/



























































































































































































































































































































































Автор: Марусик Ю.М. Ковблюк Н.М.
Теги: articulata (членистые) систематика животных специальные зоологические науки энтомология арахнология паукообразные
ISBN: 978-5-87317-772-1
Год: 2011
Похожие




Текст
РОССИЙСКАЯ АКАДЕМИЯ НАУК
Институт проблем экологии и эволюции им. А.Н. Северцова
Программа фундаментальных исследований ОБН РАН
«Биологические ресурсы России: оценка состояния
и фундаментальные основы мониторинга»
Институт биологических проблем Таврический национальный
Севера ДВО РАН универститет им. В.И. Вернадского
Ю.М. Марусик, Н.М. Ковблюк
Пауки (Arachnida, Aranei)
Сибири и Дальнего Востока России
Ответственный редактор
Член-корреспондент РАН, профессор Б.Р. Стриганова
Товарищество научных изданий КМК
Москва ♦> 2011
УДК 595.44(591.91)
ББК 28.691.82.4(25)
М29
Марусик Ю.М, Ковблюк Н.М. Пауки (Arachnida, Aranei) Сибири и
Дальнего Востока России. Москва: Товарищество научных изданий КМК.
2011.344 с.
Книга посвящена паукам, одному из самых крупных отрядов животных,
населяющих Россию к востоку от Урала. Это первое за 40 лет руководство по
изучению пауков на русском языке и первый определитель семейств пауков,
населяющих азиатскую часть России. Книга включает главы об истории изучения ара-
неофауны региона, методическую часть (методы сбора, хранения, изучения,
рисования), оригинальные политомические, дихотомические и графические
ключи для определения семейств, детальные обзоры всех 38 семейств пауков,
встречающихся в азиатской части России (морфология, перечень надвидовых
таксонов, зарегистрированных в регионе, биология, таксономия и др.),
этимологию родовых названий, словарь терминов и другие разделы. Кроме того, в
книгу включены обзоры ещё трёх семейств, нахождение которых в азиатской
части России очень вероятно, а также сведения о трёх семействах пауков,
известных из этого региона в ископаемом состоянии. Книга богато иллюстрирована
и содержит более 600 цветных и чёрно-белых иллюстраций.
Книга предназначена в первую очередь для арахнологов, но также и для
энтомологов, зоологов, экологов, специалистов в области охраны окружающей
среды, преподавателей и студентов высших учебных заведений.
На обложке: Alopecosa tanakai, фото М.М. Омелько.
ISBN 978-5-87317-772-1
© Марусик Ю.М., Ковблюк Н.М., текст,
иллюстрации, 2011
© Институт проблем экологии и эволюции
им. А.Н. Северцова РАН, 2011
© Товарищество научных изданий КМК,
издание, 2011
RUSSIAN ACADEMY OF SCIENCES
A.N. Severtsov Institute for Ecology and Evolution
Program of Fundamental Researches
«Biological Resources of Russia: State Estimation
and Fundamental Backgrounds of Monitoring»
Institute for Biological Problems V.I. Vernadsky Tavrida
of the North, FEB RAS National University
Yu.M. Marusik, M.M. Kovblyuk
Spiders (Arachnida, Aranei)
of Siberia and Russian Far East
Responsible Editor
Corresponding member of the Russian Academy of Sciences,
Professor B.R. Striganova
KM К Scientific Press Ltd.
Moscow ♦ 2011
UDC 595.44(591.91)
Marusik Yu.M., Kovblyuk M.M. Spiders (Arachnida, Aranei) of Siberia and
Russian Far East. Moscow: KMK Scientific Press Ltd. 2011. 344 p.
This book is focused on spiders occurring in Russia east of Ural Mountains. It is first
guide to study spiders in Russian written after 1971 and first identification key for
spider families occurring in the Asian part of Russia. Book contains chapters dealing
with history of studies, methods (collecting, preservation and storing, examination,
illustrating), key to families (pictorial, dichotomous and multi-entry), surveys of 38
families occurring in the Asian part of Russia (morphology, list of genera reported
from the region and their distribution, natural history, taxonomy, etc), etymology of
generic names, glossary, etc. Additional information is provided about three families
that can be found in Siberia or Far East Russia and about three families known by
fossils. Book contains over 600 colour and black-and-white illustrations.
The work will be of special interest to academics, professionals and educated
layman. It will be a useful hand-book for arachnologists, field workers and others
involved in spider-related ecological studies and conservation.
ISBN 978-5-87317-772-1
© Marusik Yu.M., Kovblyuk M.M., text,
illustrations, 2011
© A.N. Severtsov Institute for Ecology and
Evoluion,RAS,2011
© KMK Scientific Press Ltd., 2011
Посвящается Кириллу Юрьевичу Есъкову,
зачинателю планомерных исследований
пауков Сибири и Дальнего Востока
Содержание
Предисловие редактора 10
1. ВВЕДЕНИЕ 11
Цели книги 11
География 12
История изучения пауков России 13
История изучения пауков азиатской части России 15
2. МЕТОДЫ ИЗУЧЕНИЯ ПАУКОВ 19
Методы сбора 19
Минимальный набор снаряжения 19
Как узнать взрослый ли паук? 29
Как рассматривать пауков? 32
Какую использовать литературу? 44
Как определять собранных пауков? 45
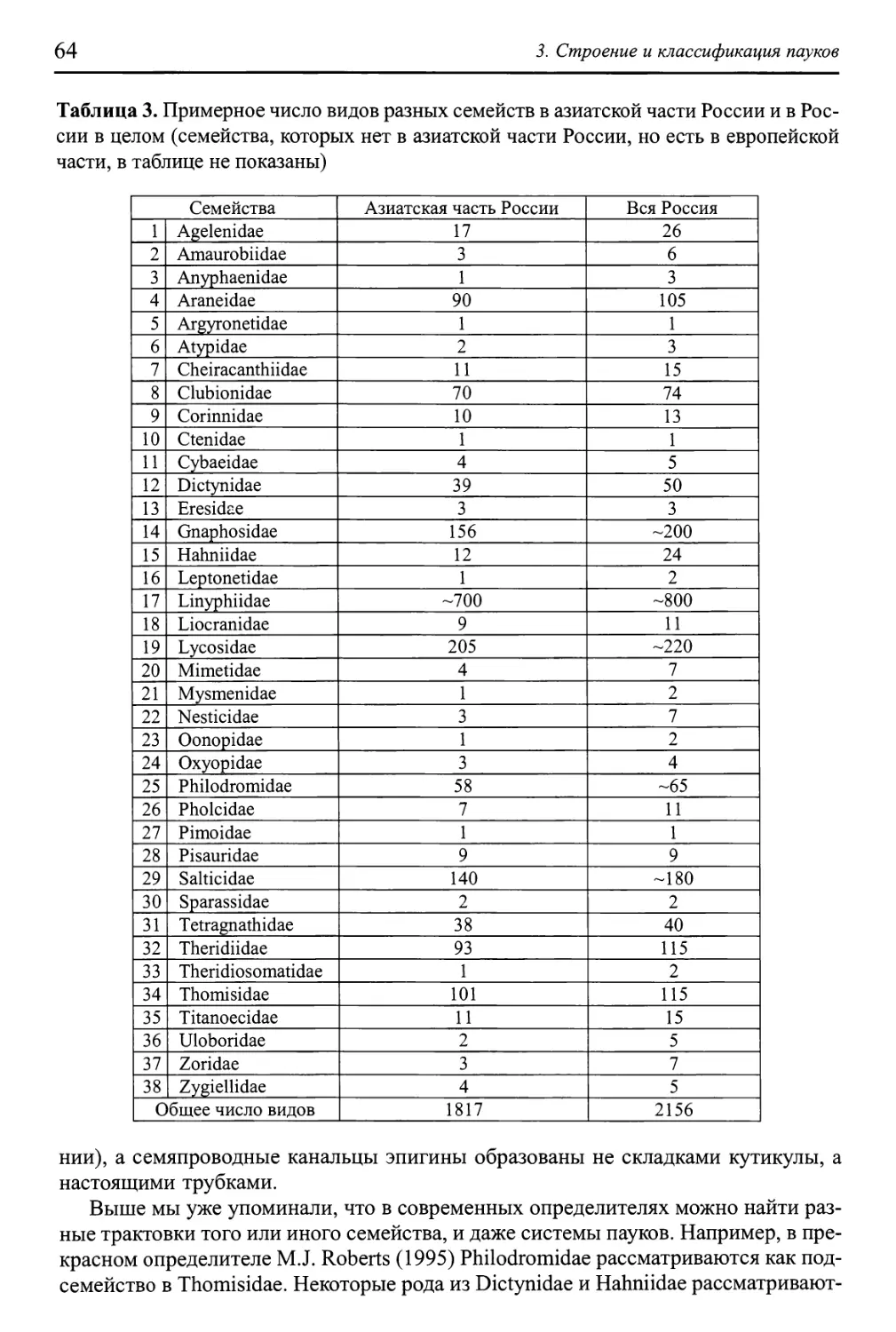
3. СТРОЕНИЕ И КЛАССИФИКАЦИЯ ПАУКОВ 47
Морфология и терминология 47
Функциональная морфология 57
Классификация пауков 60
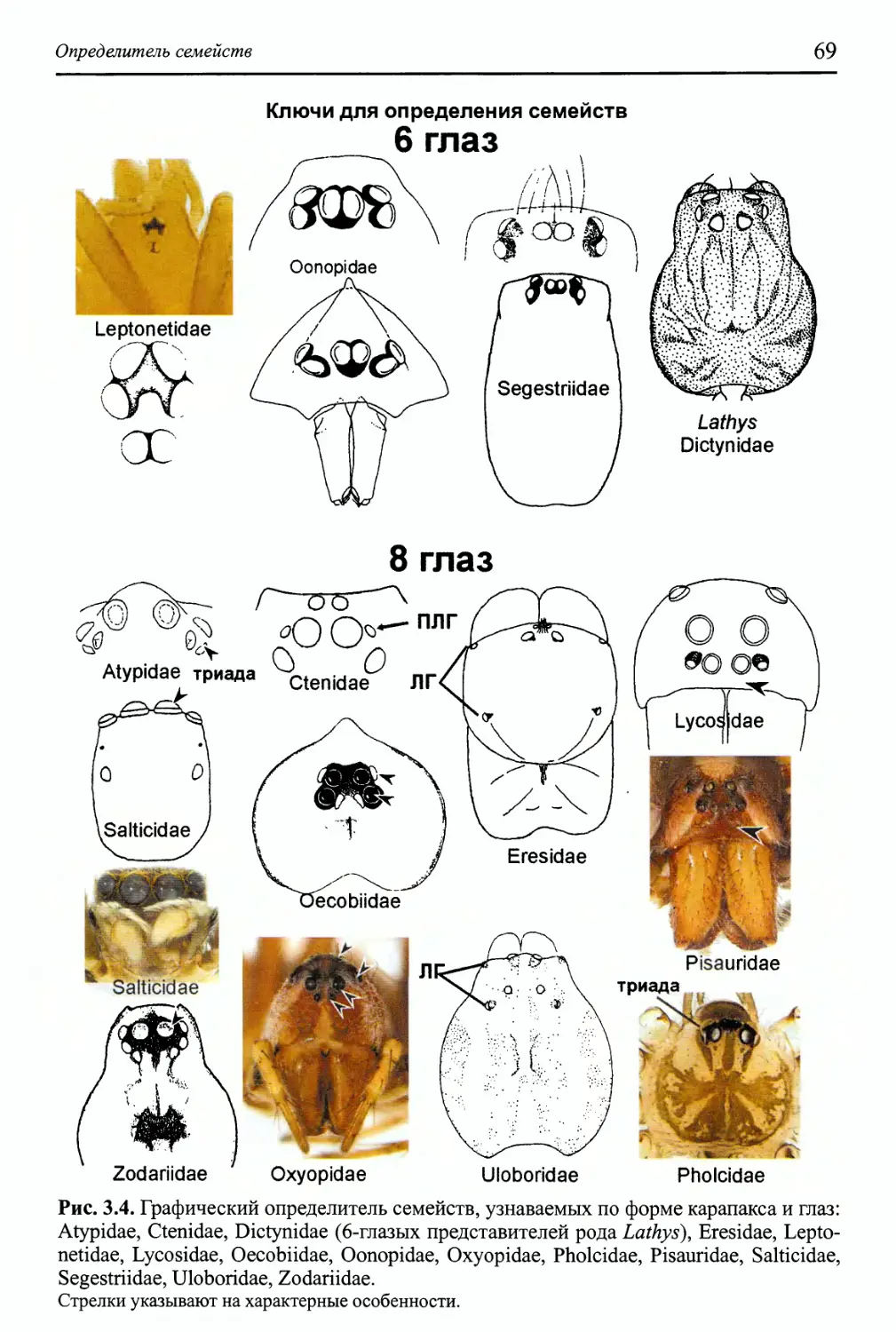
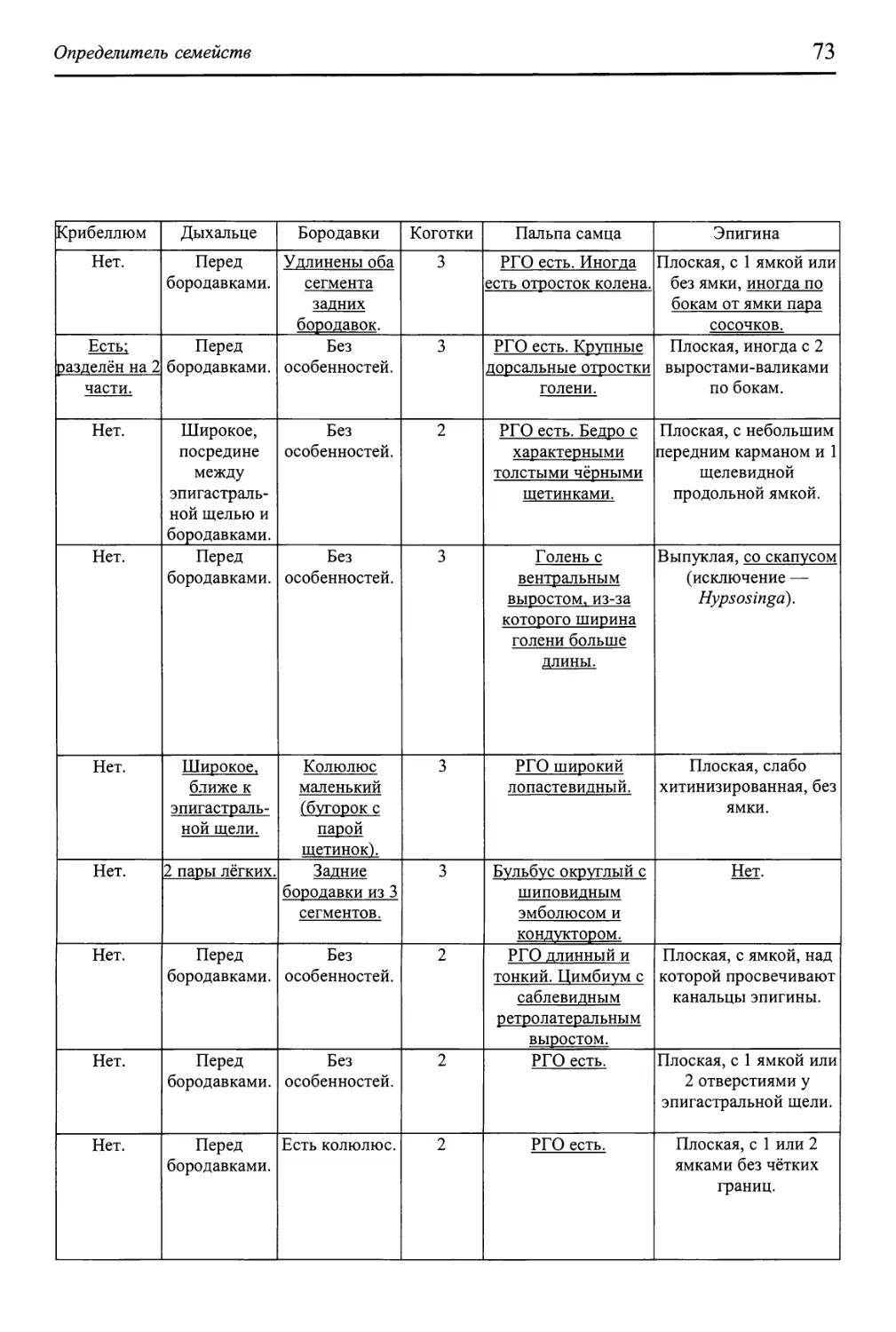
Определитель семейств 65
Как пользоваться определительными таблицами 67
Схема всего определителя 68
Ключи для определения семейств 69
4. СПЕЦИАЛЬНАЯ ЧАСТЬ 97
Схема описания семейств 97
Семейства, встречающиеся в азиатской части России 99
Семейство Agelenidae 100
Семейство Amaurobiidae 106
Семейство Anyphaenidae ПО
Семейство Araneidae 112
Семейство Argyronetidae 120
Семейство Atypidae 122
Семейство Cheiracanthiidae 126
Семейство Clubionidae 130
Семейство Corinnidae 136
Семейство Ctenidae 140
Семейство Cybaeidae 142
Семейство Dictynidae 146
Семейство Eresidae 150
Семейство Gnaphosidae 154
Семейство Hahniidae 160
Семейство Leptonetidae 164
Семейство Linyphiidae 168
Семейство Liocranidae 178
Семейство Lycosidae 182
Семейство Mimetidae 190
Семейство Mysmenidae 194
Семейство Nesticidae 198
Семейство Oonopidae 202
Семейство Oxyopidae 204
Семейство Philodromidae 206
Семейство Pholcidae 212
Семейство Pimoidae 216
Семейство Pisauridae 218
Семейство Salticidae 222
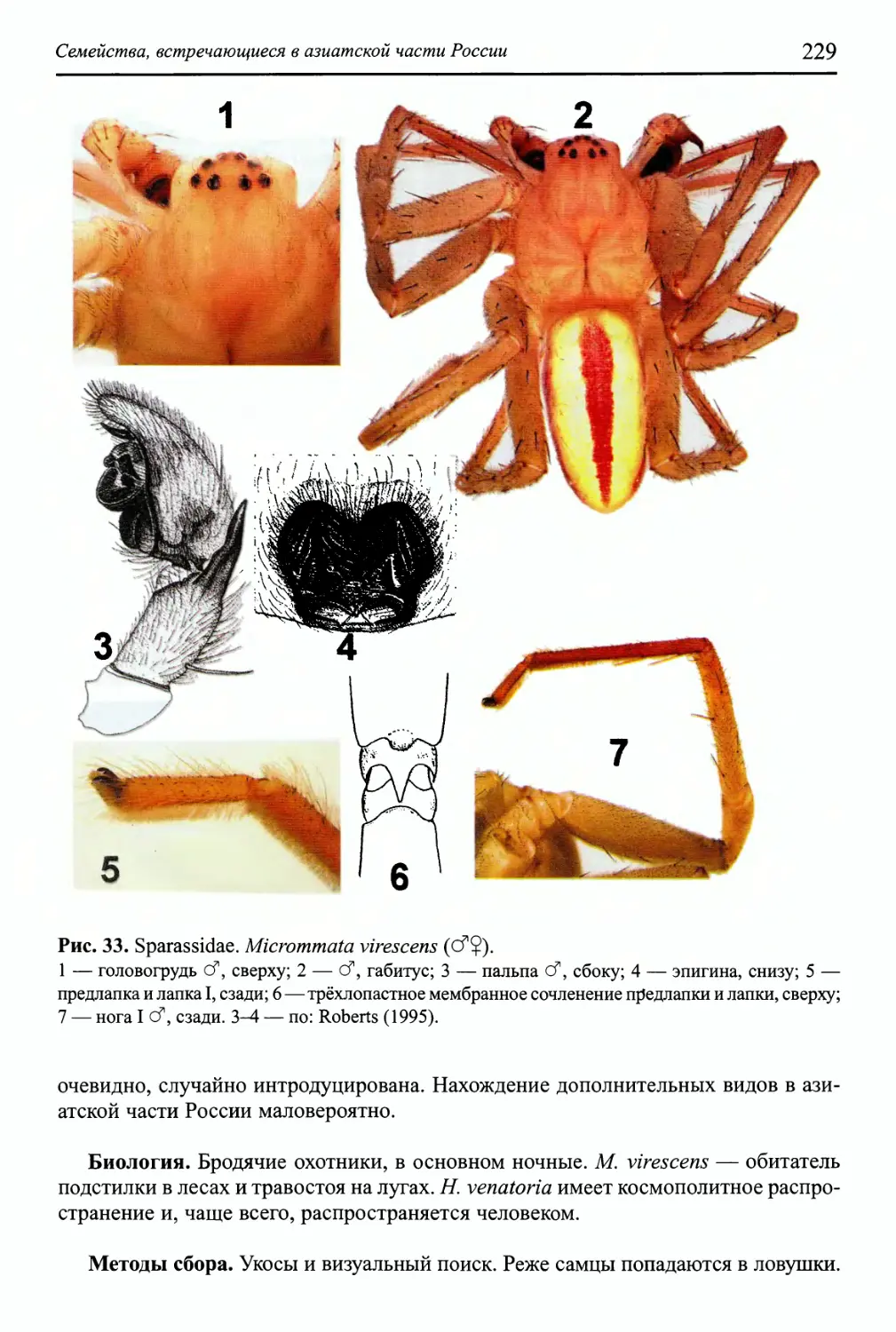
Семейство Sparassidae 228
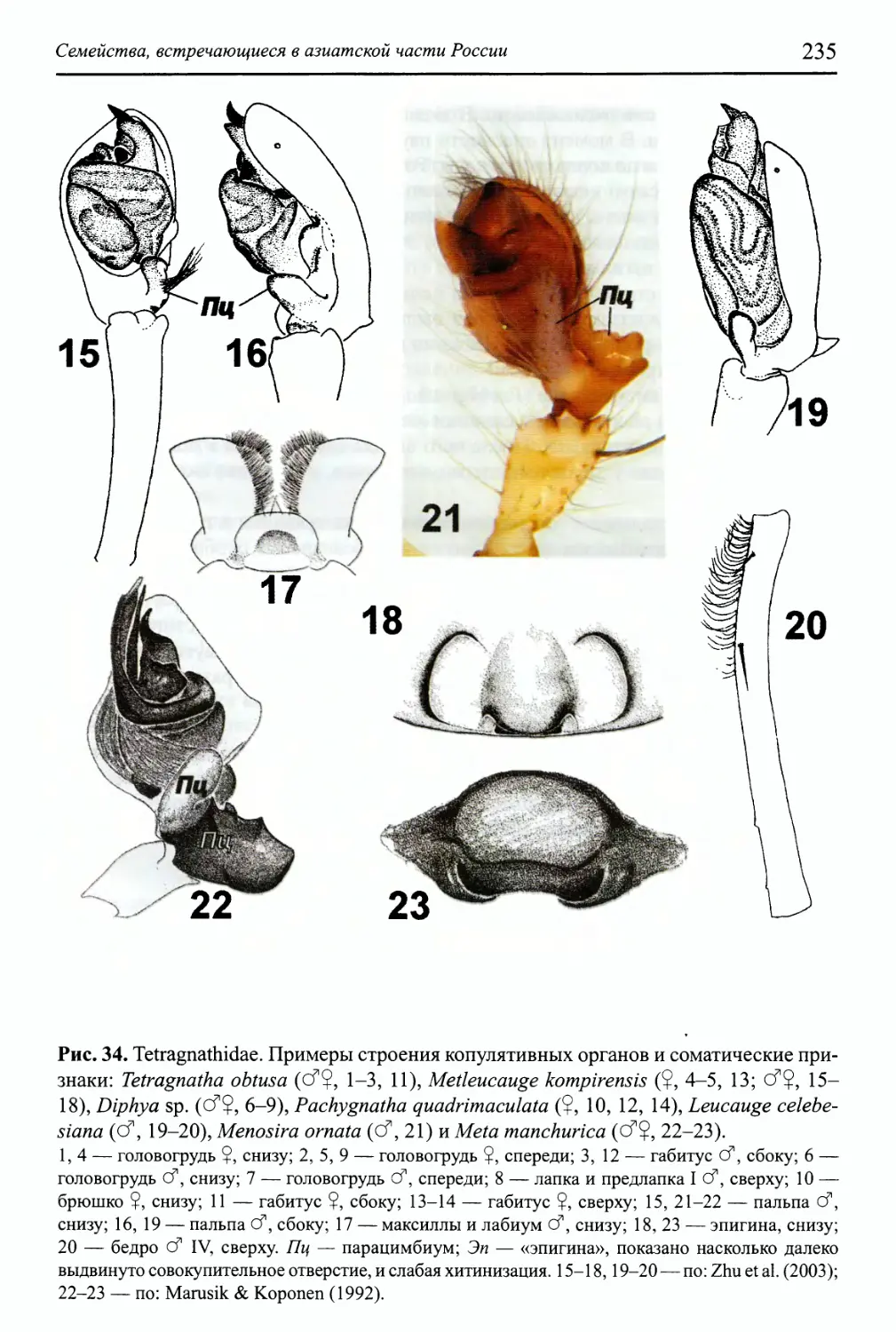
Семейство Tetragnathidae 232
Семейство Theridiidae 238
Семейство Theridiosomatidae 246
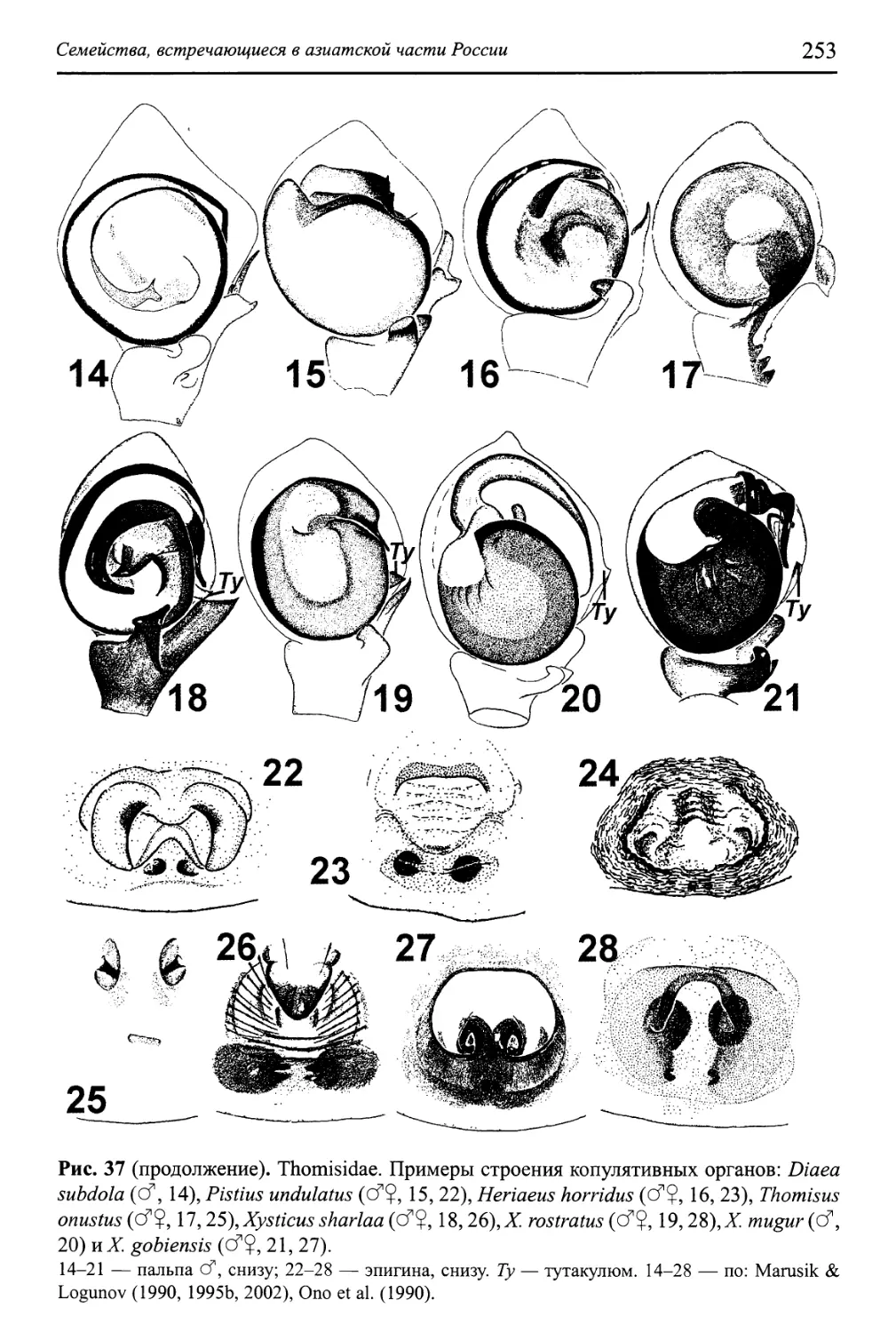
Семейство Thomisidae 250
Семейство Titanoecidae 256
Семейство Uloboridae 260
Семейство Zoridae 264
Семейство Zygiellidae 268
Семейства, нахождение которых возможно в азиатской части России 271
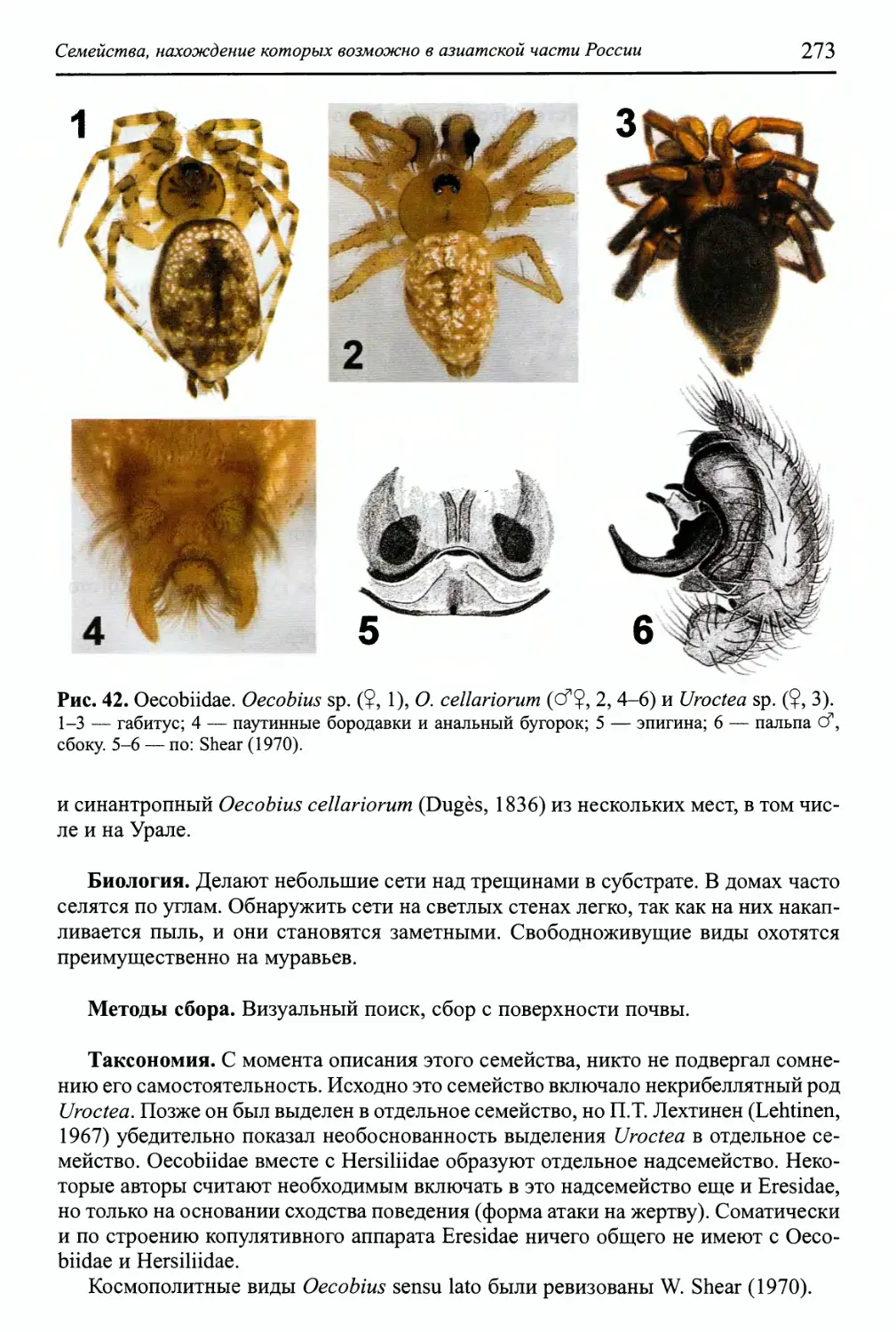
Семейство Oecobiidae 272
Семейство Segestriidae 274
Семейство Zodariidae 276
Семейства, известные из азиатской части России в ископаемом состоянии....278
Благодарности 280
ЛИТЕРАТУРА 281
ПРИЛОЖЕНИЯ 307
Приложение А. Этимология латинских названий пауков 307
Приложение Б. Список русскоязычных арахнологов-систематиков 323
Приложение В. Интернет-ресурсы, полезные для определения пауков 324
Приложение Г. Основные (настольные) книги, необходимые для изучения
и определения пауков 325
Приложение Д. Словарь 326
Указатель латинских названий 335
Content
Editor's Preface 10
1. INTRODUCTION 11
Goals 11
Geographic coverage 12
History of the study of spiders in Russia 13
History of the study of spiders in the Asian part of Russia 15
2. METHODS 19
Collecting methods 19
A minimal collecting kit 19
How to see that a specimen is adult? 29
How to observe spiders? 32
Literature to be used 44
Identifying spiders 45
3. MORPHOLGY AND CLASSIFICATION OF SPIDERS 47
Morphology and terminology 47
Functional morphology 57
Classification of spiders 60
Identification of the families 65
How to use the keys? 67
Key to the families 69
4. SPECIAL PART 97
Scheme of family description 97
Families occurring in the Asian part of Russia 99
Agelenidae 100
Amaurobiidae 106
Anyphaenidae 110
Araneidae 112
Argyronetidae 120
Atypidae 122
Cheiracanthiidae 126
Clubionidae 130
Corinnidae 136
Ctenidae 140
Cybaeidae 142
Dictynidae 146
Eresidae 150
Gnaphosidae 154
Hahniidae 160
Leptonetidae 164
Linyphiidae 168
Liocranidae 178
Lycosidae 182
Mimetidae 190
Mysmenidae 194
Nesticidae 198
Oonopidae 202
Oxyopidae 204
Philodromidae 206
Pholcidae 212
Pimoidae 216
Pisauridae 218
Salticidae 222
Sparassidae 228
Tetragnathidae 232
Theridiidae 238
Theridiosomatidae 246
Thomisidae 250
Titanoecidae 256
Uloboridae 260
Zoridae 264
Zygiellidae 268
Families those are likely to occur in the Asian part of Russia 271
Oecobiidae 272
Segestriidae 274
Zodariidae 276
Fossil families known from the Asian part of Russia 278
Acknowledgements 280
REFERENCES 281
APPENDICES 307
A. Etymology of spider names 307
Б. List of Russian speaking arachnologists 323
B. Useful internet resources 324
Г. Selected books for study and identification of spiders 325
Д. Glossary 326
Index of Latin names 335
Предисловие редактора
Пауки — многочисленная группа обитателей почвы, напочвенного и
растительного ярусов. В сообществах наземных беспозвоночных они играют существенную
роль в пищевых сетях и в определённых условиях могут контролировать обилие
других групп. Однако изучение экологии пауков и оценка их экосистемных
функций тормозится до сих пор недостатком литературы по систематике и фауне этой
группы. Настоящая книга представляет первый определитель семейств пауков
азиатской части России — Сибири и Дальнего Востока. В работе приведён детальный
обзор морфологии, биологии, ландшафтного распространения всех 38 семейств,
встречающихся на исследованной территории. Для каждого семейства пауков
составлен полный список родов с указанием их распространения. Определитель
семейств включает традиционные дихотомические таблицы, а также оригинальные
политомические ключи. Текст снабжён многочисленными иллюстрациями,
помогающими идентифицировать материал.
В книге имеется обширная вводная часть, в которой приводится история
изучения пауков в России и в Сибири, с детальной библиографией, а также география
фаунистических и таксономических исследований в азиатской России. Большую
практическую ценность для полевых работников представляет методический
раздел с подробной характеристикой методов сбора и учёта пауков разных семейств и
анализом морфологических признаков, позволяющих различать отдельные виды.
За последние 20-30 лет в азиатской России проводились активные работы по
изучению фауны пауков. Накопленный материал позволил составить данное
руководство, которое безусловно найдёт пользователей среди широкого круга арахно-
логов, энтомологов, экологов и почвенных зоологов.
Б.Р. Стриганова
1. Введение
Пауки (Aranei) — шестой по величине отряд из более чем 340 отрядов
животных. По числу видов (более 42 000 современных и ископаемых) пауки уступают
лишь четырём отрядам насекомых, а именно жукам (Coleoptera), двукрылым
(Diptera), перепончатокрылым (Hymenoptera) и чешуекрылым (Lepidoptera), а
также клещам (Acari). Обитают пауки повсеместно, вплоть до полярных пустынь, во
всех наземных биотопах и даже в пресноводных водоёмах и зонах приливов.
Пауки обладают многими качествами, существенно отличающими их от других
членистоногих. Все они облигатные хищники. Большинство пауков не
специализируются на каких-либо жертвах, и соответственно в своём распространении они
не зависят ни от кормовых растений, как фитофаги, ни от животных-хозяев, как
паразиты, ни от химического состава подстилки, как сапрофаги. Другая
существенная особенность пауков, это способность к пассивному расселению на паутинках
на огромные расстояния. Ещё одна особенность заключается в том, что пауки
имеют паутинные бородавки и могут производить паутину. Хотя многие из пауков и не
делают сетей, но все без исключения виды используют паутину для заботы о
потомстве (плетут яйцевые коконы) и для передвижения (тянут за собой
«путеводную» паутинку). Одна из главных особенностей пауков, ещё недостаточно
оцененная биологами-эволюционистами, это их копулятивный аппарат. Во всех группах
насекомых копулятивные органы и самцов и самок являются производными одних
и тех же сегментов брюшка. У пауков же совокупительные органы находятся на
разных тагмах, у самок — на брюшке, а у самцов — на педипальпах.
Копулятивный аппарат очень сложно устроен и является видоспецифичным;
совокупительные органы самцов и самок в большинстве случаев подходят друг к другу, как ключ
к замку. В связи с этим, у пауков гораздо проще (по сравнению со многими
другими группами животных) судить о том, что такое отдельный вид.
Цели книги
Идею подготовить эту книгу вызвали сетования коллег-энтомологов по поводу
того, что нет ни одного определителя пауков, даже до уровня семейств, ни по
Сибири, ни по Дальнему Востоку России. Форму изложения подсказала книга по
паукам Африки (Dippenaar-Schoeman & Jocque, 1997).
Почему выбрана азиатская часть России, а не вся страна? Дело в том, что
семейства пауков европейской части России можно определить, используя традиционный,
хотя и устаревший, «Определитель пауков европейской части СССР» В.П. Тыщенко
(1971), а также ряд англо- и немецкоязычных книг и интернет-публикаций.
Определение же некоторых дальневосточных семейств по европейским руководствам
невозможно, а ряд сибирских таксонов имеет специфичные черты. Включение всех
семейств России затруднило бы и написание ключей, и само определение. Другая
причина связана с тем, что в течение последних 25 лет пауков Сибири и Дальнего
12
L Введение
Востока России активно исследовали. В результате опубликован ряд монографий
(каталоги, описания региональных фаун, частные ревизии) и огромное количество
статей, которые до сих пор не сведены воедино. Существует лишь ряд
библиографических указателей, охватывающих литературу до 2000 г., а также общий каталог
пауков территорий бывшего СССР, где указания даны по очень широким
физико-географическим выделам и без привязки к литературе (Mikhailov, 1997,1998,1999,2000).
Основные труды первого автора книги (Ю.М.) также посвящены паукам именно
азиатской части России, да и работает он именно в этом регионе (Магадан).
При написании книги мы поставили перед собой следующие цели:
• сделать иллюстрированный определитель семейств региона;
• дать краткую морфологическую характеристику каждого из семейств;
• привести список родов, встречающихся за Уралом;
• указать, где обитают рода, известные в Сибири и на Дальнем Востоке;
• дать краткий очерк по биологии пауков каждого семейства;
• рассказать, какими методами можно собрать пауков конкретного семейства;
• привести краткие сведения об изученности каждого из семейств в мире,
России и за Уралом; привести перечень всех, либо основных, таксономических
публикаций;
• кратко указать, какие признаки используют для разграничения видов;
• очертить перспективы изучения пауков каждого семейства за Уралом.
Помимо главных целей, мы попытаемся вкратце изложить историю изучения
региона, описать методы сбора, наиболее перспективные регионы и биотопы для
коллектирования пауков, и то, как лучше рассматривать и хранить пауков.
Необходимость дополнительных вводных разделов вызвана тем, что на русском языке
никогда не выходили работы с подробным описанием того, как следует собирать
пауков. В определителе В.П. Тыщенко (1971) кратко даны самые общие
рекомендации без учёта современных возможностей. Статей, посвященных тому, как
разглядывать пауков (как рассматривать их в бинокуляр, препарировать), на русском
языке нет вовсе.
Чтобы облегчить чтение текста, все специальные термины мы вынесли в
словарик. Большинство специальных терминов снабжены английскими эквивалентами.
География
Как было сказано выше, эта книга охватывает группы пауков, которые обитают
в России к востоку от Урала (рис. 1.1). Для удобства мы называем этот регион либо
Зауральем, либо азиатской частью России. Согласно административному
(экономико-географическому) делению, этот регион состоит из Сибири (Западной,
Средней и Восточной) и Дальнего Востока.
Если делить Зауралье исходя из биогеографических или арахнологических
критериев, то чётко выделяются две области — Сибирь и юг Дальнего Востока
(регион от Амурского бассейна и южнее). Последний регион иногда называют Палеар-
хеарктикой (в него входит и прилежащий Китай, Корея, Япония), или Манчжурией
(относится к северной части Палеархеарктики). В тексте мы везде используем
термин юг Дальнего Востока.
13
Рис. 1.1. Азиатская часть России (мелкий пунктир) и наиболее изученные её регионы.
Число над чертой показывает сколько примерно видов известно сейчас, а под чертой —
сколько было известно к 1980 г.
По: Марусик (2007) с модификациями.
В пределах Сибирской части Зауралья достаточно хорошо обособлена лишь
Западная Сибирь (до Енисея) и Северо-Восток (регион к востоку от Верхоянского
хребта). Есть некоторые особенности у фаун Средней Сибири (Красноярский Край),
Якутии и южной Сибири. Эти регионы в значительной степени отличаются друг от
друга наличием или отсутствием тех или иных ландшафтов. Так в Средней
Сибири нет степных участков, в то время как на Северо-Востоке степные или степепо-
добные биотопы встречаются вплоть до 71° с.ш. Соответственно, в Туве, в Якутии
и на Чукотке есть виды (и целые группы), которые отсутствуют в Красноярском
крае, соседствующем с Тувой.
Своеобразна фауна пауков южной части Сахалина, Южных Курил и Монерона.
На этих островах есть немало видов, и даже некоторые рода, которые не
встречаются в материковой части Азии, но есть в Японии.
История изучения пауков России
Ещё три десятилетия назад пауки, как в мире, так и на территории СССР, и
России в частности, были изучены чрезвычайно слабо. К концу 70-х годов
двадцатого столетия не было мировых и региональных каталогов, существовало
ограниченное число определителей, охватывавших небольшие территории (Европейскую
часть СССР, Великобританию, Германию, Финляндию, Новую Англию (северо-
14
1. Введение
восток США)). На тот момент фауна пауков России была изучена очень слабо.
Единственный каталог был подготовлен Д.Е. Харитоновым ещё в 20-е годы прошлого
столетия и опубликован в 1932 г. К началу 80-х годов на обширной территории
России, лежащей к востоку от Уральских гор, было известно не более 500 видов
пауков - это меньше, чем было тогда известно в небольших странах -
Великобритании, Польше или Финляндии (Марусик, 2007). Помимо того, что фауна Сибири
была изучена чрезвычайно слабо, была и другая беда — очень слабо
разработанная систематика пауков, как на видовом уровне, так и на надвидовом — и во всём
мире, и у нас в частности.
Ситуация начала выправляться благодаря серии каталогов всех ныне живущих
видов пауков мира, опубликованных P.M. Brignoli (1983) и, после его смерти, N.I.
Platnick (1989, 1993). Новые таксономические сведения накапливались так
быстро, что вместо каталогов на бумажных носителях пришлось перейти на
электронный каталог, обновляемый несколько раз в год (Platnick, 2000-2010). Если в
десятилетие, предшествующее выходу каталога P.M. Brignoli, было описано 2884
новых вида, то за последующее — почти пять тысяч (4988)!
В Советском Союзе арахнологические исследования резко усилились с
выходом в свет определителя пауков европейской части СССР, подготовленного В.П. Ты-
щенко (1971). Определитель В.П. Тыщенко был первым руководством на русском
языке, благодаря которому стало возможно изучать фауну и систематику пауков.
Выход определителя закономерно способствовал появлению новых
исследователей-систематиков. Это были Е.М. Андреева, П.М. Дунин, К.Ю. Еськов, С.Л. Зонш-
тейн, А.А. Зюзин, В.И. Овчаренко, СВ. Овчинников, Н.М. Пахоруков, А.В.
Пономарёв, Л.Г. Савельева, А.В. Танасевич, Ч.К. Тарабаев, В.Я. Фет, М.Т. Штернберге
и ряд других, более молодых.
Нужно отметить, что определитель В.П. Тыщенко (1971) был не первым в
истории определителем пауков на русском языке. Первенство принадлежит С.А.
Спасскому, опубликовавшему ещё в 1925 г. «Определитель пауков Донской области»,
включающий лишь 159 видов. Эта работа и поныне важна для изучения пауков
юга России, поскольку рисунки отдельных видов до сих пор есть только здесь и
нигде более (например, Alopecosa cronebergi (Thorell, 1875)). Но для определения
пауков встречающихся в азиатской части России, этот определитель, естественно,
мало полезен. Второй попыткой создания русскоязычного определителя стала
книга Н.С. Ажегановой (1968) «Краткий определитель пауков лесной и
лесостепной зоны СССР». Но этот определитель содержал ещё меньше видов, чем работа
С.А. Спасского. Определитель В.П. Тыщенко значительно полнее, он включает 928
видов. Именно поэтому он смог стимулировать арахнологические исследования в
Советском Союзе, в том числе и в азиатской части России.
Кроме каталогов и определителей, были подготовлены сводки, обобщавшие всю
разнообразную информацию об этих животных. Единственная, но очень
качественная сводка такого рода на русском языке была подготовлена нашим выдающимся
зоологом А.В. Ивановым (1965). За рубежом было издано немало подобных
руководств, например, книги J.H. Comstock (1913), W.S. Bristowe (1939, 1941, 1958) и др.
Первым, кто начал активно изучать пауков Сибири и Дальнего Востока России,
был К.Ю. Еськов. Первая его работа по азиатской части России была
опубликована в 1979 г., а затем стало выходить по 2-3 и более работ ежегодно. Благодаря его
исследованиям и исследованиям примкнувших к нему коллег, за Уралом стало из-
15
вестно более 1800 видов вместо 500 ранее известных. За четверть столетия было
описано не менее 500 новых видов (Марусик, 2007).
В последние годы интенсивность исследований уменьшилась, и серьезно
снизился приток новых исследователей. Это связано не в последнюю очередь с тем, что
за более чем тридцать лет после появления определителя В.П. Тыщенко не вышло
ни одного определителя или справочника на русском языке, который помог бы
интересующимся пауками начать их изучение. Недостаточность определителя В.П.
Тыщенко на нынешнем этапе исследований связана с разными моментами. С одной
стороны, представления о надвидовой систематике резко изменились, число
выделяемых семейств в мировой фауне возросло с 1971 г. почти в два раза и составляет
сейчас порядка 110 (не считая полностью вымерших). Многие семейства,
фигурировавшие в определителе В.П. Тыщенко, были раздроблены, некоторые
подсемейства переброшены в другие семейства. К тому же, в европейской части отсутствует
ряд семейств, обитающих в Приморье. Поэтому определение пауков даже до
семейства представляет для начинающего арахнолога серьезную проблему.
Другая сложность для начинающих, и не только для них, заключается в том, что
огромное число публикаций по паукам Зауралья практически никак не обобщено.
Имеется всего лишь один каталог (Mikhailov, 1997), в котором приводится голый
перечень названий видов и их распространение по крупным
физико-географическим регионам. Кроме того, со времени выхода каталога К.Г. Михайлова, число
видов в регионе возросло на несколько сотен, и были обнаружены дополнительные
семейства.
История изучения пауков азиатской части России
Пионерной работой по паукам азиатской части России, была работа арахнолога
А.Э. Грубе (Grube, 1861) (см. Марусик, 2010). По материалам экспедиции на
Дальний Восток в пятидесятые годы XIX века он описал 35 новых видов пауков из
Якутии и Приамурья. Эта работа стала первой не только для России, но и вообще
первой арахнологической публикацией по паукам восточной Азии. Отрадно
отметить, что типовые материалы А. Грубэ сохранились и по сей день находятся в
музеях Санкт-Петербурга и Вроцлава.
Несколько последующих работ были выполнены британским и немецким
исследователями O.Pickard-Cambridge (1873) и L. Koch (1879a—b). Работа первого
посвящена паукам Прибайкалья, второго — паукам Западной Сибири и Приени-
сейя. На первоначальном этапе наиболее крупный вклад в изучение пауков
азиатской части России сделал польский арахнолог W. Kulczynski. Он подготовил
больше всех публикаций (Kulczynski, 1885, 1895, 1901, 1907, 1908, 1916, 1926) и
описал множество видов. Его статьи могут служить образцом и для современных арах-
нологов. Он затронул фауну и систематику пауков Камчатки, юга Красноярского
Края, Чукотки, северо-восточной Якутии и пауков-скакунчиков южной Сибири и
Дальнего Востока. Важный вклад в изучение сибирских пауков внесли француз Е.
Simon (1891) и финн Е. Odenwall (1901), описавшие ряд видов пауков из Алтая и
Прибайкалья, соответственно.
Огромный вклад в познание фауны внесли сборщики. Самыми первыми
коллекторами были Р.К. Маак и Л.И. Шренк. Р.К. Маак собирал материал в Якутии и
Приамурье, Л.И. Шренк — в Приамурье. Беспрецедентный вклад в изучение
фауны Сибири внесли ссыльные польские натуралисты Б.И. Дыбовский, А.Л. Чеканов-
16
1. Введение
ский, К. Волоссович (пауки были собраны в ходе экспедиции за трупом мамонта),
Б.К. Подгорский (Подгурский), В.К. Тачановский и вольнопоселенец А.П.(?)
Карпинский. Они собрали обширный материал на Камчатке, Чукотке, в Якутии, в
Прибайкалье. Их сборы послужили основой для статей O.Pickard-Cambridge, W. Kul-
czynski, T. Thorell. Но некоторые материалы пролежали невостребованными в
Зоологическом институте (Санкт-Петербург) до 80-х годов XX века.
Основой для публикации L. Koch (1879a) послужили материалы экспедиции
шведского исследователя А.Э. Норденшельда (А.Е. Nordenskiold).
Интересные пауки были собраны на северо-востоке Якутии Э.В. Толлем и А. Бун-
ге (1885-1886), А.А. Бирулей (1900-1902), М.И. Брусневым (1901-1903), А.В.
Колчаком и Н. Олениным (1903). Эти материалы обработал W. Kulczynski (1908).
Очень интересные сборы на Северо-Востоке России были сделаны С.А.
Бутурлиным в 1905 г. (попутно с пауками, он впервые обнаружил гнездовья розовых
чаек), экспедицией адмирала А.В. Колчака (побережье Чукотки, собирал Л.М. Ста-
рокадомский). Помимо указанных сборщиков, были и многие другие, прежде
всего в южной Сибири и на Дальнем Востоке.
После первой мировой войны темп исследований существенно снизился, хотя
сбор материала в Якутии, на Алтае, Дальнем Востоке продолжался. Между двумя
мировыми войнами по паукам Зауралья опубликовано всего несколько работ: Е. Schen-
kel (1930) и В.И. Сычевской (Sytshevskaja, 1935) по Камчатке, серия работ В.Н.
Ермолаева (Ермолаев, 1927, 1930; Ermolajev, 1928a-b, 1930, 1931, 1934a-b, 1937a-b)
по паукам Западной Сибири. Помимо этих трудов, вышло несколько статей
японских авторов (Peelle & Saito, 1932, 1933; Saito, 1932, 1933, 1934, 1935а-с) о пауках
Южного Сахалина и Курил, территорий, которые тогда принадлежали Японии.
После второй мировой войны и до конца 1970-х гг. работ стало ещё меньше. Две
очень важные работы были опубликованы A. Holm (1970, 1973). В них описаны
ранее необработанные сборы экспедиции Норденшельда и переописаны материалы,
которые ранее исследовал L. Koch (1879a). Другим исследователем, который внёс
важный вклад в исследование пауков азиатской части России, был J. Proszynski
(1979), которым описано много новых видов пауков-скакунчиков. Кроме того,
пауки Сибири и Дальнего Востока рассматривались в ревизиях пауков-бокоходов А.С.
Уточкина (1960, 1968). Несколько видов пауков-волков было описано Т.В.
Лобановой (1978) и М.Т. Штернбергсом (1979) из Саян и Прибайкалья соответственно.
Две важные работы по систематике пауков-волков подготовил А.А. Зюзин (1979а-
б). В период 1970-1980-х гг. было опубликовано более 10 фаунистических работ
М.В. Измайловой. Все её работы, включая монографию (Измайлова, 1989),
изобилуют неверными определениями, и страдают от незнания автором литературы.
Современный период мы условно отсчитываем от 1980 г., когда стал активно
публиковать свои статьи К.Ю. Еськов. Почему от Еськова? Он первым стал
работать на мировом уровне и к тому же стал разрабатывать самую сложную и богатую
видами группу в регионе. Большинство его работ посвящено систематике пауков
семейства Linyphiidae и фаунистическим исследованиям. Многие из публикаций
Еськова основаны на его же сборах. Вскоре к исследованию пауков Сибири и
Дальнего Востока подключились отечественные и зарубежные арахнологи Т.И. Олигер
(19811), П.М. Дунин (1984а-б), А.В. Танасевич (1984а), Ю.М. Марусик (1985), Т. Кго-
1 Здесь и далее указаны только первые публикации.
17
В.И. Сычевская A. Holm П.М. Дунин СВ. Овчинников
Рис.1.2. Арахнологи, внесшие существенный вклад в изучение фауны и таксономии
пауков Сибири и Дальнего Востока России: A. Grube; О. Pickard-Cambridge; L. Koch; W. Kul-
czynski; E. Simon; E. Strand; S. Saito; E. Schenkel; В.И. Сычевская; A. Holm; П.М. Дунин;
СВ. Овчинников.
Фотографий Е. Odenwall и В.Н. Ермолаева нам найти не удалось.
nestedt (1986), К.Г. Михайлов (Mikhailov, 1987), В.И. Овчаренко (Овчаренко, Мару-
сик, 1988), Д.В. Логунов (1990) и ряд других систематиков, в числе которых СН.
Данилов, СЛ. Есюнин, В.Е. Ефимик, Д.К. Куренщиков, СВ. Овчинников, СЮ.
Раков. В. Cutler, S. Koponen, R. Leech, H. Ono, N.I. Platnick, M.I. Saaristo, W. Wesoiowska.
18
7. Введение
Неоценимый вклад в изучение пауков региона внёс СИ. Головач, специалист
по двупарноногим многоножкам. Причём не столько своими сборами, сколько
помощью в издании работ отечественных авторов на английском языке в
европейских журналах. СИ. Головач помогал как с переводом, так и с публикацией работ в
западных журналах. Благодаря его усилиям вышли многие работы К.Ю. Еськова,
А.В. Танасевича, Ю.М. Марусика и К.Г. Михайлова. В 1980-е гг. требовалось
более полугода, чтобы получить разрешение из Москвы на публикацию своей статьи
в зарубежном издании. При этом необходимо было представлять, кроме
английской версии статьи, ещё и русский перевод. В 1990-е гг. СИ. Головач был в числе
создателей отечественного журнала Arthropoda Selecta и долгое время занимался
правкой английского во всех арахнологических рукописях.
Огромный вклад в изучение пауков региона внёс покойный натуралист и
профессиональный герпетолог A.M. Басарукин, который собрал обширнейшие коллекции
пауков Сахалина и Курильских островов. Д.И. Берман (Магадан) организовал самые
массовые сборы пауков в Сибири, когда в течение сезона собирались десятки тысяч
пауков в верховьях Колымы. Специалист по клопам из Якутска Н.Н. Винокуров
организовал сборы пауков по всей Якутии. Важный вклад в пополнение
коллекций пауков азиатской части России внесли сотрудники Сибирского
Зоологического музея во главе с В.Г. Мордковичем (В.В. Дубатолов, Б.П. Захаров, А.В. Барка-
лов, Р.Ю. Дудко) и сотрудники Палеонтологического института (покойный В.В. Же-
рихин, И.Д. Сукачёва) и многие другие известные и неизвестные зоологи.
Особую роль в исследовании пауков Сибири и Дальнего Востока играет
финский арахнолог S. Коропеп. В самые трудные для российской науки годы он нашёл
средства для организации совместных российско-финских экспедиций на
Полярный Урал, в Бурятию, Туву, Магаданскую область, Приморье. Помимо этого,
многие годы он добывает гранты для приезда российских коллег в Зоологический
музей Университета г. Турку (Финляндия) для работы с коллекциями. Благодаря его
непосредственному участию за десятилетие сотрудничества опубликовано более
60 совместных российско-финских работ с участием S. Коропеп, M.I. Saaristo, P.Т.
Lehtinen и более 10 российских авторов (см. Марусик, 2005). Помимо совместных
работ, вышло несколько десятков статей и две монографии (Logunov & Marusik,
2000b, 2003), выполненные российскими арахнологами в стенах Зоологического
музея в Турку.
В 2000-е гг. к таксономическим исследованиям Сибири и Дальнего Востока
подключились новые исследователи из России, ближнего и дальнего зарубежья:
Г.Н. Азаркина, В.А. Гнелица, А.В. Громов, М.М. Омелько, Л.А. Триликаускас,
D.J. Buckle, R.L. Crawford, S.Q. Li, Y.J. Song.
Совсем недавно С.Н. Данилов (2008) опубликовал каталог (вернее
аннотированный список) пауков Забайкалья.
Таким образом, видовой состав пауков Сибири и Дальнего Востока сейчас уже
выяснен в общих чертах (рис. 1.1). Бесспорно, ещё предстоят находки пауков,
новых для азиатской части России, и новых для науки. Однако, общее представление
о пауках региона (разнообразие, распределение по семействам, родственные
связи, степень эндемизма), по-видимому, останется неизменным. Это позволяет
перейти от описаний отдельных видов и составления региональных фаунистических
списков к обобщению имеющейся информации.
2. МЕТОДЫ ИЗУЧЕНИЯ ПАУКОВ
Методы сбора
Существуют стандартные методы сбора наземных беспозвоночных,
применимые и для коллектирования пауков. Это укосы энтомологическим сачком,
установка почвенных ловушек, отряхивание деревьев, сбор энтомологическим ситом, тер-
мофотоэклектором и, конечно, ручной сбор. Эти методы описаны во многих
публикациях. Здесь мы остановимся на модификациях, мало известных энтомологам
и арахнологам.
Минимальный набор снаряжения
1) Эксгаустер. При всех методах сбора желательно пользоваться эксгаустером.
Он экономит время, особенно при сборе мелких паучков, позволяет собирать
массовый материал и добывать таких пауков, которых иными способами поймать
нельзя. Устройство одного из наиболее удобных эксгаустеров показано на рисунке
2.1а-з. Внутренний диаметр эксгаустера должен совпадать или быть меньше, чем
внутренний диаметр флакона со спиртом, куда сбрасывается улов. Оптимально,
чтобы корпус был прозрачным, из пластика или стекла. В последнем случае его
концы надо обмотать прозрачным скотчем (чтобы не разбился). Наконечник
может быть сделан из стеклянной трубки. В этом случае кончик также желательно
обмотать прозрачным скотчем. Можно сделать наконечник металлическим.
Пробки могут быть корковыми, резиновыми или пластмассовыми. Всасывательную
трубку можно сделать из трубки от медицинской капельницы. Трубка может быть
жёсткой, но мундштук должен быть мягким, чтобы было удобно держать его в зубах.
Эксгаустер следует опорожнять не после поимки каждого паука, а когда наберётся
10-20 экземпляров, или когда пойман особо интересный паук. Чем жарче погода,
тем чаще следует опорожнять эксгаустер.
2) Несколько флаконов, в которые сбрасываются пауки, собранные
эксгаустером, пробиркой, или руками. Флаконов нужно несколько, чтобы не бросать много
пауков в одну пробирку и чтобы не смешивать друг с другом улов из разных
биотопов. Заполнять флаконы необходимо 70-80%-ным спиртом, примерно на
половину объема. Пробка должна легко выниматься одной рукой, но не должна сама
легко выскакивать. Она может быть резиновой, корковой или пластиковой. Флакон
может представлять собой обычную стеклянную «пенициллинку», но можно
использовать любую другую небольшую герметично закупоривающуюся ёмкость:
пластиковый бюкс из-под фотоплёнки, пробирку и т.д.
3) Сумка (рюкзак) или глубокие карманы, куда можно складывать эксгаустер и
флаконы.
20
2. Методы изучения пауков
1 (Е
В
Ж
и
и
к
Рис. 2.1. Инструменты и приспособления для сбора пауков.
а-е — детали, необходимые для сборки эксгаустера (а — стеклянная или пластиковая трубка; б —
стеклянная или металлическая трубка, через которую засасываются пауки; в — стеклянная
трубка, куда присоединяется резиновая трубка; г — пробковая или резиновая пробка, куда
вставляются трубки б и в; д — фильтр из мельничного газа; е — резиновая трубка, через которую
всасывается воздух); ж — схема снаряжённого эксгаустера; з — фотография эксгаустера; и — меч Колесова;
й — нож-копалка с закруглённым лезвием, им удобно выкапывать ямки для почвенных ловушек,
можно пользоваться и как мачете; к — сечение ножа-копалки; л-м — фотографии ножа-копалки.
Методы сбора
21
Оптимально-минимальный набор для сбора пауков разными методами
включает: 1) эксгаустер; 2) флаконы со спиртом и пустые (для живых пауков) — 10-15
штук; 3) полевая сумка (через плечо) или рюкзак; 4) сачок для кошения; 5)
раскладной зонт для просеивания подстилки и сбора с деревьев (подробнее см. ниже).
Расширенный набор может включать также матерчатые перчатки, пинцет, лист
полиэтилена, металлический прут с 2 когтями (для обдирания коры,
переворачивания камней), энтомологическое сито, спрей-репеллент для выкуривания пауков их
трещин и дупел, фонарик, фотоаппарат, GPS (для определения координат и высот),
записную книжку и карандаш, секатор, нож-копалку, и др.
Ручной сбор. Вручную собирать пауков можно с помощью пробирки или
эксгаустера. Мы предпочитаем последний инструмент. Если паук забрался в
неглубокую трещину, то поймать его можно только просунув наконечник эксгаустера в
трещину и резко вдохнув.
Наиболее оптимальное положение для сбора на почве, в травостое, на
галечниках и осыпях — на боку, когда глаза сборщика приближены к грунту на расстояние
30-50 см. Тогда видны пауки размером менее одного миллиметра.
При ручном сборе надо обязательно переворачивать камни. Чем камни
крупнее, тем больше вероятность поймать под ними интересные виды. Камни обычно
переворачивают вручную; бывает, что требуются усилия двух человек. В горах
можно ворочать камни ледорубом. Под камнями селятся самые разнообразные
пауки. При ворочании камней надо высматривать пауков не только на земле под
камнем, но и на нижней поверхности самого камня.
Сбор на скалах. В дневное время сбор на скалах эффективен только для
пауков-скакунчиков. Днём можно заметить немало сетей и убежищ, расположенных
в трещинах. Но выудить оттуда паука практически невозможно. Для
выкуривания можно использовать отпугивающие репелленты для насекомых, желательно
во флаконах, дающих мощную струю. Репеллент не должен быть опасным для
человека и убийственным для насекомых и пауков. Подобрать наиболее
эффективный не легко, но можно. После впрыскивания репеллента в трещину, пауки и
все остальные обитатели трещины спустя какое-то время начинают выбираться
наружу.
Репеллент бывает очень эффективным для сбора пауков в дуплах деревьев и на
стволах с морщинистой, но не отслаивающейся корой. После опрыскивания
поверхности, пауки начинают двигаться и становятся заметными.
Сбор с глиняных и песчаных обрывов рек, балок, оврагов. Суть метода в
постепенном обрушивании стенок. В таких местах много паучков прячется в
верхней части обрыва под растительной ветошью, а также в трещинах между
вертикальными пластами. После обрушивания вниз, через некоторое время паучки
начинают бегать и пытаться взобраться на обрыв обратно. Движение выдаёт их и
позволяет легко собрать. Таким методом можно взять многих представителей
семейств Gnaphosidae, Linyphiidae, Salticidae, Titanoecidae и других, которых трудно
добыть иначе.
22
2. Методы изучения пауков
Ручное просеивание подстилки. В местах, где есть подстилка, это самый
эффективный и уловистый метод сбора. Для этого способа достаточно иметь лист
плотного полиэтилена (50x50 см, или около того) или зонт. Сборщик садится на
колени или просто на землю, берёт рукой (желательно в перчатке) подстилку (опад,
мох) и бросает её на полог (полиэтилен, перевёрнутый зонт). Затем постепенно
разгребает постилку. Первоначально пауки впадают в оцепенение, но спустя
непродолжительное время начинают разбегаться, становясь легко заметными. Этот
метод особенно эффективен при сборе во мху или лишайниках. Обнаружить
пауков в толще этих растений невозможно, а на листе полиэтилена или ткани зонта,
они становятся заметными.
Этот способ во многом эффективнее сбора ситом, так как позволяет быстро
находить места, где концентрация пауков выше. Если в двух-трёх «жменях»
подстилки нет пауков, то следует переменить место, посмотреть более сухую или
влажную, либо более тёплую или холодную подстилку.
С помощью этого метода удалось собрать Mysmenidae практически во всех
местах Приморья, Хабаровска, Сахалина, где мы (Ю.М.) собирали. Этим же
способом собраны Mysmenidae, Leptonetidae, Oonopidae на Кавказе и других регионах,
где эти группы не были известны. Этот метод незаменим при сборе мелких пауков
семейств Linyphiidae, Hahniidae, Corinnidae, Theridiidae, Mimetidae и некоторых
других.
Энтомологическое сито (сифтер). Это устройство показано рисунке 2.2ж-й.
Есть и более простые конструкции, но они менее эффективны.
Перед началом лова мешок завязывается. Подстилка (листва, мох)
наваливается в верхний карман, прямо на сетку Когда подстилки там много, необходимо
резкими движениями трясти сетку-сито, тогда пауки и мелкие кусочки подстилки
проваливаются внутрь, а крупные листья, ветки, палочки остаются на сите. После
встряхивания, не провалившуюся подстилку выбрасывают и закладывают новую
порцию. Вываливать обогащенную подстилку с уловом для просмотра следует в
перевёрнутый зонт или на полиэтилен.
В каких условиях сито эффективнее ручного сбора? Когда лес сомкнутый и
очень тёмный, и выбирать улов, особенно мелких паучков, невозможно. В этом
случае обогащенную подстилку желательно разбирать в светлом месте. Если
время для сбора ограничено или начинается дождь, то можно быстро обогатить ситом
подстилку и сбросить улов в мешок, чтобы разобрать его уже в лабораторных
условиях. Надо помнить однако, что в пластиковых мешках пауки быстро погибают
(если жарко, то за 10-15 минут!), а неподвижные объекты очень трудно найти.
Поэтому для «высевок» предпочтительно использовать полотняные мешки.
Наилучший материал для таких мешков — мельничный газ, но годится и другая
прочная «дышащая» ткань. В других случаях (лес светлый, нет нужды торопиться)
эффективнее ручное просеивание.
Верхний обруч сифтера можно сделать из мягкой (гибкой) проволоки, или
оставить «свободный сектор». Такая конструкция очень эффективна для сбора
пауков под корой. Большие куски коры остаются на сите, а пауки проваливаются в
мешок. Это же сито можно использовать для сбора пауков в опаде, особенно если
он не глубокий, и его трудно набрать в ладонь. Верхний обруч прикладывается к
поверхности грунта, и подстилка сгребается рукой прямо в сифтер.
Методы сбора
23
Рис. 2.2. Примеры устройства почвенных ловушек и сифтера.
а-б — ловчий стаканчик с отверстиями для дренажа (а), стаканчик вкопан в грунт (б); в-г —
ловчий стаканчик в долгосрочной почвенной ловушке (в), ловушка вкопана в грунт (г); д-е —
раскладная крыша для ловушки (из листа алюминия или жести) в собранном (е) и рабочем (д)
состоянии; ж-й — схема устройства сифтера, ручки должны быть не друг над другом, а под
углом: с завязанным дном (ж); разрез, где показано, куда закладывать подстилку, и просеянная
подстилка на дне мешка (з); с развязанным дном (и); с усечённым верхним обручем (й), который
удобно приставлять к стволу дерева или к грунту (чтобы сгребать подстилку прямо в сифтер).
24
2. Методы изучения пауков
Сборы сачком. Наиболее универсален сачок диаметром 30-50 см, с жёстким
ободом и тяжёлой ручкой-древком длиной 1-1,5 м. Лучшая ткань для сачка —
мельничный газ. Верхнюю часть ткани, надеваемую на обод сачка, необходимо обшить
брезентом или другим материалом, устойчивым к разрыву (например, для
обшивания сачка очень подходит мягкая изоляция от электрического кабеля). Почему газ, а
не другая более плотная и прочная на разрыв ткань? Если газ намок (от росы или
капель дождя), он быстро высыхает. Через газовый сачок воздух легко проходит,
косить им легко, он позволяет ловить пауков даже в воде (пауков-серебрянок,
обитающих в её толще, и пауков, бегающих по водной глади). С полотняным мешком
' W
= *?з
Рис. 2.3. Примеры ловушек и методов сбора со стволов деревьев.
а — показана простая стволовая ловушка (#), и как можно использовать сачок либо сифтер с
усечённым обручем (sn); б — то же, вид сверху; в — более уловистый, но более трудоёмкий для
изготовления и установки тип ловушки; г — то же, вид сверху; д — то же, вид сверху, но стакан
заменён на мешок; е — простой сачок; ж — сачок с усечённым обручем; з-и — пример
устройства для обдирания коры, это же устройство можно применять для переворачивания камней.
Методы сбора
25
делать укосы физически тяжело, а плотный воздух в мешке сачка выталкивает
пауков, и вместо сачка пауки падают на землю. Зачем жёсткий обод и тяжёлая ручка?
Чтобы удобнее было косить по кустарникам, особенно жёстким и колючим, или по
кедровому стланику. Тяжёлая ручка пригодится и для отряхивания веток деревьев,
и как посох. Из сачка пауков удобнее выбирать не эксгаустером, а пробиркой. В
сборах, выполненных сачком, большинство особей бывает ювенильными.
Если сделать обруч сачка в виде буквы «U», на уловистости это никак не
скажется, но позволит более эффективно собирать обитателей стволов деревьев,
приставив сачок к дереву как на рисунке 2.3а-б.
Отряхивание деревьев. Наиболее просто делать отряхивание веток деревьев.
Для этого идеально подходит зонтик, обычный или раскладной (где штанга
складывается в Ъ-^ раза). Почему лучше зонт, а не специальная рамка-полог для
отряхивания? Раскладной зонт занимает меньше места, требует меньше времени для
перевода в рабочее положение, этот же зонт можно эффективно использовать для
сбора пауков и в подстилке, и на стволах деревьев. Но об этом чуть ниже. Помимо
зонта, нужно иметь ещё и палку, которой бьют по веткам. Идеально подойдёт
ручка (древко) сачка. Наиболее уловистыми являются ветки хвойных деревьев. Там
всегда больше и видов и экземпляров.
Помимо отряхивания веток, можно делать отряхивание всего дерева. Этот сбор
требует усилий как минимум двух человек и специального оборудования —
«колотушки» и «простыни». Колотушка — это тяжёлое бревно длиной в три и более
метров, которым ударяют по стволу дерева. А «простыня» раскладывается под
деревом, чтобы на неё падали пауки. Идеальной «простынёй» может служить старый
парашют. Он мало весит и легко, практически без складок, раскладывается под
деревом. Наиболее эффективно такое отряхивание утром на восходе солнца, когда
пауки ещё находятся в ночном оцепенении. Если использовать этот метод днём,
тогда многие пауки не упадут на землю, так как успеют выпустить паутину и по
ней вернуться обратно на дерево.
Сбор со стволов. Для отковыривания коры со стволов можно использовать нож-
копалку или лопатку. Собирать пауков при этом следует эксгаустером. Уловистость
метода резко возрастает, если к стволу дерева приставить «усечённый» сачок или
зонтик. При обдирании коры многие пауки выпрыгивают либо поджимают ноги и
затаиваются, и падают вниз. Если снизу подставить зонтик или сачок, тогда пауки
будут сваливаться «куда надо».
Опыт показывает, что для обдирания коры со стволов удобно использовать
металлический прут не более полуметра длиной с двумя когтями на конце. Когти
зацепляются за неровности коры, легко подцепляют отслаивающуюся кору и
отдирают её. Удобнее собирать таким способом в четыре руки. Один обдирает кору, а
другой держит зонтик (сито, сачок) и собирает пауков.
Ещё один, дополнительный способ. Он годится для сбора с гладкоствольных
пород деревьев. Большинство обитающих на коре видов пауков имеют
маскирующую окраску, и заметить их практически невозможно. Для того чтобы пауки
выдали своё присутствие, достаточно ударить по стволу чем-нибудь увесистым
(ручкой-древком сачка, тяжёлой веткой). После удара паук всегда немного смещается,
и в момент движения его можно заметить.
26
2. Методы изучения пауков
Существуют и долговременные методы сбора. О стволовых ловушках сказано в
следующем разделе, а здесь упомянем стволовую «заманиловку». Все пауки
нуждаются в укрытиях, где можно скрываться от хищников, делать убежище,
откладывать кокон. Не всегда на стволе (под корой) есть достаточно места. Это место
можно создать искусственно. Достаточно привязать к стволу свёрнутый гармошкой
непрозрачный полиэтилен, тряпку, вощёную бумагу, так, чтобы образовалось
много щелей и пустот, которые могут занять пауки под свои убежища и логова.
Проверять «заманиловку» нужно редко, можно раз в месяц.
Экзотические способы сбора пауков. Некоторые околоводные пауки, например
Theridiosoma, Pirata могут быть легко собраны, если сборщик находится по шею в
воде, а берега водоёма слегка обрывистые. В таком необычном положении
становятся легко заметными сети Theridiosoma и бегающие вдоль кромки воды Pirata.
Время сбора. Большинство арахнологов собирают в дневное время, однако
многие группы пауков могут быть собраны только ночью. В особенности это касается
южной Сибири и юга Дальнего Востока. Жаркими днями только немногие виды
активно охотятся. У многих пик активности сдвинут на ночь. Ночные укосы
позволяют собирать в массе тех пауков, которые днём встречаются единично. При этом
удобно пользоваться налобным фонариком, чтобы обе руки были свободны. В
густых зарослях и под пологом леса налобный фонарик может оказаться
незаменимым подспорьем даже днём — при разборе подстилки, например.
Очень эффективен ночной сбор пауков-кругопрядов и других тенетников.
Большинство из них днём прячутся в убежищах, а ночью перебираются в центр
паутины, и становятся заметными. Некоторых пауков, например Metleucauge (Tetragna-
thidae) можно собрать только ночью.
Самые интересные результаты даёт ночной сбор на скалах. Ночью все
тенетники и бродячие пауки выползают из трещин. Они становятся легко заметными
благодаря отбрасываемым теням. За короткое время иногда удаётся собрать сотни
пауков. Ночью много подкорных пауков выползает на поверхность коры. Норные
пауки-волки могут быть собраны тоже только ночью. Их можно заметить
благодаря отражению их глаз в свете фонаря.
На галечниках некоторые виды, особенно из пауков-волков (например, Arctosa),
днём вообще незаметны, зато ночью активны и бегают в массе. Поэтому в
экспедиционных условиях есть смысл наведываться на галечники ночью.
Опыт показывает, что тёмные сумерки и 1-2 ч после наступления темноты —
наиболее уловистое время.
Почвенные ловушки. Для пауков это, пожалуй, самый эффективный метод
сбора, позволяющий собрать самый массовый материал. Для ловушек подходят
пластиковые одноразовые стаканчики разного диаметра. Диаметр зависит от
места сборов и твёрдости субстрата. Наиболее подходящий диаметр — 5-8 см. Если
ловушек много и ставятся они временно, то для удобства их нахождения, ловушки
можно пометить лентами из пластика, ткани или бумаги. Лента-маркер вешается
на ближайший к ловушке куст или травинку. Если нет ни того, ни другого, можно
использовать колышки или отмечать начало и конец линии ловушек каменными
пирамидками.
Методы сбора
27
Наиболее практичный современный фиксатор — тосол (охлаждающая жидкость
для радиаторов автомобилей). Он относительно дёшев. В зависимости от того, в
каких условиях стоят ловушки (на жаре или в холоде, долго или краткосрочно),
можно варьировать концентрацию тосола и воды. Соотношение 1:4 может
подойти для самых разных целей, однако концентрация может быть и 100% (если
ловушки ставить на всю зиму), и 10%, если ловушки выставляются на короткое время.
Для повышения уловистости в раствор тосола следует добавить пару капель
жидкого мыла (средства для мытья посуды), чтобы уменьшить поверхностное
натяжение и тем самым ускорить утопление пауков. Кроме этого, рекомендуется
добавить уксуса (можно химического, но лучше фруктового) для запаха. Уксус
(продукт брожения) привлекает насекомых, а также и самих пауков.
Тосол можно заменить этиленгликолем или глицерином. Но эти жидкости
более дорогие. Если под рукой нет ничего из перечисленного выше, можно
использовать просто подсоленную воду. Соль не даст разлагаться паукам. Качество
материала будет при этом хуже, но это лучше, чем ничего.
Ранее в качестве фиксатора часто использовали слабый раствор формалина. Но
формалин портит самих пауков (они становятся хрупкими, теряют цвет, пальпы
вздуваются), а вдыхание его паров вредно для здоровья. Тем не менее, формалин
не высокой концентрации (2-4%) хорошо работает в ловушках. Второй соавтор
(Н.К.) с успехом применяет фиксатор такого состава: 2^4%-ный раствор
формалина; немного уксуса; несколько капель моющего средства для посуды. В жаркое
время года для защиты от полного высыхания в раствор добавляется ещё и
глицерин или этиленгликоль (примерно, 1/8-1/10 часть раствора по объёму).
Хорошие результаты даёт уксусный раствор. Можно использовать обычный
столовый 7-9%-ный уксус или разводить ледяную уксусную кислоту. Важно
помнить — «не плюй в кислоту!» — то есть, при разведении ледяной кислоты,
именно кислота льётся в воду, но не наоборот! Иначе есть опасность разбрызгивания. В
уксусный фиксатор также стоит добавить несколько капель моющего средства.
Ловушки с уксусом приходится опорожнять чаще, чем формалиновые, обычно
каждые 5-7 дней. Формалиновые ловушки могут работать до 15-20 дней. Отличие
связано с тем, что в уксусе пауки быстрее начинают мацерироваться. Если не
собирать улов более недели, то все поймавшиеся пауки могут разложиться и станут
непригодными для исследования. Формалин более практичен в жарком климате,
уксус — в более прохладном.
Чтобы стаканчики не переполнялись дождевой водой, на расстоянии 2-3 см от
поверхности можно проделать 1-2 дырки, чтобы стакан дренировался. Другой
способ — сделать крышечку над стаканом. Наиболее функциональная конструкция
показана на рисунке 2.2д-е. Такая крышка делается из листа жести или алюминия,
занимает мало места при транспортировке, легко раскладывается (разгибается) в
рабочее положение и втыкается в грунт. Крышки защитят ловушку от попадания
листьев, веточек, и сделают её менее заметной для любопытствующих.
Если предполагается использовать ловушки долго, то лучше использовать
капитальную конструкцию (рис. 2.2в-г), которая требует больше усилий для
вкапывания, но зато такую ловушку удобно опорожнять и в неё попадает меньше грязи.
Для установки ловушки, можно использовать лопатку, но удобнее — лом (если вы
передвигаетесь на своём транспорте). Лом при этом втыкается с размаха в землю,
и затем делается коническая ямка, куда удобно вставлять стаканчик. Этот метод
годится только для влажной, не каменистой почвы.
28
2. Методы изучения пауков
Для выкапывания ямок подходит и другой инструмент — нож-копалка с
полукруглым в сечении полотном (рис. 2.1й-м). Этот нож удобен и для обдирания коры,
им можно пользоваться для обрубания мелких веток.
На сильно каменистой или глинистой почве хорош для установки ловушек меч
Колесова (лесотехнический инструмент, который есть в лесничествах и
используется для посадки саженцев). Представляет собой стальной меч, длиной 40-50 см и
шириной около 10 см, на стальном стержне около 1,4-1,6 м с поперечной стальной
или деревянной ручкой (рис. 2.1и). Он весит 8-12 кг, поэтому с размаху легко
вколачивается в самую каменистую почву, буквально дробя породу; при его
проворачивании образуется круглая ямка, куда вставляется стаканчик. При некотором
навыке можно делать ямки такого диаметра, как стаканчик. При этом растительность
и почва вокруг стаканчика не повреждаются, и поэтому уловистость оказывается
гораздо выше, чем когда вокруг ловушки всё разворочено. Меч Колесова
позволяет за 1-2 часа наставить 50-100 ловушек, что очень ускоряет процесс.
Чтобы при установке стаканчика в него не насыпалась земля, ставить надо не
один стаканчик, а два, то есть вложить один стаканчик в другой. После того, как
земля вокруг стакана выровнена, верхний (в который осыпалась земля) —
вынимается, а чистый — остаётся в лунке.
В установленной ловушке тоже предпочтительно оставлять не 1, а 2 стаканчика
(один в другом). При выемке улова, когда стаканчик с раствором вынимается,
второй (нижний) в это время остаётся в земле и не даёт краям ямки осыпаться. Потом
вынутый стаканчик легко ставится обратно.
Ловушки устанавливают в линию по 10 или больше штук в контрастных
биотопах так, чтобы каждая линия облавливала только один биотоп. Расстояние между
ловушками — метр или больше. Чем больше расстояние между ловушками, тем
больше видов ловится.
Помимо простых вкапываемых ловушек, есть несколько модификаций,
позволяющих собирать пауков каменных осыпей, на водной глади, и даже на стволах
деревьев. Фиксатор во всех типах ловушек один и тот же.
Конструкция для собирания обитателей осыпей самая простая и лёгкая в
исполнении. Помимо стаканчика, необходим кусок картона (скажем 30 х 30 см), в
центре которого проделывается дырка для стакана. Картонка с ловушкой
устанавливается среди осыпи. Углы картона прижимают камнями, чтобы не унесло
ветром. Интересные результаты дают «почвенные» ловушки внутри осыпи. Такие
ловушки можно установить на целый год. Для этого нужно разобрать осыпь на
0,5-1 м, установить там несколько картонок с ловушками (фиксатор — чистый
тосол) и заложить их сверху камнями.
Водная ловушка. Для неё нужен лист фанеры или тонкий лист пенопласта. В
центре проделывается дырка для стаканчика. Чтобы конструкция не всплывала и
была вровень зеркалу воды, стакан необходимо утяжелить (свинцом,
металлическими болтами). Такие ловушки устанавливаются среди околоводной
растительности, в местах, где нет сильного волнения.
Стволовые ловушки можно делать из крупных пластиковых стаканов, но
лучше использовать плотные пластиковые мешки. Они привязываются, прикалыва-
Методы сбора
29
ются или даже приклеиваются скотчем к стволу Спичками или прутиками
делаются распорки (рис. 2.36) так, чтобы была щель не менее 1 см.
Как опорожнять ловушки? Не рекомендуется выбирать пауков в природе,
так как при этом велика опасность «пропустить» мелких пауков. Лучше
содержимое слить в пластиковую банку объёмом 1-2 л и затем спокойно разбирать
улов в лабораторных условиях. Можно вываливать пробы в пластиковые
самозакрывающиеся пакетики (Zip-lock). После того как в пакет слито содержимое
всех ловушек, следует убрать весь воздух и тщательно запечатать пакет. Есть
более лёгкий и быстрый способ. Для этого используются бязевые («почвенные»)
мешочки. Улов вываливается в мешок, при этом фиксирующую жидкость
можно сцедить в другой стакан и использовать её повторно. Затем завязанный
мешок (мешки) необходимо сложить в пластиковый контейнер, чтобы не
раздавить пробу и не дать мешку высохнуть, а потом в лабораторных условиях
начать разборку. Вместо бязевых мешочков можно использовать марлю или
широкий бинт (не менее 14 см в ширину). Отрезок бинта зажимается на отверстии
пустого стаканчика кромкой от другого стаканчика (чтобы получить такую
кромку, отрезается «верх» стаканчика, шириной 1-2 см). В результате получается
«ситечко», в которое выливается содержимое ловушки. Раствор отцеживается
для повторного использования, а улов заворачивается в «узелок» из марли и
переносится в контейнере. Прямо в поле в «узелок» надо бросить номерок или
этикетку. Материал в бязевом или марлевом мешочке можно некоторое время
хранить в холодильнике (но так, чтобы не образовался лёд, который может
повредить пауков).
Как узнать, взрослый ли паук?
Далеко не всегда в полевых условиях можно определить, взрослые ли особи
собраны. С крупными пауками таких проблем обычно не бывает. У них пальпы
самцов и эпигины самок хорошо заметны. Однако есть исключения, например, у
крупных самцов пауков-волков (Allohogna) цимбиум не сильно утолщён, и их
можно спутать с неполовозрелыми. Иногда эпигина может быть скрыта густыми
волосками, и потому не заметна. Самки Tetragnathinae не имеют эпигины,
поэтому не видно, взрослые ли они. Не имеют эпигины и Atypidae. Если в пробе есть
самцы, тогда по размеру можно представить, есть ли половозрелые самки этих
групп (они крупнее). У мелких пауков сравнительно легко опознать самцов (по
утолщённым пальпам), но самок — практически невозможно. Во многих
случаях взрослых можно отличить по размеру и более интенсивной окраске. В любом
случае, если вы только начинаете изучать пауков, то лучше брать всех подряд,
особенно мелких.
В лабораторных условиях (под бинокуляром) сложно опознать взрослых самок
лишь гаплогинных семейств (Leptonetidae, Oonopidae) и Atypidae. Если нет
самцов, то определять по самкам эти группы — мало смысла, поскольку нет ревизий и
хороших рисунков эндогины.
Нередко крупные субадультные самки Lycosidae имеют структуры,
напоминающие эпигину Если такую «эпигину» отделить, то у неё вы не обнаружите
семяпроводов и рецептакул.
30
2. Методы изучения пауков
Легко можно спутать с неполовозрелыми и взрослых самок Mysmenella (Mysme-
nidae). Эпигина очень слабо хитинизирована, не имеет ямки, а скапус можно
заметить лишь при тщательном осмотре при виде сбоку Гораздо проще рассмотреть,
нет ли хитинизированной бляшки на нижней стороне бедра. Если она есть, то
самка половозрелая.
Обращение с собранным материалом. После того как материал принесён
домой (в лагерь, лабораторию), желательно сразу же сделать этикетки, хотя бы
временные, указав географическую привязку, дату сбора, биотоп и фамилию
сборщика. Этикетка вкладывается внутрь ёмкости с пауками (ни в коем случае не
наклеивается на баночку и, тем более, не кладётся отдельно!). Этикетку лучше
всего писать тушью на кальке. Можно писать линером или карандашом. Писать
этикетку можно не только на кальке, но и на плотной бумаге. Однако, такой
вариант хуже, поскольку бумага, в отличие от кальки, размокает. В прошлые годы
многие исследователи писали этикетки на фотобумаге, поскольку она устойчива
к размоканию. Удобно сделать компьютерную этикетку. Однако
этикетки-распечатки со временем могут портиться (размокают, отпадают буквы). Несмотря ни
на что, этикетки нельзя отделять от материала или выбрасывать ни при каких
обстоятельствах!
Если нет возможности разобрать материал сразу, стоит хотя бы заменить
старый спирт новым. Если материал предстоит транспортировать на большое
расстояние, надо постараться заполнить пробирки-флаконы как можно большим
количеством спирта (чтобы меньше оставалось воздуха, пузырьки которого повреждают
пауков). Можно избежать появления пузырьков, поместив в бутылочку или
пробирку сверху над пауками большой ватный шарик. Тогда воздух останется над
ватой, и пузырьков не будет.
Где собирать пауков? Вначале о биотопах. Если цель исследования — изучить
всё многообразие видов в регионе, то, разумеется, стоит собирать везде, и
разными методами. Если же цель — найти интересных пауков, то наименее подходящие
для этого места — антропогенные луга, пастбища, сельскохозяйственные поля,
сады и огороды. Бывают, конечно, исключения: на севере немало видов, в том
числе и новых, было найдено среди строительного мусора.
Интересные результаты можно получить, собирая пауков в мало удобных
местах (и поэтому мало обследованных), на осыпях и галечниках. Тут очень высокий
уровень эндемизма. Например, на Алтае у пауков волков рода Acantholycosa он
достигает 85%. Наиболее велика концентрация пауков внизу осыпи, где она
упирается в почву. Здесь выше влажность, и встречается больше и тенетников и
бродячих пауков. Для того чтобы собрать тенетников, необходимо перевернуть массу
камней, врыться внутрь осыпи. Если поверх осыпи растут кустистые лишайники,
то среди них тоже встречаются интересные виды. Хорошие результаты дают
«почвенные» ловушки, расположенные внутри осыпи. Такие ловушки можно
установить на целый год. Уловистость их невысока, зато виды попадаются всегда очень
ценные.
На юге Дальнего Востока интересный результат может дать сбор со стволов
деревьев, и ловушками и вручную. Не исключено, что так могут быть найдены
дополнительные виды Oonopidae и даже целое семейство Segestriidae.
Методы сбора
31
В районах, где нет вечной мерзлоты, и есть пещеры, могут быть собраны новые
таксоны из Nesticidae, Leptonetidae и других семейств. Среди пещерных видов
наблюдается очень высокий уровень эндемизма.
Интересные результаты всегда дают сборы в горных тундрах, особенно в горах
Южной Сибири и Дальнего Востока.
В степной зоне, если нет каменистых склонов, то собирать вручную очень
сложно. Выручить могут лепёшки, оставляемые коровами и другими копытными. Под
ними можно найти убежища и даже логова пауков (самки вместе с коконами)
самых разных семейств.
Там, где встречаются колючие подушечные кустарники (особенно в степной
зоне), и невозможно из-за колючек собирать подстилочные формы, может помочь
секатор. Ногой (в сапоге или ботинке) приподнимается край куста и перерезается
основной ствол. Затем срезанный куст отряхивается над зонтом или
полиэтиленом. Кроме этого, необходимо рассмотреть то, что осталось на земле. Под густыми
кустами всегда повышена влажность, и там концентрируется больше насекомых и,
соответственно, пауков.
При сборе на песке, в мелких камешках или гальке имеет смысл скрести
пальцами или сгребать ладонью субстрат. Тогда мелкие паучки, лишившись убежища
или укрытия, разбегаются и становятся заметными.
Общая рекомендация: в аридных (сухих) местах пауков следует искать на
прохладных, затенённых и влажных участках. Наоборот, во влажных местах, на
болотах, пауков больше в тёплых и относительно сухих местах.
Когда собирать? О времени суток уже было сказано. Поскольку все собирают
летом, и очень мало кто осенью и ранней весной, то интересные и необычные
сборы можно сделать во внесезонье. Особенно много пауков можно поймать
поздней осенью, когда большинство Linyphiidae представлены взрослыми особями.
У некоторых видов самцы встречаются только поздней осенью и самой ранней
весной, пока ещё не полностью стаял снег.
Можно собирать и зимой, установив почвенные ловушки с крышками и залив
их 100%-ным тосолом. Опорожнить их можно ранней весной.
Наиболее перспективные регионы. Менее всего пауки изучены на северо-
западе Хабаровского края, севере Амурской области и юго-востоке Якутии. В
последнем регионе почти нет посёлков, и арахнологи там никогда не бывали. Слабо
изучены горные районы Читинской области, Восточный Саян. Не менее, а может и
более перспективным является изучение Приморья и всего Сихотэ-Алиня. Здесь
могут быть найдены дополнительные семейства, немало дополнительных и новых
родов и видов пауков.
В плане изучения видового разнообразия перспективны Камчатка и Амурская
области. По этим регионам нет даже систематических списков.
Однако необходимо подчеркнуть, что новые или редкие виды пауков могут быть
найдены в любом регионе, главное набрести или «вычислить» наиболее
необычный биотоп, или выбраться за пауками в необычное для сборов время года, или же
собирать даже в обычных биотопах, но необычным способом. Вообще, «высший
пилотаж» для арахнолога — находить новые таксоны не в тех местах, где никто
никогда не собирал, а наоборот — в регионах, которые на первый взгляд изучены
вдоль и поперёк!
32
2. Методы изучения пауков
Хранение. Оптимально хранить сборы и коллекции пауков в стеклянных
банках с притёртыми стеклянными крышками, но можно и в банках с
металлическими или пластиковыми крышками, в этом случае надо периодически проверять, не
проржавела ли или не искривилась ли крышка. Можно хранить и в пластиковых
банках, если они не протекают. В банку помещают стеклянные или пластиковые
пробирки, заткнутые ватной пробкой. Для длительного хранения пробирки ставят
вверх дном. Тогда при частичном высыхании спирта из банки, в пробирках спирт
всё же остаётся. Другой способ избежать потери материала при высыхании спирта —
это добавление в спирт глицерина (1/10-1/9 часть раствора по объёму). При
высыхании спирта, глицерин остаётся. Если в коллекции обнаружен такой подсохший
материал, достаточно просто долить новый раствор. Если же спирт высох
полностью (не был добавлен глицерин), то пауков приходится сперва несколько дней
размачивать в молочной кислоте. Лишь после этого их можно перенести в свежий
раствор спирта или спирта-глицерина. Хранить материал следует в местах, где нет
прямого солнечного света.
Этикеточные данные желательно дублировать ещё как минимум два раза — в
журналах обработки материала и в повидовой картотеке. В журнал обработки
записывается этикетка и результаты определения — количество экземпляров и
названия видов (или родов и семейств, в том случае, если материал не удалось
определить до вида). В картотеке на каждый вид заводится отдельная карточка, куда
записываются этикеточные данные всех экземпляров в коллекции. Карточки (как
и материал в коллекции) располагаются в алфавитном или систематическом
порядке. Журналы обработки и картотека очень облегчают поддержание коллекции в
порядке (так, чтобы быстро можно было найти любой экземпляр) и анализ
полученных данных (фенологии, распределения по биотопам и т.п.) при подготовке
публикаций. Полезно также иметь электронную базу данных по собственной
коллекции. Однако электронная база не должна заменять «бумажную» базу в виде
картотеки и журналов, поскольку они более надёжны (не повреждаются
компьютерными вирусами, сбоями в работе программ и т.п.).
Желательно передавать пауков, особенно тех, которыми вы не собираетесь
заниматься в ближайшее время, на хранение в центральные музеи. В России всего
несколько мест, где хорошо налажено хранение коллекций пауков. Это
Зоологический музей МГУ (куратор К.Г. Михайлов), Сибирский Зоологический музей,
Новосибирск (куратор Г.Н. Азаркина), Зоологический институт РАН
(Санкт-Петербург, куратор В.А. Кривохатский) и кафедра зоологии Пермского университета
(куратор С.Л. Есюнин).
Как рассматривать пауков?
Оптика. Наиболее удобно рассматривать пауков в бинокуляр. Это могут быть и
отечественные МБС (микроскоп бинокулярный стереоскопический) разных
модификаций, и зарубежные. Предпочтение следует отдавать тем бинокулярам,
которые дают большее увеличение.
Самое важное — это осветитель. Идеален осветитель со световодами.
Производят их за рубежом и в России, но первые очень дорогие, а вторые сложно найти,
и они весьма неудобны. Можно обойтись и обычными отечественными осветите-
Как рассматривать пауков?
33
лями, идущими в комплекте с бинокуляром. Их неудобство в том, что сложно
получить необходимую яркость освещения и сложно подвести его под нужным
углом. Кроме того, наши осветители сильно греются и испаряют спирт. Но самая
большая сложность кроется в том, что лампочки быстро выходят из строя, а
приобрести их нелегко.
Оптимизировать стандартный осветитель для МБС-9 и МБС-10 с двумя
коленами, чтобы он давал свет под большим углом, можно, убрав одно колено. Матовый
фильтр, если он стоит, следует убрать. Если есть выбор между МБС-10 и МБС-
10М, то лучше пользоваться обычным МБС-10, не модернизированным. В
подсветке к МБС-10М установлена не лампа накаливания, а галогеновая лампа. Она
ярко светит, но её свет не фокусируется в узкий пучок. А для наших целей надо
именно яркий узконаправленный пучок света, чтобы рассматривать мелкие детали
на большом увеличении.
Методика фокусировки наших бинокуляров и микроскопов: снизив
интенсивность света до минимума (диафрагмой, реостатом на трансформаторе) и двигая
лампу в патроне, надо добиться резкого изображения спирали лампы на столике
или на любой плоской поверхности, расположенной на той высоте, на которой
будут находиться в жидкости изучаемые пауки, или на зеркальце микроскопа. Для
работы восстанавливается нужная интенсивность освещения. Принцип настройки
такой же, как настройка фотоувеличителя.
Для бинокуляров МБС-10 производится двухкратный объектив-насадка. Эта
насадка одевается на объектив бинокуляра снизу и повышает увеличение в 2 раза.
В итоге максимальное увеличение для МБС-10 достигает 201х. Такая линза очень
полезна для рассматривания пауков, особенно мелких. Заказать линзу, как и сам
бинокуляр и подсветку к нему, можно через систему магазинов «Медтехника» или
непосредственно с завода-изготовителя (ОАО «Лыткаринский завод оптического
стекла», ул. Парковая, дом 1, г. Лыткарино, Московская обл., Россия, 140080).
Для рассматривания особо мелких пауков, их деталей, а также для рисования
можно использовать и просвечивающие микроскопы самых разных модификаций.
Ёмкости. Рассматривать фиксированных пауков необходимо в спирту. Можно
вместо спирта использовать и глицерин (некоторые исследователи отдают ему
предпочтение). Для большинства пауков подойдут небольшие пластмассовые или
стеклянные чашки Петри диаметром 3-5 см. Идеальная ёмкость, в которой удобно
рассматривать пауков, — «энтомологическая солонка» (рис. 2.4в-д) с закруглённым
дном.
Если рассматривать пауков много и долго, то спирт частично высыхает и «за-
жиряется», поэтому смотреть становится неудобно. Чтобы избежать этого,
необходимо подливать новый спирт, либо вообще менять содержимое солонки.
Для рассмотрения пауков с разных сторон (анфас, сбоку) удобно использовать
кусочек ваты. Если паук крупный, для этого понадобится более глубокая плошка.
На её дно можно насыпать промытый песок (есть специальные стеклянные
шарики, но найти их сложно). Паука частично нужно погрузить в песок, и тогда он
перестанет заваливаться. Есть и другой способ. Дно плошки можно залить парафином
и, после того, как он остынет, сделать в нём лунки разного размера.
Однако все эти способы не удобны при рассмотрении мелких пауков или их
частей, так как они могут затеряться в песке, светлый парафин будет отсвечивать. Если
же сделать парафин тёмным, то тёмную пальпу невозможно будет рассмотреть.
34
2. Методы изучения пауков
в
и к
л
Как рассматривать пауков?
35
Оптимальный выход из ситуации — использовать тёмный фон, а на дно
плошки положить небольшой кусочек светлой ткани размером от 5 х 5 до 10 х 10 мм.
Ткань может быть ситцевой или хлопковой (рис. 2.4в, в', л-м). Нити ткани можно
раздвинуть пинцетом или булавкой так, чтобы сделать лунку нужного размера, и
затем положить рассматриваемый объект в лунку. Если общий фон тёмный, то даже
при самом большом увеличении свет рассеиваться не будет. Нередко пальпы паука
так хорошо закрепляется на ткани, что даже при переносе плошки не меняет
своего положения. Эта техника подходит как для работы в спирте, так и в глицерине.
Для работы с просвечивающим микроскопом удобны предметные стёкла с
лункой. В этом случае можно использовать только глицерин. Покровное стекло в
большинстве случаев излишне. Чтобы расположить пальпу, эпигину или другой объект
под нужным углом и не дать ему плавать, на дно лунки необходимо поместить
буквально несколько ворсинок, выдернутых из ваты.
Инструменты. Можно использовать два пинцета, или пинцет и
препаровальную иглу Хотя бы один из пинцетов, должен иметь как можно более тонкие концы.
Самые лучшие пинцеты — глазные. Их можно найти в магазинах «Медтехники».
Стоят такие пинцеты дорого, но они удобны и будут служить годами. Оптимально
иметь набор препаровальных игл с вершиной разных форм. Иглы легко
затачиваются мелкой наждачкой, подтачиваются об серную сторону спичечного коробка.
Очень удобны иглы, заточенные в обоюдоострый стилет.
Для препарирования крупных пауков удобны одноразовые инсулиновые
шприцы с тонкими иглами. Удобство их в том, что не приходится возиться с заточкой.
Когда игла затупилась, можно взять новый шприц (они дешёвые).
Для манипулирования с самыми маленькими объектами нужны не обычные
препаровальные иглы, а очень и очень тоненькие. Для этих целей используют иглы
для рефлексотерапии (иглоукалывания) или самые тонкие энтомологические
булавки. В качестве держателя для таких игл можно использовать любые
пластиковые трубки от шариковых ручек, автоматических карандашей. Кончик такой
пластмассовой трубки поджигается спичкой или зажигалкой, и, пока он горит, в него
вставляется иголка. Как только игла стала в нужное положение, горящую трубку
надо поместить под струю воды. Пластмасса быстро застынет, и игла окажется
прочно зафиксированной в «ручке».
Игра с фоном. При работе с бинокуляром на максимальном увеличении
возникают проблемы из-за рассеивания света. Их легко избежать, если сменить фон со
Рис. 2.4. Инструменты и приспособления для работы и фотографирования: пинцеты (а-
б), «солонки» (в-е), насадка для фотографирования (й-к).
а-б — пинцеты: «глазной» пинцет, обточенный на станке (а), и классический препаровальный
пинцет (б); в-е — солонки разного размера; ж-и — бинокуляр с фотонасадкой и фотоаппаратом;
л-н — примеры изображений полученных с помощью фотонасадки и фотоаппарата.
На дне чёрной солонки (в) кусочек ткани, на котором удобно рассматривать и фотографировать
мелкие пальпы и эпигины (л-м); д — на дне солонки две микропробирки, в которых удобно
хранить отчленённые эпигины и пальпы или мелких паучков; солоноки с закруглённым дном (в-д)
удобнее пластиковой чашки Петри (е); ж — убрано одно колено у осветителя; н — под дно
прозрачной солонки подложена полоска белой бумаги для регулировки освещения (отражения).
36
2. Методы изучения пауков
светлого на тёмный. При изучении разных пауков бывает удобнее смотреть то на
тёмном, то на светлом фоне. Это зависит как от самого паука, так и от увеличения.
Например, трихоботрии легче заметить, когда фон светлый. Не обязательно делать
весь фон светлым. Можно иметь тёмный фон постоянно, а при необходимости под-
кладывать под прозрачную солонку белую клиновидную бумажку (рис. 2.4н).
Причём бывает полезно «поиграть» с положением бумажки.
Просмотр внутренних структур. Часто возникает необходимость рассмотреть
структуры на пальпе или эпигине, скрытые под другими склеритами, или же
рассмотреть в проходящем свете тёмные объекты.
Для этого можно использовать несколько методов. Один из них: поместить
объект в гвоздичное масло и, в зависимости от размера, просветление наступает
через несколько минут или часов. Но масло не всегда легко найти, к тому же оно
очень душистое.
Другой, более доступный способ — использовать или просто глицерин
(просветление идёт очень медленно), или глицерин, смешанный с молочной кислотой
(просветление занимает меньше времени). Эпигины мелких пауков хорошо
просветляются и в глицерине, и в смеси глицерина и молочной кислоты.
Вульву (эндогину) зачастую проще и быстрее рассмотреть, особенно у крупных
пауков, если предварительно «выварить» её в растворе щёлочи КОН или NaOH
(10-20%). Для этого эпигину вначале отчленяют от брюшка, а затем кладут в
слабый раствор щёлочи и греют на спиртовке (не давая закипеть) до тех пор, пока
мышцы и другие мягкие ткани не мацерируются (растворятся). Можно поместить
эпигину на несколько часов в более крепкий раствор щёлочи и не греть, чтобы
избежать возможной потери препарата из-за выплеска. Другой вариант,
позволяющий избежать выплёскивания, — поставить греться под лампой накаливания (под
настольной лампой).
После окончания мацерации эпигину вновь помещают в спирт (или сначала в
воду, потом в спирт), а затем можно переложить в глицерин. Чтобы тонкие мембра-
низированные части стали более заметными, можно поместить объект на
некоторое время в раствор Amidoschwartz (синий краситель, используется биохимиками)
в спирте. При этом слабо склеротизованные участки приобретают синий цвет. После
раствора с Amidoschwartz, эпигину переносят в глицерин. Если бросить в спирт,
окраска быстро исчезнет (краситель вымоется). Но если окраска получилась
слишком интенсивной, то эпигину можно осветлить, подержав её в спирте (степень
осветления надо контролировать под бинокуляром), и после достижения нужной
степени просветления дальше изучать в глицерине.
