/
















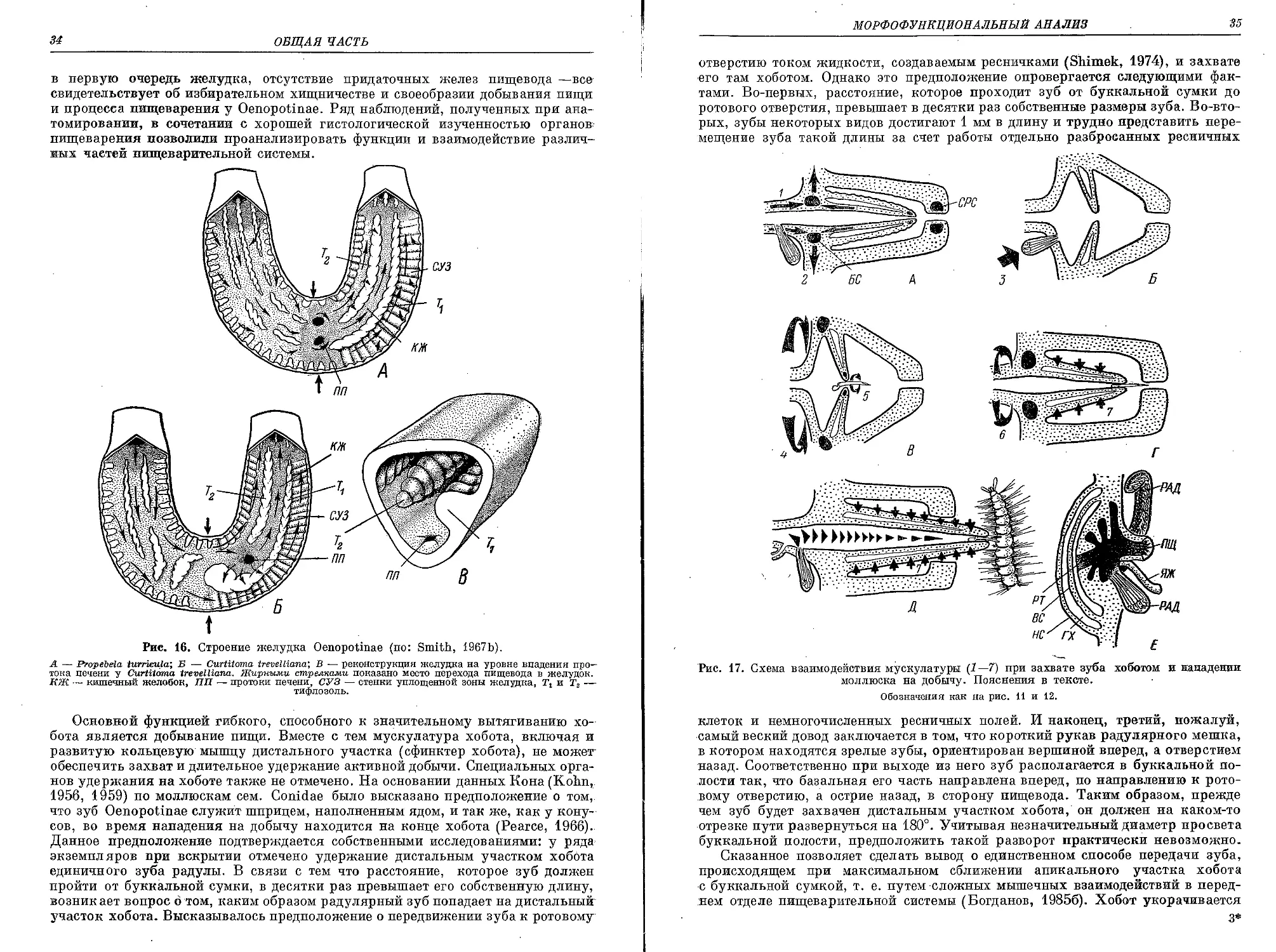






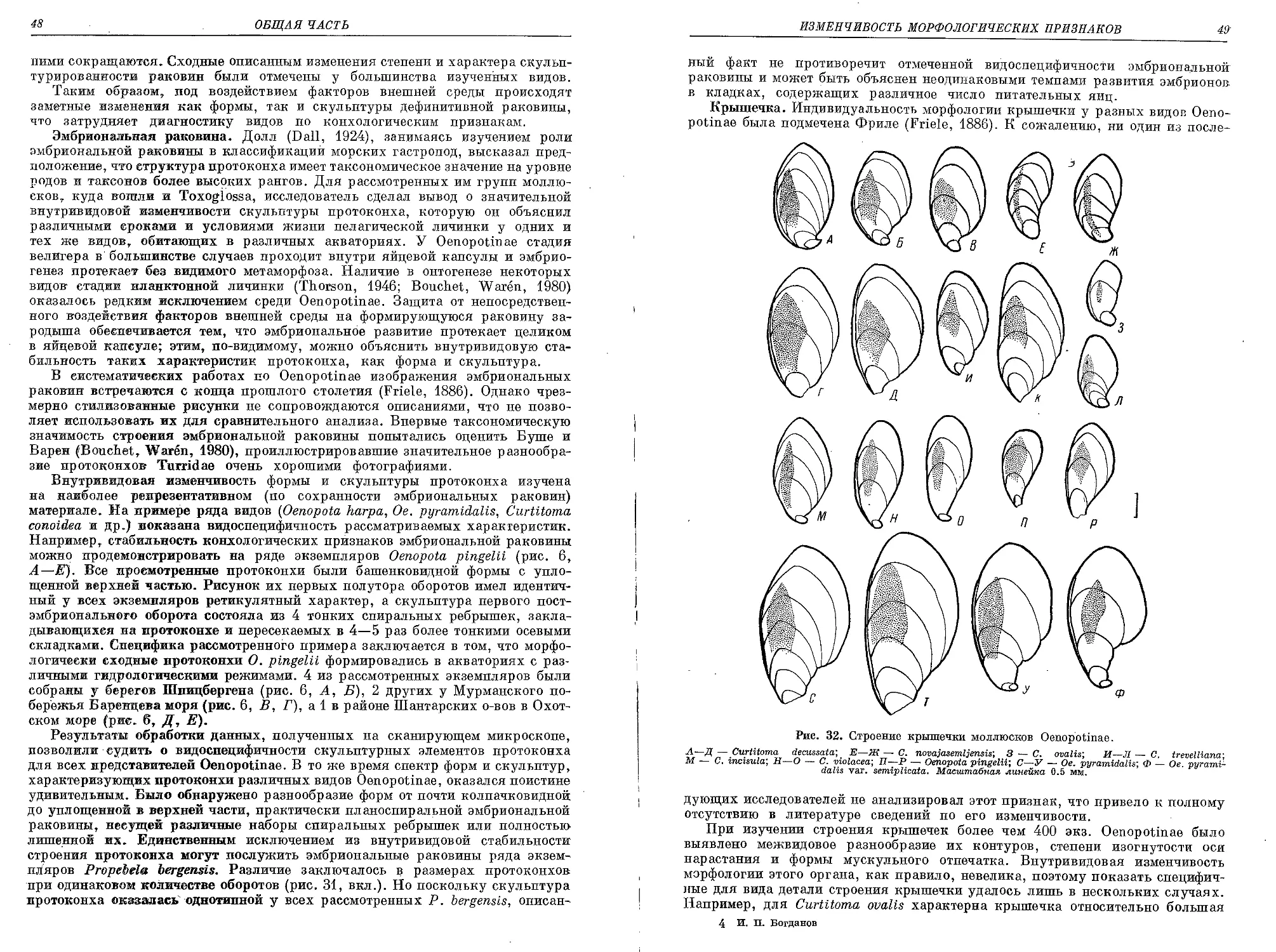
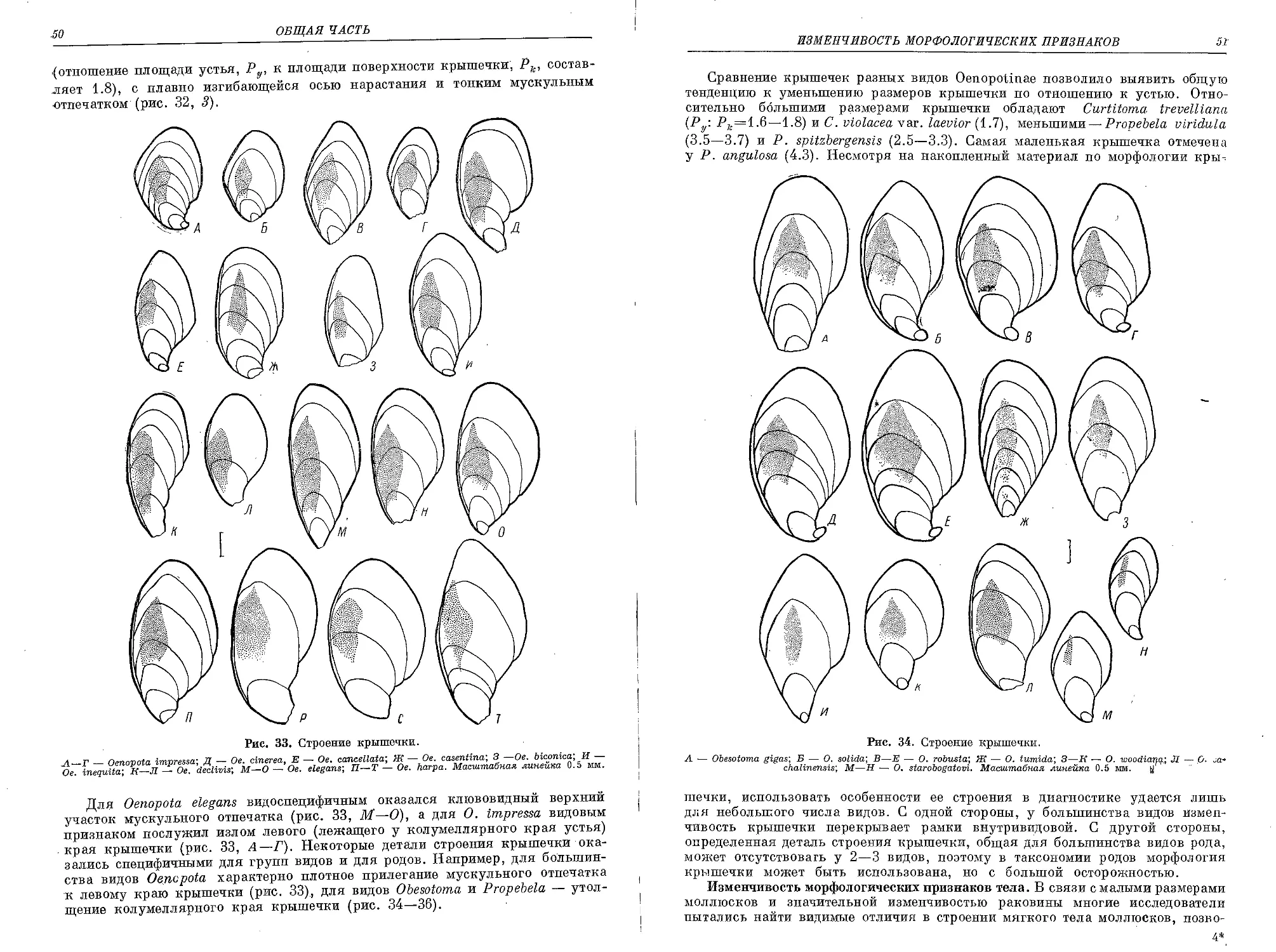



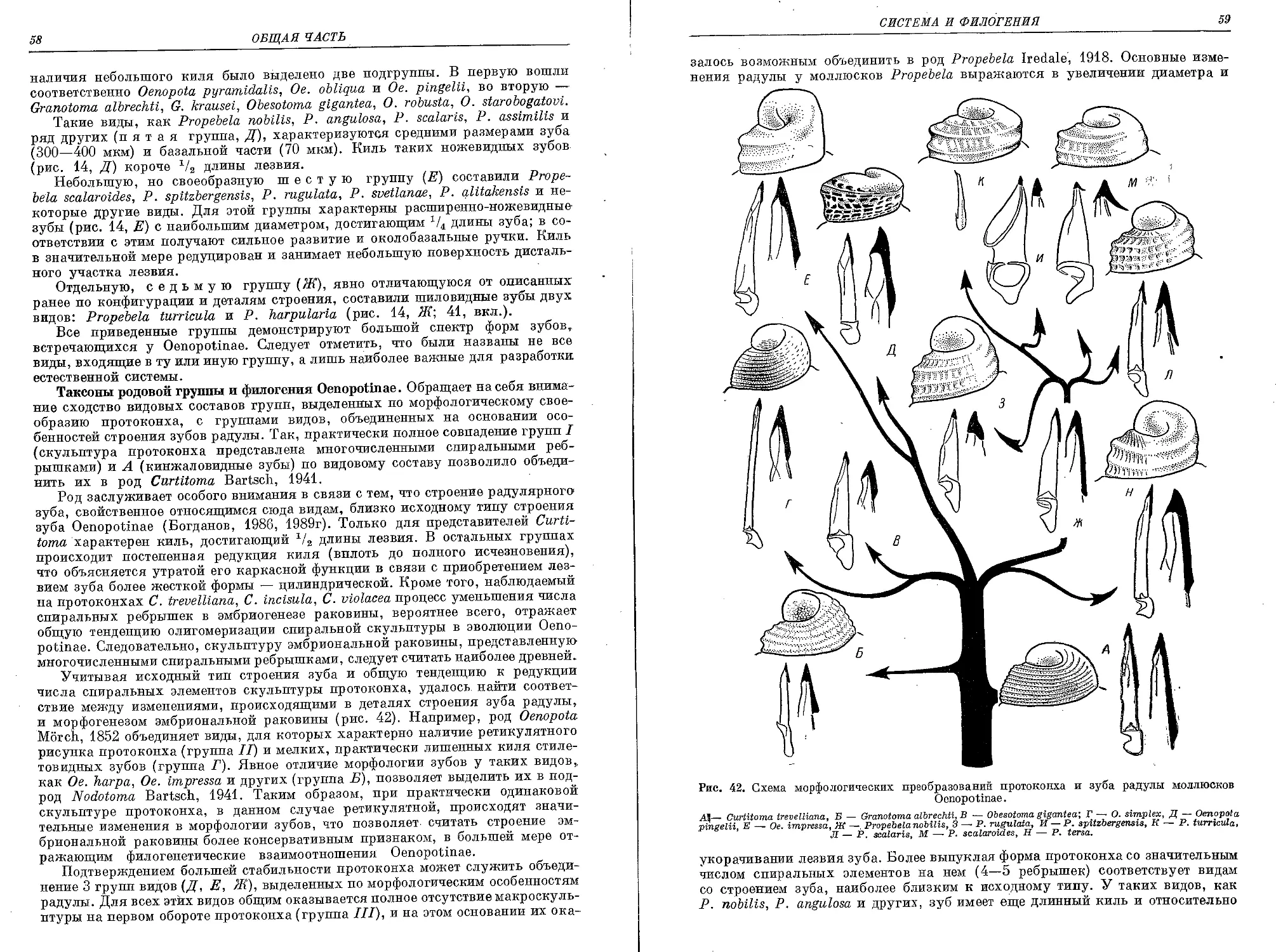
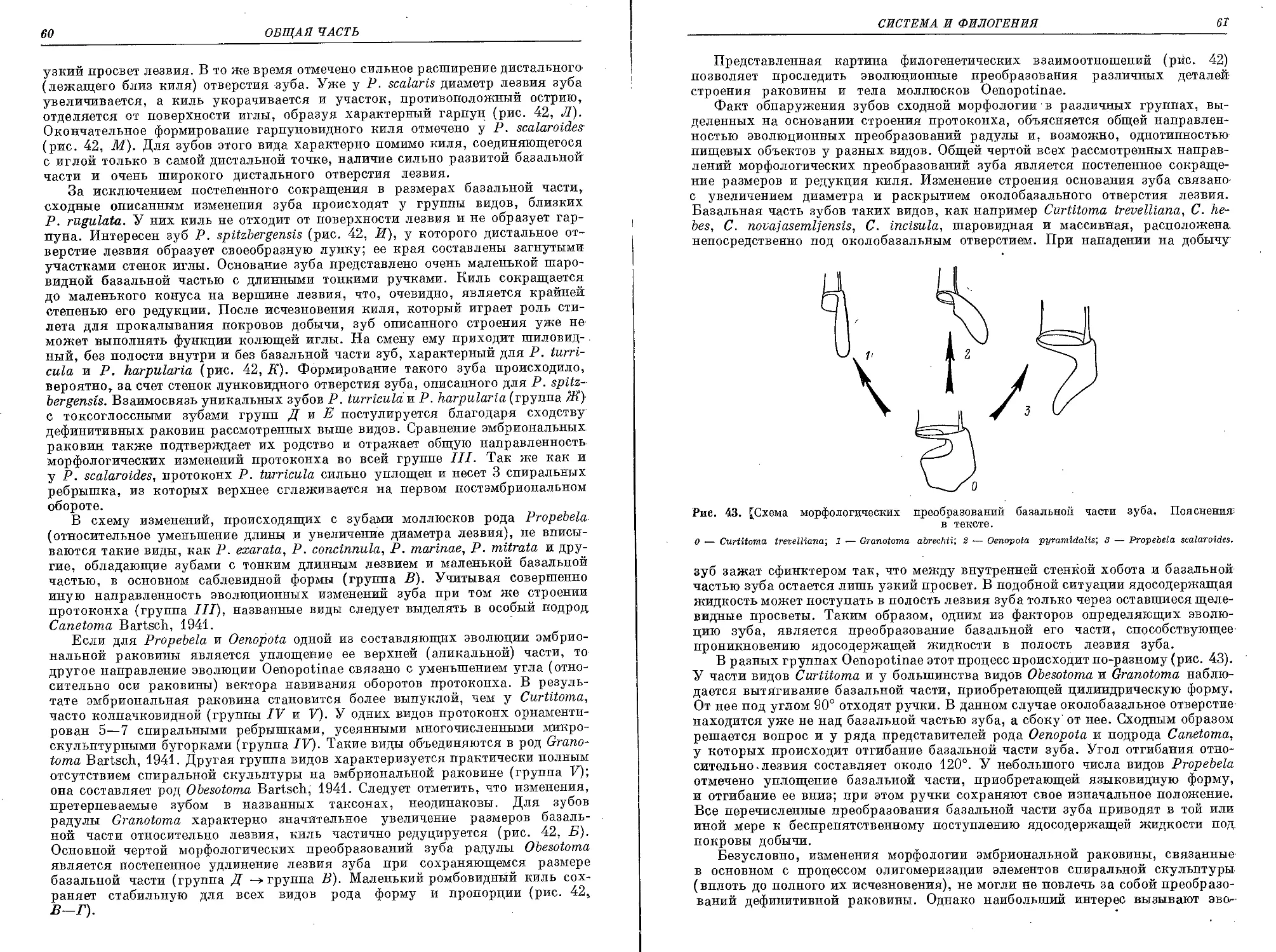


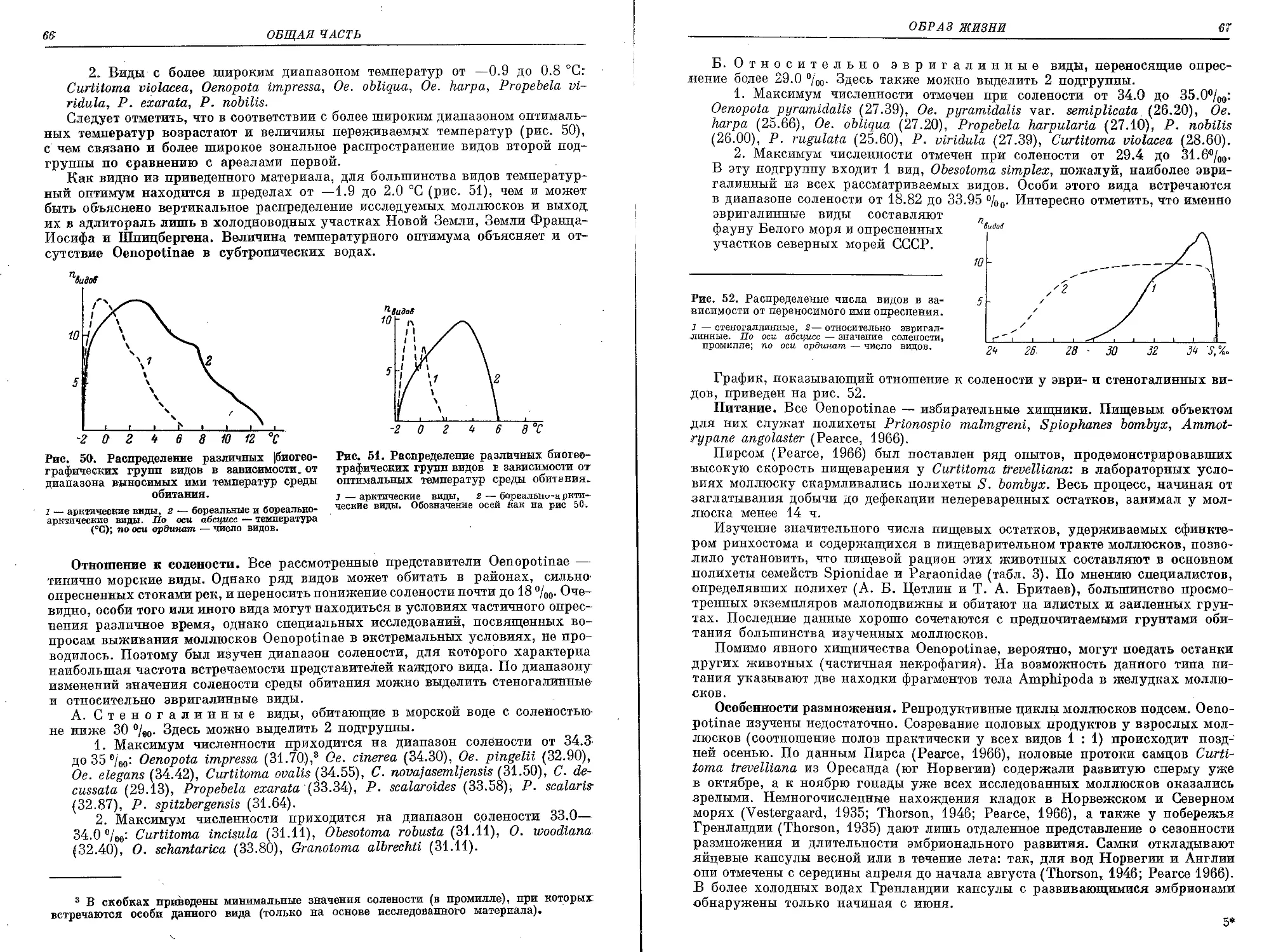
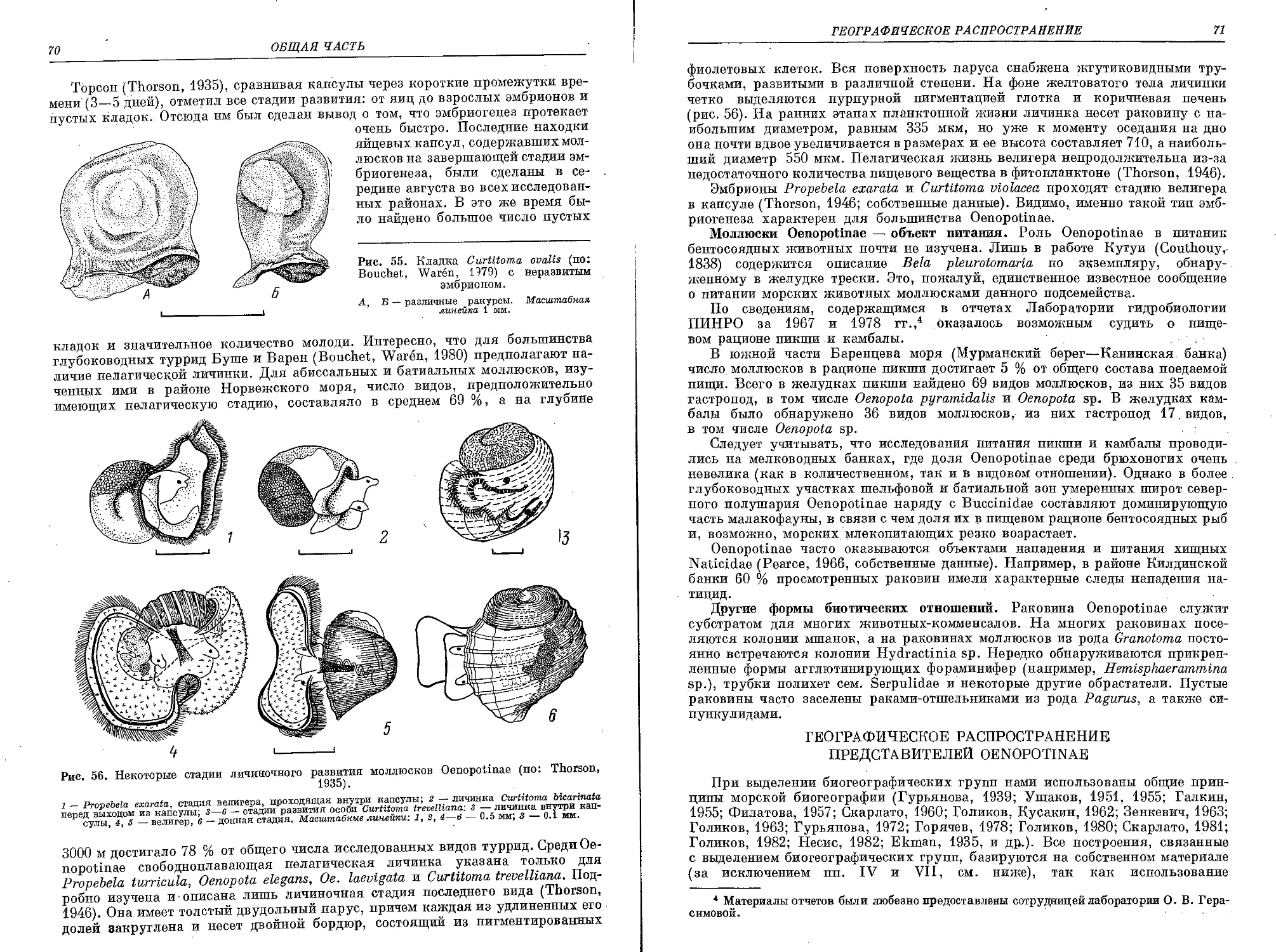




















































































Похожие




Текст
ФАуНА
СССР
МОЛЛЮСКИ
V
выпуск
з
АКАДЕМИЯ НАУК СССР
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ
ACADEMY OF SCIENCES OF THE ;U .$ .S R
ZOOLOGICAL INSTITUTE
NEW SERIES, N 142
FAUNA OF USSR
MOLLUSKS
Vol. V, N 3
I. P. BOGDANOV
MOLLUSKS OF OENOPOTINAE SUBFAMILY
(GASTROPODA, PECTINIBRANCHIA, TURRIDAE)
IN THE SEAS OF THE USSR
e
LENINGRAD
«NAUKA» PUBLISHING HOUSE
LENINGRAD BRANCH
19 90
НОВАЯ СЕРИЯ, № 142
Выпускается с 1935 г.
ФАУНА СССР
МОЛЛЮСКИ
Том V, вып. з
И. П. БОГДАНОВ
МОЛЛЮСКИ ПОДСЕМЕЙСТВА OENOPOTINAE
(GASTROPODA, PECTINIBRANCHIA, TURRIDAE)
МОРЕЙ СССР
в
ЛЕНИНГРАД
«НАУКА»
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
19 90
УДК 594.32
Богданов- И» П. Моллюски подсемейства Oenopotinae (Gastropoda, Pectinibranchia,
Turridae) морей СССР.— Л.1 Наука, 1990.—223 с. — (Фауна СССР. Нов. сер.; № 142-
Моллюски; Т. V, вып. 3).
В общей части книги рассмотрены история изучения моллюсков подсем. Oenopotinae, методы
обработки коллекционных материалов, проведен морфофункциональный анализ их строения,
исследована изменчивость морфологических признаков раковины и тела моллюсков. Кроме-
того, содержатся сведения но зоогеографии и распределению этих животных в морях СССР.
Систематическая часть книги включает определительные таблицы и диагнозы 5 родов и 78 ви-
дов Oenopotinae. Для каждого вида приведены оригинальные описания и рисунки. Библиогр-
248 назв. Ил. 475» Табл» 5.
Главный редактор серин
директор Зоологического института АН СССР
О, А. СКАРЛАТО
Редакционная коллегия:
И. М. Лихарев (отв. редактор серии), О. Л. Крыжановский (зам. отв. редактора),
С. В. Василенко, И. М» Громов, В, Ф. Зайцев, И. М. Кержнер, Л. А. Кутикова,
Г. С, Медведев, М» Е. Тер-Минасян, Н. А. Филиппова
Ответственный редактор тома
А. Н. Голиков
Рецензенты
Я. И, Старобогатов, А. А. Шилейко
1907000000-693
Б ' 042 (02)-90 557'90’ П
© Издательство «Наука», 1990
ISBN 5-02-025789-3
ПРЕДИСЛОВИЕ
Необходимость более полного и рационального использования биологиче-
ских ресурсов океана и усиление природоохранных мероприятий настоятельно
выдвигают задачу инвентаризации фауны морей СССР. Моллюски подсем. Oeno-
potinae представляют собой одну из наиболее многочисленных групп в северных
районах Тихого и Атлантического океанов и в Северном Ледовитом океане.
Обладая в большинстве случаев тонкостенной раковиной и небольшими разме-
рами, моллюски служат кормовым объектом многих бентосоядных рыб. Не-
смотря на большую значимость Oenopotinae в донных биоценозах, представи-
тели подсемейства изучены слабо. Недостаточная изученность этой Труппы
связана с мелкими размерами и значительной изменчивостью раковины. Сле-
дует также учесть, что форма и скульптура раковины для большинства специа-
листов являлись единственными признаками для дифференциации видов. Суще-
ственно и то, что после описания ряда анатомических особенностей строения
мягкого тела, в основном связанных с радулярным аппаратом, практически
никто детально не исследовал морфологические признаки тела моллюсков и
не использовал их в видовой диагностике. Имеющиеся в литературе данные
по морфологии и анатомии моллюсков изучаемой группы позволяют суДить
о значительной специализации пищедобывающего аппарата, однако механизм
его функционирования не подвергался анализу.
Настоящая работа посвящена изучению фауны и разработке системы мол-
люсков подсем. Oenopotinae. Детальное исследование морфологии раковины
и тела моллюсков в сравнительном и функциональном аспектах позволило
продемонстрировать видоспецифичность килевидного выроста дистального
участка лезвия токсоглоссного зуба радулы и обосновать использование его
формы в видовой диагностике. Обнаружено соответствие между морфологиче-
скими различиями протоконха и радулы, и как следствие — реконструирована
филогения изучаемой группы. Обширный материал позволил предварительно
оценить влияние факторов внешней среды на изменчивость морфологических
признаков у Oenopotinae, в результате чего оказалось возможным судить об их
стабильности, а соответственно и ценности для видовой диагностики. Предло-
женная в работе система группы и выделенные диагностические признаки рако-
вины делают возможным использовать представителей Oenopotinae для био-
стратиграфического анализа морских неогеновых отложений нетропических
зон северного полушария, в частности на территории СССР. Многие раковины,
обнаруженные в ископаемом состоянии, могут служить четким показателем
экологических условий в предшествующие геологические эпохи.
В процессе изучения фауны моллюсков подсем. Oenopotinae были обрабо-
таны обширные коллекции этих животных в Зоологическом институте АН СССР,
Ленинград (ЗИН АН СССР), накопившиеся более чем за столетие (1871—1988),
в Институте биологии моря ДВО АН СССР, Владивосток (ИБМ), Институте океа-
нологии им. И. П. Ширшова АН СССР, Москва (ИОАН), в Тихоокеанском ин-
ституте рыбного хозяйства и океанографии, Владивосток (ТИНРО), в его Саха-
линском отделении (СахТИНРО), а также в Зоологическом музее МГУ, Москва
(ЗМ МГУ). Всего было просмотрено более 4200 проб, содержавших свыше
12 000 экз.
6
ПРЕДИСЛОВИЕ
Все рассмотренные в настоящей работе виды обитают в бореальных и аркти-
ческих полиосоленых водах северного полушария. На севере граница распро-
странения представителей Oenopotinae доходит до. 83? с. ш. (отмечено мной).
Южными границами расселения в Тихом океане следует считать у азиатских
берегов воды, омывающие Северную Корею и о-в Хонсю, а у американских
берегов — между о-вом Ванкувер и побережьем штата Калифорния. В Атлан-
тическом океане отдельные виды встречаются вплоть до глубоководий у Испании
и Гасконского залива (у берегов Европы) и до м. Кейп-Код (у Восточных берегов
Северной Америки).
В результате обработки собственного и коллекционного материала, а также
анализа литературных данных показано, что фауна Oenopotinae Мирового
океана состоит примерно из 110 видов. Фауна Oenopotinae шельфовой зоны
морей СССР представлена 5 родами — Oenopota Morch, 1852; Curtitoma Bartsch,
1941; Propebela Iredale, 1918; Granotoma Bartsch, 1941; Obesotoma Bartsch, 1941
и включает 78 видов, из которых 47 новые для фауны СССР, а 20 видов новые
для науки.
На разных этапах работы мне была оказана существенная помощь академи-
ком М. С. Гиляровым, профессором К. В. Беклемишевым и д. б. н. Г. Б. Зеви-
ной. Ведущие специалисты ЗИН АН СССР, ЗМ МГУ и ИОАН любезно предо-
ставили материалы по исследуемой группе моллюсков. Сотрудники ЗИН
АН СССР д. б. н. А. Н. Голиков, д. б. н. Я. И. Старобогатов и к. б. н. Б. И. Си-
ренко, ЗМ МГУ к. б. н. В. Н. Горячев и ИБМ к. б. н. В. В. Гульбин консульти-
ровали меня. При работе с коллекцией ЗИН АН СССР неоценимую помощь
оказали С. И. Ягудина, М. А. Долголенко.и С. И. Стрельцова. Куратор коллек-
ции моллюсков Национального музея естественной истории США (Вашингтон)
Дж. Роузвотер (Josef Rosewater) прислал фотографии ряда голотипов, храня-
щихся в музее, и копии рисунков зубов радул типовых экземпляров видов,
описанных В. Долом и П. Бартшем.
Всем перечисленным лицам я выражаю искреннюю признательность.
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
Подотряд TOXOGLOSSA Gray, 1853
Сем. TURRIDAE Adams et Adams, 1855
Подсем. OENOPOTINAE Bogdanov, 1987
1. Род CURTITOMA Bartsch, 1941
Стр.
1. C. incisula (Verrill, 1882)...........................................................
2. C. trevelliana (Turton, 1834).........................................................
3. C. hebes (Verrill, 1880) . . .........................................................
4. C. novajasemljensis (Leche, 1878).....................................................
5. C. ovalis (Friele, 1877)..............................................................
6. C. flora (Dall, 1919).................................................................
7. C. violacea (Maghels et Adams, 1842)..................................................
8. C. conoidea (G. 0. Sars, 1878)........................................................
9. C. decussata (Couthouy, 1839).........................................................
10. C. lawrenciana (Dall, 1919).........................................................
11. C. hokkaidoensis (Bartsch, 1941).....................................................
12. C. microvoluta (Okutani, 1961).......................................................
13. C. bartschi (Bogdanov, 1985).........................................................
14. C. ptltuniensis (Bogdanov, 1985).....................................................
15. C. becklemishevt Bogdanov, 1989 .....................................................
16. C. neymanae Bogdanov, 1989 ..........................................................
84
117
119
120
121
122
123
125
127
128
130
130
131
132
132
133
2. Род GRANOTOMA Bartsch, 1941
1. G. krausei (Dall, 1886)............................................................... 134
2. G. albrechti (Krause, 1885)........................................................... 135
3. G. raduga (Bogdanov, 1985)........................................................... 137
3. Род OENOPOTA Morch, 1852
1. Подрод OENOPOTA Morch, 1852
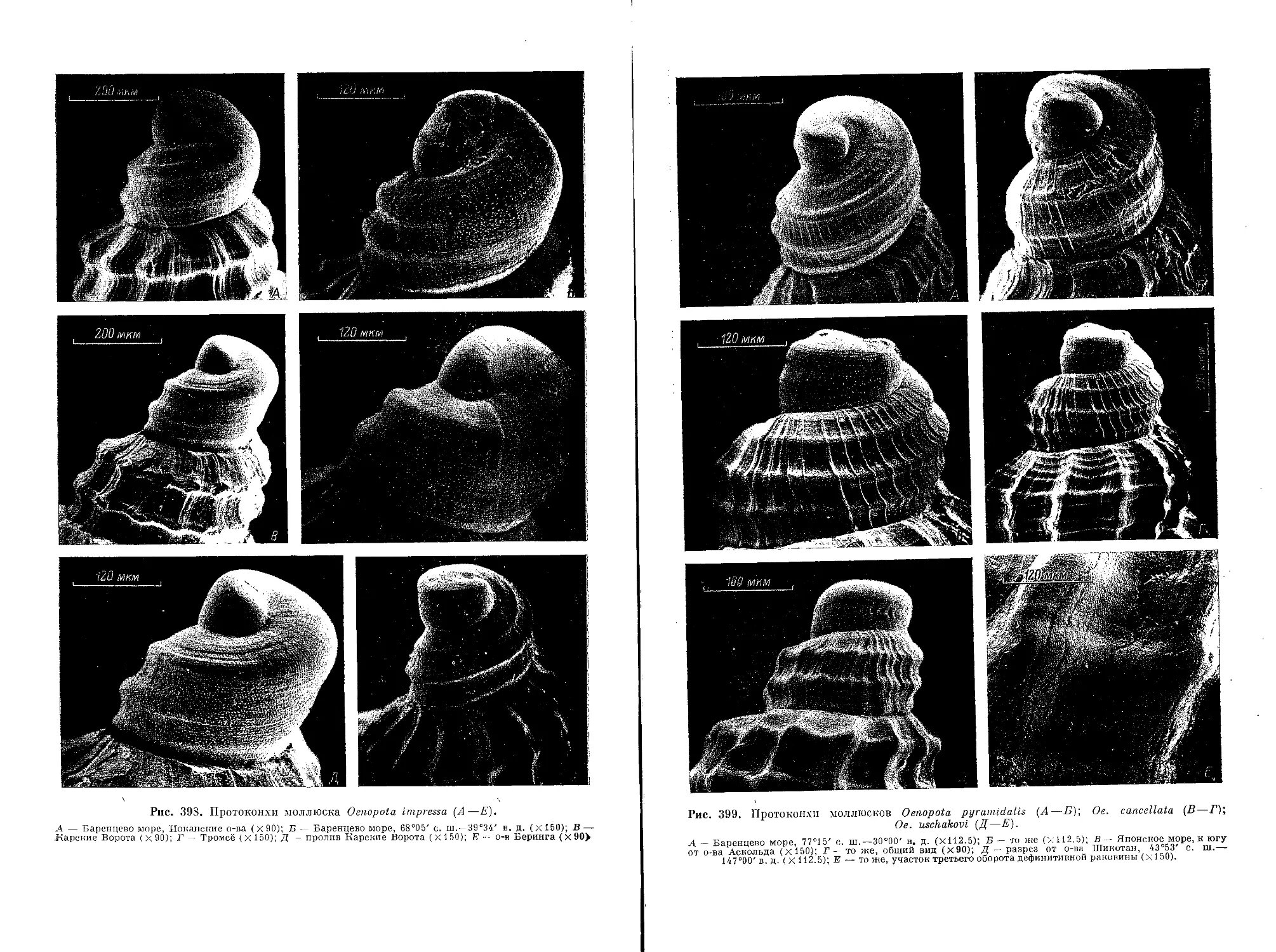
1. Oe. (Oe.) pyramidalis (Strom, 1788)................................................. 140
2. Oe. (Oe.) pingelii (Moller, 1842).................................................... 142
3. Oe. (Oe.) obliqua (0. G. Sars, 1878)................................................. 143
2. Подрод NODOTOMA Bartsch, 1941
4. Oe. (N.) impressa (Morch, 1869)...................................................... 145
5. Oe. (N.) cinerea (Moller, 1842)..................................................... 147
6. Oe. (N.) declivis (Loven, 1846)...................................................... 148
7. Oe. (-N.) elegans (Moller, 1842)...................................................... 150
8. Oe. (N.) harpa (Dall, 1884).......................................................... 151
9. Oe. (N.) cancellata (Mighels et Adams, 1840).......................... ............ 153
10. Oe. (N.) casentina (Dall, 1919)..................................................... 154
11. Oe. (N.) inequita (Dall, 1919)...................................................... 154
12. Oe. (N.) pavlova (Dall, 1919)....................................................... 155
13. Oe- (N.) uschakovi Bogdanov, 1985 .................................................. 156
14. Oe. (N.) biconica Bogdanov, 1989 ................................................... 157
15. Oe. (N.) kurilensis Bogdanov, 1989 ................................................. 157
16. Oe. (N.) elongata Bogdanov, 1989 ................................................... 158
s
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
4. Род OBESOTOMA Bartsch, 1941
1. О. japontca Bartsch, 1941 ................................................... . 160
2. О. simplex (Middendorff, 1849).................................................... 161
3. О. schantarica (Middendorff, 1849)..........................................• • . 163
4. О. gigas (Verkriizen, 1875)....................................................... 164
5. О. gigantea (Morsch, 1869)....................................................... 165
6. 0. tenuilirata (Dall, 1871)....................................................... 166
7. 0. woodiana (Moller, 1842) ....................................................... 167
8. 0. tumida (Posselt et Jensen, 1898)............................................... 168
9. 0. robusta (Packard, 1867)........................................................ 169
10. 0. tomiyaensis (Otuka, 1949)..................................................... 170
11. 0. solida (Dall, 1886) .......................................................... 171
12. 0. starobogatovi Bogdanov, sp. n................................................. 171
13. 0. sachalinensis Bogdanov, 1989 ................................................. 172
5. Род PROPEBELA Iredale, 1918
1. Подрод PROPEBELA Iredale, 1918
1. P. (P.) turricula (Montagu, 1803)............................................... 176
2. P. (P.) angulosa (G. 0. Sars, 1878)............................................. 178
3. P. (P.) nobilis (Moller, 1842)............................................... 179
4. P. (P.) assimilis (G. 0. Sars, 1878)............................................ 181
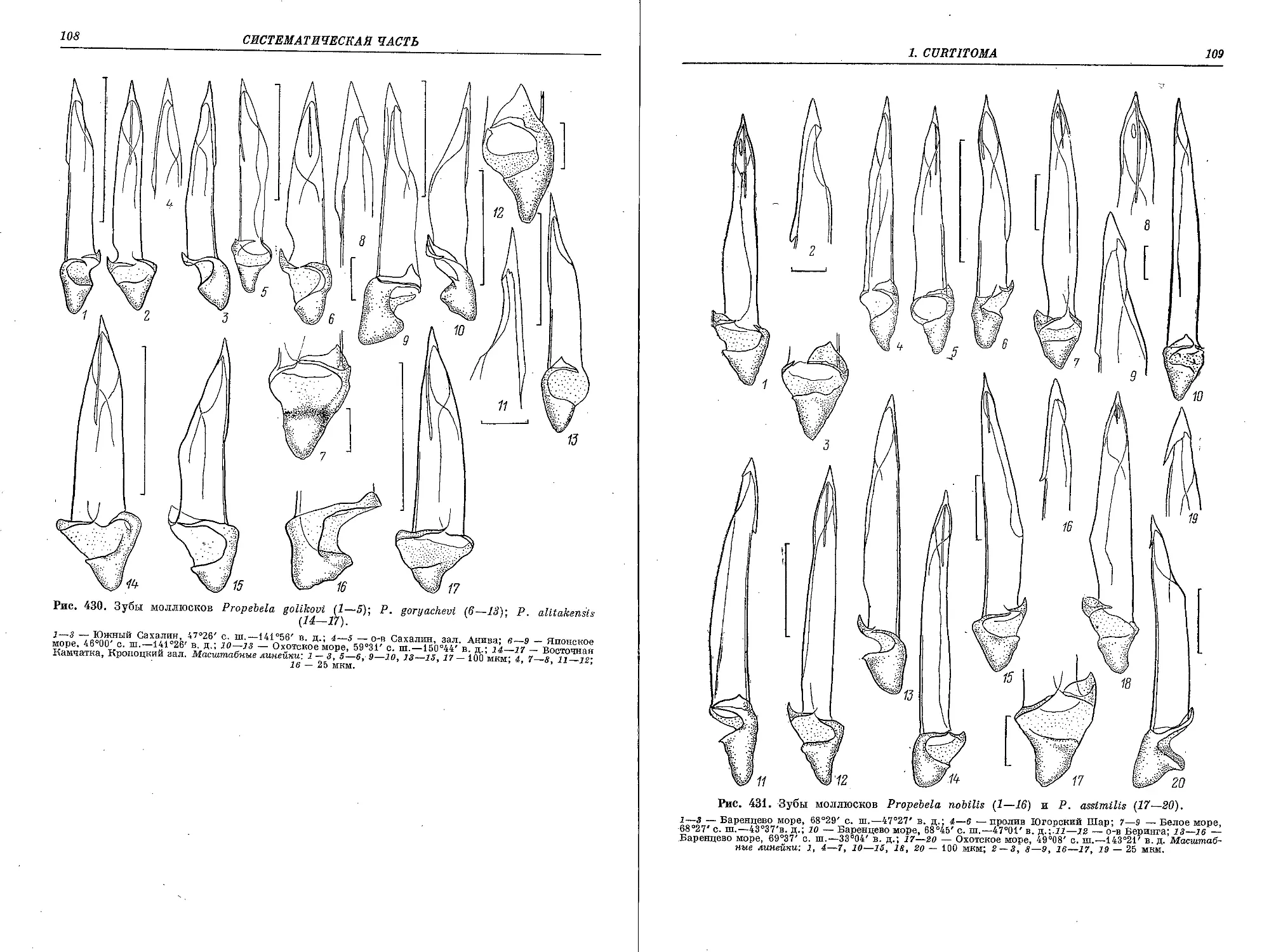
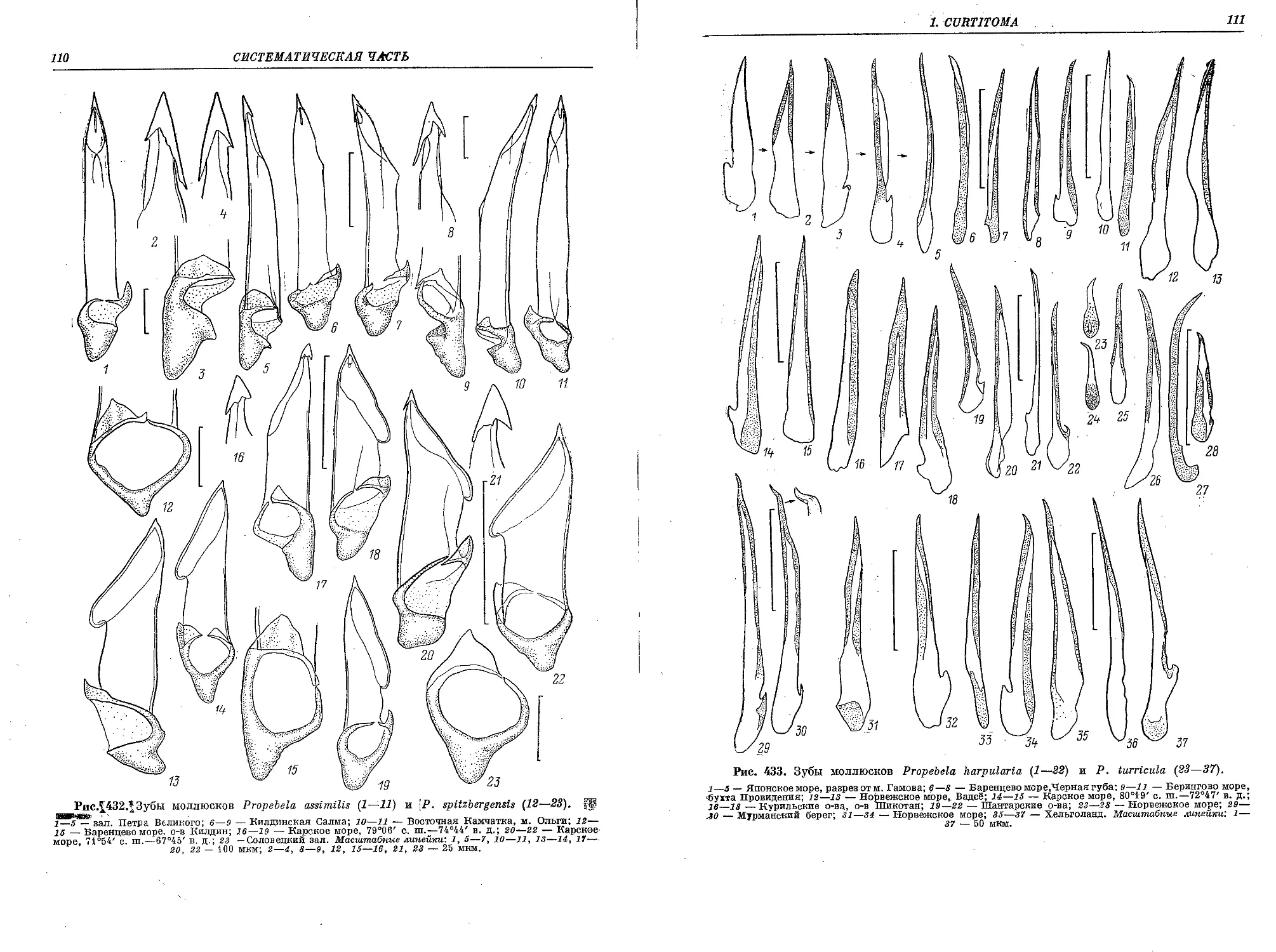
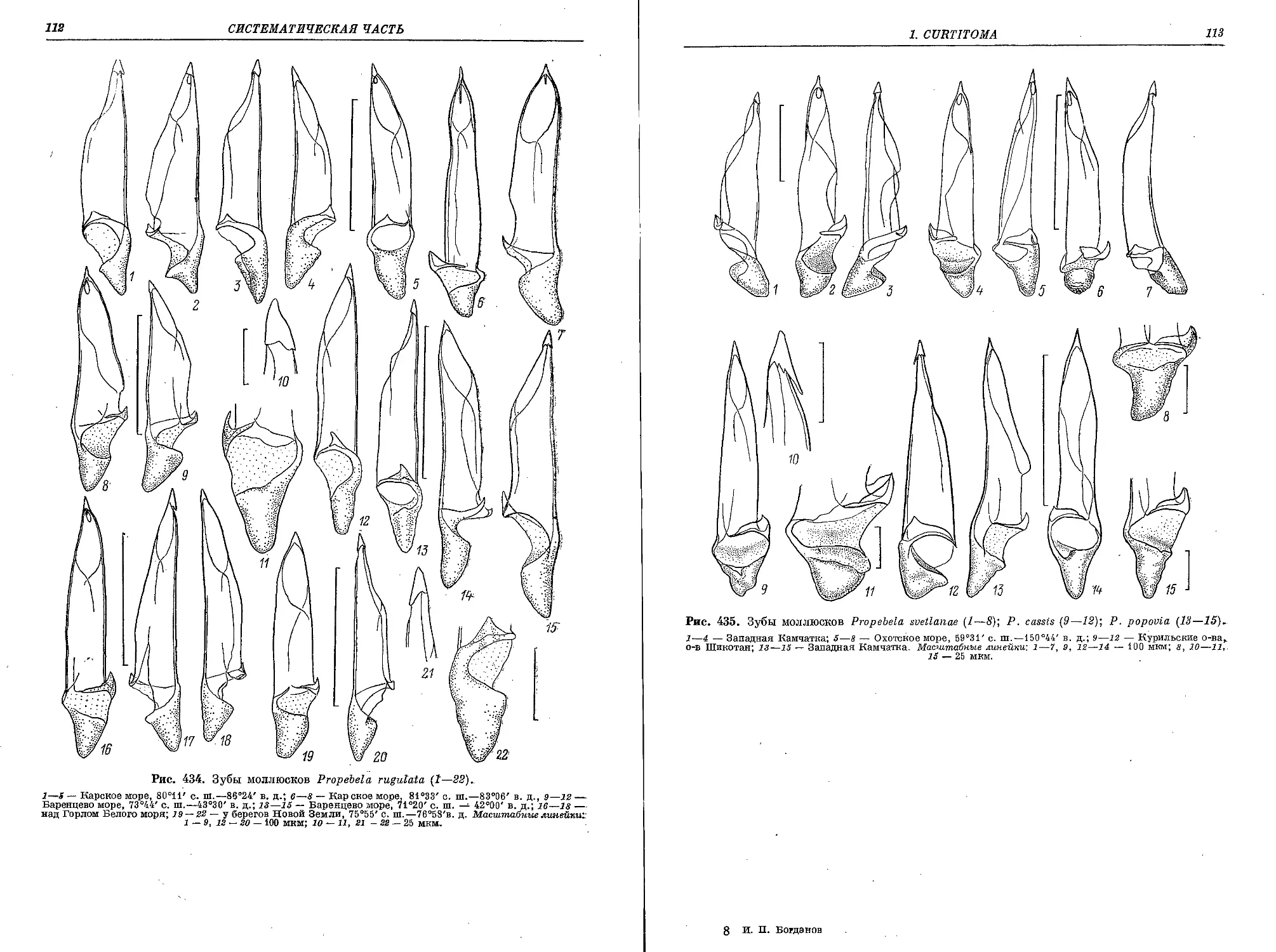
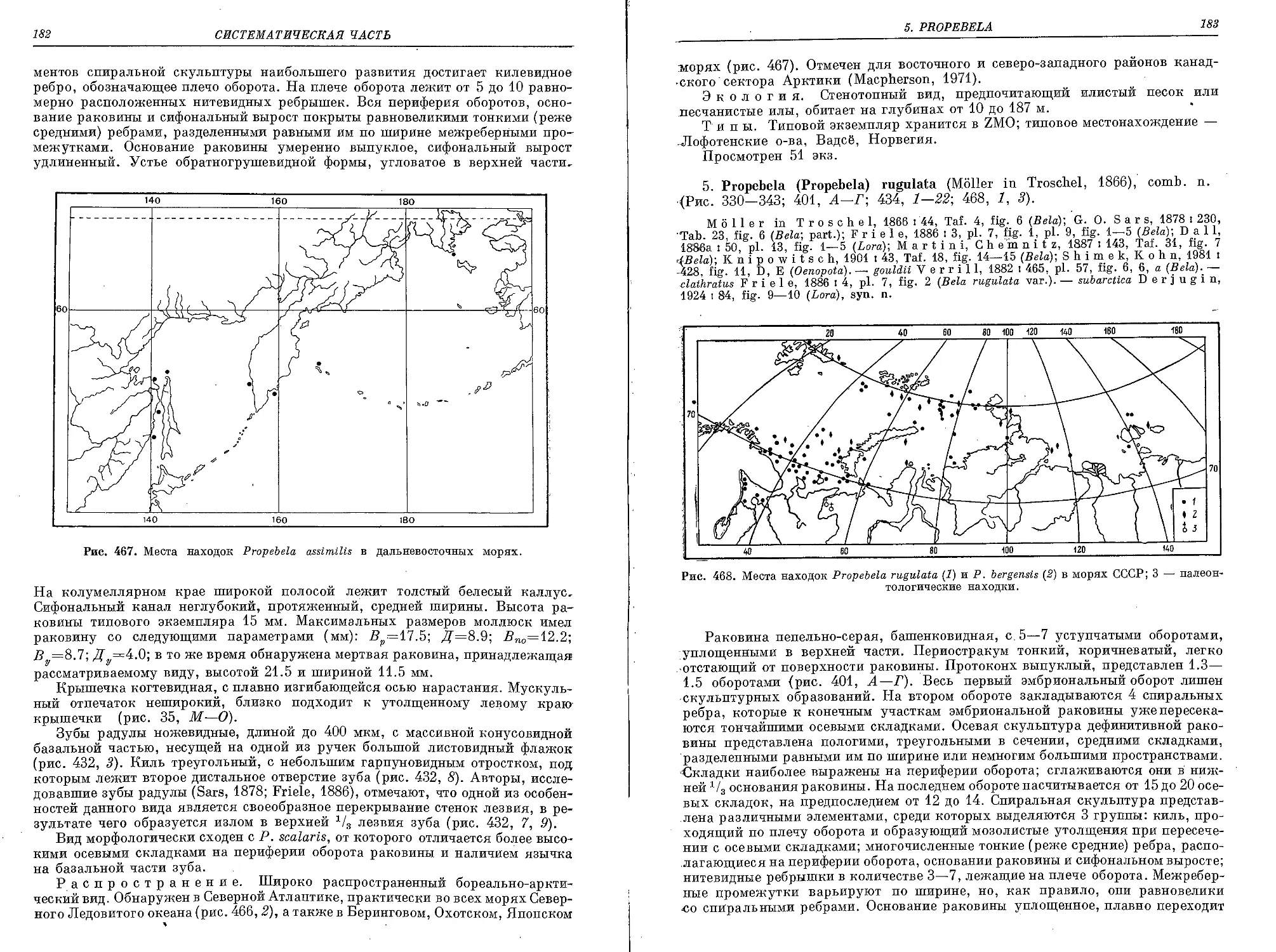
5. P. (P.) rugulata (Moller in Troschel, 1866)..................................... 183
6. P. (P.) spitzbergensis (Friele, 1886)........................................... 184
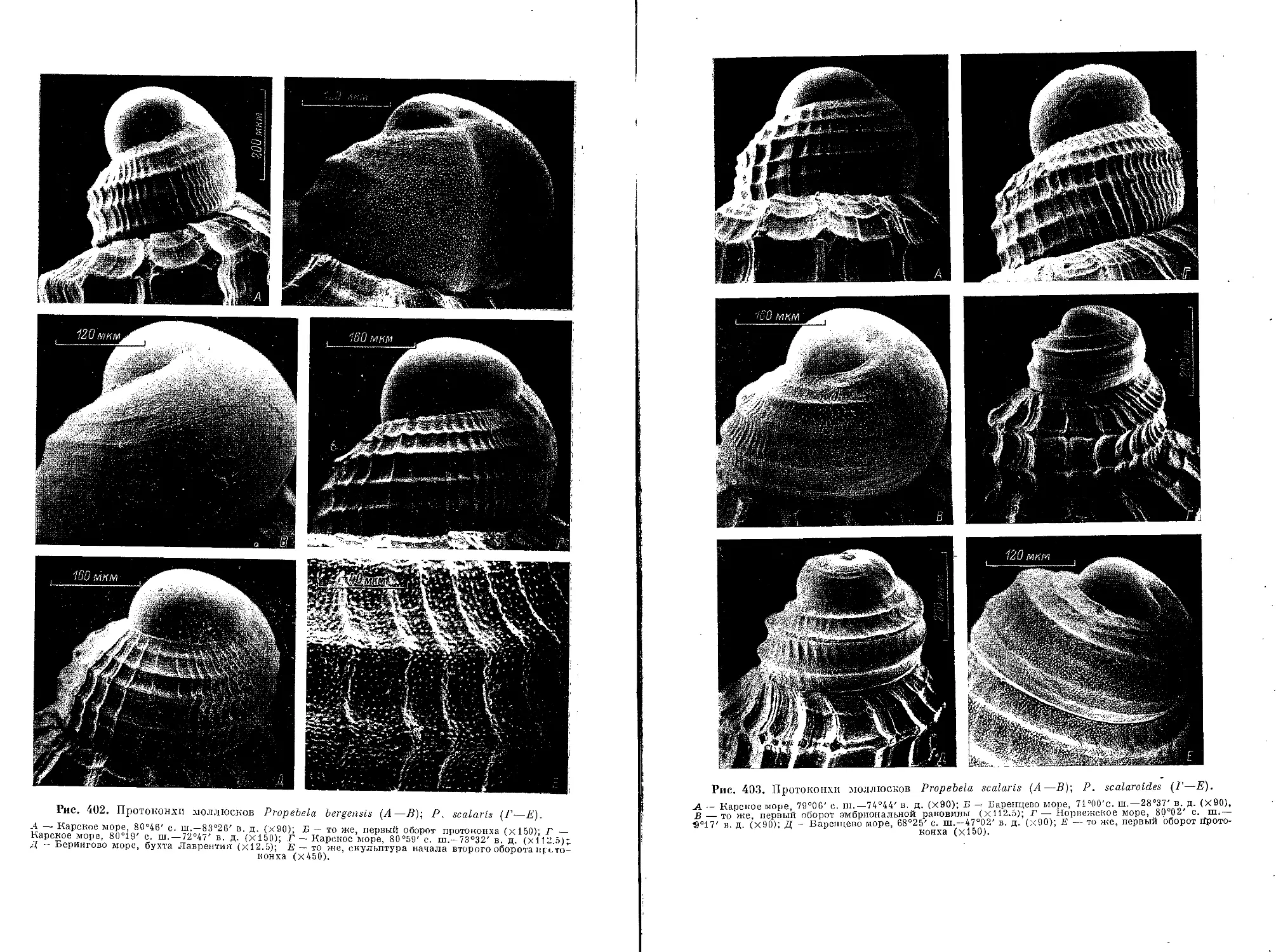
7. P. (P.) bergensis (Friele, 1886)................................................ 185
8. P. (P.) harpularia (Goiithouy, 1838)............................................ 186
9. P. (P.) popovia (Dall, 1919).................................................... 187
10. P. (P.) scalaris (Moller, 1842)................................................ 188
11. P. (P.) scalaroides (G. 0. Sars, 1878)......................................... 189
12. P. (P.) svetlanae Bogdanov, 1989 ............................................. 190
13. P. (P.) verrilli Bogdanov, 1989 .............................................. 191
14. P. (P.) cassis Bogdanov, 1989 ................................................. 191
15. P. (P.) terpeniensis Bogdanov, 1989 ........................................... 192
16. P. (P.) goryachevi Bogdanov, 1989 ............................................. 192
2. Подрод GANETOMA Bartsch, 1941
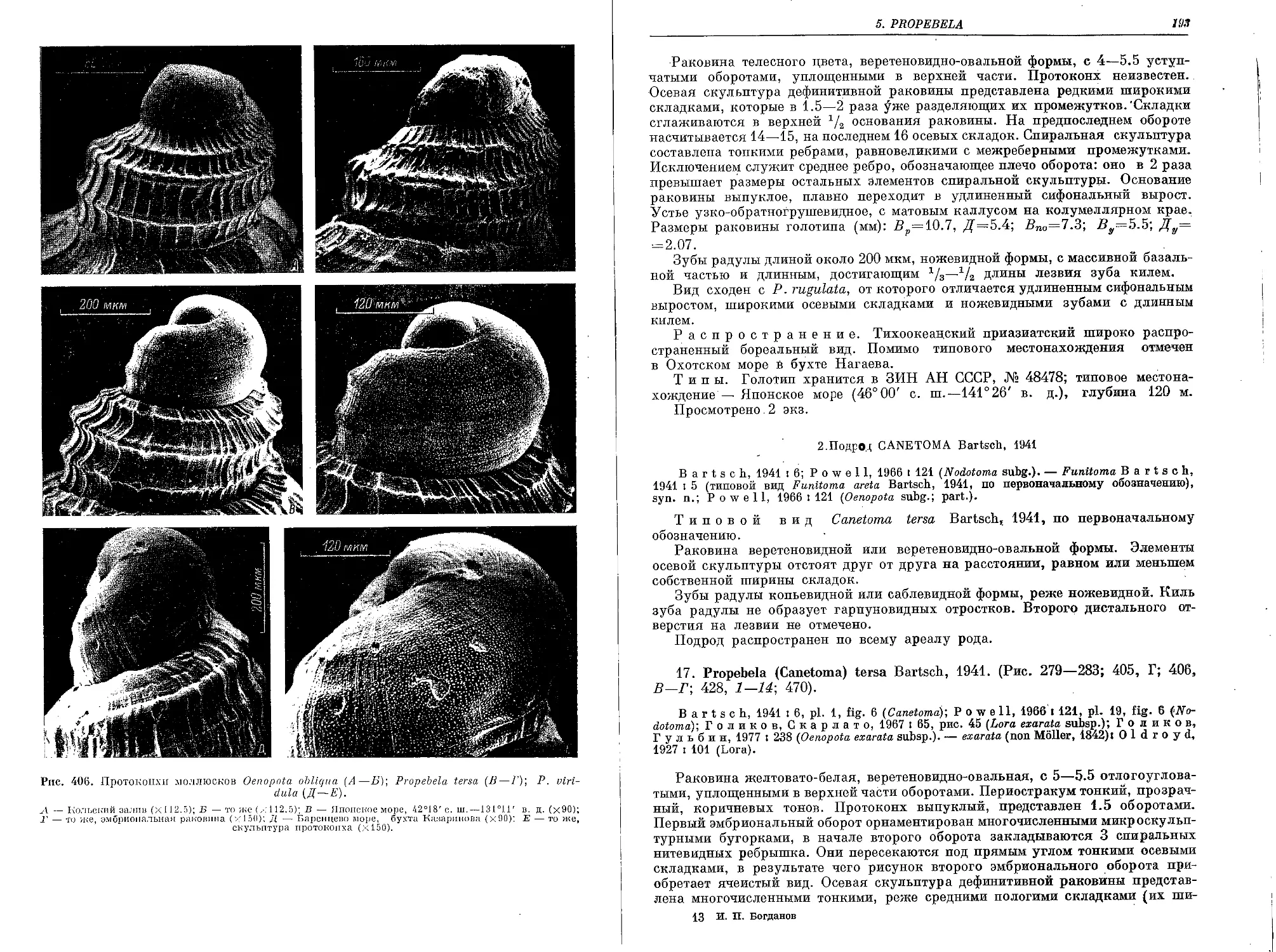
17. P. (C.) tersa Bartsch, 1941.................................................... 193
18. P. (C.) viridula (Moller, 1842)................................................ 195
19. P. (C.) exarata (Moller, 1842)................................................. 196
20. P. (C.) mitrata (Dall, 1919)...............................................‘ . 198
21. P. (C.) concinnula (Verrill, 1882) .............................................. 199
22. P. (C.) fidicula (Gould, 1850)..................................• • ............ 200
23. P. (C.) alitakensis (Dall, 1919)................................................. 200
24. P. (C.) rassina (Dall, 1919)..................................................... 201
25. P. (C.) cingulata (Golikov et Gulbin, 1977).................................. • 202
26. P. (C.) areta (Bartsch, 1941)............ . ..................................... 203
27. P. (P.) golikovi (Bogdanov, 1985) . . 204
28. P. (P.) margaritae (Bogdanov, 1985)............................................ 204
29. P. (P.) marinae Bogdanov, 1989 ................................................ 205
30. P. (P.) variabilis Bogdanov, sp. n. . ........................................ 209
ОБЩАЯ ЧАСТЬ
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Материалом для настоящей работы послужили коллекции моллюсков ЗИН АН СССР,
ИВМ ДВО АН СССР, ИОАН СССР, ТИНРО и Сах ТИНРО, ЗМ МГУ. Часть материала со-
брана автором в 1978—1983 гг. в зал. Петра Великого и в северном Приморье (Японское
море). Ниже приводится перечень сборов экспедиций и отдельных исследователей из различ-
ных районов Мирового океана, содержащих моллюсков подсем. Oenopotinae (обработаны
мной).
.. Сборы
Район работ (коллектор или экспедиция; судно; год)
Южная и юго-восточная
части Баренцева моря
Большая часть Барен-
цева моря и прилежа-
щие районы Норвеж-
ского и Гренландского
морей
Белое море
Шпицберген
Новая Земля, Земля
Франца-Иосифа, се-
верная часть Карского
моря
Юго-восточная часть
Карского моря
Море Лаптевых
Ф. Ф. Яржинский (1871), К. С. Мережков-
ский (1876), С. М. Герценштейн (1880, 1884,
1887), В. А. Фаусек (1888, 1889), Н. М. Кни-
пович (1890—1894), Г. Г. Якобсон (1893),
М. Н. Римский-Корсаков (1894), А. А. Бя-
лыницкий-Бируля (1895—1896), А. С. Бот-
кин (1896), К. Келлер (1897), А. М. Поли-
лов («Пахтусов», 1910—1913), Г. И. Горбу-
нов («Зарница», 1925), П. В. Ушаков
(«Зарница», 1927), Е. Ф. Гурьянова и
Г. С. Гурвич (ГГИ, 1928), В. В. Кузнецов
(1940), МБС («Протрактор», 1953; «Поляр-
ник», 1954; «Диана», 1955, 1956), Ю. И. Гал-
кин и Л. И. Москалев («Пр. Дерюгин»,
1958, 1959), М. В. Пропп (1959), РС-5302
(ПИНРО, 1971).
ЭНПИМ: «Мурман» (1898), «Помор» (1898—
1901), «Андрей Первозванный» (1898—1906);
МНИ: «Персей» (1924-1935); ПИНРО:
(1968—1970): РТ-61 (1968), «Водник» (1968),
«Н. Маслов» (1968—1970); «Атлантида»
(1972).
В. П. Романский («Лейтенант Скуратов»,
1908 и «Мурман», 1911), Г. С. Гурвич
(1931), «Кайра» (1934), «Онега» (1961 —
1964), ВВС ЗИН (1967, 1968, 1974, 1977).
А. А. Бируля («Бакан, 1899), А. М. Волкович
и М. К. Михайловский («Бакан», 1901),
3. Ф. Свядош («Геркулес», 1912).
Н. М. Книпович («Ермак, 1901), «Русская
полярная экспедиция» («Заря», 1900—1901),
Стрельников (1921), Г. П- Горбунов («Г. Се-
дов», 1929—1931), Л. О. Ретовский и
В. Л. Вагин (ВАИ «Ломоносов», 1931),
Г. П. Горбунов («Сибиряков», 1934),
Г. С. Сластников (ВАИ «Арктик», 1934),
Г. П. Горбунов («Садко», 1935—1937),
В. М. Колтун («Литке», 1948), Л. И. Щер-
бакова («Шторм, 1960), А. Н. Голиков
(1970), экспедиция ЗИН (1982).
В. Л. Вагин (1934), Г. С. Сластников («Арк-
тик», 1939).
«Вайгач» и «Таймыр» (1911—1914), Павленко
(«Лейтенант Дыдымов», 1915), В. Л. Ва-
гин и Н. Н. Кондаков («Русанов», 1932),
Л. О. Ретовский («Сибиряков». 1932),
Г. П- Горбунов («Садко», 1935—1937),
В. В. Макаров («Г. Седов», 1937), А. П. Анд-
рияшев («Малыгин», 1937), В. Л. Вагин
(«Литке», 1948), экспедиция ЗИН (1973).
10
ОБЩАЯ ЧАСТЬ
Район работ Сборы (коллектор или экспедиция; судно; год)
Новосибирские о-ва, Восточно-Сибирское море Северная часть Атлан- тического океана «Вайгач» и «Таймыр» (1911——1914), Ю. Д. Чи- рихин («Темп», 1934), П- В. Ушаков («Кра- син», 1935), Г. П. Горбунов («Садко», 1935—1937), А. П. Андрияшев («С. П-», 1946), экспедиция ЗИН (1973). РТ-97 (1955), В. С. Шувалов («М. Ломоно- сов», 1958), К. Н. Несис («Севастополь», 1959).
Чукотское море, Берин- гов пролив Всего около 3250 проб «Вайгач» и «Таймыр» (1911—1914), П. В. Уша- ков («Литке», 1929), МНИ (1932), П. В. Уша- ков («Красин», 1935), А. К. Леонов («Охотск», 1938), экспедиция ЗИН (1976).
Берингово море Всего около 80 проб «Вайгач» и «Таймыр» (1911—1914), А. В. Ива- нов и В. В. Макаров («Дальневосточник», 1932), Н. Н. Кондаков («Красноармеец», 1933), В. Н. Горячев («Пеламида», 1972 и «Адлер», 1973, 1979), В. Н. Лапшин («Ра- китное», 1973), А. Н. Голиков («Радуга», 1975).
Охотское море и За- падная Камчатка Курильские острова Всего около 210 проб Н. А. Смирнов и А. Г. Бегак («Командор Бе- ринг», 1907), М. Н. Павленко (1915), П. В. Ушаков («Красный Якут», 1930 и «Гагара», 1932), Е. Ф. Гурьянова (1946), КСЭ («Топорок», 1947—1949), Л. Г. Назвич («Витязь», 1951), В. Кузнецов («Витязь», 1954), А. А. Нейман («Изумруд», 1962 и «Байдар», 1964), О. Б. Мокиевский (1957), экспедиция ТИНРО—ИВМ (РТ 8-461; РТ 8-452, 1976, 1977), В. Н. Кобликов, В. Г. Аверинцев, В. А. Павлючков («По- сейдон», 1978), «Пеламида» (1985). Н. С. Спирина («Лебедь», 1954), I Курильская экспедиция ИВМ (1969), «Крылатка» (1970— 1971), «Майское» (1972), В. И. Лукин (1976), «Витязь» (1953, 1957, 1966).
Японское море Всего около 390 проб К. М. Дерюгин (1925, 1926), И. Г. Закс (1927), М. Л. Пятаков (1927, 1928), А. Н. Держа- вин (1928), Н. И. Тарасов (1928—1930), Д. М. Штенберг и А. Т. Шурин (1930), К. М. Дерюгин (1930—1933), А. В. Иванов (1931), А. В. Иванов и Д. И. Охрямкин («Россинанте», 1931), 3. И. Кобякова и А. М. Волк («Блюхер», 1933), Н. И. Тара- сов («Россинанте», 1932), Н. М. Сомова («Россинанте», 1933), А. Н. Голиков (1962, 1964, 1965), В. Л. Климова («Таманга», 1970), Белоног («Атна», 1974), В. И. Лукин («Атна, 1975), В. Н. Романов (1979), Б. И. Сиренко (1982), А. Н. Голиков (1983).
Всего около 270 проб
Примечание. В перечне приняты следующие сокращения: ВАИ — Всесо-
юзный арктический институт; ББС ЗИН — Беломорская биологическая станция
ЗИН АН СССР; ГГИ — Государственный гидрологический институт; ГОИН—Госу-
дарственный океанографический институт; МНИ — морской научный институт;
МБС — Мурманская биологическая станция; ПИНРО, ТИНРО — институты рыбного
хозяйства ц океанографии; ЭНИИМ - экспедиция для научно-промысловых иссле-
дованний Мурманска. .
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
11
По результатам обработки материалов, приведенных в перечне, была составлена карта,
отражающая степень изученности морей Полярного бассейна и северо-западной части Тихого
океана (рис. 1).
Сбор материалов осуществлялся с помощью тралов (Сигсби, Галатея, большого и малого
шлюпочных, Слизкина, зоологического и Оттертрала), драг и дночерпателей (главным
образом типа «Океан» и «Петерсена»). Небольшая часть коллекции была собрана с использо-
ванием водолазных работ.
В фондовых коллекциях ЗИН АН СССР хранятся материалы, поступившие в разные
годы от зарубежных исследователей. Г. О. Саре прислал часть определенного им материала
из вод, омывающих Западную Норвегию и Лофотенские о-ва; Г. Фриле любезно предоставил
институту часть сборов Норвежской экспедиции 1876—1878 гг.; Дж. Норберг передал сухой
материал, собранный у побережья Аляски и на севере Норвегии (Тромсё).
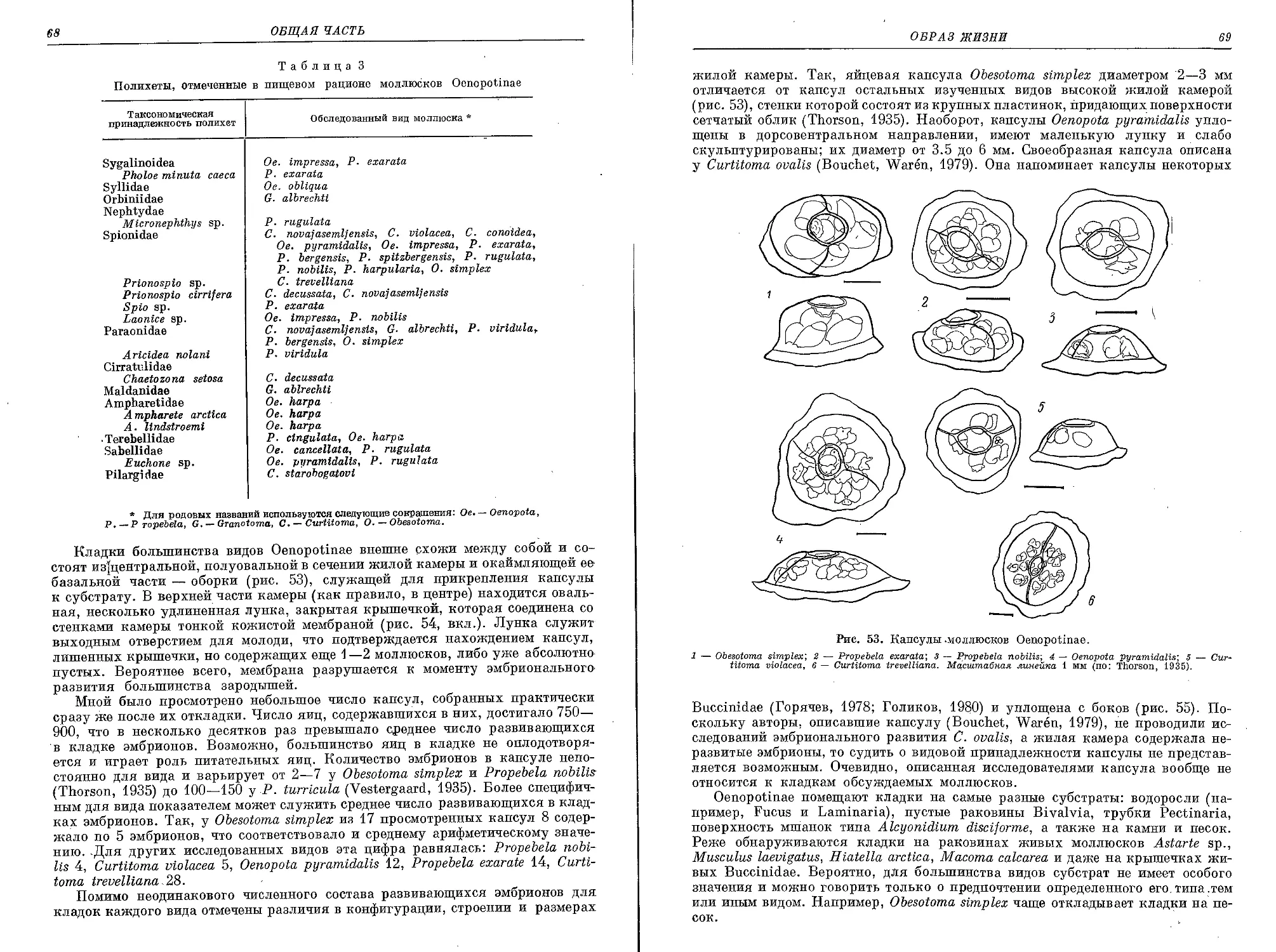
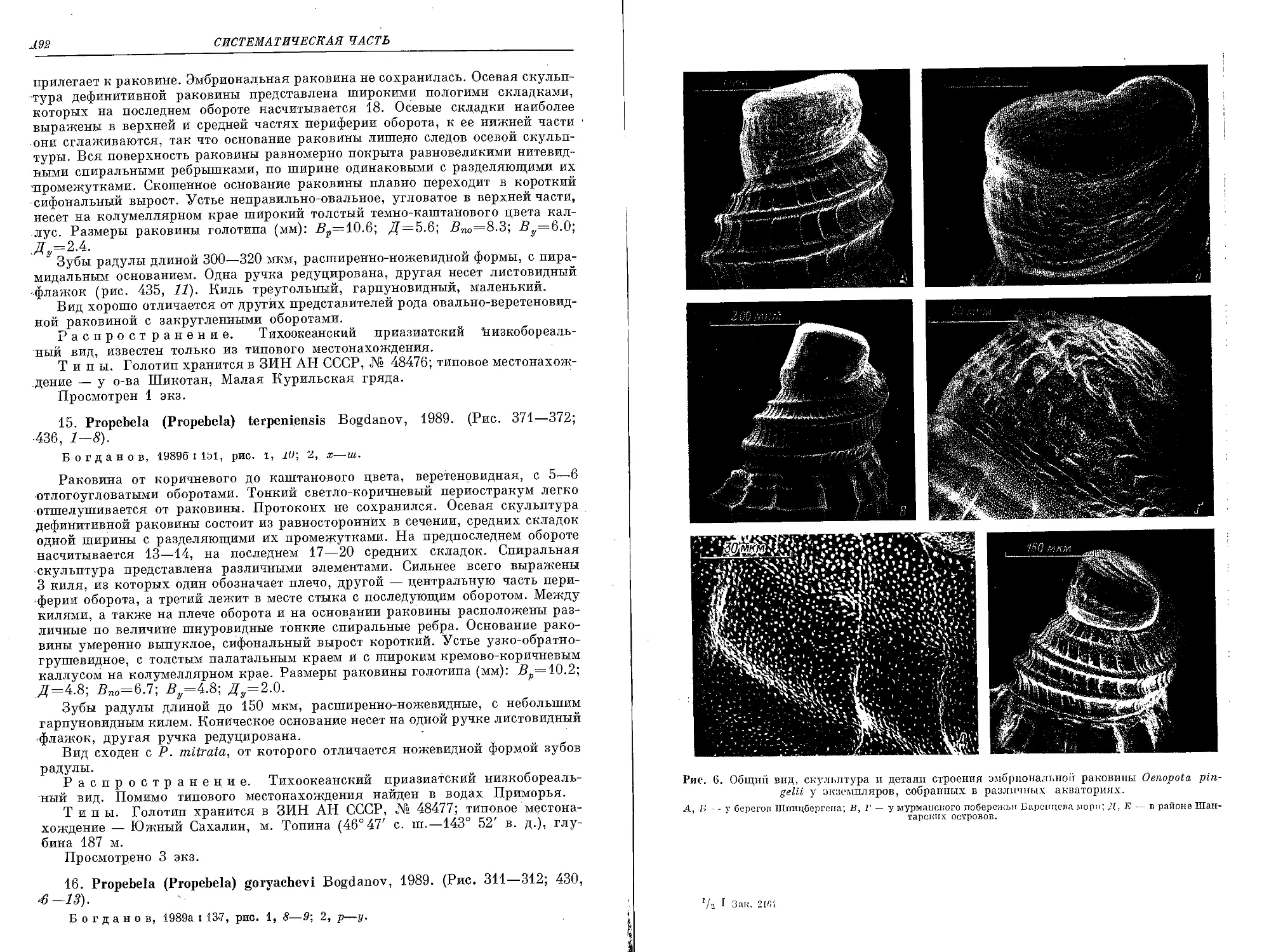
Рис. 1. Районы сбора представителей Oenopotinae, хранящихся в коллекции Зоологиче-
ского института АН СССР.
Всего просмотрено более 4200 проб, содержащих почти 12 000 экз., определенных
до вида. В каталоги отделения моллюсков Лаборатории морских исследований ЗИН АН СССР
занесено 10 860 экз. моллюсков Oenopotinae, остальные экземпляры хранятся в ЗМ МГУ
и ИВМ.
Анализ раковин моллюсков проводился под бинокуляром МБС-9. Особое внимание
уделялось изучению изменчивости осевой и спиральной скульптур дефинитивной раковины.
С этой целью для ряда экземпляров составлялось подробное описание раковины, просчитыва-
лось количество элементов спиральной скульптуры, для каждого из трех последних оборотов
считалось число осевых складок.
Анализ формы раковин проводился компараторным методом, разработанным и описан-
ным Я. И. Старобогатовым (Иззатуллаев, Старобогатов, 1984). Посредством зарисовки кон-
туров раковины данный метод позволяет наглядно сопоставить ряд параметров, характери-
зующих ее форму (таких как скорость нарастания раковины, т. е. увеличение последующего
оборота по отношению к предыдущему, шаг вдоль оси, шаг от оси, выпуклость оборотов и ряд
других). Простой и в то же время точный механизм сравнения значительного числа раковин
(нередко корродированных или обломанных) вне зависимости от возраста моллюсков явля-
ется большим преимуществом компараторного метода.
После изучения раковины из нее извлекалось тело моллюска. С этой целью приходилось
прибегать к частичному разрушению стенок раковины! на предпоследнем обороте, несколько
позади от устья, с помощью заточенной препаровальной иглы просверливалось отверстие
малого диаметра. Через него энтомологической иглой обрезался колумеллярный мускул,
и тело моллюска выдавливалось через устье. Такая методика позволяет сократить время
исследования моллюсков, извлекать их тела без существенных повреждений, сохранять рако-
вины для коллекции в практически целом состоянии и фотографировать их со стороны устья
без видимых повреждений. Фотографирование раковин производилось в Лаборатории научно-
прикладной фотокинематологии (ЛАФОКИ) АН СССР аппаратом «Hasselblad».
Изучение морфологии эмбриональной раковипы осуществлялось с помощью сканируют
щего электронного микроскопа «Hitachi S-405A» в межкафедральной Лаборатории электрон-
ной микроскопии биологического факультета МГУ. Несмотря на то что сохранность эмбрио-
нальной раковины отмечена в среднем у 1 экземпляра из 500, основную часть материала
ио скульптуре протоконха удалось получить, исследуя раковины именно взрослых моллю-
18
ОБЩАЯ ЧАСТЬ
сков. Только в двух случаях удалось сфотографировать эмбриональные раковины, извлечен-
ные из кладок. Все остальные просмотренные кладки содержали либо эмбрионы, находя-
щиеся па ранней стадии развития, либо протоконхи только начинали формироваться и
не поддавались видовой диагностике. Большинство же кладок оказались пустыми.
Прежде чем приступить к обследованию мягкого тепа моллюска, от его ноги скальпелем
отделялась крышечка для дальнейшего изучения и зарисовки. Внутреннее строение моллю-
сков исследовалось на наиболее крупных и наименее мацерированных экземплярах. На ряде
экземпляров была сделана серия гистологических срезов переднего отдела пищеварительной
системы.
Учитывая особенности строения радулярного аппарата моллюсков Oenopotinae, препа-
рирование радулярного мешка осуществлялось в глицерине, без предварительной выварки.
Заключение в незастывающую жидкость позволило исследовать зубы радулы во всех ра-
курсах. Для зарисовки радулы и дальнейшего хранения в фондовых коллекциях ЗИН пре-
парат накрывался покровным стеклом, которое окантовывалось канадским бальзамом. Всего
было изготовлено и просмотрено более 500 препаратов радул.
В последние годы рядом авторов было предложено изучать морфологию радулы мол-
люсков на сканирующем микроскопе (Kohn et al., 1972; Cerff, 1981; Reid, Lawton, 1981;
Shimek, Kohn, 1981; Сысоев, 1983, и др.). Однако незначительная толщина и прозрачность
стенок лезвия зуба позволили нам разобраться в морфологических особенностях радулы
преобладающего большинства моллюсков Oenopotinae с помощью светового ’микроскопа
при увеличениях до 400 крат. Возможности сканирующего микроскопа были использованы
в единственном случае! для выявления желобка на поверхности шиловидного зуба Propebela
turricula.
При вскрытии мягкого тела моллюсков в различных отделах пищеварительной системы
обнаруживались хеты многощетинковых червей. Часто отмечались полихеты, удерживаемые
ринхостомом моллюска. Свободный участок тела червя отделялся и, как и препараты отдель-
ных хет, передавался на определение специалистам. Просмотренный материал послужил
основой для решения вопроса о питании Oenopotinae.
При биогеографическом районировании акваторий и для выявления сходства их видового
состава использовалась общепринятая методика (Голиков, 1980; Песенко, 1982, и др.) с при-
менением уравнений Жаккара (Jaccard, 1912)
С„ 100
aj~^ + Dn+i-Cs
и Престона (Preston, 1962)
D„
/т ---------”_____
р~ Dn+Dn+i-Cs’
где D„ и Dn+i — количество видов в сравниваемых общностях, Cs — число общих видов.
Кроме того, учитывались индексы Шимкевича—Симпсона
Cs ЮО
а8 D
min
и Чекановского—Серенсена
_ 2CS• 100
+ •
Полученные результаты сводились в таблицы.
Анализ распространения видов производился на основе как собственных, так и литера-
турных данных. Рассмотрение вопроса о распространении моллюсков по глубинам, грунтам,
в зависимости от солености и температуры основывалось на этикеточных данных и записях
экспедиционных журналов. Кроме того, привлекалась обширная гидрологическая литература
по распределению течений, структурам вод и связанных с ними температурой и соленостью
в различные сезоны.
ИСТОРИЯ ИЗУЧЕНИЯ
Необходимость подробного анализа литературных данных вызвана отсут-
ствием сводки по исследованиям Oenopotinae и разрозненным состоянием имею-
щихся первоописаний, сведений по распространению и экологии этих моллю-
сков, Сочетание исторического аспекта исследования фауны с географическим
позволяет судить о степени изученности определенных акваторий и о достовер-
ности да идентификации видов.
Я плане истории изучения фауны Oenopotinae рассмотрим следующие рай-
гопы. 1. (Северо-атлантический приевропейский — охватывает акватории Север-
ИСТОРИЯ ИЗУЧЕНИЯ
13
ного и Норвежского морей, вплоть до о-вов Шпицбергена на север, до прол. Ла-
Манш и Гасконского залива — на юге; кроме того, северную и северо-восточную
части Атлантики, включая Исландию. 2. Атлантический приамериканский —
объединяет воды американского побережья от |п-ова Кейп-Код на юге до
л-ова Лабрадор на севере и воды, омывающие Гренландию. 3. Тихоокеанский
приамериканский — охватывает воды Тихого океана от штата Калифорния
до Берингова пролива, включая Алеутские о-ва. 4. Японский — к нему отно-
сятся воды, омывающие острова Японии. 5. Канадский, или американский,
сектор Арктики. 6. Евразийский сектор Арктики. 7. Дальневосточный — вклю-
чает западную часть Берингова моря, Охотское и Японское (советские террито-
риальные воды) моря.
1. Северо-атлантический приевропейский район.
Началом изучения этого района, как и исследования Oenopotinae в целом,
можно считать 1788 год, когда был описан вид Buccinum pyramidale Strom,
1788.
||Через 15 лет был описан второй вид]— Murex turricula (Montagu, 1803).
В первом и втором иллюстрированных изданиях по современным моллюскам
Великобритании и Исландии (Brown, 1827, 1844) содержится описание третьего
вида — Pleurotoma reticulata. Однако диагноз,^приведенный в работе, составлен
очень кратко, а стилизованный рисунок не отражает морфологических особен-
ностей раковины моллюска, чем, очевидно, и была вызвана многолетняя пута-
ница с его идентификацией. В 1834 г. был описан обычный для бореальных вод
вид Pleurotoma trevellianum (Turton, 1834). Рив (Reeve, 1845) описал новый для
Арктики вид Pleurotoma borealis, не вошедший, однако, в его более позднюю
сводку (Reeve, 1846).
В 1855 г. Адамсы (Adams, Adams, 1855) описали новый для арктического бас-
сейна вид Bela arctica, однако отсутствие изображения голотипа, несмотря
на наличие диагноза, не позволяет идентифицировать|этот вид. Форбс и Хэнли
(Forbes, Hanley, 1853) привели диагнозы Mangelia (Bela) turricula тз..М. (В.) tre-
velliana с детальным разбором их синонимии. В 1869 г. для фауны Шпицбергена
был описан новый вариетет Pleurotoma violacea var. gigantea Morch, 1869, воз-
веденный позже в ранг самостоятельного вида (Knipowitsch, 1901).
«Выдающейся и необходимой для точного изучения группы» (Verrill, 1882)
работой стала книга Gapca (G. О. Sars, 1878) по моллюскам, собранным у северо-
западных и северных берегов Норвегии. Приведенные в ней оригинальные под-
робные диагнозы 22 видов и 5 вариететов, отнесенных к роду Bela, сопровож-
даются данными о распространении каждого вида, приводятся глубины, на ко-
торых они были обнаружены. В монографии содержится описание 5 новых видов
рода Bela-. В. obliqua, В. angulosa, В. conoidea, В.^с alar aides, В. assimilis. Ри-
сунки, иллюстрирующие описание каждого вида, отличаются прекрасным
исполнением и в большинстве случаев,^фотографически точны.
Длительное время изучением моллюсков северо-атлантических вод зани-
мался норвежский исследователь Герман Фриле. Ряд его работ (Friele, 1873,
1874, 1879) представлен региональными сводкамиДВ ^работе 1877 года (Friele,
1877) приведены описания новых видов Pleurotoma (Bela) ovalis и Р. (В.) willei,
иллюстрированные, однако, лишь рисунками крышечки и (для первого вида)
зубов радулы. Результатом обработки материалов Норвежской северо-атланти-
ческой экспедиции 1876—1878 гг. стала монографияшо моллюскам, вторая часть
которой посвящена целиком роду Bela (Friele, 1886).. В работе приведен список
почти 30 видов и вариететов, из которых Bela schmidti, В. decussata var. finmar-
chia, В. koreni (описанная, к сожалению, по единственному ювенильному экзем-
пляру) оказались новыми для науки.
Значительное число видов (под родовым названием Bela) было описано Ло-
каром (Locard, 1892, 1896, 1897). Большинство из них, однако, относится к дру-
гим родам или сведено в синонимы к северным видам. Из собранных им в водах
Ла-Манша видов только Bela turricula и В. trevelliana относятся к Oenopotinae.
В то же время Кёлер (Koehler, 1896) описал для Гасконского залива 6 видов
14
ОБЩАЯ ЧАСТЬ
si на основании особенностей скульптуры и формы раковины повысил ранг
вариетета В. pyramidalis var. semiplicata до видового,
В начале века вышел ряд работ (Friele, Grieg, 1901; Sykes, 1906; Grieg,
1909a, 1909b; Dautzenberg, Fischer, 1912; Grieg, 1931, и др.), представляющих,
собой в основном сводки по определенным участкам Северо-атлантического евра-
зийского района. Здесь необходимо упомянуть две работы по фауне моллюсков
Исландии. Первая (Odhner, 1910) содержит список из 14 видов рода Bela,
5 из которых отмечены впервые для Исландии, а для 4 даны изображения, вто-
рая — по фауне Иса-Фиорда (Odhner, 1915) с упоминанием 13 видов рода Bela
и описанием нового вариетета В. woodiana var. lechil.
Отдельно следует остановиться на работах Варена. Часть его публикаций
представлена справочниками (Waren, 1980; 1983), содержащими краткие био-
графии выдающихся малакологов прошлого столетия, ссылки на их работы
е перво описаниями видов, указания типовых местонахождений, фотографии ти-
пов. Данные его работ, посвященных изучению глубоководной фауны Норвеж-
ского моря (Hubendick, Waren, 1974; Bouchet, Waren, 1979), вошли в совместную
с Буше публикацию (Bouchet, Waren, 1980). Книга стала одним из основных
определителей батиальной и абиссальной фауны туррид. Описания И видов,
из которых Oenopota dictyophora, Ое. convexigyra — новые для науки, включают
сведения о распространении и местах сборов, краткие дифференциальные
диагнозы и иллюстрированы хорошими фотографиями дефинитивных раковин..
2. Атлантический приамериканский район. Начало изу-
чению фауны моллюсков атлантического побережья Америки положили работы
Кутуи (Couthопу, 1838, 1839). В первой описано 3 новых вида, из которых 1 от-
несен к роду Pleurotoma (Р. bicarinata'), а 2 остальных к Fusus (F. harpularius,
F. pleurotomarius). Следует отметить, что последний вид был описан по экзем-
пляру, найденному в желудке трески. После дополнительного вскрытия более
300 рыб исследователю удалось найти еще несколько экземпляров того же вида.
Приведенные в работе диагнозы очень подробны. Впервые даны и изображения
типов, указываются места сбора и хранения типового материала. Несмотря
на то что к концу 30-х гг. XIX столетия было описано всего 6 видов Oenopotinae,
Кутуи высказывает предположение о распространении моллюсков только в мо-
рях северного полушария и об их значительном видовом разнообразии. Работа
1839 г. (Couthouy, 1839) содержит описание Pleurotoma decussata — вида, ши-
роко распространенного в северных морях. В сводке по беспозвоночным
зал. Массачусетс (Gould, 1841) в том числе и моллюскам, перечислены ранее
описанные виды и приведены рисунки, повторяющие известные изображения
типов.
Начало активного изучения фауны Oenopotinae Приамериканского района
Северной Атлантики связано с работой Мёллера (Moller, 1842), в которой содер-
жатся описания 10 новых видов, отнесенных к роду Dejrancla: D. woodiana,
D. elegans, D. cinerea, D. cylindracea, D. nobilis, D. scalaris, D. exarata, D. livida,.
D. beckii, D. viridula. Еще 2 вида были описаны им по рукописи Бека (Beck,
m.s.) — это/?, pingelit hD. vahlii. К сожалению, диагнозы всех видов были даны
кратко, без указания места хранения типовых экземпляров и не сопровождались
изображениями. В том же 1842 г. был описан широко распространенный в бо-
реальных и арктических водах вид Pleurotoma violacea (Mighels, Adams, 1842).
Первые работы по моллюскам Канадского побережья Атлантики появились
в 60-х гг. XIX в. (Walker, 1862; Packard, 1863, 1867). В них проведено детальное
изучение прибрежной фауны Лабрадора и описан новый вид Bela robusta (Pa-
ckard, 1867). Тщательный анализ имеющегося материала позволил Паккарду
обнаружить 11 видов Bela, описанных ранее из северо-восточного района Ат-
лантического океана. Однако сведение Pleurotoma harpularta в синоним к виду
Bela woodiana оказалось явно ошибочным.
В 1870 г. вышло второе, расширенное и дополненное, издание работы Гулда
(Gould, 1870), включавшее уже 8 видов, 2 из которых были описаны под родовым,
названием Pleurotoma, остальные 6 как Bela. Совершенно неоправданным ока-
ИСТОРИЯ ИЗУЧЕНИЯ
15
залось разделение видов В. vtolacea и Р. bicarinata, отнесенных к тому же к раз-
ным родам.
Сравнение 8 видов, обнаруженных у восточного побережья Северной Аме-
рики, с моллюсками европейской фауны было проведено Джефрисом (Jeffreys,
1872). На основании анализа имевшегося материала автор отметил, что у боль-
шинства видов экземпляры, собранные у европейского побережья, имели более
крупные размеры, нежели особи из приамериканских вод.
Значительный вклад в изучение Oenopotinae внес американский ученый Бе-
рилл, описавший в серии работ (Verrill, 1880, 1884; Verrill, Smith, 1884 in Ver-
rill, 1885) ряд новых приамериканских видов, встречающихся как в Атлантике,
так и в Тихом океане: Bela pygmaea, В. hebes, Lora blakei, L. subturgida, L. sub-
vitrea, L. tanneri. Наибольший интерес вызывает его публикация 1882 г. (Ver-
rill, 1882), в которой рассмотрены обнаруженные у побережья Новой Англии
18 видов и 5 вариететов рода Bela, из них В. incisula, В. concinnula, В. gouldii
оказались новыми видами, а В. concinnula var. acuta и В. decussata var. pusilia —
новыми вариететами. Детальное описание моллюсков с сопутствующим диффе-
ренциальным диагнозом иллюстрировано прекрасными рисунками раковин.
Из фаунистических работ конца прошлого века следует упомянуть сводки
по моллюскам, собранным у восточного (Posselt, 1895) и западного побережий
Гренландии (Posselt, Jensen, 1898), содержащие сведения о 18 видах .и 11 ва-
риететах, из которых Bela woodiana var. tumida, В. decussata var. inf lata и
В. pyramtdalls var. valdeplicosa описаны впервые.
Длительное время изучением фауны моллюсков северной Атлантики зани-
мался Торсон. Его работы содержат подробные данные по фауне моллюсков
у восточной (Thorson, 1934, 1944) и юго-западной части Гренландии (Thorson,
1951), а также заливов и фиордов Исландии (Thorson, 1941). Изучение Bela
vtolacea позволило автору высказать предположение о том,- что образование
вариететов bicarinata, cylindracea, brevis, beckii зависит от местообитания мол-
люсков и существенное влияние на изменчивость раковины могут оказывать
такие факторы, как глубина, течение и др.
3. Тихоокеанский приамериканский район. В 1885 г.
вышла работа Краузе (Krause, 1885) по фауне моллюсков Берингова моря.
Автор использовал как основной источник для определения моллюсков работы
по атлантическим Bela. Из 12 видов и 2 вариететов, рассмотренных в работе,
3 оказались новыми. Это эндемичные приамериканские виды Bela liitkeana,
В. metschigmensis и часто встречающийся во всем Беринговом море- вид В. al-
brechti.
