пихтовых древостоев по типам леса
Пробная площадь Порода Показатель Ярус древостоя Итого по породам
I И 1 III
Жизненное состояние древостоев
здоровые угнетенные сильно угнетенные усыхающие сухостойные здоровые угнетенные сильно угнетенные 4> S 3 § X * сухостойные здоровые угнетенные сильно угнетенные усыхающие сухостойные
1 2 3 4 5 6 7 8 9 10 И 12 13 14 15 16 17 18 19
62. 0.5 га, 1300 м, С.-З., 5°; Буковопихтовый мелкотравный II класса бонитета; Число деревьев 230, запас древесины 384 м3, состав 7ПхЗБк, сомкнутость 0.8 Пихта Бук N D Н V А L N D Н V А L 18 1 - 3 4 82 56 - 85 88 36 34 - 37 27 147 3.7 - 27 28.5 244 224 - 325 73.6 6 68 - - 32 28.7 - 220 100 13 14 5 3 4 59 47 42 36 48 25 24 24 21 22 39.4 26 7.5 3.0 7.2 175 164 168 196 73.3 15 5 65 50 - 25 23 - 52.8 9.9 188 165 - 92.5 13 41 43 ' 8 8 24 17 24 18 23 8 9 12 10 9 2.9 4.6 11.7 1.1 1.4 98 74 109 98 50.1 5 21 12 4 5 40 31 14 12 17 16 15 10 10 7 4.6 11.3 1.0 0.2 0.5 135 108 55 50 73.6 71.6 й 93.0
Итого по ярусам и древостоя в целом L 76.5 82.6 59.3 74.7
1 2 3 4 1 5 1 6 1 1 1 8
64. 0.25 га, Пих- N 27 2 - 2 -
1250 м, Ю.-З., та D 63 54 - 68 -
40°; Н 33 33 - 31 -
Пихтарник V 122 6.6 - 9.8 -
овсяницевый I А 218 236 - 239 -
класса L 95.5
бонитета;
Число
деревьев 231, Бук N - - - - -
запас древеси- D - - - - -
ны 273 м3, сос- Н - - - -
тав 9Пх1Бк, V - - - - -
сомкнутость А - - - - -
0.9 L
Итого по яру- L 95.5
сам и древос-
тоя в целом
65. 0.25 га, Пих- N 11 1 - - -
1160 м, 3., 12°; та D 94 52 - ч - -
Буково- Н 41 32 - . - -
пихтовый V 133 3.0 - - -
овсяницевый А 279 208 - - -
1а класса L 99.3
бонитета;
Число стволов Бук N 8 - - - -
203,запас D 69 - - - -
древесины 294 Н 32 - - - -
м3, состав V 39.7 - - - -
8Пх2Бк, А 220 - - - -
сомкнутость L 100
0.8
9 10 11 1 12 1 13 14 | 15 I 16 1 17 | 1 18 19
29 24 - - 1 1 11 88 24 29
46 40 - 52 34 33 20 14 19
26 24 - - 26 17 21 13 9 9
56.3 32.9 - - 2.5 0.7 9.3 18.8 1.9 4
137 139 - - - 112 144 91 76 126
86.5 43.7 86.3
3 7 6 7 1
36 40 - 29 19 28
18 22 - - - - 15 11 - 15
2.5 8.3 - - - - 2.7 1.1 - 0.4
122 135 - - - - 105 74 - -
76.9 82.6 78.4
85.5 47.8 85.6
4^
3 10 1 2 1 31 41 45 14
43 42 32 54 20 23 20 17 12
25 25 20 27 - И 13 12 10 7
5.0 16.0 0.8 5.3 - 0.2 8.2 7.9 5.7 0.5
127 146 128 225 - 82 100 91 93 80
62.9 43.0 89.0
9 3 1 2 10 23 2 3
46 49 - - 60 34 32 20 22 18
25 23 - - 24 19 17 12 11 10
16.3 5.5 - - 2.9 1.5 6.4 4.9 0.4 0.4
155 162 - - - 115 ПО 75 80 -
81.5 60.0 87.9
1 2 3 4 1 5 | 6 | 7 | 8
Итого по ярусам и древостоя в целом L 99.5
66. 0.25 га, 1200 м. С.-З., 15°; Пихтарник падубовый 1а класса бонитета; Число деревьев 133, запас древесины 340 м3, состав 9Пх1Бк, сомкнутость 0.7 Пихта N D Н V А L 21 4 1 3 - 94 52 60 109 41 33 32 36 212 11.9 3.9 38.3 228 208 240 340 84.8
Бук N D Н V А L 1 - - - 1 92 - - - 88 36 - - - 34 9.5 - - - 8.3 280 - - 53.4
Итого по ярусам и древостоя в целом L 82.9
67. 0.25 га, 950 м, 3., 8°; Буково-пихтовый мелкотравный I класса бонитета; Пихта N D Н V А L 12 - - 2 2 86 - - 92 108 34 - - 36 34 102 - - 20.4 26.5 256 - - 287 - 70.3
9 10 11 1 12 1 13 14 1 15| 161 17 1 118 19
71.2 49.1 88.5
8 7 3 2* 1 4 23 3 8
60 54 42 36 - 12 31 26 26 20
27 27 26 24 - 8 16 15 13 13
23.7 16.3 4.5 2.3 - 0.1 1.6 5.6 0.7 1.4
178 188 168 150 - 50 135 118 141 133
79.3 37.1 80.2
1 4 3 19 22 3 2
36 52 - - - 14 13 11 14 30
20 25 - 9 9 8 9 13
0.9 6.1 - - - 0.1 1.8 1.3 0.2 0.6
120 170 - - - 60 52 50 60 -
73.8 47.5 57.6
78.6 40.3 78.4
13 17 5 - - - 19 36 8 5
48 41 37 - - - 17 13 11 11
23 22 19 - - - 9 8 6 7
24.3 21.7 4.5 - - - 2.2 2.3 0.2 0.2
143 142 148 - - - 75 60 60 73
82.2 50.6 73.2
I 2 3 4 1 1 5 1 6 1 7 1 8
Число Бук N 5 - - -
деревьев 134, D 74 - - - -
запас Н 33 - - - -
древесины V 28.1 - - - -
225 м3, состав 8Пх2Бк, А L 235 - 100 - -
сомкнутость 0.7
Итого по L 75.8
ярусам и древостоя в целом
61. 0.5 га, Пих- N 22 8 1 9 13
1750 м, Ю.-З., та D 71 60 56 72 76
25-30°; Н 29 24 25 29 29
Буково- V НО 24.1 2.7 46.3 74.3
пихтовый мелкотравны й II класса бонитета; А L 211 240 254 51.6 276
Число Бук N 10 - - -
деревьев 209, D 66 - - - -
запас Н 28 - - - -
древесины V 40.1 - - - -
362 м3, состав 7ПхЗБк+Кл, А L 210 - 100 - -
сомкнутость 0.7
Клен N D - - - - -
9 10 11 1 12 13 14 | 1 15 I 16 1 17 1 18 19
3 3 - - 7 4 -
66 50 - - - - 18 20 -
27 24 - - - - 12 10 - -
11.3 6.1 - - - - 1.1 0.8 - -
210 165 - - - - 70 75 - -
89.4 57.3 94.4
84.0 51.2 74.6
10 19 5 5 6 4 19 21 5 5
48 48 40 52 56 16 16 12 12 14
23 23 22 21 22 7 8 8 10 9 4^
18.7 35.6 6.3 10.1 14.6 0.3 1.7 1.1 0.3 0.7 LA
143 167 160 216 - 66 70 5.5 65 93
55.3 47.8 52.4
22 12 7 1 3 7 4 1 1
46 34 34 38 - 24 16 18 20 16
20 21 19 19 - 17 10 10 11 8
32.6 10.4 5.5 1.0 - 1.1 0.7 0.5 0.2 0.1
155 115 120 140 - 88 62 70 75 65
85.2 69.6 91.2
2 4 1 3 5
48 44 - - 32 32 32 - - -
I 2 3 4 1 5 | 6 | 7 | 8 9 | 10 | 11 | 12 | 13 14 | 15 | 16 | 17 | 18 19
Н V А L 18 18 - - 18 3.3 5.7 - - 0.7 162 151 75.1 14 14 - 1.5 2.5 108 120 - 81.2 76.9
Итого по ярусам и древостоям в целом L 58,1 67.2 е 65.6 63.2
Примечание. N - число деревьев на пробной площади; D - средний диаметр деревьев, см; Н - средняя высота деревьев, м; V - запас деревьев, м3; А - средний возраст деревьев, лет; L - жизненное состояние, %.
Через определенное время такой подрост или дерево начинает расти нормально с характерной для вида кроной, без внешне заметных существенных признаков угнетения. Поэтому для II и III ярусов древостоя характерно наличие деревьев всех категорий жизненного состояния, в том числе здоровых деревьев.
Возраст деревьев II яруса древостоя 115-196 (225) лет (см. табл. 34). Для деревьев -этого яруса и возраста характерны более ярко выраженные конкуренция и дифференциация, чем среди деревьев I яруса. Так, среди деревьев II яруса, хотя и сохраняется тенденция к уменьшению числа стволов от здоровой категории деревьев к усыхающей и сухостойной, зачастую число стволов угнетенной категории превышает число здоровых деревьев (пробные площади 61, 62, 65, 66). Отсутствие во II ярусе отдельных типов леса, с древостоями высокой производительности I и 1а классов бонитета (пробные площади 64, 65, 66) усыхающих или сухостойных деревьев указывает на то, что конкуренция между деревьями II яруса ослабевает раньше, чем в древостоях меньшей производительности и худших условиях произрастания (пробные площади 61, 62).
Совершенно иная картина наблюдается в Ш ярусе древостоя. Возраст деревьев пихты в нем, в зависимости от типа леса и степени угнетения, колеблется от 50 (пробная площадь 66) до 144 лет (пробная площадь 64). Наличие здесь деревьев всех категорий жизненности, в том числе значительного количества усыхающих и сухостойных деревьев указывает на непрерывную жестокую борьбу и интенсивную дифференциацию среди деревьев этого яруса. Чем выше общая сомкнутость полога древостоя, тем ярче выражены эти процессы. По данным табл. 34, в III ярусе на пробной площади 66 при сомкнутости полога древостоя 0.7 насчитывалось 3 усыхающих дерева пихты, или 7.7 % от числа всех деревьев этого яруса; при сомкнутости 0.9 (пробная площадь 64) соответственно 24 усохших дерева, или 16 % от числа деревьев яруса.
На рис. 29 в виде диаграммной схемы изображено процентное соотношение числа деревьев разных категорий жизненного состояния по ярусам в пихтовом древостое. Из этой схемы видно, что для древостоя в целом (IV) распределение деревьев здоровой, угнетенной и сильно угнетенной категорий относительно равномерное, что указывает на непрерывный процесс борьбы и дифференциации среди деревьев в процессе развития древостоев. Основой и резервом древостоев являются, главным образом, деревья угнетенной и сильно угнетенной категорий, еще обладающие потенциальными возможностями для перехода в деревья здоровой категории. Анализ распределения деревьев разной категории жизненности по ярусам в спонтанном пихтовом лесу позволяет выдвинуть гипотезу о том, что потенциал развития разновозрастных древостоев во всех типах леса формируется из деревьев всех категорий жизненности. Однако их функции и рас-
Ярус древостоя
Рис. 29. Процентное соотношение числа деревьев в разных категориях жизненности по ярусам в пихтовом древостое (a, b, с, d, е - категории жизненности)
пределение по ярусам, возрастным поколениям, как было отмечено выше, существенно различаются (см. рис. 29), и это влияет на динамику и жизненное состояние ярусов, на развитие древостоев в целом. Данные, характеризующие распределение деревьев разных категорий жизненности в древостоях, могут служить основой при подборе мер ухода за насаждениями.
Классификация древостоев по их жизненному состоянию
Жизненное состояние сообществ (древостоев) - понятие, отражающее соотношение в сообществах особей (деревьев) с разным уровнем жизненности. Поэтому в основу классификации древостоев по их жизненному состоянию нами положены преобладание и роль в них особей, деревьев того или иного жизненного состояния.
Расчеты показателей жизненного состояния древостоев мы осуществляли по материалам таксации деревьев и их классификации по вышеописанной шкале на пробных площадях.
Существующие в настоящее время балльные методы оценки состояния деревьев и древостоев, по мнению Н.Я. Саликова (1979) и В.А. Алексеева (1989), не несут биологически адекватного смыслового значения. Поэтому предлагается деревьям различных категорий жизненности присваивать коэффициенты, соответствующие их состоянию, т. е. от условных чи-
еловых индексов к показателям смыслового значения. Мы также применили коэффициенты при расчетах показателей жизненного состояния древостоев. Исходя из этого, коэффициент здорового дерева приравнивается к 100 %, сухостойного - к нулю. Для деревьев промежуточных градаций указываются коэффициенты, соответствующие их жизненному состоянию к моменту перечета. По данным В.А. Алексеева, целесообразно принять, что угнетенные деревья утратили примерно третью часть жизненных потенций, сильно угнетенные - 2/3 и их состояние оценивается в 40 % от нормального. Жизненное состояние усыхающего дерева, в частности пихты кавказской, согласно нашим исследованиям, редко превышает 10 % возможного.
Мы предлагаем рассчитывать жизненное состояние одновозрастного или для определенного яруса разновозрастного древостоя, а также при необходимости, для отдельных пород в смешанном древостое по формуле:
_ 100^ + 70^+40^+101;
V - у У \ )
где Lv - относительное жизненное состояние древостоя;
И, ^2, V3, V4 - объемы древесины здоровых, угнетенных, сильно угнетенных и усыхающих деревьев лесообразователей на пробной площади или на 1 га, м3, соответственно;
100, 70, 40 и 10 - коэффициенты, выражающие жизненное состояние здоровых, угнетенных, сильно угнетенных и усыхающих деревьев, %;
И - общий запас древесины древостоя на пробной площади или на 1 га, м3, включая запас сухостойных деревьев.
Включение запаса сухостойных деревьев в оценку жизненного состояния древостоя обязательно. По нашим наблюдениям, сухостой пихты и бука высотой не менее 2/3 от высоты ствола в момент их усыхания в течение 5-7 лет после их гибели может оказывать влияние на развитие древостоя.
В разновозрастном лесу индекс жизненного состояния древостоя рассчитывается для каждого яруса отдельно и древостоя в целом. При этом расчет индекса для яруса, как отмечалось выше, производится только по запасу (объему) деревьев (формула 1). Для древостоя в целом этот показатель также вычисляется через запас, по формуле:
£ = * ^2^2 * + ^4^4 /2)
Г 7
где L - относительное жизненное состояние разновозрастного древостоя, %;
Ц, L2, Ln - показатели индексов относительного жизненного состояния древостоев ярусов, %, вычисленных по формуле 1;
И, ^2, Ип- объемы древесины соответствующих ярусов на пробной площади или на 1 га, включая запас сухостоя;
V - общий запас древесины на пробной площади, или на 1 га, включая запас сухостоя.
Для оценки древостоев по их жизненному состоянию применительно к пихтовым лесам Кавказа предлагаются следующие показатели: при индексе 100-80 % жизненное состояние древостоя или яруса древостоя оценивается как здоровое, при 79-50 % древостой или ярус древостоя считается ослабленным, при 49-30 % - сильно ослабленным, при индексе ниже 30 - разрушенным.
Определение жизненного состояния древостоев по ярусам, а также по главным лесообразующим породам в разновозрастных лесах (по формуле 1) имеет важное значение при изучении лесообразовательных процессов, строения и возрастной структуры, динамики древостоев, прогнозировании смен в лесу, позволяет учитывать особенности развития конкретных биогеоценозов.
Определение жизненного состояния сложного, многоярусного древостоя (по формуле 2) важно и целесообразно при разработке стратегии ведения хозяйства, например, для осуществления выборочной формы хозяйства. Так, в девстйвнных пихтарниках всех типов леса жизненное состояние древостоев определяется как здоровое и ослабленное с индексами от 88.5 % (пробная площадь 65) до 63.2% (пробная площадь 61, табл. 34). В таких древостоях целесообразно проведение выборочных, в частности добровольно-выборочных, рубок. В сильно ослабленных и разрушенных древостоях с индексами жизненности менее 50% добровольно-выборочные рубки мало эффективны.
Индексы жизненного состояния древостоев могут быть использованы также при выделении эталонных лесных участков, где эти индексы не должны быть ниже 80 %.
Показатели жизненного состояния древостоев нами впервые были применены также в типологических исследованиях при описании и диагностике типов лесных биогеоценозов. Так, индексы, характеризующие жизненное состояние древостоя по ярусам и лесообразователям, вычисленные по вышеописанной методике в буково-пихтовом мелкотравном типе леса II класса бонитета (пробная площадь 61), указывают, что древостой как по ярусам, так и в целом, относится к ослабленной категории. Однако сравнительно близкие значения индексов жизненности во всех ярусах -58.1, 67.2 и 65.6 % - указывают на отсутствие возрастно-восстановительных смен поколений и динамичный характер лесообразовательного процесса, достаточную устойчивость данного типа лесного биогеоценоза. Жизненность буковой части древостоя несколько выше, чем у пихтовой (91.2 против 52.4 %). Однако ценотическая позиция пихты по сравнению с буком более стабильная. Смены пихты сопутствующим видом бука не наблюдается. Состав древостоя в данном биогеоценозе 7ПхЗБк + Кл подтверждает это положение. Следовательно, в данных условиях местопроиз-
растения характеризуемый тип леса является коренным. Лесоводственные мероприятия в этом случае должны предусмотреть хозяйство на пихту.
Итак, подводя итоги исследования по данным вопросам, можно сделать следующее заключение. Изучение характера дифференциации деревьев в лесу и их классификация имеют большое значение для решения многих лесобиологических и лесохозяйственных вопросов. Дифференциация деревьев в разновозрастном и одновозрастном лесу существенно различается. Конкурентные отношения между деревьями и их дифференциация в разновозрастном лесу более сложные, чем в одновозрастном. Эти процессы во многом зависят от строения и возрастной структуры древостоев. Определение жизненного состояния и классификация деревьев осуществляются по их биолого-морфологическим и морфометрическим признакам. Расчеты жизненного состояния древостоев производятся по ярусам. В основу их положены преобладание и роль в них особей деревьев той или иной жизненности. При индексе 100-80 % жизненное состояние древостоя или яруса древостоя оценивается как здоровое, 79-50 - ослабленное, 49-30 -сильно ослабленное, ниже 30 % - разрушенное. Рекомендуемые шкалы классификации деревьев, древостоев по их жизненному состоянию и методы расчета жизненности древостоев могут быть использованы для практических и исследовательских целей. Рекомендуемая классификация деревьев позволяет также свести все многообразие категорий деревьев существующих классификаций к пяти.
ТИПЫ ПИХТОВЫХ ЛЕСОВ И ИХ ЭКОЛОГО-ГЕНЕТИЧЕСКАЯ КЛАССИФИКАЦИЯ
Типологическая классификация пихтовых лесов
Изучение лесообразовательного процесса на любом уровне невозможно осуществить в полной мере без построения четкой, объективной схемы типологической классификации лесной растительности.
Несмотря на то, что В.Н. Сукачев уделял много внимания этому вопросу, он так и не дал классификацию типов леса на биогеоценотической основе, прекрасно понимая трудности этой задачи. Он считал ее делом будущего (Рысин, 1982).
Взгляды отдельных ученых, направлений и школ по вопросам лесотипологической классификации освещались в специальной литературе неоднократно (Морозов, 1917, 1930; Сукачев, 1931-1964; Ивашкевич, 1933; Васильев, 1935; Соколов, 1936; Погребняк, 1955; Колесников, 1956, 1974; Дылис, 1964; Долуханов, 1964-1983; Махатадзе, 1965-1985; Смагин, 1965; Трасс, 1965; Голгофская, 1967, 1972; Воробьев, 1967; Васильев, 1967, 1974; Остапенко и др., 1968; Мелехов, 1968, 1980; Сочава, 1969; Александрова, 1969; Юркевич и др., 1971; Сабуров, 1971; Парфенов, 1972; Побединский, 1973; Розенберг, 1973, 1985; Назимова, 1975; Голубец, 1978; Уиттекер, 1980; Каразия, 1981; Гельтман, 1982; Василевич, 1983; Ипатов, 1983; Ды-ренков, 1985; Федорчук, 1985; Шевченко, 1985; Буш, 1985; Смологонов, 1985; Манько, 1987; Голод, 1989; Ралдугин, 1989 и другие).
Определенный вклад в развитие лесной типологии внесла рабочая секция лесной типологии Научного Совета по проблемам леса Академии наук СССР, возглавляемая долгие годы видным ученым лесотипологом Л.П. Рысиным. В работе этой секции принимали участие представители всех школ и направлений в бывшем СССР, в том числе и автор. К сожалению, плодотворная работа секции была прервана с распадом СССР.
История и современное состояние лесной типологии и их классификации в бывшем СССР подробно освещены в монографии Л.П. Рысина (1982). Для зарубежных стран эти вопросы излагаются в работах И.С. Мелехова (1980) и С.А. Дыренкова (1989). Поэтому останавливаться на них здесь нет необходимости, за исключением тех работ, которые имеют принципиальное значение для оценки современного состояния лесотипологической классификации горных лесов Кавказа и, в первую очередь тех, которые затрагивают вопросы классификации темнохвойных лесов с участием пихты кавказской. Это позволит изложить более основательно нашу точку зрения по вопросу типологической классификации пихтовых лесов.
Лесотипологические исследования на Кавказе начали развиваться в основном с начала 30-х годов, и связаны они, прежде всего, с именами
А.Г. Долуханова (1933-1989); С.Я. Соколова (1936); И.И. Тумаджанова (1934-1960); Л.Б. Махатадзе (1940-1985); П.Д. Ярошенко (1950-1962); Л .И. Соснина (1939); А.Я. Орлова (1951); Л.И. Прилипко (1954); К.Ю. Голгофской (1967-1972) и других. Несмотря на то, что типологи Кавказа вслед за В.Н. Сукачевым встали на биогеоценотические позиции и добились заметных успехов в развитии лесотипологических исследований, о полном единстве их взглядов, даже по основным вопросам, пока что говорить не приходится, и вопросы типологической классификации лесной растительности по сей день дискутируются. Дело в том, что ни одна из школ не в состоянии решить задачу разработки такой универсальной классификации, которая в желаемой степени удовлетворяла бы всем основным требуемым задачам. Соблюдение предельной естественности в рамках иерархически соподчиненных единиц одной классификационной системы фитоценозов, какой бы совершенной она не была, как справедливо отмечает А.Г. Долуханов (1971), неосуществимо.
По Р. Уиттекеру (1980), отношения иерархичности не заложены в самой природе, они достаточно условные и определены выбором подходов в классификации и путей их использования. Поэтому существование различных региональных школ и направлений лесотипологических классификаций не только закономерно, но и вполне оправдано. Видные ученые-лесоводы Г.Ф. Морозов, В.Н. Сукачев всегда были против шаблонных положений и приемов в лесной науке и практике. Это не потеряло смысла и в настоящее время при решении типологических задач.
Несомненно, лесотипологическая наука должна развиваться и впредь. Необходимы использование современных и поиски новых методических подходов к изучению типов леса, включая методы математического моделирования. Однако прогресс в теории и практике лесной типологии невозможен без единого методического подхЪда к изучению типов леса, без унификации основных типологических терминов, без сходного представления о содержании и объеме основных типологических понятий, наконец, без расчленения понятий “тип леса” и “тип лесорастительных условий”. Все это отнюдь не исключает возможности существования разных типологических направлений. Существование их необходимо для разработки более совершенных региональных классификаций, основанных на общих теоретических и методических установках.
В настоящее время, на основании работ ведущих типологов, для горных лесов Кавказа разработана эколого-генетическая (динамическая) классификация типов леса, основанная на стройном учении В.Н. Сукачева о лесной биогеоценологии.
Однако вопросы типологии и классификации пихтовых лесов в полной мере не решены. Именно с биогеоценотической, эколого-генетической позиции, нами были изучены типы пихтовых лесов и составлена их типо
логическая классификация (Бебия, 1973, 1983, 1989). Результатам этих исследований посвящена данная глава.
Наша точка зрения по основным принципам типологической классификации лесов близка и, в ряде случаев, сходна с теоретическими взглядами ведущих типологов Кавказа, прежде всего с позициями Л.Б. Махатадзе, А.Г. Долуханова, К.Ю. Голгофской.
Вместе с тем по ряду вопросов нами предлагается свое решение.
Горные леса Кавказа представляют собой особую природноэкологическую систему. Принципы выделения и классификации типов леса здесь имеют свои существенные отличительные особенности. Леса этой области характеризуются четко выраженной вертикальной зональностью. В силу этого, а также в связи с историей развития растительности данного региона, чрезвычайно богат породный состав лесов, многие виды - представители третичной реликтовой флоры. В зависимости от экспозиции,
крутизны склонов* гипсометрического положения меняются и варьируются почвообразовательные процессы, гидрологический и температурный ре-
жимы, состав атмосферы и другие экологические факторы. Все эти явления взаимосвязаны и взаимообусловлены, и они, в комплексе, определяют условия местопроизрастания, рост и развитие лесов, обусловливая многообразие типов леса, а в связи с этим сложность их классификации. Поэтому при типологии горных лесов особое значение приобретает ясность в определении содержания и объема понятия “тип леса”. Элементарная типологиче
ская единица - тип леса - это не только конкретный выдел с определенными
условиями местопроизрастания или определенными почвами и характерным для них комплексом лесной растительности. Тип леса - это, прежде всего, тип лесных биогеоценозов, по В.Н. Сукачеву (1964). Тип лесного биогеоценоза является не абстрактным понятием, а элементарной, динамически устойчивой, биологической системой, единицей учета сложного природного объекта - лесорастительного сообщества, представляющей материальную величину. Следовательно, она имеет свои размеры, которые должны измеряться единой системой мер. В противном случае понятие “тип леса” теряет практический и теоретический смысл. Нами тип леса понимается как совокупность участков леса, достаточно однородных по составу древесных пород, по другим ярусам растительности и фауне, по продуктивности, комплексу лесорастительных условий (климатических, почвенных и гидрологических), по взаимоотношению между растениями и средой, по динамическим качествам и направлению развития в пределах определенной естественно-исторической лесорастительной области и требующих при одинаковых экономических условиях однородных лесохозяйственных мероприятий.
Очевидно, что при таком определении и содержании тип леса приобретает понятие и географическое, на важность чего указывал еще Г.Ф. Мо
розов (1914). Тип леса при таком понимании становится не только элементарной таксономической, но и элементарной лесоводственной, однородной по своей природе, и хозяйственной единицей, характеризующейся конкретными качественными и количественными показателями, что, несомненно, обусловливает эффективность планирования и ведение лесного хозяйства на типологической основе.
Основными отличительными признаками типа леса мы считаем общность структуры, состава и производительности древостоев, живого напочвенного покрова и подлеска, формирования в сходных условиях местообитания, однородность лесовозобновительных процессов и динамики, реакции на антропогенные и природно-стихийные воздействия. Однако тип леса - понятие динамическое. В процессе онтоценогенетического развития он может проходить несколько полнотных или возрастных состояний - стадий, которые, по Б.П. Колесникову (1958), соответствуют понятию “тип насаждений”, по Л.Б. Махатадзе (1966) - “ассоциация”, по К.Ю. Голгофской (1967) - “тип лесного участка”, по В.Н. Сукачеву - “тип лесного биогеоценоза”.
С нашей точки зрения, наименьшая элементарная единица расчленения лесной растительности (ассоциация, тип насаждений, тип лесного участка) является не только полистным, или возрастным состоянием (стадией) развития одного лишь фитоценоза (древостоя), как это трактуется некоторыми типологами. Это, прежде всего, стадия биогеоценотического развития конкретного фитоценоза в определенных условиях местопроизрастания. Исходя из этого, наименьшую элементарную единицу лесного биогеоценоза целесообразнее называть “тип лесного биоценоза”. Активное вмешательство человека (сплошная, выборочная рубка) или природные стихийные явления (пожар, ветровал и пр.) также могут существенно изменить состояние коренного типа леса и сформировать производные типы леса. Однако на возвращение их в первоначальное состояние потребуется время значительно большее, несколько десятков лет, чем для смены одного полистного состояния древостоя другим. Например, тип леса пихтарник яс-менниковый распадается на несколько полнотных состояний: пихтарник мертвопокровный полнотой древостоя 0.9-1.0, пихтарник ясменниковый полнотой 0.6-0.8. При чрезмерно интенсивной выборочной рубке может сформироваться производный тип - пихтово-буковый разнотравный полнотой О.З-О.5.