Склериты пальпы можно легко отчленить, если предварительно её «вздуть»
(вывернуть бульбус). Для этого пальпу вначале держат в щёлочи, а затем кладут в
дистиллированную воду. Всё это нужно для того, чтобы гематодоха вздулась, и
бульбус вышел из цимбиума. Если пальпа не вздулась сразу, то процедуру нужно
повторить ещё раз.
Как рассматривать сети. Сети многих групп пауков, населяющих Сибирь и
Дальний Восток, изучены чрезвычайно слабо. Для многих родов Linyphiidae даже
не известно плетут ли они паутину. Поэтому изучение сетей может быть
перспективной областью исследования пауков.
Как рассматривать пауков?
37
Даже крупную сеть пауков-крестовиков бывает трудно рассмотреть из-за её
прозрачности. Самый простой и доступный способ сделать паутину хорошо видимой
как в природе, так и в лабораторных исследованиях — напыление. Напылять
можно любым мелким сыпучим порошком: крахмал, мука. Чем мельче, тем лучше.
Самый хороший напылитель — споры плауна. Раньше они продавались в аптеках,
как детская присыпка. Крахмал и другие вещества задерживаются на ловчих
нитях, но соскальзывают с опорных нитей. Споры же плауна, с их «ножками»,
облепляют всю паутину.
Как лучше напылять? Самый простой и эффективный способ: взять старый
носок, насыпать в него крахмал и затем потряхивать его вблизи сети. Порошок
будет просеиваться и оседать на сети. В поле такой носок лучше хранить в
полиэтиленовом мешке, чтобы порошок не размокал и не пачкал сумку (рюкзак).
Изучать сети мелких пауков-линифиид лучше в лабораторных условиях, сделав
садки разного размера, в зависимости от размера паука. При содержании пауков в
садках важно помнить, что пауки не только едят, но и пьют. В природе источником
воды для них служат, по-видимому, капельки росы. А в садок надо поставить
небольшую ёмкость с водой (например, бутылочную пробку) и следить, чтобы вода
там не высыхала.
Как приготовить иллюстрации. При изучении пауков важно уметь
зарисовывать и/или фотографировать. Если насекомых в большинстве случаев различают
габитуально, то пауков практически всех можно определить по форме копулятив-
ных органов. Если у вас есть свой рисунок непонятного паука, то можно
попытаться самостоятельно его определить, сравнивая свой рисунок с рисунками из
публикаций, или послать копию рисунка специалисту. Кроме того, рисунки могут
пригодиться для составления собственного определителя и для подготовки публикаций.
Карандашные наброски можно делать безо всяких приспособлений и устройств.
Для рисования используется простой тонко отточенный карандаш или
механический карандаш с диаметром грифеля 0,3 (оптимально) или 0,5 мм. Ниже мы
остановимся на вариантах рисования, пригодных для использования в публикациях.
Рисование с помощью окулярной сеточки и окуляр-микрометра. Это
самый простой способ, не требующий никаких приспособлений, кроме
отечественного бинокуляра МБС любой модификации. В окуляр-микрометр вставляется
сеточка, лучше сразу вместе с линейкой. Далее необходимо подобрать увеличение,
чтобы рисуемый объект входил в 5-10 клеточек (по вертикали или горизонтали).
Общий размер рисунка должен быть не менее 10 см. Затем расчертить лист бумаги
на аналогичное число клеток. Для удобства можно использовать
бумагу-миллиметровку или обычную бумагу в клеточку. Можно под обычную машинописную
бумагу подложить расчерченный лист. Далее смотрим в бинокуляр и проводим
контуры на листе бумаги, аналогично тому, как они идут в бинокуляре.
Первоначально рекомендуется сделать набросок, не детализируя. Например, если это паль-
па, нарисовать контур цимбиума, бульбуса, тегулярных отростков и эмболюса. Когда
набросок готов, можно сделать детализацию. Надо иметь в виду, что, когда вы
смотрите одним или двумя глазами, то рисунок будет выходить несколько разным.
Поэтому следует делать рисунок, используя или один или оба глаза. Сеточку лучше
ставить в окуляр, соответствующий вашему ведущему глазу. У большинства лю-
38
2. Методы изучения пауков
!4'
Рис. 2.5. Приёмы рисования с помощью окулярной сетки и микрометра,
а — вид в бинокуляр; б — контурный рисунок, сделанный по клеточкам; в — пример того, как
можно разбить одну клетку на 4 квадрата или даже 8 прямоугольников с помощью микрометра,
если в окуляр одновременно вставить и сеточку и микрометр.
дей ведущий глаз — правый. Определить ведущий глаз просто — это тот глаз,
который вы используете, когда целитесь.
Если рисуемый объект мелкий и вмещается в две-три клеточки, то следует
использовать микрометр. При совмещении сеточки и микрометра, каждый из
квадратов оказывается разбитым на 8 клеточек (прямоугольников) (рис. 2.5).
Рисовальный аппарат. Мелкие объекты удобнее рисовать с помощью
микроскопа. Для микроскопов существуют два типа рисовальных аппаратов —
рисовальная трубка или призма с зеркалом. Как их использовать, сказано в инструкции.
Рисовальный аппарат можно сделать и для бинокуляра, переделав из обычного
микроскопного (того варианта, где используется зеркало). С помощью
бинокулярного рисовального аппарата можно рисовать крупные объекты и габитус. Если при
использовании микроскопа можно свободно наводить резкость на любой слой, и
Как рассматривать пауков?
39
при этом линии не сбиваются, то при рисовании через бинокулярный рисовальный
аппарат приходится рисовать, не меняя фокус, иначе все линии сдвинутся. Для
бинокуляров некоторых зарубежных производителей (например, Motic), существуют
рисовальные аппараты, в которых на призму нанесена сеточка.
Вначале следует нанести общие линии (контуры объекта), затем постепенно
переходить к более частным. Когда набросок готов, можно убрать рисовальный
аппарат и сделать детализацию рисунка.
Масштаб. Каким бы способом не был сделан рисунок, на нём важно выставить
масштаб! Зная цену деления на сеточке окуляр-микрометра при увеличении, на
котором выполнен рисунок, и тот факт, что сторона квадрата окулярной сеточки в
бинокуляре равна 10 делениям, а на микроскопе — 5 делениям, надо рядом с
рисунком изобразить масштабную линейку определённой длины. Цена деления для
бинокуляров серии МБС при увеличении 0,6х — 0,17 мм, Iх — 0,1 мм, 2х — 0,05
мм, 4х — 0,025 мм, 7х — 0,014 мм. В зависимости от увеличения длина
масштабной линейки на рисунке может быть от 0,1 до 1 мм.
Окончательная обработка рисунка. Выше сказано, как сделать черновой
рисунок. В этом параграфе остановимся на том, как сделать этот рисунок пригодным
для публикации. Чистовой рисунок может быть: (1) сделанным тушью, или (2)
карандашным, выполненным на специальной бумаге, или (3) на обычной бумаге с
использованием карандаша и линеров. Линеры (pigment liner) — это капиллярные
ручки, надписи которыми не размываются водой и спиртом. На них обычно есть
соответствующая надпись («waterproof» и т.п.). Линеры бывают разных
диаметров. Для наших целей нужны линеры диаметром от 0,05 до 0,6 мм. Минимальный
набор — три линера с диаметром 0,1, 0,3 и 0,5 мм. Лучше использовать больше
линеров. Один из нас (Н.К.) использует линеры 6 разных диаметров и два карандаша —
тонкий и твёрдый (для контурных линий), и толстый и мягкий (для тонировки).
1. Для рисования тушью желательна специальная бумага, называемая
мелованной, или специальная калька, или бумага для черчения, или ватман. Используя све-
тостолик, переводят черновой рисунок на мелованную бумагу или кальку. Светос-
толик представляет собой ящик со стеклянной стенкой, внутри которого находятся
электрические лампочки. Яркий свет просвечивает через листы бумаги,
лежащие на стекле. Если на стекло положить оригинал (черновой рисунок), а сверху
на него — ещё один лист (для чистового рисунка), то чтобы сделать чистовой
рисунок, надо просто аккуратно обвести просвечивающий черновой вариант. Как и
при рисовании наброска, сначала необходимо провести карандашом тонкие
контурные линии, а потом обвести их тушью. Примеры рисунка тушью на рисунке
2.6г-е.
Тушью рисуют классической перьевой ручкой или специальной — рапидогра-
фом. Последние обычно дорогие, но ими удобнее рисовать. Неудобство рапидог-
рафов в том, что они быстро выходят из строя из-за забивания капилляра
высохшей тушью. Поэтому их приходится часто промывать. Как перьевые ручки, так и
рапидографы нужны разного диаметра. Минимальный набор — 2 пера — толстое
и тонкое. Толстым пером проводятся внешние контуры, а тонким — внутренние
линии. Вместо перьевой ручки или рапидографа можно использовать линеры.
40
2. Методы изучения пауков
в
б
б'
Рис. 2.6. Примеры разного типа рисунков и их редактирования: Urozelotes yutian (а, в),
Gnaphosa similis (б, е) и Pardosa palustris (г-д).
До (а-б) и после (а'-б') проведения разграничительных белых линий и нанесения
бликов; а-б — выполнены на специальной бумаге, её структура видна на рис. в (серый
режим); г-д — точечные рисунки, сделанные рапидографом (чёрно-белый режим); е —
линейный рисунок, сделан рапидографом (чёрно-белый режим).
Как рассматривать пауков?
41
Рисунок может быть исключительно контурным, или тонированным с
помощью точкования или штриховки для передачи объёма.
2. Для этого варианта необходима специальная рифлёная бумага типа Coquille-
board (рис. 2.6а-в). Найти её в России нелегко, на Западе её найти проще, но она
очень дорогая. На этой бумаге легко показывать объём рисуемых структур.
Контурный рисунок-черновик следует перенести на рифлёную бумагу с помощью того
же светостолика.
Границы склеритов лучше выделить линером, а затем перейти к штриховке.
Карандаши при таком методе лучше использовать мягкие (М; по европейской системе
обозначений — В), с более чёрным графитом, нежели обычные. В ряде случаев
можно сделать ретушь, используя мизинец, пинцет с листиком бумажки или ваткой.
Готовый рисунок можно покрыть бесцветным лаком для волос, или
специальным лаком для графических работ. Но гораздо проще рисунок сразу
отсканировать, и потом иметь дело только с цифровым изображением.
3. Самый удобный и простой способ рисования, на наш взгляд, — это
использование линеров и штриховки карандашом, а затем ретуши в графическом редакторе
(рис. 2.7а). При этом нет необходимости переносить рисунок на другую бумагу (то
есть, по сути дела, дважды делать один и тот же рисунок). Для рисования пригодна
и миллиметровка, и обычная бумага для принтеров. Если вы рисуете по сеточке, то
разлиновывать лист лучше цветными линиями (красный, зелёный, синий). Когда
черновик готов, следует оконтурить склериты линером. Более толстые склериты —
толстыми линерами, а мембранки — самыми тонкими. Когда контуры всех частей
нанесены, мягкой резинкой следует убрать карандашные линии. После этого
можно приступить к растушёвке. Не обязательно растушёвывать все части. Если цимби-
ум одноцветный, то нет смысла его «раскрашивать». Больше детализировать можно
лишь части (структуры), важные для идентификации. Растушёвка может быть не
очень детальной (тщательной), поскольку рисунок будет «доводиться» в
графическом редакторе. Можно поставить блики «штрихом» — белой краской, используемой
для исправления опечаток в документах (продаётся в канцелярских магазинах).
Доводка рисунка в графическом редакторе. После того, как готов рисунок
или готовы только контуры (штриховку ещё не делали), его следует отсканировать.
В зависимости от размера можно сканировать с разрешением 300-600 dpi (чем
крупнее рисунок, тем с меньшим разрешением). Если у вас нет цветных линий,
которые нужно убрать, тогда рисунки лучше сохранять в сером режиме (gray scale)
в формате .tif, но не jpg. Последний формат самый экономный по размеру, но при
каждом последующем сохранении качество линий и точек ухудшается, и они
превращаются в размытые полосы (линия с ореолом). Если есть цветная сетка
(клетки), то сохранять нужно в цветном режиме, с тем, чтобы затем удалить
определённый цвет. Чтобы убрать цветные линии, надо изображение разбить на три
цветовые канала RGB (red, green, blue — красный, зелёный, синий) и из трёх
результирующих картинок оставить (сохранить) ту, где нет цветных линий.
После того как изображение отсканировано, можно заняться его
редактированием. Самые распространённые и удобные редакторы — это Photoshop и PhotoPaint.
Последний продаётся в пакете с CorelDraw (редактор, необходимый для компонов-
42
2. Методы изучения пауков
400dpi a 400dpi
оригинал после размыв
серый режим Smart Blur
250d ' в 200d
ч/б ч/б
150dpi - 100dp"
ч/б ч/б
Рис. 2.7. Пример трансформации карандашного рисунка брюшка Gibbaranea omoeda в
чёрно-белый точкованный.
а — оригинал, разрешение 400 dpi; б — после небольшой размывки (Smart Blur); в-е — после
изменения разрешения в 250, 200, 150, 100 dpi и перевода в чёрно-белый режим.
Как рассматривать пауков?
43
ки снимков и незаменимый для создания карт). Оба графических редактора имеют
одинаковые возможности. У них разные интерфейсы и названия функций, однако,
добиться можно одинакового результата. Мы предпочитаем использовать Photo
Paint. К печати можно подготовить как рисунок в сером режиме, так и
классический — «точкованный».
Плохо проведённые (неудачные) линии всегда можно подправить (подтереть).
После того как рисунок приобрёл требуемый вид, его можно улучшить, проведя
белую линию вдоль границы склеритов. Линию следует проводить по контуру того
склерита, который выше (рис. 2.6а-б, а'-б'). Рисунок можно украсить бликами (рис.
2.6.а'-б'). Неудачную штриховку легко подправить с помощью размывки (Smart
Blur или Gaussian Blur), либо с помощью ретуши Effect Tool (Smudge).
Если рисунок готовится классическим, тогда после размывки (Smart Blur)
следует уменьшить рисунок до размера чуть больше того, какой вы планируете для
печати, и одновременно уменьшить разрешение до 200 dpi (при более высоком
разрешении точек получится слишком много и они будут мелкими). После этого в
Рисунке (Image), необходимо выбрать Режим (Color Mode), а там — Чёрно-Белый
(Black & White) и, наконец — Jarvis. Варианты трансформации рисунка из серого
варианта в точкованный показаны на рисунках 2.7а-е.
Фотографирование. В настоящее время можно фотографировать пауков, не
обладая даже специальной оптикой. В самом простом случае можно использовать
цифровой фотоаппарат, расположенный над объективом бинокуляра или
микроскопа. Ещё лучше, если у вас есть тринокуляр («делитель») и подходящий для него
цифровой аппарат (обычно требуется переходник). Альтернатива тринокуляру —
аппарат (фотоаппарат или специальная фотоприставка), который вставляется
вместо одного из окуляров.
Особенность фотографирования для использования иллюстраций в статьях, это
то, что лишь крупные объекты можно сфотографировать с достаточной глубиной
резкости. Мелкие пауки, отдельные пальпы и эпигины не получаются в фокусе.
Однако этот недостаток легко устраним. Необходимо сделать несколько
последовательных снимков, чуть-чуть меняя фокус. Для того, чтобы сделать из серии
фотографий один резкий снимок, существует несколько программ, которые
объединяют в одну фотографию резкие участки разных снимков (CombineZM и Helicon
Focus). На наш взгляд более удобна вторая программа, но она платная. У неё
гораздо проще меню, работает быстрее, и результат обычно лучше. Обе программы
можно найти в Интернете.
Несколько рекомендаций по редактированию. Улучшить любую фотографию
можно с помощью тех же графических редакторов, что и рисунки. Контраст
можно (лучше) усилить, затемнив тёмные участки и осветлив светлые вручную с
помощью Effect Tool (Brighten/Darken). Для тонкой ретуши следует выбирать
инструмент с нерезкими краями (нимбом). Сделать более заметными границы между
склеритами можно затемнив край верхнего склерита, и осветлив тот участок, где
нижний склерит уходит под верхний.
Поскольку фотографии печатаются в сером режиме (печать цветных снимков
очень дорогая), то снимки следует или перевести в серые, или делать пробную
печать, чтобы оценить, как видны те или иные структуры в градациях серого
цвета. В настоящее время для публикации можно сдавать в журнал цветные иллюст-
44
2. Методы изучения пауков
рации. Тогда в напечатанном виде фотографии будут чёрно-белыми (серыми), а
PDF версия вашей статьи будет иметь цветные снимки.
Какую использовать литературу?
В настоящее время существует достаточно много разных руководств, благодаря
которым можно определять до видов, родов и семейств пауков, населяющих
азиатскую часть России. Однако нет ни одного труда, где все сведения были бы сведены
воедино, и нет статей ни по одному из семейств на русском языке.
Самым главным источником, благодаря которому можно получить достаточно
полную информацию, являются периодически обновляемый каталог Н.И.
Платника (Platnick, 2010, www). Там имеются ссылки на все публикации, где есть рисунки
того или иного вида, и общие сведения об ареале. У каталога Н.И. Платника есть
свои недочёты (см. Марусик, 2009). Распространение видов дано зачастую
неверно. Если вид указан из одной точки, скажем из Калининградской области,
географически принадлежащей к Средней Европе, распространение указано как
«Россия». Список синонимов не всегда полный (часто нет данных до 1950 г.). Ссылки
на иллюстрации даны не критически. Зачастую на иллюстрациях приведены не те
виды, которые указаны в подписях.
Важен также каталог пауков бывшего СССР и три его дополнения (Mikhailov,
1997, 1998, 1999, 2000). В нём можно найти список всех пауков,
зарегистрированных к востоку от Урала, вплоть до 2000 г. Каталог К.Г. Михайлова (Mikhailov, 1997,
1998, 1999, 2000) имеет недостатки — распространение вида в пределах России
дано очень схематично, а также из каталога нельзя узнать, кто отмечал тот или
иной вид. Поэтому не всегда понятно, насколько можно доверять находкам видов в
необычных точках. Кроме того, в этом каталоге нет подробной синонимии, не
учтены новые перестановки (новые комбинации), произошедшие в 1997-2010 гг.
Однако в двух каталогах, вместе взятых, при желании можно найти почти
исчерпывающую информацию по тому или иному виду или роду.
Определители семейств, альтернативные нашему, но на английском языке,
содержатся в монографиях Roberts (1995, Средняя и Северная Европа), Dippenaar-
Schoeman & Jocque (1997, Африка), Song et al. (1999, Китай), Ubick et al. (2005,
Северная Америка), Ono et al. (2009, Япония), в обновляемом
интернет-определителе Nentwig et al. (2010, Европа). Недавно опубликован определитель семейств
пауков всего мира (Jocque & Dippenaar-Schoeman, 2006). Особенность указанных
определителей в том, что они не адаптированы к таксонам, населяющим
азиатскую часть России.
Для определения отдельных широко распространённых видов из Сибири и
Дальнего Востока можно использовать определители Roberts (1985, 1987, 1995, 1998),
Palmgren (1974a-b, 1975, 1976, 1977), Nentwig et al. (2010), Almquist (2005, 2006).
Для определения южносибирских видов незаменима книга о китайских пауках Song
et al. (1999). Для пауков, обитающих на юге Дальнего Востока, наиболее подходят
упомянутые определители Song et al. (1999) и Ono et al. (2009), а также Chikuni
(1989, Япония) и Namkung (2002, Корея). Виды, имеющие Голарктические или Си-
биро-Неарктичские ареалы, можно определять по определителям-монографиям се-
веро-американской фауны (Dondale & Redner, 1978, 1982, 1990; Platnick & Dondale,
1992; Dondale et al., 2003).
Какую использовать литературу?
45
Необходимо упомянуть, что в указанных работах, даже в самых последних, нет
единообразия в трактовке некоторых семейств. Это связано с тем, что некоторые
замечательные определители (например, Робертса и Чикуни) написаны
любителями, которые склонны придерживаться традиционных систем, но не современных.
В случае разночтения, следует ориентироваться на ту трактовку объема семейств,
которая принята в каталоге Н.И. Платника (Platnick, 2010). Это самый известный,
наиболее доступный и часто обновляемый справочник.
Подробная информация по систематике, определению, распространению и
биологии пауков азиатской части России имеется только для пауков-скакунчиков
(Logunov & Marusik, 2000b). Несколько менее подробные сведения даны в
монографии по паукам Тувы (Marusik et al., 2000). Там приведены ссылки на основные
источники для определения, сведения об ареалах и карты распространения для
более 600 видов.
Относительно подробные сведения о Linyphiidae азиатской части России
(список, синонимия, распространение по крупным выделам) можно найти в
монографии К.Ю. Еськова (Eskov, 1994).
Как определять собранных пауков?
В этом разделе мы предлагаем возможные алгоритмы определения пауков.
1). Определить семейство. Лучше начать с тех пауков (видов) которые
представлены в сборах и самцами и самками, на вторую очередь оставить виды,
представленные самцами, затем — только самками. Определение самок во многих
группах даже до семейства затруднительно (и не только для начинающих).
2). Если вид не определяется по доступным определителям, посмотреть к
какому роду он близок. Пауков из любого региона стоит «прогнать» по определителю
пауков Европы (Nentwig et al., 2010), однако идентифицировать по нему много
видов можно только в Западной Сибири.
3). Если род удалось установить (по сходству копулятивных органов), тогда
следует сверить, встречается ли такой род к востоку от Урала. Если да, тогда в
каталоге К.Г. Михайлова надо посмотреть, какие виды встречаются у нас. Есть
сложность: вид мог быть перенесён в другой род, тогда надо в каталоге N.I. Platnick (на
www) посмотреть в каких родах встречалось то или иное видовое название.
4). Посмотреть по каталогу N.I. Platnick (на www), в каких публикациях
приведены рисунки тех или иных видов, и нет ли у вас доступа к этим работам. По ним
можно попробовать определить.
5). Если вид и даже род или семейство не установлены, можно сделать
схематический рисунок пальпы и эпигины, а также внешнего вида, если на карапаксе,
брюшке или ногах есть какие-либо особенности. Если позволяют условия, можно
сделать и цифровые фотографии. Затем изображения пауков можно выслать
специалисту по той или иной группе (или нескольким сразу). Электронные адреса
русскоязычных специалистов приведены в приложении Б.
Рисунки и фотографии пригодятся для разных целей, и для создания
собственного определителя, и для последующего сличения вашей иллюстрации с
рисунками в других работах. В своё время нам (Ю.М.) удалось изучить фауну верховьев
Колымы, где обитало более 150 неописанных видов, посылая карандашные
рисунки Linyphiidae К.Ю. Еськову Если он не признавал паука по рисунку, тогда мы
посылали ему самих пауков.
46
2. Методы изучения пауков
В азиатской части России есть ещё немало неописанных видов и родов. Как
попытаться установить к какому роду относится паук? Для этого не обязательно
пересматривать все виды, относимые к тому или иному роду В первую очередь, по
каталогу N.I. Platnick (www) необходимо узнать, какой типовой вид рода. В
каталоге такие виды отмечены звёздочкой. Затем своего паука можно сравнивать с
разными типовыми видами. Разумеется, многих пауков так определить не удастся
(например, многих Linyphiidae, особенно Lepthyphantes sensu lato).
Долгий, но «спасительный» для начинающих метод определения — это
просмотр всех подряд рисунков во всех доступных (упомянутых) работах и отбор
среди этих рисунков наиболее похожих на вашего паука. Такой «тупой» метод годится
для самцов, но не для самок, так как похожие очертания эпигин встречаются у
представителей разных семейств. Отобранные похожие рисунки сравниваются с
пауком повторно и среди них выбираются самые похожие. Таким путём можно
если не определить вид, то, по крайней мере, найти виды, наиболее похожие на
ваш. При этом важно не пытаться «подогнать» ваш вид под имеющиеся рисунки.
Надо тщательно сравнить свой материал с похожими рисунками из литературы,
сверить свои экземпляры не только с рисунком, но и с описанием, причём
сконцентрироваться именно на поиске отличий, а не сходств. Тогда есть шанс не только
определить некоторые виды «тупым сравнением», но, возможно, и выявить виды,
новые для науки.
3. СТРОЕНИЕ И КЛАССИФИКАЦИЯ ПАУКОВ
Морфология и терминология
Тело паука разделено на две основные части (тагмы), головогрудь и брюшко (или
просому и опистому). Две основные части соединены узким стебельком (petiolus).
Основные части головогруди, где сосредоточены диагностические признаки, это ка-
рапакс, стернум, глаза, ротовой аппарат (хелицеры, максиллы и нижняя губа) и ноги.
Брюшко характеризуют следующие признаки, заслуживающие внимания: его
форма, верхняя и нижняя поверхности (наличие или отсутствие хитинизированных
щитков, например, скутума, на спинной или брюшной стороне), паутинные бородавки,
органы дыхания (отверстие трахей, легочные крышечки). Но особенно много
диагностических черт — в строении копулятивных органов, которые находятся на пе-
дипальпе у самца, и перед эпигастральной щелью на нижней стороне брюшка у
самки. Немало характерных черт бывает и в окраске паука (цвет, тип рисунка).
Карапакс (рис. 3.1а, ж, е) представляет собой большой склерит, покрывающий
головогрудь сверху. Его можно подразделить на головную и грудную части. У
некоторых видов это разделение чёткое, и две части разделены бороздой или
рисунком. По форме карапакс бывает круглым, овальным или грушевидным. Овальным
(яйцевидным) мы называем карапакс, где нет явственного сужения головной
части, а грушевидным — карапакс, головной отдел которого отчётливо сужен.
Головная часть несёт 6 или 8 глаз. У некоторых видов эта часть может быть
слегка приподнята над грудной или наоборот, может быть ниже грудной части
(Oonopidae, самцы Hilaira из Linyphiidae и др.). Часть видов имеют выросты
головной части (вперёд или вверх), пару ямок по бокам, заглазничные щели,
модифицированные волоски (Linyphiidae, Theridiidae). Область карапакса, которая
лежит ниже переднего ряда глаз, называется наличником или клипеусом.
В центре грудной части обычно находится так называемая медиальная щель
(борозда), она может быть поперечной или продольной. От щели внутрь
головогруди отходит вырост, к которому крепятся мышцы. Некоторые виды имеют
выросты грудного отдела (из семейств Linyphiidae, Oonopidae).
Иногда весь периметр карапакса несёт латеральные шипы (Erigone из
Linyphiidae), или весь карапкс покрыт серией точек-вдавлений (Lophomma из
Linyphiidae). Нередко карапакс покрыт густыми волосками, иногда образующими рисунок
(Salticidae). Волоски вокруг глаз могут образовывать структуры, подобные
ресницам, или бровям. У некоторых групп волоски чешуйчатые со сложной структурой
поверхности (Salticidae; Micaria из Gnaphosidae), у других утолщённые волоски-
щетинки могут иметь булавовидную форму (Ozyptila из Thomisidae).
Стернум или грудной щит (рис. 3.16) находится на нижней стороне
головогруди между тазиками. Изредка с ним сливается нижняя губа. По форме он может
быть щитовидным, округлым, овальным, почти прямоугольным или треугольным.
48
3. Строение и классификация пауков
голень,
предлапка
лапка —
коготок. хедицера
лабиум
трихоботрия
парацимбиум
тегулюм
Морфология и терминология
49
Иногда стернум имеет выросты, проходящие между тазиками; в некоторых
случаях эти выросты соединяются с аналогичными выростами карапакса.
Порой карапакс и стернум покрыты пунктировкой (точечными вдавлениями)
или вермикуляцией (извилистыми бугорками), имеют зернистую поверхность (с
точечными бугорками), или несут рисунок. Рисунок карапакса и/или стернума в
некоторых случаях бывает видо- или родоспецифичным.
Глаза (рис. 3.1 ж-з, й). Подавляющее большинство видов, населяющих
азиатскую часть России, имеет 8 глаз. Все глаза простые (не фасеточные), хотя у
некоторых могут быть сильно усложнены. Глаза располагаются на переднем крае
карапакса, хотя у ряда групп задние глаза сдвинуты кзади (Lycosidae, Salticidae, Eresidae).
У большинства групп пауков глаза расположены в два ряда. Эти ряды могут быть
прямыми, выгнутыми или вогнутыми. В отечественной арахнологии (например,
Тыщенко, 1971) изогнутость рядов глаз оценивается при виде сверху, а в
американской литературе принято оценивать, наоборот, при виде спереди (Ubick et al., 2005).
Соответственно, то, что у нас считается выгнутым рядом, для западных
исследователей кажется вогнутым. Глаза могут располагаться компактно (касаться друг
друга), либо равномерно раздвинуты на всю ширину карапакса. Боковые глаза часто
расположены рядом. Во многих случаях величина и взаимное расположение глаз,
вместе с размером наличника, могут быть использованы для определения семейств,
родов и даже видов.
В большинстве случаев глаза округлой формы, но у ряда групп некоторые глаза
могут быть угловатыми (треугольными), например у Gnaphosidae. Ещё глаза
бывают (при рассмотрении фиксированного материала) чёрными или серебристого цвета.
Считается, что чёрные глаза служат для ночного зрения, а серебристые — для
дневного. У Salticidae средние глаза переднего ряда часто ярко окрашены: могут быть
синими, зелёными, красными.
Ротовой аппарат (рис. 3.16) состоит из пары хелицер, нижних челюстей, или
максилл (=вырост тазика педипальп, =жевательная лопасть педипальп, =коксопо-
дит), непарной нижней губы (лабиум), и собственно рта. Последний прикрыт
нижней губой.
Хелицеры состоят из двух частей — собственно хелицеры (базальный членик,
paturon) и её коготка, у вершины которого открываются протоки ядовитых желез
(вырабатывающих, помимо яда, пищеварительный сок). Хелицеры бывают
разнообразной формы, и их длина всегда больше ширины. Коготок хелицеры в спокой-
Рис. 3.1. Детали строения пауков.
а — Ceratinella sp., сверху; б — Gnaphosa sp., снизу; в — брюшко Hahniidae, снизу; г —
паутинные бородавки и крибеллюм, Amaurabiidae; д — паутинные бородавки, Agelenidae; e —
головогрудь самца Tarsiphantes latithorax, сбоку; ж-з — головогрудь Lycosidae, сверху и спереди; и —
лапка IV, Theridiidae; й — головогрудь спереди, Salticidae; к-л — эпигина Neriene sp. и Oreonetides
vaginatus, снизу; м — лапка и предлапка IV, Dictynidae; н — лапка и предлапка I, Linyphiidae,
показано как измерять положение трихоботрии; о — пальпа самца Hilaira herniosa.
Сокращения: 36 — задние бородавки; ЗЛГ — задние латеральные глаза; ЗМГ — задние
медиальные глаза; Пб — передние бородавки; ПЛГ — передние латеральные глаза; ПМГ — передние
медиальные глаза; Сб — средние бородавки.
50
3. Строение и классификация пауков
ном состоянии вложен в желобок на хелицере, наподобие лезвия перочинного ножа.
Многие таксоны имеют зубчики вдоль переднего и заднего края желобка хелицер,
некоторые зубцы могут быть сильно увеличены. Иногда вместо зубчиков или в
дополнение к ним имеются модифицированные волоски. Хелицеры могут быть
сильно удлинены, тогда они обычно вытянуты несколько вперёд. Помимо зубцов,
расположенных вдоль желобка, могут быть дополнительные зубцы на передней
или боковой поверхности базального членика (Enoplognatha, Tetragnatha, Zornella).
У некоторых таксонов верхняя-боковая часть хелицеры с мощным валиком
(мыщелком) к которому крепятся мышцы (например, у Araneidae).
Хелицеры (базальные членики) могут сливаться в основании. В этом и
некоторых других случаях коготок бывает очень маленьким. Нередко по длине он
меньше ширины базального членика. Форма коготка бывает разной, от почти
треугольной до саблевидной. Обычно базальная половина коготка существенно толще
вершинной его части, которая постепенно суживается. Внутренняя часть сужающейся
половины почти всегда с мелкой пилочкой. У Tetragnatha основание коготка
может быть с выростами (зубцом или более сложными выростами).
Нижние челюсти всегда несколько длиннее своей ширины, они бывают
наклонены друг к другу или могут быть параллельными. Вершины челюстей могут быть
сужены, или закруглены по сторонам, но всегда имеют по внутреннему краю
«щётку» из специальных волосков, служащих фильтром, который защищает
небольшой рот от попадания крупных частичек пищи. У некоторых групп внутренний
или наружный-верхний край максилл видоизменён — несёт выросты (Hahniidae,
Oonopidae, Salticidae).
Нижняя губа у большинства групп свободная (не срослась со стернумом). Чаще
всего длина губы равняется или чуть меньше её ширины, но губа может быть и
удлинена. По форме она бывает полукруглой, треугольной, пентагональной, или
прямоугольной. У Linyphiidae верхняя (передняя) часть губы несколько расширена
и утолщена.
Ноги. Всегда четыре пары (рис. 3.1а), каждая из ног состоит из 7 члеников:
тазик, вертлуг, бедро, колено, голень, предлапка и лапка. В арахнологии принято
называть ноги по номерам от первой пары до четвёртой, и в тексте (при
написании) их обозначают римскими цифрами I, II, III и IV. Ноги всегда покрыты
волосками и у большинства групп несут утолщённые щетинки (=шипы). Шипы (их
число и расположение) записываются в виде разных формул, но последовательность
всегда одна — от основания членика к его вершине. На ногах всегда есть
чувствительные волоски (трихоботрии) и различные органы (тарзальный, лировидные,
щелевидные, и др.). В некоторых случаях волоски могут располагаться густо и
образовывать так называемые щётки (скопула) на нижней стороне лапок, а иногда
и предлапок. Густо расположенные волоски могут быть по всему периметру
голени (некоторые Lycosidae), в других случаях удлинённые волоски имеются только
по боками голени и предлапки. Форма и расположение шипов нередко являются
таксоно-специфичными (характерными для семейства, рода или вида). Шипы
иногда бывают очень длинными и могут быть сравнимы с длиной предлапки.
Расположение и число трихоботрии (это нитевидный чувствительный волосок,
сидящий в ямке чашевидной формы), как и шипов, нередко таксоно-специфично.
В систематике Linyphiidae используется понятие «расположение трихоботрии пред-
Морфология и терминология
51
лапки I» выраженное числом от 0 до 1. Вычисляется положение следующим
образом (рис. 3.1н): измеряется расстояние от основания предлапки до трихоботрии, и
это число делят на длину предлапки (а / Ь).
Лапка несёт на вершине два или три коготка. Все сетеплетущие группы имеют
три коготка. Два из них парные, третий расположен между ними и всегда меньше
размером. Обнаружить (рассмотреть) его бывает трудно. Большинство бродячих
пауков имеют два коготка. При этом под коготками есть пучок волосков,
называемый коготковой (подкоготковой) подушечкой (claw tuft). Коготки почти всегда
несут на себе выросты-зубчики; если их много, то коготок выглядит как расчёска. В
некоторых группах (Oonopidae, например) коготки сидят на выросте лапки,
называемом онихумом. Лапки IV у Theridiidae и Nesticidae с вентральным рядом
пильчатых щетинок (рис. 3.1 и). У пауков, имеющих крибеллюм, на предлапке IV всегда
есть каламиструм — ряд слегка изогнутых волосков, образующих
гребень-расчёску для плетения крибеллятной паутины (рис. 3.1м). Бывают каламиструмы,
состоящие не из одного ряда волосков, а из двух.
Ноги могут быть примерно равной длины, но часто бывают разной. Для
удобства изложения при описании семейств мы приводим данные в основном по
первой паре ног. Длинной ногой I мы называем ногу, которая в полтора и более раз
длиннее тела. У бродячих пауков-засадников (Thomisidae) две задние пары
существенно короче передних двух пар. У пауков-волков (Lycosidae) ноги примерно
одинаковой длины, но четвёртая пара самая длинная. Иногда используется
понятие «формула ног». Под формулой подразумевается последовательность ног, от
самой длинной до самой короткой. Например, 4:1:2:3 обозначает, что нога IV
самая длинная, а III — самая короткая. Относительная длина и толщина ног
значительно варьирует. У многих групп первая пара ног длиннее тела (иногда в 5 и более
раз). У коротконогих видов пауков ноги обычно коренастые. Передняя пара у
самцов некоторых видов (особенно у Salticidae) может быть утолщена, иметь особую
окраску, нести многочисленные железы, видоизмененные волоски.
Педипальпы (или пальпы). Это вторая пара конечностей (вслед за хелицера-
ми). Они похожи на ходильные ноги, но короче и состоят из 6 члеников: тазик
(вырост тазика образует максиллу), вертлуг, бедро, колено, голень, и лапку (пред-
лапка и лапка слиты). Лапка несёт в ряде случаев один коготок, нередко лишённый
зубчиков. Иногда коготок по толщине меньше окружающих его щетинок, и
поэтому малозаметен. У взрослых самцов всех групп пальпа видоизменена и служит
органом копуляции (об этом — в разделе про копулятивные органы). Практически
у всех групп, субадультные (предимаго) самцы имеют явственно утолщённые
лапки педипальп, но не имеют совокупительных органов.
Брюшко. Брюшко соединено с головогрудью тонким стебельком (рис. 3.1а, б).
Последний обеспечивает особую подвижность брюшка, необходимую для
плетения (ловчей паутины, коконов). Брюшко варьирует по форме. Оно бывает
сферическим, овальным, яйцевидным, каплевидным (заострённым на конце), пентаго-
нальным. Может нести угловидные (один и более) или хвостовидные выросты. В
некоторых случаях брюшко может быть уплощено или сильно удлинено.
На нижней стороне, в передней трети-четверти, имеется поперечная борозда
(щель), называемая эпигастральной (эпи — сверху, гастер — брюшко). Брюшко
52
3. Строение и классификация пауков
значительно меньше хитинизировано, нежели головогрудь, поэтому величина и
форма его может существенно варьировать. Так, у голодного паука могут быть
заметны складки (морщины), а у напитавшегося брюшко округляется и складки
разглаживаются. Брюшко всегда покрыто волосками. Волоски на верхней и нижней
стороне брюшка нередко разной формы. Иногда волоски видоизменяются в шипи-
ки, например, булавовидной формы у Ozyptila. Самцы некоторых пауков-волков
снизу на брюшке имеют шиловидные волоски. У нескольких видов пауков-крабов
взрослые самки всегда покрыты песчинками. Очевидно, при последней линьке
брюшко сверху имеет клейкий секрет, благодаря которому песчинки
приклеиваются к нему, маскируя паука.
Помимо волосков, брюшко может иметь дорсальный скутум (сильно хитинизи-
рованный участок овальной формы) или даже несколько скутумов. Помимо
дорсального, скутум может быть вентральным и даже кольцевидным (вокруг
паутинных бородавок).
У многих групп брюшко имеет пигментный или структурный рисунок. Рисунок
может быть только дорсальным, но бывает и на боках и на вентральной
поверхности. У большинства групп в передней части брюшка имеется медиальное
удлинённое пятно, обычно тёмное (но оно может быть и светлее других частей брюшка),
которое для удобства мы называем ланцетовидным (heart mark, heart spot). Часто
рисунок бывает листовидным (folium).
Брюшко несёт на себе органы дыхания и паутинные бородавки, а у самок ещё и
копулятивные органы, речь о которых пойдёт ниже.
Органы дыхания (рис. 3.1б-в). Большинство видов, населяющих азиатскую
Россию, имеет одну пару так называемых лёгких и непарное дыхальце. Два вида
(из семейства Atypidae) имеют две пары лёгких, и один вид (из семейства Oonopidae)
имеет две пары дыхалец, первая пара расположена на месте легочных отверстий, а
вторая — чуть ниже. Лёгкие представляют собой видоизменённые конечности.
Первая пара (есть у всех наших пауков, кроме Oonopidae) располагается перед эпигас-
тральной щелью. Вторая пара, если она есть, или парные дыхальца
(трансформированная в трахеи вторая пара лёгких), расположены за эпигастральной щелью.
Непарное дыхальце у подавляющего большинства видов расположено чуть
спереди от паутинных бородавок. В некоторых случаях (Argyronetidae, Anyphaenidae,
Hahniidae) дыхальце сдвинуто вперёд и располагается примерно посередине
между эпигастральной щелью и паутинными бородавками. У групп со сдвинутым
дыхальцем, последнее заметить очень легко. Дыхальце представляет собой
поперечную щель, особенно широкую у тех групп, у которых дыхальце сдвинуто вперёд от
бородавок. У групп с дыхальцем, находящимся у бородавок, оно тоже в виде щели,
но щель короче, и иногда её бывает трудно заметить.
Паутинные бородавки. Почти все наши виды имеют три пары паутинных
бородавок (видоизменённые конечности), которые обычно расположены на конце
брюшка, и могут быть видны сверху. Только у рода Zodariellum (Zodariidae) всего
одна пара паутинных бородавок. Разные бородавки называют: передними,
средними и задними (рис. 3.1г-д). У пауков, имеющих хвостовидный вырост, или у
некоторых пауков со сферическим брюшком, бородавки могут быть несколько
сдвинуты кпереди. Паутинные бородавки подвижны (имеют свою мускулатуру). Они мо-
Морфология и терминология
53
гут быть одно-, двух- и даже трёхчлениковыми. Все пары бородавок достаточно
чётко отличаются друг от друга. Они могут быть цилиндрическими,
конусовидными, и даже хвостовидными. Размер, толщина, взаимное расположение, форма
бородавок часто служат диагностическими признаками для разграничения семейств
и родов. Многие виды между передними бородавками имеют конический бугорок,
называемый колюлюсом. Иногда он может быть сильно или полностью
редуцированным. В случае неполной редукции он может быть представлен парой волосков.
У крибеллятных групп спереди от паутинных бородавок есть поперечная
пластинка, нередко разделённая пополам (крибеллюм). На крибеллюме имеется
множество маленьких паутинных желёзок, секретирующих крибеллятную паутину.
Сразу за паутинными бородавками (над ними) находится анальный бугорок.
Копулятивные органы. Копулятивные органы самок расположены в средней
части эпигастральной щели (рис. 3.16; 3.2а-б), а у самцов — на педипальпах (рис.
3.1е, о).
Копулятивный орган самок у большинства пауков представлен непарной эпи-
гиной («эпи» — над) и эндогиной («эндо» — внутри; =вульвой). У гаплогинных
пауков эпигины, как правило, нет (исключение — Pholcidae). Эпигина отсутствует
и у Tetragnathinae (Tetragnathidae). Эпигина представляет собой хитинизирован-
ную пластинку, нередко с выростами, на которой расположены ямка или два
отверстия (дырки или щели) для совокупления. Помимо отверстий и выростов, могут
иметься разного рода карманы, углубления, валики, щели. Центральная часть
эпигины лишена волосков. Эндогина (внутренняя часть эпигины) имеет пару
канальцев или щелей (=семяпроводящие канальцы), идущих от совокупительных
отверстий к рецептакулам (рис. 3.26). Рецептакулы (=семяприёмники, или спермате-
ки) — это резервуары, в которые попадают и временно там хранятся спермин.
Рецептакулы могут быть круглыми, овальными, каплевидными, фасолевидными или
удлинёнными. Удлинённые рецептакулы могут быть «свёрнутыми» в клубок и иметь
множество внутренних карманов. Если семенные канальцы длинные, то на них
нередко имеются дополнительные железы; иногда железы могут открываться
прямо в рецептакулу (Euophriinae из Salticidae). От рецептакул отходит по одному
относительно короткому резко суживающемуся оплодотворительному канальцу. По
нему сперма попадает в матку.
У гаплогинных пауков один и тот же каналец служит для поступления спермы в
рецептакулы и для вывода спермы из неё (рис. 3.2а). У этих пауков
совокупительные отверстия находятся в эпигастральной щели.
Форма выростов, ямок, карманов, щелей эпигины, и устройство эндогины в
большинстве случаев видоспецифичны. Кроме того, по форме и структуре
эпигины во многих случаях можно определить род, подсемейство и семейство пауков.
У самцов копулятивные органы находятся на апикальном членике педипальпы.
Нередко вся пальпа служит совокупительным органом, например у всех
гаплогинных пауков, а также у некоторых Salticidae, Gnaphosidae и др. У гаплогинных вся
пальпа погружается в эпигастральную щель. У других пауков бывают специальные
выросты на бёдрах, которые, так или иначе, принимают участие в копуляции.
Нередко и хелицеры участвуют в этом процессе (например, у Pholcidae и Tetragnathinae).
Собственно копулятивный аппарат самца находится на лапке педипальпы. Форма
его чрезвычайно разнообразна. В самом примитивном случае он представляет со-
54
3. Строение и классификация пауков
Гаплогинный тип Энтелегинный тип
Рис. 3.2. Схема строения эпигины у гаплогинных (а) и энтелегинных пауков (б),
строение паутины у крибеллятных (в-д) и некрибеллятных (е-ж) пауков,
в-г — крибеллятная паутинная «нить», сфотографированная сканирующим электронным
микроскопом; д — крибеллятная нить, сфотографированная в обычный микроскоп, видны опорные
нити (слева) и «путанка»; е — клейкая нить (радиальная нить) паука-крестовика с капельками
клея; ж — то же, нить помещённая на стекло, видны две опорные нити и «капелька» клея на
большом увеличении, а-г — по: Forster & Forster (1999); д-ж — по: Foelix (1982).
бой видоизменённую лапку, называемую цимбиумом (=лодочка) с выростом —
бульбусом (=луковица). В простейшем случае бульбус сферический с
конусовидным выростом — эмболюсом. Внутри бульбуса расположен канал, в который
засасывается сперма, временно там хранится, и затем извергается во время копуляции.
Эмболюс служит для введения спермы в совокупительное отверстие эпигины.
Форма цимбиума значительно варьирует у разных семейств; иногда цимбиум
существенно отличается в пределах семейства (у Theridiidae, Salticidae) и даже в
пределах рода. В некоторых семействах цимбиум имеет вырост, называемый пара-
цимбумом («пара» — около). Парацимбиум может иметь разное происхождение, и
поэтому может быть не гомологичен в разных семействах.
У большинства наших пауков бульбус представляет собой не один склерит, а
несколько. Обычно это субтегулюм (самый нижний склерит), тегулюм и эмболюс-
ный отдел. Отдельные склериты могут вторично сливаться или редуцироваться.
Каждый из склеритов может иметь свои отростки и выросты. У большинства
наших видов бульбус причленяется к цимбиуму с помощью гематодохи (мембрани-
Морфология и терминология
55
зированная трубка, свёрнутая спиралью в спокойном состоянии, и раздувающаяся
во время копуляции из-за прихлынувшей гемолимфы). В некоторых группах есть
вторая гематодоха. Среди отростков бульбуса различают эмболюс (он может иметь
несколько дополнительных выростов), медиальный (=тегулярный) отросток (=апо-
физ, =апофиза), кондуктор и некоторые другие (терминальный отросток,
субтерминальный отросток, радикс, lamella characteristica и др.).
Одинаково называемые отростки (кондуктор, медиальный, тегулярный) в
разных группах имеют разное происхождение и не гомологичны друг другу. В связи с
тем, что существует много разных выростов-отростков бульбуса, и из-за того, что
они не гомологичны, а названия отростков даны по разным критериям — одни по
положению (медиальный, терминальный), другие по месту прикрепления
(тегулярный), третьи по форме (lamella characteristica; =характерная пластинка) или по
функции (кондуктор; =проводящий) — существует большая путаница.
Медиальный отросток часто находится не посредине, а сбоку. Lamella characteristica бывает
пальцевидной, а не пластинчатой. Кондуктор в некоторых группах не направляет
эмболюс, поскольку он тонкий и мембрановидный, а защищает его (у Drassodes,
Callobius). У ряда пауков функцию направления самого себя может выполнять
эмболюс (его основание, или отростки-выросты), или направляют эмболюс другие
структуры, вплоть до желобка цимбиума.
На копулятивных органах мы остановились достаточно подробно из-за того,
что их форма и строение играет чрезвычайно важную роль в определении не
только видов, но и родов, и выше, вплоть до надсемейств и подотрядов. Помимо
собственно бульбуса и цимбиума, важную роль в определении таксонов пауков
играют отростки голени, колена, бедра, иногда и вертлуга пальпы. Специфичным (для
семейства, рода, вида) может быть число, форма, расположение таких отростков.
Размеры. Размер паука имеет значение для определения вида. Но более важен
не общий размер (=общая длина тела; измеряется от переднего края наличника до
заднего кончика брюшка, при этом паутинные бородавки не учитываются), а
длина и ширина карапакса. Очевидно, что общие размеры меняются от степени
раздутости брюшка (например, брюшко может быть сильно раздутым после еды, или у
самки перед откладкой яиц), в то время как размеры карапакса более постоянны,
они могут увеличиться только после линьки. Длина карапакса измеряется от
переднего края наличника до самой задней точки карапакса; ширина карапакса
измеряется в самом широком его месте. Ещё более существенное значение имеет размер
эпигины и пальпы. Существуют близкие виды, которые почти идентичны по
форме пальпы и эпигины, однако хорошо различаются по размерам тела, пальпы и
эпигины. Таковы, например, Parasteatoda simulans (Thorell, 1875) и P. tepidariorum
(C.L.Koch, 1841).
У некоторых видов размеры тела варьируют очень сильно, в 2-3 раза
(например, так бывает у видов рода Drassodes). Но размеры пальп и эпигин при этом
варьируют гораздо меньше, чем размеры тела. Популяции, обитающие на пределе
своих экологических возможностей (где климат не тот) или на границе видового
ареала, могут быть диморфными по размеру. Например, Gnaphosa orites Chamberlin,
1922 в лесной зоне (лесном поясе) крупная, а в горных тундрах — мелкая.
Отличаются они по размеру карапакса, эпигины и пальпы, причём переходов нет. Есть
места, где в ловушки попадаются и мелкие, и крупные особи (стык лесного пояса и
56
3. Строение и классификация пауков
горной тундры). В чём причина? Никто не изучал, но скорее всего, из-за того, что в
разных климатических условиях бывает разное число линек. Число линек не
запрограммировано жёстко. В искусственных условиях, варьируя интенсивность
питания, температуру и фотопериод, можно добиться и большего числа линек, и
меньшего. Такие работы проводили по каракурту (Эргашев, 1990). Если экземпляр не
добрал нужного размера (холодно, бескормица), то взрослеть (достигать
половозрелой стадии) приходится ускоренно, минуя одну линьку. Самцы и самки,
достигшие зрелости после меньшего числа линек, имеют меньшие размеры карапакса и
копулятивных органов.
Методика измерений. Методика измерения общей длины, а также длины и
ширины карапакса описана выше (подраздел «Размеры»). Длина сегментов ног,
пальп, бородавок всегда измеряется по дорсальной стороне, между самыми
удалёнными точками сегмента. По дорсальной стороне измерение проводится даже в
тех случаях, когда длина дорсальной стороны меньше, чем длина вентральной (так
бывает, например, с голенями пальп).
Измеряемый сегмент должен находиться в плоскости, параллельной плоскости
объектива бинокуляра (то есть, в горизонтальной). Если сегмент (нога) находятся
под углом к горизонтальной плоскости, то измерения получаются не точными из-
за оптических искажений.
Желательно проводить измерения на оторванных ногах или пальпах. Так их
легче расположить в горизонтальной плоскости. При публикации важно
оговаривать, на оторванных или на не оторванных ногах проводились измерения. На
отличия в результатах между двумя этими способами измерения сегментов ног (у
насекомых) впервые обратил внимание ещё А.А. Любищев. Если нет возможности
выполнить измерения на оторванных конечностях (из-за ценности экземпляров,
например), можно мерить сегменты, придав пауку нужное положение, опирая его
на кусочки ваты, или фиксируя его на песке или стеклянных шариках.
Если измеряется длина хелицер, то обязательно надо оговаривать, проводилось
ли измерение оторванной хелицеры или нет. Это важно, так как разные
исследователи могут измерять по-разному.
Высоту наличника лучше измерять не «в общем», а отдельно мерить
расстояние до края наличника от передних медиальных и от передних латеральных глаз.
Поскольку глаза у пауков редко бывают идеально круглыми, диаметр глаз
всегда измеряется по самому широкому диаметру линзы. Размер линзы, как правило,
меньше, чем размер пигментированного участка, на котором находится глаз.
Замерять следует именно линзу, а не пигментированный участок!
Отдельные исследователи измеряют ширину головного отдела карапакса.
Однако в большинстве случаев между головным и грудным отделом карапакса нет
чёткой границы. Поэтому этот промер получается не точным, и его лучше не
использовать.
Длина измеряемого объекта не должна превышать половину диаметра поля
зрения бинокуляра или микроскопа. Иначе могут возникнуть оптические искажения.
Степень точности измерений должна быть равна 1/20 от разности между
минимальным и максимальным значениями в выборке. Например, 10-12 мм — до 0,1 мм;
0,70-0,90 мм — до 0,01 мм. Округление проводят до ближайшего целого числа;
0,5 — до чётного. Например, значение 14,45 мм — до 14,4; а 14,55 — до 14,6.
Функциональная морфология
57
Хетотаксия. Традиционно у пауков в описаниях/переописаниях видов
приводят подробные данные о количестве и расположении шипов на ногах (хетотаксии).
Как показывает наш опыт, во многих группах пауков (Agelenidae, Amaurobiidae,
Gnaphosidae) хетотаксия подвержена высокой индивидуальной изменчивости и не
может служить для надёжного разграничения видов. Однако признаки хетотаксии
часто бывают полезны при идентификации семейств (мы используем эти признаки
в определительных ключах), а в некоторых случаях — родов и групп видов.
Функциональная морфология
Паутина. Поскольку именно паутина сделала пауков такими, какие они есть, то
остановимся кратко на свойствах паутины и на том, как пауки её используют.
Секрет, из которого создаётся паутина, производится паутинными железами,
расположенными в брюшке и открывающимися на паутинных бородавках и кри-
беллюме. Есть разные типы желёз. По форме их делят на гроздевидные (aciniform),
грушевидные (pyriform), большие ампуловидные (major amputate), малые ампуло-
видные (minor amputate), цилиндрические или трубковидные (cylindrical или tubular),
агрегированные (aggregate), лопастевидные (flagelliform или lobed), крибеллятные
(cribellate) и околокрибеллятные (paracribellate). Железы выделяют жидкий секрет
через трубочки, расположенные на вершине паутинной бородавки или на крибел-
люме. Размеры и форма паутинных трубочек специфичны для каждого типа
паутинных желёз. Секрет быстро застывает и, когда паук объединяет волокна,
вышедшие из разных трубочек, становится паутинной нитью. В зависимости от
предназначения нити, используются волокна, выходящие из разных трубок (связанных с
разными железами). Разные типы желёз имеют протоки на разных паутинных
бородавках. Ни один из видов не обладает полным набором желёз. Некоторые из
типов желёз характерны только для определённых таксонов.
Паутинные нити служат для разных целей. Это и строительство ловчей сети,
состоящей из разного типа нитей, и плетение кокона (тут тоже используются
разные нити), и сигнальная нить, и путеводная нить, которую паук тянет за собой, и
нити, которыми укутывается жертва. Кроме того, паутина используется для
плетения логовищ и для полёта.
Паутина, используемая для ловли добычи, бывает двух типов, крибеллятной и
некрибеллятной. В первом случае жертва ловится («приклеивается») благодаря
тонкой воздушной паутине, намотанной на осевое волокно (рис. 3.2в-д). Жертва,
задевая волосками и другими выростами на теле эту воздушную паутину,
зацепляется за нити. Некрибеллятные пауки ловят жертву за счёт капелек клея,
равномерно распределённых вдоль ловчей нити (у кругопрядов, рис. 3.2е-ж). Многие из
некрибеллятных пауков не используют клей для ловли жертв.
Крибеллятные сети служат долго, а сети, где используется клей, приходится
переплетать ежедневно, из-за того, что клейкое вещество подсыхает и
покрывается пылью.
Клей не всегда нужен для ловли добычи. Иногда паутина служит только
«сигнальной плоскостью». Когда жертва ступает на сеть, паук набрасывается на неё.
Нередко для повышения уловистости над плоской ловчей сетью плетётся рыхлый
«купол» (Linyphia sensu lato). Летающие насекомые, ударяясь о нити купола,
несколько оглушённые, падают на ловчую сеть, где их и подбирает паук.
58
3. Строение и классификация пауков
Далеко не все группы пауков используют ловчие сети. Те же, кто ловит добычу
с помощью сетей, используют совершенно разные конструкции. Они вкратце
рассмотрены при описании биологии разных семейств. Нередко тип сети является
характерным для отдельных родов, а иногда и видов. В ряде случаев у сетеплету-
щих пауков ловчие тенета значительно редуцируются и низводятся до нескольких
или даже до одной сигнальной нити.
Сети многих видов, обитающих в Сибири и на Дальнем Востоке, не описаны.
Не известно даже, плетут ли ловчие сети многие Linyphiidae. Нет сведений об
устройстве тенет многих Theridiidae. Между тем, устройство сетей может пролить
свет на родственные отношения многих групп пауков. Интересно также
проследить, как менялась конструкция сетей в течение эволюции отдельных групп пауков
и всего отряда в целом (на этот счёт есть разные спорные версии). Поэтому эта
область арахнологии весьма перспективна для изучения. О том, как проще
рассматривать сети, было сказано в разделе 2 (Методы изучения пауков: как
рассматривать пауков: как рассматривать сети).
Спаривание. Помимо паутины, у пауков есть и другая необычная особенность.
Это очень своеобразный совокупительный аппарат и сложное поведение,
предшествующее спариванию. Чтобы пояснить, почему так важны копулятивные органы
(и, нередко, окраска) пауков для разграничения видов и таксонов более высокого
ранга, вкратце опишем способы спаривания и поведение, ему предшествующее.
Самка сразу после имагинальной линьки способна к спариванию, к принятию и
отвержению ухаживаний. Самец же после последней линьки не может сразу
заниматься продолжением рода. Для того чтобы стать готовым к спариванию, он
должен перенести половой продукт из полового отверстия (гонопор), расположенного
спереди от средней части эпигастральной щели (там, где у самки находится эпиги-
на), в бульбус, расположенный на педипальпе. Напрямую он этого сделать не
может. Первоначально самец сплетает небольшую сеточку, называемую спермати-
ческой. Затем на неё выделяется сперма, и паук, как шприцем, засасывает её в
бульбус. Когда обе педипальпы наполнены, самец готов к спариванию.
У некоторых пауков эта процедура несколько видоизменена. Pholcus phalan-
gioides натягивает единственную нить между ногами III и на неё выделяет каплю
полового продукта. Затем берёт эту каплю хелицерами, и только потом наполняет
педипальпы (Forster & Forster, 1999).
Сближение любых хищников друг с другом всегда чревато опасностями.
Пауки — не исключение. Поэтому и у самцов и у самок возникают разные механизмы,
позволяющие распознавать друг друга, и отличать полового партнёра от
потенциальной жертвы. Поиском партнёра у пауков всегда занимается самец. У тенетных
пауков самец предупреждает самку о своём приближении особым подёргиванием
сети. Когда самец прикасается к самке, она становится неподвижной до конца
спаривания.
У пауков, лишённых сетей и не имеющих хорошего зрения (Gnaphosidae, Clubio-
nidae, Zoridae), для распознавания партнёра служат прикосновения и запах. У
пауков, обладающих хорошим зрением (пауки-волки, пауки-скакунчики), большую роль
в предкопуляционном поведении играют визуальные сигналы. У пауков-волков
очень большую роль играют и звуковые сигналы, которые издают самцы, стуча
пальпами или брюшком (Hygrolycosa rubrofasciata, см. Бенедиктов, 2006) по суб-
Функциональная морфология
59
страту (например, по сухому листику). Звуковые сигналы важны в предкопуляци-
онном поведении и у пауков-скакунчиков (например, Euophrys frontalis), у которых
самцы могут барабанить по субстрату брюшком (Maddison, 1988; Noordam, 2002).
У самцов, а иногда и у самок, многих пауков есть специальные стридуляционные
органы, которые служат для производства звуков. Эти органы могут находиться на
наружном крае базального членика хелицер (у Linyphiidae), между головогрудью и
брюшком (на их соприкасающихся поверхностях) (у Steatoda из Theridiidae) и в
других местах. Строение стридуляционных органов и производимые ими видос-
пецифичные звуки — это отдельная очень интересная область исследований в
арахнологии.
Пауки каждого семейства во время спаривания принимают специфичную позу.
Позы разных групп приведены на рис. 3.3. Сам процесс спаривания может длиться
от нескольких секунд до нескольких часов.
Для чего паукам нужны столь сложно устроенные совокупительные органы? У
гаплогинных пауков нередко вся пальпа просто вводится в эпигастральную щель
самки. Но чаще нужна жёсткая фиксация пальпы в эпигине. Для этого служат и
выросты самой пальпы, и хелицеры, которыми самец и самка сцепляются между
собой.
Cheiracanthium Thomisidae Agelenidae, Clubionidae,
Ostearius (Linyphiidae) Scytodes Araneidae, Theridiidae
Рис. 3.3. Позы при копуляции у представителей разных семейств по Forster & Forster
(1999) и Wiehle (1963). Чёрным показан самец, белым — самка.
60
3. Строение и классификация пауков
Гораздо сложнее дело обстоит у энтелегинных пауков, особенно у тех, у кого эпи-
гина устроена просто (не имеет выростов). Разные выросты пальпы самца
обеспечивают нахождение совокупительного отверстия самки. Например, отросток голени
«находит» соответствующее углубление или ямку на эпигине, затем кондуктор или
медиальный отросток, вышедший из своего обычного положения благодаря
вздутию гематодохи, ищет другую зацепку, и только один эмболюс (или эмболюс вместе
с кондуктором) вводится в эпигину. У разных пауков механизм поиска нужного
отверстия и сцепления бульбуса с нужными структурами эпигины происходит
по-своему. Механизм может различаться даже у близких групп. Иногда в спаривании и
сцеплении разных элементов участвуют и выросты голени, и даже бёдра пальпы!
Почему такой сложный механизм? Бульбус самца не имеет нервных окончаний,
и поэтому спаривание можно сравнить с открыванием замка в полной темноте, да
ещё не рукой, а протезом. Один отросток ищет дверь вообще, другой ручку, третий
замок, четвёртый замочную скважину.
Несколько слов о том, почему у некоторых групп пальпы и эпигины очень
разные по форме (Araneidae, Linyphiidae, Theridiidae), а у других, напротив, весьма
однообразные (Salticidae, Lycosidae). У первых обычно поведение ухаживания
достаточно простое. И форма копулятивного аппарата служит предохранителем от
межвидового скрещивания. У групп с однообразными пальпами и эпигинами
репродуктивную изоляцию обеспечивает не морфология, а сложное поведение
ухаживания, когда самец «танцует» перед самкой. Самцы тех групп, где ухаживание
играет важную роль, обычно ярко окрашены спереди (хелицеры, наличник,
передние ноги). Если самец принадлежит к чужому виду, самка отвергает его.
Классификация пауков
За последние четыре десятилетия (минувшие с выхода в свет определителя
В.П. Тыщенко) система пауков претерпела существенные изменения. С одной
стороны, добавилось немало новых семейств, с другой стороны, было уточнено
положение многих групп, выявлены их родственные связи. Наиболее существенный
вклад в изменение системы внёс финский арахнолог Р.Т. Lehtinen (1967). Помимо
него, свой вклад внесла и серия ревизий, выполненных американскими
исследователями N.I. Platnick, С. Griswold, новозеландским арахнологом R. Forster и
немецким арахнологом и блестящим палеонтологом J. Wunderlich. Несмотря на то, что
система пауков приобрела большую стройность и стала более логичной
(естественной), до сих пор остаётся немало спорных вопросов.
Отряд пауков делят либо на два подотряда — Mesothelae1 (=Liphistiomorphae,
членистобрюхие) и Opisthothelae2 с двумя инфраотрядами Mygalomorphae и Агапео-
morphae), либо на три подотряда — Liphistiomorphae, Mygalomorphae и Araneo-
morphae3. Первый подотряд (членистобрюхие) встречается исключительно в юго-
восточной Азии. Mygalomorphae (мигаломорфы, птицееды) и Araneomorphae (ара-
неоморфы) обитают и на территории России, в том числе в азиатской её части.
Трактовка (объём) всех трёх групп пауков (членистобрюхих, мигаломорфных и
1 Означает, что паутинные бородавки находятся посредине брюшка.
2 Означает, что паутинные бородавки находятся на конце брюшка.
3 В некоторых публикациях названия пишутся без «е» на конце, например, Araneomorpha.
Классификация пауков
61
арнеоморфных) дискуссий не вызывает. Основные морфологические отличия
между ними приведены в таблице 1. Однако существенные разногласия имеются по
системе как Mygalomorphae, так и Araneomorphae. Ниже мы подробнее
остановимся на последней группе.
Время от времени разные арахнологи предпринимали попытки создать
естественную систему пауков (Bertkau, 1878; Simon, 1892-1903; Dahl, 1913; Petrunke-
vitch, 1933; Lehtinen, 1967 и др.). Однако только две системы смогли оставить
глубокий след в надвидовой классификации пауков, а именно системы,
предложенные Е. Simon и Р.Т. Lehtinen.
Система Э. Симона была очень логичной и стройной для своего времени, и
строилась на наружно-морфологических, преимущественно соматических
признаках. Она сразу была принята арахнологическим сообществом, а в ряде случаев
используется и современными исследователями (но не систематиками). Другие
классификации, например, система А. Петрункевича, строились во многом на
анатомических признаках. Возможно поэтому, или из-за того, что схема А. Петрункевича
зиждилась на изучении сравнительного небольшого числа таксонов, она не была
Таблица 1. Основные морфологические отличия между подотрядами пауков
Подотряды
Наружная
сегментация
брюшка
Расположение
паутинных
бородавок
Количество
паутинных
бородавок
Количество
сегментов в
паутинных
бородавках
Количество лёгких
(легочных
крышечек)
Максиллы
(жевательные
лопасти тазиков
педипальп)
Хелицеры
Глаза
Представители в
азиатской части
России
Liphistiomorphae
есть
в средней части
брюшка
8 или 7
Mygalomorphae | Araneomorphae
нет
обычно в задней части брюшка
от 6 до 2 (1-3 пары)
часто более 2
4 (2 пары)
нет
нет или есть
Базальный членик направлен вперёд, а
апикальный (коготок) — назад.
сближены
нет
Atypidae
2
2-0(1 пара или нет)
есть
Базальный членик
обычно направлен вниз,
а апикальный (коготок)
— всегда вовнутрь, к
средней линии тела.
обычно расставлены (не
сближены)
все семейства, кроме
Atypidae
62
3. Строение и классификация пауков
принята. Симон же изучил огромное число групп пауков, и описал более четырёх
тысяч видов!
Однако недостатком и Симоновской системы, и всех других, было полное
игнорирование таких эволюционных явлений как параллелизм, особенно в
редукции тех или иных органов, и конвергенция. Совершенно очевидно, что при
переходе от ловли добычи с помощью сетей к бродячему образу жизни крибеллюм
перестаёт быть функциональным органом (крибелятные нити используются только для
ловчих нитей). Соответственно, крибеллюм может редуцироваться. Такой процесс
может происходить независимо в разных крибеллятных группах. Аналогичная
ситуация и с третьим коготком лапки, который используется для плетения ловчей
паутины. Потеря тенет ведёт к независимой редукции третьего коготка. Есть ещё
одна структура, которая «завязана» на ловчие сети — это паутинные железы и их
протоки, ответственные за производство паутинных волокон, используемых для
строительства тенет. Если нет сетей, то эти железы могут полностью
редуцироваться. Однако до сих пор многие арахнологи, особенно американские,
поклонники филогенетической систематики (кладистики), придают строению паутинных
бородавок сверхбольшое значение, не принимая во внимание редукцию. Есть ещё
несколько органов, устройству которых придавалось большое значение и не
учитывались явления редукции. Так, придавалось существенное значение числу ос-
тий сердца, трахейной системе и вообще органам дыхания. Но уменьшение
размера тела ведёт к неизбежному укорачиванию сердца и трахейной системы.
Соответственно, число остий уменьшается, то же самое происходит с трахеями. У
некоторых пауков полностью редуцируются лёгкие. Другой «орган», игравший важную
роль в построении классификации пауков, это форма и структура
светоотражающего слоя глаз. Однако не нужно быть физиологом, чтобы понять, что дневные и
ночные пауки должны иметь разное устройство глаз, и близкородственные
таксоны, ведущие разный образ жизни, будут различаться по этому признаку.
До работы Р.Т. Lehtinen (1967), всех аранеоморфных пауков разделяли, в
первую очередь, на крибеллятных и некрибеллятных, а эти две группы, в свою
очередь, делили на гапло- и энтелегинных. Последних подразделяли на двух- и трёх-
коготковых (см. Kaston, 1948). Строению копулятивных органов не придавали
практически никакого значения. Лехтинен же показал, что у родственных таксонов,
иногда даже внутри одного рода, крибеллюм может присутствовать или
отсутствовать (редуцироваться). То же самое применимо и к двум и трём коготкам. При
переходе к бродячему образу жизни, пауки перестают плести ловчие сети, и третий
(непарный) коготок у них редуцируется. Вдобавок к сказанному, оказалось, что
некоторые из пауков, считавшиеся гаплогинными, имеют вторично упрощённые
копулятивные органы, а другие — наоборот, исходно примитивные. Если
отталкиваться от копулятивного аппарата (не несущего адаптивной нагрузки), то
группирование семейств пауков становится гораздо проще и более естественным.
Заслуга П.Т. Лехтинена состоит в том, что он радикально перестроил систему пауков,
основываясь, в первую очередь, именно на строении копулятивных аппаратов.
Ниже мы кратко остановимся на группировке семейств, которые обитают в
Сибири и на Дальнем Востоке России (Таблица 2), поскольку названия некоторых
групп (рангом выше семейства) мы неоднократно используем в тексте.
В настоящее время среди пауков, обитающих у нас в регионе, есть две более
или менее чётко очерченные группы — гаплогинные и аранеоидные. Большинство
Классификация пауков
63
Таблица 2. Группы семейств, которые встречаются или могут быть встречены в
азиатской части России
Мигаломорф-
ные
Atypidae
Аранеоморфные
Гаплогинные
Segestriidae
Oonopidae
Leptonetidae
Pholcidae
«RTA-clade»
Крибеллятные
Amaurobiidae
Dictynidae*
Некрибеллятные
Agelenidae
Lycosidae
Pisauridae
Oxyopidae
Zoridae
Ctenidae
Argyronetidae
Cybaeidae
Hahniidae
Anyphaenidae
Cheiracanthiidae
Corinnidae
Clubionidae
Gnaphosidae
Liocranidae
Philodromidae
Sparassidae
Thomisidae
Salticidae
Zodariidae
самостоятельные
Крибеллятные
Eresidae
Oecobiidae
Titanoecidae
Uloboridae
Аранеоидные
Araneidae
Linyphiidae
Mimetidae
Mysmenidae
Nesticidae
Pimoidae
Tetragnathidae
Theridiosomatidae
Zygiellidae
Theridiidae
В азиатской части России есть и некрибеллятные таксоны.
оставшихся семейств принадлежат к так называемой «RTA-clade» (RTA — это
аббревиатура для retrolateral tibial apophysis), иначе говоря, к группе с ретролатераль-
ным отростком голени пальпы у самца. Эта группа, в отличие от гаплогинных и
аранеоидных, не является естественной. Некоторые из семейств, входящих в этот
комплекс, утратили отросток голени вторично (Lycosidae). Часть семейств,
несомненно, близко родственны между собой, но некоторые стоят особняком, например
Salticidae, Zodariidae. Крибеллятные Dictynidae по структуре пальпы не
отличаются от некрибеллятных Cybaeidae, Argyronetidae и Tegenariinae (Agelenidae).
Последняя группа имеет медиальный отросток бульбуса, которого нет у всех других
представителей этих четырёх семейств.
Среди аранеоидных семейств особняком стоят Theridiidae. В отличие от других
групп, парацимбиум у них расположен в верхней части цимбиума (а не в основа-
64
3. Строение и классификация пауков
Таблица 3. Примерное число видов разных семейств в азиатской части России и в
России в целом (семейства, которых нет в азиатской части России, но есть в европейской
части, в таблице не показаны)
Семейства
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
Agelenidae
Amaurobiidae
Anyphaenidae
Araneidae
Argyronetidae
Atypidae
Cheiracanthiidae
Clubionidae
Corinnidae
Ctenidae
Cybaeidae
Dictynidae
Eresidae
Gnaphosidae
Hahniidae
Leptonetidae
Linyphiidae
Liocranidae
Lycosidae
Mimetidae
Mysmenidae
Nesticidae
Oonopidae
Oxyopidae
Philodromidae
Pholcidae
Pimoidae
Pisauridae
Salticidae
Sparassidae
Tetragnathidae
Theridiidae
Theridiosomatidae
Thomisidae
Titanoecidae
Uloboridae
Zoridae
Zygiellidae
| Общее число видов
Азиатская часть России
17
3
1
90
1
2
11
70
10
1
4
39
3
156
12
1
-700
9
205
4
1
3
1
3
58
7
1
9
140
2
38
93
1
101
11
2
3
4
1817
Вся Россия
26
6
3
105
1
3
15
74
13
1
5
50
3
-200
24
2
-800
11
-220
7
2
7
2
4
-65
11
1
9
-180
2
40
115
2
115
15
5
7
5
2156
нии), а семяпроводные канальцы эпигины образованы не складками кутикулы, а
настоящими трубками.
Выше мы уже упоминали, что в современных определителях можно найти
разные трактовки того или иного семейства, и даже системы пауков. Например, в
прекрасном определителе M.J. Roberts (1995) Philodromidae рассматриваются как
подсемейство в Thomisidae. Некоторые рода из Dictynidae и Hahniidae рассматривают-
Определитель семейств
65
ся в Agelenidae. Это не связано с особым мнением автора, а вызвано тем, что он
арахнолог-любитель, и пользуется привычной для него старой системой. В случае
обнаружения разночтений, следует отдавать предпочтение той трактовке объёма
семейств, которая дана в каталоге Н.И. Платника (Platnick, 2010). Это самый
известный, наиболее доступный и часто обновляемый справочник. Если у вас нет
своего мнения по поводу того, кто из систематиков прав, тогда лучше рассматривать
тот или иной таксон так, как он трактуется в общедоступном справочнике. Если
вам известно, что существует несколько трактовок какого-либо семейства
современными авторами, и у вас есть своё мнение по этому вопросу, тогда отдавать
предпочтение мнению Н.И. Платника (Platnick, www) не обязательно.
Согласно последней сводке (Mikhailov, 2002) в фауне России было известно 37
семейств пауков. С учётом недавних находок дополнительных семейств Ctenidae,
Mysmenidae (Marusik & Koponen, 2000), Leptonetidae (Oliger et al., 2002), Oonopidae
(Saaristo & Marusik, 2004b), Zodariidae и Zoropsidae (Marusik, 2005) и некоторых
таксономических перестановок (перемещение в другие семейства родов Cheira-
canthium и Weintrauboa, ранее рассматривавшихся в составе Clubionidae и Linyphii-
dae), число зарегистрированных в России семейств пауков возросло до 44. Не
исключены находки в России и дополнительных семейств, например Anapidae, Fili-
statidae, Hersiliidae, Sicariidae, Segestriidae. Anapidae встречаются в Европе вплоть
до Болгарии, есть в Турции. Filistatidae известны из Крыма (наши данные) и
Закавказья, а также из Казахстана. Самая северная находка Hersiliidae — Урумчи (Синьц-
зян-Уйгурский автономный округ Китая). Популяция интродуцированного Loxosceles
laeta (Nicolet, 1849) (Sicariidae) уже многие десятилетия существует в
Зоологическом музее Университета Хельсинки (Palmgren, 1977 и S. Koponen, личное
сообщение).
В пределах части России, лежащей к востоку от Урала, в настоящее время
известны представители 38 семейств. Список этих семейств и примерное число видов,
найденных в азиатской части России и России в целом приведены в таблице 3. В
азиатской части России не найдены шесть семейств, известных в европейской
части: Dysderidae, Oecobiidae, Scytodidae, Segestriidae, Zodariidae и Zoropsidae, хотя
Segestriidae отмечены на северо-востоке Китая и в Японии, a Zodariidae известны
из сопредельных территорий Урала, Восточно-Казахстанской области и Монголии.
Для семейств с большим видовым разнообразием невозможно с точностью
указать число видов, так как есть сомнения в определениях и валидности части видов.
По большинству семейств специальных систематических работ в рамках всей
России или её азиатской части никогда не проводилось.
Определитель семейств
Сложности в подготовке определительного ключа и в его использовании.
Перечислим некоторые моменты, субъективные и объективные.
1. Тем, кто знает свою группу, не всегда легко создать ключ понятный и
удобный для начинающих или коллег, специализирующихся на изучении других
таксонов. Во многом это связано с тем, что специалист не определяет, а узнаёт своих
«подопечных». Это происходит так же, как все мы издалека узнаём своих друзей и
знакомых по походке, манере держаться, голосу и по множеству других трудно
ев
3. Строение и классификация пауков
описываемых признаков. Узнавание пауков тоже идёт «на автомате»: по габитусу,
рисунку, размеру и многим другим признакам сразу. Нередко даже сложно бывает
сформулировать сходу, почему это то семейство, а не другое. С того времени, когда
мы определяли семейства по ключу, прошло много лет, и сложности, с которыми
мы сталкивались, уже позабыты.
2. Некоторые семейства имеют уникальные черты и легко могут быть
отличимы от других буквально по одному признаку (например, Salticidae, Atypidae, Lyco-
sidae). Большинство же семейств можно опознать только по совокупности
признаков. Систематика разных групп разработана неодинаково хорошо. Так, на наш
взгляд, Theridiidae по разнообразию соматических признаков и по признакам ко-
пулятивного аппарата можно легко разбить, как минимум на 3 группы (семейства).
А семейства Pimoidae и Linyphiidae проще было бы рассматривать как одно.
Гораздо сложнее ситуация с родами. Только в некоторых семействах с родами обстоит
всё хорошо (в Salticidae, Gnaphosidae). В очень многих случаях рода явно сборные,
особенно это касается семейств Lycosidae, Theridiidae, Thomisidae, Philodromidae,
части Linyphiidae, где дать родам диагнозы практически невозможно. Поэтому мы
ограничимся только определителем семейств.
3. Некоторые хорошие (работающие) признаки сложно наблюдать. Полезный
диагностический признак — вторичная сегментация на лапках ног. Но
рассмотреть в бинокуляр, есть ли вторичная сегментация — практически невозможно.
Поэтому такие признаки в определительных таблицах мы по возможности не
упоминаем (игнорируем). Но без некоторых таких признаков обойтись невозможно.
Например, число коготков. Если паук крупный, рассмотреть коготки не составляет
труда, но у мелких пауков это достаточно сложно. Иногда однозначно можно
сказать, сколько коготков, только рассмотрев ногу в микроскоп (а не в бинокуляр). То
же самое относится и к пильчатым волоскам, встречающимся у Theridiidae и Nesti-
cidae. У некоторых групп Theridiidae они редуцированы, или выражены очень
неявно. В то же время почти все представители этого семейства отличаются габиту-
ально, но сформулировать (описать) словами это, если и возможно, то займёт
немало места. Если же, используя ключ, вы неверно интерпретировали признак, то
уйдёте «в другую степь».
4. Традиционно и в энтомологических и в арахнологических ключах
используются соматические признаки. Эти признаки используются для определения и
семейств, и родов, и видов. Особенность же пауков в том, что 90% видов (если не
больше) можно различить по форме копулятивного аппарата. Определить
подавляющие число родов (если не все 100%) можно тоже по форме пальпы самца, а
нередко и по эпигине самки. Все семейства, обитающие в нашем регионе, вполне
различимы по педипальпам самцов. Не надо даже смотреть габитус. То есть, для
определения пауков важнее не ключ, а рисунки. Ключ необходим иногда для
разграничения очень близких видов пауков, но это обычно в случае сложных групп
видов или сложных родов с достаточно однообразным копулятивным аппаратом,
но хорошими соматическими признаками (в Европе это, например, Zord).
Оптимальным было бы сочетание множества рисунков и ключей.
Если определение идёт до уровня вида, где непременно необходимо
рассмотреть пальпу самца и эпигину, то почему бы и не начинать ключ с копулятивного
аппарата, и им же заканчивать? Однако трудность в том, что копулятивный
аппарат слишком сложный. Сложный и сам по себе, и для описания словами, и для
Определитель семейств
67
рассмотрения в бинокуляр. Поэтому для определения семейств мы всё же делаем
упор на более простые для рассматривания соматические признаки.
5. Среди всего разнообразия определителей традиционно различают три
основных типа — графические, политомические и дихотомические. Графические
определители — это изображения характерных (уникальных) черт строения, по которым
можно однозначно узнать (определить) таксон. К сожалению, не все семейства
пауков азиатской части России обладают уникальными признаками; большинство
семейств можно определить только по комплексу нескольких признаков. Полито-
мический определитель представляет собой таблицу таксонов и их признаков.
Особенность политомического определителя в том, что начинать определение можно с
изучения любого признака и рассматривать признаки в любой последовательности,
отбрасывая «неподходящие» семейства до тех пор, пока не останется одно —
результат определения. При пользовании дихотомическим определителем
неизбежно приходится начинать определение с самой первой дихотомии (тезы/антитезы) и
последовательно продвигаться дальше, рассматривая признаки именно в той
последовательности, в которой построен определитель. При этом последовательность
может быть не самая удобная для данного конкретного случая, и признаки могут
быть не самые легко наблюдаемые. По нашему мнению, оптимальным является
сочетание и графических, и политомических, и дихотомических определителей.
Это позволяет в затруднительном случае «прогнать» один и тот же объект по
большему числу признаков. В итоге повышается надёжность определения.
6. В некоторых семействах содержатся отдельные необычные (нетипичные,
«маргинальные») рода, которые сильно отличаются от большинства
представителей семейства. Поэтому их идентификация затруднена. В фауне Сибири и
Дальнего Востока это Cicurina (Dictynidae), Cryphoeca (Hahniidae), Diphya (Tetragnathi-
dae), Micaria (Gnaphosidae), Phoroncidia (Theridiidae), Trachelas (Corinnidae).
Как пользоваться определительными таблицами
При определении надо обязательно сравнивать свой объект с иллюстрациями,
приведёнными в основной части книги. Определение без использования рисунков
по одному лишь тексту бессмысленно и неизбежно приведёт к ошибкам.
Иллюстрации также помогут проверить правильность своего определения (по соответствию
габитуса и всего набора признаков).
В нашем ключе на самом первом этапе мы попытались отсечь легко
определяемые семейства (те, которые определяются по глазам, бородавкам и дыхальцу) от
сложных. Это можно сделать с помощью графического определителя семейств по
уникальным чертам (рис. 3.4-3.6), а также политомического (Ключ I) или
дихотомического (Ключ II) определителей всех семейств азиатской части России.
Для Аранеоидных семейств, определение которых может вызывать трудности,
приводится дополнительный политомический определитель (Ключ III).
Для сложных семейств (тех, которые не определяются по глазам, бородавкам и
дыхальцу), мы приводим отдельный определитель (Ключ IV). Поскольку 10
трудных для определения семейств в пределах азиатской части России встречаются
только на юге Дальнего Востока, то при определении материалов из Сибири их
можно (нужно) игнорировать. Поэтому для сложных семейств сибирских пауков
мы приводим отдельный определитель (Ключ V).
68
3. Строение и классификация пауков
Схема всего определителя
Рис. 3.4-3.6. Графические определители семейств по уникальным признакам.
Ключ I. Политомический определитель всех семейств пауков азиатской части
России.
Ключ II. Дихотомический определитель всех семейств пауков азиатской части
России.
Ключ III. Политомический определитель Аранеоидных семейств.
Ключ IV. Дихотомический определитель сложных групп пауков (тех, которые
не определяются по глазам, бородавкам и дыхальцу) всего региона (Сибири и
Дальнего Востока).
Ключ V. Дихотомический определитель сложных групп пауков только Сибири.
Определитель семейств
69
Leptonetidae
Ключи для определения семейств
6 глаз
Lathys
Dictynidae
8 глаз
Г-ОТ5-
Atypidae^ триада ^аепк1ае^
Р urldae
Zodariidae
Uloboridae
Pholcidae
Oxyopidae
Рис. 3.4. Графический определитель семейств, узнаваемых по форме карапакса и глаз:
Atypidae, Ctenidae, Dictynidae (6-глазых представителей рода Lathys), Eresidae,
Leptonetidae, Lycosidae, Oecobiidae, Oonopidae, Oxyopidae, Pholcidae, Pisauridae, Salticidae,
Segestriidae, Uloboridae, Zodariidae.
Стрелки указывают на характерные особенности.
70
3. Строение и классификация пауков
A elenidae
Oecobiidae
Anyphaenidae
Agelenidae Argyronetidae
Gnaphosi ae Zodariidae Hahniidae Hahniidae
Рис. З.5. Графический определитель семейств, узнаваемых по форме паутинных
бородавок, дыхальца и брюшка: Agelenidae, Anyphaenidae, Argyronetidae, Gnaphosidae,
Hahniidae, Oecobiidae, Zodariidae.
Стрелки указывают на характерные особенности.
Определитель семейств
71
3
Mysm dae
Am
Araneidae
Theridiidae, лапка IV
gf Am
Leucauge \ \
Tetragnathldae\ \
бедро IV ^ l
Cs
Araneidae
Ara
Pb
Hs
As
Рис. З.6. Графический определитель семейств, узнаваемых по модификациям ног и их
вооружения: Araneidae, Mimetidae, Mysmenidae, Pholcidae, Pimoidae, Sparassidae, Tetragna-
thidae, Theridiidae.
Am — Araneus marmoreus, As — Araneus schrencki, Cs — Cyclosa sp., Hs — Hypsosinga sanguinea,
Pb — Pronnoides brunneus. Стрелки указвают на характерные особенности.
72
3. Строение и классификация пауков
Ключ I. Политомическии определитель всех семейств пауков азиатской
части России. Особые черты выделены
подчёркнутым шрифтом
Agelenidae
Amauro-
biidae
Anyphae-
nidae
Araneidae
Argyro-
netidae
Atypidae
Cheiracan-
thiidae
Clubio-
nidae
Corin-
nidae
Глаза
8 (4-4).
8 (4-4).
8 (4-4).
8 (4-4).
Латеральные
соприкасаются.
Медиальные
глаза образуют
квадрат.
8 (4-4).
8, компактно на
глазном бугорке.
8 (4-4).
8 (4-4).
8 (4-4).
Карапакс
От яйцевидного до
грушевидного.
Яйцевидный-
овальный.
Головной отдел слегка
приподнят.
Грушевидный.
Грушевидный.
Головной отдел у
большинства уже
груди; наличник
низкий,
сравнимый с
диаметром
передних
медиальных глаз.
Грушевидный.
Щитовидный.
Яйцевидный.
Яйцевидный.
Овально-
грушевидный.
Хелицеры
Передний край
желобка хелицер с
3-5 зубцами, а
задний-с 2-8.
Крупные, выдаются
вперёд (видны при
виде сверху).
Не
модифицированы.
Имеют
латеральный
мыщелок.
Желобок хелицер с
2 крупными
зубцами спереди и
3 — сзади.
Вытянуты вперёд;
коготки лежат на
нижней стороне
хелицер.
Иногда удлинены у
самцов.
Не
модифицированы.
Не
модифицированы.
Брюшко
Овальное.
Овальное.
Овальное.
Характерный
рисунок из 2-3
пар тёмных
крыловидных
пятен.
Разной формы:
могут быть
бугорки спереди,
выросты сзади,
заострено
спереди и сзади.
Редко со
скутумом.
Овальное.
Опушение очень
густое, особенно
вентральнр.
Овальное, с
небольшим
скутумом.
Овальное, на
конце слегка
заострено.
Овальное, на
конце может
быть слегка
заострено.
Овальное; у
самцов — с
дорсальным
скутумом.
Ноги
Без особенностей.
Длина каламиструма
не превышает
половину длины
предлапки IV.
С густой скопулой.
Голени I и/или II у
самцов часто с
мощными шипами.
Ноги 1-Й с
малочисленными
шипами, задние — с
многочисленными.
Без особенностей.
Со скопулой.
Со скопулой.
Голени и предлапки
I—II с 4 и более
парами вентральных
шипов или с
мелкими шипиками у
самцов.
Определитель семейств
73
Крибеллюм
Нет.
Есть;
разделён на 2
части.
Нет.
Нет.
Нет.
Нет.
Нет.
Нет.
Нет.
Дыхальце
Перед
бородавками.
Перед
бородавками.
Широкое,
посредине
между
эпигастраль-
ной щелью и
бородавками.
Перед
бородавками.
Широкое,
ближе к
эпигастраль-
ной щели.
2 пары лёгких.
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
Бородавки
Удлинены оба
сегмента
задних
бородавок.
Без
особенностей.
Без
особенностей.
Без
особенностей.
Колюлюс
маленький
(бугорок с
парой
щетинок).
Задние
бородавки из 3
сегментов.
Без
особенностей.
Без
особенностей.
Есть колюлюс.
Коготки
3
3
2
3
3
3
2
2
2
Пальпа самца
РГО есть. Иногда
есть отросток колена.
РГО есть. Крупные
дорсальные отростки
голени.
РГО есть. Бедро с
характерными
толстыми чёрными
щетинками.
Голень с
вентральным
выростом, из-за
которого ширина
голени больше
длины.
РГО широкий
лопастевидный.
Бульбус округлый с
шиловидным
эмболюсом и
кондуктором.
РГО длинный и
тонкий. Цимбиум с
саблевидным
ретролатеральным
выростом.
РГО есть.
РГО есть.
Эпигина
Плоская, с 1 ямкой или
без ямки, иногда по
бокам от ямки пара
сосочков.
Плоская, иногда с 2
выростами-валиками
по бокам.
Плоская, с небольшим
передним карманом и 1
щелевидной
продольной ямкой.
Выпуклая, со скапусом
(исключение —
Hypsosinga).
Плоская, слабо
хитинизированная, без
ямки.
Нет.
Плоская, с ямкой, над
которой просвечивают
канальцы эпигины.
Плоская, с 1 ямкой или
2 отверстиями у
эпигастральной щели.
Плоская, с 1 или 2
ямками без чётких
границ.
74
3. Строение и классификация пауков
Ctenidae
Cybaeidae
Dictynidae
Eresidae
Gnaphosidae
Hahniidae
Leptonetidae
Linyphiidae
Глаза
8 (2-4-2);
передние
латеральные
глаза — самые
мелкие.
8 (4-4).
8 (4-4), может
быть и 6 (2-4), но
такие не найдены
в России.
8 (4-2-2),
латеральные
глаза далеко
отстоят друг от
друга и от
средних глаз.
8 (4-4).
8 (4-4).
6 (4-2), задняя
диада сдвинута
далеко назад.
8 (4-4), формула
может быть иной
у самца.
Карапакс
Грушевидный.
Грушевидный.
Бывает разной
формы.
Прямоугольный;
головной отдел
приподнят,
наличник
низкий.
Грушевидный
или овальный.
Овальный.
Грушевидный;
наличник
высокий.
От грушевидного
до овального;
наличник
высокий. У
самцов карапакс
частое
выростами.
Хелицеры
Не модифицированы.
Не модифицированы.
У самцов часто с
вдавлением вдоль
внутреннего края и с
поперечными рёбрами
Хелицеры очень
массивные, но
короткие.
Не модифицированы.
Не модифицированы.
Слегка удлинены.
Со стридуляционными
бороздками на боковой
поверхности.
Брюшко
Овальное, на конце
заострено.
Овальное.
Овальное.
Овальное —
прямоугольное. У
самца красное с
чёрными пятнами.
Удлинённо-
овальное, у самцов
иногда со скутумом.
Овальное, с
рисунком или без.
Овальное или
каплевидное.
Овальное или
каплевидное. У
самцов и самок
иногда со скутумом
или скутумами.
Ноги
Со скопулой.
Голени и
предлапки I-
II со многими
вентральными
шипами.
Без
особенностей.
Шипов нет
(кроме Lathvs,
Devade).
Короткие,
толстые,
густо
покрыты
волосками.
Без
особенностей.
Zelotini с
гребёнкой из
вентральных
щетинок на
вершине
предлапки III.
Без
особенностей.
Длинные;
передняя пара
более чем в 3
раза длиннее
тела.
Голени с 1-2
дорсальными
щетинками.
Редко шип на
колене I.
ПредлапкаI
всегда с
трихоботрией.
Определитель семейств
75
|Крибеллюм
Нет.
Нет.
1 Крибеллюм
не
разделённый.
Cicurina без
крибеллюма.
Крибеллюм
разделён на 2
части.
Нет.
Нет.
Нет.
Нет.
Дыхальце
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
Широкое,
посредине
между
эпигастраль-
ной щелью и
бородавками.
Перед
бородавками.
Перед
бородавками.
Бородавки
Без
особенностей.
Есть колюлюс.
Без
особенностей.
Без
особенностей.
Передние
бородавки
цилиндрические
и расставлены на
диаметр (кроме
Micaria и
отчасти
Parasyrisca).
Расположены в
один
поперечный
ряд (кроме
Cryphoeca).
Без
особенностей.
Есть колюлюс.
Коготки
2
3
3
3
2
2
3
3
Пальпа самца
РГО нет.
Есть РГО. Колено с
выростом, на
котором могут быть
короткие толстые
зубчики.
Голень всегда с
отростками, если не
латеральными, то
дорсальными.
РГО нет. Тегулюм
округлый, с тонким
эмболюсом и
пластинчатым
кондуктором.
РГО есть (кроме
Callilepis).
РГО тонкий
заострённый (кроме
Cryphoeca); все
Hahniinae еще и с
выростом колена.
РГО нет. Бульбус
каплевидный, с
комплексом
отростков на
вершине.
Всегда с базальным
парацимбиумом U-
или Г-образной
формы.
Эпигина
Плоская, без ямок.
Плоская, с 1 ямкой или]
без ямки.
Плоская, с 1 или 2
ямками или без ямок.
Плоская, с 1 широкой
ямкой.
Плоская, обычно с 1
ямкой, часто с верхним
карманом или
скапусом.
Плоская, с парой
щелевидных отверстий,,
обычно с длинными
канальцами.
Нет.
Плоская или выпуклая, 1
всегда с медиальной
пластинкой.
76
3. Строение и классификация пауков
Liocranidae
Lycosidae
Mimetidae
Mysmenidae
Nesticidae
Oecobiidae
Oonopidae
Oxyopidae
Глаза
8 (4-4).
8 (4-2-2),
наличник
примерно
равен
диаметру
передних глаз.
8 (4-4).
8 (4-4).
8 (4-4).
8 (4-4). Глаза
компактно в
центре
карапакса.
L
расположены
компактной
группой.
8, в 3 ряда (2-
4-2), передние
медиальные
— самые
мелкие.
Карапакс
Грушевидный.
Грушевидный.
Яйцевидный,
высокий.
Высота больше
длины.
Грушевидный.
Сердцевидный,
длина^меньше
ширины.
Грушевидный,
грудной отдел
выше, чем
головной.У
самцов —_с
выростами.
Яйцевидный.
Наличник
высокий.
Хелицеры
Не
модифицированы.
Не
модифицированы. Может
быть
латеральный
мыщелок.
Без зубчиков.
но с
гвоздевидными
шипиками.
Не
модифицированы. Длина
хелицер меньше
высоты
наличника.
Не
модифицированы.
Зубчиков нет.
У самцов с
пальцевидными
выростами
спереди.
Не
модифицированы.
Брюшко
Удлинённо-
овальное.
Овальное.
Сферическое, с
парой бугорков.
Сферическое,
высота больше
длины.
Овально-округлое.
Овальное.
Овальное.
Удлинённое
каплевидное.
Ноги
Голени и
предлапки I—II
бывают со
многими
вентральными
шипами (2—4 и
более пар).
Со скопулой или
без.
Вентральных
шипов на голени
и предлапке I—II
от 2 до 6 пар.
Передняя
поверхность
голеней и
предлапок I—II с
рядом
чередующихся
длинных и
коротких шипов.
У самцов 1-2
мощных про-
латеральных
шипа на голени и
предлапке I. У
самок темная
склеротизованная
бляшка на бёдрах
1-П.
Лапка IV
вентрально с
пильчатыми
волосками.
Без
особенностей.
Бёдра IV
вздутые.
Шипы очень
мощные,
длинные и
оттопыренные.
Определитель семейств
а
Крибеллюм
Нет.
Нет.
Нет.
Нет.
Нет.
Нет.
Есть только
у Oecobius
si
Нет.
Нет.
Дыхальце
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
2 пары возле
эпигастраль-
ной щели;
почти
незаметны.
Перед
бородавками.
Бородавки
Без
особенностей.
Без
особенностей.
Есть колюлюс.
Есть колюлюс.
Есть колюлюс.
Задние
бородавки с
длинным и
изогнутым
апикальным
члеником.
Есть колюлюс.
Есть колюлюс.
Коготки
2
3
3
3
3
3
3
3
Пальпа самца
РГО есть.
Медиальный
отросток
характерной формы
(ширина больше, чем
высота).
РГО нет.
РГО нет.
Парацимбиум
базальный,
массивный и
хитинизированный.
РГО нет. Пальпа
крупная.
Парацимбиум очень
крупный и
оттопыренный.
РГО нет.
РГО нет. Голень
пальпы самца
крупнее бедра;
тегулюм
каплевидный, без
отростков.
Есть отростки на
голени, а иногда и на
колене.
Эпигина 1
Плоская, с 1
удлинённой ямкой.
Плоская,
разнообразной формы,
обычно с 1 ямкой и
септумом.
Слегка выпуклая, с
септумом.
Слабо склеротизована,
но со скапусом
(заметен при взгляде
сбоку).
Плоская, с
просвечивающими
рецептакулами.
Плоская, в виде
простой пластинки,
слабо склеротизована.
Нет.
Выпуклая, с
направленным вперёд
септумом, ямки нет.
78
3. Строение и классификация пауков
Philodromidae
Pholcidae
Pimoidae
Pisauridae
Salticidae
Segestriidae
Sparassidae
Глаза
8 (4-4).
8,диада
(передние
медиальные)
и 2 триады по
бокам.
8 (4-4).
8 в 2-3 ряда
(4-4 или 4-2-
2), наличник в
несколько раз
больше
диаметра
передних глаз.
8вЗряда(4-2-
2), передние
медиальные—
очень крупные,
задние меди-
альные — очень
маленькие.
6 глаз, 3
Диады.
8 (4-4).
Карапакс
От
грушевидного до
круглого,
уплощённый.
Наличник
высокий.
Округлый,
наличник
высокий,
голова
приподнята
над грудной
частью.
Грушевидный.
Грушевидный.
От сильно
вытянутого
до почти
круглого.
Яйцевидный
удлинённый.
Грушевидный.
Хелицеры
Задний край хелицер
не более чем с одним
зубчиком.
У самцов могут нести
зубчики на передней
поверхности.
Крупные, без
стридуляционных
бороздок, передний
край желобка хелицер
с 3, задний — с Ъ-А
зубчиками.
Не модифицированы.
Обычно не
модифицированы.
Могут быть сильно
удлинены.
Удлинены.
Не модифицированы.
Задний край желобка
хелицер с 3
зубчиками.
Брюшко
От палочковидного
до широко-
уплощённого.
Вытянутое (длина
больше диаметра в 2
и более раз).
Овальное.
Удлинённо-
овальное.
От овального до
вытянутого. Иногда
у самцов со
скутумом
(Chalcoscirtus).
Овальное или
яйцевидное.
Овальное, на конце
заострено.
Ноги
Бёдра
расставлены в
стороны. Шипов
много. Лапки и
дистальная часть
предлапок
передних ног со
скопулой.
Очень длинные
(передняя нога
более, чем в 4 раза
длиннее тела).
Шипов нет.
У самцов
предлапкаI в
основании
расширена и несёт
специализированные волоски.
Со скопулой и
многочисленными
шипами.
У самцов
передние ноги
могут быть
утолщены, с
густыми
волосками и
яркой окраской.
Ноги III
направлены
вперёд.
Бёдра расставлены
в стороны.
Предлапки и лапки
со скопулой.
Мембрана между -
лапкой и пред-
лапкой разделена
наЗ части.
Определитель семейств
79
Крибеллюм
Нет.
Нет.
Нет.
Нет.
Нет.
Нет.
Нет.
Дыхальце
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
Пара дыхалец,
открывающихся возле
эпигастраль-
ной щели.
Перед
бородавками.
Бородавки
Без
особенностей.
Есть колюлюс.
Есть колюлюс.
Без
особенностей.
Без
особенностей.
Без
особенностей.
Без
особенностей.
Коготки
2
3
3
3
2
3
2
Пальпа самца
РГО есть.
РГО нет. Пальпы
очень крупные.
Вертлуг с
пальцевидным
выростом. Бульбус
крупнее бедра
пальпы.
РГО нет. Цимбиум с
крупным,
коническим
выростом. Вершина
цимбиума
пальцевидная.
РГО есть.
РГО есть (кроме
Talavera). Иногда
есть отростки на
бедре, цимбиуме и
даже на вертлуге.
Голень вздутая,
крупнее цимбиума.
Тегулюм без
отростков.
РГО есть, крупный.
Эпигина
Плоская, с медиальной
пластинкой.
Медиальная пластинка
может быть вогнутой и
образовывать ямку.
Плоская, в виде
треугольной или
поперечной пластинки,
с небольшим
«скапусом».
Выпуклая, с септумом.
Плоская, с 1 ямкой,
может быть с
якоревидным
септумом.
Плоская, бывает очень
разная — с 0-2 ямками,
карманом, вырезкой
сзади.
Нет.
Плоская, без ямки.
80
3. Строение и классификация пауков
Tetragnathidae
Theridiidae
Theridioso-
matidae
Thomisidae
Titanoecidae
Uloboridae
Zodariidae
Zoridae
Zygiellidae
Глаза
8 (4-4).
8 (4-4).
8 (4-4).
Наличник
больше
передних
медиальных
глаз в
несколько раз.
8 (4-4).
8 (4-4).
8 (4-4),
латеральные
глаза далеко
отстоят друг
от друга.
8 (4-4).
Передние
медиальные
глаза —
самые крупные^
8 (4-4).
8 (4-4).
Карапакс
Грушевидный.
Сильно
варьирует.
Грушевидный.
От
грушевидного
до почти
круглого или
квадратного.
Грушевидный.
Угловато-
грушевидный.
Грушевидный.
Овальный.
Грушевидный.
Хелицеры
У большинства
удлинены, с
многочисленными
мощными шипами и с
дорсальным
выростом у самца
(все Tetragnathinae).
Сильно варьируют.
Не модифицированы.
По краям нижней
губы две круглые
ямки.
Без зубцов.
Не модифицированы.
Не модифицированы.
Коготок маленький
(меньше ширины
основного членика
хелицер).
Не модифицированы.
Не модифицированы.
Брюшко
От овального или
яйцевидного до
палочковидного.
Иногда с хвостом
(некоторые
Tetragnatha).
Сильно варьирует
от круглого до
палочковидного с
хвостом. Почти
всегда с рисунком.
Сферическое.
Обычно
пентагональной
формы, ширина
больше длины.
Часто уплощено.
Овальное. У самок
обычно
одноцветное; у
самцов иногда
дорсально с
парными белыми
пятнами.
Овальное,
заострено на
конце, передняя
треть с одним или
двумя буграми.
Овальное. У
самцов бывает
дорсальный
скутум.
Удлинённо-
овальное.
Овальное.
Ноги
Длинные, передние
ноги чаще всего в 2
и более раз длиннее
тела. Бедро IV м.б. с
рядом трихоботрий.
Ноги I—II с
хватательными
волосками (Diphya).\
Лапка IV
вентрально с
пильчатыми
волосками.
Голени III и IV
с 3—4 рядами
длинных
трихоботрий.
Бёдра расставлены в
стороны. Ноги I—II
длиннее ног III—IV.
Бёдра I всегда с
пролатеральными
шипами.
Предлапки с
дистальной
трихоботрией,
лапки без
трихоботрий.
Каламиструм почти
на всю длину
предлапки IV.
Бёдра с рядом
длинных
трихоботрий.
Без особенностей.
Голени и предлапки
I—II со многими (4 и
более пар)
вентральными
шипами.
Без особенностей.
Определитель семейств
81
Крибеллюм
Нет.
Нет.
Нет.
Нет.
Есть;
разделён на 2
части.
Крибеллюм
не
разделённый
(цельный).
Нет.
Нет.
Нет.
Дыхальце
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
Перед
бородавками.
Бородавки
Без
особенностей.
Без
особенностей.
Колюлюс есть
или
отстутствует.
Крупный
колюлюс.
Без
особенностей.
Без
особенностей.
Без
особенностей.
Всего одна
пара
бородавок,
сидят на общем
выросте.
Передние
бородавки
самцов с
щёткой чёрных
волосков.
Без
особенностей.
Коготки
3
3
3
2
3
3
3
2
3
Пальпа самца
РГО нет. Голень
длиннее своего
диаметра.
РГО нет. Голень
чашевидная,
закруглена, без
отростков. Пара-
цимбиум верхний.
РГО нет. Пальпа
очень крупная,
больше половины
карапакса.
РГО есть. Голень
с 2-3 отростками.
Цимбиум часто с
тутакулюмом.
Голень со сложно
устроенными
отростками про-,
ретролатерально и
дорсально.
РГО нет.
Парацимбиума нет.
РГО есть. Цимбиум с
тутакулюмом.
РГО есть.
РГО нет. Крупный
базальный
парацимбиум.
Эпигина 1
Редуцирована до 1
щелевидного
отверстия, вынесенного
от эпигастральной
щели назад или
простая, но с
медиальной
пластинкой.
Очень варьирует по
форме, бывают с 1
ямкой или без;
рецептакулов 1 или 2
пары.
Выпуклая, с 1 глубокой
ямкой.
Плоская, может быть
очень разной.
Плоская, без ямки, с
септумом.
Слабо хитинизирована
с двумя «сосками» или
полукруглая
хитинизированная.
Плоская, без ямок.
Плоская, с неглубокой
ямкой.
Плоская, с медиальной
пластинкой.
82
3. Строение и классификация пауков
Ключ II. Дихотомический определитель всех семейств пауков
азиатской части России
1. Две пары лёгких (перед и после эпигастральной щели); задние паутинные
бородавки 3-членистые Atypidae
- Одна пара лёгких (перед эпигастральной щелью); паутинные бородавки
всегда 2-членистые (но апикальный членик может быть незаметен) 2
2. Паутинных бородавок всего 1 пара, сидящая на общем основании... Zodariidae
- Паутинных бородавок 3 пары 3
3. Паутинные бородавки расположены в одну поперечную линию. Широкое
дыхальце находится примерно посредине между паутинными бородавками
и эпигастральной щелью Hahniidae (кроме Cryphoeca)
- Паутинные бородавки расположены не в линию 4
4. Анальный бугорок крупный, с пучком длинных волосков; апикальный
членик задних паутинных бородавок серповидно загнут внутрь; карапакс
сердцевидный и заострённый спереди (с «носиком») Oecobiidae
- Анальный бугорок не крупный, без длинных волосков; апикальный членик
задних паутинных бородавок прямой или небольшой; карапакс не
сердцевидный 5
5. Глаз 6 6
- Глаз 8 8
6. Два задних глаза соприкасаются друг с другом и отодвинуты от передних
4 глаз не менее, чем на диаметр. Мелкие (до 3 мм) и бледные гаплогинные
пауки Leptonetidae
- Все глаза собраны в более-менее компактную группу 7
7. Грудной отдел карапакса выше, чем головной отдел. У самцов карапакс
может быть с дорсальными выростами. Лапки с онихумом. Очень мелкие
(до 2 мм) и бледные гаплогинные пауки Oonopidae
- Карапакс не модифицирован. Крупные (более 6 мм) тёмные пауки, часто
с дорсальным рисунком на брюшке Segestriidae
8. Ноги очень длинные (в 4 и более раз длиннее тела), без шипов. Глаза
образуют две триады по бокам (из 3 примерно одинаковых крупных глаз),
между которыми находятся 2 маленьких глаза (передние медиальные).
