/






























































































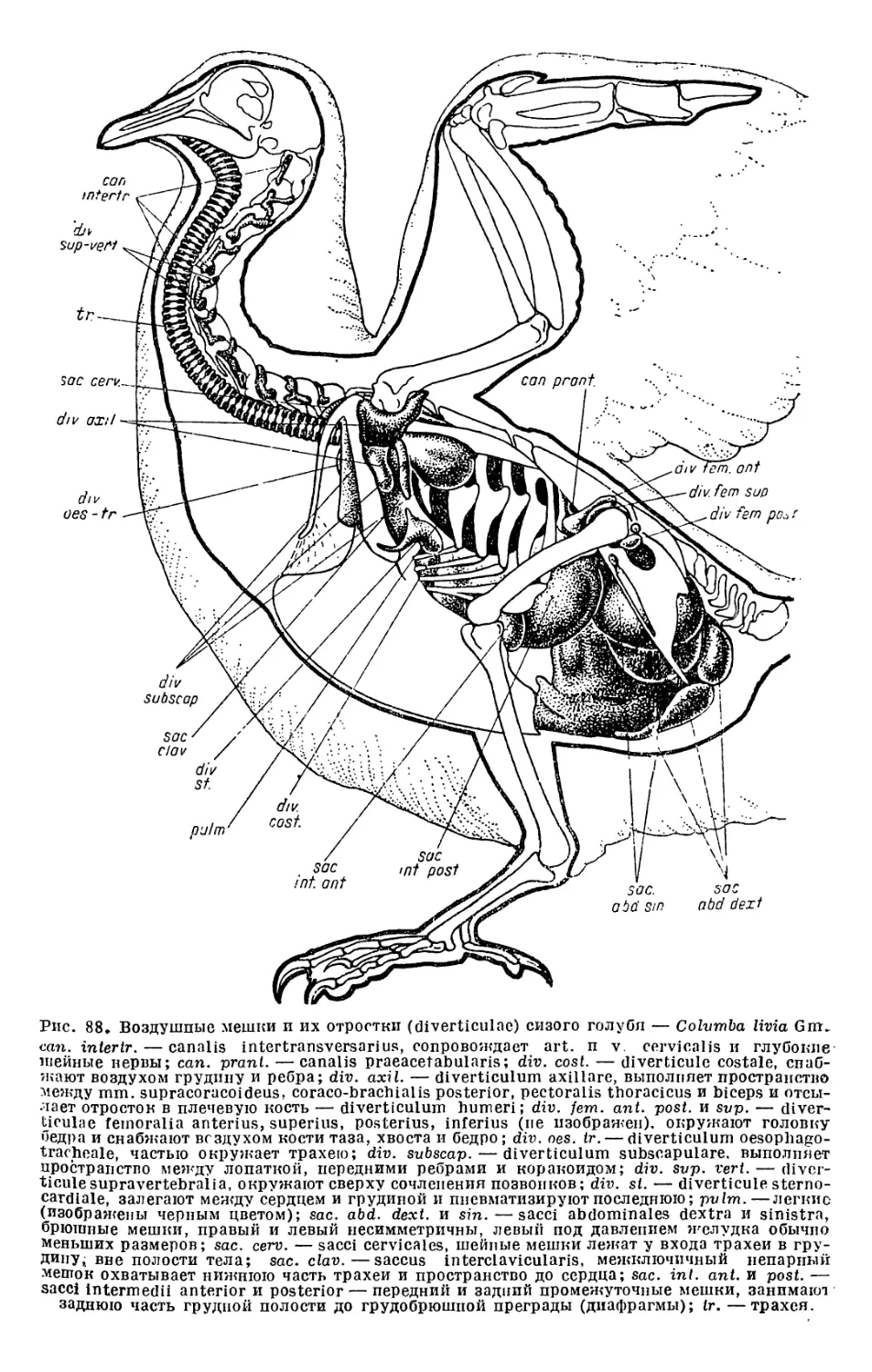



















































































































































































































































































































































































































































































Текст
A M. ШУЛЫ1ИН
ОРНИТОЛОГИЯ
ИЗДАНИЕ
ЛЕНИНГРАДСКОГО ГОСУДАРСТВЕННОГО УНИВЕРСИТЕТА
ЛЕНИНГРАД Ю4О
Л. М. ШУЛЬПИН
ОРНИТОЛОГИЯ
(СТРОЕНИЕ, ЖИЗНЬ И КЛАССИФИКАЦИЯ ПТИЦ)
ИЗДАНИЕ ЛЕНИНГРАДСКОГО
ГОСУДАРСТВЕННОГО УНИВЕРСИТЕТА
ЛЕНИНГРАД 1940
ОТ АВТОРА
Предлагаемое руководство имеет своей задачей дать основные
сведения о строении, жизни и классификации птиц, необходимые при
проработке курса орнитологии в наших университетах. Личный
интерес автора преимущественно к экологии, географическому рас-
пространению и классификации птиц выразился в более детальном
изложении этих глав, по сравнению с главами собственно морфоло-
гическими. Значительная разработанность этих вопросов для данного
класса, а также преимущественно экологическое направление спе-
циализации студентов, слушающих этот курс в Ленинградском Госу-
дарственном Университете, могут служить известным основанием
для принятого плана.
Составляя руководство, автор исходил из стремления дать книгу,
дополняющую наличную русскую литературу и в то же время даю-
щую известную систему основных знаний по данной дисциплине.
В целях более детального изложения отдельных, более важных глав
пришлось сильно сократить и даже выпустить некоторые разделы,
как, например, развитие органов, значение органов внутренней
секреции и некоторые другие, по учебным условиям не затрагиваемые
и программой лекционного курса. Кроме того, учитывая, что харак-
теристика экологии отдельных систематических групп может быть
удобно использована в имеющейся русской литературе, в системати-
ческом обзоре класса главное внимание обращено на морфологию,
в целях восполнения скудности подобных данных в русской лите-
ратуре. В связи с этим введено обсуждение филогенетического поло-
жения отдельных групп и их связей между собой, также отсутствую-
щее в доступных студенту книгах, и даны опыты построения фило-
генетических деревьев, заимствованные из специальных работ Фюр-
брингера и Пайкрафта. В систематическом обзоре морфолого-эко-
логическая характеристика доведена до семейств, в качестве предста-
вителей которых приведены наиболее обширные или интересные роды
и значительная часть видов птиц фауны Союза. Описание экологии
отдельных видов выпущено из руководства целиком, так как вклю-
чение его слишком увеличило бы объем всей книги, и отчасти соот-
ветствующие сведения могут быть найдены в уже существующей рус-
ской литературе. 1 Следует отметить также, что приложено много
1 Как продолжение этой книги, автор подготавливает сравнительно-эколо-
гический и морфологический обзор видов птиц фауны СССР, с более детальным
описанием некоторых общих явлений в жизни птиц специально территории
Союза (миграции, стационпые группировки и т. п., а также вопросы исполь-
зования и охраны).
3
усилий к тому, чтобы дополнить текст разнообразными иллюстра-
циями, не появлявшимися в наших изданиях.
При создании книги автор в значительной мере опирался на
более крупные сводки и монографии, дополняя эти сведения в неко-
торых случаях данными из литературы, посвященной более узким
и специальным вопросам. Более широко использованы недавний
обзор класса птиц, данный Штреземанном (1927—1934), известная
сводка Гадова и Зеленки (1871—1893), монументальная монография
Фюрбрингера (1888), специальные сводки Беддарда (1898), Нью-
тона и Гадова (1893), Грёббельса (1932), Ламбрехта (1936) и др.
К сожалению,многие крупные работы автор не мог иметь под руками
при составлении данной книги, что не могло не отразиться на содер-
жании настоящего руководства. В выборе классификации предпочте-
ние было отдано недавно опубликованной системе птиц Ветмора
(1929), учитывающей как современные, так и ископаемые формы и
не слишком сильно расходящейся с общепринятыми старыми клас-
сификациями. Однако в ряде случаев пришлось отступить от нее и
базироваться на других авторах. На более оригинальном материале
написаны экологические главы и распространение птиц, где автор
мог использовать результаты своих текущих исследований.
По некоторым причинам большие списки литературы по главам и
систематическим группам, составленные автором, не могли быть
помещены в это издание; некоторые указания на руководящую орни-
тологическую литературу приведены в конце книги.
Автор считает своим приятным долгом выразить благодарность
дирекции и деканату биологического факультета Ленинградского
Государственного Университета, создавшим условия для напечата-
ния настоящей книги, и профессору Д. Н. Кашкарову, содей-
ствовавшему автору советами и книгами и любезно принявшему на
себя труд по ее редактированию.
ОБЩАЯ ХАРАКТЕРИСТИКА КЛАССА ПТИЦ
Класс птиц является хорошо обособленной группой позвоночных
животных, резко выделяющейся перовым покровом тела, способ-
ностью летать (в огромном большинстве случаев) при помощи
крыльев, образуемых маховыми перьями, и гомоотермностыо, свя-
занной с большой интенсивностью жизненных процессов, свойствен-
ных организму птицы вообще.
Вместе с тем в организации птиц обнаруживается столь много
точек совпадения с организацией рептилий, что родство их и проис-
хождение из представителей класса Reptilia не вызывают сомнений.
Особенности самых разнородных сторон строения тела птиц
позволяют видеть в этой группе многочисленные признаки прой-
денной эволюции — с уровня организации рептилий на уровень,
близкий к низшим млекопитающим, но в сравнении с последней
группой развитие птиц шло совсем особым путем, особенности кото-
рого, в сущности, и подняли на высокую ступень организацию рас-
сматриваемого класса.
Все птицы в целом специализированы для обитания в воздушной
среде и передвижения по воздуху при помощи крыльев. Имеющиеся
немногочисленные исключения из этого правила могут быть рассмат-
риваемы как происшедшие вторичным путем и несут явные признаки
прежнего воздушного образа жизни.
Приспособление к полету в процессе своего развития и опреде-
лило строение представителей класса.
Передние конечности специализировались в крылья, в связи
с чем их скелет усилился в прочности, мускулатура стала более
мощной, а элементы кистевого отдела частью слились между собой,
создавая более прочную опору для маховых перьев; последние пред-
ставляют собой специализированные для данной функции контур-
ные перья, обладающие изумительной легкостью, упругостью и эла-
стичностью. В целом крыло образует весьма эффективное орудие
для поддержания тела и продвижения в воздушной среде. Функцио-
нальные задачи работы крыла повлекли за собой и изменения в строе-
нии плечевого пояса. По сравнению с грудиной рептилий, грудина
птиц сильно увеличена в размерах, образуя необходимое простран-
ство для прикрепления мощной мускулатуры. Потребность в увели-
чении поверхности грудины также повлекла за собой образование
на ней высокого костного киля, кроме птиц встречаемого лишь
у хорошо летающих млекопитающих. Образующие опору для крыла
вороньи кости (коракоиды) приобрели особую крепость и значи-
5
тельные размеры, а ключицы срослись в пружинную вилочку, до-
полнительно укрепляющую коракоиды. Крепость грудной клетки
поддерживается образованием реберных отростков (processus unci-
nati), поперечно налегающих на соседние ребра и хотя и встречаемых
еще у рептилий, но лишь у немногих форм. Таким образом передние
конечности из лазательных или бегающих конечностей рептилий
превратились в односторонне специализированный, но весьма эф-
фективный орган, даже при вторичных изменениях ни у одного из
представителей класса не возвращающийся к прежней своей ходиль-
ной функции.
Развитие весьма совершенной способности к полету подготовля-
лось общим повышением организации животного: полным отделением
венозной и артериальной кровеносных систем, лучшим использова-
нием кислорода крови и более совершенным дыханием вообще,
возникновением постоянной и высокой теплокровности, а в связи
с этим и более быстрой ассимиляции и интенсивного обмена веществ
и энергии. Интенсивная аэрация крови и высокая температура тела
создали необходимые условия для быстрого протекания нервных
реакций и явились предпосылкой для достижения повышенного пси-
хического уровня.
Естественно, что в связи с этими новыми в группе Sauropsida 1
функциями изменилась и усложнилась структура и всех остальных
органов. Легкие птицы отличаются способностью использовать кисло-
род как при вдыхании, так и при выдыхании, вследствие обра-
зования системы воздушных мешков, располагающихся между ор-
ганами, богатой сети воздушных капилляров и наличия анасто-
мозов между воздушными путями. Вместе с тем система воздушных
мешков создающая у хороших летунов высокую пневматичность
почти всех костей и даже подкожных пространств, существенно
облегчает вес тела, а располагаясь между мышцами, затрудняет
перегревание тела при сильной мускульной работе во время полета.
Быстрый обмен веществ возможен благодаря быстрому кругообо-
роту крови и следовательно относительно мощному и большому
сердцу. Температура крови у птиц в среднем составляет 42.2°,
достигая у мелких и наиболее активно жизнедеятельных форм до
44.8 и 45.5° С.
Перенос всей тяжести тела, при нахождении на земле, на задние
конечности вызвал значительное усиление их и образование харак-
терного для птиц обширного, сложного крестца (synsacrum), возник-
шего из слияния собственно тазовых позвонков с поясничными,
с частью хвостовых и последними грудными и этим укрепившего-
туловищную часть позвоночника. В задней конечности наблюдаются
слияние плюсневых костей в одну цельную кость — цевку, и образо-
вание интертарзального сочленения между голенью и одной костью;
при этом исчезает пятый луч пальцев, и тело опирается на трех-,
четырех-, редко двухпалую лапу.
Вследствие односторонней специализации передних конечностей,
1 Sauropsida составляют второй надкласс подтипа Craniata и включают
классы рептилий и птиц.
6
функцию «руки» принял на себя клюв, в связи с чем череп приобрел
особую подвижность, сохранив сочленение с шеей лишь одним мы-
щелком, причем шея значительно удлинилась. По всей вероятности
биологическая необходимость облегчения веса черепа и приближения
центра тяжести тела к середине туловища, существенная для летаю-
щего животного, вызвала редукцию зубного аппарата (сохранившегося
лишь у древних, теперь вымерших птиц) и передачу функции раз-
мельчения пищи мускульному желудку, у большинства представи-
телей класса очень объемистому и мощному. Объемистый желудок
и обширные воздушные мешки по всей вероятности и вызвали те
перемещения и изменения других внутренних органов, которые
констатируются у птиц при сравнении их с рептилиями: лопасти
печени располагаются преимущественно поперек полости тела, пра-
вый яичник и яйцевод (а иногда и правый семенник) редуцируются
п развиваются нормально лишь на левой стороне тела. Та же необ-
ходимость могла вызвать редукцию мочевого пузыря, отсутствую-
щего у птиц, и привести к выработке очень малых размеров задней
кишки, наблюдаемой в этой группе. Развитие яйца, вероятно в связи
с воздушным образом жизни животного, происходит относительно
быстро, и откладка его следует сейчас же за образованием полных
яйцевых оболочек. Явления, связанные с заботой о потомстве, вы-
ражаются у птиц разнообразными, дифференцированными и глубо-
кими приспособлениями, направленными на выращивание птенцов
и защиту их от различных внешних врагов, что в малой степени
свойственно рептилиям.
Воздушный и преимущественно дневной образ жизни требует
ориентировки главным образом зрением и слухом, и эти органы до-
стигли у птиц большого совершенства; орган обоняния менее полезен
для летающего животного и у большинства птиц развит слабо, лишь
отдельные специализированные формы представляют отклонение
в этом отношении. • .
Из органов чувств, орган зрения достиг особенно высокого разви-
тия, что повлекло за собой увеличение размеров глазных яблок и
сопровождалось значительной реконструкцией черепа, с отодвига-
нием головного мозга кзади, увеличением глазничного отдела черепа
н слиянием orbitosphenoidalia в тонкую межглазничную перегородку.
В головном мозгу особенной величины достигает мозжечок, являю-
щийся органом, координирующим сложные движения летающего
животного, но общий высокий уровень развития класса сказывается
также в значительном богатстве ассоциативных центров больших
полушарий и сложной психической жизни птиц, проявляющейся
в оригинальных брачных играх, пении и разнообразных и сложных
инстинктах вообще.
Первые представители класса Aves в ископаемом состоянии из-
вестны начиная с юрского периода. Развитие класса шло очень быст-
рыми темпами, и уже в меловой период повидимому было пред-
ставлено большинство 1 современных отрядов, и вместе с тем наибо-
1 Палеонтологически большинство отрядов птиц датируется с эоцена.
7
лее примитивные морфологические черты класса, за малым исклю-
чением, почти совершенно исчезли. В середине третичного периода
мы находим уже современные роды птиц или весьма близкие к ним
формы. В настоящее время распространение класса охватывает весь
земной шар и насчитывается около 30 000 форм птиц. Эти формы,
главным образом подвиды, группируются примерно в 10 000 видов,
2800 родов, 168 семейств. По принимаемой нами системе, ископаемые
п современные птицы могут быть подразделены на 33 отряда. Но
в то время как современные птицы могут считаться, по системати-
ческому составу, уже удовлетворительно изученными, ископаемые
известны совершенно недостаточно, и дальнейшее изучение их может
познакомить нас с немалым числом новых систематических групп.
Эволюция птиц, по достижении известного уровня, пошла по
линии преимущественно частных приспособлений к условиям место-
обитания, размножения и к особенностям питания, поэтому по плану
строения птицы представляются весьма однообразным классом:
у большинства птиц внешний облик и общий тип организации те же,
что и у вороны или вьюрка, дрозда, славки, куропатки, кулика и
утки. Наиболее уклоняющимися по организации являются: нелетаю-
щий киви, бегающие страусы, специализированные к жизни в воде
пингвины, резко выраженные хищники — крупные соколы и орлы,
летающие без устали обитатели океанов — буревестники и альба-
тросы, и мельчайшие птицы, питающиеся нектаром цветов, — ко-
лпбри.
МОРФ о л о г и я
1. ОБЩИЕ ПОКРОВЫ
Кожа. Кожа птиц состоит из двух слоев: эпителиального7
по происхождению эктодермального, слоя, или эпидермиса (epider-
mis), характеризующегося у птиц своей тонкостью в сравнении
с эпидермисом рептилий и млекопитающих, и соединительнотканного
слоя или собственно кожи (cutis, corium), возникающего из мезен-
химы. Вследствие особой тонкости эпидермиса птиц, мы не находим
у них обычного в других группах позвоночных проникновения кож-
ных сосочков в толщу эпителиальных покровов. В эпидермисе можно
различать д?а слоя клеток: верхний, тонкий, постепенно снашивае-
мый роговой слой (stratum corneum) и нижний, из живых кубических
клеток, ростковый или мальпигиев слой (stratum malpighii). В сое-
динительнотканном слое кожи различимы образованный плотной
волокнистой соединительной тканью подэпителиальный слой (stra-
tum subepitheliale) и лежащий под ним рыхлый или сетчатый слой
(subcutis). В подэпителиальном слое обильно представлены гладкие
волокна, приводящие в движение контурные перья; кроме того у птиц
с оголенными и сильно пигментированными участками кожи и мясис-
тыми выростами ее наблюдается сильная васкуляризация этого слоя
с образованием губчатых лакун. Временный застой крови в этих
лакунах вызывает набухание мясистых образований и иногда яркое
окрашивание их. Подкожный слой наиболее рыхл в своей глубокой
части, что обусловливает значительную подвижность кожи в целом.
Этот рыхлый слой является местом наибольшего накопления жировых
отложений, при упитанности птицы приводящих к образованию об-
ширных жировых прослоек (panniculus adiposus). У некоторых птиц
жировые прослойки постоянно образуются под кожей подошвы ног
и имеют здесь функцию эластичных подушек. Наконец в этом ‘же
слое лежат у крупных и хорошо летающих птиц (например многие
веслоногие) обширные воздушные пространства, связанные с воз-
душными мешками и легкими и облегчающие вес птицы.
Производными образованиями кожи являются роговые оболочки
челюстей (клюв), когти, шпоры и различные' виды перьев.
Роговой покров клюва (rhamphotheca) образуется
вследствие мощного разрастания эпидермального рогового слоя, в
некоторых местах достигающего нескольких миллиметров толщины.
Картина соотношения слоев, образующих рамфотеку, противопо-
ложна той, которую мы наблюдаем в коже тела: cutis здесь очень
9
тонок п непосредственно прилегает к надкостнице; верхние слои его
образуют многочисленные сосочки, проникающие в эпидермис и пи-
тающие его; в эпидермисе различим новый зернистый роговой слой
(stratum granulosum), лежащий непосредственно над мальпигиевым
слоем, в котором клетки переполнены зернышками кератогиалина,
промежуточной ступени кератина, образующего роговой слой рам-
фотеки. Рост этого рогового слоя невидимому во многих случаях
возобновляется периодически, вызывая
линьку рамфотеки, спадающей целиком
(тетеревиные) или — в случае образо-
вания ее отдельными роговыми пла-
стинками— по частям (чистики; рис. 1);
под этим слоем образуется новый мо-
лодой роговой покров. В других слу-
чаях однако как слущивание, так и
рост рамфотеки происходят непре-
рывно.
Переход рамфотеки в кожу головы
Рис. 1. а—клюв атлантического
тупика — Fratercula arctica L.;
пунктиром отмечены роговые пла-
стинки, развивающиеся на клюве
и около глаза к периоду размно-
жения ; сплошной линией показаны
контуры клюва во вторую половину
лета и осенью; Ъ — отпавшие рого-
вые образования; их положение на
клюве указано соответственными
буквами. По Bureau, из Stresemann.
происходит или постепенным утонче-
нием ее (без образования резкой гра-
ницы) или посредством восковицы (се-
roma) — значительного утолщения ко-
жи, то мягкой, то твердой по своему
строению и нередко ярко окрашенной.
Такая восковица характерна для днев-
ных хищников, голубей и многих по-
пугаев. В некоторых случаях она бы-
вает оперена.
Значение восковицы до сих пор
остается неясным; наиболее вероятным
объяснением является прикрывание ею
наиболее гибкого места надклювья, осо-
бенно подвижного как раз у попугаев
и отчасти у хищников.
Поверхность рамфотеки у огромного
большинства птиц является цельной,
но у трубконосых, веслоногих и неко-
торых других птиц она разбита на от-
дельные участки, как бы щитки, частью гомологизируемые головным
щиткам рептилий (Lonnberg, 1904; рис. 2). Края рамфотеки часто
несут зубцы, пластинки или тонкие насечки, облегчающие схваты-
вание и удержание добычи.
Роговой покров плюсны (podotheca). Плюсна и пальцы,
а иногда и часть голени у большинства птиц покрыты чешуйками
или роговыми пластинками, расположение которых во многих груп-
пах имеет характерный рисунок и важное систематическое значение.
В простейшем случае чешуйки однообразны и имеют вид зерен,
в более сложных они дифференцируются в пластинки и чешуйки раз-
ных типов (рис. 3). Полностью отсутствуют чешуйки лишь у неко-
торых зимородков и видов, имеющих оперенную плюсну.
10
К о г т и. Когти птиц гистологически имеют сходное строение
с рамфотекой клюва, растут беспрерывно и линяют обычно простым
постепенным обнашиванием. Развиваются когти в глубокой складке
кожи на концах пальцев, сопровождаемой впячиванием эпидермиса
в cutis; процесс образования когтя у птиц в основном представляет
сходство с процессом развития ногтя у млекопитающих.
Интересным образованием на когте является видоизмененный
в гребенку или скребок внутренний (редко внешний) край третьего
пальца (рис. 4). Он свойствен очень многим видам из различных систе-
матических групп и, возможно, употребляется для очищения кожи
Рис. 2. Роговой покров клюва птиц в сравнении с щитковаппвхМ морды у рептилий.
По своему положению отдельные части сложной рамфотеки птиц сходны с щитками
рептилий: ростральным (rostrale, г), верхнегубными (labiale, I), нижнегубными (infralabiale.
И), подбородочными (mentale, ти) и носовыми (nasale, n); 1 — голова глазчатой ящерицы —
Lacerta ocellata; 2 — змеи — Rachi delus brazili; з — слепо змейки — Glaucoma disserta;
4 —то же, вид сверху; 5 —щитохвоста — Uropeltis sp.; 6 — то же, вид сверху; 7 — ба-
клана — Phalacrocorax carbo L.; 8 — олуши — Sula bassana L.; 9 — альбатроса — Diome-
de a albatrus L. Обратите внимание па увеличение рострального щитка у Glaucoma и Uropeltis,
связанное с роющим образом жизни; это увеличение сходно с крупными размерами этого
отдела у птиц. У баклана число верхнегубпых пластинок варпирует от 3 до 4, приближаясь
к значительному количеству этих щитков у рептилий, но носовые пластинки у него исчезли
в связи с отсутствием ноздрей. Из Heilmann.
пли перьев. У некоторых видов птиц когти имеются не только на
задней конечности, но и на передней, большей частью на первом
пальце (например гуси, нанду), редко на втором (казуары, гоацин
в молодом возрасте и др.) и даже иногда на третьем (страусы). В мо-
лодом возрасте эти когти иногда помогают птенцу лазить среди веток
растений (гоацин, некоторые болотные курочки), у большинства же
птиц они имеют значение рудиментарного органа, утерявшего свое
функциональное значение, и наблюдаются лишь в эмбриональном
развитии (рис. 5). Помимо когтей, на конечностях некоторых групп
11
встречаются шпоры (calcar alae п pedis), имеющие вид острых ко-
нических образований с костяным* основанием и роговым покровом.
Общеизвестны шпоры петуха и других фазановых, сидящие на плюсне;
в других группах (пеламедеи, яканы, некоторые гуси) они образуются
на костях крыла.
Перовой покров. Перья (ptilosis), покрывающие тело
птицы, в связи с различными функциями имеют разный вид и строе-
ние, что заставляет классифицировать их в несколько типов.Наиболее
совершенное и сложное строение имеют маховые и рулевые перья,
Рис. 3. Типы роговых покровов плюсны у Passeriformes^
а — внутренняя сторона плюсны; б — наружная.
1 — пикпаспидныи тип — передняя сторона покрыта вытянутыми табличками, зад-
няя — мелкими,густо расположенными щитками с округлыми краями, или вообще лишена
щитков; тип свойствен главным образом Eurylaemidae и Cotingidae. 2 — экзаспиднын
тип — вытянутые таблички с передней стороны плюсны продолжаются па ее наружную сто-
рону и во многих случаях также и па заднюю; голая или покрытая мелкими щитками полоска
между табличками наблюдается по внутренней стороне плюсны; тип свойствен Tyrannidae
и Conopophagidae. 3 — эндаспидный тип — в противоположность второму тину, таблички
сплошь покрывают внутреннюю сторону плюсны, оставляя незаполненной полоску па на-
ружной ее стороне; тип наблюдается у Dendrocolaptidae. 4 — такзаспидный тип — передняя
сторона покрыта вытянутыми табличками, к которым прилегают по сторонам густо распо-
ложенные прямоугольные или ромбические щитки, сходящиеся своими задними краями на
подошвенной стороне плюсны; тип наблюдается у Formicariidae, Рteroptochidae, Philepittidae
и некоторых Oscines — Menuridae, Alaudidae и др. 5 — окреатный тип — передняя поверх-
ность плюсны покрыта вытянутыми табличками, которые часто сливаются в один сплошной
роговой покров, так паз. «сапожок»; к их заднему краю прилегают длинные роговые пла-
стинки, в нижней части иногда распадающиеся на щитки; тип свойствен большинству пред-
ставителей Oscines. Не изображен шестой кнемидофорный тип, в котором передняя и задняя
стороны покрыты сплошными роговыми пластинками,нередко еще сохраняющими следы слия-
ния табличек; шов между пластинками наблюдается по бокам плюсны и нередко имеет вид
оголенной или покрытой мелкими щитками полоски; этот тип свойствен питам. По Reichenow.
обусловливающие возможность полета. Менее специализированы, но
также сложно построены покровные перья, несущие разнообразные
функции: плотно покрывая друг друга наподобие черепиц, они защи-
щают тело от ветра и влаги, несут опознавательные и украшающие
отметины, а развитые пуховидные бородки, у основания лучше со-
гревают тело. По своему строению однако оба эти типа перьев близки
друг другу и объединяются под общим названием контурного пера
(pennae или plumae). Более просто построены пуховое перо (semi-
plumae) и пух (plumulae), назначение которых заключается в со-
гревании тела. Крайне специализированными являются наконец
пудреткп или порошковый пух (pulviplumae), щетпнковидные,
12
Рис. 4. Гребешки па когтях средних пальцев птиц.
а — цапли — Ardea sp. (впд сверху); b — скопы — Pandion haliaetus L. (сбоку); с — то
же (в разрезе); у т наблюдается резкий переход твердого рогового покрова когтя в более
мягкую подошвенную часть его; d — лебедя — Cygnus sp. (сбоку). По Boas, из Stresemann.
Рис. 5. Когти па пальцах крыла у гоацина и кур на эмбриональных и постэмбриональных
стадиях.
о, Ь и с — крыло птенца гоацина — Opisthocomus hoazin Miill. па различных стадиях
развития (а — снизу, b и с — сверху). Обратите внимание на длинную кисть, первоначально
превышающую по размерам предплечье, но с возрастом укорачивающуюся; па подвижность
и длину 1-го пальца; на крупные когти на 1-м и 2-м пальцах; на задержку в развитии дисталь-
ных маховых (9-го и 10-го), стоящую в связи с лазящим образом жизни; d— крыло взрослого
гоацина снизу; когти отпали, 1-й палец сильно укорочен; кисть значительно меньше пред-
плечья^ — крыло цыпленка банкивской курицы — Gallus bankiva (сцизу), с рудиментарным
когтем на 1-м пальце; f — крыло цыпленка обыкновенной курицы, более старшего возраста;
1-й палец укорочен и коготь отпал, дистальные большие и проксимальные малые маховые
отстают в развитии, как у молодого гоацина. Из Heilmann.
циализацию контурного пера,
(bj (а}
Рис. 6. Строение контурного пера.
а — общая схема, b — проксимальная
часть пера.
С — очип; Пу — дополнительное перо;
Р1.—пуховая часть опахала; U. i.—
нижнее отверстие; U. s. — верхнее от-
верстие; Ve — опахало; X— «душка»
пера, видпмая по вскрытии стенки очи-
на; I — бородки первого порядка;
II— бородки второго порядка; III —
реснички. По Boubier и Newton.
нитчатые (filoplumae) икисточковые перья, встречающиеся не у всех
видов. Все эти типы перьев представляют однако упрощение и спе-
которое можно взять за основу в на-
шем рассмотрении (рис. 6). В раз-
витом уже мертвом пере различают
ствол (scapus), несущий по краям
опахала (vexillum), проксимальную
пустую внутри и неоперенную часть
ствола — очин (calamus) и боль-
шую, дистальную, выполненную
сплошь круглыми, содержащими воз-
дух клетками — стержень (rhachis).
Стержень является основанием для
прикрепления с двух сторон бородок
первого порядка (rami), в свою оче-
редь несущих так же расположен-
ные, но более мелкие бородки вто-
рого порядка (radii). На послед-
них в свою очередь сидят микро-
скопически малые бородки третьего
порядка — реснички (radioli или
cilia), иногда снабженные крючоч-
ками (hamuli). Количество послед-
них естественно чрезвычайно ве-
лико. Даже бородки второго по-
рядка на среднем рулевом пере
американского луня (Circus hudso-
nius L.) сидят по меньшей мере
в количестве 1 250 000 штук, что
составит более 1000 м длины, если
эти бородки расположить в ряд
друг за другом.
Очин укрепляется в коже на
первом сосочке, который проникает
внутрь его, через круглое нижнее
отверстие (umbilicus inferior). Дру-
гое — верхнее отверстие (umbilicus
superior), во взрослом пере уже зам-
кнутое п имеющее вид ямки, поме-
щается в проксимальной части стер-
жня, в начале длинной бороздки,
пробегающей вдоль нижней его сто-
роны. В этой ямке у большинства птиц ответвляется так называемый
побочный стержень или дополнительное перо (hyporhachis), пред-
ставляющее уменьшенную и полупуховую копию главного пера.
Следует отметить, что перья одного и того же типа, но происхо-
дящие из разных частей тела птицы, нередко существенно отличаются
друг от друга по своей форме и строению (Chandler, 1906). Так же
отличаются деталями формы и строения аналогичные перья у пред-
ставителей разных отрядов (рис. 7).
14
Строение мелких частей пера обнаруживает удивительно тонкие
приспособления их структуры друг к другу (рис. 8). Каждая бо-
родка первого порядка (rami) имеет вид узкой пластинки, состоящей
из плотной наружной роговой оболочки (слагаемой мельчайшими,
роговыми чешуйками) и лежащих в центре клеток, наполненных
воздухом. По сторонам бородки, обращенным к другим подобным
образованиям, находятся мелкие крючкообразные выступы, в кото-
рых укрепляются основания более мелких бород второго порядка
(radii). Последние имеют различное строение в зависимости от плот-
ной или рыхлой (пуховидной) структуры пера. При пуховом строе-
нии, например в нижней части опахала,
они очень длинны и узки и состоят из
члеников, подобно усикам насекомого.
Каждый из этих члеников состоит из одной
единственной клетки и называется междо-
узлием (internodii); дистальный конец его
расширен, воспринимает проксимальный
конец следующей клетки, несет иногда
мелкие реснички и называется узлом (no-
dus). Кроме этой «членистой» части бород-
ки, объединяемой под названием перышка
(penmilum), отмечают еще расширенную
часть ее — основание (basis), которым бо-
родка прикрепляется к более крупной
части пера. В плотной части опахала, где
элементы пера тесно связаны друг с дру-
гом, бородки второго порядка дифферен-
цируются на два типа: бородки с крючоч-
ками (hamuli), сидящие на дистальной по-
верхности бородки первого порядка, и бо-
родки без крючочков, но с длинными же-
лобками, сидящие на проксимальной сто-
роне бородок. Изменения этих бородок, по
сравнению с бородками пухового пера, за-
ключаются главным образом в следующем:
Рис. 7. Строение бородок вто-
рого порядка грудного пера по-
ганки — Aechmophorus occiden-
tal's Lawr. (Colymbiformes) как
пример видоизменения бородок
контурного пера. Бородки вто-
рого порядка закручены вдоль
своей оси, что вызывает отраже-
ние света и своеобразный шелко-
вистый лоск перьев. По Chand-
ler, пз Stresemann.
удлиняется и сильно расширяется базаль-
ная часть, дорзальный край ее загибается и образует желобок (осо-
бенно велико увеличение этой базальной части у проксимальных боро-
док); перышко (pennulum) сильно укорачивается, а реснички узлов,
наоборот, сильно увеличиваются и частью загибаются в виде упругих
крючочков (hamuli). Последние развиваются только на дистальных
бородках. На проксимальных бородках сильно развиваются желобки
по краю базальной части. Ряды дистальных бородок налегают косо
на ряды проксимальных бородок и своими крючочками зацепляются
за их желобки, образуя упругое и плотное опахало (рис. 9). Разде-
ление бородок второго порядка на указанные выше типы не является
однако всюду резким и соединяется переходным строением в проме-
жуточных участках того же пера.
Развитие пера. Развитие пера столь сложного строения
не может быть рассмотрено здесь в подробностях. Более просто
15
развитие птенцового пуха (neoptilae или neossoptilae), происходя-
щее частью еще в яйце. У голубя, например, на 5-й —6-й день наси-
живания на эмбриональной коже замечаются мелкие бугорки, по-
крывающие вскоре почти все тело. Эти бугорки появляются вслед-
ствие усиленного деления клеток эпидермиса и подэпителиального
слоя, сопровождаемого богатым
(В) (£1
Рис. 8. Строение бородок второго порядка
(схематично).
А — проксимальная бородка второго по-
рядка в контурном пере; В — пуховая бо-
родка; С—полупуховая бородка; Е—
поперечный разрез бородки первого по-
рядка; D—дистальная бородка второго
порядка в контурном перс.
b — расширенное основание бородки; с —
ямка, в которой укрепляется дистальная
бородка второго порядка; ci — реснички;
ст. v. — вентрально вытянутый край бо-
родки первого порядка; dv— вентраль-
ные зубцы, сидящие в основании перыш-
ка; /. — реснички, сидящие на узлах в пу-
ховой бородке; fl — изогнутые реснички,
сидящие по дорзальному краю основания
бородок второго порядка, па перьях ту-
ловища (преимущественно груди); дг—
крючочки; 7п — желобки (изогнутый край
основания); п — узел бородки; р — пе-
рышко бородки; sd.—дистальные крюч-
кообразные выступы бородки первого по-
рядка (заусеницы); sp.—то же, прокси-
мальные выступы; sp. d.—дорзальные
зубцы, сидящие в основании перышка;
v. — косматые реснички в основании пу-
ховой бородки. По Chandler, из Boubier.
ток и постепенной редукции сосочка пульпы. Когда бородки со-
зреют, лопается роговая оболочка, кисточка бородок освобождается,
обсыхает и расправляется в гнездовой пух (рис. 10).
При развитии взрослого (окончательного или дефинитивного) пера
(teleoptilae) процесс осложняется образованием главного и побочного
стержня (рис. 10). Поэтому при образовании эпителиальных валиков на
дорзальной стороне первого сосочка обособляется комплекс клеток,
в дальнейшем входящий в состав стержня. По мере роста последнего,
на его вентральной поверхности образуются два утолщения, замыкаю-
щие между собою бороздку, заметную и на выросшем пере. Входящая
в ствол пульпа разделяется при этом на два отдела; один оказы-
притоком крови. В это же время
идет процесс впячивания кожи
по краям этого бугорка, приво-
дящий к образованию быстро ра-
стущего в длину перового сосоч-
ка, постепенно погружающегося в
кожу. Сосочек снаружи покрыт пе-
ридермой (или epitrichium — бес-
структурным роговым слоем), под
которой лежит мальпигиев слой с
намечающимся обособлением его
в три различных слоя клеток, а
под ним тонкая базальная мембра-
на, отграничивающая от этих слоев
пульпу сосочка, образованную
клетками коркового слоя и обиль-
ными питающими капиллярами.
Вместе с увеличивающимися раз-
мерами сосочка, наружный слой
клеток мальпигиевого слоя орого-
вевает и образует вокруг него ро-
говой защитный чехол. Внутрен-
ние же слои мальпигиева слоя
начинают интенсивно расти вну-
три сосочка, образуя эпителиаль-
ные валики. Основная роль в
дальнейшей дифференциации кле-
ток этих валиков принадлежит
так называемым промежуточным
клеткам, которые и образуют бо-
родки растущего пера (Schuz,R1927).
Параллельно с дифференциацией
дериватов мальпигиевого слоя на-
чинается процесс ороговения кле-
16
вается в дальнейшем заключенным в очин, другой занимает узкую
полость стержня, выходя через замыкающийся впоследствии umbi-
Рис. 9. Схематичное изображение сцепления бородок второго порядка,
г.— бородка первого порядка; d.— дистальная бородка второго порядка;
р.— проксимальная бородка второго порядка. По Pycraft, из Stresemann.
licus superior. В процессе
постепенно резорбируется,
через umbilicus inferior,
оставляя за собой ряд тон-
ких роговых колпачков,
разделенных слоями воз-
духа. Это так называемая
«душка пера», наблюдае-
мая в очине и во взрослом
состоянии и являющаяся
остатком одевавших пуль-
пу снаружи базальной мем-
браны и цилиндрических
клеток мальпигиева слоя.
Под давлением растущего
пера роговой чехол его ло-
пается, и перо постепенно
выдвигается наружу и на
чинает расправляться,про-
ходя как бы ряд стадий:
короткой кисточки, уже
частью развернутого опа-
хала и, наконец, почти
доросшего пера с остат-
ками рогового чехлика у
основания. Последние от-
падают после превращения-
пера целиком в мертвое
образование.
По своему происхо-
ждению перо выводится
из чешуи рептилий. В ис-
ороговения перового зачатка пульпа
и нижний отдел ее отступает из очина
Ram
Рис. 10. Схема развития пера.
а — начинающееся опускание обособившегося перо-
вого сосочка в глубь кожи и закладка эпителиальных
валиков,в дальнейшем образующих бородки пуха;
b — птенцовый пух в пероном сосочке; роговой че-
хол лопнул и бородки пуха расправляются; с--
контурное перо, сидящее в перовом сосочке и покры-
тое роговым чехликом; d и е —два поперечных раз-
реза пера на уровне а и Ь' рисунка с.
сог. — соедипительно-тканный слой кожи; т — оро-
говевший участок, ограничивающий «душку* пера;
р — пульпа сосочка; Rad. — бородки второго поряд-
ка; Ri.— эпителиальные валики; Ram — бородки
первого порядка; Rh — ствол пера; s. — «душка»
пера; Sh — роговой чехлик; umb. in. —нижнее от-
верстие. По Davies, из Бючли.
копаемом состоянии известны формы последних (например Еираг-
keria), судя по отпечаткам, имевшие чешую с цещпргацьным
17
современных рептилий и птиц однако не дают полных переходов между
этими типами роговых образований, хотя развиваются весьма сходно
(рис. 11). Развитие чешуй, покрывающих ноги птиц, почти, неотли-
чимое от развития чешуй рептилий, также напоминает развитие
Рис. 11. Развитие ножных чешуй и пера птиц в сравнении с развитием чешуй у рептилий (продольные разрезы).
Обратите внимание на общее сходство в развитии и близость в строении чешуй; 1—3 —развитие чешуи ужа — Tropidonotus
natrix L.; 4 —чешуя желтопузика — Ophisaurus apodus L.; 5—8—развитие чешуйки на ноге обыкновенной курицы;
9—12 — развитие пера голубя — Columba livia G-m.
с. — соединительнотканный слой кожи; е.—наружный роговой слой эпидермиса; h. — роговой покров, образуемый
мальпигиевым слоем; р.—пигментные клетки; а.—мальпигиев слой эпидермиса. Из HeilmanYi.
стержнем и боковыми насечками от него по краям, что уже напоми-
нает строение современного пера (Broom, 1913; рис. 219). Покровы*
пера. В обоих случаях эти образования закладываются как скопле-
ние эпидермальных клеток и клеток кориума. Но сосочек чешуйки
растет билатерально-симметрично, уплощается, и рост заканчивается
сплошным ороговением эпидермальных клеток, так что дефинитивная
чешуйка состоит из мезодермальной основы и эпидермоидальногр
ороговевшего покрова; чешуя никогда не линяет, а лишь постоянно
обнашивается снаружи. По наблюдениям Дэвиса (Davies, 1889),
в развивающейся чешуйке можно усмотреть
перового сосочка.
Линька и рост пера. По окон-
чании роста нового пера сосочек впадает
в недеятельное состояние, но не теряет
своей самостоятельности. . У всех птиц
периодически наступает линька, заклю-
чающаяся в выпадении старого изношен-
ного и выгоревшего оперения и выраста-
нии новых перьев. Каждое новое перо при
этом вырастает из уже существующего пе-
рового сосочка, который в период линьки
вновь пробуждается к деятельности и
образующимся новым пером выталкивает
старое (рис. 12). В зависимости от нали-
чия или отсутствия в этот период воз-
растных или половых физиологических
изменений в теле птицы, повое перо фор-
мируется или в точности подобным ста-
рому, или отличается от него, в совокуп-
ности с другими перьями образуя разные
возрастные и сезонные наряды птицы.
Помимо этого строго периодически возни-
кающего пробуждения деятельности, пе-
ровой сосочек активируется выдергива-
нием пера вне срока линьки, но при кон-
турном пере не более трех раз подряд.
После этого регенерационная способность
истощается, и наступает состояние покоя.
Только вновь возникающий период линьки
сильнее пробуждает сосочек к новой дея-
следы рудиментарного
Рис. 12. Линька перьев у ма-
гслланского очкового пингви-
на — Spheniscus magellanicus
Forster. Перо повой генерации
проникает через нижнее отвер-
стие очина в старое перо и вы-
талкивает его собой.
h — кожа; лс — мускулы, дви-
гающие перья; н.п. — новое*
перо; р.ч. — роговой чехлик ра-
стущего пера, у пингвинов про-
никающий в глубокие слои ко-
жи; с.п. —старое перо (верхний
конец его обрезан). По Lowe, из
Strescmann.
тельности.
Длительность носки пера подвержена значительным колебаниям.
Наиболее короткие сроки носятся первое птенцовое оперение, сме-
няемое часто еще в гнезде (иволга), и партии перьев, имеющие зна-
чение полового украшения. У некоторых северных куриных и гуси-
ных птиц защитный летний наряд также носится сроком не более
2—3 месяцев. Обычный однако срок носки пера равняется году.
Наиболее длинный срок носки — два года — свойствен маховым
перьям журавлей и орлов.
Отдельное Перо растет наиболее быстро, когда оно достигает
Половины своей длины, и рост значительно замедляется незадолго
перед достижением пером полных размеров. Длинные перья, соот-
19
ветственно их большей длине, растут быстрее, чем короткие перья
того же вида, но ежедневный прирост их, относительно их длины,
короче. Рост махового пера славки завирушки (Sylvia curruca L.),
длина которого равна 57 мм, серой вороны (Corvus cornix L.) — 285 мм
и журавля Bugeranus carunculatus Gm. — 480 мм, заканчивается
соответственно в 17, 32 и 54 дня. Ежедневный прирост пера у послед-
него вида достигает 13 мм, а у домашнего воробья (Passer do те sti-
lus L.) 4 мм. Смена пера часто проходит болезненно для птицы и
всегда сопровождается исхуданием. Особенно велики физиологи-
ческие затраты у тех видов, которые более или менее одновременно
теряют все маховые перья, лишаясь на некоторый период возмож-
ности летать (гуси, пастушки, журавли, фламинго и др.). Этот период
более длителен у более крупных видов. Так в группе гусиных в пе-
риод линьки мелкие утки нелетны в течение 21 дня, огарь [С as area
ferruginea Pall., весом в 1,2—1,5 кг) —35 дней, лебедь кликун (Cygnus
cygnus L., весом 8—11 кг) — 42 дней и лебедь шипун {Cygnus olor
Gm., весом 9—12 кг) — 49 дней (Heinroth, 1906).
В течение суток скорость роста также колеблется, более быстро
протекая в часы хорошего питания пера (день и начало ночи) и за-
медляясь в часы ухудшенного притока питательных материалов
(вторая половина ночи). Бородки, вырастающие в это время, остаются
на более низкой ступени дифференцирования, что вызывает на опа-
хале ясно заметные поперечные полоски, следующие друг за другом.
Исходя из этого явления, Риддл (Riddle, 1908) объясняет появление
пуховых структур в оперении вообще как следствие пониженного
питания организма в период роста этих образований. Однако пол-
ная применимость этих объяснений сомнительна.
Периодичность линьки у многих видов представляет весьма слож-
ную картину. Наиболее простым случаем является одна линька в году
после вывода птенцов (в северном полушарии — осенью), с одновре-
менной сменой мелкого и крупного пера. Более сложна линька
двойная: осенью и весной, перед гнездованием, называемая пред-
брачной, обычно ограничивающаяся сменой только мелкого пера или
части его. Наибольшую сложность представляет линька три и четыре
раза в году, с образованием специального летнего и иногда зимнего
наряда. Последние линьки всегда частичны, ограничены обычно
областью головы, шеи и туловища и известны только для уток, не-
которых куриных и рябков. Линька усложняется также захожде-
нием нарядов друг на друга, когда на птице можно найти перья
прошлого брачного наряда, перья настоящего осеннего наряда и но-
вые перья брачного наряда, что встречается у уток, чистиковыхv
гагар и других птиц. Наконец крупные птицы, особенно хищники,
меняют крупные перья весь год, постепенно, и в год обильной линьки
даже не гнездятся.
Тот или иной ход линьки обусловливает смену нарядов птицы
сезонную и возрастную (рис. 13). Как уже отчасти видно из преды-
дущего, сезонные наряды более сложны у видов с резко выраженными
вторично-половыми признаками в оперении и видов, нуждающихся
в сезонной защитной окраске или защите от холода (рис. 14). Возра-
стные же наряды наиболее постепенны у крупных, долго живущих
20
птиц. Обзор нарядов и линек представлен следующей схемой
(стр. 22, 23), заимствованной у Бианки (1913) и отчасти измененной.
Течение линьки весьма разнообразно во многих группах птиц.
Обычно линька начинается со смены маховых и рулевых перьев,
затем охватывает туловище и заканчивается на голове. Маховые
перья, если они не выпадают одновременно (см. выше), линяют
столь постепенно, что птицы не теряют способности к полету. При
этом линька одновременно
распространяется из трех
центров: большие маховые
линяют от проксимальных
к дистальным (десцендент-
но), а малые — одновре-
менно от дистальных и про-
ксимальных , двигаясь
внутрь этой части крыла
(конвергентно).У особенно
длиннокрылых видов уста-
навливаются дополнитель-
ные центры, у плохих ле-
тунов линька может итти
несимметрично на обоих
крыльях (рис. 15). Руле-
вые перья в большинстве
случаев линяют центри-
фугально, начиная от цен-
тральных рулей наружу,
но у птиц-древолазов, при
передвижении по стволу
дерева опирающихся на
удлиненные и упругие
центральные рули (дятлы,
пищухи и др.), эти перья
сменяются последними.
Центрипетально, т. е. от
краевых перьев внутрь,
линяют рулевые перья у
представителей родов Pha-
sianus, Perdix^ Alae.ctoris
Рис. 13.
Схема возрастных нарядов и линек у чаек.
Пунктирная линия изображает период роста перьев
или период линьки; сплошная линия — период носки
того или иного наряда перьев; начальным моментом
принят день вылупления (см. яйцо); спираль замы-
кается по достижении последнего взрослого наряда,
не изменяющегося (существенно) с дальнейшими
линьками.
1 — четырехлетний цикл нарядов крупного вида
северного полушария — серебристой чайки — Larus
argentatus Briinn.; 2 — четырехлетний цикл крупного
вида южного полушария — Larus dorn 'micanus Licht.
3 — трехлстний цикл вида средней величины —
Larus heermanni Cas.; 4 — двухлетний цикл мелкого
вида — обыкновенной чайки — Larus ridibundus L.;
оба последние вида рассматриваются в отношении к
северному полушарию. По Dwight, из Stresemann.
и др. Наконец у кукушек
и попугаев обычна несимметричная линька хвоста (Heinroth, 1898).
Линька птиц представляется пока мало изученным явлением,
особенно в отношении неконтурных типов пера. Пудретки в некото-
рых случаях (цапли) обладают длительным ростом, в других, наобо-
рот, в течение года имеет место несколько генераций этих перьев.
Выпадение и возобновление пуха в ряде случаев происходят в иные
сроки, чем смена контурного пера. ф
Строение второстепенных типов пера. По-
мимо рассмотренного выше контурного пера, следует рассмотреть
также строение других типов перьев. Пух (plumulae) имеет задачей
21
СХЕМА ПОСЛЕДОВАТЕЛЬНЫХ НАРЯДОВ И ЛИНЕК ПТИЦ
1) Пуховой I. Пуховидный наряд (все перья I генер.) — (у птенцовых может отсутствовать; иногда состоит из двух после-
довательных генераций; охватывает только мелкое перо)
1. Гнездовая линька — полная
2) Первый II. Гнездовой наряд (все перья II генер.)
Юношеская линька:
а) полная Ь) частичная а) полная1 Ь) частичная
3) Перц^й Ш. а) Первый простой го- Ь) Первый сложный годич- с) Первый простой полуго- d) Первый сложный полу-
зимний дичный ный дичный годичный
(все перья III генер.) (перья II и III генер.) (все перья III генер.) (перья II и III генер.)
Обнашивание и выцветание 3. Первая предбрачная линька
а) полная Ь) частичная
4) Первый IV. а) Первый простой го- Ь) Первый сложный годич- с) Первый простой полуго- d) Первый сложный полу-
брачный дичнобрачный нобрачный дичнобрачный годично брачный
(обнош. перья III генер.) (обнош. пврья II и III (все перья IV генер.) (при осложнении простого
генер.) полугод.перья III и IV ге-
нер. ; при осложнении слож-
ного полугод. — II, III и
IV генер.)
4. Первая послебрачная линька, которая бывает всегда полная2 * 4 5
5) Второй V. а) Второй простой годичный
еимний (все перья V генер.)
Обнашивание и выцветание
Ь) Второй простой полугодичный
(все перья V генер.)
5. Вторая предбрачная Л Й н ь к а
а) полная
Ъ) частичная
6) Второй
брачный
VI. а) Второй простой годично-
брачный 8
(обнош. перья V генер.)
Ь) Второй простой полугодич-
нобрачный
(все перья VI генер.)
с) Второй сложный полугодично-
брачный
(перья V и VI генер.)
6. В т о р а я послебрачная линька, которая бывает всегда полная
Далее следует повторение у одних видов V, у других V и VI нарядов,
инюгдч с небольшими в зрастными изменениями их (например, у крупных
дневных хищников). Кроме того, у стдельных видов наблюдаются следу-
ющие дополнительные наряды (римские цифры соответствуют номерам
нарядов):
Юношеский охранительный
(дополнит.)
Юношеская охранитель-
ная линька (дополнит.)
Взрослый охранительный
(дополнит.)
Взрослая охранитель-
ная линька (дополнит.)
Охранительный селезней
(дополнит.)
б. Вторая послебрачная
линька
III. d) Первый сложный
полугодичный 1 * 3
VI. с) Второй сложный
полугодичный
VII. а) Третий сложный
годичный
1 Почти полная (кроме 2 дистальных махов 1-го разр.) у молодых белых куропаток.
я Полная (включая й 2 дистальных маха 1-го разр.) у взрослых белых куропаток и других куриных.
3 Частичная линька селезней (мелкое перо).
защиту тела птицы от потери тепла. Очень короткий, слабо разви-
тый стержень его несет многочисленные, очень длинные бородки пер-
вого порядка, на которых сидят по сторонам сравнительно длинные,
почти лишенные крючочков и ресничек, мелкие бородки. Вследствие
этого, бородки пуха не связаны друг с другом и образуют на теле как
бы спутанный войлок или подшерсток, скрытый снаружи контур-
ным пером; наилучше развит этот пух у водоплавающих птиц.
Короткий очин пуха почти целиком сидит в* коже.
Простота в строении пуха рассматривалась ранее как доказатель-
ство его примитивности. Более вероятным однако является объясне-
(!а)
аь) (гь)
Рис. 14. Зимние и летние перья глухаря — Tetrao urogallus L.
1 — зимнее крупное перо, с сильно опушенным основанием опахала и
дополнительным стержнем; 2 — летнее более мелкое перо, с слабо
развитым опушением и дополнительным стержнем; а — вид сверху;
Ъ — сбоку. Оба пера сняты с задней стороны шеи самки глухаря, с оди-
наковых участков кожи. По Lonnberg, из Stresemann.
ние ее специфической ролью пуха (сохранение тепла), возможной
лишь при рыхлом строении его (Stresemann, 1927).
Птенцовый пух (neossoptilae), внешне имея большое сходство
с пухом взрослой птицы, является не чем иным как концевыми
частями бородок невыросшего контурного покровного пера, в це-
лях согревания тела птенца приобретающего пуховидное строе-
ние, Вследствие недифференцированности строящих перо клеток
эти бородки могут соединяться коротким стержнем. По мере роста
птенца, после наступающего некоторого периода покоя, перовой !
сосочек вновь, активизируется, и растущее новое перо выталкивает i
на концах своих бородок птенцовый пух (рис. 16). Соответственно
разным формам своего образования этот пух может иметь вид от-
дельных бородок первого порядка, вид взрослого пуха и вид рыхлого
с коротким стержнем пера; в последнем случае, если концевой отдел
втержня будущего взрослого пера участвует в этом образовании,
24
Рис. 15. Схема липьки маховых п рулевых перьев у представителей различных отрядов.
К. — кисть; Пп.—предплечье; Л.—левое крыло; Пр. — правое крыло. Новое перо
отмечено в виде простой вертикальной черточки, педоросшее — в виде неполной черточки;
старое неро несет снизу кружок, только что выпавшее перо отмечено буквой о.
*.— начальный период линьки маховых мексиканского большого дятла — Campophilus impe-
rialis (Gould.); 2 - период окончания липьки у домашнего воробья —Passer domesticus L.;
3 — период начала линьки у африканского сокола — Falco biarmicus tanypterus Schl.; 4 — пе-
риод окончания линьки у антильской пустельги — Cerchneis sparverius dominicensis Gm. (3 и 4 —
типичны для настоящих соколов, с центрами линьки, лежащими посредине соответствующих
отделов крыльев); 5 —линька дистальных отделов крыла у Ardea sp. проходит через одно перо
(обычно после 4-го или 5-го большого махового, считая совнутри); 6—ход линьки у кукушки —
Centococcyx affinis (Horsf.); линька асимметрична; 7 — период окончания линьки у большого
подорлика — Aquila clanga Pall.; линька начинается с трех центров в каждом отделе крыла;
8 —линька малых маховых у пеликана — Pelecanus sp.; идет несимметрично и из многих
Центров; 9, 10 — возрастные стадии и линька перьев у куриных; 9 — левая сторона — двух-
дневный фазан Виелло — Lophura ruja Raffl.; дистальные большие и малые маховые еще не
развиты; правая половина — четырехпедельпый тетерев — Lyrurus tetrix L.; эти маховые
развиваются крупнее, чем птенцовые маховые перьяио —левая и правая стороны — даль-
нейшие последовательные стадии липьки грифовой цесарки—Acryllium vullurinvm (Hardw.)
с маховыми перьями нескольких генераций; и, 12, 13 — линька хвоста у домашнего воробья—
Passer domesticus Jj.,дятла - Sphyropicus thyroideus Cass, и южпоамерикапского лупя — Circus
cynereus Vieill.; линька хвоста последнего представляет случай с чередованием липятощих
перьев. По Heinroth.
взрослое перо имеет на вершине тупо кончающийся стержень (та-
ковы, например, хвостовые перья молодых уток).1 У многих видов
контурное перо лишь постепенно в серии нарядов становится менее
рыхлым, чем в птенцовом пере.
Порошковый пух или пудретки (pulviplumulae) представляет
собой пух, специализированный н& продукции мельчайших роговых
зернышек — «пудры», размерами около 1 микрона (до 0,3 микрона).
Пудра образуется из так называемых промежуточных клеток про-
дольных валиков перового сосочка, быстро размножающихся и при
ороговении распадающихся на мельчайшие зернышки и палочки.
•Эта пудра освобождается, когда лопается роговой чехол растущего
Рис. 16. Взаимоотношения neossoptilae и teleoptilae у дневных и ноч-
ных хищных птиц.#
—дефинитивное коптурное перо сипухи—Tyto alba Scop., несущее
две генерации птенцового пуха, отделенные от пера и между собой
участками бородок первого порядка, лишенными более мелких боро-
док; 2 — рыхлое пуховое перо — mesoptilae («промежуточное перо»)
белой совы — Nyctea nyclea I»., несущее па концах бородок более про-
стой по своему строению птенцовый пух (так паз.protoptilae); 3 —
mesoptilae сапсана — Falco peregrinus Tunst., несущее на вершине
protoptilae. По Kleinschmidt, из Stresemann.
пера. Пудретки встречаются в различной стадии специализации.
Шюц (Schiiz, 1927) недавно выяснил, что способностью образовы-
вать пудру обладают очень многие птицы, не имеющие специальных
пятен порошкового пуха и развивающие ее в пуховой базальной части
пера и иногда в незначительном количестве даже в области опахала.
Рыхлое контурное оперение образует пудру в большем количестве,
чем жесткое и упругое. Собственно порошковый пух имеет вид,
изменяющийся от пуха с удлиненным периодом роста, до пуха,
1 Птенцовый пух (neossoptilae) называют также первичным или примордиаль-
ным пером— prepennae («предперо»), в отличие от собственно пуха (plumulae).
возрастная стадия которого называется preplumulae. В свою очередь neosso-
ptilae подразделяют на protoptilae — бэлее простой пух, в виде отдельных бо-
родок или пучков их (рис. 16), и mesopti'ae, обычно имеющих характер кон-
турного пера, но рыхлого строения («промежуточное перо»), хотя иногда эта
генерация напоминает больше пух, чем перо (рис. 16, 7). Обе, или первая из
этих генераций, могут полностью выпадать в развитии птицы (Ewart, 1921).
26
теряющего очин и представляющего собой видоизмененные бородки
е постоянным ростом; последние часто густо сидят на коже, образуя
пуховые пятна, располагающиеся главным образом по бокам брюха
(например цапли) и надхвостья -птицы (рис. 17).
По своим физическим особенностям продуцируемая пудра напо-
минает зерна талька и легко впитывает влагу, в чем и видят теперь
ее главнейшее назначение. Подтверждением этого соображения
ярляется широкое развитие пудреток у тех видов, у которых отсут-
ствует копчиковая железа, своим секретом также защищающая опере-
ние от смачивания’и увлажнения. Хорошо развиты пятна порошко-
вого пуха у цапель, скрытохвостов, попугаев, голубей и некоторых
других птиц. Богатое отделение пудры не остается без влияния на
окраску у некоторых видов, вызывая иногда специфический нежный
тон в окраске оперения (некоторые попугаи и голуби).
Из других типов пера следует упомянуть кисточковые
перья, окружающие выводной проток копчиковой железы (рис. 22).
Их назначение заключается в впитывании секрета железы, который
потом переносится на клюв птицы. На голове около клюва и глаз
многих птиц развиваются крупные, имеющие осязательную функ-
цию, щетинки. Около их основания обычно расположены мелкие
осязательные тельца. Особенно крупны эти щетинки, называемые
часто вибриссами, у ночных и ловящих мелких насекомых на лету
птиц (совы, козодои, киви и др.); они облегчают птицам охоту и
схватывание добычи. Крайне своеобразны нитевидные перья
(filoplumae) с длинным и тонким стержнем, почти лишенным боро-
док; они в количестве от одного до десяти сидят в окружности контур-
ного пера или пуха, образуя с ним одну перовую группу и скрываясь
обычно под покровом более длинных контурных перьев. Располо-
жение их часто подвержено особым закономерностям. За редким
исключением (нитевидные, «украшающие* в брачный период, белые
перья на шее бакланов), функция их является пока неизвестной.
Побочный (дополнительный) стержень (hyporhachis) является
частью целого пера и развивается из того же перового сосочка из
закладываемой на вентральной стороне его недифференцированной
массы промежуточных клеток. Во многих случаях он встречается
не только на кроющих контурных, но и на пуховых перьях; как
правило, дополнительный стержень бывает значительно меньше ос-
новного и имеет пуховидное строение. Значение его невидимому
заключается в уплотнении перового покрова, и в некоторых группах
он функционально замещает пух или дополняет его функцию (пин-
гвины, куры).При имеющей место линьке из зимнего в летнее перо
(тетеревиные), дополнительный стержень вылинивает вместе со всем
пером в более легкое, менее удерживающее тепло, образование
(рис. 14). У многих групп (например голуби, веслоногие, большая
часть воробьиных) побочный стержень вообще отсутствует.
Нерешенным вопросом является филогенетическое значение по-
бочного стержня. В значительном развитии его раньше видели при-
митивное свойство, исходя из того, что у некоторых древних птиц,
например казуара и эму, побочный стержень развит в одинаковой
степени с основным стержнем. Однако он совсем отсутствует в дру-
27
гих древних группах, например у страуса, нанду, киви и др. Предло-
женные гипотезы образования побочного стержня, например расще-
плением чешуи рептилий (Steiner, 1918), страдают искусственностью.
Поэтому не лишено вероятия, что побочный стержень является срав-
нительно новым образованием.
У огромного большинства видов птиц распределение перьев и
пуха не является равномерным по всему телу, а приурочено к опре-
деленным участкам, птерилиям (pterylae), между которыми нахо-
Рис. 17. Птерплоз ракоеда — Cochlearvus cochlearvus L. (I, а и b) и
солнечной цапли—Eurypyga helias Pall.
Обратите внимание па отличия в положсппи птерилий и пятен пу-
дрообразующего пуха.
а. г. — голеппал аптерия; а. т. —нижняя- аптерия; a sp. — спипная
аптерия; a. t. I. —аптерия боков тела; p.ab. — паховое пудрообразую-
щее пятно; p.cl.— околоключичное пудрообразующее пятно; р sa.—
тазовое пудрообразующее пятно; pt. al — крыловая птерилия; pt. с.—
хвостовая птерилия; pt. cap — головная птерилия; pt. ст. — голсп-
пая птерилия; pt fem. — бедренная птерилия; pt. gast.— брюшная
птерилия; pt. hum. — плечевая птерилия; pt. р. — грудная птерилия;
pt. sp. — спипная птерилия. По Nitzsch, из Stresemann.
дятся пространства кожи, не несущие .перья, — аптерии (apteriae)
(рис. 17). При этом, в пределах птерилии, перья сидят в шахматном
порядке. Распределение пуха менее правильно, и он то диффузно
рассеян по поверхности всего тела (большинство водоплавающих
птиц), то приурочен к области птерилий, то, наоборот, свойствен
только аптериям (так называемые maculae plumosae). Свойственное
немногим группам (бескилевые птицы и пингвины) равномерное рас-
пределение перьев по всему телу является вероятно вторичным
приспособлением, вследствие частью утери* способности летать, ча-
стью преимущественно водного образа жизни; некоторыми авторами
однако это свойство считается наиболее примитивным. Наличие у
страусов в прошлом аптерий устанавливается все же существованием
28
их в птенцовом оперении. Рисунок, образуемый на теле птерилиями
и. аптериями, так называемый птерилозис (pterylosis), имеет в ряде
групп существенное систематическое значение.
Наиболее обычно встречаются следующие птерилии:
1. Спинная птерилия (pteryla spinalis), непарная, идет от затылка
до хвоста, расширяется в крестце и часто охватывает здесь узкую
аптерию.
2. Плечевая птерилия (pteryla humeralis), парная, начинается не-
сколько впереди плечевого сустава и идет через проксимальную
часть плечевого отдела, обычно затем переходя вентрально в брюш-
ную птерилию; этой птерилип принадлежат так называемые лопа-
точные перья, прикрывающие на теле птицы проксимальную часть
сложенного крыла сверху.
3. Бедренная птерилия (pteryla femoralis), парная, идет вдоль
внешней стороны бедра.
4. Брюшная птерилия (pteryla gastraei), парная; крупные пте-
рилии, идущие по бокам туловища от передней области груди почти
до отверстия клоаки; иногда образуют развилок в области грудины;
между собою обе птерилии разделяются широкой apteria mesoga-
straei.
5. Головная птерилия (pteryla capitis), охватывает голову и часто
продолжается по бокам шеи.
6. Крыловая птерилия (pteryla alaris), охватывает всю область
предплечья, кисти и пальцев; к ней относятся все маховые перья
и верхние и нижние кроющие крыла.
7. Голенная птерилия (pteryla cruralis), охватывает внешнюю сто-
рону голени; у птиц с оперенной плюсной продолжается почти до
диетальныл фаланг пальцев.
8. Хвостовая птерилия (pteryla caudae), охватывает заднюю часть
тела; сюда относятся рулевые перья и верхние и нижние кроющие
их; обычно эта птерилия кпереди непосредственно переходит в спин-
ную, брюшную и бедренные птерилии.
9. Анальная птерилия (pteryla ani), вокруг анального отверстия
несет кольцо контурных перьев.
Из аптерий наибольшее значение имеют:
1. Аптерия боков шеи (apteria colli lateralis), парная, является
по существу продолжением аптерии боков тела.
2. Аптерия боков тела (apteria trunci lateralis), парная, наиболее
крупная аптерия; она обеспечивает большую подвижность крыльев
и ног и служит для помещения сложенного крыла.
3. Нижняя аптерия (apteria mesogastraei), непарная, располо-
жена по средней линии между ветвями брюшной птерилии.
4. Спинная аптерия (apteria spinale), непарная и весьма измен-
чивая, в зависимости от формы спинной птерилии.
5. Верхняя крыловая аптерия (apteria alae superiora), парная,
захватывает часть поверхности плечевого отдела и переднюю кры-
ловую перепонку.
6. Нижняя крыловая аптерия (apteria alae inferiora), парная,
захватывает нижнюю поверхность плечевого отдела передней и зад-
ней крыловой перепонки и часть предплечья.
29
7. Голенная аптерия (apteria cruralia), парная, отделяет голен-
вую птерилию от остального оперения задней конечности.
8. Головная аптерия (apteria capitis), обычно разбита на отдель-
ные участки и частью видна снаружи, частью скрыта оперением.
Перья, образующие крыло и хвост. Маховые
перья ио месту их прикрепления к скелету передней конечности
разделяются на большие маховые (remiges primariae) и малые махо-
вые (remiges secundariae или cubitales). Первые прикрепляются
к скелету 2-го и 3-го пальцев и метакарпальным костям, вторые —
к дорзальной стороне локтевой- кости, на поверхности которой они
Рис. 18. Схема прикрепления маховых перьев па скелете передней ко-
вечности.
Счет маЗтых маховых проводится от дистальных к проксимальным, счет
больших — предпочтительно от проксимальных к дистальным; т. р. —
задняя крыловая перепонка (metapatagium); рч р. — передняя крыловая
перепонка (propatagium); ъ. el. — эластическая связка (vinculum elasti-
cum). По Pycraft, из Stresemann.
вызывают экзостозы (рис. 18). Количество больших маховых,, за
исключением нелетающих форм, колеблется в пределах 10—12 перьев.
Количество малых маховых зависит от длины предплечья и поэтому
особенно велико у птиц с скользящим полетом; в общем оно ко-
леблется от 6 (колибри) до 37 (альбатрос). У последней группы
птиц потребность в увеличении планирующей поверхности вызвала
даже превращение контурных перьев заднего края плечевого отдела
в маховые перья, так называемые третьестепенные маховые (remiges,
tertiariae sive humeralis), выполняющие пространство (в виде двух
рядов) между проксимальными маховыми и стенкой тела.
С верхней и нижней сторон основания маховых перьев прикры-
ваются кроющими перьями, соответственно их расположению назы-
ваемыми кроющими больших и малых маховых
(рис. 19). На наружной поверхности крыла, нижний край образуется
30
большими кроющими (tectrices majores), затем рядом средних (сес-
trices mediae) и над ними — несколькими рядами малых кроющих
(tectrices minores). Передний край крыла покрывают более мелкие
краевые кроющие (tectrices marginales), отделяемые от малых обычно
узкой полоской аптерии. Каждому маховому соответствует по одному
перу из всех (трех) кроющих рядов перьев, но прикрепляются они
несколько* дистальнее соответствующих им нижележащих перьев,
так что поперечные ряды имеют вид сдвинутых несколько вперед и
вверх. Эта система распределения кроющих перьев нарушается
интересным, пока не вполне объясненным явлением д и аста-
та к с и и (или аквинтокубитализма) — перазвития пя-
того малого махового при сохранении связанного с ним большого
кроющего (рис. 19). Это явление свойственно как целым системати-
Рис. 19. Схема диастатаксического крыла, рассматриваемого сверху.
At. — крылышко; С. с. —carpal covert; С. г. —carpal remex; J. г. — интеркалярный
ряд; Maj. — большие кроющие; Maj. 5 — большое кроющее перо малого махового; Магд. —
краевые кроющие; Med. — средние кроющие; Мгп. — малые кроющие; Prim. — большие
маховые; Sec. — малые маховЪге; 5 — диастатаксический перерыв. По Steiner.
ческим группам (например гусиные, дневные хищники и др.), так и
отдельным формам в группах, в целом этой особенностью не обладаю-
щих (рис. 225) и следовательно не всегда может иметь существенное
таксономическое значение. У диастатаксических форм перья, распо-
лагающиеся горизонтальными рядами (параллельно оси разверну-
того крыла), делятся на два отдела: проксимальный и дистальный;
последний, по предположению Штейнера (Steiner, 1918), первона-
чально имел свое продолжение в ближайший, нижележащий, прокси-
мальный ряд, от которого он теперь сдвинут вперед, к переднему краю
крыла. Этот сдвиг, по этой гипотезе, произошел в области прикрепле-
ния пятого малого махового, вследствие чего это маховое не разви-
лось. Наиболее заметными следами этого нарушения являются раз-
личие в длине и симметричности кроющих обоих отделов одного и
того же ряда и появление перьев так называемого мштеркаляр-
ного ряда», лежащих между дистальными отделами больших и
средних верхних кроющих малых маховых. Явление эвтаксии
(или кв интокубитализ м*а — сохранения пятого махового
ЗГ
оказывается при этом вторичным, происходящим вследствие уко-
рочения предплечья и замыкания диастатаксического пропуска.
Другой особенностью среди верхних кроющих крыла являются
два пера — carpal гетех и carpal covert, лежащие в области границы
больших и малых маховых перьев и несимметричные ни одному гори-
зонтальному ряду. По объяснению Дегена (fide Stresemann, 1927)
они являются крайне редуцированными первоначальными первым
малым маховым и его большим кроющим. Редукция этих перьев
произошла по всей вероятности вследствие тех сжатий, которые
должно было испытывать перо, лежащее внутри важнейшего сгиба
крыла.
Нижние кроющие крыла располагаются подобно верхним крою-
щим. Замечательной особенностью больших и средних из них (tectri-
ces aversae) является то, что своей верхней стороной (в морфоло-
гическом отношении) они прилегают к коже и следовательно имеют
ориентировку, обратную всем прочим покровным перьям. Эта осо-
бенность объясняется закладкой их эмбрионально на верхней сто-
роне и последующим сдвигом на нижнюю сторону крыла. Свое-
образно затем наличие из средних нижних кроющих только тех,
что соответствуют шести внутренним большим маховым; основания
первых маховых оказываются лишенными их.
Как верхние, так и нижние мелкие кроющие предплечья про-
должаются на плечевой отдел, образуя здесь верхние (tectrices hume-
rales superiores) и нижние плечевые кроющие (t. h. inferiores) перья.
Эти перья часто достигают значительной величины и служат как для
увеличения поверхности крыла, так и в качестве украшац^рцих;
в первом случае особенно увеличиваются верхние кроющие, выде-
ляемые под названием верхних плечевых перьев (par^pteron),
во втором — нижние, известные как подмышечные (hypopteron).
В крыле различают еще небольшую группу перьев, сидящих
на 1-м пальце и называемых крылышком (alula или alaspuria).
Они лежат по переднему краю кистевого отдела крыла и обычно
бывают очень короткие. В крылышке различают кроющие и махо-
вые перья; число последних колеблется от 3 до 6. Значение крылышка
заключается главным образом в задержании стекания воздуха из-
под крыла при взлете, в момент которого у многих птиц оно сильно
оттопыривается и изгибается за край крыла (рис. 164).
Рулевые перья (tectrices) прикрепляются к пигостилю, т. е.
дистальным, слившимся в одну косточку, хвостовым позвонкам,
и количество их обычно соответствует числу слившихся в пигостиль
позвонков: 6—5 пар. У некоторых форм однако число их падает до
4 пар или поднимается до 12 пар, а в некоторых случаях они цочти
совсем редуцируются. Рулевые перья прикрываются в основной поло-
вине верхними и нижними кроющими хвоста (tectrices caudales
superiores et inferiores). Перья, непосредственно прилегающие к ру-
лям сверху, отличаются как большие верхние кроющие; в ряде
случаев они перерастают хвост по своим размерам (павлин).
Окраска наружных покровов. Окраска опере
ния птиц обусловлена как пигментами, так и структурными особен-
ностями пера, являясь наиболее сложной при сочетании обоих этих
32
моментов. Цвета, вызываемые пигментами, обусловлены главным
образом меланинами и липохромами. Первые имеют черноватую пли
коричневатую окраску и форму зерен пли палочек, вторые — жел-
тую, красную п иногда синюю и зеленую окраску и пли растворены
в '/Кировых капельках пли (реже) в виде хлопьев залегают в массе
рогового вещества. Образование меланина происходит пз мптохонд-
риев особых амебоидных клеток, развивающихся в перовом сосочке
пз промежуточных клеток мальппгпевого слоя. Эти амебоидные
клетки посредством длинных отростков доставляют меланиновые
зерна в клетки пера, прежде чем процесс ороговения их будет закон-
чен, Гернптцем (Gornitz, 1923) было установлено наличие двух типов
меланина: имеющего форму палочек — еумеланпна, и встречаемого
в виде круглых зерен — феомеланпна. Первый вызывает черные и
серые оттенки, второй коричневые п буроватые. Образование липо-
хромов не выяснено, по материнской субстанцией для них считается
кориосульфурин, широко представленный в жировых тканях раз-
нообразных птиц. Липохромы, встречаемые в птичьих перьях, ока-
зываются весьма разнообразными. Наиболее распространенными
являются: желтый зоофульвин (например в оперении иволги —
Oriolus oriolus L.) и красный зооэритрин (самец чечевицы — Erythrina
t’rythrina Pall.); реже встречаются липохромы зеленых тонов —
зоопразшшн (например на голове самца обыкновенной гаги — So-
muteria ntollissima L.), тураковердпн (преобладает в оперении бана-
цоедов — Turacns), фазиановердин (известен для сермунов — Itha-
ginis) и фиолетовых, например фазпанорубпн (у тех же Ithaginis).
Как указывалось, окраска пера обусловлена рядом факторов,
которые не всегда могут быть выяснены при одном макроскопиче-
ском изучении его. Наиболее обычные окраски вызываются сле-
дующими условиями (Stresemann, 1927): черная происходит вслед-
ствие густого наслоения еумелапипа; при более разреженном рас-
положении его зерен окраска получается черно-коричневой. Оттенки
от темнокрасно-коричневого до темнокорпчнево-желтого вызываются
густым залеганием феомеланпна, наоборот, оттенки светлоржав-
чато-корпчневые до глинисто-желтых обусловлены рыхлым распо-
ложением того же пигмента. Серый цвет вызывается отсутствием
пигмента в бородках первого порядка и базальных частях бородок
второго порядка при густом переполнении, еумеланином концевых
частей (pennulum) последних бородок. Тот же характер распреде-
ления пигмента, но уже феомеланпна, вызывает пустынную (тусклую
желто-бурую) окраску. Оливково-бурые и серо-бурые перья обуслов-
лены сочетанием обоих меланинов. Желтый, оранжевый и красный
тона вызываются соответствующими липохромами, обычно диффузно
пропитывающими корковый слой бородок. Белый цвет обусловлен
выполнением воздухом бесцветной роговой массы пера, главным
образом центральных клеток бородок и стержня. Голубой, в огром-
ном большинстве случаев/является цветом, вызванным структурными
особенностями, общими и большей части зеленых, синих и фиолето-
вых окрасок. Эти особенности, как правило, заключаются в значи-
тельной толщине стенок центральных клеток бородок, включающих
внутри себя пространство, наполненное воздухом; стенки этих кле-
3 Орнитология—12П9
33
ток прооодены очень оолышш числом выполненных воздухом ради
альных канальцев, в среднем слое стенок образующих густое ’.спле-
тение между собою; толщина этих канальцев стоит в определенные
отношениях к окружающей роговой массе (рис. 20). Преломление
и отражение лучей через эту мутную среду дают эффект слабогс
голубоватого блеска при общей белой окраске пера. Густые яркие
цвета получаются при скоплении пигмента на вентральной стороне
бородки, действующей как отражающая лучи ширма. Голубой, цвет
вызывается при этом скоплением здесь меланина, фиолетовый — еуме-
ланпна пли феомеланпна, в разных концентрациях. Зеленый полу-
чается комбинированием этой структуры с желтой окраской дорзаль-
ного коркового слоя бородки. Кроме того, тот или иной цветной
эффект получается комбинированием в одной бородке липохрома и
меланина пли вызывается
Рис. 20. Строение бородки голубого пера тана-
гры — Thraupis sp.
а — поперечный разрез голубой бородки первого
порядка; с — толстый роговой наружный слой;
т—толстостенные клетки с центральной возду-
хоносной полостью; р — слой пигмента;
Ъ —разрез толстостенной клетки из бородки пер-
вого порядка; t—воздухоносная центральная
полость клетки; i — стенки клетки, пронизанные
воздухоносными канальцами, образующими в
средней зоне стенки сложное сплетение. По
Knieschc и Rcnsch, из Strcseniaim.
одним из типов липохрома,
как указывалось выше.
Блестящие, призирующие
цвета птиц вызываются, глав-
ным образом структурными
изменениями бородок второго
порядка, вызывающими явле-
ние интерференции света.
Бородки оказываются сильно
расширенными и повернуты-
ми к наблюдателю своей шп‘-
рокой стороной. При этом
чрезвычайное переполнение
пигментом клеток ведет за
собой сильное утончение бес-
цветной роговой оболочки бо-
родки (вследствие растяже-
ния ее), становящейся на-
столько тонкой, что она начи-
нает ирпзировать, как стей-
ки мыльного пузыря. По из-
мерениям Ельзессера (Elsiisser, 1924), синпй цвет у блестящего
фазана (Lophophorus impeyamis Lath.) вызывается оболочками, имею-
щими 0,20 микрона толщины. Основным пигментом, выполняющим
клетки, является еумеланин и изредка феомеланин (например у фа-
занов). Альбиносы нормально иризпрующих птиц не блестят, что
очевидно объясняется отсутствием механических причин к растя-
жению оболочек клеток до необходимой тонкости их (рис.21). У птиц
с особо блестящим оперением (например блестящие фазаны, нектарки)
иризирование дает эффект Ньютоновых колец; коэффициент преломле-
ния при этом оказывается очень высок — у Lophophorus до 1,8, и
приближается к коэффициенту преломления бриллиантов (2.4)..
Окраска 'пера, по окончании периода роста являющегося мерт-
вым образованием, отражает то физиологическое состояние, в кото-
ром находился организм в период роста пера, поэтому задержка
развития пера, искусственное стимулирование деятельности перо-
34
вогб сосочка, вне обычного периода линьки и во время формиро-
вания организма, вызывает образование отличного рисунка и окраски.
В опытах Биба с Piranga erythromelaena Licht, п Doliehonys oryzi-
vorus L., имеющих резко различные по сезонам наряды, брачный и
зимний, удалось задержать осеннюю линьку. Птицы проносили
до весны яркий брачный наряд, а затем впали в линьку и из брачного
наряда перелиняли в брачный же, что указывает, что окраска опе-
рения обусловлена внутренним физиологическим ритмом организма.
Контрольный экземпляр Piranga erythromelaena Licht., допущенный
до линьки в середине зимы, перелинял, как и следовало ожидать,
в зимний наряд (Beebe, 1914). На этих яге основаниях долго рас-
тущие крупные птицы
обычно имеют хорошо вы-
раженные возрастные на-
ряды, промежуточные ме-
жду нарядом птенца и ста-
рой птицы, а молодые ра-
стущие птицы в позднее
появившихся перьях обна-
руживают более прогрес-
сивную окраску и рису-
нок.
Роговые покровы клюва
и ног, верхний слой кото-
рых снашивается посте-
пенно плп сбрасывается
периодически, нередко бы-
вают различно окрашены
1а)
по сезонам, выражая этим
различный внутренний
ритм организма. Окраска
бывает более яркой, пре-
имущественно желтой плп
красной в брачный период,
и темной или бледной —
Рис. 21. Строение бородок грудного пера норма.пню
окрашенного скворца — Shirans vulgaris L. (h) и
скворца альбиноса (а).
Клетки» дистальных бородок второго порядка у нор-
мально окрашенного скворца сильно расширены
вследствие скопления мелашша и призируют; у аль-
биноса они сохраняют типичное для дистальных бо-
родок строение, узки, несут крючочки п пе вызывают
Никакого блеска. По Renscli. из Stresemann.
зимой. Эти окраски обычно обусловлены липохромами. Изменчивая
окраска голой кожи, папрпмер у грифов Старого и Нового Света,
вызывается преимущественно или исключительно приливом плп
отливом крови.
К о ж н ы е ж е л е з ы. Кожа птиц совершенно лишена пото-
вых желез и вообще характеризуется крайней бедностью каких-
либо железистых образований. Наиболее постоянной п крупной
железой является лишь копчиковая железа (glandula uropygii),
представляющая компактное, болыйей частью явпо двудольчатое
тело, лежащее сверху на последних хвостовых позвонках у самого
основания рулевых перьев (рис. 22). Расположенный в виде выступа
выводной проток ее направлен в сторону хвоста и несет обычно на
конце группу кпеточковых перьев. Внутри железа состоит из двух
долек, слагаемых простыми радиально-расположенными каналь-
цами, впадающими в общий или двойной резервуар. Жировой секрет
35
железы вырабатывается мелкими капельками в эпителиальных клет-
ках канальцев п поступает в резервуар, при распадении образуя
однообразную зернистую массу. Выделение секрета наружу через
выводной проток происходит плп давлением клюва птицы на железу
пли действием кольцевой мускулатуры железы; выдавливающиеся
при этом капли прожиривают кпсточковые перышки, которые сма-
Рис. 22. Копчиковая железа у представителей разных отрядов.
а — оляпки — Cinclus cinclus L. (Passeriformes); Ъ—пеликана — Pelecanus sp. (Ре-
lecanifurmes) ;c—черного стрижа — Micropus apiis (L.) (Micropodii formes), в разрезе; d и e
удода — Upupa epops L. (Coracii formes). Обратите внимание на различие в строении желез:
е — с общим выводным протоком для лопастей; с — с обособленным протоком для каждой
.лопасти; ъ — с широким полем выводных протоков, изображенных в виде черных щелевид-
ных отверстий и окруженных кисточковыми перышками, обрезанными у самого основания.
— выводной проток; I. — сосок' железы; т.— собирательный резервуар железы. По
Paris и Schumacher, из Stresemann.
зывают клюв птицы. Основное Назначение секрета, по мнению неко-
торых авторов, заключается в ожирпванпп оперения и предохране-
нии его этим от намокания. В связи с этим железа наиболее хорошо
развита у водоплавающих птиц и слабо развита у. птиц с рыхлым
оперением. .В ряде групп (голуби, дрофы, некоторые попугаи) ее
функция вообще повпдпмому замещается функцией порошкового
пуха.
Зб
Кожная мускулатура. Разнообразные движения перьев
тела, выражающие обычно самочувствие и возбуждение птицы,
обусловлены кожной мускулатурой, двигающей птерилии (musculi
pterylarum) и мелкими мускулами, двигающими отдельными перьями.
Мускулы, двигающие птерилии, являются производными и тесно
связанными со скелетной мускулатурой и имеют вид тонких, отхо-
дящих от нее, лент и пучков. Наиболее значительными из них яв-
ляются: musculus subcutaneus thoracis, двигающий кожу груди,
in. s. abdominis, двигающий кожу брюха, оба отходящие от m. pecto-
ralis; к коже шеи отсылают мускульные пучки — m. sternohyoidens
и in. cucullaris; последний связан также с теменной, плечевой и спин-
ной птерплиями, которыми двигает также m. latissimus dorsi.
Рис. Диаграмма расположения перьев на коже н норовой му-
скулатуры курицы — Gallus aallus L.
а — положение рядов перовых сосочков па дорзальной (правой) сто-
роне шеи п направление залегания перьев (тонкие линии); b — му-
скульные пучки перьев, видимые сверху, по снятии кожи; с-— схема
всех мускулов, прикрепленных к одному перу (лежащему центрально).
а. I. — антеро-латеральпый ряд перьев; /. — продольный ряд перьев;
р. I.—постеро-латеральный ряд перьев; d.— мускул, опускающий
и прижимающий перо к телу, in. depressor; р. — поднимающий
перо. in. erector; г. — сближающий перья между собой, m. retractor.
По Langley.
В пределах птерилий основания контурных перьев связаны между
собой в целую систему очень короткими и топкими гладкими мускуль-
ными пучками, иннервируемыми веточками симпатического нерва
(рис. 23). Контурные перья укрепляются в коже в шахматном по-
рядке в косом положении, с вершиной, обращенной в направлении
к хвосту; внутри птерилии основание каждого пера оказывается
окруженным шестью другими очинами, которые и соединяются между
собой то косо, то перпендикулярно к очинам протянувшимися пуч-
ками. "Среди последних различают наиболее сильные пучки, сокра-
щение которых опускает перо (m. depressores), пумки, поднимающие
перо (т. ereclores), и пучки, сближающие основания перьев (т. ге-
tractores). В совокупности действие этих мускулов обусловливает,
например, поднимание и опускание хохлов, ерошенье пера, приле-
гание перьев к телу у птицы, испытывающей страх, пт. и. Отдельные
птерилии отличаются различным участием и развитием этих муску-
лов.
2. СКЕЛЕТ
В формах и свойствах скелета птиц, пожалуй, наиболее ярко
проявляются преимущественно воздушный образ, жизни представи-
37
телей этого класса и его генетическая близость к наземными пойкило-
термным пресмыкающимся. Легкость п крепость костных элементов
п резко выраженные морфологические приспособления к полету
Рис. 24. Скелет голубя, представителя одного из наиболее высоко стоящих филогенетически
отрядов птиц, в сравнении с реставрированным скелетом первоптицы — Arcliaeornis, в обилии
сохраняющим рептильные черты организации.
Сравните строение черепа, крыла, плечевого пояса п грудины, таза, плюсны и хвоста,
с. — кости запястья; cl. — ключица; со.-— коракоид; d.— пальцы; /. — бедро; /i. — малая
берцовая; h.— плечевая кость; i.—подвздошная кость; is. — седалищная кость; тле.—
кости пястья; mt. — плюсневые кости; р. — лобковая кость; ру. — пигостиль; г. — лучевая
кость; s. —лопатка; st. — грудина; tm. —плюсна или цевка, tarsometatarsus; tt. — tibio-
tarsus; и. —локтевая кость; v. — брюшные ребра; I—IV — номера пальцев. По Hellmann.
сочетаются в скелете с явным сохранением следов прошлого родства,
проявляющегося преимущественно в органах, менее затронутых
воздушным образом жизни (рпс. 24).
В целом костные ткани птиц характеризуются преобладанием
плотных структур>над губчатыми, значительной крепостью и проч-
38
Рпс. 23. Сильно пневматизированный череп тукана.
а — череп но снятии крыши клюва; Ь — продольный раз-
рез головы. По Marshall, из Hi 1 zlieinier.
ноитью, связанной с высоким содержанием солеи (наивысшим среди
позвоночных; Selenka, 1875), и исключительной пневматичиостыо,
делающей скелет весьма легким образованием.
Пневматичность создается проникновением воздуха внутрь кости,
обычно в виде выростов воздушных мешков, заполняющих внутрен-
нюю полость органа; по некоторым данным (Kennel, 1910), воздух
проникает внутрь кости даже вне воздушного мешка. Развитие пнев-
матичностп костей имеет место в эмбриональный и постэмбриональ-
ный периоды и сопровождается резорпцпей костного мозга и спон-
гпознон ткани; проникновение воздуха происходит через мелкие
отверстия (foramen pneumaticuin), образующиеся обычно в углубле-
ниях поверхности ко-
стей-.
Как правило, ко-
сти крупных птиц и
самые крупные кости
скелета вообще явля-
ются 'более пневма-
тпчнымп, причем наи-
более пневматпчным
оказывается обычно
скелет хорошо лета-
ющих птиц (например
фрегата), у которых
проникновение воз-
духа распространяет-
ся в некоторых слу-
чаях даже на конеч-
ные фаланги пальцев
(рис. 25). В теле птиц
различают три систе-
мы воздушных кост-
ных полостей, связан-
ных между собой. Воздушная система костей туловища и конечно-
стей! находится в связи с легкими, системы же черепных и нижне-
челюстных костей связаны с слуховой капсулой и носовой областью
и наполняются воздухом независимыми путями. Последние системы
наиболее широко распространены среди птиц, тогда как воздушная
система костей туловища и конечностей у ряда групп отсутствует
(например у гагар и киви), вероятно в связи с специализированным
водным пли наземным образом жизни.
Следует отметить, что развитие кости птиц гистологически проте-
кает отлично от развития кости у млекопитающих. Окостенение
у птиц происходит по поверхности хряща (перпхондрально), как
выстилка мозговой полости, а не внутри хряща (эндохондрально),
как это характерно для указанной группы (Lubosch, 1924).
Переходим к рассмотрению отдельных частей скелета.
Череп. В черепе птиц бросается в глаза своеобразная форма —
сильно суженная, вытянутая в виде клюва кпереди, огромные орбиты
с весьма тонкой межглазничпоп перегородкой и сильно вздутая череп-
39
пая коробка, сдвинутая назад за орбиты. Характерным является также
раннее слияние костей •черепа до полного исчезновения швов между
ними. По сравнению с черепом раптплпй, череп птиц отличается:
Рис. 26. Череп голуби и примитивной птицы
Archaeornis в сравнении с черепом Evparheria. саpen-
sis (Pseudosuchia, Repl ilia).
a — череп голубя; b — верхняя часть клюва утенка
красноголовой черпстп — Nyroca ferina; с — реста-
врированный череп Archaeornis; d — череп Е и par he-
.ria;e— склеротикалыюе кольцо рептилии Thalatto-
sanrus alexandrae (из триасовых отложении).
А.—угловая кость; А1.—околослезпая кость
(adlacrymale); Аг. — сочленовная кость; As. — кры-
локлпновидная кость; Во.—Основная затылочная
кость; С. — затылочный мыщелок; D. — зубная
кость; F.—лобная кость; J. — скуловая кость;
L. — слезная кость; Мл*. — верхнечелюстная кость;
W. —носовая кость; О. — всрхнсзатылочпая кость;
Ор.—заднеушпая кость; Р.—теменная кость;
Ра. — небная кость; Pf. — заднелобная кость: Рт.—
межчелюстная кость; Ро.—заглазнпчпая кость;
Pt- — крыловидная кость; Q.—квадратная кость;
Qi- — квадратпоскуловая кость; S. — чешуйчатая
кость; Sg. — надугловая кость; Sp. — пластинчатая
кость. Пз Hellmann.
значительным увеличением
мозговой коробки (рис. 26);
образованием обширных
глазничных ям, сдвигаю-
щих мозг кзади и вызы-
вающих слияние дна глаз-
ниц (orbitosphenoidalia) в
цельную, частично или
полностью окостеневаю-
] цую, межгла зничн ую пе-
регородку (septum interor-
bitale); редукцией верхней
скуловой дуги п ямы. со-
пряженной с перемеще-
нием мускулов, прикреп-
ляющихся в этой области у
рептилий па более задние
отделы черепа; изменением
переднего отдела черепа,
связанным с сильным раз-
растанием межчелюстных
костей (praemaxillare) за
счет уменьшения челю-
стных (maxillare) и в связи
с этим удлинением этого
отдела;редукцией зубного
аппарата с образованием
роговых покровов — клю-
ва; ранним замещением
примордиального (хряще-
вого) черепа костями, еще
в молодом возрасте слива-
ющимися между собой и
образующими, например
вокруг черепной полости,
одну цельную кость, и ря-
дом других, особенностей.
Вместе с, тем., по харак-
теру костных элементов,
слагающих череп, и соотно-
шениям многих из них меж-
олизок к черепу рептилий и относится к
ду собой, череп птиц очень
тому же тропибазальному и
мезокинетическому типу. Наибольшее общее сходство черепптицобна-
руживает с черепом вымерших рептилий группы Ple.rosauria (Fiirbrin-
ger, 1888). У последних череп, так же как у птиц, характеризовался
относительно крупной мозговой полостью и крупными орбитами'
40
значительной пневматнчностью костей и ранним синостозом их
между собой, удлинением предчелюстных костей и образованием
частичного (Rhamphorhynchiis) или полного (Pleranodon) роговогб
покрова клюва и т. гг., при наличии и чисто рептильных черт (верх-
них височных ям и др.). Птерозавры вели летающий образ жизни,
и это заставляет видеть в аналогичных чертах строения черепа у птиц
и рептилий приспособление к передвижению в воздухе, так как
птерозавры (судя по другим важным особенностям строения) не
родственны птицам и следовательно приобрели общие черты конвер-
гентно, как приспособление к сходному образу жизни. Значение
этих черт для полета в воздухе отмечалось выше, в общей характери-
стике. Слияние костей также может быть связано с облегчением
черепа, так как сочленение посредством швов требует большей тол-
щины костей: кости черепа частично не сливаются между собой
у страуса и пингвинов, ведущих наземный или водный, но не летаю-
щий образ жизни.
По своему развитию в онтогенезе, в черепе птиц, как и других
позвоночных, различают кости хондрального (развивающиеся из
хряща первичного черепа) и накладного плп покровного (непрефор-
мированного хрящем) происхождения. К первым в первичной череп-
ной коробке (neuralcranium) относятся: в затылочной области —
1 основная затылочная (basioccipitale), 2 боковые затылочные (ехос-
cipitalia) и 1 верхнезатылочная (supraoccipitale); в ушной области —
2 переднеушные (prooticum), 2 заднеушпые (opisthoticum) и 2 верхне-
ушные (epioticum); в глазнично-височной области — 1 основная
клиновидная (basisphenoidale), 2 крылоклиповидные (alispheuoi-
dalia), 2 глазничноклпновндные (orbithosphenoidalia), 2 переднекли-
новидные (praesphenoidalia), 1 средняя обонятельная (mesethmoi-
deum); в обонятельной области — несколько мелких, окостенений,
развивающихся из самостоятельных центров. К накладным костям
этого отдела относятся: 2 теменные (parietalia), 2 лобные (frontalia).
2 носовые (nasalia), 2 предлобные (praefrontalia), 2 чешуйча-
тые (squamosum), 1 парасфеноид (parasphenoideum) и 1 сошник
(vomer). Последние две кости закладываются тремя пли двумя
(сошник) элементами1, которые впоследствии сливаются в одну
кость.
В висцеральном отделе черепа (splanchnocranium), к которому
относятся: кости верхней челюсти — парные межчелюстные (ргае-
maxillaria) и верхнечелюстные (maxillaria); кости, развивающиеся
на небноквадратном хряще (palatoquadratum). — парные небные
(palatina), крыловидные (pterygoidea), квадратные (quadrala), ску-
ловые (jugalia пли zygomaticum) и квадратноскуловые (quadrato-
,jugalia), и кости нижней челюсти — парные угловые (angularia).
надугловые (supraangularia), сочленовные (articularia), зубные (don-
talia), подбородочные (mento-mandibularia, развиваются лишь у не-
которых дневных хищных птиц из подотряда Accipitres), пластинча-
тые (splenialia) и венечные (complementaria плп gonialia, во многих
группах отсутствующие); большинство костей являются наклад-
ными; хрящем преформпрованы лишь квадратные, сочленовные и
подбородочные кости.
4J
Переходим к более детальному рассмотрению строения черепа
(рис. 27).
В задней области черепа вокруг отверстия (foramen magnum)
мы находим у птиц обычные для позвоночных четыре затылочные
кости: нижняя пли основная (basioccipitale), две боковых
(exoccipitalia) и в е р х и я я (supraoccipilale). Основная кость у ниж-
него края затылочного отверстия несет у птиц, как и у рептилий,
единственный з а т ы л о ч н ы й мыщел о к (condylus occipita-
lis), в образовании которого у большинства птиц принимают участие
и боковые затылочные кости (см. рпс. 27). У прогрессивных групп птиц
затылочное отверстие направлено на нижнюю сторону черепа (осо-
бенно у крупноглазых видов — совы, куликов и др.), в то время как
у групп примитивных оно сохраняет положение, свойственное реп-
тилиям, т. е. направлено назад, занимая, например у гусей, почти
вертикальное положение по отношению к осп черепа.
Вследствие большой величины мозга, слуховые капсулы, поме-
щавшиеся у рептилий по бокам черепа, оказываются у птиц сдвину-
тыми далеко вниз. В образовании слуховой капсулы принимает
участие ряд костей: слуховые (periotici) — наиболее крупная передне-
ушная (prooticum). п мелкие — заднеушная (opistlioticum) и верхне-
ушная (epioticum), рано сливающиеся с соседними костями (epioti-
cum с supraoccipilale и opistlioticum с exoccipitalia) и между собой
в одну сплошную массу, создающую преимущественно дно слуховой
капсулы; затем в образовании наружных стенок ее участвуют:
сверху — чешуйчатая кость (squamosum) и верхнезатылочная (пз-
внутри), сзади — боковая затылочная (с ней сливается, образую-
щееся самостоятельно, затылочное крыло processus paroccipi-
talis), прикрывающая сзади слуховую полость, и снизу — частью
основная клиновидная кость (basisphenoidale), частью парасфеноид,
в его задней пластинчатой части (hasitemporals).
В глазнично-височной области черепа в основании лежит основная
клиновидная кость, соединяющаяся передним концом с клювом па-
расфеноида (rostrum parasphenoid ale) и иногда сочленяющаяся с кры-
ловидной костью (pterygoideum) посредством крыловидных отрост-
ков (processus pterygoideus пли processus basi plfcrygoideus).Последнее
явление характерно для ряда групп рептилий и у птиц наблюдается
преимущественно среди примитивных представителей пли сохра-
няется лишь в онтогенезе (см. рис. 31—34). Некоторые различают пер-
вичные базиптеригопдные отростки, истинно гомологичные соответ-
ствующим отросткам рептилий и свойственные представителям Ра-
laeognathae, Serpentarius и ряду других групп, и вторичные, негомо-
логичные рептилиям, располагающиеся на клюве парасфеноида зна-
чительно дистальнее и свойственные, например, куриным, утиным
и некоторым другим группам (Gaupp, 1902; Bohm, 1930). Сбоку
к основной клиновидной кости прилегают парные боковые клиновид-
ные (alisphenoidalia), образующие значительную .часть задней стенки
глазниц; по их свободному краю располагаются задние глазничные
отростки (processus orbitalis posterior). Эти отростки некоторыми
исследователями (Bittner, 1912) гомологпзируются с заднелобными
костями (postfrontale) многих рептилий, играющими важную роль
4 2
в)
Рис. 27. Черен молодого гуся — Anser anser L., сильно увеличенный.
а — сбоку; Ъ — снизу; с — сзади; d — сверху.
А— угловая кость; Аг — сочленовная кость; As — крылоклиновидная кость; Йо —
основная затылочная кость; Bsh — основная клиновидная кость; D — зубная кость; Е'—
обонятельная кость; Ео — боковая затылочная кость; F—лобная кость; 1р — межтеменная
кость; J — скуловая кость; L — слезная кость; Мх— верхнечелюстная кость; N— носо-
вая кость; О—верхнезатылочная кость; Ор— заднеушная кость; Р— теменная кость;
Ра— небная кость; Ртп — межчелюстная кость; Ро — заднеглазничная кость; Pt — крыло-
видная кость; Q — квадратная кость; Qj — квадратно-скуловая кость; It — клюв парасфе-
ноида; S —чешуйчатая кость; Sa—надугловая кость; V'—сошник. По Hellmann.
в образовании височной дуги. По мнению некоторых авторов (Huene,
Abel), височные ямы образуются этими отростками и у некоторых
современных птиц (см. ниже).
Парные передние и глазничные клиновидные кости (ргае- и orbito-
sphenoidalia) сливаются друг с другом в большей их части и образуют
значительную часть межглазшгчной перегородки (septum interorbi-
tale), частью неокостеневающей. Переднюю часть последней со-
ставляет обонятельная кость (mesethmoideum), разви-
вающаяся из нескольких центров окостенения и частью остающаяся
па стадии хряща. Эта косточка образует в н о с о в о й п о л о с т и
у одних групп птиц продольную носовую перегородку (septum nasi),
не развиваясь полностью у других групп п оставляя свободный про-
ход между носовыми полостями; это приводит к образованию сквоз-
ных (nares perviae) пли несквозных ноздрей (nares imperviae). что
является существенным систематическим признаком. Следует отме-
тить, что развитие крыловидных и глазничных клиновидных костей,
так же как образование окостенений в , обонятельной области. —
прогрессивное явление для птиц, почти не отмечаемое для рептилий
(у последних mesethmoideum полностью отсутствует). У низших
групп птиц (гусиные, фазановые и др.) носовая перегородка также
часто остается на хрящевой стадии. У всех высших птиц (надотряд
Neognathae) наблюдается также разделение обонятельной: косточки
черепно-лицевой щелью (fissura craniofacialis) на отделы
собственно носовой и межглазничный, что способствует отгибанию
верхней челюсти кверху при’ открывании клюва.
Сошник, иногда полностью отсутствующий, развивается в онто-
генезе как парная кость (у рептилий он таков и в дефинитивном со-
стоянии), впоследствии сливающаяся. Обычно сошник имеет палочко-
видную форму и соприкасается передним концом с предчелюстными
(praemaxillare) пли небными отростками ч е л ю с т н ы х
косте й (processus palatini maxillae), соединяясь задним концом
с клювом парасфеноида пли с неб и ы м и к о с т я м и
(palatina). Парасфеноид распадается на два отдела: один, возникаю-
щий из особого зачатка, есть клюв парасфеноида (rostrum parasphe-
noidale), имеющий форму заостренной балки, проникающий вперед
до носовой полости и часто сочленяющийся с крыловидными костями,
другой, возникающий из парного зачатка, есть basitemporale —
тонкая пластинка, лежащая у основания клюва и налегающая на
пижнюю поверхность основной клиновидной кости. У некоторых
групп basitemporale развивается весьма сильно и образует напра-
вленный вниз сосцевидный отросток (processus mammillaris): в этих
случаях создаются взаимоотношения костей, напоминающие поло-
жение basitemporale у Plerosauria.
Крыша черепа слагается из парных теменных, лобных, носовых,
чешуйчатых и передиелобных костей. Теменные кости (parietalia)
ограничивают заднюю часть черепной полости и нередко являются
местом прикрепления челюстной мускулатуры, образуя заметно
выступающие гребни (linea temporalis, рис. 28). Очень крупные
лобные кости (frontalia) ограничивают черепную полость сверху,
образуют крышу над глазницами и на переднем крае составляют шов
44
с костями, образующими клюв. Здесь же иногда располагаются кост-
ные наросты, являющиеся основаниями шлемов или рогов, имею-
щихся у некоторых птиц (Сasuaridae, Biicerotes), а окологлазничная
часть у водоплавающих птиц и ржанкообразных нередко несет особую
скульптуру вследствие мощного развития носовых желез, лежащих
у этой группы над орбитами. Носовые кости (nasalia) зна-
чительной частью своей поверхности образуют крышу носовой:
полости. Эта часть называется лобным отростком (processus frontalis),
высылающим кпереди два меньших отростка, охватывающих на-
ружные носовые отверстия сверху, — межчелюстной отросток (рго-
iOj
Рис. 28. Черен большого баклана — Phalacrocorax carbo L. (а) и
олуши — Sula bassana L. (b).
Обратите внимание на сложную форму задней части черепной ко-
робки и наличие гребней, к которым прикрепляется мощная че-
люстная мускулатура; к supraoccipitalc баклана прилегает ппра-
мпдообразная сесамовидная, косточка (отмечена звездочкой), увели-
чивающая область прикрепления мускулов, поднимающих че-
люсть. В черепе олуши звездочка указывает область изгиба ветви
нижней челюстц, позволяющую широкое раскрытие клюва. По
Brandt, из Stresemann.
cess us praemaxillaris) и сзади и отчасти снизу — челюстной отросток
(processus inaxillaris). Характер краев этих отростков,ограничиваю-
щих носовое отверстия, имеет существенное систематическое зна-
чение. Различают (Garrod, 1873) голоринальный тип, когда задний
край носовых отверстий закруглен, п шизоринальный, когда эти
отверстия имеют форму вытянутой кзади щели, сходящейся своими
краями кзади от. шва, между носовыми и входящими между ними
отростками предчелюстных костей (см. рис. 27). Голоринальный тип
является более первичным (Gadow, 1893) и свойствен преимуще-
ственно примитивным птицам (пингвины, буревестники,гагары и др.)
(рис. 29):
45
Рис. 29. Шизорпнальпып (а) п голори-
нальпып тип (Ь) в строении носовых по-
стен:
а — череп большой выпи — Rotavrus siel-
laris L.; b — череп большого кроншне-
па— Numenius arquata L.; звездочка —
задний кран посовых отверстий. (Ориги-
нальный рисунок.)
П р е д л о б н ы е кости (ргае-
1‘rontale) расположены по перед-
нему краю глазниц и образуют за-
щиту глаз спереди и отчасти свер-
ху. Обычно они образуют два
отростка: надглазничный (proces-
sus supraorbitalis) — по верхнему
краю глазниц, и предглазничный
(processus orbitalis anterior) — по
переднему краю глазниц, иногда
дополняемые мелкими, прилегаю-
щими костяными пластинками
(supra- или infraorbitalia), как это
свойственно некоторым рептилиям,
например динозаврам. Некоторы-
ми авторами предлобные кости
гомологпзпруются с слезными ко-
сточками (lacrymale) высших поз-
воночных.
Чешуйчатые кости (squamosum)
покрывают большую часть ушных
костей, налегают на края т е-
м е н н ы х и б о к о в ы х к л п-
н о в и д н ы х и по своему верх-
нему краю отчасти включаются в
стенку черепной полости. Нпж-
ппй отдел этой кости участвует в
образовании двух отростков, на-
правленных вниз: целиком ви-
сочного отростка (processus tem-
poralis) и вместе с крылоклпно-
впднымп (alisphenoideum) — зад-
неглаэпичного отростка (processus
orbitalis posterior плп sphenoti-
cus), возможно закладывающегося
самостоятельно и только вторично
срастающегося с черепом (Bitt-
ner, 1910). Последний отросток,
у некоторых птиц направляясь
кпереди, срастается с переднеглаз-
ничным (processus orbitalis ante-
rior, см. выше), образуя под глазом
полную подглазничную дугу (дре-
весные утки, многие попугаи и
др.). У других птиц (куры, аисты,
жаворонки и др.) за дне глазничный
и височный отростки сходятся друг
с другом, образуя замкнутое или
полузамкнутое отверстие, напо-
минающее височную яму и дугу;
46
за таковые оно и принимается некоторыми авторами (Ннепе, 19Г<:
Abel. 1919). Однако Генльман (Heilmann, 1926) отрицает гомологию
этих образований, так как у рептилий височные ямы ограничиваются
главным образом теменной костью и верхней частью чешуйчатой, не
принимающих участия в височном отверстии у птиц; при этом у птиц
яма и дуга развиваются лишь с возрастом и имеют своим назначением
защиту височного му скула, проходящего с передне-боковой поверх-
ности чешуйчатой кости (squamosum) в направлении к нпжнеп че-
люсти (рис. 30).
Рис. 30. Верхняя височная яма и дуга у птиц и рептилий.
Задняя часть черепа: 1 — Aetosaurus lerralus (Pscudosuchia. Reptilia); 2— аист попру —
Jabiru тус ter ia (Licht.); 3 — полярная гагара — Colymbux glacialis L.; 7 — фазан — Phasia-
hus colchicus L.. взрослый; 5 — фазан молодой, почти полного роста; <;—обыкновенная
курица. Обратите внимание на изменение формы яму,и -участие в ее образовании различных
элементов черепа.
As. крылоклнпопидная кость; F.—лобная кость; *Р. —теменная кость; Pf.— зад-
нелобная кость; Ро. — заднеглазничная кость: S. — чешуйчатая кость; Т. — височный
мускул. Из Hellmann.
По заднему краю чешуйчатой кости у ряда групп (преимущественно
примитивных) наблюдаются фонтапеллп, образующиеся вследствие
неполного смыкания костей вокруг хрящевого черепа.
В образовании клюва участвуют: непарная межчелюстная и пар-
ные челюстные, квадратные, крыловидные, небные и квадратно-
скуловые кости. М е ж ч е л ю с т и а я кость (praemaxilla) закла-
дывается парной, но очень скоро сливается в одну кость, в дальней-
шем принимающую вид плотного переднего тела, определяющего-
форму клюва и тремя отростками соединяющегося кзади с следую-
17
щимп костямп черепа: с лобными (посредством лобного отростка —
processus frontalis, который проникает между носовыми костями
и обычно срастается с лобными костямп), с челюстными (посредством
одноименного отростка — processus maxillaris) и с небными (также
посредством одноименного отростка — processus palatinus, уча-
ствующего в образовании костного неба).Относительно крупные раз-
меры межчелюстной кости являются характерной чертой птиц в срав-
нении с большинством рептилий. Челюстная кость (махП-
1аге) образует сравнительно небольшую часть краев надклювья,
частью вместе с челюстным отростком (processus maxillaris) преды-
дущей кости. Для этого элемента характерно образование по внутрен-
нему краю небного отростка (processus palatinus rpaxillae
или maxillo-palatinum), участвующего в образовании костяного
неба, и образование скулового отростка (processus zygomaticus),
направленного кзади и связующего эту кост^ со скуловой дугой.
Последняя или, иначе, к в а д р а т и о-с к у л о в а я кость (quad-
rato-jugale) образует как бы тонкий мостик между челюстной и
квадратной костью (quadratum) и в середине усиливается
тонкой накладной скуловой косточкой (zygomaticum пли jugale).
Этот ряд костей образует нижнюю височную дугу, характеризую-
щуюся в сравнении с гомологичной дугой рептилий тонкостью кост-
ных элементов и частью слиянием их между собой.
Квадратная кость (quadratum) имеет неправильную четырехуголь-
ную форму и служит средством прпчлененпя нижней челюсти к че-
репу и упором для квадратно-скуловой и крыловидной костей. Верх-
ний ушной отросток квадратной кости (processus oticus)
в области слуховой камеры п о д в и ж н о сочленяется с черепом
(так называемое явление стрептостилпи, характерное для класса
птиц и некоторых групп рептилий), укрепляясь связками на сустав-
ной поверхности, образуемой proticum, а иногда также и чешуйча-
той костью (squamosum). Это вызывает появление па ушном отростке
одной или двух суставных головок; наличие только одной суставной
головки является более примитивной особенностью и свойственно
например всем Palaeognathae. Другой верхний отросток (processus
orbitalis) вдается в орбиту и является местом прикрепления мощного
musculus protractor quadrati. Нижний конец квадратной кости
образует три суставные поверхности: наружную — для сочленения
с квадратно-скуловой костью, внутреннюю — для связи с крыловид-
ной костью и заднюю — для прпчлененпя нижней челюсти. Крыло-
видная кость (pterygoideum) связывает между собой задний конец
небной п только-что упомянутый внутренний отросток квадратной
кости, образуя в обопх местах сочленения. При этом у многих групп
птиц передний конец крыловидных костей на ранних стадиях онто-
генеза в виде отростка hemipterygoideum налегает на небные кости,
а впоследствии срастается с ними, образуя сочленение между основ-
ной частью pterygoideum и palatinum с приросшей к ней hemiptery-
tioideum (Pycraft, 1903).
В некоторых группах сочленение имеет место не только в ука-
занных местах, но и в средней части палочковидной по форме крыло-
видной кости с основанием клюва парасфеноида (rostrum parasphenoi-
4*
dale) или крыловидным отростком базисфеноида (processus ptery-
goideus basisphenoidale), с образованием в этих местах специальных
фасеток. Структура этих взаимоотношений крыловидной кости имеет
значительное таксономическое значение (см. ниже). Небные кости
(palatinum) в свою очередь связывают передний и задний концы
крыловидных и челюстных костей и образуют значительную часть
костного неба. С межчелюстными или верхнечелюстными костями
небные обычно образуют неподвижную связь, сзади, наоборот, со-
ставляя весьма подвижную систему.
Значение всех этих типов сочленения и связей отдельных костных
элементов очень велико, в целом создавая весьма совершенную си-
стему движений верхней челюсти, синхроничных оттягиванию
нижней челюсти. В результате это сильно увеличивает ротовую щель
без очень значительного оттягивания нижней челюсти. Движение
исходит из давления открывающейся нижней челюсти на квадрат-
ную кость, которая одновременно давит на две системы балок, ле-
жащих в одной плоскости и передающих движение клюву:’ через
квадратно-скуловую, скуловую и верхнечелюстную кости и через
крыловидную и небную кости; при этом последние скользят по пара- и
базисфеноидам и передают давление небному отростку предчелюстной
кости (processus palatinum praemaxilla). Движение клюва вверх
оказывается возможным благодаря значительной эластичности но-
совых костей и лобных отростков предчелюстных костей, без обра-
зования настоящего сочленения в этой области. Обеспечивающим
движение моментом является также образование глубокой черепно-
лицевой щели (см. выше) в области обонятельной косточки.
У некоторых птиц с сильно пневматизированным черепом (на-
пример у попугаев) в области сгиба образуется глубокая борозда,
однако не являющаяся настоящим сочленением двух костей. Таким
образом линия сгиба находится в средней части черепа, который
следовательно относится к мезокинетическому типу (Versluys,
1912), к которому и среди рептилий относятся также черепа змей и
близки черепа некоторых ящериц.
Различный тип пищи и разнообразные способы приспособления
к ней в характере добывания, дробления и удерживания вызвали
значительное разнообразие в структуре этого небно-челюстного
аппарата. Для килевых птиц Гексли (Huxley, 1867)1 и Паркером
(Parker, 1866, 1876) были отмечены четыре основных типа в его строе-
нии, имеющие до сих пор некоторое значение как таксономический
признак, хотя типы и соединяются различными переходными и укло-
няющимися структурами, обнаруженными у многих видов птиц.
Дромэогнатический тип характеризуется широким крупным сош-
ником, соприкасающимся спереди с небными отростками челюстных
костей и соединяющимся сзади с проксимальными концами небных
и дистальными крыловидных костей (рис. 31). Этот тип был уста-
новлен для скрытохвостов (Tinami formes), но по своей структуре
1 Гексли (1867) всех килевых птиц разделял на четыре группы (по его
номенклатуре — подотряды), соответственно этим типам небно-челюстного ап-
парата. В дальнейшем его система была признана искусственной, хотя, прин-
цип отчасти сохранил свое значение.
4 Орнитология—1209
49
несколько напоминает эму (Drontaeus) и отчасти других бескилевых
птиц (рис. 32, 33, 34).Сходство в соотношении небных и крыловидных
костей у этих птиц было использовано Пайкрафтом (Pycraft, 1900)
в таксономических целях: он противопоставил бескилевых птиц
вместе с Tinamijormes в качестве более древнего надотряда Palaeog-
nathae или древне-небных птиц всем остальным птицам — Neognathae
или новонебным. Основной отличительной и общей особенностью
дромэогнатического черепа и близких ему структур у различных
бескилевых является не сочленение, а срастание концов
крыловидных и небных костей между собой, причем внутренняя
Рис. 31. Дромэогнатический череп.
Тип небно-челюстного аппарата у скрытохво-
ста — Tinamus sp.; небные и крыловидные ко-
сти выделены грубыми черными контурами.
Bs. — основная клиновидная кость (Bt. —
прикрытая basitemporale); М — верхнечелю-
стная кость; Qu — квадратная кость; Ра —
небная кость; Prpt—базиптеригоидные от-
ростки основной клиновидной кости; Pt —
крыловидная кость; Rs — клюв парасфенои-
да; Votn — сошник; z — скуловая дуга. По
Gadow, из Bohm.
Рис. 32. Небно-челюстной аппарат страу-
са — Struthio camelus L. (наиболее укло-
няющийся среди бескилевых тип пеба;
сошник значительно удален от крыловид-
ных костей).
Обозначения см. рис. 31. Мр— небпыс
отростки верхнечелюстных костей.
поверхность первых, кроме того, соединяется с базиптеригоидными
отростками базисфеноида, как это имеется у рептилий (рис. 31—34).
Однако ценность этих признаков для объединения наиболее прими-
тивных птиц давно вызывает сомнение (Selenka, 1875), и в последнее
время это деление птиц Пайкрафта, вследствие наличия переходных
структур в разных группах птиц, Штреземанном (Stresemann, 1933)
рассматривается как неудачное и отбрасывается. В нашей классифи-
кации мы его сохраняем, вследствие определенного практического
удобства, которое оно представляет.
Второй тип черепа — шизогнатический, характеризуется сош-
ником, заостряющимся кпереди и раздваивающимся кзади; послед-
ним концом он обхватывает конец клюва парасфеноида. Небные
50
и крыловидные кости не приходят в соприкосновение с сошником,
но сочленяются между собой. Парные небные отростки челюстных
Рпс. 33. Небно-челюстной аппарат пан-
ду — Rhea americana L.
Обозначения см. рис. 30 и 31. По Bohm.
Рпс. 34. Небно-челюстной аппарат кпвп —
Apterix australis Shaw.
Обозначения ;см. рис. 30 и 31. По Bliom.
костей обычно лежат под небными костями п срастаются с ними, но
не сливаются ни друг с другом, ни с сошником (рис. 35). Этим типом
Сз
у
Рис. 35. Шизогнатический тип пебно-че-
люстного аппарата глухаря— Tetrao иго-
SallusL. (с рудиментарным сошником).
Обозначения см. рис. 30 и 31. По Bohm-
Рпс. 36. Шизо-десмогнатическпй пебно-челю-
стной аппарат кондора — Vultur gryphus L.
(переход к непрямому десмогнатизму; сошппк
редуцирован).
Обозначения см. рис. 30 и 31. По Bohm.
51
Рис. 37. Десмогиатический тип вебно-че-
- постного аппарата красноголовой черпе-
тп — Nyroca ferina L.
Обозначения см. рис. 31 и 32. По Bohm.
черепа обладают кулики, голуби и трубконосые и мн. др. Название
типу дано за длинные щели, образующиеся по краям сошника, между
ним и небными костями и верхне-
челюстными на всем протяжении
от хоан до конца клюва.
Третий тип — десмогнатиче-
ский, обладает узким, маленьким,
кпереди заостренным сошником,
или он даже полностью исчезает,
а небные отростки челюстных ко-
стей срастаются друг с другом в
костяную крышу неба (рис. 37).
При этом различают три рода дес-
могнатизма: прямой, когда эти
отростки прямо срастаются друг с
другом; это срастание может иметь
место полностью (гуси) или непол-
ностью (серимы) и с помощью или
без участия носовой перегородки.
В непрямом десмогнатизме слия-
ние отростков осуществляется
только этой сильно разрастаю-
щейся носовой перегородкой; в
этом случае десмогнатизм также
может быть полным (орлы, грифы,
рис. 36) или неполным (борода-
стики). Наконец в двойном десмо-
н небно-челюстные отростки и неб-
ные кости п образуют широкую твердую небную крышу, как, напри-
мер, у птиц носорогов и жиряков.
В эгитогнатпческом типе сошник туп
или вилообразен спереди и глубоко
раздваивается кзади, обхватывая клюв
парасфеноида; сошник не образует ко-
стяных соединений с носовой перего-
родкой. Небные отростки челюстных
костей расширяются на конце и не
срастаются ни между собой, ни с сош-
ником. Для небных костей характерны
так называемые мускульные отростки,
отходящие кзади и кнаружи (рис. 38).
; )тот тпп свойствен воробьиным и стри-
жеобразным. Подобно предыдущему
типу в нем также различают три рода
соотношений (Parker, 1876): полный,
неполный и составной.
Имеющиеся данные с несомнен-
ностью показывают, что из трех
типов черепа — шизо-, десмо- и эги-
тогнатического первичным является
гнатизме сходятся и сливаются
Рис. 38. Эгитогнатический типнеб-
но-челюстпого аппарата домашнего
воробья — Passer domeslicus L. Обо-
значения см. рис. 30 и 31. По
QiioBlr 4 п
52
шизогнатический, из которого посредством соединения неб-
ных костей и отростков возник десмогнатический череп и соответ-
ственным образом также эгитогнатический. Оба последних типа
небно-челюстного аппарата повидимому развивались независимо в раз-
ных систематических группах из первоначально шизогнатических
по строению птиц и следовательно не могут иметь решающего фило-
генетического значения. Соответственно таксономическое значение
этих типов не всегда согласуется с другими классификационными
признаками; являясь характерным для известных групп, в других
этот признак обнаруживает много исключений и значительное разно-
образие в самом строении
(ср. рпс. 35, 36) (Gadow,
1891; Сушкин, 1902).
Нижняя челюсть у птиц
окостеневает в онтогенезе
из одного или двух цен-
тров, с развитием в даль-
нейшем до 5 покровных
костей, обычно полностью,
без швов, срастающихся
между собой (рис. 39). Со-
членение с квадратной ко-
стью образует хондраль-
ная сочленовная кость (os
articulare), несущая на
внутренней поверхности
крупный processus mandi-
bularis. Конец клюва и
большую часть краев его
образует накладная зуб-
ная кость (dentale),
полностью срастающаяся
передним концом с одно-
именной костью противо-
положной стороны; к ней
прилегает с внутренней
Рис. 39. Нижняя челюсть: а, b — птенца северной
качурки—О се an drama leucorrhoa Vieill., с латераль-
ной (а) и медиальной (Ь) сторон, и с — хохлатого
гокко — Craxalector L., с латеральной стороны.
Ап.— угловая кость; Аг. — сочленовная кость:
С. — венечная кость; D. — зубная кость; F. т. а..—
переднее нижнечелюстное отверстие; Sa. — надугло-
вая кость; Sp.—пластинчатая; Р. ап — передняя
часть челюсти; Р.ат — сочленовная часть; Pars.pos.—
задняя часть; Рг. с. —венечный отросток — рг. со-
ronoideus; Рг. ех. — наружный отросток — рг. exter-
nus; Рг. гп.—внутренний или челюстной отросток—
рг. internus s. рг. mandibularis; Рг. pos. — задний
отросток — рг. posterior. По Pycraft и Lebedinsky, из
Stresemann.
стороны маленькая под-
бородочная косточка (os mento-mekkelianum), образующаяся хоп-
дрально у некоторых дневных хищников (Accipitres). Остальные
кости все являются покровными: наружные — угловая (angulare),
надугловая (supraangulare), и лежащие по внутренней поверх-
ности черепа — пластинчатая (operculare или spleniale) п
венечная (complimentare или goniale). Задний конец угловой
кости обычно бывает вытянут в отросток — processus angularis
posterior, к которому (как и к вышеупомянутому внутреннему от-
ростку) прикрепляются мощные пучки мускула, раскрывающего
клюв (musculus depressor mandibulae).
Интересным приспособлением в строении нижней челюсти у не-
которых рыбоядных групп (Hesperornis, Ichthyornis, Sulidae) яв-
ляется наличие сочленения или гибкого сгиба в проксимальном
53
отрезке зубной кости, дозволяющем заглатывание крупных рыб
(рис. 28).
Подъязычный аппарат птиц обычно состоит из двух непарных
элементов — entoglossum и urohyale, отсылающих кпереди парную
paraglossum, образующую скелетную основу для языка (рис. 40). От
заднего конца entoglossum отходят длинные хрящевые или окостене-
вающие двухчленные рожки (kerato-и epibranchiale), особенно длин-
ные у длипноязычных птпц и
Рис. 40. Подъязычный аппарат пурпурно-
шапочного лори — Lorius domicella L.
(Psittaciformes).
(a)—с дорзальной; (b)—вентральной и
(с)—латеральной сторон; b — entoglos-
sum; с — сочленение с paraglossum; cb —
epibranchiale; е — paraglossum; h — сим-
физ parahyalia; hb — keratobranchiale; p—
parahyalia, у лори развивающиеся в круп-
ные, сливающиеся передним концом, от-
ростки, увеличивающие область прикреп-
ления хорошо дифференцированной муску-
латуры языка;гг — слабо развитое у лори
urohyale. По Mivart, из Stresemann.
по эластичности напоминающие пру-
жину. Эти рожки обычно загиба-
ются на заднюю сторону головы,
у дятловых даже охватывая и верх-
нюю ее часть и или закругляясь
вокруг глаза или проникая далеко
вперед в полость надклювья (рис.
41).
Эти рожки, подобно аналогич-
ному аппарату у крокодилов и дру-
гих рептилий, соответствуют по
своему происхождению первой паре
жаберных дуг низших позвоноч-
ных (или третьей висцеральной
дуге). От гиоидной или второй вис-
церальной дуги остаются только
два маленьких хрящика — hyo-
е' mandibulare и stylohyale, сливаю-
щиеся впоследствии в columella
auris ушного аппарата. Проблема-
тичные остатки этой дуги будто бы
имеются также в крошечных хря-
щиках, располагающихся по краям
вышеупомянутого paraglossum или
entoglossum (так называемые para-
hyalia).
Позвоночный столб птиц мор-
фологически сходен с позвоночни-
ком рептилий, хотя и обнаружи-
вает ряд отличных черт, могущих
быть поставленными в связь с по-
летом птиц. Таковы частичное слияние позвонков между собой, по-
движность шейных позвонков, преобладающая гетероцельная форма
позвонков, позволяющая значительные изгибания позвоночника, но
не допускающая вращение, и ряд других особенностей. Количество
позвонков у птиц колеблется от 39 до 63.
Спинной позвонок состоит из тела (centrum), отходящих дор-
зально спинных дуг (neurapophysi), охватывающих с боков спинной
мозг и затем сливающихся в спинной отросток (proces-
sus spinosus) (рис. 42). Сцепление позвонков друг с другом происходит
посредством пары передних (praezygapophysi) и пары задних (p'ostzy-
gapophysi) сочленовных поверхностей косых отростков (processus
obliqui). Фасетки первых направлены вперед и вверх, а задних —
54
вниз и назад. Близ основания
сочленйвные поверхности для
первых косых отростков находятся
головки ребра (capitulum), бугорок
которого (tuberculum) прижат к
(processus transversum), сидящих
концам поперечных отростков
по бокам спинных дуг. Полу-
55
чающееся между ребром и позвонком отверстие (foramen transversum)
служит каналом, проводящим позвоночную артерию и вену и главный
ствол симпатического нерва, и сохраняется не только в области
грудных ребер, но и на шее, вследствие наличия здесь рудиментов —
pleurapophysi— исчезнувших ребер. От тела позвонка, главным
образом в шейном и грудном отделах, отходят еще вентральные,
крайне изменчивой формы, отростки (hypapophysi), то явля-
ющиеся местом прикрепления мускулов, то образующие каналы
для прохождения сонных артерий (так наз. kalapophysi).
56
Само тело свободных позвонков в разных отделах позвоночнике
столба построено различным образом. Преобладает гетероцельный
тип, с седлообразным сочленением позвонков, но у ряда групп (пинг-
вины, бакланы, попугаи и др.)широко распространен опистоцельный
тип, а в хвостовой области почти всегда имеют место процельный
или даже амфицельный и платицельный типы. Тела соседних свобод-
ных позвонков соединяются хрящевыми менисками и пробегающими
по наружной поверхности и иногда окостеневающими связками:
Шейный отдел состоит из 11—25 позвонков, в зависимости от
длины шеи. У длинношейных видов число позвонков вариирует даже
индивидуально. Два первых цозвонка специализированы в атлас
(а) (Ь) 1с)
Рис. 43. Вращасмость шеи ушастой совы —Asio otus L.
а —- смотрит назад и вверх; b — назад и вниз; с — назад. Поворот головы,
в данном случае составляющий 180°, у сов может достигать 270°. По Heinroth.
и эпистрофей, приспособленные к широким вращательным движе-
ниям и приему затылочного мыщелка (рис. 43). .Зубовидный отросток,
на котором вращается атлас, будучи по происхождению дериватом
тела атласа, полностью .срастается с эпистрофеем. Грудной отдел,
несущий ребра (из которых от 3 до 9 пар достигают грудины), обра-
зуется изменчивым числом позвонков, иногда достигающим десяти.
От 3 до 5 из них нередко сливаются между собой в одну спинную
к о с т ь (os dorsale), назначение которой, повидимому, заключается
в придании туловищу более прочной опоры и цельности, необходи-
мой в связи с интенсивной работой крыльев.
Грудные ребра птиц состоят из двух сходящихся под острым
углом отделов (рис. 24). Они полностью окостеневают и в позвоноч-
ном отделе снабжены крючковидными, отходящими назад отрост-
ками — processus uncinati; налегая на следующее позади ребро,
они способствуют большей устойчивости всей грудной клетки. Сле-
57
дует отметить, что эти отростки являются характерной особенностью
грудной клетки птиц, вне данного класса находимой лишь у кроко-
дилов и гаттерий из современных и у ряда групп из вымерших реп-
тилий.
Тазовый отдел позвоночника ш*иц замечателен образованием
огромного сложного крестца — synsacrum, состоящего из значитель-
ного числа сросшихся между собой позвонков, в совокупности обес-
печивающего опору всему телу, покоящемуся на земле только на
•двух конечностях (рис. 44). Ко-
личество позвонков в тазовом
отделе колеблется от 10 до 22,
♦распадающихся, например у
кур, на пять типов: один по-
Рис. 44. Таз и хвостовой отдел позвоночного
столба домашней курицы — Gallus gallus L.
1 — с вентральной стороны; 2 — сбоку; 3 —
то же в распиле по медиальной плоскости.
Цифры отмечают порядковые номера позвон-
ков. С. — хвостовой отдел позвоночника;
/. о. —запирательное отверстие — foramen
obturator; il. — подвздошная кость— os ilium;
is. —седалищная кость — os ischium; p— лоб-
ковая кость — os pubis; Py.—пигостиль; S.
c. — тазо-хвостовой отдел; S. I. — тазо-пояс-
ничный отдел; S. s. — тазо-крестцовый отдел;
S.t.—тазо-грудной отдел; S. I. I. — т азо-грудо-
поясничный отдел; P. p,— processus pecti-
neal is. По du Toit.
звонок — по существу груд-
ного происхождения — с сво-
бодными рудиментарными реб-
рами, четыре — без свободных
ребер, но с мощными боковыми
отростками, ди- и парапофи-
зами (последние являются руди-
ментами настоящих ребер и сли-
ваются своими утолщенными
концами между собой, с диапо-
физами и подвздошной костью),
четыре — без парапофизов и с
значительным расширением по-
звоночного канала (в связи с
поясничным вздутием спинного
мозга и выходом корешков се-
далищных нервов), два круп-
ных — с ди- и парапофизами,
несущих сочленовные поверхно-
сти— вертлужные впадины (ace-
tabula) для задних конечностей
и гомологичные крестцовым по-
звонкам рептилий (истинно-
крестцовые позвонки) и пять пе-
реходных к свободным хвосто-
вым позвонкам. Срастание всех
этих позвонков настолько полно, что следы расчленения выража-
ются только в виде межпозвоночных отверстий и поперечных
отростков. Спинные отростки позвонков сливаются-в целостный гре-
бень (crista), исчезающий однако в предвертлужном отделе. Истинно
тазовые (по происхождению) позвонки вертлужного отдела коле-
блются у птиц в числе от 1 до 3.
Хвостовой отдел позвоночника состоит из 13 позвонков, несу-
щих поперечные отростки, по происхождению соответствующие па-
рапофизам крестцовых позвонков. У бескилевых птиц и скрытохво-
стов хвостовые позвонки свободны до самого конца, но у килевых
птиц последние позвонки срастаются в цельную вертикально по-
58
Рис.. 45. Грудина а — краснозобой гагары —
Colymbus stellatus Pont, и b — большого ти-
наму ~ Tinamus major robustus Scl. ct Salv.
c — коракоид; c. s. — грудной киль; /. —
ключицы (или вилочка); т — metasternum;
р. г.—processus intermedius; р. I. а. — перед-
ний боковой отросток; р. I. р. — задние боко-
вые отростки; р. р. с.—прокоракоидный
отросток, хорошо развитый у гагар; s. s. —
spina sterni. По Eyton и Parker, из Strese-
mann.
ставленную косточку — pygostil, треугольной формы, служащую
местом прикрепления рулевых перьев.
Грудина (sternum) птиц имеет вид широкой или удлиненной пла-
стины, выпуклой вентрально, п у летающих и плавающих форм
несет посредине вертикальный грудной к и'л ь (crista или carina
sterni) (рис. 45). Развитие этого киля и его величина стоят в тесной
зависимости от мощности движении передних конечностей, так как
вдоль грудного киля прикре-
пляются частью своей длины
наиболее мощные крыловые му-
скулы. Обычно помимо киля
грудина несет спереди узкий
вырост — spina sterni, иногда
раздваивающийся на наружный
и внутренний отростки — spina
sterni externa и interna; роль
этого выступа заключается в под-
держании и укреплении креп-
кой перепонки, натянутой ме-
жду грудиной, вилочкой и ко-
ракоидами (membrana sterno-
coracoclavicularis) (рис. 46). По
обеим сторонам spina sterni на
грудине, лежат глубокие бо-
роздки сочленовных ямок для
приема коракоидов, а по боко-
вым краям грудины находятся
направленные вбок и вперед
парные передние боковые от-
ростки — processus laterales an-
leriores, ямки, принимающие
вентральные концы грудных ре-
бер, и, направленные назад,
парные задние боковые отростки
processus laterales posteriores;
последние иногда несут напра-
вленные вбок косые отростки —
processus obliqui. Задняя, наибо-
лее уплощенная часть груди-
ны, поддерживающая брюшные
органы по медиальной линии, называется metasternurn и по краям
иногда образует вторичные выступы — processus intermedii. Эта
часть грудины в онтогенезе обладает замедленным ростом п часто
развивается не в виде сплошной пластины, а в виде пластинки,
снабженной обширными вырезами (бухтами) или окнами, заполняе-
мыми иногда соединительно-тканными перепонками (фонтанелли).
Развитие грудины всегда имеет место из двух боковых хрящей,
’-оединяющихся по средней линии и совместно образующих грудной
киль (рис. 47).
Редукция полета приводит не только к исчезновению киля, но
5a
и к сокращению грудины и исчезанию spina slerni (рис. 48). По своей
форме и развитию вырезок и spina sterni грудина вообще обнаружи-
вает значительное разнообразие, стоящее в связи с различным обра-
зом жизни видов и. различным филогенетическим положением их
(рис. 49).
Плечевой пояс каждой стороны тела слагается коракоидом (со-
racoideum), лопаткой (scapula) и, за исключением бескилевых птиц,
ключицей (clavicula), которые сходятся друг с другом своими перед-
ними концами (рпс. 50). Задним концом коракоид упирается в перед-
ний край грудины. Лопатка образует сочленение с коракоидом
и посредством акромиального отростка или акромион — с ключицей;
тело лопатки лежит более
или менее параллельно
позвоночнику, налегая на
спинную сторону ребер л
прикрепляясь к ним эла-
стическими связками. Клю-
чицы каждой стороны обыч-
но срастаются своими ниж-
ними концами, образуя
характерную для класса
кость—дужку или вилочку
(furcula), которая при по-
мощи плоской вертикаль-
ной пластинки hypoclei-
dium прилегает к груд-
ному килю или spina sterni.
иногда срастаясь с ним.
Эти структурные отноше-
ния элементов плечевого
пояса в зависимости от ка-
чества полета птицы могут
существенно изменяться:
задние концы коракоидов
Рис/46.[Строение spina sterni у представителей раз-
личных отрядов.
а — серебристый фазап — Gennaeus nycthemerus^ L.
fGalliformes); b — удод— Upupa epops L. (Coracii-
jormes); c — ястреб-перепелятник — Accipiter nisusL.
( Falconiformes); d — скрытохвост — Cryptvrus nocii-
vagus (Neuwied) (Tinami formes); e — зеленый тро-
гон — Trogon viridis L. (Trogoniformes); f—серая
ворона — Corvus cornix L. (Passeriformes); cr. st.—
грудной киль; pr. I. an. — передний боковой отро-
сток грудной кости; s. а. сог. — сочленовная ямка
для коракоида; sp. с. — общий грудной вырост
(spina communis); sp. ex.—spina externa; sp. in.—
spina interna. По Fiirbringer.
то соприкасаются или срастаются, то значительно раздвигаются друг
от друга, ключицы иногда редуцируются или даже полностью исче-
зают (рис. 51), лопатка то доходит до последних грудных ребер, то
бывает очень мала, полностью срастается с коракоидом и вместо
образования острого угла с ним (у хороших летунов) составляет
тупой угол. Гомологом прокоракоида рептилий в плечевом поясе
птиц является processus procoracoidalis, образующийся на вентраль-
ной или внутренней поверхности передних концов коракоида. Верх'
ний конец коракоидов образует так называемый акрокоракоидный
отросток, несущий большую часть fossa glenoidalis для сочленения
с плечевой костью; по бокам его прилегают лопатка и ключица, сов-
местно ограничивая узкое отверстие — foramen triosseum (рис. 50).
Передняя конечность птиц слагается из плечевой (humerus)’
локтевой (ulna) и лучевой (radius) костей, костей запястья (carpus),
пястья (metacarpus) и пальцев (digiti), характерно видоизмененных
в гребущий воздух или воду аппарат — крыло. Первоначальный
60 I
Рис. 47. Развитие грудины и плечевого попса птиц (верхний ряд), в сравнении с развитием грудины у рептилий (нижний ряд).
1, 2 — образование грудины па ранних стадиях развития эмбриона курицы — Gallus даllus L.; вентральные концы ребер
сливаются в хрящевые пластинки, сближающиеся между собой; в дальнейшем латеральные части передних и задних ребер
резорбируются,а сохранившиеся вентральные части оказываются выростами грудины, разрастаются и превращаются в пе-
редние,задние и косые боковые отростки; з —дальнейшая стадия развития на эмбрионе чибиса —Vanellus vanellus L.; поло-
вники грудины и ключицы слились между собой; 4 — грудина гоаципа — Opisthocomus hoazin Mlill. незадолго до выклевы-
вания из яйца; сросшиеся ключицы (вилочка) прирастают к грудине посредством длинного отростка—hypocleidium,
по мнению Parker и Heilmann— гомологичного episternum рептилий; па дистальном конце лопатки сохраняется хрящ, го-
мологичный suprascapula рептилий; 5ns—образование грудины на первых и последних стадиях развития гаттерии —
Sphenodon punclatus Gray (Rhynchocephalia); 6 и 7—развитие грудины ящерицы — Cnemidophorus sp. (Lacertilia); гру-
дина у рептилий остается на хрящевой стадии развития; дополнительно к плечевому поясу развиваются episternum и sup-
rascapula; cl. — ключицы (вилочка); со. —коракоид; hy. —нижний отросток вилочки, hypocleidium; г. — ребра; s. — ло-
патка; ss.—suprascapula; st.—грудина. Из Heilmann. ♦
рептильные особенности здесь испытали значительные изменения
(рис. 52). При сложенном положении крыла плечевая кость приле-
гает к грудной кости и за-
нимает горизонтальное по-
ложение; с ней связаны
предплечье и кисть, рас-
полагающиеся в одной вер-
тикальной плоскости в
форме буквы Z. При кры-
ле, расправленном для по-
лета, костные элементы его
всегда образуют тупые
углы между собой, удер-
живаясь от расположения
по одной оси главным обра-
зом сухожильными тяжами
летательных перепонок —
pro- и metapatagium.
/Г/
Рис. 48. Грудина нелетающей бо-
лотной курочки — Atlaniisia го-
gersi Lowe.
Обратите внимание на малые раз-
меры грудной кости (частично
остающейся неокостеневшей—хря-
щевые участки отмечены пункти-
ром) и на незначительный труд-
ной киль, широко расставленные
коракоиды, отсутствие spina sterni
и слабую вилочку. Дальнейшие
изменения грудины в связи с поте-
рей способности к полету см. па
рис. 228. Разрез грудины по линии
ab изображен отдельно — а' Ь'.
сог. — коракоид; cr.st. — грудной
киль; cl. — вилочка; р I. а — пе-
редний боковой отросток; р. I.
р. — задний боковой отросток;
sc. — лопатка. По Lowe, из Stre-
semann.
/о7
Рис. 49. Изменения грудины и плечевого пояса у
родственных видов, в связи с различным образом
жизни.
Соотношения размеров отдельных видов пе сохране-
ны. а и Ь — грудины олуши—Sula bassana L. и
большого фрегата — Fregala aquila L. (из отряда Ре-
lecaniformes); с и d — грудины ныряющего буре-
вестника — Pelecanoides urinatrix Gm. и альбатро-
са — Diomedea exulans L. (из отряда Procellariifor-
mes). Обратите внимание на сильно выдающийся
вперёд грудной киль и длинную грудину у олуши,
стоящие в связи с типом полета этой птицы и ныря-
нием посредством падения (на полете); длинная гру-
дина защищает внутренности при плавании и ны-
рянии. в противоположность этому виду, неплава-
ющий и летающий преимущественно планирующим
полетом фрегат обладает более широкой грудиной,
причем очень мощная вилочка срастается как с груд-
ным килем, так и с акрокоракоидным отростком ко-
ракоида, чем достигается особая устойчивость пле-
чевого пояса. Среди трубконосых• ныряющий буре-
вестник обладает длинной грудиной, прямостоя-
щими коракоидами и сравнительно’ слабой вилоч-
кой, тогда как альбатрос, летающий сходно с фрега-
том, имеет почти квадратную грудину с широко рас-
ходящимися коракоидами и мощную пружинящую
вилочку. По Eyton, Milne Edwards и Forbes, из
Stresemann.
62
В соотношении и форме скелетных элементов крыла наблюдаете:1
значительное разнообразие, стоящее в связи с функцией органа
(рис. 53).
Плечевая кость является крупной трубчатой костью, несущей па
проксимальном конце эллипсоидную сочленовную поверхность —
caput humeri — для fossa glenoidalis коракоида и лопатки, а па
дистальном конце сочленовные поверхности для предплечья—trochlea
radialis и ulnaris (рис. 54). Для прикрепления мощной крыловой
мускулатуры в проксималь-
ной части кости образуются
мощные гребни: crista late-
ralis и medialis, переходящие
в соответствующие бугры —
tuberculum. Рядом с tuber -
culum inediale лежит глу-
бокая впадина — fossa pneu-
matica, с отверстием на дне
для прохода воздуха в воз-
душную полость кости. По
своему строению плечевая
кость довольно близка к
плечевой кости рептилий,
отличаясь главным образом
сильным развитием гребней,
несущих мощную крыловую
мускулатуру (рис. 217).
Из костей предплечья лок-
тевая кость больше по раз-
мерам и обычно несет следы
прикрепления к ней малых
маховых перьев. Закладыва-
ющиеся в онтогенезе двумя
рядами костные элементы за-
пястья (carpalia) в дальней-
шем сливаются дистальным
рядом с основаниями мета-
карпальных косточек; боль-
шая часть элементов про-
вис. 50. Плечевой пояс птицы.
(а) — передний копец грудины с плечевым поя-
сом орлана белохвоста—Haliaetus albicilla L.;
(b)— кости плечевого пояса воробьиной птицы
сверху и спереди.
асгос. — акрокоракоидный отросток коракоида:
асгот. — акромион; с — коракоид; cs — грудной
киль; /.—вилочка; fg. — суставная ямка для
сочленения с плечевой костью; /t.—foramen
triosseum; I.—ligamentum sternoclaviculare;
sc.—лопатка; s. co. — сочленение коракоида с
грудиной; st.—грудная кость. По Prechtl и
Holmgren, из Stresemann.
ксимального ряда также вскоре исчезает, и у взрослых птиц мы
находим только две свободных косточки: centrale radiale на лучевой
стороне и крупную кость на локтевой стороне, по происхождению
являющуюся метакарпальной косточкой пятого луча (Steiner, 1922)
(рис. 55). У нелетающих птиц сохраняется нередко лишь одна кар-
пальная косточка (рис. 56), в некоторых случаях полностью исчеза-
ющая (эму, киви).
Дистальный ряд carpalia, сливающийся с метакарпальными эле-
ментами, образует carpo-metacarpus. В последнем на эмбриональных
стадиях наблюдаются четыре metacarpalia, но первый луч почти
совсем сливается со вторым лучом, а четвертый обычно вообще
исчезает, так что у взрослых летающих птиц мы находим в этом
63
отделе только три костных элемента, срастающихся по длине или
по своим концам. При otonI второй луч оказывается наиболее кру-
пным и несет две, а третий и первый по одной фаланге пальцев.
При редукции крыла у бескилевых наиболее сохраняется именно
второй луч. В эмбриональном состоянии концевые членики первого,
второго и третьего пальцев несут когти, за немногим исключением
(рис. 56), исчезающие уже в постэмбриональном развитии.
Таз птиц имеет широкую и вытянутую форму с особенно значи-
тельным развитием подвздошной кости. Обычно лазящие и бегаю-
Рис. 51. Форма и положение вилочки (furcula) у пред-
ставителей разных отрядов.
а—V-образная вилочка серой куропатки — Perdix per-
dix L. (Galliformes)-, b—U-образпая вилочка сапсана —
Falco peregrinus Tunst. ( Falconiformes); c — несрастаю-
щаяся своими ветвями вилочка додо —Raphus cuculla-
tus L. (Didus ineptus Jj.) (Columbiformes)-, d — U-обра^
пая, срастающаяся с грудным килем вилочка розо-
вого пеликана — Pelecanus onocrotalus L. (Реlecanifor-
mes)- e— сильно отогпутая назад вилочка лебедя
кликуна—Cygnus cygnus L. (Anseri formes)' f —ви-
лочка цесарки — Guttera cristata Wagl. (Galliformes),
hypocleidium которой образует сумку, поддерживающую
.извивы трахеи; cl.—ключицы или вилочка; cr. st.—
грудной киль; hyp. — hypocleidium; s. a. cor. — сочле-
новная ямка на грудине для коракоида; st. — грудида;
tr. — трахея. По Fiirbringer.
щие формы имеют таз
более короткий и ши-
рокий, чем формы пла-
вающие^ хорошо ныря-
ющие птицы имеют осо-
бенно длинный- и узкий
таз (рис. 57). В целом
таз образуется тремя
костями; подвздошной
(ilium), седалищной (is-
chium) и лобковой (pu-
bis), сходящимися на
краях вертлужной впа-
дины (acetabulum) (р.44).
Предвертлужная часть
таза (pars glutea) уже
и как бы вогнута свер-
ху, завертлужная часть
(pars renalis) шире и
выпукла; здесь, по кра-
ям позвоночника, обра-
зуется обширное углу-
бление в подвздошной
кости (fovea renalis).
в котором лежат почки.
Подвздошная кость поч-
ти во всей своей длине
тесно срастается с позвонками, входящими в synsacrum, имея однако
иногда значительное число мелких отверстий, располагающихся
между поперечными отростками позвонков. Седалищная кость, имею-
щая также вытянутую форму, располагается по боковому краю под-
вздошной в завертлужной части и частью срастается с ней. Между
обеими костями, сразу за вертлужной впадиной, располагается об-
ширное седалищное отверстие (foramen ischiadicum).
К боковому краю седалищной в свою очередь прилегает длинная
и узкая лобковая кость, в двух местах срастающаяся с предыдущей
и таким образом оставляющая отверстия: foramen obturator и f. oblon-
gum. Задние концы лобковых костей обычно свободные (у страусов
соединяются друг с другом); эта черта, так же как отсутствие соеди-
нений дистальных концов седалищной кости, является характерным
свойством «открытого» таза птиц, редко наблюдаемым у рептилий.
64
Однако по общим соотношениям костей таз птиц весьма напоминает
таз некоторых рептилий, особенно из группы динозавров, а в своем
развитии обнаруживает черты, с несомненностью указывающие на
происхождение его из тех отношений костных элементов, которые
наблюдаются у представителей этого класса (рис. 58).
Вертлужная впадина по дну закрыта только перепонкой; по.
переднему краю костного кольца, окружающего ее, располагается
небольшой выступ — processus pectinealis, некоторыми авторами
повпдимому неправильно гомол оптируемый с praepubis рептилий.
По некоторым наблюдениям существуют половые отличия в тазе
птиц, выражающиеся у самок в большей ширине таза и в большем
Рис. 52. Скелет передней конечности первоптицы — Archaeornis siemensi Dames (Ь) и голубя —
» Columba livia Gm. (a).
c. — centrale; cr.—верхний гребень плечевой кости; dl—3 — i-й—3-й пальцы; П.—
плечевая кость; г—intermedium; тс 1—3—metacarpale 1—3; 2?.—лучевая кость;
Г. — radiate; U.—локтевая кость; U. —ulnare; 1 4- 2 — слившиеся в общую кость 1-е и 2-е
дистальные carpalia. По Heitmann.
расстоянии задних концов лобковых костей (Knobel, 1924). Однако
это наблюдение не проверено на представителях разных системати-
ческих групп.
Задняя конечность состоит из бедренной кости (femur), голени
(crus), образуемой большой и малой берцовой (tibiotarsus и fibula),
цевки, обычно называемой плюсной (tarsomet-atarsus), и пальцев.
Бедро у птиц всегда является относительно короткой трубчат&й
костью (рис. 59). На ней, почти под прямым углом к ее оси, распо-
лагается шаровидная суставная головка для соединения с тазом.
В проксимальной части кости выдаются своими размерами гребни
Для прикрепления мускулатуры: большой и малый трохантер (tro-
chanter major и minor). Дистальный конец бедра несет два мыщелка,
суставной поверхностью направленных кзади: срединный (condylus
Eiedialis) для сочленения с большой берцовой и боковой (condylus
5 Орнитология—1209 G5
Рпс. 53. Скелет передней конечности королевского пингвина —
Aptenodytes patagonica Mil. (а) и колибри— Selasphorus pla-
tycercus Swain, (b).
Обратите внимание на относительные размеры отделов конеч-
ности у этих видов в сравнении с крылом голубя (рис. 52);
кости крыла пингвина сильно уплощены; особенности функции
см. стр. 367 и 456.
г. — carpale. II.— плечевая кость; тс 1—3 — niPlacarpale 1—3;
R.—лучевая кость; s. I.—латеральная сесамовидная ко-
сточка; .s. т.—медиальная сесамовидная косточка; ph.—
Фаланги пальцев; U. — локтевая кость. По Neu и Lucas, из
Stresemann.
Рис. 54. Плечевая ;кость гуся — Anser anser L. (а и Ъ) и во-
рона — Corvus corax L. (си d).
а и с— с вентральной стороны; b и d — с дорзальной стороны:
Br. I. — место прикрепления musculus brachial is inferioris:
С—сочленовная головка — caput humeri; Cr. I.—crista
lateralis; Cr. m.—crista medialis; Ed.—processus ectepi-
condyloideus или pr. supracondyloideus lateralis; Ent. -
processus entepycondyloideus; F. p.—foramen pneumaticuni:
L. d. — следы прикрепления musculus latissimus dorsi; P'
место прикрепления musculus pectoralis; S. a. I. —sulcus an-
coneus lateralis; S. a. m. —sulcus anconeus medialis; S. ir.-~
sulcus transversus; S. c. — место прикрепления musculus supra'
coracoideus; Tb. I. — tuberculum laterale; Tb. m. — tuberculin
mediale; T. r.—trochlea radialis; T.u.— trochlea ulnaris-
, По Newton и Ga^ow.
Рис. 55. Филогенетическое
значение слияния карпаль-
ных костей.
Примитивная кисть репти-
лий состоит из трех прок-
симальных костей, двух cen-
lralia и пяти дистальных
carpalia. Уже у некоторых
динозавров 4-й и 5-й паль-
цы исчезают вместе с соот-
встств уютцп м и дистальным и
carp alia. У пер во пт и цы
Archaeornis отсутствуют не
только 4-й и 5-й пальцы с
их carpalia, по и одно ссп-
trale; l-й и 2-й дистальные
carp alia слились между со-
бой. По Паркеру, Хейль-
маппу и др., у эмбрионов
птиц. наир, гоацина, наблю-
даются эти же элементы,
по |к взрослому состоянию
из них уцелевают лишь две
самостоятельные косточки,
по этим авторам — radia-
le 4- intermedium и centra-
le-!-iilnare (см. рис. 52), по
другим — radiale 4- centrale
n inelacarpale 5-го пальца
(см. текст).
1 — пясть и запястье репти-
лии Protosa urns speneri (от-
ряд Prosauria) из верхиеперм-
ctftix отложений; 2, 3 —
левое запястье* эмбрионагоа-
ципа — Opisthocomus hoazln
Miiller с дорзальной и вен-
тральной сторон; 1, о—то
weArchaeornissiemensi Dam.;
R. —лучевая кость; U. —локтевая кость; С. — centrale; d. — пальцы; i. — intermedium;
we. — metacarpalia; r.— radiale; it.—uhiare; 1—3 — carpalia distalia. Из Heilmann.
Рис. 56. Скелет кистевого
отдела крыла казуара — Са-
suarius casuarius L.. сверху .
Первый палец, концевые фа-
ланги 3-го пальца и кар-
пальные элементы лучевой
стороны редуцированы. Уко-
роченный 2-й палец песет
коготь, dg 2 — коготь 2-го
пальца; тс. 1—3 — meta-
carpale 1-го—3-го луча;
с Уцелевший карпальный
элемент; я.—лучевая кость;
V. —локтевая кость.
(Ь}
Рис. 57. Форма таза: а — пепыряющего вида, фаэтона —
Phaelhon lepturus Daud.; b— хорошо ныряющего вида, боль-
шого баклана—• Phalacrocorax carbo L.; с — пыряющего и
более узко специализированного вида, большой поганки —
Podiceps cristatus L. Таз а и таз b принадлежат представи-
телям одного отряда Pelecani formes. По Pycraft и Dabelow.
Рис.-58. Таз рептплпй п развитие таза птиц.
а — форма таза у эмбриона прудовой черепахи — Ептуз orbicularis; b — таз молодого крокодила; с—h — последовательные ста-
дии в развитии таза У птпц; с — рогатой поганки — Podiceps aurilus L.; d и e — обыкновенной чайки — Larus ridibundus L.;
f, g> h, — курицы — Gallus gallus L. Обратите внимание па сходство первых стадий в развитии таза птиц с эмбриональной и дефини-
тивной стадиями развития таза у рептплпй; лишь в последующих стадиях седалищная и лобковая кости постепенно загибаются кзади
и занимают более пли менее параллельное седалищной кости положение, характерное для класса птиц. а. — сочленовная впадина;
/. — головка бедра; No.— nervus obturatorius; ?. —подвздошная кость; is. — седалищная кость; Р. —лобковая кость; рг. ре. —
processus pectinealis. Из Hellmann.
lateralis) для сочленения с малой берцовой костью. Переднюю поверх-
ность этого сустава прикрывает коленная чашечка (patella genu),
залегающая в бороздке между обоими мыщелками п особенно
крупная у хорошо ныряющих форм (рпс. 60).
Большая берцовая является крупной длинной костью, к которой
тесно прилегает и дистальной частью обычно сливается малая бер-
цовая, не принимающая участия в сочленении с плюсной; сочленение
с бедром также в основном ложится на два проксимальных мыщелка
Рпс. 59. Бедро и способ причлепсппл его к тазу у ныряющих п назем-
ных птиц.
С. — бедро краспозобой гагары — Со lymbus stellatiis Pont.; С я.—
положение бедра при сочленении с тазом у краснозобой гагары; II. —
бедро Hesperornis regalis Marsh.; Hr. — сочленение бедра с тазом у итого
вида;*7Ур. — сочленение бедра с тазом у среднего кронцшепа— Пите-
uius phaeopus L.; Т. — бедро тетерева — Lyrurus tetrix L.; Tt. — со-
членение бедра с тазом у тетерева; 1 —ппжпяя поверхность бедра;
2 — внутренняя поверхность бедра; at. — антптрохаптер; Ь.—таз;
с. /. — caput femori, для сочленения с тазом; с. 1. — condylus lateralis;
г. т.—condylus medialis; /. — бедро; tm — trochanter .major. По
* Heitmann.
большой борцовой и лишь частью внешнего мыщелка бодра ложится
на малую берцовую.
Сочленение голени с плюсной замечательно у птиц слиянием про-
ксимального ряда тарзальных (предплюсневых) косточек с дисталь-
ным концом большой берцовой кости и образованием ими двойных
мыщелков; дистальный ряд этих косточек в свою очередь сливается
с проксимальным концом metatarsalia или собственно плюсны
(рис. 61). Таким образом сочленение этих отделов весьма близко
напоминает то отношение, которое имеется и у рептилий, т. е. яв-
ляется интертарзальным пли межпредплюсневым, ио срастание
tarsalia с берцовой костью и плюсной здесь является полным, тогда
как у рептилий эти элементы часто сохраняют самостоятельность,
лишь связками закрепляясь по окончаниям костей обоих отделов.
•69
Глубокий желобок (fossa inlercondyloiclea), имеющийся между
дистальными мы щелка мп большой берцовой кости, продолжается
и в значительное углубление. на передней стороне ее, частью прикры-
тое костяным мостиком. Ограничивающие
ото углубление гребни (crista tibiae) про-
бегают по поверхности кости и у хорошо
ныряющих птиц продолжаются в крупный
коленный отросток (processus rotularis),
увеличивающий область прикрепления
мускулатуры лап (рис. 60).
Цевка или плюсна птиц (larsometatar-
sus) образуется в онтогенезе продольным
слиянием плюсневых костей (melalarsalia),
обычно протекающим даже без сохране-
ния швов между, ними, п вышеупомяну-
тым срастанием с дистальным рядом пред-
плюсневых косточек (рис. 62). Известное
исключение представляет лишь короткая
п широкая плюсна пингвинов и фрегатов,
прободенная . отверстиями и повпдпмому
несущая следы слияния из трех элемен-
тов (рис. 63). В образовании плюсны уча-
стия не принимает первый луч metal аг-
i-p-r
/t
rntd
t+C+I
2+3+4
mn&
oi!2
ЧЛ t+C
mt5
Pnc. GO. Голень серощекой поган-
ки — Podiceps (jrlseigena* L. (a > и
краспозобоп гагары — Colymb'its
slelldins Pont. (b).
Хорошо ныряющие виды, у которых
сильное развитие мускулатуры лап,
гл. об]), musculus gastrocnemius,
вызвало удлинение большой берцо-
вой кости в пнрамидообразпый от-
росток — processus rotularis, высту-
пающий за коленное сочленение и у
гагар функционально заместивший
коленную чашечку (см. рис. 158);
/. — бедро; /г. — малая берцовая
кость; р.—- коленная чашечка;
р. т.—processus rotularis; t.—
большая берцовая; tm. — цевка
или плюсна. По Dabelow, из Stre-
semanu.
(b)
(c)
Рис. Gl. Интертарзальпое сочленение у птиц.
Передняя сторона. (Ь)—эмбрион курицы; (с) —
первоптпца — Archaeornis siemensi Dani.; (d) — цы-
пленок курицы; Fi. —малая берцовая; Т. — боль-
шая берцовая; с — centrale; /. — fibulare; i — inter-
medium; mt 1—5 — mctatarsalia; t — tibiale; 2
3 -f- 4—дистальные tarsalia. По Heitmann.
salia, рано смещающийся кзади, сильно укорачивающийся, прйчле-
няющийся внизу tarsometatarsus и несущий короткий первый
палец (hallux).В некоторых группах птиц (например у дневных хищ-
ников) на задней поверхности плюсны развивается так называемый
hypotarsus, представляющий невидимому смещение кзади прокси-
мального отдела третьего луча metatarsalia; в этих случаях задняя
70
сторона плюсны несет резко выраженные выступы (гребни;, среди
которых залегают мощные сгибатели пальцев (рпс. 64).
Количество пальцев у птиц колеблется’от двух до пяти с наибо-
лее частым случаем в четыре пальца. Самым коротким пальцем
обычно является первый, самым длинным — третий пли четвертый.
Для многих форм характерны утеря заднего пальца п способность
второго плп четвертого постоянно пли по желанию поворачиваться
назад. В систематике наибольшее значение имеют следующие типы
Рпс. 62. Левая плюсна и пальцы, (а) —Compsognalhvs
long!pes (Theropoda, Dinosauria) из верхней юры Бава-
рии; (Ъ) —Archaeopteryx Hthographica Meyer; (с) —Ar-
chaeornis siemensi Dani.; ( d)—голубя — Columba livia Gm.
Постепенное слияние плюсневых костей в ряду предков
(s. I.) современных птиц. V Compsognathus еще сохраня-
лись 1-й п 5-й лучи, у Archaeopteryx — 4-й; Archaeorn is
metatarsalia еще не сливались между собой; тt 1—5 —
metatarsalia, от 1-го до 5-го; I—I V — 1-й — 4-й пальцы.
По Ilcilmann.
соотношения пальцев:
анизодактильный тип —
один первый палец на-
правлен назад; гетеро-
дактпльный тип — два
пальца направлены на-
зад, но один из пих
вращающийся, могу щи й
быть направлен то
Рпс. 63. Плюсна (tarso-
ineta tarsus) королевского
пингвина — Aplenodytcs
patagonica Mill. С дор-
зальной стороны. По
Watson, из Streseinann.
вперед, то назад; зигодактильный тип — два пальца, первый и
четвертый, постоянно направлены, назад (рис. 65).
3. МЫШЕЧНАЯ СИСТЕМА
Общее о б о з р е н и е. Мышечная система птиц, как и боль-
шинства высших позвоночных, настолько сложна, что может быть
описана здесь только в главнейших своих особенностях и главным
образом в элементах, имеющих отношение к основным типам дви-
жений птицы. В целом мышечная система птиц близка аналогичной
системе рептилий и отличается главным образом иным распределе-
нием и объемом мускулатуры, связанной с конечностями, разре-
шающими у птиц совершенно иные задачи движения, и отчасти
71
Рис. 64. Плюсна (цевка) и строение hypotarsus. (а,Ъ, с) —черный коршуп —- Mllrvs hor-
schvn Gm.; (d, e) —обыкновенная пустельга — Cerchneis tinuncuhis L.; (f, g)—лебедь
кликун — Cygnus cygnus L.
(a)—спереди; (b, d, f)—сзади; (с, e, g) — верхний конец сверху. Ab. II, Ab* IV —
область прикрепления и прохождения musculus abductor digit! II и m. abd. digit! IV;
Ad. II. — область прохождения m. adductor digit! II; Ad. IV — область прохождения m.
adductor digit! IV; C — суставные поверхности мыщелков для сочленения с пальцами; Ех.
Ъг. III — место прикрепления и прохождения ш. extensor brevis digiti III; Ex. br. IV —
желобок для прохождения m. extensor brevis digiti IV; Ex. d. c. — бугорки, место прикре-
пления уздечки сухожилия m. extensor digitorum communis; Ex. h. —желобок для про-
хождении сухожилия m. extensor hallucis brevis; Ex. h. br. — область прикрепления m. ex-
tensor hallicis brevis; Fl. h. br. — область прикрепления п прохождения m. flexor hallucis
brevis; Fl. perf. II — канал для сухожилия m. flexor perforates digiti II; FL prof. — каналы
для сухожилий m. flexor profundus s. perlorans; Fl. p. p. II— желобок для сухожплпят. fle-
xor perforans et perforatus digiti II; Lip. —наружный и внутренний гребнп hypotarsus’a, обра-
зующие в совокупности скользящий шарнир для проходящего пад ппми сухожилия m. ga-
strocnemius; Ну. ех.—наружный гребепь hypotarsus’a; Ну. in.—внутренний гребень
hypotarsus’a; Met. I — сочленовная поверхность для metatarsale 1-го пальца; Per. prof. —
место прохождения in. peroneus profundus; Т. а. — бугорок для прикрепления сухожилия
m. tibialis anticus; II, III, IV—мыщелки соответствующих номеров пальцев. По Qnen-
nerstedt, из Stresemann и по Сушкину.
иной мускулатурой шеи, особенно у специализированных питаю-
щихся рыбой птиц. По морфологическим особенностям мышечные
волокна птиц являются более совершенными, чем мышцы другпх
классов, в связи с высокой температурой тела, энергичным обменом
веществ и особо быстрым
движений. Мышечна я
ткань птиц является
более плотной и более
гонко волокнистой; мы-
шечные клетки очень
богаты ядрами, общая
нов ерхность которых,
вследствие их мелкости
и значительной удли-
ненности, стоит в повы-
шенных отношениях к
их объему, что вызы-
вает оживленный обмен
п напряженным характером некоторых
между веществом ядра
и клетки (Schifferde-
cker, fide Stresemann,
1927). Характерной осо-
бенностью птиц счита-
ются также ранние и
значительные окостене-
ния их сухожилий, осо-
бенно сухожилий конеч-
ности. s;
Как у других ПОЗВО-
НОЧНЫХ; мышцы ПТИЦ
иногда разделимы по
окраске на бледные (при
(а) —типы количества фаланг в пальцах; (Ъ) —типы
числа пальцев и относительного положения их (rneta-
tarsale 1-го пальца зачерчены); (а) I —наиболее обычный
тип; II — Procellariiformes, исключая род Pelecanoides;
III — стрижи — Micropus и Panyptila; IV — рябкп —
Pterocletes и козодоп — Caprimulgus; (b): I — наиболее
обычный тип анизодактильной лапы; II — отсутствие
заднего пальца, как у Rhea, Olis, Pelecanoides, Turnix,
Alcidae, Rissa; III — отсутствие 1-го пальца при зиго-
дактильном строении лапы, как у родов Sasia и Jaca-
maralcyon; IV—редукция 2-го пальца до . сохранения
лишь одной фаланги, папр. у Сеух и Alcyone; V — ре-
дукция '<-го пальца, папр. Cholornis; VI —* потеря 1-го и
2-го пальцев, папр. Struthio. По Forbes, из Stresemann.
сварении белые) и крас-
ные. Обычно это разли-
чие характерно для тех
птиц, которые часть
своей мускулатуры упо-
требляют менее часто
п напряженно; эти
мышцы оказываются
бледными, словно гголу-
прозрачными, каковы,
например, грудные мышцы многих куриных и скрытохвостов.
Гистологически бледные мускулы оказываются состоящими из срав-
нительно крупных волокон, бедных зернышками саркоплазмы (сар-
козомами) и мпофибрпллами, с рассеянными внутри клетки ядрами, в
то время как красные обладают, наоборот, темно окрашенными и
узкими волокнами, густо набиты зернышками и имеют ядра сосре-
доточенными на периферии. Установлено, что красные мускулы менее
быстро утомляются, по при неупотреблении легче ослабевают, чем
73
бледные. Последние, кроме того, на короткие сроки способны
к большей частоте сокращений.
Следует отметить широкое использование особенностей строения
мускулатуры при установлении таксономических отношений птиц,
несмотря на трудности практического изучения мышц. Отдельные
системы мускулов, цапрпмер сгибатели пальцев и мышцы гортани,
с успехом применяются для характеристики крупных систематиче-
ских групп.
Г р у и и ы м ы ш ц и м ы ш ц ы г о л о в ы. Как у других:
позвоночных, мышцы у птиц разделяются на две большие группы —
висцеральную п соматическую мускулатуру. Первая эмбриологи-
чески возникает из висцеральных пластинок целома трех первых
жаберных (глоточных) дуг. Вторая развивается из мпотомов‘ме-
диальной, париетальной стороны целома, как головной, так и туло-
вищной области.
В висцеральной мускулатуре различают три группы мышц, со-
ответственно числу висцеральных дуг. Первая развивается из бла:
стемы первой (челюстной) дуги и управляется ветвью тройничного
нерва (n. trigeminus); относящиеся сюда мускулы. (например pro-
tractor pterygoidei, ряд adduclores mandibnlae s. m. temporales)
участвуют главным образом в закрывании клюва (рис. 66). Вторая
образуется из бластемы второй дуги и управляется лицевым первом
(n. facialis); важнейшими мускулами ее являются жевательный
мускул (m. depressor mandibnlae), открывающий клюв, и так назы-
ваемый in. hyomandibularis, втягивающий язык. Третья группа,
образуемая бластемой третьей дуги, иннервируется языко-глоточным
нервом (n. glossop]iaryngeus) п состоит главным образом из т. т.
genio-hyoideus (kerato-mandibnlaris) и genio-thyreoideus, выдвига-
ющих язык из ротовой полости.
Соматическая мускулатура представлена значительно более обиль-
ными группами мышц, распадающихся на два отдела: из мпотомов
головных сомитов развиваются сложные мускулы глаз, управляе-
мые. nervus oculomolorius и n. abducens (рис. 152), а из мпотомов туло-
вища — мускулатура конечностей, хвоста, туловища п головы. По-
следний отдел управляется частью головным подъязычным первом —
л. hypoglossns (мышцы языка и верхней и нижней гортани), частью
спинными нервами (все остальные); среди последних особенно
сложна и обильна мускулатура плечевого пояса и крыла, предста-
вленная семью группами мышц, и мускулатура нижней конечности
с десятью группами, в общей сложности объединяющими много
десятков отдельных мышц.
Перейдем теперь к краткому обзору мышц и пх функций в области
шеи. туловища и конечностей (рис. 67).
Для шеи птиц характерна крайне слабая способность к боковым
сгибаниям и значительная подвижность в дорзо-вентральном на-
правлении. Наибольшая подвижность свойственна двум первым
it отчасти двум следующим шейным позвонкам, допускающим зна-
чительное вращение, которое, складываясь со сгибаниями следую-
щих отделов шеи. обусловливает поворот головы на 180°. В отно-
шении дорзо-вентральной подвижности шея разделяется иа три
74
7 — после снятия клягп;
2—после удаления mm. ad.
in and. ext. sup. и prof.;
3 — после удал on пя • mm.
ad. mand. ext. и post, и m.
pseudotcmporalis; 4 — вид
еппзу; 5 — вид еппзу по
удалении m. pter. vent.:
Ad. т. т. — т. adductor
mandibulac externus media-
ns; Ad. тц. p.‘—m. adductor
mandibulac externus profun-
dus; Ad. tn. s. — m. adductor
mandibulac externus superfi-
cialis; Ad.p.—m. adductor
mandibulac posterior; B. p.
I — pr. basipterygoideus;
B. — basitcmporale; Ch. —
хоапы; D.tn.— m. depressor
mandibulac; F. t. —for. try-
geminus; J. — jugale; L. —
лпгамепт; I. a. — m. levator
angulioris; I. po.—ligamen-
tum postorbitalc; /. s. —
lig. suborbitalc; Mand —
челюсть; Mr. — up a it рта;
Mx.—maxilla; Mx. P.—
inaxillo-palatinum; N. pt. —
n. ptcrygoideus; N.t.—ппж-
нечелюстная ветвь тройнпч-
пого перва; О — ушное от-
верстие; Ра.—palatinum;
р. pt. —m. protractor pte-
rygoidcus; Pr. a. p. — pr.
angularis posterior; Pr. a.
a. —pr. orbital is anterior;
Pr. o. p. —pr. orbi tails
posterior.; P. p. — m. pseu-
dotemporalis profundus; p.
Pt. —m. protractor ptcrygoi-
dci. P. Q.—m. protractor
quadrat!; P.s.tn. — m.pscu-
dotemporalis superior; Pt.—
ptcrygoideum; Pt. d.f Pl. I.—
дорз. и лат. части m. ptery-
goidcus dorsalis; Pt.v. I.,Pt.
v. tn. — лат. и медиальная
части m. pterygoidcus ven-
tralis; R — клюв парасфе-
ноида; R. a. —m. retractor
anguli oris; Sa—supraangu-
lare; Tp.—височная ямка;
Vrt — vnmpr
Рис. 66. Мускулатура, участвующая в движении челюстей у еппьгп — Oidcmia nigra L.
Рис. 67. Мускулатура сизого голубя — Columba livia Gin., сбоку, схематично, ai — m. ab-
ductor digiti medii; ap.—in. adductor pollicis; b— m. ilio-fibularis; bi.—m. deltoides
major; c — m. sartorius; co — in. complexus; cr—копчиковая железа; d — m.caud-
ilio-flexorius; da-—m. deltoideus minor; de — m. triceps; ea— in. patagialis (pars
bicipitis); ep — m. abductor pollicis; ex — m. extensor digiti communis; g— m. genio-
hyoideus; да — m. gastrocnemius; gl— in. genio-hyoideus, передняя часть;! i.—пъ
flexor digitoruin; if.— in. flexor collii longus; it — in. intercos tai is; ilt — m. ilio-tibialis;
il. p. — m. ilio-trochanterici; int. — m. interosseus dorsalis; is — ischium; Ic — m. lateralis
colli; lc p — m. levator costaruni posterior; Id — m. latissimus dorsi; If — m. longus colli-
anticus; Ipc — m. spinalis cervicis; Ipr — m. peroneus superficialis; m. — m. mylo cutaneus;
ma — m. mylo-hyoideus, передняя часть; mp— m. mylo-hyoideus, задняя часть; ob — ni.
obliquus externus; p — m. cucullanus, передняя часть; pb.—m. peroneus profundus; pc —
*m. pectoralis; pe — m. pubi-coccygeus; per — m. peroneus; pr— m. entepicondylo-radialis;
rca — m. rectus capitis anterior; rep. — m. rectus capitis posterior; rd. — m. extensor meta-
carpi radialis; rem. — m. rectus capitis medius; ta — m. tibialis anticuS;' te — m. temporalis
(m. masseter); t — m. trachc )-laryngeus; tr. — m. rectus abdominis analis. Из Vogt und Jung.