Таким образом, тип леса - (тип лесных биогеоценозов), в нашем понимании, включает как коренные типы, находящиеся на разных стадиях развития, так и производные, представляющие собой различные звенья восстановительных смен. Все они генетически связаны друг с другом и являются стадиями одного ряда развития на уровне единой системы в определенных условиях местопроизрастания (ТУМ). Тип лесных биогеоценозов
представляет собой типологическую гомогенную автономную экосистему, в пределах которой могут быть выделены подсистемы более низкого уровня, но характеризующиеся одним общим признаком - неспособностью к существованию и развитию вне лесного биогеоценоза (Дыренков и др., 1975).
Нами тип леса (тип лесных биогеоценозов) в таком объеме принимается за основную единицу лесной типологии в системе классификации пихтовых лесов. Она близка к понятию объема “тип леса” Б.П. Колесникова (1956) и К.Ю. Голгофской (1972), но несколько шире, чем объем таксона “тип лесного биогеоценоза” Л.В. Махатадзе (1983). Наше представление о понятии “тип леса” по объему более соответствует понятию объема “цикл типов лесных биогеоценозов” по Л.Б. Махатадзе.
Несколько иного мнения придерживается другой ведущий типолог Кавказа А.Г. Долуханов, который понятия “тип леса” и “тип лесных биогеоценозов” считает синонимами и отмечает, что тип леса - это группы близких типов лесных биогеоценозов, развивающихся в местообитаниях, экологически сходных для произрастающей растительности во всех возрастных фазах ее развития. Причем это сходство местообитаний поддерживается особенностями сложения и развития самой растительности, единообразно трансформирующей среду своего обитания. Однако в отличие от наших представлений, Д.Г. Долуханов тип лесного биогеоценоза (но не тип леса) считает низшей и в то же время основной классификационной единицей. Он не дает формулировки понятия “тип леса” и считает нецелесообразным стремиться к “укрупнению” его объема и бояться детализации.
Можно согласиться с его мнением в том, что лесоведение нуждается пе в упрощении типологических построений, а в углублении познания самих типов леса, особенностей их экологии, биологии и генезиса, хода роста и закономерностей развития древостоев, динамики их формирования и сукцессионных смен в разнообразных средах, разнообразно трансформируемых развитием биогеоценозов. Что же касается хозяйственных группировок, то, по мнению А.Г. Долуханова, они должны определяться конкретными условиями и конкретными задачами, которые подлежит решать. Все это верно, однако типологическое изучение лесов не является самоцелью. Оно необходимо не только для познания закономерностей развития лесных биогеоценозов, лесообразовательных процессов, но и для использования этих познаний в практических целях, для обоснования системы мероприятий в лесу. В этом плане чрезмерное дробление лесного биогеоценоза существенно осложняет использование типологии в лесном хозяйстве. Практически всеми ведущими типологами (Морозов, 1917; Сукачев, 1964; Мотовилов, 1955; Колесников, 1956 и др.) подчеркивается важность и необходимость ведения лесного хозяйства на типологической основе. Несмотря на это, типология при ведении хозяйства в пихтовых лесах Кавказа недостаточно используется.
Ясно, что любой вариант группировки объектов может быть оптимальным лишь для определенных конкретных целей. В связи с этим разработке лесотипологической классификации должна предшествовать по возможности четкая формулировка целей классификации и способов ее использования. Это во многом предопределяет содержание, объем классификационных единиц и их соподчинение (Федорчук, 1965). В этом плане, разумеется, наша лесотипологическая классификация не претендует на универсальность и решение общенаучных задач, а.также на роль каталога биогеоценозов. Она направлена, прежде всего, на решение практических задач. При организации и ведении лесного хозяйства в пихтовых лесах данные лесной типологии должны послужить, в первую очередь, экологической основой для принятия важнейших лесоводственных решений - выбор состава и структуры формируемых древостоев, выбор способов и технологий рубок, лесовосстановлений и оценки возможных результатов хозяйственных мероприятий. Отсюда и необходимость при лесотипологических исследованиях учета существующих и потенциальных свойств биогеоценозов.
Мы разделяем точку зрения ведущих типологов, которые производительность древостоев считают важнейшим диагностическим признаком типа леса. Лесорастительный эффект местопроизрастания наиболее полно отражается в ходе роста древесных пород (Голгофская, 1967). Исходя из этого, лесные биогеоценозы, близкие своей структурой и флористическим составом, но с древостоями разных классов бонитета, мы относим к различным типам леса. Например, пихтарник овсяницевый I класса бонитета и пихтарник овсяницевый Ш класса бонитета следует относить к разным типам леса. Эти различия в производительности древостоев обусловлены неодинаковыми лесорастительными условиями.
На Кавказе, в отдельных лесорастительных районах, пояс пихтовых лесов простирается по вертикали довольно широко, более 1000 м.
Здесь фитоценозы пихты образуют эдафически замещающий ряд, начиная от высокопроизводительных биогеоценозов (пихтарник овсяницевый 1 класса бонитета) на богатых и мощных почвах до низкополнотных (пихтарник овсяницевый III-1V классов бонитетов) на бедных, скелетных, слаборазвитых почвах. Это обусловлено различием в гидрогеоморфологи-ческих условиях их развития. В то же время, в этом высотном диапазоне при некотором сходстве почвенно-грунтовых условий климатические условия различны. В частности, с увеличением высоты над уровнем моря повышается влажность, а количество летнего тепла уменьшается, что, в свою очередь, приводит к падению производительности древостоя с 1а до IV класса бонитета (Махатадзе и др., 1977). Например, в пихтарниках Абхазии в среднегорном поясе на высоте 1000-1500 м над уровнем моря часто встречается тип леса пихтарник овсяницевый 1а класса бонитета. Однако на отметках выше 1500 м этот тип леса замещается пихтарником овсянице-
вым III класса бонитета. В данном случае имеют место высотноклиматические замещения, сходные по всем ярусам растительности, типам леса разной производительности. На это также обращали внимание А.Я. Орлов (1951). А.Г. Долуханов (1964) и др.
Большинство типологов Кавказа считают целесообразным объединить все генетически связанные эдафо-климатозамещающие типы леса в одну группу типов (типов лесных биогеоценозов). Мы также придерживаемся такого принципа и принимаем группу типов леса за следующую, после типа леса , таксономическую единицу более высокого ранга в системе лесотипологической классификации. Важнейшим условием их объединения является однородность лесообразовательных процессов и экологоморфологических особенностей субэдификаторных синузий подлеска и травяного покрова. При этом производительность древостоя в группе типов леса может колебаться в широких пределах, но в рамке тип леса она не должна превышать одного-двух классов бонитета.
По Л.Б. МЬхатадзе (1966) для хозяйственных целей типы также следует группировать в хозяйственные группы. Типы, относящиеся к одной хозяйственной группе, должны формироваться в одинаковых условиях местообитания, а также иметь сходный характер возобновления. Лишь сходство породного состава и производительности древостоев для этого недостаточно. Следовательно, хозяйственные группы не всегда могут совпадать с объединениями типов леса в группы по естественным признакам (Махатадзе, 1959).
В пихтарниках выделенные нами группы типов леса в основном соответствуют по своему объему хозяйственным группам типов леса Л.Б. Махатадзе. Для каждой из них нами предлагается определенная система мероприятий.
В данном случае экологический аспект группировки типов леса для хозяйственных целей, на наш взгляд, имеет очень важное значение. Лишь на его основе возможно многоцелевое использование, воспроизводство лесов, сохранение их экологических функций. В этом смысле выделенные нами хозяйственные группы типов леса являются эколого-хозяйственными.
Следующей единицей высшего порядка в системе классификации лесных биогеоценозов нами принята лесная формация. Смысл и объем ее сходны с понятием лесной формации Б.П. Колесникова (1956), в которую он объединил типы леса по общности главной породы.
В формацию пихтовых лесов мы включаем, помимо коренных, коротко-производные группировки, которые нередко образованы другими экологически близкими древесными породами, обычно формирующими самостоятельные лесные формации (бук, ель).
Из таксономических единиц классификации лесной растительности наивысшего ранга нами принимается “поясной комплекс формаций”. Эта
единица объединяет биогеоценотические формации пихтовых и еловых лесов, образуемых видами, близкими по биоэкологии и распространенными в едином горном, вертикально-поясном лесном поясе.
Таким образом, схема соподчинения единиц в системе нашей классификации пихтовых лесов (табл. 35) от частного к общему выглядит следующим образом: тип биогеоценоза - тип леса (тип лесных биогеоценозов) - группа типов леса (группа типов лесных биогеоценозов) - формация поясной комплекс формации.
Таблица 35
Схема соподчинения классификационных лесотипологических
единиц пихтовых лесов
Единицы классификации Тип лесного биогеоценоза Тип леса (тип лесных биогеоценозов)
Группа типов леса (группа типов лесных биогеоценозов)
Формация
Поясной комплекс формации
_____________Примеры_____________ Пихтарник овсяницевый Пихтарник овсяницевый I класса бони-
тета
Пихтарники овсяницевые
Пихтовая
Темнохвойные леса
С помощью разработанных принципов лесотипологической классификации нами составлена обобщенная схема типов леса пихтарников (табл. 40), которая приводится ниже в разделе описания типов леса.
Типы пихтовых лесов
Вопросам типологии темнохвойных, в том числе и пихтовых лесов Кавказа, посвящено много работ. Среди них'следует отметить исследования И.И. Тумаджанова (1934), Н.Я. Каца (1936). С.Я. Соколова (1936), В.А. Поварицина (1936), М.И. Сахарова (1939), А.А. Колаковского (1939), Л.И. Соснина (1939), А.Г. Долуханова (1938, 1964, 1969, 1989), А.И. Лескова (1942), В.А. Ссорина (1951), А.Я. Орлова (1951), Л.Б. Махатадзе (1965, 1966), К.Ю. Голгофской (1967), Б.Ф. Остапенко (1968), С.М. Бебия (1973, 1983, 1989) и других.
Однако среди исследователей нет единого мнения по этому вопросу. Вместе с тем региональные особенности и биогеоценотипическое разнообразие типов леса пихтарников требуют более глубокого их изучения.
В типологическом отношении темнохвойные леса Кавказа отличаются большой сложностью. Это обусловлено разнообразием лесорастительных условий и факторов среды, создающих пестроту фитоценозов. Диагностика типов леса осложняется также вследствие отсутствия четкой бонити-ровочной шкалы и таблиц хода роста, сложной структурой темнохвойных
лесов и значительной нарушенностью состояния их в результате хозяйственной деятельности человека. Именно региональные особенности и био-геоценотическое разнообразие типов леса послужили поводом противоречивого подхода большинства исследователей к определению и понятию объема типа леса.
Тип леса мы понимаем, по В.Н Сукачеву (1964), как тип лесных биогеоценозов. Правильное понимание типа леса имеет существенное значение для выявления биоэкологии породы и ведения рационального, отвечающего природе лесов, хозяйства на типологической основе. Но сложность и разнообразие типов лесов значительно затрудняют проектирование мероприятий по каждому из них. Так, в пределах ареала пихты на Кавказе среди типологического разнообразия биогеоценозов четко намечаются вертикально и регионально замещающие варианты. Нижние - до 1200 м, средние - 1200-1700 м и верхнегорные варианты с высотой 1700 м над ур. моря. Все этг* варианты существенно различаются по составу древостоя, живого напочвенного покрова, динамике и продуктивности. Поэтому, как уже отмечалось выше, типы леса целесообразно объединить в группы.
Наиболее характерными группами типов для пихтарников Кавказа являются овсяницевая, разнотравная, ожиковая, папоротниковая, разнотравно-субальпийская и группа типов с колхидским подлеском.
Группа овсяницевых пихтарников. Наиболее распространенная группа типов леса. Встречаются они на всем протяжении вертикального распространения пихтовых лесов и имеют несколько высотно замещающих типов леса как на Северном Кавказе, так и на южных макросклонах Кавказа. Занимают хорошо освещенные склоны, характеризуются высокой производительностью - до 1200 м3 на га. Древостои I, реже la, II, Ш классов бонитета.
Относящийся к этой группе тип леса пихтарник овсяницевый среднего горного пояса характеризуется высокой производительностью. Древостой I, реже II класса бонитета. Встречается на склонах до 35°.
Пихтарник овсяницевый верхнего горного пояса характеризуется средней производительностью. Древостой П-Ш класса бонитета. Встречается чаще на склонах большой крутизны, свыше 25°.
Ниже приведем описание пробной площади 12, характеризующей тип леса - пихтарник овсяницевый среднего горного пояса. Пробная площадь заложена в Рицинском госзаповеднике. Высота 1200 м над уровнем моря. Экспозиция юго-восточная, уклон местности 20-25°. Почвы бурые, лесные, мощностью 70 см. Бонитет I класса.
Древостой разновозрастный буково - елово - пихтовый, характеризуется таксационными показателями, приведенными в табл. 36.
Благоприятные условия местопроизрастания обусловили высокую продуктивность данного типа леса. Подлесок развит слабо. Встречаются единичные экземпляры черники кавказской.
Наши полевые исследования, сборы многочисленного гербария на пробных площадях, их определение послужили основанием для характеристики напочвенного покрова в различных типах леса.
Таблица 36
Характеристика древостоя по типам леса в группе овсяницевых и _______________ разнотравных пихтарников______________________
Древостой
N пр. площади Группа типов. Тип леса ярусы состав по массе средний диаметр, см средняя высота, м число стволов, шт/га Z пл. сечения, м2/га запас, кЛка полнота сом-кну-то-сть
1 2 3 4 5 6 7 8 9 10 и
1. Разнотравная I 9Пх1Бк 83 42 56 30.30 527 0.42
Пихтарник П ЮПх 39 28 36 4.84 49 0.07 -
ежевичный. Ш 7ПхЗБк 18 14 124 3.4 22 0.05 -
Класс бони- в це-
тета! лом
для
яру-
сов 9Пх1Бк - - 216 38.35 598 0.54 0.5
9. Разнотрав- I 5Пх5Бк+Е 60 38 96 26,72 455 0.44 -
ная. * 5Пх4Бк1Е
Буково- П +Ил 35 24 256 24.32 278 0.40 -
пихтовый Ш 4Пх2Бк2Е 14 13 528 8.34 55 0.14 -
ясменнико- в це-
вый. лом
Класс бони- для
тета I яру-
сов 5Пх4Бк1Е - - 880 59.38 788 0.98 0.9
+Ил
22. Разнотравная I ЮПх 75 41 105 46.48 837 0.77 -
Пихтарник П 9Пх1Бк 43 29 110 16.11 221 0.26 -
ясменнико- Ш 5Пх5Бк 20 15 105 3.22 32 0.06 -
во-кислич- в це-
ный. Класс лом
бонитета! для
яру-
сов ЮПх+Бк - - 320 65.81 1034 1.09 0.8
Продолжение табл. 36
1 2 3 4 5 6 7 8 9 10 11
12. Овсяницевая I 5Пх4Е1Бк 71 41 136 54.48 1034 0.80
Пихтарник П 5Пх2Е2Бк 41 28 116 15.16 194 0.22 -
овсянице- Ш +ИлКл
вый. Класс в це- 4ПхЗЕ2Бк 15 15 332 5.53 40 0.08 -
бонитета! лом 1Кл
для
яру- 5Пх4Е1Бк - - 564 75.17 1268 1.10 0.9
сов +ИлКл
Степень покрытия травяного покрова на пробной площади 12 составляет 0.2, доминирует овсяница горная. Встречаются ясменник душистый, кислица обыкновенная, шалфей мутовчатый, фиалка лесная, вика оранжевая, грушанка малая, купена кавказская, герань Роберта, единично -папоротник мужской. Покрытие травяного покрова неравномерное. Под сомкнутым порогом чаще встречаются ясменник, кислица. Степень покрытия их не превышает 0.1. В окнах и на освещенных участках преобладает овсяница со степенью покрытия до 0.5-0.6.
Возобновление под пологом леса хорошее. Количество крупного подроста высотбй 0.5-4 м составляют: пихты - 384 шт., ели - 1650 шт., бука - 276 шт. на гектаре. Количество мелкого подроста высотой до 0.5 м: пихты - 0.5 тыс. шт., бука - 0.3 и ели - 0.3 тыс. шт./га. Подрост на площади встречается группами в окнах. Количество всходов на пробной площади составило: пихты - 5.4 тыс. шт., бука - 0.7 тыс. шт., ели - 0.2 тыс. шт./га.
Сравнительно незначительная степень покрытия травяного покрова не может препятствовать появлению всходов и подроста. Наличие на пробной площади крупного подроста в количестве 2310 шт./га, а также деревьев третьего яруса в количестве 332 шт. подтверждает успешный процесс естественного возобновления в овсяницевых пихтарниках.
Характерно, что пихтарник овсяницевый среднего горного пояса на Северном Кавказе встречается на более пологих склонах и отличается более высокой производительностью древостоя I - 1а бонитета, чем на южном. Это подтверждается данными наших исследований (табл. 34, пробная площадь 64).
Пихтарник овсяницевый верхнего горного пояса характеризуется средней производительностью, древостой П-Ш классов бонитета, встречается чаще на склонах большой крутизны, свыше 25°. На Северном Кавказе этот тип леса отличается более низкой производительностью, древостой Ш-IV классов бонитета, среднеполнотный, возобновление слабое (Орлов, 1951).
Примечательно, что существенного различия в составе живого напочвенного покрова между этими региональными вариантами типов леса пихтарников нет.
Овсяницевые пихтарники на Северном Кавказе приурочены главным образом к центральной части пояса темнохвойных лесов, бассейнам рек Большой и Малой Лабы. К востоку от реки Лабы, как отмечает А.Я. Орлов (1951), овсяницевые пихтарники замещаются ожиковыми пихтарниками.
Группа ожиковых пихтарников. Основной доминант живого напочвенного покрова этой группы типов леса западноевропейский элемент -ожика лесная. Группа типов не имеет широкого распространения на Кавказе, но в ценотическом разнообразии типов леса она занимает определенную позицию. Распространена группа ожиковых пихтарников, в основном, по Северному макросклону Кавказа, в частности, в бассейнах рек Большой Зеленчук и Большая Лаба с высоты 1400 м над ур. моря (Орлов, 1951).
По данным А.Г.Долуханова (1989), ожиковые пихтарники встречаются также на южном склоне Кавказа, в бассейнах левых притоков среднего течения реки Ингур и некоторых правых притоков Цхенисцкали. Характерно, что в отличие от северного макросклона Кавказа, где эта группа типов леса приурочена к районам с распространением сланцев с большим содержанием железистых и марганцовистых соединений (Ссорин, 1951), в Грузии ожиковые пихтарники приурочены большей частью к склонам, состоящим из нижеюрских отложений лейаса и местами из среднеюрской порфиритовой свиты байоса (Долуханов, 1989, с. 213).
Группа образует два коренных высотно замещающих типа леса: пихтарник ожиковый среднего горного пояса II класса бонитета и пихтарник ожиковый III-1V классов бонитета верхнего горного пояса с высоты 1700 м. Для них характерны свежие суглинистые почвы средней мощности, до 60 см, подстилаемые глинистыми сланцами, реже песчаниками. Древостои бывают чисто пихтовые, иногда буково-пихтовые, а на маломощных почвах на конгломератах с примесью ели. Древостои разновозрастные с полнотой в среднегорном поясе 0.6-0.8, а в верхнегорном - 0.4-0.6.
По сравнению с овсяницевыми, ожиковые пихтарники отличаются меньшей производительностью древостоев, до 900 м3 в среднегорном поясе и до 600 м3/га в верхнегорном поясе. Естественное возобновление в них обычно слабое.
Травяной покров развит, сомкнутость его 0.3-0.8. В нем доминирует ожика лесная. Сопутствует ей овсяница горная, ясменник душистый, подлесник европейский, кислица обыкновенная, душевик крупноцветковый, герань Роберта и другие. Ближе к верхней границе леса проективное покрытие становится выше и в составе его появляются элементы субальпийского высокотравья, такие как валериана чесночниколистная, бор развесистый, смолевка многорассеченная, кочедыжник женский и другие.
Учитывая неблагоприятные условия роста близ верхней границы и слабое естественное возобновление в типах леса ожиковых пихтарников, целесообразнее допускать лишь добровольно-выборочные рубки интенсивностью в пределах годичного прироста древостоев с одновременным содействием естественному возобновлению.
Группа разнотравных пихтарников. Широко распространенная группа типов леса по всему Кавказу. К этой группе относятся следующие основные типы: ясменниковый, кислицево-ясменниковый, кислицево-подлесниковыи, пихтовые и буково-пихтовые леса. Встречаются они на свежих, хорошо дренированных, развитых бурых горно-лесных почвах. Распространены эти типы леса в основном на склонах до 30° западной и северной экспозиций. По нашим исследованиям, типы леса, относящиеся к группе разнотравных, характеризуются высокой производительностью. Древостои преимущественно I класса бонитета. Состав их может быть чисто пихтовым, а чакже с участием бука восточного, ели восточной, клена остролистного, клена высокогорного, липы кавказской, ильма, рябины обыкновенной. Полнота древостоев в среднем 0.7-0.9, иногда 1.0 (см. табл. 36).
Напочвенный покров этой группы типов леса состоит в основном из лесных растений, таких как ясменник, кислица, подлесник европейский, герань, фиалка, вороний глаз, купена, овсяница, молочай и другие. При полноте и сомкнутости древостоя меньше 0.6 могут иметь место кратковременные или длительно-производные типы леса с развитым травянистым покровом или с колхидским подлеском.
Так, по нашим исследованиям, в древостоях коренных типов разнотравной группы пихтарников при полноте 0.6 наблюдалось значительное участие в покрове ежевики кавказской и щетинистой, местами папоротников мужского и женского и др. (табл. 37). В больших окнах-прогалинах встречаются заросли бузины травянистой.
С приближением к субальпийской зоне в покрове низкополнотных древостоев начинают доминировать представители субальпийского высо-котравья и низкотравья такие, как крестовник, горечавка, душевик, гаде-лиа, борщевик, аконит и другие виды.
Следует отметить, что на прогалинах и в низкополнотных древостоях пышное развитие травяной растительности значительно препятствует появлению всходов и подроста.
В целом типы леса группы разновозрастных пихтарников характеризуются успешным естественным возобновлением. Для характеристики группы разнотравных пихтарников приводим описание трех пробных площадей.
Пробная площадь 9 заложена в Рицинском госзаповеднике в кв. 45. Высота над уровнем моря 1200 м. Экспозиция северная, уклон местности
составляет 8-15°. Почвы бурые лесные, средней мощности. Тип леса - буково-пихтовый ясменниковый I класса бонитета.
Древостой на пробной площади представлен разновозрастным буко-но-пихтовым с участием ели, ильма (см. табл. 36). Стволы деревьев пихты и бука I-П ярусов ровные, полнодревесные, хорошо очищенные от сучьев. Среди них имеются деревья исключительно высокого качества (стволы ровные, малосбежистые, высота прикрепления ветвей превышает 3/4 высо-1Ы ствола, кора гладкая). Значительное количество деревьев (880 экз/га) обусловило высокую полноту древостоя, равную 0.98, и сомкнутость полога 0.9. В силу этого деревья Ш и часть II ярусов испытывают заметное угнетение. Растут они тонкими, крона зонтикообразная. Так, при среднем диаметре деревьев Ш яруса 13 см высота их составляет 14 м. Среди них много сухих.
Количество крупного подроста незначительно и составляет 520 шт/га, однако наличие мелкого (3-10 - летнего возраста) в количестве 15.1 1ыс. шт. и всходов 18.8 тыс. шт./га подтверждает хорошие возобновительные условия в данном типе леса.
Таблица 37
Состав и обилие (по Друде) травяного покрова в группе _____________разнотравных пихтарников____________
Растения Пробные площади
9 22 1
Asperula odorata Сор. Cop. Sp-
Athyrium felix-femina - - Sol.
Calamintha grandiflpra - - Sol.
Daphne mezereum - - Sol.
Dentaria bulbifera Sol. Sol. -
Dryopteris filix-mas Sol. Sol. Cop.
Euphorbia macrocerasus Sol. , Sol. Sol.
Festuca drymeja - Sol. Sp-
Gentiana schistocalyx - Sol. Sol.
Geranium robertianum Sol. Sol. Sol.
Myosotis amoena Sol. Sol. Sol.
Oxalis acetosella Cop. sP. Sp.
Paris incompleta Sol. Sol. -
Poligonatum polyantherum Sol. Sol. -
Rubus caucasicus Sol. - Cop.
Salvia verticillata Sp. Sol. Sol.
Sambucus ebulus - Sol.
Sanicula europea - Sol. Sol.
Senecio platyphylloides - Sol. Sol.
Viola reichenbachiana Sol. Sol. -
Степень покрытия 0.1-0.2 0.2-0.3 0.7-0.8
Сомкнутость полога 0.9 0.8 0.5
Пробная площадь 22 заложена в Аибгинском лесничестве Гагринского лесхоза, кв. 54. Высота над уровнем моря 1400 м. Экспозиция северо-западная, уклон местности составляет 15°. Почвы бурые лесные суглинистые, мощностью до 80 см. Бонитет 1а класса, тип леса - ясменниково-кисличный пихтарник. Древостой разновозрастный, высокопродуктивный (см. табл. 36). Деревья пихты I яруса стройные, высота отдельных экземпляров достигает 48-50 м. Бук принимает значительное участие в Ш и меньше во II ярусах. Подлесок редкий. Представлен единичными экземплярами черники кавказской и падуба.
Несмотря на полноту древостоя, равную 1.09, и сомкнутость полога 0.8, возобновление здесь удовлетворительное. Так, общее количество крупного подроста составляет 1.4 тыс. шт., мелкого - 3.6 тыс. шт. и всходов - 50.8 тыс. шт./га.
Пробная площадь 1 заложена на лесосеке 9-летнего возраста. Место закладки - Р^ринский заповедник, район Ауадхара. Высота над уровнем моря 1650 м. Экспозиция юго-западная, уклон местности 18-20°. Почвы типичные бурые лесные, мощностью до 90 см. Бонитет 1а класса.
Древостой на пробной площади разновозрастный с незначительным участием бука (см. табл. 36). Деревья первого яруса крупномерные. Отдельные экземпляры достигают 48 м высоты при диаметре на высоте груди 152 см. Некоторые из них фаутные, суховершинные. Подлесок редкий. Состоит из черники кавказской и падуба. Низкая сомкнутость крон, равная 0.5, обусловила пышное развитие покрова. Степень покрытия покрова составляет 0.8. В силу этого на пробной площади представлен производный тип леса - ежевично-разнотравный пихтарник. Коренной тип леса на данном участке - ясменниково-кисличный пихтарник.
Возобновление неравномерное, приурочено к слабо заросшим участкам. Количество крупного подроста составляет 1.20 тыс. шт., мелкого -6.20 тыс. шт. и всходов - 12.29 тыс. шт./га.
При изучении живого покрова на пробных площадях 9, 22 и 1 была установлена корреляционная зависимость между составом, обилием, степенью покрытия травяного покрова и сомкнутостью полога древостоя. Снижение сомкнутости полога на 0.5 в разнотравных пихтарниках способствует появлению в составе покрова широкотравья, ежевики и папоротников (см. табл. 37), а также развитию подлеска из лавровишни, рододендрона понтийского, а в Аджарии и рододендрона Унгерна. Такое явление существенно влияет на процесс естественного возобновления.
К группе разнотравных пихтарников могут быть отнесены и различные эдафо-литологические варианты типов леса трахистемоновых пихтарников. Основной типообразующий эдификатор лесов реликтовый эн-дем Кавказа трахистемон восточный, который большей частью характерен для буковых лесов, чем для пихтовых. В связи с этим трахистемоновые
пихтарники имеют ограниченное распространение в Закавказье и встречаются они, главным образом, в его северо-западной части. Встречаются они и па Северном Кавказе, но в значительно меньшей степени.
Относительно биоэкологии и динамики трахистемоновых пихтарников у типологов существуют противоречивые суждения. Так, Т.И. Исаченко (1980) рассматривает трахистемоновые пихтарники в составе группы широкотравных пихтарников. По мнению А.Я. Орлова (1951), на Северном Кавказе они близки к группе папоротниково-разнотравных буково-пихтовых лесов и поэтому автор охарактеризовал их в пределах этой группы.
По данным А.Г. Долуханова (1989), в Грузии трахистемоновые пихтарники занимают слегка влажноватые экотопы, средние между группами типов леса мелкотравных и папоротниковых пихтарников. А.Г. Долуханов описал их как самостоятельную группу типов.
По нашим наблюдениям, в Абхазии типы леса трахистемоновых пихтарников приурочены, главным образом, к экотопам, близким условиям произрастания группы разнотравных пихтарников.