Нижняя губа сращена со стернумом Pholcidae
- Ноги обычной длины. Если ноги значительно длиннее тела, то на них могут
быть шипы. Нижняя губа не сращена со стернумом и глаза не образуют
триады 9
9. Глаза в 3 ряда; передние медиальные — самые крупные (в 2 и более раз
крупнее остальных глаз); задние медиальные — очень маленькие,
расположены на линии между передними латеральными и задними латеральными
глазами. Карапакс более-менее прямоугольный Salticidae
- Передние медиальные глаза обычно не самые крупные; если же их диаметр
превышает диаметр других глаз, то они не имеют цветной окраски 10
Определитель семейств
83
10. Брюшко на спинной стороне с рисунком из 2-3 пар тёмных крыловидных
пятен. Дыхальце широкое, хорошо заметное и расположено посередине
между эпигастральной щелью и паутинными бородавками. Передние
медиальные глаза в 1,5-2 раза меньше всех остальных глаз, которые примерно
равного диаметра. Бедро пальпы самца с характерными толстыми чёрными
щетинками Anyphaenidae
- Если на спинной стороне и есть подобный рисунок, то дыхальце не
посередине брюшка, а возле паутинных бородавок и плохо заметное. Бедро пальпы
самца без пучков толстых щетинок 11
11. Нижняя часть брюшка покрыта густыми длинными волосками. Дыхальце
широкое и расположено ближе к эпигастральной щели, чем к паутинным
бородавкам. Окраска брюшка одноцветная, без рисунка Argyronetidae
- Густого опушения на брюшной стороне брюшка нет. Если брюшко и
опушено, то дыхальце находится близ паутинных бородавок и на брюшке есть
дорсальный рисунок 12
12.Передние медиальные глаза — самые мелкие, не менее, чем в 2 раза меньше
всех остальных глаз и, кроме того, выдвинуты далеко вперёд от передних
латеральных глаз. Наличник высокий, во много раз превышает диаметр
передних медиальных глаз. Ноги с многочисленными длинными
оттопыренными шипами, длина которых превышает диаметр сегментов, на которых они
сидят. Эпигина выпуклая, без ямки, но с септумом, направленным вперёд
(вверх) Oxyopidae
- Передние медиальные глаза даже если и маленькие, то не выдвинуты далеко
вперёд. Шипы даже если и длинные, то не оттопыренные 13
13.Крибеллюм и каламиструм есть 14
- Крибеллюма и каламиструма нет 18
14.Карапакс прямоугольный. Задние латеральные глаза отставлены от всех
остальных глаз далеко назад (от передних латеральных — не менее чем на
десяток диаметров, т.е. примерно на 1/3 длины карапакса). У самца брюшко
дорсально ярко-красное с 2-3 парами круглых чёрных пятен. Самка дорсаль-
но пепельно-серая с проседью. Крупные, от 8 мм (самцы) до 20 мм и более
(самки) Eresidae
- Карапакс не прямоугольный, его боковые края не ровные и параллельные, а
выпуклые. Задние латеральные глаза если и отодвинуты от передних
латеральных, то на расстояние, гораздо меньшее, чем 1/3 длины карапакса 15
15. Передняя треть брюшка заметно утолщена (смотреть сбоку), иногда с
бугорками. Задние латеральные глаза заметно сдвинуты назад (по отношению к
задним медиальным). Расстояние между передними латеральными и задними
латеральными глазами — не менее 2 диаметров Uloboridae
- Брюшко примерно одинаковой толщины по всей его длине. Передние и задние
латеральные глаза сближены на расстояние, меньшее, чем 2 диаметра 16
16.Крибеллюм, если есть, то цельный (не разделён на 2 части). Мелкие пауки
(2,5-6 мм) Dictynidae
- Крибеллюм разделён на 2 части. Пауки средних размеров (больше 5 мм).... 17
84
3. Строение и классификация пауков
17.Каламиструм простирается почти на всю длину предлапки IV. У самок
брюшко обычно одноцветное, тёмное, без рисунка. У самцов брюшко дорсально
иногда с парными белыми пятнами. Если рисунок на брюшке есть, то он
всегда без ланцетовидного пятна, а только лишь с парными белыми пятнами.
Titanoecidae
- Длина каламиструма не превышает половину длины предлапки IV У самок и
самцов брюшко обычно с дорсальным рисунком, в котором присутствует
ланцетовидное пятно, но никогда не бывает парных белых пятен
Amaurobiidae
18. Брюшко обычно овальное или округлое (но есть исключения); голень пальпы
самца без РГО; эпигина обычно выпуклая (исключение — Theridiidae,
Nesticidae и некоторые Tetragnathidae) 19
См. также ключ III.
- Брюшко обычно вытянуто; голень пальпы самца обычно с РГО (исключение
— Lycosidae и Ctenidae); эпигина обычно плоская 29
19. Голень и предлапка I и II на пролатеральной поверхности с чередующимися
длинными и короткими пролатеральными шипами Mimetidae
- Голень и предлапка I и II без чередующихся длинных и коротких шипов
или вообще без латеральных шипов 20
20.Хелицеры со стридуляционными бороздами (нередко плохо заметны).
Головогрудь самцов нередко с выростами. Парацимбиум Г- или U-образный, базаль-
ный, обычно со щетинками, соединяется с цимбиумом мембраной. Брюшко
не модифицировано, овальное или каплевидное. Эпигина всегда с
медиальной пластинкой Linyphiidae
- Хелицеры без стридуляционных борозд, парацимбиум не Г-образный,
верхний или базальный, сросшийся с цимбиумом, обычно без щетинок. Брюшко
может быть с выростами 21
21. Наличник низкий (примерно равен 1-2 диаметрам передних медиальных
глаз), парацимбиум базальный (пальцевидный или лопастевидный) 22
- Наличник высокий (значительно больше, чем 2 диаметра передних
медиальных глаз), парацимбиум разной формы и положения, иногда не заметен 23
22. Головной отдел карапакса обычно не темнее остальных участков. Брюшко
овальное или каплевидное, нередко с выростами (бугорками) или заострено.
У самца часто голень I и/или II с шипами более многочисленными и
мощными, чем на других ногах. Парацимбиум пальцевидный без щетинок. Эпигина
обычно со скапусом (исключение — Hypsosinga) Araneidae
- Головной отдел затемнён; брюшко овальное; голень II у самца без особых
шипов; парацимбиум не пальцевидный; эпигина без скапуса Zygiellidae
23.Пауки мелкие (не длиннее 1,5 мм); карапакс очень высокий; у самца на
голенях и предлапках ног I есть мощный пролатеральный шип; у самки на
бёдрах I и II есть вентрально-апикальная тёмная бляшка; брюшко округлое....
Mysmenidae
- Пауки крупнее, чем 1,5 мм; самец без особых шипов на ноге I; бёдра самки
без бляшек 24
Определитель семейств
85
24.Размер 1,5-2,5 мм. Брюшко круглое. По бокам от нижней губы находится
пара округлых ямок. Бульбус круглый Theridiosomatidae
- По бокам от нижней губы нет круглых ямок, брюшко и бульбус разной формы
25
25.Бёдра IV с двумя рядами трихоботрий Leucauge (Tetragnathidae)
- Бёдра IV без пролатерального ряда трихоботрий 26
26.Максиллы длиннее нижней губы более чем в 1,5 раза, параллельны друг
другу. Хелицеры часто длинные, с зубчиками по обе стороны от коготка.
Пальпа самца может быть длиннее карапакса. Парацимбиум базальный,
массивный с двумя ветвями (Metinae) или длинный (длиннее диаметра тегу-
люма) (Tetragnathinae) или вертлуг пальпы самца с шипом (Metleucauge)
Tetragnathidae
- Максиллы длиннее нижней губы не более чем в 1,5 раза 27
27.Карапакс с продольной тёмной полосой. У самца предлапка I уплощена
и расширена в проксимальной части. Лапка IV без пильчатых волосков.
Брюшко каплевидное. Пальпа самца с сильно склеротизованным выростом
цимбиума и крюковидным парацимбиумом. Эпигина с септумом Pimoidae
- Лапка IV с серией вентральных пильчатых волосков (бывают плохо видны
или вообще отсутствуют). Максиллы могут быть наклонены друг к другу
Парацимбиум апикальный, если базальный — то крупный и сильно
оттопыренный 28
28.Максиллы не наклонены друг к другу Брюшко овально-каплевидное.
Парацимбиум базальный, большой и оттопыренный. Пальпа самки с длинным
вперёд торчащим коготком. Эпигина без ямки и трубок Nesticidae
- Максиллы наклонены друг к другу. Самец нередко со стридуляционным
аппаратом в задней части карапакса и на переднем крае брюшка. Голень
пальпы самца с закруглёнными краями, края голени плотно прилегают к
бульбусу и цимбиуму Парацимбиум верхний, пальцевидный или незаметен
(скрыт бульбусом). Если пальпа самки с коготком, то он короткий. Эпигина
без медиальной пластинки, всегда с настоящими трубками и с 1-2 парами
круглых или каплевидных рецептакул Theridiidae
29.Задние паутинные бородавки удлинены (особенно их апикальный сегмент)....
Agelenidae
- Задние паутинные бородавки если и удлинены, то их апикальный сегмент
короткий или едва заметный 30
30. Задние медиальные глаза самые крупные, значительно крупнее, чем глаза
переднего ряда. Голень пальпы самца без РГО 31
- Задние медиальные глаза если и крупнее остальных, то незначительно.
Голень пальпы самца с РГО (если без РГО, то передние паутинные бородавки
цилиндрические и раздвинуты на расстояние, равное их диаметру) 32
31. Передние латеральные глаза самые мелкие. Голени и предлапки I—II
с многочисленными (4 и более пар) вентральными шипами Ctenidae
- Глаза переднего ряда примерно одинаковые и расположены в более-менее
ровный ряд; высота наличника примерно равна диаметру глаз переднего ряда.
86
3. Строение и классификация пауков
Вентральных шипов на голени и предлапке I—II от 2 до 6 пар, но обычно
не более 3 пар Lycosidae
32. Передние паутинные бородавки раздвинуты не менее, чем на диаметр, их
базальный членик цилиндрический, а апикальный — маленький, иногда даже
незаметный (если передние бородавки не раздвинуты, то дорсальная сторона
брюшка тёмная с белыми пятнами или с металлическим блеском, задние
медиальные глаза угловатые (примерно треугольные), а эпигина без ямки)
Gnaphosidae
- Передние паутинные бородавки сближены; глаза не угловатые;
металлического блеска нет 33
33. Лапки с 3 коготками 34
- Лапки с 2 коготками 35
34. Наличник в несколько раз выше диаметра передних глаз. Колено пальпы
самца без выроста. Эпигина может быть с септумом Pisauridae
- Наличник невысокий. Колено пальпы самца с выростом. Эпигина с ямкой
или без ямки, но всегда без септума Cybaeidae
35.Края желобка хелицер без зубцов. Ноги III—IV значительно короче, чем
ноги I—II. Скопулы нет. Брюшко обычно пентагональной формы, широкое,
часто уплощено. Цимбиум обычно с ретролатеральным выростом
(тутакулюмом) Thomisidae
- Желобок хелицер с зубцами. Ноги III—IV обычно не укорочены. Лапки
и предлапки со скопулой (если скопулы нет, то есть подкоготковые
подушечки и мощные вентральные шипы на голени и предлапке 1-Й).
Брюшко более-менее удлинено. Цимбиум без тутакулюма 36
36.Нога I длиннее, чем нога IV. Цимбиум с длинным и острым саблевидным
выростом. Эпигина с округлой ямкой, над которой просвечивают
каналы семяприёмнков Cheiracanthiidae
- Цимбиум без выростов; эпигина иная 37
37.Скопула хорошо развита на лапках, а часто и на предлапках. Вентральных
шипов на голенях и предлапках I—II не более 3 пар 38
- Скопула не развита. Голени и предлапки I—II могут быть с многочисленными
(4 и более пар) вентральными шипами или с мелкими толстенькими
шипиками у самцов 40
38.3адний край желобка хелицер без зубцов или с 1 маленьким зубчиком
Philodromidae
- На заднем крае желобка хелицер зубчиков больше, чем 1 39
39. Бёдра ног расставлены в стороны примерно под прямым углом. Мембрана
между лапкой и предлапкой (смотреть на дистальном кончике предлапки)
разделена на 3 части. Скопула простирается почти на всю длину предлапки ...
Sparassidae
- Бёдра не расставлены в стороны. Мембрана между лапкой и предлапкой
обычная, не 3-лопастная. Нога IV длиннее, чем нога I. Эпигина без чётких
границ. У многих видов на тегулюме и в эпигине просвечивают тонкие
Определитель семейств
87
извилистые семенные канальцы. На предлапках скопулы или нет вообще,
или есть только на дистальной части Clubionidae
40.Карапакс с двумя чёткими тёмными продольными полосами на светлом
фоне, берущими начало от задних боковых глаз; нередко эти полосы имеют
продолжение и на дорсальной стороне брюшка. У самцов передние
бородавки снабжены щёткой чёрных волосков, направленных вовнутрь
Zoridae
- Окраска иная; бородавки без щёток 41
41. Тёмные, или брюшко светлее карапакса; рисунок если есть, то поперечный.
Бульбус без медиального отростка. Эпигина с 1 ямкой или септумом,
разделяющим 2 ямки. Рецептакулы овальные Corinnidae
- Светлоокрашенные, брюшко и головогрудь примерно одного цвета;
рисунок, если есть, то продольный. Бульбус с медиальным отростком
характерной формы (его ширина больше, чем высота). Кончик эмболюса иногда
загнут, и образует петлю, похожую по форме на «8» (иногда видна только
на большом увеличении). Ямка эпигины удлинена, имеет чёткие границы,
выглядит как продольная щель Liocranidae
88
3. Строение и классификация пауков
Ключ III. Политомическии определитель Аранеоидных семейств
Карапакс
Наличник*
Хелицеры
Максиллы/
лабиум
Брюшко
Ноги самцов
Ноги самок
Araneidae
Не
модифицирован,
плоский,часто
одноцветный.
Низкий, ~ПМГ.
Мощные,
короткие
направлены
вниз. Обычно в
основании с
латеральным
мыщелком.
-1,5
Иногда с
выростами, редко со
скутумом.
Форма разная,
иногда спереди
заострено.
Почти всегда с
рисунком.
Голени I и/или
II часто с
многочисленны
ми мощными
шипами.
Не
модифицированы, часто
с кольцами.
Linyphiidae
У самцов
нередко с
выростами,
редко с
рисунком.
Высокий,
>ПМГ.
Всегда со
стридуляцион
ными
бороздками;
иногда с
зубцами
спереди.
-1,5
Овальное,
яйцевидное;
иногда со
скутумом
(-ами) в том
числе и
вентральными.
Часто без
рисунка.
Иногда
только 1-2
дорсальных
шипа на всех
голенях.
Не
модифицированы,
редко с
кольцами.
Mimetidae
Высокий,
выпуклый, без
рисунка.
Высокий,
>ПМГ.
Ширина
одинакова по
всей длине,
вместо зубцов
гвоздевидные
шипики.
-1,5
С бугорками в
передней трети,
всегда с
рисунком.
Голени и
предлапки I и II
пролатерально
с
чередующимися длинными
и короткими
шипами.
Голени и
предлапки I и II
с пролате-
ральным рядом
чередующихся
длинных и
коротких
шипов.
Mysmenidae
Очень высокий с
поднятым
головным отделом,
у самцов высота
больше длины.
Очень высокий,
» ПМГ.
Небольшие, не
модифицированы.
-1,5
Шаровидное, с
рисунком.
Голени и
предлапки I с 1 -2
мощными
пролатеральными
шипами. Колец
нет.
Бёдра I и II с
тёмной бляшкой
вентрально. Колец
нет.
Nesticidae I
Не
модифицирован.
Без рисунка.
Высокий,
> ПМГ.
Небольшие, не
модифицированы.
-1,5; максиллы
наклонены друг
к другу.
Яйцевидное, с
рисунком или
без.
Лапка IV
вентрально с
пильчатыми
волосками.
Колец нет.
Лапка IV
вентрально с
пильчатыми
волосками.
Колец нет.
*ПМГ — передние медиальные глаза.
Определитель семейств
89
Карапакс
Наличник*
Хелицеры
Максиллы/
лабиум
Брюшко
Ноги самцов
Ноги самок
Pimoidae
Продольная
тёмная
полоса.
Высокий,
>ПМГ.
Крупные, не
модифицированы.
~1,5
Яйцевидное,
с рисунком.
ПредлапкаI
проксимально
уплощена и
расширена.
Ноги с
кольцами.
Не
модифицированы,
кольца есть.
Tetragnathidae
Не модифицирован;
головной отдел
может быть затемнён.
Высокий, > ПМГ,
или низкий ~ПМГ
Часто удлинены и
торчат вперёд, со
многими зубчиками,
иногда зубчики есть
на передней
поверхности.
>1,5
От палочковидного
до овального. Часто
серебристого или
золотистого цвета
благодаря гуанину.
Обычно не
модифицированы.
Бедро IV может быть
с пролатеральным
рядом трихоботрий
{Leucauge).
Не модифицированы,
обычно без колец.
Theridiidae
Редко с
выростами, медиальная
щель часто
глубокая, иногда
морщинистый у
самцов. С рисунком
или без.
Высокий, > ПМГ.
Не
модифицированы,
но иногда с
мощными
зубцами.
-1,5-2; максиллы
наклонены друг к
другу.
От шаровидного
до вытянутого, с
рисунком или без
(Robertus,
некоторые
Dipoena s.l.).
Лапка IV
вентрально с
пильчатыми
волосками. Иногда
с вентральными
шипиками на
ногах I и II
(Enoplognatha).
Лапка IV
вентрально с
пильчатыми
волосками. С
кольцами или
одноцветные.
Theridio-
somatidae
Высокий, с
поднятым
головным
отделом, без
рисунка.
Высокий, >
ПМГ.
Не
модифицированы.
-1,5, по
бокам от
лабиума —
округлые
ямки.
Шаровидное,
с рисунком.
Не
модифицированы,
без колец.
Не
модифицированы,
без колец.
Zygiellidae
Затемнён от
глазного поля до
медиальной
щели.
Низкий, -ПМГ
Мощные и
короткие.
Обычно в
основании с
латеральным
мыщелком.
-1,5
Овальное
уплощённое, с
рисунком
Не
модифицированы, кольца
есть.
Не
модифицированы, с
кольцами.
90 3. Строение и классификация пауков
Цимбиум
Вершина
цимбиума
Парацибиум
Голень
пальпы самца
Эпигина
Araneidae
Прямоугольный,
почти не
прикрывает
бульбус.
Широкая,
овальная.
Небольшой,
пальцевидный,
сросшийся с
цимбиумом, без
волосков.
Не
цилиндрическая, с
вентральным
конусовидным
выростом;
ширина больше
длины.
Чаще всего со
скапусом.
Linyphiidae
Иногда с
выростами,
нередко с
ретролате-
ральной
лопастью.
Широкая,
овальная.
Г- или U-
образный,
сочлённый с
цимбиумом
мембраной,с
волосками, от
небольшого
до крупного.
Часто с
отростками,
иногда с
несколькими,
длина равна
или больше
ширины.
Варьирует, но
всегда с
медиальной
пластинкой
(заметной на
виде сзади).
Mimetidae
С базальным
выростом,
сливающимся с
цимбиумом.
Широкая,
овальная.
Массивный,
сильно
склеротизо-
ванный.
Цилиндрическая, длина
больше
ширины.
Без ямки, с
септумом.
Mysmenidae
На вершине
подразделён на 2
части, широкий.
Разделена на 2
лопасти.
Нет.
Чашевидная, длина
равна ширине.
Со скапусом (он
прозрачный и
заметен только при
взгляде сбоку).
Nesticidae I
Ладьевидный.
Широкая,
овальная.
Большой,
оттопыренный, о
зубчиком.
Конусовидная,
длина больше
ширины.
Пластинчатая, с
просвечивающими
удлинёнными рецепта-
кулами.
Определитель семейств
91
Цимбиум
Вершина
цимбиума
Парацибиум
Голень
пальпы самца
Эпигина
Pimoidae
С мощным
пролатераль-
ным
выростом.
Сильно
вытянута,
пальцевидная.
Крючко-
видный,
небольшой.
Цилиндрическая, длина
равна
ширине.
С септумом.
Tetragnathidae
От ладьевидного до
длинного, иногда с
дорсальным
выростом.
Широкая и овальная,
но иногда сильно
вытянута,
пальцевидная.
Массивный
оттопыренный или
длинный
вертикальный, или
небольшой.
Конусовидная, длина
больше ширины.
Редуцирована или
плоская, редко с
выростами.
Theridiidae
Ладьевидный,
иногда с выростом.
Иногда с
выростами.
Верхний,
пальцевидный,
либо скрыт
бульбусом.
Конусовидная,
длина обычно
больше ширины,
края всегда
закруглены,
плотно прилегают
к цимбиуму и
бульбусу.
Скапуса нет.
Вульва с
настоящими
трубками, иногда
очень длинными.
Theridio-
somatidae
Ладьевидный.
Широкая,
овальная.
Небольшой,
без
волосков.
Короткая,
края
закруглены и
прилегают к
цимбиуму.
Большая, с
глубокой и
широкой
ямкой.
Zygiellidae
Ладьевидный
Широкая,
овальная.
Массивный,
слегка
оттопыренный.
От короткой до
сильно
вытянутой
(бывает более
чем в 2 раза
длиннее
цимбиума).
Без ямки, сильно
хитинизирована.
92
3. Строение и классификация пауков
Ключ IV. Дихотомический определитель сложных групп пауков
(тех, которые не определяются по глазам, бородавкам и дыхальцу) всего
региона (Сибири и Дальнего Востока)
1. Коготков 2, РГО всегда есть, эпигина плоская 2
- Коготков 3, РГО только у некоторых, эпигина разная 9
2. Бёдра торчат в стороны, перпендикулярно к оси тела 3
- Передние две пары ног направлены вперёд, задние — назад 5
3. Все ноги примерно равной величины, лапки I и II со скопулой 4
- Задние две пары ног заметно короче передних. Лапки без скопулы
Thomisidae
4. Скопула есть и на лапках и на предлапках; на заднем крае желобка
хелицер 3 зубчика Sparassidae
- Скопула есть только на дистальной части лапок; задний край желобка
хелицер без зубчиков или с 1 зубчиком Philodromidae
5. Ширина нижней губы больше её длины. Предлапки I—II вентрально с 2 или
более парами шипов 6
- Длина нижней губы значительно больше её ширины. Предлапки I—II
вентрально только с 1 парой шипов 8
6. Карапакс со светлой медиальной полосой и тёмными латеральными
полосами. Задний ряд глаз сильно выгнут. У самцов передние бородавки со щёткой
чёрных волосков, направленных вовнутрь Zoridae
- Карапакс обычно без светлой медиальной полосы и тёмных латеральных .... 7
7. Светлоокрашенные, брюшко и головогрудь примерно одного цвета;
рисунок, если есть, то продольный. Медиальный отросток характерной
формы (его ширина больше, чем высота). Эпигина с продольной щелью
и длинными рецептакулами Liocranidae
- Тёмные, или брюшко светлее карапакса; рисунок если есть, то поперечный.
Бульбус без медиального отростка. Эпигина с ямкой или септумом,
рецептакулы овальные Corinnidae
8. Нога I длиннее, чем нога IV. Пальпа самца с медиальным отростком; есть
вырост цимбиума, направленный вниз, длинный, тонкий и острый. Эпигина с
ямкой, над которой просвечивает пара или больше поперечных тёмных полос
(каналы семяприёмников) Cheiracanthiidae
- Нога IV длиннее, чем нога I. Пальпа самца без медиального отростка, и без
выроста цимбиума. Эпигина без ямки или, если ямка есть, то над ней нет
поперечных тёмных полос Clubionidae
9. Брюшко сферическое или его высота больше длины 10
- Брюшко не сферическое, а овальное или яйцевидное, его высота меньше
длины 14
10. Ноги I спереди с рядом чередующихся длинных и коротких шипов. Брюшко с
парой бугорков спереди. Желобок хелицер без зубчиков, но с гвоздевидными
Определитель семейств
93
шипиками. Парацимбиум большой, находится в основании цимбиума
Mimetidae
- Ноги без подобных шипов и брюшко без бугров, хелицеры без гвоздевидных
шипиков 11
11. Лапка IV с вентральным рядом пильчатых волосков. Голень пальпы самца
пролатерально вытянута и закруглена. Парацимбиум находится в верхней
трети цимбиума. Эпигина без медиальной пластинки Theridiidae
- Лапка IV без пильчатых волосков, парацимбиум находится в основании
цимбиума 13
13. По бокам от нижней губы 2 округлые ямки. Длина карапакса больше
высоты. Тегулюм округлый. Нога I самца без особых шипов. Эпигина с ямкой
Theridiosomatidae
- Нижняя губа без округлых ямок по бокам, высота карапакса примерно
равна длине. Голень и предлапка I самца с 1-2 толстыми шипами,
бёдра I—II самки с круглой тёмной бляшкой снизу, эпигина со скапусом
(слабо заметен) Mysmenidae
14. Медиальные и латеральные глаза раздвинуты не более чем на 1,5 диаметра.
Колено пальпы самца с отростком, на котором часто есть короткие и толстые
тёмные зубчики; РГО есть; кондуктор диктинного типа; парацимбиума нет.
Эпигина с ямкой и с трубчатыми канальцами Cybaeidae
- Медиальные и латеральные глаза обычно широко раздвинуты. Колено
пальпы самца без отростков; РГО есть только у некоторых видов;
кондуктор не диктинного типа; парацимбиум есть (бывает базальный или
верхний) 15
15.Лапка IV с вентральным рядом пильчатых волосков 16
- Лапка IV без пильчатых волосков 17
16. Самцы со стридуляционными органами на заднем крае карапакса
и вершине брюшка или со вздутой предэпигастральной областью.
Голень пальпы самца пролатерально вытянута и закруглена; парацимбиум
находится в верхней трети цимбиума. Эпигина без медиальной пластинки.
Коготок пальпы самки если имеется, то небольшой Theridiidae (часть)
- Парацимбиум базальный, очень большой и оттопыренный, заострён.
Эпигина плоская, без ямки. Пальпа самки с длинным прямым коготком
Nesticidae
17. Хелицеры со стридуляционными бороздами. Карапакс самца нередко
с выростами, или особыми волосками. Голень пальпы самца часто
с отростками; парацимбиум крупный Г- или П-образный, плоский.
Эпигина всегда с медиальной пластинкой Linyphiidae
- Хелицеры без стридуляционных борозд 18
18. Карапакс без выростов. У самцов предлапка I проксимально уплощена и
слегка искривлена. Голень пальпы самца без отростков; цимбиум со склероти-
зованным дорсальным выростом Pimoidae
- У самцов предлапка I без утолщения, цимбиум без дорсального выроста ... 19
94
3. Строение и классификация пауков
19.Наличник высокий, парацимбиум обычно большой, эпигина без выростов ....
Tetragnathidae
(часть — Dyphia, некоторые Pachygnatha, Tetragnatha)
- Наличник невысокий, парацимбиум обычно небольшой, эпигина часто со
скапусом 20
20. Хелицеры более чем в 2,5 раза превышают свою длину, с многочисленными
зубцами не только на краях желобка, но также и на передней поверхности,
а иногда и по бокам хелицеры; коготки хелицер очень длинные. Пальпа самца
длиннее карапакса; парацимбиум вертикальный, сильно вытянутый; тегулюм
сферический. Эпигины нет; половая щель отодвинута от эпигастральной
щели назад Tetragnathidae
(часть — некоторые Pachygnatha, Tetragnatha)
- Хелицеры не длиннее, чем 2 их ширины; зубцы только по краям желобка;
коготок не длиннее, чем 1,5 ширины хелицеры. Парацимбиум не
вертикальный. Эпигина есть 21
21.Максиллы в два и более раз длиннее их ширины. У самца голень II
не утолщена; пальпа длиннее карапакса. Эпигина без скапуса, но иногда
с выростом, покрытым мелкими волосками Tetragnathidae
(часть — Leucauge, Meta, Metellina, Metleucauge, некоторые Pachygnatha)
- Максиллы менее чем в два раза длиннее своей ширины. У самца голень II
обычно утолщена и с многочисленными шипами; пальпа короче карапакса.
Эпигина со скапусом, если скапуса нет, то ширина эпигины больше или
равна её длине 22
22. Брюшко самки обычно выше и шире спереди; эпигина со скапусом или
выпуклой медиальной пластинкой. У самца голени I и/или II часто с
утолщёнными шипами; парацимбиум цилиндрический короткий; медиальный
отросток с острыми на вершине изогнутыми выростами. Самец всегда меньше
самки Araneidae
- Брюшко овальное, уплощённое. Эпигина без скапуса; большая часть
медиальной пластинки погружена в эпигастральную щель. Парацимбиум толстый, не
цилиндрический; медиальный отросток без острых выростов. Самцы и самки
примерно одинаковые по размеру Zygiellidae
Определитель семейств 95
Ключ V. Дихотомический определитель сложных групп пауков
только Сибири (тех, которые не определяются по глазам, бородавкам
и дыхальцу)
1. Бёдра торчат в стороны, перпендикулярно оси тела; 2 коготка 2
- Передние две пары ног направлены вперёд, задние — назад; коготков 2
или 3 4
2. Задние две пары ног заметно короче передних; лапки без скопулы
Thomisidae
- Все ноги примерно равной величины; лапки I и II со скопулой 3
3. Скопула и на лапках и на предлапках; на заднем крае желобка хелицер
есть 3 зубчика Sparassidae
- Скопула есть только на дистальной части лапок; задний край желобка
хелицер без зубчиков или с 1 зубчиком Philodromidae
4. Хелицеры большие, направлены несколько вперёд, со многими зубцами.
Хелицеры самца могут быть с выростом спереди (на дорсальный стороне).
У самки нет эпигины Tetragnathidae
- Хелицеры иной формы; самка всегда с эпигиной 5
5. Лапки с 2 коготками, РГО есть, эпигина плоская 6
- Лапки с 3 коготками, РГО обычно нет, эпигина часто с отростками 10
6. Длина нижней губы значительно больше её ширины. Предлапки I—II
вентрально только с 1 парой шипов 7
- Ширина нижней губы больше её длины. Предлапки I—II вентрально с 2 или
более парами шипов 8
7. Нога I длиннее ноги IV. Пальпа самца с медиальным отростком;
есть длинный саблевидный вырост цимбиума. Эпигина с ямкой,
над которой просвечивают поперечные тёмные полосы (каналы
семяприёмников) Cheiracanthiidae
- Нога IV длиннее ноги I. Пальпа самца без медиального отростка
и без выроста цимбиума. Эпигина без ямки; если ямка есть,
то над ней нет поперечных тёмных полос Clubionidae
8. Карапакс со светлой медиальной полосой и тёмными латеральными
полосами, задний ряд глаз сильно выгнут. У самцов передние паутинные
бородавки со щёткой чёрных волосков Zoridae
- Карапакс без светлой медиальной полосы и тёмных латеральных 9
9. Светлоокрашенные; брюшко и головогрудь одного цвета; рисунок
продольный. Бульбус с медиальным отростком (его ширина больше, чем
высота). Эпигина с продольной щелью; рецептакулы длинные .... Liocranidae
- Тёмные, или брюшко светлее карапакса; рисунок если есть, то в виде
поперечных полос. Бульбус без медиального отростка. Эпигина
с поперечной ямкой; рецептакулы овальные Corinnidae
10. Брюшко сферическое (высота примерно равна длине и ширине) 11
96
3. Строение и классификация пауков
- Брюшко не сферическое, а овальное или яйцевидное (высота меньше
длины) 12
11. На передней поверхности голеней и предлапок I и II есть ряд
чередующихся длинных и коротких шипов. Брюшко с парой бугорков
спереди. Парацимбиум большой, в основании цимбиума Mimetidae
- Ноги I—II без пролатеральных рядов шипов. Бугорков на брюшке нет,
или если есть, то сзади. Лапка IV с вентральным рядом пильчатых
волосков. Парацимбиум незаметен Theridiidae (часть)
12. Лапка IV с вентральным рядом пильчатых волосков. Голень пальпы
самца пролатерально вытянута и закруглена; парацимбиум находится
в верхней трети цимбиума. Эпигина без медиальной пластинки, канальцы
эпигины — трубчатые Theridiidae (часть)
- Лапка IV без пильчатых волосков; парацимбиум в основании цимбиума;
эпигина с медиальной пластинкой 13
13. Наличник в 2 и более раз превышает передние глаза. Хелицеры
со стридуляционными бороздами. Парацимбиум крупный Г- или U-образный,
соединённый с цимбиумом мембраной, всегда несёт несколько волосков.
Голень пальпы самца часто с отростками. Карапакс у самцов нередко
с выростами, или особыми волосками Linyphiidae
- Наличник невысокий; хелицеры без стридуляционных борозд; парацимбиум
небольшой цилиндрический или утолщённый, причленён неподвижно;
голень пальпы самца без отростков 14
14. Брюшко самки обычно выше и шире спереди; эпигина со скапусом,
или с выпуклой медиальной пластинкой. У самца голень I и/или II часто
с утолщёнными шипами, парацимбиум цилиндрический короткий;
медиальный отросток с острыми на вершине изогнутыми выростами.
Самец всегда меньше самки Araneidae
- Брюшко овальное, слегка уплощено, с листовидным рисунком. Эпигина
без скапуса; большая часть эпигины погружена в эпигастральную щель.
Парацимбиум толстый, не цилиндрический; медиальный отросток без
острых выростов. Самец по размерам примерно равен самке Zygiellidae
4. СПЕЦИАЛЬНАЯ ЧАСТЬ
Схема описания семейств
Все семейства расположены по алфавиту для удобства их поиска. Нередко
семейства аранеоморфных пауков располагают согласно каким-либо схемам,
которые меняются. Мы же предпочли расположить строго по алфавиту, который не
подвержен переменам. Мигаломорфных пауков обычно рассматривают отдельно,
но у нас встречается только одно семейство, и мы решили дать его в общем списке.
Очень немногие группы пауков имеют русские названия, поэтому для многих
семейств мы даём транслитерированное латинское название. В англоязычных
странах, в Германии, в Японии для всех семейств имеются свои имена. Мы не стали
переводить их, но даём английские названия для всех групп (если таковые имеются).
Нередко английские названия длинные (например, daddy-long-legs — Pholcidae) или
не имеют в русском языке соответствующего эквивалента (например, money-spiders
для Linyphiidae). Английское название Oxyopidae — lynx-spiders или пауки-рыси.
Понятно (зная биологию паука), откуда пошло это название, но на наш взгляд,
проще сказать или написать «оксиопиды», нежели «пауки-рыси». К тому же, в научной
англоязычной литературе народными названиями пользуются не чаще, чем у нас.
Характеристика видового разнообразия в мире для разных семейств дана по
данным каталога Н.И. Платника (Platnick, 2009, версия 9.5, по состоянию на 30
декабря 2008 г.). Данные по России почерпнуты из наших (Ю.М.)
неопубликованных данных по состоянию на середину 2003 г. Сведения по разнообразию пауков
азиатской части России взяты по 2003 г., с учётом последующих публикаций до
2007 г. и собственным данным.
При описании тех или иных черт строения, мы основывались только на тех
таксонах, которые встречаются в нашем регионе. Описания мы пытались делать
как можно лаконичнее. Более подробно расписаны те черты, которые важны для
определения конкретного семейства, а также те признаки, которые используются
для определения родов и видов.
Диагностические признаки семейства включают сведения о размере пауков,
типе копулятивного аппарата, наличии или отсутствии крибеллюма, числе и
расположении глаз, количестве коготков на лапках. Кроме этого, указываются другие
характерные черты, которые могут быть найдены в форме карапакса или брюшка,
расположении и размере паутинных бородавок, расположении и числе дыхалец,
строении пальпы.
После диагностических признаков следует перечень таксонов (семейств и
родов), которые могут быть спутаны с представителями данного семейства, и
указания как их можно отличить — рубрика Близкие группы.
Рубрика Описание включает следующие обязательные моменты: карапакс, стер-
нум, глаза, ротовой аппарат, ноги, брюшко, паутинные бородавки, органы дыха-
98
4. Специальная часть
ния, крибеллюм-каламиструм, копулятивные органы, размер, окраска, особые
черты. Детальность их описания зависит от важности тех или иных признаков.
Карапакс мы называем грушевидным, если головная часть резко сужена по
сравнению с грудной; яйцевидным (овальным) — если головная часть не отделена
от грудной резким сужением; круглым — если ширина равна длине.
В разделе размер говорится только о размерном половом диморфизме.
Перечень родов, зарегистрированных в регионе (рубрика Рода, известные из
азиатской части России), включает даже те таксоны, которые были указаны
ошибочно (в результате ошибки определения). Такие рода сопровождаются
примечаниями. Если род представлен одним видом, то указано его название. В ряде
случаев, если в регионе обитает всего несколько видов одного семейства, то
перечислены все виды.
В рубрике Распространение указано, где обитают те или иные рода, а в ряде
случаев даны ссылки на литературу, где приводятся карты с ареалами отдельных
видов.
Рубрика Биология включает сведения о зональном и биотопическом
распространении видов, о способах и приёмах охоты, об устройстве убежищ, сетей,
коконов и другую информацию.
В рубрике Методы сбора указано, как можно собрать представителей данного
семейства.
В рубрике Таксономия рассказывается о таксономической истории таксона, с
указанием того, как он приведён в определителе В.П. Тыщенко (1971). Кроме того,
указаны спорные моменты в классификации, описано, насколько хорошо таксон
изучен в азиатской части России. Отдельно перечислены все или основные работы
по систематике конкретного семейства в регионе. Помимо этого, приведён список
наиболее важных публикаций (монографий или обзорных статей) по
сопредельным регионам.
Вслед за этим разделом, в рубрике Разграничение видов, перечислены
основные признаки, по которым можно различать виды внутри семейства.
И последняя рубрика Перспективы изучения. В ней говорится, насколько
хорошо (на наш взгляд) изучено разнообразие и таксономия группы в азиатской
части России и где можно ждать новых находок.
Семейства, встречающиеся в азиатской части России
Семейство Agelenidae C.L. Koch, 1837
Агелиниды, nayки-воронкопряды; funnelweb spiders
Рис. 4.1-14.
Крупное всесветно распространённое семейство с более чем 1100 видами и 68
родами. В России отмечено 26 видов, принадлежащих к 9 родам, а в азиатской
части России — 17 видов из 8 родов.
Диагностические признаки. Крупные, энтелегинные, некрибеллятные, 3-ко-
готковые пауки с 8 глазами в 2 ряда, задние паутинные бородавки удлинены, паль-
па с РГО.
Близкие группы. Габитуально и окраской напоминают Cybaeidae, Callobius и
Cybaeopsis из Amaurobiidae и некоторых пауков-волков Lycosidae. От Cybaeidae
отличаются удлинёнными задними паутинными бородавками. От Callobius и
Cybaeopsis отличаются отсутствием крибеллюма. От пауков-волков Lycosidae
чётко отличается гомогенными глазами в 2 ряда и удлинёнными бородавками.
ОПИСАНИЕ:
• Карапакс. От яйцевидного до грушевидного, не уплощён.
• Стернум. Щитовидный, широкий (ширина почти равна длине), у Tegena-
ria — с рисунком.
• Глаза. 8, в 2 ряда (4-4), примерно равной величины, расположены
сравнительно компактной группой (расставлены не более чем на один-два
диаметра).
• Ротовой аппарат. Не модифицирован, передний край желобка хелицер с 3-
5 зубцами, а задний с 2-8.
• Ноги. 3 коготка, с шипами, передние — всегда длиннее тела, лапки без ско-
пулы.
• Брюшко. Овальное, обычно с рисунком, но бывает и без рисунка.
• Паутинные бородавки. Передние бородавки широко расставлены; задние
бородавки значительно удлинены, апикальный членик плавно суживается, у
некоторых родов он длиннее базального членика.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Энтелегинные. Пальпа самца относительно
крупная, всегда с РГО, а у Coelotinae — с отростком колена. Кондуктор может
быть разной формы — мембрановидный {Agelena), как спиральная лента
{Paracoelotes1) или грибовидный (диктинного типа) (у Tegenaria). Эмболюс — от
короткого и толстого {Agelena) до длинного нитевидного {Paracoelotes).
Медиальный отросток бульбуса имеется. Эпигина варьирует. У Agelena это
достаточно глубокая ямка, у Coelotinae ямка может быть выражена, но
иногда нет. У самок Coelotinae эпигина всегда с парой сосочков (плоских вырос-
1В каталоге пауков мира (Platnick, www) род Paracoelotes сейчас числится под названием Pire-
neitega Kishida, 1955. Мы не принимаем эту синонимию, поскольку она сделана не по правилам
МКЗН (отдан приоритет забытому названию).
Семейства, встречающиеся в азиатской части России
101
4
6
8 7
Рис. 4. Agelenidae. Соматические признаки и габитус Tegenaria domestica (?, 1-3), Agelena
labyrinthica (4-5), Paracoelotes birulai (?, 6-7) и Alloclubionoides napolovi (cT? 8).
1 — карапакс ?, сверху; 2 — головогрудь ?, снизу; 4 — брюшко, снизу; 3,6 — паутинные
бородавки ?, снизу; 5, 7-8 — габитус, сверху.
102
4. Специальная часть
0
11
Рис. 4 (продолжение). Agelenidae. Копулятивные органы Agelena labyrinthica (cf, 9),
Alloclubionoides amurensis (C?$, 10-11), Paracoelotes sp. (?, 12) и Tegenaria domestica
(Cf?, 13-14).
9, 14 — пальпа cf, снизу; 10, 12-13 — эпигина, снизу; 11 — пальпа cf, сбоку Со — сосочки. 9,
13-14 — по: Guseinov et al. (2005); 10-11 — по: Marusik (2009b).
тов) по бокам от ямки (или медиальной пластинки). Сосочки могут быть
длинными, заострёнными на вершине {Paracoelotes), или короткими,
широкими и округлыми на вершине (Tegecoelotes).
• Размер. 6-20 мм. Самки крупнее самцов, но карапакс у самцов нередко
крупнее, нежели у самок.
• Окраска. От светло-коричневой до чёрной. Большинство видов с рисунком
и на карапаксе и на брюшке. У Tegenaria стернум всегда с тремя парами
круглых пятен, которые могут сливаться с расположенным посередине стер-
нума светлым ланцетовидным пятном.
• Особые черты. Удлинённые задние паутинные бородавки (удлинены оба
сегмента).
Семейства, встречающиеся в азиатской части России
103
Рода, известные из азиатской части России. Agelena Walckenaer, 1805; Age-
lenopsis Giebel, 1869 {A. potteri (Blackwall, 1846)); Allagelena Zhang, Zhu et Song,
2006 {A. bistriata (Grube, 1861)); Alloclubionoides Paik, 1992; Iwogumoa Kishida,
1955 (7. interuna (Nishikawa, 1977)); ICoelotes Blackwall, 1841 (C. terrestris (Wider,
1834)); Paracoelotes Brignoli, 1982; Tegecoelotes Ovtchinnikov, 1999 (T secunda
(Paik, 1971)); Tegenaria Latreille, 1804 (T agrestis (Walckenaer, 1802) и T domestica
(Clerck, 1758)).
Распространение.
Tegenaria domestica — космополитный синантропный вид, известен по всему
региону.
Т. agrestis — отмечена на юге Дальнего Востока, возможно неверно
определена.
Agelena, Paracoelotes — юг Сибири и Дальнего Востока.
Agelenopsis, Allagelena, Alloclubionoides, Tegecoelotes — юг Дальнего Востока.
Хотя первый род сравнительно недавно был интродуцирован из Северной
Америки, помимо жилых зданий, он встречается и в природных условиях.
Iwogumoa interuna — южные Курилы, Сахалин и Монерон.
Coelotes terrestris отмечен в Средней Сибири, очевидно, ошибочно, так как этот
род и вид не проходят в Азию. Возможно, это неверно определённый Paracoelotes
birulai (Ermolaev, 1926).
Биология1. Наиболее широко распространённые и известные виды — это
Agelena labyrinthica (Clerck, 1758) и Tegenaria domestica. Оба плетут воронковидную
сеть, состоящую из широкого, горизонтального щита из плотной паутины,
который к краю переходит в воронковидную трубку, где прячется паук. Первый вид
обитает среди кустов, густой травы, а второй — исключительно в помещениях,
особенно часто в тёмных подвалах. Самцы Tegenaria нередко в поисках самки
«забредают» в раковины умывальников и в ванны, откуда не могут самостоятельно
выбраться. Взрослые Tegenaria domestica встречаются в течение круглого года.
Сходную структуру имеет сеть Coelotes terrestris. Однако щит у них меньше, а
трубка, напротив, длиннее (до 20 см) и шире. Обычно Coelotinae плетут сети на
подстилке, а трубка уходит под камень или даже в землю. Виды Alloclubionoides
встречаются в затенённых местах, сети располагают у поверхности грунта, а
убежища — среди камней. Paracoelotes селятся не только под камнями, но и под
корой больших деревьев, а также в древесине лежащих трухлявых стволов.
Сети агеленид не имеют клейких субстанций, поэтому они не задерживают
упавшее насекомое, а выполняют сигнальную функцию и служат удобным субстратом,
по которому паук может беспрепятственно перемещаться, атакуя добычу. В
отличие от многих других тенетников, передвигаются они по сети не вверх ногами, а
нормально. Таким образом, охотничье поведение агеленид во многом сходно с
таковым типичных бродячих пауков.
Все изученные виды агеленид — полифаги, питающиеся разнообразными
членистоногими. Помимо мягкохитинизированных членистоногих, таких как мухи,
комары, мелкие пауки и цикадки, в диете агеленид, порой в значительном
количестве, встречаются такие хорошо защищенные или потенциально опасные насеко-
1 Раздел написан при участи Э.Ф. Гусейнова.
104
4. Специальная часть
мые как жуки, крупные прямокрылые, муравьи и даже медоносные пчелы (Nyffeler,
1982;Nentwig, 1983).
В диете Coelotes terrestris всегда доминируют жуки, что, очевидно,
обусловлено тем, что этот вид строит сети на подстилке в затенённых местах (Tretzel, 1961;
Nyffeler & Benz, 1981; Petto, 1990). Этот паук использует специализированное
хищническое поведение при охоте на крупных жуков, прокусывая сравнительно
мягкую соединительную мембрану между стернитами брюшка (Tretzel, 1961).
Спаривание, вернее ухаживание, yAgelena довольно простое. Самец,
обнаруживший сеть самки, начинает по ней постукивать, чем вводит самку в состояние транса,
после чего переносит несопротивляющуюся пассивную самку в подходящие
место и спаривается. После этого пара живёт вместе несколько недель (Jones, 1989).
Методы сбора. Лучше всего ручной сбор. Пауков легко можно заметить
благодаря их сетям. Многие виды легче собрать ночью с сетей, чем днём, когда они
прячутся в трубке, куда бывает трудно добраться.
Таксономия. Этой группе придан ранг семейства ещё в 1837 г. После этого оно
неоднократно ревизовалось (Thorell, 1870a-b; Simon, 1898; Roth, 1965; Lehtinen,
1967). Состав и рамки семейства существенно менялись, одни рода исключались
из семейства, другие, наоборот включались. К агеленидам первоначально
относили некоторых Pholcidae, Zodariidae, Gnaphosidae и даже Tetragnathidae.
Наибольшей ревизии подверглись состав и рамки семейства в работе Р.Т. Lehtinen
(1967). Он разбил таксон на два подсемейства: Ageleninae и Coelotinae. Первое
подсемейство было подразделено на четыре трибы. Многие группы (около 100
родов), ранее относимые к семейству Agelenidae, были подняты до ранга
семейства или переведены в другие таксоны. Например, до статуса семейства были
повышены Hahniidae. В семействе Agelenidae были оставлены лишь таксоны, у
которых наличествуют три общие признака: 1)3 коготка; 2) парный колюлюс; 3)
удлинённые задние паутинные бородавки. Если раньше Agelenidae сближались с Amau-
robiidae, то П.Т. Лехтинен отнёс их к самостоятельному надсемейству.
Недавно Coelotinae были перенесены в Amaurobiidae (границы которого не
менее размыты, чем Agelenidae) на основании габитуального сходства (Wunderlich,
1986). А представителей рода Alloclubionoides корейский арахнолог К.Y. Paik (1992)
вообще относил к Clubionidae!
Несмотря на неоднократную радикальную перестройку семейства, его
классификация ещё далека от удовлетворительной. Судя по строению пальпы самца,
Agelenini и Tegenariini не имеют ничего общего, и напротив, пальпа у Agelena
принципиально не отличается от таковой у Amaurobius (Amaurobiidae). Пальпы же
Tegenariini отличается от Dictyninae (Dictynidae) и Cybaeus (Cybaeidae) только лишь
наличием медиального отростка, а эмболюс и кондуктор — идентичны. Поэтому
не исключено, что семейство в обозримом будущем будет подвергнуто очередной
кардинальной перестройке.
Исходя из строения пальпы, мы считаем, что Coelotinae следует рассматривать
в Agelenidae, хотя этой группе можно придать статус семейства1.
1 Когда книга было практически завершена, Y. Nishikawa (2009) поднял статус подсемейства
Coelotinae до семейства, но в последней версии (11.5) каталога Н.И. Платника эта группа уже
перенесена в Agelenidae, хотя в прежних версиях рассматривалась в Amaurobiidae.
Семейства, встречающиеся в азиатской части России 105
Систематике агеленид региона (в основном Coelotinae) посвящены следующие
работы: Олигер (19836); Марусик, Логунов (1991); Marusik & Koponen (2000);
Овчинников (1999). Особенно важна работа последнего автора, который ревизовал
всех Coelotinae азиатской части России и описал несколько новых родов.
В последнее время описывается множество новых видов и родов агеленид из
Китая, однако обзорных работ по фауне Китая по-прежнему нет. В 2009 г. в одной
публикации (Nishikawa, 2009) было описано 44 новые вида Coelotinae/Coelotidae
из Японии.
В настоящее время этой группой в России занимается Ю.М. Марусик.
Разграничение видов. Все виды различаются формой копулятивных органов,
однако окраска и рисунок тоже могут быть полезны для разграничения видов.
Перспективы изучения. Вполне возможны находки новых и дополнительных
видов на юге Дальнего Востока.
Agelena labyrinthica (Agelenidae) in copula (фото: М.М. Омелько).
Семейство Amaurobiidae Thorell, 1870
Амауробииды; hackled-meshweb weavers
Рис. 5.1-10.
В мире известно 50 родов и 276 видов1, распространённых по всем материкам;
из них в России обитает 6 видов из 4 родов. В азиатской части России достоверно
отмечено 3 вида и 3 рода.
Диагностические признаки. Крибеллятные, энтелегинные пауки, 6-20 мм, 8
глаз в два ряда, голень пальпы самца с мощными отростками {Callobius, Cybaeopsis).
Близкие группы. Близки и напоминают Agelenidae и Cybaeidae, но
отличаются от них наличием крибеллюма. Похожи на крибеллятных Dictynidae и Titanoecidae,
но отличаются от них более крупным размером, окраской и формой копулятивных
органов.
ОПИСАНИЕ:
• Карапакс. Яйцевидный-овальный; головной отдел слегка приподнят над
грудным. Наличник невысокий.
• Стернум. Не модифицирован.
• Глаза. 8 в два ряда (4-4).
• Ротовой аппарат. Хелицеры крупные, выдаются вперёд (видны при виде
сверху).
• Ноги. 3 коготка.
• Брюшко. Овальное.
• Паутинные бородавки. Не модифицированы.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Имеются. Крибеллюм разделён на две части.
• Копулятивные органы. Пальпы самцов Callobius и Cybaeopsis с мощными
шиловидными дорсальными отростками голени. Эпигины у этих же родов
состоят из двух выростов-валиков, смыкающихся посредине и образующих
щель.
• Размер. 6-20 мм. Половой диморфизм слабо выражен.
• Окраска. Тёмная. Callobius и Cybaeopsis с рисунком на брюшке. Arctobius
светлее других родов, с характерной окраской брюшка — широкая светлая
медиальная полоса и длинное ланцетовидное пятно посреди неё. Наличие
крибеллюма и характерная окраска позволяет достоверно определять
Arctobius даже по неполовозрелым особям.
• Особые черты. Наличие крибеллюма, крупный размер, пальпа самца с
крупными дорсальными отростками голени.
Рода, известные из азиатской части России. Arctobius Lehtinen, 1967 {A. age-
lenoides (Emerton, 1919)); Callobius Chamberlin, 1947 (C. hokkaido Leech, 1971);
Cybaeopsis Strand, 1907 (C. typica Strand, 1907).
1 Согласно версии 11.5 каталога пауков Н.И. Платника.
Семейства, встречающиеся в азиатской части России
107
Распространение. Arctobius обитает по всему северу; на юг проходит по
таёжным биотопам до Монголии. Callobius и Cybaeopsis встречаются на юге Дальнего
Востока. Первый род обнаружен только на Кунашире, а второй широко
распространён на Курилах, Сахалине и Монероне.
7
3
2
8 4 10
5 6
Рис. 5. Amaurobiidae. Cybaeopsis typicus (сГ$, 1-6), Amaurobius sp. (сГ, 7-9) и Callobius
hokkaido (С?, 10).
1 — головогрудь $, спереди; 2 — предлапка IV; 3 — крибеллюм и паутинные бородавки; 4 —
эпигина, снизу; 5 — габитус ?, сверху; 6, 9 — габитус сГ, сверху; 7 — головогрудь сГ, сверху; 8,
10 — пальпа сГ, сбоку.
108
4. Специальная часть
Биология. Все виды связаны с подстилкой. Arctobius в верховьях Колымы
собран в редкостойном лиственничнике с плотным сфагновым покровом, в тополё-
во-чозениевом лесу (подстилка) и на галечниках ручьёв. На Чукотке этот род в
массе встречается среди мелких кустарничков. В Монголии несколько
экземпляров собраны в лиственничнике с Aulacomnium. Пауки, обитающие во мху и среди
кустарничков, плетут небольшие воронковидные сети. Согласно R. Leech (1972),
самки охраняют кокон сферической формы, который содержит порядка 15 яиц.
Исходя из остатков жертв, разбросанных вокруг гнезда, эти пауки питаются
муравьями и другими пауками. Особи этого вида необычно агрессивны и бросаются на
всё, что приближается к отверстию ловчей трубки (Leech, 1972).
Cybaeopsis отмечен в широком спектре биотопов с влажной подстилкой.
Единичные особи Callobius собраны на приморских скалах. Их фенология и детали
экологии неизвестны.
Методы сбора. Почвенные ловушки и ручной сбор в подстилке и под камнями.
Таксономия. Рамки семейства очерчены довольно слабо. Впервые этот таксон
выделил Т. Thorell (1869-1870), но рассматривал его как подсемейство в Agelenidae.
Позже эта группа была перенесена в Dictynidae. A. Petrunkevitch (1928) придал
этой группе ранг семейства. Первоначально в него включался и род Titanoeca Thorell,
1870 (см. например, Тыщенко, 1971). Дальневосточные представители Coelotinae
в этой работе рассмотрены в составе Agelenidae. В балтийских янтарях известно
несколько видов, относящихся к современному роду Amaurobius.
В Amaurobiidae в настоящее время выделяют порядка 10 подсемейств. Из них в
азиатской части России встречается номинативное подсемейство с двумя родами
{Callobius и Cybaeopsis) и Macrobuninae {Arctobius). R. Leech (1972) выделяет
Arctobius в отдельное подсемейство Arctobiinae.
В таксономических работах отечественных авторов рассматривался только
Cybaeopsis typica (как Callobius claustrarius, Дунин, 19846) иА. agelenoides
(Данилов, 1994). Определить все три наши вида можно благодаря работам Leech (1972),
Palmgren (1977) и Chikuni (1989).
Разграничение видов. Форма копулятивных органов и окраска/рисунок.
Перспективы изучения. Нахождение дополнительных таксонов (известных
из сопредельных регионов) маловероятно, но не исключено. В нашем
распоряжении есть самки, принадлежащие к новому виду Amaurobius из Приморья и
Хабаровского края.
Семейства, встречающиеся в азиатской части России
109
Amaurobius erberi (Amaurobiidae), самка с паучатами, Крым (фото: Н.М. Ковблюк).
Семейство Anyphaenidae Bertkau, 1878
Анифениды; tube spiders, phantom spiders
Рис. 6.1-6.
Всесветно распространённое семейство с 510 видами и 56 родами. Палеаркти-
ку населяет небольшое число видов и только один род. В России известны 3 вида
рода Anyphaena Sundevall, 1833/Голько один вид найден в азиатской части России.
Диагностические признаки. Энтелегинные пауки среднего размера (5-8 мм),
с 8 глазами и 2 коготками, дыхальце расположено посредине брюшка, пальпа
самца всегда с РГО.
Близкие группы. Несколько напоминают Clubionidae, Liocranidae, Gnaphosidae,
однако расположение дыхальца позволяет легко отличать даже неполовозрелых
Anyphaena.
ОПИСАНИЕ:
• Карапакс. Грушевидный.
• Стернум. Овальный.
• Глаза. 8, в два ряда (4-4), первый ряд выгнутый, второй почти прямой.
• Ротовой аппарат. Не модифицирован.
• Ноги. 2 коготка, густая скопула, длина передних ног чуть больше длины тела.
• Брюшко. Овальное, на конце чуть заострено.
• Паутинные бородавки. Не модифицированы.
• Органы дыхания. Пара лёгких и трахеи. Трахеи открываются дыхальцем,
расположенным посредине между эпигастральной щелью и паутинными
бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Пальпа самца с относительно крупным отростком
голени, эмболюс нитевидный, бедро в основании с пучком чёрных
волосков. Эпигина с небольшим передним карманом и щелевидной продольной
ямкой.
• Размер. 5-8 мм. Половой диморфизм слабо выражен.
• Окраска. Светло-коричневая, с характерным рисунком на брюшке (пары
«крыловидных» тёмных пятен) и на карапаксе.
• Особые черты. Положение дыхальца и специфический дорсальный
рисунок на брюшке.
Рода, известные из азиатской части России. Anyphaena Sundevall, 1833 {А.
pugil Karsch, 1879).
Распространение. Южные Курилы, юг Сахалина, остров Монерон и Приморье.
Биология. Очевидно, как и все другие виды рода, связаны с кронами деревьев.
Большинство дальневосточных особей были пойманы укосами и отряхиваньем
ветвей деревьев. Охотятся ночью. Гнёзд не плетут, только в период размножения
самки строят репродуктивные гнёзда-логова в свёрнутых листьях, которые изнутри
выстилают паутиной. Самка сидит внутри на коконе, который охраняет до вылуп-
Семейства, встречающиеся в азиатской части России
111
ления молоди. Самцы европейского вида Anyphaena accentuata (Walckenaer, 1802)
демонстрируют специфическое поведение ухаживания, благодаря которому этот
вид получил прозвище «жужжащий паук» (buzzing spider) (Bristowe, 1958). Во
время ухаживания самцы очень быстро вибрируют брюшком, которое, ударяясь о
субстрат, производит звуки, напоминающие жужжание мухи.
Методы сбора. Укосы по кустарникам, отряхивание деревьев.
Таксономия. Первоначально эта группа пауков рассматривалась в составе Clu-
bionidae. A. Petrunkevitch (1923) придал этому таксону ранг семейства. В пределах
России семейство никогда не ревизовалось. Иллюстрации единственного вида,
обитающего на Дальнем Востоке, имеются в работах Y. Chikuni (1989) и Yu.M. Marusik
(2009а).
Перспективы изучения. Находки дополнительных видов в изучаемом
регионе маловероятны.
1
Рис. 6. Anyphaenidae. Anyphaena pugil (сГ$, 1, 4-6) и A. accentuata (?, 2-3).
1 — габитус ?, сверху; 2 — головогрудь ?, спереди; 3 — брюшко ?, снизу; 4 — эпигина, снизу; 5-
6 — пальпа сГ, сбоку и снизу. 1, 4-6 — по: Marusik (2009a).
Семейство Araneidae Simon, 1895
Пауки кругопряды, пауки-крестовики, orbweb spiders
Рис. 7.1-19.
Третье по величине семейство, примерно t 2985 видами, относимыми к 167
родам. В России известно порядка 105 видов из 19 родов, а в азиатской части
России — 90 видов из 19 родов.
Диагностические признаки. Энтелегинные, некрибеллятные пауки-кругопря-
ды с 3 коготками и 8 глазами; наличник низкий; размер от 3 до 25 мм; брюшко у
большинства видов округлое, слегка или сильно налегает передней частью на
головогрудь; ноги со многими шипами; пальпа самца всегда с небольшим
неподвижным базальным парацимбиумом; эпигина у большинства самок со скапусом.
Близкие группы. Внешне (габитуально и наличием рисунка) похожи на Theri-
diosomatidae, Mysmenidae, на некоторые рода Tetragnathidae и Theridiidae и на Zygiel-
lidae. От двух первых семейств отличаются более крупными размерами, от всех
Tetragnathidae и Theridiidae — низким наличником, и от Zygiellidae
цилиндрическим парацимбиумом и отсутствием широкой продольной медиальной полосы на
карапаксе.
ОПИСАНИЕ:
• Карапакс. Грушевидный, головной отдел большинства таксонов
значительно уже грудной части; наличник низкий, сравнимый с размером глаз (кроме
Cercidia).
• Стернум. Сердцевидный или треугольный.
• Глаза. 8, латеральные сближены между собой и далеко отстоят от
медиальных, самцы Cyclosa с вынесенными вперёд передними медиальными
глазами. Передние медиальные глаза обычно расположены на примерно
одинаковых расстояниях друг от друга и от задних медиальных глаз и вместе с
ними образуют так называемый «аранеидный квадрат».
• Ротовой аппарат. Не модифицирован; хелицеры имеют латеральный
мыщелок; максиллы не удлинены (длина = 1,5 ширины).
• Ноги. 3 коготка. От коротких до очень длинных, с шипами, особенно
многочисленными и толстыми на голени I и/или II у самца (Araneus и родственные
рода). У самцов Araneus тазики I и/или II могут обладать мощным
зубовидным выростом.
• Брюшко. Разнообразно по форме, бывает овальное и яйцевидное (наиболее
широкое в передней трети), ланцетовидное, сферическое, у некоторых
видов — с выростами в передней (Araneus) или в передней и задней частях
(Cyclosa). При этом паутинные бородавки располагаются на конце брюшка,
однако могут быть сдвинуты кпереди. У одного вида, Cercidia prominens
(Westring, 1851), на дорсальной стороне брюшка имеется скутум, а спереди —
ряд шипиков.
• Паутинные бородавки. Не модифицированы.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
Семейства, встречающиеся в азиатской части России
113
1
8
1
Рис. 7. Araneidae. Araniella displicata (?, 1-2), Araneus marmoreus (cT, 3), "Eriophora"
sachalinensis ($, A), Araneus washingtoni (d\ 5), A nordmanni ($, 6-7) и Araniella у'aginumai
№8-10).
1 — головгрудь ?, спереди; 2 — ротовой аппарат ?, снизу; 3 — головогрудь сГ, сбоку; 4, 6, 9 —
эпигина, снизу; 5, 10 — пальпа сГ, снизу; 7, 8 — эпигина, сзади. Мо — медиальный отросток;
стрелкой показан шип на тазике II. 4 — по: Марусик, Кроуфорд (2006); 8-10 — по: Marusik &
Koponen (2000).
114
4. Специальная часть
11 12 13
14 15 16
17 18 19
Рис. 7 (продолжение). Araneidae. Габитус и окраска Araneus marmoreus (cf, 11), Larinio-
ides cornutus (?, 12), "Eriophora" sachalinensis (?, 13), Singa nitidula ($, 14), Araniella
displicata (cf, 15), Pronnoides brunneus (cT, 16), Cyclosa sp. (cT, 17), Hypsosinga sangui-
nea (? из Тувы, 18; сГ из Приморья, 19).
Семейства, встречающиеся в азиатской части России 115
• Копулятивные органы. Пальпа самца всегда с небольшим неподвижным
базальным парацимбиумом цилиндрической формы; голень с вентральным
выростом, благодаря которому ширина голени больше длины; бульбус
всегда с медиальным (= тегулярным) отростком, несущим хотя бы один
коготок; эмболюс короткий. Эпигина у большинства видов со скапусом
(исключение — Hypsosinga), который после копуляции иногда обламывается (у
Aculepeira, Araneus, Cyclosa, Larinia).
• Размер. 3-25 мм. Половой диморфизм выражен; самцы мельче самок, и
имеют другое вооружение ног.
• Окраска. Сильно варьирует от светлой до тёмной, с рисунком карапакса
или без, брюшко практически всегда с рисунком. Рисунок имеется и на
вентральной стороне брюшка. Рисунок чаше всего видоспецифичен, однако,
есть виды, у которых он чрезвычайно варьирует (Neoscona scylla (Karsch,
1879)). Есть виды (группа видов Araneus quadratics), у которых рисунок
одинаковый, но окраска сильно варьирует — от ярко-оранжевой до зелёной.
Araneus variegatus Yaginuma, 1960 имеет предостерегающую окраску, вент-
рал ьно брюшко с красным пятном. Укусы этого паука болезненны.
• Особые черты. Наличие колесовидных сетей; многочисленные шипы на
ногах, низкий наличник, «аранеидный квадрат» медиальных глаз, нога II у
самцов часто с мощными шипами.
Рода, известные из азиатской части России (~23).
? — недостоверные находки.
Aculepeira Chamberlin et Ivie, 1942; lAgalenatea Archer, 1951 {A. redii (Scopoli,
1763));Alenatea Songetal., 1999 {A.fuscocolorata (Bosenberg& Strand, 1906));Araneus
Clerck, 1758; Araniella Chamberlin et Ivie, 1942; Argiope Savigny et Audouin, 1826 (A.
bruennichi (Scopoli, 1772); A. lobata (Pallas, 1772)); Atea C.L. Koch, 1837; Cercidia
Thorell, 1869 (C. prominens (Westring, 1851); С levii Marusik, 1985); Chorizopes O.
Pickard-Cambridge, 1870 (C. sp.); Cyclosa Menge, 1866; Cyphepeira Archer, 1951 (C.
silvicultrix (C.L. Koch, 1845)); Eriophora Simon, 1864 (E. astridae (Strand, 1917); E.
sachalinensis (S. Saito, 1934)); Gibbaranea Archer, 1951; Hypsosinga Ausserer, 1871;
Larinia Simon, 1874; Larinioides Caporiacco, 1934; Mangora O. Pickard-Cambridge,
1889 (M acalypha (Walckenaer, 1802)); Neoscona Simon, 1864; INuctenea Simon, 1864;
Pronoides Schenkel, 1936 (P brunneus Schenkel, 1936); Singa C.L. Koch, 1836;
Stroemiellus Wunderlich, 2004 (S stroemi (Thorell, 1870))1; Zilla C.L. Koch, 1834 (Z.
diodia (Walckenaer, 1802)).
Распространение.
Aculepeira, Araneus, Araniella, Cercidia, Cyphepeira, Hypsosinga, Larinioides —
весь регион.
Argiope, Cyclosa, Gibbaranea, Larinia, Neoscona, Singa, Stroemiellus — юг
Сибири и Дальнего Востока.
Agalenatea, Mangora, Zilla — юго-запад Сибири.
Alenatea, Chorizopes, Eriophora, Pronoides — юг Дальнего Востока.
1 J. Wunderlich (2004) относит к Zygiellidae.
116
4. Специальная часть
Ареалы многих видов, встречающихся в азиатской части России, рассмотрены
в работе Marusik et al. (2000).
Биология.
Сети. Сети варьируют по размеру от десятка сантиметров (Araniella, Hypsosinga)
до нескольких метров {Araneus macacus Uyemura, 1961) и располагаются они
горизонтально или, в большинстве случаев, вертикально. Они могут располагаться
невысоко над землёй — в 5-10 см (Cercidia, Hypsosinga) или же на высоте
нескольких метров {Araneus macacus). В конструкции сети выделяют следующие типы
нитей, которые отличаются по толщине, упругости, структуре: 1) нити рамы, на
которых держится сеть; 2) радиальные нити; 3) спиральные нити (с капельками
клейкого вещества). В самой сети выделяют следующие элементы: раму, ловчую
часть, свободную зону и центр. Кроме этого, иногда встречается свободный сектор
(в котором нет спиральных нитей между двумя радиальными) и стабилимент —
зигзагообразная или прямая лента {Argiope, Cyclosa). Все виды Araneidae,
встречающиеся в азиатской части России, плетут правильную колесовидную сеть;
исключение — Stroemiellus, у которого один сектор сети свободный. Число радиальных и
спиральных нитей и размер рамы, хотя и подвержены вариациям, в большинстве
случаев видо-, а иногда и родоспецифичны. Пауки-крестовики населяют самые
разные биотопы, поэтому сети сплетаются в самом разном окружении.
Большинство стоят сети среди травы и кустарников, однако есть виды, сплетающие сети
только на деревьях, немногие виды устраивают сети в углублениях скал или среди
каменных осыпей {Aculepeira carbonarioides (Keyserling, 1892)). Некоторые
крестовики сплетают сети исключительно у воды {Araneus quadratus Clerck, 1758;
Larinioides cornutus (Clerck, 1758)). Обычно сети обновляются ежедневно
(плетётся новая ловчая спираль, если не порваны радиальные нити), обычно в сумеречное
время. Однако если погода благоприятствует, но дождь или ветер повредил сеть,
паук может плести сеть несколько раз в день. Ежедневная смена спиральных
нитей, очевидно, связана с тем, что клейкость ловчей спирали уменьшается из-за пыли
и естественного высыхания.
На берегах водоёмов, где много добычи, но мало подходящих опор для сетей
(редкие кустарники, скалы), сети могут располагаться близко друг к другу, образуя
сплошной полог, иногда в несколько слоев. Колониальные сети в азиатской части
России отмечались для "Araneus"pallasi Thorell, 1875 в Туве, на берегу озера Тере-
Холь.
Многие виды устраивают закрытое убежище недалеко от гнезда. Это могут быть
свёрнутые и сплетённые в конус листья {Araneus, Larinioides), или трубка под
корой деревьев либо в трещинах скал {Larinioides, Cyphepeira, Stroemiellus). Иногда
убежище бывает открытым, и при этом оно напоминает гнездо птицы {Aculepeira).
У некоторых видов убежищ нет, например у одного петрофильного Araneus, не
известны они у Araniella, Argiope, Cyclosa, Eriophora, Neoscona, Hypsosinga, Pro-
noides. У Cyclosa стабилимент прямой и, очевидно, служит для маскировки паука,
который сидит в центре. Стабилимент по ширине равен ширине брюшка паука, а
выросты брюшка имитируют неровности стабилимента, куда нередко вплетаются
тела жертв.
Питание. Добычу пауки-крестовики ловят благодаря ловчим сетям. Однако
жертвы пауков самостоятельно запутываются в сети крайне редко. Если летящее насе-
Семейства, встречающиеся в азиатской части России
117
комое с силой врезается в сеть, оно бывает слегка оглушённым и слегка прилипает
к сети. Чтобы добыча увязла, паук подёргивает нужные радиальные нити, чтобы за
жертву зацепились соседние ловчие нити.
Некоторые пауки сразу или после некоторого выжидания кусают жертву и
отскакивают от неё, так может повториться несколько раз, пока жертва не перестанет
трепыхаться. Иногда пауки начинают набрасывать на добычу ленты паутины и
пеленают её таким образом. Обе тактики (укусы и пеленание) могут сочетаться в
разной последовательности. Пищевой специализации у видов, населяющих
восточную Палеарктику, не наблюдали.
Забота о потомстве. Согласно В.П. Тыщенко (1971), пауки-крестовики
изготавливают 5-6 коконов и охраняют их в течение нескольких первых дней или до
смерти самки. Коконы бывают шаровидные (Araneus, Aranielld), грушевидные
(Argiope), линзовидные (Singa, Araneus variegatus Yaginuma, 1960). У последнего
вида кокон линзовидныи, поскольку устраивается под корой. Число яиц варьирует
от 15 (Cercidia prominens) до нескольких сотен.
Зимуют крестовики в виде яиц и на неполовозрелых стадиях. Иногда это могут
быть субадультные особи (Cyclosa).
Методы сбора. Укосы, отряхивание веток, визуальный поиск (крупных пауков
можно обнаружить по сетям, когда они прячутся в убежищах), некоторые виды
можно собрать вручную среди каменных осыпей (Aculepeira) или на скалах (Araneus
sp.). Cercidia попадаются только при ручном сборе в подстилке. Укосы и
визуальный поиск большиства видов наиболее эффективны ночью.
Таксономия. Ранее это семейство фигурировало также под названиями Argio-
pidae или Epeiridae. До недавнего времени в состав семейства включали и тетраг-
натидные рода Meta (см. Тыщенко, 1971) и Metleucauge, а иногда и всех Tetragna-
thidae (Levi, 1980). До сих пор дискутируется состав семейства, включать ли в него
Nephilinae1 (не встречается в России) и Zygiellinae (рассматривается здесь как
отдельное семейство).
Надвидовая систематика голарктических Araneidae разработана относительно
хорошо, прежде всего, благодаря работам М. Grasshoff (1970b-c, 1983,1986) и H.W.
Levi(1974a-b, 1977).
Систематика пауков-крестовиков азиатской части России рассматривалась в
следующих работах: Kulczynski (1885,1908); Holm (1970); Levi (1977); Бахвалов (1981);
Marusik (1986); Марусик (1989а); Marusik & Koponen (2000); Oliger et al. (2002) и
некоторых других работах.
Распространение многих видов крестовиков, населяющих Сибирь,
рассматривается в монографии по паукам Тувы (Marusik et al., 2000).
Из наиболее важных работ по сопредельным территориям следует отметить
Chikuni (1989), Dondale et al. (2003), Song et al. (1999), серию ревизий A. Tanikawa
(1989-2007) и недавнюю монографию Ono et al. (2009).
В настоящее время эта группа разрабатывается только Ю.М. Марусиком.
1 Сейчас рассматривается как отдельное семейство.
118
4. Специальная часть
Разграничение видов. Форма копулятивных органов; во многих случаях виды
хорошо различаются рисунком брюшка, как дорсальным, так и вентральным.
Самцы некоторых близких видов чётко различаются шипованием ноги I или II. В
одном случае близкие виды чётко различаются лишь размером копулятивных
органов.
Перспективы изучения. В регионе встречается несколько видов, родовая
принадлежность которых нуждается в пересмотре, например Eriophora, Larinia.
Помимо этого, имеется ряд неописанных видов из родов Aculepeira, Araneus, Atea,
Cyclosa и Larinioides. Распространение и статус некоторых видов нуждается в
уточнении. Например, самого крупного крестовика азиатской части России (и всей
России в целом) практически все исследователи традиционно относят к Araneus ven-
tricosus (L.Koch, 1879), хотя давно уже известно, что под этим названием
«скрывается» три близких вида. Из них на территории Российского Дальнего Востока
достоверно известен лишь A macacus Uyemura, 1961 (Логунов, Марусик, 2004; Marusik
et al., 2007).
Neoscona adianta (Araneidae), самка, Крым (фото: Г.А.
Прокопов).
Семейства, встречающиеся в азиатской части России
119
Argiope bruennichi (Araneidae), самка, Приморье (фото: М.М. Омелько).
Семейство Argyronetidae Thorell 18701
Серебрянки, water spiders
Рис. 8.1-6.
В мире и России известен один вид.
Диагностические признаки. Крупный размер (до 2 см), дыхальце посредине
брюшка, густое опушение нижней стороны брюшка, и обитание в воде —
позволяют легко отличить представителей этого семейства от всех других.
Близкие группы. Похожи на Cybaeidae, Amaurobiidae, Devade (Dictynidae).
Чётко отличаются расположением дыхальца и густым опушением нижней части
брюшка.
ОПИСАНИЕ:
• Карапакс. Грушевидный.
• Стернум. Не модифицирован.
• Глаза. 8 в два ряда, передние медиальные мельче задних медиальных.
• Ротовой аппарат. Хелицеры мощные; желобок хелицер несёт 2 крупные
зубца спереди и 3 — сзади.
• Ноги. 3 коготка. Передние 2 пары с малочисленными шипами, задние — с
многочисленными.
• Брюшко. Овальное, без рисунка. Нижняя часть брюшка густо опушена.
• Паутинные бородавки. Колюлюс маленький, представлен бугорком с
парой щитинок.
• Органы дыхания. Пара лёгких и трахеи с широким дыхальцем,
открывающимся посредине брюшка.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Пальпа самца с широким лопастевидным
отростком голени, бульбус диктинного типа: эмболюс, крупный и массивный
кондуктор с двумя ветвями, медиальный отросток отсутствует. Эпигина
простая, слабо хитинизирована, без ямки.
• Размер. 8-20 мм. Самцы могут быть крупнее самок, особенно размером ка-
рапакса.
• Окраска. От светло-коричневой до почти чёрной, без рисунка.
• Особые черты. Многочисленные густые волоски на брюшке
(удерживающие воздух) и широкое дыхальце, далеко отстоящее от паутинных
бородавок. Обитает в воде.
Рода, известные из азиатской части России. Argyroneta Latreille, 1804 (A.
aquatica (Clerck, 1758)).
Распространение. Вся Палеарктика. В азиатской части России встречается
везде, кроме тундровой зоны и северной части тайги.
Биология. Биология единственного вида этого семейства детально описана в
многочисленных научных и популярных изданиях, снята на видео. Первое
детальное научное описание поведения A aquatica было опубликовано В. Вагнером (1900).
Вид тесно связан с водой, пресноводными водоёмами. Убежище колоколовидной
'Н.И. Платник рассматривает таксон в составе Cybaeidae, хотя Argyronetidae имеют приоритет. По
строению копулятивных органов паук-серебрянка, на наш взгляд, гораздо ближе к Dictynidae.
Семейства, встречающиеся в азиатской части России
121
1
Рис. 8. Argyronetidae. Argyroneta aquatica (cT$).
1 — габитус, сверху; 2 — головогрудь неполовозрелого самца, спереди; 3 — брюшко
неполовозрелого самца, снизу; 4-5 — пальпа сГ, снизу и сбоку; 6 — эпигина, снизу 4 — по: Ono et al.
(2009); 5-6 — по: Roberts (1995).
формы устраивают только под водой, кокон откладывают в подводном убежище.
Самец крупнее самки.
Методы сбора. Визуальный поиск на поверхности мелких водоёмов со стоячей
водой. Пауки периодически всплывают, чтобы набрать новую порцию воздуха. Иногда
можно поймать гидробиологическим сачком или сачком из мельничного газа.
Таксономия. Рассматривается как отдельное семейство, или же как
подсемейство или триба в семействах Dictynidae (Lehtinen, 1967), Cybaeidae (см. Platnick,
2009) или Agelenidae: Cybaeinae (Grothendieck & Kraus, 1994). На наш взгляд, судя
по строению пальпы, Argyroneta наиболее близка к Dictynidae. Однако целый ряд
соматических признаков (отсутствие крибеллюма, дыхальце посредине брюшка,
густое опушение, крупный размер) позволяют рассматривать этот таксон как
отдельное семейство.
Известен всего один вид.
Перспективы изучения. Возможно, сравнительно-морфологические и
сравнительно-генетические исследования разных популяций позволят выделить подвиды.
Семейство Atypidae Thorell, 1870
Атипиды; purseweb spiders
Рис. 9.1-11.
Одно из самых небольших семейств с 3 родами и 43 видами, населяющими
Евразию, Африку и Северную Америку В России известны всего три вида рода
Atypus, два из них встречаются в Приморье.
Диагностические признаки. Крупные пауки с хелицерами, направленными
вперёд; 8 глаз на глазном бугорке, передние медиальные — самые крупные, остальные
образуют два триплета; стернум с мускульными точками; нижняя губа слита со стер-
нумом; 2 пары лёгких; задние паутинные бородавки — 3-члениковые, длинные.
Близкие группы. Не схожи ни с одной другой группой наших пауков.
ОПИСАНИЕ:
• Карапакс. Карапакс можно описать как удлинённый овал с обрубленным
передом (передний край и бока образуют прямой угол), задний край кара-
пакса равномерно закруглён.
• Стернум. Прямоугольный, с приросшей нижней губой и с 4 парами вдавле-
ний — мускульных точек (=sigilla).
• Глаза. 8, расположены на глазном бугорке; передние медиальные крупнее
других, латеральные и задние медиальные образуют глазные триплеты.
• Ротовой аппарат. Хелицеры вытянуты вперёд, коготки лежат на нижней
стороне хелицер и параллельны друг другу. Много зубчиков вдоль желобка
хелицер. Нижняя губа срослась со стернумом. Передний край максиллы
сильно удлинён.
• Ноги. 3 коготка; у самок относительно короткие и толстые, у самца
несколько длиннее тела.
• Брюшко. Овальное, с небольшим скутумом спереди (и у самцов, и у самок).
• Паутинные бородавки. Передние бородавки небольшие, тонкие, широко
расставлены. Средние бородавки короткие со скошенной вершиной, а
задние — трёхчлениковые, намного длиннее других.
• Органы дыхания. Две пары лёгких.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Пальпы самца относительно небольшие, голень
больше цимбиума, бульбус округлый с шиловидным эмболюсом и
кондуктором-мембраной. Эпигины нет, эндогина с 2 парами рецептакул.
• Размер. 10-20 мм, самцы меньше самок.
• Окраска. От коричневого до чёрного, без рисунка.
• Особые черты. Мощные хелицеры, направленные вперёд; мускульные
точки на стернуме; трёхчленные бородавки.
Рода, известные из азиатской части России. Atypus Latreille, 1804 (A. magnus
Namkung, 1986; A medius Oliger, 1999).
Распространение. Только Приморье.
Семейства, встречающиеся в азиатской части России
123
Рис. 9. Atypidae. Atypus affinis (?, 1, 5), A. medius (o\ 4, 6, 9, 11), A. muralis (cT$, 2-3, 8,
10), и Л. magnus (сГ, 7).
1 — габитус ?, сбоку; 2 — головогрудь сГ, сверху; 3^ — глазное поле, сверху; 5 — головогрудь
9, снизу; 6-7 — пальпа сГ, сбоку; 8 — вульва; 9-10 — паутинные бородавки, сбоку; 11 — габитус
СГ. 1-2, 5 — по: Wiehle (1953); 4, 6, 9 — по: Oliger (1999); 3, 8 — по: Miller (1947); 7 — по:
Логунов (1990).
Биология. A. medius собран почвенными ловушками на горном склоне южной
экспозиции в дубраве.
Биология европейских видов описана неоднократно. Очевидно,
дальневосточные виды не отличаются от них и плетут двухколенную сеть, часть которой
располагается вертикально и уходит под землю, а горизонтальная часть лежит на
поверхности, замаскированная частичками земли, кусочками опада. Когда жертва
наступает на горизонтальное колено, паук изнутри пронзает паутину коготками хели-
цер, умертвляет добычу и затаскивает в трубку
124
4. Специальная часть
Большая часть жизни паука проходит в трубке. Самцы покидают свою трубку
только для того, чтобы найти самку. Спаривание происходит внутри трубки самки.
Самец и самка сожительствуют в трубке несколько месяцев. Потом самка сама
сидит в норке с коконом и охраняет его.
Методы сбора. Почвенные ловушки (самцы). Самок можно найти только
визуальным поиском. Самки всегда находятся в убежищах, форма которых описана в
разделе «Биология». Образуют колонии — скопления десятков норок, расстояния
между которыми — от 0,5 до 2 м. То есть, если удалось найти хотя бы одну норку,
то, значит, рядом должны быть и другие. Колонии обычно находятся в густой
растительности (среди высокой травы).
Таксономия. Ранг семейства был придан этому таксону Е. Simon (1903) из-за
характерного внешнего облика. Существуют разногласия относительно состава этой
группы. Некоторые исследователи исключают род Calommata Lucas, 1837 из Aty-
pidae.
Оба вида, встречающиеся в азиатской части России, изображены в работах
Логунова (1990) и Oliger (1999).
Разграничение видов. Форма пальпы самца позволяет различить оба наших
вида.
Перспективы изучения. Многие виды встречаются очень локально, поэтому
не исключены находки других видов, известных из Китая, Кореи, или вообще
новых видов.
Логово-сеть Atypus sp. (Atypidae), Абхазия (фото: Н.М. Ковблюк).
Семейства, встречающиеся в азиатской части России 125
Atypus muralis (Atypidae), самка, Крым (фото: А.А. Надольный).
Atypus magnus (Atypidae), самец, Приморье (фото: М.М. Омелько).
Семейство Cheiracanthiidae Wagner, 18871
Хиракантиды; yellow sac spiders
Рис. 10.1-9.
Всесветно распространённое семейство, включает, по меньшей мере, 4 рода и
около 200 видов. В России известны 15 видов рода Cheiracanthium C.L. Koch, 1839,
а в азиатской части России — по меньшей мере 11 видов.
Диагностические признаки. Энтелегинные, некрибеллятные, от мелких до
крупных, 8-глазые пауки с 2 коготками, глаза широко расставлены, наличник
низкий, передние ноги заметно длиннее тела и других ног. Пальпа самца с РГО,
медиальным отростком и с тонким и острым на конце базальным выростом цимбиума,
эпигина с округлой ямкой.
Близкие группы. Габитуально, цветом и отсутствием рисунка (у большинства
видов) напоминает Haplodrassus, Parasyrisca (Gnaphosidae), Agroeca (Liocranidae),
Clubiona (Clubionidae) и Anyphaena (Anyphaenidae). От Haplodrassus отличается
нераздвинутыми передними бородавками. От Anyphaena легко отличается
^модифицированным дыхальцем. От трёх других родов, с которыми нередко путают,
Cheiracanthium уверенно отличается формой копулятивных органов (наличием
выроста цимбиума и крупной ямкой эпигины). Parasyrisca к тому же отличается
пальпой самца — с оттопыренным РГО и коротким эмболюсом, и строением
эпигины — отсутствием настоящей ямки и наличием одного или двух карманов.
ОПИСАНИЕ:
• Карапакс. Яйцевидный, без рисунка.
• Стернум. Не модифицирован.
• Глаза. 8 в два ряда (4-4), оба ряда одинаковый ширины, почти равны
ширине головной части карапакса; латеральные глаза сближены.
• Ротовой аппарат. Хелицеры удлинены, особенно у самцов, иногда с
латеральными выростами и часто с внутренней выемкой.
• Ноги. 2 коготка; светлые без колец; голени с малочисленными шипами.
Передние ноги в 1,5 и более раз длиннее тела, скопула имеется.
• Брюшко. Овальное, на конце слегка заострено.
• Паутинные бородавки. Задняя пара нередко с удлинённым апикальным
члеником, передняя пара бородавок короче задней.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Пальпы самца относительно длинные; голень у всех
видов азиатской части России с длинным и тонким РГО, раздвоенным на конце,
в дополнение к нему может быть дорсальный отросток. Цимбиум с
саблевидным ретролатеральным выростом. Ретролатеральная часть цимбиума с лопас-
тевидным выростом. Тегулюм овальный. Эбмолюс нитевидный. Медиальный
отросток длинный и очень тонкий. Эпигина у всех видов из Сибири и
Дальнего Востока России с ямкой, над которой просвечивают канальцы эпигины.
• Размер. 5-15 мм.
1 Рассматривается N.I. Platnick (2009) в составе Miturgidae.
Семейства, встречающиеся в азиатской части России
127
1
Рис. 10. Cheiracanthiidae. Cheiracanthium sp. (cT$).
1,7 — габитус сГ, сверху и сбоку; 2 — головогрудь сГ, сверху-спереди; 3 — паутинные бородавки
?, снизу; 4 — габитус ?, сверху; 5 — пальпа сГ, сбоку; 6 — эпигина, снизу; 8 — вскрытое
убежище-логово самки; 9 — логово самки. Стрелки показывают саблевидный отросток цимбиума, РГО
и характерные просвечивающие канальцы эпигины. 8-9 — по: Chikuni (1989).
128
4. Специальная часть
• Окраска. Жёлтая, светло-коричневая, иногда зеленоватая. Рисунок бывает
только на брюшке в виде ланцетовидного пятна, реже имеется «ёлочка».
• Особые черты. Саблевидный вырост цимбиума самца, первая пара ног
длиннее четвёртой. Самки сооружают полностью закрытые убежища среди
травостоя, складывая особым образом травинки (рис. 10.8-9).
Рода, известные из азиатской части России. Только Cheiracanthium C.L. Koch,
1839.
Распространение. Весь регион, кроме тундровой зоны; большинство видов
обитают в южной Сибири и на юге Дальнего Востока.
Биология1. Обитатели травостоя и кустарникового яруса. Все они — бродячие
охотники, и сетей не плетут. Пауки в дневное время скрываются в сплетённых
убежищах среди травостоя или кустарников, но могут встречаться и под камнями.
Охотятся исключительно ночью. В охотничьем поведении главную роль играют
тактильные стимулы. Пауки реагируют на добычу, если она касается их ног, и
атакуют резким наскоком (Peck & Whitcomb, 1970). Пища Cheiracanthium изучена
плохо. В агроценозах Cheiracanthium наблюдались питающимися такими
насекомыми как гусеницы, моли, тли, цикадки, а также тетраниховыми клещами (Mansour et
al., 1980). В лабораторных условиях Ch. inclusum (Hentz, 1847) охотно поедали
мух, кузнечиков, пауков, клопов, цикадок, молей и гусениц, но избегали атаковать
муравьев, некоторых жуков и паразитических ос (Peck & Whitcomb, 1970).
Половое поведение простое. Ухаживание практически отсутствует.
Дотрагиваясь до самки ногами, самец быстро забирается на неё и, отогнув её брюшко
ногами, поочерёдно вводит пальпы в эпигину Часто самцы, обнаружив гнездо суба-
дультной самки, строят рядом с ним свое гнездо и спариваются после последней
линьки самки. Спаривание обычно происходит в убежище самки.
Оплодотворённые самки через определённый период времени плетут на растениях крупное
замкнутое гнездо-логово с толстыми, жёсткими непроницаемыми стенками. Стенами
логова обычно служат свёрнутые особым образом травинки. В логове
размещается кокон и сама самка. Для некоторых видов Cheiracanthium характерна матрифа-
гия, когда вылупившаяся молодь поедает охраняющую их самку.
Некоторые виды представляют опасность для человека— Ch. punctorium (Villers,
1789) (встречается в Южной Сибири), Ch. mildei L. Koch, 1864, возможно, и
другие виды, причём (в отличие от каракурта, например) не только самки, но и в
неменьшей степени — самцы. При укусе моментально возникает очень сильная
жгучая боль в месте укуса (не зуд, как бывает при укусе гадюки; но без «запирания»
мышц, как при укусе каракурта), иррадирующая в соответствующий сегмент
конечности. Через несколько минут опухают и начинают ныть лимфоузлы на пути от
места укуса. Позже — слабый отёк в месте укуса, нарушения подвижности в месте
укуса, могут быть нарушения дыхания. Боль держится 10-20 часов, местные
симптомы исчезают через 1-2 дня (это — сводка по примерно 15 случаям укусов в
Крыму) (Н.К., собственные данные; личн. сообщ. О.В. Кукушкина).
В 2007 г. в Австрии было много случаев укусов людей Ch. punctorium. Только в
одну центральную больницу города Линц с жалобами на укус паука обратились
190 человек, однако подтверждающие это симптомы были обнаружены лишь у 8.
1 Раздел подготовлен совместно с Э.Ф. Гусейновым.
Семейства, встречающиеся в азиатской части России
129
Методы сбора. Укосы и визуальный поиск (сравнительно легко можно
обнаружить логова самок).
Таксономия. В.А. Вагнер (Wagner, 1887) описал в одной и той же работе два
новых семейства — Clubionidae и Cheiracanthidae (в оригинале окончание -idae с
одним "i")1. Первое семейство было принято арахнологами и, более того, E.Simon
(1897) подразделил его на 8 подсемейств, которым затем был придан статус
самостоятельных семейств. В то же время, Cheiracanthium был включён в состав
Clubionidae и даже рассматривался в той же трибе, что и Clubiona. Cheiracanthium
сближался с Clubiona до самого недавнего времени, пока не был переведён в
подсемейство Eutichurinae Lehtinen, 1967 семейства Miturgidae Simon, 1886 (Ramirez et al.,
1997: 45). Deeleman-Reinhold (2001) согласилась с размещением Cheiracanthium в
Eutichurinae, но перевела подсемейство из Miturgidae в Clubionidae. Главным
основанием для перевода Cheiracanthium в другое семейство и подсемейство
послужило строение задних паутинных бородовок, апикальный членик которых удлинён
(Ramirez et al., 1997: 45). Однако такое строение бородавок встречается и во
многих других семействах, например Agelenidae, Hahniidae.
Переводя Cheiracanthium из Clubionidae в другое семейство и подсемейство,
авторы (Ramirez et al., 1997), со всей очевидностью, не знали о существовании
Cheiracanthiidae, и приписали этот род в таксон, описанный гораздо позже, чем
Cheiracanthiidae. Даже если согласиться, что Eutichurus Simon, 1896 и
Cheiracanthium — родственные, то тогда подсемейство должно называться Cheiracanthiinae.
Судя по уникальному строению пальпы у Cheiracanthium и других родов этого
таксона (Summacanthium Deeleman-Reinhold, 2001, Calamopus Deeleman-Reinhold,
2001 и Calamoneta Deeleman-Reinhold, 2001), эти пауки имеют не много общего с
Eutichurus, и ещё меньше — с типовым родом Miturgidae. Кроме того, не вполне
понятны рамки Miturgidae, и относятся ли Eutichurinae к этому семейству. На наш
взгляд, логичнее согласиться с тем, что Cheiracanthium и три другие рода со
схожими пальпами следует относить к отдельному семейству. В определителе В.П. Ты-
щенко (1971) Cheiracanthium рассматривается в составе Clubionidae.
Род Cheiracanthium имеет практически всесветное распространение. Меньше
всего видов известно в Неотрописе. Возможно, он туда интродуцирован. По всей
видимости, сам Cheiracanthium может быть разбит ещё как минимум на два рода.
Виды, встречающиеся в азиатской части России, рассматривались в работах L. Koch
(1879а); Kulczynski (1885) и Holm (1973). Виды из сопредельных территорий
изображены в работах Chikuni (1989), Song et al. (1999) и Ono et al. (2009). Специально
этим родом в пределах России или восточной Палеарктики никто не занимается.
Разграничение видов. Виды различаются практически только благодаря
форме копулятивных органов. В некоторых случаях может быть использован размер
(карапакса, пальпы, эпигины), форма хелицеры самца и рисунок брюшка.
Перспективы изучения. Cheiracanthiidae — один из наименее исследованных
таксонов в азиатской части России, и крайне нуждается в ревизии.
1 Вагнер не совсем правильно образовал название семейства от рода Cheiracanthium, на самом
деле правильное написание — Cheiracanthiidae, а не Cheiracanthidae (сравни: Theridion (Theridi-
ит) — Theridiidae, Zodarion (Zodarium) — Zodariidae).
Семейство Clubionidae Wagner, 1887
Клубиониды, пауки мешкопряды; sac spiders
Рис. 11.1-12.
Согласно каталогу N.I. Platnick (2009), это семейство включает 14 родов и 555
видов. В России известен всего один род Clubiona Latreille, 1804 и 74 вида. В
азиатской части России отмечено 70 видов.
Диагностические признаки. Энтелегинные, некрибеллятные, бродячие пауки
с 2 коготками и 8 глазами, головогрудь яйцевидная, ноги с шипами, но без колец и
полос. Пальпа самца всегда с РГО, но без медиального отростка.
Близкие группы. Габитуально, цветом и отсутствием рисунка (у большинства
видов) напоминает Haplodrassus, Parasyrisca (Gnaphosidae), Agroeca (Liocraniidae),
Cheiracanthium (Cheiracanthiidae) и Anyphaena (Anyphaenidae). От Haplodrassus
отличается нераздвинутыми передними бородавками. От Anyphaena легко
отличается не модифицированным дыхальцем. От трёх других родов, с которыми нередко
путают, Clubiona достаточно чётко отличается формой копулятивных органов. В
отличие от Agroeca, у Clubiona нет продольной щели эпигины и медиального
отростка бульбуса. Parasyrisca же отличается наличием одной ямки и двух или одного
кармана эпигины, а также пальпой самца с оттопыренным РГО и эмболюсом,
скрытым под тугулюмом. Самцы Cheiracanthium обладают саблевидным выростом цим-
биума, всегда с медиальным отростком, а самки всегда имеют ямку, открытую сзади.
ОПИСАНИЕ:
• Карапакс. Яйцевидный с усечённой вершиной; медиальная борозда
продолговатая, щелевидная.
• Стернум. Овальный.
• Глаза. 8 в 2 ряда (4-4), примерно равной величины, задний ряд шире
переднего.
• Ротовой аппарат. Хелицеры от обычных до слегка удлинённых (у самцов).
• Ноги. 2 коготка, без рисунка, от коротких (короче тела) до удлинённых, с
шипами, скопула имеется. Четвёртая пара длиннее первой.
• Брюшко. Удлинённое, на конце заострено.
• Паутинные бородавки. Передние бородавки конические или
цилиндрические, сближены.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Энтелегинные. Пальпа самца не длинная, всегда с
РГО, иногда и с другими отростками голени; медиального (тегулярного) от-
Рис. 11. Clubionidae. Clubiona subtilis (сГ$, 1-2, 4-5), С. riparia (сГ$, 3, 7), С. kularensis
(СГ$, 6, 9) и С caerulescens (?, 8).
1 — головогрудь сГ, спереди; 2 — паутинные бородавки, снизу; 3 — головогрудь ?, сверху; 4 —
головогрудь О", сверху; 5 — головогрудь ?, снизу; 6-7 — пальпа сГ, снизу; 8-9 — эпигина, снизу.
6,9 — по: Marusik et al. (2002); 7 — по: Михайлов, Марусик (1995).
Семейства, встречающиеся в азиатской части России
131
1
5
6
7
132
4. Специальная часть
10 11 12
Рис. 11 (продолжение). Clubionidae. Габитус и окраска Clubiona subtilis (10, ? из
Сахалина), С. riparia (11, ? из Сахалина) и С. comta (12, сГ из Турции, вид не встречается в
азиатской части России).
ростка нет, но иногда основание эмболюса с одним или более выростом;
тегулюм часто с длинными просвечивающими семенными трубками,
свёрнутыми в петли; эмболюс от короткого до длинного. В группе видов С reclusa
бульбус всегда с кондуктором (который всегда менее хитинизирован,
нежели остальные части тегулюма). У подродаРагасШЫопа Lohmander, 1944
(типовой вид — Clubiona corticalis (Walckenaer, 1882)) пальпа — гаплогинного
типа (со вздутым тегулюмом и всеми отростками — на вершине), однако с
РГО. Эпигина разнообразной формы, но всегда плоская; иногда с ямкой, но
чаще с двумя отверстиями, расположенными вблизи эпигастральной щели.
• Размер. 3-11 мм, размерный диморфизм от слабого до умеренного.
• Окраска. Жёлтая, светло-коричневая, большинство видов без рисунка, у
некоторых есть коричневый или буроватый рисунок на брюшке.
• Особые черты. Своеобразные тонкие и сильно извитые канальцы,
просвечивающие сквозь стенки как бульбуса, так и эпигины.
Рода, известные из азиатской части России. Виды, обитающие в азиатской
части России, относятся к трём подродам: собственно Clubiona Latreille, 1804; Buclio-
па Benoit, 1977 (=Bicluona Mikhalov, 1994); Japoniona Mikhailov, 1990.
Распространение. Весь регион.
Семейства, встречающиеся в азиатской части России
133
Биология1. Биология этого семейства обнаруживает много параллельных черт
с Cheiracanthium. Как и последние, Clubiona обитают в основном на растениях и
являются бродячими хищниками, ведущими ночной образ жизни. Дневное время
суток проводят в мешковидных гнёздах, сплетённых на растениях, под корой
деревьев или просто под камнями. Питание Clubiona изучено недостаточно.
Австралийские виды были обнаружены питающимися жуками, муравьями, клопами,
пауками (Austin, 1984). Самцы часто плетут гнёзда рядом с гнёздами субадультных
самок и спариваются после линьки последних. Гнёзда сплетают только виды,
обитающие среди травостоя. Как и у Cheiracanthium, ухаживание почти не выражено.
Иногда самцы хватают хелицерами тело самок, пытающихся убежать (Bristowe,
1958). Забравшись на самку, самец загибает на бок её брюшко и вводит пальпу в
эпигину Оплодотворённые самки строят замкнутые мешковидные гнезда (откуда
и пошло английское название семейства), в которые откладывают кокон с яйцами
и охраняют его, не покидая гнезда. Охрана кладки является эффективным
способом защиты потомства Clubiona от многих паразитов и хищников, в том числе и от
особей собственного вида (Pollard, 1983). Судя по всему, как и для Cheiracanthium,
для Clubiona характерна матрифагия.
Методы сбора. Укосы, под корой деревьев, отряхивание веток, под камнями и
досками, ручной сбор и просеивание подстилки.
Таксономия. Десять лет спустя после того, как В.А. Вагнер (Wagner, 1887)
выделил семейство Clubionidae, E. Simon (1897) подразделил его на 8 подсемейств.
A. Petrunkevitch (1923) вывел из его состава Anyphaeninae, Cteninae и Heteropodinae
(=Sparassidae) и придал им статус семейств. Р.Т. Lehtinen (1967) вывел ещё три
подсемейства, придав им ранг семейств (Liocranidae, Corinnidae, Miturgidae), и ещё
один род {Micaria) перенёс в Gnaphosidae.
После такого радикального уменьшения семейства в нём остались лишь
Cheiracanthium и Clubiona, явно не родственные таксоны. По поводу принадлежности
Cheiracanthium долгое время существовали дискуссии, и его относили то к
Liocranidae, то к Eutichurinae в Miturgidae, то сближали со Sparassidae. Однако никто не
обратил внимания, что одновременно с Clubionidae было описано семейство Cheira-
canthidae Wagner, 1887.
В определителе В.П. Тыщенко (1971) Clubionidae рассматриваются в широком
смысле (как у A. Petrunkevitch (1923)) и включают Liocranidae, Corinnidae и Micaria.
Род Clubiona, как и Tetragnatha (Tetragnathidae), имеет практически всесветное
распространение. Однако есть существенная разница. Виды рода Clubiona, кроме
общего габитуального сходства, мало что объединяет. Строение бульбуса
чрезвычайно разнообразно внутри рода. В нём выделяется несколько подродов.
В настоящий момент это семейство, наряду с Salticidae, является одним из
наиболее изученных в регионе.
Виды, обитающие в азиатской части России, рассматривались в работах: L.Koch
(1879а); Kulczynski (1885, 1926); Sytshevskaja (1935); Михайлов (1992, 1994, 1995);
Mikhailov (1990,1991, 1992,1995a-b, 2003); Михайлов, Марусик (1995); Marusik et
al. (2002); Trilikauskas (2007). Множество видов дальневосточных клубион изобра-
1 Раздел подготовлен при участи Э.Ф. Гусейнова.
134
4. Специальная часть
жено в работах японских, китайских и корейских авторов, из книг которых можно
упомянуть, например, Chikuni (1989) и Song et al. (1999).
В настоящее время эта группа по-прежнему разрабатывается К.Г. Михайловым.
Разграничение видов. Виды различаются практически только благодаря
форме копулятивных органов. В некоторых случаях может быть использован размер
пауков (карапакса, пальпы, эпигины) и рисунок брюшка.
Перспективы изучения. В регионе, особенно на Дальнем Востоке,
наблюдается едва ли не максимальное в мире видовое разнообразие семейства. Не
исключены находки дополнительных, и даже новых, видов.
Clubionaphragmitis (Clubionidae), самка, Финляндия (фото: N. Fritzen).
Семейства, встречающиеся в азиатской части России
135
Clubiona neglecta (Clubionidae), самец, Финляндия (фото: N. Fritzen).
Семейство Corinnidae Karsch, 1880
Коринниды; dark sac spiders, ant-like sac spiders
Рис. 12.1-7.
Относительно крупное, всесветно распространённое семейство, с 80 родами и
956 видами. В России известно 13 видов из 3 родов, а в азиатской части России —
10 видов из 3 родов.
Диагностические признаки. Мелкие, темно окрашенные, некрибеллятные, эн-
телегинные пауки, 2 коготка, 8 глаз, передние ноги с многочисленными
вентральными шипами (4 и более пар, кроме Trachelas) или мелкими шипиками (у самцов
Paratrachelas), пальпа самца с крупным РГО. Эпигина без чётко ограниченной ямки.
Близкие группы. Внешне (габитуально и окраской) похожи на Micaria (Gna-
phosidae), но отличаются большим числом вентральных шипов на передних ногах,
крупным РГО, и ямкой эпигины без чётких границ.
ОПИСАНИЕ:
• Карапакс. Овально-грушевидный.
• Стернум. Овальный.
• Глаза. 8, в два ряда (4-4), примерно равной величины.
• Ротовой аппарат. Не модифицирован.
• Ноги. 2 коготка. Передние ноги в 1,5 и более раз длиннее карапакса, голени
и предлапки с многочисленными мощными парными вентральными
шипами {Phrurolithus) или с мелкими шипиками у самцов {Trachelas).
• Брюшко. Овальное, с рисунком; самцы Phrurolithus и Trachelas с
дорсальным скутумом.
• Паутинные бородавки. Не модифицированы, передние бородавки более хи-
тинизированы, нежели другие, колюлюс треугольный.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Пальпа самца всегда с крупным РГО, бульбус
вздутый и блестящий, без медиального отростка, эмболюс короткий. Эпигина
обычно без чёткой ямки. Нередко копулятивные отверстия забиты пробкой,
так что отверстия не видны.
• Размер. 4-6 мм. Диморфизм практически не выражен.
• Окраска. От светло-коричневой до почти чёрной.
• Особые черты. Ноги I и II с многочисленными и мощными вентральными
шипами или шипиками у самцов, отростки голени у самцов крупные.
Рода, известные из азиатской части России. Paratrachelas Kovblyuk etNadolny,
2009 {P. acuminus (Zhu et An, 1988)); Phrurolithus C.L. Koch, 1839; Trachelas L. Koch,
1872 {T. japonicus Bosenberg et Strand, 1906; T tanasevitchi Marusik et Kovblyuk,
2010).