участка: строение-первых четырех-пяти позвонков (у длинношейных
больше) позволяет свободное сгибание шеи в вентральном напра-
влении, но не дозволяет сколько-нибудь значительных изгибов
в дорзальном отношении; средний наиболее длинный отдел (5—6,
до 10 позвонков), наоборот, несгибаем вентрально, но подвижен
дорзально; наиболее короткий последний отдел из 1—4 позвонков
также сгибается легко только в вентральном направлении. Это
строение шейного отдела позвоночника связывается соответствующей
мускулатурой. Вдоль нижней стороны шеи проходят длинный
и короткий сгибатели шеи (m. longus collii anticus и т. brevis
collii), прикрепляющиеся главным образом на нижних отростках
позвонков; вдоль задней стороны шеи располагаются антагонисты
их: длинный и короткий разгибатели шеи (m. extensor collii longus
и m. spinalis cervicis), связанные главным образом с остистыми от-
ростками позвонков. Позвонки связуются между собой мелкими
musculi interspinales и intertransversarii cervicis. Кроме того, вдоль
всей шеи развиты • поднимающие голову дорзальный m. biventer
cervicis и латеральные m. m. complexus. Вдоль всей шеи тянутся
также длинные мышцы туловища — m. longissimus dorsi (Gadow, 1890).
У некоторых длинношейных, охотящихся преимущественно за
рыбой, птиц (Anhinga, Ardeae) в шейной мускулатуре наблюдаются
интересные приспособления к развитию молниеносных ударов клю-
вом (рпс. 68).
В туловищной области кроме мышц, связанных с движением ко-
нечностей, большую роль играют в связи с дыханием мышцы ребер
и брюшного пресса. Первые слагаются внешними и внутренними
межребернымп мышцами — m. т. intercostales externi и interni; из
внешних следует упомянуть слабо обособляющуюся .часть их4—
m. levatores costarum, продвигающие вперед ребра и расширяющие
этим грудную клетку. Их антагонистом являются косые брюшные
мышцы: m. т. obliquus abdominis externus и interims, прикрепляю-
щиеся на ребрах и в задней-части грудины и лобковой кости и своим
действием уменьшающие объем грудной коробки. Брюшной пресс
образуется также m. т. trans versus и rectus abdominis, укрепляю-
щимися па последних, ребрах и заднем крае грудины и дистальных
частях лобковой кости; частично они также участвуют в спадении
грудной клетки и следовательно в дыхании.
В области хвоста развита мускулатура, выполняющая его дви-
жения: на спинной стороне мускул, поднимающий хвост, — m. le-
vator coccygis, на брюшной стороне мускул, опускающий хвост, —
г1т. depressor coccygis, отгибающий его в (^торону и расправляющий
рулевые перья, — m. pubi-coccygeus externus (его антагонист —
m. pubi-coccygeus interims), мускул, приподнимающий рулевые
перья, — т. ilio-coccygeus и некоторые другие, менее важные
(рис. 69).
Более подробно следует остановиться на мускулатуре конечно-
стей. Мускулатура передней конечности у птиц очень богата и диф-
ференцирована. По характеру своего происхождения из тех или
иных отделов мускулатуры более древних Tetrapoda она может быть
сгруппирована в следующие восемь групп (см. рис. 70 и 71).
77
1. Musculus cucullaris — сравнительно слабый мускул, физиоло-
гически почти не связанный с крылом. Возникая в области головы
п дорзальной поверхности шеи, он прикрепляется главным образом
на ключице и вызывает стяжение кожи на шее и в результате
оказывает давление на зоб и топорщит перья шеи (воротник турух-
Puc. G8. Мускулатура шеи и механизм движении ее при схваты-
вании добычи у змеешейки — Anhinga anhinga L. 1 — позвоноч-
ный столб и наиболее важные сгибатели и разгибатели шеи; рез-
кий изгиб шеи наблюдается по концам удлиненного восьмого
позвонка; 2 — механизм выбрасывания шеи вперед при прока-
лывании добычи; а — при втянутой шее; b — при вытянутой
шее; fl. — часть разгибателя передней стороны шеи — m. lon-
gus collii anticus, отдпфферепцировавшаяся в самостоятельный
мускул (сгибатель краниального отдела шен); D—окостене-
вающее с возрастом сухожилие, образующее петлю на девятом
позвонке; I. с. a., I. col. ап. — главный разгибатель шеи — щ.
longus collii anticus; s— сухожпдие дорзального сгибателя
шеи — ш. spinalis cervicis; sc, sp. cerv. — сгибатель шеи — m.
spinalis cervicis; x — расстояние, на которое удлиняется шея
при сокращении m. longus collii anticus; 8 —удлиненный вось-
мой шейный позвонок. По G-arrod и Forbes, из Gadow п strese-
niann.
•
тана п райских птиц и т. п.). Мускул иннервируется шейными
нервами и веткой n. vago-accessorius.
2. Система musculi thoracici superiores, иннервируемая одно-
именными нервами. Состоит из двух групп мышц: musculi rhomboi-
dei й musculi serrati. Первые возникают на остистых отростках
позвонков и прикрепляются на дорзальных краях лопатки и клю-
чицы; их функция заключается в притяжении плечевого пояса
к позвоночнику, частью при этом несколько передвигая его кпе-
реди пли кзади. Вторые возникают на ребрах и прикрепляются на
78
лопатке, отчасти входя в состав и melapalagium; в целом они
являются антагонистами первой группы.
3. Musculus sterno-coracoideus, иннервируемый нервами loracici
inferiores; обычно незначительный по величине мускул, функциони-
рующий как приподнимающий передние ребра.
4. Система musculi brachiales superiores, иннервируемая ветками
одноименных нервов. Состоит из четырех групп мускулов, возни-
кающих в общем в дорзальной части туловища и на плечевом поясе
и прикрепляющихся по конечности на ее дорзальной стороне. Функ-
ции разнообразны: ш. latissimus dorsi приводит туловище птицы
Рис. 69. Мускулатура хвоста курицы — Gallus gallus L.
(а)—снизу, хвост отогнут кверху; (Ь)—сбоку; ad. rec. — m. adductor recticurn;
c. il. fem. — m. cand-ilio-femoralis; c. il. fl. — in. caud-ilio-flexorius; c. cl. — in. contractor
cloacalis; d. c. — in. depressor coccygis; gl. ur. —копчиковая железа; IL c. —m. ilio-coc-
cygeiis; Is — седалищная кость; lev. co.—m. levator coccygis; p. c. ex.—m. pubi-coccy-
geus externus; p. c. in. —m. pubi-coccygeus internus; sp. il. p. —spina ilium posterior; sp.
an.—m. sphincter ani; s. an. — m. suspensor ani; ir. an.—m. transverso-analis; Pit —
лобковая кость — pubis; Руд.—пигостПль? Rec.—очины рулевых перьев. По du Toil.
в горизонтальное положение во время полета; m.m. deltoidei вращают
плечо вверх и вперед и отводят его от туловища; m. scapuli-hume-
ralis подтягивают крыло к туловищу; m. т. scapular is опускают
плечо книзу и прижимают его к телу.в
5. Система musculi brachiales inferiores. иннервируемая одно-
именными нервами. Слагается тремя группами мускулов, возникаю-
щих на грудине и вентральной поверхности плечевого пояса и плеча
и заканчивающихся главным образом на вентральной поверхности
плеча и предплечья. Наиболее мощными мускулами здесь, не только
в этом разделе, но и во всем теле птицы, являются m. pectoralis и
т. supracoracoideus, составляющие по весу до х/з (колибри), 1/11
(гусь) веса всего тела. Первый тянет крыло книзу и совершает наи-
более значительную мускульную работу в теле летящей птицы, второй
79
является антагонис-
том первого, отводит
крыло п одновремен-
но вращает его. Заме-
чательным является
то, что при противо-
положной функции
оба мускула распола-
гаются одинаково на
вентральной стороне
тела и один поверх-
ностно над другим
(ср. рис. 70 п 71).
Musculus supracora-
coideus лежит в углу
между телом п килем
грудной кости и в
большей своей массе
прикрыт еще более
массивным грудным
мускулом; сухожи-
лие его, проходя дор-
зально от грудного
мускула, через fora-
men triosseum '(между
сочленовными голов-
ками плеча, корако-
ида и лопатки) выхо-
дит на дорзальную
сторону плечевой ко-
сти и прикрепляется
здесь на tuberculum
ma jus гребня этой ко-
сти (рис. 54). На этом
же гребне прикрепля-
ется масса и грудного
мускула (m. pectora-
lis). Таким образом
точки приложения
сил этих главнейших
мускулов передней
конечности почти сов-
падают, и антагони-
стическое действие их
возможно только бла-
годаря прохождению
сухожилия m. sup-
racoracoideus через
блок, дорзально ле-
жащий по отношению
80
Рис. 71. Глубокая мускулатура грудины сизого го-
лубя — Columba livia»Gm.
cor. — коракоид; corbr. br.—m. coraco-brachialis
brevis; corbr. long. — m. coraco-brachialis longus;
corst.— in. coraco-sternalis; bic.—in. biceps; /.—
вилочка; hum.—плечевая кость; ped. — грудной
мускул — m. pectoralis; sc. —лопатка; ser. m. —
m. serratus minor; spcr. — подключичный мускул —
in. supracoracoideus; st. — грудина.
По Gadow.
к грудной мышце, У птиц нелетающих, особенно бескилевых, оба
эти мускула развиты естественно весьма слабо.
Третья группа этой системы — m.m. coraco-brachiales вращают
плечевую кость спереди назад (супинируют), частью поднимают ее
н отводят от туловища.
6. Система растягивающих предплечье и кисть мускулов, иннер-
вируемых n. radialis. Слагается обычно мелкими и тонкими муску-
лами числом около двенадцати, возникающими частью еще в пре-
делах плечевого пояса и
заканчивающимися то в об-
ласти. предплечья, то в
области кисти или на фа-
лангах. Наиболее мощны-
ми являются: m. triceps
brachii, т. ectepicondylo-
ulnaris и т. ectepicondylo-
radialis, т. т. extensor
inetacarpii и нек. др.
7. Система сгибателей
предплечья п кисти, иннер-
вируемых и. medio-ulna-
ris. Эти мускулы в коли-
честве около десяти явля-
ются антагонистами ше-
стой группы. Наиболее
значительными среди них
являются: ш. biceps bra-
chii, in. brachialis infe-
rior, m. flexor carpi ulnaris
п др.
8. Несколько обособ-
ленную группу образуют
musculi propatagialis и
metapatagialis, входящие
в состав передней и задней
летательных перепонок.
Эти мускулы образуются
из частей целого ряда кры-
ловых мускулов (рис. 72).
Целесообразен теперь обзор роли этих мускулов в движении
отдельных частей конечности. По местоположению дистального при-
крепления отдельных мышц они могут быть разделены на: 1) мышцы,
движущие плечо, 2) движущие предплечье и 3) приводящие в дви-
жение кисть и пальцы.
Из мышц, движущих плечо, наибольшее значение имеют сле-
дующие мышцы.
Для подъема крыла: m. deltoides major, короткий, но очень силь-
ный мускул с широкой поверхностью прикрепления на плечевой
кости, одновременно поднимающий и супинирующий этот отдел;
supracoracoideus — значение указано выше (стр. 80); m. latissi-
<> Орнитология—1209
81
D
(6)
(4)
(5)
mus dorsi, поверхностный спинной мускул, частью возникающий
в области synsacrum.
Для опускания крыла: m. pectoralis, см. стр. 79; m. subsca-
pularis, см. стр. 79.
Для пронации крыла: m. pectoralis, m. deltoideus minor, m. sub-
scapularis.
Для супинации крыла: m. deltoides major, m. supracoracoideus,
m. scapulo-humeralis anterior, m. coraco-bracliialis posterior.
Для приведения крыла: m. scapulo-humeralis posterior.
Мышцы, движу-
щие предплечье, дей-
ствуют только в од-
ном направлении,
сгибая или разгибая
его в отношении к пле-
чевому отделу. Глав-
нейшим разгибателем
является ш. triceps,
помещающийся глав-
ным образом на дор-
зальной и ближай-
шей к телу стороне
плечевого отдела.
Наиболее важными
сгибателями являют-
ся m. biceps, лежа-
щий на вентральной
стороне плеча, и т.
brachial is inferior,
очень короткий и ли-
шенный сухожилий
мускул, соединяющий
вентральные стороны
дистальной части пле-
ча и проксимальной
части предплечья.
О ста л ьные му скулы
этого отдела мелки п
коротки; их функция ограничивается повидимому созданием упругой
связи между плечом и предплечьем в период полета.
Мышцы кисти и пальцев, помимо функции сгибателей и разги-
бателей в отношении к предыдущим отделам, играют еще значи-
тельную роль по управлению отдельными группами маховых перьев.
Развитие этих, мышц обусловливает в значительной мере особое
искусство полета у ловких летунов. Часть этих мышц возникает
в дистальной части плечевой кости (разгибатели: m. extensor meta-
carpii radialis, m. extensor digitorum communis; сгибатели: m. flexor
carpii ulnaris, m. flexor metacarpi ulnaris; m. flexor digitorum subli'
mis), часть по поверхности костей предплечья, часть в пределах
самой кисти; последние мышцы мелки и управляют главным образом
Рис. 72. Соотношения мускулов передней летательной пере-
понки — propatagium (как пример отличий в строении му-
скулатуры, используемых для систематических целей).
Левое плечо и предплечье; 1 — утки — Anas; 2 — голубя—
Columba; 3 — кукушки — Phoenicophaeus; 4 — удода —
Upupa; 5 — птицы-лиры — Мепига; 6 — враповых — Суа-
nocorax; D.—m. deltoides major; D. pl.—дельтоиднаячасть
m. propatagialis; Ld. — мускульные ленты от mm. latissi-
mus dorsi; Pt. b. — m. propatagialis brevis; Pt. I.—m. pro-
patagialis longus; T.—m. triceps; m. biceps выделен черным,
как и лепты этого мускула, отходящие к m. propatagialis.
По Fiirbringer, из Gadow и Newton.
82
первым и вторым пальцами. Соответственно плоскости движения ки-
стевого сустава, разгибатели лежат главным образом на лучевой
стороне предплечья, а сгибатели располагаются вдоль локтевой
стороны.
Из более частных приспособлений особый интерес представляет
образование сесамовидных косточек в сухожилиях сгибателей
и разгибателей крыла у птиц, использующих при полете токи воз-
духа и подолгу летающих с неподвижными крыльями (планирую-
щим полетом); это образование закрепляет крыло в растянутом
положении и облегчает мускульную работу, производимую птицей
во время полета (рис. 73).
Рис. 73. Часть передней копечпости качурки — Procellaria aequinoctialis L. с приспособ-
лением, облегчающим длительный полет с расправленпыми крыльями.
Наиболее сильный разгибатель крыла — m. extensor metacarpi radialis имеет две го-
ловки, из которых одна, более мощная, прикрепляется на processus supracondyloideus (или
рг. ectepicondyloideus) плечевой кости, другая — на сесамовидной косточке, прилежащей
к названному отростку, на которой укрепляются также сухожилия мускула, растягиваю-
щего переднюю летательную перепонку (m. propatagialis); с этим механизмом связана вто-
рая сесамовидная косточка. При расправленном крыле локтевое сочленение фиксируется
веиодвижно, причем наличие сесамовидных косточек позволяет изменения в сокращении
передней мускулатуры крыла без ослабления расправленного локтевого сочленения, а—
плечевая кость с proc, supracondyloideus lateralis; b, с — сесамовидные косточки; d, е —
головки m. extensor metacarpi radialis; / — tendo propatagialis longa, при / перекидываю-
щееся через кистевое сочленение по мостику из сесамовидной косточки; д — tendo propata-
gialis brevis; h — лучевая кость; i — локтевая кость; k, I — обе carpale; т — metacarpale.
По Reinhardt, из Stresemann.
Соотношения отдельных мускулов крыла довольно изменчивы
в различных группах птиц и широко используются в’ таксономиче-
ских целях. Наиболее употребительными являются вариации муску-
латуры, входящей в состав передней летательной перепонки (pro-
patagium) (рис. 72), используемые в систематике, начиная с иссле-
дований Гаррода (1876).
Мускулы таза и задней конечности. Обильно
расчлененная мускулатура ног лишает нас возможности описывать
более мелкие мускулы и поневоле заставляет быть схематичным.
Различают следующие десять групп этих мускулов, рассматри-
ваемых в порядке их удаления от тела к периферии конечности.
Общим руководством может служить рис. 74, представляющий
схему расположения и мест прикрепления основных мускулов ниж-
ней конечности (см. также рис. 75, 76, 77, 78).
1. Группа дорзальных musculi ilio-femorales, возникающих на
Дорзальной поверхности подвздошной кости и прикрепляющихся
*
83
Рис. 74. Схема мускулатуры ноги птиц. Мускулы отмечены сплошной черной линией, длин-
ные сухожилия — пунктиром.
О —ось тазо-бедренного сочленения; 1—m. m. ilio-trochanterici; 2 — т. ilio-femo-
ralis; 3— т. obturator; 4— т. ischio-femoralis; 5 — т. caud-ilio-femoralis; 6—пь pub'
ischio-femorails; 7—т. ilio-tibialis (или posterior externus); /а—m. ilio-tibialis anterior
(или internus); 8 — m. sartorius; 9 — m. femoro-tibialis medius; 9a — m. temoro-tibialis ex-
ternus; 11—m. ilio-fibularis; 12—m. ischio-flexorius; 13— m. caud-ilio-flexorius; И7
m. gastrocnemius; 15 — m. peroneus superficialis; 16 — m. peroneus profundus; 17 —пь P'
hialis antiOus; 18 —mm. flexores digitorum I—III; 19 — m. extensor digltorum communis
По Stolpe.
в проксимальной части бедра (рис. 74). Сюда относятся m. ilio-
femoralis externus и т. ilio-trochanterici, поднимающие и отчасти
вращающие бедро внутрь.
2. Musculus ilio-femoralis internus, возникающий на вентральном
краю подвздошной ко-
сти и оканчивающийся
на внутренней поверх-
ности бедра. Он подни-
мает, приводит и отча-
сти вращает бедро кна-
ружи.
3. Musculus ambiens
возникает на proc, ре-
ctinealis таза и прохо-
дит по передней стороне
бедра и голени. Окан-
чивается или на колен-
ной чашечке, являясь
ра згиб ате л ем голени,
или вступает в связь с
m. flexor perforati cli-
giti, являясь тогда так-
же и сгибателем второго
и третьего пальцев.
Этому мускулу непраг
вильно приписывалась
важная функция — ав-
томатическое закрепле-
ние пальцев, сжимаю-
щих ветвь во время
сна птиц (Маршаль,
1895, и др.). Однако при
более детальном иссле-
довании выя сни л о с ь
(Gadow, 1897; Brauer,
1911),что он отсутствует
у огромного большин-
ства птиц, притом пре-
имущественно древес-
ных по образу жизни
(Passeriformes, Picifor-
mes, Trogones и др.).
Лучше всего m. ambiens
развит у бегающих, пла-
вающих отрядов и днев-
ных хищных птиц. При
наличии связи этого
мускула с сгибателями
пальцев он повидимому
играет некоторую роль
Рис. 75. Мускулатура голени панду — Rhea americanu L.
Наружная поверхность левой ноги, после частичного
удаления m. peroncus super!icialis, caput externum
m. gastrocnemii, m. ilio-fibularis, m. ambiens и нек. др.
мускулов бедра; access. — part, пли т. «accessorius» му-
скула caud-ilio-flexorius; amb.—т. ambiens; с. е.
vast.— caput externum m. gastrocnemii; cd. il. fl.—
m. caud-ilio-flexorius; fem. tib. ext.—m. femoro-tibialis
externus; fl. per. II — m. flexor perforans et perforatus
digit! II; fl. per. Ill — m. flexor perforans et perforatus
digiti III; fl. pert. II — m. flexor perforatus digit! II;
fl. pert. Ill — m. flexor perforatus digiti III; fl. prof.
m. flexor profundus; gasl. — m. gastrocnemius; il. fib. - -
m. ilio-fibulares; isch. fl.—rn. ischio-flexorius; tig.
transv. — ligamentum transversum; n. isch. — nervus
ischiadicus; n. per. sup. —nervus peroneus superficialis;
per. sup.—m. peroncus superficialis; tib. ant. — m. ti-
bialis anticus. По Gadow.
85
и оканчивается на коленной чашечке пли
лени; соответственно этому он поднимает
в автоматическом сгибании пальцев при подъеме ног на ходу
птицы. Наличие этого мускула имеет существенное систематиче-
ское 1 и филогенетическое значение, так как он является наслед-
ством от рептилий (рис. 76).
4. Musculus sartorius. Возникает в дорзальной части synsacrum
в дистальной части го-
бедро и одновременно
является разгибателем
голени.
5. Группа musculi
femori - ti bi ales возни-
кает в области бедра
и подвздошной кости,
оканчивается на колен-
ной чашечке и голени.
Основной разгибатель
голени.
6. Группа musculus
obturator — m. pub-ischi-
um-f emor alis,. возни-
кают в дистальной и вен-
тральной части synsa-
crum, оканчиваются в
области бедра и иннер-
вируются nervus obtu-
rator. Функция заклю-
чается в подтягивании
п отведении бедра назад.
7. Musculu scaud-ilio-
femoralis возникает в
области хвоста и под-
вздошной кости и при-
крепляется на бедре.
Сокращения его вызы-
вают движения хвоста
косо вниз и вбок и подтягивание бедра кзади п кнаружи.
8. Группа мускулов, приводящих голень, возникающих главным
образом в области дистальной части таза и хвоста и прикрепляю-
щихся вдоль голени: m. ischio-flexorius, т. caud-ilio-flexorius, т.
ilio-fibularis и пек. др.
Обе последние группы обнаруживают значительное разнообра-
зие в местах своего прикрепления, вследствие чего важны в систе-
матическом и филогенетическом отношениях.
Рис. 76. Положение musculus ambicns у тетерева — Ly-
Turvs tetrixL. (1, 2, 3) и североамериканского аллига-
тора— Alligator missisipiensis (4,5,6), указывающее
на сходную функцию и близость этого мускула в
обопх классах.
7 и 4 — бедро с внутренней стороны; 2 п 5 — коленное
сочленение с наружной и задней сторон; 3 и 6 — с на-
ружной стороны; а — m. ambiens и его сухожилие;
/г. — малая берцовая кость; fp. — m. flexor perforatus
digiti (по мнению Гадова частично являющийся про-
изводным m. peroneus posterior рептилий); fl. — m.
femoro-tibialis; ge., gm., gi.—m. gastrocnemius oxter-
nus, medius, internus; if. — in. ilio-fibularcs; il.—
подвздошная кость; p.—лобковая кость; pa.—pa-
tella; pp. — m. peroneus posterior. По Gadow и Hell-
mann, из Hellmann.
1 Наличие или отсутствие этого мускула послужило, в частности, Гарроду
(Garrod. 1873) основанием для разделения всех птиц на два подкласса: Нота-
logonatae— обладающих этой мышцей, и Anomalogonatae—лишенных ее.
К последней группе относятся: Picifonnes, Passeriformes, Micropodiiformes,
Trogoniformes, Slrigijormes и подотряды: Podicipedes, Ardeae, Alcae. Это деле-
ние не удержалось, но для разграничения более мелких групп и определения
таксономических отношений их развитие этого мускула до сих пор имеет суще-
ственное значение.
86
В описании соотношений этих мускулов применяется формула
Гаррода, обозначающая буквами А и В наличие у musculus caud-
ilio-femoralis обособленной хвостовой и подвздошной части с соот-
ветственным прикреплением, а буквами У и X — наличие у m. caud-
ilio-flexorius обособленного прикрепления на бедре, хвосте или под-
вздошной кости (рис. 77). Наличие всех этих
образований (формула АВ ХУ) считается
наиболее примитивным явлением, из ко-
торого вследствие редукции развились
остальные комбинации. В ряде более про-
стых случаев (А: ХУ и др.) наблюдаемые
взаимоотношения в разных систематиче-
ских группах образовались почти несо-
мненно конвергентным путем.
Рис. 77. Мускулатура левого бедра канадского жу-
равля — Megalornis canadensis L., как пример положе-
ния мускулов соответственно полной формуле Гаррода
(АВХУ).
Внешняя сторона: поверхностные мускулы m. ilio-fibu-
laris и т. ilio-tibialis удалены. А — каудальная часть
ml caud-ilio-femoralis; В — подвздошная часть т. caud-
ilio-femoralis; х — musculus caud-ilio-flexorius (каудаль-
ная часть); Y — бедренная, часть («part accessorius») m.
caud-ilio-flexorius; c. il. f.— m. caud-ilio-femoralis;
c. il. fl. — m. caud-ilio-flexorius; il. fb. — m. ilio-fibu-
larls; il. fem. ex. — m. ilio-femoralis externum; il. tib. —
m. ilio-tibialis externus; is. fl. — m. ischio-f lexorius;
is. fm. — m. ischio-femoralis; N — седалищный нерв,
n. ischiadicus; p. if. — m. pub-ischio-femoralis; sart. —
m. sartorius или m. ilio-tibialis internus. По Gadow.
Рис. 78. Плюсна п нижняя
часть голени клинохвостого
орла — Aquila (Uroactus) audax
Lath.
ext. dig. com. — m. extensor
digitorum communis; ext. hall,
br. — m. extensor hallicis bre-
vis; per. sup. —m. peroneus su-
perficialis; per. prof.—m. pero-
neus profundus; tib. ant. — m.
tibialis anticus. По Milne Ed-
wards, из Gadow.
9. Группа разгибателей плюсны и пальцев, иннервируемых пер-
вой ветвью седалищного нерва.*Сюда относятся не менее восьми
мускулов, из которых наиболее крупными являются m. tibialis
anticus, m. extensor digitorum communis, m. peroneus superficialis
и нек. др. (рис. 78).
10. Группа сгибателей плюсны, пальцев и отчасти голени, иннер-
87
вируемая остальными ветками седалищного нерва. Более десяти
мускулов, главным образом развитых в области плюсны и пальцев.
Наиболее мощным является musculus gastro-cnemius, образующий
«Ахиллесово сухожилие» птичьей ноги и наиболее крупный мускул
голени. Длинные и короткие сгибатели пальцев своими сухожилиями
образуют сложные взаимоотношения, распределяясь не только в не-
сколько слоев (три), но и переплетаясь друг с другом. Наиболее
важными в систематическом отношении
//
/// IV va
. г
* V*
V!
VIII
X X!
оказались отношения друг
к другу сухожилий muscu-
lus flexor perforans pro-
fundus и m. flexor hallu-
cis. Гадовым установлено
семь типов этих отноше-
ний, называемых номерами
и применяемых для систе-
матической характеристи-
ки отдельных групп (на
рис. 79 эти отношения да-
ны с промежуточными сту-
пенями и отклонениями).
I. Оба сухожилия пере-
крещиваются, образуя про-
стой род связи —vinculum.
Этот случай наблюдается у
куриных птиц, голубей,
попугаев и др.
II. Vinculum очень ве-
лик и крепок и является
Рис. 79. Схема соотношения глубоких сгибателей
лап у птиц.
Л — сухожилие m. flexor perforans profundus; Р —-
сухожилие т. flexor hallucis; 1—4—номера
пальпев; 1 — курица — Gallus; II — киви — Apte-
ryx; III — сокола (сем. Falconidae); IV — нанду —
Rhea; Va — стриж — Micropus; Vb — момот — Мо-
motus; Vc — нолибри — Trochilus; Vd — удоды —
Upupa и Phoeniculus; VI — дятел — Picus; VII —
певчие воробьиные — Oscines; VIII — индийский
трогон — Harpactes duvauceli Temm.; IX — зимо-
родок-хохотун— Dacelo gigas Bodd.; X — южно-
американский лапчатоног — Heliornis fulica Bodd.;
XI — скопа — Pandion haliaeius L.; XII — кукуш-
ки (Cuculiformes). По Gadow и Newton.
прямым продолжением су-
хожилия m. flexor hallu-
cis; сухожилие заднего
пальца является слабыми
имеет вид боковой ветви
этого сухожилия. Тип свой-
ствен киви, пингвинам п
мн. др.
III. У многих дневных
хищников в нижней части
плюсны m. flexor делится
на две части, из которых одна проходит в задний палец, другая же
сливается с веткой m. flexor communis, проходящей во второй па-
лец. В этом случае иногда может быть значительный vinculum,
соединяющий сухожилия перед их расщеплением.
IV. У нанду, фламинго и др., где задний палец мал или отсут-
ствует, присутствуют оба. сухожидия, но вначале они сливаются
друг с другом, лишь впоследствии распадаясь перед вхождением
в пальцы.
V. У носорогов, кондоров и др. оба сухожилия лолностыо сли-
ваются перед вхождением в пальцы.
VI. У туканов и др. vinculum присутствует, но m. flexor digi-
ts
torum проходит только в третий палец; остальные пальцы снаб-
жаются трехветвистым сухожилием m. flexor
hallucis.
VIL Сухожилия пере-
крещиваются свободно,без
связи друг с другом. Сюда
относятся все воробьиные,
удоды и некоторые цапли.
В строении сухожилий
т. flexor perforans древес-
ных и отчасти даже бега-
ющих птиц обнаружено
интересное приспособление
к автоматическому удер-
живанию ветки сидящей
птицей, облегчающее на-
пряжение мускулатуры
(Schaffer, 1903).
На внутренней стороне
пальцев сухожилие этого
мускула и его сумка на
обращенных друг к другу
поверхностях покрыты сло-
ями хрящевых клеток, с
выступами на одной сто-
роне и углублениями на
другой (рис. 80). Садяща-
яся на ветку птица своим
весом вызывает вхождение
этих выступов в противо-
лежащие им углубления,
что автоматически закреп-
ляет сухожилие в одном
Рис. 80. Строение подошвенных сухожилии, облег-
чающих автоматическое сжимание ветви у домаш-
него воробья — Passer domesticus L.
/<7
положении.
6. ОРГАНЫ ДЫХАНИЯ И
ГОЛОСОВОЙ АППАРАТ
Органами дыхания у
птиц являются легкие,
стоящие в своей функции
в тесной связи с развет-
влённой системой воздуш-
ных мешков. Дыхатель-
ные пути включают в себя
также носовую и ротовую
а — сагиттальный разрез 4-го пальца; b — сумка
сухожилия ш. flexor perforans в дистальной» ее
части, с внутренней сторопы; с — разрез сухожилия
с сумкой, находящихся в состоянпп «замка» (силь-
ное увеличение); 11—V — фалапгп пальцев; el. d. —
дорзальная эластическая лента; el. v— вентральная
эластическая лепта; ех — сухожилие m. extensor;,
/. р. — сухожилие m. flexor profundus; I — хря-
щеватая выстилка подошвеппой поверхности сухо-
жилия, прекращающаяся у Г ; М — хрящеватая
выстилка подошвенной поверхности сухожилия и
внутренней поверхности его сумки; у М1 в разрезе;.
п — зубчатые возвышения па поверхности сумки;
Z — хрящеватые ребрышки на внутренней поверх-
ности сумки; у Z — в разрезе. По Shaffer.
полости, описываемые ниже (стр. 118),
верхнюю гортань, трахею, бронхи и нижнюю гортань или сиринкс.
Верхняя гортань (larynx cranialis) птиц лежит за аборальным
краем языка в виде овальной подушки с находящейся на ней про-
дольной щелью — входом гортани (ostium laryngis) (рис. 81). Гор-
89-
Ш (2) <3)
АС’.
тань у птиц образуется перстеневидным и черпаловидными хрящами.
Щитовидный и надгортанный хрящи, имеющиеся у млекопитающих,
здесь отсутствуют. Основа гортани образуется перстеневидным хря-
щем (cartilago cricoidea), состоящим из непарной вытянутой вен-
тральной части — pars ventralis, парных, почти квадратных боко-
вых частей — pars laterales и непарной дорзальной части — pars
dorsalis. Вентральная часть представляет собой обычно рано око-
стеневающую хрящевую пластинку, на внутренней поверхности ко-
торой нередко имеется продольный гребень. Верхний край дорзаль-
ного хряща по бокам несет сочленовные поверхности для соединения с
двумя парами черпаловид-
ных хрящей (cartilagines
arytaenoideae), косо залега-
ющих на оральной поверх-
ности перстеневидного хря-
ща. Эти хрящи имеют вид
трехгранных балочек, ох-
ватывающих между собой
гортанную щель; действием
лежащих на них мускулов
щель открывается и за-
крывается. Сужение щели
гортани происходит сокра-
щением musculus sphincter
laryngis (или m. arythae-
noideus transversus), коль-
цеобразно охватывающего
латеральные и дорзальные
поверхности черпаловид-
ных хрящей и дорзальный
край перстеневидного хря-
ща, а расширение — по-
средством musculus apertor
laryngis (или m. cricoary-
thaenoideus), соединяющего
медиальные края черпало-
видных балок с свободным дорзальным краем перстеневидного
хряща. Гортань в целом соединяется с другими скелетными частями
посредством musculus sternolaryngaeus и musculus laryngohyoideus.
Верхняя гортань у птиц лишена звуковых связок и не играет
существенной роли в голосовом аппарате птицы. Попадающий в гор-
тань воздух проходит первоначально через носовую полость и за-
тем через хоаны, расположенные как раз против гортанной щели.
Этот воздушный путь, прилеганием языка к орбитальному полю
и слипанием слизистых оболочек в задней части глотки, может вы-
ключать участие в нем других частей ротовой области, что имеет
особое значение для наполнения воздухом воздушных мешков, рас-
положенных в голове (см. стр. 93).
Трахея. Дыхательное горло (trachea) начинается сразу за
перстеневидным хрящем и продолжается до распадения на бронхи;
Рис. 81. Верхняя гортань королевского пингвина —
Aptenodytes patachonica Forst.
j. 2, з — с дорзальной стороны; 4— с вентраль-
ной; 5 — с латеральной; 2 — по удалении musculus
sphincter laryngis.
А. С.—черпаловидные хрящи (cartilagines агу-
thaenoideae); A L. — m. apertor laryngis; С. С.—
дорзальная часть перстеневидного хряща (cartilago
cricoidea); 8. L. —m. sphincter laryngis; T. —тра-
хеальные кольца; T. С.—вентральная и боковая
части перстеневидного хряща. По Watson, из Stre-
semann.
00
как правило, оно короче шейной части позвоночного столба и не
участвует вместе с пищеводом в его изгибах. Обычно трахея имеет
вид трубки, состоящей из костяных (редко хрящевых) колец, соеди-
ненных межкольцевыми эластичными связками; просвет трахеи
обычно уменьшается ближе к бронхам, но во многих случаях у сам-
цов уток и крохалей образует вздутия и расширения, играющие
роль резонаторов при производстве звука. У других видов трахея
значительно увеличивается в длину и образует петли, частью зале-
гающие под кожей на груди, частью проникающие внутрь киля
грудной кости (рис. 82). Распадение трахеи на короткие свободные
бронхи происходит обычно по вхождении ее в верхнее грудное от-
Рис. 82. Грудипа и плечевой пояс лебедя — Cygnus cygnus L.
Трахея глубоко врастает в киль, заворачиваясь петлей. Ьг — бронх; cl — вилочка;
сог— коракоид; cr. st. — грудной киль; sc — лопатка; tr—трахея. По Heinroth.
верстие, в редких случаях выше его. В противоположность строению
трахеи, кольца бронх обычно не замкнуты, соединяясь по медиаль-
ной стороне эластичными мембранами. В области бронх и каудаль-
ного конца трахеи у большинства птиц развивается нижняя, или
певчая гортань (syrinx), строение которой весьма разнообразно
в разных систематических группах. Этот орган является голосовым
аппаратом п у многих видов достигает значительной сложности
в своем строении (см. ниже).
Легкие и бронхи. Легкие птицы “ замечательны очень
небольшой величиной и слабой эластичностью; они заполняют дор-
зальную часть грудной полости и посредством клетчатки настолько
плотно соединяются с ребрами, что выполняют пространства между
ними (рис. 83). Нижняя поверхность легких покрыта плеврой и
оставляет между собой и грудной костью значительное простран-
91
ство — полость средостения (cavum mediastinum), занимаемую серд-
цем и воздушными мешками.
При своем вхождении в медиовентральную поверхность легких
бронхи теряют свои хрящевые полукольца и проходят в виде пере-
пончатых эластичных каналов через все легкое близ его вентраль-
ной поверхности. Начальная часть этих каналов расширяется ампуло-
образно (преддверие — vestibulum) и несет на медиодорзальноп
стороне ряд крупных отверстий (4—6), ведущих в вентральные
бронхи (entobronchii) (рис. 84). За этим расширением, основной
бронх, называемый mesobronchus, на дорзальной стороне несет
7—10 отверстий дорзальных бронхов (ectobronchii) и неопределенное
Рис. 83. Легкие и бронхи утки кряквы — Anas platyrhyncha L.
1—с дорзальной стороны; 2 — с вентральной стороны. На рис. 1 разветвления /—
7 являются дорзальными бронхами или эктобронхами;’на рис. 2 — разветвления 1—4 явля-
ются вентральными бронхами или энтобронхами. А—устье saccus cervicalis; В — устье
saccus clayicularis; С—устье saccus praethoracalis; D — устье saccus postthoracalis; К —
устье saccus abdorninalis. По Sappey, из Stresemann.
число мелких отверстий, ведущих в легочные трубочки (parabron-
chii). На эти же легочные трубочки разбиваются и дорзальные
бронхи, образуя в целом слйжно переплетающуюся систему воз-
душных путей (рис. 85), разветвляющуюся на бронхиоли,и затем
густую сеть воздушных капилляров, в которых и происходит обмен
газов (рис. 86). С этой сетью переплетается столь же сложная сеть
кровеносных капилляров. Прохождение воздуха через воздушные
капилляры происходит очевидно вследствие сокращения стенок ле-
гочных трубочек, имеющих гладкие, преимущественно кольцевые,
мускульные волокна (рис. 87).
Воздушные мешки. Упомянутый выше mesobronchus
в каудальной части легкого разветвляется на две ветви и впадает
ими в крупные воздушные мешки: брюшной (saccus abdorninalis)
02
л заднегрудной (saccus posLtlioracalis). Они являются инспиратор-
ными мешками (наполняемыми при вдохе) и играют крупную роль
в дыхании птицы (рис. 88).
Как эти, так и другие воздушные мешки птиц чрезвычайно тонки
п нежны, и стенки их представляются совершенно прозрачными
перепонками почти без уча-
стия капилляров, или нерв-
ных окончаний. Гистологи-
чески они образуются двумя
слившимися слоями — вну-
тренним, мостовидным брон-
хиальным эпителием, и на-
ружным, — имеющим харак-
тер серозной оболочки.
Остальные воздушные
мешки связаны с вентраль-
ными бронхами легких и
представляют три пары эк-
спираторных воздушных меш-
ков: шейный воздушный ме-
шок (saccus cervicalis), впа-
дающий в легкое в краниаль-
ном конце его; он образует
ряд выростов, располагаю-
щихся вдоль шеи, проника-
ющих в шейные позвонки и
их каналы и в позвоночные
отделы ребер; подключичный
воздушный мешок (saccus cla-
viculares) впадает в легкие
на вентролатеральной сторо-
не; он рано сливается в не-
парный мешок, заполняю-
щий пространство между клю-
чицами, но дает выросты в
область сердечной сумки, ме-
жду мышцами грудины в
крупные кости грудины, пле-
ча и передней конечности.
Парный предгрудный воз-
душный мешок (saccus ргае-
thoracalis) впадает в легкое
Гис. 84. Схема строения- легких и воздушных
мешков сизого голубя — Columba livia Gm.
Па левой половице схемы изображена лишь
вентральная сторона легких с экспираторными
воздушными мешками (пунктиром отмечен также
основной бронх). На правой половипс легкое
•рассматривается как бы прозрачным, причем
отмечаются лишь те воздушные пути, которые
видны с дорзальной поверхности легких и отно-
сятся к инспираторным каналам и воздушным
мешкам. Инспираторные пути и мешкп изобра-
жены черным, экспираторные—пунктиром; ах.—
diverticulum axillarc; В. — главный бронх;
С. — ventrobronchus cervicalis; са — ventrobron-
chus caudalis; cl.—ventrobronchus clavicularis;
/at.—боковая ветвь ventrobronchus caudalis;
M — mesobronchus; m. — ventrobronchus media-
ns; S. abdom.—saccus abdominalis; sb.—sacco-
bronchii; S. cerv.—saccus cervicalis; S. clav.—
saccus clavicularis; S. thor. ant. — saccus prae-
thoracalis; S. thor. post.—saccus postthoracalis;
V.—vestibulum; 1—8—дорзальные бронхи.
По Brandes, из Stresemann.
посредине его вентральной стороны;
его выросты невелики и занимают пространство в грудной полости
в области сердца, пищевода и печени.
В качестве особой системы должны быть выделены глоточно-
носовые воздушные мешки, лишь у немногих видов (напр. пели-
каны, птицы-носороги) сообщающиеся с легочными воздушными
мешками; они наполняются воздухом через глоточную полость и орби-
тальную щель. Выросты этих мешков пневматизируют кости черепа
У птиц-носорогов, туканов, сов, аистов, буревестников и др.,
93
Рис. 85. Парабропхи серого гуся — Ansar anser L. в поперечном разрезе. В правом инъе-
цированы воздушные пути, в левом сосудики artcria puimonalis. По Schulze, из Hilzheimer.
(а)
Рис. 86. Тонкое строение легкого австралийского тка-
чика — Thaeniopygia castanotis (Gould.). На препарате
видны два парабронха (Lu.), с отходящими от них
бронхиолями (Вг), соединенными друг с другом сеткой
воздушных капилляров (К); В — кровеносные сосудики.
(Ъ)
Рис. 87. Строение бронхов и
парабронхов страуса — Struthio
camelus L.
(а) —поперечный разрез двух
дорзальных бронхов с отходя-
щими от них парабронхами;
(Ь) — парабронх, вскрытый
вдоль. На внутренней поверх-
ности стенок видны ряды попе-
речных, пробегающих по кругу,
гладких мускульных пучков,
соединяющихся между собой
сплетением более тонких глад-
их мускульных волокон. По
Schulze, из Stresemann.
Рис. 88. Воздушные мешки и их отростки (diverticulae) сизого голубя — Columba lima Gm.
can. intertr. — canalis intertransversarius, сопровождает art. n v. cervicalis и глубокие
шейные нервы; can. prant. — canalis praeacetabularis; div. cost. — diverticule costale, снаб-
жают воздухом грудину и ребра; div. axil. — diverticulum axillare, выполняет пространство
между mm. supracoracoideus, coraco-brachialis posterior, pectoralis thoracicus и biceps и отсы-
лает отросток в плечевую кость — diverticulum humeri; div. fem. ant. post, и sup. — diver-
ticulae femoralia anterius, superius, posterius, inferius (не изображен), окружают головку
бедра и снабжают воздухом кости таза, хвоста и бедро; div. oes. tr. — diverticulum oesophago-
tracheale, частью окружает трахею; div. subscap.— diverticulum subscapulare. выполняет
пространство между лопаткой, передними ребрами и коракоидом; div. svp. vert. — divcr-
ticulesupravertebralia, окружают сверху сочленения позвонков; div. st. — diverticule sterno-
cardiale, залегают между сердцем и грудиной и пневматизируют последнюю; pvlm.—легкие
(изображены черным цветом); sac. abd. dext. и sin.—sacci abdominales dextra и sinistra,
брюшные мешки, правый и левый несимметричны, левый под давлением желудка обычно
меньших размеров; sac. cerv. — sacci cervicales, шейные мешки лежат у входа трахеи в гру-
дину- вне полости тела; sac. clav.—saccus interclavicularis, межключичный непарный
мешок охватывает нижнюю часть трахеи и пространство до сердца; sac. int. ant. и post. —
sacci intermedii anterior и posterior — передний и задний промежуточные мешки, занимают
заднюю часть грудной полости до грудобрюшной преграды (диафрагмы); tr. —трахея.
•проникая между пучками мышц и в кожные лопасти, иногда просто
под кожу. У многих видов выросты их проникают даже в шею,
в плечевой пояс и переднюю конечность. У некоторых аистов особые
мешки образуются под кожей шеи у ее основания и, наполняясь
воздухом, служат подушкой для массивного и длинного клюва,
покоящегося на них во время сна птицы (рис. 89).
Развитие всех этих воздушных мешков подвержено большой
изменчивости п имело бы значительное классификационное значение,
если бы не трудность изучения
их. В общем у хорсшо летающих
форм воздушные мешки и связан-
ная с ними пневматичность скелета
наиболее хорошо развитый диффе-
ренцированы, но из этого правила
имеется несколько исключений.
Наиболее пневматичный скелет,
содержащий воздух до конечных
фаланг пальцев обеих конечностей,
принадлежит птицам-носорогам и
паламедеям, не отличающимся осо-
бым искусством полета. Многие
группы отличаются не только пнев-
матичностью скелета, но и пнев-
матичностыо подкожных про-
странств, придающих подкожному
слою (subcutis) пенистый характер
(например у пеликана).
Процесс дыхания. Как
уже указывалось, воздушные
мешки, связанные с легкими, име-
ют большое значение для дыхания,
создавая большие запасы возду-
ха, помимо уже имеющегося в лег-
ких. Ветвление бронх в легких об-
разует трубчатую систему, с при-
водящей воздух функцией у дор-
зальных бронх и отводящей воздух
функцией у вентральных, что соз-
дает возможность беспрерывного
газообмена с приведением .свежего воздуха в легкие из инспира-
торных мешков в то же время, когда происходит отведение уже отра-
ботанного воздуха. Этот процесс, по теории Брандеса (Brandes, 1924),
происходит следующим образом: при расширении экзоспираторных
мешков они насасывают свежий воздух из экспираторных мешков,
заставляя его проходить предварительно через сеть воздушных ка-
пилляров, где он отдает кислород крови и получает углекислоту.
Тем временем инспираторные мешки наполняются новым свежим
воздухом. При сокращении экспираторных мешков, использован-
ный воздух выходит через преддверие и трахею наружу; отток его
в инспираторные мешки не может иметь места вследствие одновре
Рис. 89. Шейно-головной воздушный ме-
шок марабу — Leploptilos crumeniferus
Less.
Ко;ка с головы и шеи снята, тонкостен-
ный огромный мешок покрыт легкой
штриховкой. Мешок наполняется выды-
хаемым воздухом через посредство лежа-
щего под орбитой saccus orbitalis. Верх-
няя стрелка указывает на выходное в
saccus orbitalis отверстие, лежащее на вну-
тренней стороне птерпгоидной кости. По
Chapin, пз Stresemann.
менного замыкания их входных отверстий. При пырянии птицы
использованный воздух не выдыхается, а антагонистической дея-
тельностью воздушных мешков, действующих как меха, неодно-
кратно прогоняется через легкие, все с большей полнотой теряя
своп кислород. Этим считают возможным объяснить пребывание ны-
ряющих уток под водой до 15 минут, без особых затруднений в ды-
хании, и этим же, до известной степени, объясняется малая частота
дыхания у крупных птиц, составляющая у пеликана и грифа на-
пример всего 4—6 раз в минуту. Существенным для хода газообмена
в легких является также движение ребер, с которыми этот орган
тесно срастается и движениям которых следует. При каждом вздохе
поэтому боковые края легких расширяются, а медиальные части сжи-
маются; эти движения наполняют воздухом базальные части сакко-
бропхов и опустошают сжатые вентральные бронхи. Важное зна-
чение имеют также межреберные мышцы, вызывающие неравномер-
ное приближение грудины к позвоночнику и таким образом сжи-
мающие каудальную часть легкого, в то время как краниальная
может быть освобождена от давления.
Однако воздушные мешки имеют помимо указанной, связанной
с дыханием функции еще другое существенное и разнообразное зна-
чение в жизни птицы. Пневматизируя костную массу, мешки чрез-
вычайно облегчают вес птицы, что важно для летающих и отчасти
п плавающих птиц. В группе водоплавающих птиц особенной вели-
чиной отличаются брюшной и заднегрудинный мешки, объем кото-
рых регулируется в значительной степени брюшным прессом, вы-
зывая таким образом различную пловучесть тела птицы. Воздушные
мешки пеликана вмещают до 10,5 л воздуха; труп его весом в 4 кг при
заполнении воздухом этих воздушных мешков выдерживает без по-
гружения в воду нагрузку в 10,5 кг, а одна из цапель (Agamia ago-
uti Gm.) весом в 950 г выдерживала, при этих же условиях, 400 г
нагрузки, погружаясь уже от 50 г в случае ненаполнения воздухом
воздушных мешков. Важное значение воздушные мешки имеют
в охлаждении и нагревании тела. В этом отношении они возмещают
функционально отсутствующие у птиц потовые железы и особенное
значение имеют при большой мускульной работе во время полета;
роль их в нагревании периферических частей тела видна в распу-
шении и надувании мелких птиц при значительных морозах. На-
конец воздушные мешки имеют значительное биологическое зна-
чение в брачных играх и при производстве звуков как резонаторы
и украшающие образования.
Нижняя гортань — голосовой аппарат. Как
упоминалось выше, голосовым аппаратом птиц является нижняя
гортань — syrinx, образующаяся в области развилки трахеи в сво-
бодные бронхи. В связи с тем, что у многих птиц голосовые способ-
ности уже значительно вышли за пределы простых, специфических
виду, позывных криков и развились весьма дифференцированные
голосовые выражения психических ощущений, нижняя гортань
У многих групп развивается в весьма сложный орган, почти на-
столько же разнообразный по устройству, как разнообразны крики
птиц (рис. 90). Отсутствует сиринкс только у совершенно немых
7 Орнитология—1209
97
Рис. 90. Типы сиринксов и сирингиальная мускулатура.
1 — Ciconia ciconia L.; 2— Steatornis car ipensis Humb.; 3 — Aegothelesnovae-hollandiae Lath.;-
4— Thamnophilus punctaius Shaw; <5, 6*—Furnarivs rufus (Gm.); 7 —Grallaria guatema-
Lensis Prev.; 8, 9,10— Corvus corax L.; 11 — Melampitta gigantea Rotsch.; 2, 3 — бронхиаль-
ный сиринкс; 4, <5, 6, 7—трахеальный; 8, 9, io, 11—трахео-бронхиальный; 1, 10 — с дор-
зальной стороны; 6, 8 — с латеральной; 2, з, 4, 5, 7, 9,. 11—с вентральной. В рис. 9
и 11 — слева поверхностная, справа — глубокая мускулатура. I, II, III — бронх, кольца
сиринкса; Ъг.—неизмененные бронх, кольца; in — интербронхиальным апоневроз; I. s. d.—
m. laryngo-syringeus dorsalis; I. s. d. I. — m. laryngo-syringeus dorso-lateralis; I. s. v.—in-
laryngo-syringeus vent rails; I. s. v. I. m. —laryngo-syringeus ventro-lateralis; mb — мем-
браны; m. t. — мембраны барабана;. О. S. — сирингиальная косточка; р — хрящевой мо-
стик между первыми бронх, полукольцами, аналогичный pessulus; s. d. I. — m.syringeus
dorso-lateralis; st. — ni. sternotrachealis; s. v. —m. s-yringeus ventralis; s> v. I. — in. syrin-
geus ventro-lateralis; t — трахеал. кольца; t. b. — in. in. tracheo-bronchialis; tr. —барабан.
американских грифов, киви и аистов (род Ciconia). По структуре
нижней гортани различают сиринкс трахео-бронхпальный (syrinx
Iracheo-bronchialis), сиринкс трахеальный (s. Lrachealis) и сиринкс
бронхиальный (s. bronchialis). Первый тип свойствен большинству
птиц и характеризуется участием в образовании голосового аппа-
рата и соответственным изменением как трахеальных, так и брон-
хиальных колец; он описывается подробнее ниже. Трахеальный си-
ринкс свойствен ряду южноамериканских воробьиных птиц, объ-
единяемых в четыре семейства: TJendrocolaptidae^ Forrnicariidae, Pte-
roptochidae и Conopophagidae, составляющих в некоторых системах
особую группу воробьиных — Passeres tracheophonae. В этом типе
бронхи не испытывают приспособлений к произведению звуков, по-
следние же кольца трахеи (5—6) оказываются крайне утонченными,
часто неполными, скрепленными тонкой перепонкой, колеблющейся
проходящим по трахее воздухом. Голосовые мускулы располагаются
исключительно по бокам трахеи. Голос этих птиц очень громок
и прп звуке нижняя часть горла заметно раздувается. Бронхиаль-
ный сиринкс свойствен очень немногим птицам: Steatornis^ Centropus
и подсемейству Podarginae. Трахея здесь не имеет особых приспо-
соблений, но изменены верхние отделы бронх, приобретающие ко-
леблющиеся перепонки. Между этими типами сиринкса имеются
переходные образования, встречающиеся в разных систематических
группах птиц; наиболее отграничен трахеальный сиринкс.
Трахео-бронхпальный сиринкс обычно имеет следующее строение
(рис. 91). Три-шесть последних трахеальных кольца тесно соединяются
или срастаются своими краями, образуя «барабан» с крайне тонкими
стенками; к каудальному краю его прилегают бронхиальные полу-
кольца. Три первых «сирпнгпальных» полукольца отличаются своей
шириной, толщиной и неравномерным смыканием друг с другом.
Прочные эластические связки, соединяющие полукольца между собой,
между вторым и третьим, а иногда и другими полукольцами, утон-
чаются до тонкой перепонки — membrana tympaniformis externa, ниже
которой на внутренней поверхности третьего полукольца распола-
гается эластический вырост — наружная голосовая губа — labium
externum. Сходные образования развиваются и па медиальной сто-
роне бронхиальных полуколец. Здесь нижние края барабана в ме-
диальной плоскости соединяются трехгранной костяной балкой —
pessulus, по верхнему краю которой выстилающая трахею реснит-
чатая слизистая оболочка образует выдающуюся перепончатую
складку— membrana semilunaris, имеющую полулунную форму.
Прилегающие к pessulus медиальные стенки бронх образуют вну-
треннюю голосовую губу (labium internum), кзади от которой не-
обычно утонченные медиальные стенки образуют membrana tym-
paniformis interna; еще далее кзади эти стенки укрепляются
посредством эластичных волокон, образующих связку медиальных
стенок обоих бронх — bronchidesmus и ограничивающих сзади fo-
ramen interbronchiale. Играющие крупную роль в образовании
звука перепонки и губы представляют собой очень нежные образо-
вания, выстланные обычно однослойным эпителием слизистой обо-
лочки и достаточно подвижные, чтобы под влиянием мускулатуры
99
трахеи и сиринкса менять
степень своего напряже-
ния, а при прохождении
воздуха — п вибрации.
Описанный аппарат яв-
ляется местом прикрепле-
ния специальной мускула-
туры, особенно сложной и
дифференцированной у на-
стоящих певчих птиц. По
происхождению она отно-
сится к системе musculus
sterno-hyoideus и иннерви-
руется ветками подъязыч-
ного (ramus laryngeus infe-
rior и superior) и блуждаю-
м.
вш
вид спереди.
в/!е —
-Mt-----
-------В.1.
вп
--Ш.е.
щего (ramus recurrens) нер-
вов (рис. 90).Часть этой
мускулатуры связывает
трахею с ключицей и гру-
диной (m. sternotrachealis,
рис. 92), часть всецело
ограничена областью тра-
хеи и сиринкса. Первые
мускулы покидают трахею
перед распадением ее на
бронхи, и сокращение их
вызывает давление трахеи
на бронхи и вследствие
этого — вдавление внутрь
и сближение голосовых пе-
репонок в области сирин-
кса, что сужает голосовую
щель и вызывает вибрацию
перепонок при прохожде-
ние. 91. Строение трахео-брон-
хпального сиринкса.
а, а1—сиринкс домашней индей-
ки — Meleagris gallopavo L. в мо-
мент крика; Ъ, Ы —сиринкс се и;е
при свободном дыхании; с — си-
ринкс самца черного дрозда—Тиг-
dus merula L.; а1, b1, с — в разрезе,
Трахея в момент крика (а) укорачивается вследствие сокращения musculus
sterno-lrachealis и тонкие голосовые перепонки вдаются внутрь.
(С)
В—-бронхиальные полукольца; В. I, II, III — бронхиальные полукольца, вхо-
дящие в состав сиринкса; Bd.—bronchidesmus; F. ?.—foramen interbronchiale: Le.—
наружная голосовая губа; Li. —внутренняя голосовая губа; Lig. — связка, соединяющая
bronchidesmus с дорзальной стенкой трахеи; М — сирипгиальпые мышцы, располагающиеся
в области барабана; М. S.—полулунная мембрана — inembrana semilunaris; M.S.t.—
m. sterno-trachealis; M. t. e.—inembrana tympaniformes externa; M.t.i.— membrane
tympaniformes interna: P — pessulus; T — «барабан»; Tr. — трахея; Ts. — частично слив-
шиеся трахеальные кольца, соответствующие «барабану» певчих птиц. По Griilzner и Наескег,
из Stresemann.
100
musculus syrin-
Рис. 92. Плечевой пояс и трахея
серого гуся — Anser anser 1^. с дор-
зальной стороны.
С — коракоид; F — вилочка; 8 —
грудина; S. t.—musculus sterno-
tracheidis; Tr. —трахея. По Sctiio-
ler.
nun воздуха. Этим процессом обусловлен, например, главным образом
голос гусиных и куриных птиц. Ограниченный областью трахеи,
musculus tracbeo-bronchialis располагается обычно вдоль трахеи
и в разных группах видоизменением мест своего прикрепления и
распадением на самостоятельные пучки и отделы создает сложную
и дифференцированную голосовую мускулатуру. При укорачива-
нии и заканчивании на трахеальных кольцах он превращается
в muscuhis tracheal is; при отдифференцировании отделов, огра-
ниченных всецело нижней гортанью, образуются
geus (sive brevis). При наибольшем диф-
ференцировании этой мускулатуры име-
ется до 9 пар мускулов, из которых 4
являются поверхностными m. tracheo-
bronchiales (по две пары вентральных
и дорзальных) и 5 лежащих глубже —
musculi syringei (3 вентральных и 2
дорзальных). Эти мускулы изменяют
взаимоотношение бронхиальных колец
и вызывают различную напряженность
голосовых перепонок п ширину щелей,
обусловливая этим получение разно-
образных звуков. Это разнообразие зву-
ков в основном конечно зависит от диф-
ференцированной голосовой мускула-
туры. В более примитивных группах
(«бескрылые», веслоногие и др.) в обла-
сти нижней гортани мускулатура во-
обще отсутствует; у воробьиных она
проще всего у примитивных Passeres
oligomyodae с 2—3 парами мышц (цтица-
лнра) и сложна у остальных Passeres
polyrnyodae, где их не меньше семи пар.
Таким образом таксономически строе-
ние нижней гортани имеет очень боль-
шое значение и играет значительную
роль в характеристике как крупных,
так и мелких систематических групп.
Существенное значение имеет также само расположение и прикре-
пление мускулов. В группе воробьиных различают Passeres meso-
myodae с наиболее примитивным прикреплением голосовых мускулов
только на латеральных частях бронхиальных полуколец (куда отно-
сятся главным образом южноамериканские воробьиные), Passeres
anisomyodae — с прикреплением мышц на вентральных или дор-
зальных концах этих полуколец и Passeres diacromyodae — с сим-
метричным прикреплением мышц на обоих концах бронхиальных
полуколец. В последней группе различают еще 3 типа прикрепления
мускулов к. каждому полукольцу: дипломпоидный, если мышцы
прикрепляются на обоих его концах, анампоидный, если прикре-
пление имеет место только на дорзальном конце, it катамиоидный,
с прикреплением только на вентральной стороне.
.101
Рис. 93. Трахея самки цветного бекаса — llosiraiula
benghalensis L., в отличие от трахеи самца удлинен-
ная и сложенная петлей (самка этого бекаса крупнее,
ярче, окрашена и активнее в брачных играх, чем
самец. См. рис. 170).По Wood-Mason, из Stresemann.
В целом голосовой аппарат птиц оказывается построенным по
типу язычковой (органной) трубы, в которой роль нагнетающих
воздух мехов играют легкие и воздушные мешки, роль колеблю-
щихся язычков — голосовые губы и перепонки, а роль резонирующей
трубы и раструба играют трахея и ротовая полость. Подобно орган-
ной трубе, и в этом аппарате сила вдуваемого воздуха, напряженность
перепонок, ширина и длина трахеи видоизменяют звук, его интен-
сивность и тембр.Помимо строения самой нижней гортани и ее муску-
латуры, в производстве звука имеют следовательно значение форма,
длина и ширина трахеи и связанные с ней специально резонирующие
образования. С удлинением трахеи связана возможность получения
более полного п глубокого звука, и естественный отбор в этом напра-
влении привел к обра-
зованию во многих груп-
пах очень длинных тра-
хей, свернутых в петли
и залегающих то под
кожей, то врастающих
в киль грудины (рис.
93). Пневматические по-
лости в окружающих
костях при этом дей-
ствуют так же, как ре-
зонаторы. Самцы мно-
гих уток и крохалей
обладают особыми взду-
тиями трахеи или ла-
биринтами — боковыми
вздутиями барабана в
области сиринкса, на-
значение которых за-
ключается в усилении
(резонировании) опре-
деленных тонов и обра-
зовании шумов вследствие многократного отражения звука
(рис. 94). Эти образования вызывают получение голоса, совер-
шенно отличного от голоса самки, биологически соответствующего
песне певчих птиц. Наиболее распространенными резонаторами
являются все же воздушные мешки, вздувание которых близ трахеи
и гортани усиливает мощность п глубину звука. Такая функция
установлена для подключичного воздушного мешка у некоторых
птиц, а у куриных также и для шейного. У американской савки
(Oxyura jamaicensis) известно образование трахеального воздушного
мешка, надуваемого воздухом из специального отверстия в трахее.
В брачный период у многих птиц, вследствие плотного прикрывания
клюва и замыкания орбитальной щели языком, воздух попадает
в пищевод и раздувает шею; это, например,происходит в момент вор-
кования обычного сизого голубя. У дроф, кроме того/ раздувается
еще горловой мешок, сильно увеличивающийся в размерах, в других
группах имеющий значение временного хранилища пищи (рис. 95).
302
'Таким образом развитие голосового аппарата использует и захваты-
вает достаточно разнообразные органы.
Прибавление. Органы внутренней секреции —
.производные эпителия гл
О Т К II.
Из эпителия жаберной
области кишечника у по-
звоночных развиваются
различные железы, явля-
ющиеся по своей функции
органами внутренней се-
к реции.У птиц эмбриональ-
но закладывается шесть
Рис. 94. Сиринкс утки — Metopiana peposaca
Vieil., вздутый в сложный лабиринт. ь
С вентральной стороны частью ‘вскрыт; путь вы-
дыхаемого воздуха показан стрелками; аг. — ко-
стяная рамка, в которой натянута голосовая
перепонка — membrana tympaniformes; са.—
часть левого лабиринта, образующего преддверие
для воздуха, выходящего в трахею; с. — слепой
отдел в правом лабиринте; сс. — центральный
канал барабана для прохождения воздуха;
т. ty.— membrana tympaniformes; о — отвер-
стие в стенке между левой и передней камерами
лабиринта; S. Ь. — балка (pessulus) — разгора-
живающая лабиринты; t. d — правый лабиринт;
/. s. — левый лабиринт; tra. — трахея; w — вы-
ступ стенки правого лабиринта, отгораживающий
небольшой, слепо заканчивающийся отдел лаби-
ринта. Выдыхаемый воздух частью беспрепят-
ственно проходит в трахею, частью заходит в сле-
пое подразделение правого лабиринта и через
особое отверстие — в левый лабиринт, где вызы-
вает колебание голосовой перепонки, и затем вы-
ходит через узкий канал в преддверие и из него—
в центральный канал. По Gliesch, из Stresemann.
Рис. 95. Горловой мешок
дрофы — Otis tarda L., рас-
положенный под языком на
передней стороне шеи.
а—горловой мешок; Ъ —
пищевод с зобом; tr — тра-
хея. Из Coues.
вентральных (жаберных) щелей, и из них эпителий 3-й, 4-й и 6-й
щелей превращается в дальнейшем в железы внутренней секреции:
зобные (glandula thymus), паращитовидные (gl. parathyreoidea) и
так называемые эпителиальные ультимо-бронхиальные (или зажабер-
ные) тельца. В области 1-й и 2-й жаберных щелей, из выступа вен-
тральной стенки глотки, развивается крупная щитовидная железа
(gl. thyreoidea). В процессе развития все эти железы значительно
смещаются кзади и у взрослой птицы располагаются вдоль всей
103
шеи и в передней части груди, около сердца (рис. 96). В птенцовом
возрасте (главным образохм в период роста костей) они обычно раз-
вис. 96. Зобная и щитовидная железы молодого петуха—
Gallus gallus L.
Яремная вена Jvena jugularis, и блуждающий нерв, nervus
vagus, сдвинуты на спинную сторону; t — зобная железа
(thymus), разделенная на отдельные лопасти спинными
нервами; tr.—щитовидная железа (thyreoidea); р —
бронхиальные железы (parathyreoidea). По Nonidez и
Goodale, из Stresemann.
виты значительно круп-
нее, но в дальнейшем
подвергаются значи-
тельной редукции и в.
некоторых случаях ча-
стью исчезают.
Зобная железа рас-
полагается вдоль шеи,
по ходу яремной вены
п блуждающего нерва, и
разветвляющимися вет-
ками шейных нервов
вторично сегментирует-
ся на отдельные участки..
Вследствие внедрения в.
нее соединительной тка-
ни, она принимает строе-
ние альвеолярной же-
лезы, богатой кровенос-
ными сосудами. По-
окончании периода ро-
ста эта железа наиболее
сильно сокращается в-
своих размерах. На
каудальном конце ее
располагаются очень
мелкие паратиреоидные
железы и ультимоброн-
хиальное тельце, играю-
щие важную роль, как
место образования лей-
коцитов.
Щитовидная железа,
имеет вид пары кругло-
ватых телец, залегаю-
щих при входе в груд-
ную полость у сонных
артерий и каудального
конца зобной железы.
Фолликулярная парен-
хима этой железы окру-
жена тонкой соедини-
тельно-тканной капсу-
лой, от которой отходят перегородки внутрь железы, ведущие за
собой большие сосудистые стволы. В интерстициальной ткани же-
лезы разветвляется капиллярная сеть и залегают лимфатические
фолликулы. Функция щитовидной железы у птиц по видимому сов-
падает с функцией ее у млекопитающих. Она регулирует как рост
1(И
организма. так и отдельных тканей (например развитие и рост
пера).
5. КРОВЬ'II КРОВООБРАЩЕНИЕ
Разносимая ио всему телу аппаратом кровообращения кровь имеет-
чрезвычайно многостороннее значение. Помимо осуществления дыха-
ния организма, кровь разносит тканям питательные вещества, уда-
ляет ненужные вредные выделения, переносит выделяемые тканями
и органами инкреты, защищает организм от бактерий и вредных
веществ и, свертываясь, закрывает раны. Общее количество крови
в организме птицы зависит от очень многих моментов. Никогда ее
количество не бывает настолько велико, чтобы с одинаковой интен-
сивностью обеспечить питание всех органов тела; обычно наиболее
нуждающаяся система переполняется кровью, в то время как другие
снабжаются соответственно слабее, и эта картина меняется в зависи-
мости от изменений запросов отдельных органов. Сходным изменениям
подвержены и отдельные составные элементы крови.
Состав крови. Плазма крови почти бесцветна (слегка
желтоватая) и при свертывании распадается на две части: твердую —
фибрин и жидкую — сыворотку. Кроме воды, плазма содержит
белки, соли, энзимы, ряд мало исследованных продуктов внутренней
секреции и т. д. Плазма может абсорбировать значительное количество
углекислоты и служит для распределения пищи по тканям, для осво-
бождения их от ненужных продуктов обмена и т. п. Форменные эле-
менты крови представлены лейкоцитами, эритроцитами и кровяными
пластинками. Из белых кровяных клеток лейкоциты характеризуются
скоплением в плазме большого количества мелких гранул и бывают
с одним (базофильные лейкоциты) и с несколькими (оксифильные
лейкоциты) ядрами. Последние встречаются у птиц несравненно чаще,
чем у млекопитающих, и могут составлять почти половину всех лей-
коцитов вообще. Лимфоциты характеризуются отсутствием гранул
в протоплазме и бывают малой и крупной величины; по численности
они составляют половину и иногда даже до 90% всех белых кровяных,
телец. Эти белые кровяные клеткп обладают способностью изменять
свою форму и проникать амебоидными движениями через стенки кро-
веносных сосудов и между клетками тканей. Помимо содействия
абсорбированию жиров и пептонов, они способны захватывать и
переваривать посторонние тела, играя таким образом важную защит-
ную функцию в организме. Эритроциты, обусловливающие цвет
крови, имеют у птиц вид плоско двояковыпуклых и овальных телец
с крупным ядром в центре. Окраска их зависит от присутствия в них
содержащего железо белка — гемоглобина,’ который легко соеди-
няется с кислородом и углекислотой и легко отдает эти газы бедным
ими тканям. В этой дыхательной функции и заключается значение
эритроцитов в теле птицы. Хотя количество гемоглобина в крови
птиц и меньше, чем в крови млекопитающих (в сухом виде кровь
гуся содержит только 62,5% гемоглобина в сравнении с 94% и 86,5%
У человека и собаки), однако использование кислорода крови у птиц
является более совершенным, чем у этих млекопитающих. Этим ве-
роятно объясняется и меньшее количество, эритроцитов в крови птиц.
105
в сравнении с млекопитающими — всего 1,5—5,5 миллионов на
1 куб. мм крови против 2—18 миллионов у млекопитающих.
Количество и величина эритроцитов колеблются в известных
пределах в зависимости от величины птицы (обратно пропорцио-
нально) и интенсивности ее жизнедеятельности (прямо пропорцио-
нально).
Крупные птицы обладают более крупными эритроцитами п мень-
juiim числом их, что стоит в связи также и с тем, что общая дыхатель-
ная функция эритроцитов повышается пропорционально увеличению
их поверхности. Так внутри двух групп — куриных и воробьиных,
родственные представители разной величины отличаются показате-
лями, представленными в нижеследующей таблице.
1 1 1 Размер эритроцитов (в микронах) . . . Курины е *.
поредел | домашний • петух ИНДЮК
11,2 X 6,2 14 X 7 [ 15,5 X 7,5
Количество эритроцитов i 4000000 t 3300000 | 2300000 .
Воробьиные
i 1 - 1 Estrilda xubflava полевой во- робей | ворон
Размер эритроцитов (в микронах) . . . 11 X 6,2 12,5 X 6,2 15 X 6,2 .
Количество эритроцитов 5400000 5200000 3930000
Местом образования кровяных телец служат главным образом
печень, селезенка, костный мозг, ультимобронхиальные тельца и
фабрициева сумка. В эмбриональной жизни крупную роль играют
также различные скопления мезенхиматозных клеток.
Аппарат кровообращения. Кровь разносится по
телу по системам артериальных и венозных сосудов, сложно ветвя-
щихся и связанных друг с другом через посредство тончайших крове-
носных капилляров (рис. 97). Так как передача газов и питательных
веществ совершается при помощи этих последних, то стенки их чрез-
вычайно тонки и образованы лишь однослойным плоским эпителием
(эндотелием). Стенки артерий и вены, кроме эпителиального слоя и
внутренней оболочки (intima), состоят из среднего слоя гладких мышц
и эластических упругих элементов (tunica media) и наружного соедп;
нптельно-тканного слоя*, обычно с пронизывающими его эластиче-
скими волокнами (adventitia). Стенки артерий обычно подвергаются
большему давлению, чем вены, и поэтому сравнительно толще;
преимущественной особенностью вен являются клапаны, распола-
гающиеся в известных местах по ходу вен на пх внутренней поверх-
ности, препятствующие обратному току кровп.
Осуществляется движение крови сердцем, действующим как при-
сасывающий и нагнетающий насос; оно лежит в середине грудной поло-
сти и имеет форму короткого конуса, с вершиной, располагающейся ме-
106
жду долями печени. Окружающая свободно сердце околосердечная
сумка (pericardium) соединяется с позвоночным столбом, грудными
воздушными мешками и пе-
ченью.
Сердце. Сердце птиц
четырехкамерное и состоит из
двух предсердий (atrium) и
двух желудочков (ventricu-
lus) (рис. 98). В чрезвычайно
мощных мускулистых стен-
ках сердца можно различать
наружный серозный слой
(epicardium), средний мышеч-
ный (myocardium) и внутрен-
ний эндотелиальный (endo-
cardium). Полной перегород-
кой между предсердиями и
желудочками (septum atrio-
rum, s. ventriculorum) сердце
делится на две половины:
правую венозную и левую
артериальную. Однако, в эм-
бриональном периоде перего-
родка предсердий является
ситовидно - продырявленной
(так называемое овальное
окно — foramen ovale), вслед-
ствие чего происходит проток
крови. С началом легочного
дыхания отверстия замыка-
ются разрастающейся эндо-
кардиальной тканью, и сме-
шения артериальной и веноз-
ной крови уже не происхо-
дит; перегородка остается
все же тонкостенной и несет
след бывших отверстий в
форме овальной впадины
(fossa ovalis). Внутренние
подразделения сердца наме-
чаются наружными борозд-
ками: поперечной — sulcus
transversus и продольной —
sulcus longitudinalis anterior
и posterior. Соответственно
большей работе левой поло-
вины сердца, отсылающей
кровь по большому кругу
кровообращения, левый же-
лудочек объемистее, чем пра-
Рис. 97. Схема кровеносной системы птицы, с
вентральной стороны.
Артерии, за исключением art. pulmonale (отме-
чена пунктиром), поперечно исчерчены; вены —
белые, ааа — arcus ascendens aortae; a. brc. —
art. brachialis; a. desc.— aorta descendens;
A. Л— art. femoralis; a. pulm. — art. pulmo-
nale; A. sub. — art. subclavia; A. v. — art. ver-
tebralis; C. c. — art. carotis communis; C. e. —
art. carotis externa; С. г. — art. carotis interna;
с. гл. s. — vena coccygo-mesenterica; coel. —art.
coeliaca; cr.—vena femoralis; cut. abd.—vena
cutanea abdorninalis; cut. uln. — vena cutanea
ulnaris; past. — art. gastralis; hp. — art. hepatica;
hyp.— vena hypogastrica; il. — art. iliaca; il.
e.x.— vena iliaca externa; il. int. — vena iliaca
interna; isch. — art. ischiadica; Jug. d. — vena
jugularis dextra; Jug. s. — vena jugularissinistra;
L. A. — левое предсердие; L. V. —левый желу-
дочек; in. s. i. — art. mesenterica inferior; ms.
s. — art. mesenterica superior; r. — arteria и vena
renalis; R.A. —правое предсердие; rad. — art.
radialis; It. V.—правый желудочек; tib. a.—
art. tibialis anterior; tib.p.—art. tibialis posterior;
thor. — art. thoracica; Tr. aort. — truncus aortae;
uln. — art. ulnaris; V. Hr. — vena brachialis; V.
c.i., v. c. p., v. c. s. — vena cava inferior, posterior
и cranial is sinistra; v. /. — vena femoralis; V. H.
vena hepatica; v. il. c. — vena iliaca communis; v.
pulm. — vena pulmonaria; V. sub. — vena subcla-
via; v. v. — vena vertebralis. По Gadow и Newton.
107
вый, проникает и в вершину сердца и имеет более мощные стенки;
правый желудочек как бы облекает его справа и в поперечном раз-
резе имеет полулунную форму.
В правое предсердие впадают три полые вены: две передние —
vena cava cranialis dextra и v. с. c. sinistra и одна задняя — vena
cava caudalis. У примитивных птиц («бескилевые») эти вены соеди-
няются перед впадением в венозный синус (sinus venosus), как то имеет
место у рептилий, но в более высоко стоящих группах в области си-
Рис. 98. Сердце сизого голубя — Columba livia Gm., разрезанное по
медиальной линии и развернутое.
ао— arcus aortae; atr. 1.9 atr. г.—левое и правое предсердие; muse,
pap.—три папиллярных мускула левого желудочка; pulm. а. — art.
pulmonalis; It. br. ее ph.. I., tr. br. ceph. r?— правый и левый truncus
brachio-cephalicus; valv. muse. — мускулистый клапан правого атрио-
вентрикулярного отверстия, ostium atrio-ventriculare;ventr. 1.9 ventr. r. —
левый и правый желудочки. По Schimkewitsch, из Stresemann.
нуса развиваются перегородки, частью или полностью разделяющие
устья впадающих вен. Эти венозные отверстия снабжены перепонча-
тыми клапанами, препятствующими обратному течению крови из
предсердия в вены. Же лудочно-предсердное отверстие — ostium
atrium ventriculare dextrum — имеет вид косой щели. Вместо трех-
створчатого клапана млекопитающих и перепончатого клапана репти-
лий здесь существует треугольная мышечная заслонка (valvula
mtiscularis), свободным краем вдающаяся в полость желудочка и при-
крепляющаяся единственным мышечным пучком к его стенке. При вся:
кой систоле желудочков это отверстие закрывается вследствие сокра-
щения пластинки, притягивающейся к перегородке. Другое — арте-
риальное отверстие в правом желудочке (orificium arteriosum) ведет
108
в легочную артерию (arteria pulmonalis), открывается у передней стен-
ки и снабжено тремя полулунпымп.клапанамп (valvuiae semilunaris).
В левое предсердие открываются общим отверстием две. легочные
вены (venae pulmonales). Впереди итого отверстия находится утол-
щенный выступ в виде за-
слонки, свободный край
которой обращен к поло-
сти предсердия и препят-
ствует обратному току
крови в легочные вены.
Левое атрпо-вентрпкуляр-
ттое отверстие (ostium
atrioventriculare sinist-
rum), ведущее из левого
предсердия в левый желу-
дочек, закрывается дву-
створчатой или трехствор-
чатой заслонкой (valvula
hicuspidalis sive tricuspi-
dalis). Обособляющиеся на
стенках желудочка три
сосцевидные мышцы (mus-
culi papillares) отсылают
с у х о жил ьные нити (ch or d а е
lendineae) к этой заслонке.
Второе отверстие из ле-
вого желудочка, располо-
женное у продольной пе-
регородки, ведет в аорту
(truncus aortae) и снабжено
Рис. 99. Сердце лебедя шипуна — Cygnus olor L., с
более крупными сосудами.
полулунными клапанами
(valvuiae semilunaris). Са-
мо сердце птиц питается
двумя сердечными арте-
риями (arteriae coronariae
cordis) и венечной веной
(vena coronaria cordis) и
иннервируется ветвями
блуждающего и симпати-
ческого нервов.
Дуги аорты и
а р т е р и и ш е и и г о-
Область артериальной крови — светлая, область ве-
нозной крови —покрыта пунктиром, ао. d. — aorta
descendens; ar. ао. — arcus aortae; ar. brach.— arl.
brachialis; ar. car.—art. carotis; ar. mam.— art.
mammaria interna; ar. oes. as. — art. oesophagea
ascendens; ar. palm. s.—art. pulmonalis sinistra;
ar. st. cl. — art. sterno-clavicularis; ar. subcl. —
art. subclavia; ar. thor. inf. и sup. — art. thoracica
superior и inferior; ar. vert. — art. vertebralis; coel. —
art. coeliaca; gl. thyr. — щитовидная железа — gl.
tryreoidea; mes. sup.—art. incsenterica superior;
oes.—пищевод; pulm.—легкое; tr. br. ceph. - -
truncus brachioccphalicus (art. anonyma); tr. car. —
truncus caroticus; v. brach. — vena brachialis; v. cav.
.sup. d. и s. — vena cava cranialis dextra и sinistra;
v. jug. d. и v. jug. — vena jugulares dextra и sini-
stra; v. cut. abd.—vena cutaneus abdominis. Русские
названия вен и артерий см. текст, стр. 109—115.
По Gadow.
ловы (рис. 99). Из за-
кладывающихся у птиц эмбрионально шести дуг аорты, соответ-
ственных числу жаберных щелей, во взрослом организме сохраняются
только отдельные элементы. Так, arteria stapedia является остатком
ветви второй дуги; arteria carotis dorsalis — частью остатком третьей
дуги, частью остатком краниального продолжения truncus arterio-
sus (arteria carotis ventralis); arteria aortae сохраняется как правый
сосуд четвертой дуги, в отличие от млекопитающих, образуя у птиц
109
правую дугу аорты, arteria pulmonalis является остатком шестой дуги.
Остальные части их полностью исчезают, а сохраняющиеся иначе
связываются между собой.
По выходе из околосердечной сумки аорта отдает два следующие
друг за другом ствола правой и левой плече-головной артерии (arteria
brachio-cephalica dextra и sinistra), снабжающей кровью голову, шею,
грудь и крылья, после чего аорта делает изгиб вправо и назад через
правый бронх и как нисходящая аорта (aorta descendens) спускается
дорзально от пищевода. Каждая плече-головная артерия очень
быстро расщепляется на сонную и подключичную артерии (arteria
carotis dorsalis и art. subclavia), из которых первая орошает всю
область головы и шеи.
Дорзально от зобной железы, от каждой сонной артерии ответ-
вляется по тонкой позвоночной артерии (art. vertebralis), проникаю-
Рис. 100. Схема главнейших изменений сонных артерий.
Л—нормальный тип, две отдельные глубокие сонные артерии; В — обе глубокие
артерии сливаются в одну (цапля — Ardea и др.); с — то же, как и В, но проток левой
сонной артерии уменьшен (фламинго — Phoenicopterus и др.); D — левая сонная артерия
исчезла и заменена поверхностным сосудом (многие попугаи); Ао. — аорта; с. р. d. — art.
carotis profunda dextra; c. p. s. — art. carotis profunda sinistra; c. p. c. — art. carotis pro-
funda conjuncta; c.s.s.—art. carotis superficialis sinistra; su. d.— art. subclavia dextra;
su. s. — art. subclavia sinistra. По Gadow и Newton.
щей в канал поперечных отростков задних шейных позвонков и рас-
падающейся на восходящую (ramus ascendens) и нисходящую ветвь
(ramus descendens). Она орошает позвоночный столб до 5-го —6-го
грудного позвонка и проникает в полость черепа.
Собственно сонные артерии проходят на вентральной стороне шеи,
обычно в специальном канале нижних отростков позвонков. Как пра-
вило, правая и левая сонные артерии лежат рядом друг с другом,
сохраняя свою самостоятельность, но в некоторых группах (цапли,
фламинго, некоторые попугаи) они сливаются в одну • arteria carotis
conjuncta, а в ряде групп это сопровождается исчезновением одного
из стволов (рис. 100). Таким образом образуется непарная сонная
артерия, правая у некоторых дроф, левая у дятлов, воробьиных,
птиц носорогов и нек. др. Причиной нарушения этой симметрии
считается обычное расположение зоба на правой стороне, что вызы-
вает значительный отток крови послабляет правый ствол этой арте-
рии. Взамен исчезающего глубокого ствола («carotis profunda»)
110
Рис. 101. Разветвления arteria carotis externa в-
голове и шее сизого голубя — Columba livia Gm.
а — art. carotis externa; b — ar. occipitalis; c —
ушная артерия — ar. auricularis; e — лицевая арте-
рия— ar.facialis; /—артерия in. masseter; g—
артерия in. myiohyoideus; h — небная артерия;
i — носовая артерия; к — передпеглазничная ветвь
носовой артерии; I — веточки, окружающие носов» «г
отверстие; т— ar. facialis interna; п — артерия
крыши неба; о — артерия боков неба; р— артерия
спинной поверхности шеи; q — аг. mandibularis; г —
аг. pharyngea descendens; s—ar. oesophagi; t — ar.
intermandibularis; и — ar. lingualis; v — контур-
глаза; x — ушной проход; у — трахея; z — пище-
вод. Из Vocht и Yung.
иногда наблюдается поверхностная сонная артерия (carotis super-
ficialis), развивающаяся из эмбриональной arteria carotis ventralis.
На уровне первых шейных позвонков сонные артерии разделяются
на лицевую и мозговую артерии — art. carotis externa (s. facialis) и
interna (s.cerebralis) (рис. 101). Первая снабжает главным образом
язык (art. lingualis и art. intermandibularis), небо (art. palatina), че-
люсти (art. facialis), область наружного уха п горла (art. pharyngea.
descendens ) и мускулату-
ру затылка (art. occipita-
lis); вторая — мозг (art. ce-
rebralis собственно), глаз-
ное яблоко и глазницу (art.
ophthalmica), ухо (art.
stapedia) и носовую по-
лость с лобными наростами
(art. ethmoidalis).
А р т е р и и п е р е д-
н е й конечности
(крыла). Упомянутая вы-
ше подключичная артерия
(arteria subclavia) в стен-
ках тела отдает ветви arte-
ria sterno-clavicularis и
art. mammaria interna,оро-
шающие большую часть
плечевого пояса (рис. 102).
Основной ствол, входя в
конечность, ответвляет
подкрыльцовую артерию
(art. axillaris), орошаю-
щую крыло, и крупную
грудную артерию (art. tho-
racica), орошающую 2—3
ветвями грудные мышцы
и кожу. Подкрыльцовая
артерия, входя в крыло,
отдает веточку к мышцам
лопатки (art. scapularis) и в
виде плечевой артерии (art.
brachialis) проходит между мышцами в предплечье. В дальнейшем
она делится на глубокую плечевую (art. profunda brachii), луче-
вую (art. radialis) и локтевую (art. ulnaris), распадающиеся на еще
более мелкие ветви.
Разветвления нисходящей аорты. Как ука-
зано выше, загибающийся назад ствол нисходящей аорты (aorta
descendens) проходит вентрально и вдоль позвоночника. На своем
Пути она отдает значительное число парных интерсегментальных
артерий, частью небольшой величины, как то: артерии, орошающие
туловищную мускулатуру (межреберные—art. intercostales inferiores)T
половые органы (семенные артерии — art. spermatica internae), почки
ш
(почечные артерии — art. renales craniales), мускулатуру поясничной
области (поясничные — art. luinbales) и др., частью крупных по
своему калибру, как снабжающие кровью заднюю конечность —
Рис. 102. Схема артериальной системы золото-
волосого пингвина — Catarrhactes chrysocomc
Forst.
1 — аорта; 2 — truncus brachyocephalicus; 3 —
ar. subclavia; 4 — ar. vertebralis; 5 — ar. mam-
maria interna; 6* — ar. thoracica; 7 — ar. axilla-
ris-f-brachialis; 8— ar. carotis dorsalis; 9—
ar. oe-iophagea ascendens; 10 — ar. oesophagea
descendens; 11 — ar. comes nervi vagi; 12— ar.
submentalis; 13 —• ar. bronchialis; 14 —ar. inter-
costales; 15— ar. coeliaca; 16— ar. lienalis;
17 — ar. gas trails; 18 — ar. hepaticae dextrae:
19 — rarniis sinister ar. coeliaca; 20 — ramus in-
testinalis; 21 — ar. mesenterica superior; 22 —
aa. renales; 23 — aa. lumbales; 24 — ar. femora-
lis; 25 — ar. ischiadica; 26 — ar. mesenterica
inferior; 27—ar. pudenda communis; 28— ar.
coccygea media. По Watson, na Stresemann.
ла льцы. В некоторых группах
артерия теряет свою начальную
бедренной артерии.
Артерии хвостовой
седалищных артерий, нисходящая аорта резко уменьшается в вели-
бедренные и седалищные ар-
терии (art. femorales и art.
ischiaclicae). Кроме парных
ветвей, от нее отходят непар-
ные (в постэмбрпональной
жизни): чревная артерия (art.
coeliaca) и передняя и задняя
брыжеечные артерии (art.
mesenterica cranialis s. supe-
rior и art. m. caudalis s. infe-
rior), орошающие органы пи-
щеварительного аппарата.
Артерии задней
к онечност и. Каждая
задняя конечность получает
кровь из ветвей бедренной и
седалищной артерий. Первая
еще в брюшной полости де-
лится на артерии надчрев-
ную (art. epigastrica), напра-
вляющуюся к мышцам брюш-
ным и таза, и собственно бед-
ренную артерию (art. femo-
ralis), выходящую из таза
через apertura cruralis и раз-
ветвляющуюся в мышцах
бедра (исключая аддукторов).
Вторая — большая по кали-
бру — седалищная артерия,
выходящая из таза через fo-
ramen ischiadicum, в области
бедра отдает ягодичную ар-
терию (art. glutea) к аддук-
торам бедра и голени, про-
ходит в голень, орошает ее
и делится на переднюю и
заднюю большеберцовые ар-
терии (art. tibiales anterior
и posterior), орошающие све-
ими ветвями плюсну и
(например пингвины) седалищная
часть и приобретает вид ветви от
области. После ответвления
чине, но вскоре отдает еще парную, значительную по величине ветвь-
art. pudenda communis, ответвляющую свои веточки к мускулатуре
112
хвоста (musculus depressor coccygis, ischio-coccygeus, pubi-coccy-
geus), мускулатуре клоаки и ануса, фабрициевой сумке и нижним
частям яйцевода и семяпровода. Сама нисходящая аорта заканчи-
вается в виде слабой средней хвостовой артерии (art.coccygea media),
снабжающей кровью некоторые мышцы хвоста (m. levator и depressor
coccygis), копчиковую железу и окружающие кожные покровы.
Артерии пищеварительного канала, как ука-
зано выше, выходят из нисходящей аорты в виде трех непарных ство-
лов. Чревная артерия (art. соеИаса)отходитнауровнепоследних груд-
ных позвонков и, распадаясь на ветви, орошает конечную часть пище-
вода, желудок, печень, селезенку, поджелудочную железу, двенадца-
типерстную, слепые и прямую кишки. Более значительными ветвями
ее являются: arteria liepaticae, art. gastralis, art.proventriculi, art. ven-
triculi sinistra и art. gastro-duodenalis. Передняя брыжеечная арте-
рия выходит из аорты на уровне гонад, проходит в мезентериум и
орошает главным образом тонкие и слепые кишки; более круп-
ными веточками ее являются rami jejunales, art. ileo-caeco-colica,
art. colica cranialis. Задняя брыжеечная артерия покидает аорту
вблизи от бедренных артерий и питает главным образом конечный
отдел кишечника (толстую и прямую кишки); основными ее ветвями
являются art. haemorrhoidalis cranialis и art. colica caudalis.
Вены. Хотя вены обладают центростремительным потоком
крови, их целесообразно все же рассматривать подвигаясь от более
крупных к более мелким сосудам.
Впадающие в сердце вены подразделимы на вены малого и боль-
шого круга кровообращения. К первому относятся парные легочные
вепы (venae pulmonales), несущие (артериальную) кровь из легких и
при впадении в левое предсердие соединяющиеся в один ствол;
ко второму относятся три полых вены: две краниальных и одна кау-
дальная. Каждая краниальная полая вена — vena cava cranialis
sinistra и dextra —очень скоро распадается на яремную вену (vena
ugulares), собирающую кровь из шеи и головы, и подключичную вену
(vena subclavia), собирающую ее из области грудных мышц и крыльев.
Каудальная полая вена — vena cava inferior (и posterior) несет кровь
из задней части туловища (в том числе от органов брюшной полости
и тазовых конечностей) и по своему значению следовательно соответ-
ствует нисходящей аорте. Как эта вена, так и краниальные
полые вены описанным выше образом впадают в правое предсердие.
Вены шеи и головы. Яремные вены проходят в шее
сбоку от трахеи рядом с блуждающим нервом, прикрываясь тонкой
шейнокожной мышцей. В задней части шеи ответвляются позвоноч-
ные вены (vena vertebralis), сопровождающие соответствующие арте-
рии. Каждая из них подобно этим артериям распадается иа краниаль-
ную, несущую кровь из шеи и головного мозга, и каудальную ветвь,
собирающую кровь из межреберных вен. Собственно яремные вены
вблизи головы соединяются тонкой поперечной ветвью; как правило,
левая вена бывает тоньше правой и в некоторых группах (например
дятлы) почти совершенно исчезает. В области головы каждая яремная
вена распадается на vena cephalica posterior и anterior, в свою очередь
разделяющиеся на venA capitis lateralis и vena occipitalis (первая)
8 Орнитология—1209
113
и vena facialis interna и externa (вторая); из этих разветвлений наи
более крупной является лицевая вена.
Вены передней
конечности. Подклю-
чичные вены (venae subcla-
viae) служат собирающим
стволом для вен груди и
крыльев, сопровождая одно-
именные артерии. Таким об-
разом они разветвляются на
крыльцовую и наружную
грудную вены (vena axillaris
и v. thoracica externa), из
которых первая переходит в
плечевую вену (vena brachia-
lis), разделяющиеся далее
на vena profunda brachii (с
ветвями radialis и ulnaris) и
vena scapularis.
Разветвления зад-
ней полой вены. Ve-
na cava posterior проходит
вентрально от позвоночника,
вдоль нисходящей аорты, и
в области передних долей
почек разветвляется на две
vena renalis revehens commu-
nis, вскоре затем восприни-
мающие vena renalis revehens
передних долей почек. Основ-
ные стволы почечной вены в
Рис., 103. Схема сосудистой системы почек.
Артерии — черпьте; приводящие вены—темносе-
рые; отводящие — светлые; А//.—приводящая
веточка вены, лежащая на поверхности; Ао.—
нисходящая аорта; А. — апастомоз v. iliaca
communis; D.—кишечник; Eff. —веточка отводя-
щей вены вместе с артерией; К—клапан; Кар.—
капилляры; L.—печень, 'N — воротная вена
почки (почечный отдел vena hypogastrica); v.
aff.—приводящая вена верхних долей почек;
v. с. р. — нижняя полая вена; к.с.с.—vena
coccygea communis; v.c.m.—vena coccygo-me-
senterica; v.r.r.o.—выносящая вена верхних
долей почек; v.h. — vena hypogastrica; v. i. e. —
vena iliaca externa; v.r.r. — vena renalis revc-
hens communis. Кровь из области таза, хвоста
и пижнпх копечпЛтей поступает в vena hypo-
gaslrica, проходит через капилляры почек и вы-
водится через vena renalis revehens в нижнюю
полую вену; лишь при повышенном давлении
крови в воротной системе почек часть ее минует
почечпые капилляры, оттекая через анастомоз
в заднюю полую вену. По Spanner.
дальнейшем проходят по вен-
тральной поверхности по-
чек, воспринимая отводящие
мелкие вены из области зад-
них долей. •
На своем пути к почкам
задняя полая вена'восприни-
мает также парные вены от
половых органов (venae testi-
culorum или ovarii) и надпо-
чечников.
Вены задней ко-
нечности, хвоста и
почечные вены. По-
добно рептилиям, птицы об-
ладают воротной системой
почек, вследствие чего кровь из нижних конечностей, хвоста и орга-
нов области таза проходит через почки, прежде чем находит отток
через заднюю полую вену (рис. 103).
114
Из области хвоста кровь выносится парными хвостовыми венами
(vena coccygea communis), переходящими далее в тазовые вены
(v. hypogastrica) и проходящими’в почки. На уровне средних долей
последних в эти вены вливаются бедренные вены (vena femoralis
или v. iliaca externa), несущие почти всю кровь из задней конечности;
получающийся сосуд — «анастомоз» или vena iliaca communis — сое-
диняется далее с основным стволом v. renalis revehens, образуя
мощный клапан, который препятствует нормальному протоку крови
(рис. 104). Вследствие этого кровь, поступающая из задней конеч-
V.rr,
Рис. 104. Анастомоз п клапан поротной системы почек в выносящую систему vena renalis
revclicns (пли v. г. efferens).
а — фронтальный разрез через область апастомоза почки дикого лебедя—Cygnus sp.;
b — модель области анастомоза почки хохлатого жаворонка — Galerida cristata L.
А.—разрез через степки анастомоза; К1а. г.—края разреза клапана, К1. — тонкая
парусовидная стенка клапана; о — отверстие, через которое сообщаются полость анастомоза
и выносящая вена; v. И. ех. — vena iliaca externa; V. г, г. — vena renalis revehens; V.r.r.o.—
vena renalis revehens верхних долей почек, частично перерезанная; Z.v.u.— приводящая
вена нижних долей почек; W — кольцеобразное вздутие по краю клапана. На рис. а хорошо
видна вдавленпость парусовидпого клапана со стороны анастомоза. По Spanner,.
ности и таза, принуждена пройти предварительно через приносящие
ответвления обеих вен в почечную ткань; лишь пройдя последнюю,
кровь получает по выносящим венам доступ в заднюю полую вену.
Vena hypogastrica играет роль основной воротной вены почек и
воспринимает ряд более мелких вен из таза и мускулатуры бедра (vena
ischiadica, v. obturatoria), отдавая вместе с тем и наиболее много-
численные rami advehentes.
Воротная система печени. Вены пищеварительных
органов собираются в крупную воротную вену (vena portae), входя-
щую в печень двумя стволами и распадающуюся там в капиллярную
сеть воротной системы. Использованная печенью кровь выходит из
*
115
этого органа в виде двух печеночных вен (vena hepatica magna). и,
соединяясь с задней полой веной, образует нижнюю полую вену
(vena cava inferior). В левую печеночную вену впадает vena umbili-
calis, собирающая кровь с брюшных воздушных мешков и брюшины
(peritoneum), в правую — vena mesenterica, несущая кровь из тон-
ких кишек и стоящая в связи с vena coccygomesenterica, обычно
несущей кровь к почкам (рис. 103).
Селезенка и лимфатическая система. Селе-
зенка птиц сравнительно мала и лежит обычно на правой стороне
желудка, в области перехода железистого отдела в мускулистый. По
форме, окраске и функции она сходна с селезенкой рептилий.
Лимфатическая система птиц отличается от этой системы у млеко-
питающих более слабым развитием лимфатических сосудов и ред-
костью в них специальных клапанов. Лимфатические сосуды всего
тела соединяются в конечном счете в два ствола: левый и правый
грудной млечный проток (ductus thoracicus sinister et dexter), которые
под позвоночником направляются краниально к шее; они соединяются
друг с другом поперечными веточками и впадают несколько прокси-
мально от яремных вен в правую или левую краниальные полые вены.
Лимфатические сердца, существующие на эмбриональной стадии
у всех птиц, во взрослом состоянии у некоторых групп исчезают
(например у куриных), но у большинства сохраняются. Крупные
формы, например страус, обладают особенно мощными мышечными
оболочками этих сердец. Напротив, лимфатические узлы отсутствуют
у огромного большинства птиц; существуя у гусиных птиц и некото-
рых других, они обладают отличный гистологическим строением.
У гусиных они располагаются в виде двух пар возле каудальных отде-
лов яремных вен и у позвоночника возле половых желез.
Как уже указывалось (см. стр. 6), птицы обладают весьма интен-
сивным обменом веществ и, как правило, вообще характеризуются
очень энергичной жизнедеятельностью. Температура тела у птиц
в общем выше, чем у млекопитающих, и в большинстве отрядов в сред-
нем равна 42,2° С. Низкая температура свойственна примитивным
веслоногим и поганкам (39,5°), страусу (37,8°), киви (37°). В 1558 на-
блюдениях над 327 видами, сделанных A. Wetmore, температура птиц
вариировала от 38° до 44,8°, оказавшись наиболее высокой у одного из
тиранов (средняя 43,4°, высшая 44,8°), и было найдено, что у дневных
птиц температура тела значительно выше днем, чем ночью (обратно у
сов), а температура самок выше, чем самцов. Соответственно этойболь-
шой интенсивности метаболизма,.птицы обладают более быстрым кро-
вообращением и большим, относительно, объемом важнейших крове-
носных сосудов. В исследованиях Гессе (Hesse, 1921) это нашло свое
выражение в следующих наблюдениях. В соответствии с большей
потерей тепла излучением с поверхности тела у мелких птиц, чем
у крупных (за час утка теряет 6 калорий на килограмм своего веса,
голубь — 10, а воробей — 35), у мелких видов, сердце относительно
больше, чем у крупных. При одинаковой величине и весе более жи-
вой темперамент также связан всегда с более крупным сердцем.
У весящих около 200 г сороки, пустельги и чеглока вес сердца опре-
делился соответственно как 9,34°/оо? И»90/00 и 17°/оо- Соответственно
116
темпераменту и величине изменяется также и частота сердцебиения.
У мелких птиц она значительно больше. Так в ряду, образуемом
индюком в 8750 г, петухом в 1980 г, чирком в 755 г и воробьем в 30,5 г,
частота пульса определяется как 93, 312, 317 и более 460 ударов в ми-
нуту- Эти отличия сказываются и в потребностях корма, и величине,
и количестве эритроцитов, о чем см. стр. 106 и 123.
6. ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА И ПИЩЕВАРЕНИЕ
В пищеварительном аппарате, так же как и в других органах
птицы, мы находим явные черты приспособления к преимущественно
Рис. 105. Вскрытый голубь; пищеварительный аппарат.
ао— перерезанная нисходящая аорта; cl— клоака; со— слепые кишки; d. h.
желчные протоки печени; du — двенадцатпйерстная кишка ;h— печень; И — топкая кишка;
•— воронка яйцевода; ing. — зоб; I — селезенка; ое — пищевод; ov. — яичник; ov. d.
правый рудиментарный яйцевод; ov. s.—левый яйцевод; р— легкое; р. d. — дорзальная
лопасть поджелудочной железы; р. d. d — проток дорзальной лопасти поджелудочной
железы; р. v. — вентральная лопасть поджелудочной железы; р. v. d. — протоки вентраль-
ной лопасти поджелудочной железы; рг— железистый желудок; г — почки; s пижпяя
гортань; 1г — трахея; v— мускулистый желудок; v. с. — шейный отдел позвоночника.
По Шимкевпчу (с изменениями).
117
воздушному образу -жизни и специально к полету. Превращение
передних конечностей в крылья повело за собой частичное превраще-
ние челюстей в хватательный орган, каким является клюв. Вслед-
ствие необходимости облегчить скелет в связи с полетом, челюсти
птицы утеряли зубы, имевшиеся еще у археоптерикса и меловых птиц,
и задача размельчения пищи была перенесена .на желудок. В связи
с этой функцией желудок птиц более глубоко, чем у рептилий, диффе-
ренцируе'Лш на отделы, и появляется мощный мускулистый «пупок»,
форма которого намечалась уже у крокодилов (рис. 105). Вместе
с тем в пищеводе образуется расширение или зоб (ingluvies) для
временного хранения пи-
щи перед поступлением в
желудок, а в расположе-
нии других пищеваритель-
ных органов наблюдаются
некоторые изменения в
связи с сильным разви-
тием воздушных, мешков
и увеличенными размера-
ми самого желудка. Как
характерные черты пище-
варительного аппарата
птиц, необходимо также
отметить четкое выделение
двенадцатиперстной киш-
ки по сравнению с таковой
у рептилий. -
Рпс. 106. Крыша и дно (отрезанное с одной стороны
и отвернутое) ротовой полости молодой хищной
птицы.
Ch, — орбитальная щель (хоапы); G. F. — орбиталь-
ные складки; L — верхняя гортань; Ling. — язык;
Oes. — пищевод; Ph. Fr, — ряды сосочков на стен-
ках глотки; S. F.—пограничный валик орбиталь-
ного поля; Т—инфундибулярная щель. По Goep-
pert, из Stresemann.
Ротовая полость у птиц
является полостью клюва,
и роговой чехол его быстро
переходит в слизистую,
часто ярко окрашенную,
оболочку, выстилающую
полость извнутри (рис.
106). Крыша ротовой по-
лости имеет слабое углубление (соответствующее форме языка),
вдоль которого проходит узкая орбитальная щель, ведущая к хоанам.
Дно ротовой полости заполняется у многих птиц телом языка, очень
разнообразного по величине и форме в связи с характером пищи
данного вида (рис. 107); в некоторых группах язык очень мал и руди-
ментарен, теряя свое функциональное значение. Под языком у неко-
торых птиц находится сильно растяжимый мешок, образуемый стен-
ками ротовой полости, играющий то роль защечного мешка млеко-
питающих — местом временного хранения пищи (пеликан,* кед-
ровка — Nucifraga caryocathactes (L.), некоторые чистики — Aid-
dae), то роль вторичнополового органа, связанного с играми и кри-
ками птиц в период размножения (Otides). У основания языка про-
тив орбитальной щели открывается отверстие трахеи, а поверхность
сзади обоих отверстий (глотка) обычно покрыта мелкими сосочками,
вскоре переходящими в слизистую оболочку пищевода.
118
В ротовую полость открываются выводные протоки слюнных
желез, располагающихся обычно под языком, в слизистой оболочке
и на небе. Размеры и положение желез сильно вариируют, они отсут-
ствуют у веслоногих, питающихся рыбой, и хорошо развиты у зерно-
Рис. 107. Языки птиц.
а — американской славки — Oporornis agilis (Wils.) (Oscines, сем. Geolhlypidae);
b— американского чижа—Spinus tristis L. (Oscines, сем. Fringillidae); c — хохлатой
синицы — Lophophanes sp. (Oscines, сем. Paridae); d — императорского пингвина — Apte-
nodyles patagonica Mill. (Sphenisciform.es); e — канадской казарки — Bernicla canadensis L.
(Anseri formes) ; f — длинпоклювого крохаля — Merganser serra tor L.; g — багамской пищухи-
медососа— Coereba bahamensis Reich. (Oscines, сем. Coerebidae); h — медососа — Tropido-
rhynchus sp. (Oscines, сем. Meliphagidae); i — клеста — Loxia sp. (Oscines, сем. Fringillidae);
j — зеленого дятла — Picus viridis L. (Piriformis). Ho Lucas и Leibeb, пз Ililzhcimcr.
ядных птиц, нуждающихся в смачивании пищи (рис. 108). Особенно
большой величины достигают они у дятлов и некоторых ласточек
и стрижей в связи с питанием муравьями первых и постройкой гнезда
при помощи слюны у вторых (рис. 109). У большинства птиц эти
железы выделяют только слизь, муцин, иногда разной густоты
на
и клейкости, но у некоторых видов наблюдается уже в ротовой
полости частичное расщепление углеводов
Рпс. 108. Слюнные.железы утиных.
(а) — красной утки — Casarca casarca L.; голова сбоку; вскрыта
зачелюстная область; (Ь) — надклювье и верхнегортанная об-
ласть кряквы — Anas plalyrhyncha L., снизу; (с) —подклювье
и основание пищевода ее же снизу; (d)— язык и нижняя часть
гортани ее же, сверху; (е) —язык ее же, сбоку.
ао.—glandula angularis oris; cr.— gl. cricoarytaenoideae;
ma. — gl. mandibulares anteriores; mp. — gl. mandibulares po-
steriorcs; mx.—gl. maxillares; li.—gl. linguales inferiores;
Is.—gl. linguales superiores; oe.—пищевоД; pp.—gl. pala-
tinae; pt.—gl. pterygoideae. По Anthony.
в результате дей-
ствия ферментов
слюны.
Следующий за
глоткой пищевод
(oesophagus) пред-
ставляет собой
трубку, сильно ра-
стяжимую у птиц,
питающихся ры-
бой и крупными
плодами. Эта рас-
тяжимость пище-
вода является пе-
реходом к образо-
ванию зоба, мест-
ного расширения
или выроста стен-
ки пищевода, име-
ющего назначение
временно хранить
собранную пищу и
увлажнять ее (рис.
110). Обычно зоб
встречается у тех
птиц, которые мо-
гут располагать
пищей сразу в
большом количе-
стве, как, напри-
мер, дневные хищ-
ники и многие зер-
ноядные; в таких
случаях пища на-
ходится в зобу,
только пока на-
полнен желудок.
У некоторых
птиц зоб служит
для переноса птен-
цам и временного
хранения воды
(рис. 111). Как зоб,
так и пищевод ли-
шены каких-либо
желез, помимо вы-
деляющих смачи-
вающую слизь.
Желудок под-
120
разделяется на дйа, хорошо различающихся морфологически и
физиологически отдела: железистый (proventriculus) и мускуль-
ный (ventriculus) (рис. 112). Железистый желудок обычно имеет
тонкие стенки с большим количеством простых и сложных трубчатых
желез, рассеянных по складчатым внутренним стенкам желудка; часто
Рпс. 109. Поперечный разрез слюнной железы(gl. mandibularis externa)
городской ласточки — Delichon urbica L. По Antony.
сложные железы скучиваются пятнами, образуя железистые поля.
Сложные железы выделяют пепсин, и основная роль этого отдела же-
лудка заключается в расщеплении белков (протеинов) на пептоны. Од-
нако дентин, хитин и кератин, а у некоторых хищных птиц частью и
костная ткань не растворяются желудочным соком и в виде спрессо-
Рпс. 110. Форма и положение зоба у представителей разных отрядов.
1 — еловый клест — Loxia curvirostra L. (Passeriformes); 2 — сизый голубь — Columba
Iwia Gm., domestica, pull. (Columbiformes); 3—неразлучный попугайчик — Agapornis сапа
Gmel. ( Psitlaciformes).
a — пищевод; b — наполненный зоб; с — трахея; h — сердце. По Swenander, из Stre-
semann.
ванных комков извергаются через рот, как «погадки». Мускулистый
желудок имеет вид сравнительно тонкостенного мешка у хищных
и рыбоядных птиц и чрезвычайно мускулистого, толстостенного
органа у растительноядных; особенно мощна мускулатура на противо-
положных боковых стенках, радиально расходящаяся от централь-
ных сухожильных дисков (рис. 104). Сокращения этих главных н
121
(a) (bj
Рис. 111. Использование пищевода кап резервуара
для переноса воды (птенцам?). Танагра — Ccnnpso-
thraupis loricata: (а) — с сократившимися стенками
пустого пищевода; (Ь) — с растянутым пищеводом,
наполненным водой. По Вокег.
двух добавочных мускулов прессуют и перетирают пищу (зачастую
при помощи мелких камешков и особо твердых семян, заглатываемых
птицей). При этом трубчатые железы, располагающиеся в стенках
желудка, выделяют вязкий секрет, быстро сплывающийся в кожи-
стую, подобную кутикуле, оболочку, постоянно нарастающую от
стенок желудка по мере ее изнашивания (рис. ИЗ). Роль ее заклю-
чается в защите нежных стенок желудка и поддержке перетирающих
движений, в связи с чем боковые участки этой кутикулы образуют
иногда особые твердые бляшки или ороговевающие бугорки и зубцы.
В то время как у расти-
тельноядных птиц основ-
ная функция этого отдела
желудка заключается в
дальнейшем (после клюва)
раздроблении и размель-
чении пищи и разрушении
клеточных (целлулозных,
труднопер ев ариваемых)
оболочек, в тонкостенном,
легко растяжимом желудке
хищных и рыбоядных этот
отдел является преимуще-
ственно местом химиче-
ской переработки пищи,
за счет желудочных соков,
стекающих из переднего
отдела. У этой же группы
птиц нижняя часть мус-
кулистого желудка отгра-
ничивается сужением или
складкой в обособляющий-
ся пилорический отдел с не-
ясной функцией (рис. 114).
С другой стороны, у
видов, питающихся мяси-
стыми плодами и ягодами,
наблюдается уменьшение размеров желудка (и в первую очередь
мускулистого его отдела), вплоть до почти полного его псчезновения
вообще (рис. 115).
Начальный отдел кишечника представлен морфологически мало
отличающейся петлей двенадцатиперстной кишки (duodenum), обхва-
тывающей поджелудочную железу (pancreas). Выводные протоки этой
железы впадают в начальный отдел кишки, вблизи от протоков печени
п желчного пузыря; железа относительно крупнее у зерноядных и
меньше у хищных птиц (рис. 105).
Печень (hepar) у птиц имеет относительно очень крупные размеры
и развивается в виде двух лопастей, которые в связи с объемистостью
желудка и значительными размерами легких и воздушных мешков
располагаются в поперечном направлении, с наибольшей лопастью
главным образом в правой половине тела. Из двух протоков печенп,
122
впадающих в двенадцатиперстную кишку, один у большинства птиц
расширяется, образуя желчный пузырь (vesica fellea), достигающий
особенно большой величины у питающихся мясистой пищей.
Кислая среда желудка очень скоро в двенадцатиперстной кишке
меняется на щелочную, где три основных фермента поджелудочной
железы превращают крахмал пищи в легко
теины переводят в пептоны и
частью в аминокислоты, жи-
ры переводят в состояние
эмульсии, разлагая их на
жирные кислоты и глицери-
ны. Дальнейшее более пол-
ное расщепление этих про-
дуктов связано с деятельно-
стью энзимов слизистой обо-
лочки кишечника. Роль пе-
чени сводится в основном к
хранению излишних моноса-
харидов в форме гликогена
(животного крахмала), слу-
жащего резервным питатель-
ным веществом для деятель-
ности мускулатуры, и к под-
держанию постоянства соста-
ва питательных веществ в
крови.
Средний наиболее длин-
ный отдел кишечника — тон-
кие кишки (ileum) — заме-
чателен у птиц частым раз-
витием добавочных петель;
из них некоторые (например
усвояемые сахара, про-
Рис. 112. Желудок, печень и двенадцатиперстная
кишка качурки — Procellaria aequinoctialis L.
( Proce I lariiformes).
g.— мускулистый желудок; g. b. — желчный
пузырь; h. a. — печеночная артерия; L. — пе-
чень; I. h. d. — левый печеночный проток; ос s.—
пищевод; р. — поджелудочная железа; рг. — же-
лезистый желудок; h. d. — правый печеночный
(желчный) проток; v. р.—печеночная вена. По
Forbes, из Stresemann,
поджелудочная, в концевом
отрезке тонких кишек) до-
стигают особенно сложного
развития и вероятно имеют
особые функции. Тонкие киш-
ки у зерноядных форм отли-
чаются значительной длиной,
У питающихся фруктами и
мясом — укороченностыо и тонкостенностью; кроме того, крупные
формы имеют относительно более длинный кишечник, чем мелкие,
в связи с необходимостью соотношения более быстрого увеличения
объема кишки в сравнении с ее поверхностью.
Положение кишечника в полости тела весьма изменчиво и было
использовано некоторыми авторами (Гаррод, Митчелл) в качестве
систематического признака (рис. 116). Слизистая оболочка кишеч-
ника, в связи с задачей увеличения внутренней всасывающей по-
верхности, образует множество складок и выростов (рис. 117). Прямая,
сравнительно короткая кишка отграничивается от тонкой выводными
123
Рис. 113. Гистологическое строение стенок мускульного желудка хищной и зерноядной
птицы (в обоих случаях одинаковое увеличение).
(а) — вертикальный разрез стенки желудка лесного сарыча — Buteo buteo L.; (Ь)"
то же (но лишь с незначительной частью мускулистого слоя) курицы — Gallus gallus L<
а. — наружная оболочка желудка — adventitia; d. — железистый слой; тп. —муску-
листый слой; тити.—muscularis mucosae; s. — слой затвердевшего секрета, отделяемого
железами; ss. — колонки секрета; sq.—поперечные соединения колонок секрета, свиде-
тельствующие о периодическом усилении деятельности желез; tp. — tunica propria. .
Обратите внимание на разную мощность слоев секрета, tunica propria и др. и разный
тип желез —* простой кишкообразный у Buteo и собранный пучками у Gallus. По Cornselius-
(9)
₽ис. 114. Строение желудков рыбоядных птиц, являющихся представителями разных отрядов,
п “7 ираснозобой гагары — Colymbus stellatus Pontopp. ( С olymbi formes); 2— большой
поганки — Podiceps cristatus L. (Colymbiformes); 3 — черного аиста — Ciconia nigra L.
^JcoWorme8); 4— большого крохаля — Мег див merganser L. (Anseriformes); 5 — сереб-
.T /л? ^айки — Larue argentatus L. (Charadriiformes); 6 — речной крачки — Sterna hirundo
r^JVharadriiformes), сократившийся желудок; 7—малой крачки—Sterna minuta L.
^'i^radrziformes), в растянутом состоянии; 8—люрика — М ergulus alle L. (Charadriifor-
7П€8)> сократившийся желудок; 9 — его же, в растянутом состоянии; dp — железистый
отдел; ра — пилорический отдел; я —промежуточный участок. По Swenander.
Рис.. 115,. Схема постепенной редукции мускульного желудка у видов, специализирую-
щихся на питании сочными плодами растений.
Пищевод исчерчен продольно; мускульный желудок отмечен черной окраской; кишечник
исчерчен поперечно, (а) — желудок примитивного новогвинейского цветососа — Pristho-
rhamphus sp.; (b) — желудок специализированного цветососа — Dicaeum sp., превратившийся
в слепой придаток пищеварительной трубки, переваривающий животную пищу отдельно от
быстро проходящих пищеварительный аппарат ягод; (с) — желудок танагры—Euphonia
sp., превратившийся в нефункциопирующий придаток. По Desselberger, из Stresemann.
рис. 116. Главнейшие соотношения кишечных петель у птиц. Схема, вид с правой стороны.
Нисходящие отделы (считая от желудка) отмечены черными линиями, восходящие —
пунктиром. Различают закрытые петли (папр. двенадцатиперстная кишка) с нисходящим
и восходящим отделами, тесно связанными мезентерием и его сосудами, п открытые, с широ-
ким мезентерием; если нисходящий отдел лежит вправо от восходящего, петля является
«правой» (тот же duodenum и III петля, рис. с.); при положении нисходящего отдела
влево от восходящего мы имеем «левую» петлю (II, III, IV петли, рис. с.). Кишечник,
сложенный из петель, расположенных продольно оси тела, является ортоцельпым (ortho-
celous —типы a, b, с, d); при образовании из петель спирали кишечник является цикло-
цельным (cyclocoelous—е).
Различают следующие главнейшие изменения: а — isocoelous; Ь —anticoelous; с —
antipericoelous; d — isopcricoelous; а — cyclocoelous; f,g — plagiocoelous; h — telogyrous.
Тип а — свойствен большинству птиц; тпп е — большинству чаек, куликов, голубей и др.;
Ар— кукущкам, бапаноёдам и др.; h — аистам, попугаям, дневным хищникам п др.;
I, II, III, IV — номера петель. По Gadow и Newton.
{,f (2}
протоками слепых кишек (caeca), обычно парных п развитых не
у всех птиц (рис. 148). Особенно большой величины и крупного
функционального значения они достигают в некоторых группах расти-
тельноядных птиц (куриные и гусиные), у которых служат местом
переработки целлулозы в более усвояемые вещества, что достигается
обычно при помощи бактерий (Bacterium coli). В группах, где слепые
кишки отсутствуют, их роль в значительной степени повидимому
восполняется другими специальными отделами кишечника: увели-
ченной прямой кишкой или
измененным слепым отрост-
ком (diverticulum caecum
vitelli) на стенке тонкой
кишки, являющимся остат-
ком эмбрионального желточ-
ного мешка и его протока.
Расширение прямой киш-
ки в ее заднем конце пред-
ставляет собой клоаку, не-
большими складками оболоч-
ки разделяемую на три части:
coprodaeum, urodaeum и ргос-
todaeum (рис. 119). Первый
отдел сильным кольцевым му-
скулом может быть отграни-
чен от<грямой кишки и слу-
жит местом концентрации
фекальных масс; маленький,
плохо отграниченный от ргос-
todaeum, средний отдел на
выступающих внутри сосоч-
ках несет отверстия мочеточ-
ников и половых путей; по-
следний отдел запирается
мощным анальным сфинкте-
ром. На спинной стороне
proctodaeum открывается не-
большое толстостенное обра-
зование, называемое фабри-
Циевой сумкой (bursa Fabricii) » обладающее внутрисекреторной
функцией. Выяснено также, что она является местом образования
лимфоцитов и отчасти лейкоцитов. За немногим исключением
(страусы), этот орган функционирует только в молодом возрасте,
сильно редуцируясь к периоду достижения половой зрелости.
Пищеварение. Потребность в пище стоит в тесной связи
с условиями протекания обмена веществ в теле птицы, с ее величиной
и характером пищи. Так как с увеличением размеров тела вес растет
л кубических отношениях, а поверхность в квадратических, мелкие
птицы необходимо должны иметь более оживленный обмен веществ,
чтобы восполнить потерю тепла излучением с поверхности тела, чем
птицы крупные, теряющие тепла меньше относительно объема своего
(3) (4}
Рис. 117. Внутренняя поверхность кишечника.
1,2 — сорока — Pica pica L., верхний и нижний
отделы тонких кишек; 3 — курица— Gallus gallus
L., верхний отдел тонких кишек; 4 — тундровая
куропатка — Lagopus mutus Mont., основная
часть слепых кишок.
По Bujard и Schumacher, из Stresemann.
127
Рис. 118. Форма слепых кишок у представителей разных отрядов птпц.
.а — страуса—Struthio camelus L.; b— хохлатого тинаму,— Calodromas elegans d’Orb.
& G-eofr.; c — пятнистого тинаму — Nothura maculosa Temm.; d — совы — Photodilus badius
Horsf.; e —домашней курицы — Gallus gallus L.
Обратите внимание на различие слепых кишек у обоих видов тинаму. Вздутая форма
кишек Calodromas elegans невидимому может стоять в связи с накоплением переработанных
кишечником и влажных пищевых веществ, которые по выделении являются первой пищей
птенцов этого вида, обитающего сухие степи Аргентины и Патагонии.
тела. Так желтоголовый королек {Regains regulusL.), весом около 6 г,
в нормальных комнатных условиях потребляет количество пищи
в 28% веса его собственного тела, скворец же (Sturnus vulgaris L.), ве-
сомв 75 г, потребляет пищи на 12% веса тела, а домовый сыч {Athene
noctua Scop.), весом в 168 г, потребляет пищи по весу только 51/2%
веса тела. Животная пища и плоды перевариваются обычно гораздо
быстрее семян и зерен. У известного своей прожорливостью свири-
стеля ягоды проходят
весь кишечник в 8—10
минут, в то время как
зерна злаков перевари-
ваются курицей от 12 до
24 часов. Потребность
в пище значительно
возрастает при повы-
шенной трате мускуль-
ной энергии, при энер-
гичном образовании по-
ловых продуктов, при
понижении температу-
ры. Понижение послед-
ней в опытах Лапика с
IJraeginthus, маленьким
ткачиком из сем. Pl>>cei-
dae, весом в 7,5 г, с 39°С
до 16° С увеличило по-
требность птицы в пище
более чем в 3 раза
(с 6,4 до 20 калорий на
10 кв. см ее поверхно-
сти). Содержимая в не-
воле зимой в умеренной
зоне, с коротким зимним
днем в температуре 15°С,
птица не могла покры-
вать потребность тела в
пище несмотря на кор-
межку целый день и бы-
стро умирала от истоще-
ния, но в случае искус-
Рис. 119. Клоака и penis взрослого самца страуса —
Struthio camehis L.
(а) — сагиттальный разрез клоаки; (Ь) —вид вскрытой
клоаки сверху; а—penis, во втянутом состоянии вби-
рающийся в слепрй мешок b—с, в образовании которого
принимает участие bursa Fabricii; d — urodaeum; e —
koprodaeum, у страуса достигающий очень крупных раз-
меров и играющий роль временного мочевого пузыря;
/ — конус с отверстием прямой кишки; д — прямая
кишка; h — семяпровод, открывающийся выводным про-
током на сосочке h1 в urodaeum; i — мочеточник, с со-
сочком мочеточника г1. По Saint-Hilaire, из Stresemann.
ственного освещения клетки в ночные часы, птичка легко переносила
и более низкую температуру. Отсюда понятна малочисленность зи-
мовки мелких птиц в северных холодных зонах с коротким зимним
Днем и низкими температурами воздуха.
В связи с интенсивностью обмена веществ мелкие птицы гораздо
более чувствительны к голоду, чем крупные, и не выдерживают иногда
голодовки в 5—6 часов. Более крупные птицы могут обойтись
без пищи значительно дольше: 9 дней — голубь, 24 дня — белая сова,
*10 дней — орел. Длительная сопротивляемость голоду особенно
важна крупным хищным птицам, у которых добывание пищи часто
9 Орнитология—1209
129
зависит от случая, и затем перелетным птицам, пересекающим в из-
вестное время года большие водные или пустынные пространства.
В связи с этими потребностями, в организме многих птиц выработа-
лась легкая способность к ожирению, наступающему всякий раз,
когда наличная пища превышает необходимый для работы тела уро-
вень питания. Так вес домашнего гуся в 5—6 недель посредством
откорма легко может быть повышен до 50% первоначального веса
за счет накопления под кожей и в полости тела значительного коли-
чества жира. Для большинства птиц такой период ожирения наступает
периодически осенью, в связи с предстоящей зимовкой или переле-
тами, и весной на зимних квартирах, в связи с предстоящим возвра-
том на родину.
Потребность птиц в воде в разной степени выражена у дикоживу-
щих форм; большое количество пустынных и степных видов могут
обходиться без воды целыми месяцами (например джек — Chlamydolis
niacqueeni (Gray et Hard.), пустынная славка —.Sylvia папа Hempr. et
Ehr.), очевидно получая ее за счет распада жиров и углеводов пищи.
Однако в ежедневном рационе большинства других видов она необ-
ходима. При обильном питании голубь погибает от жажды на 4-н
или 5-й день. При одновременном голодании голубь погибает на
11-й день, что обусловливается расщеплением запасных белков и
жиров в теле птицы, наступающим в связи с голоданием. Это расще-
пление освобождает некоторое количество воды, которое повышает
общую сопротивляемость жажде. Сравнительная ограниченность
в потребном количестве воды у многих птиц связана с малым коли-
чеством жидких выделений в теле птицы как в виде помета, так и прп
дыхании. Потовые железы, выделяющие так много воды у млекопи-
тающих, у птиц отсутствуют.
7. ОРГАНЫ ВЫДЕЛЕНИЯ И РАЗМНОЖЕНИЯ
Половой аппарат и почки птиц в общем имеют те же основные
особенности, что и аналогичные органы рептилий. Повидимому
адаптивное значение имеют следующие отличия, однако частично по-
вторяющиеся и в некоторых группах рептилий. У всех птиц отсут-
ствует мочевой пузырь, что может быть объяснено редукцией вслед-
ствие более объемистого развития других органов птицы, вызванного
летающим образом жизни. Моча птиц имеет кашицеообразный ха-
рактер и состоит главным образом из мочевой кислоты. В половом
аппарате характерны редукция правого яичника и яйцевода и частич-
ное уменьшение семенников на этой же стороне, что может быть объяс-
нено теми же причинами (в связи с необходимостью помещения боль-
шой правой доли печени и мускулистого желудка). В строении других
^деталей этой системы птицы сближаются то с одной, то с другой из
групп рептилий.
Почки. Почки птиц лежат в обширных углублениях сложного
таза, образуемых подвздошными и отчасти седалищными костями
(рис. 120). Двумя поперечными бороздками каждая почка разде-
ляется на три доли. У некоторых птиц наблюдаются частичное слия-
ние почек по средней линии и некоторая асимметричность в распо-
130
(metanephros) и по
своему строению
Ао.
GLs.
V fem.
F.obt.
Ar. is.
V.il.int
f.is.—
V.cau.
H.
Lm.r
Pub.
CL.
-L.p.r.
V.def.
Ur.
ложении по высоте. Почки взрослых птиц по происхождению явля-
ются вторичными почками
очень близки к анало-
гичным органам у реп-
тилий (рис. 121 и 122).
Отходящие от почек
мочеточники открыва-
ются в среднем отделе
клоаки. Моча птиц, осо-
бенно богатая мочевой
кислотой, стекает по мо-
четочникам в виде бога-
той водой жидкости, но
в клоаке большая часть
воды вновь поглощает-
ся, и выделяемая на-
ружу моча имеет густой
кашицеобразный харак-
тер. Мочевая кислота
является конечным про-
дуктом азотистого' обме-
на и у птиц синтезиру-
ется из молочнокислого
аммония. Питание почек
кровью происходит по
построенной, как у яще-
риц, воротной системе
почек (рис. 103).
Половые о р г а-
п ы. Мужские половые
органы представлены
парными овальными се-
менниками, прикрепля-
ющимися к стенке тела
У верхнего конца почек.
Как правило, левый
семенник значительно
больше правого и ярче
пигментируется в слу-
чае отложения на по-
верхности зерен мела-
нина.
Величина этих орга-
нов меняется в связи с
периодами покоя и раз-
множения; в последний
период у птиц семей-
Is.
Аг. pud.
И pud.
Ar. caud.
И caud.
-/-Hee.
O.ur.
Or.u.def.
Рис.
Руд. -
120. Почки и половые органы петуха — Gallus
gallus L. в период размножения.
Ло. — aorta descendens; Ar. caud. — ar. coccygea, Ar.
cr. — ar. femoralis; Ar. is. — ar. iscliiadica; Ar. pud. —
ar. pudenda;
Cl. — клоака; F. is. —foramen ischiadicum; F. obt. —
foramen obturator; Gl. s.—надпочечные железы —
glandula suprarenalis; II.—подвздошная кость; Is.—
седалищная кость; L. a. r., L. m. r., L. p. r. — передние,
средние п задние доли почек; Or. v. def. —отверстия
семяпроводов; О. иг. —отверстия мочеточников; Pub. —
лобковая кость; Руд.—пигостиль; Rec.,—прямая
кишка; Tes. — семенники; V. caud.— vena coccygea;
V. cav.—vena cava posterior; V. def. — семяпровод;
V. fem. —vena iliaca externa (или v. femoralis); V. il.
c.—vena renalis revehens communis; V. il. int.— vena
hypogasrtica (или v. iliaca interna); V. pud. — vena
pudenda; v. r. ad. — vena renalis advehens; V. r. r.—
vena renalis revehens; Ur. — мочеточник (ureter). По
Freund, из Slreseinann.
ники весят в 300—1000
раз больше и у мелких американских славок составляют по весу
Д° 719 веса всего тела. Огромное увеличение объема их зависит от
131
интенсивного удлинения и расширения семенных канальцев (tubuli
contorti), в которых происходит развитие спермиев (рис. 123). Коли-
чество последних в ейякуляте сизого голубя достигает в это время
200 миллионов, а у петуха, после некоторого периода полового покоя,
до 4 миллиардов. Возможность такого обилия семенных нитей объяс-
няется одновременным семяобразованием во всех отделах семенни-
ков и хранением их преимущественно здесь, а не в других частях поло-
вого аппарата, хотя у некоторых форм семяпроводы (vas deferens)
I/.I/. interl.
Кар-
/1г. intral.
Gl.
Кар.
S.b.
V. aff. gl.
V. eff. gl.
V. intral.
n.
V.eff.Gl.
И interl.
S.
V. inter!.
Рис. 121. Строение почечных долек — схема.
Основные доли почек, подобно почкам рептилий, распадаются па огромное количество
микроскопических долек, в центре которых лежит широкая отводящая вепа (vena intralo-
bularis) и сопровождающая ее тонкая артерия (art. intralobularis). Артерия отсылает веточки
(vas afferens) к многочисленным почечным клубочкам (glomeruli), лежащим на периферии
почки. На поверхности долькп лежат собирающие мочевые отделения трубочки, сопровождае-
мые тонкими веточками (v. interlobularis) приводящей вены (renalis advehens). От трубочек
мочевые канальцы направляются к центральной вене, загибаются и после многих петель
заканчиваются почечными клубочками мальпигиевых телец. Капиллярная сеть располагается
между приводящими и отводящими венами, густо опутывает мочевые канальцы и восприни-
мает артериальные vasa cfferentia, исходящие из glomeruli. См. также рис. 122.
Ar. intral. — arteria intralobularis; Gl.—glomeruli; H. — мочевые канальцы; Кар.—
капилляры; Lg. — границы отдельных долек; S. — собирательные трубочки; S. Ь. — пучки
собирательных трубочек, выводящие мочу из долек; V. aff- gl- — vas afferens glomeruli;
V. eff. gl. —vas efferens glomeruli; V. interl., V. inter. —vena interlobularis* V. intral. —
vena intralobularis; V. V. interl. — разветвления venae interlobularis па поверхности почки.
По Spanner.
образуют перед впадением в клоаку небольшие расширения, до
70—120 раз увеличивающиеся в объеме в период совокуплений и по-
видимому являющиеся временным резервуаром для семени, вместо
соответствующей функции epididymis у млекопитающих.
При совокуплении самец прижимается клоакой к- клоаке самки
так, что выпрыскиваемый в urodaeum ейякулят легко попадает оттуда
в широкие устья яйцеводов. Перенос спермы облегчается значитель-
ностью и эректильностыо сосочков с выводными протоками семя-
проводов в клоаке самца: у некоторых птиц (гусиные, страусы,
кракс и тинаму) к тому же образуется мускулистый эректильный
132
вырост (penis) с бороздкой по наружной поверхности, по которой
стекает сперма (рис. 124).
Женские половые органы представлены яичником и яйцеводом.
Яичник (ovarium), в период покоя имеющий вид плотного овального
или вытянутого тела, брыжейкой (mesovarium) прикрепляется
к стенке тела против переднего конца почек. Как правило, яичник и
яйцевод отсутствуют в правой половине тела, останавливаясь в своем
развитии на позднеэмбриональных стадиях. Эта редукция характерна
для птиц п может быть объяснена огромной величиной яиц у птиц
в совокупности с необходимостью уменьшения размеров ряда внут-
Рис. 122. *ПоЕерхность почки осоеда —Pernis apivorus L. в области двух почечных долек*.
Инъецированы мочевые собирательные трубочки и канальцы и кровеносные сосуды.
Хорошо видны разветвляющиеся собирательные трубочки, крупные веточки приносящих
вен (vena renalis afferens, vena interlobularis). Vena intralobularis невидны, так как лежат
глубже верхнего слоя. Обозначения см. рис. 121. По Spanner.
ренних органов для облегчения веса тела; сильное развитие воздуш-
ных мешков, мощный мускулистый желудок, большая правая ло-
пасть печени могут механическим давлением способствовать этой ре-
дукции. Однако, как вторичное явление, нередкое у разных групп
птиц и частое (до 66% случаев) у Accipitres (главным образом у родов
Circus и Accipiter), правый яичник развивается до полного функцио-
нирования, при том же отсутствии правого яйцевода; образующиеся
в нем яйца проходят свой путь через левый яйцевод.
В период размножения яичник значительно увеличивается в раз-
мерах, приобретает гроздевидное строение й яркую желтую окраску
(рис. 125). По откладке яиц яичник быстро, через 12—20 дней, при-
нимает форму, характерную для покоящейся фазы. Столь же значи-
тельны изменения яйцевода (oviductus). В период размножения это
сильно извитая широкая трубка, дифференцированная на ряд отделов
133
Ci
и лежащая на задней стенке полости тела. Одним
концом она впадает в средний отдел клоаки, другим
свободно открывается в полость тела. В покое яйце-
вод оказывается сморщенным, тонкостенным и умень-
шенным в десятки раз до размеров мало заметного
органа.
Образование япца. Строма яичника
заключает много тысяч ооцитов, находящихся на
начальной ступени развития (рпс. 126). Ежегодно
часть их накопляет в небольшом количестве жел-
Рпс. 124. Penis нанду — Phea sp. (а) и эму — Dromaeus novae-hollan-
dia La til. (b).
a — схематически!! разрез через вентральную стенку Клоаки и penis
во втянутом состоянии; b — поперечный разрез через свободный конец
penis. Penis состоит из двух асимметричных фиброзных тел — /. Д,
сросшихся своими основаниями, и длинного и узкого слепого мешка
(или кишки), частично выворачивающегося, как палец перчатки. По
дорзальной стороне органа пробегает глубокая бороздка для стока
спермы. Невыворачивающаяся часть слепого мешка наполнена слизи-
стым секретом, выделяемым железистым эпителием стенок, что делает
орган скользким. Вследствие более сильного развития левого фиброз-
ного тела, при эрекции орган заворачивается вокруг своей осп спи-
ралью. f.fi — фиброзные тела; д. — граница между выворачивающейся
и железистой частью слепого мешка; г. М. — бороздка для переноса
спермы; о. — отверстие слепого мешка на наружном конце органа; sh.—
просвет ввороченной части органа. По Boas, из Stresemann.
ток, синтезируя его из протеинов и жировых ки-
слот, доставляемых в качестве питательного мате-
риала кровью и лимфой. С наступлением брачного пе-
риода некоторые из этих ооцитов начинают энергично
накоплять в плазме желток, увеличиваются в раз-
мерах, сначала выпячиваясь на поверхности яич-
ника, затем свешиваясь в виде стебельчатого фол-
ликула в полость тела (рис. 125). По прохождении
первого редукционного деления стенки фолликулов
лопаются, и яйцеклетка оказывается в полости тела,
где подхватывается, облекающей яичник воронкой
яйцевода.
а
Рис. 123. Сперматозоиды представителей разных отрядов.
а — зяблик —Fringilla coelebs L. (Passeriformes) ; b — иволга — Oriolus oriolus L
(Passeriformes)c — сизый голубь — Columba livia L. (Columbiformes); d — большой пе-
стрый дятел — Drgobates major L. (Piciformes). По Ballowitz, пз G-adow.
134
Количество ооци-
тов, накапливающих
значительную массу
желтка, всегда не-
сколько больше чи-
сла нормально откла-
дываемых птицей
яиц; часть их явля-
ются резервными на
случай необходимо-
сти увеличить число
яиц кладки. При пе-
реходе птицы к наси-
живанию, они под-
вергаются редук-
ции, выражающейся в
энергичном распаде
хроматинового, аппа-
рата, рассасывании
желтка и впослед-
ствии дегенерации
всей клетки.: Стенки
лопнувших фолликул
по откладке яйца
сморщиваются и так-
же быстро рассасыва-
ются без образования
желтого тела.
Попавшая в яйце-
вод яйцеклетка, под
давлением мускуль-
ных сокращений его,
проходит по всей его
длине, облекаясь по-
степенно рядом на-
ружных оболочек.
Начальный отдел яй-
цевода, воронка (in-
fundibulu m), имее т
вид широкого, край-
не тонкостенного и
нежного раструба
с многочисленными
складками, могущего
тонкими тяжами под-
тягиваться к яични-
ку (рис. 125). Ворон-
ка переходит в ко-
роткое тонкостенное
сужение (tube), рас-
Рис. 125. Яичник и яйцевод птицы в период откладки
япц — схема.
cl. — клоака; е. — белковая часть яйцевода (pars albumini-
fera); f. — стебельчатые фолликулы; inf.—воронка яйце-
вода (infundibulum); is. — перешеек (isthmus); ov. — яич-
ник; ov. d.—рудимент правого яйцевода; г. — прямая
кишка; tb. — тонкостенный отдел яйцевода (tube); vt.—
маточный отдел яйцевода (uterus); v. — мускулистый отдел
яйцевода (vagina). По Duval.
135
ширяющееся дальше в длинную, лежащую петлями, трубку, обильно
выделяющую яичный белок (pars albuminifera). Эта часть корот-
Рис. 126. Схема развития ооцита в яичнике сизого голу-
бя — Columba livia G-m.
А — ооцит диаметром в 50 микронов, сбоку; В — тот же
ооцит, сверху; С — ооцит диаметром в 600 микронов, сбо-
ку; D —ооцит диаметром в 3 мм, сбоку; Е— зрелый ооцит
диаметром в 20 мм, сбоку; F — яйцо, вынутое из яйце-
вода, сверху.
На начальных стадиях роста ооцита зародышевая бляшка
(д. V.) лежит близ анимального полюса яйца; в дальнейшем,
вместе с накоплением желтка, она оказывается перемещен-
ной ближе к центру. На более зрелых стадиях зародышевая
бляшка вновь перемещается на анимальный полюс, оставляя
след своего прохождения в виде латебры (Lat.); сростом
ооцита зародышевая бляшка сильно уплощается (так паз.
зародышевый диск — bld.). Центром образования желтка па
первых стадиях является аппарат Гольджи (у. п.), окру-
жающий центросферу с диплоидной центрозомой; в даль-
нейшем основная роль принадлежит митохондриям, пре-
вращающимся в капельки жира; вначале располагаясь в
цитоплазме в виде туманного скопления (s. с. А в дальней-
шем митохондрии распределяются на периферии (с. z.), где
гл. обр. и происходит зарождение новых жировых капель.
С началом своего роста ооцит перемещается из стромы яич-
ника на его наружную поверхность и соприкасается своей
эпителиальной оболочкой с его перитонеальным эпителием.
Место срастания обоих эпителиев образует стигму (лишен-
ную сосудов часть оболочки фолликула), которая по со-
зревании ооцита лопается, освобождая его из тканей яич-
ника; питающие яйцо оболочки (theka folliculi и theca
vasculosa) образуются вне этой стигмы, во внутренней сто-
роне ооцита.
bld. — зародышевый диск; с. с.— клоакальная халаза;
ср. — центральная протоплазма; с. st. — клоакальный ко-
нец стигмы, на котором начинается разрыв фолликулярных
оболочек; Е. А. —ось развивающегося эмбриона; д. v. —
зародышевая бляшка; L. А.—длинная ось яйца, прохо-
дящая впоследствии через халазы; Lat. — латебра; N. А.—
ось ядра; о. р. — задний край бластодермы; Р. А. — по-
лярная ось; р. р. — периферическая протоплазма; s. с. —
зона митохондриев, в которых началось образование жиро-
вых капель; sk. — стебелек фолликула; s. z. — перифери-
ческая зона митохондриев; у. п. — аппарат Гольджи. По
Bartclmez, из Stresemann.
кой перетяжкой (isth-
mus) соединяется с
толстостенной корот-
кой и широкой ча-
стью, маткой (uthe-
rus), за ней мускули-
стой трубкой, влага-
лищем (vagina), впа-
дающей в клоаку.
Большая часть
яйцевода внутри по-
крыта крупными про-
дольными складками,
в матке замещенными
многочисленными ле-
пестковидными вы-
ступами. В разрезе
яйцевод представля-
ется сложенным из
ряда слоев: снару-
жи — оболочка брю-
шины (per itoneu m).
затем идут: слой про-
дольной мускулату-
ры, слой соединитель-
ной ткани, богатый
кровеносными сосу-
дами, слой кольцевой
мускулатуры, слой
соединительной тка-
ни, толстый слой вет-
вящихся трубчатых
желез и, наконец, рес-
нитчатый эпителий с
рассеянными одно-
клеточными железа-
ми.
Уже воронка ипе-
редняя часть трубки
выделяют в виде сек-
рета , обволакиваю-
щего яйцеклетку топ-
ким слоем, очень гу-
стой слой белка, обра-
зующего на полюсах
яйца сильно скручен-
ные белковые тяжи"
халазы, плавающие
136
и/
Рис. 127. Разрез яйца птицы — схема.
al. — белок; ch. — халазы; /. а. — воздушная камера;
д. р. — зародышевая протоплазма; т. v. — желточная обо-
лочка— membrana vitellina; п.—ядро; sh. — известковая
скорлупа; s. h. т.—подскорлуповая оболочка; v.a.—
желтый (темный) желток; vf. — белый (бледный) желток*.
По Thomson.
в жидком белке (рис. 127). Их назначение заключается невиди-
мому в предохранении яйцеклетки от толчков и смягчении со-
трясения, испытываемого яйцом; кроме того, яйцеклетка свободно
поворачивается на халазах и сохраняет постоянно наиболее выгодное
при насиживании положение зародышевым диском вверх. Затем
белковая часть яйцеводов окутывает яйцо толстым слоем жидкого и
тягучего белка; последний слой образует своими волокнами как бы
сеть, наполненную жидким белком, про'боденную закрученными
халазами. Спускаясь в перешеек, яйцо покрывается скорлуповой
кожицей, впоследствии лежащей под известковой скорлупой и со-
стоящей из двух листков, между которыми на тупом конце яйца
образуется пространство, выполненное воздухом. Эта кожица состоит
из густо переплетен-
ных волокон, обвола-
кивающих яйцо всего
в 40—50% оконча-
тельного веса. Пол-
ный вес достигается
яйцом после получе-
ния слоя жидкого бел-
ка, залегающего под
упомянутой кожицей
и проникающего в
яйцо в матке уже
осмотически. Этот же
отдел яйцевода выде-
ляет белую тягучую
жидкость, содержа-
щую углекислый
кальций, затвердева-
ющую при облекании
яйца в известковую
скорлупу (рис. 128).
Отложение извести
текая различно в разные периоды образования скорлупы. Вну-
тренний слой ее, так называемый сосцевидный, состоит из кону-
совидных коротких столбиков, сливающихся кнаружи между собой
в сплошной губчатый слой, прободаемый простыми и ветвящимися
тончайшими поровыми канальцами. Снаружи скорлупа покрыта гомо-
генной эластичной кутикулой, в зависимости от рыхлости сложения
вызывающей больший или меньший блеск скорлупы; у некоторых
групп (веслоногие — Pelecanifonnes, поганки — Podicipedes, фла-
минго — Phoenicoptertformes) она рыхла, может мазать, как мел, и
достигает значительной толщины. В кутикуле открываются поровые
каналы, пронизывающие скорлупу и дающие возможность газового
обмена яйца со средой, и залегают пигменты, вызывающие разно-
образную окраску яиц. Кутикула способна при смачивании не-
сколько набухать и закрывать поровые каналы, предохраняя этим
яйцо от проникновения влаги.
Первично скорлупа яиц являлась бесцветной, и эта особенность.
тончайшими пластинками
137
сохранилась у многих современных птиц, осооенно у гнездящих-
ся в дуплах и норах. Совершенно белые яйца имеют большин-
ство сизоворонковых — Coracii/formes, голубей — Columbae и попу-
гаев — Psittaciformes. Остальные группы имеют более или менее
окрашенные яйца. В ок-
раске яиц следует раз-
личать глубокую окрас-
ку от поверхностной или
пятнистой. Первая всег-
да имеет диффузное рас-
пределение в слоях скор-
лупы и образуется по-
видимому в трубчатых
железах маточного от-
дела яйцевода. Наиболее
распространенным пиг-
ментом является голу-
бой пигмент ооциан,
одно из производных
желчного пигмента, би-
ливердина ; в немногих
других случаях были
найдены желтей оохло-
рин (казуары—Casuarii-
formes) и красный оок-
сантин (скрытохвосты—
Tinamiformes). Пятни-
стые окра ски не до статоч-
но изучены, наиболее
распространенным яв-
ляется красно-коричне-
вый оопорфирин, родст-
Рис. 128. Яйцевые оболочки.
венный пигменту крови-
оорходеину; он выраба-
(а)—схематический разрез через скорлупу, белок и
внешние слои желтка ненасижениого яйца курицы —
Gallus gallus L.; (b) —радиальный шлиф через скор-
лупу яйца страуса — Struthio camelus L. 1 — кутику-
ла; 2 — губчатый и сосцевидный слой скорлупы (а —
губчатый; Ъ — сосцевидный); 3 — по декор л уповая обо-
лочка ; 4 — слой гомогенного белка; 5—12 — слои белка
различной густоты; 13 — membrana chalazifera; 14 —
внешние слои желтка; с — сложные разветвляющиеся
канальцы в толще скорлупы, берущие начало между
сосочками сосцевидного слоя; р — поры в кутикуле, ко-
торыми заканчиваются на поверхности яйца канальцы
скорлупы. По Nathusius, из Stresemann.
тывается клетками бел-
ковой части яйцевода и
по распадении их ока-
зывается взвешенным в
клейкой белковой жид-
кости, вместе с секре-
том известковых желез,
образующих кутикулу
скорлупы. От характера
смешения разных пигментных частиц и равномерности их распреде-
ления в жидкости зависит характер рисунка и пятен на скорлупе;
закорючки и черточки есть следствие вращения яйца и сложных
движений его в яйцеводе; венчики пятен на тупом конце образуются
вследствие скопления пигмента в верхней части матки, над яйцом.
В то время как величина яйца, вообще говоря, закономерно увели-
чивающаяся с увеличением размеров тела птицы, все же обнаружи-
те
вает в этом отношении некоторые отклонения у ряда видов, в связи
с специфическими особенностями их экологии и условиями разви-
тия эмбриона, форма яйца является, повидимому, глубоким консти-
туциональным свойством п характерна для определенных системати-
ческих групп. Колебания формы имеют сравнительно узкие пределы,
хотя и встречаются даже в одной п той же кладке. В основном форма
япца определяется характером мышечных сокращений стенок яйце-
вода; при расширении пх перед движущимся яйцом и одновремен-
ном сокращении кольцевых мускулов сзади него яйцо становится
вытянутым и узкоконечным, при значительном сопротивлении стенки
яйцевода перед яйцом — яйцо становится сферическим. Обычно раз-
личают по форме шесть типов птичьих яиц: сферические, например
совы (Strigiformes), тупоэллиптические — дневные хищники (Acci-
pitres), зимородки (Haley ones'), остроэллиптические — веслоногие
(Pelecanifornies), гагары (Colymbiformes), цилиндрические — стрижи
(Мicropodiforntes), козодои (Caprimulgiformes), грушеобразные—чайки
(Lari), кулики (Limicolae), чистики (Alcae), яйцевидные — большин-
ство воробьиных (Passeriformes), куриные (Galltformes).
Главная масса яйца состоит из желтка, белка и скорлупы (рис. 127).
Собственно яйцеклетка состоит из зародышевого ядра и желтого и
белого желтка, окруженного очень тонкой желточной оболочкой
(membrana vitellina). Желтый желток очень богат жирами и состоит
из крупных, размером до 100 микронов, капель с включенными вцих
бесчисленными мельчайшими зернышками; белый желток беден жи-
рами, капли его сильно вариируют по величине, а зерна обладают
большой светопреломляющей способностью; Оба желтка распреде-
ляются в яйце чередующимися концентрическими слоями, белый
при этом окутывает желтый желток снаружи, подстилает ядро и
в виде длинной капли углубляется в центр яйцеклетки, образуя
так называемую латебру.
Оплодотворение яйца-обычно имеет место в верхнем отделе яйце-
вода. Яйцо только что отложенное обладает, как правило, уже заро-
дышевым диском, происшедшим в результате первых делений ядра
клетки.
Химический состав яйца несколько изменяется у разных форм.
У Gallus желток (в целом) состоит из 50% воды, 16% протеинов
(главным образом вителлина), 23% жира, 11% липоидов (главным
образом лецитина) и следов солей, холестероля, лутеина (вызываю-
щего желтую окраску желтка) и нек. др. Белок состоит из 85—88%
воды, 12,2% протеина, 0,5% глюкозы, следов жиров и холестероля.
Скорлупа содержит от 89 до 97% карбоната кальция, до 2% кар-
боната магнезия, до 5% фосфатов обоих элементов и до 5% органи-
ческих веществ.
Откладка яйца. Период кладки яиц, сосредоточиваю-
щийся в большинстве случаев на короткий срок, требует от самки
очень крупной затраты сил. Большинство птиц откладывает по 1 яйцу
в сутки, в очень редких случаях по 2 яйца, чаще через день или два,
иногда через 5—6 дней (большеногие куры — Megapodiidae). Так как
У многих форм кладки очень велики, птица вынуждена в короткое
время мобилизовать значительные количества питательных веществ.
13Э
Обыкновенный погоныш Porzana porzana L. нормально несет 12 яиц,
составляющих 125% веса всего тела птицы, примем 35% этого веса
приходятся на желток, особенно богатый жирами и белками. Подоб-
ные соотношения мы находим у многих уток, куриных, многих во-
робьиных. Помимо более интенсивного обмена веществ, в этот период
свойственного самке, обычно наблюдается быстрое накопление ею
жира в короткий период перед кладкой, откладываемого в печени.
Также и острая потребность организма в кальции удовлетворяется
быстрым увеличением его потребления, стимулируемым, как и в пер-
Рис. 129. Схема периферической нервной си-
стемы птицы, с вентральной стороны (а) и по-
перечный разрез через спинной мозг на уровне
выходов спинномозговых нервов (Ь).
В рис. (а) левая сторона изображает соматиче-
ские первы, правая — висцеральные. IX — п.
glossopharyngeus; X—n. vagus; С., G., Н., Р.—
его ветви в область сонных артерий, кишечника,
сердца и легких; XII. 1 — язычная ветвь, XII.
2 — ветвь в область сиринкса от nervus hypoglos-
sus; г. с. — шейное утолщение; г. I. —поясничное
утолщение спинного мозга; Sy. — симпатические
ганглии первых спинных нервов. Обозначения
рис. (b). А. II. и Р. н. — передние или вен-
тральные п задние пли дорзальные рога спин-
ного мозга, с ганглиями; Sp. G. — спинной ганг-
лий дорзального корешка спинпого нерва; Sy.
G. — симпатический ганглий; R. an., R. с. и R.
р. — передняя, соединительная (или симпатиче-
ская) и задняя ветви спинного нерва. По G-adow
и Newton.
вом случае, внутрисекретор-
ными регуляторами.
8. НЕРВНАЯ СИСТЕМА И
ОРГАНЫ ЧУВСТВ
Спинной мозг. Спин-
ной мозг (medulla spinalis)
имеет в общем цилиндриче-
скую форму, с глубокими бо-
роздами на вентральной и
дорзальной сторонах (fissura
longitudinalis ventralis et dor-
salis) (рис. 129). Он помеща-
ется в позвоночном канале и
доходит до конца его, но не
образует конского хвоста
(cauda equina), как у млеко-
питающих, переходя в ните-
образный кончик.
По длине спинного мозга
встречаются два утолщения:
шейное (intumescentia cervi-
calis), откуда выходят нервы
передней конечности (крыла),
и поясничное или крестцовое
(intumescentia lumbalis sive
sacralis), снабжающее нерва-
ми заднюю конечность. Зна-
чительность этих утолщении
зависит от величины иннер-
вируемого органа; так пле-
чевое утолщение мало у стра-
уса и велико у хорошо ле-
тающих птиц. Утолщения
спинного мозга происходят
в значительной мере за счет
увеличения серого вещества
его, но особенная величина
поясничного утолщения обу-
словлена образованием здесь
149
X/ ХП XIII XIV XV XV XIV ХШ ХП XI
Рис. 130. Плечевое сплетение и нервы левого крыла сизого голубя — Columba livia Gm.
•А — с наружной, В — с внутренней поверхности крыла; XI—XV — порядковые
номера спинных нервов; а — кожный нерв; b — нерв m. supra-coracoideus; с. d. — нервы
мелких грудных мускулов; е, f. — корешки верхнего грудного мускула; д. да — передняя
ветвь нервов предплечья; да' — n. abductor digit! medii; h — соединительная ветвь между
стволами XV и XIV нервов; i — грудные нервы; h — нерв m. coracobrachialis; I. — кожный
нерв крыла; т — верхний нерв предплечья; п — нижний нерв предплечья; п'—нерв
ш. extensor hrachii; no — нерв т. deltoides major; о — нерв т. biceps; р, — нервы т. del-
toideus; q — задняя,ветвь пижнего нерва предплечья; г, г', г1—нервы передней лета-
тельной перепонки; го — n. cubitalis; s— нерв in. pronator profundus; t — задняя петвь
переднего нижнего нерва предплечья; и — нерв m. delitoideus anterior; v — задняя ветвь
верхнего нерва предплечья; W — перв пь cubito-carpalis superficial is; W — нерв m. cubito-
carpalis profundus; x — нерв m. extensor inctacarpii; у — n. radialis; ym— нерв запястья
и кисти; z — перв m. pronator superficial; 1—нерв in. extensor digiti medii; 2— нерв
adductor pollici; 3 — нерв m. abductor poliici; 4—нерв m. interosseus. Из Vocht и Yung.
глубокой впадины (sinus rhomboidalis), раздвигающей дорзальные
столбы спинного мозга и заполненной рыхлой тканью, богатой
гликогеном; последний играет важную роль в питании нервной
энергии.
Серое вещество спинного мозга лежит внутри его и в разрезе имеет
вид расправленной бабочки. Передние (вентральные) рога его (cornu
ventrale), из которых выходят моторные корешки спинных нервов,
крупнее задних рогов (cornu dorsale), представляющих чувствитель-
ные проводящие пути. Состоящие из чувствительных волокон задние
столбы спинного мозга менее развиты, чем у рептилий и млекопитаю-
щих, что объясняется быстрым проникновением этих корешков в зад-
ние рога серого вещества; только незначительное число их подни-
мается до продолговатого мозга. Эта особенность связана с значитель-
ной локализованностью (самостоятельностью) двигательного аппарата
птиц и следовательно менее глубоким координированием движений
передней и задней конечностей, чем то наблюдается у млекопитаю-
щих и рептилий. Кроме того, и периферическая чувствительность
птиц, вследствие почти сплошного перового покрова, естественно
слабее, чем это свойственно другим высшим классам позвоночных (на-
пример млекопитающим).
Сходное явление мы наблюдаем и в передних и боковых столбах
спинного мозга, где также преобладают короткие нейроны, соединяю-
щие лишь близкие друг другу сегменты. Из длинных, идущих к го-
ловному мозгу, путей наибольшее значение принадлежит tractus
spino-cerebellaris, t. spino-bulbaris и t. spino-mesenzephalis, прово-
дящих чувство осязания, боли и тепла; из путей, идущих от голов-
ного мозга, наиболее значительны два: пучок волокон, сопрово-
ждающий tr. spino-cerebellaris по медиальной его стороне и просле-
живаемый вплоть до поясничного утолщения, и пучок, идущий от
центра равновесия в продолговатом мозгу и tectum opticum и про-
бегающий в передних боковых столбах.
Спинно-мозговые нервы. Соответственно числу
позвонков 1 от спинного мозга отходят парные спинные нервы, до-
стигающие значительного числа у длинношейных птиц, например
51 пары у страуса. В своей медиальной части они состоят из двух ко-
решков: дорзального чувствительного и вентрального моторного:
перед своим слиянием и образованием смешанного нервного ствола
дорзальный корешок образует спинной ганглий (ganglion spinale).
Смешанный нервный ствол вскоре разделяется на три ветви: ramus
anterior, управляющий кожей и мускулатурой вентральной области
и конечностей, ramus posterior, идущий к дорзальным участкам
кожи и дорзальной мускулатуре, и ramus communicans, устанавли-
вающий основное соединение с симпатическим ганглием. В области
плечевого и тазового пояса спинные нервы образуют сплетения, из
которых выходят периферические нервы, направляющиеся к той
или иной конечности.
В плечевое сплетение (plexus brachialis) соединяются вентральные
(передние) ветви 4—6 последних шейных нервов (рис. 130). Из спле-
1 Превышая однако число позвонков на одну пару нервов.
142
тения выходят дорзальные и вентральные грудные нервы (nervi
pectoralis dorsales и ventrales), управляющие мышцами ребер,
грудины и ключрцы; подлопаточные нервы (nervi subscapulares),
идущие к лопатке; крыльцовый
нерв (n. axillaris), идущий к
коже и мышце, поднимающей
плечо; лучевой нерв (n. radia-
lis), управляющий разгибате-
лями локтевого сустава, лета-
тельной перепонкой и другими
мышцами; медиальный кожный
нерв (n. cutaneus medialis), и
срединный нерв (n. medianus),
самый крупный нерв крыла,
управляющий главным образом
его сгибателями.
В области таза имеют место
трисплетения: поясничное, се-
далищное и лобковое.
В поясничное сплетение (ple-
xus cruralis) соединяются обычно
3 сцинных нерва (рис. 131).
Нервы, ( выходящие отсюда,
управляют большей частью мус-
кулатуры, идущей от таза к бе-
дру и голени и коже бедрен-
ного отдела. Наиболее мощ-
ный нерв этой группы — запи-
рательный (nervus obturator),
идущий к muscuius obturatorius
и musculus pub-ischio-femora-
lis; меньше по величине бе-
дренный нерв (nervus femora-
lis), идущий как к мышцам,
так. и к коже бедра. Седалищ-
ное сплетение (plexus ischiadi-
XXIII XXV
XXIXXII XXIV XXV!
а -
д -
-и
/Г
т‘
I ис. 131. Нервы, поясничное и седалищное
сплетения левой ноги сизого голубя —
Columba livia Gm.
XXI—XXVI — последовательные номера
стволов нервов, выходящих из спинного
мозга, а — ветвь нерва XXI; Ъ— ветвь
нерва XXII; Ь' — боковой нерв, возника-
ющий из соединения веточек а и Ь; с —
кожный нерв голени; d — нерв портняж-
ного мускула (щ. iliotibialis internus, s.
sartorius); e — нерв m. ilio-tibialis; f. g. —
корешки кожного нерва g' внутренней
поверхности голени; h. — nervus ischia-
nicus; i— нерв. m. ileo-flexor; k — нерв
jn. obliquus externus и transversus ani; I— нерв m. ischio-llexor; m — нерв in. biceps ferm>
raus; mi, Tn-—веточки перва m. biceps femoralis;n — nervus ischiadico-femoralis anterior;
° —невр m. tibialis; p— поверхностный икроножный нерв; q— глубокий икроножный
нерв; г — нерв 2-го и 3-го пальцев; s — нерв 1-го пальца; t — nervus ischiadico-femoralis
jnedius; и— нерв m. gastrocnemius; v — нерв икроножного мускула; x — n. ischiadico-
Ie,n°ralis posterior; у — нерв заднего пальца. Из Vocht и Yung.
143
Чейз) составляется 5—6 нервами; из него выходит самый крупны]
нерв тела — седалищный (nervus ischiadicus), управляющий глав
ным образом мускулатурой и кожей нижних частей конечности, о*
бедра к периферии. Основными его ветвями являются малоберцо
вып (n. fibularis) и большеберцовый (n. tibialis) нервы.
Рис. 132. Головной мозг птиц в сравнении с мозгом рептилий и млекопитающих.
7, 2, 3 — схематический продольный разрез через мозг рептилии (1), птицы (2) и
млекопитающего (3), с одинаковой штриховкой гомологичных частей мозга; 4, 5, 6 го-
ловной мозг сбоку аллигатора (4), голубя (5) и собаки (в); 7, 8 — задняя часть мозга девя-
тидневного эмбриона курицы, в разрезе и сбоку. Обратите внимание на большее общее сход-
ство мозга птицы с мозгом рептилий, чем с мозгом млекопитающих. Мозг эмбриона птиц за-
нимает промежуточное положение по отношению мозга рептилий, что особенно выражено
в небольшой величине мозжечка, ясно подразделенного на те же переднюю (а), среднюю
(т) и заднюю (р) доли, что и мозг рептилий (4).}
С. — большой мозг; СЬ. — мозжечек (а — передняя, тп — средняя и р — задняя доли);
/ — lobus flocculus; G.—полосатые тела или базальные ганглии (corpus striatum);
М.—продолговатый мозг; ol.—обонятельные доли, lobi olfactorii (в 1 — отрезанные);
ор. — зрительные бугры(thalami optici); Р. —pallium (мантияили кора большого мозга); рд-^
теменной глаз(у птиц и млекопитающих — glandula pineal is). Из Hellmann.
Лобковое сплетение (plexus pudendus) обычно развито слабой
слагается четырьмя вентральными ветвями. Нервы, выходящие
из него, с наиболее значительным хвостовым нервом (nervus cauda-
lis) иннервируют область кожи и мускулатуры хвоста и клоаки.
Головной мозг. Головной мозг птиц выделяется своей
значительной величиной в сравнении с головным мозгом, рептилий
того же роста (рис. 132). Особенной величины достигают полушария
144
переднего мозга, обычно совершенно прикрывающие сверху средний
мозг, и мозжечок, состоящий главным образом из червеобразного
отдела (vermis). Более значительная дифференцировка мозга птиц,
в сравнении с рептилиями, обусловливает более высокое развитие
большинства органов чувств и более высокий общий психический
уровень.
Увеличение размеров мозга весьма заметно и в эволюции класса
птиц, например при сравнении ископаемых и сходных с ними видов
из ныне живущих форм (рис. 133).
Головной мозг разделяется на отделы: передний, промежуточный,
средний, малый и продолговатый мозг (рис. 134).
~ г. Передний или большой мозг (cerebrum)
ереднии моз
является самым круп-
ным отделом и, подобно
большому мозгу других
позвоночных, глубоким
продольным расщепом
делится на два полу-
шария. Поверхности по-
лушарий почти лишены
борозд, гладки, серо-
красноватого цвета.
Форма полушарий, бо-
лее вытянутая у попу-
гаев п дятлов, в боль-
шинстве случаев являет-
ся шарообразной (рис,
135). В строении полу-
шарий мы находим мно-
го общих черт с строе-
нием мозга, наблюдае-
мым у пресмыкающихся.
Заслуживает быть отме-
ченным очень слабое
развитие мозолистого
тела (corpus calosum),
Рис. 133. Увеличение размеров черепной полости у
примитивных и прогрессивных видов птиц, экологи-
чески и морфологически сходных между собой.
Коптур черепа и мозговой полости: Л — Hesperornis
regalis Marsh; В — полярной гагары — Colymbus immer
Briinn.; С — Ichthyornis victor Marsh; D — пестроносой
крачки — Sterna sandvicensis Lath. По Nopcsa и Marsh,
из Lambrecht.
соединяющего оба полушария. Большая величина полушарий обу-
словлена главным образом мощным развитием полосатых тел
(corpus striatum), простирающихся в своей передней части на всю
латеральную поверхность полушарий. Наоборот, pallium (кора)
головного мозга оказывается развитой чрезвычайно слабо, на боль-
шей части своего протяжения являясь в сущности очень тонкой
оболочкой, одевающей полосатое тело: более значительно она
развита только в медио-дорзальной части полушария, срастаясь
с полосатым телом по линии слабой, параллельной оси полушария, бо-
розды — vallecula (рис. 136). Этот участок характеризуется крупными
ганглиями и может рассматриваться функционирующим как кора
мозга высших позвоночных. Из опытов однако следует, что ассоциа-
тивные функции коры могут выполняться у птиц и клетками мощно
развитого полосатого тела, что повидимому и обусловливает воз-
10 Орнитологии—1209
145
можность высокой психики у некоторых групп птиц. Затем особен-
ностью больших полушарий птиц является отсутствие аммониевых
рогов и их коммиссуры (psalterium), стоящее в связи с слабым раз-
витием обонятельного аппарата у птиц; вследствие этого и обонятель-
VH. — передний мозг; МП. — средний
мозг; НН. — малый мозг (мозжечок);
NIJ.—продолговатый мозг; Med. — на-
чало спиппого мозга; L. ol. — обонятель-
ные доли; Tr. opt. — область зрительного
бугра; Inf. —воронка нижнего мозгового
отростка; Нур. — пижпий мозговой отро-
сток — hypophysis; I—XII — черепные
нервы. По Wiedersheim, из Маршаль.
ные лопасти (lobi olfactorii) у птиц
очень малы, имеют вид небольших
яйцеобразных вздутий и часто ле-
жат на вентральной стороне полу-
шарий.
Как объем, так и детали строе-
ния больших полушарий обнару-
живают значительные изменения в
разных систематических группах,
обусловливая разный психический
уровень птиц этих групп. Наибо-
лее высоко развитым является мозг
попугаев, дятлов и воробьиных
птиц, слабее всего — мозг куриных
птиц и голубей. Различия в строе-
нии головного мозга голубя п
гуся приравниваются Эдингером
(Edinger, 1903) к различиям моз-
га кролика и собаки, а различия
между мозгом голубяи попугая —
даже собаки и обезьяны.
П р о м е ж у т о ч н ы й м о з г.
Промежуточный мозг у птиц яв-
ляется весьма укороченным от-
делом, почти целиком переместив-
шимся под большие полушария.
На нем сидят в виде возвышений
зрительные бугры (thalami optici).
слабо развитая воронка (infun-
dibulum), мозговой придаток (hy-
pophysis cerebri) и различно выра-
женная шишковидная железа (gran-
dula pinealis sive epiphysis ce-
rebri). Третий мозговой желудочек,
расположенный под зрительными
буграми, очень мал, но так же.
как у млекопитающих, делится на
отделы: дорзальный и вентральный. На ранних эмбриональных ста-
диях перед шишковидной железой закладывается париетальный ор-
ган, а в эпителии кожи над ним замечается накопление пигмента.
соответствующее теменному пятну многих рептилий; но через корот-
кое время оба эти образования исчезают. Другой особенностью,
по сравнению с мозгом рептилий, является образование ядер под
зрительными буграми в вентральной части thalamus, осуществляю-
щих передачу чувствительных возбуждений от спинного и продол-
говатого мозга к большим полушариям.
146
Средний мозг. Подобно промежуточному, средний мозг
также сдвинут на вентральную сторону и прикрыт сверху большими
полушариями. Вследствие этого объемистые tectum opticum придви-
нуты вперед, а образуемые ими по бокам крупные зрительные доли
(lobi optici) или двухолмия (corpora bigemina) передвигаются на внеш-
ние стенки промежуточного мозга и сливаются с ними. Значительное
развитие этого отдела обязано большой величине глаз птиц и поэтому
богатому развитию серого вещества, создающих мощный ассоциа-
тивный зрительный аппарат. Полость каждой зрительной доли соеди-
няется с так называемым водопроводом мозга (aquaeductus cerebri).'
ведущим из третьего в че-
твертый мозговой желудо-
чек.
Малый моз г. Ма-
лый мозг или мозжечок у
птиц очень объемист и об-
ладает очень утолщенными
стенками, так что принад-
лежащий ему четвертый
желудочек оказывается
очень суженным. В моз-
жечке различают крупное
тело (corpus), соответству-
ющее червячку (vermis)
млекопитающих, и малень-
кие боковые придатки —
боковые доли (flocculi). В
продольном разрезе моз-
жечка обнаруживается
своеобразный ветвящийся
рисунок (древо жизни —
arbor vitae), указывающий
на складчатое его строе-
ние. Тело мозжечка со-
стоит преимущественно из
так называемых спииоце-
ребеллярных волокон и образует главный орган статотонуса, т. е.
обеспечивает взаимную координацию и необходимое напряжение
мускулов, постоянно изменяющееся в зависимости от изменений
положения тела в отношении к проявляющимся в нем явлениям
тяготения.
Продолговатый мозг. Продолговатый мозг (medulla
oblongata) без резких переходов соединяется со спинным мозгом,
отделяясь от него лишь углом своего изгиба. Сверху он имеет углу-
бление — ромбоидную ямку (fossa rhomboidea), прикрытую мозжеч-
ком и являющуюся четвертым мозговым желудочком. Из продолго-
ватого мозга выходит основная масса черепных нервов, несомненно
являющихся спинно-мозговыми по своему происхождению.
Черепные н е р в ы. У птиц имеется двенадцать пар череп-
ных нервов; из них три пары (III, VI, XII) покидают продолговатый
Рис. 135. Головной мозг представителей разных
отрядов.
а — черной воропы — Corvus corone L. (Passerifor-
mes) ; b — широкохвостого лори — Lorius garrulus
L. (Psittaciformes) ; с — зеленого дятла — Picas
viridis L. (Piciformes); 1 — сверху; 2 — снизу;
3— сбоку. По Kiicnzi, из SLrcsemann.'
147
мозг вентральными корешками, являются двигательными и иннер-
вируют соматическую мускулатуру; две пары (I и 11) покидают
мозг из переднего и промежуточного его отделов и являются всецело
чувствительными нервами; и семь остальных пар выходят из продол-
говатого мозга дорзальными корешками и являются нервами пре-
имущественно сме-
шанного характера
(рис. 137 и 134).
1. Обонятельный
чувствительный нерв
(nervus olfactorius)
выходит из соответ-
ствующей обонятель-
ной доли переднего
мозга, в виде много-
численных, рыхло
друг с другом соеди-
ненных нитей, прохо-
дит в дорзомедиаль-
ную часть глазницы
и в носовую полость,
где и разветвляется в
слизистой оболочке
носовой перегородки
и дорзальной рако-
вины.
Рис. 136. Строение мантии (pallium) больших полушарий
птиц в сравнении с строением мантии рептилий и коры
млекопитающих.
7, 2, 3 — схематический поперечный разрез через мантию
больших полушарий рептилий (1), птиц (2) и млекопитаю-
щих (3); 4,5— схема нервных путей с пирамидальными
клетками в мантии птицы (4) и в коре млекопитающего
(5); 6,7 —схематический разрез через полушарие птицы
(6) и млекопитающего (7), показывающий проводящие
пути. Обратите внимание па резкое различие строения ман-
тии птиц и рептилий от строения коры млекопитающих
(у последних два мощных слоя с пирамидальными клетками,
разделенных средним слоем; у птиц пирамидальные клетки
лишь начинают появляться), на менее сложные проводя-
щие пути (4), на ограничение латеральных проводящих пуч-
ков лишь полосатым телом, в то время как у млекопитаю-
щих они возникают в коре мозга; cl. —корковый слой с
пирамидальными клетками; G. — полосатые тела (базальные
ганглии); L. — обонятельные доли; М — жировой слой;
ML — средний слой; т.—membrana, ограничивающая
мозговой желудочек; Рь Р2—два слоя пирамидальных
клеток; Т. — волокнистые проводящие пути; V.—желу-
дочки большого мозга. Из Hellmann.
II. Зрительный
чувствительный нерв
(n. opticus) начина-
ется от основания
промежуточного моз-
га и после перекреста
с другим одноимен-
ным нервом входит в
глаз, где распределя-
ется главным образом
в сетчатке.
III. Глазодвига-
тельный нерв (п. оси-
lomotorius) начинает-
ся вентрально и сзади
ножки гипофиза, вхо-
дит в глазницу зрительным отверстием (foramen opticum) и иннерви-
рует глазные мышцы’: m. rectus superior, т. rectus inferior, т. rectus
intermedins и m. obliquus inferior. Кроме того, он отдает короткую
ветвь к ресничному узлу (ganglion ciliare) и иннервирует Гардерову
железу.
IV. Блоковидный двигательный нерв (n. trochlearis) выходит
у аборального края зрительных долей, входит в глазницу вместе
с предыдущим нервом, иннервирует m. obliquus superior глаза, соеди-
148
няясь кроме того веточкой с ramus ophthalmicus тройничного
нерва.
V. Тройничный смешанный нерв (n. trigeminus) выходит двумя,
корешками, чувствительным и двигательным, из передней латераль-
ной части продолговатого мозга. Чувствительный корешок образует
полулунный или Гассеров ганглий (ganglion semilunare sive Gasseri)
п делится на ветви: глазничную (ramus ophthalmicus), верхнече-
Рис. 137. Черепные нервы левой стороны сизого голубя — Columba
livia Grin. I, — n. olfactorius; IV — n. trochlearis; V — n. trigeminus;
VII— n. facialis; VIII — n. acusticus; IX—n. glossopharyngeus;
X — n. vagus; XII — n. hypoglossus; 1,2 — первый и второй шейные
нервы; а — соединительная ветвь между n. hypoglossus и n. vagus; а —
верхнечелюстная ветвь n. trigeminus; b — нерв m. cucullaris; b' —
нерв т. temporalis; с — соединительная ветвь между n. vagus и'
первым шейным нервом; с' — n. post-orbital is; d — глазничная ветвь п.
trigeminus; е,/ — веточки этой ветви; д — Гассеровы узлы; h — ветвь,
идущая к пищеводу; i — соединительная ветвь между Гассеровым
узлом и n. facialis: h — верхняя веточка глазничной ветви; I — неб-
ная ветвь n. trigeminus; тп — соединительная ветвь между n. trigeminus
и n. facialis; п — нерв mm. mylo- и stylohyoideus; о — задняя ветвь
n. facialis; р' — нерв кожных мускулов шеи; q' — соединительная ветвь
между n. facialis и четвертым шейным нервом; г — нерв носовых отвер-
стий; s — ганглии п. glossopharyngeus; t, и — передняя и вторая ветви
этого нерва; v — соединительная ветвь с n. glossopharyngeus; х —
язычная ветвь двенадцатого нерва; у — ветвь нерва к глотке; z—
нижнечелюстная ветвь и. trigeminus. Из Vogt и Yung.
люстную (ramus maxillaris) и нижнечелюстную (ramus mandibularis).
Двигательный корешок присоединяется почти целиком к нижнечелю-
стной ветви, отдавая тоненький нерв и к глазничной веточке.
Глазничная, вероятно всецело чувствительная, ветвь, отдавая
веточку к ресничному нерву (n. ciliaris) и веточкам лобного (n. fron-
talis) и носового (n. nasalis) нервов, распределяется в носовой области,
в передней части клюва и слизистой оболочке верхней челюсти.
Верхнечелюстная чувствительная ветвь снабжает своими веточ-
ками слезную железу, конъюктиву, глазные веки, кожу в области
уздечки и значительную часть клюва.
149
Нижнечелюстная смешанная ветвь управляет musculus constrictor
dorsalis и m. adductor mandibulae и снабжает чувствительными во-
локнами слюнные железы и покровы нижней челюсти.
VI. Отводящий двигательный нерв (n. abducens) начинается от
^вентральной поверхности продолговатого мозга, проходит через
канал клиновидной кости в глазницу и разветвляется в латеральной
прямой мышце (m. rectus externus) и обеих мышцах мигательной пере-
понки (m. pyramidalis и т. quadratus).
VII. Лицевой смешанный нерв (n. facialis) начинается на лате-
ральной стороне продолговатого мозга, образует у основания ганглии
(ganglion geniculatum) и разделяется на тонкую чувствительную
переднюю (ramus anterior sive palatinus) и толстую, главным образом
двигательную, заднюю (г. posterior sive hyomandibularis) ветви.
Передняя ветвь обменивается волокнами с языкоглоточными и симпа-
тическим нервами и затем снабжает своими веточками носовую по-
лость глазницы и заднюю часть неба. Задняя ветвь управляет неко-
торыми мышцами нижней челюсти и подъязычной кости (m. dep-
ressor mandibulae, m. mylohyoideus и др.) и снабжает веточками
околоушную область. Далее она соединяется с рядом других нервов
и образует нерв — барабанную струну (chorda tympani) и др.
VIII. Слуховой чувствительный нерв (n. octaVus sive acusticus)
выходит двумя корешками из продолговатого мозга и вместе с лице-
вым нервом проникает к meatus acusticus internus. Две ветви его:
nervus vestibularis и n. cochlearis оканчиваются мелкими веточками
на разных частях слухового аппарата (внутреннего уха).
IX. Языкоглоточный смешанный нерв (n. glossopharyngeus) начи-
нается вместе с блуждающим первом в продолговатом мозгу, образует
вскоре затем подряд два ганглия (ganglion jugulare и g. petrosum),
обменивается волокнами с лицевым и блуждающим нервами и делится
на две ветви: язычную (г. lingularis) и глоточную (г. pharyngeus).