В бассейнах рек Бзыбь и ее правых притоков трахистемоновые пихтарники встречаются от 900 до 1700 м. Развиваются они как на известняковых, так и на неизвестняковых породах, в связи с чем образуют два коренных типа леса. Так, для более или менее выраженных ложбин характерен тип леса пихтарник трахистемоновый, 1-1а классов бонитета, на бескар-бонатных породах. Почвы бурые лесные, до 70 см. Древостой разновозрастный с полнотой 0.8-0.9 и производительностью до 1200 м3/га. По составу он может быть чисто пихтовым или с примесью до двух единиц бука, единично ель, клен явор, В таких древостоях много сухостоя во всех ярусах, и количество стволов подчиненных второго и третьего ярусов всегда превалирует.
Другой тип леса буково-пихтовый трахистемоновый I-II классов бонитета, развивающийся на карбонатных почвах. Древостой в этих условиях менее производительный, до 700 м3/га, с полнотой 0.7-0.8. Участие бука в нем значительно, до 4-5 единиц. Количество стволов во всех ярусах немногочисленно, среди стволов бука много фаутных, старовозрастных с раскидистой кроной.
Подлесок в трахистемоновых пихтарниках не развит. В флористическом отношении в них живой напочвенный покров может быть разнообразным, особенно в биогеоценозах на карбонатных породах. Однако в нем всегда доминирует трахистемон восточный. К сопутствующим видам относятся окопник крупноцветковый, купена многоцветковая, зубянка клубненосная, толстостенка крупноцветковая и другие, характерные для типов леса группы разнотравных пихтарников. Сомкнутость травяного покрова колеблется от 0.3 до 0.7.
В биоэкологическом понимании и ценотическом отношении рассматриваемые типы трахистемоновых пихтарников представляют определенный интерес, однако выделение их в отдельную группу типов леса с хозяйственной точки зрения вряд ли целесообразно. В этих типах, равно как и в остальных типах леса группы разнотравных пихтарников, допустимо проведение одинаковых практических мероприятий. В частности, в них можно осуществлять выборочную, а на склонах до 25° и группововыборочную форму хозяйства.
Исследователи темнохвойных лесов Кавказа А.Я. Орлов (1951) и Л.Б. Махатадзе (1966) выделяют и другую отдельную самостоятельную группу типов леса - мертвопокровные пихтарники. По мнению этих исследователей, в таких биогеоценозах отсутствие живого напочвенного покрова обусловлено недостатком почвенной влаги, а не высокой полнотой древостоя. Вместе с тем, приводимые ими отличительные особенности этих типов леса и характеристика условий их произрастания не позволяют с полным основанием отнести их к коренным типам леса, выделить в самостоятельную группу.
Следует подчеркнуть, что неблагоприятные сухие условия местообитания в этих биогеоценозах, несомненно, способствуют низкому уровню почвенной влаги, но не настолько, чтобы в таких условиях не могли произрастать хотя бы засухоустойчивые элементы. В этих условиях мезофитная порода пихта образует, к примеру, древостои высокой сомкнутости, следовательно, причины отсутствия покрова здесь и в другом.
Характерно, что по площади мертвопокровные биогеоценозы занимают обычно небольшие участки леса по склонам южной экспозиции до 40° крутизны. Для них характерны бурые лесные, слабо гумусированные, малоразвитые, щебнистые, часто скелетные почвы мощностью до 40 см и грубый лесной опад до 4-5 см толщины.
Для характеристики типа леса среднегорного мертвопокровного пихтарника по Северному Кавказу А.Я. Орлов (1951) приводит описание пробной площади 18, в древостое которой количество стволов первого, верхнего яруса составляет 370 шт., общее число их на гектаре 750. Заложена она на склоне южной экспозиции 34° крутизны.
Естественно, что сухие условия местопроизрастания и сравнительно высокая сомкнутость древесного полога, равная 0.8, не способствовали процессу разложения лесного опада, мощность которого на пробной площади составляла 4 см.
Все это и обусловливает отсутствие или слабое развитие живого напочвенного покрова в этих биогеоценозах.
Вместе с тем, в характеризуемом древостое значительное количество крупномерных стволов первого яруса, как отмечает А.Я. Орлов (с. 115), сильно фаутные, а тонкомер в большинстве случаев сильно угнетен.
Со временем в таких древостоях появление значительного количества отпада стволов, снижение общей сомкнутости полога древостоя, следовательно, и развитие живого напочвенного покрова, надо полагать, неизбежны.
Это и подтверждается тем, что на пробной площади по более освещенным местам отмечены пятна кислицы и овсяницы горной, а также единичные экземпляры вики золотистой, чины розовой, фиалки лесной, герани Роберта, купены многоцветковой, шалфея клейкого и других, то есть виды, которые в основном характерны для живого покрова группы разнотравных пихтарников.
Биоэкология и динамика таких биогеоценозов в большей степени от-ражают черты полистной стадии развития типа леса разнотравных пихтарников.
В подтверждение этого приводим описание пробной площади 15, заложенной нами в Кучба-Яштинском лесничестве Гудаутского лесхоза Абхазии (южный макросклон Кавказа) в кв. 18. Размер ее 0.25 га. Склон южной экспозиции 30° крутизны. Почвы бурые лесные, мощностью до 50 см. 11одстилка из лесного опада, грубая до 4 см толщины. Древостой по составу пихтовый с единичной примесью ели, разновозрастный, с выраженным |рехъярусным строением. Количество стволов первого яруса 176, 43% от общего их числа на гектаре. В нем значительная часть разновозрастных, фаутных, 27%. Общая сомкнутость полога 0.9. Живой напочвенный покров, равно как и подлесок, отсутствовал за исключением единичных экземпляров вики оранжевой и кислицы.
Характеризуемая пробная площадь была описана как тип леса мертвопокровный пихтарник II класса бонитета.
В 1978 году в этих лесах произошел сильный ветровал, вызвавший снижение общей сомкнутости,полога древостоя до 0.4. В 1984 году провели повторное описание данной пробной площади и обнаружили в ней развитый живой напочвенный покров, покрытие которого составляло 0.6. В нем доминировал ясменник, но значительное участие принимали также ежевика, овсяница горная, кислица, герань Роберта, шалфей клейкий, ду-шевик крупноцветковый и другие, которые, в основном, характерны для разнотравных пихтарников.
Таким образом, биогеоценозы мертвопокровных пихтарников, как показали и дальнейшие наши исследования, целесообразнее рассматривать как полнотные стадии развития отдельных типов разнотравных пихтарников.
Группа папоротниковых пихтарников. Эта группа типов леса приурочена к наиболее увлажненным местообитаниям пологих склонов. Из-за недостаточной устойчивости пихты в биогеоценозах с влажными почвами папоротниковые пихтарники не имеют широкого распространения. К этой группе относятся два типа леса: пихтарник кочедыжниковый и пихтарник страусоперовый.
Пробная площадь 6 характеризует тип леса пихтарник страусоперо-вый. Она заложена в Аибгинском лесничестве Гагринского лесхоза кв. 53. Высота над уровнем моря 1400 м. Экспозиция западная, уклон местности 5-10°. Почвы мощные бурые лесные. Бонитет 1а класса. Древостой разновозрастный пихтарник с примесью бука и клена остролистного. Несмотря на довольно высокую полноту древостоя, равную 1.11, и значительный запас древесины на гектаре - 1324 м3, сомкнутость полога составляет 0.7 (табл. 38). Обусловлено это преобладанием в древостое толстомерных стволов пихты. Крона у таких деревьев колонновидная, некоторые из них суховершинные. Подлесок развит слабо. Состоит из единичных групп черники кавказской. На прогалинах единично встречается рябина. В живом покрове, степень покрытия которого составляет 0.7-0.8, доминирует папо-ротник-страусник. Высота его достигает 1.5 м. Реже встречается папоротник мужской, кочедыжник женский, ясменник, герань Роберта, купена многоцветковая,Недотрога обыкновенная.
Таблица 38
Основные таксационные показатели древостоя _________в папоротниковых типах_________
N пр. площади Тип леса Древостой
ярусы состав по массе средний диаметр, см средняя высота, м число стволов, шт/га Е пл. сечения, м2/га запас, м3/га полнота сом-кну-то-сть
6 Пихтар- I ЮПх^Бк 79 41 130 64.35 1164 0.95
ник страу- II 8Пх2Бк 39 28 75 9.01 117 0.13 -
сопе- Ш 7Пх 23 18 ПО 4.43 43 0.06
ровый!а в це- 2Кл1Бк
класса лом
бонитета для
яру-
сов 911x1 Б к+Кл - - 315 77.79 1324 0.85 0.7
13 Пихтар- 1 ЮПх 80 42 108 53.95 1014 0.85
ник коче- П 8Пх2Бк2Пх 45 29 32 5.12 68 0.08 -
дыжнико- Ш 8Бк 18 14 132 3.26 23 0.05 -
вый I в це-
класса лом
бонитета для
яру-
сов 9Пх1Бк - - 272 62.33 1105 0.98 0.6
Возобновление наблюдается группами, приуроченными к микропо-вышенным участкам, где слабо развит папоротниковый покров. Состоит оно из пихты, бука и клена. Количество крупного подроста составляет 800 шт., мелкого - 1000 шт. и всходов - 12800 шт. на гектаре, распространение их на площади неравномерное.
Пробная площадь 13 характеризует тип леса пихтарник кочедыжниковый. Заложена она в районе Ауадхары Рицинского госзаповедника. Высота над уровнем моря составляет 1750 м. Экспозиция южная, уклон местности 20°. Почвы бурые лесные суглинистые, мощностью до 90 см. Подстилка рыхлая. Бонитет I класса. Полнота древостоя 0.98, запас, равный 1105 м3/га, свидетельствует о высокой продуктивности древостоя в данном типе леса (см. табл. 38). Однако на пробной площади деревья произрастают неравномерно. Встречаются прогалины. Перестойные деревья пихты довольно высокие. Некоторые из них достигают 50 м высоты при диаметре па высоте груди 116-120 см. Кроны рыхлые. Встречаются суховершинные экземпляры.
В подлеске единично встречаются черника кавказская и рябина.
Травяной покров мощный, степень покрытия 0.9. Высота его достигает 1.2 м. В покрове доминирует папоротник кочедыжник женский. Встречаются папоротник мужской, трахистемон восточный, ясменник, герань Роберта, кислица, овсяница, молочай, крестовник.
Естественное возобновление неудовлетворительное. Количество крупного подроста составило 1.1 тыс. шт., мелкого - 9.7 и всходов 43.0 тыс. шт. на гектаре. Казалось бы, эти цифры характеризуют успешное возобновление в данном типе леса, однако всходы и подрост встречаются группами на определенных участках. В остальных местах наблюдается полное отсутствие как всходов, так и подроста. Такой характер возобновления обусловил неравномерное распределение деревьев на площади и образование прогалин, сплошь заросших папоротниками и высокотравьем.
Таким образом, в папоротниковых пихтарниках наблюдается развитие мощного травяного покрова, препятствующего успешному возобновлению. В то же время условия роста пихты в этих местообитаниях благоприятные, что подтверждается наличием довольно крупномерных деревьев, достигающих 50 м высоты. Древостой разновозрастный, высокопродуктивный. Однако количество деревьев на гектаре сравнительно мало, в пределах 272-315 шт. (см. табл. 38).
Деревья первого яруса составляют 50-60 процентов. Среди них встречаются суховершинные. Значительное количество деревьев первого яруса обусловливает высокую полноту древостоя, иногда превышающую 1.0. В то же время из-за сравнительно малого количества деревьев всех ярусов на гектаре и неравномерного распределения их на площади сомкнутость полога всегда меньше, чем полнота древостоя. Так, на пробной площади 13
при полноте древостоя 0.98 сомкнутость полога составляет 0.6; на пробной площади 6 полнота равна 1.11, сомкнутость полога 0.7.
В силу вышеизложенного следует, что в группе папоротниковых типов пихтарников хозяйственные мероприятия следует проектировать с особой осторожностью. В этих условиях допустимы только добровольновыборочные и комплексно-выборочные способы рубок с последующим содействием процессу естественного возобновления.
Пихтарники с колхидским подлеском. В пихтарниках с подлеском из черники кавказской, падуба, лавровишни и рододендрона на южном макросклоне Кавказа распространены до 1700 м над уровнем моря, на северном макросклоне - от 800 до 1100 м. Чаще остальных встречаются буково-пихтовые леса с подлеском из черники и падуба.
Приводим описание пробной площади 8, заложенной в типе леса пихтовом черничниковом. Заложена она в Аибгинском лесничестве Гагринского лесхоза,<св. 53. Высота над уровнем моря 1250 м. Экспозиция западная, уклон составляет 30°. Почва бурая лесная, мощностью до 60 см. Бонитет П класса. Древостой с незначительным участием бука, разновозрастный. Деревья пихты сбежистые. Максимальная высота деревьев первого яруса не превышает 38 м. Распределение их на площади относительно равномерное. В составе третьего яруса преобладает бук (табл. 39).
Подлесок развитый. Доминирует черника кавказская. Единично встречается падуб. Сомкнутость яруса подлеска достигает 0.7. Высота его 1.0-1.5 м.
В травяном покрове, степень покрытия которого составляет 0.2, встречаются ясменник, овсяница, кислица, вороний глаз. На стволах перестойных деревьев и валеже растет плющ.
Возобновление удовлетворительное. Общее количество крупного подроста составляет 1.4 тыс. шт., мелкого - 4.8 тыс. шт., всходов - 40.4 тыс. шт./га. Распространение всходов и подроста на площади также относительно равномерное.
Пробная площадь 7 заложена в буково-пихтовом черничниково-падубовом типе леса в Аибгинском лесничестве Гагринского лесхоза, кв. 54. Высота над уровнем моря составляет 1500 м. Экспозиция западная, уклон местности 20°. Бонитет I класса. Почвы светло-бурые, мощностью до 60 см. Древостой буково-пихтовый, разновозрастный. Бук участвует главным образом в третьем и втором ярусах (см. табл. 39). Деревья пихты первого яруса полнодревесные. Максимальная высота их достигает 45 м.
Подлесок встречается по всей площади. Состоит он из падуба и черники кавказской. Сомкнутость яруса подлеска составляет 0.8. Высота черники достигает 1.5 м, падуба - 1 м. Травяной покров состоит из ясменника, кислицы, герани Роберта, ежевики, папоротника мужского. Степень покрытия его составляет 0.3. На валежниках и стволах перестойных деревьев
н комлевой их части встречается плющ колхидский. Возобновление удовлетворительное. Общее количество крупного подроста составляет 2.0 тыс. шт, мелкого - 4.6 тыс. шт., всходов 480 тыс. шт. на гектаре. Распределение всходов на площади относительно равномерное.
Таблица 39
Основные таксационные показатели древостоя в пихтарниках с __________________колхидским подлеском________________
N Древосто й
пр. Тип леса яру- состав по сред- сред число Е за- пол сом-
НЛО- сы массе ний няя ство- пл. пас, но- кну-
та- дна- вы- лов, сече- м3/га та то-
ди метр, сота, шт/га ния, сть
см м м2/га
i 2 3 4 5 6 7 8 9 10 и
8 Пихтарник I 9Пх1Бк 68 36 ’ 80 29.24 364 0.42
чернични- П 9Пх1Бк 45 29 150 24.17 320 0.34 -
ковый П п 8Бк2Пх 18 12 ПО 2.88 19 0.04 -
класса бони- в це-
тета лом
для
яру-
сов
9Пх1Бк - - 340 56.29 703 0.8 0.8
7 Буково- I 9Пх1Бк 76 41 90 40.38 553 0.50
пихтовый I 5Пх5Бк 43 29 145 20.96 231 0.26 -
чернични- ш 5Пх5Бк 20 5 115 3.60 25 0.04 -
ково- в це-
падубовый I лом
класса бони- для
тета .... яру-
сов
8Пх2Бк - - 350 64.94 809 0.8 0.7
19 Елово- I 5Пх5Е 67 32 24 8.38 142 0.13 -
пихтовый П 5ПхЗЕйс1
падубо- Лп+Кл 41 25 208 27.33 303 0.44 -
лавровиш- Ш ЗПх6Е1Лп
невый Ш в це- + СКлИлБ 18 10 590 14.28 107 0.23 -
класса бони- лом
тета для
яру-
сов 4Пх4Е1С1
Лп+КлБИл - - - 49.99 552 0.7 0.6
Для крупнокаменистых местообитаний, каменистых склонов известняковых гор характерны падубо-лавровишневые биогеоценозы пихтового леса. Пихта в таких местообитаниях менее устойчива. В силу этого значительное участие в древостое принимает ель восточная. Для характеристики данного типа леса приводим описание пробной площади 19.
Заложена она в Рицинском госзаповеднике. Высота над уровнем моря составляет 1000 м. Экспозиция юго-восточная. Рельеф увалистый, крутизна склона от 5 до 30° Почвы маломощные, щебнистые, хрящеватые, беспорядочно перемешивающиеся с подстилкой. Встречаются крупные обломки пород. Первый ярус древостоя состоит из пихты и ели. В остальных ярусах принимают участие сосна Коха, липа кавказская, клен, ильм и бук. Древостой низкопродуктивный. Бонитет Ш класса. Максимальная высота пихты и ели первого яруса не превышает 36 м. Деревья сбежистые. Много фаутных. Стволы плохо очищены от сучьев. Сомкнутость кроны низкая и составляет 0.6 (см. табл. 39).
Подписок сильно развит, состоит из лавровишни, падуба. Единично встречаются иглица шиповатая, черника кавказская, смородина альпийская, чубушник кавказский. Сомкнутость подлеска составляет 0.8, высота 1 м. Травяной покров незначителен. Встречаются ландыш закавказский, купена, шалфей. В тенистых местах стелется плющ, иногда взбирается на деревья.
Мощный, развитый, густой подлесок значительно препятствует успешному возобновлению. Так, на пробной площади имеется крупного подроста пихты 1.2 тыс. пгт., ели - 0.5 тыс. шт., всходов пихты - 2.1 тыс. шт., ели -3.9 тыс. пгт., других сопутствующих пород - 2.4 тыс. шт. на гектаре. Подрост на площади распределен неравномерно, встречается в виде отдельных групп.
Биогеоценозы пихтовых лесов с подлеском из рододендрона тяготеют к склонам северных румбов по неизвестняковым хребтам. В средней пихтовой зоне встречаются пихтарники с густым подлеском из рододендрона понтийского. Древостои низкобонитетные - П-Ш классов бонитета, разновозрастные. Характеризуются неудовлетворительным естественным возобновлением.
В верхней зоне в контакте с субальпийским поясом изредка может встречаться пихтарник с подлеском из рододендрона кавказского. Древостой Ш-IV классов бонитета, низкополнотный, около 0.5, разновозрастный. Участие в древостое березы, рябины, ели, клена высокогорного, бука - обычно. В подлеске густые групповые заросли рододендрона кавказского с частичной примесью черники обыкновенной. Возобновление очень слабое.
Следует отметить, что товарная ценность пихтарников с подлеском из лавровишни и рододендрона низкая. При неправильном вмешательстве
человека в этих биогеоценозах могут образоваться чистые рододендровые или лавровишневые синузии.
Пихтарники с колхидским подлеском встречаются и на Северном Кавказе, хотя в меньшей степени и не все типы леса этой группы.
По К.Ю. Голгофской (1967), типы леса рассматриваемой группы распространены здесь главным образом в бассейне реки Белой. Развиваются они на склонах различной крутизны и экспозиции на бурых лесных суглинистых почвах различной мощности. Встречаются почти по всей вертикальной протяженности пояса пихтарников. По ее данным, группа слагается из двух коренных типов леса. Колхидско-кустарниковый буко-пихтарник 1-1а классов бонитета развивается в условиях достаточного увлажнения с развитым подлеском из рододендрона понтийского, характеризуется неудовлетворительным возобновлением. Другой тип леса - колхидско-кустарниковый пихтарник III класса бонитета развивается в условиях худшего увлажнения, с развитым подлеском из черники кавказской и падуба, а также с примесью иногда лавровишни и иглицы. Характеризуется слабым возобновлением.
По данным А.Я. Орлова (1951), в пихтарниках Северного Кавказа типы леса с колхидским подлеском характерны, главным образом, для буково-пихтовых лесов. Среди них преобладают биогеоценозы с развитым подлеском из рододендрона понтийского. Естественное возобновление в этих биогеоценозах находится в прямой зависимости от степени развития подлеска, и обычно оно слабое.
Все биогеоценозы с колхидским подлеском А.Я. Орлов объединяет в две самостоятельные группы. Одна из них - группа чернично-падубовых буково-пихтовых лесов включает типы леса - падубово-буково-пихтовый и его высотно замещающий вид - чернично-буково-пихтовый. Вторая группа рододендроновых буково-пихтовых лесов включает коренной и производный типы леса с одноименными с группой названиями, но различающиеся в основном развитостью почвы. В производном типе леса она более развита, и в подлеске значительно участвует падуб.
Характерно, что в флористическом отношении состав живого напочвенного покрова биогеоценозов с колхидским подлеском пихтарников Северного Кавказа мало отличается от такового в пихтарниках южного макросклона.
Видный исследователь колхидского подлеска А.Г. Долуханов (1980) отмечает, что биогеоценозы с колхидским подлеском более характерны для буковых лесов, чем для пихтовых. Однако следует отметить, что эдифика-юрно-ценотическая роль колхидского подлеска в современных темнохвойных лесах Кавказа также весьма существенна. Он играет огромную роль в формировании и динамике развития ценозов. Отметим также, что не все элементы колхидского подлеска современных темнохвойных лесов явля
ются собственно эндемами Колхида в геоботаническом понимании. Однако эдификаторно-ценотическая позиция всех этих видов в развитии биогеоценозов темнохвойных лесов наиболее мощно выражена в Колхиде, что указывает также на сложную ценогенетическую и реликтовую черту их развития. В то же время пихтарники с колхидским подлеском характеризуются весьма своеобразными, специфическими для них чертами ценоти-ческой структуры, динамики развития, что также придает им оригинальную, отличительную особенность от темнохвойных лесов других регионов Евразии.
В этом отношении особый интерес представляют эндемичные пихтовые леса из пихты Кавакама на острове Тайвань. По данным наших наблюдений, здесь довольно широкое распространение имеют оригинальные биогеоценозы пихтовых лесов с густым, труднопроходимым “напочвенным покровом” из бамбука (Jushania niitakayamensis), высотой до 3 м. Заросли бамбука выполняют структурные функции подлеска. В таких типах леса пихта также возобновляется очень слабо. Несколько подобные пихтовые биогеоценозы встречаются и в более северных широтах, на островах Сахалин, Кунашир, Шикотан и Хоккайдо. В них нами изучены типы лесов из пихты сахалинской с густым “подлеском” из другого вида - бамбука курильского (Sasa kurilensis\ высотой до 2.5 м, которые также имеют довольно широкое распространение на всех этих островах. В эколого-ценотическом отношении оба варианта достаточно близки и можно рассматривать как широтно-зонально замещающие типы леса. Разумеется, в ценотическом и флорогенотическом отношении эти биогеоценозы пихтовых лесов существенно отличаются от биогеоцеозов темнохвойных лесов с колхидским подлеском. Однако эдификаторно-ценотическая позиция подлеска из отмеченных видов бамбука в развитии биогеоценозов из пихты Каваками и сахалинской, равно как колхидского подлеска в пихтарниках Кавказа, мощно выражена. Гипотетически это указывает на общность сложной ценогенетической и древней черты их развития.
Пихтарники с колхидским подлеском, произрастая в горных условиях, выполняют ценные почвозащитные и водоохранные функции. Поэтому в них следует проектировать мероприятия, которые должны обеспечивать естественное возобновление. В этих лесах на склонах до 30° можно допустить комплексно-выборочные рубки. Одновременно необходимо проведение мероприятий по содействию естественному возобновлению.
Группа типов разнотравных субальпийских пихтарников
Различные варианты этой группы типов произрастают у верхнего предела леса, начиная с высоты 1600 м. Многочисленные маршрутные записи, сделанные в полевых условиях этой зоны, позволили охарактеризо
вать группу типов. Для них характерны разреженные неравномерносомкнутые древостои 111—1V бонитетов. Почвы лесные, дерновые, обычно легкие или среднесуглинистые, скелетные. Древостои разновозрастные с большим количеством перестойных деревьев пихты. Стволы пихт сильно сбежистые, плохо очищенные от сучьев. Максимальная высота их не превышает 30 м. Диаметры на высоте груди достигают иногда до 100 см. В составе древостоя принимают участие ель, бук, клен, береза, рябина.
Подлесок слабо развит, в зависимости от условий местопроизрастания встречаются смородина альпийская, жимолость кавказская, рододендрон кавказский, черника кавказская, черника обыкновенная, лавровишня, волчье лыко, можжевельник и др.
Травяной покров развит. Степень покрытия его составляет 0.5-0.7. Состав разнообразный. Обычными растениями являются крестовник, аконит, чемерица Лобеля, дорикниум, валериана, колокольчик, подбел, окопник, овсяница, герань Роберта, кислица, ясменник, молочай и др.
Тяжелые лесорастительные условия (короткий вегетационный период, мощный снежный покров, низкие температуры) и развитый травяной покров обусловили очень слабое естественное возобновление. Выпас скота и заготовка жердняка пастухами также способствуют плохому восстановлению и ослаблению устойчивости этих лесов. Хотя товарная ценность древостоев низка, однако биогеоценозы этой группы типов леса, произрастая в верхней горной зоне, выполняют исключительно важные защитные функции.
В силу этого в субальпийских разнотравных пихтарниках можно проектировать только те-мероприятия, которые могут обеспечить достаточное естественное возобновление и устойчивость против неблагоприятных экологических факторов этих биогеоценозов. В них допустимы лишь санитарные рубки. На прогалинах целесообразно создавать культуры саженцами 0.7-1.0 м высоты, биогруппами на площадях 4 м2 из пород пихты, бука, кленов платанолистного и ложноплатанового, Траутфеттера, березы Медведева, глаговины и др.
Пихтарники скально-лесных комплексов. Известно, что в горных условиях вследствие сложности рельефа почвенный покров и условия местопроизрастания меняются часто на близких расстояниях. В этих неоднородных микрорастительных условиях образуются различные лесорастительные группировки.
Являясь обычными ореофитами и благодаря влажному климату, пихта кавказская и ель восточная хорошо приспособлены произрастать на каменистых субстратах, на гребнях хребтов, на маломощных скелетных почвах и образовывать такие микролесорастительные группировки. При этом пихта больше тяготеет к микроучасткам на уступах склонов, межскальным понижениям с более или менее развитыми почвами.
Скально-лесные пихтово-еловые или елово-пихтовые комплексы характеризуются неравномерно сомкнутыми низкопроизводительными рединами. Деревья значительно различаются по характеру развития, на выступах скал можно встретить отдельные деревья высотой не более 10-12 м с диаметром ствола до 70 см в спелом возрасте. В составе древостоя в виде примеси встречаются сосна Коха, осина, клен, вяз, береза.
Характерно, что в флористическом отношении скально-лесные комплексы не отличаются богатством видов, и каждый вид строго дифференцирован по экотопам. К примеру, в таких биогеоценозах по выпуклым элементам рельефа развиваются в основном литофильные элементы: вейник тростниковый, овсяница горная, по микропонижениям - ясменник, кислица и другие, то есть представители лесного разнотравья.
В результате изменения рельефа на коротком расстоянии, отдельные типы леса сменяются на протяжении незначительных площадей. В таких случаях целесообразнее выделять комплексные типы леса из разнообразных вариантов сквльно-лесных биогеоценозов.
В хозяйственном отношении скально-лесные комплексы не являются большой ценной значимостью, однако они выполняют почвозащитные функции.
Следует подчеркнуть, что выделение и изучение скально-лесных комплексов имеет важное значение в биоэкологическом и ценогенетическом понимании природы как отдельных реликтовых элементов, так и ценозов, образуемых ими.
Возможно эти ценогруппировки являются отголоском того исторического прошлого, когда в процессе эволюции реликтовые современные ле-сообразователи - пихта, ель и сосна смогли пережить и выдержать конкуренцию в борьбе за существование с другими, более молодыми конкурентоспособными видами в таких стациях в связи с их приспособленностью к этим экотопам. В этом плане скально-лесные комплексы совершенно не изучены и требуют специальных исследовании.
В заключение отметим, что результаты наших специальных типологических исследований на пробных площадях и многочисленных рекогносцировочных лесотипологических записей (более 1800) в пихтарниках большинства лесорастительных районов, а также критический анализ опубликованных литературных источников по данному вопросу позволили составить схему типологической классификации пихтовых лесов Кавказа, которая приводится ниже (табл. 40).