Семейства, встречающиеся в азиатской части России
137
1
4
Рис. 12. Coriimidae. Phrurolithus sinicus (?, 1, 4), Trachelas japonicus (сГ$, 2-3, 5,1) yl P.
claripes (?, 6).
1-2 — габитус ?, сверху; 3 — габитус сГ, сверху; 4 — головогрудь ?, сверху; 6-6 — эпигина,
снизу; 7 — пальпа самца, сбоку. 6 — эпигина после вываривания в щёлочи. Стрелка показывает
на отросток колена пальпы d\ 2-3, 5, 7 — по: Marusik & Kovblyuk (2010); 6 — по: Марусик,
Кроуфорд (2006).
Распространение.
Phrurolithus — южная Сибирь и юг Дальнего Востока.
Paratrachelas и Trachelas — только юг Дальнего Востока.
Биология. Все виды связаны с подстилкой. Сетей не плетут. Биология изучена
неудовлетворительно. Phrurolithus похожи на муравьев, встречаются в
непосредственной близости от муравейников и, вероятно, питаются муравьями.
138
4. Специальная часть
Методы сбора. Почвенные ловушки, ручной сбор и просеивание подстилки.
Таксономия. Ранее пауки этого семейства рассматривались в составе
подсемейств Corinninae и Liocraninae семейства Clubionidae. Статус семейства был
придан двум этим подсемействам П.Т. Лехтиненом (Lehtinen, 1967). Если Trachelas
традиционно рассматривался в Corinninae/Corinnidae, то Phrurolithus лишь
недавно был переведён из Liocranidae в Corinnidae (Bosselaers & Jocque, 2002). В
определителе В.П. Тыщенко (1971) оба рода рассматриваются в составе Clubionidae.
Реальное число видов в Corinnidae выше, чем указано в Platnick (2009), поскольку
Н.И. Платник не засчитал 5 родов с 44 видами, перенесённые во Phrurolithinae/
Corinnidae в работе Deeleman-Reinhold (2001), только на том основании, что ею не
был проведён кладистический анализ семейства.
Судя по строению копулятивных органов, сибирские и дальневосточные
Phrurolithus относятся, по меньше мере, к двум отдельным родам.
Виды, обитающие в азиатской части России, рассматривались в работах Danilov
(1999), Марусик, Кровфорд (2006) и Marusik & Kovblyuk (2010). Виды из
сопредельных стран изображены в работах Chikuni (1989), Song et al. (1999), Ono et al.
(2009).
Разграничение видов. Форма копулятивных органов, в некоторых случаях и
число вентральных шипов на голени и предлапке.
Перспективы изучения. Обнаружение новых или дополнительных видов
маловероятно, однако не исключено. Два рода, встречающиеся в регионе {Phrurolithus
и Trachelas), нуждаются в пересмотре их границ.
Phrurolithus festivus (Corinnidae), самка, Финляндия (фото: N. Fritzen).
Семейства, встречающиеся в азиатской части России
139
Agalenatea redii (Araneidae), самка, Ростовская область (фото: Н.М. Ковблюк).
Семейство Ctenidae Keyserling, 1877
Ктениды; tropical wolf spiders
Рис. 13.1-5.
В мире известно 477 видов, относимых к 39 родам. В России, и в Палеарктике
вообще, обитает один род. В России известен всего один вид из Приморья.
Диагностические признаки. Среднего размера (7-12 мм) энтелегинные пауки
с формулой глаз 2-4-2; с продольными полосами на карапаксе и брюшке; пальпа
самца с РГО и тупым медиальным отростком.
Близкие группы. Габитуально и окраской напоминают Zoridae, от которых
отличаются большим размером и тем, что передние латеральные глаза значительно
(в 2 и больше раз) меньше остальных.
ОПИСАНИЕ:
• Карапакс. Грушевидный.
• Стернум. Не модифицирован.
• Глаза. 8 в три ряда (2-4-2), передние латеральные глаза меньше других, а
задние медиальные — самые крупные.
• Ротовой аппарат. Не модифицирован.
• Ноги. 2 коготка. Ноги длинные у самца. Шипы многочисленные.
• Брюшко. Овальное, заострено на конце.
• Паутинные бородавки. Не модифицированы.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем у паутинных бородавок.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Энтелегинные пауки. Пальпа самца длиной как
карапакс. Цимбиум уплощён. Эмболюс длинный нитевидный. Медиальный
отросток большой, без крючка на вершине, по длине он составляет половину
длины тегулюма, с внутренней стороны с карманом. Эпигина простая, без
ямки.
• Размер. 7-12 мм. Половой диморфизм выражен.
• Окраска. Светло-коричневая, карапакс и брюшко с парой тёмных полос вдоль
медиальной полосы, карапакс дополнительно с сублатеральной
разорванной серой полосой.
• Особые черты. Передние латеральные глаза меньше, чем задние
латеральные.
Рода, известные из азиатской части России. Anahita Karsch, 1879 {A. fauna
Karsch, 1879).
Распространение. Впервые семейство отмечено Ю.М. Марусиком и С. Копо-
неном (Marusik & Koponen, 2000) с юга Приморья. За пределами этого региона
семейство в России неизвестно.
Семейства, встречающиеся в азиатской части России
141
о о
Рис. 13. Ctenidae. Anahita fauna (cT$).
1-2 — головогрудь ?, сверху и сверху-сбоку; 3 — эпигина, снизу; 4 — пальпа С?, снизу; 5 —
глазное поле, сверху. Стрелка показывает уменьшенные передние латеральные глаза.
Биология. Большинство особей были собраны нами (Ю.М.) на приморских
галечниках на юге Приморского края. Помимо прибрежных районов, большая
серия была собрана Т.П. Олигер в Лазовском заповеднике.
Методы сбора. Почвенные ловушки, ручной сбор среди приморских
галечников.
Таксономия. Е. Simon (1898) рассматривал эту группу как подсемейство в
Clubionidae. А. Петрункевич (Petrunkevitch, 1923) придал группе ранг семейства.
Р.Т. Lehtinen (1967) выделил 4 подсемейства. Таксономическое положение Anahita
не вполне ясно. Это единственный представитель семейства в Голарктике. Судя по
габитусу, окраске и форме пальпы, Anahita, возможно, относится к Zoridae.
Перспективы изучения. Нахождение дополнительных видов в регионе
маловероятно. Для определения таксономического положения рода необходимо
привлечение сравнительно-генетического метода (сравнить с представителями родов
Ctenus (Ctenidae) и Zora (Zoridae)).
Семейство Cybaeidae Banks, 1892
Цибеиды
Рис. 14.1-6.
Небольшое голарктическое семейство со 162 видами из 12 родов. В России
известно всего лишь пять видов рода Cybaeus L.Koch, 1868, из которых четыре
отмечены на Дальнем Востоке.
Диагностические признаки. Среднего размера, 8-глазые, некрибеллятные, эн-
телегинные пауки, пальпа самца с мощным выростом колена и РГО, бульбус дик-
тинидного типа.
Близкие группы. Внешне (габитуально и рисунком) похожи на Callobius,
Cybaeopsis (Amaurobiidae) и Coelotes (Agelenidae). От первых отличаются
отсутствием крибеллюма, от Coelotes — неудлинёнными бородавками, диктинидным
типом пальпы и отсутствием «сосков» эпигины.
ОПИСАНИЕ:
• Карапакс. Грушевидный.
• Стернум. Не модифицирован, щитовидный.
• Глаза. 8 (4-4), все глаза, кроме передних медиальных (самые мелкие),
примерно одинаковые по размеру.
• Ротовой аппарат. Не модифицирован.
• Ноги. 3 коготка. Передняя пара ног чуть длиннее тела, часто с кольцами.
Шипы имеются.
• Брюшко. Овальное.
• Паутинные бородавки. Колюлюс имеется. Бородавки не
модифицированы.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Пальпа самца с выростом колена и РГО, бульбус
диктинидного типа. Эпигина с ямкой, разнообразной по форме, или без ямки.
• Размер. 4-10 мм. Половой диморфизм слабо выражен.
• Окраска. От светло- до тёмно-коричневого. Карапакс и брюшко иногда с
рисунком.
• Особые черты. Мощный вырост колена пальпы самца, покрытый у многих
видов мощными (короткими и толстыми) шипиками.
Рода, известные из азиатской части России. Cybaeus L. Koch, 1868.
Распространение. Все виды известны из Приморья и Сахалинской области.
Возможно, семейство встречается и в Хабаровском крае.
Биология. Все известные нам виды встречаются исключительно в затенённых
местах с достаточно высокой влажностью. Населяют подстилку.
Жизненный цикл длится два года, по крайней мере, у единственного
исследованного вида этого рода С akiensis Ihara, 2003 в Японии (Шага, 2003). Возможно,
Семейства, встречающиеся в азиатской части России
143
/
Рис. 14. Cybaeidae. Cybaeus basarukini (сГ$? 1, 3, 5), С. kunashirensis (сГ, 2) и Cybaeus
spp. ($, 4, 6, неописанные виды).
1,2 — пальпа С?, вентрально; 3^, 6 — габитус ?, сверху; 5 — эпигина, снизу Стрелка
показывает на отросток колена пальпы о\
144
4. Специальная часть
все виды этого рода имеют два отверстия в своём убежище, которое располагается
под камнями и брёвнами.
Методы сбора. Почвенные ловушки и ручной сбор среди подстилки.
Таксономия. Ранее Cybaeus и родственные рода рассматривались в Agelenidae,
например, в определителе В.П. Тыщенко (1971). Р.Т. Lehtinen (1967) перенёс эту
группу в Dictynidae в ранге подсемейства. R. Forster (1970) придал таксону ранг
семейства. Иногда в него включают и Argyronetidae в качестве подсемейства (хотя,
так как Argyronetidae имеет приоритет над Cybaeidae, бывает, что наоборот — Су-
baeidae включают в Argyronetidae — см., например, Jocque & Dippenaar-Schoeman,
2006). В настоящее время всеми признаётся статус (ранг) этой группы как
семейства (Platnick, 2009).
Очевидно, Cybaeus заслуживает разделения на более мелкие таксоны, рода или
подрода. О гетерогенности рода свидетельствует разное строение пальпы и,
особенно, эпигины. Систематика дальневосточных видов рассматривалась всего в двух
работах: Марусик, Логунов (1991) и Oliger (1994).
В настоящее время японские виды активно ревизуют J. Ihara и другие японские
исследователи. В России этой группой занимаются М.М. Омелько и Ю.М.
Марусик.
Разграничение видов. Форма копулятивных органов; в некоторых случаях
можно использовать рисунок и окраску для разграничения близких видов.
Перспективы изучения. Несмотря на то, что известно всего 4 вида в регионе,
на самом деле разнообразие семейства гораздо выше. Просмотр
немногочисленных материалов, собранных почвенными ловушками в Приморье, позволил
выявить не менее 6 новых видов. Все виды этого рода, за редким исключением,
характеризуются очень локальным распространением.
Семейства, встречающиеся в азиатской части России
145
Cybaeus sp. (Cybaeidae), самец, Сахалин (фото: Ю.М. Марусик).
Семейство Dictynidae О. Pickard-Cambridge, 1871
Диктиниды; meshweb spiders
Рис. 15.1-18.
Всесветно распространённое семейство с 48 родами и 561 видом. В России
известно около 50 видов из 11 родов, а в азиатской части России — 10 родов и 39 видов.
Диагностические признаки. Энтелегинные крибеллятные или безкрибеллят-
ные пауки с 8 глазами, однорядным каламиструмом (если имеется), пальпы
самцов всегда с отростками голени (ретролатеральными и/или дорсальными).
Близкие группы. Наличием крибеллюма и формой тела похожи на Amaurobiidae
и Titanoecidae, однако имеют более мелкие размеры и неразделённый крибеллюм.
Близки (судя по форме пальпы) к Argyronetidae и Cybaeidae, но отличаются кри-
беллюмом и более мелкими размерами.
ОПИСАНИЕ:
• Карапакс. От грушевидного до усечённо-яйцевидного; головная часть у
большинства видов приподнята над грудной, но бывает и наоборот, грудная
приподнята над головной.
• Стернум. Примерно треугольный.
• Глаза. 8 в два ряда (4-4), в соседних регионах известны 6-глазые виды.
• Ротовой аппарат. Хелицеры вертикальные, несколько вытянуты, у самцов
часто модифицированы (с вдавлением вдоль внутреннего края, и с
поперечными рёбрами).
• Ноги. 3 коготка; у большинства видов — без шипов.
• Брюшко. Овальное.
• Паутинные бородавки. Не модифицированы.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Если имеется, то крибеллюм неразделённый.
• Копулятивные органы. Голень пальпы самца всегда с отростками, если не
латеральными, то дорсальными; кондуктор всегда двувершинный, иногда
верхняя его часть очень длинная {Lathys). Эмболюс всегда длинный. Эпиги-
на с ямкой, часто двумя, бывает без ямок вообще.
• Размер. 2,5-6 мм. Половой диморфизм слабо выражен; у самцов карапакс
может быть больше, чем у самки, но брюшко меньше.
• Окраска. От одноцветно желтоватого, до красного, зелёного или
тёмно-коричневого. Рисунок имеется на брюшке и, иногда, даже на карапаксе
(продольная светлая медиальная полоса). У видов Lathys на брюшке могут
встречаться белые гуаниновые пятна.
• Особые черты. Небольшой размер и наличие крибеллюма. Голень пальпы
самца часто с дорсальным выростом, на тупой вершине которого сидят 1-2
мощных коротких шипа. Если шипа 2, то они тесно прижаты друг к другу.
Рода, известные из азиатской части России. Archaeodictyna Caporiacco, 1928
{A. consecuta (О. Pickard-Cambridge, 1872)); Arctella Holm, 1945 (A. lapponica Holm,
Семейства, встречающиеся в азиатской части России
147
10
Рис. 15. Dictynidae. Lathys sp. (?, 1,4), Mastigusa arietina ($, 2), Argenna sp. (cT5, 3 — ~
из Большехехцирского заповедника; 10 — сГ из Тувы), ЕтЫупа wangi (сГ, 5-6), Dictyna
foliicola (СГ, 7), D. schmidti (сГ, 8-9).
1 — паутинные бородавки ?, снизу; 2-3 — брюшко ?, снизу; 5, 10 — пальпа С7, снизу; 6-7 —
пальпа О71, сбоку; 8-9 — головогрудь О71, сбоку и спереди.
148
4. Специальная часть
11 1 13
14 15 16
17
Рис. 15 (продолжение). Dictynidae. Габитус и окраска Argenna sp. ($, 11, Болыыехех-
цирский заповедник), Lathys sp. (?, 12), Emblyna borealis (?, 13), is. wangi (cT, 14), Dictyna
foliicola (cT, 15), D. тио/ог (cT, 16) и Mastigusa arietina (cT, 17-18).
11-17 — габитус, сверху; 18 — головогрудь, снизу.
Семейства, встречающиеся в азиатской части России
149
1945); Argenna Thorell, 1870 {A. patula (Simon, 1874)); Cicurina Menge, 1871 (С.
cicur (Fabricius, 1793));Devade Simon, ISS4; Dictyna Sundevall, 1833; ism WyrcaCham-
berlin, 1948; Hackmania Lehtinen, 1967 (H prominula (Tullgren, 1948)); Lathys Simon,
1884; Marylina Lehtinen, 1967 (M bicolor (Simon, 1870)) и Mastigusa Menge, 1854
(M. arietina (Thorell, 1871)).
Распространение. Archaeodictyna и Devade — юг Сибири; Argenna — юг всего
региона; Arctella, Dictyna и ЕтЫупа — весь регион, включая тундровую зону;
Hackmania — вся лесная зона; Lathys — весь регион, за исключением таёжных зон
средней и западной Сибири; Mastigusa — только Приенисейе. Наибольшее число
видов найдено на Северо-Востоке, в том числе и несколько эндемиков.
Вероятно, указания Cicurina и Marylina в южной Сибири основаны на
неверных определениях. Не исключено, что на самом деле, вместо Marylina, на юго-
западе Сибири встречается Ajmonia Lehtinen, 1967, род известный из Восточно-
Казахстанской области и Монголии (Marusik & Esyunin, 2010).
Биология. Биология очень разнообразна даже в пределах одного рода. Близкие
виды могут обитать как в подстилке, так и среди кустарников и травостоя; или, с
одной стороны, под камнями, а с другой — под корой деревьев. Многие виды
устраивают небольшие сеточки на листьях, либо в мутовках кустарников или
высохшей травы.
Коконы у Dictyna и ЕтЫупа круглые, яиц не много, около 15. В верховьях
Колымы D. arundinacea (Linnaeus, 1758) имеет 5-6 ювенильных стадий.
Методы сбора. Все методы и все биотопы.
Таксономия. Рамки этого таксона длительное время оставались неясными.
Долгое время в это семейство включали всех 3-коготковых крибеллятных пауков, не
имеющих коготковых подушечек (claw tufts). Amaurobiidae были вынесены за
пределы Dictynidae только А. Петрункевичем (Petrukevitch, 1928). Детальный
обстоятельный диагноз этого семейства дал Р.Т. Lehtinen (1967). Он же подразделил его на
несколько подсемейств и выделил множество родов. Тем не менее, надвидовая
систематика этого таксона разработана неудовлетворительно. Некоторые крупные рода,
по всей видимости, будут раздроблены на более мелкие (Dictyna, ЕтЫупа, Lathys).
Судя по строению паутинных бородавок, род Mastigusa должен быть выведен
из этого семейства и возвращён в Agelenidae.
На видовом уровне семейство относительно хорошо обработано лишь в
Северной Америке (Chamberlin & Gertsch, 1958). Таксономия сибирских и
дальневосточных видов рассматривается в работах: Kulczynski (1908, 1926); Schenkel (1930);
Дунин (19846); Марусик (1988а); Данилов (1994); Danilov (2000); Marusik & Koponen
(1998); Esyunin & Marusik (2001); Marusik & Esyunin (2010).
Разграничение видов. Форма копулятивных органов, окраска/рисунок, а иногда
и размер глаз и копулятивных органов.
Перспективы изучения. Некоторые сибирские виды описаны только по
самкам или только по самцам. Не исключены находки новых видов, прежде всего в
южных районах Сибири. Рода Dictyna, ЕтЫупа, Lathys нуждаются в пересмотре
их диагнозов.
Семейство Eresidae C.L. Koch, 1851
Эрезиды; velvet spiders
Рис. 16.1-2.
Небольшое семейство со 100 видами и 10 родами. Распространение необычно:
Евразия, Африка, и Бразилия. В России достоверно известны 3 вида, все они
встречаются и в азиатской части России.
Диагностические признаки. Крупные (8-20 мм), энтелегинные крибеллятные
пауки с прямоугольным карапаксом и 8 глазами; передняя половина карапакса
приподнята; глаза широко расставлены (боковые отстоят друг от друга на десяток
диаметров); ноги короткие и толстые. Всё тело и ноги покрыто короткими густыми
волосками. Пальпа самца без РГО.
Близкие группы. В регионе нет внешне схожих пауков.
ОПИСАНИЕ:
• Карапакс. Обратно трапециевидный или прямоугольный, передняя
половина приподнята над задней, клипеус низкий.
• Стернум. Прямоугольный; длина больше ширины в 2 раза.
• Глаза. 8, в 3 ряда (4-2-2), медиальные глаза сближены, передние
латеральные расположены по углам наличника, задние латеральные глаза далеко
отстоят от всех остальных, широко расставлены. Задние медиальные глаза
самые крупные.
• Ротовой аппарат. Хелицеры очень массивные и относительно короткие.
Нижняя губа удлинена.
• Ноги. 3 коготка, короче тела, толстые, густо покрыты волосками.
• Брюшко. Овальное — прямоугольное, с закруглёнными углами. Шире
карапакса у самки, и равно ширине карапакса у самца. Передняя часть
брюшка простирается над задней пониженной частью карапакса.
• Паутинные бородавки. Передние паутинные бородавки утолщены,
конические.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Крибеллюм хорошо выражен, разделён на 2
части.
• Копулятивные органы. Энтелегинные. Пальпа самца относительно
короткая и коренастая, без РГО. Тегулюм округлый, несёт на себе тонкий эмбо-
люс и пластинчатый кондуктор. Эпигина с ямкой, ширина которой больше
длины.
• Размер. 8-20 мм, самцы в несколько раз мельче и стройнее самок.
• Окраска. Самки чёрные с пепельным карапаксом и отчасти брюшком,
самцы с более тёмным карапаксом (меньше проседи) и ярко-красным брюшком
с 2-3 парами чёрных округлых пятен. Ноги с белыми кольцами на вершине
каждого членика.
• Особые черты. Латеральные глаза, далеко отстоящие от медиальных;
крупный размер; красная с чёрными пятнами окраска брюшка самцов.
Семейства, встречающиеся в азиатской части России
151
Рода, известные из азиатской части России. Eresus Walckenaer, 1805 (Е. kollari
Rossi, 1846; Е. cinnaberinus1 bifasciatus Ermolajev, 1937; E. granosus Simon, 1895).
Распространение. Степные районы южной Сибири и Приморье.
Биология. В южной Сибири, в Туве в частности, все особи были собраны в
полупустынных районах, на склонах гор. В Приморье немногочисленные Eresus
были собраны среди мезофитной растительности (зарастающая просека под ЛЭП).
Среди камней сеть представляет собой небольшую трубку-логово, собственно
ловчей сети не видно. Среди растительности сеть более сложная, имеется
небольшая норка в земле, небольшая трубка около 5-10 см, на поверхности переходящая
в пологовидную сеть.
Сети необычно мелкие для столь крупных пауков.
Судя по данным европейских исследователей, Eresus питается в основном
другими пауками и жуками, в том числе и жуками-скакунами. Самцы короткоживу-
щие. Самки становятся половозрелыми на третий год, откладывают один кокон.
Заботятся о нём, как и другие норные пауки, периодически поднимая к выходу,
чтобы погреть на солнце в погожий день. Отродившиеся паучата долгое время
остаются вместе с матерью, вплоть до 6-го возраста, и перезимовывают в её логове.
Отмечалось, что молодь по весне может поедать свою ослабевшую или уже
погибшую мать (так же, как у Cheiracanthium, Coelotes, Drassodes).
Методы сбора. Визуальный поиск по характерным сетям, или под камнями.
Самцы хорошо ловятся в почвенные ловушки.
1
Рис. 16. Eresidae. Eresus sp. (cT).
1 — сверху; 2 — пальпа, сбоку.
1 Rezac et al. (2008) считают, что Е. cinnaberinus — это nomen dubium.
152
4. Специальная часть
Таксономия. Первым, кто в глобальном масштабе рассмотрел семейство, был
Е. Simon (1903). Он же разделил Eresidae на два подсемейства. Р.Т. Lehtinen (1967)
в своей ревизии нисколько не изменил границы и состав семейства, но впервые
указал на возможные родственные связи. По его мнению, эти пауки близки к Zoda-
riidae. В настоящее время Eresidae сближают с Oecobiidae и Hersiliidae. Все три
семейства образуют группу, обособленную от остальных энтелегинных пауков. Хотя,
на наш взгляд, Eresidae не имеют ничего общего с Oecobiidae и Hersiliidae ни
соматически, ни в строении копулятивных органов.
Систематика рода Eresus разработана чрезвычайно неудовлетворительно, как в
рамках Европы, так и в Палеарктике в целом. В этом роде описано много видов,
причём почти все они выделены на основе окраски. Копулятивные органы при
этом зачастую вообще не описывались. Все виды, обитающие на территории СССР,
были синонимизированы с Е. cinnaberinus (Ненилин, Пестова, 1986).
Судя по строению пальпы, у нас обитает несколько видов. Очень возможно, что
Е. granosus был ошибочно определён. Этот вид описан из восточной части Китая,
и едва ли он может встречаться в степях Алтая. Недавние исследования показали,
что Е. cinnaberinus — это nomen dubium (Rezac et al., 2008), и под этим названием
в Европе отмечали три вида. Все исследованные экземпляры «Е. cinnaberinus» из
Урала и Алтая относятся к Е. kollari.
Виды, населяющие восточную часть России, рассматривались в трёх работах:
Ermolajev (1928b), Данилов (1994) и Rezac et al. (2008).
В настоящее время этот род ревизуют М. Rezac и Д.В. Логунов.
Перспективы изучения. Род Eresus крайне нуждается в ревизии. Очевидно,
дальневосточные и южносибирские популяции относятся к разным видам.
Семейства, встречающиеся в азиатской части России 153
Eresus kollari (Eresidae), самка, Чехия (фото: R. Масек).
Eresus kollari (Eresidae), самец, Чехия (фото: R. Масек).
Семейство Gnaphosidae Pocock, 1898
Гнафозиды; ground spiders
Рис. 17.1-24.
Седьмое по величине семейство, всесветно распространённое, известно 2036
видов из ПО родов. В России отмечено более 200 видов примерно из 28 родов, а в
азиатской части России — более 156 видов из 21 родов.
Диагностические признаки. Энтелегинные, некрибеллятные пауки с 8
глазами в 2 ряда; размер от мелкого до крупного; сетей не плетут, 2-коготковые;
передние паутинные бородавки цилиндрические, расставлены на диаметр или более.
Пальпа самцов, как правило, с РГО.
Близкие группы. Clubionidae, Liocranidae, Anyphaenidae, Corinnidae. От них
Gnaphosidae отличаются широко расставленными передними цилиндрическими
бородавками (исключение — муравьевидные Micarid).
ОПИСАНИЕ:
• Карапакс. Грушевидный или овальный, форма варьирует, высота низкая,
обычно с медиальной бороздой.
• Стернум. Овальный, иногда с выростами между тазиками.
• Глаза. 8, небольшие, в 2 ряда (4-4), задние медиальные глаза несколько
угловатые (треугольные).
• Ротовой аппарат. Хелицеры относительно короткие, однако у самцов
Brass odes хелицеры удлинены.
• Ноги. 2 коготка, от длинных до относительно коротких; Zelotini с гребёнкой
из вентральных щетинок на вершине предлапки III.
• Брюшко. Удлинённо-овальное, самцы иногда с маленьким дорсальным ску-
тумом.
• Паутинные бородавки. Передние бородавки цилиндрические,
параллельны друг другу, расставлены на ширину их диаметра (исключение—Micarid).
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Энтелегинные. Пальпа самца с РГО (кроме СаШ-
lepis), который обычно заострён на вершине. Эмболюс у большинства видов
тонкий и длинный, медиальный отросток имеется у большинства таксонов.
Эпигина очень разнообразна по форме, часто с верхним карманом или ска-
пусом.
• Размер. 3-20 мм. Половой диморфизм в целом слабо выражен; у самцов
Drassodes хелицеры длиннее, нежели у самок.
• Окраска. От бледно-белого до практически чёрного, большинство — без
рисунка, однако у некоторых родов (Callilepis, Micaria, Kishidaia, Sernokorba,
некоторых видов Drassodes и Gnaphosa) бывает рисунок на брюшке.
• Особые черты. Широко расставленные цилиндрические передние
бородавки, часто треугольные задние медиальные глаза.
Семейства, встречающиеся в азиатской части России
155
1
11
Рис. 17. Gnaphosidae. Соматические признаки Kishidaia albimaculata (?, 1,10), Berlandina
ubsunurica (сГ, 2, 5), Haplodrassus moderatus (сГ, З, 6), Drassodes cupreus (2, 4, 7-8).
Parasyrisca sp. ($, 9) и Micaria lenzi ((?, 11).
1-4 — паутинные бородавки, снизу; 5-7 — головогрудь, снизу; 8-9 — карапакс. сверху: ". 0-1". —
габитус, сверху- и сбоку
156
4. Специальная часть
12
13
20 21
Рис. 17 (продолжение). Gnaphosidae. Копулятивные органы Haplodrassus taepaikensis
(сГ$, 12, 16), Micaria tripunctata (сГ, 13), М. fulgens (?, 14), Drassodes cupreus (?, 15),
Zelotes longipes (?, 17), Leptodrassex memorialis (?d\ 18, 22), Haplodrassus moderatus ($,
19), Parasyrisca asiatica (cT, 20), Gnaphosa similis (cT, 21), G. muscorum (?, 23) и Drassodes
longispinus (?, 24).
12, 20-22 — пальпа сГ, снизу; 13 — пальпа сГ, сбоку; 14-19, 23-24 — эпигина, снизу 12, 16 —
по: Marusik et al. (2007); 18,22 — по: Eskov & Marusik (1995); 19,23-24 — no: Marusik & Logunov
(1995a); 21 — no: Marusik & Koponen (2000).
Семейства, встречающиеся в азиатской части России
157
Рода, известные из азиатской части России. Berlandina Dalmas, 1922; Callilepis
Westring, 1874; Drassodes Westring, 1851; Drassyllus Chamberlin, 1922; Echemus
Simon, 1878; Gnaphosa Latreille, 1804; Haplodrassus Chamberlin, 1922; Kishidaia
Yaginuma, 1960; Micaria Westring, 1851; INomisia Dalmas, 1921; Odontodrassus
Jezequel, 1965; Parasyrisca Schenkel, 1963; Phaeocedus Simon, 1893; ? Poecilochroa
Westring, 1874; IScotophaeus Simon, 1893; Sernokorba Kamura, 1992; ISosticus
Chamberlin, 1922; Tuvadrassus Marusik et Logunov, 19951; Urozelotes Mello-Leitao,
1938;Ze/otesGistel, 1848.
Распространение.
Drassodes, Gnaphosa, Haplodrassus, Micaria, Zelotes — весь или практически
весь регион.
Callilepis — практически весь регион к югу от 62°.
Parasyrisca — юг Сибири и Северо-Восток до верховий Колымы.
Urozelotes — юг Сибири и Якутия.
Drassyllus, Phaeocedus — юг Сибири и Дальнего Востока.
Berlandina, Echemus — на восток до Тувы.
Tuvadrassus — только Тува.
Kishidaia, Odontodrassus, Sernokorba — юг Дальнего Востока.
Nomisia, Scotophaeus, Sosticus, Poecilochroa — очевидно, ошибочные указания.
Наиболее важные работы, посвященные распространению гнафозид азиатской
части России: Ovtsharenko et al. (1992, 1995); Marusik et al. (2000).
Биология. Сетей не плетут. Дневные или ночные охотники. Большинство
видов связано с подстилкой, из-за чего пошло английское название "ground spiders".
Некоторые рода и виды связаны с древесной или травяно-кустарниковой
растительностью {Kishidaia, Poecilochroa, Sernokorba, Parasyrisca tyshchenkoi Ovtsharenko
et al., 1995, неполовозрелые Drassodes в таёжной зоне). Большинство видов
термофильны и заселяют теплообеспеченные сухие биотопы. Однако есть немало
видов, связанных с мезофильными местообитаниями на севере. Gnaphosa orites и G.
nigerrima — психрофильные. Первый вид является самым северным
представителем семейства на Северо-Востоке Азии. Среди подстилочных видов большинство
являются петрофилами, однако есть немногочисленные виды бриофилы.
Большинство видов — неспециализированные хищники. Известно, что Callilepis
и Micaria (Ubick et al., 2005) питаются муравьями. Только один вид, Micaria constricta
Emerton, 1894, живёт за пределами ареала муравьев. Micaria габитуально
напоминает муравьев, но этологически сходство с муравьями намного больше. Эти пауки
машут передними ногами, наподобие того, как муравьи антеннами.
Среди подстилочных видов большинство строят убежища, где самки
откладывают яйцевой кокон и охраняют его до своей естественной смерти. Ювенильные
особи, очевидно, иногда поедают мать. Зимуют неполовозрелые, вместе с
охраняющими их полуживыми самками (весной ноги самок сохраняют подвижность,
однако передвигаться они уже не могут). Коконы линзовидные (кроме Micaria),
белого или розового (у Zelotes subterraneus (C.L. Koch, 1833)) цвета.
1 Murphy (2007) считает этот род младшим синономом Haplodrassus.
158
4. Специальная часть
Пик активности большинства видов приходится на июнь и июль в таёжном
поясе, однако у некоторых видов, например Gnaphosa sticta, самцы встречаются
исключительно ранней весной, когда ещё не полностью стаял снег.
Методы сбора. Почвенные ловушки, сбор под камнями, иногда просеивание
подстилки; Kishidaia и некоторые другие рода в Южной Сибири и на юге Дальнего
Востока собираются укосами по травостою и отряхиванием ветвей.
Таксономия. Надвидовая таксономия семейства разработана недостаточно
хорошо. Это касается как деления на подсемейства, так и во многих случаях - на
рода. Рамки этого семейства существенных изменений в последнее время не
претерпели, однако в него недавно был включён род Micaria, рассматривавшийся в
определителе В.П. Тыщенко (1971) в Clubionidae. Практически все северо-амери-
канские и среднеевропейские виды, встречающиеся в азиатской части России,
хорошо изучены. В пределах Сибири ревизованы лишь Gnaphosa и Parasyrisca.
Среди крупных родов, помимо указанных выше, относительно хорошо изучены лишь
Micaria и Haplodrassus.
Наиболее важные работы, в которых рассматривается таксономия пауков
азиатской части России, следующие: Kulczynski (1885, 1908, 1926); Holm (1970); Овча-
ренко,Марусик(1988,1995); Ovtsharenkoetal. (1992,1995); Danilov (1993b, 1997b),
Eskov & Marusik (1995); Marusik & Logunov (1995a); Михайлов, Марусик (1995);
Marusik et al. (1996, 2002, 2007); Marusik & Koponen (2000, 2001a).
К важным работам, благодаря которым можно определять сибирские и
дальневосточные виды, можно отнести Platnick & Song (1986), Platnick & Shadab (1988),
Chikuni (1989), Kamura (1984, 1986, 1988, 1992, 1995, 2007 и др.), Song et al. (2004)
и Ono et al. (2009).
В настоящее время гнафозидами региона занимаются С.Л. Есюнин, Т.К. Тунё-
ва, Ю.М. Марусик, Н.М. Ковблюк, М.М. Омелько, S. Koponen.
Разграничение видов. Форма копулятивных органов, в некоторых случаях
рисунок и вооружение хелицер (Drassodes) используются для разграничения видов.
Перспективы изучения. Это семейство — одно из наиболее крупных в
азиатской части России. Практически во всех крупных родах имеются неописанные виды
(Drassyllus, Gnaphosa, Haplodrassus, Micaria, Zelotes). Некоторые рода, например
Gnaphosa, Micaria, Zelotes, явно нуждаются в пересмотре их границ, и возможно в
будущем будут подразделены на несколько родов, или, по крайней мере, подродов.
В пределах Сибири и юга Дальнего Востока возможно нахождение ряда
дополнительных родов, известных из сопредельных территорий: Coreodrassus Paik, 1984;
Fedotovia Charitonov, 1946; Leptodrassex memorialis (Spassky, 1940); Synaphosus
Platnick & Shadab, 1980; Trachyzelotes Lohmander, 1944. Находки дополнительных
видов наиболее вероятны на Алтае, в южном Забайкалье и в Приморье.
Семейства, встречающиеся в азиатской части России
159
Kishidaia albimaculata (Gnaphosidae), самка, Приморье (фото: М.М. Омелько).
Семейство Hahniidae Bertkau, 1878
Ханииды, ганииды; comb-tailed spiders
Рис. 18.1-11.
Небольшое всесветно распространённое семейство с 238 видами, относимыми
к 26 родам. В России известно 14 видов из 5 родов, а в азиатской части России —
12 видов и 4 рода.
Диагностические признаки. Мелкие 3-5 мм, 3-коготковые, 8-глазые, безкри-
беллятные пауки, паутинные бородавки расположены в один поперечный ряд, длина
их во много раз превышает ширину (кроме Cryphoeca), дыхальце сдвинуто от
паутинных бородавок ближе к середине брюшка (кроме Cryphoeca). Пальпа самца с
РГО и у всех Hahniinae с выростом колена. Эпигина плоская без ямок, вульва с
длинными семяприёмными канальцами.
Близкие группы. Внешне (габитуально и рисунком) похожи на некоторых
Lathys, Hackmania (Dictynidae), однако легко отличаются отсутствием крибеллю-
ма, расположением бородавок в поперечный ряд и широким дыхальцем, обычно
отстоящим далеко от паутинных бородавок.
ОПИСАНИЕ:
• Карапакс. Овальный, без рисунка.
• Стернум. Не модифицирован.
• Глаза. 8 (4-4), примерно одинаковой величины, передние медиальные —
самые маленькие. В соседних регионах известны 6-глазые виды.
• Ротовой аппарат. Не модифицирован.
• Ноги. 3 коготка, относительно короткие, ноги I чуть длиннее тела.
• Брюшко. Овальное.
• Паутинные бородавки. Расположены в один ряд, апикальный членик
латеральных бородавок длинный (кроме Cryphoeca).
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем вдали от бородавок (кроме Cryphoeca).
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Пальпа самца всегда с РГО, все Hahniinae ещё и с
выростом колена. Тегулюм овальный, эмболюс нитевидный длинный.
Кондуктор имеется только у Cryphoeca и некоторых Hahnia. Эпигина плоская, с
парой щелевидных отверстий, вульва с длинными семяприёмными
канальцами и одной или двумя парами рецептакул.
• Размер. 3-5 мм. Половой диморфизм практически не выражен.
• Окраска. От светло-серого или бледно-оранжевого до очень тёмного. Часть
видов с чётким рисунком на брюшке.
• Особые черты. Расположение паутинных бородавок в один поперечный ряд;
дыхальце, далеко отстоящее от паутинных бородавок.
Семейства, встречающиеся в азиатской части России
161
ю
Рис. 18. Hahniidae. Pacifantistea ovtchinnikovi ($, 1-2,7), Cryphoeca silvicola (C?¥, 3, 8,10-
11), Hahnia ononidum ($, 4—5), Neoantistea quelpartensis ($, 6), Hahnia corticicola (C?, 9).
1 — головогрудь, спереди; 2-4 — габитус ?, сверху; 5-8 — брюшко ?, снизу; 9, 11 — пальпа сГ,
сбоку и снизу; 10 — голень сГ, сбоку. Стрелка показывает положение дыхальца. 10-11 — по:
Thaler (1978).
162
4. Специальная часть
Рода, известные из азиатской части России. Antistea Simon, 1898 (A. elegans
(Blackwall, 1841)); Cryphoeca Thorell, 18701 (С silvicola (C.L. Koch, 1834)); Hahnia
C.L. Koch, 1841; Neoantistea Gertsch, 1934 (N quelpartensis Paik, 1958); Pacifanti-
stea Marusik, 2011 {P. ovtchinnikovi Marusik, 2011).
Распространение.
Antistea — на восток до Енисея;
Cryphoeca — вся Южная Сибирь и юг Дальнего Востока;
Hahnia — весь регион;
Neoantistea — юг Дальнего Востока;
Pacifantistea — Дальний Восток от Петропавловска-Камчатского до юга
Приморья.
Биология. Все виды, обитающие в регионе, связаны с почвой или собственно
подстилкой (опад, моховой покров) или с галечниками и каменными осыпями. Га-
лечниковые виды Neoantistea и Hahnia могут встречаться в огромных количествах
(десятки особей на 1 м2).
Согласно Opell & Beatty (1976), Neoantistea плетёт небольшие сети, которые
обычно не более 5 см в диаметре. Сети пологовидные, густые, располагаются в
понижениях почвы. Их трудно заметить, разве что, благодаря росе. Если
побеспокоить паука, то он скрывается в трещинах, под листья. Сетей у Antistea и Hahnia
никто не наблюдал.
Коконы у Neoantistea округлые, линзовидно-полушаровидные белого цвета.
Методы сбора. Почвенные ловушки и просеивание подстилки в лесу, среди
камней, галечников. Для Hahnia corticicola Bosenberg et Strand, 1906 и
Pacifantistea ovtchinnikovi наиболее эффективный способ сбора — сгребание (пальцами)
песка (грунта) вдоль рек и морского побережья, соответственно. После одного-
двух скребков необходимо несколько секунд подождать, и тогда лишённые
убежища пауки начинают разбегаться.
Таксономия. Долгое время эта группа рассматривалась как подсемейство Agele-
nidae. В этом же статусе таксон фигурирует в определителе В.П. Тыщенко (1971).
Р.Т. Lehtinen (1967) подразделяет Hahniidae на три подсемейства, два из которых
встречаются в азиатской части России: Cryphoecinae {Cryphoeca) и Hahniinae (все
остальные рода). J. Wunderlich (1995b) и некоторые другие исследователи
рассматривают Cryphoeca в Dictynidae. Часть исследователей полагают, что всех ха-
ниид следует считать подсемейством Dictynidae.
Надвидовая классификация разработана относительно хорошо, однако, на наш
взгляд (исходя из строения копулятивных органов), следует ожидать, что род Hahnia
будет раздроблен.
Виды, встречающиеся в азиатской части России, рассматривались в работах
Kulczynski (1926), Marusik et al. (1996) и Marusik (2011).
В настоящее время этим семейством занимается Ю.М. Марусик.
1 Судя по строению пальпы самца, этот род следует относить к Dictynidae sensu lato или Agelenidae.
Семейства, встречающиеся в азиатской части России
163
Разграничение видов. Только форма копулятивных органов; в некоторых
случаях можно использовать рисунок и окраску для разграничения не близких видов.
Перспективы изучения. В регионе имеется несколько неописанных таксонов:
новый род из Алтая, новый вид Hahnia с Сахалина и др. На наш взгляд,
заслуживает выделения в отдельный род группа видов, близких к Hahnia glacialis Soerensen,
1898, как и Н corticicola Bosenberg & Strand, 1906. Предварительные
исследования разных популяций Н ononidum Simon, 1875 указывают на то, что возможно
этот вид заслуживает разбиения на несколько викариирующих видов, или, по
крайней мере, подвидов. Несмотря на то, что семейство представлено в регионе малым
числом видов, это перспективный для изучения таксон.
Araneus pinguis (Araneidae), самка, Приморье (фото: М.М. Омелько).
Семейство Leptonetidae Simon, 1890
Лептонетиды; midget cave spiders
Рис. 19.1-6.
В мире известно 209 видов, относимых к 15 родам. Семейство имеет
исключительно Голарктическое распространение. В России семейство отмечено только в
Приморье и на северном Кавказе, всего два вида.
Диагностические признаки. Гаплогинные пауки небольшого размера с
длинными ногами, с 6 глазами, глаза расположены тремя диадами, задняя диада
находится далеко сзади за передней парой диад.
Близкие группы. Форма расположения глаз уникальна.
ОПИСАНИЕ.
• Карапакс. Грушевидный, наличник высокий.
• Стернум. Не модифицирован.
• Глаза. Шесть глаз, 4-2. Передний ряд сильно выгнут, глаза соприкасаются;
задняя пара глаз соприкасается, далеко отстоит от переднего ряда.
• Ротовой аппарат. Хелицеры слегка удлинены, с длинным и тонким
коготком, передний ряд желобка с мелкими зубчиками.
• Ноги. Длинные, передняя пара более чем в 3 раза длиннее тела.
• Брюшко. Овальное или каплевидное.
• Паутинные бородавки. Не модифицированы.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем у паутинных бородавок.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы1. Гаплогинные пауки2. Пальпа самца с
относительно длинной голенью, цимбиум не уплощён, бульбус каплевидный, с
комплексом отростков, сосредоточенных на вершине, вблизи вершины цимбиу-
ма. Эпигины нет.
• Размер. До 3 мм.
• Окраска. Бледно-белая.
• Особые черты. Пара (диада) глаз, вынесенная далеко назад.
Рода, известные из азиатской части России. "Leptoneta" Simon, 1872.
Распространение. Известен только из Лазовского заповедника. Там собрана
только неполовозрелая самка, возможно, относящаяся к новому виду (Oliger et al.,
2002). На Дальнем Востоке это самая северная находка семейства.
Биология. Единственный экземпляр собран в мелкообломочной осыпи.
Многие виды этого семейства — троглобионты, или связаны с лесной подстилкой. Леп-
1 Форма копулятивных органов нашего вида неизвестна; описание составлено по дальневосточным
видам из Японии и Кореи.
2 Недавно высказано достаточно аргументированное мнение, что Leptonetidae имеют не гапло-
гинный тип копулятивного аппарата.
Семейства, встречающиеся в азиатской части России
165
Рис. 19. Leptonetidae. Falcileptoneta ? sp. (1-2, из Приморья), К striata (3^), К aichi-
nensis (5) и К iriei (6).
1-2 — головогрудь, снизу и сверху (видны многочисленные зубцы хелицер и короткий лабиум);
3 — габитус ?, сверху; 4 — глаза ?, сверху; 5 — стернум, максиллы и лабиум, снизу; 6 — пальпа
сГ, сбоку. Стрелка показывает на далеко отстоящую диаду глаз. 3-6 — по: Ono et al. (2009).
тонетиды строят небольшие сеточки из беспорядочных нитей, или же полог очень
плотный. Пауки располагаются под сетью, вверх ногами. Если задеть сеть, пауки
падают на землю и плотно прижимают ноги к телу, чтобы стать менее заметными
(Ubick et al., 2005).
Методы сбора. Единственный экземпляр собран в мелкообломочной осыпи.
Не исключены находки и в пещерах.
Таксономия. Надвидовая систематика семейства плохо разработана. Долгое
время лептонетид объединяли с другими мелкими гаплогинными пауками Telemi-
dae, Ochyroceratidae. Впервые две последние группы были выведены из семейства
Fage (1912, 1913). Leptonetidae ревизованы только в пределах Неарктики и. отчас-
166
4. Специальная часть
ти, в Средиземноморье. Возможно, приморский вид относится к Falcileptoneta Ko-
matsu, 1970 — единственному роду, выделенному среди восточно-азиатских леп-
тонетид. Все остальные виды из Китая и Кореи относят к типовому роду Leptoneta,
хотя по строению копулятивных органов азиатские виды существенно отличаются
от западно-средиземноморских, откуда описан типовой вид. Наиболее подробную
сводку по данному семейству на Дальнем Востоке можно найти в монографии Опо
et al. (2009). Все японские виды явлются локальными эндемиками.
Разграничение видов. Только форма копулятивных органов.
Перспективы изучения. Не исключены находки нескольких видов, как в
пещерах, так и среди осыпей.
Leptoneta setulifera (Leptonetidae), самка, Китай (фото: Shuqiang Li).
Семейства, встречающиеся в азиатской части России
167
Cheiracanthium sp. (Cheiracanthiidae), самец, Приморье (фото: М.М. Омелько).
Семейство Linyphiidae Blackwall, 1859
Пауки-пигмеи, линифииды; dwarf spiders, money spiders
Рис. 20.1-23.
Это второе по величине семейство с 4359 рецентными видами и 582 родами. В
России известно около 800 видов примерно из 240 родов, а в азиатской части
России — не менее 700 видов из 221 рода (включая подрода).
Диагностические признаки. Энтелегинные, некрибеллятные, небольшие (1,2-
6 мм) пауки с 3 коготками и 8 глазами в 2 ряда; клипеус высокий; хелицеры со
стридуляционными бороздками. Самцы часто с выростами на карапаксе, пальпа
самца с базальным парацимбиумом, часто с отростками голени. Эпигина всегда с
медиальной пластинкой.
Близкие группы. Пауки, подобные линифиидам, встречаются во многих ара-
неоидных семействах (Araneidae, Theridiidae, Nesticidae, Pimoidae, Tetragnathidae)
и даже в более удалённых группах, например Uloboridae. Чаще всего за линифиид
принимают Robertus (Theridiidae) из-за отсутствия окраски и схожего габитуса. К
линифиидам некоторые исследователи относили представителей «Thymoites»
(Theridiidae), самцы которых имеют модифицированный карапакс. Линифиид всегда
можно отличить от внешне схожих таксонов по наличию стридуляционных борозд
на хелицере (не всегда легко увидеть), наличию базального U- или Г-образного
парацимбиума и медиальной пластинки эпигины.
ОПИСАНИЕ:
• Карапакс. От грушевидного до овального, клипеус высокий, самцы многих
видов с модифицированным карапаксом (выросты глазной области,
головной части сразу за глазами, выросты грудной части). Края карапакса могут
быть с зубчиками (Erigone). Заглазничная область часто со щелями и
ямками. Головная часть иногда с густыми или толстыми волосками. В редких
случаях головная часть бывает модифицирована и у самок.
• Стернум. Не модифицирован.
• Глаза. 8, в два ряда (4-4), однако формула может быть другой в случае
выростов карапакса. Боковые глаза всегда сближены. У большинства видов глаза
примерно равной величины, но могут быть и гетерогенными (передние
медиальные уменьшенные, либо задние медиальные увеличенные).
• Ротовой аппарат. Вариабельный. Хелицеры относительно длинные у всех
видов, со стридуляционными бороздками на боковой поверхности. Спереди
хелицеры могут быть с зубцом; передний край желобка хелицер иногда с
крупным (сдвоенным) зубцом. Максиллы иногда модифицированы,
наклонены друг к другу, у вершины сужены (Zornella).
• Ноги. 3 коготка. От коротких до очень длинных. Голени ног с 1-2
дорсальными щетинками. Предлапка I всегда с трихоботрией. Предлапка IV с одной
трихоботрией или без. Нога I самца в редких случаях модифицирована.
Тазик IV часто с зубчатым выростом, служащим для стридуляции.
• Брюшко. Овальное или каплевидное, чаще всего одноцветное, но бывает и
с рисунком. В редких случаях (например, у Ceratinella) есть дорсальный ску-
Семейства, встречающиеся в азиатской части России
169
Пц
П
Рис. 20. Linyphiidae. Agynetayakutsaxatilis (d\ 1-2), Zornella cultrigera (d\ 3,7), Sciastes
sp. (4), схема измерения расположения трихоботрии на предлапке I (5), A. brusnevi (d\
6), Stemonyphantes griseus (d"$, 8, 11), Oreoneta sp. (d\ 9), Tenuiphantes nigriventris (?,
10) и Perlongipalpus logunovi (?, 12).
1 — головогрудь, спереди; 2 — хелицера, сбоку; 3-4 — головогрудь, снизу и сбоку; 5 — схема
расположения трихоботрии на предлапке I; 6-9 — пальпа, сбоку; 10-12 — эпигина, снизу. Сб —
стридуляционные борозды; Фз — фронтальный зубец хелицеры; Пц — парацимбиум; Lc—lamella
characteristica. 6 — по: Marusik et al. (2002).
170
4. Специальная часть
тум. Помимо дорсального скутума бывают вентральные скутумы: непарный
перед паутинными бородавками, пара скутумов сверху-сбоку от стебелька
(иногда сливаются в один), и пара скутумов на месте лёгочных крышек
(могут сливаться с верхними). Форма скутумов отличается у самцов и самок.
• Паутинные бородавки. Не модифицированы, колюлюс имеется.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Энтелегинные пауки. Пальпа самца от короткой до
очень длинной (длиннее карапакса и тела), всегда с парацимбиумом. Пара-
цимбиум U- или Г-образный. Голень у многих с отростком или несколькими
отростками. Эпигина очень вариабельная, нередко со скапусом, или даже
двумя, но всегда трёхмерная, и всегда имеет медиальную пластинку. Вульва
без трубок, но со складками, внешне схожими с трубками.
• Размер. 1,2-6 мм, самки крупнее самцов, но нередко самцы с
модифицированным карапаксом могут быть крупнее самок.
• Окраска. От одноцветных до пёстро окрашенных. Большинство "Linyphii-
пае" и Micronetinae — с окраской брюшка, a "Erigoninae" — наоборот, с
окраской только в редких случаях.
• Особые черты. Наличие стридуляционных бороздок на хелицере (их
зачастую нелегко рассмотреть); базальный U- или Г-образный парацимбиум, при-
членённый к цимбиуму через мембрану.
Рода, известные из азиатской части России. Abacoproeces Simon, \SS4;Abiskoa
Saaristo et Tanasevitch, 2000; Acanthoneta Eskov et Marusik, 1991; Acartauchenius
Simon, 1884; Agnyphantes Hull, 1932; Agyneta Hull, 1911; Agyphantes Saaristo et
Marusik, 2004; Ainerigone Eskov, 1993; Allomengea Strand, 1912; Anguliphantes Saaristo et
Tanasevitch, 1996; Aphileta Hull, 1920; Aprifrontalia Oi, 1960; Arachosinella Denis,
1958; Araeoncus Simon, 1884; Archaraeoncus Tanasevitch, 1987; Arcterigone Eskov et
Marusik, \99A\ Arcuphantes Chamberlin et Ivie, 1943; Asiceratinops Eskov, 1992; Asio-
phantes Eskov, 1993; Asperthorax Oi, 1961; Asthenargoides Eskov, 1993; Asthenargus
Simon et Fage, 1922; Baryphyma Simon, 1884; Bathylinyphia Eskov, 1992; Bathyphan-
tes Menge, 1866; Bishopiana Eskov, 1988; Вolephthyphantes Strand, 1901; Bolyphan-
tes C.L. Koch, 1837; Carorita Duffey et Merrett, 1963; Caviphantes Oi, 1960; Centro-
merita Dahl, 1912; Centromerus F. Dahl, 1886; Ceraticelus Simon, 1884; Ceratinella
Emerton, 1882; Cnephalocotes Simon, 1884; Collinsia O. Pickard-Cambridge, 1913;
Concavocephalus Eskov, 1989; Connithorax Eskov, 1993; Crispiphantes Tanasevitch,
1992; Crosbylonia Eskov, 1988; Dactylopisthes Simon, 1884; Dactylopisthoides Eskov,
1990; Decipiphantes Saaristo et Tanasevitch, 1996; Dicymbium Menge, 1868; Diplo-
centriaHull, 1911; DiplocephalusBertkau, 1883; Diplosty la Emerton, 1882; Dismodicus
Simon, 1884; Doenitzius Oi, 1960; Drapetisca Menge, 1866; Drepanotylus Holm, 1945;
Eborilaira Eskov, 1989; Eldonnia Tanasevitch, 2008; Entelecara Simon, 1884; Epibel-
lowia Tanasevitch, 1996; Epigyphantes Saaristo et Tanasevitch, 2004; Epigytholus
Tanasevitch, 1996; Episolder Tanasevitch, 1996; Erigone Audouin, 1826; Erigonella F. Dahl,
1901; Erigonoploides Eskov, 1989; Erigonoplus Simon, 1884; Eskovia Marusik et
Saaristo, 1999; Eskovina Kocak et Kemal, 2006 (=Oinia Eskov, 1984); Estrandia Blauvelt,
1936; Evansia O. Pickard-Cambridge, 1900; Flagelliphantes Saaristo et Tanasevitch,
Семейства, встречающиеся в азиатской части России
171
13
14
15
17
Рис. 20 (продолжение). Linyphiidae. Внешний вид и карапакс "Lepthyphantes" turbatrix
(С?, 13), Lophomma vaccinii (С?, 14), Tanasevitchia uralensis (С?, 15), Perlongipalpus pini-
pumilis (cT, 16), Stemonyphantes griseus (?, 17), Saloca ryvkini (cT, 18), Scotinotylus sacer
(cT, 19), Savignia basarukini (cT, 20), Monocerellus montanus (cT, 21) и Walckenaeria
basarukini (cT, 22) и Ж golovatchi (сГ, 23).
Сб — стридуляционные борозды. 18-23 — по: Eskov & Marusik (1994).
172
4. Специальная часть
1996; Floronia Simon, 1887; Frontella Kulczynski, 1908; Frontinellina F.O. Pickard-
Cambridge, 1902; Gibothorax Eskov, 1989; Glyphesis Simon, 1926; Gnathonarium
Karsch, 1881; Gonatium Menge, 1868; Gongylidiellum Simon, 1884; Gongylidioides
Oi, 1960; Gongylidium Menge, 1868; Helophora Menge, 1866; #er6z/?/zflwtes Tanasevitch,
1992;Hilaira Simon, 1884; Я/тя/я/?/ши^ Tanasevitch, 1992;Holminaria Eskov, 1991;
Horcotes Crosby et Bishop, 1933; Hybauchenidium Holm, 1973; Hylyphantes Simon,
1884; Hypomma F. Dahl, 1886; Hypselistes Simon, 1894; Improphantes Saaristo et
Tanasevitch, 1996; Incestophantes Tanasevitch, 1992; Ipa Saaristo, 2007; Islandiana
Braendegaard, 1932;Ivielum Eskov, \9SS; Kaestneria Wiehle, 1956;KaritaTanasevitch,
2007; Kikimora Eskov, 1988; Kolymocyba Eskov, 1989; Lasiargus Kulczynski, 1894;
Lepthyphantes Menge, 1866; Leptorhoptrum Kulczynski, 1894;Lmj;/?/z/tfLatreille, 1804;
Lophomma Menge, 1868; Macrargus Dahl, 1886; Maro О. Pickard-Cambridge, 1906;
Masikia Millidge, 1984; Maso Simon, 1884; Mecynargoides Eskov, 1988; Mecynargus
Kulczynski, 1894; Megalepthyphantes Wunderlich, 1994; Mermessus O. Pickard-
Cambridge, 1899 (=Eperigone Crosby et Bishop, 1928); Mesasigone Tanasevitch, 1989;
Metopobactrus Simon, 1884; Micrargus F. Dahl, 1886; Microlinyphia Gerhardt, 1928;
Microneta Menge, 1869; Miftengris Eskov, 1993; Minicia Thorell, 1875; Minyrioloides
Schenkel, 1930 (= Baryphyma Simon, 1884); Minyriolus Simon, 1884; Moebelotinus
Wunderlich, 1995; Monocerellus Tanasevitch, 1983; Mughiphantes Saaristo et
Tanasevitch, 1999 {Suffusiphantes Saaristo et Tanasevitch, 1999; Whymperiphantes Saaristo et
Tanasevitch, 1999); Nematogmus Simon, 1884; Nenilinium Eskov, 1988; Neriene
Blackwall, 1833; Neserigone Eskov, 1992; Nippononeta Eskov, 1992; Nispa Eskov, 1993;
Notiogyne Tanasevitch, 2007; Notioscopus Simon, 1884; Obscuriphantes Saaristo et
Tanasevitch, 2000; Oculocornia Oliger, 1985; Oedothorax Bertkau, 1883; Oia Wunderlich,
1973; Okhotigone Eskov, 1993; Oreoneta Kulczynski, 1894; Oreonetides Strand, 1901;
Oryphantes Hull, 1932; Ostearius Hull, 1911; Pacifiphantes Eskov et Marusik, 1994;
Palliduphantes Saaristo et Tanasevitch, 2001; Panamomops Simon, 1884; Paraeboria
Eskov, 1990; Paraglyphesis Eskov, 1991; Parasisis Eskov, 1984; Parawubanoides
Eskov et Marusik, 1992; Pelecopsis Simon, 1864; Peponocranium Simon, 1884; Perlon-
gipalpus Eskov et Marusik, 1991; Perregrinus Tanasevitch, 1992; Perro Tanasevitch,
1992; Phlattothrata Crosby et Bishop, 1933; Pityohyphantes Simon, 1929; Pocadicnemis
Simon, 1884; Poeciloneta Kulczynski, 1894; Porrhomma Simon, 1884; Praestigia
Millidge, 1954; Procerocymbium Eskov, 1989; Pro islandiana Tanasevitch, \9$5; Pseudo-
cyba Tanasevitch, 1984; Pseudohilaira Eskov, 1990; Pseudoporrhomma Eskov, 1993;
Pseudowubana Eskov et Marusik, 1992; Saaristoa Millidge, 1978; Sachaliphantes
Saaristo et Tanasevitch, 2004; Saloca Simon, 1926; Satilatlas Keyserling, 1886; Sauron
Eskov, 1995; Savignia Blackwall, 1833; Scandichrestus Wunderlich, 1995; Sciastes
Bishop et Crosby, 1938; Scotargus Simon, 1913; Scotinotylus Simon, 1884; Semljicola
Strand, 1906; Sibirocyba Eskov et Marusik, 1994; Silometopoides Eskov, 1990; Silo-
metopus Simon, 1926; Sisicottus Bishop et Crosby, 1938; Sisicus Bishop et Crosby, 1938;
Stemonyphantes Menge, 1866; Strandella Oi, 1960; Styloctetor Simon, 1884; Tallusia
Lehtinen et Saaristo, 1972; Tanasevitchia Marusik et Saaristo, 1999; Tapinocyba Simon,
1884; Tapinocyboides Wiehle, 1960; Tapinopa Westring, 1851; Taranucnus Simon, 1884;
Tarsiphantes Strand, 1905; Tenuiphantes Saaristo et Tanasevitch, 1996; Tessamoro
Eskov, 1993; Thaleria Tanasevitch, 1984; Theoneta Eskov et Marusik, 1991; Thyreosthe-
nius Simon, 1884; Tibiaster Tanasevitch, 1987; Tibioploides Eskov et Marusik, 1991;
Tibioplus Chamberlin et Ivie, 1947; Tiso Simon, 1884; Tmeticus Menge, 1868; Tremato-
Семейства, встречающиеся в азиатской части России
173
cephalus F. Dahl, 1886; Trichobactrus Wunderlich, 1995; Trichoncus Simon, 1884;
Trichopterna Kulczynski, 1894; Troxochrus Simon, 1884; Tubercithorax Eskov, 1988;
Tunagyna Chamberlin et Ivie, 1933; Tusukuru Eskov, 1993; Typhochrestinus Eskov, 1990;
Typhochrestoides Eskov, 1990; Typhochrestus Simon, 1884; Ummeliata Strand, 1942;
Uralophantes Esyunin, 1992; Ussurigone Eskov, 1993; Uusitaloia Marusik, Koponen et
Danilov, 2001; Vermontia Millidge, 1984; Viktorium Eskov, 1988; Wabasso Millidge,
1984; Walckenaeria Blackwall, 1833; Walekenaerianus Wunderlich, 1995; Wiehlenarius
Eskov, 1990; Wubanoides Eskov, 1986; Yakutopus Eskov, 1990; Zerogone Eskov et
Marusik, 1994; Zornella Jackson, 1932.
Распространение. Ввиду большого числа родов, мы приводим только
некоторые, те, которые распространены на ограниченной территории.
Ainerigone, Dactylopisthoides, Eborilaira, Kolymocyba, Mermessus, Miftengris,
Okhotigone, Phlattothrata, Sisicottus, Tessamoro, Vermnotia, Zerogone —
Северо-Восток и/или Курилы.
Aprifrontalia, Asthenargoides, Crispiphantes, Epibellowia, Eskovina, Herbiphantes,
Himalaphantes, Neserigone, Nippononeta, Nispa, Notiogyne, Oculocornia, Oia, Oste-
arius, Pacifiphantes, Pseudoporrhomma, Sachaliphantes, Strandella, Tusukuru — юг
Дальнего Востока.
Asthenargus, Sauron, Uralophantes — Западная Сибирь.
Наиболее важные работы, посвященные распространению линифиид азиатской
части России: Eskov (1994); Saaristo & Eskov (1996); Marusik et al. (2000); Saaristo
& Marusik (2004a, c); Tanasevitch (2005, 2007a и др.).
Биология. Линифииды — наиболее широко распространённое семейство в
регионе. Они встречаются от пустынь до полярных пустынь (80°с.ш.). Населяют они
все возможные биотопы и стации, от сухих и жарких степей, до крайне холодных
ерниковых болот. Большинство линифиид живут в подстилке, во мху, под
камнями. Много видов встречается в травостое, на кустарниках, в кронах и на стволах
деревьев. Немалая доля является облигатными либо факультативными литобион-
тами. Один вид связан исключительно с затенёнными водотоками (Bathylinyphia
maior (Kulczynski, 1885)). Нередко линифииды образуют чрезвычайно плотные
популяции с концентрацией пауков, доходящей до 2000 особей на квадратный метр!
Это могут быть практически монопопуляции (образованные на 90% одним видом)
или многовидовые популяции. Первый вариант мы (Ю.М.) наблюдали на Чукотке,
в горных тундрах, в подстилке под ивой. Другая (многовидовая) популяция была
отмечена под кедровым стлаником на острове Онекотан.
Исходно все Linyphiidae — тенетники, плетущие пологовидные сети. Паук
находится на нижней стороне сети. Однако устройство сети изучено у небольшого
количества видов и родов. Наиболее хорошо известны сети Linyphia и родственных ей
родов Neriene, Microlinyphia. У них сети трёхмерные и сооружаются на высоте 10-
100 см над землёй; они удобны для наблюдения. Собственно полог состоит из
достаточно густого сплетения нитей, без определённой структуры (в отличие от,
например, кругопрядов). Несколько необычно выглядит полог у Tapinopa. Сеть у этих
пауков двумерная, и располагается чаще всего под камнями над микропонижениями.
Полог очень густо заплетён, так что паука сквозь него не видно. Сеть несколько
блестящая и больше похожа на шелковистое убежище гусениц, нежели на сеть паука.
174
4. Специальная часть
Интересно, что, несмотря на мелкие размеры, Tapinopa longidens (Wider, 1834)
обладает довольно сильным ядом. В течение нескольких минут развивается
паралич мелких мышц рядом с местом укуса (в радиусе 5-10 см), который
прекращается лишь через 3-4 часа (Н.К., собственные данные). При этом сам по себе укус не
вызывает боли и ощущается лишь как слабый укол.
Одни из наиболее необычных сетей — у Bathylinyphia maior. Они
располагаются только в сильно затенённых местах вдоль водотоков. Часть из опорных нитей
(возможно, одновременно они являются и сигнальными) могут уходить под воду!
Очевидно, эти нити закрепляются (приклеиваются), когда уровень воды ниже.
Тенета многих мелких пауков или пауков, устраивающих сети в толще
травостоя, подстилки или осыпей, очень сложно изучать. Для многих эригонин вообще
неизвестно, строят ли они сети или охотятся как бродячие пауки? Известны сети у
пауков рода Erigone. Они хорошо заметны в утреннее время, когда на них оседают
капельки росы. Располагаются они в микропонижениях (это могут быть следы от
копыт) и представляют собой простой полог 5-7 см в диаметре.
Взрослые особи у многих видов встречаются в течение всего года, однако
период, когда практически все виды представлены половозрелыми самками и самцами
приходится в азиатской части России на позднее лето - позднюю осень (с августа
и до выпадения снега). Самцы многих видов в Охотоморье и в верховьях Колымы
были собраны только в сентябре или даже в октябре, после первых снегопадов.
Весной, когда начинает таять снег, самцов этих видов собрать уже невозможно.
Из-за своего небольшого размера многие виды линифиид преадаптированы к
низким температурам (в тонких капиллярах водные растворы замерзают при
существенно более низких температурах, нежели в обычных условиях). Изучая
холодоустойчивость, нам (Ю.М.) приходилось наблюдать Anguliphantes karpinskii
(О. Pickard-Cambridge, 1873), двигающихся при температуре -11 °С. Весной (конец
марта - начало апреля) на приморских склонах в окрестностях Магадана мы
находили десятки активных видов линифиид. Если днём в это время года на почве
была положительная температура, то ночью она опускалась до -20 °С и ниже.
Коконы у линифиид бывают разнообразной формы и окраски (белые, жёлтые,
оранжеватые). Erigone имеют линзовидный кокон, сравнимый с размером
брюшка, и крепящийся к субстрату круглым пологом. У многих видов кокон крупнее, а
полог без определенной формы. Вся конструкция может быть в два раза крупнее
самки. Полог может быть тонким (похожим на бумагу) или «пушистым». В
последнем случае яйца могут быть видны сквозь него. Самки охраняют кокон, и иногда
паука можно «вычислить» благодаря наличию свежего кокона с яйцами.
Методы сбора. Все способы и все биотопы. В укосы попадаются
немногочисленные виды. Наиболее эффективны — просеивание подстилки, в том числе
моховой и лишайниковой, ручной сбор на поверхности почвы среди густого травостоя.
Множество интересных видов встречается среди камней (галечники, осыпи,
отдельно лежащие камни). Некоторые виды можно собрать только ручным сбором в
мочажинах на болотах, вдоль берегов затенённных ручьев.
Таксономия. Рамки (границы) линифиид достаточно ясны, хотя ещё недавно
эта группа рассматривалась как два семейства — Linyphiidae и Micryphantidae
(=Erigonidae) (например, в определителе В.П. Тыщенко (1971)). С другой стороны,
Семейства, встречающиеся в азиатской части России
175
из этого таксона недавно было выделено семейство Pimoidae, и периодически
обсуждается положение Stemonyphantes. Этот таксон уже рассматривается в
отдельном подсемействе Stemonyphantinae Wunderlich, 1986 и, возможно, заслуживает
выделения в отдельное семейство.
В современных границах семейства все исследователи признают два
подсемейства — Linyphiinae Blackwall, 1859 и Erigoninae Emerton, 1882. Помимо этих групп,
выделяются Dubiaraneinae Millidge, 1993; Ipainae Saaristo, 2007; Micronetinae Hull,
1920 (которых некоторые рассматривают как трибу линифиин); Mynogleninae
Lehtinen, 1967. Помимо этих таксонов, выделяют Porrhomminae (Porrhomma, Bathy-
phantes и родственные рода). Внутри как линифиин, так и эригонин существует
ещё немало таксонов, которые могут быть выделены в отдельные подсемейства
или трибы, однако надродовая систематика группы разработана недостаточно
хорошо. В азиатской части России встречаются представители пяти подсемейств:
Erigoninae, Ipainae, Linyphiinae, Micronetinae и Stemonyphantinae.
Среди Erigoninae встречается некоторое число видов с диморфными или даже
полиморфными самцами. Ди- и полиморфизм проявляется только в размере и
форме выроста головогруди у самцов. Примером таких видов может служить Oedotho-
rax gibbosus (Blackwall, 1841), который имеет крупный вырост головогруди.
Другую форму этого вида, с небольшим выростом, долго считали самостоятельным
видом или подвидом — О. tuberosus (Blackwall, 1841). В аналогичном случае, у
европейского вида Pelecopsis mengei (Simon, 1884), предполагается, что форма ка-
рапакса кодируется единственным геном (локусом) с двумя аллелями (Bryja, 2004).
Диморфный также Dactylopisthes video (Chamberlin et Ivie, 1947), а полиморфный —
Hybauchenidium aquilonare (L. Koch, 1879). У самцов последнего вида можно
проследить все переходы от невыраженного выроста головогруди до весьма высокого
(см. Holm, 1973).
Огромный вклад в изучение надвидовой таксономии этого семейства,
особенно в Северной Палеарктике, внёс К.Ю. Еськов. Когда он начал исследование лини-
фиид Сибири и Дальнего Востока, он столкнулся с сотнями новых видов,
большую часть из которых невозможно было отнести ни к одному из известных родов.
Примерно за 15 лет он описал 55 родов и 156 видов (четверть от всех видов из
азиатской части России). Огромный вклад внёс и вносит А.В. Танасевич. Им
описано из азиатской части России более 100 видов и 25 родов.
Несмотря на большое число статей, посвященных таксономии линифиид
азиатской части России, систематика этой группы далека от удовлетворительной.
Многие крупные явно полифилетичные рода до сих пор не ревизованы, например,
Centromerus, Hilaira, Lepthyphantes, Maw, Oreonetides. В регионе имеются многие
десятки неописанных видов.
Поскольку работ по систематике Linyphiidae азиатской части России
чрезвычайно много, мы перечислим только самые важные на наш взгляд: L. Koch, 1879a-
b; Kulczynski, 1885, 1908, 1926; Ermolajew, 1937a-b; Holm, 1970, 1973; Helsdingen,
1968, 1974; Еськов, 1979, 1980а-б, 1981а-б, 1984а-б, 1986, 1987а-б, 1988а-с, 1989,
1990,1991a-6;Eskov, 1986a-b, 1988,1989,1990a-b, 1991a-b; 1992,1993,1994;Eskov
& Marusik, 1991, 1992a-d; 1994, 1997; Efimik & Esyunin, 1996; Марусик, 19886,
1991 в; Marusik & Leech, 1993; Marusik et al., 1996,2001,2002; Marusik & Tanasevitch,
1998, 2003; Marusik & Saaristo, 1999; Marusik & Koponen, 2001b, 2002, 2009, 2010;
Saaristo & Eskov, 1996; Saaristo & Marusik, 2004a, с; Танасевич, 1984а-б; Танасе-
176
4. Специальная часть
вич, Еськов, 1987; Tanasevitch, 1986, 1987, 1988, 1989, 1992, 1993, 1995, 1996a-b,
2000a-b, 2005, 2006а-с, 2007a-b, 2008; Thaler et al., 1994; Saaristo & Tanasevitch,
2000; Tanasevitch & Trilikauskas, 2006. Несомненно, наиболее важная из всех
перечисленных работ — каталог линифиид северной Азии (Eskov, 1994).
Из работ по сопредельным территориям, важных для изучения сибирских и
дальневосточных Linyphiidae, можно отметить обзоры финских Linyphiidae (Palmgren,
1975, 1976), работу J. Wunderlich (1995a) по линифиидам Монголии, серию работ
A.F. Millidge (1977, 1981а—с? 1983, 1984) по линифиидам вообще и по Erigoninae
Северной Америки, в особенности. Для определения пауков Сахалина, Курил, и
Приморья важное значение имеют публикации японских авторов (Oi, 1960; Опо &
Saito, 2001; Saito, 1977, 1982a-b, 1983a-b, 1984, 1986, 1987, 1992, 1994 и др.; Saito
& Опо, 2001; Опо et al., 2009). Последнее время всё больше выходит детальных
ревизий китайских авторов, выполненных под руководством S.Q. Li (Tu & Li, 2003,
2004a-b, 2005a-b).
В настоящее время эта группа продолжает активно разрабатываться А.В. Тана-
севичем, В.А. Гнелицей, Ю.М. Марусиком, S. Koponen, D.J. Buckle, L. Tu и S. Li.
Разграничение видов. Подавляющее большинство видов различается формой
копулятивных органов, однако в некоторых случаях виды могут различаться
размером глаз, расстоянием между ними, формой и вооружением хелицер. Самцы
многих видов эригонин хорошо различаются формой карапакса.
Перспективы изучения. Linyphiidae, пожалуй, самое перспективное семейство
в плане изучения систематики и зоогеографии — здесь как нигде много новых
таксонов и нерешённых проблем. До сих пор с десяток видов остаются
известными по одному полу. Кроме того, есть немало родов, известных только по первоопи-
саниям, не всегда снабжённым качественными рисунками. Чрезвычайно слабо ли-
нифииды изучены в плане их биологии. Устройство сетей у подавляющего
большинства таксонов в азиатской части России неизвестно, равно как и у
большинства видов Европы и Северной Америки. Актуальность изучения сетей именно в
Сибири и на Дальнем Востоке связана с тем, что здесь обитает немало
эндемичных таксонов или же групп, которые в Европе и Северной Америке обитают
только в малодоступных местах.
Семейства, встречающиеся в азиатской части России
\11
Tapinopa longidens (Linyphiidae), самка, Крым (фото: Н.М. Ковблюк).
Семейство Liocranidae Simon, 1897
Лиокраниды; spiny-legged sac spiders
Рис. 21.1-6.
Небольшое, всесветно распространённое семейство со 170 видами и 29
родами. В России зарегистрировано 4 рода и 11 видов, а в азиатской части России 2
рода и 9 видов.
Диагностические признаки. Энтелегинные, некрибеллятные пауки с 2
коготками, 8 глаз в два ряда, 4-9 мм, голени и предлапки I и II с многочисленными
длинными шипами, пальпа самца с заострённым на вершине РГО, бульбус с
медиальным отростком.
Близкие группы. Внешне (габитуально и окраской) несколько похожи на
некоторых Clubiona, однако отличаются заострённым РГО, не расчленённым на части,
и высокой эпигиной с длинной продольной щелью (у Clubiona — 2 отверстия,
поперечная щель, или ямка).
ОПИСАНИЕ:
• Карапакс. Грушевидный.
• Стернум. Щитовидный.
• Глаза. 8 в два ряда (4-4), примерно одинаковой величины.
• Ротовой аппарат. Не модифицирован.
• Ноги. 2 коготка, ноги удлинённые, передние 2 пары ног с многочисленными
парными вентральными шипами на голенях и предлапках.
• Брюшко. Удлинённо-овальное.
• Паутинные бородавки. Не модифицированы, передние несколько
удлинены, цилиндрические.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Пальпа самца с РГО, бульбус Agroeca всегда с
медиальным отростком характерной формы, эпигина удлинённая.
• Размер. 4-9 мм, половой диморфизм выражен слабо.
• Окраска. Желтовато-коричневая, с рисунком или без.
• Особые черты. Ноги I—II с двумя рядами многочисленных вентральных
шипов. Кончик эмболюса иногда загнут, и образует петлю, похожую по форме
на «8».
Рода, известные из азиатской части России. Agroeca Westring, 1861 и 1Ыо-
cranum L. Koch, 1866.
Распространение. Agroeca — весь регион, кроме тундровой зоны; Ыосгапит —
западная Сибирь.
Биология1. Все виды, встречающиеся в азиатской части России — обитатели
подстилки. Некоторые виды Agroeca встречаются в лиственной подстилке, другие
обитают среди каменных осыпей.
1 Раздел подготовлен при участи Э.Ф. Гусейнова.
Семейства, встречающиеся в азиатской части России
179
Охотничье поведение и пища совершенно не изучены.
В отличие от других пауков со слабым зрением, для Agroeca характерно
сложное брачное поведение. Самцы, встретившие самок, исполняют сложный «танец»,
включающий резкие колебательные движения, поочерёдное поднятие и опускание
ног, а также очень быстрое вибрирование ногами (Bristowe, 1958).
Коконы очень характерной бокаловидной формы, и по их наличию можно
легко установить присутствие в биотопе видов Agroeca. Если кокон отложен под
камнем, то он имеет вид перевёрнутого бокала. Ножка бокала бывает очень длинной.
Иногда «бокал» маскируется налепленной на него грязью.
Методы сбора. Почвенные ловушки, ручной сбор в подстилке и среди камней.
Таксономия. Ранее пауки этого семейства рассматривались как подсемейство
в Clubionidae. Статус семейства был придан этому таксону П.Т. Лехтиненом (Leh-
tinen, 1967). До недавнего времени в состав лиокранид включали Phrurolithus,
которого сейчас относят к Corinnidae. В определителе В.П. Тыщенко (1971)
Liocranidae рассматривались в Clubionidae. Й. Вундерлих (Wunderlich, 2008c)
рассматривает эту группу как подсемейство в Zoridae. Реальное число видов в Liocranidae
ниже, чем указано в Platnick (2009), поскольку Н.И. Платник засчитал 5 родов с 44
видами, которые были перенесены из Liocranidae во Phrurolithinae/Corinnidae в
1 3
4
6
Рис. 21. Liocranidae. Agroeca cf. mongolica (cT, 1, 5, из Приморья), Agroeca sp. (?, 2, из
Тувы), A. ornata (С?, 3-4) и кокон Agroeca sp. (6).
1 — габитус сГ, сверху; 2 — эпигина, снизу; 3-4 — пальпа d\ снизу и сбоку; 5 — паутинные
бородавки, снизу; 6 — харктерная форма кокона. Стрелка указывает на медиальный отросток
характерной формы. 3-4 — по: Paquin & Duperre (2003); 6 — по: Roberts (1995).
180
4. Специальная часть
работе Deeleman-Reinhold (2001), только на том основании, что не был проведён
кладистический анализ семейства.
Liocranidae азиатской части России рассматривались в работах L. Koch (1879a);
Holm (1973); Михайлов, Марусик (1995); Danilov (1999).
Разграничение видов. Форма копулятивных органов.
Перспективы изучения. По всей видимости, в регионе встречается больше
видов рода Agroeca, часть из них, очевидно, новые для науки. Род Agroeca, его
рамки, нуждаются в пересмотре. Возможно, он будет разбит на несколько
таксонов (родов или подродов).
В настоящее время эта группа разрабатывается С.Л. Есюниным, Ю.М. Маруси-
ком и К.Г. Михайловым.
Agroeca brunnea (Liocranidae), кокон, Финляндия (фото:
N. Fritzen).
Семейства, встречающиеся в азиатской части России
181
Agroeca brunnea (Liocranidae), самец, Финляндия (фото: N. Fritzen).
Семейство Lycosidae Sundevall, 1833
Пауки-волки; wolf spiders
Рис. 22.1-21.
Всесветно распространённое семейство, четвёртое по числу видов (2339), где в
настоящее время выделяют 110 родов. В России известно, по меньшей мере, 220
видов, а в азиатской части России обитает не менее 205 видов из 16 родов.
Диагностические признаки. Энтелегинные, некрибеллятные, 3-коготковые
пауки, размер 4-30 мм; глаз 8, расположены в 3 ряда (4-2-2), задние две пары
крупнее передних в 2 и более раз, наличник низкий (меньше или чуть больше передних
медиальных глаз); пальпа самца без РГО, медиальный отросток всегда имеется.
Близкие группы. Внешне (габитуально и окраской) похожи на Zora (Zoridae),
Anahita (Ctenidae) и Pisauridae. От первых пауки-волки отличаются большим
размерным диморфизмом глаз и 3 коготками. От вторых — формой глаз, а от Pisauridae
гораздо более низким наличником. Самцы пауков-волков легко отличаются от
внешне схожих групп отсутствием РГО.
ОПИСАНИЕ:
• Карапакс. Грушевидный, головная часть уже и обычно чуть выше грудной,
медиальная бороздка всегда есть, удлинённая. Карапакс покрыт густыми
волосками. Наличник низкий (не больше 2 диаметров передних глаз).
• Стернум. Щитовидный.
• Глаза. 8, 3 ряда (4-2-2); глаза первого ряда — маленькие, расположены в
прямой ряд спереди карапакса; глаза второго и третьего рядов — очень
крупные; третья пара глаз расположена сверху карапакса; задние два ряда (2
глаза второго ряда и 2 глаза третьего ряда) образуют квадрат или трапецию.
• Ротовой аппарат. Не модифицирован. Хелицеры у крупных
представителей могут быть с латеральным мыщелком, который часто бывает красного
цвета.
• Ноги. 3 коготка, со скопулой или без, опушены, с мощными шипами,
вентральных шипов на голени I от 2 до 6 пар. Нередко у самцов передние ноги с
длинными волосками на голени, предлапке и лапке.
• Брюшко. Овальное; у самцов иногда со специализированными волосками-
шипиками на нижней стороне.
• Паутинные бородавки. Слегка удлинены и поэтому цилиндрические.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Пальпа самца без РГО; цимбиум каплевидный,
обычно шире голени, но иногда равен по ширине голени; вершина цимбиу-
ма бывает без шипов или с 1-2 или более шипами или мощными
щетинками. Субтегулюм всегда заметен, округлой формы, тегулярный (медиальный)
отросток всегда имеется, иногда смещён ретролатерально {Evippa, Xeroly-
cosa). Эмболюс лежит поперечно (исключение — Evippa, Xerolycosa, у
которых эмболюс кольцевидный), частично скрыт тегулярным отростком. Эпи-
гина разнообразной формы, чаще всего с ямкой и септумом (срединной пла-
Семейства, встречающиеся в азиатской части России
183
Рис. 22. Lycosidae. Pardosa pseudomixta (О71,1), Alopecosa cuneata (d71?, 2,5-6), Xervlycosa
mongolica (?, 3), Pardosa glacialis (?, 4), P. hyperborea (?, 7), Pfalcata (?, 8) и Mustelicosa
dimidiata (d71, 9).
1 — головогрудь сГ, спереди; 2-3 — эпигина, снизу; 4-9 — габитус, сверху.
184
4. Специальная часть
стинкой, разделяющей эпигину на две части), часто имеется верхний
карман.
• Размер. 4-30 мм, диморфизм слабый, выражается в меньшем размере
брюшка у самца.
• Окраска. От светло-коричневой до практически чёрной, обычно с
рисунком на карапаксе и брюшке, но бывает рисунок и на стернуме.
• Особые черты. Нет РГО, формула глаз, низкий наличник; самки носят
кокон, прикрепив его к паутинным бородавкам.
Рода, известные из азиатской части России. Acantholycosa F. Dahl, 1908;
Allohogna Roewer, 1955; Alopecosa Simon, 1885; Arctosa C.L. Koch, 1847; Evippa
Simon, 1882 (E. sibirica Marusik, 1995); Hygrolycosa F. Dahl, 1908 (Я. rubrofasciata
(Ohlert, 1865)); Ishicosa Roewer, I9601 (7. ishikariana (S. Saito, 1934)); Mustelicosa
Roewer, 1960 (M. dimidiata (Thorell, 1875)); Mongolicosa Marusik, Azarkina &
Koponen, 2004; Pardosa C.L. Koch, 1847; Pirata Sundevall, 1832; Sibirocosa Marusik,
Azarkina et Koponen, 2004; Tricca Simon, 1889; Triccosta Roewer, 1960 (T ipsa (Karsch,
1879)); Trochosa C.L. Koch, \S47;Xerolycosa F. Dahl, 1908.
Распространение.
Alopecosa, Pardosa, (<Tricca " — весь регион и все зоны.
Acantholycosa, Pirata, Xerolycosa, Trochosa — весь регион без тундровой зоны.
Sibirocosa — весь регион к северу от 59° с.ш. и Приморье.
Hygrolycosa — на восток до Енисея.
Allohogna, Mongolicosa, Mustelicosa — от Алтая до Забайкалья.
Evippa — Алтай и Тува.
Arctosa — юг Сибири (один вид доходит до 62° с.ш. в приенисейском регионе)
и Дальнего Востока.
Triccosta — Приморье.
Ishicosa — южные Курилы.
Наиболее важные работы (где приведены карты распространения видов),
посвященные распространению пауков-волков азиатской части России: Marusik et al.
(2000, 2004).
1 Формально рассматривается как синоним Lycosa Latreille, 1804, однако по строению копу-
лятивных органов эти рода существенно отличаются.
Рис, 22 (продолжение). Lycosidae. Копулятивные органы и нога I: Mustelicosa dimidiata
(сГ$, 10,13), Pardosa lugubris (d71,1 \), Acantholycosa sayanensis (С?, 12,15,21), Sibirocosa
sibirica (?, 14), Mongolicosa buryatica (?, 16), Tricca insignita ($, 17), Mongolicosa glupovi
($, 18), Acantholycosa altaiensis ($, 19) и Pardosa selengensis (d71, 20).
10-12 — пальпа С?,снизу; 13-14, 17-19 — эпигина, снизу; 15 — эмболюсный отдел, снизу; 16 —
эпигина, сверху; 20 — голень и предлапка I с густыми длинными волосками, сверху; 21 — пальпа
С?, сбоку. Кэ — карман эпигины; Мо — медиальный отросток; Па — палеа; Ре — рецептакула;
Се — септум; Су — субтегулюм; Эм — эмболюс;Яэ — ямка эпигины. 12, 14-16, 18-19,21 —по:
Marusik et al. (2004); 10, 13 — по: Marusik & Buchar (2004).
Семейства, встречающиеся в азиатской части России
185
10
11
12
А)
1
17
18 19
186
4. Специальная часть
Биология.
Населяют пауки-волки самый широкий спектр биотопов, встречаются и в
пустынях, и на морских пляжах, на пастбищах, лугах, по берегам рек, a Pirata может
даже легко передвигаться по воде. Длинноногие пауки родов Acantholycosa, Sibiro-
cosd, Mongolicosa населяют исключительно каменные осыпи. Только один вид Раг-
dosa lasciva L. Koch, 1879 встречается часто на стволах деревьев. Избегают пауки-
волки лишь сильно затенённые места.
В целом, пауки-волки — неспециализированные хищники. Часть видов
охотится исключительно днём (все Pardosa), другие — например, норные — только
ночью. Ночные виды могут встречаться и в гуще леса.
Все пауки-волки, населяющие Сибирь и Дальний Восток России, являются либо
бродячими, либо норными. Норных пауков-волков в регионе немного, это виды
Allohogna, Mustelicosa, "Lycosa", Xerolycosa mongolica (Schenkel, 1963). Среди
пауков-волков есть и сетеплетущие группы. Они встречаются в субтропических и
тропических районах мира.
Самки некоторых бродячих видов (Alopecosa, Arctosa, Trochosa) откладывают
коконы в особые убежища (это могут быть пустоты под камнями, либо небольшие
норки) и после откладки переходят к оседлому образу жизни, покуда не выведутся
паучата. Все виды Pardosa, Acantholycosa, Sibirocosa, Mongolicosa и некоторые Alo-
pecosa вынашивают кокон, прикрепив его к паутинным бородавкам.
Жизненный цикл у многих видов пауков-волков одногодичный. Исключение —
крупные норные пауки, и арктические виды. У некоторых наиболее северных
популяций Pardosa и Alopecosa каждая стадия (период между двумя линьками)
длится один год.
У пауков-волков сильно развито предкопуляционное поведение ухаживания,
когда самцы размахивают ногами, стучат по субстрату брюшком, педипальпами, или
даже стернумом. Кроме того, самцы некоторых видов выделяют пахучие
вещества, секретируемые первой парой ног или двумя первыми парами ног.
Форма танца видоспецифична. Некоторые виды первоначально были
выделены благодаря различиям в форме танца, и лишь затем были найдены морофологи-
ческие отличия. Благодаря удобству разведения пауков-волков, многие
исследователи используют их для изучения полового поведения (стратегии самцов, как
самки выбирают партнёра для спаривания, и многие другие аспекты).
У всех видов пауков-волков наблюдается забота о потомстве. В самом простом
случае забота сводится к охране кокона в укромном месте. При этом кокон обычно
белый и сферический. В большинстве случаев это не только охрана, так как
норные виды периодически выставляют кокон из отверстия норки, чтобы он лучше
прогрелся.
У тех видов, которые постоянно таскают кокон (линзовидный или
сферический, окрашенный или белый), отмечено, что самки могут открывать кокон и
изливать на яйца жидкость из своего ротового аппарата, после чего кокон снова
запечатывается. Наблюдались случаи, когда коконы временно погружались в воду. В
верховьях Колымы самки Pardosa algens Kulczynski, 1908 часто бегают по
поверхности наледей вместе с коконами. При этом брюшко задирается больше обычного.
Самка помогает детёнышам выбираться из кокона. Паучата сразу же
забираются на брюшко матери, и самки «убежищных» видов вновь переходят к бродячему
образу жизни, и носят на брюшке своё потомство первые несколько дней или даже
недели.
Семейства, встречающиеся в азиатской части России
187
Методы сбора. Наиболее эффективный способ — почвенные ловушки. Много
пауков можно собрать и вручную. Некоторые виды можно собрать только
вручную, например, среди осыпей, или на болотинах. Pirata piraticus (Clerck, 1758)
можно собрать троплением (ступая по болотине и надавливая ногой так, чтобы
выступала вода — тогда пауки бегут и становятся заметными). Норных пауков,
особенно самок, можно собрать только с помощью раскопок нор.
Таксономия. Это одно из наиболее чётко очерченных семейств. Тем не менее,
надвидовая классификация разработана чрезвычайно слабо и сильно запутана.
Сложность надвидовой классификации во многом связана с тем, что это
чрезвычайно однообразная группа (в соматическом плане). Предпринималось две
попытки в глобальном масштабе ревизовать надвидовые таксоны этого семейства.
Первая была предпринята C.F. Roewer (1959, 1960). Благодаря его работам число
родов увеличилось почти в два раза (с 69 до 131). Однако диагностика родов
строилась на величине и взаимном расположении глаз, числе зубчиков на хелицерах и
других соматических признаках. При этом некоторые количественные признаки
были неверно посчитаны. В результате этих работ получились искусственные рода,
например, распространённые в Зимбабве и на Ямале, или на Камчатке и в ЮАР.
Вторая попытка была ещё более неуклюжей (Guy, 1966). Эта попытка свелась лишь
к синонимизации (без просмотра материала) и понижению статуса некоторых
родов до подродов.
В 1980-1990-е гг. А.А. Зюзиным (Зюзин, 1985; Zyuzin, 1993) было продолжено
изучение надвидвой систематики пауков-волков. Им было описано два новых
подсемейства, и разработаны некоторые критерии для классификации. Большая
заслуга А.А. Зюзина состоит в расшифровке адаптивного значения некоторых
соматических признаков, используемых для классификации ликозид (Zyuzin, 1990). В
частности, ему удалось выяснить, что некоторые роющие пауки-волки отличаются
от не роющих формой карапакса (с поднятым головным отделом, но резко
уплощённым грудным отделом, что даёт возможность плотно прижимать ноги IV к
головогруди и не цепляться ими за стенки норки), наличием мощных щетинок или
шипов по краю карапакса, наличием зубцов или мощных щетинок на кончике цим-
биума (служат у неполовозрелых самцов для выкапывания норок), мощной скопу-
лой или щетинками на лапках I—II. Несмотря на то, что в идеях и подходах А.А.
Зюзина очень много нового и ценного, он лишь частично отошёл от Рёвера. Для
диагностики подсемейств, триб и видов он использовал признаки копулятивных
органов, а для диагностики подтриб и родов — исключительно соматические.
Деление семейства на подсемейства далеко от совершенства. В азиатской части
России обитают представители трёх подсемейств — Evippinae, Lycosinae и Pardo-
sinae.
На наш взгляд, большинство родов в азиатской части России (кроме недавно
ревизованных), прежде всего Pardosa, Alopecosa и Arctosa, заслуживают
разделения на многие рода. Практически все группы видов Pardosa, выделенные А.А.
Зюзиным (1979а), вполне заслуживают статуса отдельных родов. Причём сам А.А.
Зюзин отнёс одну из групп видов Pardosa не только в отдельный род Wadicosa
Zyuzin, 1985, но и в отдельное подсемейство.
К наиболее важным работам по паукам-волкам азиатской части России
относятся следующие публикации: Grube (1861); L. Koch (1879a-b); Kulczynski (1885,
188
4. Специальная часть
1907, 1908); Odenwall (1901); Holm (1970, 1973); Зюзин (1979а-б); Штернберге
(1979); Kronestedt (1986, 1990); Зюзин, Марусик (1988, 1989); Логунов (19926);
Logunov & Marusik (1995); Kronestedt & Marusik (2002); Marusik & Logunov (2002a,
2009); Marusik et al. (2004, 2010); Omelko & Marusik (2008).
Среди важных работ по сопредельным территориям можно отметить работы
Chikuni (1989); Eskov & Marusik (1995); Song et al. (1999) и серию работ Н. Tanaka
по паукам-волкам Японии.
Разграничение видов. В большинстве случаев виды различаются формой
отдельных деталей копулятивных органов. Однако нередко самцы близких видов легче
различаются другими признаками: 1) толщиной ног; 2) опушением ног; 3) шипова-
нием ног; 4) окраской ног; 5) окраской карапакса и брюшка; 6) окраской и
вооружением разных сегментов педипальпы, особенно цимбиума и голени; 7) коготками
цимбиума (числом коготков); 8) наличием особых волосков на нижней стороне
брюшка. В некоторых случаях самок близких видов можно различать шипованием
ног (особенно ноги I).
Сложности в разграничении видов связаны с тем, что у пауков-волков развито
поведение ухаживания. При распознании своего вида большую роль играет не
форма бульбуса или эпигины, а форма танца, частота ударов о субстрат (педипальпа-
ми, или брюшком, или стернумом), запахи.
Перспективы изучения. В регионе имеется немало новых, ещё не описанных
видов и родов. Пауки-волки — одна из наименее разработанных групп в мире, и
поэтому наиболее перспективная для изучения и в рамках азиатской части России,
и в более широких масштабах.
В настоящее время эта группа разрабатывается Ю.М. Марусиком, Г.Н. Азарки-
ной, Д.В. Логуновым, S. Коропеп, Т. Kronestedt и М.М. Омелько.
Семейства, встречающиеся в азиатской части России \ 89
Alopecosa tanakai (Lycosidae), самец, Приморье (фото: М.М. Омелько).
Pardosa sp. (Lycosidae), самец, Приморье (фото: М.М. Омелько).
Семейство Mimetidae Simon, 1881
Миметиды; pirate spiders
Рис. 23.1-7.
Небольшое, всесветно распространённое семейство со 153 видами и 13
родами. В России зарегистрировано 8 видов из 3 родов, из них в азиатской части
России встречается 4 вида рода Его C.L. Koch, 1837.
Диагностические признаки. Небольшие (3-6 мм), некрибеллятные, энтеле-
гинные 3-коготковые пауки, с высоким наличником, с рисунком на карапаксе и
брюшке; брюшко сферическое с парой бугорков; передние две пары ног с пролате-
ральным рядом из чередующихся крупных и мелких шипов.
Близкие группы. Внешне (габитуально и окраской) напоминают некоторых
Araneidae, Theridiidae, Linyphiidae, однако легко отличаются хватательными
шипами на передних ногах.
ОПИСАНИЕ:
• Карапакс. Яйцевидный, высокий, с рисунком, клипеус высокий.
• Стернум. Щитовидный, длина больше ширины.
• Глаза. 8, два ряда (4-4), латеральные сближены между собой, передние
медиальные глаза крупнее других.
• Ротовой аппарат. Хелицеры удлинены, с коротким коготком, примерно
равным ширине хелицеры; без зубчиков, но с гвоздевидными шипиками
(видоизменённые волоски).
• Ноги. 3 коготка. Длинные; передние ноги почти в 2 раза длиннее тела;
передние 2 пары ног с многочисленными хватательными шипами (ряд из
длинных шипов, в промежутках между которыми располагаются короткие шипы).
• Брюшко. Сферическое или яйцевидное, с бугорками, налегает на карапакс.
• Паутинные бородавки. Не модифицированы, колюлюс есть.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Пальпа самца с массивным, сильно хитинизиро-
ванным парацимбиумом в основании цимбиума; парацимбиум не
обособлен от цимбиума; бульбус сферический. Эпигина слабо выступающая, без
ямки, с септумом.
• Размер. 3-6 мм.
• Окраска. Карапакс всегда с рисунком — тёмной медиальной полосой, иногда
выраженной только в грудной части, и иногда имеются латеральные тёмные
полосы. Брюшко с нечётким рисунком.
• Особые черты. Ряд чередующихся длинных и коротких шипов на передней
поверхности голеней и предлапок I и П.
Рода, известные из азиатской части России. Его C.L. Koch, 1837.
Распространение. Весь регион без Якутии и крайнего Северо-Востока.
Семейства, встречающиеся в азиатской части России
191
Рис. 23. Mimetidae. Его sp. (сГ, 1-2 из Тувы), Его sp. (?, 3—4 из Приморья) и £. cambridgei
(5-7).
1 — головогрудь сГ, спереди; 2, 4 — нога I сГ$, сверху; 3 — габитус ?, сбоку; 5 — пальпа сГ,
сбоку; 6 — эпигина, снизу; 7 — характерная форма кокона у Его sp. 5-7 — по: Roberts (1995).
Биология. Все виды - облигатные аранеофаги (питаются только другими
пауками), что отражено в английском названии семейства. Добычей разных видов ми-
метид служат представители семейств Agelenidae, Araneidae, Dictynidae, Linyphiidae,
Philodromidae, Tetragnathidae, Theridiidae и даже другие Mimetidae!
Преимущественно миметиды специализируются на тенетных пауках. Забираясь в сеть
потенциальной добычи, миметиды имитируют трепыхание жертв или специальные
«свадебные» сигналы самцов, чтобы подманить хозяина сети поближе к себе. Ряд
длинных и коротких шипов на передних ногах служит для прочного захвата жертвы.
После захвата следует укус. Яд действует очень быстро, и жертва погибает почти
мгновенно. Высасывают жертв через небольшие отверстия, проделанные в ногах.
Коконы у Его характерной формы. Они висят на длинной нити (длина нити
больше диаметра кокона в 3 и более раз). Сам кокон сферический и несколько
каплевидный, светло-коричневый, его поверхность покрыта спиралевидными
толстыми нитями.
Установить присутствие миметид по наличию коконов легче, чем найти самих
этих пауков.
192
4. Специальная часть
Методы сбора. Только ручной сбор среди травостоя. Единичные экземпляры
попадаются при кошении и в почвенные ловушки.
Таксономия. Со времён Э. Симона (Simon, 1893) все исследователи сближают
это семейство с Araneidae и родственными группами (надсемейство Araneoidea).
Хотя они не плетут сетей (в отличие от большинства аранеоидных групп), габиту-
ально Mimetidae очень напоминают Araneidae и некоторых Theridiidae, а
строением пальпы и эпигины напоминают Linyphiidae. Тем не менее, Forster & Platnick
(1984) включили это семейство в состав надсемейства Palpimanoidea на
основании того, что они не плетут сетей, аранеофаги, а также имеют гвоздевидные (peg)
зубчики (=щетинки). Ни соматически, ни по строению копулятивных органов
Mimetidae ни в малейшей степени не напоминают Palpimanidae. Случай с включением
Mimetidae в Palpimanoidea можно было бы считать курьёзом, если бы не авторитет
Р. Форстера и Н. Платника, и то, что многие американские арахнологи до сих пор
придерживаются их точки зрения1.
Судя по всему, два вида, встречающиеся в Туве и на Дальнем Востоке, не
относятся к Его. Таксономических работ, касающихся миметид азиатской части
России, нет.
Разграничение видов. Виды различаются как формой копулятивных органов,
прежде всего формой парацимбиума и средней части эпигины, так и окраской.
Перспективы изучения. В южной Сибири и на юге Дальнего Востока имеется
несколько новых видов, по всей видимости, относящихся к неописанным родам.
1 На Международном арахнологическом конгрессе в 2007 г. и американские арахнологи
согласились, что Mimetidae следует рассматривать как аранеоидное семейство.
Семейства, встречающиеся в азиатской части России
193
Pisaura mirabilis (Pisauridae), гнездо с недавно вылупившейся молодью, Крым (фото:
Н.М. Ковблюк).
Семейство Mysmenidae Petrunkevitch, 1928
Мизмениды; spurred orb-weavers
Рис. 24.1-5.
Небольшое всесветно распространённое семейство со 107 видами и 21 родом.
В России известны всего два вида. В азиатской части России отмечен один вид.
Диагностические признаки. Мелкие, меньше 2 мм, 3-коготковые энтелегин-
ные пауки, с 8 глазами, головогрудь очень высокая, высота её почти равна длине (у
самок) или превышает длину (у самцов), у самцов голень и предлапка I с мощным
пролатеральным шипом, а бедро I и II самки с круглой склеротизованной бляшкой
в верхней части. Кругопряды.
Близкие группы. Близки и похожи на Theridiosomatidae, напоминают
некоторых Theridion со сферическим брюшком. Отличаются очень мелким размером,
окраской, наличием скапуса эпигины, тёмными склеротизированными бляшками на
бёдрах I и II у самки и мощными шипами на ноге I у самца.
ОПИСАНИЕ:
• Карапакс. Очень высокий, высота примерно равна или даже превышает
длину, клипеус высокий.
• Стернум. Не модифицирован.
• Глаза. 8 глаз, расположены компактной группой, боковые глаза
соприкасаются.
• Ротовой аппарат. Хелицеры относительно небольшие, меньше высоты
наличника.
• Ноги. 3 коготка. Относительно короткие, с немногочисленными шипами;
самки имеют небольшие округлые тёмные бляшки на вентральной стороне
бёдер I и И, а самцы — мощные пролатеральные шипы на предлапке и
голени I.
• Брюшко. Сферическое, высота больше длины.
• Паутинные бородавки. Не модифицированы, колюлюс имеется.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Энтелегинные. Пальпа самца крупная, цимбиум
подразделён на две части, эмболюс длинный. Эпигина слабо склеротизова-
на, со скапусом. Самку при беглом осмотре можно принять за
неполовозрелую из-за того, что эпигина едва заметна при виде сбоку.
• Размер. Менее 2 мм. Половой диморфизм выражен.
• Окраска. Темно окрашенные; брюшко с 4 парами белых пятен сверху и со
светлым призмовидным пятном сзади.
• Особые черты. Высокий карапакс, шипы на голени и предлапке ноги I у
самца, склеротизованная бляшка на бедре I и II у самки.
Рода, известные из азиатской части России. Mysmenella Brignoli, 1980 (М. cf.
yoW(Kraus, 1967)).
Семейства, встречающиеся в азиатской части России
195
1
Рис. 24. Mysmenidae. Mysemenella sp. (сГ?) с острова Монерон.
1,5 — габитус $ и d\ сверху; 2 — $, спереди; 3 — ?, снизу; 4 — сГ, сбоку. Стрелки показывают
хитинизированные бляшки на бёдрах I и II $, и шипы на голени и предлапке I d\
Распространение. Единственный представитель семейства известен от
Хабаровска на восток до Итурупа, на север до средней части Сахалина
(неопубликованные данные).
Биология. Многочисленные экземпляры были собраны в лесной и луговой
подстилке. Половозрелые самцы редки в июле-сентябре. На Кавказе самцы близкого
вида встречались массово в мае и июне.
Тенетник-кругопряд. CtrnMysmenella не изучены. Представители близкого рода
Mysmena строят, пожалуй, самые сложные и необычные сети среди всех пауков.
От центра сети отходят многочисленные радиальные нити паутины во всех трёх
измерениях. Между ними протянуты клейкие «спиральные» нити. Таким образом,
типичная двухмерная колесовидная сеть пауков-кругопрядов видоизменяется в
трёхмерную шаровидную (Eberhard, 1987; Miller et al., 2009). Аналогичные сети плетут
и представители северо-американских родов мизменид (Ubick et al., 2005). Сети
очень маленькие, как впрочем, и их хозяева. Функционирование сетей при ловле
добычи, поведение пауков-хозяев, а также причины столь радикальной
модификации структуры неизвестны.
196
4. Специальная часть
Методы сбора. Только ручное просеивание подстилки, ручной сбор среди
высокого травостоя, термофотоэклектор. Пауки очень мелкие; заметить их можно
только если близко склониться над грунтом или лёжа на земле.
Таксономия. Семейство никогда не ревизовалось в широких масштабах.
Первоначально этот таксон носил ранг, аналогичный трибе (Simon, 1922).
Позже Petrunkevitch (1928) поднял его до ранга подсемейства в семействе Theridiidae.
В. Гертч (Gertsch, 1960) перенёс подсемейство Mysmeninae в Symphytognathidae. A
затем этому таксону был придан ранг семейства (Forster & Platnick, 1977).
В пределах России это семейство было указано только недавно на Дальнем
Востоке (Marusik & Koponen, 2000) и на северном Кавказе (Marusik, 2005). В этих же
работах затрагивался статус дальневосточной популяции Mysmenella.
Перспективы изучения. Нахождение дополнительных видов маловероятно.
Thomisus onustus (Thomisidae), самка, Крым (фото: Г.А.
Прокопов).
Семейства, встречающиеся в азиатской части России
197
Ebrechtella tricuspidata (Thomisidae), самка, Приморье (фото: М.М. Омелвко).
Семейство Nesticidae Simon, 1894
Нестициды; cave cobweb spiders
Рис. 25.1-5.
В мире известно 206 рецентных видов, принадлежащих к 9 родам. В России
отмечено 7 видов из 4 родов. В азиатской части России обитают два вида из рода
Howaia, и новый неописанный род.
Диагностические признаки. Мелкие, до 5 мм, жёлтые с рисунком на брюшке;
лапка IV вентрально с пильчатыми волосками; лапки с 3 коготками; пальпа самца
с крупным парацимбиумом, далеко отстоящим от цимбиума, эпигина простая с
вертикально удлинёнными просвечивающими рецептакулами. Обитают в подстилке
и каменных осыпях, тенетники.
Близкие группы. Близки и напоминают Theridiidae, особенно Theridion sensu
lato. Отличаются типом рисунка, оттопыренным парацимбиумом, отсутствием ямки
эпигины и настоящих семяпроводящих канальцев, и наличем относительного
длинного прямого коготка педипальп у самки.