Целый этап в изучении Oenopotinae тесно связан с именем выдающегося
американского малаколога Уильяма|Долла. Первые его работы (Dall, 1871,
1884) содержали отдельные описания новых видов Bela, обитающих у западного
побережья северной Америки и у Аляски, таких как Bela laevigata, В. alasken-
sis (1871), В. harpa, В. murdochiana (1884). В 1886—1887 гг. вышли три работы
Долла (Dall, 1886а, 1886b, 1887), в которых он описал новые виды Bela sculptu-
rata, В. solida, В. krauset и сравнил известные приамериканские виды Bela
с европейскими. В большинстве работ по каждому виду автором дан очень точ-
ный дифференциальный диагноз. Совместно с Бартшем он описал новый вид
Bela maurellei (Dall, Bartsch, 1910). В 1919 г. вышла серия статей с описанием
приамериканской северотихоокеанской фауны рода Lora. Автором приведены
диагнозы новых видов L. babylonia (Dall, 1919с), L. flora, L.casentina, L. amiata,
L. rassina, L. galgana (Dall, 1919b) северо-западного побережья Америки. Самой
крупной его работой (Dall, 1919а) стало описание ’коллекций моллюсков
сем. Turridae, хранящихся в Национальном музее естественной истории США.
В результате им было выделено 27 новых видов рода Lora. Очень тщательные
диагнозы сопровождаются фотографиями раковин, приводятся сведения о месте
сбора типовых экземпляров и месте их хранения. Общее число видов Oenopoti-
nae, обнаруженных у северо-западного побережья Америки, не превышает 70
(Dall, 1921).
16
ОБЩАЯ ЧАСТЬ
Середина 40—60-х гг.. отмечена в отновном отдельными региональными свод-
ками (Bartsch, 1944; Burch, 1946; Bartsch, 1950; Morris, 1966), в которых лишь
упоминаются некоторые представители Oenopotinae Амеразийского района
Тихого океана.
4. Японский район. Работ японских исследователей по современ-
ным Oenopotinae очень мало. В 1951 г. для вод, омывающих о-в Хоккайдо,
было отмечено лишь 6 видов исследуемой группы (Kuroda, Kinoshita, 1951).
Ряд работ японского малаколога Хабе содержит сведения о моллюсках, отне-
сенных к роду Oenopota (Habe, Kira, 1962; Habe, Kuroda, 1962; Habe, Ito,
1965), однако существенную роль в изучении района сыграла его публикация
1958 г. (Habe, 1958) с описанием 3 новых видов: Obesotoma uchidai, О. hanasaki-
nensis, Oenopota okudai. По результатам глубоководных исследований на судне
«Сойо-Мару» была описана фауна абиссали и батиали южных Японских о-вов
(Okutani,. 1968, 1969). Из зал. Сагами описано 2 новых вида: Nematoma micro-
valuta и Funitoma delicata, обитающих на глубинах 1000—1500 м (Okutani,
1964).
5. Канадский, или американский, сектор Арктики.
Первой серьезной сводкой по фауне моллюсков американского сектора Арктики
можно считать каталог моллюсков (Bush, 1884), собранных экспедицией 1882 г.
под руководством Стирнса (W. Stearns) у побережья Лабрадора. В нем содер-
жатся сведения о 9 видах и 1 вариетете рода Bela. Впервые приведены изобра-
жения В. sarslv,. В. incisula я В. pleurotomaria из данного района, по которым
можно проводить их сравнение с европейскими видами. В 1901 г. была опубли-
кована региональная сводка по морским беспозвоночным восточного сектора
Канады (Whiteaves, 1'901), в которую вошли и результаты изучения моллюсков
Гудзонова залива (Whiteaves, 1885). Она содержит сведения о распространении
18 видов и 2 вариететов. На основании изучения значительного числа работ
предыдущих исследователей автор приводит свои суждения по синонимии ряда
видов.
Через 70 с лишним лет после публикации Долла (Dall, 1884) вышла работа
Мак-Джинити (MacGinitie, 1959) по фауне самой северной оконечности Аля-
ски — м. Барроу, которая включает 9 видов, отнесенных к родам Obesotoma,
Oenopota, Nodotoma. Описания видов сопровождаются прекрасно выполненными
фотографиями и четкими диагнозами.
По материалам, собранным в районе Чукотского поднятия у побережья
Северной Канады на станции, расположенной в 800 милях севернее Берингова
пролива, приводятся сведения о нахождении представителей Oenopotinae
в батиали. На глубинах от 540 до 1200 м было обнаружено 2 вида: Oenopota
inequita, Ое. pyramidalis (Clarke, 1963). В 1971 г. вышла работа по моллюскам
арктического сектора Канады (Macpherson, 1971), содержащая сведения о 14 ви-
дах рода Oenopota,. большинство из которых впервые обнаружены в исследован-
ном районе.
6. Евразийский сектор Арктики. Только во второй поло-
вине прошлого столетия начинается изучение северных морей России. По ре-
зультатам сборов шведской экспедиции от побережья Новой Земли до Енисея
были описаны новый вид Plearotoma novajasemljensts и вариетет Р. pyramidalis-
var. jenisseensis, распространенные в арктических водах (Leche, 1878). Для каж-
дого из И видов и 10 вариететов, обнаруженных экспедицией, приведены сведе-
ния по распространению, постанционный обзор распределения, а для ряда эк-
земпляров — промеры раковин. По материалам экспедиции на судне «Вега»
(Aurivillius, 1887) публикуются данные по фауне Bela северных морей России.
В работе содержатся сведения о 13 видах и 2 вариететах этого рода. Однако
краткие, не иллюстрированные рисунками описания не позволяют судить,
о достоверности определений ряда видов.
В конце прошлого—начале нынешнего столетий началось активное изучение-
фауны моллюсков Баренцева и Белого морей. К 1885 г. для Мурманского берега
уже известно 13 видов рода Bela (Герценштейн, 1885). Материалы, собранные
ИСТОРИЯ ИЗУЧЕНИЯ 1Г
А. Бирулей, А. Волковичем, А. Чернышевым у Шпицбергена в 1899—1900 гг.,
послужили основой для серии публикаций Книповича (Knipowitsch, 1901, 1903).
Большинство из приведенных им 12 видов и 2 вариететов отмечены впервые для
данного района. Значительную роль в познании фауны Белого моря сыграла
Соловецкая биологическая станция (Педашенко, 1897). Ряд работ, представляю-
щих, как правило, отчеты о деятельности Мурманской биостанции (Линко, 1902;
Аверинцев, 1908, и др.) или написанные по результатам экспедиций у Мурман-
ского побережья (Pfeffer, 1890; Norman, 1902; Hagg, 1905), послужили базисом
для полного описания фауны Кольского залива Дерюгиным (Дерюгин, 1915).
Работа содержит сведения по распространению и условиям обитания 15 видов
и.1 вариетета, отнесенных автором к роду Bela.
Из публикаций советских исследователей следует отметить работу Дерюгина
(1924), включающую описание нового вида Lora subarctica. Филатова и Зацепин
приводят список из 28 видов Bela для фауны Баренцева моря (Филатова, Заце-
пин, 1948).
7. Дальневосточный район. В середине прошлого столетия
были опубликованы первые сводки по фауне дальневосточных российских морей.
Миддендорф описал 2 новых вида — Pleurotoma schantarica и Р. simplex, привел
подробные диагнозы и тотальные рисунки раковин с отдельно изображенными
фрагментами скульптуры (Middendorff, 1849, 1851).
Только через 90 лет, в 1941 г., появилась публикация Бартша с описанием
8 новых видов из Японского моря (Bartsch, 1941). Работа была проделана на
основе коллекций, собранных экспедициями 1925—1933 гг. под руководством
К. М. Дерюгина.
Некоторые сведения о моллюсках Oenopotinae дальневосточных морей со-
держатся в работах Голикова с соавторами (Голиков, 1964; Голиков, Кусакин,
1978; Волова и др., 1979). В сводке по району Курильских о-вов (Голиков,
Гульбин, 1977) были описаны новые виды: Oenopota cingulata, Ое. valentlna,
Ое. kinkasanensts. Данные о нахождении Obesotoma schantarica в литоральной
зоне приведены в работе по фауне моллюсков о-ва Сахалин (Гульбин, Шуль-
мина, 1981).
Изложенный выше материал демонстрирует различную степень изученности
фауны Oenopotinae отдельных районов Мирового океана. Вследствие постоян-
ного притока сведений о нахождении новых видов и видов уже описанных,,
но ранее не обнаруженных в той или иной акватории Мирового океана,объем
обсуждаемой группы моллюсков до сих пор не был установлен.
Таблица 1
Видовой состав фауны Oenopotinae отдельных районов Мирового океана
Район Общее число видов Число эндемичных видов
Литературные данные * Собственные данные
1. Северо-атлантический приевропей- скии 41 12 4
2. Атлантический приамериканский 27 6 ?
3. Тихоокеанский приамериканский 66 — 26
4. Японский 8 — 4
5. Канадский сектор Арктики 22 1
6. Евразийский сектор Арктики 28** 36 4
7. Дальневосточный 17 68 24
* В графе «Литературные данные» приведено приблизительное число видов, в связи
с тем что, во-первых, установить правильность идентификации видов подчас не представля-
лось возможным и, во-вторых, в различных работах для одного вида часто приводились
различные названия. Здесь мы постарались сохранить позиции исследователей, в резуль-
тате чего цифры могут несколько завышать действительное число видов Oenopotinae.
** В основном сведения по Баренцеву и Белому морям.
*** в восточной части Чукотского моря.
2 И. П. Богданов
18 ОБЩАЯ ЧАСТЬ
На основе крупных региональных сводок и работ, рассмотренных выше,
а также и компилятивных сводок (Martini, Chemnitz, 1887; Kobelt, 1905; Old-
royd, 1927; Nordsieck, 1968, и др.) и списков видов (Tryon, 1884; Johnson, 1926;
Thiele, 1928; Keen, 1963, и др.) мы попытались подсчитать число видов Oeno-
potinae, составляющих современную фауну Мирового океана (табл. 1). Следует
оговориться, что для полноты картины привлекались литературные источники
даже с фрагментарным упоминанием об изучаемых моллюсках (Adams, 1840;
Carpenter, 1865; Weinkauff, 1875, 1876; Loyning, 1932; Richards, 1936; Ellis,
1960; Morris, 1960; Oskarsson, 1962, и др.).
По предварительным подсчетам, фауна Oenopotinae Мирового океана пред-
ставлена 100—110 видами. Наименее изучены район канадского сектора Арк-
тики (5) и Японский район (4).
ПОЛОЖЕНИЕ OENOPOTINAE В СИСТЕМЕ TURRIDAE
Прежде чем рассматривать вопрос таксономии Oenopotinae, необходимо уста-
новить статус исследуемых моллюсков в системе сем. Turridae.
В ряде работ, преимущественно отечественных (Голиков, Кусакин, 1962,
1978; Голиков, Гульбин, 1977; Гульбин, Шульмина, 1981, и др.), предлагается
относить их к сем. Brachytomidae Thiele, 1929. Для подсем. Brachytominae
(Thiele, 1929, 1931), входящего в состав сем. Conidae, характерно наличие
крышечки, а также радулы, которая состоит из двух рядов маргинальных зубов,
лишенных базальной мембраны. Однако в 60-х гг. нашего столетия было проде-
монстрировано независимое происхождение и различная морфология марги-
нальных зубов у вошедших в Brachytominae 8 родов (Powell, 1966). Известно
также, что образование единичных маргинальных зубов при редукции централь-
ных и латеральных и исчезновение радулярной мембраны происходят в несколь-
ких подсемействах Turridae независимо (Голиков, Старобогатов, 1975). Кроме
того, само название Brachytominae малоприемлемо (Powell, 1966; McLean,
1971) в связи с отсутствием характеристики радулы и крышечки типового вида
рода Brachytoma Swainson (Pleurotoma strombiformis Sowerby, 1839; =P. strom-
boides Sow., 1832).
Голиков и Старобогатов (1975) предложи'ли выделять в надсем. Conoidea
Rafinesque, 1815 отдельное сем. Raphitomidae Bellardi, 1875, характеризуемое
наличием только маргинальных зубов токсоглоссного строения, отсутствием
рахидальных и латеральных зубов и полной редукцией радулярной (базальной)
мембраны. Однако одной из главных характеристик подсем. Raphitominae
является отсутствие крышечки. В более поздней работе Белларди (Bellardi,
1877) предложил использовать для обозначения рассматриваемых в работе
моллюсков название Belinae Bellardi, 1874 — подсемейство, в которое входят
роды Bela, Lachesis, Clavatula, характеризующиеся наличием крышечки.
Однако у Murex nebula — типового вида рода Bela — крышечка отсутствует
(Powell, 1966), а маргинальные зубы имеют листовидное строение. Кроме того,
название Belinae, как и название Lorinae Thiele, 1925, является nomina oblita
и малопригодно из-за отсутствия четкого диагноза.
Американский исследователь Моррисон (Morrison, 1966) предложил разде-
лить всех Turridae на 3 семейства на основании особенностей строения радуляр-
ного аппарата. Выделенное им сем. Mangeliidae характеризуется отсутствием
радулярной мембраны в радулярном мешке. В это семейство им были объеди-
нены токсоглоссные моллюски с зубами, имеющими строение и используемыми
как у Conidae. Семейство Mangeliidae включает виды, представленные в основ-
ном родами Bela, Mangelia, Kurtziella, Clathromangelia, Propebela, Oenopota,
Phymorhynchus, Inquisitor, Taranis, Glyphostoma (Morrison, 1966). Однако выделе-
ние моллюсков с радулярной формулой 1 : 0 : 0 : 0 : 1 и редуцированной мем-
браной в особую группу и возведение ее в ранг семейства кажется преждевре-
менным, поскольку радулы, состоящие только из одной пары маргинальных
ПОЛОЖЕНИЕ В СИСТЕМЕ
19'
зубов в ряду, могли образовываться у Toxoglossa независимо и на разных этапах
эволюции (Powell, 1966; Shimek, Kohn, 1981).
Наиболее распространен в настоящее время взгляд на таксономический ста-
тус рассматриваемой группы видов как на представителей подсем. Mangeliinae*
Fischer, 1887 (Powell, 1942, 1966; Дмитров, 1973, и др.).
PSEUDOMELATOMINAE
ZONULISPIRINAE
BORSONIINAE
--- CLAVINAE: Calliclava
CLAVINAE: Imaclava
MANGELIINAE: Oenopota
г— STR ICTISPIR INAE
*— DAPHNELLINAE
r— CLATHURELLINAE
।—MANG-ELIINAE; Kurtziella
MITR О LUMN INA E
—1-----i------>---1-----1_____I_____L
3.0 2.5 2.0 1.5 1.0 0.5 0
Б
PSEUDOMELATOMINAE
CLAVINAE: Imaclava
CLAVINAE: Calliclava
STRICTISPIRINAE
ZONULISPIR INAE
MANG-ELIINAE: Kurtziella
MANGELIINAE: Oenopota
C LATHUR E LLIN A E
DAPHNELLINAE
M ITR О LUMN INA E
BOR SONIINAE
J_!_l_I_I_i_i i i
876543210
Рис. 2. Фенограммы подсемейств и некоторых родов Turridae, основанные на кластерном
анализе таксономических дистанций (по: Shimek, Kohn, 1981).
А — по радуле, В;— на основе геометрических характеристик раковины.
Характеризуются Mangeliinae маленькой веретеновидной, реже бикониче-
ской раковиной; радулой, состоящей из пары коротких и широких маргиналь-
ных зубов, а также отсутствием крышечки как у типового рода, так и у боль-
шинства входящих в него родов. Однако существенное отличие родов в подсем.
Mangeliinae заключается в неодинаковом строении самих зубов радулы. Изуче-
ние функциональной морфологии и эволюции токсоглоссной радулы (Shimek,
Kohn, 1981) демонстрирует большее единообразие в строении маргинальных
зубов моллюсков Oenopotinae, например с радулой некоторых Clavinae, нежели
с таковой у представителей Mangeliinae (рис. 2, А). Вместе с тем была показана
(Богданов, 1986) возможность образования зубов Oenopotinae (типичного строе-
ния) из зубов типа «hilted dagger» (см.: Powell, 1966), свойственных болыпин-
2*
20
ОБЩАЯ ЧАСТЬ
ству Mangeliinae (в частности, роду Kurtziella) и характеризующихся удлинен-
ным базальным участком и толстыми, слегка изогнутыми стенками лезвия, ко-
торые образуют дорсально открытый канал. Основные этапы эволюции марги-
нального зуба представляются в данном случае следующим образом: 1) истонче-
ние и увеличение площади поверхности стенок лезвия; 2) замыкание канала
зуба, происходящее за счет заворачивания стенок лезвия и дальнейшего их
перекрывания на дорсальной части; 3) формирование киля — дополнительного
ребра жесткости лезвия на стороне, противоположной месту перекрывания
стенок; 4) обособление базальной части и формирование «ручек» — образований
базальной части, призванных поддерживать определенный диаметр пустотелой
трубки лезвия. Иными словами, прототип зуба Oenopotinae морфологически
схож с зубами ряда Mangeliinae. Например, представители родов Bellacythara
и Euclathurella характеризуются зубами с массивной конусовидной базальной
частью с сильно развитыми ручками; тонкостенным лезвием с частично пере-
крывающимися стенками; длинным (обычно более половины длины лезвия)
килем.
Имеющиеся в настоящее время данные (Fischer, 1887; Powell, 1942, 1966;
McLean, 1971; Shimek, Kohn, 1981; Богданов, 1986, и др.) позволяют говорить
о сходстве рассматриваемой группы Oenopotinae с представителями подсем. Man-
geliinae по форме раковины (рис. 2, Б), степени развития ее осевой скульптуры,
а также по отсутствию радулярной мембраны и общности проморфологии зубов.
С другой стороны, от названного подсемейства их отличает: 1) наличие у Oeno-
potinae когтевидной крышечки, которая отсутствует у большинства Mangeliinae,
включая и изученных представителей типового рода; 2) отсутствие в половой
системе самцов изученных видов Oenopotinae мантийной поры, в то время
как у Mangelia brachystoma, представителя типового рода Mangeliinae, она есть
(Robinson, 1960). Кроме того, немногочисленные литературные данные по мор-
фологии протоконха в различных группах туррид (Powell, 1966; Bouchet,
Waren, 1980; Bernasconi, Robba, 1984, 1985, и др.) позволяют сделать вывод
о явных различиях в строении и особенностях скульптуры эмбриональных рако-
вин Oenopotinae и представителей других подсемейств Turridae.
Таким образом, несмотря на неполноту наших знаний по анатомии туррид
и морфологии их эмбриональной раковины, рассматриваемая группа видов
не может быть отнесена ни к Mangeliinae, ни к какому-либо другому подсемей-
ству Turridae и подлежит выделению ее в особое подсемейство.
Следует отметить, что признаками, характерными для подсем. Oenopotinae,
обладают не только рассматриваемые в данной работе моллюски — обитатели
Арктического бассейна и бореальных вод севера Атлантики и Пацифики,
но и 2 новозеландских рода Neoguraleus и Liracraea, у представителей которых
имеются мелкая скульптурированная раковина, протоконх, орнаментированный
тонкими спиральными ребрышками, типично токсоглоссные зубы; кроме того,
для них описана рудиментарная крышечка (Powell, 1966). К сожалению, я не
имел возможности ознакомиться с представителями названных родов и поэтому
воздерживаюсь от однозначного решения о включении их в состав подсем. Oeno-
potinae.
Вопрос о происхождении и формировании фауны моллюсков - Oenopotinae
требует как дополнительных исследований по морфологии большинства Turri-
dae, так и значительно большего числа достоверных палеонтологических нахо-
док представителей Oenopotinae. Сопоставление общего плана и деталей строе-
ния моллюсков Oenopotinae с другими представителями Turridae, проведенное
в основном на базе литературных данных (Powell, 1966; McLean, 1971; Shimek,
Kohn, 1981), свидетельствует в пользу происхождения рассматриваемых в ра-
боте моллюсков от общих с Mangeliinae предков. j
МОРФОФУНКЦИОНАЛЬНЫЙ АНАЛИЗ
21
МОРФОФУНКЦИОНАЛЬНЫЙ АНАЛИЗ
ПРЕДСТАВИТЕЛЕЙ OENOPOTINAE
Исследователи уделяли основное внимание изучению раковины и исполь-
зованию особенностей ее морфологии как единственного комплекса системати-
ческих признаков при видовой диагностике Oenopotinae. Большинство видов
описано только по дефинитивной раковине и особенностям ее скульптуры.
1АКОВИНА
Дефинитивная раковина. Моллюски Oenopotinae обладают дексиотропной
(декстральной, правозавитой) раковиной, средние размеры которой 6—10 мм.
Наибольшая высота раковины, 29 мм, отмечена для экземпляра Propebela nobilis
Рис. 3. Общий вид, основные элементы строения раковины Oenopotinae и схема измерения
ее параметров (на примере Propebela bergensis).
1 — высота раковины, 2 — высота последнего оборота, 3 — высота устья, 4 — наибольший диаметр рако-
вины, 5 — ширина устья, в — периферия оборота, 7 — плечо оборота с проходящим по нему килем, 8 —
верхний, околошовный участок оборота, 9 — средняя осевая складка, 10 — пологая широкая осевая складка,
11 — нитевидные ребрышки, 12 — тонкие ребра, 13 — средние ребра, 14 — колумеллярный край устья,
15 — палатальный край устья.
(с телом). Однако эти размеры не могут считаться максимальными, так как
в просмотренных материалах коллекции ЗИН обнаружена пустая раковина,
определенная как Oenopota harpa, со следующими размерами: высота раковины
31.4 мм, высота последнего оборота 20.0, высота устья 11.7, диаметр раковины
11.4, диаметр устья 4.5 мм. (Схема измерений раковины приведена на рис. 3).
По-видимому, это самая крупная из известных раковин Oenopotinae.
Цвет раковины не видоспецифичен, как правило, он розоватый, телесный
или светло-серый. Реже встречается песочная окраска, коричневая разных
22
ОБЩАЯ ЧАСТЬ
оттенков и у очень небольшого числа видов буро-фиолетовая. Периостракум
тонкий, часто прозрачный, иногда окрашен в более темные тона, но того же
цвета, что и раковина. Лишь у немногих видов периостракум имеет цвет, от-
личный от такового раковины. Например, раковина Curtitoma hebes почти белая
под темно-оливковым периостракумом. Периостракум, как правило, плотно
прилегает к раковине, реже он может частично отставать или отшелушиваться.
У большинства экземпляров поверхность раковины в значительной мере корро-
дирована так, что почти у 80 % просмотренных моллюсков скульптура сохраня-
ется только на последних оборотах раковины и лишь у незначительной части
моллюсков сохраняется протоконх.
Рис. 4. Основные типы формы оборотов, встречающиеся у Oenopotinae.
А — закругленные, сильно выпуклые обороты (Oenopota elegans)', Б — закругленные, умеренно выпуклые
обороты (Ое. pyramidalis); В — усеченно-конусовидные обороты (Ое. pingelii); Г — отлого-угловатые обо-
роты, тупоугловатые в верхней части (Propebela turricula); Д — уступчатые обороты, уплощенные в верхней
части (Р. rugulata); Е — уступчатые обороты, плоские в верхней части (Р. bergensis).
Форма дефинитивной раковины у разных видов варьирует от овальной
до веретеновидной или уступчатой и в основном определяет профиль оборотов.
Классификация форм оборотов (с некоторыми изменениями) дана в соответствии
с терминологией, приведенной Голиковым (1980) для раковин моллюсков рода
Bucctnum. Форма оборотов раковин Oenopotinae может быть закругленной, усе-
ченно-конусовидной, отлого-угловатой, уступчатой, в разной мере выпуклой
(рис. 4). Для оборотов с угловатым профилем выделяются: плечо оборота, около-
шовная площадка — верхний участок оборота, лежащий между плечом и верх-
ним швом оборота, и периферия, или боковой участок, оборота.
Рис. 5. Основные типы формы устья Oenopotinae.
А — широко-обратногрушевидное (Curtitoma decussata)\ Б —• узко-обратногрушевидное (Propebela svet-
lanae); В — полуовальное (Oenopota pyramidalis)-, Г — полуовальное, угловатое в верхней части (Curti-
toma trevellianaj; Д — неправильно-овальное, усеченное в верхней части (С. hebes)', Е — неправильно-
овальное, угловатое в верхней части (Obesotoma tenuilirata).
Основание раковины, как правило, округлое, в той или иной мере выпуклое,
плавно переходит в ейфональный вырост. Граница сифонального выроста
прослеживается лишь для немногих видов; например, у Oenopota harpa она
обозначена глубоким желобком.
Устье очень разнообразно по конфигурации, и его форма изменяется у раз-
ных видов от овальной до обратногрушевидной (рис. 5), что в значительной мере
объясняется профилем палатального края (в зарубежных работах — наружная
губа), скоростью навивания оборотов раковины и углом изгиба сифонального
выроста. Колумеллярный край имеет характерный S-образный изгиб и всегда
покрыт тонким каллусом, заходящим на париетальный участок последнего
оборота. Верхний участок палатального края у ряда видов Oenopotinae несет
МОРФОФУНКЦИОНАЛЬНЫЙ АНАЛИЗ
33
разной глубины анальный синус. Сифональный вырост по сравнению с таковым
большинства Turridae короткий. У группы видов, близких к Propebela turricula
и Oenopota pyramidalis, сифональный вырост удлинен и имеет легкий изгиб назад.
Сифональный канал обычно широкий и глубокий.
Скульптура раковины. Почти все исследователи считали скульптуру
раковины основным элементом в видовой диагностике обсуждаемых моллюсков.
Однако изменчивость как осевой, так и спиральной скульптуры настолько
велика, что путаница в терминологии оказалась неизбежной. Различные авторы
одни и те же элементы скульптуры обозначают как кили, ребра, наросты и т. д.
Заимствование терминологии для обозначения элементов скульптуры из работ
по другим группам моллюсков приводит к путанице в их размерных характери-
стиках. Проведенный анализ терминов дает возможность разграничить боль-
шинство типов скульптур. В настоящей работе основные элементы осевой
скульптуры именуются складками, а спиральной — ребрами и килями.
Наиболее тонкими элементами осевой скульптуры дефинитивной раковины
принято считать линии нарастания. Однако самыми мелкими осевыми элемен-
тами, играющими роль в диагностике видов, оказались узкие осевые складки.
Их характеристики следующие: ширина от 0.05 до 0.3, высота от 0.05 до 0.25 мм.
Для более крупных осевых элементов в зависимости от высоты и ширины следует
различать средние и широкие складки. Средние осевые складки характеризу-
ются шириной от 0.3 до 0.7 и высотой от 0.15 до 0.4 мм. Профиль осевых складок
разнообразен: треугольный, округлый, эллипсоидный, прямоугольный и др.
Самые крупные элементы осевой скульптуры — широкие складки (ширина
более 0.7 мм). Следует различать пологие широкие складки с высотой до 0.3
и высокие — более 0.3 мм. Существенно, что высота осевых складок неодинакова
на всем протяжении периферии оборота. В ряде случаев они сглаживаются
к основанию раковины, иногда на самом основании, реже осевые складки на
всем протяжении имеют постоянную высоту. Важной диагностической особен-
ностью является место наибольшей выраженности осевой скульптуры: оно мо-
жет быть на околошовном пространстве, чаще на периферии оборота. У Oeno-
potinae осевая скульптура обычно определяет конфигурацию и профиль пала-
тального края устья и форму линий нарастания. Осевые складки можно разде-
лить на две основные категории: сигмовидно изогнутые и прямые.
Спиральная скульптура дефинитивной раковины представлена большим на-
бором различных элементов. Самые тонкие из них обозначены нами как реб-
рышки. Ребрышки бывают двух типов: более тонкие — волосовидные и не-
сколько большего сечения — нитевидные. С ростом раковины происходит утол-
щение обоих типов ребрышек и их трансформация в ребра. В зависимости от их
ширины следует различать тонкие, средние и широкие ребра. В связи с тем что
Таблица 2
Зависимость размеров элементов.спиральной скульптуры от высоты раковины
Элементы спиральной скульптуры Размеры (ширина) элементов спиральной скульптуры, мкм
при высоте раковины от 4 до 11 мм * при высоте раковины более 11 мм
Ребрышко: волосовидное нитевидное Ребро: тонкое среднее широкое До 35 От 36 до 55 От 56 до 130 От 131 до 220 Более 221 До 39 От 40 до 65 От 66 до 170 От 171 до 250 От 250 до 600 и более
* Следует отметить, что первая графа «11 мм) характеризует в основном-
скульптурные элементы большинства моллюсков родов Curtitoma и Granotoma,
в то время как вторая (> 11 мм) — скульптуру большинства дефинитивных раковин
моллюсков Oenopota, Obesotoma, Propebela-
34
ОБЩАЯ ЧАСТЬ
одни и то же элементы спиральной скульптуры у разных видов в зависимости
от размеров раковины могут выглядеть по-разному, при описании спиральной
скульптуры помимо размерных характеристик ребер и ребрышек целесообразно
учитывать и пропорциональное их соотношение с высотой раковины (табл. 2).
У разных видов Oenopotinae элементы спиральной скульптуры имеют не
только различную ширину, но также высоту и профиль. Например, для ребер
отмечены практически все варианты перехода от выпуклого полукруглого про-
филя к уплощенному, практически прямоугольному. Ребра, имеющие прямо-
угольное сечение и разделенные узкими промежутками, получили название
шпуровидиых. Килем в настоящей работе обозначается сильно выступающее
над поверхностью раковины ребро (чаще широкое). Как правило, киль 1 и обо-
значает плечо оборота. Реже, например у Curtitoma violacea, Propebela cingulata
и у некоторых других видов, по периферии оборота раковины проходят дополни-
тельно еще 1—2 киля. Некоторые элементы осевой и спиральной скульптур
представлены на рис. 3.
Эмбриональная раковина представлена обычно полутора-двумя оборотами
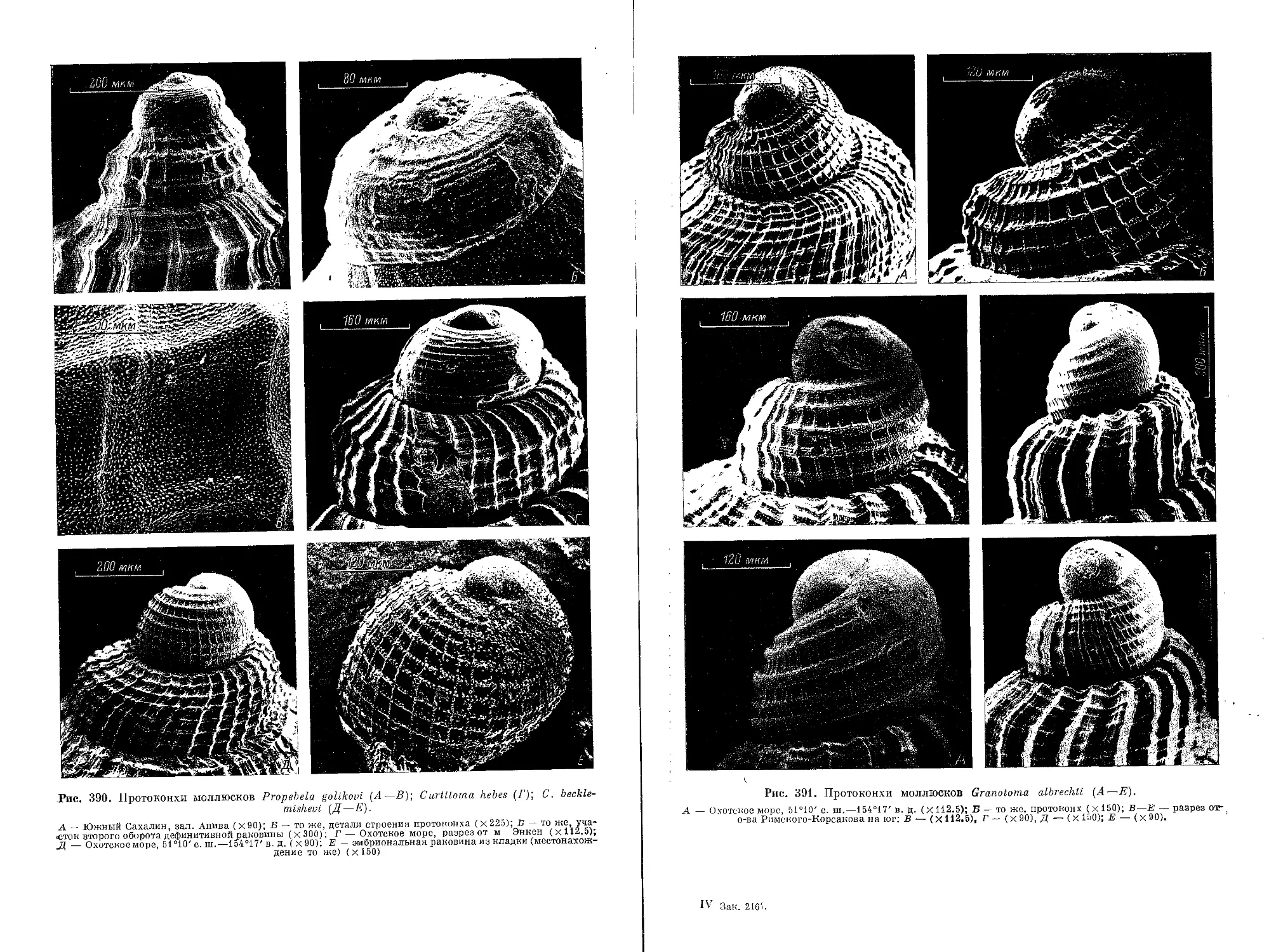
и по форме очень изменчива. Отмечены все стадии перехода от вершины рако-
вины, расположенной высоко над последующими оборотами, до погруженной
в первый полуоборот протоконха. Нами была подробно изучена скульптура
протоконхов более чем у 200 экз. раковин Oenopotinae.
У большинства видов спиральная скульптура эмбриональной раковины
представлена многочисленными волосовидными или нитевидными ребрышками.
Число их у разных видов варьирует от нескольких десятков волосовидных
ребрышек до двух тонких ребер. Закладка ребрышек у одних видов происходит
от самой макушки протоконха, у других — непосредственно в месте его перехода
в дефинитивную раковину. В таких случаях весь первый оборот эмбриональной
раковины оказывается лишенным спиральной исчерченности. Группе видов,
близких к Oenopota pingelii, свойствен ретикулятный рисунок на начальных
стадиях развития скульптуры протоконха (рис. 6, вкл.). Характерной чертой
ретикулятного рисунка является скульптура, состоящая не из осевых складок
и спиральных ребрышек, а из перемежающихся отдельных валиков и углубле-
ний (ямочек) на поверхности раковины. Направленность валиков не видоспеци-
фична, и ориентированы они могут быть под разным углом относительно оси
раковины (даже на поверхности одного экземпляра). Однако в самом ретикулят-
ном рисунке уже угадываются число и расположение будущих ребрышек.
У видов группы Obesotoma simplex протоконх лишен выраженных ребрышек,
а скульптура представлена лишь неупорядоченной тончайшей спиральной
струйчатостью. Следует отметить, что и спиральная скульптура дефинитивных
раковин у видов означенной группы всегда слабо выражена.
Осевая скульптура закладывается на эмбриональной раковине не у всех
видов Oenopotinae и отличается от осевой скульптуры дефинитивной раковины
конфигурацией и направленностью. Узкие осевые складки протоконха всегда
прямые, без каких-либо изгибов и расположены под углом к оси раковины.
На начальных оборотах дефинитивной раковины они становятся параллель-
ными оси навивания раковины, а затем приобретают окончательный облик
с сигмоидальным изгибом или изломом на околошовной поверхности. Облик
осевых складок и момент их закладки на эмбриональной раковине, вероятно,
не влияют на конфигурацию и выраженность осевой скульптуры дефинитивной
раковины.
С другой стороны, у большинства видов развитие спиральной скульптуры
дефинитивной раковины детерминировано скульптурой протоконха. Например,
у Curtitoma novaiasemljensis, С. decussata, С. conoidea многочисленные спираль-
ные ребрышки переходят в столь же неопределенное число элементов спираль-
ной скульптуры дефинитивной раковины (рис. 7, А, вкл.). У других видов
наблюдается процесс слияния ребрышек в более выраженные элементы спи-
ральной скульптуры. Так, у Curtitoma incisula и С. trevelliana на эмбриональной
раковине число спиральных ребрышек еще велико (рис. 7, Б), но постепенно
МОРФОФУНКЦИОНАЛЬНЫЙ АНАЛИЗ
85
они сливаются, образуя более толстые ребрышки (рис. 7, В), и уже на первом
постэмбриональном обороте таких ребрышек насчитывается всего 3 (рис. 7, Г).
’Столь же интересен процесс олигомеризации, проходящий на эмбриональной
раковине Curtitoma violacea. Изначально на небольшом по протяженности
отрезке протоконха закладывается множество тонких волосовидных ребрышек,
которые постепенно сближаются, образуя группы, а затем сливаются. В резуль-
тате на втором эмбриональном обороте образуется 3 небольших спиральных
киля. Два из них развиты сильнее и в дальнейшем преобладают над всей спи-
ральной скульптурой дефинитивной раковины. Один из них обозначает плечо,
другой — середину периферии. Приведенные примеры еще раз подтверждают
ведущую роль спиральной скульптуры протоконха в формировании скульптуры
дефинитивной раковины большинства видов Oenopotinae.
МОРФОЛОГИЯ МЯГКОГО ТЕЛА.
Морфология мягкого тела моллюсков была изучена более чем на 400 экз.;,
относящихся к 52 видам Oenopotinae.
Тело моллюсков расчленено на голову, ногу и висцеральный, или внутрен-
ностный, мешок и имеет окраску от телесного, нежно-розового до темно-вишне-
вого или коричневого цветов. Следует отметить, что у одних видов окраска
тела более или менее постоянна, а у других пигментация может сильно варьиро-
вать. Например, Obesotoma simplex, обитающие в дальневосточных морях, чаще
всего имеют окраску тела коричневато-вишневого цвета, в то время как для мол-
люсков этого вида, обитающих в северных морях Арктики, специфична телес-
ная, светлая окраска.
Головной отдел занимает переднюю дорсальную часть тела моллюска и
билатерально симметричен. Передняя часть головы снабжена парой конических
или почти цилиндрических щупалец, несущих чувствительные клетки. Щу-
пальца способны вытягиваться и сокращаться, но не втягиваться. Приблизи-
тельно посередине длины щупалец с наружной их стороны на округлых лопа-
стях расположены инвертированные глаза. У большинства моллюсков, оби-
тающих на глубинах более 500 м, глаза полностью редуцированы. На внешней
поверхности щупалец остается только след от них в виде слабо выраженного
утолщения. Между щупальцами на конусовидном выступе расположено отвер-
стие ринхостома.
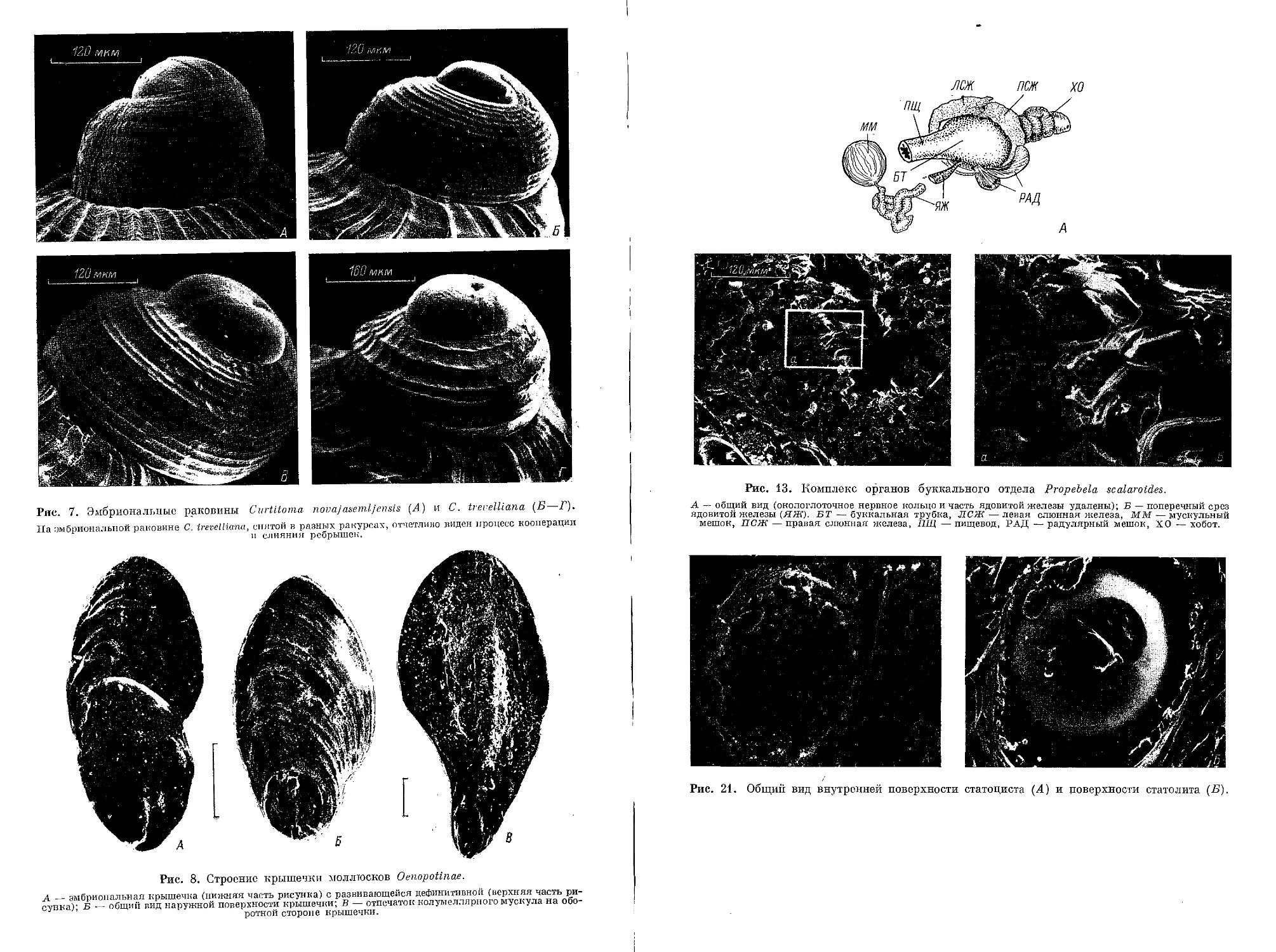
Нога, как и у большинства Toxoglossa, очень массивна. Мускулатура ноги
сильно развита; ползательная поверхность широкая, каплевидной формы.
На переднем, округлом участке ноги расположена педальная железа в форме
глубокого желобка (рис. 9).
На дорсальной поверхности заостренного участка ноги полукругом лежит
оперкулярная железа, секретирующая крышечку. У Oenopotinae размеры кры-
шечки в 2—3 раза меньше размеров устья и она безусловно является рудимен-
тарной. Крышечка листовидная, реже когтевидная, с терминально расположен-
ным ядром. Нарастание ее идет от заднего конца тела вперед по направлению
к головному отделу с постепенным изгибанием в правую сторону. Далее, при
описании крышечки, будет использован термин «ось нарастания крышечки»
(Богданов, 19836), обозначающий соответственно линию, проходящую от ядра
через все вершины следов ее нарастания. Оборотная сторона крышечки несет
след прикрепления колумеллярного мускула (рис. 8, вкл.). Отпечаток мускула
различается у разных видов по степени выраженности и форме — от округлой
до ромбовидной или клиновидной со всеми переходами (рис. 8, В, 32—36).
Мантийный комплекс органов. Мантийная полость покрыта довольно тонкой
мантией. Однако передний ее край, проходящий сразу за головой, толстостенный
и с левой стороны образует складку мантийного сифона. Сифон служит для по-
ступления воды внутрь мантийной полости. Дыхание моллюска обеспечивает
полулунно вытянутый ктенидий (рис. 9, 10), простирающийся от переднего до
заднего края мантии в левой ее части. Передний участок ктенидия лежит непо-
36
ОБЩАЯ ЧАСТЬ
средственно под основанием сифонального выроста. Характерно, что площадь
поверхности жаберных лепестков нарастает постепенно от края ктенидия, бли-
пж
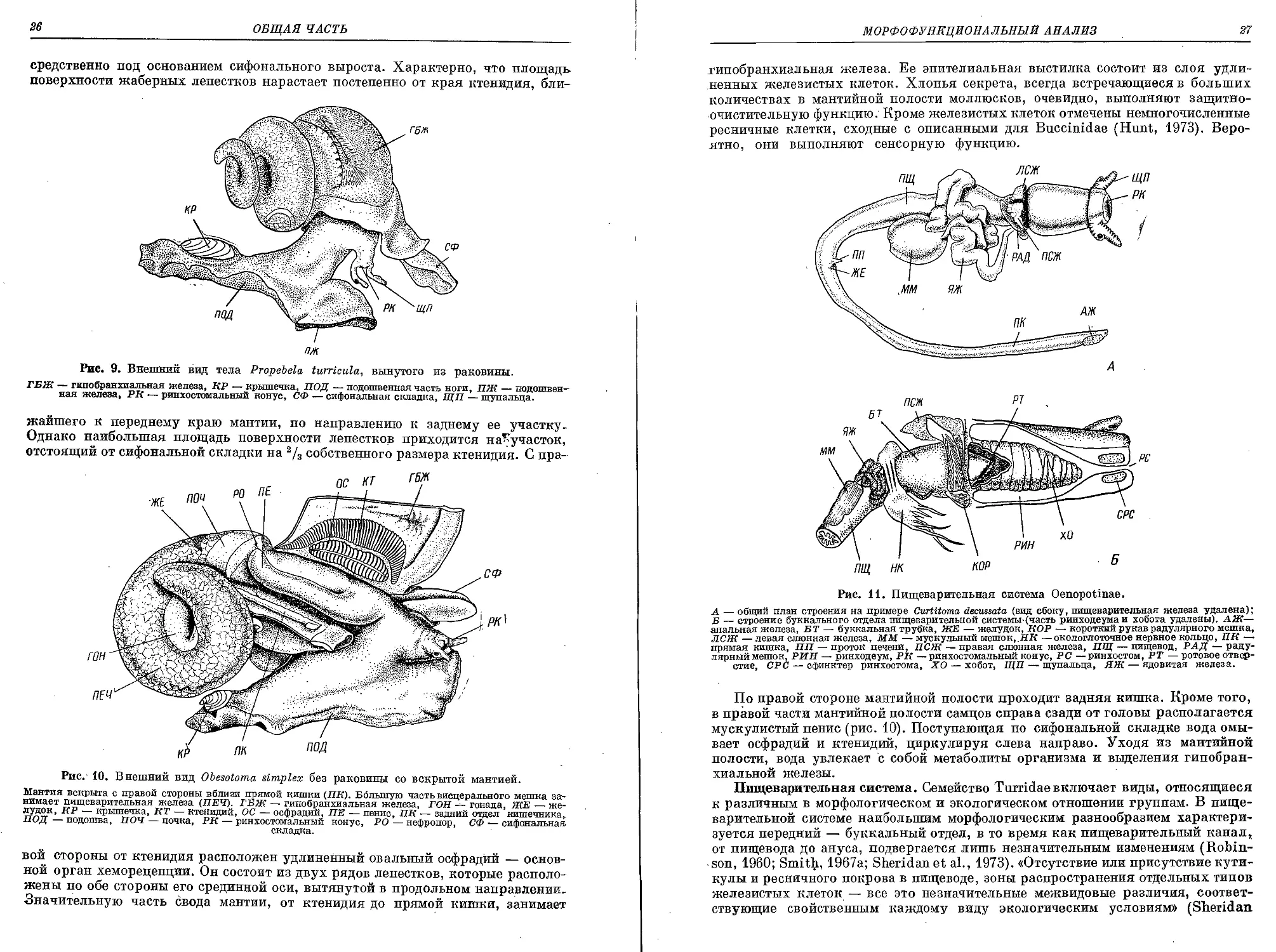
Рис. 9. Внешний вид тела Propebela turricula, вынутого из раковины.
ГБЖ — гипобранхиальная железа, RP — крышечка, ПОД — подошвенная часть ноги, ПЖ — подошвен-
ная железа, PR — ринхостомальный конус, СФ — сифональная складка, ЩП — щупальца.
жайшего к переднему краю мантии, по направлению к заднему ее участку-
Однако наибольшая площадь поверхности лепестков приходится на-участок,
отстоящий от сифональной складки на 2/3 собственного размера ктенидия. С пра-
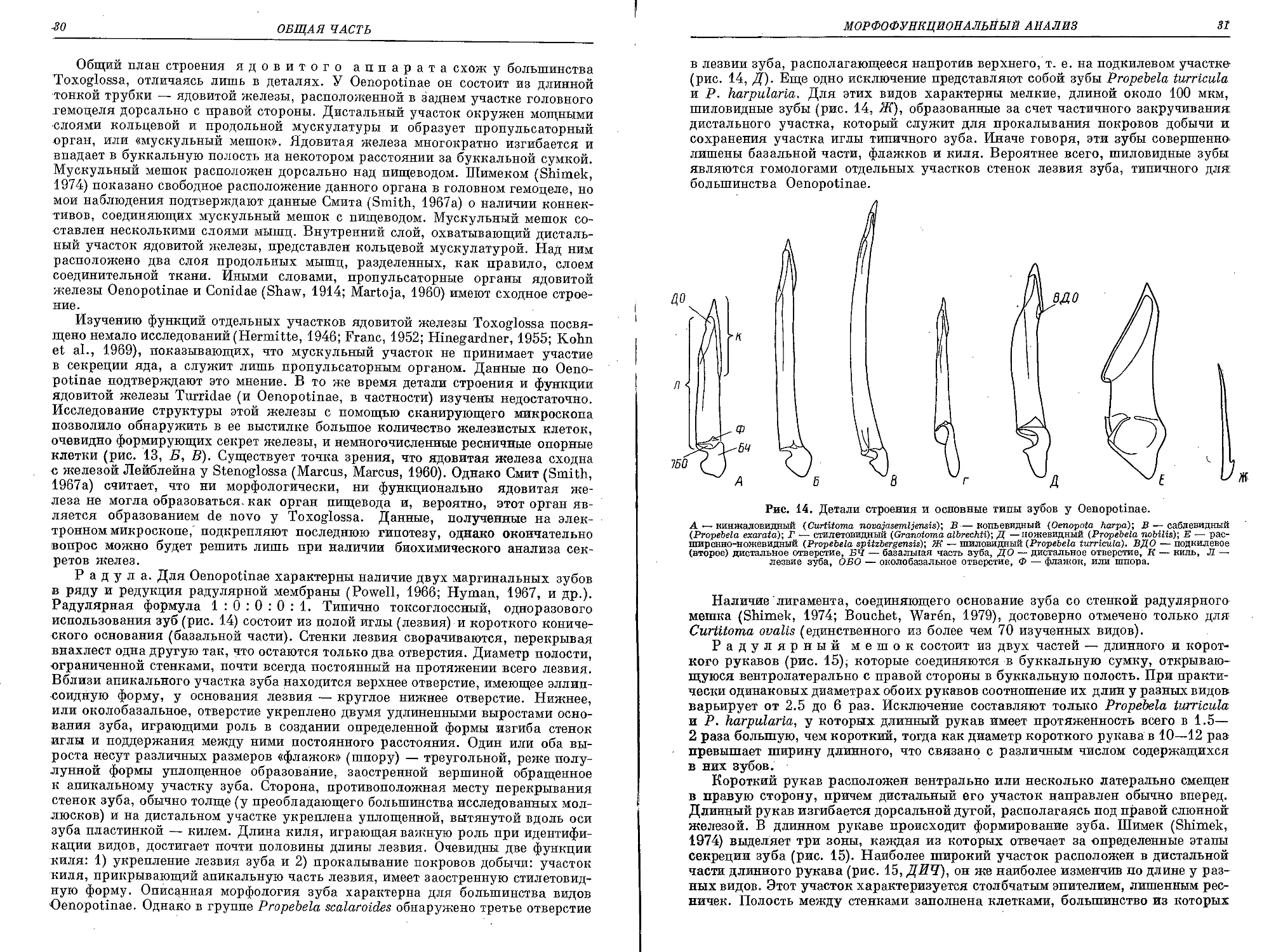
Рис. 10. Внешний вид Obesotoma simplex без раковины со вскрытой мантией.
Мантия вскрыта с правой стороны вблизи прямой кишки (ПР). Бблыпую часть висцерального мешка за-
нимает пищеварительная железа (ЛЕЧ). ГБЖ — гипобранхиальная железа, ГОН — гонада, ЖЕ — же-
лудок, RP — крышечка, НТ — ктенидий, ОС — осфрадий, ПЕ — пенис, ПН — задний отдел кишечника,
ПОД — подошва, ПОЧ — почка, PR — ринхостомальный конус, РО — нефропор, СФ — сифональная
складка.
вой стороны от ктенидия расположен удлиненный овальный осфрадий — основ-
ной орган хеморецепции. Он состоит из двух рядов лепестков, которые располо-
жены по обе стороны его срединной оси, вытянутой в продольном направлении.
Значительную часть свода мантии, от ктенидия до прямой кишки, занимает
МОРФОФУНКЦИОНАЛЬНЫЙ АНАЛИЗ
27
гипобранхиальная железа. Ее эпителиальная выстилка состоит из слоя удли-
ненных железистых клеток. Хлопья секрета, всегда встречающиеся в больших
количествах в мантийной полости моллюсков, очевидно, выполняют защитно-
очистительную функцию. Кроме железистых клеток отмечены немногочисленные
ресничные клетки, сходные с описанными для Buccinidae (Hunt, 1973). Веро-
ятно, они выполняют сенсорную функцию.
Рис. 11. Пищеварительная система Oenopotinae.
А — общий план строения на примере Curtitoma decussata (вид сбоку, пищеварительная железа удалена);
Б — строение буккального отдела пищеварительной системы-(часть ринходеумаи хобота удалены). АЖ—
анальная железа, ВТ — буккальная трубка, ЖЕ — желудок, КОР — короткий рукав радуля'рного мешка,
ЛСЖ — левая слюнная железа, ММ — мускульный мешок, НК —окологлоточное нервное кольцо, ПК —
прямая кишка, ПП — проток печени, ПСЖ — правая слюнная железа, ПЩ — пищевод, РАД — раду-
лярный мешок, РИН — ринходеум, РК —ринхостомальный конус, PC —ринхостом, РТ — ротовое отвер-
стие, СРС — сфинктер ринхостома, ХО — хобот, ЩП — щупальца, ЯЖ — ядовитая железа.
По правой стороне мантийной полости проходит задняя кишка. Кроме того,
в правой части мантийной полости самцов справа сзади от головы располагается
мускулистый пенис (рис. 10). Поступающая по сифональной складке вода омы-
вает осфрадий и ктенидий, циркулируя слева направо. Уходя из мантийной
полости, вода увлекает с собой метаболиты организма и выделения гипобран-
хиальной железы.
Пищеварительная система. Семейство Turridaeвключает виды, относящиеся
к различным в морфологическом и экологическом отношении группам. В пище-
варительной системе наибольшим морфологическим разнообразием характери-
зуется передний — буккальный отдел, в то время как пищеварительный канал,
от пищевода до ануса, подвергается лишь незначительным изменениям (Robin-
son, 1960; Smith, 1967а; Sheridan et al., 1973). «Отсутствие или присутствие кути-
кулы и ресничного покрова в пищеводе, зоны распространения отдельных типов
железистых клеток — все это незначительные межвидовые различия, соответ-
ствующие свойственным каждому виду экологическим условиям» (Sheridan
28
ОБЩАЯ ЧАСТЬ
et al., 1973). В пищеварительном тракте Oenopotinae выделяют 4 функционально-
различающихся участка (Shimek, 1974). Наиболее сложно устроен передний —
буккальный отдел, остальные 3 представлены пищеводом, желудком и кишечни-
ком с различными пищеварительными железами (рис. И, А).
Буккальный отдел пищеварительной системы начинается ринхо-
стомом, расположенным на выступающем между щупальцами ринхостомальном
конусе. Обычно он прикрыт спереди парными тонкими выростами — губами.
Для ряда видов отмечен эпителий ринхостома без ресничек (Shimek, 1974;
собственные данные), но, например, для Propebela turricula описаны ресничные
полосы непосредственно внутри ринхостома (Smith, 1967а). Ринхостом ведет
в обширную полость ринходеума (ринхоцель), окруженную сильно развитой
мускулатурой. Спереди ринхоцель ограничен очень массивной кольцевой муску-
латурой, лежащей за ринхостомом в основании щупалец и служащей сфинктером
ринходеума (рис. 11, Б). Мускулатура ринходеума состоит из сложной сети
пучков мышц, образующих стенки ринходеума и соединяющих его со стенками
тела и мускулатурой ноги. Подробно она изучена только у Oenopota levidensis-
(Shimek, 1974). Отмечено присутствие дорсальных, вентральных, дорсолатераль-
ных и латеральных мышечных валиков, создающих сложную сеть, окружающую-
ринходеум. Мускулатура ряда кольцевых складок стенок ринходеума обеспечи-
вает его расширение и сужение. При расширении происходит увеличение внут-
реннего пространства полости ринходеума, что необходимо при поглощении
пищи. Сфинктер не только закрывает отверстие, ведущее в ринходеум. Его функ-
ции многогранны: он захватывает и удерживает пищу; частично содействует
ее продвижению. Эпителий, выстилающий внутренню часть ринхостома, предста-
влен столбчатыми, сильно пигментированными клетками. Некоторые клетки
этой области секретируют кутикулу, образующую кутикулярную зону выстилки
ринхостома. По бокам от нее расположены ресничные области, образованные
также пигментированными клетками. Кроме описанных участков по бокам
ринхостома расположены группы клеток, секретирующих слизь. Эпителий,
выстилающий ринходеум, в основном также столбчатого типа. Вентральный и
латеральные участки, расположенные сразу за ринхостомом, покрыты кутику-
лой, а дорсальный участок на расстоянии х/3 от сфинктера ринходеума предста-
влен ресничной зоной. Наибольший по площади участок ринходеума, отстоящий
на х/3 от сфинктера и простирающийся почти до основания хобота, выстлан
сильно пигментированными клетками столбчатого эпителия с разбросанными
между ними многочисленными бокаловидными железистыми клетками. Реснич-
ный эпителий вновь появляется только у основания хобота.
Хобот Oenopotinae интраэмболического типа (Smith, 1967а) и состоит'
из двух стенок, между которыми расположена полость — гемоцель хобота
(рис. 12), посредством лакун соединяющаяся с головным гемоцелем. Эпители-
альная выстилка наружной стенки хобота различна по компонентам. Базальная
часть состоит из клеток столбчатого эпителия, большинство которых несет рес-
нички, между ними разбросаны железистые бокаловидные клетки. Следует от-
метить, что этот участок хобота по наличию железистых клеток соответствует
эпителиальной выстилке окружающего его участка ринходеума. Дистальный
участок лишен ресничного покрова и не содержит железистых клеток, эпите-
лий здесь кубоидальный. Этот участок хобота и вся внутренняя его стенка до
соединения с буккальной трубкой выстланы уплощенным эпителием, лишь дор-
сальная и вентральная поверхности рта представлены небольшими участками
ресничного эпителия (Shimek, 1974).
В основных чертах мускулатура интраэмболического хобота хорошо изучена
(Smith, 1967а; Sheridan et al., 1973; Shimek, 1974). Наружнаястенка его соста-
влена косой мускулатурой, в которой залегают разрозненные пучки кольцевой
мускулатуры. Для ряда видов отмечено сильное развитие кольцевой мускула-
туры дистального участка хобота, служащего своеобразным сфинктером.
Внутренняя стенка состоит из продольной мускулатуры, происходящей из:
педальной или цефалопедальной, и сливается с продольной мускулатурой, распо-
морфофункциональный анализ
39'
лагающейся снаружи буккальной трубки. Несколько радиальных мускульных
тяжей, пересекая гемоцель, соединяют наружную и внутреннюю стенки хобота
(рис. 12).
Буккальная трубка. Рот расположен в просвете хобота вблизи
его базальной части (рис. 12). У некоторых видов ротовое отверстие располо-
жено на конусовидном выросте, который является продолжением буккальной:
трубки, вдающейся в просвет хобота. Околоротовой конус, или передний отдел
буккальной трубки (собственно буккальная трубка в
Shimek, 1974), выстлан снаружи несекретирующими
клетками кубоидального эпителия (Sheridan et al.,
1973; Shimek, 1974). Ресничный покров при переходе
на внутреннюю поверхность рта постепенно редуци-
руется до отдельных разбросанных ресничных кле-
ток. Эпителий переднего отдела буккальной трубки
столбчатый, ресничный, с включением многочислен-
ных железистых клеток. Мускулатура околорото-
вого конуса представлена в основном продолжением
ретракторов хобота. Под ней расположена кольце-
вая мускулатура, наиболее развитая у основания
трубки и служащая буккальным сфинктером. Топо-
графически за буккальным сфинктером начинается
задний отдел буккальной трубки (буккальная по-
лость, — по: Shimek, 1974), выстланный простым
столбчатым эпителием. У таких видов, как Pro-
pebela turricula, Curtitoma trevelliana (Smith, 1967a),
Рис. 12. Схема строения хобота Oenopota levldensls (no: Shi-
mek, 1974).
БС — буккальный сфинктер, ВС — внутренняя стенка хобота, ГХ —
гемоцель хобота, НС — наружная стенка хобота, РТ — ротовое от-
верстие.
понимании Шимека, —
Propebela nobilis, Р. rugulata, Р. scalaroides (собственные данные), околоротовой:
конус не отмечен и местоположение ротового отверстия может быть обозначено-
приблизительно — в месте перехода базальной части внутренней стенки хобота
в задний отдел буккальной трубки.
В буккальную полость открываются протоки ядовитой и слюнных желез;
(рис. 13, А, вкл.). Часть радулярного мешка, от места впадения протоков слюн-
ных желез до буккальной полости, обозначается как буккальная сумка (Robin-
son, 1960; Smith, 1967а), состоит в основном из мускулатуры буккальной трубки
и выстлана довольно толстым слоем кутикулы (Smith, 1967а). Буккальная сумка
у разных видов может быть различной формы. В самом простом случае она пред-
ставлена узкой цилиндрической трубкой, впадающей в буккальную полость.
Усложнение или специализация буккальной сумки идет по пути расширения
протока в месте впадения в буккальную полость, так что она приобретает форму
воронки, широкой частью обращенной в буккальную полость.
Слюнные железы парные, расположены дорсолатерально в головном
гемоцеле. Правая железа, как правило, меньше левой, и проток, соединяющий
ее с буккальной сумкой, также значительно короче левого протока. Слюн-
ные железы образованы двумя типами клеток: опорными ресничными и желези-
стыми. Секрет слюнных желез выполняет одну или несколько функций. Оче-
видно, он, так же как и у Conidae, состоит в основном из протеазы, разрушающей
клетки внутри уже сформированного радулярного зуба (Marsh, 1971). Кроме
того, он может составлять один из компонентов ядосодержащей жидкости,
впрыскиваемой в добычу, или, возможно, активизирует яд по схеме, аналогич-
ной схеме превращения трипсиногена в трипсин (Shimek, 1974).
30
ОБЩАЯ ЧАСТЬ
Общий план строения ядовитого аппарата схож у большинства
Toxoglossa, отличаясь лишь в деталях. У Oenopotinae он состоит из длинной
тонкой трубки — ядовитой железы, расположенной в заднем участке головного
темоцеля дорсально с правой стороны. Дистальный участок окружен мощными
слоями кольцевой и продольной мускулатуры и образует пропульсаторный
орган, или «мускульный мешок». Ядовитая железа многократно изгибается и
впадает в буккальную полость на некотором расстоянии за буккальной сумкой.
Мускульный мешок расположен дорсально над пищеводом. Шимеком (Shimek,
1974) показано свободное расположение данного органа в головном гемоцеле, но
мои наблюдения подтверждают данные Смита (Smith, 1967а) о наличии коннек-
тивов, соединяющих мускульный мешок с пищеводом. Мускульный мешок со-
ставлен несколькими слоями мышц. Внутренний слой, охватывающий дисталь-
ный участок ядовитой железы, представлен кольцевой мускулатурой. Над ним
расположено два слоя продольных мышц, разделенных, как правило, слоем
соединительной ткани. Иными словами, пропульсаторные органы ядовитой
железы Oenopotinae и Conidae (Shaw, 1914; Martoja, 1960) имеют сходное строе-
ние.
Изучению функций отдельных участков ядовитой железы Toxoglossa посвя-
щено немало исследований (Hermitte, 1946; Franc, 1952; Hinegardner, 1955; Kohn
et al., 1969), показывающих, что мускульный участок не принимает участие
в секреции яда, а служит лишь пропульсаторным органом. Данные по Oeno-
potinae подтверждают это мнение. В то же время детали строения и функции
ядовитой железы Turridae (и Oenopotinae, в частности) изучены недостаточно.
Исследование структуры этой железы с помощью сканирующего микроскопа
позволило обнаружить в ее выстилке большое количество железистых клеток,
очевидно формирующих секрет железы, и немногочисленные ресничные опорные
клетки (рис. 13, Б, В). Существует точка зрения, что ядовитая железа сходна
с железой Лейблейна у Stenoglossa (Marcus, Marcus, 1960). Однако Смит (Smith,
1967а) считает, что ни морфологически, ни функционально ядовитая же-
леза не могла образоваться, как орган пищевода и, вероятно, этот орган яв-
ляется образованием de novo у Toxoglossa. Данные, полученные на элек-
тронном микроскопе,' подкрепляют последнюю гипотезу, однако окончательно
вопрос можно будет решить лишь при наличии биохимического анализа сек-
ретов желез.
Радула. Для Oenopotinae характерны наличие двух маргинальных зубов
в ряду и редукция радулярной мембраны (Powell, 1966; Hyman, 1967, и др.).
Радулярная формула 1 : 0 : 0 : 0 : 1. Типично токсоглоссный, одноразового
использования зуб (рис. 14) состоит из полой иглы (лезвия) и короткого кониче-
ского основания (базальной части). Стенки лезвия сворачиваются, перекрывая
внахлест одна другую так, что остаются только два отверстия. Диаметр полости,
ограниченной стенками, почти всегда постоянный на протяжении всего лезвия.
Вблизи апикального участка зуба находится верхнее отверстие, имеющее эллип-
соидную форму, у основания лезвия — круглое нижнее отверстие. Нижнее,
или околобазальное, отверстие укреплено двумя удлиненными выростами осно-
вания зуба, играющими роль в создании определенной формы изгиба стенок
иглы и поддержания между ними постоянного расстояния. Один или оба вы-
роста несут различных размеров «флажок» (шпору) — треугольной, реже полу-
лунной формы уплощенное образование, заостренной вершиной обращенное
к апикальному участку зуба. Сторона, противоположная месту перекрывания
стенок зуба, обычно толще (у преобладающего большинства исследованных мол-
люсков) и на дистальном участке укреплена уплощенной, вытянутой вдоль оси
зуба пластинкой — килем. Длина киля, играющая важную роль при идентифи-
кации видов, достигает почти половины длины лезвия. Очевидны две функции
киля: 1) укрепление лезвия зуба и 2) прокалывание покровов добычи: участок
киля, прикрывающий апикальную часть лезвия, имеет заостренную стилетовид-
ную форму. Описанная морфология зуба характерна для большинства видов
Oenopotinae. Однако в группе Propebela scalaroides обнаружено третье отверстие
МОРФОФУНКЦИОНАЛЬНЫЙ АНАЛИЗ
31
в лезвии зуба, располагающееся напротив верхнего, т. е. на подкилевом участке-
(рис. 14, Д). Еще одно исключение представляют собой зубы Propebela turricula
и Р. harpularia. Для этих видов характерны мелкие, длиной около 100 мкм,
шиловидные зубы (рис. 14, Ж), образованные за счет частичного закручивания
дистального участка, который служит для прокалывания покровов добычи и
сохранения участка иглы типичного зуба. Иначе говоря, эти зубы совершенно,
лишены базальной части, флажков и киля. Вероятнее всего, шиловидные зубы
являются гомологами отдельных участков стенок лезвия зуба, типичного для
большинства Oenopotinae.
Рис. 14. Детали строения и основные типы зубов у Oenopotinae.
А— кинжаловидный (Curtitoma novajasemljensis)', Б—копьевидный (Oenopota harpa)', В — саблевидный
(Propebela exarata); Г — стилетовидный (Granotoma albrechtiy, Д — ножевидный (Propebela nobilis)\ Е — рас-
пшренно-ножевидный (Propebela spitzbergensisy, Ж — шиловидный (Propebela turricula). ВДО — подкилевое
(второе) дистальное отверстие, БЧ — базальная часть зуба, ДО — дистальное отверстие, К — киль, Л —
лезвие зуба, ОБО — околобазальное отверстие, Ф — флажок, или шпора.
Наличие лигамента, соединяющего основание зуба со стенкой радулярного
мешка (Shimek, 1974; Bouchet, Waren, 1979), достоверно отмечено только для
Curtitoma ovalis (единственного из более чем 70 изученных видов).
Радулярный мешок состоит из двух частей — длинного и корот-
кого рукавов (рис. 15), которые соединяются в буккальную сумку, открываю-
щуюся вентролатерально с правой стороны в буккальную полость. При практи-
чески одинаковых диаметрах обоих рукавов соотношение их длин у разных видов,
варьирует от 2.5 до 6 раз. Исключение составляют только Propebela turricula
и Р. harpularia, у которых длинный рукав имеет протяженность всего в 1.5—
2 раза большую, чем короткий, тогда как диаметр короткого рукава в 10—12 раз
превышает ширину длинного, что связано с различным числом содержащихся
в них зубов.
Короткий рукав расположен вентрально или несколько латерально смещен
в правую сторону, причем дистальный его участок направлен обычно вперед.
Длинный рукав изгибается дорсальной дугой, располагаясь под правой слюнной
железой. В длинном рукаве происходит формирование зуба. Шимек (Shimek,
1974) выделяет три зоны, каждая из которых отвечает за определенные этапы
секреции зуба (рис. 15). Наиболее широкий участок расположен в дистальной
части длинного рукава (рис. 15, ДИЧ), он же наиболее изменчив по длине у раз-
ных видов. Этот участок характеризуется столбчатым эпителием, лишенным рес-
ничек. Полость между стенками заполнена клетками, большинство из которых
32
ОБЩАЯ ЧАСТЬ
является одонтобластами. Одонтобласты концентрируются на дорсальных
частях двух участков, каждый из которых секретирует один ряд зубов. Вероят-
нее всего, только лезвие зуба секретируется одонтобластами, так как базальная
часть зуба и лигамент не обнаружены в зоне одонтобластов, а отмечаются изна-
чально в средней зоне (рис. 15, СЧ) длинного рукава (Shimek, 1974). Очевидно,
что основание зуба формируется внутриклеточно в два этапа. Сначала секрети-
руется сетчатый матрикс материала зуба, после чего происходит его наполнение.
Зубы, находящиеся в дистальной части радулярного мешка, представляют
собой копии взрослых зубов с тем
Рис. И5. Схема строения радулярного
мешка и (выделяемые в нем зоны (по:
Shimek, 1974). Пояснения в (тексте.
лишь отличием, что их лезвия очень тонко-
стенны, а базальная часть сильно упло-
щена. Практически полное сходство зубов,
как формирующихся в дистальном участке
радулярного мешка, так и уже сформиро-
вавшихся, отмечено у преобладающего
большинства видов. Иная закладка зубов
у Propebela turricula и Р. harpularia. В се-
креции каждого зуба у этих видов уча-
ствует только один одонтобласт, и по мере
его роста меняется конфигурация зуба
(рис. 433, 23—28). На первых стадиях
формирования зуб представляет собой
треугольную уплощенную пластинку.
В дальнейшем апикальная часть удлиня-
ется, а края пластинки частично закручи-
ваются (рис. 433, 1—5) и зуб приобретает
окончательную шиловидную форму.
Средний участок длинного рукава
радулярного мешка самый протяжен-
ный — более 2/s общей длины рукава.
На вентральной его стенке, граничащей
-с дистальным участком, расположены клетки, которые содержат базальные
части молодых зубов, в то время как лезвия этих зубов могут находиться еще
в дистальном участке. Уже в среднем участке можно обнаружить зубы, частично
вышедшие из клеточной оболочки. Очевидно, что именно на этом участке проис-
ходит окончательное формирование как лезвия, так и основания зуба.
Проксимальный участок (рис. 15, ПЧ) характеризуется началом разрушения
клеток, формировавших зуб, и в самой его базальной части обнаруживается уже
неклеточная масса тканевых остатков. Эпителий стенок радулярной сумки на
данном участке представлен кубоидальными клетками.
Короткий рукав радулярного мешка, морфологически являющийся продол-
жением длинного рукава, функционально от него отличается. На всем протяже-
нии короткого рукава, вплоть до его дистального участка, происходит процесс
разрушения клеточного вещества в просвете лезвия зуба. Под воздействием
секрета слюнных желез зубы оказываются целиком очищенными от клеток и кле-
точных элементов. Эпителий стенок короткого рукава состоит из кубоидальных
и невысоких столбчатых клеток. Полость короткого рукава разделена на две
части поднимающейся снизу сагиттальной септой. У разных видов размеры септы
могут варьировать от маленького возвышения на дне до крупной пластинки,
перегораживающей весь короткий рукав радулярного мешка на две камеры.
Сама септа составлена клетками столбчатого эпителия, которые у основания
шире, чем в апикальной его части. Выталкивание зуба в буккальную полость
происходит за счет сокращения мускулатуры короткого рукава радулярного
мешка и стенок буккальной сумки.
Таким образом, функции радулярного мешка заключаются в формировании
зубов, их хранении и передаче в буккальную полость.
Пищевод представлен трубкой, для которой характерно наличие регу-
-лярных продольных складок на внутренней стенке. В месте соединения буккаль-
МОРФОФУНКЦИОНАЛЬНЫЙ АНАЛИЗ
33
кого отдела с пищеводом имеется эпителий нескольких типов: от простого не-
высокого ресничного с немногочисленными железистыми клетками до высокого
столбчатого с многочисленными железистыми бокаловидными клетками. Веро-
ятно, весь эпителий ресничный. В самом пищеводе число бокаловидных клеток
уменьшается, некоторые железистые клетки пищевода выделяют энзимы (Smith,
1967b). По мнению Пирса (Pearce, 1966), у Curtitoma trevelltcma черви, попадаю-
щие внутрь пищевода, подвергаются быстрому перевариванию. Возможно,
секрет железистых клеток смазывает пищу при ее продвижении на данном
участке, по также он, вероятно, участвует и в пищеварении (Shimek, 1974).
Мускулатура пищевода, как косая, так и кольцевая, расположена диффузно
и не обеспечивает активной перистальтики.
Желудок сильно упрощен по сравнению с другими неогастроподами и
представлен простой, U-образно изогнутой трубкой небольшого диаметра.
Внутренняя поверхность желудка образует два тифлозоля с расположенным
между ними кишечным желобком (рис. 16). Над тифлозолями формируется
«уплощенная зона» желудка, поверхность которой собрана в многочисленные
поперечные складки. Желудок, как и пищевод, характеризуется наличием рес-
ничного эпителия, содержащего железистые клетки. Больший тифлозоль у Pro-
pebela turricula несет преимущественно слизистые клетки (Smith, 1967b). За счет
работы ресничного эпителия в желудке образуются потоки жидкости
(рис. 16, А, Б), из которых можно выделить три основных: 1) проходящий по
левой стороне малого тифлозоля, 2) текущий между тифлозолями вдоль кишеч-
ного желобка и 3) текущий через поперечные складки уплощенной зоны. При-
нимая во внимание отсутствие у Oenopotinae зоны смешения и сортирующей зоны
и учитывая направление ресничных потоков, Смит (Smith, 1967b) пришел к вы-
воду, что пища попадает в желудок уже в значительной степени переварен-
ной. Спрессованный и слипшийся фекальный материал вдоль большого тифло-
золя поступает в кишку, в то время как переваренный материал поступает
в проток пищеварительной железы. Одно или (реже) два отверстия протока
печени открываются у начала тифлозолей, т. е. почти на границе пищевода с же-
лудком. Эпителий, выстилающий отверстие протока, составлен ресничными
клетками, но внутри собственно протока отмечены многочисленные слизистые
клетки (Smith, 1967b).
Двудольная «печень», или пищеварительная железа
занимает большую часть висцерального мешка и состоит из значительного числа
маленьких трубочек, которые, объединяясь, формируют один или два протока.
Гистология трубочек зависит от секретов, выделяемых другими железами, и
от специализации питания каждого вида (Smith, 1967b). Большая часть трубочек
составлена удлиненными пищеварительными клетками. У Propebela turricula
обнаружены также немногочисленные жгутиковые клетки, жгутики которых
направлены в просвет трубочек (Smith, 1967b).
Кишечник морфологически однороден, однако по составу эпителиаль-
ной выстилки Смит (Smith, 1967b) различает в нем четыре участка. 1. Короткий
участок, граничащий с желудком и схожий с ним по эпителиальной выстилке.
2. Протяженный участок, простирающийся вдоль колумеллярного мускула
с эпителием, состоящим из железистых и ресничных клеток. 3. Участок, топо-
графически начинающийся от колумеллярного мускула и проходящий вдоль
крыши мантийной полости до анального отверстия. Его эпителий состоит цели-
ком из коротких, густо покрытых ресничками клеток. 4. Короткий анальный
участок, выстланный многочисленными слизистыми клетками, между которыми
расположены ресничные клетки.
Функциональное назначение выделенных участков до конца не выяснено.
Возле анального отверстия в кишечник открывается анальная железа. Клетки,
выстилающие ее просвет, наполнены зелеными и черными гранулами. По-види-
мому, единственная функция анальной железы — экскреторная (Fretter, 1946).
Механизм питания. Сложное устройство переднего отдела пищеварительной
системы и резкое упрощение ее структуры от пищевода до ануса, касающееся
3 И. П. Богданов
34
ОБЩАЯ ЧАСТЬ
в первую очередь желудка, отсутствие придаточных желез пищевода —все
свидетельствует об избирательном хищничестве и своеобразии добывания пищи
и процесса пищеварения у Oenopotinae. Ряд наблюдений, полученных при ана-
томировании, в сочетании с хорошей гистологической изученностью органов
пищеварения позволили проанализировать функции и взаимодействие различ-
ных частей пищеварительной системы.
t
Рис. 16. Строение желудка Oenopotinae (по: Smith, 1967b).
А — Propebela turricula’, Б — Curtitoma trevelliana\ В — реконструкция желудка на уровне впадения про—
тока печени у Curtitoma trevelliana. Жирными стрелками показано место перехода пищевода в желудок.
КЖ — кишечный желобок, ПП — протоки печени, СУЗ — стенки уплощенной зоны желудка, Tt и Тг —
тифлозоль.
Основной функцией гибкого, способного к значительному вытягиванию хо-
бота является добывание пищи. Вместе с тем мускулатура хобота, включая и
развитую кольцевую мышцу дистального участка (сфинктер хобота), не может
обеспечить захват и длительное удержание активной добычи. Специальных орга-
нов удержания на хоботе также не отмечено. На основании данных Кона (Kohn,
1956, 1959) по моллюскам сем. Conidae было высказано предположение о том,
что зуб Oenopotinae служит шприцем, наполненным ядом, и так же, как у кону-
сов, во время нападения на добычу находится на конце хобота (Pearce, 1966).
Данное предположение подтверждается собственными исследованиями: у ряда-
экземпляров при вскрытии отмечено удержание дистальным участком хобота
единичного зуба радулы. В связи с тем что расстояние, которое зуб должен
пройти от буккальной сумки, в десятки раз превышает его собственную длину,
возникает вопрос 6 том, каким образом радулярный зуб попадает на дистальный
участок хобота. Высказывалось предположение о передвижении зуба к ротовому
МОРФОФУНКЦИОНАЛЬНЫЙ АНАЛИЗ
35
отверстию током жидкости, создаваемым ресничками (Shimek, 1974), и захвате
его там хоботом. Однако это предположение опровергается следующими фак-
тами. Во-первых, расстояние, которое проходит зуб от буккальной сумки до
ротового отверстия, превышает в десятки раз собственные размеры зуба. Во-вто-
рых, зубы некоторых видов достигают 1 мм в длину и трудно представить пере-
мещение зуба такой длины за счет работы отдельно разбросанных ресничных
Рис. 17. Схема взаимодействия мускулатуры (1—7) при захвате зуба хоботом и нападении
моллюска на добычу. Пояснения в тексте.
Обозначения как на рис. 11 и 12.
клеток и немногочисленных ресничных полей. И наконец, третий, пожалуй,
самый веский довод заключается в том, что короткий рукав радулярного мешка,
в котором находятся зрелые зубы, ориентирован вершиной вперед, а отверстием
назад. Соответственно при выходе из него зуб располагается в буккальной по-
лости так, что базальная его часть направлена вперед, по направлению к рото-
вому отверстию, а острие назад, в сторону пищевода. Таким образом, прежде
чем зуб будет захвачен дистальным участком хобота, он должен на каком-то
отрезке пути развернуться на 180°. Учитывая незначительный диаметр просвета
буккальной полости, предположить такой разворот практически невозможно.
Сказанное позволяет сделать вывод о единственном способе передачи зуба,
происходящем при максимальном сближении апикального участка хобота
с буккальной сумкой, т. е. путем сложных мышечных взаимодействий в перед-
нем отделе пищеварительной системы (Богданов, 19856). Хобот укорачивается
3*
36
ОБЩАЯ ЧАСТЬ
за счет сокращения продольной мускулатуры внутренней стенки (рис. 17, 4).
Одновременно с этим происходит сокращение продольной мускулатуры стенок
буккальной полости и дистальный участок хобота частично сближается с бук-
кальной сумкой. Благодаря сокращению мускулатуры стенок тела и, очевидно,,
мышц ноги, подходящих к базальной части хобота и ринходеума (рис. 17, А, 2)г
происходит растяжение мускулатуры буккального сфинктера. В результате
хобот приобретает коническую форму и его дистальный участок оказывается
на одном уровне с ротовым отверстием. Сокращением продольной мускулатуры
буккальная трубка укорачивается, и ротовое отверстие сближается с пищево-
дом. Следует отметить, что благодаря перечисленным мышечным действиям не’
только сокращается расстояние от радулярного мешка до дистального участка
хобота (например, у Oenopota harpa это расстояние становится равным длине-
зуба), но происходит и разворот короткого рукава радулярного мешка. В ре-
зультате он оказывается направленным вершиной назад, а отверстием вперед,
(рис. 17, Б). Соответственно зуб при выходе из радулярного мешка уже ориен-
тирован базальной частью назад и находится в положении, необходимом для
захвата его хоботом. На данном этапе зуб выталкивается из радулярного мешка
сокращением мускулатуры короткого рукава и входит в отверстие дистального-
участка хобота (рис. 17, Б). Транспортировка зуба может происходить и при
помощи ресничных полей ротового отверстия, однако их влияние несущест-
венно, так как в момент максимального сжатия расстояние от буккальной сумки
до дистального участка хобота оказывается равно, а подчас и меньше длины
самого зуба.
Здесь необходимо упомянуть о флажке, расположенном в месте перехода
базальной части зуба в лезвие. Становится очевидной роль этого образования:
при прохождении зуба через дистальное отверстие хобота происходит раздра-
жение нервных окончаний внутренней стенки апикального участка хобота шпо-
рой (флажком). В результате сфинктер хобота сокращается и захватывает зуб
(рис. 17, В, 5), при этом отверстие близ острия иглы оказывается снаружи,
хобота, а околобазальное — в просвете хобота. Хобот, несущий зуб радулы,,
может быстро вытягиваться, как только моллюск обнаружит пищевой объект.
В распрямлении хобота важную роль играют сокращение буккального сфинк-
тера, расслабление продольной мускулатуры внутренней стенки хобота и ра-
диальной мускулатуры, соединяющей наружную и внутреннюю стенки хо-
бота (рис. 17, Г).
Однако быстрота вытягивания хобота связана в основном с сокращением
кольцевой мускулатуры наружной стенки хобота, за счет чего резко возрастает
гидростатическое давление в гемоцеле хобота, ведущее к растяжению хобота
вдоль оси(рис. 17, Д, 7). Зуб, удерживаемый дистальным участком хобота, про-
бивает покровы добычи.
Умерщвление добычи. Гипотеза об умерщвлении добычи ядом,
содержащимся внутри лезвия зуба (Pearce, 1966), не объясняет механизма на-
полнения полости лезвия ядом. Для исследования этого вопроса потребовалось
прибегнуть к помощи гистологии. Просмотр серии поперечных срезов, сделан-
ных через буккальную область Propebela scalaris, выявил неравномерное разви-
тие складок стенки буккальной трубки. Они (складки) равновелики на всем
протяжении до буккальной сумки, а ниже места впадения ядовитой железы
опять переходят в одинаковой величины складки пищевода. Между буккальной
сумкой и отверстием ядовитой железы наблюдается образование ярко выражен-
ного желобка (канала), стенки которого составлены слипшимися или близко
расположенными складками (рис. 18), различными по величине и расстоянию
между ними. Вентральное расположение канала предполагает возможность
накопления в нем секрета ядовитой железы. У ряда видов, например у Grano-
toma albrechti, отверстие ядовитой железы соединяется с буккальной сумкой
рядом параллельных желобков. У некоторых видов происходит перемещение
отверстия ядовитой железы латерально вверх и вперед. Например, у Oenopota
harpa оба отверстия расположены одно над другим (рис. 18, В) и соединены ря-
МОРФОФУНКЦИОНАЛЬНЫЙ АНАЛИЗ
37
дом желобков, по которым яд может стекать из железы в буккальную сумку.
Наконец, у Propebela nobilis и Р. assimilis отмечено обособление участка буккаль-
ной полости, в результате чего проток ядовитой железы и буккальная сумка
оказываются в специальном углублении. Если яд накапливается в описанном
выше обособленном участке (желобок, канал или углубление), то он может
поступать в полость зуба либо при его выходе из радулярного мешка, либо не-
посредственно в самом мешке — в коротком его рукаве. Таким образом, пред-
положение о том, что полость лезвия является резервуаром для яда, оказыва-
ется верным, но лишь для моллюсков с длинными широкими зубами, в которых
оба отверстия достаточно малы, чтобы из них не вымывался яд при транспор-
тировке зуба и при резком выпрямлении хобота. Такие зубы встречаются только
у небольшого числа видов, например у Oenopota elegans, Oe. harpa, Propebela
scalaris, P. nobilis, Obesotoma simplex и некоторых других.
Рис. 18. Строение стенок буккальной полости.
А — поперечный срез буккальной трубки Propebela scalaris, проходящий через буккальную сумку; Б —
срез на участке между буккальной сумкой и отверстием ядовитой железы (видно формирование канала (К)
посредством сближения буккальных складок); В —продольный срез буккальной трубки Oenopota harpa
(видны многочисленные желобки, соединяющие отверстие ядовитой железы (ПЯЖ) с буккальной сумкой
(СВ)). ДР—длинный рукав радулярного мешка, ПЩ —пищевод, РТ— ротовое отверстие.
Для большинства видов Oenopotinae, у которых зубы либо очень мелкие,
либо имеют чересчур большие отверстия и яд из иглы может вымываться еще
при перемещении внутри буккальной трубки, предположение Пирса (Pearce,
1966) малоприемлемо. Наиболее логично предположить умерщвление добычи
при использовании зуба в качестве иглы шприца.
Зуб выходит из короткого рукава радулярной сумки полностью очищенным
от клёточных элементов как снаружи, так и во внутр’енней полости лезвия и
захватывается сфинктером хобота за проксимальный участок лезвия. Во время
нападения на добычу или незадолго до нанесения удара зубом происходит сжа-
тие мускулатуры дистального участка ядовитой железы, выполняющего роль
пропульсаторного органа, и яд впрыскивается в буккальную полость, где он
смешивается с секретом слюнных желез и жидкостью буккальной полости.
Создание направленного потока ядосодержащей жидкости под покровы добычи
происходит следующим образом. Задняя стенка буккальной полости за счет
сжатия мускулатуры закрывает вход в просвет пищевода (рис. 17, Е), выполняя
своеобразную функцию клапана (Shimek, 1974). Затем происходит сокращение
буккального сфинктера, в результате чего из буккальной полости секрет пере-
качивается в полость хобота. Отсюда ядосодержащая жидкость попадает
в околобазальное отверстие зуба, проходит по полости лезвия и проникает под
38
ОБЩАЯ ЧАСТЬ
покровы жертвы через дистальное отверстие зуба. Нагнетание секрета проис-
ходит за счет перистальтики, создаваемой кольцевой мускулатурой в наружной
стенке хобота. Таким образом, роль зуба сводится к прокалыванию покровов
жертвы и проведению через полость лезвия ядосодержащей жидкости. Иными
словами, зуб выполняет функцию иглы шприца, так же как и у моллюсков рода
Conus (Kohn et al., 1972).
Описанный механизм позволяет объяснить способ умерщвления добычи
моллюсками Propebela turricula и Р. harpularia. У представителей этих двух
видов зубы очень маленькие (их длина в среднем около 100 мкм), лишенные
полости внутри. Представить захват хоботом таких зубов трудно, так как
максимальная их ширина не превышает 15 мкм. Вероятно, такие зубы, причем
не один, а несколько, выбрасываются из радулярного мешка в полость буккаль-
ной трубки, где подхватываются током ядосодержащей жидкости, выпрыскивае-
мой па добычу. У рассматриваемых видов роль зуба сводится лишь к нарушению
целостности покровов жертвы, а яд воздействует контактно через ранки.
Таким образом, у большинства Oenopotinae дистальный участок хобота
несет зуб, который выполняет функции иглы шприца. В соответствии с этим,
главная функция хобота заключается в захвате зуба и нанесении им удара по
добыче. При шиловидной форме зуба меняется функция хобота. В этом случае
он не захватывает зуб, а создает целенаправленный поток ядосодержащей жид-
кости, несущей многочисленные мелкие зубы и выпрыскиваемой на добычу.
Захват добычи и пищеварение. Oenopotinae питаются пре-
имущественно полихетами (Pearce, 1966; собственные данные). Место нанесения
удара хоботом на теле червя не детерминировано. Предположение Шимека
(Shimek, 1974) об обкручивании хоботом тела полихеты при ее поглощении или
использовании зуба как якоря в теле добычи при затаскивании ее внутрь рин-
ходеума опровергается моими наблюдениями. Моллюск находит один из концов
тела парализованной полихеты и захватывает его хоботом (рис. 19, Л). Вслед-
ствие сокращения продольной мускулатуры внутренней стенки хобота проис-
ходит втягивание добычи внутрь ринходеума. Причем, учитывая подчас значи-
тельные размеры пищевого объекта, моллюск может поглощать его по частям.
Свободный участок полихеты торчит из отверстия ринхостома, зажатый и удер-
живаемый сфинктером ринходеума (рис. 19, Б). Пища, попавшая в ринходеум,
подвергается воздействию оставшейся в нем части ядосодержащей жидкости.
Как правило, уже в полости ринходеума тело червя лизируется настолько,
что превращается в однородную кашицу с разрозненными хетами. У всех ис-
следованных животных с пищевыми объектами, найденными в буккальном
отделе, было отмечено характерное вворачивание хобота внутрь буккальной
полости (Богданов, 1989в). В результате наружная стенка хобота оказывается
вплотную прилегающей к эпителию буккальной полости, а внутренняя играет
роль трубки, соединяющей полость ринходеума непосредственно с пищеводом
(рис. 19, Г, Д).
В результате вворачивания хобота происходят следующие изменения. Во-пер-
вых, увеличивается объем полости пищеварения, которая ограничена стенками
ринходеума, наружной стенкой хобота и начальными участками пищевода.
Находящаяся в камере пища подвергается воздействию ядосодержащей жид-
кости. Возможно, в лизисе принимает участие секрет бокаловидных желези-
стых клеток нижней (большей) части ринходеума и наружной стенки хобота.
Во-вторых, стенки буккальной полости не имеют мускулатуры, способной
к активным перистальтическим сокращениям. Наружная стенка хобота, во
ввернутом состоянии являющаяся стенкой буккального участка полости пище-
варения, обладает как косой, так и кольцевой мускулатурой. Таким образом,
в продвижении пищи от ринходеума к пищеводу участвует орган, способный
к активной перистальтике.
В-третьих, вворачивание хобота влечет частичное смещение радулярного
мешка к буккальному сфинктеру. В результате отверстие буккальной сумки
сплющивается, почти закрываясь. Происходит изоляция радулярного мешка,
МОРФОФУНКЦИОНАЛЬНЫЙ АНАЛИЗ
39
благодаря которой осуществляется предохранение зубов от механического
воздействия проходящей пищи. Кроме того, внутренняя стенка хобота, лежа-
щего в тесном контакте с буккальной сумкой, изолируется от дистальных острых
участков зубов.
Учитывая простое строение желудка и пищевода, можно с уверенностью
говорить о начальных этапах пищеварения в полости ринходеума. В желудке
и пищеварительной железе может происходить только окончательный лизис
и всасывание пищи. В связи с тем что железистые клетки ринходеума, пищевода
и наружной стенки хобота могут выделять небольшую долю секрета, вероятно,
основную массу пищеварительных ферментов поставляют секреты ядовитой
и слюнных желез.
Я* РАД
Рис. 19. Схема взаимодействия мускулатуры (1—6) при захвате пищи (А—Г).
Г — пучками щетинок обозначена пищеварительная камера. Д — часть буккальной трубки удалена. Обо-
значения как на рис. 11 и 12.
Кровеносная и выделительная системы ни у моллюсков Oenopotinae, ни
у Turridae в целом не изучены. На основе полученных нами данных можно кон-
статировать общую с остальными высшими Gastropoda (Stenoglossa, — in:
Fretter, Graham, 1962) схему строения кровеносной системы. Сердце примыкает
к задней левой стенке мантии и состоит из овальной формы крупного желу-
дочка и небольшого тонкостенного предсердия, сообщающихся между собой
посредством широкого атриовентрикулярного отверстия. Сердце лежит в по-
лости целома, ограниченного перикардием.
Для выделительной системы можно привести только общие сведения. Перед-
няя стенка почки образует большую часть задней поверхности мантийной по-
лости и сообщается с ней отверстием — нефропором, имеющим вид до рсо-
вентрально ориентированной щели. По краям щели развивается мускулатура,
образующая сфинктер. Слева от почки расположен перикардий, полость к ото-
40
ОБЩАЯ ЧАСТЬ
рого соединяется с полостью почки посредством реноперикардиального отвер-
стия. Наружные стенки почки пронизаны сосудами венозной системы и обра-
зуют многочисленные складки — трабекулы, вдающиеся в полость органа.
Иными словами, уровень знаний о кровеносной и выделительной системах
Turridae очень низок и требует специальных исследований. Морфологические
исследования не позволяют выявить особых межвидовых различий в строении
отдельных органов названных систем, поэтому детали их строения не исполь-
зуются в диагностике Oenopotinae ни на видовом, ни на более высоком уровнях.
Нервная система. С целью изучения нервной системы Oenopotinae было
проанатомировано 30 экз. моллюсков, относящихся к 13 видам. Окологлоточное
нервное кольцо этих животных характеризуется сильной концентрацией ганг-
лиев (рис. 20) и расположено вокруг пищевода в самом его начале, т. е. на гра-
Рис. 20. Схема строения окологлоточного нервного кольца Oenopotinae (на примере Oeno-
pota harp а).
Al—вид сзади, Б—вид справой стороны. БГ—буккальный ганглий, ГН — головной нерв, НВГ—
комиссура вентрального ганглия, ЛЦВ — левый цереброплевральный ганглий, ПГ — педальный ганглий,
ППГ — проподиальный ганглий, ПЦВ — правый цереброплевральный ганглий, СУБИ— субинтести-
нальный ганглий, СУПРИ — супраинтестинальный ганглий.
нице с буккальным отделом. Педальные, самые крупные ганглии окологлоточ-
ного кольца располагаются под буккальной трубкой. Дистальные участки
педальных ганглиев несколько обособлены и образуют так называемые про-
подиальные ганглии, от которых отходят нервные волокна, иннервирующие
ногу, а у самцов и пенис. Педальные ганглии расположены близко друг к другу
и почти сливаются в базальной части, так что соединяющие их коннективы
по существу незаметны. Верхняя, лежащая над буккально-пищеводной гра-
ницей часть нервного кольца представлена морфологически слившимися це-
реброплевральными ганглиями. У некоторых видов Oenopotinae слияние про-
изошло не до конца и ганглии выглядят гантелевидными. Переднюю, верхнюю
их часть составляет церебральный участок, иннервирующий голову, глаза,
статоцисты и хобот. Статоцисты расположены под передней частью проподиаль-
ных ганглиев на дне туловищного синуса и представлены небольшими белесыми
пузырьками с жидкостью, внутри них просматриваются известковые статолиты
(рис. 21, вкл.). От церебрального участка отходят длинные коннективы, соеди-
няющие его с буккальными ганглиями. Последние представляют собой два бобо-
видных образования, почти сросшихся на участке, противоположном отхожде-
нию церебробуккальных комиссур. Буккальные ганглии расположены вблизи
соединения короткого и длинного рукавов радулярного мешка и иннервируют
буккальный и радулярный аппараты. Плевральный участок гантелевидных
ганглиев расположен ниже и позади церебрального и отвечает за иннервацию
мантии, стенок полости тела и колумеллярного мускула. Правый и левый
цереброплевральные ганглии соединены между собой короткими комиссурами,
а с педальными ганглиями — коннективами. У большинства видов церебро-
педальные коннективы состоят из двух параллельных стволов, что свидетель-
ствует о двойственности происхождения цереброплеврального ганглия.
МОРФОФУНКЦИОНАЛЬНЫЙ АНАЛИЗ
41
Сзади от плевральной части гантелевидных ганглиев отходят четыре коннек-
тивы: две к субинтестинальному и две к супраинтестинальному ганглиям.
Субинтестинальный ганглий лежит под пищеводом, сзади от места стыка педаль-
ных ганглиев, супраинтестинальный — над пищеводом, за цереброплевраль-
ными комиссурами. От последнего отходит нерв к осфрадиальному ганглию,
который иннервирует ктенидий, осфрадий и гипобранхиальную железу. Отхо-
дящие от интестинальных к висцеральному ганглию коннективы образуют вис-
церальную петлю. От супраиытестинального ганглия коннектива идет снизу
вверх с правой стороны пищевода, от субинтестинального — с левой стороны,
сначала опускаясь вниз, а затем снова поднимаясь. Обе петли соединяются
возле желудка в правый висцеральный ганглий. Следов левого висцерального
ганглия не отмечено, очевидно, он полностью редуцирован. Правый висцераль-
ный ганглий иннервирует печень, гонаду с половыми протоками, почку, желу-
док, сердце с перикардием, аорту и жаберную вену.
Рис. 22. Схема строения половой системы самцов.
И — простата, ПЕ — пенис, ПР — проток простаты, С — семенник, СП — семенной пузырек, СПР —
семяпровод.
Половая система. Половая система самцов (рис. 22) начинается большим,
телесного цвета семенником, расположенным в верхних оборотах на колумелляр-
ной стороне висцерального мешка. Печень более темного цвета смещена к пери-
ферии завитка и частично покрывает семенник с отходящим от него гонодуктом..
Последний можно разделить на два участка: меньший — семенной пузырек и
отходящий от него длинный семяпровод. Семенной пузырек железистый и со-
стоит из клеток, содержащих в цитоплазме значительное число гранул различ-
ных размеров (Smith, 1967с). В период размножения семенной пузырек напол-
няется спермой. Семяпровод подразделяется на ренальный и паллиальный
участки: ренальный, короткий участок начинается от семенного пузырька и
внутри обильно покрыт ресничками. Снаружи он покрыт мускульными волок-
нами и имеет вид извитой трубки. В месте перехода ренального участка в палли-
альный от семяпровода к перикардиальной сумке отходит гоноперикардиаль-
ный проток, чаще его рудимент. У Curtitoma trevelliana гоноперикардиальный
проток открывается в перикардий посредством ресничной воронки (Smith,
1967с). Паллиальный участок семяпровода длинный, проходит вдоль кишеч-
ника и располагается под стенкой тела, ниже ее соединения с крышей мантийной
полости. Сначала этот участок семяпровода извит, но у задней стенки мантий-
ной полости он выпрямляется и переходит в участок семяпровода, насыщенный
железистыми клетками — простату. Обе доли простаты у Oenopotinae пол-
ностью срослись. Следует отметить, что проток, отходящий от семяпровода и
открывающийся в мантийную полость, характерный для ряда Turridae (Robin-
son, 1960), у представителей Oenopotinae не обнаружен. Простата продолжается
протоком в совокупительный орган, или пенис. Проток простаты очень тонкий
на всем протяжении и выстлан большими вакуолизированными железистыми
клетками с расположенными между ними маленькими ресничными клетками.
Перед сфинктером, закрывающим отверстие протока на конце пениса, эпите-
лий перестает быть железистым и половое отверстие выстлано только реснич-
ными клетками.
42
ОБЩАЯ ЧАСТЬ
Половая система самок (рис. 23) начинается гроздевидным яичником, лежа-
щим под печенью на колумеллярной стороне тела. Яйцевод, отходящий от
яичника, идет по внутренней поверхности висцерального мешка в направлении
Рис. 23. Схема строения половой системы самок.
ЖЖ — белковая железа, ЖСП — железа семяприемника, КС — копулятивная сумка, М — матка, ОЖ —
оболочковая^ железа, ПО — паллиальный овидукт, Я — яичник, ЯВ — яйцевод.
к колумеллярному мускулу. У Curtitoma trevelliana он короткий и выстлан рес-
ничными клетками с сильно вакуолизированной цитоплазмой и многочислен-
ными мелкими гранулами, сконцентрированными возле клеточных стенок
(Smith, 1967с). Около колумеллярного мускула начинается ренальный отдел
Рис. -24. Схема редукции (А -> В) мужской половой системы у гермафродитной особи Pro-
pebela turricula (по: Smith, 1367с). Пояснения в тексте.