В данной схеме классификации наглядно представлены сукцессионные ряды, ряды трансформации типов леса, образованные основным (коренным) типом леса и одним или несколькими производными со сменой или без смены пород. На схеме отражено также, к каким почвенным и топографическим условиям (крутизна и экспозиция склонов, высота над
Таблица 40
Типологическая классификация пихтовых лесов Кавказа
Типы условий местопроизрастания (рельеф, почва, высота над уровнем моря), индексы Группа типов леса, индекс Типы леса
коренные, индексы производные
без смены пород, индексы со сменой пород, индексы
1 2 3 4 5
I. Склоны всех экспозиций и крутизны; на склонах северных румбов до 35° крутизны; почвы мощностью до 90 см, скелетные, средние или легкие суглинки, свежие или суховатые; 900-1800 м. ТУМ-1 II. Крутые склоны южных и восточных румбов; более 30°. Почвы сравнительно маломощные, до 50 см, сильноскелетные, щебнистые, легкие суглинки, суховатые, периодически свежие. 1700-2000 м. ТУМ-П. 1. Овсяницевые пихтарники ТУМ -1 -1 То же 1. Пихтарник овсяницевый Та—I класса бонитета. Состав от 10 Пх до &Пх4Бк, ед. Е. В покрове доминирует овсяница горная. Возобновление пихтой, иногда с участием ели и бука, хорошее. Распространен во всех лесорастительных районах, за исключением 1 и 4. ТУМ -1 - 11 2. Пихтарник овсяницевый Ilin классов бонитета. Состав 10-711x0-3БК, ед Кл, Ос, Рб. В покрове преобладает овсяница. Возобновление пихтой удовлетворительное. Распространен во 2, 3, 5 лесорастительных районах. ТУМ - II - 12 Не отмечены Не отмечены 1. Сосняк овсяницевый. 5-9С5-1Пх+Е. В покрове овсяница горная с участием вейника. Возобновление пихтой с участием ели, сосной слабое. Распространен в 5 и 6 лесорастительных районах. ТУМ-1-1101 1. Рябинник разнотравный 9- 40 8Рб1-2Бк+Ос, ед. Пх, Кл. В покрове вейник, овсяница, разнотравье. Возобновление пихтой очень слабое. Распространен во 2, 3 лесорастительных районах (редко). ТУМ-11-1201
III. Крутые склоны южных и восточных румбов, 30-40°. Почвы средней мощности до 60 см, скелетные, щебнистые, суглинистые, свежие. 1400-1700 м. ТУМ-П! Ожиковые пихтарники ТУМ -III-2 1. Пихтарник ожиковый П класса бонитета. Состав 10-7Пх 0-ЗБк, ед. Е. В покрове доминирует ожика лесная. Возобновление пихтой слабое. Распространен во 2, 3, реже в 6 и 11 лесорастительных районах. ТУМ - Ш - 21 Не отмечены Не отмечены
1 2 3 4 5
IV. Крутые склоны южных румбов, более 35°. Почвы маломощные, до 45 см, скелетные, щебнистые, суглинистые, свежие, периодически суховатые. 170Q-2000 м. ТУМ - IV То же 2. Пихтарник ожиковый разнотравный Ш-IV классов бонитета. Состав 10-8ПхО-2Бк, ед. Е, Б, Рб. В покрове ожика лесная с участием овсяницы и элементов субальпийского высокотравья. Возобновление пихтой слабое. Распространен в 3, реже в 6 лесорастительных районах (редко). ТУМ-IV-22 Не отмечены • 1. Березняк вейниковый. 9Б10с, ед. Пх. В покрове доминирует вейник. Возобновление пихтой неудовлетворительное. Встречается в 3, реже в 6 лесорастительных районах. ТУМ-IV-2201
V. Склоны западных, восточных и северных румбов, до 25°. Почвы мощные, до 130 см, малощебнистые, дренированные, средние суглинки, свежие. 900-1600 м. ТУМ - V 3. Разнотравные (мелкотравные) пихтарники ТУМ-У-З 1. Пихтарник ясменниковый 1а-I классов бонитета. Состав 9-7ПХ1-ЗБк, ед. Е, Ил, Кл. В покрове доминирует ясменник. Возобновление пихтой удовлетворительное или хорошее. Распространен во 2, 3, 5, 6, 10, 11 лесорастительных районах. Возможно и в 12 районе. ТУМ -V-31 1. Буково- пихтовый ежевично-разнотравный 6-5Пх4-5Бк, ед. Ил, Кл. Полнота 0,44), 5. В покрове преобладает ежевика, папоротник. Возобновление пихтой очень слабое. Встречается в 3, 4, 5, 6, 10, 11 лесорастительных районах. ТУМ - V -311 1. Пихтово-буковый разнотравный. Бонитет II класса. Состав 8-бБк, 2-4Пх, ед. Ил, Лп. Полнота 0,3-0,5. В покрове разнотравье, степень покрытия 0,7-0,9. Возобновление пихтой очень слабое. Встречается в 4, 5 ,6, 11 лесорастительных районах. ТУМ -V-3111
1 2 3 4 5
VI. Склоны западных, восточных румбов, до 30°. Почвы сравнительно мощные, до 80 см., среднещебнистые, средние суглинки, свежие. 1500-1700 м. ТУМ-VI То же 2. Пихтарник ясменниково-подлесниковый II класса бонитета. Состав 9-6Пх1-4Бк, ед. Е, Кл, Рб. В покрове доминируют ясменник и подлесник. Возобновление пихтой удовлетворительное. Распространен во 2, 3, 5, 6 лесорастительных районах (возможно и в 12 районе). ТУМ -VI-32 1. Пихтарник крупнотравный. Состав 10-8Пх0-2Бк, ед. Е, Б, Кл. Бонитет Ш, полнота 0,3-0,5. В покрове доминирует широкотравье, степень покрытия 0,7-0,9. Возобновление очень слабое. Встречается в 4, 5, 10, 11 лесорастительных районах. ТУМ-VI-321 1. Рябинник разнотравный, бонитет Ш класса. Состав 9-8Р61-2Б, ед. Пх, Бк, Кл. В покрове разнотравье. Степень покрытия 0,7-0,8. Возобновление пихтой очень слабое. ТУМ-У1-3211
VII. Пологие склоны восточных и западных румбов, 10-25°. Почвы сравнительно мощные до 80 см, малощебнистые, легкосуглинистые, • свежие влажноватые. 900-1600 м. ТУМ-УП То же 3. Пихтарник трахистемоно-вый la—I классов бонитета. Состав 1О-7ПхО-ЗБк, ед. Е, Кл, Ил. В покрове доминирует трахистемон восточный. Возобновление пихтой очень слабое. Распространен в 4, 5 лесорастительных (возможно и в некоторых других) районах. ТУМ - VII - 33 Пихтарник ежевично-папоротниковый I—II классов бонитета. Состав 8Пх 2Бк, ед. Кл, Е, Ил. Полнота 0,3-0,5. В покрове ежевика, папоротник, степень покрытия 0,6-0,8. возобновление пихтой очень слабое. Встречается в 4, 5 лесорастительных районах. ТУМ -УП-331 Не отмечены
1 2 3 4 5
VIII. Отрицательные элементы рельефа, террасовидные уступы пологих склонов. Почвы мощные до 100 см, суглинистые, влажные. 1000-1700 м. ТУМ-УШ 4. Папоротниковые пихтарники. ТУМ-УШ-4 1. Пихтарник кочедыжниковый la—I класса бонитета. Состав 9-7Пх1-2Бк1Ил, ед. Е, Кл. В покрове доминирует кочедыжник и широкотравье. Возобновление пихтой и сопутствующими породами очень слабое. Распространен в 1, 2, 4, 5, 6, 7 лесорастительных районах. ТУМ -УШ-41 Не отмечены • Не отмечены
IX. Повышенные участки пойм, у основания склонов. Почвы мощные, до 80 см, мелкокаменистые, суглинистые, влажные. 800-1700 м. ТУМ-IX То же 2. Пихтарник страусоперо-вый, I класса бонитета. Состав 8Пх2Бк, ед. Е, Ол, Ил, Кл. В покрове доминирует папоротник страусоперовый с примесью папоротников мужского и женской), а также овсяницы, ясменника, кислицы, герани Роберта, подбела. Возобновление пихтой, буком очень слабое. Распространен в 1, 2, 3, 4, 5, 6 лесорастительных (возможно и в других) районах. ТУМ - IX - 42 Не отмечены Не отмечены
Продолжение табл. 40
1 2 3 4 5
X. Склоны северных, западных, восточных румбов до 30° крутизны. Почвы сравнительно мощные, до 80 см, скелетные, среднесуглинистые, свежие, периодически влажнова-тые. 800-1700 м. ТУМ - X 5. Пихтарники с колхидским подлеском. ТУМ-Х-5 1. Пихтарник черничниково-падубовый la—I классов бонитета. Состав 1О-6ПхО-4Бк, ед. Кл, Ил. В подлеске доминируют черника кавказская и падуб колхидский. В покрове - разнотравье Возобновление пихтой удовлетворительное или слабое. Распространен во 2, 4, 5, 7, 8 лесорастительных районах. ТУМ-Х-51 Не отмечены Не отмечены
XI. Склоны разной экспозиции, до 35° крутизны. Почвы средней мощности, до 60 см, скелетные, щебнистые, суглинистые, свежие. 600-1700 м. ТУМ-Х1 То же 2.Пихтарник рододендроновый П-Ш классов бонитета. Состав 9-6Пх1-4Бк, ед. Е. В подлеске рододендрон понтийский. Возобновление очень слабое. Распространен во 2, 4, 5 лесорастительных (возможно и в других) районах. ТУМ - XI - 52 Не отмечены 1. Заросли рододендрона понтийского. Естественное возобновление пихтой очень слабое, неудовлетворительное. ТУМ - SS XI-5201
XII. Склоны различной экспозиции и крутизны. Почвы средней мощности, до 60 см, скелетные, хрящеватые, хорошо увлажненные, среднесуглинистые, дерново-бурые. 1600-2000 м. ТУМ-XII 6. Пихтарники субальпийские разнотравные ТУМ-ХП-6 1. Пихтарник субальпийский П (Ш) класса бонитета. Состав 9-7Пх2Бк1Кл, ед. Е. В покрове доминируют элементы субальпийского высокотравья. Степень покрытия 0,6—0,8. Возобновление пихтой очень слабое. Распространен во 2, 3, 4, 5 лесорастительных (возможно и в других) районах. ТУМ - XII -61 Не отмечены 1. Субальпийский кленарник Ш класса бонитета. Состав 9КЛт1Бк, ед. Пх, Б. Полнота 0,3-0,4. В покрове густо представители субальпийского высокотравья. ТУМ -ХП- 6101
Продолжение табл. 40
1 2 3 4 5
XIII. Склоны северных, восточных, западных румбов различной крутизны. Почвы маломощные, до 40 см, скелетные, щебнистые, мелкокаменистые, увлажненные, дерново-бурые, суглинистые. 1700-2000 м. ТУМ-ХП1 То же 2. Пихтарник субальпийский Ш-IV классов бонитета. Состав 6ПхЗБк1КЛт, ед. Ос, Б, С, Рб. В подлеске рододендрон кавказский, черника обыкновенная, смородина альпийская. В покрове элементы субальпийского высокотравья, степень покрытия 0,5-0,7. Возобновление пихтой очень слабое. Распространен во 2, 3, 5 лесорастительных (возможно и в других) районах. ТУМ - ХП - 62 Не отмечены • 1. Кленарник субальпийский IV класса бонитета. Состав бКлтЗБЮс, ед. Пх, Рб. Полнота 0,3-0,4. В покрове - субальпийское высокотравье, степень покрытия 0,4-0,6. Возобновление пихтой очень слабое. Распространен во 2, 3, 5 лесорастительных районах. ТУМ - XII - 6201 2. Заросли рододендрона кавказского и субальпийского высокотравья. Возобновление пихтой очень слабое, неудовлетвори- _ тельное. ТУМ - XII - 6202 52
Примечание. ТУМ - тип условий произрастания: Пх - пихта кавказская, Е - ель восточная, Бк - бук восточный, Кл - клен остролистный, Кит ~ клен Траутфеттера, Яс - ясень обыкновенный, Ил - ильм эллиптический, Лп - липа кавказская, Рб - рябина, Ос - осина, Б -береза Литвинова, С - сосна Коха
уровнем моря и пр.) приурочен каждый сукцессионный ряд типов леса. Краткое описание почв относится, в основном, к коренным типам леса. В производных типах леса почвы могут несколько изменяться под влиянием иных ярусов растительности и продолжительности времени сукцессионных процессов.
По данной схеме можно получить краткие сведения о составе древостоя, производительности (бонитет), характере естественного возобновления по типам леса, а также о их географическом распространении (по лесорастительным районам) и другие сведения.
На схеме типологической классификации каждый приведенный горизонтальный ряд генетически связанных типов леса является в определенной степени и типом условий лесопроизрастания (ТУМ).
Использование индексации при составлении лесотипологической классификации существенно облегчает применение типологии в практических целях (при лесоустроительных работах, составлении кадастра типов леса и типов условий местопроизрастания), а также ввод типологических данных в компьютер. В схему лесотипологической классификации включены, главным образом, типы леса и их производные, которые имеют более или менее широкое распространение и представляют интерес в хозяйственном отношении. В схеме охарактеризованы 6 групп типов леса, 13 коренных и 12 производных типов леса.
СИСТЕМА МЕРОПРИЯТИЙ ПО ОПТИМИЗАЦИИ ИСПОЛЬЗОВАНИЯ ПИХТАРНИКОВ
НА ЛЕСОТИПОЛОГИЧЕСКОЙ ОСНОВЕ И ОХРАНЕ
Хозяйственные группы типов леса и системы мероприятий в них
Выше уже отмечалось, что в типологическом отношении пихтовые леса отличаются большим разнообразием и сложностью, что значительно затрудняет проектирование лесохозяйственных мероприятии по отдельным типам леса. В связи с этим, с лесоводственной и экономической точек зрения многие исследователи рекомендуют объединить типы леса в близкие в экологическом отношении хозяйственные группы, однородные или сходные по лесорастательным условиям и целенаправленности хозяйств. Идея применения лесной типологии при устройстве равнинных лесов и объединения типов леса в организационно-хозяйственные единицы - секции, позволяющие испольЛвать дифференцированный подход при решении различных лесохозяйственных мероприятий, принадлежит Г.П. Мотовилову (1955, 1957). Она основана на принципах естественной классификации типов леса. Для горных лесов Кавказа эту идею теоретически развил Л.Б. Махатадзе (1959; 1983). Основной принцип объединения типов леса в хозяйственные группы (секции) заключается в сходстве экологических (лесорастительных) условий, биоэкологических свойств древесных пород и степени их возобновляемости.
Этот принцип использован нами при объединении в хозяйственные группы генетически связанных коренных и производных типов леса, имеющих определенную целевую установку и требующих одинаковых практических мероприятий.
Нами типы леса, близкие в биоэкологическим отношении, по лесообразовательным процессам и целенаправленности хозяйства объединены в пять основных хозяйственных групп типов леса: пихтарники овсяницевые, разнотравные, папоротниковые, с колхидским подлеском и субальпийские. Для каждой группы в отдельности, на основании наших экспериментальных исследований (Бебия, 1967, 1973), разработаны системы мероприятий, способствующие повышению производительности данной категории лесов и улучшению их защитных функций.
Приводим краткую характеристику выделенных хозяйственных групп и рекомендуемых мероприятий в них.
1. Пихтарники овсяницевые. В этой группе основными целевыми установками являются: хозяйство на пихту, выращивание крупнотоварного леса, повышение производительности лесов и их защитных функций.
Для данной группы типов леса характерны, преимущественно, крутые склоны и сравнительно маломощные, средней мощности, скелетные почвы. Пихта в этих условиях недостаточно ветроустойчивая, однако древостои с высокими показателями жизненного состояния (см. табл. 34). Естественное возобновление удовлетворительное (см. рис. 11).
Допустимы добровольно-выборочные или комплексно-выборочные рубки по разработанной нами технике и технологии их проведения (Бебия, 1973). Интенсивность рубки в пределах годичного прироста древостоя. Повторяемость рубки - один раз в 15-20 лет.
Целесообразно формирование как чистых пихтовых, так и смешанных буково-пихтовых древостоев. Оптимальный состав древостоя 8-7Пх2-ЗБк, который обеспечивает наиболее высокую его производительность
2. Пихтарники разнотравные. Эта группа типов леса развивается, преимущественно, по пологим и средней крутизны склонам, на мощных, суглинистых, свежих почвах. Характеризуется высокими показателями жизненного состояния древостоев (см. табл. 34), а также хорошим естественным возобновлением (см. рис. 11). Основные целевые назначения те же, что и для предыдущей группы. Допускаются добровольно-выборочные рубки интенсивностью в пределах годичного прироста древостоя и повторяемостью один раз в 20 лет. На склонах до 20° могут быть рекомендованы также группово-выборочные рубки с окнами в 18-25 м в диаметре, округлой или овальной формы, согласно разработанной нами технике и технологии проведения рубки (Бебия, 1973). На 1 га закладываются 4-5 окон, равномерно распределенных по площади и, по возможности, приуроченных к куртинам предварительного подроста, с последующим расширением окон через каждые 12-15 лёт кольцами шириной 10-12 м. Интенсивность рубки составит до 25% запаса. Возобновительный период - 45-50 лет. В рубку назначаются все деревья I яруса, расположенные в окне. Из нижних ярусов выбираются деревья сильно угнетенной, усыхающей и сухостойной категории по разработанной нами шкале классификации деревьев (Бебия, 1999).
Для сокращения периода возобновления на лесосеке в окнах можно вводить частичные лесные культуры или провести содействие естественному возобновлению главной породы по нашей методике (Бебия, 1988). Суть этой методики излагается в следующем разделе. Для посадочного материала используется самосев (“дички”) пихты высотой 0.4-0.5 м, который под пологом и по опушкам леса всегда имеется в достаточном количестве. Метод экономичен, не требует затрат на выращивание саженцев пихты. Период возобновления может быть сокращен в два раза.
При формировании состава древостоя в смешанных буково-пихтовых древостоях участие бука целесообразно сохранять до 25-30%.
3. Пихтарники папоротниковые. Основные целевые назначения -хозяйство на пихту, формирование древостоев более высокой производи
тельности, улучшение условий лесовозобновления и сохранение защитных функции лесов.
В связи с преобладанием низкосомкнутых древостоев и очень слабым естественным возобновлением пихты и бука (см. рис. 11), допускается выборочная рубка с уборкой во всех ярусах древостоя деревьев сильно угнетенной, усыхающей и сухостойной категории жизненности.
Одновременно под пологом изреженного древостоя осуществляется содействие естественному возобновлению. Для этой цели, полосами шириной 2 м и длиной 10-15 м с разрывом в 4 м по горизонтали склона выкорчевывается почвенный покров и снимается верхний слой почвы в 5-8 см. Между очищенными полосами оставляются нетронутыми полосы шириной 3-4 м. На них укладываются выкорчеванный покров и снятая почва.
На очищенных полосах происходит возобновление, в основном лиственных пород (клен, ильм, ольха). Пихта возобновляется слабее. Однако под защитой крон лиственных пород и соответствующих рубках ухода (прореживания) в н#х пихта начинает возобновляться лучше и постепенно восстанавливать свои позиции.
Этот процесс можно ускорить путем ввода частичных лесных культур в больших окнах и прогалинах крупными саженцами лиственных пород (клен явор, ильм эллиптический, липа кавказская, ясень обыкновенный) высотой 0.5-0.7 м. Посадочные места культур обрабатываются по схеме и технологии, как на полосах для содействия естественному возобновлению.
4. Пихтарники с колхидским подлеском. Основные целевые установки - хозяйство на пихту, выращивание смешанных древостоев более высокой производительности с сохранением доминирующей позиции пихты, получение древесины, сохранение защитных функций лесов.
Рекомендуются комплексно- и добровольно-выборочные рубки интенсивностью в пределах годичного прироста древостоя.
В типах леса с вечнозеленым подлеском (падуб, рододендрон, лавровишня), в связи с низкой сомкнутостью древостоев (см. табл. 39), сильно развитым подлеском и очень слабым естественным возобновлением (см. табл. 14), допустимы лишь выборочные рубки с удалением сильно угнетенной, усыхающей и сухостойной категории деревьев.
Одновременно по всему участку целесообразно удаление подлеска 2-3 метровыми полосами по горизонтали склона. Полосы размещаются через 4-5 м по вертикали. В полосы высаживаются крупномерные саженцы лиственных пород высотой 0.5-1 м (бук, клен, липа, ильм) как быстрорастущие. Смыкание крон лиственных пород приведет к ослаблению позиции подлеска и обусловит возобновление пихты.
При формировании смешанных древостоев доля участия пихты в их составе не должна быть ниже 60 %.
5. Пихтарники субальпийские. Основное целевое назначение лесов этой группы - сохранение и обеспечение их защитной роли.
Для лесов этой группы характерны суровые экологические условия произрастания, зачастую фрагментарность их размещения. Древостои низ-кополнотные и характеризуются очень слабым естественным возобновлением. Исключительно велика их водоохранная, почвозащитная, противола-винная роль и, в первую очередь, для высокоствольного леса, произрастающего ниже.
Хозяйство в этих лесах должно быть направлено на поддержание экологической устойчивости лесов, предотвращение отрицательных антропогенных (рубки, рекреационная перегрузка), пасторальных и по возможности стихийных воздействий.
Некоторые исследователи предлагают восстанавливать субальпийские леса Кавказа исключительно путем посадок лесных культур (Махатадзе, Урушадзе, 1972). Однако достаточных практических данных, которые подтверждают эффективность таких теоретических рекомендаций по восстановлению субальпийских лесов Кавказа, пока нет. Данные наших наблюдений и многолетних стационарных исследований Абхазской лесной опытной станции по этому вопросу в Рицинском заповеднике, в урочище Ауадхара (1650 м над уровнем моря), пока не дали обнадеживающих результатов (Млокосевич, 1973; Бебия и др., 1988). Создание лесных культур в субальпийских пихтарниках сопряжено с большим риском.
Целевое назначение этой группы типов определяет особый подход к лесам этой категории. Если учесть их чрезвычайно существенное экологическое значение, станет ясна большая важность правильного ведения хозяйства в них.
В целом же без реализации предлагаемой системы мероприятий невозможно обеспечить рациональное использование и воспроизводство пихтовых лесов.
Способы рубок в пихтарниках
Произрастая в верхней горной зоне, пихтовые леса выполняют важные почвозащитные, водоохранные, климаторегулирующие, курортологические, рекреационные и целый ряд других функций. Это достаточно хорошо освещено в специальной литературе. Но следует подчеркнуть, что по режиму пользования пихтарники Кавказа отнесены к категории I группы лесов. Одновременно они являются объектами получения ценной древесины. В пихтарниках проводятся рубки главного пользования. В силу этого в деле ведения прогрессивного хозяйства в этих лесах первостепенное значение имеют способы рубок и рациональный метод технологии лесосечных
работ, обеспечивающих, в первую очередь, вышеотмеченные полезные функции.
В настоящее время лесоводственная наука достигла определенных успехов в деле решения вопроса эффективных способов рубок главного пользования для горных темнохвойных лесов.
Существенный вклад в изучение и совершенствование рубок главного пользования в горных лесах внесли исследования В.Г. Гулисашвили (1948-1971), Н.М. Горшенина (1959-1972), Б.П. Колесникова (1965), И.С. Мелехова (1966), П.И. Молоткова (1965-1967), А.В. Побединского (1967-1970) и других ученых. Изучением рубок в темнохвойных лесах Кавказа занимались К.А. Сакс (1938), А.Я. Орлов (1951), З.Я. Солнцев (1954), И.Д. Попов (1955), Б.И. Иваненко (1956), А.И. Ильин (1963), А.А. Дроби-ков (1964), К.К. Калуцкий (1967-1971), И.П. Коваль (1971), С.М. Бебия (1967-1973), И.В. Веселов (1973) и др. По мнению большинства исследователей, природе горных темнохвойных лесов Кавказа наиболее отвечает выборочная форма хозяйства. Правильность этого положения подтверждается и нашими исследованиями (Бебия, 1967-1983). Выборочной форме хозяйства для темнохвойных лесов уделяется большое внимание и за рубежом: в Германии (Kestler, 1950), Англии (Day, 1966), Югославии (Drazen, 1967), Франции (Baugemot, 1967), Австрии (Мауег, 1969). По данным С. Недялко-ва (1979), например, в разновозрастных темнохвойных лесах Болгарии производительность лесов при выборочной форме хозяйства на 44.2% выше за 100 лет, чем в одновозрастных.
Выше, при изложении результатов наших исследований по вопросам биоэкологических особенностей, плодоношения, естественного возобновления пихты кавказской, возрастного строения и структуры лесов с ее господством было установлено, что в пихтарниках целесообразнее применение добровольно-выборочных и группово-выборочных способов рубок. Однако лесоводственная эффективность указанных способов рубок будет сведена на нет, если технология лесосечных работ не обеспечит ограничения повреждаемости подроста, оставшихся деревьев на лесосеке, почвы до хозяйственно допустимой степени. К сожалению, технологический процесс лесозаготовок в горных лесах все еще далек от совершенства. Довольно сложный пересеченный рельеф местопроизрастания пихтовых лесов, крупный размер вырубаемого хлыста, порядка 7-10 м3, отсутствие техники, приспособленной для заготовки в горных условиях, обусловили сложность лесосечных работ при выборочной форме хозяйства. Этим и объясняется широкое распространение уже в течение длительного периода промышленно-выборочной рубки в пихтарниках с применением бессистемной технологии лесосечных работ. Разрешение противоречий между лесово-дственными требованиями и пользованием древесиной в горных лесах является существенным пробелом в лесном деле.
Результаты наших исследований позволили внести некоторую ясность в этот вопрос. Нами добровольно и группово-выборочные рубки были заложены для стационарных исследований в пихтарниках Кучба-Яштинского лесничества Гагринского лесхоза в квартале 18 в 1967 году (пробные площади 32, 33, 34, 35). Были разработаны техника их проведения и рациональный, широкопасечный метод технологии лесосечных работ (Бебия, 1967). В частности, техника проведения добровольно-выборочных рубок предусматривает рубки в размере среднегодичного прироста, интенсивностью не более 20 % от первоначального запаса древостоя на га. Повторяемость рубки 15-20 лет в зависимости от состояния насаждений и хода естественного возобновления. Рубки допускаются на склонах до 30° крутизны. В рубку назначаются наиболее толстомерные (старовозрастные) деревья верхнего яруса, а также фаутные деревья нижних ярусов. Группововыборочные рубки допускаются на склонах до 25° , с устойчивыми почвами и с куртинным расположением подроста. При этом спелая часть древостоя вырубается в несколько приемов (четыре-пять) окнами, диаметром 18-25 м, на протяжении двух классов возраста (40 лет).
Технология предусматривает процесс освоения лесосек с применением хлыстовой трелевки тракторами ТТ - 4, Т - 157 на склонах до 20° (на склонах большей крутизны освоение лесосек рекомендуется производить с помощью вертолетов). На лесосеке прокладываются трелевочные волоки под углом 60° к горизонталям склона через каждые 100 м. Валка деревьев осуществляется под углом 15-45° к волокам. Подтрелевка к волокам и трелевка их производится в хлыстах с обрубленными сучьями при перемещении трактора только вдоль волоков. Порубочные остатки укладываются в валы поперек волоков на расстоянии 15-20 м. Ширина валов - до 2 м, высота - 1 м. Особое внимание уделено правильной организации труда и порядку проведения технологических процессов лесосечных работ.
Исследования показали лесоводственно-эксплуатационную эффективность добровольно-выборочных и группово-выборочных рубок в пихтарниках, чем ныне применяемые в пихтовых лесах Грузии и Абхазии промышленно-выборочные рубки (табл. 41). После добровольно- и группово-выборочных рубок на лесосеке остается здоровый древостой с сомкнутостью 0.6. Установлено, что эти рубки способствуют последующему естественному возобновлению и предотвращению эрозии почвы на лесосеках.
Повышается продуктивность лесов на 11.5-14.3% (Бебия, 1967, 1972, 1973). Эти рубки были широко апробированы в производственном масштабе в пихтарниках Абхазии и Грузии.