Первая иннервирует язык и является основным вкусовым нервом птиц
и кроме того снабжает нервами m. kerato-mandibularis, глотку и
вход в гортань. Глоточная же ветвь снабжает нервными веточками
пищевод и область горловой щели.
X и XI. Блуждающий смешанный нерв (n. vagus) и прибавочный
двигательный нерв (n. accessorius) начинаются у птиц вблизи друг
от друга на латеральной поверхности продолговатого мозга, а после
выхода из черепа через яремное отверстие (foramen jugulare) сли-
ваются друг с другом. После этого ими образуется узел (ganglion
radicis vagi), отсылается ветвь (ramus accessorius externus) к кожной
мускулатуре шеи (m. cuCullaris) и коммуникационные веточки к язы-
коглоточному и подъязычному нервам. Затем вдоль сонной артерии
нерв спускается к нижней гортани и образует здесь крупный узел
(ganglion thoracicum sive nodosum). Вскоре после этого от него отхо-
дят веточки к щитовидной железе, сердцу, бронхам, желудочным
веткам симпатического нерва и крупная возвратная ветвь—ramus
recurrens, поднимающаяся вдоль пищевода и трахеи и распределяю-
щаяся веточками на их нижних стенках.
XII. Подъязычный двигательный нерв (n. hypoglossus) выходит
в вентральной задней части продолговатого мозга несколькими кореш-
150
ками, получает веточки от первого шейного и блуждающего нервов
л разделяется на две ветви — идущую к языку ramus lingual is и
идущую к нижней гортани г. laryngeus superior. Язычная ветвь упра-
вляет мышцами языка (musculi hyoglossi, m. genioglossus и др.),
по имеет и чувствительные волокна, идущие к вкусовым тельцам
кончика языка. Гортанная ветвь снабжает нервами язычно-грудную
мускулатуру (m. laryngo-liyoideus, т. sterno laryngeus, т. tracheo-
hyoideus) и мускулатуру нижней гортани.
Автономный нервный комплекс. Автономный
нервный комплекс у птиц слагается из симпатической и парасимпа-
тической систем. Симпатическая система образуется: 1) спинномозго-
вым центром, ввиде висцеромоторных столбов, лежащих в боковых
частях серого вещества спинного мозга, между шейным и крестцовым
Рис. 138. Симпатическая нервная система в области спины сизого голубя — Columba Н-
via Gm.
а — ганглии спинномозговых нервов; b — позвоночный столб; с — обрезанное ребро;
d — вентральные стволы п. sympathicus; е—дорзальный ствол n. sympathicus; / — дор-
зальный корешок спинномозгового нерва; </, I — два последних ствола плечевого нервного
сплетения; h — сердечный нерв; k — нерв внутренностей. Из Focht и Yung.
его утолщениями; 2) пограничными стволами (nervus sympaticus
vertebralis), идущими по бокам вдоль всего позвоночника и состоя-
щими из симпатических ганглиев и нервных волокон, соединяющих
их между собой (rami internodiales) и с спинномозговым центром
(rami communicans, рис. 138; 3) симпатическими волокнами, распре-
деляющимися среди внутренних органов, где местами они образуют
ганглии и сплетения (например plexus coeliacus, pl. solarus и др.).
Оба пограничных ствола соединяются друг с другом в хвостовой
области, краниально же продолжаются до симпатических ганглиев
(ganglion cervicale supremum) задних отделов продолговатого мозга,
откуда отходит вперед, вдоль сонных артерий, пара сонных нервов
(nervus carotis), снабжающих симпатическими нервами головной
отдел туловища.
В строении пограничных стволов и симпатических сплетений боль-
шое участие принимают так называемые хромофильные клетки мезо-
дермального происхождения, продуцирующие адреналин и близкие
°му гормоны. Из соединения симпатических ганглиев с тканью
151
из хромофильных клеток образуются параганглии, имеющие зна-
чение желез внутренней секреции, существенных как для самой
симпатической системы, так и для иннервируемых ею органов.
Первенствующее значение в этом отношении принадлежит надпо-
чечникам и затем каротидной железе (paraganglion caroticum), рас-
положенной у птиц вблизи ganglion nodosum и паратироидных желез;
мелкие параганглии, кроме того, рассеяны в области дуги аорты,
сердца, кишечного сплетения, шейного отдела пограничных стволов
и некоторых других органов.
Строение парасимпатической системы у птиц выяснено относи-
тельно детально только в крестцовой области. Здесь спинномозговые
центры представлены крестцовыми столбами (nucleus paracentralis
sacralis), занимающими в сером веществе спинного мозга такое же
положение, как висцеромоторные столбы, и от которых они отделены
ромбоидальной ямкой. Предположительно эта система отсылает
сосудорасширяющие волокна в область крестца и конечностей
и является таким образом антагонистом симпатической системы.
В краниальной области ее деятельность распространяется, кроме
того, и на слезные и слюнные железы.
ОРГАНЫ ЧУВСТВ
Осязательные тельца. Кожа птиц очень богата разно-
образными нервными окончаниями, особенно на оголенных от перьев
поверхностях. Простейшими из них являются интраэпителиальные
образования; нервные окончания проникают в мальпигиев слой,
ветвятся здесь и заканчиваются маленькими концевыми дисками,
тесно прилегающими к эпителиальным клеткам. Предположительная
функция их заключается в восприятии болевых и температурных
ощущений. Собственно осязательные тельца у птиц лежат под эпидер-
мисом, в соединительной ткани, и представлены главным образом
тремя типами образований: тельцами Меркеля, Грандри и Гербста
(рис. 139). В первых двух типах одни нервные окончания образуют
диск, прилегающий к крупным чувствительным клеткам, а сопут-
ствующие им свободные окончания образуют фибриллярную сеть.
Эти аппараты располагаются иногда группами и то свободно лежат
в кожной соединительной ткани (тельца Меркеля), то окружены
соединительно-тканными оболочками (тельца Грандри). Последние
найдены только в ротовой полости гусей и сов, тельца же Меркеля
широко распространены в ротовой полости самых разнообразных
птиц.
Тельца Гербста являются окончаниями безмякотных, симпати-
ческих нервных волокон, окруженных концентрически расположен-
ными соединительно-тканными оболочками-пластинками, и имеют
очень сложную структуру. Они обычно располагаются в глубоких
слоях кожи и часто в оболочках внутренних органов. В особенно боль-
шом количестве они наблюдаются у основания осязательных щети-
нок, на языке (дятлы) и клюве (гусиные) и т. п. (рис. 140).
Вкусовые органы. Вкусовые органы встречаются у птиц
в виде особых вкусовых почек, помещающихся главным образом
152
на мягком небе, в основании языка и по краям горловой щели
(рис. 141). Они состоят из групп веретенообразных, опорных и невро-
эпителиальных клеток, окруженных несколькими слоями эпители-
альных клеток и в общем имеющих форму груши. Устья их, с кон-
цевыми окончаниями вкусовых
клеток, обычно впадают в мел-
кие вкусовые ямки, на слизи-
стой поверхности ротовых обо-
лочек. Как выяснено экспери-
ментальными наблюдениями,
птицами ощущаются все четыре
вкусовых качества: соленый,
кислый, горький и сладкий; из
них последний всегда приятен
птице, а ощущение горького
вкуса иногда бывает понижено,
особенно у зерноядных, веро-
ятно вследствие привыкания к
обычно горьчащей пище.
Органы, обоняния.
Органы обоняния у большин-
ства птиц не играют большой
функциональной роли и в связи
с этим являются сравнительно
слабо развитыми. В носовой
полости можно различать пред-
дверие (vestibulum) и обонятель-
ную камеру, от латеральных
стенок которых внутрь полости
выдаются три носовые рако-
вины: верхняя (postconcha),
средняя (concha) и передняя
(praeconcha) (рис. 142). Верх-
няя, лежащая дорзально, рако-
вина имеет обычно форму бугра
и гомологична nasotur binale
млекопитающих. Она является
единственной раковиной у боль-
шинства птиц, покрытой обоня-
тельным эпителием. У некото-
рых видов она отсутствует, у
других, например у глупыша
{Fulmarus glacialis), она очень
Рпс. 139. Чувствительные тельца Грандри и
Гербста в клюве кряквы—Anas platyrhyn-
chos L.
1 — разрез через кожистый покров клюва:
сверху и впизу тельца Гербста; между ними
мелкие тельца Грандри; 2 — тельце Грандри
в разрезе, прилежащее непосредственно к
эпителию fepj; 3 — сильно увеличенный
продольный разрез тельца Грандри.
ер. — эпителий; k. — ядро по ходу топкого
безмякотного нервного волокна; nit. — чув-
ствительные диски в тельцах Грандри; пт.—
толстые мякотные нервные волокна, являю-
щиеся осевым волокном в тельцах Гербста и
образующие чувствительные диски в тельцах:
Грандри; ns — тонкие безмякотные нервные
волокна. По Botezat и Szymonowicz, из Str<-
semann.
сильно развита, завертываясь
в несколько оборотов. Средняя раковина является наибольшей по
размерам и обычно образует несколько заворотов; она гомологична
единственной раковине рептилий и основной носовой раковине
млекопитающих (maxilloturbinale), но у птиц, в огромном боль-
шинстве случаев, лишена обонятельного эпителия, как и передняя
раковина, будучи покрыта обычным респираторным эпителием
153
(рис. 143). Последняя — передняя раковина встречается только
у птиц и основной функцией имеет видимо защиту от проник-
Рпс. 140. Продольный разрез через конец клюва вальдшнепа — Scolopax rus-
ticola L. с многочисленными чувствительными клетками.
а — кость; Ъ — эпидермис; с — чувствительные клетки в ямках костной
ткани. По Leydig, из Stresernann.
новения внутрь носовой полости посторонних тел; она свешивается
с латеральной стенки непосредственно кзади от наружного носового
-отверстия. В области средней раковины дно обонятельной камеры
^сообщается проходом — хоанами с верхним отделом полости рта.
<6/
Рис. 141. Схема .двух типов вкусовых почек, наблюдающихся у птиц.
а — наиболее распространенный тип (исключая Anseri. formes, Phoenicople-
riformes и Psi Had formes); b — тип, характерный для отрядов An serif ormes и
Phoenicopteri formes.
Hz — клетки оболочки; Kg. — почечные ямки; Nz. — нервно-эпителиальпые
клетки; Р.—вкусовые поры; Slz.—опорные клетки. По Bath, из Stresernann.
454
С деятельностью носовой полости тесно связана носовая железа,
выделяющая водянистый секрет и увлажняющая чувствительный
эпителий. Эта железа весьма изменчива в размерах у разных групп
а — задняя подушка языка; ent. — os entoglossum, G. F.—орбитальная складка;
Ьаг. — верхняя гортань; М. М. — средняя раковина; N. — просвет через левое носовое от-
верстие; Ph. F.—ряд верхних сосочков глотки; R.W.—верхняя раковина (или обоня-
тельный холмик); V. М.—передняя раковина (или раковина преддверия). По Goppert,
из Streseinann.
птиц. При сильном развитии она помещается над глазами, на краю
лобных костей и у некоторых групп (гуси, дневные хищники, многие
кулики) является двупарным органом. При слабом развитии
(кукушка, козодой) она умещается
в sinus orbitalis или может совсем
отсутствовать (голубь, вальдшнеп,
сизоворонка и т. д.). Наиболее
Р,
1ис. 143. Поперечный раз-
рез через клюв кряквы —
Anas platyrhyncha L., в об-
ласти средней носовой рако-
вины.
— хоаны; с. т. — завер-
нутая спиралью средняя но-
совая раковина; Jug. —ску-
ловая кость; п — носовая
кость; pal. — небная кость;
s. п. —носовая перегородка;
Vo — веточки тройнич-
ного нерва. По Gadow.
Рис. 144. Схема носовой камеры у олу-
ши — Su la bassana L.
ch. — носовые отверстия в полость рта,
хоаны; с. ol.—редуцированная исчезнув-
шая часть носовой полости, отмеченная
пунктиром; n. ol. — обонятельный нерв—
п. olfactorius, разветвляющийся лишь в
области единственной носовой раковины,
стоящей в широком сообщении с полостью
рта; наружные носовые отверстия пол-
ностью заросли. По Gadow.
мощно’ она развита у морских ныряющих птиц (гаги, ряд нырковых
уток, чистики и др.), где секрет ее очевидно защищает оболочки от
раздражающего влияния морских солей.
155
Наружные носовые отверстия имеют у птиц различную форму и
значительное систематическое значение. Очень часто они прикры-
ваются сверху крышечкой (operculum) или густыми щетинками, защи-
щающими от проникновения посторонних веществ. У ряда водных
птиц эти отверстия совершенно зарастают (баклан, олуша), передние
носовые раковины редуцируются, и сообщение с носовой полостью
происходит только через хоаны (рис. 144).
Слабое развитие органов обоняния у птиц удовлетворительно
объясняется высоким удельным весом тех веществ, которые вызы-
вают раздражение обонятельного эпителия и следовательно длитель-
но задерживаются лишь на почве и в призем-
ных слоях воздуха. Для большинства птиц
очевидно они не могут иметь существенного
значения. Однако у немногих наземных птиц
(гусиные, кулики, киви) и у некоторых ле-
тающих групп (трубконосые, некоторые днев-
ные хищники) обоняние развито довольно
высоко, как это можно судить по объему обо-
нятельного нерва и развитию раковин и
экспериментальному привлечению на запах
американских грифов и буревестников
(рис. 145). Исключительно высокое развитие
обонятельного аппарата свойственно киви.
У хищников, сов и олуш обонятельные ощу-
щения являются дополнительными к вкусо-
вым и воспринимаются целиком или, в основ-
ном, через хоаны. Загадочным является
строение носовой полости у куриных. В на-
ружные носовые отверстия здесь открываются
два прохода, из которых передний ведет в обо-
нятельную камеру, а задний в слепую, вы-
стланную слизистой оболочкой полость, неяс-
ного значения. Плате (Plate) предполагает,
что этот орган служит для определения на-
правления ветра.
Орган слуха. Орган слуха хорошо
развит у всех птиц и по своему строению
близок к строению уха у высших рептилий
(крокодилов) и в меньшей степени млекопитающих (рис. 146). На-
ружная ушная раковина у птиц отсутствует, наружный слуховой
проход короток и барабанная перепонка обычно видна снаружи.
С поверхности проход прикрывается модифицированными короткими
перышками — кроющими ухо, и у некоторых видов (голуби, кури-
ные) отчасти небольшой поперечно поставленной’кожной складкой.
У сов наружный слуховой проход очень* объемист и сопровождается
большими кожными складками, спереди и сзади функционально
заменяющими подвижную наружную раковину.
Барабанная перепонка (membrana tympani) прикрепляется непо-
средственно к окружающим костям и отграничивает снаружи объеми-
стую полость среднего уха (рис. 147). Медиально и вентрально эта
Рис. 145. Мозг п носовые
камеры глупыша — Fu Ima-
ms glacialis L.
Правая носовая камера
вскрыта сверху; с. а. — ра-
ковина преддверия; с. та. —
средняя раковина; с. р.—
верхняя раковина, вскрытая
сверху — d, чтобы показать
ее многократную заверну-
тость; I. о. —мощно разви-
тые lobi olfactorii.no Strong,
из Stresemann.
156
1 и< • 14G. Строение уха птиц в сравнении с строением уха рептилий и млекопитающих.
ухо Рептилий; 2—птицы; 3 — млекопитающего. Слуховые косточки эмбрионов;
гтоорпиТерии* 5 — гоаципа; 6 — пустельги; 7 —лошади. Обратите внимание па сходство
одна о 1 уха ПТПЦ п Рептилий и отличие от млекопитающих. У птиц "окостеневает лишь
и CTnp$»Umella’ у млекопитающих же слуховые косточки состоят из молоточка, наковальни
1<|цепаМеЧка’ пР0Исшедших из articulare, quadratum и костной columella рептилий и птиц;
__аптиц соответствует улитке — cochlea млекопитающих;?).—полукружные каналы;
лоточрг?. о\е11а; Е-—extracolumella; I.—наковальня; I.—lagena;L.—лабиринт; М.—мо-
.$’// _ ,Л* —верхний, Pl. —нижний отросток extracolumella; Q.—quadratum; s.— sacculus;
cochlea; St. — стремя;?.—барабанная перепонка; и.—utriculus; У.»—остаток второй
'Висцеральной дуги; W — слуховой-проход.
полость узкой евстахиевой трубой (tuba Eustachii) соединяется с рото-
вой областью, а кпереди тонкой трубочкой (siphonium) соединяется
с воздушными полостями нижней челюсти (через processum mandi-
bularis internus), в обоих случаях являясь воздушным буфером для
колебаний барабанной перепонки. Перенос этих колебаний на внут-
репнее ухо осуществляется тоненьким костно-хрящевым образова-
нием — columella auris, по происхождению являющимся hyomandi-
bulare п stylohyale второй гиоидной дуги рыб. Внутренняя пластинка
этого образования — stapes
захватывает овальное окно
Рпс. 147 Схематический разрез через область
среднего уха, лабиринт и наружный слуховой
проход курицы — Gallus galbus L.
bas. — os basisphenoidale; C. —улитка — cochlea;
C. t. — слуховая полость; F. c. —fenestra rotunda;
F. v.—fenestra ovalis; L. K. — костяная кап-
сула лабиринта; L.—Hgamentum platneri; Lag.—
lagena; M. t. t. — m. tensor tympani.; Pr. ex.—
processus extracoluinellaris; Pr. i. — processus
infracolumellaris; Pr. s.— processus supracolumel-
laris; Pt.— перья, прикрывающие наружный
слуховой проход; R. — recessus stapedialis cavi
tympani; S.—sacculus; Sq.—os squamosum;
S. t. — columella; T. — барабанная перепонка —
tympanicum; T. E. — tuba Eustachii; U. —utri-
culus. По Plate, из Stresemann.
(fenestra ovalis), ведущее в на-
полненное перилимфой про-
странство внутреннего уха, а
центральная часть, имеющая
вид палочки (extracolumella),
соединяет stapes с барабан-
ной перепонкой. С послед-
ней она приходит в сопри-
косновение тремя отростками
(proc, extracoluinellaris, sup-
racolumellaris и infracolumel-
Jaris), от которых к стенкам
полости идут волокна muscu-
Ius tensor tympani, поддер-
живающие также и натяну-
тость барабанной перепонки.
Внутреннее ухо образу-
ется тремя полукружными
каналами с тремя ампулами,
вестибулярной частью, состо-
ящей из sacculus и utriculus,
и улиткой или pars basilaris
lagenae (рис. 148). С этим от-
делом, через fenestra ovalis.
приходит в соприкосновение
stapes среднего уха. Pars ba-
silaris lagena заключена в
костяную капсулу, напол-
ненную перилимфой; наиболее существенной частью ее является
натянутая в хрящевой рамке membrana basilaris, дифференци-
рованность которой определяет остроту и способность разли-
чать тона звука. Верхние слои мембраны покрываются papilla
acustica basilaris, несущей слуховые волоскп, сливающиеся выше
в membrana tectoria, гомологичную кортиевой мембране млекопи-
тающих. Все эти клеточные слои обращены внутрь ductus cochlearis.
наполненного эндолимфой, который тонким каналом — canalis reu-
nions соединяется с sacculus вестибулярной части. Как уже ука-
зывалось, слух развит у всех птиц, достигая особенной остроты у сов.
Для ряда видов последних (Strix, Asio, Aegolius) характерна би-
латеральная асимметрия в развитии наружного уха, вызванная
168
очевидно постоянным стремлением к локализации источника звукам
и отсюда более значительного функционального развития одного уха.
по сравнению с другим. Острота слуха многих птиц превышает'
остроту человеческого слуха, но объем воспринимаемых тонов и;
дифференциация их значительно ниже у птиц, чем у человека и
многих млекопитающих. Это выражается, например, в строении mem-
brana basilaris, число волокон которой у попугаев составляет всего
около 1200, в то время как у человека оно равно 13 500—24 000.
Органы зрения. Глаз птиц представляет собой орган.
А В
Рис. 148. Кожистый лабиринт сизого голубя — Со-
lumba livia Gm. А — с медиальной стороны; В — с
латеральной стороны.
аа—передняя ампулла; ае — наружная ампулла;
ар — задняя ампулла; са—передний полукружный
канал; се— наружный полукружный канал; ср —
задний полукружный канал; de — ductus endolym-
phaticus; g— нерв лагепы; I — слуховая бородавка
лагепы; ms — нервная подушка sacculus; нш — слу-
ховое пятпо — recessus utriculi; п — вход слухового
нерва; о — отолиты; та — слуховые волокна перед-
ней ампуллы; гр — слуховые волокна задней ампул-,
лы; s — sacculus; su—верхний синус; sup — задний
синус; I — лагена—lagena; и — utriculus. Ио Ret-
zius. из Vocht и Yung.
весьма совершенного стро-
ения. Только у немногих
видов этого класса органы
зрения имеют малосуще-
ственное значение в жизни
животного (например ки-
ви), являясь у огромного
большинства птиц наибо-
лее важным и совершен-
ным органом чувств вооб-
ще.
По своему строению
глаз птиц весьма сходен
с строением глаза у репти-
лий, являясь все же более
сложным образованием
(рис. 149).
В органе зрения разли-
чают глазное яблоко с
зрительным нервом и при-
даточные органы глаза
(organa oculi accessoria),
включающие веки (pal-
pebra), слезной аппарат
и двигательный аппарат
глаза. Глазное яблоко
(bulbus oculi) у птиц имеет
форму или уплощенного
в передне-заднем направлении шара, или укороченной бинокуляр-
ной Галилеевой трубки, с весьма выпуклым объективом; в послед-
нем случае, свойственном главным образом ночным птицам, глаз,
состоит как бы из трех частей: круто выпуклой кпереди,более широ-
кой и менее выпуклой кзади и соединяющего их усеченного конуса.
Подобно другим позвоночным, стенки глазного яблока образуются
тремя оболочками: наружной, средней и внутренней. В них укре-
плен преломляющий лучи хрусталик, разделяющий глаз на две
полости, наполненные водянистой влагой и стекловидным телом,
В наружной оболочке глаза (tunica fibrosa oculi externa) разли-
чают переднюю прозрачную часть — роговицу (cornea) и большую
заднюю непрозрачную — белочную (sclera). Последняя имеет хря-
щеватое и фиброзное строение с тонкими костными образованиями
159
is двух местах: подковообразной чешуйки вокруг выхода зрительного
нерва (найдена только у дятлов, воробьиных и дневных хищников)
ji кольца костных чешуек, в количестве 10—17, располагающихся
131
Рис. 149. Форма и строение глаза у птиц и рептилий.
|1родольный разрез: 1—уплощенный глаз страуса — Sthrutio camelus L., 2—полу-
гслескопический глаз орлана белохвоста — Haliaetus albicilla L.; 3 — телескопический
глаз филина — Bubo bubo L.; 4 — глаз хамелеона; 5 — глаз варана — Varanus sp.; 6—10 —
глазные гребешки киви и рептилий, сходные по своему строению; 6 — киви — Apteryx
australis Shaw.; 7 — хамелеона; 8 — веретенницы — Anguis fragilisi 9 — зеленой ящерицы —
Lacerta viridis L.; 10—варана — Varanus sp.
C—сосудистая оболочка; Co—роговица; i — ирис; L. — хрусталик; M—реснич-
ный мускул; Р — гребешок; PC — ресничный отросток; R — ретина; S — склеротическая
оболочка; Sr — окостенения склеротического кольца; п — глазная сторона; t — височ-
ная сторона глаза. По Franz, Soemmering и др., из I-lilzheimer и Hellmann.
в передней части белковой оболочки и налегающих друг на друга,
как черепицы крыши. Это склеротикальное кольцо свойственно
всем птицам (встречается и у рептилий, см. рис. .26), обладает неко-
торой, относительно друг друга, подвижностью своих элементов и
играет значительную роль в аккомодации глаза, оказывая давление
Нм)
на стенки глазного яблока и таким образом удлиняя и укорачивая
зрительную ось глаза и изменяя выпуклость роговицы. Указан-
ный выше средний! отдел яблока, имеющий вид трубки, образуется
в значительной мере этим сильно развитым склеротикальным коль-
цом. Спереди белковая оболочка переходит в роговицу, образован-
ную несколькими слоями совершенно прозрачных клеток, свободно
пропускающих лучи света. Толщина
роговицы изменчива и стоит в связи с
образом жизни птицы; она толще у
пыряющих, разыскивающих под водой
пищу, видов. Выпуклость ее почти сфе-
рпчна и при аккомодации отчасти из-
меняется.
Средняя пли сосудистая оболочка
глаза- (tunica vasculosa oculi) распола-
гается непосредственно внутрь от бел-
ковой оболочки и в области перехода
ее в роговицу загибается к глазной осп,
образуя радужину (iris), прикрываю-
щую хрусталик спереди (рис. 150).
К радужине прилегает ресничное тело
(corpus ciliare), располагающееся по
краю хрусталика. Собственно сосуди-
стая оболочка (chorioidea) выстилает за
хрусталиком всю внутреннюю поверх-
ность склеры и представляет собой очень
густое сплетение кровеносных сосудов
с многочисленными ветвящимися пиг-
ментными клетками. Свойственный! дру-
гим классам отражательный слой (tape-
tum) у птиц отсутствует (исключая козо-
доев, авдоток и страуса). Особенностью
этой оболочки у птиц (встречаемой и
у рептилий) является образование склад-
чатого выроста-гребешка (pecten), вдаю-
щегося в стекловидное тело, выполняю-
щее полость глаза. Внутрь от сосуди-
стой оболочки лежит сетчатая оболочка
(retina), сложного многослойного строе-
ние. 150. Верхняя часть ' верти-
кального разреза через глаз сизого
голубя — Columba lii'ia Gm.
а — склеротпкальнал оболочка;
b — ретина; с — сосудистая обо-
лочка— chorioidea; с —пигмент-
ная выстилка на задней стороне
ресничного отростка —corpus cilia-
ris; cl—ресничный отросток; е—
концентрические слои хрусталика;
/ — роговица; д— ирис; h — ми-
гательная перепонка; i — верхнее
векб; k — ресничный (Крамптонов)
мускул; I— наружный радиальный
слой хрусталика. Из Vocht и Yung.
пня, в которой разветвляются нерв-
ные пучки и волокна входящего в глаз зрительного нерва.
В строении глаза целесообразно различать аппарат, преломляю-
щий и проводящий световые лучи (роговица, хрусталик, стекловид-
ное тело и связанные с ними образования), и аппарат, воспринимаю-
щий раздражения, — сетчатку.
Сетчатка птиц построена сходно с сетчаткой млекопитающих
(рис. 151). Важным является распределение так называемых палочек
и колбочек, составляющих слой клеток в области наружной погра-
ничной перепонки (membrana limitans externa). У дневных птиц
в этом слое преобладают колбочки, у ночных же палочки, что стоит
И Орнитология—1209
161
в связи с преимущественным восприятием первыми элементами цве-
тов, а вторыми только светотеней и формы. Интересным является то.
совами и козодоями строением сетчатки
обладают ласточки и стрижи, зрение
что сходным с ночными
Рис. 151. Схема строения сетчат-
ки, поперечный разрез.
L. е. — membrana limitans externa,
сквозь которую проходят палочки и
колбочки; L. 1. — m. limitans inter-
na, отделяющая ретину от стекловид-
ного тела (corpus vitreum); Р. — чер-
ный пигмент у основания палочек и
колбочек; 2 — слой волокон оптиче-
ского нерва; 3 — слой внутренних
ганглиозных клеток, включенных
в нервные волокна; 4 — внутрен-
ний зернистый слой, пронизанный
нервными волокнами; 5 — средний
ганглиозный слой, состоящий из
мелких клеток, соединяющих тре-
тий и седьмой слои; 6 — внешний
зернистый слой; 7 — внешний ган-
глиозный слой, состоящий из би-
полярных клеток, продолжающих-
ся через m. limitans externa в де-
вятый слой; 9— слой палочек и
колбочек. По Gadow и Newton.
которых очевидно и днем Мало воспри-
нимает цвета. Палочки вырабатывают
большое количество красного пигмен-
та — глазного пурпура, легко обесцве-
чиваемого под влиянием света, но бы-
стро вновь вырабатываемого. Через
разрушение пурпура очевидно и проис-
ходит очувствление клеток к воздей-
ствующему раздражению. Процесс пег
редачи раздражений колбочками неиз-
вестен. В последних загадочным обра-
зованием является пузырек, помещаю-
щийся в месте сужения расширенной
части колбочки и свойственный всем
птицам. У дневных птиц этот пузырек
имеет обычно желтую или красную,
а у сумеречных — бледножелтую или
синевато-зеленую окраску. Наиболее
вероятным является функционирование
его подобно действию желтого фильтра
при фотографировании, т. е. ослабле-
ние действия синих лучей, что способ-
ствует контрастности изображения, а
в темноте кроме того устраняет хро-
матическую аберрацию. Опытом уста-
новлено, что дневные птицы с преоб-
ладанием окрашенных пузырьков над
неокрашенными, например пустельга,
зуйки и др., видят мир, подобно чело-
веку, смотрящему через красновато-
желтые очки, который воспринимает
синие и фиолетовые цвета, как слабо
насыщенные оттенки серого цвета.
Как у большинства позвоночных,
глазу птиц свойственно «слепое пят-
но», в месте выхода на сетчатку зри-
тельного нерва, не воспринимающее раз-
дражений, и желтое пятно (macula lu-
tea), дающее наиболее ясное и четкое вос-
приятие. В последнем светочувствитель-
ные (перципирующие) элементы располагаются наиболее густо.
В глазу многих птиц имеется по два и даже по три желтых пятна
(по два пятна у дневных хищников, стрижей, зимородков, по три-^
у крачек и ласточек родов Sterna и Hirundo), у некоторых же групп
желтое пятно имеет вид горизонтальной полоски (кулики, чайки,
чистики и др.). Почти всегда это пятно углубляется в неболыпу10
216
ямку (fovea). Увеличение числа желтых пятен указывает на более
совершенное зрение, на это же указывает и густота колбочек в fovea,
достигающая, например у сарыча, до 100 на 100 кв. микронов, что
в пять раз превышает количество их у человека и следовательно
в пять раз должно превышать человеческую способность различения.
В преломляющем лучи аппарате глаза центральное место зани-
мает хрусталик (lens crystallina), обычно имеющий чечевицеобраз-
ную, часто асимметричную форму. Эластичное вещество прозрач-
ного хрусталика состоит из тончайших пластинок, ориентированных
в определенных, иногда очень сложных системах. Внутренность
линзы наиболее упруга и образует ядро хрусталика (nucleus len-
tis), наоборот, края его окружены более мягким веществом — коль-
цевым валиком, отделенным от основной массы хрусталика щелью
с водянистым секретом, питающим клетки хрусталика. На кольце-
вом валике соприкасаются многочисленные отростки ресничного
тела и цинновые связки (zonula Zinnii), поддерживающие хрусталик
на месте и тесно связывающие его с оболочками глазного яблока.
Аккомодация глаза происходит главным образом изменением
формы хрусталика. Основную роль при этом играет ресничный му-
скул (musculus ciliaris), пробегающий двумя радиальными коль-
цами волокон в стенках переходной зоны роговицы и склеры.
Действие этого мускула через посредство ресничного тела и кольце-
вого валика сжимает хрусталик и этим меняет его выпуклость. Не-
которую роль при этом играет также сфинктер радужины. Этим до-
стигается аккомодация глаза, нормально установленного па дально-
зоркость, к близким объектам. Способность к этой аккомодации раз-
лична у разных видов; в то время как у баклана она в 2—3 раза пре-
вышает аккомодацию глаза у человека (40—50 диоптрии вместо
14—15), у голубей и кур она почти равна ему (8—12 диоптрий),
а у сов уменьшается в 4—7 раз (2—4 диоптрии). Некоторую роль
в аккомодации птичьего глаза играют, как указано выше, давление
склеротикального кольца и изменение длины оси глаза.
Радужина, помимо некоторого давления, оказываемого на хру-
сталик, в основном играет роль затеняющей ширмы. Опа состоит
из рыхлой соединительно-тканной стромы, спереди и сзади ограни-
ченной пограничными слоями клеток. Строма обычно бывает пере-
полнена клетками, несущими пигмент, и заключает два мускула —
ш. sphincter и m. dilatator pupillae, в зависимости от интенсивности
падающего света, рефлекторно регулирующих ширину зрачка.
Зрачек обычно бывает круглый, в редких случаях щелевидный
(Rhynchops), грушевидный ( Dryocopus) или многоугольный (пин-
гвины). Рефлекторные реакции радужины протекают у птиц чрез-
вычайно быстро, что обусловливает сохранение зрительной чув-
ствительности на ярком свету и возможность взглядов на солнце.
Вследствие изменений формы хрусталика при аккомодации, в пе-
редней камере глаза возникает переполнение ее влагой (humor
aquaeus), и часть ее, в области реснитчатого тела, проникает в ка-
меру, занятую стекловидным телом. Возникающее здесь давление
регулируется предположительно гребешком, с клетками которого
фпбрилли стекловидного тела тесно связаны. Как указывалось,
163
греоешок имеет складчатое строение и свооодным краем вдается
в полость глаза; обильные пигментом клетки, образующие его, сли-
ваются отростками в синцитии, густо переплетаемый сеточкой кро-
веносных сосудов, образующих анастомозы артерии и вены гребешка.
Помимо регуляции внутреннего давления, что может быть осуще-
ствляемо посредством богатой кровеносной сети, гребешок невиди-
мому продуцирует и питает стекловидное тело и сетчатку, лишенную
у птиц специальной артерии (arteria retinae млекопитающих).
Рпс. 152. Мускулатура и нервы глазных яблок домашнего воробья_
Passer domesticus L.
Вид сзади, на правом яблоке удален верхний прямой мускул —
in. rectus superior; С. — хиазма; b. — соединительная ветвь от ramus
ophtalmicus тройничного нерва к глазодвигательному нерву—nervus
oculomotorius; G. —гассеров ганглий — ganglion Gasscri чувствитель-
ных корешков тройничцрго нерва; I. М. —нижнечелюстная ветвь —
ramus mandibularis тройничного нерва; I. О.—верхнеглазипчная
ветвь—ramus orbitalis челюстных ветвей тройничного нерва; L.—
слезный нерв — n. lacrimalis; L. G.—слезные железы; N.—носо-
вой нерв — и. nasalis; О.—верхний косой мускул — m. obliquus
superior; Op. — ramus ophthalmicus тройничного нерва; Q. — квадрат-
ный мускул — m. quadratus; R. E. —прямой наружный мускул — m.
iectus externus; II. S.—верхний прямой! мускул — m. rectus supe-
rior; S. M. — верхнечелюстная ветвь тройпичпого нерва; S. О.—
ramus orbitalis superior верхнечелюстной ветви тройпичпого нерва;
з, 4, о. 6—третий — шестой черепные нервы. По Slonaker, из Stre-
semann.
Двигательный аппарат глаза птиц сравнительно слаб и глаза
птиц малоподвижны; это компенсируется большой подвижностью шеи
и головы. Шесть глазных мускулов, представленных уже у репти-
лий, — прямые m. т. rectus superior, inferior, lateralis, medialis
и два косых — m. m. obliquus superior и inferior вращают однако
и здесь глазное яблоко в вертикальном и горизонтальном направле-
нии (рис. 152).
Веки птиц образуются из наружного слоя кожи, мышечного слоя
и соедипительно-тканного эластичного слоя с слизистой оболочкой —
коньюктпвой (conjuctiva), которая, образуя небольшую складку,
соединяет веки с глазным яблоком. В нижнем веке кроме того имеется
1G4
соедпшггельио-тканпая или хрящевая пластинка (tarsus), служащая
для прикрепления мышцы, опускающей веко. У птиц наиболее
подвижным веком является, как правило, нижнее. Мышечный аппа-
рат этих век слагается из поперечно-полосатых мышц, поднимаю-
щих веки (m. т. levator palpebrae superior и inferior), мышцы, опу-
скающей веко (т. depressor palpebrae inferiorcs), и гладкой круговой
мышцы (т. orbicularis palpebrarum), замыкающей веки во время
сна. По краю век, вообще лишенных перьев, часто развиваются
мелкие, лишенные опахала, перышки, порой весьма сходные с рес-
ницами.
Кроме наружных век, подобно рептилиям, у птиц имеется третье
веко, или мигательная перепонка (membrana nicticans), образован-
ная двойной складкой коныоктивы и помещающаяся во внутреннем
углу глаза. Обычно опа совершенно прозрачна и служит только для
очищения роговицы. Задергивание ее на глаз происходит и при
закрытых наружных веках, действием главным образом пирами-
дальной мышцы (m. pyramidalis), расположенной па задней стороне
глаза н поддерживающей ее квадратной мышцы (ш. quadratus).
С глазом связан ряд желез, располагающихся непосредственно
у глазного яблока. Гардсрова железа (glandula Harderi) прилегает
как желтовато-белое тело на его верхней и внутренней поверхно-
стях и выделяет слизистый секрет к основанию мигательной пере-
понки, которая своими движениями и разносит его по поверхности
роговицы. Слезная железа (glandula lacrimal is) располагается во
внешнем углу глаза п выделяет водянистый секрет под верхнее
веко. Омыв роговицу, секрет попадает в открывающийся во вну-
треннем углу глаза слезный проток (ductus lacrimal is), ведущий в но-
совую полость, в область ее средней раковины.
Вследствие бокового положения глаз у птиц, впечатление, по-
лучаемое каждым глазом, в значительной мере независимо, и птица
очевидно в состоянии уделять некоторое время напряженное
внимание двум противоположно размещенным предметам. Даже
у сов, с глазами, сдвинутыми кпереди, бинокулярное зрение
весьма несовершенно, и глубина рассматриваемого объекта уста-
навливается частью на основании опыта, частью попеременным
рассматриванием тем и другим глазом. Таким образом у большин-
ства видов зрительное поле весьма велико и достигает почти 360Л
Частичное сближение оптических осей повидпмому возможно как
кпереди, так и кзади. У домашнего воробья оптическая ось, распо-
ложенная к медиальной линии тела под углом в 65°, может быть
приближена к ней на 25°, что однако не дает еще ясного биноку-
лярного зрения. Произвольное частичное сближение оптических
осей кзади известно у очень многих птиц (кулики, вороны, козодой
и др.), что дает возможность контролировать объекты, находящиеся
сзади птицы, не совершая движения головы.
Э _ к о л_____________о г__________и я
Развитие любой формы жизни протекает в тесной зависимости
внутренних, конституциональных структур п процессов от окру-
жающей среды; свойства этой среды в значительной мере определяют
и общий облик животного, характер движений, развитость органов
чувств, периодичность жизненных отправлений и многие другие
стороны организации животного и его жизнедеятельности. Вместе
с конституциональными особенностями живого вещества, среда вхо-
дит в эволюционный процесс как формирующий фактор, постоянно
являясь, в известной своей констелляции, непременным условием
самого существования данной жизненной формы и определенных
присущих ей жизненных отправлений.
Конкретной среДой для каждой формы птиц является биотоп
и населяющий его комплекс животных и растений — биоценоз.
Изучение экологии и распространения птиц указывает на чрезвы-
чайную важность бптопа и биоценоза, как факторов, определяющих
распределение, облик и поведение птицы. Ареал огромного боль-
шинства птиц определяется соответствующими виду биотопическимп
условиями, облик п способы движений вида стоят в соответствии
с характером биотопа, питание и фенология показывают тесную
и многостороннюю связь с другими организмами данного биоценоза
и установившимся в этих условиях климатическим режимом.
Разные виды обнаруживают при этом различную степень этой
зависимости, разную пластичность и приспособляемость, по общая
подчиненность известным бпотопическим условиям остается не-
сомненной.
Среди различных сторон зависимости от среды, на первом месте
необходимо поставить общий характер движений птиц, общий облик
пли хабитус птиц, как следствие определенной адаптации к тому
или иному биотопу п способу добывания пищи, и самый характер
питания н овладения пищевым материалом.
1. ОБИТАЕМАЯ СРЕДА И ОСНОВНЫЕ СПОСОБЫ ПЕРЕДВИЖЕНИЯ
Современные данные морфологии, экологии и палеонтологии
птиц делают наиболее вероятным происхождение птиц от рептилии,
приспособленных к древесному образу жизнп. За это говорят как
приспособление первичных птиц Archaeopteryx- п Archaeornis к ДРе'
весному образу жизнп, так и преобладание среди птиц древесного
образа жизни вообще и несомненная вторичность глубоких присно-
166
соблений к беганию или плаванию. Таким образом есть основания
рассматривать все разнообразие движений птиц как происшедшее
из основных движении, свойственных лазящим рептилиям. Тако-
выми были, повидимому, бег, ползание на четырех конечностях и
прыжки, совершаемые с ветки на ветку. Из этих прыжков предполо-
жительно развился сначала планирующий, а потом и активный
способ полета, с прогрессивной специализацией в этом направлении
передних конечностей. В связи с стремлением использовать для
поисков пищи не только поверхность листвы и ветвей, но п стволы
деревьев, из прыжков специализировалось лазание по стволам с опо-
рой на хвост, как это характерно для дятлов, пищух и других птиц.
Вместе с тем вторичный переход к движению по земле сопрово-
?кдался выработкой приспособлений к бегу или шаганию но отно-
сительно ровной плоскости, а переход к жизни в воде или на берегу
привел к выработке плавающих движений с дальнейшей специали-
зацией способности к нырянию в поисках подводной пищи. Каждый
из этих типов движения от форм мало специализированных в даль-
нейшем приводил к образованию форм односторонне специализи-
рованных, терявших способность к ряду других движений, как,
например: фрегат — к передвижению по земле и плаванию, пин-
гвины — к летанию, гагары — к бегу, хождению или прыганию
и т. д.
В вопросе о происхождении способов движений птиц основную
трудность представляет объяснение перехода от пятипалой ноги
древесных рептилий к четырехпалой ноге древесных птиц с противо-
поставляющимся задним пальцем, так как в настоящее время мы
не находим необходимой серии переходов между ними. Возможный
путь этого перехода представляют способы движения древесной
рептилии Polychrus marmoratus из семейства Iguanidae. Обладая
пятипалыми лапами, эта рептилия в ряде движений своих конеч-
ностей чрезвычайно напоминает соответствующие движения ног
птицы (рис. 153). При прыжке с ветки на ветку Polychrus marmo-
ratus поворачивает задние ноги вперед и наиболее длинными 2-м,
3-м и 4-м пальцами обхватывают ветку, причем 1-й палец оказы-
вается сзади и как бы противопоставляется остальным; 5-й палец
при этом оказывается нефункциопирующпм. При более постоянном
повороте задних ног вперед, как это имело место у динозавров и
должно было иметь место у предков птиц, 5-й палец вообще терял
оы свое значение при беге и прыжках по веткам и вследствие неупо-
требления должен был бы исчезнуть, а 1-й — укрепиться в своем
положении противоположности остальным пальцам. Функциони-
рование 5-го пальца у современных древесных рептилий, медленно
лазящих вдоль веток, объясняется тем, что он играет роль заднего
пальца вследствие большого поворота всей ноги наружу, под боль-
шим углом к осп тела, как это вообще свойственно рептилиям.
Преобладающим типом движения древесных птиц является пры-
гание с ветки на ветку обеими ногами сразу. Упомянутый выше тип
ноги большинства воробьиных с тремя пальцами, направленными
вперед, и одним назад является очевидно вполне соответствующей
этой задаче структурой, так как он свойствен более 90% всех лес-
167
пых птиц. В других группах однако распространен другой тип
строения пог, с 4-м или 2-м пальцем (реже), направленным назад,
так что нога получает большую силу и цепкость за ветви деревьев,
что можно рассматривать как более высокую ступень специализации
в этом направлении. Из этого типа движения несомненно развилась
и способность к ползанию по стволам деревьев, цепляясь за кору
крупными и острыми когтями лап и передвигаясь вверх короткими
прыжками, опираясь при этом дополнительно на жесткие, упругие
и загнутые в вентральном
направлении на конце ру-
левые перья. Представите-
лями этого движения явля-
ются дятлы, но конвергентно
этот же или сходный способ
лазания развился пво многих
семействах воробьиных, па-
пример у пищух, поползней,
некоторых древолазов (Den-
drocolaptidae), гавайских цве-
точниц (Jlelerorhynchus из се-
мейства Drepanididae), ткачп-
ковых (Phormoplectes preussi
Sjost. н др.из семейства Plocei-
dae) и неворобышых птиц из
семейства Phoeniculidae. Не-
которые из этих видов в усло-
виях скалистого ландшафта
перешли и к жизни на ска-
лах и в ряде случаев (Ticho-
drama murarut L., Sitta neu-
mayer Michah. н др.) всецело
приспособились к лазанию
по каменистым поверхно-
стям, не изменив по существу
структуры задних конечно-
стей. Как эта группа, так в
представители семейства Sd-
ином направлении, так как
Рис. 153. Употребление пальцев йог игуаной —
Polychrus marmoratus при движении по дере-
вьям: 1 — при лазании; 2 — при прыжке с ветки
па ветку. По Bdker.
tidae вообще, развились несколько в
они не пользуются хвостом в качестве опоры и в состоянии лазить
в любом направлении по вертикальной плоскости, в том числе и
вниз головой. Возможность этого дается очень сильной мускула-
турой ног и способностью весьма значительного отведения ноги
в сторону; при этом положении птпца цепляется верхней ногой
поперек ствола дерева и в то же время подпирает себя ниже нахо-
дящейся ногой, упирающейся в кору дерева.
Уклоняющуюся форму лазящего способа движения представляют
стрижи, на ногах которых все четыре пальца направлены вперед
п снабжены очень острыми когтями. При помощи этого аппарата
они подвешиваются на вертикальных стенах, скалах и т. д., цепляясь
всеми четырьмя когтями и опираясь на всю длину короткой плюсны-
168
Большинство лесных птиц, спускаясь на землю, передвигаются
по ней, прыгая короткими прыжками, пе умея шагать плп бегать.
Исключение из этого составляют • только крупные птицы, которые
пе могут прыгать в силу механических причин — значительно боль-
шей затраты мускульной энергии на прыжок, чем на шаг, чтр очень
существенно для тела крупной птицы вследствие возрастания его
массы пропорционально кубу, тогда как соответствующая попереч-
нику мускула сила растет только как квадрат ого длины. Птицы же,
эволюционировавшие среди открытого ландшафта — в степях, пу-
стынях и болотах, и передвигающиеся по земле, как правило, не
прыгают, а ходят или бегают, независимо от их размеров и массы.
Иногда при этом разный способ движения может быть свойствен
членам узкой систематической группы в зависимости от образа
их жизни. Так из семейства враповых (Corvidae) лесные сойка
(Garrulus) и кукша (Cractes) прыгают, а свойственные открытому
ландшафту грач (Тгурапосога.г)^ галка (Coloeus) и саксаульная
сопка (Podoces) только шагают пли бегают. В некоторых группах
однако способность к тому пли иному движению весьма консерва-
тивна, и при всех условиях они сохраняют тот же способ движения;
так голуби и в лесу только бегают пли ходят, а соловьи (Eritha-
cinae) и в степях и в пустынях только прыгают (например чеканы).
Необходимо отметить также, что мелкие наземные птицы, живу-
щие па почве, усеянной мелкими препятствиями, как, например,
па лесной, также преимущественно передвигаются по ней прыжками.
Передвижение птиц по земле шагами плп бегом весьма сходно
с тем же процессом у человека. При этом нога, отталкивающаяся
от земли, вначале своей осью подается несколько назад, при одно-
временном сгибе пнтертарзалыюго и нпжнеплюсиевого сочленения.
Передвигаясь относительно другой ноги и опускаясь на землю, опа
вытягивается в этих суставах и сначала касается земли концами
пальцев. Опускаясь па всю ступню, эта йога становится опорной,
в то время как соседняя начинает проделывать только-что пройден-
ную фазу рассматриваемой конечности. В следующие моменты опор-
ная нога образует уменьшающийся тупой угол между плоскостями
пальцев и плюсны, и вместе с тем увеличивается угол между бедром
и голенью, что передвигает тело вперед. Это движение приводит
плюсну в положение острого угла к плоскости стопы, что влечет за
собой сокращение сгибателей пальцев и сопровождается подъемом
стопы сзади, что вновь толкает тело птицы вперед. В этот последний
момент другая нога касается земли и начинает становиться опорной
ногой.
Необходимость согласования центра тяжести тела с переменой
опорного значения конечности вызывает боковые качания тела,
компенсирующие эти изменения. Как выяснено Штольпе (Slolpe,
1932), протекание этих движений обусловливается главным образом
строением коленного сустава. Сгибание коленного сустава опираю-
щаяся ноги передвигает этот сустав несколько наружу, так что
оеДро и тело переносятся на сторону опорной ноги, но антптрохантер
Допускает отведение бедра лишь на определенное короткое рас-
стояние. Следствием этого при сгибе колена опорной ноги яв-
169
ляется неизбежный перенос заднего конца тела на сторону пере-
носимой ноги, в то время как передний конец с центром тяжести
иереносптся на противоположную сторону.
Чем короче плюсны и чем ниже лежит центр тяжести тела,
а также чем больше длина тела относительно его ширины, тем боль-
шего размаха достигают боковые колебания тела, необходимые для
сохранения равновесия. Наиболее хорошо это4 выражается в пере-
валистых движениях, свойственных уткам и гусям. У первых, как
у особенно хорошо ныряющих птиц, движение на земле затруд-
няется, кроме того, иным строением ноги, более приспособленной
it’ гребным движениям в воде, чем хождению по суще. Так как ноги
у них отставлены далеко назад, то при движении по земле им при-
ходится держать тело выпрямленным круто вверх, достигая этим
перемещения центра тяжести на плоскость опоры. Но центр тя-
Гис. 154. Иоло/кснис тела и пог при плавании: (а) —чайки; (Ъ) —утки; (с)—гуся;
(d) —нырковой утки; (е) —баклана; (f) —гагары. По Dabclow.
жести у этих птиц находится очень высоко, что затрудняет баланси-
рование тела во время движения. Большая устойчивость дости-
гается опорой не только на пальцы, но и на всю плюсну (чистики)
или па хвост (пингвины). Последние при передвижении короткими
шажками для удержания равновесия размахивают и вытягивают
назад тяжелые крылья пли делают неуклюжие прыжки; в случае
особой спешки для них ’ характерно особое скользящее движение
лежа па брюхе, с отталкиванием от льда или снега быстрыми дви-
жениями крыльев.
Наиболее специализированными типами передвижений птиц яв-
ляются плавание (и ныряние) и полет, в которых некоторые группы
достигли изумительного совершенства. Поэтому целесообразно более
подробное рассмотрение этих способов передвижения.
Плавание и ныряние п т и ц. Использование водной
среды для обитания у многих видов птиц связано с развитием спо-
собности к плаванию, потребовавшей выработки ряда приспособле
ний. Из 33 отрядов птиц эти приспособления развились у отрядов-
ЧТО
Рис. 155. Груд-
ная кость хорошо
ныряющего пред-
ставителя подот-
ряда чистиков
(Alcae), тонко-
клювой кайры —
Uria aalge Pont.
По Eyton, из Stre-
semann.
Ilesperornithijormes, Ichthyornilhijormes, Spheniscijorntes, Procellari-
iformes, Colymbifonnes, Pelecanijormes, Phoenicopterifornies, Anseri-
jormes и подотрядов Lari, Alcae n Hetiornithes, жизнь которых тес-
нейшим образом связана с водной стихией. Но кроме того способ-
ность плавать более или менее широко свойственна и сухопутным
видам в силу их малого удельного веса. Однако нормально исполь-
зуют эту возможность только представители следующих (в основном
большей частью сухопутных) подотрядов: Opisthoconii, Haiti и Li-
micolae.
Приспособления к плаванию. Воз-
можность пловучестп тела птиц обусловлена малым
удельным весом, вследствие обширных воздушных
мешков, пневматичностп костей, пухлости и лег-
кости перового покрова, способного задерживать
внутри себя большое количество воздуха. Роль по-
следнего фактора ясно показывают измерения Хейп-
ротом (Heinroth, 1924—1932) удельного веса селезня
кряквы, который у оперенной птицы составлял 0,6,
а у ощипанной 0,91. Оперенный селезень, весом в
1337 г, вытесняет 2060 куб. см воды, ощипанный
же, весом всего в 1270 г, вытесняет 1390 куб: см,
что указывает, что перовой покров его, весом всего
только в 67 г, задерживал до 650 куб. см воздуха.
Эта причина является основной, почему любая
птица, попавшая в воду, в состоянии держаться
па ее поверхности, пока перо ее полностью не про-
питается водой.
У птиц, более глубоко приспособленных к пла-
вающему образу жизни, существенно изменяется
форма тела. Она удлиняется, приобретает валько-
ватую форму, несколько уплощенную в дорзовен-
тральном направлении (рис. 154). Такая форма бо-
лее благоприятствует движению тела в воде, чем
корпус наземных птиц с выпуклой грудью. Вслед-
ствие уплощения тела, происходит перемещение
центра тяжести в дорзальном направлении и кзади,
что влечет за собой малую погружаемость в воду
грудной части и более значительную погружаемость
задней части, особенно во время гребных движений ног, проталки-
вающих тело вперед. При этом, как правило, удлиняется грудная
клетка и далее кзади прикрепляются и сильно удлиняются задние
ребра, защищая внутренности от давления воды (рис. 155).
Оперение, одевающее тело, также испытывает значительные изме-
нения: контурное перо становится более густым и жестким, на
нижней поверхности тела исчезают аптерии, и перо оказывается
покрывающим густо всю поверхность тела, наиболее тесно соприка-
сающуюся с водой. Под контурным пером развивается очень густой
и теплый слой пуха, а под кожей образуется толстый жировой слой*
прекрасно защищающие организм от охлаждения водой. Необхо-
димость сделать перо несмачиваемым привело к развитию особенно
17 L
крупных по размерам копчиковых желез, секрет которых заботливо
разносится птицами по
поверхности оперения, ожирпвая его.
Из органов движения наиболее зна-
чительные изменения испытывают ноги
и крылья. Пальцы ног приобретают пла-
вательные лопасти, развитие которых идет
в общем по двум направлениям: 1) с об-
разованием па пальцах кожистых оторо-
чек или более упругих пластинок, как у
плавунчиков из куликов, лысухи из ку-
рочек п поганок или 2) с образованием
кожистых перепонок между пальцами,
охватывающих передние пальцы, иногда
вместе с задними, что свойственно гуси-
ным, чистикам, чайкам, веслоногим, га-
гарам и пек. др. (рис. 156). В целях
уменьшения трения, при перемещении ла-
пы сзади вперед пальцы складываются
и представляют сравнительно малую по-
верхность; это же значение очевидно имеет
и уплощение с боков плюсны у гагар и
поганок, по форме сходной с лезвием но-
жа, вследствие чего нога с наименьшим
трением разрезает воду. Движения от-
дельных лап у большинства плавающих
птиц независимы и только при особой
быстроте эти движения объединяются для
получения большего толчка. Так, напри-
мер, поступают при взлете пеликаны.
Спокойно плавающие гусиные часто пря-
чут одну лапу в оперение и передвига-
ются движениями только одной ноги.
Крыльями при плавании пользуются
в сущности только одни пингвины. Это
вызвало глубокие изменения в их строе-
нии, сопровождающиеся потерей способ-
ности к полету. Прекрасно развитые кры-
лья - ласты полностью взяли на себя
функцию лап, которые уже не играют
роли в продвижении тела, вытягиваются
назад и содействуют лишь изменению
направления движения. Пингвины пла-
вают, глубоко погружаясь телом в
воду, н не в состоянии при плавания
([)
?nes); (d) — большой
чина—Phalaropus sp.
Рис. 156. Строение лап у плавающих представителен
различных отрядов.
(а)—пеликана—Pelecanus sp. (Pelecani formes); (b)
пингвина ( Sphenisciformes) ; (c) — чайки (Charadriifor-
поганки — Podiceps cristatus L. (С о lymbi formes) (e) —плавун-
(Chara dr ii formes); (f) — лысухи — Fulica (lira L. (Grui formes) •
Из Reichcnow.
172
менять степень погружения, как некоторые другие группы. Из
других водоплавающих только бакланы и змеешейки при пла-
вании . окунают крылья в воду и иногда возможно помогают ими
при плавании; вследствие этого крылья постоянно намокают и тре-
буют длительной сушки на земле. Остальные водные птицы при
плавании держат крылья сложенными и или приподнятыми над
уровнем воды (например пеликаны) или спрятанными в удлиненные
контурные перья боков тела, которые образуют род защищенного
от воды кармана и предохраняют крыло от намокания. У нырцо-
вых уток крылья не намокают даже при глубоком и длительном ны-
рянии, при котором для передвижения утки пользуются одними
ударами лап.
Ныряние. Подобно тому как среди огромного количества
птиц, могущих держаться на воде, хорошо плавать могут сравни-
тельно немногие группы, так и среди собственно водоплавающих
птиц способность нырять развилась лишь у особенно искусно пла-
вающих и специально к нырянию приспособленных форм. Основное
приспособление в нырянии направлено на преодоление весьма
упругой среды, что требует значительной силы и соответствую-
щего ее приложения. Это достигается в общем двумя путями: или
падением птицы с высоты, с использованием той силы, которую дает
скорость движения падающего тела, или получением мощных толч-
ков крыльями или лапами, отталкивающимися в воде, сочетаемым
с параллельным увеличением удельного веса тела.
Первый способ естественно не дает возможности проникнуть
па значительную глубину, по используется довольно широко раз-
личными, в том числе и неплавающими формами, как то: скопой из
хищников, различными зимородками, фаэтонами, олушами, некото-
рыми пеликанами и бакланами из веслоногих, некоторыми крач-
ками (Sterna), чайками (Lams leucopterus Faber, и Rissa) и тру-
бконосыми (Pujfinus pujfinus L. и Pujfinus gravis O’Reilly). Необ-
ходимая для проникновения в воду скорость развивается или про-
стым падением или очень крутым скользящим полетом.
Второй способ, как указывалось, сопровождается развитием спе-
циальных приспособлений к произвольному увеличению удельного
веса тела и задержке дыхания под водой. Это достигается особенно
крупными размерами инспираторных воздушных мешков, из ко-
торых давлением брюшного пресса по желанию может быть удален
воздух. Кроме того, пневматичность скелета у хорошо ныряющих
птиц уменьшается вместе с специализацией всего тела в этом на-
правлении. Она уже очень мала у поганок, крохалей, бакланов и
пингвинов п полностью исчезает у гагар и тупиков (Fratercula)
пз чистиковых птиц. Некоторым доказательством того, что это явле-
ние связано именно с нырянием, является различие в пневматич-
постп ныряющих п неныряющих представителей веслоногих; по-
еледние, как, например, фрегат и пеликан, в противоположность
первым, отличаются исключительной пневматичностыо своего ске-
лета. Увеличение удельного веса достигается также у птиц неболь-
шой величины (поганки, нырковые утки, лысухи) произвольным
прижатием пера к телу и удалением таким образом заключенного
173
в перовом покрове воздуха. По наблюдениям Штуббса (Stubbs.
1.910), эффект этого действия у малой поганки (Podiceps ruficollis
Pall.) увеличивает ее удельный вес с 0,66 до 0,84. Эта же способность
невидимому является основной причиной наблюдаемой у поганок
то легкой пловучести почти всего тела на поверхности воды, то по-
гружения его целиком jb воду, с выставлением на поверхность только
части шеи и головы, подобно общеизвестному положению пери-
скопа.
Продолжительность пыряния обычно сравнительно коротка и
констатирована у морянки в 70 сек., у Oideniia perspicillata L. до
60 сек. Однако ряд видов (Oidemia jusca L., Colynibus и Mergas}
свободно остаются под водой иногда до 2 мин., и в условиях опыта,
без вреда для птицы, домашняя кряква смогла пробыть под водой
от 11 до-17 мин., что указывает на возможность весьма длительной
задержки дыхательных движений птицей. Возможность этого объяс-
няется механизмом дыхания птицы, который располагает как боль-
шим запасом воздуха в воздушных мешках, так и возможностью
при погружении много раз перегонять воздух из задних'мешков
в передние и обратно, каждый раз используя тот же воздух все
с большей полнотой. Важным моментом является при этом сравни-
тельное богатство крови ныряющих птиц (установлено на утках),
обусловливающее большое накопление кислорода в крови во время
вдыхания. Отношение количества крови (вес) к весу тела особенно
резко разнится в положительную сторону для утки при сравнении
с курами. У первых это отношение выражается как 1 к 14—21, у вто-
рых, как 1 к 21 до 46. Кроме того, накопление углекислоты в крови
у уток не возбуждает дыхательный центр, что имеет место у
млекопитающих, а лишь парализует его. Затем в кровеносном
аппарате ныряющих гусиных, гагар и бакланов наблюдается силь-
ное увеличение размеров нижней полой вены (vena cava inferior),
у крохалей, например, в 6 раз превышающей калибр аорты. По пред-
положению Гратиолет (Gratiolet, 1867, по Stresemann, 1931), здесь
скопляется и застаивается кровь, приносимая из задней половины
тела посредством vena cava posterior и обеих печеночных вен.
если этого требуют обстоятельства дыхания; в таких случаях
в легкие поступает кровь только из передней половины туловища
п крыльев, гонимая через сердце пз обеих верхних полых вен (vena
cava cranialis). Надо думать, что этот процесс дает возможность пра-
вильнее использовать дыхательные возможности крови при за-
держке дыхательных движений.
Развитие этих приспособлений и мощные гребные движения перед-
них пли задних конечностей под водой дают возможность некоторым
видам, ныряя, достигать значительных для птиц глубин. Обычная
глубина разыскивания пищи под водой для нырковых морскпх
уток (турпаны, гага, морянки) составляет от 3 до 6 м, но иногда они
проникают до 8—10 м, а бакланы и пингвины могут спускаться
ловидимому еще глубже. Пингвины заглатывают мелкие камни,
необходимые для правильного хода пищеварения, доставая их на
дне водоемов, так что в обледенелой Антарктике ряд данных о гео-
логической природе морского дна был получен только изучением
174
тех галек, которые находили в желудке добытых в этих местах
пингвинов (Allen, 1928).
Дьюаром (Dewar, 1924) было установлено полезное правило для
ориентировочного определения глубины, на которую погружается
птица. Из многочисленных наблюдении над нырковыми утками и
поганками, с измерениями глубины ныряния и его продолжитель-
ности, выяснилось, что время, проводимое птицей под водой, уве-
личивается пропорционально глубине, достигаемой птицей, и что
время, затрачиваемое птицей на поиски пищи па дне, обычно равно
количеству времени, затрачиваемому на погружение на глубину
одного фатома (.-= 183 см), у большинства птиц равному 10 сек.
Таким образом из продолжительности пребывания птицы под водой
можно вывести достигаемую ими в данном случае глубину. Для
поганки, например, кормившейся в морском прибрежье и нырявшей
в течение 27—28 сек., 32—35 сек. и 35—36 сек., вычисленная глу-
бина в 3,2 м, 4,1 м и 4,5 м оказалась соответствующей действи-
тельной. При этом рекомендуется первые 20 сек. считать соответ-
ствующими одному фатому глубины, так как 10 сек. уходят на
поиски пищи, а каждые следующие 10 сек. относить за соответствую-
щее число фатомов. По Дьюару, это правило имеет приложимость
к обычному нырянию в поисках корма как для нырковых уток и по-
ганок, так и для бакланов, гагар и чистиков. Для лысухи ука-
занное отношение изменяется, так как этот вид не тратит время
па розыски корма, а, спустившись на дно, схватывает клювом рас-
тение и сейчас же всплывает вверх; поэтому первые 10 сек. целе-
сообразно считать, как и последующие, за 180 см глубины.
Движение под водой, как указывалось, совершается при помощи
гребных движений ногами или крыльями. Можно выделить четыре
основных способа таких движений (Stresemann, 1931).
1. Большинство ныряющих птиц пользуются для продвижения
только толчками ног и крылья в это время держат скрытыми под
контурными перьями. Таковы все гагары, поганки, лысухи, кро-
хали и из нырковых уток роды Nyroca и Bi^iura.
2. Движение вперед достигается гребными толчками пог, но
в качестве руля служат также, полураскрытые крылья. Таковы
бакланы и змеешейки.
3. Как крылья, так и ноги служат для движения вперед и изме-
нения направления. Сюда относится ряд морских уток, как гаги,
турпаны, каменушка, морянка и утка-пароход (Tachyeres).
4. Движение вперед происходит только при помощи крыльев, ноги
же служат только в качестве рулевого аппарата. Таково движение
под водой чистиковых, пингвинов и ныряющих буревестников
(Piiffiniis, Pelecanoides и др.) (рис. 157).
Движение при этом ног и крыльев может быть рассмотрено обо-
собленно.
Большинство видов получают необходимую силу, для движения
вперед посредством одновременного толчка обеими ногами, что
ооусловливается рядом соответствующих приспособительных черт
в строении тела и самих конечностей. Этим видам свойствен очень
узкий длинный таз, что сближает относительное положение ног
175
друг к другу il дает возможность общего сближенного действия их
снизу пли сзади тела. Кроме того, как правило, бедро оказывается
относительно уменьшенным по длине, что содействует сближению
конечностей, а голень сильно увеличенной, что имеет своим след-
ствием более удаленное кзади положение интертарзального со-
членения вытянутых пог и таким образом возможность гребных
движений сзади тела; удлиненность голени увеличивает вместе с тем
область прикрепления двигательной мускулатуры лап и увеличи-
Рпс. 157. Двп/кенпя плавающего баклана — Phalacrocorax
carbo L. По Dabelow.
вает их длину. При этом большая берцовая у этих видов значительно
вращается относительно заметно удлиненной малой берцовой кости.
Движения ног при нырянии осуществляются повидимому двумя
различными способами. Ныряющие веслоногие отводят ноги под
туловищем вперед и одновременно отталкиваются обеими лапами
с перепонками, расправленными в одной плоскости; внутренние
пальцы ног при этом почти соприкасаются, друг с другом, и так
как перепонка соединяет все четыре пальца ног, то получается
очень значительная по размерам отталкивающая плоскость. Боль-
шая берцовая испытывает при этом вращение внутрь относительно
малой берцовой кости; бедро в это время тесно прилегает к телу п
положением антитрохантера удерживается от отведения, однако
при- посредстве голени оно играет значительную роль в движения
176
ноги и несет сильную мускулатуру. Сильно развитая чашечка не
препятствует относительно свободным движениям колена в сагит-
тальной плоскости.
Гагары, поганки и нырцовые утки отводят при нырянии лапы
вперед по бокам тела и затем боковыми движениями их, направлен-
ными друг к другу кзади от туловища, отталкиваются и получают
движение вперед. Наиболее резко этот тип движения выражен у по-
ганок и гагар, имеющих весьма специализированное строение зад-
них конечностей. Большая берцовая кость у этих видов образует
направленный проксимально длинный пирамидальный отросток
(processus rolularis). у гагар функционально полностью поглощаю-
Рис. 158. Фирма тела и леван задняя конечность фазана —- Phusluи us colchirus L. и гагары —
Colymbus sp. —наземной неплавающей птицы и птицы, хорошо плавающей н ныряющей, но
неспособной к передвижению ио земле.
(а)—туловище гагары со спины (по удалении кожи); (Ь)п (с) —леван задняя конеч-
ность фазана и гагары, сбоку; обратите внимая не на соотношение величины отделов ноги
и мускулов; (d) и (?) —туловище фазана и гагары, после удаления кояш, сбоку; / — бе-
ДРо; р — передний конец голени; рг — processus rolularis; I — голень (tibioiarsus); tin—
плюсна (larso-Hielalarsns). Ho Jleiliiiaiin.
Щий чашечку. Образование его вызывается потребностью удлинить
мощные мускулы — сгибатели и разгибатели питертарзального со-
членения, прикрепляющиеся вдоль голени. Вместе с тем этот отро-
сток ограничивает сгибаемость коленного сустава в сагиттальной
плоскости, и при разгибе в колене большая берцовая соответствую-
щей мускулатурой (musculus femoro-tibialis и in. ilio-tibialis) вра-
щается наружу, противоположно тому, что наблюдается у бакланов;
в этом же направлении вращается и плюсна относительно дисталь-
ного конца этой же кости. Бедро у этих форм является наиболее
укороченным и двигающимся только в трансверсальной плоскости
тела, сравнительно незначительно продвигая нижние отделы ноги
вперед и назад. Таким образом и бедро и голень оказываются с весьма
ограниченной свободой движений, прилегают тесно к телу и пол-
ностью обтягиваются кожей, как бы включаясь в состав тела
(рис. 158). По выражению Хейнрота (Heinrotb, 1924—1932), ноги
Г- Орнитология—1209 №
этих форм оказываются заключенными как бы в узкую юбку, позво-
ляющую двигаться только плюсневому отделу ноги, выходящему
из тела возле самого хвоста. Следовательно голень не может быть
оттянута в сторону от тела, как у большинства птиц, подвижна
/д;
Рис. 159. Движения ныряющей черпошейпоп
поганки — Podiceps nigricollis Brehm.
(а) — фаза отталкивания; (Ь) — фаза приве-
дения ног. Палево вид сбоку, направо —спе-
реди и сверху; кружки указывают положение
тазо-бедрепного, коленного й интертарзалъ-
ного суставов. По Frank и Neu, из Stresemann.
в сущности только вперед и взад
и несколько вращается вокруг
своей длинной оси. Вследствие
такой конструкции конечности
к передвижению на суше эти
формы почти совершенно неспо-
собны.
Анализом, проведенным кп-
норапидсъемкоп, движения зад-
них конечностей у поганок
рисуются в следующем виде
(рис. 159): вначале пнтертар-
зальное сочленение сильно сги-
бается, плюсна передвигается
вперед и прижимается к телу,
обращаясь узким задним краем
вниз; при этом сложенные
пальцы оборачиваются концами
вперед. Затем голень поворачи-
вается на 90° кнаружи и пере-
водит в это же положение
плюсну, задний край которой
теперь оказывается обращенным
вверх; в этот же момент начи-
нают расправляться плаватель-
ные пластинки, также обра-
щенные наружу вверх. Далее
бедро и голень резко отводятся
кзади, и расправленные пла-
стинки отталкиваются от воды,
описывая концами пальцев род
эллипса. При этом движения
лопастей обеих ног приходятся
почти друг против друга и кзади,
как бы отталкивая позади тела
широкий столб воды. Вслед за
этим голень отводится в преж-
нее положение, постепенно вра-
щаясь несколько внутрь, интер-
тарзальное сочленение вновь
сгибается, и плюсна поворачивается задним краем книзу в исход-
ное положение (Frank u. Neu, 1929, fide Stresemann, 1931).
Скорость движения под водой этим способом передвижения выяс-
нена недостаточно. У лучших ныряльщиков, как, например, гагар?
она превосходит во всяком случае скорость птицы, плывущей на
поверхности воды. У одной из мелких поганок? поганки черношей'
178
ной {Podiceps nigricollis Brehm), она установлена в 0,8 м пли 0,3 м
в секунду под влиянием каждого гребного движения. У гагар эта
скорость должна быть значительно большей.
На пути приспособления к нырянию этим способом, посредством
гребных движений пог, лежат однако механические препятствия,
в силу которых ныряющий вид в процессе эволюции также не может
достичь большой величины, как и летающие птицы. Поэтому вряд
ли вообще могли существовать на земле ныряющие виды больше Не-
sperornis — наиболее крупного вида, нырявшего таким же способом.
Так как увеличение задних конечностей в той же пропорции, как и
увеличение тела птицы, превращает конечность в слишком длинный
рычаг, чтобы он был способен экономно давать нужный эффект в со-
противляющейся его движениям водной среде, то с увеличением
размеров тела мы закономерно наблюдаем относительное к телу
уменьшение длины задних конечностей. Так, в отношении к длине
туловища, длина бедра у самой маленькой поганки, поганки малой
[Podiceps rujicollis (PaU .), 120—200 г весом] составляет 39,6%, а у по-
ганки большой {Podiceps cristatus L., 900—1250 г весом) всего лишь
25,1%. Подобные соотношения свойственны и другим группам тех
же специализаций.
Из птиц’, пользующихся гребными движениями крыльев при ныря-
нии, чистики и нырковые утки не расправляют крыльев под водой, как
во время полета в воздухе. Движение вперед получается главным обра-
зом ударами нерасправленных, согнутых в кистевом сгибе крыльев, то
оттягиваемых от тела, то вновь прижимаемых к нему. Такие движения
особенно характерны для нырковых уток, прибегающих одновременно
к помощи и гребных движений ног. Кайры при нырянии пользуются
крыльями более широко и в некоторых случаях почти летят под водой,
более широко расправляя крылья и делая ими в воде более значи-
тельные взмахи. Движения обоих крыльев при этом всегда являются
одновременными. Наибольшей свободой движений крыльями, превра-
щенными в ласты, располагают пингвины, могущие плыть как одно-
временными, так и разновременными взмахами крыльев и совер-
шающие при этом до 3—4 ударов крыльями в минуту.
Исключая пингвинов, приспособления крыльев к движениям
под водой сравнительно невелики и заметны только у более специа-
лизированных форм. Как правило, кости предплечья и плечевого
отдела у них являются сильно уплощенными дорзовентрально, чтобы
облегчить проведение крыла вперёд. В большинстве случаев крылья
У этих форм меньше по величине, вследствие большего удобства
таких плоскостей в плотной среде, причем крыло является заострен-
ным, весьма компактным, с очень жесткими маховыми перьями.
Сила, затрачиваемая организмом па движение крыльями в воде,
очень значительна и, в противоположность обычным условиям по-
лета, велика не только при опускании крыльев, но и при поднима-
нии их. Вследствие этого мускулы, поднимающие крыло, особенно
musculus supracoracoideus, сильно увеличиваются в размерах и
становятся почти равными по массе musculus pectoralis пли даже
превышают их (особенно у пингвинов). Это развитие мускулатуры
сопровождается в свою очередь удлинением грудной кости и уве-
179
личением гребня грудины, к которым прикрепляется эта муску-
латура.
Процесс уменьшения крыльев и специализация к водному образу
жизни в ряде групп заходят настолько далеко, что ряд видов уже
потерял способность летать или обладает весьма несовершенным
полетом. Среди буревестниковых на последней стадии оказались
представители рода Pelecanoid.es, способные только к короткому
трепещущему полету. Среди нырковых уток, утка-пароход [77/-
chyeres cinereus (Gm.)] совсем потеряла способность к полету. Край-
ней специализации и потери способности к полету достигли пин-
гвины. Редукция крыльев наблюдается и у ряда представителен
группы, ныряющей при помощи гребных движений ног, но здесь
это вызвано как крайней специализацией к водному образу жизни,
так и сравнительной безопасностью от врагов. В процессе эволю-
ции крылья оказались просто ненужными в жизни вида. Такой про-
цесс наблюдается в группе бакланов (Nannopterum harrisi RoLsch.),
поганок (CeMropelind). чистиков (Pingiiinus impennis L.) п в группе,
сходной с гагарами. — Hesperornithifonnes (Hesperornis).
Благоприятствующим моментом этой эволюции является проти-
воположность требований к строению птицы со стороны условий
жизни в воде и в воздухе. Развитие способности к плаванию и ны-
рянию требует уплощения и удлинения туловища, возможность
увеличения’ удельного веса, уменьшения размеров крыльев, что
противоречит условиям полета в воздухе.
Применение крыльев для движения под водой дает скорость,
близкую к Toil, которая развивается птицами гребными движениями
лап и, как и в последнем случае, превышает скорость движения
по поверхности воды. Исключительном скоростью движения обла-
дают пингвины, которые, по определению Мэрфи (Murphy, 1915)4
достигают в некоторых случаях 10 м в секунду. О силе движения,
развиваемой этими птицами, легко судить по прыжкам, которые
они делают из воды на высоту 2 м, стремясь вскочить на морской
лед, круто обрывающийся в глубокую воду.
П о л е т птиц'
Особенности строения крыла, обеспечивающие способность
к полету, весьма разнообразны. Они выражаются: в характере
сочленений отдельных костей передней конечности (см. стр. 63),
в мощности п расположении мускулатуры (см. стр. 78—83), в форме
крыла и относительных размерах его к массе тела и соотно-
шении частей крыла между собой, а также в структуре составляющих
его маховых и кроющих перьев. Последняя группа особенностей
может быть рассмотрена в связи с полетом в настоящей главе.
Образующие поддерживающие плоскости маховые перья при-
крепляются к поверхности локтевой и кистевых костей крыла п
по мере’удаления от основания крыла увеличиваются в крепости,
эластичности и обычно величине. Как мы видели ранее (см. стр. 30),
они разделяются на малые и большие маховые перья. На разверну-
том крыле малые маховые оказываются прикрепленными очинами
180
более или менее перпендикулярно к локтевой кости, большие же
маховые, по мере удаления от проксимальной части крыла, оказы-
ваются прикрепленными под острым углом к кости, п этот угол все
уменьшается в наиболее дистальных перьях; последнее (плп пер-
вое) большое маховое прикрепляется вдоль конечной фаланги 2-го
пальца и образует таким образом прямолинейное продолжение длин-
ной осп крыла. На 2-м и 3-м пальцах крыла, обладающих некоторой,
хотя и слабой, подвижностью, укрепляются четыре первых ма-
ховых, в то время как остальные шесть располагаются поперек
метакарпальных костей крыла, прикрепляясь к обеим их ветвям.
Лишь в редких случаях количество больших маховых превышает
десять штук, зато количество малых маховых весьма изменчиво,
стоит в связи с длиной костей предплечья и у особо длиннокрылых
птиц может достигать тридцати семи штук.
Все маховые расположены черепицеобразно, таким образом, что
наружное узкое опахало пера прикрывает широкое внутреннее
опахало соседнего, что имеет большое значение при полете. При
давлении воздуха на плоскость распростертого крыла снизу, вну-
тренние опахала плотно пригибаются, так что получается сплошная
поверхность: при давлении же сверху, внутренние спахала отги-
баются, так что между маховыми получаются зазоры, и этим умень-
шается сопротивление крыла воздуху. Просветы, образуемые очи-
нами, у. основания перьев прикрываются верхними и нпжнимн крою-
щими крыла; при этом, в связи с задачей укрепления слабо при-
крепленных малых маховых (укрепленных в надкостнице только
концами очинов), большие кроющие перекрещивают принадлежа-
щее им маховое, в то время как в дистальной части крыла они лежат
параллельно стержням больших маховых. Остальная поверхность
скелета крыла прикрывается средними и мелкими, кроющими, ко-
торые закругляют передний край его и создают ему легко обтекае-
мую форму, напоминающую в поперечном разрезе форму поддержи-
вающих поверхностей у современных самолетов.
Маховое перо является в высокой степени эластичным образо-
ванием, допускающим значительный изгиб и вместе с тем связность
составляющих его бородок. Нормально перо несколько согнуто
в сагиттальном и поперечном направлении, так что является выпук-
лым на спинной стороне. В полете, под давлением воздуха, перо
выпрямляется и может даже несколько изгибаться в противополож-
ном направлении. Наружное и внутреннее опахала, будучи почти
симметричны друг другу во внутренних малых маховых, в напра-
влении к дистальному концу крыла, становятся все более несим-
метрично развитыми на маховых перьях, при этом наружное опахало
сильно суживается, а внутреннее сохраняет прежние размеры пли
несколько расширяется. При этом у птиц, обладающих парящим
полетом, на дистальных маховых происходит в вершинной их по-
ловине сужение обоих опахал, вследствие чего на распростертом
крыле они выступают по краю крыла в форме широко растопырен-
ных пальцев и называются пропеллерными маховыми. Устойчивость
опахал давлению воздуха обеспечивается структурой бородок I и
" порядка. Бородки 1 порядка расположены по отношению к оси
.181
давления своей узкой стороной и вследствие этого особенно упруги,
кроме того, при выпрямлении пера в воздухе они располагаются
по отношению к стержню под более острым углом и теснее приле-
гают друг к другу. Под влиянием того же процесса происходит лег-
кий поворот по осп широких базальных частей бородок II порядка,
вследствие чего опахало принимает более плотную структуру и
крепче происходит сцепление микроскопических роговых крю-
чочков.
Форма крыла, так же как и строение маховых перьев, обусловли-
вает особенности полета птиц. В этом отношении можно различать
несколько основных типов крыловой поверхности, соединенных
между собой однако достаточно обильными переходами.
1. Острым крылом, при умеренно длинном предплечье, обладают
ласточки, стрижи, некоторые соколы, щурки, крачки, многие ку-
лики. Их полет отличается большой быстротой и допускает как
гребной способ полета, так и скольжение и иногда парение.
2. Широким коротким крылом обладает большинство воробьиных.
Им свойствен преимущественно так называемый трепещущий волно-
образный полет, а более крупным формам — и скользяще-гребной
полет, не отличающиеся значительной скоростью, но довольно эко-
номные по затрате мускульной энергии.
3. Широким длинным крылом с резко выступающими пропел-
лерными маховыми обладают крупные хищные, многие веслоногие,
аисты и цапли и др. Этот тип крыльев обычно связан с способностью
парить, но допускает и скользяще-гребной полет.
4. Широким коротким крылом при круто согнутых маховых
обладают многие куриные птицы, трехперстки, болотные курочки
и др. Они обладают быстрым, ио очень неловким полетом, и быстро
истощают имеющийся запас энергии. Птицы летят преимущественно
скользяще-гребным полетом, но прибегают и к скользящему полету.
3. Очень узким длинным крылом, при весьма удлиненном
предплечье, обладают трубконосые птицы, отличающиеся весьма со-
вершенной способностью к скользящему и парящему полету.
Однако не следует думать, что форма крыла всегда определяет
способ и искусство полета. Нередко совершенно одинаковая форма
крыла сопровождается различными способностями птиц к полету,
вследствие большего пли меньшего развития мускулатуры или иных
свойств. Существенно важной для развития того или иного типа
полета является также величина птицы, так как это определяет
механические свойства полета. Чем крупнее птица, тем она легче
летает, парящим п скользяще-гребным полетом, так как с увеличе-
нием массы тела уменьшается относительное лобовое сопротивление.
Но это же свойство (вес птицы) уменьшает способность к трепещу*
щему полету, так как последний требует затраты большого коли-
чества мускульной энергии, у крупных птиц всегда относительно
меньшей, чем у мелких, согласно с общим биологическим законом
о зависимости силы животного от его величины.
Механические основы полета птиц. Механи-
чески функция крыльев в простейшем полете птиц ближе всего
может быть уподоблена движению поддерживающих плоскостей
182
планера в воздухе. Исследования полета аналогичных аппаратов
установили, что при таком движении плоскости, при котором ее по-
верхность будет образовывать острый угол с осью движения (в вер-
тикальном разрезе), образующееся сопротивление воздушной среды
будет оказывать давление на плоскость, почти перпендикулярное
ей, и обладать определенной подъемной силой. Эта подъемная сила
изменяется в грубых чертах прямо пропорционально площади пло-
скости п зависит от синуса вышеуказанного угла наклона плоскости
(или угла атаки) п квадрата скорости движения; вместе с тем она
обратно пропорциональна тяжести движущегося тела (ср. рис. 160).
Увеличение угла атаки (в известных, ограниченных размерах) есте-
ственно ведет как к увеличению подъемной силы, так и к уменьшению
скорости движения. Таким образом, следовательно, увеличение скоро-
сти движения может быть получено уменьшением угла атаки. В свою
очередь, от величины угла атаки зависит положение центра давле-
ния (пли точки опоры о воздух). При угле атаки, близком к 90° (или
при вертикальном, к осп движения, положении плоскости), давле-
ние является наибольшим в центре плоскости и равномерно убывает
к краям, так как здесь обтекающий воздух оказывает меньшее со-
противление. При уменьшении угла атаки, точка опоры передви-
гается кпереди и тем больше, чем меньше угол атаки, что и вызывает
наибольшее развитие подъемной силы в передней части плоскости.
При этих же условиях точка опоры тем ближе передвигается к пе-
реднему краю плоскости, чем больше скорость ее движения. По-
следние положения объясняют, почему продолговатые плоскости
более удобны для полета: величина подъемной силы зависит от
отношения формы плоскости к оси движения, в случае же располо-
жения длинной стороны перпендикулярно к оси движения, плоскость
получает большее давление снизу. Опытом также установлено, что
вогнутые книзу поверхности обладают большей подъемной силой,
чехМ плоские, причем такие поверхности сохраняют свою подъемную
силу и при несколько наклонной вперед хорде. Утолщение передней
стороны плоскости при этом совершенно’ незначительно увеличивает
лобовое сопротивление и весьма благоприятно для лучшего исполь-
зования подъемной силы; задний край, наоборот, в случае отсут-
ствия утолщения и наличия отгибаемости, создает большую устой-
чивость движения, облегчая стекание воздуха и препятствуя его
вибрации. Другими средствами устойчивости является положение
центра тяжести плоскости между ее передним ребром и точкой опоры
и наличием позади главной плоскости дополнительной, играющей
роль стабилизатора. Большую роль в полете играет также общая
обтекаемость форм движущегося тела, снижающая лобовое сопро-
тивление п позволяющая воздуху без образования вредных зави-
хрений стекать с поверхности тела. Наиболее обтекаемые формы
обладают плавными закругленными контурами, суженными кпереди
и кзади.
Отмеченные особенности находят полное приложение к строению
летящей птицы. Несущие плоскости у птицы представлены кры-
льями, удовлетворяющими вполне требованиям механики: они про-
долговаты и расположены длинной стороной перпендикулярно к оси
183
полета, поверхность их несколько вогнута снизу, передний край
утолщен и удобно обтекаем, задний край эластичен п может вслед-
ствие этого допускать дорзовентральные изгибы. Общая форма ле-
тящей птицы с вытянутым вперед клювом, плотно прилегающим
к телу оперением, суживающимся в большинстве случаев хвостом
весьма удобно обтекаема. Вытянутое туловище, хвост, а иногда и
длинные выставляемые назад ноги могут играть роль стабилиза-
тора. Другим условием устойчивости у птиц является расположение
центра тяжести тела значительно ниже основной плоскости опоры,
что достигается высоким прикреплением крыльев к телу п положе-
нием тяжелых органов — грудной мускулатуры, пищеварительных
органов и т. д. — книзу от этой плоскости. В строении крыльев,
как несущих плоскостей, у птицы имеется ряд преимуществ перед
летальными аппаратами человека. Главнейшие из них — это спо-
собность по произволу уменьшать и увеличивать их площадь боль-
шим плп меньшим расправлением крыла. Этим птица меняет крыло-
вую нагрузку, т. е. отношение веса тела к площади крыла, что
имеет большое значение в механике полета. Чем больше крыловая
нагрузка, тем больше скорость движения птицы. Изменением кры-
ловой нагрузки таким образом птица может отчасти управлять своей
скоростью и в доступных ей рамках всегда соблюдать мехаппческп-
онтимальные условия. Возможность перемещать крылья относи-
тельно тела более вперед и кзади позволяет перемещать центр тя-
жести тела соответственно требованиям скорости движения и меха-
нике поворотов. В сложных случаях птица использует и другие
части тела, голову, шею, ноги, при их помощи сохраняя равновесие
и требуемую ловкость изменения пути движения. Хотя уменье
использовать свои органы на полете в значительной степени зало-
жено в организме птицы, совершенство этих движений достигается
у большинства летунов длительным периодом обучения и после-
дующей практикой.
Способы полета. Обратимся теперь к рассмотрению самих
способов полета. Наблюдаемый нами полет птиц бывает двух видов:
это или передвижение в воздухе без движения (плп почти без дви-
жения) крыльями, застывшими в одном положении (и лишь слегка
меняющими крыловую нагрузку, частичным уменьшением их пло-
щади), плп передвижение при помощи ударов крыльями, которыми
птица как бы поддерживает себя в воздухе. Первый удобно мо-
жет быть назван п л а н и р у ю щ и м пли парящим по-
ле т о м и объединяет все те случаи, когда животное скользит
вниз в воздухе, «парит», летая кругами, пли даже поднимается
вверх силой воздушного течения без движений крыльями. Второй
называется г р е б н ы м или (лучше) пропел л и р у ю щ и м п о-
л е т о м и объединяет разнообразные движения, сопровождаемые
маханьем крыльями, как. например, плавный гребной полет во-
рона, быстрый полет утки, фазана, трепещущий полет колибри
и т. д. Такие частные особенности в пределах обоих типов полета
обусловливаются конечно более частными моментами в механике
полета. Развитие способности к тому плп иному типу полета у мно-
гих форм птиц достигает, значительной специализации, при которой
J84'
может иметь место ограниченная способность пользоваться другим
типом полета. В других случаях однако способности к разнообраз-
ному полету сохраняются, и различные способы движения вклю-
чаются в полет вида как разные фазы, приспособленные к выполне-
нию топ или иной специальной задачи в жизни животного.
Ниже описываются лишь основные способы полета.
Л. II л а н и р у ю щ и й п о л е т
I. Скользя 1ц и и п о л е т наблюдается обычно у птиц
выше галки величиной и является весьма распространенным видом
полета. У огромного большинства видов это одна из фаз обычных
способов передвижения в воздухе. Начальная сила движения вперед
получается или в результате отталкивания при взлете пли разви-
вается в гребном полете. При этом полете птица скользит по наклон-
ной плоскости, почти не двигая более
пли менее расправленными крыльями.
Механика этого полета полностью сов-
падает с изложенными выше закономер-
ностями в движении плоскостей пла-
нера. Приложение этих закономерно-
стей к скользящему полету птицы мы
видим на прилагаемом рисунке (рис.
160); АВ представляет наклонную пло-
скость скольжения, проводимую через
центр тяжести тела; С’—плоскость рас-
правленного крыла, образующая с АВ
угол а, пли угол атаки. Направление
действия обра зующейся подъемной
силы указано компонентом 5У, про-
рис. 160. Схема взаимодействии сил
при скольжении птицы.
1В — плоскость скольжения цен-
тра тп жести тела; С — плоскость
расправленного крыла: SG—на-
правление действия силы тяжести:
S У — направление подъемной силы :
SV — сила, движущая птицу впе-
ред; SZ — сила лобового сопротив-
ления ; а — угол атаки. См. текст.
Но Lorenz.
тпвоположной по значению силе тя-
жести SG. Сила, движущая птицу вперед, отложена па АВ в виде
слагающей SV силы тяготения SG. Противодействующей ей силой
является слагающая SZ подъемной силы 5У, выражающей вели-
чину лобового сопротивления, испытываемого птицей. Как мы ви-
дели выше, 5У — подъемная сила, изменяется пропорционально
площади крыльев, квадрату скорости движения и синусу угла атаки.
Сила, движущая вперед, или быстрота скольжения, следовательно,
обратно пропорциональна площади крыльев, синусу угла атаки
и величине лобового сопротивления. Таким образом мы видим,
что предпосылки для достижения большей скорости движения при
отом полете и пловучести в воздухе противоречивы друг другу, и со-
вершенствование того или иного качества неизменно снижает воз-
можности противоположного. Процесс специализации в том или
Другом направлении осложняется кроме того некоторыми механи-
ческими неудобствами, определяющими грань возможностям поль-
зоваться тем или иным видом планирующего полета. Так лобовое
сопротивление, изменяющееся прямо пропорционально плоскости
сопротивления, при уменьшении величины птицы становится важ-
ным критическим фактором. При увеличении птицы, плоскость со-
1.85
противления ее тела увеличивается в квадрате, а вес в кубе, следо-
вательно лобовое сопротивление относительно уменьшается. Такны
образом с уменьшением величины птицы лобовое сопротивление резко
возрастает, что затрудняет скольжение небольших птиц и препят-
ствует скольжению мелких видов. Угол наклона плоскости сколь-
жения по мере уменьшения размеров тела становится по необходи-
мости все более крутым и у мелких воробьиных приближается к 45е.
что биологически оказывается неприемлемым. Вес тела при своем
увеличении также становится критическим для возможностей по-
лета, так как он не должен переходить определенных отношений
к поверхности крыла и при значительном увеличении потребовал
Рис. 1G1. Положение крыльев и маховых у парящего и скользящего
сипа —Gyps fulvus llalb.
(а) —парящий против ветра, с набором высоты; (Ъ) —без изменения
высоты; (с) —парящий по ветру, с потерей высоты; (d,e,f) — скользя-
щий, с различным положением крыльев. По Ahlborn, из Stresernann.
бы развития огромных крыльев, биологически и механически для
птицы невозможных. Это объясняет, почему летающие птицы не пре-
вышают размеров кондора и альбатроса и почему более крупные
формы, как, например, страус, являются нелетающими.
2. П а р я щ и и п о л е т наблюдается обычно у крупных птпп
и заключается в том, что птица скользит без взмахов крыльями,
не теряя высоты или даже поднимаясь вверх (рис. 161). Необходимая
энергия для полета этим способом извлекается из восходящих воз-
душных течений, которые весьма нередки и возникают несколькими
способами. Главнейшими из них для парения сухопутных птин
являются течения термического происхождения. Поверхность земли
освещенная солнцем, нагревается сильнее атмосферы; от нее воздух
прилегающих слоев расширяется и, сделавшись легче вышележащих
слоев, поднимается вверх. Такие течения особенно сильны в странах
с теплым климатом и открытым ландшафтом, там, где сильна инсо-
Л 86
ляция ii интенсивен нагрев почвы. Нередко эти течения вследствие
своей мощности поднимаются до больших высот. Находясь в таком
течении, птица плавно и ровно скользит в поднимающейся среде
и вследствие этого поднимается вместе с ней. При этом птицы обычно
описывают круги, при достаточной шпроте течения совсем не выходя
из его пределов и все время поднимаясь медленным и ровным тем-
пом. Другого типа течения получаются при ветре, вследствие различ-
ной скорости горизонтального движения воздуха на разной высоте.
Ближе к земле, от трения об ее поверхность и различные предметы,
движение воздуха является более замедленным и сопровождается
образованием вихрей как с вертикальной, так и горизонтальной
осью вращения. Последние как бы катятся по поверхности земли,
причем с передней надветренной стороны их воздух поднимается,
а с задней подветренной — опускается. Площади восходящих те-
чений в этих потоках сравнительно невелики, а вследствие горизон-
тального перемещения вихря они все время перемещаются в одном
направлении. Хищные птицы, пользующиеся этими потоками для
парения, все время описывают круги, стараясь возможно чаще пере-
летать восходящее течение, которое их поднимает. Наконец восходя-
щие течения воздуха образуются также при столкновении ветра
с каким-либо вертикальным препятствием, как то: склон горы, об-
рыв, опушка леса, от которого ветер отражается отчасти вверх.
Эти течения не перемещаются и, смотря по расположению преграды,
могут распространяться на значительное пространство. Такими те-
чениями особенно часто пользуются у берегов моря чайки и в горах
хищники.
Сходно с этими течениями воздуха образуются восходящие токи
на море, где ветер отражается от надветренной стороны волны. По-
лучающиеся обильные, но невысокие воздушные течения исполь-
зуются для полета длиннокрылыми трубкопосымп птицами — бу-
ревестниками и альбатросами. Запасшись энергией, они могут на
время подняться значительно выше в воздух, но потом вновь сколь-
зят к поверхности воды, вновь поднимаются кверху и т. д.
Как уже указывалось, парящим полетом пользуются главным об-
разом крупные птицы с большими и длинными или широкими крылья-
ми п сравнительно низкой крыловой нагрузкой; в силу этих осо-
бенностей они обладают весьма пологим углом плоскости скольжения.
В. Г р е б п о й п л п н р оно л л п р у ю щ и й и о л с т
1. Трепещущий полет является наиболее активным
типом полета, требующим большой затраты мускульной энергии.
Им пользуются все птицы во время взлетов и приземления, а мелкие
11 во время полета вдаль, в горизонтальном направлении. Движения
крыльев при этом не только поддерживают птицу в воздухе, но и дают
значительный толчок вперед и допускают весьма крутые подъемы;
размах крыльев при этом всегда более значителен и делается с ви-
димым усилием. Поднимающаяся птица держит тело круто припод-
нятым, так что взмахи крыльев происходят почти в горизонтальной
плоскости, вследствие чего тело поднимается круто кверху. При
187
иарата к парящему полету и
Рис. 162. Взмах крыла во время тре-
пещущего полета. Стрелками указано
распределение давления воздуха на
дорзальную сторону больших махо-
вых. По Lorenz.
летом поднимается вверх и в
опускании крыла оно поворачивается передним краем вперед,
как бы загребая воздух и в общем своим действием весьма напоми-
ная работу воздушного винта. При подъеме крыла дистальная часть
его отворачивается таким образом, что его плоскость образует с плос-
костью малых маховых тупой угол градусов в 100—130. при этом
большие маховые разъединяются и, получая давление теперь с верх-
ней стороны крыла, являются таким образом и во время подъема плос-
костью, содействующей движению тела вперед (рис. 162).
Затрата мускульной энергии на этот тип полета настолько ве-
лика, что крупные птицы, вообще относительно массы своего тела
гораздо более слабосильные, чем мелкие, взбегают пользоваться
этим полетом сколько-нибудь долгое время. У наиболее крупных
видов, таких, как орлы, грифы, альбатросы, эта способность вообще
мало развита, вследствие специализовапиости их летательного ап-
слишком большой тяжести их тела.
Мелкие птицы, наоборот, широко
пользуются этим полетом и в силу .ме-
ханических причин, при полетах
вдаль главным образом, принуждены
пользоваться именно трепещущим по-
летом. Наиболее распространен среди
них так называемый волнообразный
полет, состоя! цп it из отдельных
сменяющих друг друга траэкторий.
При этом птица быстрыми ударами
крыльев (трепещущий полет) подни-
мается косо вверх и, развив необхо-
димую скорость, прижимает к телу
крылья и опускается далее по по-
логой траэкторви подобно летящей
нули. Утратив необходимую ско-
рость, она опять трепещущим но-
ющем повторяет тот же маневр. Этот
способ полета экономит затрачиваемую им работу, а при длитель-
ности полета зачастую и единственно возможен, так как вполне го-
ризонтальное передвижение трепещущим полетом крайне невыгодно,
вследствие очень большого лобового сопротивления (косо к осп дви-
жения поставленное тело) и малой скорости, только и возможной при
таком полете. Избегая большого лобового сопротивления и в волни-
стом полете, птицы в снижающейся частя траектории не держат
крылья распростертыми, а прижимают их к телу; отсутствие работы
в этот период компенсирует повышенную затрату энергии в на-
чале описываемой траектории.
2. Скользя щ е-г р е б н о й иоле т. Этот тип полета свой-
ствен всем крупным и средним по величине птицам во время пх
горизонтального передвижения вдаль. 13 это время птица предста-
вляется наблюдателю равномерно, и обычно неторопливо, машущей
крыльями, а тело ее расположенным почти в горизонтальном поло-
жении (рис. 163). Как это было выяснено наблюдениями Лилиенталя
и Лоренца (Lorenz, 1933), этот тип полета представляет собой соче-
1R8
танпе скольжения с активными взмахами, поддерживающими тело
в воздухе, или, в формулировке Лоренца, скольжение с возмеще-
нием высоты. Протекание этого движения представляется в следую-
щем виде: при опускании крыльев с силой Ъниз, давление, встречае-
мое разными частями их, неравномерно; малые маховые, имеющие
меньший размах движения, испытывают мепьшее давление, большие,
описывающие большую дугу, — более значительное. Так как наи-
более дистальные маховые при расправленном крыле находятся
под углом к оси движения, а точка опоры располагается у перед-
него края крыла, то давление воздуха вызывает частичный поворот
кисти передним краем вниз и отгиб кверху заднего края плоскости
больших маховых. В результате этого тело получает как подъем
тела кверху, так в некоторую поступательную энергию. Начинаю-
щийся вслед за этим взмах крыльями переводит кисть в обратное
положение и активно увеличивает угол атаки дистальной части крыла.
Встречным давлением воздуха крылья, без особой затраты мускуль-
ной энергии со стороны птицы, поднимаются кверху и становятся
в начальное положение. Во время этого взмаха крыльями птица
естественно несколько опускается книзу, скользя в то же время
весьма полого вперед, по
следующий же удар 1
крыльями вниз подни- __---
мает ее на прежнюю вы-
соту и помимо уже имею- - V^C_jL-
щейся скорости сколь-
жения дает ДОПОЛНИ- Гис- схема Г1а».п.:м11це-1’|)ебп<»го полета. По Lorenz’
тельный толчок вперед.
Малые маховые, при этом мало меняющие угол атаки п мало откло-
няющиеся от горизонтального положения, играют главным образом
роль поддерживающих плоскостей, большие же маховые выполняют
всю активную работу по подъему тела и отчасти продвижению его
вперед. Скорость, даваемая этим полетом, может быть очень значи-
тельной, н большинство наиболее быстро летающих птиц достигает
значительной скорости как раз именно в этом полете. Некоторые из
них произвольно могут регулировать быстроту движения, прибегая
к более сильным и глубоким ударам. При этом после каждого удара
крыльями вниз птица часто уменьшает их площадь, притягивая их
к телу, что увеличивает крыловую нагрузку и уменьшает лобовое
сопротивление, значительно увеличивающее скорость. Эта способ-
ность более широко свойственна тем птицам, которые обладают не
слишком большими или не слишком маленькими крыльями и имеют
Достаточно сильную крыловую мускулатуру.
Вкратце следует рассмотреть также те движения, которые свой-
ственны птице при стремлении подняться в воздух или опуститься
нв землю плп тот пли иной насест.
Взлет, так же как и посадка, по-разному осуществляется
птицами в зависимости от их способности к тому пли иному типу
полета. Мелкие птицы, в совершенстве владеющие трепещущим по-
петом, наиболее легко взлетают, будучи способны нередко почти
вертикально подниматься вверх и даже задерживаться в воздухе
I *чо
на одном месте на короткое время. Более крупным птицам подъем
с места трепещущим полетом стоит очень значительных усилий,
и они предпочитают помогать ему толчками ног, несколькими прыж-
ками, причем подъем вертикально вверх для них совсем недоступен,
и они лишь постепенно взбирают в высь. Бегающие птицы, как жу-
равль, дрофа и др., предпочитают при этом брать разбег, после ко-
торого они постепенно поднимаются в воздух. Наиболее труден старт
для водоплавающих птиц с тяжелым телом и маленькими крыльями.
Как правило, они первоначально плывут, затем начинают ударять
Рпс. 1G4. Взлет орла беркута — Aquila chrysaetos L.
1 —птица с силой отталкивается от скалы, взвивает крылья вверх, в почти верти-
кальную плоскость. и начинает расправлять кистевое, сочленение; 2, 3 и 4 — подъем в
воздухе. Обратите внимание на положение маховых перьев и крылышка. По Knight,
из Stresemann.
крыльями по воде; при постепенном ускорении, тело все выше выходит
из воды, п при достижении минимальной скорости, необходимой для
скользяще-гребного полета, птицы совсем отделяются от воды. Взлету
крупных птиц сильно помогает ветер; против ветра многие из них прямо
с места могут взлететь скользяще-гребным полетом. Крупные птицы,
присаживающиеся на возвышенных местах, слетают, бросаясь вниз,
и, развив этим достаточную скорость, переходят на скользяще-греб-
ной или скользящий полет (например,сразу начиная парить) (рис. 164).
Посадка большинства птиц сопровождается встречным тре-
пещущим движением крыльями, т. е. по своему направлению обрат-
ным тому, которое развивалось птицами при подъеме. Мелкие птицы
этим способом получают вполне достаточное торможение, так что
посадка для них не представляет особой трудности. Более крупные,
190
однако, садясь на возвышенные предметы, опускаются сначала
ниже их п погашают слишком большую скорость скользящим дви-
жением вверх перед посадкой. Таким же способом, в прин-
ципе, садятся многие птпцы, например вороны, на землю. Они летят
некоторое время над самой землей, потом слегка взлетают и опу-
скаются на землю, стараясь повернуть тело против направления дви-
жения и погашая еще не израсходованную скорость несколькими
прыжками по земле. Торможение часто еще имеет место задолго до
посадки; оно достигается или изменением положения тела поперек
к оси полета пли косо,плп изменением положения крыльев, превращая
их как бы в парашют и опуская их концы, что делает «парашют»
более выпуклым и более эффективно тормозящим.
При посадке на воде крупные водоплавающие птицы тормозят
скольжением по самой поверхности воды, чем они напоминают по-
садку гидросамолетов.
Основные направления в специализации
полета. Как уже указывалось, морфологические и механические
свойства той или иной птицы в значительной мере предопределяют
характер ее полета, как и вообще возможный объем доступных ей
движений в воздухе. Вместе с тем выяснено, что использование ме-
ханической силы ветра в том пли ином его выражении требует опре-
деленной структуры летающих поверхностей и аппаратов, причем
эффективное использование какого-либо одного свойства воздушной
среды или аппарата неизбежно понижает приспособленность этого
аппарата к эффективному использованию других свойств и снижает
в этом направлении качество полета. Из этого следует, что перво-
классная способность к полету означает высокое использование ле-
тательных свойств в каком-либо одном направлении, всегда сочетаю-
щееся с наличием недостатков в полете другими способами. По
мнению Лоренца (Lorenz, 1933), может быть установлено пять основ-
ных направлений специализации полета, причем между ними воз-
можны и существуют различные переходные состояния.
В кратком виде предлагаемая им схема заключается в следующем.
А. Развитие в направлении быстрого, сильного и активного по-
лета, способного бороться с сильным ветром. Предпосылками для
этого полета являются: 1) наличие относительно высокой крыловой
нагрузки, предрасполагающей к значительной скорости движения
при скользящем и скользяще-гребном полете; 2) малое лобовое
сопротивление, увеличивающее скорость движения; 3) короткие,
упругие маховые перья, и 4) возможность широкого размаха крыла.
Первое, второе и третье качества (взятые вместе) обычно свя-
заны с узкой и острой формой кры%а, с большими маховыми, стоя-
щими в соприкосновении друг с другом своими опахалами до самой
вершины. Эти качества соединяются с возможностью широкого раз-
маха преимущественно двумя способами: или сравнительно неболь-
шая крыловая площадь способна совершать большой размах вслед-
ствие длинной плечевой кости, что свойственно, например, многим
трубконосым птицам, или небольшое, узкое и острое крыло обладает
очень быстрыми ударами и таким образом сравнительно мало теряет за-
пое энергии. Последний способ, например, свойствен нырковым уткам.
19L
В. Развитие в направлении более верткого и легкотормозимого
полета. Предпосылками являются: 1) наличие низкой крыловой на-
грузки: 2) незначительный размах обоих крыльев, и 3) наличие боль-
ших рулевых перьев. Эти качества легко соединимы друг с другом
обычно с значительным расширением проксимальной части крыла..
Типичным полетом этого рода Лоренц считает полет турако (Тп-
racos); в этом же направлении развито крыло сойки, кукши, лесных
ястребов и др.
С. Развитие в направлении к быстрому ускорению полета (к бы-
строму набиранию начальной скорости полета) посредством трепе-
щущего способа продвижения в воздухе. Предпосылками являются:
1) сильно выгнутые дистальные маховые (выпуклое крыло с пропел-
лерными большими маховыми); 2) короткое крыло, и 3) способность
к затрате большого количества мускульной силы, хотя бы на корот-
кий промежуток времени.
Типичным представителем птицы с таким полетом является
фазан.
D. Развитие способности к трепещущему полету на одном месте
(с задержкой в воздухе на одном месте). Предпосылками являются:
1) маленькое, острое п узкое крыло, способное к очень быстрым дви-
жениям и поэтому сравнительно экономно затрачивающее необхо-
димую энергию; 2) значительная сила и выносливость мускулатуры
(в связи с возможно малыми размерами тела); 3) возможно большее
число больших маховых, расположенных перпендикулярно к осп
крылового удара, и 4) мускулатура, поднимающая крыло, приблизи-
тельно пропорциональная мускулатуре, опускающей крыло. Наи-
более развитым в этом направлении типом является колибри.
Е. Развитие в направлении достижения длительного (безустан-
ного) поискового полета и планирования. Предпосылками являются:
1) плавное (сравнительно медленное) снижение при скольжении,
и 2) большой размах обоих крыльев и следовательно связанная
с этим значительная способность к боковой устойчивости. Это до-
стигается как уменьшением крыловой нагрузки (например сарыч),
так и сравнительным уменьшением лобового сопротивления, например
увеличением абсолютной величины тела, что мы наблюдаем у грифов.
Механические задачи того или иного полета далеко не всюду
позволяют развитие переходных типов. В большинстве случаев эти
переходные типы достигаются довольно разнообразно, в зависимости
от величины птицы и ее систематического положения. Совместимость
в полете одного вида качеств разных направлений возможна, по мне-
нию Лоренца, следующим образом.
АсВ. Быстрый (скоростной) *полет с поворотливостью. Здесь
с требованиями скоростного полета совместима только одна пред-
посылка поворотливости — длинные рули, остальные же предпосылки
(А1 и Bl, А4 и В2) прямо противоположны друг другу. Возможность
сочетания длинного хвоста с скоростным полетом, с задачей увели-
чения поворотливости птицы, мы находим у ласточек, фрегатов,
рябков и нек. др. Длинные хвосты обычно при этом вытянуты в одну
или две острых косицы, обеспечивающие лучшее стекание воздух'8
и устраняющие таким образом образование вихрей и вибрацию перьев.
192
В большинстве случаев все же там, где при скоростном полете нужна
значительная поворотливость, она возмещается способностью к тре-
пещущему полету на месте.
АсС. Быстрый полет со способностью к ускорению полета трепе-
щущими движениями. Единственной. предпосылкой последнего, по
мнению Лоренца, вполне соединимой с быстрым полетом, является
большая мускульная сила (С4). Эта сила дает голубям и некоторым
попугаям, вообще более приближающимся к типу А, способность
к проявлению большего ускорения в определенные моменты полета
и к получению нужной скорости при старте. Как правило, быстро
летающие длиннокрылые птицы, с относительно слабой мускулату-
рой, опускающей крылья, и длинным плечевым отделом, с большим
трудом взлетают с ровного места при тихой погоде (например фре-
гат, многие буревестники). Необходимость малого лобового сопро-
тивления в типе (А2) стоит в противоречии с выпуклыми пропеллер-
ными маховыми (С1), важными для получения большой начальной
скорости; вредное действие последних для развития большой ско-
рости парализуется в некоторых случаях (например перепелятник)
частичным сгибом крыла в кисти и подгибанием пальцев, что делает
поверхность крыла цельной и заостренной.
AcD. Быстрый полет со способностью трепещущего полета на месте.
Быстро летающие птицы с острым, коротким, узким крылом часто
могут прекрасно трепетать на одном месте (ласточки, голуби, неко-
торые попугаи), так как подобный тип крыла является одной из пред-
посылок обоих типов полета (А4 и D1). Однако некоторые виды с таким
же крылом вряд ли способны’ к проявлению этого качества, как,
например, стрижи, щурки, чеглоки и т. д. Вообще, кромё желатель-
ности высокой крыловой нагрузки, ни одна из предпосылок к бы-
строму полету с коротким крылом не противоречит условиям полу-
чения трепещущего полета, да и эффект высокой крыловой нагрузки
может быть снижен развитием соответствующей мускульной силы.
Только быстрые летуны с удлиненным плечевым отделом не могут
развивать трепещущий полет на месте.
АсЕ. Быстрый полет со способностью к длительному полету и пла-
нированию. Совмещение обеих способностей возможно при условии,
что медленность снижения при скольжении, необходимом для плани-
рования, достигается не столько снижением крыловой нагрузци,
сколько уменьшением лобового сопротивления. Это может быть
в результате более совершенно обтекаемой формы тела и несущих
плоскостей, что имеется, например, у альбатроса, и из хищников
У коршуна, или увеличением абсолютной величины тела, как в при-
мерах крупных орлов и различных грифов. Альбатросы могут быть
рассматриваемы как в равной мере удовлетворяющие задачам обоих
типов полета. Среди птиц с длительным, так называемым поисковым
полетом, как, например, чайки, встречается способность противо-
стоять сильным ветрам увеличением крыловой нагрузки частичными
’ гибами крыла.
ВсС. h оворотливость со способностью быстрого увеличения на-
чальной скорости трепещущим полетом вообще не противоречат
Друг другу и очень часто оказываются соединенными в полете вида.
1 а О рнптологи я—1209
193
Птицы, удовлетворяющие только первому типу полета, вообще по-
видимому редки.
BcD. Поворотливость сэ способностью трепещущего полета па
одном месте. Это качество, как замечает Лоренц, теоретически вполне
допустимо, практически невидимому никогда не совмещается водном
виде в сколько-нибудь значительном развитии, вследствие того, что
биологически они друг друга замещают.
ВсЕ. Поворотливость со способностью к длительному полету и пла-
нированию. Оба качества могут быть совмещены друг с другом при
очень низкой крыловой нагрузке и следовательно при условии,
когда птицам не приходится бороться с сильным ветром. Таков
полет многих сов (преимущественно лесных).
CcD. Способность к быстрому развитию большой начальной ско-
рости со способностью трепещущего полета на месте. Эти условия по
непонятным причинам почти несоединимы.
СсЕ. Способность к быстрому развитию большой начальной ско-
рости со способностью к длительному полету и планированию. Сов-
мещение этих качеств никогда не достигает значительного совер-
шенства уже потому, что длинные и большие крылья и значительная
величина тела препятствуют возможности развития большой ско-
рости непосредственно с места. Примером совмещения все же этих
качеств, по Лоренцу, является серая неясыть, которая хотя в основ-
ном летает по типу сарыча, имеет однако выпуклые, пропеллерные
маховые (короткие’и ограниченные очень небольшой частью крыла).
Более сильное развитие пропеллерных маховых наблюдается у ястре-
биной совы.
DcE. Способность к трепещущему полету на месте со способно-
стью к длительному полету и планированию. К этой комбинации
способны повпдимому только мелкие птицы последнего типа, как.
например, обыкновенная чайка и др., не достигающие однако боль-
шого совершенства в трепетании на месте в воздухе.
Этот обзор, конечно, не может претендовать на исчерпывающую
полноту, но удовлетворительно систематизирует различные укло-
нения в характере и сочетании возможностей в полете.
2. ОБЗОР ПРИСПОСОБЛЕНИЙ К ХАРАКТЕРУ МЕСТООБИТАНИЯ И
СПОСОБУ ДОБЫВАНИЯ ПИЩИ
В процессе эволюции птицы приспособились к весьма разнооо-
разной обстановке и дали удивительное изобилие систематических
форм. Но наиболее древняя для птиц среда — лесная растительность
оказалась очевидно наиболее благоприятной, так как здесь мы на-
ходим наибольшее разнообразие биологических типов и особенное
богатство видов. Наименее заселенной оказалась водная среда
а к жизни в земле, в почве, не перешел ни один из известных предста-
вителей этого класса.
Помимо тех глубоких изменений в организации животного, ко-
торые требовались приспособлением к полету, процесс эволюД^
птиц, как это уже указывалось ранее, не сопровождался резкими из-
менениями формы животных внутри самого класса. По развит*111
194
всех существенных птичьих особенностей, изменения в организме
птиц имели в сущности поверхностное идиоадаптивное значение,
вызывавшееся приспособлением к различным частным способам до-
бывания пищи и к качествам ее, а также стремлением к самосохра-
нению и размножению. Результатом этого является изобилие типов
окрасок и цветов, разнообразие в строении клювов и лап и т. д,,
при сравнительном однообразии внутренней глубокой организа-
ции. Если бы можно было из свойств всех отдельных видов синте-
зировать обобщенный тип птицы, этот тип слабо отличался бы от
произвольно взятого вида класса птиц. Если бы то же самое мы сде-
лали с млекопитающими и рептилиями, обобщенный тип их отли-
вался бы очень глубокими и многообразными чертами от каждого
любого вида этих классов.
Таким образом, следовательно, в сравнении например с млеко-
питающими, приспособления птиц оказались более односторонними
п в целом им свойственно меньшее разнообразие организации и ме-
нее глубокое использование всех сред, за исключением воздушной.
При всех этих особенностях нельзя однако отрицать, что каждая
птица несет все же штамп той среды, которой опа свойственна, п что
каждый узко взятый тип среды сходно модифицирует в одном напра-
влении всю населяющую его фауну птиц в целом. Так как система-
тический состав фауны в значительной мере определяется историче-
ским расселением видов, а разобщенность материков является глу-
боким препятствием для этого процесса, то родство форм отдельных
фаун далеко не однородно и определяется в значительной мере
характером современной и прошлой связи этих территорий между
собой. Среда же, даже в отдаленных участках суши, часто явля-
ется весьма сходной и по существу создает фауне одпе и те же
условия существования. Так, пустыни мы имеем в Азии, Африке,
Австралии, Северной и Южной Америке; скалистые и низменные
берега моря окаймляют все континенты; лес в разных модифика-
циях рассеян почти по всем странам. Это сходство в условиях су-
ществования фауны при различии ее систематического состава,
обусловленном историческими условиями, вызвало в эволюции птиц
многочисленные проявления конвергенции в виде выработки в раз-
ных систематических группах сходных типов адаптации.
Наиболее резкими примерами такой конвергенции являются
такие парные группы, как дневные хищники и совы, буревестники
11 чайки, пингвины и многие чистики, куропатки и скрытохвосты
и мн. др. (рис. 165). Все это группы, далеко удаленные друг от друга ио
своему родству, но внешне имеющие глубокие черты сходства, вслед-
ствие обитания в сходных условиях и выполнения сходной пли оди-
наковой роли в природе. Другим примером является приспособле-
ние типа «древолаза», например дятла, которое несмотря на широкое
распространение собственно дятлов появилось и в других группах,
особенно в семействе древолазов (Dendrocolaptidae)^ семействе
пищух (Certhiidae), семействе поползней USittidae), семействе тка-
ииков (Ploceidae) и др.
Таким образом развитие приспособлений, повторяющихся в фау-
11 ах при сходных условиях, является широко распространенным
195
сути
tv называемая система является ио
жизненных форм, как они понимаются
•
(2/
скопы —
zpylrrnen-
явлением. Беглый обзор показывает, что при всем их разнообразии
эти приспособления сравнительно легко группируются в ограничен;
нос число основных адаптивных типов. Возникает вопрос, не це-
лесообразно ли было бы объединить эти типы в какую-либо систему,
облегчающую обзор в помогающую осмысливать экологию весьма
многочисленного видами класса? Опыт такой системы разрабаты-
вается й настоящее время, но здесь мы можем воспользоваться
только самыми общими результатами, преследуя лишь цель ориен-
тировочного обзора.
дела классификацией
тер в экологии расте-
ний. Принципы уста-
новления ее однако
необходимо видоиз-
менять • от класса к
классу. вследствие
известной специфика-
ции жизненных от-
правлений разных си-
стематических групп.
Если, например, в
группе млекопитаю-
щих в качестве ос-
новных признаков в
группировке адапта-
ций наиболее удобен
характер питания в
совокупности с ха-
рактером занимаемо-
го жилья, для птин
более удобна груп-
пировка их по ха-
рактеру движений в
совокупности со спо-
собом и местом ра-
зыскивания нищи. Принимая это, мы основываемся на предпо-
ложении, что главнейшие локомоторные приспособления вида и его
хабптус определяются способом разыскивания и добывания пищи,
как основным жизненным мотивом, формирующим как облик, так
и жизнь животного п, в известной мере, избираемость тех или иных
стации. Остальные приспособления вида, какой бы важный характер
для существования вида они не имели, все же являются более част-
ными и, как правило, формируются эволюцией зависимо от первого
момента.
Устанавливаемая согласно этому принципу экологическая клас-
сификация птиц конечно не лишена затруднительных случаев. Эво-
люция пе может быть стиснута в произвольные узкие рамки и со-
здает много уклоняющихся от основных штампов форм. - Переход*
ные положения здесь так же обильны, как в генетической класси-
фикации, основанной на кровном родстве. Существенным является
Рис. 165. Явление конвергенции в строении лап
Pavdion haldwtvs L. (]) и рыбной совы — Ketupo
sis L- (2).
Обратите внимание на сходство чушуйчатых покровов, форму
ногтей и оборотнопалость 4-го пальца, возникшие в связи с
сходным способом добывания пищи. Среди дневных хищни-
ков скопа представляет единственный вид, обладающий
оборотнопалостью 4-го пальца, как то свойственно совам.
По Blanford, из SLreseniann.
19G
также то. что многие формы оказываются обладающими изумитель-
ной приспособляемостью, как при наличии узких структурных
особенностей, так и при отсутствии их.
В этой классификации целесообразно применение трех понятий:
<<тпп», «класс» п «форма». «Тип» устанавливается по о с и о в и о м у
характер у движений в поисках пищи, и это понятие ближе
всего отвечает основным жизненным форхмам растений в интерпре-
тации их Раункпером; количество типов естественно очень невелико.
По отношению к птицам, первые попытки общей классификации их
исходили как раз из этих экологических групп или категорий,
п мы можем отчасти заимствовать оттуда их номенклатуру. «Форма»
является группой видов, сходных друг с другом по хабитусу, дви-
жениям и способу добывания пшцп настолько, что отличия в этом
отношении между видами и родами малы или неуловимы; различия
выражаются главным образом в склонностях к разным (но близким)
биотопам (стациям) и самому составу пищи. Таковы, например, объ-
единяемые в одну «форму» все представители родов llotacilla, Calo-
bates, Budytes, Dendronanthus. Henicurus п пек. др. Сюда, как пра-
вило, входят целиком целые genum geographicum Рента (Rensch,
1929). Наконец «класс» является совокупностью ряда форм, сход-
ных по тем же признакам, но п более резко различающихся одна
от другой. Таков, например, класс жаворонков, объединяющий жа-
воронков собственно, коньков, трясогузок и т. д. Классы нередко
будут совпадать с границами отдельных семейств и отрядов, но в
других случаях обнаруживают более разнообразный ассортимент
систематических форм. Я предпочитаю называть классы произвольно
избираемой терминологией, учитывающей хабитус и способ движе-
ний, им свойственные.
Можно установить семь адаптивных типов:
1. Водоплавающие.
2. Бродные-полуводные.
3. Бегающпе-наземные.
4. Скачущпе-наземно-кустарные.
5. Шественные-наземно-древесные.
6. Лазящпе-кустарно-древесные.
7. Охотящиеся на лету.
Перейдем теперь к обзору этих типов и составляющих их классов
11 тех общих приспособлений, которые их характеризуют. Специаль-
ное внимание уделяется приспособлениям, связанным с добыванием
ппщи, и приспособлениям в строении клюва, лап и хвоста (рис. 166,
Тип водоплавающих объединяет птиц, проводящих всю млн
оолыпую часть жизни на воде и добывающих пищу плавая пли ныряя
за ней. Если в известные периоды своей жизни некоторые виды, вхо-
дящие в тип, и кормятся на земле, их связь с водой и приспособле-
ния к жизни на ней, как, например, у гусей, все же совершенно
очевидны.
„ Населяемые этим типом водные биотопы разнообразны: это при-
брежные области моря, озера, реки и различные более мелкие во-
доемы. Птицы этой группы обладают вытянутым челнообразным
197
21
Рис. 166. Кеновы птиц. Г—большой крохаль — ATerfims mergari.se г L.: 2— розовый
пеликан — Pelccanus onocrolalus L.; J — цапля ракоед — Cancroma cochlearia L.; 4 —фла*
мппго — phoenicoplerus roseus L.; a — водорез — Rhynchops nigra L.; 6' —малая колпица-
Platalea rninor 'I emm. w Schleg.; 7 —кулик лопатепъ —Ew'ynorhynchus pygmaevs L.J $ "
большая белая ржанка — Chionis alba (G-m.); 9 —ипатка —Lunda cirrhata Pall.; 10 — бо-
родавчатый ибис — Geroniicus papillosus (Temm.); 11—бекас-отшельппк — Capella sob-
taria (Hodgs); 12 — кривоносый зуек — Anarlvynchus frontalis Q. & G.; 13 — марабу —
toptilvs sp.; 14 — орлипоклювая колибри — Evtoxeres aquila Bourc.; 15 — султаискпп ДП*
тел — Chrysocolaptes stricklandi (Lay); 16 — мечеклюв — Docimastes ensifer (Boiss.); К 7
африканский аист-разипя — Anaslomvs lamelligerus Temm.; 18 — колибри — Avocelhuty
recur?'irostra (Sw.); 19—белобрюшка — Phaleris psittaculus Pall.; 20 — зубчатоклювы»
голубь — Didvncvlus slrigiroslris Jerd.; 21 — шпрококлювка — Platyrhynchus inystacetf
(Aieill.) (Passeriformes, сем. 7'yrannidae).
телом, несущим под кожей, особенно на нижней стороне, значитель-
ный слой жира в целях предохранения тела от охлаждения. В строе-
ние. 167. Строение ног некоторых птиц. 1—морская сорока — llucmatepus ostralegus L.;
травник — Tringa tetanus L.; 3 — якана; 4—дневная хищная птица; 5—франко-
,п — Francolinus sp.; б — дрозд; 7 — водяной пастушок — Hallus aquaticus L.; 8 — черная
'Рачка — Hydrochelidon nigra L.; 9—голубь; 10 — сова; 11—травник — Tringa tetanus
ь.; 12 — Pteroplochos niegapodius Kittl.; 13 — попугай; 14 — зимородок — Alcedo atthis L.;
гр - ильмовый стриж; 16 —обыкновенный козодой — CaprimuIgus europaeus L.; 17 —дя-
,ел зеленый — Picas canus Gm.; 18 — черный стриж — Micropus apus L.; 19 — колибри —
Eriocnemis luciani (Bourc.); 20— саджа — Syrrhaptes paradoxus (Pall.).
ниц ног, хвоста, органов дыхания и пр. имеется много приспособи-
тельных черт, связанных с способностью плавать и нырять (см.
стр. 171—180).
199
Рис. 168. Хвосты некоторых птиц. 1—дре-
волаз — Schizura desmursi Gay.; 2 — петухо-
хвостый тиран — Alpctiurus tricolor (Vicill.);
3 — попугай ь-рошка — Nasiterna pusio Scl.;
1 — средни и дптел — Dryobates тле dins (L.).
Преобладающей пищей этой группы являются рыба, водные ра-
кообразные, моллюски, насекомые и их личинки, в значительно
меньшей степени также водная и прибрежная растительность, ее
листья, клубни и семена. В зависимости от характера этой нищи
и особенностей самих птиц видоизменяются и способы добывания
ее. Питающиеся рыбой пингвины, многие чистики, крохали, гагары,
некоторые поганки и бакланы ныряют в воду и догоняют ее под
водой, хватая клювом. Острые края клюва, направленные назад,
острые пилообразные роговые зазубрины на краях его (крохали)
пли острый крючок на конце клюва помогают удержанию скользкой
и вертлявой добычи, is7 пингвинов эту роль кроме того играет значи-
тельный по размерам язык, как
л небо, сплошь усаженный силь-
ными, направленными назад,
роговыми сосочками, при по-
мощи которого птицы ориенти-
руют добычу в клюве таким
образом, чтобы она легче -
рыба, например, передним кон-
цом! вперед — могла просколь-
знуть в глотку. Змеешейка, до-
гоняя рыбу сходным! образом,
однако не хватает ее в клюв,
а протыкает ее острым! тонким
клювом, как стилетом, и, появ-
ляясь с добычей на поверхность
воды, резким! взмахом! головы
скидывают рыбу с клюва вверх
п вновь ловит в открытую пасть.
При этом, охотясь под водой,
птица все время держит голову
и клюв в свободном положении,
примерно так, как копьемета-
тель держит копье, прежде чем
он заметит добычу. Приблизмв-
с силой выбрасывает голову впе-
ред, с удивительной точностью попадая в свою цель. Эти действия
возможны благодаря специальным! видоизменениям! скелета шеи
и развитию соответствующей мускулатуры. Клювы некоторых
чистиковых птиц поражают своей своеобразной, сплющенной с бо-
ков, формой с острым! крючком на конце.. Таковы, например, роды
Lunda, Fratercula н А1са„ Этот клюв является повидпмому приспо-
соблением к разрыванию грунта, в котором эти хорошо ныряющие
•морские формы могут находить любимых ими рыб и моллюсков.
Таким! способом! повидимому ловит пескороек (Ammodytes) обыкно-
венная гагарка. При этом схваченная поперек рыбка при помощи
языка продвигается глубже в расщеп клюва и здесь зажимается
языком против неба, что дает возможность поймать птице еще не-
сколько рыбок, прежде чем направиться к своим птенцам. В этой
же группе чистиков многочисленные мелкие виды питаются главных
шись на нужное расстояние, птица
200
образом планктоном и мелкими ракообразными. Клюв их коротоку
широк у основания и заканчивается без крючка или особого острия.
Они склевывают животных с поверхности или ловят их в воде.
У Phaleris клюв при этом вздернут кверху и имеет вид заостренной
лодочки. Так как пища некоторых видов чрезвычайно мелка, у них
образуется обширный подгорловой мешок, в котором скапливается
добываемая пища (см. стр. 118). К сходному образу жизни при-
способлен один из родов трубконосы.х птиц — Pelecaiioides, пре-
красно ныряющий и для своей систематической группы имеющий
чрезвычайно маленькие крылья. В своих движениях и способе
добывания пищи он приобрел почти полное сходство с этими чи-
стиками.
Иное направление в этом типе свойственно уткам, ныркам
п отчасти лысухе. Главным объектом питания их являются
беспозвоночные, частью добываемые пырянием (см. стр. 170), частью
Рис. 169. Клюв широконосой утки — Spatula, clypeuta L. По
Brehm, из Hilzheimer.
доставаемые из воды на более мелких местах длинной шеей и клю-
вом. Целый ряд форм при этом, не изменяя существенно строение
клюва, перешел к питанию растительными веществами: семенами,
листьями водяных растений, клубеньками. Питающиеся донными
моллюсками морские гаги и турпаны ныряют на глубину до 6—10 м
и имеют обычно крепкий грубый клюв с хорошо развитым уплощен-
ным ноготком на конце. Близки к ним сходные по биологии пресно-
водные гоголи и чернети. Питающиеся нежными личинками на-
секомых и растительностью, они имеют клюв более тонкий и снаб-
женный по краям тонкими роговыми пластинками. У питающейся
мелкими моллюсками и ракообразными широконоски этот тип клюва
достигает наиболее своеобразного строения. На внутренней поверх-
ности краев надклювья у этого вида свисают вниз узкие и частые
роговые пластинки, по своему расположению напоминающие строе-
ние так называемого китового уса у китообразных (рис. 169). Про-
тив окончаний этих пластинок на краю подклювья также имеются
роговые насечки, и сходные пластинки п щетинки, направленные
в бок, несут края мясистого и крупного языка, выполняющего цо-
201
лость клюва. Взаимодействие этих пластинок образует род сита,
способного процеживать воду, по задерживать различные, даже
весьма мелкие организмы. Кормящаяся таким способом в грязи
утка, как говорят, «щелокчет»: действующий как мощный поршень
язык всасывает воду через чуть приоткрытый клюв, а затем при за-
крытом клюве вновь выжимает ее через ситовидный аппарат, па ко-
тором остаются идущие в пищу мелкие животные п семена. С подоб-
ным функционированием клюва координированы и другие особен-
ности его строения, как, например, очень значительная его чув-
ствительность (см. стр. 153), а также расположение хоан далеко кзади,
у основания клюва, что защищает их при помощи языка от про-
никновения воды и грязи. Увеличение чувствительности и поверх-
ности клюва некоторых уток достигается также разрастанием ро-
гового покрова его в особые лопасти, как, например, у австралий-
ской утки Malacorhynchos.
Совсем особым путем приспособления пошли пеликаны, снабжен-
ные огромным горловым мешком на длинном клюве, которым они
подхватывают в воде мелкую рыбу. При этом подклювье принимает
форму сачка, очевидно действующего наиболее эффективно для
данной цели. Этот способ охоты будет более добычливым при воз-
можности преследовать стайную рыбу, и пеликаны постоянно на-
блюдаются на общественных охотах, когда они стаями загоняют
рыбу в мелкие заливы и на мели, где ее легче вычерпывать. Обще-
ственные охоты за стайными рыбами устраиваются и многими дру-
гими формами этого типа: крохалями, бакланами, змеешейкой
и мн. др.
Переходное положение к другим типам занимают «гуся» и пла-
вунчики. Первые, хотя проводят большую часть жизни па воде,
кормятся главным образом на берегу и на суше, преимущественно
нежной зеленью. Грубо зазубренные края клюва, приходящиеся
друг против друга, и большой острый ноготок являются приспособле-
нием к этому типу питания. Ряд упомянутых видов этого класса —
Channel, Ansercmas, Cereopsis выработали уже дальнейшие при-
способления к наземной жизни: вырезанные плавательные пере-
понки. более грубый клюв и др. Плавунчики представляют собой
мелких куличков, перешедших к образу жизни мелкой уточки. Они
уже не столько береговая птица, сколько птица открытой воды,
мелких луж и озерков. На пальцах ног у них образовались широкие
лопастные перепонки, и они прекрасно плавают, собирая корм —
водяных блох и личинок насекомых — в верхних слоях воды своим
тонким, как пинцет, клювом. При этом очень часто плавунчик бы-
стро вращается па поверхности воды и этим создает движение воды,
поднимающее ближе к поверхности планктонные организмы, здесь
и вылавливаемые этой птичкой.
В тип б р о д н ы х - п о л у в о д и ы х птиц можно отнести длинно-
ногих, связанных с болотами и побережьями птиц, типичными пред-
ставителями которых являются цапли, аисты, журавли, большин-
ство куликов, многие болотные курочки и др.
Как правило, для них типичны длинные плюсны (нередко п го-
лени частью являются обнаженными от оперения); пальцы их обычно
202
тонки и длинны и лишены полных перепонок, хотя и имеются от-
дельные исключения. Эти птицы кормятся шагая или бегая по бе-
регу водоемов, пли среди густой травы пли камышевых зарослей,
и добывают пищу то подстерегая ее, то активно разыскивая в укром-
ных уголках, то предпринимая к этому специальные уловки. Наи-
более типичные для этого типа «формы» имеют обычно длинную
вытянутую шею и длинный заостренный клюв. Последний однако,
как и в предыдущей группе, испытывает особенно крупные изме-
нения в зависимости от характера пищи и способа ее добывания.
Добывающие рыбу пли крупных беспозвоночных имеют обычно
крупный и острый, слегка конический клюв с эластическими и по-
тому способными к некоторому расширению ветвями подклювья
и весьма растяжимым пищеводом. Такой клюв типичен для цапель,
аистов и теневой птицы. Некоторые виды однако приобрели весьма
расширенный и вздутый клюв, иногда очень крупный по размерам,
как, например, китоглав (Balaeniceps) (рис. 245) пли ракоед (Сап-
сто! па).
Другие формы, питающиеся мелкими беспозвоночными и среди
них главным' образом червями или личинками, как, например,
ибисы и многие кулики, имеют клюв тонкий и длинный, часто за-
гибающийся книзу. Этим клювом птицы прободают мягкую влажную
почву и, так как роговой покров клюва чрезвычайно богат чувстви-
тельными клетками, ощущают и вытаскивают из почвы личинок
насекомых и червей. Наиболее интересно в этом отношении устроен
клюв у бекасов '(Capella), у которых он способен открываться на
конце под землей, без расширения клюва у основания (рис. 140).
Весьма своеобразные изменения клюва мы наблюдаем у некото-
рых куликов (рпс. 166). Распространенная у нас на севере камне-
шорка (Arenaria inlerpres L.) разыскивает свой корм под камешками,
щепочками, растениями на морском побережье, поднимая и пере-
вертывая их, и клюв ее является соответственно измененным ору-
дием: он резко вздернут кверху, несколько уплощен сверху и весьма
крепок. Другой наш кулик (Ibidorhyncha struthersi Vig.), живу-
щий по галечникам горных рек Тянь-шаня, имеет клюв круто за-
гнутым книзу: он разыскивает им пищу в щелях среди камней у бе-
регов реки. Новозеландский Anarhynchus frontalis Q. & G. также
переворачивает камешки в поисках пищи, но он приподнимает их не
зверх. а вбок, и его клюв постоянно шилообразно загнут вправо.
Впды рода Haematopus приспособлены к питанию главным образом
ракушечными моллюсками, раковины которых они весьма искусно
вскрывают ударами своего долотообразного клюва. Последний
весьма уплощен с боков и заострен на конце и может служить в дан-
ном случае не только долотом, но и рычагом, разворачивающим
крышки раковины. В ином направлении развит клюв кулика ло-
патня (Eurynorhynchus pygmaeus L.); он сильно уплощен сверху вниз
п расширен на конце в маленькую ложечку, которой эта птичка хва-
тает в мелких водоемах мельчайших беспозвоночных. Совершенно
но этому же типу построен клюв у колпицы (Platalea из отряда Ci-
coniifonncs), функционирующий таким же образом. У других форм
однако, несмотря на микроскопические размеры объектов питания,
203
клюв имеет характер пинцета, которым например ходулочник {1Ц-
/nantopus) и шилоклювка (Recurvirostra) весьма ловко таскают
пищу из мелкой воды. Крайне оригинальным в этой группе птиц
является фламинго {Phoenicoplerus), представляющий пример экстрен-
ного приспособления неумеющей нырять птицы к питанию мелкими
водными животными. Чрезвычайно длинные ноги и шея дают ему
возможность глубоко заходить в воду, где он щелокчет грязь клювом,
сходным с клювом многих уток. В противоположность последним,
более подвижным, в клюве фламинго является однако надклювье,
и клюв изогнут колеичато, так что птица опускает его в воду верх-
ней стороной вниз. Основной пищей фламинго являются мелкие
ракообразные Artemia salina, мелкие моллюски {Cerithium.) и
черви.
Промежуточное положение к следующему типу занимает группа
пастушков, водяных курочек и якан. Для большинства из них ха-
рактерно обитание среди болотной растительности, где они кормятся
как животным, так и растительным кормом. Сжатое с боков тело
характерно для тех форм, которые живут в густых- зарослях. Специ-
фической чертой являются тонкие и длинные пальцы ног, дающие
возможность бегать по болотистому грунту. В этом отношении наи-
более специализированы различные яканы (отряд (Jharadriijorn^s}
с чрезвычайно длинными пальцами ног. вследствие чего тяжесть
тела распределяется на столь большую площадь, что птички могут
свободно бегать по плавающим листьям водяных кувшинок п другой
плавающей растительности. Клюв у большинства этих форм корот-
кий, приспособленный к склевыванию пищи с поверхности различ-
ных растений пли щипанию их листьев. *
Следующим крупным адаптивным тимом являются бегающие-
наземные птицы, держащиеся в разнообразных, но преимуще-
ственно сухих, наземных местообитаниях и приспособленные к лег-
кому передвижению по земле. Представителями типа являются:
дрофы, страусы и их ближайшие родственники, рябки, фазановые
птицы, большеногие куры, скрытохвосты, трехперстки, серимы, сек-
ретари; из куликов—авдоткп, бегунки п др.; из голубей—венце-
носные голуби; из воробьиных — птица-лира, жаворонки, трясогузки,
коньки и мн. др. Все эти виды быстро бегают по земле и сравнительно
редко поднимаются на деревья, даже если они и обитают в древес-
ных участках. Преобладающей пищей являются главным образом
растительные вещества — семена, молодые побеги, почки, клубни
и т. д., хотя многие не брезгают не только насекомыми и другими
беспозвоночными, но даже мелкими позвоночными животными.
Из приспособлений к питанию этой группы следует отметить
развитие способности у многих форм к разгребанию земли и назем-
ного сора, в поисках пищи, что свойственно многим фазановым,
большеногим курам, мадагаскарским пастушковым куропаткам,
птице-лире и др. Некоторые мелкие воробьиные (например Pteroptt’
chos) ворошат наземные листья, с силой отбрасывая их особыми двц'
жениями ног и перебегая по листьям. Наконец у других форм ра3'
рывание мусора и земли происходит посредством клюва. У кеклика
(Alaectoris) и блестящего фазана .(Lophophorus) несколько крючко-
204
ватый клюв специально служит орудием вырывания из земли пита-
тельных луковиц и клубней различных лилейных растений. Жура-
вли вырывают клубни растений обычно в мягкой болотной и луговой
почве и в состоянии совершать это своим прямым и сильным клювом.
Большинство птиц этой группы, питающихся растительными объек-
тами, удовольствуется однако просто сбором семян и листочков с по-
верхности земли или склевыванием из цветочных головок и обла-
дает простым, несколько вздутым,, типа кур, клювом. Собирающие
насекомых птицы обычно имеют более тонкий и слабый клюв, прямой
и заостренный. Птицы, копающиеся в земле или в пыли, нередко
имеют ноздри защищенными мягкими крышечками, или специально
разрастающимися покровами (например у кагу — Rhinoch cietus), или
густо разрастающимися перышками (Thinocorys и др.). Наконец
у питающихся крупными плодами клюв нередко бывает сильным
и крючковатым, иногда слабо уплощенным (эму, казуар и др.).
В строении ног следует отметить укорочение пальцев у видов,
живущих на твердой степной почве, уменьшающее вредное трение
при беге птиц. У некоторых форм, особенно живущих на песке, на-
пример у дрофы красотки (Chlamydotis), подушечки пальцев упло-
щаются и расширяются, что делает опору пальцев на сыпучем грунте
более прочной. Наоборот, у форм, живущих на каменистом или твер-
дом грунте, иногда наблюдается слияние пальцев между собой,
у саджи (Syrrhaptes) приводящее, например, к образованию кро-
шечной лапки, с плоской мозолистой ступней и широкими, как ко-
пытца, когтями сросшихся пальцев. Саджи прекрасно, очень быстро,
бегают по твердому щебнистому грунту.
Небольшой «класс» птиц в этой группе представляют формы,
подобные киви (Apteryx} и вальдшнепу (Scolopax). Это обитатели
сырых зарослей с обильным валежником, в котором, как и в мягкой
почве, они могут разыскивать свой корм из червей и личинок, вы-
таскивая их длинным клювом из мягкой земли. Интересно, что оба
•этих представителя приобрели сходный облик сходный тип при-
способления совершенно разными путями. Вальдшнеп ориентируется
главным образом осязанием и зрением (рис. 140), которые у него
очень хорошо развиты, киви же преимущественно обонянием, вос-
полняющим недостатки слабого зрительного аппарата.
Особую группу по характеру приспособлений представляют се-
римы (Сапата. Chunga) и птица-секретарь. Это быстро бегающие
степные птицы, с длинной голенью и короткими пальцами ног,
преследующие преимущественно рептилий и особенно змей. Послед2
ние схватываются пальцами ног и у секретаря с силой отбрасы-
ваются на землю, что нередко вызывает смерть животного. Своеоб-
разной чертой в связи с этим является наличие отличного от осталь-
ных, острого, кругло загнутого когтя на внутреннем пальце, которым
хватается и придерживается рептилия, а у серим, кроме того, боль-
шого пучка щетинообразно стоячих у основания клюва перьев, за-
щищающих повидимому лицо от опасных укусов животного.
Следует упомянуть также, что многим из птиц этого типа свой-
ственно стремление при появлении врага лечь на землю и, вытянув
по земле шею или прижав голову к туловищу, застыть в таком со-
206
стоянии. Это поведение, в совокупности с имеющейся обычно охра-
нительной окраской, совершенно скрывает птицу.
Особый тип приспособлений представляют ск а ч у щи е - на-
земно-кустарные птицы. Это обычно мелкие птицы, проводя-
щие большую часть времени на земле и среди валежника, где они
главным образом и разыскивают пищу, но скрывающиеся в случае
опасности в гущу кустарников и на деревья, среди которых они также
могут передвигаться с достаточной ловкостью. Основной пищей этой
группы, как и предыдущей, являются семена, личинки и взрослые на-
секомые, живущие не земле и в мертвой лесной подстилке, иногда и
зеленые части растений. Значительная часть этой группы птиц неспо-
собна к бегу только вследствие указанных выше (в разделе о движе-
нии) механических причин. Представителями этой группы являются:
крапивники (JTroglodytidae)' муравьеловки (F'ortnicariidae)ч многие
Pteroptochidae, Conopophagidae, земляные ракши (Bracliypteracii-
dae), ряд кукушек (например Centro pus), часть соловьиных, зави-
рушки (Prunellidae), ряд попугаев, вьюрков и мн. др. Значительное
число форм этой группы характеризуется очень крупными задними
конечностями, иногда непропорционально крупными в отношении
к телу, обычно коротким хвостом, иногда заметными приспособле-
ниями к обитанию в тени, под пологом леса, что выражается в отно-
сительно крупных глазах, темной окраске, коротких тупых и окру-
глых крыльях, в очень громком и сильном голосе. Но в отношении спо-
соба хватания пищи эта группа сходна с последующими и лишена
каких-либо только в ее пределах встречающихся приспособлений.
Следует упомянуть, что и в этой группе мы встречаем развитие круп-
ных гребущих листву задних конечностей, свойственных иногда
совсем небольшим птичкам.
Близким типом приспособлений является группа ш е с т в е н ных-
наземно-древесных птиц, включающая таких птиц, как краксы.
многие голуби, многие врановые, удоды,некоторые носороги, скворцы,
представители семейства Icteridae и ряд попугаев (Pezoporus и др.).
Обычно это птицы крупные или средней величины, передвигающиеся
по земле шагами и разыскивающие пищу главным образом на земле,
но значительную часть времени проводящие и гнездящиеся на де-
ревьях. По питанию это частью всеядные, частью зерноядные и на-
секомоядные формы. Некоторые из них питаются и плодами, собирая
их даже с веток деревьев.
i Наиболее обширным и численно богатым типом являются лазя-
щие-кустарно-древесные птицы. Сюда относится преобладаю-
щее наше птичье население лесов и кустарных зарослей, а частью и ска-
листых местообитаний. Основными классами организации здесь явля-
ются: сойки, кукушки, славки, попугаи, древолазы, мухоловки, ту-
каны, сорокопуты, синицы, дроздовые и мн. др. Пищей группы явля-
ются насекомые и их личинки, собираемые на коре, листьях и цветах
растений, взрослые насекомые, ловимые в воздухе, ягоды и плоды-
семена, нектар и пыльЦа цветов, а у некоторых и птенцы и яйца дрУ'
гих птиц, разыскиваемые при кочевках по лесу. Из особых приспо-
соблений к добыванию пищи отметим наиболее интересные. В группе
мухоловов мы находим преимущественно формы с относительно вер-
206
тпкальноп посадкой, подстерегающие добычу сидя спокойно па сучке,
иногда даже в тени дерева. При появлении поблизости удобной
жертвы, птичка взлетает в воздух и, ловко управляя своим полетом,
хватает насекомое в воздухе и вновь садится на старое место. Боль-
шинство видов этих форм обладает уплощенным, расширенным у осно-
вания клювом, ширина которого еще увеличивается сильно развитыми
приротовыми упругими щетинками. У многих клюв на конце снаб-
жен крючком, а при ловле крупных и жестких насекомых клюв яв-
ляется более объемистым, высоким и сильным. Ловящие бабочек
жакамары (сем. Galbulidae) имеют длинный и упругий клюв, схва-
тывающий насекомое, как сильный пинцет. Птицы, очищающие от
насекомых листву и мелкие веточки деревьев, обладают обычно
сильными и цепкими конечностями и маленьким шиловидным пли
слегка изогнутым клювом. Древолазы и группы, переходные к ним,
как указывалось, отличаются очень сильными конечностями и не-
редко жестким хвостом, служащим опорой. В зависимости от спе-
циализации древолаза, клюв вариирует от тонкого и гибкого, про-
никающего в трещины коры, до массивного долота, отбивающего
от дерева, в поисках личинок в древесине, крупные щепки.
Особо крупную группу приспособлений представляют птицы,
потребители пыльцы и нектара цветов. В этом участвуют семей-
ства: колибри ( Trochilidae), щеткоязычные попугаи (Trichoglossi-
dae), медососы (Meliphagidae). иектарницы (Nectariniidae), гавай-
ские цветочницы (Drepanididae). белоглазки (Zosteropidae), Di-
caeidae, Coerebidae и отдельные представители других семейств.
Приспособление к характеру и форме цветов здесь достигает уди-
вительного разнообразия и специализации. Питающиеся нектаром
имеют обычно длпннцй и тонкий клюв, иногда, сообразно форме
цветка, своеобразно изогнутый; питающиеся пыльцей имеют клюв
обычно более грубой формы. Неменьшая специализация свойственна
языку, который у сосущих нектар сворачивается в одну или две
трубочки, образованные нередко особо разрастающимися бахром-
ками языка. У питающихся пыльцей язык покрыт щетинками, пли
имеет форму ложечки, или рассечен на бахромки, напоминающие
кисточку. Все эти приспособления очевидно вырабатывались в глу-
бокой связи с эволюцией соответствующих групп растительности
и остаются не без влияния на них. Орнитофилия пли опыление ра-
стений птицами имеет существенное значение в тропиках, и в Бра-
зилии например число опыляемых птицами семейств растений соста-
вляет до 39% всех семейств, опыляемых при помощи каких бы то
ни было животных (Porsch, 1929).
Но не менынее разнообразие в строении клюва и языка мы на-
ходим у птиц зерноядных. Здесь птицы, питающиеся мелкими семе-
нами, вытаскиваемыми из окружающих их оболочек, имеют клюв
обычно узко заостренный и несколько вздутый у основания. Питаю-
щиеся семенами толстыми, с твердой оболочкой, имеют очень круп-
ный конический клюв, п внутренние поверхности челюстей нередко
ооразуют специальные ямки, иногда с своеобразной насечкой, ме-
'Кду которыми зажатое семя разрушается мощной мускулатурой
клюва. Особенно интересен клюв клеста, концы челюстей которого
207
перекрещиваются и заходят друг за друга. Различные виды клестов
являются специализированными к разным видам хвойных деревьев,
семенами которых они питаются, вскрывая клювом чешуйки шишек
и искусно отворачивая их особым поворотом клюва. Значительную
специализацию мы находим также в строении клюва и других орга-
нов у таких прекрасно лазящих древесных птиц, как попугаи.
Последний адаптивный тип — птицы, охотящиеся на лету,—
также представляет комплекс весьма разнообразных адаптаций,
основным свойством которых является значительное искусство по-
лета, во время которого многие птицы не только высматривают свою
пищу, но и добывают ее. Преобладающими в этой группе являются
хищные и насекомоядные птицы.
Наиболее массовое значение имеют следующие классы: класс
чаек, в который могут быть объединены, кроме собственно чаек,
большинство буревестников и водорезы. Это преимущественно длин-
нокрылые птицы, прекрасно летающие, нередко способные к весьма
эффективному парящему и планирующему полету. Для большинства
птиц характерна удивительная выносливость к продолжительности
полета. Все они связаны также и с водой, на которой могут плавать
и отдыхать, но лишь в редких, исключительных случаях способны
нырять. В связи с способностью плавать все они обладают дере-
пончатопалыми плюснами. Их пищу составляют плавным образом
плавающие по поверхности животные, различные отбросы и живот-
ные. держащиеся на побережье. Более континентальные формы из
них являются почти всеядными.
.Соответственно способу добывания шнци — выхватыванию ее из
воды или подбиранию на берегу — клюв у них в большинстве слу-
чаев сильный, загнутый и заостренный на корце, обычно значитель-
ной длины. Наиболее специализированным и оригинальным клювом
обладает водорез (род Rlu/nchops). Его клюв сильно сжат с боков и
напоминает по форме лезвие ножа, при этом подклювье значительно
(1—2 см) выдается за конец надклювьяипо бокам исчерчено тонкими
бороздками, невидимому являющимися местом скопления чувстви-
тельных клеток. Эта форма летает над самой водой, производя весьма
незначительные движения крыльями и держа клюв на 1/3 опущенным
в воду. Прикосновение к мелким рыбкам и ракообразным вызывает
хватательные движения клюва. Так как этот способ питания неза-
висим от освещения, водореза приходится наблюдать на таких охот-
ничьих полетах как в дневное, так и в ночное время.
Класс ныряющих с воздуха объединяет также птиц, связанных
с водной стихией и обладающих тем же поисковым полетом, как пре-
дыдущая группа. Сюда могут быть объединены весьма разнообраз-
ные по систематическому положению виды: фаэтон, крачки, ряд чаек
(Rissa, Lotus leucopterus Faber), некоторые буревестники (Puffin^
pujfinus L. и P. gravis O’Rielly), олуши (Sula}, даже пеликаны (P^
lecanus occidentalis L., P. oc. thagus Mol.).
Типичным для всех этих форм, как указывалось, является вы-
слеживание добычи предварительно в воздухе, на спокойном полете.
Высмотрев ее,, птицы кидаются вниз, полусложив крылья, развивая
этим значительную скорость, вследствие которой они с силой про*
нпкают в воду и хватают здесь клювом добычу. Такие крупные
формы, как олуши, могут при этом развивать очень значительную
силу для весьма глубокого ныряния. У всех этих форм мы находим
плотную, сравнительно короткую, шею и остро кончающийся клюв,
Облегчающий этим погружение под воду. Близки к этим формам фре-
гаты й буревестники, не ныряющие в воду, но отнимающие добычу
у других форм, нападая на них сходным образом в воздухе.
Среди хищных птиц, по способу питания почти целиком входя-
щих в этот адаптивный тип, можно наметить несколько адаптивных
классов. Массовыми и характерными из них являются, например,
группы грифов, сарычей и ястребов. Первые (включая грифов Ста-
рого и Нового Света) формой и строением крыльев приспособлены
к длительному парению высоко в воздухе и обслеживаншо огром-
ных районов. С огромной высоты они высматривают наличие где-
пибудь павших животных, к которым очень быстро собираются
в степных и горных районах целыми группами. Это типичные не-
крофаги, и бить живую добычу они не умеют. Вследствие этого,
в противоположность остальным хищным птицам, их плюсны слабы,
приспособлены более для передвижения по земле, чем для схваты-
вания, и пальцы вооружены лишь короткими и слабо загнутыми
когтями. Но клюв, приспособленный к разрыванию мяса, крупен
и круто- загнут. Характерной особенностью большинства из них
является также оголенность от перьев лицевых частей головы,
а частд и всей шеи. Это дает им возможность даже через малые есте-
ственные отверстия и разрывы в коже крупных павших животных
проникнуть внутрь трупа, а частью и освобождает от накопления
грязных остатков среди оперения.
Группа сарычей, куда могут быть зачислены, помимо собственн >
сарычей (Buteo, Archibuteo и др.), также большинство орлов и дру-
гие близко родственные роды, также обладает широкими и мощ-
ными крыльями, но их способность к парению ограниченнее и вместе
с тем способность к активным видам полета и ловкость более разно-
сторонни, чем у грифов (см. стр. 186). Эти формы в большинстве своем
высматривают добычу на лету, а затем бросаются на нее и ловят
ее на земле (в редких случаях и на лету). Они характеризуются нор-
мальным строением лап хищной птицы, с круто загнутыми и острыми
когтями и умеренно широкими крыльями. Группа ястребов объеди-
няет преимущественно лесных птиц, обладающих очень вертким
и ловким полетом и способных преследовать добычу и на лету. Для
них крайне характерны подстерегание добычи сидя где-либо среди
густых веток дерева и внезапный налет на жертву и преследование
ее среди веток деревьев и кустов. Основной добычей ястребов, в за-
висимости от величины птицы, являются различные воробьиные,
куриные и врановые птицы. Особую группу представляют луни,
Держащиеся открытых ландшафтов и сравнительно медленным по-
петом облетающие свою охотничью территорию, летая над самой
землей. Это охотники на мелких грызунов и мелких полевых птиц,
с тРУДОм излавливающие летящую добычу и, подобно ястребам,
норовящие невзначай броситься на нее из-за прикрытия. Наи-
пучшпх летунов среди хищников, по силе, быстроте и выносливости
И Орнитология—1209
209
полета, представляют соколиные, в зависимости от величины спе-
циализирующиеся на охоте то за крупными утками и другими пти-
цами, то за воробьиными и различными летающими насекомыми.
В группе последних форм интересное приспособление, связанное
с добыванием пищи, представляет способность задерживаться в воз-
духе и быстрыми взмахами крыльев поддерживать тело на одном
месте, что используется для выслеживания и более тщательного
рассмотрения предполагаемой жертвы. Помимо этих основных групп,
мы находим у хищных немало отклонений и узких специализаций,
как, например: охотящийся на летучих мышей М achaeramphiis.
питающийся почти исключительно моллюском Ampullarius свое-
образный род Roslrainus, охотящийся главным образом на гнезда
и яйца птиц Icliaetus, и мн. др. Совы, во многих отношениях предста-
вляющие группу конвергентную дневным хищникам, отчасти ха-
рактеризуются теми же приспособлениями (см. стр. 449).
В этот тип необходимо отнести и обширную группу ласточек,
а также близких экологически им стрижей, Artamidae, Pseudoclu-
lidonidae, тиркушек и козодоев. Это все птицы, специализированные
на охоте за насекомыми, которых они ловят на лету, исключительно
преследуя их в воздухе. Для них характерны острые длинные крылья,
удивительно быстрый и легко управляемый полет, широкая пасть
при коротком клюве, обильные щетинки у его основания. Внутри
этой группы мы находим уже более детальную специализацию, на-
пример у стрижей — к улавливанию насекомых более высоких слоев
воздуха, у береговых ласточек — насекомых, летающих над водой,
у козодоев — ночных насекомых и т. п. Несколько обособленное
место занимают щурки (Meropidae'), преследующие главным образом
перепончатокрылых, которых они ловят длинным и острым клювом
и, несмотря на их жало, поедают.
3. РАЗМНОЖЕНИЕ И ИНДИВИДУАЛЬНОЕ РАЗВИТИЕ
Процесс размножения вызывает в жизни птиц развитие разно-
образных и по своему биологическому смыслу весьма глубоких
приспособлений, обеспечивающих воспитание и выживание потом-
ства. Вследствие важности этого процесса в жизни животных, мы
остановимся на изложении явлений, связанных с ним, более подробно.
Достижение половозрелост и. Как правило, на-
ступление половозрелости птицы зависит от ее величины: чем птица
крупнее, тем позднее развивается в ней способность производить
половые продукты. Самки мелких ткачиков способны к оплодотво-
рению уже в возрасте 6 месяцев, и начало гнездования в этом воз-
расте вероятно для многих воробьиных сходного роста в тропиче-
ских и теплых странах. Огромное большинство воробьиных однакс
становятся половозрелыми 8—12 месяцев. Крупные представители
Passeres, например род Corvus, достигают половозрелости только ко
второму году жизни; в этот же срок начинают гнездится мелкие
крачки и чайки, мелкие дневные хищники, утки и нырки. К концу
третьего года жизни приступают к размножению крупные чайки,
гагары, орлы, крупные веслоногие и гусиные (гаги). На четвертом-
210 I
пятом году жизни начинает гнездиться страус. От этой схемы отдель-
ные группы и виды обнаруживают иногда отклонения. Как правило,
самки созревают в половом отношении несколько раньше самцов
п иногда на год, полгода раньше приступают к размножению.
Обнаруживающаяся у многих птиц к концу первого года спо-
собность к размножению застает их весной в молодом пере, развив-
шемся задолго до проявления половых стремлений. Поскольку ряду
этих видов весенняя линька вообще не свойственна, птицы присту-
пают в этот год к размножению в молодом наряде, нередко резко
отличающемся от наряда взрослой половозрелой птицы. Из птиц
нашей фауны это явление свойственно чечевице [Erythrina erythrina
(Pall.)], горихвостке {Phoeniciirus phoenicurus L.), сынехвостой
славке [Tarsiger cyanura (Pall.)], перепелятнику (Accipiter nisus L.)
и вероятно многим другим. На следующий год эти птицы наде-
вают уже взрослый наряд, в большинстве случаев с более или
менее резко выраженными вторичными половыми признаками. У не-
которых видов развитие вторично половых признаков наступает,,
в противоположность указанной группе птиц, очень рано, уже
в рыхлом птенцовом пере, но обычно бывает ограничено лишь раз-
личием в окраске маховых и рулевых перьев. Таким образом, на-
пример, может быть установлен пол у птенцов дубоноса {Coccothrau-
stes coccothraustes L.), находящихся еще в гнезде, или у птенцов
восточноазиатского каменного дрозда {Manticola solitaria magna La
Toushe).
Половой диморфизм и ф о р м ы «б рак а». Половой
диморфизм представляет у птиц широко распространенное явление,
нередко принимающее резко выраженную форму. Степень развития
вторично-половых отличий зависит обычно от характера связей по-
лов в период размножения и тех средств, которые стоят на службе
полового отбора. Связь полов у огромного большинства птиц пред-
ставляет форму моногамии, весьма изменчивой по продолжитель-
ности. У многих уток заинтересованность пары друг в друге про-
должается только от первых брачных игр до первого снесенного яйца
пли выклюнувшегося птенца. У многих воробьиных, наоборот, пара
не только вместе выкармливает птенцов, но выводит в году до двух-
трех выводков. На небрачный период пары обычно расстаются друг
с Другом, но, в связи с присущей большинству видов способностью
возвращаться на место бывшего гнездования, между ними часто
вновь устанавливается заинтересованность и <<брак» на новый гнездо-
вой период. Такие случаи с наибольшей частотой установлены для
крупных хищных птиц, аистов, гусей и т. д. У сипухи {Tyto alba,
^сор.) был установлен случай «длительного брака», продолжавшийся
не менее восьми лет. У многочисленной группы птиц вторично-поло-
вые отличия хотя иногда бывают очень резко выражены, в массе
все же не столь значительны, как при иных формах «брака». В пре-
делах очень многих групп, например у пингвинов, гагар и поганок,
буревестников, некоторых веслоногих, зимородков, щурок, стрижей
11 вертишейки, вторично-половые различия вообще отсутствуют.
Полигамия встречается у птиц как в виде полиандрии (много-
мужества), так и полигинии (многоженства). Пример первой формы
211
мы находим у плавунчиков, трехперсток, цветного оекаса и скрыто-
хвостов, пример второй формы — в дикой фауне — у павлина, вдо-
вушек (из ткачпков) и повпдимому у некоторых дроф. В условиях
одомашнения эта форма распространена у обыкновенного петуха,
индейки и утки. Для всех этих случаев характерен чрезвычайно рез-
кнй половой диморфизм с более яркой, пышной окраской самцов
в случае полигинии и с более яркой раскраской самок в случае
полиандрии (рис. 170). При последней форме на самцов падают также
все заботы по устройству гнезда, высиживанию и воспитанию птенцов.
Не менее резко половой диморфизм выражен также в группе соб-
ственно «безбрачных» птиц, у которых встреча самца с самкой имеет
кратковременный характер, и между полами не устанавливаются
сколько-нибудь длительные привязанности. Самцы в этой группе
окрашены гораздо ярче самок и не принимают ни малейшего участия
пе только в гнездовых делах, но и в самке после происшедшего со-
вокупления. Примерами таких видов являются глухарь, тетерев,
ряд американских тетеревиных и повидимому некоторые дрофы.
Отличие брачного наряда от небрачного, носимого в осенне-
зимний период, достигается в некоторых группах птиц (многие во-
робьиные, например вьюрок, овсянки, подорожники, многие жа-
воронки и др.) обносом кончиков перьев, имеющих вид каемок,
лишенных пигмента и скрывающих под собой собственно брачную
окраску. Параллельно с сменой нарядов у многих птиц происходит
изменение окраски роговых покровов и кожных выростов, к брач-
ному периоду обычно приобретающих яркие цвета: желтый, голубой,
красноватый (клюв и ноги у зяблика, вьюрка, многих уток); кожистые
лопасти [розетки над глазами у тетеревиных, горловые лопасти
у сатиров (Tragopan)\ к весне обычно увеличиваются в размерах
и интенсивнее окрашиваются. Сообразно с различием ролей, играе-
мых полами в выращивании потомства, длительность носки брачных
нарядов и время их смены различны по полам. Самцы уток, например,
не принимающие участия в насиживании, линяют уже в июне, самки
же, по взматереиии птенцев — в июле или августе. Также отличен
ход линек у самцов, и самок белой куропатки.
Как уже указывалось, форма развития вторично-половых отли-
чий весьма разнообразна и в значительной части связана с характе-
ром брачных игр. То и другое взаимно обусловливают друг друга,
притом обычно в брачных играх только и раскрывается функциональ-
ное значение этих отличий. Хотя отдельные стороны этих отличий
друг с другом связаны, обзор их целесообразно осуществлять исходя
из отдельных признаков.
Наиболее постоянным вторично-половым признаком является
различие в величине. У огромного большинства видов самцы немного
больше самок, иногда с четкими различиями только в средних цифрах*
Значительное превышение самца над самкой свойственно немногим
птицам. Наиболее заметно оно у глухаря, мускусной утки и дрофы,
самцы которых весят 4,5—7 кг, 4 кг и 11—16 кг, а самки только
2,5—5 кг, 2кг и 5—6 кг. Количественно незначительным исключенном
из этого правила являются обратные соотношения. Самка больше
самца преимущественно у видов, в которых самец принимает на себя
212
?пс. 170. Цветной бекас — Rostralula benghalensis (L.), самка (правая фигура) окрашена более ярко и крупнее, чем самец (левая фигура).
Jlo Bannermann,
у перепелятника, например, самец весит
Рис. 171. Вторично-половые отличия в Форме и рыл а у
чибиса — Vane llu s vanellus L.
Верхнее крыло типично для самца, нижнее — для
самки. Значительное удлинение и расширение четвер-
того-девятого больших маховых вызывают у самца во
время воздушных брачных игр характерный и далеко
слышимый шум. По Frohawk, из Stresemann.
заботы по высиживанию яиц и вождению птенцов. Таковы трех-
перстки, скрытохвосты, киви и казуары. То же явлений невидимому
свойственно яканам и ряду — может быть даже большинству —
куликов. У ряда групп дневных и ночных хищников самки также
более крупны по размерам, что, возможно, связано с выкармли-
ванием птенцов первое время самцами. Благодаря меньшей величине,
последний более ловок и добывает вместе с тем более мелкую добычу,
более ценную для молодых птенцов, чем добыча крупная. Различия
между самцом и самкой в этих группах иногда очень значительны.
108—178 г, а самка 220—
320 г (Stresemann, 1928).
Различия в окраске
также широко распро-
странены и выражаются
либо в альтернативной
раскраске (например у
клеста — самец красный,
самка — зеленая), либо
в постепенных количе-
ственных изменениях,
связанных с большей
концентрацией тех же
пигментов у особей муж-
ского пола. Эти раз-
личия проявляются так-
же в развитии дополни-
тельных пигментов, об-
разовании структурных
изменений в пере, вызы-
вающих блеск, и появ-
лении вместо однообраз-
ной диффузной окраски
более или менее слоя;-
ного рисунка. Отличия
в окраске могут охватывать не только перо, но также окраску клюва,
кожных выростов и радужину глаза (например у скворца, некото-
рых куриных и др.) и т. д.
В случаях особо резкого полового диморфизма наблюдается раз-
витие украшающих пучков перьев, иногда значительной длины п
пышности. Такие перья замечательны например для райских птиц-
Обычно эти перья представляют собой измененные и специализиро-
ванные обычные контурные перья туловища. Так у павлина верхние
кроющие хвоста перерастают хвост и развиваются в длинные укра-
шающие и расправляемые веером перья «ложного хвоста». У ко-
зодоя Macrodipteryx longipennis Shaw, пара внутренних маховых
вырастает в длинные гибкие перья. У райских мухоловок (р°Д
Tschitrea) также непомерно переразвиваются центральные рулевые
перья. У чибиса Vanellus oanellus L. крылья самца являются более
широкими, чем у самки, в связи с сложными эволюциями в воздухе-
которые проделывает в брачный период птица (рис. 171). Достаточно
214
хорошо известно также образование выростов кожи в виде лопастей
на горле и своеобразных роговых бровей и гребней, столь обычных
па голове у куриных птиц.
Некоторые изменения наблюдаются даже в строении скелета.
Так у попугаев обнаружены половые отличия в форме таза, более
широкого у самки, вероятно в связи с развитием яйца. У хищных
птиц найдены половые отличия и в строении других частей скелета;
особенно значительны здесь различия в форме грудины, более узкой
и длинной у самцов. Наиболее удивительны различия в форме клюва
многих птиц-носорогов и особенно у птицы гуйи (Heteralocha аси-
tirostris Gould.) из Новой Зеландии, у самки которой клюв раза
в два длиннее клюва
самца и вместо пря-
мой формы резко за-
гнут и утончается
книзу (рис. 172). Раз-
личия в форме клюва
у этого вида вызы-
вают и различный спо-
соб питания, хотя пти-
цы держатся конечно
в одной стации. .
Указанные анато-
мические особенности
хотя и представляют
вторично-половые от-
личия, однако не стоят
в связи с брачными
играми, а являются,
так сказать, консти-
туциональными вто-
рично-половыми от-
личиями. Из анатоми-
ческих особенностей
Рис. 172. Гуйя— Неteralocha aculirostris G-ould. (пилений
рисунок — самка, верхний — самец). По Evans.
строение голосового
аппарата наиболее явно стоит на службе полового отбора. По сравне-
нию с голосом рептилий голос птиц несравненно более дифференциро-
ван, и у многих видов развился целый лексикон звуков, соответст-
вующих различным биологическим моментам. Общеизвестная способ-
ность большинства птиц к пению является часто вторично-половой
особенностью, обусловленной очевидно более дифференцированным
строением нижней гортани у самцов и более энергично функциони-
рующей мускулатурой ее. Эта особенность наблюдается и у непевчих
видов, самцам которых присущи своеобразные брачные крики. У них
часто наблюдается удлинение трахеи, свертывание ее в петли, удли-
нение бронхов, вздутие их и трахеи и образование особых боковых
нздутиц-лабиринтов. Последние наиболее сложные образования изве-
стны главным образом для утиных и являются причиной своеобразных,
иногда едва слышных, звуков, издаваемых самцом при брачных
играх.
215
Брачные игры. Процесс спаривания предваряется п со-
провождается брачными играми, смысл которых частью заключается
в установлении знакомства и тесных психических уз между птицами.
частью в приведении птицы в состояние готовности к копуляция.
Длительность периода этих брачных игр бывает различна: у круп-
ных птиц этот период обычно бывает дольше, у мелких — короче;
кроме того, на длительность их влияет
Рис. 173. Отдельные моменты из брачных игр боль-
ших поганок — Podiceps cristatus L.
1 — «кошачья поза»; играющие на воде поганки
сближаются друг с другом, растопырив перья ворот-
ника и хохлы п издавая при этом своеобразные гром-
кие крики; 2 — «пингвиповый тапец»; в наиболее на-
пряженные моменты брачных игр оживленно плаваю-
щие поганки ныряют, вырывают пучки водных расте-
ний и, подплывая друг к другу грудью, поднимаются
с воды почти вертикально, держа зелень в клювах.
По Huxley, нз Stresernann.
совместная' или раздельная
жизнь полов во внебрач-
ное время и сжатость сро-
ка размножения климати-
ческими условиями, что
наблюдается например в
полярной области. У круп-
ных птиц брачные игры
начинаются весной очень
рано и первоначально
имеют значение только
скрепить «знакомство» птиц
между собой и создать из-
вестную привязанность;
это может иметь место прп
половых органах, находя-
щихся в состоянии пол-
ного покоя. Только впо-
следствии, ближе К пе-
риоду спаривания, игры
увеличиваются в интенсив
ности и напряженности к
приобретают задачу при-
ведения в эротическое со-
стояние как самки, так в
самца. Предполагается,
что физическая готовность
к копуляции происходит
через психическую готов-
ность, обусловливающую,
например у самки, про-
цесс овуляции.
Брачные игры вырабатываются постепенно из обычных движении
организма, в процессе отбора специализирующихся и приобретающих
особую напряженность. У самца эти движения имеют значение глав-
ным образом показа, выставки свойственных ему украшений, импо-
нирующих самке, движения же самок носят «кокетливый» характер
(рис. 173, 174-, 175, 176, 177). Наибольшее воздействие на психику
•получается при этом как при ярких, блестящих окрасках и сложных
рисунках, так и контрастности раскраски и неожиданности, внезап-
ности изменения формы, цвета и звука. Поэтому мы видим в брачных
играх примеры обнаружения вдруг ярких пятен, внезапные изме-
нения очертаний тела, издавание новых сильных звуков, в обычное
время несвойственных данному виду. При этом самцы всегда пн*
216
стинктивно показывают себя самкам «в самом лучшем светеж От
дельные моменты этих брачных игр имеют своим прямым назначе-
нием пробудить инстинкт гнездостроения, выкармливания птенцов,-
вызвать потребность в копуляции. Так как брачные игры выраба-
тываются очевидно действием полового отбора, они развиваются
в весьма специфичные виду формы поведения и окраски, весьма раз-
личные в большинстве случаев даже у близких видов и иногда в от-
дельных популяциях того же вида. Напряженность этих игр требует
затраты значительной энергии, которая обеспечивается обычно упи-
танностью самцов перед брачным периодом.
Формы, которые принимают брачные игры, у различных видов
весьма разнообразны. У видов с одинаковой окраской обоих полон
/
Рис. 174. Брачные игры синих пингвинов — Pygoscelis adeliae Ilomb. ct Jac.
Самка разгребает в земле гнездовую ямку и поджидает самца. При приближении послед-
него птицы особыми движениями и криками как бы приветствуют друг друга (этот момент
и изображен на фотографии). После заключения прочного знакомства, самец начинает помо-
гать постройке гнезда, подтаскивая к нему камешки. По Lewick, из Stresemann.
обычно и участие полов в играх является одинаковым, у видов с раз-
личной окраской полов роль их в играх оказывается различной.
Некоторым опытом классификации брачных игр может являться
группировка их по тем органам, на восприятие которыми они рас-
считаны (Thomson, 1923).
1. Ориентированные на врсприятие органами зрения: а) экспози-
ция отличительных особенностей окраски и формы тела, обычно свя-
занных с соответствующими импонирующими движениями, как, на-
пример, распускание хвоста павлином, прыжки и движения токую-
щего тетерева и т. д.; б) «подсказывающие» движения, направленные
на пробуждение инстинкта гнездостроения и т. д., выражающиеся
в одаривании самцом самки веточками и листочками, в кормлении
самцом самки и т. д.
217
2. Ориентированные на восприятие органами слуха: сюда отно-
сятся пение птиц, воркование и произведение
других, аналогичных
по значению, звуков;
кроме того, произве-
дение звуков хло-
паньем крыльев, сту-
ком по дереву (дят-
лы), движением пе-
рьев хвоста на лету
(бекасы) и крыла (го-
голь) и т. д.
3. Ориентирован-
ные на восприятие
органами осязания:
сюда относимы обще-
известные «поцелуи»
голубей и других
птиц, поглаживание
и охорашивание опе-
рения другого пола
Рис. 175. Брачные игры соснового тетерева — Dendroaa-
pus obscuras richardsoni Dougl.
Токующая птица сильно раздувает шейные воздушные меш-
ки, причем перья разворачиваются в виде двух белых розе-
ток по бокам шеи, среди которых выступают пурпурно-крас-
ные оголенные участки кожи. Помимо образования украше-
ний, надутые воздушные мешки играют роль резонаторов,
усиливая протяжный брачный крик, свойственный птице.
По Brooks, из Bent.
И Т. П.
У многих ВИДОВ
брачные игры однако
носят комплексный
характер и входят
одновременно во все
указанные рубрики.
У птиц, не гнездящихся общественно, с брачными играми самца
обычно бывает связана и борьба за территорию — некоторый уча-
сток площади, куда не допускаются другие самцы и самки того же
ЗРпс. 176. Токование полынного тетерева (или острохвоста) — Centrocercus urophasianus Bon-
1—в начальный момент тока: птица начинает падувать шейные воздушные мешки-
расправляет веером хвост, приподнимает сложенные крылья; 2 — в разгар тока: шейвис |
мешки сильно вздуваются, сложенные крылья принимают почти вертикальное положение-
По Horsfall, из Stresemaim.
218
вида и в пределах которого в дальнейшем будет строиться гнездо.
Положение и значительность избираемой территории определяются
экологией и величиной птицы; за удержание ее в брачный период
самцы того же, а иногда и близких видов вступают между собой в оже-
сточенные драки, а оживленное пение их является как бы указанием,
что данный участок уже занят.
Основное биологическое значение
этих территорий заключается в обес-
печении будущего выводка достаточ-
ным запасом корма, собираемого ро-
дителями поблизости гнезда.
Выбор гнездовой территории, так
же как й места самого гнезда, у пе-
релетных птиц падает обычно на сам-
ца, раньше самки возвращающегося с
зимовки. Там, где самец не принимает
участия в воспитании потомства, эта
забота естественно падает на самку.
Постройка гнезда в большинстве слу-
чаев проводится обоими, с преиму-
щественной ролью самки, которая
сплетает гнездо и выстилает его
нежными веществами, в то время как
самец главным образом собирает и
таскает материал. Чем сложнее вооб-
ще постройка, тем более возведение
ее падает на кого-либо одного из
пары. В таких случаях самцы часто
не принимают участия в постройке
гнезда и только сопровождают самку
в ее сборах гнездового материала.
Гнездостроен и е. В не-
которых случаях стремление самцов
строить гнездо не находит себе удо-
влетворения в одной постройке и
вызывает постройку еще нескольких
гнезд, обычно недоконченных, но
Употребляемых в качестве спальных
гнезд самим самцом. Наиболее из-
кестна в этом отношении гнездовая
Рис. 177. Токующий сепын журавль—
Megalornis grus L. ilo HcinroLh.