ГП — гоноперикарднапьный проток, ПЕР — перикардиальная сумка, СПМ — семяприемник. Остальные
обозначения как на рис. 22 и 23.
яйцевода, проходящий вплотную с перикардием, и у ряда туррид (но не у Oeno-
potinae) дает от себя проток к околосердечной сумке. Ренальный отдел сосед-
ствует с белковой железой, после которой начинается паллиальный отдей яйце-
вода. Эпителий белковой железы состоит из ресничных клеток с большими сли-
зистыми клетками в промежутках (в основном на дорсальной стороне). Палли-
альный отдел начинается значительным расширением яйцевода, от которого
ИЗМЕНЧИВОСТЬ МОРФОЛОГИЧЕСКИХ ПРИЗНАКОВ 43
отходит проток железы семяприемника. Проток выстлан ресничными клетками,
и в нем обнаружены прикрепленные к стенке спермин (Smith, 1967с), что указы-
вает на его функционирование в качестве семяприемника. Железа семяприем-
ника расположена на дорсальной поверхности белковой железы, и ее эпители-
альная выстилка состоит из больших железистых клеток, секрет которых со-
действует сохранению находящейся внутри спермы (Smith, 1967с). Оболочковая
(или скорлуповая) железа расположена на незначительном расстоянии от семя-
приемника и лежит на правой стороне мантийной полости, под кишечником.
Латеральные стенки железы развиты сильнее, чем дорсальная и вентральная,
так что в поперечном сечении ее просвет выглядит как дорсовентральная щель.
Оболочковая железа открывается в копулятивную сумку, выстланную реснич-
ным эпителием, содержащим как слизистые, так и железистые клетки. Копуля-
тивная сумка открывается в мантийную полость на уровне анального отверстия
или несколько впереди него.
Oenopotinae раздельнополы, но у некоторых особей Propebela turricula
Отмечен гермафродитизм (Smith, 1967с), что у Toxoglossa встречается очень
редко. У ряда экземпляров данного вида Смит (Smith, 1967с) обнаружил раз-
витие обеих половых систем, но простата была частично редуцирована (рис. 24,
Л). Значительное число самок имело помимо нормально развитой женской
половой системы редуцированную мужскую, которая была представлена буль-
бой, оставшейся от семенника, и отходящим от нее тонким протоком к муску-
листому пенису (рис. 24, Б). Наибольшая атрофия мужской половой системы
отмечена для самок, у которых рядом с белковой железой был обнаружен руди-
мент семенника и короткий участок семяпровода; пенис практически редуци-
рован (рис. 24, В). Смит (Smith, 1967с) считает, что гермафродитизм Propebela
turricula — приспособление к сохранению вида с небольшим ареалом и пола-
гает, что вся популяция должна быть гермафродитной. Однако в случае с Pro-
pebela turricula (все другие исследованные нами виды оказались раздельно-
полыми) лучше говорить не о гермафродитизме, а о протерандрии: у молодых
животных развивается мужская половая система, а у взрослых — женская,
что не раз отмечалось и у других Pectinibranhcia.
ИЗМЕНЧИВОСТЬ МОРФОЛОГИЧЕСКИХ1 ^ПРИЗНАКОВ
Дефинитивная раковина. Исторически описание и диагностика Oenopotinae
базировались преимущественно на морфологии дефинитивной раковины, зна-
чительная изменчивость которой и послужила основной причиной слабой
разработки систематики группы. Для выявления размаха внутривидовой из-
менчивости раковины и элементов ее скульптуры было проведено детальное
изучение нескольких групп видов, сходных по морфологии дефинитивной
раковины. Первую группу образовали Obesotoma simplex и О. schantarica,
принимаемые большинством исследователей за один вид. В другую группу
вошли все описанные подвиды и вариететы вида Curtitoma violacea. В третью
группу были объединены моллюски с оспариваемой видовой принадлежностью,
а именно: Propebela rugulata, Р. nobilis, Р. gouldii, Р. spltzbergensls и Р. Ьег-
gensis. Многие исследователи (Friele, 1886; Kobelt, 1905, и др.) названные
виды считали подвидами Р. rugulata. Выбор названных 8 видов был обусловлен,
во-первых, различными взглядами исследователей на их статус; во-вторых,
необходимостью найти конхологический признак, наименее подверженный
изменчивости и четко характеризующий тот или иной вид; в-третьих, большим
объемом материала в коллекциях ЗИН.
Для изучения явления, описанного уже в конце прошлого века .(Verrill.,
1882), которое заключается в том, что морские моллюски высоких широт,
в частнбсти многие виды Oenopotinae, обладают как высокими (стройными),
так и низкими (приплюснутыми) раковинами, был применен компараторный
метод их сравнения (Иззатуллаев. Старобогатов, 1984).
а
ОБЩАЯ ЧАСТЬ
После просмотра и зарисовки с помощью рисовального аппарата контуров
раковины более чем 400 экз. моллюсков из группы Р. rugulata удалось выде-
лить 9 форм. Термин «форма» обозначает здесь определенные изменения про-
порций раковины, зависящие в основном от скорости навивания оборотов (шаг
вдоль оси, — Raup, 1967) и легко выявляемые компараторным методом. Срав-
нение стройности раковин проводилось без предварительного учета видовой
принадлежности моллюсков, т. е. одновременно на всех 5 видах группы. В ре-
зультате при наложении каждой последующей формы на предыдущую (от более
удлиненной к более укороченной) был обнаружен плавный переход между
основными тремя формами (рис. 25). Следует отметить, что ни одна из основных
Рис. 25. Применение колшараторггого метода при сравнении раковин и изучении моллю-
сков группы Propebela rugulata.
В центре показаны три основные формы. Цифры обозначают каждую из обнаруженных форм: от наиболее
удлиненной (2) к самой укороченной (9).
форм не соответствовала какому-либо определенному виду. Например, рако-
вины P.nobilis вписались в 5 форм, из которых 2 были основными, а Р. rugu-
lata во все 9. Последний факт, очевидно, может служить объяснением выде-
ления различными авторами многочисленных вариететов Р. rugulata. Таким
образом, действие компараторного метода при сравнении раковин морских
моллюсков ограничено, и в отличие от ряда пресноводных моллюсков (Андре-
ева, Старобогатов, 1981; Иззатуллаев, Старобогатов, 1984, и др.) форма.рако-
вины у представителей Oenopotinae не может служить четким таксономическим
признаком.
Несколько иные результаты были получены при обследовании более чем
300 раковин Obesotoma simplex и О. schantarica. Компараторным методом срав-
нения было выделено 10 форм раковин. Несмотря на большую изменчивость
«стройности» раковины, удалось выделить 2 группы, соответствующие рас-
ИЗМЕНЧИВОСТЬ МОРФОЛОГИЧЕСКИХ ПРИЗНАКОВ
45
сматриваемым видам: в 4 формы вписывались все раковины О. schantarica,
а в 6 — О. simplex (рис. 26). Несмотря на то что виды не обладали общими
формами, диапазон внутривидовой изменчивости раковины оказался чрезмерно
велик. Например, форма раковины, идентичная О. simplex, была отмечена
у О. gtgantea.
В группе Curtitoma vtolacea (в отличие от рассмотренных' ранее) наблю-
далась различная степень стройности раковины, но независимые формы выде-
лить не удалось. Иными словами, все просмотренные экземпляры (около 270),
относящиеся к различным вариететам — от самого стройного С. vtolacea var.
laevior до типичной формы, различались только шагом вдоль оси раковины.
Различия формы раковин у одного вида удалось соотнести с гидрологи-
ческими характеристиками среды обитания. В литературе высказывалось мне-
ние о взаимосвязи существования различных вариететов вида с глубиной или
широтностью его местонахождения (Friele, 1886), предполагалось также, что
степень стройности раковины зависит от экологических условий, в которых
раковины температурного фактора (Голиков, 1980). Данная точка зрения
оказалась приемлемой и для Oenopotinae. Из всех изученных факторов (грунт,
соленость, содержание кислорода, глубина и температура) изменения послед-
него коррелировали в наибольшей степени с изменениями скорости навивания
оборотов раковины. Выяснилось, что с уменьшением температуры (сравнение
моллюсков осуществлялось на материале из акваторий, где сезонные колеба-
ния гидрологических условий минимальны) высота раковины и ее относитель-
ная толщина уменьшаются. Так, наиболее стройная раковина для Propebela
nobtits была отмечена в водах побережья Мурманска (среднегодовая темпера-
тура 6 °C), а самая приземистая — в море Лаптевых (£=1 °C). Подтверждением
влияния гидрологических условий, в первую очередь температурного фактора,
на скорость навивания оборотов послужил ряд находок. Например, в одной
беломорской пробе были обнаружены раковины, вписывающиеся в одну и
ту же форму, но относящиеся (по морфологии зубов радулы) к разным видам:
Р. nobtlis и Р. spttzbergensts (рис. 27, вкл.).
Статистический анализ параметрических характеристик был проведен
на 3 наиболее многочисленных в изученном материале видах. Для Curtitoma
novajasemljensis, С. trevelliana, С. decussata удалось показать изменения ско-
рости навивания оборотов раковины в зависимости именно от температуры
(рис. 28). При сопоставлении изменений параметров раковины (например,
высоты и наибольшего диаметра) было выяснено, что в различных акваториях
их изменение происходит разными темпами. Так, по достижении одного и
того же диаметра, равного-3.5 мм, высота раковины у С. novajasemljensis со-
ставила в наиболее прогреваемых районах Восточно-Сибирского моря 7.0 мм
(рис. 29), в то время как у особей, обнаруженных в Карском море (при темпе-
ратуре 1.1 °C), высота раковины была на 1.4 !им меньше (5.6 мм). Таким об-
разом, в различных акваториях раковины одного вида могут иметь различную
форму, что обусловлено влиянием на нее факторов внешней среды, и в первую
очередь температуры. В более теплых акваториях чаще встречаются моллюски
с более стройной раковиной. Недостатком изучения изменчивости формы ра-
ковины статистическим методом является необходимость большой выборки
ОБЩАЯ ЧАСТЬ
моллюсков различных возрастных групп из сопоставимых по гидрологическим:
характеристикам проб. К сожалению, для большинства видов это требование
не может быть удовлетворено, в связи с тем что Oenopotinae, как правило,
не образуют скоплений. Кроме того, этот метод менее нагляден и ошибка изме-
рений, особенно для мелких экземпляров, может затушевывать небольшие
изменения формы раковины, выявляемые компараторным методом.
Для ряда видов был отмечен половой диморфизм, проявляющийся в отно-
сительно более широкой раковине у самок, чем у самцов. Например, в пробе,
содержащей 12 экз. С. trevelliana одинакового размера, было отмечено 5 рако-
вин с более широкими оборотами, чем у остальных 7 (рис. 30, вкл.). Вскрытие
моллюсков выявило соотношение самцов и самок соответственно 7 : 5. Уве-
личение последнего и (реже) предыдущих оборотов раковины и проявление
ее полового диморфизма наблюдаются только у взрослых особей после на-
Рис. 28. Зависимость изменений
параметров раковины Curtitoma
trevelliana от температуры среды
обитания.
По оси ординат — высота раковины,
по оси абсцисс — наибольший ее диа-
метр.
Рис. 29. Зависимость изменений параметров раковины
Curtitoma novajasemljensis от температуры среды оби-
тания.
По оси ординат — высота раковины, по оси абсцисс — наиболь-
ший ее диаметр. А — Восточно-Сибирское море (t=3 °C), Б —
Новая Земля (2=1 °C); В — Новая Земля (2=0 °C); Г — Кар-
ское море (2=—1.1 °C).
ступления половой зрелости. Использование компараторного метода позволил»
отметить еще одно интересное проявление полового диморфизма, выражающееся
в различной степени отгибания колумеллярного края сифонального выроста.
Так,, например, для самцов Propebela nobilis характерен более сильный отгиб
колумеллярного края сифонального выроста в сторону от оси раковины, чем
для самок.
Некоторые изменения формы раковины могут быть связаны с поврежде-
ниями (как правило, обломами) палатального края устья, которые часто обна-
руживаются у обитающих на литорали и в сублиторали моллюсков; реже
подобные изменения связаны с возрастными перестройками. Так, у старых
особей Р. nobilis встречается утолщение палатального края устья (в 3—4 раза
превышающее толщину стенок раковины), которое в результате приобретает
вид ребристого мозолистого нароста. Утолщение стенок устья влечет за собой
не только выгибание наружу массивного палатального края, но и изгибание
назад сифонального выроста. Описанные изменения безусловно связаны с про-
должающейся секрецией раковины, происходящей на фоне сильного замедле-
ния роста моллюска.
Все сказанное демонстрирует зависимость формы раковины от различных
биотических и абиотических факторов и свидетельствует о сильной ее измен-
чивости, что затрудняет использование данного признака при идентификации
видов.
ИЗМЕНЧИВОСТЬ МОРФОЛОГИЧЕСКИХ ПРИЗНАКОВ 47
Более стабильным конхо логическим признаком оказалась скульптура
раковины. На видах тех же трех групп была исследована стабильность скульп-
турных элементов раковины. Для группы Curtitoma violacea были отмечены
всевозможные варианты скульптуры раковины и составлен вариационный ряд
от практически полного отсутствия скульптурных элементов (что свойственно
большинству моллюсков типичной формы) до экземпляров с развитыми двумя
килями (вариетет bicarinata) и далее к форме, названной нами nordica (для нее
характерно появление помимо килей ярко выраженных тонких осевых скла-
док). Это подтвердило полиморфность С. violacea, отмеченную при изучении
форм раковины. Например, в Баренцевом море у о-ва Виктория было обнару-
жено несколько экземпляров, у которых скульптура верхних 3.5—4 оборотов
оказалась характерной для формы «bicarinata» и состояла из многочисленных
узких осевых валиков и двух спиральных килевидных ребрышек. На пред-
последнем обороте раковины скульптура становилась менее рельефной, а к по-
следнему обороту она практически полностью сглаживалась, оставляя лишь
еле намеченные спиральные ребрышки, и соответствовала скульптуре типич-
ной «violacea».
Для группы Propebela rugulata размах внутривидовой изменчивости круп-
ных осевых и спиральных скульптурных образований оказался в большинстве
случаев перекрывающим межвидовой. Так, у Р. nobilis, Р. rugulata и Р. spitz-
bergensis имеются как средние, так и тонкие осевые складки различной ширины
и высоты. Столь сильная изменчивость крупных элементов скульптуры, отме-
ченная у видов рассматриваемой группы, делает их практически непригодными
для диагностики. Единственное достоверное отличие по скульптуре между
названными видами заключается в развитии на киле Р. nobilis, обозначающем
плечо оборота, дополнительных 3—6 тонких ребрышек. Они резко увеличивают
поверхность киля, который на последних оборотах сглаживается, расширяясь
до размеров осевых складок. Все просмотренные экземпляры Р. rugulata и
Р. spitzbergensis характеризовались резко обозначенным развитым килем,
лишенным дополнительных спиральных образований.
Широкие пологие осевые складки, расположенные на околошовном про-
странстве Obesotoma schantarica и сглаживающиеся к последней трети пери-
ферии оборота, служат хорошим отличием этого вида от О. simplex. Иначе
говоря, у разных видов Oenopotinae внутривидовой размах изменчивости
скульптуры неодинаков и может перекрывать внутривидовой.
Попытка соотнести нестабильность скульптуры раковины внутри вида
с изменениями гидрологии среды его обитания позволила сделать ряд пред-
варительных замечаний. Так, было подмечено, что степень развития скульп-
туры внутри вида наиболее тесно связана с изменениями солености — фактора,
действующего в сочетании с температурой. Так, в Белом море при солености
26—28 %0 осевая скульптура преобладающего большинства раковин Curtitoma
violacea представлена только линиями нарастания. В Баренцевом море при
продвижении от м. Нордкап на север в Арктику, |т/е. из вод с более высокой
{34.5 %о) в воды с более низкой (32.6 %0) соленостью, выраженность тонких
осевых складок заметно ослабевает. Сходное соответствие орнаментированности
раковины изменениям солености удалось показать для южных и средних Ку-
рильских о-вов, где в пробах из вод с высокой соленостью осевая скульптура
раковин С. violacea состояла из резко обозначенных тонких складок, а при
понижении солености они сглаживались.
Ряд изменений скульптуры раковины в различных акваториях наблюдается
у Propebela nobilis. При выходе из Белого моря (соленость 28 %0) и продвиже-
нии в более теплые участки Баренцева моря с более высокой соленостью (33—
34 %0) осевые складки на раковинах этого вида становятся более выпуклыми,
а расстояния между ними увеличиваются. При дальнейшем продвижении на
север к Шпицбергену и в сторону о-ва Новая Земля наблюдается опреснение
воды до 30—31 %0, одновременно с этим отмечается сглаживание скульптуры
раковины, осевые складки становятся более пологими, а промежутки между
48
ОБЩАЯ ЧАСТЬ
ними сокращаются. Сходные описанным изменения степени и характера скульп-
турироваиности раковин были отмечены у большинства изученных видов.
Таким образом, иод воздействием факторов внешней среды происходят
заметные изменения как формы, так и скульптуры дефинитивной раковины,
что затрудняет диагностику видов по конхологическим признакам.
Эмбриональная раковина. Долл (Dall, 1924), занимаясь изучением роли
эмбриональной раковины в классификаций морских гастропод, высказал пред-
положение, что структура протоконха имеет таксономическое значение на уровне
родов и таксонов более высоких рангов. Для рассмотренных им групп моллю-
сков, куда вошли и Toxoglossa, исследователь сделал вывод о значительной
внутривидовой изменчивости скульптуры протоконха, которую он объяснил
различными сроками и условиями жизни пелагической личинки у одних и
тех же видов, обитающих в различных акваториях. У Oenopotinae стадия
велигера в большинстве случаев проходит внутри яйцевой капсулы и эмбрио-
генез протекает без видимого метаморфоза. Наличие в онтогенезе некоторых
видов стадии планктонной личинки (Thorson, 1946; Bouchet, Waren, 1980)
оказалось редким исключением среди Oenopotinae. Защита от непосредствен-
ного воздействия факторов внешней среды на формирующуюся раковину за-
родыша обеспечивается тем, что эмбриональное развитие протекает целиком
в яйцевой капсуле; этим, по-видимому, можно объяснить внутривидовую ста-
бильность таких характеристик протоконха, как форма и скульптура.
В систематических работах по Oenopotinae изображения эмбриональных
раковин встречаются с конца прошлого столетия (Friele, 1886). Однако чрез-
мерно стилизованные рисунки не сопровождаются описаниями, что не позво-
ляет использовать их для сравнительного анализа. Впервые таксономическую
значимость строения эмбриональной раковины попытались оценить Буше и
Варен (Bouchet, Waren, 1980), проиллюстрировавшие значительное разнообра-
зие протоконхов Turridae очень хорошими фотографиями.
Внутривидовая изменчивость формы и скульптуры протоконха изучена
на наиболее репрезентативном (по сохранности эмбриональных раковин)
материале. На примере ряда видов (Oenopota harpa, Oe. pyramidalts, Curtitoma
conoidea и др.) показана видоспецифичность рассматриваемых характеристик.
Например, стабильность конхологических признаков эмбриональной раковины
можно продемонстрировать на ряде экземпляров Oenopota pingelii (рис. 6,
А—Е). Все просмотренные протоконхи были башенковидной формы с упло-
щенной верхней частью. Рисунок их первых полутора оборотов имел идентич-
ный у всех экземпляров ретикулятный характер, а скульптура первого пост-
эмбрионального оборота состояла из 4 тонких спиральных ребрышек, закла-
дывающихся на иротоконхе и пересекаемых в 4—5 раз более тонкими осевыми
складками. Специфика рассмотренного примера заключается в том, что морфо-
логически сходные протоконхи О. pingelii формировались в акваториях с раз-
личными гидрологическими режимами. 4 из рассмотренных экземпляров были
собраны у берегов Шпицбергена (рис. 6, А, Б), 2 других у Мурманского по-
бережья Баренцева моря (рис. 6, В, Г), а 1 в районе Шантарских о-вов в Охот-
ском море (рис. 6, Д, Е).
Результаты обработки данных, полученных на сканирующем микроскопе,
позволили судить о видоспецифичности скульптурных элементов протоконха
для всех представителей Oenopotinae. В то же время спектр форм и скульптур,
характеризующих протоконхи различных видов Oenopotinae, оказался поистине
удивительным. Было обнаружено разнообразие форм от почти колпачковидной
до уплощенной в верхней части, практически планоспиральной эмбриональной
раковины, несущей различные наборы спиральных ребрышек или полностью
лишенной их. Единственным исключением из внутривидовой стабильности
строения протоконха могут послужить эмбриональные раковины ряда экзем-
пляров Propebela bergensis. Различие заключалось в размерах протоконхов
при одинаковом количестве оборотов (рис. 31, вкл.). Но поскольку скульптура
протоконха оказалась' однотипной у всех рассмотренных Р. bergensis, описан-
ИЗМЕНЧИВОСТЬ МОРФОЛОГИЧЕСКИХ ПРИЗНАКОВ
49
ный факт не противоречит отмеченной видоспецифичности эмбриональной
раковины и может быть объяснен неодинаковыми темпами развития эмбрионов,
в кладках, содержащих различное число питательных яиц.
Крышечка. Индивидуальность морфологии крышечки у разных видов Oeno-
potinae была подмечена Фриле (Friele, 1886). К сожалению, ни один из после-
Рис. 32. Строение крышечки моллюсков Oenopotinae.
А—Д — Curtitoma decussata; Е—Ж — С. novafasemljensis; 3 — С. ovalis; И—Л — С. trevelliana;
М — С. incisula; Н—О — С. violacea; П—Р — Oenopota pingelii; С—У — Oe. pyramidalis; Ф — Oe. pyrami-
dalis var. semiplicata. Масштабная линейка 0.5 мм.
дующих исследователей не анализировал этот признак, что привело к полному
отсутствию в литературе сведений по его изменчивости.
При изучении строения крышечек более чем 400 экз. Oenopotinae было
выявлено межвидовое разнообразие их контуров, степени изогнутости оси
нарастания и формы мускульного отпечатка. Внутривидовая изменчивость
морфологии этого органа, как правило, невелика, поэтому показать специфич-
ные для вида детали строения крышечки удалось лишь в нескольких случаях.
Например, для Curtitoma ovalis характерна крышечка относительно большая
4 И. П. Богданов
50
ОБЩАЯ ЧАСТЬ
(отношение площади устья, Р , к площади поверхности крышечки, Рк, состав-
ляет 1.8), с плавно изгибающейся осью нарастания и тонким мускульным
отпечатком (рис. 32, 3).
Рис. 33. Строение крышечки.
Л—Г — Oenopota impressa", Д — Ое. cinerea, Е — Ое. cancellated, Ж — Ое. casentincr, 3 —Ое. biconica", И —
Ое. inequita; К—JI — Ое. declivis*, М—О — Ое. elegans; П—Т — Ое. harpa. Масштабная линейка 0.5 мм.
Для Oenopota elegans видоспецифичным оказался клювовидный верхний
участок мускульного отпечатка (рис. 33, М—О), а для О. impressa видовым
признаком послужил излом левого (лежащего у колумеллярного края устья)
края крышечки (рис. 33, А—Г). Некоторые детали строения крышечки ока-
зались специфичными для групп видов и для родов. Например, для большин-
ства видов Oenopota характерно плотное прилегание мускульного отпечатка
к левому краю крышечки (рис. 33), для видов Obesotoma и Propebela — утол-
щение колумеллярного края крышечки (рис. 34—36).
ИЗМЕНЧИВОСТЬ МОРФОЛОГИЧЕСКИХ ПРИЗНАКОВ
Сравнение крышечек разных видов Oenopotinae позволило выявить общую
тенденцию к уменьшению размеров крышечки по отношению к устью. Отно-
сительно большими размерами крышечки обладают Curtitoma trevelliana
(Р : Pfe=1.6—1.8) и C. vtolacea var. laevior (1.7), меньшими — Propebela viridula
(3.5—3.7) и P. spitzbergensis (2.5—3.3). Самая маленькая крышечка отмечена
у Р. angulosa (4.3). Несмотря на накопленный материал по морфологии кры--
Рис. 34. Строение крышечки.
А — Obesotoma gigas; Б — О. solida; В—Е — О. robusta.', Ж — О. tumida’, 3—В — О. woodiar^ Л —- О. за*
chalinensis', М—Н — О. starobogatovi. Масштабная линейка 0.5 мм. у
гаечки, использовать особенности ее строения в диагностике удается лишь
для небольшого числа видов. С одной стороны, у большинства видов измен-
чивость крышечки перекрывает рамки внутривидовой. С другой стороны,
определенная деталь строения крышечки, общая для большинства видов рода,
может отсутствовать у 2—3 видов, поэтому в таксономии родов морфология
крышечки может быть использована, но с большой осторожностью.
Изменчивость морфологических признаков тела. В связи с малыми размерами
моллюсков и значительной изменчивостью раковины многие исследователи
пытались найти видимые отличия в строении мягкого тела моллюсков, позво-
4*
52
ОБЩАЯ ЧАСТЬ
лившие бы легко идентифицировать виды. Например, при описании моллю-
сков Pleurotoma turricula и Р. trevelliana в диагнозе (Jeffreys, 1867) приводились
такие детали, как форма ноги, относительная длина щупалец, расположение
и цвет глаз и]т. д., т. е. признаки, подверженные не только значительной внутри-
Рис. 35. Строение крышечки.
А—В — Propebela nobilis; Г—Е — Р. rugulata; Ж —- Р. spitzbergensis; 3—К — Р. bergensis; Л — Р. spit-
zbergensis; М^О — Р. assimilis; П—С — Р. scalaris; Т — Р. fidlcula; У—Ф — Р. turricula. Масштабная
линейка 0.5 мм.
видовой изменчивости, но и морфология которых в значительной мере зависит
от фиксатора и момента фиксации. В то же время привлечение отдельных
структур тела или определенных систем его органов для идентификации видов
успешно практикуется в таксономии многих групп моллюсков. Как правило,
существенные для видовой диагностики изменения связаны с органами половой
(Голиков, 1963, 1980; Горячев, 1978; Шилейко, 1984, и др.) или пищевари-
тельной (Галкин,'1955, и др.) системы. Как в общем плане, так и в деталях
строения половая система Oenopotinae не подвержена сколько-
нибудь заметным межвидовым изменениям. Отмечены незначительные раз-
ИЗМЕНЧИВОСТЬ МОРФОЛОГИЧЕСКИХ ПРИЗНАКОВ
53
личия в расположении железы семяприемника у самок — сверху или сбоку
от белковой железы, а у самцов — в степени редукции гоноперикардиального
протока. Мужской и женский копулятивные аппараты не имеют никаких до-
полнительных образований, морфология которых могла бы облегчить видовую
диагностику, и различаются у отдельных особей лишь размерами, что, как
правило, зависит от степени половозрелости моллюска.
Рис. 36. Строение крышечки.
•А—В — Propebela angulosa', Г—Е — Р. exarata\ Ж—М — Р. viridula; Н^П — Р. variabilis; Р — Р. mit-
rata', С — Р. marinae. Масштабная линейка 0.5 мм.
Значительная межвидовая и внутривидовая изменчивость наблюдается
в строении пищеварительной системы. Часть сравнительно-
морфологических замечаний об изменчивости ринходеума, буккального аппа-
рата и желудка приведена в разделе «Морфофункциональный анализ». Сле-
дует отметить морфологическое разнообразие ядовитого аппарата, состоящего
из ядовитой железы и ее пропульсаторного органа (мускульного мешка).
Существенные различия в длине и диаметре ядовитой железы оказались неза-
висимыми ни от возраста моллюсков, ни от их видовой принадлежности. Так,
например,, у нескольких экземпляров Curtitoma trevelliana одного пола и воз-
раста (все изученные моллюски были из одной пробы и имели одноразмерные
раковины) длина ядовитой железы менялась в пределах 25 % относительно
среднего значения. Сравнение формы мускульных мешков показало их силь-
ную внутривидовую изменчивость.
Радула. Наиболее надежным с точки зрения видовой диагностики ока-
зался радулярный аппарат. В 1853 г. впервые было высказано предположение
о значимости морфологии радулы при определении видовой принадлежности
моллюсков рода Bela (Forbes, Hanley, 1853). Видоспецифичность радулы была
54
ОБЩАЯ ЧАСТЬ
проверена на 8 видах рода Bela Трошелем (Troschel, 1866), который выявил
и проиллюстрировал сложное строение зубов радулы. Для описания раз-
личных участков зуба им были введены такие обозначения, как лезвие, руко-
ять, лист (наиболее широкая часть основания, от которой отходит лезвие)
и др. Часть из предложенных им обозначений используется и поныне.
Строение зубов 19 видов рода Bela из Норвежского моря описано Сарсом1
(G. О. Sars, 1878). Берилл, продемонстрировав значительную изменчивость-
формы раковины моллюсков рода Bela (Verrill, 1882), указал на видоспецифич-
ность морфологии зубов радулы исследованных им видов. Таксономическая
ценность морфологии зубов была наглядно показана Фриле (Friele, 1886).
Исследователь выделил группы видов как по морфологическим особенностям
раковины, так и по конфигурации зубов. Например, Bela spitzbergensis была
описана на основании отличий в морфологии зуба, несмотря на сходство с В. ru-
gulata в строении раковины. Автор отмечал различия в деталях строения зу-
бов, объясняя их внутривидовой изменчивостью. С другой стороны, для группы
Язидов, близких В. trevelliana, характерно очень сходное строение зубов радулы..
Идентифицировать виды этой группы автор считает возможным только по.
раковине,
Инымй Словами, использование морфологических особенностей строения
зубов радулы было предложено для диагностики Oenopotinae уже во второй:
половине прошлого столетия, однако в определении таксономического веса:
этого признака не было достигнуто согласия.
Мной специально была изучена видоспецифичность зубов на более чем
20 видах Oenopotinae. Детальный анализ изменчивости радулы показал,, что-
при относительно постоянной морфологии зуба его параметры у преобладаю-
щего большинства видов могут сильно варьировать. Так, например, у Pro-
pebela nobills, несмотря на постоянную конусовидную конфигурацию основания
зуба, пропорции его могут изменяться у разных экземпляров, характеризуя'
различные формы: от высококонической до почти равносторонней в сечении.
Наиболее показательным из всех размерных характеристик зуба оказалось,
отношение его общей длины к длине базальной части. Именно по этому кри-
терию удалось легко .различить виды, слабо дифференцируемые по раковине..
Например, для Р. rugulata отношение длины зуба к его основанию составляет
в среднем 3.09 (при наибольшем значении 3.2), тогда как для Р. nobilis мини-
мальное отношение составляет 4.3 при среднем 4.7. Еще большие различия^
отмечены между зубами Obesotoma schantarica со средним значением отношения,,
равным 3.8 (максимальное 4.2), и О. simplex со средним показателем 9.0 (ми-
нимальное 7.3).
Детальный анализ общей морфологии радулы позволил выявить ряд функ-
ционально важных деталей зуба, практически не подверженных внутривидовой
изменчивости. Например, у Propebela exarata, Р. viridula и ряда других видов
специфичной оказалась форма флажка (рис. 428). Oenopota impressa и Propebela
scalaroides хорошо идентифицируются по базальной части зуба, которая в ниж-
ней части уплощена (рис. 432). Однако наряду с общей морфологией зуба наи-
более важное значение для систематики и понимания эволюции рассматривае-
мой группы имеет киль. Относительные его размеры и форма оказались практи-
чески не подверженными внутривидовой изменчивости.
Таким образом, наибольшую ценность для видовой диагностики моллюсков
Oenopotinae представляют особенности строения спиральной и (в меньшей
степени) осевой скульптуры как дефинитивной, так и эмбриональной раковины,,
а также общая морфология и детали строения зубов радулы..
СИСТЕМА И ФИЛОГЕНИЯ
55
СИСТЕМА И ФИЛОГЕНИЯ
В настоящее время имеется несколько точек зрения относительно числа
родов, составляющих рассматриваемую группу токсоглоссных моллюсков.
Окончательное разрешение этого вопроса осложняется еще и тем, что дли-
тельное время существовала путаница в синонимии самих родовых названий.
Только на первых этапах изучения Oenopotinae менее чем за 30 лет было пред-
ложено несколько родовых названий, таких как Defrancia (non Bronn, 1825)
Millet, 1826; Pleurotomoides Bronn, 1831; Pleurotomina Beck, 1847; Bela Gray,
1847; Ischnula Clarke in Gray, 1847; Lora Gistel, 1848; Oenopota Morch, 1852;
Onopota Adams et Adams, 1858 и ряд других.
Синонимия родовых названий подробно рассмотрена в ряде работ (Adams,
Adams, 1858; Dall, 1918; Bartsch, 1941; Powell, 1942, 1966). Ниже остановлюсь
лишь на названиях, длительное время и наиболее часто употреблявшихся
в литературе.
BELA Gray, 1847
Грей (Gray, 1847а) опубликовал некоторые данные из монографии Лича,
содержавшие список и предложения по классификации британских моллюсков.
В списке моллюсков 6 видов были обозначены родовым названием Bela. В связи
с этим ряд исследователей предлагали установить авторство названия рода как
Bela Leach in Gray, 1847. Однако Грей (Gray, 1847а, 1847b), кроме списка мол-
люсков, не приводит больше никаких сведений из рукописи Лича. Типовым
видом Bela., по последующему обозначению (Gray, 1847b), служит Murex nebula
Montagu, 1803. Поэтому следует считать авторство рода как Bela Gray, 1847.
М. nebula обладает типичной для Mangelia раковиной (Bartsch, 1944; Powell,
1966) и не имеет ничего общего с рассматриваемой в работе группой.
LORA Gistel, 1848
Родовое название Lora употреблялось многими авторами (вслед за Dall,
1886b, 1918) вместо Bela. Название Lora предложено (Gistel, 1848) как заме-
щающее для обозначения моллюсков Defrancia Millet, 1826 (типовой вид De-
francia viridula Fabricius, 1760). Однако D. viridula Moller, 1842 резко отли-
чается от описанного Фабрициусом вида (Morch, 1852), а именно мёллеровский
вид большинством исследователей принимался за типовой. Кроме того, родо-
вые названия Defrancia Millet, 1826 и Defrancia Moller, 1842 являются млад-
шими гомонимами Defrancia Bronn, 1825, сведенного в свою очередь в синоним
к Pleurotomoides Bronn, 1831.
OENOPOTA MSrch, 1852
G середины XX столетия в публикациях прочно укрепилось родовое на-
звание Oenopota как замещающее для Bela и Lora. Практически всех Oenopo-
tinae до настоящего времени продолжают описывать как представителей одного
рода Oenopota, совершенно не принимая во внимание два обстоятельства:
1) наличие описания рода Propebela Iredale, 1918, отличающегося от Bela
(=Oenopotd) формой раковины и угловатостью оборотов; 2) существование
предложения на основании различий в формах раковин, а также морфологи-
ческих особенностей осевой и спиральной скульптуры разделить все виды
на 12 родов, из которых Turrttomella, Nodotoma, Funitoma, Cestoma, Granotoma,
Nematoma, Curtitoma, Venustoma, Canetoma, Obesotoma выделены впервые
(Bartsch, 1941). , .
Значительный вклад в изучение таксономии Turridae внес американский
малаколог Пауэлл (Powell, 1942, 1951, 1960). В работе 1966 года (Ppwell,
1966) он, относя изучаемых моллюсков, к подсем. Mangeliinae, предлагает, объ-
-единить их в 4 рода:- Oenopota (с подродом Funitoma}, Propebela, Obesotoma
56
ОБЩАЯ ЧАСТЬ
и Nodotoma. Последняя работа, хотя и является в настоящее время основным
источником определения надвидовых таксонов сем. Turridae, не положила,
конец разногласиям в понимании числа родов Oenopotinae.
Следует отметить, что до настоящего времени диагностика родов и подро-
дов базировалась только на морфологических особенностях дефинитивной
раковины. Проведенные исследования позволили установить, что при выде-
лении таксонов родового ранга в подсем. Oenopotinae главенствующую роль
играют морфологические особенности протоконха и строения зубов радулы.
Эмбриональная раковина. Для различных видов было отмечено сходство
ряда признаков (касающихся в основном формы и скульптуры) протоконха,
на основании которых удалось разделить весь просмотренный материал на 5'
более или менее обособленных групп.
В первую группу (7) вошли моллюски, для которых характерно на-
личие множества (подчас неопределенного числа) спиральных ребрышек на
слегка выпуклой эмбриональной раковине. Это Curtitoma novajasemljensis,
С. decussata, С. conoidea. Скульптура их эмбриональных раковин представлена
10—30 спиральными ребрышками, без видимых изменений переходящих в реб-
рышки дефинитивной раковины. У С. incisula, С. trevelliana, С. vtolacea и ряда
других также отмечена закладка большого числа ребрышек, но еще на эмбрио-
нальной раковине происходит процесс их объединения, а затем и слияния
в 3—4 тонких ребра, дающих начало элементам спиральной скульптуры де-
финитивной раковины.
Главной отличительной чертой второй группы моллюсков (II) служит
наличие ретикулятного рисунка на первом обороте протоконха, в зависимости
от степени развития которого следует выделять внутри группы две подгруппы.
К первой из них отнесены виды с сильно развитым ретикулятным рисунком,,
покрывающим весь первый оборот эмбриональной раковины: Oenopota cinerea,
Ое. declivis (рис. 37, А, вкл.), Ое. pingelii и др. Названные виды характеризу-
ются наличием уплощенной площадки на апикальном участке эмбриональной
раковины, которая образуется за счет навивания ее первого оборота в одной
плоскости. Иными словами, направление вектора навивания первого оборота,
эмбриональной раковины составляет 90° к оси раковины. Вторая подгруппа,
характеризуется постепенным исчезновением ретикулятного рисунка на про-
токонхе Ое. pyramidalis и Ое. cinerea до практически полного его стирания
(у Ое. impressa и Ое. elegans). Одновременно с ослаблением рисунка на первом:
обороте протоконха происходят изменения скульптуры последнего его полу-,
оборота и первых оборотов дефинитивной раковины. Так, если в первой под-
группе поздний эмбриональный и первые дефинитивные обороты характери-
зуются наличием 4—6 спиральных ребрышек, то для второй подгруппы
типична спиральная скульптура, состоящая из 3—4 ребрышек, а в слу-
чае с Ое. impressa даже из 2 (рис. 37, Б, вкл.). Значительными оказываются
и изменения формы протоконхов второй подгруппы относительно первой,
заключающиеся в еще большем отклонении вектора навивания протоконха.
В ряду Ое. delcivis Ое. harpa Ое. impressa его угол по отношению к оси
раковины возрастает от 90 до 120°. Однако еще внутри кладки вектор нави-
вания оборотов протоконха постепенно изменяет направление и становится
параллельным вектору навивания оборотов дефинитивной раковицы (уже
к моменту выхода моллюска из кладки). Описанное изменение не влечет за
собой излома оси раковины, однако приводит к погружению вершины прото-
конха в первый полуоборот эмбриональной раковины (рис. 37, Б).
Третья группа видов (III) сходна со второй по происходящему в ней
процессу уменьшения числа спиральных элементов скульптуры протоконха.
Так, для большей части видов, а именно Propebela angulosa (рис. 38, А, вкл.),
Р. scalaris, Р. rugulata, Р. bergensis, Р. exarata, Р. assimilis и ряда других,
характерна закладка на поздней стадии эмбрионального развития раковины
(1.3—1.5 оборота) от 3 до 5 спиральных ребрышек. На протоконхах Р. turri-
cula, Р. harpularia, Р. scaldroides (рис. 38, Б) изначально закладываются также.
СИСТЕМА И ФИЛОГЕНИЯ
57
3 спиральных ребрышка, из которых верхнее редуцируется уже к концу пер-
вого оборота дефинитивной раковины, а оставшиеся 2 получают преимуществен-
ное развитие по сравнению с остальными (постэмбриональными) спиральными
элементами.
Отличительной чертой рассматриваемой группы служит сильное развитие
на первом обороте протоконха микроскульптурных элементов, состоящих
из отдельно разбросанных бугорков, при полном отсутствии макроскульптур-
ных образований. Изменения формы протоконха происходят в данной группе
от выпуклого у Р. nobilis, Р. bergensis, Р. scalaris по пути постепенного упло-
щения к почти планоспиральному первому обороту эмбриональной раковины
у Propebela harpularia и Р. scalaroides (рис. 38, Б). Таким образом, наблю-
дается постепенное увеличение угла между вектором навивания эмбриональных
оборотов и осью раковины приблизительно от 70 до почти 90°.
Четвертая, малочисленная, но своеобразная группа (IV) объединяет
Granotoma albrechti, G. krausei и G. raduga. Их протоконхи характеризуются
наличием четко обозначенных, полукруглых в сечении 5—7 спиральных реб-
рышек, пересекаемых в 2—3 раза более тонкими прямыми, слегка наклонен-
ными вперед, очень узкими осевыми складками. Элементы как осевой, так
и спиральной скульптуры протоконха густо покрыты микроскульптурными
бугорками, практически отсутствующими в промежутках между ребрами
(рис. 39, вкл.).
Представители данной группы хорошо обособляются от остальных изучен-
ных мной видов наличием частичной гетерострофии: ось навивания колпачко-
видного протоконха несколько наклонена относительно оси навивания дефи-
нитивной раковины, что особенно заметно благодаря спиральным ребрышкам,
идущим не параллельно, а под значительным углом ко шву. В результате
нижние 2—3 ребрышка протоконха прикрываются по мере роста раковины
первым дефинитивным оборотом.
Виды последней, пятой группы (7) характеризуются уменьшением
угла между вектором навивания оборотов протоконха и осью раковины, т. е.
по сравнению с другими группами эмбриональные раковины таких видов, как
Obesotoma tenuilirata, О. gigantea, О. simplex, выглядят более выпуклыми, кол-
пачковидными. Отличительной чертой группы, к которой относятся перечис-
ленные виды, является практически полное отсутствие скульптуры на прото-
конхе: еле заметная спиральная струйчатость пересекает редкие следы нара-
стания на протяжении всей поверхности эмбриональной раковины (рис. 40,
вкл.).
Радула. По сходству общего облика и конфигурации зубов с учетом деталей
их строения, в первую очередь формы киля, все исследованные виды Oenopo-
tinae объединяются в несколько групп.
В первую группу (Л) входят Curtitoma novajasemljensis, С. trevelliana,
С. incisula, С. ovalis и другие, зубы которых отличаются малыми размерами,
массивной базальной частью и килем, достигающим (а подчас превышающим)
1/2 длины лезвия (кинжаловидные зубы, рис. 14, А). Второй группе (Б)
свойственны большие копьевидные зубы, достигающие (а иногда превышаю-
щие) 1 мм в длину, с широким просветом лезвия и массивным основанием
(рис. 14, Б). Дистальный участок лезвия снабжен относительно крупным ки-
лем, достигающим 1/5 общей длины копьевидного зуба. В эту группу входят
Oenopota elegans, Ое. cinerea, Ое. harpa, Ое. cancellata, Ое. impressa и ряд других.
К этой группе близки виды, составляющие третью группу (В), — Pro-
pebela exarata, Р. tersa, Р. variabilis, Obesotoma simplex, обладающие саблевид-
ными длинными зубами (до 1.2 мм), но со значительно меньшим диаметром
лезвия (рис. 14, В). У них слабее развит киль, а базальная часть зуба значи-
тельно уступает в размерах, составляя от 1/8 до 1/13 общей его длины.
Тонкие маленькие стилетовидные зубы (рис. 14, Г) с длиной лезвия, всего
в 2—3 раза превышающей размеры базальной части, составили следующую,
четвертую группу (Г), в которой на основании полного отсутствия или
58
ОБЩАЯ ЧАСТЬ
наличия небольшого киля было выделено две подгруппы. В первую вошли
соответственно Oenopota pyramidalis, Ое. obliqua и Ое. pingelii, во вторую —
Granotoma albrechti, G. krauset, Obesotoma gigantea, 0. robusta, 0. starobogatovi.
Такие виды, как Propebela nobilis, P. angulosa, P. scalaris, P. assimilis и
ряд других (пятая группа, Д), характеризуются средними размерами зуба
(300—400 мкм) и базальной части (70 мкм). Киль таких ножевидных зубов
(рис. 14, Д) короче 1/2 длины лезвия.
Небольшую, но своеобразную шестую группу (Е) составили Prope-
bela scalaroides, Р. spitzbergensis, Р. rugulata, Р. svetlanae, Р. alitakensis и не-
которые другие виды. Для этой группы характерны расширенно-ножевидные
зубы (рис. 14, Е) с наибольшим диаметром, достигающим 1/i длины зуба; в со-
ответствии с этим получают сильное развитие и околобазальные ручки. Киль
в значительной мере редуцирован и занимает небольшую поверхность дисталь-
ного участка лезвия.
Отдельную, седьмую группу (Ж), явно отличающуюся от описанных
ранее по конфигурации и деталям строения, составили шиловидные зубы двух
видов: Propebela turricula и Р. harpularia (рис. 14, Ж; 41, вкл.).
Все приведенные группы демонстрируют большой спектр форм зубовт
встречающихся у Oenopotinae. Следует отметить, что были названы не все
виды, входящие в ту или иную группу, а лишь наиболее важные для разработки
естественной системы.
Таксоны родовой группы и филогения Oenopotinae. Обращает на себя внима-
ние сходство видовых составов групп, выделенных по морфологическому свое-
образию протоконха, с группами видов, объединенных на основании осо-
бенностей строения зубов радулы. Так, практически полное совпадение групп I
(скульптура протоконха представлена многочисленными спиральными реб-
рышками) и А (кинжаловидные зубы) по видовому составу позволило объеди-
нить их в род Curtitoma Bartsch, 1941.
Род заслуживает особого внимания в связи с тем, что строение радулярного1
зуба, свойственное относящимся сюда видам, близко исходному типу строения
зуба Oenopotinae (Богданов, 1986, 1989г). Только для представителей Curti-
toma характерен киль, достигающий 1/2 длины лезвия. В остальных группах
происходит постепенная редукция киля (вплоть до полного исчезновения),
что объясняется утратой его каркасной функции в связи с приобретением лез-
вием зуба более жесткой формы — цилиндрической. Кроме того, наблюдаемый
на протоконхах С. trevelliana, С. incisula, С. violacea процесс уменьшения числа
спиральных ребрышек в эмбриогенезе раковины, вероятнее всего, отражает
общую тенденцию олигомеризации спиральной скульптуры в эволюции Oeno-
potinae. Следовательно, скульптуру эмбриональной раковины, представленную
многочисленными спиральными ребрышками, следует считать наиболее древней.
Учитывая исходный тип строения зуба и общую тенденцию к редукции
числа спиральных элементов скульптуры протоконха, удалось найти соответ-
ствие между изменениями, происходящими в деталях строения зуба радулы,
и морфогенезом эмбриональной раковины (рис. 42). Например, род Oenopota
Morch, 1852 объединяет виды, для которых характерно наличие ретикулятного
рисунка протоконха (группа II) и мелких, практически лишенных киля стиле-
товидных зубов (группа Г). Явное отличие морфологии зубов у таких видов,,
как Ое. harpa, Ое. impressa и других (группа Е), позволяет выделить их в под-
род Nodotoma Bartsch, 1941. Таким образом, при практически одинаковой
скульптуре протоконха, в данном случае ретикулятной, происходят значи-
тельные изменения в морфологии зубов, что позволяет считать строение эм-
бриональной раковины более консервативным признаком, в большей мере от-
ражающим филогенетические взаимоотношения Oenopotinae.
Подтверждением большей стабильности протоконха может служить объеди-
нение 3 групп видов (Д, Е, Ж), выделенных по морфологическим особенностям
радулы. Для всех этих видов общим оказывается полное отсутствие макроскуль-
птуры на первом обороте протоконха (группа III), и на этом основании их ока-
СИСТЕМА И ФИЛОГЕНИЯ
59
залось возможным объединить в род Propebela Iredale, 1918. Основные изме-
нения радулы у моллюсков Propebela выражаются в увеличении диаметра и
Рис. 42. Схема морфологических преобразований протоконха и зуба радулы моллюсков
Oenopotinae.
А|— Curtitoma trevelliana, Б — Granotoma albrechti, В — Obesotoma gigantea; Г — О. simplex, Д — Oenopota
pingelii, E — Oe. impressa, Ж —. Propebela nobilis, 3 — P. rugulata, И — P. spitzbergensis, К •— P. turricula,
JI — P. scalaris, M — P. scalaroides, H — P, tersa.
укорачивании лезвия зуба. Более выпуклая форма протоконха со значительным
числом спиральных элементов на нем (4—5 ребрышек) соответствует видам
со строением зуба, наиболее близким к исходному типу. У таких видов, как
Р. nobilis, Р. angulosa и других, зуб имеет еще длинный киль и относительно
60
ОБЩАЯ ЧАСТЬ
узкий просвет лезвия. В то же время отмечено сильное расширение дистального
(лежащего близ киля) отверстия зуба. Уже у Р. scalaris диаметр лезвия зуба
увеличивается, а киль укорачивается и участок, противоположный острию,
отделяется от поверхности иглы, образуя характерный гарпун (рис. 42, Л).
Окончательное формирование гарпуновидного киля отмечено у Р. scalaroldes
(рис. 42, М). Для зубов этого вида характерно помимо киля, соединяющегося
с иглой только в самой дистальной точке, наличие сильно развитой базальной
части и очень широкого дистального отверстия лезвия.
За исключением постепенного сокращения в размерах базальной части,
сходные описанным изменения зуба происходят у группы видов, близких
Р. rugulata. У них киль не отходит от поверхности лезвия и не образует гар-
пуна. Интересен зуб Р. spitzbergensis (рис. 42, И), у которого дистальное от-
верстие лезвия образует своеобразную лунку; ее края составлены загнутыми
участками стенок иглы. Основание зуба представлено очень маленькой шаро-
видной базальной частью с длинными тонкими ручками. Киль сокращается
до маленького конуса на вершине лезвия, что, очевидно, является крайней
степенью его редукции. После исчезновения киля, который играет роль сти-
лета для прокалывания покровов добычи, зуб описанного строения уже не
может выполнять функции колющей иглы. На смену ему приходит шиловид-
ный, без полости внутри и без базальной части зуб, характерный для Р. turri-
cula и Р. harpularia (рис. 42, К). Формирование такого зуба происходило,
вероятно, за счет стенок лунковидного отверстия зуба, описанного для Р. spitz-
bergensis. Взаимосвязь уникальных зубов Р. turricula и Р. harpularia (группа Ж)
с токсоглоссными зубами групп Д и Е постулируется благодаря сходству
дефинитивных раковин рассмотренных выше видов. Сравнение эмбриональных
раковин также подтверждает их родство и отражает общую направленность
морфологических изменений протоконха во всей группе III. Так же как и
у Р. scalaroldes, протоконх Р. turricula сильно уплощен и несет 3 спиральных
ребрышка, из которых верхнее сглаживается на первом постэмбриональном
обороте.
В схему изменений, происходящих с зубами моллюсков рода Propebela
(относительное уменьшение длины и увеличение диаметра лезвия), не вписы-
ваются такие виды, как Р. exarata, Р. concinnula, Р. martnae, Р. mitrata и дру-
гие, обладающие зубами с тонким длинным лезвием и маленькой базальной
частью, в основном саблевидной формы (группа В). Учитывая совершенно
иную направленность эволюционных изменений зуба при том же строении
протоконха (группа III), названные виды следует выделять в особый подрод
Canetoma Bartsch, 1941.
Если для Propebela и Oenopota одной из составляющих эволюции эмбрио-
нальной раковины является уплощение ее верхней (апикальной) части, то
другое направление эволюции Oenopotinae связано с уменьшением угла (отно-
сительно оси раковины) вектора навивания оборотов протоконха. В резуль-
тате эмбриональная раковина становится более выпуклой, чем у Curtitoma,
часто колпачковидной (группы IV и F). У одних видов протоконх орнаменти-
рован 5—7 спиральными ребрышками, усеянными многочисленными микро-
скульптурными бугорками (группа IV). Такие виды объединяются в род Grano-
toma Bartsch, 1941. Другая группа видов характеризуется практически полным
отсутствием спиральной скульптуры на эмбриональной раковине (группа V);
она составляет род Obesotoma Bartsch, 1941. Следует отметить, что изменения,
претерпеваемые зубом в названных таксонах, неодинаковы. Для зубов
радулы Granotoma характерно значительное увеличение размеров базаль-
ной части относительно лезвия, киль частично редуцируется (рис. 42, Б).
Основной чертой морфологических преобразований зуба радулы Obesotoma
является постепенное удлинение лезвия зуба при сохраняющемся размере
базальной части (группа Д -> группа В). Маленький ромбовиднйй киль сох-
раняет стабильную для всех видов рода форму и пропорции (рис. 42,
В—Г).
СИСТЕМА И ФИЛОГЕНИЯ
61
Представленная картина филогенетических взаимоотношений (рис. 42)
позволяет проследить эволюционные преобразования различных деталей
строения раковины и тела моллюсков Oenopotinae.
Факт обнаружения зубов сходной морфологии в различных группах, вы-
деленных на основании строения протоконха, объясняется общей направлен-
ностью эволюционных преобразований радулы и, возможно, однотипностью
пищевых объектов у разных видов. Общей чертой всех рассмотренных направ-
лений морфологических преобразований зуба является постепенное сокраще-
ние размеров и редукция киля. Изменение строения основания зуба связано-
с увеличением диаметра и раскрытием околобазального отверстия лезвия.
Базальная часть зубов таких видов, как например Curtitoma trevelliana, С. he-
bes, С. novajasemljensis, С. incisula, шаровидная и массивная, расположена
непосредственно под околобазальным отверстием. При нападении на добычу
Рис. 43. £Схсма морфологических преобразований базальной части зуба. Пояснения
в тексте.
О — Curtitoma trezeUiana\ 1 — Granotoma abrechti; 2 — Oenopota pyramidalis; 3 — Propebela scalaroldes.
зуб зажат сфинктером так, что между внутренней стенкой хобота и базальной
частью зуба остается лишь узкий просвет. В подобной ситуации ядосодержащая
жидкость может поступать в полость лезвия зуба только через оставшиеся щеле-
видные просветы. Таким образом, одним из факторов определяющих эволю-
цию зуба, является преобразование базальной его части, способствующее
проникновению ядосодержащей жидкости в полость лезвия зуба.
В разных группах Oenopotinae этот процесс происходит по-разному (рис. 43).
У части видов Curtitoma и у большинства видов Obesotoma и Granotoma наблю-
дается вытягивание базальной части, приобретающей цилиндрическую форму.
От нее под углом 90° отходят ручки. В данном случае околобазальное отверстие
находится уже не над базальной частью зуба, а сбоку' от нее. Сходным образом
решается вопрос и у ряда представителей рода Oenopota и подрода Canetoma,
у которых происходит отгибание базальной части зуба. Угол отгибания отно-
сительно . лезвия составляет около 120°. У небольшого числа видов Propebela
отмечено уплощение базальной части, приобретающей языковидную форму,
и отгибание ее вниз; при этом ручки сохраняют свое изначальное положение.
Все перечисленные преобразования базальной части зуба приводят в той или
иной мере к беспрепятственному поступлению ядосодержащей жидкости под,
покровы добычи.
Безусловно, изменения морфологии эмбриональной раковины, связанные
в основном с процессом олигомеризации элементов спиральной скульптуры
(вплоть до полного их исчезновения), не могли не повлечь за собой преобразо-
ваний дефинитивной раковины. Однако наибольший интерес вызывают эво--
63
ОБЩАЯ ЧАСТЬ
impressa
harpa
casentina
pavlova
elegans
•declivis
cancellate
biconica
mequita
sachalinensis
kurilensis
elongate
uschakovi
obliqua ---
pyramidalis
pingelii —
Рис. 44. Филогенетические отношения ви-
дов рода Oenopota.
obusta
woodiana
tumida
turricula
neymanae
harpularia
violacea
schantarica
simplex------
gigantea-----
gigas ------'
starobogatovi
tenuilirata -
japonica---
tomiyaensis
solida-----
Рис. 45. Филогенетические отношения ви-
дов рода Obesotoma.
trevelliana
fiora
spitzbergensis
rugulata
bergensis
assimilis
scalans
anguiosa
svetlanae
popovia
terpeniensis
verrilli
nobilis
goryachevi
novajasemljensis
hebes
mcisula
becklemishevi
bartschi
piltuniensis
conoidea
ovalis
decussata
lawrenciana
microvoluta'
hokkaidoensis
raduga
Рис. 47. Филогенетические отноше-
ния видов родов Granotoma и Cur-
titoma.
Рис. 46. Филогенетические отноше-
ния видов рода Propebela.
ОБРАЗ ЖИЗНИ
63-
люционные изменения, происходящие не со спиральной, а с осевой скульпту-
рой дефинитивной раковины. Изучение филогенетических взаимоотношений
видов выявило тенденцию к постепенному сглаживанию линий нарастания
в скульптуре дефинитивной раковины с последующим исчезновением на них
следов анального синуса. Например, у Curtitoma trevelliana, С. conoidea, С. in-
cisula и большинства других видов Curtitoma серповидный анальный синус
ярко выражен. Преобладающее число видов Granotoma и Oenopota и некоторые
Obesotoma имеют сглаженный серповидный синус, но у О. simplex и Oenopota
impressa он окончательно исчезает и линии нарастания выглядят почти пря-
мыми.
У всех представителей рода Propebela анальный синус отсутствует, а линия
нарастания эволюционирует от прямой к преломленной в верхней части. При-
чем излом происходит в направлении навивания оборотов раковины. Сходные
изменения, связанные со стиранием анального синуса, трактуются как про-
грессивные для большинства представителей ряда подсемейств Turridae (Po-
well, 1966). Морфологические изменения дефинитивной раковины у эволюци-
онно молодых видов сопровождаются увеличением размеров раковины и ис-
чезновением анального синуса.
Эволюционные изменения морфологии крышечки касаются уменьшения
относительных ее размеров, более резкого изгибания оси нарастания (наблю-
дается на начальной стадии роста), сужения отпечатка колумеллярного мус-
кула и смещения его от левого края крышечки в центр.
Обобщая рассмотренные данные, можно сделать вывод, что наиболее арха-
ичными особенностями Oenopotinae, по-видимому, являются: 1) короткий
массивный зуб небольшого диаметра с уплощенным основанием, широкими
короткими ручками и длинным килем, равным или превышающим 1/2 длины:
лезвия; 2) протоконх выпуклой формы, скульптура которого состоит из много-
численных спиральных ребрышек; 3) наличие на линиях нарастания следов^
анального синуса; 4) относительно крупная крышечка с плавно изгибающейся
осью нарастания.
С учетом архаичности тех или иных признаков и направленности их эволю-
ционных преобразований были построены схемы филогенетических взаимоот-
ношений исследованных видов (рис. 44—47). Однако отсутствие палеонтоло-
гических данных не позволяет рассмотреть вопрос о филогенетических взаимо-
отношениях видов в историческом аспекте.
ОБРАЗ ЖИЗНИ МОЛЛЮСКОВ OENOPOTINAE
Распределение по глубинам. Oenopotinae имеют очень широкий диапазон
глубин распространения: верхней границей служит литораль, а нижней ба-
тиаль (Bouchet, Waren, 1980). Отдельные представители подсемейства отмечены
на глубине до 4743 м, а по данным А. В. Сысоева (устн. сообщ.), — ив ультра-
абиссали на глубинах свыше 7 тыс. м.
Анализ распределения исследованных видов1 позволяет выделить несколько
основных групп.
А. Стенобатные циркумлиторальные (здесь и далее
терминология по: Голиков, 1982) виды, поднимающиеся в адлитораль. Отме-
чены на глубинах до 39 м, но чаще всего встречаются на глубинах 2—10 м:
Propebela areta, Р. cingulata, Obesotoma starobogatovi, О. simplex, Curtitoma no-
vajasemljensis.
Б. Эврибатные элиторальные виды. В этой, наиболее-
представительной по числу видов группе выделяют 3 подгруппы в соответствии
с глубинами, характеризующимися пиком численности моллюсков.
1 Сведения о распространении моллюсков Oenopotinae приводятся только для видов,,
которые представлены достаточной для достоверности данных выборкой.
64
ОБЩАЯ ЧАСТЬ
1. Пик численности приходится на глубины 50—80 м. К этой подгруппе
относятся Curtitoma hebes (15—494),2 Granotoma albrechti (3.5—900), Pro-
pebela alitakensis (28—180), P. turricula (10—260).
2. Пик численности приходится на глубины 15—НО м. В эту подгруппу
входят Oenopota harpa (6—310), Ое. impressa (5—500), Propebela nobtits (7—
515), P. virtdula (7—780), Obesotoma japonica (8—380), Curtitoma violacea
(1.5-669).
3. Пик численности размыт — приходится на глубины 20—200 м. Эта
подгруппа объединяет виды, практически равномерно распространенные по
всей глубине: Oenopota pyramidalis (14.5—400), Ое. elegans (7—660), Ое. ci-
nerea (28—422), Obesotoma schantarica (5—330), 0. robusta (29—860), Prope-
bela exarata (4—320), P. tersa (60—900), P. scalaris (1.5—660), P. rugulata
(9.5—500), P. scalaroides (3—430), Curtitoma decussata (1.5—820), C. trevel-
liana (3—430). Для последнего вида отмечается 2 пика численности: один
на глубине 40—80, другой на глубине 160—220 м.
В. Эврибатные сублиторальные виды, обитающие на глу-
бинах от 2 до 500, но с пиком численности на 20—50 м: Propebela harpularia,
Р. spitzbergensis, Obesotoma gigantea, О. woodiana, Oenopota pingelii, Oe. obli-
•qua, Curtitoma laivrenciana.
Г. Эврибатные батиальные виды, обитающие на глубинах
250—2500 м: Propebela abyssicola (250—1181), Curtitoma ovalis (1900—2300),
C. microvoluta (1076—1400). Указать пик численности для отдельных предста-
вителей выделенной группы не представляется возможным из-за малочислен-
ности материала.
Литературные данные позволяют расширить знания о батиметрических
границах распространения ряда видов. Так, верхней границей распространения
Curtitoma ovalis следует считать глубины 557—699, а нижней 2775 м (здесь и
ниже сведения взяты из: Bouchet, Waren, 1980). Для Oenopota elegans нижней
границей распространения является 1301, а для Curtitoma violacea 932 м. Од-
нако существенных поправок в выделение групп (осуществленное на основе
распределения видов по глубинам) литературные данные не вносят. Общая кар-
тина вертикального распределения моллюсков подсем. Oenopotinae, основан-
ная на собственном материале, представлена на рис. 48.
Распределение по грунтам. Несмотря на подвижный образ жизни рассматри-
ваемых хищных гастропод, можно говорить о явной приуроченности большин-
ства видов к определенным типам грунтов и выделить следующие группы.
А. Эвритопные виды, встречающиеся на всех типах грунтов: от ска-
листого и каменистого до чисто илистого. Однако в этой группе можно выделить
4 подгруппы.
1. Виды, чаще всего обитающие на мягких илистых грунтах с при-
месью более крупных частиц (галька, гравий, ракуша): Oenopota declivts,
Ое. elegans, Curtitoma violacea, C. decussata, C. conoidea, C. incisula, C. ova-
lis, C. trevelliana, Propebela rugulata, P. spitzbergensis.
2. Виды, предпочитающие песчанистые грунты: Oenopota pyramidalis,
Oe. pyramidalis var. semiplicata, Propebela viridula, P. exarata, P. nobilis,
P. scalaris, P. tersa, Obesotoma simplex, O. gigantea, O. schantarica, O. robusta,
Granotoma albrechti.
3. Виды, предпочитающие илистый песок и песчаный ил: Curtitoma he-
bes, Obesotoma starobogatovi, О. solida, Oenopota harpa, Oe. impressa, Oe. obli-
qua, Oe. pingelii, Propebela scalaroides, P. harpularia, P. assimtlis, P. alita-
kensis, P. angulosa.
4. Вид, предпочитающий глинистые грунты: Obesotoma woodiana.
Б. Стенотопные виды, место обитания которых приурочено к оп-
ределенному типу грунта.
2 В скобках указаны минимальная и максимальная глубины нахождения живых особей
данного вида в исследованном материале.
ОБРАЗ ЖИЗНИ
65
1. Виды, обитающие на чистых илах: Oenopota cinerea, Curtitoma nova-
jasemljensis.
2. Виды, обитающие на чистом песке: Propebela turricula, Р. areta, Obe-
sotoma japonica.
Соотношение числа видов, предпочитающих тот или иной тип грунта, отра-
жено на рис. 49. Наиболее многочисленную группу составляют виды, которые
обитают преимущественно на илистых
грунтах. Доля находок моллюсков на
скалах и ракушечниках невелика, что,
вероятно, связано с распространением
полихет, которыми они питаются.
Отношение к температуре. Термо-
патия видов Oenopotinae неодинакова.
Достаточно сказать, что некоторые мол-
люски были обнаружены при темпера-
Рис. 48. Распределение числа видов в зависи-
мости от глубины их обитания.
По оси абсцисс — число видов, по оси ординат —
глубина в метрах.
туре, достигающей 15 °C, тогда как большая часть видов встречается только
в водах, температура которых не превышает 2 °C. В зависимости от диапазона
оптимальных для вида температур можно выделить следующие группы.
А. Стенотермный, умеренно-тепловодный атлан-
тический вид: Propebela turricula.
Б. Стенотермные, холодноводные арктические
виды, обитающие в диапазоне температур от —1.9 до 6.5 °C. В этой группе
в зависимости от наибольшей частоты встречаемости в определенном диапазоне
температур выделяются следующие подгруппы.
П6идов
20 Y
Рис. 49. [Распределение видов по предпочитаемым грунтам.
По оси абсцисс —• тип грунта: 1 — скалы, камень; 2 — гравий,
галька, ракуша; з — песок; 4 — илистый песок; 5 — ил с камнями,
ил с галькой; 6 — чистый ил. По оси ординат — число видов, пред-
почитающих данный тип грунта.
1. Виды, пик численности которых приурочен к температурам от —1.8
до —0.8 °C: Curtitoma novajasemljensis, С. trevelliana, Oenopota elegans,
Oe. cinerea, Propebela spitzbergensis.
2. Виды, пик численности которых приходится на диапазон температур
от —0.8 до 1.0 °C: Curtitoma incisula, Obesotoma woodiana, О. robusta, О. sta-
robogatovi.
3. Виды промежуточной подгруппы, встречаются чаще всего при тем-
пературе от —1.98 до 0 °C: Curtitoma decussata, Propebela scalaroides, P. sca-
laris, P. rugulata.
В. Эвритермные умеренно-холодноводные бо-
реально-арктические виды, обитающие в диапазоне температур от
—2.0 до 12.8 °C. При выделении оптимальных температур обитания настоящая
группа может быть разделена на две подгруппы.
1. Виды с узким диапазоном оптимума от 0.3 до 1.4 °C: Oenopota pyra-
midalis, Ое. pyramidalis var. semiplicata, Propebela harpularia, Obesotoma
schantarica, Granotoma albrechti.
5 И. П. Богданов
6g
ОБЩАЯ ЧАСТЬ
2. Виды с более широким диапазоном температур от —0.9 до 0.8 °C:
Curtitoma violacea, Oenopota impressa, Oe. obliqua, Oe. harpa, Propebela vi-
ridula, P. exarata, P. nobilis.
Следует отметить, что в соответствии с более широким диапазоном оптималь-
ных температур возрастают и величины переживаемых температур (рис. 50),
с чем связано и более широкое зональное распространение видов второй под-
группы по сравнению с ареалами первой.
Как видно из приведенного материала, для большинства видов температур-
ный оптимум находится в пределах от —1.9 до 2.0 °C (рис. 51), чем и может
быть объяснено вертикальное распределение исследуемых моллюсков и выход
их в адлитораль лишь в холодноводных участках Новой Земли, Земли Франца-
Иосифа и Шпицбергена. Величина температурного оптимума объясняет и от-
сутствие Oenopotinae в субтропических водах.
Рис. 50. Распределение различных [биогео-
графических групп видов в зависимости, от
диапазона выносимых ими температур среды
обитания.
1 — арктические виды, г — бореальные и бореально-
арктические виды. По оси абсцисс — температура
(°С>; по оси ординат — число видов.
Ге
Рис. 51. Распределение различных биогео-
графических групп видов г зависимости от
оптимальных температур среды обитания.
] — арктические виды, 2 — бореальни-а ркти-
ческие виды. Обозначение осей как на рис 50-.
Отношение к солености. Все рассмотренные представители Oenopotinae —
типично морские виды. Однако ряд видов может обитать в районах, сильно'
опресненных стоками рек, и переносить понижение солености почти до 18 °/00. Оче-
видно, особи того или иного вида могут находиться в условиях частичного опрес-
нения различное время, однако специальных исследований, посвященных во-
просам выживания моллюсков Oenopotinae в экстремальных условиях, не про-
водилось. Поэтому был изучен диапазон солености, для которого характерна
наибольшая частота встречаемости представителей каждого вида. По диапазону
изменений значения солености среды обитания можно выделить стеногалинные
и относительно эвригалинные виды.
А. Стеногалинные виды, обитающие в морской воде с соленостью'
не ниже 30 °/в0. Здесь можно выделить 2 подгруппы.
1. Максимум численности приходится на диапазон солености от 34.3
до 35 e/00: Oenopota impressa (31.70),3 Oe. cinerea (34.30), Oe. pingelii (32.90),
Oe. elegans (34А2), Curtitoma ovalis (34.55), C. novajasemljensis (31.50), C. de-
cussata (29.13), Propebela exarata (33.34), P. scalaroides (33.58), P. scalaris
(32.87), P. spitzbergensis (31.64).
2. Максимум численности приходится на диапазон солености 33.0—
34.0 °/ee: Curtitoma incispla (31.11), Obesotoma robusta (31.11), О. woodiana
(32.40), О. schantarica (33.80), Granotoma albrechti (31.11).
3 В скобках приведены минимальные значения солености (в промилле), при которых
встречаются особи данного вида (только на основе исследованного материала).
ОБРА 3 ЖИЗНИ
67
Б. Относительно эвригалинные виды, переносящие опрес-
нение более 29.0 °/00. Здесь также можно выделить 2 подгруппы.
1. Максимум численности отмечен при солености от 34.0 до 35.0°/ю:
Oenopota pyramidalis (27.39), Oe. pyramidalis var. semiplicata (26.20), Oe.
harpa (25.66), Oe. obliqua (27.20), Propebela harpularia (27.10), P. nobilis
(26.00), P. rugulata (25.60), P. viridula (27.39), Curtitoma violacea (28.60).
2. Максимум численности отмечен при солености от 29.4 до 31.6'°/0в.
В эту подгруппу входит 1 вид, Obesotoma simplex, пожалуй, наиболее эври-
галинный из всех рассматриваемых видов. Особи этого вида встречаются
в диапазоне солености от 18.82 до 33.95 %0. Интересно отметить, что именно
эвригалинные виды составляют
фауну Белого моря и опресненных
участков северных морей СССР.
Рис. 52. Распределение числа видов в за-
висимости от переносимого ими опреснения.
1 — стеногаллинные, 2— относительно эвригал-
линные. По оси абсцисс — значение солености,
промилле; по оси ординат — число видов.
График, показывающий отношение к солености у эври- и стеногалинных ви-
дов, приведен на рис. 52.
Питание. Все Oenopotinae — избирательные хищники. Пищевым объектом
для них служат полихеты Prionospio malmgreni, Spiophanes bombyx, Ammot-
rypane angolaster (Pearce, 1966).
Пирсом (Pearce, 1966) был поставлен ряд опытов, продемонстрировавших
высокую скорость пищеварения у Curtitoma trevelliana: в лабораторных усло-
виях моллюску скармливались полихеты S. bombyx. Весь процесс, начиная от
заглатывания добычи до дефекации непереваренных остатков, занимал у мол-
люска менее 14 ч.
Изучение значительного числа пищевых остатков, удерживаемых сфинкте-
ром ринхостома и содержащихся в пищеварительном тракте моллюсков, позво-
лило установить, что пищевой рацион этих животных составляют в основном
полихеты семейств Spionidae и Paraonidae (табл. 3). По мнению специалистов,
определявших полихет (А. Б. Цетлин и Т. А. Бритаев), большинство просмо-
тренных экземпляров малоподвижны и обитают на илистых и заиленных грун-
тах. Последние данные хорошо сочетаются с предпочитаемыми грунтами оби-
тания большинства изученных моллюсков.
Помимо явного хищничества Oenopotinae, вероятно, могут поедать останки
других животных (частичная нек-рофагия). На возможность данного типа пи-
тания указывают две находки фрагментов тела Amphipoda в желудках моллю-
сков.
Особенности размножения. Репродуктивные циклы моллюсков подсем. Oeno-
potinae изучены недостаточно. Созревание половых продуктов у взрослых мол-
люсков (соотношение полов практически у всех видов 1 : 1) происходит позд-
ней осенью. По данным Пирса (Pearce, 1966), половые протоки самцов Curti-
toma trevelliana из Оресанда (юг Норвегии) содержали развитую сперму уже
в октябре, а к ноябрю гонады уже всех исследованных моллюсков оказались
зрелыми. Немногочисленные нахождения кладок в Норвежском и Северном
морях (Vestergaard, 1935; Thorson, 1946; Pearce, 1966), а также у побережья
Гренландии (Thorson, 1935) дают лишь отдаленное представление о сезонности
размножения и длительности эмбрионального развития. Самки откладывают
яйцевые капсулы весной или в течение лета: так, для вод Норвегии и Англии
они отмечены с середины апреля до начала августа (Thorson, 1946; Pearce 1966).
В более холодных водах Гренландии капсулы с развивающимися эмбрионами
обнаружены только начиная с июня.
5*
68
ОБЩАЯ ЧАСТЬ
Таблица 3
Полихеты, отмеченные в пищевом рационе моллюсков Oenopotinae
Таксономическая
принадлежность полихет
Обследованный вид моллюска *
Sygalinoidea
Pholoe minuta caeca
Syllidae
Orbiniidae
Nephtydae
Micronephthys sp.
Spionidae
Oe. impressa, Р- exarata
Р. exarata
Ое. obliqua
G. albrechti
Prionospio sp.
Prionospio cirrifera
Spio sp.
Laonice sp.
Paraonidae
Artcidea nolani
Cirratulidae
Chaetozona setosa
Maldanidae
Ampharetidae
Ampharete arctica
A. lindstroemi
Terebellidae
Sabellidae
Euchone sp.
Pilargidae
P. rugulata
C. novajasemljensis, C. violacea, C. conoidea,
Oe. pyramidalis, Oe. impressa, P. exarata,
P. bergensis, P. spitzbergensis, P. rugulata,
P. nobilis, P. harpularia, 0. simplex
C. trevelliana
C. decussata, C. novajasemljensis
P. exarata
Oe. impressa, P- nobilis
C. novajasemljensis, G. albrechti, P. viridula,
P. bergensis, 0. simplex
P. viridula
C. decussata
G. ablrechti
Oe. harpa
Oe. harpa
Oe. harpa
P. cingulata, Oe. harpa
Oe. cancellata, P. rugulata
Oe. pyramidalis, P. rugulata
C. starobogatovi
* Для родовых названий используются следующие сокращения: Ое. — Oenopota,
Р.—Propebela, G.— Granotoma, С. —Curtitoma, О. — Obesotoma.
Кладки большинства видов Oenopotinae внешне схожи между собой и со-
стоят из|центральной, полуовальной в сечении жилой камеры и окаймляющей ее
базальной части — оборки (рис. 53), служащей для прикрепления капсулы
к субстрату. В верхней части камеры (как правило, в центре) находится оваль-
ная, несколько удлиненная лунка, закрытая крышечкой, которая соединена со
стенками камеры тонкой кожистой мембраной (рис. 54, вкл.). Лунка служит
выходным отверстием для молоди, что подтверждается нахождением капсул,
лишенных крышечки, но содержащих еще 1 —2 моллюсков, либо уже абсолютно
пустых. Вероятнее всего, мембрана разрушается к моменту эмбрионального'
развития большинства зародышей.
Мной было просмотрено небольшое число капсул, собранных практически
сразу же после их откладки. Число яиц, содержавшихся в них, достигало 750—
900, что в несколько десятков раз превышало среднее число развивающихся
в кладке эмбрионов. Возможно, большинство яиц в кладке не оплодотворя-
ется и играет роль питательных яиц. Количество эмбрионов в капсуле непо-
стоянно для вида и варьирует от 2—7 у Obesotoma simplex- и Propebela nobilis
(Thorson, 1935) до 100—150 у Р. turricula (Vestergaard, 1935). Более специфич-
ным для вида показателем может служить среднее число развивающихся в клад-
ках эмбрионов. Так, у Obesotoma simplex из 17 просмотренных капсул 8 содер-
жало по 5 эмбрионов, что соответствовало и среднему арифметическому значе-
нию. .Для других исследованных видов эта цифра равнялась: Propebela nobi-
lis 4, Curtitoma violacea 5, Oenopota pyramidalis 12, Propebela exarate 14, Curti-
toma trevelliana. 28.
Помимо неодинакового численного состава развивающихся эмбрионов для
кладок каждого вида отмечены различия в конфигурации, строении и размерах
ОБРАЗ ЖИЗНИ
69
жилой камеры. Так, яйцевая капсула Obesotoma simplex диаметром 2—3 мм
отличается от капсул остальных изученных видов высокой жилой камерой
(рис. 53), стенки которой состоят из крупных пластинок, придающих поверхности
сетчатый облик (Thorson, 1935). Наоборот, капсулы Oenopota pyramidalis упло-
щены в дорсовентральном направлении, имеют маленькую лунку и слабо
скульптурированы; их диаметр от 3.5 до 6 мм. Своеобразная капсула описана
у Curtitoma avails (Bouchet, Waren, 1979). Она напоминает капсулы некоторых
Рис. 53. Капсулы-моллюсков Oenopotinae.
1 — Obesotoma simplex; 2 — Propebela exarata; 3 — Propebela nobilis; 4 — Oenopota pyramidalis; 5 — Cur-
titoma violacea, 6 — Curtitoma trevelliana. Масштабная линейка 1 мм (no: Thorson, 1935).
Buccinidae (Горячев, 1978; Голиков, 1980) и уплощена с боков (рис. 55). По-
скольку авторы, описавшие капсулу (Bouchet, Waren, 1979), не проводили ис-
следований эмбрионального развития С. avails, а жилая камера содержала не-
развитые эмбрионы, то судить о видовой принадлежности капсулы не представ-
ляется возможным. Очевидно, описанная исследователями капсула вообще не
относится к кладкам обсуждаемых моллюсков.
Oenopotinae помещают кладки на самые разные субстраты: водоросли (на-
пример, Fucus и Laminaria), пустые раковины Bivalvia, трубки Pectinaria,
поверхность мшанок типа Alcyonidium disciforme, а также на камни и песок.
Реже обнаруживаются кладки на раковинах живых моллюсков Astarte sp.,
Musculus laevigatas, Hiatella arctica, Macoma calcarea и даже на крышечках жи-
вых Buccinidae. Вероятно, для большинства видов субстрат не имеет особого
значения и можно говорить только о предпочтении определенного его. типа .тем
или иным видом. Например, Obesotoma simplex чаще откладывает кладки на пе-
сок.
70
ОБЩАЯ ЧАСТЬ
Торсон (Thorson, 1935), сравнивая капсулы через короткие промежутки вре-
мени (3—5 дней), отметил все стадии развития: от яиц до взрослых эмбрионов и
пустых кладок. Отсюда им был сделан вывод о том, что эмбриогенез протекает
очень быстро. Последние находки
яйцевых капсул, содержавших мол-
\\ люсков на завершающей стадии эм-
бриогенеза, были сделаны в се-
редине августа во всех исследован-
7 ных районах. В это же время бы-
/ ло найдено большое число пустых
Рис. 55. Кладка Curtitoma ovalis (по:
Bouchet, Waren, 1379) с неразвитым
эмбрионом.
А, Б — различные ракурсы. Масштабная
линейка 1 мм.
кладок и значительное количество молоди. Интересно, что для большинства
глубоководных туррид Буше и Варен (Bouchet, Waren, 1980) предполагают на-
личие пелагической личинки. Для абиссальных и батиальных моллюсков, изу-
ченных ими в районе Норвежского моря, число видов, предположительно
имеющих пелагическую стадию, составляло в среднем 69 %, а на глубине
Рис. 56. Некоторые стадии личиночного развития моллюсков Oenopotinae (по: Thorson,
1935).
1 — Propebela exarata, стадия велигера, проходящая внутри капсулы; 2 — личинка Curtitoma bicarinata
перед выходом из капсулы; 3—6 — стадии развития особи Curtitoma trevelliana: 3 — личинка внутри кап-
сулы, 4, 5 — велигер, 6 — донная стадия. Масштабные линейки: 2, 2, 4—d — 0.5 мм; з — 0,1 мм.
3000 м достигало 78 % от общего числа исследованных видов туррид. Среди Oe-
nopotinae свободноплавающая пелагическая личинка указана только для
Propebela turricula, Oenopota elegans, Oe. laevigata и Curtitoma trevelliana. Под-
робно изучена и описана лишь личиночная стадия последнего вида (Thorson,
1946). Она имеет толстый двудольный парус, причем каждая из удлиненных его
долей закруглена и несет двойной бордюр, состоящий из пигментированных
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
71
фиолетовых клеток. Вся поверхность паруса снабжена жгутиковидными тру-
бочками, развитыми в различной степени. На фоне желтоватого тела личинки
четко выделяются пурпурной пигментацией глотка и коричневая печень
(рис. 56). На ранних этапах планктонной жизни личинка несет раковину с на-
ибольшим диаметром, равным 335 мкм, но уже к моменту оседания на дно
она почти вдвое увеличивается в размерах и ее высота составляет 710, а наиболь-
ший диаметр 550 мкм. Пелагическая жизнь велигера непродолжительна из-за
недостаточного количества пищевого вещества в фитопланктоне (Thorson, 1946).
Эмбрионы Propebela exarata и Curtitoma violacea проходят стадию велигера
в капсуле (Thorson, 1946; собственные данные). Видимо, именно такой тип эмб-
риогенеза характерен для большинства Oenopotinae.
Моллюски Oenopotinae — объект питания. Роль Oenopotinae в питании
бентосоядных животных почти не изучена. Лишь в работе Кутуи (Couthouy,
1838) содержится описание Bela pleurotomaria по экземпляру, обнару-
женному в желудке трески. Это, пожалуй, единственное известное сообщение
о питании морских животных моллюсками данного подсемейства.
По сведениям, содержащимся в отчетах Лаборатории гидробиологии
ПИНРО за 1967 и 1978 гг.,4 оказалось возможным судить о пище-
вом рационе пикши и камбалы.
В южной части Баренцева моря (Мурманский берег—Канинская банка)
число моллюсков в рационе пикши достигает 5 % от общего состава поедаемой
пищи. Всего в желудках пикши найдено 69 видов моллюсков, из них 35 видов
гастропод, в том числе Oenopota pyramidalis и Oenopota sp. В желудках кам-
балы было обнаружено 36 видов моллюсков, из них гастропод 17. видов,
в том числе Oenopota sp.
Следует учитывать, что исследования питания пикши и камбалы проводи-
лись на мелководных банках, где доля Oenopotinae среди брюхоногих очень
невелика (как в количественном, так и в видовом отношении). Однако в более
глубоководных участках шельфовой и батиальной зон умеренных широт север-
ного полушария Oenopotinae наряду с Buccinidae составляют доминирующую
часть малакофауны, в связи с чем доля их в пищевом рационе бентосоядных рыб
и, возможно, морских млекопитающих резко возрастает.
Oenopotinae часто оказываются объектами нападения и питания хищных
Naticidae (Pearce, 1966, собственные данные). Например, в районе Килдинской
банки 60 % просмотренных раковин имели характерные следы нападения на-
тицид.
Другие формы биотических отношений. Раковина Oenopotinae служит
субстратом для многих животных-комменсалов. На многих раковинах посе-
ляются колонии мшанок, а на раковинах моллюсков из рода Granotoma посто-
янно встречаются колонии Hydractinia sp. Нередко обнаруживаются прикреп-
ленные формы агглютинирующих фораминифер (например, Hemisphaerammina
sp.), трубки полихет сем. Serpulidae и некоторые другие обрастатели. Пустые
раковины часто заселены раками-отшельниками из рода Pagurus, а также си-
пункулидами.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
ПРЕДСТАВИТЕЛЕЙ OENOPOTINAE
При выделении биогеографических групп нами использованы общие прин-
ципы морской биогеографии (Гурьянова, 1939; Ушаков, 1951, 1955; Галкин,
1955; Филатова, 1957; Скарлато, 1960; Голиков, Кусакин, 1962; Зенкевич, 1963;
Голиков, 1963; Гурьянова, 1972; Горячев, 1978; Голиков, 1980; Скарлато, 1981;
Голиков, 1982; Несис, 1982; Ekman, 1935, и др.). Все построения, связанные
с выделением биогеографических групп, базируются на собственном материале
(за исключением пп. IV и VII, см. ниже), так как использование
4 Материалы отчетов были любезно предоставлены сотрудницей лаборатории О. В. Гера-
симовой.
72
ОБЩАЯ ЧАСТЬ
литературных данных осложняется постоянно встречающимися неточностями
в определении большинства видов.
На основании проведенных исследований можно выделить следующие типы
ареалов Для моллюсков Oenopotinae, распространенных только в холодных и
умеренных водах северного полушария.
I. Тихоокеанские широко распространенные бо-
реальные виды (7): Curtitoma lawrenctana, Granotoma albrechti, G. krausei,
Oenopota pavlova, Oe. casentina, Oe. inequita, Obesotoma solida. Обитают в низко-
и высокобореальных районах у берегов Азии и Америки; распространены от
Южного Приморья и о-ва Хонсю у берегов Азии и от Ванкувера у берегов Аме-
рики до южной части Чукотского моря.
II. Тихоокеанские приазиатские широко распростра-
ненные бореальные виды (9): Curtitoma пеутапае, С. becklemishevi,
Oenopota kurilensts, Obesotoma schantarica, О. sachalinensis, Propebela margaritae,
P. areta, P. marinae, P. goryachevi. Обитают в низко- и высокобореальных
районах у берегов Азии.
III. Тихоокеанские приазиатские низкобореаль-
ные виды (15): Curtitoma microvoluta, С. hokkaidoensis, Oenopota uschakovi,
Oe. biconica, Oe. Candida, Oe. elongata, Obesotoma japonica, O. tomijaensis, Pro-
pebela verrilll, P. terpeniensis, P. cassis, P. cingulata, P. golikovi, P. exquisita,
P. venusta. Распространены от северной Кореи и о-ва Хонсю до севера Татар-
ского пролива в Японском море и от п-ова Босо на тихоокеанском побережье
Японии до Южно-курильского мелководья и южной части о-ва Итуруп; в Охот-
ском море у берегов Сахалина (на мелководье до 30—40 м) от зал. Анива до
вал. Терпения.
IV. Тихоокеанские приамериканские низкобо-
реальные виды 5 (приводятся только на оснований литературных данных)
(4): Oenopota pytisa, Ое. surana, Obesotoma dtegensis, О. sixta. Распространены
у берегов Британской Колумбии и Калифорнии.
V. Тихоокеанские широко распространенные вы-
сокобореальные виды (7): Curtitoma flora, Propebela mitrata, P. fi-
dtcula, P. variabilis, P. alitakensis, P. popovia, P. rassina. Обитают в высоко-
бореальных районах у побережий Азии и Америки и встречаются в прилежащих
акваториях.6
VI. Тихоокеанские приазиатские высокобореаль-
ные виды (4): Curtitoma bartschi, С. piltuniensis, Granotoma raduga, Propebela
svetlanae. Расселены от м. Терпения и о-ва Итуруп на север до Берингова про-
лива и прилежащих акваторий.
VII. Тихоокеанские приамериканские высокобо-
реальные виды 5 (приводятся только на основании литературных данных)
(9): Granotoma lutkeni, Obesotoma chiachiana, О. quadra, О. kyskana, О. miona,
Oenopota althorpensts, Propebela tenuissima, P. alaskensls, P. monterealis. Распро-
странены у берегов Аляски и Британской Колумбии до Ванкувера.
VIII. Амфибореальные широко распространенные
виды тихоокеанского происхождения (7): Curtitoma in-
cisula, С. hebes, Oenopota cancellata, Obesotoma tenuilirata, 0. robusta, Propebela
concinnula, P. tersa. Отмечены в бореальных водах Тихого океана и в высоко-
бореальных водах американского побережья Северной Атлантики.
IX. Атлантические евразийские бореально-суб-
тропические виды, заходящие в арктические воды (2): Propebela turri-
cula, Р. bergensis. В субтропиках встречаются только на больших глубинах в ба-
тиали.
6 Суждения о родовой принадлежности приведены только на основе морфологических
особенностей дефинитивной раковины.
6 Например, представители Р. fidicula обнаружены в Чукотском море у берегов
о-ва Врангеля, и этот вид можно считать тихоокеанским высокобореально-арктическим.
Но выделение нового типа ареала мы считаем преждевременным в связи с недостаточной
изученностью распространения группы в Чукотском море.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
73
Х. С; Широко распространенные бореально-аркти-
ческие виды, преимущественно атлантического происхож-
дения (9): Curtitoma trevelliana (рис. 57), Oenopota. elegans, Ое. declivis,
Ое. obliqua, Ое. cinerea, Obesotoma woodiana, Propebela exarata, P. rugulata,
P. scalaris. Распространены в высокобореалышх районах Северной Атлантики
и циркумполярно в Арктическом бассейне.
XI. Атлантические высокобореальные виды (2): Cur-
titoma avails, Propebela angulosa. Распространены от Шотландии и Северной
Ирландии у берегов Европы и от м. Код у берегов Америки до юго-западной
части Баренцева моря, западных берегов Шпицбергена и южной части Гренлан-
дии. С. ovalis может быть охарактеризована еще и как высокобореально-бати-
альный вид.
XII. Широко распространенные бореально-арк-
тические виды, преимущественно тихоокеанского проис-
хождения (10): Curtitoma violacea, Oenopota harpa, Oe. pyramidalis, Oe. pin-
gelii, Oe. impressa, Obesotoma simplex, Propebela viridula, P. harpularia, P. no-
bilis, P. asstmilis. Распространены циркумполярно в Арктическом бассейне и
отмечены в Беринговом, Охотском, Японском морях, а также в Северной Ат-
лантике.
XIII. Тихоокеанский бореально-арктический вид
тихоокеанского происхождения (1): Curtitoma decussata.
Распространен в низко- и высокобореальных водах Тихого океана и доходит
в Полярном бассейне до восточных берегов Новой Земли.
XIV. Евразийские арктическо-бореальные виды в ы-
сокобореально-атлантического про и схождения (4):
Curtitoma conoidea, С. novajasemljensis, Obesotoma gigas, Propebela spltzbergen-
sts. Распространены в западной части Арктического бассейна и встречаются
в высокобореальных водах Атлантического океана.
XV. Арктические, преимущественно евразийские виды (4).
Распространены только в Северном Ледовитом океане: Propebela scalaroides,
Obesotoma gigantea — на запад от п-ова Таймыр; Obesotoma starobogatovi, О. tu-
mida в Восточно-Сибирском море, море Лаптевых, а также в южной части Кар-
ского моря, практически отсутствуют в Баренцевом море (рис. 58).
Представленная классификация демонстрирует соответствие распростране-
ния видов Oenopotinae ареалам других групп морских беспозвоночных (Уша-
ков, 1951; Гурьянова, 1955; Скарлато, 1956; Кусакин, 1956, 1958; Скарлато,
1960; Голиков, Кусакин, 1962; Голиков, 1963; Голиков, Кусакин, 1978; Голи-
ков, 1980; Скарлато, 1981, и др.).
74
ОБЩАЯ ЧАСТЬ
Весь ареал подсемейства был разбит на произвольно выбранные мелкие ре-
гионы. Значительное число широко распространенных видов не позволяет
в полной мере использовать уравнение Престона (Preston, 1962), показывающее
степень различия видового состава сравниваемых выделов (табл. 4, нижняя
часть). Выяснение степени сходства видового состава сравниваемых районов
осуществлялось по формулам Жаккара (Jaccard, 1912), Шимкевича—Симпсона
и Чекановского—Сёренсена (Песенко, 1982). Результаты сравнений представ-
лены в табл. 4 и 5.
Рис. 58. Места нахождения Obesotoma starobogatovi.
Как видно из приведенных в таблицах данных, зал. Петра Великого, воды
Северного Приморья и Татарского пролива, мелководье заливов Анива и Тер-
пения, воды Южных Курильских о-вов могут быть объединены в Северояпон-
скую низкобореальную подобласть Тихоокеанской бореальной области. Юж-
ная граница подобласти сильно размыта у северных и восточных берегов о-ва
Хоккайдо, так как там проходит холодное течение Оя-Сио (Kuroda, Kinoshita,
1951). Северная ее граница проходит в районе м. Терпения и сейерной части
о-ва Итуруп.
В состав фауны Северояпонской низкобореальной подобласти входит 39 ви-
дов Oenopotinae, из них 34 % эндемичные для подобласти, 32 тихоокеанские
широкобореальные, 29 бореально-арктические тихоокеанского происхождения и
5 % тихоокеанские высокобореальные виды, заходящие за северную границу
подобласти. В районе от Северного Приморья до зал. Петра Великого обнару-
жено 33 вида, из которых 36.3 % — тихоокеанские широкобореальные, 33.3
тихоокеанские приазиатские низкобореальные, 30.3 % бореально-арктические
тихоокеанского происхождения. В число приазиатских низкобореальных вхо-
дят 3 эндемичных вида (9 %), а именно: Propebela verrilli, Oenopota biconica,
Oe. elongata. В заливах Анива и Терпения обнаружено 24 вида, из которых
46 % бореально-арктические тихоокеанского происхождения, 29 тихоокеан-
ские широкобореальные, 21 тихоокеанские низкобореальные, 4 % тихоокеан-
ские высокобореальные виды, заходящие в зал. Терпения. В число низкоборе-
альных входит 1 вид (4 %), эндемичный для данного района: Propebela terpenten-
sis. У Южных Курильских о-вов эндемичных видов не обнаружено, а по видо-
вому составу этот район наиболее близок району Северного Приморья.
Изложенный материал, включая сопоставление районов (сходство и разли-
чие видовых составов и степень их эндемизма), позволяет выделить в пределах
Североядонской низкобореальной подобласти две провинции: Приморскую и
Южиосахалиискую.
Таблица 4
Сравнение видового состава Oenopotinae в различных частях ареала подсемейства. В верхней части таблицы
по Чекановскому—Сёренсену, в нижней по Престону
Район исследования
Q СО L© •<? L© 00 чН СО L© тН 00 1© 00 нН СО СО СО Nf СО СО © гч g< И § S 3 £ L© СО О *<? © СО СО 00 CD чн k© th CO CO CO <? © 1© CM 1© S CM CO GM CM GM CM tH GM CM CM Sf CD CO Sf 1© CO CO CO Sf CO b- CO CO . U -!1 P n d
Q l— 0 -cf M t>-in О СЧ in CO О Ш XH О in xf 0 SP MO CO -гч „ M ta S' “ ® О Й 00 ед О E ф и HLO>-Dh^OOJCOs?04?hOl'jOlOO’HL.OCOHC4 r_!O Д D И CM CO v- bl bl bl H CM CM CM OO © © St* Л CC О Ю St1 LQ © CD О Ф u U & И E
Ч b- b> b- CD 1© <? CO CD sj< GM О CO 00 О О и О 00 О 00 О СО О Ю £ ~ ° со1 об О 00 СМ'тН <? CM ’Н CM CO LO CD О к© со О о об 6н °? °? ИЙЕч Д’ • тН СО тн тн тн тн тН т-l CM CM СО k© CM СО "SJ1 1© *<J< СО 1© Ю Ф О О Е 5 Е* ГС >>Ч К
Ч £ 2 Е I Д Obl©LOOCDb-C©b-0*sFOOb-OOOcC)CDkO-HCM О th СМ ф’о^СО ь g о u6'^Oo6ot^t^l6t><D^t<o^OsrGM^t^THCM^0^l?^ Е и 2 -Q ® S CO GM GM СО GM GM GM GM sf i© L© с© ‘•З* 1© СО к© CD k© М* ООО Н Ж О 5 о - я о а г - р<
Q СО О тгн L© тн СО 1© b- 1© CD СМ CM тн СМ СМ СО О 00 СО СО 00 с© оо CDOtHOthOCM’iOCM CM CM k© ь- CM b-' COOHCO^^1^0^^ ев E 2 H C^] И H H H H н H co co H CMCM co <? 1© CO О О О О 0 * Q 0
Q CO 0 CO О CO CM H 00 *<JH О CM 0 co 0 00 00 co CD © CD CO CO CO 1 £ £ | S § £ сбюбюоббтнм^ббнсбнсо©^^^^^0^ X‘ о S « Й2 CMCM CM CM CM CM TH CM CO CO Ю CM s? CO Ю CO CD ©©©©© E .© 2* >*
Р co 0 cm co co ь- i© 0 м< ь, 00 1© co 00 t- o oo ooooocoiD =2o ®«co<o®oo CO CO н CO co CM CM CM co co co co M* *<r co ю OOOOOO Ea£§o£ei , cc | 2 0 a
Ч CM CO CO H CM LO L.O L.O 0 00 CM C5 LO О О О CO CO 03 Q3 L-O co CM V oicDcO^^OMOOLOUOLOoioodCcoS^^^^^^^ CM *<Л CM CM CM н CM CM CM *<t< L© CM Sf LO CO ООФООФО ~ - «Що 1 4 ЕП И О —О
t> co ю co co co 00 co l© см и cm l© cm co 00 t- l© co cd cm co l© So , “ e 66666coo'}6coi6^6h66w^l^^^l^1^ °3>ao 1е5- CO CO CM CM GM CM CM CM CM CO *<Л t- CO L© b- ОФОФООФО =л^>.<© 0
ч EC *h (-3 Д и oooob-ocMi>-i©-<tcDLOL©oocM^i©cocoLOcocoa2LOc© a S<2 0 ' S^g.«Ag CM co CM CO CM CO CM CM CO CM CO CD <? b, ФОООООООО сз£гсо&°£ П й1 E JS 0 utL
ч . * в w w 2 b LO О *<Л см CO О 00 О t> t> TH CO О _ CD н >1© b- <? I© b- CO CO 00 тбббнбббббббб® I^OOOOcOlOLQcOLOt-b. ps g-o g О _ CM *<л GM <? co CO CM CM CM CO CO co <? ФФООООООФО ‘“pt? a< О £
ч 1 " " J iu Ы н L и 3 CM 00 <? sf LO CO L© 0 <? LO 0 CM sf TH CD О CD '•& lu© OO H Ю CD S 4 £ = &S, CO CO CO Г~ 1П CO.in Г' 1Л Ю 1Л 10 OOOOOOOOOOO “^язЕГц) 3
<3 отноооооосооо^о Г"-—. оооооонсмооь-мсо !$22“£e|h Сз 00 L© *-H 1© *<J* L© st* b- CO " ® « J M l“* ^s?COs?4?sJ<COCOST<4?*<fH ООФФОООООООО ' °3
<3 — 1 - ° ЦМ a-> ' 2 CMCMCMtHIDOCDtHCDO 1© О th CO Sj* L© CM CM t-ч CM CD b- Q3 4 1 5 -°u® didcdad^idd^cdin^”^.®^1^.1^'?^ 6L|aSB® l>\t*sWLOLOOOcr>r- ФООФФОООООООО я Q 03 0 0 M «
ч д * “ 0 tO co s 0 :c 0 c cic o я сю я9 g ® -E cj t; и H t s ® Jt' _• •• • c so 00 t- m m co m m cn m in -cf co co S.3SSr’S,>' lOCOOCDOOCDO-ICOh' - u ь n. 0 (1) 1 b- CO L© CD b- b- co t> Г- О О О О CD О О О О О О О О О . нЦ)5*аои| Й C C S M Г7 ”
Q Qj & W CJ e« sfCDCMOCDCMOcO ОСМСМтнООЬ-фООФСОО^^сМО 5 о bs 1 Q ^.X^.X^^X’.X^OOr-iOr-^LOLOlDlOCOLOCO^CDcD ЙЩ '£ ' >• 00 i-O CO l© CO CO н 1© • • » • • • • • * ♦ * * _♦ • aS H S “’I CO GM CD CO b-b- b. br 0 О О О О © О 0 OO О О 0 © Ф a £ И • N 0. J2
Q CD ~ Q, 0 - H О © CO 00 © и oc> CD н b- TH CD СЗ О О CO тН н О b- CM CD S|sgg < i> xf co co t> 00 co О О О О О О 0 О О О О О О О О О Эёрйёпй
Ч Ost*DDNOHOCOOXDOblSt*LCOO o6i--o6iddd£“^®^“^«^-?c’?14':!:?';,^ir! §£§„«> b-co co co 00 00 0 ©©© О ©©©©©©©©©©© © oaa^oyj S Ф « 0 t 1
ч 5* »Й 1 У =* M* © ST 00 00 _ H b-L© CD CO co TH © © ©CM 00 H SJH CM co b- 00 -СЙсГ ^«<^©00^^ Ф « НЧ отоомоом ©©©d©d©©©©©©©©©©©© 5^2 .22м "i ! ГС и 1-1 uC
ч .— 0 | jj< РЧ Фф s CDOOCDGM ФнОнфЛЬОнО^ФСО^^ФФОСЧ « *,a £ g" Й X^n^^M00bCW*Wl/WLO4Wm^LO©LOLC: ОС)5“^ИФ OOCOCDCD Ф©ОФФФФФФО©ФФО©ФООО n/S“b^a «-»a> c. q ф
Q w 1 2H b-CDCO^ СМЮСОтНСОФСОтн^ФтНтНСОСОСМСМ©1©тНОО £-2 2^ '^jWbCOCDbC01DLO^bSt*L©lOM*M*COSt*Sf*COLOst* >& Л о о ns ё см gm ь- cm _ a _ rvQ 0 5 b- LO b- 0 О ©ОФ ОФ О О© О © © 0 0 © 0 © ©© Ейхо Era к § 1 s 5 § я
о го CJ ” Я C co CO 1©1©1©00tHL©CDI©C©^4COCD4hC©©-'HGMGMCO00tH 5 £ 1 § S s cororldddddccddoddddccddddd KagSaa® 15 ° §O<«g
о < -s°S О 0 00 M О in ч-< -rt -гЧ CO co О 'Ct* 00 CO ^4 Sf <M in in О 1П CO n-gg1®» ^int^coi^oocoi^cococo^ooin^comin^i^ioint^co a , s g g gi0 S'- d d d d d d d d d d d d dd d d d d d d dd ®„§аий «nocogs?