Таблица 41
Лесоводственно-эксплуатационная характеристика _____________разных способов рубок____________
Наименование показателей Способ рубки
промышленновыборочный (средние данные из 3 пробных площадей) добровольновыборочный группововыборочный
Полнота древостоя до рубки 0.9 0.8 0.8
Полнота после рубки 0.3 0.6 0.6
Интенсивность рубки по запасу, % / м3 45.3/411 14.6/106 16.8/16
В том числе дровяного, % - 30.0 31.3
Количество дровяных деревьев до рубки, % 12.3
Дровяных деревьев после рубки 41.0 - -
Диаметр срубленных деревьев, см 92 (60-172) 28 (12-88) 42(12-148)
Уничтожено деревьев при лесосечных работах по запасу, % 12.0 1.0 0.9
Число уничтоженных деревьев, % 18.7 2.5 2.7
Повреждено оставшихся Деревьев, % 33.6 12.1 6.7
Уничтожено подроста высотой 0.5 м и выше, % 82.1 7.3 10.0
Нарушена поверхность почвы, % 75.9 10.8 6.0
Потери почвы через 2 года после рубки 326 - 36
Примечание. Данные на 1 га
С 1968 года в темнохвойных лесах Абхазии и Грузии стали допускать только добровольно- и группово-выборочные рубки (Правила рубок главного пользования в лесах Грузинской ССР, 1968, 1980), в которых были учтены разработанные нами техника и технология их проведения. Нами в 1998 году был также разработан “Лесной Кодекс Республики Абхазия”, в котором добровольно-выборочные и группово-выборочные способы рубок рассматриваются в качестве основных для пихтовых лесов.
Опыт рационального способа восстановления изреженных пихтовых лесов
Известно, что выборочная форма хозяйства рассчитана на естественное воспроизводство лесов. Однако проведенные исследования показали, что естественное возобновление вырубок в пихтовых лесах зачастую зави
сит от высоты над уровнем моря, типа леса, а также способов и технологии рубок. В отдельных трудновозобновляющихся типах леса (папоротниковых, с кустарниковым подлеском, субальпийских) процесс естественного возобновления растягивается на длительный период. Не возобновляются длительное время и лесосеки, где была нарушена технология лесозаготовок и отсутствует благонадежный подрост. На таких лесосеках в целях сокращения срока восстановления, сохранения защитных свойств лесов целесообразно после рубок проводить лесовосстановительные мероприятия.
Классические схемы создания лесных культур пихты на вырубках, как показал многолетний производственный опыт в пихтарниках Северного Кавказа, не всегда дают положительные результаты (Мальцев, 1979). К тому же, такие культуры - дорогостоящие. Посевы семян пихты непосредственно на лесокультурную площадь также оказались ненадежным способом лесовосстановления. Они страдают от фауны, заглушаются быстрорастущим на лесосеке высокотравьем, и требуют в течение ряда лет трудоемкого агротехнического ухода.
Под пологом естественных насаждений, по опушкам леса пихты кавказской наблюдается обилие самосева разных пород, различного возраста и жизненности. Такой “лесной питомник” можно использовать для получения посадочного материала главных лесообразующих пород. Метод экономичен, не требует затрат на выращивание растений. Самосев можно использовать для подсадки низкополнотных, изреженных древостоев, для восстановления больших окон и прогалин в лесу. Для этой цели нами в 1967-1970 годах были заложены опытные участки в различных лесорастительных условиях на разной высоте над уровнем моря.
Участок I. Южный склон Гагринского хребта, 500 м над уровнем моря, склон 25-30° крутизны. Древостой с сомкнутостью полога 0.4-0.5, III класса бонитета, состав ЗД5Гр2Бк. Почвы перегнойно-карбонатные до 50 см.
Участок II. Расположен в Рицинском заповеднике, на высоте 950 м над уровнем моря. Площадь обезлесенная от прошедшего в 1959 году верхового пожара. Склон юго-восточной экспозиции, 20-30° крутизны. Почвы бурые лесные, до 60 см глубины.
Участок III. Расположен в урочище Ауадхара Рицинского заповедника, на высоте 1650 м над уровнем моря, в верхнем поясе произрастания пихтовых лесов. Рельеф сравнительно ровный, плато бывшего русла речки. Неподалеку отдельными куртинами произрастают: клен Траутфетгера, бук, а по ущелью - ольшаники. Заросли последних развиваются в местах увлажнения, образованного проточными водами. Почвы аллювиальные, мощные до 90 см.
Участок IV. Расположен в Кучба-Яштинском лесничестве Гагринского лесхоза. Высота 1200 м над уровнем моря. Рельеф ровный. Тип леса- буково-пихтовый мелкотравный (ясменниковый) I класса бонитета.
Сомкнутость полога древостоя 0.2-0.3. Почвы бурые лесные, мощные до 110 см. Лесосека промышленно-выборочной рубки, двухлетней давности.
Посадочный материал - самосев пихты, выкапывался из-под полога леса с небольшим комом. Растения брали 5-10-летнего возраста, высотой 0.2-0.4 м, хорошего качества. Эти растения высаживались в рединах древостоев, на открытых площадях с размещением 2.5x2.5 м на постоянное место. Приживаемость посадок достигала 80-96%. Дальнейший уход заключался в приствольном рыхлении в течение трех лет и прокосе в междурядьях до пяти лет.
В целом наши многолетние наблюдения за опытными посадками по использованию самосева пихты для восстановления коренных древостоев показали эффективность этого способа. По нашим данным (Бебия и др., 1970, 1988), хороший рост и развитие посадок самосева пихты наблюдаются в поясе 500, 900 и 1200 м над уровнем моря. Так, на участке I пихты в биоэкологическом везрасте 25-27 лет в среднем достигли высоты Юм, диаметра у корневой шейки 14 см. Эти показатели в поясе 950 м (участок П) составили соответственно 12 м и 15 см. На высоте 1200 м над уровнем моря (участок Ш) высота посадки пихты составила чуть меньше 11.4 м, диаметр - 17 см, текущий прирост по высоте 50-78 см. Хвоя на деревьях держится 5-6 лет, наблюдается начало дифференциации деревьев по возрасту.
Значительно хуже растут и развиваются посадки самосева пихты на открытой площади на высоте 1650 м (участок IV), хотя эта высота относится к поясу пихтовых лесов. Посадки страдают здесь от мощного снежного покрова, достигающего более 10 м толщины. Отрицательное влияние на рост и развитие растений оказывают также: наличие поздних весенних и ранних осенних заморозков; зимние морозы, достигающие -19 °C; зимнее объедание корней, стволов, ветвей, хвои кротами и полевками. Растения постоянно ломаются, обнаруживают кущение. В возрасте 25 лет растения не превышают 1.5 м высоты и 5.6 см диаметра. Прирост по высоте составил 10-12 см.
Безусловно, результаты наших исследований по восстановлению из-реженных пихтовых лесов не могут быть окончательными. Исследования в этом направлении необходимо продолжать. Однако опыт использования самосева для восстановления лесов достаточно эффективен и может быть широко использован в производстве.
Созологическая характеристика пихтовых лесов
Проблема охраны природы, нормализации и оптимизации взаимодействия человека, общества и природы в глобальном, региональном и ло
кальном масштабах выдвигается на первое место среди наиболее актуальных проблем современности.
И сегодня, когда на территории стран бывшего СССР складывается новая система общегуманистических принципов, роль разумного природопользования, направленного на поддержание структурного разнообразия и оптимального функционирования природных систем, возрастает чрезвычайно сильно. В этом плане особая ответственность ложится на власти и население тех регионов, в которых сосредоточены уникальные природные комплексы, представляющие общечеловеческую ценность. Несомненно, к числу таких регионов относится Кавказ. В силу ряда геологических и исторических предпосылок на территории Кавказа, и в первую очередь Колхиды, сложилась своеобразная, рефугиальная система природных комплексов, нигде более в мире не встречающихся. Одним из важнейших компонентов этих природных комплексов являются пихтовые леса.
Говоря о пихтарниках Кавказа как об уникальном природном комплексе, необходимо в первую очередь отметить следующее.
Реликтовые девственные пихтовые леса Кавказа пока еще произрастают на достаточно больших площадях и могут представлять автономные экологические единицы, обеспечивающие естественное саморегулирование системы. Поэтому они являются хранителями и источниками генетического фонда разнообразия живого мира. Эти леса являются также важнейшими источниками информации, отражающими реальную динамику природных процессов и явлений в лесу, что открывает возможность не только для широких общебиологических заключений, но и для разработки систем практических мероприятий по ведению рачительного лесного хозяйства, для принятия решений в сфере отношения общества и природы.
И сегодня, когда площади пихтовых лесов в мире катастрофически сокращаются, когда целиком род пихта находится,на грани регрессии, проблема сохранения и рационального использования пихтарников Кавказа становится крайне актуальной.
Еще в 1972 году в югославском городе Любляне на заседании отделения 1.1 “Девственный лес” Международного союза лесных исследовательских организаций (IUFRO), посвященном специально проблеме пихты, было принято обращение ко всем странам Европы и отдельно ко всем лесным учреждениям Югославии (по числу и величине существующих девственных лесных резерватов Югославия занимает ведущее место в Европе) принять срочные меры для сохранения остатков девственных пихтарников. В этом отношении девственные пихтарники Кавказа представляют, несомненно, огромную ценность как в региональном, так и в глобальном масштабе.
В то же время большую тревогу вызывает состояние этих лесов, снижение продуктивности и защитных функций, исчезновение или сокра
щение ареала многих ценных видов и форм растений и обеднение их генофонда в результате хозяйственной деятельности человека и, прежде всего, рубок главного пользования.
К настоящему времени в разных странах ряд охраняемых территорий охватывает около 135 тыс. га лесов пихты кавказской (табл. 42).
Однако границы этих охраняемых пихтарников в большинстве искусственные, разрозненные, существенно различаются размерами, назначением, установленным режимом и не всегда охрана в них может считаться удовлетворительной. Они не охватывают весь спектр коренных природных комплексов в пределах ареала пихты кавказской. Все это не может создавать гарантию сохранения генофонда ценных видов растений и растительных сообществ.
Все нарастающее хозяйственное использование уникальных сообществ реликтовой флоры нарушает узкий диапазон экологических условий, к которым они приспособлены. А это может привести к необратимым изменениям их оригинального флористического состава.
Таблица 42
Охраняемые территории, охватывающие участки лесов
с участием пихты кавказской
Страна Заповедник Общая площадь, га В том числе площадь лесов с участием пихты
тыс. га %
Абхазия Рииннский Псху- 16289 10483 64.3 (70.3)
Гумистинский 40818 8917 21.8(23.5)
Грузия Алгетский 6000 1369 22.8(27.1)
Боржомский 17948 399 2.2 (2.6)
Россия Кавказский биосферный заповедник 263277 110400 41.9 (68.0)
Тебердинский 84996 3500 4.1 (22.5)
Турция Бейкоз 465 - -
Кекез 324 - -
Каздажи 240 - -
Примечание. В Турции общая площадь пихтовых лесов (пихт кавказской, Борнмюлле-ра, киликийской, троянской) 183880 га, доля заповедных 0.33%. Примерно треть площади пихтарников занимают леса из пихты кавказской, но в них нет заповедных территорий (Gelik, 1989). В скобках указаны проценты от покрытой площади лесов. Данные таблицы составлены по материалам книги “Заповедники Кавказа” (М., Мысль, 1990)
Самый крупный среди охраняемых территорий - Кавказский биосферный заповедник, в котором пихтовые леса занимают до 110.4 тыс. га, примерно 68% покрытой лесом площади заповедника. Однако после распада СССР система охраны в нем существенно изменилась не в лучшую
сторону. Работы по мониторингу и изучению природы в целом сведены до минимума.
На территории других заповедников ситуация еще более сложная. Все они не в состоянии выполнять в должной мере свои заповедные функции. Общим и существенным бедствием всех охраняемых территорий стало сильное антропогенное давление. К примеру, Рицинский заповедник с 1996 года преобразован в Рицинский национальный парк. На части его территории уже допущены рубки главного пользования. Альпийский пояс, прилегающий к территории национального парка, не включен в его состав. Из-за отсутствия защитной буферной зоны субальпийские леса национального парка и альпийские луга находятся под чрезмерной нагрузкой пастьбы скота. Пасторальные факторы здесь существенно влияют отрицательно на охрану ценных видов флоры и динамику растительных сообществ.
Ясно, что если не будут приняты меры по спасению еще сохранившихся девственных пихтарников Кавказа, то их ждет та же печальная участь, что и девственных лесов Европы, о которой с большой тревогой сообщается в работах видных исследователей (Henkel, 1954; Godwin, 1975; Mayer, 1975; Schmidt-Vogt, 1977; Leibundgut, 1982 и другие).
Проблема сохранения девственных пихтарников Кавказа как в заповедниках, так и вне их территорий, стала одной из наиболее актуальнных научных и практических задач современности.
Видовой и фитоценотический принцип охраны реликтовых пихтарников Кавказа
Как отмечает В.Е Соколов (1986), заповедники - интенсивно развивающаяся специфическая' форма научной и соответственно хозяйственной деятельности человеческого общества, функционально необходимый элемент в системе природопользования. Одна из генеральных функций заповедников заключается в резервировании и сохранении генетического фонда природы (Краснитский, 1983). Однако с резким ростом технического прогресса, урбанизации и рекреации на Кавказе осуществление полноценной охраны генофонда разнообразного живого мира в пихтарниках становится все более проблематичным. При нынешнем темпе и характере иррационального природопользования и мощном антропогенном, хозяйственном давлении в заповедниках происходят изменения сформировавшихся за многие годы биогеоценотических комплексов, появляются вторичные и обедненные низкопродуктивные сообщества. Существенно ухудшаются условия обитания и исчезают отдельные виды ценных растений, которые наука еще не успела изучить. Потеря многих из них явится невосполнимым уроном.
Достаточно отметить, что из 62 видов деревьев и кустарников, произрастающих в пихтовых лесах Кавказа, 9.7% отнесены к категории редких и исчезающих и были занесены в Красную Книгу СССР. Это: Betula те-grelica, Ruscus colchica, Quercus pontica, Buxus colchica, Staphylea colchica, Taxus baccata.
Проведенные нами исследования показали, что в Красную книгу СССР не вошли еще много ценных растений пихтовых лесов, заслуживающих статуса редких, исчезающих и подлежащих охране и заповедованию. Поэтому мы отдельным списком приводим ниже еще 13 видов растений в целях включения их в региональную Красную книгу.
Список растений, подлежащих включению в региональную Красную книгу
l. Acer sosnowskii. Североколхидский (абхазский), скально-лесной, известняковый, меЛфильный, эндем. В пихтарниках Гагринского хребта встречается в виде единичной примеси.
2. Castanea sativa. Реликтовый южноевропейско-кавказско-малоазиатский, лесной, мезофильный. В пихтарниках встречается изредка. На Кавказе из-за бессистемных рубок естественные каштанники практически вырублены.
3. Corylus colchica. Северо-колхидский, субальпийский, известняковый, литофильный, эндем. В субальпийских пихтарниках изредка.
4. Daphne caucasica. Редкий общекавказский, лесной, мезофильный. В пихтарниках очень редко. Биоэкология вида мало изучена.
5. Daphne pseudosericea. Редкий северо-колхидский, известняковый, литофильный, эндем. Для пихтовых лесов малохаракгерный, изредка встречается в нижней части пояса пихтарников.
6. Rhododendron ungemii. Южно-колхидский, лесной, мезофильный, эндем.
7. Sorbus migarica. Довольно редкий северо-колхидский, скальнолесной, известняковый, эндем. Встречается в субальпийских пихтарниках.
8. Ulmus glabra. Западно-европейский, лесной, реликтовый. В пихтарниках в виде примеси до 1500 м над уровнем моря. Из-за голландской болезни вид стал редким.
9. Blechnum spicant. Вид с сильно дезъюнктивным ареалом в пределах Евразии. Реликтовый, лесной, мезофильный. В пихтарнике редко.
10. Colchicum speciosum. Общекавказский, лесной, высокогорный и луговой, мезофильный. В пихтарниках редко. Чрезмерное использование в практических целях резко сократило популяции вида.
11. Corydalis caucasica. Колхидский, лесной, мезофильный, эндем.
12. Pachyphragma macrophyllum. Колхидский с широкими кавказскими, ирригациями, лесной, мезофильный, реликтовый, эндем. Вид больше характерен для лиственных лесов. В пихтарниках встречается реже.
13. Polystichum woronowii. Редкий, колхидско-гирканский, лесной, мезофильный. Для пихтарников мало характерен.
Осуществление охраны редких и исчезающих видов, приведенных в списках, невозможно без сохранения среды их обитания - тех фитоценозов, в которых они существуют или которые образуют.
Безусловно, заповедники играют существенную природоохранную роль, однако они не в состоянии охватить все ценотическое разнообразие растительности, учесть все региональные особенности, включить все редкие и подлежащие охране сообщества. Поэтому выделение и охрана таких сообществ и вне территории заповедников очень важны (Рысин, 1984).
До недавнего времени во всем мире в системе охраны растительного мира приоритетом считалась охрана генофонда природной флоры, особенно редких и исчезающих видов. И только недавно возник вопрос о сохранении сообществ в целом как среды обитания отдельных видов растений. Опыт показывает, что долговременное сохранение того или иного вида возможно лишь при сохранении большей части его ценопопуляций в пределах естественного ареала, при условии сохранения тех исторически, относительно сложившихся сообществ, в которых вид произрастает. В свою очередь, растительное сообщество может сохраниться только при условии стабильности всех компонентов биогеоценоза. Такой системный биогеоце-нотический подход в наиболее полной мере отвечает современным положениям о развитии природы (Степанова, 1982; Парфенов, 1983; Прилипко, 1984; Рысин и др., 1985; Седельников, 1988). Наряду с понятием “генофонд” стало утверждаться и понятие “ценофонд” как характеристика фи-тоценотического разнообразия и богатства растительного покрова. Однако, как совершенно справедливо отмечает В.П. Седельников (1988), для растительных сообществ не разработано четкой общепризнанной категориальной шкалы редкости.
Наиболее полный перечень признаков для определения значимости редкости сообществ приводится в шкалах редкости фитоценозов, предлагаемых С.М. Стойко (1982), М.П. Шиловым (1982), В.П. Седельниковым (1988). С учетом мнения этих исследователей для пихтарников Кавказа для выделения сообществ, подлежащих охране, мы предлагаем следующую шкалу фитосозологической оценки.
1. Природоохранное значение - значимость сообществ в стабилизации природных процессов в целом горных экосистем, отдельных урочищ, ландшафтов. К таким сообществам следует относить субальпийские пихтарники, сообщества на площадях постлавинных участков, на участках источников минеральных вод.
2. Научное значение - как эталон, характеризующий биогеоценозы пихтовых лесов, куда можно отнести высокопродуктивные эталонные лесные участки, эталоны коренных типов лесных биогеоценозов; сообщества, включающие виды с пониженным эколого-биологическим потенциалом на границе ареала пихты, а также встречающиеся интразонально; биогеоценозы коренных или условно-коренных типов с присутствием подлежащих охране реликтовых и эндемичных видов; сообщества, отражающие историю формирования растительного покрова; уникальные культурфитоценозы из аборигенных или интродуцированных видов, имеющие научно-экспериментальное значение.
3. Народнохозяйственное значение - как эталон высокопродуктивных биогеоценозов; как источник ценного растительного сырья; как источник интродуцентов; как местообитание ценопопуляций хозяйственно важных уничтожаемых видов; как объект эстетико-воспитательного, рекреационного значения.
Такая фитооозологическая оценка позволяет наиболее полно выявить и обеспечить охрану ценных видов и сообществ растений на экосистемном уровне.
С учетом приводимых показателей фитосозологической оценки современное рациональное природопользование в пихтарниках Кавказа должно обеспечить сохранение как отдельных видов растений, так и целых природных комплексов, особенно эталонных. В этом отношении особый интерес представляют пихтарники, произрастающие на известняковых массивах Гагринского хребта в Абхазии и горных систем Фишт, Оштен на Северном Кавказе. Именно здесь сконцентрированы уникальные и вместе с тем наиболее ранимые объекты природы. Они отличаются наибольшим богатством флоры пихтовых биогеоценозов (Бебия и др., 1987; Алтухов и др., 1989). В них произрастает большая часть эндемичных, реликтовых, а также редких и исчезающих видов флоры пихтарников Кавказа. В этих лесах сосредоточены оригинальные, большей частью эндемичные, очень редкие биогеоценозы. Это пихтарники, которые в определенных экологических условиях формируют субэдификаторные синузии из: Pachyphragma macrophyllum, Trachystemon orientate.
Пихтарники с доминированием в покрове Senecio correvonianus - абхазского, альпийско-известнякового, литофильного эндемика, встречаются только в верхней части пояса пихтовых лесов Гагринского хребта. Оригинальные пихтарники с подлеском реликтового рододендрона (Rhododendron ungemii) произрастают лишь на юго-восточной границе ареала пихты в Аджарии и Лазистане (Турция). Ясно, что биогеоценозы таких сообществ должны быть включены в категорию редких и подлежат охране.
Не только присутствие третичных реликтов, но и образование особых, своеобразных сообществ с участием пихты вдали от ее основного ареала представляет большой научный интерес для познания истории формирования растительности Западного Кавказа. Еще В.П. Малеев (1939) подчеркивал важность и необходимость всяческой охраны местопроизрастания таких сообществ. По имеющимся данным (Малеев, 1939; Орлов, 1951; Алтухов и др., 1989), а также по нашим наблюдениям, на Северном Кавказе, пихтовые сообщества, оторванные от своего сплошного распространения, встречаются - около Горячего Ключа, по реке Чепси и Аюк, по р. Большая Собачка, на северном склоне Шабановского перевала, севернее станицы Тхамаха по р. Шебеш, по р. Чепси на склонах г. Поднавислой, у Хребтовой щели на возвышенности у долины р. Коурзе, по северному склону г. Щетка и г. Чубатой, по Афипсу и первых Волчьих ворот. Самым крайним пунктом пихтарников на южном склоне является местообитание в урочище Верхний Дефан под г. Лысой.
А.М. Амирханов и др. (Заповедники Кавказа, 1990) также отмечают произрастание пихты на территории Северо-Осетинского заповедника, но местонахождение такого пункта, крайне отдаленного от восточной границы сплошного ареала, не уточняется. Безусловно, все местонахождения крайних точек сообществ с участием пихты должны подлежать охране.
Перед лесным хозяйством стоит важнейшая задача - повысить продуктивность и улучшить качественный состав лесов (Атрохин, 1967). Для этого необходимы эталонные образцы, которых следует придерживаться при организации и ведении хозяйства (Лосицкий и др., 1980). С нашей точки зрения, эталонными являются насаждения коренных типов леса, которые по составу, продуктивности и качеству наилучшим образом отвечают целям хозяйствования, т. е. в спелости дают древесину требуемых сортиментов, эффективно выполняют водорегулирующие, почвозащитные и другие полезные функции, наиболее полно используют плодородие почвы, обладают наивысшим годичным приростом древесины в данных условиях местопроизрастания и наиболее устойчивы против вредных факторов. Индекс жизненного состояния таких древостоев по нашим расчетам не менее 80 %.
По имеющимся данным (Ушатин, 1962), в естественных биогеоценозах Северного Кавказа встречались уникальные, самые высокопроизводительные в масштабах Европы пихтарники с запасом до 2500 м3/га. По результатам наших исследований (Бебия и др., 1985), запас высокопродуктивных пихтовых древостоев, которые могли бы служить эталонами, в Абхазии составляет 1684 м3/га.
В табл. 43 приведены основные показатели эталонного пихтарника трахистемонового 1а класса бонитета, произрастающего в Аибгинском лесничестве Гагринского лесхоза на высоте 1200 м над уровнем моря. Экспозиция южная, склон 30° крутизны.
Таксационные показатели эталонного пихтового древостоя (пихтарник трахистемоновый 1а класса бонитета)
Таблица 43
Ярус Состав Полнота Сомкнутость D ср., см Коэффициент вариации, % Н ср.,м Коэффициент вариации, % Возрастная стадия Класс возраста Число деревьев Запас
шт/га % м3/га %
I ЮПх 1.0 0.4 98 26 44 8.3 Перестойная XVII и более 12 2.6 418 25
Старая XII-XVI 24 5.2 460 27
Спелая ГХ-ХП 60 13.0 620 37 ю
и 8Пх2Бк 0.2 0.2 32 47 22 35 Приспевающая VI-VIII 60 13.0 144 8.5 о ьэ
Средневоз-
растная Ш-V 100 22.0 36 2.0
ш 8Пх2Бк - 0.1 10 20 9 11 Молодая 1-П 204 44.2 9 0.5
Как видно из табл. 43, нетронутый рубкой пихтарник характеризуется разновозрастностью, сложной структурой с вертикальной сомкнутостью полога. В таких древостоях представлены деревья всех возрастных групп в соотношении, обеспечивающем непрерывное их существование. Значительный запас I яруса, установление среднего возраста не по числу стволов, а по запасу (сумма площадей сечений) создавали ложное представление о сплошной перестойности разновозрастных пихтарников и их неминуемой гибели. Этот ошибочный тезис, как уже отмечалось выше, служил главным аргументом при назначении рубок высокой интенсивности, которые привели к снижению производительности пихтовых лесов на значительных площадях как на Северном, так и на Южном макросклоне Кавказа (Иваненко, 1956; Беленко, 1964; Дробиков, 1965; Гулисашвили, 1965; Коваль, 1977; Бебия, 1973 и др.).
Лучшей моделью для организации непрервыного и равномерного пользования лесом могут служить разновозрастные, многоярусные эталонные насаждения, максимально использующие пространство, солнечную энергию и почвенное плодородие. Эталонные древостои площадью не менее 3-5 га надо сохранять для каждой природной зоны, каждого высотного пояса и типа леса. Они будут образцами оптимального состава, возрастной структуры, строения, устойчивости и производительности. Изучение их позволит правильно регулировать возобновление вырубок, целенаправленно осуществлять реконструкцию малоценных насаждений и формировать высокопродуктивные. Эталоны имеют важное значение и как хранилища генофонда древесных пород, поэтому они должны выделяться и сохраняться не только в заповедниках, но и в лесах хозяйственного значения. Это особенно важно в связи с понижением общей продуктивности древостоев из-за усиления хозяйственной деятельности главным образом рубок. Так, еще в 40-50-е годы в Абхазии также встречались пихтарники с запасом до 2500 м3/га (Малеев, 1936). Теперь таких массивов нет даже в Рицинском заповеднике.
Охрана редких сообществ является дополнительной, но весьма существенной мерой, которая наряду с заповедниками и эталонными участками позволит охарактеризовать весь ценофонд пихтарников Кавказа.
С целью сохранения многообразия отмеченных природных комплексов в пихтарниках требуется.
I. По южному макросклону:
а) прекращение всех видов хозяйственной деятельности, в первую очередь рубок, на территории Рицинского национального парка до проведения его функционального зонирования;
б) расширение границы РНП на северо-западе с включением в его территорию всего массива Гагринского хребта, относя его в зону охраняемой территории.
в) присоединение к территории РНП всей площади, примыкающей к нему, альпийского пояса и высокогорий. Территория РНП в таких границах примкнется с северной стороны к Кавказскому биосферному заповеднику (КБЗ), что будет иметь чрезвычайно важное экосистемно-природоохранное значение, в первую очередь для пихтарников, что особенно важно. Кроме того, территория РНП охватит весь спектр верхне-высотных природных поясов региона и обеспечит равновесие и естественное саморегулирование в природных комплексах.
П. По Северному Кавказу мы полностью разделяем точку зрения сотрудников КБЗ и других исследователей, которые предлагают расширить границы заповедника, проводя их на северном макросклоне Главного Кавказского хребта в районе горных массивов Фишта, то есть от Черкесского перевала на северо-запад в верховье р. Чегурсанки, далее на север в верховья ручья Водопадного, включая гору Фишт, озеро Псенодах, к верховью притоков р. Цице, включая г. Оштен, и далее на северо-восток до границ заповедника в раЖже приюта по р. Армянке. Этот участок площадью 3.5 тыс. га, как отмечают М.Д. Алтухов и С.А. Литвинская (1989), должен охраняться как область питания пресных вод, как крупнейший в бывшем СССР карстовый массив, как один из центров формирования и сохранения представителей третичной флоры, уникальных пихтарников на известняковых массивах.
III. Проведение систематизации и составление кадастра всех редких сообществ и эталонных участков на всем ареале распространения пихты, с целью их дальнейшей охраны и организации постоянного контроля за состоянием и динамикой сообществ.
Суммируя все изложенное, следует подчеркнуть, что охрана генофонда и ценофонда пихтарников Кавказа, поставленная на научную основу, позволит не только сохранить их как основную трофическую часть биоты, но и значительно повысить степень рационального использования. Использование пихтарников и их сохранность следует при всем противоречии рассматривать как составные части единой проблемы и решать ее нужно комплексно на уровне единой системы.
ЗАКЛЮЧЕНИЕ
Особенностью настоящей работы является то, что вопросы лесообразовательного процесса, лесопользования и созологии пихтарников Кавказа впервые рассматриваются в единой системе.