ОПИСАНИЕ:
• Карапакс. Грушевидный, не модифицированный, клипеус высокий.
• Стернум. Щитовидный.
• Глаза. 8, в два ряда, латеральные сближены между собой. Передние
медиальные мельче других.
• Ротовой аппарат. Хелицеры не модифицированы, с 2-3 зубчиками вдоль
переднего края желобка. Максиллы над нижней губой наклонены друг к другу.
• Ноги. С 3 коготками, одноцветные, с малочисленными шипами, покрыты
ровными рядами очень характерных толстых щетинок. Лапка IV вентрально
с пильчатыми волосками. Педипальпы самок с крупным коготком.
• Брюшко. Овально-округлое.
• Паутинные бородавки. Немодицифцированы, колюлюс развит.
• Органы дыхания. Пара лёгких, трахейное дыхальце открывается у
паутинных бородавок.
• Крибеллюм-каламиструм. Отсутствуют.
• Копулятивные органы. Энтелегинные пауки. Пальпа самца без отростков
голени; парацимбиум крупный, оттопыренный; эмболюс длинный
нитевидный. Эпигина простая, в виде пластинки с просвечивающими удлинёнными
рецептакулами; семяпроводы неразвиты.
• Размер. До 5 мм. Половой диморфизм слабо выражен.
• Окраска. Светлые, брюшко чаще всего с рисунком из 4 пар тёмных пятен.
Белых гуаниновых пятен нет.
• Особые черты. Оттопыренный парацимбиум. Ноги покрыты стройными
рядами ровных и одинаковых по высоте щетинок. Педипальпа самки с
длинным коготком.
Рода, известные из азиатской части России. Howaia Lehtinen et Saaristo, 1980
(некоторые считают его синонимом Nesticella Lehtinen et Saaristo, 1980).
Семейства, встречающиеся в азиатской части России
199
Рис. 25. Nesticidae. Howaia brevipes (сГ$, 1-2, 4-5) и Н kerzhneri (сГ, 3).
1 — габитус $, сверху; 2 — габитус сГ, сбоку; 3-4 — пальпа сГ, снизу; 5 — эпигина, снизу 3 -
Марусик (1987а).
Распространение. Известны два вида, обитающие на юге Дальнего Востока:
Н. brevipes (Yaginuma, 1970) (Курилы и Сахалин) и Н kerzhneri Marusik, 1987
(Амурская область, Хабаровский край и Приморье).
Биология. Подавляющее большинство представителей семейства —
обитатели пещер и крупнообломочных осыпей и россыпей. Но в Палеарктике известны
два современных рода, обитающие вне пещер, Howaia и Nesticella. Ископаемые
пауки из Балтийских янтарей Epopino Petrunkevitch, 1942 также вели внепещер-
ный образ жизни. Экология пауков-нестицид, и Howaia в частности, не изучены.
Howaia обитают как в затенённой подстилке, так и на лугах. Иногда встречаются
среди мелкообломочных осыпей. Коконы круглые, белого цвета. Самки, очевидно,
встречаются в течение всего года. Самцы обычно более редки, нежели самки,
встречаются в июле-сентябре.
200
4. Специальная часть
Методы сбора. Разгребание или просеивание подстилки, среди осыпей.
Таксономия. Впервые этот таксон (Nesticeae) был выделен Е. Simon (1894) в
подсемействе Tetragnathinae семейства Argiopidae (=Araneidae). Ранг семейства этой
группе придал М. Dahl (1926). Помимо 206 рецентных видов известно 11
ископаемых видов из 4 родов. Большинство видов обитают в Голарктике. Надвидовая
систематика голарктических и ориентальных видов относительно хорошо
разработана благодаря работе Lehtinen & Saaristo (1980). J. Wunderlich (1986) и некоторые
другие исследователи рассматривают Howaia и Nesticella как синонимы. В работе
Marusik & Guseinov (2003) обосновывается самостоятельность этих родов.
Виды и рода нестицид достаточно хорошо различаются формой парацимбиу-
ма. Два дальневосточных вида рассматривались в работах Марусика (1987а; Ма-
русик, Кровфорд, 2006). Иллюстрации Н brevipes имеются также в японских,
китайских и корейских определителях.
Разграничение видов. Только форма копулятивных органов.
Перспективы изучения. В Приморье обитает неописанный род. Возможно
обнаружение в пещерах Дальнего Востока рода Cyclocarcina Komatsu, 1942, широко
распространённого в пещерах Японии. Не исключены находки и дополнительных
видов Howaia.
Семейства, встречающиеся в азиатской части России
201
Pirata piratoides (Lycosidae), самка, Приморье (фото: М.М. Омелько).
Семейство Oonopidae Simon, 1890
Оонопиды; dwarf hunting spiders; goblin spiders
Рис. 26.1-8.
Сравнительно небольшое всесветно распространённое семейство с 512
видами, относимыми к 75 родам. В России известно два рода и два вида. На востоке
России отмечен один вид, из монотипичного рода.
Диагностические признаки. Мелкие гаплогинные пауки, 6 глаз расположены
компактной группой, бёдра задних ног несколько вздуты (скакательные).
Близкие группы. Форма расположения глаз уникальна.
ОПИСАНИЕ:
• Карапакс. Грушевидный с высокой грудной частью (выше головной), у самца
модифицирован (с двумя выростами дорсально).
• Стернум. Удлинённо-сердцевидный.
• Глаза. 6, расположены компактной группой.
• Ротовой аппарат. Максиллы удлинены, в 2 раза длиннее нижней губы. Хе-
лицеры маломощные, у самцов с пальцевидными выростами.
• Ноги. 2 коготка, сидящие на онихуме. Передние ноги чуть длиннее тела.
Нога III у самца почти равна длине тела. Бёдра задних ног несколько вздуты
(скакательные).
• Брюшко. Овальное.
• Паутинные бородавки. Колюлюс небольшой, задние и передние
бородавки одинаковой величины, цилиндрические.
• Органы дыхания. Легочные крышки имеются, но лёгкие редуцированы.
Трахеи не объединены и открываются дыхальцами чуть ниже лёгких.
Отверстия дыхалец практически незаметны.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Гаплогинные. Голень пальпы самца крупнее
бедра, по размеру примерно равна тегулюму; тегулюм каплевидный.
Копулятивные органы самки без заметных выростов. Самку можно принять за
неполовозрелую.
• Размер. 1,5-1,8 мм. Половой диморфизм не выражен.
• Окраска. Без рисунка, бледно-жёлтая.
• Особые черты. 6 глаз в компактной группе, грудь выше головы, вздутые
бёдра IV.
Рода, известные из азиатской части России. Ferchestina Saaristo et Marusik,
2004 (К storozhenkoi Saaristo et Marusik, 2004).
Распространение. Только южное Приморье. В Азии это самая северная
находка семейства.
Биология. Единственный вид, известный из Приморья, собран фумигацией
стволов деревьев.
Методы сбора. Фумигация стволов, обдирание коры.
Семейства, встречающиеся в азиатской части России
203
Рис. 26. Oonopidae. Ferchestina storozhenkoi (cT$).
1 — головной отдел ?, сверху; 2 — вершина хелицер С?, спереди; 3 — карапакс и хелицеры сГ,
спереди; 4 — карапакс сГ, сбоку; 5 — пальпа ?, сбоку; 6 — стернум, лабиум и максиллы сГ, снизу;
7 — «эпигина»; 8 — пальпа сГ, сбоку 1-8 — по: Saaristo & Marusik (2004b).
Таксономия. Впервые на самостоятельность этой группы указал Е. Simon (1893).
В этой же работе он привёл ключ для всех известных ему родов. Семейство
изучено чрезвычайно слабо. Единственная ревизия пантропических родов была
предпринята М. Сааристо (Saaristo, 2001). До настоящего времени нет обзоров даже
европейских оонопид. Судя по всему, семейство включает много больше видов,
нежели известно сейчас. Так, предварительный неполный обзор австралийской
фауны позволил выявить более 200 новых видов и родов (М. Harvey & М. Saaristo,
личное сообщение). Разнообразие этого семейства в границах бывшего СССР
также много выше ранее известного. В каталоге 1997 г. (Mikhailov, 1997) указано
всего два рода и три вида. По нашим данным, это семейство включает не менее 10
видов из более, чем 6 родов. Единственная работа по паукам Oonopidae азиатской
части России опубликована Saaristo & Marusik (2004b). Единственный
представитель фауны Дальнего Востока принадлежит всесветно распространённому
подсемейству Orchestininae Chamberlin et Ivie, 1942.
В настоящее время это семейство (в пределах СНГ) изучает Ю.М. Марусик.
Разграничение видов. Форма копулятивных органов, возможно и форма кара-
пакса у самца.
Перспективы изучения. Не исключены находки дополнительных видов в
Приморье.
Семейство Oxyopidae Thorell, 1870
Оксиопиды, пауки-рыси; lynx spiders
Рис. 27.1-7.
Небольшое всесветно распространённое семейство с 426 видами и 9 родами. В
России известен один род с 4 видами, три из которых отмечены в азиатской части
России.
Диагностические признаки. Среднего размера энтелегинные пауки с 8
характерно расположенными глазами, образующими три ряда (2-4-2), лапки с 3
коготками, ноги с мощными шипами, брюшко каплевидное.
Близкие группы. Формула глаз уникальна.
ОПИСАНИЕ:
• Карапакс. Яйцевидный, с высоким клипеусом.
• Стернум. Щитовидный, в задней части удлинён и разделяет тазики IV.
• Глаза. 8, расположены в три ряда (2-4-2), второй ряд сильно выгнут.
Передние медиальные глаза наиболее мелкие.
• Ротовой аппарат. Не модифицирован.
• Ноги. 3-коготковые, ноги относительно длинные, передняя пара
приблизительно в 1,3-2,0 раза длиннее тела. Шипы очень мощные.
• Брюшко. Удлинённое каплевидное.
• Паутинные бородавки. Не модифицированы, средняя пара наименьшая по
размеру, колюлюс имеется.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Энтелегинные. Пальпа самца с хорошо развитыми
отростками голени, иногда имеются отростки и на колене; цимбиум
заострён на вершине, длинные волоски закрывают тегулюм. Эпигина с септумом,
направленым вверх (вперёд), ямки нет.
• Размер. 5-10 мм. Половой диморфизм слабо выражен.
• Окраска. Все зауральские виды тёмноокрашенные, в спирту рисунок
бывает не выражен. В общих чертах рисунок состоит из медиальной светлой
полосы на карапаксе и брюшке; ланцетовидное пятно на брюшке тёмное.
• Особые черты. Расположение глаз уникально.
Рода, известные из азиатской части России. Oxyopes Latreille, 1804 (О. hetero-
phthalmus Latreille, 1804; О. licenti Schenkel, 1953; О. ramosus (Martini et Goeze, 1778)).
Распространение. Род распространён по всей Сибири и Дальнему Востоку. О.
licenti замещает О. ramosus к востоку от Алтая. О. heterophtalmus доходит на
восток до Прибайкалья.
Биология. Населяют травостой, активные дневные охотники, сетей не плетут,
хотя согласно исследованиям Р. Джексона, паутина играет существенную роль при
ловле добычи. Коконы округлые, прикрепляются самками среди листьев, иногда
вокруг сплетается небольшая сеть. Самка охраняет кокон. В целом биология мало
изучена.
Методы сбора. Укосы. Поздней весной и в начале лета самок можно собирать
вручную с коконов на траве. В почвенные ловушки попадаются очень редко.
Семейства, встречающиеся в азиатской части России
205
Рис. 27. Oxyopidae. Oxyopes ramosus (С??, 1-2, 4, 6) и О. /zcewtf (С?$, 3, 5, 7).
1-2 — головогрудь ?, спереди и сверху; 3 — габитус о", сверху; 4 — колено и голень пальпы сГ,
сбоку; 5-6 — эпигина, снизу; 7 — пальпа сГ, спереди. 4-6 — по: Marusik et al. (1996); 7 — по:
Ono et al. (2009).
Таксономия. Семейство в широких масштабах никогда не ревизовалось. По
мнению П.Т. Лехтинена (Lehtinen, 1967), Oxyopidae следует рассматривать в
составе надсемейства Pisauroidea. H. Homann (1975) и СЕ. Griswold (1993)
сближают оксиопид с иными семействами.
Два сибирских викариирующих вида рассматривались в работе Марусика с
соавторами (Marusik et al., 1996). В настоящее время пауками-оксиопидами
восточной Палеарктики занимается С.Л. Есюнин и Т.К. Тунёва (Есюнин, Тунёва, 2009).
Разграничение видов. Форма копулятивных органов и окраска.
Перспективы изучения. Не исключены находки дополнительных видов,
прежде всего на юге Дальнего Востока и в южной Сибири.
Семейство Philodromidae Thorell, 1870
Пауки-бокоходы; small huntsman spiders
Рис. 28.1-19.
Относительно небольшое, практически всесветно распространённое семейство
с 27 родами и 531 видом. В России известно 5 родов и более 60 видов, из них в
азиатской части России отмечено 58 видов из тех же 5 родов.
Диагностические признаки. Энтелегинные, некрибеллятные пауки среднего
размера (3-10 мм) с 8 глазами в два ряда, ноги примерно равной длины, бёдра
расставлены в стороны, лапки передних ног со скопулой, пальпа самца с РГО, эпи-
гина с более или менее выраженной медиальной пластинкой.
Близкие группы. Габитуально схожи со Sparassidae и некоторыми Thomisidae
(особенно с Diaea и Oxytate). От первых отличаются меньшим размером и
отсутствием зубчиков на внутренней стороне хелицер, кроме того, тем, что между
лапками и предлапками нет трёхчленной мембраны. От пауков-крабов (Thomisidae)
чётко отличаются примерно равной длинной всех ног и наличием скопулы.
ОПИСАНИЕ:
• Карапакс. От грушевидного до практически круглого (ширина = длине),
слегка или сильно уплощённый (особенно у дендробионтных Philodromus).
• Стернум. Щитовидный, задняя часть разделяет тазики IV.
• Глаза. 8 в два ряда (4-4), практически равной величины, оба ряда выгнуты
(при виде сверху), боковые глаза значительно удалены друг от друга (более
чем на 2 диаметра).
• Ротовой аппарат. Не модифицирован. Задний край хелицер не более чем с
одним зубчиком.
• Ноги. 2 коготка, все пары ног примерно равной длины, бёдра расставлены в
стороны (под углами, близкими к прямым по отношению к оси тела), шипы
многочисленные, лапки и дистальная часть предлапок передних ног со
скопулой.
• Брюшко. Форма варьирует, но сзади всегда резко сужается, яйцевидная,
овальная, овально-удлинённая (Tibellus), иногда пентагональная (у
Philodromus).
• Паутинные бородавки. Не модифицированные, конические, колюлюса нет.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Пальпа самца всегда с РГО; отростков голени
исходно два, но вентральный может быть сильно или полностью
редуцированным. Бульбус иногда без выростов (некоторые Thanatus), но чаще всего с
мембранизированным выростом (кондуктором). Пальпа некоторых
Philodromus может нести дополнительные выросты. Эпигина плоская, чаще
всего с хорошо ограниченной медиальной пластинкой. У Tibellus эта часть сильно
уменьшена по высоте. У некоторых Philodromus верхняя часть медиальной
пластинки вогнута и образует ямку.
Семейства, встречающиеся в азиатской части России
207
1
8
10
Рис. 28. PModromidae. Внешний вид Thanatus formicinus (?, 1,4, 8), Philodromus cortici-
nus (С?, 2,9), P margaritatus (d\ 3,5), Tibellus oblongus (C?$, 6-7) и Philodromus alascensis
(¥, 10).
1-2 — головогрудь, спереди; 3 — головогрудь, сверху; 4-7, 9-10 — габитус, сверху; 8 — пред-
лапка и лапка I. Стрелками показана скопула.
208
4. Специальная часть
• Размер. 3-10 мм, размерный диморфизм выражен не сильно.
• Окраска. От светлой до тёмной. Иногда практически без рисунка, но чаще
всего с рисунком на карапаксе и брюшке. Ланцетовидное пятно на брюшке
всегда имеется. Половой диморфизм в окраске (рисунке) у некоторых групп
(Tibellus) не выражен, однако у большинства групп — отчётливый.
• Особые черты. Уплощённое тело, бёдра расставлены в стороны, ноги
примерно равной длины, скопула на лапках передних ног.
Рода, известные из азиатской части России.
Apollophanes О. Pickard-Cambridge, 1898; Artanes Thorell, 18691; Ebo Keyserling,
1884; Philodromus Walckenaer, 1826; Thanatus C.L. Koch, 1837; Tibellus Simon, 1875.
Распространение. Все рода распространены по всей Сибири и Дальнему
Востоку, за исключением Ebo, который известен только в южной Сибири.
Наиболее важные работы, посвященные распространению филодромид
азиатской части России: Logunov (1996a); Efimik (1999); Marusik et al. (2000).
Биология. Филодромиды сетей не плетут, ведут бродячий образ жизни.
Большинство наших видов — дневные хищники. Населяют широкий спектр биотопов
во всех природных зонах, и, в отличие от других бродячих видов (пауки-волки,
гнафозиды), освоили все ярусы. Причем один и тот же вид может охотиться в
совершенно разных условиях. Philodromus alascensis Keyserling, 1884 и Thanatus
bungei (Kulczynski, 1908) встречаются и на осыпях, и на почве, и среди травостоя,
и, кроме того, на кустах и деревьях, а первый вид обычен ещё и в теплицах (на
северо-востоке Азии) и в других строениях. Однако большинство видов
адаптировано к определённым условиям. Так удлинённые Tibellus и некоторые Philodromus
обитают только среди травостоя. Часть видов Philodromus обитают только на
стволах деревьев. Artanes marusiki Logunov, 1997 обитает на скалах и активен в тёмное
время суток. Thanatus kolymensis Marusik, 1991 — облигатный литобионт,
встречается среди каменных осыпей и россыпей в горных тундрах.
Самки многих Philodromus и Thanatus откладывают линзовидный кокон и
прикрепляют его к субстрату (камням, сухим листьям, коре деревьев) с помощью
тонкого полупрозрачного полога и охраняют его, сидя поверх полога. Кокон крупный,
чуть меньше размера тела, а полог часто превышает размах ног.
Самки Tibellus, напротив, размещают свои коконы на травинках или невысоких
кустарниках, и вокруг кокона сплетают плотный белый полог. Похожие коконы и у
хортобионтных (обитающих в травостое) Philodromus. Зимуют филодромиды, судя
по всему, только неполовозрелыми.
Методы сбора. Все способы и все биотопы. Виды, встречающиеся на скалах,
проще всего собирать ночью. Подкорные виды собираются обдиранием коры.
Самок Tibellus можно обнаружить благодаря белым коконам среди травостоя. Самки
всегда охраняют их.
1 Название использовалось Д.В. Логуновым (Logunov, 1997) при описании нового вида, однако в
работе не был приведён анализ рода и перечень видов, которые к нему относятся.
Семейства, встречающиеся в азиатской части России
209
11
14 15
17
18
Рис. 28 (продолжение). Philodromidae. Примеры строения копулятивных органов:
Philodromus vinokurovi (С?, 11), .Р utotchkini (cf, 12), P. mysticus (cf, 13), Apollophanes
macropalpus (С?$, 14, 17), Thanstus kolymensis (cf, 15), Tibellus asiaticus (0*2, 16, 19) и
Philodromus aryy (?, 18).
11-16 — пальпа сГ, снизу; 17-19 — эпигина, снизу 1-12, 14-15, 17-18 — по: Марусик (1991 г).
210
4. Специальная часть
Таксономия. Е. Simon (1895) включил этот таксон в ранге подсемейства в
Thomisidae. Многие исследователи придерживались такой же трактовки этой группы
пауков до 70-х и даже 90-х годов XX столетия. Филодромиды рассматриваются как
подсемейство в определителе В.П. Тыщенко (1971) и в достаточно свежем
определителе M.J. Roberts (1995). Первым, кто указал на принципиальные отличия
между Philodromidae и Thomisidae, был Н. Homann (1975). Эти группы пауков
существенно отличаются морфологией глаз, числом хромосом, личиночным
развитием. Со своей стороны можем добавить, что два семейства существенно
различаются и морфологией копулятивных органов. J. Coddington и H.W. Levi (1991)
сближают филодромид со Sparassidae и Selenopidae.
Надвидовая классификация группы разработана недостаточно. Если рода^о/-
lophanes, Thanatus и Tibellus очерчены достаточно хорошо, то Philodromus
нуждается в серьёзном пересмотре. Для отдельных видов и групп видов внутри
Philodromus было предложено несколько родов, по меньшей мере 4: Artanes Thorell,
1869, Horodromoides Gertsch, 1933, Philodromoides Scheffer, 1904 и Rhysodromus
Schick, 1965.
Несмотря на то, что филодромидам посвящено несколько специальных
таксономических работ (Марусик, 1991r;Logunov, 1996;Efimik, 1999), изучена эта группа
ещё недостаточно хорошо. Даже в хорошо изученном роде Thanatus есть ещё
некоторое количество новых видов. Слабее других изучен Philodromus.
Работы, посвященные изучению систематики филодромид азиатской части
России: Kulczynski (1885; 1908); Данилов (1991); Марусик (1991г); Логунов (19926);
Ляхов (1992); Logunov (1996а, 1997); Efimik (1999); Szita & Logunov (2008).
В настоящее время эта группа разрабатывается Д.В. Логуновым.
Разграничение видов. Для определения видов используют почти
исключительно форму копулятивных органов, хотя в некоторых случаях «работает» хетотаксия
(число и расположение шипов) на цимбиуме и голени, размер тела, длина ног.
Большинство видов хорошо различаются рисунком тела, хотя самки хортобионтных
Philodromus после откладки и высиживания кокона часто меняют свою окраску.
Перспективы изучения. Как уже было сказано выше, род Philodromus
нуждается в пересмотре. В регионе имеются ещё неописанные виды Thanatus. Вполне
вероятно нахождение новых для науки и для региона видов Philodromus, особенно
в Приморье.
Семейства, встречающиеся в азиатской части России
211
Philodromus sp. (Philodromidae), самка, Приморье (фото: М.М. Омелько).
Семейство Pholcidae C.L. Koch, 1851
Фольциды; daddy-long-leg spiders
Рис. 29.1-5.
Относительно крупное, всесветно распространённое семейство с 1000 видов и
85 родами. В России зарегистрировано 11 видов из 3 родов, а в азиатской части
России — 7 видов и 2 рода.
Диагностические признаки. Гаплогинные пауки, с 8 глазами и 3 коготками, с
круглым карапаксом, удлинённым брюшком и чрезвычайно длинными ногами (в 5
и более раз длиннее тела). Пальпа самца очень толстая, с голенью и бульбусом,
превышающими по длине бедро пальпы.
Близкие группы. От других длинноногих пауков Bathylinyphia (Linyphiidae),
Tetragnatha (Tetragnathidae) отличается отсутствием шипов на ногах, круглым
карапаксом и наличием триад глаз.
ОПИСАНИЕ:
• Карапакс. Практически округлый, наличник высокий, голова несколько
приподнята над грудной частью. Часто с рисунком.
• Стернум. Плоский и округлый, сросшийся с нижней губой.
• Глаза. 8, расположены компактной группой, 2 передних медиальных —
самые мелкие, и по бокам от них — 2 триплета с более крупными глазами.
• Ротовой аппарат. Хелицеры маленькие, частично сросшиеся; у самцов
модифицированы и могут нести несколько зубчиков на передней поверхности.
Нижняя губа срослась со стернумом.
• Ноги. 3 коготка. Чрезвычайно длинные, даже бедро превышает длину тела.
Вся передняя нога в 5-9 раз длиннее тела. Шипов нет. В момент опасности
ноги легко аутотомируются (паук отбрасывает их сам).
• Брюшко. Удлинённо-овальное, длина более чем в два раза превышает
ширину. Часто с рисунком.
• Паутинные бородавки. Передние бородавки толстые и цилиндрические,
слегка расставлены, с коротким колюлюсом между ними. Задние бородавки
мельче, конической формы.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Пальпа самца короткая и очень широкая, голень
часто толще бёдер ходильных ног и по размеру превышает бедро пальпы.
Вертлуг с пальцевидным выростом. Сам бульбус также крупнее бедра
пальпы. Несмотря на гаплогинность, эпигина имеется — в виде треугольной или
поперечной пластинки с небольшим «скапусом».
• Размер. 4-10 мм, с расставленными ногами — до 100 мм. Самцы мельче
самок, но их ноги заметно длиннее.
• Окраска. Бледно окрашены, зачастую с характерным (видоспецифичным)
рисунком на карапаксе и брюшке.
• Особые черты. Сверхдлинные ноги, очень крупные пальпы самцов.
Семейства, встречающиеся в азиатской части России
213
Рис. 29. Pholcidae. Pholcus phungiformes (cT$).
1-2 — головогрудь $, сверху и спереди; 3 — головогрудь О4, сбоку; 4 — габитус О4, сбоку; 5 — $,
снизу
214
4. Специальная часть
Рода, известные из азиатской части России. Pholcus Walckenaer, 1805; Psilo-
chorus Simon, 1893 (P simoni (Berland, 1911)).
Распространение. Pholcus — Улан-Уде, Магадан (недавно интродуцированы,
синантропы), юг Читинской области (Сохондинский заповедник), и весь юг
Дальнего Востока (Сахалин, Южные Курилы, Хабаровский край и Приморье).
Psilochorus simoni — отмечен в Западной Сибири (возможно, неверно
определён или интродуцирован).
Биология. Большинство наших видов населяют жилые дома (и не могут жить
за их пределами) и другие человеческие постройки (на юге Дальнего Востока),
обычны в естественных и искусственных пещерах. В природных условиях
обитает, судя по всему, один вид P. zichyi Kulczynski, 1901. Его мы находили в самых
разных условиях в каменных осыпях в центральной Монголии, Сохондинском
заповеднике, на южных каменистых склонах в Приморье и даже среди приморских
галечников в Приморье, причём в больших количествах (Marusik & Koponen, 2000).
Все фольциды сетеплетущие, сети их рыхлые, бесформенные, в постройках и
пещерах всегда располагаются по углам вблизи потолка. При прикосновении к сети,
пауки начинают её сильно трясти, очевидно, чтобы стать менее заметными.
Установлено, что помимо добычи, попавшейся в сети, фольциды могут питаться и
другими пауками, забираясь в их сети и приманивая подёргиванием сети,
имитирующим трепыхание жертвы.
Копуляция происходит на сети и длится несколько часов (Jones, 1989). Другие
исследователи указывают, что копуляция длится до 6 минут (Forster & Forster, 1999).
Особенность копуляции заключается в том, что, как и у всех гаплогинных пауков,
пальпы вводятся не поочерёдно, а сразу две, одновременно! Общая ширина двух
пальп нередко превышает ширину брюшка.
Кокон круглый; покровы его очень тонкие, сетковидные, не скрывающие яиц.
Самка носит кокон в хелицерах.
Методы сбора. Сети большинства видов можно обнаружить визуально. Самих
пауков собирать проще всего вручную. P. zichyi встречается под камнями, в том
числе и среди приморских галечников. Большинство наших видов синантропы.
Таксономия. Эта группа пауков давно имеет ранг семейства, и трактовка Pholci-
dae сколько-нибудь существенно не изменилась со времён описания. Е. Simon (1893)
трактовал это семейство как энтелегинное, а Р. Lehtinen (1967) — как гаплогинное,
и сближал его с наиболее примитивными аранеоморфными пауками — Hypochilidae.
Позже их рассматривали как родственных к Filistatidae (крибеллятное семейство),
или Plectreuridae-Diguetidae. На наш взгляд, это гаплогинное семейство, но с
чрезвычайно сложно устроенным копулятивным аппаратом, не характерным для
других групп. Кроме того, большинство видов семейства имеет 8 глаз, а не 6, как у
всех остальных гаплогинных семейств (Oonopidae, Dysderidae, Leptonetidae и др.).
Кроме того, у самки всегда имеется эпигина (хитинизированная генитальная
пластинка). Существует мнение, что Pholcidae — это настолько своеобразная линия
эволюции пауков, что они должны быть выделены в отдельный подотряд (Старо-
богатов, 1985), однако этот подотряд плохо обоснован и не признан арахнологами.
Семейства, встречающиеся в азиатской части России
215
Тем не менее, у фольцид и вправду есть ряд уникальных черт, которые могут быть
использованы для обоснования подотряда, в частности особый план строения ко-
пулятивного аппарата у самцов, способ наполнения бульбуса семенем, способ
плетения кокона.
Pholcidae подразделяется на несколько подсемейств. В России встречаются
представители только номинативного подсемейства. Систематика видов азиатской
части России рассматривалась в работах Т.И. Олигер (Олигер, 19836; Oliger, 1998) и
отчасти в работе Marusik & Koponen (2000).
В настоящее время обзор фольцид азиатской части России делает Д.В.
Логунов.
Перспективы изучения. Pholcidae азиатской части России никогда не
ревизовались. Из сопредельных территорий (Китай, Корея) описано немало видов, и
поэтому не исключено, что на юге Дальнего Востока будут найдены новые и
дополнительные виды. Нам известны несколько видов из Приморья и Хабаровского края,
которые формально ещё не отмечены на территории России.
Pholcus phalangioides (Pholcidae), самка, Крым (фото: Н.М. Ковблюк).
Семейство Pimoidae Wunderlich, 1986
Пимоиды; large hammock-web spiders
Рис. 30.1-9.
Небольшое семейство, ограниченное в распространении только Голарктикой и
Юго-Восточной Азией. Известно 33 вида, относимые к 4 родам. В России
известен только один вид из Сахалина и Монерона.
Диагностические признаки. Энтелегинные, некрибеллятные, аранеоидные,
небольшие или среднего размера (6-8 мм) пауки с 3 коготками и 8 глазами в 2 ряда,
передние медиальные глаза самые крупные. Брюшко с чётким рисунком. Самцы с
модифицированой предлапкой I (уплощена в основании), цимбиум треугольной
формы при виде сбоку (за счёт конического выроста), вершина цимбиума пальцевидная.
Близкие группы. Близки к Linyphiidae, особенно к Stemonyphantes, отличаются
уплощённой в основании предлапкой I у самцов, коническим, сильно хитинизиро-
ванным выростом цимбиума, и пальцевидной вершиной цимбиума. Самки не имеют
чётких отличий, кроме как отсутствие стридуляционных бороздок на хелицерах.
ОПИСАНИЕ:
• Карапакс. Грушевидный, наличник относительно низкий (1,4-1,5
диаметров передних медиальных глаз).
• Стернум. Длина больше ширины, с выростом между тазиками IV.
• Глаза. 8 глаз; передние медиальные — самые крупные, остальные пример-
• но одинаковой величины, боковые глаза соприкасаются; передние
медиальные глаза сближены — расстояние между ними меньше, чем их диаметр.
• Ротовой аппарат. Хелицеры крупные, без стридуляционных бороздок
(только у Weintrauboa, у других родов — есть); передний край желобка хелицер с
3 зубчиками, задний край с 3^4 зубчиками. Максиллы в 2 раза длиннее
нижней губы.
• Ноги. С шипами и тёмными кольцами, у самцов длиннее, чем у самок; пред-
лапка I у самцов модифицирована — уплощена в основании, слегка
искривлена и несёт специализированные волоски.
• Брюшко. Овальное, с чётким рисунком.
• Паутинные бородавки. Не модифицированы, колюлюс имеется.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Энтелегинные пауки. Пальпа самца относительно
крупная; голень без отростков; цимбиум с крупным, сильно хитинизирован-
ным коническим ретролатеральным выростом; вершина цимбиума
пальцевидная; парацимбиум крючковидный, с крупным основанием. Эмболюс
длинный, нитевидный. Эпигина простая, с септумом.
• Размер. 6-8 мм, самки крупнее самцов.
• Окраска. Карапакс с широкой тёмной медиальной полосой, брюшко с
рисунком.
• Особые черты. Модифицированные ноги I у самцов; конический вырост
цимбиума; отсутствие стридуляционных бороздок на хелицерах.
Рода, известные из^ азиатской части России. Weintrauboa Hormiga, 2003
(W. insularis (Saito, 1935)).
Семейства, встречающиеся в азиатской части России
217
1 2
Ч
9
6 7
Рис. 30. Pimoidae. Weintrauboa insularis (cT$).
1,5 — пальпа сГ, сверху; 2, 4 — пальпа сГ, сбоку; 3 — эпигина, снизу; 6 — предлапка I сГ; 7 —
нога I сГ; 8-9 — габитус ? и сГ, сверху Яг/ — парацимбиум. 1-2, 6-7 — по: Hormiga (2003); 3-5 —
по: Hormiga (2008); 8-9 — по: Chikuni (1989).
Распространение. Единственный вид известен с юга Сахалина и Монерона.
Хотя в каталоге Н.И. Платника (Platnick, 2009) для России указаны два вида рода
Weintrauboa, на самом деле у нас обитает только один вид (см. Hormiga, 2008).
Биология. Биология нашего вида неизвестна. Пологовидные сети близкого вида
W. contortipes (Karsch, 1881) изображены в работе Shinkai & Takano (1984: 27). Сети
имеют наклон около 45°, расположены невысоко над почвой, по всей видимости,
вверх и вниз от сети отходят поддерживающие нити.
Методы сбора. Неизвестно. Вероятно, почвенные ловушки и ручной сбор.
Таксономия. Семейство ревизовал в мировом масштабе Г. Хормига (Hormiga,
1994,2003). Судя по всему, разнообразие семейства выше и распространение шире.
Один монотипический род, Nanoa Hormiga, Buckle et Scharff, 2005, из Неарктики
недавно перенесён в другое семейство (Wunderlich, 2008a). В России известен
всего один вид. Этот вид рассматривается всего в трёх публикациях: Chikuni (1989) и
Hormiga (2003, 2008).
Перспективы изучения. Нахождение дополнительных видов маловероятно.
Семейство Pisauridae Simon, 1890
Пизауриды; nursery web spiders, raft or fishing spiders (Dolomedes)
Рис. 31.1-9.
Относительно небольшое семейство с 339 видами и 53 родами. В России
отмечены 9 видов из двух родов1, все они встречаются и в азиатской части России.
Диагностические признаки. Крупные (10-20 мм), энтелегинные, некрибел-
лятные пауки, с 8 глазами, задние глаза крупнее передних, ноги длинные с
шипами, скопула имеется, пальпа с РГО.
Близкие группы. Габитуально и отчасти формулой глаз напоминают Lycosidae,
однако чётко отличаются высоким наличником, в несколько раз превышающим
передние глаза. Кроме того, самцы пизаурид всегда с РГО, в то время как его нет у
пауков-волков.
ОПИСАНИЕ:
• Карапакс. Грушевидный, иногда с обрезанной вершиной. Наличник много
' больше диаметра передних глаз.
• Стернум. Не модифицирован.
• Глаза. 8 глаз, в 2-3 ряда (4-4 или 4-2-2), задний ряд можно считать либо
сильно выгнутым, либо рассматривать как два отдельных ряда. Размер
передних глаз в 1,5-2 раза меньше задних.
• Ротовой аппарат. Не модифицирован.
• Ноги. 3 коготка, длинные и относительно толстые, со многими шипами,
скопула имеется.
• Брюшко. Удлинено, постепенно сужается к концу.
• Паутинные бородавки. Не модифицированы.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Пальпа самца с РГО, у Pisaura он стандартной
формы — длинный отросток, сужающийся к вершине. У Dolomedes основание
РГО находится ниже вершины голени, сам РГО пластинчатый, у некоторых
видов сильно варьирует по форме. Эпигина у Pisaura с якоревидным септу-
мом. У Dolomedes эпигина представлена одной ямкой.
• Размер. 10-20 мм.
• Окраска. В целом пауки тёмные, Pisaura со светлой медиальной полосой
на карапаксе, a Dolomedes с двумя субмаргинальными светлыми полосами
на карапаксе и на брюшке.
• Особые черты. Крупные пауки, передние глаза меньше задних, наличник
высокий; самки носят кокон в хелицерах.
Рода, известные из азиатской части России. Dolomedes Latreille, 1804; Pisaura
Simon, 1885.
1 По мнению некоторых исследователей, оба рода принадлежат разным семействам.
Семейства, встречающиеся в азиатской части России 219
Рис. 31. Pisauridae. Pisaura ancora (сГ$, 1-3), P. lama (?, 4), Dolomedes fimbriatus (?, 5-
6), D. bukhkaloi (cT, 7) и D. senilis (cT?, 8-9).
1,5 — головогрудь ?, спереди; 2, 7 — пальпа сГ, снизу; 3 — пальпа сГ, сбоку; 4, 9 — эпигина,
снизу; 8 — отросток голени сГ, сбоку. 2-А — по: Логунов (1990); 7-9 — по: Марусик (1988а).
220
4. Специальная часть
Распространение. Dolomedes — весь регион, кроме тундровой зоны; Pisaura -
юг Сибири и Дальнего Востока.
Биология. Dolomedes населяет исключительно околоводные биотопы, где есть
открытая вода, хотя бы временно, a Pisaura — обитатели травостоя и кустарников.
Пизауриды — довольно активные дневные охотники с хорошо развитым
зрением. Для некоторых видов Dolomedes достоверно известно, что они могут питаться
небольшими рыбками, и даже приманивают их особым постукиванием передними
ногами по водному зеркалу.
Поведение ухаживания у Pisaura очень своеобразное: самцы приподносят
«свадебный» подарок в виде жертвы, замотанной в паутину. Недавно установлено, что
подарок, напоминающий по форме кокон, имеет и стимулирующий характер.
Причём, чем светлее (белее) паутина, в которую завёрнут подарок, тем он
привлекательнее (Stalhandske, 2002). Иногда самка поедает самца после совокупления. В
экспериментальных условиях только 40% самцов без подарка смогли спариться с
самками, в то время как с подарком — 90%.
Коконы у пизаурид очень крупные, самки охраняют и постоянно таскают их,
зажав в хелицерах. Перед вылуплением молоди коконы крепятся к растениям
(травостой, низкие кусты), затем кокон чуть приоткрывается и вокруг него сплетается
так называемая выводковая (nursery) сеть. Молодь Dolomedes после покидания
выводковой сети держится чаще всего вдали от воды, и ведёт себя как представители
рода Pisaura.
Методы сбора. Почвенные ловушки, укосы, ручной сбор. Самок легко
обнаружить благодаря защитным логовам, которые они устраивают для своих коконов.
Самки всегда находятся вблизи логова. Dolomedes можно обнаружить визуально
вдоль берегов болот и озёр, когда они охотятся. Pisaura легко можно собрать
ночью с помощью укосов.
Таксономия. Границы семейства очерчены неудовлетворительно.
Первоначально это семейство было подразделено на три таксона, которым позже был придан
ранг подсемейств, в том числе и Dolomedinae и Pisaurinae. P.T. Lehtinen (1967)
повысил ранг первого таксона до семейства, но другие арахнологи не поддержали
его мнение. P. Sierwald (1990) сделала попытку ревизовать семейство, однако не
было найдено ни одной синапоморфии для всех родов. Даже выводковая сеть (из-
за которой пошло английское название) отмечена всего лишь у 9 родов. Судя по
строению пальпы, Dolomedes и несколько внеголарктических родственных родов
вполне заслуживают выделения в отдельное семейство Dolomedidae, как считает
Р.Т. Lehtinen (1967).
Пауки этого семейства рассматривались в работах Bonnet (1929); Марусик
(1988а); Логунов (1990); Renner (1988). Пауки-пизауриды смежных территорий
рассматриваются в работах Chikuni (1989), Song et al. (1999) и Ono et al. (2009).
Разграничение видов. Только форма копулятивных органов (но эпигины
Dolomedes довольно сходны друг с другом).
Перспективы изучения. Возможно, ревизия пауков рода Dolomedes позволит
выявить дополнительные виды, в том числе и новые для науки.
Семейства, встречающиеся в азиатской части России
221
Dolomedes sp. (Pisaundae), самка с коконом, Приморье (фото: М.М. Омелько).
Семейство Salticidae Blackwall, 1841
Пауки-скакунчики, сальтициды; jumping spiders
Рис. 32.1-22.
Это самое крупное семейство пауков с 5202 рецентными видами и 563 родами.
Распространено оно всесветно. В России известно более 180 видов из более чем 50
родов, а в азиатской части России — более 140 видов, по меньшей мере, из 31 рода.
Диагностические признаки. Энтелегинные, некрибеллятные пауки, с 2
коготками и 8 глазами, расположенными в три ряда (4-2-2), глаза переднего ряда много
крупнее глаз второго ряда, карапакс высокий, прямоугольный, ноги относительно
короткие. Пальпа самца с РГО (нет только у Talavera), медиальный отросток
отсутствует.
Близкие группы. Благодаря крупным глазам переднего ряда, Salticidae не
могут быть спутаны ни с одной из других групп.
ОПИСАНИЕ:
• Карапакс. От прямоугольного до почти круглого (Rhene) при виде сверху,
иногда длина превышает ширину в 2 и более раз (Myrmarachne). Высота
большая, задний скат карапакса бывает очень крутым. Иногда головная часть
приподнята над грудной. Передняя часть бывает украшена яркими
волосками (иногда образующими рисунок) или имеет поверхностную окраску.
Верхняя часть также часто с рисунком.
• Стернум. Разной длины, от щитовидного до удлинённого.
• Глаза. 8 в 3 ряда (4-2-2), передние медиальные очень крупные, передние
латеральные меньше, передний ряд глаз занимает всю ширину карапакса.
Глаза второго ряда очень маленькие. Вокруг передних глаз могут быть ярко
окрашенные «реснички».
• Ротовой аппарат. Хелицеры от обычных до сильно удлинённых (самцы
Myrmarachne и Salticus). Задний край желобка с одним, несколькими или многими
зубцами. Нижняя губа и максиллы нередко модифицированы у самцов.
• Ноги. 2 коготка, у большинства относительно короткие (передние ноги
обычно короче тела). Иногда передние ноги самца бывают сильно утолщены,
несут густые уплощённые волоски, благодаря чему нога кажется ещё шире.
Нередко первые ноги ярко окрашены и с рисунком.
• Брюшко. От овального до удлинённо-овального, часто чуть заострено на
конце. Чаще всего с рисунком, иногда очень ярким. Самцы некоторых родов
с дорсальным скутумом, у самцов одного вида (Euophrys frontalis) есть и
вентральный скутум.
• Паутинные бородавки. Не модифицированы, цилиндрические, часто
торчат из-под брюшка.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Коцулятивные органы. Энтелегинные. Пальпа самца относительно
короткая (не длиннее головогруди), толстая. РГО имеется у всех, кроме Talavera.
Семейства, встречающиеся в азиатской части России
223
11
Рис. 32. Salticidae. Внешний вид Euophrys frontalis (С?, 1,7,10), Asianellus kazakhstanicus
(С?, 2, 8), Philaeus chrysops (О71, 3), Phlegra fasciata (?, 4), Sitticus mirandus (cf, 5, 11), 5.
eskovi (cf, 6) и Myrmarachne formicaria (cf, 9).
1-6, 9 — габитус, сверху; 7-8, 10-11 — головогрудь, спереди. Euophrys frontalis: 1 и 7 — светлая
форма; 10 — тёмная форма. 1,6 — по: Марусик, Кроуфорд (2006); 2-3. 5. 8. 11 — по: Marusik et
al. (2007).
224
4. Специальная часть
Иногда имеются дополнительные отростки голени. Бывают отростки и на
бедре и даже на вертлуге. Цимбиум иногда с выростами в базальной части,
иногда верхняя часть удлиняется {Yllenus). Бедро, голень и цимбиум могут
нести ряды модифицированных «окрашенных» волосков. Тегулюм разный
по форме, апофизов (подвижных отростков), как правило, нет; но кроме эм-
болюса иногда имеется один отросток. Отросток может быть выростом те-
гулюма {Pellenes) или выростом основания эмболюса (Aelurillus). Эмболюс
от длинного и тонкого, до короткого. У Heliophanus и некоторых других групп
эмболюс сросся с тегулюмом. Эпигина очень вариабельная — может быть
без ямки, с ямкой, с двумя ямками, иногда с нижним карманом, или
вырезкой.
• Размер. 2,5-12 мм. Самки обычно крупнее самцов, но у Myrmarachne
самцы кажутся длиннее из-за длинных хелицер, направленных вперёд.
• Окраска. Значительно варьирует, от бледной до тёмной. Большинство
видов с рисунком. Рисунок создаётся волосками, структурной окраской и
пигментами. Самцы чаще всего ярче окрашены, нежели самки, однако иногда
самцы бывают одноцветно чёрными, а самки имеют рисунок {Chalcoscirtus).
• Особые черты. Огромные передние медиальные глаза; передвижение
прыжками.
Рода, известные из азиатской части России. Aelurillus Simon, 1884; Asianellus
Logunov et HQciak, 1996; Carrhotus Thorell, 1891; Chalcoscirtus Bertkau, 1880; Den-
dryphantes C.L. Koch, 1837; Euophrys C.L. Koch, 1834; Evarcha Simon, 1902; Har-
mochirus Simon, 1885; Helicius Zabka, 1981; Heliophanus C.L. Koch, 1833; Marpissa
C.L. Koch, 1846; Mendoza Peckham et Peckham, 1894; Myrmarachne MacLeay, 1839;
Neon Simon, 1876; Pellenes Simon, 1876; Philaeus Thorell, 1869; Phintella Strand,
1906; Phlegra Simon, 1876; Plexippoides Proszynski, 1984; Pseudeuophrys F. Dahl,
1912; Pseudicius Simon, 1885; Rhene Thorell, 1869; Salticus Latreille, 1804; Sibianor
Logunov, 2001; Sitticus Simon, 1901; Synageles Simon, 1876; Synagelides Strand, 1906;
Talavera Peckham et Peckham, 1909; Tuvaphantes Logunov, 1993; Yaginumaella
Proszynski, 1979; Yllenus Simon, 1868.
Распространение.
Chalcoscirtus, Dendryphantes, Euophrys, Heliophanus, Sitticus — весь регион
вплоть до тундровой зоны.
Aelurillus, Asianellus, Evarcha, Pellenes, Phlegra, Neon, Salticus, Sibianor,
Talavera — весь регион до 65° с.ш.
Carrhotus, Marpissa, Philaeus, Pseudeuophrys, Pseudicius, Synageles — южная
Сибирь и юг Дальнего Востока.
Yllenus — южная Сибирь.
Harmochirus, Helicius, Mendoza, Myrmarachne, Phintella, Plexippoides, Rhene,
Synagelides, Yaginumaella — юг Дальнего Востока.
Tuvaphantes — только Тува.
Наиболее важные работы (где приведены карты распространения видов),
посвященные распространению пауков-скакунчиков азиатской части России: Logunov
& Marusik (2000b, 2003) и Logunov & Kronestedt (2003).
Семейства, встречающиеся в азиатской части России
225
12
16
1
1
20
21
Рис. 32 (продолжение). Salticidae. Примеры строения копулятивных органов: Yllenus
kalkamanicus (cf$, 12, 14), Sitticus mirandus (?, 13), Evarcha michailovi ($, 15), Rhene
atrata (cf$, 16-17), Yaginumaella medvedevi (C?$, 18, 21), Sitticus fas ciger (cf, 20) и
Heliophanus baicalensis (?, 22).
12 — пальпа сГ, сбоку; 13-16,19,21 —эпигина, снизу; 17-18,20,22 — пальпасГ, снизу. 12-14 —
по: Marusik et al. (2007); 22 — по: Marusik & Cutler (1989).
226
4. Специальная часть
Биология. Salticidae населяют в азиатской части России все природные зоны и
продвигаются на север вплоть до острова Врангеля (-71,5° с.ш.). Встречаются они
во всех биотопах и всех стациях. Хотя большинство видов населяют травостой и
кустарники, есть и подстилочные (в лесу) виды {Neon, некоторые Euophrys, Synage-
lides, Yaginumaella), и дендробионты (некоторые Sitticus), и петрофилы
(большинство Chalcoscirtus, некоторые Heliophanus, Sitticus). Большинство видов на севере
населяют наиболее теплообеспеченные биотопы, однако есть и исключения. Так,
хотя большинство Chalcoscirtus — термофильные виды, С. alpicola (L. Koch, 1876)
и С. hyperboreus Marusik, 1991 селятся среди мхов и лишайников на северных
склонах и в горных тундрах.
Судя по всему, большинство наших видов — неспециализированные хищники.
В тропиках и субтропиках встречается немало форм, специализированных на
питании термитами, москитами, и даже пауками, например Portia, Cyrba. Для вида
Cyrba, встречающегося в Закавказье, характерна полифагия, но большую часть его
диеты составляют пауки рода Oecobius (Oecobiidae) (Guseinov et al., 2004; Mapy-
сик, 2006).
До недавнего времени считалось, что все Salticidae охотятся (находят жертв)
благодаря глазам, устроенным наиболее совершенно во всём отряде. Однако
оказалось, что представители выше упомянутого рода Cyrba представляют
исключение. Выискивают свою жертву они с помошью первой пары ног, которую
используют как усики, ощупывая пространство впереди себя.
Для всех скакунчиков характерна забота о потомстве. Многие виды
откладывают небольшое число яиц. Самка сплетает логово без какого-либо выхода, в
котором откладывает один кокон. Несколько исследованных гнёзд Chalcoscirtus в
верховьях Колымы содержали 4-6 яиц. Чуть большее число яиц (8-11) было в коконах
Euophrys proszynskii Logunov, Cutler et Marusik, 1993.
Этологии сальтицид посвящена специальная работа Д.Б. Ричмана и P.P.
Джексона (1991), опубликованная на русском языке.
Методы сбора. Все способы и все биотопы. Логова некоторых видов {Sitticus)
располагаются в метёлках тростника. Иногда бывает по нескольку логовов на
одной метёлке. Некоторых Pellenes можно заметить благодаря белым логовам среди
травы и мелких кустарничков. Многие виды встречаются под корой деревьев,
среди осыпей и под камнями. В таких местах (особенно на хорошо прогреваемых
склонах) иногда образуются целые колонии, когда под одним плоским камнем
располагаются десятки «домиков» одного или нескольких видов. Наскальные, и
вообще петрофильные, виды легко обнаружить визуально, но активны такие пауки,
только когда светит солнце.
Таксономия. Среди семейств, богатых видами, как у нас, так и в мире вообще,
Salticidae — наиболее хорошо очерченное семейство. Есть некоторые сомнения в
статусе тропических Lyssomaninae, однако в целом представители семейства легко
опознаются благодаря очень крупным передним медиальным глазам и чуть более
мелким передним латеральным. Хотя границы семейства ясны, родственные связи
этой группы остаются неясными. Сальтицид сближали и с Thomisidae, и с Gnapho-
sidae, Clubionidae и некоторыми другими семействами.
Семейства, встречающиеся в азиатской части России
227
Пауки, населяющие Сибирь и Дальний Восток России, относятся к нескольким
подсемействам: Aelurillinae, Dendryphantinae, Euophryinae, Heliophaninae, Hyllinae,
Mogrinae, Myrmarachninae и Plexippinae. Все эти подсемейства относительные
молодые (не встречаются в балтийских янтарях, возраст которых составляет около 50
млн. лет).
Salticidae, пожалуй, самая изученная группа пауков азиатской части России. Это
касается и надвидовой и видовой классификации, а также распространения. В
отличие от других групп со многими видами и родами, здесь практически не
осталось «проблемных» родов.
Хотя первые работы с описанием скакунчиков азиатской части России были
написаны A. Grube (1861) и L. Koch (1879b), зачинателем изучения этой группы
следует считать W. Kulczynski (1895), который подготовил специальную работу,
посвященную этому региону. После практически 70-летнего промежутка, работы
по систематике сальтицид азиатской части России были активизированы другим
польским исследователем J. Proszynski. Он переописал ряд малоизвестных видов,
описанных ещё А. Грубе, и описал немало новых видов, преимущественно из
Дальнего Востока (Proszynski, 1971, 1979). Благодаря этим работам и активному
содействию самого Е. Прушиньского, систематикой пауков азиатской части России
занялись П.М. Дунин (1984а), С.Н. Данилов, Д.В. Логунов и Ю.М. Марусик.
Скакунчикам Сибири и Дальнего Востока России посвящено так много работ, что здесь
мы перечислим только основные: Kulczynski (1895, 1908); Proszynski (1971, 1979,
1987); Дунин (1984а); Марусик (1988а, 1990, 1991а-б); Marusik & Cutler (1989);
Логунов (1991); Logunov (1992a-b, 1993a-c, 1996b, 1998, 1999); Логунов, Марусик
(1991); Logunov & Wesolowska (1992, 1995); Danilov & Logunov (1994); Logunov et
al. (1993, 1999); Logunov & Marusik (1994b, 1999a-b, 2000a-b, 2003); Logunov &
HQciak (1996); Logunov & Rakov (1996); Logunov & Kronestedt (1997, 2003); Rakov
(1997); Azarkina (2004). К обобщающим работам можно отнести каталог пауков-
скакунчиков Северной Азии (Logunov & Marusik, 2000b). Важной работой, без
которой нельзя обойтись при изучении пауков азиатской части России и других
регионов, является периодически обновляемый электронный каталог Е. Прушинського
(http://salticidae.org/salticid/main.htm).
В настоящее время изучением систематики пауков-скакунчиков азиатской
части России и других регионов продолжают активно заниматься Д.В. Логунов и
Г.Н. Азаркина.
Перспективы изучения. Несмотря на то, что это семейство относится к
наиболее изученным в регионе, тем не менее, не исключены находки дополнительных
новых видов. Сравнительное изучение разных популяций широко
распространённых видов может выявить новые для науки виды.
Семейство Sparassidae Bertkau, 1872
Гигантские бокоходы, large huntsmen spiders
Рис. 33.1-7.
В мире известно 1056 видов, относимых к 83 родам. В России и азиатской
части России известно всего два вида из двух родов.
Диагностические признаки. Крупные и очень крупные энтелегинные пауки,
два коготка, ноги расставлены в стороны, предлапки и лапки со скопулой.
Близкие группы. Близки и напоминают Philodromidae, легко отличаются
более крупным размером и наличием зубчиков на заднем крае хелицер.
ОПИСАНИЕ:
• Карапакс. Грушевидный.
• Стернум. Не модифицирован.
• Глаза. 8 в два широких ряда. Передний ряд выгнутый, задний почти
прямой. Глаза примерно одинаковой величины.
• Ротовой аппарат. Не модифицирован. Задний край желобка хелицер с 3
зубчиками.
• Ноги. 2 коготка; длинные, все расставлены в стороны примерно под
прямым углом к оси тела, примерно одинаковой длины; предлапки и лапки со
скопулой. Мембрана между лапкой и предлапкой разделена на 3 части, что
обеспечивает большую подвижность.
• Брюшко. Овальное, заострено на конце.
• Паутинные бородавки. Не модифицированы. Колюлюса нет.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся дыхальцем у
паутинных бородавок.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Энтелегинные. Пальпа самца крупная, с мощным
отростком голени. Эмболюс длинный, нитевидный или утолщённый. Эпи-
гина очень крупная, сильно хитинизирована, без ямки.
• Размер. 7-30 мм. Половой диморфизм имеется.
• Окраска. От коричневого (Heteropoda) до салатового или розового
(Micrommata). Самцы Micrommata с красными полосами (медиальной и
латеральными) на брюшке.
• Особые черты. Крупный размер; ноги (бёдра), расставленные в стороны;
скопула на лапках и предлапках; голень пальпы самца с единственным, но
очень крупным и хитинизированным отростком.
Рода, известные из азиатской части России. Heteropoda Latreille, 1804 (Я.
venatoria (Linnaeus, 1767)); Micrommata Latreille, 1804 (M. virescens (Clerck, 1758)).
Распространение. В России известно всего два вида, оба встречаются в
азиатской части России. Н venatoria известна всего по одной самке, собранной на
морском побережье в Приморье (Marusik & Kuzminykh, 2010), а М. virescens
распространена по всей южной Сибири и на юге Дальнего Востока России. Н venatoria,
Семейства, встречающиеся в азиатской части России
229
1
-if
t^&^l^^*
~4—
Рис. 33. Sparassidae. Micrommata virescens (сГ$).
1 — головогрудь сГ, сверху; 2 — сГ, габитус; 3 — пальпа сГ, сбоку; 4 — эпигина, снизу; 5 —
предлапка и лапка I, сзади; 6—трёхлопастное мембранное сочленение предлапки и лапки, сверху;
7 — нога I сГ, сзади. 3-4 — по: Roberts (1995).
очевидно, случайно интродуцирована. Нахождение дополнительных видов в
азиатской части России маловероятно.
Биология. Бродячие охотники, в основном ночные. М. virescens — обитатель
подстилки в лесах и травостоя на лугах. Н. venatoria имеет космополитное
распространение и, чаще всего, распространяется человеком.
Методы сбора. Укосы и визуальный поиск. Реже самцы попадаются в ловушки.
230
4. Специальная часть
Таксономия. Долгое время это относительно крупное семейство было
совершенно неизученным, как на видовом, так и на надвидовом уровне. Е. Simon (1897)
рассматривал эту группу как подсемейство (Sparassinae) в Clubionidae. Т.Н. Jarvi
(1912, 1914) попытался ревизовать этот таксон в мировом масштабе и придал ему
ранг семейства. Во второй половине 20-го века велась дискуссия, как правильно
называть это семейство — Sparassidae или Heteropodidae. Под вторым названием
эта группа фигурирует в монографиях по Африке (Dippenaar-Schoeman, Jocque,
1997) и Советскому Союзу (Mikhailov, 1997). Последние годы это семейство
активно разрабатывает P. Jager вместе соавторами в рамках Старого Света. Описано
немало новых родов и видов из Средней Азии, Юго-Восточной Азии, Японии.
Перспективы изучения. Нахождение дополнительных видов в регионе
маловероятно.
Micrommata virescens (Sparassidae), самец, Прибайкалье (фото: В.Г. Шиленков).
Семейства, встречающиеся в азиатской части России
231
Yaginumaella sp. (Salticidae), самка, Приморье (фото: М.М. Омелько).
Семейство Tetragnathidae Menge, 1866
Тетрагнатиды; long-jawed spiders (Tetragnathinae),
silver marsh spiders (Leucauginae)
Рис. 34.1-23.
Крупное, всесветно распространённое семейство с 48 родами и 937 видами. В
России известно, по меньшей мере, 39 видов из 6 родов. В азиатской части России
встречаются 37 видов.
Диагностические признаки. Некрибеллятные, 8-глазые пауки; брюшко чаще
всего удлинённое, с серебристым или золотистым пигментом; ноги и пальпы
длинные, максиллы в полтора и более раз превышают по длине нижнюю губу
Близкие группы. Metinae (Meta, Metellina, Metleucauge) могут быть спутаны с
некоторыми Araneidae, Linyphiidae и Zygiellidae. От всех отличаются
удлинёнными максиллами. Самцы дополнительно отличаются от Araneidae очень длинными
передними ногами и удлинённой голенью пальпы; от Linyphiidae — неподвижно
приросшим оттопыренным или прямым парацимбиумом; от Zygiellidae —
отсутствием медиального отростка и округлым тегулюмом.
ОПИСАНИЕ:
• Карапакс. От коротко-грушевидного до удлинённо-грушевидного, есть
несколько видов с ромбовидным карапаксом; наличник невысокий
(исключение — некоторые Pachygnatha).
• Стернум. Удлинённый.
• Глаза. 8, в два ряда (4-4), латеральные часто сближены.
• Ротовой аппарат. Максиллы удлинены, как минимум в 1,5 раза длиннее
нижней губы; ширина максилл в 1,5-2 раза меньше длины. Хелицеры у
большинства видов удлинены, с многочисленными мощными шипами и с
дорсальным выростом у самца (все Tetragnathinae). Коготок хелицер у
основания иногда с зубчиком.
• Ноги. 3 коготка. Передние ноги длинные, чаще всего в 2 и более раз
длиннее тела. У Leucauge базальная половина бедра IV с двумя рядами трихобот-
рий (диагностический признак рода).
• Брюшко. Овальной, яйцевидной формы, иногда чуть заострено на конце, у
многих видов (Tetragnatha) длина превышает ширину в 2 и более раз.
Иногда с хвостовидным выростом.
• Паутинные бородавки. Не модифицированы.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками; иногда дыхальце может быть
несколько отнесено от бородавок.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Пальпа самца с базальным крупным
парацимбиумом, он может быть вертикальным или оттопыренным (кроме Leucauge и
Metleucauge). Голень у всех видов (кроме некоторых Pachygnatha) в несколько
раз длиннее своего диаметра. Тегулюм обычно крупный, шаровидный или
слегка овальный, с просвечивающим широким семенным канальцем. Тегу-
Семейства, встречающиеся в азиатской части России
233
люм несёт только эмболюс (обычно длинный) и кондуктор, обычно
служащий опорой-стержнем для длинного эмболюса (исключение Meta,
основание эмболюса у которой с выростами). У Metleucauge вертлуг пальпы самца
удлинён и несёт шип (диагностический признак рода). Эпигина у всех видов
Tetragnathinae редуцирована до простого щелевидного отверстия,
вынесенного далеко за пределы эпигастральной щели по направлению к паутинным
бородавкам. У представителей других подсемейств эпигина сравнительно
простая, плоская и без выростов, всегда с медиальной пластинкой.
• Размер. 3-15 мм.
• Окраска. Большинство видов имеют серебристый или золотистый пигмент
на брюшке. Вентрально окраска брюшка чаще всего — чёрная (кроме
Pachygnatha). Некоторые дальневосточные виды Tetragnatha имеют зеленоватую
окраску.
• Особые черты. Удлинённые максиллы.
Рода, известные из азиатской части России: Diphya Nicolet, 1849
(собственные данные); Leucauge White, 1841; Meta C.L. Koch, 1836; Metellina Chamberlin et
Ivie, 1941; Metleucauge Levi, 1980; Pachygnatha Sundevall, 1823; Tetragnatha Latreille,
1804.
Распространение.
Tetragnatha, Pachygnatha — весь регион; последний род не известен лишь в
верховьях Колымы и на Индигирке (регионе с экстраконтинентальным климатом).
Diphya, Leucauge, Meta и Metleucauge — юг Дальнего Востока.
Metellina — Западная Сибирь, но, судя по всему, один вид встречается и в
Приморье.
Ареалы более 10 видов тетрагнатид, населяющих азиатскую часть России,
приведены в работе Marusik et al. (2000).
Биология. В нашем регионе тетрагнатиды встречаются во всех зонах, вплоть
до 71° с.ш. Населяют широкий спектр биотопов с умеренной и высокой
влажностью.
Все виды — кругопряды; исключение составляют лишь взрослые Pachygnatha,
которые вторично утратили способность плести сеть. Сети у всех видов
правильные, но с более редкими и относительно нежными нитями спирали, нежели у
пауков-крестовиков (Araneidae). Центр сети чаще всего свободный (не заплетённый).
Питаются они преимущественно комарами и другими амфибиотическими
насекомыми. В экспериментальных условиях установлено, что при обилии комаров
Tetragnatha перестаёт плести сети, и охотится подобно паукам засадникам.
Сети большинства видов устраиваются среди травостоя, кустарников и в
нижних частях крон деревьев. Исключение — лишь Pachygnatha, которые сплетают
сети у поверхности почвы, и Meta — облигатный троглобионт. Metellina и
Metleucauge устраивают сети практически всегда в затененных местах. Сети многих
видов Tetragnatha и все Metleucauge располагаются у воды и практически всегда
горизонтально.
Tetragnatha может быть встречена на сети в дневное время, но обычно сидит на
близлежащих травинках, прижавшись к ним и вытянув ноги вдоль травинки. За-
234
4. Специальная часть
4 6
1
2
5
3
8 О
1
13 14
Семейства, встречающиеся в азиатской части России
235
22
23
Рис. 34. Tetragnathidae. Примеры строения копулятивных органов и соматические
признаки: Tetragnatha obtusa (cf$, 1-3, 11), Metleucauge kompirensis ($, 4-5, 13; сГ$, 15-
18), Diphya sp. (cf$, 6-9), Pachygnatha quadrimaculata ($, 10, 12, 14), Leucauge celebe-
siana (cT, 19-20), Menosira ornata (cT, 21) и Meta manchurica (C?$, 22-23).
1,4 — головогрудь ?, снизу; 2, 5, 9 — головогрудь ?, спереди; 3, 12 — габитус сГ, сбоку; 6 —
головогрудь 0\ снизу; 7 — головогрудь С?, спереди; 8 — лапка и предлапка I d\ сверху; 10 —
брюшко ?, снизу; 11 — габитус ?, сбоку; 13-14 — габитус ?, сверху; 15, 21-22 — пальпа сГ,
снизу; 16, 19 — пальпа сГ, сбоку; 17 — максиллы и лабиум 0\ снизу; 18, 23 — эпигина, снизу;
20 — бедро С? IV, сверху. Пц — парацимбиум; Эп — «эпигина», показано насколько далеко
выдвинуто совокупительное отверстие, и слабая хитинизация. 15-18,19-20 — по: Zhu et al. (2003);
22-23 — по: Marusik & Koponen (1992).
236
4. Специальная часть
метить паука при этом очень сложно. В дневное время их легко собирать с
помощью сачка, укосами. В момент опасности пауки могут опускаться на воду, и
способны передвигаться по поверхности воды (Foelix, 1982). Metleucauge охотятся
(выбираются в центр сети) исключительно ночью. Днём они прячутся в сплетении
корней, и найти их очень сложно, хотя сети встречаются довольно часто. Сети
Metellina могут располагаться вертикально. Эти пауки, как и Leucauge, легко
собираются укосами на лугах и опушках.
Meta сплетает сети исключительно в пещерах, гротах, среди
крупнообломочных осыпей, но несколько экземпляров были собраны в брошенных домах под
строительным мусором на острове Шикотан и среди поросшего мхом бурелома в
тёмном лесу на острове Сахалин.
Спаривание у Tetragnathinae {Pachygnatha, Tetragnatha) происходит в
необычной манере. Самцы сближаются с самками «лицом» к «лицу» и захватывают хели-
церами коготки хелицер самки. После чего вводят свои пальпы в половую щель,
но не поочерёдно, как у других энтелегинных пауков, а сразу две пальпы
одномоментно.
Tetragnatha откладывают линзовидные коконы на травинки и прикрепляют их
тонким пологом золотистого цвета. Зимуют неполовозрелые особи, которые
осенью встречаются до первых морозов. Изучена холодоустойчивость зимующих
особей у Т. externa (Linnaeus, 1758) (самый распространённый вид нашей фауны,
встречающийся от Индии до 70° с.ш.) (Россолимо, Марусик, 1989). Она достигает -22°С.
Коконы у Meta белые, каплевидные, много крупнее самого паука, крепятся к
«потолку» несколькими нитями. Заметить присутствие Meta гораздо легче
благодаря коконам, нежели по сетям или самим паукам.
Методы сбора. Укосы {Tetragnatha, Leucauge, Metellina), почвенные ловушки и
ручной сбор на поверхности грунта {Pachygnatha). Metleucauge проще всего
собрать ночью, когда пауки забираются в центр сети. Meta можно собрать только в
пещерах и гротах, несколько особей собраны среди развалин деревянных
строений.
Таксономия. В азиатской части России обитают представители трёх
подсемейств: Tetragnathinae {Pachygnatha, Tetragnatha), Metinae {Meta, Metellina,
Metleucauge) и Leucauginae {Leucauge).
Хотя семейство Tetragnathidae было выделено сравнительно давно, ещё в 1866 г.,
благодаря работам Е. Simon (1892 и др.) его долгое время рассматривали как
подсемейство в Araneidae. Ведущий специалист по паукам кругопрядам мира Н. W. Levi
рассматривал эту группу как подсемейство, по крайней мере до 1981 г.
Подсемейство Metinae до самого недавнего времени большинством авторов включалось в
Araneidae, причём зачастую даже без выделения в отдельное подсемейство (см.,
например, определитель В.П. Тыщенко, 1971). P.M. Brignoli (1983) рассматривал и
Metinae и Tetragnathinae в статусе отдельных семейств. Если вопрос о том,
включать ли Metinae в Tetragnathidae, или придать ему статус отдельного семейства —
дискуссионный, то в настоящее время никто не оспаривает, что Metinae не
родственны Araneidae.
Несколько представителей дальневосточной фауны до недавнего времени
рассматривали в роде Dyschiriognatha Simon, 1893, однако сейчас доказано, что они
Семейства, встречающиеся в азиатской части России
237
имеют мало общего с типовым видом этого рода, и были перенесены в Pachygnatha
(Zhu et al., 2003).
Tetragnatha — один из немногочисленных родов пауков, имеющих всесветное
распространение. Ранее выделялись несколько родов внутри Tetragnatha, однако
была показана несостоятельность таких таксонов. Из-за отсутствия хитинизиро-
ванной эпигины Tetragnathinae иногда считают (или просто называют) «полуэнте-
легинными». «Полу-» — из-за того, что самцы имеют такие же пальпы, как у энте-
легинных пауков.
Два других рода {Pachygnatha, Leucauge) также имеют широкое
распространение. Первый известен по всему Старому Свету (без Австралии) и в Неарктике, а
второй — пантропическое распространение. Если Pachygnatha можно считать
относительно «хорошим» родом (из-за однообразия внешнего вида и копулятивных
органов), то единственной чертой, объединяющей виды рода Leucauge, является
наличие ряда трихоботрий на бедре IV. Копулятивные органы в этом таксоне
чрезвычайно разнообразны, и он, на наш взгляд, несомненно, должен быть разделён на
несколько родов.
Таксономии пауков тетрагнатид азиатской части России посвящено всего три
работы: Strand (1907); Marusik & Koponen (1992); Kurenshchikov (1994).
По сопредельным регионам подготовлены замечательнае сводки в Китае (Zhu
et al., 2003) и в Японии Ono et al. (2009). Род Tetragnatha Японии и всей
юго-восточной Азии ревизован в серии работ японской исследовательницы Т. Окума
(Okuma, 1988-1989).
Разграничение видов. В пределах родов Tetragnatha и Pachygnatha виды
проще всего различаются не копулятивными органами, а вооружением хелицер (в том
числе и наличием/отсутствием зубчика на коготке хелицер) и, нередко, окраской
карапакса {Pachygnatha), стернума {Т externa (Linnaeus, 1758) и Tpinicola L. Koch,
1870) и брюшка. В ряде случаев имеет значение форма (профиль) брюшка.
Представители других подсемейств различаются почти исключительно формой пальп и
эпигин.
Перспективы изучения. Вполне возможно обнаружение дополнительных
видов на юге Дальнего Востока, новых как для региона, так и для науки. На наш
взгляд, под названием Pachygnatha clercki Sundevall, 1823 в регионе скрывается
несколько близких видов.
Семейство Theridiidae Sundevall, 1833
Теридииды; cobweb spiders, comb-footed spiders
Рис. 35.1-35.
Пятое по величине семейство в мире, всесветно распространено, насчитывает
109 родов и 2293 вида. В России известно около 115 видов более чем из 32 родов,
а в азиатской части России — около 93 видов из 29 родов.
Диагностические признаки. Энтелегинные, некрибеллятные пауки, от
мелкого до среднего размера, с 8 глазами, 3 коготка, на лапках IV — серия пильчатых
загнутых щетинок, наличник высокий.
Близкие группы. Габитуально и окраской схожи с Mysmenidae, Nesticidae,
Theridiosomatidae, некоторыми Araneidae, Linyphiidae, Tetragnathidae и Uloboridae.
В большинстве случаев, Theridiidae можно отличить по пильчатым щетинкам
на лапках IV (подобные есть только у Nesticidae). Самцов Theridiidae можно
однозначно отделить от сходных таксонов по наличию верхнего парацимбиума и по
закруглённой на вершине голени пальпы. Самок отличить сложнее, особенно Rober-
tus, которые очень напоминают некоторых Linyphiidae (но у Robertus нет стридуля-
ционных борозд по наружному краю базального сегмента хелицер). Подробнее о
различиях близких аранеоидных групп смотри определительную таблицу (Ключ III).
ОПИСАНИЕ:
• Карапакс. Форма сильно варьирует, от круглого и овального до
грушевидного. Высота от низкой до очень высокой. У самцов может быть с
выростами. Клипеус иногда очень высокий, может быть вертикальным или пологим,
у одного вида Rhomphaea с рострумом. Иногда грудная часть может быть с
ямкой или длинными бороздами.
• Стернум. Сильно варьирует по форме.
• Глаза. 8, в два ряда (4-4), латеральные соприкасаются (исключение — Latro-
dectus и Episinus).
• Ротовой аппарат. Варьирует, у всех групп максиллы наклонены друг к
другу, хелицеры могут быть короче наличника (Dipoena и родственные рода), у
самцов некоторых родов (например, Enoplognatha, Steatoda) хелицеры
удлинены и несут мощные зубцы спереди, или даже по всему периметру
хелицеры (у Enoplognatha monstrabilis Marusik & Logunov, 2002).
• Ноги. 3 коготка, лапки IV с рядом слегка загнутых зазубренных (пильчатых)
щетинок, служащих для плетения сетей. Иногда эти щетинки сильно
редуцированы. Шипы имеются или отсутствуют. У самцов Enoplognatha и
некоторых других родов на ногах вентрально могут быть шиловидные выросты
кутикулы.
• Брюшко. Очень разнообразной формы, от сферической и овальной до
каплевидной и пентагональной, иногда с выростами в передней или задней
части. Длина выростов значительно варьирует, от коротких (короче тела, Phoron-
cidia), до очень длинных (длиннее тела, Rhomphaea). У самцов Theridion
sensu lato и Enoplognatha предэпигастральная область удлинена, «вздута» и
возвышается над постэпигастральной частью.
• Паутинные бородавки. Не модифицированы, колюлюс имеется или
отсутствует.
Семейства, встречающиеся в азиатской части России
239
Рис. 35. Theridiidae. Robertus ungulatus ($, 1), Steatoda bipunctata ($, 2), Enoplognatha
sp. (cf5 3), Phycosoma mustelina ($, 4, 7), Thymoites oleatus ($, 5), Enoplognatha serra-
tosignata ($, 6), Theridion sibiricum ($, 8) и Enoplognatha gramineusa (cf 5 9).
1,4 — головогрудь ?, спереди; 2 — паутинные бородавки ?, снизу; 3 — стридуляционный орган
С?5 снизу; 5-6 — эпигина, снизу; 7-8 — головогрудь ?, снизу; 9 — передняя часть головогруди С?,
снизу.
240
4. Специальная часть
13
14
15
16
17
Рис. 35 (продолжение). Theridiidae. Внешний вид: Ohlertidion ohlerti (cf$, 10-11), Thy-
moites oleatus ($, 12), Steatoda bipunctata (?, 13), Enoplognatha gramineusa (cf,14), Phyco-
soma mustelina ($, 15), Robertus ungulatus ($, 16), Theridion sibiricum ($, 17), Rhomphaea
hyrcana (cf, 18) и Phoroncidia pilula (?, 19).
10, 12-13, 15-16 — габитус ?, сверху; 11, 14 — габитус сГ, сверху; 17, 19 — габитус ?, сбоку; 18 —
габитус 0\ сбоку.
Семейства, встречающиеся в азиатской части России
241
Рис. 35 (продолжение). Theridiidae. Примеры строения копулятивных органов и
соматические признаки: Enoplognatha serratosignata (сГ,-20), Heterotheridion nigrovariegatum
(ti\ 21), Robertus kastoni (сГ$, 22, 28, 31), "Theridion" palmgreni (cT, 23), Enoplognatha
monstrabilis (O71, 24-25, 33-35), Rhomphaea hyrcana (cT$, 26, 30), Arctachaea nordica (O71,
27) и Robertus sibiricus (cT?, 29, 32).
20, 31-32 — пальпа С?, сбоку, виден пальцевидный парацимбиум; 21 — цимбиум без бульбуса,
показан кармановидный парацимбиум; 22 — вершина цимбиума с пальцевидным парацимбиу-
мом; 24 — шипики на предлапке I у С?; 25 — нога I С?; 26-27 — пальпа С?, снизу; 28-30 —
эпигина, снизу; 33-35 — хелицера спереди, сбоку и сзади. Стрелка указывает на пальцевидный
(20,22) и кармановидный (21) парацимиум. 22,28-29, 31-32 — по: Eskov (1987): 23 — по: Мару-
сик, Целлариус (1986); 24-25, 33-35 — по: Marusik & Logunov (2002a); 26 — по: Логунов, Мару-
сик(1990).
242
4. Специальная часть
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Энтелегинные. Пальпа самца варьирует по
размеру, иногда по длине может превышать головогрудь; голень чашевидная, рет-
ролатеральная часть голени несколько вытянута и закруглена, без апофизов.
Цимбиум с верхним парацимбиумом, который может быть двух типов: 1)
пальцевидный, иногда заострённый на конце (Enoplognatha, Robertus, Cru-
stulina, Steatoda и др.); 2) своего рода карман внутри цимбиума {Theridion
sensu lato). Цимбиум может быть с апикальными выростами, форма его очень
вариабельна. Бульбус тоже разный по форме, с разным числом отростков.
Эмболюс от длинного нитевидного, до короткого. Эпигина очень варьирует
по форме, бывают с ямкой или, наоборот, с выростом; скапуса нет. У
большинства видов эпигина плоская. Вульва у большинства групп с настоящими
трубками, иногда очень длинными. Рецептакул обычно 2, но у всех родов,
близких к Dipoena, их 4.
• Размер. 2-15 мм, в большинстве случаев размерный половой диморфизм
имеется.
• Окраска. Очень разнообразная, от одноцветных и без рисунка {Robertus),
до очень вариабельных по цвету и рисунку в пределах одного вида {Rugathodes
nigrolimbatus (Yaginuma, 1972)).
• Особые черты. Самцы многих видов имеют стридуляционные органы на
заднем крае карапакса (серия гребней) и вершине брюшка (сильно хитини-
зировано, с зубчиками, о которые трутся гребни карапакса). Лапки IV вент-
рально с рядом слегка загнутых зазубренных щетинок, голень пальпы самца
закруглена ретролатерально, верхний парацимбиум, вульва с трубками.
Рода, известные из азиатской части России. Arctachaea Levi, 1957 {A. nordica
(Chamberlin et Ivie, 1947)); Canalidion Wunderlich, 2008 (C. montanum (Emerton,
1882)); Chrysso O. Pickard-Cambridge, 1882; Crustulina Menge, 1868; Cryptachaea
Archer, 1946 (C riparia (Blackwall, 1834)); Dipoena Thorell, 1869 {D. torva (Thorell,
1875)); Enoplognatha Pavesi, 1880; Episinus Walckenaer in Latreille, 1809; Euryopis
Menge, 1868; Heterotheridion Wunderlich, 2008 (Я. nigrovariegatum (Simon, 1873));
Lasaeola Simon, 1881; Latrodectus Walckenaer, 1805 {L. tredecimguttatus (Rossi, 1790));
Neottiura Menge, 1868; Ohlertidion Wunderlich, 2008 {O. ohlerti (Thorell, 1870)); Pai-
discura Archer, 1950 {Ppallens (Blackwall, 1834)); Parasteatoda Archer, 1946 (=Achae-
aranea ad partem); Phoroncidia Westwood, 1835 (i^ pilula (Karsch, 1879)); Phycosoma
O. Pickard-Cambridge, 1879; Phylloneta Archer, 1950; Platnickina Ko£ak et Kemal,
2008 {=Keijia Yoshida, 2001); Rhomphaea L. Koch, 1872 {R. sagana (Donitz et Strand,
1906)); Robertus O. Pickard-Cambridge, 1879; Rugathodes Archer, 1950; Steatoda Sun-
devall, 1833; Stemmops O. Pickard-Cambridge, 1894 {S. nipponicus Yaginuma, 1969);
Takayus Yoshida, 2001; Theridion Walckenaer, 1805; Thymoites Keyserling, 1884; Ya-
ginumena Yoshida, 2002 {Y. castrata (Bosenberg et Strand, 1906)).
Распространение.
Arctachaea, Canalidion, Enoplognatha, Euryopis, Ohlertidion, Phoroncidia, Phylloneta,
Robertus, Rugathodes, Steatoda, Theridion, Thymoites — по всему региону, но на
севере большинство родов встречается только в Якутии и Магаданской области.
Семейства, встречающиеся в азиатской части России
243
Crustulina, Neottiura, Parasteatoda, Takayus — южная Сибирь и юг Дальнего
Востока. Синантропные Parasteatoda могут встречаться и севернее.
Dipoena, Lasaeola, Paidiscura — только в южной Сибири до Прибайкалья.
Heterotheridion, Latrodectus — только на юго-западе.
Episinus — Западная Сибирь и юг Дальнего Востока.
Chrysso, Platnickina, Phycosoma, Rhomphaea, Stemmops, Yaginumena — только
юг Дальнего Востока.
Единственная работа, где приведены карты с распространением отдельных
видов теридиид, была опубликована Ю.М. Марусиком с соавторами (Marusik et al.,
2000).
Биология. Как и морфология, биология пауков этой группы чрезвычайно
разнообразна. Они населяют все типы биотопов и все ярусы, от подстилки до крон
деревьев. Несколько видов Steatoda являются синантропами. Кроме Steatoda, в
жилищах часто поселяются Parasteatoda. На север теридииды продвигаются вплоть
до тундровой зоны. При продвижении на север, видовой состав очень обедняется.
Наибольшее разнообразие в пределах азиатской части России наблюдается на юге
Дальнего Востока.
В сравнении с другими семействами, среди теридиид высока доля видов,
обитающих на деревьях и кустарниках. В этом плане они сравнимы только с пауками-
крестовиками.
Подавляющее большинство наших видов плетут паутину, однако у Euryopis,
Dipoena и родственных родов сеть редуцировалась до нескольких сигнальных
нитей. Эти пауки охотятся исключительно на муравьев. Видимо поэтому на севере,
где мало муравьев, встречается всего один вид Euryopis. Крайне редуцированы
сети и у Episinus. Помимо упомянутых родов, при случае (если попадут в сеть)
используют в пищу муравьев и представители многих других родов.
Сети Parasteatoda, Theridion очень прочные и могут удерживать очень крупную
добычу, во много раз превышающую размер паука. Особенно жёсткие и прочные
сети у каракуртов Latrodectus, которые могут ловить даже мелких ящериц! В ловле
добычи иногда принимают участие клейкие нити, но часто для этого служат и
неклейкие. У части сетеплетущих видов сеть плоская (пологовидная), располагается
над поверхностью, от неё книзу отходят клейкие нити, служащие для поимки
ползающих насекомых. Если сеть располагается высоко над землёй, то она обычно
трёхмерная. Летающее насекомое врезается в верхнюю часть сети (рыхлое и
беспорядочное сплетение нитей) и, оглушённое, падает на густую пологовидную
паутину, расположенную внизу, где паук и умерщвляет добычу. На сетях некоторых
видов теридиид подвешены камешки на паутинках (у Latrodectus, отдельных
видов Theridion). Их функция непонятна; возможно, они обеспечивают поддержание
нужной формы сети. Многие сетеплетущие виды имеют убежище. Оно может быть
разной формы. Часто сети камуфлируются листиками, а иногда — остатками жертв.
Устройство сети может сильно меняться с возрастом, отличаться у взрослых и
молодых, у молодых разных возрастов. У каракурта, например, сети молодых
особей похожи на напёрсток, подвешенный над землёй на паутинках, а сети
взрослых — классическая для теридиид конструкция с пологом, с «растяжками» сверху
и снизу полога, с логовом в глубине сети.
244
4. Специальная часть
Некоторые виды Enoplognatha (у нас это Е. margarita Yaginuma, 1964) охотятся
как засадники, располагаясь в цветках растений.
После оплодотворения самка откладывает несколько коконов. Коконы обычно
сферические, у большинства групп белые, однако могут быть и зеленоватыми,
голубыми или иного цвета. У некоторых видов могут быть выросты на сферическом
коконе. Коконы могут иметь гладкую поверхность (Theridion) или могут быть
покрыты толстым рыхлым слоем паутины (Steatoda). Иногда кокон крепится к
бородавкам, и тогда самки могут переносить его.
Самки охраняют кокон. У нескольких видов отмечено, что самки могут
кормить своё потомство. В тропиках есть социальные виды теридиид. У некоторых
видов ослабевшая и перезимовавшая самка, очевидно, служит первой едой для
своего потомства.
Методы сбора. Все методы и все биотопы, в том числе и в домах. Много
интересных видов можно собрать отряхиванием ветвей, особенно хвойных пород
деревьев.
Таксономия. Theridiidae, несомненно, — одно из наиболее сложных для
изучения семейств. Трудности связаны со сложным и разнообразным строением пальп
у самцов. До настоящего времени неизвестны чёткие рамки этой группы.
Единственный признак, объединяющий теридиид, это пильчатые щетинки на лапках IV.
Однако этот признак есть не у всех (у некоторых таксонов, не плетущих сетей, они
редуцированы), а с другой стороны присущ также и Nesticidae. Морфологически и
этологически это семейство чрезвычайно гетерогенно. Из Theridiidae
периодически выделяют некоторые таксоны в отдельные семейства (например, Synotaxidae)
или переводят отдельные рода в другие семейства, в том числе и новые, например
Comaroma Bertkau, 1889 переведена в Anapidae; Cepheia Simon, 1894 в Synaphridae;
Cyclocarcina Komatsu, 1942 и Eidmannella Roewer, 1935 в Nesticidae, a Trogloneta
Simon, 1922 в Mysmenidae. Причем некоторые рода, вынесенные из Theridiidae,
относятся к гаплогинным семействам! Длительное время существовала практика
отнесения новых непонятных родов, не «проходящих» в Araneidae, Linyphiidae и
некоторые другие (когда ещё не были известны Anapidae, Mysmenidae, Theridioso-
matidae), в Theridiidae.
На наш взгляд, в обозримом будущем из состава семейства будут выведены
рода, близкие к Euryopis и Dipoena. Эту группу называют Hadrotarsinae, хотя есть и
другие названия для этой группы, например Euryopinae или Dipoeninae.
О неразработанности этой группы свидетельствует и отношение числа видов к
числу родов — 26 (на один род приходится в среднем 26 видов), в то время как в
целом по отряду это соотношение составляет всего 11.
Наибольший вклад в изучение этого семейства внёс H.W. Levi; помимо ревизий
неарктических и неотропических родов, он ревизовал все известные в мире рода
(Levi & Levi, 1962). До настоящего времени ни одно семейство с числом видов
более 1000 не ревизовано в таком аспекте. На наш взгляд, много, если не
большинство, родов синонимизировано Г.В. Леви неправомочно.
Недавно была предпринята попытка пересмотреть надвидовую систематику этой
группы с помощью кладистических методов в мировом масштабе (Agnarsson, 2004).
Однако сколь-нибудь существенного прогресса, кроме ревалидизации некоторых
родов, эта попытка не принесла.
Семейства, встречающиеся в азиатской части России
245
Среди родов, встречающихся в азиатской части России, наиболее слабо до
последнего времени бы разработан род Theridion, но недавно J.Wunderlich (2008b)
восстановил валидность нескольких синонимизированных ранее родов и описал
несколько новых.
Таксономических работ, касающихся Theridiidae азиатской части России, очень
немного, это: L. Koch (1879); Kulczynski (1908); Holm (1970, 1973); Марусик, Цел-
лариус (1986); Марусик (1988а); Логунов (19926); Marusik & Logunov (2002a) и
некоторые другие.
По сопредельным территориям имеются обстоятельные ревизии Yoshida (2003);
Zhu (1998) и обзоры Song et al. (1999), Ono et al. (2009). Многие голарктические и
сибиро-американские виды могут быть определены благодаря работам H.W. Levi
(1951, 1954, 1957а-Ьидр.).
Разграничение видов. В большинстве случаев виды уверенно различаются
формой копулятивных органов, однако очень многие виды можно легко отличить
по рисунку брюшка и даже карапакса. Помимо этого, некоторые виды можно
диагностировать по форме брюшка и карапакса. Самцы Enoplognatha хорошо
различаются формой хелицер.
Перспективы изучения. В регионе, особенно на юге Дальнего Востока,
имеется немало неописанных видов и даже родов. Возможны находки
дополнительных видов, описанных из сопредельных регионов. Пауки-теридииды — одна из
наименее разработанных групп в мире, и поэтому наиболее перспективная для
изучения и в рамках азиатской части России, и в более широких масштабах.
В настоящее время эта группа разрабатывается Ю.М. Марусиком и А.В.
Громовым.
Семейство Theridiosomatidae Simon, 1881
Теридиосоматиды; ray spiders
Рис. 36.1-6.
В мире известно 77 видов, относимых к 12 родам. В России отмечены два вида,
а в азиатской части России — один.
Диагностические признаки. Менее 3 мм, брюшко сферическое, длинные три-
хоботрии на голени III и IV; по краям у основания нижней губы две круглые ямки.
Близкие группы. Похожи на Mysmenidae и некоторых Araneidae и Theridiidae
{Theridion sensu lato, Parasteatoda, Dipoend). Отличаются наличием ямок по бокам
от нижней губы.
ОПИСАНИЕ:
• Карапакс. Грушевидный, не модифицированный; клипеус больше передних
медиальных глаз в несколько раз.
• Стернум. Щитовидный.
• Глаза. 8, в два ряда, латеральные сближены между собой, все примерного
одинакового размера.
• Ротовой аппарат. Хелицеры относительно короткие. По бокам от нижней
губы имеется пара круглых отверстий желёз.
• Ноги. Относительно короткие, с 3 коготками, голени III и IV с 3-4 рядами
длинных трихоботрий, которые более чем в два раза превышают диаметр
голени.
• Брюшко. Сферическое.
• Паутинные бородавки. Не модифицированы, колюлюс крупный.
• Органы дыхания. Пара лёгких, трахейное дыхальце открывается у
паутинных бородавок.
• Крибеллюм-каламиструм. Отсутствуют.
• Копулятивные органы. Энтелегинные пауки. Пальпа самца очень
крупная, почти округлая, по ширине больше половины карапакса. Парацимбиум
базальный, маленький. Эпигина крупная с глубокой ямкой.
• Размер. 1,5-3 мм. Половой диморфизм выражен.
• Окраска. Брюшко с окраской.
• Особые черты. Круглые отверстия желез по краям нижней губы.
Рода, известные из азиатской части России. Theridiosoma О. Pickard-Cam-
bridge, 1879.
Распространение. В России и СНГ известен только один род Theridiosoma с
двумя видами. В азиатской части России известен только один вид Т epeiroides
Bosenberg et Strand, 1906 из юга Дальнего Востока.
Биология. Обитают исключительно в затенённых биотопах с высокой
влажностью. Кругопряды. В «рабочем» состоянии сеть слегка коническая, паук сидит вне
сети, напротив центра, и натягивает нить, прикреплённую к центральной части.
Семейства, встречающиеся в азиатской части России
247
1 2
Рис. 36. Theridiosomatidae. Theridiosoma epeiroides (сГ$).
1 — габитус ?, сверху; 2 — ? снизу, видны ямки по бокам от нижней губы; 3 — медиальный
отросток бульбуса, снизу; 4 — пальпа С?, снизу-спереди; 5 — пальпа С?, спереди; 6 —
характерная форма кокона. Стрелка указывает на ямку сбоку от нижней губы. 3-5 — по: Марусик (1989а);
6 —по: Roberts (1995).
Сеть Т. gemmosum (L. Koch, 1877) детально описана Wiehle (1931). Коконы
золотисто-коричневые, округлые, диаметром около 3 мм, прикрепляется к поверхности
нитью до 25 мм. Коконы встречаются чаще, нежели сами пауки.
Форма сети и процесс её постройки у Т. epeiroides описаны ^работе Shinkai &
Shinkai (1985), опубликованной на японском языке. Начальные стадии сооружения
сети ничем не отличаются от таковых у пауков-кругопрядов, и строящаяся сеть
всегда имеет густо заплетённый центр. После того, как сделана ловчая спираль,
центр удаляется, к месту слияния радиусов крепится нить, с помощью которой
сеть удерживается в вогнутом состоянии. Когда жертва задевает сеть, паук
отпускает крепёжную нить, сеть несколько выстреливает и провисает, благодаря чему
жертва быстрее запутывается.
248
4. Специальная часть
Методы сбора. Визуальный поиск сетей или коконов в затемнённых влажных
местах.
Таксономия. Надвидовые таксоны этого семейства в мировом масштабе
ревизованы в работе Д. Коддингтона (Coddington, 1986). Theridiosoma относится к
номинативному подсемейству. Диагностические рисунки дальневосточного вида
приведены в работах Марусика (1989а), Chikuni (1989), Namkung (2002) и Опо et al.
(2009).
Разграничение видов. Только форма копулятивных органов.
Перспективы изучения. По всей видимости, на юге Приморья обитает ещё
один вид, относимый к Wendilgarda Keyserling, 1886 (у нас имеется два субадульт-
ных самца). Нахождение дополнительных таксонов маловероятно.
Pardosa "lugubris" (Lycosidae) in copula, Приморье (фото: М.М. Омелько).
Семейства, встречающиеся в азиатской части России
249
Marpissa pulla (Salticidae), самец, Приморье (фото: М.М. Омелько).
Семейство Thomisidae Sundevall, 1833
Пауки-бокоходы, пауки-крабы; crab spiders
Рис. 37.1-28.
Шестое по величине семейство в мире, всесветно распространено,
насчитывает 174 рода и 2085 видов. В России известно, по меньшей мере, 115 видов из 16
родов, а в азиатской части России — 101 вид из 16 родов.
Диагностические признаки. От мелких до крупных, 2-коготковые, некрибел-
лятные, энтелегинные пауки, 8 глаз в 2 ряда, ноги с многочисленными шипами, без
скопулы; пальпа самца с 2 и более отростками голени. У большинства видов длина
брюшка равна или меньше ширины.
Близкие группы. Внешне несколько напоминают Philodromidae, другую
группу пауков бокоходов, однако Thomisidae отличаются тем, что у них нет скопулы, и
задние две пары ног заметно короче передних.
ОПИСАНИЕ:
• Карапакс. Форма варьирует, от грушевидной до овальной или почти
круглой, или квадратной. У некоторых родов (Bassaniana, Coriarachne) уплощён.
У отдельных таксонов покрыт булавовидными щетинками. Некоторые виды
с глазными выростами. Наличник высокий, вертикальный, но может быть и
пологим. У Heriaeus карапакс покрыт торчащими волосками, иногда
волнистыми.
• Стернум. Сердцевидный.
• Глаза. 8, в два ряда (4-4), боковые глаза часто расположены на общем
бугорке, который часто белого цвета, иногда заострённый. Размер глаз может быть
одинаковым, но у многих видов передние латеральные — крупнее других.
• Ротовой аппарат. Хелицеры короткие и толстые в основании. Зубцов вдоль
желобка нет.
• Ноги. 2 коготка. Передние две пары ног длиннее последних двух. Ноги
относительно толстые, длина передних может быть длиннее тела. Ноги с
шипами; всегда есть несколько пролатеральных шипов на бёдрах I, особенно
много шипов на предлапках. Иногда часть шипов булавовидные. Скопулы и
подкоготковых подушечек нет. У Heriaeus ноги покрыты торчащими
изогнутыми или даже волнистыми волосками.
• Брюшко. Варьирует по форме от круглого и овального до
удлинённо-овального, округло-трапециевидного и угловатого. Углы могут быть боковыми или
дорсальными. Наиболее широкая часть брюшка — задняя треть. У Heriaeus
брюшко покрыто торчащими волнистыми волосками, часто белыми.
• Паутинные бородавки. Не модифицированы.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Пальпы самца с РГО, голенных отростков от двух
до трёх, цимбиум у большинства видов с ретролатеральным выростом
(называемым тутакулюмом), бульбус (тегулюм) дисковидный, с тегулярными
Семейства, встречающиеся в азиатской части России
251
1
7 8
10 11
Ту
Рис. 37. Thomisidae. Внешний вид и копулятивные органы: Oxytate striatipes (С?, 1),
Lysiteles coronatus ($, 2), Diaea suspiciosa ($, 3), Synema globosum ($, 4, 12), Misumena
vatia (O71, 5), Tmarus horvathi (?, 6), Ozyptila scabricula ($, l),Xysticus dzhungaricus (cf, 8,
10) и Jf. cowcreto (С??, 9, 11, 13).
1-9 — габитус; 10-11 — пальпа сГ, снизу; 12 — головогрудь ?, спереди; 13 — эпигина, снизу
Ту — тутакулюм.
252
4. Специальная часть
отростками (1-3) или без них. Эмболюс может быть длинным и начинаться
вблизи голени, или коротким и начинаться в верхней части тегулюма. У
большинства видов эмболюс относительно толстый; у одних таксонов эмболюс
утончается к вершине, у других — наоборот утолщается {Xysticus nenilini).
Эпигина варьирует по форме, может быть с отчётливой ямкой, иногда ямка с
выростами; у части видов на месте ямки находится выпуклость, которая
возвышается над остальной частью эпигины. У ряда таксонов ямки нет, зато
есть скапус, или своего рода септум. У многих групп эпигина без чётких
форм.
• Размер. 3-14 мм. Размерный диморфизм от нечёткого, до очень явного. В
случае сильного полового диморфизма размер самцов может существенно
варьировать {Thomisus).
• Окраска. Сильно варьирует от бледно-белой, до жёлтой, зеленоватой, или
почти чёрной с красными пятнами. Рисунок имеется как на карапаксе, так и
на брюшке. Рисунок брюшка может быть чётким или размытым. Некоторые
цветочные виды могут быть полиморфными по окраске и рисунку. Окраска
(цвет) у особей отдельных видов может резко меняться в течение
нескольких дней, в зависимости от цвета субстрата {Misumena vatia).
• Особые черты. Тутакулюм (вырост цимбиума), бёдра ног направлены в
стороны, пентагональная форма брюшка.
Рода, известные из азиатской части России. Bassaniana Strand, 1928 {В. deco-
rata (Karsch, 1879)); Coriarachne Thorell, 1869 (C. depressa (C.L. Koch, 1837)); Diaea
Thorell, 1869 {D. subdola O. Pickard-Cambridge, 1885); Ebelingia Lehtinen, 2005 {E.
kumadai (Ono, 1985)); Ebrechtella F. Dahl, 1907 {E. tricuspidata (Fabricius, 1775));
Heriaeus Simon, 1875; Lysiteles Simon, 1895; Misumena Latreille, 1804 (M vatia (Clerck,
1758)); Oxytate L. Koch, 1878 {O. striatipes L. Koch, 1878); Ozyptila Simon, 1864;
Pistius Simon, 1875; IRuncinia Simon, 1875; Synema Simon, 1864; Thomisus Walcke-
naer, 1805; Tmarus Simon, 1875; Xysticus C.L. Koch, 1835.
Распространение.
Misumena, Ozyptila, Xysticus — весь регион, вплоть до тундровой зоны.
Coriarachne, Ebrechtella, Heriaeus, Pistius, Synema, Thomisus, Tmarus — юг
Сибири и Дальнего Востока.
Bassaniana, Ebelingia, Diaea, Oxytate — только юг Дальнего Востока.
Lysiteles — Алтай и юг Дальнего Востока (дизъюнктивный ареал).
?Runcinia — только Западная Сибирь.
Ареалы более 30 видов томизид, населяющих азиатскую часть России,
приведены в работе Marusik et al. (2000).
Биология. Пауки-бокоходы по типу охоты относятся к бродячим-засадникам.
Населяют они самые разные биотопы и стации. Немногочисленные виды
населяют кроны деревьев {Lysiteles, Pistius, некоторые Tmarus, Xysticus vachoni Schenkel,
1963), стволы {Xysticus sibiricus Kulczynski, 1908, Bassaniana, Coriarachne),
большинство видов населяют кустарники, травостой, подстилку, есть несколько видов,
связанных с камнями (галечники, осыпи и россыпи). Многие виды охотятся,
поджидая жертву в цветах. Причём, если некоторые группы специализированы для
Семейства, встречающиеся в азиатской части России
253
Рис. 37 (продолжение). Thomisidae. Примеры строения копулятивных органов: Diaea
subdola (d\ 14), Pistius undulatus (сГ$, 15, 22), Heriaeus horridus (сГ$, 16, 23), Thomisus
onustus (cT$, 17,25),Xysticussharlaa(cT$, 18,26),X rostratus (cT$, 19,28),X mugur(C?,
20) и X gobiensis (cT?, 21, 27).
14-21 — пальпа О*, снизу; 22-28 — эпигина, снизу Ту — тутакулюм. 14-28 — по: Marusik &
Logunov (1990, 1995b, 2002), Ono et al. (1990).
254
x «4. Специальная часть
охоты на цветах, например Ebrechtella, Misumena, Synema, то Xysticus albidus Grese,
1909 одинаково часто встречается и на цветах (в солнечную погоду) и в подстилке,
и даже среди камней на галечниках.
В пределах семейства коконы варьируют по форме от плоского до
шаровидного, их покровы могут быть «пушистыми» или как бумага. Коконы Xysticus линзо-
видные, к субстрату не прикрепляются, но всегда охраняются самкой. Дендроби-
онтный X. sibiricus помещает коконы в вертикальные щели древесины или под
корой. Размер кокона 8-10 мм. Многие виды размещают коконы под камнями.
Число яиц в коконе может сильно варьировать, так у X. albidus оно колеблется
от 15 до 54 (среднее число 29), а у X. sibiricus — 40-66.
Методы сбора. Все способы и биотопы.
Таксономия. Долгое время семейство Thomisidae имело более широкие рамки
и включало в себя подсемейство Philodrominae, которое сейчас всеми относится к
самостоятельному семейству, причём неродственному Thomisidae. Обе группы
рассматривались в одном семействе и в капитальной монографии Р.Т. Lehtinen (1967)
и в определителе В.П. Тыщенко (1971). В настоящее время надвидовая
таксономия пауков-бокоходов активно разрабатывается П.Т Лехтиненом (Lehtinen, 2002,
2004; Marusik et al., 2005).
В пределах северной Голарктики, Thomisidae — одно из наиболее хорошо
изученных крупных семейств. Однако это относится только к видовому уровню. Два
самых крупных рода, Ozyptila и Xysticus, нуждаются в пересмотре их границ и
дроблении на более мелкие таксоны. Xysticus внешне (соматически) очень
однородный род, однако копулятивные органы самцов существенно различаются у
разных видов. Предпринималось несколько попыток реанимировать ранее синони-
мизированные рода, например Psammitis Menge, 1868 (Ono, 1978). Однако эти
попытки были мало аргументированными и, более того, в работе X. Оно (Опо, 1978)
не рассматривались даже типовые виды родов, а лишь умозрительные концепции
родов, без указания видов, которые к ним относятся. Более обоснованная попытка
разбить Xysticus по крайней мере на два рода была предпринята Й. Вундерлихом
(Wunderlich, 1987). Он отнёс все виды без медиального отростка к Psammitis Menge,
1868. Однако бульбус в пределах этого таксона оказалася вновь чрезвычайно
разнообразным и, более того, многие виды имели выросты тегулюма, которые Й. Вун-
дерлих почему-то не считал апофизами. На наш взгляд, всё дело в том, что Xysticus
включает около десяти чётко очерченных таксонов, и только для некоторых уже
имеются названия: Bassaniodes Pocock, 1903, Proxysticus Dalmas, 1922, Psammitis
Menge, 1868, Spiracme Menge, 1876.
Аналогична ситуация и с родом Ozyptila, к которому относят всех мелких пауков-
бокоходов с булавовидными щетинками на ногах, карапаксе, и иногда на брюшке.
Первые виды томизид из азиатской части России были описаны ещё в 1861 г.
(Grube, 1861). Позже виды азиатской части России рассматривались в следующих
таксономических работах: Kulczyhski (1885,1908,1926); Simon (1895); Grese (1909);
Уточкин (1968); Holm (1970); Hippa et al. (1986); Марусик (1988а, 19896); Marusik
& Chevrizov (1990); Логунов (1990, 1992а-б); Ono et al. (1990); Danilov (1993a);
Logunov & Marusik (1994a, 1998); Logunov (1995); Марусик, Логунов (1995);
Azarkina & Logunov (2001); Logunov et al. (2001); Marusik & Logunov (2002b);
Семейства, встречающиеся в азиатской части России
255
Seyfulina & Mikhailov (2004); Marusik (2008); Marusik & Omelko (2008); Marusik et
al. (2010). Распространение многих видов пауков-бокоходов в Сибири
рассматривается в монографии по паукам Тувы (Marusik et al., 2000).
Разграничение видов. В большинстве случаев виды отличаются формой копу-
лятивных органов, в некоторых случаях может быть использована окраска (тип
рисунка) брюшка и головогруди и хетотаксия.
Перспективы изучения. Как уже говорилось выше, имеется немало
нерешённых проблем, касающихся надвидовой систематики пауков-бокоходов. Несмотря
на хорошую изученность Thomisidae азиатской части России, в регионе есть ещё
ряд неописанных видов Xysticus sensu lato и Ozyptila sensu lato, прежде всего в
Приморье.
Эта группа разрабатывается Д.В. Логуновым и Ю.М. Марусиком.
Oxytate striatipes (Thomisidae), самец, Приморье (фото: М.М. Омелько).
Семейство Titanoecidae Lehtinen, 1967
Титаноециды; rock weavers
Рис. 38.1-10.
В мире известно 5 родов и 45 видов, из них в России отмечены 2 рода и 12
видов, а в азиатской части России известны 2 рода и 11 видов. Семейство имеет
практически всесветное распространение, но большинство видов известно из Го-
ларктики.
Диагностические признаки. Крибеллятные тенетники, 3-коготковые пауки
среднего размера 5-12 мм, 8 глаз, крибеллюм разделён на 2 части, каламиструм
почти на всю длину предлапки, голень пальпы самца с многочисленными
отростками, в том числе и пролатерально, эмболюс длинный нитевидный.
Близкие группы. Похожи на Dictynidae и Amaurobiidae, однако чётко
отличаются своеобразной формой отростков голени на пальпе самца. От диктинид
отличаются крибеллюмом, разделённым на 2 части (у диктинид крибеллюм обычно
цельный). Самки этого семейства легко отличаются от Amaurobiidae одноцветным
брюшком и плоской эпигиной без явственных выпуклостей (лопастей) и без
медиальной пластинки.
ОПИСАНИЕ:
• Карапакс. Грушевидный.
• Стернум. Овальный.
• Глаза. 8, в 2 ряда (4-4), примерного одинакового размера.
• Ротовой аппарат. Не модифицирован.
• Ноги. 3 коготка, длина сравнима с размером тела, предлапки с
предвершинной трихоботрией, лапки без трихоботрий.
• Брюшко. Овальное.
• Паутинные бородавки. Не модифицированы.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Крибеллюм разделённый; каламиструм
длинный, почти на всю длину предлапки.
• Копулятивные органы. Голень пальпы самца со сложно устроенными
отростками про-, ретролатерально и дорсально; эмболюс длинный, может быть
частично скрыт тегулюмом. Тегулюм с одним единственным отростком в
нижней части. Эпигина в виде пластинки, без ямки, но со слабо
выраженным септумом.
• Размер. 5-12 мм. Половой диморфизм слабо выражен. У самок брюшко
несколько больше.
Рис. 38. Titanoecidae. Titanoeca sibirica (сГ$, 1, 3^, 7-8), Nurscia albofasciata (cT, 2) и
Titanoeca eca (сГ?, 5, 9-10).