ч 3 й>и-„о ©LCLCst*COHQinOst*J\IOSf*0'?COCOl-0©LOOOa3PO © « » 0 5 B B и L© b- 00 00 00 b- 00 co CO © I© b- L© *cf< L© ЧГ1 CO Ю M* CO 1© >L© H & 7 k3 1 К 0©00©©0©©©CD©©©©0©©0©©©© - = 1 H Q «Р
ищи О и * О ОН «СО ТГЮО Г-00 О фнеч МН >Н^£ dr г Ji S г 1-ч«М’-'Лсог~а>л-^-*»-<«-'н-Ч1-чн — 1—Ji * v J QQQQQQQQQQC)C3QC)QC)|^C)C)CjC)C)CjC) Я & a n S р, -УФ Св Ф I 3 О к Е <©
a
I
R
Ю
сб
l©
cd
tf
S
R
\O
сб
сб
ч 1>ОМСОМС:фЬ.^01СОС^тнФФтсОФФ>.С0^0 ^ОС^^С^О^^ОчОФСО-чОФСССОФО^Ч^гч СМч^СМСМСМ’чЧ'ччсМСМС01©С©СОѩѩѩ[>.ООС©с©[>.ООЕ-
ч 00 О О *<? 00 О >© О СМ СМ О CM О О О СМ С© С© [>• СО Г"; о qq СМ С©С©СЭС0С01ЛС01Л1©ччОС01©'^ОСМс©’чЧ[?ч.С0С0^СМ00 СМ*<?-чЧСОСМСМСМСМСОСО1©С©СО,<?С©[?ч00Ф1>.[>.1>-’^ <?
q СО СО СО GM GM GM СМ СО СО 1© t— t- t— О» Ю С© 1©‘с© О0 м <? о t^c©t?^t<ododo6odt^i?^i©cMc©cMcMcM,<?c6uoc6’H'5-<odi© СМ СО СМ СМ чЧ чЧ чЧ 'гЧ см СМ *<? С— со С— С— t> 1© С© 1© С© 00 со <?
q софоффф^^^^смоффччоссмф^сооо С© С© чЧ чЧ чЧ ^ч СУЗ ©3 ©3 СО С© С-- чЧ [-• СО СО О CO 00 С© СМ С© чН 00 CO^CMCOCOCOCMCMCMSFUOUOCOUOOOOOOOO*^ СО Sf< <?
оосососососососососооосоооосо^ оз со оз <? oocdcdcdcocdcdcdcocdoocdooooo^^LOcdcdc© СМ СМ чЧ чЧ чЧ чЧ чЧ •’И чЧ со <? см со <? *<С <? 1© см со со <?
*<? СО чЧ sf со см со *<у <? 00 со 00 t> Г"- <? Г-; О О СО ОЗ 1© ^co^^^^cocMco^^tr^coodtr^tr^^tr^Z^oocoirSod *<? СО -чЧ sf <? СО CM СО sj< м< [ч, со 00 Г- Г-' <? Г- CM Sf со см см
сГ DDOCb-b-cocob-b-ocooccooco^OLOc^-cooo чЧ чЧ 1© d чЧ чЧ СО СО •’Ч чЧ* d СО ч-ч 1© СО L© 00 чч О 00 1© СО СМ* СМ •^^CMLO^^COCO^^LOOO^t-OOt^-k© 1© СМ 1© <? <?
<5 <©c©oc©c©c©cococ^t>-t>-ocoococo oot4.i©oooo cdcdocdcdcdcdcdcdcdcdocdocdcdSidoo^ocdo CM^CMCMCMCM’4-<C©CMCM*<t<C©COc©r-t>- со см со *<? СО 1© L©
c©ocMcoooc©i©oo*<fi©ocMcocM*<t<^oooocMo-'^coc> coocMcot^cocotr^ait^ocMcocM^SocMLOcModoocoo СО •<? см СО СМ СО СМ СМ СМ со 1© Г- со Г— 03 Lrt^COCM^CO*^!©
Ч «©«©«©^«©СО^ОСМиОООЗф^^оО^-ччСОСОЕ^СОфСМ о6с©ОО^о6с©<ОзО-чЧ[?^Оо6с>СММС0ОЗо6чЧОЗСМС©1©о6 СМ *<? СМ со СМ СО СМ СО со 1© t— 1© [•— *<}<СМСМСМ-чЧ^С0С0С0
Q С© О СО СМ чЧ t>- СМ О СМ О СО СМ Ф СМ чч 1© !>• 03 СО 03 С© О СМ • обос©СМо6чЧ-чЧ1©’<-чОС©*-3<1©СО’чЧ1>^о6г,*Ф^обг>03-чЧ СМ С© СО L© со <? М< СО *<? 1© 1© 00 1© 1© со см см см см -чн см со
Q OCOOOOOOOOtr^LOCOC©^ СЗФ!>ОО1П^!>00М;Ь; dcoLodddodiO'Ccicocioq г^сообсосоччобоз^оосб' t> С© 00 С© I"- 1© t> С© С© 1© L© чЧ ч-Ч чЧ чЧ CM -Ч< -rt TH '-н
4 ^COC©^^^COOO^t^O^CM^CMCMG>C©CO*<t<CO^OOOO r^co^t^cMt?^i©c©[>.c60’<!r<cdod-’H,M<c©t>-c©><-<'IHr^oic© i© со м< sf *чГ со со <? *<? l© со sf <? >© со <? со см см
Q COC©OCOOOOOi©t-’-r-0<£tO*<?<l©l©CMOOt-’-Ot-’-C3*<f<LOC''- ^c©o<^ododcMo6o6i©^Mi©od*<fc6c©*<f-^4oic6’^t?^oo6 OO^UOLOOOCSOC©!?- СМСО’^'^СМ’^СМ’ЧН'^СМ'^СМ’^
4 1© С> СО СО 1© 00 СО СО OC©sfOa3srOOC^OOOC©L©oOi© 1>оод>н'гч1>о6-^Н’^’^Ф05од1©*<^’чн*<?’чназс©1Лсмс6,<^ 00 <? С© СО СО 00 Q© СО 00 С© СМ со тН СМ СМ 'Т-i СМ •’ГН чн тН
Q i©c©e©i©'<rcMoc©^ooi©c>i©coc©cMoooc©[>.oi©c© С© С©0<^СМодоСМ^и1ЛОо6с>,<?1ЛС©-<?С)сОС©’чНСМ’сОС© Г-СМ Г-Г"-00 00 t-СО С© 1© СМ С© TH -ГН тН СМ -гч СМ -ч-к -^ч тН
q” ОС©ОООООО 00L©4tfC©C>C©CMC©C©C©Sf«CT3*<t<001©t- ОС©0 1©ОООМОС©’оо’г^Ос^Г'ѩѩ'^С^сМСОС©СМсд 00 sf Г- Ь- оо 03 С© 1© СО СМ С© тН чгЧ -ГЧ чгЧ тН СМ тН тН
Q смо*<?1Лоо 0001Л'<Г’^чфсмаз<озс©ОФс©с©01Лсз одос©юОО^од1?^е6с6од1>с6смсмс©с©с6с©сзсмосм 00 “xf £>• t— *^*<?*<?,<?СМСО'^Ч'^ч,чи •’гч 'гч тч -г-ч тН •’гч
q i©c©‘<rocM^oocMc©’4-ie©4^4-jOcoc©'^4-4t>..<jHococoi© О С© О СМ 1© М Ф ©3 t>- 00 тч С© !>• О 00 С© •чн С© СО 1© тч ©i СО СЗ^ ФСО 03 t- С© Ю *<? CO СМ х© СМ >гЧ ГЧ -^ч <гЧ -<r-i СМ -гч чгЧ
q ©3 С© С© OC30CO’^<CMCMSj<C©1'tf’-r4C©COCOOOC©C©l©t> фс©с©>-чсм oo^cd^^cdcdcsc^Ldost^iOidLocdcM'-tf oo*<fc©t- 00 00 С© 1© L© СО СМ тЧ 'ГЧ Ч~ч чгч чгч 'ГЧ т-ч тч
q СМсСОЗ^^Е^’гчЮСМ^ОСМСМСМОччОО^СМкЛкЛС©^!© ѩѩОсМ-^1>ООСООО'<ГОСМСзч-<0[?-СМ’чЧ'<Г1ЛСООЗ’чЧСО t- С© СО L© с© -<JH 1© vfH СО CM 1© CM CM -ЧЧ CM чч -^ч тН
q С© О ОЗФГ^ОООСМ0 1ЛСО^ООС©*<?СОО'МНОО’ЧЧС©00'^ с©ОмсмоосМ’ччс©1>1>-ччс©аз[?^-ччазососМ’ччсоаз'гн СО С© 1© с© 1© 1© «4< со СМ СМ *<? тЧ тч тч -^ч тЧ -гЧ
q О 0'с?’ччО’ччОСМСМС31ЛФОЗСОСМС©1>СМ’чЧСЗСМСМСМ о^ооз-<?с)ео1Л,<?азоз1<?ФсО’ччсмфсм1мч^)©о6-ччсм С© СМ СМ СМ СО СМ СМ чч -чч CM CM SJ* тЧ ^ч СМ СМ СМ ЧЧ тЧ СМ чч СМ СМ
Q C00030C©C©OCMCMOCOt>-OOl©44J©OOCOC©CMC©l©t> ^cdocмoдc©o6^c©^oдoзt?dt-<fo6cмt>l©, сб-ччсзсм*^ СО1©С©С©С©1©С©*с}|Ю,<}<С©1©чЧчЧчЧ--ч--ччЧ СМ чЧ тЧ -чЧ
онмп'#ле['»с:очмп'г чсз«оч,1Л<с>с»соС5Чччмчччччтче1етеде1е1 QQQQQQQQQQQQQCjCieiClCliqQiqCiCliq
1 Обозначения районов исследования как в табл.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
77
Берингийская (Алеутская) высокобореальная подобласть Тихоокеанской
бореальной области включает в себя остальные, выделенные в Тихом океане
районы (2114—Z?19). Всего для подобласти нами отмечено 54 вида, из которых 14
(26 %) эндемичны. Общее соотношение видов следующее: 35 % тихоокеанские
широко распространенные бореальные, 35 тихоокеанские высокобореальные
(с учетом эндемичных), 17 бореально-арктические тихоокеанского происхожде-
ния, 9 бореально-арктические атлантического происхождения, 4 % арктиче-
ско-бореальные.
На самостоятельность Охотского моря как подразделения Берингийской
высокобореальной подобласти указывают многие исследователи (Гурьянова,
1935; Галкин, 1953; Голиков, 1963; Голиков, Кусакин, 1971, 1978, и др.), вы-
деляющие его в Охотоморскую провинцию. В Охотском море из обнаруженных
нами 23 видов- 52 % составили тихоокеанские широко распространенные бо-
реальные, 31 бореально-арктические тихоокеанского происхождения, 17 %
тихоокеанские высокобореальные приазиатские виды (вид Propebela svetlanae
-эндемичен для района).
Воды у Юго-Восточной Камчатки, Средних и Северных Курильских о-вов
населены 23 видами Oenopotinae, из которых 53 % тихоокеанские широко рас-
пространенные бореальные, 26 бореально-арктические тихоокеанского проис-
хождения, 21 % тихоокеанские высокобореальные, включая эндемичный вид
Curtitoma bartschi. У берегов Аляски, по материалам из коллекции ЗИН
АН СССР и по литературным данным, известно 30 видов, из которых 30 % ти-
хоокеанские высокобореальные (20 % эндемичны для района), 27 бореально-
арктические тихоокеанского происхождения, 17 тихоокеанские широко распро-
страненные бореальные и 7 % бореально-арктические атлантического проис-
хождения. Результаты обработки материала позволяют говорить как об отдель-
ных регионах Берингийской высокобореальной подобласти о трех провинциях:
Охотоморской, Алеутской и Курило-Ка м ч а т с к о й
(последняя объединяет Средние и Северные Курильские о-ва с Юго-Восточной
Камчаткой).
Вопрос о северной границе Берингийской высокобореальной подобласти
решается различными исследователями неоднозначно. Некоторые относят се-
верную часть Берингова моря к Арктической области (Виноградов, 1948;
Галкин, 1955; Нейман, 1960; Голиков, 1963, 1980, и др.). Другие проводят гра-
ницу между Тихоокеанской бореальной и Арктической областями в районе Бе-
рингова пролива или несколько к северу от него (Ушаков, 1953; Скарлато,
1981, и др.), что отвечает и моим результатам обработки материала по Oenopo-
tinae.
Арктическую область населяют 36 видов Oenopotinae, состав которых раз-
нообразен: 78 % бореально-арктические виды (39 % тихоокеанского и 39 %
атлантического-происхождения), 14 арктическо-бореальные виды арктического
происхождения, 8 тихоокеанские широко распространенные бореальные все-
ленцы, 8 составляют эндемичные виды и 3 % (1 вид) высокобореально-аркти-
ческие атлантического происхождения.
Своеобразным по фаунистическому составу районом оказалось Чукотское
море, включая и о-в Врангеля, где обнаружено 19 видов. Из них 52 % бореально-
арктические тихоокеанского происхождения, 18 % составили тихоокеанские
широко распространенные бореальные виды, 18 бореально-арктические ат-
лантического происхождения, 6 арктическо-бореальные и 6 % (1 вид Obesotoma
tumida) арктический эндемик, обнаруженный только в Чукотском море. Как
видно из сравнения видового состава различных участков Мирового океана,
фауна Чукотского моря и о-ва Врангеля имеет большое число видов, общих
с Беринговым морем. Следуя мнению ряда авторов (Андрияшев, 1939; Ушаков,
1940; Гурьянова, 1951; Ушаков, 1952, и др.), мы предлагаем выделять в преде-
лах Арктической области Чукотско -Американскую провинцию.
Районы у Западной и Восточной Гренландии, Северной Исландии, северная
часть Баренцева моря, а также Карское море и море Лаптевых сходны по фау-
Рис. 59. Схема биогеографического районирования на основе анализа распространения Oenopotinae.
-3 — Тихоокеанская бореальная область: 1 — Северояпонская низкобореальная подобласть, 2,3 — Берингийская высокобореальная подобласть (2 — Охотоморская
провинция); 4, 5 — Арктическая область; 4 — Чукотско-Американская провинция, 5 — Западно-Сибирская провинция; в — Атлантическая бореальная область.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
79
нистическому составу, и их следует выделить вЗападно-Сибирскую
(Гурьянова, 1939, 1951) провинцию Арктической области. Всего для провинции
отмечен 31 вид, из которых 39 % атлантические бореально-арктические виды,
32 тихоокеанские бореально-арктические, 13 арктическо-бореальные, 10 аркти-
ческие, 6 % атлантические высокобореально-арктические приевропейского
происхождения. 1 арктический вид — Obesotoma gigantea эндемичен для про-
винции.
Атлантическая бореальная область, в состав которой вошли 35 видов Oe-
nopotinae, ограничена у берегов Америки: на юге м. Код и водами к северу от
Ньюфаундленда; у берегов Европы: проливом Ла-Манш, Северо-Западным
Шпицбергеном и мелководьем у Новой Земли. В состав фауны области входит
37 % атлантических бореально-арктических видов, 29 тихоокеанских боре-
ально-арктических, 11 арктическо-бореальных, 11 % амфибореальных при-
американских широко распространенных видов тихоокеанского происхожде-
ния. 2 вида, или 6 %, являются эндемичными и встречаются только в батиали.
Фауна моллюсков Северной Атлантики очень сходна во всей области, од-
нако в ней можно выделить два подразделения: евразийский и амеразийский.
Фауна моллюсков Западного Шпицбергена, Северной Норвегии и южной части
Баренцева моря насчитывает 26 видов, из которых 42 % атлантические боре-
ально-арктические, 35 тихоокеанские бореально-арктические, 15 арктическо-
бореальные, 8 % атлантические приевропейские высокобореально-арктические.
В Белом море фаунистический состав тот же, что и в Баренцевом, но насчитывает
всего 17 видов, из которых 47 % атлантические бореально-арктические, 35 ти-
хоокеанские бореально-арктические, 12 арктическо-бореальные, 6 % атланти-
ческие приевропейские высокобореально-арктические. Фауна Oenopotinae аме-
риканского побережья Северной Атлантики известна только по литературным
источникам. Она близка по видовому составу фауне южной части Баренцева
моря (табл. 4 и 5) и насчитывает 14 видов, из которых 42 % — атлантические
бореально-арктические, 29 тихоокеанские бореально-арктические и 29 % —
амфибореальные приамериканские широко распространенные виды тихоокеан-
ского происхождения.
Изложенный материал демонстрирует большое сходство фаунистического
состава Атлантической бореальной и Арктической областей, что свидетельствует
о возникновении бореально-арктических видов атлантического происхождения
в районе, лежащем на стыке двух областей. Биогеографическое районирование
моллюсков подсем. Oenopotinae представлено на рис. 59.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Как уже было отмечено в Общей части, многие авторы использовали для опи-
сания моллюсков широко варьирующие признаки, например окраску тела,
форму и строение щупалец или соотношение отдельных параметров раковины,
в частности отношение общей высоты раковины к высоте последнего оборота
или к наибольшему ее диаметру. В работе предпринята попытка избежать в опи-
саниях употребления признаков, изменчивость которых превышает границы
внутривидовой, и даются только диагностические признаки.
В диагнозе видов для удобства чтения используется широко распространен-
ный в иностранной литературе термин «анальный синус», обозначающий вы-
емку верхней части палатального края устья. (Безусловно, данный термин сле-
дует понимать как указание на топографическую близость синуса палатального
края устья раковины к анальному отверстию моллюска). При описании зубов
радулы используются два термина: «флажок» и «шпора» для обозначения одного
и того же образования, а именно уплощенного выроста ручек базальной части
зуба. Под «флажком» следует понимать широкое протяженное образование,
обычно полукруглой или треугольной формы. Термин «шпора» используется
только для обозначения очень маленького равнобедренно-треугольного образо-
вания на ручке.
В ряде случаев при сравнении экземпляров из евразийских окраинных морей
с моллюсками, обитающими у американских берегов, я использовал неопубли-
кованные материалы зарубежных коллег с их любезного разрешения. Так, при
исследовании ряда видов, описанных Доллом (W. Н. Dall), но не сопровожден-
ных иллюстрациями, вспомогательными данными послужили фотографии голо-
типов, присланные из Национального музея естественной истории США Джо-
зефом Роузвотером (J. Rosewater). Этот исследователь предоставил для исполь-
зования копии зарисовок зубов радул некоторых видов, описанных Доллом и
Бартшем (Р. Bartsch). Авторство части препаратов неизвестно, но больший
объем работы сделан Дж. Моррисоном (J. Morrison). В названных и сходных
случаях обязательно делаются оговорки об источниках рассматриваемых дан-
ных.
В специальной части используются следующие сокращения.
При описании параметрических характеристик раковины:
Вр — общая высота раковины;
Впо— высота последнего оборота;
Вр — высота устья;
Д — наибольший диаметр (или ширина) раковины;
Ду — ширина устья.
При указании музеев, институтов и других мест хранения типовых экземпля-
ров:
ЗИН АН СССР — Зоологический институт АН СССР, Ленинград;
Amh. Coll. — Amherst College, Massachusetts, USA;
BMNH — British Museum (Natural History), London;
BSNH — Boston Society of Natural History;
GITU — Geological Institute of Tokyo University;
IRSN — Institut Royal des Sciences Naturelies, Brussels;
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
81
MNHN — Museum National d’Histoire Naturelle, Paris;
SMNH — Swedish Museum of Natural History, Stockholm;
Stuttg. RC — Stuttgart Royal Cabinet (Stuttgarter Mus. fur Naturk.);
USNM — United States National Museum of Natural History, Washington;.
ZMB — Zoological Museum, Bergen;
ZMC — Zoological Museum, Copenhagen;
ZMO — Zoological Museum, Oslo=Museum Christiania.
Сем. TURRIDAE Adams et Adams, 1855
Подсем. OENOPOTINAE Bogdanov, 1987
Богданов, 1987:35.— Raphitominae (non Bellardi, 1875; part.): Голиков,
Старобогатов, 1975 : 214. — Mangeliinae (part.): Fischer, 1887 : 58Г, Morri-
son, 1966:2; Powell, 1966:97; McLean, 1971:128; Shimek, Kohn,
1981 : 428. — Lorinae Thiele, 1925 : 92 (nomen oblitum). — Brachytominae (part.):
Thiele, 1929 : 362; Голиков, Гу ль бин, 1977 : 235 (Brachytomidae)', Голи-
ков, Кусакин, 1978 : 203 (Brachytomidae').
Раковина маленькая (высота в среднем 14—16 мм), овально-веретеновидной
или башенковидной формы, одноцветная, с медленно нарастающими оборо-
тами, разделенными прижатым швом. Последний оборот занимает от х/2 до
2/3 высоты раковины. ПрОтоконх сосцевидной (реже бочонковидной) формы,
как правило, орнаментирован 2—20 спиральными ребрышками. Спиральная
и у большинства видов осевая скульптуры дефинитивной раковины хорошо
развиты. Палатальный край устья (наружная губа) всегда очень тонкий,
ломкий. Сифональный вырост короткий, сифональныи канал умеренной глу-
бины, относительно широкий. Крышечка когтевидной или каплевидной формы,
с терминально расположенным ядром.
Радула состоит из пары маргинальных зубов (радулярная формула
1 : 0 : 0 : 0 : 1) типично токсоглоссного строения: округлая или цилиндриче-
ская базальная часть несет полое лезвие, укрепленное в дистальной части
килем. Базальная мембрана отсутствует.
Представители подсемейства распространены циркумполярно в Арктике
и в бореальных водах Тихого и Атлантического океанов. В ископаемом состоя-
нии известны начиная с миоцена (Япония).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (4). Раковина тонкостенная, маленькая (высота до 5—11 мм). Эмбриональная
раковина орнаментирована многочисленными (более 5) нитевидными спи-
ральными ребрышками.
2 (3). При пересечении осевой и спиральной скульптур образуются узлы,
придающие поверхности раковины сходство с орнаментацией кукуруз-
ного початка. Киль менее х/4 длины лезвия зуба радулы ......
.....................................................2. Granotoma Bartsch
3 (2). Спиральная скульптура дефинитивной раковины представлена тонкими
ребрами и при пересечении с осевой скульптурой не образует узелков,
придающих поверхности раковины сходство с орнаментацией кукурузного
початка. Киль, как правило, занимает около х/2 длины лезвия . . .
................................................... 1. Curtitoma Bartsch
4 (1). Раковина толстостенная, крупная (высота до 20—30 мм). Спиральная
скульптура эмбриональной раковины состоит из 2—4, редко 5 ребрышек.
5 (6). Сигмоидальный изгиб на элементах осевой скульптуры в различной мере
выражен и соответствует форме анального синуса. Эмбриональная рако-
вина уплощена в верхней части, или ее макушка погружена в первый
полуоборот протоконха; орнаментирована в различной степени выражен-
ным ретикулятным рисунком ............................3. Oenopota Morch
6 И. П. Богданов
•82
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
'6 (5). Сигмоидальный изгиб на элементах осевой скульптуры отсутствует, реже
слабо выражен. Эмбриональная раковина практически всегда выпуклая
в верхней части, первый ее оборот не орнаментирован ретикулятным ри-
сунком и лишен осевой скульптуры.
7 (8). Раковина овально-веретеновидной или овальной формы. Спиральная
скульптура дефинитивной раковины, как правило, слабо выражена, без
четкого киля на плече оборота. Спиральная скульптура на эмбриональ-
ной раковине практически полностью отсутствует..........................
.............................................4. Obesotoma Bartsch
'8 (7). Раковина башенковидной или веретеновидной формы. Среди элементов
спиральной скульптуры дефинитивной раковины наблюдается преиму-
щественное развитие киля, обозначающего плечо оборота. Спиральная
скульптура эмбриональной раковины представлена 2—5 ребрышками,
которые закладываются на втором обороте протоконха......................
............................................. 5. Propebela Iredale
1* Род CURTITOMA Bartsch, 1941
Bartsch, 1941 : 6. — Nematoma Bartsch, 1941 : 6 (типовой вид Nematoma
hokkaidoensis Bartsch, 1941, по первоначальному обозначению), syn. n.— Widalli Bog-
danov, 1986 : 46 (Oenopota subg., типовой вид Pleurotoma trevellianitm Turton, 1827, по пер-
воначальному обозначению), syn. n. — Oenopota (part.): Morch, 1852 : 73; N ord-
si e c k, 1968 : 157; Powell, 1966 : 121; Bouchet, W a г ё n, 1980 : 66. — Bela
{non Gray, 1847; part.): G. 0. Sars, 1878 : 222; Friele, 1886 : 2; Martini,
Chemnitz, t887 : 137.
Типовой вид Cirtitoma hecuba Bartsch, 1941 = BeZa incisula Verrill,
1882, по первоначальному обозначению.
Раковина маленькая, ее высота, как правило, не превышает 11 мм, овально-
веретеновидная, тонкостенная. Эмбриональная раковина выпуклая, орнамен-
тирована многочисленными нитевидными спиральными ребрышками, которые
уже на последнем полуобороте протоконха могут сливаться в 3—5 шнуро-
видных ребрышка. Осевая скульптура дефинитивной раковины представлена
узкими, реже средними осевыми складками, иногда только линиями нараста-
ния. Спиральная скульптура дефинитивной раковины представлена много-
численными тонкими ребрами. Сифональный вырост короткий. Крышечка когте-
видной формы, ось нарастания имеет сильный излом на начальных стадиях
роста. Мускульный след либо тонкий, расположенный в срединной части,
либо широкий, не прилегает к левому краю крышечки. Зубы радулы кинжало-
видного, реже стилетовидного типа, с хорошо развитым килем.
Род распространен в арктических и бореальных водах северного полу-
шария. В ископаемом состоянии известен с плиоцена (Англия, Исландия).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (4). Раковина состоит из уступчатых оборотов, уплощенных в верхней части.
2 (3). Осевая скульптура представлена тонкими валиками, равновеликими
спиральным ребрышкам на всей поверхности последнего оборота и осно-
вания раковины. Валики не дают резкого сигмоидального излома . . .
4. С. novajasemljensis (Leche)
3 (2). Осевая скульптура представлена пологими осевыми складками, раз-
витыми только в верхней части оборотов. Верхняя часть складок, распо-
ложенных на плече последнего оборота, дает резкий сигмоидальный излом
........................................... 1. С. incisula (Verrill)
4 (1). Раковина состоит из закругленных или отлогоугловатых оборотов.
5 (12). Спиральная скульптура раковины получает преимущественное разви-
тие над элементами осевой скульптуры по крайней мере на ранних дефи-
нитивных оборотах.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
83
6 (9). Обороты раковины закругленные, умеренно выпуклые, раковина оваль-
ная. Осевая скульптура не развита.
7 (8). Зубы радулы мелкие (до 140 мкм в длину), с длинным килем, дости-
гающим х/2 длины лезвия, и с массивным основанием.......................
..................................... 14. С. piltuniensis (Bogdanov)
8 (7). Зуб радулы средних размеров (до 245 мкм), с небольшим килем, зани-
мающим до х/4 общей длины лезвия, и с тонким цилиндрическим основа-
нием ........................................13. С. bartschi (Bogdanov)
9 (6). Раковина веретеновидная, состоит из оборотов отлогоугловатой формы.
Элементы осевой скульптуры представлены в различной мере развитыми
складками.
10 (11). Спиральная скульптура одинакова на периферии оборота и на осно-
вании раковины, кили отсутствуют................16. С. neymanae Bogdanov
11 (10). Среди элементов спиральной скульптуры наибольшее развитие полу-
чают два киля, один из которых обозначает плечо оборота, другой —
нижнюю треть его периферии .............................................
................ 7. С. violacea (Mighels et Adams) (var. bicarinata)
12 (5). Элементы спиральной скульптуры равновелики с осевыми складками
или развиты слабее их.
13 (20). Обороты раковины отлогоугловатой формы, тупоугловатые в верхней
части. Зубы радулы кинжаиовидной формы.
14 (15). Устье узко-обратногрушевидной формы, округлое основание раковины
плавно переходит в удлиненный сифональный вырост........................
.................................... 15. С. becklemishevi Bogdanov
15 (14). Устье полуовальной формы, угловатое в верхней части, сифональный
вырост короткий, основание раковины скошенное.
16 (17). Практически прямые осевые складки равновелики на всей поверхности
оборота и равны по величине пересекающим их спиральным ребрам . .
................................................... 3. С. hebes (Verrill)
17 (16). Осевые складки отсутствуют на основании раковины, развиты на пери-
ферии и приобретают резкий сигмоидальный изгиб на плече оборота.
18 (19). Устье по высоте составляет более половины общей высоты раковины.
Зубы радулы кинжаловидные, с квадратным основанием, киль достигает
х/2 длины лезвия ...............................2. С. trevelliana (Turton)
19 (18). Устье небольшое, около ~4S общей высоты раковины. Зубы радулы
удлиненно-кинжаловидные, с цилиндрическим основанием и небольшим
килем .................................................6. С. fiora (Dall)
20 (13). Раковина состоит из закругленных, умеренно-выпуклых оборотов.
Зубы радулы стилетовидные.
21 (22). Устье сильно вытянуто, превышает V2 общей высоты раковины . . .
.... 12. С. microvoluta (Okutani)
22 (21). Устье меньше или (редко) равно г/2 общей высоты раковины.
23 (26). Зубы радулы менее 100 мкм, с очень коротким лезвием, часто не пре-
вышающим длину основания зуба. Киль маленький: V5—1/7 общей длины
зуба.
24 (25). Осевые складки узкие, многочисленные (от 24 до 36 на последнем
обороте), имеют плавный сигмоидальный изгиб ............................
..................................................................................9. С. decussata (Couthouy)
25 (24). Осевые складки пологие, немногочисленные (18—22), имеют резкий
сигмоидальный перелом в верхней четверти периферии оборота . . .
............................................. 10. С. lawrenciana (Dall)
26 (23). Зубы радулы, как правило, значительно длиннее 100 мкм, с небольшим
основанием и в 4—5 раз превышающим его длину лезвием. Киль упло-
щенный, превышает 1/i общей длины зуба.
27 (30). Осевая и спиральная скульптуры равновелики.
.28 (29). Раковина овальной формы. Основание зуба радулы несет лигамент
. ................................................ . 5. С. ovalis (Friele)
6*
84
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
.29 (28). Раковина веретеновидной, реже веретеновидно-овальной формы. Осно-
вание зуба радулы лишено лигамента ......................................
.......................................7. С. violacea (Mighels et Adams)
30 (27). Осевая скульптура преобладает над спиральной.
31 (32). Осевые складки тонкие, многочисленные (20—31), промежутки между
ними в 2—3 раза шире самих складок .... 8. С. conoidea (G. О. Sars)
32 (31). Осевые складки немногочисленные (от 14 до 20 на последнем обороте),
широкие, пологие, в 4—5 раз превышают ширину разделяющих их про-
межутков ...................................11. С. hokkaidoensis (Bartsch)
1. Curtitoma incisula (Verrill, 1882), comb. n. (Рис. 63—65; 386, В—E;
407, 21—23; 408, 1-6; 439, 7).1
Verrill, 188,2 : 461, pl. 43, fig. 12, pl. 57, fig. 14 (Bela)-, Bush, 1884 : 238, pl. 9,
fig. 10 (Bela)-, Krause, 1885 : 279, Taf. 18, fig. 13 (Bela)', Martini, Chemnitz,
1887 : 149, Taf. 31, fig. 15 (Bela)-, К о b e 1 t, 1905 : 257, Taf. 84, fig. 19 (Bela decussata
subsp.); О 1 d г о у d, 1927 : 108 (Bela)-, Macpherson, 1971 : 118, pl. 7, fig. 8, map 52
(Oenopota). — hecuba Bartsch, 1941 ; 6, pl. 1, fig. 3, syn. n.; Powell, 1966 : 121,
pl. 19, fig. 8 (Kodotoma).— impressa (non Morch, 1869)1 V e r r i 11, 1880 : 365 (Bela).—
decussata (non Couthouy, 1839): Tryon, 1884: 217, pl. 29, fig. 65 (Bela; part.).
Раковина маленькая, веретеновидно-овальная, сероватая или белая, обычно
с рыжеватым налетом на завитке, с 5—5.5 уступчатыми оборотами, уплощен-
ными в верхней части. Периостракум очень тонкий, зеленовато-белый или рыже-
ватый, легко отстающий. Протоконх слегка выпуклый, представлен 1.5 оборо-
тами, орнаментированными многочисленными спиральными нитевидными реб-
рышками, которые уже к началу второго эмбрионального оборота сближаются
и в дальнейшем сливаются в 3 нитевидных ребрышка (рис. 386, В—Е). Осевая
скульптура закладывается только на постэмбриональных оборотах и состоит
из средних пологих осевых складок, стирающихся в нижней части периферии
оборота и исчезающих в верхней 1/4 основания раковины. На плече оборота
осевые складки приобретают резкий серповидный изгиб, в результате чего
создается впечатление о смазанности всей скульптуры на околошовной поверх-
ности. Пространства, разделяющие осевые элементы, всегда в 2—6 раз тоньше
самих складок, которых на последнем и предпоследнем оборотах насчитывается
от 14 до 20. Спиральная скульптура представлена в разной мере развитыми
тонкими шнуровидными ребрами. Они более широкие на периферии оборота
и сужаются на основании раковины. Промежутки, разделяющие их, всегда
одинаковы по величине и в 3—10 раз £же самих ребер.
Основание раковины всегда прямое, с небольшой выемкой в месте перехода
в короткий сифональный вырост. Устье широко-обратногрушевидное, углова-
тое в ве'рхней части, на колумеллярном крае несет широкую полосу белесова-
того каллуса. В верхней части палатального края устья расположен глубокий
синус. Сифональный канал неглубокий, но широкий.
Максимальные размеры экземпляра из типовой серии, обнаруженного у бе-
регов Новой Англии (Verrill, 1882) (мм): Вр=8; Д—^.Ь; Впо=6; Вр=4.5.
В Японском море обнаружена раковина высотой 10.5 мм.
Крышечка когтевидная, почти ромбовидная; ось нарастания имеет резкий
изгиб на ранней стадии ее формирования. Мускульный след тонкий, смещен
в левую четверть крышечки (рис. 32, М).
Радула состоит из зубов длиной 130—150 мкм типичной для рода кинжало-
видной формы. Киль достигает 1/2 длины лезвия и имеет некоторое отхождение
от стенок лезвия на противоположной дистальному участку зуба поверхности
(рис. 408, 3). Основание зуба массивное, ручки развиты, на одной из ручек
расположена низкая широкая шпора. Исследованные тихоокеанские моллюски
имели немного более удлиненное лезвие зуба, чем на рисунках, сопровождаю-
1 Рис. 60—406 даны на вклейке.
1. CURTITOMA
85
Рис. 407. Зубы моллюсков Curtitoma trevelliana (1—7); С. novajasemljensis (8—13); С. beckle-
mishevi (14—20); C. incisula (21—23).
1—2 — Гренландское море; 3—4— Земля Франца-Иосифа; 5—7 — Баренцево море, Мурманский берег;
8 — море Лаптевых; 9—11 — Новая Земля, Горбовы острова; 12 — Новая Земля, зал. Медвежий; 13 — Бе-
лое море; 14—16 — Охотское море, 51°10' с. ш.—154°17' в. д.; 17—20 — Западная Камчатка; 21—22 —
о-в Сахалин, зал. Анива; 23 — Чукотское море. Масштабные линейки: 1—8, 10—14, 16—18, 21—23 —
100 мкм; 9 — 10 мкм; 1S, 19—20 — 20 мкм.
86
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 408. Зубы моллюсков Curtitoma incisula (1—6) и С. violacea (7—29).
1 — о-в Сахалин, м. Чихачева; 2—4 — Японское море; 5—в — Чукотское море; 7—9 — Карское море;
J0—12 — Баренцево море, 70°11' с. ш.—46°00' в. д.; 13—15 — о-в Парамушир; 1в—19 — Баренцево море,
Поморский рейд; 20—22 — Северная Земля, прол. Вилькицкого; 23—24 — Баренцево море, Мурманский
берег; 25—26 — Карское море, о-в Визе; 27—29 — северная часть Карского моря. Масштабные линейки:
1, 5—17, 19—29 — 50 мкм; 2—4, 13 — 25 мкм.
1. CURTITOMA
87
Рис. 409. Зубы моллюсков Curtitoma hebes (1—6); С. avails (7—9); С. bartschi (10—17)-
С. flora (18—19); С. violacea (20—22). ’
™9«OTOK2£,sJ°Pe’ РазРез от м- Энкен; 4—в — Японское море, Татарский пролив- 7_9 — Гренландское
Море, глубина 2045 м; 10—11 — о-в У рул, близ м. Китового; 12—15 — Охотское море- 16_17 — Японское
^Ж,’о?'в У°мерон; У~19 — Камчатка, м. Хребтовый, го—22 — зал. Петра Великого, 42°24'^? ш —
101 28 в. д. Масштабные линеики: 1—3, 5—6, 20—21— 100 мкм; 7—12, 15—19 — 50 мкм- 4 13—14 22
25 мкм. * ’ ’
88
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 410. Зубы моллюсков Curtitoma conoidea (7—15) и С. decussata {16—33).
1—3 — мурманский берег; 4—5 — Земля Франца-Иосифа; в—10 — Мурманский берег; 11—15 — Карское
море- 16—17 — о-в Сахалин, м. Елизаветы; 18—21 — Восточно-Сибирское море; 22—26 — Северная земля;
ZI—30 _зал Петра Великого; 31—33 — Берингов пролив. Масштабные линейки: 1—5, 7—13, 17—
33 — 50 мкм; в, 14—16 — 25 мкм.
1. CURTITOMA
89
Рис. 411. Зубы моллюсков Curtitoma decussata (1—8); С. lawrenciana (9—13); С. пеутапае
(14—18); С. piltuniensis (19—82); Granotoma raduga (23—27).
i__2__о-в Беринга; 3—i — Карское море; 5—в — о-в Биннета, 76°38' с. ш.—149°42 в. д.; 7—3 Бе-
рингово море, бухта Эмма; 9—10 — Западная Камчатка; 11—13 — Японское море, разрез от м. Среднего,
^4__2Q__Японское море, Советская гавань; 17—18 — Западная Камчатка; 19—22 — Восточный Сахалин*
зал Пильтун: 23—27 — бухта Русская, Восточная Камчатка. Масштабные линейки: 1—9, 13—15, 19—21,
23—24, 26—27 —t 50 мкм; 10—12, 16—18 22, 25 — 25 МКМ.
90
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 412. Зубы моллюска Granotoma albrechti (1—22).
1—4 — Камчатка, бухта Русская; 5—7 — Чукотское море, 68°10' с. ш.—176°50' з. д.; 8—13 —северо-за-
падная часть Японского моря, 46°30' с. ш.—138°25' в. д.; 14—18 — Камчатка, траверз бухты Моржовой;
J9—22 — Татарский пролив. Масштабные линейка: 1—7, 11—гг — 50 мкм; 8—10 — 25 мкм.
1. CURTITOMA
91
Рис. 413. Зубы моллюсков Granotoma krausei (1—16).
1—2 — о-в Сахалин, пос. Катангин; 3—4 — Восточная Камчатка; 5—7 — охотоморское побережье о-ва
Сахалин; 8—11 — Татарский пролив, 50°02' с. ш,—140°46' в. д.; 12—16 — Шантарские о-ва. Масштабные
линейки: 1—-3, 5—8, 11—12, 15— 50 мкм; 4, 9—10, 13—14, 16 — 25 мкм.
92
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 414. Зубы моллюсков Oenopota cinerea (1—12) и Ое. impressa (13—23).
1—2 — Мурманский берег; 3 — Баренцево море, 70°08' с. ш.—38°55' в. д.; i—9 — восточная часть Карского-
моря, 77°52' с. ш.—99°23' в. д.; 10—12 — Баренцево море, 69°37' с. ш.—34°09' в. д.; 13—15 — Берингово-
море, 58°31' с. ш.—175°58' з. д.; 16—1а — Баренцево море, Мурманский берег; 19—23 — море Лапте-
вых. Масштабные линейки: 1—3, 3—10, 13—13, 17—19,21 —100 мкм; 3—7, 11—12, 15—16, 20, 22,
23 — 25 мкм.
1. CURTITOMA
93-
Рис. 415. Зубы моллюсков Oenopota impressa (1—12) и Ое. [harpa (13—23).
I__3 — Камчатка, траверз бухты Моржовой; 4—7 — Баренцево море; 8—10 —о-ва Новая Земля; 11—12 —
Новосибирские о-ва; 13 — Берингов пролив; 14 —море Лаптевых; 15—17 — Баренцево море; 18—19 —
Японское море- 20—22 — Баренцево море. Масштабные линейки: 1—5, 8—10,12, 14, 19, 22 — 100 мкм; 6—
7, 11, 13, 15—18, 20—21 — 25 мкм.
24
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 416. Зубы моллюсков Oenopota harpa (1—5); Ое. elegans (6—15)’, Ое. declivis (16—24).
1—S — Баренцево море; 4— Японское море; 5— Баренцево море; 6—8—Баренцево море, Ура-губа;
—11 — Мурманский берег; 12—15 — Белое море; 16—п — Баренцево море, о-в Колгуев; 18—20 — Кар-
ское море, 79°16' с. ш.—100°04' в. д.; 21—24 — Карское море, 77°47' с. ш,—100°19' в. д. Масштабные ли-
нейка: 1—3, 6—8, 11, 13—14, 16—18, 21—23 — 100 мкм; 4—5, 9—10, 12, 15, 19—20, 24 — 25 МКМ.
1. CURTITOMA
9$
Рис. 417. Зубы моллюсков Oenopota declivis (1—13) и Ое. cancellata (14—27).
1—2 — Баренцево море, Мурманский берег; 3—6 — Килдинский пролив; 7, 10—11 — Мурманский берег
69’21' с. ш. — 35’03' в. д.; 8—9 - Мурманский берег, о-в Еретики; 12—13 — Килдинская Салма; 14—19 —.
зал. Петра Великого, разрез от о-ва Аскольд; 20—23 — зал. Петра Великого, разрез отм. Поворотного; 24_
21 — зал. Петра Великого, отм. Среднего. Масштабные линейки.: 1—2, 4—6, 12—15, 18, 22—26 *— юо’МКМ-
S. 7—11, 16—17, 19—21, 27 — 50 МКМ.
96
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 418. Зубы моллюсков Oenopota cancellata (1—5); Ое. kurilensis (6—12); Ое. uschakovi
(13—16); Ое. pavlova (17—22).
1—S — Японское море, у о-ва Аскольд; ff—7 — Курильские о-ва, 46°59' с. ш,—152°11' в. д.; в—10 —
о-в Симушир; 11—12 — о-в Уруп, м. Китовый; 13—16— о-в Итуруп, 44°22' с. ш.—147°09' в. д.; 17—19 —
Японское море, зал. Петра Великого; 20—22 — о-в Сахалин, траверз м. Лопатина. Масштабные линейки:
1—2, 6—7, 9—14, 17—22 — 100 мкм; 3—5, 3, 15—16 — 25 мкм.
1. CURTITOMA
97
Рис. 419. Зубы моллюска Oenopota pyramidalis (1—94).
льная, ча°ть Баренцева моря; 5—8 — у западного побережья о-вов Новая Земля; 9—13 —
Мурманский берег, 14—16 — Карское море; 17—19 — Норвежское море; 20—23 —,о-в Визе, Карское море;
24—27 — Белое море; 28—29 — Баренцево море; 30—34 — Охотское море. Масштабные линейки.-. 1—6,
9—14—ts, 20—21, 23, 24—34 — 50 мкм; 7—8, 12—13, 19, 22 — 25 мкм.
7 И, П, Богданов
98.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 420. Зубы моллюсков Oenopota obliqua (1—10) и Propebela viridula (11—24).
1—з — Баренцево море, 79°17' с. ш.—44°35' в. д.; 4—5 — Баренцево море, Черная губа; в—10 — Карское
море, 79°27' с. ш.—76°40' в. д.; 11—13 —Маточкин Шар, 73°15' с. ш.—54°05' в. д.; И—15 — Баренцево
море, 68°05' с. ш.—39°33' в. д.; 16—18 —Японское море, Татарский пролив; 19—21 — Баренцево море,
69°39' с. ш.—59°25' в. д.; 22—24 —о-ва Новой Земли, 72°25' с. ш.—52°32' в. д. Масштабные линейки:.
1—4, 6—12, 14—17, 21—24 — 100 мкм; 5, 13, 18—20 — 25 мкм.
1. CURTITOMA
99
12
Рис. 421. Зубы моллюсков Obesotoma sachalinensis (1—10); О. gigas (11—14); О. woodiana
(15—18). '
1—г — Японское море, бухта Роджерса; з—7 — Японское море, 42°51' с. ш.—133°54' в. д.; 8—10 — юж-
ная часть Охотского моря; 11—14 — Земля Франца-Иосифа, о-в Хейса; 15—16 —Баренцево море, 69°46*
•г. та.—54°39'' в. д.; 17—18 — Белое море. Масштабные линейки.: 1—3, 5, 8—9, 11, 14—16 — 100 мкм‘.
4, 6—7, 10, 12—13, 17—18 — 25 МКМ.
7*
100
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 422. Зубы моллюсков Oenopota casentina (1—3); Oe. inequita (4—9); Oe. biconica (10—
18); Oe. elongata (19—23); Oe. pingelii (24—SO).
1—3 — Новосибирские о-ва, 44°42' с. ш.—147’56' в. д.; 4—в — Японское море, Де-Кастри, о-в Южный; 7—
9 — Японское море, 49’02'с.ш.—140’21' в. д.; 10—13 — Японское море, зал. Ольги; 14—18 —Северные Ку-
рильские о-ва; 19—23 — Японское море, 42°17' с ш.—в. д.; 24—26 — Карское море, 80’14' с. ш.—
80’12' в. д.; 27—30 — море Лаптевых, 76’47' с. ш.—117’24' в. д. Масштабные линейки: 1—3, 7, 10, 13, 14,
17, 19—20, 24—26, 28—30 — 100 мкм; 4—6, 8—9, 11—12, 13—16, 18, 21—23, 27 — 25 МКН.
1. CURTITOMA
101
Рис. 423. Зубы моллюсков Obesotoma woodiana (1—11); О. ’.tumida (12—16); О. robusta
(17—28).
I—в — Баренцево море, 68°48' с. ш.—43°32' в. д; 4—6 — Баренцево море, 68°28' с. ш.—42°46' в. д.; 7—
II — Баренцево море, 69°38' с. ш.—58°42' в. д.; 12—16 — Северный Ледовитый океан, 76°31' с. ш.—
118°46' в. д,; 17—19 — Японское море, Де-Кастри; 20 — 21 — Японское море, зал. Петра Великого; 22—25—
Японское море, зал. Чихачева, о-в Устричный; 26-—27 — Японское море, от о-ва Аскольда; 23 — о-в Саха-
лин, м. Елизаветы. Масштабные линейки; 1—3, 5-—6, 9, 12—15, 17,19—24, 26 — 27 — 100 мкм; 4, 7 —
3, 10—11, 16, 18, 25, 28 — 25 мкм.
102
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 424. Зубы моллюсков Obesotoma schantarica (1—5); О. faponica (6 7); О. solida (8 8},
О. tomiyaensis (10—12); О. simplex (13—20).
1- о-в Сахалин, м. Елизаветы; 2-5 - Охотское море, Шантарские о-ва и
Петра Великого у о-ва Русского; 8-9 - зал. Петра Великого; 10-12 - Японское
20 — Баренцево море, Корабельная бухта. Масштабные линеики.: 1-3,5-12, 15, 20 1UU мкм, 4,
1. CURTITOMA
IOS
Рис. 425. Зубы моллюсков Obesotoma gigantea (I—10) и О. starobogatovi (11—22).
1__з __Карское море, о-в Визе; 4—10 — Земля Франца-Иосифа, о-в Виктория; 11—13 — море Лаптевых,.
71°58' с. ш.—132°16' в. д.; 14 -16 — море Лаптевых, м. Ляховской; 11—20 - Карское море, 73°36 с. ш.—
78°22' в д.; 21—22 — Новосибирские о-ва. Масштабные линейки: 1—1, 10—13, 13—11, 20—22 — 100 мкм;
8—9, 14, 18—19 — 25 мкм.
104
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 426. Зубы моллюсков Obesotoma starobogatovi (1—4)', О. tenuilirata (5 9)’, Curtitoma
hokkaidoensis (10—12)-, Propebela marinae (13—22).
1—4 — Карское море; 5—9 — о-в Сахалин, м. Елизаветы; 10—12 — Охотское море; 13—17 — Охотское
море 41)°24' с. ш,—148°46' в. д.; 18—22 — Курильские о-ва, 44°21' с. ш.—148°24' в. д. Масштабные линеики-.
1—2, 4—6, 9—11, 15, 19—20 — 100 мкм; 3, 7—8, 12—14, 10—18, 21—22 — 25 МКМ.
1. CURTITOMA
105
Рис. 427. Зубы моллюсков Propebela fidicula (1—7); Р. margaritae (8—9); Р. concinnula
(10—16); Р. mltrata (17—20).
1 3 . Восточная Камчатка, Кроноцкий зал.; 4—7 — о-в Врангеля, 8—9 — Восточная Камчатка, о-в Ка-
рагинский; ю—13 — Охотское море, 56°09' с. ш.—144°30' в. д.; 74—16 — Западная Камчатка: 17-—20 —
Курильские о-ва. Масштабные линейки: 1—3, 5—10, 12, 14—15, 18—19 — 100 мкм; 4, 11, 13, 16—17, 20 —
25 мкм.
106
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 428. Зубы моллюсков Propebela tersa (1—14) и Р. exarata (15—26).
1—4 — Тихий океан, 38°57' с. ш.—143°23' в. л.; 5—7 — зал. Петра Великого; 8—11 — о-в Беринга; 12—
14 — зал. Петра Великого; 15—18 — Баренцево море, 69°12' с. ш.—46°48' в. д.; 19—22 — Мурманский бе-
рег; 23—26 — Кольский зал. Масштабные линейки: 1, 4 — 6, 1»—11, 16—18, 21—22, 25 — 100 мкм; 2—3,
7—9, 12—15, 19—20, 23—24, 26 — 25 МКМ.
1. CURTITOMA
107
Рис. 429. Зубы моллюска Propebela voriabilis (1—24).
1—5 - о-в Беринга; 6-11 - о-в Медный; 12-17 - о-в Беринга; 13-22 -Командорские о-ва о-вАрий
Камень- 23—24 — о-в Беринга. Масштабные линеики: 1—2, в, 9—10,13 — 14, 17, 19 — 20, 23 24 1U0 мкм,
у 15-.16г is> 21—22 — 25 мкм.
108
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 430. Зубы моллюсков Propebela
golikovi (1 5); Р. goryachevi (6—13); Р. alitakensis
(14—17).
? ш-~141Л58' в- д-; ^-5 - о-в Сахалин, зал. Анива; в-9 - Японское
море, 46 00 с. ш,—141 26' в. д., 10—13 — Охотское море, 59°31' с. ш.—150°44’ в. д • 14—17 — Восточная
Камчатка, Кроноцкий зал. Масштабные линейки' 1 — з, 5—6, 9—10, 13—15 17 — 100 мкм' 4 7____8 п____w
16 — 25 мкм. ’ ’ * ’
1. CURTITOMA
109
Рис. 431. Зубы моллюсков Propebela nobilis (1—16) и Р. assimilis (17—20).
iemnr Баренцево море, 68°29'с. ш.—47°27' в. д.; 4—в —пролив Югорский Шар; 7—9 —Белое море,
Ь8 27 с. ш,—-43 37 в. д.; 10 — Баренцево море, 68°45' с. ш,—47°01' в. п.;-11—12 — о-в Беринга; 13—16 —
.Баренцево море, 69 J37 с. ш.—33°04' в. д.; 17—20 — Охотское море, 49°08' с. ш,—143°21' в. д. Масштаб-
ные линеики: 1, 4—7, 10—15, 13, 20 — 100 мкм; 2 — з( 8—9, 16—17, 19 — 25 мкм.
по
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис.^432.^3убы моллюсков Propebela assimilis (1 —11) и spitzbergensis (13—33).
7^^зал. Петра Великого; в—9 — Килдинская Салма; 10—11 — Восточная Камчатка, м. Ольги; 12—
15 — Баренцево море, о-в Килдин; 16—19 — Карское море, 79°06' с. ш.—74°44' в. д.; 20—22 — Карское
море 71°54' с ш.—67°45' в. д.; 23 —Соловецкий зал. Масштабные линейка: 1, 5—7, 10—11, 13—14, 11—
20, 22 — 100 мкм; 2—4, 8—9, 12, 15—16, 21, 23 — 25 мкм.
1. CURTITOMA
111
Рис. 433. Зубы моллюсков Propebela harpularia (1—22) и P. turricula (23—37).
1—S — Японское море, разрез от м. Гамова; в—8 — Баренцево море,Черная губа; 9—11 — Берингово море,
‘бухта Провидения; 12—13 — Норвежское море, Вадсё; 14—15 — Карское море, 80°19' с. ш.—72°47' в. д.;
1в—18 — Курильские о-ва, о-в Шикотан; 19—22 — Шантарские о-ва; 23—28 — Норвежское море; 29—
ЛО — Мурманский берег; 31—34 — Норвежское море; 35—37 — Хельголанд. Масштабные линейки: 1—
37 — 50 мкм.
иг
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 434. Зубы моллюсков Propebela rugulata (1—22’).
1—5 — Карское море, 80°11' с. ш.—86°24' в. д.; 6—8 — Кар ское море, 81°33' с. ш.—83°06' в. д., 9—12 —
Баренцево море, 73°44' с. ш.—43°30' в. д.; 13—15 — Баренцево море, 71°20' с. ш. — 42°00' в. д.; 13—18 —
над Горлом Белого моря; 19 — 22 — у берегов Новой Земли, 75°55' с. ш.—76°58'в. д. Масштабные линейки'.
1—9, 12 —20 —100 мкм; 10 — 11, 21 - 22 — 25 мкм..
1. CURTITOMA
113
Рис. 435. Зубы моллюсков Propebela svetlanae (1—S); P. cassis (9—12)’ P. popovia (13—15).
1—4 — Западная Камчатка; 5—8 — Охотское море, 59°31' с. ш.—150°44' в. д.; 9—12 — Курильские о-ва,,
о-в Шикотан; 13—15 — Западная Камчатка. Масштабные линейки: 1—7, 9, 12—14 — 100 мкм; 8, 10—11,
16 — 25 мкм.
8 И. П. Богданов
114
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 436. Зубы моллюсков Propebela terpeniensis (1—8); Р. verrllli (9—15); Р. ^cingulata
(16—19). '
£—s — Японское море, у о-ва Аскольд; в—а — Охотское море, восточный берег Южного Сахалина; 9—12 —
Западная Камчатка; 13—15 •— Приморье; 15—19 — о-в Монерон. Масштабные линейки." 2—3 6—7 9
11, 15—1в, 18 — 100 мкм; 1, 4—5, 8, 10, 12, 13—14, 17, 19 — 25 мкм. ’
1. CURTITOMA
115
Рис. 437. Зубы моллюсков Propebela angulosa (1—9) и Р. scalaris (10—22).
1—3 — Баренцево море, 69°02' с. ш.—37°27' в. д.; 4—7 — Баренцево море, 80°10' с. ш.—33°59' в. д ; 8—
9 — Мурманский берег; 10—13 — Баренцево море, 68°05' с. ш_39°34' в. д.; 14—17 — Карское море,
79°06' с. ш.—74’44' в. д.; 18—20 —о-ва Новая Земля, губа Мелкая; 21 — 22 —Новая Земля, в проливе
о-вов Баренца. Масштабные линейки: 1, 3—4, в—12, 14, 1в—18, 21, 22 — 100 мкм; 2, 5, 13, 15, 19—20 —-
25 мкм.
8*
Ц6
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 438. Зубы моллюсков Propebela scalaris (1—9) и Р. scalaroldes (10—18).
1—2—над Горлом Белого моря; 3—4 — Баренцево море, Мурманский берег; 5—9— Карское море,
..78°08' с. ш.—73°47' в. д.; 10—13 — Баренцево море, 68°28' с. ш.—35°12' в. д.; 14—15 — Баренцево море,
у о-ва Килдин; 16—18 - Баренцево море, Мурманский берег. Масштабные линейки: 1, 3—6, 9—10, 13—15,
17 — 100 мкм; 2, 7—8, 11—12, 16, 18 —25 мкм.
.щих первоописание (Verrill, 1882, pl. 57, fig. 14), но строение лезвия и базаль-
ной части ро всех случаях оказалось одинакоьым.
Вид хорошо отличается от других предстаьителей рода сьоеобразной ры-
, резкой анального синуса, сохраняющейся на осеьой скульптуре раковины р Риде
резкого сигмоидального излома иерхней части осеных складок.
Распространение. Широко распространенный амфибореальный рид
. тихоокеанского происхождения. Обитает и бореальных родах Тихого океана и
в нысокобореальных родах американского побережья Атлантики. Обнаружен
у берегов Ньюфаундленда, Западной Гренландии (Posselt, Jensen, 1898), в Бе-
ринговом проливе (Krause, 1885), в Тихом океане в зал. Св. Лаврентия, у Ко-
. мандорских о-вов, у северо- и юго-восточных берегов Камчатки, а также в Охот-
ском море у Курильских о-вов, у берегов Приморья и в Татарском проливе
, в Японском море .(рис. 439,7).
1. CUHTtTOMA
117
вида на-
Экология. Обитает на глубинах от 15 до 494 м, однако существуют
указания на нахождения моллюсков на глубинах до 914 м (Verrill, 1882).
Наиболее частые находки особей данного вида приходятся на глубины 15—
100 м. Обитает преимущественно на заиленных грунтах (илистый' песок, ил
с примесями гальки, гравия, ракуши и т.
грунтах (20 %), и на галечнике (до 9 %
от общего числа просмотренных моллюс-
ков). Оптимальная температура сущест-
вования для моллюсков данного
ходится в границах от —-1.78 до 2.15 °C.
Вид стеногалинный, встречается в диапа-
зоне солености от 32.94 до 33.89 °/00, только
1 экз. был найден при солености 31.11 °/00.
Типы. Место хранения типов Bela
incisula неизвестно; типовое местонахо-
ждение — Лабрадор. Голотип, С. hecuba,
хранится в USNM, № 428871; типовое
местонахождение — Японское море.
Просмотрено 186 экз.
2. Curtitoma trevelliana (Turton, 1834),
comb. n. (Рис. 7, Б—Г; 30; 57; 72-77;
.386, А—Б; 407, 1-7).
д.), реже встречается на песчаных
Рис. 439. Места находок Curtitoma
incisula (2) и С. hepes (2).
Turton, 1834 i 351 (Pleurotoma); For-
bes, 1850 : 227 (Bela); Forbes, Hanley,
1853 : 452, pl. 112, fig. 1,2 [Mangelia (Be-
la)}; Adams, Adams, 1858 : 92 (Bela);
J effreys, 1867 : 398 (Pleurotoma); 1872 : 222,
pl. 91, fig. 8 (Pleurotoma); G. O. Sars, 1878:
235, tab. 6, fig. 13; tab. 9, fig. 4 (Bela); Tryon,
1884 : 221, pl. 33, fig. 65 (Bela; part.); Герцен-
штейн, 1885:690 (Pleurotoma); Friele, 1886:
11, pl. 8, fig. 7—10, pl. 10, fig. 10—11 (Bela);
Martini, Chemnitz, 1887:146, Taf. 31,
Fig. 11 (Bela); Locard, 1892:54 (Bela); 1897:
231 (Bela); Knipowitsch, 1901:44 (Bela);
К о b e It, 1905 : 266, Taf. 85, Fig. 18—20 (Bela);
О I dr о yd, 1927 : 123 (Lora); Pearce, 1966:81,
fig. 1 (Lora); .Bouchet, Waren, 1980:75,
fig. 155—156, 264 (Oenopota); Waren,
1983 : 167, pl. 8, fig. 7 (Oenopota). — reticulata
Brown, 1827, pl. 48, fig. 29, 30 (Pleurotoma;
nomen oblitum); 1844:8, pl. 5, fig. 29—30
(Pleurotoma); Sykes, 1906:177 (Lora); Daut-
zenbetg, Fischer, 1912:46 (Bels); Mac-
pherson, 1971:121, pl. 7, fig. 13, map 54
(Oenopota). — finmarchia Friele, 1886:12, pl. 8, fig. 11—13 (Bela decussata var.).—
decusstjta (non Couthouy, 1839): Reeve, 1846:38, pl. 19, fig. 159 (Pleurotoma). — viri-
dula (non Moller, 1842): Tros die 1, 1866:19, tab. 4, fig. 2 (Bela).
Раковина, как правило, кремовых тонов, овально-веретеновидной формы,
с 5—6 уступчатыми оборотами. Периостракум тонкий, желтоватый, полупро-
зрачный, плотноприлегающий. Эмбриональная раковина представлена 1.5—
1.7 оборотами выпуклой формы, орнаментированными 15—20 нитевидными спи-
ральными ребрышками (рис. 7, Б; 386, А—Б). При переходе на дефинитивную
раковину спиральные ребрышки сближаются, а затем сливаются в тонкие
ребра (рис. 7, Г). Средние размеры протоконха: ширина 290—310, высота 190—
200 мкм. Осевая скульптура дефинитивной раковины изменчива и представлена
у разных экземпляров средними или узкими складками. Иногда осевые складки
могут отсутствовать на последнем обороте и его скульптура представлена не-
большими осевыми валиками. Прямые на периферии оборота раковины осевые
118
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
складки приобретают на плече оборота сигмоидальный изгиб; в верхней части
основания они сглаживаются. Пространства, разделяющие осевые складки,
также сильно варьируют в размерах: от 1/2 до в 2—3 раза превышающих по ши-
рине собственно складки. На последнем обороте раковины насчитывается от 14
до 25 осевых складок, на предпоследнем от 13 до 19. Спиральная скульптура
представлена различными элементами: от волосовидных ребрышек до тонких
ребер. Она покрывает всю поверхность раковины, однако четкой закономер-
ности в чередовании элементов спиральной скульптуры, как правило, не наблю-
дается. Основание раковины сильно уплощенное, без границы переходящее
в короткий сифональный вырост. Устье узко-обратногрушевидное, несет
на колумеллярном крае тонкий матовый каллус. Сифональный канал широкий,
неглубокий.
Наиболее крупный экземпляр, обнаруженный у Мурманского побережья,
имеет следующие параметры (мм): В =10.6; Д=5.2; Впо—1Л\ В =5Д-, Д,=
=2.3.
Крышечка когтевидная, полуовальная, с относительно полого изгибаю-
щейся осью нарастания. Мускульный отпечаток занимает срединную часть ле-
вой ее половины (рис. 32, И—Л).
Изображения зубов радулы (G. О. Sars, 1878 : Tab. 9, fig. 4; Friele, 1886 :
pl. 10, fig. 10—11) полно отражают особенности радулы. Зубы кинжаловидной
формы (407, 7—7), киль достигает х/2 длины лезвия (реже превышает); базаль-
ная часть неправильно-полукруглой формы, одна из ручек несет уплощенный
флажок.
Вид схож по форме раковины с С. violacea и С. flora, от которых отличается
кинжаловидной формой зуба. От С. hebes отличается более стройной раковиной
с хорошо развитой осевой скульптурой, представленной тонкими осевыми
складками.
Изменчивость. Впервые на значительную изменчивость вида указал
Герценштейн (1885), обнаруживший 3 основные вариации: 1) экземпляры с тон-
кой осевой скульптурой, не преобладающей над спиральной и образующей по-
добие сети (схожей с описанной — Forbes, Hanley, 1853); 2) осевые складки
едва выражены, а спиральная скульптура явно преобладает; 3) наибольшая
по числу особей группа моллюсков (соответствующая описанию — G. О. Sars,
1878) с преобладающими в верхней части оборота выраженными осевыми склад-
ками. В исследованном материале описанные вариации встречаются наиболее
часто.
Замечания. Употребление младшего синонима — Pleurotoma trevel-
liana для обозначения вида обоснованно предлагают использовать Буше и
Варен (Bouchet, Waren, 1980) в связи с тем, что данное название широко рас-
пространено в литературе. Название Р. reticulata является nomen oblitum.
Так, в период с 1827 по 1904 г. только в 3 из 78 вышедших работ используется
название «reticulata» (по: Dautzenberg, Fischer, 1912).
Распространение. Атлантический широко распространенный бо-
реально-арктический вид, обитает в высоко- и низкобореальпых районах Се-
верной Атлантики, в Арктику заходит до Новосибирских о-вов. Отмечен в Ка-
надском секторе Арктики, у Западной и Восточной Гренландии, у берегов
Ньюфаундленда и Лабрадора, Новой Англии, Новой Шотландии в Девисовом
проливе и у м. Код, у Исландии, Шотландии, в Гасконском зал., у берегов
Англии и Норвегии. Обнаружен в Баренцевом, Белом, Карском морях и в море
Лаптевых; многочисленны/его находки у берегов архипелага Земли Франца-
Иосифа, о-вов Северной Земли и Новосибирских о-вов.
Экология. Обитает от литорали до 430 м, отмечено нахождение отдель-
ных особей на глубине до 1447 м (Голиков, 1964). В Баренцевом море наиболь-
шая частота встречаемости приходится на глубины 120—220 м, в то время
как в районе о-вов Новая Земля он встречается только на глубинах 0—40 м.
В юго-восточной части Карского моря пик численности моллюсков приходится
на глубины 40—60 м. В Гасконском заливе моллюски обнаружены на глубине
1. CURTITOMA
119
'900 м (Locard, 1897). Преобладающее большинство находок экземпляров вида
связано с заиленными грунтами (ил с камнями, илистый песок, чистый ил —
более 90 %), хотя некоторые представители были встречены и на песчанистых
грунтах. Температурный оптимум находится в границах от —1.8 до —1.0 °C,
отмечены живые моллюски в акваториях с придонной температурой 2.8 °C.
Предпочитаемая соленость от 33.8 °/00 и выше, однако 2 экз. были найдены при
меньшей солености: один при 32.97, другой при 31.02 %0.
Типы. Типы, Pleurotoma, trevelliana, хранятся в BMNH (Oldroyd, 1927).
Лектотип (Waren, 1983) хранится в USNM, № 190860; типовое местонахожде-
ние — Скарборо, Великобритания. Место хранения типов, Р. reticulata,
неизвестно; типовое местонахождение — Greenock (Шотландия).
Просмотрено более 1250 экз.
3. Curtitoma hebes (Verrill, 1880), comb. n. (Рис. 66—68; 390, Г- 409,1—в’,
439, 2).
Verrill, 1880 : 367 (Bela)-, 1882 : 459, pl. 57, fig. 7 (Bela)-, Tryon, 1884 : 217,
pl. 29, fig. 69 (Bela decussata var.); Martini, Chemnitz, 1887 ! 162, Taf. 33, fig. 3
.(Bela)-, К о b e 1 1, 1905 : 257, pl. 84, fig. 5 (Bela decussatasubsp.).
Раковина белесая, веретеновидная, с 4—5 закругленными, сильновыпук-
лыми оборотами, причем верхние из них, как правило, обломаны. Тонкий, про-
зрачный, от темно-оливкового до зеленовато-белого цвета периостракум плотно
прилегает к раковине. Протоконх умеренно выпуклый, представлен 1.3—
1.5 оборотами, несущими до 14 нитевидных спиральных ребрышек (рис. 390, Г).
Осевая скульптура закладывается только на ранних постэмбриональных
оборотах. Осевая скульптура дефинитивной раковины составлена многочислен-
ными валиками и тонкими складками, причем сечение последних бывает от полу-
овальной до уплощенно-прямоугольной формы. Четкой детерминации в рас-
положении элементов осевой скульптуры на поверхности раковины не наблю-
дается. Как складки, так и валики равномерно развиты на плече и периферии
оборота и на основании раковины. На околошовной поверхности они при-
обретают сигмоидальный изгиб. На последнем обороте раковины насчитывается
от 32 до 48 различных элементов осевой скульптуры. Элементы спиральной
скульптуры па околошовном пространстве представлены многочисленными
(8—12) равновеликими с разделяющими их промежутками нитевидными реб-
рышками. Периферию оборота и основание раковины покрывают тонкие шнуро-
видные ребра, по ширине в 1.5—4 раза превышающие межреберные простран-
ства. Пересечение практически равновеликих элементов осевой и спиральной
скульптур придает поверхности раковины сетчатый рисунок.
Основание раковины слабовыпуклое, плавно переходит в короткий сифо-
нальный вырост. Устье неправильно-овальной формы, уступчатое в верхней
части. Тонкий белесоватый каллус лежит узкой полосой на колумеллярном
крае. В верхней части палатального края сипус не выражен. Сифональный
канал широкий, относительно глубокий, короткий.
Размеры типового экземпляра (мм): Вр=8.0; Д=5.0; 2?ПО=6.35; ^=5.0;
Ду=1.8 (Verrill, 1882); максимальные размеры 5^=9.0; Д=5.5 отмечены для'
особи из типовой серии.
Зубы ножевидной формы, с массивной, сильно уплощенной базальной
частью и килем, длина которого превышает 1/2 длины лезвия (рис. 409, 1—6).
Лезвие относительно большого диаметра. Шпора очень узкой полосой лежит
па одной из ручек.
Вид схож по раковине с С. novajasemljensis, от которого отличается более
округлыми оборотами раковины, неправильно-овальным, усеченным в верх-
ней части устьем и наличием тонких осевых складок.
Распространение. Амфибореальный широко распространенный вид
тихоокеанского происхождения. Обитает в бореальных водах Тихого океана
120
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
и в высокобореальных водах американского побережья Атлантики. Помимо
типового местонахождения вид отмечен у западного побережья Камчатки и
у берегов о-ва Сахалин в Охотском море, у Южных Курильских о-вов и в Япон-
ском море в Приморье и у западных берегов о-ва Сахалин (рис. 439, 2).
Экология. Обитает на глубинах от 60 до 925 м, наиболее часто встре-
чается в диапазоне глубин 120—200 м. Все имеющиеся экземпляры собраны
с песчанистых, илистых и илисто-песчанистых грунтов. Отмечен только
в водах с океанической соленостью (не ниже 33.3 °/00) при температуре
от — 0.3 до 1.0 °C.
Рис. 440. Места находок Curtitoma novajasemljensis (7) и С. ovalis (2); 3 — палеонтологиче-
ские находки.
Типы. Место хранения типового экземпляра не указано; типовое место-
нахождение — Ньюпорт, у берегов штата Родайленд.
Просмотрено 35 экз.
4. Curtitoma novajasemljensis (Leche, 1878), comb. n. (рис. 7, A; 69—70;:
387, А —Б; 407, 8-13; 440, 7).
Leche, 1878 1 53, Tab. 1, fig. 15 (Pleurotoma); Герцеиштейн, 1885:690-
(Pleurotoma); Martini, Chemnitz, 1887 : 165, Taf. 33, fig. 16 (Bela); К о b e 11,
1905 1 242, Taf. 85, fig. 13, 14 (Bela); О 1 d г о у d, 1927 : 111 (Lora); Macpherson,.
' 1971 1 119, pl. 7, fig. 14, map. 52 (Oenopota).
Раковина маленькая, овальная, желтовато-пепельного цвета, с 4—6 уступ-
чатыми оборотами, уплощенными в верхней части, из них только последние
2—2.5 оборота сохраняют скульптуру, остальные корродированы. Периостра-
кум тонкий, прозрачный, бледно-оливкового цвета, плотно прилегающий.
Протоконх сосцевидной формы, представлен 1.5 оборотами выпуклой формы
(рис. 387, А, Б). Спиральная скульптура протоконха представлена много-
численными волосовидными ребрышками. На последних стадиях развития
протоконха закладывается осевая скульптура, которая сначала не превышает
размеры следов нарастания раковины (рис. 387, J5), но затем приобретает раз-
меры тонких складок. Осевая скульптура дефинитивной раковины-представлена
тонкими многочисленными (до 60 на последнем обороте), упорядоченными ва-
ликами. Последние одинаково развиты как на плече и на периферии оборота,
так и в верхней 1/2 основания раковины. Полное стирание осевых валиков;
отмечается лишь на сифональном выросте. Спиральная скульптура представ-
лена многочисленными тонкими ребрами, равновеликими с осевыми валиками..
Так как промежутки, разделяющие элементы и осевой и спиральной скульптуры,.
1. CURTITOMA
1S1
в 1.5—2 раза шире валиков и ребер, то последние при пересечении придают
поверхности раковины ярко выраженный сетчатый рисунок. Основание ра-
ковины слабовыпуклое, без четкой границы переходит в короткий сифональ-
ный вырост. Устье полуовальной формы, угловатое в верхней части. Колумел-
лярный край несет узкую полосу тончайшего каллуса. Анальный синус на
палатальном крае устья практически отсутствует. Сифональный канал протя-
женный, глубокий.
Размеры голотипа (мм): _вр=8.0; Д=4.5; /?г,=4.0 (Oldroyd, 1927). Из рай-
она Карского моря обнаружена раковина со следующими параметрами: Вр=
= 11.6; Д=7.8; 5„0=8.4; ^=6.2; Ду=3.2.
Крышечка когтевидная, с плавно изгибающейся осью нарастания. Мус-
кульный отпечаток представлен тонкой линией, проходящей вдоль оси кры-
шечки (рис. 32, Е-—Ж).
Зубы радулы ножевидные, до 150 мкм в длину (рис. 407, 8—13), с массив-
ным основанием неправильно-овальной формы. Лезвие удлиненное, неболь-
шого диаметра. Киль достигает х/2 длины лезвия.
Вид отличается от других видов рода уступчатыми оборотами и мелкой
решетчатой скульптурой раковины.
Р аспр остр анение. Арктическо-бореальный, преимущественно евра-
зийский вид. Обитает в северных морях СССР и у берегов Норвегии в Баренце-
вом море (рис. 440, 1), отмечен в северной части канадского сектора Арктики
(Macpherson, 1971).
Экология. Обитает на глубинах от 3 до 450 м, однако наибольшее
число находок связано с глубинами 5—70 м. Отмечено явное предпочтение
илистых и заиленных грунтов и низких температур от —1.6 до —1.0 °C, хотя
встречается в диапазоне от —2.0 до 1.1 °C. Моллюски обитают в основном при
солености выше 34 °/00, однако обнаружены некоторые особи и при солености
около 20.41 °/00 (в море Лаптевых).
Типы. Место хранения типового материала неизвестно. Высказывалось
предположение о хранении его в ЗИН АН СССР (Oldroyd, 1927), однако в кол-
лекции он не обнаружен. Типовое местонахождение — Маточкин Шар, Новая
Земля.
Просмотрено 848 экз.
5. Curtitoma ovalis (Friele, 1877), comb. n. (Рис. 71; 409, 7—9; 440, 2—3).
F r i e 1 e, 1877 : 9, fig. 5, 5a [Pleurotoma (Bela)]', 1886 : 14, pl. 8, fig. 21, 22, pl. 10, fig. 19
(Bela)', Martini, C he mnitz, 1887 : 166, Taf. 33, fig. 15 (Bela)', Kobelt, 1905 :
258, pl. 84, fig. 15, 16 (Bela decussata var.); Bouchet, Waren, 1979 : 227, fig. 15, 53
(Oenopota); 1980:68, fig. 144, 258—259 (Oenopota).— exigua Jeffreys, 1883 : 399,
pl. 44, fig. 10 (Pleurotoma); Tryon, 1884 : 216, pl. 33, fig. 73 (Bela).
Раковина маленькая, овальная, очень тонкостенная, белая, с 3.5—4 за-
кругленными, умеренно выпуклыми оборотами. Периостракум очень тонкий,
прозрачный. Протоконх небольшой, 840—880 мкм, выпуклый, представлен
1.5—1.7 умеренно выпуклыми оборотами, покрыт многочисленными нитевид-
ными спиральными ребрышками. Осевая скульптура в виде тонких валиков
закладывается на ранних стадиях развития постэмбриональной раковины
(Bouchet, Waren, 1980). Осевая скульптура дефинитивной раковины представ-
лена многочисленными осевыми валиками, разделенными в 2—3 раза пре-
вышающими их собственную ширину промежутками. Валики одинаково вы-
ражены на всей поверхности раковины, исчезая лишь в нижней 1/,1 основания
раковины. Осевая скульптура имеет плавный, распространяющийся на всю
поверхность периферии оборота (вдоль оси навивания раковины) сигмоидаль-
ный изгиб. На последнем обороте насчитывается до 40, на предпоследнем
до 32 осевых валиков. Спиральная скульптура состоит из равновеликих на всей
поверхности оборотов, основания раковины и сифонального выроста нитевид-
122 СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ных спиральных ребрышек, равных по ширине разделяющим их межреберным
пространствам. В связи с тем что выраженность ребрышек такая же, как и
у осевых складок, пересечение скульптурных элементов придает поверхности
раковины сетчатый рисунок. Основание раковины уплощенное, плавно пере-
ходит в короткий сифональный вырост. Устье полуовальной формы, с тончай-
шим прозрачным каллусом на колумеллярном крае. Палатальный край несет
небольшую вырезку анального синуса. Сифональный канал .широкий и глу-
бокий.
Высота раковины типового экземпляра 4.6, ширина 2.1 мм. Максимальная
высота раковины 4.8 мм (место сбора: 80°01' с. ш.—03°19' в. д.).
Крышечка небольших размеров, с плавно изгибающейся осью. Мускуль-
ный отпечаток в виде узкой полоски, занимает срединную часть крышечки
(рис. 32, 3).
Зубы радулы длиной 100—120 мкм, стилетовидной формы, с коротким ос-
нованием и длинным, до х/2 длины лезвия килем (рис. 409, 7—9). Можно отме-
тить полную морфологическую идентичность исследованных зубов с изображе-
нием, приведенным для типового экземпляра (Friele, 1877 : fig. 5, а).
Вид более всего сходен с С. novajasemljensis, от которого отличается закруг-
ленными, умеренно выпуклыми оборотами раковины и стилетовидной формой
зуба.
3 амечания. Редкость данного вида в коллекциях привела к тому, что
многие авторы сводят его в синонимы к ряду сходных по раковине видов (на-
пример, С. hebes, С. novajasemljensis и др.), однако характерное строение зубов
радулы и тонкое строение скульптуры помогают легко распознать этот вид.
Распространение. Атлантический, преимущественно евразийский
высокобореальный вид. Обитает в батиальной зоне к западу от Британских о-вов
(Sykes, 1906), в Норвежском море (Friele, 1886) и у Шпицбергена (рис. 440, 2).
Экология. Обитает на глубинах от 557 до 2 775 м в основном на заилен-
ных грунтах при отрицательных температурах и при солености, близкой 34—
35 %0.
Типы: Лектотип (Bouchet, Waren, 1979) Pleurotoma ovalis и 12 паралекто-
типов хранятся в ZMB; типовое местонахождение — северная Атлантика
(63°22' с. ш.—05°29' з. д., глубина 2 222 м). Голотип, Р. exigua, находится
в BMNH, N 85.11.5.4580; типовое местонахождение расположено к северу от Геб-
ридских о-вов (59°5Г с. ш,—08°18' з. д., глубина 1 020 м).
Просмотрено 7 экз.
6. Curtitoma fiora (Dall, 1919), comb. n. (Рис. 78; 409, 18—19).
Dall, 1919b : 302 (Lora)', О 1 d г о у d, 1927 I 103 (Lora).
Раковина зеленовато-белая, удлиненно-веретеновидная, толстостенная,
с 6—6.5 закругленными, умеренно выпуклыми оборотами. Периостракум отно-
сительно толстый, рыжевато-коричневый. Эмбриональная раковина корроди-
рована, ее скульптура не изучена. Осевая скульптура дефинитивной раковины
составлена широкими пологими складками, разделенными в 3—5 раз меньшими
промежутками. Складки сглаживаются к нижней части периферии оборота,
а наибольшая их выраженность приходится на околошовную поверхность, где
они приобретают сигмоидальный изгиб. На последнем обороте раковины на-
считывается до 17 складок, на предпоследнем до 16. Спиральная скульптура
представлена тонкими шнуровидными ребрами, разделенными в 5—7 раз более
узкими межреберными промежутками. На периферии последних дефинитивных
оборотов наблюдается закладка тонких нитевидных ребрышек между шнуровид-
ными ребрами по 1 в каждом межреберном промежутке. Ребрышки, лежащие
на околошовной поверхности, как правило, в 1.5—2 раза тоньше ребер перифе-
рии оборота. Основание раковины уплощенное, плавно переходит в короткий
сифональный вырост. Устье неправильно-овальной формы, угловатое в верх-
1. CURTITOMA
123
ней части. Белый каллус широкой толстой полосой покрывает колумеллярный
край устья. Палатальный край в верхней своей части несет неглубокий синус.
Сифональный канал неглубокий, широкий, относительно короткий.
Размеры типового экземпляра (мм): 5р=9.75; Д=4.0; /?по=6.0; /?3,=4.0.
-Исследованный экземпляр из Охотского моря имеет высоту раковины 11.4 мм.
Зубы радулы до 120 мкм в длину, удлиненно-кинжаловидные, с цилиндри-
ческим основанием и небольшим килем (рис. 409, 18—19); одна из небольших
ручек несет листовидную шпору.
Рис. 441. Места находок Curtitoma violacea в северных морях СССР; 1 — палеонтологи-
ческие находки.
Вид наиболее сходен с С. trevelliana, от которого отличается высоким за-
витком и рыжеватой окраской раковины, а также стилетовидной формой
зубов.
Распространение. Тихоокеанский широко распространенный вы-
сокобореальный вид. Обнаружен в районе Алеутских о-вов и Аляски (Oldroyd,
1927), также известен из Охотского моря (м. Хребтовый).
Типы. Голотип хранится в USNM, № 220399а; типовое местонахождение —
о-в Адах, Алеутские о-ва.
Просмотрен 1 экз.
7. Curtitoma violacea (Mighels et Adams, 1842), comb. n. (Рис. 83—96; 387,
B—E; 388, A—E; 408, 7—29; 409, 20—22; 441; 442).
Mighels, Adams, 1842 : 51, pl. 4, fig. 21 (Pleurotoma}; Stimpson, 1851 : 49
{Mangelid}; Adams, Adams, 1858 : 92 [Pleurotoma (Bela)]; Troschel, 1866 : 43,
pl. 4, fig. 1 (Bela); Gould, 1870 : 353, fig. 622 (Bela); G. O. Sars, 1878 : 238, Tab. 17,
fig. 2, Tab. 9, fig. 8 (Bela); V e r r i 1 1, 1882 I 481, pl. 57, fig. 16 {Bela bicarinata var.); Pos-
selt, Jensen, 1898 : 155 (Bela bicarinata var.); О 1 dr о у d, 1927 : 121 (Bela);
Thorson, 1944 i 109 (Bela); Филатова, Зацепин, 1948 : 390, табл. 48, рис. 19
(Lora); Bouchet, Waren, 1980 : 74, fig. 145, 261 (Oenopota). — bicarinata Cou-
th о u y, 1838 r 104, pl. 1, fig. 11 (Pleurotoma, nom. praeocc., non Wood, 1828); Gould,
1841 : 281, fig. 186 (Pleurotoma); Stimpson, 1851 : 49 (Mangelia); Adams, Adams,
1858 : 95 (Defrancia); Gould, 1870 : 349, fig. 618 (Pleurotoma); G. O. Sars, 1878 : 237,
Tab. 9, fig. 7, Tab. 16, fig. 12 (Bela); Tryon, 1884 : 214, pl. 27, fig. 26, pl. 28, fig. 32, 34
(Bela); Friele, 1886 : 15, pl. 8, fig- 18, pl. 10, fig. 15—16 (Bela); Martini, Chem-
nitz, 1887 : 152, Taf. 32, fig. 5 (Bela); Dall, 1889 : 100, pl. 50, fig. 15 (Mangelia); К o-
b e 1 1, 1905:261, Taf. 83, fig. 15 (Bela); Dautzenberg, Fischer, 1912 : 44
(Bela); Oldroyd, 1927 : 110 (Bela); Macpherson, 1971 : 113, pl. 7, fig. 6, map. 50
(Oenopota); Hubendick, Waren, 1974 : 31, fig. 231 (Lora); Abbott, 1974 : 286,
Fig. 3388 (Oenopota); Голиков, Гульбин, 1977:237 (Oenopota).— cylindracea
Moller, 1842:86 (Defrancia); M б г c h, 1868:215 [Pleurotoma (Bela) violacea var.];
Posselt, Jensen, 1898 t 157 (Bela bicarinata var.); К о b e 1 t, 1905 : 262 (Bela bi-
124
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
carinata subsp.); Thorson, 1944:110 (Bela violacea var.).— beckii Moller, 1842 : 87
(Defrancia); Posselt, Jensen, 1898 : 157 (Bela bicarinata var.); Thorson, 1944 :
112 (Bela violacea var.). — laevior G. 0. Sars, 1878 : 239, Tab. 17, fig. 3 (Bela violacea
var.); Krause, 1885:277 (Bela violacea var.).— rufescens G. 0. Sars, 1878:238,
Tab. 16, fig. 11 (Bela bicarinata var.). — geminolineata Friele, 1879": 277 (Bela bicarinata;
var.); 1886 : 16, pl. 8, fig. 19 (Bela bicarinata var.); Martini, Chemnitz, 1887 : 235,
Taf. 42, fig. 8 (Bela bicarinata var.); К о b e 1 1, 1905 : 262, Taf. 83, fig. 19 (Bela bicarinata
subsp.). — nodulosa Krause, 1885 : 277, Taf. 18, fig. 4, 12 (Bela violacea var.). — minus'
cularia L о с a r d, 1897 : 259, pl. 12, fig. 28—30 (Bela).
Раковина от пепельного цвета до вишневого, овально-веретеновидная,
с 5—6 оборотами отлогоугловатой формы, тупоугловатыми в
верхней части.
Рис. 442. Места находок Curtitoma vio-
lacea в дальневосточных морях СССР.
Периостракум тонкий, прозрачный, желто-
ватый. Эмбриональная раковина (рис. 387,
388) сильновыпуклая, представлена 1.3—
1.4 оборотами. Спиральная скульптура
появляется в конце первого оборота про-
токонха и представлена многочисленными
волосовидными ребрышками. Однако уже
начало второго эмбрионального оборота
обозначено всего 3 ребрышками, которые
переходят на дефинитивную раковину.
Верхнее из них постепенно редуцируется,
а 2 остальных трансформируются в кили.
Осевая скульптура закладывается только
на дефинитивной раковине и варьирует
у разных экземпляров от линий нараста-
ния до тонких складок, но обычно пред-
ставлена многочисленными (до 45 на по-
следнем обороте) небольшими осевыми ва-
ликами, разделенными в 1.5—4 раза более
широкими промежутками. Осевые элемен-
ты на плече оборота имеют сигмоидальный
изгиб; на верхней х/3 основания рако-
вины они сглаживаются. Спиральная
скульптура представлена многочисленными
тонкими, реже нитевидными ребрами.
Межреберные промежутки составляют
х/2—1/i ширины разделяемых ими ребер.
Преобладающее развитие на поверхности
раковины или по крайней мере на ранних
оборотах получают 2 киля. Один киль
обозначает плечо, другой — нижнюю
треть периферии оборота. У многих эк-
земпляров на последнем обороте можно
видеть третье килевидное ребро, обозна-
чающее границу периферии оборота и ос-
нования раковины, т. е. проходящее по
месту стыка предыдущего оборота с после-
дующим. Основание раковины, как правило,
слабовыпуклое, почти уплощенное, без видимой границы переходящее в корот-
кий сифональный вырост. Устье обратногрушевидной формы, с белесым или
кремового цвета каллусом на колумеллярном крае. Анальный синус неглубо-
кий. Сифональный канал глубокий, как правило, узкий.
Крышечка когтевидной, удлиненной формы, с резко изгибающейся осью.
на начальных стадиях роста. Мускульный отпечаток ромбовидный, узкой поло-
сой лежит в правой половине крышечки (рис. 32, Н—О).
Зубы радулы маленькие, от 70 до 135 мкм, стилетовидные, с непостоянной
формы основанием й длинным (более х/2 длины лезвия) килем (рис. 408—409),
1. CURTITOMA
125
Особенности зубов радулы постоянны у всех форм и вариететов и соответствуют?
имеющимся изображениям (G. О. Sars, 1878; Friele, 1886).
Вид хорошо дифференцируется по веретеновидной раковине и, как правило",
прослеживающимся на ней 2 спиральным килям.
Изменчивость. Значительный полиморфизм раковины, наличие мно-
гочисленных форм (см. Общую часть) и изменчивость скульптурных элементов
привели к выделению большого числа подвидов и вариететов. Например, эк-
земпляры, описанные как violacea, отличаются от bicarinata отсутствием ярко
выраженных 2 килей на последних дефинитивных оборотах. Отличие формы
laevior (рис. 92) от bicarinata заключается в большей высоте раковины при оди-
наковой ширине, иными словами, в большей стройности. Следует считать все
эти подвиды и вариететы одним видом в связи с наличием переходных форм
и однотипностью морфологии зубов радулы.
Распространение. Широко распространенный бореально-арктиче-
ский вид тихоокеанского происхождения. Встречен во всех арктических морях,
в бореальных водах Тихого и Атлантического океанов, обитает у побережья
Норвегии, Шпицбергена, в Баренцевом и Белом морях, в водах архипелагов
Новой Земли и Земли Франца-Иосифа, у о-вов Северной Земли, в море Лапте-
вых у Новосибирских о-вов и в Чукотском море у о-ва Врангеля, в Беринговом
море у берегов Восточной Камчатки, повсеместно в Охотском море и у берегов
Приморья и в Татарском проливе в Японском море. Кроме того, вид отмечен
в канадском секторе Арктики (Macpherson, 1971), в Лабрадоре, у берегов Бри-
танской Колумбии, Ньюфаундленда, Новой Англии, в зал. Массачусетс, у бе-
регов Западной и Восточной Гренландии, Западной Ирландии и Исландии.
Экология. Обитает от литорали до 869 м, однако раковины находили
на глубине до 2505 м (Bouchet, Waren, 1980). Наибольшая его встречаемость
отмечена на 6—80 м в дальневосточных водах и 20—НО м в Северном Ледовитом
океане. У архипелага Земли Франца-Иосифа заходит в литоральную зону,
максимум его численности отмечен на глубинах 0—20 м. Встречается практи-
чески на всех типах грунта, реже на глинистых, наиболее часто обитание вида"
связано с илистыми и заиленными грунтами. Моллюски встречаются при соле-
ностях от 28.6 до 35.0, в основном при 33.8—35.0 %0. Вид относительно эври-
термный, встречается при температурах от —2.1 до 7.9 °C. Следует отметить
скоррелированность нахождения различных инфранодвидовых форм с темпера-
турным оптимумом. Так, наибольшая частота встречаемости типичной формы
violacea отмечена при температурах от 0 до 2.0 °C, в то время как форма bicari-
nata встречается в основном при диапазоне температур от —1.8 до 0 °C.
Типы. Типы Pleurotoma violacea (без номеров) находятся в Amh. Coll.,
Великобритания; типовое местонахождение — Casco Вау, Мэн. Типовой эк-
земпляр Р. bicarinata, по-видимому, хранится в BSNH; типовое местонахожде-
ние — воды зал. Массачусетс (east of Nahant). Типы Defrancia cylindracea,
D. beckii должны находиться в ZMC; однако на запрос от 25.02.1979 г. Хопнер.
(G. Норпег) ответил, что они не обнаружены в музее; типовое местонахожде-
ние — побережье Гренландии. Место хранения типов Bela violacea var. laevior
и В. bicarinata var. rufescens, вероятно, в ZMO; типовое местонахождение —
северо-восток Норвежского моря. Тип В. bicarinata var. nodulosa хранится
в Stuttg. ВС; типовое местонахождение — зал. Св. Лаврентия. Голотип В. mi-
nuscularia — в MNHN (Bouchet, Waren, 1980); типовое местонахождение —
юго-восточная часть Бискайского залива (43°01' с. ш.—09°38' з. д., 2018 м)..
Просмотрено более 1340 экз.
8. Curtitoma conoidea (G. О. Sars, 1878), comb. n. (Рис. 97—103; 389, Г—Е‘,
410, 1—15; 443, 2).
G. О. Sars, 1878 i 236, tab. 16, fig. 14 (Bela)-, Tryon, 1884 : 221, pl. 29, fig. 61
(Bela)’, Герценштейн, 1885 : 691 (Pleurotoma); Friele, 1886 : 13, pl. 8, fig. 14
(Bela decussata var.); Martini, Chemnitz, 1887 : 155, Taf. 32, fig. 8 (Bela); К o-
b e 1 1, 1905 : 259, pl. 83, fig. 13 (Bela decussata subsp.); Thorson, 1944 : 118 (Bela
decussata var.).
126
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Раковина белая, веретеновидной формы, с 5.5—7 закругленными, умеренно
выпуклыми оборотами. Периостракум тонкий, плотно прилегающий, обычно
светло-коричневый. Протоконх выпуклый, сосцевидной формы, представлен
1,4—1.5 сильновыпуклыми оборотами. Спиральная скульптура первого эмб-
рионального оборота слагается из рядов микроскульптурных бугорков. У начала
второго оборота протоконха появляются и довольно быстро развиваются 8—
9 тонких ребер, которые дают начало спиральной скульптуре дефинитивной
раковины. Элементы осевой скульптуры закладываются на последних этапах
развития протоконха (рис. 389, /’). На дефинитивной раковине осевая скуль-
птура представлена многочисленными сигмовидными тонкими складками, наи-
более выраженными в верхней х/2 оборота и сглаживающимися в нижней 1/,1
периферии оборота или на основании раковины. Складки разделены в 4—5 раз
Рис. 443. Места находок Curtitoma decussata (1) и С. conoidea (2) в Северных морях СССР;
3 — палеонтологические находки.
более широкими промежутками. На последнем обороте насчитывается от 22
до 36 осевых складок, на предпоследнем от 16 до 24. Спиральная скульптура
состоит из многочисленных нитевидных ребрышек, равновеликих на всей по-
верхности оборотов, на основании раковины и на сифональном выросте. Разде-
ляющие ребрышки промежутки одинаковой с ними ширины. Основание рако-
вины уплощенное, плавно переходящее в сифональный вырост средних разме-
ров. Устье чаще всего полуовальное, угловатое в верхней части. Колумелляр-
ный его край несет широкую полосу тонкого белесого, реже кремового цвета
каллуса. В верхней части палатального края лежит глубокая врезка анального
синуса. Сифональный капал протяженный, неглубокий, но широкий. Высота
раковины типового экземпляра 15 мм (раковина составлена 7 оборотами).
Наибольший исследованный экземпляр имел следующие размеры (мм): =-11.8;
Д=6.2; £по=8.6; ^=6.3; Д^=2.8.
Крышечка когтевидная, с резким изломом оси нарастания на ранних ста-
диях роста. Ромбовидный мускульный след занимает большую часть левой
стороны крышечки.
Зубы радулы длиной от 90 до 120 мкм, стилетовидной формы с цилиндриче-
ским основанием, несущим на одной из двух ручек маленькую равнобедренно-
треугольную шпору (рис. 410, 6). Киль достигает х/3 длины лезвия.
Вид сходен с С. decussata, от которого отличается высоким завитком и морфо-
логией зубов радулы, в основном цилиндрической формой базальной части
зуба, а также тем, что у С. conoidea шпора отходит почти от основания зуба,
в то время как у С. decussata она расположена на срединном участке
одной из ручек.
1. CURTITOMA
12T
Распространение. Евразийский арктическо-бореальный вид вы-
сокобореально-атлантического происхождения. Помимо Норвежского моря
отмечен повсеместно в Баренцевом и Белом морях, в северо-восточной части
Карского моря и западной части моря Лаптевых (рис. 443, 2).
Экология. Обитает на глубинах от 1.5 до 820 м, у берегов Северной
Шотландии найдена раковина (без тела) на глубине 1008 м (Odhner, 1915),
однако наиболее часто встречается на
глубинах 20—120 м. Большинство мол-
люсков собрано на илистых и заилен-
ных грунтах. Предпочитают соленость
более 34 ,°/00.
Типы. Типовой материал, вероятно,
хранится в ZMO; типовое местонахожде-
ние — Вадсё, Норвегия.
Просмотрено 153 экз.
9. Curtitoma decussata (Couthouy, 1839),
comb. n. (Рис. 104—108; 389, A—В; 410,
16—33; 411, 1—8; 443, 2, 3; 444).
Couthouy, 1839:183, pl. 4, fig. 8
(Pleurotoma); Gould, 1841 : 280, fig. 185
(Pleurotoma); Stimpson, 1851 : 49 (Man-
gelta); Gould, 1870 : 354, fig. 623 (Bela);
V err ill, 1882:479, pl. 43, fig. 13, (Bela);
Tryon, 1884 : 217, figs. (Bela, part.); F r i e 1 e,
1886 : 12, pl. 8, fig. 11, pl. 10, fig. 12 (Bela,
part.); Martini, Chemnitz, 1887 : 153,
Taf. 32, fig. 6 (Bela); Thorson, 1951 : 61
(Bela); Macpherson, 1971 : 115, pl. 7,
tig. 9, Map. 51 (Oenopota). — tenuicostata [M.
S a r s, 1869 : 259 (Pleurotoma, nomen nudum)]:
G. O. S a r s, 1878 : 237, Tab. 17, fig. 1 (Bela);
V e r r i 1 1, 1882 : 481 (Bela decussata var.);
Tryon, 1884 : 217, pl. 28, fig. 35, 36 (Bela
decussata-vax., part.); Martini, Chemnitz,
1887:150, Taf. 32, fig. 1, 2 (Bela); D a u t-
zenberg, Fischer, 1912 : 49 (Bela); Thor-
son, 1944 : 116 (Bela); Mac Ginitie,
1959 : 134 pl. 16, fig. 4 (Oenopota); Bou-
chet, Waren, 1980 : 73, figs. (Oenopota). —
willei, Friele, 1886:13, pl. 8, fig. 16,17,
pl. 10, fig- 14 (Bela tenuicostata var.); К o-
b e 1 1, 1905 : 254, pl. 84, fig. 6 (Bela).
Рис. 444. Места находок С. decus-
sata в дальневосточных морях.
Раковина белая, овальная, реже
овально-веретеновидная, с 5—6 закруг-
ленными, сильновыпуклыми оборотами. Периостракум очень тонкий, легко-
отстающий, светло-оливкового цвета. Эмбриональная раковина (рис. 389, А—В)
практически сходна с таковой у С. conoidea, но отличается от нее менее выпук-
лыми оборотами. Осевая скульптура дефинитивной раковины представлена
многочисленными валиками или тонкими осевыми складками, разделенными
в 1.5—3 раза более широкими промежутками. Наиболее выражены элементы
осевой скульптуры в верхней части периферии оборота. На основании рако-
вины, как правило, сохраняются следы осевых складок. Элементы осевой скуль-
птуры сигмоидально изогнуты. На последнем обороте их насчитывается от 25
до 34, на предпоследнем от 18 до 23. Спиральная скульптура равновелика на
всей поверхности раковины и представлена многочисленными тонкими ребрыш-
ками. Межреберные промежутки в 1.5—2 раза у-же их. Основание раковины
уплощенное, плавно переходит в короткий сифональный вырост. Устье широко-
обратногрушевидной формы, несет на колумеллярном крае тонкую полосу
138
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
белого каллуса, на палатальном — довольно глубокий анальный синус. Сифо-
нальный канал протяженный, широкий, средней глубины. Высота раковины
типового экземпляра, Pleurotoma decussata, 8.7, ширина 3.7 мм. Наибольшие
размеры (5р=12.5; Д=6.0, мм) указаны для раковины из зал. Массачусетс
(Verrill, 1882).
Крышечка когтевидная, с резко изгибающейся осью на ранних этапах фор-
мирования. Ромбовидный мускульный след расположен вблизи левого края
крышечки (рис. 32, А—D).
Зубы радулы стилетовидной формы, с очень коротким лезвием, иногда не
превышающим длину основания зуба (рис. 411, 1—2). Киль, составляющий
х/4 длины лезвия, занимает дистальную часть зуба. Характер зубов С. decussata
позволяет говорить об их морфологической идентичности с зубами Bela tenui-
costata (Friele, 1886, Tab. 10, fig. 13) и отнести их к одному виду.
Вид сходен с С. lawrenciana морфологией зубов радулы и формой раковины,
но отличается от него наличием многочисленных тонких осевых складок.
Замечания. Значительная изменчивость размеров осевой скульптуры
при небольших размерах раковины повлекла многочисленные ошибки в сино-
нимии вида. Bela tenuicostata и ее вариетет willet на основании сходства морфо-
логических особенностей раковины и зубов радулы следует считать младшими
синонимами С. decussata, в то время как все тихоокеанские виды, такие как
С. hebes, С. lawrenciana и другие, являются номинативными.