Развитие теории лесообразовательного процесса (ЛОП) началось сравнительно недавно, и полного представления о ней не сформировалось. Нами ЛОП понимается как процесс возникновения, развития, разрушения и смены лесных сообществ во времени в результате взаимовлияния биотических, абиотических факторов и антропогенного воздействия. Основные закономерности ЛОП пихтовых лесов установлены при изучении их генезиса, современного распространения, природных условий произрастания; при исследований морфо-биоэкологических особенностей пихты кавказской, процессов возобновления лесов, их строения, возрастной структуры и развития, биогеоценотических особенностей и динамики лесных сообществ, а также реакции их на проведение хозяйственных мероприятий.
Проведенные исследования свидетельствуют об автохтонном горном происхождении пихтовых лесов Кавказа. В конце плиоцена здесь в средне-верхне-горном поясе произрастали обширные леса из пихты кавказской, с примесью бука восточного, ели восточной и других представителей ценотического состава современных пихтарников.
Отличительной особенностью современных пихтарников Кавказа является относительное флористическое богатство их состава (207 видов). Среди них 82 % являются элементами средиземногорной флористической области, т. е. выходцы из высокогорной флоры; реликты составляют 38 %, в том числе 90 % древесных видов. Позиция 85 % реликтов в ценотическом составе коренных типов леса устойчивая. Встречаемость их составляет 17-100 %, фитоценотический индекс превышает 34. Каждый их них является характерным и постоянным, как минимум, для двух типов леса. Лишь около 15 % реликтов относятся к факультативным видам, но и они обнаруживают постоянную связь с коренными фитоценозами, каждый из них характерен, по крайне мере, для одного типа леса.
Высокая степень преемственности ценотического состава между плиоценовыми и современными пихтарниками, достаточно устойчивая позиция реликтовых видов в составе последних, сравнительно высокое содержание в них колхидских (36 %) и общекавказских (23 %) эндемичных элементов, а также, в силу естественно-исторических причин, отсутствие существенных трансформаций антропогенных воздействий на спонтанное развитие пихтовых лесов в историческом прошлом свидетельствуют не только о древнем автохтонном формировании их основного флористического ядра, но и позволяет утверждать, что девственные пихтарники здесь
сохранили черты реликтовости. Это придает им особую уникальность в региональном и глобальном масштабе и создает необходимость охраны.
Основные морфо-биологические и экологические черты пихты кавказской освещались и раньше в работах ряда исследователей. Но проведенные более глубокие исследования, в том числе с использованием экспериментальных опытов, позволили впервые отметить сравнительно низкую морфологическую пластичность данного вида (в природе известны лишь одна его форма и одна разновидность) и выдвинуть предположение о том, что пихта кавказская представляет собой более старую ветвь в своей секции, а также о более или менее завершенном процессе формирования и обособления ее как самостоятельного вида. Эти же морфо-био- и экологические ее черты можно рассматривать как проявление общебиологической закономерности, отражающей эволюционную специализацию вида и приспособления его к произрастанию в изолированном от других видов пихт Колхидском рефугиуме.
МатериальЯ исследования свидетельствуют о том, что современное распространение пихтовых лесов на Западном Кавказе обусловлено геологическими и историческими причинами, климатическими, прежде всего гидротермическими, условиями, биоэкологическими особенностями пихты, а также влиянием антропогенных воздействий. Характер распространения пихты определяется комплексным воздействием этих ведущих факторов на развитие лесов в конкретных условиях местообитания.
Стратегия вида пихты в пределах ее ареала достаточно устойчивая, даже на границе ареала. Являясь доминантам - эдификатором растительных сообществ, пихта наиболее свободна от приспособления к условиям межвидовой конкуренции в сообществах. Об этом свидетельствуют, прежде всего, ее высокие показатели встречаемости во всех типах лесов (100 % и фитоценотического индекса (5400). Во всех типах леса возобновление идет пихта по пихте, смены ее сопутствующими породами субэдификато-рами не наблюдается.
Почва, составляя неотъемлемую часть леса как биогеоценоза, находится в постоянном единстве и взаимодействии с лесом на уровне лес -почва - лес. Под пихтовыми лесами формируются, главным образом, бурые лесные кислые и бурые лесные псевдооподзоленные почвы. Отличительной особенностью их является слабая выраженность дифференциации по генетическим горизонтам профиля. Оба типа почвы достаточно плодородны для произрастания высокопроизводительных (до 2000 кбм/га) пихтовых, буково-пихтовых древостоев.
В семяношении пихты урожайные годы наблюдаются через 1-2 года. В разновозрастных пихтарниках семяносят деревья I и П яруса. Деревья Ш яруса плодоносят единично. Нетронутые рубками разновозрастные пихтовые древостои в количественном и качественном отношениях вполне
достаточно обсеменяются для беспрерывного возобновления. На 1 га урожаи семян пихты достигает 149 кг, или 1062 тыс. шт. полнозернистых. Исходя из этого, при рубках в этих лесах следует ориентироваться, главным образом, на естественное восстановление главной породы без дополнительных затрат на искусственное лесовосстановление.
Наилучшее естественное возобновление наблюдается в разнотравных, с подлеском из черники кавказской и овсяницевых пихтарниках, удовлетворительное - в пихтарниках с подлеском из падуба. Пихтарники ожиковые, папоротниковые, с подлеском из лавровишни, рододендрона характеризуются слабым возобновлением. В пределах группы типов леса лучшее формирование возобновления наблюдается при сомкнутости полога 0,6 для групп типов овсяницевых, ожиковых, с подлеском из черники кавказской и падуба, 0,5-0,6 для разнотравных. В папоротниковой группе типов лучшее возобновление можно ожидать при сомкнутости полога древостоя 0,6. Эти показатели необходимо учитывать при проведении рубок в пихтарниках.
В естественных окнах пихта хорошо возобновляется куртинами. В пределах группы типов леса процесс возобновления зависит, главным образом, от величины и формы окон, от крутизны и экспозиции склона. В овсяницевых и разнотравных пихтарниках лучше возобновляются окна средних размеров, диаметрами от 18 до 24 м.
Материалы изучения девственных пихтовых лесов Кавказа свидетельствуют о том, что древостои во всех группах типов леса характеризуются абсолютно-разновозрастной структурой и сложным ступенчатым вертикальным строением полога. Насаждения отличаются большой амплитудой колебания возраста (до 450-500 лет), высоты (до 60 м) и диаметра (до 2 м). Кривые распределения этих показателей многовершинны. Реже встречаются относительно разновозрастные древостои, для которых в ряду распределения также характерно значительное колебание возраста (61-120 лет). Но основная масса стволов (70-80 %) сосредоточена в двух - трех классах возраста. Распределение стволов по возрасту близко к нормальному.
Отличительной особенностью абсолютно-разновозрастных насаждений пихты кавказской является непрерывный характер возрастной динамики, в процессе которой происходит смена отдельных старовозрастных деревьев или групп деревьев более молодыми. В насаждениях, как правило, всегда представлены все возрастные группы древостоя и подрост, благодаря чему обеспечивается устойчивость леса. Возрастно-восстановительный тип динамики путем смены отдельных отмирающих поколений новыми, более молодыми для девственных насаждений пихты кавказской не является характерным. Эти выводы чрезвычайно важны и должны быть учтены при разработке практических мероприятий, при назначении рубок в лесу. Они опровергают ошибочный тезис о якобы развитии девственных разно
возрастных насаждений пихты кавказской до "перестойного" состояния. Постоянный процесс естественного возобновления, сложная возрастная структура древостоев и непрерывный характер их развития являются важнейшими факторами основополагающих приспособлений на уровне сообществ, которые обеспечивают пихтарникам Кавказа длительное процветание.
Для лесотаксационных работ и лесоводственных целей сложные разновозрастные древостои оказалось наиболее целесообразно разделять на три яруса, не включая подроста. Первый, верхний ярус состоит из старовозрастной и спелой части древостоя. Второй, средний ярус - из средневозрастной и приспевающей части. Третий ярус представлен молодыми деревьями. При таксации насаждений таксационные показатели следует устанавливать для каждого яруса в отдельности. Такой метод таксации имеет важное значение для проектирования хозяйственных мероприятий и организации рационального пользования лесов.
Ведение лесного хозяйства в пихтарниках Кавказа на лесотипологической основе все <ще остается важнейшей и актуальной задачей современного лесоводства. Нами, на основе анализа и обобщений большого фактического материала личных исследований, в том числе на типологических стационарных пробных площадях, разработана впервые для пихтарников Кавказа наиболее полная классификация типов леса. В ее основу положен эколого-генетический принцип. С учетом пространственно-временных факторов развития лесных сообществ выявлены этапы развития сообществ от общего к частному (от коренного типа леса к производному) и от простого к сложному (восстановление в исходный тип леса). Этапами развития служат генетически связанные биогеоценозы (ассоциации, пол-нотные стадии), которые вместе с коренным образуют единую биогеоцено-тическую систему - тип леса (тип лесных биогеоценозов). Составлена система соподчинения типов леса и типа условий местопроизрастания. Совокупность близких в биоэкологическом, ценотическом отношениях и генетически связанные эдафо- и климатозамещающие типы леса нами объединены в естественные группы типов леса. Это пихтарники: овсяницевые, ожиковые, разнотравные, папоротниковые, с вечнозеленым колхидским подлеском, с листопадным колхидским подлеском, субальпийские и скально-лесные комплексы.
С лесоводственной и экономической точек зрения типы леса, близкие в биоэкологическом отношений, однородные или сходные по лесообразовательным условиям, целенаправленности хозяйств и требующие проведения одинаковых практических мероприятий, объединены в пять основных хозяйственных групп типов леса. Это - пихтарники овсяницевые, ожиковые, разнотравные, с подлеском из колхидских элементов и субальпийские. Для каждой секции предлагается система мероприятий, обеспечивающих рациональное использование данной категории лесов, повыше
ние производительности .р улучшение защитных функции. Такой комплексный и системный подход к объединению типов леса позволяет вести практические мероприятия в более крупном масштабе.
Биоэкологическце особенности пихты кавказской, характер естественного возобновления, строение и возрастная структура древостоев, важные защитные функции пихтовых лесов обусловливают эффективность выборочной формы хозяйства в них. Добровольно-выборочные, комплексновыборочные и группово-выборочные способы рубок наиболее отвечают природе этих лесов и в лесоводственно-эксплуатационном отношении являются более эффективными, чем применяемые в настоящее время промышленно-выборочные и постепенные рубки. Лесоводственно-экономическая эффективность этих рубок, с применением разработанной нами техники и технологии лесосечных работ, выражается в сохранении защитных функций лесов, сохранении лесной среды, обеспечении естественного возобновления главной породы без дополнительных затрат, обеспечении равномерного и постоянного пользования лесом с повышением общей его продуктивности на 11-14 %,повышением производительности труда на лесосечных работах на 8-9 % по сравнению с бессистемным методом освоения лесосек при промышленно-выборочных способах рубок.
Пихтовые леса, произрастая в верхней горной зоне на значительных площадях (320 тыс. га) Западного Кавказа, выполняют важные первостепенные почвозащитные, водоохранные, климаторегулирующие, курортологические и другие полезные функции. Они являются хранителями уникального генофонда растений, редких природных комплексов, источниками ценных видов для интродукции. Получение древесины в этих лесах должно носить подчиненный характер.
В настоящей работе в основу решения задач сохранения богатейшего разнообразия растительного мира пихтовых лесов положен принцип охраны генофонда и ценофонда реликтовых, эндемичных, редких, исчезающих видов растений и эталонных сообществ. Во флоре пихтарников Кавказа 6 видов древесных растений из 62 (10 %) отнесены к категории редких, исчезающих и занесены в Красную книгу СССР. Установлено, что еще 13 видов растений являются редкими и исчезающими, подлежат охране и занесению в региональную Красную книгу.
В качестве некоторых практических мер по охране генофонда ценных видов растений и природных комплексов предлагаются:
а) расширение границ Рицинского национального парка с включением в зону его охраняемой территории всего массива Гагринского известнякового хребта и присоединением всей площади альпийского пояса и высокогорий, примыкающей к нему;
б) расширение границ Кавказского биосферного заповедника за счет присоединения к нему известняковых горных массивов Фишт - Оштен,
общей площадью 3,5 тыс. га. В таких границах охраняемые территории Кавказского биосферного заповедника и Рицинского национального парка примкнут друг к другу и станут целостной обширной площадью, что чрезвычайно важно в экосистемно- природоохранном отношении, в первую очередь для пихтарников Западного Кавказа;
в) необходимо проведение систематизации и составление кадастра всех редких сообществ и эталонных участков в пределах всего ареала распространения пихты, с целью их дальнейшей охраны, организации постоянного контроля за состоянием и динамикой сообществ.
Настоящая работа имеет определенную методическую ценность. Нами предлагается новая шкала лесоводственной классификации подроста, шкала оценки естественного возобновления, новый метод классификации деревьев в разновозрастном лесу и оценка жизненного состояния древостоев, биогеоценотический подход в охране ценного разнообразия видов и сообществ лесов.
Ценнейшее, величественное наследие природы - девственные пихтарники, к сожалению, могут уйти в прошлое безвозвратно. Очень скоро остатки настоящих спонтанных пихтовых лесов можно будет наблюдать и изучать только в немногих заповедниках и специальных резерватах. Поэтому, выполняя настоящую работу, автор стремился добиться и другой немаловажной цели - побудить специалистов к изучению коренных пихтовых лесов везде, где они еще сохранились.
СПИСОК ЛИТЕРАТУРЫ
Агеенко А.С. Пихта Майра на острове Сахалине. //Вопросы геог. ДВ, Хабаровск : Изд. Приамурского филиала ГО СССР, 1963. Сб. 6. С. 202-204.
Александрова В.Д. Классификация растительности. М.-Л., 1964. 275 с.
Александрова М.С. Интродукция хвойных в ГБС АН СССР. //В кн. : Интродукция древесных растений. М., Наука, 1980. С. 48-65.
Алексеев В.А. Текущий прирост - важнейший признак жизнеспособности подроста ели. //Лесное х-во. 1973. № 6. С 15-19.
Алексеев В.А. Световой режим леса. Л., 1975. 225 с.
Алексеев ВАДиагностикаживненного состояния деревьев //Лесоведение, 1989. №4. С. 51-57.
Алексеев С.В., Молчанов А.А. Выборочные рубки в лесах Севера. М., 1954. 146 с.
Алисов Б.П. Климат СССР. М.: Высш. Школа, 1969. 104 с.
Алтухов М.Д., Литвинская С. А. Охрана растительного мира на Северо-Западном Кавказе. Краснодар, 1979.189 с.
Альбов Н.М. Леса Абхазии. Одесса, Тип.“Славянка”, 1882. 19 с.
Альбов НМ. Очерк растительности Колхиды. //Земледелие, 1896. № 1. С. 13-47.
Альпы-Кавказ. Современные проблемы конструктивной географии горных стран. Науч, итоги франко-советских полевых симпозиумов в 1974 и 1976 гг. М., 1980. 328 с.
Ареалы деревьев и кустарников СССР. Л.: Наука, 1977. Том 1. 164 с.
Арнольд Ф.К. Русский лес. С.-Петербург, 1891. 707 с.
Атрохин В.Г. Биологические основы формирования высокопродуктивных насаждений. М.: Лесная пром., 1967. 180 с.
Базумова Г.Г. Морфологические показатели вегетативного побега хвойного подроста. И Стационарные исследования в лесах Сихотэ-Алиня. Владивосток, 1977. С. 52-58.
Баранов В.И. Этапы развития флоры и растительности в третичном периоде на территории СССР. М.: Высш, школа, 1952. 364 с.
Баранов В.И., Смирнов М.Н. Пихтовая тайга на предгорьях Алтая. !Плр. Пермского биол. науч.-исслед. инет., 1931-1932. Т. 4. С. 1-96.
Баранов Н.И., Григорьев К.И. К вопросу о строении и росте ельников Севера. //В кн.: Географические вопросы лесного хозяйства. М. 1955. Вып. V. С. 16-24.
Бахсолиани Т.Г. Характер развития естественного возобновления и травяного покрова типов леса бассейна р. Кодор. //Тр. Тбил. ин-та леса. М.: Лесн. Пром-ть, 1968. Т. XVII. С. 113-135.
Бебия С.М. Рубки главного пользования в горных лесах Абхазии. //Науч.-техн. конф. Абхазской НИЛОС, Сухум: Алашара, 1967. С. 24- 28.
Бебия С.М. Выживаемость подроста пихты кавказской в окнах группово-выборочных рубок. //Лесоведение, 1972. № 2. С. 51-57.
Бебия С.М. Группово-выборочные рубки в пихтовых лесах Грузии, техника и рациональная технология их применения. //Тр. Тбил. ин-та леса. Тбилиси, 1972. Т. 19-20. С. 96-111.
Бебия С.М. Дифференциация деревьев в разновозрастных лесах и их классификация. //Материалы науч. Сессии Совета Бот. Садов Закавказья. Сухум, 1973. С. 74-75.
Бебия С.М. Пихтовые леса Абхазии и основы ведения хозяйства в них. Автореферат канд. диссерт. Львов, 1973. 27 с.
Бебия С.М. Естественное возобновление в буковых и пихтовых лесах Абхазии. //Сообщ. АН ГССР, Тбилиси. 1978, Т. 89, № 2. С. 453-455.
Бебия С.М. К вопросу типологии лесов южного склона западной части Большого Кавказа. //В кн.: Современные проблемы лесной типологии. Львов, 1983. С. 73-75.
Бебия С.М. Выборочное хозяйство и повышение продуктивности в горных лесах. //Гр. Сух. бот. сада, Тбилиси, 1984. Вып. 28. С. 130-137.
Бебия С.М/ Биоэкологические и лесоводственные особенности пихты кавказской.. //Журнал Folia Dendrologica. Братислава, 1987. № 16. С. 159-178.
Бебия С.М. Леса Абхазии и пути их улучшения. //Тез. докл. научн.-техн. конф. Сухум, 1988. С. 7-10.
Бебия С.М. Особенности флоры пихтарников Кавказа. //В кн. Проблемы флористики и систематики растений Кавказа. Сухум, 1991. С. 13-19.
Бебия С.М. К истории и географии пихтовых лесов Кавказа. //СБ СР. М.: 1994, Вып. 2. С. 65-67.
Бебия С.М. Проблемы и перспективы интродукции пихты кавказской. //В кн.: Биологическое разнообразие. Интродукция растений. С.-Петербург, 1995. С. 31-132.
Бебия С.М. Оценка успешности естественного возобновления в пихтарниках Кавказа. //Лесное хоз-во, 1997. № 6. С. 23-25.
Бебия С.М. Дифференциация деревьев в лесу, их классификация и определение жизненного состояния древостоев. //Материалы науч, сессии посвященной 90-летию А.А. Колаковского. Сухум, 1999. С. 44-65.
Бебия С.М., Кобахия А.З. Способы восстановления изреженных буково-пихтовых лесов Абхазии. //Тезисы докладов 3-й науч.-техн. конф. Абхазской НИЛОС, Сухум: Алашара, 1970. С. 3-5.
Бебия С.М., Кобахия А.З. Сохраняемость подроста при различных способах рубок в горных буково-пихтовых лесах Абхазии. Сухум: Алашара, 1972. С. 81-88.
Бебия С.М., Каландия М.Д. Особенности строения возрастной структуры пихтовых лесов Абхазии и их значение для ведения лесного хоз-ва. //В кн. Леса Абхазии. Сухум : Алашара, 1972. С. 57-68.
Бебия С.М., Тугуши К.Л., Лейба В.Д. Значение эталонных насаждений для моделирования высокопродуктивных лесов. //Лесное хоз-во, 1985. №9. С. 32-36.
Бебия С.М., Колаковский А.А., Адзимба З.И., Долуханов А.Г. и др. Пицунда-Мюссерский заповедник. М.: Агропромиздат, 1987. 190 с.
Бебия С.М., Млокосевич Б.В., Кобахия А.З., Лейба В.Д. Опыт восстановления изреженных лесов самосевом. ИТез. докл. научно-технической конференции. Сухум, 1988. С. 19-20.
Белоновская Е.А., Гребенщиков О.С., Давыдова М.В. Биота экосистем Большого Кавказа. М.: Наука, 1990. 275 с.
Берг Л.С. Труды по теории эволюции. (1922-1930). Л.: Наука, 1977.388 с. Биосфера, эволюция, пространство, время. М.: Прогресс, 1988. 463 с.
Бобров Е.Г. Лесообразующие хвойные СССР. Л. 1978. 189 с.
Буш К.К. Актуальные проблемы лесной типологии. НВ кн. Современные проблемы лесной типологии. М.: Наука, 1985. С. 20-24.
Буш Н.А. Ботанико-географический очерк Кавказа. М.-Л.: АН СССР, 1935. 107 с.
Быков Б.А. О вертикальной поясности в связи с общим законом зональности. И Вестник АН Каз. ССР, 1954. № 8. С. 46-56.
Вальтер Г. Растительность земного шара. Эколого-физиологическая характеристика. М., 1974. Т. 2. Леса умеренной зоны. 424 с.
Василевич В.И. Статистические методы в геоботанике. Л.: Наука, 1969.232 с.
Василевич ВИ Очерк теоретической фитоценологии Л.: Наука, 1983.247 с.
Васильев Н.Г. Пихта цельнолистная в лесах Южного Приморья. //Лесн. Хоз-во. 1958. № 4. С. 13-17.
Васильев Н.Г., Колесников Б.П. Чернопихтово-широколиственные леса Южного Приморья. М.-Л., 1962. 147 с.
Васильев Я.Я. Взаимоотношения между пихтой и елью в Европейской части СССР. //Сов. бот., 1935. № 2. С. 68-74.
Веселов И.В. Смешанные леса из пихты .и бука на Северном Кавказе и их биологическая продуктивность. Краснодар, 1973. 211 с.
Воробьев Д В. Методика лесотипологических исследований. Киев, 1967. 388 с.
Воробьев Д.П. Дикорастущие деревья и кустарники Дальнего Востока. Л.,1968. 278 с.
Воронов АГ. Место биогеографии в системе наук и ее будущее. //В кн.: Теоретические и прикладные аспекты биогеографии. М.: Наука.1982. С. 4- 22.
Воронов П.В. Управление ростом и развитием деревьев. М.-Л.: Гослесбумиздат, 1954. 112 с.
Воропанов П.В. Ельники Севера. М.-Л.: Гослесбумиздат, 1950. 180 с.
Вульф Е.В. Историческая география растений. М.-Л.: Изд. АН СССР, 1944. 546 с.
ТгтЯЗЛ. Борьба за существование как фактор эволюции. Л., 1976.166 с.
Гаузе Г.Ф. Экология и некоторые проблемы происхождения видов. //В кн.: Экология и эволюционная теория. Л.: Наука. 1984. С. 7-108.
Гвоздецкий Н.А. О высотной зональности как основной закономерности ландшафтной дифференциации горных стран. //В кн.: Вопросы ланшафтоведения. Алма-Ата, 1963. С. 14-22.
Гвоздецкий Н.А. Кавказ. М., 1963. 262 с.
Гельтман В.С. Географический и типологический анализ лесной растительности Белоруссии. Минск, 1982. 256 с.
География тесных ресурсов земного шара. М.: Изд. Иностранной литературы, 1960. 665 с.
Герасимов М.В. Пихта кавказская. М.-Л.: Гослесбумиздат, 1948.176 с.
Гигаури Г.Н. Закономерности строения, роста и развития буковых и хвойных лесов Грузии и ведения хоз-ва в них. Автореферат дис. д-ра с-х наук, Тбилиси, 1973. 52 с.
Гигаури Г.Н. Некоторые особенности возрастного развития темнохвойных и буковых лесов Грузии. ИТр. Тбилисского ин-та леса. Тбилиси, 1974. Т. XXI. С. 201- 207.
Глазов Н.М. Возрастные структуры девственных лесов. //Лесн. хоз-во 1979. №6. С. 47-48.
Гниденко В.И. О взаимоотношении бука, пихты, ели и динамики естественного формирования смешанных древостоев. //Лесоводство и агролесомелиорация. Киев, 1967. Вып. 12. С. 57-64.
Голгофская К.Ю. Типы буковых и пихтовых лесов бассейна р. Белой и их классификация. /Пр. Кавказского гос. заповедника. 1967. № 9. С. 157-284.
Голгофская К.Ю. Опыт составления типологической классификации лесной растительности Кавказского заповедника. //Проблемы типологии и классификации лесов. Свердловск, 1972. С. 167-176.
Голгофская К.Ю. Использование принципов географо-генетического направления при типологической классификации и районировании горных лесов. //В кн. Эколого-географические и генетические принципы изучения лесов. Свердловск, 1983, С. 60-63.
Голгофская К.Ю. Использование типов леса в лесном хоз-ве и лесоустройство горных лесов. //В кн. Проблемы использования типов леса в лесном хоз-ве и лесоустройстве. Свердловск, 1986. С. 42-46.
Голод Д.С. Парфенов В.И., О световом режиме в еловых фитоценозах. 1/Богганикъ исследования. Минск, 1966. Вып. 8. С. 122-130.
Голубец М.А. Ельники Украинских Карпат. Киев, 1978. 264 с.
Горчаковский П.Л. Пихтовая тайга Среднего Урала. //Зап.
Уральского отдела Географ, общества СССР, 1954. Вып. 1. С. 12-77.
Грант В. Эволюционный процесс. М.: Мир, 1991. 448 с.
Гребенщиков О.С. Опыт климатической характеристики основных растительных формаций Кавказа //Бот. Журнал, 1974. № 2. Т. 59. С. 161-176.
Гроссгейм А.А. Анализ флоры Кавказа. //Тр. бот. ин-та Азерб. филиала АН СССР. 1936. Т. 1. С. 1- 260.
Гроссгейм А.А. К географии хвойных лесов Кавказа. //Тр. Азерб. гос. ун-та. Баку, 1939. Вып. 1. С. 10-56
Гроссгейм А. А. Растительный покров Кавказа. М.: МОИП. 1948. 267с.
Гулисашвили В.З. Некоторые экологические особенности ели восточной и пихты кавказской и климатические границы их распространения. //Сооб. Груз. Филиала АН СССР, 1940. Т. 1 № 1. С. 55 - 62.
Гулисашвили В.З. Распространение лесообразующих хвойных пород в Закавказье и взаимоотношение между ними. //Бот. жур. 1951 № 3. С. 277-287.
Гулисашвили В.З. Горное лесоводство. М.-Л.: Гослесбумиздаг, 1956.354 с.
Гулисашвили В.З. Природные зоны и естественно-исторические области Кавказа. М.: Наука, 1964. 327 с.
Гулисашвили В.З. Лесное хоз-во в горных лесах СССР. //В кн.: Лесное хоз-во и лесная промышленность СССР. Докл. к VII Международному лесному конгрессу. М.: Лесная пром., 1972. С. 172-182.
Гулисашвили В.З. Практика ведения выборочного хозяйства в горных лесах. М., 1972. 96 с. *
Гулисашвили В.З. Внутривидовые взаимоотношения и регулирование численности особей в растительных сообществах. //Сов. вопр. лесоведения и лесной биоценологии. М.: Наука, 1974. С. 233- 245.
Гулисашвили В.З., Махатадзе Л.Б., Прилипко Л.И. Растительность Кавказа. М.: Наука, 1975. 233 с.
Гумбольдт А. География растений. М.-Л., 1936. 230 с.
Давыдов МВ. Карпатские пихтарники. //Лесное хоз-во. 1956, № 5. С. 29 - 31.
Данилов Д.Н. Плодоношение пихты сибирской. //Бюлет. МОИП, 1951. Т. 56, Вып. 1.С. 90-93.
Дарвин Ч. Происхождение видов. М.-Л, 1937. 762 с.
Дерябин Д.Н., Букштынов А.Д. Лесоводственное значение хвойного подроста. М.: Лесн. пром., 1970. 93 с.
Докучаев В.В. Учение о зонах природы. М.: Географгиз, 1948. 64 с.
Долуханов А.Г. Геоботанический очерк лесов ущелья р. Чхалты. ИГ?. Тбил. бот. ин-та. 1938. Т. 5. С. 1 -103.
Долуханов А.Г., Сахокия М.Ф. Опыт геоботанического районирования Закавказья. //Сообщ. АН ГССР. 1941. Вып. 4. С. 341 - 348.
Долуханов А.Г. О некоторых закономерностях формирования и смен основных формаций лесной растительности Кавказа. //Тр. Тбил. бот. ин-та. 1958. Т. 19. С. 71 -121.