Деятельность крапивника. Вероятно
°лизкие к этим стимулы вызывают образование особых построек
австралийскими птицами беседочницами, строящими из гнездо-
вого материала небольшие галереи, украшенные цветами и ка-
кпми-нибудь блестящими вещами, среди которых самец с самкой
проводят брачные игры. Украшение гнезд блестящими надкрыльями
пауков и какими-либо другими предметами довольно широко рас-
пространено среди птиц (сорока, сойка, поползни и др.).
Местонахождение гнезда и материал, употребляемый для него,
чрезвычайно разнообразны и находятся в теснейшей связи с эко-
219
логией птицы. Обычно материал для гнезда собирается там же.
где птица кормится, и составляется, так сказать, всякими подруч-
ными веточками, былинками и т. п., изменяющимися сообразно
роду постройки и стадии ее выполнения. У многих птиц при этом
выработалась весьма узкая приспособленность к характеру мате-
риала и положению гнезда, что может являться фактором, огра-
ничивающим их распространение. У других, наоборот, выработалась
значительная приспособляемость, дающая возможность помещать
гнездо в различной обстановке
Рис. 178. Лепное гнездо рыжего починка —
Fumarins г it f us Gin.
1 — гнездо, построенное почти цетином из глины;
па гнезде его строитель. Все птицы около 75 г,
вес гнезда около 3—4,5 кг; 2—распиленное
и строить его из довольно разно-
образного материала. Поэто-
му целесообразно различать
птиц стено- и эвринидов. При-
мером последних может яв-
ляться например сокол сап-
сан, строящий гнездо и на
скале, и на ветках дерева, и
на земле; примером первых
является королек, гнездя-
щийся всегда среди мелких
веточек свешивающихся сни-
зу пышных еловых ветвей-
лап. Эти же свойства сказа-
лись и в характере гнезд раз-
личных отрядов птиц. При-
способления к частным усло-
виям экологии отдельных ви-
дов чрезвычайно осложнили
эволюцию птичьего гнезда и
явились причиной широкой
конвергенции в гнездострое-
нии отдельных видов птпц
различных таксономических
групп. Так, например, по-
требность гнездиться в пор-
ках или пустотах свойственна
представителям отрядов-
пингвинов, трубконосых, веслоногих, гусиных, чистиков, голубей,
гнездо, показывающее строение его из двух ка-
мер — гнездовой и прихожей. По JDoello Jurado,
из SLrescinann.
п» пугаев, сизоворонок, сов, дятлов и многим воробьиным, гнездя-
щимся кроме того и многими другими способами.
Не вдаваясь в детали, обзор типов гнезд можно сделать следую*
щим образом. Наиболее простым гнездом является ямка в земле,
выложенная несколькими травинками, что мы находим у мно-
гих куликов, чаек и куриных птиц. Более совершенные формь
наблюдаются у бакланов, цапель, некоторых чаек, которые вокру1
яиц укладывают валик веточек и иногда создают громоздкую по-
стройку, помещая ее и на ветвях деревьев. Получающееся улучше-
нием этой формы—-гнездо в виде чаши, с плотно сплетенными краями
и дном, у наших птиц наилучшего совершенства достигает, ДО
жалуй, у зяблика. Стремление защитить гнездо сверху привело к о°'
разованию гнезд-домиков, с входом сбоку, как, например, гнезда
220
пеночек и долгохвостой синицы, а стремление при этом сделать
гнездо недоступным даже для легкого врага, главным образом раз-
ных лазящих хищников, создало висячий тип гнезда, укрепленный
на кончиках веток деревьев, как, например, у иволги или ремеза.
В другом направлении шло развитие гнездостроения у других птиц.
То же основное стремление защитить гнездо от врага, сделать его
возможно более незаметным, создало птиц дуплогнездников, частью
гнездящихся в различных готовых пустотах, частью самостоятельно
выдалбливающих дупла (дятлы, некото-
рые синицы). Та же потребность вырабо-
тала способность представителей разных
отрядов рыть норы, главным образом клю-
вом, и на конце их устраивать гнездовые
камеры-расширения. Еще большего искус-
ства в лепной работе достигают птицы,
делающие гнезда из земли и глины. Упо-
требление этого материала может варии-
ровать от применения земли в небольшом
количестве, для укрепления основания
гнезда и стенок его (как у большинства
дроздов), до лепки гнезда целиком из
глины, как делает скалистый поползень
пли птица-печник (Furnarius, рис. 178).
У ласточек и стрижей в этой лепке боль-
шую роль играет слюна, к этому периоду
выделяемая в большом количестве сильно
увеличенными железами, крепче скрепля-
ющая комочки грязи, веточки, травинки
пт. д., а у саланган ставшая почти исклю-
чительным материалом для гнездострое-
ння.
Из наиболее интересных приспособле-
ний гнезд к тем или иным условиям сре-
ды следует упомянуть гнездо фламинго,
построенное из грязи в виде башенки, с
Рис. 179. Гнездо поползня —
Neosi.Ua chrysinptera Lath.
Птица кормит птенцов, сидящих
в гнезде. Гнездо строится из ку-
сочков коры, располагаемых
своей длинной осью параллель-
но стволу дерева и укрепляе-
мых паутиной. Благодаря та-
кому строению и охранитель-
ной окраске гнездо почти неви-
димо. По G-ibbert, из Stre.se-
inanii.
потном на верхнем конце и являющееся *
приспособлением против наводнений тех мелей, на которых дер-
жатся фламинго. Ходулочник, делающий гнездо на плоских берегах,
также надстраивает гнездо, сделанное из веток, в период наводнений.
Ьольшинство мелких птиц, вьющих открытое гнездо, убирает гнездо
снаружи мелкими лишаями, кусочками коры и т. п., делая его чрез-
вычайно похожим на нарост на коре или стволе дерева (рис. 179).
В целях защиты гнезд от мелких хищников многие птицы в тропиках
строят гнезда совсем рядом с гнездами ос, или устраивают его на
Деревьях, обитаемых жалящими муравьями, или выдалбливают по-
мещение для него в термитниках. Мелкие птицы, кроме того, часто
Уездятся вблизи гнезд крупных и сильных птиц, могущих защитой
своего гнезда обеспечить сохранность гнезду мелкой птицы, распо-
ложенному поблизости. В наших условиях к этому средству прибе-
гают например воробьи, гнездящиеся в гнездах орлов, орланов и
221
скоп, и парочки ушастых поганок, гнездящиеся под защитой коло-
ний чаек плп крачек (см. об этом также на стр. 301).
Процесс постройки гнезда мало исследован. Ласточки начинают
лепить гнезда с создания опоры его дну и затем выводят стенки,
постепенно накладывая комочек грязи за комочком и скрепляя их
вкладываемыми соломинками и перьями. Береговые ласточки, щурки
п зимородки роют своп норы клювом, лапками откидывая землю
назад. Большинство птиц, устраивающих гнездо опирающимся на
какой-либо предмет, сначала создают основу дна гнезда, затем воз-
водят боковые стенки и после этого выкладывают гнездо мягким
материалом и обвивают его веществами, имитирующими окружаю-
щую обстановку. Самый процесс свивания сопровождается харак-
терными движениями всего тела, во время которых птица глубоко
сидит в гнезде и напряженно и быстро дрожит (Schlosser, 1927). Это
дрожащее впрессовывание туловища имеет очевидно значение смять
гнездовый материал по форме тела и теснее стрясти и уложить отдель-
ные травинки. У черного дрозда этот процесс продолжается точно че-
тыре секунды, после чего самка приподнимается из лотка напять се-
кунд и затем вновь опускается, повторяя те же движения. При каждом
впрессовывании птица поворачивается примерно на 35° по сравне
нию с прежним положением и после 2—3 поворотов в одну сторону
поворачивается в сторону противоположную. Такое впрессовывание
возобновляется неоднократно и подряд, не слетая с гнезда, может
продолжаться до 30 раз. Очевидно эти движения то в ту, то в другую
сторону и создают ту плотную, «свитую» ткань стенок гнезда, кото-
рая поражает у многих видов своей ажурностью и вместе с тем проч-
ностью постройки. От времени до времени птица, кроме того, стре-
мится клювом укрепить и вплести травинки в стенку гнезда, особенно
при сопротивлении отдельных менее гибких и длинных травинок.
Как правило, гнездо строится главным образом по утрам, когда как
само гнездо, так и собираемый для гнезда материал влажны от росы
и сравнительно более податливы. При еще более сложных гнездах
птицы протыкают стенки гнезда клювом с травинкой, продергивают
последнюю через стенку и неоднократным повторением этого ма-
невра постепенно укрепляют бока гнезда и делают их более плот-
ными (рис. 180). Славка портниха в состоянии одним волокном сшить
3—4 пары противостоящих отверстий на краях свернутых листьев,
внутри которых она делает гнезда, и при этом укрепить края волокна
в предохранение от проскальзывания его. Висячие гнезда начинаются
с предварительного оплетания конца ветви длинными волокнами
и укрепления их на ней; свисающие концы этих волокон в дальнею
шем являются первыми основаниями для стенок гнезда.
Длительность постройки гнезда даже в сравнительно сложных
случаях обычно невелика, несмотря на работу над гнездом только
по несколько часов в день. Даже при приготовлении нор, береговая
ласточка, например, в состоянии вырыть за двое суток ход более 60 см
длиной. Самка дерябы приготовляет гнездо за 7—8 дней и даже за
3 дня. Сложная, шарообразная, убранная кусочками лишайников
снаружи п выстланная перьями (случается до 2000 перьев в одном
гнезде) постройка долгохвостых синиц заканчивается парочкой
-222
в 18 дней, причем само гнездо готого в сущности уже на 9-й
день.
Кладка яиц. Количество яиц, сносимых в гнездо видом,
различно у разных групп птиц и часто представляет собой система-
тический признак, очевидно выработавшийся в связи как с физиоло-
гическими, так и экологическими особенностями вида или система-
тической группы. Здесь очевидно сказывается способность вида про-,
тпвостоять различным враждебным факторам, или возможность ком-
пенсировать мдлое число яиц неоднократным гнездованием. Во мно-
гих случаях причины, обусловливающие установившийся размер
кладки, не являются вполне понятными.
Рис. 180. Отдельные моменты в плетении гнезда ткачпком — Quelea quelea L.
А 2 — способы завязывания лубяных волокон, употребляемые птицей; 3 — вьющая
гнездо птица; взаимодействие в постройке лап и клюва; 4, 5, 6 — последовательные стадии
в завязывании волокна вокруг ветви. Стрелка указывает направление перемещения во-
локна птицей; а — место щипка волокна клювом. По Friedemann, из Stresemann.
Только одно яйцо в кладке имеют многие крупные дневные хищ-
ники, крупные чайки, большинство чистиков и все трубконосые.
Затем отдельные виды других отрядов: козодоев, стрижей, воробьи-
ных, голубей, скрытохвостов, дроф и куликов.
Два яйца в кладке свойственны большинству голубей, колибри,
журавлям и гагарам; кроме того многим козодоям, пингвинам, чи-
стикам и киви.
Три яйца — нормальное число в кладке рябков и большинства
паек. Четыре яйца нормально свойственны трехперсткам и боль-
шинству куликов. Пять-шесть яиц нормально свойственны мелким
Дневным хищникам, многим совам, многим воробьиным умеренных
зон.
Наибольшее количество яиц имеют в кладке представители трех
отрядов: куриных, гусеобразных и воробьиных. Из первых серая
кУропатка несет до 18—22 яиц, из вторых чирок-свистунок несет
Д° 16 яиц, из воробьиных особенно плодовиты синицы: большая
223
песет .до 12—15 штук, долгохвостая до 16—19 штук. Это количество
япц в кладке подвержено значительным колебаниям п зависит от
весьма разнообразных обстоятельств. К внутренним определяющим
причинам относится возраст, с определенного момента вызывающий
уменьшение половой продукции. У курицы, например, уменьшение
количества яиц падает в период со второго года до восьми лет
в определенной пропорции, равной 88% каждого предыдущего года
(Brody, 1924). Ворони мохноногий канюк, имеющие среднюю кладку
в 3—4 яйца, к старости несут только по одному яйцу.*В свою очередь
и молодые птицы, при длительном развитии, несут яиц меньше, чем
взрослые особи.
Из внешних факторов существенное значение имеет обилие пищи
к началу ^размножения и в период его. В годы, обильные мышевид-
ными грызунами, наблюдаются, например, почти двойные по размерам
кладки у болотных сов, мохноногих канюков и белых сов и гнездо-
вание их иногда в неурочное время (осенью, ранней весной и т. д.).
Кроме того, в отношении размеров кладки можно подметить еще
некоторые закономерности. Крупные виды кладут яйца меньше,
чем их меньшие по размерам родственники. Виды, несущие крупные
яйца, имеют их обычно в меньшем количестве, чем родственные впды
с яйцами меньших размеров. Большинство птенцовых птиц, распро-
страненных в тропиках п в умеренной зоне, имеет разный размер
кладок:-в тропиках меньше, в умеренной зоне больше. Еще большее
число яиц несут те же виды на субарктических широтах. Различие
при этом может быть очень значительным: в тропиках многие во-
робьиные несут только по 2—1 яйцу, в умеренной зоне 5—6, север-
нее до 7—8 яиц. Примером может служить каменка, несущая в се-
верной Африке 5 яиц, в Германии преимущественно 6, в Гренландия
7—8 и даже до 9. Вероятной причиной этого явления считается влия-
ние северного, более длинного, чем в тропиках, дня. Продолжитель-
ный период освещения дает возможность более длительного питания,
а следовательно и большей продукции яиц. В силу этих же причин
возникает и более легкая возможность выкормить увеличенный вы-
водок. С другой стороны, возможно, что это свойство выработалось
в борьбе за существование, как приспособление в борьбе с суровыми
условиями жизни в северных широтах и как средство, только вос-
полняющее потери, несомые видом в период длинных и опасных
перелетов.
Интересной особенностью тех видов, которые несут 4 яйца п
больше в кладке, является способность нести большее число япц.
чем нормально, в случае удаления части кладки. При этом разные
впды реагируют по-разному, но в общем, при кладке в 4—5 яиц.
удаление!—2 яиц часто не вызывает докладывания яиц до полного
числа, удаление же 3—4 яиц вызывает описываемое явление. Повп-
димому птица испытывает некоторый дискомфорт при'насиживании
1—2 яиц (возможно через наседное пятно), что стимулирует психпк)
и деятельность полового аппарата; при этом докладка может иметь
место только в случае удаления части яиц в первые дни по окончании
кладки, пока яичник не испытывал еще ретрогрессивных изменений
По окончании докладки новым удалением части яиц можно вызвать
224
этот процесс заново. Таким образом удалось у болотной камышевки
(Acrocephalus paluslris Bechst.) довести число отложенных яиц до 11.
Штиве довел галку до откладки 15 яиц, а рекордистами в этих опытах
явились вертишейка, за 62 дня отложившая 62 яйца, и золотой
дятел (Colaptes auratus L.), снесший 71 яйцо. Это же качество в сущ-
ности использовано человеком при разведении яйценоских домаш-
них птиц, выведенные расы которых в состоянии нести в год до 200—
300 янц, а в течение своей жизни дать более 1000 яиц. Рекордистами
являются пз кур белый леггорн, несущий до 357 яиц в год (365 дней),
п из уток белый индийский бегун, снесший 365 яиц за 365 дней.
В случае удаления пз гнезда всех яиц, процесс иногда может итти
тел» же порядком, особенно если имело место удаление еще незакон-
ченной кладки. При удалении же кладки спустя несколько дней по
ее снесении, птица может вновь отложить кладку, что обычно сопро-
вождается постройкой нового гнезда и в сущности вновь возникаю-
щими брачными играми. Снесение новой кладки в этом случае зави-
сит в значительной степени от продолжительности этих игр и срока
постройки гнезда. У голубей это имеет место на 7-й—10-й день,
у черной вороны через 15 дней, у дерябы на 10-й день. У более круп-
ных птиц этот срок более длинен. При разрушении гнезда с уже на-
сиженной кладкой, восстановление наступает значительно позднее;
здесь необходим гораздо более длительный путь развития ооцитов,
к этому сроку уже значительно ретрогрессировавших и частью впав-
ших в состояние покоя. По существу эти вторичные кладки яиц пред-
ставляют собой явление, аналогичное нормальным вторым кладкам,
широко распространенным у многих птиц теплых широт. Оно свой-
ственно в нашей фауне воробьям, многим вьюрковым, дроздам, слав-
ковым и другим птицам.
Нормальная вторая кладка (в противоположность вынужден-
ным) начинается обычно по выводе потомства. При этом птпца вновь
переживает брачные игры, строит новое гнездо и т. д., как в пре>-
дыдущем случае. Наличие этих вторых кладок определяется главным,
образом сроком, необходимым на воспитание потомства, и в случае'
быстрого развития птенцов и длинного богатого кормом лета птица
может иметь еще третью и даже четвертую кладки (например во-
робьи) (рис. 181). У некоторых видов при этом происходит интересное
захождение одного цикла на другой: так самец дроздовидной камы-
шевки по выводе птенцов принимает на себя все заботы по выращи-
ванию выводка, в то время как самка строит второе гнездо и кладет
новую кладку, а у водяной камышницы, например, птенцы первого
вывода помогают родителям в выкармливании птенцов второй кладки.
Н а с и жива н и е. У птиц с мало выраженным половым димор-
физмом в насиживании принимают обычно участие оба пола, в дру-
гих случаях — тот пол, который имеет более однообразную и менее
заметную окраску. Прп участии обоих полов обычно устанавливается
смена их в насиживании, причем ведущая роль принадлежит то
самцу, то самке. В некоторых случаях это имеет повидимому при-
способительное защитное значение, как, например, у страуса, где
верный самец насиживает ночью, а бурая самка насиживает днем.
Время смены часто является строго фиксированным и имеет место
/
Орнитология—1209 225
два раза в сутки; сама смена при этом сопровождается особыми це-
ремониями, тихими криками и приносом к гнезду нового строитель-
ного материала. В редких случаях насиживание бывает совместным,
как днем, так и ночью, что имеет место например у мелких ткачиков.
При участии в насиживании какого-либо одного пола, другой не-
редко помогает этому процессу приносом корма для насиживающей
птицы. При отсутствии этой помощи мелкие птицы периодически
отлучаются от гнезда на короткий срок, чтобы перехватить немного
пищи. У американского крапивника это отсутствие однако не про-
должается долее 6 минут каждый раз. Более крупные птицы часто
Рис. 181. Диаграмма гнездового периода 85 пар домовых крапивников — Troglodytes aedw
ЛЧеШ.» гнездившихся в окрестностях Орнитологической станции в Огио (США) в 1927 г.
Черные линии указывают время п длительность гнездового периода, начиная от от-
кладки 1-го яйца до вылета птенцов из гнезда; перерыв линии пунктиром означает прежде-
временное прекращение гнездового периода; белые кружочки — нахождение гнезда в долине,
черные кружочки — гнездование на возвышенности (больше 274 м над ур. м.). Диаграмма
показывает значительную индивидуальную вариацию в начале гнездового периода у разных
пар с амплитудой от 10 мая до 31 августа; к вторичному гнездованию переходят лишь птицы,
рано приступившие к гнездованию вообще. По Baldwin and Bowen, из Stresernann.
приспособляются при этом к очень длительному воздержанию от
поисков пищи, существуя исключительно за счет тех жировых от-
ложений; которые накапливаются (специально) в большем коли-
честве к периоду размножения. Этот пост у обыкновенной гаги про-
должается до 28, а у эму даже до 60 дней полного воздержания от
пищи.
У огромного большинства птиц самый процесс насиживания,
сущность которого заключается в передаче кладке теплоты тела
родителя и поддержании этой теплоты необходимый срок, осуще-
ствляется с помощью наседных пятен, образующихся на теле взрос-
226
At.
l е.
!.th.
t.p.
Рис.
пятна и их кровяное питание у
Ath.e."
-М.а.
LS2. Паседпьте __ __ . _____ _______
обыкновенной чайки — Lar us ridibundus L.
Кожа левой половины туловища отпрепарирована и от-
вернута в сторону (пунктирный контур); fascia superfi-
cialis показана косой штриховкой, паседпые пятна —
черным цветом, птерилип — пунктиром. Прп I и II по-
казаны отверстия для выхода наружной грудной арте-
рии, питающей главным образом грудные наседные
пятна, под кожу. Л. i.—подкожная грудная арте-
рия — arteria subcutanca (thoracis) плп аг. incubatoria;
A. th. e', A. th. e''—ветви наружной грудной арте-
рии— аг. thoracica externa; М.а.—musculus subcu-
laneus abdominis; M. th.—m. subcutaneus tlioracis;
t. p. — главная ветвь arteria pelvica interna (являю-
щаяся ветвью arteria femoralis). По Lange, из Strese-
mann.
лой птицы к этому периоду (рис. 182). Те части кожи птицы, которые
приходят в соприкосновение с кладкой, испытывают при этом со-
стояние, напоминающее воспаление. Подкожная клетчатка сильно
разрыхляется, повышается ее васкуляризация, исчезает к этому сроку
обычный подкожный
слой жира, в некото-
рых группах сосудистая
ткань при этом сильно
разрастается, приобре-
тает губчатое строение
и переполняется лим-
фой. Расположенные на
этом участке перовые
сосочки и связанные с
ними гладкие перовые
мускулы полностью ис-
чезают. Температура те-
ла на этих участках
оказывается несколько
повышенной, а в сово-
купности с более повы-
шенной температурой
того пола, который уча-
ствует в насиживании,
достигает заметного
превышения над сред-
ней температурой тела
данного вида. Развитие
наседных пятен подчи-
няется в основном гор-
мональному стимулу и
обнаруживается неред-
ко ранее откладки яиц;
прп этом они обнару-
живаются иногда у еще
неполовозрелых особей,
появляясь например в
летний период у обык-
новенных чаек-одноле-
ток [Lotus ridibundus
L.). Размеры и место-
положение этих пятен
заметно вариируют в
зависимости от характера птерплозиса в данной систематиче-
ской группе. В большинстве случаев пятно располагается в об-
ласти брюшной аптерии, отграниченной по бокам боковыми брюш-
ными птерилиями (pteryla gastrei), но иногда кроме того развивается
в области боковых аптерий, между главной и боковой ветвями ниж-
ней птерилии, и оказывается таким образом разбитым на несколько
участков. Далеко не всегда величина наседного пятна соответствует
•227
ванном к гнездовому периоду среди
Рис. 183. Императорский пингвин — Aptenodytes
iorsteri Gray, согревающий птенца на расправлен-
ных плавательных перепонках, па льду Антарк-
тики. По Wilson, из Stresernann.
неличные кладки, очевидно требуя от времени до времени перемеще-
ния яиц или тела птицы: но это соответствие в величине пятен и
в количестве яиц обычно имеется у птиц с небольшой кладкой.
()тсутствуют наседные пятна только у гусеобразных и некото-
рых веслоногих, у которых водный образ жизни мог вызвать
исчезновение их, в связи с необходимостью более совершенного
отепления нижней поверхности брюха, постоянно соприкасающе-
гося с водой. У гусеобразных этот недостаток возмещается образо-
обычного пуха особого длин-
ного п иначе окрашенного
пуха, который высиживаю-
щая яйца птица выщипывает
у себя на груди и брюхе и
употребляет на выстилку
гнезда и прикрытие яиц на
время покидания гнезда.
Плохо проводя тепло, оче-
видно этот пух создает усло-
вия, сходные с эффектом тер-
мостата. Из веслоногих олу-
ши согревают яйца главным
образом плавательными пе-
репонками, которые они рас-
правляют на яйцах, опуска-
ясь на гнездо. У пингвинов,
высиживающих их единствен-
ное яйцо в стоячем положе-
нии, наоборот, оно иногда
поддерживается на перепон-
ках лап. У этого удивитель-
ного но приспособлениям
отряда кожа тела перед кло-
акальным отверстием обра-
зует род кармана, прикры-
ваемого сверху брюшной
кожной складкой, в которой
обычно поддерживается м
высиживается яйцо и согре-
вается птенец в его более раннем возрасте (рис. 183).
Насиживание кладки у большинства птиц начинается после от-
кладки последнего яйца, так что вначале отложенные яйца, при
больших кладках, могут находиться в состоянии покоя до 20 и
30 дней. Так поступают все гусеобразные, куриные, кулики и боль-
шинство воробьиных. Меньшая группа птиц начинает насиживание
с первого отложенного яйца (например совы, стрижи, дневные хищ-
ники, попугаи, цапли, аисты, поганки, гагары, веслоногие, журавли
п др.), некоторые занимают промежуточное положение, весьма не-
плотно насиживая первое яйцо (голуби), или приступают к насижи-
ванию незадолго до окончания кладки (дятлы, страусы, некоторые
пастушки). Во многих из этих случаев то или иное доведение можно
228
Рие. J84. Полупловучее гнездо малой поганки —
Podiceps riificollis no'caehollandiae Steph.
(’верху гнездо с япцами, старательно укрытое птицей
водяными растениями; внизу —то те гнездо, остав-
ленное открытым. По Burrell, пз Stresemann.
рассматривать как приспособление к тем или иным особенностям
гнездования. Так, насиживание с первого япца у цапель, нередко
устраивающих гнезда с очень неглубоким гнездовым лотком на
кустах, сохраняет яйца от вываливания при качании веток ветром.
Насиживание по окончании кладки, у птиц с большим количеством
яиц, приводит к более или менее одновременному вылуплению птен-
цов, что облегчает заботу
о прокормленип их и за-
щите, а у выводковых птиц
является и необходимой
предпосылкой их воспита-
ния.
При малой величине
кладок разновременность
вывода птенцов не имеет
особых неудобств, при
крупных кладках в неко-
торых случаях это вызы-
вает значительную разно-
возрастность птенцов и у
хищников, цапель и аистов
обычно приводит к гибели
младших птенцов еще до
вылета из гнезда; у пога-
нок каждый из высижи-
вающих супругов вслед-
ствие этого часто удоволь-
ствуется выводом только
двух первых птенцов, по-
кидая после этого гнездо
и остальных, еще не вы-
лупившихся птенцов.
У птиц, насиживающих
с начала кладки, затруд-
няется воровство яиц и ра-
зорение гнезд со стороны
мелких вредных птиц,
вроде сойки, сороки и др.,
нему часто подвергаются
птпцы, оставляющие гнездо
открытым на более или менее длинный промежуток времени. Как
в той, так и другой группе, в процессе эволюции, высиживающие
птицы и яйца приобретают различную покровительственную окраску
(см. стр. 300), а у гусиных и поганок развивается способность, оста-
вляя гнездо, прикрывать яйца пухом (первые) или зелеными вод-
нымц растениями (вторые), вытащенными пз воды поблизости от
г«езда (рис, LSI).
Использование для инкубации яиц те п-
4 ° й сред ы. Чрезвычайно своеобразным способом высиживания
ЯИЧ обладают большеногие куры Малайского архипелага и Австра-
229
пии. в наиболее простых случаях напоминая поведение большинства
рептилии. Однако развился этот способ повпдимому совершенно не-
зависимо от последних. Большеногие куры совершенно освободились
от высиживания яиц теплотой своего тела, вместо этого закапывая
яйца в землю, где птенцы развиваются под влиянием теплоты почвы,
держащейся на уровне 25—30° С. Таким образом поступает Eulipoa
waUacei (Gray), закапывающая свои япца на глубину 35—60 см,
в темный вулканический песок побережий Молукских островов.
Вйды рода Megapodius или вырывают в рыхлой земле яму до глубины
в 60—100 см, откладывают на дне ее яйца и закрывают выброшенной
землей, или, сгребая ногами, собирают большую кучу лесного му-
сора, внутрь которого и закапывают япца. Наибольшей сложности
этот инстинкт достигает у родов Leipoa, Aepypodius и Talegalltis.
Они вырывают ногами в песчаной почве довольно обширные ямы и
заполняют их специально собранными различными растительными
остатками (листьями, веточками). Под влиянием сырости, в этих
остатках постепенно развивается гниение, продуцирующее теплоту
в 30—36°. Спустя несколько месяцев по устройстве этой ямы, сор-
ные куры вновь разыскивают ее и за несколько дней до снесения
яйца вырывают в середине этой гнпющей массы небольшую камеру;
в дальнейшем в ней откладываются, с интервалом в несколько дней,
от 6 до 8, иногда даже до 20 яиц, распределенных обычно в несколько
этажей и прикрываемых сверху п между собой песком, смешанным
с гниющим веществом. Рыхлость этой смеси обеспечивает доступ
воздуха к развивающемуся яйцу. В устройстве этих куч самке у не-
которых видов помогает и самец; целесообразно отметить также,
что яйца зарываются поодиночке, в вертикальном положении, тупым
концом кверху, и что расположение их по 3—б штук в каждом этаже
на некотором расстоянии друг от друга очевидно указывает на извест-
ную систематичность в роющей деятельности птицы.
Развитие птенцов в отложенных яйцах происходит очень медленно,
не менее 42 и даже 70 дней, вероятно в зависимости от температуры
гниющих остатков. В яйцах, отложенных непосредственно в песок,
развитие происходит частью за счет теплоты самого песка, нагре-
того лучами тропического солнца, поддерживающейся на высоте 30°,
частью за счет теплоты, развиваемой самим эмбрионом и сохраняю-
щейся в яйцевой камере вследствие плохой теплопроводности песка.
В строении самого яйца мы находим существенные приспособления
к длительному развитию. Яйцо очень крупно (12—17% веса тела,
в то время как у одинаковой по величине курицы яйцо весит только
4%) и чрезвычайно богато желтком (до 220% веса белка, в то время
как у кур только 62%). Поэтому вылупляющиеся птенцы хорошо
развиты, покрыты длинным пухом и обладают уже значительно
развитыми маховыми перьями. По вылуплении они самостоятельно
прокладывают себе дорогу через кучу наружу и сейчас же могут хо-
рошо подлетывать. Спустя некоторое время они повидимому при-
соединяются к своим родителям.
Временное зарывание яиц в песок, обычно на дневные часы, от-
мечается у некоторых куликов. Однако среди этой группы к более
широкому использованию почвенной теплоты для высиживания яиц
230
перешел только один крокодилов сторож (Plizvianiis aegypints L.),
который зарывает свою кладку яиц на глубину 10 см в нагретый
солнцем песок речных островов. Птицы однако следят за этим ме-
стом п, как говорят, в жаркие часы дня ложатся здесь на песок,
чтобы может быть ослабить накаляющее действие прямых солнечных
лучей. Детали поведения этого вида однако недостаточно известны.
Основные черты в эмбриональном разви-
тии птиц (рис. 185). Как уже указывалось (см. стр. 139), опло-
дотворение яйца происходит в верхнем отделе яйцевода. Отклады-
ваемое яйцо нередко
находится уже в на-
чальных стадиях дро-
бления зародышевого
диска. Дробление
яйца птицы происхо-
дит по меробластиче-
скому тппу, сходному
с тппом дробления,
наблюдаемым у реп-
тилий. Зародышевый
диск охватывается
процессом деления
только в центральной
его части, ближе к
периферии лишь на-
мечая распадение
плазмы на крупные
бластомеры. Эти крае-
вые клетки стоят в
сообщении с питаю-
щим желтком и обра-
зуют промежуточную
зону между дробя-
щейся и инертной
частью желтка; по-
Рис. 185. Развитие куриного яйца.
(а) —яйцо перед насиживанием; (Ъ) — яйцо в середине
второго дня насиживания; (с) — яйцо в конце пятого дня
насиживания; (d)—яйцо на седьмой день насиживания.
1 — воздушная камера; 2 — халазы; 3 — белок; 4 — под-
скорлуповая оболочка; 5 —area ораса, окружающая кажу-
щуюся более темной area pellucida; 6 — аллантоис, лежа-
щего па левом боку эмбриона, окруженного амниотической
оболочкой; 7 — area vasculosa; 8 —аллантоис с его сосу-
дами на фоне желточного мешка; границы аллантоиса выде-
ляются благодаря огибанию его края проходящими в его
стенках сосудами; эмбрион просвечивает через оболочки.
По Кегг, из Stresernann.
степенным дроблением бластомеров эта зона все более отодвигается в
сторону периферии и увеличивает таким образом площадь зародыше-
вого диска. Вследствие быстрого роста в стороны, центральная часть
диска вскоре утончается и образуется только одним просвечиваю-
щим слоем клеток, в то время как краевые его части являются много-
слойными и непросвечивающпми. Это является причиной образо-
вания. спустя сутки с начала дробления, area pellucida (пли про-
зрачное поле) и окружающей ее area ораса (или темное поле).
Эмбрион в дальнейшем образуется всецело в области area pellu-
cid а. в области же area ораса образуются органы временные,
исчезающие в течение или к концу эмбриональной жизни. Однослой-
ная часть диска, являющаяся первичной эктодермой, приподнимается
над подстилающим ее белым желтком, образующаяся полость дроб-
ления (так наз. бластоцель) наполняется жидкостью, и создаются
условия для развития гаструляцип. При этом, в области заднего
23L
края промежуточной зоны образуется бластопор, и свободный
край диска с боков и сверху медленно начинает вворачиваться и
расти внутрь полости дробления. Мигрирующие и растущие внутрь
клетки образуют энтодерму, заполняют полость и входят в сопри-
косновение с внутренними боковыми краями ее, к этому времени
утолщающимися скоплением эктодермальных клеток в зародышевые
валики. Параллельно с этим процессом происходит интенсивное раз-
множение клеток дорзальной губы бластопора, развивающееся в пе-
реднем направлении п приводящее к образованию так называемой
многослойной первичной полоски в медиальной и задней частях дор-
зальной стенки гаструлы. Вдоль этой полоски в скором времени
образуется желобок, разделяющий полоску на две продольные пер-
вичные складки. Перед передним концом (Гензеновским узлом) этих
складок на продолжении первичного желобка в скором времени об-
разуется новая короткая полоска, которая является зачатком хорды
и мезодермы и дает стимул к образованию нервных валиков па
лежащей над ней нервной пластинке.
Параллельно с ростом этих образований удлиняется и принимает
более грушеобразную форму area pellucida, суживающаяся к зад-
нему концу эмбрионального диска. I
В области первичней бороздки также выделяются мезодермаль-
ные клетки, которые мигрируют в пространство между двумя пер-
вичными листками и расселяются к периферии, образуя при этом
сосудистую кровеносную ткань. Образование этой ткани суще-
ственно отстает от центробежного распространения экто- и энто-
дермы, вследствие чего принято различать на этих стадиях в area
ораса три зоны: area vasculosa — в пределах распространения эле-
ментов мезодермы, area vitellina interna — в области наличия только
двух зародышевых листков, и area vitellina externa — в области
открытого желтка п краевой зоны зародышевого диска. Центро-
бежный рост всех трех слоев идет однако очень быстро, и к седьмому
дню они покрывают уже всю область желтка. Слой мезодермальных
клеток очень скоро расщепляется на два слоя: самато- и спланх-
ноплевру, между которыми возникает целомическая полость.
Тем временем медуллярные складки, разрастаясь кзади, захва-
тывают в свою область п первичную бороздку, при этом медулляр-
ный желобок замыкается, начиная спереди, в трубку.
Обособление эмбриона происходит как в силу его быстрого роста*
так и вследствие углубления той полулунной борозды, которая
с самого начала образовывалась впереди головного отростка. Послед-
няя, углубляясь под головной отдел, продолжается в дальнейшем и
по бокам выступающего тела эмбриона, а впоследствии вместе с бо-
роздкой, углубляющейся кзади от эмбриона, отграничивает от желтка
и хвостовой отдел, в конечном счете обусловливая связь продолго-
ватого тела эмбриона с желточным мешком только посредством
узкого желточного прохода. Обособляющийся эмбрион, располо-
женный первоначально параллельно короткому диаметру яйца, на-
чинает несколько перемещаться своей осью ближе к продольной осн
яйца, головой к его тупому концу, изгибается вдоль своей оси я
ложится на желток своей левой стороной. С течением времени пи-
232
тательный желточный ма-
териал затрачивается иа
рост эмбриона, тело его
все более увеличивается,
тогда как желточный ме-
шок все более уменьша-
ется. К моменту вылупле-
ния сильно уменьшивший-
ся в размерах желточный
мешок вбирается внутрь
брюшной полости, стенки
его замыкаются за ним в
области пупка, а оболочки
мешка съеживаются и от-
брасываются как беспо-
лезные организму. Остатки
желточного стебелька, ме-
жду пупком и пищевари-
тельным каналом, сохра-
няются у многих, преиму-
щественно выводковых
птиц в виде diverticulum
caecum vitelli — короткого
отростка, напоминающего
рудиментарную слепую
кишку и находящегося на
стенке тонкой кишки.
Одновременно с потребле-
нием желтка происходит
расходование и белочной
массы яйца, становящейся
весьма тягучей в начале
эмбрионального развития;
за несколько дней до вы-
лупления белок полностью
используется, так что эм-
брион со своими оболоч-
ками занимает
полость яйца.
Вис. 186. Развитие зародышевых
[молочек эмбриона
Ins gallus L.;
ИЯПП “'H’CpiDlU ДСП!» 11 <* L-
<b) — на девятый день па-
|р н <—на дменаддахып
тппл.пЛ?и??Пваппп- AU- — аллан-
-• - полость аллантоиса:
внутренний листок аллан-
: Ат.С. —
1ППоп‘г“ ««шниии; j4.O. V. - СОе-
иочтЖ амниотической и серозной
ПОЛНОСТЬЮ
курицы, Gal-
последовательные
стадии; схема.
па четвертые! день наспжп-
1Рп,иванпя; (с)—на двенадцатый
?iuiA3»-r
тп • xpCHilllU di 11UTUК UJJLJlilH-
„jea; Am.— амнион; Ат.С. —
,!& .амнпона; A.S. V. — сое-
тип»"*'" «““«этической н серозной
S эктодерма; Е.Е.В.С.
ЕсТ
Мез
Ent
АН +Ат.
Ur -
A.S.V -п
D.5.N.5
Ser
Ат
Eiw.S -
Ат С
АН !~
Ат^Ё^\
Е.Е.В.С.
АН./-^
оболочек; D.S.N.S. — выход желточного мешка в ну-
— экстра-эмбриональпап полость тела; Etw.— белот?:
энтодерма; Mes.—мезодерма; M.R.— мезодермальное
_ _ ~ _ ЭМбрИОпальнЬ1н
Y.S.S. — перего-
колкпА~~'Лелковый мешок; Ent. —--™——, ------ -----------------
nvnnv. TiV- — membrana vitellina; Ser. — серозная оболочка; Umb.
к, иг. — стебелек аллантоиса (urachus); Y.S.—желточный мешок;
родни желточного мешка. По Lillie, из Slrcsemann.
233
Подобно рептилиям и млекопитающим, птицы обладают в эмбрио-
нальном состоянии двумя зародышевыми оболочками: амнион и ал-
лантоис (рпс. 186). Амнион является обширной оболочкой, образую-
щейся только эктодермальным и саматоплевральным листком и воз-
никающей, следовательно, после расщепления мезодермы. Перво-
начально эти два слоя образуют полулунную складку впереди головы;
которая растет и постепенно наволакивается на голову эмбриона.
В это же время подобная складка образуется сзади хвоста, и своим
продолжениями обе складки связываются по бокам. В дальнейшем
эмбрион как бы погружается в эти складки, которые сливаются над
ним в сплошной покров. Таким образом эмбрион оказывается заклю-
ченным в двойной мешок, внутренний пз которых называется амнион
и оказывается лежащим в полости, называемой амниотической и
вскоре наполняющейся жидкостью — liquor amnii, представляющей,
по содержанию солей и белков, наиболее благоприятную среду для
нежных тканей зародыша. Этот внутренний мешок имеет эктодер-
мальную стенку, направленную внутрь, и саматоплевральную, на-
правленную наружу; обратное отношение пластов наблюдается
в серозной оболочке, которая, оставаясь в частичном соединения
с амнионом над спинной стороной эмбриона, сильно разрастается
вокруг желточного мешка, прилегая своей наружной стенкой к жел-
точной оболочке, и в конце концов выстилает внутреннюю поверх-
ность подскорлуповой оболочки.
Аллантоис, образуется как вырост пищеварительного канала, не-
посредственно впереди анального отверстия. Наружная стенка его
образуется спланхноплеврой, а внутренняя — «энтодермой. Он быстро
разрастается в крупный мешок, приходящий в соприкосновение
с внутренними оболочками яйца, где срастается с внутренней по-
верхностью серозной оболочки и в процессе роста обволакивает
почти всю ее подскорлуповую поверхность. Стенки аллантоиса бо-
гато снабжаются кровью в связи с его задачей как органа дыхания,
осуществляемого через поры в скорлупе яйца, и органа выделения.
К концу периода насиживания, когда у эмбриона уже начинают
функционировать легкие, аллантоис сморщивается и отбрасывается
вместе со скорлупой, и лишь его узкий и вытянутый стебелек сохра-
няется некоторое время как urachus на внутренней поверхности
брюшной стенки птенца.
В эмбриональном развитии птиц имеется много общего с разви-
тием рептилий, как это легко видеть из сопоставления соответствую-
щих стадий развития этих животных (рис. 187). В целях более кон-
кретного ознакомления с эмбриональными стадиями развития птенца:
привожу хронологический обзор эмбрионального развития обыкно-
венной курицы.
Первый день. 1-й—8-й час насиживания. На-
чало дробления сильно вариирует в зависимости от длительно-
сти нахождения яйца в яйцеводе птицы. Обычно первые
роздкп появляются уже в желтке, поступающем в маточньй
«отдел яйцевода, и отложенное яйцо имеет зародышевый диск ДО
4 мм ширины. В этот период охлаждение яйца вызывает только
приостановление развития, повышение же температуры вь1П1е
234
нормальной температуры насиживания может быть губительным для
жизни.
В этот период образуются описанные выше area pellucida и ораса,
образуются зародышевая полоска, бластопор п зародышевые валики
и первые рассеянные клетки
мезодермы, приуроченные к
задней части area pellucida,
несколько мутнеющей к этому
времени (эмбриональный щи-
ток).
8-й—12-й час. Прими-
тивная полоска увеличива-
ется до трети длины area
pellucida, которая становится
овальной. Происходит энер-
гичное образование клеток
мезодермы.
12-й—16-й час. На при-
митивной полоске, увеличи-
вающейся до двух третей те-
перь уже грушеобразной area
pellucida, появляется первич-
ный желобок.
16-й—20-й час. Впереди
первичной полоски образу-
ется темная черта, обозна-
чающая зачаток хорды, раз-
деляющий мезодерму на пра-
вую плевую половины. Экто-
дерма над хордой утолщается
в медуллярную пластинку,
образующую вскоре медул-
лярные утолщения и лежа-
щий между ними желобок.
Впереди этих образований в
•жтодерм начинает выпячи-
ваться полулунная головная
складка и намечается место
°УДУщей амниотической
складки.
20-й—24-й час. Полу-
лунная-головная складка бы-
СтР° увеличивается в разме-
рах и приподнимается над
Ориональным диском; тем
вРеменем медуллярные утол-
щения соприкасаются друг
с Другом в области ме-
Рпс. 187. Эмбриональные стадии развития курицы
(правая колонка) в сравнении с соответствую-
щими стадиями развития ящерицы (левая ко-
лонка).
Обратите внимание на возрастающую диверген-
цию на последних стадиях. По Keibel и Selenka,
из Hellmann.
^Уллярного желобка и, начиная с переднего конца, обращают
его в мозговую трубку. Открытые концы медуллярных утол-
235
щений захватывают при этом передний отдел первичной бо-
роздки, задняя часть которой в дальнейшем исчезает. В этот же период
происходят расщепление мезодермы на наружный и внутренний
листки? л образование вторичной (целомической) полости тела.
Это расщепление однако не захватывает всей мезодермы и ограни-
чивается только боковыми ее частями, так называемыми боковыми
пластинками, не затрагивая часть, тяготеющую к хорде, так называе-
мые позвоночные пластинки. В последних очень скоро появляются
поперечные бороздки, дробящие их на кубические отделы-сомиты.
Первая пара сомитов появляется в 20-м часу жизни на уровне
переднего конца первичной полоски, следующие появляются между
ней и полоской, так что последняя все время отодвигается кзади,
и удлинение эмбриона имеет место между сомитами и полоской.
Образование и рост сомитов, в дальнейшем появляющихся в ко-
личестве более 50 пар, имеют место в течение нескольких первых
дней. Та часть эмбриона, которая находится впереди первого сомита,
соответствует в дальнейшем голове, а остальные —2 шее, телу и
хвосту. В период всех этих изменений area ораса распространяется
постепенно по поверхности желтка, и появляется уже значительно
сосудистое ноле вокруг эмбриона.
Второй день. Медуллярные складки, закрываясь, быстро
превращаются в трубку; передняя часть ее впереди первого сомита
образует четыре последовательных вздутия мозговых пузырей; из
стенок переднего мозгового пузыря образуются выросты глазных
пузырей. В конце будущей головы намечаются слуховые ямки.
Число сомитов за этот день увеличивается от 4—5 до 15. Одно-
временно происходит образование сердца и главнейших кровенос-
ных сосудов из спланхнического листка мезодермы на вентраль-
ной стороне будущей глотки. При образовании последней, соеди-
няются медиовентральные выступы спланхнических листков обеих
сторон в области S-образного изгиба головной складки и таким
образом отграничивают снизу начальный отдел кишечной трубки.
Мезодермальные стенки этих выступов, вентрально от образую-
щейся глотки, утолщаются и сливаются, образуя внутри пустоты
в виде двух трубок. Кпереди и кзади каждая трубка продолжается
в крупные сосуды — первую дугу аорты кпереди и желточные вены
кзади. Вскоре медиальная перегородка, разделяющая эти трубки,
резорбируется, образуя зачаток сердца, которое начинает пульси-
ровать, сначала очень редко и медленно. Обе дуги аорты, в ко-
торые сердце гонит кровь, вскоре загибаются кверху по бокам
гсишечной трубки и отсюда идут по направлению к хвосту, отда-
вая по пути пару желточных артерий, разветвляющихся 'в пре*
делах area vasculosa; по краям последней к этому времени обра-
зуется также sinus или vena terminals, через которую и обильна
ветви других вен кровь собирается в две желточные вены, впа-
дающие в венозный задний отдел сердца. Пз других образован^
следует отметить появление в области 6-го—S-го сомита заклад^
первичных почек (Вольфовых протоков), начало сгиба головы в^
реди от более оформляющейся хорды и быстрое развитие амниоти<
ской складки, уже прикрывающей спереди головной отдел эзмбриона.
236
В трети й день бластодермальный диск захватывает почти
половину поверхности желтка и, так как белок уже значительно
уменьшился в количестве, сосудистая сеть area vasculosa прибли-
жена к скорлупе. Помимо сосудистой системы желточного мешка,
в теле эмбриона развилась собственная сеть. Пара передних и
задних кардинальных вен собирает кровь в теле и гонит ее через
правый и левый Кювьеровы протоки в сердце. Пара первичных аорт
соединилась теперь в одну дорзальную аорту, которая в области
шеи, вместо одиночных правой и левой дуг, пополняется следу-
ющими 2—С) парами дуг. Однако вследствие неодновременного
образования и превращений их, в каждый момент они нс пред-
ставлены более чем 3—4 парами. В дальнейшем первая, вторая
п пятая дуги очень скоро исчезают; третья пара продолжается
в шею il в голову в виде внутренних и наружных arteria carotis и
отсылает также arteria subclavia в передние конечности; четвер-
тая дуга правой стороны превращается в восходящую дугу боль-
шой аорты, а левая ее ветвь исчезает; шестая или последняя пара
преобразуется потом в легочные артерии, из которых правая
некоторое время связывается еще со стволом аорты посредством
ductus Botalli. Одновременно с этими изменениями происходит пре-
вращение сердца из простой трубки в четырехкамерный орган.
При этом сердечная трубка S-образно изгибается, передняя часть
сгиба вздувается п разделяется перегородкой, растущей с внутрен-
ней стенки на правый и левый желудочки. Процесс перестройки
сердца затягивается на несколько дней; на пятый день сходная пе-
регородка разделяет заднюю или венозную часть сердца па правое
и левое предсердия, причем полное разделение их происходит не
ранее чем на двенадцатый день, вследствие сообщений предсердий
между собой через foramen ovale. На пятый день перегородка же-
лудочков продолжается в bulbus arteriosus и подразделяет его на
truncus arteriosus и truncus pulmonalis. В связи с развитием легких,
появляются п легрчные вены, связующие легкое с левым предсерг
Дпем сердца. К концу пятого дня большинство важнейших артерий
в вен тела в основном приобретает свое дефинитивное развитие.
Помимо этих важных изменений в развитии кровеносной системы,
в третий день дифференциация широко захватывает и большинство
Других органов. Сильно разрастаются пузыри головного мозга, а
задний мозг распадается на мозжечек и продолговатый мозг.
" связи с глазными пузырями образуются хрусталики. Намечаются
обонятельные ямки. Вдоль нервной трубки появляются зачатки че-
репных и спинных нервов. Заканчивается образование задней и пе-
редней кишек; вторая при этом подразделяется на пищевод, желудок
11 Двенадцатиперстную кишку, первая на кишечник и клоаку. В виде
вентрального выроста пищеварительной трубки впереди желудка
закладываются парные легкие. Подобные же выросты из двенадцати-
верстной кишки образуют печень и поджелудочную железу; значи-
тельно прогрессирует развитие Вольфовых тел и их протоков.
В этот период эмбрион несколько скрючивается вдоль своей оси
в ложится на желток левой Стороной своего тела.
I е т в е р т ы й день знаменуется появлением зачатков пе-
237
редких и задних конечностей, пока имеющих вид уплощенных кони
ческпх выростов. Амнион теперь полностью окутывает эмбрион i
лежит почти непосредственно под скорлуповой оболочкой яйца
Голова эмбриона подвернута более чем под прямым углом книзу
хвост загибается кзади и внутрь. Головные и мозговые пузырв
чрезвычайно увеличились в размере, на голове могут быть разли
чены клюв и зачатки обонятельных и слуховых органов. В полости
тела уже заложены почки, мочеточники и гонады. Аллантоис вы-
ступает теперь как грушеобразный мешок, стенки которого сопро-
вождаются сосудиками, отходящими от желточных вен и дорзаль-
ной аорты.
В пятый день бластодерм уже оказывается охватывающш!
весь желток, a area vasculosa покрывающей около двух третей его
поверхности. Аллантоис уже служит теперь дыхательным органом п
стоит в соприкосновении с внутренней поверхностью яичной скор
лупы над правой стороной эмбриона. Конечности эмбриона несколько
удлинились, но все чрезвычайно сходны между собой. Отдельные,
в будущем костные, элементы начинают приобретать хрящевое строе
ние (череп, челюсти, позвонки и др.).
Начиная с шестого дня, в эмбрионе, до сего времени
очень сходном с соответствующими стадиями развития других Ат-
niota, начинают появляться характерные черты, принадлежащие
классу птиц. Появляются зачатки зоба и слепых кишек, замечаются
различия в структуре передних и задних конечностей, желудок
дифференцируется на мускулистый и железистый отделы, челюсти
приобретают форму клюва и проявляется птичье расположение че
репных элементов. Мочеточники вторичных почек приобретают са
мостоятельное сообщение с клоакой.
На седьмой день полость амниона приобретает значи-
тельные размеры, сопровождаясь увеличением амниотической жид-
кости; амнион начинает медленно и ритмично пульсировать, вслед-
ствие чего эмбрион медленно покачивается то в ту, то в другую сто-
рону. Аллантоис к этому времени распространяется очень широко
под скорлуповой оболочкой п наполняется жидкостью. Area vascu-
losa тем временем лишается своего терминального синуса вследствие
рассасывания, а желточные артерии и вены связуются с эмбрионол
общим стволом, становясь всего лишь ветвями брыжеечных сосудов.
После кратковременного увеличения в объеме желточного мешка
(вследствие абсорбирования быстро уменьшающегося количества
белка) желточный мешок вскоре опять опорожняется и на один*
надцат ый день собирается в многочисленные внутренне
складки. Кишечник тем временем сильно увеличивается в размер?
и частью свертывается в изгибы и петли.
К пятнадцатому дню аллантоис выполняет почти весь
внезародышевой целом и лежит непосредственно под скорлупок*
будучи отделен от скорлуповой оболочки тонкой серозной обо*
лочкой и остатками желточной, с которыми он срастается. Ра3‘
ветвленная, сеть сосудов сопровождает его стенки. На девя1'
надцат ый день яичный белок совершенно исчезает, и
точный мешок вбирается в брюшную стенку эмбриона.
238
У самого эмбриона на седьмой день непропорционально'
крупная голова перестает расти быстрее, чем остальное тело, по-
является зачаток языка на дне ротовой полости и закрываются жа-
берные щелп на ясно образующейся шее.. Уже на восьмой
день появляются костные пятна в области будущего клюва, на
двенадцатый ден.ь клюв принимает более определенную
форму и покрывается роговой оболочкой. На тринадцатый
день могут быть замечены когти на пальцах ног и иногда на
пальцах крыла и чешуйки на передней поверхности плюсны и паль-
цев. На шестнадцатый день эти образования являются
более твердыми и с более значительным покрывающим их слоем
рога. Перовые сосочки начинают появляться на поверхности кожи
на в о с ь м о й день, будучи особенно заметными по средней
лпнпп спины и на бедрах; на тринадцатый день эти со-
сочки располагаются на большей части тела, значительно увеличи-
ваются и частью имеют уже характер пеньков с просвечивающим
пигментом. За день или за два до вылупления роговые чехлпки на них
лопаются, и птенец уже в яйце оказывается покрытым длинным пер
впчным пухом. Хрящевой скелет заканчивается образованием в зна-
чительной части уже на тринадцатый день, и к этому же
периоду происходит достаточная дифференциация мускулатуры. Око-
стенение, напротив, имеет место лишь в немногих органах и является
весьма незначительным даже при вылуплении цыпленка. На че-
тырнадцатый день эмбрион лежит уже в общем вдоль длин-
ной оси яйца, головой к тупому концу, почти касаясь клювом внут-
ренней стенки воздушной камеры в конце яйца. На двадцатый
день клюв, вооруженный яйцевым зубом, прорывает эту пере-
понку, и птица начинает дышать воздухом, содержащимся в этой
камере. Легкие начинают функционировать, и организм перестает
снабжать кровью аллантоис, который вследствие этого сморщивается
п дегенерирует, а пупочное отверстие закрывается (рис. 188).
Постэмбриональное развитие. Вылупляющиеся
птенцы почти у всех птиц требуют известной заботы взрослых и более
пли менее продолжительное время воспитываются ими. Необходи-
мость этой заботы и длительность ее в значительной мере опреде-
ляются тем состоянием, в котором птенец появляется из яйца; обычно
°Н бывает более развит при более длинном эмбриональном периоде
, снабженном большим запасом питательного вещества,
тип развития вероятно являлся первичным для птиц,
наиболее близким к тому процессу, который мы видим у рептилий и
который требует сравнительно малое участие родителей в воспитании
птенцов. Этот тип индивидуального развития, однако, имеет ряд
°собых физиологических и биологических трудностей: требует боль-
шей затраты питательных веществ на каждое яйцо, ведет таким об-
разом к откладке крупных яиц и следовательно меньшему их числу,-
нынуждает откладку яиц через значительный промежуточный срок,
требует большей затраты энергии на насиживание. В процессе эво-
Л1°Ции для многих птиц оказалось более выгодным укоротить эмбрио-
нальный период и удлинить период выкармливания птенцов. Отдель-
ЙЬ1е группы достигли при этом различных ступеней развития по
и при яйце
Последний
239
этому пути, вследствие чего мы находим серию переходов между тем
и другим способами развития. Крайние группы этого ряда принято
характеризовать названием выводковых и птенцовых птиц. Под
выводковыми (nidifugae) понимаются птпцы, птенцы которых сразу
iro выводе из яйца в состоянии самостоятельно перемещаться и добы-
вать корм (рис. 189). Таковыми являются обычно отряды, большую
часть жизни проводящие на земле или у воды. У птенцовых птпц
(nidicolae) птенцы вылупляются недоразвитыми, часто голые, сле-
пые, с неразвитыми конечностями, длительный период не могут по
кинуть гнезда и самостоятельно добывать корм (рис. 190). К этой
группе относится большинство древесных птиц.
Количественно большая часть птиц легко входит в ту или внук
группу, но имеются также отряды, занимающие промежуточное по.
Риг. 188. Птенец серой цапли —• Ardea cinerea L. перед вылуплением.
Положение птенца 'в период проклевывания скорлупы. Па конце надклювья вндеь
яичный зуб, положение пальцев ног таково, что они легко могут быть введены в образующее^
отверстие и содействовать дальнейшему вскрытию скорлупы. (Сильно увеличено). По НОг
stein, из Stresemann.
ложение. Наиболее резко выраженными выводковыми птицами яв
ляются киви и большеногие куры. Птенец первого, за исключением
величины, почти ничем не отличается от взрослой птицы; птенць
вторых сразу по вылуплении могут быть вполне самостоятельный
и в состоянии даже летать. Высоко развитой выводковостыо об.#
дают также гусиные и куриные птицы, в меньшей степени кулики
Чайки и чистики, гагары и поганки занимают промежуточное поло
жение, но по общему развитию птенцов все же ближе к выводков^
птицам: первые выкармливаются родителями некоторое вре#
в гнезде, вторые нежны и неспособны к значительной перемен
места. Неопределенное положение занимает гоацин, выкармливав
мый взрослыми птицами, но могущий уже в раннем возрасте лазить
по веткам и даже плавать. В полной мере птенцовыми являют^
уже пингвины и трубконосые. Дальнейшую стадию возрастаю^
птенцовости мы имеем в отрядах аистовых и дневных хищников с птен
2-Ю
цами беспомощными, но появляющимися из яйца уже зрячими.
Наиболее типично птенцовыми являются веслоногие, совы, стрижи,
зимородки, сизоворонки, удоды, кукушки, дятлы и воробьиные.
Сравнительно быстрый рост птенцов обусловливается богатым
притоком пищи, получаемой птенцами особенно обильно при вы-
Орнитология—1209
24 L
Рис. 190. Пример птенцовой птицы:
птенец птицы-носорога — Сгапог-
rhinus cassidix Temm. (Coraciifor-
mes), просящий есть. По I-Ieinrich,
из Stresemann.
кармливании их родителями. Первые дни по вылуплении птенец
особенно нуждается в согревании, но в состоянии обходиться без
нищи, благодаря запасу яичного желтка в полости тела. Последний
прп вылуплении составляет от 1/3 до \/7 первоначального количества
желтка в яйце и вообще его тем больше, чем больший процент желтка
(‘оставляет вес свежего яйца. У цыпленка он потребляется главным
образом в первые три дня, исчезая в организме на шестой день;
перед исчезновением он отчасти накапливается в печени в качестве
запасного питательного материала. Количество потребляемой птен-
цом пищи чрезвычайно велико, и у птенцовых птиц первые дни может
превышать ежедневно вес самого тела.
Вследствие этого и прирост идет
быстрым темпом и в первые дни мо-
жет составлять до 40—45% первона-
чального веса тела (рис. 191). У круп-
ных птенцовых птиц в период роста на
прирост идет около всего количе-
ства пищи; так у орлана белохвоста
для увеличения веса тела на 170 г не-
обходимо около 500 г мясной и рыбной
пищи. Характер роста птенцов тйкпм
образом зависит от количества пищпп
потому быстрее у птенцовых птиц, чем
у выводковых, самостоятельно разыски-
вающих и собирающих свою пищу.
Притом, чем меньше абсолютный вес
взрослой птицы, тем быстрее он дости-
гается растущим молодым организмом.
Вес выклюнувшегося птенца составляет
около 2/3 веса свежего яйца; остальное
теряется испарением в процессе разви-
тия эмбриона. Через 8 дней молодой
крапивник весит, подобно старому, уже
8 г, в то время как чеглок только
за 26 дней достигает веса в 220 г.
а фаэтон только на сорок первый
день весит, как и взрослый, 570 г. Существенное значение при этом
имеет конечно то, что величина яйца относительно веса тела гораздо
больше, как правило, у мелких птиц по сравнению с более круп-
ными. Крапивник при вылуплении весит уже г/8 веса взрослой птицы,
а фаэтон всего только 71з (Stresemann, 1928).
Крупное значение имеет также различный темп развития, вообще
присущий данному виду или группе. Так, например, черный дятел
насиживает 14 дней, а по вылуплении уже на десятый день весит
в шестнадцать раз больше, чем при вылуплении. Упомянутый же
выше фаэтон насиживает 28 дней и на десятый день по вылуплении
весит не больше, чем в три с половиной раза превышая только что
выклюнувшегося птенца.
Длительность пребывания в гнезде птенцовых птиц зависит в зна-
чительной мере от того состояния, в котором они вылупляются из
242
Рпс. 191. Возрастные стадии большого баклана — Phalacrocoravtcarbo L.
возп7/11ЦО 11 еще слепой птенец; возраст— 12 часов; (Ь) —возраст— 10 дней; (с) —
раст— 15 д11еи; (а) —возраст— 25 дней; (в)—летный птенец, молодой наряд; (/)—взрос-
лая птица в брачном наряде. По Ileinroth.
яйца, а также и от размеров окончательно выросшей птицы. Про-
должительность пребывания в гнезде птенца непропорционально
увеличивается у крупных птиц, главным образом в зависимости от
скорости роста перьев, относительный прирост которых значительно
уменьшается с увеличением пх абсолютной длины. Эта же причина
вызывает длительный рост и длиннокрылых мелких птиц, в сравне-
нии с близкими к ним по величине короткокрылыми воробьиными.
Последние в среднем проводят в гнезде столько же дней, сколько
насиживается кладка (т. е. около 12—14 дней); деревенская же ла-
сточка находится в гнезде до 21 дня, а стриж даже до 35—42 дней.
Влияние величины особенно заметно на хищниках: в то время как
птенцы перепелятника сидят в гнезде 28—30 дней,-беркут покидает
гнездо только спустя 90 дней гнездовой жизни. Период же насижи-
вания у этих видов составляет 31—35 и 44 дня. Столь длительное
пребывание в гнезде может быть обусловлено, положим, также и
сравнительной труднодоступностыо и защищенностью птенцов от
врагов. Последний фактор резче всего сказывается у дуплогнездни-
ков, как правило, долго находящихся в гнезде. Так крупный вя-
хирь, гнездящийся открыто, сидит в гнезде всего 21 день, в то время
как значительно более мелкий клинтух находится в. гнезде около
28 дней. Очень долго находятся в гнезде и мелкие синицы: большая
до 23 дней. .
Соответственно влиянию этого фактора, птицы, гнездящиеся на
земле if находящиеся в наибольшей опасности, оставляют гнездо
очень рано, например, полевой жаворонок на девятый день, соло-
вей на одиннадцатый и т. п.
Поскольку степень развития выклюнувшегося птенца весьма
различна, столь же различным оказывается и пуховой покров его
в разных систематических группах. Все выводковые вылупляются
покрытыми пухом и' иногда (большеногие куры) даже с короткими
маховыми перьями; птенцовые, как правило, появляются из яиц
голыми или покрытыми очень редкими и рассеянными пушинками, и
только позднее на них развивается свежий пуховой покров или сразу
рыхлое контурное оперение (mesoptylae). Основное значение птенцо-
вого пухового покрова заключается очевидно в выполнении трех за-
дач: 1) защищать тело от охлаждения, соответственно чему птенцовый
пух обычно обладает длинными пушистыми бородками и соответ-
ственно росту тела обладает иногда длительным периодом роста,
причем появляется нередко на теле двумя генерациями (см. стр. 26);
2) обеспечивать птенцу незаметность для возможных врагов — в связи
с чем окраска пуха птенцов выводковых птиц обычно соответствует
окружающему фону и сверху обычно темнее, а снизу светлее, иЗ) обес-
печивать птенцу отличение родителями от сходных видов, почему пу-
ховая окраска птенцов обычно имеет видовые признаки. У некоторых
видов пушинки частью своеобразно видоизменяются в украшающие
образования. У ‘некоторых дуплогнездников особенно длинные пу-
шинки, повидимому, являются дополнительными* элементами орга-
нов осязания.
Пуховой пбкров птенца обычно очень скоро замещается контур-
ным оперением, обычно очень рыхлой структуры и более примитпв-
244
ной окраски, чем взрослое контурное перо. Продолжительность
носки его колеблется от двух недель до одного года. Более долго
носятся маховые и рулевые перья; обычно в молодом возрасте они
имеют отличную форму от дефинитивного пера: более заострены,
частью совсем недоразвиваются и образуют крыло более тупое и
округлое, чем у взрослой птицы.
Развитие тела и отдельных органов у птенцов происходит с раз-
личной быстротой в разных группах, в зависимости от важности
того или иного органа в соответствующие этапы жизни. Так нырко-
рые утки, находящие в воде достаточную безопасность и обильный
корм, быстро научаются плавать, но очень поздно приобретают спо-
собность летать; маховые у них начинают только что появляться
на крохотных крыльях в то время, когда они почти совсем выросли
п уже одеты плотным контурным пером. В противоположность им,
у куриных очень рано развивается крыло: у большеногих кур частью
уже в яйце, у павлина через неделю по вылуплении. Соответственно
этому и у других птиц наблюдаются ^различия в росте отдельных ча-
стей тела^ Однако отдельные органы несмотря на их важность могут
медленно приобретать свойственную им форму. Так длинный клюв
кроншнепа и горбатый клюв фламинго лишь постепенно достигают
своей дефинитивной формы, хотя функционируют и в период птенцо-
вой жизни уже как специализированные органы; в этом случае
функция как бы формирует орган, обладающий уже тенденцией к раз-
витию в определенном направлении.
По вылуплении, птенцы, вследствие несовершенств терморегуля-
ционных способностей их организма, нуждаются в частом согрева-
нии, выполняемым обычно тем полом, который высиживал яйца.
Период такого согревания более значителен у птенцовых птиц, чем
у выводковых, и длительность его зависит от роста птенцов, его
пухового покрова и климата страны, обитаемой видом. С течением
времени родители все менее и менее уделяют внимание этому согре-
ванию, пока наконец птенцы настолько подрастут, что уже не могут
спрятаться, под взрослой птицей. У близкой к нашим овсянкам
Melospiza melodia (Wils.) длительность обогревания, по данным* Нис
(Nice, 1933—1934), менялась по вылуплении следующим образом
(в процентах, к общему времени наблюдения за гнездом).
Дли- тель- ность наблю- дений в ча- сах Д И и
1 2 3 4 5 6 7 8 о 10 п
Май — .39 ? 50,9 47,1 40,0 62,0 42,1 22,9 9 4,7
Июль 38 64 59,6 56,4 54,2 14,4 5,2 — 7,3 | — — —
Средний вес птенца .... 1,7 3,5 (М ю 6 8,4 9,8 12,6 15.8' ? 16,8 184
Выкармливание и воспитание птенцов протекает весьма разно-
травно у птиц с различной экологией. Основная тяжесть этой ра-
245
•боты ложится на ту птицу, которая высиживала яйца, но в боль-
шинстве случаев здесь помогает и другой партнер, также выкар-
мливая птенцов или принося только пищу, которую основной вос-
питатель распределяет среди птенцов. В ряде случаев другой парт-
нер принимает на себя заботу только о части выводка.
Только птенцы некоторых выводковых птиц в состоянии без
особого обучения сразу самостоятельно отыскивать пищу и личным
опытом научаться выбору ее. В большинстве случаев птенцы выкар-
мливаются родителями, что сопровождается во многих случаях
образованием разнообразных приспособлений. В простейших слу-
чаях, как это делают куриные, взрослые отыскивают пищу и осо-
быми звуками сзывают молодежь к лакомому кусочку, который
они сами расклевывают. Пастушковые и крачки при этом подносят
корм птенцам в клюве. Сидящим в гнезде птенцам хищники при-
носят добычу в когтях ir разрывают ее перед птенцами на кусочки.
Другие приносят пищу в зобу и или (как например аисты) отры-
гивают ее на край гнезда, где Ьна уже склевывается птенцами, или
ко мят птенцов непосредственно из зоба и пищевода. Влрюследнем
случае птенец часто глубоко запускает клюв в глотку взрослой
птицы и принимает здесь отрыгиваемую пищу (например пеликаны,
бакланы) или клювы птиц скрещиваются, и корм из клюва одной
птицы языком передвигается в клюв птенца (альбатросы, колпицы).
Иногда отрыгивание пищи происходит прямо в рот птенцу обхва-
тывающему клювом голову взрослой птицы (козодои, стрижи):
колибри, таким же .образом кормящие птенцов нектаром, погру-
жают длинный клюв глубоко в пищево’д птенцов. Последние спо-
собы питания явились по всей вероятности, начальными ‘стадиями
для развития рефлекса у птенцов держать клюв открытым при поя-
влении взрослой птицы. Широко открытый клюв, писк при этом
и своеобразное дрожание головы характерны у ожидающих пищу
птенцов всех воробьиных, дятлов, кукушек, удодов и некоторых
других. Этот рефлекс возникает уже у самых слабых птенцов, не-
редко еще слепых, при малейшем раздражении птенца сотрясением
или иногда затенением. Родители всовывают пищу в глотку птенцу
клювом всю целиком или отрыгивают ее из зоба и кормят малень-
кими порциями. Условным раздражителем для родителей является
в свою очередь яркая окраска полости рта птенца, обычно оран-
жевого и ьт ясного цвета, иногда с резко контрастирующими пят-
нами или блестящими образованиями на роговых сосочках неба
к по углам клюва (рис. 192). Эти же особенности облегчают и ориен-
тировку кормящих птиц в темных гнездах, дуплах и норах. Харак-
терно .для этих птенцов также образование особых вздутий и утол-
щений по углам клюва, обладающих особенной чувствительностью.
Конечно способ кормления птенцов может меняться в зависи-
мости от возраста их. Кормление птенцов нередко не ограничивается
периодом гнездовой жизни . и продолжается некоторое время по
вылете птенцов, особенно у более крупных птиц. Приобретение
навыков к самостоятельному разыскиванию и добыванию пищи
происходит главным образом подражанием действиям взрослых
птиц. Дневные хищники проводят длительную выучку, приучаясь
246
самостоятельно схватывать добычу на лету, после предварительных
уроков в подхватывании убитых животных, бросаемых им роди-
телями.
Пища птенцов,, помимо различий, обусловливаемых способом
жизни взрослых птиц, обычно изменяется в процессе роста самих
птенцов. Первоначально она имеет более нежный и легко перева-
риваемый характер и лишь постепенно приближается к составу
пищи взрослых. Зачастую корм, передаваемый птенцам, предвари-
тельно обрабатывается и раз-
мягчается в зобу взрослых птиц.
У голубей сами стенки зоба
испытывают жировое переро-
ждение и образуют творожистую
массу, которой первое время п
выкармливаются птенцы. Коли-
чество приемов пищи и вели-
чина их весьма изменчивы, в
зависимости от рода пищи, ско-
рости роста птенца и способов
самого кормления. Большин-
ство мелких птиц кормится ро-
дителями через очень короткие
перерывы, так что за день у
синиц насчитывается, напри-
мер, до. 350—400 прилетов к
гнезду; крупные кормятся с бо-
лее значительными перерывами
и иногда не более 2—3, даже
1 раза на день, как, например,
фаэтон, потребляющий зато пи-
щи за раз до -г/з веса всего тела
птенца. Так как неблагоприят-
ная погода часто затрудняет
выкармливание птенцов, то в
большинстве случаев наблю-
дается способность их выдержи-
Рпс. 192. Небные розетки птенца мадагаскар-
ской кустарниковой кукушки—Coua cristata L.
Белый центр и края розеток резко выделя-
ются на фоне красной окраски слизистой
оболочки; с возрастом розетки исчезают. По
Bluntschli.
вать довольно длительный пе-
риод голодания, у стрижа могущий продолжаться даже свыше 20 дней.
Помимо кормления птенцов на родителей падают и другие, свя-
занные с выращиванием потомства заботы. Во избежание слишком
большого загрязнения гнезда экскрементами, у птенцов открыто
гнездящихся крупных видов существует рефлекс выпрыскивания
экскрементов в сторону через край гнезда, что обеспечивает чистоту
гнездовой подстилки. У мелких птиц, преимущественно гнездя-
щихся в дуплах, этот рефлекс не развит, и родители сами уносят
экскременты из гнезда, выбрасывая их на некотором расстоянии
°т него. Эта очистка облегчается тем, что экскременты оказываются
облеченными весьма тягучим и прочным секретом, образуя как бы
мешочек. Птица захватывает нередко этот мешочек в момент дефе-
кации, когда птенец принимает своеобразную, направленную аналь-
247
ным отверстием вверх, позу; анальное отверстие при этом обычно
оказывается отороченным' венцом перышек, отлично окрашенных
от соседнего оперения, что облегчает ориентировку взрослой птицы
в темноте гнездовой камеры.
Помимо этого, взрослые птицы защищают иногда птенцов от
солнца, расправляя над ними крылья, обмахивая их, прикрывая
иногда веткой с зелеными листьями, защищают их от врагов. В связи
с последней задачей замечательно развитие ряда инстинктов, как то:
Рис. 193. Гнездовая колония альбатросов — Diomedea immutabilis Rotsch. на острове Лан-
зан. По Palmer.
способность притворяться больной,. с целью отведения врага от
гнезда, временное подавление инстинкта самосохранения и пр.
(см. стр. 299).
Выводковые птицы, как правило, сейчас же по выводе уводят
птенцов от гнезда и ведут слегка кочующую жизнь поблизости,
в довольно узком районе, более или менее сплоченной семьей. Птен-
цовые покидают гнездо обычно после того как приобретут способ-
ность перепархивать, и, пока они подкармливаются родителями,
также продолжают жить сплоченной семьей.
Длительность такой семейной кочующей жизни определяется
различными моментами, среди которых важнейшим является на-
личие у вида второй и третьей кладок. В последнем случае роди'
тели вскоре изгоняют птенцов с территории гнездования и предо'
248
ставляют их всецело самостоятельному существованию. Птенцы
держатся более или менее сплоченным выводком и вскоре соеди-
няются с другими выводками в общие стайки, предпринимающие
и миграции, без участия старых экземпляров. У выводковых при-
вязанность родителей к птенцам может быть очень длительной и
продолжаться, например у гусей, почти целый год.
Недостаток места дл*я гнездования и беззащитность птиц иногда
являются побудительной причиной образования массовых гнездо-
вых колоний, насчитывающих нередко до нескольких десятков
тысяч экземпляров. Более часто такие колонии образуют водные,
преимущественно морские птицы, как, например, чистиковые, пинг-
вины, трубконосые (рис. 193), веслоногие, но такт^е и многие назем-
ные, как, например: щур-
ки, стрижи, ласточки
(рис. 194), некоторые тка-
чики (рис. 195), грачи и
др. Иногда защиты такой
колонии ищут мелкие дру-
гие, также колониальные
виды, как, например, уша-
стая поганка, гнездящая-
ся вколониях мелких чаек.
Гнездование такими спло-
ченными поселениями вы-
зывает образование спе-
цифических инстинктов.
Обычно колония зани-
мается из года в год и на
территории своего ядра
является наиболее пере-
населенной; здесь птицы
располагаются гнездить-
ся нередко на очень близ-
Рис. 194. Гнездовая колония ласточек — Pelroche-
lidon lunifrons Say.
Ретортообразпые гнезда состоят почти исключи-
тельно из пропитанной слюной и высохшей грязи:
гнезда укреплены на склонах скалы. По Storer, из-.
Stresemann.
ком расстоянии друг от
Друга, и весной, при занятии колонии, между претенден-
тами на место для гнезда разыгрываются ожесточенные драки.
Развитие колониальности гнездования в отдельных случаях при-
вело к образованию общественных построек, как, например,
У некоторых ткачей, строящих общую крышу, прикрывающую от-
дельные гнезда (рис. 195). Защита гнезд в колониях происходит
сразу массой птиц и естественно является более интенсивной, чем:
защита гнезда одной парой. Как правило, однако высиживание яиц.
11 кормление птенцов не являются общими, и птицы прекрасно узнают
свое гнездо, руководствуясь главным образом памятью на место
11 отчасти индивидуальными особенностями в окраске яиц. и птен-
Ц°в, вариирование которой особенно велико именно у общественно
^ездящихся птиц. У пингвинов однако развита наклонность вы-
сиживать чужие яйца, стащенные у соседей, и у них же и некото-
рых крачек (например Sterna sandwicensis Lath.), после некоторого
ПоДрастания птенцов и сбивания их в стаи, наблюдается обществен-
ное выкармливание птенцов, без различения уже на своих и чужих.
Наиболее высокого развития достигает все же общественная жизнь
пингвинов, у которых колонии выставляют своих сторожей, суще-
ствуют общественные игры и наблюдается большее развитие ин-
стинктов, направленные на общую пользу колонии.
Птицы-паразиты. Птицы, по праву называющиеся пара-
зитами, подкладывающие яйца в гнезда другим видам, довольно
широко распространены и многочисленны и включают не только
общеизвестную кукушку,
но и многих других пред-
ставителей этого семейства,
Рпс. 195. Гнездовая колония общественного ткани-
на — Philetairus socius Lath. По Stark, из Strese-
iriann.
а также американских
скворцов (Icteridae), тка-
чикрв (Ploceidae), медо-
ука зчиков (Indicatoridai]
и один вид уток
ronetta). Развитие этой
удивительной особенности
в данных систематических
группам происходило по-
видимому независимо, хотя
и сходными путями, вы-
зывая очень близкое по-
ведение и морфологиче-
ские приспособления. Бо-
лее детально это явление
было прослежено однако
только у обыкновенной ку-
кушки.
Самцы п самки куку-
шек не образуют постоян-
ных пар в период размно-
жения, встречаясь только
для совокупления; каждый
из них обладает известной
территорией, защищаемой
от других кукушек того
же пола, куда они возвращаются ежегодно, подобно большин-
ству птиц. В пределах своего района каждая самка разыскивает
гнезда предпочтительно тсго вида воробьиных птиц, в гнезде кото-
р. г она сама вывелась, и лишь в случае недостатка последних она
подкидывает яйца в гнезда других видов, близких им по биологии.
В основной массе яйца откладываются в гнезда с неполными и све-
жими кладками, но вследствие недостатка гнезд частью и с наси-
женными яйцами, или брошенными и пустыми. Таким образом часть
яиц пропадает, часть выкидывается или забрасывается вместе с клад-
кой приемными родителями, и вылупляется, как показывают ана-
лизы находок, только около 60% всех яиц. Соответственно с этими
условиями, у кукушек выработалась способность производить зна-
чительное количество яиц (до 26), откладываемых с промежутками
.250
по меньшей мере в двое суток, что обеспечивает также необходи-
мое время для поисков следующих гнезд. Как полагают, стимулом
к развитию фолликула в яичнике самки является находка кукуш-
Рпс. 196. Величина яйца в отношении к величине тела у выводковой птицы — кулика (а),
птенцовой птицы — дрозда (Ь) и птицы, подкидывающей яйца в гнезда других птиц — ку-
кушки (с). Для сравнения подобраны птицы приблизительно одинаковой величины. По
Zimmer, из Ililzlieimer.
кой готового гнезда или строящей гнездо пары птичек. Созреваю-
щее через 4—5 дней яйцо откладывается поэтому в большинстве
случаев в еще свежую кладку. Установлено также, что в некоторых
случаях кукушка уничтожает насиживаемые
кладки, и этим побуждает их владельцев к
новой кладке, совпадающей по времени с со-
зреванием ее яйца.
Как показывают имеющиеся наблюдения,
среди самок кукушек обычно существует изве-
стная специализация к откладке яиц в гнезда
определенного вида или группы видов, с кото-
рыми у них наблюдается и наибольшее сходство
в окраске яиц. Только в случае недостатка по-
стоянных хозяев, кукушка данной биологиче-
ской расы использует гнезда и других видов.
Во многих географических районах эта спе-
циализация однако нерезка, вследствие лег-
кой смешиваемости популяций по причине рых-
лых брачных связей между самцами и самками
кукушек. Как правило, общая приспособлен-
ность к роду хозяев сказывается не только
в окраске яиц кукушки, но также и в их очень
малой величине (х/зз веса тела, в то время как
У одинакового ростом рябинника —Turdus
pilaris L. яйцо весит’ 1/15 веса тела (рис. 196) и в особой крепости
скорлупы, препятствующей расклевыванию яиц мелкими выну-
жденными. хозяевами.
Для эмбрионального развития кукушенка достаточна сравни-
тельно очень короткая для величины птицы инкубация. Первое
Рис. 197. Птенец кукуш-
ки — Cuculus canorus L.
пытается выбросить яйцо >
из гнезда.
Возраст около 50 часов.
По Hcinroth, из SLrcse-
mann.
251
время птенец совершенно гол, и кожа его обладает резкой чувстви-
тельностью к прикосновению посторонних объектов.’ Это свойство
является основанием беспокойного поведения птенца в гнезде в слу-
чае нахождения в нем яиц и других птенчиков. Если последние
при движении птейца очутятся у него на спине, особенно в неболь-
шом углублении в тазовой области, кукушенок сейчас же судорожно
вытягивается на ногах и стремится выбросить эту ношу через край
гнезда (рис. 197). Эта реакция и приводит в большинстве случаев
к гибели всех сводных братьев кукушенка еще в первые дни по
вылуплении.
Растет птенец очень быстро, вызывая своими просьбами о пище
энергичную деятельность приемных родителей. В отличие от дру-
гих птиц, молодой кукушек ок совершенно нем и не трясет
крыльями, в чем видят приспособление к способу выкармливания
кукушки, так как незнакомый крик мог бы пугать приемных роди-
телей, а движения крыльев могли бы разрушить слабое гнездо мел-
ких птичек. Только уже по достижении почти полного роста моло-
дой кукушенок переходит к самостоятельному образу жизни и,
в частности, совершенно независимо от старых кукушек или прием-
ных родителей совершает перелет на зимовку в южные страны.
4. КЛИМАТИЧЕСКИЕ ФАКТОРЫ И ВЫЗЫВАЕМЫЕ ИМИ ЯВЛЕНИЯ
В ЖИЗНИ ПТИЦ
Развитие каждого вида происходит в определенной среде, к усло-
виям которой, в процессе своей эволюции, вид приспособляется
настолько, что нормальные, средние, к с лебанг я климата не сказы-
ваются на благосостоянии популяции, или переносясь видом «без-
болезненно», или складываясь особенно благоприятно для него.
Большинство типично пустынных птиц в своей жизни подвергается
экстремным климатическим влияниям не потому, что они не в силах
избежать их, а потому, что они свободно выносят или даже пред-
почитают эти условия. Они преуспевают лучше всего в пустыне-
Наличие этой приспособленности объясняет, почему во многих слу-
чаях резкие, на наш взгляд, проявления климатических условия
не вызывают со стороны птиц особых реакций. Ряд особенностей
при этом делает птиц очень легко переносящими эти условия: весьма
совершенная теплокровность и терморегулирование, прекрасные
изолирующие качества перового покрова и способность к значи-
тельным миграциям, которые повидимому и дали птицам преиму*
щество в борьбе за существование и содействовали всесветному рас'
пространению этого класса.
• Однако указываемая приспособленность относится главным оо-
разом к средним климатическим условиям того или иного место-
обитания и значительно менее совершенна к резким отклоне-
ниям от этих средних условий. Наблюдение показывает, чт0
большинство птиц весьма чувствительно к непривычным для но*
отклонениям климатических влияний. Эта чувствительность н°'
вышается при недостаточном питании, большой затрате энергии я
других неуравновешенных физиологических состояниях организм?1
252
Гак чувствительность повышена в период линьКи, в период после
высиживания яиц и выкармливания птенцов, истощающих взрослую
птицу. Велика чувствительность яйца и птенца птицы к длитель-
ным нарушениям оптимальных условий. Яйцо не выносит нагре-
вания выше 46°, а в конце периода насиживания понижения темпе-
ратуры ниже 15°; птенец в первый период своей жизни является
в сущности хладнокровным организмом, и теплокровный и термо-
регулирующий механизм устанавливается постепенно,* по мере раз-
вития и роста молодой птицы. Известна также относительно малая
сопротивляемость влияниям климата молодых жйвотных вообще.
Из этого следует, что климат в жизни, размножении и распро-
странении птицы имеет весьма существенное значение и опреде-
ляет течение многих жизненных процессов. Они успешнее всего
протекают при некотором, специфическом для каждого вида, соче-
тании и размахе климатических факторов, создающих для вида
так называемую «зону комфорта», отклонение от которой в ту или
другую сторону одинаково приводит к неблагоприятным или гибель-
ным последствиям.
На организм климат оказывает воздействие всей совокупностью
создающих его факторов, но среди них можно выделить наиболее
важные, основные — свет, тепло, влажность,t движения воздуха,
осадки, и второстепенные, насколько известно, оказывающие менее
существенное воздействие. После рассмотрения влияния отдельных
факторов мы рассмотрим влияние климата в совокупности, по тем
периодическим сезонам, которые он вызывает.
Свет. Соотношение длительности дня и ночи под разными
широтами земли, как известно, различно и подвержено определен-
ной закономерности. На экваторе день и ночь круглый год длятся
по 12 часов, в то время как за полярным кругом летом солнце целые -
месяцы не заходит за горизонт, а зимой целые месяцы не появляется
из-за горизонта. В средних широтах разница менее значительна,
но все же летом день- значительно превышает длительность ночи,
а зимой, наоборот, ночь превышает день. Общее значение этого
явления сказалось и в животном мире, вызвав развитие специали-
зированных как дневных, так и ночных животных. Среди птиц резко
преобладает количество первых, в то время как вторые даже в тро-
пиках представлены главным, образом с.овами и козодоями. Однако
к ночному образу жизни перешли и отдельные представители дру-
гих отрядов: охотящийся на летучих мышей Мachaerhamphus (из
Дневных хищных птиц), питающиеся главным образом червями и
личинками киви, ряд куликов (вальдшнеп, бекасы, авдотка и др.),
питающиеся рыбой и земноводными некоторые цапли (особенно
кваква, выпи, Cochlearius и др.), некоторые растительноядные попу-
раи (Stringops), морские, питающиеся планктоном, мелкие виды
тРубконосых (качурки, некоторые буревестники и др.) и водорез
(Rhynchops)‘из подотряда чаек. Кроме того, к ночному образу жизни,
хотя бы частично, переходят некоторые другие птицы,* например
пресноводные утки, гуси, а во время периодических перелетов дея-
тельный образ жизни ведет ночью, хотя и не кормится, большин-
ство как мелких, так и крупных птиц.
263
Виды, ведущие ночной образ жизни, обладают целым рядом
приспособлений к разыскиванию пищи в условиях чрезвычайно
ослабленного освещения. Особенно*это сказывается на строении
глаз, отличающихся у ночных птиц повышенным количеством пало-
чек в сетчатке глаза, богатством их зрительным пурпуром, способ
ноитыо к особенно быстрой адапцип при сравнительно слабой акко-
модации. У сов глаза при этом обладают особенно своеобразной
формой (см. рис. 149). В совокупности это строение обеспечивает до-
статочно резкое зрение в условиях темноты, но вместе с тем делает
эти формы особенно чувствительными и к дневному освещению.
Приспособляемость отдельных форм к этому весьма различна: в то
время как из сов сипуха {Tijto alba Scop.) и мохноногий сыч [Лф-
Hus junereus (L.)] вылетают на охоту только после значительного
потемнения, ястребиная сова [Surnia ulula (L.)] ведет в значитель-
ной мере дневной образ жизни.
Ночные виды отличаются также особой тонкостью слуха, и совы
обладают даже весьма своеобразной наружной ушной раковиной.
В связи с ориентировкой их по слуху стоит и особая мягкость пх
оперения, в том числе п маховых перьев, непроизводящих шума
при полете. У ночных же видов мы находим и наилучшим образом
развитый обонятельный аппарат, особенную роль играющий в поисках
пищи у киви, у глупыша [Fuhnarus glacialis (L.)] и других трубко-
носых птиц. У других видов большую роль играет сильно развитое
чувство осязания. Так им в значительной степени руководствуется
вальдшнеп при разыскивании личинок и червей в мягкой земле, водо-
рез (род. Rhynchops) (см. стр. 208), кормящиеся ночью утки и др.
Периоды активности ночных и дневных птиц противоположны:
у первых активность пробуждается с наступлением сумерек пли
ночи и заканчивается с утренними сумерками; дневные просыпаются
от сна с окончанием утренних сумерек и восходом солнца п засы-
пают с заходом его. Некоторые птицы проявляют наибольшую дея-
тельность в переходный период сумерек и в- светлые лунные ночи,
как, например, водорез и некоторые козодои, но собственно суме-
речных видов не существует. Пробуждаемость видов к активности
наступает в определенной последовательности. Фотометрическим
методом было установлено, что каждому виду соответствует типич-
ная для него иЦтенсивность света, при которой происходит его про-
буждение, измеряемая единицами света (люксами). Так весной чер-
ный и певчий дрозды пробуждаются к активности при освещении
в 0,1 люксов, кукушка — 1, большая синица — 1,8, славка Черно-
головка — 4, зяблик — 12 и домашний воробей — 20 люксов. При-
мерно этой же интенсивности света соответствует и время впадения
в сон у соответствующих видов. Однако, как мы видели, длина дня
и ночи зависит от географической широты и сезона, что весьма суще-
ственно изменяет поведение и жизнь птиц. В летнее время в высо-
ких широтах птицы активны значительно более длинный срок, чем
под тропиками, и у некоторых видов в любое время суток можно
слышать пение и наблюдать их разыскивающими корм. Потрео-
ность во сне здесь очень значительно сокращается и различно вира'
жена в зависимости от погоды и периода размножения. У больший-
251
ства воробьиных при этом.период покоя приходится на вечерние часы
(18—23 ч.), и к полночи большинство особей уже активны, и всюду
слышится пение (Palmgren, 1932). В этих условиях круглосуточного
освещения интенсивность света теряет уже свое значение условного
раздражителя, и потребность покоя пли сна сокращается до необхо-
димого минимума п передвигается на тот период суток, который
наиболее неблагоприятен для самочувствия и активности птицы.
В средних широтах таким периодом является ночь, в высоких широ-
тах — повышенная сухость и тепло после полуденных часов. Чем
резче выражены эти последние качества, тем резче выражен п пе-
риод покоя в суточной жизни птиц, при прохладной же п сырой
погоде этот период покоя является более слабым и расплывчатым.
Наиболее резко выраженный цикл под 70s с. ш. имеет зяблик, удо-
влетворяющийся в июне—июле всего лишь 5 часами сна в сутки.
На потребности во сне сказывается и возбудимость птицы. В раз-
гар периода размножения потребность во сне у птиц особенно пони-
жена, а в период перелета многие птицы способны обходиться почти
без сна, летя ночью и кормясь и отдыхая днем.
Различия в продолжительности активности в тропиках и на се-
вере должны иметь значительное влияние на жизненные явления
птиц, но пока они недостаточно выяснены. Повидимому одним из
следствий этого различия в продолжительности активности является
меньшая половая производительность птиц юга: у значительного
числа широко * распространенных видов в тропиках мы находим
вкладке всего лишь 2—3 яйца, в то время как в средних широтах
.они или их родичи несут по 5—6 яиц, а на севере число яиц дости-
гает 7—9 (см. примеры на стр. 224). Вместе с тем отсутствие света
зимой в полярной области и краткость зимнего дня в таежной полосе
должны являться важным фактором в распределении птиц на зи-
мовке и их экологии. В таежной полосе это вызывает малочислен-
ность птиц на зимовке, на севере же вообще могут существовать
зимой только те виды, которые в состоянии приспособиться к усло-
виям пониженного освещения. Неожиданным образом эта приспо-
собляемость у ряда видов довольно велика. Так в Гренландии под
ш. зимуют не только белые куропатки и кречеты, но и неко-
торые воробьиные — ворон, чечетка, пуночка, хотя солнце зимой
не появляется вообще в течение двух месяцев. Из чистиков, толсто-
клювая кайра [Uria lomvia (L.)], тупик [Fratercula arclica (L.)] и
чистик [Cepphus grylle mandti (Licht.)] зимуют даже в некоторые
зимы под 80° на севере Шпицбергена, тогда как в летнее время они
живут в условиях круглосуточного дня.
Изменениям в длине дня в настоящее время приписываются
'также стимуляция развития гонад у птиц и следовательно сроки
гнездования; эти же причины, по мнению Роуэна (Rowan, 1925).
вызывают сроки прилета птиц в наши широты с далеких южных
зимовок (см. стр. 284).
Тепло. Лучистая энергия солнца является одним из основ-
ных факторов жизни на земле и главным образом через то тепло.
к°торое получает наша планета. Температурный режим играет роль
основного условия в жизни растений и хладнокровных животных
255.
и уже через посредство этих организмов, создающих для птиц укры-
тие и пищу, влияет на их жизнь. Не менее велико и непосредствен-
ное влияние температуры, хотя теплорегулирующий механизм тела
птиц обеспечивает известную выносливость к отклонениям темпе-
ратуры воздуха от нормальной для вида. В опытах Кенди (Кеп-
deigh, 1934), с выживаемостью домашнего воробья в разных темпе-
ратурных условиях выяснилось, что верхней критической для
вида температурой тела является 46,7°С, а низшей J 21°, 23,9°С.
за пределами которых жизнь птицы прекращается. Значительно уже.
конечно, .зона температурного оптимума животного. В отношении
к температуре внешней среды мы имеем среди птиц, как и других жи-
вотных, разные степени тепло- и хладолюбивости, стено- и эвритер-
мии. Приспособляемость птиц к температурным условиям может
быть весьма значительна. Императорский пингвин размножается на
льду Антарктики зимой, при температуре,, нередко понижающейся
в это время др —50° С. В области Верхоянского полюса холода
с температурами до —70° С зимуют черноклювые глухари, кукшп.
темноголовые синички (Poecile cincta kolymensis But.) и некоторые
другие виды. В оцытах Роуэна (Rowan, 1925) североамериканские
вьюрки [Junco hyemalis (L.)] великолепно прожили зиму в вольере
при температуре, спускающейся до —40° С. С друге й стороны, в пу-
стынях Средней Азии мы имеем довольно разнообразную фауну,
птиц, живущую при температуре воздуха, доходящей до —{—35° С.
и температуре почвы, поднимающейся до {-70° С. Известна разно-
образная и довольно богатая фауна холодных плоскогорий Тибета
и Памира с средними температурами летом до -[-14° и зимой до —18".
указывающими на весьма суровые континентальные условия, кото-
рые могут выносить местные птицы. Эти резкие границы выносимых
температур возможны только благодаря развитию специальных пр
способлений в организме птицы. Среди последних особую роль играет
теплокровность с высокими температурами крови (см. стр. 116)
и обладающий прекрасными изоляционными свойствами перовой
покров. Пронизанный большим количеством воздуха, легкий и упру-
гий, он задерживает рассеивание, путем излучения, тепла, выра-
батываемого телом. Наибольшей плотности этот покров достигает
в зимнее время, накануне которого он вырастает на смену более
рыхлому летнему перу. Зимнее оперение у зимующих на севере ви-
дов отличается большей длиной и густотой и большим количеством
пуховых перьев. Наиболее теплое оперение свойственно видам,
живущим в холодном климате. Общеизвестны высокие согревающе
качества гагачьего пуха, ценимого особенно для авиационных шле-
мов и курток, требующих высокую легкость и повышенные согре-
вающие качества. Высоко ценятся также перовые и пуховые шкур^
северных гагар, кайр, лебедей и гусей. Это теплое зимнее оперение
у некоторых птиц, например у тетеревиных, заменяется на лето
более легким пером, лишенным базальных пуховых частей. Суш6'
ственной защитой от холода является также толстый подкожный
жировой слой, накапливаемый осенью к зиме многими северными in*
цами. С этой же задачей стоцт в связи и наблюдающееся увеличение
размеров птиц в холодных континентальных странах в сравнении
•256
с особями того же вида или их близкими’родичами из более южных
и мягких в климатическом отношении районов. Эта закономерность,
известная под названием правила Бергмана, обеспечивает бо^ее
выгодное в холодном климате соотношение между поверхностью
тела, излучающего тепло и массой его: увеличение размеров тела
уменьшает это соотношение. Помимо размеров, зимующие на севере
птицы, как, например, тетеревиные, в сравнении с фазановыми, обла-
дают более округлым компактным телом, более плотной и короткой
шеей п иногда более укороченными конечностями и клювом. При-
мерами, подтверждающими правило Бергмана, являются белая
куропатка {Lagopus lagopus L.) с самыми крупными расами на Ново-
сибирских островах и в степях Казахстана и самыми мелкими в сред-
ней части Европейской СССР и юго-восточной Сибири, или толсто-
клювая кайра [Uria lomoia (L.)], в сравнении с тонкоклювой \Uria
aalge (Pontop.)]. Усиленный расход тепла в холодном климате ком-
пенсируется также большим потреблением пищи, в сравнении с по-
треблением ее в теплое время года (или в теплых странах).
Иного характера приспособления к перенесению высокой тем-
пературы: это более редкий, короткий, менее пуховой покров, частое
наличие значительных оголенных участков кожи (цесарки, грифы,
аисты п др.), весьма нередкое отложение черного пигмента в коже,
особенно в случае оголенности частей тела (как ширмы от вредного
действия прямых солнечных лучей), и повышение теплоотдачи
испарением с поверхности легких и воздушных мешков. Как известно
(см. стр. 35), птицыне обладают потовыми железами, и заменяющим
их деятельность является дыхание с широко раскрытым клювом,
что часто можно видеть в жаркую погоду, например, у ворон.
Как уже указывалось, температура птенцов значительно отли-
чается от таковой у взрослых. В начале .своей жизни птенцы по
своим температурным реакциям определенно являются пойкилотерм-
ными (холоднокровными) животньйий, и температура их стоит в пря-
мой связи с температурой гнезд$Н она повышается при нагревании
их взрослой птицей и понижается при отсутствии ее. Постепенно,
независимо от окружающих условий, температура тела птенцов
повышается, например, у американского домового крапивника,
в среднем на 0,3° С в день, и к возрасту в девять дней способность
к температурному контролю (поддержанию собственной температуры
на известном уровне) более или мёнее устанавливается. Установление
этой способности обусловлено увеличением общей продукции теплоты
в теле животного, уменьшением отношения поверхности тела к его
массе п связано с развитием перьев, установлением функционирования
воздушных мешков и развитием соответствующего нервного и гормо-
нального контроля. Однако, нагревание тела свыше нормальной
температуры для вида быстро влечет за собой гибель птенца, причем
летальные температуры нагревания оказываются относительно оди-
наковыми для всех возрастов и составляют для крапивника 46,6° С.
большинства птиц мы видим те или иные приспособления в пове-
дении, обеспечивающие птенцу в гнезде защиту от перегревания
и губительного действия прямых солнечных лучей (рис. 198 и 199).
детальные температуры охлаждения зависят от возраста, и у кра-
17 Орнитология—1209
257
швнпка смерть наступает перед установлением температурного
юнтроля при температуре в 8,3° С, а после установления его — при
; Рис. 198. .Гнездо уУравьеловки — Thamnomanes caesins glavcus .Cab.
Гнездо защищено .сверху от солнца и. дождя листьями, которые птица подбирает с земли
и укрепляет над гнездом. По Beebe, из Stresernann.
температуре в 15,6° С. Таким-образом с увеличением возраста птицы
степень^ ^охлаждения тела без летального исхода уменьшается и
' 35' наименьшей является.
как мы видели, у взрос
лой птицы (при темпе
ратуре в 21,9° С).
По отношении? к теп
пературе воздуха темпе
р а тур а яйца увеличь
вается с течением пику
бации за счет теплоты
образуемой самим эм
брионом. Выше уже ука
зывалось, что у неко
торых зформ (большено-
гих ,кур и нырковой
утки; Охуига) эта-/те-
плота может быть очень
значительной. Однако в
большинстве случаев
Рис. 199. Самка ястреба перепелятиица — Accipiler
nisus L.; и жаркие часы дня стоит над гнездом, защи-
щая птенцов от прямых солнечных лучей широко рас-
правленными крыльями. no^Owen, из Streseipann.
она «слишком мала и не
может длительно под-
держиваться без при-
ведения тепла извне.
Сообразно общей закономерности, наблюдаемой у теплокровны*
животццх,. заключающейся в том, что молодые - их. стрдип более
258
выносливы к холоду (таким образом животное в своем развитии про-
ходит как бы стадию хладнокровного существа), известна способ-
ность яиц выносить очень значительное охлаждение. У кур яйца
дают здоровых, цыплят, если в свежем состоянии они будут под-
вергнуты замораживанию даже до — 4° и — 6° С и хранимы в та-
ком виде некоторое время. Известно, что близкий к белой ку-
ропатке пютландский граус {Lagopus scoticus Lath.) неоднократно
в период кладки яиц захватывается весенними морозами, и много
раз находили его яйца полностью замерзшими, и тем не менее
пз них в дальнейшем выводились • цыплята. У мелких воробьи-
ных это свойство повидимому менее выражено. С продвижением
развития t эмбриона в яйце охлаждение его становится все более
вредным п если не сразу губит зародыш, то вызывает ослабление
его жизненности и создает‘условия для гибели его в дальнейшем.
Все же при охлаждении до 15 — 21° С эмбрионы всех стадий северо-
американского домашнего крапивника (Troglodytes aedon Vieill.)
выживают при длительности охлаждения до 16 и изредка даже до
24—30 часов. Прп этом охлаждение в первый период инкубации
(у Troglodytes aedon Vieill. до 8-го дня) вызывает в дальнейшем
оттяжку в вылуплении, в конце же инкубации это охлаждение даже
не сказывается на ней (Kendeigh, 1934).
Таким образом уже в самой организации яйца и птенца мы нахо-
дим приспособления к вышеуказанным перерывам в инкубации,
на случай спугивания птицы с гнезда хищником, ненастной погоды
пли длительного отсутствия птицы в поисках пищи. Однако, как
мы видели, эти приспособления имеют предел, компенсируемый
обычно соответствующим поведением насиживающей птицы, как то:
большой привязанностью птицы к гнезду в последние периоды
насиживания, более частым возвращением родителей к птенцам
н согреванием их в первые периоды жизни и т. п. Надо думать,
нто и характер соответствующих эмбриональных приспособлений
различен в зависим(хзти от экологии птицы, о чем однако наши све-
дения пока весьма незначительны.
Из прямых температурных влияний па взрослую птицу наиболь-
шее значение, повидимому, имеет характер ночных температур в связи
с длиной ночи, а также дневные максимальные температуры воз-
духа. Роль первых видна из того, что на охлаждение птицы реаги-
руют повышением метаболизма, большим потреблением пищи и боль-
шей подвижностью, что невозможно в ночные часы. Этот фактор
является основной причиной, ограничивающей число зимующих
нтиц в высоких и средних широтах, если это не компенсируется
возможностью отыскивать пшцу в темноте (см. выше). Высокие
температуры воздуха оказывают вредное влияние не только пря-
А1ьш повышением температуры тела, ио и (в большей степени) воз-
действуя на активность птицы, видоизменяя течение обмена веществ
Организме. Наблюдениями установлено, что у домашнего воробья об-
Мен веществ достигает минимума при температуре воздуха в 33,9° С,
каковую можно рассматривать как критическую. Вместе с умень-
шением метаболизма уменьшается п активность, так как малейшее
Усилие причиняет нежелательный подъем тепературы тела. Если
259
такая температура и выносится птицей более или менее длительный
период, вследствие испарения влаги с поверхности легких и воздуш-
ных мешков и связанного с этим охлаждения тела, то условия суще-
ствования индивидуума все же являются тяжелыми.
Кроме только что описанных зависимостей в жизни птиц от
температурных условий большую роль играют и другие отклоне-
ния от привычной виду нормы. При этом важны не только абсолют-
ные величины этих отклонений, но и продолжительность этих откло-
нений, а также приуроченность их к. тому или иному моменту жиз-
ненного цикла вида. Резкие уклонения от нормы имеют особо ката-
строфический характер.
Влажность воздуха. Влажность воздуха может быть
рассматриваема в двух отношениях: абсолютного количества водя-
ных паров, содержащихся в воздухе, ийи так называемой абсолют-
ной влажности, и количества их к тому напряжению водяных паров,
которое могло бы насытить воздух при данной температуре, так
называемой относительной влажности. Последнее понятие является
более важным в биологическом отношении, так как относительная
влажность обусловливает возможность и величину испарения.
Как правило, относительная влажность, при неизменном количестве
паров в воздухе, варипруёт обратно температуре: когда температура
поднимается, относительная влажность уменьшается, и обратно,
при понижении температуры относительная влажность увеличи-
вается. Однако в природе постоянно наблюдаются и иные условия,
при которых низкая влажность сочетается с низкой температурой
и высокая влажность — с высокой температурой.
Прямое влияние этого фактора сравнительно невелико и в боль-
шинстве случаев с трудом отделимо от влияния температуры. Нор-
мально та или иная влажность усугубляет или ослабляет действие
температуры. При низкой температуре высокая относительная влаж-
ность может быть весьма вредным фактором, так как она повышает
теплопроводность воздуха и увеличивает отдачу тепла телом через
его поверхность и особенно через легкие и воздушные мешки. Таким
•образом низкая температура более легко переносима при низкой
же влажности, т. е. более сухом воздухе. В свою очередь при высо-
кой температуре воздуха высокая относительная влажность препят-
ствует быстрому испарению с поверхности воздушных путей и тела
и охлаждению этим тела, и комбинация этих факторов в целом может
иметь*весьма губительное влияние на животное; следовательно
высокая температура воздуха переносится лучше также при большей
сухости воздуха.
Значительная влажность воздуха также вредна и для эмбриональ-
ного развития в яйце. Прц наблюдениях над развитием яиц .белых
леггорнов выяснилось, что 80-процентная влажность при инкубация
вызывает более быстрый рост и потребление кальция, но причиняет
значительную смертность перед самым вылуплением; 40-процентная
влажность хотя и задерживает рост, все же не отражается существенно
на смертности эмбрионов. Это указывает, что и при развитии яйца
должна быть специфическая виду относительная влажность воздух*
которая может и инстинктивно поддерживаться взрослыми птен-
260
цами. Это значение, например, может иметь свойственное многим
видам подкладывание в гнезда зеленых листьев, как, например, у
осоедов и других дневных хищников, или у ласточки Progne purpurea
Cate sb.
О значительном влиянии повышенной влажности на окраску
оперения, очевидно через метаболизм организма, можно судить по
совпадению интенсивности окраски географических рас одного вида
с обилием осадков в различных районах области распространения
его (Gornitz, 1923; Серебровскпй, 1925).
Тот же эффект экспериментально был получен Бибом (W. Beebe.
1907) у горлинки Scardafella inca Less, при содержании ее в перена-
сыщенной влажностью атмосфере. С каждой последующей линькой
новое оперение все более и более темнело, пока меланин не заместил
полностью все те части пера, которые раньше были белыми.
О с а д к и. Осадки имеют крупное значение в жизни птиц, не
тблько через посредство растительности или животных, служащих
им пищей, но и прямым воздействием на организм птицы. Их влия-
ние различно, в зависимости от их формы, длительности и сочетае-
мости с другими факторами, из которых особенно важны темпера-
тура и ветер.
К частичному перенесению жидких осадков оперение тела птгц
приспособлено значительной жесткостью и черепитчатым перекры-
тием перьев, а также постоянной смазанностыо его выделениями
копчиковой железы или обсыпанностыо пудрой пудреток. Эта сма-
занность пера /Кировым секретом особенно значительна у водопла-
вающих птиц, что и облегчает им ненамоканпс тела, несмотря па
длительное нахождение в воде.
В силу этих причин небольшой и несильный дождь не может
иметь значительного влияния на тело взрослой птицы. Однако дожди
затяжные в конце концов вызывают намокание пера, что может
привести к очень большой потери тепла и гибели птицы. Еще более
губительные последствия может иметь ливень, который все птицы
стремятся избежать, укрываясь под густую листву, ветви деревьев
птому подобную защиту; сильный ливень вызывает намокание даже
водоплавающих птиц настолько, что они тонут.
Молодые птицы еще более подвержены намоканию, чем старые,
Как в силу более рыхлого пера, так и из-за неуменья найти соответ-
ствующее укрытие. Как яйца, так и молодые птенцы обычно защи-
щаются от дождя взрослыми птицами, прикрывающими их своим
телом. Однако, если они все-таки успеют намокнуть, смерть их от
°хлаждения наступает в очень скором времени. Поэтому «вымокание
птенцов, в частности различных куриных птиц — тетеревов, куро-
ааток, фазанов и т. д., имеющих охотничье значение, с давних пор
признается как весьма важный фактор в уменьшении численности
популяций этих видов. В случае намокания гнезда и кладки яиц
атица часто бросает это гнездо. Овлажнение поверхности яйца вредно
Действует на будущего эмбриона, вследствие чего обычно и не реко-
мендуется обмывание яиц при вкладывании их в инкубатор.
Не менее сильную роль часто играют наводнения, образующиеся
в Результате дсждей, особенно сказывающиеся на птицах, гнездя-
261
щихся на земле и возле водоемов,. В горных странах лцвни вызы-
вают значительный подъем воды в реках и разлитие их по долине,
что губит весьма обширное птичье население. В СССР это явленно
особенно свойственно Амуро-Уссурийскому краю с его летними
дождями, что повидимому и вызывает сравнительную бедность
здешней болотной и водной фауны. Вообще же известно, что сильные
дожди в гнездовой сезон могут иметь столь сильное влияние па
птичью популяцию, что восстановление ее численности будет воз-
можно только спустя несколько лет.
Далее, очень велико прямое влияние града на птиц,: в котором
комбинируется действие ливня с механической силой отдельных
градин. Известны случаи, когда птицы собирались тысячами после
прохождения полосы такого града, причем действие его сказывалось
не только на мелких птицах, но и на крупных: градом были уби-
ваемы дрофы-и другие достаточно крупные птицыо , j
Прямым губительным влиянием обладает также ожеледь. По
отношению к крупным открыто живущим птицам, например дро-
фам, она может вызвать образование ледяных пластинок на смо-
ченных перьях тела, что чрезвычайно отяжеляет и истощает птицу
и кроме того быстрее ведет к более глубокому промоканию оперения.
На мелких птиц вредно действует образование-ожеледи на ветках
деревьев, затрудняющее не только разыскивание пищи, но и само
движение в кроне растения. В группе тетеревиных мы имеем неви-
димому специальное приспособление к этому явлению в виде роговых
бахромок, образующихся к зиме по краям пальцев, что дает птицам
возможность более прочно держаться на таких ветках и вместе
с тем облегчает и передвижение по более или менее рыхлому снегу
(Формозов, 1932).
Прямое влияние, сходное с влиянием сильного дождя, оказывает
также мокрый снег, падающей хлопьями и тающий на теле, смачи-
вающий оперение. Кроме этого, снег оказывает косвенное влияние
затрудняя добывание пищи. Его значение при этом особенно велико,
если он ложится глубоким и, под влиянием ветра, весьма уплотня-
ющимся слоем. Такой покров совершенно скрывает корм, делает
его труднодоступным даже разгребающим движениям ног и вслед-
ствие этого является одниэд: из основных факторов распределения
многих птиц на зимовке и их зимнего поведения. Так наличие я
величина снежного покрова определяют распределение и встречае-
мость на зимовке перепелов, хохлатых и черных жаворонков и дру*
гих птиц в наших степях. Вместе с тем толстый и рыхлый снежный
покров является необходимым условием для успешной зимовки многих
тетеревиных в наших лесах, так как, Зарываясь в него, они проводят
здесь ночи в сравнительном тепле, будучи защищены от ветра в перво-
ды сильных морозов нашей зимы. Однако оттепель может вызвать
образование тонкой ледяной корочки — наста на поверхности снега,
п тогда место спасения от стужи может сделаться для этих птпП
суровой тюрьмой и могилой.
Жидкие осадки могут оказывать и другое, .более косвенное влия-
ние на фауну птиц. Дождь, в особенности в;совокупности с низкой
температурой, может понизить активность насекомых и‘ за трудна
262
птицам питание. Частые дожди способны очищать воздух от насе-
комых и вызывать голодовку у стрижен, ласточек и мухоловок. Недо-
статок осадков и вызываемая этим засуха не только иссушают расти-
тельность, но сказываются и на птичьем населении. Известно, что
в пустынных районах в сухие годы многие птицы даже и не присту-
пают к размножению, а гонады их остаются неразвитыми. Но даже
ив влажных районах очень сухое лето вызывает сокращение периода
гнездования и ускоряет наступление липьки у воробьиных птиц.
Умеренная влажность, вызванная достаточными осадками, в степях
изоне лесной растительности, как правило, благоприятствует обпль-.
ному размножению. Но бывает и наоборот.* мышеядные дневные и
ночные хищники (юлее обильно, размножаются в сухпе годы, когда
засуха вызывает обильный урожай мышевидных грызунов; в эти
же периоды в тропиках рыбоядные водоплавающие, птицы раз-
множаются^ и расселяются усиленнее, вследствие появления боль-
шого количества отмелей и берегов, на которых они е успехом
могут ловить рыбу, и образования более мелких водоемов, в которых
водные животныеf концентрируются в большом количестве.
Давление воздуха. Благодаря своему преимущественно
воздушному образу жизни птицы обладают широкой приспособляе-
мостью к различиям атмосферного давления, хотя повидимощу спо-
собны ощущать даже небольшие его изменения. Современные подъемы
на высочайшие горные вершины (Гималаи) обнаружили птиц даже
на высотах в 6000 м, куда, в царство льда, снега и голого камня,
они поднимались (клушица — Pyrrhocorax pyrrhocorax L.), следуя
за альпинистами в надежде на лагерные объедки. Нормально рас-
пространение птиц кончается на высотах альпийских лугов. Альпий-
ские и субальпийские высоты, как правило, обладают в гнездовое
время своим особым птичьим населением, но значительное число
зтпх форм , с наступлением зимы, а иногда и непогоды, спускается
значительно ниже, даже к подножию гор, что говорит за очень высо-
кую приспособленность птиц к колебаниям атмосферного давления.
Только немногие из этих форм, вроде улларов (видов рода Тetraogallus).
возможно приспособлены к узким границам атмосферного давления.
Помцмо собственно горных видов, легко выносят изменения дав-
ления и многие равнинные формы, во время перелета поднимающиеся
на высоты в 4000 — 5000 м и переваливающие Альпы, Тянь-шань.
Памир, Тибет, и т. д.‘ Даже такие плохие детуны п низинные формы,
как коростель и болотные курочки, были находимы на этих высотах.
Но независимо, от. гор некоторые птицы во время перелета поднима-
ются иногда на значительные высоты, как, например, журавлц, гуси
11 т. д,5 констатированные астрономами и летчиками на высотах
в 3000—5000.м. ’ ' :
Таким образом легкая приспособляемость птиц к изменениям
Давления несомненна. Она обусловлена по всей вероятности объеми-
стыми воздушными ^мешками, которые дают возможность смягчать
резкие изменения давления и создавать в дыхательном, аппарате
условия, наиболее благоприятные организму.
При этой широкой выносливости к изменениям давления изуми-
тельной является, чувствительность птиц к небольшим изменениям
2>3
давления, вызываемым прохождением циклонов, гроз и т. д. С дав-
них пор известна способность птиц предчувствовать перегну погоды,
что отражается в поведении, (купанье в пыли), активности и крикли-
вости. Так как основную роль играет, повидимому, чувствитель-
ность к изменениям давления, необходимо допустить наличие у птиц
особого механизма, воспринимающего эти изменения. По гипотезе
Гизлера (1905), таким механизмом являются очины контурных/глав-
ным образом маховых перьев, пустые внутри, соединенные с чувстви-
тельным перовым сосочком отверстием, заткнутым живой тканью.
По Гизлеру, эти очины можно уподобить строению коробки ане-
роида. Изменяющееся давление воздуха передается чувствительным
окончаниям перового сосочка, что вызывает зуд, стимулирующий
у птиц* потребность купаться.
Ветер. Ветер оказывает на птиц двоякое влияние: термическое
и механическое. Обычно ветер оказывает охлаждающее влияние на
тело птицы. Он раздувает и ерошит перья, покрывающие тело,
и увеличивает циркуляцию воздуха вокруг него. Это сказывается
благоприятно при высоких температурах воздуха, вследствие охла-
ждения поверхности тела и усиления испарения воды через ротовую
полость, влекущее за собой также ослабление внутренних темпе-
ратур. Но это же влияние ветра при низкой температуре воздуха
может быть весьма неблагоприятным вследствие переохлаждения
тела. Так как сильный ветер обычно сопровождается дождем или
снегом, термическая роль ветра может быть очень значительна и
в сильной степени сказывается на жизненной стойкости индивидуума.
Механическая роль вехра определяется его силой и также раз-
лична в зависимости от сопровождающих обстоятельств. Слабый
ветер только на плохих летунов оказывает вредное влияние и сле-
довательно легко переносится большинством даже мелких птиц.
При более сильном ветре мелкие птицы в большинстве случаев ищут
защиты среди каких-либо густых зарослей. Сильный ветер не играет
роли только для крупных птиц с сильным активным полетом, а ура-
ганный ветер может быть используем только особенно искусными
и сильными летунами, например альбатросами, буревестника мп к
т. д., и все же требует-от них много ловкости и энергии. Ветер
вообще играет важную роль для взлета многих крупных тяжелых
видов и необходим для полета парящих форм. Для последних обра-
зование, направление п сила ветра имеют большое значение (см.
стр. 186).
Существенную роль играет ветер в жизни птиц во время пере-
лета . Очень сильный ветер многие птицы предпочитают пережидать,
держась более или менее укрытых мест. Если сила ветра допускает
перелет вида, он летит при ветре любого направления. Неоднократно
высказывавшиеся взгляды за наибольшую благоприятность про-
тивного или попутного ветра на подтверждаются фактами. Реакция
птиц на ветер обусловливается исключительно его механической
силой в отношении к данному виду, сложившимся исторически
направлением перелета и силой стимула, движущего птицу. В соот-
ветствии с этими обстоятельствами поведение птиц будет различно
и на разных отрезках их пути будет протекать при различных уело-
264
виях. Говоря обобщенно, наиболее благоприятными для перелете!
являются слабые или умеренные ветры\ при сильных же ветрах —
попутный и Даже противный ветер, несмотря на необходимость затра-
ты большей энергии на борьбу с ним; при сильных. ветрах небла-
гоприятны боковые ветры, так как они вызывают относ птиц в сто-
рону от нормальной для вида перелетной трассы. Насколько велико
может быть дрейфование птиц в сторону от прямого пути, показывает
обилие залетов птиц на острова, лежащие вне нормальной линии
пролета. На Канарских островах в составе фауны имеются 16 регу-
лярно пролетающих видов, которые по существу являются регулярно
повторяющимися массовыми залетами, обязанными дрейфующему
действию ветров. Подобные факты заставляют предполагать значи-
тельную смертность некоторых популяций в море во, время се-
зонных перелетов, вследствие заноса масс птиц в сторону от пра-
вильной, обеспечивающей отдых и корм дороги.
Влияние климата в целом. Уже рассматривая отдель-
ные факторы, мы указывали, что многие из них не действуют изолиро-
ванно, а имеют место в связном комплексе факторов, причем зна-
чение фактора изменяется в зависимости от характера данного ком-
плекса. Влияние климата и есть воздействие такого комплекса,
складывающегося в характерные типы в определенных ландшафтах
н зонах земли. Климат накладывает определенный отпечаток на
фауну птиц данного ландшафта, определяя экологический состав-
фауны, степень ее оседлости, периодические сроки отдельных жизнен-
ных явлений и в значительной степени само поведение птицы.
Обычно мы наблюдаем известную приспособленность вида к кли-
мату своего ареала, выработавшуюся за длительный путь эволюции
вида; эта приспособленность наиболее значительна в тех районах,
где птица гнездится. В районах гнездования мы наблюдаем наиболее
благоприятные условия для существования вида, в пределах ареала;
вне этих районов в комплексе климатических факторов могут быть
такие изменения, которые уже не гарантируют преобладающего
благополучия данной популяции. Хотя причины той или иной
картины расселения и распределения вида по площади очень сложны
к помимо климатических причин важную роль играют наличие
соответствующих стаций, конкуренция и факторы исторического
порядка, исключительно важная роль климата не подлежит со-
мнению. Мы можем различать в отношении к виду несколько кате-
горий климатических влияний. Под зоной комфорта следует рас-
сматривать такое состояние климатических элементов, которое бла-
гоприятствует развитию самых сложных и глубоких жизненных про-
цессов вида, как, например, размножение. Как правило, это влия-
ние выражается в том, что оно не подавляет жизнедеятельности,
физиологической ритмики животных и вследствие этого практически
не ощущается ими. Вне зоны комфорта лежит зона диском-
форта, в которой неблагоприятные климатические влияния уже на-
чинают ощущаться видом и на которые вид реагирует изменением
своего поведения, и наконец можно различать зону границ вынос-
ливости, в которой существование вида оказывается уже необеспе-
ченным и которую он всячески стремится избежать.
265
В целом, наиболее благоприятными для жизни птиц -явля-
ются очевидно условия влажного тропического .климата, в кото-
рых мы находим наиболее богатые, наиболее разнообразные по
составу фауны. Огромное большинство птиц ведет здесь оседлый
образ жизни, если и передвигаясь в поисках пищи/то всегда в пре-
делах очень узкого района. В этом климате размножение птицы
обусловливается преимущественно внутренними, .физиолргическимп.
а не внешними причинами. Большинство видов в целом размножается
круглый год, хотя особи этих видов и имеют йекоторую периодич-
ность в размножении; эта периодичность у разных,особей наступает
в разное время, благодаря чему яйца птенцов ^ вылетевших моло-
дых птиц можно встретить здесь в любое время йода* У некоторых
видов однако [например у Tschitrea rufocinerear^Caib.) в Африке]
взрослые особи находятся повидимому беспрерывно: в состояния
размножения, судя по развитию гонад у добываемых/экземпляров
(Ghapin, 1934). Неразмножающиеся особи разные видов, часто
соединяются в общие стайки и кочуют в пределах; очень;.узкого,
района, постоянно возвращаясь в район бывшего гнездовья.Только
у немногих видов кочевки имеют более обширныйнхарактер:-, это
обычно плодоядные формы и виды, связанные с цветением какого-
либо дерева; и размножение у этих форм наступает в период и в.районе
более массового цветения этих деревьев.
Иная картина наблюдается в странах с более или, денее резко
выраженными периодическими изменениями климата^. В тропиках
это находит свое выражение в смене периодов дождей с периодами
засух. Таких периодов бывает два или четыре в течение года, с раз-
ными сроками ,цх начала и конца к северу и югу от- эц>ватора.
К периоду засухи деревья теряют свою листву, вся трава засыхает,
водоемы пересыхают, и в совокупности вся природа .производит
на птичье население эффект, сходный с эффектом зимы.в умеренных
и субполярных зонах земного шара. С возвращением периодов
дождей быстро оживает растительность и возобновляется обильная
жизнь насекомых. Эти сезонные изменения вызывают появление
сезонных изменений и в жизни орнитофауны. Для большинства
птиц период дождей является периодом размножения, после кото-
рого, с наступлением засухи, птицы начинают кочевать или отле-
тают к югу от экватора, где под аналогичными широтами в это'р момент
начинается период дождей, где птица и переживает неблагоприятное
для нее время года. Необходимо иметь в виду, что такимюбразом
резко выраженную сезонность в жизни jjtj ц и появление.перелетов
мы имеем уже под 10—15° шпроты. Далее к северу различия сезо-
нов все более углубляются, и зимний период становится все более
трудно переносимым на месте сезоном года. Явленш перелета захва-
тывают огромный процент видов фауны, перелет — его пути ..и осу-
ществление являются,все более сложными, а в ж; они птиц выра-
батываются и дополнительные биологические черты, обеспечи-
вающие с наименьшим риском переживание «мертвого» годового
сезона.
Из приспособлений, имеющих такой сезонный, вызванный клима-
тическими условиями, характер, заслуживают рассмотрения сл^
266
дующие:• изменения в питании, осеннее ожирение птиц, развитие
инстинкта приготовления запасов, впадение во временную спячку,
развитие кочующего или бродячего образа жизни, развитие перелета.
Смей а питания. В условиях изменчивой в климатическом
отношении природы имеется очень мало специализированных на
один род пищи видов птиц, придерживающихся его в течение круглого
года. Как правило, мы имеем изменение режима питания соответ-
пвенно изменению сезонов года, протекающего для большинства
впдов совершенно закономерно (рис. 200). Наиболее резко это выра-
жено у птиц умеренных и приполярных широт, летом питающихся
насекомыми, их личинками и другими беспозвоночными, осенью
нередко кормящихся ягодами и различными плодами и зймой,
?ис. 200. Состав пищи взрослых американских сорок —Pica pica hudsonia Sab. по мебйцам
года и в обобщенной годовой пропорции (правая колонка).
Диаграмма осповапа на результатах изучения содержимого 313 желудков. По Kalmb&ch.
в случае зимовки в более или менее северных широтах, питающихся
'сменами и зернами различных растений. Эта приспособляемость
ооеспечивает виду выживание в этих суровых условиях.
Осеннее ожирение широко распространено у птиц
Умеренных и северных зон и является подготовкой как для пере-
несения низких температур зимой (вследствие плохой теплопро-
водности жира), так. и 'в качестве запаса питательных веществ на
^учай временной бескормицы и как запас энергии для предстоящего
многим из них длительного перелета. Способность птиц этой фауны
Скоплять жир является во многих случаях изумительной; жир
чозкет располагаться толстым слоем под всей поверхностью кожи.
й значительными скоплениями отлагаться среди внутренних брюш-
аи* органов. Быстрота его накопления может . быть весьма зна-
^тельна: мелкие птицы, начинающие жиреть по окончании линьки,
За одну неделю из худого экземпляра могут превратиться в чрез-
2G7
вычайно жирную птицу. Серый гусь в одомашненном состоянии
может за 4—6 недель увеличиться в весе на 2 кг или на 50 % перво-
начального веса. При значительном потреблении этого жира, в резуль-
тате большой мускульной работы во время перелета, птица вновь
тощает, но во время остановок может вновь за несколько дней, при
хороших условиях питания, набрать новый запас жира и пуститься
в путь с обеспеченным запасом питания для мускульной энергии.
Эти отложения жира главным образом стимулируются внутри-
секреторной деятельностью организма.
Инстинкт собирания запасов пищи пробу-
ждается обычно у ряда видов в осеннее время, в период обильного
урожая семян и плодов различных растений. По всей вероятности
эта способность развилась из свойства, присущего и в период лет-
него образа жизни, как, например, многим сорокопутам, насажи-
вающим излишнюю добычу на шипы, колючки, обломанные веточкг.
в районе своей обычной охоты. В случае отсутствия обычной добычи
сорокопут повидимому использует эти запасы. В осеннее время
собирание запасов свойственно многим врановым (кедровке, сойке,
кукше), дятлам [Melanerpes jormicivorus (Swains)], поползням,
некоторым синицам (Poecile atricapillus L.) и некоторым совам
(Glaucidiurn passerinum L.). Кедровка при этом срывает кедровые
шишки, раздалбливает чешуи, достает «орешки» и набирает их
в большом количестве (8—10 шт.) в горловой растяжимый мешок
под языком и затем рассовывает их в трещины коры, под корни,
в ямки во мху и т. д. Сходным образом поступает сойка с дубовыми
желудями. Калифорнийский дятел [Melanerpes formicivorus (Swains)]
прячет дубовые желуди поодиночке в выдолбленные им самим ямки
в коре дерева, накапливая запасы более чем в 1000 желудей на
одном дереве. В случае отсутствия урожая желудей он удовлетворяет
свой инстинкт приготовления запасов, пряча в приготовленные
ямки мелкие камешки (рис. 201). Синицы прячут семечки за чешуйки
коры, мелкие совы собирают в дупла запасы пойманных ими мелких
птичек, -мышевидных грызунов и т. д., потребляя их в периоды
бескормицы. Отыскивание этих запасов не обязательно является
сознательным актом, хотя память у птиц, в особенности у врано-
вых, развита весьма высоко. Способность приготовлять запасы
исторически вырабатывалась для обеспечения кормом на небла-
гоприятный период всей популяции вида в целом; так как орехи пря-
чутся птицами в привычные для них местечки, где они обычно разыски-
вают свою пищу, то и в дальнейшем они вновь натыкаются на спрятан-
ное, хотя данный запас мог быть сделан й другим индивидуумом.
Развитие этих привычек очевидно подобно таким же привычкам
многих млекопитающих.
Впадение во временную спячку (оцепенение).
Крайне интересным, но мало исследованным является временное
впадение в спячку, наблюдавшееся достоверно только у ласточек
(Lorenz, 1932; Dupond, 1932). Это явление издавна служило пред'
метом легендарных рассказов о зимней спячке ласточек и ДРУГ11Х
птиц и сопровождалось невероятными баснями, вследствие чего
уже давно было подвергнуто сомнению и забыто.
268
Однако недавно .оно наблюдалось с полной достоверностью
Лоренцом в долине Дуная в конце сентября 1932 г., при установив-
шемся продолжительном ненастье. Деревенские ласточки, незадолго
перед тем начавшие интенсивный пролет, были замечены скопляв-
шимися массами около трех домов. Здесь ласточки залезали в щели
и пустоты, прижимаясь друг к другу и образуя целые скопления.
При попытке их взять обнаружилось, что они находятся в более
глубоком и крепком сне, чем это нормально для них даже в ночных
условиях: сбор ласточек снаружи этих скоплений даже не будил
следующие и более глубокие слои этих птиц. Однако при схваты-
вании эти ласточки сейчас же пробуждались и, не проявляя каких-
либо внешних признаков болезненного состояния^ л$вко и бодро
улетали. Очевидно этот
сон не был подобен сну
больных птиц, впадаю-
щих в него и в дневные
часы, с трудом поднима-
ющих веки и при этом
обычно йроявляющих
вялость и распущен-
ность оперения. Мы мо-
жем видеть в этом яв-
лении приспособитель-
ную реакцию на нена-
стную дождливую по-
году, очищающую воз-
дух от летающих на-
Рис. 201. Пищевые запасы «дятла сборщика» — Mela-
nerpes formicivorus Swains.
секомых, — постоянной
пищи ласточек. Соби-
раннем в группы в за-
щищенных местах, с
плотным прилеганием
Фуг к другу, птицы
(1)—желуди, защемленные птицей в кору отмершей
дубовой ветви; (2)—камешки, воткнутые дятлом в
дырки ствола дерева, приготовленные им для собирания
желудей; сбор камешков осуществляется деятельной
птиЩей повидимохму при отсутствии желуде и. По Ritter,
, из Stresemann.
лучше экономят вну-
треннее тепло, а впадением в сон понижают обмен веществ, что
в целом лучше благоприятствует переживанию неблагоприятной'
ПОГОДЫ.
К сожалению это интересное явление мало выяснено не только
с физиологической, но и с экологической стороны.
Кочующий или бродячий образ ж и з н и в осен-
Нее п зимнее время развивается, как правило, у большинства видов
Умеренных и высоких широт; некоторые при этом являются пере-
летными птицами и удаляются далеко на юг. Строго говоря, на
ЭТцх шпротах оседлых видов почти не существует. Наибольшей
оседлостью обладают куриные, все же предпринимающие зимой
х°тя бы небольшие кочевки из глубины леса ближе к речным доли-
Нам (рябчик), из одних типов леса в другие (глухарь). Неулетевшие
Далеке на юг птицы в большей или меньшей мере ведут зимой бро-
дачнй образ жизни, передвигаясь по стране в поисках более кормных
Мест- Эти движения в некоторых случаях носят повидймому совер-
269
шенно беспорядочный характер (свиристели, снегири), хотя в боль-
шинстве случаев имеют все же явно южное направление. Птицы
постепенно стайками . спускаются к югу, задерживаясь на более
высоких или низких широтах наличием подходящего корма или
вследствие мягкости погоды. Таким образом миграции этих птиц,
размах и направление их определяются кормом и суровостью погоды
и дают возможность в нужной степени избежать неблагоприятных
условий зимнего времени. Сходная картина наблюдается и в тро-
пиках, с периодическим сухим временем года; значительная часть
птиц и здесь ведет бродячий образ жизни.
Перелет с несомненностью развился пз этих сравнительно
коротких кочевок и представляет теперь стимулируемое весьма
глубоким инстинктом явление ежегодного передвижения птиц на
значительное, иногда захватывающее десятки тысяч километре;
расстояние. Переходные, положения, связывающие резко выражен-
ный дальний перелет с короткими кочевками, имеются в обилии
и в настоящее время, не только в пределах крупных систематических
групп, но даже в пределах вида. Широким применением кольце-
вания было выяснено, что у многих видов в пределах одной и той
же популяции могут встречаться особи как оседлые, так и слабо
кочующие, а также и перелетные на сравнительно значительное
расстояние. Так, например, чибис в Англии частью остается на
зимовку, частью зимует в северной Франции, частью отлетает на
юг Испании. Как правило, наибольшей склонностью к оседлости
обладают старьте особи, а более значительный перелет совершают
сравнительно молодые индивидуумы. Однако весьма распростра-
нены явления и весьма дальнего перелета всей популяции вида
в целом.
Так как перелет представляет очень крупное и комплексное
явление в жизни птиц, он заслуживает обособленного и более деталь-
ного рассмотрения.
ПЕРЕЛЕТ ПТИЦ
Под перелетом следует понимать периодически, соответственно
со сменой теплого и холодного или сырого и сухого сезонов года,
повторяющееся перемещение популяции вида из области гнездовья
в область, где вид живет весь период неблагоприятных для него
условий на родине. Размах таких перемещений, как уже указы-
валось, может быть очень велик, причем области зимовки (как обычно
называют районы, где перелетный вид живет более или менее оседло
вне времени гнездования), то могут непосредственно соприкасаться
с областью гнездовья, то отделяться от нее значительным простран-
ством, через которое вид только пролетает, не задерживаясь на
длительный период. Наиболее длинными пролетными путями обла-
дают -главным образом северные кулики и некоторые крачки, и*
которых пестрохвостый вртренник — Limosa lapponica (L.),
голь — Tringa erytHropus (Pall.), некоторые песочники и ржанки
и многие другие (рис. 202), летуя в области северных тундр и побе-
режий. арктпки, } зимуют в умеренной зоне южного полушария-
270
ложной Австралии, Тасмании, Новозеландии, южной Африке
,и Патагонии. Область пролета их по кратчайшему расстоянию
может составлять 10—15 тыс. километров, пролетаемых видом:
в течение 1,5—2 месяцев каждую весну и осень.
Пролетные пу-
ти. Кац протяженно,
так и характер области
пролета ^данной попу-
ляции обусловливаются
в значительной степени
формой гнездового и
зимнего ареала и эколо-
гией вида. Если протя-
жение области зимовки
по широте соответствует
более или менее протя-
жению гнездового ареа-
ла, а пространство, ле-
жащее между ними, не
представляет особых
преград для пролета
птицы, мы имеем обычно
перелет,так называемым
широким фронтом, т. е.
пролетающие птицы рас-
сеиваются на всем этом
промежуточном про-
странстве; вся популя-
ция вида в целом как
бы перемещается в на-
правлении зимовок, не.
Рис. 202. Распространение и миграции американских
особен длиннохвостой крачки — Sterna paradisaea Briinn.
Черные точки — места гнездования; треугольники —
места зимовок; белые кружочки — места находок в период
пролета; стрелки — направление п пути пролета основ-
ной массы, особей/та зимовку; крест с .точкой — места
кольцевддия; крест — места находок окольцованных
. птиц.
После гнездования в Канаде п Гренландии птицы переле-
тают Атлантический океап к берегам Европы и следуют
на зимовки вдоль берегов Африки и Южной Америки.
Наиболее удаленные друг от друга точки гнездования и
зимовки этих крачек отстоят па 11 000 миль; так как
крачки летят кружным путем, вероятный путь этих птиц
составляет ежегодно около 25 000 миль. По Lincoln.
меняя относительного
положения своих частей
(рис. 203).
При значительно бо-
-iee ограниченном про-
тяжении по широте
зимнего ареала в срав-
нении с гнездовым, ле-
тящие птицы при при-
ближении, гг к зимовке
очевидно, сгущаются на
°°лее ( узкой террито-
рии, и область пролета принимает характер воронки пли может
сужаться до сравнительно узцого пролетного потока. При более
ограниченных экологических /требованиях птиц пролет может кон-
центрироваться на немногие участках и вести ^образованию резко
выраженных пролетных путей, которыми следует перемещающаяся
популяция. Как правило, однако эти пролетные пути представляют
ппшь наибольшие сгущения летящей птицы, рассеивающейся в мень-
271
шем количестве и на остальной площади, лежащей между зимними
п летними ареалами. Обусловленные экологическими причинами,
пролетные пути следуют протяжению определенного ландшафта:
так приморские и береговые птицы следуют обычно береговой линии
морей, придерживаясь ее основного направления, птицы речные
следуют рекам, горные птицы часто следуют горным цепям, а птицы
лесные скопляются преимущественно в более удобно, относительно
направления движения,
Рис. 203. Гнездовой и зимний ареалы п пролетные
пути американской овсянки — Zonotrichia ^uerula
(Nutt.).
Грубый пунктир ограничивает область массового про-
лета этого вида, местами образующую полосу не шире
500 миль. Тонкий пунктир включает район более или
менее регулярного пролета. Точки отмечают места
случайных или спорадических встреч. По Lincoln.
расположенных лесных
районах (рис. 204).
Направление перелета
о бу с л ов лено отно ситель-
ными положениями гнездо-
вого и зимнего ареалов и
необязательно идет в се-
веро-южном направлении.
Речные и озерные птицы
часто зимуют у побережий
незамерзающего моря под
той же широтой, откуда
они улетают, и пути пере-
лета их следовательно дол-
жны лежать в широтном
направлении. Крупную
роль в определении этих
путей играет также исто-
рический • момент — про-
цесс их образования. Из
совпадения географически
вероятного расселения ви-
да с обычным путем его
пролета было выведено
правило (Seebohm, 1892:
Menzbier, 1898) о повто-
рении путями пролета
путей расселения видов.
Вследствие именно этих
причин некоторые виды
летят на зимовки в во-
сточном или западном на-
правлении, как, например, многие Г американские по происхожде-
нию виды в тундрах у северо-восточной Сибири, расселившиеся Д°
Таймыра и летящие осенью в восточном направлении, или, наобо-
рот, азиатские виды в Аляске, улетающие в западном направлении.
Но расселение может иметь место не только на гнездовье, но и на
зимовке, следствием чего могут образоваться иногда парадоксальны?
по направлению пути, как, например, перелет амурского кобчика
(Erythropus ainurensis Radde), летующего в южной половине восточ-
ной Сибири и летящего через собственно Китай, Индию и Индии-
•ское море в южную п восточную Африку.
272
2---------------------------3 -............................................. 4 llllllllllllllllil 5
Рис. 204. Главнейшие области зимовок птиц Евразии и пути перелета к ним: 1 —западноафриканские зимовки и пути тГерелета к ним;
2 — восточноафриканские зимовки п пути перелета к ним; 3 — передне азиатские и индийские зимовки и пути перелета к пим; 4 — пути вер-
тикальных миграций и зимовки гималайских птиц; .5 —ппдо-малайские и восточноиндийские зимовки и пути перелета к ним; в — амери-
канские зимовки и пути перелета к пим. Густота линий указывает степень интенсивности пролета в данном направлении; попа ряду с этим
в любой местности могут быть отмечены и другие единичные типы перелета. Схема по Boubier, с изменениями.
О возможных перемещениях зимнего ареала свидетельствует
также распространение некоторых видов, например, корольков
{Regulus azoricus Seebohm и teneriffae Seebohm) на Азорских остро-
вах и Regulus clarci в Гондурасе, близко родственных Regulus regu-
lus L. и Regulus satrapa Licht., распространенных в настоящее
время значительно севернее этих широт и не встречающихся здесь
даже в период осенне-зимних кочевок. Нахождение этих южных
форм может быть объяснено только былым, более южным, распро-
странением северных видов и областью перелетов их некогда во
всяком случае до более южных широт.
Однако не следует думать, что пролетный путь может лежать
только в области, в которой когда-то лежал гнездов ,й или зимний
ареал вида. Пролетные пути могут образоваться и независимым,
т. е. вторичным способом, посредством открытия новых удобных
мест зимовок. Повидимому основным условием образования этих
новых путей является расселение вида в новые районы, где он встре-
чает близкие по экологии виды, обладающие иными пролетными
путями и зимовками: стайки этих видов, улетая осенью, увлекают
с собой особей расселяющегося вида и вместе с ними возвращаются
на следующий год; постоянное повторение этого явления и вызывает
образование нового пролетного пути у расселившегося вида. Такими
пролетными путями п видим му обладают летящие через Среднюю
Азию восточносибирские горные коньки (Anthus japonicus Temm.
etSchl.), пролетающие через восточную Азию острохвостые кулики
(Heteropygia acuminata Horsf.), зимующие в Британии рогатые жаво-
ронки (Eremophila alpestris L.) со Скандинавского полуострова и
мнтие другие виды, ист >рич< ски чуждые для этих областей про-
лета. Эта особенность может быть более легко выражена у видов,
находящихся в стадии интенсивного расселения и своими передо-
выми пунктами далеко удаляющихся от нормальных зимовок вида.
Птицы далеко не исключительно следуют установившимся исто-
рически традициям, но могут довольно сложным образом видоиз-
менять пролетный путь, соответственно экологическим или физи-
ческим запросам. Это следует из факта существования искривлений
линии пролета и так называемых петлеобразных пролетных путей.
Пролетные потоки птиц во многих случаях огибают моря или гор-
ные хребты и пустыни, пролегая по более удобным для вида местам
й далеко не всегда следуя к, атчайшему расстоянию между зимов-
к°й и гнездовым ареалом. Упомянутый выше амурский кобчик
следует по крутой кривой на свои зимовки в Африке, не сокращая
расстояния перелетом через Центральную Азию. Довольно боль-
шое число видов при перелетах весной придерживается отлич-
ного пути, чем осенью, в совокупности за год описывая как бы род
эллипса (петлю) в своих странствованиях между зимним и летним
ареалом. Классическим примером такого перелета является американ-
ский подвид золотистой ржанки (Charadrius dominicus Mull.), гнез-
ДУющей в тундре Северной Америки и на Канадском архипелаге
(рис. 205). Осенью большая часть этих ржанок улетает в восточном
направлении к северным берегам Лабрадора и затем летит прямо
к югу через Бермудские и Малые Антильские острова в Гвиану и
18 Орнитилоги я—120 9
273
Рис. 205. Распространение и миграции золотистой
ржанки — Charadrius dominicus (Miill.).
Североамерикапский подвид — С. d. dominicus
(Miill.), гнездящийся в тундрах Канады п па Канад-
ском архипелаге (черные точки); осенью отлетает к
берегам Лабрадора и затем прямым путем достигает
зимовок в Бразилии и северной Аргентине. Весной
птицы возвращаются другим, более коротким путем,
через долину Миссисипи. Гнездящийся в Сибири и
па Аляске подвид — С. d. fulvus (Gm.) (крестики);
частью зимует на островах Гавайских, Маркизских
и Поумоту в Тихом океане и возвращается обратно
тем же самым путем. По Lincoln.
затем в пампасы 71ргентины и Бразилии. Весной на этом пути вид
не встречается, но зато обнаруживается сильный пролет его из
Южной Америки вдоль Панамского перешейка и затем долиной
Миссисипи прямо к северу. В нашей фауне такой пример предста-
вляет восточносибирская черная казарка [Branta bernicla nigricans
Lawr.), гнездящаяся в приморской тундре от устья Лены на западе
до залива Коронации в Се-
верной Америке на восто-
ке. С зимовок на Печилий-
ском заливе и Желтом
море птицы летят весной
к области гнездовья, рас-
сеиваясь повидимому ко-
нусом (воронкой), соответ-
ственно положению гнез-
дового ареала, но наи-
большее сгущение наблю-
дается по трассе, ведущей
через Средний Амур и
Среднюю Лену под Якут-
ском. Осенью на средней
Лене черная казарка отсут-
ствует, но наблюдается
интенсивный пролет ее в
западном отрезке Охот-
ского моря и на нижнем
Амуре. Очевидно основ-
ные массы птиц избирают
различные пролетные пути
для весеннего и летнего
периода. Различные петле-
образные пути известны не
только для различных ку-
ликов, уток, гусей, но и
для некоторых мелких
птиц, и обусловливаются
повидимому р а з личными
причинами. Как правплог
весенний путь является
более прямым и коротким
и отвечает более сильно вы-
раженному весной стрем-
лению быстрее долететь до мест гнездования. Отклоняющийся
осенний путь объясняется экологическими причинами и предваритель-
ным перед отлетом скоплением птиц на лежащих в стороне кормо-
вых угодьях, где птицы откармливаются перед отлетом и, улетая
отсюда в южном направлении, естественно следуют уже друг011
дорогой. Для мелких птиц в различии этих путей имеет повидимом)
некоторое значение также ветер, вызывающий различное отклони
ние летящих стай в разные стороны по сезонам.
274
Несмотря на колоссальность расстояния, пролетаемого многими
птицами, все больше накапливается данных, указывающих на
значительное постоянство пролетных путей, характерных для вида.
Это постоянство распространяется не только на популяцию в целом,
но и на отдельных особей. Кольцеванием было установлено, что
через Гельголанд неоднократно пролетали черные дрозды (Turdus
merula L.), окольцованные здесь же в предыдущие сезоны. Одна
из американских овсянок [Zonotrichia albicollis (Gm.)], окольцо-
ванная в 1916 г. на зимовке в Георгии, на юге Соединенных Штатов,
в 1600 км от ближайших мест гнездования этого северного вида,
вновь добывалась здесь в 1917, 1920 и 1921 гг. Нормально держась
в стайках, этот вид был представлен здесь каждый год экземпля-
рами, окольцованными здесь же ранее и очевидно частью совместно
и регулярно странствовавшими по одной и той же дороге. То же
самое было констатировано для многих других видов. Это пока-
зывает, что верность месту гнездования, как правило, свойственная
большинству видов, является очевидно частью верности большего
числа особей всем ее миграционным путям в целом, включая вер-
ность как месту родины, так и месту зимовки.
Течение перелета и его экологические
у с л о в и я. Как в период пролета, так и на зимовке вид придер-
живается тех же или сходных условий местности, в которых мы
находим его и в лучший период жизни. Экологические связи однако
становятся рыхлее, и формы лесные могут удовольствоваться кустар-
никами, виды густых болотных зарослей — любыми типами водое-
мов. Как правило, перелетный путь проходит преимущественно
через удобные виду ландшафты или он опускается на отдых или
остановку только в тех участках маршрута, где расположены соот-
ветствующие стации. В еще большей степени эти экологические
требования сказываются в распределении вида на зимовках. Сход-
ным образом сохраняется приверженность к тому же типу питания.
Этот известный консерватизм в требованиях к условиям мест-
ности и питания является причиной фенологической приурочен-
ности перелета вида к состоянию ландшафта пути пролета. Как
правило, птицы пролетают в то время, когда находимые ими по
пути условия оказываются сходными с летними условиями на их
родине. В наиболее резкой форме это свойственно поздно приле-
тающим птицам с наиболее глубоко развитым перелетным инстинк-
том. Вместе с тем птицы прилетают на родину к тому сроку, когда
наблюдается уже в природе развитие необходимых для их жизни
условий — привычной пищи и укрытия. Соловей появляется в тот
период, когда на кустарниках уже разворачиваются листья, камы-
шевки — когда подрастает молодой камыш, ласточки — в период
появления летающих в воздухе насекомых. Таким образом
всюду наблюдается постоянный спектр прилета, растягивающийся
обычно на 1,5—2 месячный период. Под Ленинградом первыми,
около 18 марта, появляются передовые грачп, затем передовые
’Кворцы (26 марта), 1—10 апреля начинается пролет лебедей, приле-
тают полевые и лесные жаворонки, пустельги, зяблики, коноплянки,
Дорябы и др. На 11—20 апреля приходится массовый прилет гусей,
*
273
чаек, уток, многих куликов (кроншнепы, дупелп, бекасы, вальдш-
непы п др.), журавлей, певчих дроздов и других воробьиных. Около
30 апреля впервые раздается песнь пеночки теньковки, 1 мая —
песнь лесного конька, 9 мая — кукушки, 12 мая — соловья. К концу
мая прилетают самые поздние виды: стрижи, иволги, сизоворонки.
Еще более рано, в феврале — марте, идет в основном прилет птиц па
южной Украине. На глубоком севере, на Таймыре, первые птицы
появляются только 6 мая (пуночка — Passerina nivalis L.). Гуси
(черные казарки — Branta bernicla nigricans Lawr.) появляются
только в начале июня, и лишь со второй половины этого месяца
начинается размножение птиц (кладка, насиживание и т. д.).
Птицы, поздно прилетающие, как правило, рано и отлетают,
так что осенний спектр отлета оказывается обратным весеннему.
Под Ленинградом его картина, например, развертывается следующим
образом: около 25 августа отлетают стрижи {Micropus apus L.),
около 11 сентября начинается пролет дупелей {Capella media Inth.).
16-го — журавлей {Grus grus L.), 17-го — славок черноголовой
{Sylvia atricapilla L.), 20-го — сарычей {Buteo buteo L.), 21-го —
пеночек теньковок {Phylloscopus collybitus Vieil.), 24-го — ласточек
{Hirundo rustica L.), белых трясогузок (Motacilla alba L.), 25-ro —
коньков {Anthus trivialis L.), 26-ro — зябликов {Fringilla coelebs L.).
и в это же время кончается отлет черноголовой и сарычей. 28 сен-
тября начинают отлетать жаворонки {Alauda arvensis L.), малиновки
{Erithacus rubecula L.), 4 октября — певчие дрозды {Turdus erice-
iorum Turton) и белобровки (Turdus musicus L.), 11-го начинают
отлетать грачи {Trypanocoraxfrugilegus'Li.), 20-го — чайки {Larussyp)>
21-го — лебеди {Cygnus cygnus L.). В то же время к 18 октября
заканчивается отлет зябликов {Fringilla coelebs L.), теньковок.
23-го — малиновок, 29-го — лебедей. Около 12 ноября исчезают
последние чайки. В октябре и ноябре появляются и различные
зимующие у нас северные птицы (свиристели, щуры, снегири и др.)-
Некоторые виды однако не поспевают вывести детей в короткий
срок при позднем прилете, несмотря даже на более длинный северный
день и вследствие этого оказываются уклоняющимися от этого правила.
Таков, например, черный стриж {Micropus apus L.), задерживаю-
щийся в Финляндии иногда до сентября, в то время как в северной
Италии он отлетает уже в конце июля. Известное исключение пред-
ставляют повидимому также кукушки, невоспитывающие самостоя-
тельно птенцов, начинающие отлетать уже вскоре по окончания
кладки яиц самками. Так, например, хохлатая кукушка {Clamator
glandarius L.) пз Египта уже отлетает в течение июня несмотря
на прилет свой туда в начале февраля. Это имеет также своим след-
ствием то, что молодые кукушки летят на зимовку уже совершенно
самостоятельно, обычно вне связи как со своими настоящими, так
и приемными родителями.
Так как поступательное движение весны развертывается мед-
леннее в сравнении с возможной скоростью перелета, особенно хоро-
ших летунов, мы обычно наблюдаем определенную закономерность
в порядке прилета особей северных и южных популяций того ?ке
вида. Вначале обычно появляются местные особи данного вида и
276
лишь в дальнейшем появляются особи, гнездящиеся далее на севере;
пролет последних может тянуться очень долго; так, например,
под 43° с. ш. в Южно-Уссурийском крае гнездящиеся личинкоеды
(Pericrocotiis divaricatus Raffl.) появляются уже 5—10 мая, а про-
лет гнездящихся севернее особей продолжается даже до 31 мая.
Более длинен этот период у видов с более обширным распростра-
нением по мерт диану, например жаворонков, многих коньков н
других птиц, но в большинстве случаев эти северные п южные попу-
ляции представлены уже разными подвидами. Это явление связано
в свою очередь с длительной задержкой гнездящихся на севере
птиц на зимовке, где под экватором, в Африке например, можно
встретить перепела еще в половине апреля.
Весьма распространенной особенностью сезонных миграции яв-
ляется также разделение самцов п самок с молодыми на отдельные
стаи и пролет в разные сроки. При этом обычно молодые птицы
спускаются дальше всего на зимовке; почтп так же далеко и
отчасти или всегда смешанно с ними оседают на зимовку самки и
ближе всего к гнездовой области зимуют старые самцы. При частич-
ной зимовке вида на местах гнездовья остаются здесь всегда самцы,
притом всегда более старые экземпляры. Это явление широко свой-
ственно различным утиным, многим хищникам и значительному
числу воробьиных. Наиболее вероятным основанием для этого
является различный ход развития стимулирующих перелет влия-
ний в организме птицы, в зависимости от пола п возраста. Сроки
пролета также вариируют, как указано, в связи с полом и возрастом.
Распространеннейшим явлением служит ранний прилет самцов
по сравнению с самками, часто опережающий последних на 4—
12 дней. Биологические значение этого факта обычно видят в закреп-
лении самцами сразу по прилете гнездовой территории, что обеспе-
чивает выкармливание будущего потомства достаточным количеством
корма(см. стр. 216). Однако более ранний прилет самцовможет обусло-
вливаться также более близким расположением их зимовок. Позднее
всего прилетают на родину молодые птицы, которые нередко, в случае
наступления половой зрелости спустя год и больше от рождения,
весной летят медленно и часто оседают, не долетев даже до области
гнездовья вида. Так разнообразные молодые кулики, гнездящиеся
на севере, в большом количестве проводят лето в степях и болотах
степной полосы, так и не долетая в первый год жизни до мест гнездо-
вья. Молодые некоторых видов, например белых аистов и некоторых
прачек, первое лето вообще повидимому не покидают области зимовки.
Разновременный отлет, по полам и возрастным группам к югу часто
сопровождается разновременным наступлением линьки. Большин-
ство молодых мелких птиц перед отлетом перелинивает только мелкое
неро, после чего сейчас же начинает мигрировать, в то время как
старые птицы могут иметь еще несколько выводков п перед отлетом,
кроме мелких, меняют и крупное оперение. Вследствие этого у очень
многих видов молодежь отлетает значительно раньше старых, само-
стоятельно странствуя к югу или почти без сопровождения старых
Птиц. Однако в ряде случаев взрослые перед отлетом вообще не
проходят линьки п отлетают раньше молодых (например сорокопут-
277
жулан); также рано улетают почему-либо не гнездившиеся экзем-
пляры или особи, не прпнпмающпе участия в кормлении и вождении
птенцов, как, например, самки многих северных куликов.
Образ жизни во время перелета и зимовки варипрует у разных
видов. Более поздно прилетающие и рано улетающие формы менее
склонны к кочевкам во время пролета, быстрее и с большей целе-
устремленностью добираясь до зимних пли летних квартир. Неко-
торые однако склонны и ’в это время к значительным кочевкам,
в сущности ведя бродячий образ жизни и на зимовке, что наблю-
дается у некоторых хищников и стрижей. Большинство птиц однако
в области зимовки хотя бы часть времени ведет всецело оседлый
образ жизни, во время которого птицы отдыхают и накапливают
новые силы к предстоящему полету, а частью и претерпевают осен-
нюю плп весеннюю предотлетную линьку. Образ жизни в значитель-
ной мере определяется также экологическими запросами вида.
Птицы, находящие пищу в изобилии и в концентрированном виде,
как, например, розовые скворцы в области Синда, встречаются
большими стаями и почти оседлы, вследствие чего они обладают
очень небольшим зимним ареалом; принужденные разыскивать
пищу на большом расстоянии, например трубконосые, обычно
кочуют всю’зиму в пределах большой области и обладают, в сравне-
нии с местами гнездовья, обширными зимними ареалами. Как в обла-
сти зимовки, так отчасти и на пролете мигрирующие популяция
подчиняются влияниям сезонного климата и иногда, соответственно
смене сухпх и дождпвых времен года, перемещаются на значительной
территории.
Скорость перелета определяется главным образом ско-
ростью полета и частотой и продолжительностью остановок. Послед-
ние подвержены большой изменчивости не только по видам, но я
по сезонам. Уже указывалось, что весной птицы летят с большей
быстротой:, чем осенью, причем поздно прилетающие особенно спо-
собны покрывать большие пространства в короткие сроки. Восточно-
сибирский веретенник, пролетающий до зимовок в Ново-Зеландип
и Тасмании по прямой до 12 000 км, покрывает это расстояние осенью
за 2—3 месяца, а весной за 1—1,5 месяца. Вследствие этого неуди-
вительно, что многие кулики весной почти не наблюдаются во многих
пунктах их пролета, — настолько короткое время проходит волна
мигрирующей популяции этих видов.
Фенологическими наблюдениями удалось также установить, что
весной, по мере приближения к местам гнездовья, скорость пере-
лета у многих видов значительно увеличивается. Так, по сводке
Кука (Cooke, 1915), в Северной Америке средняя скорость продви-
жения мигрирующих птиц на пространстве от Нового Орлеана до
южной Манитобы составляет 23 английских мили в день; 16 видон
при этом уже с южной Мпннезоты развивают скорость до 40 миль
в день. С южной Манитобы до озера Атабаска у 12 видов скорость
повышается до 72 миль в день, у летящих далее 5 видов она повы-
шается к Большому Невольничьему озеру до 116 миль, а на Аляск}
они прибывают со скоростью 150 миль в день (рис. 206). Скорость
и ход прилета совершенно очевидно следуют движению весенних
278
Рпс. 20G. Миграции черношапочпой американской слав-
ки — Dendroica striata (Forst.),
Изо хропьГ соединяют пупкты с одновременным прилетом
птиц весной; с продвижением к северу птицы летят бы-
стрее, что выражается более широким расстоянием ме-
жду изохропами. С 30 апреля по 10 мая птицы летят в
среднем со скоростью 30 миль в день, в период же 25—
30 мая — нс менее 200 миль в день. По Lincoln.
изотерм. Сводками Кайгородова для Европейской части СССР
установлено, что движение популяции кукушки весной (изохроны
первого кукования) совпадает с ходом апрельских изотерм и соста-
вляет в среднем 80 км в день; для грача эта связь намечается с изо-
термами марта и составляет около 55 км в день. Лист же летит со
скоростью 60 км в день.
О средней скорости
перелета отдельных ви-
дов можно судить в ря-
де случаев по окольцо-
ванным птицам, добы-
вавшимся вскоре по
кольцевании их во вре-
мя миграций. Так обык-
новенные ГО piIXBO CTKII,
окольцованные в Герма-
нии, осенью неодно-
кратно добывались в
Средиземноморье и Аф-
рике. Наибольшую бы-
строту дал экземпляр,
успевший за 6 дней по-
крыть расстояние по
прямой в 1000 км, что
составляет до 167 км в
день. Другой экзем-
пляр, улетевший уже за
2070 км, находился в
полете уже 47 дней, что
доставляет до 44 км в
день. Из этих и подоб-
ных им данных следует,
что птицы не обладают
постоянной скоростью
полета, но по всей веро-
ятности сочетают дли-
тельный полет с проме-
жутками отдыха. О том,
что многие виды в состоянии покрывать, не отдыхая, колоссальные
расстояния, известно из нормального нахождения многих птиц
на пролете на островах, весьма удаленных от ближайшей береговой
линии, и из нередких залетов даже мелких птиц на удаленные океани-
ческие островки. Наибольший интерес вызывает перелет золотистой
ржанки (Charadrius dominicus Mull.) на зимовку на Гавайские
острова, расположенные в 3300 км от ближайшего берега (Алеут-
ских островов). Было подсчитано, что при скорости полета, равной
26 м в секунду (обычной скорости куликов средней величины),
ржанка должна лететь это пространство 35 часов и сделать до 252 000
взмахов крыльями. Но на этих островах зимует в большом коли-
честве целый ряд других азиатских и американских куликов, как
279-
Numenius tahitiensis Gm., Heleractitis incanis Gm. и brevipes Vieilk
и др., одинаково с ржанкой нормально не спускающихся на воду,
что показывает, что это отнюдь не является исключительным свой-
ством одного вида. Не менее удивительную способность показывают
различные случаи залетов. В декабре 1927 г. из Англии штормовым
ветром была занесена на американскую сторону Атлантического
океана (Ньюфаундленд) стая в сотню чибисов, как указывалось,
нормально зимующих к юго-западу не далее Пиренейского полу-
острова. Принимая во внимание силу ветра, можно считать, что
птицы перелетели расстояние в 3500 км за 24 часа, что кажется
почти невероятным для этого, не отличающегося хорошими качествами
полета, вида. На какие подвиги способны еще худил е летуны, пока-
зывают залеты на Бермуды (1000 км, от берега Северной Америки
в Атлантическом океане) коль бри (Trochilus kolibris L.), коростеля
(Crex crex L.) и жаворонка (Alauda arvensis L.); последние два
вида должны были пролететь из Европы не менее 6000 км, чтобы
попасть на эти острова.
Подобные примеры показывают, что большинство птиц могут
лететь с очень значительной скоростью, выдерживая весьма дли-
тельное напряжение полета. Поэтому сравнительную медленность
движения их по пролетным путям можно объяснить значительными
задержками в пути, временной стационарностью или возможными
кочевками в сторону. Что такое поведение является вполне законо-
мерным, следует» из известно!! правильности продвижения птиц
во время перелета. Так белый аист, улетая из Германии во второй
половине августа, прилетает в южную Африку в ноябре п на зимовке,
в области Капа, появляется в декабре. Колючехвостый стриж (Hirun-
dapus caudacutusIj^Xh.^ улетая из Восточной Сибири в начале августа,
весь август и сентябрь держится в восточном Китае, только в декабре
появляется в южной Австралии, где держится до февраля, затем
странствует в Тасманию, где дер?кится до марта или середины апреля.
Последний вид особенно указывает на широкие кочевки по пути
и общую медленность и «календарность» продвижения по маршруту,
несмотря на удивительную быстроту (до 170 км в час) и выносливость
полета, которые при большей целеустремленности и скорости полета
могли бы за несколько дней перенести птицу из области гнездовья
до южных пунктов зимовки.
Высота перелета. До недавнего времени считалось, что
птицы во время сезонных перелетов стремятся подняться на боль-
шую высоту, что и является причиной редкости наблюдений пере-
лета некоторых видов. Однако благодаря развитию' воздушных
сообщений выяснилось, что это не соответствует действительности.
Лишь в крайне редких, исключительных, случаях приходилось
констатировать летящие стаи журавлей на высотах до 5000 м над
землей. Нормально крупные птицы не поднимаются во время про-
лета выше 1500 м, предпочитая вообще держаться ниже 1000 м над
землей. Мелкие же птицы нормально летят ниже 300 м, во время
ветра спускаясь к самой земле, вследствие меньшей скорости здесь
ветра, благодаря трению о поверхность почвы. В пределах этих
величин высота полета колеблется от вида к виду и стоит в связи
280
с привычным способом полета и поведения птицы. Так крупные
хищники летят на значительной выс<те, луни над самой землей,
жаворонки, трясогузки также не поднимаются высоко в воздух,
тогда как журавли нормально летят на высотах около 1000 м над
землей. Указываемые данные относятся к полету относительно земли,
а не уровня моря, так как на высоких плоскогорьях п над цепями
гор многие птицы бывают принуждены нормально лететь на высо-
тах в 4000 п 5000 м. Перелет в ночное время в большинстве случаев
не отличается от перелета, наблюдающего ся днем, но пногда (утки,
мелкие птицы) протекает на меньших высотах.
Необходимо отметить, что время суток, избираемое для пере-,
лета, во многих случаях отличается значительным постоянством
и зачастую не соответствует обычным периодам активности птицы.
Кроме собственно ночных птиц, ночью в большом количестве летят
мелкие воробьиные птицы, кулиги, утки и птицы многих других отря-
дов. Лишь в дневные часы наблюдается лет дневных хищников. В ос-
новном это объясняется временем кормежки птицы и возмо?кностью
временных голодовок. Мелкие птицы с весьма живым обменом веществ
не могут голодать даже одних сут. к, вследствие этого они днем
кормятся, очень короткое время проводят во сне, а значительную
часть ночи и сумерки продолжают перелет. Крупные дневные хищ-
ники или кормятся по пути, медленно продвигаясь к югу, или совсем
воздерживаются временами от пищи, настойчиво продолжая пролет
п отдыхая во сне в ночное время.
Таким образом наиболее активный пролет большинства видов
наблюдается в ночные часы. Однако в разгар весеннего пролета
многие виды, предпочитающие лететь в ночное время, в большом
количестве продолжают лететь и днем.
Физиологические условия п е р е л е т а. Из наи-
более загадочных с физиологической стороны вопросов миграций
птиц следует считать вопрос о том, откуда берется энергия в орга-
низме птицы для преодоления часто весьма длительных перелетов и
огромных расстояний. Что именно вызывает и стимулирует начало
и течение перелета и, наконец, каким образом птицы в состоянии
следовать своему пролетному пути, сохраняя-на тысячи километров
не только правильное направление, но и следуя во многих случаях
через точно определенные участки.
Энергия перелета. Рассмотрение каждодневной деятель-
ности птиц в летний период их жизни и грубая оценку затрачиваемой
пмп энергии обнаруживают факт весьма значительной близости
между количеством движения птиц летом и количеством его, затра-
гиваемым в среднем в период перелета. Наблюдениями установлено.
Что самка обыкновенной горихвостки за 16 часов 30 минут 419 раз
пормила птенцов, принося корм из окрестностей гнезда. Если мы
примем расстояние, на котором собиралась пища, равным только
оО м, что несомненно ниже имеющегося отношения в действитель-
ности, то птица оказывается вынужденной сделать не менее 42 км
За День в поисках пищи, т. е. очень близко к указанной выше цифре
в 44 км средней скорости пролета, установленной по данным кольце-
пания. Гораздо большее расстояние должна налетать деревенская
/81
ласточка, кормящая птенцов также до 400 раз в день, но каждый
раз летающая значительно дальше и дольше, чем горихвостка.
Невероятно большой должна показаться также работа стрижа,
с визгом носящегося в воздухе долгий летний день, а иногда и ночью,
и летающего со скоростью никак не меньше 64 км в час; 900 км рас-
сматриваются как очень низкая оценка для налетанного стрижем
расстояния в течение целого дня. Таким образом с полным правом
энергия, развиваемая видом в период перелета, может рассматри-
ваться как «деятельность каждодневной жизни, внезапно направлен-
ная на определенную цель» (Nicholson, 1927). Однако далеко не
все виды в период летней жизни проявляют такую же подвижность,
как в период перелета (например большинство мелких зерноядных
птиц, перепел, кукушка и мн. др.), п организм должен подготовиться
к затрате большого количества энергии во время миграции. Основные
запасы при этом, повидпмому,создаются в виде жира, который накап-
ливается у перелетных птиц еще в большем количестве и с большей
быстротой, чем у птиц, зимующих в условиях значительного холода
(см. стр. 267). Накопление жира контролируется органами внутренней
секреции, предположительно гонадами, стимулирующими отложе-
ние жира в процессе их редукции (Groebbels, 1932), но в целом осу-
ществление этого контроля еще не выяснено. Соответственно быстрой
потребляемости жира в процессе мускульной работы, во время
остановок и отдыха происходит вновь быстрое его накопление,
так что птица в состоянии таким образом восстановить утраченные
запасы после особенно тяжелого перелета.
Дополнительными источниками энергии являются для организма
птицы отложения гликогена в печени, мускулатуре, в поясничном
утолщении спинного мозга и запасной белок. Однако запасы этих
веществ быстро резорбируются, и значительность их для организма
в период перелета не является достаточно ясной.
Стимулы перелета. Мы уже указывали, что «перелетный
инстинкт» с различной глубиной развит у птиц и что у типично пере-
летных птиц время его пробуждения не обнаруживает зависимости
от непосредственного влияния внешних, например климатических
факторов, влияние которых несомненно на кочевку зимующих у нас
птиц. Но даже рассмотрение поведения кочующих птиц заставляет
предполагать присутствие в их организме в определенное время осо-
бого расположенияк перелету. В то время как в последней
группе это расположение разрешается в передвижении популяция
под влиянием плохой погоды, недостатка корма и т. д., в группе
собственно перелетных, птиц перелет возникает уже в силу этого
внутреннего физиологического стимула. Интересным моментом явля-
ется также то, что это предрасположение может быть различно ра3'
вито не только у близких видов, но даже особей одного и того#6
вида (см. сказанное о миграциях чибиса на стр. 270), что указывает
на значительную изменчивость в некоторых условиях этого физио-
логического состояния. Все это заставляет рассматривать «перелет-
ный инстинкт» как весьма сложное и комплексное явление, в про-
явлении которого играют взаимодействующую роль как период^
282
ческая деятельность желез внутренней секреции, так и достаточно
развитый у птиц психический момент.
Существование такого внутреннего импульса хорошо доказывают
содержимые в неволе комнатные птицы, постоянно ко времени про-
лета особей того же вида па воле впадающие в особое состояние бес-
покойства: птицы «бьются» в клетке по ночам, несмотря на то, что
они могут быть совершенно изолированы от криков своих сотова-
рищей и внешних условий погоды вообще. Регистрацией подвиж-
ности птиц в это время при помощи самопишущих аппаратов удалось
установить некоторые любопытные особенности их поведения. У пере-
летных птиц могут быть различаемы два типа жизнедеятельности:
суточный и годовой. Последний распадается на две перелетных фазы,
разделяемых фазами покоя. Сопоставление течения перелетных фаз
у комнатных птиц с пролетом особей этого же вида на воле выявило
общее совпадение сроков «беспокойства» у подопытных и свободных
птпц в близлежащих районах, при этом беспокойство опытных птиц
заметно растягивалось п на тот срок, пока особи того же вида еще
продолжали перелет далее к северу (весной) или к югу (осенью),
несмотря на то, что в месте экспериментирования этих птиц уже
ле оказывалось. Этот факт очевидно указывает на то, что длитель-
ность «пролетной фазы» потенциально определяется не влиянием
местных условий и, повпдимому, не мускульной усталостью, а
определенным сроком времени, в период которого вероятно ра-
ботает гормональное влияние. На воле этот процесс конечно
осложняется психологической связью (тяготением птиц к опре-
деленным знакомым и родным местам), могущей оказывать реши-
тельное влияние на поведение птицы. Суточный тип жизнедея-
тельности, как показывают те же исследования, в период вне-
пролетнып заключается в активности днем и сне ночью, в период
же пролета проявляется у птиц активность и в ночные часы. Как
выяснилось, в последнем случае переход от дневной активности
к ночной сопровождается коротким перерывом активности, более
значительным у славок, при затемнении и перед рассветом, и совсем
кратким, уже в густых сумерках, у дроздов и соловьиных птиц.
Этот суточный ритм удерживается в том же состоянии даже при
содержании птицы при постоянном затемнении, что показывает,
что ритм может существовать и независимо от условий освещения.
Но вместе с тем оказалось, что после 13-дневного содержания в таких
условиях, при дальнейшем искусственном освещении клетки ночью
11 затемнении днем, ритм может быть передвинут и даже разбит на
более короткие сроки в 6 и 9, 6 и 6 часов. Последний результат
указывает на то, что ритм устанавливается под влиянием осве-
щения. Но из первого опыта видно, что после этого в организме
устанавливается определенная физиологическая периодичность, под-
держивающаяся и в отсутствии освещения. Этот опыт таким образом
показывает нам, какова зависимость и как может передвигаться
Цикл при изменении внешних условий. Несомненно, что сходным
образом изменяется и внутренне обусловленный годовой цикл
при отклоняющихся от нормы колебаниях сезонных внешних факто-
ров. В известных, каждый раз однако довольно узких, границах
293
погода может замедлить развертывание цикла плп ускорить его,
Передвигание и превращение циклов мы наблюдаем у видов, пере
селяемых (натурализуемых) из северного полушария в южное, как.
например, у черного дрозда, щегла, зеленушки и других видов
в Австралии, зяблика в южной Африке и т. д. Пока однако не уста-
новлено на опыте влияние этих перемещений на ход и развитие самою
перелета. Указанное выше изменение суточного цикла может еже-
годно осуществляться при перелете далеко на юге зимующих видов,
попадающих постепенно осенью из северной области с коротким
днем в южную область с длинным днем и относительно короткой
ночью. Влияние этого момента однако также не исследовано.
Совпадение периодической деятельности некоторых органов,
способных к внутренней секреции, с периодичностью перелета
вызвало предположение о наличии тесной зависимости между деятель-
ностью этих органов и развитием стремления к перелету. Наиболь-
шее значение в стимулировании перелета Роуэном (Rowan, 1926.
1927, 1929, 1931, 1932), Шильдк ахером (Schildmacher, 1933-
1934) и некоторыми другими авторами придавалось гормонам
гонад, а Вагнером (Wagner, 1930) — щитовидной железе.
Так как птица обычно наблюдается в состоянии перелета в период
развития и редукции гонад (см. стр. 131), а оседлость ее падает на пе-
рис д их наибольшего или наименьшего развития, предст. яло доказать,
что I зменение развития гонад может автоматически вызвать и появле-
ние стремления к перелету. Этого Роуэну удалось достичь полу-
чением преждевременного развития гонад, в результате постепенного
увеличения освещаемости американских вьюрков {J unco hiemalis L.).
содержимых зимой в комнатных условиях. Находившиеся в состоя-
нии редукции гонады птиц, в результате такого освещения, в тече-
ние 5—6 недель достигали максимальной величины, птица начи-
нала интенсивно петь и проявляла обычный весной половой интерес.
Контрольные птицы, не подвергавшиеся этим условиям, не обна-
руживали при этом никаких существенных изменений в своем пове-
дении. В то же время удалось искусственным уменьшением света
у птиц с начавшимся нормальным развитием гонад в марте и апреле
вызвать обратное развитие гонад вплоть до доведения их до зим-
него неактивного состояния. Когда эти подопытные птицы вместе
с контрольными были выпущены зимой на волю, то оказалось,
что в окрестностях остались держаться только птицы с неразви-
тыми гонадами или с полным развитием гонад, большинство же
прочих, с гонадами в состоянии интенсивного роста или сокращения,
исчезло, очевидно обнаружив наклонность к перелету. Это дало
Роуэну основание рассматривать перелет как явление, стимули-
руемое гормонами, выделяемыми половыми железами в период
своего развития или редукции, в свою очередь стоящего в зависи-
мости от количества часов активности, которыми может располагать
птица. Местом образования этих гормонов Роуэн считал интер-
стициальную ткань как в яичниках, так и семенниках, которая
по его мнению, наилучше развйта как раз в растущих или умень-
шающихся гонадах.
Шильдмахер пытался подтвердить влияние половых гормонов
284
на пролет иначе поставленными опытами. Он вспрыскивал в осен-
нее время в грудные мускулы самок обыкновенной горихвостки
(Phoenicurus phoenicurus L.) препарат женского полового гормона
прогинон, с целью ослабить или совсем загасить стремление к пере-
лету. Однако примененные слишком большие дозы этого гормона
не вызвали полного угасания перелета, как предполагается, вслед-
ствие того, что они переводили стремление к перелету, в обратное
по значению стремление, наблюдаемое в весеннее время. По мнению
автора, опыты все же доказывают, что осенний перелет есть след-
ствие уменьшения полового гормона, так как введение полового
гормона все же ослабляло перелет, причем введение гормона тре-
бовалось в большем количестве к концу осени, когда инстинкт
перелета развит особенно сильно.
Однако все эти опыты не лишены существенных возражений.
Биссоннетт (Biss nnette, 1931), повторивший опыты Роуэна над
скворцом (Sturnus vulgaris L.) с сходным результатом, не нашел
оснований для признания промежуточной ткани гонад, как органа,
пр дуцирующего специальный гормон, и вообще склонен рассматри-
вать совпадение в развитии гонад с проявлением перелетного инстинк-
та как обусловленное общим посторонним им стимулом. Такой
стимул, по предположению Штреземанна, может исходить от перед-
них долей гипофиза, которые есть основание рассматривать как
периодически действующую полигормональную железу. Шокертом
(Schockaert) было показано, что инъекция ее гормона резко уско-
ряет процесс сперматогенеза у уток. С биологической стороны
выводы Роуэна и Шильдмахера мало приемлемы вследствие нали-
чия большого количества противоречащих их положениям фактов.
Если в ряде случаев мы действительно наблюдаем соответствие
между развитием гонад и стадией перелета (например у Pericrocotus
roseus divaricatus Raffl., Emberiza spodocephala Pall., Emberiza
fiureola Pall, и др.), то с другой стороны известно, что поздно при-
летающие виды уже в начале перелета имеют сильно набухшие
гонады. У некоторых видов это набухание гонад происходит задолго
До начала перелета. Непонятным является также сильное развитие
инстинкта перелета у молодых птиц несмотря на стационарность
в развитии их гонад и вообще слабую развитость их. Также не
согласуется с этой гипотезой факт довольно однообразного процесса
созревания гонад при весьма различном выражении явлений про-
ста нередко даже у одного вида. Что свет является не основным
фактором, следует как из указанных выше наблюдений над комнат-
ными птицами, так и фактов перелетов птиц, зимующих в южном
полушарии. Созревание гонад у этих видов начинается в феврале —
апреле, когда здесь устанавливается или уже развивается осень,
с Уменьшающимся количеством дневных часов, а не увеличиваю-
щимся, как требует гипотеза Роуэна. Кроме того, не менее выра-
женным перелетом обладают многие ночные виды, для которых свет
следовательно должен был бы оказывать обратное влияние. Затем
значительная часть зимовок лежит в полосе тропиков, где отно-
пгения дневных часов к ночным постоянны круглый год и где свет
таким образом также не мог бы оказывать требуемого влияния.
285
Все эти соображения заставляют рассматривать стимуляцию пере-
лета как явление более сложное, чем это предполагается изло-
женными гипотезами.