Распространение. Тихоокеанский бореально-арктический вид ти-
хоокеанского происхождения. Распространен в Тихом океане, в Японском,
Охотском морях и повсеместно в Беринговом море (рис. 444), найден в Чукот-
ском море, море Лаптевых, в Восточно-Сибирском и Карском морях. Отмечен
в канадском секторе Арктики, от п-ова Фокс до Гудзонова зал. в Лабрадоре,
у берегов Новой Шотландии, зал. Массачусетс и у берегов Западной и Восточной
Гренландии и Северо-Западной Исландии. Найден у берегов Шпицбергена,
Финмаркена и о-вов Новая Земля.
Экология. Обитает на глубинах от 1.5 до 820 м, в Северной Атлантике
отмечен на глубине до 2 582 м (Thorson, 1944). Однако наибольшее число нахо-
док связано с глубинами 20—120 м. Моллюски предпочитают илистые и заилен-
ные грунты, на всех остальных типах грунтов было обнаружено менее 20 %
исследованных особей. Вид встречен при температурах от —1.98 до 4.0 °C,
однако температурный оптимум для него составляет от —1.8 до —1.0 °C. Как
правило, моллюски обитают в водах с высокой океанической соленостью
34.5—35.0 °/00, тем не менее отмечено 2 находки в Восточно-Сибирском море
при солености 29.13 и 24.76 %0.
Типы. Типы Pleurotoma decussata хранятся в музее BSNH; типовое место-
нахождение — зал. Массачусетс. Синтипы Bela tenuicostata — в ZMO (Bouchet,
Waren, 1980), типовое местонахождение — Lofoten, Норвегия. Синтипы
Р. wlllei — в ZMB; типовым местонахождением считается Западный Шпицбер-
ген, глубина 1203 м (Bouchet, Waren,. 1980).
Просмотрено 368 экз.
10. Curtitoma lawrenciana (Dall, 1919), comb. n. (Рис. 109—111; 411, 9—13;
445).
Dall, 1919a : 43, pl. 15, fig. 6 (Lora; pro tenuilirata sensu Krause, 1886). — nazanensis
Dall, 1919a : 45, pl. 15, fig. 8 (Lora), syn. n.; Mac Giniti e, 1959 : 134, pl. 16, fig. 3
(Oenopota), — tenuilirata (non Dall, 1871): Krause, 1886 : 274, pl. 18, fig. 8 (Bela); Ma c-
<1 i n i t i e, 1959 : 132, pl. 16, fig. 1 (Obesotoma).
Раковина телесного цвета, веретеновидная, с 4.5—5.5 оборотами отлогоуг-
ловатой формы. Периостракум тонкий, коричневатых или рыжеватых тонов.
Протоконх не исследован. Осевая скульптура дефинитивной раковины пред-
ставлена многочисленными пологими складками, разделенными в 4—6 раз
1. CURTITOMA
129
более узкими, чем их ширина, промежутками. Складки ярко выражены в верх-
ней части оборота и сглаживаются на основании раковины. Видоспецифичным
признаком является характерная сигмовидная изогнутость элементов осевой
скульптуры. Изгиб приходится на плечо оборота, в результате на околошовной
поверхности складки наклонены вперед, а на периферии оборота назад.
На последнем обороте раковины насчиты-
вается от 20 до 26 складок, на предпо-
следнем от 16 до 22. Спиральная скульп-
тура представлена преимущественно шну-
ровидными, средними ребрами, наиболь-
шая их ширина приходится на центральную
часть периферии оборота. Межреберные
промежутки в 6—10 раз уже шнуровид-
ных ребер. Околошовная поверхность
покрыта волосовидными ребрышками. Ос-
нование раковины уплощенное, граница
его перехода в короткий сифональный
вырост, как правило, обозначена слабо.
Устье узко-обратногрушевидной формы,
с широкой полосой толстого бежевого цве-
та каллуса на колумеллярном крае и не-
большой выемкой анального синуса на па-
латальном. Сифональный канал узкий,
глубокий. Высота раковины, описанной
Краузе, 15.5 мм. Наибольший известный
экземпляр имеет размеры (мм): В =17.4;
Д=8.8 (MacGinitie, 1959).
Крышечка с плавно изгибающейся осью
нарастания. Мускульный отпечаток эллип-
соидной формы, занимает большую часть
левой стороны крышечки, но не прилегает
к левому краю.
Зубы радулы длиной до 140 мкм, сти-
летовидной формы. Одна из ручек сильно
редуцирована, другая несет треугольную
уплощенную шпору (рис. 411, 10). Киль
достигает 1/,1 общей длины зуба.
Вид сходен с С. decussata, от которого
отличается наличием относительно немно-
гочисленных (18—22) пологих средних
осевых складок. Рис. 445. Места находок Curtitoma
Замечание. Долл (Dall, 1919а) lawrenciana.
описал сходный по морфологии рако-
вины вид Lora nazanensis, отличающийся формой устья (полуовальной, углова-
той в верхней части) и более выраженными осевыми складками. Мной найдены
все переходы от формы, вписывающейся в Lora nazanensis, до характерной для
С. lawrenciana. Практически полное соответствие морфологии зубов у всех
изученных экземпляров и рисунки радул Моррисона (присланные Дж. Роузво-
тером) позволяют говорить об идентичности долловских видов nazanensis
и lawrenciana не только по скульптуре раковины, но и по морфологии зубов
радулы.
Распространение. Тихоокеанский широко распространенный бо-
реальный вид. Обитает в низко- и высокобореальных районах у берегов Азии
и Америки и выходит в Северный Ледовитый океан. Многочисленны находки
в Чукотском и Беринговом морях, а также у берегов Восточной и Южной Кам-
чатки. Кроме того, вид отмечен у Южных Курильских о-вов и в Приморье
(Японское море).
9 И. П. Богданов
130
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Экология. Обитает на глубинах от 4 до 380 м, наибольшая встречае-
мость приходится на глубины 20—45 м.
Типы. Типовой экземпляр Lora lawrenciana хранится в Stuttg. RC; типо-
вое местонахождение — Plover Вау. Голотип L. nazanensis хранится в USNM,.
№ 220327; типовое местонахождение — Nazan Вау, о-в Атка, Алеутские о-ва..
Просмотрено 75 экз.
11. Curtitoma hokkaidoensis (Bartsch, 1941), comb. n. (Рис. 117—120; 426,.
10—12).
Bartsch, 1941 : 9, pl. 1, fig. 1 (N ematoma)', О k u t a n i, 1964 : 431, pl. 7, fig. 9?
(N ematoma); 1966 : 24, pl. 2, fig. 8, 9 (N ematoma); Powell, 1966 : 121, pl. 19, fig 10 (A'o-
dotomd}.
Раковина белая, слегка коричневатая в верхней части оборотов, веретено-
видная, с 4.5—5.5 закругленными, умеренно выпуклыми оборотами. Перио-
стракум тонкий, прозрачный, легко отстающий от поверхности раковины.
Верхние обороты, как правило, корродированы, и скульптура эмбриональной
раковины не сохраняется. Осевая скульптура дефинитивной раковины пред-
ставлена пологими средними складками, которые имеют на околошовной по-
верхности сигмоидальный изгиб, а на периферии оборота отклонены назад..
Наиболее выражены складки в центральной части периферии оборота; сглажи-
ваются в двух направлениях: наверху — возле шва, внизу — в верхней х/3 ос-
нования раковины. Разделены они промежутками, в 1.5 раза меньшими или
реже равными по ширине складкам. Численность осевых складок в значитель-
ной мере варьирует у различных экземпляров, и на последнем обороте рако-
вины их насчитывается от 16 до 24, на предпоследнем от 14 до 18. Спиральная,
скульптура равновелика на всей поверхности оборотов, основания раковины:
и на сифональном выросте и представлена многочисленными тонкими нитевид-
ными ребрами. Межреберные пространства немногим у'же самих ребер или равны
им по ширине. Основание раковины выпуклое, сифональный вырост удлинен-
ный. Устье широко-обратногрушевидной формы, с широкой, относительно глу-
бокой выемкой анального синуса на палатальном крае. Колумеллярный край<
несет полосу тонкого, телесного цвета каллуса. Сифональный канал протяжен-
ный, широкий, умеренной глубины. Высота типового экземпляра 9.3 при ши-
рине 5.3 мм. Высота раковины наибольшего исследованного экземпляра 11.6 мм..
Зубы радулы до 160 мкм, стилетовидные, с коротким конусовидным основа-
нием, с 2 тонкими ручками, одна из них вооружена очень мелкой шпорой;
(рис. 426, 10—12). Киль уплощенный, превышает J/4 общей длины зуба.
Вид сходен с С. microvoluta, от которого отличается более уплощенной ра-
ковиной, небольшим сифональным выростом и осевыми складками, имеющими-
более сильный сигмоидальный изгиб.
Распространение. Тихоокеанский приазиатский низкобореальный
вид. Встречен от зал. Сагами до берегов о-ва Хоккайдо, у южных берегов о-ва
Сахалин, у берегов Приморья и в Татарском проливе.
Экология. Обитает на глубинах от 160 (Приморье) до 1400 (зал. Са-
гами) м, в основном на заиленных грунтах, реже с примесью гальки и песка.
Тип ы. Типовой экземпляр хранится в USNM, № 204951; типовое место-
нахождение — к юго-востоку от о-ва Хоккайдо (Albatross, st. 5036), глу-
бина 848 м.
Просмотрено 7 экз.
12. Curtitoma microvoluta (Okutani, 1964), comb. n. (Рис. 116).
Okut ani, 1964 : 431, pl. 6, fig. 10 (N ematoma); 1966 : 24, pl. 2, fig. 10—11 (Nematoma).-
Раковина светло-коричневая, веретеновидная, очень тонкостенная, с 3.5—
4.5 закругленными, умеренно выпуклыми оборотами. Периостракум тонкий,,
прозрачный, легко отстающий. Верхние обороты раковины сильно корродиро-
1. CURTITOMA'
131
ваны, протоконх неизвестен. Осевая скульптура дефинитивной раковины пред-
ставлена выпуклыми, треугольными в сечении, средними складками, исчезаю-
щими лишь в нижней трети основания раковины и сигмоидально изогнутыми
на околошовной поверхности. Разделены складки в 2 раза более узкими, чем
их ширина, промежутками. На последнем обороте раковины насчитывается
от 16 до 18 осевых складок, на предпоследнем от 12 до 14. Спиральная скуль-
птура состоит из многочисленных, равновеликих на всей поверхности раковины
тонких нитевидных ребер, разделенных вдвое меньшими по ширине межребер-
ными промежутками. Основание раковины уплощенное, сифональный вырост
.длинный. Устье полуовальное, угловатое в верхней части (там, где расположен
глубокий вырез анального синуса). Розоватый каллус лежит широкой полосой
на колумеллярном крае. Сифональный канал протяженный, глубокий, очень
•широкий. Отношение высоты раковины к ширине (ВрхД) у моллюсков, собран-
ных в зал. Сагами, было следующим (мм): 9.8x4.5 и 10.0x4.9. Высота рако-
вины, найденной в Северном Приморье, 9.7 мм. Крышечка и радула не иссле-
дованы.
Вид хорошо отличается от остальных представителей рода сильно вытяну-
тым устьем узко-обратногрушевидной формы, превышающим х/2 общей высоты
'раковины.
Распространение. Тихоокеанский приазиатский низкобореальный
вид. Встречен в зал. Сагами, у о-ва Мияке (34°05.5' с. ш.—139°29.3' в. д.)
и в Северном Приморье у о-ва Аскольд.
Экология. К сожалению, имеются данные только по вертикальному рас-
пределению. В зал. Сагами вид был обнаружен на глубине 710—770, у о-ва Ми-
яке на 1230—1350, у о-ва Аскольд на 269—340 м.
Т и п ы. Типовой экземпляр хранится, вероятно, в Токийском университете
или в RFRL, Tokay; типовое местонахождение — зал. Сагами, 35°05.5' с. ш.—
139°29.3' в. д., глубина 710—770 м.
Исследован 1 экз.
13. Curtitomabartschi (Bogdanov, 1985) comb. n. (Рис. 113—115; 409,16—17).
Богданов, 1985a : 452, рис. 1, ф—ц, рис. 2, 6 (Oenopota).
Раковина светло-коричневая, овально-веретеновидная, очень тонкостенная,
с 4—5 закругленными, слабовыпуклыми оборотами. Периостракум тонкий,
прозрачный. Эмбриональные обороты не сохранились. Осевая скульптура дефи-
нитивной раковины состоит из многочисленных валикообразных линий нара-
стания, которые пересекаются тонкими волосовидными ребрышками, так что
поверхность периферии оборота приобретает струйчатый характер. Спиральные
ребрышки многочисленны, равновелики и равноудалены друг от друга на пери-
ферии оборотов, основании раковины и сифональном выросте. Основание рако-
вины прямое, незаметно переходящее в сифональный вырост. Устье полуоваль-
ной формы, несет на колумеллярном крае тончайший беловатый каллус. В верх-
ней части палатального края расположена небольшая выемка анального синуса.
Сифональный канал широкий, глубокий. Размеры голотипа (мм): В =9.6;
Д=5.1; Впо=1.1- /?у=5.9; Ду=2.4.
Зубы радулы кинжаловидные (рис. 409, 16—17), небольших размеров:
в длину достигают 245 мкм; средняя ширина лезвия 19 мкм; базальная часть
зуба округлая.
Вид сходен с С. hebes, от которого отличается более удлиненной формой
раковины, слабо развитой осевой скульптурой й широким округлым основанием
зуба радулы.
Распространение. Тихоокеанский приазиатский высокобореаль-
;ный вид. Найден у берегов о-ва Уруп в северной и западной частях Охотского
моря.
9*
132
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Типы. Голотип, хранится в ЗИН АН СССР, № 37977; типовое местона-
хождение — у побережья о-ва Уруп близ м. Китового, глубина 150 м, на песча-
нистом грунте с частыми выходами скал.
Просмотрено 6 экз.
14. Curtitoma piltuniensis (Bogdanov, 1985), comb. n. (Рис. 112; 411, 19—22)..
Богданов, 1985a : 451, рис. 1, и—м; 2, 3 (Oenopota).
Раковина сероватая, почти белая, овальная, тонкостенная, с 3.5 закруглен-
ными, умеренно выпуклыми оборотами. Периостракум пепельного цвета, очень-
тонкий, прозрачный. Эмбриональные обороты не сохранились. Осевая скульп-
тура дефинитивной раковины представлена сигмоидально изогнутыми линиями
нарастания. Спиральная скульптура как на периферии, так и на основании ра-
ковины состоит из тонких нитевидных равновеликих ребрышек, расстояния
между которыми равны им по ширине. Умеренно выпуклое основание рако-
вины плавно переходит в короткий прямой сифональный вырост. Устье полу-
овальной формы, с широкой полосой плотного матового каллуса на колумел-
лярном крае. В верхней части палатального края залегает неглубокая вырезка,
анального синуса. Сифональный канал глубокий и очень широкий (ширина,
около 1.5 мм). Размеры голотипа (мм): В =7.7; Д=5.2; 5ПО=6.5; В =5.2;
Ду=2.7.
Зубы радулы маленькие, длиной до 140 и шириной до 10.8 мкм, стилетовид-
ные; базальная часть вытянутая, узкая (рис. 411, 19—22).
Вид морфологически сходен с Lora quadra Dall, 1919, но отличается более-
вытянутым основанием и устьем, а также слабо развитыми элементами осевой и
спиральной скульптур. Qi Bela decussata var. inflata Posselt et Jensen, 1898 отли-
чается более низким завитком и слабо развитой осевой скульптурой. Радула
схожа с Oenopota pyramidalis (Strom, 1782), но виды сильно различаются
формой и скульптурой раковины.
Распространение. Тихоокеанский приазиатский высокобореаль-
ный вид. Известен только из типового местонахождения.
Типы. Голотип (единственный экземпляр) хранится в ЗИН АН СССР;
№ 37981; типовое местонахождение — Восточный Сахалин, пос. Пильтун,
глубина 5 м.
15. Curtitoma becklemishevi Bogdanov, 1989. (Рис. 60—62; 390, Д—Е; 407„
14-20).
Богданов, 1989а : 134, рис. 1, 1—2; 2, а—в.
Раковина светло-серая, тонкостенная, овально-веретеновидная, с 4—4.5 обо-
ротами отлогоугловатой формы. Периостракум тонкий, сероватый. Эмбриональ-
ная раковина выпуклой формы, ее скульптура состоит из многочисленных
(более 15) спиральных нитевидных ребрышек. Осевая скульптура дефинитив-
ной раковины представлена линиями нарастания и тонкими осевыми складками,,
выраженными в различной мере. Чаще всего осевые складки слабо развиты.
Как складки, так и линии нарастания имеют на околошовной площадке сигмо-
идальный изгиб, соответствующий по форме изгибу анального синуса. Спираль-
ная скульптура представлена на дефинитивной раковине шнуровидными спи-
ральными ребрами, равномерно расположенными на всей поверхности рако-
вины. Основание раковины плавно переходит в удлиненный сифональный вы-
рост. Устье обратногрушевидной формы, с тонким, слегка бежеватым каллусом
на большей части колумеллярного края. Наибольшая высота раковины до-
10 мм. Размеры голотипа (мм): 5р=7.7; Д=3.9.
Зубы радулы кинжаловидной формы, длиной до 300 мкм, несут большой,
до 1/2 длины лезвия киль. Основание короткое, массивное, шпора треугольной
формы (рис. 407, 14—20).
2. GRANOTOMA
133
Вид конхологически сходен с С. hebes, от которого отличается округлым ос-
нованием раковины, формой устья и удлиненным сифональным выростом.
Распространение. Тихоокеанский приазиатский широко распро-
страненный бореальный вид. Помимо типового местонахождения известны на-
ходки в Татарском проливе и у берегов Приморья.
Типы. Голотип хранится в ЗИН АН СССР, № 48467; типовое местонахож-
дение — Охотское море, 51°10' с. ш.—154°17' в. д., глубина 583 м.
Просмотрено 14 экз.
16. Curtitoma neymanae Bogdanov, 1989. (Рис. 79—82; 411, 14—18).
Богданов, 19335 : 147, рис. 1, 1—3; 2, а—г.
Раковина светло-бежевая или кремовая, веретеновидная, тонкостенная,
с 4.5—5 оборотами отлогоугловатой формы. Очень тонкий, прозрачный перио-
стракум плотно прилегает к раковине. Эмбриональная раковина не изучена.
Осевая скульптура дефинитивной раковины представлена сигмоидально изо-
гнутыми тонкими осевыми складками, наиболее выраженными в верхней части
оборота. Они постепенно сглаживаются на периферии и практически исчезают
на основании раковины. Промежутки, разделяющие тонкие пологие осевые
складки, превышают их по ширине в 2—3 раза. Спиральная скульптура дефи-
нитивной раковины представлена развитыми в различной мере элементами.
На периферии оборота лежат 3 спиральных тонких ребра, одно из которых про-
ходит по плечу оборота, другое обозначает нижнюю треть периферии. Третье
ребро лежит на стыке с последующим оборотом, в результате чего создается впе-
чатление о наличии двойного шва между оборотами. Между 3 описанными тон-
кими ребрами, а также на околошовной площадке развиваются многочисленные
равновеликие шнуровидные ребрышки. Основание раковины и сифональный
вырост орнаментированы развитыми в различной степени тонкими спиральными
ребрами, как правило, равными по величине 3, описанным для периферии обо-
рота. Слабовыпуклое основание раковины плавно переходит в короткий сифо-
нальный вырост. Устье неправильно-овальное, усеченное в верхней части, несет
на колумеллярном крае толстый матовый каллус. Наибольшая высота рако-
вины 7 мм.
Зубы радулы маленькие, их длина не превышает 100 мкм, стилетовидные
(рис. 411, 14—18). Базальная часть тонкая, цилиндрическая; шпора малень-
кая, треугольная.
Вид наиболее сходен с инфраподвидовой формой С. violacea, var. bicarinata,
от которой отличается более удлиненной раковиной с пологими тонкими осе-
выми складками; наличием на околошовной площадке равновеликих нитевид-
ных ребрышек, одинаковыми по величине элементами спиральной скульптуры
как на периферии, так и на основании раковины.
Распространение. Тихоокеанский приазиатский широко распро-
страненный бореальный вид. Встречен в Северном Приморье и у берегов Запад-
ной Камчатки.
Типы. Голотип хранится в ЗИН АН СССР, № 48468; типовое местона-
хождение — Японское море, зал. Чихачева, близ пос. Де-Кастри, глубина
0.1 м.
Просмотрено 5 экз.
2. Род GRANOTOMA Bartsch, 1941
Bartsch, 1941 : 5; Богданов, 1986 : 45 {Oenopota subg.). — Oenopota (part.):
Morch, 1852:73; Dall, 1918:329; Powell, 1966:121; Bouchet, Waren,
1980 : 66. — Bela (non Gray, 1847): Dall, 1887 : 300 (part.).
Типовой вид Bela krausei Dall, 1886b, по первоначальному обозна-
чению.
134
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Раковина небольшая, средняя ее высота 5—8 мм, овально-веретеновидной
формы, очень тонкостенная. Эмбриональная раковина выпуклая, с частично
наклоненной относительно оси навивания дефинитивной раковины осью про-
токонха. Скульптура эмбриональной раковины представлена 5—7 нитевидными
спиральными ребрышками одинаковой величины, которые разделяются про-
межутками, в 4—5 раз превышающими ширину ребрышек. Элементы спираль-
ной скульптуры пересекаются в 5—7 раз более тонкими осевыми складками, в ре-
зультате чего рисунок, покрывающий поверхность протоконха, приобретает
ячеистый вид. Вся поверхность эмбриональной раковины усеяна многочислен-
ными микроскульптурными бугорками, причем основными местами их кон-
центрации являются элементы осевой и спиральной скульптур.
Значительное развитие на дефинитивной раковине элементов осевой и спи-
ральной скульптур приводит к тому, что ширина промежутков между ними зна-
чительно (в 4—10 раз) сокращается. В результате рисунок скульптуры рако-
вины становится сходным с орнамёнтацией кукурузного початка. Сифональный
вырост короткий и широкий. Крышечка когтевидная, с плавно изгибающейся
осью нарастания; мускульный след чаще всего расположен вблизи ее левого
края.
Зубы радулы стилетовидные, как правило, не превышают 150 мкм в длину.
На базальной части зуба развивается только 1 ручка, несущая шпору.
Род распространен в бореальных водах Тихого океана (единичные экземп-
ляры отмечены также в Чукотском море). Достоверные палеонтологические
находки неизвестны.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Осевые складки широкие, пологие, не более 15 на последнем обороте
раковины .......................................3. G. raduga (Bogdanov)
2 (1). Осевые складки средние, уплощенные, многочисленные (от 19 до 44 на
последнем обороте).
3 (4). Осевые складки трапециевидные или квадратные в сечении. Зубы радулы
с характерным гарпуновидным килем...................1. G. krausei (Dall)
4 (3). Осевые складки полуовальные в сечении. Зубы радулы с килем, не обра-
зующим гарпуновидный вырост.....................2. G. albrechti (Krause)
1. Granotoma krausei (Dall, 1886). (Рис. 128-135; 393, B—E-, 413, 1—16;
446, 2).
Dall, 1886b : 301, pl. 4, fig. 4. (Bela); 1919a : 46, pl. 15, fig. 3 (Lora); Oldroyd,
1927 : 107, pl. 7, fig. 7 (Lora); Bartsch, 1941 : 5, pl. 1, fig. 9; Powell, 1966 : 121,
pl. 19, fig. 11 [Oenopota (Funitoma)].
Раковина белая, веретеновидной формы, с 5—6 закругленными, умеренно
выпуклыми оборотами. Периостракум тонкий, полупрозрачный, легкоотстаю-
щий, зеленовато-желтый или коричневатый. Протоконх сильновыпуклый.
Первый его оборот лишен макроскульптурных образований и равномерно по-
крыт микроскульптурными бугорками. В начале второго оборота протоконха
закладывается 6 спиральных нитевидных ребрышек, из которых 3 нижних одно
за другим покрываются последующим оборотом. На элементах как осевой,
так и спиральной скульптуры эмбриональной раковины происходит концент-
раций микроскульптурных бугорков (рис. 393, Д), сохраняющаяся и на пост-
эмбриональных элементах скульптуры. Осевая скульптура дефинитивной ра-
ковины представлена многочисленными, прямоугольными в сечении, тонкими
складками, равновеликими на всей поверхности оборотов и основания раковины.
Промежутки, разделяющие их, в 2—4 раза уже складок; последние имеют на
околошовной поверхности сигмоидальный изгиб. На последнем обороте насчи-
тывается от 26 до 38 осевых складок, на предпоследнем от 20 до 28. Спиральная
скульптура представлена тонкими шнуровидными ребрами, равновеликими
2. GRANOTOMA
135
практически на всей поверхности раковины, за исключением сифонального вы-
роста. Последний покрыт близко расположенными многочисленными нитевид-
ными ребрышками. Ребра разделены в 3—4 раза более узкими, чем их ширина,
межреберными промежутками. Одноразмерность элементов осевой и спираль-
ной скульптур придает поверхности раковины ячеистый вид. Основание рако-
вины умеренно выпуклое, граница его перехода в сифональный вырост средних
размеров всегда отмечена желобком. Устье
узко-обратногрушевидной или полуоваль-
ной формы, его верхняя часть угловатая.
Колумеллярный край покрыт тонким, те-
лесного цвета каллусом, в верхней части
палатального края расположен довольно
глубокий анальный синус. Сифональный
канал глубокий, средней ширины. Раз-
меры типового экземпляра (мм): JBjt)=9.0;
Д=3.2; В =АД. Наибольшие .размеры
раковины были отмечены для экземпляра,
найденного к юго-востоку от о-ва Ас-
кольда на глубине 80 м. Его параметры
следующие (мм): 73р=1О.О; Д=4.8; Вп0=
=6.8; ^=5.3; Д>2.1.
Зубы радулы стилетовидной формы
(рис. 413), с широкой базальной частью
и характерным гарпуновидным
Флажок петлевидной формы, расположен
на единственной развитой ручке. Отмеча-
ется практически полное сходство иссле-
дованного мной материала с изображени-
ями зубов, полученными от Моррисона
(исследован экземпляр, определенный
Доллом, номер по каталогу 224024), за
исключением более уплощенной базальной
части у последних.
Вид сходен с G. albrechti, от которого
отличается удлиненной раковиной с менее
рельефной скульптурой; морфологией зу-
бов, в основном гарпуновидной формой
киля.
Распространение. Тихооке-
анский широко распространенный бо-
реальный вид, обитающий в низко- и вы-
сокобореальных районах у берегов Азии
и в высокобореальных районах у бере-
гов Америки. Обнаружен в Японском
море в зал. Петра Великого, в Татарском проливе, в Охотском море у берегов
Южной Камчатки, в Беринговом море в бухте Провидения и у о-ва Беринга
(рис. 446, 2). У берегов Америки распространение не изучено, указан для вод,
омывающих Аляску.
Экология. Обитает на глубинах от 17 до 140 м на заиленных и песчани-
стых грунтах.
Типы. Типы хранятся в USNM (в работах Долла номера голотипа нет);
типовое местонахождение — Порт-Этчез, Аляска.
Просмотрено 43 экз.
Рис. 446. Места находок Granotoma
albrechti (7) и G. krausei (2) в мо-
рях СССР.
килем.
2. Granotoma albrechti (Krause, 1885), comb. n. (Рис. 39; 122—127; 391,
A—E; 392, Д—Е; 393, А—Б; 412, 1—22; 446, 7).
136
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Krause, 1885 : 276, pl. 18, fig. 3, 11 (BeZa); Dall, 1886b : 300, pl. 4, fig. 1 (BeZa);
Martini, Chemnitz, 1887 : 233, Taf. 42, fig. 3 (BeZa); Oldroyd, 1927 : 106, <
pl. 11, fig. 5 (Bora); Голиков, Гульбин, 1977 : 236 (Oenopota).
Раковина беловатая, реже со слабым зеленоватым оттенком, овально-вере-
теновидной формы, с 4—5 закругленными, умеренно выпуклыми оборотами.
Периостракум тонкий, легкоотстающий, коричневатый или желтовато-серый,
полупрозрачный. Эмбриональная раковина выпуклой сосцевидной формы
(рис. 391). Первый оборот протоконха практически весь лишен макроскульп-
турных элементов, но равномерно покрыт микроскульптурными бугорками.
Уже на исходе первого эмбрионального оборота отмечается закладка спираль-
ных нитевидных ребрышек, которые на втором обороте протоконха резко
увеличиваются в размерах. На втором обороте происходит закладка осевых
складок, которые в 4—7 раз тоньше пересекаемых ими спиральных элементов
скульптуры протоконха. С момента закладки нитевидных ребрышек и осевых
складок все микроскульптурные бугорки концентрируются на элементах макро-
скульптуры, в результате чего последние приобретают гранулированный вид.
Осевая скульптура дефинитивной раковины представлена средними складками,
чаще полуовальными или имеющими в сечении вид равнобедренного треугольника.
Промежутки, разделяющие их, в 3—7 раз уже самих складок. Выраженность
складок практически одинакова на всей поверхности оборотов и на основании
раковины. На околошовной поверхности складки имеют сигмоидальный изгиб.
На последнем обороте раковины насчитывается от 20 до 38 складок, на пред-
последнем от 17 до 34. Спиральная скульптура состоит из многочисленных тон-
ких ребрышек, равновеликих на всей поверхности раковины и разделенных
в 4—5 раз более узкими, чем их ширина, межреберными промежутками. Одно-
размерность осевой и спиральной скульптур придает поверхности раковины
ячеистый вид. Основание раковины сильно уплощенное, плавно переходящее
в средних размеров сифональный вырост. Устье полуовальной, реже непра-
вильно-овальной формы, угловатое в верхней части. Тонкий белый каллус
покрывает широкой полосой колумеллярный край устья; в верхней части пала-
тального края расположен неглубокий синус. Сифональный канал широкий
и глубокий. Размеры голотипа (мм): 75^=10, Д=Ъ мм; длина зуба 0.15 мм.
Максимальные размеры раковины (мм) — Бр=10.2; Д=5.5; 5mo=7.2; В,=Ъ.8;
Ду=2.3 отмечены для экземпляра из Охотского моря. В литературе (Dall,
1886b) приводится максимальная высота раковины — 11.2 мм — для моллюска
из района Берингова пролива (Port Clarence).
Крышечка когтевидная, яйцевидной формы, с тонкой полосой мускульного
следа, проходящей вдоль плавно изгибающейся оси нарастания.
Зубы радулы стилетовидной формы (рис. 412), с редуцированным килем
на дистальном участке лезвия. Единственная развитая ручка базальной части
несет маленькую треугольную шпору.
Вид наиболее сходен с G. krauset, от которого отличается более приземистой
формой раковины и зубами радулы с цилиндрической формой основания.
Распространение. Тихоокеанский широко распространенный бо-
реальный вид, обитающий в низко- и высокобореальных районах, преимущест-
венно у берегов Азии. Обнаружен в Северном и Южном Приморье, у Японского
и Охотоморского побережий о-ва Сахалин, в северной части Охотского моря
и у Средних Курильских о-вов. В Беринговом море встречается от Восточной
Камчатки до Берингова пролива. Найден в Чукотском море близ о-ва Врангеля
(рис. 446, 1).
Экология. Обитает на глубинах от 3 до 900 м, однако максимум числен-
ности приходится на глубины 40—120 м. Моллюски встречены на всех типах
грунтов, но более 60 % находок пришлось на песчанистые грунты. Обнаружен
при температурах от —1.23 до 11.2 °C. Представители вида встречаются в диа-
пазоне солености от 32.82 до 34.14°/00, не найдены в водах, опресненных ниже
31.11 »/00.
3. OENOPOTA
137
Типы. Голотип (коллекционный номер не указан) хранится в Stuttg. RC;
типовое местонахождение — Plower Вау (Берингов пролив).
Просмотрено 354 экз.
3. Granotoma raduga (Bogdanov, 1985), comb. n. (Рис. 121; 392, A—Г; 411,:
23—27).
Богданов, 1985a : 451, рис. 1, д—з; 2, 2 (Oenopota);
Раковина кремового цвета, тонкостенная, овально-веретеновидная, с 3—,
4 закругленными, сильновыпуклыми оборотами с канальчатым швом. Перио-
стракум тонкий, прозрачный. Протоконх округлой формы. Первые полобо-’
рота протоконха покрыты многочисленными, беспорядочно расположенными
микроскульптурными бугорками. Последующие участки протоконха имеют
ячеистый вид благодаря равномерно расположенным элементам осевой и спи-
ральной скульптур (рис. 392, А). Осевая скульптура дефинитивной раковины
состоит из почти прямых, слегка наклоненных назад средних складок» сигмо-
, видно изогнутых на околошовной поверхности. Складки наиболее выражены
на периферии оборота и сглаживаются в нижней 1/3 основания раковины.
На околошовной площадке, где спиральная скульптура выражена слабо, за-
метны многочисленные волосовидные утолщения линии нарастания. Складки
разделены равными им по ширине пространствами. На последнем обороте их
насчитывается от 19 до 22, на предпоследнем от 14 до 18. Не периферии оборо-
тов и основании раковины спиральная скульптура сильно развита и имеет вид
уплощенных равновеликих тонких ребер, ширина которых в 2—2.5 раза пре-
вышает ширину разделяющих их промежутков. Основание раковины выпук-
лое. На прямом сифональном выросте спиральные ребрышки многочисленны,
равновелики и равноудалены, они примерно в 2 раза меньше ребрышек перифе-
рии оборота. Устье широКо-обратногрушевидной формы, с толстым матовым
каллусом на почти прямом колумеллярном крае. Сифональный канал узкий
и глубокий. Размеры голотипа (мм): В =10.3; Д=5.9; 5ИО=7.6; В =5.8;
Д#=2.7 мм.
Зубы радулы маленькие, их длина достигает у голотипа 130 мкм, узкие
(ширина лезвия около 7.5 мкм), стойким цилиндрическим основанием (рис. 411,
23—27). Развита только 1 ручка, несущая маленькую треугольную шпору.
Вид хорошо отличается от других видов рода приземистой формой и изящ-
ной скульптурой дефинитивной раковины.
Распространение. Тихоокеанский приазиатский высокобореаль-
ный вид. Обитает в Охотском море у берегов Восточного Сахалина и в Беринго-
вом море у берегов Восточной Камчатки.
Экология. Отмечен на глубинах от 115 до 925 м, на илистых и заилен-
ных грунтах.
Типы. Голотип хранится в ЗИН АН СССР, № 37975; типовое местонахож-
дение — бухта Русская, Восточная Камчатка, глубина 115 м, на мягких илах.
Просмотрено 8 экз. (в том числе голотип).
3. Род OENOPOTA Morch, 1852
Mor ch, 1852 : 73; Adams, Adams,. 1858 : 654 (part.); Dall, 1918 : 329
(part.; =Bela, Lora)‘, 1919a : 40 (part.); Bartsch, 1941:3 (part.; =Pleurotomoides);
Powell, 1966 : 121 (part.); Bouchet, Waren, 1980 : 66 (part.); Богданов,
1986 : 45 (Oenopota subg.). — Bela (non Gray, 1847): G. 0. S a r s, 1878 : 222 (part.);
F r i e 1 e, 1886 : 2 (part.); Martini, Chemnitz, 1887 : 137 (part.); Dall, 1918 :
318. — Lora (non Gistel, 1848): Dall, 1918 : 328; 1919a : 39 (part.); Oldroyd, 1927 : 95
(part.); Bartsch, 1941 : 3; Powell, 1966 : 143.
Типовой вид Fusus pleurotomarius Couthouy, 1838=7?uccmum pyrami-
dale Strom, 1788, по последующему обозначению (Dall, 1918).
138
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Раковина веретеновидной формы, средних или крупных размеров (до 31 мм
высотой), как правило, толстостенная. Эмбриональная раковина уплощена
в верхней части, или ее макушка погружена в первый полуоборот протоконха.
Характерной чертой эмбриональной раковины служит в различной мере выра-
женный ретикулятный орнамент скульптуры первого оборота протоконха.
Число элементов спиральной скульптуры последнего оборота протоконха ко-
леблется у разных видов от 2 до 4, изредка до 5 ребрышек. Осевая скульптура
закладывается на постэмбриональном обороте в виде тонких осевых складочек.
На дефинитивной раковине она представлена средними и широкими осевыми
складками. Спиральная скульптура дефинитивной раковины развита значи-
тельно слабее осевой и, как правило, в сечении имеет прямоугольную форму.
Основание раковины выпуклое, сифональный канал вытянут.
Крышечка имеет каплевидную форму, ось нарастания чаще всего без резкого
изгиба на ранних стадиях формирования. Левый край крышечки очень мощный,
утолщенный. Мускульный отпечаток лежит в срединной части или прилегает
к левому краю, как правило, заострен в верхней части.
Род распространен в бореальных и арктических водах северного полушария.
В ископаемом состоянии представители рода известны с плиоцена (Япония,
Англия, Исландия).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (6). Зубы радулы мелкие, не более 130 мкм, стилетовидной формы, киль
редуцирован.
2 (3). Раковина с усеченно-конусовидными оборотами. Равновеликие эле-
менты осевой и спиральной скульптур при пересечении образуют узелки,
придающие раковине чешуйчатый рисунок ...............................
...................................... 2. Oe. (Oe.) pingelii (Moller)
3 (2). Раковина с закругленными или отлогоугловатыми оборотами. Скульп-
тура не придает раковине чешуйчатого рисунка.
4 (5). Раковина с отлогоугловатыми оборотами, уплощенными в верхней части.
Осевая скульптура представлена средними складками, имеющими на
периферии оборота наклон назад и преломляющимися на плече оборота
.......................................3. Oe. (Oe.) obliqua (G. О. Sars)
5 (4). Раковина с закругленными, умеренно выпуклыми оборотами. Осевая
скульптура представлена в различной мере выраженными складками,
имеющими плавный сигмоидальный изгиб в верхней Ч2 периферии обо-
рота ..................................1. Oe. (Oe.) pyramidalis (Strom)
6(1). Зубы радулы более 140 мкм, часто очень крупные (200—800 мкм),
копьевидной формы, киль ярко выражен, занимает до длины лезвия
зуба.
7 (8). Осевая скульптура выражена слабее спиральной .................
......................................10. Oe. (N.) casentina (Dall)
8 (7). Осевая скульптура преобладает над спиральной.
9 (18). Осевая скульптура раковины представлена широкими, полуовальными
в сечении складками, превышающими в 3—-7 раз разделяющие их про-
межутки.
10 (15). Спиральная скульптура представлена сплющенными шнуровидными
ребрами, разделенными узкими желобками.
11 (12). Осевые складки расположены далеко друг от друга, на последнем
обороте их не более 11.....................14. О. (N.) biconica Bogdanov
12 (И). Осевые складки расположены близко друг к другу, на последнем
обороте их более 18.
13 (14). Осевые складки пересечены 2—4 очень глубокими желобками. Зубы
радулы изогнутые...........................4. Oe. (N.) impressa (Morch)
14 (13). Осевые складки не перерезаны спиральными глубокими желобками.
Зубы радулы прямые........................8. Oe. (N.) harpa (Dall)
3. OENOPOTA
139
15 (10). Спиральная скульптура представлена тонкими округлыми многочис-
ленными ребрышками с практически равными им по ширине межребер-
ными промежутками.
16 (17). Осевые складки сглаживаются на периферии оборотов. Зубы радулы
не превышают в длину 150 мкм, киль превышает V3 длины лезвия ....
..........................................И. Oe. (N.) inequita (Dall)
17 (16). Осевые складки сглаживаются на основании раковины. Зубы радулы
в длину превышают 250 мкм, киль занимает менее 1/3 длины лезвия . . .
..........................9. Oe. (N.) cancellata (Mighels et Adams)
18 (9). Осевые складки меньше или равны по ширине разделяюшим их проме-
жуткам.
19 (24). Осевые складки выражены не только на периферии оборотов, но и на
основании раковины.
20 (21). Основание раковины округлое. Обе ручки небольшой конической
базальной части зуба радулы несут листовидные флажки.....................
........................................ 5. Oe. (N.) cinerea (Moller)
21 (20). Основание раковины скошенное. Только 1 из 2 ручек массивной ба-
зальной части несет шпору, как правило, треугольной формы.
22 (23). Обороты раковины закругленные. Весь первый оборот эмбриональной
раковины покрыт ярко выраженным ретикулятным рисунком ....
...................................... 6. Oe. (N.) declivis (Loven)
23 (22). Обороты раковины отлогоугловатой формы. На первом обороте эмбрио-
нальной раковины сохраняются только следы ретикулятного рисунка
.............................................. 7. Oe. (N.) elegans (Moller)
24 (19). Осевые складки сглаживаются на периферии оборотов или на основа-
нии раковины.
25 (30). Осевые складки на околошовной поверхности имеют ярко выраженный
сигмоидальный изгиб.
26 (29). Спиральная скульптура равновелика на всей поверхности оборотов
и на основании раковины.
27 (28). Обороты раковины закругленные, умеренно выпуклые. Скошенное
основание раковины плавно переходит в сифональный вырост. Зубы
радулы с массивной базальной частью и шлемовидным дистальным участ-
ком киля ................................15. Oe. (N.) kurilensis Bogdanov
28 (27). Обороты раковины отлогоугловатой формы. Вытянутый сифональ-
ный вырост четко отграничен от основания раковины. Зубы радулы с от-
логоконусовидной базальной частью и тонким игловидным дистальным
участком киля.................................12. Oe. (N.) pavlova (Dall)
29 (26). Спиральная скульптура представлена на одном обороте различными
элементами: от нитевидных ребрышек до шнуровидных ребер с различной
шириной разделяющих их промежутков......................................
.....................................16. Oe. (N.) elongata Bogdanov
30 (25). Осевые складки прямые на околошовной поверхности, на ранних обо-
ротах несут лишь след сигмоидального изгиба........................ . .
....................................,13. Oe. (N.) uschakovi Bogdanov
1. Подрод OENOPOTA Morch, 1852
Раковина веретеновидной или удлиненно-веретеновидной формы. Состав-
лена, как правило, закругленными или усеченно-конусовидными оборотами.
Осевая скульптура представлена многочисленными складками, имеющими
в верхней части ярко выраженный сигмоидальный изгиб. Ось нарастания,
как правило, резко изгибается на ранних стадиях формирования крышечки.
Зубы радулы маленькие, до 130 мкм, стилетовидной формы; киль редуци-
рован.
Подрод распространен на всем ареале рода.
140
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
1. Oenopota (Oenopota) pyramidalis (Strom, 1788). (Рис. 191—208; 397,
Д—Е; 399, А—Б; 419, 1—34; 447).
Strom, 1788 : 296, pl. 1, fig. 22 (Buccinum); Stimpson, 1851 : 49 (Mangelia);
T г о s c h e 1, 1866 : 43, Tab. 4, fig. 3 (Bela); G. 0. Sars, 1878 : 222, Tab. 16, fig. 3,
Tab. 8, fig. 8 (Bela); Tryon, 1884 : 215, pl. 28, fig. 40, 47, pl. 34, fig. 71, pl. 30, fig. 92
(Bela); Г ер ценит e йн, 1885 : 685 (Pleurotoma); Friele, 1886 : 22, pl. 8, fig- 25,
pl. 10, fig. 21 (Bela); Martini, Chemnitz, 1887 : 159, Taf. 32, fig. 14, 15 (Bela);
Posselt, Jensen, 1898 : 161 (Bela); Knipowitsch, 1901 : 47 (Bela); К o-
b e 11, 1905 : 259, pl. 83, fig. 22, 23 (Bela); S c h 1 e s c h, 1924 : 327, Taf. 11, fig. 9—10
(Lora); Филатова, Зацепин, 1948 : 389, табл. 98, рис. 14 (Bela); MacGinitie,
1959 : 136, pl. 16, fig. 12—13, pl. 17, fig. 6—7 (Oenopota; part.); Powell, 1966 : 121,
pl. 9, fig. 1 (Oenopota); Macpherson, 1971 : 121, pl. 7, fig. 12, map. 53 (Oenopota). —
pleurotomaria Couthouy, 1838 : 107, pl. 1, fig. 9 (Fusus); Adams, Adams, 1858 : 92 (Bela);
Gould, 1870 : 355, fig. 625 (Bela); Verrill, 1882 : 478 (Bela); Bush, 1884 : 238,
pl. 9, fig. 7 (Bela).— vahlii Beck inMoller, 1842 : 86 (Defrancia); Knipowitsch,
1901 : 47 (Bela pyramidalis var.). — semiplicata G. O. Sars, 1878 : 222, Tab. 16, fig. 4
(Bela pyramidalis var.). — jenisseensis L e c h e, 1878 : 56, Tab. 1, fig. 17 (Pleurotoma pyra-
midalis var.); Martini, Chemnitz, 1887 : 165, Taf. 33, fig. 8, 9 (Bela). —- rufa
(non Montagu, 1803): Gould, 1841 : 290, fig. 192 (Fusus); Forbes, Hanley, 1853 :
454, pl. 112, fig. 3—5, pl. TT, fig. 4 [Mangelia (Bela); part.]; J effreys, 1867 : 392 (Pleu-
rotoma; part.). — simplicata (non Locard, 1895): Koehler, 1896 : 140, pl. 5, fig. 2 (Bela).
Рис. 447. Места находок Oenopota pyramidalis (1) и Oe. pyramidalis var. semiplicata (2) в мо-
рях СССР; 3 — палеонтологические находки.
Раковина от буровато-коричневого до телесного цвета, удлиненно-веретено-
видная, с 6—8 закругленными, умеренно выпуклыми оборотами. Периостракум
плотный, не прозрачный, коричневатый. Протоконх неправильно-цилиндриче-
ской формы, орнаментирован 3—5 спиральными тонкими ребрами. На первом
эмбриональном обороте сохраняются следы ретикулятного рисунка, на втором
обороте закладывается осевая скульптура в виде тонких осевых складочек,
расположенных параллельно оси протоконха (рис. 397, Д—Е). На дефинитив-
ной раковине осевая скульптура представлена средними или широкими склад-
ками, наиболее рельефными в верхней части периферии оборотов и сглаживаю-
щимися на основании раковины. Многочисленные осевые складки (от 16 до 19
на последнем обороте) имеют ярко выраженный сигмоидальный изгиб. Они раз-
делены равными по ширине или чуть более узкими промежутками. Спиральная
скульптура представлена различными элементами: от волосовидных ребрышек
до узких шнуровидных ребер, весь набор которых может присутствовать на
раковине одного экземпляра. Четкой детерминированности в расположении
различных элементов спиральной скульптуры не отмечено. Основание рако_
вины выпуклое, место перехода в удлиненный сифональный вырост всегда четк0
3. OENOPOTA
141
обозначено. Устье полуовальной, реже широко-обратногрушевидной формы
с полосой белесого или кремового каллуса на колумеллярном крае и глубокой
выемкой синуса, занимающего верхнюю х/3 палатального края. Сифональный
канал узкий, глубокий. Наибольший экземпляр из просмотренной коллекции
:имел высоту раковины 15.8 мм. В литературе указывается на еще большие раз-
меры; так, в канадском секторе Арктики обнаружена раковина высотой 20
,(Macpherson, 1971), в Иса-Фьорде 23 мм (Odhner, 1915). .
Крышечка со значительно утолщенным левым краем. Мускульный отпеча-
ток занимает большую часть левой ее стороны, но расположен относительно
.далеко от края (рис. 32, С—Ф).
Зубы радулы стилетовидные, характерной для вида формы (рис. 419),
с небольшим коническим основанием и легкой изогнутостью околобазальной
г/з лезвия зуба. Развита одна из ручек (Troschel, 1866; G. О. Sars, 1878; соб-
ственные данные). Морфология и параметры отдельных частей зубов могут
сильно варьировать. Однако специфичные черты — отогнутость цилиндриче-
ской базальной части, выпуклость стенок дистального участка лезвия, пре-
имущественное развитие одной из ручек, несущей маленькую шпору, по-
стоянны для выделяемых по конхологическим особенностям варие-
тетов.
Вид наиболее сходен с Ое. pingelii, от которого отличается сильно изогну-
тыми широкими осевыми складками и тонкой спиральной скульптурой.
Изменчивость. Исследованный материал позволяет говорить о зна-
чительной изменчивости моллюсков. В коллекции ЗИН АН СССР имеются все
переходные формы от типичной к вариететам semiplicata и jenisseensis. Первая
характеризуется менее стройной раковиной и осевой скульптурой, стираю-
щейся уже в нижней х/4 периферии оборота; вторая — очень стройной высокой
раковиной и небольшим числом (до 12) осевых складок. Найдена новая форма,
характеризующаяся значительной выпуклостью основания раковины и стер-
тостью осевых складок на поздних дефинитивных оборотах (рис. 205—208);
она распространена в Карском море и холодноводных участках Барен-
цева.
Распространение. Бореально-арктический широко распростра-
ненный вид. Встречен у берегов Аляски, м. Барроу, у арктического побережья
Канады от Гудзонова залива до пролива Принца Уэльса, Гренландии, Ислан-
-дии, в Лабрадоре, у Северной и Южной Норвегии, Шпицбергена, Мурманского
побережья до Финмаркена и у берегов Новой Земли в Баренцевом море (рис. 447).
'Описан из Белого моря (Герценштейн, 1885), Карского и Восточно-Сибирского
-морей (Odhner, 1915), Берингова пролива (Clarke, 1963), в Беринговом море
от о-ва Нунивак до зал. Пьюджет-Саунд (Verrill, 1882). Известен для глубинных
вод Бискайского залива (Koehler, 1896).
Экология. Отмечен на глубинах от 5 (Белое море) до 330 м, в литера-
туре называется верхняя граница 2 м (Иса-Фьорд), нижняя — до 1 410 м (Би-
скайский залив) (Koehler, 1896) и до 1 959 м (Девисов пролив) (Odhner, 1915),
возможно до 2 010 м (Clarke, 1963). Наибольшая частота встречаемости на глу-
бинах 20—150 м. Вид обитает практически на всех грунтах с предпочтением
песчанистых (около 60 %). Представители вида были встречены в диапазоне
температур от —0.4 до 8.2 °C, в основном при 0.6—1.4 °C, при солености от
. 27.39 др 34.8, преимущественно при 34.4—34.8 0/00.
Типы. О типах Buccinum pyramidale никаких сведений в литературе не об-
наружено. Вероятно, они хранятся в ZMC, однако наш запрос (25.02 1979 г.)
не привел к их обнаружению. Типовое местонахождение — Северное море.
Голотип Fusus pleurotomarius хранится в BSNH; типовое местонахождение —
зал. Массачусетс, глубинные воды. Типы Defrancia vahlii и Bela pyramidalis
var. semiplicata, по-видимому, хранятся в ZMO. Место хранения типов Pleuro-
Лота pyramidalis var. jenisseensis неизвестно.
Просмотрено 290 экз. вида и 92 экз., относящихся к различным инфра-
£ подвидовым формам.
148
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
2. Oenopota (Oenopota) pingelii (Moller, 1842), comb. n. (Рис. 6; 209—210;
422, 24—30-, 448).
Moller, 1842 2 : 86 (Defrancia)', Adams, Adams, 1858 : 92 (Bela)', Morch, 1868 ’
215 [Pleurotoma (Bela)]; G. 0. S a r s, 1878 : 223, Tab. 16, fig. 5 (Bela)', Verrill, 1882 : 464,
pl. 43, fig. 5 (Bela)', Tryon, 1884 : 217, pl. 29, fig. 64 (Bela)', Friele, 1886 : 22, pl. 8,
fig. 26, a, pl. 10, fig. 20a (Bela)', Martini, Chemnitz, 1887 : 156, Taf. 32, fig. 10 (Bela)',
Posselt, J ensen, 1898 : 160 (Bela)', О d h n e r, 1910 : 12, Taf. 1, fig. 17 (Bela)', 1915 :
220 (Bela)', S c h 1 e s c h, 1924 : 328 (Lora).
Раковина светло-коричневая, высокая, веретеновидная, с 6—7 усеченно-
конусовидной формы оборотами. Периостракум плотный, полупрозрачный,,
коричневатый. Эмбриональная раковина цилиндрической формы, уплощенная
в верхней части. Первый эмбриональный оборот несет ярко выраженный рети-
Рис. 448. Места находок Oenopota pingelii в Северных морях СССР. '
кулятный рисунок (рис. 6), второй — 4 спиральных ребрышка различной ве-
личины. Спиральная скульптура пересечена равномерно расположенными тон-
кими осевыми складками, придающими протоконху сетчатый рисунок. Осевая
скульптура дефинитивной раковины представлена пологими, сигмовидно изо-
гнутыми средними складками, выраженными на периферии оборотов и сглажива-
ющимися на основании раковины. Разделяющие их промежутки, как правило,,
в 2—3 раза уже самих складок. На последнем обороте раковины насчитывается
от 20 до 36 осевых складок, на предпоследнем от 18 до 23. Спиральная скульп-
тура состоит из равновеликих средних шнуровидных ребер, разделенных в 1.5—
2 раза более узкими межреберными пространствами. Максимального развития
достигает ребро, обозначающее нижнюю х/3 периферии оборота и придающее
ему усеченно-конусовидную форму. Вниз от этого ребра, к сифональному вы-
росту, и вверх, ко шву, наблюдается постепенное сокращение в размерах эле-
ментов спиральной скульптуры. При пересечении с осевыми складками происхо-
дит утолщение спиральных ребер, в результате чего последние приобретают
структуру четок. Слабовыпуклое основание плавно переходит в вытянутый
сифональный вырост, покрытый нитевидными ребрышками. Устье узко-об-
ратногрушевидной формы с тонким коричневым каллусом на колумеллярном
крае и довольно глубоким анальным синусом, занимающим верхнюю х/3 пала-
тального края. Сифональный канал неглубокий, средней ширины. Размеры
2 В работе Мёллера (Moller, 1842) автором этого и ряда других названий указан Бэк
(Beck), и в литературе широко принято авторство Beck in Moller. Однако поскольку нет ни-
каких свидетельств того, что Мёллер приводит данные Бэком диагнозы этих видов, автором
должен считаться Мёллер.
3. OENOPOTA 143
•.наибольшего в коллекции экземпляра (обнаружен у Мурманского побережья)
следующие (мм): 5р=10.6; Д=4.3; 2?ПО=6.4; £^=4.6; Дд=Л.1. Наибольшие
из описанных размеров раковины — высота 13.7 при ширине 5.6 (Thorson,
1941) и £?р=16 мм (G. О. Sars, 1878).
Крышечка каплевидной формы, со слабо изгибающейся осью нарастания.
Мускульный отпечаток занимает срединную часть крышечки (рис. 32, П—Р).
Зубы радулы мелкие (не более 150 мкм), со слабо развитым килем и с длин-
ным цилиндрическим основанием, несущим только 1 ручку с маленькой шпорой
;(рис. 422, 27). В отличие от рисунков, приведенных в работе Фриле (Friele,
1886, Tab. 10, fig. 20), зубы исследованных моллюсков имели более удлинен-
ные лезвие и базальную часть при тех же пропорциях зубов.
Вид хорошо отличается от сходных по форме раковины видов своеобразной
скульптурой, состоящей из равновеликих осевых складок и спиральных ребер,
образующих при пересечении толстые узлы-четки.
Распространение. Широко распространенный бореально-арктиче-
ский вид, отмечен у берегов Западной и Юго-Восточной Гренландии, Исландии,
Шпицбергена, восточного Финмаркена, у Мурманского побережья Баренцева
моря и в Белом море, в северной части Карского моря и западной части моря
Лаптевых (рис. 448), обнаружен в Охотском море и в зал. Св. Лаврентия.
Кроме того, вид обитает у американских берегов Северной Атлантики: у Новой
Шотландии, в водах Лабрадора.
Экология. Обитает на глубинах от 3 до 774 м (Thorson, 1944), встречаясь
в основном на 20—25 м, практически на всех типах грунтов, предпочитая,
однако, илистый песок или песчанистый ил, в диапазоне солености от 32.8
.до 35.0 °/00, с пиком численности при 34.3—35.0 °/00.
Тип ы: Типы, по-видимому, хранятся в ZMO; типовое местонахож-
дение — воды, омывающие Гренландию.
Просмотрено 58 экз.
3. Oenopota (Oenopota) obliqua (G. О. Sars, 1878). (Рис. 211—213; 406, A—Б;
-420, 1—10; 449).
G. О. S a r s, 1878 : 226, Tab. 16, fig. 6, Tab. 8, fig. 11 (Bela); Tryon, 1884 : 219, pl. 28,
Tig. 50 (Bela); Герценштейн, 1885 : 687 (Pleurotoma); Friele, 1886 : 7, pl. 7, fig. 15—
17, pl. 10, fig. 1—2 (Bela); Martini, Chemnitz, 1887 : 146, Taf. 31, fig. 12 (Bela);
Posselt, Jensen, 1898 : 165 (Bela); К о b e 1 1, 1905 : 250, Taf. 83, fig. 4 (Bela); Mac-
pherson, 1971 : 119, pl. 7, fig. 3, map 53 (Oenopota).
Раковина белая, небольшая, веретеновидной формы, с 5—6 оборотами от-
. логоугловатой формы, тупоугловатыми в верхней части. Периостракум тонкий,,
желтоватый или светло-коричневый, легко отстающий от поверхности раковины.,
Протоконх сосцевидной формы, представлен 1.5—1.7 уступчатыми оборотами,
орнаментированными 3 спиральными ребрышками (рис. 406, А—Б). Осевая
скульптура дефинитивной раковины представлена тонкими складками, наибо-
лее выраженными в центральной части периферии оборота и сглаживающимися
в нижней половине основания раковины. Осевые складки имеют на плече обо-
рота слабо выраженный сигмоидальный изгиб, на периферии оборота они ско-
тттр.ны назад и разделены в 2—4 раза более широкими промежутками. На послед-
нем обороте раковины насчитывается от 16 до 22 осевых складок, на предпослед-
нем от 13 до 15. Спиральная скульптура состоит из тонких ребер и нитевидных
ребрышек. Наиболее развиты среди элементов спиральной скульптуры 2 тон-
ких ребра: одно из них проходит по плечу, другое по срединной части перифе-
рии оборота. Остальные элементы спиральной скульптуры получают развитие
преимущественно под ребром, отмечающим середину периферии оборота,
и не имеют упорядоченного расположения. Они разделены различной ширины
межреберными промежутками: от х/2 до 3/2 ширины самих ребер. Основание ра-
ковины практически прямое, плавно переходит в короткий сифональный вы-
рост. Устье неправильно-овальной формы, угловатое в верхней части; на колу-
144
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
меллярном крае несет кремового цвета каллус, в верхней части палатального
края — довольно глубокий анальный синус. Сифональный канал неглубокий,,
относительно широкий. Высота раковины типового экземпляра 9 мм. Наиболь-
шие размеры раковины из коллекции ЗИН АН СССР (мм) — /?р=8.5; Д=3.85,-:
7?ПО=5.6; /?3,=3.8; Д =1.6 — отмечены для моллюска из Баренцева моря.
Зубы радулы маленькие, длиной 100—120 мкм, стилетовидные, с очень тон-
ким цилиндрическим основанием (рис. 420, 1—10). Одна из ручек редуциро-
вана, другая развита и несет маленькую шпору. Длина киля составляет 1/,1 об-
щей длины зуба радулы. Исследованные мной зубы морфологически сходны
с изображениями, приведенными для североатлантических моллюсков
(G. О. Sars, 1878 : Tab. 8, fig. 11; Friele, 1886 : pl. 10, fig. 1—2), но отличаются
меньшей отогнутостью базальной части.
Рис. 449. Места находок Oenopota obliqua в морях СССР.
Вид сходен с Ое. cinerea, от которого отличается прямым основанием рако-
вины, плавно переходящим в сифональный вырост, и стилетовидной формой
зубов.
Распространение. Атлантический широко распространенный вы-
сокобореально-арктический вид. Отмечен в северном и северо-западном рай-
онах канадского сектора Арктики, у берегов Западной Гренландии, Исландии,
у Норвежского побережья, в Кольском заливе, в Белом море, у о-вов Новой
Земли, у о-вов Земли Франца-Иосифа, в центральной части Карского моря-
и в море Лаптевых (рис. 449).
Экология. Обитает на глубинах от 6 до 350 м, чаще всего встречается
на 10—50 м, отмечен на глубинах до 395—406 м (Odhner, 1915). Обитает на всех
типах грунтов, предпочитая илистые и заиленные (более 60 % всех находок).
Моллюски встречаются при температурах от —1.83 до 4.2 °C, в основном при
отрицательных температурах. Большинство животных отмечено в акваториях
с высокой соленостью 34—35 °/00, но встречаются и при более низких ее зна-
чениях — до 27.2 °/00.
Типы. Типовой экземпляр находится в ZMO; типовое местонахождение —
Хаммерфест, Норвегия.
Просмотрено 108 экз.
2. Подрод NODOTOMA Bartsch, 1941
Bartsch, 1941 : 5 (=Bela, Lora', part.); Powell, 1966 : 122 (Obesotoma subg.; part.)..
Типовой вид Pleurotoma impressa^M.6rch, 1869, по первоначальному
обозначению.
3. OENOPOTA
145
Раковина веретеновидной или кубаревидной формы, как правило, с отлого-
угловатыми или уступчатыми оборотами. Осевая скульптура представлена
в основном немногочисленными складками, имеющими в верхней части слабый
сигмоидальный изгиб (или изгиб практически отсутствует). Крышечка с плавно
изгибающейся осью нарастания.
Зубы радулы крупные, до 1 мм в длину, копьевидной формы, с хорошо
развитым килем.
Подрод распространен по всему ареалу рода.
4. Oenopota (Nodotoma) impressa (Morch, 1869),3 comb. n. (Рис. 37, 5;
161—167; 398, A—E; 414, 13—23; 415, 1—12; 450; 451).
Morch, 1869 : 21 [Pleurotoma (Ischnula)]; Leche, 1878 : 54, pl. 1, fig. 16 (Pleurotoma);
Tryon, 1884 : 220, pl. 33, fig. 69 (Bela); Krause, 1885 : 278 (Bela); Герценштейн,
1885 : 686 (Pleurotoma); F r i e 1 e, 1886 : 18, pl. 8, fig. 1, 2, pl. 10, fig. 9 (Bela); Knipo-
w its c h, 1901 : 45 (Bela); К о b e 1 1, 1905 : 237, Taf. 84, fig. 11—13 (Bela turricula subsp.);
Oldroyd, 1927 : 95 (Lora); Bartsch, 1941 : 5, pl. 1, fig. 2 (Nodotoma); M ac Gini-
t i e, 1959 : 137, pl. 16, fig. 14,15 (Nodotoma); Powell, 1966 : 121, pl. 19, fig. 5 (Nodotoma).—
sarsli V e r r i 1 1, 1880 : 364 (Bela; pro cancellata sensu G. O. Sars, 1878); 1882 : 484 (Bela);
Bush, 1884 : 237, pl. 9, fig. 8 (Bela); Tryon, 1884 : 218, pl. 28, fig. 49 (Bela); Martini,
Chemnitz, 1887 : 147, Taf. 31, fig. 13 (Bela); Posselt, Jensen, 1898 : 165 (Bela);
К о b e 1 1, 1905 : 245, pl. 28, fig. 21 (Bela); Dautzenberg, Fischer, 1912 : 48
(Bela). — cancellata (non Mighels et Adams, 1840): G. O. Sars, 1878 : 224 Tab. 23, fig. 3,,
Tab. 8, fig. 9 (Bela).
Рис. 450. Места находок Oenopota impressa в северных морях СССР.
Раковина серая, веретёновидная или веретеновидно-овальная, с 6—7 обо-
ротами отлогоугловатой формы, тупоугловатыми в верхней части. Периостра-
кум плотный, полупрозрачный, пепельного цвета, легкоотстающий от поверх-
ности раковины. Протоконх цилиндрической формы, макушка погружена
в первый полуоборот эмбриональной раковины (рис. 398). Первый оборот орна-
ментирован только микроскульптурными бугорками, но уже к началу второго
эмбрионального оборота закладываются 2 тонких спиральных ребра, одно из ко-
торых обозначает середину периферии, другое — ее верхний участок. Осевая
скульптура дефинитивной раковины представлена близко расположенными
средними складками, промежутки, разделяющие их, составляют 1/4—1/7 ши-
рины складки. Сигмоидальный изгиб в верхней части элементов осевой скульп-
туры намечен слабо. Складки заходят на основание раковины, и их следы про-
слеживаются даже на сифональном выросте. На последнем обороте их насчиты-
3 Авторство вида часто указывается как «Beck in Morch»; см. примет, на с. 142.
Ю И. П. Богданов
146
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
вается от 16 до 20, на предпоследнем от 13 до 18. Спиральная скульптура пред-
ставлена широкими ребрами: 2 или 3 на периферии оборота, 4 или 5 на плече
оборота и 4 или 5 на основании раковины. Межреберные промежутки более чем
в 10 раз уже ребер, в результате чего создается впечатление спиральной скульп-
туры, состоящей из глубоких узких желобков.
Основание раковины выпуклое, четко отграничено желобком от сифональ-
гного выроста средних размеров. Последний украшен 5—7 шнуровидными сред-
ними ребрами. Устье обратногрушевидной
формы, угловатое в верхней части. Каллус
кремового цвета, толстой полосой лежит
на колумеллярном крае. Сифональный
канал неглубокий, средней ширины. Вы-
сота раковины типового экземпляра 15 мм
при ширине 7 мм. Наибольший экземпляр
был найден в Беринговом море; параметры
его раковины (мм): 2?p=12.4; Д=6.1;
7?no=8.8; jBj,=6.3; ^^=2.6. Наибольшая
раковина (без тела) высотой 13.2 мм най-
дена у Мурманского побережья.
Крышечка когтевидной неправильно-
овальной формы, с часто расположенными
линиями нарастания. Мускульный отпеча-
ток ромбовидной формы, расположен чуть
правее оси нарастания (рис. 33, А— Г).
Зубы радулы длиной до 450 мкм, копь-
евидной формы, со слегка изогнутым лез-
вием, киль протяженный, превышает раз-
меры дистального отверстия зуба. Базаль-
ная часть желтого цвета, пирамидальной
формы, несет 2 развитые ручки, одна из
которых вооружена небольшим узким
флажком, другая — широким пологим
(рис. 414, 22—23). Наблюдается полное
сходство морфологии зубов у исследован-
ных нами моллюсков с изображениями,
приводимыми для Bela impressa (Friele,
1886 : pl. 10, fig. 9), «2?. cancellata»
(,G. 0. Sars, 1878 : Tab. 8, fig. 9) и В. sar-
sii (Friele, 1886 : pl. 10, fig. 7—8).
Вид похож по раковине на Ое. harpa, от
которого легко отличается широкими осе-
Рис- ^451. Места находок Oenopota выми складками, пересеченными очень
impressa в дальневосточных морях глубокими 2-4 желобками, а также изо-
гнутостью зубов радулы.
Распространение. Широко распространенный бореально-аркти-
ческий вид. Обнаружен у берегов Норвегии и Шпицбергена, у южных берегов
Баренцева моря, у архипелага Земля Франца-Иосифа, а также в Карском море,
море Лаптевых (рис. 450). Повсеместно обнаружен в Чукотском, Беринговом,
Охотском морях, а также в Приморье и Татарском заливе в Японском море
(рис. 451). В Тихом океане у американских берегов обнаружен от м. Барроу
до о-ва Кадьяк (Аляска).
Экология. Обитает на глубинах от 5 (у Земли Франца-Иосифа) до 500 м,
наибольшее количество находок приходится на глубины 12—100 м. Представи-
тели вида встречены на всех типах грунтов, однако прослеживается явное пред-
почтение ими илистых и песчаных грунтов (более 85 % всех находок). Моллюски
обитают в диапазоне температур от —1.82 до 10.2 °C, в основном предпочитая
3. OENOPOTA
147'
температуры от —1.8 до 0.8 °C, при солености 31.70—35.04 °/00, предпочитая
33.4-34.8 °/00.
Типы. Типовая серия Pleurotoma, impressa хранится в ZMC; типовое ме-
стонахождение — Bellsound (Норвегия). Тип Bela sarsii находится в ZMO,
типовое местонахождение Тромсё (Норвегия).
Просмотрено 237 экз.
5. Oenopota (Nodotoma) cinerea (Moller, 1842). (Рис. 136—139; 394, A—E;
414, 1—12; 452).
Moller, 1842 : 86 (Defrancia); M б r ch, 1868 : 215 [Pleurotoma (Bela)]; Jeffreys, 1876:
330 (Bela)-, G. 0. Sars, 1 878 : 227, Tab. 23, fig. 4 (Bela); Tryon, 1884 : 218, pl. 28, fig. 48
(Bela); Friele, 1886 : 9, pl. 7, fig. 23a, pl. 10, fig. 6 (Bela); Martini, Chemnitz,.
1887 i 156, Taf. 32, fig. 9 (Bela); Posselt, Jensen,' 1898 : 150 (Bela); К о b e 1 t,
1905 : 250, pl. 83, fig. 10 (Bela); Odhner, 1910 : 11, pl. 1, fig. 19 (Bela); Thorson,
1951 : 60 (Bela); Macpherson, 1971 : 114, pl. 7, fig. 5, map 50.
Рис. 452. Места находок Oenopota cinerea в морях СССР.
Раковина зеленовато-белая, веретеновидная, с 5.5—6.5 оборотами отлого-
угловатой формы, тупоугловатыми в верхней части. Периостракум тонкий, жел-
товатый или светло-коричневый, легкоотстающий. Протоконх цилиндрической
формы, с уплощенной верхней площадкой (рис. 394). Первые 1.2—1.3 эмбрио-
нальных оборота несут складки, придающие протоконху ретикулятный рису-
нок. Уже на последних этапах развития первого эмбрионального оборота про-
исходит обособление 4—5 нитевидных спиральных ребрышек, которые на вто-
ром обороте занимают всю его поверхность (ретикулятный рисунок исчезает).
На втором эмбриональном обороте закладывается осевая скульптура в виде
узких складок, постепенно достигающих размеров спиральных ребер, так что
к первому постэмбриональному обороту они становятся равными, а затем и боль-
шими по высоте, чем элементы спиральной скульптуры. На более поздних обо-
ротах дефинитивной раковины осевая скульптура представлена пологими
средними складками, наиболее развитыми на плече оборота и редко доходя-
щими до нижней части основания раковины. На периферии оборота складки
имеют небольшой наклон назад, а на плече оборота — сигмоидально изогнуты.
Разделены они промежутками, как правило, равными собственной ширине.
На последнем обороте раковины насчитывается от 17 до 22 осевых складок,
на предпоследнем от 15 до 19. В верхней, большей части периферии оборотов,
спиральная скульптура представлена тонкими нитевидными ребрами, разде-
ленными вдвое более узкими межреберными промежутками. В нижней части
периферии оборотов, на основании раковины и частично на сифональном вы-
росте скульптура состоит из шнуровидных широких ребер, разделенных про-
10*
148
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
межутками, которые более чем в 10 раз у'же самих ребер. На периферии послед-
него оборота насчитывается 2—3 широких ребра, на основании раковины около
6, на сифональном выросте 2—4. Основание раковины выпуклое, плавно пере-
ходит в удлиненный сифональный вырост. Устье полуовальной формы, с неболь-
шой вырезкой анального синуса в верхней х/5—1/6 палатального края; с толстым
жремового цвета каллусом на колумеллярном крае. Сифональный канал корот-
кий, относительно глубокий, широкий. Типовой экземпляр имеет раковину
высотой 6.5 мм. Наибольший экземпляр, обнаруженный в восточной части
Баренцева моря, имеет следующие параметры (мм): 2?p=14.2; Д=6.8; 7?гао=9.4;
72^=7.0; ^=3.0. Известна раковина высотой до 22 мм (Odhner, 1915).
Крышечка когтевидная, с плавно изгибающейся осью нарастания. Левый
край крышечки утолщенный, мускульный отпечаток не прилегает к немуа
вверху заострен (рис. 33, Д').
Зубы радулы длиной до 600—700 мкм, копьевидные, прямые, с относительно
небольшим коническим основанием, обе ручки несут почти одинаковые по раз-
мерам листовидные флажки (рис. 414, 6). Киль в 1.5—2 раза больше дисталь-
ного отверстия зуба. Морфология исследованных зубов соответствует изобра-
жениям в литературе (Friele, 1886 : pl. 10, fig. 6).
Вид сходен с Oe. elegans, от которого отличается более уплощенной формой
раковины и характерной формой эмбриональной раковины, первый оборот
которой целиком покрыт ретикулятным рисунком.
Распространение. Атлантический высокобореально-арктический
вид. Отмечен в канадском секторе Арктики от зал. Маккензи до о-ва Элсмира,
у берегов Южной и Западной Гренландии, Исландии между о-вами Фарое
и Шотландией, около Северной Норвегии и Финмаркена, у Шпицбергена, в юж-
ной части Баренцева моря и в восточной части Карского моря; встречен у Ново-
сибирских о-вов (рис. 452).
Экология. Обитает на глубинах от 4.5 м в канадском секторе Арктики
.(Macpherson, 1971) до 761 м у Шпицбергена (Thorson, 1941); мною отмечен
на глубинах от 28 до 422 м. Встречается преимущественно на илистых и заилен-
ных грунтах при температуре от —1.69 до 1.6 °C, однако наибольшее число
-экземпляров отмечено при температуре от —1.6 до —1.1 °C при солености
,;34.3—34.67 °/00.
Типы. Типовые экземпляры хранятся в ZMC; типовое местонахождение —
.воды, омывающие Гренландию.
Просмотрено 28 экз.
6. Oenopota (Nodotoma) declivis (Loven, 1846). (Рис. 37, A; 140—146; 395,
A—E; 416, 16—24; 417, 1-13; 453, 2).
Loven, 1846 i 145 (Tritonium); J effreys, 1867 : 330 (Pleurotoma); G. O. Sars,
1.878 : 224, Tab. 16, fig. 10 (Bela); Tryon, 1884 : 218, pl. 28, fig. 38 (Bela); Герцен-
ш т e й h, 1885 : 686 (Pleurotoma); Friele, 1886 : 8, pl. 7, fig. 20—22 (Bela cancellata var.);
‘Martini, Chemnitz, 1887 : 157, Taf. 32, fig. 11 (Bela); Posselt, J ensen, 1898 :
152 (Bela); К о b e 1 t, 1905 : 249, Taf. 83, fig. 5—9 (Bela cancellata subsp.); Thorson,
1935 : 36 (Bela cancellata var.); 1944 : 121 (Bela); Macpherson, 1971 : 115, pl. 7, fig.
10, map 51 (Oenopota). — cancellata (non Mighels et Adams, 1840): Troschel, 1866 : 42,
Taf. 4, fig. 9 (Bela); Friele 1886:8, pl. 7, fig. 18—19, pl. 10, fig. 3—5 (Bela).
Раковина серого цвета, тонкостенная, удлиненно-веретеновидная, с 5.5—
'6.5 закругленными, умеренно выпуклыми оборотами, слегка угловатыми в верх-
ней части. Периостракум тонкий, желтоватый, прозрачный, плотноприлегаю-
щий. Эмбриональная раковина представлена 1.5—1.7 округлыми оборотами,
причем весь первый оборот покрыт ретикулятным рисунком, в котором наме-
чены будущие 5—6 спиральных ребрышек, проявляющихся только на втором
•обороте протоконха (рис. 395). Осевая скульптура закладывается на втором
обороте в виде тонких складочек и получает окончательное развитие только
и а первых оборотах дефинитивной раковины (рис. 395, Г). Осевая скульптура
3. OENOPOTA
149
поздних оборотов дефинитивной раковины представлена тонкими рельефными
складками, треугольными в сечении, разделенными в 2—2.5 раза более широ-
кими промежутками. Складки выражены практически одинаково на поверхно-
сти последнего оборота и на всем основании раковины; на околошовной поверхно-
сти они приобретают незначительный сигмоидальный изгиб. На последнем обо-
роте раковины насчитывается от 18 до 22 осевых складок, на предпоследнем от
14 до 18. Спиральная скульптура состоит из шнуровидных средних ребер, за-
нимающих большую поверхность оборотов раковины, основание и сифональный
вырост. Шнуровидные ребра разделены в 4—6 раз более узкими промежутками.
На околошовной поверхности спиральная скульптура представлена 5—9 тон-
кими волосовидными ребрышками. Основание раковины слабовыпуклое, си-
-фональный вырост средних размеров. Устье неправильно-овальное, угловатое
в верхней части. Белый или кремового цвета каллус широкой полосой покры-
вает колумеллярный край устья. В верхней части палатального края располо-
Тис. 453. Места находок Oenopota elegans (1) и Oe. declivis (2) в морях СССР и в сопредель-
ных водах (по собственным данным).
жена неглубокая вырезка анальног синуса. Сифональный канал короткий,
широкий, умеренной глубины. Наибольшая высота раковины — 20 мм указана
для моллюска из вод Северной Норвегии (G. О. Sars, 1878).
Крышечка каплевидной формы, с очень слабо изогнутой осью нарастания.
Широкий мускульный след прилегает к левому краю крышечки (рис. 33, К—Л).
Зубы радулы копьевидные, длиной 300—500 мкм, основание несет 2 ручки,
только на одной из них развит уплощенный флажок (рис. 417, 11). Морфология
исследованных мной зубов четко отвечает имеющимся в литературе описаниям
и рисункам (Troschel, 1866; Friele, 1886).
Вид сходен с Oe. elegans, от которого отличается более округлыми оборотами,
незначительной шириной осевых складок и ярким ретикулятным рисунком
эмбриональной раковины.
Распространение. Атлантический, широко распространенный вы-
сокобореально-арктический вид. Распространен в высокобореальных районах
Северной Атлантики, повсеместно в Баренцевом море и в северной части Кар-
ского моря (рис. 453, 2). Отмечен у берегов Западной Гренландии, Исландии,
Финмаркена (Thorson, 1944), а также в канадском секторе Арктики: от о-ва Соме-
рсет и Бассейна Фокс до о-вов Белчер в Гудзоновом заливе (Macpherson, 1971).
Экология. Обитает на глубинах 15—860 м, однако в большей массе
встречен на глубинах 120—220 м, предпочитает заиленные грунты и низкие
(от —1.8 до —0.5 °C) температуры.
Типы. Место хранения типового материала неизвестно; типовое место-
нахождение — Финмаркен, Северная Норвегия.
Просмотрено 186 экз.
150
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
7. Oenopota (Nodotoma) elegans (Moller, 1842). (Рис. 147—154; 396,4—Z7;
416, 6—15; 453, 7).
Moller, 1842:86 (Defrancia); Jeifreys, 1867 : 331 (Pleurotoma); G. 0. Sars,
1878 .: 225, Tab. 16, fig. 15, Tab. 8, fig. 12 (Bela); Martini, Chemnitz, 1887 : 141,
Taf. 31, fig. 5 (Bela); Posselt, Jensen, 1898 : 151 (Bela); К о b e 1 t, 1905:247,
Taf. 83, fig. 2 (Bela cancellata subsp.); MacGinitie, 1959 : 135, pl. 16, fig. 7,8;
Bouchet, Waren, 1980 : 73, fig. 140, 266. — cancellata (non Mighels et Adams,
1840): Tryon, 1884 : 218, pl. 29, fig. 53 (Bela; part.).
Раковина пепельно-серая, удлиненно-веретеновидная, с 6.5—7.5 оборотами
отлогоугловатой формы, тупоугловатыми в верхней части. Периостракум тон-
кий, полупрозрачный, желтоватый. Протоконх бочонковидной формы, пред-
ставлен 1.5—1.7 угловатыми оборотами. На первом эмбриональном обороте со-
храняются следы ретикулятного рисунка (рис. 396). С середины этого оборота
намечается закладка 3 тонких спиральных ребер, из которых верхнее редуци-
руется к моменту формирования первых дефинитивных оборотов. Иногда закла-
дывается 4 ребра, но в таких случаях нижнее закрывается последующим оборо-
том. Оставшиеся 2 ребра получают развитие на дефинитивной раковине и обо-
значают один срединную часть периферии оборота, другой — его плечо. Осевая
скульптура дефинитивной раковины представлена треугольными в сечении,
покатыми тонкими или средними складками, на периферии оборота слегка от-
клоненными назад и приобретающими легкий сигмоидальный изгиб на плече
оборота. Складки практически одноразмерны на всей поверхности последнего
оборота и на основании раковины, их следы заметны и на сифональном выросте.
На последнем обороте раковины насчитывается от 18 до 24 осевых складок,
на предпоследнем от 16 до 20. Элементы спиральной скульптуры можно разде-
лить на 2 группы. На плече оборота расположены многочисленные (до 12) ни-
тевидные или волосовидные ребрышки. На периферии оборотов, основании ра-
ковины и сифональном выросте лежат различной ширины шнуровидные ребра,
превышающие ширину разделяющих их промежутков в 3—7 раз. Элементы
спиральной скульптуры представлены, как правило, определенным типом ребер
(чаще всего средними ребрами), однако у некоторых экземпляров отмечено чере-
дование элементов. Так, между средними шнуровидными ребрами могут закла-
дываться 1—3 волосовидных ребрышка (рис. 149—150), в результате чего спи-
ральная скульптура становится неупорядоченной. Основание раковины слабо-
выпуклое, сифональный вырост укороченный. Устье обратногрушевидной
формы, с широкой полосой матового, телесного цвета каллуса на колумеллярном
крае. В верхней части палатального края лежит неглубокая вырезка анального
синуса. Сифональный канал протяженный, умеренной ширины и глубины.
Высота раковины типового экземпляра 5.5 мм. Наибольшие размеры имеет ра-
ковина моллюска, обнаруженного у Мурманских берегов (мм): В =17.6; Д=
= 7.0; В„о=10.5; £^=7.7; Ду=3.3.
Крышечка каплевидная, с почти прямой осью нарастания. Заостренный
в верхней части мускульный отпечаток плотно прилегает к утолщенному ле-
вому краю крышечки (рис. 33, М—О).
Зубы радулы копьевидной формы, с массивной округлой базальной частью и
длинным килем, вдвое превышающим размеры дистального отверстия лезвия
(рис. 416, 6—15}. Флажок небольшой. Исследованные зубы сходны с изображен-
ными у Сарса (G. О. Sars, 1878 : Tab. 8, fig. 12).
Вид хорошо отличается от других представителей рода угловатостью основа-
ния раковины. ОтОе. declivls отличается укороченным сифональным выростом и
широким основанием раковины. Важным отличием вида является рудиментар-
ный ретикулятный рисунок протоконха.
Распространение. Атлантический широко распространенный вы-
сокобореально-арктический вид. Обнаружен у берегов Исландии, Западной
Гренландии, Шпицбергена, Северной и Южной Норвегии, в Баренцевом море
и повсеместно в Карском море (рис. 453, 7). Отмечен в Бискайском заливе
(Bouchet, Waren, 1980).
3. OENOPOTA
151
Экология. Обнаружен на глубинах от 21 до 860 м, имеются сведения
о его нахождении на глубинах до 1 203 м (Голиков, 1964). Наибольшее число
моллюсков отмечено на глубинах 120—220 м. Вид предпочитает заиленные и
илистые грунты с примесями камней, гравия, гальки и ракуши (более 75 %
находок). Моллюски встречаются в диапазоне температур от —1.8 до 3.6 °C и
солености от 32.76 до 35.04 °/00, предпочитая, однако, температуры от—1.8 до
—0.8 °C и соленость 34.4—35.0 °/оо-
Типы. Типовой экземпляр хранится в ZMC; типовое местонахождение —
воды Западной Гренландии.
Просмотрено 159 экз.
Рис. 454. Места находок Oenopota harpa в северных морях СССР и сопредельных водах;
' 1 — палеонтологические находки.
8. Oenopota (Nodotoma) harpa (Dall, 1884). (Рис. 155—160; 397, A—Б', 415,
13—22; 416, 1—5; 454; 455).
Dall, 1884 : 523 (Bela); 1886b : 300, pl. 4,. fig. 2 (Bela)-, Martini, Chemnitz,
1887 : 236 (Bela); Oldroyd, 1927 : 106 (Lora); MacGini tie, 1959 : 134, pl. 6, fig. 5,
>6. — plicifera Schmidt, 1872 : 57, Tab. 4, fig. 4, 5 (Mangelia; nom. praeocc., non Wood,
1848); Leche, 1878 : 58 (Pleurotoma); Aurivilli us, 1887 : 352 (Bela); К о b e 11,
1905 : 266, Taf. 84, fig. 22,23 (Bela); S c h 1 e s c h, 1924 : 328, Taf. 11, fig. 13 (Lora), -schmid-
.ti F r i e 1 e, 1886 : 10, pl. 8, fig. 6 (Bela; nom. n. pro plicifera); Martini, Chemnitz,
1887:231, Taf. 42, fig. 1 (Bela); Knipowitsch, 1901:47 (Bela). ]
Раковина белая или светло-коричневая, удлиненно-веретеновидная, с 6.5—
7.5 отлогоугловатыми оборотами, реже закругленными, умеренно выпуклыми.
Периостракум толстый, непрозрачный, коричневатый. Эмбриональная рако-
вина цилиндрической формы, со слабыми следами ретикулятного рисунка на
нервом эмбриональном обороте. Спиральная скульптура протоконха представ-
лена 3 тонкими спиральными ребрами. Осевая скульптура дефинитивной рако-
вины состоит из треугольных в сечении, широких складок, ярко выраженных как
на периферии оборота, так и на основании раковины. На околошовной поверх-
ности складки имеют значительный сигмоидальный изгиб, разделены они про-
межутками в 5—7 раз более узкими, чем сами складки. На последнем обороте
раковины насчитывается от 18 до 23 складок, на предпоследнем от 15 до 18.
Спиральная скульптура состоит из шнуровидных уплощенных широких ребер,
разделенных значительно более узкими межреберными промежутками. В ре-
зультате несоответствия ширины ребер и межреберных промежутков создается
впечатление о спиральной скульптуре, состоящей как бы из узких желобков.
Основание раковины умеренно выпуклое, с четкой границей, обозначенной
желобком в месте перехода в удлиненный сифональный вырост. Устье непра-
152
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
вильно-овальной формы, угловатое в верхней части, с небольшой выемкой аналь-
ного синуса на палатальном крае и массивной широкой полосой кремового цвета
каллуса на колумеллярном крае. Сифональный канал глубокий, средней ши-
рины. Размеры голотипа (мм): 2?р=17.О; Д=8.0; /?по=12.5;р?у=10.0; Др=3.5.
Наибольший экземпляр (с телом) из Баренцева моря имел раковину с парамет-
рами (мм): Вр=21.3; Д=12.0; 5ПО=18.3; /?3,=13.5; Др=5.0, наибольшая по раз-
мерам раковина (без тела), найденная
у о-ва Колгуев, имела параметры (мм):
Вр=31.4; Д=11.4; £„о=20.0; 5у=11.7;
я^.
Крышечка когтевидная, неправильно-
овальная, со значительно изгибающейся
осью нарастания. Мускульный отпечаток
может быть заостренным в верхней части
(рис. 33, П—Р, Т) или закругленным
(рис. 33, С). Он может плотно прилегать
к левому краю крышечки или распола-
гаться в срединной ее части.
Зубы радулы копьевидной формы, дли-
ной до 1 мм, с конусовидным оранжевым
основанием и большим, в 2—3 раза пре-
вышающим дистальное отверстие зуба
килем (рис. 415, 13—22). Лезвие зуба
слегка изогнуто. Одна из ручек несет
равнобедренную шпору. Исследованные
зубы имеют полное сходство с изображе-
ниями радулы паратипа (№223402; USNM),
присланными Дж. Роузвотером.
Вид сходен с Oe. impressa,, от кото-
рого отличается более округлыми оборо-
тами, осевыми толстыми складками, не
прорезанными спиральными глубокими
желобками, и конусовидным основанием
зуба.
Распространение. Широко,
распространенный бореально-арктический
вид тихоокеанского происхождения. Оби-
тает циркумполярно в Арктическом бас-
сейне и в морях Северной Атлантики и Се-
верной Пацифики. Найден в канадском
Рис. 455. (Места находок Oenopota секторе Арктики у м. Барроу, в водах
harpa в дальневосточных морях. Британской Колумбии, у западных и
восточных берегов Шпицбергена, в Нор-
вежском, Баренцевом, Карском морях и в море Лаптевых (рис. 454), а также
в Чукотском, Беринговом и Охотском морях (рис. 455).
Экология. Обитает на глубинах от 6 до 324 м, встречается в основном
на 15—110 м на заиленных с примесями камней, гальки, ракуши и песка грунтах
(более 55% находок), в меньшей степени — на других типах грунтов. Моллюски
обитают в диапазоне температур от —2.0 до 9.11 °C, преимущественно от —1.96
до 2.0 °C, при солености от 25.66 (в Белом море) до 35.04 %0, предпочитая мор-
скую соленость 33.6—34.9 %0.
Типы. Голотип Bela harpa хранится в USNM, № 40959; типовое местона-
хождение — о-в Нунивак,. Аляска. Типовой экземпляр В. schmidti хранится
в ZMO; типовое местонахождение — Берингово море.
Просмотрено 297 экз.
3. OENOPOTA
153
9. Oenopota (Nodotoma) cancellata (Mighels et Adams, 1840),- comb. n.
(Рис. 168-173; 399, В—Г; 417, 14-27; 418, 1—5).
Mighels, Adams, 1840 : 50 (Fusus); 1842 : 52, pl. 4, fig. 18 (Fusus); Gould, 1870 :
335, fig. 625 (Bela); Verrill, 1882 : 475, pl. 43, fig. 10—11, pl. 57, fig. 13 (Bela); Tryon,
1884 : 218, pl. 29, fig. 67 (Bela; part.); Martini, Chemnitz, 1887 : 149, Taf. 31, fig. 16
(Bela); К о b e 1 1, 1905 : 246, Taf. 83, fig. 1 (Bela).
Раковина белая, розоватая или коричневатая, веретеновидная, с 7—8 за-
кругленными выпуклыми оборотами. Периостракум плотный, легкоотстаю-
щий, светло-коричневый. Протоконх выпуклый, первый его оборот цилиндриче-
ской формы, украшен ретикулятным рисунком (рис. 399, В—Г). Спиральная
скульптура закладывается на второй половине эмбрионального оборота и со-
стоит из 3 тонких ребер. Осевая скульптура закладывается на втором эмбрио-
нальном обороте в виде тонких осевых складочек, имеющих наклон влево. Осе-
вая скульптура дефинитивной раковины представлена сильновыпуклыми, полу-
круглыми в сечении, средними (реже широкими) складками, равновеликими
с разделяющими их промежутками. Складки наиболее выражены в нижней
половине периферии оборотов и практически сглаживаются около шва и на
нижней 1/3 основания раковины. Элементы осевой скульптуры имеют значитель-
ный наклон назад и лишь в самой верхней части сигмоидально изогнуты. На
последнем обороте раковины насчитывается от 18 до 22 осевых складок, на пред-
последнем от 16 до 19. Спиральная скульптура на поверхности оборотов и
в верхней половине основания раковины представлена тонкими волосовидными
ребрышками, равновеликими между собой и с разделяющими их промежутками.
В нижней части основания раковины и на сифональном выросте спиральная
скульптура имеет вид тонких ребер, которые в 2—3 раза превышают по ширине
разделяющие их промежутки. Основание раковины умеренно выпуклое, си-
фональный вырост удлиненный. Устье узко-обратногрушевидной формы,
угловатое в верхней части; несет на колумеллярном крае кремового цвета
каллус, на палатальном — небольшую вырезку анального синуса. Сифональный
канал короткий, относительно широкий, неглубокий. Наибольшая высота
раковины 20, при диаметре 7.75 мм (Verrill, 1882).
Крышечка средних размеров, каплевидная, с плавно изгибающейся осью
нарастания. Мускульный отпечаток шлемовидной или ромбовидной формы,
расположен в левой части крышечки, но не прилегает к левому ее краю
(рис. 33, Е).
Зубы радулы копьевидной формы, длиной до 500 мкм, с массивным основа-
нием, несущим на одной из ручек заостренную шпору. Киль удлиненный,
с шлемовидной колющей Частью (рис. 417, 17). Имеющиеся изображения зубов
(Verrill, 1882 : pl. 57, fig. 13) и исследованные мной радулы внешне иден-
тичны .
Вид сходен с Oe. biconica, от которого отличается более выпуклыми оборотами
и прямым основанием раковины, менее развитой спиральной скульптурой и
узко-обратногрушевидной формой устья.
Распространение. Амфибореальный широко распространенный
вид тихоокеанского происхождения, отмечен в бореальных водах Тихого океана
и в высокобореальных водах американского побережья Атлантики. Помимо
типового местонахождения обитает в заливах Каско, Массачусетс (Gould, 1870),
у берегов Новой Шотландии ив Лабрадоре, обнаружен также в зал. Петра Вели-
кого в Японском море.
Экология. Найден на глубинах от 12 до 132 м, отмечен до 176 м (Ver-
Till, 1882), на илистых и заиленных грунтах при температуре от —0.6 до 8.8 °C и
солености 31.98—33.27 °/00.
Типы. Типовой материал, вероятно, хранится в BSNH, вид описан по
экземпляру, найденному в желудке пикши; из вод Новой Англии.
Просмотрено 34 экз.
154=
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
10. Oenopota (Nodotoma) casentina (Dall, 1919), comb. n. (Рис. 177—178;:
422, 1~3).
Dall, 1919b : 302 (Лога); Oldroyd, 1927 : 105 (Lora).
Раковина серого цвета, веретеновидная, с 5—6 оборотами отлогоугловатой
формы, тупоугловатыми в верхней части. Периостракум тонкий, полупрозрач-
ный, желтоватый. Верхние обороты корродированы, протоконх не сохранился.
Осевая скульптура дефинитивной раковины представлена на ранних оборотах
тонкими пологими складками, разделенными в 1.5—2 раза более широкими
пространствами. К последнему обороту складки становятся столь пологими, что
практически не выдаются над поверхностью раковины. На предпоследнем обо-
роте насчитывается около 16 складок. Все элементы осевой скульптуры приобре-
тают на плече оборота незначительный сигмоидальный изгиб. Спиральная
скульптура состоит из многочисленных, неупорядоченных в расположении,,
очень пологих и всегда шнуровидных средних и тонких ребер. Как правило,
в верхней части оборотов преобладают тонкие ребра, а в нижней части и на ос-
новании раковины — средние. Межреберные промежутки представляют собой
желобки, которые в 2—3 раза у^же самых тонких элементов спиральной скульп-
туры. Основание раковины слабовыпуклое, сифональный вырост относительно
короткий. Устье полуовальное, угловатое в верхней части, несет на колумел-
лярном крае узкую полоску тонкого каллуса кремового цвета. Вырезка аналь-
ного синуса слабо различима в верхней части палатального края устья. Сифо-
нальный канал короткий, глубокий и широкий. Размеры голотипа (мм): В =9.0;.
Д=5.0; /?ио=7.0. Раковина высотой 12.8 мм найдена у восточного побережья
о-ва Сахалин (47°52' с. ш.—-144°38' в. д.).
Крышечка когтевидная, с плавно изгибающейся осью нарастания. Ланцето-
видный мускульный след лежит в срединной части крышечки (рис. 33, Ж).
Зубы радулы длиной до 150 мкм, копьевидные, с высококоническим основа-
нием и длинным килем, в 2—4 раза превышающим размеры дистального отвер-
стия зуба (рис. 422, 1—3). Одна из ручек несет маленькую шпору.
Вид сходен с Ое. inequita, от которого отличается угловатыми оборотами,,
отсутствием выраженных пологих осевых складок и высококонической формой,
базальной части зуба радулы.
Замечания. Исследованные экземпляры практически не отличаются
от типового (фотография голотипа была предоставлена Дж. Роузвотером).
Распространение. Тихоокеанский широко распространенный бо-
реальный вид, обитает в высоко- и низкобореальных районах у берегов Азии и.
Америки. Обнаружен у восточных берегов Южного Сахалина и у Южных Ку-
рильских о-вов, и в зал. Монтерей, Калифорния.
Типы. Голотип хранится в USNM, № 209257; типовое местонахождение —
Point Pinos Light, зал. Монтерей, Калифорния.
Просмотрено 5 экз.
11. Oenopota (Nodotoma) inequita (Dall, 1919). (Рис. 175—176; 422, 4—9).-
Dall, 1919a : 44—45, pl. 16, fig. 9 (Lora); Oldroyd, 1927 : 96 (Lora); Clarke,
1963 : 98.
Раковина желтовато-белая, веретеновидная, с 6—6.5 оборотами отлогоугло-
ватой формы, тупоугловатыми в верхней части. Периостракум тонкий, полу-
прозрачный, коричневатый. Верхние обороты, как правило, сильно корродиро-
ваны. Скульптура протоконха неизвестна. Осевая скульптура дефинитивной ра-
ковины представлена пологими широкими складками, разделенными равными
им по ширине или в 1.2—1.5 раза меньшими промежутками. Складки наиболее-
выражены в верхней части оборотов, в верхней ]73 оборота они имеют также сла-
бый сигмоидальный изгиб. В нижней части оборота складки начинают сглажи-
ваться и исчезают уже на границе с основанием раковины. На последнем и пред-
последнем оборотах насчитывается от 16 до 20 осевых складок. Спиральная
3. OENOPOTA
155
скульптура состоит из многочисленных, различных по ширине и не упорядочен-
ных в расположении средних и узких шнуровидных ребер. Межреберные про-
межутки очень узкие, имеют вид желобков. Ребра нижней 1/3 основания и си-
фонального выроста расположены на большем расстоянии друг от друга —
межреберные промежутки здесь подчас равны по ширине ребрам. Основание
раковины сильно уплощенное, плавно переходит в удлиненный сифональный
вырост. Устье узко-обратногрушевидное, с неглубокой вырезкой анального
синуса в верхней части палатального края и с кремового цвета тонким каллу-
сом на колумеллярном крае. Сифональный канал протяженный, относительно
глубокий. Размеры голотипа (мм): Вр=11.0; Д=4.0; Впо—7.0.
Крышечка средних размеров, каплевидная, с плавно изгибающейся осью
нарастания. Ланцетовидный мускульный отпечаток расположен вблизи слабо
утолщенного левого края крышечки, но не прилегает к нему (рис. 33, И).
Зубы радулы маленькие, длиной до 150 мкм, копьевидные, с закругленно-
коническим основанием, несущим на одной из ручек заостренный треугольный,
на другой — полуовальный флажки (рис. 422, 4—9). Киль вдвое превышает раз-
меры дистального отверстия зуба. Исследованный материал морфологически
сходен с изображениями зубов радулы голотипа (рисунки Моррисона).
Вид похож на Ое. casentina, от которого отличается округлыми оборотами и
широкими пологими осевыми складками.
Изменчивость. Изображение голотипа в отличие от исследованных
мной моллюсков характеризуется более выраженным плечом оборота и скошен-
ным основанием раковины.
Распространение. Тихоокеанский широко распространенный бо-
реальный вид, обитает в низко- и высокобореальных районах у берегов Азии и
Америки. Встречен от зал. Пловер в Беринговом море до Аляски у американс-
кого побережья и в Японском море у азиатских берегов.
Типы. Голотип хранится в USNM, № 222238; типовое местонахождение
Plover Вау, Берингово море.
12. Oenopota (Nodotoma) pavlova (Dall, 1919), comb. n. (Рис. 184—186;
.397, В—Г; 418, 17-22).
Dall, 1919a : 43, pl. 14, fig. 9 (Lora).
Раковина белая, веретеновидная, с 6—6.5 закругленными, умеренно выпук-
лыми оборотами. Периостракум очень тонкий, плотно прилегающий, кремового
цвета. Протоконх усеченно-конусовидной формы, представлен 1.6—1.9 оборо-
тами; начальные этапы его развития сопровождаются появлением слабого рети-
кулятного рисунка. На втором эмбриональном обороте развивается килевидное
ребрышко, которое проходит по плечу оборота (рис. 397, В—Г). Осевая скульп-
тура дефинитивной раковины представлена пологими средними складками, наи-
более выраженными в верхней части оборота и имеющими на околошовной по-
верхности (верхняя 1/3 оборота) сигмоидальный изгиб. Складки сглаживаются,
не достигая основания раковины. Они разделены в 2 раза более узкими про-
странствами, чем сами складки. На последнем обороте насчитывается от 18 до
22 осевых складок, на предпоследнем от 16 до 18. Спиральная скульптура пред-
ставлена многочисленными, равновеликими на всей поверхности раковины шну-
ровидными ребрышками, которые превышают по ширине разделяющие их про-
межутки в 2 раза. На первых 2—3 постэмбриональных оборотах отмечено явное
преобладание 2 килевидных ребрышек над остальными элементами спиральной
.скульптуры. Они разделяют периферию оборота на практически равные 3 части.
•Однако уже на четвертом дефинитивном обороте ребрышки сглаживаются.
Основание раковины уплощенное, плавно переходящее в удлиненный сифональ-
ный вырост. Устье обратногрушевидной формы, с неглубоким вырезом анального
синуса в верхней 1/4—х/5 палатального края и с тонким кремового цвета каллу-
<сом на колумеллярном крае. Сифональный канал широкий, неглубокий. Раз-
156
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
меры типового экземпляра (мм): 7?р=18.5; Д=9.0; 7?ПО=12.5. Максимальные-
размеры раковины в исследованном материале составляли 12.7 мм.
Крышечка с плавно изгибающейся осью нарастания. Ланцетовидный мус-
кульный отпечаток расположен вблизи утощенного левого ее края.
Зубы радулы копьевидные, очень изящные, длиной до 150 мкм, с округло-
конусовидным основанием, большим флажком на одной из ручек и длинным,
в 4—5 раз превышающим дистальное отверстие зуба килем (рис. 418,
17—22).