1 — габитус 9, сверху; 2 — габитус О*, сверху; 3 — предлапка и лапка IV, ?; 4-5 — пальпа о\
снизу; 7-8 — голень пальпы О*, сбоку и сверху; 9 — пальпа сГ, сверху; 10 — эпигина, снизу. 5, 9-
10 — по: Eskov & Marusik (1995); 4, 7-8 — по: Marusik (1995).
Семейства, встречающиеся в азиатской части России
257
1
258
4. Специальная часть
• Окраска. Карапакс красноватый (у свежесобранных экземпляров) или
коричневый. Брюшко однотонное у самок, у самцов одноцветно-чёрное или
чёрное с белыми парными пятнами дорсально.
• Особые черты. Наличие разделённого крибеллюма; самки с однотонным
брюшком; самцы с тёмным брюшком и у некоторых видов дорсально с
парными белыми пятнами. Пальпа самца с несколькими сложно устроенными
выростами голени, в том числе и дорсальными.
Рода, известные из азиатской части России. Nurscia Simon, 1874 (N. albofas-
ciata (Strand, 1907)) и Titanoeca Thorell, 1870 (10 видов, с учётом неопределённых).
Распространение. Nurscia — только юг Дальнего Востока, Titanoeca — весь
регион, кроме тундровой зоны.
Биология. Большинство видов связаны с отдельно лежащими камнями,
галечниками, осыпями, однако один и тот же вид может обитать как среди камней, так и
в подстилке, например Т. nivalis Simon, 1874 и Т. sibirica L. Koch, 1879. Пауки
сплетают небольшие неправильные сети, некоторые участки которых могут
напоминать отдельные сектора сети пауков-кругопрядов. Сам паук при этом сидит в
убежище (под камнем, листиком).
Самки встречаются в течение всего года (поздней осенью очень
малочисленны), а самцы — только летом. Зимуют немногочисленные самки и неполовозрелые
особи разных стадий. Судя по промерам карапакса, Т. nivalis в верховьях Колымы
имеет пять личиночных стадий. Кокон округлый и содержит 20-42 яиц. Самки и
отдельные неполовозрелые особи, обитающие среди галечников, сплетают
зимовочный кокон-убежище, который, очевидно, предохраняет от заливания талой
водой.
Методы сбора. Почвенные ловушки (куда ловятся в основном самцы), ручной
сбор в подстилке и среди камней.
Таксономия. Самостоятельность семейства обоснована П.Т. Лехтиненом (Leh-
tinen, 1967). До этого таксон рассматривался в Amaurobiidae (см., например, Ты-
щенко, 1971). Виды, обитающие в Сибири и на Дальнем Востоке,
рассматривались в работах L. Koch (1879), Holm (1973), Дунин (19846), Данилов (1994) и Marusik
(1995).
Разграничение видов. Форма копулятивных органов и окраска/рисунок.
Перспективы изучения. Не исключены находки дополнительных видов
Titanoeca в юго-западной Сибири (в сопредельных регионах обитают виды, не
найденные в Сибири), а также на Дальнем Востоке. В нашем распоряжении имеется
самка с Кунашира, которая отличается от всех известных нам видов очень
крупными размерами.
Семейства, встречающиеся в азиатской части России
259
Titanoeca sp. (Titanoecidae), самец и самка, Крым (фото: Н.М. Ковблюк).
Семейство Uloboridae Thorell, 1869
Улобориды; hackled-orb web spiders
Рис. 39.1-11.
В мире известно 266 видов, относящихся к 18 родам. В азиатской части России
отмечены два вида из двух родов, а во всей России — 5 видов из 3 родов.
Диагностические признаки. Менее 6 мм, удлинённое овальное брюшко, ноги
3-коготковые, имеются крибеллюм и каламиструм, бёдра с длинными трихоботри-
ями.
Близкие группы. Похожи на некоторых Theriididae, Linyphiidae, Araneidae,
однако легко отличаются наличием крибеллюма, каламиструма и формой глаз.
ОПИСАНИЕ:
• Карапакс. Угловато-грушевидный.
• Стернум. Не модифицирован.
• Глаза. 8 глаз в два выгнутых ряда, глаза далеко отстоят друг от друга;
латеральные глаза не сближены.
• Ротовой аппарат. Не модифицирован.
• Ноги. 3 коготка, бёдра с рядом длинных трихоботрий. Передние ноги
относительно длинные (длиннее тела), остальные короче. Иногда с кольцами.
• Брюшко. Овальное при виде сверху, заострено на конце, передняя треть с
одним или двумя буграми (иногда не выражены), направленными вверх.
• Паутинные бородавки. Не модифицированы, базальные членики
несколько удлинены.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем у паутинных бородавок.
• Крибеллюм-каламиструм. Имеются. Крибеллюм неразделен. Каламиструм
однорядный.
• Копулятивные органы. Энтелегинные пауки. Пальпа самца относительно
мелкая. Парацимбиума нет, заметных отростков бульбуса тоже. Эпигина слабо
хитинизирована и выглядит как два соска (Uloborus), или полукруглая без
заметных отверстий (Octonoba).
• Размер. 3-6 мм. Половой диморфизм выражен.
• Окраска. Чёткого рисунка обычно нет; окраска от белой, кремовой или
светло-серой до почти чёрной. Иногда на карапаксе — чёткие продольные
тёмные полосы.
• Особые черты. Крибеллятные кругопряды. Сети практически всегда со ста-
билиментом, колесовидные.
Рода, известные из азиатской части России. Octonoba Opell, 1979 и Uloborus
Latreille, 1806.
Распространение. Uloborus имеет практически всесветное распространение.
Транспалеарктический вид U walckenaerius Latreille, 1806 известен из степей
южной Сибири и на юге Дальнего Востока. Octonoba yesoensis (Saito, 1934) имеет
Семейства, встречающиеся в азиатской части России
261
Рис. 39. Uloboridae. Uloborus walckenaerius (С?$, 1, 4-5, 9-11) и Octonoba yesoensis
(С??, 2-3, 6-8).
1 — головогрудь ?, сверху; 2 — головогрудь субадультного С?, сверху; 3—4 — паутинные
бородавки и крибеллюм, снизу; 5-6 — габитус ?, сбоку; 7 — пальпа С?, снизу; 8-9 — эпигина, снизу; 10 —
пальпа С?, сбоку; 11 — предлапка и лапка IV. 7-10 — по: Yoshida (2009).
262
4. Специальная часть
Кавказско-Дальневосточный дизъюнктивный ареал. В азиатской части России
известен на юге Дальнего Востока к востоку от Читинской области.
Биология. Оба вида, встречающиеся в азиатской части России — кругопряды.
Сети небольшие, располагаются горизонтально. Сети О. yesoensis описаны Мару-
сиком (19876). Этот вид сооружает сети исключительно в затенённых местах. Сети
со спиральным стабилиментом в центре, реже стабилимент прямой, иногда вовсе
отсутствует. U. walckenaerius в степной зоне делает сети на открытых местах
среди кустарника или вблизи скал и обрывов. На Дальнем Востоке этот вид
встречается только в затенённых местах. В отличие от других кругопрядов, сети плетут
(переплетают ловчую спираль) не каждый день, а только после того как она
порвётся (уловистость крибеллятной нити не уменьшается со временем). Ювениль-
ные особи младших возрастов у О. yesoensis плетут сеть в виде полога, без
явственных радиальных и спиральных нитей.
Из-за редукции ядовитых желёз, после поимки добыча тщательно укутывается
в паутину, после чего паук кусает жертву и вводит пищеварительные ферменты
внутрь.
Коконы у наших видов уплощённые, звёздчатые, сероватого цвета.
Методы сбора. Укосы, отряхивание, визуальный поиск сетей.
Таксономия. Таксономическое положение Uloboridae — предмет дискуссий.
Первоначально их рассматривали совместно с другими пауками-кругопрядами, до
тех пор, пока пауки не были разделены на крибеллятных и некрибелятных.
Некоторые исследователи (Lehtinen, 1967; Тыщенко, 1986) считают крибеллятных
кругопрядов не родственными другим кругопрядам (Araneidae, Tetragnathidae, Theri-
diosomatidae, Mysmenidae и др.). Североамериканские авторы и ряд европейских
объединяют всех кругопрядов. Исходя из строения копулятивных органов,
расположения глаз и структуры кутикулы, на наш взгляд, Uloboridae не родственны
другим кругопрядам.
Относительно хорошо улобориды изучены в Неарктике (серия работ В. Opell),
и на Дальнем Востоке, в Японии, Китае и Корее (работы Н. Yoshida).
Оба рода, встречающиеся в азиатской части России, относятся к
номинативному подсемейству (Uloborinae). Виды, встречающиеся в пределах бывшего
Советского Союза (известно всего 6), специально не изучались. О. yesoensis из
Амурской области и Приморья рассматривалась в работе Марусика (19876).
В настоящее время семейством в пределах Палеарктики никто специально не
занимается.
Разграничение видов. Только форма копулятивных органов.
Перспективы изучения. В Приморье и на южных Курилах возможно
обнаружение дополнительных видов из рода Octonoba. Возможно степные и
дальневосточные популяции U. walckenaerius не конспецифичны.
Семейства, встречающиеся в азиатской части России
263
Pardosa hyperborea (Lycosidae), самец, Финляндия (фото: N. Fritzen).
Семейство Zoridae F.O. Pickard-Cambridge, 1893
Зориды; spiny leg spiders
Рис. 40.1-8.
Семейство с небольшим числом видов (77), распространённых по всему миру.
Выделяют 14 родов. В России зарегистрировано 7 видов одного рода Zora C.L. Koch,
1847, а в азиатской части России — всего три вида.
Диагностические признаки. Небольшие, некрибеллятные, 2-коготковые эн-
телегинные пауки с 8 глазами, расположенными в 2 ряда. Пальпа самца с РГО и
медиальным отростком, эпигина с неглубокой ямкой.
Близкие группы. Внешне (габитуально, окраской, расположением глаз)
напоминают некоторых Lycosidae и особенно Anahita (Ctenidae), однако отличаются
меньшими размерами и расположением глаз. Вооружением ног
(многочисленными вентральными шипами на ногах I и II) похожи на Corinnidae и Liocranidae, но
отличаются от них рисунком карапакса, а также расположением и размером глаз.
ОПИСАНИЕ:
• Карапакс. Овальный, головная часть чуть уже грудной, наличник низкий.
• Стернум. Щитовидный.
• Глаза. 8, два компактных ряда (4-4), передний ряд прямой - слегка
выгнутый, задний — сильно выгнут. Передние глаза мельче задних; наличник
низкий. Медиальные глаза двух рядов почти соприкасаются.
• Ротовой аппарат. Не модифицирован.
• Ноги. Передние ноги длиннее тела в 1,5 и более раз, голени и предлапки I-
II вентрально с двумя рядами многочисленных шипов.
• Брюшко. Удлинённо-овальное.
• Паутинные бородавки. Не модифицированы у самок; у самцов передние
бородавки со щёткой чёрных волосков, направленных к оси тела. Задние
бородавки длиннее передних.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Пальпа самца с невысоким РГО, тупым на
вершине; бульбус с тонким крючковидным медиальным отростком и длинным
нитевидным эмболюсом. Форма РГО необычная — отросток не плоский, как у
большинства других пауков, а имеет сложную трёхмерную форму, из-за чего
под разными ракурсами один и тот же отросток выглядит очень по-разному.
Такое строение РГО затрудняет определение видов у зорид. Эпигина с
неглубокой ямкой, парой круглых рецептакул и относительного толстыми
канальцами.
• Размер. 4-7 мм, половой диморфизм выражен слабо.
• Окраска. От светлой до тёмной. Самцы темнее самок. Рисунок очень
характерный: карапакс с широкой светлой медиальной полосой, парой тёмных
полос по бокам от неё, кроме того, имеется пара субмаргинальных светлых
Семейства, встречающиеся в азиатской части России
265
Рис. 40. Zoridae. Zora parallela ($, 1, 3), Z spinimana (C?$, 2, 4-8).
1 — головогрудь ?, спереди; 2 — головной отдел карапакса ?, сверху; 3 — паутинные бородавки
сГ, снизу; 4 — габитус ?, сбоку-сверху; 5 — пальпа С?, сбоку; 6 — отросток голени пальпы С?,
сбоку; 7 — эпигина после мацерации, сверху; 8 — С?, тазики IV. 5-7 — по: Miller (1947); 8 — по:
Heimer & Nentwig (1991).
полос и ещё одна пара тёмных маргинальных (итого на карпаксе 3 светлые и
4 тёмные продольные полосы).
• Особые черты. Два ряда шипов вентрально на передних 2 парах ног, форма
и расположение глаз, рисунок карапакса.
Рода, известные из азиатской части России. Zora C.L. Koch, 1847 (Z nemoralis
(Blackwall, 1861), Z. cf. nemoralis, Z spinimana (Sundevall, 1832)).
Распространение. Весь регион, кроме тундровой зоны.
Биология. Обитатели подстилки в широком спектре биотопов с умеренной и
высокой влажностью. Избегают лишь сухих мест и сильно затенённых участков.
Самки встречаются в течение всего года. Самцы относительно редки.
Zora — активные дневные охотники. Самки крепят белый уплощённый кокон
на камни или сухие листья, и охраняет его, распластавшись поверх кокона.
266
4. Специальная часть
Методы сбора. Почвенные ловушки и ручной сбор среди подстилки.
Таксономия. Длительное время голарктический род Zora доставлял немало
неприятностей для классификации пауков (Lehtinen, 1967). Его относили то к Lycosidae
(схожий габитус и окраска), то к Clubionidae или Liocraniinae (отсутствует скопула)
и, наиболее часто, к Ctenidae (наличие множества вентральных шипов на
передних голенях и предлапках) (Lehtinen, 1967). Впервые ранг семейства этому
таксону придал Р.Т. Lehtinen (1967), однако границы его чётко не обозначил. Часть
родов была впоследствии выведена из семейства, например Zoropsis Simon, 1878. Не
исключено, что и некоторые другие рода из Австралазии и Неотрописа,
числящиеся в семействе, будут выведены из Zoridae. С другой стороны, это семейство
может пополниться родами, ныне относимыми к Ctenidae, например Anahita. В
определителе В.П. Тыщенко (1971) Zora рассматривается в отдельном подсемействе
Zorinae семейства Clubionidae. Й. Вундерлих (Wunderlich, 2008c) рассматривает
это семейство более широко и включает в него Liocranidae как подсемейство
Liocraninae. Виды азиатской части России в таксономических работах не
изучались. Семейством никто в настоящее время не занимается.
Разграничение видов. Форма копулятивных органов довольно однообразна,
проще всего виды различаются рисунком карапакса, наличием особых
утолщённых волосков на тазиках IV (у Z spinimana), формой РГО.
Перспективы изучения. На большей части Сибири (от Тувы до Магаданской
области) обитает ещё не описанный вид, близкий к Zora nemoralis. He исключены
находки дополнительных видов, в том числе и новых для науки, в Южной Сибири
и малоизученных регионах, например в Северном Приамурье.
Семейства, встречающиеся в азиатской части России
267
Pisaura mirabilis (Pisauridae), самка, Финляндия (фото: N. Fritzen).
Семейство Zygiellidae Simon, 1929
Зигиеллиды
Рис. 41.1-8.
Небольшое всесветно распространённое семейство, включающее не менее 159
видов из 7 родов, в том числе один ископаемый род. В России обитает 5 видов из 3
родов, а в азиатской части — 4 вида из 3 родов.
Диагностические признаки. Энтелегинные, некрибеллятные, с 8 глазами, 3-
коготковые пауки-кругопряды мелкого и среднего размера, брюшко слегка
приплюснуто, пальпа самца с крупным парацимбиумом.
Близкие группы. Габитуально и рисунком напоминают Enoplognatha (Theri-
diidae), Metleucauge, Meta sensu lato (Tetragnathidae) и некоторых Araneidae. От
первых легко отличаются низким клипеусом; кроме того, самцы зигеллид не имеют
модифицированных хелицер, а эпигина самки всегда с медиальной пластинкой (нет
у теридиид). От Metleucauge и других Tetragnathidae отличаются существенно
меньшими размерами и более короткими ногами; самцы не имеют шипа на вертлуге
пальпы, а у самок медиальная пластинка большей частью спрятана в эпигастраль-
ную щель (находится на поверхности брюшка у Metleucauge и других
Tetragnathidae). От Araneidae отличаются затемнённой головной частью. Ширина колена пальпы
у самцов не больше длины, парацимбиум не пальцевидный, а эпигина без скапуса.
ОПИСАНИЕ:
• Карапакс. Грушевидный, невысокий, наличник низкий, без густого
опушения.
• Стернум. Не модифицирован.
• Глаза. 8 в два ряда (4-4), боковые соприкасаются друг с другом, клипеус
низкий.
• Ротовой аппарат. Не модифицирован, длина максилл не превышет 1,5 их
ширины.
• Ноги. 3 коготка. Относительно длинные и толстые.
• Брюшко. Овальное, слегка уплощено.
• Паутинные бородавки. Не модифицированы.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Пальпа самца с парацимбиумом, ширина которого
больше длины, медиальный отросток имеется, тегулюм часто с выростом,
кроме эмболюса имеется крупный округлый кондуктор, а у некоторых
таксонов и удлинённый терминальный отросток. Ширина голени не больше
длины членика. Эпигина простая, трёхмерная, с медиальной пластинкой, без
выростов.
• Размер. 4-8 мм, размерного диморфизма нет.
• Окраска. Карапакс чаще всего с рисунком, брюшко всегда с листовидным
рисунком.
• Особые черты. Кругопряды со свободным сектором сети, вдоль которого
проходит сигнальная нить.
Семейства, встречающиеся в азиатской части России
269
Рис. 41. Zygiellidae. Parazygiella dispar (С?$).
1-2 — головогрудь $, спереди и снизу; 3 — габитус ?, сверху; 4 — брюшко $, снизу; 5, 8 —
пальпа сГ, снизу и сбоку; 6-7 — эпигина, снизу и сзади. Мо — медиальный отросток. 5-8 — по:
Levi (1974a).
Рода, известные из азиатской части России. Parazygiella Wunderlich, 2004
(Р dispar (Kulczynski, 1885)); Zygiella F.O. Pickard-Cambridge, 1902 (Z atrica (C.L.
Koch, 1845); Z x-notata (Clerck, 1758)); Yaginumia Archer, 1960 (Y. sia (Strand, 1906);
[Stroemiellus Wunderlich, 2004 (S. stroemi (Thorell, 1870)1].
1 Мы рассматриваем этот род в Araneidae
270
4. Специальная часть
Распространение.
Parazygiella — Дальний Восток на север до северного Охотоморья.
Находки двух видов Zygiella на Дальнем Востоке сомнительны и нуждаются в
подтверждении, равно как и находка Parazygiella montana (C.L. Koch, 1839) в
Средней Сибири.
Yaginumia sia формально не отмечена в России, но нам она известна с южных
Курил.
Биология. Parazygiella dispar в Охтоморье, на Сахалине и Курилах —
массовый, но локальный вид. Встречается на приморских скалах. В северо-западной
Кайаде этот вид обычен на скалах, но встречается и в лесу на стволах деревьев.
Убежища устраивает в трещинах скал или под корой. Размер экземпляров может
существенно варьировать.
Известно, что Yaginumia sia в Японии обычна в населённых пунктах, сети
строит у воды, под перилами мостов, на скамейках.
Методы сбора. Визуальный поиск среди скал и на коре деревьев.
Таксономия. Положение и статус таксона составляет предмет дискуссий.
Впервые отдельный статус этой группе придал Е. Simon (1929), называя её Zygiellaea.
Долгое время эта группа, как род Zygiella, рассматривалась в Araneidae. H.W. Levi
(1980) рассматривал этот род в подсемействе Metinae. P.Т. Lehtinen et al. (1979) и
Marusik et al. (2000) рассматривали Z stroemi (Thorell, 1870) в Araneidae, а
остальные виды рода — в Tetragnathidae. Griswold et al. (1998) и Scharff & Coddington
(1997), напротив, весь род рассматривают в Araneidae. Недавно J. Wunderlich (2004)
выделил эту группу в отдельное семейство и разбил Zygiella на несколько родов.
Одновременно (хронологически даже чуть раньше) китайские исследователи
описали подсемейство Guizygiellinae Zhu et al., 2003 в составе Tetragnathidae,
представители которого ранее рассматривались в Zygiella.
В определителе В.П. Тыщенко (1971) фигурирует как род пауков-крестовиков.
В каталоге N.I. Platnick (2009) разные рода этого семейства фигурируют в Araneidae
и Tetragnathidae.
Stroemiellus Wunderlich, 2004 (S. stroemi (Thorell, 1870)) мы рассматриваем как
род Araneidae.
Таксономия видов, встречающихся в Сибири и на Дальнем Востоке России,
рассматривалась только в работах Kulczynski (1885) и Levi (1974а). Все виды,
встречающиеся в России и странах СНГ, ревизовал H.W. Levi (1974a).
Разграничение видов. Виды различают только по форме копулятивных
органов. Самки различаются лишь формой и пропорциями медиальной пластинки.
Перспективы изучения. Нахождение новых и дополнительных таксонов в
азиатской части России маловероятно.
Семейства, нахождение которых возможно
в азиатской части России
Семейство Oecobiidae Blackwall, 1862
Экобииды; star-legged spiders
Рис. 42.1-6.
Всесветно распространённое небольшое семейство со 107 видами и 6 родами.
В России известен один синантропный вид рода Oecobius.
Диагностические признаки. Мелкие, энтелегинные, крибеллятные пауки с 8
глазами (4 чёрных круглых, 4 серебристых полукруглых), карапакс сердцевидный,
брюшко овальное, задние паутинные бородавки с длинным серповидно
изогнутым апикальным члеником. Анальный бугорок с венчиком волосков.
Близкие группы. Форма головогруди, глазная формула, крупный анальный
бугорок с венчиком волосков и наличие крибеллюма делают семейство непохожим
на остальные группы.
ОПИСАНИЕ:
• Карапакс. Сердцевидный, длина меньше ширины, плоский, с рисунком.
• Стернум. Сердцевидный, длина меньше ширины.
• Глаза. 8, компактная группа с двумя рядами (4-4), иногда глаза
расположены почти в центре карапакса; передние латеральные и задние медиальные
глаза полукруглые серебристые (белые), остальные — круглые и чёрные.
• Ротовой аппарат. Хелицеры короткие, их коготок тоже, зубчиков нет. Мак-
силлы наклонены друг к другу, почти соприкасаются.
• Ноги. 3 коготка, ноги чуть длиннее тела, коренастые, с кольцами.
• Брюшко. Овальное.
• Паутинные бородавки. Передние и медиальные короткие и утолщённые,
задние с длинным и изогнутым апикальным члеником, анальный бугорок с
венчиком длинных волосков. Задние бородавки торчат из-под брюшка.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Есть только у рода Oecobius sensu lato, у
самцов каламиструм может отсутствовать.
• Копулятивные органы. Энтелегинные. Пальпа самца без РГО, цимбиум
обычной формы, бульбус округлый с торчащим медиальным отростком.
Эпигина в виде простой пластинки, слабо склеротизована.
• Размер. 2-3 мм.
• Окраска. Бледные, с тёмным рисунком на карапаксе и отдельными точками
на брюшке.
• Особые черты. Форма карапакса, глазная формула, венчик волосков на
анальном бугорке.
Распространение. Есть невысокая вероятность того, что синантропные виды
этого семейства будут найдены в Приморье или в других регионах. Космополит-
ный Oecobius navus Blackwall, 1859 известен из соседней Японии. Кроме этого, в
Корее и Китае известны крупные некрибеллятные Uroctea Dufour, 1820. Однако
вероятность их нахождения у нас очень мала. В России известен космополитный
Семейства, нахождение которых возможно в азиатской части России
273
1
Рис. 42. Oecobiidae. Oecobius sp. (?, 1), О. cellariorum (cT$, 2, 4-6) и Uroctea sp. (?, 3).
1-3 — габитус; 4 — паутинные бородавки и анальный бугорок; 5 — эпигина; 6 — пальпа сГ,
сбоку 5-6 — по: Shear (1970).
и синантропный Oecobius cellariorum (Duges, 1836) из нескольких мест, в том
числе и на Урале.
Биология. Делают небольшие сети над трещинами в субстрате. В домах часто
селятся по углам. Обнаружить сети на светлых стенах легко, так как на них
накапливается пыль, и они становятся заметными. Свободноживущие виды охотятся
преимущественно на муравьев.
Методы сбора. Визуальный поиск, сбор с поверхности почвы.
Таксономия. С момента описания этого семейства, никто не подвергал
сомнению его самостоятельность. Исходно это семейство включало некрибеллятный род
Uroctea. Позже он был выделен в отдельное семейство, но П.Т. Лехтинен (Lehtinen,
1967) убедительно показал необоснованность выделения Uroctea в отдельное
семейство. Oecobiidae вместе с Hersiliidae образуют отдельное надсемейство.
Некоторые авторы считают необходимым включать в это надсемейство еще и Eresidae,
но только на основании сходства поведения (форма атаки на жертву). Соматически
и по строению копулятивного аппарата Eresidae ничего общего не имеют с
Oecobiidae и Hersiliidae.
Космополитные виды Oecobius sensu lato были ревизованы W. Shear (1970).
Семейство Segestriidae Simon, 1893
Сегестрииды; tubeweb spiders
Рис. 43.1-5.
Небольшое всесветно распространённое семейство с тремя родами и 111
видами. В России отмечен один вид из рода Segestria.
Диагностические признаки. Гаплогинные, 6-глазые, темно окрашенные, 3-ко-
готковые пауки, с третьей парой ног, направленной вперёд.
Близкие группы. Формой тела и отчасти окраской напоминают некоторых
Gnaphosidae, Clubionidae, однако наличие всего 6 глаз, направленная вперёд
третья пара ног, и пара дыхалец, расположенных вблизи лёгких, позволяют легко
узнавать этих пауков.
ОПИСАНИЕ:
• Карапакс. Яйцевидный, удлинённый, клипеус низкий.
• Стернум. Удлинённо-овальный, с меж-тазиковыми выростами.
• Глаза. 6, 3 диады.
• Ротовой аппарат. Хелицеры, максиллы и нижняя губа удлинены. Коготок
хелицер небольшой.
• Ноги. 3 коготка, третья пара ног направлена вперёд.
• Брюшко. Овальное или яйцевидное.
• Паутинные бородавки. Короткие, не модифицированы.
• Органы дыхания. Пара лёгких и пара дыхалец, открывающихся чуть ниже
лёгких.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Гаплогинные, голень самца вздутая, крупнее цим-
бимума, тегулюм как луковица, с длинным постепенно суживающимся
слегка изогнутым нерасчленённым эмболюсом. Семяпроводы есть. Эпигины нет.
• Размер. 6-15 мм, самцы мельче самок.
• Окраска. Тёмные, иногда с рисунком на брюшке, вентрально брюшко
светлее.
• Особые черты. 3 диады глаз, третья пара ног направлена вперёд; парные
дыхальца, расположенные вблизи лёгких.
Распространение. Единственный вид и род Segestria Latreille, 1804 (S. senoculata
(Linnaeus, 1758)) известен в европейской части России вплоть до Урала. Вполне
возможно его нахождение в прилегающих районах западной Сибири. Кроме того,
виды этого же рода (вероятно, ошибочно отнесённые к Ariadna Audouin, 1826)
известны в Японии, Корее и Китае. Мы (Ю.М.) находили неописанный вид в
окрестностях Пекина, под корой деревьев. Не исключено, что при изучении фауны
стволов деревьев в Приморье, либо на южном Сахалине или Курилах, будут найдены
представители этого семейства.
Биология. Сети плетут среди камней или под корой деревьев.
Методы сбора. Вручную среди камней и под корой.
Семейства, нахождение которых возможно в азиатской части России
275
1
4 5
Рис. 43. Segestriidae. Segestria senoculata (сГ$) из Турции.
1 — сГ, габитус; 2 — головогрудь сГ, сверху; 3 — брюшко ?, снизу; 4 — пальпа сГ, сбоку;:
головогрудь 2, снизу.
головогрудь ?, снизу
Таксономия. Длительное время эта группа считались подсемейством Dysderi-
dae C.L. Koch, 1837. Самостоятельность этого таксона была впервые доказана Forster
& Platnick (1985). В сколь-нибудь широких масштабах группа никогда не
ревизовалась. В настоящее время нет арахнологов, специализирующихся в изучении рецен-
тных Segestriidae.
Семейство Zodariidae Thorell, 1881
Зодарииды; ant-eating spiders
Рис. 44.1-6.
Относительно крупное, всесветно распространённое семейство с 875 видами и
74 родами. В России достоверно известно два вида из рода Zodarion Walckenaer,
1826 s.l. и один вид рода Zodariellum Andreeva et Tyshchenko, 1968.
Диагностические признаки. Энтелегинные, некрибеллятные, мелкие или очень
мелкие, 3-коготковые, 8-глазые пауки, передние медиальные глаза намного
крупнее других. Коготки хелицер меньше ширины хелицер. Пальпа самца с изогнутым
РГО.
Близкие группы. Форма тела, окраска, всего 1 пара паутинных бородавок
(задние и средние отсутствуют), сидящих на общем основании, крупные передние
глаза в противовес остальным мелким глазам, делают зодариид непохожими на
других пауков.
ОПИСАНИЕ:
• Карапакс. Грушевидный, относительно плоский, наличник низкий.
• Стернум. Щитовидный.
• Глаза. 8, формула необычная: 2 передних медиальных — самые крупные,
остальные мелкие, компактно располагаются сзади за передними
медиальными.
• Ротовой аппарат. Хелицеры относительно короткие, с маленьким коготком
(меньше ширины основного членика хелицер), максиллы наклонены друг к
другу.
• Ноги. 3 коготка, ноги длинные и тонкие.
• Брюшко. Овальное, слегка сплюснутое.
• Паутинные бородавки. Всего одна пара, сидят на общем выросте. Колю-
люс редуцирован до нескольких щетинок.
• Органы дыхания. Пара лёгких и трахеи, открывающиеся одним
дыхальцем перед паутинными бородавками.
• Крибеллюм-каламиструм. Нет.
• Копулятивные органы. Энтелегинные. Пальпа самца с длинным
изогнутым кзади РГО, цимбиум с тутакулюмом (ретролатеральным выростом). Эм-
болюс длинный, нитевидный.
• Размер. 2,5-4 мм. Самцы чуть меньше самок.
• Окраска. Одноцветные бордовые или с рисунком на брюшке и иногда на
карапаксе.
• Особые черты. Бордовая окраска, передние медиальные глаза намного
крупнее всех остальных; РГО завёрнут назад.
Распространение. Один вид {Zodariellum nenilini (Eskov, 1995)) известен на
южном Урале. Этот же вид встречается в Восточно-Казахстанской области и в
Монголии, поэтому не исключено, что он будет найден в Западной Сибири или на
Алтае. Всего в Монголии встречается три вида рода Zodariellum.
Семейства, нахождение которых возможно в азиатской части России
277
1
Рис. 44. Zodariidae. Zodariellum schmidti (?, 1), Z. nenilini (C?$, 2,4-5) и Zodarion cyprium
(сГ, з, 6).
1-2 — габитус $, сверху; 3 — головогрудь d\ сверху; 4 — пальпа сГ, снизу; 5 — эпигина, снизу;
6 — брюшко о", снизу 1-2, 4-5 — по: Marusik & Koponen (2001).
Биология. Известно, что пауки этого семейства и подсемейства являются об-
лигатными мирмекофилами, причём разные виды зодариид специализируются на
разных видах муравьев. Среди близких видов встречаются как специалисты
(охотящиеся на один вид муравьев), так и генералисты (охотятся на несколько видов
муравьев).
Виды рода Zodarion Walckenaer, 1826 строят на камнях куполовидные
«домики», которые снаружи инкрустируют (маскируют) мелкими камешками. В таком
«домике» самка делает кокон и охраняет его.
278
4. Специальная часть
Методы сбора. Почвенные ловушки и ручной сбор среди камней.
Таксономия. Относительно недавно сделана мировая ревизия родов этого
семейства (Jocque, 1991). Все виды, обитающие в России, принадлежат к
номинативному подсемейству. Монгольские и восточно-казахстанские виды были
ревизованы Marusik & Koponen (2001c). На территории СНГ никто специально этим
семейством не занимается. В Европе систематику и этологию этой группы исследует
Stano Pekar из Чехии.
Zodarion sp. (Zodariidae), гнездо, Крым (фото: Н.М. Ковблюк).
Семейства, известные из азиатской части России
в ископаемом состоянии
В таблице 3 приведено 38 семейств, зарегистрированных в азиатской части
России, однако на этой территории отмечено больше семейств. Речь идёт о трёх
ископаемых семействах. Это Juraraneidae Eskov, 1984 (Juraraneus Eskov, 1984, J.
rasnitsyni Eskov, 1984), Lagonomegopidae Eskov et Wunderlich, 1995 (Lagonomegops
Eskov et Wunderlich, 1995, L. sukatchovae Eskov et Wunderlich, 1995); Mecico-
bothriidae Holmberg, 1882 {Cretohexura Eskov et Zonstein, 1990; C. coylei Eskov et
Zonstein, 1990). Помимо этих семейств, отмечены ещё и Segestriidae (со знаком
вопроса: ювенильные экземпляры) из таймырских янтарей (Eskov & Wunderlich,
1995).
Из Juraraneidae известен только один вид по отпечатку из Бурятии (Eskov, 1984).
Датируется средней юрой (-165 млн. лет). Не исключено, что в дальнейшем это
название будет синонимизировано с Araneidae.
Lagonomegopidae были найдены в таймырских янтарях, датируемых меловым
периодом (-82-86 млн. лет), более древних, чем балтийские. Эта группа
принадлежит надсемейству Palpimanoidea. Все виды этого семейства имеют очень
крупные задние медиальные глаза. По мнению К.Ю. Еськова и его
коллег-палеонтологов, это семейство было меловым аналогом Salticidae. После первой находки на
Таймыре, представители ещё четырёх родов этого семейства были
зарегистрированы в янтарях из Канады, США, Бирмы, Иордании и Испании. Всего описано 6
видов. В кайнозойских отложениях не найдено ни одного представителя этой
группы. Скакунчики же ни разу не отмечались в мезозое, но зато представлены во всех
хорошо изученных отложениях кайнозоя (балтийские и более молодые янтари).
Mecicobothriidae — мигаломорфное семейство, отмечено в Забайкалье. В
нынешнее время это семейство (9 видов из 4 родов) известно исключительно из
Нового Света (Северной и Южной Америки). В ископаемом состоянии известно 2
вида из одного рода из Азии (Забайкалье и Монголия), датируемые меловым
периодом (65-140 млн. лет).
Благодарности
Авторы очень благодарны Э.Ф. Гусейнову (Институт зоологии АН
Азербайджана, Баку) за участие в написании очерков по биологии отдельных семейств; S.
Коропеп (Зоологический музей, Университет Турку, Финляндия) за
предоставленное оборудование для фотографирования пауков и возможность работать с
коллекциями.
В процессе подготовки рукописи разного рода консультации, связанные с
этимологией родовых названий, нам оказывали А.В. Танасевич (Москва), Don Cameron
(Ann Arbor, USA), P.Т. Lehtinen (Турку, Финляндия) и Michael I. Saaristo. Особенно,
мы признательны Д.В. Логунову (Манчестер) и К.Г. Михайлову (Москва) за
разностороннюю помощь при подготовке монографии. Мы также признательны СЮ.
Стороженко (Биолого-почвенный институт ДВО РАН, Владивосток), который дал
толчок к написанию определителя. Brandi Fleshman (Fairbanks, Alaska, USA)
отредактировала язык в английской версии аннотации.
Натурные фотографии нам любезно предоставили А. А. Надольный, М.М. Омель-
ко, Г.А. Прокопов, В.Г. Шиленков, N. Fritzen, R. Macek и Shuqiang Li.
Г.Н. Азаркина, Д.А. Милько (Бишкек, Киргизия), Т. Kronestedt (Stockholm,
Sweden), N. Tsurusaki (Tottori, Japan), W. Wesolowska (Wroclaw, Poland) помогли
нам в поиске фотографий арахнологов, внёсших существенный вклад в изучение
пауков азиатской части России.
Г.Н. Азаркина (Новосибирск), М.М. Омелько (Владивосток), А.А. Надольный
(Таврический национальный университет, Симферополь), О.В. Кукушкин (Кара-
дагский природный заповедник НАНУ, Феодосия) смотрели рукопись на разных
этапах подготовки; П.Е. Гольдин (Таврический национальный университет,
Симферополь) смотрел приложение А (этимология). Кроме того, рукопись читали
студенты А.Р Мустафаев и В.Г. Маннаппова (Таврический национальный
университет, Симферополь).
Работа первого автора (Ю.М.) была частично поддержана грантами РФФИ №№
09-04-01365 и 11-04-01716-а.
Работа второго соавтора (Н.К.) была частично поддержана Карадагским
природным заповедником НАН Украины.
ЛИТЕРАТУРА
Ажеганова Н.С. 1968. Краткий определитель пауков (Aranei) лесной и лесостепной зоны
СССР // Определители по фауне СССР, издаваемые Зоол. ин-том АН СССР. Л.:
Наука. Вып. 98. 147 с.
Бахвалов В.Ф. 1981. Новые виды пауков-кругопрядов (Aranei, Araneidae) из Сибири и
Советского Дальнего Востока// Энтомол. исслед. в Киргизии. Фрунзе: Илим. Вып. 14.
С.142-150.
Бенедиктов А.А. 2006. Кто поёт, когда насекомые молчат? // Химия и жизнь. Вып.7. С.63.
Вагнер В.А. 1900. Водяной паук (Argyroneta aquatica CI.), его индустрия и жизнь как
материал сравнительной психологии // Бюл. Моск. о-ва испыт. прир. Нов. сер. Т. 14.
С.61-174, Pl.Ia.
Данилов С.Н. 1991. Новый вид пауков рода Tibellus (Aranei, Philodromidae) из Забайкалья
// Зоол. ж. Т.70. Вып.8. С.139-141.
Данилов С.Н. 1994. Крибеллятные пауки (Aranei, Cribellatae) Забайкалья // Энтомол.
обозр. Т.73. Вып.1. С.200-209.
Данилов С.Н. 2008. Каталог пауков (Arachnida, Aranei) Забайкалья. Улан-Удэ: Изд-во
БНЦСОРАН. 108 с.
Дунин П.М. 1984а. Материалы к изучению фауны пауков Дальнего Востока (Arachnida,
Aranei). I. Семейство Salticidae // Фауна и экол. насекомых юга Дальнего Востока.
Владивосток: ДВЦ АН СССР. С. 128-140.
Дунин П.М. 19846. Материалы к изучению фауны пауков Дальнего Востока (Arachnida,
Aranei). II. Секция Cribellatae // Фауна и экол. насекомых юга Дальнего Востока.
Владивосток: ДВЦ АН СССР. С. 141-146.
Ермолаев В.Н. 1927. Новый вид p. Coelotes Blackwall (Araneae, Agalenidae) из Западной
Сибири // Ежег. Зоол. муз. АН СССР. 127(1926). С.347-355.
Ермолаев В.Н. 1930. К вопросу о распространении в Сибири тарантула Hogna (Trochosa)
singoriensis (Laxm.) // Русск. энтомол. обозр. Т.24. Вып.3-4. С.212-217.
Еськов К.Ю. 1979. Три новых вида пауков семейства Linyphiidae (Aranei) из Сибири //
Фауна и экол. паукообразных. Тр. Зоол. ин-та АН СССР. Т.85. С.65-72.
Еськов К.Ю. 1980а. Два новых вида пауков рода Maro (Aranei, Linyphiidae) из Сибири //
Зоол. ж. Т.59. Вып.7. С. 1100-1103.
Еськов К.Ю. 19806. Систематические замечания о пауках рода Hummelia (Aranei,
Linyphiidae) с описанием нового вида // Зоол. ж. Т.59. Вып. 11. С. 1743-1746.
Еськов К.Ю. 1981а. Пауки из родов Eboria, Latithorax, Rhaebothorax и Typhochrestus
(Aranei, Linyphiidae) в фауне Сибири // Зоол. ж. Т.60. Вып.4. С.496-505.
Еськов К.Ю. 19816. Таксономия пауков рода Hilaira (Aranei, Linyphiidae) // Зоол. ж.
Т.60. Вып. 10. С. 1486-1493.
Еськов К.Ю. 1984а. Новые и малоизвестные роды и виды пауков (Aranei, Linyphiidae) с
Дальнего Востока // Зоол. ж. Т.63. Вып.9. С.1337-1344.
Еськов К.Ю. 19846. Таксономия пауков рода Oreonetides (Aranei, Linyphiidae) // Зоол. ж.
Т.63. Вып.5. С.662-670.
Еськов К.Ю. 1986. О пауках рода Рею Tanasevitch, 1985 (Aranei, Linyphiidae) // Зоол. ж.
Т.65. Вып.П.С.1738-1742.
282
Литература
Еськов К.Ю. 1987а. Новые данные о пауках рода Hilaira (Aranei, Linyphiidae) фауны
СССР // Зоол. ж. Т.66. Вып.7. С. 1020-1031.
Еськов К.Ю. 19876. Пауки неарктических родов Ceraticellus и Islandiana (Aranei,
Linyphiidae) в фауне Сибири и Дальнего Востока // Зоол. ж. Т.66. Вып. 11. С. 1748-1752.
Еськов К.Ю. 1988а. Пауки рода Wabasso (Aranei, Linyphiidae) в фауне СССР // Зоол. ж.
Т.67. Вып.1.С.136-140.
Еськов К.Ю. 19886. Пауки родов Mecynargus, Mecynargoides gen.n. и Tubercithorax gen.n.
(Aranei, Linyphiidae) в фауне СССР // Зоол. ж. Т.67. Вып.12. С.1822-1832.
Еськов К.Ю. 1988с. Семь новых монотипических родов пауков семейства Linyphiidae
(Aranei) из Сибири // Зоол. ж. Т.67. Вып.5. С.678-690.
Еськов К.Ю. 1989. Новые монотипические роды пауков семейства Linyphiidae (Aranei)
из Сибири. Сообщение 1 // Зоол. ж. Т.68. Вып.9. С.68-78.
Еськов К.Ю. 1990. Новые монотипические роды пауков семейства Linyphiidae (Aranei)
из Сибири. Сообщение 2 // Зоол. ж. Т.69. Вып.1. С.43-53.
Еськов К.Ю. 1991а. К таксономии и зоогеографии пауков родов Маго и Oreonetides
(Aranei, Linyphiidae) // Зоол. ж. Т.70. Вып.4. С.45-54.
Еськов К.Ю. 19916. Пауки рода Savignya (s. str.) (Aranei, Linyphiidae) в фауне Дальнего
Востока и Центральной Азии // Зоол. ж. Т.70. Вып.5. С. 140-144.
Есюнин С.Л., Тунёва Т.К. 2009. Обзор пауков-рысей группы heterophthalmus рода Oxyopes
(Aranei, Oxyopidae) Палеарктики // Зоол. ж. Т.88. Вып.2. С. 164-175.
Зюзин А.А. 1979а. Таксономическое изучение палеарктических пауков рода Pardosa
C.L. Koch (Aranei, Lycosidae). I. Таксономическая структура рода // Энтомол. обозр.
Т.58. Вып.2. С.431-447.
Зюзин А.А. 19796. Заметки о двух редких видах рода Acantholycosa F. Dahl (Aranei,
Lycosidae) фауны СССР // Зоол. ж. Т.58. Вып.11. С. 1736-1738.
Зюзин А.А. 1985. Критерии рода и подсемейства в систематике пауков семейства
Lycosidae (Aranei) с описанием нового рода и двух новых подсемейств // Фауна и
экол. пауков СССР. Тр. Зоол. ин-та АН СССР Т. 139. С.40-51.
Зюзин А.А., Марусик Ю.М. 1988. Новый вид пауков рода Acantholycosa (Aranei,
Lycosidae) из Восточной Сибири // Зоол. ж. Т.67. Вып.7. С. 1083-1085.
Зюзин А.А., Марусик Ю.М. 1989. Новый вид пауков рода Pardosa (Aranei, Lycosidae) из
Магаданской области // Энтомол. обозр. Т.68. Вып.2. С.432^434.
Иванов А.В. 1965. Пауки, их строение, образ жизни и значение для человека. Л.: Изд-во
Ленинградск. ун-та. 304 с.
Измайлова М.В. 1989. Фауна пауков юга Восточной Сибири. Иркутск: Изд-во Иркутск.
ун-та. 182 с.
Лобанова ТВ. 1978. Новые виды пауков семейства Lycosidae (Aranei) // Таксономия и
экол. членистоногих Сибири. Новосибирск: Наука, Сиб. отд. С.6-9.
Логунов Д.В. 1990. Новые данные о пауках семейств Atypidae, Pisauridae и Thomisidae
фауны СССР // Членистоногие и гельминты. Новосибирск: Биол. ин-т СО АН СССР.
С.33-43.
Логунов Д.В. 1991. Пауки семейства Salticidae (Aranei) Тувы. 1. Шесть новых видов из
родов Sitticus, Bianor и Dendryphantes II Зоол. ж. Т.70. Вып.6. С.50-60.
Логунов Д.В. 1992а. Обзор пауков рода Tmarus Simon, 1875 (Araneae, Thomisidae)
фауны СССР с описанием нового вида // Сиб. биол. ж. Вып.1. С.61-73.
Логунов Д.В. 19926. К фауне пауков Болынехехцирского заповедника (Хабаровский
край). I. Семейства Araneidae, Lycosidae, Philodromidae, Tetragnathidae, Theridiidae,
Литература
283
Thomisidae // Сиб. биол. ж. Вып.4. С.56-68.
Логунов Д.В., Марусик Ю.М. 1990. Пауки рода Argyrodes (Aranei, Theridiidae) СССР //
Зоол. ж. Т.69. Вып.2. С. 133-136.
Логунов Д.В., Марусик Ю.М. 1991. Переописание и морфологические различия Bianor
aurocinctus (Ohlert) и В. aemulus (Gertsch) (Aranei, Salticidae) // Сиб. биол. ж. Вып.2.
С.39-47.
Логунов Д.В., Марусик Ю.М. 1992. Пауки рода Phoroncidia (Aranei, Theridiidae) СССР
// Фауна и экология пауков, скорпионов и ложноскорпионов СССР. Тр. Зоол. ин-та
АН СССР. Т.226 (1990). С.91-97.
Логунов Д.В., Марусик Ю.М. 2004. Отряд Агапеае - пауки // Биоразнообразие Сохон-
динского заповедника. Членистоногие. Новосибирск-Чита. С.41-80.
Ляхов О.В. 1992. Ebo distinctivus sp.n. (Агапеае, Philodromidae) с Алтая // Зоол. ж. Т.71.
Вып.7. С. 147-149.
Марусик Ю.М. 1985. Ревизия пауков рода Cercidia Thorell (Aranei: Araneidae) с
описанием нового вида // Вестн. ЛГУ. Вып.З. С. 3-7.
Марусик Ю.М. 1987а. Три новых вида пауков семейства Nesticidae (Aranei) фауны СССР
// Зоол. ж. Т.66. Вып.З. С.461-463.
Марусик Ю.М. 19876. Систематика и биология паука-кругопряда Octonoba yesoensis
(Aranei, Uloboridae) // Зоол. ж. Т.66. Вып.4. С.613-616.
Марусик Ю.М. 1988а. Новые виды пауков верховий Колымы // Зоол. ж. Т.67. Вып. 10.
С. 1469-1482.
Марусик Ю.М. 19886. Три новых вида пауков семейства Linyphiidae (Aranei) с Северо-
Востока СССР // Зоол. ж. Т.67. Вып.12. С.1914-1918.
Марусик Ю.М. 1989а. Новые данные о фауне и синоними пауков СССР (Arachnida,
Aranei) // Фауна и экология пауков и скорпионов. М.: Наука. С.39-52.
Марусик Ю.М. 19896. Два новых вида рода Xysticus и замечания по синонимии пауков-
бокоходов (Aranei, Thomisidae, Philodromidae) Сибири // Зоол. ж. Т.68. Вып.4. С. 140-
145.
Марусик Ю.М. 1990. Пауки рода Chalcoscirtus (Aranei, Salticidae) фауны СССР 1 // Зоол.
ж. Т.69. Вып.6. С.45-57.
Марусик Ю.М. 1991а. Пауки рода Chalcoscirtus (Aranei, Salticidae) фауны СССР. 2 //
Зоол. ж. Т.70. Вып.1. С. 19-31.
Марусик Ю.М. 19916. Пауки рода Chalcoscirtus (Aranei, Salticidae) фауны СССР. 3 //
Зоол. ж. Т.70. Вып.2. С.22-29.
Марусик Ю.М. 1991 в. Новые данные о пауках подсемейства Linyphiinae (Aranei,
Linyphiidae) Восточной Сибири // Зоол. ж. Т.70. Вып.6. С.61-67.
Марусик Ю.М. 1991г. Пауки-бокоходы семейства Philodromidae (Aranei) Восточной
Сибири // Зоол. ж. Т.70. Вып. 10. С.48-58.
Марусик Ю.М. 2005. 60 лет Сеппо Копонену - большому другу российской
арахнологии // Arthropoda Selecta. T.13. Вып.З. С.171-177.
Марусик Ю.М. 2006. Пауки-скакунчики - двойное исключение // Природа. Вып.2. С.85.
Марусик Ю.М. 2007. Пауки (Arachnida: Aranei) азиатской части России: таксономия,
фауна, зоогеография. Автореф. дис. на соиск. уч. степ. д. биол. н. СПб.: СПбГУ. 36 с.
Марусик Ю.М. 2009.10 лет самому востребованному арахнологическому каталогу Plat-
nick N.I. 2000-2009. The world spider catalog, versions "0"-9.5. New York: American
Museum of Natural History, online at http://research.amnh.org/entomology/spiders/catalog/
index.html // Arthropoda Selecta. T.18. Вып. 1-2. С. 115-117.
284
Литература
Марусик Ю.М. 2010. Имена в российской арахнологии. Адольф Эдуард Грубе (Adoplh
Eduard Grube) 1812-1880 // Arthropoda Selecta. T.19. Вып.2. С.109-111.
Марусик Ю.М., Кроуфорд Р.Л. 2006. Пауки (Aranei) острова Монерон // Растительный и
животный мир острова Монерон (Материалы Международного сахалинского
проекта). Владивосток: Дальнаука. С. 171-195.
Марусик Ю.М., Логунов Д.В. 1991. Пауки надсемейства Amaurobioidea (Aranei)
Сахалина и Курильских островов // Зоол. ж. Т.70. Вып.9. С.87-94.
Марусик Ю.М., Логунов Д.В. 1995. Малоизвестные виды пауков семейств Salticidae и
Thomisidae (Aranei) Дальнего Востока СССР // Энтомологические исследования на
Северо-Востоке СССР. Владивосток: ДВО АН СССР, 2: 131-140.
Марусик Ю.М., Целлариус А.Ю. 1986. Новый вид пауков рода Theridion (Aranei, Theri-
diidae) из Карелии // Зоол. ж. Т.65. Вып.П. С.1736-1738.
Михайлов К.Г. 1992. Пауки рода Clubiona Latreille 1804 (Aranei, Clubionidae)
Советского Союза (фаунистический обзор) // Фауна и экол. пауков, скорпионов и ложноскор-
пионов СССР. Тр. Зоол. ин-та АН СССР. Т.226. С.60-69.
Михайлов К.Г. 1994. Bicluona Michailov, subgen.n., новый подрод пауков рода Clubiona
(Aranei, Clubionidae) из Восточной Азии // Зоол. ж. Т.73. Вып.П. С.52-57.
Михайлов К.Г. 1995. Выделение внутриродовых группировок у пауков рода Clubiona
(Aranei, Clubionidae): типологическая процедура // Зоол. ж. Т.74. Вып.4. С.70-81.
Михайлов К.Г. 1996. Фауна пауков территории бывшего СССР // Успехи соврем, биол.
Т. 116. Вып.2. С.231-238.
Михайлов К.Г, Марусик Ю.М. 1995. Пауки Северо-Востока СССР. Семейства
Clubionidae, Zoridae, Liocranidae и Gnaphosidae (род Micaria) (Arachnida, Aranei) //
Энтомологические исследования на Северо-Востоке СССР. Владивосток: ДВО АН СССР.
Вып.2. С.90-113.
Ненилин А.Б., Пестова М.В. 1986. Пауки семейства Eresidae фауны СССР // Зоол. ж.
Т.65. Вып.П. С.1734-1736.
Овчаренко В.И., Марусик Ю.М. 1988. Пауки сем. Gnaphosidae (Aranei) Северо-Востока
СССР (Магаданская обл.) // Энтомол. обозр. Т.67. Вып.1. С.204-217.
Овчаренко В.И., Марусик Ю.М. 1995. Дополнительные данные о пауках семейства
Gnaphosidae (Aranei) Северо-Востока Азии // Энтомологические исследования на
Северо-Востоке СССР. Владивосток: ДВО АН СССР. Вып.2. С. 114-130.
Овчинников СВ. 1999. К надвидовой систематике пауков подсемейства Coelotinae
(Агапеае, Amaurobiidae) бывшего СССР // Tethys Entomological Research, Алматы.
T.l.C.63-80.
Олигер Т.И. 1981. К фауне пауков (Агапеае) Лазовского государственного заповедника
им. Л.Г. Капланова // Пауки и насекомые Дальнего Востока СССР. Владивосток: Биол.-
почв. ин-т ДВНЦ АН СССР. С.3-10.
Олигер Т.И. 1983а. Новые виды пауков семейств Lycosidae и Araneidae из Лазовского
заповедника // Зоол. ж. Т.62. Вып.2. С.303-305.
Олигер Т.И. 19836. Новые виды пауков семейств Pholcidae, Clubionidae и Agelenidae из
Лазовского заповедника // Зоол. ж. Т.62. Вып.4. С.627-629.
Олигер Т.И. 1985. Новые виды и новый род пауков-пигмеев (Aranei, Micryphantidae) из
Приморья // Энтомол. обозр. Т.64. Вып.З. С.645-648.
Олигер Т.И. 1991. Новый вид пауков ^omAraneus группы diadematus (Aranei, Araneidae)
из Приморья // Зоол. ж. Т.70. Вып.8. С. 141-143.
Россолимо Т.Е., Марусик Ю.М. 1989. Холодоустойчивость некоторых видов пауков
Магаданской области // Зоол. ж. Т.68. Вып.З. С.136-139.
Литература
285
Ричман Д.Б., Джексон P.P. 1991. Этология пауков-скакунчиков // Сиб. биол. ж. Вып.4. С.ЗЗ-
41.
Садыков А.С., Ахунов А.А., Салихов Ш.И. 1985. Яд каракурта. Ташкент: Издательство
ФАН. 168 с.
Спасский С. А. 1925. Определитель пауков Донской области. Новочеркасск: Типогр.
Знание. 62 с, 2 табл.
Старобогатов Я.И. 1985. Систематическое положение и система отряда Пауков (Aranei-
formes) // Фауна и экология пауков СССР. Тр. Зоол. ин-та АН СССР Т. 139. С.4-16.
Танасевич А.В. 1982. Новые род и виды пауков семейства Linyphiidae (Aranei) из Боль-
шеземельской тундры // Зоол. ж. Т.61. Вып.10. С.1501-1508.
Танасевич А.В. 1983. Новые роды и виды пауков семейства Linyphiidae с полярного
Урала // Зоол.ж. Т.62. Вып.2. С.215-221.
Танасевич А.В. 1984а. Новые виды пауков рода Agyneta Hull, 1911 (Aranei, Linyphiidae)
из Сибири и Средней Азии // Науч. докл. высш. шк. Биол. н. Вып.5. С.47-53.
Танасевич А.В. 19846. Новые и малоизвестные пауки семейства Linyphiidae (Aranei) из
Болынеземельской тундры // Зоол. ж. Т.63. Вып.З. С.382-391.
Танасевич А.В. 1985. К изучению пауков (Aranei) Полярного Урала // Фауна и экол.
пауков СССР Тр. Зоол. ин-та АН СССР. Т. 139. С.52-62.
Танасевич А.В., Еськов К.Ю. 1987. Пауки рода Lepthyphantes (Aranei, Linyphiidae) в
фауне Сибири и Дальнего Востока // Зоол. ж. Т.66. Вып.2. С. 185-197.
Тыщенко В.П. 1971. Определитель пауков Европейской части СССР // Определители по
фауне СССР, издаваемые Зоол. ин-том АН СССР. Л.: Наука, Лен.отд. Вып. 105. 281 с.
Тыщенко В.П. 1986. Новое подтверждение конвергентного происхождения круговых
ловчих сетей у крибеллятных и некрибеллятных пауков // Докл. АН СССР. Т.287.
Вып.5. С.1270-1273.
Уточкин А.С. 1960. Пауки рода Synaema группы globosum в СССР // Зоол. ж. Т.39. Вып.7.
С.1018-1024.
Уточкин А.С. 1968. Пауки рода Xysticus фауны СССР (определитель) (учебное
пособие). Пермь: Пермск. гос. ун-т. 73 с.
Штернберге М.Т. 1979. Новые виды пауков семейства Lycosidae (Aranei) из Забайкалья //
Latvijas Entomologs. Вып.21. С.86-90.
Эргашев Н.Э. 1990. Экология ядовитых пауков Узбекистана. Ташкент: ФАН. 189 с.
Agnarssonl. 1996. Islenskar konguaer. Reykjavik: Fjolrit Nattufraedistofnunar. 175 p.
Agnarsson I. 2004. Morphological phylogeny of cobweb spiders and their relatives (Araneae,
Araneoidea, Theridiidae) // Zool. J. Linn. Soc. Vol.141. P.447-626.
Almquist S. 2005. Swedish Araneae, part 1: families Atypidae to Hahniidae (Linyphiidae
excluded) // Insect Syst. Evol., Suppl. No.62. P. 1-284.
Almquist S. 2006. Swedish Araneae, part 2: families Dictynidae to Salticidae // Insect Syst.
Evol., Suppl. No.63. P.285-601.
Austin A.D. 1984. Life history of Clubiona robusta L. Koch and related species (Araneae,
Clubionidae) in South Australia // J. Arachnol. Vol.12. No.l. P.87-104.
Azarkina G. 2004. New and poorly known Palaearctic species of the genus Phlegm Simon,
1876 (Araneae, Salticidae) // Rev. arachnol. T.14. P.73-108.
Azarkina G.N., Logunov D.V 2001. Separation and distribution of Xysticus cristatus (Clerck,
1758) andX. audax (Schrank, 1803) in eastern Eurasia, with description of a new species
from the mountains of central Asia (Aranei: Thomisidae) // Arthropoda Selecta. Vol.9.
No.2. P.133-150.
286
Литература
Baert L. 1994. Walckenaeria koenboutjei, a new Siberian erigonid spider (Araneae, Linyphi-
idae) // Bull. Inst. r. Sci. nat. Belg., Entomol. T.64. P.5-7.
Bertkau P. 1878. Versuch einer naturlichen Anordnung der Spinnen, nebst Bemerkungenzu-
einzelnen Gattungen // Arch. Naturg. Bd.44. S.351^410.
Bonnet P. 1929. Sur une nouvelle espece de Dolomedes (araneide) de la region de Г Amour
(Siberie orientale) // Bull. Soc. ent. France. P.267-269.
Bosselaers J., Jocque R. 2002. Studies in Corinnidae: cladistic analysis of 38 corinnid and
liocranid genera, and transfer of Phrurolithinae // Zool. Scr. Vol.31. P.241-270.
Brignoli P.M. 1983. A catalogue of the Araneae described between 1940 and 1981. Manchester
Univ. Press. 755 p.
Bristowe W.S. 1939. The comity of spiders. London: Ray Society. Vol.1. P. 1-228.
Bristowe W. S. 1941. The comity of spiders. London: Ray Society. Vol.2. Rxiv + 229-560p.
Bristowe W.S. 1958. The World of Spiders. London: Collins, xii + 304 p.
Bryja V. 2004. Male trimorphism in Pelecopsis mengei (Erigoninae, Linyphiidae, Araneae) //
F. Samu, Cs. Szinetar (eds.). European Arachnology 2002. Budapest: Plant Protection
Institute & Berzsenyi College. P.73-79.
Cameron H.D. 2005. An etymological dictionary of North American spider genus names // D.
Ubick, P. Paquin, RE. Cushing & V Roth (eds.). Spiders of North America: an identification
manual. American Arachnological Society. P.274-330.
Chamberlin R.V, Gertsch W.J. 1958. The spider family Dictynidae in America north of
Mexico // Bull. Amer. Mus. Nat. Hist. Vol.116. P.l-152.
Chikuni Y. 1989. Pictorial encyclopedia of spiders in Japan. Tokyo: Kaisei-sha Press. 310 p.
[на японском]
Coddington J. A. 1986. The genera of the spider family Theridiosomatidae // Smithson. Contrib.
Zool. No.422. P.l-96.
Comstock J.H. 1913. The spider book. N.Y.: Doubleday Page & Company. 722 p.
Dahl F. 1913. Vergleichende Physiologie und Morphologie der Spinnentiere unter besonderer
Berucksichtigung der Lebensweise. 1. Die Beziehungen des Korperbaues und der Farben
zurUmgebung. Jena. 113 S.
Dahl M. 1926. Spinnentiere oder Arachnoidea. Springspinnen (Salticidae) // Die Tierwelt
Deutschlands. Jena. Lfg.3. 55 S.
Danilov S.N. 1993a. Crab spiders (Aranei: Thomisidae, Philodromidae) of Transbaikalia. 1 //
Arthropoda Selecta. Vol.2. No.l. P.61-67.
Danilov S.N. 1993b. Spiders of the genus Micaria (Araneae, Gnaphosidae) from Siberia //
Ann. Naturhist. Mus. Wien. Bd.94/95B. P.427-431.
Danilov S.N. 1997a. New data on the jumping spiders from South Siberia (Aranei
Salticidae) // Arthropoda Selecta. Vol.6. No. 1/2. P. 113-116.
Danilov S.N. 1997b. New data on the spider genus Micaria Westring, 1851 in Asia (Aranei
Gnaphosidae) // Arthropoda Selecta. Vol.5. No.3/4. P.113-116.
Danilov S.N. 1999. The spider family Liocranidae in Siberia and Far East (Aranei) // Arthropoda
Selecta. Vol.7. No.4. P.313-317.
Danilov S.N. 2000. New data on the spiders of the family Dictynidae (Araneae) from Siberia //
Ecologia, Bratislavia. Vol.19 (Suppl.3). P.37-44.
Danilov S.N., Logunov D.V 1994. A faunistic review of the jumping spiders (Aranei Salticidae)
of Transbaikalia // Arthropoda Selecta. Vol.2 (for 1993). No.4. P.25-39.
Deeleman-Reinhold C.L. 2001. Forest spiders of South East Asia: with a revision of the sac
and ground spiders (Araneae: Clubionidae, Corinnidae, Liocranidae, Gnaphosidae, Prodi-
domidae and Trochanterriidae). Leiden: Brill. 591 p.
Литература
287
Dippenaar-Schoeman A.S., Jocque R. 1997. African Spiders: An Identification Manual // Plant
Protection Res. Inst. Handbook. Pretoria. No.9. 392 p.
Dondale CD., Redner J.H. 1978. The insects and arachnids of Canada, Part 5. The crab spiders
of Canada and Alaska, Araneae: Philodromidae and Thomisidae // Research Branch,
Agriculture Canada. Publ. No. 1663. 255 p.
Dondale CD., Redner J.H. 1982. The insects and arachnids of Canada. Part 9. The sac spiders
of Canada and Alaska. Araneae: Clubionidae and Anyphaenidae // Research Branch,
Agriculture Canada. Publ. No. 1724. 194 p.
Dondale CD., Redner J.H. 1990. The insects and arachnids of Canada. Part 17. The wolf
spiders, nurseryweb spiders, and lynx spiders of Canada and Alaska (Araneae: Lycosidae,
Pisauridae, and Oxyopidae) // Research Branch, Agriculture Canada. Publ. No. 1856. 383 p.
Dondale CD., Redner J.H., Paquin P., Levi H.W. 2003. The insects and arachnids of Canada.
Part 23. The orb-weaving spiders of Canada and Alaska (Araneae: Uloboridae, Tetragna-
thidae, Araneidae, Theridiosomatidae). Ottawa: NRC Research Press. 371 p.
Dong S.J., Zhu M.S., Yoshida H. 2005. Twelve new species of the family Uloboridae
(Arachnida: Araneae) from China // Acta arachnol. Vol.54. P.81-92.
Eberhard W. G. 1987. Web-building behavior of anapid, symphytognathid and mysmenid spiders
(Araneae) // J. Arachnol. Vol. 14. P.339-356 .
Efimik VE. 1999. A review of the spider genus Tibellus Simon, 1875 of the East Palaearctic
(Aranei, Philodromidae) // Arthropoda Selecta. Vol.8. No.2. P. 103-124.
Efimik V.E., Esyunin S.L. 1996. A new subgenus and a new species of Walckenaeria Blackwall,
1833, from the Urals with remarks on the distribution of some unicornis-gvoup species in
the Palearctic (Aranei Linyphiidae) // Arthropoda Selecta. Vol.5. No. 1-2. P.63-73.
Ermolajew W. 1928a. Eine neue Aranea-Art vom Altai // Zool. Anz. Bd.77. H.9-10. S.209-
213.
Ermolajew W. 1928b. Materialen zur Spinnenfauna Westsibiriens I—II // Arch. Naturgesch.
Bd.92A. H.7. S.97-111.
Ermolajev W. 1930. Beschreibung einer neuen sibirischen Spinnengattung aus der Familie
Linyphiidae // Zool. Anz. Bd.90. H.7-8. S.216-221.
Ermolajew W. 1931. Zur Frage der Nordgrenze des Verbreitungsgebietes der Barenspinne in
Nordwestasien // Zool. Anz. Bd.95. H.l-2. S.62-64.
Ermolajew W. 1934a. Materialen zur Spinnenfauna Westsibiriens. III. Die Spinnen der Stadt
Tomsk // Folia zool. hydrobiol. Vol.7. No.l. S. 130-147.
Ermolajew W. 1934b. Materialen zur Spinnenfauna Westsibiriens. IV Notizen iiber die
Spinnenfauna der Krautdecke in der Waldsteppe des Rayon Kamenj (West-Sibirien) //
Folia zool. hydrobiol. Vol.7. No.l. S. 147-148.
Ermolajew W. 1937a. Beitrage zur Spinnentier-Fauna der Stadt Tobolsk (West-Sibirien) //
Festschr. Strand. Bd.2. S.519-524.
Ermolajew W. 1937b. Beitrag zur Kenntnis der altaischen Spinnen // Festschr. Strand. Bd.3.
S.596-606.
Eskov K.Y. 1984. A new fossil spider family from the Jurassic of Transbaikalia (Araneae:
Chelicerata) //Neues Jahrbuch fur Geologie und Palaontologie, Monatshefte. P. 645-653.
Eskov K.Yu. 1986a. A new species of the Mediterranean spider genus Erigonoplus Simon
1884 from northeastern Siberia (Arachnida: Araneae: Linyphiidae) // Senckenberg. biol.
Bd.66. H.4-6. P.333-337.
Eskov K.Yu. 1986b. On Veles Pakhorukov 1981 and Wubanoides n.gen., two Siberian linyphiid
genera (Arachnida: Araneae: Linyphiidae)// Senckenberg. biol. Bd.67. H.l-3. P. 173-182.
288
Литература
Eskov K.Yu. 1987. The spider genus Robertus O.Pickard-Cambridge in the USSR, with an
analysis of its distribution (Arachnida: Araneae: Theridiidae) // Senckenberg. biol. Bd.67.
H.4-6. P.279-296.
Eskov K.Yu. 1988. The spider genera Savignya Blackwall, Diplocephalus Bertkau and
Archaraeoncus Tanasevitch (Aranei, Linyphiidae) in the fauna of Siberia and the Soviet
Far East // Folia Entomol. Hung. T.49. P. 13-39.
Eskov K.Yu. 1989. New Siberian species of erigonine spiders (Arachnida, Aranei, Linyphiidae)
// Spixiana. Bd.ll. H.2. P.97-109.
Eskov K.Yu. 1990a. On the erigonine spider genera Dactylopisthes Simon, 1884 and Dactylo-
pisthoides gen. nov. (Arachnida, Araneae: Linyphiidae) // Reichenbachia. Bd.28. Nr.l.
P. 1-5.
Eskov K.Yu. 1990b. The spider genus Collinsia O. Pickard-Cambridge, 1913 in the fauna of
Siberia and the Soviet Far East (Arachnida: Araneae: Linyphiidae) // Senckenberg. biol.
Bd.70. H.4-6. P.287-298.
Eskov K.Yu. 1991a. New linyphiid spiders from Siberia and the Far East. 1. The genus
Holminaria gen. nov. (Arachnida, Araneae: Linyphiidae) // Reichenbachia. Bd.28. Nr.2.
P.97-102.
Eskov K.Yu. 1991 b. New linyphiid spiders from Siberia and the Far East. 2. The genus Paragly-
phesis gen. nov. (Arachnida, Araneae: Linyphiidae) // Reichenbachia. Bd.28. Nr.2. P. 103-
107.
Eskov K.Yu. 1992. A restudy of the generic composition of the linyphiid spider fauna of the
Far East (Araneida: Linyphiidae) // Ent. scand. Vol.23. P. 153-168.
Eskov K.Yu. 1993. Several new linyphiid spider genera (Aranei, Linyphiidae) from the Russia
Far East // Arthropoda Selecta. Vol.2. No.3. P.43-60.
Eskov K.Yu. 1994. Catalogue of the linyphiid spiders of northern Asia (Arachnida, Araneae,
Linyphiidae). Sofia-Moscow: PENSOFT Publ. 144 p.
Eskov K.Yu., Marusik YM. 1991. New linyphiid spiders (Aranei, Linyphiidae) from East
Siberia // Korean Arachnol. Vol.6. No.2. P.237-253.
Eskov K.Yu., Marusik Yu.M. 1992a. On the mainly Siberian genera Wubanoides, Parawuba-
noides gen.n. and Poeciloneta (Araneidae: Linyphiidae) // Arthropoda Selecta. Vol. 1. No. 1.
P.21-38.
Eskov K.Yu., Marusik Yu.M. 1992b. The spider genus Centromerus (Aranei, Linyphiidae) in
the fauna of Siberia and the Russian Far East, with an analysis of its distribution //
Arthropoda Selecta. Vol.1. No.2. P.33-46.
Eskov K.Yu., Marusik Yu.M. 1992c. On the Siberio-Nearctic spider genus Silometopoides
(Araneida: Linyphiidae) // Reichenbachia. Vol.29. Nr.l9. P.97-103.
Eskov K.Yu., Marusik Yu.M. 1992d. On Tunagyna and Thaleria, two closely related Siberio-
Nearctic spider genera (Aranei, Linyphiidae) // Ent. Scand. Vol.22. P.405^416.
Eskov K.Yu., Marusik Yu.M. 1994. New data on the taxonomy and faunistics of North Asian
linyphiid spiders (Aranei Linyphiidae) // Arthropoda Selecta. Vol.2 (for 1993). No.4. P.41-
79.
Eskov K.Y, Marusik Y.M. 1995. On the spiders from Saur Mt. range, eastern Kazakhstan
(Arachnida: Araneae) // Beitr. Araneol. Bd.4 (fur 1994). P.55-94.
Eskov K.Yu., Marusik Yu.M. 1997. A new species of erigonine spider genus Erigonoplus
(Araneida, Linyphidae) from south Siberia // Arthropoda Selecta. Vol.6. No. 1-2. P.91-93.
Eskov K.Y, Wunderlich J. 1995. On the spiders of the Taimyr ambers, Siberia, with the
description of a new family and with general notes on the spiders from the Cretaceous resins //
Beitr. Araneol. Bd.4 (fur 1994). P.95-107.
Литература
289
Eskov K.Y., Zonstein S.L. 1990. First Mesozoic mygalomorph spiders from the Lower
Cretaceous of Siberia and Mongolia, with notes on the system and evolution of the infraorder
Mygalomorphae (Chelicerata: Araneae) //Neues Jahrbuch fur Geologie und Palaontologie,
Abhandlungen. Bd.178. P.325-368.
Esyunin S.L., Marusik Yu.M. 2001. A new species of the genus Devade Simon, 1884 from
Mongolia, with a notes on D. tenella (Tyshchenko, 1965) (Aranei: Dictynidae) // Arthropoda
Selecta. Vol.9. No.2. P. 129-131.
Esyunin S.L., Efimik V.E. 1996. Catalogue of the spiders (Arachnida, Aranaei) of the Urals.
Moscow: KMK Scientific Press Ltd. 229 p.
Fage L. 1912. Etudes sur les araignees cavernicoles. I. Revision des Ochyroceratidae (n. fam.) //
Arch. zool. exper. gen. T.10. No.5. P.97-162.
Fage L. 1913. Etudes sur les Araignees cavernicoles. II. Revision des Leptonetidae // Arch.
zool. exper. gen. T.10. No.5. P479-576.
Foelix R.F. 1982. Biology of spiders. Cambridge: Harvard Univ. Press. 306 p.
Forster R.R. 1970. The spiders of New Zealand. Part III // Otago Mus. Bull. Vol.3. P. 1-184.
Forster R.R., Forster L. 1999. Spiders of New Zealand and their worldwide kin. Univ. Otago
Press, vii + 270 p.
Forster R.R., Platnick N.I. 1977. A review of the spider family Symphytognathidae (Arachnida,
Araneae) // Amer. Mus. Novit. No.2619. P. 1-29.
Forster R.R., Platnick N.I. 1984. A review of the archaeid spiders and their relatives, with
notes on the limits of the superfamily Palpimanoidea (Arachnida, Araneae) // Bull. Amer.
Mus. nat. Hist. Vol.178. P. 1-106.
Forster R.R., Platnick N.I. 1985. A review of the austral spider family Orsolobidae (Arachnida,
Araneae), with notes on the superfamily Dysderoidea // Bull. Amer. Mus. nat. Hist. Vol. 181.
P. 1-230.
Gertsch W.J. 1960. Descriptions of American spiders of the family Symphytognathidae //
Amer. Mus. Novit. No. 1981. P. 1-40.
Grasshoff M. 1970a. Die Gatrung Kilima n. gen. (Arachnida: Araneae: Araneidae) // Sencken-
berg. biol. Bd.51.S.119-128.
Grasshoff M. 1970b. Die Tribus Mangorini. I. Die Gattungen Eustala, Larinia s. sir., Larinopa
n. gen. (Arachnida: Araneae: Araneidae-Araneinae) // Senckenberg. biol. Bd.51. S.209-
234.
Grasshoff M. 1970c. Die Tribus Mangorini. II. Die neuen Gattungen Siwa, Paralarinia, Fara-
dja, Mahembea und Lariniaria (Arachnida: Araneae: Araneidae-Araneinae) //
Senckenberg. biol. Bd.51. S.409-423.
Grasshoff M. 1983. Larinioides Caporiacco 1934, der korrekte Name fur die sogenannte
Araneus cornutus-Gmppe (Arachnida: Araneae) // Senckenberg. biol. Bd.64. S.225-
229.
Grasshoff M. 1986. Die Radnetzspinnen-Gattung Neoscona in Afrika (Arachnida: Araneae) //
Zool. Wetensch. Bd.250. S. 1-123.
Grese N. 1909. Die Spinnen der Halbinsel Jamal // Annu. Mus. Zool. Acad. Sci. S.Petersb.
T.14. S.325-331.
Griswold C.E. 1993. Investigations into the phylogeny of the lycosoid spiders and their kin
(Arachnida: Araneae: Lycosoidea) // Smithson. Contrib. Zool. No.539. P. 1-39.
Griswold C.E., Coddington J.A., Hormiga G., Scharff N. 1998. Phylogeny of the orb-web
building spiders (Araneae, Orbiculariae: Deinopoidea, Araneoidea) // Zool. J. Linn. Soc.
Vol.123. P. 1-99.
290
Литература
Grothendieck К., Kraus О. 1994. Die Wasserspinne Argyroneta aquatica: Verwandtschaft
und Spezialisation (Arachnida, Araneae, Agelenidae) // Verh. naturwiss. Ver. Hamburg
(N.F.). Bd.34. S.259-273.
Grube E. 1861. Beschreibungen neuer, von den Herren L. v. Schrenk, Maack, С v. Ditmar u. a.
im Amurlande und in Ostsibirien gesammelter Araneiden // Bull. Acad. Imp. Sci. St.-Peters-
bourg. Ser.3. T.4. No.3. P. 161-180.
Guseinov E.F., Cerveira A.M., Jakson R.R. 2004. The predatory strategy, natural diet, and life
cycle of Cyrba algerina, an araneophagic jumping spider (Salticidae: Spartaeinae) from
Azerbaijan // N. Z. J. Zool. Vol.31. P.291-303.
Guseinov E.F., Marusik Y.M., Koponen S. 2005. Spiders (Arachnida: Aranei) of Azerbaijan
5. Faunistic review of the funnel-web spiders (Agelenidae) with the description of a new
genus and species // Arthropoda Selecta. Vol.14. No.2. P. 153-177.
Guy Y. 1966. Contribution a Г etude des araignees de la famille des Lycosidae et de la sous-
famille des Lycosinae avec etude speciale des especes du Maroc // Trav. Inst, scient. cherif
(Zool.). T.33. P. 1-174.
Heimer S., Nentwig W. 1991. Spinnen Mitteleuropas: Ein Bestimmungsbuch. Berlin: Verlag
Paul Parey. 543 S.
Helsdingen P. J., van. 1968. Comparative notes on the species of the Holarctic genus Stemony-
phantes Menge (Araneida, Linyphiidae) // Zool. Meded. Leiden. Bd.43. P. 117-139.
Helsdingen P.J., van. 1974. The affinities of Wubana andAllomengea with some notes on the
latter genus (Araneae, Linyphiidae) // Zool. Meded. Leiden. Bd.46. P.295-321.
Hippa H., Koponen S., Oksala I. 1986. Revision and classification of the Holarctic species of
the Ozyptila rauda group (Araneae, Thomisidae) // Ann. zool. Fenn. Vol.23. P.321-328.
Hippa H., Oksala I. 1985. A review of some Holarctic Agyneta Hull s.str. (Araneae,
Linyphiidae) // Bull. Brit, arachnol. Soc. Vol.6. Pt.7. P.277-288.
Holm A. 1970. Notes on spiders collected by the "Vega" expedition 1878-1880 // Entomol.
Scand. Vol.1. No.3. P. 188-208.
Holm A. 1973. On the spiders collected during the Swedish expedition to Novaya Zemlya and
Yenisey in 1875 and 1876 // Zool. Scr. Vol.2. No.3. P.71-110.
Homann H. 1975. Die Stellung der Thomisidae und der Philodromidae im System der Araneae
(Chelicerata, Arachnida) // Z. Morph. Bd.80. S. 181-202.
Hormiga G. 1994. A revision and cladistic analysis of the spider family Pimoidae (Araneoidea:
Araneae) // Smithson. Contrib. Zool. No.549. P. 1-104.
Hormiga G. 2003. Weintrauboa, a new genus of pimoid spiders from Japan and adjacent
islands, with comments on the monophyly and diagnosis of the family Pimoidae and the
genus Pimoa (Araneoidea, Araneae) // Zool. J. Linn. Soc. Vol.139. P.261-281.
Hormiga G. 2008. On the spider genus Weintrauboa (Araneae, Pimoidae), with a description
of a new species from China and comments on its phylogenetic relationships // Zootaxa.
Vol.1814. P. 1-20.
Ihara Y 2003. Cybaeus akiensis n. sp. (Araneae: Cybaeidae) from western Honshu, Japan,
with some notes on its biology // Acta Arachnol. Vol.52. P.51-57.
Jarvi Т.Н. 1912. Das Vaginalsystem der Sparassiden. I // Ann. Acad. Sci. Fenn. Bd.4. S.l-
131.
Jarvi Т.Н. 1914. Das Vaginalsystem der Sparassiden. II // Ann. Acad. Sci. Fenn. Bd.4. S.132-
248.
Jocque R. 1991. A generic revision of the spider family Zodariidae (Araneae) // Bull. Amer.
Mus. nat. Hist. Vol.201. P. 1-160.
Литература
291
Jocque R., Dippenaar-Schoeman A.S. 2006. Spider families of the World. Tervuren: Royal
Museum for Central Africa. 336 p.
Jones D. 1989. A Guide to spiders of Britain and Northern Europe. Revised ed. London:
Hamlyn. 320 p.
Kamura T. 1984. Notes on Japanese gnaphosid spiders (I) // Atypus. No.85. P. 1-8.
Kamura T. 1986. Notes on Japanese gnaphosid spiders (II) // Atypus. No.87. R9-20.
Kamura T. 1987a. Notes on Japanese gnaphosid spiders (III): Callilepis nocturna (Linnaeus)
and Zelotes potanini Schenkel // Atypus. No.89. P. 1-6.
Kamura T. 1987b. Two new species of the genus Zelotes (Araneae: Gnaphosidae) from Japan //
Akitu (n. ser.). Vol.85. P. 1-7.
Kamura T. 1987c. Three species of the genus Drassyllus (Araneae: Gnaphosidae) from Japan //
Acta arachnol. Vol.35. P.77-88.
Kamura T. 1987d. Redescription of Odontodrassus hondoensis (Araneae: Gnaphosidae) //
Proc. jap. Soc. syst. Zool. Vol.36. P.29-33.
Kamura T. 1988. A revision of the genus Gnaphosa (Araneae: Gnaphosidae) from Japan //
Akitu. Vol.97. P. 1-14.
Kamura T. 1990. Notes on Japanese gnaphosid spiders (IV): One newly recorded species and
two little-known species of Japan // Atypus. No.95. P.32-38.
Kamura T. 1992a. Notes on Japanese gnaphosid spiders (V). Three rare species in Japan //
Atypus. No. 100. P. 17-22.
Kamura T. 1992b. Two new genera of the family Gnaphosidae (Araneae) from Japan // Acta
arachnol. Vol.41. P. 119-132.
Kamura T. 1995a. A newly recorded spider from Japan, Phaeocedus braccatus (L. Koch)
(Araneae: Gnaphosidae) // Acta arachnol. Vol.44. P.43^6.
Kamura T. 1995b. A new species and a newly recorded species of the genus Haplodrassus
(Araneae: Gnaphosidae) from Japan // Acta arachnol. Vol.44. P. 123-127.
Kamura T. 1998. Taxonomic notes on Gnaphosidae (Araneae) from Japan // Acta arachnol.
Vol.47. P. 169-171.
Kamura T. 2001. A new genus Sanitubius and a revived genus Kishidaia of the family
Gnaphosidae (Araneae) // Acta arachnol. Vol.50. P. 193-200.
Kamura T. 2007. Spiders of the genus Haplodrassus (Araneae: Gnaphosidae) from Japan //
Acta arachnol. Vol.55. P.95-103.
Kaston B.J. 1948. Spiders of Connecticut // Bull. Conn. St. geol. nat. Hist. Surv. Vol.70. P.l-
874.
Kluge N.J. 2007. Case 3371. Araneidae Clerck, 1758, Araneus Clerck, 1758 and Tegenaria
Latreille, 1804 (Arachnida, Araneae): proposed conservation // Bull, of Zool. Nomencl.
Vol.64. No.l.P.15-18.
Koch L. 1879a. Arachniden aus Sibirien und Novaja Semlja eingesammelt von der schwedischen
Expedition im Jahre 1875 // Kongl. Svenska Vet.-Akad. Handl. Bd.16. H.5. S. 1-136.
Koch L. 1879b. Uebersicht der von Dr. Finsch in Westsibirien gesammelten Arachniden //
Verh. zool.-bot. Ges. Wien. Bd.28 (1878). S.481^90.
Kronestedt T. 1986. Studies on species of Holarctic Pardosa groups (Araneae, Lycosidae).
III. Redescriptions of Pardosa algens (Kulczynski), P. septentrionalis (Westring), and P.
sodalis Holm // Ent. scand. Vol.17. P.215-234.
Kronestedt T. 1990. Separation of two species standing as Alopecosa aculeata (Clerck) by
morphological, behavioural and ecological characters, with remarks on related species in
thepulverulenta group (Araneae, Lycosidae) // Zool. Scr. Vol.19. P203-225.
292
Литература
Kronestedt Т., Marusik Yu.M. 2002. On Acantholycosa solituda (Levi & Levi) and A. sterneri
(Marusik) (Araneae, Lycosidae), a pair of geographically distant allied species // Acta
arachnol. Vol.51. No.l. P.63-71.
Kulczynski W. 1885. Araneae in Camtschadalia a Dre B. Dybowski collectae // Pam. Wydz.
matem.-przyr. Akad. Umiej. Krakow. Т.Н. P. 1-60.
Kulczynski W. 1895. Attidae musei zoologici Varsoviensis in Siberia orientali collecti // Rozpr.
spraw. wydz. mat. przyrod. Akad. umiej. Cracov. T.32. P.45-98.
Kulczynski W. 1901. Arachnoidea // G. Horvath (Hrsg.). Zoologische Ergebnisse der dritten
asiatischen Forschungsreise des Grafen Eugen Zichy. Budapest, Leipzig. Bd.2. S.311-
369.
Kulczynski W. 1907. Fragmenta arachnologica V. IX. Araneae in Terra Tshuktshorum a Cel.
Podhorski lecitae // Bull. Acad. Sci. Cracovie. CI. Sci. math, narur. T.6. P.585-595.
Kulczynski W. 1908. Araneae et Oribatidae expeditionum Rossicarum in insulas Novo-Sibiricas
annis 1885-1886 et 1900-1903 susceptarum // Mem. Acad. Imp. Sci. St.-Petersbourg.
Ser.8. CI. phys.-math. T.18. No.7. P. 1-97.
Kulczynski W. 1916. Araneae Sibiriae Occidentalis Arcticae // Mem. Acad. Imp. Sci. Petrograd.
Ser.8. CI. phys.-math. T.28. No.ll. P. 1^4.
Kulczynski W. 1926. Arachnoidea camtschadalica // Annu. Zool. Mus. AN USSR. T.27. No. 1.
P.29-72.
Kurenshchikov D.K. 1994. The spider genus Tetragnatha Latreille, 1804, from the southern
Far East of Russia (Aranei Tetragnathidae) // Arthropoda Selecta. Vol.3. No. 1-2. P.57-64.
Leech R.E. 1972. A revision of the Nearctic Amaurobiidae (Arachnida: Araneida) // Mem.
ent. Soc. Canada. Vol.84. P. 1-187.
Lehtinen P.T. 1967. Classification of the Cribellate spiders and some allied families, with
notes on the evolution of the suborder Araneomorpha // Ann. Zool. Fennici. Vol.4. P. 199-
467.
Lehtinen P.T. 2002. Generic revision of some thomisids related Xo Xysticus C.L.Koch, 1835
and Ozyptila Simon, 1864 // S. Toft, N. Scharff (eds.). European Arachnology 2000:
Proceedings of the 19th European Colloquium of Arachnology. Aarhus Univ. Press. P.315-
327.
Lehtinen P.T. 2004. Taxonomic notes on the Misumenini (Araneae: Thomisidae: Thomisinae),
primarily from the Palaearctic and Oriental regions // D.V Logunov, D. Penney (eds.).
European Arachnology 2003. Proc. 21st Europ. Colloq. Arachnol., St.-Petersb., 4-9 Aug.
2003. Moscow: KMK Sci. Press Ltd. P. 147-184.
Lehtinen P.T, Koponen S., Saaristo M.I. 1979. Studies on the spider fauna of the southwestern
archipelago of Finland 2. The Aland mainland and the island of Eckero // Mem. Soc.
Fauna Flora fenn. Vol.55. No.l. P.33-52.
Lehtinen P.T, Saaristo M.I. 1980. Spiders of the Oriental-Australian region. II. Nesticidae //
Ann. Zool. Fenn. Vol.17. P.47-66.
Levi H.W. 1951 New and rare spiders from Wisconsin and adjacent states // Amer. Mus.
Novit. No.l501.P.l^ll.
Levi H.W. 1954. Spiders of the genus Euryopis from North and Central America (Araneae,
Theridiidae) // Amer. Mus. Novit. No. 1666. P. 1^18.
Levi H.W. 1957a. The spider genera Enoplognatha, Theridion, and Paidisca in America north
of Mexico (Araneae, Theridiidae) // Bull. Amer. Mus. Nat. Hist. Vol.112. No.l. P.l-123.
Levi H.W. 1957b. Spiders of the new genus Arctachaea (Araneae, Theridiidae) // Psyche.
Vol.64. No.3. P.102-106.
Литература
293
Levi H.W. 1974a. The orb-weaver genus Zygiella (Araneidea, Araneidae) // Bull. Mus. Сотр.
Zool. Vol.146. No.5. P.267-290.
Levi H.W. 1974b. The orb-weaver genera Araniella and Nuctenea (Araneidea, Araneidae) //
Bull. Mus. Сотр. Zool. Vol.146. No.6. P.291-316.
Levi H.W. 1977. The American orb-weaver genera Metepeira, Kaira, and Aculeipeira north
of Mexico (Araneae: Araneidae) // Bull. Mus. Сотр. Zool. Vol.148. No.5. P.185-238.
Levi H.W. 1980. The orb-weaver genus Mecynogea, the subfamily Metinae and the genera
Pachygnatha, Gleognatha andAzilia of the the subfamily Tetragnathinae north of Mexico //
Bull. Mus. Сотр. Zool. Vol.149. P. 1-74.
Levi H.W., Levi L.R. 1962. The genera of the spider family Theridiidae // Bull. Mus. сотр.
Zool. Harv. Vol.127. P. 1-71.
Logunov D.V. 1992a. Definition of the spider genus Talavera (Araneae, Salticidae), with a
description of a new species // Bull. Inst. roy. sci. natur. Belg. Entomol. T.62. P.75-82.
Logunov D.V 1992b. The spider family Salticidae from Tuva. II. An annotated check-list of
species // Arthropoda Selecta. Vol.1. No.2. P.47-71.
Logunov D.V. 1993 a. Notes on the penicillatus species group of the genus Sitticus Simon, 1901
with a description of a new species (Araneae, Salticidae) // Genus. Vol.4. No.l. P. 1-15.
Logunov D.V. 1993b. Notes on two salticid collections from China (Araneae Salticidae) //
Arthropoda Selecta. Vol.2. No.l. P.49-59.
Logunov D.V. 1993c. New data on the jumping spiders (Araneae Salticidae) of Mongolia and
Tuva // Arthropoda Selecta. Vol.2. No.2. P.47-53.
Logunov D.V. 1995. Contribution to the northern Asian fauna of the crab spider genus Xysticus
C.L. Koch, 1835 (Aranei Thomisidae) // Arthropoda Selecta. Vol.3. No.3^1. P.lll-118.
Logunov D.V 1996a. A critical review of the spider genera Apollophanes O.P.-Cambridge,
1898 and Thanatus C.L.Koch, 1837 in North Asia (Araneae, Philodromidae) // Rev.
arachnol. Т.Н. Fasc.3. P.133-202.
Logunov D.V. 1996b. A review of the genus Phlegra Simon, 1876 in the fauna of Russia and
adjacent countries (Araneae: Salticidae: Aelurillinae) // Genus. Vol. 7. P.533-567.
Logunov D.V. 1997. Taxonomic notes on some Central Asian philodromid spiders (Aranei
Philodromidae) // Arthropoda Selecta. Vol.6. No. 1-2. P.99-104.
Logunov D.V. 1998. Pseudeuophrys is a valid genus of the jumping spiders (Araneae,
Salticidae) // Rev. arachnol. T.12. Fasc.ll. P.109-128.
Logunov D.V. 1999. Redefinition of the genera Marpissa С L. Koch, 1846 and Mendoza
Peckham & Peckham, 1894 in the scope of the Holarctic fauna (Araneae, Salticidae) //
Rev. arachnol. T.13. Fasc.2. P.25-60.
Logunov D.V, Cutler В., Marusik Yu.M. 1993. A review of the genus Euophrys C.L. Koch in
Siberia and the Russian far East // Ann. Zool. Fennici. Vol.30. P.101-124.
Logunov D.V, He^ciak S. 1996. Asianellus, & new genus of the subfamily Aelurillinae (Araneae,
Salticidae) // Ent. Scand. Vol.26. P. 103-117.
Logunov D.V, Kronestedt T. 1997. A new Palearctic species of the genus Sitticus Simon, with
notes on related species in the floricola group (Araneae, Salticidae) // Bull. Br. arachnol.
Soc. Vol.10. Pt.7. P.225-233.
Logunov D.V, Kronestedt T. 2003. A review of the genus Talavera Peckham and Peckham,
1909 (Arareae, Salticidae) // J. nat. Hist Vol.37. P. 1091-1154.
Logunov D.V, Marusik Yu.M. 1994a. A faunistic review of the crab, spiders (Araneae,
Thomisidae) from the mountains of South Siberia // Bull. Inst. Roy. Sci. Natur. Belgique,
Entomol. T.64. P. 177-197.
294
Литература
Logunov D.V., Marusik Yu.M. 1994b. New data on the jumping spiders of the Palaearctic
fauna (Aranei, Salticidae) // Arthropoda Selecta. Vol.3. No. 1-2. P.101-115.
Logunov D.V., Marusik Yu.M. 1995. Spiders of the family Lycosidae from the Sokhondo
reserve, Chita Area, East Siberia (Arachnida: Araneae) // Beitr. Araneol. Bd.4 (fur 1994).
P. 109-122.
Logunov D.V., Marusik Yu.M. 1998. A new species of the genus Xysticus from the mountains
of South Siberia and Mongolia (Araneae, Thomisidae) // Bull. Br. arachnol. Soc. Vol.11.
Pt.3. P. 103-106.
Logunov D.V, Marusik Yu.M. 1999a. New species and records of the jumping spiders from
the Russian Far East (Araneae, Salticidae) // Acta Arachnologica. Vol.48. No.l. P.23-29.
Logunov D.V, Marusik Yu.M. 1999b. A brief review of the genus Chalcoscirtus Bertkau,
1880 in the fauna of Central Asia and the Caucasus (Araneae, Salticidae) // Arthropoda
Selecta. Vol.7. No.3. P.205-226.
Logunov D.V, Marusik Yu.M. 2000a. Miscellaneous notes on Palaearctic Salticidae
(Arachnida: Aranei) // Arthropoda Selecta. Vol.8. No.4. P.263-292.
Logunov D.V, Marusik Yu.M. 2000b. Catalogue of the jumping spiders of northern Asia
(Arachnida, Araneae, Salticidae). Moscow: KMK Scientific Press Ltd. 299 pp.
Logunov D.V, Marusik Yu.M. 2003. A revision of the genus Yllenus Simon, 1868
(Arachnida, Araneae, Salticidae). Moscow: KMK Scientific Press Ltd. 167 pp.
Logunov D.V, Marusik Yu.M., Rakov S. 1999. A review of the genus Pellenes in the fauna of
Central Asia and the Caucasus (Araneae, Salticidae) // J. Nat. Hist. Vol.33. No.l. P.89-
148.
Logunov D.V, Marusik Yu.M., Trilikauskas L. 2001. A new species of the genus Xysticus
from South Siberia (Araneae, Thomisidae) // Reichenbachia. Bd.34. P.33-38.
Logunov D. V, Rakov S.Y 1996. A review of the spider genus Synageles Simon, 1876
(Araneae, Salticidae) in the fauna of Central Asia // Bull. Inst. Roy. Sci. Nat. Belg., Entomol.
T.66. P.65-74.
Logunov D.V, Wesolowska W. 1992. The jumping spiders (Araneae, Salticidae) of
Khabarovsk Province (Russian Far Easr) // Ann. Zool. Fennici. Vol.29. P. 113-146.
Logunov D.V, Wesolowska W. 1995. New data on some poorly known Palaearctic species of
Sitticus (Araneae: Salticidae) // Genus. Vol.6. No.2. P. 163-175.
Maddison W.P. 1988. A common method of sound production by courting jumping spiders
(Araneae, Salticidae) // J. Arachnol. Vol.16. P.267-269.
Mansour F., Rosen D., Shulov A. 1980. Biology of the spider Chiracanthium mildei
(Arachnida: Clubionidae) // Entomophaga. Vol.25. No.3. P.237-248.
Marusik Yu.M. 1986. The orb-weaver genus Larinia Simon in the USSR // Spixiana. Bd.9.
H.3. P.245-254.
Marusik Yu.M. 1995. A review of the spider genus Titanoeca from Siberia (Arachnida: Araneae:
Titanoecidae) // Beitr. Araneol. Bd.4 (fur 1994). P. 123-132.
Marusik Yu.M. 2005. New family and new interesting records of spiders (Aranei) from the
European part of Russia // Arthropoda Selecta. Vol.14. No.l. P.89-91.
Marusik YM. 2008. Synopsis of the Ozyptila rauda-group (Araneae, Thomisidae), with
revalidation of Ozyptila balkarica Ovtsharenko, 1979 // Zootaxa. Vol.1909. P.52-64.
Marusik Yu.M. 2009a. A check-list of spiders (Aranei) from the Laso Reserve, Maritime
Province, Russia // Arthropoda Selecta. Vol.18. No. 1-2. P.95-109.
Marusik Yu.M. 2009b. First description of the male ofAlloclubionoides amurensis (Ovtchinni-
kov, 1999) (Aranei: Amaurobiidae) //Arthropoda Selecta. Vol.18. No. 1-2. P.51-55.
Литература
295
Marusik Yu.M. 2011. A new genus of hahniid spiders from the Far East Asia // Zootaxa.
No.2788. P.57-68.
Marusik Yu.M., Azarkina G.N., Koponen S. 2004. A survey of East Palaearctic Lycosidae
(Aranei). II. Genus Acantholycosa Dahl, 1908 and related new genera // Arthropoda Selecta.
Vol.12. No.2. P.101-148.
Marusik Yu.M., Buchar J. 2004. A survey of East Palaearctic Lycosidae (Aranei). 3. On the
wolf spiders (Araneae, Lycosidae) collected in Mongolia by Z. Kaszab in 1966-1968 //
Arthropoda Selecta. Vol.12. No.2. P.149-158.
Marusik Yu.M., Chevrizov B.P. 1990. Three new crab spiders from the Asian part of the
USSR//Reichenbachia. Bd.27. H.15. P.89-93.
Marusik Yu.M., Cutler B. 1989. Description of the males of Dendryphantes czekanowskii
Proszynski andHeliophanus baicalensis Kulczynski (Araneae, Salticidae)//Acta Arachnol.
Vol.37. No.2. P.51-55.
Marusik Yu.M., Esyunin S.L. 1998. A new species of the spider genus Pelecopsis Simon, 1864
(Aranei, Linyphiidae) from South Siberia // Arthropoda Selecta. Vol.6. No.3^1. P. 105-108.
Marusik Yu.M., Esyunin S.L. 2010. On the northernmost Ajmonia Caporiacco, 1934 (Aranei:
Dictynidae: Dictinynae) // J. Nat. Hist. Vol.44. No.5-6. P.361-367.
Marusik Yu.M., Fritzen N.R., Song D.X. 2007. On spiders (Aranei) collected in central Xinjiang,
China // Arthropoda Selecta. Vol.15. No.3. P.259-276.
Marusik Yu.M., Gnelitsa V A., Koponen S. 2006. A survey of Holarctic Linyphiidae (Araneae).
1. A review of the Palaearctic genus Notioscopus Simon, 1884 // Bull. Brit, arachnol. Soc.
Vol.13. Pt.8. P.321-328.
Marusik Yu.M., Guseinov G. 2003. Spiders (Arachnida, Aranei) of Azerbaijan. I. New families
and genera records // Arthropoda Selecta. Vol. 12. No.l. P.29^16.
Marusik Yu.M., Hippa H., Koponen S. 1996. Spiders from the Altai area, South Siberia //
Acta Zoologica Fennica. Vol.201. P. 11^45.
Marusik Yu.M., Koponen S. 1992. A review of Meta (Araneae, Tetragnathidae) with description
of two new species // J.Arachnol. Vol.20. No.2. P. 137-143.
Marusik Yu.M., Koponen S. 1998. New and little known spiders of the subfamily Dictyninae
(Araneae, Dictynidae) from South Siberia // Entomol. Probl. Vol.29. No.2. P.79-86.
Marusik Yu.M., Koponen S. 2000. New data on spiders (Aranei) from the Maritime Province //
Arthropoda Selecta. Vol.9. No.l. P.55-68.
Marusik Yu.M., Koponen S. 2001a. A new species of the genus Gnaphosa (Araneae, Gnapho-
sidae) from Mongolia and new species records from east Palaearctic // Acta Arachnol.
Vol.50. No.2. P.135-144.
Marusik Yu.M., Koponen S. 2001b. Revision of the Holarctic spider genus Procerocymbium
Eskov, 1989 (Araneae: Linyphiidae) // Acta Arachnol. Vol.50. No.2. P. 145-156.
Marusik Yu.M., Koponen S. 2001c. Spiders of the family Zodariidae from Mongolia
(Arachnida: Araneae) // Reichenbachia. Bd.34. P.39^18.
Marusik Yu.M., Koponen S. 2002. Vermontia, a linyphiid genus new to Palaearctic and new
records of spiders from northeast Asia (Araneae) // Bull. Br. arachnol. Soc. Vol.12. Pt.4.
P.159-165.
Marusik Yu.M., Koponen S. 2009. A new species of Uusitaloia Marusik, Koponen & Danilov
(Araneae: Linyphiidae: Erigoninae), a genus through to be monotypic, from the Wrangel
Island NE Russia // Acta Entomologica Fennica. Vol.20. P. 18-21.
Marusik Yu.M., Koponen S. 2010. A review of the Holarctic genus Tmeticus Menge, 1868
(Araneae, Linyphiidae), with a description of a new genus // ZooKeys. No.59. P. 15-37.
296
Литература
Marusik Yu.M., Koponen S., Danilov S.N. 2001. Taxonomic and faunistic notes on linyphiids
of Transbaikalia and south Siberia // Bull. Brit, arachnol. Soc. Vol.12. Pt.2. P.83-92.
Marusik Yu.M., Koponen S., Vinokurov N.N., Nogovitsyna S.N. 2002. Spiders (Aranei) from
northernmost forest-tundra of northeastern Yakutia (70°35'N, 134°34'E) with description
of three new species // Arthropoda Selecta. Vol. 10. No.4. P.351-370.
Marusik Yu.M., Kovblyuk M.M. 2010. The spider genus Trachelas L.Koch, 1872 (Aranei:
Corinnidae) in Russia // Arthropoda Selecta. Vol.19. No.l. P.21-27.
Marusik Yu.M., Kuzminykh G.V 2010. On two spider genera new to Russia (Aranei:
Corinnidae, Sparassidae) // Arthropoda Selecta. Vol.19. No.2. P.97-100.
Marusik Yu.M., Leech R. 1993. The spider genus Hypselistes, including two new species,
from Siberia and the Russian Far East (Araneida: Erigonidae) // Can. Entomol. Vol. 125.
P.1115-1126.
Marusik Yu.M., Lehtinen P.Т., Kovblyuk M.M. 2005. Cozyptila, a new genus of crab spiders
(Araneae, Thomisidae, Thomisinae, Coriarachnini) from the western Palaearctic //
Arthropoda Selecta. Vol.13. No.3. P.151-163.
Marusik Y.M., Logunov D.V. 1990. The crab spiders of middle Asia, USSR (Aranei,
Thomisidae). 1. Descriptions and notes on distribution of some species // Korean Arachnol.
Vol.6. No.l. P.31-62.
Marusik Yu.M., Logunov D.V. 1995a. Gnaphosid spiders from Tuva and adjacent territories,
Russia (Arachnida: Araneae: Gnaphosidae) // Beitr. Araneol. Bd.4 (fur 1994). P. 177-210.
Marusik Y.M., Logunov D.V. 1995b. The crab spiders of Middle Asia (Aranei, Thomisidae), 2 //
Beitr. Araneol. Bd.4 (fur 1994). P. 133-175.
Marusik Yu.M., Logunov D.V. 1998. Taxonomic notes on the Evarchafalcata species complex
(Aranei, Salticidae) // Arthropoda Selecta. Vol.6. No.3^1. P.95-104.
Marusik Yu.M., Logunov D.V. 2002a. New faunistic records for the spiders of Buryatia
(Araneae), with a description of a new species from the genus Enoplognatha (Theridiidae) //
Arthropoda Selecta. Vol.10. No.3. P.265-272.
Marusik Yu.M., Logunov D.V 2002b. New and poorly known species of crab spiders (Aranei:
Thomisidae) from South Siberia and Mongolia // Arthropoda Selecta. Vol. 10. No.4. P.315—
322.
Marusik Yu.M., Logunov D.V. 2009. New faunistic records of spiders collected from the
mountain Altai (Arachnida, Aranei) // Arthropoda Selecta. Vol.18. No.3^1. P.145-152.
Marusik Yu.M., Logunov D.V, Koponen S. 2000. Spiders of Tuva, South Siberia. Magadan:
IBPN FEB RAS. 252 p.
Marusik Y.M., Omelko M.I. 2008. First description of the female ofOzyptila utotchkini
(Araneae: Thomisidae) // Zootaxa. Vol.1823. P.65-68.
Marusik Y.M., Omelko M.I., Koponen S. 2010. A survey of the East Palaearctic Lycosidae
(Aranei). 5. Taxonomic notes on the easternmost Palaearctic Pirata species and on the
genus Piratosa Roewer, 1960 // Arthropoda Selecta. Vol.19. No.l. P.29-36.
Marusik Yu.M., Ryabukhin A.S., Kuzminykh G.V. 2010. New data on spiders and harvestmen
(Arachnida: Aranei & Opiliones) from Western Koryakia, Kamchatka Peninsula //
Arthropoda Selecta. Vol.19. No.4. P.227-236.
Marusik Yu.M., Saaristo M.I. 1999. Review of East Palaearctic species of the genus Minicia
Thorell, 1875 with descriptions of two new genera (Aranei: Linyphiidae: Erigoninae) //
Arthropoda Selecta. Vol.8. No.2. P.125-130.
Marusik Yu.M., Tanasevitch A.V. 1998. Notes on the spider genus Styloctetor Simon, 1884
and some related genera, with description of two new species from Siberia (Aranei:
Linyphiidae) // Arthropoda Selecta. Vol.7. No.2. P. 153-159.
Литература
297
Marusik Yu.M., Tanasevitch A.V. 2003. Two new erigonine spiders (Aranei, Linyphiidae)
from mountains of South Siberia // Arthropoda Selecta. Vol.11. No.2. P. 159-165.
Marusik Yu.M., Tanasevitch A.V, Kurenshchikov D.K., Logunov D.V. 2007. A check-list of
the spiders (Araneae) of the Bolshekhekhtsyrski Nature Reserve, Khabarovsk Province,
the Russian Far East // Acta Arachnologica Sinica. Vol.16. No.l. P.37-64.
Mikhailov K.G. 1987. Contribution to the spider fauna of the genus Micaria Westring, 1851
of the USSR. I (Aranei, Gnaphosidae) // Spixiana. Bd.10. H.3. P.319-334.
Mikhailov K.G. 1990. The spider genus Clubiona Latreille 1804 in the Soviet Far East, 1
(Arachnida, Aranei, Clubionidae) // Korean Arachnol. Vol.5. No.2. P. 139-175.