Столь же недостаточной представляется п попытка связать пере-
лет с деятельностью щитовидной железы. Скармливание птицам,
находившимся в состоянии покоя, препарата этой железы в опытах
Вагнера вызывало будто бы у них типичное беспокойное состояние
перелета. Этот результат однако трудно связать, поскольку известна
способность этих желез усиливать обмен веществ, с наблюдающейся
склонностью перелетных птиц к значительному ожирению.
Как уже указывалось, из наблюдений перелетных птиц в ком-
натных условиях вытекает, что «состояние перелета» продолжается
определенное время, весьма сходное как в опытных условиях, так
и в естественных. Это дает основание полагать, что именно эта, обу-
словленная деятельностью невыясненных гормонов, срочность опре-
деляет время остановки птицы в области гнездовий и зимовок.У опыт-
ных, знакомых со своим пролетным путем и зимними и гнездовыми
квартирами особей это свойство дополняется активным выбором
знакомых мест и своего рода психической привязанностью к при-
вычной обстановке. Однако как в том, так п в другом случае внеш-
ние факторы могут влиять на конкретный результат оседания во
время перелета, причем действие этих факторов сильнее для моло-
дых и одиночно летящих птиц, чем для старых и летящих стаями.
Длительное ненастье и сильные противные ветры могут задержать
птиц, вследствие чего потребность осесть может появиться раньше,
чем птица достигнет пункта своего назначения. Пока неясно, на-
сколько старая птица может противодействовать этому влиянию,
но есть основание думать, что нередко наблюдаемое массовое осе-
дание птиц на гнездовье по южному краю ареала, где нормально
вид редок, как, например, гнездование годами шилохвости (DajiU
acuta L.) в южнойУкраине,или зимовки севернее нормальной области
зимовок могут объясняться как раз этими причинами. Столь же
возможен, при благопрятных ветрах и т. п., перелет птицами области
гнездования, особенно молодыми или заблудившимися. В таких
случаях птица может осесть за 100—500 км севернее своего ареала,
что и приводит к образованию случаев летних и весенних залетов,
иногда сопровождаемых гнездованием и весьма нередких во многих
фаунах. К таким случаям повидимому можно отнести например не-
однократные летние находки серого галликрекса {Gallicrex cine-
reus) на Дальнем Востоке.
Интересными опытами, поставленными с целью выяснения спо-
собности птицы потерять стремление к перелету при ее попадании
на место родины, являются эксперименты Штиммельмайера (Stun'
melmayer, 1932) с пеночкой-весничкой {Phylloscopus trochilus W
и обыкновенной горихвосткой {Phoenicurus phoenicurus L.). Птицы
были пойманы осенью на месте гнездовья и содержались всю зпзО
в клетке. Весной, в период пролета этих форм и интенсивного бес
покойства данных экземпляров, они были выпущены на месте гнез-
довья. Птицы остались на этом месте, что указывает, если в опьпе
не было ошибки, на большую роль «чувства родины» у старых осооеп.
.286
Крайне интересной, но маловыясненной особенностью является
очевидное расселение птиц под влиянием стимула перелета. Рас-
селение многих птиц происходит главным образом явно в напра-
влении пролета, причем происходит очень быстро, иногда приводя
к образованию парадоксально длинных пролетных путей. Так
северная пеночка (Phylloscopus borealis Blas.), зимующая на Малай-
ском архипелаге, расселялась через северную Сибирь и север Евро-
пейской части СССР до Скандинавии, весничка же, наоборот, из
Европейской части СССР расселялась опять-таки по северу до Ана-’
дыря, улетая зимовать в Африку. Сходно с ней расселялись
турухтан (Pavoncella pugnax L.), гаршнеп [Lymnocryptes minima
(Brunn.)], дупель [Capella media (Lath.)] и многие другие виды.
Есть основание предполагать, что эта волна настойчиво продви-
гающихся пионеров составляется молодыми особями с удлиненным
сроком действия стимула перелета, вследствие чего они стремятся
все дальше и дальше лететь по своему маршруту и оседают таким
образом на новых для вида местах. Происходит ли таким образом
передвижение всего ареала вместе с зимовками в одном направле-
нии (т. е. с продвиганием зимовок к северу), или расширение аре-
ала к северу сопровождается одновременно с расширением его
нк югу, сейчас трудно решить утвердительно. Вероятно это может
протекать различно у разных видов. Однако параллельное рас-
ширение и зимнего ареала и таким путем чрезвычайное удлинение
пролетных путей очень вероятны для многих птиц, в частности для
гнездующих на севере куликов и крачек, имеющих огромные про-
зетные пути, до 12000—17000 км длиной, и для некоторых хищ-
ников [например Erythropus amurensis (Radde)].
Известным подтверждением этого явления служит факт перелета
северных популяций вида на зимовку южнее той области, где зимуют
особи из более южных частей гнездового ареала. Наилучше извест-
ный случай такого рода имеется в подвидах Paserella iliaca (Merrem.)
(рис. 207), гнездовой п зимний ареалы которых располагаются по
горным цепям северо-западного побережья Соединенных Штатов
н Аляски, от острова Кадьяк до Калифорнии. Гнездящиеся в наи-
оолее южной части гнездового ареала Р. iliaca juliginosa Ridgw.
оседлы, гнездящиеся севернее ее — Р. il. townsendi (Audub.) зи-
муют сразу к югу от гнездового ареала вида, а гнездящиеся на даль-
нем севере расы— Р. il. unalaschensis (Gm.), insularis Ridgw., sinuosa
зимуют все вместе наиболее далеко к югу от области гнездования
вида.
Замечательным фактом является то, что расы, отстоящие от осед-
ной копуляции на равном расстоянии, зимуют в одной области,
а не в разной, как поступают расы, находящиеся в ином отноше-
нии к этой же оседлой популяции.
Пока необъясненным является оседание некоторых видов, напри-
мер Botaurus stellaris L, Coturnix communis L. и др., на гнездова-
ние в области их зимовок, что нормально не свойственно огромному
°ольшинству видов; неясны ни причины оседания, ни механизм
°°разования его.
Ориентация птиц во время пролета, способность их
287
находить правильный путь на гнездовье пли зимовку, становится
все более загадочной и сложной, по мере того как более детально
выясняется картина миграций птиц в целом. Имеющиеся наблю-
дения и опыт заставляют принимать как наличие зрительного ориен-
тирования, путем запечатления определенных зрительных впеча-
тлений (иначе говоря, памяти), так и наличие пока непонятного
и ближе не анализированного чувства направления, обеспечиваю-
Рпс. 207. Гнездовые и зимние ареалы западноамерпкан-
ских подвидов овсянки — Passerella iliaca Mcrrein.
Распространение па гнездовье очерчено сплошной ли-
нией; места зимовок заполнены пуантелыо. 1 — Passe-
rella iliaca unalaschensis (Gm.); 2 — P. i. insularis
Ridgw.; «3 — P. i. sinuosa; 4 — P.i. annedens Ridgw.;
’> — P. i. townsendi (And.); 6 — P.i. fuliginosa Ridgw.
По Swarth, из Lincoln.
щего правильность об-
щего направления пе-
релета.
Чувство на-
п р а в л е и и я. Необ-
ходимость принятия су-
ществования этого чув-
ства следует из фактов
физической невозмож-
ности многих видов
ориентироваться по ка-
ким - либо приметам
местности. В таком по-
ложении в особенности
находятся морские пти-
цы , преимущественно
ра зличные трубконосые.
Многие из этих форм
гнездятся на уединен-
ных океанических остро-
вах, кочуют же на ши-
роком пространстве оке-
анов, скопляясь зимой
иногда на огромном
расстоянии от своей
родины. Таков, напрп-
мер, серый буревест-
ник (Puff in us gravis
О. Reylli), гнездующий
на островах, лежащих
возле Новой Зеландии,
а зимующий в массе
у калифорнийских ое-
регов. Ни течение, ни цвет морской воды, ни тем более бере-
говая линия не могут служить этим птицам руководством для отыски-
вания затерянных на пространстве тихоокеанских вод островков,
служащих для гнездовья, так как это типичные птицы открытого
свободного моря. В сходных условиях несомненно оказываются
многие зимующие на море птицы, как то: чпетики, пингвины, гагары-
казарки, нырковые утки и крохали, держащиеся нередко вдалп
от берегов в течение значительной части года. Немало также и назем-
ных птиц, совершающих обширные перелеты через морские пре*
странства и с удивительной точностью попадающих на остров3?
288
где они гнездятся или зимуют. Таковы, например, стрижи Micro-
pus unicolor Jercl. и Micropus murinus Brehm, гнездящиеся на Ма-
дейре и зимующие в Африке, с пространствами открытого моря
на пути в 500 км, пли приводившиеся уже примеры с золотистой
ржанкой (Charadrius dominions чМй11.) и аляским кроншнепом (Nu-
menius tahitiensis Gm.), зимующими на островах Полинезии и
Гавайских, рассеянных друг от друга на сотни и тысячи кило-
метров и от места родины — Аляски и северо-восточной Азии — уда-
ленных не менее, чем на 3300 км. Сходный пример представляют
две перелетные кукушки Новой Зеландии, зимующие на Соломо-
новых островах [Chalcites lucidus (Gm.)] или на островах Полине-
зии к востоку до острова Товарищества и до Маркизовых островов
[Urodynamis taitiensis (Sparrm.)]. Мадагаскарские кукушки (Сиси-
lus poliocephalus Lath.), щурка (Merops superciliosus L.) и сизово-
ронка (Eurystomus glaucurus Mull.) перелетают через 200—300 км
Мозамбикский пролив и зимуют в восточной Африке. Наличия по-
добной же способности требуют и факты перелета молодых птиц
на зимовку без руководства старых, что свойственно видимо многим
птпцам. Наиболее изумительными примерами этого могут являться
подкладывающие яйца кукушки, молодые которых улетают с родины
обычно поодиночке и определенно без руководства как настоящими,
так п приемными родителями. Интересно, что и возвращение в гнез-
довую область происходит невидимому поодиночке, и при всем том
у кукушек наблюдаются не только подвиды, но и мелкие экологи-
ческие расы, различие между которыми выражается в приспо-
соблении цвета их яиц к цвету яиц тех птиц, которые обычно в дан?
ной местности являются для них приемными родителями. Что это же
свойство присуще и молодым, обычно странствующим в компании
со старыми, явствует из опытов Тинемана (Stresemann, 1933) с моло-
дыми белыми аистами (Ciconia ciconia L.). Несколько штук этих
птиц были пойманы, окольцованы и задержаны в неволе, пока все
старые аисты не покинули страну. После этого, с значительным
запозданием против их отлета, молодые птицы были выпущены
п, как впоследствии оказалось, птицы улетели на зимовку в общем
правильном направлении.
За наличие чувства направления говорит способность многих
морских птиц ориентироваться в густом тумане, лежащем на де-
сятки километров, а также странствование большинства мелких
дневных птиц ночью, в том числе и в темные безлунные ночи, когда
об ориентировке зрительным путем для большинства дневных птиц
конечно не может быть и речи.
Зрительное ориентирование. О том, что птицы
в состоянии также ориентироваться по памяти, развитой у них
весьма высоко, следует из наблюдений над почтовыми голубями.
Они явно руководствуются приметами местности, а при попада-
нии в новую местность стремятся ориентироваться, летая широ-
кими кругами и разыскивая знакомые уже черты района. Схватив
особенности своего положения, они продолжают лететь далее я пра-
вильном направлении уже с очевидной уверенностью. Память голу-
°ей позволяет им возвращаться к старой голубятне, при попадании
Орнитология—1209 «89
в окрестности ее даже после двухлетнего отсутствия. Эта способ-
ность ориентирования в разной степени конечно свойственна от-
дельным особям и может быть повышена дрессировкой и селекцией.
Некоторые наблюдения позволяют заключить, что нормально голуби
пытаются ориентироваться и в совершенно незнакомой местности,
правильный пролет через которую им позволяет чувство напра-
вления. Основным импульсом при этом является очевидно стремле-
ние не пропустить местоположение голубятни, вследствие чего
голуби прекращают полет или легко теряются в тумане и в темноте.
Положение голубя при этом удачно сравнивалось с положением
в тумане человека, который хотя мог хорошо знать местоположение
дома и иметь компас, но тем не менее не мог бы ориентироваться,
вследствие отсутствия указаний на местности и возможности опре-
делить, как далеко он зашел от своего дома.
О наличии хорошо развитого зрительного ориентирования среди
диких птиц говорит и та точность, с которой большинство старых
птиц возвращается с зимовки на место своего гнездовья, чтобы
занять здесь вновь ту самую территорию, на которой они ранее
гнездились. Эта способность была подтверждена последние годы
обильными результатами по кольцеванию. Так из 26 экземпляров
обыкновенной горихвостки (Pkoenicurus phoenicurus L.), окольцо-
ванной в Германии на месте гнездовья, в последующие годы 21 экзем-
пляр был найден на том же самом месте, несмотря на то, что птицам
приходилось зимовать по меньшей мере под северными широтами
в Африке (Судан и Абиссиния). Способность возвращения на старое
место гнездовья, надо полагать, является еще более сильно выражен-
ной у крупных птиц, более способных удерживать правильное напра-
вление в силу лучшей борьбы с действием ветра. Далеко летящие
птицы при этом обладают повидимому наиболее четкой ориенти-
ровкой и способностью находить места гнездовья. О том, что эта
острота ориентировки распространяется не только на места гнез-
довья, но и на места зимовок и вероятно и путь пролета, следует
из результатов кольцевания зимних птиц в Георгии (Соединенные
Штаты), приведенных выше (см. стр. 275).
Способность ориентировки можно проверить у птиц, пользуясь
ярко выраженным стремлением большинства из них в период гнез-
дования вернуться к гнезду, в случае удаления их в другое место.
Наиболее интересные опыты в этом отношении были произведены
над двумя морскими крачками — Anous stolidus L. и Sterna jus-
cata L., образующими массовые гнездовья на островах главным
образом тропического пояса. Взрослые крачки этих видов лови-
лись и кольцевались на маленьком островке Тортугас, у входа
в Мексиканский залив, и затем увозились пароходом в клетках
в направлении главным образом к порту Гальвестон, лежащему
в этом же заливе, и к мысу Гаттерас, расположенному на побе-
режье к северо-востоку ст Флориды.
К месту гнездования с первого направления вернулись крачки,
указанные в нижеследующей таблице.
Из двух крачек Sterna, выпущенных в море у мыса Гаттерас
(на расстоянии 1368 км от гнездовья), обе вернулись обратно через
290*
Расстояние от гнездовой колонии, на которой были выпущены птицы Число выпущен- ных птиц Из них вернулось па место гнездо- вания Выясненная продолжитель- ность поисков ими мест гнездовья
832 км 4 Anous 6 Sterna 3 Anous ! 5 Sterna । 3 суток 22,5 час. . ! 7 » 21 »
1050 » 2 Anous । 2 Anous ! 11 » 13 » 17 » ’ 2- »
1267 » 4 Anous 1 Anous 6 » 2 »
6 Sterna 2 Sterna 7 » 6 » 12 »
4 суток 23 часа. Последний опыт особенно интересен тем, что в район
выпуска эти крачки нормально не попадают, так как он лежит вне
границ ареала этих форм, и следовательно птицы должны были
ориентироваться здесь в совершенно незнакомых условиях. Из рас-
смотрения количества времени, затраченного этими крачками на воз-
вращение, можно полагать, что птицы стремились к цели, устана-
вливая в большинстве случаев сразу же относительно правильное
направление полета, но розыски гнездовий сопровождались поле-
тами и в боковых направлениях в целях ориентировки. В дальней-
шем этот же тип опытов был повторен с многими другими видами
птиц (Hirundo rustica L., Riparia riparia L., Sturnus vulgaris L.,
Sterna macriira Naum, и др.) с более или менее сходными резуль-
татами. При этих опытах следует ожидать, что при выпуске птиц
В районах их более частых кочевок и на маршруте обычных их пере-
летов возвращение будет происходить более быстро, с меньшим
процентом потерь, чем в иных случаях, но это не было еще про-
верено.
Существенную роль в выборе правильного направления и более
верной ориентировки играет сбивание большинства видов птиц
в период пролета в стаи. Связность этих стай изменчива от вида
к виду и сезона к сезону. Как правило, она наиболее значительна
именно на период интенсивного пролета. При образовании этих
°тай из старых и молодых птиц, опыт старых особей может способ-
ствовать более правильной ориентировке молодых птиц и прово-
дить первое знакомство их с пролетной дорогой более верным и
надежным способом. Кроме этого, из взаимодействия особей всей
стаи вообще происходит нивелирование уклонений инстинкта и
суммируется выбор пути, более правильного и более массово свой-
ственного виду. У наиболее склонных к стайности видов все от-
клонения в развитии стимула пролета, как в виде ускорения, так
и запаздывания, могут не развиваться до преждевременной реали-
зации в сравнении со всей массой особей, вследствие привязан-
ности этих экземпляров к стае. Таким образом перелет в стаях
может вести к наиболее правильному и жизненному для вида резуль-
тату, в то время как перелет отбившихся одиночек может давать
маибольшее число уклонений с пути и потери где-нибудь в дороге.
291
5. ПРИСПОСОБЛЕНИЯ, СВЯЗАННЫЕ С СТРЕМЛЕНИЕМ
К САМОСОХРАНЕНИЮ
Смерть является судьбою всякого живого существа, но несмотря
на это стремление жить может рассматриваться одним из основных
свойств организма, так как, за исключением патологических слу-
чаев, каждое животное всеми доступными ему средствами стре-
мится избежать ее приближения. Условия жизни однако играют
очень важную определяющую роль в длительности индивидуаль-
ной жизни и одни организмы ставят в условия постоянной опасности,
другие* наоборот, относительно освобождают от постоянной воз-
можности гибели, отсрочивая на более длинный период наступле-
ние смерти. В условиях ежеминутной возможной гибели живут
мелкие птицы, стоящие в начале трофических (пищевых) цепей,
окруженные не только враждебной общей средой, но имеющие п
массу живых врагов. Крупные хищные птицы в значительной мере
свободны от последних.
Длительность индивидуальной жизни обусловливается двумя
основными причинами: конституциональными особенностями, по-
тенциально благоприятствующими долголетию, и биоценотическпмп,
конкретно определяющими возрастной состав общей популяции
вида.
Продолжительность жизни. Данные о потенцио-
нальном долголетии птиц естественно весьма отрывочны и пока
не могут рисовать исчерпывающе возможные пределы жизни. В основ-
ном они составляются рекордами выживаемости птиц в клеточ-
ных условиях и случаями добычи окольцованных птиц через зна-
чительный промежуток после их закольцевания. Более достовер-
ные из этих данных о продолжительности жизни некоторых птиц
сгруппированы в следующем списке. Большинство данных отно-
сится к содержанию в неволе; цифра, сопровождаемая звездочкой,
получена из материалов по кольцеванию.
Страус (Struthio camelus L.)......................
Киви (Apteryx australis mantelli Baril.)..........
Пингвин (Spheniscus demersus L.J .................
Альбатрос (Diomedea exulans L.J...................
Чайка серебристая (Larus argentatus Brunn.) . . .
Ч .йка обыкновенная (Larus ridibundus* L.) . . . .
Олуша (Sula bassana L.J...........................
Пеликан (Pelecanus onocrotalus L.) ...............
Лебедь-шипун (Cygnus olor Gmel.j..................
Лебедь малый (Cygnus bewickii Jarrell^............
Гусь серый (Anser anser L., domesticus)...........
Канадская казарка (Branta canadensis la.) ....
Кряква (Anas plathyrhyncha L.)....................
Чирок трескунок (Anas querquedula L.).............
Шилохвость (Dafila acuta L.)......................
Фламинго (Phoenicopterus roseus Pall.^............
Белый аист (Ciconia ciconia L.J...................
Черный аист (Ciconia nigra L.)....................
Каравайка (Plegadis falcinellus L.)...............
Серая цапля (Ardea cinerea L.)....................
Рыжая цапля (Ardea purpurea L-7...................
Чибис (Vanellus oanellus L.)......................
40
20
12
46
44, 15*, 13*
21, 13*, 12*
23
52
21
24
65
29, 33
22, 10x/o*
41
13*
27
12, 24, 10*
10, 30
9, 10*
22, 11, 20. 60, 153A*
10, 23, 13*
13, ll3/4*, 1Г/2*
292
Веретенник чернохвостый (Limosa limosa L.J . . .
Кулик сорока (Haematopus ostralegus L.J........
Дрофа (Otis tarda L.J..........................
Журавль серый (Megalornis grus L.).............
Павлин (Pavo cristatus L.).....................
Курица (Gallus gallus L., domesticus)..........
Рябок белобрюхий (Pterocles alchata L.)........
Сизый голубь (Columba livia Lt.)...............
Королевский гриф (Sarcoramphus papa 1117 • • • •
Черный гриф [Aegypius monachus (L.)]...........
Бородач (Gypaetos barbatus L.J.................
Орлан-белохвост (Haliaetus albicilla L.).......
Сарыч большой (Buteo buteo L.).................
Филин (Bubo bubo Lt.)..........................
Неясыть серая (Strix aluco Lt.)................
Попугай (Ara macao L.).........................
Попугай серый (Psittacus erythacus L.).........
Ворон (Corvus corax L.J........................
Галка (Coloeus monedula Lt.) .... *............
Сорока (Pica pica L.)..........................
Иволга (Oriolus oriolus L.)....................
Скворец (Sturnus vulgaris L.)..................
Скворец розовый (Pastor roseus Pall J..........
Зеленушка (Chloris chloris Lt.)................
Коноплянка (Acanthis cannabina L.).............
Канарейка (Serinas canaria Lt.)................
Зяблик (Fringilla coelebs L.)..................
Воробей домашний (Passer domesticus Lt.).......
Желтая овсянка (Emberiza bruniceps Brand J . . .
Хохлатый жаворонок (Galerida cristata LJ . . . .
Белая трясогузка (Motacilla alba L.)...........
Большая синица (Parus major L.)................
Черный дрозд (Turdus merula L.)................
Певчий дрозд (Turdus philomelos Lt.)...........
Соловей (Luscinia megarhynchos Lt.)............
Деревенская ласточка (Hirundo rustica L.J . . . .
12*
30
27
42
более 19
30
17
22, 29, 28, 35
более 52
более 32
23
48
23, 18, 25, 15x/o*
34, 53. 68
27
64
50
69, 50
И
21
8*
7, 9, 10
8, И, 12
12
17, 20
17, 20
И
11—18, 14
13
13
более 10
9
11, 20, 10*
17, 83/4*
10, 9, 25
9
Менее достоверные данные указывают на способность попугаев до-
стигать 80—102 лет, стервятника (Neophron percnopterusLt.) 101 года,
белоголового сипа (Gyps fulous Gm.) 117 лет, канадской казарки
(Branta canadensis L.) больше 100 лет.
Таким образом в целом необходимо признать, что птицы могут
обладать значительной долговечностью и возраста в 50 60 лет
вполне могут достигать крупные птицы, а мелкие 20—35 лет. Как
правило, невидимому внутри родственной группы-крупные формы
вообще более долговечны, чем мелкие, но что возраст все же не
всецело пропорционален росту, видно из сопоставлений со страусом,
который, хотя весьма легко одомашняется и давно разводится в не-
воле, однако не живет дольше 40 лет, да и возраста в 20 22 года
достигает сравнительно редко. Большинство приведенных выше
Цифр представляет все же редкие явления, более редкие явления,
чем наличие столетнего человека. Это легко констатируется напри-
мер на курах, которые в огромной массе погибают уже ранее восьми-
летнего возраста и 24—30 лет достигают в исключительно редких
случаях. Возможно, что эта смертность связана также с повышен-
ной половой продуктивностью, стоящей, ^ак полагают, в обрат-
ном отношении к продолжительности жизни. У кур, при повышен-
293
ной продуктивности яиц, половая продукция почти полностью
угасает после 8 лет жизни. Но имеющиеся данные указывают, что
при нормальном укороченном половом цикле половая способность
сохраняется еще в очень значительном возрасте. Так известна пара
филинов, в возрасте 68 и 53 лет регулярно размножавшаяся и за
35 лет имевшая потомство в 93 птенца. Содержавшаяся в неволе
самка африканского орла [Aquila тарах (Тетш.)] еще в возрасте
более 30 лет положила яйцо; 24-летние садовая славка и жаворо-
нок пели до последнего дня жизни.
Из наблюдений на воле известно сообщение Стинструпа о мор-
ской чайке (Laras marinus L.), воспитанной из гнезда на Фарерских
островах и выпущенной на волю. Птица была настолько ручная,
что ела пищу из рук своего воспитателя. С течением времени она
спарилась с другой чайкой и гнездилась на близлежащих скалах,
сохраняя доверчивость, в противоположность своей подруге. Эта
птица еще гнездилась на воле в 63-летнем возрасте.
Биоценотические факторы, ограничивающие долголетие птицы,
представлены как неблагоприятными . климатическими факторами,
так и разнообразными врагами, из которых главнейшую роль играют
хищники. Смертность от этих причин наиболее значительна в раннем
возрасте, вследствие неопытности молодых птиц и их физической
слабости. Общие эмпирические данные в ^отношении мелких пев-
чих птиц указывают на смертность в периоде от откладки яйца до
вылета молодых приблизительно в количестве 60—64% (в среднем),
т. е. гнездо покидает только х/3 возможных птенцов; из этой послед-
ней цифры только около половины сохраняется к следующему
гнездовому периоду. Если число яиц, откладываемых за лето, равно 5(|
то к осени популяция вида увеличивается примерно на 5/3, а к сле-
дующей весне только на 5/6 в отношении к первоначальному насе-
лению. Если мы имеем вид, не испытывающий существенного при-
роста популяции и не проявляющий тенденцию к расселению и рас-
ширению или сужению ареала (что, теоретически говоря, никогда
не имеет места), то из этих расчетов можно вывести, что вероят-
ный средний возраст половозрелой особи этого вида равен 2,8 года.
В этих условиях количество выживающих молодых птиц в конеч-
ном счете должно быть равно погибающим половозрелым птицам.
Эти соображения могут быть полезны для определения степени
отхода особей в.изучаемых популяциях. В действительности однако
популяция никогда не остается на одном уровне, а постоянно
испытывает значительные колебания численности, в благоприятные
годы расселяясь и расширяя свой ареал, в неблагоприятные годы
разреживаясь и сокращая область своего обитания.
Общая правильность приведенных выше данных о ходе смерт-
ности указывается между прочим и данными кольцевания. Мэги
(Magee, 1928) в течение 6 лет прикармливал мексиканских чечевид
[Erithrina mexicana (Mull.)] и регулярно отлавливал и кольцевал
их. Результаты, полученные им за эти годы, приведены в ниже-
следующей таблице.
Если принять во внимание, что подкармливаемые птицы весьма
прочно держатся в районе кормежек и, как обычно для мелких
294
Год кольцева- ния Количество окольцованных птиц Количество птиц, пойманных повторно
1923 1924 | 1925 | 1926 1927 | 1928 | 1-е полугодие
1922 254 33 19 5 3 2 1
1923 1092 — 71 41 22 13 1
1924 1043 — — 87 44 26 8
1925 1510 — — — 182 62 27
1926 1084 — — — — 122 28
1927 1441 — — — — — 89
птиц, не стесняются многократно иттп в ловушку, можно с боль-
шой вероятностью принять, что большинство особей, не попадав-
шихся в ловушки вторично, погибли от тех или иных причин. Тогда
из приведенных данных следует, что средний возраст особи данной
популяции составляет около 2 лет 9 месяцев, что подтверждают
приводившиеся выше расчеты Буркитта (Burkitta, 1926). Из сос-
тава в 621 повторно отмеченных чечевиц лишь по одной особи имели
не менее 8 и 7 лет и четыре — не менее 6 лет (Stresemann, 1929).
Сходные результаты рисуются данными кольцевания и для
других видов. Так в отношении обыкновенной чайки {Larus ridi-
bundus L.) и белого аиста {Ciconia ciconia L.) полученные цифры
приведены в нижеследующей' таблице.
Названпн видов Коли- чество добы- тых околь- цован- ных птиц Количество добытых экземпляров
возраст в годах
1 2 3 4 5 6 7 8 9 10
larw.s ridibundus 140 1 105 1 20 | 8 1 1 ' 2 4 — — — 1 -
| Ciconia ciconia 94 54 10 10 7 6 1 2 1 2 1 4
Приспособления к самосохранению. Этот
жесткий естественный отбор у птиц, как и прочих живых существ,
выработал разнообразные приспособления к достижению возможно
большей выживаемости. Среди них приспособления к защите от
врагов отличаются рядом специфических особенностей: они напра-
влены к тому, чтобы или обмануть глаза врага, или, в редких слу-
чаях, его обонятельное чутье, или устрашить его неожиданными
Для него звуками и движениями, или отогнать его применением
тех или иных орудий защиты. Из указанной выше закономерности
° преобладающей гибели особей в раннем возрасте следует, что
наибольшего развития приспособления к самосохранению достигают
именно в этом возрасте, когда, кроме того, слабы и возможные актив-
ные средства защиты. В общем это положение действительно имеет
место, хотя тщательному исследованию оно еще недостаточно
подвергалось (стоит вспомнить широкое распространение охра-
нительной окраски яиц, преобладающей охранительной окраски
295
птенцов, смену защитного молодого наряда при возмужании птицы
на заметные и яркие наряды, свойственные большинству видов
этого класса).
Структурные и цветовые приспособления, связанные с самосо-
хранением, развиваются в теснейшей зависимости от психических
рефлексов организма, так что поведение вида оказывается координи-
рованным с той функцией, которую несет указанное выше приспособ-
ление. Во многих случаях это, связанное например с охранительной
окраской, поведение может затормаживать весьма распространен-
ный среди различных животных рефлекс бегства. Что и последний
развивается как приспособление к защите, видно из того, что он
совершенно отсутствовал у многих форм, прежде их столкновения
с человеком. Так, по описаниям старых путешественников, на многих
впервые открытых островах, не посещавшихся человеком, птицы про-
являли вначале полную безбоязненность и подпускали к себе человека
вплотную. Полную безбоязненность по отношению к человеку прояв-
ляют и сейчас многие обитатели островов Галапагосского архипелага,
мало посещаемого человеком. Как правило, однако рефлекс бегства
свойствен почти всем взрослым птицам (и некоторым молодым),
причем дистанция, на которой он возникает, тем больше, чем круп-
нее птица и чем она заметнее (белые птицы осторожнее темноокра-
шенных).
Почти каждая птица при нападении на нее активно защищается
доступными средствами, но специальные приспособления в этом
направлении развиты не столько против внешних врагов, сколько
против индивидуумов того же вида. Большая часть этих приспо-
соблений выработалась в процессе драки самцов или самок в период
тока или ухаживанья. Таковы, например, шпоры — костные и рого-
вые острые наросты на плюсне у куриных, на сгибе крыла у пела-
медей, шпорцевых чибисов и пастушков, которыми птицы в состоя-
нии остро поранить друг друга, или воротники из длинных перьев
у турухтанов, ослабляющие силу удара клювом, или удары крыль-
ями у гусей, лебедей и других крупных птиц. При личной защите
большинство птиц пользуется теми орудиями, которые употребля-
ются ими для добывания пищи: хищники и совы — острыми ког-
тями (и отчасти клювом), цапли и аисты — ударами клюва по воз-
можности в глаза нападающего, секретарь — ударами ног, боль-
шинство воробьиных — щипля клювом. Страус ударом ноги в со-
стоянии перебить голень человека. Во многих случаях эта защита
сопровождается устращакЛцими реакциями: гуси злобно шипят,
вертишейка шипит и странно ворочает вытянутой шеей и головой,
напоминая змею, синицы шипят, сидя в дупле, если их потрево-
жить во время насиживания яиц, буревестники и качурки изрыгают
и выплевывают на нападающего вонючую и маслянистую жидкость,
молодые хищники забрызгивают экскрементами, выпь и филин, под-
нимая перья, увеличивают размеры тела и принимают угрожающую
позу.
Более пассивные способы защиты сводятся главным образом
к охранительной окраске, широко свойственной как птенцам, яйцам
так и многим взрослым птицам. Как правило, наличие ее связан^
296
с рефлексом затаиванья, при котором заметившая опасность птица
или прижимается к субстрату и вытягивается вдоль него, или за-
стывает в обнаруженной позе, пока приближающаяся опасность
не вызовет реакции бегства. Иногда нормальная защитная поза
имитирует какой-нибудь предмет,
так, например, высиживающий яйца
козодой —Nyctibius griseus Gm. сидит
почти вертикально с вытянутой шеей,
вследствие чего он становится до
чрезвычайности схожим с основанием
обломанной ветви (рис. 208). Не ме-
нее оригинальную позу занимает при
этом и исполинский белоног [Podar-
gusstrigoides (Lath.)] (рис. 209).
Наиболее часто охранительная
окраска под цвет субстрата развива-
ется у тех взрослых птиц, которые,
или будучи деятельны ночью, день
проводят во сне, расположившись
где-нибудь в укромном уголке, или
принимают участие в длительном
высиживании яиц на сравнительно
открытом месте, или вследствие сла-
бости крыльев мало могут иметь
шансов на своевременное укрытие от
вра^а. Примерами таких форм могут
являться козодои, удивительно мало
заметные на фоне своей среды, вслед-
ствие сходства своей темной пестрой
окраски с нею. Так обыкновенный
Рис. 208. Насиживающий серый испо-
линский козодой — Nydibhis дг Isens
Gm.
Единственное яйцо кладки отложено
в углубление, образовавшееся па об-
ломанном стволе дерева вследствие
вышивания древесины. Насиживаю-
козодой {Caprimulgus europaeus L.)
и индийский козодой (Caprimulgiis
indicus jo taka Temm. et Schleg.) по-
хожи на цвет бурой коры или на бу-
рый лесной покров почвы, пустынный
щие птицы, а впоследствии также и
птенцы принимают вертикальное по-
ложение тела с вытянутой шеей, что
делает их чрезвычайно похожими на
обломанный сук; сходство увеличи-
вается охранительной буро-пестрой
окраской, причем птица почти полно-
стью закрывает глаза. Окружающая
обстановка—тропический лес (острова
Тринидад). По Muir, из Stresernann.
козодой {Caprimulgiis aegyptius Licht)
совершенно сливается своей окраской
с песчано-бурой окраской песка пу-
стыни, а африканский скалистый
козодой {Caprimulgus ruficollis
lemm.) —с окраской скалистой почвы
обитаемых им мест Центральной
Африки. Охранительно окрашены буро-рыжеватые самки видов
тетеревиных, самки и самцы вальдшнепа, бледнобурые авдотки,
многие жаворонки, из лесных плохо летающих птиц: пищухи, не-
которые дятлы, большинство пеночек. Охранительным свойством
обладают не только окраски, совпадающие по цвету с фоном, но и
Разбивающие, благодаря своей пестроте, облик птицы, затушевы-
вающие контур животного. Этим свойством очень многие птицы ин-
стинктивно пользуются. Известным охранительным свойством могут
297
обладать и сравнительно однообразно, но под тон среды окрашенные
птицы, как, например, чайки, в большинстве своем стально-серые
сверху и белые снизу. Однако в некоторых случаях охранительная
окраска может являться ложной, т. е. образующейся не под дей-
ствием естественного отбора. Таков, например, общий блеклый, пес-
чано-бурый тон окраски пустынных птиц, который вызывается физи-
ческим действием пустынного солнца (выцветанием).
Птицам в малой степени свойствен миметизм, и наиболее частые
случаи этого подражания направлены главным образом на имити-
рование хищных птиц. Такими примерами являются, например,
многие кукушки, обладающие в ряде форм [Cuculus canorus L,
Cuculus optatus Gould., Hiercoccyx jugax Horsf., Hilrococcyx varins
'(Vahl.) и др.] сходством с небольшими ястребами родов Accipiter,
Astur [особенно Л.
badius (Gm.)]%HBiiffl-
stur. Вероятной вы-
годой этого является
то, что благодаря
этому сходству они
чаще спугивают мел-
ких птиц, облетая
различные стации, и
обнаруживают этим
большее количество
гнезд, в дальнейшем
используемых . для
подкладки своих яиц.
Другие кукушки сб-
ладают удивитель-
ным сходством в окра-
ске с теми видами
птиц, которым они
подбрасывают свои
яйца. Так Surniculus
Рис. 209. Насияшвающий исполинский белопог —'Podargus
strigoides Lath.
Обладающая охранительной окраской птица принимает
позу, почти параллельную ветви дерева, застывая с вытя-
нутой шеей и силыю^прищуривая глаза. По Irby, из Strc-
semann.
lugubris (Horsf.) в совершенстве имитирует дронго — Dicrurus ater
(Негш.), а одна из Eudynamys подражает в окраске молодым птенцам
майны—Eulabes javanensis (Osbeck.), в гнездах которых они преимуще-
ственно воспитываются. Имеются однако примеры и других подража-
ний. На островах Малайского и Папуасского архипелагов виды рода
Мimeta (один из родов сем. иволг — Oriolidae) до деталей имитирую1
окраску и величину видов рода Philemon (из медососов—Meliphagii^
причем совпадение соответствующих пар видов разных родов вы-
ражается не только в сходной общей окраске и поведении, но
даже общей структуре наростов на клюве и характере отдельных
цветовых пятен. Почти каждый вид рода Philemon имеет соответ-
ствующего, сходно с ним окрашенного спутника из рода MimM
•связь обоих родов оказывается столь постоянной, что имелась воз-
можность, по открытии своеобразного нового вида того же рода
М imeta на острове Жилоло, предсказать наличие здесь же и вида
рода Philemon, что дальнейшие исследования блестяще подтвер'
298
дили. В данном случае ясно, что подражающей птицей является
Mmeta, так как это сравнительно слабая птица и обладает окраской,
уклоняющейся от нормальной в группе иволг; Philemon же яв-
ляется сильной, крикливой и задиристой птицей, так что 2—3 эк-
земпляра их легко сгоняют ворону или ястреба, пытающихся, на-
пример, присесть на дерево, занятое этим видом.
Наиболее совершенным случаем подражания в классе птиц счи-
тается подражание Tylas edwardi Hartl, виду Xenopirostris polleni
(Sches.). Последний вид относится к сорокопутам (сем. Laniidae) и
является сильной и крупноклювой птицей, с характерной для со-
рокопутов окраской, но несколько диморфичной: может быть окра-
шен снизу то в белый, то в бурый цвет. Tylas edwardi Hartl, явля-
ется видом и родом с несколько неопределенным систематическим
положением и считается наиболее близким к роду Hypsipetes^ отно-
симым то к дроздам (Turdidae)^ то к Pycnonotidae. Однако в противо-
положность Hypsipetes, имеющим незаметную однотонную окраску,
Tylas «перо в перо» сходна с Xenopirostris в обеих ее цветовых фа-
зах и отличается лишь несколько по величине и иной форме лап и
клюва. К сожалению как в этом, так и в других случаях экология
этих видов недостаточно известна, чтобы мы могли ясно представлять
пользу такого подражания.
* Интересным является далее случай подражания одного хищника
другому, сравнительно мало опасному для мелких птиц, которого
поэтому они мало боятся. Широко распространенный в Южной Аме-
рике ястреб (Accipiter pileatus Temm.) в области, тяготеющей в Рио-
Жацейро, принимает окраску местного насекомоядного вида Наг-
рад diodon Temm., безвредного для мелких птиц. Сходство здесь
распространяется как на взрослое, так и на юношеское перо, кото-
рые весьма различны. Подобный же характер, по своему зна-
чению, носит подражание хищного Spizaetus lanceolatus, обитающего
на о. Целебес, местному осоеду—Pernis celebensis Wai., или южно-
американской каракары — Jbycter americanus (Bodd.) местному
краксу из рода Ortalis.
Активная защита у птенцов выражена значительно слабееДсм.
указанные выше факты), а во время гнездовой жизни в большинстве
случаев вообще отсутствует; этот недостаток восполняется как охра-
нительной окраской, так и соответствующими реакциями родителей
птенцов. Из действий последних замечательна «притворяемость боль-
ными», свойственная взрослым многих форм как при беспомощных
птенцах, так и в последний период насиживания. При этом рефлекс
негства является в значительной мере заторможенным; птицы пры-
гают поблизости от врага с полураспущенными крыльями и жалоб-
ными криками и почти даются в руки. Однако стремление поймать
птицу приводит к большей активности бегства, что во многих слу-
чаях заканчивается отвлечением внимания врага и отведением его
°т гнезда или птенца на безопасное расстояние. Беспомощные птенцы
птенцовых птиц приобретают охранительный наряд большей частью
только ко времени покидания гнезда, выводковые же носят его с мо-
^онта появления из яйца. Обычно этот наряд является пестрым, те-
мной окраски и соответствует субстрату; таку болотных куликов он
299
бурых и коричневатых тонов, соответственно цвету почвы, у крачеь
же светлый, желтоватый или беловатый и соответствует цвету песка
или гальки. Только птицы хорошо защищенные, например обществен-
но-гнездящиеся, имеют окраску птенцов нейтральную или весьма за-
метную, играющую роль сигнала для родителей при отыскивании ими
своих птенцов. При защитной окраске птенцам часто свойственны
и защитные позы. Птенец Nyctibius (см. выше) также прямо сидш
в гнезде, как и взрослые птицы. Подросшие птенцы рыжей цапли
(Ardea purpurea L.), находясь в гнезде, вытягивают кверху рыжую
шею с поднятым кверху клювом и застывают в таком положении.
Широко известна аналогичная поза молодых и старых выпей. В слу-
"*"436 опасности, птенцы куликбв, дроф,
куриных прижимаются и вытягиваются
на земле. Окраска яиц птиц является
в большинстве случаев защитной и гар-
монирует в той или иной степени с окру-
жающим фоном. Первичный цвет яиц
был белый, который и удерживается
сейчас у тех видов, у которых кладка
Спрятана от взоров в дупла или норки
(дятлы, зимородки, сизоворонки, щуркп
и др.). Если птицы устраивают гне^а
более или менее открыто, то выраба-
тываются соответствующие защитные
реакции у родителей: кладка насижи-
вается начиная с первого отложенного
яйца, и поэтому нормально гнездо не
оставляется с открытой кладкой. Так
делают голуби, колибри, совы, цапли
и поганки. Иногда до инкубации и во
время перерывов ее яйца покрываются
защитно окрашенным пухом, выщипан-
ным из груди (гусеобразные), листьями
(некоторые куриные) или водяными ра-
стениями, вытащенными из воды по со-
Рис. 210. Насиживающая желто-
брюхая мухоловка — Мгсгоеса fla-
vigaster Gould. Гнездо настолько
мало, что сидящая птица полно-
стью прикрывает его своим распу-
шенным оперением. Гнездо содер-
жит только одно яйцо. По Harvey,
ш из Stresemann.
седству с гнездом, как делают поганки. В случае вторичного пере-
хода к гнездованию в защищенных местах защитная окраска обычно
ослабляется или теряется. Это хорошо видно на окраске яиц мно-
гих пеночек, гнездящихся в гнездах-домиках, на горных вьюрках
(Leucosticte, Montifringilla), на топориках (Lunda и Fratercula) из
группы чистиков, гнездящихся в норах. Ослабленная окраска яиц
свойственна большинству синиц, гнездящихся в дуплах.
Примерами защитной окраски является окраска яиц куликов,
чаек, тетеревиных, козодоя, жаворонков, варакушки, соловьев и
мн. др.
Действие естественного отбора, направленное на выработку за-
щитных приспособлений, захватывает и весь процесс гнездостроения
и выкормки птенцов. Как правило, строя гнездо, птица держится
очень скрытно, и в период инкубации самец держится от него в не-
котором отдалении и нередко мало заметен. Дятел, выдалбливаю-
300
щпй дупло для гнезда, уносит стружки от своей работы, дабы не
выдать его местонахождение. Помет птенцов мелкие птицы обычно
подхватывают в момент дефекации и уносят далеко о! гнезда. Само
гнездо у активно мало защищенных видов устраивается по возмож-
ности малозаметным (например гнездо Microeca из сем. Мuscicapidae,
рис. 210), помещаясь под защитой ветвей, листьев, устраиваясь
в виде домика или тщательно маскируясь, как, например, гнезда
зяблика и долгохвостой*синицы, украшенные кусочками лишайника,
взятого с того же дерева. Очень часто гнездо устраивается висячим
на конце длинной ветви, иногда, как, например, у ремеза, подвеши-
ваясь над водой и на столь тонких ветвях, что враг не в состоянии
до него добраться. Вход в гнездо при этом затрудняется сужением
входного отверстия до незначительной величины.
Особ,ое значение эти меры защиты имеют в жарких тропиках,
где рептилии, главным образом, ящерицы и змеи, являются одними
из очень распространенных грабителей гнезд. На следующей таб-
лице изображены, по данным Пека (Peck, 1910), процентные отно-
шения видов птиц, имеющих различные типы гнезд в разных усло-
виях; с одной стороны, в штате Иова в Северной Америке, на
40—45° с. ш., и с другой стороны, в области Британского Гон-
дураса, в тропиках центральной Америки.
Из таблицы ясно видно, насколько повышается процент особо
защищенных гнезд в тропиках, в сравнении с умеренными шпротами
1 । Районы Опирающиеся нижней частью, сверху откры- тые гнезда Гнезда п дуп- лах Открытые сверху, подве- шенные гнезда Гнезда с боко- вым входом
Штат Иова. 65 21 8 6
Британский Гондурас . . . 43 32 10 15
В связи с наличием этих же врагов, в тропиках очень распро-
странено гнездование мелких птиц в колючих кустах или употре-
бление для построек колючих веточек, через которые многим
фагам сравнительно трудно добраться до гнезда.
Защитное значение имеет также стремление многих птиц к стай-
1ости или общественному гнездованию. Последнее свойство создает-
я вообще вследствие различных причин: недостаточности удобных
1ест для гнездования, как, например, для гнездящихся на скалах
истиков и некоторых чаек, или прямой потребности в защите своих
оплеменников, вследствие той выгоды, которую дает виду раз-
евающаяся в гнездовых колониях взаимопомощь (пингвины). При
впадении какого-нибудь хищника на птицу или гнездо в колонии,
округ него быстро собирается целая стая птиц, начинающих устра-
!ающе нападать на него и громко кричать. Прямыми наблюдениями
становлено, что каждая птица отражает опасность для своего
лизлежащего гнезда, но усилия многих птиц сливаются в общую
оллективную защиту, обычно весьма эффективную. Так на севере
олониальные чайки легко отгоняют даже песца и крупных хищных
301
бургомистров. Отдельные беззащитные виды очень часто ищут сосед-
ства такой колонии, которая защитой своих гнездовий защищает и их
гнезда. Так на наших болотах мелкая ушастая поганка (Podi-
ceps nigricollis Brehm.) гнездится почти исключительно маленькими
колониями среди крупных колоний чаек — Lotus i idibundus L. и minu-
tus (Pall.), а в тундрах севера известна предпочитаемость мелких ка-
зарок [Branta ruficollis Pall., Anser albijrcms (Scop.)] гнездиться по-
близости от гнезд сапсана или белой совы. Возможно, что этими же
потребностями объясняется наблюдаемая в различных странах склон-
ность воробьев гнездиться в нишах стенок крупных гнезд хищни-
ков. В тропических странах очень многие мелкие птицы устраи-
вают гнезда в непосредственном соседстве с постройками ос, владельцы
которых не обращают внимания на соседних квартирантов, но, за-
щищая свой мед и личинок, способны отогнать и весьма крупных
врагов, против которых гнездящиеся в соседстве птицы без своих
жалящих союзников почти беззащитны. Высокое развитие обще-
ственных инстинктов у пингвинов приводит к появлению обще-
колониальной заботы о птенцах, к усыновлению покинутых птенцов
вследствие гибели их родителей, к известному разделению колонии
на птиц, кормящих птенцов, и птиц, охраняющих их от нападения
хищных морских птиц.
Склонность к общественности проявляет большинство птиц и вне
периода гнездования, причем у некоторых видов, например грачей,
скворцов, куликов и т. д., стаи составляются преимущественно из
особей того же вида, в других же случаях являются смешанными
из представителей различных групп. Так, например, в наших ле-
сах постоянно встречаются стайки синичек, совместно с большими
малым дятлом, пищухой, поползнем и другими видами. В тропиках
известны смешанные стаи мелких воробьиных, кочующих в поисках
пищи, и главным образом походных муравьев и др. Виды, состав-
ляющие стаи, на разный лад, разными способами разыскивают нуж-
ную им пищу, и выгода, извлекаемая ими в таком общении, заклю-
чается в предупреждении о неожиданной опасности. Появление
хищника сейчас же вызывает у заметивших его особей тревожный
крик и реакцию бегства и моментально отражается на поведении
прочих птиц. Вследствие этого, тревожный крик, как сигнал об
опасности, свойствен почуй всем видам птичьей фауны (в том числен
нестайным видам), и значение его понимается не только особямя
данного вида, но и другими видами, даже других классов. Так тре-
вожный крик Buphaga ofricana L. в Африке заставляет сейчас же
насторожиться пасущихся буйволов, а испуганный крик воробьев
на гумне заставляет и курицу укрыться от ястреба в безопасное место.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
Современное географическое распространение птиц сложилось
под влиянием ряда факторов, среди которых такие факторы, как
экологические особенности этой группы (способность птиц к даль-
ним перелетам и др. черты, см. стр. 253—287), пройденный путь
эволюции и относительная древность птиц (с юры, многие современ-
ные роды — с олигоцена и миоцена, см. стр. 333—348), а также поло-
жение первичного очага происхождения отдельных видов — должны
были играть и играют особенно крупную .роль. Помимо этого важ-
ное значение для расселения птиц имела история отдельных ча-
стей суши и из более современных факторов — влияние человека,
преобразующего своей деятельностью природу. Таким образом,
понимание современного ареала вида и распространения класса
птиц в целом возможно только при учете очень широкого комплекса
факторов и их взаимодействия между собой. Это недостижимо в
кратком обзоре, и задачей настоящей главы является лишь указание
на некоторые общие явления в распространении птиц и обзор фауны
но отдельным районам земного шара.
По отношению к населяемой ими стации 1 отдельные виды птиц
могут являться относительно стено- или эвристационными формами.
Примерами первых могут являться связанные с еловым лесом: чиж
(Spinus spinus L.), кедровка (Nucifraga caryocathactes L.), королек
(Regulus regulus L.), синица-московка [Periparus ater (L.)] и мн. др.;
среди птиц стеностационные виды более многочисленны в группе
зерноядных и в группе лесных птиц, например, более часты в отно-
шении к хвойным типам леса. Примерами эвристационных форм
могут являться: ворона (Corous cornix L.), пустельга (Tinun-
^lus tinunculus L.), кряква (Anas plathyrhyncha L.) и др. Обычно
всей своей организацией, поведением и периодикой жизненных про-
цессов каждый вид приспособлен к обитаемой им стации и к от-
дельным условиям (факторам), которые в совокупности образуют
сРеду для существования животного. В отношении видов стенеста-
дионных наличие соответствующей стации часто является основным
Моментом, определяющим характер их распространения (или распре-
деления) в той или иной стране. Однако для других видов ближай-
шее изучение показывает, что они еще не заняли полностью со-
ответствующих им стаций, а находятся в процессе заселения их;
1 Йод стацией понимается выделяющийся какими-либо признаками участок
остности, относительно однородный на всей своей территории.
303.
в этих случаях ареал вида в данной стране находится очевидно в тес-
ной зависимости от положения в ней первичной области расселения
вида, пли очага его возникновения (первичного обособления). Лишь
с течением времени вид полностью заселит подходящие для него
условия местности, и исторический фактор образования ареала
отступит на второй план перед влиянием современных факторов в рас-
селении животного. В дальнейшем вид может оказаться в неблаго-
приятных условиях, популяция его начнет сокращаться п может
разбиться на ряд маленьких островков, уцелевших в наиболее благо-
приятных для вида участках прежнего ареала. На этой стадии раз-
вития ареала вида также может отсутствовать закономерная связь
распространения животного с территорией распространения под-
ходящей для него стации.
Наблюдение показывает, что в комплексе факторов, слагающих
стацию, наибольшее значение для птиц имеет растительность, со-
здающая обычно защитную обстановку, условия для гнездования п
кормовые ресурсы, если не в прямом значении, так в косвенном,
питая животных, идущих в пищу насекомоядным и хищным птицам.
Роль каждого из этих факторов в отдельности может иметь решающее
значение в ограничении ареала вида; значение последнего фактора
подтверждают большинство специализированных по питанию видов,
как, например, многие виды колибри, клесты (Loxia), арчевый дубо-
нос (Mycerobas carneipes Hodgs) и мн. др., своим ареалом тесно свя-
занные с ареалами определенных видов растений; примерами зна-
чения второго фактора могут являться, например, формы ремезов
(Remiza), связанные в своем распространении с ивой, тополем плп
камышом, из опушения семян которых они строят гнездо, гоголь
(Bucephala clangula L.), гнездящийся только в дуплах, беркут
(Aquila chrysaetus L.), требующий для гнезда или наличие дерева
или уступа скалы, и мн. др. Однако приспособляемость вида к из-
менениям типичной для него стации иногда может быть очень зна-
чительной, примером чему может служить, например, обыкновен-
ный деятел, в сущности лесная птица, но в качестве особой формы -
Dryobates major leucopterus Sev., проникающий на гнездовье в средне-
азиатские песчаные пустыни и гнездящийся здесь в низеньких дерев-
цах саксаула, а за неимением их — даже в отвесах песчаных бар'
ханов.
Замечательной чертой в распространении многих птиц является
сравнительно небольшая область гнездования при обширности топ
территории, на которой данные виды могут быть встречены на кочев-
ках, перелете или на зимовках. Это объясняется, повидимому, осо-.
бой требовательностью птиц в период вывода молодых, вследствие,
повышенной чувствительности молодых птиц к влияниям климати-
ческих факторов и нуждаемости в определенных видах корма. Та-
ким образом в ограничении ареала видов существенное значение (
в известные периоды их жизни имеют климатические факторы. 11
границы области гнездования могут совпадать с ходом изотерм?
изогиет и другими климатическими показателями (Серебровскпй’
1926). Так, например, выяснено, что северная граница гнездового
ареала Troglodytes aedon aedon Vieill. определяется низкими ноч-
304
пымн температурами, а южная — высокими дневными температурами
(Kendeigli, 1934). Однако «существенное значение в ограничении
указанных ареалов имеет также своеобразный «инстинкт родины» —
иначе говоря, стремление гнездиться там же, где данный экземпляр
вырос или гнездился раньше (что констатировано для многих видов
птиц): это явление заставляет рассматривать этот класс как отно-
сительно консервативный но способности к расселению и служит
нередко единственным объяснением наличия крошечных ареалов
у таких хорошо летающих видов птиц, какими являются, например,
стрижи п ласточки. Несмотря на наличие этого стремления у многих
видов животных, в известных рамках необходимо признать способ-
ность особей к активному выбору для себя соответствующей среды
п отчасти района для гнездования. Так, например, розовый скворец
свободно и почти ежегодно внутри своего ареала меняет места
своего гнездования, выбирая участки, сильно зараженные кубышками
саранчевых, служащих ему для выкармливания птенцов; арчевый
дубонос в западном Тянь-шане годами гнездится то в субальпийской,
то ниже — в древесно-кустарниковой зоне; многие утки, в зависи-
мости от степени местных разливов, легко меняют места гнездовья
и в некоторых случаях задерживаются на гнездовье южнее нормаль-
ной области гнездования.
Основным стимулом к расселению животных является перена-
селение данной территории и, так сказать, «давление», которое
в результате этого оказывают более сильные и верные месту родины
пли кочевки индивиды в отношении более слабых. Возможно однако,
вто па этой основе у многих видов уже выработался механизм, за-
ранее (до собственно перенаселения) вызывающий процесс расселе-
ния. Установлено как кольцеванием, так и простым наблюдением,
что молодым птицам свойственны в большей мере, чем старым, по-
иски новых мест обитания, широкие откочевки и оседание в- сто-
роне от родины. Расселяющимися, занимающими впервые новую
Для вида территорию, являются по преимуществу молодые птицы.
Кроме того, для птиц может быть констатирована специфическая спо-
собность к расселению ио пути пролета, подмеченная у многих видов
и выражающаяся в наиболее энергичном расселении по линии про-
лета, с каждым годом все далее и далее от былой границы ареала.
Вероятной причиной этого является стимулирование пролета деятель-
ностью гонад, вызывающих состояние пролета все более длительное
время, вследствие чего вид расширяет область своего распространения
все далее по линии пролета. Таким образом идет расселение, например
к западу, овсянки дубровника (Emberiza aureola Pall.), чечевицы {Ery-
erythrina Pall.). северной пеночки (Phylloscopus borealis Blas.) и
Др., а к востоку пеночки-веснички {Phylloscopus trochilus L.), глупой
ржанки {Charadrius morinellus L.), турухтана [Pavoncella pugnax(Ij.)\
11 Других видов, достигающих теперь из первоначальной и вероятно
недавней стадии на меридиане Енисея уже берегов Берингова моря.
Замечательной чертой такого расселения является во многих слу-
чаях отсутствие или редкость расселения в сторону, «в бока», от
основного направления пролета, несмотря на наличие соответствую-
Щих стаций и других необходимых виду условий.
20 Of] НТО.к I•ИН- 1209
305
Быстрота расселения варпирует в зависимости от вида и геогра-
фических условий. В среднем, очевидно в связи со способностью
к перелетам, птицы расселяются быстрее, чем другие группы по-
звоночных животных, как об этом можно судить по составу фауны
океанических островов. Многие мелкие и слабые птички, как, на-
пример, вьюрки, белоглазки (Zosterops}, болотные курочки (Porzm]
п др. были в состоянии
Рис. 211. Расселение скворца — Sturnus vulgaris L.
в Северной Америке, начиная с 1896 по 1926 г.
Поселение возникло' от 160 птиц, выпущенных в
1890—1891г. в Центральном парке IIыо-Иорка. Гра-
ницы гнездового ареала к определенным годам отме-
чены сплошными или пунктирными линиями. Вне
ареала 192G г. лежат изолированные находки птиц,
преимущественно в зимнее время. По Cooke, из
Stresemann.
прочно, а иногда и неодно-
кратно, заселять острова,
удаленные за 2000—4000км
от ближайшего континен-
та. Весьма быстрым ока-
зался процесс расселения
у многих видов евразиат-
ских птиц г щегла [С ardue-
lis carduelis (L.)], домаш-
него воробья {Passer do-
mesticus L.), зяблика (Frin-
gilla coelebs L.), скворца
(Sturnus vulgaris L.) и дру-
гих видов, завезенных ко-
лонистами из Европы в Но-
вую Зеландию, Австралию,
южную Африку и Соеди-
ненные Штаты. Домашний
воробей за восемьдесят лет
сумел широко распростра-
ниться по Северной Аме-
рике и стать одним пз
серьезнейших врагов сель-
ского хозяйства. Столь же
быстрое распространение
обнаружил за последние
три десятилетия обыкно-
венный скворец, ввезен-
ный в конце прошлого сто-
летия в район Нью-Йорка
(рис. 211). Из наблюдений
следует, что расселение
вида идет успешнее в том
случае, когда проникно-
вение вида в новую область происходит более значительными
численно группами и от времени до времени повторяется, так
сказать, поддерживая [новую колонию. Малочисленные и эпизо-
дические залеты обычно неэффективны, за исключением происхо-
дящих в наиболее благоприятных условиях. Расселение, как
правило, идет в пределах привычной для вида среды и нередк0
участками, создающими более благоприятные условия. Так, напри;
мер, дубровник (Emberiza aureola Pall.), гнездящийся в Восточной
Сибири в довольно широком комплексе стаций,- но наиболее изооп-
306
лующий в .луговой пойме, исторически недавно расселился по всей
Западной Сибири п большей части Европейской СССР, причем
встречается здесь только в луговой пойме; есть основа-
ния считать, что расселение по другим стациям, вне основной, опти-
мальной для вида, представляет в большинстве случаев вторичный
этап. Однако известны примеры расселения с переходом и в другие
среды, в случае их общего соответствия экологическим требованиям
вида.
С процессом расселения обычно стоит в связи и процесс эволю-
ции вида. Дивергенция признаков протекает наиболее успешно
прп географической изолированности отдельных частей общей попу-
ляции или, во всяком случае, меньшем общении ее частей между
собой. Как правило, видообразование протекает быстрее у оседлых
видов и в популяциях, разделенных какими-либо преградами. По-
следними являются, смотря по экологии вида, более значительные
морские преграды, высокие горы и вообще резко отличающиеся и
обширные по площади стации. В тропическом влажном климате,
благоприятствующем развитию оседлости среди птиц, существен-
ной преградой являются даже реки, как, например, Амазонка и
ее притоки, и различные высотные зоны. Чем более вид проявляет
стремление жить в глубине леса, тем более эффективными для него
являются подобные преграды. Длительное существование вида
в разных высотных зонах влечет обычно за собой расщепление его
на разные высотные расы, с дальнейшей эволюцией до значения ви-
дов и более крупных систематических единиц. В Андах Колумбии
(Южная Америка) этот процесс может быть прослежен невидимому
на разных этапах (Chapman, 1917). Так Troglodytes musculus Naum..
kgelaius icterocephalus L. и Zenaida rujicauda Bp. свойственны уме-
ренной, субтропической п тропической высотным зонам, будучи пред-
ставлены в них каждый по крайней мере двумя близкими подвидами.
Другие же горные виды [Penelope montagni (Bp.), Andigena hypoglau-
cus Gould и др.] близки соответственным видам предгорной зоны
[Penelope cristata L., Andigena nigrirostris Waterh.), но отличаются
резко, не образуя морфологических переходов. Этот же процесс обна-
ружен и на других горных системах. Для южной Венецуэлы, шапри-
мер, установлено происхождение местной овсянки (Emberizoides duidae
Chapm.), свойственной субтропической зоне горного хребта Дуида,
°Т близкого вида — Emberizoides sphenurus herbicola (Vieill.). рас-
пространенного в окружающей тропической зоне (Chapman, 1931).
Следами этого процесса можно считать приуроченность близких
видов к двум или трем различным высотным зонам в горных систе-
мах нашего Союза. Так, например, па южном Сихотэ-Алине, в зоне
*пироколиственного леса, живут болотная гаичка (Poecile palustris
С) и полосатая мухоловка (Hemichelidon griseisticta Pall.), заме-
чаемые в вышележащей зоне хвойной тайги близкими видами: черно-
головой гаичкой (Poecile atricapillus L.) и сибирской мухоловкой
vleinicheldon sibirica Pall.). На Алтае наблюдается еще большая
Дифференциация: Poecile palustris L. обитает в предгорной полосе
п некоторых горных ущельях, Р. atricapillus L. широко распростра-
нена выше в горах и еще выше распространена Poecile cinctc
*
397
Pall.. занитющая полосу кедровой тайгп, окаймляющую верхнюю
лесную зону. Однако в данном случае, в отличие от вышеуказанных
промеров, взятых пз тропической зоны, происхождение близких
Друг другу видов вряд ли мыслимо протекавшим непосредственно
друг от друга и в данном месте, так как все они уже хорошо обособ-
лены и несомненно прошли весьма длительный путь расселения;
однако общий характер пх эволюции вероятно проходил сход-
ным путем. Влпзкую.картину в распределении родственных видов и
подвидов мы часто наблюдаем п в соседних друг другу шпротных
б по гео гр а фп че скп х зр и а х.
Как правило, чем чаще сменяются поколения, тем больвю имеют
шансов проявиться мутационные изменения, а чем меньше популя-
ция, тем быстрее они распространяются среди данного населения.
В связи с этим, на островах, а также по хребтам и вершинам гор,
в случае оседлости видов или отсутствия интенсивного пролета
через данную территорию, процесс видообразования обычно идет
быстрее, чем в относительно монотонных материковых условиях.
Существенную роль играют также внешние раздражители, особенно
при новом воздействии их, как, например, при расселении вида в
новые для пего, климатически отличающиеся районы, ставящие вид
в иные условия. Для птиц имеет большое значение также сегрега-
ция популяций не только в гнездовой период, но и в период кочевки
л пролета: образование новых пролетных путей, как правило, ве-
дет за собой систематическое отчлепение той популяции, которая
с этим новым пролетным путем связана.
Имеются отдельные попытки определить возраст подвидовых
отличий некоторых видов, исходя из сопоставления заселения ви-
дов каких-либо географических ландшафтов с вероятным возра-
стом их. Таким образом норвежский подвид варакушки (Cyunosylvui
svecica gaetkei Kleinsch.) устанавливается существующим не больше
<8000—10 000 лет; подвид хохлатого жаворонка Нильской дельты
(Galerida cristala nigricans Brehm.) ие больше 5000—И ООО лет (Mo-
reau, 1932); шотландский вид белой куропатки (Lago/nis scoticux
Lath.) — не больше 30 000 .лет и т. п. Однако эти цифры все же
весьма гадательны и могут быть оспариваемы. Скорость дифферен-
циации вида и обособления систематических рас очевидно весьма
изменчива, как изменчивы экология, наследственная конституция п
окружающие физические условия.
Хотя расселение большинства видов птиц, исключая некоторых
хищных и подкладывающих япца в чужие гнезда, может пттп без
тесной связи одного вида с другим, все же, в силу общей зависи-
мости от окружающей, главным образом, растительной среды, рас-
селение отдельных видов находится обычно в известной координа-
ции с целым комплексом видов. Эти комплексы видов являются
закономерными компонентами биоценозов и разделяют нередко пк
судьбу, как то: перемещения, деградацию п исчезновение пли, на-
оборот, дальнейшую дифференциацию. Пернатые компоненты ряда
биоценозов, свойственных какой-либо стране, рассматриваются как
орнитологическая фауна данной местности и в зависимости от ланд-
шафта и соотношения с окружающими фаунами несут характерные
308
особенности и признаки. Состав, расселение и иные закономерности
в сложении фаун п являются главным образом предметом изучения
оринтогеографни.
Из общих закономерностей в распределении птиц необходимо
отметить следующие: количество видов резко увеличивается в на-
правлении от полярных зон к экватору и также от холодных и жар-
ких пустынь к сырому тропическому лесу. Однако в обратной про-
порции очень часто . наблюдается изменение в численности вида;
в тропическом лесу виды гораздо малочисленное особями, чем в ле-
сах умеренной зоны. Количество видов зависит также от относитель-
ной длительности существования и спокойного развития данной
фауны. При значительности этих сроков видообразование сильно
повышает количество выделившихся видов в пределах рода и ро-
дов в пределах семейств с одновременным процессом глубокой диф-
ференциации экологических особенностей видов и более всесторон-
него использования окружающих условий. Наиболее резко это
выражено в фауне Южной Америки, наиболее богатой птицами
вообще. Здесь в пределах горной страны Эквадор, по площади
равной Испании, насчитывается более 1200 гпездятцпхся видов,
а в пределах всей Южной Америки насчитывается больше 2500 ви-
дов (не считая многочисленных подвидов), тогда как в гораздо бо-
лее значительной по площади территории Палеарктики насчитывают
только 1100 видов. Параллельно с этим, для этих крупных древних
фаун характерно также сохранение и древних эндемиков, изолиро-
ванных видов, обособленных родов, семейств и других групп, не имею-
щих близкого родства или с родством в столь же обособленных ви-
дах, весьма удаленных по распространению. Еще в большем! степей и
зтпм свойством сохранения древних эндемов обладают острова,
особенно удаленные от берегов, пли океанические, на которых они
уцелевают б.чагодаря отсутствию конкуренции.
Стремление охватить все разнообразие животного мира, и птиц
в Частности, и сгруппировать многочисленные фауны в легко, обо-
зреваемые группы давно выдвинуло идею зоогеографического райо-
нирования фаун. В. основном это районирование как раз проводи-
лось на птицах, и наиболёе общепринятым является деление Склэ-
тера. предложенное в 60-х годах и липть (‘легка измененное Уол-
лесом (1876). Обычным принципом такого районирования фаун яв-
ляется выделение географических территорий, характеризующихся
распространением здесь эндемичных форм и, не принимая во внима-
ние изменения фауны, могущих быть в высотных поясах или. ио раз-
личным элементам ландшафта. Построенная по такому принципу
зоогеографическая карта основных делений суши земного шара при-
ведена на стр. 310. Однако построение зоогеографических карт воз-
можно в из несколько иных принципов, позволяющих строгое и по-
следовательное подчинение от мельчайших фаунистических подраз-
делений. до высших. За первичную ориитогеогрцфическую группи-
ровку может быть принята группировка птиц, связанных со стацией.
1100 в пределах однородной стации распределение птиц является
ввпболее равномерным. Нанесение на карту таких группировок дает
|,аА[ наноолее верную картину распределения птиц по данной террп-
309
Рис. 212. Зоогеографическая карта мира.
I. Пеогея; No 1—чилийская подобл.; Л’е 2— бразильская подобл.; Ne 3 — амазонская подобл.; Хе 4— £убапдская
подобл.; Хе .5— центральноамериканская подобл.; А’е в — антильская подобл. II. Нотогея: N 1 — австралийская подобл.;
X 2—папуасская подобл.; X 3— новозеландская подобл. III. Арктогеп: восточная обл. (О); эфиопская обл. (А);
голарктическая обл. (П). Восточная область: 01 — переднеппдпйская подобл.; 02 — малайская подобл.
Эфиопская область: А1 — западноафриканская; Л2 — посточпо-тояшоафрпканскал; АЗ — мадагаскарская.
Голарктическая область: палсарктика (Ир), псарктпка (Нп) и арктика (Па). Палеарктика: Пр1 — евро-
пейско-рибпрснап подобл.; Пр2—средпземноморспо-псррднсаппатскап подобл.; ТТрЗ — црптральпоазпатская подобл.;
it тАоетпчп(>пзипт('1;;1г1 подобл. ТТ оярктпка: JTni — сепороамсрпка некая подобл.; ТГп2 — сопорекап подоб.т. ТТо Newton,
Магепч, Cbnpln тт др.
тории. При учете распространения на больших территориях эти
группировки приходится систематизировать и обобщать. Вследствие
мозаичности сочетания разнообразных стаций в пределах той пли
иной местности, в одно целое приходится объединять и различные
стадионные группировки птиц с разной фауной, но с одним пре-
обладающим территориально типом их. Достаточно резкое различие
в этих преобладающих типах дает возможность различать зоогеогра-
фические участки. Сходным образом, по более общим признакам,
участки группируются в округа, округа в провинции, провинции
в области, с установлением между ними иногда дополнительных,
переходных единиц.
Установление таких территориальных группировок видов все же
весьма усложняется проникновением разнородных элементов друг
в друга и высотной зональностью. Более высокие пояса гор в южных
шпротах северного полушария несут более северные группировки
видов (по своим родственным связям), или оставшиеся здесь от не-
когда располагавшихся здесь северных биоценозов, отошедших потом
в области равнины к северу, пли проникшие сюда активным рас-
селением и постепенно сложившиеся в особые группы форм. Также
п на равнине: проникающие, например, в пустыню реки ведут за
собой чуждые пустыне биоценозы, а более или менее обширные
участки степи среди лесов могут вести за собой и степных животных,
чуждых лесу. Вследствие этого, любые зоогеографические деления
обычно отграничены нерезко, а своими элементами взаимно прони-
кают друг в друга, образуя внутри одной области как бы островки
фауны другой, и местами создают подлинное смешение разных фауни-
стических элементов. Смешанность черт часто увеличивается значи-
тельным активным приспособлением видов к чуждым для них усло-
виям существования.
Такая смешанность черт естественно наиболее выражена по
краям какой-либо территории, причем разные края ее будут но-
сить разный смешанный характер. Так фауна сахаро-азиатских
пустынь на юго-западных своих окраинах воспринимает много
эфиопских форм, на юго-восточных — индийских, на северо-западе —
средиземноморских, на востоке — китайских; однако основной кон-
тингент форм, специально приспособленных к типичным пустынным и
степным условиям и широко распространенный, остается тем же самым.
При районировании фауны птиц руководствуются только гнез-
дящимися птицами, так как экологические требования птиц в этот
период наиболее определенны, и они ведут оседлый образ жизни.
Щнако и другие категории птиц имеют известное значение для
характеристики местности. Вследствие большей оседлости, в тро-
пиках лучше оказываются характеризованными птицами не только
стации, но и переходные зоогеографические территории, по при-
чине длительного приспособления к ним специальных форм. Так
лесостепь в северном полушарии состоит в сущности из лесных пли
степных форм, не имея, строго говоря, своих видов; но переходная
полоса от саванн к тропическому лесу в Африке имеет свои формы
п изобилии, несмотря на общее ландшафтное сходство с северной
лесостепью.
3! L
Накопляющиеся данные все более указывают на известный парал-
лелизм в распределении птиц с распределением высшей раститель-
ности. Зто может быть объяснено несомненной зависимостью птиц
от типов растительности, а также, во многих” случаях, легкостью
расселения при содействии ветра. Близкие сроки эволюции и рас-
селения по земному шару высших растении и класса птиц также
создают предпосылку общего сходства в распространении этих орга-
низмов. Однако растительность гораздо детальнее отражает гео-
графические судьбы .ландшафтов, ибо она во много раз богаче, разно-
образнее специализирована, более прямым образом зависит от свойств
климата и почвы тт вообще быстрее видоизменяется. Так флора
южной оконечности Африки выделяется но причине богатства энде-
миками и оригинальности в особое* капе,кое царство, что не может
быть подтверждено птицами, хотя и имеющими сходные ио пропс-’
хождению элементы (например род Promerops)\ причиной этого
является то, что канская флора приурочена к очень ограниченной
территории и экологически однообразна. Отсутствие горных поясов
и лесной растительности в совокупности с указанной особенностью
флоры не дает для развития и снецнал пзацшг орнитофауны еоответ-
ствующей среды.
При выделении и обозначении’ орннтогеографнческпх единиц
приходится учитывать площадь относительно однородной фауны,
ее фаунистическую ограниченность, степень древности родственных
связей и характер взаимоотношений г другими, странами. Чисто
формальное статистическое сравнение фаун но численности эндеми-
ков неприменимо, так как обилие видов определяется в значитель-
ной степени экологической обстановкой: пустыня столь же древ-
няя, как тропический лес, всегда будет населена гораздо более
малым числом видов, вследствие ее. однообразия и скудности кор-
мовых ресурсов. Также умеренные п полярные с,трапы обладают
по экологическим условиям более бедной в видовом отношении
фауной. Также и полной равноценности зоогеографических единиц
в территориальном отношении невозможно добиться, вследствие раз-
нообразия стран п их животного мира.
Нижеследующий обзор зоогеографических областей по необхо-
димости краток и поверхностен, В системе зоогеографических деле-
ний приняты во внимание взгляды Ньютона, Чапина. Склетера.
Мерриэма, Уоллеса, Северцева, Сушкина и др., но внесены некото-
рые видоизменения там, где они представлялись необходимыми.
Области северного полушария рассматриваются более подробно,
чем южного, причем по отношению к ним осуществлены указанные
выше принципы группировки фаун, повлекшие за собой установле-
ние областей и подобластей в новых границах, схематично отра-
женных на зоогеографической карте Евразии (рис. 213).
Весь наземный животный мир земного шара может быть разде-
лен на три фаунистических царства: арктогейское. иеогейское и
нотогейское, характеризуемые значительной: изолированностью в
своем развитии, разными преобладающими систематическими груи'
нами и разным морфологическим уровнем эволюции, которого в
массе достигло пернатое население на той или другой территории.
3J2
Наиболее обширным царством является арктогепское, включающее
в себя и всю территорию СССР.
Арктогейское царство охватывает всю Евразию, Африку, Ии-
домалайскпй архипелаг и Северную Америку. Все эти территории
в течение третичного и четвертичного периодов находились в об-
щении между собой и настолько глубокой зависимости, что фауна
отдельных областей сложилась под существенным влиянием сосед-
них стран. Крупные климатические изменения этих стран явились
причиной глубоких перемещений фауны, обширных вымираний древ-
313
mix групп п мощного расцвета новых, более высоко морфологически
стоящих. Все основные, более высоко приспособленные и жизне-
способные группы возникли повидпмому на этой территории п
уже впоследствии, по установлении связей с южными, изолирован-
ными материками, стали проникать в южном направлении и теснпть
более древнюю фауну.
В пределах арктогейского царства можно различать области
аркто-альпийскую, голарктическую лесную, область североамери-
канских степей и пустынь, область сахаро-азиатских степей и пус-
тынь, эфиопскую и восточную или‘индийскую.
А р к т о - а л ь и и и с к а я область разделяется на две под-
области: арктическую и альпийскую.
Арктическая подобласть является циркумполярной,
охватывающей бассейн Ледовитого океана, арктические острова и
тундры Евразии и Северной Америки. Южная граница ее проходит по
краю северного редколесья; этот последний образует переходную к
лесной полосе, так называемую лесотундру, в которой область пред-
ставлена языками или вкраплениями тундры. По горным системам,
на более или менее значительной высоте, островки арктической под-
области далеко проникают к югу, переходя постепенно в альпий-
скую подобласть и далее к югу вливаясь в альпийские зоны только
отдельными своими элементами.
Основными стациями подобласти являются разные типы тундр,
арктических лугов, озер, каменистые россыпи, морские побережья
и море, покрытое большую часть года если не ледовым покровом,
то плавающим льдом.
Фауна подобласти сравнительно небогата числом видов, но нередко
изумительно богата численностью особей. В связи с длительной
холодной зимой все птицы являются перелетными или хотя бы ко-
чующими; гнездование многих форм является однако эндемичным
для области. Наиболее характерные виды и роды: 'Colymbus adamsi
Gray, 1 С, immer Brunn., C, stellatus Pontop.; Fulmarus; Plotus;
Uria loinvia L.; Cepphus; Fratercula; Lunda; Cerorhinca; Phalens;
Aethia; Lams glaucus Brunn., L. leucopterus VieiL; Хеша; Rhodo-
stethia; Pagophila; Sier cor arias, ряд видов; Arenaria; Squatarola;
Charadrius dominions Mull., Ch. pluvialis L.; Eudromias morinellus
(L.); Limosa lapponica, (L.); Crocethia; Limonites minuta (Leisl.).
L. ntficollis (Pall.); Heteropygia; Crymophilus; Chen; Branla, ряд
видов; Clangula; Arctonetta; Somateria; Erionetta; Heniconetta; P№
lacrocorax, ряд видов; Lagopus mutus Mont.; Archibuteo (частью):
Hierojalco, ряд видов; Nyctea; Plectrophenax; Calcarius lapponicus^\
An thus cervinus Pall, и нек. др.
Гнездящаяся фауна птиц проникает местами далеко к северу,
так на земле Гранта до 82—83° с. ш. еще гнездятся из гусиных
Branta bernicla L., Somateria spectabilis (L.) и Clangula hyemalis (14
из чаек — Stercorarius parasiticus L. и Sterna paradisea Brunn.; пз
1 Русские названия видов н указания па систематическое положение
а также родов if семейств, следует искать в систематической части (стр. 349—«н 1»
в целях краткости эти сведения опущены из настоящей главы»
314 /
куликов — Phalaropus julicarius (L.), Calidris canulus L., Crocethia
alba (Pall.). Arenaria inter pres (L.); из куриных — Lagopus iniitus
rupestris Gm.; из воробьиных — Pledrophenax nivalis L.; из сов —
Nyclea scandiaca L. На земле Франца Иосифа, под 8Г с. ш. и север-
нее‘найдены еще па гнездовье: Branta bernicla. glaucogaster Brehm.,
Fuhnarus glacialis L., Lams hyperboreus Gm., Pagophila eburnea
(Phipps.), Stercorarius parasiticus (L.), Plotus alle (L.). Cepphus grylle
mandti (Licht.) и вероятно ряд других видов..
Кочующие глупыши (Ful/nams glacialis L.) наблюдались в море
еще под 85° с. ш.
Генетически фауна арктической подобласти ведет свое начало из
следующих источников: наиболее древнего элемента, связанного
с морскими берегами и побережьями внутренних водоемов, и более
молодых — выходцев из лесной зоны, альпийской зоны гор и не-
когда лежавших значительно дальше к северу степных ландшафтов.
Значительная часть этих выходцев с юга приспособилась к арктиче-
ской обстановке в период ледниковой эпохи. Наибольшее богатство
область обнаруживает на‘Чукотском полуострове, на Аляске и Бе-
ринговом море, повидимому являющихся и территорией наиболее
древнего развития ее биоценозов.
А л ь п и й с к а я и о д о б л а с т ь охватывает фауну птиц,
сложившуюся в условиях альпийских зон горных систем северного
полушария. В северных широтах эта фауна близка к фауне аркти-
ческой подобласти, но к югу она становится более специфичной,
обнаруживая вместе с тем родственные связи с фауной, населяю-
щей нижние зоны гор. Преобладают виды, по образу жизнп свя-
занные со скалами, альпийскими лугами, снежниками и карлико-
выми кустарниками. Наиболее характерные роды и впды: Anthus
spinoletta (L.), A. roseatus Bly th.; Eremophila alpestris (L.) (частью);
Leucosticte (ряд видов); M ontif ringilia (ряд (видов); Acanthis flavi-
rostris L. (частью); Prunella (ряд видов); Phoenicurus erythrogastra
(Guld.); Tichodroma nniraria L.; Pyrrhocorax graculus L., P. pyrrho-
corax L.; Tetraogallus; Ylhaginis; Lerwa; Lagopus mutus L.(частью);
Eudroinias morinelliis (L.).n др.
Генетически фауна подобласти преимущественно сложилась из
первоначально лесных и кустарных горных видов, колонизовавших
бесснежные высоты и в дальнейшем распространившихся в этой
среде по другим горным системам. Однако систематическая обособ-
ленность многих форм настолько резка, что прямые исходные формы
Для ряда альпийских видов не могут быть указаны. Сходным пу-
тем проникли в альпийскую зону также виды из-степей и болот-
ных группировок. Более поздним является проникновение в эту
подобласть собственно арктических элементов.
Голарктическая лесная область охватывает весь
северный и умеренный лесной пояс северного полушария. На се-
вере она связана переходной зоной лесотундры с арктической обла-
стью, проникая в тундру разреженными лесами и кустарными за-
рослями; на юге область отделяется от пустынно-степных областей
или переходной зоной лесостепи, пли островками располагается по
г°рам, частью же постепенно переходит в тропические леса эква-
315
термальных областей. Сообразно своему огромному протяжению
и значительному охвату северного полушария по широте — это
самая крупная биогеографическая область; она разделяется па
шять подобластей: подобласть фауны северных хвойных лесов (ши-
рокой зоной от Скандинавского полуострова, через Сибирь", до
Камчатки и Анадыря и в Северной Америке — Аляска, Капа да и
часть Соединенных Штатов); подобласть фауны европейских широко-
лиственных лесов (большая часть Европы, часть берегов Средизем-
ного моря, Малой Азии, Кавказа я др.), подобласть фауны восточно-
азиатских широколиственных лесов (главным образом от Даурии,
Амура и Сахалина на севере, через Японию и лесной Китай до при-
мерно водораздела к югу от Янг-тзе-цзяна) и подобласть фауны
широколиственных и южных хвойных лесов Соединенных П1татов
с двумя резко характеризованными и разорванными провинциями:
провинция фауны тихоокеанской лесной области Северной Аме-
рики (от Ситки к югу, захватывая леса к западу от Скалистых гир
и к югу до Калифорнии) и провинция фауны аллеганских широко-
лиственных лесов Соединенных Штатов (ют области Великих озер
на север до тропической Флориды на тоге и прерий на западе).
В целом область характеризуется большим количеством энде-
мичных родов, но почти лишена эндемичных семейств (преимуще-
ственно этой области свойственны семейства Bombycillidae, частью
Prunellidae и Tetraonidae).
Фауна области относительно небогата и явно обеднена под влия-
нием обширного плейстоценового оледенения, развивавшегося на
значительной части этой территории и вызвавшего вымирание ра-
нее более богатой фауны частью тропического и субтропического
характера и значительные перемещения сохранившихся ее элемен-
тов. Вследствие этих же причин отдельные составляющие ее фауны
легко вступали в связь с соседними областями и заимствовали от
них значительное количество форм.
Л о д о б л а с т ь ф а у п ы с сворных х в о й и ы х лесов
на юге переходит в биоценозы широколиственных лесов разного
состава пли почти непосредственно граничит со степью. В южных
своих частях естественно она имеет более разнообразный видонов
состав, чем в северных. Южнее общей своей границы отдельные
языки и островки этой подобласти приурочены к южнее лежащим
горным системам: Пиренеям, Альпам, Тянь-тпаню, Гималаям, горам
Сычуани п Ганьсу западного Китая, Сперра-Невада и Скалистым
горам п Аллеганам Северной Америки. Основными стациями под-
области являются: разные типы хвойного леса, участки мелколист-
венных лесов, травянистые и моховые лесные болота, берега рек п
водоемов, возникшие под влиянием человека поля и др.
Большинство птиц подобласти перелетны, другие кочуют в пре-
делах ее же границ. Состав фауны сравнительно небогат. Из энде-
мичных и наиболее характерных родов и видов следует упомянуть.
Colymbus arcticus L., С\ pacijicus Lawr.; Podic&ps auritus L., P. grisei-
gena (Bodd.); Glottis nebularius (Gunn.); Pseudo to tanus guttifer Nordnt.;
Heteractitis; Melanonyx; Bucephala; Lagopus lagopus (L.) (частью).
Tetrao; Falcipennis; Canachites; Dendrogapus; Scotiaptex: Surnu^
316
Gluacidium passerinum L.. Gl'. gnome; Archibuteo lagopus L.; Picoides:
Rombycilla; Poecile Ulrica pillus L.. P. cinctus Pall.; Periparus ater L.;
Regains (частью); An thus gustavi Svvinh.; Acanthis linaria L.; Frin-
gilla niontif ringilia L.; Pyrrhula pyrrhula L.; Loxia\ Pinicola, Crac-
tes\ Nucijraga, Spinas (частью); Zonotrichia^ Junco hyeinalis, Carpv-
dncus. ряд видов; Troglodytes (частью): Tardus eunomus (Pall.),
T. naumanni Temin.. T. obscuras Gm.. 7’. musicus L.; Ilylocichla.
(diciae (Baird.) и др.
Подобласть разделяется па два отдела: палеарктический в Ста-
ром Свете п иеарктическпй в Новом Свете с тяготеющими к ним и
частью изолированными высокогорными поясами, проникающими
к югу до Пиренеев. Альп. Карпат. Кавказа. Тянь-шаня. западного
Китая и Гималаев в Евразии п гор Сперра-Невада, Скалистых и
Аллеганских в Америке.
По своему происхождению фауна подобласти составляется глав- '
ным образом из древних фаунистических элементов, населявших
когда-то хвойные леса северных прптихоокеанскпх стран, элементов
хвойных зон горных систем северной Азии и выходцев из зоны ши-
роколиственных лесов. Влияние плейстоценового оледенения ска-
залось .между прочим ив значительных перемещениях этих биоцено-
зов. в связи с которыми хвойная зона северного происхождения
(ставшая сборной по составу) распространилась на весь север
Евразии и север Америки, образовав колонии и на горах, .лежащих
южнее. Наиболее древними и богатыми но фаунистическому составу
районами области являются Канадская тайга, Охотский округ и
хвойный пояс Гималаев и западного Китая.
И о д о б л а с т ь фа у н ы е в р о н е й с к и х ш п р о к о-
л н с т в е и н ы х .лесов граничит с северной лесной подобластью
довольно широкой переходной полосой, тянущейся от Финского
залива примерно но линии, проходящей через Ярославль, Пермь,
Номень. Новосибирск. Конфигурация остальных границ еще более
рожна. вследствие проникновения в глубь массивов этихЛесов уча-
стков степей (Венгрия, Украина), образования между степной
областью и лесами широкой переходной полосы лесостепи, сохра-
нонпя на горах высотных зон северной подобласти и островного
сохранения самих широколиственных лесов, местами в этих же и
более южных горных районах, к востоку до Тянь-шаня и Гималаев,
11 мощной изменяющей деятельности человека.
Подобласть образуется двумя провинциями разного историче-
ского значения: провинцией собственно широколиственных лесов и
Провинцией горного Средиземноморья. До ледникового периода
°ба эти района хотя н взаимодействовали друг с другом, были хо-
роню обособлены и резко фаунпстическп характеризованы, но пере-
мещения, вымирание if перестройка биоценозов под влиянием плей-
стоценового оледенения были здесь столь велики, что обе провинции
в настоящее время глубоко взаимодействуют между собой своими’
авт°хтонными элементами и территориально во многих районах
е тРУдом разграничиваемы. Исторически и фаунально к провинции
11п,роколиствеиных лесов принадлежат преимущественно биоценозы
°)ковых, дубовых и смешанных широколиственных лесов, сырых
317
лугов п травянистых болот и т. п., а к средиземноморской — био-
ценозы скал, вечнозеленых кустарников, горных сосновых лесов и
ксерофи^гных склонов. Горные системы Средиземного моря в значи-
тельной степени участвовали в образовании альпийской фауны,
впоследствии частью широко расселившейся, частью пополненной
элементами других биогеографических районов. Основной водо-
раздел между обеими провинциями проходит по Пиренеям, южным
склонам Альп, отрезает к югу Балканский полуостров, Закав-
казье, Копет-даг, западный Тянь-шань и южные склоны западных
Гималаев. Однако элементы и биоценозы широколиственных лесов
проникают глубоко в эти системы, а отдельные средиземноморские
виды расселились далеко к северу, до Англии и Дании, и к востоку
в Центральную Азию.
Характерными, частью эндемичными формами подобласти яв-
ляются: Picus viridis L.; Dendrocoptes medius (L.); Alaectoris; Phusid-
nus, ряд форм; Columba palumbus L., C. oenasL., C. fusca Pall.-, Slrep-
topelia turturL., Aegypius monachus (L.); Gyps ftilous (Habl.); Gypat-
tus barbatus (L.); Pernis apioorus L.; Merula; Monticola, saxatilis L.:
Prunella modular is L.; Lanius minor Gmel., L. collurio L., L. senator
L.; Lophophanes cristatus L.; Remiz, ряд видов; Sitta tephronobi
Sharpe, S. canadensis kruperi Poel.; Sylvia hortensis Gm., 5. myslaceti
Men., S. deserticola Tristr.; Chersophilus duponti (Vieill.); Diplootocus
moussieri (Olph.-Gal.); Acrocephalus strepera Vieil.; Miliaria; Eniberizo
stew ar ti Blyth., E. cirlus L., E. cinerea Strickl., E. hortulana L., E. buc-
hanani Blyth.; Serinus citrinella L.; Rhodopechys; Oraegithus: Cardm-
Us; Luscinia megarhynchos Brehm и L. philomela L.; Oenanthe pleschanhi
(Lep.), 0. hispanica. (L.) и др.; Irania; Scotocerca; Cyanopica cyami
coocki Bp. и мн. др.
Генетически фауна подобласти слагается главным образом из
древних автохронных элементов горного Средиземноморья, остатков
древней фауны третичных лесов Европы подтропического типа,
и пришлых элементов восточноазпатского происхождения, про-,
никших сюда вместе с упомянутой выше инвазией с востока. Обед-
нение фауны за ледниковый период здесь было наиболее сильным
из всех широколиственных подобластей северного полушария.
В период оледенения фауна сохранялась в немногих районах —
убежищах, из которых впоследствии она вновь распространилась
по большой территории, сохраняя однако местами следы своей преж-
ней приуроченности. Более северные и адаптивные формы широко-
лиственной провинции уцелели еще по южной таежной полосе, со-
храняя ненарушенными связи с восточноазиатской фауной, не-
смотря на преобладающе таежный характер местности. Формы среди-
земноморской провинции приняли большое участие в заселении
пустынь и степей, особенно в их тугайной (‘приводной) раститель-
ности.
Подобласть фауны восточноазпатских ши-
роколиственных лесов граничит с северной тайгой бо-
лее резко, чем подобная область в Европе, в меньшей степени об-
разуя переходную смешанную полосу. В пределах Манч?курии и
Приамурья эта подобласть занимает южные и нижние склоны гор,
318
долины рек н более или менее значительные равнины, которые не
многочисленны вследствие горного характера местности. Наиболее
адаптивные форпосты этой фауны проникают к северу по долинам
рек и южным склонам сопок. В направлении к подступающей с за-
пада степи границы более стушевываются и отчасти носят лесостен-
ной характер, при котором островки лесной растительности несут
фауну рассматриваемой подобласти, а степь — фауну пустынно-степ-
ной области. На территории западного Китая граница обусловлена
стыком с горными островками северной лесной подобласти. В напра-
влении к югу фауна все более обогащается подтропическими и тропи-
ческими элементами и с значительной постепенностью сливается
с тропической фауной южного Китая.
Основными стациями подобласти являются широколиственные,
смешанные и более однообразные дубовые леса, высокотравные
луга, травянистые болота, водоемы, морские каменистые побережья
п поля; влияние человека местами (восточный Китай, значительная
часть Кореи и Японии) очень велико и совершенно уничтожило
былую местную фауну.
Фауна подобласти довольно богата и обнаруживает значитель-
ное родство с индомалайскими формами. Перелет у большинства
птиц развит слабо, и зимовки обычно лежат близко.
Наиболее характерными и частью эндемичными формами яв-
ляются: Ciconia boyciana Swinh.; Dendronessci; Ery thro pus aniuren-
sis Radcle; Butastur indicus (Gmel.); Ketupa doeriesi; Accipiter virga-
lus Temm. & Laug.; Grus japonensis Mull., G. vipio Pall.; Aegia litis
placidusGray; Hirundapus caudacula Lath.; Dryobates japonicus (Seeb.);
hngipicus nanus (Vig.), J. kizuki Temm.; Corvus leoailanti Less.;
Emberiza tristrami Swinh., E. jankowskii Tacz., E. elegans Temm.,
E. chrysophrys Pall., E. rulilaPaW., E. yessoensis Sw.; Eophona inigra-
toria Hart., E. personata Temm. & Schleg.; Chloris sinica L.; Sturnia
stamina Pall.; Spodiopsar cineraceus Temm.; Polyomyias muginiaki
Temm.; Zanthopygia; Anthus menzbieri Shulp.; Horeites 'cantans Temm.;
Phylloscopus plumbeitarsus Swinh., Ph. occipitalis Bly th.; Acrocephalus
bistrigiceps Swinh.; Pseudaedon sibilans Swinh.; Larvivora cyane (Pall.);
Cyanopica cyana Pall.; Turdus hortuloruin Scl.,T. pallidas Gm.; Mon-
ticola gularis (Sw.) и мн. др.
Генетически подобласть образуется обильными, значительной
Древности, автохрониыми элементами и пришельцами из индокитай-
ских тропиков. Эта фауна на значительной части территории не
испытала больших плейстоценовых потрясений и перенесла их без
значительных перемещений. Ряд данных указывает на то, что об-
щий состав фауны здесь сохраняет те типы и соотношения, которые
сУЩествовалп в более северных зонах в период перед развитием оле-
денения. В дальнейшем эта фауна частью была сдвинута к югу,
пастью сохранилась на более или менее прежних местах, частью
°ыла уничтожена или влилась отдельными элементами в образо-
вавшийся обширный пояс северных хвойных лесов. Интересным
пРедставляется также приспособление многих ее элементов к суро-
вым таежным условиям Алтая, Саян, Камчатки, где широколиствен-
ен растительность исчезла; птицы, экологически изменившись,
319
вошли в другой биоценоз. Таковы в Саянах: Larvivora. Pseudaedon.
Лcrbivocula и др.: на Камчатке: Coccothraustes. Hemichelidon grisei-
slicta (Swinh.) п др.
Входящие в подобласть острова (южный Сахалин, Япония,* Риу-
Кпу, Формоза и Боппп), имея тот же общин характер, сохраняют
более древнюю эндемичную фауну, особенно острова более мелкие и
расположенные южнее. На Риу-Кпу живет примитивная сойка —
Lalocitla. своеобразный родич седого дятла Sapheopipo и эндемичный
род Icoturus из сем. Sylviidac. На Бонине пз трех древних эндемиков —
Chaunoproclus ferreirostris (Vig.), Hapalopteron familiare Kittl. и
Aegilhocichla lerreslris l\i Lil. Chaunoproclus является ближайшим
родичем североамериканского кардинала. Хотя основная фауна
Формозы совпадает с китайской, однако здесь находятся: Oriolus
Irailli (Vig.), Lioptila auricularia (Swinh.), Actino dura egertoni (Gould.),
Cyanops nucha I is (Gould.), Hierophasis swinhoei (Gould.), Syrinaticus
mikado и др., ближайшие родственники которых живут не ближе
нижней Индии. Это указывает на то, что пижнеиндийский элемент
в фауне Китая раньше был более богат и что и в южном Китае основ-
ная часть фауны за плейстоцен сместилась несколько к югу, как то
наблюдается и в северной части подобласти (см. выше).
П о д о б л а ст ь ф а у и ы широко л и с т в е и н ы х п
то ж и ы х х в о й н ы х л е с о в Соединенных Штатов, кйк указыва-
лось, состоит из двух провинций. Провинция аллеганских широко-
лиственных лесов занимает всю восточную часть Соединенных Шта-
тов к северу до области Великих озер и более монотонных хвойных
лесов Канады, к югу до побережья Мексиканского залива. На западе
провинция граничит с прериями, образуя по ландшафту картину,
сходную с Евразией по экологическим условиям. Эта зона лесо-
степи представляет собой переходный район к области американ-
ских степей. По характеру биоценозов провинция близко напоми-
нает восточвоазиатскую подобласть, также представляя постепенный
переход от умеренных по составу лесов на* севере к биоценозам тро-
пического характера на юге. Влияние человека очень велико и ме-
стами уничтожило местную фауну. Фауна провинции сравнительно
богата, хорошо обособлена от предыдущих подобластей и обнару-
живает большое количество неотропических по происхождению форм.
Значительное число видов являются перелетными, с зимовками
в северных частях Южной Америки.
Из характерных и частью эндемичных форм следует упомянуть:
Rallus virginianus L.; Porzana Carolina (L.); Colinus virginianus (L.);
Zjenaidiira macroura L.; Baeolophus bicolor L.; Penthestes carolinensis
(And.); Polioptila caerulea L.; Meleagris gallopavo silveslris Vieilld
Dolichonyx oryzivorus L.; Philochela minor (Gmel.); Lanivireo flaw
irons (Vieil.); L. solitarius (Wols.); Passerculus princeps Magh.;^*
serherbulus henslowi (And.), P. maritimus (Wils.); Pipilo erythrophlw
mils L.; Cardinalis cardinalis L.; Guiraca caerulea (L.); Passerina cyanM
(L.) и мн. др.
Генетически фауна провинции слагается: из немногих эндемич-
ных форм; форм, по происхождению общих с восточноазиатской под-
областью; форм, проникающих из северной подобласти, и форм
320
неотропнческих, в значительной степени находящих здесь свою
северную границу. Из последних многие являются общими формами
для этой и тихоокеанской лесной провинции (см. ниже). По про-
исхождению как эта, так и следующая провинции представляют
весьма давно (в начале неогена?) обособившуюся часть умеренного
лесного пояса, развившуюся в значительной мере самостоятельно,
хотя и сохранившую еще живые генетические связи с восточной
Азией. Обширное оледенение Северной Америки не сказалось губи-
тельно на фауне вследствие возможности открытого отступления
к югу. Кроме того, фауна имела возможность значительного обо-
гащения выходцами из неотропической области, что особенно резко
выделяет эти провинции из других подобластей Голарктики.
Разъединение обеих провинций путынностепной областью, особенно
широкой на юге, с различным типом климатических условий на
обеих сторонах материка явилось условием для обособления обеих
провинций.
Провинция фауны тихоокеанского побережья Северной Америки
занимает узкую полосу по берегу Тихого океана, на севере гранича
с Ситкинским округом неарктической таежной провинции, на юге
и востоке вступая в соприкосновение с прилежащими районами об-
ласти степей и пустынь Северной Америки. Географически весь
район представляет горную страну, покрытую преимущественно
вечнозелеными хвойными, но южного типа, лесами и различными
типами лиственного леса. Основными биотопами являются: горные
хвойные леса, лиственные леса по долинам рек, каменистые склоны
с кустарниками, горные реки, кустарные заросли и др. Характерны
Для провинции следующие формы: Icterus bullocki Swains.; Hespe-
riphona vespertina (Coop.); Nucifraga Columbiana (Wils.); Spicella
breweri Cass.; Oreospiza chlorura (Towns.); Chamaea; Psaltiparus]
Zamelodia melanocephala (Swains); Calamospiza melanocorys (Stejn.);
Bendroica auduboni (Towns.); Dendroica townsendi (Towns.); My ade-
pts townsendi (And.); Ixoreus naevius и мн. др. Некоторые из ука-
занных выше видов отмечают наличие древнего эндемизма в про-
винции, с весьма изолированным систематическим положением. Из
зтого центра много видов повидимому расселилось в соседние рай-
°ны Калифорнии и Скалистых гор.
Обеим провинциям широколиственных и южных хвойных лесов
дверной Америки свойственно большое количество неотропических
во происхождению форм и форм древних североамериканских, но
стоящих в генетической связи с неотропиками. Часть из них ука-
зана выше. Из подобных групп наиболее интересны семейства Geoth-
typidae и Odontophoridae, а также представители южноамерикан-
ских по происхождению семейств: Tyrannidae и Trochilidae.
Область североамериканских степей и пу-
с т и н ь занимает всю центральную и южную часть Северной Аме-
рики и обладает очень сложными очертаниями, вследствие про-
никновения в нее северных фаун длинными языками и островами
п° системам Скалистых гор и Сиерра-Невада. Благодаря протяже-
нию по широте на 40°, северные ее районы сильно отличаются от
10}кных и должны быть поделены на несколько расположенных по
Орнитология—1209
321
долготе (и широте) округов. По общим физико-географическим усло-
виям область близко напоминает экстратропические пустыни и
степи Старого Света и сухие горы средиземноморской провинции.
Основными стациями области являются различные типы степей,
пустынь, пустынных гор, заросли кустарников, можевеловые леса
и различные типы преимущественно стоячих водоемов.
Характерными и частью эндемичными для области являются:
Pelecanus erythrorhynchus Gmel.; Limosa fedoa L.; Steganopus tricolor
(Vieil.); Centrocercus urophasianus (Bp.); Grus mexicana (Miill.); Speo-
tyto cunicularia hypogaea Bp.-, Falco mexicanus Schleg.; Anthus spra-
guei (And.); Chondestes grammacus (Say); Pipilo maculatus L.-, Ammod-
ramus bairdi-, Rhynchophanes mccowni-, Calcarius ornatus Towns.; Tym-
panuchus cupido L. и мн. др.
Характерные элементы области по происхождению являются или
древними переселенцами из лесных районов Северной Америки, или
потомками общих с Евразией болотных береговых типов, вроде CJia-
raridus nivosus, либо специализированными выходцами из неотрошг
ческой Америки. Хорошая характеризованность пустынностепной
фауны указывает на значительную древность ее обособления. Важ-
ным является далее отсутствие таких широко распространенных
пустынно степных форм Старого Света, как представители подотрядов
Otides, Pterocletes и др.
Область фауны сахаро-азиатских степей
и пустынь занимает обширнейшую часть территории Старого
Света как по долготе, так и по широте, и благодаря этому значительно
отличается на своих крайних границах. На севере пределы области
взаимодействуют с лесной областью, образуя переходную лесостеп-
ную полосу, а местами островками илй языками проникая в горные
районы средиземноморской провинции или восточноазпатской ши-
роколиственной подобласти. На юге Сахары область постепенно пе-
реходит в саванны Африки, а на юго-востоке то выходит на берег
моря, то граничит с индийскими сухими тропическими лесами, то
постепенно переходит в высокогорные пустыни Тибета, граничащие
с наиболее юговосточными и южными островками голарктической
фауны. Фауна пустынь и степи обычно сравнительно бедна и на зна-
чительные территорий весьма однообразна.
Основными стациями области являются различные типы степей
и пустынь, заросли кустарников и- малочисленные внутренние
водоемы.
Характерными и отчасти эндемичными для области являются:
М armaronetta angustirostris Menetr.; Oxyura leucocephala (Scop.);
Casarca ferruginea (Pall.); Charadrius alexandrinus L., Ch. leschend-
ulti Less.; Cursorius cursor (Lath.); Chlamydotis-, Pterocles arenarius
(Pall.), P. alchata L.; Syrrhaptes; Buteo rufinus Cretz.; Caprimutyfls
aegyptius Licht.; Alauda, ряд форм; Calandrella; Melanocorypha; Saw
lauda; Alaemon-, Rhamphocorys-, Pterocorys-, Oenanthe deserti Temm.,
O. isabellina (Cretz.), 0. moesta Licht.; Ammomanes deserti Licht., A
phoenicura Frankl.; Galerida*, Sylvia папа (Hempr. & Ehr.), S. curructi
L., ряд подвидов; Anthus campestris L.; Bucanetes-, Passer simple
Licht., P. ammodendri Gould., P. griseigularis Sharps, P. enigmaticus
322
Sar., P. moabiticus Tristr.; Lanius pallidirostris Cass., L. isabellinus
Hempr. et Ehr.; Crateropus squamiceps (Cretz.), C. caudatus (Drap.),
C. fulvus (Desf.); Panur us biarmicus L.; Corvus riificollis Less., род
Podoces и мн. др.
В пределах этой обширной территории могут быть выделены че-
тыре провинции: провинция степей и северных полупустынь, провин-
ция среднеазиатских пустынь, провинция южных пустынь и провин-
ция высокогорных тибетских пустынь. Первая провинция захва-
тывает всю пустынностепную территорию от Венгрии (и островками
до Испании и Алжира) на западе, до Пекина на востоке и к югу до
Усть-урта, Балхаша, восточного Тянь-шаня и Ордоса и характе-
ризуется из птиц такими формами, как Otis, Saxilauda, Pterocorys,
Syrraptes paradoxus Pall, и др., естественно, как и последующие
провинции, распадаясь на ряд главным образом по долготе ориенти-
рованных округов. Участки этой провинции языками и островками
проникают местами, преимущественно по горам, и в южнее лежа-
щую провинцию среднеазиатскую. Последняя занимает северную
зону Сахары, северную Персию, равнинную Среднюю Азию, восточ-
ный Туркестан и южную Гоби до Алашаня. Характерными формами
являются: Chlamydotis, Lanius pallidirostris Cass., Sylvia папа.
Hemp. & Ehrenb., Passer simplex Licht., P. ammodendri Gould и др.
Провинция южных пустынь занимает южную Сахару, Аравию, южные
пустыни Персии, Белуджистана и Синда и частью разбита на отдель-
ныео.стровки. Из характерных форм упомянем Еироdotis arabs L., ряд
форм Crateropus, Ammomanes, Rhamphocorys, некоторые Pterocles и др.
Тибетская провинция занимает огромное плоскогорье Тибета,
проникая в восточный Памир и отдельными элементами в централь-
ный Тянь-шань. Из характерных форм наиболее замечательны:
Pseudopodoces, Syrrhaptes tibetanus Gould., Melanocorypha maxima
Gould., Megalornis nigricollis Przew., Charadrius mongolus (atrifrons)
Pall., Ibidoryncha struthersi Vig., Lams bruneicephalus Jerd. и др.
Генетически фауна этой области находится в значительной связи
с фауной восточноафриканской области, но обладает также своими
Древними эндемичными родами {Otis, Chlamydotis, Podoces, Pseudo-
podoces, Saxilauda, Melanocorypha, Bucanetes, и др.). Распростра-
нение видов этих родов указывает на вероятность нескольких центров
в образовании пустынностепной фауны. Таковыми являются повп-
димому западная часть Гоби, Персия и западная Сахара. Образо-
вание пустынной фауны происходило под воздействием соседних
лесных и горных фаун, давших много приспособившихся к пустыне
и степи форм. В разных пустынных очагах таковыми были отличные
исходные формы, вследствие чего фауна пустынь обнаруживает
нередко значительное родство с ближайшими фаунами: средиземно-
морской горной провинции, фауной горного Китая, саваннами
Африки и др.
Переходим теперь к рассмотрению южных областей, следуя де-
лениям Уоллеса и других авторов (рис. 212).
Э ф и опекая область охватывает всю Африку к югу
от Сахары, включая юго-западный угол Аравии, Мадагаскар и острова
Сокотра, Занзибар, Асценсион и Елены.
* 303
Зоогеографические подразделения ее были в последнее время
разработаны Чапином (Chapin, 1932), и хотя в этой обработке от-
сутствует необходимая характеристика делений, мы будем следовать
ей. Всю эфиопскую область можно разделить на три подобласти:
западноафриканскую, восточно- и южноафриканскую и мадагаскар-
скую. В западноафриканскую подобласть входят основные массивы
тропических сырых лесов и прилежащие к ним части саванн, фауна
галерейных лесов которых близка к основной фауне тропического
леса. Эта подобласть характеризуется родами: Tigriornis,Pteronetta,
Dryotriorchis, Urotriorchis, Phasidus, Himantornis, Psittacus, Cerato-
gymna, Trachylaemus, Gymnobucco, Melignomon, Melichneutes, Ver-
reauxia, Ixonotus, Lobotos, Stiphrornis, Dyaphorophyia, Stizorhina,
Fraseria, Picathartes, Pseudochelidon, Malimbus, Brachycope, Pholidor-
nis, Parmoptila, Spermospiza и др. Она подразделима на провинцию
гвинейских лесов с округами Верхне- и Нижне-Гвинейским и про-
винцию гвинейских саванн с округами Верхне-Гвинейским, саванн
Убанги-Уелле, саванн южного Конго и саванн Уганда Униоро.
Фауна сырых тропических лесов Африки обнаруживает зна-
чительную близость к фауне подобных же лесов индийской области
и некогда несомненно стояла в территориальной связи с ними. Сокра-
щение африканских тропических лесов до сравнительно незначитель-
ной территории обусловлено повидимому общим увеличением су-
хости климата этого континента, что доказывается рядом физио- и
биографических фактов и между прочим прерывчатым распростра-
нением птиц тропического леса и подобным же распространением
птиц, свойственных горным поясам. Близость этой фауны к фауне
тропических лесов индийской области ставит вопрос о возможности
иного районирования фаун обеих этих областей.
Подобласть восточной и южной Африки охватывает всю запад-
ную Африку полукольцом и занимает всю остальную часть конти-
нента эфиопской области. На севере, по краю южной Сахары она
граничит с характеризованной выше областью сахаро-азиатских
пустынь и отрезает к эфиопской области также юго-западный угол
Аравии. По преимуществу вся эта территория представляет саванны,
а на юго-западе и пустыни. В составе подобласти различают три
провинции: суданскую с округами пустынным и саванн, северо-
восточноафриканскую с округами Абиссинского нагорья и Сомали,
восточно-южную с округами восточноафриканского нагорья Родезии,
восточноафриканской низменности, юго-восточного влажного и юго-
западного пустынного; с предпоследним округом стоит в связи рас-
сматриваемый как особый округ — высокогорные пояса Централь-
ной Африки с подразделениями на Камерунский и Восточноафри*
канский районы.
Восточная и южная подобласть Африки характеризуется сле-
.дующими родами птиц: Struthio, Microparra, Grus, Pterocles, Acryl-
Hum, Vultur, Otogyps, Sagittarius, Gallirex, Gymnoschizorhis, Tra~
chyphonus, Rhinopomastus, Chloropeta, Eurocephalus, Urolestes, Corvur
tur, Amydrus, Cosmopsarus, Chalcopsar, Textor, Dinemellia, PloceipaS'
ser, Rhynchostrutus, Promerops, Salpornis, Argya, Brady pterus
и мн. др.
324
По составу фауны эта подобласть близка к фауне саванн Индии
п также сравнительно недавно стояла в связи с ними. Обширность
территории этой подобласти является сравнительно новым явлением,
возникшим в результате развития более сухого климата. Несомненно,
однако, что этой подобласти принадлежат и наиболее древние степ-
ные и пустынные очаги в развитии фауны, расположенные частью на
юге, частью на северо-западе подобласти. Эти очаги явились топ
колыбелью, в которой первоначально развивались группы степных
и пустынных птиц, впоследствии заселившие частью и более север-
ные степи и пустыни.
Это широкое расселение некоторых африканских по происхож-
дению групп, а также весьма значительное родство с фауной индий-
ской области являются причиной сравнительно слабой характери-
зованности континентальной Африки древним эндемизмом. Наибо-
лее интересными в этом отношении формами являются семейства:
Coliidae, Musophagidae, Phoeniculidae, Numididae (с одним представи-
телем в Марокко), Sagittarius, Balaeniceps, Scopus, Smithornis, Pseu-
Aocalyptomena, Pseudochelidon, Promerops, Parmoptila, Picathartes.
Struthio по происхождению также вероятно относится к этой терри-
тории, хотя еще в плиоцене он был широко распространен в саха-
ро-азиатских пустынях к северу до Херсона и Забайкалья. Харак-
терными для континента являются также следующие группы
и роды: цесарки с родами Numida, Acrylium, Guttera, Agelastes,
Phasidus; голошейный турач — Pternistes, венценосный журавль —
Balearica; из хищников Melierax и Helotarsus*, из попугаев Psitta-
cus и Poicephalus', из бородастиков Trachyphonus, Lybius, Tricho-
laema, Barbatula; из трогонов Apaloderma*, из мухоловок Brador-
nis, Batis, Platysteira, Prionops, Laniarius', из скворцов Buphagus,
Lamprocolius, Lamprotornis. Кроме того, в Африке же может счи-
таться лежащим центр видового разнообразия следующих семейств:
Otididae, Pteroclidae, Aegypiinae, Indicatoridae, Capitonidae, Mero-
pidae, Ploceidae, Alaudidae и Nectariniidae.
Мадагаскарская подобласть составляется Мадагаскаром с при-
лежащими (но на значительном расстоянии) островами Маврикия,
Реюнион, Сейшельскими, Коморскими и др. Сам Мадагаскар представ-
ляет плато, покрытое сухими лесами и саваннами, по краю которого
расположены зоны сырого тропического леса. Флора и фауна весьма
своеобразны и хранят большое количество эндемиков, из которых
многие весьма древнёго возраста. Мадагаскар отделился от Африки
невидимому в раннетретичное (может быть и раньше) время, и фауна
его развивалась изолированно. Соединение с Индией, предполагав-
шееся ранее, теперь считается сомнительным. Фауна птиц Мада-
гаскара составлялась вероятно главным образом заносным путем,
хотя некоторые древние формы (например вымершие недавно пред-
ставители нелетающей страусообразной группы Aepyornithijormes
!1Нек- друг.), возможно, являются остатками материковой фауны.
Муть проникновения и родина древних эндемиков неясны. К тако-
ВЫм относятся роды Monias и Mesoenas, составляющие эндемичный
^Дотряд, семейства Brachypteraciidae и Leptosomatidae (из отряда
^0Taciiformes), семейство Philepittidae, близкое к питтам, семейство
325
Vangidae, близкое к сорокопутам, и роды Coracopsis, Coua, Falculia,
Нуposit la и др.
Широко распространенные и частью характерные для Африки
семейства: Coliidae, Musophagidae, Bucerotidae, Otididae, Picidae и
Capitonidae — на Мадагаскаре совершенно отсутствуют. Обильный ряд
форм сравнительно недавно иммигрировал на Мадагаскар и частью
еще не образовал своих видов. Из африканских по происхождению
таковыми являются: Scopus, Threskiornis, Thalassornis, Agapornis,
Polyboroides, Vinago, Oena, Corvus albus и др. Индийского происхож-
дения род Alectroenas, виды Ninox, Otus rutilus (Pucher.), Cuculus
poliocephalus Lath., Copsychus, Hypsipetes и др.
Тяготеющие к Мадагаскару острова, вследствие своей удален-
ности и давности обособления, населены хотя и бедной, но почти
сплошь эндемичной фауной, частью также неясного происхождения.
Таковы, например, нелетающий очень крупный голубь додо—Raphus
cucullatus L. и крупная нелетающая болотная курочка — Aphanyp-
teryx, истребленные вскоре после открытия этих островов (острова
Маврикия). Более молодые эндемики являются преимущественно
индийского происхождения, например скворец — Fregilupus, личин-
коед— Oxynotus, попугаи — Mascarinus и Palaeornis, стриж—Calio-
calia и др.
Индийская пли восточная область охваты-
вает весь Индостан (без Синда), южные склоны Гималаев, Индо-
китай и южный Китай и Малайский архипелаг к востоку до (вклю-
чая) Явы, Бали, Борнео п к северу от них — Филиппины. Поло-
жение границ на западе нуждается в ревизии, так как саванны
Индостана обнаруживают значительную близость к фауне соответ-
ствующих ландшафтов восточной Африки. Но фауна тропических
лесов Индостана явно тяготеет к восточной Индпи, так же, как и
южные склоны Гималаев. Цейлон по своей фауне примыкает к юж-
ному концу Индостана, сохраняя однако в горных лесах несколько
эндемических видов и два рода — Elaphrornis (из Tim alii dae) и Stur
nornis (из Sturnidaep
В целом для области характерно изобилие куриных, особенно
роскошных по окраске фазановых форм, с родами: Tragopan, Gen-
naeus, Polyplectron, Pavo, Argusianus, Gallus, обилие эндемичных
мелких куропаток, как то: Arboricola, Caloperdix, Rollulus. Из
попугаев наиболее обильны представителями роды Palaeornis и Psil-
tacinus, из Caprimulgiformes род Batrachostomus, из бородастиков
роды Cyanops, Psilopogon, Calorhamphus, из трогонов Pyrotrogon,
пз нектарниц Arachnotera. Campophagidae и Bicaeidae имеют в этой
области центр своего видового разнообразия и особенно обильно
представлены формами родов Pericrocotus, Bicaeum и Prionochih^'
Вследствие несколько неясного зоогеографического положения
Передней Индии, в области можно принять лишь две хорошо раз-
личающиеся подобласти — индийскую и малайскую, с отнесением
к последней всего Малайского архипелага, Филиппин*и Малакского
полуострова.
Наиболее характерными для индийской подобласти являются
роды: Perdicula, Microperdix, Gennaeus, Crocopus, Thaumatibis, Gra-
326
phocephalus, Oryceros, Gecinulus, Brachypternus, Hydrornis, Digenea,
Volvocivora, Molpastes, Pellorneurn, Schaeniparus, Lioptila, Uro-
cichla, Franklinia, Machlolophus, Aegithaliscus, Urocissa и др.
Наиболее характерные элементы подобласти свойственны гор-
ным и более северным лесным районам. Для подобласти характерно
также значительное родство с фауной Китая, Японии и отчасти юго-
восточной Сибири.
Малайская подобласть, сходно с тропической Южной Америкой,
является богатейшим фаунистическим районом, с обильным древ-
ним и молодым эндемизмом. До плейстоцена значительная часть
входящих в подобласть островов составляла одно целое, вследствие
чего фауна сохраняет в основном по всей территории весьма близкий
характер. Все острова покрыты роскошным тропическим лесом
с хорошо выраженной областью вечновлажных горных лесов
в более высоких зонах. Для всей подобласти характерно изобилие
видов и родов в группах кукушек, дятлов, Capitonidae и тималий.
Для Явы эндемичны Crocias, Stasiasticus и Psaltria\ для Борнео —
Lobiophasis, Pityriasis, A llocotops, Chlainydochaera, Heterococcyx,
Chlorocharis и др.; для Бали — Leucopsar\ для Филиппин — Sar-
cophanops, Dasylophus, Lepidogrammus, Pithecophaga, Sarcops, Good-*
fellowia, Rhabdornis, Zosterornis, Ptilocichla, Dasycrotopha, Hypo-
cryptadius и др. Кроме собственно индийских и малайских форм,
на территории малайской подобласти расселилось в настоящее время
значительное число форм австралийской области, хотя пограничная
«Уоллесова линия» между островами Бали и Ломбок п Борнео и
Целебес и сейчас сохраняет некоторое значение фаунистического
водораздела. На Филиппины характерно проникновение бореаль-
ных по происхождению форм — Loxia п Pyrrhula.
Неогейское царство охватывает весь континент Южной
Америки к севёру вплоть до подножия Мексиканского плоскогорья
и включая Антильские острова. Являясь по площади примерно
равной Африке и располагаясь в сходных широтах, Южная Аме-
рика на много богаче ее фаунистически. В пределах Южной Америки
насчитывается не меньше 2500 гнездящихся видов («кругов форм»),
пз которых огромный процент являются эндемичными району,
образуя в значительной степени замкнутые и изолированные систе-
матические группы. Вся фауна Южной Америки характеризуется
сравнительно низким морфологическим уровнем развития, прибли-
жаясь в этом отношении к фауне Австралии и Новой Зеландии или
проявляя еще более примитивные черты.
Эти особенности материка объясняются длительным изолирован-
ным развитием его животного мира, вследствие длительного периода
обособленности Южной Америки от Северной, по крайней мере по
плиоцен. Значительная связь обоих континентов осуществлялась
невидимому через архипелаг островов, что и дало возможность
представителям ряда северных групп проникнуть в неотропическую
область, например: Corvidae, Motacillidae, Alaudidae, Turdidae,
Fringillidae, Hplcyonidae, Geothlypidae, Odontophoridae, и обратно,
ю'Кным группам проникнуть на север, например: Tirannidae, Tro-
chilidae, Cracidae, Vireonidae, Troglodytidae и Mimidae.
327
Неогейское царство обычно рассматривается состоящим из одной
неотропической области, разделяемой на 4 или 6 подобластей. В
целом всей области свойственно не менее 24 эндемичных семейств,
частью одновременно являющихся и единственными семействами
отдельных отрядов; если же сюда присоединить и семейства, отдель-
ные представители которых проникают и в Северную Америку, но
главным образом свойственны неотропической области, то число
семейств повысится до 35. Эндемичны Ю. и Ц. Америке: Rheidae,
Tinamidae, Cracidae, Opisthocomidae, Thinocoridae, Psophiidae, Cari-
amidae, Palamedeidae, Eurypygidae, Steatornithidae, Rhamphastidae,
Galbutidae, Momotidae, Bucconidae, Trochilidae, Todidae, Coerebidae,
Pteroptochidae, Formicariidae, Dendrocolaptidae, Phytotomidae, Cotin-
gidae, Pipridae и Tyrannidae.
Вместе с тем характерным является отсутствие некоторых широко
распространенных и частью проникающих в Северную Америку
групп, как, например, Tetraonidae, Syl^iidae и Laniidae, илигруищ
широко распространенных в тропиках Старого Света, как Phasia-
nidae, Aegypiinae, Sturnidae, Bucerotidae, Meropidae, Coraciidae,
Nectariniidae, Muscicaipidae, Campophagidae, Pycnonotidae и др.
• В качестве отдельных подразделений неотропической области
рассматриваются: чилийская подобласть, охватывающая южную
половину Южной Америки до 30° ю. ш. на восточном и 5° ю. ш.
на западном берегах с значительным продолжением к северу по аль-
пийской зоне Анд; бразильская подобласть, занимающая область
бразильских саванн и кампосов; амазонская подобласть, охватываю-
щая низменность реки Амазонки и Гвиану; субандская подобласть,
охватывающая значительную часть Колумбии, Венецуэлы и Галапа-
госские острова; центральноамериканская подобласть, занимающая
весь перешеек между Северной и Южной Америкой, и антильская
подобласть, охватывающая Антильские и Багамские ‘острова. В ха-
рактеристику подобластей мы однако не можем входить подробно, за
недостаточностью новых систематизированных данных. Чилийская
подобласть характеризуется умеренным и холодным климатом, степ-
ными и пустынными ландшафтами и широколиственными лесами
по западным склонам южных Анд. Характерны для подобласти не
менее 5 гнездящихся видов пингвинов, ржанка — Chionis, зобатые
песочники — Attagis и Thinocorys и ряд представителей семейства
Pteroptochidae и Phytotomidae. Ряд форм, преимущественно свойствен-
ных чилийской области, проникает и в бразильскую, вообще имею-
щую переходный характер, таковы, например, нанду— Rhea, серимы---
Cariamidae и Palamedeidae. Амазонской, центральной и колумбий-
ской подобластям свойственно наибольшее число характерных для
Южной Америки групп. Особый интерес представляют относимые
к последней подобласти Галапагосские острова, расположенные на
экваторе в 100 км от берега. Животный мир этих островов, вулкани-
ческих . по происхождению, впервые навел Ч. Дарвина на мысль
о способе эволюции организмов. Здесь обильно представлена энде-
мичная и развившаяся на островах из неизвестного корня группа
вьюрков — Geospizinae, в особенностях которых еще легко усмо-
треть тот путь адаптивной радиации, который привел к образованию
328
многочисленных видов этой группы. Из других эндемиков островок
интересны Nesomimus^ близкий к Mimus, и Nesopelia, близкая к Ze-
naida, и сейчас обитающих на противолежащих берегах Южной
Америки.
Подобласть Антильских островов представляет собой древний
архипелаг, вследствие промежуточного положения между континен-
тами Северной и Южной Америки, населенный выходцами с них
обеих. Северное влияние сказывается в наличии здесь таких пред-
ставителей, как эндемичный вид Loxia. Из древних эндемиков архи-
пелага интересны резко обособленное семейство Todidae и роды:
Dulus, PhoenicophilusMimocichla, Microligea^ Ferminia, Torreornis.
Starnoenas, Prionoteles, Teninotrogon и др.
Нотогейское царство включает в себя Австралию,
Новую Зеландию и все острова, начиная от уоллесовой линии к во-
стоку и юго-востоку, почти вплоть до берегов Южной Америки.
Этот преимущественно островной мир хранит наиболее примитив-
ную фауну и замечателен, подобно неогеи, исключительно высоким
процентом древнего эндемизма. В целом же фауна сравнительно небо-
гата, так как большинство островов очень мелки и удалены друг
от друга, а более крупные острова, как Новая Зеландия или Австра-
лия, лежат в умеренном климате и последняя на большей части
пространства представляет собой пустыню.
Для всего царства в целом характерны крупные нелетающие род-
ственники страуса — эму и казуары, весьма разнообразные райские
птицы и беседочницы — Paradiseidae, чрезвычайно изобильные фор-
мами попугаи, голуби, зимородки и медососы. Из попугаев подсе-
мейства Cyclopsittacinae Nasiterninae ограничены вообще этой
территорией, в то время как подсемейства Cacatuinae, Platycerci-
пае и Loriinae приурочены к ней за очень немногим исключением.
Из голубей особенно богато представлены плодоядные голуби —
Carpophaga и Ptilinopus и своеобразные венценосные голуби (Goura)
и наземные — Caloenas и Otidiphaps. Из зимородков наиболее бо-
гаты роды: Clytoceyx, Tanysiptera, Symci, Melidora, Saurotnarptis,
Сеух и Alcyone. Здесь же главным образом распространены ласточ-
ковые сорокопуты — Artamidae. Вместе с тем замечательно для
всего царства отсутствие широко распространенных в остальном
Старом Свете настоящих вьюрков, дятлов и грифов.
Нотогея разделяется на австралийскую, папуасскую с Поли-
незией и новозеландскую подобласти.
Австралийская подобласть охватывает материк с Тасманией и бли-
жайшими прилегающими мелкими островами, но вероятно за выче-
том значительной части горного Квинсленда, населенного главным
образом попуасскими птицами. За исключением восточной горной
части, одетой умеренными и тропическими лесами, большая часть
Австралии представляет пустынную степь, частью поросшую кустар-
никами. Фауна Австралии сравнительно небогата (666 гнездящихся
видов, относимых к 286 родам и 99 семействам), изобилует древними
и молодыми эндемиками (треть всех родов и 70%зарегистрирован-
ных видов). Наиболее характерно обилие видов из семейства Ме-
uphagidae — медососов, попугаев, голубей и примитивных ткачи-
329-
нов. Из эндемичных родов наиболее замечательны: Dromaius, Мепига,
Atrichornis, а также Strep era, Cereopsis, Biziura, Malacorhynchus,
Anseranas, Stictonetta, Chenonetta, Chenopsis, Pedionomus и мн. др.
При отнесении Квинсленда к австралийской подобласти, Австралия
разделяется на три округа: Торресовский, Кассовый и Эремийский
(пустынный), одновременно этим отличая и основные элементы
фауны: первый — богатый новогвинейскими (папуасскими) фор-
мами, проникшими на территорию Австралии сравнительно недавно;
Кассовый — представляющий наибольшее сосредоточение древних
типов, и Эремийский — населенный измененными формами пер-
вых двух округов, а по растительности наиболее древний среди
прочих округов. Несмотря на значительность территории этого
округа, вследствие пустынности, он сравнительно беден птицами.
Наиболее характерны для него: Leipoa ocellata Gould. (Megapodiidae),
Eupodotis, Oedicnemiis, Eremiornis carteri North., Ashbyia lovensis
(Sylviidae),- Lacustroica whitei Camp. (Meliphagidae) и др. Более
молодые эндемики Австралии обнаруживают наибольшую близость
к птицам Новой Гвинеи и Индии.
Новозеландская подобласть, помимо островов Новой Зеландии,
включает острова Чатам, Ауклэнд, Норфольк, Мэкквари и др. Не-
смотря на весьма южное положение, Новая Зеландия покрыта лесной
растительностью, притом носящей преимущественно подтропиче-
ский и умеренный характер. Высокие горы островов имеют хорошо
выраженный альпийский пояс. Вследствие своей изолированности
и весьма древнего отделения от материка, Новая Зеландия полу-
чила весь свой высший животный мир заносным путем, причем он
развился здесь далее вне острой конкуренции. По этой причине
Новая Зеландия весьма богата древними эндемиками и изолированно
стоящими систематическими группами. Особенно поражает большое
количество потерявших способность летать видов, что оказалось
возможным благодаря отсутствию наземных млекопитающих. Наибо-
лее вероятным является происхождение этих нелетающих форм от
летавших видов, прошедших всю редукцию крылового аппарата
в пределах самой Новой Зеландии. Таковыми являются несколько
видов киви (Apteryx), недавно вымершие разнообразные предста-
вители отряда Dinornithiformes, из попугаев Strigops, из гусей вымер-
ший гигантский гусь — Cnemiornis, из болотных курочек Go№
ralus и вымершая Aptornis и др. Из других древних эндемиков сле-
дует упомянуть попугая — Nestor, куличка — Anarhynchus и Thy
nornis, примитивных воробьиных Acanthisitta, Xenicus, Traversia,
из Oscines ряд родов с изолированным или неясным систематическим
положением, например Turnagra, Heteralocha, Creadion, Callaeas,
Clitonyx, Prosthemadera, Pogonornis, Myiomoira и др.
Входящие в состав новозеландской области острова получили
свою, в общем весьма бедную, фауну невидимому из Австралии,
через посредство Новой Зеландии, и также характеризуются своим
эндемизмом. Наиболее замечательными из них являются нелетающие
пли почти нелетающие виды: болотная курочка — Cobalus modesty
Hutt, на Чатаме и утка — Nesonetta aucklandica Gray и крохаль-
Mergus australis (Hombr. & Jacq.) на Ауклэндских островах.
330
Папуасская подобласть охватывает Новую Гвинею, Малые Зонд-
ские острова, Целебес, Молукские, архипелаг Бисмарка, Соломоновы
и другие, входящие в собственно папуасскую провинцию, и острова
Полинезии, входящие в полинезийскую провинцию той же подобласти.
Папуасская провинция представляет наиболее богатый остров-
ной мир в пределах всей потогеи. Входящие в нее острова срав-
нительно крупны и близко расположены друг к другу и в значи-
тельной мере покрыты роскошными низменными и горными лесами
с богатым и разнообразным животным миром.
По составу фауны этот островной мир несет вместе с тем переход-
ные черты от еще более богатей малайской фауны на западе, к срав-
тельно бедной фауне Полинезии и уклоняющейся фауне Австралии,
но обладает и значительным числом древних и новых эндемиков.
Наиболее характерны длй провинции и богато представлены здесь
группы: Paradiseidae, Laniidae, Muscicapidae, Meliphagidae, Dica-
eidae, Campophagidae, Pittidae, Halcyones, Cuculi, Columbae, Psittaci,
Megapodiidae и Casuarii. Из малайских форм однако здесь уже
отсутствуют представители семейств Trogonidae, Eurylaemidae, Capi-
lonidae, Bucerotidae и за малым исключением отсутствуют дятлы.
Почти все острова, особенно более крупные, хранят немало древ-
них эндемиков. Наиболее замечательны из них: на Малых Зонд-
ских островах — Alopecoenas hoedti (Schl.), Pachycephala nudigula
Hart, (семейство Pachycephalidae), Lophozosterops dohertyi Hart.; на
Тиморе — Buttikoferella*, на Целебесе — poj\biMegacephalon, Arami-
dopsis, Meropogon, Cissura, Malia, Geomalia, Heinrichia, Cataponera,
Coracornis, Enodes, Scissirostrum, Streptocitta', на Южно-Молукских
островах (Серан, Буру) — Tephrozosterops, Madanga*, на Северо-Молук-
скпх островах — Segnioptera, Lycocorax, Habroptila, Melitograis, Neos-
colopax-, на Новой Гвинее — Paramythia, Goura, Anuraphasis, Salvado-
nna, Oreostruthus, Daphaenositta и др.; на архипелаге Бисмарка —
bkranostreptus, Ortygocichla*, на Соломоновых островах—Micro-
goura, Edithiornis, Mochthopoeus.
Отдельные острова проявляют различную близость друг к другу,
представляя немало головоломных проблем по истории и эволю-
ции фауны. Как общую закономерность можно отметить наиболь-
шую близость к малайской фауне фауны Малых Зондских остро-
ги и Целебеса; к Полинезии проявляют близость острова Соломо-
новы и Бисмарка, к Австралии — южная часть Гвинеи.
Полинезийская провинция охватывает мелкие, рассеянные в юж-
ной половине Тихого океана острова к северу до группы Гавайских
островов. Заселение этих вулканических островов могло происхо-
дить только заносными, хорошо летающими птицами, вследствие
чего орнитологическая фауна бедна и богата эндемизмом. Наибо-
лее широко распространены голуби — Gallicolumba, Ptilinopus, Glo-
hcera и Phlogoenas, болотная курочка — Porzana, из сов род Strix,
нз зимородков род Halcyon, из стрижей Callocalia, из мухоловок
"hipidura и Myiagra,m личинкоедовЛа/я^е, из толстоголовых соро-
копутов Pachycephala, из белоглазок Zosterops, из медососов Myzo-
те1а, из ткачей Erythrura, из скворцов Aplonis, из славковых Асго-
cephalus -и Ротагеа.
Из групп островов наиболее оригинальной фауной обладают:
Новая Каледония с древними эндемиками — Rhinochetus (подотряд
Rhinocheti), Tricholimnas (Ralli), Phaenorrhina и Drepanoptila (Du-
culinae), Megalurulus ( Timaliidae) и Physocorax (Corvidae); на
Фиджи из древних эндемиков имеются: V itia, Trichocichla, Lampro-
На, Muscylva и Pyrrhulopsis', на Самоа — Didunculus, Pareudias№\
на Каролинских — Metabolus', на Баобельтаоб — Megazoster ops; на
Палау — Psamathia, на острове Зайпан — Cleptornis.
Наиболее оригинальной и вместе с тем сравнительно богатей
фауной отличаются Гавайские острова, выделяемые иногда также
в самостоятельную провинцию. Наиболее экологически разнооб-
разным и вместе с тем эндемичным семейством являются представи-
тели группы гавайских цветочниц — Drepanididae, занимающей
изолированное положение среди воробьиных. Из других эндемиков
замечательны: Phaeornis, Moho и Chaetoptila (Meliphagidae). Свое-
образным далее является отсутствие широко распространенных
в Полинезии представителей отрядов голубей, попугаев и зимород-
ков. Северное положение и относительная близость к Америке
объясняют наличие здесь представителей родов, чуждых остальной
Полинезии, как, например, роды Corvus, Asio, Ruteo, Gallinuh
Fulica и Nycticorax.
Антарктическая область охватывает побережье Антаркти-
ки и рассеянные в южном полярном море острова Южно-Шотландскпе,
Оркнейские, Южной Георгии, Буве, принца Едварда, Крозет, Кергуе-
ленские, Макдональда, Хеачд, св. Павла, Ново-Александровскиепдр.
Все птицы, обитающие здесь,-за исключением Anthus antarctic^
Cab., живущего на болотах южной Георгии, связаны в своем пита-
нии с морем и населяющими его животными. Наиболее характерны
для Антарктики пингвины, ржанка Chionis и разнообразные труб-
коносые птицы (особенно роды М acronectes, Prion, Halobaena, Pagoi-
гота). Чайки, крачки и утки играют здесь второстепенную роль;
из них в полярном бассейне распространены: Stercorarius skua intt
cormicki Saund., Sterna vittata Gm., St. virgata Cab., Anas georgica
Gm., An. eatoni Sharp, и An. eatoni drygalskii Reich.
Строго антарктическими являются только пингвины: Арtenodf
tes forsteri Gray и Pygoscelis adeliae (Hombr. & Jacq) и поморник Ster
corarius skua maccormicki Saund. Последние летом проникают иногда
до 84°, 26' ю. ш., но основная масса обычно не идет южнее кое-где
обнажающихся побережий антарктического континента. Почти не
выходят за пределы ледяного пака качурки — Pagodroma п№
(Gm.) и Thalassoeca antarctica (Gm.).
Большинство обитателей антарктического бассейна явно авто-
хтонного происхождения. Интересным является здесь наличие
биполярных элементов, т. е. свойственных также и арктической
области. Таковы Stercorarius skua Brunn., представленные здесь осо-
бой формой, и близкий к северному глупышу род М acronectes.
Наиболее вероятным является проникновение этих видов с се-
вера в ледниковую эпоху, когда границы полярных областей было
значительно расширены и миграции полярных птиц должны был»
быть более длинными.
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ КЛАССА
ПТИЦ
Вопрос о происхождении класса птиц находится в более благо-
приятном положении, чем вопрос о происхождении многих других
систематических групп животных, вследствие находок в ископае-
мом состоянии промежуточной формы между птицами и рептилиями
(рис. 214). Archaeopteryx и очень близкий к нему Archaeornis из
юрских отложений Баварии обладают настолько смешанными чер-
тами обоих классов, что мнения палеонтологов о систематическом
положении этих форм расходятся: Дамес (Dames, 1884, 1897) счи-
тал, что Archaeopteryx является уже настоящей птицей и значительно
удален от места ответвления птиц от класса рептилий, Хейльман
(Heilmann, 1926) же рассматривает эту форму как теплокровную
п оперенную, подобно птицам, рептилию (рис. 215).
Безусловно птичьими чертами в строении археоптерикса яв-
ляются перовой покров и отчасти строение плечевого пояса, таза и
конечностей, но строение черепа и особенно несущие зубы челюсти,
Длинный хвостовой отдел позвоночника, брюшные ребра, свободные
тарзальные элементы несут еще явные рептильные признаки. Птичьи
черты, по своему значению, все же перевешивают в характеристике
археоптерикса и в систематическом отношении являются более важ-
ными признаками, чем его рептильные особенности. Некоторая часть
последних в свою очередь встречается в других группах птиц, в ка-
честве отдельных удержанных признаков, иногда менее резко выра-
женных. В целом, археоптерикс (вместе с археорнис) все же настолько
отличается от прочих групп, что заслуживает выделения в самостоя-
тельный подкласс, равноценный массе всех прочих птиц. Это отлпчие
было зафиксировано впервые Геккелем (Hackel, 1866), установившим
подклассы Saururae для юрских птиц и Ornithurae для позднейших
11 в том числе современных птиц.
Вследствие важности представителей Saururae в вопросе о про-
хождении птиц, рассмотрим подробнее их особенности (рис. 24).
Череп, сохранившийся лишь у археорниса, имел еще рептильную
форму, но отличался уже большей тонкостью и легкостью своего
кроения. Вследствие малой величины мозговой коробки лобные
Хтп отличаются очень незначительной величиной в сравнении
1 современными птицами. Наоборот, глаза были очевидно очень
кРУпны, судя по величине орбит, и снабжены кольцом в 10—12
(клеротикальных пластинок. Спереди глазницы были отграничены
вред глазничной ямки посредством os adlacrymale, отсутствующей
* с°временных птиц, что указывает на акинетичность черепа, в про-
333
Рис. 214. Перионтица Сименса — Ardiaeornis siemensi Dames.
Отпечаток в известковых сланцах верхнеюрского периода (титон) из Блуменберга б-лВ8
Ейхштетта в Германии. Оригинал сохраняется в Берлинском музее. По Heiimai
Рис. 215. Первоптица — Archaeornis siemensi Dames.
Реставрация Хейльманна. Верхний рисунок — самец, внизу самка. По Hellmann-
тпвоположность всем современным птицам. Крупная предглазнич-
пая ямка отодвигает далеко вперед положение голоринального носо-
вого отверстия. Межчелюстная кость относительно короче, а челю-
стная длиннее, чем у современных птиц; обе они несли 13 острых ко-
нических зубов примерно одинаковой величины и сидевших в отдель-
ных альвеолах (так называемый тормодонтный тип). Существование
этих зубов, расположенных вплоть до переднего конца челюстей,
исключает присутствие рамфотеки и подчеркивает рептильный ха-
рактер черепа (рис. 26).
Позвоночник состоял из 50 позвонков (приблизительно), именно
10—11 шейных, 11—12 грудных, 2 поясничных, 4—6 крестцовых
и 20—21 хвостовых. Длина хвоста у археорнис составляет 17—18 см.
Шейные и грудные позвонки повидимому являлись амфицельными,
но точно это, за качеством сохранности, не удается установить.
Крестцовые позвонки повидимому не срастались в сложный кре-
стец (synsacrum), вследствие чего подвижность туловищных поз-
вонков была более значительной, чем у современных птиц. Хвосто-
вые позвонки к концу хвоста становятся все меньше, напоминая ту
картину, которую мы находим у «бескилевых», Hesperornis и скрыто-
хвостов, у которых однако это свойство повидимому является вто-
ричным, коррелятивно связанным с полной редукцией или слабостью
полета. Ребра были очень тонки и лишены processus uncinati, ко-
торые могли оставаться на хрящевой стадии и поэтому не сохра-
ниться. Грудные отделы ребер неизвестны, но грудная часть обоих
найденных экземпляров вообще не сохранилась. Между тазом п
плечевым поясом однако рассеяны брюшные ребра, в количестве
двенадцати пар у археорнис, очевидно являвшиеся еще наследством
от рептилий, у которых они довольно широко распространены.
В плечевом поясе обнаруживаются некоторые отличия между
обоими ископаемыми видами (рис. 216). Саблеобразная, типичная
для птиц лопатка соединяется с коракоидом у археоптерикса сино-
стотически, а у археорниса подвижно; коракоид у первого вида,
•сходно с современными «бескилевыми», представляет прямоуголь-
ную широкую плитку, в то время как у археорниса приближается
к форме балки, типичной для гребных крыльев. Прокоракоид неви-
димому оставался на хрящевой стадии. Ключица чрезвычайно
сильна, U-образна, с обеими половинками, полностью сросшимися,
без медиального апофиза. У археоптерикса она была сильнее п
кругла в поперечнике, в то время как у археорниса оказывается
несколько сплющенной с боков.
Передние конечности представлены хорошо развитыми крыльями,
от которых даже сохранились отпечатки вполне нормально развитых
маховых перьев. Плечевая кость (рис. 217) равна 63 мм длины и снао-
жена довольно сильно развитым гребнем crista lateralis, указываю-
щим на значительное развитие крыловой мускулатуры; однако в це-
лом плечевая кость слабее, чем у хорошо летавших птиц и рептилий.
Radius и ulna развиты, как у типичных летающих птиц. В кистевом
сочленении на лучевой стороне сохранились следы двух свободных
карпальных костей, число которых однако должно быть большим
в связи с большей подвижностью сочленения, чем у современных
336
птиц. Кисть состоит из трех свободных удлиненных метакарпальных
костей, из которых первая только в треть длины остальных; прокси-
мальные концы их несколько уплощены с боков и возможно непод-
вижно прилегали друг к другу. Три свободных, совершенно не срос-
шихся пальца состоят из 2, 3 и 4 фаланг, несут большие и круто
загнутые когти. Вся кисть в целом стояла несомненно еще на весьма
низкой, напоминающей рептилий, ступени развития.
Сохранность таза оставляет желать много лучшего, но все же
отвечает на основные филогенетические вопросы. Соединение таза
Рис. 216. Грудина и плечевой пояс первоптицы — Archaeornis siemensi
Dames в сравнении с грудиной ближайших рептилий и примитивных
птиц.
А — грудппа Archaeornis; С —грудина Carcosaurus из меловых отло-
жений; Е— грудина Euparkeria; Н — грудина Hesperornis regalis
Marsch; cl. — ключицы; Co. — коракоид; ер. — episternum; h. —
плечевая кость; р.—precoracoideum; S.—лопатка; St.—грудина.
Обратите внимание на форму коракоида у Archaeornis, близкую кора-
коиду рептилий; по внутреннему краю его сохранились края отвер-
стий, прободающих коракоид и прекоракоид у рептилий. И у прими-
тивных птиц (Hesperornis) еще сохраняются рептильные черты в форме
коракоида и в наличии несрастающихся ключиц. По Nopsca, Broom,
Marsch и Hellmann, из Hellmann.
f позвоночником очевидно было весьма слабым (рис. 218). Подвздош-
ная кость срасталась с позвоночником не синостотически, а синде-
смотически, причем только с 4—6 позвонками, а не с 10—22, как
• современных птиц. В связи с этим, подвздошная кость отличалась
очень небольшой величиной, и предкрестцовый отдел значительно
превышал посткрестцовый. Прямое соприкосновение подвздошных
костей не имело места. Соединение с другими тазовыми костями
оыло опять-таки только синдесмотическим и происходило только
По краям вертлужной впадины, дно которой, как у птиц, оставалось
втянутым мембраной. Направленные назад седалищные кости не
°°разовывали между собой симфиза, еще существовавшего у Pseu-
337
dosuchia, что могло иметь место в связи с увеличением размеров
яиц и твердостью их скорлупы. Края седалищных костей отчасти,
возможно, оставались на хрящевой стадии. Лобковые кости, как
у всех птиц, направлены уже назад и по длине превосходят седалищ-
ные. У археорнис дистальные концы их возможно оставались сво-
бодными, но у археоптерикса они сливались в симфиз подобпо
тому, как это было свойственно еще псевдозухиям.
Задние конечности несут в общем облике птичьи черты, но отли-
чаются в ряде существенных моментов. Проксимальная головка
бедра не составляет прямой угол со стволом кости, а образует угол
Рис. 217. Плечевая кость Archaeornis и Archaeopteryx в сравнении с плечевой
костью птерозавра и птиц.
Правая плечевая кость: 1 — Rhamphorhynchzis koheni (Plerosduria) из верх-
неюрских отложений Баварии; 2 — Archaeornis siemensi Dam.; 3 — Archaeo-
pteryx lithographica Meyer; 4 — Ichthyornis dispar Marsch; 5 — обыкновенная
курица: 1 и 2 — с дорзальной стороны; з, 4, 5 — с вентральной стороны.
Узкая форма кости Archaeornis и Archaeopteryx, небольшая величина сочлене-
ний с предплечьем и сравнительно слабый верхний гребень (тс.) указывают на
плохие летательные способности животного. По Plieninger, Petronievics and
Woodward, Marsch и Hellmann, из Hellmann.
около 130°, из чего следует, что коленное сочленение было направ-
лено значительно вбок, как это имеет место у многих рептилии,
и само движение ног должно было протекать отлично от современ-
ных птиц. В тарзальном сочленении сохранились от рептилий fibulare.
tibiale и intermedium, входящие в состав проксимальных tarsalia.
Дистальные tarsalia утеряли свою самостоятельность и слились
с metatarsalia. Таким образом мы имеем здесь интертарзальнып
тип сочленения. В отличие от птиц, у археорнис малая берцовая
не редуцирована дистально, а входит в соприкосновение с fibulare,
что также должно было влиять на характер движений. Метатар'
зальные кости 2-го, 3-го и 4-го пальцев в отличие от большинства
птиц полностью отделены друг от друга, хотя тесно соприкасаются
и лежат в одной плоскости. На задней стороне к ним прилегает
короткое metatarsale 1-го пальца, как у птиц, направленного назад-
338
От пятого луча рептилий здесь уже не осталось никаких следов.
Количество фаланг пальцев редуцировано до 2, 3, 4и 5, снабженных
сильными и крутыми когтями (рис. 61 и 62).
Рис. 218. Таз первоптицы Archaeopteryx в сравнении с тазом ближайших рептилий и
(]) более прогрессивных птиц.
pter]/Y Таз Orniihosuchus Woodwardi, сбоку; (2)—то же, сверху; (3)—таз Archaeo-
Pato erCfKy; (4) —то же, сверху; (5) — таз Apatornis, сбоку; (6) — цыпленок павлина,
ds. —Хтп ™s’ сб0«У; (7 ) — таз курицы — Callus gallus L., сбоку; (8) — то же, сверху.
Удалит₽3альпая поверхность подвздошной кости; i — подвздошная кость; is.—
Обратит?3*1 кость; %—foramen obturator; Р.—pubis; pr. ре.—processus peclinealis.
количрпт Внпмание па прогрессивный в ряду приведенных видов процесс увеличения
на поет а П03в0нк0в» сливавшихся в synsacrum, и увеличения и удлинения тазовых костей,
степенный поворот в положении лобковых костей спереди к положению параллельно
седалищным костям и на увеличение слияния костей. Из Heilinann.
Не может быть никаких сомнений, что крылья и хвост юрских
Птиц несли развитые маховые и рулевые перья и птицы были покрыты
*
339
перьями и на большей части тела, за исключением головы и плюсны.
Маховые перья археорнис, в количестве не менее 10 больших п
значительно большего числа малых, обладали узкими внешними
и широкими внутренними опахалами, что характерно для крыльев
с летающей функцией. Наибольшая длина 5 и 6 больших маховых
указывает на округлость крыла и приспособленность его к полету
через короткие расстояния. Маховые укреплялись на метакарпаль-
ных отделах и основных фалангах 2-го и 3-го пальцев, в то время
как 1-й палец был совершенно свободен от маховых, и alula очевидно
отсутствовали. Некоторые из маховых достигали по меньшей мере
13 см длины. Рулевые, обладавшие сходной структурой, попарно
располагались по краям хвостовых перьев, мржет быть за исклю-
чением 5 проксимальных позвонков; таким образом в хвосте было
не меньше 40 рулевых, достигавших в средней части хвоста 65 мм
в длину.
Анализ рассмотренных особенностей позволяет видеть в юрских
птицах еще плохих летунов, ведших преимущественно, древесный
образ жизни. Птица лазила преимущественно в сплетении ветвей,
цепляясь не только пальцами ног, но и когтями крыльев, и реша-
лась перелетать только короткие расстояния, преимущественно с бо-
лее высокого пункта, вниз, к основанию дерева. По размерам тела
(величиной с Сороку) и величине и форме несущей крыловой поверх-
ности юрская птица должна была обладать слабым скользяще-греб-
ным полетом, незначительная сила которого видна из довольно
рыхлого соединения костей плечевого пояса и строения грудного
отдела позвоночного столба. Как на земле, так и по ветвям птица
передвигалась очевидно на всех четырех конечностях, сходно с яще-
рицами, как это следует из отставленных вбок по бокам тела колен-
ных сочленений. Присутствие брюшных ребер при этом повидимому
указывает, что брюхо могло часто находиться в соприкосновении
с грунтом или корой дерева и требовало более существенной, чем
наружный покров, защиты. Наиболее полную гипотетическую кар-
тину среды, в которой обитали юрские птицы, и характеристику их
образа жизни дает Хейльманн (Heilmann, 1926).
«Представим себе,что мы находимся на одном из больших островов Юрского
моря. Теплый и влажный бриз обдувает наши лица; мы видим перед собой
темносииее море, разнообразящееся островами, уходящими за горизонт. Волны
разбиваются вдали от берега о коралловый риф, на котором греются на солнце
огромные морские рептилии. Прилив сейчас на исходе...
На земле, почва покрыта мелкими папоротниками и мхами; здесь еще не
имеется травы. Бриз слабо колышет большие листья низких, похожих на
пальмы деревьев на соседних холмах, но они являются саговниками, а не паль-
мами. Низкий кустарник составляет передний план небольшой рощицы, в на-
правлении к которой мы прокладываем наш путь; позади же многочисленные
древовидные папоротники поднимают кверху их изящные пышные листья.
В некотором отдалении, несколько похожих на араукарий хвойных поднимаются
лад ландшафтом, который выглядит подобным парку.
Общий вид не так странен, как можно было бы ожидать, но тем не менее
несколько монотонен, ибо мы не видим ни единого цветка где бы то ни было,
ни роя вьющегося около них насекомых. Здесь имеются, правда, некоторые
осы и немногие неуклюжие бабочки, но они не обладают органами для выса-
сывания меда.,
340
Прибой воли и рев ветра — единственные звуки, достигающие уха, в ином
отношении природа почти нема. Нет шума и гама чаек по берегам, рептилии
немы, по тонкое слабое стрекотание кузнечиков уже слышно...
Внезапно оперенное существо снимается с вершины древовидного папорот-
ника, плавно планируетв воздухе, несомое расправленными крыльями, и пытается
поймать стрекозу. Но последняя уклоняется ловким поворотом, и птица про-
должает скользить немного, прежде чем она способна остановиться. Наконец
ей удается повернуть, она хлопает маленькими крыльями и пытается под-
няться несколько выше в погоне за ее сверкающей добычей. Ее движения выгля-
дят слабыми и неловкими, очевидно требующими значительных усилий, и оказы-
ваются безуспешными. Полет стрекозы легок и проворен; с легкостью она
надувает тщетные усилия птицы и очевидно рассматривает это преследование
не более как занятный спорт.
Немного спустя, однако, мы видим, как стрекоза парит в воздухе, выше
вершин деревьев, оставляя птицу тяжело хлопать крыльями вниз к корням
ствола саговника и быстро взлезать к вершине. При ближайшем рассмотрении
эта птица оказывается видом, напоминающим Археорнис. Крылья частично
сложены и цепляются когтями трех свободных пальцев за грубую древесную
нору. Хотя тело покрыто перьями, вид голой головы и шеи производит впечат-
ление, что животное является рептилией, маскирующейся в чужие перья. Что
касается головы, то кажется едва ли возможным, чтобы она принадлежала
птице.
Мы не могли бы назвать переднюю час^ь зубастых челюстей клювом, ибо
па них отсутствует сплошной роговой покров и они покрыты толстой кожей,
укрытой крупными щитками, как на голове ящерицы. Ноздри открываются
спереди морды, как у рептилий, ирис глаз сверкает кораллово-красным и ушное
отверстие хорошо заметно. Голая грубая кожа сера, перья на шее черные с бе-
лым, в то время как остальное перо бледно-коричневое.
Тем временем Археорнис поднялся по стволу и прыгает на больших перистых
листьях; наконец он комфортабельно уселся на средней жилке одного из них п
начал издавать ряд грубых хриплых криков. Момент спустя, другой Археорнис
скользит в воздухе и садится поблизости к нему. У этой новой птицы голые
части головы и шеи красноваты, ее оперение густого стального цвета и более
роскошно. Это очевидно самец, который сейчас же начинает прыгать около пер-
вой птицы. Он наклоняет низко голову, приподнимает красивые голубоватые
перья спины и распускает перья длинного хвоста, как бы показывая всю их
красоту. После этого он поднимает голову кверху и надувает мощный горловой
мешок, который постепенно приобретает яркий красный оттенок...
Но в этот самый момент идиллия приходит к концу. Второй самец опу-
скается с треском и пытается схватить самку за шею... Это возбуждает пыл
первого самца. С поднятыми перьями и открытой пастью он бросается на своего
соперника и они сцепляются в драке. Они схватили друг друга зубами и пока
пх перья развеваются во всех направлениях, они находят дорогу до кожи и мяса
ДРУГ друга их длинными и острыми когтями крыловых пальцев. Кровь течет
обильно, и, как бесформенный ком перьев, два соперника падают с дерева па
землю, где борьба продолжается среди листьев папоротника.
Оставленная самка является конечно теперь заинтересованным зрителем,
смотрящим вниз с листьев саговника. То один, то другой самец появляется на
поверхности, каждый пытается освободить себя и обратиться к самке с но-
вым ухаживанием; они забыли обо всем вокруг них.
Вдруг мы замечаем изящное маленькое существо, движущееся с быстротой
стрелы через открытый подлесок. С маленькими передними лапками, прижа-
тыми к груди, головой и передней частью туловища, вытянутыми вперед, п
хвостом, приподнятым кверху, оно кажется летящим по воздуху, несомым силь-
ными задними конечностями. Это Компсогнатус (Comps о gna thus) в поисках
добыли.
Как только маленький динозавр достигает катающегося клубка перьев
сРЭДи папоротников, он останавливается на момент разобраться в происходя-
щем. Клубок теперь превращается в двух самцов, один из которых лежит бес-
сильным на земле, в то время как другой поднимается на ноги, тряся своими
перьями.
Тем временем самка на дереве заметила проворную рептилию и разра-
жается резкими пронзительными криками предупреждения. Но ее самец внизу
341
кажется "не обращает на это внимание, напоенный победой и гордостью; он
поднимает голову, чтобы объявить о победе всему миру, когда внезапно Компсо-
гнатус прыгает на него, и крик победителя замирает в приглушенном вере-
щанье.
Одолев птицу, маленький динозавр садится на задние ноги (по способом,
совершенно отличным от кенгуру). Он деликатно и изящно сложен, с роговыми
зернистыми щитками, которые на спине становятся несколько больше и сли-
ваются друг с другом, образуя как бы хорошо заметный продольный гребень.
Ноги его очень похожи на ноги птицы; тремя пальцами обеих передних лапок
он держит теперь добычу и пожирает...»
Как указывалось выше, несмотря на наличие смешанных черт
п противоречивость в мнениях специалистов, юрских птиц прихо-
дится относить в класс птиц, рассматривая лишь в качестве наибо-
лее примитивного и самостоятельного подкласса. Собственно пере-
ходное звено — Proavis пли «предптицу» приходится создавать
гипотетически, исходя из анализа тех морфологических особенно-
Рис. 219. Реставрация скелета — Orni lliosuchus Woodxvardi.
Слева — S — отпечатки чешуи с центральным стержнем и боковыми насечками, напо-
минающими перо. По Hellmann.
степ, которые свойственны, с одной стороны, юрским п современным
птицам, с другой стороны, ближайшим к ним группам рептилии.
Из последних наиболее близки к птицам и являются вероятно исход-
ной группой для них так называемые псевдозухии (Pseudosuchiay
отряд вымерших рептилий, наиболее известный из триаса, но обо-
собившийся повидимому уже с пермского возраста. В этой группе
находят целый ряд родов, в которых различные исследователи
видят предков таких групп, как динозавры, птерозавры и кроко-
дилы. Ряд мелких родов из этого отряда, особенно Ornithosuchiis
(рис. 219) и Euparkeria, обнаруживают наибольшую морфологи-
ческую близость и к юрским птицам. Наиболее выделяет их в этом
отношении наличие следующих особенностей: мощно развитые зад-
ние конечности, могущие всецело являться опорой для тела, хотя
бы временно, как у современных ящериц Chlamydosaurus] более
слабые передние конечности, имеющие значение схватывающего
органа и иногда совершенно не принимающие участия в движении,
длинный хвост, челюсти, снабженные сидящими в альвеолах зубами.
Ряд особенностей указывает на большую близость в строении и
в других органах. В плечевом поясе имелись ключицы, кисть перед*
них конечностей состояла по меньшей мере из трех лучей (I—Ш)-
В тазовом поясе лобковые кости были направлены вперед и вниз,
342
а седалищные назад; те и другие сливались в попарном симфизе;
тазовые кости срастались с очень малым числом крестцовых позвон-
ков. Задняя конечность была типичной пятилучевой конечностью
рептилий, без противопоставления 1-го пальца. У Euparkeria однако
1-й палец был уже слебее развит, чем 3-й, что указывает на более
приподнятое положение тела при движении. Брюшная область была,
очевидно в связи с преобладающим передвижением на задних ко-
нечностях, укреплена брюшными ребрами. Череп являлся акинети-
ческим, с неподвижной квадратной костью и большими преорби-
тальными ямками. Наружные покровы состояли из чешуй очень
оригинального строения. У Euparkeria эти чешуи имеют вытяну-
тую форму и прободены осью, от которой по бокам ответвляются
короткие бороздки, так что чешуйка напоминает строение пера.
Рпс. 220. Реставрация—Ornilhosuchus Woodward!. По Heilmann.
Внешний вид ближайшего к птицам Ornithosuchus, в реконструк-
ции Хейльманна, избражен на прилагаемом рисунке (рис. 220).
Ход эволюции этих мелких рептилий в ту стадию, которую мы
находим в археоптериксе и в еще более поздних и рецептных пти-
цах, различными авторами представляется в разном виде. Боль-
шинство [в том числе Гексли (Huxley, 1868, 1870); Марш (Marsch,
1880); Абель (Abel, 1911); Биб (Beebe, 1915) и др.] считают более
вероятным приобретение предком птицы вначале способности пры-
гать и лазить по стволам и ветвям деревьев, и уже только в про-
цессе этой древесной жизни у них развились и удлинились крыловые
перья, образовав первичное крыло (рис. 221). Меньшее число авторов
[среди них Фюрбрингер (Furbringer, 1888); Брум (Broom, 1913);
Нопкса (Nopcsa, 1907,1923) и др. ] допускают развитие птиц из наземных,
бегающих на двух ногах предков, поднявшихся в воздух в силу той
помощи, которую должны были оказывать новообразующиеся крылья
па сильном бегу или при прыжках в воздух. По нашему мнению,
последняя гипотеза является весьма спорной и маловероятной.
• 343
Последнее время значительное внимание привлекло предполо-
жение Биба (Beebe, 1915) о четырехкрылой стадии в эволюции птиц,
основанное на наблюдении крупных зачатков перьев па задней
Рис. 221. Гипотетический предок птиц — предптица, Proavis, по представлениям
Хейльманна. По Heilmann.
стороне бедра у эмбрионов и птенцов некоторых голубей и других
птиц, что рассматривалось как рудимент некогда более сильно раз-
витого здесь ряда маховых перьев (рис. 222). Подкреплением для
этой гипотезы является нахождение в области задней конечности
аналогичных по функции перьев на отпечатке у археоптерикса.
Согласно этому воззрению, предок птицы жил среди лесной обста-
344
новей и, прыгая с дерева не дерево, постепенно приобрел способ-
ность парашютировать благодаря развитию удлиненных перьев, по
Рис. 222. Рудименты тазового крыла у птенцов птиц.
(а) и (Ь) —четырехдневный птенец белокрылой горлицы — Me lope На asialira; (с) и
(d) — трсхдневный и несколько старше птенец сизого домашнего голубя; (в) и (/) — птенцы
Фазана и хохлатого баклана — Phalacrocorax graculus L., без признаков раппего развития
перьев в области бедренной птерилии; д — бедренная птерилпя зеленого дятла — Picas
tiridis L. По мнению Хейльмапна, «тазовое крыло» птиц есть лишь ненормально ускоренное
развитие перьев, без признаков их дальнейшей деградации и, следовательно, нс рудимент
далекого прошлого. По Beebe и Heilmann, из Heilmann.
задним краям обеих конечностей и хвосту, построенному, как у ар-
хеоптерикса (стадия Tetrapteryx, рис. 223). Но в процессе эволюции
функция крыла у передних конечностей развилась далее, а задние
конечности стали спе-
циализироваться в ином
направлении и утеряли
парашютирующее зна-
чение; археоптерикс
представляет уже ста-
цию значительной ре-
дукцией этого заднего
летательного аппарата;
в дальнейшем через ста-
дию -Оуор^п/яконструк-
Ция птицы приобрела
°°лее современный об-
лик. Однако последую-
щим авторам (Heilmann,
1926) не удалось найти
Рис. 223. Предполагаемая четырехкрылая стадия Те tra-
pteryx в ряду предков птиц, в сравнении с первоптпцен
Archaeornis.
(а) — Telrapleryx; (b) —Archaeornis. По Beebe и Hell-
mann, из Heilmann.
I
345
подтверждений существованию у эмбрионов рудиментов истинно
маховых перьев в области бедра, и гипотеза поэтому остается на
умозрительной стадии.
Филогенетически в эволюции птиц большой интерес представ-
. ляют еще меловые птицы (из Канзаса Северной Америки) Ichthyornis
п Hesperornis, главным образом вследствие наличия зубов в их
челюстном аппарате. Обе группы обладали уже весьма совершен-
ным строением и весьма близко напоминали, с одной стороны, чаек
(Ichthyornis), с другой — современных гагар. Ichthyornis прекрасно
летал, что доказывает строение скелета его крыла и грудины с боль-
шим килем; Hesperornis утерял способность летать повидимому вто-
рично, вследствие односто-
ронней специализации (рпс.
224). Вследствие наличия
признаков зубов у ископае-
мых преставителей некото-
рых других отрядов, частью
даже более поздних по возра-
сту, в последнее время суще-
ствует тенденция рассматри-
вать наличие зубов у иско-
паемых форм как некоторую
филогенетическую стадию,
проходившуюся рядом отря-
дов независимо. Согласно
этой точки зрения, Ichthyor-
nis и Hesperornis размеща-
ются, как отряды, среди со-
временных отрядов птиц,
располагаясь в соседстве
Colymbijormes и Lari и Limi-
colae. По другой точке зрения
они являются одним из трех
224. Реставрация скелета Hesperornis regalis НЭДОТрЯДОВ ПТИЦ, ПриурОЧбН
Marsch из верхнего мела Канзаса. По Marsch, нь!М ВСвпелО К мелОВОМУ
из Stresemann. м *
времени и уже в то время
.достигшим значительного разнообразия в своих адаптациях (см.
ниже).
Второй надотряд птиц, представленный из современных форм
преимущественно нелетающими видами (кроме скрытохвостов), вслед-
ствие глубоких различий между отрядами, представляет, возможно,
полифелитически возникшую группу. Большинство авторов схо-
дится в своих взглядах на потерю этими видами способности летать,
вследствие как специализации к бегающему способу передвижения,
так и по причине достигнутого в связи с этим передвижением крупного
роста. По их мнению, отдельные отряды этой группы независимо
потеряли способность летать и приобрели современные сходные черти
вторичным путем, как следствие приспособления к сходному образу
жизни. Немногие авторы (например Lowe, 1931), наоборот, считают
их неспособность летать первичным явлением й всю группу Ratitae —
:346
возникшей монофилитически. Повидимому более правильным яв-
ляется все же рассматривать весь подотряд Palaeognathae как объеди-
нение сравнительно древних форм, дошедших до нашей эпохи только
nlo Coracias
% Bucerotes
% Halcyones
Cotharls
Psitlaciformes
'ifialli
Irogoruformes
ColiifSoffrmes
Oiciformes passenformes
Vo Columbae
V0CucuLi
Micropodiuormes
% Opislhocomi
Colymbiformes^Hesperorms)
Sphentsciforme-
Dinormthiformes
/Epyorn 11h i form es
n/0 Pierer eles
n!o
VoLinicol#
ornilhi-
formes
Turnici-
formes.
%An/nmae
Vо An seres
1 Mill 2 » J Ж
Capnmulyi formes
Stripformes
%Ciconiap
Pelecomformes
VoMusopbogil л/
c Meapodudoe
Tinamuormes
Rheifo
Slrulhioniformes
Casuarii-
formes
n/0Crues
n/0Ralli
VoRhinochsli
% Europoyat
% Mesoenades
n/o Canamidse
Pro cellar: i formes
'Apteryyiformes
^Ilc- 225. Филогенетические отношения отрядов и некоторых подотрядов класса птиц по
представлениям Пайкрафта.
Номенклатура систематических групп, применяемая Пайкрафтом, заменена номенклатурой,
^меняемой в настоящей книге (см. «Краткий систематич. обзор кл. птиц», стр. 349—510).
2аллельная ветвям штриховка — группа PataeofiFnathae; косая штриховка — диаста-
^сические формы; клетчатая штриховка—эвтаксические формы; белые участки среди
'Риховкц — наличие в данной группе как эвтаксическпх, так и диастатаксических форм.
По Pycraft.
Немногих представителях и филогенетически представляющих более
Примитивную из рецентных видов ступень в эволюции изучаемого
^асса.
Филогенетические отношения в пределах отряда Новонебных
1TlI4 {Neognathae) также во многих случаях не являются достаточно
347
ясными. В нижеследующем систематическом обзоре взаимоотноше-
ния отдельных групп рассматриваются вкратце на основе воззрений
Гадова (Gadow, 1893), Фюрбрингера (Fiirbringer, 1888), Ветмора
(Wetmore, 1929) и Штреземанна (Stresemann, 1933), причем в основу
положена система Ветмора (1929), измененная лишь в немногих
деталях.
К сожалению, имеющиеся более новые таксономические системы
лишены попыток представить взаимоотношения форм посредством
филогенетического дерева. Более обоснованные попытки такого рода
принадлежат Пайкрафту (Paycraft, 1900) и Фюрбрингеру (Fiirbringer,
1888). Здесь приводятся обе эти схемы с целью отразить существую-
щие взгляды (рис. 225 и 226). Расхождения данных рисунка и форму-
лированной в тексте точки зрения по поводу положения той или
иной группы могут наглядно представить различие мнений на исто-
рический процесс эволюции птиц как в целом, так и в отношении
отдельных мелких групп.
Osccnes
Jracheophones
35
Pterocletess.
„Eurypygaet
JWicropodilform es
Striglforrnes
TodL.
Coractiformes
GO
65
Cathartae
,Gastornithes"
Anhimae-PaLamedeifonnes~
Aepyornithiformes.
6
CasuarLlformes„^_
StruthLordform es
Archaeopterygiformes.
105
/ \HalcyoncformBs.
_ .Caprlm и Lgiform es
Coraciae
Pels rgo - Herod u
__ Petecantformes
„ • i.
JjjCLnes
----1---Otigomyodi
.Jroh^U
.....-L......MtcropodU
.EuryLaemi
PtCLformes.
Meropes
CucuLl___
Musophage..^
_ Passeriformes
. Pico-Pas seres*
_CaULformes
__TrogonlfOrjnes
Hal clones.
Leo -Passeriformes
EueuUformes^.
Opisthocomt...
Galli..
Turniciformes.
J?rues*..
Psitta ciformes'.
Julia*»:..
Jchthyornithcformes. _
Tlnarntformes.
„ Apteryfftformes"^.
„ A lectorom lthes\. L
Рис. 226. Филогенетическое развитие класса птиц по представлениям М. Фюрбрппгера.
Названия отрядов и подотрядов, в тех случаях, когда их объем совпадает с принятом
далее системой Ветмора, Даны по этой системе* (см. «Краткий систематич. обзор кл. птиц»,
стр. 349—510); в других случаях сохранена номенклатура Фюрбрингера, причем названия
ввяты в кавычки. Из них упомянуты: отряды — Pelargornithes, Alectorornithes и Coracornithes,
подотряды — Palamedeiformes, Anseriformes, Pоdicipitiformes, C iconiiformes, Charadriiformes,
Ralliformes, Apterygiformes, Pico-Passeriformes, Halcyoniformes и Coraciiformes; надсемейства
(«gens») — Gastornithes, Hesperornithes, Pelargo-Herodzi, Jchthyornithes, Laro-Limicolae, Eury-
pygae, Grues, Fulicariae, Hemipodii, Pico-Passeres и Todi; объем (состав) этих групп ясен из
схемы. Цифрами обозначены семейства (и в некоторых случаях подсемейства и роды); отме-
чены: 1 — Archaeopterygidae; 2 —Hesperornithidae; 3 — EnaHornilhid^e; 4 — Ichlhyorni-
thidae; 5 — Struthionidae; 6 — Rheidae; 7 — Casuariidae; 8 — Dromaedidae; 9 — Dromorni-
thidae; 10 —Apterygidae; 11 — Dinornithidae; 12 — Tinamidae; 13 — Sphenisci dae; 14 — Pro-
cellariidae; 15 — Colymbidae; 16 — P о dicepe di dae; 17 — Phaelhontidae; 18 —Pelecanidac;
19 — Pelagornithidae; 20 — Sulidae; 21 — Phalacrocoracidae; 22 — Anhingidae; 23 — Frc-
gathidae; 24 — Ardeidae; 25 — Scopidae; 26 — Balaenicipetidae; 27 — Ciconiidae; 28 — Thres-
kiomithidae; 29—Phoenicopteridae; 30 — Palaelodidae; 31—• Anhimidae; 32— Mergidae;
33 — Anatidae; 34 — Anseridae; 35 — Cygnidae; 36 — Cnemiornithinae (по Фюрбрингеру,
курпцые гуси и нелетающие вымершие гуси Новой Зеландии); 37 — Cathartidae; 38 — Тега-
Accbpltres
____faiconcformes
.........Phoanicopteriformes
taro-Lim ico Lae ’
.. _CLeo niiformes
-Anseres
___Colymblfomes*
Hesperornithes
______„Jchthyorntthes"
Charadriiformes"
____________Coracornithes’'
. . Procellariifomes
CiconiLformes"
-----------------------„Pedlcipltiformes"
Ansertformes
. ..SpheniscLformes
___Rheiformes
thornithidae ♦ 39 — Sagittarii dae; 40 — Pandioninae; Лмрйгтвэ Guvo-Falco-
43 — Vulturinae (по Фюрбрингеру, 40—43 являются подсемействами семейства Gwpo a*ico~
nidae, по своему объему равными подотряду Accipitres нашей системы), Elphasidninae-
Megapodiidae; 46 — Phasianidae; 47 — Numidirtae; 48 - Mele^^rniL7
50— Tetraonznae; 51—род Дгдна; 52— Opisthowrudae; оЗ Ре^пп^интеоу вымеиший
cidae; 55 - Mesoenatidae; 56 - Rallidae; 57 - Aytornithidae (по^ Фюрбрингеру, вымецшия
род Aptornis); 58 — Helzornithzdae; 59 — Rhinochelidae; 60 — El77ypv9}^J,idle, 67 __ ™hin&-
62 _ Psophiidae; 63 - Aramidae; 64 — Gruidae; 65 - Ohdae; 66 ^^n^nididae- 71 7^
coridae; 68 — Bwrhinidae; 69 — Dromadidae; 70a — Glareohdae, 70b Chzonzaia^, 71,7
и 7 3 — три подсемейства — Charadriinae, Scolopacinae и ^hyn9hael^rid^7л — Surrha/ntzdae-
сером в пределах семейства Charadriinae; 74 — Laridae; 73 -/Ыае,76
77—Raphidae; 78 — Columbidae; 79 — Musophagidae^ 80 — Сисипичаемые фюп-
82 — род Strigops; 83, 84 — два подсемейства — Asionino? п.,^д^а± cip^imulgidae-, —
брингером в пределах сов; 85 — Steatornithidae; 86 — Poda^ozdac^^о. S — L^toSmatidae-
Micropodidae; 89 - Trochilidae; 90 - Coliidae; 91 - . 9Л - То^^-98 —
y.3 — Coracizdae; 94 — Alcedznzdae; 95 — Meropidae; 96 — . 102 — Bucconidae;
Upupidae; 99 — Phoeniculidae; 100 — Bucerotidae; igl - ^^^^idae- 107 — Pici-
103 — Rhaxnpha&txdae; 104 — Indxcatoridae; R>5 — Capitonidae, 106 yng , 34g_510)
dae; 108 — Eunjlaemidae; 109 — Atrichornithidae; jio - S^nzirzdae (см. стр.
По Furbringer.
краткий систематический обзор класса
птиц
[Положение в системе: второй класс надкласса SAuropsidA (первый
ласс — REPtiliA), пятый класс подтипа черепных — CrAniotA;
hniotA — третий подтип типа ChordAtA; ChordAtAj— седьмой тип
’АЗДЕЛА EumEtAZOA; EuiDEtAZOA — ВТОрОЙ РАЗДЕЛ ПОДЦАрСТВА MEtAZOA.]
ПОДКЛАСС I: SAUKUBAE (=АВСПАЕОК-
NITHES). —ЯЩЕРОХВОСТЫЕ ПТИЦЫ
Вымершие, известные по двум ископаемым экземплярам и от-
печатку одного пера, юрские птицы, характеризующиеся обильными
общими чертами с рептилиями. Диагностически важны: длинный
хвост, с большим числом уменьшающихся к концу хвоста позвонков,
несущих по бокам парные рулевые перья; наличие брюшных ребер,
несрастание метакарпальных костей и трех пальцев крыла и воору-
жение их когтями при наличии хорошо развитых маховых перьев;
шдичпе в челюстях конических зубов, сидящих в обособленных
альвеолах (подробности организации см. стр. 333).
Подкласс обнимает единственный отряд Archaeopterygifornies
единственным семейством Archaeopterygidae. Известны три вида:
bcliaeopteryx lithographica Meyer (перо), A. niacrura Owen и Archaeor-
siemensi Dames, из верхней юры (тптон) Баварии.
ПОДКЛАСС II: OKNITHUKAE (=NEOK-
NITHES).—ВЕЕРОХВОСТЫЕ
Обширный, обнимающий и всех современных птиц, подкласс,
ископаемом состоянии представленный начиная с меловых отло-
жений. Ornithurae являются, невидимому, прямыми потомками Sau-
чгм, развившимися из последних уже к концу юрского периода; 1
деловое время они обладали еще отчасти весьма примитивными для
Руппы птиц чертами, напр. зубами, ио были представлены уже как
етающпми, снабженными килем и пигостилем, птицами, так и целе-
йшими, с редуцированной передней конечностью, килем грудины
пигостилем.
1 Однако, по некоторым воззрениям, Archaeopteryx п Archaeornis представ-
ит ветвь, уклонившуюся в сторону, так что прямые предки современных
пщ остаются неизвестными.