Вид сходен с Ое. elegans, от которого отличается менее округлым основанием
раковины, более тонкой осевой скульптурой и мелкими зубами с конусовидной
базальной частью. От большинства видов рода отличается скульптурой прото-
конха только с одним развитым килевидным ребром.
Изменчивость. Моллюски из коллекционных материалов ЗИН АН
СССР обладают более стройными раковинами по сравнению с типовым экземпля-
ром.
Распространение. Тихоокеанский широко распространенный бо-
реальный вид, обитающий в низко- и высокобореальных районах у берегов
Азии и Америки. Помимо типового местонахождения отмечен в северной части
Японского моря и у берегов о-ва Сахалин.
Типы. Голотип хранится в USNM, № 226200; типовое местонахождение —
о-ва Прибылова, Берингово море.
Просмотрено 13 экз.
13. Oenopota (Nodotoma) uschakovi Bogdanov, 1985. (Рис. 189—190; 399,
Д—Е; 418, 13-16).
Богданов, 1985а : 450, рис. 1, а—г; 2, 1.
Раковина телесного цвета, тонкая, удлиненно-веретеновидная, с 5.3 за-
кругленными, умеренно выпуклыми оборотами, с прижатым швом. Периостра-
кум тонкий, желтоватый. Протоконх выпуклой формы, представлен 1.5 уступ-
чатыми оборотами. Первый эмбриональный оборот несет следы ретикулятного
рисунка, на втором обороте развиваются 3 тонких спиральных ребра, из кото-
рых одно обозначает плечо оборота, другое — нижнюю часть периферии и на-
ходится на стыке с последующим оборотом, третье ребро обозначает середину
периферии оборота (рис. 399, Д). Узкие осевые валики появляются на последней
1/,1 эмбриональной раковины. Осевая скульптура на протоконхе и первых 2 де-
финитивных оборотах состоит из почти прямых, параллельных оси нарастания
раковины узких складок, по ширине вдвое меньших, чем разделяющие их про-
межутки. На более поздних оборотах осевые складки приобретают легкий сиг-
моидальный изгиб на околошовной поверхности. Осевые складки отчетливее
выражены на периферии оборота и сглаживаются к его основанию. На послед-
нем обороте раковины насчитывается 19 складок, на предпоследнем 18. Среди
элементов спиральной скульптуры первых постэмбриональных оборотов выде-
ляются 3 более высоких ребрышка, являющихся продолжением эмбриональных,
и многочисленные волосовидные ребрышки, имеющие постэмбриональную за-
кладку. На поздних дефинитивных оборотах разниц,. их размерах стирается,,
и они становятся равновеликими и равноудаленными еренно выпуклое осно-
вание раковины плавно переходит в удлиненный сифона ьный вырост и покрыто
такими же по величине ребрышками, как и периферия последнего оборота.
Устье неправильно-овальной формы, с тонким белым каллусом на колумелляр-
ном крае. Сифональный канал широкий и глубокий. Размеры голотипа (мм):
5р=10.4; Д=4.8; 5„О=7.7; В,=ЪЛ\ Др=2.3.
Зубы радулы крупные, длиной до 300 мкм, копьевидной формы, с массивным
коническим основанием, несущим 2 тонкие ручки. На одной из ручек располо-
жена треугольная шпора. Киль в 2.5 —3 раза превышает размеры дистального,
отверстия лезвия.
3. OENOPOTA
157-
Виц наиболее сходен с Ое. cancellata, от которого отличается низкими и менее
изогнутыми осевыми складками, массивным основанием зуба радулы и цилинд-
рической формой оборотов эмбриональной раковины.
Распространение. Тихоокеанский приазиатский низкобореаль-
ный вид. Встречен у Южных Курильских островов и в южной части Охотского,
моря.
Типы. Голотип хранится в ЗИН АН СССР, № 37974, типовое местона-
хождение — зал. Рока, о-в Итуруп (44°24'2'' с. ш.—147°09'1" в. д., глубина
160 м.)
Просмотрено 3 экз.
14. Oenopota (Nodotoma) biconica Bogdanov, 1989. (Рис. 174; 422, 10—18)-
Богданов, 1989а : 134, рис. 1, 3; 2, в—е.
Раковина серого цвета, веретеновидная, почти биконическая, с 5.5—6 обо-
ротами, толстостенная. Периостракум светло-серый, слегка желтоватый, лег-
коотстающий. Эмбриональная раковина не изучена. Осевая скульптура дефини-
тивной раковины состоит из толстых пологих, сигмоидально изогнутых осевых
складок, которые значительно превышают по ширине разделяющие их проме-
жутки. По высоте осевые складки наиболее выражены в верхней части перифе-
рии оборота и постепенно сглаживаются ко шву, так что на основании раковины
отсутствуют даже следы складок. Спиральная скульптура представлена различ-
ными по ширине средними ребрами, прямоугольными в сечении. Между сред-
ними ребрами закладываются тонкие, однако все межреберные промежутки оди-
наковы по ширине. Скошенное основание раковины плавно переходит в корот-
кий сифональный вырост. Устье полуовальной формы, с толстым матовым, слег-
ка розоватым каллусом на колумеллярном крае. Размеры типового экземпляра
(мм): Вр=10.0; Д=4.8; Впо=7.1; Ву=5.1; Ду =2.3.
Крышечка с плавно изгибающейся осью нарастания, проходящей вдоль.,
утолщенного левого края. Мускульный отпечаток довольно узкой полосой ле-
жит по правую сторону от оси нарастания (рис. 33, 3).
Зубы радулы мелкие, длиной 140—160 мкм, копьевидной формы, с массив-
ным округлым основанием и длинным килем, в 2—2.5 раза превышающим дис-
тальное отверстие зуба (рис. 422, 15).
Вид наиболее сходен с Ое. cancellata, от которого отличается исчезающими на;
скошенном основании раковины осевыми складками, наличием средних шнуро-
видных спиральных ребер, различных по величине. Скошенность основания'
придает раковине биконическую форму.
Распространение. Тихоокеанский приазиатский низкобореаль-
ный вид, известен только из зал. Ольги, Японское море.
Типы. Голотип хранится в ЗИН АН СССР, № 48469; типовое местонахож-
дение — зал. Ольги, Японское море, глубина 22 м.
Просмотрен 1 экз.
15. Oenopota (Nodotoma) kurilensis Bogdanov, 1989. (Рис. 179—183; 418.
6—12).
Б отдано в, 19896 : 150, рис. 1, 3—4; 2, д—з.
Раковина, как правило, оливкого цвета, веретеновидная, с 4.5—5.5 закру-
гленными, умеренно выпуклыми оборотами, очень тонкостенная. Периостракум
очень тонкий, прозрачный, плотно прилегает к раковине. Эмбриональная рако-
вина не изучена. Осевая скульптура дефинитивной раковины слагается из часто
расположенных, сигмовидно изогнутых средних складок, равновеликих по вы-
соте и ширине. Наибольшее развитие осевые складки получают на околошовной
пространстве, на периферии оборота они постепенно сглаживаются. У ряда
просмотренных экземпляров уже на периферии последнего оборота происходит
практически полное исчезновение осевых складок, на других раковинах сгла-
U58
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
живание складок приурочено к основанию раковины (рис. 182—183). Спираль-
ные ребра многочисленны, расположены равномерно по всей поверхности рако-
нины, они практически одноразмерны как между собой, так и с разделяющими
их межреберными промежутками. Основание раковины плавно переходит
в средних размеров сифональный вырост. Устье неправильно-овальной формы,
усеченное в верхней части. Тонкий каллус более светлых тонов, чем раковина,
покрывает большую часть колумеллярного края устья. Сифональный канал
широкий и относительно глубокий. Средние размеры раковины 10—12 мм, раз-
меры раковины голотипа (мм): Z?p=11.8; Д=5.5; Впо=10.6; Ву=6.3; Д —2.5.
Зубы радулы копьевидные, длиной 200—250 мкм, с массивной базальной
настью и шлемовидным дистальным участком киля (рис. 418, 8). Треугольная
шпора расположена на одной из ручек.
Вид сходен с Oe. cancellata, от которого отличается тонкостенной раковиной,
многочисленными осевыми складками меньших размеров, но с более выражен-
ным сигмоидальным изгибом, а также массивной, неправильной формы базаль-
ной частью зуба.
Распространение. Тихоокеанский приазиатский широко распро-
страненный бореальный вид, встречается в водах, омывающих Южные, Средние
,и Северные Курильские острова.
Типы. Голотип хранится в ЗИН АН СССР, № 48470; типовое местонахож-
дение — Охотское море, 46°59' с. ш.—152°11' в. д., глубина 11 м.
Просмотрено 38 экз.
16. Oenopota (Nodotoma) elongata Bogdanov, 1989. (Рис. 187; 188; 422,
19—23).
Богданов, 1989а : 137, рис. 1, 4—5', 2, ш—к.
Раковина кремового цвета, удлиненно-веретеновидная, с 4.5—5 закруглен-
ными, умеренно выпуклыми оборотами. Периостракум тонкий, прозрачный,
плотноприлегающий. Протоконх неизвестен. Осевая скульптура дефинитивной
раковины слагается из большого числа средних складок (до 20 на предпоследнем
•обороте и до 24 на последнем). Элементы осевой скульптуры, повторяя профиль
палатального края устья, имеют в верхней части сигмоидальный изгиб. Складки
равновелики как на околошовной поверхности, так и на периферии, однако
на основании раковины они сглаживаются. Элементы спиральной скульптуры
представлены в основном двумя типами ребер. На периферии оборота располага-
ются 3—4 округлых в сечении, спиральных шнуровидных ребра. Между ними,
а также на всей околошовной поверхности и на основании раковины располо-
жены практически одинаковые по величине многочисленные тонкие спиральные
ребра, ширина которых уменьшается с переходом на сифональный вырост.
Основание раковины округлое, сифональный вырост длинный (у голотипа
2.7 мм), составляет более 1/4 всей высоты раковины. Устье обратногрушевидное.
Как правило, весь колумеллярный край покрыт толстым матовым каллусом.
Сифональный канал глубокий. Размеры голотипа (мм): 2?gJ=8.0; Д=3.7; Вп0=
= 5.9; 7?2,=4.6; Д =1.7. Наибольшая высота раковины 8.3 мм.
Зубы радулы длиной до 176 мкм имеют ножевидную форму (рис. 401).
Коническое основание (рис. 422., 21—23) на одной из ручек несет треугольную
шпору. Киль превышает 1/4 общей длины зуба.
Вид наиболее сходен с Oe. uschakovi, от которого отличается округлым основа-
нием раковины, длинным сифональным выростом, а также наличием сигмо-
видного изгиба осевых складок.
Распространение. Тихоокеанский приазиатский низкобореаль-
пый вид. Известен только из типового местонахождения.
Типы. Голотип хранится в ЗИН АН СССР, № 48471; типовое местонахожде-
ние — Японское море (42°17' с. ш.—131°11' в. д.), на глубине 1 076—1 400 м
на илистом с конкрециями грунте.
Просмотрено 2 экз.
4. OBESOTOMA
159>
4. Род OBESOTOMA Bartsch, 1941
Bartsch, 1941 : 4; Powell, 1966 : 121; Богданов, 1986 : 45 (Oenopota'
subg.). — Oenopota (part.): Morch, 1852 : 73; В о u c h e t, Waren, 1980 : 66. — Bela:
(non Gray, 1847; part.): G. 0. S a r s, 1878 : 222; Friele, 1886 : 2; Martini, Chem-
nitz, 1887 : 137-
Типовой вид Obesotoma japonica Bartsch, 1941, по первоначальному
обозначению.
Раковина овально-веретеновидной, реже овальной формы, больших размеров,
(в среднем 17—23 мм), как правило, толстостенная, Эмбриональная раковина
выпуклая, слабо орнаментирована или совсем лишена скульптуры. Осевая
скульптура дефинитивной раковины варьирует у разных видов от линий нараста-
ния до широких пологих складок. Сигмоидальный изгиб на элементах осевой
скульптуры отсутствует или слабо выражен. Спиральная скульптура всегда
слабо развита. Основание раковины скошено, сифональный вырост небольших
размеров. Крышечка с плавно изгибающейся или практически прямой осью,
нарастания. Мускульный отпечаток занимает срединную часть крышечки.
Зубы радулы копьевидной или саблевидной формы, с относительно неболь-
шим ромбовидным килем.
Род распространен по всему ареалу подсемейства. В ископаемом состоянии
представители рода известны для северной части Тихого океана начиная с плио-
цена (Камчатка, Япония, Калифорния), для северной Атлантики — с плейсто-
цена (Англия, северо-восток европейской части СССР).
ЕГАВЛИЦА^ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (4). Осевая скульптура представлена только линиями нарастания.
2 (3). Раковина приземистая, отношение высоты раковины к высоте последнего'
оборота от 1.19 до 1.25. Зубы радулы саблевидной формы..................
.........................................2. О. simplex (Middendorff)
3 (2). Раковина стройная, отношение высоты раковины к высоте последнего»
оборота от 1.30 до 1.43. Зубы радулы копьевидной формы..................
............................................. 5. О. gigantea (Morch)
4 (1). Осевая скульптура представлена в той или иной мере развитыми осе-
выми складками.
5 (10). Раковина очень крупная, средние размеры 16—20 мм.
6 (7). Обороты слабовыпуклые, в результате чего завиток приобретает кони-
ческую форму. Зубы радулы мелкие, длиной менее 150 мкм . . . .
............................................. 1. О. japonica Bartsch
7 (6). Обороты умеренно выпуклые, завиток уступчатой формы. Зубы радулы,,
как правило, превышают 200 мкм в длину.
8 (9). Спиральная скульптура на околошовной поверхности представлена
в 1.5—2 раза более тонкими ребрами, чем на периферии оборота, в резуль-
тате на раковине всегда четко определяется плечо оборота. Киль зуба
радулы всегда более общей длины зуба....................................
...................................... 13. О. sachalinensis Bogdanov
9 (8). Спиральная скульптура практически на всей поверхности раковины
представлена одноразмерными ребрышками. Плечо оборота не обособлено..
Киль зуба радулы всегда менее Ve общей длины зуба.......................
...................................... 3. О. schantarica (Middendorff}
10 (5). Раковина средних размеров (10—13 мм), как правило, не более 16 мм
высотой.
11 (12). Зубы радулы ножевидной формы ..................................
..............................12. О. starobogatovi Bogdanov sp. n..
12 (11). Зубы радулы копьевидной формы.
13 (20). Раковина составлена закругленными оборотами, в разной степени:
выпуклыми, четкой границы между плечом и периферией оборота не-
наблюдается.
160
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
14 (17). Осевая скульптура представлена очень пологими складками, как пра-
вило, исчезающими уже на нижней г13 периферии оборота.
15 (16). Спиральная скульптура представлена округлыми тонкими ребрами.
Основание раковины четко отграничено резким изгибом от вытянутого
сифонального выроста...............................4. О. gigas (Verkriizen)
16 (15). Спиральная скульптура представлена широкими шнуровидными реб-
рами. Основание раковины плавно переходит в сифональный вырост
.................................................6. О. tenuilirata (Dall)
17 (14). Осевая скульптура представлена высокими широкими складками,
сглаживающимися только на нижней половине основания раковины.
18 (19). Раковина почти овальная, сифональный вырост короткий. Базальная
часть зуба радулы округлая......................10. О. tomiyaensis (Otuka)
19 (18). Раковина веретеновидная, сифональный вырост удлиненный. Базаль-
ная часть зуба радулы высококоническая...............11. О. solida (Dall)
20 (13). Раковина составлена отлогоугловатыми или уступчатыми оборотами,
имеющими четкую границу между плечом и периферией.
21 (22). Раковина составлена уступчатыми оборотами, уплощенными в верх-
ней части. Осевые складки исчезают на периферии оборота. Околодисталь-
ная часть лезвия зуба большего диаметра, чем остальная часть лезвия
......................................... 8. О. tumida (Posselt et Jensen)
22 (21). Раковина составлена отлогоугловатыми оборотами. Осевые складки
не исчезают на периферии оборота. Лезвие зуба одного диаметра на всем
протяжении.
23 (24). Плечо занимает верхнюю 1li оборота раковины. Осевые складки
у взрослой раковины многочисленны — более 18 на последнем обороте.
Зубы радулы большие, в длину превышают 20'0 мкм..........................
......................................................................................7. О. woodiana (Moller)
24 (23). Плечо занимает от V2 до 2/3 всего оборота раковины. Осевые складки
у взрослой раковины немногочисленны, менее 18 на последнем обороте.
Зубы радулы небольшие, в длину менее 150 мкм.............................
.................................................................................. 9. О. robusta (Packard)
1. Obesotoma japonica Bartsch, 1941. (Рис. 259—261; 424, 6—7).
В artsh, 1941 : 7, pl. 1, fig. 11; Powell, 1966 : 121, pl. 19, fig. 13; Голиков,
Гул ьбин, 1977 : 239 (Oenopota).
Раковина телесного цвета, веретеновидная, с 5—7 закругленными, умеренно
выпуклыми оборотами. Периостракум кожистый, непрозрачный, плотноприле-
гающий. Скульптура эмбриональной раковины не исследована. Осевая скульп-
тура дефинитивной раковины представлена пологими широкими складками,
в верхней части имеющими незначительный сигмоидальный изгиб. Складки
выражены только на периферии оборота и разделены равновеликими им по ши-
рине промежутками. По мере навивания оборотов раковины осевая скульптура
ослабевает и сглаживается, так что у некоторых экземпляров на последнем обо-
роте складки слабо различимы. Их число на последнем и предпоследнем обо-
ротах колеблется от 14 до 16. Спиральная скульптура представлена многочис-
ленными, равновеликими на всей поверхности раковины тонкими ребрами, раз-
деленными вдвое более узкими межреберными промежутками. Основание рако-
вины уплощенное, сифональный вырост удлиненный. Устье полуовальной
формы, с широкой полосой толстого кремового цвета каллуса. Сифональный
канал протяженный, широкий, глубокий. Высота раковины типового экземпляра
124.9, ширина 12.5 мм. Наибольшие размеры раковины (мм) отмечены для мол-
люска, найденного у о-ва Шикотан на глубине 8 м: В =28.0; Д=13.0; £?по=
21.0; ^=16.0; Д^=5.0.
Зубы радулы копьевидной формы, с коническим основанием и маленьким
^ромбовидным килем, длиной не более 150 мкм.
4. OBESOTOMA
161
Вид сходен с О. robusta, от которого отличается большими размерами рако-
вины, отсутствием угловатости оборотов и большим числом равномерно распо-
ложенных тонких шнуровидных спиральных ребер.
Распространение. Тихоокеанский приазиатский низкобореаль-
ный вид, обитает в Японском море и у берегов Малой Курильской гряды.
Экология. Обитает на глубинах от 8 до 380 м, отмечен до 700 м (Голи-
ков, Гульбин, 1977). Распространен в основном на глубинах 15—110 м, преиму-
щественно на чистом песке.
Типы. Голотип хранится в USNM, № 204967; типовое местонахождение —
Японское море, глубина 763 м.
Просмотрено 9 экз.
Рис. 456. Места находок Obesotoma simplex в северных морях СССР и в сопредельных водах.
2. Obesotoma simplex (Middendorff, 1849). (Рис. 214—218; 400, Д-, 424,
13-20- 456; 457).
Middendorff, 1849 I 119 (Pleurotoma}', 1851 t 223, Taf. 12, fig. 15, 16 (Pleurotoma}',
G. O. Sars, 1878 I 239, Tab. 17, fig. 4, Tab. 23, fig. 11, Tab. 9, fig. 9 (Bela', = laevigata); Гер-
ценштем, 1885 I 692 (Pleurotoma)', Odhner, 1910 I 11 (Bela violaceae var.); Oldroyd,
1927 1120 (Lora); MacGinitie, 1959 I 133, pl. 16, fig. 2 ( = laevigata); S h i-
m ek, Kohn, 1981 I 428, fig. 11, A (Oenopota). — laevigata Dall, 1871 1 98, pl. 16, fig. 7
(Bela)', 1886b I 300, pl. 3, fig. 7 (Bela). — moerchi L e c h e, 1878 1 57, pl. 1, fig. 18 (Pleurotoma
violacea var.); Oldroyd, 1927 I 112 (Lora). — schantarica (non Middendorff, 1849)l Try-
fl n, 1884 1 214, pl. 30, fig. 96, 97, pl. 29, fig. 56 (Bela-, part.); Голиков, Гульбин,
1977 I 242 (Oenopota', part.).
Раковина, как правило, сиреневого или фиолетового цвета, овально-веретено-
видная, с 4—5 закругленными, умеренно выпуклыми оборотами. Периостра-
кум очень тонкий, полупрозрачный. Завиток составляет 1/5—1/7 высоты рако-
вины. Верхние обороты дефинитивной раковины, как правило, корродированы.
Протоконх сильновыпуклый, представлен 1.5—1.7 закругленными, умеренно
выпуклыми оборотами. Эмбриональная раковина практически лишена, скульп-
турных элементов, если не считать слабую спиральную струйчатость (рис. 400,
Д). Осевая скульптура дефинитивной раковины как таковая отсутствует. Иногда
линия нарастания сопровождается незначительными осевыми утолщениями.
Спиральная скульптура представлена многочисленными волосовидными реб-
рышками, равновеликими между собой и с разделяющими их межреберными
промежутками (на всей поверхности раковины). Основание раковины уплощен-
ное, плавно, без видимой границы переходящее в удлиненный сифональный
вырост. Устье полуовальной формы, с широкой полосой толстого кремового
цвета каллуса на колумеллярном крае. Сифональный канал широкий, относи-
11 И. П. Богданов
162
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
тельно глубокий. Максимальная высота раковины 15 мм отмечена для моллюска
из Карского моря (73° 36' с. ш.—78° 22' в. д.).
Крышечка когтевидная, с плавно изгибающейся осью нарастания, широкий:
мускульный отпечаток лежит в срединной части крышечки.
Зубы радулы саблевидной формы, с отогнутым основанием. Базальная
часть зуба овальной формы, несет 2 ручки, вооруженные флажками, один из-
которыхтреугольной, другой — полулунной формы. Киль небольших размеров,,
ромбовидный, ненамного превышает дистальное отверстие зуба.
Рис. 457. Места находок Obesotoma simplex
в дальневосточных морях СССР.
Рис. 458. Места находок Obesotoma schan—
tarica в морях СССР.
Вид похож на О. schantarica, от которого отличается отсутствием широких,
осевых складок и саблевидной формой зубов.
Изменчивость. Вид очень изменчив как по форме (стройности или
уплощенности), так и по окраске раковины. На этом основании одни авторы
сводили номинальные виды в синонимы к О. simplex, другие дробили вид. От-
личия, найденные Доллом при описании Bela laevigata (в основном параметричес-
кие), несущественны и вписываются во внутривидовую изменчивость В. simplex.
К примеру, экземпляры из канадского сектора Арктики сравнивались с моллюс-
ками, обнаруженными у Восточной Гренландии, и оказались сходными (Мас-
Ginitie, 1959 : 133). Подробно вопрос изменчивости раковины О. simplex рас-
смотрен в Общей части.
4. OBESOTOMA
163
Распространение. Широко распространенный бореально-аркти-
ческий вид, обитает циркумполярно в Арктическом бассейне (рис. 456), отмечен
в Беринговом, Охотском и Японском морях (рис. 457) и в Северной Атлантике.
Отмечен для вод западного побережья Гренландии, севера и востока Исландии
{Thorson, 1944), а также у берегов Аляски (Dall, 1886а).
Экология. Обитает от литорали до 180 м, предпочитая глубины 5—
40 м; на всех типах грунтов, в основном на песчанистых (более 75 % находок).
Моллюски встречаются чаще всего при температурах от —0.6 до 0.8 °C, хотя
диапазон температур, при которых найдены моллюски, составляет от —1.47
до 5.03 °C. Вид относительно эвригалинный, обитает в воде со значениями соле-
ности 18.82—33.95, в основном при 29.4—31.6 °/оо-
Типы. Типовой экземпляр Pleurotoma simplex должен был храниться
в ЗИН АН СССР, но, по-видимому, утерян, так как мы его не смогли обнару-
жить; типовое местонахождение — Охотское море. Тип Bela laevigata находится
в USNM; типовое местонахождение — St. Michael, Нортон Саунд, Аляска.
Место хранения типов Р. violacea var. morchi установить не удалось; типовое
местонахождение — Маточкин Шар, Новая Земля.
Просмотрено 245 экз.
3. Obesotoma schantarica (Middendorff, 1849). (Рис. 219—220; 424, 1—5;
458).
Middendorff, 1849 i 118 (Pleurotoma)', 1851 : 223, Taf. 12, fig. 17—19 (Pleurotoma);
Tryon, 1884 i 214, pl. 34, fig. 76 (Bela; part.); H a b e, I t o, 1965 t 81, pl. 30, fig. 16; P o-
w e 1 1, 1966 1 121; Голиков, Гульбин, 1977 I 242 (Oenopota; part.); Петров,
1982 i 61, табл. 9, рис. 20 (Oenopota).
Раковина пепельно-серая, овально-веретеновидная, очень толстостенная,
с 5—6 закругленными, умеренно выпуклыми оборотами. Периостракум тол-
стый, кожистый, легкоотстающий, коричневатых тонов. Верхние обороты ра-
ковины сильно корродированы. Протоконх неизвестен.
Осевая скульптура дефинитивной раковины представлена широкими поло-
гими складками, разделенными в 1.5—2 раза более узкими промежутками.
Складки немного более рельефны на периферии оборота, чем на основании ра-
ковины и сифональном выросте. Они имеют плавный сигмоидальный изгиб, рас-
пространяющийся практически на всю поверхность периферии оборота. На пос-
леднем и предпоследнем обороте раковины насчитывается около 14—16 осевых
складок. Спиральная скульптура состоит из многочисленных нитевидных ребры-
шек, реже тонких ребер, равновеликих на всей поверхности оборотов, основа-
ния раковины и на сифональном выросте. Промежутки, разделяющие ребра,
равны им по ширине или немногим уже их. Основание раковины сильно упло-
щенное, сифональный вырост короткий. Устье полуовальной формы, угловатое
в верхней части, с широкой полосой толстого белого каллуса на колумеллярном
крае. Палатальный край толстостенный, с незначительной выемкой анального
синуса в верхней части. Сифональный канал короткий, очень широкий и глубо-
кий. Самый крупный экземпляр с высотой раковины 28 мм был обнаружен
в зал. Петра Великого (Японское море).
Крышечка очень плотная, коричневая, с сильно изгибающейся осью нараста-
ния. Мускульный отпечаток расположен в срединной части крышечки, широкий.
Зубы радулы копьевидной формы, длиной до 250 мкм, с практически равным
дистальному отверстию ромбовидным килем. Высококоническое основание от-
клонено от оси зуба (рис. 424, .3).
Вид сходен с О. sachalinensis, от которого отличается отсутствием угловатости
оборотов и мелкими зубами с отогнутым основанием, несущим только 1 флажок.
Замечания. В связи со значительной путаницей в диагностике вида,
имеющейся в литературе, многие данные по распространению и экологии вида
оказываются сомнительными.
Распространение. Тихоокеанский приазиатский .широко распро-
страненный бореальный вид, обитает в низко- и высокобореальных районах
11»
164
систематическая часть
у берегов Азии. Вид отмечен в зал. Петра Великого и в Татарском проливе?
в Японском море, в Охотском море, у Курильских о-вов и берегов Восточной
Камчатки, в северной части Берингова моря и у о-ва Врангеля (рис. 458).
Экология. Обитает на глубинах от 14.5 до 400 м, в основном на песча-
нистых грунтах, реже с примесями ила, гальки, гравия, при температурах от
—1.47 до 12.4 °C и солености 33.51—34.26 °/00.
Типы. Типовой экземпляр хранится в ЗИН АН СССР, № 21721; типовое-
местонахождение — район Шантарских о-вов, Охотское море.
Просмотрено 85 экз.
Рис. 459. Места находок Obesotoma gigantea (1) и О. gigas (2); 3 — палеонтологические на-
ходки.
4. Obesotoma gigas (Verkriizen, 1875),4 comb. n. (Рис. 221; 421, 11—14;.
459, 2).
V erkuzen, 1875 1 239, Taf. 8, fig. 6, 7 (Pleurotoma (Bela)]; Martini, Che m-
n i t z, 1887 ! 160, Taf. 32, fig. 16 (Bela); К о b e 1 t, 1905 t 264, Taf. 84, fig. 12 (Bela schanta-
rica subsp.). — schantarica (non Middendorff, 1849)t T г у о n, 1884 i 214, pl. 33, fig. 70 (Bela;
part.); Friele, 1886 1 17, pl. 8, fig. 20, pl. 10, fig. 17, 1.8 (Bela; part.).
Раковина светло-коричневая, овально-веретеновидная, с 4—6 закруглен-
ными, умеренно выпуклыми оборотами. Периостракум тонкий, полупрозрачный,,
зеленовато-коричневый. Верхние обороты окатаны, форма и скульптура прото-
конха неизвестна. Осевая скульптура дефинитивной раковины представлена,
очень пологими тонкими складками, неравномерно расположенными на поверх-
ности оборота и разделенными пространствами, равными им по ширине или.
в 2—4 раза большими. Складки одинаковы по величине как на поверхности обо-
рота, так и на основании раковины. Число их непостоянно у различных экземп-
ляров, но, как правило, превышает 20 на последнем и предпоследнем оборотах.
Спиральная скульптура представлена равновеликими на всей поверхности
последнего оборота и основания раковины тонкими шнуровидными ребрами,
разделенными вдвое более узкими межреберными пространствами. Основание
раковины сильновыпуклое, резко отграничено от укороченного сифонального
выроста. Устье полуовальной формы, с широкой полосой тончайшего телесного
цвета каллуса на колумеллярном крае. Палатальный край составлен очень-
тонкой стенкой и несет в верхней своей 1/7—4/8 части неглубокую вырезку
анального синуса. Сифональный канал широкий, глубокий, средней длины-
Размеры типового экземпляра (мм): 2?p=12.0; Д=8.0; Z?2,=10.0; Д^=5.5.
4 Авторство вида часто указывается как «Beck in Verkriizen»; см. примем, на с. 142..
4. OBESOTOMA
165
Крышечка крупная, как правило, с прямой осью нарастания, проходящей
по срединной части. Мускульный отпечаток небольшой, заостренный в верхней
части, лежит в центре крышечки (рис. 34, А).
Зубы радулы копьевидной формы, длиной свыше 300 мкм, с небольшим кону-
совидным основанием, несущим на каждой из ручек по флажку: один из них
треугольной, другой — полуовальной формы (рис. 421, 13). Киль превышает
в 2—3 раза размеры дистального отверстия зуба. Диаметр дистального участка
лезвия зуба несколько больший, чем в околобазальной части. Для
исследованных экземпляров свойственны те же морфологические особенности зубов
радулы, что изображены Фриле для Bela schantartca(FAQ\Q, 1886 : pl. 10, fig. 17,
18). Данный факт свидетельствует о неправильной трактовке многими европей-
скими исследователями вида О. schantarica.
Вид сходен с О. gigantea, от которого отличается низким завитком, полуо-
вальной формой устья и формой зубов радулы, в основном закругленной базаль-
ной частью и протяженным килем.
Распространение. Евразийский арктическо-бореальный вид высо-
кобореального атлантического происхождения, отмечен в водах Северной Норве-
гии, у Шпицбергена, в Баренцевом и Белом морях (рис. 459, 2).
Типы. Место хранения типового экземпляра не указано (возможно, ZMO);
типовое местонахождение — Вадсё, Норвегия.
Просмотрено 8 экз.
5. Obesotoma gigantea (Morch, 1869), comb. n. (Рис. 222—225; 425, 1—10',
459, 1, 3).
Morch, 1869 i 22 [Pleurotoma (Bela) violacea var.]; L e c h e, 1878 I 57, Tab. 1, fig. 19
(Bela violacea var.); Aurivillius, 1887 t 348 (Bela violacea var.); Posselt, Jen-
sen, 189.8 i 158 (Bela violacea var.; part.); Knipowitsch, 1901 I 48, Taf. 18, fig. 10—11
(Bela); К о b e 1 t, 1905 I 263, Taf. 84, fig. 20, 21 (Bela bicarinata subsp.); D an t z e nb erg,
Fischer, 1912 i 43, pl. 1, fig. 1, 2 (Bela); Qdhner, 1915 I 221 (Bela); Harmer, 1915:
291, Tab. 32, fig. 24 (Bela). — woodiana (non Moller, 1842) i Knipowitsch, 1901 ! 49,
Taf. 18, fig. 12, 13 (Bela).
Раковина кремовая или светло-коричневая, веретеновидная, с 5.5—6.5 обо-
ротами отлогоугловатой формы, тупоугловатыми в верхней части. Периостра-
кум очень тонкий, коричневый, полупрозрачный, легкоотстающий. Верхние
обороты дефинитивной раковины, как правило, корродированы, скульптура про-
токонха неизвестна. Осевая скульптура дефинитивной раковины представлена
неупорядоченными в расположении на поверхности оборота линиями нарастания.
Последние наиболее заметны в верхней части периферии оборотов, хотя просле-
живаются и на основании раковины. Спиральная скульптура состоит из много-
численных нитевидных ребрышек, равновеликих на всей поверхности раковины
как между собой, так и с разделяющими их межреберными промежутками. Ос-
нование раковины умеренно выпуклое, плавно переходящее в удлиненный си-
фональный вырост. Устье широко-обратногрушевидной формы, с широкой по-
лосой тонкого, кремового или коричневатого цвета каллуса на колумеллярном
крае. Сифональный канал узкий, глубокий. Параметры самого крупного, до-
стоверно идентифицированного экземпляра (мм): Z?p=19.0; Д=10.0; Z?no=12.5;
^=12.0; найден у Маточкина Шара, Новая Земля (Leche, 1878). В литературе
указывается на обнаружение моллюсков с высотой раковины до 22.5—24.6 мм
(Odhner, 1915).
Крышечка каплевидная, с практически прямой осью нарастания. Мускуль-
ный отпечаток небольшой.
Зубы радулы очень маленькие, длиной 180—220 мкм, копьевидной формы,
с высококоническим основанием, несущим на обеих ручках флажки, один из
которых треугольной, другой — полуовальной формы (рис. 425, 1—10). Киль
маленький, ромбовидный.
Вид хорошо отличается от других представителей рода формой зубов с вы-
сококоническим основанием.
166
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Распространение. Арктический евразийский вид, отмечен, в Иса-
Фьорде, Исландия; у островов архипелагов Шпицберген, Земля Франца-Иосифа,
в центральной части Карского моря и в юго-западной части моря Лаптевых
(рис. 459,1). Есть указания на нахождение вида у западных берегов Гренландии
Рис. 460. Места находок Obesotoma
tenuilirata в морях СССР.
(Odhner, 1915).
Экология. Обитает на глубинах
от 9 до 280 м, в основном на глубинах
от 12 до 40 м, преимущественно на песча-
ных грунтах, реже с примесями ила.
Типы. Место хранения типов не
указано (возможно, они находятся в IRSN);
типовое местонахождение — Hornsund,
Западный Шпицберген.
Просмотрено 47 экз.
6. Obesotoma tenuilirata (Dall, 1871),
comb. n. (Рис. 40; 231-235; 400, E; 426,
5-9-, 460).
Dall, 1871 t 98 (Bela laevigata var.); Ver-
rill, 1882 i 463 (Bela)-, Krause, 1885 : 274, Taf.
18, fig. 8 (Bela)-, Dall, 1919a 1 42, pl. 15,
fig. 4 (Lora)', Oldroyd, 1927 I 118 (Lora). —
simplex (non Middendorff, 1849): Verrill, 1880:
367 (Bela).
Раковина светлая, телесного цвета, ве-
ретеновидно-овальной формы; с 3—4 за-
кругленными выпуклыми оборотами. Пери-
остракум плотный, желтоватый или корич-
невый, легкоотстающий. Верхние обороты
раковины, как правило, окатаны. Про-
токонх сосцевидной формы, представлен
1.5 сильновыпуклыми оборотами, покры-
тыми тончайшей спиральной струйчатостью
(рис. 400, Е). Осевая скульптура дефини-
тивной раковины представлена очень по-
логими, незначительно возвышающимися
над поверхностью раковины средними
складками, различными только в верхней
части периферии оборота. Они разделены
в 5—7 раз более узкими, чем их ширина,
промежутками и на околошовной поверх-
ности имеют слабый сигмоидальный изгиб. На последнем обороте раковины
насчитывается от 20 до 24 осевых складок, на предпоследнем от 19 до 22. Спи-
ральная скульптура представлена тонкими и средними ребрами, чаще шнуро-
видными, нежели нитевидными. Межреберные промежутки всегда в 3—5 раз
уже ребер. Последние равновелики практически на всей поверхности раковины,
за исключением околошовного пространства, где элементы спиральной скульп-
туры в 1.5—4.7 раза тоньше остальных ребер. Основание раковины слабовыпук-
лое, плавно переходящее в небольшой сифональный вырост. Устье неправиль-
но-овальной формы, угловатое в верхней части, с небольшой выемкой анального
синуса в верхней 1/7—1/8 палатального края и узкой полосой белого матового
каллуса на колумеллярном крае. Сифональный канал короткий, неглубокий,
очень широкий. Наибольшие размеры раковины из типовой серии (мм): В =
23.3; Д=10.5; £по=17.0.
Зубы радулы , копьевидной формы, с массивным округлым основанием.
Киль длинный, на проксимальном участке гарпуновидный. Одна из ручек
несет маленькую треугольную шпору (рис. 426, 7).
4. OBESOTOMA
167
Вид сходен с О. simplex, от которого отличается тонкостенной раковиной и
копьевидными зубами.
Распространение. Амфибореальный широко распространенный
вид тихоокеанского происхождения. Отмечен в бореальных водах Тихого океана
и в высокобореальных водах американского побережья Северной Атлантики.
Вид встречен в Беринговом море от о-вов Шумагина, Аляска, до Алеутских
о-вов и на север в Арктику до м. Барроу, повсеместно в Охотском море в зал.
Петра Великого и в Татарском проливе в Японском море (рис. 460). Отмечены
также находки вида в Чукотском море у о-ва Врангеля (собств. данные) и в зал.
Св. Лаврентия (Verrill, 1882).
Экология. Обитает на глубинах от 9 до 210 м преимущественно на или-
стых грунтах.
Типы. Голотип хранится в USNM, № 226186; типовое местонахождение —
Fort St. Michael, Нортон Саунд, Аляска.
Просмотрено более 80 экз.
7. Obesotoma woodiana (Moller, 1842), comb. n. (Рис. 236—240; 421, 15—18;
423, 1—11; 461).
Moller, 1842 I 86 (Defrancia); L e c h e, 1878 i 54 (Pleurotoma); Posselt, Jen-
sen, 1898 i 145, Taf. 1, fig. 3 (Bela); К о b e 1 1, 1905 I 244, Taf. 84, fig. 1—3 (Bela turricula
subsp.); Schlesch, 1924 i 326, Taf. 11, fig. 2—3 (Lora); Oldroyd, 1927 t 111 (Lora).
Раковина телесного цвета, веретеновидная, с 5—6 оборотами отлогоуглова-
той формы, тупоугловатыми в верхней части. Периостракум плотный, кожистый,
серого цвета. Верхние обороты раковины, как правило, корродированы. Эм-
бриональная раковина не исследована. Осевая скульптура дефинитивной ра-
ковины представлена прямыми, очень пологими средними складками, наиболь-
шая выраженность которых приходится на плечо оборота. В верхней половине
основания раковины складки сглаживаются. Промежутки в 4—6 раз уже ши-
рины складок. На последнем обороте раковины насчитывается от 16 до 24 эле-
ментов осевой скульптуры, на предпоследнем от 14 до 19. Спиральная скульп-
тура. на самых ранних дефинитивных оборотах представлена 2 килевидными реб-
рышками, из которых одно отмечает плечо, другое — нижнюю 1/3 периферии
оборота. На последних оборотах у большинства экземпляров сохраняется киле-
видное ребро, проходящее по плечу оборота, немногим превышающее размерами
остальные элементы спиральной скульптуры. Последние представлены поло-
гими, не рельефными тонкими (реже средними) ребрами, расстояние между кото-
168
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
рыми составляет х/5—% их собственной ширины. Основание раковины выпуклое,
сифональный вырост удлиненный. Устье широко-обратногрушевидной формы,
с широкой полосой белесого каллуса на колумеллярном крае. Сифональный
канал протяженный, узкий, неглубокий. Высота раковины типового экземпляра
6 мм. Наибольший экземпляр (высота раковины 16.2 мм) найден в Баренцевом
море (рис. 240).
Крышечка каплевидной формы, с практически прямой осью нарастания.
Мускульный отпечаток занимает центральную часть крышечки (рис. 34, 3—К).
Зубы радулы крупные, длиной 200—400 мкм, копьевидной формы, с откло-
ненным назад основанием, несущим на одной из ручек уплощенную треуголь-
ную шпору; дистальная часть длинного киля имеет шлемовидную форму
(рис. 423, 7).
Вид похож на О. robusta, от которого отличается более уступчатыми оборо-
тами, низким завитком, большими зубами радулы с массивным основанием и
шлемовидным дистальным участком киля.
Распространение. Атлантический высокобореально-арктический
вид. Обитает в юго-восточной части Баренцева моря, у о-вов Земли Франца-
Иосифа, у южной оконечности о-вов Шпицбергена (рис. 461).
Экология. Встречается на глубинах от 5 до 240 м, в основном от 20
до 60 м; на илистом песке и глине (до 60 % находок), реже на других типах
грунтов при температуре от —0.96 до 6.54 °C и солености, превышающей
32.47°/00, преимущественно от 33.3 до 34.3 °/00.
Типы. Типовой экземпляр хранится в ZMC; типовое местонахождение —
Арктика.
Просмотрено 111 экз.
8. Obesotoma tumida (Posselt et Jensen, 1898), stat, et comb. n. (Рис. 241 —
242; 423, 12—16).
Posselt, J ensen, 1898 i 145, Taf. 1, fig. 3, a (Bela woodiana var.).
Раковина телесного с желтизной цвета, кубаревидная, с 4—4.5 уступча-
тыми оборотами, уплощенными в верхней части. Периостракум тонкий, легко
отшелушивающийся, светло-коричневый. Верхние обороты раковины сильно
корродированы, протоконх не сохранился. Осевая скульптура дефинитивной
раковины состоит из пологих средних складок, разделенных вдвое меньшими
промежутками. Элементы осевой скульптуры выражены в верхней части обо-
рота и полностью сглаживаются уже в нижней 1/3 периферии оборота. На по-
следнем и предпоследнем оборотах насчитывается около 22—28 складок. Спи-
ральная скульптура на ранних дефинитивных оборотах представлена 3—5 рав-
ноудаленными (межреберные промежутки в 1.5—2 раза шире ребер) тонкими
ребрами, лежащими на периферии оборота. На поздних дефинитивных оборо-
тах встречаются различные элементы спиральной скульптуры: без определен-
ного порядка чередующиеся средние и тонкие ребра. У одних экземпляров
численно преобладают тонкие нитевидные ребра, у других — шнуровидные
средние. Межреберные промежутки на последнем обороте составляют 1/2'—
1/в ширины самих ребер. Основание раковины выпуклое, сифональный вырост
прямой, короткий. Устье полуовальной формы, угловатое в верхней части.
На колумеллярном крае лежит широкая полоса белого каллуса. Сифональный
канал короткий, широкий, относительно глубокий. Наибольший изученный
экземпляр имел высоту раковины 13 мм (рис. 241).
Крышечка каплевидная, с практически прямой осью нарастания. Мускуль-
ный отпечаток булавовидный, занимает срединную часть крышечки и лежит
непосредственно под осью нарастания (рис. 34, Ж).
Зубы радулы длиной от 250 до 600 мкм, копьевидной формы, с небольшой,
отклоненной назад базальной частью и тонким килем, несколько превышающим
в длину дистальное отверстие зуба. Характерная особенность зубов радулы дан-
4. OBESOTOMA
169
ного вида состоит в некотором увеличении, диаметра окрлодистальцой части
лезвия относительно околобазальной (рис. 423, 12.—13).
Вид сходен с О. woodiana, от которого отличается уступчатыми оборотами,
уплощенными в верхней, части, осевыми складками, стирающимися на .пери-
ферии оборота, и утолщенными в ди-
стальной части лезвия ,зубами.
Распространение. Арктиче-
ский вид, отмечен в Восточно-Сибирском
море и море Лаптевых; найден также
в водах Западной Гренландии.
Типы. Типовой материал хранится,
вероятно, в ZMC; типовое местонахожде-
ние — воды Западной Гренландии.
Просмотрено 4 экз.
9. Obesotoma robusta (Packard, 1867),
comb. n. (Рис. 243—248; 423,17—28', 462).
Packard, 1867 i 232, pl. 7, fig. 12
(Bela). — raricostulata Golikov в Го ли-
ков,; Ска p л ат о, 1984 1 432, рис. 12 (Oeno-
pota), syn. n.
Раковина различных оттенков корич-
невого цвета, часто коричнево-вишневая,
уступчатая, веретеновидно-овальная, тол-
стостенная, с 5—6 отлогоугловатыми
оборотами, тупоугловатыми в верхней ча-
сти. Периостракум плотный, полупрозрач-
ный. Верхние обороты практически всегда
сильно корродированы. Эмбриональная
раковина не изучена. Осевая скульптура
дефинитивной раковины состоит из широ-
ких складок, треугольных в сечении. Наи-
более выражены они на плече оборота,
к верхнему и нижнему швам сглажива-
ются. На основании раковины прослежи-
ваются лишь следы складок. На послед-
нем обороте раковины насчитывается от
16 до 18 складок, на предпоследнем от
14 |до 16. Среди элементов спиральной
скульптуры выделяется тонкий киль, кото- Рис. 462. Места находок Obesotoma
рый проходит по плечу оборота и дает мозо- robusta в морях СССР,
листые утолщения при пересечении с осе-
выми складками. Над килем, на околошовной поверхности, расположено до
12 тонких волосовидных ребрышек. Ниже киля располагаются средние шнуро-
видные ребра. Они практически равновелики на периферии оборотов, на осно-
вании раковины и в верхней части сифонального выроста. Разделяют их,
как правило, равные по ширине ребрам промежутки. Осевые складки и спи-
ральные ребра обычно светлее разделяющих их промежутков, вследствие чего
все элементы скульптуры выглядят очень рельефно. Основание раковины уме-
ренно выпуклое, сифональный вырост средних размеров. Устье широко-обратно-
грушевидной формы, угловатое в верхней части. Телесного цвета каллус лежит
толстым слоем на колумеллярном крае. Сифональный канал глубокий, узкий.
Высота раковины типового экземпляра 4.5 мм при ширине 2.8 мм. Самый круп-
ный экземпляр, обнаруженный в зал. Чихачёва (пос. Де-Кастри), имел высоту
раковины 15.2 мм.
Крышечка крупная, когтевидная, с сильно изгибающейся на ранних этапах
формирования осью нарастания. Мускульный отпечаток широкий, ромбовид-
170
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ный, занимает большую часть левой стороны крышечки, однако не достигает
утолщенного его левого края (рис. 34, В—Е).
Зубы радулы копьевидной формы, очень маленькие, их длина не превышает
150 мкм. Основание зуба отогнуто назад, на одной из ручек расположена очень
тонкая треугольная шпора (рис. 423, 28); киль ромбовидный.
Вид сходен с О. woodiana, от которого отличается высоким завитком, поло-
гими широкими осевыми складками и небольшими зубами радулы с маленьким
ромбовидным килем.
Распространение. Амфибореальный широко распространенный
вид тихоокеанского происхождения. Помимо Лабрадора распространен в бо-
реальных районах Тихого океана, преимущественно у берегов Азии. Отмечен
в зал. Петра Великого и северной части Японского моря, у Охотоморского по-
бережья о-ва Сахалин, берегов Западной и Восточной Камчатки, о-ва Беринга,
в северной части Берингова моря и у о-ва Врангеля в Чукотском море (рис. 462).
Экология. Встречается на глубинах от 7 до 660 м в основном до 200 м,
на песчанистых и илистых грунтах при температурах от —1.78 до 1.75 °C и со-
лености 31.11—33.53 °/оо-
Типы. Типовой материал Bela robusta хранится в BSNH; типовое место-
нахождение — Caribou Isl., Лабрадор (вид описан по пустым раковинам).
Голотип О. raricostulata хранится в ЗИН АН СССР, № 33929; типовое место-
нахождение — восточное побережье Южного Сахалина, глубина 149 м.
Просмотрено 52 экз.
10. Obesotoma tomiyaensis (Otuka, 1949), comb. в. (Рис. 255—256; 424,
10—12).
Otuka, 19491308, pl. 13, fig. 17 (Lora).
Раковина телесного цвета, овальная, реже овально-веретеновидная, очень
толстостенная, с 5 закругленными, сильновыпуклыми оборотами. Периостра-
кум тонкий, полупрозрачный, коричневатый, легкоотстающий. Скульптура
протоконха не сохранилась. Осевая скульптура дефинитивной раковины со-
стоит из полуовальных в сечении, широких складок, разделенных на послед-
нем обороте практически равными по ширине, а на предыдущих в 1.5—2 раза
более широкими промежутками. Складки прямые, имеют незначительное откло-
нение вперед на околошовной поверхности. В верхней 1/3 основания раковины
они сглаживаются. На последнем обороте раковины насчитывается от 15 до
17 складок, на предпоследнем около 14. Спиральная скульптура представлена
средними или узкими шнуровидными ребрами, как правило, равновеликими
с разделяющими их промежутками. Элементы спиральной скульптуры равно-
велики на всей поверхности раковины, за исключением околошовного про-
странства, где они несколько тоньше и представлены нитевидными ребрышками.
Основание раковины выпуклое, сифональный вырост короткий. Устье полу-
овальное, несет на колумеллярном крае широкую полосу толстого, кремового
или розового цвета каллуса. Сифональный канал короткий, узкий. Высота
раковины голотипа 14.0 мм при ширине 7.0 и высоте устья 7.5 мм.
Зубы радулы маленькие, длиной до 130 мкм, копьевидные, с закругленной
цилиндрической базальной частью, лезвием малого диаметра и небольшим
ромбовидным килем (рис. 424, 10—12).
Вид наиболее сходен с О. solida, от которого отличается коротким сифональ-
ным выростом, очень толстыми и широкими осевыми складками и округлым
основанием зубов радулы.
Распространение. Тихоокеанский приазиатский низкобореальный
вид. Помимо типового местонахождения (палеонтологического) отмечен только
в северной части Японского моря.
Типы. Голотип хранится в GITU, № Y-0020; типовое местонахождение —
Tomiya, Minato.machi, плиоценовые отложения Японии.
4. OBESOTOMA 171
Просмотрено 2 экз. Для 1 экз. на этикетке не указаны координаты место-
нахождения. 2-й экземпляр обнаружен у о-ва Монерон1(46° 07' с. ш.—144° 45'
в. д.) на глубине 67 м на песчаном грунте с камнями при придонной темпера-
туре 5 °C.
И. Obesotoma solida (Dall, 1886), comb. n. (Рис. 249—254; 257—258; 424,
8—9).
Dall, 1886b i 301, pl. 3, fig. 4 (bora); Oldroyd, 1927 I 116 (bora).
Раковина телесного цвета, овально-веретеновидная, с 5—5.5 закругленными,
иногда тупоугловатыми в верхней части, умеренно выпуклыми оборотами.
Периостракум плотный, коричневатый. Скульптура эмбриональной раковины
не изучена. Осевая скульптура дефинитивной раковины состоит из практи-
чески прямых широких складок, наиболее выраженных на периферии оборота
и сглаживающихся, как правило, уже в верхней 1/3 основания раковины. Осе-
вые складки в 3—4 раза шире разделяющих их промежутков. На последнем
обороте насчитывается от 13 до 16 складок, на предпоследнем от 12 до 14. Спи-
ральная скульптура представлена многочисленными шнуровидными средними
ребрами, равновеликими на периферии оборотов, основании раковины и сифо-
нальном выросте. Промежутки, разделяющие их, всегда в 2—3 раза уже самих
ребер. На околошовной поверхности ребра более тонкие, но также шнуровид-
ные. Основание раковины умеренно выпуклое, плавно переходит в удлиненный
сифональный вырост. Устье полуовальное, угловатое в верхней части; колу-
меллярный край несет широкую, относительно толстую полосу белесого кал-
луса. Сифональный канал глубокий, средней ширины. Наибольшие размеры
раковины типовой серии (мм): Вр= 3.0; Д=7.0; Z?no=8.0. В зал. Петра Вели-
кого нами найдена раковина высотой 18.0 мм.
Крышечка когтевидная, с плавно изгибающейся осью нарастания, желтая.
Мускульный отпечаток ромбовидный, занимает практически всю левую поло-
вину крышечки и прилегает к ее левому, утолщенному краю (рис. 34, Б).
Зубы радулы копьевидные, длиной до 130 мкм, лезвие очень малого диа-
метра. Основание зуба высококоническое, отогнуто назад, на одной из ручек
несет равностороннюю треугольную шпору. Киль ромбовидный, маленький
(рис. 424, 8).
Вид морфологически сходен с О. robusta, от которого отличается округлыми
оборотами раковины и широкими осевыми складками, а также высококони-
ческой базальной частью и тонким лезвием зуба.
Распространение. Тихоокеанский широко распространенный бо-
реальный, преимущественно приазиатский вид. Помимо шельфов Аляски и за-
падной части Алеутских о-вов отмечен в зал. Петра Великого и зал. Чихачева
(пос. Де-Кастри), Японское море.
Экология Обнаружен на глубинах от 5 до 380 м, на илистом песке
или песчанистом илу.
Типы. Типовая серия хранится в USNM; типовое местонахождение —
побережье о-ва Кыска, Аляска.
Просмотрено 43 экз.
12. Obesotoma starobogatovi Bogdanov, sp. n. (Рис. 58; 226—230; 400, A—Г-
425, 11—22; 426, 1—4).
Раковина белого, реже кремового цвета, овально-веретеновидная, с 4—5 обо-
ротами отлогоугловатой формы; покрыта тонким полупрозрачным, плотнопри-
легающим периостракумом. Верхние обороты, как правило, сильно корроди-
рованы. Эмбриональная раковина сосцевидной формы, несет до 10 пологих
нитевидных ребрышек, густо усеянных микроскульптурными бугорками
(рис. 400, А—Г). Осевая скульптура дефинитивной раковины представлена по-
172 СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
логими, слегка выступающими над поверхностью раковины тонкими складками,
наиболее выраженными на плече оборота. Складки на периферии оборота рас-
полагаются параллельно оси раковины и залегают на основании раковины.
На плече оборота они преломляются и на околошовной пространстве имеют
направленность влево. Элементы спиральной скульптуры представлены ните-
видными ребрышками, которые (как и разделяющие их промежутки) равно-
велики на всей поверхности раковины. Основание раковины умеренно выпуклое.
Сифональный вырост короткий. Устье узко-обратногрушевидное, с тонким
матовым каллусом на колумеллярном крае. Сифональный канал открытый,
неглубокий. Размеры голотипа (мм): Вр=6.6-, #=4.0; 2?ПО=4.8; Ву—3.5; Ду=
= 1.7. Наибольший экземпляр, найденный в море Лаптевых у о-ва Малый
Ляховский на глубине 3.5 м, имел размеры (мм): В =11.8; #=6.0; 2?ПО=8.3;
Ву=6.0; #,=2.6.
Крышечка каплевидной формы, со слабо изогнутой осью нарастания. Му-
скульный отпечаток в виде тонкой полосы, топографически расположен не-
посредственно под осью (рис. 34, М—Н).
Зубы радулы ножевидной формы, с массивным конусовидным основанием,
несущим на одной из ручек полуовальный флажок (рис. 425, 14—19). Неболь-
шой киль занимает самую дистальную часть лезвия зуба (рис. 425,18).
По раковине вид наиболее сходен с О. colpoica, от которого отличается более
выпуклыми оборотами, обратногрушевидной формой устья и наличием осевых
складок. От О. gigantea отличается по форме зубов радулы (рис. 425) — более
массивной базальной частью и широким околобазальным отверстием лезвия.
Распространение. Арктический евразийский вид. Распространен
в. западной части Восточно-Сибирского моря, в море Лаптевых и Карском море
(рис. 58).
Экология. Обитает на глубинах от 3 до 40 м, в основном на глубинах
6,5—20 м; встречается только на песчанистых, илистых и глинистых грунтах.
Отмечен при температурах от —1.02 до 2.09 °C и солености 29.22—31.61 °/оО.
Типы. Голотип хранится в ЗИН АН СССР, № 48472; типовое местонахож-
дение: Новосибирские о-ва (74° 29' 5” с. ш.—140° 29' в. д.), глубина 20.5 м,
песчанистый грунт.
Просмотрено 257 экз.
13. Obesotoma sachalinensis Bogdanov, 1989. (Рис. 262—264; 421, 1—10).
Богданов, 19896 I 150, рис. 1, 5—6\ 2, и—м.
Раковина светло-коричневая, овально-веретеновидная, с 4—4:5 закруглен-
ными, умеренно выпуклыми оборотами. Периостракум тонкий, от белого до бу-
роватого цвета, плотноприлегающий. Эмбриональные обороты, как правило,
корродированы. Осевая скульптура дефинитивной раковины представлена по-
логими широкими складками, которые разделены различными по ширине про-
межутками; ослабевает в нижней части периферии оборота и уже на основании
раковины следов осевых складок не видно. Спиральная скульптура представ-
лена многочисленными тонкими ребрами, которые шире разделяющих их про-
межутков. В верхней части оборота, близкой ко шву, ребра тоньше, чем на пе-
риферии. Основание умеренно выпуклое, сифональный вырост короткий.
Устье полуовальной формы, палатальный его край покрыт тонким светло-
коричневого цвета каллусом. Сифональный канал протяженный, узкий. Раз-
меры раковины голотипа (мм): 5р=9.2; #=5.4; Впо=7.3; Вр=5Л; Ду=2.3.
Наиболее крупная раковина высотой 20.2 мм обнаружена у побережья о-ва Са-
халин.
Крышечка крупная, когтевидная, с плавно изгибающейся осью нарастания.
Крупный, практически ромбовидный мускульный отпечаток расположен вблизи
утолщенного левого края крышечки (рис. 34, #).
5. PROPEBELA
173
Зубы радулы копьевидной формы, длиной 400—500 мкм. Основание зуба
высококоническое, закругленное, несет 2 флажка, из которых один треугольной,
.другой — полуовальной формы (рис. 421, 7, 10).
Вид похож на О. schantarica, от которого отличается угловатостью оборотов,
«более крупными зубами с массивным основанием и длинным килем.
Распространение. Тихоокеанский приазиатский широко распро-
страненный бореальный вид. Отмечен в северной части Японского моря и юж-
ной части Охотского моря.
Экология. Обитает на глубинах от 5 до 150 м на илисто-песчаных грун-
тах.
Типы. Голотип хранится в ЗИН АН СССР, № 48473; типовое местонахож-
„дение — южная часть Охотского моря, глубина 146--—150 м.
Просмотрено 12 экз.
5, Род PROPEBELA Iredale, 1918
I г е d а 1 е, 1918 I 32; Bartsch, 1941 t 4 (part; =Bela, Lora)', Powell, 1966 I 120;
Богданов, 1986 1 45 (Oenopota subg.)-Oenopota (part.)i Morch, 1852 I 73; Bo u-
•c h e t, Ware n, 1980 t 66 (= Turritoma, Turritomella). — Turritomella Bartsch, 1941a:
143 (nom. n. pro Turritoma Bartsch, 1941, типовой вид Turritoma exquisita Bartsch, 1941, no
первоначальному обозначению).— Bela (non Gray, 1847; part.)! G. O. Sars, 18781222;
F r i e 1 e, 1886 i2; Martini, Chemnitz, 1887 1 137. — Pleurotomina (non Beck, 1847)1
'M б r c h, 1852 : 73.
Типовой вид Murex turricula Montagu, 1803, по первоначальному
обозначению.
Раковина крупная, высотой до 20—25 мм, веретеновидной формы, с отлого-
угловатыми или уступчатыми оборотами, в основном толстостенная. Протоконх
представлен 1.5—2 оборотами слабовыпуклой или уплощенной формы, причем
первый оборот лишен макроскульптурных элементов, которые закладываются
на втором эмбриональном обороте в виде 3—4, реже 5 спиральных ребрышек.
Осевая скульптура закладывается на первом дефинитивном обороте. Скульп-
тура дефинитивной раковины представлена средними и широкими осевыми
складками. Сигмоидальный изгиб отсутствует на складках, которые всегда
прямые или изогнутые в верхней части в левую сторону. Спиральная скульп-
•тура представлена различными по величине элементами, но, как правило, на-
< блюдается преимущественное развитие киля, обозначающего плечо оборота.
'Основание раковины пологое, умеренно выпуклое, устье (за редким исключе-
. ми ем) угловатое в верхней части.
Крышечка когтевидная, с плавно изгибающейся осью нарастания. Мускуль-
ный отпечаток узкий относительно ширины крышечки.
Зубы радулы от ножевидной до саблевидной и шиловидной формы.
Род распространен в бореальных и арктических водах северного полуша-
рия. В ископаемом состоянии известен в Тихом океане с миоцена (Япония),,
в Атлантике — с плейстоцена (Англия, Исландия).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (4). Зубы радулы шиловидной формы, базальная часть не развита.
2 (3). Основание раковины скошенное, завиток высокий, более 1!ъ общей вы-
соты раковины. Осевые складки прямые . ............................
1. Р. (Р.) turricula (Montagu)
3(2). Основание раковины округлое, завиток невысокий, менее V3 общей
высоты раковины. Осевые складки чуть скошены вперед на плече обо-
рота .................................8. Р. (Р.) harpularia (Couthouy)
4 (1). Зубы радулы иной формы, базальная часть развита.
5 (18). Зубы радулы копьевидной или саблевидной формы.
6 (9). Элементы осевой скульптуры имеют преимущественное развитие над
спиральными ребрами. Раковина не имеет ячеистого вида.
174
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
7 (8). Раковина составлена закругленными, умеренно выпуклыми оборотами,,
киль отсутствует. Зубы радулы саблевидные, лезвие зуба всегда изогнуто,,
округлая базальная часть несет на каждой из ручек по флажку ... .
................................ 30. Р. (С.) variabilis Bogdanov, sp. n.
8 (7). Раковина составлена отлогоугловатыми оборотами, плечо оборота
обозначает в той или иной степени развитый киль. Зубы радулы копьевид-
ные, одна ручка базальной части зуба редуцирована, другая несет упло-
щенный флажок ............................18. Р. (С.) viridula (Moller)»
9 (6). Осевая и спиральная скульптуры равновелики и при пересечении при-
дают раковине ячеистый вид.
10 (17). Зубы радулы саблевидной формы.
11 (12). Устье значительно превышает V2 высоты раковины. Ячеистый рисунок
раковины ярко выражен на ранних оборотах и ослабевает к последнему
обороту раковины............................29. Р. (С.) marinae Bogdanov
12 (11). Устье меньше или (редко) равно 1/2 высоты раковины. Ячеистый рису-
нок не ослабевает на последнем обороте раковины.
13 (14). Раковина состоит из уступчатых оборотов, уплощенных в верхней
части. Сглаженные средние осевые складки в 1.5—2 раза меньше разде-
ляющих их промежутков ..........................20. Р. (С.) mitrata (Dall)»
14 (13). Раковина состоит из отлогоугловатых оборотов. Средние осевые складки
высокие, рельефные, как правило, шире разделающих их промежутков.
15 (16). Осевые складки полукруглые в сечении. Базальная часть зуба радулы
цилиндрическая, языковидного выроста не имеет........................
................................... 21. Р. (С.) concinnula (Verrill),
16 (15). Осевые складки треугольные в сечении. Базальная часть зуба радулы
пирамидальной формы, имеет языковидный вырост...........................
....................................... 19. Р. (С.) exarata (Moller)»
17 (10). Зубы радулы копьевидной формы .... 17. Р. (С.) tersa Bartsch
18 (5). Зубы радулы ножевидной или расширенно-ножевидной формы.
19 (20). Осевая скульптура представлена тонкими складками, равновеликими'
по высоте и ширине со спиральными ребрами ..............................
................................... 28. Р. (С.) margaritae (Bogdanov)»
20 (19). Осевая скульптура представлена в основном широкими и средними
складками, явно преобладающими над элементами спиральной скульп-
туры (исключения могут составлять лишь 1—3 высоких киля).
21 (38). Зубы радулы расширенно-ножевидной формы. Лезвие большого диа-
метра и с широким дистальным отверстием зуба.
22 (23). Обороты закругленные, сильновыпуклые. Раковина овально-веретено-
видная......................................14. Р. (Р.) cassis Bogdanov
23 (22). Обороты раковины уступчатой или отлогоугловатой формы. Раковина,
веретеновидная или башенковидная.
24 (27). Спиральная скульптура представлена различными элементами, среди;
которых широкие ребра (более 3) равновелики по высоте осевым складкам.
25 (26). Осевые складки средние, не более чем в 3—4 раза превышают ширину
разделяющих их промежутков. Киль, обозначающий плечо оборота,
развит. Базальная часть зуба округлая, дистальное отверстие зуба не-
превышает х/3 длины лезвия, киль с гарпуновидным выростом ....
...................................... 15. Р. (Р.) terpeniensis Bogdanov
26 (25). Осевые складки широкие, в 6—7 раз шире разделяющих их проме-
жутков. Киль не развит. Базальная часть зуба высококоническая, ди-
стальное отверстие зуба превышает V2 длины лезвия, киль без гарпуно-
видного выроста.................................9. Р. (Р.) popovia (Dall)'
27 (24). Спиральные элементы представлены, как правило, тонкими ребрами,
несравнимо меньшей величины, чем осевые складки. Развитие получает
только 1 киль, обозначающий плечо оборота, который может быть равно-
велик элементам осевой скульптуры.
28 (31). Осевые складки в 2—4 раза шире разделяющих их промежутков..
5. PROPEBELA
175
.29 (30). Осевые складки прямые, идут параллельно оси навивания раковины.
Киль зуба радулы маленький, занимает самую апикальную часть лезвия
............................................ 12. Р. (Р.) svetlanae Bogdanov
30 (29). Осевые складки отклонены назад на периферии оборота и проходят
под углом к оси навивания. Киль зуба радулы длинный, достигает х/2
длины лезвия ................................23. Р. (С.) alitakensis (Dall)
-31 (28). Осевые складки, как правило, уже разделяющих их промежутков.
32 (33). Раковина крупная, средние размеры 18—20 мм. Гарпуновидный
вырост большой, составляет более чем V2 длины киля зуба радулы. Ба-
зальная часть зуба уплощенная, отогнута книзу.............................
................................11. Р. (Р.) scalaroldes (G. О. Sars)
33 (32). Раковина небольшая, средние размеры 8—10 мм. Гарпуновидный
вырост киля зуба радулы маленький или отсутствует. Базальная часть
зуба конусовидная.
34 (35). Дистальное отверстие зуба лунковидное, образовано завернутыми
стенками лезвия, в результате чего создается впечатление канта, обрам-
ляющего весь периметр отверстия ..........................................
................................... 6. Р. (Р.) spitzbergensis (Friele)
- 35 (34). Стенки дистального отверстия зуба радулы не заворачиваются и не
образуют кант.
- 36 (37). Раковина тонкостенная, состоит из отлогоугловатых оборотов . . .
............................................. 7. Р. (Р.) bergensis (Friele)
- 37 (36). Раковина, как правило, толстостенная, состоит из уступчатых обо-
ротов, уплощенных в верхней части ........................................
.......................... 5. Р. (Р.) rugulata (Moller in Troschel)
- 38 (21). Зубы радулы ножевидной формы; дистальное отверстие зуба, как
правило, не превышает х/3 длины лезвия. Последнее всегда малого диа-
метра.
- '39 (42). Раковина высококоническая, завиток очень высокий, более 0.45 от
общей высоты раковины. Отношение высоты раковины к высоте устья
всегда не менее 2.4 (в среднем 2.53).
40 (41). Осевые складки прямые. Слегка изогнутое лезвие зуба несет на дисталь-
ном участке маленький, не достигающий 1/в длины зуба треугольный
гарпуновидный киль...........................13. Р. (Р.) verrilli Bogdanov
41 (40). Осевые складки скошены на околошовной поверхности. Лезвие зуба
радулы прямое, несет на дистальном участке удлиненный киль, дости-
гающий х/4 общей длины зуба ...............2. Р. (Р.) angulosa (G. О. Sars)
-42 (39). Раковина в той или иной мере веретеновидная, завиток невысокий,
менее 0.40 от общей высоты раковины. Отношение, высоты раковины
к высоте устья не превышает 2.3.
• 43 (48). Раковина составлена закругленными отлогоугловатыми оборотами,
усеченными в нижней четверти.
44 (45). Раковина маленькая (средние размеры 5—6 мм), очень толстостенная,
ее основание скошенное, сифональный вырост короткий. Устье полуоваль-
ное. Осевые складки отклонены назад ... 26. Р. (С.) areta (Bartsch)
45 (44). Средние размеры раковины более 8 мм, она составлена стенками
нормальной толщины, основание раковины закругленное, сифональный
вырост большой. Устье обратногрушевидное. Осевые складки прямые,
параллельны оси раковины.
46 (47). Среди элементов спиральной скульптуры развиты 2 киля, равные
по размерам осевым складкам. Зубы радулы маленькие, длиной до 150 мкм,
базальная часть массивная, составляет более г/3 общей длины зуба. Киль
маленький, треугольный, под ним не образуется дополнительного дисталь-
ного отверстия..................25. Р. (С.) cingulata (Golikov et Gulbin)
47 (46). Развит только 1 киль, обозначающий плечо оборота и значительно
уступающий в размерах осевым складкам. Длина зубов более 300 мкм,
базальная часть составляет менее V4 от размеров зуба. Под гарпуновидным
176
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
килем образуется дополнительное отверстие..........................
...........................................4. Р. (Р.) assimilis (G. О. Sars)
48 (43). Раковина составлена уступчатыми или отлогоугловатыми оборотами,
тупоугловатыми в верхней части и не усеченными в нижней четверти.
49 (56). Осевая скульптура прямая, без резких изломов, сохраняется на осно-
вании раковины,-
50 (53). Элементы осевой скульптуры представлены широкими осевыми склад-
ками, которые в 2—4 раза превышают по ширине разделяющие их про-
межутки.
51 (52). Спиральная скульптура на периферии оборота и в верхней части
основания раковины представлена средними шнуровидными ребрами,
которые превышают ширину разделяющих их промежутков в 2 раза . . .
..............................................22. Р. (С.) fidicula (Gould)
52 (51). Спиральная скульптура представлена на всей поверхности раковины
тонкими нитевидными ребрами, равновеликими с разделяющими их про-
межутками ....................................24. Р. (С.) rassina (Dall)
53 (50). Элементы осевой скульптуры представлены тонкими или средними
складками, разделенными в 2—3 раза более широкими промежутками.
54 (55). Киль, проходящий по плечу оборота раковины, очень широкий и несет
на своей поверхности 5—7 тонких ребрышек. Киль зуба радулы занимает
до 1/3 длины лезвия.............................3. Р. (Р.) nobilis (Moller)
55 (54). Киль плеча оборота раковины тонкий, без дополнительной скульптуры.
Киль зуба радулы маленький, занимает менее Ve длины лезвия . . .
..............................................10. Р. (Р.) scalaris (Moller)
56 (49). Осевая скульптура развита на периферии оборота, дает резкий излом>
вперед на плече и исчезает на основании раковины.
57 (58). Раковина толстостенная, каштанового цвета. Киль, обозначающий
плечо оборота, не развит или только намечен. Сифональный вырост не-
большой. Одна из ручек базальной части зуба радулы несет маленькую
шпору .....................................27. Р. (С.) golikovi (Bogdanov)
58 (57). Раковина тонкостенная, телесного цвета. Киль сильно развит, высту-
пает над поверхностью раковины в такой же степени, как и осевые складки.
Сифональный вырост удлиненный. Одна из ручек базальной части зуба
радулы несет большой, треугольной формы флажок...........................
.................................... 16. Р. (Р.) goryachevi Bogdanov
1. Подрод PROPEBELA Iredala, 1918
Раковина башенковидной, реже веретеновидной формы. Элементы осевой
скульптуры, как правило, отстоят друг от друга на расстоянии, большем или
равном собственной ширине складок.
Зубы радулы ножевидной, расширенно-ножевидной или шиловидной формы.
На киле лезвия зуба радулы имеются гарпуновидные выросты. На лезвии зуба
образуется дополнительно второе дистальное отверстие.
Подрод распространен по всему ареалу рода.
1. Propebela (Propebela) turricula (Montagu, 1803). (Рис. 368—370; 404,,
Д-Е; 405, А; 433, 23—37; 463, 2).
Montagu, 1803 I 262, pl. 9, fig. 1 (Murex)i Gould, 1841 1 292, fig. 193 (Fusus);-
Brown, 1844 I 7, pl. 5, fig. 51—52 (Fusus); Reeve, 1846 l 39, pl. 19, fig. 162 (Pleurotoma);.
Forbes, Hanley, 1853 I 450, pl. Ill, fig. 7, 8, pl. TT, fig. 2 [Mangelia (Bela)]; Jef-
freys, 1867 i 395 (Pleurotoma; part.); Gould, 1870 i 351. fig. 620 (Bela); Martini,
Chemnitz, 1887 I 138, Taf. 31, fig. 1 (Bela); L о с a r d, 1892 i 54, fig. 42 (Bela); 1897 I 230-
(Bela); К о b e 1 1, 1905 I 234, Taf. 82, fig. 1 (Bela); Dautzenberg, Fischer, 1912 I 41
(Bela; part.); Iredale, 1918 i 32; Oldroyd, 19271115 (Lora); Powellr
1966 i 120, pl. 19, fig. 3;' Bouchet, Waren, 1980 I 72, fig. 154, 267 (Oenopota)..
5. PROPEBELA
177'
Раковина розоватая, удлиненно-веретеновидная, башенковидная, с 6—7
(иногда до 8.5) уступчатыми оборотами, уплощенными в верхней части. Перио-
стракум тонкий, прозрачный, плотноприлегающий. Протоконх слабовыпуклый,
первый оборот слегка возвышается над вторым эмбриональным оборотом, не-
сущим 3 спиральных тонких ребра (рис. 404, Д—Е). Верхнее ребро постепенно
редуцируется на дефинитивных оборотах. Осевая скульптура дефинитивной
раковины представлена ярко выраженными, полукруглыми в сечении, прямыми
складками, разделенными равными или в 1.2—1.5 раза более широкими про-
странствами. Складки имеют на плече оборота небольшой изгиб вперед. Они
практически одноразмерны на всей поверхности оборота, незначительно сгла-
живаются на основании раковины, достигая сифонального выроста. На послед-
нем обороте насчитывается от 14 до 20 складок, на предпоследнем от 12 до 18.
Спиральная скульптура представлена в основном двумя элементами: нитевид-
ными ребрышками и тонкими ребрами. Причем ребрышки занимают верхнюю
часть оборота, а ребра — нижнюю. Наиболее выражены ребра на основании;
раковины. Среди элементов спиральной скульптуры ранних дефинитивных обо-
ротов наиболее выделяются 2 килевидных ребра, одно из которых проходит
по плечу, обозначая его, другое — в нижней х/3 периферии оборота. У боль-
шинства экземпляров на последних оборотах сохраняется только пологий,,
слабо выраженный киль, покрытый 4—5 нитевидными ребрышками. Нижнее
килевидное ребро не развивается и становится одноразмерным (реже чуть более
широким), с тонкими ребрами. Основание раковины слабовыпуклое, сифональ-
ный вырост удлиненный. Устье узко-обратногрушевидной формы, с широкой
полосой плотного белого каллуса на колумеллярном крае. Сифональный канал
протяженный, узкий, неглубокий. Максимальная высота раковины 13.3 мм от-
мечена мной для моллюска из Баренцева моря. В литературе указано на на-
хождение моллюска, высота раковины которого 15.0 мм при ширине 5.0 мм
(Locard, 1892).
Крышечка маленькая, когтевидная, с резко изгибающейся на ранних стадиях
роста осью нарастания. Эллипсоидный мускульный отпечаток узкой полосой
лежит в левой части крышечки (рис. 35, У—Ф).
Зубы радулы шиловидные, как правило, не длиннее 100—120 мкм. Мной
найдены все формы зубов, описанные ранее в литературе (Forbes, Hanley,.
1853 : pl. ТТ, fig. 2; Troschel, 1866 : Taf. 4, fig. 8; Friele, 1886 : Tab. 9, fig. 25).
Вид сходен с P. harpularia', от которого отличается более высоким завитком
и менее округлым основанием.
12 и. П. Богданов
:178
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Распространение. Атлантический евразийский бореально-суб-
тропический вид, заходит в арктические воды. Обитает в высоко- и низкоборе-
альных районах европейской части Северной Атлантики, заходит в Арктику
до о-вов Новой Земли и о-ва Визе в Карском море (рис. 463, 2). Отмечен для за-
падной и восточной части Англии и Уэльса, на всех побережьях Ирландии,
Шотландии и у Исландских берегов, а также в скандинавских водах. Ряд нахо-
док сделан в Средиземном море у. западных и юго-западных берегов Португа-
лии (460—620 м), на севере Испании (608 м).
Экология. Встречен на глубинах от 10 до 260 м, наибольшая частота
встречаемости приходится на 40—80 м. Имеется сообщение о нахождении вида
недалеко от западной части Марокко на глубине 1900 м (Locard, 1892). Чаще
всего особи данного вида обитают на песчанистых грунтах, реже с примесью
камней и ракуши, только 1 раз отмечено нахождение вида на плотном илу.
Типично стеногалинный океанический вид. Минимальная соленость, при кото-
рой он встречается, составляет 33.23 %0. Данных по температуре накоплено
недостаточно для определения диапазона переносимых температур, однако
:можно уверенно говорить, что максимальная температура обитания не превы-
шает 5.2 °C.
Типы. Типовой экземпляр хранится в Royal Albert Museum, Exeter,
.Англия; типовое местонахождение — Сандвич, Кент, Великобритания.
Просмотрено 17 экз.
'2. Propebela (Propebela) angulosa (G. О. Sars, 1878), comb. n. (Рис. 38, A;
:313—315; 437, 1—9).
G. O. S a r s, 1878 : 227, Tab. 16, fig. 16 (Bela); Герценштейн, 1885 I 687 (Pleuro-
:\toma); Martini, Chemnitz, 1887 I 142, Taf. 31, fig. 6 (Bela); Knipowitsch,
1901 i 42 (Bela); К о b e 1 1, 1905 ! 248, Taf. 83, fig. 3 (Bela cancellata subsp.); О d h n e r,
1910 i 12, pl. 1, fig. 11 (Bela); Schlesch, 1924 t 328 (Lora); Thorson, 1941 I 100-
.(Bela,). — cancellata (non Mighels et Adams, 1840) I T г у о n, 1884 I 218, pl. 28, fig. 46 (Bela;
part.).
Раковина белая, удлиненно-веретеновидная, тонкостенная, с 6—7 оборотами
.отлогоугловатой формы, тупоугловатыми в верхней части. Периостракум про-
зрачный, тонкий, кремовых тонов. Протоконх сильновыпуклый, представлен
1.5 отлогоугловатыми оборотами. Первые 3/4 эмбрионального оборота лишены
.скульптуры, далее на периферии оборота появляются 4 спиральных ребра,
равноудаленных друг от друга. Осевая скульптура дефинитивной раковины
^представлена тонкими складками, разделенными в 1.5—2 раза более широкими
промежутками. На периферии оборота складки имеют незначительный наклон
назад, а на плече оборота приобретают слабую сигмоидальную изогнутость.
Наиболее выражены складки в верхней части оборота, в нижней 1/3 основания
раковины они сглаживаются. На последнем обороте насчитывается от 14 до 15
Лосевых складок, на предпоследнем от И до 12. Спиральная скульптура пред-
оставлена на обороте раковины в основном тонкими ребрами и нитевидными реб-
рышками, чередующимися друг с другом. Они разделены, как правило, в 2—
2.5 раза более широкими межреберными промежутками. На основании рако-
вины и на сифональном выросте спиральные ребрышки равновелики как между
.собой, так и с разделяющими их промежутками. Основание раковины уме-
ренно выпуклое, плавно переходит в удлиненный сифональный вырост. Устье
узко-обратногрушевидной формы, с тонким полупрозрачным кремового цвета
каллусом на колумеллярном крае. В верхней 1/4 палатального края расположен
неглубокий анальный синус. Сифональный канал неглубокий, узкий. Высота
раковины типового экземпляра 12.5 мм. Наибольший экземпляр в коллекции
.'ЗИН АН СССР был найден у Мурманского побережья; высота его раковины
11.9 мм.
Крышечка очень маленькая, когтевидная, с плавно изгибающейся осью
щарастания. Мускульный отпечаток занимает большую часть поверхности левой
5. PROPEBELA
179”
стороны крышечки и почти вплотную прилегает к левой, несколько утолщенной
ее стороне. Форма мускульного отпечатка варьирует у различных экземпляров
от ланцетовидного до трапециевидного1 (рис. 36, А—В).
Зубы радулы ножевидной формы, длиной до 150—170 мкм (у типового эк-
земпляра радула длиной 190 мкм), с конусовидным основанием, несущим на од-
ной из ручек маленькую треугольную шпору (рис. 437, 2). Киль удлиненный,.,
достигает 1/4 длины зуба. Зубы, исследованные мной, сходны с изображением,,
приведенным для типового экземпляра (G.O. Sars, 1878 : Tab. 8, fig. 10).
Вид хорошо отличается от других представителей рода высоким завитком
и смазанной верхней, лежащей на околошовной площадке частью осевых скла-
док.
Рис. 464. Места находок Propebela nobilis в морях СССР; 5 — палеонтологические на.ходки.
Распространение. Атлантический высокобореальный вид. Рас-
пространен от м. Код у берегов Америки и южной части Гренландии до юго-
западной части Баренцева моря и западных берегов Шпицбергена, включая
Шотландию и Северную Ирландию. Отдельные экземпляры найдены в Белом
море.
Экология. Обитает на глубинах до 280 м, верхней границей распро-
странения может служить находка у берегов Исландии на глубине 23 м (Thor-
son, 1941). Встречается преимущественно на песчанистых грунтах.
Типы. Типовая серия хранится в ZMO; типовое местонахождение —
Вадсё, Норвегия.
Просмотрено 38 экз.
3. Propebela (Propebela) nobilis (Moller, 1842). (Рис. 316—321; 431, 1—16;
464; 465). .
Moller, 1842 t 85 (Defrancia); Morch, 1852 I 73 (Pleurotomina); 1877 I 439 (Pleuro-
toma turricula var.); L e c h e, 1878 I 51 (Bela turricula var.; part.); G. O. Sars, 1878 I 228,.
Tab. 16, fig. 19, 20, Tab. 8, fig. 14 (Bela); Tryon, 1884 I 219, pl. 28, fig. 41 (Bela; part);
Krause, 1885 t 275 (Bela); Герценштейн, 1885 I 689 (Pleurotoma); Friele, 1886t-
5, pl. 7, fig. 8, pl. 9, fig. 13—-15 (Bela); Martini, Chemnitz, 1887 i 145, Taf. 31, fig. 10
(Bela); Posselt, J ensen, 1898 i 147 (Bela); К о b e 1 t, 1905 t 237, Taf. 82, Fig. 4 (Bela..
turricula subsp.); Schlesch, 1924 I 327, Taf. 6, fig. 13—14, Taf. 11, fig. 14 (Lora); О 1 d-
r о у d, 1927 i 109, pl. 7, fig. 9 (Lora); Филатова, Зацепин, 1948 t 389, табл. 48,.
рис. 13 (Lora); Oskarsson, 1962 I 150, fig. 141 (Lora); Powell, 1966 t 121. — brevi-
spira L б у n i n g, 1932 1 15, pl. 2, fig. 26 (Bela nobilis var.).— turricula (non Montagu,
1803) l П e т p о в, 1982 I 63, рисунки (Oenopota).
Раковина пепельно-серая, реже светло-коричневая, башенковидная, круп-
ная, с 7—8 уступчатыми оборотами, плоскими в верхней части. Периостракум
12*=
JL80
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
тонкий, легкоотстающий, прозрачный, всегда коричневатый. Эмбриональная
раковина выпуклая, скульптура протоконха не сохраняется у взрослых особей.
Осевая скульптура дефинитивной раковины представлена высокими средними
складками, разделенными в 2—3 раза более широкими промежутками. Осевые
складки одинаково выражены как на плече оборота, так и на его периферии,
сглаживаются они только в верхней трети основания раковины. На последнем
-обороте раковины насчитывается от 16 до 19 складок, на предпоследнем от 13
до 16. Спиральная скульптура представ-
лена средними спиральными ребрами и
равновелика на всей поверхности рако-
вины, включая и ее основание. Межребер-
ные пространства в 1.5—2 раза уже ре-
бер. Особого развития среди элементов
спиральной скульптуры достигает киль,
обозначающий плечо оборота. Киль ши-
рокий и пологий, каких-либо утолщений
при пересечении с элементами осевой
скульптуры не образует. Характерной
чертой вида служит наличие на поверх-
ности киля 5—7 тонких спиральных ребер.
Основание раковины умеренно выпук-
лое. Граница его, перехода в удлиненный
сифональный вырост всегда четко обозна-
чена. Устье узко-обратногрушевидное,
с широкой толстой полосой белесоватого
каллуса на колумеллярном крае. Сифо-
нальный канал широкий, умеренно глу-
бокий. Высота раковины типового экзем-
пляра 7.5 мм. Максимальные размеры
раковины отмечены у моллюска из Ба-
ренцева моря (мм): 2?p=29.0; Д=13.9;
2?ПО=19.3; 2?у=14.1; /7^=5.0. размеры ра-
ковины, описанной под названием brevi-
spira, составляют (мм): 7?р=18.0; Д=
= 10.0; 5^=11.0; ^=4.0 (Loyning, 1932).
Крышечка неправильно-полуовальной
формы, с плавно изгибающейся осью нара-
стания, левый край ее сильно утолщен.
Мускульный отпечаток, обычно заострен-
ный в верхней части, занимает большую
часть левой стороны крышечки (рис. 35,
Рис. 465. Места находок Propebela Л-—В).
nobilis в дальневосточных морях. Зубы радулы большие, длиной до
400-500 мкм, ножевидной формы, с мас-
сивным конусовидным основанием, несущим на одной из ручек листовидный
флажок (Troschel, 1866 : Taf. 4, fig. 4; G. О. Sars, 1878 : pl. 8, fig. 14; собствен-
ные данные). Длина киля в 2—3 раза больше дистального отверстия зуба;
в непосредственном соседстве с килем расположено третье отверстие зуба,
оно небольших размеров и имеет вид удлиненной щели (рис. 431, 8).
Вид сходен с Р. rugulata, от которого отличается наличием на киле дополни-
тельных 5—7 спиральных ребрышек и относительно длинным килем на зубе
радулы.
Распространение. Широко распространенный бореально-аркти-
ческий вид, обитает в Чукотском море, у берегов арктической Аляски, Грен-
ландии, Норвегии, в бореальных водах обнаружен у Алеутских, Командорских,
Курильских о-вов и у о-ва Сахалин, в Японском море отмечен в зал. Петра
5. PROPEBELA
181
Великого и в Татарском проливе (рис. 465). Вид найден в' Баренцевом море
в юго-восточной части и у берегов Шпицбергена, в Белом море повсеместно,
у восточных берегов Новой Земли, у Новосибирских о-вов и в западной части
моря Лаптевых (рис. 464). Отмечен в Лабрадоре (Thorson, 1951).
Экология. Обнаружен на глубинах от 7 до 515 м, в основном на 10—
100 м; указывается нахождение на глубинах до 1 710 м (Koehler, 1896). Оби-
тает на всех типах грунтов, в основном на песчанистых, при температурах от
—2.0 до 12.8 °C и солености 26—34.8 °/00, обычно при 33.2—34.6 °/00.
Типы. Типовой экземпляр Dejrancia nobilis хранится в ZMC; типовое ме-
стонахождение — побережье Западной Гренландии. Тип Bela nobilis var.
brevispira хранится в ZNO; типовое местонахождение — о-в Врангеля (75° 56'
с. ш.—162° 57' в. д.), глубина 53 м.
Просмотрено более 560 экз.
Рис. 466. Места находок Propebela spitzbergensis (7) и Р. assimilis (2) в северных морях СССР;
3 — палеонтологические находки.
4. Propebela (Propebela) assimilis (G. О. Sars, 1878), comb. n. (Рис. 322—
327; 431, 17—20; 432, 1—11; 404, А—Б; 466, 2—3; 467).
G. O. S a r s, 1878 I 231, Tab. 23, fig. 8 (Bela}; F r i e 1 e, 1886 I 5, pl. 9, fig. 9, 10 (Bela
.rugulata vex.); Martini, Chemnitz, 1887 I 143, Taf. 31, fig. 8 (Bela); Posselt,
J e n sen, 1898 I 149 (Bela exarata var.); К о Ь e 1 1, 1905 : 235, Taf. 82, fig. 2 (Bela turricula
subsp.); Dautzenberg, Fisher, 1912 I 42 (Bela turricula var.). — turricula (non Mon-
tagu, 1803) I T г у о n, 1884 I 219, pl. 28, fig. 44 (Bela; part.); Macpherson, 1971 I 123,
pl. 7, fig. 11, map 54 (Oenopota). — scalaris (non Moller, 1842)i Bouchet, Waren, 1980 t
73, fig. 149, 150 (Oenopota; part.).
Раковина телесного цвета, башенковидная, с 6—7 уступчатыми оборотами,
уплощенными в верхней части. Периостракум тонкий, коричневатый, плотнопри-
легающий. Протоконх выпуклой формы, представлен 1.7 оборотами. Первый
эмбриональный оборот закругленный, лишен какой-либо скульптуры. Второй
•оборот отлогоугловатый, несет 3 спиральных тонких ребра, из которых верхнее
редуцируется уже на первом дефинитивном обороте, 2 остальных обозначают
соответственно плечо и середину периферии оборота. Осевая скульптура
дефинитивной раковины представлена ярко выраженными, полу-
круглыми в сечении, средними складками. Наибольшего развития
складки достигают в верхней части периферии оборота, на основании
раковины они сглаживаются. Складки разделены почти равными или немногим
меньшими по ширине пространствами. На последнем обороте раковины насчи-
тывается от 14 до 17 осевых складок, на предпоследнем от 13 до 14. Среди эле-
182
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ментов спиральной скульптуры наибольшего развития достигает килевидное
ребро, обозначающее плечо оборота. На плече оборота лежит от 5 до 10 равно-
мерно расположенных нитевидных ребрышек. Вся периферия оборотов, осно-
вание раковины и сифональный вырост покрыты равновеликими тонкими (реже
средними) ребрами, разделенными равными им по ширине межреберными про-
межутками. Основание раковины умеренно выпуклое, сифональный вырост
удлиненный. Устье обратногрушевидной формы, угловатое в верхней части.
Рис. 467. Места находок Propebela assimilis в дальневосточных морях.
На колумеллярном крае широкой полосой лежит толстый белесый каллус.
Сифональный канал неглубокий, протяженный, средней ширины. Высота ра-
ковины типового экземпляра 15 мм. Максимальных размеров моллюск имел
раковину со следующими параметрами (мм): 2?р=17.5; Д=8.9; 7?ПО=12.2;
Ву=8.7; Д^=4.0; в то же время обнаружена мертвая раковина, принадлежащая
рассматриваемому виду, высотой 21.5 и шириной 11.5 мм.
Крышечка когтевидная, с плавно изгибающейся осью нарастания. Мускуль-
ный отпечаток неширокий, близко подходит к утолщенному левому краю
крышечки (рис. 35, М—О').
Зубы радулы ножевидные, длиной до 400 мкм, с массивной конусовидной
базальной частью, несущей на одной из ручек большой листовидный флажок
(рис. 432, 3). Киль треугольный, с небольшим гарпуновидным отростком, под
которым лежит второе дистальное отверстие зуба (рис. 432, 8). Авторы, иссле-
довавшие зубы радулы (Sars, 1878; Friele, 1886), отмечают, что одной из особен-
ностей данного вида является своеобразное перекрывание стенок лезвия, в ре-
зультате чего образуется излом в верхней 1/3 лезвия зуба (рис. 432, 7, 9).
Вид морфологически сходен с Р. scalarts, от которого отличается более высо-
кими осевыми складками на периферии оборота раковины и наличием язычка
на базальной части зуба.
Распространение. Широко распространенный бореально-аркти-
ческий вид. Обнаружен в Северной Атлантике, практически во всех морях Север-
ного Ледовитого океана (рис. 466, 2), а также в Беринговом, Охотском, Японском
5. PROPEBELA
183
морях (рис. 467). Отмечен для восточного и северо-западного районов канад-
•ского сектора Арктики (Macpherson, 1971).
Экология. Стенотопный вид, предпочитающий илистый песок или
песчанистые илы, обитает на глубинах от 10 до 187 м.
Типы. Типовой экземпляр хранится в ZMO; типовое местонахождение —
.Лофотенские о-ва, Вадсё, Норвегия.
Просмотрен 51 экз.
5. Propebela (Propebela) rugulata (Moller in Troschel, 1866), comb. n.
(Рис. 330-343; 401, А—Г; 434, 1-22; 468, 1, 3).
Moller in Troschel, 1866 i 44, Taf. 4, fig. 6 (Bela); G. 0. Sars, 1878 i 230,
Tab. 23, fig. 6 (Bela; part.); Friele, 1886 1 3, pl. 7, fig. 1, pl. 9, fig. 1—5 (Bela); Dall,
1886a ! 50, pl. 13, fig. 1—5 (Lora); Martini, Chemnitz, 1887 I 143, Taf. 31, fig. 7
dBela); Knipowitsch, 1901 t 43, Taf. 18, fig. 14—15 (Bela); Shimek, Kohn, 1981 i
-428, fig. 11, D, E (Oenopota). — gouldii V e r r i 11, 1882 1 465, pl. 57, fig. 6, 6, a (Bela)..—
clathratus Friele, 1886 t 4, pl. 7, fig. 2 (Bela rugulata var.). — subarctica Derjugin,
1924 i 84, fig. 9—10 (Lora), syn. n.
Рис. 468. Места находок Propebela rugulata (1) и P. bergensis (2) в морях СССР; 3 — палеон-
тологические находки.
Раковина пепельно-серая, башенковидная, с. 5—7 уступчатыми оборотами,
уплощенными в верхней части. Периостракум тонкий, коричневатый, легко
•отстающий от поверхности раковины. Протоконх выпуклый, представлен 1.3—
1.5 оборотами (рис. 401, А—Г). Весь первый эмбриональный оборот лишен
скульптурных образований. На втором обороте закладываются 4 спиральных
ребра, которые к конечным участкам эмбриональной раковины уже пересека-
ются тончайшими осевыми складками. Осевая скульптура дефинитивной рако-
вины представлена пологими, треугольными в сечении, средними складками,
разделенными равными им по ширине или немногим большими пространствами.
Складки наиболее выражены на периферии оборота; сглаживаются они в ниж-
ней 1/3 основания раковины. На последнем обороте насчитывается от 15 до 20 осе-
вых складок, на предпоследнем от 12 до 14. Спиральная скульптура представ-
лена различными элементами, среди которых выделяются 3 группы: киль, про-
ходящий по плечу оборота и образующий мозолистые утолщения при пересече-
нии с осевыми складками; многочисленные тонкие (реже средние) ребра, распо-
лагающиеся на периферии оборота, основании раковины и сифональном выросте;
нитевидные ребрышки в количестве 3—7, лежащие на плече оборота. Межребер-
ные промежутки варьируют по ширине, но, как правило, они равновелики
по спиральными ребрами. Основание раковины уплощенное, плавно переходит
184
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
в удлиненный сифональный вырост. Устье обратногрушевидной формы, угло-
ватое в верхней части, несет на колумеллярном крае широкую полосу белого
или кремового цвета каллуса. Сифональный канал протяженный, неглубокий,,
средней ширины. Наиболее крупный экземпляр в нашей коллекции имел
следующие параметры (мм): Вр=13.0; #=5.8; Впо=1.1; А?/=5.9; Д =2.1.
Крышечка когтевидная, с сильно изгибающейся осью нарастания. Секиро-
видный мускульный отпечаток расположен вблизи утолщенного левого края
крышечки (рис. 35, Г — Е).
Зубы радулы расширенно-ножевидной формы, с массивным коническим осно-
ванием, несущим только 1 ручку с небольшой треугольной шпорой(рис. 434, 22),
вторая ручка рудиментарна-(рис. 434, 11). Киль представляет собой маленькое-
утолщение апикального участка лезвия (рис. 434,10), под которым расположено
небольшое лунковидное второе дистальное отверстие. Отмечается полное сход-
ство исследованных радул с изображениями зубов моллюсков, собранных в Нор-
вежском море (Troschel, 1866 : Taf. 4, fig. 6; Friele, 1886 : pl. 9, fig. 1—5).
Вид сходен с P. assimilis, от которого отличается сильным развитием спи-
ральной скульптуры и зубами радулы с маленьким килем на дистальной части
зуба.
Замечания. Первоописание очень короткое и не позволяет четко иден-
тифицировать вид; то же касается описания и рисунков зубов радулы. По сути
можно говорить о том, что вид описан только по радуле. Следует отметить, что
радула Bela gouldii (описанная и изображенная) идентична рисункам В. rugu-
lata (Troschel, 1866; собственные данные).
Распространение. Атлантический широко распространенный вы-
сокобореально-арктический вид, обитает у берегов Норвегии — Бодо, Тромсё,
Хаммерфест (Friele, 1886), а также в большинстве районов Баренцева и Белого
морей, отмечен в Карском море и море Лаптевых (рис, 468, 1). Обнаружен
у американских берегов в заливах Мэн, Массачусетс, Кейп-Код.
Экология. Обитает на глубинах от 9.5 (в Белом море) до 500 м, преиму-
щественно на 20—100 м, на песчанистых с примесями ила или заиленных грун-
тах (до 70 % всех находок) при температурах от —1.8 до 6.2 °C и солености
от 25.6 (в Белом море) до 35.0 °/00.
Типы. О типе Bela rugulata сведений нет. Место хранения типового эк-
земпляра В. gouldii неизвестно; типовое местонахождение — зал. Мэн. Типо-
вое местонахождение Lora subarctica — Баренцево море (место хранения не ука-
зано).
Просмотрено 165 экз.
6. Propebela (Propebela) spitzbergensis (Friele, 1886), stat, et comb. n.
(Рис. 344-347; 432, 12-23; 466, 1).
Friele, 1886 I 4, pl. 7, fig. 3, 4, pl. 9, fig. 6, 6, a{Belarugulata var.); Knipowitsch,
1901 i 43 {Bela rugulata var.); О dhner, 1915 1 211 (Bela rugulata var.; part.); О 1 dro у d,
1927 t 117, pl. 7, fig. 10 {Lora rugulata subsp.).
Вид по форме и скульптуре раковины сходен с Р. rugulata. Морфологичес-
ким отличием является большее развитие киля, обозначающего плечо оборота,
и более рельефная спиральная скульптура (Friele, 1886 : 4). Высота раковины
голотипа 19.0 мм при ширине 10.0 мм.
Крышечка когтевидная, удлиненная, ось нарастания слабо изгибается.
Мускульный отпечаток в виде тонкой полоски расположен поблизости от утол-
щенного левого края (рис. 35, Ж, Л).
Зубы радулы расширенно-ножевидные, длиной до 200 мкм, с массивным
основанием, несущим на одной из ручек большой листовидный флажок
(рис. 432, 23). Киль маленький, шлемовидный, занимает апикальный участок зуба
(рис. 432, 16, 21). Дистальное отверстие зуба лунковидное, образовано завер-
нутыми стенками лезвия, в результате чего создается впечатление каймы, об-
рамляющей весь периметр отверстия.
5. PROPEBELA
185
Вид сходен по форме раковины с Р. rugulata и Р. nobilis, от которых отли-
чается широко-ножевидной формой зуба с характерно окаймленным дистальным
•отверстием лезвия.
Распространение. Евразийский арктическо-бореальный вид высо-
кобореально-атлантического происхождения. Обитает у западных, восточных
и северных берегов Шпицбергена, в Баренцевом море у Мурманского побережья,
у о-ва Килдин, в Карском море в основном в центральной и северной его ча-
стях, в Белом море у Соловецких о-вов и в море Лаптевых у северного побе-
режья Новосибирских о-вов (рис. 466, 7).
Экология. Встречается на глубинах от 7 до 450 м, в основном на 20—
120 м; преимущественно на илистых и заиленных грунтах (более 80 % всех
находок). Обитает при температурах от —1.8 до 1.2 °C, в основном в диапазоне
от —1.6 до —1.0 °C, и солености 31.64—35.03 ?/00, преимущественно 34.5—
:35.0 •>/„„.
Типы. Типовая серия хранится в ZMO; типовое местонахождение —
Magdaline Вау, Шпицберген.
Просмотрено 74 экз.
7. Propebela (Propebela) bergensis (Friele, 1886), comb. n. (Рис. 31, A—Б;
348-352; 401, Д—Е; 402, A—B; 468, 2).
Friele, 1886 I 4, pl. 7, fig. 5, pl. 9, fig. 7—8 (Bela rugulata var.); Bouchet, Wa-
T e n, 1980 I 73, figs. (Oenopota; — detegata, furfuraculata). — detegata L о с a r d, 1897 I 256,
pl. 21, fig. 12—14 (Bela). — furfuraculata L о с a r d, 1897 I 254, pl. 21, fig. 9—11 (Bela).