Долуханов А.Г. Вопросы естественной классификации лесных ценозов. //Тр. Тбил. бот. ин-та. 1959. Т. 20. С. 289 - 301.
Долуханов А.Г. Темнохвойные леса Грузии. Тбилиси: Мецниереба. 1964. 127 с.
Долуханов А.Г. Вопросы типологии горных лесов в связи с природной относительностью непрерывности их растительного покрова. //Тр. МОИП. М.: Наука, 1970. Т. 38. С. 24 - 33.
Долуханов А.Г. Колхидский подлесок. Тбилиси: Мецниереба. 1980. 262 с.
Долуханов А.Г. О некоторых дискуссионных вопросах лесной типологии и особенностях типологического изучения горных лесов. //В кн.: Современные проблемы лесной типологии. М.: Наука, 1985. С.113-119
Долуханов АТ. Растительность Грузии. Тбилиси: Мецниереба. 1989.238 с.
Дорофеев П.И. Развитие третичной флоры СССР по данным палеокарпологических исследований. Автореферат диссерт. На соискание уч. ст. доктора наук. Л., 1964. 43 с.
Дорофеев П.И. О некоторых проблемах истории флоры. //Бот. жур. 1965. Т. 50, № 11. С. 1509 - 1522.
Дорофеев П.И. Третичная флора бассейна Омолоя. //В кн.: История флоры и растительности Евразии. Л.. 1972. С. 41 - 112.
Дробиков А.А. Изменение радиационного режима при рубке леса в пихтовых древостоях. //Сб. науч. тр. ВНИИ лесоводства и механ. лесн. х-ва, 1972. Вып. 11. С. 15-27.
Дыренков С.А., Чертов О.Г. Лесная типология в СССР и за рубежом. //В кн. Лесоведение и лесоводство. М., 1975. Т. 1. С. 190 - 253.
Дыренков С.А. Роль признаков растительности и местообитаний в лесной типологии. //Лесоведение, № 4. 1982. С.З 1-37.
Дыренков С.А. Структура и динамика таежных ельников. Л., 1984.174 с.
Завадский К.М. Вид и видообразование. Л., 1968. 404 с.
Заклинский Н.С. Леса Абхазии. М., 1931. 211 с.
Заповедники Кавказа. М.: Мысль, 1990. 336 с.
Захаров С.А. К характеристике высокогорных почв Кавказа. Изв. Константиновского межевого ин-та., 1914. Вып. 5. 368 с.
Захаров С.А. Опыт классификации почв Закавказья. /ГУр. почв, сектора Груз, филиала АН СССР. 1935. Т. 1. С. 3 - 32.
Злобин Ю.А. Оценка качества ценопопуляций подроста древесных пород. //Лесоведение, 1976. № 6. С. 72 -79.
Зонн С.В. Горно-лесные почвы Северо-Западного Кавказа. М.: Изд-во АН СССР, 1950. 334 с.
Иванов Н.Г. Реакция подроста пихты кавказской на условия освещения под пологом буково-пихтовых лесов. кн. Проблемы лесоведения в горных лесах. М., 1984. С. 53 - 61.
Иванов Д.А., Косович Н.Л. О работе ассимиляционного аппарата древесных пород. //Журн. Русск. ботан. о-ва. 1932. Т. 17, № 1.
Ивашкевич Б.А. Девственный лес, особенности его строения и развития. //Лесное х-во и лесн. пром., 1929, № 11. С. 40 -47: № 12. С. 41 -46.
Ильинская И.А. К систематике ископаемых Faqaceae СССР. //В кн. Систематика и эволюция высших растений. Л.: Наука, 1980. С. 20 - 29.
Ипатов В.С. Дифференциация древостоя. //Вест. ЛГУ. Сер. биол., 1969. Вып. 3. С. 43 - 54.
Кайрюкштис Л. Научные основы формирования высокопродуктивных елово-лиственных насаждений. М., 1969. 208 с.
Каппер О.Г. Хвойные породы. М.-Л.: Гослесбумиздат, 1954. 304 с.
Каразия С.П. Типологическая классификация лесов Литвы - основа специализированного хозяйства. Каунас, 1981. 17 с.
Карпов В.Г. Экспериментальная фитоценология темнохвойной тайги. Л. 1969. 335 с.
Касимов В.Д. Классификация елового подроста под пологом леса. //Лесное х-во, 1960. № 7. С. 9 -11.
Катрушенко И.В. О фитосинтетической адаптации многолетней хвои подроста ели Picea abies к свету. //Бот. журн. 1965. Т. 50, № 8. С. 1119-1120.
Кеппен Ф. Географическое распространение хвойных деревьев в Европейской России и на Кавказе. СПб., 1885. 634 С.
Кларк Г Доисторическая Европа. Изд. иностр, лит-ры. М., 1953. 330 с.
Коваль И.П. Многоцелевое использование горных лесов Северного Кавказа. //Лесное х-во, 1977. № 1. С. 36- 40.
Коваль И.П. Средосберегающая технология освоения лесных ресурсов горных территорий. //Сб. науч. тр. ВНИИЛМ. М., 1987. С. 85 -94.
Коваль И.П. О стратегии лесопользования в горных лесах. //Лесное хоз-во Северного Кавказа. М., 1996. Вып. 22. С. 6 -11.
Колаковский А.А. К истории флоры колхидского рефугиума. //В кн. Академику В.Н. Сукачеву к 75-летию со дня рождения. М.-Л.: Изд. АН СССР, 1956. С. 275 - 285.
Колаковский А.А. Ботанико-географическое районирование Колхиды. /Пр. Сух. бот. сада. Сухум, 1958. Вып. И. С. 124 -166.
Колаковский А.А. Растительный мир Колхиды. Изд.МГУ. 1961. 459 с.
Колаковский А.А. Ботанико-географическое районирование Колхиды. //Тр. Сух. бот. сада. Сухум, 1968. С. 141-196.
Колаковский А.А. Вертикальная поясность лесной растительности Колхиды в третичное время. //Тр. Тбил. ин-та леса. 1974. Т. 21. С. 98 -115.
Колаковский А.А., Шакрыл А.К. Сарматская флора Абхазии. Шр. Сух. бот. сада. 1976. Вып. 22. С. 98 - 148.
Колесников Б.П. Кедровые леса Дальнего Востока. //Тр. ДВФ АН СССР, 1956. Т. И (IV). С. 1 - 260.
Колесников Б.П. Состояние советской лесной типологии и проблемы генетической классификации типов леса. //Изв. Сибир. отд. АН СССР, 1958, №2. С. 109- 122.
Колесников Б.П. Генетический этап в лесной типологии и его значение. //Лесоведение, 1974. № 2. С. 3- 20.
Колесников в.П. Проблемы охраны растительного мира. //Журн. Общ. биологии 1976. Т. 37, № 5. С. 635 - 647.
Калуцкий К.К. Буковые леса Северного Кавказа и основные вопросы ведения х-ва в них. Автореф. докт. диссертации. Воронеж, 1972. 42 с.
Комаров В.Л. Меридиональная зональность организмов. //В кн.: Дневник 1-го Всерос. съезда рус. бот. в Петерб. 1921. № 3. С. 27 - 28.
Комаров В Л. Происхождение растений. М.: Изд. АН СССР. 1961.190 с.
Комендар В.И. Форпосты горных лесов. Ужгород, 1966. 205 с.
Комин Г.Е. Смены поколений в разновозрастных лесах и возможности их прогноза. //Лесоведение, 1982. № 4. С. 49 - 55.
Комин Г.Е., Семечкин И.В. Возрастная структура древостоев и принципы ее типизации. //Лесоведение, 1970. № 2. С. 24 -33.
Комисаров Д.А., Штейнвольф ЛЛ. Интенсивность фотосинтеза подроста ели в разных экологических условиях. //Сб. “Свет, режим, фотосинтез и прод. леса. М.: Наука, 1967.
Кондратюк Е.Н. Основные этапы истории формирования и развития флоры хвойных на территории Украины. //В кн. Природная обстановка и фауны прошлого. Киев. Изд. Наукова думка, 1965. С. 124 - 142.
Корчагин А.А. Происхождение хвойных лесов Евразии. 1Г£ез. докл. по секции географ, наук. Л., 1945. С. 13 -15
Корчагин А.А. Строение растительных сообществ. НВ кн. Полевая геоботаника. Л., 1976, Т. 5. С. 1 - 320.
Крамер П.Д., Козловский Т.Т. Физиология древесных растений. М.: Лесная пром. 1983. 462 с.
Краснитский AM Проблемы заповедного дела М.: Лесная пром. 1983.192 с.
Краснов АН. Южная Колхида и ее место среди других субтропических областей земного шара. //Русск. Субтропики. 1912, № 8. С. 1- 425.
Криницкий Г.Т. Об определении жизнеспособности подроста древесных растений биоэлектрическим методом. /Мзь. Вузов. Леей. Журн. 1984, №4. С. 22-25.
Криштофович А.Н. Эволюция растительного покрова в геологическом прошлом и ее основные факторы. //В кн. Материалы по истории флоры и растительности СССР. 1946. Вып. 2. С. 21 - 86.
Крылов Г.В., Марадудин И.И., Михеев Н.И., Казакова Н.Ф. Пихта. М.: Агропромиздат. 1986. 239 с.
Кузнецов Н.И. Принципы деления Кавказа на ботанико -георафические провинции. //Зап. АН по физико-математ. отдел., 1909. Т. 24, вып. 1.С. 1-174.
Лавренко Е.М. История флоры и растительности СССР по данным современного распространения растений. //В кн.: Растительность СССР. М.-Л., 1938. Т. 1.С. 235-296.
Лархер В. Экология растений. М.: Мир, 1978. 384 с.
Лесков А.И. Верхний предел лесов в горах Западного Кавказа. И Бот. журн. СССР. 1932. Т. 17, № 2. С. 227-259.
Ломов В.М. Типы строения и особенности таксации насаждеий пихты Сев. Кавказа. Автореф. канд. диссерт. Воронеж, 1971. 28 с.
Лосицкий К.Б., Чуенков В.С. Эталонные леса. М., 1980. 182 с.
Малеев В.П. Флора и растительность Абхазии. И В кн. Абхазия, геобот. и лесовод, очерк. М.-Л.: Изд. АН СССР, 1936. С. 1-45.
Малеев В. П. Растительность причерноморских стран. / Эвксинская провинция Средиземноморья/, ее происхождение и связи. И Тр. Боган, ин-та АН СССР. Геоботаника. 1940. Сер. 3. Вып. 4. С. 135-249.
Малеев В.П. Третичные реликты во флоре Западного Кавказа и основные этапы четвертичной истории его флоры и раст-ти. И В кн.: Матер, по истории флоры и растительности СССР, 1941. Вып. 1. С. 61-144.
Малеев В. П. Основные этапы развития растительности Средиземноморья и горных областей юга СССР / Кавказа и Крыма/ в четвертичный период. // Тр. Никитского бот. сада. Симферополь, 1948. Т. 25, вып. 1-2. С. 3-28.
Малышев А.И. Стратегия и тактика охраны флоры. / Бот. журн. 1980. Т. УШ, №6. С. 875-881.
Мальцев М. П. О некоторых лесоводственных особенностях пихты кавказской в молодых культурах на вырубках. // Сб. науч, трудов ВНИИЛМ, М. 1979. Вып. 14. С. 11-20.
Манько ЮЛ Пихтово-эловыелеса северного Сихоте-Алиня. Л., 1967.224 с.
Манько Ю.И. Ель Аянская. Л., 1987. 280 с.
Марадудин И.И. Естественное возобновление пихты сибирской под пологом насаждений на Салаире. И В кн. Продуктивность и
восстановительная динамика лесов Зап. Сибири. Новосибирск: Наука, 1971. С. 195-200.
Марков К.К. Палеогеография. М.: МГУ, 1960. 267 с.
Матюшкин Е.Н. Региональная дифференциация лесной фауны палеоарктики в прошлом и настоящем. И В кн. Теоретические и прикладные аспекты биогеографии. М.: Наука, 1982. С. 59-80.
Махатадзе ДБ. Темнохвойные леса Кавказа. М.: Лесная пром-ть, 1966.159 с.
Махатадзе Л.Б., Урушадзе Т.Ф. Типологическая классификация лесов Кавказа как основа ведения хозяйства. И Лесоведение, 1977, № 2. С. 3-15.
Махатадзе Л.Б. Организация хозяйства на лесотипологической основе в горных лесах Кавказа. // В кн.: Современные проблемы лесной типологии. М.: Наука, 1985. С. 120-126.
Махатадзе ЛЬ., Попе» ИД Типы леса Закавказья. М: Лесн. пром-ть, 1965.132 с.
Маценко А£. Обзор рода Abies Mill.. И В кн.: Ботанические материалы гербария Бот. ин-та им. Комарова АН СССР, 1963. Т. ХХП. С. 33-42.
Маценко A.fi. Пихты восточного полушария. // В кн.: Флора и систематика высших растении. Тр. БИН АН СССР. М.-Л.: Наука, 1964. Серия 1, вып. 13. С. 3-103.
Медведев Я.С. Растительность Кавказа. Опыт ботанической географии Кавказского перешейка. Тифлис, 1919. 485 с.
Мелехов И.С. Рубки главного пользования. М.. Лесная пром-ть, 1966. Втор. изд. 374 с.
Мелехов И С. Динамическая типология леса //Лесн. хоз-во, 1968. № 3. С. 15-21.
Мелехов И.С. Лесоведение. М., 1980. 408 с.
Мигунова Е.С. К теории лесообразовательного процесса. И В кн.: Теория лесообразовательного процесса. Красноярск, 1991. С. 98-101.
Мильков Ф.Н. Природные зоны СССР. М.: Мысль, 1977. 293 с.
Мильков Ф.Н., Гвоздецкий Н.А. Физическая география СССР. М.: Мысль, 1969. 461 с.
Мишин В. Реликтовые полидоминантные сообщества с буком и их значение для объяснения происхождения развития и ценологической дифференциации современных буковых лесов Балканского п-ва. // Тр. Тбил. ин-та леса. Тбилиси, 1974. С. 116-127.
Млокосевич Б.В. К вопросу акклиматизации древесных пород в горных условиях Абхазии. И Научно-техн. конф. Абхазской НИЛ ОС. Сухум: Алашара, 1967. С. 15-19.
Молоткова И.И. Закономерности распределения деревьев по толщине в разновозрастных пихтовых насаждениях Закарпатья. И Изв. Высш. учеб, заведен. Лесн. журн. 1966, № 1. С. 42-48.
Молчанов А.А. Гидрологическая роль леса. М.: АН СССР, 1960. 487 с.
Морозов Г.Ф. Лес как явление географическое. И Матер, по изуч. русского леса. С. - Петербург, 1914. Вып. 1. С. 121.
Морозов Г.Ф. Учение о лесе. 7-е изд. М.-Л., Гослесбумиздат. 1949. 455 с.
Мотовилов Г.П. Роль лесной типологии в лесном хозяйстве. М, 1955.49 с.
Назимова ДИ Горные темнохвойные леса Западного Саяна. Л., 1975. 120 с.
Недялков С. Некоторые особенности устройства лесов с выборочной формой хозяйства. //Лесн. хоз-во. 1979. № 3. С. 60-62.
Нейштадт М.И. История лесов и палеогеография СССР в голоцене. М.: Изд-во АН СССР, 1957. 404 с.
Нейштадт М.И. Особенности развития лесов на территории СССР в голоцене. И В кн.: Современные проблемы геог. М., 1964. С. 207-214.
Некрасова Т.П., Рябиков А.П. Плодоношение пихты сибирской. Новосибирск: Наука, СО, 1978. 150 с.
Нестеров В.Г. Общее лесоводство. М.-Л.: Гослесбумиздат, 1954. 656 с.
Нешатаев Ю.Н. Пихта сибирская на западной границе своего ареала. И Бот. журн. 1963. Т. 48, № 1. С. 96-98.
Овсянкин В.Н. Пихтовые леса Северного Кавказа. // Тр. Всесоюзного заочного лесотехнического ин-та. М.: Изд. высш, и сред, спец, образования РСФСР, 1964. № 8. С. 33-57.
Орлов А. Я. Темнохвойные леса Северного Кавказа. М.: Изд-во АН СССР, 1951.256 с.
Остапенко Б.Ф. Диагностика и хозяйственная группировка типов леса северного склона Большого Кавказа // Тр. Харьк. СХИ, 1968. Т. 72. С. 111-257.
Парфенов В.И. Обусловленность распространения на границах ареалов. Минск, 1980. 205 с.
Пианка Э. Эволюционная экология. М.: Мир, 1981. 399 с.
Плотников В.В. Эволюция структуры растительных сообществ. М.: Наука, 1979. 275 с.
Побединский А.В. Лесная типология - основа лесохозяйственных мероприятий. //Лесн. хоз-во. 1973. № 9. С. 16-20.
Побединский А.В. Рубки главного пользования. М.: Изд-во “Лесн. пром-ть”. 1980. 191 с.
Поварницын В.А. Типы лесов Абхазии. И В кн. Абхазия, геобот. и лесовод, очерк. М.-Л.: Изд. АН СССР, 1936. С. 47-65.
Поварницын В.А. Леса из сибирской пихты в СССР. И В кн.: Акад. В. Н. Сукачеву к 75 лет. со дня рождения. М.-Л.: Изд. АН СССР, 1956. С. 408-424.
Погребняк П.С. Основы лесной типологии. 2-е изд. Киев. 1955. 456 с.
Попов К.П. К уточнению восточной границы ареала Picea orientalis и Abies nordmanniana на Большом Кавказе. //Боган, журн. 1982. Т. 67,№4. С. 507-513.
Попов М.Г. К вопросу о происхождении тайги. И Сб. статей по результатам исследований в области лесного хозяйства и лесной промышленности в таежной зоне СССР. М.-Л.: Изд-во АН СССР, 1957. С. 19-28.
Правила рубок главного пользования в горных лесах Сев. Кавказа М., 1993. 20 с.
Правила рубок главного пользования и лесовосстановительных рубок в лесах ГССР. Тбилиси, 1980. С. 20.
Придал М.В. Фенотипический анализ популяций пихты Нордманна в Кавказском заповеднике. //Лесоведение, 1977, № 6. С. 70-79.
Прилипко Л.И. О необходимости и критериях выделения редких и эталонных растительных сообществ для охраны. // В кн.: Охрана природы Грузии. Тбилиси, 1984. Вып. XII, С. 91-101
Прасолов Л.И. Горно-лесные почвы Кавказа. И Тр. Почвенного ин-та им. В.В. Докучаева. М., 1947. Т. XXV. С. 5-28.
Протопопов В.В. Средообразующая роль темнохвойного леса. Новосибирск: Наука, 1975. 328 с.
Райт В.Р. Введение в лесную генетику. М.: Лесная промышленность. 1978. 470 с.
Растительность европейской части СССР. Л.,: Наука, 1980. 429 с.
Рикли М. Материалы к географии и истории флоры Кавказа и Высокой Армении. //Естествознание и география. М. № 8/10. С. 126-144.
Ричардс П.Р. Тропический дождевой лес. М.: Изд-во Иностр, лит-ры. 1961.448 с.
Розенберг В.А. Принципы и основные единицы классификации еловых и пихтовых лесов Дальнего Востока. И Второе Всесоюзное совещание по лесной типологии. / Тез. докл./. Красноярск, 1973. С. 95-97.
Ролдугин И.И. Еловые леса Тянь-Шаня. / Флора, классификация и динамика/. Алма-Ата: Наука Каз. ССР, 1989. 304 с.
Рысин Л.П. Влияние лесной растительности на естественное возобновление древесных пород под пологом леса. //В кн.: Естественное возобновление древесных пород и количественный анализ его роста. М.: Наука, 1970. С. 7-53.
Рысин Л.П. Лесная типология в СССР. М.: Наука, 1982. 217 с.
Рысин Л.П. Краковская Декларация о спасении Европейских лесов. (5 - 7. VI. 89 г.). И Лесоведение, 1990. N 5. С. 76-78.
Сабан Я. А. Экология горных лесов. М.: Лесная пром-ть, 1982. 168 с.
Сабашвили М.Н. Почвы Грузинской СССР. Тбилиси, 1965. 549 с.
Савченко АЛЛ. Возобновление пихтовых лесов. М.: Лесн. пром-ть. 1970.
80 с.
Саликов Н.Я. Критерий жизненности насаждений. И Веста, с-х науки. 1979. № 8. С. 120-123.
Сафаров И.С. Редкие и исчезающие виды дендрофлоры Восточного Закавказья и их охрана. //Боган, журнал, 1986. Т. 71, № 1. С. 102-107.
Сахаров М.И. Типы леса в верховьях реки Мзымта. И Тр. Кавказ, залов., 1939. Вып. 2. С. 83-125.
Северцев АН Главные направления эволюционного процесса М, 1967. 202 с.
Семечкин И.В. Динамика возрастной структуры древостоев и методы ее изучения. И В кн.: Вопросы лесоведения. Красноярск, 1970, Т. 1. С. 422-446.
Семечкин И.В. Поколение деревьев - естественная элементарная единица при изучении строения древостоев в статике и динамике. // В кн.: Итоги изучения лесов Дальнего Востока. Владивосток, 1967. С. 117-125.
Серебряков И.Г. Экологическая морфология растений. М., 1962. 378 с.
Сидельников В.П. Высокогорная растительность Алтай-Саянских гор. области. Новосибирск: Наука. 1988. 222 с.
Синнот Э. Морфогенез растений. М., 1963. 603 с.
Синская Е.И. Основные черты эволюции лесной растительности Кавказа в связи с историей видов. И Бот. журнал, 1933. № 3. С. 89-102.
Смит У.Х. Лес и атмосфера. М.: Прогресс, 1985. 429 с.
Смолоногов Е.П. Генетическая классификация типов леса и восстановительно-возрастная динамика кедровых лесов Урало-Сибирской тайги. Автореф. докт. диссерт. Свердловск, 1987. 47 с.
Соколов В.Е. Заповедники СССР: их цели и задачи. // В кн.: Итоги и перспективы заповедного дела в СССР. М.: Наука, 1986. С. 5-13.
Соколов Н.С. К истории растительности Большого Кавказа. И Веста. МГУ, сер. геогр., 1962. Вып. 5, С. 40-44.
Соколов С.Я. Классификация типов лесов Абхазии. И Абхазия, геобот. и лесовод, очерк. М.-Л.: Изд. АН СССР, 1936. С. 203-212.
Солберг О., Солберг Д. Популяционная биология и эволюция. М.: Мир, 1982.488 с.
Солнцев Г.К. Совершенствование лесохозяйственного производства Сев. Кавказа. И Сб. науч. тр. НИИ горлесэкол. М., 1996. Вып. 22. С. 3-5.
Соснин Л.И. Типы лесов Кавказского госзаповедника. И Тр. Кавказ, госзаповедника. М., 1939. Вып. 2. С. 5-82.
Сосновский Д.И. Основные формы растительного покрова Кавказа в их географическом размещении. И Сов. бот., 1947. Т. 15, №6. С. 318-328.
Сочава В.Б. К фитосоциологии темнохвойного леса. И Журн. Рус. ботан. о-ва. 1930. Т. 15, № 1-2. С. 7-41.
Сочава В.Б. Позиция некоторых реликтов древней флоры в современных фитоценозах Кавказа. Доки. АН СССР, 1947. Т. 56, № 3. С. 313-316.
Сочава В.Б. О происхождении буковых лесов Кавказа. И Известия АН СССР, сер. биологическая, 1949. Т. 2. С. 224-236.
Спурр С.Г., Барнес Б.В. Лесная экология. Пер. с 3-го английского изд. М.: Лесная пром-ть, 1984. 479 с.
Ссорин В.А. Пихтовые леса Верхней Сванетии. И Боган, журн. 1951. Т. 36, № 5. С. 468-496.
Станюкович К.В. Растительность гор СССР. Душанбе: Изд. Дониш, 1973.411 с.
Степанов Б.П. Охрана редких и находящихся под угрозой исчезновения объектов растительного мира: структура проблемы и координация научно-исследовательских работ. И В кн.: Охрана растительных сообществ и находящихся под угрозой исчезновения экосистем. М., 1981. С. 3-5.
Стойко С.М. Категоризация редких, уникальных и типичных фитоценозов и их интегральная созологическая оценка. II Материалы I Всесоюзн. конференции по охране редких сообществ. М., 1982. С. 5-7.
Столяров Д.П.; Кузнецова В.Г. Разновозрастные ельники и ведение хозяйства в них. М., 1979. 168 с.
Сукачев В.Н. О влиянии интенсивности борьбы за существование между растениями на их развитие. И ДАН СССР, 1941. Т. 30. № 8. С. 752-755.
Сукачев В.Н. Динамика лесных биогеоценозов. И В кн.: Основы лесной биогеоценологии. М., 1964. С. 458-486.
Основные понятия лесной биогеоценологии. II В кн.: Основы лесной биогеоценологии. М.: Наука, 1964а. С. 5-49.
Сукачев ВН. Проблемы фитоценологии. Избран, труды. Л., 1975. Т. 3.539 с.
Сукачев В.Н., Зонн С.В. Методические указания к изучению типов леса. М., 1961. 144 с.
Сябряй С.В. Происхождение горно-таежных лесов Карпат. И Бюл. МОИП. Отдел, биолог. 1992. Т. 97, Вып. 3. С. 92-97.
Тахтаджян А.Л. К вопросу о происхождении умеренной флоры Евразии. И Бот. журн. 1957. Т. 42. № И. С. 1635-1653.
Теория лесообразовательного процесса. Тезисы докладов. Красноярск, 1991. 186 с.
Тимофеев В.П. Роль елового подроста в восстановлении вырубок. И Лесн. х-во, 1968. № 12. С. 9-15.
Тимофеев-Ресовский Н.В. и др. Краткий очерк теории эволюции. М., 1977.297 с.
Тимухин Н.Т., Горчарук Л.Г. Особенности современного почвообразования в буково-пихтарниковых биогеоценозах Западного
Кавказа. И Почвенно-биогеоценологические исследования на Северо-Западном Кавказе. Пушкино, 1990. С. 5-17.
Ткаченко М.Е. Первобытные и выборочные леса Севера России. И Лесовод., 1929, № 10-11. С. 12-31.
Толмачев А.И. К истории возникновения и развития темнохвойной тайги. М.-Л.: Изд-во АН СССР, 1954.156 с.
Толмачев А.И. Роль миграции и автохтонного развития в формировании высокогорных флор земного шара. // Проблемы ботаники, 1960. Т.5.С. 18-31.
Толмачев А.И. Основные пути формирования растительности высокогорных ландшафтов северного полушария. И Боган, журн. 1948. Т. 33, №2. С. 161-180.
Толмачев А.И. Современные проблемы географии растений. II Вести. Ленинградского у-та. 1968. № 9. С. 24-29.
Трасс Х.Х. О значении доминантов растительных сообществ для классификации растительного покрова. // В кн.: Проблемы современной ботаники. М.-Л., 1965. Т. 1. С. 247-250.
Третьяков НВ. Закон единства и строения насаждений. М-Л., 1927. С. 113.
Тумаджанов И.И. К постплиоценовой истории лесной растительности Сев. Кавказа. //Тр. Тбил. бот. ин-та. 1955. Т. 17. С. 161-221
Тумаджанов И.И. Опыт дробного геоботанического районирования Северного склона Большого Кавказа. Тбилиси: Изд. АН ГрССР, 1963. 241 с.
Тумаджанов И.И. Основные черты истории и географии лесной растительности Большого Кавказа в плестоцене и голоцене. И Изв. АН СССР. Сер. Геогр., 1973. № 2, С. 34-43.
Уиттекер Р. Сообществам экосистемы. Пер. с англ., М., 1980. 327 с.
Уранов А.А. Жизненное состояние вида в растительном сообществе. И Бюллет. МОИП. Отд. биол.. 1960, Т. XV, вып. 3. С. 5-11.
Урушадзе Т.Ф. Почвы горных лесов Грузии. Тбилиси: Мецниереба, 1987. 243 с.
Успенский Е.И., Максимов Г.Н., Чапурин В.А. Оценка жизненного состояния елового подроста. И Науч. докл. высш. шк. Биологические науки, 1974. С. 63-66.
Ушатин П.Н. Основы организации лесного х-ва в горных лесах СССР. М., 1962. 89 с.
Фалалеев Э.Н. Пихтовые леса Сибири и их комплексное использование. М.: Лесная пром-ть, 1964. 165 с.
Федоров Ан. А. История высокогорной флоры Кавказа в четвертичное время, как пример автохтонного развития третичной флористической основы. И Матер, по четвертич. периоду СССР. 1952. Вып. 3. С. 49-87.
Федоров Ан. А. Флора Юго-западного Китая и ее значение для познания растительного мира Евразии. И Комаровские чтения, БИН АН СССР, 1957. Вып. 10.