349
Морфологически подкласс характеризуется срастанием метакар.
пальных костей и иным развитием пальцев: второй является наибо-
лее длинным пальцем, в то время как третий б. м. редуцировани
тесно прилегает ко второму. Брюшные ребра отсутствуют. Хвост
образуется не более чем 13 позвонками, причем 5 или 6 последниу
позвонков б. ч. срастаются в отдельную косточку (пигостиль), bj
которой укрепляются основания хвостовых перьев, располагающиеся
в виде веера.
НАДОТРЯД 1: ODONTHOGNATHAE — ЗУБОЧЕЛЮ-
СТНЫЕ
Ископаемые птицы меловых отложений Северной Америки п
Европы, характеризующиеся сохранением некоторых, впоследствии
исчезающих, рептильных черт. Диагностически важно наличие на-
стоящих зубов, сидящих вдоль краев нижней челюсти и на верхне-
челюстной кости (maxilla), в то время как межчелюстная кость была
лишена их. Замещение зубов происходило в вертикальном напра-
влении, как у крокодилов и динозавров. Характерно также отсут-
ствие соединений между дистальными концами седалищной п под-
вздошной костей.
Эта группа охватывала вероятно большую часть птиц мелового
периода, но в ископаемом состоянии представлена только двумя
отрядами, с представителями, ведшими водный образ жизни.
ОТРЯД 1: HESPERORNITHIFORMES. — ГЕСПЕРОРНИСЫ
Ископаемые зубатые птицы, характеризующиеся зубами, сидя-
щими в бороздках, длинной, широкой и плоской (без киля) гру-
диной, весьма редуцированными передними и сильными плаватель-
ными задними конечностями.
В эту группу объединяется ряд ископаемых меловых птиц, из-
вестных частью по весьма недостаточным остаткам, распределенных
в дв? семейства: Hesperornithidae и Enaliomithidae.
Семейство 1: Hesperornithidae
Объединяет наиболее хорошо известных представителей группы:
Hespernis regalis Marsch, Н. crassipes Marsch (рис. 227) и другие виды
из меловых отложений Канзаса,а также род Hargeria. По своим адап-
тивным особенностям эти птицы очень близко напоминают совре-
менных гагар, что обычно объясняется конвергенцией в приспо-
соблении к сходному образу жизни. Характерны мощные задние
конечности с четырьмя пальцами, несущими плавательные пластинки,
как у видов р. Podiceps. Передние конечности очевидно не функцио-
нировали, судя по сохранившейся тонкой и маленькой плечевой
кости. Таз отличался значительной массивностью, длиной и узостью.
Позвоночник состоял из гетероцельных позвонков, а в хвостовом
отделе был лишен пигостиля. Строение тела выявляет в птицах этой
группы мощных ихтиофагов, типа современных гагар. В отлпчпсот
350
последних, ветви нижней челюсти соединялись на конце хрящей, что
в совокупности с особым устройством квадратомандибулярного сочле-
нения позволяло значительное расширение цлюва, сходное с соответ-
Рис. 227. Hesperornis regalis Marsch.
Реставрация Хейльмаппа. По Heil шапп.
ствующим приспособлением некоторых веслоногих. Череп, обладаю-
щий общей конфигурацией и строением черепа гагар, отличался,
очень маленькой черепной полостью, но более значительной полостью
для обонятельных долей мозга. Hesperornis crassipes Marsch по длине
превышал сто сантиметров, т. е. был больше самых крупных совре-
менных гагар.
351
Семейство 2: Enaliornithidae
Нижне- и среднемеловые птицы, по характеру приспособлений
напоминающие гагар и гесперорнисов, но с амфицельными грудными
позвонками и менее полным срастанием метатарзальных костей.
Известны роды: Enaliornis, Neogaeornis, Baptornis, Eupterornis (палео-
эоцен), описанные частью по весьма фрагментарным остаткам.
Систематическое положение отдельных родов и всего семейства
спорно; при некоторых глубоко примитивных особенностях есть
черты весьма большого сходства с современными гагарами и поган-
ками.
ОТРЯД 2: ICHTIiyORNITHIFORMES. — ИХТИОРНИСЫ
Ископаемые птицы среднего и.верхнего мела Северной Америки
(Канзас), напоминавшие чаек п бакланов и обладавшие сильным
полетом. Характерны амфицельные позвонки и мощный грудной
киль. Строение передних конечностей и грудины указывает на зна-
чительное совершенство полета. Таз мал и широк. Задние конеч-
ности с совершенна сросшимися metatarsalia, напоминающими чаек.
Имеется пигостиль. Ветви нижней челюсти без крепкого симфиза,
сходно с Hesperornis.
Известно лишь одно семейство Ichthyornithidcie, с призна-
ками отряда. Наилучшей сохранностью обладают Ichthyornis victor
Marsch и dispar Marsch и некоторые другие виды того же рода. Кроме
того известен Apatornis celer Marsch и сомнительный по системати-
ческому положению род Cimolopteryx.
НАДО ТРЯД 2: PALAEOGNATHAE.—ДРЕВНЕНЕБНЫЕ
ПТИЦЫ
Преимущественно нелетающие птицы южного полушария, в це-
лом характеризующиеся обилием древних примитивных черт. Общим
.анатомическим признаком является примитивное строение неба:
сохранение свойственных рептилиям крупных первичных базипте-
ригоидных отростков основной клиновидной кости (processus basi-
рterygoi dei); тесное срастание (швом) передних частей птериго-
идов с небными косточками; крупный, занимающий значительную
часть крыши черепа — сошник (у более прогрессивных птиц сош-
ник мал, первичные базиптеригоидные отростки отсутствуют п,
в связи с этим, соединение р terygoi deum с palalinum достигается
сочленением, с образованием между ними сочленовных головок
(см. стр. 49).
Другими примитивными чертами (однако, прокидывающимися
п среди новонебных птиц, Neognathae} являются: наличие длинного,
вворачивающегося, как у рептилий, penis’a, остановка в развитии
колец крупных дыхательных путей на хрящевой стадии, отсут-
ствие у большинства представителей группы ппгостпля. Квадрат-
ная кость укреплена в черепе малоподвижно и лишь одной сустав-
ной головкой; орбитальная щель имеет сердцевидную или овальную,
а не щелевидную форму.
352
Многие другие своеобразные особенности, общие большинству
отрядов этой группы, образовались повидимому конвергейтно,
в силу сходных условий жизни. Изучение морфологии членов группы
указывает на значительную обособленность входящих в нее отря-
дов и независимое происхождение их от общего ствола птиц, на раз-
ных, но всякий раз весьма древних и близких ступенях его разви-
тия. Наиболее примитивными формами следует считать эму и страуса,
наиболее прогрессивными — скрытохвостов.
В целом вся группа ведет происхождение от летающих форм,
но большинство видов в процессе эволюции утеряли это свойство,
и к настоящему времени способность летать сохранили только скрыто-
хвосты. Основными причинами этой редукции повидимому являются:
Рис. 228. Грудины нелетающих птиц.
1 — казуар — Casuarins sp.; 2 — нанду — Rhea sp.; 3 — киви — Apteryx sp.; 1 и 2 —
с вентральной стороны; 3—с дорзальной стороны.
cs. — слившиеся между собой коракоид и лопатка, так паз. coracoscapula. По Owen,
из Stresernann.
частью развитие организации бегающего типа, происходившее па-
раллельно с увеличением размеров тела, в силу чего способность
полета могла теряться механически вследствие крупной величины
п тяжести животных; частью вследствие эволюции видов на удален-
ных от континента островах, лишенных существенных врагов для на-
земных птиц. Происхождение нелетающих форм из летающих доказы-
вается строением передней конечности по типу настоящего крыла
с маховыми, большими кроющими и т. п., несмотря на значительную
редукцию отдельных элементов. Прекращение функционирования
крыла повлекло за собой уменьшение грудной мускулатуры, в свою
очередь вызвавшее редукцию грудного киля (рис. 228). Параллельно
шло уменьшение и ослабление плечевого пояса, укорочение коракоида
и лопатки, синостотическое срастание их под тупым углом, исчез-
новение акрокоракоидного отростка лопатки, ключиц, исчезнове-
ние и уменьшение крыловой мускулатуры, уменьшение и печезно-
23 Орнитология—1209
353
вение ппевматпчностп костей (за исключением крупной бедренной
кости сильно развитых ног) и т. д. Потеряв функцию аппарата,
поддерживающего тело в воздухе, маховые и рулевые перья видо-
изменились в украшающие перья; это сопровождалось исчезнове-
нием различий в структуре пера между базальной и верхушечной
его частями и исчезновением разделения перового покрова на контур-
ное, пуховое и другие типы и исчезновением аптерий, еще замет-
ных на коже эмбриона. Рассученность пера делала ненужным ожи-
ривание его, вследствие чего подверглась редукции и копчиковая
железа.
Надотряд Palaeognathae разделяется на семь отрядов: Struthioni-
jorm.es, Rheijormes, С asiiariijormes, Aepyornithijormes, A pt er у formes.
Dinornithijormes и Tinamijormes.
ОТРЯД 1: STRUTH1ONIFORMES. — СТРАУСЫ
Крупнейшие из современных птиц, обитатели степей и полу-
пустынь Африки и Аравии. Вследствие недоразвития мускулатуры
и костных элементов крыла — нелетающие, превосходно бегаю-
щие птицы с двупалыми ногами (сохранились только третий и чет-
вертый пальцы). Тело покрыто черно-бурым или бурым оперением,
за исключением оголенной шеи и головы. По питанию являются
преимущественно растительноядными.
Единственное семейство Struthionidae, с одним рецентным видом,
страус — Struthio camelus L., известным в шести подвидах и еще
в плиоцене распространенным в южной Европе и пустынно-степной
Азии. Держится парами и группами, по природе являясь преиму-
щественно моногамом. Оба пола участвуют в высиживании и вожде-
нии птенцов, но основные заботы ложатся на самца. Гладкие, белые,
цвета слоновой кости, яйца откладываются в углубление почвы,
выскребанное самцом, в количестве до 12—15 штук, достигая весом
каждое 1600 г. Инкубация длится 42 дня. Птенцы выводковые,
воспитываются преимущественно самцом.
Взрослым птицам свойствен половой диморфизм, повидимому
стоящий в связи с хорошо выраженными брачными «танцами» и
криками самца; последний отличается от самки белыми перьями
крыла и хвоста, весьма пушистыми и своеобразно завитыми, и
черной окраской остального оперения. Вследствие спроса на эти перья
в галантерейной торговле страус ревностно преследуется, во многих
районах прежнего ареала уже истреблен и с прошлого столетия
разводится в полудомашнем состоянии в южной и северной Африке,
Калифорнии, Аризоне и некоторых других местах с промышленной
целью (страусовые фермы). Разводится в зоопарке Аскания Нова
(СССР), но пока без промыслового использования. Молодые имеют
диффузно пеструю окраску.
По своей организации страусы являются одними из самых свое-
образных и примитивных современных птиц, утерявших способность
к полету, гл. обр. в связи со своим гигантским ростом (до 260 см)
и весом (70—90 кг). Благодаря бегающему способу передвижения,
задние конечности развиты очень сильно, и двупалость есть явное
35.4
вторичное приспосооление к оегу, аналогичное процессу редукции
боковых пальцев в ряду предков лошади. Передние фаланги пальцев,
укорочены и наиболее длинный (третий) палец несет толстый, тупой
и широкий коготь, напоминающий копытце, в то время как на вто-
ром (четвертом) пальце коготь почти отсутствует. На бегу шаг птицы
достигает 4 м длины! Узкий, но сильный таз сохраняет примитив-
ные черты в виде выреза между седалищно-подвздошными костями,
(fissura ilio-ischiadica) и прочность симфиза лобковых костей, так
паз. таз закрытого типа. Редукция летательных способностей свя-
зана с значительной дегенерацией всего плечевого пояса п грудины.
Последняя, имея уплощенный (плоскодонный — отсюда название Ratl-
ine, применяемое к нелетающим представителям Palaeognathae и
установленное Мерремом, 1813) характер, ’лишенадаже следов киля,
но в задней части имеет широкое возвышение (proluberantia sterni),
на которое птица опирается, лежа на земле. Слившиеся между собой
коракоид и лопатка (korako-scapula) очень коротки, коракоид сильно
уплощен и расширен, крупный processus procoracoldeus лопатки
отграничен от коракоида покрытым мембраной окном; этот отросток
является вторичным образованием, аналогичным procoracoid’y реп-
тилий, и принимает, вместе с коракоидом, основное прикрепление
сохраняющейся еще крыловой мускулатуры (гл. обр. musculus sup-
racoracoideus и subcoracoideus), покидающей почти целиком место
прежнего прикрепления посредине грудины. Musculus supracoracoi-
deus, поддерживающий плечевую кость, вследствие более тупого угла
сочленения костей в плече, скользит к головке humerus в плоской
ложбине передней поверхности конца коракоида, -без образования
акрокоракоидным отростком известного блока, характерного для
летающих птиц. Вилочка уже утеряна; в крыле дистальные отделы
сильно укорочены в отношении к плечевой кости; карпальные эле-
менты локтевой стороны исчезли, но пальцы сохраняют нормальную
для Ornithurae структуру; первый и второй (редко и третий) пальцы
при этом на конце несут круто загнутые когти, что является при-
митивной чертой. Среди крыловых перьев можно различить 16 боль-
ших, 20—23 малых маховых и 4 пера, входящих в состав alula, но
опахалы их симметричны и рассучены, как и перья, покрывающие
все тело. В противоположность остальным Palaeognathae, страус еще
сохранил пигостиль, образованный 4 позвонками. Огромное коли-
чество рулевых (50—60) служит только для украшения и имеет
тот же рассученный характер.
Крайне своеобразным и примитивным является у страуса строе-
ние пищеварительного аппарата. Обычной пищей страуса являются
плоды пустынно-степных ксерофитов с толстой оболочкой, прогла-
тываемые целиком. В связи с этим клюв очень широк, а пищевод
растяжим. Твердые оболочки плодов перетираются в мускулистом
шелудке, обладающем крайне мощной мускулатурой и сильными,
перетирающими дисками. Между объемистым железистым и неболь-
шим мускулистым отделом включается обширный гладкостенный
отдел, подобно длинному мешку свешивающийся дорзально от
мускулистого желудка и служащий для временного хранения пищи.
Кишечник сравнительно прост, но чрезвычайно длинен, в 20 раз
♦
355
превышая длину туловища и достигая более 14 м длины. Особенно
длинна при этом прямая кишка, собранная в многочисленные извивы.
Сравнительно короткие слабые кишки (70 см) заставляют рассмат-
ривать удлиненную прямую кишку как место главнейшего расщеп-
ления трудно усвояемой целлюлозы, происходящего, как известно,
при деятельном участии бактерий. Этой чертой, курьезным образом,
страус вновь напоминает некоторых копытных (лошадей). В об-
ласти клоаки замечателен весьма обширный и растяжимый сорго-
daeum, являющийся по функции мочевым пузырем, накапливающим
мочу перед дефекацией.
Среди особенностей, связанных с размножением, следует отме-
тить видоизменение перьев крыла и хвоста в украшающие, отличаю-
щиеся особой пышностью и закрученностью; подобно остальному опе-
рению тела, они обладают весьма простыми и недифференцированными
далее бородками I порядка. Во время токования, сопровождаемого
очень своеобразными позами и маханьем развернутых крыльев (что
вероятно и является причиной меньшей редукции передних конеч-
ностей, чем у австралийских бескрылых птиц), самцы издают гром-
кий ревущий звук, сильно раздувая шею. Тем не менее сиринкс
весьма примитивно развит, голосовые мускулы отсутствуют и имеются
лишь внутренние перепончатые складки. Самки страуса немы.
Следует заметить, что строение неба у страуса уклоняется от струк-
туры, наблюдаемой у прочих Palaeognathae. Сошник не приходит
в соприкосновение с крыловидными костями, будучи вынесен далее
вперед. Движение верхней челюсти осуществляется только палоч-
ковидными небными костями, спереди упирающимися на небные
отростки верхнечелюстных костей, а сзади сливающимися с крыло-
видными костями. Последние (см. рис. 32), передавая движение
квадратной кости, скользят, упираясь на базиптеригоидные отро-
стки, вдоль клюва парасфеноида и передвигают этим небные кости
вперед и назад. Движение сошника осуществляется уже небными
отростками верхнечелюстных костей, входящих в соприкосновение
с ним вентрально от rostrum parasphenoidale.
Еще в историческое время страус был распространен очень ши-
роко, повидимому проникая на север до Сирии, Палестины, Аравии
и восточной Персии. Древние письменные источники упоминают
страуса в качестве птицы, привозившейся как подарок из Турке-
стана в Китай, что может указывать на обитание его в начале нашей
эры где-то поблизости от Средней Азии. Из плиоценовых и плейсто-
ценовых отложений находки страуса известны с южной Украины
и Забайкалья, так что распространение его в Евразии надо признать
весьма обширным в период существования так наз. Гиппарионовой
фауны. В ископаемом состоянии известно до 11 видов страусов.
ОТРЯД 2: RHEIFORMES. — НАНДУ
Крупные обитатели степей и полупустынь Южной Америки.
Подобно страусам, нелетающие, бегающие (с трехпалыми ногами)
птицы, с длинными волнистыми маховыми и кроющими крыла,
играющими существенную роль в брачных церемониях, но без хво-
356
дней
длится
Рис. 229. Найду — Rhea americana L.
По Evans.
стовых (рулевых) перьев. Преимущественно травоядные, ощипы-
вающие с земли нежные зеленые листочки, птицы.
Единственное семейство Rheidae с двумя рецептными видами —
Rhea americana L. и R. pennata Orb. (рис. 229), распространенными
от Патагонии до саванн Бразилии и высокогорных степей Перу.
Оба пола окрашены одинаково, в общем однообразной гл. обр.
бурой окраски, держатся группами, самец во главе нескольких
самок. Все заботы по высиживанию и воспитанию птенцов ложатся
на самца, который приготовляет гнездо в виде ямки в земле; самка
несет от 10 до 15 яиц, весом до 660 г.
Птенцы выводковые.
Мягкие рыхлые перья нанду, ли-
шенные добавочного стержня, хотя не
отличаются красотой страусовых
перьев, ценятся в торговле; ради них,
мяса и яиц нанду промышляются и во
многих местах уже истреблены. Успешно
акклиматизированы в Аскания Нова.
Внешне нанду наиболее близки
австралийскому эму, но как от него,
так и от страуса отличаются весьма су-
щественными особенностями. Пальцы
трехпалых сильных задних конечностей
заканчиваются сжатыми с боков ког-
тями. Более широкий чем у страуса
таз лишен симфиза лобковых костей,
но образует симфиз лобковых и седа-
лищных костей каждой стороны и зам-
кнутую incisura ischiadica. Крылья
стоят на гораздо менее значительной
стадии редукции, вероятно в связи с
активной ролью, играемой ими в брач-
ных играх. Кисть длиннее пред-
плечья, а в целом оба отдела длиннее
плечевого отдела, обратно тому, что наблюдается у страуса. Сохра-
няются все карпальные элементы, первый палец значительной ве-
личины и на конце несет острый коготь и пять перьев, составляющих
alula. Сохраняются 12 больших и 16 малых маховых, последние
перекрываются более длинными украшающими большими кроющими.
Соответственно более дифференцированному крылу, coracoscapula
обязует более острый угол, но вилочка отсутствует, a processus
procoracoideus менее развит. Грудина имеет более треугольную форму
с выступом, напоминающим protuberantia страуса. Рулевые совер-
шенно отсутствуют и хвостовые позвонки не образуют пигостиля.
В строении небного аппарата наблюдается значительное совпа-
дение с строением неба у казуаровых. Сошник отодвинут далее
кзади и соединяется швом как с небными, так и крыловидными ко-
стями, в то же время вилкообразно охватывая rostrum. Таким об-
разом движение quadratum передается *по осп черепа на верхне-
и нижнечелюстные кости не только через цепь крыловидных и неб-
357
ных костей, но и через сошник. Базиптеригоидные отростки здесь
являются как опорой всего небного аппарата в целом, так и специаль-
ным шарниром в движении крыловидных костей (рис. 33).
Пищеварительный аппарат менее специализирован, чем у страуса,
в связи с более нежной пищей. Мускулистый желудок значительно
слабее и лишен перетирающих пластинок; кишечник короче, не пре-
вышает восьмикратную длину туловища, но относительно короткая
задняя кишка восполняется крайне длинными слепыми кишками
с признаками спирального клапана, в целом столь же обширными,
как сам кишечник. Роль мочевого пузыря на себя принимает здесь
вместительная и существующая в течение всей жизни фабрициева
сумка.
Следует также отметить значительное развитие сиринкса, наи-
более выраженного у нанду среди всех нелетающих Palaeognathae:
он снабжен парой настоящих трахео-бронхиальных мускулов, mem-
brana tympaniformes и pessulus, с соответствующим образом видо-
измененными последними трахеальными и начальными бронхиаль-
ными кольцами. Голос самца, в связи с этим, состоит уже из двух
тонов.
Положение нанду в системе, будучи достаточно обособленным,
показывает вместе с тем смесь примитивных с сравнительно прогрес-
сивными для Palaeognathae особенностями, которые связывают его,
с одной стороны, с страусом, с другой стороны, с казуарами и осо-
бенно эму, и затем с скрытохвостами, с которыми они разделяют и со-
временную родину. В некоторых отношениях нанду можно рассма-
тривать как примитивных, стоящих на уровне эму и потерявших
способность летать, скрытохвостов. В ископаемом состоянии нанду
известен только из Южной Америки, не древнее плиоцена, откуда
из Патагонии описан весьма близкий Heterorhea dabbenei Rov.
ОТРЯД 3: CASUARIIFORMES. — КАЗУАРООБРАЗНЫЕ
Крупные нелетающие птицы Австралийской области, с глубокой
редукцией передних конечностей, и без рулевых перьев, с трехпа-
лыми бегательными ногами, с перовым покровом из двуветвистых
перьев [побочный ствол (стержень) столь же хорошо развит, как
и основной].
Казуары представлены двумя, хорошо характеризованными се-
мействами: Casuariidae и Dromaedidae. Для обоих семейств харак-
терен небный аппарат, напоминающий строение неба нанду, но с еще
более мощным сошником, сдвинутыми кзади базиптеригоидными^от-
ростками и крупными небными отростками верхнечелюстных костей,
срастающихся как с сошником (что не наблюдается у нанду), так
и с межчелюстной костью. Будучи сравнительно всеядными и пло-
доядными птицами, казуары имеют слабый и маленький мускули-
стый желудок и сравнительно короткий и широкий кишечник с ма-
ленькими слепыми кишками. Фабрициева сумка не функционирует
во взрослом состоянии. В крайне редуцированных передних конеч-
ностях характерна короткость дистальных отделов крыла, в целом
по размерам меньше плечевой кости, и слияние карпальных костей
358
с метакарпальными (с полной редукцией первого пальца). В задних
конечностях особой длиной отличается плюсна, много превышающая
размеры бедра; пальцы несут сильные сжатые с боков когти. Таз
весьма объемистый и широкий, характеризуется срастанием на кон-
цах седалищной и подвздошной костей. Далее характерна грубая
зернистость и густозеленая пигментация скорлупы яиц.
Семейство 1: Casuariidae. — Казуары
Один род Casuarius, обильный видами и подвидами на Новой
Гвинее, соседних частях Австралии и островах Ару, Дерам и др.
Рис. 230. Однолопастной казуар —Casuarius ипаррсп-
diculatus Bly th. По Evans.
(рис. 230). Обитатели густых тропических лесов, питающиеся глав-
ным образом опадающими плодами деревьев. Тело, заметно сжатое
Рис. 231. Крыло казуара — Casuarius casuarius L. с вну-
тренней стороны; маховые (с. г.) превратились в ого-
ленные стержни. Но Pycraft, из Streseinanii.
с боков, весом до 80—100 кг, покрыто черным волосовидным опере-
нием, на шее и голове имеются голые ярко окрашенные мясистые
359
части и костяной шлем. Клюв сжатый с боков. Внутренний палец
ног б длинным острым когтем, употребляющимся в качестве защиты.
Маховые, в виде шести голых стержней, сидят на крайне редуци-
рованных крыловых костях, почти незаметных в оперении тела
(рис. 231). Преимущественно моногамы; родительские заботы лежат
гл. обр. на самце. Самка кладет 3—5 изумрудно-зеленых яиц.
В ископаемом состоянии известен 1 вид из плейстоцена Ав-
стралии.
Семейство 2: Dromacdidae. — Эму
Один род Dromaius с единственным рецептным видом — D. по-
vaehollandiae Lath, в Австралии и Тасмании. Обитатель кустарни-
ковых песчаных пустынь и степей, питающийся гл. обр. плодами
и ягодами. По общему хабитусу напоминает нанду, с уплощенным
сверху вниз клювом, оперенной головой, общим серо-бурым опере-
нием, без «шлема» на голове, общим весом достигая 45—55 кг. Махо-
вые (7 больших и 10 малых) с побочным стержнем, очень коротки
и недифференцированы. Все крыло не более 20 см длины и мало за-
метно в оперении тела.
Преимущественно моногамы, с частичным участием и самки в вос-
питании птенцов. Кладка до 9 яиц, весом каждое до 600 г. Инкуба-
ция длится 56 дней. Птенцы выводковые, с темным продольнополо-
сатым рисунком по светлому фону.
В сравнении с казуаром, явно специализированным к жизни
в чаще, эму представляется более генерализованной птицей. Отно-
сительно более крупное чем у казуара крыло находится на еще боль-
шей ступени редукции, так как сохранило лишь один палец, воору-
женный когтем, и единственную metacarpale. Несмотря на это,
в плечевом поясе сохранились рудименты ключицы, и коракоиды
соприкасаются у своего основания. В связи с более грубой раститель-
ной пищей эму имеет более длинный кишечник — 330—360 см,
в то время как у казуаров он не превышает 153 см и чрезвычайно
широк (пища — мясистые плоды). Замечательно для эму значитель-
ное развитие обонятельных долей и бугров, указывающее на род-
ство с бескрылыми. Своеобразным также является эластический со-
едините льно-тканный трахеальный мешок, надуваемый в брачный
период воздухом, выходящим из трахеи через отверстие неполных
колец на вентральной стороне и вызывающим громкие барабанные
звуки.
В ископаемом состоянии известно 4 вида эму из плейстоцена Ав-
стралии и один мелкий вид вымер в историческое время на острове
Кенгуру.
Наиболее близким родством казуаровых следует считать нанду-
В ископаемом состоянии близкая форма Genyornis newtoni Stirl. &
Zeitz., раза в два превышавшая по размерам казуаров, известна
из плейстоцена южной Австралии. Несколько сомнительно родство
также весьма крупной формы Dromornis australis Owen из третичных
отложений Австралии, выделяемой некоторыми в особое семейство
Dromornithidae,.
360
ОТРЯД 4: AEPYORNITH1FORMES. — МАДАГАСКАРСКИЕ
СТРАУСЫ
Вымершие гигантские страусоподобные птицы, высотой до 3 м
известные из плейстоценовых отложений Мадагаскара, эоцена и оли
гоцена Египта и северной Африки. При общем сходстве с страусами
отличаются чрезвычайно мощными
трехпалыми задними конечностями
и очень широкой грудной костью;
судя по маленькой сохранившейся
плечевой кости, ненайденное крыло
должно было стоять на глубокой
ступени редукции.
Найденные части скелетов и
осколки скорлупы яиц относят к
одиннадцати видам и пяти родам
одного семейства • Aepyornithidae.
Наилучше известный мадагаскарский
Aepyornis (рис. 232), повидимому по-
добно казуарам, являлся лесной
птицей и возможно исчез уже в
историческое время, вместе с возра-
ставшим обезлесением острова. Судя
по структуре поверхности теменных
костей, у некоторых видов Aepyornis
на голове был украшающий хохол из
крупных перьев. Замечательны хо-
рошо сохранившиеся яйца Aepyornis,
достигавшие 9—12,5 кг весом, более
.чем в 5—7 раз превышающие по объ-
ему яйца страуса (рис. 233). Известны
роды: Stromeria, Mullerornis, Aepyor-
nis, Eremopezus и Psainmornis.
Отношение мадагаскарских стра-
усов к другим отрядам недостаточно
ясно, несмотря на хорошую сохран-
ность ряда находок. Ближайшими
формами рассматриваются моа, но
Рис. 232. Скелет Aepyornis maximus Is.
G-eoffr., из плейстоцена юго-западного
Мадагаскара. Рядом (правая фигура)
скелет страуса — Struthio camelusL. По
Monnier, из Stresernann.
сходство, возможно, вызвано конвер-
генцией в связи с гигантским ростом
птицы (мощные задние конечности и
таз). Своеобразными являются гру-
дина с широко расставленными коракоидами, значительный плече-
вой пояс, сильно пневматизированные бедренные кости, значитель-
ная короткопалость и размеры яиц.
Есть указания на недавнее существование этих птиц па Мада-
гаскаре, так как были находимы кости с следами обработки их чело-
веком. Поэтому с мадагаскарскими страусами связывают рассказы
арабских сказок о гигантских птицах «рок», переносивших в своих
когтях слонов, отнесенные еще^Марко-Поло и др. к Мадагаскару.
361
ОТРЯД 5: APTERYGIFORMES. — БЕСКРЫЛЫЕ
Нелетающие, частью уже почти истребленные птицы Новой Зе-
ландии, с крайне редуцированным крылом, четырехпалыми ногами.
Рис. 233. Яйцо Aepyornis (? maximus Is. Geoff г.) из плейстоценовых отложе-
ний Лмбовомбе, Мадагаскар; рядом яйцо курицы. Пз Lambrecht.
‘С сходным с типом казуара, но уклоняющимся строением небно-
челюстных костей. Сошник срастается с небными и птеригоиднымп
костями, не сочленяясь с небными отростками челюстных костей.-
Базиптеригоидные отрост-
ки хорошо развиты. Перья,
покрывающие тело, лише-
ны побочных стержней.
Единственное семейство
Apterygidae — киви, с од-
ним рецентным родом Apte-
ryx, с тремя видами на
обоих островах Новой Зе-
ландии (рис. 234). Обита-
тели густых сырых лесов,
добывающие пищу (червей
и личинок насекомых)
длинным клювом в мягкой
земле. Величиной с круп-
Гпс. 234. Киви — Apleryx australis Shaw. По Evans. ПЫХ Кур, НВ СИЛЬНЫХ Н0-
гах, с рыхлым бурым
оперением, почти совершенно скрывающим крошечные крыловые
косточки.
Моногамы. Самка больше самца (2,5 и 1,5 кг), родительские за-
362
боты лежат гл. обр. на самце. Кладка помещается в небольших
норах и состоит пз одного, редко двух, очень крупных (450 г) яиц,
весом до !/5 веса матери. Птенцы выводковые, по типу п окраске
оперения весьма сходные с взрослыми птицами.
По своему строению киви представляет весьма специализиро-
ванную, занимающую нишу в биоценозе, аналогичную нише на-
шего вальдшнепа, форму, с приспособлениями, развившимися на
.весьма примитивной основе. Пневматизация скелета полностью
отсутствует, и наблюдается сравнительно сокращенное развитие
воздушных мешков. Ноги очень мощны, приспособлены к раз-
гребанию земли. Первый палец удержался как почти не функ-
ционирующий придаток. Крупный, но узкий таз сохраняет при-
митивную incisura ischiadica, но не образует симфиза. лобковых
костей; дистальные концы ischium сильно расширены. В передней
конечности скелет чрезвычайно короток, дистальные отделы его
в целом вдвое меньше плечевого отдела и находятся в состоянии по-
стоянного сгиба .(под углом в 45е). Кпсть сохраняет лишь одну (редко
две) carpale и 2—3 элемента второго луча, на конце вооруженного
острым когтем и несущего 13 слабо отдифференцированных коротких
маховых перьев. Однако прекрасно сохраняется еще, м. б. благо-
даря постоянному сгибу, pro- и metapatagium. В плечевом поясе
отсутствует вилочка, coracoscapula изогнута под углом всего в 130°,
грудина характерна своей шириной и обширными вырезками по зад-
нему краю. Хвостовые позвонки образуют еще небольшой пигостиль
(пз 2—3 позвонков), но рулевые перья уже исчезли.
В поисках пищи киви руководится гл. обр. обонянием, что до-
казывается сильно развитыми bobi olfactorii, обширными обоня-
тельными полостями и проникновением обонятельного нерва в них
через ситовидную пластинку. Ноздри находятся близ самого конца
длинного клюва, что среди птиц свойственно только киви и явно
стоит в связи со способом добывания пищи. Клюв при этом окружен
длинными осязательными вибриссами. Маленькие глаза находятся
на пути к редукции, pecten уже почти исчез, lobi optici мозга очень
малы. Птица становится активна только в густых сумерках.
Пищеварительный аппарат отличается значительной длиной (до
150 см), с довольно длинными, но узкими слепыми кишками, напо-
минающими caeca скрытохвостов. Мускулистый желудок соответ-
ственно роду пищи слаб, язык несмотря на длинный клюв — очень
короток.
Следует отметить весьма примитивный сиринкс, лишенный му-
скулов, но обладающий membrana tympaniformes. Трахея в целом,
подобно предыдущим отрядам, остается на хрящевой стадии раз-
вития. Крик киви — короткий и резкий свист.
В отличие от всех прочих Palaeognatliae обладает копчиковой
железой.
По систематическому положению бескрылов следует рассматри-
вать как резко уклонившуюся в сторону, специализированную ве-
точку того ствола, который привел к образованию эму и ответвил
нанду. Ряд особенностей, вероятно конвергентного значения, сбли-
жает бескрылов также с примитивными пастушковыми и скрытохво-
363
стыми. В ископаемом состоянии известны два очень близких рода
Pseudapteryx и Metaptepyx; последний из плейстоцена Австралии.
ОТРЯД G: DINORN1THIFORMES. — МОА
Крупные вымершие страусообразные птицы Новой Зеландии^
характеризующиеся полным отсутствием крыльев, крайне мощным
Рис. 235. Скелет и левая задняя конечность
представителен отряда Dinornithi formes: а —
Euryapteryx elephantopus Owen., из плейстоцена
Южного острова Новой Зеландии; Ъ — Dinornis
giganteus Owen., из плейстоцена Северного острова
Новой Зеландии. Высота скелета 154 см. По Owen,
из Stresemann.
развитием четырехпалых ног
и двойными, с побочным
стержнем перьями, покры-
вавшими тело.
Единственное семейство
Dinornithidae — моа, в кото-
ром известно пять родов с
двумя десятками видов, пред-
ставленных нередко почти
полными скелетами из плей-
стоценовых отложений Но-
вой Зеландии и частью еще
живших на островах перед
колонизацией их европей-
цами. Различают роды: Di-
nornis, Megalapteryx, Anoina-
lopteryx, Emeus и Euryapteryx
(рис. 235). Преимущественно
крупные формы, от метра до
3,3 м высоты.. Повидимому
это были лесные птицы, в
значительной степени питав-
шиеся корнями папоротников
и других растений, которые
добывались разгребанием зем-
ли лапами.
По своему строению моа
обнаруживают тесное родство
с бескрылыми, с сходными
чертами с эму и отчасти с
А еру orn is; последнее сходство
имеет преимущественно кон-
вергентный характер. Заме-
чательны весьма сильные зад-
ние конечности с особенно
мощными и укороченными
плюснами, несущими так-
же сравнительно короткие
пальцы, массивное тело и толстая довольно длинная шея. Череп
сравнительно мал, с коротким тупым или загнутым крючком книзу
клювом. Строение небного аппарата, глазниц и обонятельных поло-
стей указывает на родство с киви, несмотря на другую форму че-
репа. Также весьма близка, снабженная вырезками, грудина, не
364
сохранившая даже сочленовной ямки для плечевой кости, но несу-
щая два-три полных ребра. Таз, по морфологии будучи сходен с та-
зом киви, отличается значительной шириной и мощностью, напоми-
ная в этом отношении таз Aepyornis. Строение и окраска перьев
весьма сходны с таковыми же у эму. Трахеальные кольца, в проти-
воположность остальным Palaeognathae, окостеневали. Кроме ске-
лета и перьев найдены многочисленные остатки яиц, не превышав-
ших по весу 2,2 кг, с разветвленными порами в толще скорлупы.
ОТРЯД 7: TINAMIFORMES. — СРЫТОХВОСТЫ (ТИНАМУ)
Внешне весьма сходные с куропатками, наземные, летающие?
мелкой и средней величины птицы Южной и центральной Америки-
В строении черепа характерно соприкосновение по средней ли-
нии небных костей и срастание их задними концами с сошником
и крыловидными костями (дромеогнатический тип неба). Послед-
ние, скользя по типичным базиптеригоидным отросткам, двигают
надклювье одновременно через сошник и небные кости, срастающиеся
спереди с неоными отростками
верхнечелюстных костей. Грудина
несет крупный киль; коракоид и
лопатка разъединены.
Скрытохвосты илитинамы пред-
ставлены лишь одним семейством
Tinamidae, с 5 родами: Tinainus,
Nothocercus, Crypturellus, Rhyncho-
tus, Calopezus, насчитывающими
около 50 видов (рис. 236). Они оби-
тают степи, кустарники и леса и
по своей биологии во многих отно-
шениях сходны с наземными ку-
риными. Характерна сравнительно
длинная и тонкая шея, с тесно при-
легающим оперением, и округлое
туловище с очень коротким хвос-
том, обычно скрытымв очень густом
Рис. 236. Большой скрытохвост — Rhyn-
chotus rujescens (Temm.). По Evans.
п длинном оперении надхвостья. Подобно куриным, пища гл. обр.,
но не исключительно, растительная. Самки обычно больше самцов.
Забота о потомстве ложится гл. обр. на самцов, и обычно наблю-
дается полиандрия: самки откладывают в углубления на земле от
1 до 10 яиц, распределяя их между отдельными самцами. Яйца за-
мечательны своей блестящей, одноцветной, но разнообразной окрас-
кой (от желтых и зеленых тонов до черной окраски) и значительно
более крупными размерами, чем, напр., яйца куриных. Инкубация
длится 19—20 дней. Птенцы выводковые, воспитываемые у большин-
ства видов самцом.
Строение скрытохвостов имеет многочисленные особенности, де-
лающие систематическое положение их довольно загадочным. Наи-
более вероятным является близкое родство их с нанду, но они об-
ладают также особенностями, общими им с трехперстками, пастуш-
366
новыми и куриными. По ряду признаков их можно рассматривать
как специализированную примитивную группу, пошедшую по «ку-
риному» адаптивному пути развития и конвергентно выработавшую
сходные особенности. На длительный путь эволюции указывают
значительные структурные отличия, имеющиеся между отдельными
родами группы. В ископаемом состоянии известны только пз плио-
цена Южной Америки (род Т inamisornis).
Из анатомо-морфологических особенностей внимание привле-
кают к себе следующие элементы. Грудина очень длинна, крайне
узка и снабжена очень длинными и узкими, пробегающими парал-
лельно, боковыми отростками, отчасти напоминая грудину куропа-
ток. Мощные и бледные, как у кур, грудные мускулы прикрепляются
как к гребню, так и к боковым отросткам, сохраняя соотношение
частей, близкое к куриным. Плечевой пояс прекрасно развит и снаб-
жен крепкой вилочкой. Крылья коротки, широки и выпуклы, с 10
большими маховыми. 4—5 грудных позвонков сливаются между
собой, таким образом и в этой особенности напоминая куриных.
Хвост однако, как указывалось, не развит, хотя 8—12 коротких
рулевых сидят на редуцированном, составленном 2—3 позвонками,
пигостиле. Таз очень широк, с глубокой и широкой incisura isch'ia-
dica, и весьма схож с тазом киви.
В пищеварительном аппарате замечательно наличие объемистого
зоба. Слепые кишки относительно короче, чем у куриных, но у Са-
lopezus своеобразно видоизменены и объемисты, в связи с ролью их
в период вождения птенцов (см. стр. 128). Далее замечательны при-
сутствие маленькой, обычно слабо оперенной, копчиковой железы
и примитивное развитие у большинства видов сиринкса. Дополни-
тельный (побочный) стержень в пере, хотя присутствует у большин-
ства видов, находится явно в состоянии исчезновения и дегенерации.
Наблюдается своеобразное сцепление опахалов всех перьев, осо-
бенно маховых. Среди всех Palaeognathae единственно скрытохвосты
располагают ясно выраженным птерилозом.
НАДОТРЯД 3: NEOGNATHAE. — НОВОНЕБНЫЕ
ПТИЦЫ
Огромное большинство современных и ископаемых птиц, в целом
характеризующихся более прогрессивным строением, чем птицы
двух предыдущих надотрядов. Небно-челюстной аппарат не дромеог-
натического или близкого к нему строения, а шизогнатического
и видоизмененных из него десмогнатического пли эгитогнатического
типов.
Надотряд объединяет 23 отряда.
ОТРЯД 1: SPHENISCIFORMES. — ПИНГВИНЫ
Средней и крупной величины (от 4 до 33 кг), нелетающие, с крьг
льями, видоизмененными в ласты, плавающие и ныряющие птицы
антарктических и субантарктических вод. Вместе с холодными те-
366
чениями проникают к северу до южных берегов Австралии, Новой
Зеландии, юго-западной Африки (до Китовой бухты), западных бе-
регов Южной Америки и даже Галапагосских островов. Весьма
своеобразные внешне птицы, обликом напоминают северных чисти-
ков, обладают вертикальной посадкой вальковатого туловища, ко-
роткими четырехпалыми, с плавательными перепонками лапками,
коротким хвостом, толстой шеей и обычно острым сильным клювом.
По питанию пингвины преимущественно ихтиофаги или потребители
головоногих и планктонных ракообразных. Пингвины гл. обр.
прибрежные птицы, на крайнем юге удовлетворяющиеся иногда даже
побережьями ледяного пака, на двухметровую высоту которого
они в состоянии выпрыгивать прямо из воды; обычно же живут по
каменистым берегам, гнездясь колониально и откладывая яйца в ще-
лях скал, россыпях, или делают гнезда, собранные из приморской
гальки. Гнездящийся на льду императорский пингвин согревает яйца и
птенца на перепонках лап, под защитой свешивающейся вниз складки
кожи брюха. В кладке, в зависимости от размеров птицы, 1—2 (до 3)
белых яйца. Оба родителя не отличаются окраской, обладают гром-
ким и грубым голосом, участвуют в высиживании и воспитании птен-
цов, иногда «общественно» выкармливая молодежь. Инкубация длится
от 31 (мелкие) до 53 (крупные) дней. Птенцы вылупляются слепыми
и развиваются медленно; спускаются на воду только .приобретя де-
финитивное перо, что у мелких происходит в возрасте 3, крупных —
4, 5 месяцев. Пингвины составляют единственное семейство Sphe-
niscidae с примерно 13—17 видами, относимыми к родам: Apteno-
dytes, PygosceliSj Eudyptes, Megadyptes, Eudyptula. и Spheniscus.
Биологически пингвины представляют собой глубоко специали-
зованную группу, с крайне оригинальным комплексом приспособле-
ний. Вальковатая форма тела, с короткой толстой шеей при мощных
эластических ластах-крыльях делает из них самых искусных плов-
цов (см. стр. 172). Крылья, при плавании вращающиеся почти винто-
образно в плече и почти несгибаемые в своей плоскости, замечательны
уплощением всего скелета и закреплением сочленений в полурастяну-
том состоянии. Последнее достигается в локтевом суставе образова-
нием двух сесамовидных косточек, включенных в сухожилие muscu-
lus triceps, а в кистевом составе — сильно увеличенной ulnare;
В обоих случаях окостенения исключают возможность значительного
сгиба и вместе с действием мускулатуры делают крыло весьма упру-
гим к давлению среды иа лучевую сторону его при плавании. Соот-
ветственно этой задаче, фаланга первого пальца подверглась полной
редукции, метакарпальные элементы почти полностью слились,
а фаланги 2-го и 3-го пальцев относительно укрупнились. Вслед-
ствие относительной неподвижности дистальных отделов крыла му-
скулатура их подверглась деградации и сокращению. Большинство
сгибателей и разгибателей кисти и предплечья сохранились только
на стадии сухожилий. Совсем отсутствуют обычные у птиц сгибатель
11 пронаторы предплечья: musculus biceps и musculus pronator su-
blimis nlprofundus. Мощно развита лишь мускулатура, вращающая
крыло в целом, особенно главный разгибатель предплечья muscu-
lus triceps, вместе с musculus latissimus dorsi, осуществляющий
3G7
гребущее движение крыла кзади. Musculus supracoracoideus, отво-
дящий крыло вверх, одновременно приводит к погружению туловища
под воду и вследствие этого развит исключительно сильно и превы-
шает по массе musculus pectoralis. Соответственно развитию этой
мускулатуры, грудная клетка весьма прочна, а грудной киль хорошо
развит. Имеющиеся восемь грудных ребер несут очень крупные pro-
cessus uncinati; особой крепостью обладают коракоиды с отростками
(procoracoid, acrocoracoid) ключицы и сильно расши-
ряющиеся кзади лопатки. Как маховые, так и их
кроющие полностью исчезли (см. дальше), и все
крыло оказывается покрыто тесно прилегающими че-
шуевидными перышками, весьма напоминающими
внешне чешуйчатый покров рептилии (рис. 237).
Необходимая водной хорошо ныряющей птице
подвижность туловища достигается легкой сгибае-
мостыо шейного отдела в сагиттальном направлении
и туловищного в боковом, сопровождаемом мощным
развитием мускулатуры. Разгибатель позвоночника
musculus biventer cervicis очень длинен; сильно
развит сгибатель шеи m. longus colli anticus с слу-
жащими ему для прикрепления гипапофизами перед-
них грудных позвонков. Подвижность грудного от-
дела позвоночника обусловлена как сжатостью этих
позвонков с боков, так и опистоцельным строением
пх, начиная с третьего грудного позвонка, выражен-
ным в наиболее полной форме в сравнении с дру-
гими водоплавающими. В целях той же подвиж-
ности, synsacrum, состоящий из 12—14 позвонков,
не срастается полностью с тазовым поясом, а оста-
Рис. 237. Перо с
внутренней по-
верхности крыла
золотоволосого
пингвина— Eudy-
ptes chrysocome
Forst. В основа-
нии опахала вид-
ны остатки рого-
вой сумки, в ко-
торой перо глубо-
ко сидит в коже
(и с которой ча-
стью срастается).
По Studer, uaStre-
semann.
вляет щель между ним и подвздошной костью, затя-
нутой широкой связкой.
Короткие задние конечности при плавании от-
тягиваются назад и играют лишь роль руля, заме-
щая этим короткий, хотя и жесткий хвост. Основ-
ную роль ноги играют для передвижения на суше,
которое совершается на пальцах, то неуклюжим ша-
гом, то короткими прыжками; при спешке же птица
ложится на брюхо и отталкивается от среды (снега)
быстрыми ударами крыльев и ног. Стоя, птица опира-
ется на всю, очень короткую и широкую плюсну, три
metatarsalia которой срослись только по своим концам, разделяясь
глубокими бороздами и частью отверстиями для прохождения кро-
веносных сосудов. В строении ног замечателен также сильный muscu-
lus ambiens, сухожилие которого проходит над коленкой на поверх-
ности (в борозде) очень крупной коленной чашечки, назначение ко-
торой прикрыть весьма выпуклое у стоящей птицы коленное сочле-
нение. Все четыре пальца ног направлены вперед, по первый палец
очень мал и б. ч. свободен, остальные соединяются плавательной
перепонкой.
Устойчивости на земле птицы помогает также хвост из 16—-20
368
относительно коротких перьев, на которые птицы опираются в стоя-
чем положении. Эти рули укреплены на типичном пигостиле и имеют
оригинальное строение: вентральная бороздка на упругом и широ-
ком стержне очень широка и U-образна, так что по бокам ее обра-
зуются выпуклые и жесткие валики, весьма повышающие упру-
гость пера.
Как у нелетающей птицы, удельный вес тела очень высок, до
0,98, вследствие чего при плавании птица глубоко погружается в воду.
Пневматичность скелета совершенно отсутствует, и отростки воздуш-
ных мешков не заходят даже в крупные кости; вследствие этого на
черепе и челюстях частично сохраняются швы отдельных сливаю-
щихся между собой костей. Все тело окутано мощным подкожным
жировым слоем, в целях защиты от охлаждения.
Подобно перьям крыла и хвоста, адаптивно видоизменено и все
остальное оперение, равномерно покрывающее все тело, так что ап-
терии совершенно отсутствуют. Контурное перо имеет широкий,
сильно сплющенный сверху вниз и эластичный ствол и также упло-
щенные бородки; бородки II порядка нормально развиты обычно
только в основании пера, в дистальной же его части они теряют
сцепленность друг с другом и имеют вид длинных жестких ресни-
чек. Вследствие этого, оперение приобретает обычно грубый воло-
совидный и чешуевидный характер. Каждое перо искривлено таким
образом, что между ним и кожей находится тонкий слой воздуха,
в котором расправляется пуховидный дополнительный стержень
пера. Собственно пух и какие-либо другие виды перьев полностью
отсутствуют. Все перья своими стержнями очень глубоко косо сидят
в коже, тесно соединены с ороговевающими стенками перьевых
сосочков и связаны мощной кожной мускулатурой. Птицы смазы-
вают свое оперение жировым выделением крупной копчиковой же-
лезы, вокруг парных выводных отверстий, несущей двойной ворот-
ничек перьев.
Как уже частью упоминалось, крайне своеобразно протекает
линька. Новое перо образуется и растет глубоко в субкутикулярном
слое кожи и появляется наружу одетое длинным роговым чехлом,
который на вершине охватывает очин старого пера; при этом вер-
шина нового пера проникает также в очин старого через его нижнее
отверстие. Это тесно связывает оба поколения перьев, а так как
линька приходится на очень сжатый срок (у некоторых видов —
Ю дней) и одновременно охватывает значительные участки тела, то
освобождение старых перьев происходит сразу целыми партиями,
вследствие некоторого сцепления перьев одной генерации между
собой. Таким образом старое перо сваливается с тела целыми лоску-
тами, не без участия соответствующих движений тела и клюва птицы;
старое перо уносит вместе с собой и роговой чехол нового вырос-
шего пера. В период линьки пингвины все время остаются на суше,
проводя время стоя, и совершенно не едят; в это время антарктиче-
ские пингвины иногда совершенно заносятся снегом. Молодые пинг-
вины перед получением первого дефинитивного оперения сменяют
два последовательных пуховых наряда.
Не менее значительны приспособления в пищеварительном ап-
369
парате. Рамфотека клюва у ряда пингвинов является сложной,
разбитой на куски; наружные носовые отверстия отсутствуют.
Обонятельные ощущения воспринимаются повидимому через хоаны,
так как lobi olfactorii и обонятельный эпителий развиты хорошо^
Мощно развитые носовые железы помещаются во впадинах над
глазницами. Язык и небо густо покрыты большими, направленными
назад, ороговевшими сосочками, способными удерживать скользкую
добычу. Сильно растяжимый пищевод ведет в очень объемистый
железистый желудок, с отверстиями желез, сосредоточенными в обо-
собленный участок (juga). Мускулистый желудок мал. Кишечник
очень длинен и, включая duodenum, сложен многочисленными пет-
лями; слепые кишки, наоборот, очень малы. Фабрициева сумка со-
храняет значительную величину всю жизнь и повидимому периоди-
чески изменяется в своих размерах
Многочисленные особенности пингвинов позволяют их рассматри-
вать как весьма специализированную группу, ближайшее родство
которой намечается только в отношении к трубконосым. С ними их
сближают: шизогнатизм черепа (но парный сошник, в отличие от
трубконосых, не срастается с небными костями и совершенно от-
сутствуют базиптеригоидные отростки), голориния, непрободенные
ноздри, сложная рамфотека, наличие трахейной перегородки и ряд
сходных черт в строении пищеварительного аппарата, мускулатуры п
в индивидуальном развитии.Наиболее замечательны следующие мо-
менты : несмотря на различия в функциональном употреблении крыла,
мускулатура крыла и плечевого пояса очень сходна, особенно сбли-
жаясь с ныряющим и плохо летающим Pelecanoides. В обеих группах
наблюдается редукция musculus biceps brachii. У ископаемого пин-
гвина Palaeeudyptes antarcticus Huxley плечевая кость была от-
носительно длиннее к размерам крыла, чем у современных форм.
Выступающие по локтевому краю крыла рецентных форм, особенно
сильно уплощенные перья по положению соответствуют маховым
перьям и насчитываются в количестве 36 штук; по некоторым на-
блюдениям, эти перья после первой линьки становятся короче, что
может указывать на процесс филогенетической редукции их. Харак-
терное для пингвинов неполное срастание метатарзальных костей
является повидимому псевдопримитивным свойством, так как все
раннетретичные пингвины отличаются от рецентных более совер-
шенным срастанием этих элементов и много более длинной и узкой
плюсной вообще. Для обеих Сравниваемых групп характерны также
медленность индивидуального развития и два последовательных
птенцовых пуховых наряда. Некоторые черты сходства пингвинов
с веслоногими и гагарами обусловлены повидимому всецело конвер-
гентными явлениями или признаками, унаследованными от общего
предка. Рассмотренное дает основание видеть в пингвинах ветвь,
развивающуюся с трубконосыми из общего источника, но различ-
ными путями,причем в пингвинах более специализированную и остав-
шуюся вместе с тем на более примитивной общей стадии развития.
По некоторым взглядам (Geoffroy St. Hilaire J. и Le Maout,
1855; Stejneger, 1885; Menzbier, 1887), в последнее время особенно
защищаемым Р. R. Lowe (1933, 1939), пингвины настолько обо-
370
соблены, что должны быть выделены в качестве самостоятельного
подкласса. По мнению Lowe, специализация пингвинов к водному
образу жизни выработана на очень примитивной основе, непосред-
ственно из особенностей строения общего предка птиц; по этому
взгляду, пингвины и их предки никогда не летали.
По своему распространению и вероятно происхождению пингвины
тяготеют к южному полюсу и полушарию. В ископаемом состоянии
известны третичные и четвертичные пингвины с наиболее древними
и уже нелетавшими формами, относимыми к эоцену и миоцену, об-
наруженные на Новой Зеландии, Южно-Шотландских островах
и в Патагонии. Наиболее крупные из них, напр. Anthropornis пог-
denskjoldii Wyman, превышали вдвое современных самых крупных
пингвинов Aptenodytes forsteri Gray, весящих 32 кг. Мелкие пингвины
повидимому не были меньше современных Spheniscus demersus L.,
ростом в 50 см. Всего известно в ископаемом состоянии около 35 ви-
дов, относимых к 23 родам.
ОТРЯД 2; PROCELLARIIFORMES. — ТРУБКОНОСЫЕ
Разнообразной величины, внешне сходные с чайками, прекрасно
летающие, длинно- и узкокрылые, с загнутым крючком на конце
надклювьем и ноздрями в особых характерных трубочках, птицы
морей всего света. Некоторые прекрасно ныряют, многие являются
сумеречными и ночными птицами. Пища состоит гл. обр. из голово-
ногих, ракообразных, медуз и рыб, добываемых в воде или собирае-
мых с поверхности моря. Гнездятся колониально, на морских остро-
вах. Гнездо устраивается на земле, в щелях между камнями, или само-
стоятельно вырытых норах; кладка только из 1 белого яйца. Оба
пола окрашены одинаково, преимущественно в темную окраску.
Инкубация от 38—40 (мелкие) до 60 дней (у крупных форм). Птенцы
вылупляются слепыми и обладают медленным эмбриональным и пост-
эмбриональным развитием: 61 день у весящих 40 г Hydrobates ре-
lagicus, до 4 месяцев у весящих 7,5 кг альбатросов. Характерен
способ защиты птенцов и взрослых птиц этой группы—выплевыванием
на врага едкой и пахучей жидкости. Единственное семейство Рго-
cellariidae с примерно 130 видами, относимыми к 22 родам и распре-
деляемым по 4 подсемействам.
Подсемейство Diomedeinae— альбатросы, с родами: Diomedea
(рис. 238), Тhalassogeron, Phoebetria; подсемейство Procellariinae —
буревестники, с родами: Мacronectes, Fulmarus, Priocella, Daption,
Procellaria, Puffinus, Pachyptila и др.; подсемейство Hydrobatinae —
качурки, с родами: Hydrobates, Oceanodroma, Fregetta и др.; подсе-
мейство Pelecanoidinae — нырковые буревестники с единственным
родом Ре lecano ides.
В морях СССР распространены сравнительно немногие формы.
Из главнейших следует упомянуть: из альбатросов — зимующий
У Д.-В. побережий белоспинный альбатрос — Diomedea albatrus
Pall.; из буревестников — кочующий на Черном море малый буре-
вестник — Pujfinus puffinus Brunn., зимующие у Д.-В. побережий
тонкоклювый и бурый буревестники — Puffinus tenuirostris (Temm.)
371
и P. griseus (Gm.); гнездящийся в арктических и субарктических
морях глупыш — Fulmarus glacialis (Ь.);из качурок, гнездящихся
местами у Д.-В. побережий, вилохвостая качурка — Oceanodroma
monorhis (Swinh.), северная — О. leucorrhoa (Vieill.) и сизая ка-
чурка — О. furcata (Gm.).
Биологически трубконосые представляют собой резко харак-
теризованную пелагическую группу, гл. обр. тяготеющую к океа-
ническим просторам южного полушария. В этой группе мы находим
наибольшую приспособленность к летающему образу жизни над во-
дой, хотя один род с 5—6 видами уклонился в направлении преиму-
щественно плавающего и ныряющего образа жизни. Полет большин-
ства видов отличается удивительной легкостью и способностью к дли-
тельному скольжению и парению, особенно у крупных видов. В связи
с этим крупным птицам
Рис. 238. Странствующий альбатрос — Diomedea ехи-
lans L. По Evans.
свойственны высокая пнев-
матичность скелета, мощ-
ная крыловая и грудная
мускулатура. Парящие
крупные виды отличаются
особенно длинным плечом
и предплечьем, вследствие
чего при постоянном числе
больших маховых (11), ко-
личество малых маховых
сильно увеличивается, до-
стигая у альбатроса (раз-
мах крыла 3,5 м) 37 -}- спль-
но увеличенные кроющие
плеча и собственно плече-
вые, выполняющие про-
странство между маховы-
ми и телом птицы. Мел-
кие птицы, летающие над
волнами более трепещу-
щим или порхающим полетом, отличаются особенным удлине-
нием кисти и (меньше) предплечья. У видов с длительным парящим
полетом развит особый механизм, закрепляющий проксимальные
отделы крыла в расправленном положении и этим облегчающий
производимую мускулами работу (см. рис. 73). При этом на
лучевой стороне локтевого сочленения развиваются две сеса-
мовидные косточки, из которых внутренняя, тесно прилежащая
к processus supracondyloideus lateralis дистального конца плечевой
кости, при полном расправлении крыла передвигается дистальнее
и запирает вращение ulna в достигнутом положении. Вместе
с тем, соседняя ей внешняя сесамовидная косточка является
местом прикрепления одной из головок сильнейшего разгибателя
кисти musculus extensor metacarpi radialis и сухожилий, натя-
гивающих propatagium: tendo propatagialis longa и brevis, что
позволяет перенос мускульных тяжей от плеча к кисти без затра-
гивания собственно локтевого сочленения и следовательно может
372
приводить к изменению положения кисти и пальцев при фикси-
рованном соотношении проксимальных отделов крыла. Преимуще-
ственно статическая задача крыловой мускулатуры, в поддержании
крыла в расправленном состоянии, выражается также в развитии
главным образом приключичной части musculus pectoralis, подтя-
гивающей крыло вперед (в связи с чем мощное развитие U-образных
ключиц и передний синостоз их с грудиной), и резкой вообще диффе-
ренциации musculus pectoralis на два слоя: приводящий крыло к телу
(вниз) и удерживающий крылья в расправленном положении. Рез-
кое отклонение от этих особенностей проявляет лишь Pelecanoi-
des, внешне напоминающий чистика и сходный с ним и по строению
узкой длинной грудины, сжатых ключиц, узкого таза; вследствие
употребления крыльев гл. обр. для «полета под водой», весь скелет
их сильно уплощен и укорочен, приобретая и ряд особенностей, на-
поминающих пингвинов. Однако некоторые из типичных буревест-
ников, напр. Puffinus, также способны к коротким полетам под во-
дой, при нырянии с воздуха, без существенных структурных изме-
нений.
Все трубконосые прекрасно плавают. Передние пальцы ног соеди-
нены перепонками, задний палец очень мал или отсутствует. Харак-
терной чертой в строении конечностей является мощное развитие
crista tibiae, вытянутой в широкий выступающий за колено отросток.
У многих однако задние конечности сравнительно малы и коротки.
Некоторые мелкие качурки пользуются ногами для торможения
своего полета, происходящего над самой водой, если они замечают
плавающую пищу, причем на лету они погружают ноги в воду с рас-
ставленными плавательными перепонками (напр. Fregetta)] у этих
видов конечность значительно длиннее, особенно голень и плюсна,
пальцы сильнее и вооружены плоскими когтями, передняя же сто-
рона плюсны гладка или покрыта косо лежащими поперечными
щитками.
Туловищное оперение состоит как из контурных, так и пухо-
вых перьев; последние покрывают тело всюду и имеют дополни-
тельные стержни. Смазка оперения может происходить из большой,
снабженной перовой кисточкой у выходного отверстия, копчиковой
железы.
В связи с характером питания замечательно весьма высокое раз-
витие органов обоняния, выражающееся в очень крупной величины
lobi olfactorii и, одетых обонятельным эпителием, сильно закручен-
ных раковинах носовых полостей. В связи с этим очевидно носовые
отверстия помещаются на концах трубочек, образованных удлине-
нием хрящевой стенки носовых полостей, одетых рамфотекой. Раз-
меры и форма этих трубочек вариируют повидимому соответственно
роли обоняния в жизни птицы. Наилучше обоняние развито у Ful-
marus, хуже всего у Pelecanoides. Как у большинства морских птиц,
У трубконосых имеются большие носовые железы, помещающиеся
в ямках крыши глазных орбит.Снабженный большим крючком клюв
трубконосых характеризуется сложной рамфотекой. У ряда форм,
питающихся планктоном, замечательно образование роговых пла-
стинок по краям и внутренней поверхности надклювья, сочетаемых
с расширенной формой клюва и весьма растяжимой кожей подклю-
373
вья, функционирующей как горловой мешок (рис. 239). Шизогна-
тический тип строения неба, свойственный трубконосым, у Diomedea
имеет тенденцию к неполному десмогнатизму. У многих форм на-
блюдаются различной величины ложные базиптеригоидные отростки.
Пищевод обладает сильной растяжи-
мостью и ведет в огромный желези-
стый желудок, в сравнении с кото-
рым мускулистый желудок мал, сдви-
нут на сторону и относительно слаб.
Кишечник сравнительно короток, а
слепые кишки крайне малы или от-
сутствуют.
По строению трубконосые оказы-
ваются стоящими, ближе всего к пинг-
винам. Кроме уже указанных осо-
бенностей (см. стр. 370), они прояв-
ляют сходство с ними в одинаковом
числе шейных позвонков, парных
сонных артериях и кардинальных
венах, свободных грудных позвон-
ках и др. и у Pelecanoides в значи-
тельной длине гипапофиз на груд-
ных позвонках, в связи с мощным
развитием сгибателя шеи (musculus
longus collii anticus), и нек. др. Ряд
общих черт из уже .указанных связы-
вает трубконосых также с веслоно-
гими, но это родство более отдаленно,
и разделение групп произошло, по
эоцена, причем трубконосые остались
на более примитивной стадии разви-
тия. Столь напоминающие трубконо-
сых внешне чайки представляют
совершенно обособленно развиваю-
щуюся группу, что указывается напр. резко различным строе-
нием органов, функционально связанных с сходным образом жизни
(мускулатура крыла, органы пищеварения) и различиями в индиви-
дуальном развитии.
Древнейшие ископаемые трубконосые известны из эоцена Африки
(Gigantornis), верхнего олиго цена Франции (Puffinus и Hydrornis)
и миоцена Северной Америки (Puffinus и Fulmarus), позднее встре-
чаются почти исключительно современные роды (исключая Ploto-
tornis).
ОТРЯД 3: COLYMBIFORMES. — Г АГ APO ОБРАЗНЫЕ
Мелкой и средней величины, плавающие и ныряющие птицы,
с далеко кзади поставленными ногами, что не допускает передви-
жение по земле. Внешне характерны: довольно длинное туловище,
длинная шея, острый, умеренной длины клюв с простой рамфоте-
Рпс. 239. Клюв питающегося планкто-
ном ширококлювого буревестника —
Pachyplila Jorsteri Lath.
Обратите внимание па мясистый язык,
на роговые пластинки, свешивающиеся
по краям клюва п функционирующие
сходно с роговой бахромой клюва уток,
и на сильно растяжимое дно подклю-
вья, образующее подъязычный (горло-
вой) мешок. По Wilson, пз Strese-
mann.
мнению Гадова (1893), не позднее
374
кой, четырехпалые лапы с сильно сплющенными с боков плюснами,
передние пальцы которых или соединены перепонкой или оторочены
упругими плавательными пластинками, толчками которых, произво-
димыми почти сзади тела, птицы передвигаются в воде. Хвост коро-
ток и у поганок почти отсутствует. Дефинитивный пух равномерно
распределяется по всему телу и подобно контурному перу несет до-
полнительный стержень.
Анатомически характеризуются шизогнатизмом, при отсутствии
базиптеригоидных отростков, голорпнией, отсутствием носовой пе-
регородки; в строении конечностей и поясов характерны U-образная
вилочка с hypocleidium, разъединенные коракоиды и мощное развитие
пирамидообразных processus rotularis больших берцовых костей.
В пищеварительном аппарате замечательны обособление пилори-
ческого отдела мускулистого желудка, довольно длинный кишеч-
ник, функционирующие, но небольшие, слепые кишки. Имеется
оперенная копчиковая железа.
Довольно сходные по биологии птицы, объединяемые в эту группу,
все же легко разделимы на два подотряда: Colymbi — гагары и Ро-
dicipedes — поганки.
ПОДОТРЯД 1: COLYMBI. — ГАГАРЫ
Крупной величины, питающиеся рыбой, искусно ныряющие, пе-
репончатопалые, обитатели морских вод и крупных озер арктиче-
ской и бореальной зон северного полушария. Специализированные
водоплавающие формы, вылезающие (так как ходить не могут) на
сушу только к гнезду, устраиваемому из водных растений на краю
берега водоема. Оба пола, почти неотличимые по окраске, принимают
участие в воспитании птенцов. Кладка из двух (редко трех) отно-
сительно крупных, оливково-бурых с черными пятнами яиц. Инку-
бация длится 28 дней. Птенцы выклевываются покрытые темноокра-
шенным пухом, весьма подвижны и быстро начинают плавать. Перед
получением контурного оперения перелинивают два пуховых на-
ряда, развивающихся из тех же перовых сосочков.
Подотряд образует единственное семейство Colymbidae и один
род Colymbus, составленный 5 видами, представленными и в фауне
СССР. Наиболее известны: Colymbus arcticus L. —чернозобая га-
гара, широко распространенный бореальный вид; С. stellatus Роп-
lopp. — краснозобая гагара, арктический и субарктический вид;
С. immer Brunn. — полярная гагара, северная Европа и восточная
Америка; С. adamsi Gray — белоклювая гагара, арктические побе-
режья Азии.
Ископаемый род Colymboides известен с верхнего эоцена Фран-
ции и занимает промежуточное положение между этим и следующим
отрядом. Виды, относимые к рецентному роду Colymbus, известны
начиная с олигоцена. (Особенности строения см. Podicipedes.)
ПОДОТРЯД 2: PODICIPEDES. — ПОГАНКИ
Мелкие и средней величины, с своеобразным рыхлым оперением,
птицы преимущественно внутренних водоемов всей земли. Питаю-
щиеся водными беспозвоночными и рыбками, специализированные
376
плавающие и ныряющие птицы, устраивающие из водных растений
пловучие гнезда, помещаемые среди болотной растительности. Пальцы
ног оторочены плавательными пластинками и на конце несут плоские
ногти. Одинаково окрашенные, оба пола участвуют в воспитании
потомства. Кладка из 4—6 белых яиц, в защиту от врагов прикры-
ваемых птицей при сходе с гнезда водной растительностью. Инку-
бация продолжается 20—25 дней. Вылупляющиеся птенцы одеты
пушком, обычно черной основной окраски, с белыми продольными
полосками. Птенцы первое время согреваются и сохраняются ро-
дителями среди оперения и под крыльями, но вскоре сходят и на воду.
Один птенцовый пуховой наряд.
Известно около 25—30 видов, группируемых в 4 рода единствен-
ного семейства Podicipedidae. Роды: Podiceps, Aechmophorus, Ро-
dilymbus и Centropelma. Представлены в СССР 5 видами, из которых
наиболее известны и широко распространены: чомга — Podiceps
cristatus (L.), серощекая поганка — Podiceps griseigena (Bodd.).
красношейная поганка — Podiceps auritus (L.) и черношейная по-
ганка —Podiceps nigricollis Brehm.
В ископаемом состоянии известны лишь современные роды, на-
чиная с олигоцена.
Родственные взаимоотношения Colymbi и Podicipedes явствуют
из большого количества общих признаков, хотя некоторые отличия
также довольно существенны. Наиболее важные из последних сопо-
ставлены в этой таблице.
Colymbi
Шейные позвонки — 14—15
Грудина узкая и длинная
Грудные позвонки свободны, с
_]_-образными, довольно длин-
ными гип апофизами
Patella отсутствует и замещает-
ся всецело processus rotularis
Fibula достигает интертарзаль-
ного сочленения
Носовые железы крупные, лежат
в ямках крыши орбит
Пилорический отдел желудка
слабо выражен
Musculus ambiens есть
Сиринкс трахео-бронхиальный с
симметричной мускулатурой
Сонные артерии парные
Маховые — 11
Podicipedes
17—21
Сравнительно широкая и укоро-
ченная
3—4 позвонка слились в os dor-
sale; с простыми гипапофизамп
Крупная, пирамидообразная
Не достигает этого сочленения,
не доходя до него на одну треть
tibia
Малы, не образуют ямок
Резко выражен и обычно набит
заглоченными птицей перьями
в защиту кишечника от мелких
костей
Отсутствует
Бронхо-легочный с асимметрич-
ной мускулатурой
Только левая
12
376
Рулевые перья — 16—18
Спинная аптерия ограничена
только шеей
Передние пальцы ног соединены
общей перепонкой
Крайне редуцированы
Ограничена только спиной
Несут плавательные пластинки
заканчиваются широкими ног-
тями
Эти отличия указывают на достаточно давнее расхождение обеих
групп и несколько различные пути их эволюции после этого, причем
поганки достигли большей специализации. Из географического рас-
пространения группы можно заключить с большой вероятностью,
что в то время как гагары развивались гл. обр. в северном полуша-
рии, тяготея к арктическому бассейну и, в стационном отношении,
к морям и их побережьям (широкое расселение ряда видов на внутрен-
них пресных водах — вторично), поганки эволюционировали на
суше, вырабатывая приспособления к биотопам различных вели-
чин и особенностей, и широко расселились по земному шару. Взаи-
моотношения обеих групп с другими отрядами весьма темны (отда-
ленное родство может быть намечаемо только в отношении к Alcae)^
и в целом Colymbijormes занимают изолированное положение среди,
самых примитивных отрядов новонебных птиц.
ОТРЯД 4: PELECANIFORMES (STEGANOPODES).—
ВЕСЛОНОГИЕ
Весьма разнообразные по величине и форме (напр. пеликан.,
баклан, фрегат), плавающие и (обычно) хорошо летающие птицы,
распространенные по пресным и морским водоемам почти всей земли.
Из внешних признаков характерны сложная рамфотека и связанность
всех четырех пальцев коротких ног сплошной плавательной пере-
понкой. При сходном типе питания (pjbi6bi) замечательны различным
направлением специализации в ее добывании, изменяющейся от пре-
следования добычи нырянием (бакланы) и выхватыванием пз воды
при стремительном падении с воздуха (олуши, фрегаты, фаэтоны)
до ловли на мелководье объемистым клювом, выхватывая ее как
сачком (пеликаны). По индивидуальному развитию птенцовые, вы-
лупляющиеся слепыми с медленным развитием птицы.
Анатомически эта группа характеризуется комбинацией следую-
щих признаков: десмогнатизмом с изменчивостью в сторону шизо-
гнатизма и при отсутствии базиптеригоидных отростков; в пище-
варительном аппарате общими являются: отсутствие ротовых желез,
сильная растяжимость пищевода, обширность железистого желудка
и незначительность мускулистого при наличии изменчивого пилори-
ческого отдела; слепые кишки сильно укорочены; в оперении харак-
терны: диастатаксическое крыло с 11 маховыми, развитость пуха,
покрывающего все тело, отсутствие или слабость развития побоч-
ного стержня в контурном пере, птенцовый наряд из praeplumulae,
а не ргаереппае (предшественников контурного пера); большая коп-
чиковая железа лишена сосцевидного выводного протока, открываясь
изменчивым числом отверстий, окруженных кисточковыми перьямп.
377
Отряд подразделим на четыре подотряда: Phaethontes, Pelecani,
Fregatae и Odontopteryges, из которых последний представлен вымер-
шей формой с проблематичным систематическим положением.
Систематическое положение веслоногих в целом определяется их
родственными взаимоотношениями с трубконосыми, аистовыми и при-
митивными хищными птицами. «Их родство с Procellaritformes про-
является не столько в абсолютном сходстве многих признаков, сколько
в том, что многие особенности Pelecaniformes могут быть выведены
прямо из соотношений, имеющихся у трубконосых; другими словами:
веслоногие оказываются выше специализированными из одинаковой
основы с трубконосыми. Обе группы повидимому прошли длинный
параллельный путь развития» (Gadow, 1893). Что касается родства
с Ciconivformes и Cathartae, то морфологический анализ показывает
значительно более раннее отщепление Pelecaniformes от общего ствола,
но на стадии, когда ряд выработавшихся признаков мог унаследо-
ваться этими группами одновременно. Такими напр. являются:
десмогнатическое строение неба, выклевывающиеся голыми птенцы,
отсутствие боковой шейной аптерии, соотношения ножной мускула-
туры, слабая развитость или отсутствие мускулатуры, сиринкса и
мн. др. Столь отличная, удаляющая друг от друга эти группы,
внешняя морфология во многих отношениях есть следствие сильной
специализации и резко отличного образа жизни, приобретенного за
период длительной эволюции.
ПОДОТРЯД 1: PHAETHONTES. — ФАЭТОН
Небольшие, внешне напоминающие крачек, прекрасно летающие
птицы тропических морей, с преобладающей белой окраской, сильно
удлиненными центральными рулевыми и заостренным к концу клювом.
Рпс. 240. Краспоклювый фаэтон—Phaelhon aethereus L. По Evans.
Фаэтоны преимущественно прибрежные птицы, питающиеся ры-
бой, головоногими и ракообразными, добываемыми подобно крач-
кам нырянием с воздуха, после высматривания их на полете (рис.
240). Оба пола мало отличимы. Птицы даже на гнездовье редко сое-
диняются в группы. Гнезда помещаются обычно в щелях скал или
378
иод кустарником на голой земле, на уединенных берегах и островах.
Кладка только из 1 яйца, с коричневато-красными пестринами.
Инкубация продолжается 28 дней. Птенец вылупляется покрытый
густым белым пухом и развивается очень медленно (63 дня). Из-
вестны только 3 вида единственного рода и семейства: Phaethon,
семейство Phaethontidae.
Морфологически фаэтоны в группе Pelecaniformes занимают изо-
лированное положение; от прочих подотрядов их отличают: отсутствие
перегородки между ноздрями (т. е. nares perviae), довольно крупные
щелеобразные носовые отверстия, наличие сошника, имеющегося
также п у фрегатов, относительно большой язык, более примитивное
строение мускулатуры ног (формула Гаррода: AXY), пятнистая
окраска яиц и другие признаки, стоящие в связи с типом адаптации
птицы. Приспособительный отпечаток несут: заостренный клюв,
тонко иззубренный по краям для удержания добычи, малый по
размерам пилорический отдел желудка, высокая пневматичность
кожи на нижней стороне туловища, ослабляющая силу удара в воду
при нырянии с воздуха, и другие особенности. Маленькие слабые
задние конечности едва служат для движения по земле и отличаются
как утерей musculus ambiens, так и отсутствием обычного прохо-
ждения musculus iliofibularis через сухожильную петлю,что наблю-
дается еще только у стрижей.
Среди всех Pelecaniformes фаэтоны обладают наибольшим коли-
чеством примитивных особенностей. В ископаемом состоянии они
известны из нижнего эоцена (Prophaeton}.
ПОДОТРЯД 2: PELECANI. — ПЕЛИКАНОВЫЕ
Крупной и средней величины водоплавающие птицы морских по-
бережий и крупных внутренних водоемов. Характеризуются увели-
ченным числом шейных позвонков (17—20; у фрегатов и фаэтойов
14—15), редуцированными до узкой щели или совершенно заросшими
носовыми отверстиями и ясно выраженной носовой перегородкой,
слабым развитием носовых желез,умещающихся в ростральной части
орбит, отсутствием сошника, чрезвычайно редуцированным и ве-
роятно нефункционирующим уже языком и другими особенностями.
В экологическом отношении для подотряда характерны гнездова-
ние колониями, одноцветная окраска яиц, выкармливание птенцов
из сильно растяжимого пищевода. Некоторые из этих признаков
однако свойственны и фрегатам.
Различают шесть семейств: Pelecanidae, Cyphornithidae^ Pela-
gornithidae, Sulidae, Phalacrocoracidae и Anhingidae, из которых два
семейства представлены только вымершими формами.
Семейство 1: Pelecanidae. — Пеликаны
Крупные птицы с длинным, оригинального устройства клювом.
Надклювье плоское и на конце образует крутой крючок, а кожа ме-
жду ветвями подклювья образует легко растяжимый горловой ме-
шок. Свойственны крупным, преимущественно тепловодным бас-
сейнам Старого и Нового Света. Единственный род Pelecanus с 8 ви-
379
дамп, из которых два: пеликан розовый — Pelecanus onocrotalus L.
и серый — Pelecanus crispus Bruch, распространены в СССР, в Арало-
Каспийском и Черноморском бассейнах.
Пеликаны представляют собой крайне своеобразно приспособлен-
ную группу птиц. Обладая весьма пневматичным скелетом и даже раз-
ветвленной под кожей, среди корней перьев, системой воздушных
пузырьков, они отличаются высокой пловучестыо, неспособны ны-
рять и, несмотря на объемистость тела, прекрасно летают как сколь-
зяще-гребным, так и отчасти парящим полетом. Преимущественно
это стайные птицы, добывающие рыбу загоняя ее с шумом всем об-
ществом на мелкие места, или в заливы, и выхватывая из воды клю-
вом, как сачком. При этом крайне эластичные ветви нижней челюсти,
под влиянием сокращения musculus sterno-hyoideus и musculus ge-
nio-glossus, тянущих к голове конец челюсти, сильно изгибаются,
образуя почти круглый обруч, на котором подвешен широкий гор-
ловой мешок. Отдельные пеликаны (напр. Pelecanus occidentalis thagus
Mol. западной Америки) однако приобрели способность нырять за
рыбой и с воздуха, как олуши.
Гнездятся пеликаны колониально, гл. обр. в обширных болоти-
стых зарослях, на островах или уединенных берегах, устраивая
объемистые гнезда из тростника, морской травы и т. п. Кладка со-
стоит из 2—3 желтоватых яиц с рыхлым меловым слоем на поверх-
ности. Инкубация длится 36 дней. Птенцы вылупляются слепые и го-
лые, впоследствии одевающиеся пухом.
В своем строении пеликаны обнаруживают наибольшую близость
к олуше, хотя отдельные черты соединяют их только с фаэтонами
(утеря musculus ambiens; короткая малая берцовая, дистально не
доходящая до сочленения; незаросшие носовые отверстия и пр.).
Из характерных черт, кроме упомянутых (клюв и пр., см. выше),
необходимо отметить наименьшее в подотряде число шейных позвон-
ков—17; слияние грудных позвонков в обширную os dorsale; очень
большое, 20—24, число рулевых перьев в коротком округлом хвосте
и крупную копчиковую железу, снабженную 6—9 отверстиями.
По своим особенностям пеликаны могут рассматриваться односторонне
специализированными формами на наиболее примитивной для под-
отряда основе. В ископаемом состоянии настоящие пеликаны из-
вестны с верхнего эоцена (роды Pelecanus и Protopelicanus).
Семейство 2: Cyphornithidae
Сходные с пеликанами формы, с переходными признаками к олу-
шам и отчасти к бакланам. Известны два миоценовых рода, Cyphor-
nis и Palaeochenoides.) сохранившиеся в весьма фрагментарных
остатках.
Семейство 3: Pelagornithidae
Представлены ископаемым Pelagornis^ из миоцена Франции. Ги-
гантские пеликаны с плечевой костью длиной в 58 см, по ряду при-
знаков стоявшие уже ближе к олушам, чем к пеликанам. Сюда же
относят последнее время роды Argillornis и Lip to mis из эоцена —
миоцена Европы и Патагонии.
380
Семейство 4: Sulidae. — Олуши
Крупные, преимущественно белые или чернобелые, с коническим
заостренным на конце и иззубренным на краях клювом и острыми
длинными крыльями, птицы морских и пресноводных побережий
Старого и Нового Света. Единственный род Sula, с 7 видами (рис. 241).
Олуши прекрасно летают, способны к длительному парению,
весьма легки и пловучи, вследствие чего добывают пищу бросаясь
с воздуха в воду, чем приобретают достаточную силу для проникно-
вения на значительную глубину и плавания под водой. Резкость
удара об упругую среду отчасти парализуется очень густым опере-
нием и особенно значительной пневматичностью кожи в форме боль-
ших подкожных воздушных мешков и воздушных пузырьков, распо-
лагающихся между основаниями очинов перьев. С этим же способом
добывания пищи находит-
ся в связи длинная, защи-
щающая внутренности от
удара, грудная клетка и
кость. Своеобразна кон-
струкция нижней челюсти
с легкой сгибаемостью вет-
вей ее в области spleniale
и заметным скольжением
наружу их основания в
месте сочленения с quad-
ratum, при открывании
клюва; эта конструкция
допускает широкое рас-
ширение пасти при про-
глатывании крупной до-
бычи. Гнездятся олуши
обычно колониально на
Рпс. 241. Олуша — Sula bassana L. По Evans.
скалах и откосах мор-
ских побережий, образуя многотысячные поселения, и в сухом
климате (запад Южной Америки, юго-западная Африка и др.) яв-
ляются главнейшими продуцентами высокоценного гуано. Гнездо
устраивается из морской травы. Кладка состоит из 1, редко 2 зеле-
новатых яиц. Инкубация продолжается 42 дня. Птенцы вылупляются
слепыми, голыми, с черной кожей и медленно развиваются. Птенцо-
вый пуховой наряд белый.
По систематическому положению олуши занимают промежуточ-
ное положение между пеликанами и бакланами, обнаруживая однако
наибольшее родство с последними. Их соединяют сними одинаковое
число шейных позвонков (18), заросшие наружные носовые отверстия,
сходное строение задних конечностей. С пеликанами им общи: от-
сутствие собственной мускулатуры у сиринкса, наличие образуемого
катапофизами канала и высокая пневматичность подкожных про-
странств. Из специфических черт, кроме вышеупомянутых, следует
отметить наличие 16 рулевых в хвосте, сохранение только правой
сонной артерии, пять отверстий в копчиковой железе.
381
В ископаемом состоянии олуши, относимые к роду Sula, известны
с олигоцена [Sula ronzoni (Gerv.), S. loxostyla Cope и др.]. Переход-
ное положение к бакланам занимают роды Miosula (миоцен), Elopte-
гух (верхний мел), Eostega и Aetiornis (эоцен).
Семейство 5: Phalacrocoracidae. —Бакланы
Средние и крупные, длинношейные, с длинным жестким хвостом
и длинным крючковатым клювом, прекрасно ныряющие темноокра-
шенные птицы, распространенные почти по всему свету. Предста-
влены только 2 родами: Nannopterum и Phalacrocorax с примерно
26 видами. Из последнего рода в СССР широко *распространены: Ph.
carbo L. — баклан большой; Ph. aristotelis (L.) — баклан длинно-
носый, свойственный берегам Черного моря; Ph. pygmaeus (Pall.) —
малый баклан, обычный в Средней Азии; Ph. filamentosus (Temm.
&Schleg.) — японский баклан; Ph. pelagicus Pall. —тихоокеанский
баклан и Ph. bicristatus Pall. — краснолицый баклан, свойственные
морским Д.-В. побережьям.
Бакланы специализированы на добывании пищи, обычно рыбы,
под водой, в совершенстве ныряя и схватывая ее вооруженным крюч-
ком клювом. Обычно являясь колониально гнездящимися птицами,
они держатся в период размножения предпочтительно на островах
или около скал. Гнезда устраиваются из морской травы и сучьев
на скалах и деревьях; оба пола, одинаково окрашенные, принимают
участие в воспитании птенцов. Кладка состоит из 4—6 голубоватых
яиц. Инкубация длится 23 дня. Птенцы выклевываются слепыми,
голыми, с черной кожей и в дальнейшем характеризуются медлен-
ным развитием. Молодежь выкармливается, как у пеликанов и олуш,
из пищевода родителей, куда они засовывают голову и клюв. Пухо-
вой птенцовый наряд черный или серый. ч
По систематическому положению бакланы близки змеиным пти-
цам, разделяя вместе с ними ряд общих адаптивных особенностей:
большой удельный вес (вследствие почти полной апневматичностп
скелета), прекрасную способность нырять при помощи одновремен-
ных толчков широких лап, причем крылья несколько оттопыриваются
от тела и вместе с длинным жестким хвостом из 12—14 рулевых слу-
жат для изменения направления движения. В строении задних ко-
нечностей характерна длинная fibula, доходящая до плюсны и поз-
воляющая вращение большой берцовой кости относительно малой при
гребных движениях ног. В связи с необходимостью большей гибко-
сти ныряющей птицы, грудные позвонки у обеих групп опистоцельны
и несут длинные гипапофизы для прикрепления мощной шейной му-
скулатуры, а таз отличается относительной узостью и длиной. Харак-
терно также образование сесамовидной косточки в области затылка,
имеющей форму стилета и сочленяющейся с supraoccipitale. Она обра-
зуется из окостенения соединительно-тканной прослойки между мощ-
ными мускулами, закрывающими клюв (musculus adductor mandi-
bulae) и прикрепляющимися в области затылка, и служит для увели-
чения их основания и развития ими большей силы. Эта мускулатура
дает возможность удерживать клювом сильных рыб и развита зна-
382
чительно мощнее у бакланов, чем у змеешеек. У обеих групп копчи-
ковая железа несет лишь по два отверстия. Особый интерес среди
бакланов представляет галапагосский баклан— Nannopterum harrisi
Rotsch., с сильно редуцированным крылом и уже потерявший вся-
кую способность летать.
В ископаемом состоянии бакланы известны с олигоцена (роды
Oligocorax, Miocorax, Paracorax, Australocorax и Phaldcrocorax).
Семейство 6: Anhingidae. — Змеиные птицы, змеешейки
Крупные, очень длинношейные и длиннохвостые птицы с штык о-
образным острым клювом, свойственные пресным водам тропического,
пояса Старого и Нового Света.
Единственный род Anhinga (= Plotus) с 3 видами (рис. 242).
Змеиные птицы замечательны односторонней специализацией
в преследовании рыбы, ко-
торую они осуществляют под
водой или близ поверхности,
плавая с почти целиком по-
груженным в воду телом и вы-
ставленной длинной шеей.
При этом голова с острым
клювом носится шеей с свое-
образными движениями, слов-
но копье в руке человека,
и при приближении к добыче
резким движением выкиды-
вается вперед и с молние-
носной быстротой накалы-
вает рыбу на клюв, после
чего, уже на поверхности во-
ды, рыба сбрасывается с клю-
ва вверх, вновь ловится и про-
глатывается головой вперед.
Этот способ охоты возможен
вследствие особой конструк-
ции удлиненной до 19—20 по-
звонков шеи, имеющей резкий
изгиб посредине (см. рис. 68,
стр. 78). Благодаря особому
Рис. 242. Чернобрюхая змеешейка — Plotus те-
lanogasler (Gm.). По Evans.
строению сочленовных поверх-
ностей позвонков, передняя ее часть, кончая 8-мпозвонком, сгибается
только вентрально, задняя же часть только дорзально, 8-й позвонок
при этом сильно удлинен. Разгибатель шеи—musculus spinalis cervicis,
прикрепляющийся дорзально на 2-м—4-м позвонках, проходит сухо-
жилием через петлю у 9-го позвонка и связывается с последующими
позвонками; сгибатель шеи, мощный musculus collii anticus, возни-
кает на длинных гипапофизах грудных позвонков и заканчивается
гл. обр. на 8-м—10-м позвонках. Мощным сокращением обоих этих
мускулов изгиб шеи на 8-м позвонке резко выпрямляется, и шея удли-
383-
няется, что и развивается в необходимую силу для штыкового удара
клювом. Внутренние края узкого клюва тонко зазубрены для удер-
жания скользкой добычи.
Обычной стацией змеиных птиц являются реки и крупные внутрен-
ние водоемы. Гнездятся они, устраивая гнездо, подобно некоторым
бакланам, на кустах пли деревьях, и несут от 3 до 5 яиц. Вылупляю-
щиеся слепыми и голыми, птенцы впоследствии одеваются в белый
пух. В богатых кормом местах змеиные птицы ведут стайный образ
жизни.
От близких к ним Бакланов змеиные птицы отличаются, помимо
указанного, рядом других своеобразных черт. Обширный желези-
стый желудок имеет железистые поля концентрированными в спе-
циальных слепых выростах желудка. Пилорический отдел особенно
велик, причем отверстие его в двенадцатиперстную кишку защи-
щается густым скоплением волосовидных отростков, защищающих
также и вход в пилорический отдел со стороны собственно желудка.
В строении задне!! конечности замечательно сухожилие musculus
ambiens, проходящее в глубоком желобке на коленной чашечке,
а не пробуравливающее ее, как это свойственно бакланам. Сонные
артерии в противоположность последним также представлены только
левым стволом. Характерна одновременная линька всех маховых,
так что птица, подобно утиным, временно теряет способность летать.
В ископаемом состоянии семейство известно, начиная с эоцена
(род Proto plotus). Виды, относимые к современному роду, известны
из плейстоцена Венгрии, Австралии и Мадагаскара.
ПОДОТРЯД 3: FBEGATHAE. — ФРЕГАТЫ
Крупные, очень длиннокрылые, вилохвостые, с длинным крючко-
ватым клювом, птицы прибрежий морей тропического пояса. Одни
из лучших и выносливейших летунов, охотящиеся за летучими ры-
бами и беспозвоночными, выплывающими на поверхность, причем
бросаются на добычу как бы ныряя за ней в воздухе, схватывая,
прежде чем она вновь уйдет в воду. Постоянно преследуют в море
других рыбачащих птиц, заставляя их бросить пойманную добычу,
которую и подхватывают на лету. Вследствие маленьких с редуци-
рованными плавательными перепонками ног, спускаются на воду
только как исключение и не способны к значительному передвиже-
нию по земле. Отдыхают и гнездятся (колониями) на островах, на
скалах и на деревьях, делая гнезда из сухих сучьев. Оба пола при-
нимают участие в воспитании птенцов.Кладка из одного яйца желто-
ватой окраски. Птенец выклевывается слепым и голым, вскоре об-
растает белым пухом, но развивается медленно.
Известны лишь одно семейство Fregathidae и один род Fregata
с пятью видами (рис. 243).
В структурном отношении фрегаты представляют собой край-
нюю специализацию к воздушному образу жизни и по выносливости
полета уступают только альбатросам. Резко выражены адаптивные
особенности превосходного летуна: исключительная пневматичность
скелета, длинные крылья (в противоположность крупным трубко-
384
носым птицам, с длинной кистевой частью и предплечьем), широкая
п короткая грудная кость с высоким килем и слившимися с ним
и акрокоракоидным отростком ключицами. Крыловая мускулатура
с musculus pectoralis, дифференцированным на два слоя, и сесамовид-
ной косточкой — местом
прикрепления разгибателя
кисти — musculus extensor
metacarpi radialis. Харак-
терны гетероцельность
грудных позвонков и ко-
роткий п широкий таз.
Изолированное положе-
ние фрегата в пределах от-
ряда мотивируется рядом
оргинальных черт. Наибо-
лее своеобразно строение
плюсны, очень короткой и
широкой, с поверхностны-
ми продольными борозда-
ми, указывающими на со-
став ее из трех metatar-
, salia. Является ли это
примитивной чертой или
новым приспособлением, рпс 243 Больпюй фрсгат _ Pregala aquila Б По
В частности выражением Evans,
тенденции переместить тя-
жесть сидящего тела возможно ближе к опоре и облегчить схваты-
вание опоры пальцами, как думают некоторые, пока неясно. Ряд осо-
бенностей фрегата, по всей вероятности конвергентных, объеди-
няет его с фаэтоном: наличие сошника, 14—15 шейных позвонков,
парные сонные артерии и нек. др.
ПОДОТРЯД 4: ODONTOPTERYGES
Крупные, структурными чертами более всего напоминающие
олуш и фрегатов, ископаемые эоценовые птицы с длинным зубчатым
клювом. Зубцы были образованы рамфотекой и по строению напоми-
нали зубчики клюва крохалей (Mergus, Anseres}. Известны только
одно семейство Odontopterygidae п один род и вид Odontopteryx to-
liapica Owen. Сомнительное положение занимает Pseudodontornis
longirostris (Spulski), известный по фрагментам черепа неизвестного
возраста.
ОТРЯД 5: CICONIIFORMES. — ЛИСТООБРАЗНЫЕ
Разнообразные по величине, длинношейные, длинноногие и обычно
длинноклювые болотные птицы, распространенные почти по всей
земле. Биологически характеризуются животноядностыо и птенцо-
востыо. Анатомически—десмогнатизмом и исчезающими вторич-
ными базпптеригоидными отростками, наличием пилорического от-
25 Орнитология—1209
386
дела и очень малыми размерами слепых кишек. Задний палец б. ч.
хорошо развит, крыло является диастатаксическим.
Отряд объединяет пять довольно обособленных групп, рассма-
триваемых нами как подотряды: Ardeae^ Scopides, Balaenicipites.
Ciconiides и Threskiornithides. Их систематическое положение опре-
деляется родством с веслоногими, фламинго и отдаленными связями
с Accipitres, обсуждаемым в отдельных подотрядах.
ПОДОТРЯД 1: ARDEAE. — ЦАПЛИ
Разнообразной величины, очень длинношейные (19—20 шейных
позвонков) птицы, с прямым острым на конце (б. ч.) и слегка зазу-
бренным на краях клювом и гребенчатым краем когтя среднего пальца.
Обитатели болот всего света, исключая субполярных холодных
областей.
Цапли являются преимущественно ихтиофагами, подстерегаю-
щими добычу стоя на вахте в удобном месте водоема и внезапно
схватывающими ее резким выбросом вйеред длинной шеи и клюва;
иногда на охоте они тихо расхаживают или преследуют бегом усколь-
зающую добычу. В связи с этим, в строении шеи наблюдается особен-
ность, напоминающая соответствующий аппарат змеиных птиц:
дистальная часть шеи с 6-го позвонка сгибается только вентрально,
а проксимальная сгибается дорзально; для обеспечения верности
и быстроты удара клювом/шея почти лишена сгибаемости в сторону
и в спокойном состоянии и на полете укладывается S-образно, при-
тягиваясь к телу и торча вперед клювом. Во время сна клюв сверну-
той таким образом шеи защемляется между кистевым сгибом крыла
и грудью. В связи с обитанием в болотах, птицам свойственны очень
длинные передние и относительно очень длинный и низко располо-
женный задний пальцы. В пищеварительном аппарате характерно
наличие длинного заостренного языка, растяжимого пищевода, боль-
шого пилорического отдела желудка и непарной очень маленькой сле-
пой кишки. Вследствие затруднений боковых движений щей, коп-
чиковая железа рудиментарна, и ее функция восполняется пятнами
пудреток, расположенных в передней части груди, на надхвостье
и в паховой области.
По времени деятельности значительная часть цапель является
ночными или сумеречными птицами. Гнездование обычно является
.колониальным на деревьях, кустах или в тростниках, в гнездах,
устраиваемых из ветвей и грубых стеблей растений. Кладка из 3—3
(редко 7, у тропических видов — 2) голубоватых,зеленоватых и олив-
ковых яиц, у некоторых родов покрытых пятнами. Инкубация, в за-
висимости от размеров тела птицы, продолжается от 17 до 30 дней.
Вылупляющиеся беспомощными птенцы имеют только один пухово'й
наряд (состоящий из ргаереппае) серой или светлокорпчневоп окраски
и выкармливаются вначале из пищевода*
В настоящее время насчитывается около сотни видов цапель,
распределяемых по 20 родам и двум семействам: Ardeidae и Cochlea-
riidae\ последнее заключает только один род Cochlearius с двумя ви-
дами, свойственными тропической Южной и Средней Америке, и ха-
386
растеризуется широким плоско-выпуклым клювом, редуцированным
языком, непрободенными ноздрями и более короткой шеей. Семей-
ство Ardeidae характеризуется указанными выше признаками от-
ряда. Различают роды: Botaurus^ Ixobrychus, Nycticorax, Butorides,
Ardea, Phoyx, Ardeola, Bubiilciis, Egretta и др.
В СССР широко распространены серая цапля — Ardea cinerea L.
п выпь — Botaurus stellaris (L.). Преимущественно на юге, по водое-
мам зоны степей и отчасти широколиственных лесов, распростра-
нены: рыжая цапля — Ardea purpurea L., белая цапля — Egretta
alba (L.), кваква — Nycticorax nycticorax (L.), обыкновенный вол-
чок — Ixobrychus miuutus (L.). На южные окраины Союза заходит
ряд мелких и субтропических цапель, из которых наиболее инте-
ресны: чепура нужда — Egretta garzetta (L.), коровья цапля — Ви-
oulcus ibis (L.), желтая цапля — Ardeola ralloides (Scop.), амурская
зеленая кваква — Butorides javanica amurensis Schrenck. и амурский
каштановый волчок — Ixobrychus eurythnius (Swinh.).
Систематическое положение цапель внутри отряда, через Scopus
wBalaeniceps. легко связуется с аистами, а вне отряда проявляет опре-
деленную близость к веслоногим, с которыми их особенно объединяют:
сложный hypotarsus, ортоцёльный тип расположения кишечника,
строение желудка, окраска яиц, способ выкармливания птенцов
п т. д., опуская уже своеобразную конструкцию шейного аппарата,
сходную с таковой у Anhinga, но вызванную повидимому конверген-
цией. Вследствие этого ответвление цапель от общего ствола Cico-
niiformes следует рассматривать наиболее глубоким., В ископаемом
состоянии цапли известны начиная с нижнего олигоцена (род Рго-
herodius); род Ardea известен с среднего миоцена, род Butorides
с плейстоцена. Из третичных родов также известны роды Goliathia,
Ardeacites, Eoeeornis и Botaurites.
Характерные морфолого-анатомические особенности подотряда ?
кроме указанных, заключаются также: в весьма значительных разме-
рах сошника (наиболее крупного во всем отряде), прободенности
ноздрей и интерорбитальной перегородки, трахеобронхиальном си-
ринксе, 11 маховых и б. ч. 12 рулевых, редком пухе, свойственном
только аптериям, и, в связи с этим, значительном развитии побочных
стержней на контурных перьях, отсутствии musculus ambiens
и нек. др.
[ПОДОТРЯД 2: SCOPIDES. — МОЛОТОГЛАВЫ
Небольшая, отчасти напоминающая квакву, коричневато-бурая,
с умеренной длины клювом и лапами, птица, свойственная леси-
стым побережьям рек тропической Африки и Мадагаскара. Известно
лпшь одно семейство Scopidae с единственным видом Scopus umbretta.
Gmelin (рис. 244)/
По строению эта форма занимает промежуточное положение
между цаплями и аистами, но отчасти, как и по биологии, отличается
своеобразными чертами. Птица живет парами и питается рыбками,
лягушками и т. п., преследуя их на бегу или расхаживая, как аист.
Отдыхает обычно на деревьях или в гнезде, представляющем объеми-
*
387
печи с несколькими камерами и
Рис. 244. Молотоглав — Scopus гстЪгеИа G-m. Пз
Evans.
Рис. 245. Китоглав — Balaeniceps rex
Gould. По Evans.
стую постройку на дереве из сучьев и стеблей, сложенных в форме
одом. Кладка состоит пз
3 или больше яиц, бе-
лого цвета. Инкубация
длится 21 день. Птенцо-
вый пуховой наряд
(ргаереппае) серый. В
морфологии молотогла-
ва наиболее своеобраз-
ными являются: клюв
прямой, твёрдый, сильно
сжатый с.боков, с борозд-
ками, пробегающими от
ноздри до коготкообра#
но загнутого на конце
надклювья, и наличие
только 10 маховых пе-
рьев в крыле. Из черт,
общих с цаплями, сле-
дует отметить: сложный
hypotarsus, низко поса-
женный задний палец,
гребенчатость когтя
среднего пальца, си-
ринкс, снабженный мус-
кулатурой, положение шейной аптерии, отсутствие musculus ambiens.
С другой стороны, .Scopus близки аистам по малому числу позвон-
ков (16), непрободенным ноздрям,
сплошной интерорбитальной перего-
родке, редуцированному языку, от-
сутствию пудреток и некоторым дру-
гим особенностям. Мускулатура мо-
лотоглава вообще объединяет черты
обеих этих групп.
ПОДОТРЯД 3: BALAENICIPETES. —
КИТОГЛАВЫ
Крупная птица папирусовых бо-
лот верховьев Нила и Конго, с очень
большим, широким, на конце крюч-
коватым клювом. Единственное се-
мейство Balaenicipitidae с 1 видом
Balaeniceps rex Gould, (рис. 245).
Китоглав, подобно Scopus, так-
же занимает промежуточное поло-
жение между аистами и цаплями,
проявляя наибольшую близость к
аистам. По морфологическим особенностям китоглав может рассма-
триваться как примитивный аист, сохранивший еще много общих
388
черт с цаплями (п Scopus) и экологически специализированный в том
же направлении. В связи с обитанием в залитых водой болотах,
птице свойственны длинные пальцы ног. По походке и повадкам ки-
тоглав напоминает аиста, но преследует основную свою пищу —
рыб Protopterus и Polypterus— подобно цаплям, и летит, как послед-
ние, притянув шею к туловищу. Гнездо устраивается в болотных
зарослях; кладка состоит из 2 белых яиц; птенцы одеты .серым пу-
хом с розоватым налетом.
В морфологии птицы, как уже отмечено, перевес берут черты аиста.
В этом отношении особенно должны быть упомянуты: 16—17 шейных
позвонков, наличие носовой и межглазпичной перегородок, малень-
кий язык, распределение пуха как по птерилиям, так и по аптериям,
весьма укороченный побочный стержень в пере, отсутствие шейной
аптерии, аистовый тип щиткования плюсны и нек. др. С другой
стороны, копчиковая железа редуцирована и на спине расположены
два больших пудрообразующих пятна; более сходны с цаплями
строение пищеварительного канала с огромным железистым желуд-
ком и своеобразным пилорическим отделом его, строение таза и дру-
гие признаки. Ряд этих признаков общи также китоглаву и Scopus.
ПОДОТРЯД 4: CICONIAE. — АИСТЫ
Большие птицы, с крупным, заостряющимся к концу клювом,
населяющие травянистые болота и сырые луга Старого и Нового
Света. Из внешних признаков для аистов особенно характерно строе-
ние ног и клюва; задний палец короток и прикрепляется выше пе-
редних, когти тупы и лишены на переднем среднем пальце гребня,,
покров плюсны из шести сторонних щитков, на надклювье отсутствует
продольная бороздка. Биологически это птицы более сухих (чем
это характерно для цапель) местообитаний, с характерной спокойной
расхаживающей походкой, прекрасно летающие и способные к дли-
тельному парящему полету. Пища — мелкие рептилиц, амфибии,
насекомые, моллюски; обычно собираются с земли. Держатся и гнез-
цятся отдельными парами, устраивая обширные гнезда из ветвей,
на деревьях, скалах, как исключение на земле в болотах. Некоторые
легко становятся захребетниками человека. Кладка состоит из 3—5
эелых яиц. Инкубация длится около ЗО^цней. Птенцы являются зря-
чими с первого дня, покрытыми редким коротким белым пухом (ргае-
реппае), впоследствии дополняемым более длинным и густым ргае-
plumulae. Корм родителями приносится на край гнезда.
Известно около 20 видов, распределяемых по 10 родам и относимых
< одному семейству Ciconiidae. Роды: Ciconia, Sphenorhynchus.
^eptoptilos, Ephippiorhynchus, Ibis, Jabiru и др. В СССР встречаются:
эелый аист — Ciconia ciconia (L.), (рис. 246), исчезающий в настоя-
дее время амурский аист — Ciconia boyciana Swinh. и наиболее
лпроко распространенный черный аист — Ciconia nigra (L.).
В ископаемом состоянии аисты известны с нижнего олпгоцена
Palaeoephippiorhynchus), современные роды появляются уже с верх-
чего олигоцена (Leptoptilos) и плиоцена (Ciconia). Из ископаемых
юдов интересны: Propelargus, Pseudotantalus (миоцен Европы, бли-
383
зок к рецептному американскому Ibis), Amphipelargus, Palaeope-
1 argils и нек. др.
Морфолого-анатомические особенности аистов выявляют как спе-
цифические черты, так и разнообразные отношения родства с ца-
плями (через Balaeniceps и Scopus), ибисами, Pelecaniforines, Phoeni-
copteriformes и Cathartae, так что филогенетически аисты являются
очень интересной центральной группой отряда, сохранившей черты,
в значительной части уже утерянные близкими родичами, более
специализированными и пошедшими по боковой, сильно отклоняю-
щейся линии развития. Из своеобразных черт замечательно сильное
развитие воздушных мешков
носоглоточной системы, спус-
кающихся у большинства ви-
дов далеко вниз под кожей
шеи, но не вступающих в
связь с мешками легочной
системы. Они наполняются
при выдыхании через saccus
orbitalis, расположенный под
глазницами, и служат повщць
мому гл. обр. как эластич-
ная подушка для клюва во
время сна. При этом голова
втягивается в плечи, а клюв
торчит вперед, покоясь на
надутой воздухом передней
стороне шеи. Особенно ве-
лика эта подушка у марабу,
в связи с его огромным клю-
вом, который и на лету птица
принуждена нести, втянув
шею, как цапля, в то время
Рпс. 2'16. Бепьгп аист —^icouia cicghIu L. По как друГие аПСТЫ ЛеТВЮТ С
вытянутой шеей. Характерно
далее отсутствие голосовых мускулов в сиринксе, у большинства
видов лишенном и мембран, так что аисты большей частью немы п
восполняют этот недостаток особым щелканьем клюва. Отдельные
виды имеют однако более совершенный голосовой аппарат, у марабу
(трахеальный сиринкс) напоминающий даже сиринкс примитивных
воробьиных, у самца Ibis с сложенной петлями трахеей. В связи
с парящим полетом грудная клетка более прочна, вилочка тесно
прилегает к килю и у ряда видов срастается с ним, a musculus ре-
cloralis дифференцируется в два слоя. В пищеварительном аппа-
рате должно быть отмечено наличие железистых полей в первом
и пилорического отдела во втором (мускулистом) разделах же-
лудка. Имеются очень маленькие парные слепые кишки.
Из особенностей, выявляющих родственные взаимоотношения,
достойны внимания: связующие с ибисами густая оперенность шеп
и упрощенный hypotarsus, связующие с цаплями (промежуточных
Balaeniceps и Scopus опускаем) прободенность ноздрей и голорийпя,
390
наличие лишь одной вырезки на грудине; связующие с фламинго
наличие рудиментарного penis’a п расчлененности шейных воздуш-
ных мешков на отдельные камеры.
ПОДОТРЯД 5: THRESKIORNITHIDES. — ИБИСЫ
Рис. 247. Колпица — Platalea leuco-
rodia L. По Evans.
Птицы средней величины, с гибким, изогнутым книзу и за-
остренным к концу или ложкообразным клювом, населяющие бо-
лотистые прибрежья тропической и умеренной зон. Обычно подвиж-
ные и стайные ибисы разыскивают пищу, состоящую из червей,
моллюсков, ракообразных и насекомых, или ковыряясь длинным
клювом в мягкой почве или грязи водоемов, или (колпицы) вытря-
сывая ее из пл а на мелководье характерными боковыми движениями
широкого и чувствительного клюва.
Из внешних морфологических черт ха-
рактерны наличие длинных бороздок,
пробегающих от ноздрей по поверхно-
сти надклювья, почти до конца его,
связанность пальцев короткими пере-
понками и обычная оголенность кожи
на лицевых частях и шее. При полете
вытягивают шею и плюсны. Гнездятся
б. ч. колониально, устраивая гнезда из
веток и стеблей, на деревьях, кустах,
в тростниках и на скалах. Кладка
обычно из 2—3 голубоватых яиц, иногда
покрытых пятнами. Инкубация длится
21 день. Птенцы, покрытые белым, ко-
ричневатым или черным пухом, вы-
кармливаются из глотки и пищевода
родителей.
В подотряде единственное семейство
Tkreskiornithidae с тридцатью (прибли-
зительно) видами, распределяемыми по
Ю родам. Внешне резко различающиеся
формой клюва, колпицы и ибисы легко
скрещиваются между собой и приносят
плодовитое потомство. Известны роды: Threskiornis, Plegadis,
llagedashia, Nipponia, Platalea, Ajaja и др. В СССР встречаются
нормально только три вида: каравайка — Plegadis falcinellus (L.),
японский или китайский ибис — Nipponia nippon (Тешш.) и кол-
пица— Platalea leucorodia L. (рис. 247).
В ископаемом состоянии ибисы встречаются начиная с олиго-
Цеца (род Ibidopsis). Из ископаемых родов известны: Ibidopsis,
ibidopodia, Pro tibis.
Морфологически ибисы обнаруживают наибольшее родство с
инетами, но проявляют большую примитивность, что не может найти
своего выражения в линейном расположении вследствие относи-
тельно прямых связей между цаплями, Balaeniceps, Scopus и Cico-
niae. Ближайшей формой из аистов является род Ibis (=Tantalus).
391
Очевидно ибисы представляют собой боковую, рано отделившуюся
и более специализированную ветвь общего ствола.
Основные мо’рфолого-анатомические отличия от аистов заклю-
чаются в следующем: шизориния (Platalea занимает переходное
положение, у которой кроме того более сильно развиты и носовые
железы), грудина с четырьмя вырезками (а не с двумя) и вилочка
не достигает грудины, нижняя челюсть длинным крючкообразным
processus angularis posterior (что отличает также и от прочих под-
отрядов), более полный ассортимент ножных мускулов, контурные
перья с побочным стержнем, только 11 маховых (а не 12) и другие
признаки. Из других особенностей важно отметить варьирование
формы трахеи, у колпиц разветвляющейся на бронхи уже в области
шеи и уложенной петлями в грудине, отсутствие голосовой муску-
латуры, отсутствие пудреток, оперенность копчиковой железы, ре-
дуцированность языка и пр.
ОТРЯД 6: PHOEN1COPTERIFORMES. — ФЛАМИНГО
Крупные, весьма длинноногие и длинношейные, с перепончато-
палыми лапами и коленообразно изогнутым клювом, красные или
беловато-розовые птицы мелких лагун и соленых водоемов Старого
(без Австралийской области) и Нового Света. Только одно семей-
ство Phoenicopteridae с двумя родами Phoenicopterus и Phoenicon-
naias и четырьмя видами.
Фламинго представляет собой весьма своеобразно специализи-
рованных птиц, свойственных преимущественно теплому поясу.
Это стайные птицы, держащиеся обычно на мелководье, богатом
илом и кишащем мельчайшими беспозвоночными. Длинные ноги
дают возможность птице глубоко, по брюхо, заходить в воду и,по-
грузив голову на дно, вылавливать из взмученной воды питательные
частицы и животных. Это происходит при помощи ситовидного аппа-
рата клюва, по принципу строения напоминающего клюв уток, по
с чрезвычайно мясистым и мощным языком и вследствие этого вы-
пуклым объемистым подклювьем и плоским надклювьем; края
обоих, как и край языка, усажены роговыми пластинками и зуб-
чиками, при закрытом клюве образующими ситовидный аппарат.
При опущенной на дно водоема голове, благодаря форме клюва,
подклювье оказывается сверху надклювья и образует неподвижную
часть. При кормежке надклювье приоткрывается, действием языка
всасывается вода с мелкими животными, а по закрывании клюва
выжимается через ситовидный аппарат, оставляя питательный ма-
териал, отобранный весьма чувствительным языком и клювом, на
их внутренней поверхности. Основной пищей являются мелкие
ракообразные (Artetnia salina), черви (Tubijex) и моллюски [Се-
riihium, Cardium и др.). Гнездятся фламинго колониально на впо-
следствии обсыхающих мелях, строя гнездо из цла в воде, в виде
усеченного конуса, высота которого определяется глубиной мелко-
водья (до 40—50 см). В лоток па его вершине откладываются 1—2.
относительно крупных, белых, с рыхлой меловой поверхностью яйца.
Оба взрослых принимают участие в воспитании птенцов, выкле-
392
вывающихся через 30—32 дня зрячими и весьма активными. Пухо-
вой наряд снежно-белый, замещаемый через четыре недели серым.
Спустя 4 дня птенцы покидают гнездо и бегают поблизости, причем
выкармливаются родителями отрыгнутой пищей около двух недель:
в дальнейшем они кормятся самостоятельно, подобно взрослым.
Вначале мягкий и прямой, напоминающий молодую колпицу, клюв
вскоре горбится и постепенно приобретает размеры, форму и изгиб,
характерные для взрослых птиц.
Своеобразное сочетание различных особенностей, как говорит
Гадов, «делало фламинго со времен Линнея истинным яблоком
раздора между орнитологами. Одни принимали его за нечто вроде
лебедя, с ногами аиста и лапами гуся, другие считали за аиста
с преобразованным утиным клювом, третьи поднимали его до уровня
группы, равнозначной всем аистообразным и всем гусеобразным,
п рассчитывали этим разрешить трудность задачи». Более детальное-
исследование показало наибольшее родство фламинго все же*
с аистами и ибисами со столь значительными уклонениями, что
введение его в группу, общую с ними, крайне затрудняет их дефи-
ницию, а с другой стороны, вполне оправдывает выделение его
в самостоятельный отряд. Родство фламинго с аистами следует из
весьма сходного строения грудины и грудной клетки, ^аза, кры-
ловой мускулатуры с musculus pectoralis, дифференцированным на
два слоя, в связи с сходным, хотя и не парящим, типом полета. На
лету фламинго вытягивает шею и ноги, принимая благодаря их
длине совершенно своеобразный вид. Так же сходны: строение муску-
латуры ног (хотя есть и своеобразные черты), строение шейных воз-
душных мешков, кишечник, рудиментальный penis, диастатакспче-
ский тип крыла, 12 маховых, оперенная копчиковая железа, птерк-
лозис, характер окраски и многие другие особенности, общие им
пгусеобразным (десмогнатизм, голориния, прободенные ноздри и др.).
Некоторые черты объединяют фламинго в первую очередь с иби-
сами и более узко — с колпицами (развитость носовйх желез, на-
личие затылочных фонтанеллей, мощно развитый processus angu-
laris posterior нижней челюсти, первоначальная форма клюва и др.).
Общие черты с гусеобразными следует рассматривать как развившиеся
конвергентно, так напр. строение языка, лап, недоразвитость зад-
него пальца и у отдельных видов его исчезновение, 19 шейных по-
звонков, число рулевых (12—16), функционирующие слепые кишки,
выводковость и одновременную линьку маховых, делающие фла-
минго неспособным к полету на двух-, трехнедельный период. Kai:
эти, так и другие подобные черты легко объясняются приспособле-
нием к типу питания и движения. Так сходное в принципе с гусе-
образными строение клюва образовано однако иным участием отдель-
ных костей; играющая у последних большую роль praemaxillare,
у фламинго очень мала, и надклювье образовано гл. обр. maxil-
lare. В строении ног развитие перепонок объясняется тем, что фла-
минго прекрасно плавает и бродит по вязкому илу. Следует отметить
наконец некоторые своеобразные черты фламинго: вторичные ба-
зиптеригоидные отростки сохраняются всю жизнь; сиринкс бронхо-
трахеальный (голос фламинго напоминает гусиный); большие воз-
393
душные полости под кожей, весьма мощный мускулистый желудок,
отделенный от железистого длинным участком, постоянное расшире-
ние пищевода в нижней его части (выполняющее функцию времен-
ного вместилища пищи?), щпткованпе плюсны спереди и сзади
поперечными табличками п нек. др. Часть этих особенностей может
рассматриваться как унаследованная от веслоногих.
В ископаемом состоянии виды современных родов обильно
встречаются начиная с верхнего олигоцена. Наиболее ранние п
сильно измененные представители этой же группы известны даже
с мела (Scaniornis и Parascaniornis). Наиболее интересны филоге-
нетически виды рода Elornis из олигоцена Франции и Англии с не-
сколько укороченными ногами и ряд видов рода Palaelodus, с верх-
него олиго цена по миоцен Европы, относимые иногда в особое се-
мейство Palaelodidae. Последний отличался еще более короткой
голенью п плюсной и значительно более длинными пальцами.
В отличие от современного Phoenicopterus, metatarsalia были сильнее
сплющены с боков, a hypotarsus построен сложнее, так что эти
формы явно стояли ближе к аистам, чем рецентные роды-и виды.
ОТРЯД 7: ANSERIFORMES. — ГУСЕОБРАЗНЫЕ
Отряд объединяет водных, гл. обр. растительноядных птиц, есте-
ственно распадаясь на два подотряда: Anseres и Anhimae. Обеим
этим, довольно резко различающимся по биологии, группам свой-
ственна следующая комбинация признаков, одновременно отличаю-
щая их и от всех остальных птиц: выводковость, десмогнатическое
строение неба с хорошо развитыми вторичными базиптеригоиднымп
отростками и наличие двух пар стерно-трахеальных мышц (musculus
ypsilo-lrachealis и musculus sterno-trachealis).
Следующие морфолого-анатомические черты также общи обоим
подотрядам. В строении скелета: сходное число шейных позвонков
(у Anseres 16—25, у Anhimae 19—20), грудная кость с вырезом
с каждой стороны, разделенные базальные концы коракоидов,
U-образная вилочка, сходное число ребер, голоринальный тип но-
совых костей, отсутствие носовой перегородки, наличие длинного
загнутого кверху processus angularis posterior. В кровеносной си-
стеме характерно наличие парных сонных, артерий. В пищевари-
тельном аппарате наблюдается сходный тип строения кишечника,
функционирующие слепые кишки. В перовом покрове: строение птен-
цового пуха, редуцированность побочного стержня в контурном
пере п в связи с этим обильно покрывающий все тело пух, отсутствие
шейной аптерии, одинаковое число (11) больших маховых, диастата-
ксический тип крыла и в общем согласное число рулевых перьев
(у Anseres 14—24, у Anhimae 12—14).
ПОДОТРЯД 1: ANHIMAE. — ПАЛАМЕДЕИ
Крупные, внешне напоминающие куриных, голенастые птицы
прибрежий стоячих вод Южной Америки. Характерно грузное оваль-
ное туловище, длинная шея пз 19—20 позвонков; ' четырехпалые,
ЗЭ4
лишенные плавательных перепонок, ноги, длинный хвост, большие
широкие крылья с шпорами на кисти и крючковатый куриный клюв
с восковицей. Пищу добывают, как гуси, сщипывая нежную болот-
ную растительность. Гнездо в виде большой кучи камыша устраи-
вается в мелкой воде. Кладка состоит из 2—6 желтовато-белых япц.
Инкубация длится 6 недель. Птенцы выводковые, весьма напоминаю-
щие по поведению и окраске гусят.
Только одно семейство Anhimiclae с двумя родами Anhima и
Chauna и тремя видами (рис. 248).
Паламедеи представляют собой крайне своеобразных птиц, обна-
руживающих прямую близость только к гусиным птицам и отличаю-
щихся от них рядом примитивных и специализированных черт.
Гадов рассматривает их как древнейших, еще живущих членов
общего ствола гусеобразных
птиц, ответвившихся даже еще
раньше, чем ясно наметилось
разделение Ciconiijorines и Anse-
rijorrnes. Поэтому отдельные не-
ясные черты сходства, которые
они проявляют также с фла-
минго и куриными, могут от-
части рассматриваться как на-
следство пройденной былой, еще
мало дифференцированной ста-
дии развития Anseriformes.
Гадов указывает, что из 40
наиболее важных морфолого-
анатомических признаков па-
ламедеи совпадают или близки
с Anseres примерно в 30, в
том числе в строении неба, ко-
нечностей, грудины, таза, пи-
щеварительном аппарате, мускулатуре сиринкса и др. (см. выше).
Из отличий этой группы наиболее существенны: отсутствие processus
uncinati, penis’а, подкожного жирового слоя и аптерий. Ряд наи-
более выделяющихся особенностей явно имеет адаптивный характер.
Напр., сильные довольно длинные ноги с покровом из шестиуголь-
ных щитков, распространяющимся отчасти и на голень, и с длинными
пальцами, что легко ставится в связь с полуболотным образом
жизни; задний палец при этом посажен очень низко и вооружен
длинным когтем. Скелет отличается крайней пневматичностыо вплоть
до фаланг пальцев и дополняется весьма разветвленной системой
воздушных мешков, из которых особенно замечательна система мел-
ких пузырей, распространенная под кожей и придающая ей эмфизе-
матозный характер. Это свойство очевидно связано с прекрасным
полетом паламедеи, который в брачный период сопровождается
длительным парением на большой высоте и характерными криками
(сиринкс у паламедеи трахео-бронхиальный). В пищеварительном
аппарате характерен объемистый железистый желудок с большими
полями желез, сильный, но маленький мускулистый желудок и круп-
Рпс. 248. Рогатая паламедея — Anhhna сог-
nuta. По Brehm, из Ililzheimer.
395
ные, своеобразной формы, слепая и толстая кишки. Своеобразным
является очень густой перовой и пуховой покров (причем перья
лишены, как правило, побочных стержней), распределенный равно-
мерно по всему телу. Мощные шпоры, помещающиеся на метакар-
пальных костях (I п II), служат птице очевидно для защиты.
ПОДОТРЯД 2: ANSERES. — ГУСИНЫЕ
Разнообразной величины, плавающие, с характерным строением
клюва птицы всего света. Клюв характеризуется прямой формой, мяг-
ким и чувствительным роговым покровом, оканчивающимся на вер-
шине твердым коготком, и наличием по краям клюва роговых пласти-
нок или зубцов. Вследствие своей обширности (более 250 видов)
группа с трудом поддается общей биологической характеристике.
В большинстве своем гусиные обитают пресные воды, болота, но также
и морские побережья, прекрасно плавают и б. ч. ныряют. В связи
с этим для большинства характерна округлая или вытянутая форма
тела, соединенные плавательными перепонками лапы, густое, упру-
гое контурное оперение, смазываемое выделениями крупной копчи-
ковой железы. Ряд форм преимущественно кормится на берегу,
срывая или скусывая клювом части растений, но большая часть
добывает корм в воде, то роясь в илу и высеивая или вылавливая
клювом питательные растения и мелких животных, то преследуя
рыбок и крупных водных насекомых. Сообразно этому наблюдается
значительное разнообразие в строении клюва, с грубыми зубцами
и большим ноготком у пасущихся на земле, широкого и плоского
с пластинками по краю у питающихся ’мелкими водными обитате-
лями, узкого и длинного с острыми зубчиками у преследующих
рыбу. Крупные виды обычно не ныряют и в связи с этим обладают
наиболее длинной шеей (у лебедей 23—25 шейных позвонков), пре-
следующие пищу под водой имеют наиболее укороченную шею
(16—17 позвонков); большинство же кормится на мелких местах
и погружается в воду весьма неглубоко. В связи с характером пищи,
клюв и язык отличаются высокой чувствительностью и богаты ося-
зательными тельцами» (тельца Грандри). Относительно хорошо также
развито обоняние, судя по крупным носовым полостям. Морские
виды имеют обычно очень крупную носовую железу, располагаю-
щуюся по верхнему краю орбиты, секрет которой повидимому ней-
трализует раздражающее действие соленой воды. Пищеварительный
аппарат отличается мощным мускулистым желудком и длинными
слепыми кишками у растительноядных форм; последние малы у ры-
боядных.
Вне периода размножения гусиные являются стайными птицами,
а в брачный период преимущественно моногамы, но основные за-
боты по выводу птенцов ложатся гл. обр. на самку. Яйца откла-
дываются обычно в ямку на земле под защитой растительности,
в дальнейшем выкладываемую специальным пухом самки, линяю-
щей к этому периоду. Этим пухом, обычно близким по окраске
к фону окружающей местности, яйца прикрываются во время отсут-
ствия матери. Некоторые виды однако гнездятся в скалах, дуплах?
396
пногда чужих гнездах, Heteronetta и отчасти Erisniatura иногда
подкладывают яйца в чужие гнезда. Кладка состоит из желтоватых,
голубоватых или зеленоватых яиц и весьма изменчива по величине:
обычно 4—6 яиц у гусей, 8—12 у уток, колеблясь в зависимости
от величины птицы и условий гнездования. Инкубация сравнительно
коротка у гнездящихся на земле: 22—26 дней (утки), 28 (гуси),
35 (лебеди), но значительно дольше у гнездящихся в дуплах: .у ма-
ленькой утки Aix — 31 день,’ у крупной утки Cairina — 35 дней.
Выклевывающиеся зрячими и покрытыми густьп® пухом птенцы
весьма активны с первого дня, плавают и ныряют, и только водятся
н защищаются, а не выкармливаются родителями. Единственный
птенцовый пуховой наряд вариирует от чисто белого до темнобурого
с светлыми пятнами. Последующие перья взрослых нарядов разви-
ваются из того же перового сосочка.
Вследствие своей распространенности, массовости и значитель-
ных размеров птицы этого отряда имеют»крупную экономическую
ценность как объект охоты и разнообразного хозяйственного исполь-
зования (см. стр. 521—532).
В целом весьма гомогенная и довольно однообразная по органи-
зации группа, с трудом подразделима на группы меньшего значе-
ния. Обычно различают четыре семейства:
Семейство 1. Mergidae — крохали, с узким клювом с круто за-
гнутым коготком на конце, с твердыми острыми, направленными
назад, зубчиками по краю клюва, с родами: Mergus, Merganser,
Lophodyt.es, Mergellus.
Семейство 2. Anatidae — утки, с широким клювом, несущим по
краям роговые пластинки, и коротким ноготком, с родами: Merga-
netta, Salvadorina, Erisniatura, Somateria, Heniconetta, Oidemia,
Histrionicus, Thachyer.es, Biicephala, Nyroca, Metopiana, Netta, Spa-
tula, Heteronetta, Anas, Cairina, Aix, Nettapus, Bentrocygna и др.
.Семейство 3. Anseridae — гуси, с широким и высоким клювом,
с широким крупным ноготком и грубо зубчатыми краями клюва.
Роды: Tadorna, Casarca, Anser, Eulabeia, Philacte, Melanonyx, Cyg-
nopsis,Branta, Rujibrenta (рис. 249), Cereopsis (рис. 251), Anseranasu др.
Семейство 4. Cygnidae лебеди, с голой уздечкой, длинной
шеей, с широким клювом с грубыми пластинками по краям. Роды:
Coscoroba, Cygnus (рис. 250).
Виды из всех этих семейств весьма обильно представлены в СССР.
Наиболее распространенные: в семействе Mergidae — крохалей:
луток — Mergellus albellus (L.), большой крохаль — Mergus mer-
ganser Ъ. и’длинноносый крохаль — Mergus 'serrator (L.), распро-
страненные через весь Союз и свойственные гл. обр. рекам и круп-
ным водоемам. Из семейства Anatidae — уток: чирок-свистунок —
Querquedula crecca (L.), чирок-трескунок — Querquedula querquedula
(L.), клохтун — Querquedula jormosa (Georgi), широконоска — Spa-
tula clypeata (L.), кряква — Anas platyrhyncha L., серая утка —
Anas stepera L., шилохвость — Anas acuta L., свиязь — Marcca
penelope (L.), белоглазый нырок — Nyroca rufa (L.), красноголовый
нырок — Nyroca ferina (L.), хохлатая чернеть — Nyroca fuligula
(L.), морская чернеть — Nyroca marila (L.), гоголь — Bucephala
397
clangula (L.), морянка — Clangula hyemalis (L.), каменушка — Hi-
strionicus histrionicus (L.), синьга — Oidemia nigra (L.), турпан —
Oidemia fusca (L.), мандаринка — Aix galericulata (L.) и савка —
Erismatura leucocephala (Scop.)- Большинство из этих видов на гне-
здовье свойственны пресным во-
Pnc. Краспозобал казарка — Пи fi-
bre н la ruficollis Pall, llo Evans.
дам и распространены через весь
Союз пли значительную часть его.
Арктическим морским побережьям
свойственны гл. обр. гаги: обыкно-
венная — Somateria mollisima. (L.),
гребенушка — Somateria spectabilis
•(L.), очковая — Somateria fischeri
(Brandt) и малая гага — Henico-
netta stelleri (Pall.).
Из семейства Anseridae — гу-
сей: серый гусь — Anser anser (L.),
белолобый гусь — Anser albifrons
(Scop.), пискулька — Anser ery-
thropus (L.), гуменник — Melano-
nyx fabalis (Lath.). Более редки и
узко распространены : сухоиос —
Cygnopsis cygnoides (L.), горный
белошей — Philacte canagicus (Se-
гусь — Eulabeia. indica (Lath.),
wast.), белый гусь — Anser hyperboreus Pall., черная казарка —
Branta bernicla (L.), белощекая казарка — Branta leucopsis (Bechsl.).
краснозобая казарка — Rufibrenta ruficollis (Pall.), пеганка — Ta-
dorna tadorna(L.) и крас-
ная утка — Tadornafer-
ruginea (Pall.). Из ce- <
мейства Cygnidae — ле-
бедей, в СССР встреча-
ются гл. обр.: кликун —
Cygnus су gnus* (Li.}, ши-
пун — Cygnus olor (Gm.)
и малый лебедь — Cyg-
nus minor (Pall).
По систематическому
положению гусиные яв-
ляются изолированной
группой, обнаруживаю-
щей близкое родство
только с Anhimae, че-
рез эту последнюю свое-
образную группу форм
Рис. 250. Малый лебедь — Cygnus bewickii Jarr. По
Evans.
Anseres связывается и с Ciconiiformes. Различные водоплавающие-
веслоногие, гагары, поганки, чайки чужды гусеобразным.
В морфологической характеристике гусиных, кроме указанного,
следует подчеркнуть следующие черты. Большинство видов, входя-
щих в эту группу, обладает сильным, быстрым п выносливым поле-
том. Однако есть и нелетающие (Tachyeres cinereus Gm.) или почтя
398
нелетающие (Nesonetta auclandica Gray.) виды, характеризуемые зна-
чительной редукцией крыла. При плавании необходимая сила раз-
вивается толчками ног, но крупные морские утки Somateria, Oidemia
помогают движению кроме того и крыльями. Задний палец у всех
видов очень укорочен и высоко прикреплен, но, несмотря на. это.
у хорошо ныряющих видов он также снабжен особой кожистой ло-
Рпс. 251. Скелет куриного гуся — Cereopsis novae-hollandiae Lath, (левая фигура) и скелет
гигантского гуся — Cnemiornis calcitrans Owen., утерявшего способность летать, из плейстон
Цепа Южного острова Повой Зеландии (правая фигура). Высота скелета Cnemiornis 91,5 ехц
оба изображения уменьшены в одинаковых отношениях. По Owen, из Stresernann.
пастью, что применяется как диагностический признак при разде-
лении группы на подсемейства. Характерно наличие musculus
ambiens.
Большое разнообразие проявляется в строении трахеи. У ряда
лебедей, имеющих громкий, трубный, музыкальный голос, трахея
образует в грудине петлю, проникая в нее вентрально от вилочки,
У Anseranas трахея лежит двойной петлей на поверхности грудного
мускула; самцам большинства видов свойственны расширения тра-
хеи ц вздутия в виде костяного лабиринта, являющиеся причиной
появления у самца в брачный период отличного, нередко весьма
399
музыкального голоса; строение трахеи имеет поэтому существенное
таксономическое значение. Наконец весьма оригинальной чертой
является наличие длинного penis’а, выворачивающегося сходно с ре-
nis’oM нанду {Rhea). Его сохранение возможно обусловлено тем,
что спаривание у этих форм обычно происходит на воде.
В ископаемом состоянии некоторые пз современных родов уток
(Anas, Fuligula) известны с верхнего олиго цена, а гуси (Anser) и
лебеди (Cygnus) с миоцена. Из более древних интересна находка
в верхнем эоцене Франции Romainvillia, близкой к современной
Dendrocygna, в настоящую эпоху не выходящей за пределы тропи-
ков, и наиболее примитивной утки GallorniS' straeleni Lambr., извест-
ной пз верхнего мела Франции.
Особый интерес представляет также нелетающий гусь из плей-
стоцена Новой Зеландии — Cnemiornis calcitrans Owen, весьма близ-
кий австралийскому роду Cereopsis (рис. 251). Он обладал весьма
редуцированным крылом и уже плоской грудной костью.
ОТРЯД 8: FALCONIFORIIES. — ДНЕВНЫЕ ХИЩНЫЕ ПТИЦЫ
Разнообразные по величине, хищные, с крюч -ообразно загнутым
клюром и восковицей у основания его, обычно с острыми загнутыми
когтями, птенцовые птицы различных сред почти всего света. Группа
распадается на три подотряда: Cathartae, Sagittarii и Accipitres,
сохраняя однако общими значительное число и биологических осо-
бенностей. В целом характеризуются: десмогнатизмом (проходя ши-
зогнатическую стадию в индивидуальном развитии и нередко удер-
живая ее и в взрослом состоянии), голоринальным типом носовых
костей, metasternum, на обоих краях несущей вырезы или фонта-
иелли (однако у многих видов полностью зарастающие), наличием
musculus ambiens, двух сонных артерий, мясистым крупным языком,
весьма расширенным пищеводом с резко выраженным зобом и весьма
слабым и растяжимым мускулистым желудком и редуцированными
(в разной степени) слепыми кишками. В наружных покровах харак-
терно, как правило, наличие побочного стержня (исключая С а-
thartae и Pandion), равномерное распределение пуха как на птерп-
лиях, так и на аптериях, с образованием местами пудреток у неко-
торых' родов. Копчиковая железа хорошо развита и обычно бывает
оперена. Диастатаксическое крыло, как правило, несет И (редко 10)
маховых. Количество рулевых перьев 12, редко 14.
По индивидуальному развитию это птенцовые, вылупляющиеся
.зрячими, но с медленным развитием птицы. Птенцовый пух, обычно
белый или серый, вначале состоит из ргаереппае, в дальнейшем до-
полняемым и перерастающим позднее развивающимся praeplumulac.
Птенцы выкармливаются кусочками пищи, разрываемой родителями
на гнезде и подаваемой им в клюве. Грифы выкармливают птенцов
пз зоба.
По систематическому положению дневные хищники ведут свое
происхождение от птиц, близких апстообразным и веслоногим. Это
родство проявляется гл. обр. через примитивную группу CdthaTtae,
АОО
в то время как Accipilres представляют собой сравнительно новую
и конечную ветвь, наиболее отошедшую от предков и богатую фор-
мами.
ПОДОТРЯД 1: CATHARTAE. — АМЕРИКАНСКИЕ ГРИФЫ
Крупные, с оголенной от перьев головой и удлиненным крючко-
ватым клювом, птицы Южной п южной части Северной Америки.
Биологически замещают грифов Старого Света, подобно им питаясь
преимущественно падалью и беспомощными позвоночными живот-
ными. Обитают горы, степи, отчасти леса, прекрасно летают, обла-
дая парящим полетом, легко передвигаются по земле. Гнездятся
на скалах, деревьях, часто в дуплах. Кладка из 2—3 чисто белых
или покрытых красновато-коричневыми пятнами яиц. Птенцы вы-
клевываются зрячими, но ха-
рактеризуются длительным раз-
витием. В морфолого-анатоми-
ческом отношении американские
грифы образуют обособленную
н весьма примитивную группу,
проявляющую существенные
черты сходства вне отряда с
апстообразнымп и веслоногими
птицами (см. стр. 376 и 390). Са-
thartae, несмотря на наличие мно-
гих черт специализации, оста-
лись как бы на более низкой
ступени развития и наименее
выработали в своих представи-
телях хабптус и повадки днев-
ной хищной птицы. Они пре-
красно бегают по земле благо-
даря сравнительно длинным но-
гам п напоминающему куриных
строению лап, с длинным средним
п укороченным задним пальцем
и мало искривленными когтями.
Впротивоположность остальным
Рис. 252. Кондор — Vvltur gryphus L., сам-
ка. По Evans.
дневным хищникам, американские грифы сохранили хорошо разви-
тое обоняние (с крупными носовыми камерами и мощными lobi
olfactorii), играющее существенную роль при разыскивании добычи.
Пз примитивных черт ими удержаны: еще функционирующие всю
жизнь базиптеригоидные отростки; лишенная голосовых мускулов
нижняя гортань с полностью замкнутыми бронхиальными кольцами
вследствие чего птицы немы; пилорический, снабженный нитевидными
выростами, как у Anhinga, отдел желудка. Кроме того, характерно
отсутствие надбровного отростка praefrontale, отсутствие носовой
перегородки, наличие на metasternum грудины двух пар вырезок,
отсутствие слепых кишек, отсутствие дополнительного стержня
в оперении и оголенность копчиковой железы.
20 Орнитология—1209
491
Подотряд разделяется на два семейства: современных американ-
ских грифов, Cathartidae, с родами: Vultur^ Sarcorarnphus, Gymno-
gyps, Coragyps и Cathartes (рис. 252) и вымершее семейство Тети-
thornithidae с родом Teratomis пз плейстоцена Калифорнии и сомни-
тельного по систематическому положению рода Tapinopus из эоцена
Франции.
Семейство Cathartidae также представлено в ископаемом состоя-
нии видами рецептных родов Sarcorhamphus, Gymnogyps, Vultui\
Coragyps и Cathartes (с миоцена — плиоцена) и вымершими Phasing
gyps. Palaeogyps.) Cathartidarum и Plesiocathartes из отложений позд-
нее эоцена. Из них наиболее интересен последний, известный пз
верхнего олигоцена Франции, указующий на распространение этого
семейства ранее и в Европе.
ПОДОТРЯД 2: SAGITTARII. — ПТИЦЫ СЕКРЕТАРИ
Рис. 253. Секретарь — Sagittarius serpen tar ius
Miller. По Brehm, из riilzheimer.
Крупная, внешне напоминающая серим, длинноногая п длинно-
хвостая птица степей Африки. Единственное семейство Sagitta-
riidae с видом Sagittarius ser-
pentarius Miller (рис. 253).
Своеобразно измененный п
специализированный хищ-
ник, преследующий преиму-
щественно рептилий (особен-
но змей) и насекомых и уби-
вающий их ударом о землю.
Прекрасно летает, но редко
пользуется крыльями, перед-
вигаясь гл. обр. бегом на
своих длинных, с короткими
пальцами и круто загнутыми
когтями, ногах. Гнездится
устраивая объемистые гнезда
из ветвей и стеблей на низ-
ких деревьях и кустах. Клад-
ка из двух белых яиц, с весь-
ма медленным развитием.
Инкубация длится около 42
дней: становится летным пте-
нец только спустя четыре ме-
сяца.
В морфологическом отно-
шении, помимо своеобраз-
ной специализации, секре-
тарь удержал хорошо выра-
женные примитивные чертыг
ставящие его в промежуточное положение между предыдущим и
последующим подотрядом. Ему еще свойственны функционирующие
во. взрослом состоянии базиптерпгоидные отростки, хотя строение
ноздрей (imperviae), трахео-бронхиальный сиринкс, оперенная коп-
402
чпковая железа уже сближают его с Acciptlres. Характерно строение
грудины с сужающимся кзади metasternum, лишенным вырезов,
и срастание грудного киля с вилочкой, строение таза, а также уко-
рененность носовых камер и отсутствие раковины преддверия.
В ископаемом состоянии известны крайне близкие Amphiser-
pentarius robustus Miln. Edwards и A. Schlosseri Gail, (из верхнего
олигоцена Франции), отличающиеся более толстыми и короткими
плюснами.
ПОДОТРЯД 3: ACCIP1TRES. — НОРМАЛЬНЫЕ ХИЩНЫЕ ПТИЦЫ
Весьма разнообразные по величине, форме и образу жизни днев-
ные хищники, характеризующиеся несквозными ноздрями, руди-
ментарными базиптеригоидными отростками, трахео-бронхиальным
сиринксом, свободной, связанной лишь связками с грудиной ключи-
цей, короткими слепыми кишками и оперенной копчиковой железой.
Характерно далее наличие надбровного отростка praefrontale. В мор-
фологии форм наиболее разнообразны приспособления, связанные
с добыванием пищи, отражающиеся па способах полета, посадки,
движений по земле, строения клюва и лап. За редким исключением
(Gypohierax, Daptrius), питаются животной пищей, то добываемой
на лету (сокола, ястреба, орлы, сарычи и др.), то схватываемой
с земли (луни, сарычи и др.), с ветвей деревьев (Kaupifalco, Micra-
slur) или разыскивая павших животных (грифы, стервятники и др.).
Гнездятся на земле, на скалах, на деревьях, иногда в дуплах, не-
редко занимая чужие гнезда. В брачный период большинству видов
свойственны игры в воздухе, сопровождаемые своеобразным поле-
том. Кладка от 1 (крупные виды) до 6 (мелкие) белых пли голубо-
ватых, часто покрытых коричневатыми и сероватыми пятнами яиц.
Инкубация в зависимости от роста птицы длится 28—55 дней.
Птенцы выкармливаются обычно при участии обоих родителей, но
растут очень медленно и покидают гнездо у крупных видов только
на 3-м—4-м месяце.
Весьма богатая видами и родами группа (около 240 видов, рас-
пределяемых в сотню родов) разделима на два семейства.
Семейство 1: Accipitridae ( = Aquilidae). — Орлиные
С небной поверхностью рогового надклювья без продольного
гребня (иногда с возвышением в задней части, но резко ограничен-
ным спереди). Характерно положение границы оперения на боках
нижней челюсти в виде острого угла (в некоторых случаях оперение
не заходит на бока нижней челюсти, тогда темя является голым
или покрыто пухом). Анатомические черты: наличие hemiptery-
goid’a, сиринкс без резко выраженных membrana tympaniformis
externa и с голосовым мускулом, укрепляющимся на втором брон-
хиальном кольце. Принимаются подсемейства и роды:
Подсемейство 1. Elaninae, с родами: Elanus, Chelictinia п Ла-
thaerhamphus.
Подсемейство 2. Perninae — осоеды, с родами: ElanoideSy Avi-
tfda, Pernis, Chondr ohier ax и др.
403
Подсемейство 3. Miloinae— коршуны, с родами: Harpagus,
Г linia, Rostrhamus, Milons, llaliastur п др.
Подсемейство 4. Accipitrinae — ястребы, с родами: Accipiter, Ме-
Чегах, Urotriorchis п др.
Подсемейство 5. Buteoninae— сарычи, с родами: Buteo, Ви-
tnstur, Harpia, Pitecophaga, Aquila и др.
Подсемейство 6. Aegipiinae — грифы, с родами: Aegypius, Gyps,
Pseudogyps, Neophron, Gypohierax, Gypaetus и др.
Подсемейство 7. Circinae — луни, с родами: Circus, Geranospiza,
Gymnogenys.
Подсемейство 8. Circaetinae — змееяды, с родами: Circaetus,
Dryotriorchis, Eutriorchis и-др.
Подсемейство 9. Pandioninae — скопы, с родом Pandion.
В СССР это семейство представлено многими видами. Наиболее
важны: тетеревятник — Astur gentilis L. (=Л. palniubarius L.), пере-
пелятник — Accipiter nisus (L.), луговой лунь — Circus pygargus (L.),
полевой лунь — Circus cyaneus (L.), болотный лунь — Circus aeru-
ginosas (L.), коршун черный — Milons korschun, орлан-белохвост —
Haliaelus albicilla (L.), беркут — Aquila chrysaetus (L.), могильник —
Aquila heliaca Sav., степной орел — Aquila nipalensis Hodgs, под-
орлик — Aquila clanga Pall., обыкновенный канюк — Buteo buteo
(L.), степной канюк — Buteo rujinus (Cretzschm.), мохноногий ка-
нюк — Buteo lagopus (Pont.), змееяд — Circaetos gallicus Gmel.,
осоед — Pernis apivorus L., скопа — Pandion haliaetus (L.), стервят-
ник — Neophron perenopterus (L.), сип — Gyps fulous (НаЫ.), чер-
ный гриф — Aegypius monachus (L.), ягнятник — Gypaetus barba-
tus (L.).
Семейство 2: Falconidae. — Соколиные
С небной поверхностью рогового надклювья с резким продоль-
ным гребнем, который постепенно понижается кпереди. Характерно
также положение границы оперения на боках нижней челюсти в виде
прямой или слабо изогнутой линии, направленной вперед и вниз
(в случае же, если оперение не заходит на бока нижней челюсти,
темя остается покрытым перьями). Анатомические черты: сиринкс
с большими membrana tympaniformis externa, несущей голосовой
мускул, нижнечелюстными фонтанеллями и os dorsale. Принимаются
следующие подсемейства и роды:
Подсемейство 1. Ilerpetothcrinae, с родами: Micrastur, Негре-
thotheres.
Подсемейство 2. Polyborinae, каракары, с родами: JJaptrius,
Aliloago, Senex, Polyborus и др.
Подсемейство 3. Poliohieracinae, карликовые соколы, с родами:
S piziapteryx, Poliohierax, Mier oilier ах и др.
Подсемейство 4. Falconinae, настоящие соколы, с родами: Сет-
chneis, Aesalon, Erythropus, Hypotriorchis, Falco, Hierojalco и др-
В СССР встречаются представители только последнего подсе-
мейства. Наиболее широко распространены: сапсан — Falco pereg-
rin us Tunst, балобан—Falco cherrug Gray, чеглок — Hypotriorchis
401
subbuteo L., дербнпк — Aesalon columbarius (L.), пустельга — Cerdt-
neis tinunculus (L.) п кобчик Erythropus vesper linns (L.). Более
редки кречеты — Hierojalco gyrfalco L., и лишь степной области свой-
ственна степная пустельга — Cerchneis naumanni (Fleisch).
В ископаемом состоянии подотряд известен с нижнего эоцена
(род Lithornis). Современные роды (Aquila, Haliaetus. Buteo) по-
являются частью с эоцена, частью с миоцейа. Из вымерших родов
наиболее интересны: Palaeocircus, Teracus, Aquilavus, Palaeogyps,
Palaeohierax и нек. др.
ОТРЯД 9: GALLIFORMES. — КУРООБРАЗНЫЕ
Наземные и древесные, преимущественно растительноядные птицы
с шизогнатическим, голоринальным черепом, эвтаксическпм кры-
лом, с сохранением побочного стержня в оперении, мышечной фор-
мулой ABXY, с присутствующим expansor secundarium и большим
зобом.
В состав отряда входят два следующие подотряда.
ПОДОТРЯД 1: GALLI. — КУРЫ
Наземные и древесные птицы, с сильными четырехпалыми ногами,
короткими широкими крыльями, выводковые и преимущественно
растительноядные, распространенные почти по всей земле. Харак-
терна комбинация следующих анатомических особенностей: наличие
processus obliquus на processus lateralis posterior грудной кости и
вторичных базиптеригоидных отростков в небном аппарате черепа,
большой зоб, очень мускулистый желудок и длинные слепые кишки.
По своей экологии это довольно разнообразная группа, тяго-
теющая в основном к наземному образу жизни. Главные заботы по
воспитанию птенцов ложатся на самку, которая нередко значи-
тельно меньше самца.
Куриные представляют собой хорошо отграниченную группу
большого возраста, обнаруживающую близость, кроме гоацина,
только к скрытохвостам и отчасти к трехперсткам, но в обоих слу-
чаях эта близость может быть обусловлена широкой конвергенцией,
вызванной сходным образом жизни. Последнее в особенности ка-
сается общего хабитуса, строения ног, крыльев, морфологии клюва
и образа жизниг В отношении к этим группам наиболее характерными
чертами являются все же следующие: в отличие от скрытохвостов
небный аппарат построен по шизогнатическому типу с слабым па-
лочковидным сошником и коротким клювом парасфеноида, небные
косточки сочленяются с крыловидными костями, но не охватывают
rostrum, сдвигаясь несколько вбок. Птеригоид скользит при дви-
жении по клюву парасфеноидной кости, упираясь в соответственные
выступы со скользящей поверхностью, — вторичные базиптерпгоид-
ные отростки. В области таза incisura ischiadica частичным сраста-
нием ilium и ischium превращена в foramen ischiadicum. Трахеаль-
ные кольца окостеневают, а сиринкс обладает значительно большей
сложностью (чем у Тinamtformes). Иным закономерностям подвер-
406
жено распределение пуха, вообще малочисленного, встречаемого
только на аптериях, а не на птерилиях, что наблюдается у скрыто-
хвостов; отсутствуют также намечающиеся у последних пудретки.
Также характерным является щиткование спереди плюсны в два
раза, в то время как у Tinamtforines щитки располагаются в один
ряд. Все эти особенности указывают на иной путь развития Galli,
ведущих филогенетически свое начало м. б. и из ветвей, близ-
ких к месту ответвления скрытохвостов. От трехперсток куриные
отграничиваются присутствием os dorsale, иным строением meta-
sternum грудины, отсутствием spina externa и interna, наличием
зоба и рядом других признаков.
Обособление куриных вероятно падает уже на конец мела, так
как характерные формы подотряда известны даже из нижнего эоцена
(род Palaeophasianiis, из Северной Америки).
В отряде насчитывается несколько сотен современных и 6—7 де-
сятков ископаемых видов, распределяемых в 86 родов и 4 семейства.
Семейство 1: Cracidae. — Кракссвые
Преимущественно древесные, с длинным хвостом и круто загну-
тыми когтями на пальцах, б. ч. гнездующие на дереве, куры Южной
и Центральной Америки.
Характерна высокая пнев-
матичность скелета, уко-
роченность отростков и вы-
резов грудины, удлинен-
ность трахеи у самцов и
расположение ее петлями,
дегенерировавший побоч-
ный стержень в оперении
и сохраняющийся еще pe-
nis. Кладка состоит из
2—3 относительно круп-
ных яиц. Инкубация длится
22—29 дней. Ряду родов
свойственно уклонение в
строении черепа в виде не-
полного десмогнатизма.
Рис. 254. Хохлатый Ираке - Стах aleetor L. По ПринимаютсяРОДЫ: СгаХ,
Evans. Ortalis, Penelope, Oreopha-
sis, Mitu и др. (рис. 254).
В ископаемом состоянии известен лишь род Ortalis (Ort. phen-
gitis Wetm.) из нижнего плиоцена Небраски, указывающий на не-
давнее более обширное распространение этой группы к северу.
Семейство 2: Gallinuloididae
Ископаемый вид Gallinuloides wyomingensis Eastm. из среднего
эоцена Северной Америки со смешанными признаками ряда семейств
куриных (Cracidae, Tetraonidae). Наиболее близок повидимому
к роду Сгах или Ortalis.
406
Семейство 3: Megapodiidae.— Большеногие куры
Наземные, темно окрашенные куры Австрало-Малезпи с силь-
ными гребущими ногами и замечательным размножением. Очень
крупные яйца откладываются
в согребаемыс птицами боль-
шие кучи лесного мусора и
гумуса и согреваются тепло-
той тления его, или яйца
согреваются теплотой'вулка-
нпческого песка (см. стр.
229). Птенцы вылупляются
без участия взрослых, уже
покрытые перьями и способ-
ные подлетывать. При сход-
стве грудины с грудиной
краксовых их отличает от-
сутствие извивов трахеи, пра-
вой сонной артерии, пневма-
тпчностп костей и penis’а.
Принимаются 7 родов:
Megapodius, Eulipoa, Leipoa,
Talegalla, Мacrocephalon, Alec-
iura, Aepypodius (рис. 255).
Рис. 255. Кустарпап индейка—Aleclura la-
thami Lath. По Evans.
В ископаемом состоянии известен лишь один род Chosornis (Ch.
praeteritus De Vis) из плейстоцена Австралии.
Семейство 4: Phasianidae. — Фазановые
Основная масса куриных птиц, весьма разнообразных по окраске
п слЬженпю. Характерно прикрепление заднего пальца на ноге
выше уровня прикрепления передних, наличие -глубоких вырезов
в грудине и длинных боковых отростков ее, наличие обеих сонных
артерий, оперенность копчиковой железы, длинный побочный стер-
жень в оперении, отсутствие penis’a. Гнездо устраивается па земле,
п сравнительно небольшие яйца насиживаются только самкой.
Многочисленные представители семейства группируются в че-
тыре подсемейства:
Подсемейство 1. Numidinae — цесарки, с африканскими родами:
Numida, Guttera, Phasidus и др.
Подсемейство 2. Meleagrinae — индейки, с 2 видами родов: Ме-
leagris и Agriocharis в Центральной Америке и Соединенных Штатах.
Подсемейство 3. Phasianinae — настоящие куры, распространенные
почти по всей земле и наиболее многочисленные, с родами: Colinus,
Odontophorus и др. (американские по своему распространению роды,
выделяемые часто в особое подсемейство Odontophorinae)^ Lerwa,
Ammoperdix, Tetraogallus, Tetraophasis, Alectoris, Francolinus, Per-
dix, Coturnix, IthaginisjbTragopan, Lophophoras, Crossoptilon, Gen-
naeus, Gallus, Pucrasia, Phasianus, Chrysolophus, Polyplectron, Argu-
sianus, Pavo и др.; с многочисленными представителями в фауне
407
СССР , как то: перепел — Coturnix coturnix (L.), серая и бородатая
куропатки'— Perdix perdix L. п P. daurica (Pall.), каменная куро-
патка или кеклик — Alectoris kakelik (Falk.), уллар — Tetraogallus
caucasicus (Pall.) и близкие виды, кавказский фазан — Phasiamis
Рпс. 25G. Дикуша — Falcipennis falcipennis (Hartl.) (ппжппй рисунок — самец, верхний —
самка). По Jamashina.
colchicus L. п др. Более ограниченно распространены турач —
Francolinus orientalis (L.), горная курочка — Ammoperdix griseogu-
laris (Brandt), немой перепел — Coturnix japonica Temm. & Schleg.
Подсемейство 4. T atraoninae — тетеревиные с евразиатскпмп
ii североамериканскими родами: Telrao, Lyrurus, Lagopus, Tetrastes,
Bonasa, Bendragapus, Tympanuchus, Falcipennis (рис. 256), Centro-
cercus и др., из которых в СССР широко распространены гл. обр-
408
лесные п имеющие оольшое охотопромысловое значение впды г
рябчик — Tetrastes bonasia (L.), тетерев-косач — Lyrurus tetrix (L.).
обыкновенный и каменный глухарь — Tetrao urogallus L. и T. par-
virostris Bon. и тундряные и болсГгные белая и тундряная куропатки—
Lagopus lagopus (L.) и L. mutus (Mont.). Более узко распространены
кавказский тетерев — Lyrurus mlokosiewiczi (Tacz.) и дикуша —
Falci pennis jalcipenn.is (Hartl.) (см. стр. 521—532).
В ископаемом состоянии, как указывалось, семейство появляется
с нижнего эоцена (Palaeophasianus meleagroides). Современные роды
известны с миоцена (Melecigris) или нижнего плиоцена (Phasianus,
Gallus и др.). Из ископаемых родов наиболее интересны: Archeo-
phasianus, Palaeortyx, Palaeocry ptonyx, Taoperdix, Palaeoperdix, Pa-
ropavo, Palaeotetrix и др.
Рис. 257. Гоации — Opisthocomus hoatzin Miilk.
По Evans.
ПОДОТРЯД 2: OPISTHOCOML — Г0АЦ1ШЫ
Сходные с краксами, древесные четырехпалые птицы тропиче-
ской Южной Америки, с своеобразными глубокими приспособле-
ниями к обитанию в затоп-
ляемых лесах и мангровых
зарослях. Известно только
одно семейство Opisthocomi-
dae с единственным видом
Opisthoconius hoatzin Muller
(рис. 257). При общем сход-
стве с куриными, обильны
отличия, вызванные уклоня-
ющимся образом жизни. Под
влиянием давления мощно
развитого зоба, служащего
основным органом для пере-
тирания грубых листьев де-
ревьев, которыми питается
птица, киль грудины исчезает
спереди и сохраняется толь-
ко в задней ее части. Гру-
дина почти без вырезов,
срастается с корацоидами и
вилочкой и грудные муску-
лы несет гл. обр. в своей
задней части (рис. 258). Мус-
кулистый желудок крайне мал
и тонкостенен, а слепые ки-
шки укорочены. Филогенети-
чески интересны: отсутствие
вторичных базиптеригоидных
отростков, свойственных бли-
жайшим родственникам, по-
кров плюсны- маленькими щитками и распределение пуха не только
по аптериям, но и между перьями птерилий. Интересной особен-
ностью является образование у птенцов в первые недели жизни
409
острых подвижных когтей на концах 1-го и 2-го пальцев крыла,
при помощи которых птенец цепляется, лазая среди спутанных
ветвей обитаемых нм зарослей.
Гоацин почти не летает и не спускается на землю. Гнездо устраи-
вается постоянно над водой из рыхло сложенных веточек и несет
от 2 до 3 пятнистых яиц. Птенцы выкармливаются из зоба, подобно
голубям. Долго находящиеся в гнезде птенцы, одетые бледноко-
рпчневым пухом, являются в сущности промежуточными по разви-
тию между птенцовыми и выводковыми птицами, так как в случае
опасности спасаются бегст-
вом, лазая среди ветвей вы-
шеописанным способом, или
спрыгивают в воду, ныряют
и уплывают при помощи дви-
жений крыльев и ног. По ми-
новении опасности они вновь
влезают в гнездо. Вследствие
такого образа жизни у птен-
цов маховые развиваются мед-
ленно, но в дальнейшем, с по-
терей во взрослом состоянии
способности лазить и пла-
вать при помощи крыльев,
рост и смена маховых во
время линьки наблюдаются
как у куриных. Эвтаксиче-
ское крыло гоацина несет де-
рне. 258. Грудипа и плечевой попе гоацина —
-Opislhocomus hoatzin Mull.. с вентральной и дор-
зальной сторон.
Обратите внимание на редуцированный спереди
киль под влиянием давления мощно разросше-
гося зоба. По Huxley, пз Stresemann.
сять маховых, перья тела
снабжены длинным побочным
стержнем. Длинный хвост,
позволяющий лазящей птице
лучше балансировать, со-
стоит из десяти р’улевых.
Крайне своеобразный образ жизни и уклоняющиеся от куриных,
имеющие очевидно адаптивный характер, черты явились причиной
спорного положения гоацина в системе. Вследствие конвергенции,
приобретенные особенности напоминают, помимо куриных, также
мало специализированных кукушек, бананоедов и голубей. Наи-
более вероятным считается теперь рассматривать гоацина ранней
боковой ветвью куриных, еще на весьма примитивной основе пере-
шедшего к односторонней и исключительной специализации. В иско-
паемом состоянии гоацин известен из плейстоцена (?) Бразилии
и восточного Перу; кроме того близкий (но несколько сомнитель-
ный по родству) род известен из эоцена — олигоцена Франции:
Filholornis, с тремя видами.
ОТРЯД 10: TURNICIFORMES. — ТРЕХПЕРСТКИ
Мелкие, внешне напоминающие перепелов, степные и луговые
птицы тропических и субтропических стран Старого Света. Питаются
преимущественно семенами растений. Самки окрашены ярче сам-
410
цов п больше их размерами; подобно самцам многих куриных,
отличаются драчливостью и склонны к полиандрии. Кладка со-
стоит из 2 (Ortyxelus) — 4 пятнистых япц, но сопровождается по-
вторными кладками (известны случаи сноса за 55 дней 33 яиц),
которые распределяются между самчпками, выполняющими все обя-
занности по высиживанию и воспитанию птенцов. Инкубация длится
12—13 дней. Птенцы выклевываются покрытые пухом и характери-
зуются быстрым развитием: на 10-й день они уже подлетывают и
надевают птенцовый перовой наряд, сменяемый на наряд взрослых
на 5-й—6-й неделе. В конце 4-й недели они уже прогоняются вы-
полнявшими родительские обязанности самцами, а на четвертом
месяце становятся половозрелыми (тропические виды).
Трехперстки известны примерно в количестве 27 видов, относи-
мых к двум семействам.
Семейство 1. Pedionomidae, с единственным видом Pedionomus
torquatus Gould, свойственным Австралии (рис. 259). Характери-
зуется наличием заднего
пальца, дпастатаксическим
крылом, с первыми тремя
наружными малыми махо-
выми более укороченными
и слабыми, чем прокси-
мальные, сходно с тем, что
наблюдается у куриных.
Семейство 2. Turnicidae
с родами Turnix и Orty-
xelus, характеризуемые от-
сутствием заднего пальца
п эвтаксическим крылом.
Из этого семейства в наши
пределы, в Уссурийский
крап, заходит распростра-
нение Turnix tanki Ыап-
Рис. 259. СтспноИ странник — Pedionomus torquatus
Gould. По Evans.
jordi Blyth.
По систематическому положению трехперстки представляют за-
гадочную группу, весьма обособленную, хотя и обладающую чер-
тами разных отрядов: куриных, пастушковых, голубей, рябков и
ржанковых. В биологическом отношении эта группа аналогична
куриным, с тем же типом движений, с коротким, округлым и выпук-
лым крылом (с 10 маховыми) и теми же особенностями полета. Однако
грудина имеет более пастушковый характер и сзади имеет лишь
одну пару весьма глубоких вырезок, processus obliqui отсутствуют,
вилочка без крупной hypocleidium, а короткая и неперфорирован-
ная spina communis не образует поверхностей для сочленения с ба-
зально-медиальными концами коракоидов, так что последние оста-
ются разделенными, не перекрещиваясь, как это имеет место и у мно-
гих куриных. Своеобразным является также образование очень
крупной, до слияния 9 позвонков, os dorsale. В пищеварительном
аппарате, несмотря на сходный пищевой режим, характерны отсут-
ствие зоба и наличие значительной растяжимости переднего отдела
411
пищевода, что стоит, между прочим, в связи с брачными играми,
так как пищевод сильно раздувается при производстве характерных
звуков, издаваемых в этот период. Мускулистый желудок силен
и слепые кишки хорошо развиты. Замечательно строение черепа
с эгитогнатическим по форме сошником, широким и тупым спереди,
вильчатым кзади, прилегающим к длинному и пневматизированному
парасфеноиду. Processus palatini maxillae весьма слабы и крючко-
образны. Базиптеригоидные отростки возникают на basisphenoidale,
что напоминает отношение, наблюдаемое в палеогнатическом че-
репе. В строении костей, образующих носовую камеру (nares per-
viae), наблюдается значительное сходство с строением их у голубей.
Также своеобразно строение нижней гортани. Трахеальные кольца
остаются на хрящевой стадии. Контурное оперение с длинными
побочными стержнями. Эта комбинация особенностей заставляет
рассматривать трехперсток как весьма древнюю группу, удержав-
шую черты многих групп, в дальнейшем разошедшихся и утеряв-
ших прямую связь друг с другом. За это говорит и глубокая диффе-
ренциация семейств. Австралийский Pedionoinus отличается от соб-
ственно трехперсток сплошь примитивными чертами; к указанным
особенностям необходимо прибавить сохранение обеих сонных
артерий и более полную мускульную формулу для задних конеч-
ностей (ABXY). Между обоими семействами наблюдается развитие
в разных направлениях, так что их связь мыслима только через
общего предка. Это доказывается тем, что у Pedionoinus из обеих
сонных артерий левая является более слабой (у Turnicidae отсут-
ствует правая), а из ножных мускулов формулы Гаррода склонен,
к редукции отдел А (а не В, отсутствующий у тех же трехперсток).
ОТРЯД 11: GRUIFORMES. — ЖУРАВЛЕОБРАЗНЫЕ
Этот крупный отряд, включающий одиннадцать подотрядов,
имеет несколько сборный характер и представляет значительное
разнообразие как биологических, так и анатомических особенностей
у включенных в него форм. Преимущественно это наземные птицы,
хотя отдельные группы и виды и гнездятся и садятся для отдыха на
деревья. Число общих им признаков сравнительно ограничено, но,
несмотря на отклонения в различных особенностях, все включенные
сюда группы обнаруживают наибольшее родство именно друг к другу,
а не к другим соседним отрядам. Журавлеобразные могут быть оха-
рактеризованы как преимущественно болотные и наземные птицы,
с шизогнатическим черепом (с тенденцией к десмогнатизму у Rhino-
cheti и Cariamae), лишенным базиптеригоидных отростков, с гру-
диной, лишенной spina interna, и разделенными базальными концами
коракоидов, с трахео-бронхиальным сиринксом и с весьма просто
построенным пуховым нарядом. Зоб отсутствует. Для входящих
сюда подотрядов характерна вся комбинация этих признаков, а не
каждый признак в отдельности. 1
1 В классификации этого и следующего (Charadriiformes} отрядов я придер-
живаюсь более консервативной системы Ветмора, считая, что предложения
Р. R. Lowe (1931) должны быть предварительно подвергнуты основательному
412
В отряде Graijormes различаем 11 подотрядов: Mesoenades,
Ralli, Heliornithes, Rhinocheti, Eurypygae, Cariamae, Phororhaci,
Diatrymae, Psophiae, Grues, Glides,
ПОДОТРЯД 1: MESOENADES. — МАДАГАСКАРСКИЕ ПАСТУШКИ
Мелкие, по общему облику напоминающие повидимому земляных
кукушек, длиннохвостые и короткокрылые птицы саванн Мадага-
скара. Обитают преимущественно ксерофильные леса, проводя все
время па земле, быстро бегая на сильных довольно длинных ногах,
редко прибегая к употреблению крыльев. Питаются насекомыми,
гл. обр. муравьями. Гнездятся на земле и на кустах. Кладка состоит
из 2—3 темных пятнистых яиц. Птенцы выводковые, одетые черным
пухом. Известно единственное семейство Mesoenatidae с 3 видами
родов Mesoenas (= Мesites) и Monias,
Эта маленькая групка птиц долго блуждала в системе между
отрядами голенастых, воробьиных и куриных птиц, но наиболее
близка повидимому к пастушковым птицам. Вне сомнения это оста-
ток весьма древней и поэтому теперь хорошо изолированной группы,
благодаря островному распространению находящейся в процессе
редукции крыльев. Вероятно, наиболее правильно будет рассматри-
вать Mesoenades как одну пз древних боковых ветвей генерализован-
ного еще ствола Gruiformes, Из анатомических черт замечательны сле-
дующие особенности: череп шизогнатический, без базиптеригоидных
отростков и боковых затылочных фонтаиеллей, по строению небных
элементов явно журавлиного типа. Грудина типа трехперсток, без
processus obliqui с очень длинными proc, lateralis anterior, с кото-
рыми сочленяются 3 пз 5 достигающих грудины ребер. Очень длин-
ные коракоиды весьма слабы и соединяются с лопаткой под более
тупым углом (58°), упираясь в грудину на уровне 3-го—4-го грудин-
ного ребра. Вилочка совершенно исчезла. Характерна очень круп-
ная spina communis. Грудной киль сильно отодвинут кзади. Сиринкс
с парным musculus tracheo-bronchialis. В пищеварительном аппарате
отсутствует зоб, но мускулистый желудок значителен и очень длинны
слепые кишки. Весьма своеобразно распределены по телу пять
пятен пудреток, в связи с чем вероятно и стоит отсутствие копчи-
ковой железы. Контурные перья лишены побочных стержней. Крыло
с 10 маховыми перьями эвтаксического типа.
За древность группы говорит также сходство ряда особенностей
с другими наиболее малочисленными п изолированными группами
отряда, особенно Eurypygae и Rhinocheti, с которыми Mesoenades
разделяет наличие пудреток, образование шеи из семнадцати по-
звонков и другие признаки.
ПОДОТРЯД 2: RALLI. — ПАСТУШКИ
Мелкой и средней величины, несколько уплощенные с боков,
короткохвостые, невысокие на ногах птицы болотных и луговых
критическому разбору, в виду резко отличной трактовки им этих групп. Оба
указанных отряда (исключая подотряды Ralli и Heliornithes, выделяемые им
В особый отряд Ralliformes} объединяются им в один отряд под названием
Telinatoformes.
413
п отчасти водных местообитаний, распространенные почти по всей
земле. Обычно плохо летающие, но прекрасно бегающие-и искусно
пробирающиеся среди густой травянистой растительности, нередко
плавающие формы (в этих случаях с своеобразной кожистой ото-
рочкой по краям пальцев). Клюв сжатый с боков, с сквозными
ноздрями. Питаются преимущественно семенами, зеленью и бес-
позвоночными (черви, моллюски, насекомые). Нередко более активны
в сумеречное и ночное время. Гнездятся на земле плп на поваленном
тростнике, устраивая гнезда из стеблей трав. Кладка состоит из
3—12 яиц. Оба пола принимают участие в воспитании птенцов.
Птенцы выводковые, покрытые густым черным или темнокоричне-
вым пухом, с поздно развивающимися маховыми. Подобно жу-
равлям, пастушки одновременно меняют маховые перья и временно
становятся неспособными к полету. Многие островные формы па-
стушков утеряли способность летать и представляют собой виды,
находящиеся в различной стадии редукции крыльев с сопутствую-
щими изменениями в других различных органах.
Подотряд состоит примерно из 150 видов, объединяемых в одно
семейство Rallidae с тремя подсемействами.
Подсемейство 1. Rallinae, с родами: Gallira.llus, Crex, Porzana,
Rallus, Atlantisia и др.
Подсемейство 2. Gallinulinae, с родами: Gallinula, Gallicrex, Por-
phyria, Notornis и др.
Подсемейство 3. Fulicinae, с родом Fulica.
В СССР известно около 11 видов, из них наиболее распростра-
ненными являются: лысуха — Fulica atra L., коростель — Crex
crex (L.), пастушок — Rallus aquaticus L. и обыкновенный пого-
ныш — Porzana porzana (L.). Более узко распространены: сул-
танка — Porphyria poliocephalus (Lath.), камышница — Gallinula
chloropus (L.), большой погоныш — Porzana paykulli (Ljungh.), ма-
лый погоныш — Porzana рагу a (Scap.), курочка крошка — Porzana
pusilia (Pall.) п нек. др.
По систематическому положению пастушковые представляют
весьма цельную, хорошо отграниченную и внутри себя довольно одно-
образную группу. Наибольшая близость ими обнаруживается в отно-
шении к журавлиным и лапчатоногим, причем в отношении к первым
во многом они стоят глубже и являются более примитивными. Мно-
гие характеризующие их особенности могут быть сведены к приспо-
соблениям, связанным с своеобразным образом жизни и слабым
полетом, таковы, напр., тонкие, постоянно разделенные коракоиды.
U-образная, не достигающая грудины и лишенная hypocleidium
вилочка, отсутствие spina interna sterni, весьма длинные и отста-
вленные вбок processus lateralis posterior, отсутствие os dorsale
(обеспечивающее большую подвижность позвоночному столбу), длин-
ные и сильные ноги с относительно длинными пальцами и всегда
присутствующим задним пальцем, круглое, тупое крыло, умеренно
длинная шея из 14—15 позвонков. Другие особенности менее объ-
яснимы приспособительными чертами. В строении таза характерна
узкая и высокая предацетабулярная часть и обычно короткая, ши-
рокая постацетабулярная часть; foramen ischiadicum уменьшен до
414
круглого отверстия, что в целом может стоять в связи с их бегающим
наземным образом жизни. Ножные мускулы сохраняются в полном
комплекте. В пищеварительном канале характерно отсутствие зоба,
наличие очень маленького, но сильного мускулистого желудка,
функционирующие длинные слепые кишки, особенно у растительно-
ядных Fulica, Gallinula и других форм. Имеются обе сонные арте-
рии. Сиринкс типично трахео-бронхиальный. В наружных покровах
своеобразно мягкое, рыхлое, менее тесно сцепленное крючочками
контурное оперение, несущее весьма редуцированный побочный
стержень. Пух весьма просто построенный распределяется по всему
телу. Крыло у летающих форм диастатаксическое с 11 маховыми,
у нелетающих превращающееся нередко в эвтакснческое с редуци-
рованным числом маховых (до 9—8). Рулевых, обычно очень ко-
ротких и мягких, 12—14. На первом пальце в крыле развит коготь,
играющий некоторую роль при передвижении и лазании птенцов.
В ископаемом состоянии пастушковые известны с эоцена Европы
и Северной Америки (роды Eocrex, Palaeor alius, Gypsornis. Querey-
ryllus и др.). Наиболее интересны из ископаемых: Gypsornis cuvieri
М. Edwards пз верхнего эоцена Франции, сходный с южноамери-
канской Aramides, затем Aptornis dejossor Owen пз плейстоцена Но-
вой Зеландии, нелетающая гигантская форма (до 80 см высоты),
с сросшимися коракоидами и грудиной и весьма короткими и силь-
ными плюснами и нек. др. Современные роды известны начиная
с плиоцена (Rallus, Gallinula и др.).
ПОДОТРЯД 3: HELIORNITHES. — ЛАПЧАТОНОГИ
Мелкие, внешне напоминающие небольших поганок, птицы за-
болоченных прибрежий рек тропических лесов, прерывисто распро-
страненные в отдельных областях Южной Америки, Африки и
Индий. С длинным туловищем, покрытым плотным, как у уток,
оперением, относительно длинной шеей (на 2 позвонка длиннее шеи
пастушков), маленькой головой и узким удлиненным клювом. Ха-
рактерны острые крылья, длинный жесткий хвост и длинные пальцы
ног с кожистыми боковыми лопастями, подобно лапам лысухи.
Прекрасно плавают и ныряют, могут бегать по земле и недурно,
хотя и низко, летают.
Пища животная и растительная. Биология известна слабо. У не-
которых гнездо строится из веток в кустах над водой и откладывается
до 5—7 яиц, типа пастушковых. Птенцовые, у Ileliornis вылупляю-
щиеся даже голыми. Пуховой наряд серо-коричневый сверху, белый
снизу, с темной грудной полоской и длинным хвостом, как у утят
Anseres.
Известно только одно семейство 1leliornithidae с 3 видами, отно-
симыми к разным родам: Ileliornis julica Bodd. (Южная Америка),
Podica senegalensis Vieill. (Африка) и Helio pais persona la Gray (Ниж-
няя Индия, Суматра) (рис. 260).
Систематическое положение лапчатоногов загадочно и спорно,
в виду наличия смешанных черт, указывающих на родство с раз-
ными группами (Eurypygae, Mesoenades, Ralli). Большинство авто-
415
ров сближает их с пастушковымп, а Гадов рассматривает как и
выше специализированных по сравнению с ними. Несомненными
являются глубокая древность этих форм и односторонняя специали-
зация в направлении, сходном с биологией поганок. Строение че-
репа, отсутствие os dorsale, длинные слепые кишки, когти на пер-
вом пальце крыла, птерилозис, сиринкс, основные скелетные осо-
бенности сходны с таковыми, наблюдаемыми у пастушковых. Расши-
Рпс. 2G0. Сенегальский лапчатоног — Podica senegalensis Vieill. По Bannermann.
ренпе задней части грудины и вытянутая форма грудной клетки,
почти сплошное оперение брюшной стенки, сильно укороченные
плюсны, жесткий и по форме напоминающий баклана хвост, пластин-
чатые оторочки по краям пальцев, плотное оперение почти с полной
редукцией дополнительного стержня могут быть поставлены в’связь
с водным образом жизни. Мускулатура ног очень сильна, особенно
отличается сильным развитием musculus biceps, и прогрессивнее,
чем у пастушковых. Резко отличаются также от Ralli тем, что вилочка
синостотпчески сливается с грудиной. Лобковые и седалищные
кости отчасти срастаются. Крыло эвтаксическое с 10 маховыми
перьями.
ПОДОТРЯД 4: RHINOCHETJ. — КАГУ
Величиной и хабптусом напоминающая квакву, плохо летающая
птица болотистых лесных участков Новой Каледонии. Rhinochelus
jiibatusVerr. &Des Murs. (рис. 261) — единственный вид сем. Rhino-
chetidae.
Стальносерой общей окраски, с умеренной длины ногами, клю-
вом и хвостом и большим белым хохлом на затылке. Питается пре-
имущественно улитками, червями и насекомыми, частью доставае-
мыми клювом из глубины почвы. Гнездо устраивается из веток п
листьев на земле; в кладке только одно пятнистое яйцо типа пастуш-
416
заставляют рассма-
Рис. 261. Кагу— Rhinochetus jubatus J.
Verr. & Des Murs. По Evans.
ковых. Инкубация длится 36 дней. Птенцовые, но при вылуплении
покрытые пухом, по окраске .напоминающим пух бекасов.