Раковина белая, веретеновидная, очень тонкостенная, с 5—5.5 оборотами
отлогоугловатой формы, тупоугловатыми в верхней части. Периостракум тон-
кий, полупрозрачный, коричневатый. Протоконх сильновыпуклый, диаметром
850—1000 мкм (Bouchet, Waren, 1980), представлен 1.5—1.7 оборотами. Первый
эмбриональный оборот лишен макроскульптурных образований, но густо по-
крыт микроскульптурными бугорками. На втором обороте происходит закладка
4 нитевидных спиральных ребрышек. Практически одновременно с ними за-
кладываются и элементы осевой скульптуры, в результате чего второй оборот
протоконха приобретает ячеистый вид (рис. 401, Е). Осевая скульптура дефини-
тивной раковины представлена тонкими складками, треугольными в сечении,
разделенными в 2—3 раза более широкими промежутками. Складки выражены
одинаково на всей поверхности оборотов и основания раковины. На последнем
обороте насчитывается от 14 до 20 осевых складок, на предпоследнем от 13 до 16.
Спиральная скульптура на плече оборота представлена 2—4 нитевидными реб-
рышками, на остальной поверхности раковины — тонкими ребрами. Наиболь-
шее развитие среди элементов спиральной скульптуры отмечено для килевид-
ного ребра, проходящего по плечу оборота. Межреберные промежутки на пери-
ферии оборотов в 3—5 раз шире ребер, на основании раковины и на сифональ-
ном выросте, как правило, равны им. Основание раковины уплощенное, сифо-
нальный вырост небольшой. Устье узко-обратногрушевидной формы, с тонким
белесоватым или розоватым каллусом на колумеллярном крае. Сифональный
канал протяженный, узкий.
Крышечка маленькая, когтевидная. Ромбовидный мускульный отпечаток
немного смещен в левую сторону от срединной части крышечки (рис. 35, 3—К).
Зубы радулы той же конфигурации, что и у Р. rugulata.
Вид сходен с Р. rugulata, от которого отличается небольшой раковиной с тон- .
кой решетчатой скульптурой и высоким сильновыпуклым первым оборотом
протоконха.
Замечания. Основанием для выделения нового вариетета (Friele,
1886) послужили следующие морфологические особенности Р. bergensis: 1) зави-
ток длиннее устья и 2) большее число осевых складок, чем у типичной формы.
‘Следует отметить, что отдельные экземпляры отвечают указанным требованиям,
;но у некоторых моллюсков высота завитка может быть равна высоте устья.
186
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Очевидно, что данный признак (как и у большинства других видов) обладает-
большим диапазоном изменчивости.
Распространение. Атлантический евразийский бореально-суб-
тропический вид, заходящий в арктические воды, распространен в западной
части Арктического бассейна и встречается в бореальных водах Северной
Атлантики (рис. 468,2). Отмечен в батиали (Locard, 1897; Bouchet, Waren, 1980).
Типы. Синтипы Bela rugulata var. bergensis (Bouchet, Waren, 1980) хра-
нятся в ZMB и в BMNH; типовое местонахождение — Sognefjord, Берген,.
180 м. Голотипы В. detegata и В. furfuraculata хранятся в MNHN (Bouchet,
Waren, 1980). Типовое местонахождение В. detegata — южное побережье Пор-
тугалии, глубина 532 м; В. furfuraculata — западное побережье Марокко, глу-
бина 1 900 м.
Просмотрено 27 экз.
8. Propebela (Propebela) harpularia (Couthouy, 1838), comb. n. (Рис. 355—
367; 404, В—Г; 433, 1-22; 463, 1, 3).
Couthouy, 1838 i 106, pl. 1, fig. 10 (Fusus); Gould, 1841 i 291, fig. 191 (Fusus);:
Stimpson, 1851 I 48 (Mangelia); Adams, Adams, 1858 1 92 (Bela); Gould, 1870 г
353, fig. 621 (Bela); G. O. Sars, 1878 : 234, Tab. 16, fig. 17 (B ela); V err ill, 1882 i 473,
pl. 43, fig. 14, pl. 57, fig. 9 (Bela)-, Tryon, 1884 i 219, pl. 29, fig. 51 (Bela); Герцен-
штейн, 1885 i 687 (Pleurotoma); F r i e 1 e, 1886 i 20, pl. 4, fig. 23, 24 (Bela); Martini,
Chemnitz, 1887 t 139, Taf. 31, fig. 2 (Bela)-, Dall, 1889 i 98, pl. 50, fig. 17 (Bela); P о s-
s e 11, Jensen, 1898 t 144 (Bela); К о b e 1 t, 1905 I 243, Taf. 82, fig. 21 (Bela); Grieg,.
1909b I 26 (Bela); Harmer, 1915 i 287, Tab. 32, fig. 15 (Bela); Oldroyd, 1927 i 112:
(Lora); Grant, Gale, 1931 I 524 (Lora); Morris, 1960 : 220, Tab. 39, fig. 6 (Lora);
1966 t 104, Tab. 40, fig. 4 (Lora); Macpherson, 1971 1 117, map 52 (Oenopota); Голи-
ков, Гульбин, 1977 ! 239 (Oenopota). — rosea M. S a r s in L о v e n, 1846 I 144 (Trito-
nium); J effreys, 1867 i 396 (Pleurotoma turricula var.); G. O. S a r s, 1878 1 234, Tab. 23,
fig. 10 (Bela harpularia var.); V e r r i 1 1, 1882 t 485 (Bela); Martini, Chemnitz,.
1887 t 139, Taf. 31, fig. 3 (Bela); К о b e 1 t, 1905 t 238, Taf. 82, fig. 5 (Bela turricula subsp.);.
Oldroyd, 1927 I 120 (Lora). — harpularioides Golikov et Fedjakov in Голи-
ков, 1987 t 124, рис. 69 (Oenopota), syn. n.
Раковина, розоватая или коричневатая, веретеновидно-овальная, реже ве-
ретеновидная, с 5—6 оборотами отлогоугловатой формы, тупоугловатыми
в верхней части. Периостракум очень тонкий, неплотно прилегающий, кремо-
вого или желтовато-коричневого цвета. Протоконх представлен 1.8 оборотами,,
причем первый эмбриональный оборот навивается практически планоспирально.
Он лишен макроскульптуры, но густо усеян микроскульптурными бугорками.
На втором эмбриональном обороте закладываются 3 спиральных ребрышка,-
получающие дальнейшее развитие на дефинитивной раковине. Осевая скульп-
тура дефинитивной раковины представлена средними складками, округлыми
в сечении и часто равновеликими с разделяющими их промежутками. Складки
наиболее выражены на периферии оборота, сглаживаются они на основании
раковины; на плече имеют небольшой наклон вперед. На последнем обороте-
насчитывается от 13 до 20 складок, на предпоследнем от 12 до 18. Спиральная
скульптура состоит из многочисленных тонких или средних шнуровидных ре-
бер, разделенных в 2—3 раза более узкими межреберными промежутками.
Элементы спиральной скульптуры одноразмерны почти на всей поверхности
оборота и основания раковины, лишь иногда большее развитие получают 3 ребра
(эмбриональной закладки): одно проходит по плечу, другое — по срединной
части периферии оборота. Третье ребро лежит на стыке оборотов и видно только
на последнем обороте, где оно отмечает верхнюю границу основания раковины.
Основание раковины выпуклое, сифональный вырост средних размеров. Устье-
обратногрушевидной формы, с белесоватым (реже бежевым) каллусом на колу-
меллярном крае. Анальный синус отсутствует. Сифональный канал средней
ширины, глубокий. Наиболее крупная раковина обнаружена в южной части
Баренцева моря (мм): Вр—12.4; Д=б.8; Впо—1Л; Ву—бЛ); Ду—2Л. В литера-
туре содержатся сведения о нахождении моллюска с параметрами раковины.
(мм): Вр=17.0; Д=7.5; Вио=11.5; Ву=9.0; #у=3.0 (Verrill, 1882).
5. PROPEBELA
187
Зубы радулы шиловидные (рис. 433, 1—22), различной, конфигурации. На
ранних стадиях формирования они более листовидные, чем на поздних (рис. 433,
1—5, стадии формирования зуба в длинном рукаве радулярного мешка). Часть
описанных стадий отмечена и другими исследователями (G. О. Sars, 1878 :
Tab. 9, fig. 3; Friele, 1886 : pl. 9, fig. 23—24).
Вид похож на P. turricula, от которого отличается более округлыми обо-
ротами, слабой угловатостью их плеча и чуть скошенными осевыми складками.
Распространение. Широко распространенный бореально-аркти-
ческий вид, обитает в высокобореальных районах Северной Атлантики, отме-
чен в Лабрадоре, у берегов Ньюфаундленда, у п-ова Новая Шотландия, в зал.
Массачусетс, от п-ова Кейп-Код до о-ва Лонг-Айленд, у северных и восточных
берегов Исландии, у северных и южных берегов Норвегии; в Арктическом
'бассейне обнаружен у берегов Северной Земли, Земли Франца-Иосифа, Шпиц-
бергена, в Баренцевом и Белом морях (рис. 463, 1), а также в море Лаптевых
и Восточно-Сибирском море. В дальневосточных морях обитает в зал. Провиде-
ния в Беринговом море, у Курильских и Шантарских о-вов в Охотском море,
у о-ва Монерон и в зал. Петра Великого в Японском море.
Экология. Встречается на глубинах от 5 до 500 м, у восточного по-
бережья Северной Америки до 693 м (Thorson, 1944), однако наибольшая частота
встречаемости приходится на глубины 20—100 м. Представители вида обитают
на всех типах грунтов, предпочитая, однако, мелкофракционные: 70 % особей
встречено на песчанистых и илистых грунтах. Диапазон температур, при ко-
тором были обнаружены моллюски, составил от —1.8 до 7.5 °C, наиболее
часто животные встречаются при температурах от —1.0 до 2.0 °C. Большинство
моллюсков встречается при солености 34.4—35.0 °/00. Минимальная соленость
обитания 27.1 °/00.
Типы. Голотип Fusus harpularius хранится в BSNH; типовое место-
нахождение — зал. Массачусетс (описан по экземпляру, обнаруженному в же-
лудке рыбы). Типовой экземпляр Tritonium roseum хранится в ZMO; типовое
местонахождение — Арктика (Oldroyd, 1927). Голотип Oenopota harpularioides
хранится в ЗИН АН СССР, № 35054; типовое местонахождение — Кандалакш-
ский залив, Белое море, глубина 52 м.
Просмотрено 402 экз.
9. Propebela (Propebela) popovia (Dall, 1919), comb. n. (Рис. 373; 435,
13—15).
Dall, 1919a I 42, pl. 14, fig. 8 [Lora)’, Oldroyd, 1927 I 117 [Lora).
Раковина белая, веретеновидная, с 5—6 оборотами отлогоугловатой формы,
тупоугловатыми в верхней части. Периостракум тонкий, полупрозрачный.
•Эмбриональная раковина не изучена. Осевая скульптура дефинитивной рако-
вины представлена полукруглыми в сечении, широкими складками. Они ярко
выражены на всей поверхности оборота и незначительно сглаживаются на осно-
вании раковины. На последнем и предпоследнем оборотах раковины насчиты-
вается по 14—16 осевых складок, разделенных в 1.5—2 раза меньшими про-
странствами. Среди элементов спиральной скульптуры наблюдается четкая
дифференцировка. Так, в верхней части плеча'оборота и на нижней части осно-
вания раковины, как и на сифональном выросте, спиральная скульптура пред-
ставлена нитевидными ребрышками. На периферии оборота (и прилегающих
участках) расположено по 4—5 средних шнуровидных ребра, между которыми
.залегает по 1—2 тонких ребра. В результате поверхность раковины приобре-
тает гофрированный вид. Межреберные промежутки всегда значительно тоньше
самих ребер. Основание раковины умеренно выпуклое, плавно переходит в не-
большой сифональный вырост. Устье узко-обратногрушевидной формы, с ши-
рокой полосой белого матового каллуса на колумеллярном крае. Сифональный
•.канал узкий, неглубокий. Размеры раковины типового экземпляра (мм): Вр=
188
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
=13.0; Д=5.5; Z?no=8.0. Исследованный мной моллюск имел раковину высо-
той 8.7 мм.
Зубы радулы расширенно-ножевидные, с относительно небольшим кони-
ческим основанием, несущим на одной ручке листовидный флажок, другая ручка
рудиментарна. Киль небольших размеров, занимает апикальный участок лезвия,
дистальное отверстие зуба широкое и протяженное, может достигать х/2 общей
длины зуба (рис. 435, 13—15).
Вид сходен с Р. terpentensis, от которого отличается более пологими обо- '
ротами и всегда чередующимися элементами спиральной скульптуры: между
каждыми 2 ребрами залегает шнуровидное ребрышко.
Распространение. Тихоокеанский широко распространенный вы-
сокобореальный преимущественно приамериканский вид. Встречен у американ-
ского побережья от Бристольского залива в Беринговом море до зал. Монтерей,.
Калифорния, а также у берегов Западной Камчатки.
Типы. Голотип хранится в USNM, № 220883- типовое местонахождение —
Бристольский залив, Берингово море.
Просмотрен 1 экз.
10. Propebela (Propebela) scalaris (Moller, 1842), comb. n. (Рис. 375—383;
402, Г—E; 403, Л—В; 437, 10—22; 438, 1—9; 469, 1).
Moller, 1842185 (Defrancia); Leche, 1878 l 52 (Pleurotoma turricula var.);
G. O. S a r s, 1878 1 229, Tab. 23, fig. 5, Tab. 8, fig. 16 (Bela); Verrill, 1882 г 471, pl. 57,
fig. 12, 12a (Bela); Dall, 1886a 1 153, pl. 7, fig. 8 (Bela); Friele, 1886 i 6, pl. 7, fig. 9—14,
pl. 9, fig. 16-—22 (Bela); Martini, Chemnitz, 1887 t 161, Taf. 33, fig. 1 (Bela); Pos-
selt, Jensen, 1898 I 149 (Bela exarata var.); К о b e 1 t, 1905 I 239, Taf. 82, figs. (Beltz
turricula subsp.); Dautzenberg, Fischer, 1912 : 42 (Bela turricula var.); Old-
royd, 1927 i 96, pl. 7, fig. 8 (Lora); Филатова, Зацепин, 1948 i 389, табл. 98,
рис. 17 (Lora); Bouchet, Waren, 1980 I 73, fig. 151, 268 (Oenopota; part.). —• americana
Packard, 1867 t 285, pl. 7, fig. 11 (Bela); Tryon, 1884 I 220, pl. 33, fig. 64 (Bela); К o-
b e 1 1, 1905 1 244, Taf. 82, fig. 13 (Bela). — ecarinata G. O. S a r s, 1878 : 229, Tab. 16, fig. 9!
(Bela scalaris var.). — abyssicola Friele, 1886 t 6, pl. 7, fig. 12—13, pl. 9, fig. 21—22 (Belct
scalaris var.).
Раковина белая или розоватая, удлиненно-веретеновидная или башенковид-
ная, с 6—7 уступчатыми оборотами, уплощенными в верхней части. Периостра-
кум тонкий, легкоотстающий, желтоватый. Протоконх выпуклый, представлен
1.7—2 оборотами. Первый оборот украшен микроскульптурными бугорками,
на втором эмбриональном обороте закладываются и получают развитие 5 спи-
ральных тонких ребер. Приблизительно через 1/4 оборота после закладки спи-
5. PROPEBELA
. 189'
ральных ребер появляются осевые складки, которые по величине (ширине и вы-
соте) равновелики с элементами спиральной скульптуры. Благодаря равномер-
ности расположения складок и ребер второй оборот протоконха приобретает-
сетчатый рисунок. Осевая скульптура дефинитивной раковины представлена:
полукруглыми в сечении, тонкими складками, отделенными друг от друга в 2—
3 раза более широкими промежутками. Осевые складки одноразмерны на всей
поверхности последнего оборота и на основании раковины. В верхней части
(на плече оборота) они имеют небольшой изгиб вперёд. На последнем обороте
раковины насчитывается от 18 до 20 осевых складок, на предпоследнем от 14
до 18. Спиральная скульптура на плече оборота представлена 4—7 нитевидными
ребрышками; на периферии, основании раковины и на сифональном выросте
лежат многочисленные равновеликие тонкие ребра, разделенные равными им
по ширине межреберными промежутками. Киль на поздних дефинитивных обо-
ротах слабо выражен и практически не преобладает над элементами спиральной
скульптуры периферии оборота. Основание раковины умеренно выпуклое, сифо-
нальный вырост удлиненный. Устье обратногрушевидной формы, угловатое
в верхней части, несет на колумеллярном крае широкую полосу кремового цвета,
каллуса. Сифональный канал протяженный, относительно неглубокий, широ-
кий. Высота раковины типового экземпляра 6.5 мм. Наибольшие размеры рако-
вины (мм): Бр=24.2; Д=11.5; БПО=16.2; 7^=12.5; ^=5.0. Известна рако-
вина, высота которой 25.0 мм при ширине 11.0 мм (Verrill, 1882).
Крышечка крупная, когтевидная. Мускульный отпечаток с заострен-
нойверхней частью, расположен недалеко от утолщенного левого края крышечки
(рис. 35, П—С).
Зубы радулы длиной 250—300 мкм, ножевидные, с массивным основанием,
несущим на одной из ручек уплощенный флажок. Лезвие зуба имеет характер-
ный изгиб назад. Киль маленький, шлемовидный, образует гарпуновидные
выросты (рис. 438, 2, 7—8). За счет своеобразного заворачивания стенок лезвия
зуба и большого размера дистального отверстия создается впечатление излома
верхней 2/з лезвия (отмечено и у Фриле: Friele, 1886: pl. 9, fig. 17, 20).
Вид сходен с Р. assimilis, от которого отличается тонкими осевыми складками
и более высоким завитком, а также гарпуновидным килем зуба радулы.
Распространение. Широко распространенный бореально-арктичес-
кий вид, вероятно, атлантического происхождения. Обитает в высокобореаль-
ных районах Северной Атлантики и циркумполярно в Арктическом бассейне,,
заходит в бореальные районы Тихого океана. Отмечен в Беринговом море до
о-ва Ситка, Аляска, в Северном Ледовитом океане в море Лаптевых, в северной"
части Карского моря, повсеместно в Баренцевом море (рис. 469, 7) и у берегов
Западной Норвегии. У атлантического побережья Северной Америки описано
нахождение от п-ова Кейп-Код до Лабрадора.
Экология. Обитает на глубинах от 1.5 до 660 м, встречаясь в основном
на 10—190 м; практически на всех типах грунтов, предпочитая песчанистые,
при температурах от —1.9 до 5.5 °C (в основном от —1.4 до —0.6 °C); предпочи-
тает океаническую соленость 34.3—35.0 °/00.
Типы. Типовой экземпляр Defrancia scalaris хранится в ZMC; типовое
местонахождение — воды Западной Гренландии. Типы Bela americana, по-види-
мому, хранятся в BSNH; типовое местонахождение — Square Island, п-ов Лаб-
радор.
Просмотрено 65 экз.
И. Propebela (Propebela) scalaroides(G. О. Sars, 1878), comb. n. (Рис. 38, Б;
384—385; 403, Г—Е; 438, 10—18; 469, 2).
G. О. Sars, 1878 I 231, Tab. 23, fig. 7 (Bela); Tryon, 1884 t 226, pl. 28, fig. 39 (Bela-
tirricula subsp.); Friele, 1886 t 5, pl. 7, fig. 7, pl. 9, fig. 11—12 (Bela rugulata var.); M a r-
tini, Chemnitz, 1887 i 158, Taf. 32, fig. 12 (Bela); К о b e 1 t, 1905 i 240, pl. 82, fig. 19-
(Bela turricula subsp.); Odhner, 1915 t 211 (Bela rugulata var.); Schlesch, 1924 t 326,.
Taf. 11, fig. 4 (Bela rugulata subsp.).
190
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Раковина, как правило, белая удлиненно-веретеновидная, с 6.5—7.5 обо-
ротами отлогоугловатой формы, тупоугловатыми в верхней части. Перио-
•стракум тонкий, коричневых оттенков, плотноприлегающий. Протоконх вы-
пуклый, представлен 1.8 оборотами, причем первый оборот навивается практи-
чески планоспирально и лишен скульптурных элементов. На втором эмбрио-
нальном обороте закладываются 3 ребрышка, из которых верхнее редуцируется
уже на первом постэмбриональном обороте (рис. 403, Д). Осевая скульптура
закладывается только на дефинитивной раковине и представлена тонкими склад-
ками, разделенными в 1.5—2 раза более широкими промежутками. Складки
наиболее выражены в верхней части оборота и сглаживаются на основании рако-
вины, исчезая в его нижней 1/3. На последнем обороте насчитывается от 16
.до 26 складок, на предпоследнем от 13 до 22. Спиральная скульптура на плече
оборота представлена 5—7 нитевидными ребрышками, плечо оборота обозначает
•тонкое килевидное ребро. На периферии оборотов, на основании раковины
и на сифональном выросте расположены многочисленные, равновеликие с раз-
деляющими их промежутками тонкие ребра. Основание раковины умеренно
выпуклое, сифональный вырост вытянутый. Устье неправильно-полуовальной
•формы, угловатое в верхней части; палатальный край несет широкую полосу
толстого белого каллуса. Сифональный канал протяженный, широкий, уме-
ренно глубокий. Высота раковины голотипа 23 мм, максимальные размеры рако-
вины в коллекции ЗИН АН СССР (мм): В =26.5; Д=11.5; Z?no=18.6; В =
= 13.5; Д=ЪЛ.
Зубы радулы расширенно-ножевидной формы, с массивным основанием, ди-
стальная часть которого отогнута вниз. Лезвие очень большого диаметра
•{более 1/4 общей длины зуба). Характерная черта вида, отмеченная на рисунках
Capca(G. О. Sars, 1878 : Tab. 8, fig. 15), — форма киля: он практически целиком
отходит от апикального участка лезвия и образует-далеко выступающий гарпун
(рис. 438, 11, 16).
Вид хорошо отличается от других представителей подсемейства по морфоло-
гическим особенностям зуба радулы — гарпуновидным килем и отогнутой ба-
зальной частью основания зуба.
Распространение. Арктический евразийский вид. Помимо побе-
режья Финмаркена и Лофотенских о-вов отмечен у Шпицбергена, северного
побережья Исландии (Иса-Фьорд), в Баренцевом море и северной части Кар-
ского моря (рис. 469, 2).
Экология. Обитает на глубинах от 3 (Земля Франца-Иосифа) до 426 м,
встречаясь в основном на 150—.300 м; на всех типах грунтов, предпочитая
илистые и заиленные, при температурах от —1.7 до 3.2 °C, преимущественно
в диапазоне от —1.5 до —0.3 °C, и солености 33.58—34.94 °/00.
Тип ы. Типовые экземпляры хранятся в ZMO; типовое местонахождение —
западный Финмаркен.
Просмотрено 32 экз.
12. Propebela (Propebela) svetlanae Bogdanov, 1989. (Рис. 328—329;
•435, 1—8).
Богданов, 19896 I 151, рис. 1, 7—8; 2, н—р.
Раковина телесного цвета, веретеновидная, с 4.5—5 уступчатыми оборотами,
уплощенными в верхней части. Периостракум тонкий, коричневатый, полу-
прозрачный. Протоконх не изучен. Осевая скульптура дефинитивной раковины
представлена средними складками, в 3—4 раза более тонкими, чем разделяющие
их промежутки. Осевые складки одинаковы по высоте на всей периферии обо-
рота, уменьшаются в размерах на плече оборота и сглаживаются на основании
раковины. На предпоследнем обороте насчитывается 12—14, на последнем
15—17 таких складок. Спиральная скульптура представлена различными
по ширине тонкими шнуровидными ребрами. Они, как правило, превышают
5. PROPEBELA
19T
по размерам разделяющие их промежутки. Основание раковины умеренно
округлое, без резких границ переходит в короткий сифональный вырост.
Устье узко-обратногрушевидное, с толстым коричневатым каллусом на всей
колумеллярной поверхности. Сифональный канал узкий, умеренно глубокий.
Размеры раковины голотипа (мм): Вр=1.3; Д=3.9; j?no=5.6; Z?g,=4.0; Д^=1.7..
Высота раковины наибольшего экземпляра 10.8 мм.
Зубы радулы длиной от 140 до 280 мкм, расширенно-ножевидной формы,,
с массивным конусовидным основанием, которое несет на одной из ручек упло-
щенный треугольный флажок. Треугольный киль занимает самую апикальную-
часть зуба (рис. 435, 7). Под килем расположены 2 дистальных отверстия
зуба: одно — оставшееся за счет неполного заворачивания стенок лезвия,,
другое — вновь образовавшееся, лунковидное.
Вид сходен с Р. assimilis, от которого отличается низким завитком раковины
и расширенно-ножевидными зубами с маленьким треугольным килем.
Распространение. Тихоокеанский приазиатский высокобореаль-
ный вид. Помимо бухты Нагаева известен только у побережья Западной Кам-
чатки.
Типы. Голотип хранится в ЗИН АН СССР, № 48474; типовое местонахож-
дение — бухта Нагаева (59° ЗГ с. ш.—150° 44' в. д.), Охотское море, глубина.
10 м.
Просмотрено 2 экз.
13. Propebela (Propebela) verrilli Bogdanov, 1989. (Рис. 353—354; 436,.
9—15).
Богданов, 1989а I 137, рис. 1, 6—7; 2, л—п.
Раковина светло-коричневая, удлиненно-веретеновидная, с 7—8 отлогоугло-
ватыми оборотами. Периостракум тонкий, коричневатый, плотноприлегающий.
Протоконх неизвестен. Осевая скульптура представлена редко расположен-
ными средними складками, треугольными в сечении. Промежутки, разделяю-
щие складки, в 2—4 раза шире их. На последнем и предпоследнем оборотах на-
считывается по 11—12 осевых складок. Спиральная скульптура сильно варьи-
рует в размерах, наибольшее развитие получает киль, проходящий по плечу
оборота. Основание раковины скошенное, орнаментировано только элементами?
спиральной скульптуры. Устье узко-обратногрушевидное, с тонким матовым
каллусом на колумеллярном крае. Сифональный канал узкий, относительно
глубокий. Размеры раковины голотипа (являющейся одновременно и наиболее-
крупной) (мм): _5p=11.9; Д=4.6; 2?„О=7.2; 5^=4.7; Ду=1.8.
Зубы радулы длиной до 220 мкм, ножевидной формы, с конусовидным осно-
ванием и небольшим треугольным гарпуновидным килем (рис. 436, 13). Лезвие-
зуба слегка изогнуто.
По форме раковины вид похож на Р. angulosa, от которого отличается пря-
мыми (не скошенными на околошовной площадке) осевыми складками и малень-
ким треугольным килем на дистальном участке лезвия зуба. От остальных видов:
рода отличается высоким завитком.
Распространение. Тихоокеанский приазиатский низкобореаль-
ный вид, обитает в Приморье.
Типы. Голотип хранится в ЗИН АН СССР, № 48475; типовое местонахож-
дение — Приморье, разрез от м. Пельма, 47° 4Г с. ш.—139° 16' в. д., глубина-
31 м, ил с примесью гальки и гравия.
Просмотрено 3 экз.
14. Propebela (Propebela) cassis Bogdanov, 1989. (Рис. 374; 435, 9—12).
Богданов, 19896 I 151, рис. 1, 9; 2, с—ф.
Раковина светло-каштанового цвета, овально-веретеновидная, с 5 закруглен-
ными сильновыпуклыми оборотами. Периостракум тонкий, прозрачный, плотно-
J92 СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
прилегает к раковине. Эмбриональная раковина не сохранилась. Осевая скульп-
тура дефинитивной раковины представлена широкими пологими складками,
которых на последнем обороте насчитывается 18. Осевые складки наиболее
выражены в верхней и средней частях периферии оборота, к ее нижней части
они сглаживаются, так что основание раковины лишено следов осевой скульп-
туры. Вся поверхность раковины равномерно покрыта равновеликими нитевид-
ными спиральными ребрышками, по ширине одинаковыми с разделяющими их
^промежутками. Скошенное основание раковины плавно переходит в короткий
сифональный вырост. Устье неправильно-овальное, угловатое в верхней части,
несет на колумеллярном крае широкий толстый темно-каштанового цвета кал-
лус. Размеры раковины голотипа (мм): 2?р=10.6; Д=5.6; Z?no=8-3; В = 6.0;
Х=2.4.
Зубы радулы длиной 300—320 мкм, расширенно-ножевидной формы, с пира-
мидальным основанием. Одна ручка редуцирована, другая несет листовидный
• флажок (рис. 435, 11). Киль треугольный, гарпуновидный, маленький.
Вид хорошо отличается от другйх представителей рода овально-веретеновид-
ной раковиной с закругленными оборотами.
Распространение. Тихоокеанский приазиатский ’низкобореаль-
ный вид, известен только из типового местонахождения.
Типы. Голотип хранится в ЗИН АН СССР, № 48476; типовое местонахож-
дение — у о-ва Шикотан, Малая Курильская гряда.
Просмотрен 1 экз.
15. Propebela (Propebela) terpeniensis Bogdanov, 1989. (Рис. 371—372;
436, 1—8).
Богданов, 19890 1151, рис. 1, JV; 2, х—ш.
Раковина от коричневого до каштанового цвета, веретеновидная, с 5—6
отлогоугловатыми оборотами. Тонкий светло-коричневый периостракум легко
отшелушивается от раковины. Протоконх не сохранился. Осевая скульптура
дефинитивной раковины состоит из равносторонних в сечении, средних складок
одной ширины с разделяющими их промежутками. На предпоследнем обороте
насчитывается 13—14, на последнем 17—20 средних складок. Спиральная
скульптура представлена различными элементами. Сильнее всего выражены
3 киля, из которых один обозначает плечо, другой — центральную часть пери-
ферии оборота, а третий лежит в месте стыка с последующим оборотом. Между
килями, а также на плече оборота и на основании раковины расположены раз-
личные по величине шнуровидные тонкие спиральные ребра. Основание рако-
вины умеренно выпуклое, сифональный вырост короткий. Устье узко-обратно-
грушевидное, с толстым палатальным краем и с широким кремово-коричневым
каллусом на колумеллярном крае. Размеры раковины голотипа (мм): В =10.2;
,Д=4.8; 5ИО=6.7; ^=4.8; Д>2.0.
Зубы радулы длиной до 150 мкм, расширенно-ножевидные, с небольшим
гарпуновидным килем. Коническое основание несет на одной ручке листовидный
флажок, другая ручка редуцирована.
Вид сходен с Р. mitrata, от которого отличается ножевидной формой зубов
радулы.
Распространение. Тихоокеанский приазиатский низкобореаль-
ный вид. Помимо типового местонахождения найден в водах Приморья.
Типы. Голотип хранится в ЗИН АН СССР, № 48477; типовое местона-
хождение — Южный Сахалин, м. Топина (46° 47' с. ш.—143° 52' в. д.), глу-
бина 187 м.
Просмотрено 3 экз.
16. Propebela (Propebela) gorvachevi Bogdanov, 1989. (Рис. 311—312; 430,
•6-13).
Богданов, 1989а 1 137, рис. 1, 8—9; 2, р—у.
Рис. 6. Общий вид, скульптура и детали строения эмбриональной раковины Oenopota pin-
gelii у экземпляров, собранных в различных акваториях.
А, Б - - у берегов Шпицбергена; В, Г — у мурманского побережья Баренцева моря; Д, Е — в районе Шан-
тарскпх островов.
!/е -f Зак. 21П1
Рис. 7. Эмбриональные раковины Curtitoma novajasemljensis (Л) и С. trevelliana (В—Г).
На эмбриональной раковине С. trevelliana, снятой в разных ракурсах, отчетливо виден процесс кооперации
и слияния ребрышек.
Рис. 8. Строение крышечки моллюсков Oenopotinae.
А — эмбриональная крышечка (нижняя часть рисунка) с развивающейся дефинитивной (верхняя часть ри-
сунка); Б — общий вид наружной поверхности крышечки; В — отпечаток колумеллярного мускула на обо-
ротной стороне крышечки.
лсж псж хо
Рис. 13. Комплекс органов буккального отдела Propebela scalaroid.es.
А — общий вид (окологлоточное нервное кольцо и часть ядовитой железы удалены); Б — поперечный срез
ядовитой железы (ЯЖ). БТ — буккальная трубка, ЛСЖ — левая слюнная железа, ММ — мускульный
мешок, ПСЖ — правая слюнная железа, ПЩ — пищевод, РАД — радулярный мешок, ХО — хобот.
Рис. 21. Общин вид внутренней поверхности статоциста (4) и поверхности статолита (Б).
Рис. 30. Раковины самки (Л) и самца (Б)
Curtitoma trevelliana из одной пробы.
Рис. 27. Propebela nobilis (Л) и Р. spitzber-
gensis (Б) из района Воронки Белого моря,
вписывающиеся в одну форму. Пояснения
в тексте..
Рис. 31. Разноразмерные эмбриональные раковины Propebela bergensis, обнаруженные
в южной части Баренцева моря.
А — аномальная, Б — типичная форма.
Рис. 37. Эмбриональные раковины Oenopota declivis (Л) и Oe. impressa (Б).
Рис. 38.
Эмбриональные раковины
Propebela а п i> и losa (/I) и Р. scalaroldes (Б).
Рис. 39. Эмбриональная раковина Granotoma albrechti (Л) и участок первого оборота’ прото-
коиха с многочисленными неупорядоченными микроскульптурными бугорками (Б).
Рис. 40. Эмбриональная раковина Obesotoma tenuilirata.
.1 — общий вид, Б — участок скульптуры первого оборота протоконха.
Рис. 41. Зубы радулы Propebela turricula (Б, В) и детали их строения (4, Г).
Г __ на заднем плане по центру виден желобок, образованный завернувшимися стенками зуба.
Рис. 54. Фрагменты клад-
' ки Сurtitoma decussata.
Изображения получены с по-
мощью сканирующего мик-
роскопа. На правом рисунке
видна кожистая мембрана
крышечки.
11 *' 'ЛГЛ “ •—*>
Рис. 60—71. Раковины моллюсков рода Curtitoma.
60—62 - С. becklemishevi: 60— Вр=7.7 мм, Охотское море, 51°36'с. ш.—154°46'в. д., глубина 418 м; 61 —
Вр— 7.7, Японское море, 45°12' с. ш,—137°27' в. д., 583 м; 62 — В^=6.9, 51 °10' с. ш.—154°17'Jb. д., 580 л<;
63—65 — С. incisula: 63 — Вр=8.1, зал. Петра Великого, разрез от м. Среднего, 88 м; 64 — Вр=5.5, Охот-
ское море, о-в Сахалин, траверз м. Лихачева, 60 м; 65 — Вр=6.0, Татарский пролив, 47°39' с. ш,—
139с15' в. д., 205 м; 66—68 — С. hebes: 66 — Вр=7.0, Японское море, зал. Петра Великого, 180 м; 67 —-
Вр=7<5, Охотское море, о-в Сахалин, разрез от пос. Пограничный, 200 м; 68 — В^=6.8, Японское море, раз-
рез от м. Энкен, 155 м; 69—70 — С. novajasemljensis: 69 — 0^=4.9, море Лаптевых, о-в Котельный, 8 м; 70 —
Вр=33, Карское море, 76°38' с. ш.—78°51' в. д., 62 м; 71 — С. ovalis, Bp=i.8, 80°01' с. ш.—03°19' в. д.,
2380 м.
Рис. 72—82. Раковины моллюсков рода Curtitoma.
72—77 — С. trevelliana: П — Вр=6.9, 81°35' с. ш,—75°57' в. д., глубина 225 м; 73 — Вр=8.1, 73°38' с. ш.—
27°14' в. д., 422 м; 74 — Вр=8.3, Карское море, 76°36' с. ш.—74°48' в. д., 157 м; 75 — Вр=6.1, Баренцево
море, 74°45' с. ш.—17°20' в. д., 168 м; 76 — В^=10.0, 69°00' с. ш,—43°00' в. д., 70 м; 77 — Вр=11.8,
78°00'с. ш,—12°13' в. д., 245 м; 78— С. fiora, Вр=11.4, Охотское море, м.Хребтовый, 28 м; 79—82 — С. пеу-
тапае: 79 — Вр=7.0, Западная Камчатка, 12 м; 80 — Вр=6.4, Японское море, Советская гавань, 8 м; 81—
Вр—в.Э, Японское море, Де-Кастри, 0.1 м; 82 — Вр=6.7, Западная Камчатка, 8 м.
Рис. 83—94. Раковины моллюска Curtitoma violacea.
S3 —-Ву>=5.9. Японское море, зал. Петра Великого, глубина 210—170 м; 84 —В^=7.1, Баренцево море
70’39' с. ш.-53°42' в. д., 41 м; 85 — £^=5.7, 73’18' с. ш.—55’53' в. д., 121 м; 86 — В^=7.4, 77°50' с. ш.—
104’07' в. д., 207 м; 87 - Bp=7.0, 79°24' с. ш,—70’48' в. д., 526 м; 88 — В/)=6.8, о-в Сахалин, траверз оз.
Айнское, 40 м; 89 — В1О=6.4, Пенжинский залив, 60’32' с. ш.—155’23' в. д., 40 м; 90 — В^=7.0, Южная,
Курильская гряда, 25 м; 91 — В^=9.8, Японское море, зал. Петра Великого, 12 м, 92 — 13^=7.6, Земля.
ФраНца-Иосифа, Британский канал, 80’24' с. ш,—52’10' в. д., 15 м; 93 — Вр=1.5, Баренцево море, Помор-
ский рейд, 17.5 м; 94 — Вр=5.1, Новая Земля, губа Черная, 25 м.
П Зак. 2164.
Рис. 95—105. Раковины моллюсков рода Curtitoma.
95—96 — С. violacea: 95 — Вр=8.1, 70°11' с. ш.—46’00' в. д., глубина 84 м; 96 — Вр=10.6, Му рманский'бв'
per, 17 м; 97—103 — С. conoidea: 97 — Вр=&.7, 80°00'с. ш.—102’05'в. д., 350 м; 98 — Вр=8.9, 80°45’с. nr.~
29’57' в. Д., 445 м; 99 — Bp=91, 78°31 с. Ш,—66’44' в. д., 385 м; 100 — Вр=9.2, 80’1' с. ш.—86°24’ В.Д.,
250 м; 101 — Bp=9.S, море Лаптевых, 77’10' с. ш,—120°50' в. д., 220 м, 102 — Вр=И.2, 69°33' с. ш.—
32’52' в. д., 260 м; 103 — Bj,=10.1, 69’05' с. ш.-37’35* в. д., 178 м; 104—105 - С. decussata: 104 — Вр—
=8.3, 72’39'с. ш,—64’18'в. д., 62 м; 105 — Вр=7 7, 81’35'с. ш.—75’57' в. д., 225 м.
Рис. 106—115. Раковины моллюсков рода Curtitoma.
106—108 — С. decussata: 106 — Вр=9.2, 79°16'с. ш.—80°31' в. д., глубина 80 м; 107 — Бр—7.5, 75’25’с. ш.=-«
81’27' в. д., 24 м; 108 — Вр=5.9, 77’27' с. ш.— 86°15' в. д., 112 м; 109—111 — С. lawrenciana: 109 Вр=
=7.1, о-в Беринга, 54 м; 110 — Вр—7.5, Восточная Камчатка, Кроноцкий залив, траверз м. Ольги, 148 м]
111 — Вр—9.2, Западная Камчатка, 51°25' с. ш.—156’27' в. д., 54 м; 112 — С. piltuniensis: Вр=7.6, Восточ-
ный Сахалин, зал. Пильтун, 5 м; 113—115 — С. bartschi: 113 — Вр=9.6, о-в Уруп, близ м. Китовый, 150 MJ
114 — Вр=7.4, Охотское море, 414 м; 115 — Вр=9.5, Охотское море, 345 м.
Рис. 116—124. Раковины моллюсков родов Curtitoma и Granotoma.
116 — С. microvoluta: Вр==ЭЛ, Японское море, от о-ва Аскольда, глубина 269 -340 м; 117—120 — С. hok-
kaidoensis: 117 — Bp=it.i, 56°09' с. ш.—144°30' в. д., 307 м; 118 — Вр—9.д, Приморье, разрез отм. Пещер-
ного, 160 м; 119 — Вр=6Л, южная часть Охотского моря, 216 м; 120 — Вр=6.5, Татарский пролив, 200 м;
121 —{G. raduga,. голотип, Вр=6.6, Восточная Камчатка, бухта Русская, 111 м; 122—124 - G. albrechti:
122 — Вр=11.2, о-в Уруп, м. Китовый, 214 м; 123 -- Вр=7.5, Охотское море, 24 м; 124 — Вр=10.0, Западная.
Камчатка, 51°25' с. ш.—156°27' в. д., 56 м.
Рис. 125—135. Раковины моллюсков рода Granotoma.
125—127 — G. albrechti: 125 — Bp=7.i, о-в Беринга, глубина 60 м; 126 — В^=7.0, Охотское море, м. Лан-
тарский, 62 м; 127 — 13^=6.8, 50°02' с. ш,—140°46' в. д., 70 м; 128—135 — G. krausei: 128 - 13^=8.3, Охот-
ское море, 53°05' с. ш.—144°10' в. д., 70 м; 129 — Вр=9.6, северо-западная часть Японского моря,
47°04' с. ш.—139°29' в. д., 70 м; 130 — 13^=10.1, Японское море, 50°02' с. ш.—140°46' в. д., 70 м; 131 —
Бр=8.7, Охотоморское побережье восточного Сахалина, 70 м; 132 — В^=8.2, Татарский пролив,
50°03' с. ш.—142°01' в. д., 94 м; 133 — Вр=10.7, о-в Сахалин, пос. Пильтун, 5 м; 134 — Bp=11.2, Японское
море, 40°14' с. ш.—143°21' в. д.; 135 — J3p==l 18, о-в Беринга, траверз р. Фонтанка, 87 м.
Рис. 136—146. Раковины моллюсков рода Oenopota.
136—139 — Ое. cinerea: 136 — Вр=16.0, прол. Вилькицкого, 77*52' с. ш.—99*23' в. д., глубина 51 м; 137 —
Вр=14.1, Баренцево море, 70*08' с. ш.—38°55' в. д., 195 м; 138 — Вр=17.0, Баренцево море,
69804* с. Ш.—36°09' в. д., 85 м; 139 — Вр=11.6, 69*37' с. ш.—34*09' в. д., 230 м; 140—146 — Ое. declivis:
140 — Вр=12.6, Баренцево море, Кильдинский пролив, 100 м; 141 — Вр=11.9, Северная Земля.
79*16'с. ш,—100°04' в. д., 145 м; 142 — Вр=9.6, Мурманский берег, 42 м; 143 — Вр=10.1,Баренцево море,
о-в Колгуев, 24 м; 144 — Вр=10.8, прол. Вилькицкого, 77*47' с. ш.—100*19' в. д., 100 м; 145 — Вр=12.5,
Баренцево море, Ура-Губа, 98 м; 146 — Вр=13.7, Баренцево море, 69*46' с. ш.—46*12' в. д., 77 м.
Рис. 147—157. Раковины моллюсков рода Oenopota.
<47—154 — Ое. elegans: 147 — Вр=ПЛ, Баренцево море, 69°12'с. ш.—36°10' в. д., глубина 180 м; 148 —
Вр=17.1, Килдинская салма, 110 м; 149 — Вр=16.4, Баренцево море, о-в Килдин, 100 м; 150 — Вр=18.0,
Мурманский берег Баренцева моря, 98 м; 151 — Вр=18.6, Баренцево море, 69°12' с. ш.—35°13' в. д., 80 м;
152—Вр=10.8, Баренцево море, Ура-Губа, 219-204 м; 153 — Вр—21.0, Карское море, 77°27' с. ш.—
82°35' в. Д., 57 м; 154 — Вр=18.3, Килдинский пролив, 100 м, 155—157 — Ое. harpa: 155 — Вр=19.5, Мур-
Шанский берег, 68°56' с. ш.—37°26' в. д., 155 м; 150 — Вр=28.0, юго-восточная часть Баренцева моря^
69о20' с. ш.—56°00' в д., 18 м; 157 — Вр=17.0, море Лаптевых, 28—30 м.
Рис. 158—167. Раковины моллюсков рода Oenopota.
158—160 — Oe. harpa: 158 — Вр=9.1, Берингово море, о-в Беринга, глубина 67 м; 159 — В^=12.7, о-в Са-
халин, траверз м. Лопатина, 250 м; 160 — Вр=8.8, Новосибирские о-ва, 77°16' с. ш.—145°24' в. д., 45 м;
161—167 — Oe. impressa: 161— Вр=7.4, Новосибирские о-ва, 77°16' с- ш.—145°24' в. д., 45 м; 162 — Вр=
= 10.3, Баренцево море, Мурманский берег, 18 м; 163 — Bp=11.9, Финмаркен, 13 м; 164 — Вр=14.1, Ба-
ренцево море, Иоканские о-ва, 40 м; 165 — Ву;=6.4, Охотское море, о-в Сахалин, м. Елизаветы, 100 м; 166 —
JB=6.5, Баренцево море, Мурманское побережье, 93 м; 167 — В^=8.0, о-в Сахалин, Охотоморское по-
бережье, 180 м.
Рис. 168—178. Раковины'моллюсков рода Oenopota.
168—173 — Oe. cancellata: 168 — Вр = 12.1, Японское море, разрез от м. Поворотного на юг, глубина 128 м;
169 — Вр=12.5, зал. Петра Великого, от м. Среднего, 12 м; 170 — Вр=10.3, зал. Петра Великого, от м. Сред-
него, 12 м; 171 — Вр=10.1, зал. Петра Великого, у о-ва Акольд, 132 м; 172 — Вр=14.2, зал. Петра Вели-
кого, у о-ва Аскольд, 132 м; 173 — В? = 10.3, Японское море, о-в Аскольд, 79 м; 174 — Ое. Ысопгса: Вр~
= 10.0, Японское море, зал. Ольга, 22 м; 175—176 — Oe, inequita: 175 — Вр = 12.0, Японское море, зал.
Петра Великого, 67.5 м; 176 — В^=11.6, Японское море, зал. Лихачева, о-в Южный, 5 м; 177—178 —
Oe casentina: 177 — Вр=12.8, восточное побережье южного Сахалина, 47°52' с. ш.—144°38' в. д.. 134 м;
178 — В^=10.3, Охотское море, 44°42' с. ш.—147°56'в . д., 291 м.
Рис. 179—190. Раковины моллюсков рода Oenopota.
479—183 — Ое. kurilensis: 179 — В?=9.5, Курильские о-ва, о-в Симушир, глубина 43 м; 180 — Вр=11.8,
Охотское море, 46°59' с. ш.—152°11' в. д., Им; 181 — Вр=13.3, Курильские о-ва, о-в Шикотан, 151 м; 182 —
Sp=ll.l, Курильские о-ва, о-в Уруп, м. Китовый, 83 м; 183 — Вр=11.2, Курильские о-ва, о-в Уруп, м. Ки-
товый, 83 м; 184—186 — Ое. pavlova: 184 — Вр=7.7, о-в Сахалин, траверз р. Малка, 60 м; 185 — Вр=12.7,
-о-в Сахалин, траверз м. Лопатина, 250 м; 186 — Вр=12.4, Японское море, 42°28' с. ш.—132°50' в. д., 160 м,
487—188 — Ое. elongata: 187 — Вр=8.0, Японское море, 42°17' с. пг.—131°1Г в. д., 1076—1400 м; 188 —
Вр=8.3, Японское море, 42°18' с. ш.—131°24' в. д., 1370 м; 189—190 — Ое. uschakovi: 189 — Вр=10.4, о-в
Итуруп, зал. Рока, 44°24' с. ш.—147°09' в. д., 160 м; 190 — Вр—7.4, Охотское море, Южные Курильские
о-ва, 130 м.
Рис. 191—202. Раковины моллюска Oenopota pyramidalis.
-191 — Вр=10.3, пролив Югорский Шар, глубина 44 м; 192 — В^=12.2, Татарский пролив, 49’02' с. ш.—
140’21' в. д., 44 м; 193 — Вр=15.8, Баренцево море, Долгая Губа, 13 м; 194 — Вр=17.7, Баренцево море,.
'74°80' с. ш.—17°30' в. д., 230' м; 195 - Bp=10.4, Баренцево море, 71 °16' с. ш,—25’34' в. д., 315 м; 196 —
Вр=16.9, Карское море, 77°11' с. ш.—96’15' в. д., 80 м; 197 — Вр=13.4, Карское море, о-в Визе,
"79°27' с. ш.-76°40' в. д., 34 м; 198 — Вр=14.5, Баренцево море, 68°39' с. ш.—38’40' в. д., 18 м; 199 — Вр=
=12.0, Баренцево море, 3 м; 200 — Вр2=3.2, южная часть Баренцева моря, 68’30' с. ш.—42’20' в. д., 61 MJ.
201 — Вр—11.2, Мурманский берег, 40 м, 202 — Вр=11.3, Соловецкие о-ва, 23 м.
Рис. 203—213. Раковины моллюсков рода Oenopota.
•203—208 — Ое. pyramidalis: 203 — Вр=11.6, Баренцево море, 71°03' с. ш.—35°00' в. д., глубина 165м; 204 -
Вр=12.5, восточная часть Карского моря, 51 м; 205 — Вр —15.8, Земля Франца-Иосифа, о-в Греэм-Белл»
5 м; 206 — Вр~П.З, Карское море, о-в Визе, 79°27' с. ш.—76°40' в. д., 34 м; 207 — Вр = 14.2, Земля Франца-
Иосифа, о-в Грезм-Белл, 5 м; 208 — Вр=19.0, Баренцево море, Мурманский берег, 77 м; 209—210 — Ое. pin—
gelii: 209 - Вр=10.7, Баренцево море, 69°27' с. ш.—32°24' в. д., 97 м; 210 — Вр = 10.4, море Лаптевых,.
76°47' с. ш.—117°24' в. д., 43 м; 211—213 — Ое. obliqua: 211 — Вр=8.1, Баренцево море, 79°18' с. ш.—
44°35' в. д., 147 м; 212 — В^=9.5, Баренцево море, Черная губа, 8 м; 213 — Вр=8.1, Карское море, о-в Визе»
79°27' с. ш.—76°40' в. д., 34 м.
Рис. 214—225. Раковины моллюсков рода Obesotoma.
214—218 — О. simplex-. 214 — 13^=9.3, юго-восточная часть Баренцева моря, глубина 62 м; 215 — Вр=9.0,
«ого-восточная часть Баренцева моря, 62 м; 216 — Вр = 7.2, Охотское море, зал. Александры, 8 м; 217 —
Вр—Э.4, Баренцево море, Корабельная бухта, 6—10 м; 218 - 23^=11.1, Баренцево море, Корабельная бухта,
"6—10 м; 219—220 — О. schantarica: 219 - голотип, Вр=1, Охотское море, Щантарские о-ва; 220 — Вр=
=28.0, Японское море, зал. Петра Великого, к югу от о-ва Русского, 5 м; 221 — О. gigas: В^=14.0, Земля
•Франца-Иосифа, о-в Хейса, 20 м; 222—225 — О. gigantea: 222 — Вр = 16.9, Земля Франца-Иосифа, о-в Вик-
тория, 30—40 м; 223 — В^=17.5, Карское море, 80°11' с. ш.—75°02' в. д., 32 м; 224 — Вр=15.1, Земля
•Франца-Иосифа, о-в Виктория, 30—40 м; 225 — В^=13.6, Земля Франца-Иосифа, о-в Виктория, 30—40 м.
Рис. 226—236. Раковины моллюсков рода Obesotoma.
226—230 — О. starobogatovi: 226 — Вр=в.6, восточная часть Карского моря, глубина 11.7 м; 227 —2-Вр—
=6.3, Новосибирские о-ва, о-в Котельный, 8 м; 228 — Вр=4.9, море Лаптевых, м. Песчаный, 10 м; 2291—
Вр=6.8, Новосибирские о-ва, о-в Котельный, 17 м; 230 — Вр=8.4, Новосибирские о-ва, о-в Котельный, 82м;
231—235 — О. tenuilirata: 231 — Вр=9.0, Охотское море, 55’42' с. ш.—154’58' в. д., 66 м; 232 — Вр=8.3^
Татарский пролив, 9 м; 233 — Вр=8.9, северо-западная часть Японского моря, 47’10' с. ш.—139’30' в. д.,
234 — Вр=12.5, Западная Камчатка, 52’44' с. ш.—155’10' в. д., 120 м; 235 — Вр=9.Ь, о-в Сахалином. Бел-
линсгаузена, 121 м; 236 — О. woodiana: Вр=16.0, Баренцево море, 69’26' с. ш.—60’00' в. д., 14 м.
Рис. 237—248. Раковины моллюсков рода Obesotoma.
237—240 — О. woodiana: 237 — Вр=12.4, Норвежское море, 75’10' с. ш.—16’39' в. д., 200 м; 238 — Вр=
=12.3, Баренцево море, 69’38' с. ш.—58’42' в. д., 24 м; 239 — Вр=12.0, Баренцево море, 68’28' с. ш.—
42’46' в. д., 59 м; 240 — Вр=16.2, Баренцево море, 70’05' с. ш,—47’36' в. д., 77 м; 241—242 — О. tumida'-
241 — Вр=13.0, Северный Ледовитый океан, 76’31' с. ш.—118’46'в. д., 64 м; 242 — Вр=11.1, Новосибирски?
о-ва, 77’44' с. ш. -145’32' в. д., 26 м; 243—248 — О. robusta: 243 — Вр=9.7, зал. Лихачева, о-в Устричный,
7 м; 244 — Вр=8.3, Японское море, 42’17' с. ш.—131’11' в. д., 925 м; 245 — Вр=10.0, Восточная Камчатка,
Кроноцкий зал., бухта Ольги, 148 м; 246 — Вр=11.9, Японское море, зал. Лихачева, о-в Устричный, 7,М4
247 — Вр=13.0, зал. Петра Великого, от о-ва Аскольда на юг, 269—349 м; 248 — В^=15.2, Японское море,
Де-Кастри, бухта Южная, 10 м.
Рис. 249—259. Раковины моллюсков рода Obesotoma.
249—254, 257—259 — О. solldtr. 249 — Вр=13.5, зал. Петра Великого, к югу от о-ва Аскольда, глубина
262 м; 250 — Вр=14.0, зал. Петра Великого, от м. Среднего, 121 м; 251 - Вр=13.1, зал. Петра Великого,
42°24' с. ш,—131 °28' в. д., 210 м; 252 — Во==11.4, зал. Петра Великого, от м. Среднего, 121 м; 253 — В? =
= 13.8, зал. Петра Великого, разрез от о-ва Русского на юг, 330—380 м; 254 — Вр=18.0, зал. Петра Вели
кого, от м. Среднего на юг, 88 м; 257 — Вр=11.7, Японское море, Де-Кастри, у о-ва Клыкова, 15 м; 258 —
Вр=12.4, зал. Петра Великого, с северной стороны о-ва Аскольд, 20—24 м, 259 — Вр=23.0, зал. Петра Ве-
ликого, на юг от о-ва Русского, 330—380 м; 255—256 — О. tomiyaensis: 255 — Вр=12.0, ?; 256 — Вр=11.3,
у о-ва Монерон, 46°07' с. ш.—141°45' в. д., 67 м.
Рис. 260—271. Раковины моллюсков родов Obesotoma и Propebela.
260—261 — О. japonica: 260 — Bjo=24.1, о-в Сикотан, бухта Сикотан, 8 м, 261 — Bjo=27.0, зал. Петра Ве-
ликого, о-в Петров, 42°51' с. ш.—133°54' в. д., 35 м; 262—264 — О. sachalinensis: 262 — Вр—20.2, Охото-
морское побережье о-ва Сахалин; 263 — Вр=17.0, Японское море, бухта Роджерса, 5 м; 264 — Вр=16.5,
южная часть Охотского моря, 146—150 м; 265—271 — Р. viridula: 265 — Вр=11.5, Баренцево море,
68°05' с. ш.—39°33' в. д., 17—19 м; 266 — В^=11.3, Новая Земля, Маточкин Шар, 73°15' с. ш.—54°05' в. д.,
8 м; 267 — Вр=8.5, Татарский пролив, 50 м; 268 — 13^=10.0, Баренцево море, 69°35' с. ш.—50°05' в. д.,
70 м; 269 — Вр—7.7, Охотское море, зал. Пильтун, 35 м; 270 — Вр=9.7, Чукотское море, 70°44' с. ш.—
170°40' з. д., 34 м; 271 — В^=10.8, Баренцево море, 68°48' с. ш.—43°10' в. д., 53 м.
Ш Зак. 2164
Рис. 272—283. Раковины моллюсков рода Propebela.
272—273 — Р. viridula: 272 — Вр=13.0, Баренцево море, 72°25' с. ш.—52°32' в. д., глубина 37 м; 273 —
Вр=7.9, 69°39'с. ш.—59°25' в. д., 50 м; 274—275 — Р. marinae: 274 — Вр—10.7, Курильские о-ва»
44°21' с. ш.—148°24' в. д., 414 м; 275 — Вр=7.3, Охотское море, 49°24' с. ш.—148°46' в. д., 1076 м; 276—
278 — Р. exarata: 276 — Bjo=10.9, Баренцево море, Кольский зал., 56 м; 277 — Вр=10.8, Карское море»
о-в Визе, 34 м; 278 — Bjo=10.1, Баренцево море, Килдин, Могильная Губа, 20 м; 279—283 — Р. tersa: 279 —
В^=6.4, о-в Беринга, 89 м; 280 — Вр=10.7, Курильские о-ва, о-в Шумшу, 38°57' с. ш.—143°23' в. д., 350—
340 м; 281 — В^=10.9, зал. Петра Великого, на юг от о-ва Карамзина, 67 м; 282 — В?=11.5, зал. Петра
Великого, у о-ва Русского, 240 м; 283 — В^=9.1, Японское море, 42°18* с. ш.—131°11' в. д., 925 М.
Рис. 284—294. Раковины моллюсков рода Propebela.
284 — Р. mitrata: Вр—12.0, Курильские о-ва, 51°44' с. ш.—155°09' в. д., 120 м; 285—288 — Р. concinnula’
285 — В^=8.2, восточное побережье южного Сахалина, 46°50' с. ш.—143°53' в. д., 183 м; 286 — В^=7.7’
о-в Беринга, траверз р. Фонтанка, 100 м; 287 — Вр=12.2, Западная Камчатка, 51°25' с. ш.—156°27' в. д."
42 м; 288 — 13^=12.5, Западная Камчатка, 51°25' с. ш.—156°27' в. д., 42 м; 289—293 — Р. margaritae: 289 —
Вр=5.8, Японское море, от м. Гамова на юг, 190 м; 290 — Вр=9.2, Восточная Камчатка, м, Ольги, 200 м!
291 —голотип,- Bj0=12.4, о-в Беринга, 55°23' с. ш.—165°23' в. д., 100 м; 292 — Вр=9.1, Восточная Кам-
чатка, м. Ольги, 200 м; 293 — Вр=8.2, Командорские о-ва, о-в Медный, 200—150 м; 294 — Р. fidicula: Вр—
= 11.5, о-в Врангеля, 16 м.
Рис. 295—306. Раковины моллюсков рода Propebela.
295—300 — Р. uarlabilis: 295 — 5^=10.4, Командорские о-ва, о-в Беринга, глубина 100 м; 296 — Вр=9.5,
Командорские о-ва, о-в Беринга, 100 м; 297 — Вр = 13.9, Командорские о-ва, о-в Медный, 98 м; 298 — Вр=
=11.3, Командорские о-ва, о-в Беринга, 100 м; 299 — Вр=13.1, Командорские о-ва, о-в Арий Камень, 60 м;
300 — В^=12.4, Командорские о-ва, о-в Медный, 120—130 м; 301—302 — Р. golikovi; 301 — Вр=7.0, Япон-
ское море, м. Ракума, 26 м; 302 -- Вр=7.4, Японское море, м. Ракума, 25 м; 303—304 — Р. areta: 303 —
Вр=5.6, Японское море, о-в Монерон, 30 м; 304 — В^=5.3, о-в Маканруши, м. Верхний, 44—50 м; 305—
306 — Р. alitakensls: 305 — Вр=7.3, Восточная Камчатка, Кроноцкий зал., бухта Ольги, 148 м; 306 — Вр =
= 7.0, Восточная Камчатка, Кроноцкий зал., бухта Ольги, 148 м.
Рис. 307—317. Раковины моллюсков рода Propebela.
307 — Р. rassina: Вр = 10Л, о-в Беринга, 70 м; 308—310 — Р. cingulata: 308 — Вр=8.1, Японское море, зал
Ольги, Тихая гавань, 8.5 м; 309 — 13^=8.6, о-в Сахалин, зал. Анива, 16 м; 310 — Bp = S.3, о-в Шикотан,
бухта Анама, 8—7 м; 311—312 — Р. goryachevi: 311 — В^=10.7, Японское море,46°00' с. пг.—141 °26' в. д.,
120 м; 312 — Ву,=6.8, Охотское море, бухта Нагаева, 59°31' с. ш. —150°44' в. д., 10 м; 313—315 — Р. ап-
gulosa: 313 — Вр=11.9, Баренцево море, Мурманский берег, 178 м; 314 — В^=11.4, Баренцево море,
69°02' с. ш.—37°27' в. д., 184 м; 315 — Вр=10.4, Баренцево море, 80°10' с. ш.—33°56' в. д., 280 м; 316—
317 — Р. nobilis: 316 — Вр=15.8, Баренцево море, 68°45' с. ш.—47°01' в. д., 55 м; 317 — 5^=11.3, Соло-
вецкий зал.
Рис. 318—329. Раковины моллюсков рода Propebela.
318—321 — Р- nobilis: 318 — Bp==16.5, Баренцево море, 69°37' с. ш.— 33°04' в. д., глубина 135 м; 319 —
Вр=28.2, Белое море, 68°27' с. щ,—43°37' в. д., 30 м; 320 — 5^=19.0, Баренцево море, 71°30’ с. ш.—
51°07' в. д., 61 м; 321 — В?=17.0, Баренцево море, 68°00' с. ш.—47°36' в. д., 46 м; 322—327 — Р. assimilis
322 — 5^=15.2, западный берег о-вов Новой Земли, 13 м; 323 — 5^=14.0, Восточная Камчатка, 324 —
Вр=13.5, Охотское море, 49°08' с. ш,—143°21' в. д., 25 м; 325 - Вр=14.0, Татарский пролив, 60 м’; 326 -
Вр=14.3, Мурманский берег, Еретики; 327 — 5^=9.8, Баренцево море, 68°25' с. ш.—39°52' в. д., ИО-
121 м; 328—329 — Р. svetlanae 328 — Вр=7.3, Охотское море, бухта Нагаева, 59°ЗГ с. ш.—150°44' в. д.»
10 м; 329 — В;о=10.8, Охотское море, Западная Камчатка.
Рис, 330—341. Раковины моллюска Propebela rugulata.
330 — Вр=10.6, Карское море, 77°57' с. ш.—75°10' в. д., 310 м; 331 — Вр=13.2, Пуховый зал.; 332 — Вр=
=13.8, море Лаптевых, П°Ц’ с. ш.—105°02' в. д., 189 м; 333 — Вг>=12.9, Баренцево море, 79°36 с. ш.—
53°59' в. д., 297 м; 334 — Вр=9.1, Баренцево море, 7О'?39' с. ш,—53°42' в. д., 41 м; 335 — Вр—10.7, Северный
Ледовитый океан, 81°33' с. ш.—83°06' в. д., 300 м; 336 — В^р=10.9, Земля Франца-Иосифа, о-в Хейса, 28—
46м; 337 — Вр=12.5, 80°11' с. ш,—86°24' в. д., 250 м; 338— Вр=11.5, Новая Земля, вход в зал. Медвежий,
70—64 м; 339 — В/)=14.3, Новосибирские о-ва, 75°37® с. ш.—146°45' в. д., 11.5 м; 340 - Вр=15.0, Норвеж-
ское море, 67°65' с. ш.—11°21' в. д., 200 м; 341 — В^=15.0, над Горлом Белого моря, 81 м.
Рис. 342—352. Раковины моллюсков рода Propebela.
342—343 — Р- rugulata: 342 — 5^=10.3, 78°55' с. ш.—76°58' в. д., глубина 115 м; 343 — Вр=11.4, Карское
море, 78°20'с. ш.—79°10' в. д., 175 м; 344—347 — Р. spitzbergensis: 344 —Вр=7.0, Карское море,
71°54' с. ш.—67°45' в. д., 40 м; 345 — Вр=1.2, Новосибирские о-ва, 77°47' с. ш.—143°35' в. д., 45 м; 346 —.
Вр—ЗЛ, Баренцево море, 70°15' с. ш.—41°20' в. д., 125 м; 347 — В?=11.2, Соловецкий зал.; 348—352 —
Р. bergensis: 348 — Вр=8.в, Баренцево море, 73°30' с. ш.—27°30' в. д., 310 м; 349 — Вр=8.1,
4М°35' с. пг.—75°57' в. д., 225 м; 350 — Bp=8.S, Баренцево море, 71°31' с. пг.—34°59' в.д., 120 м; 351 — Вр~
=7.8, 80°46' с. ш,—83°26' в. д., 290 м; 352 — В?=8.0, 78°00' с. ш.— 76°00' в. д.
Рис. 353—363. Раковины моллюсков рода Propebela.
353—354 — Р. verrilli: 353 — 73^=11.9, Приморье, разрез отм. Пельма, глубина 31 м; 354 — _В^=9.9, При-
морье, разрез от м. Пельма, 31 м; 355—363 — Р. harpularia: 355 — В^=8.2, у о-ва Визе, 37 м; 356 — Вр=
=6.4, Берингово море, зал. Провидения, бухта Эмма, 18 м; 357 — Вр=7.1, Баренцево море, бухта Казари-
нова, 17—9 м; 358 — 13^=9.5, Баренцево море, 69°19' с. ш.—34°00' в. д., 56 м; 359 — 13^=8.5, Баренцево-
море, Губа Ярнышная, 50—36 м; 360 — 13^=11.4, Баренцево море, Порчниха, 13 м; 361 — Вр=9.8, Финмар-
хен; 362 — Вр=10.9, Шантарские о-ва, 2.5 м; 363 — Вр=8.8, Баренцево море, о-в Колгуев, 69°15' с. ш.—
48°08' в. д., 26 м.
Рис. 364—374. Раковины моллюсков рода Propebela.
364—367— Р. harpularia: 364 — Вр=9.1, Японское море, разрез от м. Гамова, глубина 64 м; 365 — Вр=
==8.7, Новосибирские о-ва, о-в Большой Ляховский, 31 м; 366 — Вр=8.1, о-в Монерон, 17 м; 367 — Вр=
=12.1, о-в Монерон, 52 м; 368—370— Р. turricula: 368 — Вр=13.3, Баренцево море, 68°17' с. ш.—48°31' в. д.,
44 м; 369 — Вр=13.1, Хельголанд; 370 — Вр=12.0, Баренцево море, 69°00' с. ш.—48°00' в. д., 18 м; 371—
372 — Р. tetpeniensis: 371 — Вр=10.2, южный Сахалин, зал. Терпения, м. Тонина, 46°47'5" с. ш.—
143°52'3" в. д., 187 м; 372 — Вр=8.3, разрез отм. Ливенорна, 46°26' с. ш,—143°37' в. д., 51 м; 373 — Р. ро-
povia: Вр=8.7, Западная Камчатка, 85 м; 374— Р. cassis: Вр=10.6, Малая Курильская гряда, о-в Шикотан.
Рис. 375—385. Раковины моллюсков рода Propebela.
375—383 — Propebela scalaris: 375 — 13^=17.3, Баренцево море, 68°05' с. ш.—39°34' в. д., глубина 17—
19 м; 376 — £р=16.0, Баренцево море, Мурманский берег, 105 м; 377 — Вр=16.3, Новая Земля, Губа Мел-
«ая, 6.5 м; 378 — Bp=14.0, Баренцево море, Териберка, 12 м; 379 — Вр=14.1, Баренцево море, 77 м; 380 —
Вр=11.1, Карское море, 79°06' с. ш.—74°44' в. д., 307 м; 381 — В?=13.8, Баренцево море, 71°00'с. ш. —
28°37' в. д., 250 м; 382 — Bp=li.9, у о-ва Килдин, 67 м; 383 — В^=26.5, северная часть Норвежского моря;
384—385 — Р. scalaroides: 38i - Вр=26.0, Баренцево море, Мурманский берег; 385 — Вр =22.8, Баренцево
море, 69°28' с. ш.—35°12' в. д., 168 м.
Рис. 386. Протоконхи моллюсков Curtitoma trevelliana (А—Б) и С. incisula (В~Е).
А — Карское море, 79°38' с. ш. —74°24' в. д. (увел, х 150); Б - Карское море, 81 °00' с. ш. —75°12' в. д.
(Х112.5); В — Восточная Камчатка, Кроноцкий зал. (х 112.5); Г — Охотское море, разрез от м. Энкега
<X 112.5); Д — Командорские о-ва, северо-западная оконечность о-ва Беринга (х 112.5); Е — о-в Сахалин^
м. Безымянный (х 112.5).
Рис. 387. Протоконхи моллюсков Curtitoma novajasemljensis (А, Б) и С. violacea (В—Е).
Л — Карское море, 73°28' с. ш,—80°28' в. д. (х 150); Б - детали строения протоконха (X 300); В — Карское
зморе, у о-ва Визе, 81°35' с. ш,—75°57' в. д. (х 150); Г -- Мурманский берег (X 150); Д — Карское море
у о-ва Визе (х 150); Е — Южно-Курильский пролив, 45°55' с. ш.—146°08' в. д. (х 112.5).
Рис. 388. Протоконхи моллюска Curtitoma violacea.
А — Шпицберген, 76°50' с. ш.~25°30' в. д. (х 112.5); Б — участок второго оборота протоконха (Х45О>£
В — о-в Визе 81°35' с. ш.—75°57' в. д. (X 112.5); Г — Карское море, 82°09' с. ш.—83°08' в. д. (х90); Д —
эмбриональная раковина из кладки (Х112.5); Е — о-в Визе (Х90).
Рис. 389. Протоконхи моллюсков Curtitoma decussata (А—В) и С. conoidea (Г—Е).
А Карское море, 79°01' с. ш.—84°30' в. д. (х 112.5); В — участок скульптуры второго оборота эмбрио-
нальной раковины (Х300); В—Шпицберген, 80°46' с. ш,—29°57' в. д. (х 112.5): Г — 81°35' с ш —
75°57' в. д. (X 112.5); Д — 80°1Г с. ш,— 86°24' в. д. (X 150); Е — 79°00' с. ш. 83 = 12' в. д. (х 112 5)
Рис. 390. Протоконхи моллюсков Propebela golikovi (А—В)\ Curtitoma hebes (Г); С. beckle-
mishevi (Д—Е).
А -- Южный Сахалин, зал. Анива (Х&О); Б — то же, детали строении протоконха (Х225); Б — то же уча-
сток второго оборота дефинитивной раковины (х 300); Г — Охотское море, разрез от м Энкен (Х112.5);
Д — Охотское море, 51°10'с.ш.—154°17'в. д. (х90); Е — эмбриональная раковина из кладки (местонахож-
дение то же) (X 150)
''С': /<КМ1
А — Охотское море, 51 °1 О' с. ш.—154°17' в. д. (х 112.5); Б — то же, протоконх (х 150); В—Е — разрез от
о-ва Римского-Корсакова на юг; В — (х 112.5), Г — (х 90), Д — (х 150); Е — (х 90). 1
IV Зак. 2161.
Рис. 392. Протоконхи моллюсков Granotoma raduga (А—Г) и G. albrechti (Д~Е).
А Тс£по«ское море’ 42°18' с- ш.—131’11' в. д. (X 150); Б — Охотское море, 51’10' с. ш.-154’17' в д.
<х750); В — то же, общий вид (х 90); Г — то же, микроскульптура первого оборота протоконха (х 750k
Д - зал. Петра Великого, 42°24' с. ш.-131°28' в. д. (Х112.5); Е — та же проба (х 150)).
Рис. 393. Протоконхи моллюсков Granotoma albrechti (А—Б) и G. krausei (В—Е).
А — западное побережье Южного Сахалина, 47°18' с. ш,—141°55' в. д. (х 112.5); Б — то же, участон
скульптуры первого оборота протоконха (X 750); В — зал. Петра Великого, 42°24' с. ш.—131°28' в. д
(X 90); Г - зал. Петра Великого, разрез от м. Гамова на юг (X 90); Д — то же, участок первого оборота эм-
бриональной раковины (х450); Е — то же, участок третьего оборота дефинитивной раковины (Х150).
Рис. 394.
Протоконхи моллюска Oenopota cinerea (А—Е).
Л — Карское, море, 81 °11' с. ш.—75°20' в. д. (X 150); Б — то же, общий вид (Х90); В — Баренцево море,
73°28'с.ш.—34°57' в. д. (х225); Г — т те, общий вид (X 90); Д —море Лаптевых, 77°25' с. ш.—
118°17' в. д. (;<90); Е — Северный Ледовитый океан, 82°09' с. ш.--83°08' в. д. (х90).
Рис. 395. Протоконхи моллюска
Oenopota declivis (А—Е).
Л. — Баренцево море, Килдинский пролив (х 90); Б — то же, участок верхнего оборота протоконха (х 225};
В — участок второго оборота протоконха, Килдинская Са.тма (х 450); Г — Карское море, 79°57' с. ш.—
75°03' в. д. (X112.5); Д — Карское море, 79°38' с. ш.—74°42' в. д, (Х112.5); Е — Баренцево море,
69°23' с. ш,—32°65'в. д. (Х150).
Рис. 396. Протоконхи моллюска Oenopota elegans (А—Е).
А — Мурманский берег (X 150); Б — Карское море, 76°41' с. ш,— 88°10' в. д. (х 112.5); В—Е — Охотское
море, северо-восточная часть о-ва Сахалин; В (х 150), Г (Х90), Д (Х112.5), Е (х 750)
Рие. 397. Протоконхи моллюсков Oenopota harpa {А—Б'}, Ое. pavlova (В—Г); Ое. pyrami-
dalis (Д—Е).
-А - Карское море, о-в Визе, 79°27' с. ш.—76°40' в. д. (Х112.5); Б — Баренцево море, 68°15' с. ш,—
48°>.г>' в. д. (х 150); В — разрез от м. Анива, 45°44' с. ш.—143.°56' в. д. (X 90); Г — то же, участок скульп-
туры эмбриональной раковины (X 450); Д — Карское море, 79°27' с. ш,—76°40' в. д. (X 90); Е — Северная
Земля, 76°59' с. ш.—100°20' в. д. (х90).
Рпс. 393. Протоконхи моллюска Oenopota impressa (А—Е).
А — Баренцево море, Иоканские о-ва (х 90); Б - Баренцево море, 68°05' с. ш.-39°34' в. д. (х 150); В —
Жарение Ворота (X 90); Г — Тромсё (X 150); Д - пролив Карские Ворота (X 150); Е о-в Беринга (х 90>
Рис. 399. Протоконхи моллюсков Oenopota pyramidalis (А—Б)-, Ое. cancellata (В—ГУ
Ое. uschakovi (Д—Е).
А — Баренцево море, 77°15' с. ш,—30°00' в. д. (Х112.5); Б - то же (X 112.5); В Японское море, к югу
от о-ва Аскольда (Х150); Г - то же, общий вид (Х90); Д — разрез от о-ва Шикотан, 43°53' с. ш.—
147°00' в. д. ( X 112.5); Е — то же, участок третьего оборота дефинитивной раковины (х 150).
Ри::. 400. Протоконхи моллюсков Obesotoma starobogatovi (А—Г); О. simplex (Д); О. tenui-
lirata (Е).
"г море Лаптевых, о-в Котельный (х 150); Б — море Лаптевых, Вилькпн знак ( х 150); В — то же (х 150)’
J - до же, участок скульптуры второго оборота эмбриональной раковины (хЗОО)- Д — Восточная Кам-
чатка, зал. Олюторский (Х90); Е — Японское море, 464)0' с. ш,—142°П'в д. 1x90)
Рис. 401. Протоконхи моллюсков Propebela rugulata (A -J'y. Р. bergensis (Д—Е),
А — Карское море, 80°46' с. ш.—83°26'' в. д. (Х112.5); Б — то же (Х90); В — Гренландское море,
.S0°02' с. ш. — 9°17' в. д. (х 112.5); Г — Карское море, 81°35' с. ш. — 7-5°57' в. д. (х 112.5); Д — Маточкин Шар
(Х112.5); Е — Карское море (х90).
Рис. 402. Протоконхи моллюсков Propebela bergensis (Л—В)\ Р. scalaris (Г—Е).
А — Карское море, 80°46' с. ш,—83°26' в. д. (Х90); Б — то же, первый оборот протоконха (Х150); Г —
Карское море, 80°19' с. ш. — 72°47' в. д. (Х150); Г — Карское море, 80°59' с. ш.—73°32' в. д. (Х112.5);.
Д — Берингово море, бухта Лаврентия (Х12.5); Е — то же, скульптура начала второго оборота прсто-
конха (х450).
Рис. 403. Протоконхи моллюсков Propebela scalaris (А—В); Р. scalaroides (Г—Е).
Л - Карское море, 79’06' с. 111.-74’44' в. д. (Х90); Б - Баренцево море, 71°00'с. ш,—28’37'в д. (Х90),
_В — то же первый оборот эмбриональной раковины (Х112.5); Г — Норвежское море, 80 02 с. пь —
S°17' в. д (X90): Д - Баренцево море, 68’25' с. ш.-47°02' в. д. (х90); Е — то же, первый оборот Прото-
кокка (Х150).
Рис. 404. Протоконхи моллюсков Propebela assimilis (А—Б); Р. harpularia (В—Г); Р. tur-
ricula (Д—Е).
А — Карское море, 80°19' с. ш,—72°47' в. д. (Х112.5); Б — Мурманский берег (Х112.5); В — Баренцев»
море, Черная Губа (х90); Г — то же, детали строения первого оборота протоконха (х225); Д — у северных.
берегов^Шотландии (х225); Е — то же, общий вид (Х112.5).
Рис. 405. Протоконхи моллюсков Propebela turricula (Л); Р. areta (Б)-, Р. exarata (В);
Р. tersa (Г); Р. cingulata (Д—Е).
А — у о-ва Колгуева, 69°15' с. ш.— 48°08' в. д. (х90); Б - о-в Беринга (Х112.5); В — Баренцево море.
71 02 С. ш,—49°54' в. д. (Х90); Г — Татарский пролив, 48°25' с. ш.—142°04' в. д (Х112.5); Д — о-в Са-
халин, разрез от пос. Пильтун (Х90); Е — Японское море, зал. Петра Великого (х90)
Рис. 406. Протоконхи моллюсков Oenopota. obliqua. (А—Б}; Propebela tersa (В — Г); Р. viri-
dula (Д—Е).
Л — Кольский залив (X 112.5); Б — то же (X 112.5); В — Японское море, 42°18' с. ш.—131 °11' в. д. (х90);
Г — то же, эмбриональная раковина (х 150); Д — Баренцево море, бухта Казаринова (Х90); Е — то же,
скульптура протоконха (Х150).
5. PROPEBELA
11)3
Раковина телесного цвета, веретеновидно-овальной формы, с 4—5.5 уступ-
чатыми оборотами, уплощенными в верхней части. Протоконх неизвестен.
Осевая скульптура дефинитивной раковины представлена редкими широкими
складками, которые в 1.5—2 раза ^же разделяющих их промежутков.'Складки
сглаживаются в верхней 1/2 основания раковины. На предпоследнем обороте
насчитывается 14—15, на последнем 16 осевых складок. Спиральная скульптура
составлена тонкими ребрами, равновеликими с межреберными промежутками.
Исключением служит среднее ребро, обозначающее плечо оборота: оно в 2 раза
превышает размеры остальных элементов спиральной скульптуры. Основание
раковины выпуклое, плавно переходит в удлиненный сифональный вырост.
Устье узко-обратногрушевидное, с матовым каллусом на колумеллярном крае.
Размеры раковины голотипа (мм): 2?р=10.7, Д=5.4; 2?пО=7.3; Вд=Ь.Ъ', Ду—
=2.07.
Зубы радулы длиной около 200 мкм, ножевидной формы, с массивной базаль-
ной частью и длинным, достигающим 1/3—1/2 длины лезвия зуба килем.
Вид сходен с Р. rugulata, от которого отличается удлиненным сифональным
выростом, широкими осевыми складками и ножевидными зубами с длинным
килем.
Распространение. Тихоокеанский приазиатский широко распро-
страненный бореальный вид. Помимо типового местонахождения отмечен
в Охотском море в бухте Нагаева.
Типы. Голотип хранится в ЗИН АН СССР, № 48478; типовое местона-
хождение — Японское море (46°00' с. ш.—141°26' в. д.), глубина 120 м.
Просмотрено 2 экз.
2.Подрод CANETOMA Bartsch, 1941
Bartsch, 1941 i 6; Powell, 1966 i 121 (Nodotoma subg.). — Funitoma Bartsch,
1941 i 5 (типовой вид Funitoma areta Bartsch, 1941, по первоначальному обозначению),
syn. n.; Powell, 1966 I 121 (Oenopota subg.; part.).
Типовой вид Canetoma tersa Bartsch, 1941, по первоначальному
обозначению.
Раковина веретеновидной или веретеновидно-овальной формы. Элементы
осевой скульптуры отстоят друг от друга на расстоянии, равном или меньшем
собственной ширины складок.
Зубы радулы копьевидной или саблевидной формы, реже ножевидной. Киль
зуба радулы не образует гарпуновидных отростков. Второго дистального от-
верстия на лезвии не отмечено.
Подрод распространен по всему ареалу рода.
17. Propebela (Canetoma) tersa Bartsch, 1941. (Рис. 279—283; 405, Г; 406,
В-Г, 428, 1—14-, 470).
Bartsch, 1941 t 6, pl. 1, fig. 6 (Canetoma)', Powe 11, 1966 t 121, pl. 19, fig. 6 (No-
dotoma)', Голиков, Скарлато, 1967 i 65, рис. 45 (Lora exarata subsp.); Голиков,
Гульбин, 1977 t 238 (Oenopota exarata subsp.). — exarata (non Moller, 1842): О 1 d г о у d,
1927 t 101 (Lora).
Раковина желтовато-белая, веретеновидно-овальная, с 5—5.5 отлогоуглова-
тыми, уплощенными в верхней части оборотами. Периостракум тонкий, прозрач-
ный, коричневых тонов. Протоконх выпуклый, представлен 1.5 оборотами.
Первый эмбриональный оборот орнаментирован многочисленными микроскульп-
турными бугорками, в начале второго оборота закладываются 3 спиральных
нитевидных ребрышка. Они пересекаются под прямым углом тонкими осевыми
складками, в результате чего рисунок второго эмбрионального оборота при-
обретает ячеистый вид. Осевая скульптура дефинитивной раковины представ-
лена многочисленными тонкими, реже средними пологими складками (их ши-
13 И. П. Богданов
IM
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
рина в 2—3 раза превышает собственную высоту), разделенными, как правило,
равными им по ширине промежутками. Осевые складки равновелики на плече
и на периферии оборота и сглаживаются в верхней 1/4 основания раковины.
На околошовной пространстве элементы осевой скульптуры слегка скошены
вперед. На последнем обороте раковины насчитывается от 26 до 30 осевых скла-
док, на предпоследнем от 18 до 22. Спиральная скульптура равновелика по вы-
соте осевым складкам или (реже) развита сильнее и представлена на периферии
оборота и на основании раковины средними шнуровидными ребрами. Расстоя-
ние между ребрами равно их ширине. По плечу оборота, обозначая его, прохо-
дит киль, в 1.5 раза превосходящий по величине спиральные ребра периферии
оборота и в 4—6 раз — ребрышки околошовного пространства. Основание ра-
ковины умеренно выпуклое, как правило, плавно переходящее в небольшой
сифональный вырост. Устье узко-обратногрушевидное, с тонким белесым кал-
Рис. 470. Места находок Propebela tersa в морях СССР.
лусом на колумеллярном крае. Сифональный канал глубокий, широкий. Вы-
сота раковины типового экземпляра 7.0 мм при ширине 4.0 мм. Наибольшие
параметры раковины отмечены для моллюска из зал. Петра Великого (мм):
£р=11.5; Д=5.7; 5ЭТО=7.9; Ву=5.7; Д^=2.3.
Крышечка каплевидной формы, с практически прямой осью нарастания.
Мускульный отпечаток занимает срединную часть крышечки.
Зубы радулы длиной 350—500 мкм, копьевидной формы, со слегка отогнутой
назад базальной частью, несущей характерный язычок (рис. 428, 9). Оба флажка
полуовальной формы.
Вид сходен с Р. exarata, от которого отличается низким плечом и выпуклыми
оборотами раковины, а также копьевидной формой зубов.
Р а с пр остранение. Амфибореальный широко распространенный
вид тихоокеанского происхождения. Обитает у берегов Приморья и у о-ва Моне-
рон в Японском море, у северо-восточных берегов о-ва Сахалин, у Средних и
Южных Курильских о-вов и у Западной Камчатки в Охотском море. Отмечен
в зал. Массачусетс.
Экология. Обитает на глубинах от 60 до 900 м, преимущественно в диа-
пазоне 65—110 м, отмечена находка раковины на глубине 1045 м (Голиков,
Гульбин, 1977). Моллюски предпочитают песчанистые грунты (до 70 % всех
находок), реже илистые. Вид обнаружен при солености 32—34 °/о0.
Типы. Место хранения типового экземпляра USNM, № 428877; типовое
местонахождение — Приморье, Японское море.
Просмотрено 66 экз.
5. PROPEBELA
195
18. Propebela (Canetoma) viridula (Moller, 1842), comb. n. (Рис. 265—273;
406, Д-Е; 420, 11—24; 471; 472).
Moller, 1842 t 87 (Defrancia; non Fabricius, 1760);5 Morch, 1852 i 73 (Oenopota);
G. 0. S a r s, 1878 I 235, Tab. 16, fig. 7, 8, Tab. 9, fig. 5 (Bela); T г у о n, 1885 t 223, pl. 27,
fig. 20 (SeZa); Martini, Chemnitz, 1887 I 154, Taf. 32, fig. 7 (Bela)', К о b e 1 t, 1905 t
'255, pl. 84, fig. 8 (Bela decussata subsp.). — arctic a Adams, Adams, 1855 t 121 (BeZa);
Macpherson, 1971 i 111, pl. 7, fig. 2, map 49 (Oenopota-, part.). — kobelti Verkriizen,
1875 i 238 (Bela); К о b e 1 1, 1876 t 178, pl. 4, fig. 5 (Bela); F r i e 1 e, 1886 t 13, pl. 8, fig. 15
(Bela); К о b e 1 t, 1905 i 254, Taf. 83, fig. 12 (Bela decussata subsp.). — borealis (non Reeve,
1846)t.S c hl e s c h, 1924 I 327, Taf. 7, fig. 1, 2 (Lora).
Раковина от пепельно-серого до светло-каштанового цвета, овально-веретено-
видная, с 5—6 оборотами отлогоугловатой формы, тупоугловатыми в верхней
части. Периостракум тонкий, коричневый, реже ржаво-бурый, плотно прилегает
к поверхности раковины. Протоконх выпуклый, представлен 1.8 умеренно вы-
Рис. 471. Места находок Propebela viridula в северных морях СССР.
пуклыми оборотами. Первый эмбриональный оборот покрыт многочисленными
микроскульптурными бугорками, на втором обороте происходит закладка
2 тонких спиральных ребер. Осевая скульптура дефинитивной раковины состоит'
из широких пологих складок, склаживающихся в верхней х/4 основания рако-
вины. В верхней части оборота на околошовной поверхности осевые складки
получают небольшой изгиб вперед. Пространства, их разделяющие, в 2—3 раза
уже самих складок. На последнем обороте раковины насчитывается от 18
до 25 складок, на предпоследнем от 16 до 20. Спиральная скульптура представ-
лена равновеликими на всей поверхности раковины средними и тонкими реб-
рами. На периферии оборота они, как правило, шнуровидные. В результате
создается впечатление об их преобладании над элементами спиральной скульп-
туры плеча оборота и основания раковины. Межреберные промежутки однораз-
мерны и равны по ширине самим ребрам. Основание раковины уплощенное,
плавно переходит в средних размеров сифональный вырост. Устье полуоваль-
ное, угловатое в верхней части. Широкий толстый каллус обычно коричневатых
оттенков. Сифональный канал широкий, неглубокий. Высота раковины типо-
вого экземпляра 4.3 мм. Максимальные размеры раковины отмечены для мол-
люска из Баренцева моря (мм): Z?p=14.3; Д=7.0; 2?ПО=Ю.З; В~=1.7; Д#=З.О.
Крышечка когтевидная, с сильно изгибающейся осью нарастания. Му-
скульный отпечаток варьирующей конфигурации, занимает срединную часть
6 Ряд авторов середины прошлого столетия вслед за Мёллером считали данный вид си-
нонимом Trttonium viridulum Fabricius, 1760, однако Мёрч, изучавший типовые материалы,
указал различие названных видов.
' 13*
196
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
крышечки и Отстоит на большом расстоянии от ее левого утолщенного края
(рис. 36, Ж—М).
Зубы радулы длиной 150—250 мкм, копьевидной формы, с относительно не-
большой закругленной базальной частью, которая несет на одной из ручек
уплощенный флажок, другая ручка в значительной мере редуцирована (рис. 420,
Рис. 472. Места находок Propebela
viridula в дальневосточных морях
СССР.
11—24). Киль в 2—2.5 раза превышает
размеры дистального отверстия лезвия.
Вид сходен с Р. variabilis, от которого'
отличается более низким завитком, часто
расположенными средними осевыми склад-
ками и копьевидной формы зубами.
Распространен и е. Широко
распространенный бореально-арктический
вид, обнаружен в Арктическом бассейне,,
в Северной Атлантике, в Беринговом,
Охотском и Японском морях (рис. 472).
Встречается в южной части Баренцева мо-
ря, (у южных берегов Новой Земли и
в Белом море (рис. 471). Отмечен в ка-
надском секторе Арктики и у берегов^
Западной Гренландии.
Экология. Обитает на глубинах
от 0 до 780 м, преимущественно на 10—
100 м; на всех типах грунтов, в ochobhoml
на песчанистых (около 70 % находок), при
температурах от —1.64 до 7.6 °C и со-
лености 27.39—34.87 °/00.
Типы. Типовые экземпляры Defran-
eta viridula, очевидно, хранятся в ZMO;
типовое [местонахождение — воды Запад-
ной Гренландии. Место хранения Bela
arctica неизвестно; типовое местонахож-
дение указывается очень широко — арк-
тические моря (Macpherson, 1971). Место
хранения В. kobelti не указано (вероят-
нее всего, ZMO); типовое местонахожде-
ние — воды Северной Норвегии.
Просмотрено 311 экз.
19. Propebela (Canetoma) exarata (Mol-
ler, 1842), comb. n. (Рис. 276-278; 405,
В; 428, 15—26; 473).
Moller, 1842 1 85 (Defrancia)', Morch, 1852 t 74 (Pleurotomina); Troschel,
1866 i 43, pl. 4, fig. 5 (Bela); G, O. S a r s, 1878 1 232, Tab. 16, fig. 18, Tab. 9, fig. 1 (Bela);
Герценштейн, 1885 I 689 (Pleurotoma); Friele, 18&6 t 19, pl. 8, fig. 24 (jSefa); Mar-
tini, Chemnitz, 1887 i 140, Taf. 31, fig. 4 (Bela); Posselt, Jensen, 1898 i 146.
(Bela); Knipowitsch, 1901 i 44, pl. 18, fig. 16—17 (Bela); К о b e 11, 1905 s 236,
Taf. 82, fig. 3 (Bela turricula subsp.); Thorson, 1935 1 39, fig. 40 (Bela); 1941 t 103 (Bela);
Филатова, Зацепин, 1948 I 389, табл. 98, рис. 16 (Lora). — mitrula L о v e n, 1846 t
144 (Bela); G. O. S a r s, 1878 i 233, Tab. 23, fig. 9, Tab. 9, fig. 2 (Bela); Martini, Chem-
nitz, 1887 i 158, Taf. 32, fig. 13 (Bela); К о b e 1 t, 1905 ! 241, Taf. 82, fig. 12 (Bela turricula
subsp.); Thorson, 1941 t 104 (Bela).
Раковина белая, реже пепельная, высокая, башенковидная, тонкостенная,
с 5.5—6.5 оборотами отлогоугловатой формы, уплощенными в верхней части.
Периостракум очень тонкий, неплотно прилегающий. Протоконх представлен
2 оборотами, причем первый эмбриональный оборот выпуклой формы и лишен
макроскульптурных образований (рис. 405, В). Второй оборот угловатый, ор-
5. PROPEBELA
197
наментирован 3 спиральными тонкими ребрами. Непосредственно перед дефи-
нитивными оборотами происходит закладка осевых складок в виде едва намечен-
ных утолщений, заметных только в межреберных участках. Осевая скульптура
дефинитивной раковины представлена многочисленными средними складками,
имеющими небольшой изгиб вперед на плече оборота. В сечении осевые складки
имеют вид равнобедренного треугольника. Складки разделены равными им
по ширине, реже меньшими промежутками. На последнем обороте раковины
насчитывается от 23 до 25 складок, на предпоследнем от 16 до 21. Спиральная
скульптура на периферии оборота и на основании раковины представлена тон-
кими, как правило, шнуровидными спиральными ребрами, разделенными в 1.5—
2 раза более широкими межреберными промежутками. Киль, проходящий по
плечу оборота, незначительно превышает высоту ребер периферии оборота.
На плече оборота залегают многочисленные нитевидные спиральные ребрышки,
равновеликие с разделяющими их промежутками. Основание раковины слабо-
выпуклое, без резкой границы переходящее в средних размеров сифональный
вырост. Устье узко-обратногрушевидной формы, с тонким коричневатым кал-
лусом на колумеллярном крае. Высота раковины типового экземпляра 4.5 мм.
Максимальные размеры (мм): Вр=12.9; Д=5.6; ВПО=8.2; В^=5.6; Д&=2.2.
Крышечка каплевидной формы, с практически прямой осью нарастания на
поздних этапах формирования. Мускульный отпечаток неопределенной кон-
фигурации, расположен в срединной части крышечки (рис. 36, Г—Е).
Зубы радулы длиной до 800 мкм, саблевидной формы, с небольшой пира-
мидальной базальной частью, имеющей языковидный вырост и несущей на од-
ной из ручек характерный для вида листовидный флажок (рис. 428, 26). Лезвие
небольшого диаметра, всегда изогнуто.
Вид сходен с Р. tersa, от которого отличается более высоким завитком и,
главное, саблевидной формой зубов.
Замечания. Ныне можно говорить, что моллюсков, известных под
названием «mitrula», следует относить к рассматриваемому виду. Во-первых,
мной найдены все переходные формы как по раковине, так и по скульптуре.
Во-вторых, при исследовании коллекционного материала ЗИН АН СССР и при
изучении литературных данных [G. О. Sars, 1878 : Tab. 9, fig. 1 (exarata), fig. 2
(mitrula)] отмечена полная идентичность морфологических особенностей строе-
ния зубов радулы Р. exarata и Bela mitrula.
Распространение. Атлантический высокобореально-арктический
вид. Распространен в высокобореальных районах Северной Атлантики и в Ар-
ктике. Описан из вод Лабрадора, у о-ва Новая Англия, Восточной и Западной
198-
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Гренландии, в Иса-Фьорде у берегов Исландии, Западной Ирландии, Нор-
вегии. Обитает у берегов Шпицбергена,.в юго-восточной и центральной частях
Баренцева моря, в центральной и северной частях Карского моря, отмечен в Бе-
лом море и в центральной части моря Лаптевых, а также у берегов' Новоси-
бирских о-вов (рис. 473). •
Экология. Обитает на глубинах от 4 до 320 м, в литературе имеются
данные о нахождении вида у западных берегов Ирландии на глубине 2.214 м
(Odhner, 1915). Моллюски предпочитают песчаные грунтьв(более 70 % всех
находок), но встречаются и на илистых. Диапазон температур, при которых най-
дены живые моллюски, находится в пределах от —1.76 до 6.85 °C, однако чаще
всего они встречаются при температурах от —1.2 до 1.6 °C. Вид обитает при
солености 33.34—34.90, в основном 34.4—34.8 °/00.
Типы. Типовой экземпляр Defrancia exarata хранится в ZMO; типовое
местонахождение — воды Западной Гренландии. Типы Bela mitrula хранятся,
вероятно, в SMNH; типовое местонахождение — воды Северной Норвегии.
Просмотрено более 770 экз.
20. Propebela (Canetoma) mitrata (Dall, 1919), comb. n. (Рис. 284; 427,
17—20).
Dall, 1919a : 43, pl. 16, fig. 1 (Lora); О 1 d г о у d, 1927 I 119 (Lora).
Раковина серая, башенковидная, с 6—6.5 уступчатыми оборотами, упло-
щенными в верхней части. Периостракум плотный, коричневатый, легкоот-
стающий. Верхние обороты обычно окатаны, скульптура эмбриональной рако-
вины неизвестна. Осевая скульптура дефинитивной раковины представлена
прямыми узкими складками: рельефными на ранних оборотах и более пологими
на поздних дефинитивных оборотах. Складки выражены одинаково на всей по-
верхности оборота и сглаживаются в нижней части основания раковины. Про-
межутки, разделяющие осевые элементы, на ранних дефинитивных оборотах
всегда превышают ширину складок, на поздних — в 1.5—2 раза уже их. На по-
следнем обороте раковины насчитывается от 24 до 26 складок, на предпоследнем
от 16 до 18. Спиральная скульптура представлена на ранних дефинитивных обо-
ротах тонкими ребрами, на поздних — средними шнуровидными ребрами. По-
следние разделены в 1.5 раза меньшими по ширине межреберными промежут-
ками. Ребра многочисленны и практически равновелики на всей поверхности
последнего оборота, за исключением более сильного — килевидного ребра, про-
ходящего по плечу оборота. Отсутствие заметного преобладания элементов осе-
вой скульптуры над таковыми спиральной придает поверхности раковины (при
пересечении ребер со складками) сетчатый рисунок. Основание раковины уме-
ренно выпуклое, сифональный вырост относительно короткий. Устье полу-
овальное, угловатое в верхней части. Матовый белого цвета каллус широкой
полосой лежит на колумеллярном крае устья. Сифональный канал очень ши-
рокий, глубокий. Размеры раковины голотипа (мм): 2?р=24.0; Д=11.0; Впо=
=16.0. Наибольший из просмотренных экземпляров имел раковину высотой
12 мм.
Крышечка крупная, когтевидная на ранних стадиях роста и практически
яйцевидная на поздних. Ось нарастания резко изогнута. Бобовидный мускуль-
ный отпечаток занимает большую часть левой стороны крышечки, но не приле-
гает к утолщенной левой ее стороне (рис. 36, Р).
Зубы р адулы длиной до 700 мкм, саблевидные, с маленькой базальной частью,
имеющей характерный вырост — язык (рис. 427, 17). Флажок широкий, невы-
сокий. Киль маленький, несколько превышает в длину размеры дистального
отверстия зуба. По общей морфологии исследованные радулы соответствуют
изображениям зубов на рисунках Моррисона.
Вид похож на Р. exarata, от которого отличается сглаженными средними осе-
выми складками и уплощенным вытянутым треугольным флажком зуба радулы.
5. PROPEBELA 799.
Замечания. Исследованные моллюски отличаются от типового экзем-
пляра большей уплощенностью верхней части оборота и как следствие — бо-
лее башенковидной формой раковины.
Распространение. Тихоокеанский широко распространенный вы-
сокобореальный вид. Распространен вдоль американского побережья от Бе-
рингова пролива до о-вов Шумагина, Аляска; отмечен у берегов Курильских
о-вов и у охотоморского побережья о-ва Сахалин.
Типы. Голотип хранится в USNM, № 87857; типовое местонахождение —
Порт Кларенс, Аляска, Берингов пролив.
21. Propebela (Canetoma) concinnula (Verrill, 1882), comb. n. (Рис. 285—288;
427, 10—16).
Verrill, 1882 I 468, pl. 43, fig. 15, pl. 57, fig. 11 (Bela)', Tryon, 1884 t 220, pl. 29,
fig. 54 (Bela', part.); Martini, Chemnitz, 1887 1 148, Taf. 31, fig. 14 (Bela)', Kobel t,
1905 1 242, Taf. 82, fig. 20 (Bela). — acuta Verrill, 1882 1 470, pl. 57, fig. 10 (Bela contig
nula var.). — exarata (non Moller, 1842)i Verrill, 1880 I 366 (Bela).
Раковина белая, веретеновидная, с 5.5—6.5 отлогоугловатыми оборотами,
тупоугловатыми в верхней части. Периостракум толстый, зеленоватый или ко-
ричневатый, легкоотстающий. Эмбриональная раковина окатана, скульптура
протоконха неизвестна. Осевая скульптура дефинитивной раковины состоит из
многочисленных, полуовальных в сечении, средних складок, разделенных рав-
ными по ширине или немногим более узкими промежутками. На околошовной
поверхности складки изогнуты вперед; в нижней х/3 основания раковины они
сглаживаются. На последнем обороте раковины насчитывается от 20 до 26 скла-
док, на предпоследнем от 19 до 21. Спиральная скульптура представлена на
плече оборота 4—6 нитевидными ребрышками, на периферии оборота и на ос-
новании раковины — средними шнуровидными ребрами, разделенными рав-
ными им по ширине промежутками. Элементы спиральной скульптуры при пе-
ресечении с осевыми складками образуют утолщения в виде узелков. Вследствие
одноразмерности складок и средних ребер поверхность периферии оборота ра-
ковины приобретает ячеистый вид. Основание раковины уплощенное, плавно
переходит в удлиненный сифональный вырост. Устье обратногрушевидной
формы, усеченное в верхней части. Белый каллус широкой полосой лежит на
колумеллярном крае. Сифональный канал относительно мелкий и узкий. Сред-
ние размеры раковины из типовой серии (мм): 5^=10.0; 27=4.5; В^=Ь.Ь; наи-
большие: 2?p=11.5; 27=5.25; В^=Ь.0; 27^=2.0. Наиболее крупный экземпляр
с высотой раковины 12.5 мм отмечен для побережья Западной Камчаткй.