Федоров Ан. А. О флористических связях Восточной Азии с Кавказом. // В кн.: Материалы по истории флоры и растительности СССР. М.-Л.: Изд-во АН СССР, 1958. Вып. 3. С. 230-248.
Федорчук В.Н., Дыренков С.А. Выделение и распознавание типов леса. Методы исследования. Л., 1975. 35 с.
Фирсова В.П., Ржанникова Г.К. Почвы Южной тайги Урала и Зауралья. И Тр. инст-та экологии раст. и животных. Свердловск, 1972. Вып. 85. С. 3-87.
Фридланд В.М. Почвы высокогорий Кавказа. // Генезис и геог. почв. М.: 1966. С. 43-82.
Хильми Г.Ф. Теоретическая биофизика леса. Изд-во АН СССР, 1957. 208 с.
Хохряков АП. Анализ флоры Колымского нагорья. М.: Наука, 1989.152 с.
Хуторцев И51. Семеношение пихты кавказской на Северо-Западном Кавказе. //Тр. СКЛОС. Майкоп, 1975. Вып. 12. С. 159-170.
Цельникер Ю.Л. Адаптация лесных растений к затенению. И Бот. журн. 1968. № 10, т. 53. С. 1478-1491.
ЧочиеваКИ Ископаемые Тахосйасеае Колхиды. Тбилиси: Мецниереба, 1985. 92 с.
Шавнин А.Г. Строение, рост и особенности таксации ельников Приморского края. И Разновозрастные леса Сибири, Дальнего Востока и Урала. Красноярск. Изд-во СО АН СССР, 1967. С. 81-92.
Шатилова И.И. История развития позднеплиоценовой растительности Западной Грузии. Тбилиси^ 1984. 106 с.
Шввденко АИ. Пихтовые леса Украины. Львов: Высшая школа, 1980.192 с.
Шилов М.П. Экофонд, генофонд, ценофонд. // В кн.: Охрана раст. сообществ редких и находящихся под угрозой исчезновения экосистем. М., 1982. С. 16-18.
Шмалыаузен И.И. Интеграция биологических систем и их саморегулирование. //Бюлл. МОИП,отд. биологии, 1961. Т. 66, вып. 2. С. 104-134.
Шоо Р. История растительности Венгрии. И Доки. зарубежных ученых на съезде ВБО. Л., 1958. С. 71-78.
Юркевич И.Д., Гельтман В.С. Соотношение понятия “лесная ассоциация” и “типлеса”. //Боган, журн., 1970. Т. 55, № 1. С. 34-41.
Юркевич И.Д., Голод Д.С., Парфенов В.И. Типы и ассоциации еловых лесов. Минск. 1971. 351 с.
Юрцев БА Вопросы происхождения темнохвойной тайги в свете новейших палеоботанических исследований. //Бот. журн., 1972. № 11. С. 1455-1469.
Alexander R.R. Initial partial cutting in Old-growth Spruce-Fir. USDA Forest Serv. Research Paper RM-76. Colorado, 1971. P. 20.
Axelrod D. I. Tertiari floras and topographic histori of the Snake River Basin, Idaho. И Geological Societi of America. 1968. Bulletin, 79. P. 713-734.
Axelrod Daniel I. Environment of the Middle Eocen (45 Ma) Thunder Mountain Flora, Central Idaho. //Nationale Geographic Research. 1990. Vai. six, P. 355-363.
Bean W.J. Trees et Shrubs hardy in The British Isles. Eighth ed Vol. I. London: John Murray, 1980. 845 p.
Chamberlain Ch. J. Gimnosperms: Structure and evolutions. N.-Y., Chicago Press, 1935. - 484 p.
Clements F.E. Plant Succession and Indicators. New Jork, 1928.
Cooper F. The ecology of Fire. H Sci. Amer., 1961. Vol. 204, N 4. P. 215-241.
Davis PH Flora cf Turkey and the Aegean Islands. Edinburgh, 1965. Vol. 1.567 p.
Dangler A. Waldbau auf Okologischer Grundlage. Hamburg und Berlin: Verlag Paul Parey, 1982. Zw. Bd. 280 S.
Eis S. Cone production of Douglas - fir and grand fir and its climatic requirements. // Can. J-Forest Res, 1973,3 N 1. P. 60-70.
Endo K. Forest Soils of the Warm Temperate Zone in Japan. И Soils as Site Factor to Forests of the Temperate and Cold Zones. Proc. Symp. Zvolen, 1977. Vol. 14. P. 111-119.
Farjon Aljos. Pinaceae. Konigstein. Koeltz Sci. Books. 1990. ХП - 330 p.
Flora of Taiwan. Taipei, Taiwan, ROC, 1994. Sec. ed. Vol. I. 648 p.
Franklin J.F. Ecology of noble fir. I I Abstract, Trul Fir Symposium -Seattle, Washington, 1981. P. 7-8.
Gaussen H. Les Gymnosperms actualles et fossiles. // Trav. Lab. Forest. Toulouse: 2, 1,1, 7. 1964. S. 321-480.
Gelik O., Ozkan N. The Turkish Forestry. General Directorate of Forestry. Publ. number: 673, Serial number: 30. Ankara, 1989. 126 p.
Godwin H. History of the natural forest of Britain: establishment, dominance and destruction. // Phill. Trans. Roy. Soc. London, 1975. 271. P. 47-67.
Greguss P. Fossil Gymnospermen Woods in Hungary from the Permien to the Pliocene. Budapest: Akad. Kiad6,1967. 137.
Henkel W. Das Tarmenproblem. //Forst und Jagd. 1954, Jg. 4, Hft. 1. S. 20-22.
International Dendrological Researth Institute, IncfIDRI). News Brief. Medfield, 1994. N 6. P. 2-8.
Yang Jeng-Chuan, Pan Fuh-Jiunn. The current status of native woody vegetation in Taiwan. // Temperate Trees under Threat. International Dendrolodical Society. Taipei, 1996. P. 89-120.
Jain K.K. A taxonomic revision of the Himalayan firs. И Indian Forester. 1975. Vol. 101,N3.PP. 199-204.
Kaji M. Studies on the ecological geography of subalpine conifers. Distribution patem of Abies martiesii in relation of the effect of climate in the postglacial warm period. // Bull. Tokyo Univ. Forest. 1982. N 72. P. 31-120.
Kolhs J. Struktur und Entwicklungsdynamik im subalpinen Fichten-Wald Schloflberg I Linz. Wien, 1974. 130 s.
Leibundgut H. Ober Zweck und Methodik der Struktur und Zuwachsanalise von Urwdldem. H Schweiz. Z. Forstwesen, 1959. Bd. 110. S. 111-124.
Leibundgut H. Ober die Dynamik europaischer Urwdlder. // Allgemeine Forst Zeitschrift. 1978. N 24. S. 686-690.
Leibundgut H. Europdische Urwdlder der Bergstufe. Bern, Stuttgart: Verlag Paul Haupt. 1982. 308 S.
Liu Tang-Shiu. A Monograph of the Genus Abies. Taipei, China, 1971. 608 p.
Mayer H. T^nnenreiche Walder am Sudabfall der Mittleren Ostalpen. Munchen Basel Wien: BLV Verlagsgesellschaft, 1969. 259 S.
Mayer H. Die Tanne - ein unentbehrlicher Okologischer Stabilisator des Gebirgs-Waldes. Jahrbuch des Vereins zum Schutze der Alpenpflanzen und -Tiere. Munchen, 1975. 30 S.
Mesterhazy Zsolt. Conifer Treasury of the World. Budapest, 1995. 546 p.
Mirow N.T. The Genus Pinus. The Ronald Press Company. New York, 1967. 602 p.
Mitchell A.E. The genus Abies in Scotland..H Trans. Bot. Soc. Edinburgh, 1971. Vol. 41, pt. 2. P. 201-212.
Mittfeld J. Die Europdischen und Mediterranen Abies-Arten. II In.: Die Pflanzenareale: 1,2. Jena, 1926. S. 22-29.
Nakamura T. Succession and Differentiation of Forest Communities in the Subalpine Region of Mt. Fuji. //Bull. Tokyo Univ. For. 1992. 87. P. 159-173.
Owens, John W. Sexual reproduction of Abies amabilis. //Canad. J. Bot; 1977. Vol. 55, N. 21. P. 2653-2667.
Rubner K. Die Pflanzen geographischen Grundlagen des Waldbaues.. 5. Aufl. Radebeul, Berlin, 1960. 620 s.
Rushforth K. D. Conifers. London: Chistopher Helm, 1987. 232 p.
Sarvas R. Der nordische Urwald. //Schweiz. Zeitschr. Forstwes., 1959. Bd. 3. S. 319-345.
Schmidt-Vogt H. Die Fichte. Hamburg, Beilin, Veriag Paul Parey, 1977. Bd. 1. 677 S.
Simpson G. G. This view of life. N. Y., 1964. 308 p.
Smith G. D. Soil Classification. 7-aproximotion. Soil Survey Staff, USA, 1960.
Soljanik Ivan. О kavkaskoj jeli (Abies nordmanniana) njezina svojstva mogucnosti i potrebe prosirvania u. Jugoslaviji. .//Sumarski, 1963. List, god. 87, 1-2, C. 23-36.
Strasburger E. Lehrbuch der Botanik. Stuttgart-New York, Gustav Fisher Verlag, 1983. 32 Auflage. 1161 S.
Szafer W. Flora pliocenska z Krosienko nad Dunajcem. Krakdw, 1947. 213 s.
Szafer W. Pliocenska flora okolic Czorsztyna i jej stosunek do plejstocenu. //Inst. Geolog. Prace, Warszawa, 1954. Tom. XI. s. 1-238.
Taverner R. and Smith G. The concept of Braunerde (Brown Forest Soils) in Europae and the Unitied States. Adv. Argon, 1957. 9. P. 217-289.
Thomasius H., Hartig M. Erfahrungen mit dem Anbau von Abies Nordmanniana (Stev.) Spach in den Waldern der DDR. //Follia dendrologica. 1985. N 12, S. 177-188.
Traquillini W. Phisiological ecology of the Alpen Timberline. B.-Helderberg; W. Y.: Spriger, 1979. 131 p.
Tschermak L. Die naturalische Verbreitung der Baumarten des Waldes im Alpenvorland Ober- und Niederdstereichs. //Cbl. ges. Forstwesen, 79. 1962. N3. S. 113-131.
Unsere Umwelt. Wald. Waldschadenserhebung. Verlag GmbH. Grufelfing. 1984. S. 68-77.
Vyskot M. Tchechoslowakische Urwaldreservate als Lehrobjekte. // ADgemeine Forst Zeitschrift. 1978. N 24. S. 696-697.
Walter H. Vegetationszonen und Klima. Jena: VEB Gustav Fischer Verlag, 1970. 244 S.
Wieser RE. Zu Fichien-Typen. //Allg Forst Zeitschrift, 14.1959. N14. S. 40Ф406.
Zukrigl K. Der Rotwald. als Forschungsobjekt. // Allgemeine Forst Zeitschrift. 1978. N 24. S. 700-702.
THE FIR FORESTS OF CAUCASUS
Prof. Bebya Sergej
Botanical Institute of the Abkhazian Academy of Sciences, Sukhum
SUMMARY
The study of virgin forests and especially Abies nordmanniana (Stev.) Spach has a special place in the practice of botanical and forestry investigations. These investigations give an enormous real material for comprehending the regularity of the forming and dynamics of the plant cover of the whole Caucasus. In this sence the explanation of the origin and evolution of Abies forests and the studying of the forest-formation process in them, to which the present work is devoted, mean a great scientific and practical interest.
In florocenogenetic respect Abies forests of Caucasus excels in their extraordinary originality and peculiarity. They are one of the most productive and richest forest comunities of Europe. In them a lot of terterialy relic and endemic plants have preserved. They form characteristic fitolandscapes similar in a well known degree to those dominating here for the last geological periods.
However the accelerated tempo and scales of exploitation have lead to a significant disorder and diminishing of productivity of Abies nordmanniana forests, and to the decrease of their area as well as to weakening the cenotic position of many relics and endemics in them.
Solving the problem of preservation, renewal and rational using of Abies forests has become one of the actual tasks in modem science of forestry and practice.
The monograph is a complete work fulfilled under consideration of several sciences: paleogeography, floristics, geobotanics, forest science, forestry and sozology. For the first time it touches all aspects of forest-formation process of Abies forests of Caucasus on the level of a single system, states and scientifically grounds the reactions of forests to people’s economic activity. For the first time Abies forests have been studied all around on the level of individual entire formation. Genesis, history of forming and regularities of modem spread of Abies forests are ascertained. It has been made a new, more perfect sketch-map of forest division into districts of the Abies nordmanniana’s area. Morpho-biological, ecological, forestry and phitocenotic peculiarities have been studied with the help of experimental observations. Pecularities of seed-bearing and natural renewal of Abies in forest biogeocenotics are ascertained. The main regularities of the age structure, composition and dynamics of stands, character of
differentiation of the trees in the forest are ascertained.
It is determined the strategy of the species Abies nordmanniana within the limits of its natural habitat. It is worked out a new, for Abies forests more entire regional ecogenetic classification of forest types, system of the co-ordination of the type of condition of the growing place and forest types. It is worked out the system of measures, on forest-typological base, for optimal using and reproduction of Abies forests. New methods are worked out as well as a new technology of voluntarily and group-selective fellings of trees. In the work it is given, for the first time, a sozological appreciation of Abies forests. It is worked out a categorical scale of the rerities of fitocenotics and described a network of natur-protection objects in Abies forests.
There are ascertained 13 species of plants beeing subject to protection and to bringing in the regional Red Book.
The results of all these investigations form a fundamental, bioecological basis of working out the system of controlling with forest forming processes in order to make rational forestry in Abies forests and in their preservation.
At the end of the work you can find main conclusions and recomendations. They are as follows.
The made investigations prove the autochthonous mountain origin of Abies forests of Caucasus. At the end of Pliocene here in the middle-upper mountain belt extensive Abies nordmanniana forests grew, with the admixture of Fagus orientalis, Picea orientalis and other species of cenotical compound of modem Abies forests.
The relative floristic exuberance of their compound (207 species) is concerned to be the distinguishing pecularity of modem Abies forests of Caucasus. Among them 82 % are elements of the mediterrane-mountain* floristic region, i.e. immigrants of high-altitude flora. Relics compose 38 %, among them 56 % woody species. In cenotical compound of indigenous types of forests the position of 85 % of relics is stable. Their presence amounts to 17-100 %, phytocenotical index outnumbers 34. Every of them is typical and permanent , like minimum, for two types of forest. 15% of relics only concern optional species, but they have permanent connection with indigenous phytocenotics. Every of them is typical for one of the type of forest at the least.
The high degree of continuity of cenotical compound between Pliocene and modem Abies forests, sufficiently stable position of relic species in the compound of modem Abies forests, comparatively high content in their Colchidean (36%) and common Caucasian (23%) endemic elements prove not only the ancient autochthonous forming of their main floristic kernel, but also allow to approve that the virgin Abies forests have preserved the relic characteristics.
♦A new "mediterrane-mountain region" was assigned for the first time by Mr Alfred Kolakowskij in 1956. It includes territories of mountain regions of the globe only.
It attaches them a peculier uniqueness in regional and global scale as well as necessity of preservation.
The investigation of morpho-biological and ecological peculiarity has given the possibility of supposition that Abies nordmanniana is a more ancient evulionary branch. These morpho-biological and ecological peculiarities can be considered as the manifestation of common-biological regularity directed to the evolutionary specialization of species and accomodation to growing in Colchidean refugium, isolated from other Abies species.
Investigation materials show that modem spreading of Abies forests in West-Caucasus is conditioned by geological and historical reasons, climatic, first of all hydrothermic conditions, bioecological peculiarities of Abies as well as by anthropogenic influences. Character of the spreading of Abies is determined by complect influence ff these main factors on the development of forests in concrete conditions of location.
Strategy of species of the Abies nordmanniana is sufficiently stable within its natural habitat, and even on the boarder of its natural habitat. Being a dominating edificator * of plant communities, Abies is the most free from adaptation to the conditions of interspecies compatition in the communities. It is proved first of all by its high indices of presence in all types of forest (100 %) and by the phytocenotic index (5400). In all types of forest the renewal is going on from Abies to Abies, its change with accompanying species cannot be observed.
Soil, forming the integral part of forest like biogeocenose, is in a permanent unity and interaction with the forest on the level of forest-soil-forest. Under Abies forests are being formed mainly brown forest acid and brown forest pseudopodzol soils. The weak expression of the differentiation according to genetic horisons of profile is to be considered its distinctive peculiarity. Both types of soil are enough fertile for growing of high productive (to 2000 m3/ha) Abies and Fagus-Abies stands.
In the seed-bearing of Abies good years have been observed in 1 to 2 year’s time. In Abies forests of different age trees of I-st and П-nd layer bear seeds. Trees of III -rd layer bear seeds alone. Spontaneous Abies stands of different age give quite enough seeds both in quantity and quality for a continuous renewal. On 1 ha the seed yield of Abies reaches 49 kgs or 1062 thousand pcs of good quality. Consequently at felling trees in these forests you must base your orientation mainly on natural revival of the main species without additional expenses of artificial reforestration.
The best natural renewal is observed in mixtaherbosum Abies forests with an undergrowth of Vaccinium and in Festucosum Abies forests, and a satisfactory one - in Abies forests with an undergrowth of Ilex aquifolium. At the
same time a weak renewal is characteristic of the Luzulosum and Filicosum Abies forests with an undergrowth of Laurocerasus and Rododendron. Within the group of forest types better forming of renewal is observed at 0,6 cover of stand for groups of Festucosum and Luzulosum types with an undergrowth of Vaccinium and Hex aquifolium, and 0,5-0,6 for mixtaherbosum ones. In the Filicosum group of types better renewal can be awaited at 0,6 cover of stand. These indices must be bore in mind at felling trees in Abies forests.
In natural windows Abies renews well in flower-beds. Within the group of forest types the process of renewal depends mainly on the size and form of windows, on the steepness and layout of the slope. In Festucosum and Mixtaherbosum Abies forests windows of middle size, with a diameter from 18 to 24 m renew better.
The materials of the study of virgin Abies forests in Caucasus show that in all groups of forest types the structure of absolutely different age and complex stepped vertical composition of the cover are characteristic of the stands. The woods surpass by their big amplitude of wavering of age (up to 450-500 years), height (up to 60 m) and diameter (up to 2 m). The crooked line of these indices are multipeaky. Rarely can be found stands of relatively different age, of which a considerable wavering of age (61-120 years) is also characteristic. However the main mass of trunks (70-80 %) is concentrated in two or three classes of age. The distribution of trunks according to the age is near to normal.
The distinguishing peculiarity of the Abies nordmanniana woods of absolutely different age is the continuous character of age dynamics, in the process of which the change of the individual old trees or group of trees is going on by young ones. In die woods, in general, allways all age groups of stand and saplings are present, owing to what the stability of the forest is assured.The agerenewing type of dynamics by means of changing individual dying generations by younger ones is of the virgin woods of Abies nordmanniana not characteristic.These conclusions are extremely important and must be taken into consideration when working out the practical measures, when appointing the fellings in the forest. They refute the misconclusion of the supposed development of virgin woods of Abies nordmanniana of different age to a "reconstructed" condition. The continual process of natural renewal, complex age structure of stands and continuous character of their development are the most important factors of essential adaptations on the level of assotiations, which provide for the long flourishing of the Abies forests of Caucasus.
For forest surveying works and for purposes of forestry it has seemed to be the most expedient to share the complex stands of different age into layers tiers, the saplings not included. The first, upper layer consists of the old-aged and ripe part of the stand. The second, middle layer consists of the middle-aged and ripening part. The third one is characterized by young trees. When surveying forests, the estimation indices must be stated separately for every layer.This
method of forest surveying is very important in case of projecting economic measures and organizing the rational use of the forests.
Conducting forestry in the Abies forests of Caucasus on forest typological basis is still the most important and actual task of the modem forestry. On the base of analysis and generalization of big actual material of personal investigations, including those made on typological stationary test places, we have worked out for the first time the most complete classification of types of forest for the Abies forests of the Caucasus. It is based on ecological-genetic principle. Taking into account the space and time factors of development of forest assotiations, phases of development of assotiations have been revealed from common to particular (from indigenous type of forest to derived one) and from simple to difficult (reforestration into initial type of forest). As phases of development serve the genetically tied up biogeocehotics (assotiations, fullness of stands), which form together with the indigenous one a united biogeocenotic system - type of forest (type of forest biogeocenotics). We have united the aggregate of, in bioecological, cenotical respects near and genetically tied up edapho and climate substituting types of forest into natural groups of forest types. They are Abies forests: Festucosum, Luzulosum, Mixtaherbosum, Filicosum, with evergreen Colchideaq undergrowth, with deciduous Colchidean undergrowth, Subalpine and Rocky-forest complexes.
From point of view of forestry and economics the types of forest, which are in bioecological respect near, homogeneous or similar according to forest-forming conditions, purposefullness of forestry and demanding identical practical measures, are united in five basic economical groups of types of forest or sections. They are Abies forests Festucosum, Luzulosum, Mixtaherbosum, with undergrowth of Colchidean elements and subalpine. For every section a system of measures is offered, which results rational use of the given category of forests, rise in the productivity and improvement of protective functions.This complex and systematizing approach to the unification of forest types allows taking practical measures on a larger scale.
The bioecological pecularities of Abies nordmanniana, character of natural renewal, composition and age structure of stands, important protective functions of Abies forests presuppose the effectiveness of the selective forestry in them.The voluntarily selective and group-selective way of fellings suit best the nature of these forests and in forestry-expluatation respect they are more effective than the industry-selective and gradual fellings of nowadays.The forestry-economic effectiveness of these fellings, with the edoption of the by us worked out technics of their realization and technology of forest felling works express itself in the preservation of the protective functions of forests, in the preservation of forest habitat, in protecting the natural renewal of the main species without additional expenses, in securing the even and continual use of forest with increasing its
general productivity to 11-14 %, by comparision with the systemless method of the occupation of felling areas at the industry-selective fellings.
Abies forests, growing in the upper mountain zone on considerable areas (320 thousand ha) of West-Caucasus, fulfil important paramount soil- and water-protective, climate-regulating functions and those of convalescence, etc. They are the keepers of unique genofond of plants, rare nature complexes, and the source of valuable species for introduction. The receipt of wood must have a subordinated character in these forests.
In the present work the decision of the tasks to preserve the richest flora’s variety of Abies forests underlies the principle of the protection of genofond and cenofond of relic, endemic, rare, disappearing plant species and standard assotiations. In the flora of Abies forests of Caucasus 6 species of arborescent plants from 62 ones (10 %) are taken into the rare, disappearing category and entered in the Red Book of the USSR. It has been ascertained, that there are 13 further rare and disappearing plant species and they must be preserved and taken into the regional Red Book, too.
In the conclusion of the work you can find a number of recommandations of several practical steps for the preservation of genofond of valuable plant species and nature complexes.
The majestic phenomenon of nature - the virgin Abies forests, regrettably, may disappear off the face of the earth forever and in a very short time you can observe and study the remainders of present spontaneous Abies forests in several national parks and special reserves only. Therefore it is necessary to study the indigenous forests everywhere where they still exist.
236
СОДЕРЖАНИЕ
ПРЕДИСЛОВИЕ.............................................. 3
ВВЕДЕНИЕ................................................ 7
ГЕНЕЗИС ПИХТОВЫХ ЛЕСОВ И ИХ РАСПРОСТРАНЕНИЕ НА КАВКАЗЕ 13
Происхождение и систематика рода пихты............. 13
Кавказ как рефугиум реликтовой дендрофлоры и позиция пихты в ней.............................. 23
ПРИРОДНЫЕ УСЛОВИЯ И ЗАКОНОМЕРНОСТИ СОВРЕМЕННОГО РАСПРОСТРАНЕНИЯ ПИХТОВЫХ ЛЕСОВ КАВКАЗА.................. 33
Природные условия произрастания пихтовых лесов.............................................. 33
Рельеф и геологическое строение.................... 33
Климат............................................. 36
Почвы.............................................. 37
Основные закономерности широтного и высотного распространения пихтовых лесов..................... 42
Видовой состав пихтовых лесов и взаимоотношение пихты с сопутствующими породами.................... 46
Лесорастительное районирование области распространения темнохвойных лесов................. 50
ОСНОВНЫЕ ЧЕРТЫ МОРФОЛОГИИ, БИОЛОГИИ, ЭКОЛОГИИ ПИХТЫ КАВКАЗСКОЙ И ЕЕ ЛЕСОВОДСТВЕННЫЕ ОСОБЕННОСТИ............. 56
Морфология, биология и экология пихты.............. 56
Некоторые особенности семяношения пихты кавказской. 63
Естественное возобновление пихты под пологом леса.. 70
Возобновление пихта в естественных окнах........... 79
Лесоводственная классификация подроста пихты....... 87
Шкала оценки естественного возобновления в пихтарниках... 94
237 ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ СТРОЕНИЯ И ВОЗРАСТНОЙ СТРУКТУРЫ ПИХТОВЫХ ЛЕСОВ................................. 99
Классификация лесов по их происхождению и влиянию экзогенных факторов....................... 100
Строение и возрастная структура пихтовых лесов.... 104
Онтогенез пихты кавказской и возрастная динамика древостоев............................... 115
Ярусное строение пихтовых древостоев.............. 131
Дифференциация деревьев в лесу и их классификация... 134
Закономерности дифференциации деревьев в разновозрастном лесу............................ 137
Классификация деревьев по их жизненному состоянию... 138
Классификация древостоев по их жизненному состоянию. 148
ТИПЫ ПИХТОВЫХ ЛЕСОВ И ИХ ЭКОЛОГО-ГЕНЕТИЧЕСКАЯ КЛАССИФИКАЦИЯ........................................... 152
Типологическая классификация пихтовых лесов....... 152
Типы пихтовых лесов............................... 159
СИСТЕМА МЕРОПРИЯТИЙ ПО ОПТИМИЗАЦИИ ИСПОЛЬЗОВАНИЯ ПИХТАРНИКОВ НА ЛЕСОТИПОЛОГИЧЕСКОЙ ОСНОВЕ И ОХРАНЕ...... 186
Хозяйственные группы типов леса и система мероприятий в них...............'................. 186
Способы рубок в пихтарниках....................... 189
Опыт рационального способа восстановления изреженных пихтовых лесов.................................... 192
Созологическая характеристика пихтовых лесов...... 194
Видовой и фитоценотический принцип охраны реликтовых пихтарников Кавказа............................... 197
ЗАКЛЮЧЕНИЕ.............................................. 205
ЛИТЕРАТУРА.............................................. 211
SUMMARY................................................. 230
Рис. 1. Типично разновозрастные пихтарники
Рис. 2. Пихтарники трахистемоновые
Рис. 3. Дифференциация деревьев и самоизреживание древостоя пихты кавказской
Рис.4. Высокопроизводительный древостой пихты кавказской
Рис.5. Взятие пробы древесины на дрань часто вызывает стволовую гниль
Рис. 6. Шишконошение пихты кавказской
. IV
Рис. 7. Двухвершинность пихты кавказской
Рис. 8. Пихтарники, пройденные опытными добровольно-выборочными рубками
Рис. 9. Окно группово-выборочных рубок в буково-пихтовом лесу
Рис. 10. Подрост пихты в окне группово-выборочных рубок через три года
Рис. 11. Разновозрастный древостой пихты сахалинской с напочвенным покровом сазы курильской на о. Кунашире
Монография
Сергей Михайлович Бебия
ПИХТОВЫЕ ЛЕСА КАВКАЗА
Редактор Благодатова Н.Д.
Оригинал-макет подготовил Бебия С.М.
Верстка Сырейщиков Д.Г.
Книга издана в рамках Проекта Глобального экологического фонда "Сохранение биоразнообразия Российской Федерации"
Дизайн обложки Желт Дебреци и Ли Айли
Фото автора
Дизайн цветных иллюстрации Имре Ретезар, черно-белых иллюстраций Сырейщиков Д.Г.
Лицензия ЛР № 020718 от 02.02.1998 г.
Лицензия ПД № 00326 от 14.02.2000 г.
Подписано к печати 30.01.2002 г.
Бумага 80 г/м2 "Снегурочка"
Объем 17,0 п.л.Тираж 500 экз.
Формат 60x88/16
Ризография Заказ № 59
Издательство Московского государственного университета леса. 141005. Мытищи-5, Московская обл., 1-я Институтская, 1, МГУЛ. Телефон: 588-54-15, 588-57-62, 588-53-48. Факс: 588-51-09 E-mail: izdat@mgul.ac.ru