Изолированный род с ближайшим систематическим родством с сол-
нечной «цаплей» Южной и Центральной Америки — Eurypyga helias,
с которой он имеет много общих признаков: сходно построенное небо,
обилие пудреток в оперении, наличие os dorsale, глубокие височные
впадины, структурные особенности нижней челюсти, весьма корот-
кие слепые кишки, характер распределения пуха, голую копчиковую
железу, одинаковое число маховых (10) и ряд биологических
черт (пища, поведение и др.).
Несмотря на крайнюю географическую удаленность обеих форм,
морфологические и биологические особенности
тривать оба эти вида как сравни-
тельно близко родственные спе-
циализированные пережитки гене-
рализированного ствола журавле-
образных птиц. Особенно интерес-
ным является сохранение сходных
признаков даже в окраске крыльев
и позах возбуждения и ухажива-
ния за самкой.
Из приспособительных черт
своеобразны крупные трубчато-
образно завернутые крышечки ноз-
дрей, образующие защиту ноздрей
при копании клювом в почве. Воз-
можно, что в связи с этим же стоит
развитие носовой перегородки. Из
других характерных черт, отли-
чающих от Eurypyga, следует упо-
мянуть: наличие только 16 шейных
позвонков, грудина более сходная с грудиной журавлей, без выреза
по заднему краю, редуцированная spina externa, наличие шейной
аптерии, крупно развитый побочный стержень в контурном пере
и эвтаксическое крыло.
Древность обеих форм сказывается также в сохранении ими при-
знаков, характерных для Ciconiijormes, причем Rhinochetus наиболее
сближается с Scopides, a Eurypyga с Nycticorax (Ardeae). Сближаю-
щими моментами одновременно являются как хабитус форм, так и
некоторые морфолого-анатомические черты. Однако последние имеют
бесспорно меньший вес, чем признаки родства в пределах Grui-
jorrnes, и должны рассматриваться только как обусловленные удер-
жанием этими изолированными и древними формами значительного
числа примитивных особенностей.
ПОДОТРЯД б: EURYPYGAE. — СОЛНЕЧНЫЕ ЦАПЛИ
Единственное семейство Eurypygidae и один вид — Eurypyga
helias Pallas (рис. 262), напоминающий небольшую цаплю, свой-
ственный тропическим болотным лесам Центральной и Южной Аме-
27 Орнитология—1209 417
рики. С тонкой длинной шеей, умеренной длины, тонким и прямым
клювом и ногами, с крайне пестрым оперением, напоминающим по
пестроте ночных бабочек. Характерно расправление крыльев и
хвоста веерообразно при возбуждении. Питается гл. обр. насеко-
мыми, склевываемыми с ветвей и листьев внезапными ударами
клюва, не несходно с цаплями. Гнездо устраивается из веток, грязи
и сухих листьев на
Рпс. 262. Солнечная цапля — Eurypyga helias Pall. По
Evans.
земле или низких вет-
вях. Кладка состоит
из 2 яиц типа пастуш-
ковых. Инкубация
длится 27 дней. Птен-
цы почти выводко-
вые, покидающие
гнездо спустя не-
сколько дней по вы-
луплении. Пуховой
наряд красновато-бе-
лый с темными про-
дольными и попереч-
ными полосками.
Ближайшая кагу древняя форма, анатомически характеризуе-
мая шизоринальным черепом без затылочных фонтанеллей, пробо-
денными ноздрями, грудиной, сходной с грудиной многих пастуш-
ковых, с вырезами в ее задней части, частью слитыми (3) грудинными
позвонками (os dorsale), 18 позвонками, составляющими шею, крайне
короткими слепыми кишками, многочисленными пудретками (но
с постоянным ростом составляющего их пуха в отличие от кагу),
диастатаксическим крылом и др.
ПОДОТРЯД 6: CARIAMAE. — СЕРИМЫ
Крупные, внешне весьма напоминающие африканских секрета-
рей или наших авдоток и джеков, быстро бегающие птицы степей
Южной Америки. Хвост, шея и ноги длинные, в основании надклювья
пучок торчащих вверх густых щетиновидных перьев, прикрываю-
щих ноздри. Пища состоит гл. обр. из змей, ящериц и насекомых,
убиваемых ударом плюсны (коготь второго пальца загнут, как
у хищной птицы). Гнездо устраивается из веток, помещается обычно
на густых кустах или низких деревьях. Кладка из двух белых с ко-
ричневато-красными пятнами яиц. Птенцовые, но вылупляющиеся
с открытыми глазами и покидающие гнездо ранее, чем поднимутся
на крылья. Пуховой наряд коричневато-желтого цвета.
Сходство с секретарями, долгое время признававшееся обуслов-
ленным родством, основано на конвергенции; из современных птпд
ближайшими систематически являются трубачи (Psophiae), а пз
вымерших очень близки гигантские, утерявшие способность ле-
тать, Piatryma и Phororhacos (эоцен Северной Америки и миоцен
Патагонии).
Серимы представлены современным семейством Cariamidae с 2>ви-
дами разных родов: Cariama (рис. 263) и Chunga и ископаемым се-
418
мейством Hermosiornidae^ с родами Procariama и Hermosiornis из
*нижнего плиоцена Южной Америки.
В целом подотряд анатомически характеризуется шизогнатиче-
ским голоринальным черепом с тенденцией к десмогнатизму и без
затылочных фонтанеллей.
Характерно наличие os
uncinatum. Слепые кишки
длинные, носовая перего-
родка отсутствует, над-
хвостовая железа голая.
Крыло эвтаксическое с 10
маховыми. Контурные пе-
рья снабжены побочными
стержнями.
ПОДОТРЯД 7: PHORORHACI
Гигантские ископаемые
птицы Южной Америки,
относимые главным обра-
зом к миоцену. При об-
щем внешнем сходстве с
серимами отличались по-
видимому полной потерей
способности к полету и
огромным массивным че-
репом, с вздутым крюч-
коватым клювом, в целом
достигавшим у Phororhacos
longissimus даже 3/4 метра
длины. Различают два се-
мейства : Phororhacidae и
Brontornithidae. В пер-
вом наиболее известны
Рпс. 263. Серпма — Cariama cristata L. По Evans.
роды Phororhacos и Pelecyornis, более
фрагментарно сохранились Prophororhacus и Callornis. В семействе
Brontornithidae различают роды: Brontornis, Liornis^Moreno-Мегсе-
ratia^ Physornis, Lophiornis, Ameghinia и Staphylornis, часть которых
сомнительна. В отдельное семейство Opisthodactylidae, относимое
иногда в эту группу, выделяют Оpisthodactylus patagonicus Am.,
известный по весьма фрагментарным остаткам.
ПОДОТРЯД 8: DIATRYMAE
Гигантские ископаемые эоценовые птицы Америки и Европы,
по образу жизни, повидимому, напоминавшие серим, а в анатоми-
ческих чертах обнаруживающие значительное сходство с предста-
вителями Phororhaci, что обычно объясняется конвергенцией. Ске-%
лет нижних конечностей весьма массивен и напоминает моа, кости
крыла крайне редуцированы, и птицы очевидно не летали. Голори-
нальный и десмогнатический череп длинен, с сжатым с боков и высо-
ким клювом, с массивной нижней челюстью и широким затылком.
419
Известно лишь одно семейство Piatrymidae с родами Piatryma и
Omorhamphus. Наиболее полно известен скелет Piatryma steini Matt/
et Grang. из эоцена Уайоминга, достигавшая 2 м высоты (рис. 264).
Единственным европейским прёдставителем этой, в основном аме-
риканской группы являтся Piatryma sarasini Schaub из нижнего
эоцена Швейцарии. В последнее время родство Piatryma с серимами и
журавлями подвергается сомнению, и делаются попытки связать их
с цаплями.
В эту же группу иногда относят в виде особого семейства Gastor-
Remiornis, Pasornis и Macrornis,
представляющих гигантских,
б. ч. фрагментарно сохранив-
шихся эоценовых птиц Евро-
пы.
nitidae, ряд родов: Gastornis,
Matt.&
Рис. 264.
Grang. По
Реставрация Diatryma steini
Matthew and Granger, из Heilmann.
ПОДОТРЯД 9: PSOPHIAE. -
ТРУБАЧИ
. Крупные, обликом напо-
минающие цесарку на более
длинных ногах, стайные, пло-
хо летающие птицы тропиче-
ских лесов Южной Америки.
Характерны округлый склад
тела, длинные ноги и шея
(17—18 позвонков), очень ко-
роткий клюв и хвост. Пища
преимущественно раститель-
ная, собираемая с земли.
Гнездо устраивается в дуп-
лах или на крупных ветвях.
Кладка состоит из 6—10 бе-
лых яиц. Птенцы выводко-
вые, покрытые светлокорич-
невым пухом. Ближайшими
журавли и серимы, но по
отклоняются и от них и со-
родственниками трубачей являются
многим особеностям птицы резко
храняют глубоко примитивные черты. Анатомически характери-
зуются шизогнатическим голоринальным черепом без затылочных
фонтанеллей и носовой перегородки, наличием ряда (5) надглаз-
ничных косточек (подобно Tinamiformes), частичным слиянием спин-
ных позвонков (os dorsalej, грудиной без вырезов и восемью груд-
ными ребрами, оперенной надхвостовой железой. Крыло эвтакои-
ческое с 10 большими маховыми, рулевых 10. Известно только
одно • семейство Psophiidae с несколькими видами рода Psophia
(рис. 265).
ПОДОТРЯД 10: GRUES. — ЖУРАВЛИ
Крупные и средней величины голенастые бегающие птицы,
с удлиненным туловищем, шеей и клювом, связанные обычно с бо-
лотами и степями почти всех частей света. Биологические осо-
420
бенности существенно разнятся по семействам;Характеризуются в це-
лом шизогнатическим шизоринальным черепом, без носовой перего-
родки и базиптеригоидных отростков. Затылочные фонтанелли от-
сутствуют. Сохраняются обе сонные артерии. Нижняя гортань тра-
хео-бронхиального типа. Крыло диастатаксическое, контурные перья
в побочными стержнями, пух распределяется как по аптериям, так и
птерилиям, пудретки отсутствуют, в хвосте двенадцать рулевых
перьев. Копчиковая железа оперена.
Подотряд составляется двумя семействами: Aramidae и Gruidae.
Семейство 1: Aramidae. — Арамы
С единственным видом Aramus scolopaceus Gmel., распространен-
ным по лесным болотам Южной и Центральной Америки и Фло-
риды. Напоминают обликом
крупного пастушка с умерен-
но длинной шеей и слегка
загнутым клювом. Питаются &
почти исключительно улит-
ками рода Ampullarius. В вос-
питании птенцов принимают
одинаковое участие оба ро-
дителя. Объемистое гнездо,
свитое из веток и стеблей,
устраивается на земле или
на дереве. Кладка состоит
из 5—8 пятнистых яиц. Птен-
цы выводковые, покрытые
черным пухом, долго вы-
кармливаемые родителями в
гнезде.
По своему строению Ara-
nas является несомненным
журавлем, хотя с еще не-
развитым внешне журавли-
ным обликом и односторонне
специализированным. Морфо-
лого-анатомические черты вы-
ражают преимущественно рис. 265. Трубач — Psophia crepitans L. По
специализацию формы, чем Evans.
ее примитивность. Харак-
терны длинные пальцы ног, развившиеся очевидно как при-
способление для передвижения по болотистой почве и удержания
раковины моллюска в момент освобождения его из раковины, при
употреблении в пищу. Шея в сравнении с настоящими журавлями уко-
рочена и состоит только из 17 позвонков; из них последний несет сво-
бодные ребра. Грудных позвонков 7. Вилочка отличается отсутствием
hypocleidium. Spina externa очень мала. Больших маховых только
десять. Трахея самцов, как обычно и у журавлей, удлинена и свер-
нута в длинную петлю, располагающуюся однако выше вхождения
421
трахеи в грудину, что является примитивным признаком, так как
примерно в таком же виде свойственно и самкам ряда видов настоя-
щих журавлей (Sarcogeranus, Anthropoides, Tetrapteryx и Balearica).
В ископаемом состоянии семейство известно из среднего мио-
цена Небраски (Северная Америка), в форме Aramornis longurio
Wetm. Другими близкими формами являются Euryonotus brachy-
pterus и Е. argentinus Mercer, из миоцена Аргентины.
Семейство 2: Gruidae. —Настоящие журавли
Крупные, широко распространенные птицы, с особенно длинной
шеей (19—20 шейных позвонков) и плюснами, но с пальцами умерен-,
ной длины. Характерны для большинства форм удлиненность трахеи
у обоих полов и прохождение ее петлями в киле грудной кости.
Рис. 26G. Венценосный журавль — Balearica pa-
vonine, L. По Evans.
в связи с чем стоит гром-
кий трубный голос этих птиц.
Главным образом степные и
болотные, хорошо бегающие
птицы. В отличие от сходных
внешне аистов, никогда не
садятся на деревья и кусты.
Пища гл. обр. растительная,
частью животная, иногда вы-
капываемая клювом из зем-
ли. Гнездо в виде подстилки
из стеблей трав, устраива-
ется на земле. Кладка со-
стоит из 1—2 пятнистых яиц.
Инкубация у крупных видов
длится до 34 дней. Птенцы
выводковые, покрытые жел-
товатым пухом. Для семей-
ства характерна одновремен-
ная линька маховых, делаю-
щая их временно неспособны-
ми к полету. У крупных ви-
дов поэтому она происходит
не каждый год (через год).
За исключением Balearic
журавлям свойственно по 11
маховых перьев. Характерны
для семейства далее особен-
ности, связанные с хорошо развитым полетом этих в большинстве
случаев перелетных птиц. Грудина узка и длинна, без вырезов по
заднему краю, с семью грудинными ребрами. Вилочка у перелетных
видов синостотически срастается с килем.
В ископаемом состоянии известны с среднего миоцена Европы:
Palaeogrus princeps Portis и др. (Probalearica, Paragrus, Protogrus,
Pliogrtis и др.).
Подотряд состоит из одного семейства Gruidae с 7 родами и 16—
422
19 видами. Роды: Grus, Sarcogeranus, Anthropoides, Balearica(piic. 266),
Antigone и др.
В СССР распространены девять видовиз них гл. обр. серый
журавль — Grus grus (L.) и в степях юга — Anthropoides virgo (L.).
Более узко распространены или редки даурский журавль —
Grus vipio Pall., черный журавль — Grus monacha Temm., манджур-
ский журавль — Grus japonensis (Mull.), стерх,— Sarcogeranus leuco-
geranus Pall, и нек. др.
ПОДОТРЯД 11: OTIDES. — ДРОФЫ
Крупные и средней величины, обликом напоминающие кур,
быстро бегающие и летающие птицы степей и пустынь Старого Света.
Характерны умеренно длинные шея и трехпалые ноги при округлом
плотном туловище. Пища преимущественно растительная, допол-
няемая насекомыми, ящерицами и грызунами в летнее время. Гнездо
устраивается на земле в ямке,
в густой траве. Кладка состоит
из 1—5 пятнистых яиц. Инку-
бация длится у дрофы 30 дней.
Самцы п самки постоянных пар
у некоторых видов повидимому
не образуют,^ и забота о птен-
цах ложится на самку. Самка
по размерам значительно мень-
ше самца. Птенцы выводковые,
покрытые очень густым корот-
ким пухом желтоватой общей
окраски с черным рисунком.
Около 30 известных видов
объединяются в единственное се-
мейство Otidae с принимаемыми
родами: Otis, Eupodotis, Chla-
mydotis , Tetr ах и др.
В СССР в степях и пусты-
нях распространены только три
вида: дрофа — Otis tarda L.
(рис. 267), стрепет — Tetrax tet-
Рпс. 267. Дрофа — Otis tarda L. По Evans.
rax L. и вихляй (или джек) — Chlamydotis macqueeni Gray.
Дрофы представляют достаточно цельную и хорошо отграни-
ченную группу. Хотя через посредство Burhinus они и связуются
с Charadrivformes, но основное родство их с журавлями представляется
несомненным. Череп голоринальный, без носовой перегородки и
затылочных фонтанеллей, но с прободенной межглазничной перего-
родкой. Шея образуется 16—18 позвонками. В грудине отсутствуют
spina sterni, и задний край имеет две пары вырезок, как у куликов.
В пищеварительном аппарате своеобразно строение весьма длинных
слепых кишек, имеющих сильно расширенной среднюю часть с про-
дольными складками слизистой оболочки и многочисленными округ-
лыми железками, а в концевом отделе усеянную ворсинками, по-
423
добно строению кишечника. Повидимому в этом проявляется при-
способление к преобладающей растительной пище. Сиринкс трахео-
бронхиальный, но голосовые мускулы или отсутствуют или слабо
развиты. В связи с этим повидимому пищевод у Otis tarda L. обла-
дает зобообразным расширением, а у некоторых видов и значительной
растяжимостью во время, токования, когда птица принимает стран-
ные позы и взъерошивает свое опе.рение. В это время он раздувается
до огромной величины, и играет роль резонатора, усиливающего
издаваемые птицей 'звуки; у обыкновенной дрофы для этой цели слу-
жит также особый горловой мешок, открывающийся под языком.
Характерным является также наличие рудиментарного penis’a
(сходно с Burkinas'). В оперении следует отметить замеча-
тельную розовую окраску пуха, покрывающего только аптерии;
такой же цвет свойствен и базальным пуховым частям контурного
пера, снабженного большим дополнительным стержнем. Копчиковая
железа, в связи с обитанием в сухом климате, совершенно отсутствует
и замещается обильным пудрообразованием самого оперения. Крыло
диастатаксическое. Мышечная формула Гаррода ВХУ.
В ископаемом состоянии виды рецентного рода Otis известны
начиная с верхнего миоцена. К среднему эоцену относится очень
близкий и древнейший род подотряда: Palaeotis (Р. Weigelti Lamb.).
ОТРЯД 12: CHARADRIIFORMES. — РЖАНКО ОБРАЗНЫЕ
Отряд объединяет преимущественно водных и береговых птиц
типа ржанки, чайки и чистика. Количество общих им признаков
невелико, но как и в составе других сложных объединений, напр.
Ciconiiformes, входящие в отряд группы обнаруживают наиболее
тесное родство между собой, в целом заметно обособляясь от осталь-
ных отрядов. Ближайшими родственными группами данному отряду
являются лишь Gruiformes и Turnicijonnes, с которыми родство
обнаруживается гл. обр. в подотрядах Jacanae и Thinocori. Обосо-
бление этих отрядов от общего ствола происходило повидимому
очень близко друг к другу.
Общими признаками отряда являются: шизогнатический, преиму-
щественно шизоринальный череп, с хорошо развитым сошником;
metasternum с одной или двумя парами вырезок, обычно весьма
коротких и ограниченных только задним краем грудины, spina
interna sterni отсутствует; вилочка U-образная; прокоракоидный
отросток обычно хорошо развит; processus ectepicondyloideus у летаю-
щих форм хорошо развит, шейных позвонков 15—16; обе сонные
артерии развиты; как правило, птицы, входящие, в отряд, имеют
диастатаксическое крыло, с 11 маховыми перьями, из которых на-
ружное обычно сильно укорочено. Птенцы вылупляются зрячими,
покрытыми пухом и обычно выводковые, хотя нередко длительно
выкармливаются в гнезде родителями.
Исключая глубокие ледяные пустыни Арктики и Антарктики,
этот отряд распространен решительно по всей земле.
В состав отряда входят подотряды: 1 — Jacanae, 2 — Thinocori,
3 —Limicolae, 4 — Lari, 5 — Alcae.
424
ПОДОТРЯД 1: JACANAE. — ЯКЛНЫ
Мелкие, сходные с пастушками, птицы тропических озер и болот.
Характерны высокие ноги с очень длинными и тонкими четырьмя
пальцами и когтями, дающими птице возможность бегать по плаваю-
щим листьям водных растений и зыбучей болотной грязи. По питанию
сходны с пастушками.
Гнезда устраиваются на
земле или у водоемов,
на плавающих листьях
водных растений. Клад-
ка состоит из 4, редко
5, пестрых или одно-
цветных яиц. Самец
меньше самки и несет
все заботы по воспита-
нию птенцов. Птенцы
выводковые, умеющие
плавать, покрытые пе-
стрым пухом.
По морфолого-анато-
мическим особенностям
яканы близки пастуш-
кам и куликам, имея
смешанные черты. Пер-
вых напоминает строе-
ние таза, имеющиеся у
некоторых кожистые ли-
цевые лопасти, пище-
варительный аппарат,
вторых — строение груд-
ной клетки, шизори-
ния, наличие вторич-
ных б а зипте р игои дных
отростков, образование
шпор на сгибе крыла
и другие особенности.
Преобладание все же
ржанковых черт в ана-
томическом строении за-
Рпс. 268. Африканская якана — Actophilornis afri-
canus Gm. на плавающих листьях растений. По Ban-
nermann.
ставляет видеть в них примитивную ветвь общего ствола ржан-
кообразных, приобретающих черты пастушковых конвергентным
путем. Из своеобразных черт необходимо отметить еще почти пол-
ную редуцированность слепых кишек, полную мускульную формулу
ног (АВХУ), но без специального длинного сгибателя заднего пальца,
указывающего на развитие специализированной конечности якан из
ноги нормального типа. Задний палец весьма длинен, прикреплен
очень низко и несет длинный, несколько вздернутый кверху, коготь.
На первом пальце крыла имеется короткий коготь. Сиринкс с одной
парой трахео-бронхиальных мускулов. Из Limicolae ближайшими
42а
формами являются повидимому примитивные Charadriidae (Rhynchaea
и др.).
Известно 11 видов единственного семейства Jacanidae, распреде-
ляемых по родам Jacona, Actophilornis, Hydrophasianus и др. (рис. 268).
ПОДОТРЯД 2: THINOCORL — ЗОБАТЫЕ БЕКАСЫ
Мелкие, внешне напоминающие рябков, быстробегающие, остро-
и длиннокрылые обитатели степей и полупустынь Патагонии и суб-
альпийской зоны южноамериканских Анд. Характерен округлый
склад тела, низкие, короткопалые плюсны, короткий куриный клюв
с ноздрями, прикрывающимися кожистыми крышечками, умеренной
длины хвост. Питаются преимущественно семенами растений, в связи
'с чем стоит наличие обширного зоба, мощного мускулистого желудка,
длинной прямой и слепых кишек. Яйца откладываются в углубление
почвы, кладка состоит из 4 грушевидных пестрых яиц, типа ржанок.
Имеется только одно семейство Thinocoridae с четырьмя видами
двух родов Thinocorus и Attagis.
Зобатые бекасы представляют интереснейший случай конвер-
генции с рябками и отчасти трехперстками. Систематическое .по-
ложение их долгое время вследствие этого было неясно, и только
изучение анатомического строения Thinocorus п Attagis выяснило
ближайшее родство их с ржанковыми (и особенно Glareolidae).
Отклонения в строении этой группы от типичных ржанок, вызванные
приспособлением к питанию семенами и жизнп в горных степях,
настолько значительны, что зобатых бекасов следует признать под-
отрядом значительного возраста, ответвившегося еще от весьма
генерализированных предков современных семейств Limicolae. По ске-
летным особенностям туловища Thinocoridae вполне укладываются
в рамки Limicolae, но сходно с тем, что наблюдается у Turniciformes.
череп неполно эгитогнатический с очень широким и сзади раздвоен-
ным сошником, полной носовой перегородкой и голоринальным
строением носовых костей. Базиптеригоидные отростки однако
отсутствуют и processus palatini maxillae коротки и утолщены.
Носовые железы хорошо развиты, но лежат в отличие от куликов
в углублениях крыши глазниц. Особенности пищеварительного аппа-
рата уже были упомянуты. Мускульная формула ног полная
(АВХУ). Сиринйс с одной парой.трахео-бронхиальных мышц. Копчи-
ковая железа оперена. Вероятно в связи с обитанием в холодном
климате, аптерии покрыты густым пухом черного цвета.
ПОДОТРЯД 3: LIMICOLAE. — КУЛИКИ
Мелкие и средней величины голенастые птицы, б. ч. длинно-
ногие, длинноклювые, острокрылые, с умеренной величины хвостом.
Обычно болотные или равнинные формы с общим космополитическим
распространением. Пища преимущественно состоит из беспозвоноч-
ных (черви, моллюски, ракообразные), отчасти насекомых. Гнездо
устраивается обычно на земле, в ямке. Кладка состоит из 1—4 пяг
нистых яиц.' Птенцы выводковые, одетые пестроокрашенным пухом.
42В
Филогенетические связи, вследствие включения сюда нескольких
уклоняющихся групп, разнообразны. Ближайшее родство наблю-
дается с чайками, чистиками и дрофами. Анатомически семейства
куликов объединяются шизогнатизмом черепа с раздвоенным сзади
сошником, отсутствием spina interna, несоприкасающимися базаль-
ными частями коракоидов, оперенной копчиковой железкой с одним
протоком, присутствием всегда musculus ambiens, сложным hypo-
tarsus. Многие признаки, общие для большинства форм, отсутствуют
у отдельных уклоняющихся родов и групп. К таким особенностям
надо отнести преобладающее у большинства форм шизоринальное
строение носовых костей, значительное развитие носовых желез,
metasternum с. двумя парами неглубоких вырезок, вилочка с hypo-
cleidium. Хороцю развитые слепые кишки, дистатаксическое крыло
с 11 маховыми, 12 рулевых. Контурное перо с побочным стержнем.
Подотряд разделяют обычно на пять семейств: Burhinidae, Dro-
madidae, Glareolidae, Chionididae и Charadriidae.
Семейство 1: Burhinidae.— Авдотки
Трехпалые, сумеречные и ночные, с очень крупными глазами
кулики степей, пустынь, пустошей и побережий моря жаркого и
умеренного пояса. Известно 9 видов, относимых к трем родам Burhi-
Рпс. 269. Сенегальская авдотка — Burhinus senegallensis Swains. По Bannermann.
nus, Orthorhamphus и Esacus (рис. 269). Характерны: голориния,
отсутствие носовой перегородки, затылочных фонтанелл и базипте-
ригоидных отростков. У сухопутных форм оперение обильно отде-
ляет пудру.
427
Преимущественно • парные или держащиеся одиночками птицы.
Яйца откладываются в ямку в песке, в количестве 1—2 пестрых яиц,
и отличаются от яиц большинства куликов овальной, а не грушевид-
ной формой. Инкубация у обыкновенной авдотки длится 26 дней.
В отличие от остальных куликов обнаруживают значительную
близость к Olides следующими признаками: внешние черты сходства,
отсутствие заднего пальца, мускулатура ног (ВХУ), голориния,
весьма рудиментарный penis, сиринкс, лишенный мускулов, и др.,
за что некоторыми авторами (Garrod, Forbes, Furbringer, Lowe)
объединяются с дрофами.
В ископаемом состоянии известен близкий ’род Milnea gracilis
Lyd. из верхнего олиго цена Франции и очень крупный вид Bathor-
nis veredus Wetm. из олигоцена Колорадо. *
В СССР только один представитель — обыкновенная авдотка —
Burkinas oedicnemus (L.), распространенный гл. обр. в пустынях и
степях Средней Азии и по пескам крупных южных рек Украины
(Днепр, Дон и др.).
Семейство 2: Dromadidae. — Рачьи ржанки
Единственный вид Dramas ardeola Paykull, несколько напоми-
нающий кулика-сороку, но более длинноногий, четырехпалый,
черный с белым кулик, распространенный по песчаным побережьям
западной части Индийского океана и Красного моря. Замечательно
колониальное гнездование в вырытых в песке самими птицами длин-
ных (1—1,20 м) и глубоких (50 см) норах. Кладка состоит из одного
очень большого белого яйца, весом в г/4 часть птицы. Птенцовые.
Птенцы одеты одноцветно серым пухом. Питаются крабами и моллюс-
ками. Характеризуются шизоринальным без базиптеригоидных от-
ростков и затылочных фонтанеллей черепом, очень крупными носо-
выми железами, гребенчатостью когтя среднего пальца и неболь-
шими плавательными перепонками.
Систематически стоят повидимому ближе всего к Glareola, Cursu-
rius и отчасти Recurvirostra.
Семейство 3: Glareolidae. —Тиркушки
Ржанкообразные птицы степей пустынь и песчаных берегов рек
Старого Света. Внешне довольно разнообразная группа. Собственно
тиркушки характеризуются на лету обликом ласточек, укороченным
клювом с широким разрезом рта, ловят насекомых обычно в воздухе
и гнездятся часто колониально. Пустынные (Cursorius, Peltohyas,
Rhinoptilus) отличаютсй длинными плюснами, бледной под цвет
почвы окраской и корм собирают с земли. Крокодилов сторож —
Pluvianus aegyplius, z&№4ST&№n комменсализмом с крокодилом, на
коже которого он разыскивает пищу (насекомых и червей) и зары-
ванием яиц (3—4) в горячий речной песок, предоставляя инкубацию
их действию солнца. Яйца группы характеризуются более крупной
формой, чем то свойственно большинству куликов. Инкубация
у Glareola длится 17—18 дней. Птенцы выводковые, пуховой наряд
428
окрашен под цвет почвы с сложным черным рисунком. Анатомически
характеризуются шизоринией с тенденцией у некоторых к голори-
нии, отсутствием базиптеригоидных отростков и затылочных фонта-
неллей. В противоположность остальным куликам ноздри несквозные
и замечательны крупные носовые железы. Плюсны спереди и сзади
покрыты поперечными щитками и средний палец имеет гребенчатый
коготь.
Известно около 25 видов, распределяемых по родам: Glareola^
Galachrysia, Pluvianus, Cursorius и др.
В СССР представлены степной тиркушкой — Glareola nordmanni
Fisch.-Wald., луговой — Gl. pratincola (L.) и пустынным бегунком —
Cursorius cursor (Lath.).
Семейство 4: Chionididae. — Белые ржанки
По величине и хабитусу напоминающие голубей, снежно-белые
птицы морского побережья островов Антарктики. Характерно плот-
ное округлое туловище, короткие плюсны и клюв, тяжеловатый
полет и неспособность плавать. Роговой покров клюва образует
над ноздрями особую чешуевидную крышечку. Пища гл. обр. из
слизняков, рачков, морской травы, собираемых на скалах в период
отлива. Гнездо устраивается в щелях скал; кладка из 2—3 с голубым
фоном пятнистых яиц. Инкубация длится 28 дней. Пуховой наряд
коричневый с нежным черным рисунком.
Только два монотипных рода: Chionis и Chionarchis.
Белые ржанки из Limicolae наиболее близки к так наз. куликам-
сорокам (род Haematopus), но представляют все же обособленную
более примитивную группу, многими исследователями рассматри-
ваемую как стоящую на полпути между куликами и чайками.
Кроме своеобразной ноздревой покрышки, очень характерны
также сильно вздутые lacrymalia и nasalia, отчасти прикрывающие
глубокие вдавления, образующиеся мощными носовыми (надбров-
ными) железами. Череп характеризуется широким сошником, но
лишен базиптеригоидных отростков, затылочных фонтанеллей и но-
совой перегородки. Processus angularis mandibulae заострен, длинен
и загнут книзу. Базальные концы коракоидов почти соприкасаются,
вилочка без hypocleidium, задний край грудины с одной парой
вырезок. Слепые кишки относительно очень длинны. Оперение очень
густо и жестко. Задний палец функционирует.
Семейство 5: Charadriidae. — Ржанковые
Основная масса куликов (более 100 видов), весьма разнообразных
по облику, размерам и повадкам, почти всесветного распростране-
ния. Вследствие разнообразия, с трудом характеризуемы биоло-
гически. Морфологически объединяются шизоринальным черепом,
С базиптеригоидными отростками и затылочными фонтанеллями, но
без носовой перегородки. В сиринксе для многих видов характерно
недоразвитие musculus tracheo-bronchialis.
Семейство распадается на 12 подсемейств, из которых одно (Pres-
byornithinae) включает только ископаемые формы. Различают под-
429г
семейства: Rostratulinae, Haematopodinae, Vanellinae, Charadriinae,
Thringinae, Arenariinae, Scolopacinae, Eroliinae, Ibidorhynchinae, Re-
cur virostrinae, Presbyornithinae и Phalaropinae. Классификация ржан-
ковых однако явно недостаточно разработана.
Подсемейство 1. Rostratulinae— цветные бекасы с родами: Rostra-
tula и Nycticryphes.
Подсемейство 2. Haematopodinae — кулики-сороки, с единст-
венным родом Haematopus.
Подсемейство 3. Vanellinae — чибисы, с родами: Chettusia, Vanel-
lus, Microsarcops^ Lobivanellus, Hoplopterus и др.
Рис. 270. Шллоклювка — Recurvirostra * avocetta L. По Bannermann.
Подсемейство 4. Charadriinae — ржанки, с родами: Squatarolo,
Phwialis, Charadrius, Eupoda, Eudromiqs и др.
Подсемейство 5. Tringinae— улиты, с родами: Numenius, Li-
mosa, Tringa, Pseudototanus, Xenus, Actitis, Heteroscelus и др.
Подсемейство 6. Arenariinae — камнешарки, с родами: Aphriza и
Arenaria.
Подсемейство 7. Scolopacinae — бекасы, с родами: Capella, Sco-
lopax, Philohela, Lymnocryptes и др.
Подсемейство 8. Eroliinae — песочники, с родами: Calidris, Сто-
cethia, Eurynorhynchus, Erolia, Liinicola, Tryngites, Philomachus и др-
Подсемейство 9. Ibidorhynchinae — серпоклювы, с единственным
видом рода Ibidorhyncha.
Подсемейство 10. Recurvirostrinae — шилоклювки, с родами: Hi-
mantopus, Recurvirostra (рис. 270).
430
Подсемейство 11. Presbyornithinae, с единственным (ископаемым)
представителем Presbyornis prevetus Wetm., сходным с родом Recur-
virostra, из эоцена Колорадо.
Подсемейство 12. Phalaropinae— плавунчики, с родами: Phala-
ropus, Steganopus^TL Lobipes.
Из СССР известно около 75 видов, из них до 70 видов, гнездя-
щихся в наших пределах. Наиболее распространены: тулес —
Squatarola squatarola (L.), европейская золотистая ржанка — Р1и-
vialis apricarius L., американо-сибирская золотистая ржанка —
Pluvialis dominions (Mull.), хрустан—Eudromias morinellus (L.),
галстучник — Charadrius hiaticula L., малый зуек — Charadrius
dubius Scop. и другие ржанки, чибис — Vanellus vanellus (L.), камне-
шарка — Arenaria interpres (L.), кулик-сорока— Haematopus ostralegus
L., большой кроншнеп — Numenius arquata (L.), средний кроншнеп—
Numenius phaeopus (L.), большой веретенник — Limosa limosa (L.),
малый веретенник — Limosa lapponica (L.), мородунка — Xenus cinerea
(Guld.), черныш — Tringa ochropus L., фифи — Tringa glareola L.,
перевозчик — Actitis hypoleucos (L.), большой улит — Tringa nebu-
laria (Gunn.), щеголь — Tringa erythropus (L.), травник — Tringa
totanus (L.), турухтан — Philomachus pugnax (L.), песчанка — Cali-
dris alba (Pall.), кулик воробей — Erolia minuta (Leisl.), белохвостый
песочник — Erolia temmincki (Leisl.), чернозобик — Erolia alpina
(L.), грязовик —Limicola falcinellus (Pontopp.), гаршнеп —Limno-
cryptes minima (Brunn.), вальдшнеп—'Scolopax rusticola L., бе-
кас — Capella gallinago (L.), дупель — Capella media (Lath.), кру-
глоносый плавунчик — Phalaropus lobatus (L.) и мн. др.. Роды с огра-
ниченным распространением в пределах Союза: Eupoda, Chettusia,
Lobivanellus, HimantQpus, Recurvirostra, Macrorhamphus, Arquatella,
Ancylochilus, Heteractitis, Pseudototanus, Eurynorhynchos, Hetero-
pygia, Rostratula и др.
В ископаемом состоянии кулики появляются с эоцена, в родах,
близких современным {Presbyonis, см. выше). Рецентные роды появ-
ляются часто уже с среднего и конца эоцена {Tringa, Philoheld),
с олигоцена (Vanellus) и позднее.
ПОДОТРЯД 4: LARI. — ЧАЙКИ
Разнообразные по величине птицы, представляемые общеизвест-
ными типами чаек и крачек (мартышек), по берегам и островам
рек и различных водоемов, распространенные по всей земле. Внешне
характеризуются удлиненным туловищем, длинными острыми
крыльями, перепончатопалыми лапами, с острым или характерно
загнутым крючком на конце клювом. Преимущественно рыбоядные
и насекомоядные, плавающие, но ныряющие только с воздуха.
Гнездо устраивается из стеблей трав на земле, на скалах (редко на
кустах). Кладка состоит из 1—4 грушевидных пятнистых яиц,
типа ржанковых. Часто колониально гнездящиеся, стайные птицы.
Птенцы выводковые, но долго кормимые родителями. Птенцовый
пуховой наряд напоминает наряд ржанок.
Родственные отношения довольно тесны с ржанковыми и чисти-
ковыми птицами; для чаек характерны следующие анатомические
431
признаки: шизогнатический и шизоринальный череп с хорошо раз-
витым сошником, без базйптеригоидных отростков, затылочных
фонтанеллей и носовой перегородки, большие хорошо развитые
носовые (надбровные) железы. Шейных позвонков 15. Грудина и
скелет плечевого пояса» как у ржанок, но вилочка* без hypocleidium.
Базальные концы коракоидов сходятся сзади spina externa sterni.
Грудные позвонки, как у А 1сае и ржанок, — опистоцельные. Сиринкс
типично трахео-бронхиальный с*хорошо развитыми мускулами. Пе-
редние пальцы связаны плавательной перепонкой; задний палец
мал или отсутствует.
Представляя цельную, довольно хорошо отграниченную группу
и резко отличаясь от куликов обликом и внешними признаками,
по своему строению чайки однако весьма близки к ним и особенно
семейству ржанковых и должны рассматриваться как побочная
прогрессивная и специализированная ветвь последних. Отличия от
чистиковых гораздо более значительны, так что внешнее сходство
с ними может быть было приобретено конвергентным путем.
В ископаемом состоянии семейство известно начиная с нижнего
эоцена [Halcyornis toliapicus Konig) и вообще в .эоцене представлено
уже рядом родов: Limosavis, Palaeotringa. Современные роды (sensu
latQ) появляются с олигоцена (Larus).
Отряд охватывает около 140 видов и разделяется на четыре
семейства: Laridae, Stercorariidae, Sternidae и Rynchopidae.
Семейство 1: Laridae. —Настоящие чайки
Сложены по типу обыкновенной чайки — Lams ridibundus L.
с светло окрашенным оперением, с характерной формы сильным
Рис. 271. Черноголовый хохотун-—Larus ichthyaetus Pall. По Evans.
крючковатым клювом с цельной (сплошной) рамфотекой. Характерны:
мускульная формула ног АХУ; слепые кишки рудиментарны. Насе-
ляют различные типы водоемов, подбирая пищу гл. обр. с воды или
432
земли, ь кладке ооычно z—J яйца, распространены по всей земле,
за исключением тропического пояса. Принимаются роды: Larus,
Rissa, Rhodostetia, Pagophila, Хета и др. (рис. 271). Представлены
20 видами в СССР, с наиболее обычными напр. в полярных морях:
Larus hyperboreus Gm., L. leucopterus Vieill., Rissa. trydactyla
(L.), Pagophila eburnea (Phipps) и др.; на пресных водах Larus mi-
nutus Pall., L. ridibundus L., L. canus L.; на южных морях Larus
ichthyaetus (Pall.), L. genei Breme и др. Монотипные роды в СССР;
Хета sabini (Sab.), Rhodostethia rosea (Megill.).
Семейство 2: Stercorariidae.— Поморники
Обликом схожи с чайками, но темной или бурой окраски, с удли-
ненными центральными рулевыми, с крючковатым клювом, с рам-
фотекой, разделенной на 4 части, с мышечной формулой АХУ, сдлин-
ными слепыми кишками. Всеядные и гл. обр. хищные птицы, добы-
вающие пищу гл. обр. отнимая ее у других птиц. В кладке только
2 яйца. Только один род Stercorarius с биполярным распростра-
нением. В СССР гл. обр. поморник длиннохвостый — Stercorarius
parasiticus (L.), поморник короткохвостый — Stercorarius cepphus
(Brunn.) и поморник средний — Stercorarius pomarinus Temm.
Семейство 3: Sterni dae. — Крачкп
Сложены по типу обыкновенной крачкп — Sterna hirundo L.,
внешне напоминая ласточек, с вилообразным или вырезанным хвостом
н узкими длинными крыльями. Клюв, утончающийся к вершине,
с прямым острым концом. Питаются мелкой рыбой и насекомыми,
бросаясь часто за добычей с воздуха и как бы ныряя за ней. Кладка
из 1—3 яиц. Слепые кишки рудиментарны. Мускульная формула
АВХУ. По распространению почти космополиты.
Известно 42 вида, относимых к 10 родам: Sterna, Gelochelidon,
Gygis, Anous, Chlidonias, Hydroprogne и др.
В СССР представлены гл. обр. обыкновенной крачкой — Sterna
hirundo L., малой — St. albifrons, чегравой — St. tschegrava Lepech.,
длиннохвостой — St. paradisea Brunn., белокрылой — Chlidonias
leucoptera (Temm.), черной — Ch. nigra (L.), белощекой — Ch. hib-
rida (Pall.) и др.
Семейство 4: Rynchopidae. — Водорезы
Сложены по типу крачек, черной с белым окраски, с уплощенным
с боков подклювьем, значительно выступающим за конец надклювья
и играющим роль чувствительного органа при отыскивании пищи,
собираемой с поверхности воды. Кладка из 3—5 яиц. Мускульная
формула АВХУ. В отличие от остальных семейств musculus ambiens
отсутствует. Слепые кишки рудиментарны. Зрачок в виде щели.
Распространены в тропическом поясе, преимущественно в устьях рек;
летают над водой с погруженным в воду подклювьем. Только один
род Rhynchops с тремя видами.
28 Орнитология—1209
433
ПОДОТРЯД б: ALCAE. — ЧИСТИКИ
Рис. 272. Бескрылый чистик—Pinguinus irnpennis L.
(крупная фигура) и люрик—Plotus alle L. Обратите вни-
мание па относительную величину крыла у крупного,
потерявшего способность летать, вида; бескрылый чи-
стик вымер около 1844 г. По Thomson.
Мелкой и средней величины морские, хорошо ныряющие (гребя
крыльями под водой) и плавающие птицы Ледовитого и северных
частей Атлантического и Тихого океанов. С удлиненным туловищем,
далеко отодвинутыми назад перепончатопалыми лапами, короткими
острыми крыльями и разнообразным по форме клювом. Пища состоит
гл. обр. из ракообраз-
ных и рыб. Гнездятся
обыкновенно массовыми
колониями на скалистых
берегах. Очень круп-
ные яйца (1—2) сно-
сятся без собственно
гнезда в трещины скал,
ниши россыпей, откла-
дываются на скалистых
уступах или в вырытых
самими птицами норах.
Инкубация длится 30
дней у кайр, гнездя-
щихся открыто, и 40
дней у топориков, гнез-
дящихся в норах. Вы-
водковые, но с длитель-
ным выкармливанием
птенцов родителями. Пу-
ховой наряд по окрас-
ке сходен с окраской
взрослой птицы и со-
стоит из перьев двух ге-
нераций: пухового и по-
лупухового; первый но-
сится только короткий
срок и перелинивает во
второй еще в гнезде.
Известно около 30
видов, относимых к 14
родам единственного се-
мейства Alcidae. Роды:
Plotus, Pinguinus (вы-
мер), Alca, Uria, Cepphus, Pseuduria, Brachyramphus, Synthlibor-
hamphiis, Aethia, Fratercula, Lunda, Cerorhinca и др. (рис. 272).
Основная масса видов Alcidae распространены в северной части
Тихого океана и Беринговом море, и в СССР встречаются предста-
вители всех современных родов. Наиболее широко распространены
однако только следующие виды: длинноклювая кайра — Uria aal^
(Pontopp.), короткоклювая кайра — Uria lomvia (L.), тупик —Fra-
tercula arctica (L.), чистик — Cepphus grylle (L.) и нек. др.
По систематическому положению чистики близки чайкам и кули-
434
кам и являются повидимому ответвившимися от предков современ-
ных Limicolae и Lari в период их разделения. В современных формах
они представляют собой глубоко специализированную группу, в от-
дельных представителях, напр. бескрылом чистике — Pinguinus im-
pennis, утерявших даже способность летать вследствие редукции
крыла п приспособления его к передвижению под водой.
Как в морфологическом, так и биологическом и географическом
отношении в этой группе наблюдается значительная конвергенция
в развитии с пингвинами, черты которых, в менее узко и высоко
специализированной форме, группа и отдельные виды повторяют.
Из приспособительных особенностей следует отметить весьма длин-
ную п относительно узкую грудную кость, с далеко назад заходя-
щими 7—8 грудными ребрами, защищающими тело от водяного
давления; грудные позвонки свободны и, как у пингвинов (а также
Lari и Liniicolae), опистоцельны, что обеспечивает большую гибкость
туловища. Передние позвонки с большими гипапофизами, несущими
мощный сгибатель шеи, musculus collii longus anticus. Скелет крыла
несколько уплощен и укорочен, особенно в предплечье, в связи
с употреблением крыла для движения под водой. Крыло острое,
с очень жесткими 11 маховыми. В задних конечностях задний палец
мал или отсутствует, а передние соединены плавательной перепон-
кой. Плюсна коротка. Мускульная формула более прогрессивная
(АВХ, AX), musculus ambiens отсутствует. Некоторые виды, нахо-
дясь на земле, сидят почти в вертикальном положении па всей
плюсне и с трудом передвигаются на твердой поверхности. Хвост
очень короток, пз 12—16 рулевых. Носовые железы велики. Форма
клюва в ряде видов стоит в связи с характером пищи: у питаю-
щихся планктонными рачками он короток, нередко вздернут, у ловя-
щих рыбу — удлинен, у некоторых сплющен с боков, возможно
в связи с разрыванием песка при добыче моллюсков и держащихся
на дне рыб, а также рытьем нор. Язык мясистый, вытянутый, иногда
с щетинками у основания. У планктоноядных (Plotus, Aethia и
вероятно других близких родов) в период кормления птенцов разви-
вается объемистый горловой (подъязычный) мешок. Слепые кишки
обычно рудиментарны. Контурное оперение жесткое, с крупным
побочным стержнем. Пух очень густой, распределяется равномерно
по всему телу.
Интересны приспособления в форме и окраске яиц. У колониально
гнездящихся на уступах скал видов яйцо резко грушевидной формы,
что сохраняет его от падения (скатывания) с узкого уступа, и весьма
разнообразной пятнистой окраски, облегчающей отыскивание своих
яиц птицей. У птиц, гнездящихся в щелях скал и в норах, яйца
овальные и нередко совершенно лишены какой-либо окраски. Птенцы
видов, гнездящихся на уступах, являются также более выводко-
выми, чем вылупляющиеся в защищенных местах (россыпях, норах).
Лпнька всех маховых наступает одновременно, так что птицы вре-
менно теряют способность к полету.
Из особенностей, указывающих на родство, необходимо отметить
еще: шизогнатический, шизорпнальный череп, без базиптеригоид-
ных отростков, иногда затылочных фонтанеллей и носовой перего-
*
435
родкп, как л у чаек. Базальные концы коракоидов разобщены.
У ряда видов рамфотека периодически линяет, спадая отдельными
кусками, что стоит в связи с специально брачной раскраской. Пте-
рилозис в общем типа Lari и Limicolae.
В ископаемом состоянии известны начиная с эоцена. Наиболее
замечательны: Mancalla calif or niense Lucas, нелетающий гигантский
вид (высота около 70 см), близкий к роду Uria, из плиоцена Кали-
форнии; Nautilorius и Hydrolherikornis пз эоцена Северной Америки,
с относительно более (?) длинными libiolarsus, чем современные
виды.
ОТРЯД 13: COLUMBIFORMES. — ГОЛУБЕОБРАЗНЫЕ
Отряд объединяет голубей и рябков. Это преимущественно расти-
тельноядные, наземные или древесные птицы с следующими общими
морфолого-анатомическими признаками: череп шизогнотический и
шизоринальный, с носовой перегородкой и обычно с базиптеригопд-
ными отростками, но без сошника (исключая Didunculus); шейных
позвонков 14—16; metasternum с двумя парами вырезок; базальные
концы коракоидов разобщены; плечевой отдел крыла относительно
короток, с очень сильным проксимальным отрезком и глубокой
коракопдноплечевой ямкой; processus ectepicondyloideus перемещен
несколько в проксимальную сторону; hypotarsus сложный; носовые
железы слабы или отсутствуют; сонные артерии нормальные; в пи-
щеварительном аппарате характерно наличие большого зоба; крыло
обычно диастатаксическое с 10—11 большими маховыми; копчиковая
железа мала или отсутствует, функционально’ замещаясь пудро-
образованием растущими перьями; нижняя аптерия велика, про-
должаясь от зоба до анального отверстия.
Из двух подотрядов, составляющих эту группу, большинство
анатомов рассматривает Pterocletes как более примитивных. Основа-
нием для этого является большее количество примитивных черт,
как напр.: полная мускульная формула (АВХУ), наличие побочного
стержня в пере, длинные слепые кишки и ряд других признаков.
ПОДОТРЯД 1: PTEROCLETES. — РЯБКИ
Мелкие, быстро летающие, обликом напоминающие голубей птицы
сухих степей и пустынь Африки и южной Азии. Характерен корот-
кий клюв; маленькие, густо оперенные, иногда с слитыми пальцами,
лапки; длинные острые крылья; заостренный хвост.
Питаются семенами пустынных растений, собирая их на земле,
передвигаясь своеобразным семенящим бегом. Преимущественно стай-
ные птицы. Самец отличается более яркой окраской. Гнездятся па
земле, откладывая 2—4 пятнистых, овальной формы, яйца в углубле-
ние почвы. Птенцы выводковые, с густым коротким пестрым пуховым
нарядом, окрашенным под цвет почвы. Гнездясь нередко в глубине
безводных мест пустыни, ежедневно летают на водопой, причем пьют,
как голуби, всасывая воду клювом и принося птенцам воду в зобу п
в смоченном оперении.
436
Только одно семейство Syrrhaptidae, с примерно 30 видами двух
родов Pterocles и Syrrhaptes. В фауне СССР представлены гл. обр.
следующими видами: саджа — Syrrhaptes paradoxus (Pall.), черно-
брюхий рябок — Pterocles arenarius (Pall.) и белобрюхий рябок —
Pterocles alchata, (L.) (рис. 273).
По своему строению, помимо своего родства с голубями, рябки
обнаруживают значительную близость к куликам, особенно ржан-
ковым, и должны быть рассматриваемы как группа, ответвившаяся
поблизости возникновения Liinicolae и вместе с тем оставшаяся на
более примитивной стадии в сравнении с голубями, параллельно
которым они в основном развивались. Вследствие своеобразного
образа жизни легко выявляются приспособительные и специализи-
рованные черты: напоминающий ржанок и отчасти голубей сильный
скользяще-гребной полет; острые длинные крылья с 11 маховыми
перьями; плотное весьма густое оперение с почти редуцированным до-
полнительным стержнем; короткие ножки, с замечательным у Syrrhap-
Рпс. 273. Саджа — Syrrhaptes paradoxus Pall. По Evans.
les слиянием пальцев и образованием общей подотекп с широкими ту-
пыми когтями, напоминающими копытца; задний палец у Pterocles
слаб и высоко поставлен, а у Syrrhaptes совсем отсутствует. По неко-
торым сведениям, некоторые виды разгребают землю, как куры,
разыскивая семена. Скелет отличается сильной пневматизирован-
ностыо. Наружные носовые отверстия с оперенной кожистой кры-
шечкой (operculum) в защиту от проникновения пыли. Клюв формою
весьма напоминает клюв куриных. Объемистый зоб представляет
собой расширение вентральной и латеральной стенок пищевода
(но не двойной вырост. пищевода, как у голубей). Мускулистый
желудок очень крепок, а слепые кишки очень длинны (в отличие
от голубей) и сопровождаются образованием многочисленных про-
дольных складок по внутренней поверхности. Кишечник в общем
близок к кишечнику голубей, но имеет более примитивный характер
петель. Весьма объемист желчный пузырь. Копчиковая железа очень
мала и оголена от перьев, что восполняется обильным пудрообразо-
ванием пуховыми частями перьев.
Из выявляющих родство особенностей следует отметить, что
череп, грудина, плечевой пояс, таз, мускулатура очень близки к та-
ковым голубей. Сиринкс отличен, с двумя парамп трахео-бронхиаль-
ных мускулов и симметричными стернотрахеальными мускулами.
437
В птерплозпсе наблюдается отсутствие шейных аптерий и наличие
(хотя и редкого) пуха на аптериях. В отличие от птенцовых голубей,
рябки выводковые, и пуховой наряд птенцов напоминает пуховой
наряд ржанок.
В ископаемом состоянии известны лишь представители современ-
ного рода Pterocles (Pt. validus, larvatus и sepultus Milne-Edwards)
пз Франции, возрастом не старше верхнего эоцена.
ПОДОТРЯД 2: COLUMBAE. — ГОЛУБИ
Древесные и наземные, представляемые типом сизого голубя,
в большинстве хорошо летающие птицы, распространенные почти
по всей земле, с наибольшей численностью и разнообразием на Ма-
лайском архипелаге и в Австралийской области. Пища разыски-
вается на земле пли склевывается с веток и вершин деревьев. Вне
периода гнездования обычно стайные птицы. Гнездятся обычно на
деревьях, устраивая просвечивающие, небрежно свитые из мелких
веток гнезда или гнездясь в дуплах и на земле. Кладка состоит пз
2, редко 1 (у крупных форм), белых глянцевитых яиц. В воспитании
птенцов принимают участие оба родителя. Птенцы выклевываются
слепыми, покрытыми волосовидным нежным редким пушком, и
выкармливаются из зоба, первое время особым «молочком» — про-
дуктом жирового перерождения эпителия, выстилающего стенки
зоба, а в дальнейшем набухшими в зобе семенами. Развитие харак-
теризуется сравнительно коротким эмбриональным (инкубация длится
14 дней у мелких и до 30 дней у крупных видов) и длинным постэмб-
риональным периодом. Юношеская линька осенью является полной
линькой.
Известно около 500 видов, с трудом группируемых в семейства и
подсемейства. Принимаемая классификация имеет ориентировочный
характер.
Семейство 1: Rapliidae (Dididae). —Дронты
Только два рода и вида: дронт — Raphus cucullatus L. (—Didus
ineptus L.) и пустынник — Pezophaps solitaria Gmelin с Маскаренскпх
островов (о. Маврикия и о. Родригец), истребленные к семнадцатому
веку (рис. 274). Очень крупные, нелетающие, наземные птицы,
с редуцированным грудным килем, spina sterni, вилочкой и крылом
и в отличие от остальных голубей без базиптеригоидных отростков.
Семейство 2: Columbidae. — Настоящие голуби
Хорошо летающие формы с сохраняющимися базиптеригоиднымп
отростками. Подразделимы на 9 подсемейств. 1
1 Некоторые авторы (см. папр. Peters, 1937) подразделяют эту группу
только па четыре подсемейства: Treroninae, Columbinae, Gourinae и Diduncu-
linae.
438
Рис. 274. Скелет Дронта —Haphvs ciiciillalus L., из
плейстоценовых отложен пг острова Маврикия. По
Lambrecht.
Подсемейство 1. Didunculinae—зубиатоклювые голуби. Един-
ственный вид Didunculus strigirostris Peale на островах Самоа.
Подсемейство 2. Duculinae— плодоядные голуби. Роды: Du-
cula, Myristicioora, Phae-
riorhina и др.
Подсемейство 3. Ptili-
nopodinae. Роды: Ptilino-
pus, Alectroenas и др.
Подсемейство 4. Trero-
ninae. Роды: Treron, Vi-
llage, Crocopus и др.
Подсемейство 5. Geo-
trygoninae. Роды: Goura
(рис. 275), OtidiphapsyiRp.
Подсемейство 6. Pha-
binae. Роды: Phaps, Geo-
phaps, Calopelia и др.
Подсемейство 7. Colu-
mbinae'— настоящие голу-
би. Роды: Columba, Edo-
pistes и др.
Представлены в СССР
лесными: вяхирем — Co-
lumba palumbus L., клинту-
хом — С. oenas L., и жи-
вущими на скалах и у че-
ловеческих жилищ сизым
голубем — Columba livia
Gm. (родоначальник до-
машних рас голубей) и
каменным голубем — С. ги-
pestris Pall.
Подсемейство 8. Peristerinae. Роды: Peristera, Chamaepelia, Scar-
dafella и др.
Подсемейство 9. Columbigallinae— горлицы. Роды: Streptopelia,
Turtur, Zenaidura, Onopopelia и др.
В СССР представлены гл. об. обыкновенной горлицей — Strep-
topelia turtur (L.), большой горлицей — Streptopelia orientalis (Lath.)
п узко распространенными южными горлицами: кольчатой — Strep-
topelia decaocto (Friv.), малой — Stigmatopelia cambayensis (Gm.)
и нек. др.
Систематическое положение голубей Гадов резюмирует следующим
образом: «Ближайшими родственниками голубей являются рябки.
Обе группы вместе коренятся в примитивных Limicolae, из которых
голуби развились в растительноядных древесных птиц. По их общему
строению они находятся только на средней стадии развития, но в от-
ношении птенцовостп, строения зоба, оперения, нижней гортани,
способности к полету, географического распространения и числа
видов они достигли очень высокой специализации. Они являются
конечной веткой на периферии филогенетического дерева птиц».
43 9
В ископаемом состоянии голуби известны с верхнего олигоцена
[Gerandia calcaria Miln.-Edw.) Современные роды появляются с верх-
него плиоцена (Columba).
В морфолого-анатомическом отношении голуби имеют, как ука-
зывалось, много черт, общих с рябками. Не останавливаясь на этих
общих чертах (см. стр. 436), отметим только отличия: metasternum
обычно с парой крупных латеральных и парой небольших медиаль-
ных вырезок (из которых последние часто замещены фонтанеллями
и в общем весьма напоминают грудину рябков); аберрантные голуби
Raphus, Pezophaps и Ridiinculus в этом представляют отличия.
Коракоидные ямки разобщены; spina interna крупна и на вершине
часто вырезана; spina externa очень мала и сильно вариирует; ви-
лочка V-образна,почти без
Рис. 275. Вепцсноспьти голубь — Goura coronata L.
Цо Evans.
hypocleidium и yDuculinae
часто даже не соединяется
посредине; 3—5 спинных
позвонков сливаются в os
dorsale; задний палец при-
крепляется почти на уро-
вень передних; мускуль-
ная формула у большин-
ства АВХУ; клюв харак-
терной формы, с мягкой
восковицей, прикрываю-
щей обычно щелевидные
отверстия; носовые железы
отсутствуют; зоб представ-
ляет собой двухстороннее
выпячивание пищевода;
желудок и кишечник при-
способлены или к твердой
пище из семян (в этих слу-
чаях мускулистый желу-
док очень силен, его
стенки иногда сопровождаются образованием костных бляшек,
кишечник более или менее длинен) и зерен или к питанию фрук-
тами и пропуску их семян (в таких случаях мускулистый желу-
док очень слаб, а кишечник широк и короток); слепые кишки
рудиментарны или совсем отсутствуют; положение musculus sterno-
trachealis несимметрично на обеих сторонах вследствие сдвигания
трахеи в сторону зобом (б. ч. влево); нижняя гортань характер-
ного строения: последние (три) трахеальные кольца, несущие го-
лосовые мембраны, очень тонки, вентрально соединяются костным
или хрящевым мостиком, а на дорзальной стороне весьма утон-
чаются или исчезают; голосовые мускулы прикрепляются непо-
средственно к мембранам. Контурные перья характерны очень ши-
роким и плоским (утолщенным) стержнем, перед вершиной вдруг
сужающимся, и очень обширными пуховыми краями перьев, зани-
мающими значительную часть их длины; возможно в связи с послед-
ним явлением побочный стержень и пух отсутствуют; последний
440
представлен местами нитевидными образованиями. Птенцовый пух
очень простого строения. Крыло б. ч. диастатаксическое за исклю-
чением ряда уклоняющихся родов.
ОТРЯД 14: CUCULIFORMES. — КУКУШКИ
Мелкой и средней величины птицы, наземные пли древесные,
распространенные почти по всей земле; характеризуются десмогна-
тическим голоринальным черепом без базиптеригоидных отростков,
но с носовой перегородкой, с двумя парами вырезок по заднему краю
грудины (у некоторых родов однако зарастающих во взрослом со-
стоянии). Сиринкс обычно трахео-бронхиальный. Сонные артерии
парные. Носовые железы отсутствуют. Ноги обычно зигодактиль-
ного (или переходного к нему) типа, с сохраняющимся musculus
ambiens. Крыло эвтаксическое с 10 большими маховыми; рулевых
перьев 10, очень мелкий и спорадичный пух распределяется только
по аптериям; имеется боковая шейная аптерия. Птенцовые; птенцы
выклевываются слепыми и голыми.
В целом группа хорошо изолирована и только через посредство
бананоедов приближается к гоацину, которого напоминает харак-
тером индивидуального развития, гнездостроением, а из анатоми-
ческих черт характером metasternum, отсутствием spina interna и
базиптеригоидных отростков, числом рулевых и некоторыми дру-
гими особенностями. Однако отличия в целом настолько велики,
что рядом анатомов (Фюрбрингер, Гадов, Беддард) отрицается связь
Opisthocomus даже с генерализованными Cucultformes.
Отряд распадается на два резко характеризованных подотряда:
Musophagi и Cuculi.
ПОДОТРЯД 1: MUSOPHAGI. — БАНАНОЕДЫ
Древесные птицы, по поведению, а отчасти и внешне, напоми-
нающие наших соек, свойственные тропическим лесам Африки.
Питаются исключительно плодами и почками деревьев, держась
почти всегда на вершинах их. Гнездо устраивается из мелких веточек
в развилках ветвей и по рыхлости постройки напоминает гнездо
голубей. Кладка состоит из 2, относительно весьма крупных, белых
С розоватым или желтоватым налетом яиц. Оба пола окрашены
одинаково и в равной степени участвуют в воспитании птенцов. Инку-
бация длится 18 дней. Выклевывающиеся птенцы вначале голы, затем
покрываются очень густым и коротким^черным пухом и развиваются
очень медленно. В случае тревоги рано покидают гнездо и лазают по
ветвям сходно с гоацином, цепляясь как лапами, так и хорошо
развитыми когтями первого пальца в крыле. К семи неделям начи-
нают перепархивать, сохраняя еще однако птенцовый пуховой наряд.
Взрослые птицы окрашены обычно в яркие цвета, из которых заме-
чателен красный цвет, продуцируемый содержащим медь пигментом
турацином, свойственным, насколько известно, только этой группе
441
птиц. Пигмент легко растворим в воде, и птицы теряют его после
каждого сильного дождя.
В составе подотряда только одно семейство Мusophagiclae, с при-
мерно сорока видами, относимыми к 8 родам: Turctcus, Gallirex,
Musophaga, Corythaeola, Crinijer и др. (рис. 276).
В систематическом отношении бананоеды равноценны кукушкам,
отличаясь от них большим количеством специализированных черт.
Разобщение обеих групп должно
иметь однако солидный возраст.
Происхождение бананоедов наме-
чается сходством с гоацином, где-то
по близости от группы курообраз-
ных, хотя некоторые авторы объяс-
няют это сходство как возник-
шее всецело конвергентным путем
(см. выше).
Из анатомических особенно-
стей бананоедов необходимо отме-
тить отсутствие сошника в черепе п
наличие постоянно os uncinatum,
свойственной только немногим ку-
кушкам. Носовые отверстия в про-
тивоположность таковым у куку-
шек, щелеобразны и значительно
сдвинуты вперед. Режущие края
клюва обычно зазубрены. Meta-
sternum постоянно имеет две пары
Рис. 276. Зелспоспппнып турако — Gal-
lirex chlorochlamys Shelley. Ho Evans.
вырезок, базальные части кора-
коидов скрещиваются, ключица U-образпа и без hypocleidium; гру-
дины достигают пять пар ребер. На ногах 4-й палец не оборачивается
назад, хотя значительно отводится в сторону. В связи с питанием
преимущественно фруктами, кишечник короток и широк, приспо-
собленный, подобно кишечнику некоторых голубей, к прохожде-
нию косточек, и без слепых кишек. В оперении характерны корот-
кость спинной птерилии и наличие побочных стержней. Копчико-
вая железа является оперенной.
ПОДОТРЯД 2: CUCULI. — КУКУШКИ
Древесные, наземные п кустарниковые птицы, обычно хорошо
летающие, длиннохвостые, распространенные почти по всей земле,
но с наибольшим обилием и разнообразием в тропической зоне.
Группа в указанных пределам ведет весьма разнообразный образ
жизни, от форм, проводящих всю жизнь в кронах деревьев, до форм,
обитающих в кустарниковой степи и быстро бегающих по земле.
Пища почти у всех видов животная, состоит преимущественно пз
насекомых и их личинок, гусениц, а у крупных бегающих форм
также из рептилий и мелких позвоночных. Общеизвестное свойство
обыкновенной кукушки подкладывать яйца в гнезда других видов
птиц свойственно многим родам, паразитирующим на различных вп-
442
дах и группах воробьиных птиц (см. стр. 250); большинство однако
строит собственные гнезда из веток и стеблей, помещая их среди веток
деревьев и на земле, и несет от 2 до 6 одноцветных белых, голубова-
тых и зеленоватых яиц, высиживаемых обоими родителями. Птенцы
выклевываются слепыми и голыми и в индивидуальном развитии не
покрываются пухом, спустя некоторое время одеваясь птенцовым
пером, развивающимся из длинных пеньков; некоторые предвари-
тельно бывают одеты волосовидным покровом (trichoptili), представ-
ляющим собой видоизмененные концы этих пеньков.
Подотряд состоит только из одного семейства Cuculidae с двумя
стами видов, распределяемых на 47 родов. Имеющиеся группи-
ровки родов в подсемейства являются спорными, так как анатоми-
ческое строение
многих н особенно
уклоняющихся ви-
дов неизвестно.
Различают 5 под-
семейств.
Подсемейство 1.
Crotophaginae с ро-
дами Guira и Сго-
tophaga, замеча-
тельными - очень
крупной величи-
ной яиц (до 17 и
:}5% веса самки) и
совместным выси-
живанием своих
кладок нескольки-
ми самками в од-
ном гнезде.
Подсемейство 2.
Diplopterinae, с ро-
дами Taper а (=
Diplopterus) и Dro-
Рис. 277. Земляная кукушка — Carpococcyx radiatus (Тсшш.).
По Evans.
тососсух.
Подсемейство 3. Neomorphinae. с родами:Neomorphus, Gcococcyx,
Сагрососсух (рис. 277), Cenlropus, Saurothera и дрл
Подсемейство 4. Coccystinae, с родами: Clanialor, Urodynaniis,
Eudynamis, Scythrops и др.
Подсемейство 5. Cuculinae с родами: Cuculus, Hierococcyx, Chry-
sococcyx. Heterococcyx и др.
В СССР известно 5 видов кукушек. Наиболее широко распро-
странены: обыкновенная — Cuculus canorus L. и глухая — С. ор-
latus Gould. Юго-восточной Сибири свойственны еще: кукушка ма-
лая — Cuculus poliocephalus Lath., индийская — С. micropterus Gould,
и ширококрылая — Hierococcyx jugax (Horst.).
В ископаемом состоянии кукушки известны с верхнего эоцена
Франции, где была найдена Dynamopterus velox Miln.-Edw., близкая
к рецептной малайской — Eudynamis.
443
В систематическом отношении, вне родства с бананоедами, ку-
кушки являются хорошо изолированной группой, что однако может
быть обусловлено недостаточным знанием многих уклоняющихся
родов. Антомическп они характеризуются наличием в черепе сош-
ника, но обычно сильно редуцированного, и отсутствием os uncina-
tum, за исключением уклоняющихся родов Scythrops и Piaya. Вы-
резки па metasternum часто превращаются в фонтанеллп или пол-
ностью зарастают, коракоиды разобщены, вилочка V-образна и
с длинным, прилегающим к грудному килю, hypocleidium (за исклю-
чением того же рода Scythrops), только три или четыре пары ребер
достигают грудины. Режущие края челюстей (за исключением Scy-
throps) без зазубрин. В пищеварительном аппарате интересен как бы
волосистый покров внутренних стенок желудка, обусловленный
однако не органическим строением его, а проникновением в кути-
кулу острых волосков гусениц, которыми многие кукушки предпо-
чтительно питаются, и застреванием их здесь. Слепые кишки, как и
кишечник, довольно длинны. Нижняя гортань чрезвычайно вариирует
в пределах группы, у некоторых родов является чисто бронхиальной
и у Crotophaga близко напоминает сиринкс Steatornis\ преобладает
однако типично трахео-бронхиальный сиринкс, с одной парой го-
лосовых мускулов. В контурном оперении побочные стержни очень
редуцированы или отсутствуют. Копчиковая железа оголена.
ОТРЯД 15: PSITTACIFORMES. — ПОПУГАИ
Преимущественно древесные, представляемые общеизвестными
формами попугаев, с вздутым массивным клювом и зигодактильными
(лазательными) ногами, птицы тропического и умеренного поясов
(исключая Голарктики). Большинство попугаев являются лесными
формами, обитающими в кронах деревьев, где птицы лазают по вет-
вям, цепляясь как ногами, так и клювом, в поисках пищи, состоя-
щей преимущественно из семян и плодов, а в некоторых группах —
из цветочного нектара и насекомых. Весьма склонные к стайному
образу жизни, попугаи нередко и гнездятся колониально, или одна
пара поблизости от другой. Гнездом служит дупло дерева, обычно
самостоятельно выдалбливаемое птицей, или иногда пустоты в тер-
митниках и, у единичных видов, в норах, в щелях скал, без гнезда
на земле и в массивном общественном гнезде, типа ткачей, само-
стоятельной постройки. Кладка состоит из 2—5 белых, гладких и
округлых яиц. В воспитании птенцов принимают участие оба пола
или одна самка. Инкубация длится от 18 до 30 дней в зависимости
от величины птицы. Птенцы выклевываются слепыми и обычно
голыми, но вскоре покрываются редким пушком (ргаереппае), и
некоторое время спустя этот наряд заменяется вторым пуховым
нарядом отличной окраски. Птенцы развиваются очень медленно и
у крупных видов не покидают дупла до 2—3 месяцев. Выкармли-
ваются птенцы содержимым зоба, передаваемом в клюве.
В настоящее время известно около семисот форм попугаев, отно-
симых примерно к сотне родов. Классификация группы не разра-
ботана. Вследствие разнообразия переходов между отдельными груп-
444
нами п сравнительной незначительности структурных отличий при-
нимают только одно семейство Psittacidae, разбиваемое на 10 под-
семейств. 1
Подсемейство 1. Lorrinae. Роды: Lorius, Eos, Trichoglossus
и Др.
Подсемейство 2. Cyclopsittacinae. Единственный род Cyclop-
sitta. о
Подсемейство 3. Micropsittinae— дятловые попугаи. Род Mi-
cropsitta (=N asiterna).
Подсемейство 4. Kakatoeinae. Роды: Kakatoe, Licmetis и др.
Подсемейство 5. Strigopinae. Род Strigops (рис. 278).
Подсемейство 6. Nestorinae — кеа. Род Nestor.
Подсемейство 7. Platy cercinae. Роды: Platy cercus, M elopsit ta-
cos, Pezoporus, Geop-
sittacus и др.
Подсемейство 8. ч
Palaeornithinae—вое- '
ковоклювые попу-
гаи. Роды: Loriculus,
Palaeornis, Agapornis
и др.
Подсемейство 9.
Psittacinae. Роды:
Psittacus ,Poicephalus,
Coracopsis.
Подсемейство 10.
Conurinae. Роды:
Amazona, Ara, Ara-
tinga, Enicognathus и
др.
Систематически ПО- ₽пс. 278. СовипыП попуган — Slrigops habroplilus Gray.
По Evans.
пугай представляют
резко отграниченную
п цельную группу, судя по распространению и анатомической обо-
собленности, весьма значительного возраста. В ископаемом состоя-
нии они однако редки ранее плиоцена; из более ранних наиболее
интересны Archaepsittacus verreauxi Mil.-Edw. пз верхнего олиго-
цена Франции, близкий западноафриканскому Psittacus erythacuslj.,
iiAratinga fratercula Wetmore из среднего миоцена Северной Америки,
близкая Aratinga carolinensis L.
Систематически более близки попугаям только кукушки и сизо-
воронковые, но, судя по характеру сходных признаков, ответвление
попугаев может быть мыслимо только от общего ствола этих групп,
прежде чем он распался на отдельные отряды. Вследствие этого и
в результате глубокой специализации и эволюции попугаев, общие
всем этим группам признаки создают уже впечатление достаточно
отдаленного родства.
1 Питерс (Peters, 1937) в последнее время выделяет лишь шесть подсемейств:
Strigopinae, Nestorinae, Loriinae, Micropsittinae, Kakatoeinae n Psittacinae.
445
Морфолого-анатомические особенности попугаев во многих: слу-
чаях носят явно приспособительный характер, выявление чего облег-
чается их своеобразной биологией. Наиболее распространенной пи-
щей попугаев являются семена и плоды растений, твердые оболочки
которых клюв попугаев выработался разрушать. Вероятно эта задача
и была первичной задачей в эволюции клюва этого отряда. Верх-
няя челюсть, вытянутая крючком над нижней, очещэ подвижна,
образуя настоящее сочленение как с частью лобной кости, так и
в области небных костей, вследствие, чего может значительно отги-
баться вверх. Область этого сочленения прикрыта сверху мягкой
вздутой восковицей, нередко покрытой перышками. Относительно
массивная нижняя челюсть соединяется с черепом мощной мускулату-
рой и благодаря косому вперед повороту сочленовной головки quad-
ratum может несколько двигаться вперед и назад и способна к неболь-
шим боковым движениям. Из челюстной мускулатуры особенно сильно
развит у многих форм musculus adductor mandibulae externus, кото-
рый прикрепляется и на костных подглазничных отростках, образуе-
мых post- и praefrontale, у многих видов образующих полное костное
полукольцо. Строение небно-челюстного аппарата десмогнатическое.
без базиптеригоидных отростков и сошника (вероятно в связи с сво-
бодой движения верхней челюсти); небные кости при этом очень
велики и в проксимальной их половине превращены в почти верти-
кально располагающиеся пластинки; все это создает весьма сильными
прочный аппарат, позволяющий у крупных форм раздавливать весьма
крепкие плоды. У этих форм дистальная часть небной поверхности
надклювья, там, где кончается усеченный конец подклювья, покрыта
тонкими, поперечно пробегающими рубчиками, подобно поверхности
напильника, и при содействии языка служит для удержания захва-
ченных плодов. Мускулистый язык большинства попугаев на своем
конце с вентральной стороны снабжен как бы ногтем, а с дорзаль-
ной — чувствительным эпителием, с щетинками у питающихся пыль-
цой и соком цветов, или с ложкообразным углублением у питающихся
семенами. Благодаря его подвижности плод удерживается и уклады-
вается в нужном положении и силой давления челюстей раздавлп?
вается. Размельчение пищи также осуществляется клювом, с чем
.вероятно стоит в связи сравнительная слабость мускульного же-
лудка; оно обеспечивается разнообразным движением челюстей,
могущих в совокупности производить как бы глодающие и пере-
жевывающие движения. Попугаи часто отрыгивают пищу из
объемистого зоба (имеющегося у всех) и вновь размельчают ее
клювом. В пищеварительном аппарате замечательны далее наличие
небольшого промежуточного отдела между мускулистым и желе-
зистым желудками, значительная длина кишечника и полное
отсутствие слепых кишек и (у большинства) желчного пузыря.
В строении конечностей обращают на себя внимание следующие
черты: плюсна очень коротка и вместе с тем толста и сильна, с широ-
ким сложным hypotarsus’ом; ее сочленение с голенью допускает
значительное вращение, осуществляемое внутрь мощным musculus
peroneus profundus. Musculus ambiens у большинства родов отсут-
ствует. Положение пальцев является типично зигодактильным, прп-
446
чем 4-it очень длинен. Когти круто загнуты, за исключением Pezo-
porus. преимущественно бегающего по земле. Каждая нога отли-
чается значительной самостоятельностью движений и употребляется
для подношения пищи клюву. Вместе с тем при лазании у многих
форм клюв помогает подтягивать тело. Интересным является та’кже
опистоцельный тип грудных позвонков, делающий тело более гибким-.
За исключением немногих форм, попугаи хорошо летадот скользяще-
гребным полетом. Крыло у попугаев диастатаксическое, имеет 10 боль-
ших и 8 малых маховых перьев; в связи с крылом особенно мощно
развит musculus supracoracoideus. Из особенностей строения попу-
гаев, которые в настоящий момент трудно поставить в связь с их
образом жизни, замечательны следующие черты. Шейных позвон-
ков 14, как исключение 13 и 15; в последнем случае (Strigops) сво-
бодное ребро 15-го позвонка очень длинно и несет короткий гру-
динный отдел. Грудина и плечевой пояс представляют значительные
вариации. Metasternum б. ч. с одной парой фонтанеллей, реже с па-
рой вырезок, иногда зарастают и фонтанелли. Киль обычно высок,
но у плохо* летающих Strigops он имеет вид низкого валика. Spina
externa sterni обычно длинна и вилкообразна. Spina interna отсут-
ствует. Вилочка весьма изменчива, независимо от способностей к по-
лету. Обычно она V-образна и слаба, всегда без hypocleidium, не-
редко с ба колена ее не соединяются и более или менее редуцированы.
Верхний гребень плечевой кости велик и напоминает по форме гре-
бень humerus у Columbtformes. Весьма изменчиво положение сонных
артерий, представленных обычно или двумя глубокими артериями
пли левой коллатеральной, правой глубокой. Последний случай
свойствен всем попугаям Северной и Южной Америки, что может
указывать на образование ими замкнутой родственной группы (Фюр-
брингер), и этот же тип свойствен африканским попугаям типа Psitta-
cus и части австрало-папуасских родов. Напротив, парные глубокие
сонные артерии свойственны всем Loriinae и многим родам индо-
австралийской области. Разнообразие изменчивости сонных арте-
рий именно в Старом Свете заставляет предполагать происхожде-
ние их именно здесь, а не в Америке (Фюрбрингер), что подтвер-
ждается и зоогеографическим изучением этой группы. Своеобразно
строение непарной нижней гортани, не имеющей membrana tympa-
niformis interna и pessulus и представленной только одной парой
наружных голосовых связок и специальными мускулистыми расши-
рителями (один) и замыкателями (два) ее, в то время как musculus
sternotrachealis сильно редуцирован или отсутствует. В оперении
замечательно обилие у всех попугаев продуцирующего пудру пуха,
концентрирующегося главным образом в области надхвостья и по
бокам туловища. В связи с этим может стоять недоразвитие копчи-
ковой железы у ряда родов, хотя у большинства она хорошо развита
и оперена. В оперении присутствуют и крупные побочные стержни
и обыкновенный пух, распределяющийся по всему телу.
Попугаи являются одними из самых развитых в психическом
отношении птиц, что выражается в значительной величине больших
полушарий, развитии их височных и лобных долей и наилучшем
среди всех птиц развитии коры большого мозга. Это сопровождается
447
хорошим развитием ассоциативных способностей. В связи с боль-
шой способностью к имитации звуков, благодаря мускулистости и
подвижности языка, попугаи легко выучиваются повторять слова и
правильно ассоциировать их произношение с соответствующими мо-
ментами их опыта.
ОТРЯД 16: STRIG1FORMES. — СОВЫ
Преимущественно ночные, хищные птицы, с коротким крючко-
ватым, одетым восковицей, клювом, с оборотнопалой лапой (4-й па-
лец может поворачиваться назад), с направленными характерно
вперед глазами, окруженными дискообразно расположенными перь-
ями и крайне мягким пушистым оперением. Распространены почти по
всей земле. Пища животная. Гнездо обычно без какой-либо под-
стилки на земле, в дупле, в чужих гнездах. Кладка состоит из 1—
8 белых шаровидных яиц. В воспитании птенцов принимают уча-
стие оба пола. Инкубация длится от 25 (мелкие) до 34 (крупные
виды) дней. Птенцы вылупляются слепыми, но покрытыми белым
пухом (ргаереппае), сменяемым затем полупуховым нарядом (из
mesoptilae), носимым короткий период.
Известно около четырехсот форм сов. Отряд делят на два се-
мейства.
Семейство 1: Tytonidae. — Сипухи
С маленькими орбитами, пневматизированной межглазничной пе-
регородкой, одной парой вырезок грудины, непневматизированноп
вилочкой и гребенчатым когтем сред-
него пальца,. Известны два рода:
Tyto и Heliodilus. В западных ча-
стях СССР распространена относи-
мая сюда сипуха — Tyto alba (Scop.)
(рис. 279).
Рис. 279. Сипуха — Tyto alba (Scop.).
По Evans.
Семейство 2: Strigidae. — Нормаль-
ные совы
С большими орбитами, двумя па-
рами вырезок грудины и пневматизи-
рованной вилочкой.
Сюда относятся роды: Kelupa,
Bubo. Asio, Otus, Nyctea, Surnia,
Ninox, Athene, Glaucidium. Speotyto,
Strix, Scotiaptex и др.
В СССР представлены: неясытью
серой — Strix aluco L., длиннохво-
стой— S. uralensis Pall., лапланд-
ской — Scotiaptex nebulosa Forst,
ушастой совой — Asio otus (L.), бо-
лотной — Л. flainmeus (Pontopp.),
448
мохноногим сычом — Aegolius funereus (L.), сычиком воробьиным —
Glaucidium passerinum (L.), домовым сычом •— Athene noctua (Scop.),
ястребиной совой — Sarnia ulula (L.), белой совой — Nyctea scan-
diaca (L.), совкой—Otus scops (L.),. филином— Bubo bubo L., го-
лолапым филином — Ketupa blakistoni Seeb., иглоногой совкой —
Xinox scutulata Raffles и др.
В систематическом отношении совы представляют собой группу,
конвергентным путем выработавшую многие особенности дневных
хищных птиц, в связи с сходным хищным образом жизни. Ближай-
шими родственниками их являются однако только Caprimulgiformes
(особенно Podargus и отчасти Steatornis), в более отдаленном род-
стве стоят также Coraciiformes. В сравнении с этими группами совы
являются более примитивными подряду признаков (наличие шизо-
гиатизма, полного сошника, базиптеригоидных отростков, вырезки
па грудине, наличие слепых кишек) и должны рассматриваться
отделившимися от общего ствола вскоре за ответвлением кукушек и
попугаев. В ископаемом состоянии совы известны начиная с эоцена
(род Minerva, Protostrix). Рецентные роды Bubo, Asio, Strix появ-
ляются уже с олигоцена, но ископаемые находки по совам вообще
очень бедны.
Подобно попугаям, многие морфологические особенности сов
легко могут быть поставлены в связи с их образом жизни, и в целом
вся группа может рассматриваться как весьма специализированная
п биологически параллельная дневным хищникам. В противопо-
ложность последним, они активны гл. обр. в ночное и сумеречное
время, хотя отдельные виды могут быть преимущественно деятельны
днем, как напр. Surnia, Nyctea, некоторые Glaucidium. В связи с бо-
лее ограниченными возможностями, которые представляет ночь для
ловли добычи, совы проявляют значительно меньшее разнообразие
в способах и предметах охоты, гл. обр. питаясь мелкими наземными
млекопитающими, мелкими птпцами, летающими в воздухе насеко-
мыми и (редко) рыбами и лягушками. Отсюда значительная одно-
родность в приспособлениях и цельность всей группы.
Добываемая пища обычно хватается лапами, вооруженными
острыми круто загнутыми когтями. Четвертый палец оборотнопал.
Как пальцы, так особенно плюсна нередко бывают оперены, но у до-
бывающих рыбу {Ketupa, Scotopelia) плюсны вытянуты и голы
и формой напоминают лапы скопы, а у ловящих насекомых (Ninox)
покрыты длинными жесткими щетинками (видоизмененными
перьями), облегчающими удержание мелкой и скользкой добычи.
Разрывается добыча клювом, сходным с клювом хищников, но более
укороченным, и с выпуклой, обычно оперенной, восковицей. Ориен-
тировка в поисках добычи происходит обычно по слуху, в связи
с чем стоит не только тонкое его развитие, но образование круп-
ных ушных раковин, асимметрия в развитии черепа (см. стр. 158)
п развитие подвижного лицевого опахала из перьев, помогающего
активно улавливать звуки. Тонкость звуковых восприятий обеспечи-
вается весьма легким неслышным полетом, обусловленным как
особым строением маховых перьев, так и общей мягкостью всего
оперения, вызванной необычайной длиной свободного pennulum бо-
29 Орнитология*—1209
449
родок П порядка. Большую роль в ориентировке играет также острое
зрение направленных вперед телескопообразных по форме крупных
глаз. Глаза сов обладают весьма совершенным (для птиц) телеско-
пическим зрением, захватывающим около 60° всего поля, но наличие
только боковых fovea и неподвижность глаз должны восполняться
большой подвижностью головы, поворот которой допускается строе-
нием позвоночника на 270°. В связи с питанием необходимо отметить
мясистую и крупную форму языка, отсутствие зйба, неперевари-
ваемость костей желудочным соком, вследствие чего несъедобные
части через короткое время извергаются через клюв ввиде погадок,
длинные и своеобразно вздутые на конце слепые кишки, присут-
ствие желчного пузыря.
Из своеобразных морфологических черт следует отметить шизо-
гнатический тип строения неба, с сохранением сошника (кроме
Speotyto) и функционирующих базиптеригоидных отростков. Rostrum
parasphenoidei очень короток. Строение носовых костей голорпналь-
ного типа. Шейных позвонков 14, грудные позвонки гетероцельные.
не образующие os dorsale. Коракоиды соприкасаются базальными
частями грудных ребер. Metasternum с двумя парами вырезок (с од-
ной парой у Tyto). В строении лап замечательны сильная укорочен-
ность основных фаланг 3-го и 4-го пальцев (сходно с тем, что на-
блюдается у Pandion), упрощенный hypotarsus и отсутствие muscu-
lus ambiens (в противоположность Accipitres, имеющих сходный тин
ноги и не лишенных musculus ambiens). У большинства родов ниж-
няя гортань бронхиального типа с большой наружной голосовой
мембраной и одной парой голосовых мускулов. Этот орган сильно
вариирует, что имеет существенное значение в связи с ночным обра-
зом жизни птиц. Сонные артерии всегда парные. Контурное перо
крупное, с широкой пуховой базальной частью и вследствие этого
без дополнительного стержня и обыкновенного пуха. Крыло диаста-
таксическое, с 11 маховыми. Копчиковая железа оголена.
ОТРЯД 17: CAPRIMULGIFORMES. — КОЗОДОЕОБРАЗНЫЕ
Ночные птицы, внешне напоминающие крупных ласточек, с сла-
быми ногами, длинными крыльями, весьма широким разрезом клюва
и покровительственной, напоминающей цвет коры дерева или мерт-
вый покров почвы, окраской. Оперение мягкое, как у сов. Дневные
часы проводят спокойно сидя на ветвях деревьев, пнях и поверх-
ности земли, сливаясь с ней своей окраской. С трудом двигаются
короткими шажками по земле, вследствие короткой плюсны, исклю-
чая бегающего, с длинными плюснами, Nyctidronias. Гнездятся без
гнезда, откладывая яйца на землю, или в уклоняющихся группах —
в дуплах, на пнях, на коре ветвей, иногда устраивая незатейли-
вое гнездо. Яйца одноцветные или пестрые, иногда защитной окраски.
Оба пола участвуют в воспитании птенцов. Кладка состоит из 1—2.
редко 3—4 яиц. Инкубация у европейского козодоя длится 16—
17 дней. Птенцы вылупляются зрячими п покрыты коротким густым
пухом (praepennae). Выкармливаются из глотки взрослых, охваты-
вая своим клювом клюв взрослой птицы. Питаются гл. обр. насе-
комыми, ловимыми в воздухе широким клювом, подобно ласточкам.
450
Уклоняющиеся группы питаются мелкими позвоночными (амфибиями)
п сидячими насекомыми, склевываемыми с ветвей (Podargidae), и
плодами деревьев (Steatornis).
Отряд разделяется на пять хорошо охарактеризованных се-
мейств.
Семейство 1: Steatornitilldae. — Жиряки
Единственный род и вид Steatornis caripensis Humb., распростра-
ненный в области горных тропических лесов Южной Америки.
Замечательны массовые гнездовья (и обитание) в глубоких темных
пещерах, из которых птицы вылетают только ночью, питаясь в это
время сочными плодами высоких деревьев, скусываемыми на лету
с кончиков веток. Косточки плодов извергаются в виде погадок,
которые служат гнездовой подстилкой. Кладка состоит только из
I—2 белых яиц. Птенцы выклевываются голыми, лишь позднее
одеваясь темным пухом, и удивительно жирны.
Морфологически жиряки отличаются от остальных козодоев бо-
лее плотным оперением, сильным, загнутым в длинный крючок на
конце, клювом, окруженным длинными, направленными вперед,
щетинками, оголенной вплоть до пяточного сустава маленькой ла-
пой. Череп очень близок к черепу сов (напр. Bubo bubo L.), но с двой-
ным десмогнатизмом с базиптеригопдными отростками. Носовая
перегородца, как у сов и Podargus, окостеневает и также более,
чем у Caprimulgidae, развито лобночелюстное сочленение. Нижняя
гортань типично-бронхиальная.
По целому ряду признаков жиряки занимают промежуточное
положение между совами и козодоями и очевидно представляют
собой пережиток весьма древней группы. Ряд черт указывает однако
на одновременную сильную специализацию (десмогнатизм черепа,
мышечная формула XV и др.).
Семейство 2: Aegotlielidae. — Совиные лягушкороты
Единственный род Aegotheles с примерно двенадцатью видами,
распространенными гл. обр. в Новой Гвинее, Австралии, на Север-
ных Молукских островах и в Новой Каледонии. По вертикальной по-
садке напоминает сов (причем сидящая птица обхватывает ветвь паль-
цами, чтс несвойственно большинству козодоеобразных). Днюет и
гнездится в древесных дуплах, где без всякого гнезда откладывает
до .4 белых яиц.
Aegotheles наиболее близок к Podargidae, ио помимо особых биоло-
гических черт его отличает значительно более слабый клюв с носо-
выми отверстиями ближе к концу клюва и длинными осязательными
щетинками, направленными вперед. По этим признакам и отсутствию
пудреток этот род сближается с Steatornis, Характерной чертой яв-
ляется отсутствие слепых кишек.
Семейство 3: Podargidae. — Совпные козодоп
Крупные, по облику близкие козодоям, обитатели преимущест-
венно тропических лесов Малайского архипелага и Австралийской
области. Пищу ловят как на лету, так и собирают повидимому
л
45 L
с ветвей. Устраивают гнездо из веточек или мха, лишайников п
листьев в развилках ветвей деревьев. Кладка из 1—3 япц. Птенцы
одеты белым пухом.
Известны только два рода — Podargus и Batrachostomus с двумя
десятками видов.
В морфологическом отношении Podargidae характеризуются силь-
ным, довольно массивным клювом, крючкообразно загнутым на конце
вниз, с щелевпднымп ноздрями, прикрытыми кожей у основания
клюва. Череп десмогиатпческпй, ио без базиптеригопдных отростков.
Metasternum с двумя парами вырезок. Сиринкс бронхиальный.
Замечательны большие пятна порошкового пуха, расположенные
в области надхвостья. Лапа способна обхватывать ветки; четвертый
палец ее состоит из 5 фаланг и способен оборачиваться назад, как
у сов.
Семейство 4: Nyetibiidae. — Исполинские козодои
Единственный род Nyctibius. с шестью формами, свойственный
тропическим частям Южной и Центральной Америки. Замечательно
гнездование на обломанных ветвях деревьев и пнях, с откладкой
единственного яйца в ямку из древесной трухи, причем высиживаю-
щая птица занимает почти вертикальное положение, напоминая фор-
мой и окраской обломанный сук. В нормальном положении Nycti-
bius сидит, как большинство древесных птиц, поперек ветвей. Мор-
фологически Nyctibius занимает промежуточное положение между
Podargidae и Caprimulgidae. Строение пальцев, грудины, порошко-
вого пуха подобно первым, но клюв по своему строению ближе
к клюву настоящих козодоев, отличаясь однако боковым зубцом
с краю надклювья и своеобразно отогнутым концом подклювья.
В противоположность Podargidae, прокоракоидный отросток тонок,
хотя сохраняет те же соотношения. Характерна окраска ириса:
желтая или оранжевая.
Семейство 5: Caprimulgidae. — Настоящие козодои
Небольшие козодои, представляемые обычным европейским козо-
доем, Caprimulgus europaeus L., распространенные почти по всей
земле. Свойственны различным типам стаций, питаются исключи-
тельно насекомыми, ловимыми на лету. Пальцы неспособны к
обхвату веток, потому и на дереве птицы сидят всегда вдоль ветвей.
Кладка из 1—2, окрашенных покровительственно яиц, откладывае-
мых без гнезда на земле. Птенцы покрыты пестрым, обычно бледным
пухом, быстро покидают место вывода, хотя держатся поблизости.
Известно около сотни форм, относимых к родам: Chordeilcs,
Caprimulgus, Phalaenoptilus, Nyctidromus и др.
В СССР представители только одного рода Caprimulgus.
Наиболее широко распространен обыкновенный козодой — Сар-
riniulgus europaeus L. (рис. 280). Кроме того, пустыням Средней Азии
свойствен буланый козодой — Caprimulgus .aegyptius Licht., а юго-
восточной Сибири — индийский козодой —Caprimulgus indicus Lath.
Морфологически эта группа характеризуется коротким тонким
и слабым строением клюва, с носовыми отверстиями, помещающи-
452
мися на концах мягких коротких трубочек. Череп шизогнатический
или десмогнатический с базиптеригоидными отростками. Язык ко-
роток, треугольный по форме. Metasternurn только с одной парой
вырезок. Нижняя гортань трахео-бронхиального типа. Пудреткп
отсутствуют, побочный стержень в пере рудиментарен. Четвертый
палец ног неспособен оборачиваться, состоит только из 4 фаланг,
коготь среднего пальца имеет гребенчатый край. Плюсна частью опе-
рена, частью покрыта поперечными пластинками.
Морфологически козодоеобразные более близки только совам,
вслед за которыми их ветвь очевидно и отделилась от общей крупнел
ветви класса птиц; родство с стрижеобразными и сизоворонками
является более отдаленным и обусловленным только близостью
корней этих групп между собой. В целом группа, даже включая
такие переходные, древние «звенья», как Steatornis, Aegotheles,
Podargus и др., представляется весьма цельной и резко отграничен-
ной.
Морфолого-анатомическими признаками отряда являются сле-
дующие. Череп шизогнатический или десмогнатический, голори-
нальный, с сошником,
носовой перегородкой,
с функционирующими
базиптеригоидными от-
ростками. Исключение
представляет семейство
Podargidae с десмогна-
тпческим черепом без
функционирующих ба-
зиптеригоидных отро-
стков. Восковица от-
сутствует. Processus an-
nularis mandibulae очень
короток. Щейных позвонков 14 (редко 13—15), грудные поз-
вонки свободные и гетероцельные (у Steatornis опистоцельные);
кольцо атласа неполно на дорзальной стороне, как у сов. Meta-
sternum с 1 или 2 парами вырезок. Имеется только 4 грудных
ребра. Основания коракоидов разобщены. Spinae sterni отсутствуют
или spina externa рудиментарна. Плюсна обычно очень коротка.
Hypotarsus имеет канал. Из ножных мускулов отсутствует musculus
ambiens. Язык укорочен и более или менее рудиментарен, зоб от-
сутствует. Кишечник сравнительно короток с длинными вздутыми
па конце, как у сов, слепыми кишками. Желчный пузырь имеется.
Нижняя гортань весьма изменчива в этой группе, как и у сов, и
вариирует от типично-бронхиальной у Steatornis и Podargus до трахео-
бронхиальной у Caprimulgidae. Сонные артерии обычно парные.
Кожные покровы очень тонки. Мягкое контурное перо такого же
строения, что и у сов, но с побочными стержнями. Пух покрывает
только аптерии. Копчиковая железа мала и оголена, а у Podargidae
и Nyctibius совершенно отсутствует, вероятно в связи с наличием
У этих форм пудреток. Крыло диастатаксическое, обычно с 10 боль-
шими маховыми. Хвост с 10 рулевыми.
Рис. 280. Обыкновенный козодоп — Caprimvlgiis eura-
liaeus L. с широко раскрытым клювом, в позе защиты.
По I-Ieinroth, из HilHieimcr.
453
ОТРЯД IS: MICROPO 1)1 FORMES. — СТРИЖЕОБРАЗНЫЕ
Острокрылые, быстро летающие мелкие птицы, представленные
стрижами и колибри, составляющими резко характеризованные под-
отряды. Общими признаками обеих групп, отличающими от других
птиц, являются: строение крыла из очень укороченной плечевой
части и очень длинной кистевой,* несущей 10 больших маховых,
с наиболее длинными дистальными махами. Малые маховые очень
укорочены и малочисленны (G—11). В строении крыла наиболее
своеобразен весьма укороченный и толстый плечевой отдел с мощным
condylus radialis, глубокой и широкой fossa olecrani, с processus
ectepicondyloideus, смещенными в проксимальную сторону, и с очень
крупной п искривленной crista superior humeri. Грудина без вы-
резов, но иногда с 1—2 фонтанеллями по заднему краю и с очень
короткими spina externa и interna. Грудных ребер только четыре.
Вилочка U-образная с коротким hypocleidium. Череп эгитогнатиче-
ский, голоринальный, с носовой перегородкой. Шейных позвонков
13—14. В очень маленьких й сравнительно слабых задних конечно-'
стях отсутствуют musculus ambiens и отделы В и Y формулы Гар-
рода. Из ‘сонных артерий сохраняется только левая (исключая
Cypseloides). Слепые кишки, за редким исключением, полностью
редуцированы. В оперении контурные перья еще сохраняют побоч-
ный стержень. Копчиковая железа оголена.
Птицы строят гнезда. Яйца белого цвета, овальной формы, напо-
минающей яйца козодоев. Птенцы выклевываются слепыми и долго
выкармливаются.
Из сравнения обеих групп следует, что колибри более специали-
зированы п занимают более высокую морфологическую ступень,
чем стрижи. На это указывает напр. потеря ими пуха, редукция
побочных стержней, потеря musculus sternotrachealis, приобретение
зоба и изумительно цветистой раскраски самцов. У стрижей вместе
с тем мы еще встречаем формы с двумя сонными артериями, и грудина
нередко несет фонтанеллп, что указывает в отношении этих призна-
ков на известную примитивность. В эмбриональном состоянии ко-
либри имеют клюв весьма короткий и напоминающий клюв стрижен.
Систематическое положение отряда вызывало много споров.
Большинство авторов высказывалось за родство . стрижеобразных
с козодоями (Huxley, 1867; Hartert, 1892; Gadow, 1892; Beddard, 1898;
Thompson, 1901, и др.) пли воробьиными (Shufeldt, 1885; Furbringer,
1888, 1902; Harting, 1901, и др.). Последнее время этот вопрос был
предметом специального исследования П. Р. Лоу (Р. В. Lowe, 1939),
который нашел стрижеобразных наиболее близкими примитивным
воробьиным и решительно отвел родство с козодоеобразиыми. По
мнению Лоу, в отряд Passeriformes в качестве подотрядов должны
быть включены кроме собственно воробьиных (Passeres) и рогоклю-
вов (Eurylaemi) также дятлы (Pici) и между ними — обособленные
друг от друга стрижи и колибри. Это предложение хорошо обос-
новано автором, но уже не могло быть использовано соответствую-
щим образом в настоящем обзоре.
454
ПОДОТРЯД 1: MICROPODIL — СТРИЖИ
Внешне сходные с ласточками, лучшие летуны среди птиц, рас-
пространенные почти по всей земле. Пищу, состоящую из мелких
насекомых, особенно мелких жучков (напр. у обыкновенного стрижа
н Европе, из некоторых Halticine, Psyliodes chrysocephalus, видов
Tachyporus и Aphrophora} ловят исключительно на лету коротким,
но с глубоким широким расщепом, клювом. По земле большинство
не в состоянии двигаться и прицепляется только к поверхности
высоких предметов: скалам, стволам деревьев и т. д.; в связи с этим
у большинства все четыре пальца ног обращены вперед и снабжены
острыми когтями; птицы прицепляются ими, опираясь на всю плюсну.
У некоторых видов однако 1-й или 4-й пальцы противостоят осталь-
ным, пли только 1-й палец повернут назад, что позволяет садиться
на ветки подобно прочим птицам. Гнездятся нередко колониально,
устраивая гнездо в дуплах высоких деревьев, щелях скал, под крышей
и т. д.; ряд видов строят гнезда прикрепляя их к ветвям, Micropus
Aorws Heugl. роет норки в песчаных склонах, некоторые виды занимают
чужие гнезда. Гнездо строится из собираемых на лету в воздухе
растительных пушинок, перьев и соломинок, скрепляемых клейкой
слюной, отделяемой сильно* увеличивающимися на этот период слюн-
ными железами. У саланган слюна преобладает в постройке гнезда,
а у Callocallia francica (Gm.) гнездо целиком состоит из слюны.
У прикрепляющих гнездо к ветвям деревьев [Hemiprocne) гнездо
очень мало и мелко и в состоянии поддерживать только 1 яйцо; птица
высиживает его сидя рядом на ветке над гнездом и в защиту от
выпадения яйца от ветра прилепляет его слюной к гнезду. В кладке
только 2 (редко 1—3) яйца. Оба родителя выкармливают птенцов,
которые вылупляются после (б. ч.) 18 дней насиживания слепыми
н большей частью (при гнездовании в дуплах, щелях и т. д.) го-
лыми; только выращиваемые в открытых гнездах на деревьях (Сур-
siurus, Hemiprocne) появляются на свет густо опушенными. Пост-
эмбриональное развитие птенцов весьма длительно.
Подотряд заключает только одно семейство Micropodidae с сот-
ней форм, распределяемых по родам: Callocalia, Chaetura, Cypseloi-
des, Micropus, Aeronautes, Cypsiurus и др. В СССР наиболее распро-
странен черный стриж — Мicropus apus (L.) и белопоясничный стриж
Micropus pactficus (Lalli.). Кроме того, в более южных районах
встречаются: белобрюхий стриж — Micropus melba (L.), малый
стриж — Micropus ctffinis (Gray) и колючий стриж — Hirundapus
caudacutus. (Lath.).
Морфологически стрижи характеризуются эгитогнатическпм чере-
пом с коротким, но глубоко расщепленным клювом, трахео-брон-
хиальной нижней гортанью с одной парой бронхиальных мускулов
и musculus sterno-trachealis, наличием только 13 шейных позвонков
(у Chaetura — 14),е характерным строением пальцев (см. выше), ба-
зальные фаланги которых редуцированы. В сравнении с колибри,
стрижи имеют относительно маленькую по размерам печень, но
сохраняют желчный пузырь. Крыло эвтаксическое с 8—9 малыми
маховыми, за исключением родов Streptoprocne и Cypseloides, пмею-
456
щих диастатакспческое крыло с 10—11 малыми маховыми. Первый
палец крыла вооружен когтем (отсутствует у Hemiprocne). Пух встре-
чается только на аптериях, в контурном пере побочный стержень
есть и длинен.
Ископаемые стрижи известны начиная с эоцена — олигоцепа
(роды Cypselavus, Aegialornis). С верхнего олпгоцена появляются уже
рецентные роды — Cypselus и Callocalia.
ПОДОТРЯД 2: TROCHILI. — КОЛИБРИ
Мелкие и мельчайшие птицы, от мелкой ласточки до шмеля ро-
стом, обычно сверкающие яркими ирпзирующими окрасками, оби-
тающие Южную и Северную Америку, с огромным преобладанием
в тропиках этой части света. Мельчайшие виды весят до 1,6—1,8 г
и несут яйца, равные примерно 0,2 г. Характерен сильный трепе-
щущий жужжащий полет с столь быстрыми взмахами крыльев (30—
50 взмахов в секунду), что на полете у птички заметны только два
туманных пятна по. бокам тела на месте крыльев. Четырехпалые
ножки очень малы и слабы, плюсна только в половину длины сред-
него пальца, задний палец направлен назад. Все колибри являются
древесными обитателями, питающимися преимущественно нектаром
цветов и находимыми в цветах мелкими насекомыми. Поведение
большинства видов сходно с поведением бабочек бражников: пере-
мещаясь в воздухе настолько быстро, что они почти не заметны для
глаза, они «застывают», быстро жужжа крыльями, перед цветком
и тонким заостренным клювом высасывают из него нектар. Малень-
кая величина и быстрота движений связаны с исключительно высо-
ким обменом веществ. Для мелких видов (напр. весящего 3 г Chlo-
restes caeruleus) констатировано 5—6 дыханий в секунду. В неволе
колибри потребляют за сутки двойной вес питательной жидкости,
в сравнении с весом тела. Питаясь гл. обр. нектаром цветов, колибри
выработали тонкие приспособления к его добыванию, и многие виды
тесно специализированы к определенным видам растений (рис. 281).
Гнездятся колибри, устраивая маленькое чашкообразное гнездо,
помещаемое в развилке мелких веток или прикрепляемое на концах
длинных листьев деревьев. Гнездо вьется из мха, кусочков лишай-
ников и растительного пуха. Все заботы о потомстве лежат только
на самке, в связи с чем самки обычно окрашены бледнее самца,
а самцам свойственны различные брачные украшения, в виде хохлов,
воротничков, длинных хвостовых перьев и т. д. Самцы весьма драч-
ливы, но не поют. Голос колибри состоит из коротких свиристящих
звуков. Кладка состоит из двух матово-белых яиц. Инкубация длится
14—19 дней. Птенцы вылупляются слепыми и голыми, с длинным
и редким ржавчатым пухом только на спине. Птенцы выкармливаются
цветочным нектаром, вкачиваемым им прямо в пищевод через длин-
ный клюв. Развиваются птенцы медленно. •
Колибри представляют весьма цельную и однообразную по внут-
ренней организации группу, недопускающую дробления на несколько
семейств. Единственное семейство Trochilidae состоит почти из 600
форм, относимых к 120 родам, проявляя таким образом крайнее
456
разнообразие в окраске, украшениях, формах хвоста и клюва. Оби-
лие всевозможных переходов и отклонений затрудняет дробление
даже на подсемейства. Последних различают два.
Подсемейство 1. Phaethornithinae. Тускло окрашенные колибри,
обитающие преимущественно под пологом леса и питающиеся гл. обр.
насекомыми и паучками. Передние пальцы в основной половине
срослись. Средние рулевые хвоста длиннее крайних. Крыло диаста-
таксическое. Роды: Phaethornis, Threnetes, Eutoxeres^ Doryfera и др.
Подсемейство 2. Trochilinae.
Преимущественно ярко окра-
шенные колибри, питающиеся
цветочным нектаром и держа-
щиеся на ярком свету. Перед-
ние пальцы свободны до осно-
вания. Средние рулевые сильно
удлинены. Крыло эвтаксиче-
ское. Наибольшее число родов:
Archilochus, Florisuga, Oreotrochi-
lus, Selasphorus, Lanipornis и
мн. др.
Морфологические особенно-
сти колибри в наружных чер-
тах специализации изобильны,
но во внутренней организации
они довольно однообразны и до-
статочно близко примыкают к
стрижам, так что ряд специа-
листов их рассматривает только
как семейство. Из явно адап-
тивных черт отметим: клюв, ва-
риируя в длине и форме, обыч-
но приспособлен к типу основ-
ных питающих птичку цветов.
У некоторых видов клюв пре-
вышает длину тела. Обычно
клюв тонок и заострен на кон-
це с завернутыми внутрь, не-
редко с тонкой насечкой, ре-
жущими краями. Соответственно функции клюва язык узкий, спо-
собный далеко вытягиваться (с длинными заворачивающимися
вокруг головы подъязычными рожками), в дистальной части расще-
пленный на две узкие, несколько закрученные полоски, совокупным
действием образующие трубочку. Взрослые без зоба, но у птенцов
образуется маленький шарообразный зоб. Мускулистый желудок
очень маленький. Слепые кишки отсутствуют за исключением корот-
ких рудиментов у некоторых родов. В строении крыльев указанные
выше черты укорочения плеча и предплечья и удлинения кисти до-
ходят здесь до своего крайнего выражения. Грудина относительно
много больше, чем у прочих птиц. В связи с типом полета, musculus
supracoracoideus также массивен, как и musculus pecloralis. Относе-
457
Рис. 281. Колибри — Archilochus colubrislj.^
момент опыления цветка Macranlhera Lecontei.
1, 2, 3 — цветы*этого растения. Пестик созре-
вает раньше тычинок и по оплодотворении вя-
нет и засыхает, в то время как тычинки до-
стигают полного роста и созревают. Пыльца
осыпается на затылок сосущей птички и при-
липает к нему в виде желтого пятна; при по-
сещении цветка с зрелым пестиком послед-
ний приходится рыльцем против затылка и
соприкасается с пыльцой. По Pickens, из
Stresernann.
телыю длинное, острое крыло весьма плоско, с 10 большими махо-
выми и только 6 малыми маховыми. Дистальные маховые весьма
упруги, иногда с весьма утолщенным стержнем, прикрепляющимся
на относительно сильно расширенных и удлиненных фалангах
2-го пальца. С интенсивным обменом стоит в связи весьма крупное
сердце, у мелких видов составляющее до 25°/00 веса тела. Из других
особенностей необходимо отметить эгитогнатическое строение кост-
ного неба; число шейных позвонков варпирует между 14 и 15; ниж-
няя гортань с двумя парами трахео-бронхиальных мускулов, muscu-
lus sterno trachealis отсутствует; трахея ветвится выше торакаль-
ного отверстия; побочный стержень в пере обычно очень укорочен,
пух совсем отсутствует; крыло у большинства эвтаксического типа,
за исключением более примитивной группы тускло окрашенных ко-
либри, у которых оно диастатакспческое, коготь на первом пальце
крыла отсутствует; нога анизодактильного типа.
ОТРЯД 19: COLIIFORMES.-ПТПЦЫ-МЫШП, 1 МЫШАНКИ
Мелкие, величиной с крупных вьюрков, по движениям весьма
напоминающие синиц, кустарниковые птицы Африки. Подобно си-
ницам, они странствуют мелкими стайками по лесам и кустарникам,
лазая и вешаясь в разнообразных позах в густых древесных кронах
и ветвях кустарников. При общей коричневатой, с мелким волни-
стым рисунком, окраске, характерен очень длинный ступенчатый
хвост, хохол’из рассученных перьев на голове, конический слабо
загнутый, напоминающий вьюрков клюв и весьма подвижные сво-
бодные до основания пальцы ног, 1-й и 4-й пальцы которых пово-
рачиваются и вперед и назад. Пища состоит из мелких плодов, ягод
и почек. Гнездо строится в виде чаши в ветвях деревьев и кустар-
ников из стеблей и листьев. Кладка заключает 2—5 белых или пят-
нистых яиц. Птенцы с длительным развитием, первоначально голы,
затем с редким и короткм пуховым нарядом. Выкармливаются,
подобно певчим птицам, всовыванием корма птенцам в клюв, в связи
с чем последние имеют яркую окраску полости рта (черную, с оран-
жево-желтым языком).
Отряд составляет единственное семейство Coliidae с 6 видами рода
Colius (рис. 282).
Морфологически птицы-мыши занимают относительно изолиро-
ванное положение. Наибольшее сходство по внутренней организации
они обнаруживают с стрижами и африканскими козодоями, несмотря
на совершенно различный образ жизни и питание. Характер сбли-
жающих признаков дает основание видеть в них группу, близкую
к еще генерализованному стволу козодоев и стрижей и развивавшуюся
очевидно в виде самостоятельной ветви. По происхождению эта
группа должна стоять также недалеко от корней группы трогонов.
Из морфолого-анатомических особенностей Colius следует отметить
следующие. Череп десмогнатический, но с непрямым десмогнатизмом
1 Название дано как за общую окраску, так и за свойство лазить вдоль
ветвей, опираясь па всю плюсну.
45 s
п рудиментарным сошником; строение носовых костей голорпналь-
лого типа, носовая перегородка имеется; базиптерпгопдные отростки
полностью отсутствуют; шейных позвонков 13; грудина с двумя
парами глубоких вырезов свое-
образнойформы и крупной spina
externa; коракоиды разобщены
своими основаниями, вилочка
1:-образная с крупным hypoclei-
dium; ноги, как указано выше,
пампродактильного типа, един-
ственного в классе птиц; когти
длинны и остры, плюсна спере-
ди покрыта пластинками, уве-
личивающимися в размерах к
дистальному концу плюсны; мы-
шечная формула АХУ; muscu-
lus ambiens отсутствует. Ниж-
няя гортань трахео-бронхиаль-
ного типа с двумя парами го-
лосовых мышц; голос птиц
стрекочущий и свистящий, в
последнем случае сходный с го-
лосом крупных ржанок. Сон-
ная артерия сохраняется толь-
ко левая. Мускулистый желу-
док слаб, кишечник короток и
широк, слепые кишки отсутст-
вуют. Печень также мала, но
снабжена желчным пузырем.
Бросается в глаза значительная
толщина кожи. Птерилозис за- Рпс 28, Мы1ШШ1>.а _ CoHtw sll,alus Gm.
мечателен шириной СВОИХ пте- По Evans.
рилий. Контурные перья ко-
ротки и рыхлы, но с крупными побочными стержнями. Пуховые перья
отсутствуют. Крыло эвтаксическое с 10 большими маховыми. Хвост
из 10 рулевых. Копчиковая железа оперена.
ОТРЯД 20: TROGONIFORMES. — ТРОГОНЫ
Величиной с дрозда или кукушку, ярко окрашенные и обычно
длиннохвостые лесные птицы тропического пояса. Характерен ко-
)откий, у основания широкий клюв, с коротким крючком на конце,
жруженный направленными вперед крупными щетинками; плюсна
коротка и частью оперена, с 1-м и 2-м пальцами, направленными назад,
острыми крупными когтями; хвост нередко с своеобразными выре-
(анными на концах рулевыми; роскошная общая окраска, сверху
л. обр. зеленая, иризирующая, снизу красная или желтая, с разно-
>бразной дополнительной расцветкой. Птицы держатся в густых
'роппческпх лесах, где с веток деревьев короткими взлетами бро-
аются и ловят, подобно мухоловкам, пролетающих насекомых, или
459
подобным же способом скусывают на лету мелкие плоды с веток де-
ревьев, вновь после этого возвращаясь на старое место. Гнездятся
в дуплах деревьев или роют пустоты в крутых откосах и в тер-
митниках. Кладка из 2—4 белых глянцевитйх яиц высиживается
обоими родителями. Долго разви-
тие. 283. Квезал — Pharomachrus то-
cinno De la Llave. По Evans.
вающиеся птенцы по вылуплении
первоначально голы.
Только одно семейство Trogoniiw
с 50 формами, относимыми к 13 ро-
дам. Распространены прерывисто: в
наибольшем количестве (ок. 30 форм)
в Центральной и Южной Америке —
роды: Trogon, Pharomachrus и др.
(рис. 283); затем около 16 форм в юго-
восточной Азии, на Больших Зонд-
ских и Филиппинских островах --
роды: Harpactes и Apalharpactes, на-
конец 3 вида в Африке — роды: Ара-
loderma и Heterotrogon.
Морфологически трогоны пред-
ставляют весьма изолированную
группу, с оригинальным смешением
систематически важных черт, при-
сущих разным группам примитивных
Coraciomorphae. Более определенное,
хотя и достаточно отдаленное родство
они обнаруживают только с козо-
доями, Colius, и отчасти с Coraciae.
По примитивности многих свойствен-
ных им признаков (шизогнатизм, с
сохраняющимися базиптеригоиднымп
отростками, с крупным сошником,
крупным побочным стержнем в пере
и др.) они могут быть поставлены
в системе только на один уровень
с совами и ведут свое отщепление ве-
роятной общего ствола с этими пти-
цами и козодоями.
В морфологической характери-
стике трогонов необходимо отмс-
тить следующие черты: череп ши-
зогнатический с базиптеригоиднымп
отростками и крупным сошником;
, строение носовых костей голорп-
нального типа; носовая перегородка имеется; шейных позвонков 1э;
грудина напоминает грудину сизоворонок с двумя парами очень
глубоких вырезов и вилкообразной длинной spina externa; кора-
коиды соприкасаются; вилочка U-образная, с liypocleidium; нога
(см. выше) гетеродактильного типа, единственного во всем классе
птиц, с своеобразным строением сгибателей пальцев. Musculus
460
anibiens отсутствует. Сонная артерия сохраняется только левая.
Язык трехсторонней формы, короткий, заостренный спереди. Зоба
нет. Мускулистый желудок очень велик, но тонкостенен. Слепые
кишки коротки. Нижняя гортань трахео-бронхиального типа (голос
свистящий). Кожа замечательна своей тонкостью и нежностью,
перья держатся весьма слабо и при прикосновении спадают целыми
партиями. Птерилозис замечателен отсутствием бифуркации спин-
поп птерплпи. Пух отсутствует, зато кроющие перья весьма рыхлой
п мягкой структуры и с очень крупными побочными стержнями.
Крыло эвтаксическое, с 10 большими маховыми. Рулевых перьев
двенадцать. Копчиковая железа оголена.
Трогоны представляют собой весьма интересную зоогеографиче-
скую группу, ведущую свое происхождение вероятно из Старого
Света, несмотря на наибольшее разнообразие сейчас именно в Южной
Америке. Большинство американских трогонов ярко и пестро окра-
шены, причем самки мало уступают в окраске самцам. У некоторых
видов однако самки окрашены значительно скромнее самцов, сбли-
жаясь в этом отношении с молодыми птицами. У большинства же
азиатских трогонов окраска весьма тусклая, причем самец напоми-
нает скромно окрашенную самку американских птиц, а самка — его
птенца. Африканские птицы уже несколько более сближаются с аме-
риканскими птицами. Сходное распределение признаков дает иззуб-
ренность клюва, свойственная взрослым особям большинства аме-
риканских птиц; у их молодых края клюва гладкие или с короткой
выемкой перед концом клюва, причем это свойственно всем возрас-
там азиатских трогонов. С этим интересно сопоставить то, что аме-
риканские виды, в отличие от трогонов Старого Света, почти исклю-
чительно питаются плодами и ягодами, срывая их так же, как
трогоны ловят насекомых; последние совершенно не употребляют
какой-либо растительной пищи. Интересным является также, что
несмотря на полную п глубокую прерывчатость распространения,
трогоны сохранили полное единство облика, а африканские Apalo-
dernia несомненно являются близкими родственниками американ-
ского рода Trogon. На былую связь в распространении трогонов
Старого и Нового Света на территории Европы указывает нахождение
их в ископаемом состоянии во Франции, относимое к олигоценовому
времени — Paratrogon gallicus Miln.-Edw., близкий к роду Trogon.,
и м. б. эоценовые находки видов рода Archaeotrogon. j
ОТРЯД 21: CARACIIFORMES. — РАКШЕОБРАЗНЫЕ
Разнообразная по биологии группа птиц, объединенная следую-
щими общими признаками: череп десмогнатический, голорпнальный,
с носовой перегородкой и без базиптеригоидных* отростков (послед-
нее исключая Buc^rotes)] шейных позвонков 14—15; metasternym
с одной или двумя парами вырезок; вилочка U-образная. Располо-
жение пальцев анизодактильного типа. Мускульная формула АХУ
(у Halcoynes АХ). Musculus ambiens отсутствует.
Группа составляется семью небольшими, резко характеризован-
ными подотрядами: Halcyones^ Todi, Mornoti^ Coraciae^ Upupae,
461
M eropes и Bucerotes, обнаруживающими несомненное родство между
собой. Все эти группы развивались повидпмому в виде отдельных
самостоятельных ветвей, но с различным обособлением друг от друга.
Наиболее низкое положение занимают по особенностям их органи-
зации Coraciae, наиболее высокое U pupae, обнаруживающие при-
знаки, сближающие эту ветвь уже с Passeres. С другой стороны.
Coraciae обнаруживают признаки родства с Cuculijornies и Capri-
mulgiformes.
ПОДОТРЯД 1: CORACIAE. — РАКШИ
Внешне напоминающие некоторых вороньих птиц (гл. обр. галок),
ярко окрашенные древесные или. наземные птицы Старого Света.
Для древесных форм характерен сильный, в брачный период кувыр-
кающийся в воздухе, полет и грубый, напоминающий карканье,
голос. Наземные виды перелетают только на короткое расстояние,
передвигаясь по звмле гл. обр. прыжками. Питаются различными
членистоногими, гл. обр. насекомыми, у древесных форм нередко
ловимыми на лету, у наземных собираемыми с земли и выгребаемыми
пз почвы лапами. Гнездятся в дуплах или земляных норах, отклады-
вая- (без. подстилки) 3—6 белых глянцевитых округлых яиц (Со-
racias). Инкубация у обыкновенной сизоворонки длится 19 дней.
Птенцы вылупляются голыми, позже покрываются длинными пень-
ками молодых перьев, поздно лопающихся. Птенцового пухового
наряда нет. Развитие птенцов медленное.
Подотряд состоит из трех семейств: Leptosomalidae, Brachypteru-
ciidae и Coraciidae, объединяемых следующими морфологическими
признаками. Череп с очень длинным processus postfrontalis, дости-
гающим скуловой дуги; замечательна очень крупная и вытянутая
os lacrymale; сошник рудиментарен; шейных позвонков 13—14; гру-
дина с 1 илд 2 парами глубоких вырезов, с 4—5 грудными ребрами,
с длинной вилкообразной spina externa, но без spina interna; осно-
вания коракоидов разобщены; вилочка U-образная, с hypocleidiuni.
Ноги синдактильные с сросшимися базальными фалангами передних
пальцев (исключая Leptosomus, у которого 4-й палец может отворачи-
ваться в сторону). Мускульная формула АХУ. Сонные артерии
парные (у Leptosomus глубокие артерии заменены вторичными по-
верхностными). Нижняя гортань трахео-бронхиального типа. Язык
длинный без роговых бахромок. Зоб отсутствует, слепые кишки
длинные, имеется желчный пузырь. Кроющие перья несут крупные
побочные стержни, пух отсутствует. Крыло обычно диастатаксн-
ческое, с 11 большими маховыми. Рулевых 12. Копчиковая железа
оголена; у Leptosomus она отсутствует и замещена парными пятнами
порошкового пуха, расположенными в поясничной области.
•
Семейство 1: Leptosomatidac. — Кирумбо
Единственный вид — Leptosomus discolor Herm., распростра-
ненный на Мадагаскаре и Коморских островах. Коротконогая, от-
носительно крупная древесная ракша с очень резким половым ди-
462
морфизмом. Обитают лесные опушки, пцтаются преимущественно
гусеницами, гнездятся в дуплах деревьев. Замечательны воздушные
игры, сопровождаемые свистом, напоминающим крик хищных птиц.
Характерно положение щелевидных ноздрей под прикрытием рого-
вой крышечки на середине клюва, бронхиальный сиринкс, наличие
пудреток, далеко отворачивающийся 4-й палец, в то время как 2-it
и 3-й пальцы срослись у основания. Наличием пудреток, яркой фио-
летовой иризирующей раскраской и другими особенностями кирумбо
напоминают бапаноедов и некоторых кукушек.
Семейство 2. Bracliypteraciidae. — Земляные ракши
Короткокрылые наземные птицы Мадагаскара с удлиненными
плюснами, разгребающие почву лапами при разыскивании пищи.
В связи с этим своеобразно расположение когтей: на 1-м и 4-м паль-
цах они повернуты наружу, а на 2-м и 3-м — внутрь. Гнездятся в зем-
ляных норах. Крылья округлые, слабые, некоторые виды с очень
длинным хвостом и сильно вариирующим по величине клювом. Из-
вестно 5 видов родов: Brachypteracias, Geobiastes, Atelornis и Urate-
lornis.
Семейство 3: Coraciidae. — Древесные ракши (сизоворонки)
Ярко окрашенные, но без металлических оттенков в оперении...
древесные птицы почти всего Старого Света. Носовые отверстия
у основания клюва (морфологические признаки, основные для всего
отряда, см. выше). Гнездятся в дуплах или норах.
Известно около 30 форм, относимых к родам Coracias и Eurysto-
nius, В СССР представлены: сизоворонка — Coracias garrula L.
п восточный широкорот — Eurystomus orientalis L.
Систематическое положение подотряда рассматривается обычно
как наиболее низкое во всем отряде, обнаруживающее признаки
родства с кукушками и козодоями, а в пределах отряда — с Hal-
cyones. Наиболее примитивные черты во всей группе свойственны оче-
видно кирумбо.
В ископаемом состоянии представлено лишь семейство Coraciidae
видом Geranopterus alatus Miln.-Edw., из верхнего эоцена Франции,
наиболее близким повидимому к эфиопскому Eurystomus glaiicurus
Mull.
ПОДОТРЯД 2: HALCYONES. — ЗИМОРОДКИ
Обычно мелкие, лесные и прибрежные птицы с удлиненным нено-
вом, укороченным хвостом и яркой раскраской, распространенное
почти по всем частям света. Первоначально лесные, питающиеся
насекомыми птицы, из которых развился независимо ряд родов ихтио-
фагов, ловящих рыбу подобно крачкам, т. е. ныряющих за ней с воз-
духа, но предварительно нередко караулящих ее сидя на ветке над
водой. Гнездятся в большинстве случаев в норах, самостоятельно
вырытых в земляных откосах, иногда в дуплах деревьев или пустотах
построек термитов и муравьев. Кладка из 2—7 белых глянцовитыхг
463
насчитывают около 20
Рис. 284. Ракетохвостый зи-
мородок — Tanysiptera dea
L. По Wallace, из Evans.
почти сферических яиц откладывается без подстилки, в дальнейшем
накапливающейся из погадок птички. Инкубация длится 21—24 дня.
Птенцы вылупляются голыми; впоследствии они покрываются пень-
ками птенцового пера, поздно разворачивающегося. Пуховой наряд
отсутствует.
В единственном, входящем в подотряд семействе Alcedinidae
родов. Принимают два подсемейства: подсе-
мейство Alcedininae с родами: Alcedo (рис.
284), Alcyone, Ceryle и др.; подсемейство
Haley oninae с широким клювом, с родами:
Clytoceyx, Dacelo, Melidora, Halcyon и др.
В СССР представлен гл. обр. обыкно-
венный или голубой зимородок — Alcedo
atthis (L.). В южные пограничные районы
заходят: белогрудый красноносый зиморо-
док — Halcyon smyrnensis (L.), ошейниковый
красноносный зимородок — Halcyon pileala
Bodd. и малый пегий зимородок — Ceryle
rudis (L.).
Морфологически -зимородки характери-
зуются следующими признаками. Череп без
сошника; os lacrymale расширена и удли-
нена; клюв у питающихся насекомыми и
иногда мелкими рептилиями крупен и срав-
нительно широк, у Melidora с крючком на
надклювье, а у Clytoceyx короткий, широкий
и вздутый, повидимому служащий для вы-
капывания земляных червей; у питающихся
рыбой клюв сплющенный с боков, по общей
форме сходный с клювом дятла; шейных-
позвонков 14—15; metasternum с двумя па-
рами глубоких вырезов. Spina externa длинна
и вилкообразна, spina interna отсутствует,
коракоиды разобщены, вилочка U-образная,
без hypocleidium. Проксимальные, шейные
и спинные позвонки с хорошо развитыми ги-
папофизами. Слабые маленькие ноги синдак-
тильные, с сросшимися 3-м и 4-м пальцами
почти по всей длине и 2-м и 3-м па льцами у осно-
вания; у Сеух и Alcyone все дистальные фаланги 2-го пальца отсут-
ствуют. Мускульная формула ног АХ. Язык очень короток, кишечник
длинен, слепые кишки рудиментарны, не функционирующие. Ниж-
няя гортань трахео-бронхиального типа с двумя парами голосовых
мускулов. Сонные артерии парные. Пух имеется, густой на апте-
риях, более спорадичный на птерилиях. В контурном пере побоч-
ный стержень отсутствует. Крыло у первичных форм преимуще-
ственно диастатаксическое, у питающихся рыбами гл. обр. эвта-
ксическое. Больших маховых 11, но у Alcedo 10. Рулевые большей
частью очень короткие, в количестве 12 (редко 10). Копчиковая
железа оперена (исключая род Tanysiptera\
464
Систематически зимородки довольно тесно примыкают к ракшам,
представляя вместе с тем сильно специализированную группу. Не-
сколько более отдаленное родство они обнаруживают к щуркам
и момотам.
ПОДОТРЯД 3: MEROPES. — ЩУРКИ
стайные птицы. Из насекомых
на мух. ос и пчел, проглаты-
Рис. 285. Золотистая щурка — Merops
apiaster L. По Evans.
Ярко и пестро окрашенные, ловящие на лету насекомых сходно
с ласточками, птицы Старого "Света. Характерны длинное вытянутое
тело, заостренный саблевидный клюв, острые крылья (исключая
Nyctiornis) и длинный хвост. Преимущественно степные или лесо^-
степные, колониально гнездящиеся и
с особенным предпочтением охотятся
ваемых целиком, не откусывая жа-
ло. Полы окрашены одинаково и
принимают сходное участие в воспи-
тании птенцов. Кладка в 6 белых яиц
откладывается в самостоятельно вы-
рытоп длинной норе, обычно в пес-
чаных откосах и обрывах. Птенцы
выклевываются голыми; впоследствии
подобно зимородкам покрываются
пеньками птенцового пера.
Подотряд состоит только из одно-
го семейства Meropidae, насчитыва-
ющего около 40 форм, относимых к
6 родам. Роды : Merops (рис. 285),
Dicrocercus. Melittophagus. Nyctiornis
и др.
В СССР распространены: золо-
тистая щурка — Merops apiaster L.
и зеленая щурка — Merops persicus
Pall.
Морфологически щурки харак-
теризуются следующими особенно-
стями. Череп с рудиментарным сошником; шейных позвонков 14;
грудина с двумя парами глубоких вырезок, с 4 парами грудных ребер
п длинной spina communis; коракоиды скрещиваются. Ноги синдак-
тпльиые, того же типа, как у Haley ones, возможно являясь приспособ-
лением к разрыванию земли, при постройке нор. Musculus ambiens от-
сутствует. Мускульная формула АХУ. Слепые кишки длинные, функ-
ционирующие, имеется желчный пузырь. Сохраняется только левая
сонная артерия (но парные у Nyctiornis'). Пух отсутствует. Контур-
ное перо несет рудиментарный побочный стержень. Крыло эвтапев-
ческое с 11 большими маховыми (у Merops и Nyctiornis 10). Рулевых
перьев 12. Копчиковая железа оголена.
Щурки обнаруживают ближайшее родство с зимородками и,
с другой стороны, с удодами, значительно далее отстоя от момотов,
несмотря на внешнее значительное сходство. По общему уровню
своей организации щурки уже приближаются к удодам, значительно
Удаляясь от более примитивных сизоворонок.
31» Орнитология—1209
4G5
ПОДОТРЯД 4: MOMOTL — ЛОМОТЫ
Внешне близко напоминающие щурок, по поведению соответ-
ствующие сизоворонкам, пестро окрашенные, с длинными хвостами
и грубо зазубренным
Рис. 286. Момот—
H/.s brasi lipnsis Lath. IJo
Evans.
по режущим краям клювом, лесные птицы
Центральной и Южной Америки. Крылья
короткие, округленные (4-й — 6-п маховые
самые длинные).JB xBOCTe замечательно строе-
ние длинных центральных рулевых перьев,
опахало которых таким образом (в изве-
стном возрасте) ощипывается птицей, что
концевые части перьев остаются связанными
с основным опахалом только одним стерж- •
нем (ракетообразные рулевые). Характерным
является выщипывание тех частей пера, ко-
торые уже развиваются суженными. Плюсна
относительно длинна. Полы окрашены оди-
наково. Пища гл. обр. насекомые, преиму-
щественно жуки, частью фрукты. Кладка из
4 белых глянцевитых яиц откладывается в
самостоятельно вырытых норах.
Анатомически характеризуются следую-
щими признаками. Череп с рудиментарным
сошником: шейных позвонков 15; грудина с
двумя парами вырезок, частью замкнутых в
фонтаиелли:* Синдактилия ног слабо выра-
жена. Мускульная формула АХУ, musculus
ambiens отсутствует. Язык длинен, бахром-
чат на конце. Мускулистый желудок масси-
вен, кишечник относительно длинен. Жел-
чный пузырь имеется. Слепые кишки отсут-
ствуют. Побочный стержень в контурном
пере рудиментарен, а пух отсутствует. Кры-
ло эвтаксическое с 11 большими маховыми.
Копчиковая железа оголена или с рудимен-
тарными перышками.
Только одно семейство Momotidae с 20 видами и 7 родами. Роды:
Momotus. В aryphi hen gus, Urospatha, Hylomanes и др. (рис. 286).
Ближайшими родичами момотов являются территориально близ-
кие тоди, несмотря на резкое различие в облике и поведении обеих
групп. Более отдаленно родство с Coraciae и Halcyones. По проис-
хождению момотов некоторые рассматривают (Гадов) как неотро-
пическую боковую ветвь С or aciif ormes, берущую свое начало как раз
между обеими вышеупомянутыми группами Coraciae и Halcyones.
ИОДОТРЯД 5: TODI. — ТОДИ (ПЛОСКОКЛЮВЫ)
Мелкие, ярко-зеленые, с коротким хвостом, с длинными ногами
и уплощенным тонко зазубренным на краях клювом, кустарниковые
птички Антильских островов. Пища — насекомые, собираемые с вет-
466
вей или ловимые r воздухе. сходно с мухоловками. Кладка из 3—4
белых, округлых яиц помещается в самостоятельно вырытую норку
в откосах и берегах рек, гнездовая камера которой выстилается ко-
решками, мохом и растительным вой-
локом. Птенцы выклевываются голыми.
Только одно семейство Todidae с пятью ви-
дами одного рода Todus (рис. 287). Систе-
матически близки зимородкам и момотам;
морфологически характеризуются непол-
ным десмогнатизмом и отсутствием сош-
ника. Межглазнпчная перегородка с круп-
ными фонтанеллями. Грудина сходна с
грудиной зимородков, с двумя парами вы-
резок; spina externa длинна и вилкообраз-
на. Основания коракоидов разобщены.
Шейных позвонков 15. Ноги синдактиль-
ные, даже с частичным срастанием дисталь-
ных фаланг > пальцев. Мускульная фор-
мула АХУ, musculus ambiens отсутствует.
Язык длинный, заостренный, кишечник
Рис. 287. Плоскоклюв—Todi/s
viridis L. По Evans.
очень короток. Слепые кишки короткие.
на конце колбообразно
вздутые, подобно caeca Strigiformes. Ниж-
няя гортань трахео-бронхиального типа с частью срастающимися кон-
цами, с одной парой голосовых мускулов. Крыло эвтаксическое с 10
большими маховыми. Рулевых двенадцать. Копчиковая железа опе-
рена. Пух отсутствует, побочный стержень в пере рудиментарен.
Тоди обнаруживают близкое родство только к момотам, но и в отно-
шении к ним все же занимают совершенно обособленное положение.
ПОДОТРЯД 6: UPUPAE. — УДОДЫ
Ярко окрашенные, мелкие, древесные и наземные птицы Африки
с Мадагаскаром и Евразии. Характерен длинный, обычно саблеоб-
разно-загнутый клюв, относительно длинный хвост, короткие, но
сравнительно сильные ноги с слабо выраженной синдактилией (сра-
стаются только основные фаланги 3-го и 4-го пальцев). Гнездятся
в древесных дуплах (Upupa. также в щелях скал и пустых норках).,
строят гнездо, но высиживает яйца только самка, в то время как самец
ее кормит. Кладка из 3—7 одноцветно-белых плп зеленовато-голубых
яиц. Инкубация у Upupa длится 16 дней. Выкармливаются пищей,
всовываемой птенцам в клюв; в связи с этим последние, так же, как
и воробьиные, имеют ярко окрашенную ротовую полость и толстые
чувствительные вздутия по углам клюва.
Интересным защитным образованием является образование в круп-
ном копчиковой железе у насиживающих самок вонючей черно-корич-
невой смазки, выпрыскиваемой из железы прп защите.
Подотряд образуется двумя семействами: Upupidaen Phoeniculidae.
Семейства 1: Upupidae. — Настоящие удоды
G единственным родом Upupa и десятью формами, распростра-
ненными в Евразии и Африке. Хвост прямой, короче крыла. Харак-
467
терем пестрой ркраскоп и длинным приподнимающимся хохлом на
голове. Питается преимущественно насекомыми, их личинками и чер-
вями. разыскивая пх на земле и вытаскивая клювом пз почвы. Птенцы
первоначально одеты густым серовато-белым пухом. В СССР эта
группа представлена обыкновенным удодом — U pupa epops L., ши-
роко распространенным во всей степной и лесостепной полосах и юж-
ных горных районах.
Семейство 2: Plioeniculidae. — Древесные удоды
Включает три рода: Phoeniculus, Scoptelus и Phino pomas tits,
с двадцатью пятью формами, распространенными в тропической
Африке. Характерен ступенчатый, длинный, с крыло пли длиннее,
хвост. Преимущественно птицы тропического леса, разыскивающие
пищу—различных членистоногих — в коре деревьев и ветвей, лазая,
подобно пищухам, по стволам, опираясь на хвост. Хвост линяет,
как у дятлов, центрпфугально, начиная с предпоследней пары.
Птенцы вместо пуха покрыты трпхоптплямп. образующимися пепь-
ками первого птенцового контурного пера.
Морфологически удоды стоят очень близко к птицам носорогам,
представляя в сравнении с ними более прогрессивную группу,
в ряде признаков своей организации весьма близко приближаю-
щуюся к воробьиным. Из особенностей строения следует отметить
следующие: в черепе сошник не сохраняется; шейных позвонков 14;
первый позвонок (атлас) свободен; metasTernum только с одной парой
вырезок, иногда зарастающих до образования фонтанеллей; Сохра-
няется только левая сонная артерия. Язык очень мал. Слепые кишки
отсутствуют. Нижняя гортань трахео-бронхиального типа, но без
голосовых мускулов. Мускульная формула ног АХУ, musculus
ambiens отсутствует. Flexor liallucis longus совершенно независим
от flexor perforans digilornm, так же как у настоящих Passeres.
Обыкновенный пух у взрослых отсутствует. Пуховой наряд птенцов
является видоизмененными окончаниями контурных перьев (как
у Pusseres). Имеется шейная и спинная аптерии. Phoeniculus отли-
чаются наличием редуцированного побочного стержня в контурнохм
вере. Крыло эвтакспческое. с 10 большими маховыми перьями.
Рулевых перьев десять. Копчиковая железа с общим выводным про-
током.
ПОДОТРЯД 7: ВUCEROTES — ПТИЦЫ-НОСОРОГИ
Крупные и средней величины, темные или пестро окрашенные
(черные с белым и желтым) птицы тропиков Старого Света, с огром-
ным клювом п сильно пневматизированными костнороговыми наро-
стами на нем и голове (рис. 288). Преимущественно древесные, разы-
скивающие пищу в кронах деревьев, вне брачного периода стайные
птицы. Только род Biicorvus ведет наземный образ жизни и, в противо-
положность остальным, передвигающимся на. земле и ветвях прыж-
ками, носорогам, легко ходит и бегает по земле. Полет тяжелый,
скользяще-гребной, прерываемый у некоторых короткими периодами
46S
скольжения. Питаются фруктами, плодами и ягодами, отчасти насе-
комыми, a Bucorvus п рептилиями и даже павшими животными.
Крайне своеобразным является гнездование в больших дуплах,
входное отверстие которых суживается самцом и самкой при помощи
глины или помета птиц: после этого самка проскальзывает в дупло,
где замуровывается вплоть до оставления небольшого отверстия
для кончика клюва (рис. 289). Самка несет от 1 до 4 белых матовых
яиц и высиживает их будучи кормпмой самцом, передающим ей
пищу через оставленное отверстие. Корм — полупереваренные в же-
лудке самца плоды извергаются им облеченные в оболочку пз выде-
лений стенок желудка и в таком виде передаются самке, а впослед-
Рнс. ‘288. Клювы различных птиц носорогов.
(и) — щитоклюва— Bhinoplax vigil Forst.; (b)— африканского носорога— Bycuuislps
cylindricus Тепин.; (с) — рогатого ворона — Bucorvus abyssinicus Bodcl.; (d)— ново-
гвинейского складчаторога — IlJiynlidoceros plicatus (Forst.). Из Reichenovv.
ст вин и птенцам. Инкубация продолжается у Lophoceros 28 дней.
Самка покидает дупло не ранее 3 недель по вылуплении птенцов и за
период насиживания претерпевает полную линьку мелкого и круп-
ного пера. В дальнейшем самец с самкой совместно выкармливают
птенцов, покидающих дупло спустя 1—2 месяца по вылуплении.
Птенцы первоначально совершенно голы, по с весьма пневматизиро-
ванным подкожным слоем (в защиту от охлаждения?); покидая
гнездо, они бывают очень жирны. При выкармливании их корм
вкладывается в раскрытый (как у воробьиных) клюв птенцов.
В группе различают только 1 семейство Bucerolidae с 70 формами,
распределяемыми на 16 родов. Роды: Bucorvus, Bycanistes, Buceros,
Gyinnolaenius и др.
Морфологически птицы-носороги стоят довольно близко к удодам,
представляя по сравнению с ними более примитивную группу;
происхождение их приходится искать вместе с Upupae поблизости
Halcyones п в соседстве вероятно с одновременно ответвившимися
Meropes. К последним птицы-носороги стоят по своей внутренней
организации ближе, чем к Halcyones, что дает основание предпола-
гать и возможность их совместного ответвления.
Из особенностей организации птиц-носорогов необходимо отме-
тить следующее. Череп десмогнатическцй с базиптерпгоидными от-
Рмс. 289. Гомрай — Dichoceros Licornis L. Самец
кормит самку, замурованную в дупле. По Newton.
ростками, однако не дости-
гающими птеригоидов; сош-
ник отсутствует, режущие
края клюва зазубрены; шей-
ных позвонков 14—15, пер-
вые два позвонка слились;
metasternum с одной парой
маленьких вырезок. Имеется
длинная spina communis ster-
ni. Основания коракоидов со-
прикасаются. С грудиной со-
членяются 8 пар ребер. Ви-
лочка U-образная, chypoclei-
dium. Конечности пневмати-
зированы до дистальных фа-
ланг, но осевой скелет* не-
пневматичен. Передние паль-
цы ног срастаются: 2-йи3-й
на 3/2—1 фалангу, 3-й и 4-п
на 2—3 фаланги. Исключение
представляет Bucorous с очень
длинной плюсной и относи-
тельно короткими пальцами.
Мускульная формула ног
АХУ; musculus ambiens от-
сутствует. СУХОЖИЛИЯ ПП1-
sculus flexor hallucis п mu-
sculus flexor profundus соединяются между собой (через vincu-
lum) в дистальных частях плюсны (в отличие от Upupae). Ниж-
няя гортань трахео-бронхиальная с одной парой голосовых мус-
кулов. Имеется только левая сонная артерия (исключая Висог
viis. у которых она парная). Язык очень короткий. Зоб отсутствует.
Слепые кишки отсутствуют. Обыкновенный пух и побочные стержни
полностью отсутствуют. Крыло эвтаксическое с 10 маховыми. Руле-
вых перьев десять. Копчиковая железа оперена.
В ископаемом состоянии Bucerotidae (несколько сомнительные)
известны из верхнего эоцена Франции (Cryptornis antiquus Miln.-
Edw., близкие к роду Lophoceros).
ОТРЯД 22: PICI FORMES. — ДЯТЛООБРАЗНЫЕ
Древесные птицы мелкой и средней величины, характеризуемые
следующими общими признаками: строение носовых костей голо-
рпнального типа, с сохраняющейся носовой перегородкой. Бази-
470
птеригоидные отростки отсутствуют. Глубокие височные ямы. Шейные
позвонков 14; спинные позвонки свободны; грудина с двумя парами'
вырезов (у Pici иногда превращающихся в фонтанелли), с крупной
spina externa (spina interna отсутствует); пять полных ребер, частью
сочленяющихся с длинным processus lateralis anterior sterni; кора-
коиды разобщены; вилочка U-образная, без hypocleidium; имеющаяся
сесамоидная os humero-scapulare значительна; musculus deltoidens
достигает значительной длины, сопровождая почти всю плечевую
кость. Ноги зигодактильного типа. Hypotarsus сложный, в простей-
шем случае с тремя каналами; musculus ambiens отсутствует. Нижняя
гортань только с одной парой трахео-бронхиальных мускулов.
В птерилозисе характерны узость птерилий и значительность апте-
рии. Обыкновенный пух отсутствует. Крыло тупое, эвтаксическое,
с 10 большими и 10—12 малыми маховыми. Зоб отсутствует.
Биологически все птицы этой группы характеризуются гнездо-
ванием в дуплах или в норах и несут одноцветно белые яйца. Птенцы
вылупляются слепыми и голыми. Птенцовый пуховой наряд отсут-
ствует; исключение представляет дятел — Centurus aurijrons (Wa'gl).
Отряд разделяется на четыре подотряда; Galbulae, Rhaniphasli,
Capitonides и Pici.
Систематически отряд обнаруживает многие черты сходства
с примитивными воробьиными — Eurylaeini и Clamatores, вместе
с последними ведя свое происхождение от сравнительно примитив-
ных Coraciijormes, поблизости Halcyones и Momoti.
ПОДОТРЯД 1: GALBULAE. — ЯКАМ АРЫ
Мелкие, ярко окрашенные обитатели густых тропических лесов
Южной п Центральной Америки, подстерегающие пролетающих
насекомых, спокойно сидя на ветке, п затем взлетающие за ними по-
добно мухоловкам. Гнездятся, устраивая гнезда на конце самостоя-
тельно вырытой в землистых откосах длинной норы. Кладка состоит
из 2 — 4 кругловатых блестящих яиц.
Различают два семейства.
Семейство 1: Galbulidae.—Настоящие якамары1 (Блестянки)
Внешне напоминающие щурок, с металлически блестящим опе-
рением, обычно длинноклювые, относительно длиннохвостые и ко-
ротконогие птички. Питаются преимущественно бабочками, пресле-
дуя их подобно мухоловкам.
Известно около 20 форм, относимых к 7 родам. Роды: Brachygalba,
Jacmnaralcyon, Jacanierops, Galbula и др.
Из морфологических признаков характерен клюв, с резким раз-
витием граней, чем он напоминает клюв Picidae, также сходна и уко-
рененность крайней пары рулей. Вилочка повидимому без hypoclei-
dium. Хвост ступенчатый, но центральные две пары рулей равны
между собой.
1 Якамары — местное бразильское название.
471.
Семейство 2: Bueeonidae. — Пуховки 1 (леипвки)
Мелкие. внешне напоминающие лесных зимородов пли мелких
кукушек, птицы теневых .лесных участков, питающиеся гл. обр. жу-
ками п прямокрылыми. Кладка только из 2 яиц. Клюв умеренной
длины, сжатый, иногда сжатый с боков, часто с крючком на конце,
без заметных граней, с щетинками у основания. Хвост малоступен-
чат, пли прямой. Общая окраска обычно невзрачная, часто с пят-
нами и пестринками, что напоминает молодые наряды многих во-
робьиных. От Galbulidae отличаются также деталями (строения)
черепа и наличием меж-
ключичного выроста в
вилочке. Известно око-
ло 40 форм, относимых
к 13 родам: В песо. Ма-
lacoplila, Chelidoplera я
др. (рпс. 290).
Морфологически яка-
ма ры представляют со-
бой группу наиболее
примитивных дятлооб-
разных птиц, пз всего
от р я да п р о я вл я ю щп х
наибольшее родство с
Caracii- и (даже) Сиси-
liformes. Из особенно-
стей внутренней орга-
низации их отличают от
остальных Picijornies гл.
обр. очень крупные про-
коракоидные отростки,
ф у н к ц и о н и р у ю щ в е
длинные, вздутые колбо-
образно на конце, сле-
пые кишки и парные
сонные артерии — всё признаки, свойственные гл. обр. более низко
стоящим отрядам. Однако по общему строению грудины, плечевого
пояса, зигодактильному типу ноги (с утерей первого пальца у Jacu-
niaralcyon, сходно с Picoides} и типу сгибателей пальцев это уже
весьма близкая к дятлам группа. Из других особенностей следует
упомянуть: десмогнатическое строение костного неба, с парным
сошником (?), но сильно редуцированным. Spina extern а вило-
образна на конце. По строению musculus propatagialis обнаружи-
вается близость к Merops й Upupa, сходство с которыми имеется в
в других чертах. В отличие от остальных отрядов Piciformes желчный
пузырь не обнаружен. Язык очень длинен, тонок, заострен на конце.
Короткие мускулы 4-го пальца очень сильны. Мускульная формула
1 Название пуховки происходит поводимому от свойства птиц в спокой-
ном положении приподнимать контурное оперение, так что фигура птицы при-
нимает форму пухового шара или пуховки.
472
варпирует: АХУ пли АХ. Покровные перья с весьма редуцирован-
ными стержнями, а у М alacoplila и совсем без побочных стержней.
Больших маховых 10, рулевых 12. Копчиковая железа оголена.
ПОДОТРЯД 2: RHAMPHASTL — ПЕРЦЕЯДЫ (ТУКАНЫ)
Подвижные древесные птицы тропических лесов Центральной
и Южной Америки с огромным ярко и пестро окрашенным клювом
и пестро окрашенным оперением. Держатся в кронах деревьев, пи-
таясь фруктами, иногда членистоногими, япцами и птенцами. Ок-
раска полов одинакова. Гнездятся в дуплах деревьев, откладывая
без гнезда 1—4 относитель-
но маленьких с матовой
скорлупой япца. Птенцы
с очень медленным разви-
тием, у Rhaniphastos. про-
зревающие только в воз-
расте 3 недель.
Из других биологиче-
ских черт интересно поло-
жение, занимаемое тука-
ном во ci/е, с клювом, при-
крытым спинными перья-
ми,]! высокоподнятым и за-
ложенным целиком на спи-
ну хвостом, что является
невидимому приспособле-
нием к ночевкам в дуплах.
Единственное семейство
Rhamplutstidae с 60 форма-
ми, относимыми к 7 родам:
Rham,phastos,A ndigena, Ple-
roglossus, Selenidera и др.
(рис. 291).
Систематически перцея-
ды являются повидимому
ближайшими родичами
С a piton idae, несмотря на со-
вершенно ОТЛИЧНЫЙ внеш- Рпе. -291. Тукан — nhainphaslos ar'u'l Vii;. По Evaib.
нпй облик.
Из морфологических особенностей наиболее яркой является
форма клюва, уплощенная с боков, с зазубренным режущим краем:
Носовые отверстия лежат по его заднему краю. Не взирая на зна-
чительную величину, клюв очень легок благодаря сильной пневма-
тизированностп его скелета. Уздечка и окружность глаз оголены от
перьев. Череп десмогнатпческнй. Язык длинный, ороговевший, густо
бахромчатый по краям и на конце. Мускулистый желудок слаб и
объемист. Кишечник же относительно короток и широк, слепые
кишки отсутствуют. Замечателен очень длинный кишкообразный
желчный пузырь. Правая лопасть печени значительно больше левой.
473
В плечевом поясе характерные короткие ключицы, не соединяющиеся
в вилочку. Крылья тупые, сравнительно слабые. Только левая сон-
ная артерия (исключая Rhamphastos dicoldrus L.). В нижней гортани
3—4 трахеальные кольца сливаются в костный барабан. Копчиковая
железа оперена. В крыле 11 маховых, из которых дистальные очень
укорочены. Рулевых 10, при линьке у многих видов сменяемых в цен-
трппетальном порядке. Хвост обычно крупный и прямой пли слегка
.ступенчатый.
ПОДОТРЯД 3: CAPITONIDES
Мелкой и средней величины насекомоядные и фруктоядные птицы,
распространенные в лесных тропиках почти всего света. Характе-
ризуются следующими признаками. Череп эгптогнатический, сошник
спереди расщеплен, вилочка, если и имеется, лишена hypocleidium.
Spina ex Lerna sterni длинна, но не вилкообразна. Доли печени почти
равны по величине. Желчный пузырь длинен, короткие мускулы
4-го пальца очень слабы пли отсутствуют. Мускульная формула
АХУ. Побочный стержень в контурном пере развит. Копчиковая
железа оперена.
По многим признакам птицы этой группы представляются при-
митивными дятлами, менее специализированными или специализиро-
ванными в ином биологическом направлении. Подотряд разделяется
на два семейства: Indicatoridae и Capitonidae.
Семейство 1: Indicatoridae. — Медоуказчики
Мелкие, величиной с воробья пли скворца, невзрачно окрашен-
ные птицы, внешне и по повадкам напоминающие вертишеек, рас-
пространенные в тропиках Старого Света, с наибольшим преоблада-
нием в Африке. Мало подвижные и мало заметные, они обращают
на себя внимание только короткими криками, да своеобразными
повадками, давшими им их народное название. Обычной пищей явля-
ются насекомые и их личинки, собираемые с веток и коры деревьев:
у некоторых видов однако основной пищей являются личинки и ку-
колки пчел (и вероятно ос). Очевидно в связи с тем, что ме до указ-
чики, вооруженные лишь коротким, сравнительно слабым клювом,
не в состоянии проникнуть в пчелиные постройки, у них развилась
способность привлекать беспокойным поведением и криками к этим
ульям и гнездам человека, чем в частности славятся два вида рода
Indicator [7. indicator (Gm.) и variegata Less.]. Эта способность
развилась повидимому из сожительства (явление так паз. ком-
менсализма) с медоедом, Mellivora са pensis. по отношению к кото-
рому птицы проявляют то же поведение. Однако, с этими же криками
возбужденная птица может привести следующего за Heit человека
и к леопарду, змее и т. п., а за медоеда нередко принимает и собаку.
Другой род Melichneutes привлекает повидимому своих помощ-
ников, производя особый шум суженными и измененными крайними
рулевыми. Бросающуюся в глаза значительную толщину кожи медо-
указчиков ставят в связь с защитой против пчелиного жала. В брач-
(4 /4
ный период медоуказчикп производят барабанную трель ударами
клюва по сухим веткам деревьев, подобно тому, как это делают дятлы.
Все члены этой группы не строят собственных гнезд, подкидывая
свои оелые, слабо глянцовитые япца в гнезда видов семейства Pici-
dae и Capitonidae. В группе различают около 12 видов, относимых
к 4 родам: Indicatin' (ареал прерывчат: Африка, Гималаи, Малакка,
Суматра, Борнео), Melignomon и Мelichneutes (область Конго и
Камерун в Африке) и Prodotiscus (Африка).
Систематически медоуказчикп представляют группу, близко род-
ственную бородастикам и дятлам, но в оригинальной смеси этих об-
щих черт. Вследствие этого их приходится рассматривать как само-
стоятельную генетически веточку, более примитивную и более
сохранившую черты общих предков. Из морфологических особенно-
стей следует отметить: череп эгптогнатпческпй, с широко расставлен-
ными processus palatini maxillae; щелевидные носовые отверстия
лежат у самого конька клюва и открываются вверх, а не вбок;
ключицы соединяются в вилочку; грудина имеет значительное сход-
ство с грудиной Jynx, крыло острое, с 2 маховыми самыми длинными,
но число больших маховых только 9, иногда с первым — рудимен-
тарным: хвост обычно прямой, из 12 рулевых (у Prodotiscus — 10).
В нижней гортани несколько трахейных колец образуют костяной
барабан.
Семейство 2: Capitonidae. — Бородастики
Ярко и пестро окрашенные птицы лесных тропиков, с толстым
сильным клювом, окруженным обильными длинными щетинками.
Обитают преимущественно опушки лесов и рощи древесной расти-
тельности в стели, лазая среди веток деревьев. Питаются главным
образом ягодами и мясистыми плодами, а также насекомыми и их
личинками, доставаемыми преимущественно из-под коры пли гни-
лой древесины. Большинство издает сильный звенящий крик, напо-
минающий крик некоторых дятлов. Гнездятся в дуплах стволов
деревьев и их ветвей, самостоятельно выдалбливаемых мощным клю-
вом подобно дятлам. Некоторые однако устраивают гнезда в термит-
никах, a Trachyphonus вырывает косые узкие норы в ровной твердой
почве, выкидывая землю ногами. Кладка состоит из 2—5 яиц, весьма
схожих с яйцами дятлов. Инкубация у Tricholaenia, leuconielan Bodd.
длится 14 дней.
Известно около 140 видов, относимых к 18 родам, с центром рас-
пространения в Африке. Роды: Tricholaenia, Gymnobucco, Megalaenia,
Cyanops, Xantholaenia и др. .
Морфологически бородастики характеризуются следующими чер-
тами: череп эгитогнатическнй с тенденцией к десмогнатпзму, в связи
вероятно с несколько вздутой формой клюва; последний прямой
пли слегка изогнутой формы, с одним или несколькими зубцами по
краю надклювья; носовые отверстия лежат у конька клюва и обычно
прикрыты щетинками; крылья тупые, короткие; хвост короткий,
113 10 рулевых, крайняя пара-их обычно укорочена; ключицы не
соединяются в вилочку; контурное перо несет побочный стержень;
475
язык длинный с роговыми краями и с вильчатым или бахромчатым
концом; правая доля печени немного больше левой; желчный пузырь
очень велик; кишечник весьма объемист, но короток; нижняя гор-
тань без костяного барабана, образуемого трахейными кольцами.
ПОДОТРЯД 4: PICI —ДЯТЛЫ
Обширная группа небольших древесных лазящих птиц, с ха-
рактерным долотообразным сильным клювом и (обычно) жесткими
рулевыми перьями, на которые опираются птицы, лазящие по коре
дерева. Распространены по всей земле, исключая безлесные про-
странства, Нотэгею и Мадагаскар. Пища добывается преимущественно
из-под коры деревьев и состоит гл. обр. из насекомых и их личинок.
Кроме того птицы кормятся собирая насекомых с ветвей, истребляя
муравьев и их куколки («яйца») и потребляя различные древесные се-
мена (семена хвойных, орехи, желуди ит.д.). Оба пола окрашены
большей частью различно. В брачный период свойственна так наз. «ба-
рабанная трель» — дребезжание сухого сука под влиянием ударов
клювом, заменяющее брачное пение.Гнездятся обычно выдалбливая
самостоятельно дупло в подгнившем дереве. Кладка из 2 (в тропиках)
до 8 и 12 (Dryobates и Jynx) белых, слабо блестящих яиц, отклады-
вается на дно дупла без подстилки. Инкубация коротка, у Dryocopus
martins только 13 дней; насиживают оба пола, самец преимущественно
ночью. Птенцы вылупляются голыми (исключение Centurus aurijrons
Wagl., у которого птенцы слабо опушены). Развитие птенцов быстрое.
Различают два семейства: Jyngidae и Piciclae.
Семейство 1: Jyngidae. — Вертишейки
С единственным родом Jynx и двумя видами в Африке и Евразии.
Рулевые перья мягкие, клюв слабый, редко употребляемый в ка-
честве долота. Гнездятся преимущественно в чужих дуплах. Кончик
языка гладкий, без щетинок. Ножки подъязычной кости заходят
в левый sinus orbital is. Musculus tracheohyoides прикрепляется
в нижней части трахеи. Сошник отсутствует (так -же, как у р. Sn-
sia, возможно относящегося к этому же семейству).
В СССР по всей лесной зоне широко распространена обыкновен-
ная вертишейка — Jynx tarquilla L.
Семейство 2: PiciЛае. — Дятлы (настоящие дятлы)
С долотообразным клювом. Кончик языка несет щетинки. Йошки
подъязычной кости, в случае большой длины их, заходят в правый
sinus orbitalis или заворачиваются за правую орбиту.
В этой группе известно около 500 форм, распределяемых на
48 родов. Систематика дятлов недостаточно разработана и отличается
искусственностью.
Принимают следующие подсемейства: Piciinininae, с родами: /JJ-
ciunnus, Nesoctites, Verreauxia и Sasia; Psilor/iinae, с родами: ChrU'
socolaptes^ Brachypternus. Dinopiuin, и др.; Picinae. с, родами: Picus-,
47G
('hloronerpes, Cola pies, Geocolaptes. M iilleripicus и др.; Dryobali-
mie. (‘ родами: Dendrodromas. Dryobales, Leucontopicus, Hypo picus,
Jimgipicus. Sphyra picus. Picoides, Sapheopipo, Dryocopus н др.
В СССР представлены 15 видами; из них наиболее распростра-
нены: большой пестрый дятел — Dryobales major (L.), белосппниый
дятел — Dendrodromas leucotos (Bechst.), малый дятел — Leuconto-
picus minor (L.), желна — Dryocopus martins (L.). трехпалый дятел —
Picoides tridactylus (L.), седой дятел — Picas canus Gm. Более узкие
ареалы занимают: зеленый дятел — Picus viridis L., чешуйчатый
дятел — Picus squama tus Gould, белокрылый дятел — Dryobales leucop-
terns (Salvad.), сирийский дятел—Dryobates syriacus (Hemp, el
Ehrenb.). полосатый дятел — Dendrodromas liljordi (Sharp, et Dress.),
средний дятел — Dendrocopies medius (L.). малый и большой остро-
крылые дятлы — Jyngipicus kizuki (Temin.) и J. nanus (Vig.).
Морфологически дятлы представляют собой группу с ярко выра-
женными приспособлениями в строении клюва, языка и хвоста к ори-
гинальному образу жизни. Клюв прямой или, иногда, слабо изогну-
тый, обычно с резко выраженными гранями, конец его б. ч. долото-
образен. Роговое вещество рамфотеки лежит очень твердым и тол-
стым слоем. Кости черепа, особенно клюва и крыши его, весьма
крепки, небно-челюстной аппарат малоподвижен и межглазничная
перегородка очень тверда. Язык очень длинен, с длинными ножками
подъязычной кости, загибающимися на крышу черепа и заходящими
в надклювье или заворачивающимися вокруг глаза; конец языка
обычно заострен п несет направленные назад щетинки. Благодаря
зтпм щетинкам и липкости язык действует то как липкий прутик,
то как гарпун, протыкающий добычу, то как тонкий крючок, вслед-
ствие длины могущий проникать в узкие ходы живущих в коре и
древесине насекомых. С функцией языка связаны очень объемистые
нижнечелюстные слюнные железы, выделяющие весьма липкий
секрет. Далее своеобразно специализирован хвост, состоящий из
12 рулей, крайняя пара которых сильно редуцирована и иногда
(у карликовых дятлов) совсем отсутствует. У Verreau.ria плюется
только 8 рулевых перьев. Рули играют роль опорного органа для
тела птицы, в связи с чем очень жестки и упруги, а стержни несут
глубокую вентральную бороздку (как у пингвинов). Хвост мягок
только у карликовых дятлов (Picumninae) и Jynx, благодаря чему
эти виды часто сидят поперек веток, как большинство воробьиных.
Некоторые дятлы однако уклонились от древесного образа жизни
п приспособились к жизни в открытой степи, прыгая по земле в по-
исках насекомых, гл. обр. муравьев, каковы напр. некоторые виды
Colaptes в Южной Америке и Geocolaptes в южной Африке, для гнезда
роющие норы в береговых обрывах. Некоторые дятлы гнездятся
в постройках термитов (Campethera nivosa Swains.) или древесных
муравьев (Мicropternus}.
Из морфологических особенностей дятлов необходимо отметить
следующие черты: череп шизогнатический:; сошник сильно редуци-
рован; слезная кость (os lacryrnale) отсутствует, функционально
замещаясь os eclelhmoideum; os jugale также отсутствует (Кра-
совский, 1936); крыловидные кости своеобразной формы; в гру-
477
динз ^Jjina fek'ierna длинна и впльЧАТЗ; вйлйЧка U-образна. без
flypocfeidium. nepfcktii (задний палец) оЧёйь мал, часто не функ-
ционирует, у Некоторых родов* (Plcoides, Sasici. Dinopium) сов-
сем отсутствует. Мускульная формула AXV или АХ. Очень ма-
лсйькйе йосовые железы помещаются в задней части орбиты. Сонная
ар^фтгй Сохраняется только левая, в шейном отделе, проходящая
частью в костном канале, образуемом 4 позвонками. Трахеоброн-
хиальная нижняя. Гортань без костного барабана, с одной парой боко-
вых голосовых мускулов. Слепые кишки отсутствуют» Желчный
пузырь длинен и кишкообразен. Копчиковая железа имеет двудоль-
ную форму и снабжена на конце перьевой кисточкой (исключая
ряд родов с оголенной железой). Побочные стержни рудиментарны
или во всяком случае очень малы. Полупух и контурные перья отде-
ляют йудру (в особенности у Mulleripiciis). Юношеская линька (в про-
тивоположность остальным подотрядамPiciformes) полная; внутренние
большие маховые птенцового крыла у многих родов остаются весьма
маленькими и не функционируют. При линьке хвоста последними
вменяются центральные рулевые. Птенцы в период пребывания
в дупле имеют роговые пяточные наросты (помогающие взби-
раться по стенкам дупла?), свойственные также туканам и бюрода-
стикам.
Систематически дятлы имеют ближайших родственников. в> боро-
да стиках, сходных не толы-со обилием общих анатомических черт,
но и некоторых биологических. Однако вследствие достаточной
обособленности той п другой группы целесообразнее их рассматри-
вать как подотряды. Вместе с Capitonides дятлы также близко род-
ственны Coraciae. п ответвление их общими или близко соприкасаю-
щимися стволами имеет место вероятно поблизости ветвей момотов
и зимородков. Наличие некоторых общих черт с примитивными
воробьиными Ewrylaemi и Мепигае объясняется ответвлением послед-
них поблизости ют группы дятловых. Но кроме того,, имеющиеся
у всех дятлообразных мелкие черты сходства с Clainatores и Oscines
у дятлов выражены особенно сильно, что указывает на достижение
ими из всей группы наиболее высокого морфологического уровня,
соответствующего и наиболее прогрессивному отряду класса птиц —
воробьиным.
Из современного распространения дятлов, гл. обр.. отсутствия
в Австралии, Новой Гвинее и на Мадагаскаре, вытекает, что они яв-
ляются сравнительно поздно развившейся и расселившейся группой.
В связи с обилием дятлов в Южной Америке и наличием только здесь
примитивных дятлообразных — Galbulae и Rhainphastir родиной Pici
вероятнее всего нужно считать тропическую Южную, и Центральную
Америку. В ископаемом состоянии однако дятлы известны начиная
с верхнего олигоцена Франции (Palaeopicus, archiaci и consobriniis
Miln.-Edw., близкие к рецептному Picus).
ОТРЯД 23: PASSERIFORMES. — ВОРОБЫШООБРАЗНЫЕ
Основная масса видов птиц, примерно 3/5. всех видов птиц вообще,
-в количестве почти 18 000 форм пли 6000 видов. Преимущественна
478
мелкие и средней величины древесные и кустарные птицы,- хота разни "
образце приспособления столь велико, что. за исклКУЧеНйе^ водной
среды, все остальные типы стаций имеют своих многочисленных
обитателей и специализированные виды.
Общие признаки группы заключают следующие особенности:
череп эгитогнатическпй (исключая некоторых представителей се-
мейства Dendrocola ptidae и Conopophagidae), без базиптеригоидных
отростков; шейных позвонков 14 (исключая Eurylaenii с 15 позвон--
ками); грудина с 1 парой вырезок, иногда превращающихся в фон--
танелли; в этом признаке уклоняются однако Pteroptochidaein Сопо-
pophagida'e с 2 парами вырезок и род Мепига. лишенный вырезок
вообще; прокоракоидный отросток мал. Spina externa sterni длинна
и вилкообразна на конце (исключая некоторых Cotingidae, Eurylaenii
и Philepittd). Spina interna sterni отсутствует; вилочка имеет расши-
ряющиеся проксимальные концы (сходно с Pici, Capitonides nRham-
phasti) и имеет liypocleidium (исключая опять-таки род Мепига);
у Atrichornis ключицы не соединяются в вилочку. Пять (у Согсогах 6)
полных ребер. Передние пальцы свободны до основания (исключения
есть среди Clamatores и Eurylaemi). Назад направлен только один
(первый) палец, более сильный, чем передние, и прикрепляющийся
на одном с ними уровне. Hypotarsus сложный. Musculus ambiens
отсутствует. Мускульная формула ног АХУ (у Dicriirus AX). Muscu-
lus flexor hallucis longus не соединяется (посредством vinculum)
с musculus flexor digitorum profundus (perforans) (исключая Eury-
la.emi). Сонная артерия сохраняется только левая. Слепые кишки
рудиментарны и превращены в лимфо-эпителиальные органы. Крыло
эвтакспческое, с 10 (или 9-j-1) большими маховыми; у многих,
преимущественно примитивных форм однако имеется и одиннадца-
тое редуцированное дистальное маховое. Малых маховых 9 (у Me
лига 10). Обычное число рулевых 12, но нередки уклонения, у Menu.
16, у Stipiturus и др. 6. Линька больших маховых происходит десп
дентно, малых — конвергентно к середине, рулевых перьев цен'
фугально (Cerlhia. представляет исключение, меняя рулевые,
дятлы). Побочный стержень в пере или полностью отсутствуй
очень мал {Oscines и сем. Acanthisitlidae). Пух покрывает
аптерии и обычно спорадичен, пышно развит он только у
Пудреткп встречаются лишь у Artamus (в виде четыре х
Копчиковая железа всегда присутствует и оперена. 7
Из биологических особенностей можно указать на птен’
относящихся сюда представителей. При этом птенцы
слепыми и несут обычно только на верхней стороне
(praepennae). Однако птенцы видов, гнездящихся в
дах, бывают полностью оголены, а также и некото1
открыто {Pycnonotus, Lanius и Dicrurus). Инкубаи
от 11 дней у мелких форм до 21 дня у крупных (j
по этим особенностям представляет Мепига, наг
дней и имеющая птенцов густо опушенных. Все
ливают птенцов всовывая им пищу в открыты
ротовая полость птенцов бывает обычно я
иногда сигнальные пятна.
Г(С
,ен>
три-
как
гт или
только*
Ci/wto-
до в ость .всех
вылупляются
редкий, пушок
закрытых гиез-
рые гнездящиеся
4яя продолжается
лором).. Отклонение
ляжмвающая 35—40
/ воробьиные выкарм-
Д шиов.,. в связи с чем
pFo «окрашена, и . несет
479
Классификация отряда, особенно Oscines, находится в мало раз-
работанном. в известных отделах прямо в хаотическом, состоянии.
Это обусловлено частью анатомическим однообразием группы, частью
неизученностыо строения, частью широкой распространенностью яв-
лений глубокой конвергенции. Крупные подразделения группы осно-
вываются гл. обр. на строении нижней гортани, щитковании плюсны
и соотношениях (связях) пальцев, как наименее изменчивых под
влиянием образа жизнп.
В составе отряда различают четыре подотряда: Eitrylaemi, Clama-
tores, Мепигае. и Oscines.
ПОДОТРЯД 1: EURYLAEMI. — РОГОКЛЮВЫ
Внешне напоминающие некоторых сизоворонок, переходная к ним
группа лесных тропических птиц Индо-Малайской области и Африки.
Клюв короткий, плоский п широкий с немногими щетинками у осно-
вания. Крыло короткое и округленное. Держатся преимущественно
в кронах густого леса. Гл. обр. насекомоядные, отчасти плодоядные
«формы. Гнезда устраиваются в виде висящей на ветвях сумки с бо-
ковым входом; предпочти-
Рпс. 29?. Зеленый рогоклюв — Са liiptomena viridis
Balli. По Evans.
тельио помещаются на све-
шивающихся над водой
ветвях. Яйца обычно пят-
нистые (у Smithornis одна-
ко чисто белые).
Единственное семей-
ство Eurylaemidae с 20
видами родов: Енгу lacmus,
С a ly р to тепа. Рsendee а 1ур-
tomaena. Smithornis и др.
(рис. 292).
Резко отличающаяся от
остальных воробьинооб-
разных группа птиц с сле-
дующими характерными
особенностями: сухожилия
сгибателейпальцев: muscu-
lus flexor hall и cis longus и
inusculus flexor digitorum
prof and us соединены между
собой сухожильной лентой
(vinculum) (как у сизоворонок); шейных позвонков 15; spina externa
slerni с закругленным концом, задняя сторона плюсны покрыта шести-
сторонними щитками (а не табличками, как у остальных Passerifor-
mes), хорошо развито лобно-носовое сочленение клюва; 2-й и 4-й
пальцы ног срастаются с 3-м пальцем на 1-й и 2-й проксимальных
фалангах; трахео-бронхиальная нижняя гортань без голосовых му-
скулов Дли лишь с одним слабым мускулом мезомнопдного типа; сон-
ные артерии парные; главной артерией ног является arteria ischia-
dica (так наз. гомеомерный тип). Больших маховых 10 1, у Calyplo-
тепа 9 1. Рулевых 12.
486
Рогоклювы представляют по существу переходную группу между
сизоворонковыми и дятлообразными и настоящими воробьиными.
По строению неба, лобно-носового сочленения, некоторым чертам
птерилозиса, числу позвонков они сближаются более с первыми
группами, очевидно сохраняя в этом примитивные черты.
ПОДОТРЯД 2: CLAMATORES. — КРИЧАЩИЕ ВОРОБЬИНЫЕ
Примитивные воробьиные, с иижпей гортанью трахео-бропхиаль-
пого или бронхиального типа, с мезомиопдным или анизомиоидпым
типом голосовой мускулатуры. Связки глубоких сгибателей пальцев
(см. Eurylaenii) пе имеется.
В эту группу относятся 11 семейств, с почти 2000 видов, распро-
страненных гл. обр. в Южной (и отчасти Северной) Америке и тро-
пиках Старого Света. По типу нижней гортани группируются иногда
в две группы: с трахео-бронхпальной гортанью — Ilaploophonae,
с семействами Acanthisittidae, Philepitlidae и Pittidae в Старом
Свете и семействами Tyrannidae, Pipridae, Cotingidae, Phytotoniidae
в Новом Свете, и с трахеальной гортанью— Tracheophonae, с семей-
ствами Conopophagidae, Pteroptochidae, Formicariidae и Dendroco-
laptidae. свойственными Новому Свету.
Семейство 1: Acanthisittidae
Мелкие, относительно длинноногие, по облику напоминающие
питт, а по повадкам крапивников, лесные птицы Новой Зеландии
(п соседних островов) с трахео-броихпалытым сиринксом и пшзорп-
нальным строением носовых
костей. Имеется только одна
пара голосовых мускулов
мезомиоидиого типа. Крылья
короткие, тупые, с 10 боль-
шими маховыми. Гнездятся в
полудуплах. Яйца чисто бе-
лого цвета. Роды: Accinlhi-
sitta, Xenicu9& Traversia.
Семейство 2: Philepittidae
Мелкие наземные лесные
птицы Мадагаскара, обликом
напоминающие питт, с тра-
хео-бропхиальиыммезомноид-
ным сиринксом, верхнее брон-
хиальное кольцо которого
сильно вытянуто. Spina ex-
terna ster-ni мала и невилко-
Рпс. 293. Мадагаскарская пптта — Philepilta
jala Bodd. Но Evans.
образна. Крыло с 10 боль-
шими маховыми (и длинным дистальным). Рулевых 12. Покров
плюсны такзасштдного типа. Яйца чисто белые.
Известен только один род Philepitta с двумя видами (рис. 293).
31 Орнитология—1209
481
Семейство 3: Pittidae. — Питты
Мелкие наземные, ярко и пестро окрашенные, лесные птицы
Зондских островов, Индии, Австралии и Африки, с трахео-бронхиаль-
ным мезомиоидным сиринксом, верхнее кольцо которого имеет нор-
мальное строение. Грудина с одной парой очень глубоких вырезов
и уклоняющейся от остальных Passeriformes формой metasternum,
с очень крупной и вилкообразной spina externa sterni. Число шейных
позвонков вариирует между 15 и 14. Крыло с 10 большими маховыми
(и длинным дистальным). Рулевых
12. Покров плюсны кнемидофорного
типа. Пища из насекомых и червей
разыскивается на земле. Птицы пе-
редвигаются большими прыжками,
Рис. 294. Короткохвостая питта — Pitta
brachyura L. По Evans.
Рис. 295. Ласточке хмштьш тиран —
Milvulns tyrannuus (”.. По Evans.
сходно с дроздами. Полет очень слаб; Гнездо открытое, помещаю-
щееся на земле. Яйца пятнистые, с коричневыми и красными пе-
стринками по белому фону.
Только один род Pitta, с единственным видом в Африке и 40 фор-
мами в остальной части ареала (рис.- 294).
Семейство 4: Tyrannidae. — Тираны
Мелкие, гл. обр. древесные, внешне часто напоминающие мухо-
ловок и сорокопутов, в основном насекомоядные птицы Америки
(исключая Phaeomyias и др., питающихся ягодами). Сиринкс трахео-
бронхиальный мезомиоидного типа. Membrana tympaniformes interna
с натягивающим хрящиком. Главнейшая артерия ног — седалищ-
482
ная. Щиткованпе плюсны экзаспидпого типа. Концы маховых перьев
нередко сужены, являясь причиной своеобразных звуков, издаваемых
при брачных церемониях. Гнезда разнообразного типа, с преобла-
данием открытых, чашкообразных. Яйца большей частью пятнистые.