Зубы радулы длиной до 600 мкм, изогнутые, саблевидной формы, соответ-
ствуют описанию Веррила (Verrill, 1882). Киль относительно небольшой, в 1.5—
2 раза превышает длину дистального отверстия зуба. Базальная часть конусо-
видная, закругленная; флажок маленький', полуовальный.
Вид сходен с Р. mitrata, от которого отличается высоким завитком; скульп-
турой раковины, образующей толстые узлы в месте пересечения осевых и спи-
ральных элементов; отсутствием языка на базальной части зуба.
Распространение. Амфибореальный широко распространенный
вид тихоокеанского происхождения. Отмечен в приазиатских бореальных во-
дах Тихого океана и в высокобореальных приамериканских водах Атлантики.
Встречен от Новой Англии до п-ова Новая Шотландия, обитает у берегов
Ньюпорта, у м. Код, в зал. Массачусетс; в зал. Каско, а также в Тихом океане
у о-ва Беринга и у берегов Западной Камчатки.
Типы. Место хранения типов Bela concinnula и В. concinnula var. acuta
не указано; типовое местонахождение — Мартас Виньярд, Лабрадор.
Просмотрено 12 экз.
BOO
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
22. Propebela (Canetoma) fidicula (Gould, 1850). comb. n. (Рис. 294; 427и
1-7).
Gould, 1850 1 142 (Fusus); 1852 I 233, fig. 284 (Fusus); Tryon, 1884 I 222, pl. 32,
fig. 17 (Bela); Arnold, 1903 i 209, pl. 7, fig. 10 (Bela); Oldroyd, 1927 I 102 (Lora);
Shimek, Kohn, 1981 I 428, fig. 11, В (Oenopota). — amiata Dall, 1919b : 303 (Lora),
syn. n.
Раковина белая, башенковидная, с 4—5 уступчатыми оборотами, уплощен-
ными в верхней части. Периостракум тонкий, полупрозрачный, коричневатый,
плотноприлегающий. Верхние обороты исследованных экземпляров корроди-
рованы, скульптура протоконха неизвестна. Осевая скульптура дефинитивной
раковины состоит из полукруглых в сечении, средних складок, разделенных
в 4—5 раз более узкими промежутками. Осевые складки наиболее выражены на
периферии оборота, на плече получают наклон вперед; сглаживаются в нижней
трети основания раковины. На последнем обороте насчитывается от 17 до 19 осе-
вых складок, на предпоследнем около 14. Спиральная скульптура на плече обо-
рота представлена нитевидными ребрышками (от 5 до 7). На периферии оборота
и в верхней части основания раковины залегают средние шнуровидные ребра,
разделенные в 2—3 раза более узкими межреберными пространствами. Нижняя
половина основания раковины и сифональный вырост украшены тонкими шну-
ровидными ребрами. Основание раковины умеренно выпуклое, плавно переходит
в удлиненный сифональный вырост. Устье узко-обратногрушевидной формы,
с тонкой полосой кремового цвета каллуса на колумеллярном крае. Сифональ-
ный канал глубокий, относительно широкий. Высота раковины типового эк-
земпляра 11.3 мм при ширине 5.6 мм. Наибольшая из ископаемых раковин
имела параметрь! (мм): Z?p=10.0; Д=5.6; Вп0=8.5; Ву=6.2; Ду=2А. Наиболее
крупный из исследованных мной экземпляров имел раковину с размерами (мм):
5р=12.4; Д=5.6; 5я0=8.5; ^=6.2;- Ду=2.4.
Крышечка каплевидной формы, с практически прямой (на поздних стадиях
формирования) осью нарастания. Мускульный отпечаток в виде тонкой полосы,
занимает срединную часть крышечки (рис. 35, Т).
Зубы радулы длиной до 250—300 мкм, ножевидные. Небольшое коническое
основание несет на одной из ручек уплощенный листовидный флажок (рис. 427,
4). Киль не превышает размеров дистального отверстия зуба и имеет гарпуно-
видный отросток.
Вид сходен с Р. concinnula, от которого отличается низким завитком, очень
пологими осевыми складками и ножевидными зубами.
Распространение. Тихоокеанский высокобореально-арктический
вид. Вдоль американского побережья отмечен от Алеутских о-вов до зал. Бел-
лингем, Пьюджет-Саунд, обитает в Чукотском море у о-ва Врангеля и у берегов
Восточной Камчатки. Найден в плиоценовых (Сан-Педро) и плейстоценовых
(Санта-Барбара) отложениях Калифорнии (Arnold, 1903).
Типы. Типовой экземпляр Busus fidicula хранится в USNM; типовое ме-
стонахождение — Пьюджет-Саунд. Голотип Lora amiata хранится в USNM,
№ 214219; типовое местонахождение — Belkoffski, Аляска.
Просмотрено 2 экз.
'23. Propebela (Canetoma) alitakensis (Dall, 1919), comb. n. (Рис. 305—306;
430, 14—77).
Dall, 1919a t 43, pl. 15, fig. 7 (Lora).
Раковина желтовато-серая, веретеновидная, с 5—7 сильновыпуклыми обо-
ротами отлогоугловатой формы. Периостракум тонкий, прозрачный, легкоот-
стающий. Верхние обороты раковины сильно корродированы, скульптура про-
токонха не исследована. Осевая скульптура дефинитивной раковины состоит из
ярко выраженных, сильновыпуклых, полукруглых в сечении, средних складок,
имеющих незначительный изгиб в верхней части и слегка отклоненных назад
5. PROPEBELA
801
на периферии оборота. На периферии оборота складки наиболее выражены, по-
степенно ослабевают на основании раковины и сглаживаются на границе с си-
фональным выростом. Пространства, разделяющие складки, равны им по ши-
рине. На последнем и предпоследнем оборотах раковины насчитывается; от 12
до 14 складок. Спиральная скульптура четко дифференцирована. На плече обо-
рота расположены многочисленные нитевидные ребрышки. На периферии обо-
рота, на основании раковины и на сифональном выросте она представлена в раз-
личной мере выраженными ребрами. Как правило, между каждым шнуровид-
ным средним ребром залегают 1—2 тонких ребра. Межреберные пространства
практически всегда больше или равны по ширине ребрам. Основание раковины
умеренно выпуклое, четко отграничено от небольшого сифонального выроста.
Устье узко-обратногрушевидное, с полосой белого матового каллуса на колу-
меллярном крае. Сифональный канал глубокий, средней ширины. Размеры ра-
ковины голотипа (мм): Вр=20.0; Д=9.0; 2?яо=14.О. В нашем распоряжении
были раковины высотой 7.0—7.3 мм.
Зубы радулы длиной до 140 мкм, расширенно-ножевидные, с массивным
пирамидальным основанием, несущим на одной из ручек широкий листовидный
флажок. Киль длинный, достигает 1/2 длины лезвия.
Вид хорошо отличается от других представителей подрода расширенно-
ножевидными зубами с длинным килем.
Распространение. Тихоокеанский высокобореальный вид. У аме-
риканских берегов распространен от о-ва Унимак на восток до зал. Алитак,
о-в Кодьяк, у азиатских берегов найден только в Кроноцком заливе.
Экология. Обитает на глубинах от 28 до 180 м, преимущественно на
заиленных грунтах.
Типы. Голотип Lora alitakensis, хранится в USNM, №222088; типовое ме-
стонахождение — о-в Унимак, Алеутские острова.
Просмотрено 2 экз. из бухты Ольги, Кроноцкий залив, Восточная Камчатка,
глубина 148 м.
24. Propebela (Canetoma) rassina (Dall, 1919), comb. n. (Рис. 307).
Dall, 1919a i 303 (Лога); Oldroyd, 1927 I 117 J Lora)-, Boss et al., 1968:273
(Lora).
Раковина белая или желтовато-серая, башенковидная, с 6—6.5 оборотами
отлогоугловатой формы, тупоугловатыми в верхней части. Периостракум тон-
кий, полупрозрачный, телесного цвета. Эмбриональная раковина не сохрани-
лась. Осевая скульптура дефинитивной раковины состоит из полукруглых
в сечении, средних складок, ширина которых равна ширине разделяющих их
промежутков. Складки выражены в равной степени на всей поверхности пери-
ферии оборота и на основании раковины. На последнем обороте их насчитывается
от 16 до 20, на предпоследнем от 14 до 16. Спиральная скульптура состоит из
многочисленных тонких нитевидных ребер, разделенных несколько меньшими
или равными им по ширине межреберными промежутками. На плече оборота
спиральные ребра более узкие, чем на периферии оборота и основании рако-
вины. Основание раковины сильно уплощенное, без ярко выраженной границы
переходит в относительно короткий сифональный вырост. Устье узко-обратно-
грушевидное, с широкой полосой толстого кремового цвета каллуса на колу-
меллярном крае. Сифональный канал средней глубины и ширины. Размеры ра-
ковины типового экземпляра (мм): 2?p=10.0; Д—5.5; ВпО=7.0. Исследованная
раковина имела высоту 10.7 мм. Радула не изучена.
Вид отличается от других представителей подрода широкими высокими осе-
выми складками, значительно превышающими элементы ' спиральной скульп-
туры.
Распространение. Тихоокеанский высокобореальный вид. По-
мимо типового местонахождения обнаружен только у о-ва Беринга.
208
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Типы. Голотип хранится в USNM, № 214224; типовое местонахождение —
юго-западная часть о-ва Гагемейстера, Бристольский залив.
Исследована единственная раковина (без тела), найденная у берегов о-ва
Беринга на глубине 70 м.
25. Propebela (Canetoma) cingulata (Golikov et Gulbin, 1977),’ comb. n.
(Рис. 308—310; 405, Д—E\ 436, 16—19-, 474).
Голиков, Гульбин, 1977 I 243, рис. 16, 18 (Oenopota).
Рис. 474. Места находок Propebela cingulata.
Раковина светло-коричневого цвета, с более бледными ребрами и столбиком,
стройная, башенковидная, с 5.5—6 оборотами отлогоугловатой формы; в верх-
ней и нижней частях обороты тупоугловатые. Периостракум коричнёва’тый,
полупрозрачный. Протоконх представлен 1.5 оборотами, причем первый оборот
лишен макроскульптуры и покрыт мелкими микроскульптурными бугорками
(рис. 405, Д—Е). Начиная со второго оборота, происходит закладка спираль-
ной скульптуры, представленной на эмбриональной раковине 3 тонкими реб-
рами. Верхнее из них при переходе на дефинитивную раковину полностью реду-
цируется, а 2 оставшихся преобразуются в килевидные ребра. Осевая скульп-
тура дефинитивной раковины представлена полукруглыми в сечении, сред-
ними складками, одинаково выраженными практически на всей поверхности
раковины: на плече и периферии оборота и на основании раковины. Проме-
жутки несколько превышают ширину самих складок. На последнем и предпо-
следнем оборотах насчитывается от 12 до 15 осевых складок. Спиральная скульп-
тура верхних оборотов состоит из 2 килевидных ребер, которые на последних
дефинитивных оборотах превращаются в настоящие кили. Один киль проходит
по плечу оборота, обозначая его, второй — по нижней 1/3 периферии оборота.
На последнем обороте обнаруживается третье килевидное ребро, проходящее
в верхней части основания раковины и лежащее, как правило, на границе
между основанием и периферией оборота. Все пространство между килями и
килевидными ребрами покрыто многочисленными нитевидными ребрышками,
равновеликими с разделяющими их промежутками. Основание раковины уме-
ренно выпуклое, сифональный вырост небольшой. Устье неправильно-овальной
формы, усеченное в верхней части; на колумеллярном крае несет тонкую по-
лосу кремового цвета каллуса. Сифональный канал узкий и глубокий. Раз-
меры раковины голотипа (мм): 2?р=8.5; Д=3.7; 5л0=5.7; 5^=4.0; Д =1.3.
5. PROPEBELA
303
Наибольшие размеры имел экземпляр, найденный у о-ва Шикотан, высота его
раковины 9.3 мм при ширине 3.8 мм.
Зубы радулы мелкие, длиной до 140 мкм, ножевидной формы. Основание
большое, коническое, на одной из ручек несет листовидный флажок. Киль ма-
ленький, треугольный, с намечающимся гарпуновидным отростком (рис. 436,
19).
Вид сходен с Р. rassina, от которого легко отличается по наличию на пери-
ферии оборотов 2 килей, равновеликих по высоте с осевыми складками.
Распространение. Тихоокеанский приазиатский низкобореаль-
ный вид. Помимо типового местонахождения отмечен в зал. Анива, о-в Саха-
лин, в Приморье и у о-ва Монерон, Японское море.
Экология. Встречен на глубинах до 39 м, в основном до 10 м; преиму-
щественно на песчаных грунтах.
Типы. Голотип хранится в ЗИН АН СССР, № 28266; типовое местона-
хождение — у о-ва Шикотан, глубина 7—8 м.
Просмотрено 19 экз.
26. Propebela (Canetoma) areta (Bartsch, 1941), comb. n. (Рис. 303—304;
405, Б).
Bartsch, 1941 i 11, pl. 1, fig. 8 (Funitoma)', Powell, 1966 I 121, pl. 19, fig. 4,
{Oenopota (Funitoma)!
Раковина от светло-каштанового до коричнево-вишневого цвета, овально-
веретеновидная, с 5—6 отлогоугловатыми, в верхней и нижней частях тупоугло-
ватыми оборотами. Периостракум тонкий, легкоотстающий, прозрачный. Про-
токонх представлен 2 оборотами, причем первый эмбриональный оборот уме-
ренно выпуклый, лишен макро- и микроскульптурных образований. На вто-
ром обороте закладываются 3 спиральных ребра, из которых верхнее остается
слабо развитым и постепенно исчезает, а оставшиеся 2 ребра к концу второго
эмбрионального оборота приобретают килевидную форму. Осевая скульптура
дефинитивной раковины представлена ярко выраженными средними склад-
ками, полукруглыми в сечении. Они отклонены назад и разделены равными
им по ширине пространствами. Складки более выражены на периферии оборота,
сглаживаются в нижней части основания раковины. На последнем обороте на-
считывается около 14 складок, на предпоследнем от 11 до 13. Спиральная скульп-
тура представлена на ранних оборотах 2 килевидными ребрышками (продолже-
ние эмбриональных), на поздних дефинитивных оборотах — средними ребрами,
равновеликими на всей поверхности раковины и разделенными в 2 раза более
узкими межреберными промежутками. Основание раковины сильно уплощен-
ное, и колумеллярный край сифонального выроста составляет с ним практи-
чески прямую линию. Устье неправильно-овальное, усеченное. В верхней
части палатального края лежит неглубокий анальный синус. Колумеллярный
край устья покрыт узкой полосой тонкого каллуса розоватого или вишневого
цвета. Сифональный канал короткий, широкий, неглубокий. Высота раковины
типового экземпляра 8.5 мм при диаметре 3.7 мм. Максимальные размеры ра-
ковин, исследованных мной экземпляров, не превышали 5.9 мм.
Вид отличается от остальных представителей подрода маленькой, очень тол-
стостенной раковиной и протоконхом, первый оборот которого абсолютно ли-
шен скульптурных элементов.
Замечания. Исследованные экземпляры имели более выраженные
(в большей мере выступающие над поверхностью раковины) осевые складки,
чем у типового экземпляра. Возможно, это объясняется возрастными разли-
чиями.
Распространение. Тихоокеанский приазиатский широко распро-
страненный бореальный вид. Обитает у Средних и Северных Курильских о-вов.,
у Охотоморского побережья о-ва Сахалин, в Японском море отмечен у о-ва Мо-
нерон и у берегов Приморья.
304 СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Экология. Обитает на глубинах от 0 до 50 м преимущественно на пес-
чаных и скалистых грунтах.
Типы. Типовой экземпляр хранится в USNM, № 428857; типовое место-
нахождение — Японское море.
Просмотрено 22 экз.
27. Propebela (Canetoma) golikovi (Bogdanov, 1985), comb. n. (Рис. 301—
302; 390, A—B; 430, 1-5).
Богданов, 1985a 1 452, рис. 1, c—y', 2, -5 (Oenopota).
Раковина от каштанового до светло-коричневого цвета, веретеновидная,
толстостенная, с 5.5—6 отлогоугловатыми оборотами. Периостракум тонкий,
слегка матовый, легкоотстающий. Протоконх представлен 1.5 оборотами выпук-
лой формы, причем первый эмбриональный оборот погружен во второй за счет
практически планоспирального навивания эмбриональной раковины. Неболь-
шая часть второго оборота протоконха несет многочисленные волосовидные
спиральные ребрышки, которые при переходе на дефинитивную раковину
сливаются в 2 тонких спиральных ребра. Осевая скульптура состоит из прелом-
ленных в верхней части, узких складок, резче всего выраженных на периферии
оборота, сглаживающихся на основании раковины. Осевые складки разделены
вдвое более широкими промежутками. На последнем обороте насчитывается
14, на предпоследнем 12 складок. Спиральная скульптура на последних обо-
ротах представлена различной величины тонкими ребрами с одинаковыми меж-
реберными промежутками. Сильнее всего развиты ребра, являющиеся продол-
жениями эмбриональных; одно из них обозначает плечо оборота, другое —
нижнюю х/3 его периферии. На околошовной площадке спиральная скульптура
в виде нитевидных ребрышек, развита слабо. Основание раковины выпуклое,
со спиральной скульптурой, чуть слабее выраженной, чем на периферии по-
следнего оборота. Устье узко-обратногрушевидной формы, с тонким матовым
каллусом на колумеллярном крае. Сифональный канал узкий, глубокий. Раз-
меры раковины голотипа (мм): Вр=8.1; Д—3.8; Впо—Ъ.8’, Ву=^.2', Дд=1.§,
Зубы радулы маленькие, длиной до 140 мкм, ножевидные, с длинным килем,
в 1.5^-2 раза превышающим размеры дистального отверстия лезвия. Базальная
часть небольшая, конусовидная, на одной из ручек несет маленькую шпору.
•Вид сходен с Р. alitakensis, от которого отличается маленькой раковиной,
менее выпуклыми оборотами и более часто расположенными осевыми складками,
а также ножевидной формой зубов. Характерная черта вида — беловатая ок-
раска высоких осевых и спиральных элементов скульптуры.
Распространение. Тихоокеанский приазиатский низкобореальный
вид. Встречен у южного и западного побережий о-ва Сахалин и в Северном
Приморье.
Экология. Обитает на глубинах от 15 до 46 м на илистых и илисто-
песчаных грунтах.
Типы. Голотип хранится в ЗИН АН СССР, № 37984; типовое местона-
хождение— прибрежье о-ва Сахалин (47° 25'5” с. ш.—141° 55'5” в. д.) глу-
бина 32 м.
Просмотрено 17 экз.
• 28. Propebela (Canetoma) margaritae (Bogdanov, 1985)f comb. n. (Рис. 289—
293; 427, 8-9).
Богданов, 1985a I 451, рис. 1, н—p, 2,. 4 (Oenopota).
Раковина серая, удлиненно-веретеновидная, толстостенная, с 5—7 оборо-
тами отлогоугловатой формы. Периостракум тонкий, сероватый, легкоотстаю-
щий. Эмбриональная раковина представлена 1.7 оборота, ее скульптура плохо
сохранилась. Осевая скульптура дефинитивной раковины представлена высо-
5. PROPEBELA
205
кими тонкими складками, ширина которых в 3—4 раза меньше разделяющих
их промежутков. Складки прямые, в верхней части слегка наклонены вперед,
они ярко выражены на периферии оборота и сглаживаются к его основанию.
На последнем обороте раковины насчитывается до 25, на предпоследнем до
20 складок. Спиральная скульптура на первом постэмбриональном обороте со-
стоит из 2 нитевидных ребрышек, с ростом раковины их количество увеличива-
ется и к последнему обороту они становятся равновелики (и по ширине, и по
высоте), сильнее прочих остается развитым только ребро, обозначающее плечо
оборота. Основание раковины округлое, покрыто спиральными ребрами, рав-
ными по величине таковым периферии оборота. Сифональный вырост удли-
ненный. Устье узко-обратногрушевидной формы, колумеллярный край покры-
вает узкая полоса толстого беловатого каллуса. Сифональный канал узкий и
глубокий. Размеры раковины голотипа (мм): В =12.4; Д=4.9; Вка=7А;В =
= 5.2; Д,=2.0.
Зубы радулы маленькие, длиной до 150 мкм, ножевидной формы. Длина
киля не превышает размеров дистального отверстия лезвия. Базальная часть
высококонической формы, небольшой полуовальный флажок развит только
на одной ручке (рис. 427, 8—9).
Вид сходен с Р. concinnula, от которого отличается более тонкими осевой и
спиральной скульптурами и ножевидной формой зубов.
Распространение. Тихоокеанский приазиатский широко распро-
страненный бореальный вид. Помимо типового местонахождения отмечен у бе-
регов о-ва Медного, у Восточной Камчатки и в Японском море в зал. Петра
Великого.
Типы. Голотип хранится в ЗИН АН СССР, № 37982; типовое местона-
хождение — о-в Беринга, на траверзе р. Фонтанка (55°22'5" с. ш.—
165°53' в. д.), глубина 100 м, мелкопесчаный илистый грунт.
Просмотрено 8 экз.
29. Propebela (Canetoma) marinae Bogdanov, 1989. (Рис. 274—275; 426,
13—22).
Богданов, 19896 i 152, рис. 1, 11—13; 2, щ—я.
Раковина телесного цвета, овально-веретеновидная, с 4.5—5 уступчатыми
оборотами, уплощенными в верхней части. Тонкий полупрозрачный кремового
цвета периостракум легко отстает от поверхности раковины. Эмбриональная
раковина не изучена. Осевая скульптура дефинитивной раковины представлена
непостоянным числом пологих тонких складок, сглаживающихся к нижней
части периферии оборота. Спиральная скульптура состоит из многочисленных
средних ребер, различных по высоте и ширине. На первых 3—4 дефинитивных
оборотах в местах пересечения осевой и спиральной скульптур, образуются
узелки, благодаря чему верхние обороты приобретают ячеистый рисунок, ос-
лабевающий к последнему обороту раковины. Умеренно выпуклое основание
раковины плавно переходит в вытянутый сифональный вырост. Устье узко-
обратногрушевидное, угловатое в верхней части, с тонким матовым каллусом
на колумеллярном крае. Размеры раковины голотипа (мм): В=10.7; Д=5.7;
5л0=8.4; ^=6.4; Ду=2.5.
Крышечка небольшая, каплевидная, со слабо изгибающейся осью нараста-
ния. Мускульный отпечаток имеет ромбовидные очертания, как правило, он
ромбовидной формы и расположен в центральной части крышечки (рис. 36, С).
Зубы радулы длиной 400—550 мкм, саблевидные, с небольшим полуоваль-
ным основанием, несущим на ручках 2 флажка, один треугольной, другой полу-
овальной формы. Киль длинный, в 2—3 раза превышает размеры дистального
отверстия зуба (рис. 426, 13, 18).
Вид сходен с Р. viridula, от которого отличается слабым развитием осевых
элементов скульптуры и саблевидной формой зубов с 2 флажками на ручках.
2Q6
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Р аспространение. Тихоокеанский приазиатский широко распро-
страненный бореальный вид. Помимо типового местонахождения отмечен в рай-
оне Курильских о-вов (44°2Г с. ш.—148°24' в. д.) и в Японском море
(42°58' с. ш. — 134°07' в. д.).
Экология. Встречен на глубинах от 300 до 1 076 м на заиленных грун-
тах.
Т и и ы. Голотип хранится в ЗИН АН СССР, № 48479; типовое местонахож-
дение — Охотское море (49°24' с. ш.—148°46' в. д.), глубина 1 076 м, грунт
серо-зеленый ил, придонная температура 2.46 °C, соленость 34.25 °/00.
Просмотрено 3 экз.
Рие. 475. Места находок Propebela variabilis в морях СССР.
30.. Propebela (Canetoma) variabilis Bogdanov, sp. n. (Рис. 295—300; 429r
7-24; 475).
Раковина от светло-коричневого до различных оттенков розового цвета, ве-
ретеновидно-овальная, с 5—6 закругленными, умеренно выпуклыми оборотами.
Периостракум тонкий, плотноприлегающий. Протоконх не сохранился. Осевая,
скульптура дефинитивной раковины состоит из широких пологих осевых скла-
док, которых на предпоследнем обороте насчитывается 14—16, на последнем
около 20. Спиральная скульптура представлена средними шнуровидными, пря-
моугольными в сечении ребрами. По ширине они в 4—7 раз превышают разде-
ляющие их промежутки. Спиральные ребра практически равновелики на всей
поверхности раковины, но несколько утончаются на околошовной поверхности.
Основание раковины выпуклое, устье обратногрушевидной формы, с толстым:
розоватым каллусом на колумеллярном крае. Сифональный канал узкий и
глубокий. Размеры раковины голотипа (мм): 7??=11.3; Д=5.1; Вп0=7.9; В =
=5.2; Ду=2.3. Высота раковины наиболее крупного экземпляра 13.9 мм.
Крышечка когтевидная, удлиненная, с плавно изгибающейся .осью нараста-
ния и с утолщенным левым краем. Мускульный отпечаток ромбовидной или за-
кругленно-ромбовидной формы, в верхней части всегда заостренный, топогра-
фически расположен под осью крышечки (рис. 36, Н—П).
5. PROPEBELA
207
Зубы радулы длиной до 800 мкм, саблевидные, с небольшим округлым ос-
нованием, несущим по маленькому флажку на каждой из ручек- Киль удлинен-
ный, в 1.5—2 раза превышает размеры дистального отверстия зуба. Лезвие
зуба всегда изогнуто (рис. 429).
По раковине вид похож на Р. harpularia, от которого отличается саблевид-
ными зубами радулы.
Замечания. Изменчива как форма раковины, так и ее скульптура.
Сильнее всего варьирует форма раковины, что связано с различной степенью
выпуклости оборотов и основания раковины. Что касается изменчивости скульп-
туры, то известны экземпляры с ярко выраженными элементами осевой и спи-
ральной скульптур (рис. 295—296). Вместе с тем найдены раковины, у которых
•спиральная ребристость на последних дефинитивных оборотах отсутствует,
а осевые складки едва выступают над поверхностью раковины (рис. 300). Самым
надежным диагностическим признаком вида остается стабильная морфология
зубов радулы.
Распространение. Тихоокеанский высокобореальный вид. По-
мимо типового местонахождения отмечен в Беринговом проливе и у берегов
Курильских о-вов (рис. 475).
Типы. Голотип хранится в ЗИН АН СССР, № 48480; типовое местона-
хождение — о-в Беринга, Командорские о-ва, глубина 100 м.
Просмотрено 37 экз.
ЛИТЕРАТУРА
Аверинцев-С. Зоологические работы Мурманской Биологической станции за 1904—1908 гг. //
Тр. Ими. СПб. О-ва Естествоисп. 1908. Т. 39, вып. 1, № 8. С. 288—328.
Дмитров О. В. Турриды позднего эоцена — раннего миоцена юга СССР. М., 1973. 212 с.
Андреева С. И., Старобогатов Я. И. Новые виды моллюсков семейства Pyrgulidae (Gastro-
poda, Pectinibranchia) из Аральского моря//Зоол. журн. 1981. Т. 60, вып. 1.
С. 29—35.
Андрияшев А. П. Очерк зоогеографии и происхождения фауны рыб Берингова моря и со-
предельных вод. Л., 1939. 185 с.
Богданов И. П. Влияние солености и температуры на скульптуру и форму раковины брюхо-
ногих моллюсков рода Oenopota Morch, 1852 // Науч. докл. высш. шк. Биол. науки.
Ift83a. С. 5.
Богданов И. П. Значимость признаков в диагностике моллюсков рода Oenopota (Gastropoda,
Turridae) // Моллюски: Систематика, экология и закономерности распространения.
Сб. 7. Л., 19836. С. 51—52.
Богданов И. П. Новые виды брюхоногих моллюсков рода Oenopota (Gastropoda, Turridae)
из дальневосточных морей СССР // Зоол. журн. 1985а. Т. 64, вып. 3. С. 448—453.
Богданов И. П. Морфофункциональный анализ строения переднего отдела пищеварительной
системы моллюсков рода Oenopota (Gastropoda, Turridae) // Зоол. журн. 19856. Т. 64,
вып. 11. С. 1632—1639.
Богданов И. П. К построению естественной системы моллюсков рода Oenopota (Toxoglossa,
Turridae) // Зоол. журн. 1986. Т. 65, вып. 1. С. 36—46.
Богданов И. П. Комплексный подход к изучению таксономических признаков морских
гастропод на примере моллюсков подсемейства Oenopotinae subfam. nov. // Моллюски:
Результаты и перспективы их исследований. Сб. 8. Л., 1987. С. 35—37.
Богданов И. П. Пять новых япономорских видов Oenopotinae (Gastropoda, Turridae) //
Зоол. журн. 1989а. Т. 68, вып. 9. С. 134—138.
Богданов И. П. Семь новых видов Oenopotinae из Охотского моря // Зоол. журн. 19896.
Т. 68, вып. 11. С. 147-152.
(Богданов И. П.) Bogdanov I. Р. Peculiar feeding mechanism of the Oenopotinae molluscs
(Gastropoda, Turridae) // Conchiglia. 1989b. Anno 21, N 246—249. P. 39—46.
(Богданов И. П.) Bogdanov I. P. Morphological transformations in radula and protoconch
of Oenopotinae Bogdanov, 1987 // Conchiglia. 1989г. Anno 21, N 233—236. P. 35—47.
Виноградов Л. Г. О зоогеографическом районировании дальневосточных морей//
Изв. ТИНРО. 1948. Т. 28. С. 162—167.
Волова Г. Ю., Голиков А. Н., Кусакин О. Г. Раковинные брюхоногие моллюски залива
Петра Великого. Владивосток, 1979. 210 с.
Галкин Ю. И. Материалы к характеристике фауны моллюсков залива Анива (Охотское
море) // Тр. Зоол. ин-та АН СССР. 1953. Т. 13. С. 242—249.
Галкин Ю. И. Брюхоногие моллюски трохиды дальневосточных и северных морей СССР.
Л., 1955. 132 с. (Опред. по фауне СССР, изд. Зоол. ин-том АН СССР; № 57).
Герценштейн С. Материалы к фауне Мурманского берега и Белого моря // Тр. Имп. СПБ
Общ. Естеств. 1885. С. 635—814.
Голиков А. Н. Брюхоногие моллюски рода Neptunea Bolten. М.; Л., 1963. 217 с. (Фауна СССР.
Нов. сер.; № 85. Моллюски; Т. 5, вып. 1).
Голиков А. Н. Брюхоногие и лопатоногие моллюски (Gastropoda et Scaphopoda) северной
части Гренландского моря и районов к северу от Шпицбергена и Земли Франца-
Иосифа // Тр. Аркт. и Антаркт. НИИ. 1964. Т. 259. С. 340—354.
Голиков А. Н. Моллюски Buccininae Мирового океана. Л., 1980. 465 с. (Фауна СССР.
Нов. сер.; № 121. Моллюски; Т. 5, вып. 2).
Голиков А. Н. О принципах районирования и унификация терминов в морской биогеогра-
фии Ц Морская биогеография. М., 1982. С. 94—98.
Голиков А. Н. Моллюски Белого моря. Л., 1987. 328 с. (Опред. по фауне СССР, изд. Зоол. ин-
том АН СССР; № 151).
Голиков А. Н., Гульбин В. В. Брюхоногие переднежаберные моллюски (Gastropoda, Рго-
sobranchiata) шельфа Курильских островов. II. Отряды Hamiglossa—Homoestropha//'
Фауна прибрежных зон Курильских островов. М., 1977. С. 172—286.
Голиков А. Н., Кусакин О. Г. Фауна и экология брюхоногих моллюсков (Gastropoda, Pro-
ЛИТЕРАТУРА
809
sobranchia) литорали Курильских островов // Исслед. дальневост, морей СССР. 1962.
Вып. 8. С. 248—346.
Голиков А. Н., Кусакин О. Г. Фауна и распределение раковинных брюхоногих моллюсков
на литорали морей СССР // Моллюски: Пути, методы и итоги их изучения. Сб. 4. Л.,
1971. С. 27-29.
Голиков А. Н., Кусакин О. Г. Раковинные брюхоногие моллюски литорали морей СССР.
Л., 1978. 256 с. (Опред. по фауне СССР, изд. Зоол. ин-том АН СССР; № 116).
Голиков А. Н., Скарлато О. А. Моллюски залива Посьет (Японское море) и их экология //'
Тр. Зоол. ин-та АН СССР. 1967. Т. 42. С. 5-154.
Голиков А. Н., Скарлато О. А. Раковинные брюхоногие и двустворчатые моллюски шельфа
южного Сахалина и их экология // Биоценозы и фауна шельфа южного Сахалина.
Л., 1984. С. 368—487. (Исслед. фауны морей; Т. 30 (38)).
(Голиков А. Н., Старобогатов Я. И.) Golikov A. N., Starobogatov Y. I. Systematics of pro-
sobranch gastropods // Malacologia. 1975. Vol. 15, N 1. P. 185—232.
Горячев В. H. Брюхоногие моллюски рода Neptunea Roding Берингова моря. М., 1978..
90 с.
Гульбин В. В., Шульмина М. В. Брюхоногие моллюски литорали острова Сахалин. Влади-
восток, 1981. С. 62-74 (Сб. работ АН СССР ДВНЦ; № 24).
Гурьянова Е. Ф. К зоогеографии дальневосточных морей// Изв. АН СССР. Отд. математ..
и естеств. наук. 1935. № 7. С. 1229—1235.
Гурьянова Е. Ф. К вопросу о происхождении и истории развития фауны Полярного бас-
сейна Ц Изв. АН СССР. Сер. биол. 1939. № 5. С. 679—704.
Гурьянова Е. Ф. Бокоплавы морей СССР и сопредельных вод (Amphipoda—Gammaridae).
Л., 1951. 1029 с. (Опред. по фауне СССР, изд. Зоол. ин-том АН СССР; № 41).
Гурьянова Е. Ф. Схема зоогеографического деления донной фауны Курило-Сахалинского-
района // Атлас океанографических основ рыбопоисковой карты южного Сахалина—
южных Курильских островов. Л., 1955. Т. 1. С. 87—88.
Гурьянова Е. Ф. Зоогеографическое районирование моря. Фауна Тонкинского залива и
условия ее существования//Исслед. фауны морей. 1972. Т. 10, вып. 18. С. 8—21.
Дерюгин К. М. Фауна Кольского залива и условия ее существования // Зап. Акад. наук.
.. Физ.-мат. отд-ние. 1915. Т. 34, сер. 8, № 1. 929 с.
Дерюгин К. М. Баренцево море по Кольскому меридиану (33°30' в. Д.) И Тр. Сев. н.-про-
мыслов. эксп. М.; Л., 1924. Вып. 19. 102 с.
Зенкевич Л. А. Биология морей СССР. М., 1963. 739 с.
Иззатуллаев 3. И., Старобогатов Я. И. Род Melanopsis (Gastropoda, Pectinibranchia) и его-
представители, обитающие в водоемах СССР // Зоол. журн. 1984. Т. 63, вып. 10. С.
1471—1483.
Кусакин О. Г. К фауне и флоре осушной зоны острова Кунашир // Тр. пробл. и тематич..
совещ. Зоол. ин-та АН СССР. 1956. Т. 6. С. 98—115.
Кусакин О. Г. Сезонные изменения на литорали южных Курильских островов // Вестн. ЛГУ..
Сер. биол. 1958. № 3, вып. 1. С. 116—130.
Линко А. К. Мурманская биологическая станция и Санкт-Петербургское Общество Есте-
ствоиспытателей. Отчет лаборанта за лето 1900, 1901 и 1902 гг. // Тр. Имп. СПб.
О-ва Естествоисп. 1902. Т. 33, вып. 1. С. 1—28.
Нейман А. А. Количественное распределение бентоса в восточной части Берингова, моря //:
Зоол. журн. 1960. Т. 39, вып. 9. С. 3—6.
Несис К. Н. Зоогеография Мирового океана: сравнение зональности пелагиали и регио-
нального членения шельфа (по головоногим моллюскам) // Морская биогеография.
М., 1982. С. 114—134.
Педашенко А. Д. Отчет о состоянии и деятельности Соловецкой биологической станции
в 1897 г. Ц Тр. Имп. СПб. О-ва Естествоисп. 1897. Т. 28, вып. 1, № 7. 34 с.
Песенко Ю. А. Количественные методы фаунистических исследований. М., 1982. 287 с..
Петров О. М. Морские моллюски антропогена северной части Тихого океана. М., 1982.
144 с. (Тр. Геол, ин-та АН СССР; Вып. 357).
Скарлато О. А. К биогеографии дальневосточных морей Советского Союза на примере дву-
створчатых моллюсков // Тр. пробл. и тематич. совещ. Зоол. ин-та АН СССР. 1956..
Вып. 6. С. 83—92.
Скарлато О. А. Двустворчатые моллюски дальневосточных морей СССР (отряд Dysodonta).
М.; Л., 1960. 150 с. (Опред. по фауне СССР, изд. Зоол. ин-том АН СССР; № 71).
Скарлато О. А. Двустворчатые моллюски умеренных вод северо-западной части Тихого
океана. Л., 1981. 480 ;.
Сысоев А. В. Морфология радулярных зубов некоторых Turridae (Gastropoda, Toxoglossa)
северо-западной части Тихого океана // Зоол. журн. 1983. Т. 62, вып. 11. С. 1621 —
1628.
Ушаков П. В. Донное население как показатель теплых и холодных течений в Чукотском
море Ц Проблемы Арктики. 1940. Т. 7—8. С. 21—25.
Ушаков П. В. О морской донной фауне в районе южных Курильских островов // Докл.
АН СССР. 1951. Т. 30, № 1. С. 125—128.
Ушаков П. В. Чукотское море и его донная фауна// Крайний Северо-Восток Союза ССР.
М.; Л., 1952. Т. 2. С. 3-78.
Ушаков П. В. Фауна Охотского моря и условия ее существования. М., 1953. 459 с.
14 И. П. Богданов
210 ЛИТЕРАТУРА
Ушаков П. В. О значении пролива Лаперуза в формировании фауны юго-восточной части
Охотского моря Ц Докл. АН СССР. 1955. Т. 105, № 6. С. 1371—1374.
Филатова 3. А. Зоогеографическое районирование северных морей по распределению дву-
створчатых моллюсков Ц Тр. ИОАН СССР. 1957. Т. 23. С. 195—215.
Филатова 3. А., Зацепин В. И. Класс Gastropoda — брюхоногие моллюски // Определитель
флоры и фауны северных морей СССР. М., 1948. С. 358—401.
Шилейко А. А. Наземные моллюски подотряда Pupillina фауны СССР (Gastropoda, Pulmo-
. nata, Geophila). Л., 1984. 399 с. (Фауна СССР. Моллюски; Т. 3, вып. 3).
Abbott В. Т. American seashells. 2nd ed. New York. 1974. 633 p.
Adams С. B. Descriptions of thirteen new species of New England shells// Bost. J. Nat. Hist.
.1840. Vol. 3, N 3. P. 318-332.
Adams H., Adams A. The genera of recent Mollusca; arranged acording to their organization.
London. Vol. 2. 1854. P. 1-92; 1855. P. 93-284; 1858. P. 541—660.
Arnold R. The paleontology and stratigraphy of the marine Pliocene and Pleistocene
of San Pedro, California // Mem. Calif, acad. Sci. 1903. Vol. 3. P. 9—420.
Aurivillius C. W. S. Arktiska Hafmollusker. II Placophora och Gastropoda // A. E. Norden-
skiold. Vega-Expedioneus Vetenskapliga lakttagelser. Stockholm, 1887. S. 311—383.
Bartsch P. The nomenclatorial status of certain northern turritid molluscs // Proc. Biol. Soc.
Wasch. 1941. Vol. 54. P. 1—14.
Bartsch P. Turritomella, new subgeneric name for Turritoma Bartsch, 1941 Ц Nautilus. 1941a.
Vol. 54. P. 143.
Bartsch P. Some notes upon West American Turrid molluscs // Proc. Biol. Soc. Wasch. 1944.
Vol. 57. P. 25-30.
Bartsch P. New West American turrids // Nautilus. 1950. Vol. 63, N 3. P. 87—99.
Bellardi L. I Molluschi de i terreni terziarii del Piemonte e della Liguria, II Gasteropoda (Pleu-
rotomidae). Torino, 1877. Vol. 29. 360 p.
Bernasconi M. P., Robba E. The Pliocene Turridae from Western Liguira. I. Clavinae, Turrinae,
Turriculinae, Grassispirinae, Borsoniinae, Clathurelinae // Bull. Mus. Reg. Sci. Nat.
Torino. 1984. Vol. 2, N 1. P. ,257—358.
Bernasconi M. P., Robba E. The miocene species of the genus Spirotropis (Turridae, Gastropoda)
in Europe//Bull. Mus. Reg. Sci. Nat. Torino. 1985. Vol. 3, N 1. P. 203—220.
Boss K. J. J., Rosewater J., Ruhoff F. The zoological taxa of William Healey Dall// U. S.
Nat. Mus. Bull. 1968. N 287. 427 p.
Bouchet P., Waren A. The Abyssal molluscan fauna of the Norwegian Sea and its relation
to other faunas//Sarsia. 1979. Vol. 64. P. 211—243.
Bouchet P., Waren A. Revision of the North-East Atlantic bathyal and abyssal Turridae (Mol-
lusca, Gastropoda) // J. Moll. stud. 1980. Suppl. 8. 119 p.
Brown Th. Illustrations of the recent conchology of Great Britain and Ireland. 1st ed. Smith
Elder and Co. Edinbourgh, 1827. 144 p.
Brown Th. Illustrations of the recent conchology of Great Britain and Ireland. 2nd ed. London,
1844. 144 p.
Burch J. O. Distributional list of the West American marine mollusks from San Diego, Califor-
nia to the Polas Sea, pt II, Gastropods//Proc. Conch. Club South. California. 1946.
Vol. 2, N 62. P. 17—42.
Bush K. J. Catalogue of Mollusca dredged on the coast of Labrador by the expedition of Stearn //
Proc. U. S. Nat. Mus. 1884. Vol. 6. Washington. P. 236—247.
Carpenter P. P. Diagnoses specierum et varietatum novarum molluscorum, prope sinum Puge-
tianum a Kennerlio Doctore, nuper decesso, collectorum // Proc. Acad. Nat. Sci. Phila-
delphia. 1865. P. 58—69.
Cerff F. C. Scanning electron microscopy of the radulae of two morphs of Cerithidea decollate
(Linn.) // 20 Annu. Conf. Electron. Microsc. Soc. South. Afr. Durban. 2—4 Dec. 1981.
Proc. Vol. 11. Johannesburg, 1981. P. 95—96.
Clarke A. H. Arctic archibental and abyssal molluscs. II. Molluscs dredged from drifting station
Charlie (Alpha)// Nat. Mus. Canada Bull. 1963. Vol. 185, N 7. P. 90—109.
Couthouy J. P. Descriptions of new species of Mollusca and shells and remarks on several Polypi
found in Massachusetts Bay//Bost. J. Nat. Hist. 1838. Vol. 2, N 1. P. 53—111.
Couthouy J. P. Monograph on the family Osteodesmacea of deshayes, with remarks on two spe-
cies of Patelloidea, and descriptions of new species of marine shells a species of Anculotus,
and one of Eolis // Bost. J. Nat. Hist. 1839. Vol. 2, N 2. P. 129—189.
Dall W. H. Descriptions of sixty new forms of molluscs from the west coast of North American
and the North Pacific Ocean, with notes on others already described // Amer. J. Conchol.
1871. Vol. 7, N 2. P. 93—160.
Dall W. H. New or specially interesting shells of the Point Barrow expedition // Proc. U. S.
Nat. Mus. 1884. Vol. 7, N 33. P. 523—526.
Dall W. H. Contributions to the natural history of the Commander Islands. N 6: Report on Be-
ring Island Mollusca collected by Mr. Nicholas Grebnitzki//Proc. U. S. Nat. Mus.
' 1886a. Vol. 9. P. 209—219.
Dall W. H Report on the Mollusca, pt 1 // Bull. Mus. Gompar. Zool. 1886b. Vol. 12, N 6.
P. 171—318.
ЛИТЕРАТУРА
SIT
Dall W. H. Supplementary notes on some species of mollusks of the Bering Sea and vicinity //
Proc. U. S. Nat. Mus. (1886) 1887. Vol. 9, Sign. 19-20. P. 297-310.
Dall W. H. Shell-bearing marine mollusks and brachiopods of the south-eastern coast of the Uni-
ted States// Proc. U. S. Nat. Mus. 1889. N 37. 221 p.
Dall W. II. Notes on the nomenclature of the mollusks of the family Turridae Ц Proc. U. S. Nat.
Mus. 1918. Vol. 54. P. 313—333.
Dall W. H. Description of new species of mollusks of the family Turritidae from the west coast
of America and adjacent regions// Proc. U. S. Nat. Mus. Washington. 1919a. Vol. 56,
N 2288. P. 1-86.
Dall W. H. Descriptions of new species of mollusca from the North Pacific Ocean in the collec-
tion of the United States National Museum//Proc. U. S. Nat. Mus. 1919b. Vol. 56
N 2295. P. 293—371.
Dall W. H. New shells from the north-west coast // Proc. Biol. Soc. Wash. 1919c. Vol. 32.
P. 249-252.
Dall W. H. Summary of the marine shell-bearing mollusks of the north-west coast of America,,
from San Diego, California, to the Polar Sea, mostly contained in the collection
of the U. S. N. M., with illustrations of hitherto unfigured species // Bull. U. S. Nat. Mus.
1921, N 112. P. 1-216.
Dall W. H. Notes on molluscan nomenclature // Proc. Biol. Soc. Wash. 1924. Vol. 37. P. 87—90.
Dall W. H., Bartsch P. New species of shells collected by Mr. John Macoun at Barkley Sound,.
Vancouver Island British Columbia//Mem. Canad. Dep. of Mines. 1910. N 14. 22 p.
Dautzenberg Ph., Fischer H. Mollusques provenant des compagnes de 1’Hirondelle et de la Prin-
cesse-Alice dans les mers du Nord // Res. Camp. Sci. sur son Yacht Albert 1 de Monaco.
1912. Vol. 37. 629 p.
Ekman S. Tiergeographie des Meeres. Leipzig, 1935. 542 S.
Ellis D. V. Marine infaunal benthos in North America // Arctic Inst, of North Amer. Tech.
Paper. Montreal. 1960. N 5. 53 p.
Fischer P. Manuel de conchyliologie et de paleontologie conchyliologique on histoire naturelie'
des mollusques vivants et fossiles. Paris, 1887. 1369 p.
Forbes E. Report on the investigations by British Marine zoology by means of the dredge. Pt I.
The Infralittoral. London, 1850. P. 192—263.
Forbes E., Hanley S. A history of British Mollusca and their shells. London, 1853. Vol. 3. 616 P,
Franc A. Notes ecologiques et anatomiques sur Philbertia purpurea (Montagu) // Bull. Mus..
Nat. Hist. Nat. Paris. 1952. Ser. 2, N 24. P. 302—305.
Fretter V. The pedal sucker and anal gland of some British stenoglossa // Proc. Malac. Soc.
London, 1946. Vol. 27. P. 126—130.
Fretter V., Graham A. British prosobranch molluscs, their functional anatomy and ecology //
Roy. Soc. London. 1962. 755 p.
Friele H. Oversigt over de i Bergens Omegn forekommende skaldaekte Mollusker // Kristiania
vidensk.-selsk. Forhandl. 1873. 24 S.
Friele H. Oversigt over de i Bergens Omeng forekommende skaldaekte Mollusker// Forhandl,
i Vidensk. Kristiania. 1874. P. 289—312.
Friele H. Preliminary report on the Mollusca // Nytt Mag. Nat. Christiania. 1877. Bd 23. S. 1 —
10.
Friele H. Catalog der auf der norvegischen Nordmeer-expedition bei Spitzbergen gefundenen '
Mollusken // Jb. Dtsch. Malakozool. Ges. 1879. Bd 1, Ges. 6. S. 264—286.
Friele H. Mullusca II // The Norvegian North-Atlantic Expedition 1876—1878. 1886. Vol. 16,
P. 1—44.
Friele H., Grieg J. A. Mollusca III. Christiania, Grondahl and sons bogtr. 1901: Den Norske-
Nordhavs-Exp. 1876—78 //Zoologi. 1901. Bd 7. 129 S.
Gistel J. Naturgeschichte des Thierreichs fiir hohere Schulen. Stuttgart, 1848. Bd 16, H. 216.
4 S., 32 Taf.
Gould A. Report on the invertebrate of Massachusetts. Cambridge,' 1841. 373 p.
Gould A. Proceedings of the Boston Society of Natural History. (1849) 1850. Vol. 3. P. 136—145,
Gould A. United States exploring expedition. During the years 1838—1842 under the command
of C. Wilkes. Philadelphia, 1852. Vol. 12: Mollusca and Shells. P. 507.
Gould A. Report on the invertebrate of Massachusetts. 2ed ed. comprising the mollusca. Boston,
1870. 524 p.
Grant U. S., Gale H. R. Catalogue of the marine pliocene and pleistocene Mollusca of Cali-
fornia and adjacent regions//Mem. San-Diego Soc. Nat. Hist. 1931. Vol. 1. P. 1—1036.
Gray J. E. On the classification of British Mollusca, by W. E. Leach, M. D. // Ann. Mag. Nat.
Hist. 1847a. Vol. 20. P. 267—273 (October).
Gray J. E. A list of the genera of recent Mollusca, their synonyma and types// Proc. Zool.
Soc. London. 1847b. Vol. 15. P. 129—219.
Grieg J. Invertebres du fond// Due d’Orleans. Croisiere Oceanographique accomplie a Bord
de la Belgica dans la Mer du Gronland 1905. Bruxelles, 1909a. 63 S.
Grieg J. Brachiopods and molluscs // Report of the second Norwegian Arctic Expedition
in the «Fram» 1898—1902. 1909b. N 2. 41 p.
Grieg J. Supplement to the Brachiopoda, Scaphopoda and Lamellibranchiata // Reports
on the scientific results of the «Michael-Sars» North Atlantic deep-sea Expedition 1910. .
1931. Vol. 5, N 4. P. 3—5.
14*
212
ЛИТЕРАТУРА
Habe Т. The fauna of Akkeshi Bay. XXV. Gastropoda//PAMBS. 1958. N 8. P. 1—40.
Habe T., Ito K. Shells of the world in color. Vol. 1: The Northern Pacific. Hoikusha, 1965. 176 p.
Habe T., Kira T. Shells of the Western Pacific in color. Hoikusha, 1962. Vol. 1. 224 p.
Habe T., Kuroda T. Coloured Illustrations of the shells of Japan. Hoikusha, 1962. Vol. 2. 182 p.
Hagg R. Mollusca und Brachiopoda gesammelt von der Schwedischen Zoologischen Polar-
Expedition nach Spitzbergen, dem nordostlichen Gronland und Jan Mayen im Jahre
1900//Ark. Zool. Stockholm, 1905. Bd 2, N 13. 137 S.
Harmer F. W. The pliocene Mollusca of Great Britan, being supplementary to S. V. Wood’s
monograph of the Crag Mollusca // Paleontol. Soc. London. 1915. Vol. 68. P. 201—302.
Hermitte L. C. D. Venomous marine mollusks of the genus Conus // Trans, roy. Soc. trop. Med.
Hyg. 1946. Vol. 39. P. 485-512.
Hinegardner R. T. The anatomy and histology of the venom apparatus in several gastropods
of the genus Conus// M. S. Thesis Univ. South. Calif. 1955. P. 47—50.
HubendickB., Waren A. Smasnackor fran svenska Vastkusten. 6. Slaktena Emarginula, Lacuna,
Natica, Lora // Arstr. Goteborgs Naturhist. Mus. 1974. P. 25—32.
Hunt S. Fine structure of the secretory epitelium in the hypobranchial gland of the prosobranch
gastropod mollusc Buccinum undatum L. // J. Mar. Biol. Assoc. U. K. 1973. Vol. 53,
N 1. P. 59—71.
Hyman L. H. The Invertebrates: Mollusca. I. Me Graw-Hill Book Company. New York. 1967.
Vol. 6. 792 p.
Iredale T. Molluscan nomenclatural problems and their solution // Proc. Malac. Soc. London.
1918. Vol. 13. P. 28-40.
Jaccard P. The distribution of the flora in the alpine zone//New. Phitol. 1912. Vol. 11.
P. 37—50.
Jeffreys J. G. British conchology, or an account of the Mollusca which now inhabit the British
Isles and the surrounding seas. Vol. 4: Marine shells, in continuation of the Gastropoda
as far as the Bulla family. London, 1867. 487 p.
Jeffreys J. G. The mollusca of Europe compared with those of eastern North America // Ann.
Mag. Nat. Hist. 1872. Ser. 4, N 10. P. 237—247.
Jeffreys J. G. On the Mollusca procured during the Cruise of H. M. S. «Triton» between the Heb-
rides and Faroes in 1882 // Proc. Zool. Soc. London. 1883. P. 389—399.
Johnson C. W. A list of the molluscs collected by Mr. Owen Bryant along the coast of Labrador,
New Faundland and Nova Scotia // Nautilus. 1926. Vol. 40. P. 21—25.
Keen A. M. Marine molluscan genera of Western North America. Stanford, California, 1963.
126 p.
Knipowitsch N. M. Zoologische Ergebnisse der Russischen Expeditionen nach Spitzbergen.
Mollusca und Brachiopoda. I. Uber die in den Jahren 1899—1900 in Gebiete von Spitz-
bergen gesammelten recenten Mollusken und' Brachiopoden // Ann. Mus. Zool. St. Pe-
tersb. 1901. Bd 6. 105 S.
Knipowitsch N. M. Zoologische Ergebnisse der Russischen Expeditionen nach Spitzbergen.
Mollusca und Brachiopoda//Ann. Mus. Zool. St. Petersb. 1903. Bd 8. S. 133—143.
Kobelt W. Beitrage zur Arctischen Fauna//Jb. Dtsch. Malakozool. Ges. 1876. Bd 3. S. 61 —
76, 165-180.
Kobelt W. Iconographie der Schalentragen der europaischen Meeres-Conchylien, Fam. Pleuro-
tomidae. Weisbaden. 1905. S. 219—406.
Koehler R. Resultats scientifiques de la Campagne du «Caudan» dans le Golfe de Gascogne
Aout-Sept. 1895. Paris, 1896. 271 p.
Kohn A. J. Piscivorous gastropods of the genus Conus// Proc. Nat. Acad. Sci. Wash. 1956.
Vol. 42. P. 168—171.
Kohn A. J. The ecology of Conus in Hawaii // Ecol. Monogr. 1959. Vol. 29. P. 47—90.
Kohn A. J., Saunders P. R., Wiener S. Prelimenary studies on the venom of the marine snails
Conus// Ann. N. Y. Acad. Sci. 1969. Vol., 90. P. 706—725.
Kohn A. J., Nybakken J. W., Van Mol J.-J. Radula tooth structure of the gastropod Conus
imperialis elucidated by scanning electron microscopy // Science. 1972. Vol. 176. P. 49—
51.
Krause A. Ein Beitrag zur Kenntniss der Mollusken-Fauna des Beringsmeeres// Arch. f. Nat.
1885. Bd 51. N 1. S. 256—302.
Kuroda T., Kinoshita T. A Catalogue of Marine Molluscan Shells of Hoccaido // Bull, of Hok-
kaido Reg. Fish. Res. Labor. Fish. Agency. 1951. N 2. 40 p.
Leche W. Ofversigt ofver de af Svenska Expeditionerna till Novaja Semlja och Jenissej 1875
och 1876 Insamlade Hafs-Mollusker//Kongl. Svenska Vetensk.-Acad. Handl. 1878.
Vol. 16, N 2. S. 1—86.
Locard A. Les Coquilles marines des Cotes de France. Baillere. Paris, 1892. 384 p.
Locard A. Mollusques testaces et Brachiopodes // Resultats scientifiques de la compagne
du «Caudan»// Ann. Univ. Lyon. 1896. P. 129—242.
Locard A. Mollusques testaceas. I. Expeditions Scientifiques du Travailleur et du Talisman.
Masson. Paris, 1897. 516 p.
Loven S. Index Molluscorum litora Scandinaviae occidentalia habitantium // Ofvers. Kongl.
Svenska Vetensk. Acad. Handl. Stockholm. 1846. Vol. 3, N 5. P. 135—161.
Loyning P. Loricata and Gastropoda from the Siberian Arctic Ocean. Norwegian North Polar
Expedition with the «Mand» // Sci. Res. 1932. Vol. 5, N 14. P. 3—19.
ЛИТЕРАТУРА
213
MacGinitie N. Marine Mollusca of Point Barrow, Alaska Ц Proc. U. S. Nat. Mus, 1959. Vol. 109,
N 3412. P. 59—208.
Macpherson E. The marine Molluscs of Arctic Canadian. Prosobranch gastropods, chitons and
scaphopods / Nat. Mus. of Canada. Ottawa, 1971. 149 p.
Marcus E., Marcus E. On Hastula cinerea//Bol. Fac. Filos. Cienc. Univ. Sao Paulo. Zool.
1960. N 23. P. 26—65.
Marsh H. The foregut glands of vermivorous Cone shells//Austral. J. Zool. 1971. Vol. 19,
N 4. P. 313—326.
Martini A., Chemnitz J. H. Systematischen Conchylien-Cabinet von Martini und Chemnitz.
Die Familie Pleurotomidae. Conch. Cab. Nurnberg, 1887. Vol. 4, N 2. 248 p.
Martoja M. L’appareil venomeux de Conus mediterraneus//Ann. Sci. Natur. Zool. 1960.
Ser. 12, vol. 2. P. 43—57.
McLean S. H. J. A revised classification of the family Turridae with the proposal of new subfa-
milies, genera and subgenera from the Eastern Pacific// Veliger. 1971. Vol. 14, N 1.
P. 114—130.
Middendorff A. T. Beitrage zu einer malacologia Rossica. St. Petersbourg, 1849. Abt. 2, 3.
S. 1-187.
Middendorff A. T. Mollusken. Reise in den ausserstern Norden und Osten Sibiriens wahrend
der Jahre 1843 und 1844. St. Petersbourg, 1851. Bd 2. S. 164—464.
Mighels J. W., Adams С. B. The descriptions of new species of molluscs// Proc. Bost. Soc.
Nat. Hist. 1840. Vol. 1. P. 1—50.
Mighels J. W., Adams С. B. Description of twenty-four species of the shells of New England //
Bost. J. Nat. Hist. 1842. Vol. 4, N 1. P. 37—54.
Moller H. P. C. Index Molluskorum Groenlandiae // Naturhist. Tidsskr. 1842. Bd 4. P. 76—97.
Montagu G. Testacea, or natural history of British shells. London, 1803. 291 p.
Morch O. A. L. Catalogus conchyliorum quae reliquit D. Alphonso D’Aguirra et Gadea comes
de Yoldii. Hafniae, 1852. 170 p.
Morch O. A. L. Fauna Molluscorum Islandiae // Vid. Medd. Natur. For., Kjobenhavn. 1868.
P. 185-227.
Morch O. A. L. Catalogue des mollusques du Spitzberg, recueillis par le Dr. H. Kroyer pendant
le voyage de la corvette la Recherche en juin 1838 // Mem. Soc. malacol. Belguique.
1869. Bd 4. P. 7—32.
Morch O. A. L. The Greenland Fauna. IV. Edited by Dr. Robert Brown. London, 1877. Appen-
dix. S. 435—442.
Morris P. A. A field guide to the shells of our Atlantic and Gulf Coasts. Cambridge (Mass.),
1960. 236 n.
Morris P. A. A field guide to shells of the Pacific coast and Hawaii. Boston, 1966. 297 p.
Morrison J. P. E. On the families of Turridae // Ann. Rep. Amer. Malacol. Union. 1966. Vol. 32.
P. 1-2.
Nordsieck F. Die europaische Meeres-Gehauseschnecken (Prosobrancia). Stuttgart, 1968. 273 p.
Norman A. M. Notes on the natural history of East Finmark. Mollusca // Ann. Mag. Nat. Hist.
1902. Ser. 7, vol. 10. P. 43—53.
Odhner N. Hj. Marine Mollusca of Ireland in the collections of the Swedish State Museum //
Arch. f. Zool. Stockholm, 1910. Bd 7, N 4. 31 S.
Odhner N. Hj. Die Molluskenfauna des Eisfjordes. Stockholm, 1915. Bd 54, N 1. 274 p.
Okutani T. Report on the archibenthal and abyssal gastropod mollusca mainly collected from Sa-
gami Bay and adjacent waters by the R. V. Soyo-Maru during the years 1955—1963 //
Jour. Facul. Sci. Univ. Tokyo. Sec. II. 1964. Vol. 15, N 3. P. 371—447.
Okutani T. Archibenthal and Abyssal'mollusca collected by R. V. «Soyo-Maru» from Japansee
waters during 1964 //Bull. Tokai Reg. Fish. Res. Lab. Japan. 1966. N 46. P.
1—32.
Okutani T. Bathyal and Abyssal Mollusca trowled from Sagami Bay and the South off Boso Pe-
ninsula by R. V. Soyo-Maru 1965—1967 // Bull. Tokai Reg. Fish. Res. Lab. 1968. Vol. 56.
P. 7-54.
Okutani T. Archibenthal and abyssal Mollusca collected by the R. V. Soyo-Maru from Japansee
waters during 1964// Bull. Tokai Reg. Fish. Res. Lab. 1969. Vol. 57. P. 4—12.
Oldroyd I. S. The marine shells of the west coast of North America// Stanford Univ. Publ.
Ser. Geol. Sci. 1927. Vol. 2, N 1. P. 1-300.
Oskarsson I. Skeldirafauna Islands. 2. Seasniglar med skei (Gastropoda, Prosobranchia, Tecti-
branchia). Reykjavik. Ungef. Prentsmidjan Leiftur. 1962. 167 S.
Otuka Y. Fossil mollusca and rocks of the Kiyosumi Group exposed at Minato Machi, Chiba
Prefecture and its environs//Jap. J. Geol. Geogr. 1949. Vol. 21, N 14. P. 295—309.
Packard A. S. List of animals dredged near Caribon Isl., Southern Labrador//Canad. Natu-
ralist. 1863. Vol. 8, N 6. P. 401—429.
Packard A. S. Observations on the glacial phenomena of Labrador and Maine, with a view
of the Recent invertebrate fauna of Labrador // Mem. Bost. Soc. Nat. Hist., n. s. (1866).
1867. Vol. 1, N 2. P. 210-303.
Pearce J. B. On Lora trevelliana (Turton) (Gastropoda, Turridae) // Ophelia. 1966. Vol. 3.
P. 81—91.
Pfeffer G. Die Fauna der Insel Jeretik, Port Wladimir, an der Murmanskiiste. 1. Teil// Jb.
214
ЛИТЕРАТУРА
Posselt И. J. Ostgronlandske Mollusker // Medd. om Gronland. Kjobenhavn. 1895. Bd 19.
S. 61—94.
Posselt H. J., Jensen A. S. Gronlands Brachiopoder og Bloddyr in Conspectus Faunae Groen-
landicae // Medd. om Gronland. Kjobenhavn. 1898. Bd 23. S. 1—298.
Powell A. W. B. The New Zealand recent and fossil Mollusca of the family Turridae with general
notes on turrid nomenclature and sistematics // Bull. Auckl. Inst. Mus. 1942. N 2.
P. 1—188.
Powell A. W. B. Antarctic and subantarctic mollusca: Pelecypoda and Gastropoda // Discovery
Rep. 1951. Vol. 26. P. 46—197.
Powell A. W. B. Antarctic and subantarctic Mollusca// Rec. Auckl. Inst. Mus. 1960. Vol. 5,
N 3, 4. P. 158—160.
Powell A. W. B. The molluscan families Speightiidae and Turridae // Bull. Auckl. Inst. Mus.
1966. N 5. 184 p.
Preston F. W. The cannonical distribution of commoness and rarity // Ecology. 1962. Vol. 43,
N 3. P. 185—215.
Raup D. M. Geometric analysis of shell coiling // J. Paleontol. 1967. Vol. 41, N 1. P. 43—65.
Reeve L. A. Conchologia Iconica. 3. Mangelia. London. 1845. 252 p.
Reeve L. A. Conchologia Iconica 1. Pleurotoma. London. 1846. 369 p.
Reid E. T., Lawton J. R. Scanning electron microscopy as an aid to limped taxonomy, biology
and ecology//20. Annu. Conf. Electron Microsc. Soc. South. Afr., Durban. 2—4 Dec.
1981. Proc. Vol. 11. Johannesburg, 1981. P. 119—120.
Richards H. G. Recent and pleistocene marine shells of James Bay//Amer. Mid. Nat. 1936.
Vol. 16, N 2. P. 528—545.
Robinson E. Observations on the toxoglossan gastropod Mangelia brachystoma (Phillipi) //
Proc. Zool. Soc. London. 1960. Vol. 135. P. 319—338.
Sars G. O. Mollusca regions arctical Norvegiae. Bidrag til Kundskaben om Norges Arktiske-
Fauna // Christiania. 1878. Bd 1. S. 1—466.
Sars M. Fortsatte bemerkinger over det dyriske livs udbredning i havets dybder// Forhandl.
Vidensk. Kristiania. (1868) 1869. P. 246—275.
Schlesch H. Zur Kenntnis der pliocanen Cragformation von Hallbjahrnarstadur, Tjornes, Nord-
island und ihrer Mollusken-Fauna // Abh. Arch. Molluskenk. 1924. Bd 1, H. 3. S. 309—
369.
Schmidt M. F. Wissenschaftliche Resultate der zur Aufsuchung eines angekundigten Mammuth
cadavers von der Kaiserlichen Akademie der Wissenschaften an dem unteren Jenissei
ausgesandten Expedition//Mem. Acad. Imp. Sci. St. Petersbourg, 1872. Vol. 7, N 1.
P. 1—168.
Shaw H. Anatomy of Conus tulipa and textile//Quart. J. micr. Sci. 1914. Vol. 60..
P. 40—49.
Sheridan R., Van Mol J.-J., Bouillon J. Etude morphologique du tube digestif de quelques
Turridae (Moll.—Gastr.—Prosobr.—Toxogl.) de la region de Roscoff // Cahiers Biol,
mar. 1973. Vol. 14, N 2. P. 159—188.
Shimek R. L. The Morphology of the Buccal Apparatus of Oenopota laevidensis (Gastropoda,.
; Turridae) // Ztschr. Morph. Tiere. 1974. Bd 80. S. 59—96.
Shimek R. L., Kohn A. J. Functional morphology and evolution of the toxoglossan radula//
Malacologia. 1981. Vol. 20, N 2. P. 423—438.
Smith E. H. The proboscis and oesophagus of some British Turridae // Trans. Roy. Soc. Edinb.
1967a. Vol. 67. P. 1—22.
Smith E. H. The neogastropod stomach, with notes on the digestive diverticula and intestine //'
Trans. Roy. Soc. Edinb. 1967b. Vol. 67. P. 23-42.
Smith E. H. The reproductive system of the British turridae (Gastropoda, Toxoglossa) //
The Veliger. 1967c. Vol. 10, N 2. P. 176—182.
Stimpson W. Shells of New England, a revision of the synonymy of the testaceous mollusks,
of New England. Boston, 1851. 58 p.
Strom H. Beskrivelse over norske insecter// Kong. Dan. Vidensk. Selsk. Skrift. 1788. Vol. 3..
P. 296—297.
Sykes E. R. On the Mollusca procured during the «Porcupine» expeditions 1869—1870. Supple-
mental notes. Pt 3 // Proc. Malac. Soc. London. 1906. Vol. 7. P. 173—190.
Thiele J. Handbuch der Zoologie: Solenogastres—Mollusca—Echinodermata. Berlin; Leipzig,
1925. Bd 5. S. 1—260.
Thiele J. Arktische Loricaten, Gastropoden, Scaphopoden und Bivalven//Fauna Arctica..
Jena, 1928. Bd 5, N 2. S. 563-632.
Thiele J. Handbuch der systematischen Weichtierkunde. Jena, 1929. S. 1—376.
Thiele J. Handhuch der systematischen Weichtierkunde. Stuttgart, 1931. S. 377—778.
Thorson G. Marine mollusks It The Scoresby Sund committees 2nd East Greenland Expedition,
in 1932 etc. // Medd. om Gronland. 1934. Bd 104, N 17. 8 p.
Thorson G. Studies on the egg-capsules and development of Arctic marine prosobranchs // Medd.
om Gronland. 1935. Bd 100, N 5. S. 1—71.
Thorson G. Marine Gastropoda Prosobranchiata// The Zoology of Iceland. Copenhagen, 1941.
Vol. 4, N 60. 150 p.
Thorson G. The zoology of east Greenland. Marine Gastropoda Prosobranchiata//Medd...
om Gronland. 1944. Bd 121, N 13. 180 p.
ЛИТЕРАТУРА
315
Thorson G. Reproduction and larval development of Danish marine bottom invertebrates //
Medd. Komm. Havunders. 1946. Ser. Plankton, Vol. 4, N 1. 523 p.
Thorson G. Scaphopoda, Placophora, Solenogastres, Gastropoda, Prosobranchiata, Lamelli-
branchiata//Medd. om Gronland. 1951. Bd 81, N 2. 177 p.
Troschel F. H. Das Gebiss der Schnecken. Berlin, 1866. Bd 2. 48 S.
Tryon G. W. Manual of conchology: Conidae, Pleurotomidae. London, 1884. Vol. 6. 413 p.
Turton W. Description of some new and rare British species of shells // Lond. Mag. Nat. Hist.
1834. Vol. 7. P. 350—353.
Verkriizen T. A. Bericht uber einen Schale-Ausflug im Sommer 1874 // Jb. Dtsch. Malakozool.
Ges. 1875. Bd 2. P. 229-240.
Verrill A. E. Notice of recent additions to the marine invertebrata of the northeastern coast
of America, with descriptions of new genera and species and critical remarcs on others //
Proc. U. S. Nat. Mus. 1880. Vol. 3. P. 356—405.
Verrill A. E. Catalogue of marine mollusca. Added to the fauna of New England during the past
ten years // Trans. Con. Acad. 1882. Vol. 5. P. 447—587.
Verrill A. E. Second catalogue of mollusca recently added to the fauna of the New England coast
and adjacent parts of the Atlantic// Trans. Con. Acad. 1884. Vol. 6. P. 139—294.
Verrill A. E. Third catalogue of mollusca recently added to the fauna of New England coast
and the adjacent parts of the Atlantic// Trans. Con. Acad. 1885. Vol. 6, pt 2. P. 395—
452.
Vestergaard K. Uber den Laich und die Larven von Scalaria communis (Lam.), Nassaris pyg-
maeus (Lam.) und Bela turricola (Mont.) // Zool. Anz. Leipzig, 1935. Bd 109. S. 7—8.
Walker D. Notes on the zoology of the late arctic expedition under Captain Sir F. L. McClin-
tock // R. N. J. Roy. Dublin Soc. 1862. Vol. 3. P. 61—77.
Waren A. Marine Mollusca described by John Gwyn Jeffreys, with the location of the type ma-
terial // Spec. publ. Conch. Soc. of Great Britain and Ireland. 1980. 60 p.
Waren.A. Marine mollusca described by W. Turton and W. Clark// J. Conch. 1983. Vol. 31,
. . N 3. P. 161—171.
Weinkauff H. C. Uber eine kritische Gruppe des genus Pleurotoma Lam.] sehsu stricto // Jb.
Dtsch. Malakozool. Ges. 1875. Bd 2. S. 285—292.
Weinkauff H. C. Beitrage zur Klassification der Pleurotomen // Jb. Dtsch. Malakozool. Ges.
1876. Bd 3. 203 S.
Whiteaves J. F. List of marine invertebrates from Hudson’s Straite // Bell. Roy. Rep. on Hud-
son’s Bay. Geol. Survey Canad. 1885. Vol. 4. P. 58DD—60DD.
Whiteaves J. F. Catalogue of the marine invertebrata of Eastern Canada // Geol. Survey Ca-
nada. 1901. N 722. 271 p.
SUMMARY
The Oenopotinae subfamily mollusks form one of the most numerous groups of benthos
animals in the northern regions of the Pacific and Atlantic Oceans as well as of the Polar Ocean.
78 Oenopotinae species belonging to 5 genera, namely Curtitoma, Oenopota (nominate subg.
and subg. Nodotoma,), Obesotoma, Granotoma, Propebela (nominate subg. and subg. Ca-
netoma) have been discovered in the seas of the USSR. 20 hitherto unknown species and 47
new for the fauna of the USSR species have been described. The subfamily whs studied on the ba-
sis of the collection at the Zoological Institute of the Academy of Sciences of the USSR. The total
of more than 12 000 specimens were studied and defined up to the species.
The detailed study of shells and bodies of mollusks made it possible to reveal the most
species-specific characters for the group. Of the shell elements, the highest stability is characte-
ristic of the form, size and protoconch sculpture (fig. 6). The morphological analysis of the major-
systems of organs has shown that interspecific differences lie mostly in the digestive system.
The study of radula morphology proved some functionally important parts of the tooth practi-
cally not liable to changes, while the keel, strengthening the distal portion of the tooth blade
(fig. 14) turned out to be the most essential for the species diagnostics, phylogeny and evolution
of the group as a whole. The morphological changes which reflect the evolutionary development
of the Oenopotinae, are shown to be connected with the oligomerization of the elements of the pro-
toconch spiral sculpture, with the stepwise obliteration of the anal sinus along the growth lines
of the definitive shell and with the keel reduction on the distal part of the radular tooth (fig. 42)
(Богданов, 1989 г). The changes in the radular tooth structure appeared to be correlated with
the morphogenesis of the protoconch. Evolutionary transformations in the morphology of the cap
are connected with lessening of its relative size, sharper curve of the growth line and narrowing
of the columellar muscle imprint (fig. 32—36). The characters supporting the evolutionary con-
cept appeared to be of major significance for the supraspecific taxonomy. Morphological pecu-
liarities of the radula and protoconch served in general for the distinguishing of genera and sub-
genera. A series of chaiacters, such as the presence of a claw-shaped cap, specific sculpture
on the protoconch, absence of the mantle pore in the male genital system confirm the attitude,
towards the species studied as representatives of a separate subfamily of Oenopotinae.-
OENOPOTINAE Bogdanov,’ 1987
The shell is small (14—16 mm on the average), oval-spindleshaped or bench-formed, always;
monochromatic, with slowly growing whorls, separated by a pressed seam. The last whorl
occupies from % to 2/s of the shell height. The protoconch is nipple-shaped (rarely barrel-shaped)-
and is ornamented, as a rule, by 2 to 20 spiral ribs. The spiral sculpture and (in most species)-
the definitive one are well developed in the definitive shell. The palatal edge of the opening
(the outer lip) is always very thin and fragile. The siphonal process is short and the siphonal canal
is moderately deep and relatively wide. The cap is claw- or drop-shaped with the nucleus located
terminally.
The radula consists of a pair of marginal teeth (the radular formula is 1 : 0 : 0 : 0 : !)
of typically toxoglossal structure with rounded or cylindrical basal part, which bears a hollow
blade strengthened by the keel in its distal portion. The basal membrane is absent.
It should be noted that the Oenopotinae mollusks have a lot in common with representatives-
of Mangeliinae subfamily, the form and sculpture of shell and general promorphology of radular
teeth supporting the idea. They might have probably had a common ancestor.
The study presents the outcomes of morphological and functional analysis of the structure
and organization of mollusks’ bodies. The study of musculature and epithelial lining of the ali-
mentary system made possible to open up the secrets of the feeding mechanism of the Oenopo-
tinae mollusks (Богданов, 1989b). It is shown that the tooth performs the role of a hollow syringe
needle during the attack on the prey as in Conidae mollusks. The tooth is passed from the radular-
sac during the maximum closeness between the distal part of proboscis and the buccal sac
(fig. 17). The initial stages of digestion take place in the anterior section of the alimentary sy-
stem. In mollusks with the food in the rhynchodaeum, the turning of the proboscis into the buc-
cal cavity has been noted. As a result, a sharp increase in the volume of the digestive cavity
takes place, the latter being limited by the rhynchodaeum walls, the outer proboscis wall and
by the initial portion of the oesophagus (fig. 19). The stomach has a simple structure and is sligh- -
tly separated from the oesophagus.
SUMMARY
217
The questions of biogeography, ecology and distribution of the Oenopotinae mollusks
are discussed in the paper. For each of 78 species keys with pictures of radular teeth, photoes
of the definitive shell and protoconch accompanied by the information on distribution and type
material are given.
Diagnoses of new species
Obesotoma starobogatovi sp. n. (fig. 58; 226—230; 404—405 part.; 432 part.).
The shell is most similar to that of O. colpoica, differs by more convex whorls and
by the presence of thin axial ribs. It differs from O. gigantea by the form of radular teeth
(fig. 404): i. e., by a more massive basal part and a wide basal opening of the blade. — Western
part of the East Siberian, Laptev and Kara Seas.
Holotype: Novosibirskiye Ostrova (74°29.5' N—140°29z E) depth 20.5 m; preserved
at the Zoological Institute, Leningrad, No 48472. — Dimensions of the holotype shell are
(in mm): height 6.6, width 4.0, body whorl height 4.8, aperture height 3.5, aperture width 1.7.
Propebela (Canetoma) variabilis sp. n. (fig. 295—300; 408; 475).
The shell is similar to that of P. harpularia, but can be easily distinguished by a sabre-
shaped radular teeth. — Bering strait, off the shores of Komandor and Kuril Is.
Holotype: Bering I., Komandor Is. depth 100 m; preserved at the Zoological Insti-
tute, Leningrad, No 48480. — Dimensions of the holotype shell are (in mm): heigt 11.3, width
5.1, body whorl height 7.9, aperture height 5.2, aperture width 2.3.
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ МОЛЛЮСКОВ1
abyssicola, Bela scalaris var. 64, 188
acuta, Bela concinnula var. 15, 199
alaskensis, Propebela, Bela 15, 64, 72
albrechti, Granotoma, Bela, Lora, Oenopota
15, 31*, 36, 57, 58, 59*, 61*, 64-66,
72, 90*, 134, 135
alitakensis, Propebela (Canetoma), Lora 58,
64, 72, 108*, 175, 200, 204
althorpensis, Oenopota 72
americana, Bela 188, 189
amiata, Lora 15, 200
angulosa, Bela cancellata 178
angulosa, Propebela, Bela, Lora, Pleurotoma
13, 51, 53*, 56, 58, 59, 64, 73, 115*,
178, 191
arctica, Bela, Oenopota 13, 195, 196
areta, Propebela (Canetoma), Funitoma, Oe-
nopota (Funitoma) 63, 65, 72, 175, 193,
203
assimilis, Bela exarata var., Bela rugulata
var., Bela turricula var. 181
assimilis, Propebela, Bela 13, 37, 52*, 56,
58, 64, 73, 109*, 110*, 176, 181, 184,
189, 191
babylonia, Lora 15
bartschi, Curtitoma, Oenopota 72, 77, 83,
87*, 131
beckii, Bela bicarinata var., Bela violacea
var. 15, 124
beckii, Defrancia 14, 124, 125
becklemishevi, Curtitoma, Oenopota 72, 83,
85*, 132
Bela 13—18, 53—55, 82, 133, 137, 159, 173
Belinae 18
Bellacythara 20
bergensis, Bela rugulata var. 185
bergensis, Propebela, Oenopota 21*, 22*,
43, 48, 52*, 56, 57, 72, 175, 183*, 185
bicarinata, Bela, Defrancia, Mangelia, Oe-
nopota, Pleurotoma 14, 15, 70*, 123, 125
bicarinata, Curtitoma violacea var., Bela
violacea vht. 47, 83, 133
biconica, Oenopota (Nodotoma) 50*, 72,
74, 100*, 138, 153, 157
blakei, Lora 15
borealis Reeve, Pleurotoma 13
borealis Schl., Lora 195
brachystoma, Mangelia 20
Brachytoma 18
Brachytomidae 18, 81
Brachytominae 18, 81
brevis, Bela violacea var. 15
brevispira, Bela nobilis var. 179
Buccinum 22
cancellata auct., Bela 145, 148, 150, 178
cancellata, Oenopota (Nodotoma), Bela 50*,
57, 72, 95*, 96*, 139, 153, 157, 158
cancellatus, Fusus 153
Candida, Oenopota 72
Canetoma 55
Canetoma (subg.) 60, 61, 62*, 193
casentina, Oenopota (Nodotoma), Lora 15,
50*, 72, 100*, 13'8, 154, 155
cassis, Propebela 72, 113*, 174, 191
Cestoma 55
chiachiana, Obesotoma 72
cinerea, Oenopota (Nodotoma), Bela, Def-
rancia, Pleurotoma (Bela) 14, 50*, 56,
57, 64—66, 73, 92*, 139, 144, 147
cingulata, Propebela (Canetoma), Oenopota
17, 24, 63, 72, 114*, 175, 202
clathratus, Bela rugulata var. 183
Clathromangelia 18
Clavatula 18
Clavinae 19
colpoica, Oenopota 172
concinnula, Propebela (Canetoma), Bela 15,
60, 72, 105*; 174, 199, 200, 205
Conidae 18, 29, 30
Conoidea 34
conoidea, Bela decussata var., Bela decussata
125
conoidea, Curtitoma, Bela, Pleurotoma 13,
24, 48, 56, 63, 64, 73, 84, 88*, 125, 126*
Conus 38
convexigyra, Oenopota 14
Curtitoma 55, 58, 60, 61, 62*, 63, 81, 82
cylindracea, Bela bicarinata var., Bela vio-
lacea var., Pleurotoma (Bela) violacea
var. 14, 15, 123
cylindracea, Defrancia 123, 125
declive, Tritonium 148
declivis, Bela cancellata, Bela cancellata var.
148
declivis, Oenopota (Nodotoma), Bela, Pleu-
rotoma 50*, 56, 64, 73, 94*, 95*, 139,
148, 150
1 Курсивом выделены синонимы, жирным шрифтом — названия родов и таксономи-
ческих единиц выше рода, жирными цифрами — страницы с описаниями, звездочкой —
страницы с иллюстрациями к описаниям.
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
219
decussata auct., Bela, Pleurotoma 84, 117 .
decussata, Curtitoma, Bela, Mangelia, Oe-
nopota, Pleurotoma 14, 22*, 24, 27*, 45,
49*, 56, 64—66, 73, 83, 88*, 89*, 126*,
127, 129
Defrancia 14, 55
delicata, Funitoma 16
detegata, Bela 185, 186
dictyopnora, Oenopota 14
diegensis, Obesotoma 72
ecarinata, Bela scalaris var. 188
elegans, Bela cancellata 150
elegans, Oenopota (Nodotoma), Bela, Def-
rancia, Pleurotoma 14, 22*, 37, 50*,
-56, 57, 64—66, 70, 73,‘94*, 139, 148,
149*, 150, 156
elongata, Oenopota (Nodotoma) 72, 74, 100*,
139 158
Euclathurella 20
exarata auct., Lora, Bela 193, 199
exarata, Bela turricula 196
exarata, Propebela (Canetoma), Bela, Def-
rancia, Lora, Plerotoma 14, 53*, 54, 56,
57, 60, 64, 66, 68, 69*, 70*, 71, 73, 106*,
174, 194, 196, 198
exigua, Bela, Pleurotoma 121, 122
exquisita, Propebela, Turritoma 72, 173
fidicula, Propebela (Canetoma), Bela, Fu-
sus, Lora, Oenopota 52*, 72, 105*, 176,
200
finmarchia, Bela decussata var. 13, 117
flora, Curtitoma, Lora 15, 72, 83, 87*, 118,
122
Funitoma 55, 193
furfuraculata, Bela 185, 186
galgana, Lora 15
geminolineata, Bela bicarinata, Bela bicari-
nata var. 124
gigantea, Bela bicarinata, Bela violacea var.,
Pleurotoma violacea var. 13, 165
gigantea, Obesotoma, Bela 45, 57, 58, 59*,
. 64, 73, 79, 103*, 159, 164*, 165, 172
gigas-, Bela schantarica 164
gigas, Obesotoma, Bela, Pleurotoma (Bela)
51*, 73, 99*, 160, 164
Gly phostoma 18
golikovi, Propebela (Canetoma), Oenopota
72, 108*, 176, 204
goryachevi, Propebela 72, 108*, 176, 192
gouldii, Propebela, Bela 15, 43, 183, 184
Granotoma 55, 60, 61, 62*, 63, 71, 81, 133
hanasakinensis, Obesotoma 16
harpa, Oenopota (Nodotoma), Bela, Lora
15, 21, 22, 31*, 36, 37*, 40*, 48, 50*,
56—58, 64, 66, 67, 73, 93*, 94*, 138,
146, 151
harpularia, Propebela, Bela, Lora, Oenopota,
Mangelia 14, 31, 32, 38, 56—58, 60,
64, 65, 67, 73, 111*, 173, 177*, 186, 207
harpularioides, Oenopota 186, 187
harpularius, Fusus 14, 186, 187
hebes, Bela decussata, Bela decussata var. 119
hebes, Curtitoma, Bela 15, 22*, 61, 64, 72,
83, 87*, 117*, 118, 119,122, 128, 131,133
hecuba, Curtitoma, Nodotoma 82, 84, 117
hokkaidoensis, Curtitoma, Nematoma, No-
dotoma 72, 82, 84, 104*, 130
impressa auct., Bela 84
impressa, Bela turricula 145
impressa, Oenopota (Nodotoma), Bela, Lora,
Pleurotoma (Ischnula) 50*, 54, 56—58,
59*, 63, 64, 66, 73, 92*, 93*, 138, 144,
145, 152
incisula, Bela decussata 84
incisula, Curtitoma, Bela, Oenopota 15, 16,
24, 49*, 56—58, 61, 63—66, 72, 82, 84,
85*, 86*, 117*
inequita, Oenopota (Nodotoma), Lora 16,
50*, 72, 100*, 139, 154
inflata, Bela decussata var. 15, 132
Inquisitor 18
Ischnula 55
japonica, Obesotoma, Oenopota 64, 65, 72,.
102*, 159, 160
jenisseensis, Bela 140
jenisseensis, Pleurotoma pyramidalis var. 16,
140
kinkasanensis, Oenopota 17
kobelti, Bela 195, 196
kobelti, Bela decussata 195
koreni, Bela 13
krausei, Granotoma, Bela, Lora, Oenopota
(Funitoma) 15, 57, 58, 72, 91*, 133, 134,
135*, 136
kurilensis, Oenopota (Nodotoma) 72, 96*,
139 157
Kurtziella 18, 20
kyskana, Obesotoma 72
Lachesis 18
laevigata, Oenopota, Bela 15, 70, 161, 163
laevior, Curtitoma violacea var., Bela violacea
var. 45, 51, 124, 125
lawrenciana, Curtitoma, Lora 64, 72, 83,
89*, 128, 129*
lechi, Bela woodiana var. 14
levidensis, Oenopota 28, 29*
Liracraea 20
livida, Defrancia 14
Lora 15, 55, 137
Lorinae 18, 81
Jutkeana, .Bela 15
lutkeni, Granotoma 72
Mangelia 18, 55
Mangeliidae 18
Mangeliinae 19, 20, 55, 81
margaritae, Propebela (Canetoma), Oenopota
72, 105*, 174, 204
marinae, Propebela (Canetoma) 53*,.60, 72,
104*, 174, 205
maurellei, Bela 15
metschigmensis, Bela 15
microvoluta, Curtitoma, Nematoma 16, 64,
72, 83, 130
minuscularia, Bela 124, 125
miona, Obesotoma 72
280
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
mitrata, Propebela (Canetoma), Lora 53*,
60, 72, 105*, 174, 192, 198, 199
mitrula, Bela 196—198
mitrula, Bela turricula 196
moerchi, Lora, Pleurotoma violacea var. 161,
163
monterealis, Propebela 72
murdochiana, Bela 15
nazanensis, Lora, Oenopota 128—130
nebula, Murex 18, 55
Nematoma 55, 82
Neoguraleus 20
neymanae, Curtitoma 72, 83, 89*, 133
nobilis, Pleurotoma turricula var. 179
nobilis, Propebela, Bela, Defrancia, Lora,
Pleurotomina, Pleurotoma 14, 21, 29, 31*,
37, 43—47, 52*, 54, 57, 58, 59*, 64, 66—
68, 69*, 73, 109*, 176, 179, 185
Nodotoma 16, 55, 56
Nodotoma (subg.) 58, 62*, 144
nodulosa, Bela violacea var. 124, 125
novajasemljensis, Curtitoma, Bela, Lora,
Oenopota, Pleurotoma 16, 24, 31*, 45,
46*, 49*, 56, 57, 61, 63, 65, 66, 73, 82,
85*, 119, 120*, 122
Obesotoma 16, 50, 55, 60, 61, 62*, 63, 82, 159
obliqua, Oenopota, Bela, Pleurotoma 13,
58, 64, 66, 67, 73, 98*, 138, 143, 144*
Oenopota 16, 18, 55, 58, 60, 61, 62*, 63, 8.1,
09 147 IRQ
Oenopota’(subg.) 62*, 82, 139, 173
okudai, Oenopota 16
Onopota 55
ovalis, Bela decussata var. 121
ovalis, Curtitoma, Bela, Oenopota, Pleuro-
toma (Bela) 13, 31, 49*, 57, 64, 66, 69,
70*, 73, 83, 87*, 120*, 121
pavlova, Oenopota (Nodotoma), Lora 72,
96*, 139, 155
• Phymorhynchus 18
piltuniensis, Curtitoma, Oenopota 72, 83, 89*,
132
pingelii, Oenopota, Bela, Defrancia, Lora,
Pleurotoma (Bela) 14, 22*, 24, 48, 49*,
56, 58, 59*, 64, 66, 73, 100*, 138, 141,
142
Pleurotoma 14
pleurotomaria, Bela 16, 71, 140
pleurotomarius, Fusus 14, 137, 140
Pleurotomina 55, 173
Pleurotomoides 55
plicifera auct., Mangelia 151
plicifera, Bela, Lora, Pleurotoma 151
popovia, Propebela, Lora 72, 113*, 174, 187
Propebela 18 , 50, 55 , 59—61, 62*, 63, 82, 173
Propebela (subg.) 62*, 176
pusilia, Bela decussata var. 15
pygmaea, Bela 15
pyramidale, Buccinum 13, 137, 140
pyramidalis, Oenopota, Bela, Lora, Mangelia,
Pleurotoma 16, 22*, 23, 48, 49*, 56,
58, 61*, 64, 65, 67, 68, 69*, 71, 73, 97*,
132, 138, 140
pytisa, Oenopota 72
quadra, Obesotoma, Lora 72, 132
raduga, Granotoma, Oenopota 57, 72, 89*,
134, 137
Raphitomidae 18
Raphitominae 18, 81
raricostulata, Oenopota 169, 170
rassina, Propebela (Canetoma), Lora 15, 72,
176, 201, 202
reticulata, Bela, Lora, Oenopota, Pleurotoma.
13, 117—119
robusta, Obesotoma, Bela 14, 51*, 58, 64—
66, 72, 101*, 160, 161, 168, 169, 171
rosea, Bela, Lora 186
rosea, Bela harpularia var., Pleurotoma tur-
ricula var. 186
roseum, Tritonium 186, 187
rufa, Mangelia (Bela), Pleurotoma 140
rufescens, Bela bicarinata var. 124, 125
rufus auct., Fusus 140
rugulata, Propebela, Bela, Lora, Oenopota
22*, 29, 43, 44*, 47, 52*, 54, 56, 58r
59*, 60, 64, 65, 67, 73, 112*, 175, 180,.
183, 184, 185, 193
sachalinensis, Obesotoma 51*, 72, 99*, 159,
163, 172
sarsii, Bela 16, 145, 147
scalaris auct., Oenopota 181
scalaris, Bela turricula, Pleurotoma turricula
var. 188
scalaris, Propebela, Bela, Defrancia, Lora,.
Oenopota 14, 36, 37*, 52*, 56—58, 59*r
60, 64—66, 73, 115*, 116*, 176, 182,.
188
scalaroides, Bela rugulata, Bela rugulata var.,.
Bela turricula 189
scalaroides, Propebela, Bela 13, 29, 30, 54,.
56—5,8, 59*, 60, 61*, 64—66, 73, 116*,.
175, 189
schantarica auct., Bela, Oenopota 161, 164
schantarica, Obesotoma, Bela, Oenopota,
Pleurotoma 17, 43, 44, 45*, 47, 54, 64—
66, 72, 102*, 159, 162*, 163, 173
schmidti, Bela 13, 151, 152
sculpturata, Bela 15
semiplicata, Bela pyramidalis var. 14, 49*,.
64, 65, 67, 140*, 141
simplex auct., Bela 167
simplex, Bela violacea var. 161
simplex, Obesotoma, Bela, Lora, Oenopota,
Pleurotoma 17, 24, 25, 26*, 37, 43, 44,
45*, 47, 54, 57, 59*, 63, 64, 67, 68, 69*,
73, 102*, 159, 161, 167
sixta, Obesotoma 72
solida, Obesotoma, Bela, Lora 15, 51*, 64,
72, 102*, 160, 170, 171
spitzbergensis, Bela rugulata var., Lora ru-
gulata 184
spitzbergensis, Propebela 31*, 43, 45, 47,
51, 52*, 54, 58, 59*, 60, 64—66, 73,
110*, 175, 181*, 184
starobogatovi, Obesotoma 51*, 58, 63—65,.
73, 74*, 103*, 104*, 159, 171
Stenoglossa 30, 39
strdmbiformis, Pleurotoma 18
stromboides, Pleurotoma 18
subarctica, Lora 17, 183, 184
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
821
subturgida, Lora 15
subvitrea, Lora 15
surana, Oenopota 72
svetlanae, Propebela 22*, 58, 72, 77, 113*,
175, 190
tanner!, Lora 15
Taranis 18
tenuicostata, Bela decussata var. 127
tenuicostata, Bela, Oenopota, Pleurotoma 127,
128
tenuilirata auct., Bela, Obesotoma 128
tenuilirata, Bela laevigata var. 166
tenuilirata, Obesotoma, Bela, Lora 22*, 57,
72, 104*, 160, 166
tenuissima, Propebela 72
terpeniensis, Propebela 72, 74, 114*, 174,
188, 192
tersa, Lora exarata, Oenopota exarata 193
tersa, Propebela (Canetoma), 'Canetoma, No-
dotoma 57, 59*, 64, 72, 106*, 174, 193,
197
tomiyaensis, Obesotoma, Lora 72, 102*, 160,
170
Toxoglossa 19, 25, 30, 48
. trevelliana, Curtitoma, Bela, Lora, Oenopota
22*, 29 , 33, 41, 42, 45 , 46*, 49*, 51—
54, 56—58, 59*, 61*, 63—65, 67, 68,
- 69*, 70*, 73*, 83, 85*, 117, 123
trevellianum, Pleurotoma 13, 82, 117
tumida, Bela woodiana var. 15, 168
tumida, Obesotoma 51*, 73, 77, 101*, 160,
168
turricula auct., Bela, Oenopota 179, 181
turricula, Propebela, Bela, Lora, Mangelia
(Bela), Murex, Oenopota, Pleurotoma 13,
22*, 23, 26*, 28, 29, 31*, 32, 33, 38,
42*, 43, 52*, 56, 58, 59*, 60, 64, 65,
. 68, 70, 72, 111*, 173, 176, 187
turriculus, Fusus 176
Turridae 18, 20, 30, 41, 56, 63, 81
Turrltoma 173
Turritomella 55, 173
uchidai, Obesotoma 16
uschakovi, Oenopota (Nodotoma) 72, 96*,.
139, 156, 158
Vahlii, Bela pyramidalis var. 140
vahlil, Defrancia 14, 140
valdeplicosa, Bela pyramidalis var. 15
valentina, Oenopota 17
variabilis, Propebela (Canetoma) 53*, 57,.
72, 107*, 174, 196, 206
venusta, Propebela 72
Venustoma 55
verrilli, Propebela 72, 74, 114*, 175, 191
violacea, Bela bicarinata var. 123
violacea, Curtitoma, Bela, Lora, Mangelia,.
Oenopota, Pleurotoma 14, 15, 24, 25, 43,
45, 47, 49*, 56, 58, 64, 66—68, 69*,
71, 73, 84, 86*, 87*, 118, 123, 124*
viridula auct., Bela 117
viridula, Bela decussata 195
viridula, Propebela (Canetoma), Bela, Def-
rancia, Oenopota 14, 51, 53*, 54, 55, 64,
66, 67, 73, 98*, 174, 195, 205
Widalli 82
willei, Bela, Pleurotoma 13, 127, 128
willei, Bela tenuicostata var. 127
woodiana auct., Bela 165
woodiana, Bela turricula 167
woodiana, Obesotoma, Bela, Defrancia,
Lora, Pleurotoma 14, 51*, 64—66, 73z
99*, 101*, 160, 167, 169, 170
СОДЕРЖАНИЕ
Предисловие ................................................................ 5
Систематический указатель видов ........................................... 7
Общая часть .............................................................. 9
Материал и методы исследования.......................................... 9
История изучения....................................................... 12
Положение Oenopotinae в системе Turridae............................... 18
Мбрфофункциональный анализ представителей Oenopotinae ................. 21
Раковина ........................................ : . ............. 21
Дефинитивная раковина (21). — Эмбриональная раковина (24).
Морфология мягкого тела ............................................ 25
Головной отдел (25). — Нога (25). — Мантийный комплекс органов (25).—
Пищеварительная система (27). — Механизм питания (33). — Крове-
носная и выделительная системы (39). — Нервная система (40). — Поло-
вая система (41).
Изменчивость морфологических признаков..................................43
Дефинитивная раковина (43). — Эмбриональная раковина (48). — Кры-
шечка (49). — Изменчивость морфологических признаков тела (51).
Система и филогения................................................... 55
Образ жизни моллюсков Oenopotinae .................................... 63
Распределение по глубинам (63). — Распределение по грунтам (64). —
Отношение к температуре (65). — Отношение к солености (66). — Пита-
ние (67). — Особенности размножения (67). — Моллюски Oenopotinae —
объект питания (71). — Другие формы биотических отношений (71).
Географическое распространение представителей Oenopotinae . . . . . е . « 71
Систематическая часть ..................................................... 80
Таблица для определения родов......................................... 81
1. Род Curtitoma.................................................... 82
2. Род Granotoma .................................................. 133
3. Род Oenopota ................................................... 137
4. Род Obesotoma .................................................. 159
5. Род Propebela .................................................. 173
Литература . . , , ,...................................................... 208
Summary . ................................................................ 216
Указатель латинских названий моллюсков.................................... 218
CONTENTS
Preface .........................................................................
Systematic list
General ..............................................................•..........
Material and research methods................................................
Background of the study......................................................
Taxonomical position of Oenopotinae in Turridae family.......................
Morphological and functional analysis of Oenopotinae specimens...............
Shell ....................................................................
Definitive shell (21). — Protoconch (24).
Soft body morphology......................................................
Head (25). — Foot (25). — Mantle set morphology (25). — Alimentary
system (27).- — Feeding mechanism (33). — Blood conveying and excre-
tory systems (39). — Nervous system (40). — Genital system (41).
Variability of morphological characters .....................................
Definitive shell (43). — Protoconch (48). — Operculum (49). — Body (51).
System and phylogeny ........................................................
Mode of life.................................................................
Distribution in depths (63).— Distribution on sea bed soils (64). —
Response to temperature (65). — Response to salinity (66). — Nutrition
(67). — Peculiarities of reproduction (67). — Oenopotinae as feeding objects
(71). — Other forms of biotic relations (71).
Geographical distribution of Oenopotinae specimens...........................
Systematics ....................................................................
Defining table of genera.....................................................
Curtitoma genus .................................... .....................
Granotoma genus ..........................................................
Oenopota genus
Obesotoma genus...........................................................
Propebela genus ..........................................................
References ..............................................................
Summary................................................................
Index of names........................................................... > . «
5-
7
9
9’
12
18
21
21
25-
43
55-
63
71
80
81
82
133
137
159
173
208
216-
218-
Научное издание
Игорь Петрович Богданов
МОЛЛЮСКИ ПОДСЕМ. OENOPOTINAE МОРЕЙ СССР
Фауна СССР
Утверждено к печати
Зоологическим институтом
Академии наук СССР
Редактор издательства С. Н. Моисеенко
Технический редактор О. Б. Мацылееич
Корректоры Г. А. Александрова и Г. Н. Мартьянова
ИБ № 44453
Сдано в набор 06.12.89. Подписано к печати 09.10.90.
Формат 70х108*/1«- Бумага типографская № 2.
Гарнитура обыкновенная. Печать высокая.
Усл. печ. л. 19.6 4- 4.9 вкл. Уел. кр.-от. 24.5.
Уч.-изд. л. 26.71. Тираж 900. Тип. зак. № 2164.
Цена 5 р. 30 к.
Ордена Трудового Красного Знамени издательство
«Наука». Ленинградское отделение.
199034, Ленинград, В-34, Менделеевская пин., 1.
Ордена Трудового Красного Знамени Первая
типография издательства «Наука».
199034, Ленинград, В-34, 9 пиния, 12.