Mikhailov K.G. 1991. The spider genus Clubiona Latreille 1804 in the Soviet Far East, 2
(Arachnida, Aranei, Clubionidae) // Korean Arachnol. Vol.6. No.2. P.207-235.
Mikhailov K.G. 1992. The spider genus Clubiona Latreille, 1804 (Arachnida, Aranei,
Clubionidae) in the USSR fauna: a critical review with taxonomical remarks // Arthropoda
Selecta. Vol.1. No.3. P.3-34.
Mikhailov K.G. 1995a. New or rare Oriental sac spiders of the genus Clubiona Latreille 1804
(Aranei Clubionidae) // Arthropoda Selecta. Vol.3. No.3^. P.99-110.
Mikhailov K.G. 1995b. Erection of infrageneric groupings within the spider genus Clubiona
Latreille, 1804 (Aranei, Clubionidae): a typological approach // Arthropoda Selecta. Vol.4.
No.2. P.33-48.
Mikhailov K.G. 1997. Catalogue of the spiders of the territories of the former Soviet Union
(Arachnida, Aranei). Moscow: Zoological Museum of the Moscow State University.
416 p.
Mikhailov K.G. 1998. Catalogue of the spiders (Arachnida, Aranei) of the territories of the
former Soviet Union. Addendum 1. Moscow: KMK Scientific Press Ltd. 50 p.
Mikhailov K.G. 1999. Catalogue of the spiders (Arachnida, Aranei) of the territories of the
former Soviet Union. Addendum 2. Moscow: Zoological Museum, Moscow State University.
39 p.
Mikhailov K.G. 2000. Catalogue of the spiders (Arachnida, Aranei) of the territories of the
former Soviet Union. Addendum 3. Moscow: Zoological Museum, Moscow State University.
33 p.
Mikhailov K.G. 2002. The spider fauna of Russia and other post-Soviet republics: a 2000
update // S. Toft, N. Scharff (eds.). European Arachnology 2000. Proceedings of the 19th
European Colloquium of Arachnology, Aarhus 17-22 July 2000. P255-259.
Mikhailov K.G. 2003. The spider genus Clubiona Latreille, 1804 (Aranei: Clubionidae) in the
fauna of the former USSR: 2003 update // Arthropoda Selecta. Vol.11. No.4. P.283-317.
Miller F. 1947. Pavouci zvirena hadcovych stepi u Mohelna // Arch. Sv. Vyzk. ochr. prirod.
kraj. zem. Morav. Vol.7. P. 1-107.
Miller J.A., Griswold C.E., Yin CM. 2009. The symphytognathoid spiders of the Gaoligong-
shan, Yunnan, China (Araneae, Araneoidea): Systematics and diversity of micro-
orbweavers // ZooKeys. Vol. 11. P.9-195.
Millidge A.F. 1977. The conformation of the male palpal organs of Linyphiid spiders and its
application to the taxonomic and phylogenetic analysis of the family (Araneae:
Linyphiidae) // Bull. Brit, arachnol. Soc. Vol.4. Pt.l. P. 1-60.
Millidge A.F. 1981a. The erigonine spiders of North America. Part 3. The genus Scotinotylus
Simon (Araneae: Linyphiidae) // J. Arachnol. Vol.9. No.2. P. 167-214.
Millidge A.F. 1981b.The erigonine spiders of North America. Part 5. The genus Satilatlas
(Araneae: Linyphiidae) // Bull. Amer. Mus. Nat. Hist. Vol.170. P.242-253.
298
Литература
Millidge A.F. 1981c. A revision of the genus Gonatium (Araneae: Linyphiidae) // Bull. Brit.
arachnol. Soc. Vol.5. Pt.6. P.253-277.
Millidge A.F. 1983. The erigonine spiders of North America, Part 6. The genus Walckenaeria
Blackwall (Araneae, Linyphiidae) // J. Arachnol. Vol.11. No.2. P. 105-200.
Millidge A.F. 1984. The erigonine spiders of North America. Part 7. Miscellaneous genera
(Araneae, Linyphiidae) // J. Arachnol. Vol.12. No.2. P. 121-169.
Murphy J. 2007. Gnaphosid genera of the world. St Neots, Cambs: British Arachnological
Society. Vol.1. P.i-xii, 1-92; Vol.2. P.i-ii, 93-605.
Namkung J. 2002. The spiders of Korea. Seoul: Kyo-Hak Publishing Co. 648 p.
Nentwig W. 1983. The prey of web-building spiders compared with feeding experiments
(Araneae: Araneidae, Linyphiidae, Pholcidae, Agelenidae) // Oecologia (Berlin). Vol.56.
No.l.P.132-139.
Nentwig W., Gloor D., Hanggi A., Kropf С 2010. Spiders of Europe. Version 10.2010: http://
www.araneae.unibe.ch/
Nishikawa Y. 2009. A new genus and 44 new species of the family Coelotidae (Arachnidae,
Araneae) from Japan // H. Ono (ed.). The Spiders of Japan with keys to the families and
genera and illustrations of the species. Kanagawa: Tokai Univ. Press. P.51-70.
Noordam A.P. 2002. Abdominal percussion and ventral scutum in male Euophrys frontalis
(Araneae: Salticidae) // Ent. Ber., Amst. Vol.62. No.l. P.17-19.
Nyffeler M. 1982. Field studies on the ecological role of the spiders as insect predators in
agroecosystems (abandoned grassland, meadows, and cereal fields) report. Ph.D. thesis.
Zurich: Swiss Federal Inst, of Tech. 174 p.
Nyffeler M., Benz G. 1981. Okologische Bedeutung der Spinnen als Insektenpradatoren in
Wiesen und Getreidefeldern // Mitt. Dt. Ges. Allg. Angew. Ent. Bd.3. H.l-3. S.33-35.
Odenwall E. 1901. Araneae nonnulae Sibiriae Transbaicalensis // OfVers. Finska Vet.-Soc.
Forhandl. Bd.43. P.255-273, 1 tab.
Oi R. 1960. Linyphiid spiders of Japan // J. Inst. Polytech. Osaka Cy Univ. Vol. 11(D). P. 137-244.
Okuma С. 1988а. A revision of the genus Tetragnatha Latreille (Araneae, Tetragnathidae) of
Asia, Part I // J. Fac. Agr. Kyushu Univ. Vol.32. P.165-181.
Okuma С 1988b. A revision of the genus Tetragnatha Latreille (Araneae, Tetragnathidae) of
Asia, Part II // J. Fac. Agr. Kyushu Univ. Vol.32. P. 183-213.
Okuma C. 1989. Distribution and subdivisional classification of the spider genus Tetragnatha II
Y. Nishikawa, H. Ono (eds.). Arachnological Papers Presented to Takeo Yaginuma on the
Occasion of his Retirement. Osaka: Osaka Arachnologists' Group. P.53-62.
Oliger T.I. 1993. A new Hypsosinga Ausserer, 1871 (Aranei Araneidae) from the Maritime
Province, Russia // Arthropoda Selecta. Vol.2. No.3. P.65-66.
Oliger T.I. 1994. A new species of the spider genus Cybaeus L. Koch, 1868, from the Petrov
Is., Sea of Japan // Arthropoda Selecta. Vol.3. No. 1-2. P.51-55.
Oliger T.I. 1998. The spider genus Pholcus Walckenaer, 1805, in the Maritime Province,
Russian Far East (Aranei: Pholcidae) // Arthropoda Selecta. Vol.7. No.2. P. 111-115.
Oliger T.I. 1999. A new Atypus from the Russian Far East (Aranei: Atypidae) // Arthropoda
Selecta. Vol.7. No.3. P.201-204.
Oliger T.I., Marusik Yu.M., Koponen S. 2002. New and interesting records of spiders (Araneae)
from the Maritime Province of Russia // Acta Arachnologica. Vol.51. No.2. P.93-98.
Omelko M.M., Marusik YM. 2008. A survey of the east Palearctic Lycosidae (Aranei). 4. On
two somatically similar species of Alopecosa from the Russian Far East // Arthropoda
Selecta. Vol.16 (for 2007). No A P.237-243.
Литература
299
Ono H. 1978. Thomisidae aus dem Nepal-Himalaya. I. Das Genus Xysticus С L. Koch 1835
(Arachnida: Araneae) // Senckenberg. biol. Bd.59. S.267-288.
Ono H. (ed.). 2009. The spiders of Japan with keys to the families and genera and illustrations
of the species. Kanagawa: Tokai Univ. Press. 739 p.
Ono H., Marusik Yu.M., Logunov D.V. 1990. Spiders of the family Thomisidae from Sakhalin
and the Kurile Islands // Acta Arachnol. Vol.39. No.l. P.7-19.
Ono H., Matsuda M, Saito H. 2009. Linyphiidae, Pimoidae // H. Ono (ed.). The Spiders of
Japan with keys to the families and genera and illustrations of the species. Kanagawa:
Tokai Univ. Press. P.253-344.
Ono H., Saito H. 2001. New species of the family Linyphiidae (Arachnida, Araneae) from
Japan // Bull. Natn. Sci. Mus., Tokyo. Ser.A. Vol.27. No.3. P. 159-203.
Opell B.D. 1979. Revision of the genera and tropical American species of the spider family
Uloboridae // Bull. Mus. сотр. Zool. Harv. Vol.148. P.443-549.
Opell B.D. 1984. Phylogenetic review of the genus Miagrammopes (sensu lato) (Araneae,
Uloboridae) // J. Arachnol. Vol.12. P.229-240.
Opell B.D., Beatty J.A. 1976. The Nearctic Hahniidae (Arachnida, Araneae) // Bull. Mus.
Сотр. Zool. Vol.147. No.9. P.393^133.
Ovtsharenko VI., Platnick N.I., Marusik Yu.M. 1995. A review of the Holarctic ground spider
genus Parasyrisca (Araneae, Gnaphosidae) // Amer. Mus. Novit. No.3147. P. 1-55.
Ovtsharenko VI., Platnick N.I., Song D.X. 1992. A review of the North Asian ground spiders
of the genus Gnaphosa (Araneae, Gnaphosidae) // Bull. Amer. Mus. Natur. Hist. Vol.212.
P. 1-88.
Paik K.Y. 1992. A new genus of the family Clubionidae (Arachnida, Araneae) from Korea //
Korean Arachnol. Vol.8. No.l. P.7-12.
Palmgren P. 1974a. Die Spinnenfauna Finnlands und Ostfennoskandiens. IV Argiopidae,
Tetragnathidae und Mimetidae // Fauna Fennica. Vol.24. 70 S.
Palmgren P. 1974b. Die Spinnenfauna Finnlands und Ostfennoskandiens. V Theridiidae und
Nesticidae // Fauna Fennica. Vol.26. 54 S.
Palmgren P. 1975. Die Spinnenfauna Finlands und Ostfennoskandiens. VI. Linyphiidae 1 //
Fauna Fennica. Vol.28. 102 S.
Palmgren P. 1976. Die Spinnenfauna Finnlands und Ostfennoskandiens. VII. Linyphiidae 2 //
Fauna Fennica. Vol.29. 126 S.
Palmgren P. 1977. Die Spinnenfauna Finnlands und Ostfennoskandiens. VIII. Argyronetidae,
Agelenidae, Hahniidae, Dictynidae, Amaurobiidae, Titanoecidae, Segestriidae, Pholcidae
und Sicariidae. Anhang: Bestimmungstabelle der in Finnland representierten Spinnen-
familien und kommentiertes Register der Teile I—VIII der Spinnenfauna // Fauna Fennica.
Vol.30. 50 S.
Paquin P., Duperre N. 2003. Guide d'identification des araignees de Quebec // Fabreries,
Suppl. No. 11. P. 1-251.
Parker J.R. 1993. Names of spiders // Br. arachnol. Soc, Members' Handbook. Rl—17.
Peck W.B., Whitcomb W.H. 1970. Studies on the biology of a spider Chiracanthium inclusum
(Hentz) // Bull. Arkansas Agri. Expt. Sta. No.753. P. 1-76.
Peelle M.L., Saito S. 1932. Spiders from the southern Kurile Islands. I. Araneida from Iturup //
J. Fac. Sci. Hokkaido. Ser.6, Zool. Vol.2. No.2. P. 83-96.
Peelle M.L., Saito S. 1933. Spiders from the southern Kurile Islands. II. Araneida from Shiko-
tan // J. Fac. Sci. Hokkaido. Ser.6, Zool. Vol.2. No.3. P. 109-123.
Petto R. 1990. Abundance and prey of Coelotes terrestris (Wider) (Araneae, Agelenidae) in
hedges // Bull. Brit, arachnol. Soc. Vol.8. Pt.6. P.185-193.
300
Литература
Petrunkevitch A. 1923. On families of spiders // Ann. N. Y. Acad. Sci. Vol.29. P.145-180.
Petrunkevitch A. 1928. Systema Aranearum // Trans. Conn. Acad. Arts Sci. Vol.29. P. 1-270.
Petrunkevitch A. 1933. An inquiry into the natural classification of spiders, based on a study
of their internal anatomy // Trans. Conn. Acad. Arts Sci. Vol.31. P.303-389.
Pickard-Cambridge O. 1871. Arachnida // Zool. Rec. Vol.7. P.207-224.
Pickard-Cambridge O. 1873. On some new species of Araneida, chiefly from Oriental Siberia //
Proc. zool. Soc. Lond. P.435^152.
Platnick N.I. 1974. The spider family Anyphaenidae in America north of Mexico // Bull. Mus.
сотр. Zool. Harv. Vol.146. P.205-266.
Platnick N.I. 1989. Advances in Spider Taxonomy 1981-1987: A Supplement to Brignoli's A
Catalogue of the Araneae described between 1940 and 1981. Manchester Univ. Press. 673 p.
Platnick N.I. 1993. Advances in spider taxonomy 1988-1991, with synonymies and transfers
1940-1980. New York. 846 p.
Platnick N.I. 2009. The world spider catalog, version 9.5. American Museum of Natural History,
online at: http://research.amnh.org/entomology/spiders/catalog/index.html
Platnick N.I., Dondale CD. 1992. The insects and arachnids of Canada. Part 19. The ground
spiders of Canada and Alaska. Araneae: Gnaphosidae // Research Branch, Agriculture
Canada Publ. No.1875. 297 p.
Platnick N.I., Shadab M.U. 1988. A revision of the American spiders of the genus Micaria
(Araneae, Gnaphosidae) // Amer. Mus. Novit. No.2916. P. 1-64.
Platnick N.I., Song D.X. 1986. A review of the zelotine spiders (Araneae, Gnaphosidae) of
China // Amer. Mus. Novit. No.2848. P. 1-22.
Pollard S.D. 1983. Egg guarding by Clubiona cambridgei (Araneae, Clubionidae) against
conspecific predators // J. Arachnol. Vol.11. No.3. P.323-326.
Proszynski J. 1971. Redescriptions of the A.E. Grube's Siberian species of Salticidae (Aranei)
in the collection of the Wroclaw Zoological Museum // Ann. Zool. T.28. No. 11. P.205-
226.
Proszynski J. 1979. Systematic studies on East Palaearctic Salticidae, 3. Remarks on Salticidae
of the USSR // Ann. Zool. T.34. No. 11. P.299-369.
Proszynski J. 1987. Atlas rysunkow diagnostycznych mniej znanych Salticidae 2. Siedlce:
Zeszyty Naukowe Wyzszej Szkoly Rolniczo-Pedagogicznej. 177 p.
Proszynski J. 2010. Monograph of the Salticidae (Araneae) of the World: http://salticidae.org/
salticid/main.htm
Rakov S.Y. 1997. A review of the spider genus Evarcha Simon, 1902 in Middle Asia (Aranei
Salticidae) // Arthropoda Selecta. Vol.6. No. 1/2. P. 105-112.
Ramirez M.J., Bonaldo A.B., Brescovit A.D. 1997. Revision del genero Macerio у comentarios
sobre la ubicacion de Cheiracanthium, Tecution у Helebiona (Araneae, Miturgidae,
Eutichurinae) // Iheringia (Zool.). Vol.82. P.43-66.
Renner F. 1988. Wiederbeschreibung von Dolomedes strandi Bonnet und Anmerkungen zur
Taxonomie sibirischerDolomedes-Arten (Araneae: Pisauridae) // Stuttgarter Beitr. Naturk.
Ser.A (Biol). Bd.427. S. 1-6.
Rezac M., Pekar S., Johannesen J. 2008. Taxonomic review and phylogenetic analysis of
central European Eresus species (Araneae: Eresidae) // Zool. Scripta. Vol.37. P.263-287.
Roberts M.J. 1985. The spiders of Great Britain and Ireland. Vol.1: Atypidae to Theridio-
somatidae. Colchester, England: Harley Books. 229 p.
Roberts M.J. 1987. The spiders of Great Britain and Ireland Vol.2. Linyphiidae. Colchester,
England: Harley Books. 204 p.
Литература
301
Roberts M.J. 1995. Collins field guide: spiders of Britain & Northern Europe. London: Harper
Collins. 383 p.
Roberts M.J. 1998. Spinnengids. Baarn, Netherlands: Tirion. 397 p.
Roewer C.F. 1959. Araneae Lycosaeformia II (Lycosidae) // Explor. Pare natn. Upemba Miss.
G. F. deWitte. T.55. S.l-518.
Roewer C.F. 1960. Araneae Lycosaeformia II (Lycosidae) (Fortsetzung und Schluss) // Explor.
Pare natn. Upemba Miss. G. F. de Witte. T.55. S.519-1040.
Roth V.D. 1965. Genera erroneously placed in the spider families Agelenidae and Pisauridae
(Araneida: Arachnida) //Ann. ent. Soc. Am. Vol.58. P.289-292.
Saaristo M.I. 2001. Dwarf hunting spiders or Oonopidae (Arachnida, Araneae) of the
Seychelles // Insect Syst. Evol. Vol.32. P.307-358.
Saaristo M.I., Eskov K.Yu. 1996. Taxonomy and zoogeography of the hypoarctic erigonine
spider genus Semljicola (Araneida, Linyphiidae) // Acta Zool. Fennica. Vol.201. P.47-69.
Saaristo M.I., Marusik Yu.M. 2004a. Revision of the Holarctic spider genus Oreoneta Kulczyn-
ski, 1894 (Arachnida; Araneae; Linyphiidae) // Arthropoda Selecta. Vol. 12 (for 2003).
No.3^1. P. 185-220.
Saaristo M.I., Marusik Yu.M. 2004b. Ferchestina, a new genus of oonopid spiders from Russian
Far East (Aranei, Oonopidae) // Arthropoda Selecta. Vol. 13. No. 1-2. P.51-54.
Saaristo M.I., Marusik Yu.M. 2004c. Two new petrophilous micronetine genera Agyphantes
gen. n. and Lidia gen. n. (Araneae, Linyphiidae, Micronetinae) from eastern Palearctic
with descriptions of two new species // Bull. Brit. Arachnol. Soc. Vol.13. Pt.3. P.76-82.
Saaristo M.I., Tanasevitch A.V 2000. Systematics of the Bolyphantes-Poeciloneta genus-group
of the subfamily Micronetinae Hull, 1920 (Arachnida: Araneae: Linyphiidae) //
Reichenbachia. Bd.33. P.255-265.
Saito H. 1977. A new spider of the genus Porrhomma (Araneae: Linyphiidae) from caves of
Tochigi Prefecture, Japan // Acta arachnol. Vol.27. P.48-52.
Saito H. 1982a. Notes on Japanese Linyphiidae, I // Acta arachnol. Vol.31. P. 17-26.
Saito H. 1982b. Notes on linyhiine [sic] and erigonine spiders from Hokkaido, Japan // Kishidaia.
No.49. P.8-21.
Saito H. 1983a. On Japanese species of Gnathonarium (Araneae: Linyphiidae) // Atypus.
No.83. P.3-6.
Saito H. 1983b. Description of a new soil dwelling linyphiine spider of the genus Syedra
(Araneae: Linyphiidae) from Japan // Edaphologia. Vol.29. P. 13-16.
Saito H. 1983c. [Notes on linyphiine and erigonine spiders from Hokkaido, Japan II] // Insect,
Utsunomiya. No.34. P.50-60 [на японском].
Saito H. 1984. Descriptions of four new species of the soil dwelling linyphiine spiders (Araneae:
Linyphiidae) from the Kyushu and Kanto Districts // Edaphologia. Vol.31. P. 1-8.
Saito H. 1986. New erigonine spiders found in Hokkaido, Japan // Bull. natn. Sci. Mus. Tokyo.
Ser.A. Vol.12. P.9-24.
Saito H. 1987. On some linyphiid spiders added to the spider fauna of Japan // Heptathela.
Vol.3. No.2. P.l-13.
Saito H. 1992. Two new linyphiine spiders (Araneae: Linyphiidae) from Japan // Edaphologia.
Vol.48. P. 1-5.
Saito H. 1994. Descriptions of four new species of the soil dwelling linyphiine spiders (Araneae:
Linyphiidae) from the Kyushu and Kanto Districts // Edaphologia. Vol. 31. P. 1-8.
Saito H., Ono H. 2001. New genera and species of the spider family Linyphiidae (Arachnida,
Araneae) from Japan // Bull. natn. Sci. Mus. Tokyo. Ser.A. Vol.27. P. 1-59.
302
Литература
Saito S. 1932. Description of two new spiders of Araneida from the northern Kurile Islands //
Trans. Sapporo nat. Hist. Soc. Vol.12. No.2-3. P. 100-104.
Saito S. 1933. Araneida from the northern Kurile Islands, with descriptions of two new species //
Bull. Biogeogr. Soc. Japan. Vol.4. No.2. P. 122-131.
Saito S. 1934. A supplementary note on spiders from southern Saghalin, with descriptions of
three new species // Trans. Sapporo nat. Hist. Soc. Vol. 13. P.326-340.
Saito S. 1935a. Spiders from the northern Kurile Islands II // Trans. Sapporo nat. Hist. Soc.
Vol.14. No. LP. 100-104.
Saito S. 1935b. Further notes on spiders from southern Saghalin. (The third supplement) //
Trans. Sapporo Nat. Hist. Soc. Vol. 14. No.l. P.51-54.
Saito S. 1935c. Further notes on spiders from southern Saghalin, with descriptions of three
new species // Annot. zool. Japon. Vol. 15. P.58-61.
Scharff N., Coddington J. A. 1997. A phylogenetic analysis of the orb-weaving spider family
Araneidae (Arachnida, Araneae) // Zool. J. Linn. Soc. Vol. 120. P.355^134.
Schenkel E. 1930. Die Araneiden der schwedischen Kamtchatka-Expedition 1920-1922 //
Ark. Zool. Bd.21A. H.15. S.l-33.
Seyfulina R.R., Mikhailov K.G. 2004. Three new species of genus Xysticus C.L. Koch, 1835
(Aranei: Thomisidae) from the Amur area // Arthropoda Selecta. Vol. 12 (for 2003). No.3-
4. P.251-254.
Shear W.A. 1970. The spider family Oecobiidae in North America, Mexico, and the West
Indies // Bull. Mus. сотр. Zool. Vol.140. P. 129-164.
Shinkai A., Shinkai E. 1985. The web-building behavior and predatory behavior of Theridio-
soma epeiroides (Araneae: Theridiosomatidae) and the origin of the ray-formed web //
Acta arachnol. Vol.33. P.9-18.
Shinkai E., Takano S. 1984. A field guide to the spiders of Japan book. Japan, Tokai University
Press. 204 p.
Sierwald P. 1990. Morphology and homologous features in the male palpal organ in Pisauridae
and other spider families, with notes on the taxonomy of Pisauridae (Arachnida: Araneae) //
Nemouria (Occas. Pap. Delaware Mus. nat. Hist.). Vol.35. P. 1-59.
Simon E. 1881. Les arachnides de France. Paris. T.5. P. 1-180.
Simon E. 1891. Liste des Arachnides recueillis par Rabot dans la Siberie occidentale en 1890 //
Bull. Soc. Zool. Fr. T.16. P. 107-109.
Simon E. 1892. Histoire naturelle des araignees. Paris. T.l. P. 1-256.
Simon E. 1893. Histoire naturelle des araignees. Paris. T.l. P.257^188.
Simon E. 1894. Histoire naturelle des araignees. Paris. T.l. P489-760.
Simon E. 1895. Histoire naturelle des araignees. Paris. T.l. P.761-1084.
Simon E. 1897. Histoire naturelle des araignees. Paris. T.2. P. 1-192.
Simon E. 1898. Histoire naturelle des araignees. Paris. T.2. P.193-380.
Simon E. 1901. Histoire naturelle des araignees. Paris. T.2. P.381-668.
Simon E. 1903. Histoire naturelle des araignees. Paris. T.2. P.669-1080.
Simon E. 1922. Description de deux arachnides cavernicoles du midi de la France // Bull. Soc.
ent. France. P. 199-200.
Simon E. 1929. Les arachnides de France. Paris. T.6. Pt.3. P.533-772.
Song Y.J., Li S.Q., Marusik Y.M. 2005. Redescription on Dicymbium facetum (L. Koch)
(Araneae, Linyphiidae) with first report on its male // Acta Zootaxon. Sin. Vol.31. No.2.
P.330-334.
Song D.X., Zhu M.S., Chen J. 1999. The spiders of China. Hebei Science & Technology
Press. 640 p.
Литература
303
Song D.X., Zhu M.S., Zhang F. 2004. Fauna Sinica: Invertebrata Vol. 39: Arachnida: Araneae:
Gnaphosidae. Beijing: Science Press. 362 p.
Stalhandske P. 2002. Nuptial gifts of male spiders function as sensory traps // Proc. Roy. Soc.
Biol. sci. Vol.269. No. 1494. P.905-908.
Strand E. 1907. Sud- und ostasiatische Spinnen. I // Abh. naturforsch. Ges. Gorlitz. Bd.25.
H.l.S.107-215, 1 Taf.
Sytshevskaja VI. 1935. Etude sur les Araignees de la Kamtchatka // Folia zool. hydrobiol.
Vol.8. No.l.P.80-103.
Szita E., Logunov D. 2008. A review of the histrio group of the spider genus Philodromus
Walckenaer, 1826 (Araneae, Philodromidae) of the eastern Palaearctic region // Acta zool.
Acad. Sci. Hungar. T.54. No.l. P.23-73.
Tanasevitch A.V. 1986. New and little-known species of Lepthyphantes Menge 1866 from the
Soviet Union (Arachnida: Araneae: Linyphiidae) // Senckenberg. biol. Bd.67. H. 1-3. P. 137-
172.
Tanasevitch A.V. 1987. New species of Lepthyphantes Menge 1866 from the Soviet Far East,
with notes on the Siberian fauna of this genus (Aranei, Linyphiidae) // Spixiana. Bd.10.
H.3. P.335-343.
Tanasevitch A.V. 1988. Some new Lepthyphantes Menge (Aranei, Linyphiidae) from Mongolia
and the Soviet Far East // Folia entomol. hung. T.49. P. 185-196.
Tanasevitch A.V. 1989. A review of the Palaearctic Poeciloneta Kulczynski (Aranei,
Linyphiidae) // Spixiana. Bd.ll. H.2. P. 127-131.
Tanasevich A.V. 1990. Zoogeography of the genus Lepthyphantes in the USSR (Araneae,
Linyphiidae) // Acta zool. fenn. Vol. 190. P.357-362.
Tanasevich A.V. 1992. New genera and species of the tribe Lepthyphantini (Aranei
Linyphiidae Micronetinae) from Asia (with some nomenclatorial notes on linyphiids) // Arthropoda
Selecta. Vol. 1. No. 1. P.39-50.
Tanasevitch A.V. 1993. Another new species of Lepthyphantes Menge from the Russian Far East
(Arachnida: Araneae: Linyphiidae: Micronetinae) // Reichenbachia. Bd.30. H.l. P. 1-3.
Tanasevitch A.V. 1995. Two new genera of the family Linyphiidae from Tuva, South Siberia,
Russia (Arachnida Aranei Linyphiidae) // Arthropoda Selecta. Vol.4. No.3^. P.65-69.
Tanasevitch A.V. 1996a. New species of genus Incestophantes Tanasevitch, 1992 from southern
Siberia and the Far East, with notes on systematics of this genus (Arachnida: Araneae:
Linyphiidae: Micronetinae) // Reichenbachia. Bd.31. P. 113-122.
Tanasevitch A.V 1996b. Reassessment of the spider genus Wubanoides Eskov, 1986 (Arachnida:
Araneae: Linyphiidae) // Reichenbachia. Bd.31. P. 123-129.
Tanasevitch A.V. 2000a. On some Palaearctic species of the spider genus Agyneta Hull, 1911,
with description of four new species (Aranei: Linyphiidae) // Arthropoda Selecta. Vol.8.
No.3. P.203-213.
Tanasevitch A.V 2000b. New species of the family Linyphiidae from south Siberia, Russia
(Arachnida: Araneae) // Reichenbachia. Bd.33. P.243-253.
Tanasevitch A.V. 2005. New or little-known species of Agyneta and Nipponeta from Asia
(Aranei: Linyphiidae) // Arthropoda Selecta. Vol.13. No.3. P.165-170.
Tanasevitch A.V 2006a. New or little-known Maw O.P.-Cambridge from Siberia and the
Russian Far East (Aranei: Linyphiidae: Micronetinae) // Arthropoda Selecta. Vol. 14. No.3.
P.259-268.
Tanasevitch A.V. 2006b. On some Linyphiidae of China, mainly from Taibai Shan, Qinling
Mountains, Shaanxi Province (Arachnida: Araneae) // Zootaxa. Vol. 1325. P.277-311.
304
Литература
Tanasevitch A.V. 2006c. Five new Linyphiidae spiders from the Russian Far East, with notes
on synonymy (Arachnida: Aranei) // Arthropoda Selecta. Vol. 15. No.l. P.29-38.
Tanasevitch A.V. 2007a. New linyphiid taxa from Siberia and the Russian Far East, with notes
on the genera Notioscopus Simon and Carorita Duffey et Merrett (Aranei: Linyphiidae) //
Arthropoda Selecta. Vol.15. No.2. P. 141-152.
Tanasevitch A.V. 2007b. On a small linyphiid spider collection from Simushir Island, Kurile
Islands, Russia, with notes on Stemonyphantes sibiricus Grube (Aranei: Linyphiidae) //
Arthropoda Selecta. Vol.15. No.3. P.255-258.
Tanasevitch A.V 2008. New records of linyphiid spiders from Russia, with taxonomic and
nomenclatural notes (Aranei: Linyphiidae) // Arthropoda Selecta. Vol. 16. No.2. P.115-
135.
Tanasevitch A.V, Koponen S. 2007. Spiders (Aranei) of the southern tundra in the Russian
plain // Arthropoda Selecta. Vol. 15. No.4. P.295-345.
Tanasevitch A.V, Trilikauskas L.A. 2006. A new species of the genus Savignia Blackwall
from the Khabarovsk Province, Russian Far East (Aranei: Linyphiidae: Erigoninae) //
Arthropoda Selecta. Vol.14. No.3. P.269-270.
Tanikawa A. 1989. Japanese spiders of the genus Larinia Simon (Araneae: Araneidae) // Acta
arachnol. Vol.38. P.31-47.
Tanikawa A. 1992a. A revisional study of the Japanese spiders of the genus Cyclosa (Araneae:
Araneidae) // Acta arachnol. Vol.41. P. 11-85.
Tanikawa A. 1992b. A revision of the Japanese spiders of the genus Metleucauge Levi, 1980
(Araneae: Tetragnathidae) // Acta arachnol. Vol.41. P. 161-175.
Tanikawa A. 1993. A new species of the genus Meta (Araneae: Tetragnathidae) from Japan //
Acta arachnol. Vol.42. P.97-101.
Tanikawa A. 1995a. A revision of the Japanese spiders of the genus Araniella (Araneae:
Araneidae) // Acta arachnol. Vol.44. P.51-60.
Tanikawa A. 1995b. Two new species of the spider genus Diphya (Araneae: Tetragnathidae)
from Japan and Taiwan, with notes on the known species // Acta arachnol. Vol.44. P. 101—
111.
Tanikawa A. 1998. A revision of the Japanese spiders of the genus Neoscona (Araneae:
Araneidae) // Acta arachnol. Vol.47. P. 133-168.
Tanikawa A. 1999. Japanese spiders of the genus Eriovixia (Araneae: Araneidae) // Acta
arachnol. Vol.48. P.41-48.
Tanikawa A. 2000. Japanese spiders of the genus Eriophora (Araneae: Araneidae) // Acta
arachnol. Vol.49. P. 17-28.
Tanikawa A. 2001. Twelve new species and one newly recorded species of the spider genus
Araneus (Araneae: Araneidae) from Japan // Acta arachnol. Vol.50. P.63-86.
Tanikawa A. 2007. An identification guide to the Japanese spiders of the families Araneidae,
Nephilidae and Tetragnathidae. Arachnological Society of Japan. 121 p.
Thaler K. 1978. Die Gattung Cryphoeca in den Alpen (Arachnida, Aranei, Agelenidae) //
Zool. Anz. Bd.200. S.334-346.
Thaler K., Helsdingen P., Deltshev C. 1994. Vikariante Verbreitung im Artenkomplex von
Lepthyphantes annulatus in Europa und ihre Deutung (Araneae, Linyphiidae) // Zool. Anz.
Bd.232. S.lll-127.
Thorell T. 1869. On European spiders. Part I. Review of the European genera of spiders,
preceded by some observations on zoological nomenclature // Nova Acta reg. Soc. sci.
Upsaliae. T.7. No.3. P. 1-108.
Литература
305
Thorell Т. 1870а. Remarks on synonyms of European spiders. Part I. Uppsala. 96 p.
Thorell T. 1870b. On European spiders //Nov. Act. reg. Soc. sci. Upsaline. T.7. No.3. P.109-
242.
Tretzel E. 1961. Biologie, Okologie, und Brutpflege von Coelotes terrestris (Wider) (Ara-
neida, Agelenidae), II: Brutpflege // Zeits. Morph. Okol. Tiere. Bd.50. S.375-542.
Trotta A. 2005. Introduzione ai ragni italiani (Arachnida Araneae) // Memorie della Societa
Entomologica Italiana. Vol.83. P.3-178.
Trilikauskas L. 2007. A new species of the genus Clubiona Latreille, 1804 (Araneae:
Clubionidae) from the Russian Far East // Acta Biol. Univ. Daugavp. Vol.7. P. 1-3.
Tu L.H., Li S.Q. 2003. A review of the spider genus Hylyphantes (Araneae: Linyphiidae) from
China // Raffles Bull. Zool. Vol.51. P.209-214.
Tu L.H., Li S.Q. 2004a. A preliminary study of erigonine spiders (Linyphiidae: Erigoninae)
from Vietnam // Raffles Bull. Zool. Vol.52. P.419^133.
Tu L.H., Li S.Q. 2004b. A review of the Gnathonarium species (Araneae: Linyphiidae) of
China// Rev. Suisse Zool. ТЛИ. Р.851-864.
Tu L.H., Li S.Q. 2005a. A new species of the genus Hylyphantes (Araneae: Linyphiidae) from
Sichuan Province, China // Acta zootaxon. sin. Vol.30. P.62-64.
Tu L.H., Li S.Q. 2005b. Chinese linyphiid spiders housed at Dr Schenkel's collection in Basel
(Araneae, Linyphiidae) // Acta zootaxon. sin. Vol.30. P.861-862.
Ubick D., Paquin P., Cushing P.E., Roth V 2005. Spiders of North America: an identification
manual. American Arachnological Society. 377 p.
Vellard J. 1924. Etudes de zoologie. I. Araneidae // Arch. Inst. Vital. Brasil. Vol.2. P. 1-32, 121—
170.
Wagner W. 1887. Copulationsorgane des Mannchens als Criterium fur die Systematik der
Spinnen // Horae Soc. Entomol. Ross. T.22. P.3-132.
Wesolowska W., Marusik Yu.M. 1990. Notes on Heliophanus camtschadalicus Kulczynski,
1885 (Aranei, Salticidae) and the related species // Korean Arachnol. Vol.6. No.l. P. 91-
100.
Wiehle H. 1931. Spinnentiere oder Arachnoidea. Araneidae // Die Tierwelt Deutschlands.
Jena. Lfg.27. S.l-136.
Wiehle H. 1953. Spinnentiere oder Arachnoidea (Araneae) IX: Orthognatha-Cribellatae-Hap-
logynae-Entelegynae (Pholcidae, Zodariidae, Oxyopidae, Mimetidae, Nesticidae) // Die
Tierwelt Deutschlands. Jena. Lfg.42. S. 1-150.
Wiehle H. 1963. Spinnentiere oder Arachnoidea (Araneae). XII. Tetragnathidae-Streckspinnen
und Dickkiefer // Die Tierwelt Deutschlands. Lfg.49. S.l-76.
Wunderlich J. 1980. Uber europaische Symphytognathidae (Arach.: Araneae) // Verh. naturw.
Ver. Hamb. (N.F.). Bd.23. S.259-273.
Wunderlich J. 1986. Spinnenfauna gestern und heute: Fossile Spinnen in Bernstein und ihre
heute lebenden Verwandten. Wiesbaden: Quelle & Meyer. 283 S.
Wunderlich J. 1987. Die Spinnen der Kanarischen Inseln und Madeiras: Adaptive Radiation,
Biogeographie, Revisionen und Neubeschreibungen. Langen, West Germany: Triops Verlag.
435 S.
Wunderlich J. 1995a. Linyphiidae aus der Mongolei (Arachnida: Araneae) // Beitr. Araneol.
Bd.4 (fur 1994). S.479-529.
Wunderlich J. 1995b. Beschreibung einer bisher unbekannten Art der Gattung Cryphoeca
Thorell 1870 aus der Turkei (Arachnida: Araneae: Dictynoidea: ?Dictynidae) /7 Beitr.
Araneol. Bd.4 (fur 1994). S.719-721.
306
Литература
Wunderlich J. 2004. Fossil spiders in amber and copal // Beitr. Araneol. Bd.3(A-B). P.l-
1908.
Wunderlich J. 2008a. Descriptions of fossil spider (Araneae) taxa mainly in Baltic amber, as
well as on certain related extant taxa // Beitr. Araneol. Bd.5. P.44-139.
Wunderlich J. 2008b. On extant and fossil (Eocene) European comb-footed spiders (Araneae:
Theridiidae), with notes on their subfamilies, and with descriptions of new taxa // Beitr.
Araneol. Bd.5. P. 140-469.
Wunderlich J. 2008c. On extant and fossil spiders (Araneae) of the RTA-clade in Eocene
European ambers of the families Borboropactidae, Corinnidae, Selenopidae, Sparassidae,
Trochanteriidae, Zoridae si. 1., and of the superfamily Lycosoidea // Beitr. Araneol. Bd.5.
P.470-523.
Yoshida H. 1982. Spiders from Taiwan II. Three species of the genera Hyptiotes and
Miagrammopes (Araneae: Uloboridae) // Proc. jap. Soc. syst. Zool. Vol.22. P. 18-20.
Yoshida H. 2003. The spider family Theridiidae (Arachnida: Araneae) from Japan.
Arachnological Society of Japan. 224 p.
Yoshida H. 2009. Uloboridae, Theridiidae, Ctenidae // H. Ono (ed.). The Spiders of Japan
with keys to the families and genera and illustrations of the species. Kanagawa: Tokai
Univ. Press. P. 142-147, 356-393, 467^68.
Zhu M.S. 1998. Fauna Sinica: Arachnida: Araneae: Theridiidae. Beijing: Science Press, xi +
436 p.
Zhu M.S., Song D.X., Zhang J.X. 2003. Fauna Sinica: Invertebrata Vol. 35: Arachnida: Araneae:
Tetragnathidae. Beijing: Science Press, vii + 418 pp.
Zyuzin A.A. 1990. Studies on burrowing spiders of the family Lycosidae (Araneae). I.
Preliminary data on structural and functional features // Acta Zoologica Fennica. Vol. 190.
P.419-422.
Zyuzin A.A. 1993. Studies on the wolf spiders (Araneae: Lycosidae). I. A new genus and
species from Kazakhstan, with comments on the Lycosidae // Mem. Queensl. Mus. Vol.33.
No.2. P.693-700.
Приложения
Приложение А
Этимология латинских названий пауков
Этимология латинского названия отряда Пауки.
Почему пишут то Aranei (в основном русскоязычные авторы), то Агапеае? Ага-
nei — это множественное число от Araneus Clerck, 1758. Агапеае —
множественное число от Aranea Linne, 1758. В латинском языке araneus (мужской род)
означает просто «паук». Aranea (женский род) первоначально означало «сеть паука», но
позже стало обозначать и самого паука. Два родовых названия {Araneus Clerck и
Aranea Linne) долго конкурировали до тех пор, пока Международный комитет по
зоологической номенклатуре в 1948 г. не отдал приоритет названию Клерка.
Недавно (Kluge, 2007) было обращение в Международную комиссию по
зоологической номенклатуре с предложением изъять родовое название Aranea Linne, 1758, то
есть сделать его недействительным. Это предложение было принято (Bulletin of
Zoological Nomenclature, 2009, 66(2): Opinion 2224).
Два названия отряда также долго конкурировали друг с другом. В 1996 г. на
арахнологическом конгрессе в Женеве после голосования было рекомендовано
использовать название Агапеае (представитель от России К.Г. Михайлов голосовал
против, за Aranei). Рекомендация, в отличие от решения Международной
комиссии по зоологической номенклатуре, обязательной силы не имеет. Правила же
Международного кодекса зоологической номенклатуры не распространяются на
названия отрядов. Поскольку валидным названием признано Araneus, a Aranea вообще
изъято, то логичнее и грамматически правильнее использовать название Aranei.
Этимология родовых названий.
Родовые и видовые названия пауков, обитающих в азиатской части России,
создавались и зарубежными и отечественными систематиками в течение примерно
250 лет. Эти названия имеют самое разное происхождение, порой причудливое и
трудно понимаемое в наше время.
Этимология обычно объясняется при введении названия для нового рода или
вида, однако, в старых (да и во многих современных) работах таких объяснений не
приводится. В зоологической номенклатуре допускаются названия,
представляющие собой любые произвольные сочетания букв. Поэтому расшифровка
этимологии многих названий сильно затруднена и не всегда возможна.
Подробную этимологию, включая оригинальное написание на греческом или
латыни можно найти у J.R. Parker (1993) (для пауков, обитающих в
Великобритании), а также у H.D. Cameron (2005) (для родов пауков, обитающих в Северной
Америке). Нередко трактовка слов существенно отличается в этих источниках. Судя
по детальной аргументации и эрудиции, более тщательно с этимологией
разобрался H.D. Cameron (2005).
Изучение этимологии не сводится к простому листанию латинского либо
греческого словаря и поиску подходящих, созвучных слов. Зачастую необходимо
внимательно смотреть первоисточник, знать этимологию других названий того же
автора, язык на котором говорил и писал автор и историю того времени, когда созда-
308
Приложения
валось название. Помимо этого нужно знать первоисточники, с которыми работал
тот или иной арахнолог, какими книгами он увлекался. Есть несколько американо-
сибирских родов, названия которым дали битвы Наполеона в Европе, например
Talavera (битва при Talavera). Ещё при изучении происхождения названия важно
знать, как в разных странах произносились, и соответственно транскрибировались
греческие названия. Совсем нелишне иметь представление и о мировых религиях.
Немало названий взято из древнегреческой и древнеримской мифологии, Ветхого
Завета, Авесты (священной книги зороастризма).
R.V. Chamberlin, описавший очень много таксонов, в том числе и тех, которые
водятся в России, использовал слова американских индейцев гошутов. При этом
многие из этих названий имеют гибридную этимологию: одно слово (корень) — из
гошутского, другое — из латыни или греческого.
Е. Simon дал названия очень многим таксонам пауков, когда он был ещё совсем
молодым (в 16 лет), например, Neoscona Simon, 1864. Тогда, будучи ещё
школьником, он не знал греческий и латынь в полной мере и допускал много ошибок при
образовании названий пауков. В зрелые годы Е. Simon допускал также немало
грамматических и даже орфографических ошибок (Cameron, 2005). Кроме Э. Симона,
С.А. Walckenaer, знавший латынь и греческий не очень хорошо, при образовании
названий также допустил много ошибок.
Некоторые авторы просто смотрели в словарь в поисках подходящих слов,
никоим образом не связанных по смыслу с описываемым пауком. Е. Simon и C.R.
Crosby & S.C. Bishop описали как никто много родов и для того, чтоб не попасть в
омонимию, выискивали любые слова из греческого и латинского словаря, которые
к описываемому таксону не имели никакого отношения. Именно так, например,
появились названия Horcotes Crosby & Bishop, 1933 и Tmarus Simon, 1875. Другие
искали подходящее слово в произведениях античных авторов. Так появилось
название Dipoena Thorell, 1869 (по имени одного из критских скульпторов, имя
которого фигурирует у Плиния). Всё это затрудняет восстановление значения разных
названий.
Нередко в разных группах пауков существуют свои традиции образования над-
видовых таксонов. Например, при образовании родовых названий у пауков-волков
есть традиция к названию какого-либо хищного животного добавлять окончание —
-osa, по аналогии с первым названием Lycosa (от lycos — волк), например, Pardosa
(от леопарда), Arctosa (от медведя). C.F. Roewer в двух работах описал сразу
несколько родов, названия которых производятся от хищников: Dingosa (динго),
Hyaenosa (гиена), Mustelicosa (ласка), Lynxosa (рысь). Аналогичную этимологию
имеют Crocodilosa Caporiacco, 1947 (крокодил) и Alopecosa Simon, 1885 (лиса). В
Linypiidae и некоторых других семействах принято давать окончание -neta
(латинизированное греческое слово «прядильщик, пряха»). Кроме того, в Linypiinae очень
традиционное окончание -yphantes (греческое слово xxpavrnc; — ткач). Крупный
род Lepthyphantes сейчас интенсивно разбивается на более мелкие, и названия этим
родам даются чаще всего по названию типового вида с добавлением окончания —
phantes (например, Tenuiphantes, от tenuis — название типового вида).
Аналогичное окончание есть и у пауков-скакунчиков, например Dendryphantes, Tuvaphantes.
Этимология родовых названий отечественных авторов самая разная. Это и
патронимы (Bishopiana, Crosbylonia, Eskovia, Holminaria, Ivielum, Nenilinium, Tanase-
vitchia, Uusitaloia, Viktorium, Wiehlenarius), и гибридные названия, производные от
Приложение А. Этимология латинских названий пауков
309
названий родов и географических названий (Arcterigone, Asiceratinops, Asiophantes,
Kolymocyba, Pacifiphantes, Sibirocosa, Tuvaphantes), реже описательные названия
(Concavocephalus, Perlongipalpus, Procerocymbium). Для некоторых родов из
Курил и Сахалина К.Ю. Еськов использовал слова аборигенного народа — айнов.
Нередко рода называются созвучно к близким либо похожим таксонам (Hilaira —
Pseudohilaira. Glyphesis — Paraglyphesis. Tapinocyba — Pseudocyba, Tibioplus —
Tibioploides). Часть родовых названий составлена из названий двух близких родов:
Eborilaira (Eboria & Hilaira), Bathylinyphia (Bathyphantes & Linyphia). Самое
курьёзное название имеет сказочную этимологию: Kikimora palustris — кикимора
болотная.
Этимология видовых названий.
При описании новых видов отечественные авторы часто дают им названия,
производные от географических названий (например, yeniseicus, sibiricus, kolymensis,
tuvinensis, sachalinensis, wrangeliana), или патронимы, в честь сборщиков (ryvkini,
zherikhini, dudkoi, glupovi) и коллег (например, nenilini, tyshchenkoi, basarakuni,
holmi). Часть названий — описательные и указывают на морфологические
особенности (rubens — красный, barbatus — бородатый, minaretifer — несущий минарет)
или приуроченность к тем или иным биотопам (pinipumilis — от Pinispumila, larice-
torum — от Larix, sphagnicolus — от Sphagnum, petrophila — любящая камни) и
природным (широтным и высотным) зонам (alpina, borealis, hyperboreus, arcticus).
Нередко видовые названия, образованные от одних и тех же географических
названий, пишутся по-разному, например alaskensis (Agyneta) и alascensis (Philo-
dromus), jeniseicus (Bathyphantes) wyeniseica (Micaria). To же самое относится и к
патронимам, например mikhailovi (Zelotes) и michailovi (Evarcha).
Расшифровка этимологии некоторых родовых названий пауков из
азиатской части России.
В этом разделе использованы сокращения:
а. — английское слово (корень);
л. — латинское слово (корень);
лл. — два латинских слова или корня;
г. — греческое слово или корень;
гг. — два греческих слова или корня;
м. — мифологическое;
мет. — метафорическое;
п. — патроним;
р. — русское слово (корень).
Abiskoa — от названия типового вида abiskoensis, типовое местообитание
которого Abisko, Швеция.
Acantholycosa — от г. шип (колючка) и Lycosa: паук-волк с шипами
(многочисленными).
Acanthoneta — от гг. шип и -neta (прядильщик): прядильщик с шипами
(многочисленными).
Achaearanea — замещающее название для Achaea (преоккупировано в Lepi-
doptera). От греческих слов Achaea (область в Греции) и агапеа (паук).
310
Приложения
Aculepeira — от л. шип и г. область в Греции Эпеирос. Название образовано
добавлением к названию типового вида aculeata родового названия Epeira. Такой
способ образования родовых названий был обычен для Chamberlin & Ivie.
Aelurillus — от г. дикий кот: движущийся подобно коту.
Agalenatea — вероятно, от Agalena (одно из написаний Agelena) и Atea (род
Araneidae).
Agelena — от г. группа, живущие вместе: много сетей в одном месте.
Agelenopsis — от Agelena и г. подобный: паук внешне очень похожий на Agelena.
Agnyphantes — гг. первый корень очевидно от неизвестный и ткач.
Agroeca — от г. поле или земля (agro) и живущий (oecos).
Agyneta — от г. без самки; самка была описана через несколько лет после
самца. Имеется и другая трактовка происхождения названия: г. Agy- слабые
конечности, и вторая часть родового названия Microneta (-neta — прядильщик).
Agyphantes — от Agyneta и Lepthyphantes.
Ainerigone — от р. айны (название народа) и Erigone: род обитает там, где
раньше жили айны.
Alenatea — производное от названия похожего рода Agalenatea.
Alloclubionoides — от г. другой/около и родового названия Clubiona. Род
первоначально отнесён к Clubionidae, но сейчас рассматривается в Coelotinae.
Allohogna — от г. другой/около и родового названия Hogna: род, близкий к Hogna.
Allomengea — от г. другой/около и родового названия Mengea: похожий, но не
такой, как Mengea.
Alopecosa — от г. Alopec- (лиса) и -osa (обычное родовое окончание для пауков-
волков): «паук-лиса».
Amaurobius — от гг. тёмный и живёт: живущий в темноте или незаметный
(невыразительный).
Ambanus — от маньчжурского «амбань» (вельможа, сановник). Очевидно из-за
крупного размера. Синоним Alloclubionoides.
Anahita — от имени древней иранской богини Anahita в Авесте (священная книга
зороастризма).
Anelosimus — от г. невиданный.
Anguliphantes — от л. angulatus (типовой вид) и Lepthyphantes (род, в котором
первоначально рассматривался типовой вид).
Antistea — от л. быть выдающимся (о дыхальце?), надзиратель или учитель
(священник).
Anyphaena — от гг. без и ткач: не плетущий сетей.
Aphileta — от гг. недружественный, нелюбимый.
Apollophanes — от гг. Apollo (м.) и -phanes, обычное окончание многих
собственных имён, например, Aristophanes: «паук-Аполлон».
Aprifrontalia — этимология первого корня неясна, очевидно название связано с
выростом головной части у самца.
Arachosinella — этимология неясна, возможно название связано с Arachosia
(Anyphaenidae).
Araeoncus — от гг. тонкий и опухоль: о выросте головной части.
Araneus — л. паук.
Araniella — л. уменьшительное от паук: «паучок».
Приложение А. Этимология латинских названий пауков
311
Archaeodictyna — от г. древний и Dictyna.
Archaraeoncus — от г. древний (архаичный) и Araneoncus.
Arctachaea — замещающее название для Achaea; от гг. Арктика и Achaea
(область в Греции).
Arctella — от гг. Арктика и уменьшительный суффикс -ella. Род описан из
Лапландии.
Arcterigone — от Арктика и Erigone.
Arctobius — от гг. Арктика и -bios (живущий).
Arctosa — от г. или л. медведь или Большая Медведица (северное созвездие) и
-osa (обычное родовое окончание для пауков-волков): «паук-медведь» или
«северный паук-волк».
Arcuphantes — от arcuatulus (типовой вид) и г. ткач.
Argenna — 1) от г. белый: белое поле вокруг эпигины, или 2) от г. медленный.
Argiope — г.м. имя нимфы, но возможно и от другого мифологического
персонажа.
Argyrodes — л. серебряный.
Argyroneta — от л. серебряный и г. плести: «плетущий серебро».
Artanes — возможно, от названия древнего города или царской династии в
Армении.
Asianellus — от Asia и -llus (окончание у близкого рода Aelurillus).
Asiceratinops — от Азия и Ceratinops.
Asiophantes — от Азия и -yphantes (ткач): «ткач-азиат».
Asperthorax — от лл. грубый и грудь (о структуре грудной части карапакса).
Asthenargoides — от Asthenargus и л. суффикс -oides (похожий).
Atea — этимология не понятна. Атеа — небольшая область в северо-восточной
Испании.
Atypus — 1) от г. ужасный, или 2) от гг. нет и типичный: «необычный».
В alius — г. прыгающий вокруг, танцующий: «танцор».
Baryphyma — от гг. большой (массивный) и вырост: о выросте карапакса.
Bassaniana — замещающее название для Bassania (город Elbasan в
современной Албании).
Bathylinyphia — от Bathyphantes и Linyphia: из-за совмещения признаков двух
родов.
Bathyphantes — от гг. глубокий и ткач: плетущий паутину в глубине зарослей.
Berlandina — п. L. Berland, известный французский арахнолог.
Bicluona — название образовано от Clubiona перестановкой первых двух слогов.
Bishopiana — п. Sherman Bishop, известный американский арахнолог.
Bolephthyphantes — от двух родовых названий Bolyphantes и Lepthyphantes.
Bolyphantes — от гг. от сети и ткач: «ткач сетей».
Bucliona — мететеза от Clubiona.
Callilepis — от гг. красивый и чешуйка: красивый узор (рисунок) из чешуек.
Callobius — от названий двух родов Callioplus и Amaurobius.
Canalidion — от canal и Theridion.
Carorita — п. Mrs Carol Rita Duffey.
Carrhotus — п. от имени возничего колесницы участвовавшего в олимпийских
гонках.
Caviphantes — от гг. ямка (впадина, пещера) и ткач: «пещерный ткач».
312
Приложения
Centromerus — от гг. жало или острие и часть: об эпигине с острым выростом
(скапусом).
Ceraticelus — производное от Ceratinella и -elus — подобный.
Ceratinella — от гг. сделанный из рога и уменьшительного суффикса: о скутуме
на брюшке. Исходно название дано как уменьшительное от Ceratinops Menge, 1868.
Cercidia — от г. ткацкий челнок: форма брюшка. Это замещающее название для
Cerceis (преоккупировано в ракообразных) — от м. имени морской нимфы
Цирцеи — дочери Океана и Тетис.
Chalcoscirtus — от гг. медный, бронзовый и прыгать, Scirtus — м. имя сатира из
сопровождения Диониса: «бронзовый сатир».
Cheiracanthium — от гг. рука и колючка: о выросте цимбиума, «колючерукий».
Chorizopes — от гг. разделённый и глаза.
Chrysso — г. от прилагательного «золотистый» (о цвете типового вида).
Cicurina — от л. домашний, ручной, скучный, неинтересный: «домашний паучок».
Clubiona — 1) от гг. слава и жизнь; или 2) от гг. хорошо известный и делать
беспорядки; или 3) от келья, малая комната: самки некоторых видов заточают себя
в убежище, где находится кокон: «отшельник».
Cnephalocotes — от гг. мягкий как шерсть и голова: «мягкоголовый».
Coelotes — от г. полость, ниша: паук, делающий или занимающий полости (под
камнями), «полостник».
Collinsia — п. Collins, возможно в честь W.P. Collins, специалиста по
диатомовым водорослям.
Concavocephalus — от лл. вогнутый и голова: карапакс с углублением между
задними медиальными глазами и наличником, «вогнутоголовый».
Connithorax — замещающее название для Conithorax (от гг. конический и грудь):
«кониче скогрудый».
Coriarachne — от гг. Coris (клоп, постельный клоп) и паук: «плоский паук».
Crispiphantes — от л. кучерявый (волнистый) и г. ткач: «кучерявый ткач».
Crosbylonia — п. Cyrus Richard Crosby (1879-1937), известный американский
арахнолог.
Crustulina — л. раковина, или твёрдый: из-за твёрдого брюшка.
Cryphoeca — от гг. скрытый и дом, или жить: «скрытоживущая».
Cryptachaea — от г. скрытый и Achaearanea.
Cybaeopsis — от Cybaeus и -opsis (подобный).
Cybaeus — от г. Cybaea, торговое судно.
Cyclosa — от г. круг: о сети.
Cyphepeira — от л. кубок, чаша (о форме скапуса) и Epeira.
Dactylopisthes — от гг. палец и сзади: из-за выроста карапакса.
Dactylopisthoides — уменьшительное от Dactylopisthes.
Decipiphantes — от decipiens — л. обманщик (типовой вид) и Lepthyphantes (род,
в котором первоначально рассматривался типовой вид).
Dendryphantes — от гг. дерево и ткач; паук, обитающий на деревьях.
Diaea — 1) г. через или в течение и ручей; 2) замещающее название для Diana
(римская богиня).
Dictyna — 1) г. сеть (dictis); или 2) г.м. в честь критской богини Диктинны (имя
означает «плетущая сети») — покровительницы охоты и рыболовства, любимой
спутницы Артемиды; по другой версии — это второе имя (прозвище) Артемиды
Приложение А. Этимология латинских названий пауков
313
(покровительницы охоты с помощью сетей); 2) л.м. в честь богини Diana —
латинский эквивалент Артемиды.
Dicymbium — от гг. или лл. дважды и цимбиум: о большом выросте голени
пальпы самца, создающем видимость двух цимбиумов.
Diplocentria — от гг. двойной и шпора: о голени пальпы самца.
Diplocephalus — от гг. двойной и голова, о форме головогруди: «двуглавый».
Diplostyla — от гг. двойной и вырост, стилет: два выроста эпигины.
Dipoena — от Dipoinos, греческий скульптор.
Dismodicus — 1) от лл. нет и обычный: «необычный»; 2) гг. два и шишка
(колесо): голова с двумя округлыми выростами: «шишкоголовый».
Doenitzius — п. W. Donitz, немецкий арахнолог.
Dolomedes — г. хитрый, коварный.
Drapetisca — от г. беглый, убегает недалеко в сторону: «беглец».
Drassodes — г. хватать (цеплять).
Drassyllus — от г. хваткий (цепкий) в действии, уменьшительное название от
Drassodes.
Drepanotylus — от гг. серп (или крюк) и выпуклость (вздутие).
Ebelingia — п. Karl Ebeling, финский исследователь, частично спонсировавший
поездки П.Т. Лехтинена в Юго-Восточную Азию.
ЕЬо — 1) возможно произвольная комбинация букв, созвучная Odo (тевтонское
имя); 2) сокращённое (ник) от Ebur, имя, обычное в средневековой Германии.
Eboria — от л. Eboracum, название города Йорка в Англии.
Eborilaira — от Eboria и Hilaira. Eboria — от л. Ebor — область York в Англии.
Echemus — м. Эхемус, царь Аркадии.
Eldonnia — замещающее название для Eldonia (преоккупировано), от р. «елда»,
хотя название дано из-за крупной и сильно склеротизованной эпигины
(напоминающей «елду»).
ЕтЫупа — от embolus и Dictyna: самцы имеют утолщённый эмболюс.
Enoplognatha — от гг. вооруженный и челюсти: о хелицерах самца.
Entelecara — от гг. совершенный (полный) и голова, о карапаксе самца: «пре-
красногол овый».
Eperigone — от г. сверху и Erigone.
Epibellowia — от эпигина и a. bellows (кузнечные меха), из-за того, что эпигина
крепится к брюшку структурой, напоминающей гармошку.
Epigyphantes — от epigynatus (типовой вид) и Lepthyphantes (род в котором
первоначально рассматривался типовой вид).
Epigytholus — от лл. эпигина и купол: о куполовидной форме эпигины.
Episinus — от г. вредный.
Episolder — согласно пояснению автора, от «по-видимому, слившихся частей
эпигины».
Eresus — от г. «я налажу (перемонтирую)»: возможно о том, что сеть липкая.
Erigone — м. дочь Икара, перенесённая в небо как созвездие Дева: о
распространении с помощью паутины по воздуху.
Erigonella — уменьшительное от Erigone.
Erigonidium — от Erigone и г. подобный: похожий на Erigone.
Erigonoploides — от Erigonoplus и подобный.
314
Приложения
Erigonoplus — от Erigone и -oplus (несущий, вооружённый), возможно о
выросте головной части у самца.
Eriophora — гг. несущий шерсть, о густых волосках, покрывающих тело.
Его — м. от имени Эрос (бог любви): из-за того, что пауки этого рода любят (в
качестве пищи) других пауков (есть и другие, менее интересные варианты).
Eskovia — п. Кирилл Юрьевич Еськов (р. 1956), известный российский
арахнолог, палеонтолог, писатель. Описал типовой вид рода.
Eskovina — замещающее название для Oinia Eskov, 1984.
Estrandia — п. Embrik Strand, известный арахнолог и энтомолог, работавший
долгое время в Риге.
Euophrys — от гг. красивый, хороший, настоящий (ей-) и брови: о волосках,
окантовывающих передние медиальные глаза.
Euryopis — от гг. широкий и спина: широкое брюшко.
Evarcha — от гг. хороший и правитель.
Evippa — очевидно, от гг. "eu" (=ev — настоящий, хороший) и лошадь: очень
быстро бегают.
Ferchestina — от Far East и Orchestina: Orchestina с Дальнего Востока.
Flagelliphantes — от flagellifer (типовой вид) и Lepthyphantes (род в котором
первоначально рассматривался типовой вид).
Floronia — от л. цветок, живущий среди цветов.
Frontella — очевидно производное от Frontina Simon, 1884.
Frontinellina — производное от Frontina Simon, 1884.
Gibbaranea — от лл. горб (бугор) и паук: о двух конических выростах на
брюшке.
Gibothorax — от лл. горб (бугор) и грудь: о вздутом грудном отделе карапакса.
Glyphesis — от г. выемка (зазубрина) на стреле: о форме эпигины.
Gnaphosa — от г. шерстяной, инструмент для пыток (мет.): «власяница».
Gnathonarium — г. челюсть: о мощных хелицерах.
Gonatium — г. колено: об увеличенном колене пальпы самца.
Gongylidiellum — от Gongylidium, уменьшительное.
Gongylidioides — от Gongylidium и г. -oides (подобный).
Gongylidium — от гг. округлый и маленький: «шарик».
Hackmania — п. Walter Hackman, финский арахнолог, автор монографии по
паукам Ньюфаундленда.
Hahnia — п. Carl Wilhelm Hahn (1786-1836), известный немецкий арахнолог.
Halorates — г. море: живущий около моря.
Haplodrassus — от гг. простой и зажимать (хватать).
Heliophanus — от гг. Солнце и лампа; можно найти только когда светит солнце:
«подсолнечник».
Helophora — от гг. коготь (ноготь, гвоздь) и нести: о длинной эпигине.
Herbiphantes — от л. трава и г. -yphantes (ткач): собирается укосами в траве.
Heriaeus — возможно 1) п. от ХогрШх;, один из древнейших аттических
трагиков или 2) город (Heria) в Испании.
Heteropoda — от гг. разные и нога: «разноногий».
Heterotheridion — от г. разный (другой) и Theridion.
Hilaira — 1) п. Etienne Geoffrey Saint Hilaire (1772-1844), французский зоолог-
эволюционист; 2) г. слегка блестящий.
Приложение А. Этимология латинских названий пауков
315
Himalaphantes — от Гималаи и Lepthyphantes. типовой вид описан из Гималаев.
Holminaria — п. Аке Holm, известный шведский арахнолог.
Horcotes — г. давать клятву, офицер принимающий клятву. Произвольно взятое
название.
Howaia — в честь Liz Но Wai из Фиджи, которая помогала в сборах пауков Р.Т.
Lehtinen.
Hybauchenidium — от гг. горб, шея и уменьшительного окончания: «мелочь
горбатая».
Hygrolycosa — от гг. влажный и Lycos а: паук-волк, который обитает в сырых
местах.
Hylyphantes — от гг. материал и ткач.
Нуротта — от гг. под и глаза: о глазах, расположенных под выростом на кара-
паксе самца, «насупленный».
Hypselistes — от г. высокий (о высоте над уровнем море): обитающий в горах,
«горец».
Hypsosinga — от г. высокий и родового названия Singa: Singa с высоким
наличником.
Improphantes — от improbulus — дерзкий, нагловатый (типовой вид) и Lepthyz
phantes (род, в котором первоначально рассматривался типовой вид).
Incestophantes — от видового названия incestus (непристойный) и Lepthyphantes
(род, в котором первоначально рассматривался типовой вид).
Ipa — п. в честь Toimo Ilmari Saaristo. Ipa — прозвище производное от Ilmari.
Ishicosa — от названия типового вида ishikariana, и окончания -cosa.
Islandiana — от Исландия (типовой вид из Исландии).
Ivielum — п. Wilton Ivie, известный американский арахнолог.
Iwogumoa — японское название для пауков Coelotinae, где iwo — дом (простое
жилище), gumo — паук.
Japoniona — от japonica (типовой вид) и Clubiona.
Kaestneria — п. A.G. Kaestner (1719-1800), немецкий арахнолог.
Karita — согласно автору — произвольная комбинация букв, созвучно с Саго-
rita, родом, в котором ранее рассматривался типовой вид.
Keijia — п. Keiji Yoshida, отец японского арахнолога Н. Yoshida.
Kikimora — м. кикимора, типовой вид palustris: «кикимора болотная».
Kishidaia — п. К. Kishida, известный японский арахнолог.
Kolymocyba — от Колыма и Aulacocyba (близкий род).
Larinia — 1) г. толстый; 2) от римского города Larinium.
Larinioides — от Larinia и -oides (подобный).
Lasaeola — возможно, от г. лохматый.
Lasiargus — г. лохматый и г.м. Argus (персонаж с несчётным количеством глаз).
Lathys — 1) замещающее созвучное название для Lethia, возможно от
«скрываться от преследования»; или 2) от г. Lathyrus — растение вика.
Latrodectus — гг. кусающий скрыто.
Lepthyphantes — от гг. тонкий (маленький, хрупкий) и ткач. Большинство родов
выделенных из Lepthyphantes происходит от названия типового вида и
окончания -phantes, например Anguliphantes, Decipiphantes.
Leptoneta — от г.л. тонкий (маленький, хрупкий) и прясть.
Leptodrassex — от Leptodrassus и суффикса -ех (бывший).
316
Приложения
Leptorhoptrum — г. дверной молоток: возможно, о мощных хелицерах.
Linyphia — от гг. лён и ткать.
Lophomma — от гг. бугорок и глаза: о приподнятой головной части карапакса у
самца.
Lycosa — г. мет. волк: охотящийся как волк (1ус — волк, но lycos — рвущий
(терзающий как волк — мет.). Название идёт ещё от Аристотеля.
Lysiteles — имя героя из пьесы «Trinummus» («Три монеты») римского
комедиографа Тита Мамкция Плавта (л. Titus Maccius Plautus).
Macrargus — от г. большой, крупный и Argus. Argus — м.г. персонаж с
несчётным количеством глаз.
Mangora — этимология неясна (Cameron, 2005).
Маю — п. Publis Vergillius Maro (Вергилий, 70-19 гг. до н.э.), поэт, автор
«Энеиды».
Marpissa — от г. хватать — набрасываться.
Masikia — от типового местообитания типового вида, реки Masik, остров Banks,
Северо-Западные Территории, Канада.
Maso — 1) от л. самец, или г. жевать; 2) п. С. Papirius Maso, упоминался в
работах Плиния и Цицерона.
Mecynargoides — от Mecynargus и г. -oides (подобный).
Mecynargus — от г. удлиняться и Argus, типовой вид longus.
Megalepthyphantes — от г. большой и Lepthyphantes.
Mesasigone — от л. Средняя Азия (mesasiatic = среднеазиатский), и -gone — от
Erigone.
Meta — 1) л. конический: о брюшке; 2) м.п. Meta, первая жена Эгея.
Metellina — 1) от л. металлический (металл): об окраске брюшка; 2) двойное
уменьшительное (-ella и -ina) от Meta.
Metleucauge — от Meta и Leucauge: пауки этого рода совмещают признаки двух
родов.
Metopobactrus — гг. лоб и дубина (палка, бревно), дубиноподобный вырост лба:
« дубиноголовый».
Micaria — от 1) г. блестящий; 2) замена одной буквы в Macaria, преокупирован-
ного в Lepidoptera.
Micrargus — от гг. маленький и Argus. Argus — м.г. персонаж с несчётным
количеством глаз.
Microlinyphia — от г. маленький и Linyphia.
Micrommata — от гг. маленький и глаз.
Microneta — от гг. маленький и сеть (ткать): «маленький ткач».
Miftengris — от «миф тенгр» (= голова рыбы), название Северного Сахалина в
языке нивхов.
Minyrioloides — уменьшительное от Minyriolus.
Minyriolus — г. мелкий и л. уменьшительный суффикс -ol л. крошечный:
«малюсенький».
Misumena — от гг. ненависть (гнев) и ярость.
Moebelotinus — род, близкий к Moebelia и Scotinotylus.
Mongolicosa — от Монголия и Lycosa: «монгольский паук-волк».
Monocerellus — от лл. один и рог; об особом выросте карапакса из слившихся
щетинок: «единорог». С уменьшительным суффиксом — «маленький единорог».
Приложение А. Этимология латинских названий пауков
317
Mughiphantes — от mughi (типовой вид) и г. -yphantes (ткач).
Mustelicosa — от ласка (Mustela) и окончание -cosa, типичное для
пауков-волков: «паук-ласка».
Mygalomorphae — от гг. полевая мышь и форма (габитус): из-за того, что
покрыты густыми волосами и живут в норах.
Myrmarachne — от гг. муравей и паук: «паук-муравей».
Mysmenella — уменьшительное от Mysmena, хотя Mysmena мельче, чем Mys-
menella.
Nenilinium — п. Андрей Борисович Ненилин, известный советский арахнолог.
Neoantistea — г. новый и Antistea.
Neon — п. Neon, историческая личность, упоминающаяся в хрониках Полибия
(Polibius).
Neoscona — 1) гг. грамматически неверное сочетание слов плести и тростник:
сети среди тростника; 2) от гг. новый и пыль.
Neottiura — от гг. гнездо и охранять: самка таскает с собой кокон и охраняет его.
Neriene — м. имя жены Марса.
Neserigone — от nescius (л., неизвестный) и Erigone.
Nippononeta — от л. Nippon (Япония) и Meioneta.
Nispa — «старший» на языке айнов.
Notiogyne — от Notioscopus и epigyne.
Notioscopus — 1) от г. сырой (влажный) и л. вид (внешний); обитает в
заболоченных местах; 2) заметный и наблюдатель (о необычном выросте на головогруди
самца).
Nuctenea — от гг. ночь и плести: «ночной ткач».
Obscuriphantes — от obscurus (типовой вид) и г. -yphantes (ткач).
Octonoba — согласно автору таксона, это простая комбинация букв, хотя
очевидно что происходит от названия типового вида octonarius (состоящий из 8 — о 8
бугорках на брюшке у типового вида).
Oculocornia — от лл. глаза и рог: задние медиальные глаза сидят на отдельных
бугорках.
Oecobius — от гг. дом и живущий: «домовой».
Oedothorax — от гг. вздутие (выпуклость) и грудь: о форме карапакса самца.
Ohlertidion — от ohlerti (типовой вид) и Theridion.
Oia — п. Rishiri Oi, известный японский арахнолог.
Oinia — п. Rishiri Oi, известный японский арахнолог.
Okhotigone — от Охотского моря и Erigone.
Oreoneta — от гг. гора и прядильщик: встречается в горах.
Oreonetides — от Oreoneta и сын: «сын горной прядильщицы».
Oryphantes — вероятно от г. гора и ткач: встречается в горах.
Ostearius — 1) л. привратник, портье (в Англии часто встречается в
помещениях); 2) от г. шустрый (быстрый).
Oxyopes — 1) от острый и «такие глаза»; острое зрение: «остроглазый»; 2) от гг.
острый (колючий) и ноги: о мощных шипах на ногах.
Ozyptila — от гг. острый (колючий) и мягкое перо: о булавовидных щетинках.
Pachygnatha — от гг. толстый и челюсти.
Pacifantistea — от Pacific (Тихий океан) и Antistea: указывает на
распространение (от Камчатки до Хоккайдо) и родственные связи.
318
Приложения
Pacifiphantes — от Pacific (Тихий океан) и Bathyphantes (близкий род):
указывает на распространение рода по обе стороны от Тихого океана, на Дальнем Востоке
и в Канаде.
Paidiscura — очевидно производное от Paidisca (синоним Thymoites) и г.
охранять.
Palliduphantes — комбинация из pallidus (типовой вид) и Lepthyphantes.
Panamomops — от гг. все и безупречный: о глазах, «глазастый».
Paraclubiona — от г. para- (за, около) и Clubiona.
Paracoelotes — от г. para- (за, около) и Coelotes.
Paraeboria — от г. para- (за, около) и Eboria.
Paraglyphesis — от г. para- (за, около) и Glyphesis.
Parasisis — от г. para- (за, около) и Sisis — п. от имени Sisis, отец короля Малой
Армении Антипатра (упоминается у Страбона).
Parasteatoda — от г. para- (за, около) и Steatoda.
Parasyrisca — от г. para- (за, около) и Syrisca — л. сирийская женщина,
«сирийка».
Paratrachelas — от г. para- (за, около) и Trachelas.
Parawubanoides — от г. para- (за, около) и Wubanoides.
Parazygiella — от г. para- (за, около) и Zygiella, замещающее название для Zygia
(«ярмо», эпитет богини Геры, покровительницы браков).
Pardosa — от гг. леопард и -osa, типичное окончание для пауков-волков:
пятнистый как леопард, «паук-леопард».
Pelecopsis — 1) каска и лоб; о задних глазах, сидящих на возвышенности; 2) от
гг. чёрный и вид (внешний).
Pellenes — 1) замещающие название для Peles, Peles — римский бог,
покровитель пастухов и скота. Чтобы сделать название как можно более созвучным был,
очевидно, выбран город Pellene в греческой области Achaea; 2) от г. землистый
(глиняный): о цвете (окраске).
Peponocranium — от гг. арбуз и череп (голова): о форме выроста карапакса
самца, «арбузоголовый».
Perlongipalpus — от слов через-длинная-пальпа: пальпа самца необычно
длинная, «сверх-длиннопальпый».
Perregrinus -замещающее название для Peregrinus (странник, пришелец со
стороны): «странник».
Perm — замещающее название для Рего (этимология неизвестна).
Phaeocedus — от гг. серый и похороны: о траурной окраске, «траурный».
Philaeus — г.м. сын Аякса, одного из персонажей Одиссеи.
Philodromus — от гг. любящий и бродить: «бродяга».
Phintella — производное от Phintia и уменьшительного суффикса -ella.
Phlattothrata — забавное заимствование из английского перевода «Лягушек»
Аристофана: «phlattothrattophlattothrat» — это пародия Эвридипа (герой комедии)
на высокий поэтический стиль Эсхила (другого героя комедии); это длинное слово
можно перевести на русский как «ля-ля-ля» («траля-ля-ля»).
Phlegm — от местности в Греции (упоминается Геродотом), название города
явно производно от «горелый», что возможно указывает на вулкан: о пепельной
окраске, «погорелец».
Pholcus — от г. косоглазый (или взгляд украдкой) либо кривоногий.
Приложение А. Этимология латинских названий пауков
319
Phoroncidia — от гг. нести, колючка и уменьшительного суффикса: о мелком
размере и выростах на брюшке (у некоторых видов выросты заострённые).
Phrurolithus — от гг. страж (охранник) и камень: охраняющий камень.
Pirata — г. пират: от того, что автор вида поймал Araneus piraticus (типовой вид
рода) бегущим по воде среди тростника: «пират».
Pisaura — от л. города в Италии Pisauram (сейчас Pesaro), либо от реки Pesaro.
Pistius — от г. верующий.
Pityohyphantes — от гг. сосна и сеть: встречается на соснах, «сосновый ткач».
Platnickina — п. известного американского арахнолога Norman I. Platnick,
замещающее название для Keijia.
Pocadicnemis — от гг. волос и нога: о волосатых ногах (длинные волоски на
ногах), «мохноногий».
Poecilochroa — от гг. пятнистый (многокрасочный, пёстрый) и кожа:
«пятнистый».
Poeciloneta — от гг. пятнистый (многокрасочный, пёстрый) и прядильщик.
Porrhomma — от гг. далеко и глаза: глаза далеко отстоят друг от друга.
Praestigia — от л. впереди и г. шило: о выросте в передней части карапакса у
самцов.
РгосегосутЫит — от лл. выдающийся (=выступающий далеко) и цимбиум: с
выростом цимбиума.
Proislandiana — л. перед и Islandiana.
Pseudeuophrys — от г. ложный и Euophrys.
Pseudicius — от г. ложный и Icius.
Pseudocyba — от г. ложный и Tapinocyba.
Pseudohilaira — от г. ложный и Hilaira.
Pseudoporrhomma — от г. ложный и Porrhomma.
Pseudowubana — от г. ложный и Wubana.
Rhene — возможно м. от Rhene — имя нимфы (наяды).
Rhomphaea — от г. меч (особый тип широкого меча): о вытянутом брюшке, «ме-
чебрюхий».
Robertus — п. в честь Robert Jocelyn, старшего сына О. Pickard-Cambridge.
Rugathodes — от л. с морщинами (бороздками), производное от Rugatha:
«морщинистый».
Saaristoa — п. в честь известного финского арахнолога Michael I. Saaristo.
Sachaliphantes — от Сахалин и Lepthyphantes.
Saloca — от г. возбуждённый.
Salticus — от г. прыгать или танцевать: попрыгунчик, скакунчик.
Satilatlas — возможно, производное от м. Atlas, Atlantis.
Sauron — от имени персонажа из «Братство Кольца» Дж.РР Толкина.
Savignia — п. в честь M.J.C.L. de Savigny (1777-1851).
Scandichrestus — от Scandinavia и Typhochrestus.
Sciastes — очевидно от эпитета Аполлона, данного ему поэтом Ликофроном
(Lycophron). Смысл неясен.
Scotargus — от гг. тёмный и медленный: «сонный».
Scotinotylus — от гг. тёмный и зёрнышко (мозоль): о выросте на карапаксе
самца, «мозолеголовый».
Segestria — от л. покрытие или мантия.
320
Приложения
Semljicola — от названия острова Новая Земля (semlja латинизированное) и
л. -cola (обитающий).
Sibianor — от Сибирь и рода Bianor.
Sibirocosa — от Сибирь и Lycosa: «сибирский паук-волк».
Sibirocyba — от Сибирь и Tapinocyba (типовой вид рода Sibirocyba
первоначально описан в Tapinocyba).
Silometopoides — от Silometopus и г. подобный.
Silometopus — от гг. широкий-вздёрнутый и лоб (о выросте карапакса у самца).
Singa — от г. причинять вред (вредить).
Sisicottus — от Sisis (отец Антипатра, царя Малой Армении) и -cottus. Вместе,
по-гречески — «несуществующий».
Sisicus — м. Сизис, сын Антипатра, царя Малой Армении.
Sitticus — очевидно п. в честь Publius Sittius, друга Цицерона, окончание
созвучно Salticus.
Sosticus — г. способный спасти или сохранить.
Steatoda — от г. жирный (много жира), упитанный.
Stemmops — от гг. гирлянда (венок, корона) и глаза: глаза расположены кругом.
Stemonyphantes — от гг. нить и ткач.
Strandella — п. Embrik Strand, известный арахнолог и энтомолог, работавший
долгое время в Риге.
Styloctetor — от гг. заострённый инструмент и штукатур: об отростке на голени
пальпы самца.
Suffusiphantes — от suffusus (типовой вид) и Lepthyphantes.
Synageles — от гг. вместе и «на свету (солнца)».
Synagelides — производное от Synageles.
Synema — гг. кроваво-красный.
Takayus — от курорта в Японии, Takaya.
Talavera — вероятно, от местности в Испании, где произошло одно из
известных сражений 28 января 1809 г., в котором участвовал герцог А.У. Веллингтон,
глава союзных войск в битве при Ватерлоо.
Tallusia — п. в честь двух сестёр из финской семьи Tallus: Ulla и Kirsti, с
которыми были знакомы оба автора (Lehtinen и Saaristo).
Tanasevitchia — п. Андрей Викторович Танасевич (р. 1956), известный
российский арахнолог. Описал типовой вид рода.
Tapinocyba — от гг. скромный и куб: о небольшом выросте карапакса самца.
Tapinocyboides — напоминающий Tapinocyba.
Tapinopa — от гг. низкий и лицо: невысокий клипеус.
Taranucnus — 1) от кельтского божества, латинизированное имя которого Taranis
(бог грома); 2) от г. крюк.
Tarsiphantes — от л. tarsi (лапки) и г. -phantes (ткач).
Tegecoelotes — от Tegenaria и Coelotes (похож на тот и другой род).
Tegenaria — от средневекового латинского слова, означающего «делающий
матрасы»: «матрасник».
Tenuiphantes — от tenuis (типовой вид) и Lepthyphantes (род в котором
первоначально рассматривался типовой вид).
Tessamoro — «приморцы» (обитатели морского побережья) на языке айнов.
Приложение А. Этимология латинских названий пауков
321
Tetragnatha — от гг. четыре и челюсти: о удлинённых максиллах и длинных
хелицерах.
Thaleria — п. Konrad Thaler (1940-2005), знаменитый австрийский арахнолог.
Thanatus — от 1) г. смерть или 2) м. перевозчик через реку Стикс.
Theoneta — комбинация из двух родовых названий: Theonina и Microneta.
Theridion — от г. крошечный зверёк.
Theridiosoma — от Theridion и г. soma (тело): тело как у Theridion.
Thomisus — этимология не совсем ясна, поскольку название образовано не по
правилам, как и многие другие родовые названия у С.А. Walckenaer. Вероятно, от г.
плеть, хлыст: автор полагал, что паук ловит добычу набрасывая нить на жертву.
Thymoites — имя собственное из «Илиады» Гомера, брат короля Приама.
Thyreostenius — от гг. щитовидная дверь и сила.
Tibellus — 1) по созвучию с macellus, одним из видов, включённым в род; 2) от
г. щуковидный (или флейтовидный), как берцовая кость (о форме тела).
Tibiaster — от tibia: самец имеет сильно расчленённую голень пальпы.
Tibioploides — от Tibioplus и г. подобный.
Tibioplus — от л. tibia (голень) и вооруженный: о большом отростке голени
пальпы самца у типового вида.
Tiso — от г. кто-либо (любой).
Titanoeca — от гг. белый (белая земля, известняк) и обитать: обитающий в
известняке.
Tmarus — от названия горы в Греции, Tomaros, упоминается Вергилием.
Tmeticus — от гг. рубить, автор (Menge) интерпретировал как «с острым зубом».
Trachelas — не вполне понятна этимология: 1) эпитет (прозвище) императора
Константина Великого (274-337), означает «бычья шея»; 2) о выпуклом карапаксе.
Trachyzelotes — от г. шершавый и Zelotes: о щётке из шипов на переднем крае
хелицер.
Trematocephalus — от гг. дырка и голова: о форме карапакса самца.
Tricca — от г. волос: возможно о волосатых пальпах.
Triccosta — производное от Tricca.
Trichobactrus — от Trichoncus и Metopobactrus.
Trichoncus — от гг. волос и опухоль.
Trichopterna — от гг. волос и пятка (ступня): очевидно из-за наличия трихобот-
рии на предлапке IV.
Trochosa — 1) от г. бегущий быстро и -osa (как у Lycosa). 2) от г. колесо или шар
(мяч): о форме кокона.
Troxochrus — 1) от гг. грызун (или долгоносик) и цвет: грудная часть сильно
толстая (грубая); 2) от гг. есть (глодать) и шкура (кожа).
Tubercithorax — от гг. бугорок и грудь: головогрудь с небольшим выростом.
Tuberta — от г. вздутие.
Tunagyna — от гошутского (североамериканские индейцы Gosiute) «прямой» и
эпигина: о длинной прямой эпигине.
Tusukuru — от айнского «шаман».
Tuvadrassus — от Тува и Drassus.
Tuvaphantes — от Тува и -phantes (ткач).
Typhochrestinus — производное от Typhochrestus.
Typhochrestoides — производное от Typhochrestus.
322
Приложения
Typhochrestus — 1) первая часть не имеет смысла, chrestus — полезный; 2) от гг.
волосатый и нуждающийся (в чём-то): о карапаксе без волосков, «лысый».
Uloborus — от г. смертельный, убийственный укол. Забавно, что как раз
Uloboridae, и Uloborus в частности, лишены развитых ядовитых желез.
Ummeliata — замещающее название для Hummelia: п. David Hummel,
шведский энтомолог.
Urozelotes — Cameron (2005) не смог найти этимологию первого слога (иго).
Автор рода (Mello-Leitao) также не поясняет этимологию. Вероятно, от немецкого
суффикса — иг, означающего «древний, примитивный» или, что более вероятно,
от г. иго или иг (хвост) и Zelotes: Zelotes с хвостом (о торчащих бородавках).
Ussurigone — от р. Уссури и Erigone.
Uusitaloia — п. в честь финского акаролога М. Uusitalo.
Vermontia — от штата Вермонт в США.
Viktorium — п. Виктор Петрович Тыщенко (1937-1986), выдающийся
советский энтомолог и арахнолог.
Wabasso — из «Песни о Гайавате» Лонгфелло, означает Север.
Walckenaeria — п. в честь С.А. Walckenaer (1771-1852), известного
французского арахнолога.
Walckenaerianus — от Walckenaeria: пальпа и головогрудь подобны у
Walckenaeria.
Weintrauboa — п. в честь американского зоолога R.L. Weintraub.
Whymperiphantes — от whymperi (типовой вид) и Lepthyphantes.
Wiehlenarius — п. в честь Н. Wiehle (1884-1966), выдающего немецкого
арахнолога.
Wubanoides — от Wubana (по-гошутски «волос на вершине», о длинных
щетинках на карапксе) и подобный.
Xerolycosa — от г. сухой и Lycosa: паук-волк из сухих мест.
Xysticus — от латинского слова, заимствованного из греческого и означающего
«атлет».
Yaginumaella — п. Takeo Yaginuma, известный японский арахнолог.
Yaginumena — п. Takeo Yaginuma, известный японский арахнолог.
Yaginumia — п. Takeo Yaginuma, известный японский арахнолог.
Yakutopus — от Якутия и Notioscopus.
Zelotes — от 1) г. с усердием (рвением), поклонник, подражатель, или 2) Simon
the Zealot (Зелот), один из апостолов (учеников) Иисуса Христа.
Zerogone — от «zero» и Erigone: неизвестен самец, и непонятны родственные
связи.
Zilla — от имени собственного из Ветхого Завета, Zillah в английском варианте
библии, и Zilla — в немецком.
Zora — от г. чистый (не смешанный с чем-либо).
Zornella — п. в честь Anders Zorn (1860-1920), известного шведского
художника. Цимбиум самца с 3 крупными щетинками, напоминающими кисти.
Приложение Б. Список русскоязычных арахнологов-систематиков 323
ПРИЛОЖЕНИЕ Б
Список русскоязычных арахнологов-систематиков
ФИО
Азаркина Галина
Николаевна
Гнелица Валерий
Анатольевич
Громов Александр
Викторович
Гусейнов Эльчин
Фазилевич
Данилов Сергей
Николаевич
Есюнин Сергей
Леонидович
Зонштейн Сергей
Львович
Ковблюк Николай
Михайлович
Логунов Дмитрий
Викторович
Марусик Юрий
Михайлович
Михайлов Кирилл
Глебович
Овчаренко Владимир
Иванович
Олигер Татьяна
Ивановна
Омелько Михаил
Михайлович
Пономарёв Александр
Викторович
Танасевич Андрей
Викторович
Триликаускас
Лаймонас
Альбертович
Тунёва Татьяна
| Константиновна
E-mail
urmakuz@gmail.com
gnelitsa@mail.ru
alexander_gromov@yahoo. com
apsheron@list.ru
hun-vey-bin@rambler.ru
sndanilov@list.ru
Sergei.esyunin@psu.ru
znn@post.tau.ac.il
kovblyuk@mail.ru
dpavuk@ngs.ru
dmitri.v.logunov@manchester.ac.uk
yurmar@mail.ru;
yurmar@newmail.ru
mikhailov2000@gmail.com
ovtshare@amnh.org
ns_zap@mail.ru
omelkom@gmail.com
ponomarev 1952@mail.ru
and-tan@mail.ru
laimont@mail.ru
gnaphos@yandex.ru
Основные интересы
Salticidae; все группы на
Алтае
Linyphiidae
Theridiidae; весь регион
Agelenidae;
поведение
Corinnidae, Micaria,
Liocranidae; все группы в
Забайкалье
все группы; Урал и
прилежащие регионы
мигаломорфные пауки и
Filistatidae Палеарктики
Gnaphosidae; Крым,
Восточное Средиземноморье,
Кавказ
Salticidae, Lycosidae,
Philodromidae, Thomisidae,
Zodariidae
все группы;
весь регион
Clubionidae, Corinnidae,
Cheiracanthiidae, Liocranidae
Gnaphosidae
все группы;
Нижне-Свирский заповедник
(Ленинградская область)
Lycosidae юга Дальнего
Востока; Cybaeidae,
Gnaphosidae
все группы; европейская
Россия, Кавказ, Западный
Казахстан
Linyphiidae
все группы;
Приамурье, Алтай и Горная
Шория
Gnaphosidae
324
Приложения
Приложение В
Интернет-ресурсы, полезные для определения пауков
Каталог пауков мира — Platnick N.I. 2011. The world spider catalog, version 11.5. American
Museum of Natural History: http://research.amnh.org/entomology/spiders/catalog/
index.html
База данных К.Г.Михайлова «Пауки России и сопредельных стран [до 2000 г.]»: http.7/
www.zin.ru/biodiv/aranei.asp
База данных по распространению пауков в Европе — Helsdingen P., van. 2010. Fauna
Europaea: Araneae. Version 2.2: http://www.faunaeur.org/
Каталог и определитель пауков-скакунчиков (Salticidae) мира — Proszynski J. Monograph
of the Salticidae (Araneae) of the World: http://salticidae.org/salticid/main.htm
Определитель пауков Европы — Nentwig W., Gloor D., Hanggi A., Kropf C. Spiders of
Europe. Version 10.2010: http://www.araneae.unibe.ch/
Статьи по паукам, опубликованные в Русском Артроподологическом Журнале «Arthro-
poda Selecta», в свободном доступе в формате PDF: http://arthropodaselecta.
britishspiders.org.uk
Статьи по паукам, опубликованные в журнале Acta Arachnologica (Arachnological Society
of Japan), в свободном доступе в формате PDF: http://www.jstage.jst.go.jp/browse/asjaa/
_vols
Все работы по паукам, опубликованные в журналах Американского Музея Естественной
Истории (Нью-Йорк), в свободном доступе в формате PDF: http://digitallibrary.amnh.
org/dspace/
Все работы, опубликованные в американском Journal of Arachnology, в свободном доступе
в формате pdf: http://www.americanarachnology.Org/JOA_online.html#issues
Список публикаций и коллекция рисунков по паукам Китая — Endemic spiders in China:
http://www.amaurobiidae.com/araneae/
Периодическое издание Smithsonian Contributions to Zoology — все работы в свободном
доступе в формате PDF: http://www.sil.si.edu/SmithsonianContributions/Zoology/
index.cfm
Журнал Bulletin of the Museum of Comparative Zoology — некоторые (но не все) работы
по паукам в свободном доступе в формате PDF: http://www.mcz.harvard.edu/
Publications/search_pubs.html?publication=bullmcz
Форум русскоязычных арахнологов «RusArachnology»: http://groups.google.com/group/
RusArachnology
Приложение Г. Основные (настольные) книги
325
Приложение Г
Основные (настольные) книги,
необходимые для изучения и определения пауков
Иванов А.В. 1965. Пауки, их строение, образ жизни и значение для человека. Л.: Изд-во
Ленинградск. ун-та. 304 с.
Сейфулина P.P., Карцев В.М. 2010. Пауки средней полосы России. Атлас-определитель
с обзором биологии пауков. М.: Фитон+. 608 с.
Тыщенко В.П. 1971. Определитель пауков Европейской части СССР // Определители по
фауне СССР, издаваемые Зоол. ин-том АН СССР. Л.: Наука, Ленинградск. отд.
Вып.105. 281 с.
Almquist S. 2005. Swedish Araneae, part 1: families Atypidae to Hahniidae (Linyphiidae
excluded) // Insect Syst. Evol., Suppl. 62. P. 1-284.
Almquist S. 2006. Swedish Araneae, part 2: families Dictynidae to Salticidae // Insect Syst.
Evol., Suppl. 63. P.285-601.
Chikuni Y. 1989. Pictorial encyclopedia of spiders in Japan. Tokyo: Kaisei-sha Press. 310 p.
[на японском].
Dondale CD., Redner J.H. 1978. The insects and arachnids of Canada, Part 5. The crab spiders
of Canada and Alaska, Araneae: Philodromidae and Thomisidae // Research Branch,
Agriculture Canada, Publ. No. 1663. 255 p.
Dondale CD., Redner J.H. 1982. The insects and arachnids of Canada. Part 9. The sac spiders
of Canada and Alaska. Araneae: Clubionidae and Anyphaenidae // Research Branch,
Agriculture Canada, Publ. No. 1724. 194 p.
Dondale CD., Redner J.H. 1990. The insects and arachnids of Canada. Part 17. The wolf
spiders, nurseryweb spiders, and lynx spiders of Canada and Alaska (Araneae: Lycosidae,
Pisauridae, and Oxyopidae) // Research Branch, Agriculture Canada, Publ. No. 1856.
383 p.
Dondale CD., Redner J.H., Paquin P., Levi H.W. 2003. The insects and arachnids of Canada.
Part 23. The orb-weaving spiders of Canada and Alaska (Araneae: Uloboridae, Tetragna-
thidae, Araneidae, Theridiosomatidae). Ottawa: NRC Research Press. 371 p.
Eskov K.Yu. 1994. Catalogue of the linyphiid spiders of northern Asia (Arachnida, Araneae,
Linyphiidae). Sofia-Moscow: PENSOFT Publ. 144 p.
Heimer S., Nentwig W. 1991. Spinnen Mitteleuropas: Ein Bestimmungsbuch. Berlin: Verlag
Paul Parey. 543 S.
Jocque R., Dippenaar-Schoeman A.S. 2006. Spider families of the World. Tervuren: Royal
Museum for Central Africa. 336 p.
Logunov D.V., Marusik Yu.M. 2000. Catalogue of the jumping spiders of northern Asia
(Arachnida, Araneae, Salticidae). Moscow: KMK Scientific Press Ltd. 299 p.
Marusik Yu.M., Logunov D.V., Koponen S. 2000. Spiders of Tuva, South Siberia. Magadan:
IBPN FEB RAS. 252 p.
Mikhailov K.G. 1997. Catalogue of the spiders of the territories of the former Soviet Union
(Arachnida, Aranei) // Sbornik trudov Zoologicheskogo muzeya MGU. Vol.37. 416 p.
Namkung J. 2002. The spiders of Korea. Seoul: Kyo-Hak Publishing Co. 648 p. [на корейском]
Ono H. 2009. The Spiders of Japan with keys to the families and genera and illustrations of the
species. Kanagawa: Tokai Univ. Press. 739 p. [на японском]
Palmgren P. 1974a. Die Spinnenfauna Finnlands und Ostfennoskandiens IV. Argiopidae,
Tetragnathidae und Mimetidae // Fauna Fennica. Vol.24. S.l-70.
326
Приложения
Palmgren P. 1974b. Die Spinnenfauna Finnlands und Ostfennoskandiens. V. Theridiidae und
Nesticidae // Fauna Fennica. Vol.26. S.l-54.
Palmgren P. 1975. Die Spinnenfauna Finlands und Ostfennoskandiens. VI. Linyphiidae 1 //
Fauna Fennica. Vol.28. S. 1-102.
Palmgren P. 1976. Die Spinnenfauna Finnlands und Ostfennoskandiens. VII. Linyphiidae 2 //
Fauna Fennica. Vol.29. S. 1-126.
Palmgren P. 1977. Die Spinnenfauna Finnlands und Ostfennoskandiens. VIII. Argyronetidae,
Agelenidae, Hahniidae, Dictynidae, Amaurobiidae, Titanoecidae, Segestriidae, Pholcidae
und Sicariidae. Anhang: Bestimmungstabelle der in Finnland representierten Spinnenfa-
milien und kommentiertes Register der Teile I—VIII der Spinnenfauna // Fauna Fennica.
Vol.30. S. 1-50.
Platnick N.I., Dondale CD. 1992. The insects and arachnids of Canada. Part 19. The ground
spiders of Canada and Alaska. Araneae: Gnaphosidae // Research Branch Agriculture
Canada Publ. No. 1875. 297 p.
Roberts M.J. 1985. The spiders of Great Britain and Ireland. Vol.1. Atypidae to Theridio-
somatidae. Colchester, England: Harley Books. 229 p.
Roberts M.J. 1987. The spiders of Great Britain and Ireland. Vol.2. Linyphiidae. Colchester,
England: Harley Books. 204 p.
Roberts M.J. 1995. Collins Field Guide: Spiders of Britain & Northern Europe. London: Harper
Collins. 383 p.
Roberts M.J. 1998. Spinnengids. Baarn, Netherlands: Tirion. 397 S.
Song D.X., Zhu M.S., Chen J. 1999. The spiders of China. Hebei Science & Technology
Press. 640 p.
Song D.X., Zhu M.S., Zhang F. 2004. Fauna Sinica: Invertebrata Vol. 39: Arachnida: Araneae:
Gnaphosidae. Beijing: Science Press. 362 p. [на китайском]
Ubick D., Paquin P., Cushing P.E., Roth V. 2005. Spiders of North America: an identification
manual. American Arachnological Society. 377 p.
Zhu M.S., Song D.X., Zhang J.X. 2003. Fauna Sinica: Invertebrata Vol. 35: Arachnida: Araneae:
Tetragnathidae. Beijing: Science Press, vii + 418 p. [на китайском]
Приложение Д. Словарь
327
Приложение Д
Словарь
ad partem — лат., от части, частично.
-ае — окончание, которое во многих латинских словах указывает на множественное число.
Агапеае — лат. название отряда Пауки. Название было впервые введено в
номенклатуру К. Линнеем в 1758 г. в 10-м издании «Системы Природы...» (Linnaeus С. 1758.
Systema Naturae. Ed. X. Holmiae. 821 p. (Araneae, p. 619-624)). Используется наряду
с названием Aranei, введённым в употребление К. Клерком на год раньше. См. также
Приложение А. Этимология названий.
Aranei — лат. название отряда Пауки. Название было впервые введено в номенклатуру
Клерком в 1757 г. в работе «Пауки Швеции...» (Clerck С. 1757. Svenska spindlar (Aranei
Suecici). Stockholmiae, 154 p.). Используется наряду с названием Агапеае,
введённым в употребление К. Линнеем на год позже. См. также Приложение А.
Этимология названий.
cf. (сокращение от лат. confinis / conformis — соседний / похожий) — близкий вид.
Например, A us cf. bus — какой-то вид рода A us, близкий к A us bus.
Cribellatae (лат.), cribellate (англ.) — крибеллятные, пауки с крибеллюмом.
Dionycha (двух-коготковые) — пауки с двумя коготками на лапках ног.
Ecribellatae (лат.), ecribellate (англ.) — некрибеллятные, пауки без крибеллюма.
Entelegynae (лат.), entelegyne (англ.) — см. энтелегинные пауки.
Haplogynae (лат.), haplogyne (англ.) — см. гаплогинные пауки.
lamella characteristica (лат.) — отросток бульбуса (радикса), имеющий видоспецифич-
ную форму. В строгом смысле этот отросток имеется только у Micronetinae
(Linyphiidae), однако термин используется и для не гомологичной структуры у Ipainae
(Linyphiidae) и у Linyphiinae. Обычно имеет плоскую форму, но бывает и толстым.
RTA — аббревиатура для retrolateral tibial apophysis (англ.) — ретролатеральный го-
ленный отросток пальпы самца (РГО).
RTA-clade — большая группа энтелегинных семейств пауков, которые имеют
ретролатеральный отросток на голени пальпы у самца (РГО). Однако в эту группу входят и
Lycosidae, хотя у них нет отростка голени, но не входят Linyphiidae (часть из которых
имеют отростки голени) и Titanoecidae (тоже имеют отростки голени).
sensu lato (лат.) — в широком смысле (в широкой трактовке).
s.l. (лат.) — сокращение для sensu lato — в широком смысле (в широкой трактовке).
sensu stricto (лат.) — в узком смысле (в узкой трактовке).
s.s. (лат.) — сокращение для sensu stricto — в узком смысле (в узкой трактовке).
Trionycha (трёх-коготковые) — пауки с тремя коготками на лапках ног.
Австралазия — Австралия, Новая Зеландия и юго-восточная Азия.
апофиз (англ. apophysis/apophyses) — вырост или отросток пальпы самца (может быть
на разных частях пальпы — на бульбусе, цимбиуме, голени, колене, бедре), иногда
подвижный.
апикальный (от лат. apex — верхушка) — вершинный, находящийся на вершине.
аранеоидные — семейства, близкие к Araneidae, в азиатской части России — это Aranei-
dae, Linyphiidae, Mimetidae, Mysmenidae, Nesticidae, Pimoidae, Tetragnathidae, Theridio-
somatidae, Theridiidae и Zygiellidae.
аранеоморфные — все высшие пауки (относящиеся к подотряду Araneomorphae). У
нас — это все семейства, кроме Atypidae.
328
Приложения
аранеофаг (от лат. araneus — паук + греч. phagos — пожиратель) — питающийся
пауками.
аридный (от лат. aridus, англ. arid — сухой) — засушливый.
аутотомия (от греч. autos — сам + греч. tome — отсечение) — отбрасывние
(отламывание) своих ног самим пауком; защитная реакция, наблюдаемая при резком
раздражении, например, при схватывании хищником. У молодых пауков утраченные ноги
обычно восстанавливаются после следующей линьки. Восстановившиеся ноги
(регенераты) отличаются от нормальных размером, окраской, хетотаксией.
базальный (англ. basal, от греч. basis — основа, основание) — находящийся в
основании.
бедро (лат. и англ. femur/femora) — пятый сегмент ноги (если считать с дистального
конца) и четвёртый (с дистального конца) сегмент пальпы; обычно это самый
мощный сегмент конечности.
бриофил (от греч. bryon — мох + греч. philus — любящий) — предпочитающий мох.
бульбус (от лат. bulbus — луковица) — копулятивный орган на лапке педипальпы
самца.
вентральный (лат. ventralis, англ. ventral) — расположенный снизу, на брюшной
стороне.
вертлуг (лат. и англ. trochanter) — сегмент ноги или пальпы, следующий за тазиком
(второй с проксимального конца).
видоспецифичный — характерный только для конкретного вида.
викарирование (от лат. vicarius — заменяющий, заместитель) — замещение одного
вида другим, но близкородственным, в разных географических регионах.
вогнутый ряд (англ. recurved) — о форме рядов глаз; это такой ряд глаз, в котором
медиальные глаза стоят позади от латеральных; оценивается при виде сверху, хотя в
западной литературе — при виде спереди.
вульва (лат. и англ. vulva) — внутренняя часть эпигины. То же самое, что и эндогина.
выгнутый ряд (англ. procurved) — о форме рядов глаз; это такой ряд глаз, в котором
медиальные глаза стоят впереди латеральных; в нашей традиции оценивается при
виде сверху, хотя в западной литературе — при виде спереди.
габитуальный — относящийся к габитусу.
габитус (лат. и англ. habitus — внешность, наружность) — внешний вид, облик (форма
тела).
гаплогинный (англ. haplogyne; от греч. haplos — одиночный + греч. gyne —
женщина) — особый тип копулятивного аппарата, когда у самки нет эпигины.
Гаплогинные (лат. Haplogynae, англ. haplogyne) — пауки, у которых эпигина имеет
только одну пару канальцев (протоков), которые служат и как осеменительные, и как
оплодоторительные. Самцы имеют просто устроенные пальпы (исключение — Phol-
cidae), пальпа без гематодохи. Эпигина, вернее генитальная пластинка, без ямок и
щелей. В азиатской части России — это только Pholcidae и Oonopidae, но, кроме них,
могут быть найдены Segestriidae, Sicariidae и Scytodidae.
гематодоха (англ. haematodocha; от греч. haimatos — кровь) — приёмник гемолимфы,
мешковидный орган, соединяющий бульбус с цимбиумом. В покое свёрнут, а во
время спаривания раздувается под давлением гемолимфы.
глазное поле — передняя часть карапакса, на которой расположены глаза; это может
быть прямоугольник, углами которого служат латеральные глаза.
Голарктика (англ. Holaractic; от греч. holos — весь + arktikos — северный,
находящийся с той же стороны, где созвездия Большой и Малой Медведицы вместе с По-
Приложение Д. Словарь
329
лярной звездой) — биогеографическая область, охватывающая всё северное
полушарие примерно севернее 30° с.ш.; состоит из двух подобластей — Неарктики и
Палеарктики.
голень (лат. и англ. tibia/tibiae) — третий с конца сегмент ноги и второй (с дистального
конца) сегмент пальпы.
гомологичный (от греч. homologia — соответствие) — для органов и структур,
имеющих общее происхождение.
гонопор (от греч. gonos — семя + poros — проход, отверстие) — половое отверстие
самца.
гуанин — светлый (белый, жёлтый, зеленоватый) пигмент.
гуаниновый — из пигмента гуанина, расположеного отдельными пятнами, чаще всего
белого или серебристого цвета.
дендробионтный (от греч. dendron — дерево + biontos — живущий) — обитающий на
деревьях.
дизъюнктивный (от англ. disjunction — разобщение, обособление) — ареал
разорванный, не сплошной. Например, виды с кавказско-дальневосточным дизъюнктивным
ареалом обитают на Кавказе и на Дальнем Востоке, но отсутствуют между ними —
в Сибири, в степной зоне и прочих регионах.
диктинный — тип пальпы, характерный для семейства Dictynidae и некоторых других,
когда кондуктор грибовидный, отходит от основания эмболюса, эмболюс длинный,
основание эмболюса большое и округлое (рис. 15.5,10).
дистальный (от лат. distare — отстоять, находиться на расстоянии) — находящийся
дальше от места прикрепления органа (ноги, например).
дорсальный (лат. dorsalis, англ. dorsal) — расположенный сверху, на спинной стороне.
дыхальце — щель, через которую поступает воздух в трахеи.
ерник — густой кустарник (обычно в таёжной и тундровой зоне) высотой 30-100 см.
ЗЛГ — задние латеральные (^боковые) глаза.
ЗМГ — задние медиальные (^средние) глаза.
имаго (от лат. imago — образ, вид) — взрослая (половозрелая) стадия развития. В этой
стадии пауки, как правило, не линяют и не растут (исключение — членистобрюхие
пауки и птицееды).
каламиструм (лат. и англ. calamistrum) — ряд изогнутых волосков или щетинок на
предлапке IV, который служит для прядения (расчесывания) особого типа паутины.
карапакс (лат. и англ. carapace) — склерит, закрывающий головогрудь сверху; на нём
расположены глаза.
карман эпигины — склеротизованный неподвижный участок эпигины,
ограничивающий её ямку; может быть верхним, нижним или боковыми.
клипеус (лат. и англ. clypeus) — наличник, область карапакса между его передним
краем и передними глазами.
коготковые подушечки (англ. claw tufts) — пучки волосков под коготками ног у
некоторых пауков.
коксоподит — то же самое, что и максиллы.
колено (лат. и англ. patella/patellae) — сегмент ноги или пальпы, расположенный
между голенью и бедром.
колюлюс (лат. и англ. colulus) — непарный вырост между передними паутинными
бородавками; иногда редуцирован до двух волосков. По всей видимости, представляет
собой рудимент крибеллюма. А.В. Иванов (1965) и В.П. Тыщенко (1971) пишут —
колюлус (сложнопроизносимое сочетание).
330
Приложения
кондуктор (лат. и англ. conductor — сопровождающий, проводник) — вырост бульбу-
са, служащий для направления эмболюса во время копуляции или для защиты эмбо-
люса в состоянии покоя. Используется для негомологичных структур.
конспецифичный (англ. conspecific) — относящийся к тому же виду.
копуляторные каналы (англ. insemination ducts, entrance ducts) — каналы в эндоги-
не самки, ведущие от копулятивного (совокупительного) отверстия к рецептакуле
(сперматеке). То же самое, что и осеменительные канальи
крибеллюм (лат. и англ. cribellum) — ситовидная пластинка; поперечная пластинка
спереди от паутинных бородавок со многими паутинными трубочками, служащая
для прядения особого типа паутины.
крибеллятные (лат. Cribellatae, англ. cribellate) — пауки, имеющие крибеллюм.
криптическая окраска (от греч. kryptos — тайный, скрытный) — маскирующая окраска.
лабиум (лат. и англ. labium) — нижняя губа (непарный склерит, расположенный между
тазиками педипальп).
лапка (лат. и англ. tarsus/tarsi) — последний сегмент ноги или пальпы.
латеральный (лат. lateralis, англ. lateral) — расположенный сбоку, боковой.
лёгкие (англ. book lungs) — лёгкие пауков; органы дыхания, представляющие собой
видоизменённые конечности; располагаются снизу брюшка спереди от эпигастраль-
ной щели (бывают и сзади от эпигастральной щели, например, у Atypidae). Снаружи
видны не сами лёгкие, а лёгочные крышечки (англ. book lung opercula),
прикрывающие их.
литобионт (англ. lithobiont; от лат. lithos — камень + biontos — живущий) —
обитающий среди камней; здесь применяется для пауков, обитающих исключительно среди
камней, в отличие от петрофилов («любящих камни»).
максиллы (лат. и англ. maxilla/maxillae) — нижние челюсти, образованные выростами
(жевательными лопастями) тазиков педипальп.
медиальная пластинка эпигины (англ. median plate) — средняя часть эпигины,
обособленная щелью от боковых частей.
медиальная щель (или борозда) (англ. median furrow) — продольная щель (борозда)
вдоль грудной части карапакса, иногда представляет собой поперечное вдавление.
Служит для прикрепления мышц.
медиальный (лат. medialis, англ. medial) — средний, срединный.
медиальный отросток (англ. median apophysis) — отросток (вырост) тегулюма;
иногда называется тегулярным отростком. Может быть расположен как посредине, так и
сбоку. У амауробиоидных пауков обычно с крючком на вершине. Используется для
негомологичных структур.
мезофильный (от греч. mesos — средний, промежуточный + греч. philus — любящий) —
умеренно влажный (о битопе, или о предпочитаемом биотопе).
мигаломорфные (лат. Mygalomorphae) — пауки, входящие в отдельный
одноимённый подотряд (инфраотряд). В фауне России встречается два семейства, а в азаитс-
кой части только одно семейство — Atypidae.
мирмекофил (англ. myrmecophilous; от греч. myrmex — муравей + греч. philus —
любящий) — любящий (поедать) муравьев.
МКЗН — Международный кодекс зоологической номенклатуры.
монотипичный (англ. monotypic, monobasic; от греч. monos — один, единственный +
греч. typos — образец) — род или семейство с одним видом.
Приложение Д. Словарь
331
мускульные точки — хитинизированные бляшки на верхней части брюшка,
служащие для прикрепления мышц.
мыщелок — бугорок на наружной (ретро-латеральной) поверхности в основании хели-
церы.
надвидовая систематика (англ. supraspecific systematics) — систематика,
рассматривающая таксоны от уровня подрода и выше (род, триба, подсемейство и т.д.).
наличник (лат. и англ. clypeus) — область карапакса между передним его краем и
передними глазами.
Неарктика (англ. Nearctic; от греч. neos — новый, находящийся в Новом Свете + греч.
arktikos — северный, находящийся с той же стороны, где созвездия Большой и
Малой Медведицы вместе с Полярной звездой) — подобласть Голарктики,
находящаяся в Северной Америке (большая часть Мексики, США и Канада).
некрибеллятный (лат. Ecribellatae; англ. ecribellate) — без крибеллюма.
Неотропис (англ. Neotropis; от греч. neos — новый, находящийся в Новом Свете +
греч. tropikos — тропический) — биогеографическая область, приходящаяся на
Южную и Центральную Америку.
нижняя губа (лат. и англ. labium) — непарный склерит, расположенный между
тазиками педипальп.
номинативный (англ. nominative) — «давший название», применяется для подвидов и
более высоких категорий, например вид Dictyna tyshchenkoi состоит из двух
подвидов: D. tyshchenkoi tyshchenkoi и D. tyshchenkoi wrangeliana. Первый из подвидов —
номинативный. Подсемейство Tetragnathinae является номинативным для Tetragnathi-
dae (и наоборот).
облигатный (от лат. obligatus — обязательный, непременный) — обязательный
(например, облигатный литобионт — обитающий исключительно среди камней).
онихум (лат. и англ. onychium) — вырост лапки ноги, на котором сидят коготки; служит
для обеспечения большей подвижности коготков и обеспечивающий лучшее
сцепление с субстратом. Из наших пауков имеется у Oonopidae.
оплодоторительные каналы (англ. fertilization ducts) — каналы в эндогине самки,
ведущие от сперматеки к матке.
осеменительные каналы (англ. insemination ducts, entrance ducts) — каналы в
эндогине самки, ведущие от копулятивного (совокупительного) отверстия к рецептакуле
(сперматеке). То же самое, что и копуляторные каналы.
отросток (англ. apophysis/apophyses, outgrowth, process) — вырост какой-либо части
пальпы самца. Чаще термин «apophysis» применяют для длинных и подвижных
(сидящих на мембранках) выростов, a «outgrowth» — для коротких и неподвижных
выступов.
Палеарктика (англ. Palaearctic; от греч. palaios — древний, относится к Старому
Свету + греч. arktikos — северный, находящийся с той же стороны, где созвездия
Большой и Малой Медведицы вместе с Полярной звездой) — область Голарктики,
занимающая всю северную часть Евразии и северную Африку.
пальпа (лат. palpus/palpi; англ. palp или palpus) — то же, что и педипальпа —
конечность, расположенная между хелицерами и ногами.
пантропический (англ. pantropical; от греч. pan — всё + греч. tropikos —
тропический) — распространённый по всему тропическому поясу.
парацимбиум (лат. и англ. paracymbium/paracymbia; от греч. para — возле, около,
рядом + греч. cymbium — лодочка) — склерит (отросток, вырост), причленённый
332
Приложения
или отходящий от цимбиума. Используется для негомологичных структур. Парацим-
биум может быть базальный и верхний. Нижний (базальный) парацимбиум
находится в основании цимбиума (например, у Linyphiidae, Nesticidae). Верхний
(апикальный) парацимбиум находится в верхней части цимбиума (например, у Theridiidae).
пармула (лат. и англ. parmula) — отросток заднего края эпигины.
педипальпа (англ. pedipalp) — то же, что и пальпа — конечность, расположенная
между хелицерами и ногами.
петрофил (~ный) (англ. petrophilous; от греч. petra — скала, утёс, камень + греч.
philus — любящий) — «любящий камни»; применяется для пауков, которые часто
(преимущественно) встречаются среди камней. В отличие от литобионтов, такие пауки
могут обитать и в местах без камней.
ПЛГ — передние латеральные (=боковые) глаза.
ПМГ — передние медиальные (=средние) глаза.
предкопуляционный (англ. precourtship) — период, предшествующий копуляции
(«ухаживание»). Самцы многих видов в этот период устраивают «танцы»,
«демонстрации» или другие действия, чтобы расположить к себе самку и не быть принятыми за
добычу.
предлапка (лат. и англ. metatarsus/metatarsi) — предпоследний сегмент ноги.
проксимальный (от лат. proximus — ближайший) — находящийся рядом с местом
прикрепления органа (ноги, например).
пролатеральный (англ. prolateral; от лат. pro спереди + лат. lateralis — боковой) —
расположенный сбоку-спереди.
психрофильный (англ. psychrophilous; от греч. psychria — холод + греч. philus —
любящий) — «любящий холод»; применяется для пауков, обитающих в холодных
условиях.
радикс (англ. radix; от лат. radix — корень) — отросток или часть тегулюма, который
несёт помимо эмболюса и другие отростки. Не гомологичен у разных групп.
Синоним — эмболюсный отдел.
РГО (англ. RTA — retrolateral tibial apophysis) — ретролатеральный отросток голени
пальпы самца.
ретролатеральный (англ. retrolateral; от лат. retro сзади + лат. lateralis — боковой) —
расположенный сбоку-сзади.
рецентный (англ. recent; от лат. recenter — недавно, только что) — современный
(противопоставляется вымершим/ископаемым).
рецептакула (лат. и англ. receptaculum/receptacula) — часть репродуктивного органа
самки, обычно округлой формы, которая служит резервуаром, куда попадает сперма
после копуляции.
рострум (лат. и англ. rostrum — нос, клюв) — вырост наличника или глазной области
(если он направлен вперёд).
РТА-клада — то же, что и RTA-clade.
свободный сектор сети (англ. free sector) — участок сети, в котором нет ловчих
(спиральных) нитей.
семяприёмник — то же самое, что и сперматека. А.В. Иванов (1965) и В.П. Тыщенко
(1971) пишут этот термин как «семеприёмник».
септум (лат. и англ. septum — перегородка) — неподвижный вырост эпигины,
разделяющий её на две половины (левую и правую) и не имеющий свободной вершины (в
отличие от подвижного скапуса).
Приложение Д. Словарь
333
скапус (лат. и англ. scapus, scape) — вырост эпигины, крепящийся к её передней части
(реже к задней), свободный на вершине. Часто имеет кармашек (ямку) на вершине.
Иногда скапус может обламываться (Araneidae). Используется для негомологичных
структур.
склерит (англ. sclerite; от греч. scleros — твёрдый) — сильно хитинизированный
участок паука, чаще всего чётко обособленный от других; применяется для структур,
расположенных на головогруди или пальпе самца (не обязательно сильно хитинизиро-
ванных).
склеротизованный (англ. sclerotised; от греч. scleros — твёрдый) — сильно
хитинизированный.
скопула (лат. и англ. scopula — веник, щётка) — густые волоски на нижней стороне
лапки (иногда, и предлапки), формирующие своего рода щётку.
скутум (лат. и англ. scutum/scuta — щиток) — склеротизованный участок брюшка.
соматический (англ. somatic; от греч. soma — тело) — применяется ко всем частям
тела, кроме копулятивных органов (педипальпа самца, эпигина) и полового
отверстия самца.
сперматека (англ. spermatheca/spermathecae; от греч. sperma — семя + греч. theke —
ящик, хранилище; лат. receptaculum seminis — хранилище семени) —
семяприёмник.
стабилимент (англ. stabilimentum) — зигзагообразная, прямая или спиралевидная лента
на колесовидной ловчей сети у некоторых крестовиков Araneidae (Argiope, Cyclosa)
и Uloboridae (Octonoba).
стернум (лат. и англ. sternum) — непарный склерит, расположенный между тазиками
ног.
стридуляционные — «стрекочущие», служащие для производства звуков.
стридуляционные бороздки/желобки (англ. stridulating furrows/files/ridges) —
бороздки-гребни, расположенные параллельно друг другу; могут быть на боковой
стороне хелицер (Linyphiidae), на заднем крае карапакса (Theridiidae) и на других
участках тела. Служат для издавания звука. По гребням паук проводит выростами
тазиков или других частей. Трение выростов о бороздки производит звук. Используется
для негомологичных структур.
субадультный (лат. subadultus; англ. subadult) — предимагинальный; возраст (стадия),
предшествующая половозрелой. На этой стадии у самцов уже есть вздутия на конце
педипальп, и у многих самок есть выпуклости над эпигастральной щелью.
субмаргинальный (англ. submarginal) — обычно о полосе, расположенной сразу над
полосой, проходящей по самому краю карапакса (или дорсальной стороны брюшка).
субтерминальный отросток (англ. subterminal apophysis) — расположенный у
вершины, но не на ней, используется для отростка у Araneidae лежащего чуть ниже
эмболюса.
тагма — крупный обособленный отдел тела; у пауков две тагмы — головогрудь и
брюшко.
тазик (лат. и англ. соха/сохае) — самый проксимальный сегмент конечности (ноги или
пальпы).
таксон (англ. taxon/taxa) — здесь используется для систематических групп разного
ранга (видов, групп видов, подродов, родов, семейств и т.д.).
тегулюм (англ. tegulum; от лат. tegulum — крыша) — основная часть бульбуса, к
которой крепятся все отростки и от которой отходит эмболюс.
334
Приложения
тегулярный отросток (англ. tegular apophysis) — отросток, отходящий от тегулюма.
Синоним — медиальный отросток. Используется для негомологичных структур.
терминальный (англ. terminal; от лат. terminalis — конечный, концевой) —
расположенный на вершине (конце) чего-либо. Используется для называния отростков пальпы у
некоторых пауков.
термофильный (англ. termophilous; от греч. thermos — тёплый + греч. philus —
любящий) — теплолюбивый.
трихоботрия (лат. и англ. trichobothrium/trichobothria) — чувствительный волосок,
чётко отличается от всех других тем, что он очень тонкий, торчит перпендикулярно
поверхности членика, выходит из круглой ямки.
тутакулюм (лат. и англ. tutaculum) — обособленная часть цимбима, расположена ретро-
латерально, служит для поддеражания эмболюса в спокойном состоянии, или может
служить для направления эмболюса к отверстию в эпигине. Характерен для
семейства Thomisidae, но встречается и в других группах.
хетотаксия (лат. и англ. chaetotaxy; англ. spination) — число и расположение шипов на
ногах.
цимбиум (англ. от лат. cymbium — лодочка) — верхний членик педипальпы (лапка) у
самцов, к которому крепится бульбус.
шип (англ. spine, macroseta/macrosetae) — утолщённая щетинка (волосок). В
энтомологии шипом называют только неподвижные выросты, но в арахнологии не так.
эмболюс (лат. и англ. embolus; от греч. embolus — металлический нос судна, таран) —
часть совокупительного органа самца, на котором находится отверстие, ведущее в
семенной каналец пальпы самца. Через отверстие спермин засасываются в пальпу, и
впрыскиваются в совокупительное отверстие самки.
эндогина (лат. endogyne; англ. endogyna; от греч. endon — внутри + греч. gyne —
женщина) — внутренняя часть эпигины, в случае гаплогинных пауков — это и есть весь
копулятивный орган самки. То же самое, что и вульва.
энтелегинный (англ. entelegyne; от греч. enteles — законченный, совершенный + греч.
gyne — женщина) — тип устройства копулятивного аппарата, когда у самки есть
эпигина.
Энтелегинные (лат. Entelegynae; англ. entelegyne) — энтелегинные (то есть, не гапло-
гинные) аранеоморфные пауки. Пауки, у которых эпигина имеет две пары канальцев
(протоков) — осеменительные или копуляторные (ведут от совокупительного
отверстия к сперматеке) и оплодоторительные (ведут от сперматеки к матке). Самцы
имеют сложно устроенные пальпы со множеством склеритов (субтегулюм, тегулюм, и
др.), пальпа всегда имеет гематодоху. Эпигина у этих пауков, как правило, с
выростами, углублениями, часто с заметными совокупительными отверстиями. В азиатской
части России — это все аранеоморфные семейства, кроме Pholcidae и Oonopidae.
эпигастральная щель (англ. epigastral furrow; от греч. epi — сверху, над + греч.
gaster — желудок, брюхо, пузо) — поперечная щель (борозда), которая находится
снизу брюшка в передней его части. Спереди-посредине от неё находятся копуля-
тивные органы самки и половое отверстие самца, а по сторонам от них — лёгочные
крышки.
эпигина (лат. и англ. epigyne; от греч. epi — сверху, над + греч. gyne — женщина) —
копулятивный орган самки, находится в средней части эпигастральной щели, имеет
совокупительные отверстия или щели, часто с выростами; эпигина всегда более хи-
тинизирована, нежели соседние участки.
ювенильный (англ. juvenile, immature; от лат. juvenilis — юный) — неполовозрелый.
Указатель латинских названий
Названия ранга отряда выделены жирным шрифтом; названия группы
семейства — обычным шрифтом, группы рода и вида — курсивом. Для родовых
названий в скобках указана принадлежность к семейству, для видовых названий через
запятую указана родовая принадлежность. Жирным выделены страницы, где
начинается описание семейств.
Abacoproeces (Linyphiidae) 170
Abiskoa (Linyphiidae) 170, 309
acalypha, Mangora 115
Acantholycosa (Lycosidae) 30, 184, 186, 309
Acanthoneta (Linyphiidae) 170, 309
Acari 11
Acartauchenius (Linyphiidae) 170
accentuata, Anyphaena 111
Achaearanea (Theridiidae) 242, 309
Aculepeira (Araneidae) 115, 116, 117, 118, 310
acuminus, Paratrachelas 136
Aelurillinae 227
Aelurillus (Salticidae) 224, 310
affinis, Atypus 123
Agalenatea (Araneidae) 115, 310
Agelena (Agelenidae) 100-104, 310
Agelenidae 49, 57, 63, 64, 65, 70, 72, 85,100,
101, 102, 104, 106, 108, 121, 129, 142, 144,
149, 162, 191
Ageleninae 104
Agelenini 104
agelenoides, Arctobius 106, 108
Agelenopsis (Agelenidae) 103, 310
Agnyphantes (Linyphiidae) 170, 310
agrestis, Tegenaria 103
Agroeca (Liocranidae) 126, 130, 178, 179, 180,
310
Agyneta (Linyphiidae) 169, 170, 309, 310
Agyphantes (Linyphiidae) 170, 310
aichinensis, Falcileptoneta 165
Ainerigone (Linyphiidae) 170, 173, 310
Ajmonia (Dictynidae) 149
akiensis, Cybaeus 142
alascensis, Philodromus 207, 208, 309
alaskensis, Agyneta 309
albidus, Xysticus 254
albimaculata, Kishidaia 155
albofasciata, Nurscia 256, 258
Alenatea (Araneidae) 115, 310
algens, Pardosa 186
Allagelena (Agelenidae) 103
Alloclubionoides (Agelenidae, Amaurobiidae)
101, 102, 103, 104,310
Allohogna (Lycosidae) 29, 184, 186, 310
Allomengea (Linyphiidae) 170, 310
Alopecosa (Lycosidae) 14, 183, 184, 186, 187,
308,310
alpicola, Chalcoscirtus 226
altaiensis, Acantholycosa 184
Amaurobiidae 57, 63, 64, 72, 84, 100, 104, 106,
107, 108, 120, 142, 146, 149, 256, 258
Amaurobius (Amaurobiidae) 104, 107, 108, 310,
311
Ambanus (Agelenidae, Amaurobiidae) 310
amurensis, Alloclubionoides 102
Anahita (Ctenidae) 140, 141, 182, 264, 266, 310
Anapidae 65, 244
ancora, Pisaura 219
Anelosimus (Theridiidae) 310
Anguliphantes (Linyphiidae) 170, 174, 310, 315
Antistea (Hahniidae) 162, 310, 316
Anyphaena (Anyphaenidae) 110, 111, 126, 130,
310
Anyphaenidae 52, 63, 64, 70, 72, 83, 110, 111,
126, 130, 154
aquatica, Argyroneta 120, 121
Aphileta (Linyphiidae) 170, 310
Apollophanes (Philodromidae) 208, 209, 210,
310
Aprifrontalia (Linyphiidae) 170, 173, 310
aquilonare, Hybauchenidium 175
Arachosinella (Linyphiidae) 170, 310
Araeoncus (Linyphiidae) 170, 310
Aranea 307, 309
Araneae 307, 326
Araneill,307, 326
Araneidae 50, 60, 63, 64, 71, 72, 84, 88, 90, 94,
96, 112, 113, 114, 116, 117, 168, 190, 191, 192,
200, 232, 233, 236, 238, 244, 246, 260, 262,
268, 269, 270, 279, 326, 332
Araneomorphae 60, 61, 326
Araneus (Araneidae) 71, 112, 113, 114, 115,
116, 117, 118,307,310,317,327
Araniella (Araneidae) 113, 114, 115, 116, 117,
311
Archaeodictyna (Dictynidae) 146, 149, 311
336
Указатель латинских названий
Archaraeoncus (Linyphiidae) 170, 311
Arctachaea (Theridiidae) 241, 242, 311
Arctella (Dictynidae) 146, 149, 311
Arcterigone (Linyphiidae) 170, 309, 311
Arctobiinae 108
Arctobius (Amaurobiidae) 106, 107, 108, 311
Arctosa (Lycosidae) 26, 184, 186, 187, 308, 311
Arcuphantes (Linyphiidae) 170, 311
Argenna (Dictynidae) 147, 148, 149, 311
Argiope (Araneidae) 115, 116, 117, 311, 332
Argiopidae 117, 200
Argyrodes (Theridiidae) 311
Argyroneta (Argyronetidae) 120, 121, 311
Argyronetidae 52, 63, 64, 70, 72, 83, 120, 121,
144, 146
Ariadna (Segestriidae) 274
arietina, Mastigusa 147, 148, 149
Artanes (Philodromidae) 208, 210, 311
arundinacea, Dictyna 149
aryy, Philodromus 209
Asianellus (Salticidae) 223, 224, 311
asiatica, Parasyrisca 156
asiaticus, Tibellus 209
Asiceratinops (Linyphiidae) 170, 309, 311
Asiophantes (Linyphiidae) 170, 309, 311
Asperthorax (Linyphiidae) 170, 311
Asthenargoides (Linyphiidae) 170, 173, 311
Asthenargus (Linyphiidae) 170, 173, 311
astridae, Eriophora 115
Atea (Araneidae) 115, 118, 310, 311
atrata, Rhene 225
atrica, Zygiella 269
Atypidae 29, 52, 61, 63, 64, 66, 69, 72, 82, 122,
123, 124, 326, 329
Atypus (Atypidae) 122, 123, 311
baicalensis, Heliophanus 225
Ballus (Salticidae) 311
Baryphyma (Linyphiidae) 170, 172, 311
basarukini, Cybaeus 143
basarukini, Savignia 171
basarukini, Walckenaeria 171
Bassaniana (Thomisidae) 250, 252, 311
Bassaniodes (Thomisidae) 254
Bathylinyphia (Linyphiidae) 170, 173, 174, 212,
309,311
Bathyphantes (Linyphiidae) 170, 175, 309, 311,
316
Berlandina (Gnaphosidae) 155, 157, 311
Bicluona (Clubionidae) 132, 311
bicolor, Marylina 149
bifasciatus, Eresus 151
bipunctata, Steatoda 239, 240
birulai, Paracoelotes 101, 103
Bishopiana (Linyphiidae) 170, 308, 311
bistriata, Allagelena 103
Bolephthyphantes (Linyphiidae) 170, 311
Bolyphantes (Linyphiidae) 170, 311
borealis, Emblyna 148, 309
brevipes, Howaia 199, 200
bruennichi, Argiope 115
brunneus, Pronoides 71, 114, 115
Bucliona (Clubionidae) 132, 311
bukhkaloi, Dolomedes 219
bungei, Thanatus 208
buryatica, Mongolicosa 184
caerulescens, Clubiona 130
Calamoneta (Cheiracanthiidae, Miturgidae) 129
Calamopus (Cheiracanthiidae, Miturgidae) 129
Callilepis (Gnaphosidae) 75, 154, 157, 311
Callobius (Amaurobiidae) 55, 100, 106, 107,
108, 142,311
Calommata (Atypidae) 124
cambridgei, Его 191
Canalidion (Theridiidae) 242, 311
carbonarioides, Aculepeira 116
Carorita (Linyphiidae) 170, 311
Carrhotus (Salticidae) 224, 311
castrata, Yaginumena 242
Caviphantes (Linyphiidae) 170, 312
celebesiana, Leucauge 235
cellariorum, Oecobius 273
Centromerita (Linyphiidae) 170
Centromerus (Linyphiidae) 170, 175, 312
Cepheia (Synaphridae) 244
Ceraticelus (Linyphiidae) 170, 312
Ceratinella (Linyphiidae) 49, 168, 170, 312
Cercidia (Araneidae) 112, 115, 116, 117, 312
Chalcoscirtus (Salticidae) 78, 224, 226, 312
Cheiracanthidae 129
Cheiracanthiidae 63, 64, 72, 86, 92, 95, 126,
127, 129, 130, 133,322
Cheiracanthium (Cheiracanthiidae) 65, 126,
127, 128, 129, 130, 133, 151,312
Chorizopes (Araneidae) 115, 312
chrysops, Philaeus 223
Chrysso (Theridiidae) 242, 243, 312
cicur, Cicurina 149
Cicurina (Dictynidae) 67, 75, 149, 312
cinnaberinus, Eresus 151, 152
claripes, Phrurolithus 137
claustrarius, Callobius 108
clercki, Pachygnatha 237
Clubiona (Clubionidae) 126, 129, 130, 132,
133, 178,310,312
Clubionidae 58, 63, 64, 65, 72, 87, 92, 95, 104,
110, 111, 126, 129, 130, 132, 133, 138, 141,
154, 158, 179, 226, 230, 266, 274, 310, 322
Cnephalocotes (Linyphiidae) 170, 312
Coelotinae 100, 103, 104, 105, 108, 310
Coelotes (Agelenidae, Amaurobiidae,
Coelotidae) 103, 104, 142, 151, 312, 317
Coleoptera 11
Указатель латинских названий
337
Collinsia (Linyphiidae) 170, 312
Comaroma (Anapidae) 244
comta, Clubiona 132
Concavocephalus (Linyphiidae) 170, 309, 312
concretus, Xysticus 251
Connithorax (Linyphiidae) 170, 312
consecuta, Archaeodictyna 146
constricta, Micaria 157
contortipes, Weintrauboa 217
Coreodrassus (Gnaphosidae) 158
Coriarachne (Thomisidae) 250, 252, 312
Corinnidae 22, 63, 64, 67, 72, 87, 92, 95, 133,
136, 137, 138, 154, 179,264,322
Corinninae 138
cornutus, Larinioides 114, 116
coronatus, Lysiteles 251
corticalis, Clubiona 132
corticicola, Hahnia 161, 162, 163
corticinus, Philodromus 207
coylei, Cretohexura 279
Cretohexura (Mecicobothriidae) 279
Cribellatae 326, 329
Crispiphantes (Linyphiidae) 170, 173, 312
Crocodilosa (Lycosidae) 308
cronebergi, Alopecosa 14
Crosbylonia (Linyphiidae) 170, 308, 312
Crustulina (Theridiidae) 242, 243, 312
Cryphoeca (Hahniidae) 67, 75, 82, 160, 161,
162,312
Cryphoecinae 162
Cryptachaea (Theridiidae) 242, 312
Ctenidae 65, 69, 74, 84, 85, 140, 141, 182, 264,
266
Ctenus (Ctenidae) 141
cultrigera, Zornella 169
cuneata, Alopecosa 183
cupreus, Drassodes 155, 156
Cybaeidae 63, 64, 74, 86, 100, 104, 106, 120,
121,142, 143, 144, 146,322
Cybaeinae 121
Cybaeopsis (Amaurobiidae) 100, 106, 107, 108,
142,312
Cybaeus (Cybaeidae) 104, 142, 143, 144, 312
Cyclocarcina (Nesticidae) 200, 244
Cyclosa (Araneidae) 71, 112, 114, 115, 116,
117, 118,312,332
Cyphepeira (Araneidae) 115, 116
cyprium, Zodarion 277
Cyrba (Salticidae) 226
Dactylopisthes (Linyphiidae) 170, 312
Dactylopisthoides (Linyphiidae) 170, 312
Decipiphantes (Linyphiidae) 170, 312, 315
decorata, Bassaniana 252
Dendryphantes (Salticidae) 224, 308, 312
Dendryphantinae 227
depress a, Coriarachne 252
Devade (Dictynidae) 74, 120, 149
Diaea (Thomisidae) 206, 251, 252, 253, 312
Dictyna (Dictynidae) 147, 148, 149, 312, 313,
330
Dictynidae 49, 63, 64, 67, 74, 83, 104, 106, 108,
120, 121, 144, 146, 147, 148, 149, 160, 162,
191,256,328
Dictyninae 104
Dicymbium (Linyphiidae) 170, 313
Diguetidae 214
dimidiata, Mustelicosa 183, 184
Dingosa (Lycosidae) 308
diodia, Zilla 115
Dionycha 326
Diphya (Tetragnathidae) 67, 80, 233, 235
Diplocentria (Linyphiidae) 170, 313
Diplocephalus (Linyphiidae) 170, 313
Diplostyla (Linyphiidae) 170, 313
Dipoena (Theridiidae) 89, 238, 242, 243, 244,
246,308,313
Dipoeninae 244
Diptera 11
Dismodicus (Linyphiidae) 170, 313
dispar, Parazygiella 269, 270
displicata, Araniella 113, 114
Doenitzius (Linyphiidae) 170, 313
Dolomedes (Pisauridae) 219, 220, 313
Dolomedidae 220
Dolomedinae 220
domestica, Tegenaria 101, 102, 103
Drapetisca (Linyphiidae) 170, 313
Drassodes (Gnaphosidae) 55, 151, 154, 155,
156, 157, 158,313
Drassyllus (Gnaphosidae) 157, 158, 313
Drepanotylus (Linyphiidae) 170, 313
Dubiaraneinae 175
Dyschiriognatha (Tetragnathidae) 236
Dysderidae 65, 214, 275
dzhungaricus, Xysticus 251
Ebelingia (Thomisidae) 252, 313
Ebo (Philodromidae) 208, 313
Eboria (Linyphiidae) 309, 312, 317, 313
Eborilaira (Linyphiidae) 170, 173, 309, 312
Ebrechtella (Thomisidae) 252, 254
eca, Titanoeca 256
Echemus (Gnaphosidae) 157, 312
Ecribellatae 326, 330
Eidmannella (Nesticidae) 244
Eldonnia (Linyphiidae) 170, 312
elegans, Antistea 162
Emblyna (Dictynidae) 147, 148, 149, 313
Enoplognatha (Theridiidae) 50, 89, 238, 239,
240, 241, 242, 244, 245, 268, 313
Entelecara (Linyphiidae) 170, 313
Entelegynae 326, 333
Epeiridae 117
338
Указатель латинских названий
epeiroides, Theridiosoma 246, 247
Eperigone (Linyphiidae) 172, 313
Epibellowia (Linyphiidae) 170, 173, 313
Epigyphantes (Linyphiidae) 170, 313
Epigytholus (Linyphiidae) 170, 313
Episinus (Theridiidae) 238, 242, 243, 313
Episolder (Linyphiidae) 170, 313
Epopino (Nesticidae) 199
Eresidae 49, 63, 64, 69, 74, 83,150, 151, 152, 273
Eresus (Eresidae) 151, 152,313
Erigone (Linyphiidae) 47, 168, 170, 174, 310,
313,315,316,320,321
Erigonella (Linyphiidae) 170, 313
Erigonidae 174
Erigonidium (Linyphiidae) 313
Erigoninae 170, 175, 176
Erigonoploides (Linyphiidae) 170, 313
Erigonoplus (Linyphiidae) 170, 314
Eriophora (Araneidae) 113, 114, 115, 116, 118,
314
Его (Mimetidae) 190, 191, 192, 314
eskovi, Sitticus 223
Eskovia (Linyphiidae) 170, 173, 308, 314
Eskovina (Linyphiidae) 170, 314
Estrandia (Linyphiidae) 170, 314
Euophryinae 227
Euophrys (Salticidae) 59, 222, 223, 224, 226,
314,318
Euryopinae 244
Euryopis (Theridiidae) 242, 243, 244, 314
Eutichurinae 129, 133
Eutichurus (Miturgidae) 129
Evansia (Linyphiidae) 170
Evarcha (Salticidae) 224, 225, 309, 314
Evippa (Lycosidae) 182, 184, 314
extensa, Tetragnatha 236, 237
falcata, Pardosa 183
Falcileptoneta (Leptonetidae) 165, 166
fasciata, Phlegra 223
fasciger, Sitticus 225
fauna, Anahita 140, 141
Fedotovia (Gnaphosidae) 158
Ferchestina (Oonopidae) 202, 203, 314
Filistatidae 65, 214
flmbriatus, Dolomedes 219
Flagelliphantes (Linyphiidae) 170, 314
Floronia (Linyphiidae) 172, 314
foliicola, Dictyna 147, 148
formicaria, Myrmarachne 223
formicinus, Thanatus 207
frontalis, Euophrys 59, 222, 223
Frontella (Linyphiidae) 172, 314
Frontinellina (Linyphiidae) 172, 314
fulgens, Micaria 156
Gibbaranea (Araneidae) 42, 115, 314
gibbosus, Oedothorax 175
Gibothorax (Linyphiidae) 172, 314
glacialis, Hahnia 163
glacialis, Pardosa 183
globosum, Synema 251
glupovi, Mongolicosa 184, 309
Glyphesis (Linyphiidae) 172, 309, 314, 317
Gnaphosa (Gnaphosidae) 40, 49, 55, 154, 156,
157, 158,314
Gnaphosidae 21, 47, 49, 53, 57, 58, 63, 64, 66,
67, 70, 74, 86, 104, 110, 126, 130, 133, 136,
154, 155, 156,226,274,322
Gnathonarium (Linyphiidae) 172, 314
gobiensis, Xysticus 253
golovatchi, Walckenaeria 171
Gonatium (Linyphiidae) 172, 314
Gongylidiellum (Linyphiidae) 172, 314
Gongylidioides (Linyphiidae) 172, 314
Gongylidium (Linyphiidae) 172, 314
gramineusa, Enoplognatha 239, 240
granosus, Eresus 151, 152
griseus, Stemonyphantes
Guizygiellinae 270
Hackmania (Dictynidae) 149, 160, 314
Hadrotarsinae 244
Hahnia (Hahniidae) 160, 161, 162, 163, 314
Hahniidae 22, 49, 50, 52, 63, 64, 67, 70, 74, 82,
104, 129, 160, 161, 162
Hahniinae75, 160, 162
Halorates (Linyphiidae) 314
Haplodrassus (Gnaphosidae) 126, 130, 155,
156, 157, 158,314
Haplogynae 326, 327
Harmochirus (Salticidae) 224
Helicius (Salticidae) 224
Heliophaninae 227
Heliophanus (Salticidae) 224, 225, 226, 314
Helophora (Linyphiidae) 172, 314
Herbiphantes (Linyphiidae) 172, 173, 314
Heriaeus (Thomisidae) 250, 252, 253, 314
herniosa, Hilaira 49
Hersiliidae 65, 152,273
heterophthalmus, Oxyopes 204
Heteropoda (Sparassidae) 228, 314
Heteropodidae 230
Hetewtheridion (Theridiidae) 241, 242, 243, 314
Hilaira (Linyphiidae) 47, 49, 172, 175, 309,
312,314,318
Himalaphantes (Linyphiidae) 172, 173, 315
hokkaido, Callobius 106, 107
Holminaria (Linyphiidae) 172, 308, 315
Horcotes (Linyphiidae) 172, 308, 315
Horodromoides (Philodromidae) 210
horridus, Heriaeus 253
horvathi, Tmarus 251
Howaia (Nesticidae) 198, 199, 200, 315
Указатель латинских названий
339
Hyaenosa (Lycosidae) 308
Hybauchenidium (Linyphiidae) 172, 175, 315
Hygrolycosa (Lycosidae) 58, 184, 315
Hyllinae 227
Hylyphantes (Linyphiidae) 172, 315
Hymenoptera 11
hyperborea, Pardosa 183
hyperboreus, Chalcoscirtus 226, 309
Hypochilidae 214
Hypomma (Linyphiidae) 172, 315
Hypselistes (Linyphiidae) 172, 315
Hypsosinga (Araneidae) 71, 73, 84, 114, 115,
116,315
hyrcana, Rhomphaea 240, 241
Improphantes (Linyphiidae) 172, 315
Incestophantes (Linyphiidae) 172, 315
inclusum, Cheiracanthium 128
insignita, Tricca 184
insularis, Weintrauboa 216, 217
interuna, Iwogumoa 103
Ipa (Linyphiidae) 172, 315
Ipainae 175, 326
ipsa, Triccosta 184
iriei, Falcileptoneta 165
Ishicosa (Lycosidae) 184, 315
ishikariana, Ishicosa 184
Islandiana (Linyphiidae) 172, 315, 318
Ivielum (Linyphiidae) 172, 308, 315
Iwogumoa (Agelenidae) 103, 315
japonicus, Trachelas 136, 137
Japoniona (Clubionidae) 132, 315
jeniseicus, Bathyphantes 309
jobi, Mysmenella 194
Juraraneidae 279
Juraraneus (Juraraneidae) 279
Kaestneria (Linyphiidae) 172, 315
kalkamanicus, Yllenus 225
Karita (Linyphiidae) 172, 315
karpinskii, Anguliphantes 174
kastoni, Robertus 241
kazakhstanicus, Asianellus 223
Keijia (Theridiidae) 242, 315
kerzhneri, Howaia 199
Kikimora (Linyphiidae) 172, 309, 315
Kishidaia (Gnaphosidae) 154, 155, 157, 158, 315
kollari, Eresus 151, 152
kolymensis, Thanatus 208, 209, 309
Kolymocyba (Linyphiidae) 172, 173, 309, 315
kompirensis, Metleucauge 235
kularensis, Clubiona 130
kumadai, Ebelingia 252
kunashirensis, Cybaeus 143
labyrinthica, Agelena 101, 102, 103
laeta, Loxosceles 65
Lagonomegopidae 279
Lagonomegops (Lagonomegopidae) 279
lama, Pisaura 219
lapponica, Arctella 146
Larinia (Araneidae) 115, 118, 315
Larinioides (Araneidae) 114, 115, 116, 118, 315
lasciva, Pardosa 186
Lasaeola (Theridiidae) 242, 243, 315
Lasiargus (Linyphiidae) 172, 315
Lathys (Dictynidae) 69, 74, 146, 147, 148, 149,
160,315
latithorax, Tarsiphantes 49
Latrodectus (Theridiidae) 238, 242, 243, 315
lenzi, Micaria 155
Lepidoptera 11,309,315
Lepthyphantes (Linyphiidae) 46, 171, 172, 175,
285, 308, 310, 311, 312, 313, 314, 315, 319
Leptodrassex (Gnaphosidae) 156, 158, 315
Leptoneta (Leptonetidae) 164, 166, 315
Leptonetidae 22, 29, 31, 63, 64, 65, 69, 74, 82,
164, 165,214
Leptorhoptrum (Linyphiidae) 172, 316
Leucauge (Tetragnathidae) 85, 89, 94, 232, 233,
235,236,237,316
Leucauginae 232, 236
levii, Cercidia 115
licenti, Oxyopes 204, 205
Linyphia (Linyphiidae) 57, 172, 173, 309, 311,
316
Linyphiidae 16, 21, 22, 31, 36, 45, 46, 47, 49,
50, 58, 59, 60, 63, 64, 65, 66, 74, 84, 88, 90, 93,
96, 97, 168, 169, 171, 173, 174, 175, 176, 190,
191, 192, 212, 216, 232, 238, 244, 260, 322,
326,331,332
Linyphiinae 170, 175, 326
Liocranidae 63, 64, 76, 87, 92, 95, 110, 126,
133, 138, 154, 178, 179, 180, 264, 266, 322
Liocraninae 138, 266
Liocranum (Liocranidae) 178
Liphistiomorphae 60, 61
lobata, Argiope 115
logunovi, Perlongipalpus 169
longidens, Tapinopa 173
longipes, Zelotes 156
longispinus, Drassodes 156
Lophomma (Linyphiidae) 47, 171, 172, 316
Loxosceles (Sicariidae) 65
lugubris, Pardosa 184
Lycosa (Lycosidae) 184, 186, 308, 309, 314,
316,318,320
Lycosidae 29, 49, 50, 51. 60, 63, 64, 66. 69, 76,
84, 86, 100, 182, 183, 184. 218. 264. 266, 322,
326
Lynxosa (Lycosidae) 308
Lysiteles (Thomisidae) 251, 252, 316
340
Указатель латинских названий
Lyssomaninae 226
macacus, Araneus 116, 118
Macrargus (Linyphiidae) 172, 316
Macrobuninae 108
macropalpus, Apollophanes 209
magnus, Atypus 122, 123
maior, Bathylinyphia 173, 174
major, Dictyna 148
manchurica, Meta 235
Mangora (Araneidae) 115,316
margarita, Enoplognatha 244
margaritatus, Philodromus 207
marmoreus, Araneus 71, 113, 114
Maw (Linyphiidae) 172, 316
Marpissa (Salticidae) 224, 316
marusiki, Artanes 208
Marylina (Dictynidae) 149
Masikia (Linyphiidae) 172, 316
Maso (Linyphiidae) 172, 316
Mastigusa (Dictynidae) 147-149
Mecicobothriidae 279
Mecynargoides (Linyphiidae) 172, 316
Mecynargus (Linyphiidae) 172, 316
medius, Atypus 122, 123
medvedevi, Yaginumaella 225
Megalepthyphantes (Linyphiidae) 172, 316
memorialis, Leptodrassex 156, 158
Mendoza (Salticidae) 224
mengei, Pelecopsis 175
Mermessus (Linyphiidae) 172, 173
Mesasigone (Linyphiidae) 172, 316
Mesothelae 60
Meta (Tetragnathidae) 94, 117, 232, 233, 235,
236,268,316
Metellina (Tetragnathidae) 94, 232, 233, 236, 316
Metinae 85, 232, 236, 270
Metleucauge (Tetragnathidae) 26, 85, 94, 117,
232,233,235,236,268,316
Metopobactrus (Linyphiidae) 172, 316
Micaria (Gnaphosidae) 57, 67, 75, 133, 136,
154-158,309,316,322
michailovi, Evarcha 225, 309
Micrargus (Linyphiidae) 172, 316
Microlinyphia (Linyphiidae) 172, 173, 316
Micrommata (Sparassidae) 228, 229, 316
Microneta (Linyphiidae) 172, 310, 316, 319
Micronetinae 170, 175, 326.
Micryphantidae 174
Miftengris (Linyphiidae) 172, 173, 316
mikhailovi, Zelotes 309
mildei, Cheiracanthium 128
Mimetidae 22, 63, 64, 71, 76, 84, 88, 90, 93, 96,
190-192, 326
Minicia (Linyphiidae) 172, 316
Minyrioloides (Linyphiidae) 172, 316
Minyriolus (Linyphiidae) 172, 316
mirandus, Sitticus 223, 225
Misumena (Thomisidae) 251, 252, 254, 316
Miturgidae 126, 129, 133
moderatus, Haplodrassus 155, 156
Moebelotinus (Linyphiidae) 172, 316
Mogrinae 227
mongolica, Agroeca 179
mongolica, Xerolycosa 183, 186
Mongolicosa (Lycosidae) 184, 186, 316
Monocerellus (Linyphiidae) 172, 316
monstrabilis, Enoplognatha 238, 241
montanum, Canalidion 242
montanus, Monocerellus 171
Mughiphantes (Linyphiidae) 172, 317
mugur, Xysticus 253
mundus, Zelotes 40
muralis, Atypus 123
muscorum, Gnaphosa 156
Mustelicosa (Lycosidae) 183, 184, 186, 308, 317
mustelina, Phycosoma 239, 240
mysticus, Philodromus 209
Mygalomorphae 60, 61, 317, 329
Mynogleninae 175
Myrmarachne (Salticidae) 222-224, 317
Myrmarachninae 227
Mysmena (Mysmenidae) 195, 317
Mysmenella (Mysmenidae) 30, 194-196, 317
Mysmenidae 22, 30, 63-65, 71, 76, 84, 88, 90,
93, 112, 194, 238, 244, 246, 262, 326
Mysmeninae 196
Nanoa (Pimoidae, Pumiliopimoidae) 217
napolovi, Alloclubionoides 101
navus, Oecobius 272
Nematogmus (Linyphiidae) 172
nemoralis, Zora 265, 266
nenilini, Xysticus 252
nenilini, Zodariellum 211, 276
Nenilinium (Linyphiidae) 172, 308, 317
Neoantistea (Hahniidae) 161, 162, 317
Neon (Salticidae) 224, 226, 317
Neoscona (Araneidae) 115, 116, 308, 317
Neottiura (Theridiidae) 242, 243, 317
Nephilinae 117
Neriene (Linyphiidae) 49, 172, 173, 317
Neserigone (Linyphiidae) 172, 173, 317
Nesticeae 200
Nesticella (Nesticidae) 198-200
Nesticidae 31, 35, 63, 64, 66, 76, 84, 85, 88, 90,
93, 168,198,238,244,326,331
nigerrima, Gnaphosa 157
nigriventris, Tenuiphantes 169
nigrolimbatus, Rugathodes 242
nigrovariegatum, Heterotheridion 241, 242
nipponicus, Stemmops 242
Nippononeta (Linyphiidae) 172, 173, 317
Nispa (Linyphiidae) 172-173, 317
Указатель латинских названий
341
nitidula, Singa 114
nivalis, Titanoeca 258
Nomisia (Gnaphosidae) 157
nordica, Arctachaea 241, 242
nordmanni, Araneus 113
Notiogyne (Linyphiidae) 172, 173, 317
Notioscopus (Linyphiidae) 172, 317, 321
Nuctenea (Araneidae) 115,317
Nurscia (Titanoecidae) 256, 258
oblongus, Tibellus 207
Obscuriphantes (Linyphiidae) 172, 317
obtusa, Tetragnatha 235
Ochyroceratidae 165
Octonoba (Uloboridae) 260-262, 317, 332
Oculocornia (Linyphiidae) 172, 173, 317
Odontodrassus (Gnaphosidae) 157
Oecobiidae 63, 65, 69, 70, 76, 82, 152, 226,
272, 273
Oecobius (Oecobiidae) 77, 226, 272, 273
Oedothorax (Linyphiidae) 172, 175, 317
ohlerti, Ohlertidion 240, 242
Ohlertidion (Theridiidae) 240, 317
Oia (Linyphiidae) 172, 173, 317
Oinia (Linyphiidae) 170, 317
Okhotigone (Linyphiidae) 172, 173, 317
oleatus, Thymoites 239, 240
ononidum, Hahnia 161, 163
onustus, Thomisus 253
Oonopidae 22, 29, 30, 47, 50-52, 63-65, 69, 76,
82,202,203,214,327,330,333
Opisthothelae 60
Orchestininae 203
Oreoneta (Linyphiidae) 169, 172, 317
Oreonetides (Linyphiidae) 49, 172, 175, 317
orites, Gnaphosa 55, 157
ornata, Agroeca 179
ornata, Menosira 235
Oryphantes (Linyphiidae) 172, 317
Ostearius (Linyphiidae) 172, 173, 317
ovtchinnikovi, Pacifanistea 161, 162
Oxyopes (Oxyopidae) 204, 205, 317
Oxyopidae 63, 64, 69, 76, 83, 97, 204, 205
Oxytate (Thomisidae) 206, 251, 252
Ozyptila (Thomisidae) 47, 52, 251, 252, 254,
255,317
Pachygnatha (Tetragnathidae) 94, 232, 233,
235-237,317
Pacifiphantes (Linyphiidae) 172, 173, 309, 317
Pacifanistea (Hahniidae) 161, 162, 317
Paidiscura (Theridiidae) 242, 243, 318
pallasi, 'Araneus" 116
pollens, Paidiscura 242
Palliduphantes (Linyphiidae) 172, 318
palmgreni, "Theridion" 241
Palpimanidae 192
Palpimanoidea 192, 279
palustris, Kikimora 309, 314
palustris, Pardosa 40
Panamomops (Linyphiidae) 172, 318, 132
Paraclubiona (Clubionidae) 132, 318
Paracoelotes (Agelenidae) 100-103, 318
Paraeboria (Linyphiidae) 172, 318
Paraglyphesis (Linyphiidae) 172, 309, 318
parallela, Zora 265
Parasisis (Linyphiidae) 172, 318
Parasteatoda (Theridiidae) 55, 242, 243, 246, 318
Parasyrisca (Gnaphosidae) 75, 126, 130, 155—
158,318
Paratrachelas (Corinnidae) 136, 137, 318
Parawubanoides (Linyphiidae) 172, 318
Parazygiella (Zygiellidae) 269, 270, 318
Pardosa (Lycosidae) 40, 183, 184, 186, 187,
308,318
patula, Argenna 149
Pelecopsis (Linyphiidae) 172, 173, 318
Pellenes (Salticidae) 226, 224, 318
Peponocranium (Linyphiidae) 172, 318
Perlongipalpus (Linyphiidae) 169, 171, 172,
309,318
Perregrinus (Linyphiidae) 172, 318
Perro (Linyphiidae) 172, 318
Phadocedus (Gnaphosidae) 157, 318
phalangioides, Pholcus 58
Philaeus (Salticidae) 223, 224, 318
Philodromidae 63, 64, 66, 78, 86, 92, 95, 191,
206,210,228,250,322
Philodromoides (Philodromidae) 210
Philodromus (Philodromidae) 206, 207, 208,
209,210,309,318
Phintella (Salticidae) 224, 318
Phlattothrata (Linyphiidae) 172, 173, 318
Phlegra (Salticidae) 223, 224, 318
Pholcidae 53, 63, 64, 69, 71, 78, 82, 97, 104,
212,214,215327,333
Pholcus (Pholcidae) 58, 213, 214, 318
Phoroncidia (Theridiidae) 67, 238, 240, 242, 319
Phrurolithinae 138, 179
Phrurolithus (Corinnidae) 136-138, 319
phungiformes, Pholcus 213
Phycosoma (Theridiidae) 239, 240, 242, 243
Phylloneta (Theridiidae) 242
pilula, Phoroncidia 240, 242
Pimoidae 63, 64, 66, 71, 78, 85, 89, 91, 93, 168,
174,216,326
pinicola, Tetragnatha 237
pinipumilis, Perlongipalpus 171, 309
Pirata (Lycosidae) 26, 184, 186, 187. 319
piraticus, Pirata 187, 319
Pireneitega (Agelenidae) 100
Pisaura (Pisauridae) 218-220. 319
Pisauridae 63, 64. 69. 78. 86. 182, 218
342
Указатель латинских названий
Pisaurinae 220
Pisauroidea 205
Pistius (Thomisidae) 252, 253, 319
Pityohyphantes (Linyphiidae) 172, 319
Platnickina (Theridiidae) 242, 243, 319
Plectreuridae 214
Plexippinae 227
Plexippoides (Salticidae) 224
Pocadicnemis (Linyphiidae) 172, 319
Poecilochroa (Gnaphosidae) 157, 319
Poeciloneta (Linyphiidae) 172, 319
Porrhomma (Linyphiidae) 172, 175, 319
Porrhomminae 175
Portia (Salticidae) 226
potteri, Agelenopsis 103
Praestigia (Linyphiidae) 172, 319
Procerocymbium (Linyphiidae) 172, 309, 319
Proislandiana (Linyphiidae) 172, 319
prominens, Cercidia 112, 115, 117
prominula, Hackmania 149
Pronoides (Araneidae) 115, 116
proszynskii, Euophrys 226
Proxysticus (Thomisidae) 254
Psammitis (Thomisidae) 254
Pseudeuophrys (Salticidae) 224, 319
Pseudicius (Salticidae) 224, 319
Pseudocyba (Linyphiidae) 172, 309, 319
Pseudohilaira (Linyphiidae) 172, 309, 319
pseudomixta, Pardosa 183
Pseudoporrhomma (Linyphiidae) 172, 319
Pseudowubana (Linyphiidae) 172, 319
Psilochorus (Pholcidae) 214
pugil, Anyphaena 110, 111
punctorium, Cheiracanthium 128
quadratus, Araneus 115, 116
quadrimaculata, Pachygnatha 235
quelpartensis, Neoantistea 161, 162
ramosus, Oxyopes 204, 205
rasnitsyni, Juraraneus 279
reclusa, Clubiona 132
Rhene (Salticidae) 222, 224, 229, 319
Rhomphaea (Theridiidae) 238, 240-243, 319
Rhysodromus (Philodromidae) 210
riparia, Clubiona 130, 132
riparia, Cryptachaea 242
Robertus (Theridiidae) 89, 168, 238-242, 319
rostratus, Xysticus 253
RTA-clade 63, 326, 331
rubrofasciata, Hygrolycosa 58, 184
Rugathodes (Theridiidae) 242, 319
Runcinia (Thomisidae) 252
ryvkini, Saloca 171, 309
Saaristoa (Linyphiidae) 172, 319
sacer, Scotonotylus 171
sachalinensis, Eriophora 113-115
Sachaliphantes (Linyphiidae) 172, 173, 319
sagana, Rhomphaea 242
Saloca (Linyphiidae) 171, 172, 319
Salticidae 21, 47, 49-51, 53, 54, 60, 63, 64, 66,
69, 78, 82, 133, 222, 223, 226, 227, 279, 322
Salticus (Salticidae) 222, 224, 319
sanguinea, Hypsosinga 71, 114
Satilatlas (Linyphiidae) 172, 319
Sauron (Linyphiidae) 172, 173, 319
Savignia (Linyphiidae) 171, 172, 319
sayanensis, Acantholycosa 184
scabricula, Ozyptila 251
Scandichrestus (Linyphiidae) 169, 172, 319
schmidti, Dictyna \A1
schmidti, Zodariellum 277
schrencki, Araneus 71
Sciastes (Linyphiidae) 172, 319
Scotargus (Linyphiidae) 172, 319
Scotinotylus (Linyphiidae) 171, 172, 319
Scotophaeus (Gnaphosidae) 157
scylla, Neoscona 115
Scytodidae 65, 327
secunda, Tegecoelotes 103
Segestria (Segestriidae) 274, 275, 319
Segestriidae 30, 63, 65, 69, 78, 82, 274, 275,
279, 327
selengensis, Pardosa 184
Selenopidae 210
Semljicola (Linyphiidae) 172, 320
senilis, Dolomedes 219
senoculata, Segestria 274, 275
Sernokorba (Gnaphosidae) 154, 157
serratosignata, Enoplognatha 239, 241
sharlaa, Xysticus 253
sia, Yaginumia 269, 270
Sibianor (Salticidae) 224, 320
sibirica, Evippa 184
sibirica, Sibirocosa 184
sibirica, Titanoeca 256
sibiricum, Theridion 239-240
sibiricus, Robertus 241
sibiricus, Xysticus 252
Sibirocosa (Lycosidae) 184, 186, 309, 320
Sibirocyba (Linyphiidae) 172, 320
Sicariidae 65, 327
Silometopoides (Linyphiidae) 172, 320
Silometopus (Linyphiidae) 172, 320
silvicola, Cryphoeca 161, 162
silvicultrix, Cyphepeira 115
similis, Gnaphosa 40, 156
Simithidion (Theridiidae) 320
simoni, Psilochorus 214
simulans, Parasteatoda 55
Singa (Araneidae) 71, 114, 115, 117, 320
sinicus, Phrurolithus 137
Указатель латинских названий
343
Sisicottus (Linyphiidae) 172, 173, 320
Sisicus (Linyphiidae) 172, 320
Sitticus (Salticidae) 223-226, 320
Sosticus (Gnaphosidae) 157, 320
Sparassidae 63, 64, 71, 78, 86, 92, 95, 133, 206,
210,228,230
Sparassinae 230
spinimana, Zora 265, 266
Spiracme (Thomisidae) 254
Steatoda (Theridiidae) 59, 238-240, 242-244,
320
Stemmops (Theridiidae) 242, 243, 320
Stemonyphantes (Linyphiidae) 169, 171, 172,
174,216,320
Stemonyphantinae 175
sticta, Gnaphosa 158
storozhenkoi, Ferchestina 202, 203
Strandella (Linyphiidae) 172, 173, 320
striata, Falcileptoneta 165
striatipes, Oxytate 251, 252
stroemi, Stroemiellus 115, 269, 270
Stroemiellus (Araneidae) 115, 116, 269, 270
Styloctetor (Linyphiidae) 172, 320
subdola, Diaea 252, 253
subterraneus, Zelotes 157
subtilis, Clubiona 130, 132
Suffusiphantes (Linyphiidae) 172, 320
sukatchovae, Lagonomegops 279
Summacanthium (Cheiracanthiidae) 129
suspiciosa, Diaea 251
Symphytognathidae 169
Synageles (Salticidae) 224, 320
Synagelides (Salticidae) 224, 226, 320
Synaphosus (Gnaphosidae) 158
Synaphridae 244
Synema (Thomisidae) 251, 252, 254, 320
Synotaxidae 244
taepaikensis, Haplodrassus 156
Takayus (Theridiidae) 242, 243, 320
Talavera (Salticidae) 79, 222, 224, 320
Tallusia (Linyphiidae) 172, 320
tanasevitchi, Trachelas 136
Tanasevitchia (Linyphiidae) 171, 172, 308, 320
Tapinocyba (Linyphiidae) 172, 309, 318, 320
Tapinocyboides (Linyphiidae) 172, 320
Tapinopa (Linyphiidae) 172, 173, 320
Taranucnus (Linyphiidae) 172, 320
Tarsiphantes (Linyphiidae) 49, 172, 320
Tegecoelotes (Agelenidae) 102, 103, 320
Tegenaria (Agelenidae) 100-103, 320
Tegenariinae 63
Tegenariini 104
Telemidae 165
Tenuiphantes (Linyphiidae) 169, 172, 308, 320
tepidariorum, Parasteatoda 55
terrestris, Coelotes 103, 104
Tessamoro (Linyphiidae) 172, 173, 320
Tetragnatha (Tetragnathidae) 50, 80, 94, 133,
212, 232, 233, 235-237, 321
Tetragnathidae 26, 53, 63, 64, 67, 71, 80, 84, 85,
89, 91, 94, 95, 104, 112, 117, 133, 168, 191,
212, 232, 236, 238, 262, 268, 270, 326, 330
Tetragnathinae 29, 53, 80, 85, 200, 232, 233,
236, 237, 330
Thaleria (Linyphiidae) 172, 321
Thanatus (Philodromidae) 206, 207, 208, 210,
321
Theoneta (Linyphiidae) 172, 321
Theridiidae 22, 47, 49, 51, 54, 58-60, 63, 64,
66, 67, 71, 80, 84, 85, 89, 91, 93, 96, 112, 129,
168, 190-192, 196, 198, 238, 244-246, 268,
322,326,331,332
Theridion (Theridiidae) 129, 194, 198, 238-
244-246, 321
Theridiosoma (Theridiosomatidae) 26, 246-248,
321
Theridiosomatidae 63, 64, 80, 85, 89, 91, 93,
112, 194, 112, 194, 238, 244, 246, 262, 326
Thomisidae 47, 51, 63, 64, 66, 80, 86, 92, 95,
206, 210, 226, 250, 251, 254, 255, 333
Thomisus (Thomisidae) 252, 253, 321
Thymoites (Theridiidae) 168, 239, 240, 242, 321
Thyreosthenius (Linyphiidae) 172, 321
Tibellus (Philodromidae) 206-210, 321
Tibiaster (Linyphiidae) 172, 321
Tibioploides (Linyphiidae) 172, 309, 321
Tibioplus (Linyphiidae) 172, 309, 321
Tiso (Linyphiidae) 172, 321
Titanoeca (Titanoecidae) 172, 321, 208, 256,
258, 321
Titanoecidae 21, 35, 63, 80, 84, 106, 146, 256,
326
Tmarus (Thomisidae) 25, 252, 282, 308, 321
Tmeticus (Linyphiidae) 172, 321
torva, Dipoena 242
Trachelas (Corinnidae) 67, 136-138, 321
Trachyzelotes (Gnaphosidae) 158, 321
tredecimguttatus, Latrodectus 242
Trematocephalus (Linyphiidae) 172, 321
Tricca (Lycosidae) 184, 321
Triccosta (Lycosidae) 184, 321
Trichobactrus (Linyphiidae) 172
Trichoncus (Linyphiidae) 172, 321
Trichopterna (Linyphiidae) 172
tricuspidata, Ebrechtella 252
Trionycha 326
tripunctata, Micaria 156
Trochosa (Lycosidae) 184, 186, 321
Trogloneta (Mysmenidae) 244
Troxochrus (Linyphiidae) 173. 321
Tubercithorax (Linyphiidae) 173. 321
tuberosus, Oedothorax 175
344
Указатель латинских названий
Tuberta (Hahniidae) 321
Tunagyna (Linyphiidae) 173, 321
turbatrix, "Lepthyphantes" 171
Tusukuru (Linyphiidae) 173, 321
Tuvadrassus (Gnaphosidae) 157, 321
Tuvaphantes (Salticidae) 224, 308, 309, 321
Typhochrestinus (Linyphiidae) 173, 321
Typhochrestoides (Linyphiidae) 173, 321
Typhochrestus (Linyphiidae) 173, 322
typica, Cybaeopsis 106, 108
tyshchenkoi, Dictyna 330
tyshchenkoi, Parasyrisca 157
ubsunurica, Berlandina 155
Uloboridae 63, 64, 69, 80, 83, 168, 238, 260,
262, 322, 332
Uloborinae 262
Uloborus (Uloboridae) 260, 261, 322
Ummeliata (Linyphiidae) 173, 322
undulatus, Pistius 253
ungulatus, Robertus 239, 240
uralensis, Tanasevitchia 171
Uralophantes (Linyphiidae) 173, 322
Uroctea (Oecobiidae) 272, 273
Urozelotes (Gnaphosidae) 157, 322
Ussurigone (Linyphiidae) 173, 322
utotchkini, Philodromus 209
Uusitaloia (Linyphiidae) 173, 308, 322
vaccinii, Lophomma 171
vachoni, Xysticus 252
vaginatus, Oreonetides 49
variegatus, Araneus 115, 117
vatia, Misumena 251, 252
venatoria, Heteropoda 228, 229
ventricosus, Araneus 118
Vermontia (Linyphiidae) 173, 322
video, Dactilopisthes 175
Viktorium (Linyphiidae) 173, 322
vinokurovi, Philodromus 209
virescens, Micrommata 228, 229
Wabasso (Linyphiidae) 173, 322
Wadicosa (Lycosidae) 187
Walckenaeria (Linyphiidae) 171, 173, 322
Walckenaerianus (Linyphiidae) 173, 322
walckenaerius, Uloborus 260-262
wangi, Emblyna 147, 148
washingtoni, Araneus 113
Weintrauboa (Pimoidae) 65, 216, 217, 322
Wendilgarda (Theridiosomatidae) 248
Whymperiphantes (Linyphiidae) 172, 322
Wiehlenarius (Linyphiidae) 172, 322
wrangeliana, Dictyna 330
Wubanoides (Linyphiidae) 172, 322
x-notata, Zygiella 269
Xerolycosa (Lycosidae) 182-184, 186, 322
Xysticus (Thomisidae) 251-255, 322
Yaginumaella (Salticidae) 224-226, 322
yaginumai, Araniella 113
Yaginumena (Theridiidae) 241, 243, 322
Yaginumia (Zygiellidae) 269, 270, 322
Yakutopus (Linyphiidae) 173, 322
yakutsaxatilis, Agyneta 169
yeniseica, Micaria 309
yesoensis, Octonoba 260-262
Yllenus (Salticidae) 224, 225,
Zelotes (Gnaphosidae) 40, 156-158, 309, 322
Zelotini 74, 154
Zerogone (Linyphiidae) 173, 322
zichyi, Pholcus 214
Zilla (Araneidae) 115,322
Zodariellum (Zodariidae) 52, 276, 277
Zodariidae 52, 63, 65, 70, 80, 82, 104, 129, 152,
276
Zodarion (Zodariidae) 129, 276, 277
Zodarium (Zodariidae) 129
Zora (Zoridae) 66, 141, 182, 264-266, 322
Zoridae 58, 63, 64, 80, 87, 92, 95, 140, 141,
179, 182,264,266
Zorinae 266
Zornella (Linyphiidae) 50, 168, 169,173, 322
Zoropsidae 65
Zoropsis (Zoropsidae) 266
Zygiella (Zygiellidae) 269, 270
Zygiellaea 270
Zygiellidae 63, 64, 80, 84, 89, 91, 94, 96, 112,
115,232,268,326
Zygiellinae 217
Книги Товарищества научных изданий КМК
БИОЛОГИЯ
СЕРИЯ «ОПРЕДЕЛИТЕЛИ ПО ФЛОРЕ И ФАУНЕ РОССИИ»
Деревья и кустарники зимой. 2-е изд. [Вып.9]. Е.Т. Валягина-Малютина. 2007. 268 с.
Формат 170 х 240 мм. Те. перепл. — Цена 220 руб. — Булавоусые чешуекрылые
Восточной Европы. [Вып.8]. А.Л. Львовский, Д.В. Моргун. 2007. 443 с, 8 цв.вкл.
Формат 170 х 240 мм. Те. перепл. — Цена 400 руб. — Флора Северо-Западного Кавказа.
[Вып.7]. А.С. Зернов. 2006. 664 с. Формат 170x240 мм. Те. перепл. —Цена 500руб. —
Флора Нижнего Поволжья. Том 1 (споровые, голосеменные, однодольные). [Вып.6].
А.К. Скворцов (ред.). 2006. 435 с. Формат 170 х 240 мм. Те. перепл. — Цена 400 руб.
Планируется: Е.Т. Валягина-Малютина. Деревья и кустарники средней полосы
европейской части России (иллюстрированный определитель).
ПРОЧИЕ ОПРЕДЕЛИТЕЛИ ПО ФЛОРЕ И ФАУНЕ
Атлас молоди пресноводных рыб России. А.П. Макеева и др. 2011. 383 с, ил.
Формат 215 х 300 мм. В обл. — Цена 700 руб. — Краткий определитель мохообразных
Подмосковья. Е.А. Игнатова и др. 2011. 272 с, ил., 48 цв. вкл. Формат 165 х 240 мм.
В обл. — Цена 350 руб. — Иллюстрированные определители свободноживущих
беспозвоночных евразийских морей и прилегающих глубоководных частей
Арктики. Том 2. Немертины, головохоботные, малощетинковые черви, пиявки,
погонофоры, эхиуры, сипункулы, форониды, плеченогие. В.И. Сиренко (ред.). 2010. 186
с, ил. Формат 170 х 240 мм. Те. перепл. — Цена 300 руб. — Определитель
зоопланктона и зообентоса пресных вод Европейской России. Том 1. Зоопланктон. В.Р.
Алексеев (ред.). 2010. 495 с, ил. Формат 170x240 мм. Те. перепл. — Цена 600 руб. —
Флора и фауна Белого моря. Иллюстрированный атлас. А.Б. Цетлин и др. (ред.).
2010. 471 с, цв. фотоатлас. Формат 175 х 245 мм. Те. перепл. — Цена 1500 руб. —
Растения Российского Западного Кавказа. Полевой атлас. А.С. Зернов. 2010. 448
с, цв. фотоатлас. Формат 155 х 220 мм. Те. перепл. — Цена 600 руб. — Стрекозы
Восточной Европы и Кавказа: Атлас-определитель. В.Э. Скворцов. 2010. 623 с, ил.
Формат 205 х 145 мм. Тв. перепл. — Цена 400 руб. — Определитель наземных
моллюсков лесостепи Правобережного Поволжья. Т.Г. Стойко, О.В. Булавкина. 2010.
96 с, ил., 17 цв.табл. Формат 165 х 235 мм. В обл. — Цена 220 руб. —
Иллюстрированные определители свободноживущих беспозвоночных евразийских морей и
прилегающих глубоководных частей Арктики. Том 1. Коловратки, морские пауки,
ракообразные: усоногие, тонкопанцирные, эуфаузииды, неполнохвостные, мизи-
ды, капреллиды. Б.И. Сиренко (ред.). 2009. 189 с, ил. Формат 170 х 240 мм. Тв.
перепл. — Цена 300 руб. —Атлас дикорастущих растений Ленинградской области.
2010. 664 с, цв. фотоатлас. Бум. мелов. Формат 130 х 170 мм. Тв. перепл. — Цена
600руб. — Определитель сосудистых растений Оренбургской области. З.Н. Ряби-
нина, М.С. Князев. 2009. 758с, ил. Формат 170x240мм. Тв. перепл. —Цена 600руб.
— Растения средней полосы Европейской России. Полевой атлас. И.А. Шанцер. 3-
е изд. 2009. 470 с, цв. фотоатлас. Бум. мелов. Формат 130 х 170 мм. Тв. перепл. —
Цена 500 руб. — Флора водоемов Волжского бассейна. Определитель
сосудистых растений. Л.И. Лисицына и др. 2009. 219 с, ил. Формат 143 х 210 мм. В обл. —
Цена 150 руб. — Определитель грибов России. Порядок Афиллофоровые. Вып.З.
И.В. Змитрович. 2008. 278 с. Формат 145 х 218 мм. Тв. перепл. — Цена 300 руб. —
Определитель сосудистых растений Соловецкого архипелага. К.В. Киселёва и др.
2004. 175 с, цв. фото. Формат 170 х 240 мм. Тв. перепл. — Цена 220 руб. —
Иллюстрированное руководство для ботанических практик и экскурсий в Средней
России. В.Э. Скворцов. 2004. 506 с. Формат 170 х 240 мм. Тв. перепл. — Цена 300 руб. —
Иллюстрированный определитель растений Средней России. Том 3. И.А. Губанов
и др. 2004. 520 с. Формат 210 х 295 мм. Тв. перепл. — Цена 350 руб. Том 2
(распродан; готовится 2-е издание) — Том 1. 2002. 526 с. с портр. — Цена 350 руб. — Флора
мхов средней части Европейской России. Том 2. М.С. Игнатов, Е.А. Игнатова. 2004.
С. 609-944. Бум. мелов. Формат 195x270 мм. Тв. перепл. — Цена 300 руб. Том 1. 2003.
С. 1-608, илл. — Цена 500руб. — Определитель грибов России. Дискомицеты. Вып.1.
Копротрофные виды. В.П. Прохоров. 2004. 255 с. Формат 145 х 218 мм. Те. перепл.
— Цена 200 руб.
Планируется: Ан.А. Шилейко, Т.С. Рымжанов. Фауна наземных моллюсков Казахстана и
сопредельных территорий.
СЕРИЯ «РАЗНООБРАЗИЕ ЖИВОТНЫХ»
Повелители бездны [Вып.5]. АН. Островский. 2009. 216 с, 16 с. цв. вкл. Формат 145
х 205 мм. — Цена 220 руб. — Мамонт [Вып.З]. А.Н. Тихонов. 2005. 90 с, цв. вкл.
Формат 145 х 205 мм. — Цена 100 руб. — Гидра: от Абраама Трамбле до наших дней
[Вып.1]. С.Д. Степаньянц и др. 2003. 101 с. + цв.вкл. Формат 145 х 205 мм. — Цена
100руб.
Планируется: А.А. Островский. Подводный мир (в печати).
УЧЕБНИКИ ДЛЯ ВУЗОВ
Общая арахнология. Краткий курс. Часть 1. Введение. Малые отряды. К.П
Михайлов. 2011. 65 с. Формат 145 х 203 мм. В обл. — Цена 60руб. — Протистология:
Руководство. К. Хаусман и др. Пер. с англ. 2010. 495 с, ил. Формат 170 х 240 мм. Те.
перепл. — Цена 600 руб. — Жизнь на дне. Био-экология и био-география бентоса.
И.А. Жирков. 2010. 453 с, ил. Формат 170 х 240 мм. Те. перепл. — Цена 600 руб. —
Морская биогеоценология. И.В. Бурковский. 2006. 285 с. Формат 170 х 240 мм. Те.
перепл. — Цена 220 руб. — Основы микологии (морфология и систематика грибов
и грибоподобных организмов). Л.В. Гарибова, С.Н. Лекомцева. 2005. 220 с. Формат
170 х 240 мм. Те. перепл. — Цена 220 руб. — Малый практикум по зоологии
беспозвоночных. Часть 1. 2-е изд. И.А. Тихомиров и др. 2008. 304 с, 14 ч/б вкл. Формат 170
х 240 мм. Те. перепл. — Цена 250 руб. — Основы биогеографии. В.Г. Мордкович.
2005. 236 с, 1 цв. вкл. Формат 170 х 240 мм. Те. перепл. — Цена 220 руб.
Планируется: И.А. Тихомиров. Малый практикум по зоологии беспозвоночных (часть 2).
СЕРИЯ «СОВРЕМЕННАЯ ОТЕЧЕСТВЕННАЯ БИОЛОГИЯ»
Экология и биогеография. Избранные работы. Ю.И. Чернов. 2008. 580 с. Формат
170 х 240 мм. Те. перепл. — Цена 500 руб. — Избранные труды. Н.В. Кокшайский.
2008. 410 с. с портр. Формат 170 х 240 мм. Те. перепл. — Цена 400 руб. —
Избранные труды. В.В. Кучерук. 2006. 523 с. с портр. Формат 170 х 240 мм. Те. перепл. —
Цена 300 руб. — Избранные труды. Е.Н. Матюшкин. 2005. 658 с. с портр. Формат
170 х 240 мм. Те. перепл. — Цена 400 руб. — Избранные труды по эволюционной
биологии. А.П. Расницын. 2005. /V + 347 с. с портр., 16 фототаблиц. Формат 170 х 240
мм. Те. перепл. — Цена 250 руб. — Избранные труды. Организм, геном, язык. В.М.
Медников. 2005. 452 с. с портр. Формат 170 х 240 мм. Те. перепл. — Цена 300 руб.
Планируется: СМ. Разумовский. Избранные труды.
НАУЧНО-ПОПУЛЯРНЫЕ ИЗДАНИЯ
Невероятная зоология. В.Н. Танасийчук. 2-е изд. 2011. 372 с, в обл. Формат 140 х
213 мм. — Цена 220 руб. — Моя биосфера: научно-художественное изложение
биосферных знаний. ПИ. Флёрова. 2010. 247 с, ил. Формат 170 х 240 мм. Те. перепл. —
Цена 300 руб. — Киты и история китобойного промысла: взгляд из Японии. М.
Комацу С. Мисаки. 2005. 142 с, в те. перепл., цв. вкл. Формат 145 х 215 мм. — Цена
200 руб.
Планируется: В.Н. Танасийчук. Цокотуха ли муха.
СПРАВОЧНЫЕ ИЗДАНИЯ, ИСТОРИЯ БИОЛОГИИ
Столовые корнеплоды в России. В.И. Леунов. 2011. 272 с, ил., цв. книга. В те.
перепл. Формат 210 х 290 мм. — Цена 900 руб. — Промысловые, перспективные для
промысла и кормовые беспозвоночные Российских морей. Э.Н. Егорова, Б.И.
Сирен ко. 2010. 285 с, 8 вкл., 16 цв. вкл. В те. перепл. Формат 170 х 240 мм. — Цена 350
руб. — Методы исследования сообществ микроартропод. М.Б. Потапов, Н.А.
Кузнецова. 2011. 84 с, ил., 4 цв. вкл., в обл. Формат 145 х 215 мм. — Цена 60 руб. —
Методические подходы к экологической оценке лесного покрова в бассейне
малой реки. Л.Б. Заугольнова, Т.Ю. Браславская (отв. ред.). 2010. 383 с, ил., 14 цв. вкл.,
в те. перепл. Формат 170 х 240 мм. — Цена 500 руб. — Биологическая систематика:
эволюция идей. И.Я. Павлинов, Г.Ю. Любарский. Сб. тр. Зоол. музея МГУ. Том 51.
2011. 667 с, в те. перепл. Формат 170 х 240 мм. — Цена 600 руб. — Как невозможно
жили мы. В.В. Меншуткин (сост.). 2011. 219 с, 11 с. вкл., в обл. Формат 140 x200 мм.
— Цена 270 руб. — Каталог типовых образцов сосудистых растений Восточной
Азии, хранящихся в Гербарии Ботанического института им. В.Л. Комарова (LE).
часть 2 (Китай). А.Е. Грабовская-Вородина (отв. ред.). 2010. 517 с, в те. перепл.
Формат 150 х 210 мм. — Цена 500 руб. — Культурные растения Главного
ботанического сада им. Н.В. Цицина Российской академии наук: 60 лет интродукции. А.С.
Демидов (отв. ред.). 2011. 511 с, 50 цв. вкл., в те. перепл., суперобл. Формат 170 х
240 мм. — Цена 700 руб. — Растительные ресурсы России. Дикорастущие
цветковые растения, их компонентный состав и биологическая активность. Том 3. А.Л.
Вуданцев (отв. ред.). 2010. 601 с, в те. перепл. Формат 170 х 240 мм. — Цена 600
руб. — Большая энциклопедия цветочных многолетников. М.М. Диев. 2011. 515 с,
цв. фотоатлас. Формат 210 х 295 мм. — Цена 1500 руб. — «Философия зоологии»
Жана-Батиста Ламарка: взгляд из XXI века. AM. Шаталкин. 2009. 606 с, ил., в те.
перепл. Формат 150 х 220 мм. — Цена 500 руб. — Справочник по семеноведению.
В. Вроувер, А. Штелин. Пер. с. нем. 2010. 694 с. в те. перепл. Формат 210 х 290 мм. —
Цена 1100 руб. — Эволюция и систематика: Ламарк и Дарвин в современных
исследованиях. Сб. тр. Зоол. музея МГУ. Том 50. А.В. Свиридов, А.И. Шаталкин (отв.
ред.). 2009. 366 с, 40 с. вкл., в те. перепл. Формат 150 х 220 мм. — Цена 300 руб. —
Растительные ресурсы России. Дикорастущие цветковые растения, их
компонентный состав и биологическая активность. Том 2. А.Л. Вуданцев (отв. ред.). 2009.
513 с, в те. перепл. Формат 170 х 240 мм. — Цена 600 руб. — История
Зоологического музея МГУ: Идеи, люди, структуры. Г.Ю. Любарский. 2009. 744 с, ил., в те.
перепл. Формат 210 х 290 мм. — Цена 900 руб. — Вид и видообразование. Анализ
новых взглядов и тенденций. (Тр. Зоол. ин-та РАН, Прил. № 1). 2009. 298 с, в те.
перепл. Формат 210 х 267 мм. — Цена 500 руб. — Международный кодекс
ботанической номенклатуры. 2009. 282 с, в те. перепл. Формат 145 х 220 мм. — Цена 300
руб. — Виды и формы хвойных, культивируемые в России. Часть 2. Picea, Thuja.
Д.Л. Матюхин и др. 2009. 288 с, бум. мелов., цв. фото. Те. пер. Формат 170 х 240 мм.
— Цена 700 руб. — Виды и формы хвойных, культивируемые в России. Часть 1.
Juniperus, Cephalotaxus, Taxus, Torreya. 2-е изд. Д.Л. Матюхин и др. 2009. 259 с,
бум. мелов., цв. фото. Те. пер. Формат 170 х 240 мм. — Цена 700 руб. — Учёные в
вихре времени. Воспоминания об орнитологах, защитниках природы и других
натуралистах. Е. Новак. Пер с нем. 2009. 473 с, в те. перепл. Формат 150 х 207 мм.
— Цена 500 руб. — Каталог б йоты Беломорской биологической станции МГУ. А.В.
Чесунов и др. (ред.). 2008. 384 с, в те. перепл. Формат 170 х 240 мм. — Цена 500 руб.
— Страна ББС. Е. Каликинская. 2008. 534 с, в те. перепл. Формат 170 х 245 мм. —
Цена 500 руб. — Каталог чешуекрылых (Lepidoptera) России. СЮ. Синев (ред.).
2008. 424 с, в те. перепл. Формат 220 х 290 мм. — Цена 620 руб. — Растительные
ресурсы России. Дикорастущие цветковые растения, их компонентный состав и
биологическая активность. Том 1. А.Л. Вуданцев (отв. ред.). 2008. 421 с, в те.
перепл. Формат 170 x240 мм. — Цена 500 руб. — Введение в палеоэнтомологию. В.В.
Жерихин и др. 2008. 371 с, в те. перепл. Формат 170 х 240 мм. — Цена 300 руб. —
Мировая коррелятивная база почвенных ресурсов: основа для международной
классификации и корреляции почв. Пер. с англ. 2007. 278 с, в обл. Формат 145 х
215 мм. — Цена 150 руб. — Мозаика судеб биофаковцев МГУ 1930-1960 годов
поступления. Том II. 1951-1960-е годы. Л.И. Лебедева (сост.).2007. 640 с, те. перепл.
Формат 145 х 220 мм. — Цена 300 руб. — Том I. 1930-1950-е годы. 2007. 479 с, те.
перепл. Формат 145 х 220 мм. — Цена 200 руб. — Ваш любящий Валя. Валентин
Александрович Догель (1882-1955). Письма домой. СИ. Фокин (ред.). 2007. 266 с, +
40 с, ил., в обл. Формат 145 х 210 мм. — Цена 150 руб. — Эрнст Майр и
современный эволюционный синтез. Э.И. Колчинский. 2006. 149 с, ч/б вкл., в обл. Формат
145 х 215 мм. — Цена 100 руб. — Александр Николаевич Формозов: Жизнь
русского натуралиста. А.А. Формозов. 2006. 208 с, в обл. Формат 135 х 203 мм. — Цена 100
руб. — Пока горит свеча... Очерки по истории кафедры зоологии беспозвоночных МГУ.
2-е изд. В.В. Малахов. 2006. 153 с, бум. мелов., в обл. Формат 145 х 210 мм. — Цена
120 руб. — Конспект фауны земноводных и пресмыкающихся России. СП.
Кузьмин, Д.В. Семенов. 2006. 139с, в те. перепл. Формат 145x210мм. — Цена 150руб.—
Список птиц Российской Федерации. Е.А. Коблик и др. 2006. 281 с, бум. мелов.,
печать двухцветная, в обл. Формат 145 х 215 мм. — Цена 220 руб. — Биологический
факультет МГУ. А.И. Нетрусов и др. (ред.). 2005. 242 с, в те. перепл. Формат 170x240
мм. — Цена 220 руб. — Эволюционные факторы формирования разнообразия
животного мира. Э.И. Воробьева, Б.Р. Стриганова (ред.). 2005. 308 с, в те. перепл.
Формат 170 х 240 мм. — Цена 220 руб. — Каталог типовых образцов сосудистых
растений Восточной Азии, хранящихся в Гербарии Ботанического института им. В.Л.
Комарова (LE). часть 1 (Япония и Корея). В.И. Грубое (ред.). 2004. 188 с, ил. Формат 140
х 205 мм. В обл. — Цена 80 руб. — Биология гидротермальных систем. А.В. Гебрук
(ред.). 2002. 543 с. с цв. вкл., в те. перепл. Формат 210 х 260 мм. — Цена 400 руб.
Планируется: В.Б. Голуб, М.Н. Цуриков, А.А. Пронин. Коллекции насекомых: сбор,
обработка и хранение материала. — В. Хенниг. Филогенетическая систематика (пер. с англ.).
НАУЧНЫЕ МОНОГРАФИИ
Новости систематики высших растений. Том 42. Н.Н. Цвелёв (отв. ред.). 2011. 250
с. Формат 145 х 213 мм. Те. перепл. — Цена 300руб. — Ботанические сады в
современном мире: теоретические и прикладные исследования. А.С.Демидов (отв. ред.).
2011. 765 с, ил. Формат 210 х 295 мм. Те. перепл. — Цена 900 руб. —
Восстановление и мониторинг природной флоры. Б.Р. Стриганова, А.А. Маслов (отв. ред.). 2010.
116с, ил. Формат 145 х 210 мм. В обл. — Цена 100 руб. — Онтогенетическая
систематика и новая модель эволюции Bilateria. А.В. Мартынов. 2011. 286 с, ил., 16 цв.
вкл. Формат 170 х 240 мм. Те. перепл. — Цена 350 руб. — Конспект фауны жуков
южной тайги Западной Сибири (в бассейне Нижнего Иртыша). СП. Бухкало и др.
2011. 267 с. Формат 145 х 215 мм. В обл. — Цена 180 руб. — Орхидные северо-
запада Европейской России (Ленинградская, Псковская, Новгородская области).
П.Г. Ефимов.'2011. 211 с, цв. фотоатлас. Формат 170 х 240 мм. Те. перепл. — Цена
600 руб. — Сравнительная анатомия, структурные преобразования и адаптивная
эволюция аппарата двуногой локомоции птиц. А.В. Зиновьев. 2010. 285 с, ил., 2 цв.
блока. Формат 210 х 290 мм. Те. перепл. — Цена 500 руб. — Особенности
экспонирования коллекций декоративных растений. Вып. 2. А.С. Демидов (отв. ред.). 2011.
128 с, ил. Формат 140x213 мм. В обл. — Цена 100 руб. — Серебряноборское
опытное лесничество: 65 лет мониторинга. Б.Р. Стриганова, А.А. Сирин (отв. ред.). 2010.
260 с, ил., 8 цв. вкл., 6 карт. Формат 170 х 240 мм. Те. перепл. — Цена 400 руб. —
Тараканы. Биология, экология, санитарно-эпидемиологическое значение,
контроль численности синантропных тараканов. В.П. Дремова, Н.А. Алешо. 2011. 306
с, ил. Формат 145x210 мм. В обл. — Цена 300 руб. — Панцирные клещи Монголии.
Б. Баяртогтох. 2010. 371 с, ил. Формат 21 Ох 290мм. Те. перепл. — Цена 500руб. —
Миграция воробьиных птиц: остановки и полёт. Н.С. Чернецов. 2010. 173 с, ил.
Формат 170 х 240 мм. Те. перепл. — Цена 220 руб. — Новости систематики низших
растений. Том 44. А.Д. Потёмкин (отв. ред). 2010. 378 с, ил. Формат 150 х 220 мм.
Те. перепл. — Цена 300 руб. — Диатропика, эволюция и систематика. Ю.В.
Чайковский. 2010. 407 с, ил. Формат 150 х 215 мм. Те. перепл. — Цена 400 руб. — Роденто-
логия. П.А. Пантелеев. 2010. 221 с, ил. Формат 145 x220 мм. Те. перепл. — Цена 360
руб. —Ускользающий мир: экологические последствия утраты местообитаний. И.
Хански. Пер. с англ. 2010. 340 с, ил. Формат 170 х 240 мм. Те. перепл. — Цена 400
руб. — Калужская флора. ИМ. Решетникова и др. 2010. 548 с, ил., 212 с. цв. вкл.
Формат 170 х 240 мм. Те. перепл. — Цена 1000 руб. — Сообщества и популяции
животных: экологический и морфологический анализ (Тр. Ин-та систематики и
экологии животных СО РАН. Вып. 46). В.Н. Большаков (отв. ред.). 2010. 256 с, ил.
Формат 150х 220 мм. Те. перепл. — Цена 300 руб. — Структура и динамика
зоопланктона Рыбинского водохранилища. В.И. Лазарева. 2010. 183 с, ил. Формат 150 х 220
мм. Те. перепл. — Цена 300 руб. — Растительность полуострова Камчатка. В.Ю.
Нешатаева. 2009. 537 с, ил., 16цв.вкл. Формат 170 x240 мм. Те. перепл. —Цена 600
руб. — Бета-казоморфины и их роль в регуляции поведения. В.А. Дубынин, А.А.
Каменский. 2010. 306 с, ил. Формат 145 х 221 мм. Те. перепл. — Цена 300 руб. —
Историческая экология озёрных зооценозов. Н.Н. Смирнов. 2010. 225 с, ил.
Формат 145 х 221 мм. Те. перепл. — Цена 220 руб. — Экология кустарниковой улитки
Fruticicola fruticum. Д.В. Зейферт, И.М. Хохуткин. 2010. 91 с, ил. Формат 140x203
мм. В обл. — Цена 100 руб. — Новости систематики высших растений. Том 41. Н.Н.
Цвелёв (отв. ред.). 2009. 341 с. Формат 145x213 мм. Те. перепл. — Цена 300 руб. —
Северный олень. Управление поведением и популяциями. Оленеводство. Охота.
Л.М. Баскин. 2009. 284 с, ил. Формат 21 Ох 290мм. В обл. — Цена 350руб. — Новости
систематики низших растений. Том 43. АД. Потёмкин (отв. ред). 2009. 409 с, ил.
Формат 150х 220 мм. Те. перепл. — Цена 300 руб. — Гомологическая изменчивость
морфологических структур и эпигенетическая дивергенция таксонов: основы
популяционной мерономии. А.Г. Васильев, И.А. Васильева. 2009. 511 с, ил. Формат
150x220 мм. Те. перепл. — Цена 500руб. —Жуки-листоеды подсемейства Alticinae
Индокитая. Л.Н. Медведев. 2009. 224 с, ил. Формат 170 х 240 мм. На англ. яз. Те.
перепл. — Цена 300 руб. — Флора и фитогеография печеночников Камчатки и
прилегающих островов. В.А. Бакалин. 2009. 367 с, ил., 24 цв.вкл. Формат 170 х 240 мм.
Те. перепл. — Цена 400 руб. — Змеи Кавказа. Б. С. Туниев и др. 2009. 223 с, ил., 80 цв.
вкл. Формат 220 х 290 мм. Те. перепл. — Цена 900руб. — Личинки жуков-листоедов
России. Ю.М. Зайцев, Л.Н. Медведев. 2009. 246 с, ил. Формат 170 х 240 мм. Те.
перепл. — Цена 300 руб. —Динамика населения диких копытных России: гипотезы,
факторы, закономерности. А.А.Данилкин. 2009. 310 с, ил. Формат 170x240 мм. Те.
перепл. — Цена 300 руб. — Структура и функции лесов Европейской России. 2009.
389 с, ил. Формат 145 х 220 мм. Те. перепл. — Цена 300 руб. — Acrothoracica,
сверлящие ракообразные. Г.А. Колбасов. 2009. 452 с, ил. Формат 145 х 220 мм. Те.
перепл. — Цена 500 руб. — Конспект флоры Кавказа. Том 3(1). А.Л. Тахтаджян (отв.
ред.). 2008. 469 с. Формат 170 х 240 мм. Те. перепл. — Цена 600 руб. —
Сравнительная карпология семейства Urticaceae. Т.Н. Кравцова. 2009. 368 с, ил., 32 с. прил., 2
цв.вкл. Формат 170 х 240 мм. Те. перепл. — Цена 500 руб. — Животные глинистой
полупустыни Заволжья (конспекты фаун и экологические характеристики). А.А.
Тишков (отв. ред.). 2009. 164 с, ил. Формат 145 x220 мм. Те. перепл. — Цена 220 руб.
Аннотированный список жесткокрылых насекомых Центральной Мещеры. В.Б.
Семёнов. 2009. 168 с. Формат 140 х 200 мм. В обл. — Цена 100 руб. — Высшие бази-
диомицеты лесных и луговых систем Жигулей. В.Ф. Малышева, Е.Ф. Малышева.
2008. 342 с, ил., 8 с. цв. вкл. Формат 145 х 220 мм. Те. перепл. — Цена 300 руб. —
Новости систематики высших растений. Том 40. Т.В. Егорова (отв. ред.). 2008. 365
с. Формат 148 х 213 мм. Те. перепл. — Цена 300 руб. — Экосистема эстуария реки
Невы: биологическое разнообразие и экологические проблемы. 2008. 477 с.
Формат 170 х 240 мм. Те. перепл. — Цена 500 руб. — Пойменные луга Северной
Монголии. Часть 1. (Тр. Совм. Росс.-Монг. компл. биол. экспед. Том 49). 2008. 240 с, 16
цв.вкл., 2 цв. карты. Формат 170 х 240 мм. Те. перепл. — Цена 300 руб. — Звери
Алтая. Часть 1. Крупные хищники и копытные. Г.Г. Собанский. 2008. 414 с, 16 цв.вкл.
Формат 170 х 240 мм. Те. перепл. — Цена 400 руб. — Растения Центральной Азии.
Вып.146. В.И. Грубое (отв. ред.). 2008. 223 с. Формат 164 х 238 мм. В обл. — Цена
220 руб. — Виды и сообщества в экстремальных условиях. Сборник,
посвященный 75-летию академика Юрия Ивановича Чернова. А.Б. Бабенко и др. (ред.). 2009.
494 с, ил., 16 цв. вкл. Бум. офсетн. Формат 170 х 240 мм. Те. перепл. — Цена 500 руб.
— Экология океанического обрастания в пелагиали. И.Н. Ильин. 2008. 241 с, ил.
Формат 145 х 220 мм. Те. перепл. — Цена 220 руб. — Семязачаток цветковых
растений. И.И. Шамров. 2008. 350 с, ил., 8 ч/б вкл. Бум. офсетн. Формат 170 х 240 мм. Те.
перепл. — Цена 300 руб. — Цитологические карты политенных хромосом и
некоторые морфологические особенности кровососущих мошек России и
сопредельных стран. Атлас. Л.А. Чубарева, Н.А. Петрова. 2008. 135 с. + 218 с. ил. Бум. офсетн.
и мелов. Формат 170 х 240 мм. Те. перепл. — Цена 400 руб. — Сцелиониды Палеар-
ктики. Подсемейство Scelioninae. СВ. Кононова, М.А. Козлов. (Определители по
фауне, издав. Зоол. ин-том РАН, вып. 172). 2008. 489 с, ил. Формат 170 х 240 мм. Те.
перепл. — Цена 600 руб. — Новости систематики низших растений. Том 41. МЛ.
Андреев (отв. ред.). 2007. 355 с, ил. Формат 150 х 220 мм. Те. перепл. — Цена 220
руб. —Демографическая структура и популяционная динамика певчих птиц. В.А.
Паевский. 2008. 235 с, ил., цв.вкл. Формат 170 х 240 мм. Те. перепл. — Цена 300 руб.
— Хомячки рода Phodopus: систематика, филогеография, экология, физиология,
поведение, химическая коммуникация. Н.Ю. Феоктистова. 2008. 414 с, ил., цв.вкл.
Формат 150 х 215 мм. Те. перепл. — Цена 500 руб. — Эволюция экосистем Европы
при переходе от плейстоцена к голоцену (24 - 8 тыс. л.н). А.К. Маркова и др. 2008.
556 с, цв., ил. Формат 170 х 240 мм. Те. перепл. — Цена 500 руб. — Пространствен-
но-этологическая структура популяций грызунов. B.C. Громов. 2008. 581 с, ил.
Формат 150 х 220 мм. Те. перепл. — Цена 500 руб. — Веслоногие ракообразные отряда
Harpacticoida Белого моря: морфология, систематика, экология. П.Н. Корнев, Е.С.
Чертопруд. 2008. 379 с, ил., с портр. Формат 170 х 240 мм. Те. перепл. — Цена 300
руб. —Абрикос в Москве и Подмосковье. А.К. Скворцов, Л.А. Крамаренко. 2007. 188
с, ил, 32 цв. вкл. Формат 145 х 220 мм. Те. перепл. — Цена 200 руб. — Активный
связный мир. Опыт теории эволюции жизни. Ю.В. Чайковский. 2008. 726 с, ил.
Формат 150 х 220 мм. Те. перепл. — Цена 350 руб. — Исследования по ихтиологии
и смежным дисциплинам на внутренних водоёмах в начале XXI века (к 80-летию
профессора Л.А. Кудерского). 2007. 645 с, ил., с цв. портр. Формат 170 х 240 мм.
Те. перепл. — Цена 500 руб. — Флора сосудистых растений Таймыра и
сопредельных территорий. Часть 1. Аннотированный список флоры и её общий анализ. Е.Б.
Поспелова, И.Н. Поспелов. 2007. 457 с, ил. + 16 с. цв.вкл., вложенный лазерный диск.
Формат 170 х 240 мм. Те. перепл. — Цена 400 руб. — Зимовка и
холодоустойчивость муравьев на северо-востоке Азии. Д.И. Берман и др. 2007. 261 с, ил. + 28 с.
цв.вкл. Формат 170 х 240 мм. Те. перепл. — Цена 300 руб. — Паразитизм как форма
симбиотических отнощений. В.А. Ройтман, С.А. Беэр. 2008. 310 с, ил. Формат 170
х 240 мм. Те. перепл. — Цена 300 руб. — Синаптонемный комплекс - индикатор
динамики мейоза и изменчивости хромосом. Ю.Ф. Богданов, О.Л. Коломиец. 2007.
358 с, ил. Формат 170 х 235 мм. Те. перепл. — Цена 300 руб. — Пространственная
экология почвенных животных. АД. Покаржевский и др. 2007. 174 с, ил. Формат
170 х 240 мм. Те. перепл. — Цена 220руб. — Коллекция культур базидиомицетов LE
(БИН). Каталог штаммов. Н.В. Псурцева и др. 2-е изд. 2007. 116 с, цв. и ч/б вкл. 16 с.
Формат 165 х 235 мм. В обл. На англ. яз. — Цена 120 руб. — Сосудистые растения
Карельского перешейка (Ленинградская область). А/О. Доронина. 2007. 574 с, ил.
Формат 170 х 240 мм. Те. перепл. — Цена 500 руб. — Экосистема малой реки в
изменяющихся условиях среды. А.В. Крылов, А.А. Бобров (отв. ред.). 2007. 372 с,
ил. + 8 с. цв. вкл. Формат 170 х 240 мм. Те. перепл. — Цена 400 руб. — Дыхание
почвенных беспозвоночных. Ю.Б. Вызова. 2007. 328 с. Формат 150 х 220 мм. Те.
перепл. — Цена 220 руб. — Новости систематики высших растений. Том 39. Т.В.
Егорова (отв. ред.). 2007. 370 с. Формат 148 х 213 мм. Те. перепл. — Цена 220 руб. —
Теоретические и практические проблемы изучения сообществ беспозвоночных:
памяти Я.И. Старобогатова. А.И. Кафанов (отв.ред.). 2007. 306 с. с портр. Формат
145 х 220 мм. Те. перепл. — Цена 220 руб. — Растения Центральной Азии. Вып.16.
В.И. Грубое (отв. ред.). 2007. 135 с. Формат 164 х 238 мм. В обл. — Цена 150 руб. —
Паразиты рыб озера Байкал (фауна, сообщества, зоогеография, история
формирования). О. Т. Русинек. 2007. 571 с, ил. + 24 с. цв. вкл. Формат 170 х 240 мм. Те.
перепл. — Цена 500 руб. — Новости систематики низших растений. Том 40. МЛ.
Андреев (отв. ред.). 2006. 339 с, ил. Формат 150 х 220 мм. Те. перепл. — Цена 220
руб. — Состояние и проблемы продукционной гидробиологии. К 100-летию проф.
Г.Г. Винберга. 2006. 329 с, ил., портр. Формат 150 х 220 мм. Те. перепл. — Цена 220
руб. — Микокалициевые грибы Голарктики. А.Н. Титов. 2006. 296 с, ил. + 40 с. цв.
вкл. Формат 170 х 240 мм. Те. перепл. — Цена 220 руб. — Биология морских
нематод. А.В. Чесунов. 2006. 367 с, ил. Формат 170 х 240 мм. Те. перепл. — Цена 220 руб.
— Пресноводные раковинные амебы. Ю.А. Мазей, А.Н. Цыганов. 2006. 300 с, ил.
Формат 170 х 240 мм. Те. перепл. — Цена 220 руб. — Позвоночные животные
северо-востока центрального региона России. В.А. Зайцев. 2006. 513 с, ил., цв. вкл.
Формат 170 х 240 мм. Те. перепл. — Цена 300 руб. — Семейство гвоздичные во
флоре Кыргызстана. ГА. Лазьков. 2006. 272 с, ил. Формат 170 x240 мм. Те. перепл.
— Цена 220 руб. — Новости систематики высших растений. Том 38. ТВ. Егорова
(отв. ред.). 2006. 377 с. Формат 148 х 213 мм. Те. перепл. — Цена 220руб. —
Адаптации хальцидоидных наездников к паразитированию на ложнощитовках в
условиях различных широт. Е.С. Сугоняев, Н.Д. Войнович. 2006. 263 с. Формат 170 х 240
мм. Те. перепл. — Цена 220 руб. — Гепатоидные кожные железы млекопитающих.
С.А. Шабадаш, О.Ф. Чернова. 2006. 217 с. Формат 170 х 240 мм. Те. перепл. — Цена
220 руб. — Эволюция биосферы и биоразнообразия. К 70-летию А.Ю. Розанова.
СВ. Рожнов (отв. ред.). 2006. 600 с. с портр. Формат 170 х 240 мм. Те. перепл. —
Цена 400 руб. — Флора субарктических гор Евразии и высотное распределение её
видов. В.Б. Куваев. 2006. 568 с, ч/б вкл. Формат 170 х 245 мм. Те. перепл. — Цена
400 руб. — Пластинчатоусые жуки подсемейства Scarabaeinae фауны России и
сопредельных стран. О.Н. Кабаков. 2006. 374 с, цв. вкл. Формат 170 х 240 мм. Те.
перепл. — Цена 400 руб. — Атлас-определитель усоногих раков надсемейства
Chthamaloidea Мирового океана. О.П. Полтаруха. 2006. 198 с. Формат 170 х 240
мм. Те. перепл. — Цена 200 руб. — Растения Центральной Азии. Вып.15. В.И. Грубое
(отв. ред.). 2006. 143 с. Формат 164 х 238 мм. В обл. — Цена 150 руб. —
Млекопитающие Вьетнама. Г.В. Кузнецов. 2006. 420 с. Формат 170 х 240 мм. Те. перепл. — Цена
400 руб. — Нематоды надсемейства Drilonematoidea - паразиты дождевых
червей. С.Э. Спиридонов, Е.С. Иванова. 2005. 296 с. Формат 170 х 240 мм. Те. перепл. —
Цена 220 руб. — Бделлоидные коловратки фауны России. Л.А. Кутикова. (Тр. Зоол.
ин-та РАН, т.305). 2005. 315 с. Формат 170 х 240 мм. Те. перепл. — Цена 220 руб. —
Кариотипы паразитических перепончатокрылых. В.Е. Гохман. 2005. 185 с, бум.
офсетн. и мелов. Формат 150 х 220 мм. Те. перепл. — Цена 120 руб. — Ископаемые
цветковые растения. Том 4. Nyctaginaceae - Salicaceae. Л.Ю. Буданцев (ред.). 2005.
466 с, бум. офсетн. и мелов. Формат 228 х 295 мм. Те. перепл. — Цена 600 руб. —
Китайская восковая пчела на Дальнем Востоке России. В.Н. Кузнецов. 2005. 111 с,
бум. мелов., цв. фото. Формат 148 х 215 мм. В обл. — Цена 100 руб. — Животное
население почв бореальных лесов Западно-Сибирской равнины. Б.Р. Стригано-
ва, ИМ. Поряди на. 2005. 234 с. Формат 170 х 240 мм. Те. перепл. — Цена 220 руб. —
Лишайники - индикаторы радиоактивного загрязнения. Л.Г. Бязров. 2005. 476 с.
Формат 150 х 220 мм. Те. перепл. — Цена 220 руб. — Полорогие А.А. Данилкин.
(серия «Млекопитающие России и сопредельных регионов»). 2005. 550 с, цв. вкл.
Формат 170 х 240 мм. Те. перепл. — Цена 400 руб. — Земноводные российского
Дальнего Востока. С.Л. Кузьмин, И.В. Маслова. 2005. 434 с, цв. вкл. Формат 170 х 240 мм.
Те. перепл. — Цена 300 руб. — Трансформационная типологическая систематика.
Б.П. Захаров. 2005. 164 с. Формат 145 х 210 мм. В обл. — Цена 100 руб. — Ветвисто-
усые ракообразные отряда Ctenopoda мировой фауны (морфология, систематика,
экология, зоогеография). Н.М. Коровчинский. 2004. 410 с. Формат 170 х 240 мм. Те.
перепл. —Цена 300руб. — Атлас волос млекопитающих. Тонкая структура остевых
волос и игл в сканирующем электронном микроскопе. О.Ф. Чернова, Т.Н. Целико-
ва. 2004. 429 с, бум. мелов. Формат 170 х 240 мм. Те. перепл. — Цена 400 руб. —
Зоогенная дефолиация и лесное сообщество. Е.Н. Иерусалимов. 2004. 263 с.
Формат 148 х 213 мм. Те. перепл. — Цена 150 руб. — Морфогенез и эволюция. В.Г.
Черданцев. 2003. 360 с. Формат 145 х 205 мм. Те. перепл. — Цена 200 руб. — Голово-
хоботные черви Мирового Океана (Определитель морской фауны). А.В. Адрианов,
В.В. Малахов. 1999. 328 с, бум. мелов. Формат 205 х 285 мм. В обл. — Цена 220 руб.
Планируется: Птицы России и сопредельных регионов. Веслоногие, аистообразные, фла-
мингообразные. — Л.Г. Вартапетов (отв. ред.). Птицы Сибири: структура и динамика
фауны, населения и популяций (в печати). — Л.А. Кудерский. Избранные труды. Том 1.
Исследования по ихтиологии, рыбному хозяйству и смежным дисциплинам (в печати). —
Б. Баяртогтох. Фауна и экология панцирных клещей Монголии (в печати).
Заказать эти и другие издания (география, история, медицина) изд-ва КМК можно по
адресу: 123100 Москва, а/я 16 изд-во КМК, Михайлову Кириллу Глебовичу
Комп. почта: mikhailov2000@gmail.com
Интернет: http://avtor-kmk.ru (аннотаиии изданиях <-,'-
Факс: (495) 629-4825
Тел. (495) 692-5894 раб.
Научное издание
МАРУСИК Юрий Михайлович, КОВБЛЮК Николай Михайлович
Пауки (Arachnida, Aranei) Сибири и Дальнего Востока России.
Москва: Товарищество научных изданий КМК. 2011. 344 с.
При участии ИП Михайлова КГ.
Верстка: Св.В. Найденко, К.Г. Михайлов
Для заявок:
123100, Москва, а/я 16, Издательство КМК
электронный адрес mikhailov2000@gmail.com
http://avtor-kmk.ru
Подписано в печать 25.07.2011. Тираж 400 экз.
Формат 70x100/16. Объём 22 п.л. Уч.-изд.л. 28,6. Бумага мелованая.
Отпечатано в ООО «Галлея-Принт», Москва, 5-я Кабельная ул., 56