Известно более 700 форм многих родов: Alecturus, Tyrannus,
Myiarchus, Onychorhynchus, Elaenia, Pyrocephaliis, Oxyruncus, Xol-
mis и др. (рис. 295). 1
Семейство 5: Pipridae. — Манакипы
Рис. 296. Мапакпп— Pipra men ta-
ils Scl. По Evans.
Мелкие древесные, напоминающие по повадкам синиц, гл. обр.
насекомоядные пт^ры Южной и Центральной Америки. Вне брач-
ного периода ведут стайный кочующий
образ жизни. Гнездо устраивается от-
крыто. Яйца б. ч. пятнистые. Трахео-
бронхиальный сиринкс без натягиваю-
щего хрящика для внутренней голосо-
вой мембраны. Плюсны относительно
длинные, 2-й и 4-й пальцы срослись па
протяжении полутора фаланг (кроме
Piprites}, главная артерия ног — бед-
ренная, щиткование плюсны экзаспид-
ного типа. Роды: Pipra, Piprites, Chiro-
xiphia, Manacus, Machaerop terns, Neopel-
ma и др. (рис. 296).
Семейство 6: Cotingidae. -- Котинги
Древесные, обычно ярко окрашенные птицы Южной и Централь-
ной Америки, весьма разнообразные по хабитусу и повадкам. Гнез-
Рис. 297. Звонарь — Procnias niveus Bodd. Птица обладает
звучным голосом; у кричащей птицы мясистый полый вы-
рост приподнимается и торчит в виде рога кверху (правая
фигура). По Evans.
дятся в дуплах или
устраивают открытые
гнезда, помещаемые
среди ветвей дере-
вьев. Яйца пятни-
стые, разнообразной
окраски. Питаются
главным образом яго-
дами, плодами и от-
части насекомыми.
Нижняя гортань тра-
хео - бронхиального
типа, без хрящика
внутренней голо совой
перепонки, с измен-
чивой (двупарной)
мускулатурой. Пре-
обладает катакромиоидный тип. Некоторые с очень громким металлп-
чески-звучащим голосом. Основная артерия ног — arteria femoralis.
1 Обычно это огромное семейство разделяют на ряд подсемейств, напр.:
Fluvicolinae, Tyranninae, Myiarchinae, Platyrinchinae, Euscarthminaey Serpo-
phaginae и Elaeniinae.
483
(Гетеромерный тип). Щиткование плюсны пе экзасппдного типа
(преимущественно ппкнасппдного типа). Своеобразны видоизменен-
ные маховые, играющие роль в брачных играх.
Систематически близки к Pipridae.
Известно около 170 видов. Роды: Phoenicircus, Ampelion, Cotinga,
liupicola, 1 Attila, Tityra, Pyroderus, Cephalopterus и др. (рис. 297).
Семейство 7: Pliytotomidae. — Траворезы
Мелкие, южноамериканские птицы с характерно зазубренным
по краям вьюрковым клювом и весьма укороченным п широким
кишечником. Питаются листочками, почками и плодами. По пове-
дению напоминают вьюрков. Вне брачного периода держатся стаями.
Гнезда устраиваются открытые. Яйца пятнистые. С трахео-брон-
хпальпым сиринксом и основной артерией пог — седалищной.
(Гомэомерный тип.) Известно только два вида одного рода Phy to tom а.
Семейство 8: Conopopliagidae. — Гусешщседы
Мелкие лесные, внешне напоминающие соловьев (и зорянок)
птички Южной Америки, живущие в густой растительности, питаю-
щиеся преимущественно мелкими жучками и передвигающиеся
сильными прыжками. Яйца пятнистые. Череп шизогнатический.
Носовые отверстия без специальной крышечки. Грудина с 2 парами
вырезов. Нижняя гортань трахеального типа, без трахео-бронхиаль-
ных мускулов, но с коротким processus vocalis. Щиткование плюсны
якзасппдного тина.
Два рода: Conopophaga и Corythopsis.
Семейство 9: Pteroptocliidae
Наземные, гл. обр. живущие в густых кустарниках, быстро
бегающие и скачущие птицы Южной Америки. Ноги длинные и
сильные и некоторыми видами употребляются для разгребания
наземного покрова в поисках пищи. Крылья очень короткие, округ-
лые, почти не употребляемые для передвижений. Питаются пре-
имущественно насекомыми, пауками п улитками. Гнездо обычно
помещается в коротких норках. Кладка из 2 белых япц. Череп
:)гптогнатпческпй. Грудина с двумя парами вырезок. Нижняя
гортань трахеального типа, без или с одной парой трахео-бропхпаль-
пых мускулов п с хорошо развитым processus vocalis, к концу кото-
рого прикрепляется musculus sterno trachealis. Носовые отверстия
прикрыты большой крышечкой (operculum). Щиткование плюсны
такзаспидного типа.
Различают около 30 форм, относимых к родам: Pterop tachos,
S су taiopus, Eugr^lla, Scdorchilus, Aerop ter nis п др.
1 Род Rupicola нередко выделяется в особое семейство Rupicolidae.
44
Семейство 10: Formicariidae. — Муравьеловки
Мелкие, гл. обр. питающиеся насекомыми (особенно муравьями),
преимущественно лесные птицы Южной Америки, известные постоян-
ным спутничеством походным колоннам муравьев и термитов. Дер-
жатся частью на* деревьях, частью на земле, гнездо строят открытое,
чашкообразное, на земле или в кустах. Яйца б. ч. пятнистые. Строе-
ние носовых костей шпзоринального типа. Нижняя гортань трахе-
ального типа, с одной парой трахео-бронхиальных мускулов (у неко-
торых редуцированы) и у некоторых родов с развитым processus
vocalis (напр. Chamaeza). Покров плюсны такзаспидного типа.
Крыло коротко и сильно округлено. Своеобразна значительная
пушистость и длина оперения надхвостья.
Обширная группа видов (около 350) многих родов: Taraba,
Grailaria, Thamnophilus, Мугтоthernia, Hypocnemis, Chamaeza, For-
micarius и др.
Семейство 11: Dendrocolaptidae. — Древолазы
Обширная группа видов различного экологического облика,
то сходные с дятлами, то сходные с дроздами, жаворонками, камен-
ками, пищухами и др. Распространены по Южной и Центральной
Америке. Сообразно этому разнообразию гнездятся то в дуплах,
то в закрытых гнездах, то в небольших норках в земле; всегда однако
гнездо закрытого типа и яйца постоянно одноцветные, белые,
иногда зеленоватые или голубоватые. Пища состоит преимущественно
пз насекомых, пауков и т. п.^и иногда ягод. Нижняя гортань трахе-
ального типа, с двумя парами трахео-бронхиальных мускулов
и хорошо развитым processus vocalis. Щиткование плюсны эттда-
спидного типа.
Известно около 700 форм пз родов: Geositla, Fumarins, С indo des,
Synallaxis, Philydor и мп. др.1
ПОДОТРЯД 3: MEXURAE. — ПРИМИТИВНЫЕ ВОРОБЬИНЫЕ
Небольшая группа форм, свойственная Австралии. Нижняя
гортань диакромиоидного типа. Покров плюсны такзаспидного
типа, с косым положением боковых чешуй. Оба входящие в подотряд
семейства по ряду признаков (строение кишечника, плечевых муску-
лов и др.) близки к дятлообразным и ведут свое происхождение
повидимому от- группы, общей с этим отрядом. В отношении к Oscines
Мепигае занимает не только обособленное, но в морфологическом
отношении^!! глубоко примитивное положение.
Семейство 1: Atrichornitliidae
Мелкие,^кустарниковые, внешне напоминающие крупных камы-
шевок, почти нелетающие, но хорошо лазающие и бегающие птицы
Австралии, только с двумя парами голосовых мускулов нижней гор-
1 Некоторые авторы разделяют это семейство на два: Furnariidae (с подсе-
мействами: Furnariinae, Synallaxinae, Margarornithinae, Phzlydorinae и Scle-
rurinae) п семейство Dendrocolaptidae собственно.
485
тайп п редуцированными ключицами (сходно с Capitonides). Крылья
очень короткие, круглые, с 10 большими маховыми. Рулевых перьев
12. Щелевидные носовые отверстия прикрыты крышечкой.
Известно только два вида одного рода Atrichornis (= Atrichia).
Семейство 2: Menuridae.—Птицы-лиры
Внешне напоминающие фазанов, с хвостом в форме лиры и силь-
ными гребущими землю (как куры) ногами, лесные птицы Австралии.
•Обитают участки с густым подсе-
Рис. 298. Птица-лира — Menura supcrba
Davies. По Evans.
дом и зарослями папоротников.
Питаются насекомыми, червями,
улитками и отчасти семенами,
собирая их из разрываемой нога-
ми мертвой лесной подстилки и
употребляя крылья только для
редких коротких взлетов или под-
держания равновесия. Подобно
некоторым куриным, самцы птиц-
лир устраивают своеобразные брач-
ные игры на небольших точках
и замечательны исключительной.
способностью подражания в криках
п пении. Самки строят на земле
или в разветвлении ветвей и ство-
лов низко на дереве большое гнез-
до, с крышей и боковым входом.
КлаД^а состоит только из 1 круп-
ного яйца мелкопятнистой (по
светлому фону) окраски. Инкуба-
ция длится 35—40 дней и осуще-
ствляется только самкой. Птенцы
покрыты очень густым и длинным
серым или черным пухом (ргае-
реппае) и развиваются крайне
медленно.
Череп неполно эгитогнатиче-
ский; processus palatini Maxillae
не окостеневают; сошник спереди
закруглен. Грудина с очень малень-
кой и неглубокой боковой вырезкой. Вилочка развита, но без hypo-
cleidium. Нижняя гортань с тремя парами трахео-бронхиальных
мускулов. Слепые кишки хотя сильно редуцированы, повидимому
все же функционируют. Крыло с 10 1 большими и 10 малыми
маховыми. Рулевых перьев 16.
Один род Menura с двумя видами (рис. 298).
ПОДОТРЯД 4: OSCINES. — ПЕВЧИЕ ВОРОБЬИНЫЕ
Основная масса воробьиных птиц, весьма разнообразная по
экологии и внешнему виду. В этой группе насчитывается около
4000 видов, подразделяемых почти на 50 семейств. Обтцсм особеп-
486
ностьто их является трахео-бропхиальный тип гортани с дпакромио-
идным прикреплением пяти, семи и больше пар голосовых трахео-
бронхиальных мышц, 3 что является условием развития в этой
группе, как правило, весьма богатого и разнообразного голоса
с появлением многих первоклассных певцов. Нижние кольцы тра-
хеит срастаются в костный барабан. Покров плюсны преимущественно
•окреатного типа (редко такзаспидного типа у отдельных форм
Muscicapidae, Alaudidae и Laniidae).
Анатомическая характеристика семейств затруднительна вслед-
ствие сравнительного однообразия строения группы и гл. обр.
из-за ее неразработанности. Вследствие этого в настоящем случае
приходится ограничиваться только внешне-морфологическими приз-
наками. Выяснение происхождения группы, по мнению Гадова,
следует ожидать от изучения австралийских воробьиных птиц, в
общем обладающих более низкой морфологической организацией,
но не от Мепигае, являющихся повидимому боковой ветвью
воробьинообразных.
В ископаемом состоянии известны уже с верхнего эоцена, откуда
описаны (из Франции): Palaegithalus cuvieri Gerv. (сравниваемая,
из современных, то с Sitta, то с Sylvia) и Laurillardia longirostris
Milne-Edwards (сходная с Promerops из Meliphagidae)\ однако более
детальное положение обеих этих форм среди Passeriformes спорно,
так как они могут относиться и к Clamalores. Более определенно
положение Palaeospiza из верхнего миоцена Колорадо (Северной
Америки) (см. ниже). Рецептные роды Motacilla и Lanius появляются
уже с верхнего олигоцена, в миоцене появляются роды F ringilia
п Corvus, а с плиоцена Alauda, Sitta, Anthus и мн. др.
Семейство 1: Alaudidae. — Жаворонки
Мелкие, наземные, быстро бегающие, преимущественно степные
и пустынные птицы Старого Света и Северной Америки. Гнездят-
ся па земле, устраивая чашкообразиое гнездо в углублении почвы.
Яйца густо пятнистые, с коричневыми, серыми или краснова-
тыми пестринками по светлому фону. Самцам свойственна песня на
лету, причем птицы часто задерживаются с песней в воздухе на од-
ном месте трепещущими движениями крыльев, иногда звучно уда-
ряя их друг о друга (Mirafra). Питаются насекомыми, семенами
и зелеными частями растений.
Морфологически характеризуются такзаеппдным типом щитко-
вапия и латнплантарным типом строения плюсны (округленным
задним краем плюсны), с относительно короткими пальцами, но
длинным и прямым задним когтем. Клюв тонкий, иногда почти ко-
нический. Крыло острое с 10 большими маховыми. Общая окраска
однообразно пестрая. Только одна линька в течение года. Интересно
сохранение у жаворонков в нижней гортани рудиментарного pes-
1 Однако у ряда форм, относимых к Oscines, обнаружено весьма упрощен-
ное строение нижней гортани, напр. южноафриканской Sphenoeacus (Sylvii-
dae), австралийских Grallina (Тimaliidae) и Plilotis (Meliphagidae}, новозеланд-
ской Prosthemadera (Meliphagidae).
487
stilus, единственно во всем подотряде. Центр распространения
находится в Африке. Различают около 26 родов: Certhilauda,
Alaemon, Chersophilus, Eremophila, Mirajra, Ammomanes, Galerida,
Lullula, Alauda, Calandrella, Saxilauda, Melanocorypha и др.
Обильно представлены в фауне СССР. Наиболее распростра-
нены: полевой жаворонок — Alauda arvensis L., лесной жаворонок —
Lullula arborea (L.) и хохлатый жаворонок — Galerida crista la (L.).
С областью зоны степей и пустынь связаны большая п малая калан-
дры — Melanocorypha calandra (L.) и М. bimaculyta (Menetr.), черный
жаворонок — Saxilauda yeltoniensis (Forst.), короткопалый и малый
жаворонки — Calandrella brachydactyla (Leisl.) п C. minor (Cab.)
и нек. др. Области северных и альпийских тундр свойствен Егето-
phila alpestris (L.).
Семейство 2: Palaeospizidae
Единственный пока известный ископаемый вид Palaeospiza bella
Allen пз верхнего миоцена Колорадо (Северная Америка), с призна-
ками, частью южноамериканских Colingidae, частью представителей
Oscines. По мнению Ветмора (Wetmore, 1925), занимает промежуточ-
ное положение между кричащими воробьиными и воробьиными
с острым краем задней стороны плюсны (акутиплаптарный тип
плюсны), оставляя в стороне жаворонков.
Семейство 3: llirundinidae. — Ласточки
Мелкие, острокрылые, прекрасно летающие птицы, распростра-
ненные почти по всей земле. Питаются, подобно стрижам, ловя
насекомых в воздухе и неустанно гоняясь за ними преимущественно
в открытых местах. Гнезда устраиваются пз грязевых комочков,
соломинок, перьев и т. и., слепляемых слюной, иногда почти цели-
ком из грязи, и помещаются на скалах, деревьях, постройках чело-
века. Некоторые роют норы в откосах пли гнездятся в дуплах и
щелях скал. Часто колониальны. Яйца белые или с красноватыми и
бурыми пятнами по белому фону. Самцу свойственна щебечущая песня.
Морфологически характеризуются тонким вытянутым туловищем,
уплощенной головой, коротким широким с большим разрезом клю-
вом, очень маленькими слабыми ножками, вилообразным пли выре-
занным хвостом. Больших маховых 9. Только одна линька в году.
Анатомически интересны крупные размеры слюнных желез, осо-
бенно в брачный период. Известно около 125 форм, относимых
к 13 родам, с центром распространения в Африке. Роды: Ripariaу
Ptyonoprogne, Hirundo, Progne, Delichon и др.
В СССР гл. обр. распространены: деревенская ласточка пли
косатка — Hirundo rustica (L.), городская ласточка — Delichon urbica
L. и береговая ласточка — Riparia riparia (L.)
Семейство 4: Campephagidae. — Лпчинкоеды
Мелкие, обычно длиннохвостые, по повадкам напоминающие то>
мухоловок, то сорокопутов, птицы, преимущественно тропических
стран восточного полушария. Питаются насекомыми, собирая их
с ветвей или листьев пли взлетая за ними в воздух, как мухоловки.
Гнезда устраиваются из тончайших веточек, лишайника и паутины
488
в виде маленьких чаш, помещаемых высоко на ветвях деревьев,.
Яйца пестро пятнистые по белому или цветному фону.
Морфологически своеобразно строение стержней перьев над-
хвостья, в базальной части весьма жестких и затем вдруг стано-
вящихся мягкими. Клюв обычно уплощенный, нередко узкий,
с слабыми щетинками. Ножки маленькие, короткие, хвост преиму-
щественно ступепчат или закруглен. 2-е—5-е большие маховые
паи длинные. Известно около 200 форм, относимых к родам, с центром
распространения в Индо-Австралийской области. Роды: Graucalus,
Campephaga, Lalage, Syntmorphus, Pericrocotus и др.
В СССР только 1 вид— Pericrocotus roseus divaricatus RaffI., захо-
дящий к нам в Уссурийский и Амурский края пз Китая и Японии .
Семейство 5: Biachypodidae (Pycnonotidae). — Коротконогие дрозды
(бюльбюли)
Величиной с дроздов, по повадкам напоминающие славок, лес-
ные птицы тропиков п субтропиков Старого Света. Преимущественно
держась в кронах деревьев, реже в кустарниках, коротконогие
дрозды или бюльбюли питаются гл. обр. насекомыми и ягодами.
Чашкообразное гнездо помещается среди веток деревьев. Яйца б. ч.
пятнистые, красноватыми пестринками по белому фону. Прекрасные
певцы, нередко не уступающие по красоте песни дроздам.
Морфологически для бюльбюлей характерно мягкое, несколько'
рыхлое, более.длинное на надхвостье оперение, с обильными, рас-
сеянными, нитчатыми перьями, особенно на затылке и спине. Плюсны
очень коротки, крылья закруглены, с 4-м—6-м махами самыми
длинными, хвост небольшой, прямой или округленный. Известно'
около 260 форм,.относимых к 30 родам, с центром распространения
в Индийской области. Роды: Chloropsis, Hypsipetes, Hemixus, Crini-
ger, Trachycomus, Bleda, Jxocincla, Joie, Rubigula, Pycnonotus, Irena,
Microscelis и др.
Семейство 6: Motacillidae.—Трясогузки
Мелкие, подвижные, наземные, быстро бегающие птички, дер-
жащиеся гл. обр. на лугах, в степи, на побережьях рек и озер почти
всей земли. Питаются мелкими насекомыми, преимущественно
склевывая их с земли и стеблей растений, п иногда мягкими семе-
нами. Гнездо устраивается в виде чашки пз стеблей и волос или
в углублении земли или каких-нибудь пустотах среди камней, под
крышами зданий и т. д, Яйца преимущественно мелко и густо пят-
нистые с бурыми, коричневатыми и красноватыми пестринами.
Некоторым родам свойственна простенькая песня на крыльях в воз-
духе, другие поют сидя на кустах и возвышениях.
Морфологически характерны: тонкая форма тела, острые кры-
лья с 9 большими маховыми, тонкий клюв, слабо изогнутый, длин-
ный коготь заднего пальца. В отличие от напоминающих их жаво-
ронков — плюсна акутиплантарного типа. Хвост обычно удлиненный,
tia конце почти прямой. Группа состоит примерно из 120 форм, разделяе-
мых на 8 родов, с центром распространения в Евразии и Африке. Роды:
Dendronanthus, Calobates, Budytes, Motacilla, Anthus, Macronyx и др.
489
В СССР широко распространены: белая трясогузка — Motacilla
adba L., желтая трясогузка — Budytes Hava L., лесной конек— Ап-
.Ihus trivialis L., луговой конек — Anthus pratensis L., полевой конек —
Anthus campestris (L.) и нек. др. В горных районах широко распро-
странены горная трясогузка —• Calobates cinerea TunsI. в горный
конек — Anthus spinoletta L.
Семейство 7: Cinclidae. — Оляпки
Мелкие, по форме и повадками напоминающие крапивников,
птицы быстрых горных ручьев и речек горных стран Евразии и
системы Анд запада Америки. Отличаются способностью нырять
под воду мелких речек п бегать здесь по дну в поисках мелких вод-
ных насекомых, их личинок и некоторых беспозвоночных. Объеми-
стые гнезда из мха и листьев в виде шара с боковым входом устраи-
ваются в камнях, пещерах и скалах,- предпочтительно поблизости
водопадов. Яйца белые с красноватыми пестринками.
Морфологически характеризуются сильными, с длинными плюс-
нами, ногами, узким прямым клювом с крышечкой над ноздрями,
коротким вздернутым кверху хвостом, коротким округлым крылом.
Аптерии покрыты густым длинным пухом. Известно около 20 форм
единственного рода Cinclus. В СССР распространены: обыкновенная
оляпка — Cinclus cinclus L. и черная оляпка — Cinclus pallasi Temm.
Семейство 8: Troglodytidae. —Крапивники
Очень мелкие, крайне подвижные птички, держащиеся всегда
в густой чаще, особенно в кустарниках. Питаются мелкими насе-
Гис. 299. Крапивник — Troglodytes trog-
lodytes L. По Evans.
комыми, склевываемыми с расти-
тельности и земли. Гнездо устраи-
вается из мха, листьев и стеблей
растений в виде шара с боковым
входом и помещается обычно в ча-
ще ветвей. Яйца белые с красно-
ватыми пестринами. Оба пола
отличаются неутомимым, крайне
звонким пением.
Морфологически характерны
тупые округлые крылья с наиболее
длинными 3-м—5-м большими ма-
ховыми, обычно короткий, из
12 рулевых, закладываемых вверх
(на спину), хвост, сравнительно
длинные плюсны. Особенно харак-
терна общая коричневая окраска
перьев с черными и бурыми попе-
речными полосками и пестринами.
Известно много форм, относимых
к 18 родам; все, кроме 3 родов, распространены в Северной п
Южной Америке. Роды в Евразии: Troglodytes, Pnoepyga и Spe-
4.И)
laeornis. П.рпшщаются роды: М icrocerculus, Henicorhina, Troglodytes,
Leucole pis, Cistothorus и др. (рис. 299).
В СССР распространен единственный вид: обыкновенный кра-
пивник — Troglodytes troglodytes (L.).
Семейство 9: Mimidae. — Пересмешники
Напоминающие дроздов величиной и посадкой, древесные и
кустарные птицы Южной и Северной Америки. Питаются насеко-
мыми и их личинками, собираемыми с веток и на земле. Гнездятся
устраивая открытое чашкообразное гнездо. Яйца напоминают яйца
дроздов, голубые или беловатые, одноцветные или покрытые пестрин-
ками. Некоторые виды отличаются прекрасным пением и удивитель-
ной способностью подражать другим птицам и различным звукам.
Морфологически характеризуются тупым крылом из 10 больших
маховых, с 3-м—5-м маховыми самыми длинными, сильными плюс-
нами, спереди покрытыми поперечными табличками, и хвостом из
J2 рулевых.
Известно много форм, относимых к 13 родам. Роды: Donacobius,
Me.lanotis, Taxostoma, Mimus, Cinclocerthia, Phamphocinclus и др.
Семейство 10: Turdidae. —Дрозды
Преимущественно мелкие, древесные и наземные, передвигающиеся
по земле скачками, птицы, распространенные почти по всей земле.
Весьма разнообразны по экологии, частью являясь лесными пти-
цами, частью кустарными, скалистыми, степными и пустынными.
Пища часто разыскивается иа земле и состоит преимущественно из
насекомых и их личинок и, временами, различных ягод и плодов.
Гнездятся в большинстве строя открытое чашкообразное гнездо,
помещаемое на земле в кустах или на деревьях; другие формы гнез-
дятся в дуплах, пустотах скал, постройках и даже норах. Яйца раз-
нообразны. Самцы в большинстве прекрасно поют.
Из морфологических признаков характерно для большинства
родов слияние щитков плюсны, за исключением 2—3, расположенных
выше основания пальцев. Передние пальцы, особенно внутренний
и средний, глубоко расщеплены. Крыло заостренное, длиннее хвоста,
с 1-м маховым очень укороченным и 2-м длиннее 6-го махового.
Молодые птицы носят пестрый наряд, отличный от взрослого. Эта
группа весьма обильна видами и родами. Различают роды: Turdus,
Oenanthe, Saxicola, Cercomela, Myiophonus, Monticola, Diplootocus,
Chaimarrornis, Sicilia, Grandala, Erythropygia, Erithacus, Phoeni-
curus, Rhyacornis, Cyanosylvia, Icoturiis, Calliope, Tarsiger, Larvivora,
Pseudaedon, Luscinia и др.
В СССР наиболее распространены: певчий дрозд—Tardus ericeloruin
Furt., черный дрозд — Merula merula L., каменка — Oenanthe oenan-
the (L.), луговой чекан — Saxicola rubetra (L.), горихвостка обык-
новенная — Phoenicurus phoenicurus (L.), зорянка — Erithacus rube-
cula L., варакушка — Cyanosylvia svecica (L.), соловей западный —
Luscinia megarhynchos Brehm и соловей восточный — Luscinia luscinia
49 L
(L.). Из представителен других родов в СССР встречаются: каменный
дрозд — Monticola saxatilis (L.), синяя птица — Myiophonus caeru-
leus (Scop.), синехвостка— Tarsiger cyanurus (Pall.), красношейка —
Calliope calliope Pall, и мп. др.
Семейство II: Zeledoniidae. — Водяные дрозды
Единственный вид Zeleclonia coronata Rigw. в горах Коста Piikiu
Семейство 12: Timaliidae. — Тималпи
Мелкие, величиной с славок и дроздов, кустарные и наземные-
птицы, распространенные в тропиках и субтропиках Старого Света.
Питаются различными насекомыми, пауками, улитками и т. тг.,
собирая их препмуществеи-
Рпс. 300. Тпмалшг — Timalia maculata Temm. По
Evans.
ио на земле, частью на вет-
вях п листьях. Гнездятся
строя открытое чашкооб-
разное гнездо. Япца одно-
цветно голубые или белые
или покрытые разнообраз-
ными пестринками.
Морфологически харак-
теризуются б. ч. коротки-
ми тупыми крыльями, с 10
большими маховыми, на-
личием 12 рулевых, вну-
тренний и задний пальцы
йог равной величины. Ноз-
дри часто прикрыты пе-
рышками п щетинкамиг
хотя не скрыты ими со-
вершенно. В углу клюва
хорошо развиты щетинки.
Окраска птенцов сходна
с окраской взрослой сам-
ки.
Весьма многочисленная видами и родами группа. Различают
роды: Timalia, Orthonyx, Pycnoptilus, Eniciiriis, Crateropus, Ponia-
torhinus, Dryonastes, Turdinus, Leiothrix, Garrulax и др.
В Таджикистан, на самый юг СССР к нам заходит Garrulax
lineatum (Vig.), относимая в эту группу.
Семейство 13: Sylvildae. — Славковые
Мелкие, кустарные и древесные птички, распространенные почти
по всему Старому Свету, своей подвижностью и пением весьма
оживляющие наши леса и заросли. Питаются преимущественно
мелкими насекомыми и ягодами. Гнездо разнообразной постройки,
то открытое чашковпдное, то шарообразное с боковым входом,
492
помещаемое гл. обр. в кустах и иногда на дереве или земле. Яйца
разнообразны, обычно пятнисты.
Из морфологических признаков следует указать наличие 10 боль-
ших маховых, 10 или 12 рулевых, открытые, неприкрытые перышками
ноздри, обычно тонкий и узкий, б. м. шиловидный клюв. Полы
обычно окрашены одинаково, птенцы окрашены сходно с взрослой
самкой, ио обычно в более яркие цвета.
Весьма. многочисленная родами и видами группа. Различают
роды: Camaroplera, Macrosphenus, Hylia, Stiphrornis, Acanthiza,
Horeites, Cettia. Scotocerca, Prinia, Phragmaticola, Lusciniola, Eremo-
mela, Sylvia, Cislicola, Phylloscopus, Regulus, Hippolais, Acrocephalus,
Locustella и др.
В СССР весьма многочисленны и составляют основное население
.лесной и кустарной растительности; наиболее обыкновенные пеночки:
весничка — Phylloscopus trochilus (L.), теньковка — Phylloscopus
collybita (Vieill.); славки: садовая — Sylvia borin (Bodd.), Черно-
головка — S. atricapilla (L.), серая — S. communis Lath., зави-
рушка — S. curruca (L.); камышевки: дроздовидная — Acrocephalus
arundinaceus (L.), барсучек — A. schoenobaenus (L.) и мн. др. Из
представителей других родов упомянем: широкохвостую камы-
шевку — Cettia cetli (Магш.), обыкновенного сверчка — Locustella
naevia (Bodd.), толстоклювую камышевку — Phragmaticola aedon
(Pall.), пеночку-пересмешку — Hippolais icterina (Vieill.).
Семейство 11: Muscicapidae. — Мухоловки
Мелкие древесные птички Старого Света, населяющие обычно
леса, перелески и сады и добывающие насекомых в воздухе, взле-
тая за ними коротким характерным полетом с насеста и вновь воз-
вращаясь на старое место. Преимущественно держатся в кронах
деревьев и любят опушки и прогалины; помимо указанного способа,
собирают также насекомых с листьев и ветвей, что однако менее
характерно. Гнездятся устраивая гнезда на деревьях открыто среди
ветвей, преимущественно у их основания плп в дуплах и полудуплах.
Гнездо строится из мелких веточек и стеблей растений, выстилаемых
перьями и растительным пухом. Яйца одноцветны пли пятнисты
с красноватыми и коричневатыми пестринками по голубому или
белому фону.
Из морфологических черт наиболее характерными чертами явля-
ются: обычно уплощенный сверху вниз широкий клюв с более или
менее обильными и сильными щетинками по краям клюва. Ножки
маленькие и относительно слабые. Крыло тупое, с 10 большими
маховыми, из которых 3-е и 4-е или 4-е и 5-е наиболее длинны, а
1-е короче половины самого длинного. Птенцовый наряд резко
пятнистый.
Своими менее типичными формами семейство тесно соприкасается
с славками Sylviidae, а отчасти и с сорокопутами Laniidae, так что
положение многих родов пока является спорным.
В семействе различаются до 750 форм и почти 90 родов, из кото-
рых приведем роды: Muscicapa, Microeca, Alseonax, Hemic lielido п,
Hedymela, Polyomyias, Zanthopygia, Siphia.
493
Рис. 301. Райская мухоловка — Tschilrea pa-
radisi L. По Evans.
В СССР широко распространены: серая мухоловка — Muscicapa.
striata (Pall.), пеструшка — Hedymela hypoleuca PalL, малая мухо-
ловка — Siphia parva Bechst. В Сибири широко распространены г
сибирская мухоловка — Hemichelidon sibirica Gm., ширококлювая
мухоловка — Arizelomyia daurica Pall, и др. Замечательны по своей;
красоте заходящие в южные районы Союза из субтропиков райская
мухоловка — Tschitrea paradisi
(L.) (рис. 301), синяя мухолов-
ка — Cyanoptila cyanomelaena
Temm. и белобровая мухоловка
Zanthopygia zanthopygia Hay.
Семейство 15: Pachycephali-
dae. — Толстоголовые copc-
копуты
Преимущественно мелкие.,
напоминающие обликом то со-
рокопутов, то мухоловок, а по
поведению отчасти синиц и дят-
лов, птицы Восточной п Австра-
лийской области. Как правило,
древесные и кустарные птицы,
питающиеся насекомыми, иногда
мелкими плодами. Некоторые
весьма искусно отделяют клю-
вом кору деревьев, разыскивая
под ней насекомых. Гнезда от-
крытые, чашкообразные, строят-
ся среди ветвей. Яйца напоми-
нают яйца сорокопутов.
Из внешних признаков ха-
рактерен сильный с крючком на
конце клюв, с носовыми отверстиями, лежащими в ямках, с мягкими,
краями и частично обросшими перышками. Больших маховых 10,
из них 1-е в две трети длины 2-го. Хвост в две трети длины крыла,
обычно закругленный или вырезанный. Половой диморфизм выра-
жен, в оперении характерно наличие желтой окраски.
Известно около 120 форм, относимых к родам: Pachycephala,
Pitohui, Eopsaltria, Hyloterpe, Pachycephalopfis, Pachycare и др.
Семейство 16: Prionopidae. — Лесные сорокопуты
Мелкие, напоминающие обликом сорокопутов, птицы Африки
и Передней Азии. Обычно древесные и кустарные формы, с преоб-
ладающей темной окраской оперения, питающиеся насекомыми,
улитками и плодами. Гнездо рыхлое, открытое, среди веток дерева
или кустов, или в дуплах и щелях скал. Яйца обычно пятнистые.
Внешне характеризуются сильным на конце загнутым клювом, с вы-
резкой на режущих краях надклювья перед вершиной. Бока плю-
494
сны покрыты многими мелкими щитками, несливающимися в общие"
пластинки. В оперении обычны коричневый, черный, серый и бе-
лый цвета.
Известно около 90 форм, относимых к 5 родам: Nilaus, Еигосе-
phalus, Prionops.) Sigmodus, Hypocolius.
Семейство 17: Vangidae
Небольшое семейство мелких и средней величины птиц, обли-
ком и поведением напоминающее сорокопутов, но св клювом разно-
образного строения. Преимущественно кустарные, отчасти назем-
ные птицы, свойственные только Мадагаскару.
Из морфологических черт характерны: покров плюсны из боко-
вых щитков; крыло короткое, с 4—5 или 4—6 маховыми наиболее
длинными; оперение надхвостья отличается особенной длиной п
мягкостью. Принимаются роды: Vanga, Xenopirostris, Aerocharisr
Oriolia, Ar lamia, Cyanolanius и др.
Семейство 18: Iraniidao. — Сорокопуты
Мелкие древесные и кустарные птицы, распространенные почти
по всей земле, исключая Южную Америку. Населяют преимуще-
ственно лесные опушки,
редколесье, кустарные за-
росли ит. п., напоминая
по поведению мухоловок.
Пищу — преимущественно
различных насекомых —
добывают взлетая и ловя
в воздухе или подбирая
с земли. Крупные виды
ловят также мелких гры-
зунов, птичек, таскают
птенцов. Характерна спо-
собность некоторых видов
накалывать добычу па ко-
лючки растений, запасая
ее впрок. Открытые чашко-
образные гнезда помеща-
ются на деревьях или ку-
старниках. Яйца пятни-
стые, разнообразной ок-
Рпс. 302. Большой сорокопут — Lantus ехсиЪПог L.
Ho Evans.
раски.
Из морфологических черт характерна сильная форма клюваг
несколько сжатая с боков, с крючком на верхней челюсти и глубо-
кой выемкой перед ним. Плюсны довольно коротки, одетые спереди
поперечными табличками. Крыло довольно тупое с 10 большими
маховыми. Оперение мягкое, у крупных форм серое с черным и
белым, у мелких разноцветной, иногда весьма пестрой окраски.
Известно около 180 форм, относимых к 20—25 родам. Роды:
Lanins, Urolestes, Мalaconolus, Laniarius, Chaiinonotiis ii’np. (рис. 302)fc.
495
В СССР группа представлена широко распространенными: боль-
шим сорокопутом—Lanins excubilor L., жуланом — Lanins col-
Inrio L., рыжехвостым сорокопутом — Lanins cristains L. В южные
районы СССР заходят: красноголовый сорокопут—Lanins sena-
tor L., длиннохвостый сорокопут —Lanins cry Uironolus (Vig.) и др.
Семейство 19. Artamidae. — Ласточковые сорокопуты
Мелкие, внешностью и поведением напоминающие ласточек,
но родственные сорокопутам, пндо-австралпйскпе птицы. Преи-
мущественно обитают степные равнины, луга, берега рек и т. п.
Питаются насекомыми, гоняясь за ними на лету, подобно ласточкам.
Вне брачного периода обычно держатся большими стаями. Чашко-
образное гнездо устраивается в дуплах плп на. ветвях деревьев из
рыхло сплетенных стеблей, мелких корешков, листочков и мха.
Япца сходны по окраске с яйцами сорокопутов. В кладке 2—4 яйца.
Из морфологических черт характерны длинные, доходящие до
конца хвоста пли длиннее, крылья с 9-J-1 большими маховыми, из
которых 2-е и 3-е наиболее длинны. Хвост короткий и обычно прямой
плп вырезанный. Ножки очень малы и слабы. Клюв короткий,
-сжатый с боков к вершине, с немногими щетинками у основания.
Характерно наличие маленьких пятен пудреток по бокам тела,
на надхвостье и в пахах. Известно около 20 видов единственного
рода Artanins.
Семейство 20: Pseudochelidonidae
Мелкие, внешностью напоминающие ласточек и Artamidae,
быстро летающие птицы западной Центральной Африки, населяющие
гл. обр. берега рек. Питаются насекомыми, подобно ласточкам,
добываемым на лету. От сходных Artamidae отличаются отсутствием
пудреток, очень коротким и широким у основания клювом и бле-
стящей зелеповато-черной окраской с красным клювом и ногами.
Только один вид: Psendochelidon eurystomina Hartl, с подвидами.
Семейство 21: Bombycillidae. — Свиристели
Мелкие, несколько напоминающие поведением мухоловок, лес-
ные птицы Евразии и Северной и Центральной Америки. Держатся
преимущественно в кронах деревьев, летом питаются насекомьрии,
в остальное время преимущественно ягодами, в поисках которых
широко кочуют. Гнездятся на деревьях, устраивая чашкообразное
гнездо из мелких веточек, стеблей и лишайников. Яйца пятнистые.
Морфологически характеризуются коротким широким клювом,
с носовыми отверстиями, частью прикрытыми мелкими перышками
и короткими щетинками; ноги короткие, крыло с 10 маховыми.
Хвост относительно короткий, прямой.
Это семейство является сравнительно сборной группой форм,
частью с сомнительным систематическим положением. Различают 10—
12 форм, относимых к 5 родам: Bombycilla, Phainoptila, Phainope-
pla, Ptilogonys, Dnlns.
496
В СССР распространены лишь два вида, свойственные области
хвойных лесов: обыкндвенный свиристель — Bombycilla garrulus (L.)
п японский свиристель — Bombycilla japonica (Sieb.).
Семейство 22: Sturnidae. —Скворцы
Мелкие, подвижные, наземно-древесные, бегающие обычно по
земле в поисках пищи, нередко ярко металлически окрашенные,
птицы всего Старого Света. Питаются насекомыми, их личинками,
червями, ягодами. Гнездятся преимущественно в дуплах, скалах,
строят гнездо из стеблей трав и кладут от 4 до 7 обычно одноцветно
голубых, иногда испещренных красноватыми пятнами яиц. Преиму-
щественно птицы лесостепи, опушек лесов, приречной раститель-
ности.
Из морфологических черт характерна короткость 1-го махового,
не достигающего половины длины 2-го махового и обычно имеющего
совсем маленькие размеры. В крыле 10 маховых с наиболее длин-
ными 2-м—4-м маховыми. Клюв обычно прямой, заостренный,
без щетинок у основания; у некоторых родов однако нескоцько
вздутый, вороньей формы. Плюсны сильные.
Известно около 200 видов, относимых к 37 родам с центром распро-
странения в Индийской и Эфиопской областях. Роды: Cosmopsarus,
Sturnus, Spodiopsar, Sturnopastor, Creatophora, Pastor, Sturnia,
Gracupica, Acridotheres и др.
В СССР широко распространен обыкновенный скворец — Stur-
nus vulgaris L. и в области степей и пустынь розовый скворец —
Pastor roseus Pall. На окрайнах проникают на нашу территорию:
серый скворец — Spodiopsar cineraceus (Temm.), малый скворзц —
Sturnia sturnina (Pall.) и индийская майна — Acridotheres tristis L.
Семейство 23: Oriolidae. — Иволги
Мелкие, величиной с скворца или дрозда, ярко окрашенные
лесные птицы Старого Света. Держатся всегда в густых кронах
деревьев, питаются насекомыми, преимущественно гусеницами, и
отчасти плодами. Висячее, в виде чаши, гнездо помещается в раз-
вилке ветвей дерева, прикрепляясь к ним лубяными волокнами и
гибкими травинками. Яйца розовато-белые с красновато-коричне-
выми пестринами. Птицы издают прекрасные флейтовые крики.
Морфологически характеризуются сравнительно тонким вытя-
нутым клювом, с открытыми ноздрями и без.щетинок у основания;
крыло с 10 большими маховыми, с 3-м—5-м наиболее длинными;
плюсна относительно крупная; хвост прямой; в оперении преоб-
ладает ярко-желтая и черная окраска.
Известно около 70 форм, распределяемых в ряд родов с центром
распространения в области Малайского архипелага. Роды: Oriolus.
Mimeta, Sphecotheres и др.
В СССР только представители рода Oriolus*. обыкновенная ивол-
га — Oriolus oriolus (L.), туркестанская иволга — Oriolus kundoo
Sykes и индийская иволга — Oriolus indicus'iQv^.
32 Орнитология—1909
437
Семейство 24: Dicruridae. — Дронго
Величиной с дрозда, древесные, по повадкам напоминающие
крупных мухоловок, птицы тропиков и субтропиков Старого Света.
Держатся в кронах деревьев.
Рис. 303. Дронго — Dissemurvs paradiseus L.
По Evans.
временами взлетая за пролета-
ющими насекомыми. Подобно
щуркам, ловят пчел и ос, загла-
тывая их вместе с жалом. Из-
редка питаются плодами. Гнез-
до устраивается чашкообразно,
висячим между развилками вет-
вей. Яйца пятнистые, напоми-
нают яйца сорокопутов. Сам-
цам свойственна довольно слож-
ная песнь и способность под-
ражания.
Морфологическими особен-
ностями являются: сильный,
напоминающий по форме ворон,
клюв, обычно с густыми, напра-
вленными вперед щетинками v
основания; тупое крыло с
4-м—5-м маховыми самыми длин-
ными; короткие плюсны; хвост
из 10 рулевых преимуществен-
но вилкообразной или лирооб-
разной формы; однотонная чер-
ная, реже серая, несколько блестящая окраска. Известно около
70 форм, относимых к родам: Dicrurus, Chaetorhynchus, Chibia. Edo-
lius, Dicranostreptus, Ehringa и др. (рис. 303).
Семейство 25: Cracticidae
Величиной с дроздов, внешне напоминающие сорокопутов и
врановых, древесные и наземные птицы Австралии и Папуасии.
Из морфологических черт характерен длинный прямой и силь-
ный клюв с свободными щелеобразными носовыми отверстиями.
Первое маховое сильно укорочено, 3-е и 4-е пли 4-е и 5-е большие
маховые самые длинные. Общая окраска оперения напоминает
ворон; птенцовый наряд коричневый с поперечным рисунком.
Известно немного форм, относимых к родам: Cracticus, Strepera.
Pityriasis, Gymnorhina и др.
Семейство 26: Corvidae. — Врановые
Вариирующие по величине от размеров скворца до ворона, частью
наземные, частью древесные, преимущественно всеядные птицы,
распространенные почти по всей земле. По экологии весьма измен-
чивы. Древесные формы передвигаются преимущественно прыжками
498
и питаются часто плодами, ягодами и крупными семенами; назем-
ные шагают или бегают, собирая как животный, так и растительный
корм с земли. Гнездятся обычно на деревьях, иногда на скалах,
постройках и в дуплах, устраивая массивное гнездо из ветвей, стеб-
лей и мха, иногда с крышей сверху. Некоторые виды колониальны.
Яйца обычно пятнистые, с бурыми пестринками по зеленоватому
фону. В психическом отношении одни из наиболее развитых птиц.
Морфологически характеризуются сильным, слабо изогнутым
или прямым клювом с густо оперенными ноздрями; уздечка опере-
на щетинкообразными перьями; плюсны сильные; крыло тупое,
иногда округлое с первым маховым длиннее кроющих. Хвост прямой.
Известно около 300 форм, распределяемых почти в 40 родов.
Роды: Corvus, Gymnocorax, Corvultur, Coloeus, Pica, Dendrocitta,
Aphelocoma, Cyanocitta, Cyanopica, Cracles, Garrulus, Lalocitta, Cyano-
cephalus, Nucifraga, Pyrrhocorax, Podoces и др.
В СССР широко распространены: ворона серая и черная —
Corvus cornix L. и С. corone L., ворон — Corvus corax L., грач —
Trypanocorax jrugilegus L., галка — Coloeus monedula (L.), сорока —
Pica pica (L.), кедровка — Nicijraga caryocathacles (L.), сойка —
Garrulus glandarius (L.), кукша — Cractes injaustes (L.). Из интерес-
ных форм, но редких, следует отметить также: голубую сороку —
Cyanopica cyanus (Pall.), саксаульную сойку — Podoces pandtri
Fisch., клушицу — Pyrrhocorax pyrrhocorax (Ь.)-и альпийскую галку—
Pyrrhocorax graculus (L.).
Семейство 27: Para disei dae.—Райские птицы
Ярко окрашенные, весьма подвижные, древесные и наземные
птицы Новой Гвинеи, Австралии и Северных Молукских островов,
замечательные своеобразием своих брачных игр. По преимуществу
являются птицами тропического леса, разыскивающими пищу —
плоды, насекомых и их личинок — среди ветвей и листьев древесных
крон. Чашкообразное открытое гнездо устраивается среди ветвей
деревьев. Яйца, .по 2 в кладке, весьма разнообразны по окраске.
Характерен резкий половой диморфизм с очень яркой окраской
самцов и обильными украшающими перьями, придающими птицам
не только разнообразную окраску, но и изменчивую форму. Самцы
собираются в брачный период на ток в кроне какого-либо излюб-
ленного дерева, принимая здесь разнообразные позы и щеголяя
яркими красками и формами своего оперения и издавая громкие
звучные крики. Обычный же голос, каркающий и грубый, напоминает
ворон. Группа родов проводит брачные игры на земле, устраивая
«беседки» и украшая их и расположенные около них точки яркими
цветами и камешками (эти роды выделяют иногда в особое под-
семейство Ptilonorhynchinae).
По морфологическим признакам райские птицы тесно примыкают
к воронам, отличаясь от них гл. обр. своеобразием нарядов и окраски
у самцов. Уздечка однако не покрыта щетиновидными перышками,
как у ворон, а покрыта гладким, коротким, бархатистым оперением.
Крыло, как правило, с наиболее длинными 4-м и 5-м большими
*
499
маховыми. Украшающие перья самцы носят в течение 3/4 года. У неко-
торых родов трахея очень удлинена и проникает в области вилочки
под кожные покровы тела, располагаясь на груди в виде длинной
извитой петли.
Известно около 120 форм райских птиц, относимых к 33 родам.
Роды: Paradisea, Ptilonorhynchus, Chlamydera, Manucodia, Phony-
gammus, Parotia, Lophorina, Astrapia, Semioptera, Pteridophora,
Cicinnurus, Seleucides и др.
Семейство 28: Paradoxornithidae. — Камышевы© синицы
Мелкие (до величины дрозда)
мущественно тростниковым или
Рис. 304. Усатая синица—Panurus biarmicus
(L). По Evans.
подвижные, свойственные преи^
бамбуковым зарослям Евразии
птицы, искусно лазящие среди
густых растений. Питаются пре-
имущественно семенами и мел-
кими насекомыми.
Из морфологических призна-
ков характерны: высокий и у
многих форм короткий, сжатый
с боков клюв с сильно изогну-
тым надклювьем и волнообраз-
ным режущим краем клюва;
носовые отверстия прикрыты
короткими перышками; крыло
очень коротко и округло, 5-е—
7-е маховые самые длинные.
Известно около трех десят-
ков видов, относимых к 6—8
родам: Conostoma, Paradoxor-
nis, Suthora, Panurus и др. В
СССР по озерам степей ши-
роко распространена усатая
синица — Panurus biarmicus (L.)
(рис. 304). В Уссурийский край
заходит Suthora webbiana Gray.
Семейство 29: Paridae. — Синицы
Мелкие, весьма подвижные, преимущественно древесные птички,
распространенные почти по всей земле. Держатся главным образом
в кронах деревьев,, где,* лазая по ветвям и свободно подвешиваясь
даже к тонким веточкам, разыскивают корм, состоящий преиму-
щественно из насекомых, их личинок и яиц, а в зимнее время отчасти
и из семян. Некоторые виды, напр. ремеза, держатся большей
частью в кустарниках и тростниках. Гнездятся в дуплах деревьев,
устраивая рыхлые гнезда из животного и растительного пуха и
перьев: некоторые строят весьма искусные закрытые гнезда, сви-
вая их из растительного пуха с боковой трубкой для входа и помещая
над водой на свешивающихся ветвях или среди тростинок (ремеза),
500
или помещая в развилках отзолов и крупных ветвей дерева и при-
готовляя из мха, лишайников, перьев и т. д., напр. Aegithalos.
Все синицы несут большое количество яиц (до 8—12), обычно
белых, с мелкими красноватыми
Рис. 305. Большая синица — Parus major L.
По Evans.
пестринками.
Из морфологических приз-
наков отметим: клюв короткий
и прямой, иногда заостренный;
носовые отверстия прикрыты ще-
тинками или перышками; плюс-
ны сильные, с относительно ко-
роткими, срастающимися у са-
мого основания пальцами; кры-
лья короткие, тупые, с 10 боль-
шими маховыми. Оперение мяг-
кое и особенно длинное на над-
хвостье. Известно около 300 ви-
дов синиц, отнесенных к 17—21
родам: Различают роды: Parus,
Cyanistes, Lophophanes, Poecile,
Periparus, Chamaea, 1 Aegitha-
los, Remiz, Leptopoecile, Lopho-
basileus, Psaltiparus и др.
(рис. 305).
В СССР весьма распростране-
ны: большая синица — Parus
major L., зеленая лазоревка — Cyanistes caeruleus L., белая Лаза-
ревна — Cyanistes су anus Pall., московка — Periparus ater L., хохлатая
синица—Lophophanes cristatus L., гайка черноголовая — Poecile
atricapillus L., гайка болотная — Poecile palustris L., длиннохво-
стая синица — Aegithalos caudatus (L.). Более узко распространены
ремез — Remiz pendulinus (L.) и близкие виды, буроголовая гайка —
Poecile cinctus Bocld. и нек. др.
Семейство 30: Si ttidae. — Поползни
Мелкие лесные (и скалистые) птицы, по движениям напоминаю-
щие частью синиц, частью дятлов, распространенные в Евразии,
на Зондских островах, в Австралии и Северной Америке. Держатся
преимущественно в кроне дерева, лазают в любом положении
по ветвям и стволу, кормясь частью насекомыми, частью плодами
(орехами, ягодами и т. д.), раздалбливая их, подобно дятлам, клю-
вом. Гнездо устраивается в дупле, причем отверстие дупла сужается
до размеров птицы, замазываясь глиной (Sitta). Яйца белые с крас-
новатыми и коричневатыми пестринками.
Из морфологических признаков характерен прямой или слегка
вздернутый кверху клюв, сильные плюсны с отцосительно длинным
задним пальцем и крупными круто загнутыми когтями, тупые округ-
лые крылья, короткий прямой хвост. •
1 Нередко выделяют в особое семейство Chamaeidae (крапивниковых синиц).
501
Поползней известно около 60 форм, относимых к родам: Dendro-
phila, Sitta, Daphaenosilta и. др.
В СССР распространены два вида: наиболее широко распро-
страненный — обыкновенный поползень — Sitta europaea L. и южные:
поползень Крюпера — Sitta krilperi Pelz. и скалистый—Sitta neu-
mayer Michah.
Семейство 31: Hyposittidae. — Мадагаскарские поползни
В это семейство выделяется единственный вид Hypositta согаТ
lirostris Newt., распространенный на Мадагаскаре, отличающийся
коротким клювом, загнутым на конце крючком и с зарубинкой на
краю надклювья, и другими признаками.
Семейство 32: Certhiidae. — Пищухи
Мелкие, напоминающие по форме и поведению маленьких дятлов,
лесные или горные
Рис. 30G. Пищуха — Cer-
thia familiaris L. По
Evans.
птицы почти всего света, исключая Южную
Америку. Подобно дятлам, птички лазают по
стволам деревьев, опираясь на жесткие руле-
вые перья, и длинным изогнутым клювом до-
стают из трещин коры мелких насекомых и ли-
чинок, составляющих их пищу. Один укло-
няющийся вид и род обитает скалы и, лазая
по их поверхности, помогает движениям взма-
хами крыльев. Гнездятся в дуплах, щелях и
пустотах за отставшей корой или (Tichodroma)
в щелях скал. Яйца напоминают яйца по-
ползня.
Из морфологических черт своеобразно строе-
ние длинного хвоста, служащего опорой (у
Tichodroma хвост короткий^, длинные, с ост-
рыми когтями пальцы ног, изогнутый тонкий
саблевидный клюв. Крылья коротки и тупы.
В семействе известно около 40 форм, отно-
симых к родам: Certhia, Salpornis, Tichodroma,
Rhabdornis, Climacteris (рис. 306).
В СССР распространены только три вида:
широко расселившаяся обыкновенная пищу-
ха — Certhia jamiliaris L. и свойственные юж-
ным окрайнам страны: гималайская пищуха —
Certhia himalayana Vig. и стенолаз — Tichodro-
ma muraria (L.).
Семейство 33: Nectariniidae.— Нектарпицы
Мелкие, очень подвижные, по повадкам напоминающие мелких
синиц, птицы тропиков Старого Света. Нектарницы являются дре-
весными и кустарными птицами, как бы замещающими в Старом
Свете отсутствующих здесь колибри. Подобно последним, они пи-
502
гаются нектаром цветов и мелкими насекомыми, находимыми в цветоч-
ных чашечках, но в отличие от колибри не задерживаются перед ними
в воздухе, а лазают по веткам и подвешиваются к цветам, как синицы,
разыскивая насекомых также на листьях и взлетая за ними иногда
в воздух. Полет нектарниц менее быстр и напоминает полет славок
и синиц. Гнезда устраиваются из стеблей растений в виде закрытого
мешочка с боковым входом, подвешиваемого к концам ветвей деревьев
и кустарников; стенки сна-
ружи убираются мхом и ку-
сочками коры, внутри гнез-
до выстилается растительным
пухом. Яйца пятнистые,
весьма разнообразные по
окраске. Обычен резко выра-
женный половой диморфизм,
с яркой, металлически ири-
зирующей окраской самцов.
Из морфологических осо-
бенностей замечательна из-
менчивость формы клюва,
специализированной, как у
колибри, к форме чашечек
цветов определенных видов
растений, в отношении к
которым нецтарницы явля-
ются важными опылителями. Язык очень длинен и узок, с про-
дольным желобком, также напоминает язык колибри. Крыло ту-
пое с 10 большими маховыми, из которых 3-й—5-й образуют вер-
шину крыла.
Нектарницы известны в количестве около 250 форм с наибольшим
распространением в Эфиопской области. Чаще принимаются роды:
Arachnoterax Antherptes, Cinnyris, Nectarinia, Drepanorhynchus
n др. (рис. 307).
Рис. 307. Великолепная нектарнпца — Cinnyris
splendidus Shaw. По Evans.
Семейство 34: Meliphagidae. — Медососы
Обычно мелкие, древесные и кустарные птицы Австрало-Папуас-
ской области, Малайских островов и южной Африки, по облику
и повадкам напоминающие то синиц, то славок, то дроздов. Медо-
сосы питаются, преимущественно нектаром цветов и мелкими насе-
комыми, находимыми в цветочных чашечках, а также мелкими
мягкими плодами. Значительное количество видов связано с распро-
странением определенных эвкалиптов, к питанию нектаром которых
эти виды приспособлены. Гнездятся обычно в ветвях деревьев,
устраивая открытое чашкообразное гнездо. Яйца.пятнистые, крас-
ными пестринками по белому или розоватому* фону.
Из морфологических признаков характерны: узкий небольшой
клюв с тонкой насечкой по режущим краям у конца его, узкий
длинный язык, расщепленный и часто бахромчатый на конце, с частич-
но свернутыми в трубочку краями, служащий для всасывания
503
нектара. Тупое крыло с 10 большими маховыми и довольно длинные
и сильные плюсны.
Медосссы известны в количестве около 280 видов, распределяемых
почти на 40 родов. Наиболее резко выраженными являются роды:
Philemon, Meliphaga, Prosthemadera (рис. 308), Manorhina, Руспо-
pygius, Myzomela, Lich-
mera, Meliornis, Prome-
rops и др.
Семейство 35: Dicaeidae
Преимущественно
мелкие лесные и ку-
старные птицы, обычно
напоминающие синиц
по повадкам, свойствен-
ные Индии, Малайскому
архипелагу и Австра-
лии. Питаются гл. обр.
насекомыми, пауками,
мягкими семенами и нек-
таром. Гнезда закры-
тые, в виде грушеобраз-
Рис. 308. Тун — Prosthemadera novae zelandiae Gm. По
Evans.
ных построек, подве-
шенных на концах ветвей или (Pardalotus) в дуплах и пустотах п
даже норках. Яйца обычно белые, в редких случаях* пятнистые.
Морфологически характеризуются коротким, прямым, шило-
образным, у основания нередко расширенным клювом; края над-
клювья иногда пильчаты; язык обычно расщеплен или бахромчат.
Из десяти больших маховых первое сильно укорочено и обычно
скрыто кроющими. Хвост короткий и прямо обрезанный. Известно
около 100 форм, относимых к родам: Dicaeuni, Oreocharis, Urocharis.
Pristorhamphus, Pardalotus, Paramythia.
Семейство 36: Zcsteropidae. — Белоглазки*
Мелкие, по облику и повадкам напоминающие пеночек, весьма
подвижные древесные и кустарные птички тропиков и субтропиков
Старого Света. Кормятся гл* обр. насекомыми, отчасти семенами
и ягодами, обыскивая ветви и листья растений. Чашкообразные
гнезда помещаются в развилках ветвей и украшаются снаружи
мхом и лишайниками, а внутри выстилаются волосом. Яйца белые
или голубые, обычно в числе 3—5 штук. Самцам свойственна до-
вольно сложная песенка.
Из морфологических особенностей для большинства белогла-
зок характерно тонйое кольцо вокруг глаза из коротких белых
перышек; преобладающая окраска зеленая или желтая; клюв тон-
кий, шиловидный, с щелевидными ноздрями, с слабо развитыми
щетинками у основания; язык с гладкими краями, вилообразно
разветвляется на конце; крыло с 9 большими маховыми перьями..
504
Белоглазок насчитывают около 150 форм, относимых к родам:
Zosterops, Hypoctyptadius, Lophozosterops и др. В СССР встречается
только 1 вид, краснобокая белоглазка — Zosterops erythropleurus
Swinh., заходящий своим ареалом к нам в Уссурийский край и
на Амур.
Семейство 37: Vireonidae
Мелкие, кустарные и лесные птицы Северной и отчасти Южной
Америки, напоминающие обликом мелких сорокопутов или круп-
ных славок. По питанию являются, насекомоядными, в северных
широтах ежегодно мигрирующими видами. Гнездо висячее, при-
крепляющееся в развилках веточек деревьев и кустарников; клад-
ка состоит из 4—5 пятнистых яиц. Большинство видов являются
прекрасными певцами.
Из морфологических черт характерен напоминающий сороко-
путов клюв, на конце загибающийся резким крючком; частью слив-
шиеся основания пальцев ног; , короткие тупые крылья, с 10 боль-
шими маховыми. Преобладающая окраска однообразно-зеленая,
без пятен. Принимаются роды: Vireo, Pachysylvia, Vireolanius и др.
Семейство 38: Prunellidae. — Завирушки
Единственный род Prunella, свойственный всей Евразии. Мел-
кие птички, по окраске несколько напоминающие самок домашнего
воробья, ведущие частью наземный, частью кустарный образ жизни.
Клюв /твердый, постепенно утончающийся к концу, с округлым
коньком надклювья. Плюсны сравнительно короткие, с передней
стороной, покрытой пластинками. Третье большое маховое самое
длинное. Хвост довольно короткий, прямой или вырезанный. Птен-
цовый наряд пятнистый, сходно с дроздами. Кормятся преиму-
щественно на земле насекомыми и семенами. Чашкообразные откры-
тые гнезда помещаются обычно в кустах или щелях камней и скал.
Кладка состоит из 4—6 голубых яиц.
В СССР встречается несколько видов. Более широко распро-
странены: обыкновенная завирушка — Prunella modularis (L.), аль-
пийская завирушка — Prunella collaris (Scop.), черногорлая зави-
рушка — Prunella atrogularis (Brandt), лесотундровая завирушка —
Prunella montanella (Pall.) и др.
Семейство 39: Compsothlypidae. — Американские славки
Мелкие подвижные, обычно ярко и пестро окрашенные птички
Южной и особенно Северной Америки. По внешнему облику напо-
миная наших славок и некоторых пеночек, они весьма разнообраз-
ны по своей экологии: некоторые как бы замещают в Северной
Америке наших славок, другие ведут себя как мухоловки, третьи —
как коньки, Mniotilta лазает по коре дерева подобно пищухе;
ряд .форм напоминают по движениям синиц и т. д. . Питаются
преимущественно насекомыми,' мелкими личинками, ягодами и мяг-
кими семенами. Гнезда б. ч. чашкообразные, иногда шарообразные,
как у пеночки, помещаются преимущественно на деревьях или
в кустарных зарослях. Яйца напоминают яйца пеночек и синиц,.
605
белого цвета с красноватыми пестринками, откладываются обычно
в количестве 2—6. Из морфологических признаков характерно
наличие только 9 больших маховых; тупое крыло, с наиболее длин-
Рис. 309. Черно-белая американская славка — Mnio-
tvlta varia L. По Evans.
ными 3-м и 4-м маховыми,
редко 2-м и 3-м, слабый
тонкий клюв, обычно яркая
пестрая окраска.
В семействе различают
около 300 видов, распре-
деляемых по 30 родам.
Роды: Rhodmocichla, Geoth-
lypis, Dendroica, Oporornis,
Mniotilta и др. (рис. 309).
Семейство 40: Thraupi-
dae. — Танагры
Мелкие, обычно ярко
окрашенные, внешне и по
повадкам весьма напоми-
нающие вьюрков, лесные
птицы Южной и Северной
Америки. Кормятся преимущественно на земле насекомыми и мяг-
кими семенами, которые в отличие от вьюрковых не очищают от
кожуры. Гнезда устраиваются обычно в кустах в виде открытой
чашки из стеблей расте-
ний, мха и т. п. Яйца
пятнистые, весьма напо-
минают яйца овсянок.
Клюв танагр обычно ко-
нический, но слабее клю-
ва вьюрков, с несколько
загнутым на конце над-
клювьем и более откры-
тыми носовыми отвер-
стиями. Больших махо-
вых, девять, из них 2-е—
3-е пли 1-е—2-е наибо-
лее длинные, 1-е маховое
длиннее малых маховых.
' В семействе разли-
чают около 400 форм,
разделяемых на 62 рода:
Euphonia, Thraupis^ Pi-
ranga, Cissopis, Orchesti-
Рис. 310. Бразильская тапагра — Rhymphocoelus brasi-
lius L. По Evans.
ciis и др.
Семейство 41: Tersinidae
По окраске напоми-
нающие танагр, а по внешности — ласточек, мелкие ярко окра-
шенные птицы тропической Южной Америки. Единственный род —
506
Tersina ( = Procnias). Клюв очень короток и уплощён сверху, с очень
широким основанием, носовые отверстия открываются на конце очень
коротких трубочек. Крылья длинные и острые, хвост прямой. Птицы
гнездятся в дуплах деревьев или в норках в земле и кладут 3—4
чисто белых яйца.
Семейство 42: Ploceidae. —Ткачикп
Мелкие,, по внешности и повадкам напоминающие воробьев,
являющихся членами этой же группы, лесные, лесостепные и степ-
ные птицы Старого Света. Кормятся преимущественно семенами,
освобождая клювом ядрышко от одевающих его оболочек, но в гнездо-
вое вр^мя поедая и выкармливая птенцов также насекомыми и их
личинками. Преимущественно стайные и часто колониально гнездя-
щиеся птицы. Гнездо устраивается на дереве или у степных видов,
в высокой траве или кустарниках в виде шарообразной, рыхлой
постройки из стеблей травянистых растений, с входом сбоку; неко-
торые строят более сложные гнезда, создавая большие обществен-
ные постройки на ветках деревьев, с сотнями гнезд под одной общей
крышей и входным отверстием снизу. У многих видов наблюдается
резкий половой диморфизм, с более яркой окраской самца и обычно
двойной линькой оперения его в течение года. Яйца то одноцвет-
ные, то пестрые, с густыми коричневатыми или бурыми пестрин-
ками по белому, голубоватому или коричневатому фону.
По внешним морфологическим признакам — строению клюва,
крыла, плюсны — ткачики весьма близки вьюркам. Наиболее суще-
ственные отличия заключаются в строении рогового и костного неба,
нижней гортани (сирпнкса) и полной осенней линьке молодых (моло-
дые вьюрковые сменяют осенью лишь мелкое перо); большинству
ткачиковых свойственны также десять больших маховых перьев, из
которых первое часто очень мало. Биологически характерны устрой-
ство закрытых сверху гнезд*(и частое помещение гнезд в дуплах
и других укрытиях), колониальность гнездования и стайный образ
жизни нередко в течение круглого года. По развитию нижней гор-
тани, голоса и постройке гнезда ткачи в сравнении с вьюрками
стоят на более нпзкой ступени развития. Отличают около 600 видов
ткачиков, относимых к 75 родам с центром распространения в Эфиоп-
ской области. Вне тропиков и субтропиков ткачики очень малоч
численны. В связи с человеком почти по всему миру, расселяется
лишь обыкновенный домашний и полевой воробей.
К этому семейству 1 относятся роды: Textor, Passer, Dinemellia,
Histurgops, Plocepasser, Sporopipes, Malimbus и др.
В СССР из этой группы широко распространены: домашний
воробей — Passer domesticus (L.), полевой воробей — Passer топ-
tanus (L.) и более узко (в южных горных и степных районах) испан-
ский воробей — Passer hispaniolensis (Temm.)’, саксауловый воро-
бей— Passer ammodendri Gould., каменный воробей — Petronia pet-
1 Сушкин (1927) различает в этом семействе шесть подсемейств: ВиЬаЛог-
nithinae, Р loco passer inae, Passerinae, Sporopipinae, Ploceinae и Estrildinae,
607
ronia; затем снежные вьюрки — Montif ringilia nivalis L., в южных
горных странах, и короткопалые воробьп — Carpospiza brachydactyU
Вр., в пустынных горах Средней Азии и Закавказья.
Семейство 43: Fringillidae. — Вьюрки
Мелкие птички с коническим клювом, представляемые обще-
известными видами, как^чиж, щегол, чечетка, зяблик и др., распро-
страненные по всей земле, исключая Австрало-Папуасскую область.
Весьма разнообразные по экологии, будучи свойственны самым
разнообразным местообитаниям. Преимущественно зерноядные пти-
цы, часто и птепцов выкармливающие семенами пз зоба. В брачным
период однако большинство становится насекомоядными видами.
Вне брачного периода обычно стайные птицы. Гнездо устраивается
в виде открытой кверху чашки, из мелких стеблей, веточек, кореш-
ков, волос и перьев, часто украшенных снаружи мхом и лишайни-
ками. Кладка обычно из 3—5 пятнистых яиц. Часто наблюдается
резкий половой диморфизм: самцы окрашены ярче и обычно обла-
дают красивой песней.
Из внешних морфологических признаков вьюрков для боль-
шинства видов важнейшим признаком является наличие только
9 больших маховых перьев; по.многим внешним особенностям эта
группа близка ткачам (см. это семейство). Вьюрки являются одним
пз самых богатых видами и подвидами семейств, с наибольшим
распространением в северной умеренной зоне. Насчитывается всего
около 1100 видов вьюрков, относимых примерно к 150 родам: Frin-
gilla, Leacosticte, Mycerobas, Eophona, Spizella, Passerella, Janco,
Poospiza, Melospiza, Poocetes, Pinicola, Coccothraustes, Hesperiphona,
Emberizoides, Embernagra, Geospiza, S рог ophila, Loxia, Emberiza, Melo-
p.hus, Plectrophenax, Calcarius, Phrygilus, Spiza, Gubernatrix, Pyrrhula,
Erythrina, Erythrospiza, Rhodopechys, Rhodospiza, Uragus, Chloris,
Serinas, Oraegithus, Spinas, Cardaelis п др.
Некоторыми авторами семейство вьюрковых разделяется на
два семейства, собственно вьюрковых — Fringillidae и овсянковых —
Emberizidae (роды Calcarius, Zonotrichia, Emberiza и др.). Другие
авторы рассматривают эти подразделения только как подсемейства,
иногда выделяя еще ряд подсемейств, напр; Coccothraustinae, Рут-
rhulinae, Sporophilinae и др. Сушкин (Sushkin, 1928), принимая
указанные семейства, подразделяет первое на подсемейства: Fringil-
linae (род Fringilla), Cardaelinae (роды Acanthis, Spinas, Pyrrhula
и др.) и Cardinalinae (роды Richmondena, Sporophila и др.). По мне-
нию этого автора, оба эти семейства относятся даже к разным над-
семействам: к надсемейству Fringilloidei, куда входят также семейства
Ploceidae и Drepanididae, и к надсемейству Emberizoidei, куда отно-
сятся также семейства Thraupidae, Icteridae и Coerebidae.
В СССР представители семейства весьма многочисленны. Наи-
более известны из собственно вьюрков: щегол — Cardaelis carduelis
(L.), чиж — Spinas spinas (L.), коноплянка — Acanthis cannabina
(L.), чечетка — Acanthis jlammea (L.), зеленушка — Chloris chloris
(L.), дубонос — Coccothraustes coccothraustes (L.), снегирь — Pyrrhula
508
\pyrrhula (L.), чечевица — Erythrina erythrina (Pall.), щур—Pinicola
'micleator (L.), еловый клест —Loxia curvirostra L., зяблик — Frin-
«illa coelebs L., северный выорок — Fringilla montifringilla L. и
mi. др., обычно менее широко распространенные.
Из овсянок следует отметить: обыкновенную овсянку — Emberiza
dtrinella L., дубровника — Emberiza aureola Pall., седоголовую
овсянку — Emberiza spodocephala Pall., садовую овсянку — Emberiza
liortulana L., овсянку-крошку — Emberiza pusilia Pall, и из других
родов: пуночку — Plectophenax nivalis L. и лапландского подо-
рожника — Calcarius lapponicus L.
Семейство 44: Drepanididae. — Гавайские цветочницы
Мелкие, обычно однообразно в зеленые тона окрашенные, под-
вижные древесные и кустарные птички Гавайских островов. По
повадкам напоминают мел-
ких синиц или славок,
особенно хлопотливо пере-
летающих около цветов
пли почек деревьев. Пи-
таются пыльцей, нектаром
цветов, мелкими насеко-
мыми.
Морфологически для
птиц этой группы харак-
терно строение языка как
у медососов, как приспо-
собление к сосанию нек-
тара, при весьма разнооб-
разной форме клюва, в не-
которых родах напоминаю-
щего даже клюв клеста.
Больших маховых 10, из
них 1-е сильно укорочено.
Почти все представители
этой группы обладают ре*з-
Рпс. 311. Мамо — Drepanis pacifica Gm. По Wilson
and Evans, из Evans.
ким и весьма неприят-
ным запахом, не зависящим от способа питания. Принимаются
роды: Drepanis\ Psittirostra, Hemignathus. Loxops и др. (рис. 311),
Семейство 45: Coerebidae. — Цветочницы
Очень мелкие, обычно ярко й пестро окрашенные, по повадкам
напоминающие мелких синиц, птички тропических частей Южной
и Центральной Америки. Обитают преимущественно на опушках,
в Садах и перелесках, держатся в кронах, где разыскивают корм
среди мелких ветвей, листьев и цветов. Питаются мелкими насе-
комыми, нектаром и пыльцей цветов и мягкими мелкими плодами
и ягодами. Гнезда строятся шарообразными из тонких стеблей,
с входом сбоку, и подвешиваются обычно на концах свешивающихся
609
ветвей. Яйца (2—4 в кладке) пятнистые с красно-коричневыми
пестринками по красновато-белому или голубоватому фону.
Из морфологических черт характерно строение языка, расще-
пляющегося на конце пли образующего кисточку. Клюв по форме
’ Гис. 312. Сахарная птица—Coereba, jlaveola Мах.
• По Evans.
весьма изменчив. Крыло
образуется 10 больши-
ми маховыми, из кото-
рых 1-е крайне дэедуци-
ровано.
Известно около 100
форм, относимых к 9 ро-
дам: Coereba (рис. 312),
Conirostrum, Oreomanes..
Hemidacnis и др.
Семейство 46: Icteri-
Американские
иволги
Внешностью и повадками напоминающие скворцов и .иволг,
пестро и нередко ярко окрашенные птицы Южной и Северной Аме-
рики. Весьма разнообразны по экологии, то являясь лесными пти-
цами, то степными и полевыми, птицами болотных зарослей и т. д.
Питаются преимущественно насекомыми и их личинками, иногда
плодами, отдельные роды — преимущественно семенами и зернами.
Очень часто являются колониальными и стайными птицами. Гнез-
дятся устраивая то длинные' мешковидные висячие гнезда, при-
Рис. 313. Балтиморская иволга — Icterus ballimore L. По Evans.
крепленные к концам ветвей, с боковым входом, то гнезда типа
иволг, или открытые гнезда в траве, на земле. Так наз. воловьи
птицы ( Molothrus) подкидывают яйца в гнезда других птиц, частью
видов той же группы Icteridae. Яйца разнообразные,, всегда густо
покрытые пестринами.
Известно около 190 видов, распределяемых на 25 родов. Роды:
Zarhynchus, Ostinops, Cassicus, Icterus, Agelaius, Molothrus, DoU-
chonyx, Sturnella и др.
ыо
ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ ПТИЦ
Птицы являются одним из наиболее распространенных и в видо-
вом отношении богатых классов позвоночных животных. Заселяя
самые разнообразные типы местности и встречаясь в значительном,
иногда массовом, числе, они играют очень крупную роль в эконо-
мике природы. Своей деятельностью птицы влияют на численность,
других животных, поедая одних, конкурируя с другими, являясь,
пищей третьих; они влияют на растительность, частью поедая зеле-
ные части растения и семена, частью распространяя последние и со-
действуя таким образом расселению растений. У берегов Финского за-
лива один квадратный километр обычного смешанного леса населяют
в среднем около 600 пар гнездящихся птиц (Palmgren, 1928), утраиваю-
щихся в численности после появления птенцов. Но наиболее пора^
жают.обилием птиц «птичьи горы». Подсчет кайр (Uria lomvid), гнез-
дящихся на западных берегах Новой Земли, дал ориентировочную
цифру в. четыре миллиона экземпляров. Так как средний вес кайр
равен 2,5 кг, то живой вес всей массы новоземельскпх кайр должен
составлять десять миллионов килограммов. Если предположить,
что одна кайра для своего пропитания извлекает из моря 250 г
рыбы в сутки п принять, что кайра живет на Новой Земле в средней
4 месяца ежегодно, мы должны сделать вывод, что все население
кайр за сутки добывает из моря около одного миллиона килограмм
рыбы, или за 120 суток — сто двадцать тысяч тонн этого живого
вещества! (Портенко, 1931).,
Значение птиц в жизни человека также велико и. разнообразно.
Оно может быть сведено к определенным типам: птйцы могут быть
полезны или вредны человеку в ёго сельскохозяйственной, лесо-
культурной пли какой-либо иной деятельности: они используются
как промысловый пли охотничий (спортивный) объект; они издавна
культивируются в домашнем хозяйстве; наконец птицы важны для
человека как переносчики болезней и паразитов, опасных как для
него, так и его подсобных животных. Под этими разделами значение
птиц и рассматривается ниже.
В своей сельскохозяйственной деятельности человек встречает
со стороны птиц как помощь, в форме истребления различных вре-
дящих посевам и плодам насекомых, истребления семян сорных ра-
стений ит. п., так и вред, в виде уничтожения полезных человеку
животных, зерновых культур, плодов, порчи древесины деревьев
и т. п. Эта роль птиц лишь в редких случаях является определенно
йолезной пли несомненно вредной, так как птицы различно реа-
511
гируют на окружающую обстановку и пищу в зависимости от их
изменения, сезона, условий года, наконец возраста самого живот-
ного и его состояния. В большинстве случаев птица различными
сторонами своей жизни приносит человеку одновременно и пользу
и вред, так что суждение о ее экономическом значении может быть
выведено лишь на основании детального ознакомления с жизнью
животного и сравнения указанных сторон ее деятельности. Слож-
ность этих отношений легко может быть показана на некоторых при-
мерах. Клест-еловик (Loxia curvirostra L.) обитает хвойные, преи-
мущественно еловые, леса Евразии и питается почти исключительно
семенами ели и отчасти сосны, которые созревают в шишках, растре-
скивающихся и высыпающйх семена на землю. Ведя преимуще-
ственно стайный и кочующий образ жизни, клест быстро реагирует
на неурожай семян ели (который бывает весьма изменчив) и пере-
летает из района в район, в более кормные места, обследуя огромные
территории; кормясь, клест срывает и не вполне спелые шишки,
зажимает их лапами и затем искусными поворотами своеобразного
клюва (см. рис. 207) вскрывает чешуи и достает семена; обычно
использовав лишь часть семян шишки, клест бросает ее и прини-
мается за. другую, снимая такйм образом с дерева весьма значитель-
ную часть урожая. Семенами ели питается целый ряд других живот-
ных, и в том числе особенно белка, используемая человеком как
пущной зверек. Деятельность многочисленных клестов очевидно
вредна для человека, так как сужает кормовую базу ценного зверька
и к тому же уменьшает потенциальную возможность расселения
ценного дерева. Однако при детальном изучении обнаруживается
(Формозов, 1934), что, сбрасывая шишки с семенами на землю, клест
оказывает и благодеяние для белки, так как лежащие в сырости на
земле шишки дольше сохраняют семена (вследствие смыкания чешуи)
и смогут быть использованы белкой тогда, когда другие обычные
древесные корма ее уже использованы. Таким образом клест содей-
ствует промышленнику, в известной мере способствуя жизни
зверька — объекта его охоты, в тяжелые для животного периоды.
Практически эта польза Сможет быть существеннее для человека,
чем вред от частичного поедания семян, идущих в пищу тому же
зверьку, и уменьшения возможностей для самосева ели. Не менее
сложны отношения сибирской кедровки (Nucifraga caryocathactes
macrorhynchus Brehm), распространение которой в Сибири в значи-
тельной мере связано с распространением сибирского кедра (Pinus
cembra sibirica Маут.), семенами которого она питается. В этом отно-
шении кедровка является серьезным конкурентом ряда промысло-
вых животных, в том числе соболя и белки, в жизни которых семена
кедра играют очень важную роль. Но в то же время кедровка, бла-
годаря своему стремлению устраивать запасы орехов и искусному
полету, является несомненно важнейшим разносчиком семян кедра
и весьма существенно содействует его распространению, невозмож-
ному, вследствие тяжести плодов, без помощи животных. Наконец
кедровка играет отрицательную роль для человека, поедая мелких
лесных птиц и их яйца (например синиц), несомненно полезных
в жизни леса уничтожением многих вредных насекомых. Если мы
512
отметим теперь, что массовые миграции кедровки, предпринимаемые
ею время от времени из-за местных неурожаев кедровых и еловых
семян, могут быть использованы человеком для прогнозов о числен-
ности и условиях жизни некоторых промысловых животных [Формо-
зов (Formosof) 1933], мы получим столь сложную картину по-
ложительного и отрицательного значения этой птицы для человека,
что суждение о ее деятельности вряд ли может быть категорическим
и универсальным.
Почти таково же положение с нашими пестрыми дятлами (Dryo-
bates major L.), которые, с одной стороны, повреждением кэры и
древесины деревьев,
поеданием семян и
орехов приносят вред,
с другой, уничтоже-
нием различных ко-
роедов и других дре-
весных насекомых
приносят несомнен-
нуюпользу (рис.314).1
Вы да л б лива я дупл о
для своего гнезда,
дятлы повреждают де-
рево, понижают его со-
противляемость гриб-
ным инфекциям и раз-
носят последнюю и в
то же время создают
места гнездовья для
различных насекомо-
ядных дуплогнездни-
ков, в большинстве
своем весьма полез-
ных для леса. Если
принять во внимание,
что последние виды
при отсутствии дят-
линых дупл оказыва-
ются редкими в лесу
и что пара дятлов в. среднем выдалбливает в районе своего гнез-
дования 10—12 более или менее законченных гнезд, используя из
них лишь одно (Берлепш, 1900), легко уяснить, что и эта роль
дятлов весьма немаловажна.
Во многих случаях эта последняя и вредная деятельность не
Рис. 314. Наросты древесины, вызванные иа дереведят-
лами, вследствие разрушения коры и луба.
(а) — продольный разрез древесины сосны в области ч<коль-
цевания» дерева дятлом, невидимому iDryobates major L.
Многим дятлам свойственно продалбливание коры дерева
весной, в виде кольца углублений, через которые птица
пьет сладкий сок, поднимающийся весной по стволу. Удары
клювом наносятся наискось, что облегчает истечение сока,
и повторяются нередко из года в год па том же самом месте;
(Ъ)ствол сосны, покрытый кольцами разрастания дре-
весины, вызванными «кольцеванием» дерева дятлом —
Sphyrapicus varius G-rael. По Fuchs и Title! из Stresemann^
1 Так, например, выяснено (Положенцов и Кнорре, 1936), что в Бузулукском
бору большой пестрый дятел уничтожает летом многих вредных насекомых
и в том числе майских жуков, но не менее 9—10 месяцев в году питается семе-
нами сосны и других, преимущественно хвойных, пород деревьев. По произве-
денным подсчетам за год, каждый дятел уничтожает более 157 500 семян сосны,
что конечно неблагоприятно влияет на возобновление этого- ценного дерева
в степных лесах.
33 Орнитология—1209 * 513
сочетаются одновременно, а перемежаются во времени. Так многие
насекомоядные птпцы, весьма полезные летом, осенью на кочевках
и перелетах появляются в садах человека и становятся весьма вред-
ными уничтожением различных ягод, плодов и фруктов и вызывают
организацию мешкотной или дорогостоящей защиты этих культур
человеком.
Из этих примеров ясно, что полезность птицы является весьма
изменчивым свойством, и следовательно отношение человека к ее
деятельности казалось бы должно вариировать сообразно обстоя-
тельствам. При малом хозяйственном освоении того или другого
участка природы даже потенциально вредные птицы в сущности
являются безразличными, и вред становится тем ощутимее, чем осно-
вательнее человек использует то или иное богатство природы. При
этом в практике человека была бы желательной возможность регу-
лировать или использовать этот вред или пользу, привлекая их на
полезный период, удаляя при других обстоятельствах. Однако тех-
нически это мало достижимо, почему приходится довольствоваться
суммарным определением полезности вида и при активном отноше-
нии к нему в общем заботиться о нем, несмотря на отдельные
отрицательные стороны, наблюдающиеся в его деятельности. Сле-
дует притом еще раз подчеркнуть, что и кажущийся вред может
иметь положительное значение. Так хищники, охотящиеся за куро-
патками, естественно ловят главным образом слабых и больных
животных и этим способствуют уменьшению эпизоотий и большей
выносливости всей популяции в целом.
Выяснение полезности птиц обычно достигается отстрелом жи-
вотных и определением содержимого желудков, а также наблюде-
нием над поведением и питанием птиц в природе и выборочным корм-
лением в неволе с последующим определением количества съеден-
ной пищи (Rorig, 18). 1 Подобное изучение ведется с пятидесятых
годов прошлого столетия (Jenks, 1858; Treadwell, 1858) и в ряде
стран осуществляется с большой планомерностью. Массовость ма-
териала (по многим видам количество исследованных желудков исчи-
сляется сотнями), захват всех сезонов и возрастов и отстрел птиц
в различных стациях, обитаемых ими, очень существенны для полу-
чения более объективной картины значения птицы в хозяйстве че-
ловека. Этой методикой было выяснено, что семья скворцов за день
может уничтожить более 350 гусениц, саранчуков и улиток; кукушка
в час истребляет иногда до ста гусениц; горихвостка в лето съедает
до миллиона насекомых, а королек в течение года потребляет до
четырех миллионов еще более мелких насекомых (Коротнев, 1931). Вы-
считано, что в долинах рек Сакраменто и Сан Джоакин в Калифорнии,
площадью в 11000000 акров, на каждые два акра культурной земли
приходится по одной паре луговых жаворонков (Sturnella magna L.).
1 Для хищных птиц и сов также широко применяется изучение погадок,
в которых остатки пищи зачастую легко определимы.-Так, например, Пидо-
пличка (1930, 1932, 1937 гг. и др.) исследовал более 3900 погадок сов, собран-
ных более чем в 400 пунктах; в этих погадках были обнаружены остатки
84 897 грызунов и различных мелких беспозвоночных. Материал исследований
Утендорфера (Uttendorfer, 1924) по питанию птиц состоял из 31 267 погадок.
514
Так как в среднем каждая пара производит четырех птенцов, каждый
из которых весит около 30 г пока он в гнезде и поедает ежедневно
пищи, по весу равной в среднем половине его собственного веса,
то вся популяция луговых жаворонков обеих долин должна скарм-
ливать птенцам'ежедневно не менее 343,5 т насекомых и их личинок.
Подобными же подсчетами выяснено (Мекленбурцев, 1935), что розо-
вый скворец — Pastor roseus L., съедающий за день до 200 г саранчи
в день, большой гнездовой колонией уничтожает до 100 т саранчи
в течение одного месяца.
Столь же большим оказывается и вред птиц, когда дело идет о мно-
гочисленных и широко распространенных птицах. Подсчитано, что
в одной Великобритании потери урожая, приносимые домашним
воробьем (Passer domesticus L.), составляют почти баснословную
цифру в 8000 000 фунтов стерлингов. Большой баклан (Phalacrocorax
carbo L.), гнездящийся в Астраханском заповеднике в дельте Волги
в количестве около 4800 пар, съедает в год, по некоторым подсчетам,
до 4132,8 т рыбы, что составляет около 1,3% от годового улова
рыбы дельтовыми промыслами Волги (Сушкина, 1932). 1
Детальными наблюдениями в общем выяснено, что большинство
птиц приносит больше пользы, чем вреда, и во многих случаях вполне
заслуживает хозяйственной заботы о них и стремления увеличить
их численность. Ценность их особенно увеличивается тем фактом,,
что в случае начинающегося массового размножения каких-либо
насекомых ими начинают кормиться даже птицы, нормально эти
виды не потребляющие в пищу; совокупная деятельность этих птиц
снижает численность вредителей и уменьшает их вредное действие.
Поэтому многие и видят основную полезность насекомоядных птиц
в том, что они постоянно сдерживают размножение насекомых на
некотором низком уровне и предотвращают таким образом массовые
«напасти» их на наши сельскохозяйственные и лесные культуры.
Вкратце обзор наиболее полезных и вредных птиц может быть
проведен следующим образом.
К полезным птицам, уничтожающим вредных грызунов (мышей,
полевок, сусликов и др.) в полях и лугах, можно отнести, например,
следующих птиц: белый аист — Сiconia ciconia L., ворон — Corvus
corax L., ворона серая — Corvus cornix L., ворона черная — Corvus
corone L., лесной канюк — Buteo buteo L., канюк мохноногий — Buteo
lagopus L., степной орел — Aquila nipalensis, ряд луней — Circus
cyaneus L., Circus macrourus L., пустельга обыкновенная — Cerchneis
tinunculus L., болотная сова — Asia jlammeus L., сова ушастая —
Asia otus L., сыч — Athene noctua L. и целый ряд других птиц.
К птицам полезным уничтожением вредных насекомых в лесу
относятся: ряд синиц — Parus major L., Periparus ater L., Cyanistes
1 По другим подсчетам (Дюнин, 1936), количество бакланов в дельте Волги
определяется в 16 000—18 000 голов, причем лишь на территории Астраханского
заповедника, где в 1933 г. было около 8000 голов, баклан уничтожает не менее
4788 т в год. Однако экономическая ценность баклана определяется не только
влиянием его на рыбное хозяйство человека. Поселяясь колониями, баклан
губит и портит*рощи, располагающиеся по течению рек, весьма ценные для чело-
века в безлесной местности.
515
c^eruleus L., Poecile atricapillus L., Poecile palustris L., Lophopha-
nes cristatus L., пищуха — Certhia jamiliaris L., поползень — Sitta
europaea L., ряд славок — Sylvia atricapilla L., Sylvia hortensis L.,
Sylvia communis L., Sylvia curruca L., ряд мухоловок — Muscicapa,
striata, Hedymela hypoleuca Pall., Siphia parva L., ряд *пеночек — Phyl-
loscopus trochilus L., Phylloscopus collybita L., Phylloscopus sibilatrix
L., Phylloscopus nitidus viridanus L., Hippolais icterina L., желто-
головый королек — Regulus regulus L., крапивник — Troglodytes
troglodytes L., ряд дроздов — Turdus viscivorus L., Turdus pilaris L.,
Turdus ericetorum Turt., горихвостка — Phoenicurus phoenicurus L.,
малиновка — Erithacus rubecula L., кукушка — Cuculus canorus L.,
иволга — Oriolus oriolus L., козодой — Caprimulgus europaeus L.,
вертишейка — Jynx torquilla L., ряд дятлов—Picus viridis L.,
Picus canus L., Dryobates major L., Dendrodromas leucotos L.,
Leucontopicus minor L., зяблик—Fringilla coelebs L.. чиж — Spinus
spinus L., сорока — Pica pica L.,‘сойка — Garrulus glandarius L.,
совка сплюшка — Otus scops L. и мн. др.
На полях и лугах уничтожают много вредных насекомых следую^
щие виды: обыкновенный скворец — Sturnus vulgaris L., ряд тря-
согузок— Budytes jlava L., Budytes citreola Pall., некоторые ов-
сянки — Emberiza citrinella L., Emberiza.aureola Pall., жаворонки —
Alauda arvensis, Galerida cristata L., Calandrella cinerea L., Calandrella
minor, некоторые коньки — Anthus campestris L., Anthus praten-
sis L., сизоворонка — Coracias garrula L., удод — Upupa epops L.,
щурка — Merops apiaster L., обыкновенная и степная пустельга —
Cerchneis tinunculus L., Cerchneis naumanni L., кобчик —Erythropus
vespertinus L., грач— Trypanocorax jrugilegus L., поблизости рек
чайки — Larus ridibundus L., Larus canus L. и другие птицы.
Из птиц, приносящих вред, отметпм лишь ястреба тетеревят-
ника — Astur gentilis L., перепелятника — Accipiter nisus L., бо-
лотного луня — Circus aeruginosus L. и филина.— Bubo bubo L.,
почти всегда охотящихся за теплокровными позвоночными, обитаю-
щими лесные, болотные или открытые стации. Многие другие виды
повинны в преследовании и уничтожении ценных для человека /ки-
вотных, но большинство из них одновременно приносят и пользу
поеданием вредных насекомых, моллюсков, сорняков и т. п.
Полезная деятельность птиц вызывает стремление увеличить их
численность и привлечь для гнездовья в наши сады, парки и лесные
насаждения.
Приемы привлечения птиц на гнездовье различны и основаны на
знании особенностей их образа жизни и поведения. 1 Значительное
число полезных птиц является дуплогнездниками и очевидно они
нуждаются в дуплах для помещения гнезда, причем величина этих
дупл, а также и характер летного отверстия должны соответствовать
привычкам птицы. Например маленькие синички {Cyanistes cyanus
Pall., С. caeruleus L., P. atricapillus L. и др.,) не гнездятся в больших
дуплах. Чтобы добиться гнездования в искусственной дуплянке
1 В виду важности широкого распространения сведений о* способах при-
влечения птиц, полезно остановиться на этом вопросе несколько детальнее.
516
определенного вида, необходимо дуплянку или скворечник помещать
в тех условиях, которые данный вид предпочитает; для синицы-
лазоревки, например — в лиственных участках парка, для мос-
ковки — в хвойной части или среди елей и т. д.
Наиболее простым и вполне удобным способом приготовления
этих гнездовий является сколачивание их в виде продолговатого
ящика, как делаются скворечники. Они изготовляются из слегка
оструганных и распиленных досок толщиной в 2—2,5 см, совершенно
сухих и лишенных посторонних запахов и большого количества
смолы. Дощечки должны быть пригнаны друг к другу возможно
плотнее, не оставляя щелей, и сколачиваются гвоздями или свин-
чиваются шурупами. Летное отверстие делается в верхней трети
передней стенки с таким расчетом, чтобы кошки не могли лапой
через отверстие добраться до гнезда. Крышку скворечника нужно
делать с небольшим выступом над передней стенкой,’как бы навесом,
и не приколачивать, а привинчивать 3—4 винтами, так как большин-
ство птиц любит с каждой кладкой строить новое гнездо, и может
случиться, что спустя несколько лет дупло будет настолько забито
гнездовой трухой, что это будет препятствовать поселению некоторых
видов птиц; кроме того, в гнездовой трухе всегда разводится очень
много птичьих паразитов, которые чрезвычайно беспокоят птиц,
а иногда и побуждают птицу бросить гнездо или ведут к гибели
птенцов. Поэтому полезно от времени до времени осматривать скво-
речники и, отвинчивая крышку, удалять старые сор и труху, а иногда
и окуривать дупло (конечно, не в гнездовое время) серными парами.
Задней своей стороной скворечник крепко прикрепляется (винтами
и гвоздями) к более длинному бруску дерева или к палке, которая
уже приколачивается к стволу дерева, поверхности здания и т. п.
Для большинства мелких птиц скворечники следует вешать на вы-
соте 3—5 м от земли, и лишь для скворцов, сизоворонок, пустельги
и т. д. рекомендуется помещать их еще выше, на высоте 5—8 м.
К общим правилам развески скворечников относятся: скворечник
должен при всех условиях находиться преимущественно на под-
ветренной стороне, чтобы ветер возможно меньше задувал в него;
положение скворечника должно быть вертикальным или передней
стороной слегка наклонным вниз, но ни в коем случае не .вверх,
так как в этом случае внутрь будет попадать дождь, а птенцам будет
трудно выбираться из гнезда; скворечник должен быть прибит
очень прочно к стволу, чтобы он не качался, что пугает птицу;
скворечники надо развешивать еще до прилета птиц, с ранней весны
или даже осени, чтобы можно было проверить их устойчивость и спо-
собность противостоять дождям; вешать скворечники надо не гуще,
чем по одному на дереве средней величины, а при малом количестве
скворечников и большом парке, еще реже, стараясь более равно-
мерно распределить их по этой территории.
Величина скворечника, как указывалось, должна изменяться
сообразно величине птицы, для которой он делается. Обычно раз-
личают пять основных размеров скворечников, которые приведены
на прилагаемой табличке (даны внутренние размеры, в милли-
метрах).
617
Глубина скворечника от крышки до дна. . . .
Ширина скворечника между стенками..........
Ширила летного отверстия...................
Расстояние верхнего края лстпого отверстия
от крышки..................................
I II III IV V
233 233 336 420 420
95 95 125 180 2 0
28 32 46 60 85
55 50 60 60 60 i
Из них I тип приспособлен преимущественно для мелких синиц:
лазоревки {С у artistes caeruleus L.), пухляка {Poecile atricapillus L.),
болотной гаечки {Poecile palustris L.), пищухи {Certhia familiaris L.);
II тип — для большой синицы {Parus major L.), поползня {Sitta
europaea L.), горихвостки {Phoenicurus phoenicurus L.), мухоловки-
пеструшки {Hedymela hypoleuca Pall.), вертишейки {Jynx torquilla L.),
малого пестрого дятла {Leucontopicus minor L.); Ill тип — для
скворца {Sturnus vulgaris L.), большого пестрого дятла {Dryobates
major L.) и аналогичных им по величине; IV тип — для зеленого
дятла {Picus viridis L.), седого дятла {Picus canus Gm.) и удодов
{Upupa epops L.), и V тип — для клинтуха {Columba oenas L.),
сизоворонки {Coracias garrula L.), пустельги {Cerchneis tinuncu-
lus L.), галки {Coloeus monedula L.), небольших сов, черного дятла
{Dryocopus martius L.), лутка {Mergellus albellus L.), гоголя {Bu-
cephala clangula L.) и нек. др. Следует указать, что многое в посе-
лении птицы зависит от соответствия местоположения скворечника
ее привычкам, от обилия корма поблизости, отсутствия конкуренции,
многочисленности птиц этого вида. Поползень охотно занимает и
сравнительно большие дупла, искусно сужая их входные отверстия
при помощи глины; скворцы селятся иной раз в дуплах, в которые
они явно с трудом протискиваются, ит. д., но указанные размеры
в среднем наиболее соответствуют указанным видам.
Необходимость приготовления таких маленьких скворечников, как
I и II типы, частью объясняется тем, что более крупные и сильные
птицы, например галки, скворцы, большие синицы и воробьи,
легко .отнимают скворечники у более мелких и слабых птиц, если им
только подходит по величине их летное отверстие.
Некоторые из указанных птиц явно предпочитают более естествен-
ный характер дупл и лучше поселяются лишь в дуплянках, т. е.
гнездовых домиках, сделанных не из сколоченных досок, а в обруб-
ках дерева, высверленного или выдолбленного внутри. Таковы дятлы,
иногда галки, клинтухи, сизо1 оронки и вообще более крупные птицы.
В таких случаях, чтобы привлечь их, приходится предлагать им
долбленые домики, соответствующие указанным выше размерам.
Наиболее неохотно поселяются в искусственных жилищах крупные
дятлы, в большинстве случаев способные самостоятельно выдолбить
для себя дупло, соответственно своим вкусам.
Наконец следует указать, что некоторые дуплогнездники, на-
пример серая мухоловка, гнездятся лишь в полудуплах, иначе
говоря, в очень мелких дуплах с широким летным ^отверстием. При-
518
готовить их можно в виде тех же скворечников, но сильно укорочен-
ных (15—18 см), и в качестве летного отверстия следует сделать
широкий вырез под крышкой, высотой в 5—7 см и шириной в 10—
15 см,.
Привлечение мелких дуплогнездников в наши сады и парки яв-
ляется наиболее распространенным и разработанным способом при-
влечения птиц вообще. Несомненно, этот способ привлечения птиц
может быть значительно расширен. В Карелии и в Приалтайской
степи издавна ставят на озерах большие «скворечники» для поселе-
ния гоголя, которого используют в промысловом отношении, отбирая
часть его кладки. Вероятно такие дуплянки можно ставить и для
утки мандаринки [Dendronessa galericulata (L.)] Уссурийского края
и для некоторых крохалей. Для красной утки (Casarca ferruginea
Pall.), гнездящейся в норах, в Дании вырывают в холмах у берега
моря длинные ходы с отнорками, в которых эти утки поселяются
целыми колониями, и также частично используются для сбора яиц
(Naumann, 2-е изд.). Для гаги в Исландии и Норвегии издавна при-
готовляют удобные места для гнездования и охраняют их, весьма
эффективно используя эту ценную птицу (см. ниже). Мелкие хищ-
ники: обыкновенные и степные пустельги (Cerchneis tinunculus L. и
С. naumanni Fleisch) и кобчики [Erythropus vespertinus (L.)], ценные
истреблением вредных грызунов и саранчи, охотно гнездятся в дуп-
лянках и с пользой могут быть привлечены в нуждающиеся в них
степные места.
В отношении свободно гнездящихся птиц, строящих свои гнезда
открыто среди ветвей кустарников, прибегают в парках и садах
к обрезке кустарников и ветвей деревьев, чтобы вызвать образование
мутовок, густых скоплений веток и обильное омертвение их, среди
которых птицы гнездятся с большей охотой, чем среди сравнительно
разреженных зарослей. Той же цели добиваются связывая пучки
ветвей кустов между собой, получая этим участки кустарных зарос-
лей с более густым ветвлением. Еще лучшие результаты получаются
специальными посадками более густых и удобно для птиц ветвящихся
кустарников, располагаемых группами, в виде живых изгородей.
Наиболее ценны для этого густые посадки молодых елок, боярыш-
ника, калины, шиповника, ежевики и мн. др. При сочетании этих
зарослей с посадками густых деревьев (ель, на юге — пирамидаль-
ный тополь и др.) и соответствующей подрезке можно добиться очень
густых зарослей, богато населяемых мелкими птичками. Очень ценно
включить в эти заросли кусты и деревья, покрывающиеся осенью
ягодами: жимолость, рябина, черемуха, калина, бузина и нек. др.,
так как ягоды осенью и отчасти зимой составляют для многих птиц
очень важный корм, привлекающий помимо местных птиц нехмало
и зимующих видов: свиристелей, щуров, снегирей и т. п.
Сходного характера мероприятия проводимы и в отношении по-
левых и отчасти водных птиц, но они сложнее, более трудоемки и
дороги (см. ниже). Целесообразнее подумать о пернатой фауне при
производстве разных хозяйственных мероприятий. Так, выкорче-
вывая лес под пашню, можно оставлять небольшие заросли кустар-
ников, в которых будут находить себе укрытие куропатки, тетерева,
619
а зимой и осенью перелетные птицы. В степи и лесостепи следует
особенно бережно относиться к зарослям степных кустарников по
оврагам и небольшим колкам еще существующей древесной расти-
тельности. Также в отношении болот и озер. В низовьях Сыр-дарьи
из-за немногих гектаров посева иногда безрассудно осушают сотни
гектаров озер и камышевых зарослей; через несколько лет почва под
посевами на осушенном дне озера истощается, пашня забрасывается,
а население теряет немалый заработок от охоты на тех гусей, уток и
пеликанов*, которые гнездились на этих озерах ранее или останавли-
вались здесь во время пролета. Поэтому забота об охране болотной
и водоплавающей птицы, как нашего природного богатства, должна
стоять не менее высоко, чем забота и о многих других формах сель-
ского и лесного хозяйства. В западных странах, чтобы увеличить
количество дичи, иногда прибегают к засевам болот специальным
видом болотного злака, так называемым канадским рисом, который
любят пресноводные утки. Такие опыты делались и у нас, и нет сом-
нения, что в бедных дичыо местах и этот способ ценен для увеличения
нашей болотной и озерной фауны.
Привлекая птиц, приходится бороться и с их врагами. Из них
особенно опасны кошки и бродячие собаки, и борьба с ними должна
заключаться в отпугивании их, а в отношении кошек и в создании
препятствий в охоте за гнездами. Для этого, помимо ранее указан-
ных предосторожностей, прибегают к обкручиванию на высоте 2 м
ствола дерева со скворечником несколькими рядами колючей про-
волоки, через которую кошка не может пробраться. Вредных хищ-
ных птиц (см. ниже) необходимо отстреливать, а гнезда их в пределах
охраняемого района систематически разорять.
Не менее существенной является иногда и забота о зимующих
у нас птицах. Для многих йз них суровая и длительная зима является
настоящим критическим периодом года, так как пища скудна и для
многих, кроме того, укрыта снегом. Гибель птиц от бескормицы зимой
несомненно уменьшает численность многих видов к летнему периоду
и таким образом ослабляет их возможное полезное значение в этот
период года. В такие суровые зимы весьма целесообразна в садах и
парках подкормка птиц, для чего прикрепляют у дерева им на помощь
(так, чтобы птицам было удобно слетать к нему) небольшой столик
или дощечку и высыпают на нее крошки и кусочки сухого хлеба, или.
лучше семена конопли, льна и разных злаков. Птицы скоро замечают
этот корм и регулярно появляются здесь в часы кормежки.
В противоположность привлечению полезных птиц, борьба с вред-
ными птицами гораздо менее распространена, так как обычно свя-
зана с довольно крупными расходами. Наиболее интенсивным спо-
собом борьбы является разорение гнезд вредных птиц, затем отстрел
их, особенно хищных видов. Для последних применяется также
устройство ловушек или капканов, пристраиваемых к клеточкам,
в которых помещается приманная птица; практикуется также охота
с приманным филином. Последняя основана на той ненависти, ко-
торую вызывают в дневных птицах ночные совы; филин помещается
на открытом месте и привлекает внимание пролетающих хищников,
которые подлетают к нему и могут быть убиты охотником из засады.
520
Во многих странах население ознакамливается с ролью хищников
специальными брошюрами, и ранее практиковалась выдача премий
за представленные лапы добытых вредных птиц.
Промысел и охота на птицу издавна играли значительную роль
в жизни человека. Судя по сохранившимся орудиям лова, промысел
птиц сетями был развит еще у жителей свайных построек Швейца-
рии, относимых к неолиту. У туземного населения многих стран
до сих пор весьма распространено использование птиц на мясо, на
шкурку и перья, идущие на украшение. По словам летописцев,
в эпоху древней Руси охота на крупных птиц и собирание яиц со-
ставляли один из видных промыслов населения на территории Евро-
пейской части СССР. В странах с более обедненной природой это
использование птиц как пищевого продукта отходит на второй план,
и охота на птицу приобретает преимущественно спортивное значение.
При этих условиях человек стремится не столько получить выгоду
добычей мяса или какого-либо иного продукта, сколько в процессе
охоты извлечь эстетическое наслаждение или удовлетворение физ-
культурного порядка. Тем не менее, промысловое значение охоты
на птицу сохраняется еще во многих странах и в том числе у нас
в СССР, особенно в зоне тундры, лесных пространствах севера и
(на водоплавающую птицу) в степных просторах юга.
Использование промыслово-охотничьей птицы происходит по
нескольким направлениям: дичный промысел, использование на
перо и пух, промысел яиц и наконец спортивное направление, вклю-
чающее не только охотничьих птиц собственно, но и птиц комнат-
ных, содержимых в неволе из-за их красоты или приятного пения.
Особняком стоит использование гуано — помета птиц, в огромном
количестве накапливающегося на месте массовых гнездовий, в усло-
виях особенно сухого климата.
Дичный промысел наиболее развит в районах, обладающих мас-
согыми гнездовьями птиц, главным образом в северных и южных
умеренных и субполярных широтах. В СССР подобный промысел
развит преимущественно в тундре и таежной зоне и местами в зоне
степи и пустыни (западносибирские озера, фазан на Сыр-дарье и др.).
В тундровой зоне издавна существует промысел на линную птицу,
главным образом гусей (рис. 315, 316), заготавливаемых впрок ту-
земным населением в вечной мерзлоте; производится отлов сильями
белых куропаток {Lagopus lagopus L.) и на побережьях с птичьими
базарами добываются кайры (главным образом Uria lomvia L.),
заготавливаемые для корма собак на зиму. В таежной зоне лежит
основная область заготовки экспортной лесной дичи, среди которой
первое место занимают рябчик (Tetrastes bonasia L.), тетерев (Lyru-
rus tetrix L.) и белая куропатка; меньшее значение имеют глухари
(Tetrao urogallus L. и Т. par vir os tris Bon.), утки и гуси.
В основном дичный промысел имеет потребительское значение
для местного охотничьего населения, составляя обычно важное под-
спорье в хозяйстве человека. Как товарный продукт, дичь исполь-
зуется лишь в районах, тяготеющих к железным дорогам, обеспе-
чивающим вывоз дичи в города и за границу. По условиям этого
транспорта, вывоз дичи возможен лишь в замороженном состойнйп
521
и осуществляется преимущественно зимой; это определяет промыс-
ловую ценность боровой дичи (наземных птиц, главным образом
тетеревиных), добываемой поздней осенью и зимой, и малую исполь-
зуемость в товарном отношении водоплавающих птиц, исчезающих
1?ис. 315. Загон линных белолобых казарок — Anser albifrons (Scop.) в сети промышленни-
ками в низовье реки Индигирки.
Сети расставлены на земле в виде широкой воронки, раскрытой в сторону воды. Стадо
линных гусей окружается на воде охотниками в легких лодках и медленно гонится к сетям,
в которых и запирается. По Zenzlnov.
Рис. 31G. Избиение линных белолобых казарок, после загона их в сети.
.Лтицв схватываются за шею пониже головы, и несколькими поворотами туловища
в воздухе шея ломается. По Zenzinov.
пз промысловых районов до наступления морозов. Для использо-
вания птицы в промысле важны также относительно крупная ве-
личина ее и легкость добывания различными самоловами (силками,
слопцами, тенетами и т. п.) или стрельбой пз ружей. Последние
годы, в связи с развитием консервной и коптильной промышленности,
начали вовлекаться в промысел и запасы дичи южных районов
(области пролета и зимовок), и при этом стали использоваться такие
виды, как пеликаны, бакланы, цапли, чайки и поганки; но пока
заготовки этих птиц еще не достигли широких масштабов.
Общая сумма дичного промысла определяется различно, от
37 000 000 штук (Туркин, 1902) до 70 000 000 (Соловьев, 1926) и
свыше 100 000 000 штук (Бутурлин, 1933), добывавшихся ежегодно
в довоенное время. В денежном выражении это оценивается по тогдаш-
ним заготовительным ценам в размере от 7 000 000 до 20000 000 руб.,
из которых почти на 2 500 000 руб. дичи экспортировалось за
границу.
Вне СССР дичный промысел более значительно развит в Канаде
и на пролетных водоплавающих птиц в Голландии, Франции и дру-
гих странах Европы, а также в Китае. В южных странах он носит
большей частью подсобное значение. Более значителен лишь про-
мысел пингвинов н$ островах к югу от Тасмании и Новой Зеландии,
добываемых здесь в количестве до 150 000 штук ежегодно (Mawson,
1921).
Необходимо отметить, что этот промысел во всех странах земетно
падает в своем значении, вследствие сокращения первобытных тер-
риторий, резкого уменьшения запасов птицы, замены дичных про-
дуктов продуктами домашнего птицеводства и других отраслей сель-
ского, хозяйства и т. п. Истребление дачи неумеренной охотой осо-
бенно наглядно сказалось в Северной Америке. В 1864 г. некоторые
торговцы Нью-Йорка продавали от 150 000 до 200 000 штук дичи
за полгода и в то же время отдельные торговцы получали от постав-
щиков зараз до 20 т лугового тетерева [Tympanuchus cupido (L.)], что
составляет около 20 000 штук этой птицы (Palmer, 1899). В настоящее
время эта птица сохранилась в совершенно ничтожном числе в одном
из восточных штатов и вряд ли может быть сохранена от вымирания.
Знаменитый странствующий голубь {Ectopistes migratoriiis L.) еще
в 1878 г. был описан гнездящимся в штате Мичиган огромной коло-
нией, покрывавшей 100 000 акров; здесь было заготовлено не менее
1 500 000 голубей убитыми и 80 532 голубя живыми. Теперь этот
голубь совершенно вымер в природе и недавно последний экземпляр
погиб и в неволе. Подобные случаи известны в истории многих куль-
турных стран. В СССР запасы дичи еще весьма велики и в промысло-
вом отношении несомненно еще не используются в достаточной степени.
Промысел пера и пуха в известной мере связан с дичным промыс-
лом, так как попутно используются и различные отходы этого про-
мысла. Однако до недавнего времени существовала и самостоятель-
ная охота за птицами, специально с целью добычи крыльев, хвостов
и вообще красивых перьев, употреблявшихся главным образом для
женских украшений. Самостоятельным промыслом является также
сбор гагачьего пуха.
523
Самим промысловым населением перо добытых птиц широко ис-
пользуется для приготовления перин и подушек; для этой цели идет
главным образом мягкое и мелкое перо уток и гусей. Туземное на-
селение Сибири, Канады, Гренландии, Аляски с давних пор исполь-
зует также шкурки гагар, поганок, лебедей, гусей и других водо-
плавающих птиц для приготовления или красивой отделки верхней
одежды. На севере Западной Сибири из лебединого меха иногда де-
лались малицы. Переработка птичьих шкурок в меха существовала
в довоенное время в Европе и в некотором количестве осуществляется
в настоящее время и в СССР. Наилучшие меховые шкурки полу-
чаются от гагар, поганок, лебедей, пеликанов, бакланов, гусей,
уток и некоторых других птиц, имеющих сравнительно крупные
размеры. Для получения меха употребляется преимущественно зим-
няя и позднеосенняя птица, находящаяся в свежем брачном наряде.
Шкурки различают на перовые (сохраняющие перо на коже) и пухо-
вые, с выщипанным пером; и те.и другие обычно имеют красивый
внешний вид и не требуют подкраски. По носкости перовые шкурки
гагар и поганок не уступают шкуркам белки, горностая и лисицы;
пуховые шкурки гуся близки по носкости к шкуркам сурка и прочнее
шкурок крота, а пуховые шкурки уток в этом отношении схожи
с шкурками кролика и зайца (Успенский, 1934). Весьма ценным
качеством птичьего меха являются его легкость и теплота, вследствие
чего он особенно пригоден для одежды летчиков; вообще же птичий
мех идет на приготовление детских шубок, женских шляп, муфт,
воротников и пригоден для подбивки пальто.
О размерах этого промысла в прошлом и в настоящее время можно
судить по следующим цифрам. На Обдорской ярмарке в 1877—1878 г.
было продано 3000 лебяжьих шкурок, в 1881 г. — 5175; шкурок
гагар и специально снятых красивых шеек их в 1858 г. было продано
2611, в 1870 г.—800 и в 1877—1878 г.—2000. На Констан-
тиновской ярмарке в б. Акмолинской области было продано в 1894 г.
800 лебяжьих шкурок и 8000 шкурок гагар (Силантьев, 1898).
До войны в Европе, главным образом в Голландии, ежегодно пере-
рабатывалось свыше 10 000 лебяжьих шкурок и свыше 50 000 шкурок
домашних гусей. В начале текущего столетия шкурки гагар и по-
ганок в огромном количестве (не менее 500 000 ежегодно) вывози-
лись из России, преимущественно из Западной Сибири, в Герма-
нию по цене до 1,5 марки за штуку (Успенский, 1934). В настоящее
время считается вполне реальной добыча в СССР шкурок птиц на
сумму в 2 000 000 руб. (Спангенберг, 1934).
Кроме Европы и СССР, птичьи меха добывались в Канаде, За-
падных штатах Америки, северной и южной Африке. Еще более
широкое распространение имела в недавнем прошлом торговля кра-
сивым пером, крыльями и хвостами птиц, употреблявшимися в ши-
роких размерах для украшения шляп и других частей одежды. У евро-
пейских народов это стало входить в моду в мужском костюме с XI Пв.
и в женском с XVI в. и в той или иной форме продолжалось до первых
лет настоящего столетия. Особенно ценились перья страусов, нанду,
марабу, эгретки цапель и перья различных райских птиц и много-
численных видов колибри. Очень скоро однако были найдены спо-
524
собы использования и более скромно окрашенных пли рассученных
перьев, и в промысел были вовлечены в сущности все виды птиц,
особенно крупных и средних размеров, могущих быть добываемыми
массами. До войны Россия занимала первое место по доставке па
западноевропейский рынок продуктов этого промысла, оценивав-
шихся приблизительно в 1 500 000 рублей и находивших сбыт для
переработки в «фантазийное перо» в Германии (на 700 000 руб.),
во Франции (500 000 руб.) и Австро-Венгрии (300 000 руб.) (Шнитке,
1925). Особенно пострадали от этого промысла Среднее и Нижнее
Поволжье (Житков, 1914), Украина, Западная Сибирь и Средняя
Азия. Есть сведения об отправке из Архангельска в Европу в 1900-х
годах партии одних крыльев белых куропаток весом в 10 т (Oldys,
1894). Заметное уменьшение количества птиц, вследствие такого уни-
чтожения их, вызвало в Америке, Европе, а затем и в России широ-
кое движение в защиту птиц и борьбу с модой на птичьи украшения.
В результате спрос на перья прекратился, промысел упал, и в на-
стоящее время в большинстве стран птицы находятся под охраной
закона, не разрешающего охоту в летнее время и вообще массовое
истребление птиц для целей моды. Следует отметить, что в СССР
заготовки птиц, главным образом сорочьих шкурок, еще практико-
вались в 1925—1927 гг. (вывезено 180000 сорок на сумму в 36 000руб.;
Залевский, 1935).
О размерах торговли перьями ярких экзотических птиц легко
судить по предложению на лондонских аукционах в 1913 г.: 32 000
перьев райских птиц, 27 000 затылочных перьев венценосных голу-
бей и 4000 подхвостовых перьев марабу. Стоимость одного экземпляра
красивейших райских птиц достигала для Paradisea apoda L. до 240
марок, Paradisea raggiana Scl. до 160 марок, a Manucodia jobiensis
Salvad. даже до 400 марок (Neunzig, 1931).
Из объектов этого промысла следует остановиться на промысле
белых цапель в целях получения эгреток, добыче страусовых перьев
н гагачьего пуха. Из белых цапель (род Egrettd) особенно ценится
большая снежно-белая цапля, в состав брачного наряда которой
входят длинные (до 48—50 см), сильно рассученные плечевые перья,
вырастающие обычно в количестве 30—50 штук и называемые в тор-
говле эгретками. Цена на эти перья, снятые со свежего экземпляра,
доходила в Астрахани в 1912 г. до 600 руб. за фунт, что составляет
наряд приблизительно 25—30 экземпляров (Житков, 1914). Интен-
сивное преследование этих цапель по всей Европе и в России привело
к резкому сокращению их колоний и местами к полному исчезнове-
нию их; только, в сущности, развитие движения охраны природы
спасло эту птицу от полного истребления; в СССР эта цапля нахо-
дится под строгой охраной закона. Промысел сохранился главным
образом в Южной Америке, где в Венецуэле практикуются специаль-
ная охрана массовых гнездовий этой птицы и сбор выпадающих
в послебрачный период перьев. Общая ценность этого промысла
в одном штате Апуре достигала в 1912—1914 гг. 171 555— 98 967 фун-
тов стерлингов. В Тунисе в недавнее время были проведены инте-
ресные опыты по одомашнению цапель, для чего молодыми они вы-
бирались из гнезд, помещались в большие вольеры, включавшие
525
участки оолота с деревьями, где впоследствии и размножались.
(Neunzig, 1931).
Промысел страусов для добычи перьев существовал еще в глу-
бокой древности. Преследование сильно сократило численность этих
птиц и вызвало попытки к одомашнению, которые увенчались успе-
хом. В настоящее время страусовые фермы существуют не только
в Африке (родине страуса), но также в Австралии, Аргентине, Бра-
зилии, Флориде, Аризоне и Калифорнии. Численность домашних
страусов только в одной Британской южной Африке годами дости-
гает 776 313 штук (1913 г.), обнаруживая однако с каждым годом
заметное падение (в 1926 г. — 104 578 штук); в Соединенных Штатау
Америки численность страусов в 1914 г. достигала 10 000 штук.
Ценные перья получаются от страуса обстрижкой, производимой
каждые 7—8 месяцев. Экспорт перьев из южной Африки — основ-
ной области культуры страуса — достигал по ценности в 1913 г.
2 953 587 фунтов стерлингов, но к 1925 г. упал до 203 976 фунтов.
Наибольший интерес в промысловом отношении представляет
для нас сбор'гнездового пуха обыкновенной гаги {Somateria molis-
simaL.), широко распространенной по морским побережьям и остро-
вам Арктики. Эта крупная утка, под покровительством человека
способная гнездиться массами на небольшой территории, утепляет
гнездо пухом, выщипанным из груди, отличающимся пышностью,
исключительной легкостью и упругостью; гагачий пух незаменим
там, где требуется особенно легкая и утепленная одежда. Одно гнездо
дает 20—30 г чистого пуха (Формозов, 1930), килограмм которого
расценивается сейчас около 600 руб. Таким образом одна гнездя-
щаяся самка дает весьма значительный доход, тем более ценный,
что в области своего распространения гага довольно нетребова-
тельна к условиям гнездования и в сущности не требует никакой
заботы, кроме охраны от хищников. В Исландии, славящейся своим
гагачьим хозяйством, гаги образуют густые многочисленные насе-
ления. «Мы высадились на скалистый, изъеденный волнами берег, —
пишет Шепард, путешествовавший по северо-западной окраине этого
острова, — повсюду были гаги и их гнезда; большие бурые самки
сидели на гнездах в огромных количествах и на каждом шагу выска-
кивали из-под наших ног. Лишь с большим трудом нам удавалось,
не наступать на гнезда. На противоположном берегу, как раз над
уровнем прилива, тянулась очень толстая стена из больших камней,
вышиной около трех футов. Из низа стены, с обеих сторон было
вынуто через один по камню, так что образовался ряд квадратных
помещений, служащих птицам гнездами. Почти каждое помещение
было занято, и пока мы шли вдоль берега, из них вылетала целая
вереница гаг, одна за другой. Поверхность воды была совершенно
белая, от самцов, приветствовавших своих буроватых подруг гром-
ким и шумным криком. Даже дом человека был настоящее чудо.
Окружающие его земляные стены и отверстия окон были заняты
гагами; вокруг дома, на земле, бахромой сидели те же птицы. Мы
могли их видеть также на торфяных скатах крыши, а одна гага си-
дела на скребке у порога. На лужайках, обращенных к морю, дерн
был снят квадратными кусками, величиной около 18 дюймов, и каж-
526
дая из образовавшихся ямок была занята гагами. Этой птицей были
наводнены ветряная мельница, все надворные постройки, все бугры,,
все скалы и трещины. Гаги были повсюду. Многие из них оказались
такими ручными, что мы могли гладить их на их гнездах, а хозяйка
сказала нам, что вряд ли на всем острове найдется хоть одна наседка,
которая не позволила бы ей взять из гнезда яиц без страха и сопро-
тивления» (Формозов, 1930). Собранный пух очищается вручную или
специальными машинами, сортируется и поступает в продажу. Ма-
ленькая Исландия вывозит ежегодно 2400—2800 кг гагачьего пуха.
Норвегия 1200—1600 кг, южная Гренландия 1200—2000 кг. Со-
временный промысел в СССР очень мал (не свыше 500 кг), хотя не?
сомненным является, что при хозяйственном подходе к гаге, соот-
ветствующей охране гнездовий и привлечении ее, гага представляла’
бы ценную статью экспорта как для наших европейских побережий,
так вероятно и для отдельных районов азиатского сектора Арктики.
Когда-то наша страна даже стояла на первом месте по добыче га-
гачьего пуха, вывозя его (по данным Германа, относящимся к 1804—
1805 гг., и расчетам Формозова, 1930) по 30 000—35 000 фунтов и со-
бирая вообще около 120 000—140 000 фунтов, общей ценностью не
менее чем на 12 000 000 руб. по современным ценам. Развитие и ин-
тенсификация промысла гагачьего пуха является одной из важных
задач в освоении нашего севера.
Не последнюю роль в использовании перьев птиц играло ранее
приготовление писчих перьев, распространенных с VI и особенно
с XVII вв. и употреблявшихся до XIX в., до появления металли-
ческих перьев. В основном для этой цели употреблялись гусиные
перья, преимущественно домашних птиц. Меньшее значение, но
сохранившееся до настоящего времени, имеет приготовление из
очинов поплавков, кистей и щеточек, зубочисток, мушек для рыбо-
ловных крючков и различных украшений в виде цветов, бабочек
и т. п., распространенное у некоторых народов.
Промысел птичьих яиц является одним из самых распространен-
ных видов использования дикой птицы, издревле осуществляемый
хищнически и не только специально промышленниками, но зачастую
попутно, при производстве каких-либо других работ, особенно рыб-
ной ловле. Более регулярно используются массовые гнездовые ко-
лонии чаек, кайр, бакланов, чибисов, цапель и других птиц, гнезда
которых легко находимы, скучены .на небольшом пространстве,
сравнительно легко достигаемы; попутно собираются яйца и всех
других более крупных птиц, гнезда которых окажутся поблизости.
В сборке яиц принимают широкое участие подростки и женщины;
кладка обычно выбирается целиком, а сильно насиженные яйца
зачастую разбиваются. Урон, наносимый дикой птице в результате
такого промысла, несомненно очень велик и может быть является
главнейшей причиной общего оскудения крупной птицы, отмечае-.
мого для многих районов.
Соловьев (1926) оценивает ежегодную сборку яйца дикой птицы
по всему СССР в количестве не менее 60 000 000 штук. Сбор яиц
кайр только одной первой кладки на Новой Земле может составлять
2 000 000 экземпляров (Портенко, 1931). В районе Чанских озер
527.
в Западной Сибири каждым крестьянским хозяйством собирается
ежегодно для еды две-три сотни яиц (в среднем) водоплавающих
птиц; в 1930 г., когда средний сбор яиц составлял лишь 145 яиц
на семью, общее потребление района (47 000 хозяйств) должно было
(•оставлять около 9 000 000 яиц, не считая еще многих тысяч погу-
бленных, сильно насиженных кладок (Жданович, 1931). По весьма
ориентировочным подсчетам, сбор яиц чаек на берегах Онежского
и Кандалакшского заливов составлял до войны не менее 1 000 000 эк-
земпляров.
Основная масса собираемых яиц употребляется населением для
личного потребления, так как по вкусу яйца даже рыбоядных птиц
почти неотличимы от яиц кур и гусей. На севере яйца идут также
в пищу собакам и служат для привады песцов и других промышляе-
мых хищников. В конце прошлого столетия на Командорских остро-
вах ежегодно собирались массами яйца и, нагруженные в бочки, от-
правлялись в Сан-Франциско и Петропавловск для розничной про-
дажи. До 1922 г. норвежцы вывозили яйца кайр с Новой Земли для
целей мыловаренного производства.
Крупный промысел на яйца птиц существует и во многих других
местах земного шара. Сбор яиц кайр на Фораллонских островах
в Калифорнии еще в 1850—1856 гг. оценивался в 100 000—200 000
долларов в год. В южном полушарии интенсивно используются
различные трубконосые птицы (Palmer, 1899), (рис. 317).
В .организации яичного промысла отрицательной стороной яв-
ляются бесхозяйственное разорение гнездовых колоний, многократ-
ный сбор яиц в них, уничтожение насиженных яиц и небрежность
в обращении с огнем, что на южных озерах вызывает пожары ка-
мышей, которые губят огромные массы птицы. В целях сохранения
наших дичных богатств промысел яиц может быть допускаем лишь
на действительно массовых гнездовьях, с выбором (и лишь частич-
ным) только яиц первой кладки, с одновременной организацией
охраны гнездовий от хищников и браконьеров, с заботой о сохране-
нии гнездовий от степных и лесных пожаров.
Переходя к обзору охотничье-спортивного значения птиц, необ-
ходимо подчеркнуть, что для обширных районов СССР и всего мира
оно не уступает значению промысла дичи, а иногда и превышает
его по своей общей ценности.
Объект этой охоты обычно .остается в личном потреблении охот-
ника и числендо добывается, как правило, в сравнительно неболь-
шом количестве. Этот вид* охоты является способом использования
досуга, укрепляющим здоровье, развивающим выносливость, силу,
меткость, ориентировку в природе и знание и понимание ее. Цен-
ность этой охоты не столько в добыче мяса и шкурок, сколько в той
закалке, в том воспитании человека, которое создается в процессе
отыскивания, наблюдения и преследования птицы. Число лиц, за-
нимающихся подобной охотой, очень велико. По подсчетам Соло-
вьева (1926), из наиболее вероятной цифры охотников в СССР, рав-
ной 2 000 000 человек, не менее 500 000 составляют любителей-
спортсменов и около 1 250 000 человек занимаются охотой в смешан-
ных целях, т. е. отчасти под влиянием тех же стимулов. Спортивное
£38
значение охоты еще более значительно в других странах и, например
в Соединенных Штатах Америки, число спортсменов достигает
5 000 000 человек.-
Спортивная охота использует более широкий ассортимент видов,
включая и значительную часть тех видов, которые являются про-
мысловыми. Существенными являются не только величина и массо-
вость того или иного животного, но и характер возможных способов
охоты. Возможность охотиться с собакой, отыскивающей затаив-
шуюся птицу и после выстрела приносящей ее хозяину, возмож-
ные. 317. Сборы япц альбатроса — Biome dea immulabilis Botsch. па островах Лайзан.По
Palmer.
ность стрелять с места, из засады, трудность и эффектность стрельбы
на лету — все это методы, в значительной мере определяющие ис-
пользуемые для охоты виды. Спортивная охота является почти все-
цело охотой ружейной, в противоположность промысловому исполь-
зованию птиц, основанному в значительной мере на применении раз-
личных самоловов.
В СССР наиболее ценимыми объектами спортивной охоты яв-
ляются утки, куриные и кулики. Из первых особенно ценны широко
распространенные: кряква (Anas plathyrhyncha L.), серая утка
(Anas strepera L.), шилохвость (Anas acuta L.), чирок-свистунок
(Nettion crecca L.j и чирок-трескунок (Querquedula circia L.), более
доступные обычным способам охоты и обладающие особенно вкус-
ным мясом. Менее используются обычно: свиязь (Anas penelope L.),
34 Орнитология—1209
529
широконоска (Anas clypeata L.), хохлатая чернеть (Nyroca cristata
L.), морская чернеть (Nyroca marila L.), гоголь (Bucephala clan-
gula L.), из гусей — серый гусь (Anser anser Ь.)и гуменники (An-
ser fabalis) и в еще меньшей степени крохали, лебеди, казарки и
пеганки. Из видов, имеющих крупное, но относительно локальное
значение, надо указать: красноголовую чернеть (Nyroca jerina L.),
клоктуна [Nettion jonnosum (Georgi)], белоглазого нырка [Nyroca
nyroca (Guld.)] и белолобую казарку (Anser albifrons Scop.). На уток
обычно охотятся: с подхода или подъезда на лодке к кормящимся
стаям; из засады на зорях во время перелета с жировочного водоема
на другой, для отдыха; с манной «криковой» уткой на селезней,
засев поблизости в укрытое место; с собакой на затаивающихся
молодых уток и другими способами. На гусей охотятся главным об-
разом из засады, подкарауливая птиц, летящих на кормежку или
обратно.
Из куриных птиц наиболее ценны в спортивной охоте тетерев
косач, рябчик и серая куропатка; в меньшей степени развита охота
на глухаря, белую куропатку и перепела. Локальное, но крупное
значение имеет охота на кеклика (Alaectoris kakelik Falk.) и фазана
(Phasianus colchicus L.). Наиболее распространена охота на токах:
на тетерева — обычно из шалаша, выстроенного у токовища, на
глухаря — с подхода к «поющей» птице, на рябчика, живущего в
однобрачии, — подманивая его «на пищик», которым подражают
его голосу. Кроме того, охотятся с собакой по рослым выводкам,
подманивают зимой к чучелам, выставленным на кормовых березах
(тетерев), скрадывают на кормежке.
Из куликов, типично спортивных птиц по характеру своего исполь-
зования, распространена охота на вальдшнепа (Scolopax rusticola
L.), бекаса (Capella gallinago Briss.) и дупеля (Capella media L.),
в меньшей степени на гаршнепа (Lymnocriptes minima Brunn.),
турухтана (РаооцсеИа pugnax L.), чернохвостого веретенника (Li-
mosa limosa L.), большого кроншнепа (Numenicus arquatus L.), и
чибиса (Vanellus vanellus L.); остальные кулики обычно исполь-
зуются лишь попутно, вследствие своей слишком малой величины,
или осторожности и малочисленности. Охотятся на этих птиц, по-
мимо обычного скрада или подхода, преимущественно с легавой со-
бакой, делающей над птицей стойку; на дупеля и турухтана рас-
пространена охота также на токах и на вальдшнепа — на тяге, под-
карауливая птицу в вечерних или утренних сумерках совершающей
характерные брачные перелеты.
Меньшее охотничье значение по своей малой распространен-
ности или локальности имеют голуби, рябки, поганки и различные
журавлеобразные. Из голубей более ценны: вяхирь (Columba рalum-
bus L.), клинтух (Columba oenas L.) и дикий сизый голубь (Columba
livia Briss.), добываемые обычно осенью, во время их кормежек на
полях или перелетах. Рябки, представленные у нас главным обра-
зом тремя видами, саджей (Syrrhaptes paradoxus Pall.), чернобрю-
хим рябком (Pterocles arenarius Pall.) и белобрюхим рябком (Ptero-
cles alchata L.), являются излюбленным объектом охоты в Средней
Азии, но не распространены в СССР за ее пределами. Эта группа
ЬЗО
птиц добывается интереснейшей охотой на водопоях, к которым
птицы прилетают в определенное время суток большими, иногда
громадными стаями. Из журавлеобразных наибольшее значение
имеют дрофиные: стрепет (Tetrax tetrax L.), дрофа (Otis tarda L.)
и отчасти джек (Chlamydotis macqueeni Gray.), являющиеся излюб-
ленным объектом степной охоты. Эти весьма осторожные птицы добы-
ваются обычно с подъезда, при медленном приближении к птице в про-
цессе объезда ее по кругу, или нагоном на спрятавшегося охотника. Из
того же отряда более часто добывается также лысуха (Fulica atra
L.) и изредка, более или менее случайно, коростель (Crex crex L.),
курочки (род Porzana), журавли- (Megalornis) и др.; исключитель-
ная скрытность этих видов, живущих в густой болотной раститель-
ности, или крайняя осторожность мешают их использованию. Что же
касается поганок и лысух, недостаточная спортивная охота на них
объясняется частью скрытным образом жизни видов и исключитель-
ной способностью к нырянию, частью некоторыми предрассудками
охотников.
В настоящее время трудно подсчитать количество птиц, добывае-
мых подобной спортивной охотой. Ойо несомненно составляет значи-
тельную часть общей добычи птиц в СССР, оценка которой уже была
приведена выше. В других странах продукты спортивной охоты со-
ставляют подавляющую часть общего числа добываемых птиц.
Широкое развитие спортивной охоты привело во многих странах к
необходимости мероприятий по охране птицы, выразившихся в уста-
новлении коротких сроков охоты, отнесении их лишь на осень и
зимний период, с запрещением охоты не только в период вывода
птенцов — летом, но й весной, в период, подготовительный к раз-
множению. К другим мероприятиям относятся: установление опре-
деленных лимитов отстрела птиц на охоте, допускаемого для охот-
ника, организация заказников для восстановления запасов дичи и
более постоянных заповедников, призванных сохранить участки
природы в относительно первобытном состоянии. В СССР для ох-
раны птиц применяется большая часть этих мероприятий и широко
осуществляются организация охотничьих хозяйств и закрепление
угодий за определенными коллективами охотников. В число хозяй-
ственных мероприятий по охоте входят: отстрел вредных хищников,
подсадка кормовых растений (для уток например — подсевы канад-
ского риса— Zizania aquatica, ценных видов рдестов — Potamo-
geton pectinatus и различных других видов), устройство защитных
полос из кустарников и других растений для гнездования птиц и
укрытия от непогоды, иногда зимняя подкормка и т. п. За границей
практикуется также обогащение фауны акклиматизацией новых
охотничьих видов птиц; так в Северной Америке удачно акклимати-
зированы чуждые местной фауне серая куропатка (Perdix perdix L.),
обыкновенный фазан (Phasianus colchicus L.; Phillips, 1928); в Шот-
ландии реакклиматизирован глухарь (Tetrao urogallus L.) и не-
которые другие виды (Ritchie, 1920). С этими мероприятиями ко-
нечно сочетаются и общие усилия, направленные к защите и охране
птиц от истребления, распространяемые на все группы птиц. Цен-
ность этих усилий велика и для СССР, несмотря на сравнительное
*
531
богатство нашей природы. К настоящему времени и на нашей тер-
ритории вымерли или сильно сократились в численности целый ряд
видов птиц, некогда являвшихся обильными. На Командорских
островах у восточных берегов Камчатки, вместе с известной мор-
ской коровой {Rhytina stelleri) вымер большой и неплохо летавший
стеллеров баклан {Phalacrocorax perspicillatus Pall.), почти исчез
из северо-восточных тундр Сибири белый гусь {Chen hyperboreus
Pall.), еще не так давно встречавшийся здесь огромными стаями;
вымерла хохлатая пеганка {Pseudotadorna cristata Kur.,) встречав-
шаяся ранее в Южно-Уссурийском крае. Немало есть и птиц, уже
сейчас настолько немногочисленных, что если не принять соот-
ветствующих мер, они наверняка последуют за предыдущими. Так
очень редки стали в СССР: китайский ибис {Nipponia nippon Temm.),
китайский белый аист {Ciconia boyciana Swinh.), стерх {Sarcoge-
ranus leucogeranus Pall.), журавль-монах {Grus monachus PalL),
даурский журавль {Grus vipio) и нек. др. Это все крупные,
очень осторожные птицы, которые исчезают просто потому, что
не терпят соседства человека и при малейшей тревоге бросают
свои насиженные места. Гораздо больше таких видов, которые исче-
зают пока-что только местами, по преимуществу там, где в отно-
шении их резче сказалась деятельность человека. Так несмотря
па охрану, пока еще малочисленны на Украине белые цапли {Eg-
retta alba L.), исчезают стрепета [Tetrax tetrax L.), исчезли во многих
районах на гнездовье серые гуси {Anser anser L.), крайне редки
стали в устьях рек Черного моря пеликаны {Р elecanus onocrotalus L.).
Немногими колониями сохранились еще в Казахстане красивые
фламинго {Phoenicopterus roseus Pall.), стали редки на мурманском
берегу обыкновенные гаги и т. д. Исчезновение этих птиц конечно
стоит в связи как с спортивным преследованием их, так и слишком
интенсивным промыслом, и поэтому. является итогом всякой охоты
и деятельности человека вообще.
По существу лишь одним из разделов охот спортивного исполь-
зования птиц является ловля мелких птиц, главным образом раз-
личных воробьиных, для содержания их в неволе. Распространение
этого обычая приурочено преимущественно к городам п в СССР не
имеет столь большого значения, как в других странах. Тем не менее
для известных групп населения эта «охота» доставляет много удо-
вольствия и вызывает к жизни специальный небольшой промысел.
В СССР наиболее излюбленными объектами содержания в неволе
являются: чиж {Spinus spinus L.), щегол {Carduelis carduelis L.),
снегирь {Pyrrhula pyrrhula L.), реполов {Acanthis cannabina L.),
овсянка {Emberiza citrinella L.), зяблик {Fringilla coelels L.) и боль-
шая синица {Parus major L.y. Менее распространены, требуя за
собой более тщательного ухода: восточный соловей {Luscinia philo-
mela L.), лесной жаворонок {Lululla arborea L.)t полевой жаворонок
{Alauda arvensis L.), певчий дрозд {Turdus ericetorum Turton.),
черный дрозд {Turdus merula L.), славка Черноголовка {Sylvia
atricapilla L.), зорянка (Erithacus rubecula L.), московка {Parus
ater L.), гаичка {Poecile atricapillus L.), лазоревка {Cyanistes coeru-
leus L.), скворец {Sturnus vulgaris L.) и ряд других. Из привозных
532
и разводимых в неволе наичаще встречаются: канарейка (Serinus
canaria L.), волнистый попугайчик {М elopsithacus itndulatus Shaw.)
и нек. др. Кроме того, в неволе держат перепелов {Coturnix cotur-
nix L.) и кекликов {Alaectoris kakelik Falk), ценя последних за драч-
ливость самчиков между собою. Содержание всех этих птиц свя-
зано с развитием птицеловства. Добывают мелких птиц насто-
роженными сетками (понцами, лучком), самоловами (чопцы, силки),
ловлей на птичий клей и другими способами.
В других странах этот «спорт» является более широко развитым
и использует огромное количество ярко и красиво окрашенных
тропических видов: различных попугаев, ткачиков, вьюрковых,
танагр и многих насекомоядных. В начале нашего столетия в Со-
единенные Штаты ежегодно импортировалось не менее 300 000 кле-
точных птиц (Oldys, 1899). В Англии, Шотландии, Бельгии, Гер-
мании и некоторых других странах в массе разводятся канарейки
для широкой торговли ими. В Граце (Австрия) некоторые села спе-
циально промышляют разведением канареек и славятся высокими
качествами этих птиц (особенно канарейки Андреасбурга). Орга-
низация специальных обществ любителей комнатной птицы и вы-
ставок сопутствует этому занятию.
Следует хотя бы кратко упомянуть о том, что человек издавна
использует некоторых птиц в качестве своих помощников, «ловчих
птиц», при охоте на других животных. Для этой цели приучают
обычно ястребов-тетеревятников {Astur gentilis L.), ястребов-пере-
пелятников {Accipiter nisus L.), орлов-беркутов {Aquila chrysaetosL.),
соколов-сапсанов {Falco peregrinus Tunst.) и кречетов {Hierofalco
gyrfalco L.), которые вынимаются из гнезд маленькими птенцами,
воспитываются в неволе и дрессируются для охоты на фазанов,
серых куропаток, кекликов, тетеревов (тетеревятник, сапсан),
перепелов (перепелятник), лисиц, зайцев, даже дроф и джейранов
(орлы) и т. п. Эта охота повидимому существовала у народов
Азии уже за тысячу лет до нашей эры, а в средние века, при-
мерно до конца XVIII века, была широко распространена
и у европейских народов. В настоящее время эта охота более
широко практикуется лишь у степных народов Средней и Цен-
тральной Азии, хотя местами еще сохраняется и в других стра-
нах, например в Европе (Англия, Германия и др.).
Сходным образом исполь?уется баклан {Phalacrocorax carbo L.),
дрессируемый главным образ м в Китае и Японии для охсты за
рыбой, которая отбирается у птицы, как только последняя по-
явится из воды с добычей; чтобы рыба не могла быть проглочен-
ной, на шею баклана надевается узкое кожаное кольцо, недо-
пускающее расширение пищевода.
Наконец особый тип промыслового использования птиц пред-
ставляет эксплоатация гуано (птичьего помета), осуществляемая
в некоторых засушливых тропических областях земного шара. От-
ложения гуано встречаются на месте массовых гнездовий птиц почти
во всех широтах, но обычно в небольших количествах и низкого
качества. Гуано, имеющее коммерческую ценность, ограничено
исключительно засушливыми областями тропиков и приурочено
533
главным образом к островкам западного берега Южной Америки
(Чинча, Гуанап®, Лобос и др.). Менее богатые залежи находятся на
островах Галапагосских, Лайзан и некоторых других островах,
в южной части Тихого океана, расположенных возле Нижней Кали-
форнии, Венецуэлы, Колумбии, Экуадора, Боливии, Австралии
(о. Сван), южной и западной Африки (Асценсион, Ишабоэ, залив
Алгоа и др.) и на Аравийском побережье (Palmer, 1899). Гуано упо-
требляется как удобритель, и его ценность заключается в богатстве
соединениями азота и фосфатов. 1 В сухих тропиках помет птиц
быстро высыхает и с течением времени не претерпевает значитель-
ных изменений, в то время как в влажных и более холодных обла-
стях быстро наступающее гниение приводит к потере почти всех
органических соединений, а щелочи и фосфаты вымываются дож-
дями. Основными производителями гуано на Перуанском побережье
(острова Чинча и др.) являются бакланы и олуши, образующие
колосальные гнездовые колонии. 2 Кроме того, крупное значение
здесь же и в других местах имеют альбатросы и другие трубконосые,
пеликаны, чайки и крачки и местами пингвины. Ценность гуано,
как удобрителя, была известна перуанцам более четырех столетий
тому назад. В перйод существования государства инков гуановые
острова и гнездовые колонии их тщательно охранялись, и истреб-
ление птиц в гнездовый сезон каралось смертью. Впервые сведения
о гуано были привезены в Европу А. Гумбальдтом (1804), но их
ценность была осознана лишь в сороковых годах этого столетия.
Признание гуано ценным удобрителем повлекло за собой интенсив-
ную эксплоатацию Перувианских островов и поиски других отло-
жений; в пятидесятых годах экспорт составлял около 200 000 т
и вскоре достиг общей суммы вывоза в 5 000 000 т. Первое время
запасы перувианского побережья казались неисчислимыми, так
как местами отложения гуано на островах Чинча залегают на глу-
бину в 30 м, и были подсчитаны в количестве 12 376 100 т. Однако
вскоре интенсивный вывоз стал сильно истощать эти запасы, что
повлекло за собой постепенное угасание этой торговли. Использо-
вание гуано еще более сократилось с введением искусственных удоб-
рителей. 3
1 По своему богатству азотистыми соединениями перувианское гуано счи“
тается более чем в тридцать три раза более эффективным удобрителем, чем навоз
скотного двора; особенная ценность гуано заключается в благоприятном для
абсорбции растениями сочетании химических соединений (Murphy, 1936).
2 Основными производителями в настоящее время являются прежде всего
Phalacrocorax bougainoillii (Less.) и затем Sula oariegata (Tschudi) и Pelecanus occi-
dentalis thagus Mol. Ряд ранее важных видов — ныряющие буревестники —
Pelecanoides garnodii (Less.) и др., крачка—Lar osier па inca (Less.) и пингвин —
Spheniscus humboldti Меуеп теперь потеряли свое значение (Murphy, 1935).
3 Последние десятилетия использование перувианского гуано вновь воз-
растает, в результате более разумного и хозяйственного использования совре-
менных колоний. За пятилетия с 1909 по 1934 г. добыча гуано развивалась сле-
дующим образом (в метрических тоннах): 1909—1914 гг.—50 916, 1914—1919 гг.—
63 866, 1919—1924 гг. — 75 280, 1924—1929 гг. — ИЗ 749, 1929—1934 гг. —
121 131, а в 1934 г. добыча превысила 160 000 т (Coker, 1935). Добыча сильно
колеблется также по годам, вследствие периодического изменения в численности
птиц, благодаря меняющимся метеорологическим и гидрологическим условиям
района (Murphy, 1936).
534
Переходим к обзору использования домашних птиц в хозяйстве
человека. В этом отношении основное значение принадлежит курам,
в меньшей степени уткам и гусям и третьестепенное и частью деко-
ративное значение имеет разведение индеек, цесарок, лебедей, раз-
личных фазанов и некоторых других красивых птиц. Несколько
особняком стоит голубеводство, имеющее частью чисто любитель-
ский, частью важный военный интерес.
Ближайшим диким видом птиц к нашей курице является банкив-
ский петух {Gallus gallus L., G. bankiva L.), распространенный no
всей Индии и на Индо-Малайском архипелаге; он рассматривается
прародителем домашних кур, одомашненным по всей вероятности
в весьма отдаленное время в пределах Индии. Куры упоминаются
уже в «Ведах» древних индусов, составление которых относится
к 1500—2000 лет до нашей эры. В Европу куры попали значительно
позднее, так как из ранних авторов о них упоминает лишь Аристо-
фан, живший за 400—500 лет до нашего летоисчисления. Позднее
куры были расселены очень широко, причем образовали большое
количество разнообразных пород, используемых человеком для раз-
личных целей.
Сельскохозяйственное значение курицы очень велико. Яйца и
мясо птицы очень питательны и вследствие легкой переваривае-
мости особенно ценны для больных; пух и перо находят себе широ-
кое применение для набивки подушек, украшения и т. д. (см. выше);
птичий помет является прекрасным удобрителем, в этом отношении
превышая в 3—4 раза конский помет; наконец птица потребляет
большое количество вредных в хозяйстве насекомых и червей. В от-
ношении разведения, курица представляет исключительные удоб-
ства; птица быстро достигает половой и хозяйственной зрелости,
несет много яиц и вследствие этого может быть быстро размножена;
удовлетворяясь вообще сравнительно небольшим количеством корма,
курица к тому же может быть вскормлена на малоценных отбросах
сельского хозяйства, в виде неполноценного зерна и других отхо-
дов; часть времени куры могут быть содержимы вообще на свободе
и почти не требуя за собой ухода; содержание птицы возможно в
дешевых постройках и доступно при небольших средствах. Куро-
водство в сущности возможно повсеместно, начиная от крайнего
севера до крайнего юга; продукты птицеводства всегда находят
спрос и очень удобны1 для сбыта (Иванов, 1931). Вследствие этого,
данная отрасль сельского хозяйства считается одной йз наиболее
перспективных и как в мелких формах, так и в виде крупных хозяйств
получила широкое распространение как у нас, так и за границей.
В России до конца прошлого столетия птицеводство (в основ-
ном куроводство) являлось мелкой отраслью хозяйства, пре-
имущественно подсобного значения. Вывоз продуктов (главным
образом яиц) начался лишь в восьмидесятых годах, достигнув в
предвоенный период значительных размеров, на сумму свыше
100 000 000 руб. в год. В 1911 г. производство продуктов птицевод-
ства в Европейской части России оценивалось в сумме'320 000 000
руб., и экспорт составлял к общему вывозу продуктов животновод-
ства около 40%; в 1913 г. эта цифра поднялась до 45%. После рево-
535
люцпи птицеводство стало быстро возрастать начиная с 1923 г.
В 1928 г. заготовка яиц достигла 17 800 вагонов, из которых 9 115
вагонов было экспортировано.
С развитием коллективизации и совхозного строительства в
СССР создаются чрезвычайно благоприятные условия для расши-
рения и укрупнения птицеводства. Являясь доступным самому мел-
кому хозяйству, производство только при условии крупного хозяй-
ства может использовать все новейшие достижения науки и стать
особенно выгодным. Специализация хозяйств в том или ином на-
правлении, создание больших инкубаторов и севооборотов, обеспе-
чивающих птицу кормами и выгулами, обеспеченность подготовлен-
ным техническим и высококвалифицированным руководящим персо-
налом, естественно, доступны только крупным организациям. В на-
стоящее время такие крупные хозяйства уже созданы и продолжают
создаваться. Число яйцемест в инкубаториях СССР растет быстрыми
темпами. Если в 1929 г. инкубаторов имелось всего на 260 000 яйце-
мест, то уже в 1931 г. их число поднялось до 21 500 000 яйцемест,
и в 1932 г. достигло 42 000 000 яйцемест. Проводится большая ра-
бота по селекции птицы, по организации племенных рассадников,
подготовке кадров птицеводов различной квалификации, на кото-
рых и ложится эта чрезвычайно важная и ответственная задача по
развитию нашего птицеводства.
Все породы кур можно разделить на следующие типы и группы
(Иванов, 1931).
ТИПЫ
I. Бойцовый
II. Яйценоский
ГРУППЫ И ПОРОДЫ
| Породы
| Малайские бойцовые
) Индийские бойцовые или азиль
| Английские бойцовые старого типа
Английские бойцовые нового типа
V Корнуэльско-индийские бойцовые
( 1. ГРУППА СРЕДИЗЕМНОМОРСКАЯ
Породы
Итальянские (ливорнские, легхорны)
Анконские
Кастильские
Минорки
Испанские черные белолицые
Андалузские ’
2. ГРУППА СРЕДНЕЕВРОПЕЙСКАЯ
Породы
Гамбургские
Кампинские
Польские
Русские простые
Русские ушанки
Русские гилянские
Русские орловские
Русские павловские
3. ГРУППА ЮГО-ВОСТОЧНАЯ ЕВРО-
ПЕЙСКАЯ
Породы
Голошейпые или семиградские
536
III. Мясной
IV. Общепользовлтельный
V. Декоративный
1. ГРУППА АНГЛИЙСКАЯ
Породы
Доркинги
Суссенекие
2. ГРУППА ФРАНЦУЗСКАЯ
Породы
Брестские
Ляфлеш
Гу даны
Крэвкеры
Мантские
3. ГРУППА БЕЛЬГИЙСКАЯ
Породы
Куку-де-малинь (кукушечные)
Брекель
Брабантские
4. ГРУППА АЗИАТСКАЯ
Породы
Кохинхины
Брамы
Лангшаны
1. ГРУППА АНГЛИЙСКАЯ
Породы
Орпингтон
2. ГРУППА ФРАНЦУЗСКАЯ
Породы
Фавероль
3. ГРУППА АМЕРИКАНСКАЯ
Породы
Доминикские
Плимут-рок
Виандот
Род-айленд
Породы
Бесхвостые
Курчавые
< Шелковые
Иокогама
Феникс
Бентамки или карликовые
Бойцовый тип кур создан человеком путем подбора и тренировки?
в связи с издревле существующим развлечением — петушиными
боями. Птицы этого типа отличаются небольшой головой с коротким
острым клювом, длинной мускулистой шеей, крепкой грудью и не-
сколько вертикальным положением туловища; петухи отличаются
большой храбростью, выносливостью и силой. Наиболее древними
породами являются малайские и индийские бойцовые куры, выведен-
ные повидимому в южной Азии.
537
Яйценоский тип кур по своему происхождению является евро-
пейским типом, по мнению некоторых исследователей, возникшим
из итальянских кур. Это птицы средней величины, отличающиеся
высокой производительностью (130—200 и выше яиц в год). Среди-
земноморская группа пород выведена из итальянских и испанских
кур, улучшенных в Англии и Америке. Из них наибольшей
известностью пользуются легхорны и минорки. Первые получили
свое название от города Ливорно в Италии, откуда они были
впервые вывезены. По яйценоскости эти куры занимают одно из
первых мест, давая от 120 до 200 яиц в год и в отдельных
линиях даже до 300—315 яиц, 65—70 г весом. Характерна также
ранняя половозрелость и носка яиц уже с 6—7-месячного возраста.
По окраске это преимущественно куры белой, куропатчатой или чер-
ной масти, с высоким, прямостоящим и листовидным гребнем, с бе-
лыми ушными мочками и пышным вертикальным хвостом. Вес ку-
рицы достигает 2 кг, петуха — 2,5 кг. Это одна из наиболее распро-
страненных пород, в Соединенных Штатах Америки составляющая
до 80% всего поголовья кур. В СССР легхорн особенно рекомен-
дуются для южных яйцепромышленных районов. Минорки близки
к легхорнам и отличаются главным образом иным цветом кожи и ног
(белой или серой кожей, вместо желтой, и телесным цветом ног,
вместо желтого или черного). Это более крупные куры, весом, до
•3—3,5 кг (сГсГ), и преимущественно темной окраски, выведенные из
кур о. Минорка. По хозяйственным качествам минорки славятся
величиной своих яиц, достигающих 75—85 г весом, хорошей носко-
стью (до 180 яиц в год) и скороспелостью. Рекомендуются для хозяй-
ственной полосы СССР. Существенное значение имеют и другие по-
роды этой группы кур.
Среднеевропейская группа пород объединяет главным образом
гамбургских, польских и русских кур. Первые обладают преимуще-
ственно пятнистой окраской, розовидным гребнем и серой кожей;
хозяйственном отношении они хуже средиземноморских пород,
так как мелки (cfd* 2—2,5 кг, 99 до 1—1,5 кг) -и несут небольшие
яйца. Польские куры, характеризующиеся густым хохлом перьев
на голове, являются скорее декоративной породой, хотя до появ-
ления итальянских кур в Западной Европе считались лучшими не-
оушками. «Русские куры» являются в сущности собирательным на-
званием для ряда наших местных пород, до сих пор недостаточно
изученных как в отношении происхождения, так и хозяйственных
качеств. Среди них находятся породы, сходные с гамбургскими (на-
пример южнорусские ушанки, белые, палевые, ситцевые и др.);
другие напоминают азиатских кур (орловские куры, юрловские голо-
систые и нек. др.). Это преимущественно небольшие куры, не всегда
с хорошей яйценоскостью, но с большой выносливостью, нетребова-
тельностью и неприхотливостью в отношении корма, ухода и содер-
жания. Наиболее выделяются своими качествами ушанки, с харак-
терными украшениями на голове в виде густых бак, прикрывающими
ушные мочки. Они достигают значительного веса (сГсГ 3—3,3 кг,
99 2—2,5 кг), несут довольно много яиц (до 200 штук в год)
и сравнительно не боятся морозов. Затем следует отметить орлов-
.538
ских кур, достигающих 4—4,5 кг весом (с?сГ до 2,5—3,3 кг) и
несущийся до глубокой осени, и оригинальную породу голосистых
кур, выведенных в с. Юрлове Ливенского района (ЦЧО) любите-
лями петушиного пения.
В последнюю группу яйценоских кур — юго-восточную евро-
пейскую — относят голошейных или семиградских кур, встречаю-
щихся и в южных районах СССР. Семиградские куры лишены опе-
рения на шее и отличаются ярко-красной окраской кожи ее. Это
хорошие несушки,, дающие до 180 довольно крупных яиц в год,
скороспелые и хорошо откармливающиеся.
Мясной тип кур отличается обычно крупными размерами, скоро-
спелостью, быстрой откармливаемостью и очень нежным, сочным и
вкусным мясом; способность к носке яиц у них обычно понижена.
Их значение в куроводстве СССР пока очень невелико. Из наиболее
ценных пород следует отметить доркингов, имеющих по пять паль-
цев на ногах. Они являются умеренными несушками, но обладают
отличными мясными качествами; вес петухов достигает 4—4,8 кг,
кур 3—3,2 кг. Из французской группы пород особенно ценны брест-
ские куры, сохраняющие значительную яйценоскость, ляфлеш,
дающие во Франции лучших по качеству каплунов и пулярдок, и
гуданы, напоминающие польских кур крупным хохлом на голове
и помимо вкусного мяса хорошо несущиеся (до 120—150 яиц в год).
Из азиатских пород, вообще говоря более примитивных по мясным
качествам, наиболее выделяются лангшаны, очень крупные (cfd до
4—4,4 кг, 99 до 3,2—3,6 кг), выносливые и довольно хорошие не-
сушки. Довольно хороши также брамы, одни из наиболее крупных
кур (cTcf до 4,4—4,8 кг, 99 Д° 3,6—4 кг) и своеобразные кохинхины,
отличающиеся массивностью (cfcf до 5,2, 99 Д° 4,4 кг весом), корот-
ким хвостом и очень густой оперенностью ног.
Общепользовательный тип сочетает хорошую яйценоскость с
значительными мясными качествами, выносливостью и неприхот-
ливостью. Из породистых кур этот тип является одним из наиболее
широко распространенных. Куры английской породы орпингтон
дают хорошее мясо, быстро откармливаются, несут до 160 яиц в год
и хорошо выносят сырые и холодные местности. Выведенные во
Франции фавероль — скороспелы, дают отличное мясо и довольно хо-
рошие несушки (до 120—130 яиц в год); вследствие своей выносли-
вости, они хорошо разводятся в СССР даже в средней и северной
полосах. Из американских пород крупные плимут-рок (Jd* до 4,3 кг,
99 ДО 3,7 кг) дают до 120—130 яиц весом в 60—70 г и отличаются
очень вкусным мясом, выносливостью и неприхотливостью. Близ-
кие к ним виандот несут до 170 яиц меньшей величины и выделяются
скороспелостью и хорошей откармливаемостью. Для условий СССР
также очень хороши род-айленд, прекрасно сочетающие в себе раз-
личные хозяйственные качества.
Декоративный тип кур не имеет хозяйственного значения и раз-
водится главным образом любителями. Из многочисленных пород,
относящихся в эту группу, следует отметить своеобразных феник-
сов, разводимых в Японии, петухи которых имеют необычайно
длинный хвост, достигающий в рекордных случаях больше 6 м
539
(Beebe, 1921). Замечательны шелковые куры, покрытые шелкови-
стыми нитевидными перьями и имеющие темнофиолетового цвета ко-
жу и ноги. Своеобразны богатая представителями группа бентамок
или карликовых кур, оригинальные бесхвостые куры, курчавые,
падуанские, иокогама и мн. др.
Переходя к другим видам домашней птицы, необходимо остано-
виться на индейках, цесарках, утках и гусях и отдельных породах
голубей. Индейки происходят от североамериканского дикого вида,
одомашненного мексиканцами еще в глубокой древности; предпола-
гают, что в образовании культурных пород этого вида принимали
участие разные формы Meleagris gallopavo L. — мексиканская, се-
вероамериканская из восточных штатов и может быть некоторые
другие. В Европу индейка ввезена в 1530 г. Индейки дают превос-
ходное белое и сочное мясо и несут до 30—40 яиц в год. Они отлично
разводятся в странах с умеренным климатом, не перенося сырости
и больших холодов. Курочки начинают нестись в возрасте 9 меся-
цев и являются прекрасными наседками. Из пород индеек ценны
крупные бронзовые разновидности весом до 15 кг (самцы, самки до
8,3 кг), а также черные, белые, красные и аспидные или голубые
птицы, частью ценимые и как декоративные породы. В СССР
индейки разводятся почти повсеместно, причем, уступая иностран-
ным породам в величине, по качеству мяса, плодовитости и выносли-
вости они превосходят их.
Цесарки имеют своей родиной Африку, где они*были одомаш-
нены за несколько столетий до нашей эры. Исходной формой яв-
ляется Numida meleagris L., которая и сейчас еще широко распро-
странена в этой стране. Цесарки дают вкусное мясо, напоминающее
мясо фазанов, и несут до 60—100 довольно мелких яиц в год. От-
личаясь нетребовательностью на корм, они легко разводимы летом
на полувольном содержании и в СССР особенно пригодны для южной
части страны. Известно несколько пород цесарок — белые, голубые
п серые, из которых некоторые на юге Франции разводятся с про-
мышленной целью.
Среди уток мы встречаем два одомашненных вида: обыкновенную
крякву (Anas plathyrhyncha L.), прирученную, повидимому неза-
висимо, в древности в Китае и Риме, и мускусную утку или шипуна
(Cairina moschata L.), в диком состоянии распространенную в Аме-
рике; оба вида бедны породами; из крякв наибольшее хозяйствен-
ное значение имеют руанские утки, пекинские и индийские бегуны.
Первые, выведенные во Франции, интересны крупной величиной
(до 4—4,8 кг), вкусным мясом, хоре шей носкостью (до 100 яиц в
год) я выносливостью. Пекинские происходят из Китая и отли-
чаются бело-кремовой окраской, вздернутым хвостом и хорошей вы-
носливостью и неприхотливостью. По качеству мяса и носкости они
близки к предыдущим. Индийские бегуны повидимому выведены
в Индии и затем улучшены в Англии. Это мелкие утки, весом до
1,8—2 кг, отличающиеся значительной носкостью, обычно до 200
яиц в год. Среди других пород, преимущественно мясных по сво-
ему назначению, следует отметить английских эйльсбюри, голубых
шведских уток, английских орпингтон и американскую породу
540
каюга, особенно ценимую гастрономами. Мускусные утки разводятся
главным образом в Бразилии и ценятся за хорошие мясные качества
и крупную величину (до 4 кг и более); птицы обладают оригиналь-
ным мускусным запахом.
Домашние гуси разводятся исключительно для получения мяса,
гусиного жира, пуха, пера и отчасти пуховых шкурок. Существую-
щие породы ведут свое происхождение от двух диких форм серого
гуся (Anser anser L.) и сухоноса [Cygnopsis cygnoides (L.)], одомаш-
ненных в глубокой древности. Из иностранных пород пользуются
известностью тулузские, эмденские, померанские и китайские
гуси, из гусей СССР — главным образом тульские, арзамасские,
холмогорские и ленточные. Тулузские гуси выведены во Франции
и являются самой крупной породой (до 16 кг), с большими жиро-
выми мешками на животе и подбородке. Эмденские гуси происходят
из Германии, отличаются меньшей величиной, но более скороспелы
и подвижны. Померанские гуси наименее улучшены в сравнении
с предыдущими. Китайские гуси выведены в Китае из сухоноса
и характеризуются более тонкой и гибкой шеей и вздутием у осно-
вания клюва; они отличаются большой выносливостью, неприхот-
ливостью и довольно хорошей носкостью при посредственном каче-
стве мяса. Тульские гуси очень неприхотливы, мясисты и жирны,
но небольшой величины (6—6,4 кг). Более крупные арзамасские
гуси обладают преимущественно белым оперением. Наиболее круп-
ные из русских гусей — холмогорские (до 10 кг) отличаются шишко-
образным вздутием на лбу и представляют повидимому помесь туль-
ских гусей с китайскими; они прекрасно откармливаются, выно-
сливы и нетребовательны к корму и хорошо несутся. Ленточные гуси
возникли в районе Дуная и северного берега Черного моря и по-
мимо мясных качеств ценятся и как декоративная птица, благодаря
своеобразной курчавости или удлиненности контурных перьев.
Помимо хозяйственно ценных видов, широко разводятся в пар-
ках и садах Европы также различные другие птицы, назначение
которых чисто декоративное. Таковы павлины, многочисленные
виды фазанов [Phasianus reevesi Gray, Chrysolophus pictus L., Chry-
solophus amherstiae Leadb., Crossoptilum auritum (Pall.), Gennaeus
nyctemerus L. и мн. др.], лебедей (Cygnus cygnus L., Cygnus olor Gm.,
Cygnus melanocoryphus Mol., Chenopsis atrata Lath.) и уток (J)en-
dronessa galericulata L., Lampronessa sponsa L. и др.)*
Домашний голубь, ‘образующий многочисленные породы, про-
исходит от дикого скалистого голубя (Columba livia Gm.), широко
распространенного в горах и у нас в СССР. В Египте изображения
голубя встречаются на стенах древних храмов п гробниц, относя-
щихся к 3000 лет до нашей эры. Отсюда голубь распространился
в Грецию, Рим, Сирию, Персию и Индию, где однако возможно и
независимое приручение этой птицы. Долгое время у многих на-
родов голуби являлись предметом культа, что объясняет долго со-
хранявшееся предубеждение против употребления голубей в пищу.
Впоследствии внимание было обращено как на эту сторону исполь-
зования голубей, так и на дрессировку их для установления почто-
вой связи.
541
Всех домашних голубей можно разбить на три группы пород:
декоративные, мясные и почтовые (Черепенин, Назаров, Орлов.
1937). Первые отличаются пестрой окраской, нередко своеобразной
формой тела, размерами и развитием украшающих перьев и широко
распространены у любителей голубеводства. Из относящихся к этой
группе пород дутыши или зобастые голуби возникли в Голландии
и Бельгии и замечательны способностью, воркуя, исключительно
сильно раздувать свой зоб. Трубачи, выведенные в Средней Азии,
обладают весьма продолжительным воркованьем, по звуку напо-
минающим барабанный бой. Трубастые или павлиньи голуби, про-
исходящие из Индии, отличаются очень большим числом рулевых
перьев, достигающим сорока; токующие птицы’ расправляют хвост
веером, как павлин, раздувают зоб и своеобразно дрожат. Тонные
голуби, также выводимые из Индии, ценятся за склонность много,
быстро и сильно летать, обычно стайками и нередко на большой
высоте. Из них турманы или катуны оригинально кувыркаются
в воздухе, хлопающие голуби издают ударами крыльев особый гром-
кий звук во время полета, а кружастые подолгу вьются в воздухе.
Замечательные чубатые голуби или якобины, происходящие из Азии,
отличаются своеобразным воротником или капюшоном из перьев,
облегающим шею и голову. Миловидные мелкие голубки — чайки,
также азиатские по происхождению, характеризуются «манишкой»
из курчавых, веерообразно-расходящихся на груди перьев, кожи-
стой горловой складкой и коротким толстым клювом. Наконец в эту
же группу декоративных голубей входят различные цветные голуби,
в остальном близкие к обыкновенному сизому голубю.
Мясные породы голубей, *к которым относятся исполинские или
римские и куриные голуби, отличаются главным образом крупной
величиной и значительным весом. Исполинские голуби были вы-
ведены на юге Франции улучшением крупных итальянских сизых
голубей и по внешности напоминают последних, отличаясь большими
размерами (размах 105 см, вес самца до 1250 г). Куриные голуби
происходят из Азии и в Европу попали около 300 лет тому назад;
короткое широкое туловище, длинная шея, длинные толстые ноги
и вздернутый хвост придают им значительное сходство с курами.
Эти голуби особенно ценятся своим вкусным мясом. По этому послед-
нему качеству к этой группе близки турецкие голуби, относимые
также к почтовым голубям (см. ниже) и откармливаемые для по-
лучения вкусного мяса.
Мясные качества голубей были оценены еще римлянами, и вгна-
стоящее время в Западной Европе и Соединенных Штатах Америки
голубей разводят с промышленной целью для откорма. Особенно
развито мясное голубеводство во Франции, Англии, Бельгии и от-
части в Германии и Италии, где широко используется племенной
материал. В СССР заготавливались Союзптицепродуктом и другими
организациями главным образом домашние голуби и дикие сизые
скалистые голуби. Мясные породы голубей (римские и турецкие)
разводятся в настоящее время на Украине, Кавказе, Урале й ме-
стами в Средней Азии (Никитин, 1934). Следует заметить, что по
качеству и составу своего мяса голуби не уступают^курам, давая
542
даже большее количество белков и обладая менее водянистым мял
сом.
Что касается почтовых голубей, то они разделяются на две
большие группы пород: породы восточные и породы европейские
или бельгийские. Восточные породы были отчасти выработаны уже
в древности, так как применение голубей в качестве средства связи
наблюдалось уже 2000 лет тому назад, в Египте и Персии. Отли-
чаясь хорошей способностью ориентировки на местности, восточные
голуби несколько тяжеловесны и не могут подолгу летать. С ожив-
лением интереса к голубиной почте, главным образом с коммерче-
скими целями, в XVII в. началось улучшение этих пород скрещи-
ванием с более легкими породами, в частности чайками и турманами.
В результате, в Голландии и Бельгии (и отчасти Англии, Франции и
Германии) были выведены европейские породы, отличающиеся как
хорошей памятью местности и ориентировкой, так и способностью
пролетать большие расстояния без отдыха. К восточным породам
относятся карьер, дракон, багадет, скандерун, польские и турец-
кие голуби, внешне характеризуемые голым окологлазным коль-
цом и бородавчатой восковицей. Карьер, выведенный в Малой Азии
и Египте, является родоначальником английских гонцов, отличаю-
щихся весьма значительной способностью к ориентировке и острым
зрением. Более вынослив в полете небольшой дракон, выведенный
в Англии скрещиванием карьера с турманом, и более крупный скан-
дерун. Происходящий из Азии багадет выделяется весьма быстрым
полетом, дикостью и неуживчивостью. Менее определенны в своих
признаках польские и турецкие голуби. В настоящее время евро-
пейские или бельгийские голуби интенсивно вытесняют применение-
восточных пород для установления почтовой связи.
Лучшими голубями бельгийского происхождения считают ант-
верпенских, люттихских и брюссельских почтарей, по внешности
отличающихся цветом оперения и отчасти формой клюва. При об-
щем сходстве с обыкновенным сизым голубем, почтари выделяются
широкой мускулистой выпяченной грудью, короткой сильной шеей,
мускулистыми крепкими крыльями, небольшим окологлазным коль-
цом, крупной восковицей и сильным и толстым клювом.
Использование почтовых голубей в настоящее время ограничи-
вается почти исключительно военными целями, хотя оно могло бы
найти широкое применение и в хозяйственной жизни, осуществляя,
например, связь удаленных промыслов, лесо- и торфоразработок,
отдельных предприятий и т. д. Применение голубей в армии нача-
лось преимущественно с франко-прусской войны 1870—1871 г.,
для связей между крепостями, фортами и войсковыми частями.
В дальнейшем появляются специальные подразделения военного
почтового голубеводства. В условиях современного оснащения ар-
мии техническими средствами голубиная связь признается целесо-
образной для связи десантов, рейдирующей конницы и частей, от-
резанных или окруженных противником, со своими войсками; для
связи кораблей и подводных лодок, вышедших в открытое море,
*с берегом; для связи с разведывательными частями и частями при-
крытия; для доставки донесений с самолетов или аэростатов п для
543
.дублирования технических средств связи (Черепенин, Назаров,
Орлов, 1937). Предварительно голуби тренируются для работы в той
или иной местности, так как возврат птицы на привычную им голу-
бятню или передвижной голубиный пост в главном основан на зри-
тельном ориентировании и запоминании местности (см. стр. 289).
Радиус работы голубей при передвижной голубятне составляет
30—40 км. При работе в совсем незнакомой местности обычно тре-
буется дрессировка в течение двух-трех дней. Дальняя связь при
длительной дрессировке может осуществляться на 300—400 км и
даже до 1000 км. Хорошо прирученные голуби могут летать и в ноч-
ное время, но при этом голубятня должна быть освещена пзвпутри.
Скорость лета почтовых голубей равна 60—70 км в час, при высоте
над землей 300—350 м в хорошую и 100—150 м — в плохую по-
году.
Передаваемую депешу или «голубеграмму» вкладывают в особый
футлярчик из алюминия или ствол гусиного пера, прикрепляемый
к плюсне голубя. Продолжительность иочтовой службы голубя до-
стигает десяти лет.
Переходим к краткой характеристике санитарного значения птиц
в жизни человка. Это значение изучено совершенно недостаточно
и по современным данным представляется гораздо менее значитель-
ным, чем роль млекопитающих. Однако существенность его все же
не подлежит сомнению.
Вместе с млекопитающими многие дикие виды птиц являются
временными или постоянными хозяевами паразитов, которые при
известных условиях вызывают опасные заболевания у соприкасаю-
щегося с ними человека. Так, например, переносчик таежного энце-
фалита на нашем Дальнем Востоке — клещ обитает не только на
млекопитающих, но и на ряде видов птиц, могущих в широкой сте-
пени способствовать распространению этой болезни (Павловский).
Клещевой возвратный тиф, свойственный в СССР южным районам,
особенно Средней Азии, переносится клещем Ornithodorus papillipes,
находимым и на птицах. Японская речная лихорадка, вирус кото-
рой обнаружен в клеще Trombicula, связана с наличием ряда видов
млекопитающих и птиц, на которых этот клещ живет (Elton, 1931).
Несомненным является участие птиц в подобном распространении
и многих других опасных болезней. Длинный ряд возбудителей бо-
лезней у домашних птиц переносится при участии диких видов птиц,
особенно филогенетически и экологически близких им. Таковы,
например, холера птиц, чума птиц, тиф птиц, дифтерит, оспа птиц
и нек. др. (Сизов, 1931). Наблюдающийся в птичниках на индей-
ках и курах типун вызывается круглым червем, от которого в при-
родных условиях часто гибнут также скворцы и грачи (Ellon and
Buckland, 1928). Домашний воробей является носителем и распро-
странителем клещей Germanyssus gallinae и Uermanyssus avium
доставляющих очень много беспокойства и заболеваний домашней и
комнатной птице (Ewing, 1911), а первый иногда нападает п на чело-
века (Braun, 1908).
Птицы, живущие в норах грызунов, а также вероятно хищники,,
питающиеся последними, являются зачастую переносчиками блох
.544
(Сергеев, 1936), чем в широкой степени могут способствовать распро-
странению чумных эпидемий.
Как более детальный пример значения птиц в вызывании болез-
ней у людей, можно привести попугайную болезнь или пситтакоз.
Эта болезнь выражается у попугаев в затрудненном дыхании, на-
сморке, грудном хрипе и поносе; птица сидит на жердочке с полу-
закрытыми глазами и взъерошенными перьями. В разгар болезни
появляются конвульсии, и птица обычно умирает. Возбудитель
пситтакоза до сих пор не найден, так же, как не наблюдалось заболе-
ваний попугаев на воле; очевидно редкая в естественных условиях
болезнь вспыхивает и быстро распространяется среди скученных
птиц в клетках, содержимых без достаточного ухода.
При контакте с больным попугаем легко заболевают люди пожи-
лого возраста и иногда молодежь. Болезнь начинается болью су-
ставов, слабостью и сильной жаждой; затем начинаются сильные
головные боли, сопровождаемые часто бредовым состоянием, появ-
ляется сильный кашель, лихорадочное состояние и упадок сердеч-
ной деятельности. Кризис, а иногда и смерть наступают на 12-й—
13-й день после начала болезни. Смертность от этой болезни у людей
вообще составляет от 20 до 40% всех заболеваний ею. При благо-
приятном исходе температура к концу третьей недели спадает, но
человек становится вполне работоспособным не ранее, чем через
2—3 месяца после начала болезни.
Распространение этой болезни ограничено пока странами Ев-
ропы и Америки. В 1892 г. в Париже было констатировано 49 забо-
леваний, из которых 33 закончились смертью. В 1929 г. в Германии
было 215 случаев заболеваний попугайной болезнью, и из них 45
со смертельным исходом. В этот же год болезнь посетила Францию,
Италию, Швейцарию, Чехию, Голландию, Польшу, Данию, Шве-
цию, Алжир, Египет и Канаду, причем смертность во всех этих стра-
нах достигала до 20% заболеваний. В отдельных случаях болезнь
поражала целые семьи и людей лишь кратковременно входивших
в соприкосновение с больной птицей. Свое происхождение болезнь
ведет из Южной Америки (Grzimek, 1933).
РУКОВОДЯЩАЯ ЛИТЕРАТУРА
Литература по орнитологии очень велика, особенно по фауни-
стике и экологщт, так как эта область принадлежит к одному
из наиболее разрабатываемых отделов зоологии. В год появляется
от 1000 до 1500 статей и книг, частью рассеянных по весьма разно-
образным изданиям. Однако обзор этой литературы, в большей ее
части, сравнительно легко доступен, благодаря обилию специальных
журналов, регистрирующих выходящие работы и рецензирующих их.
В целях облегчения интересующимся возможности углубления све-
дений, почерпнутых из настоящей работы, ниже даются краткие
указания основных сводок, обзоров и исследований по отдельным
крупным разделам данной дисциплины. По недостатку места, принт
лось ограничиться несколько субъективным выбором лишь глав-
|/в35 Орнитология-*1209
545
нейших источников. Дополнительные ёсылки, нередко более де*
тальные, могут быть получены из работ авторов, указанных в тек-
сте: руководствуясь годом издания работы, ее легко отыскать в еже-
годниках основного каталога зоологической литературы («Zoolo-
gical Record», London) или в других обзорных или специальных
журналах, указанных ниже.
Систематический обзор класса птиц в целом наиболее полно
представлен на русском языке в 4-м изд. А. Б р э м а, Жизнь жи-
вотных, где отдел птиц составляет 4 тома, обработанных В. М ар-
шале м, Ф. Хемпельманом и О. Цур-Штрассеном
(изд. Девриен, СПб., 1914). Более сжато обзор дан М. А. М е н з б и*
р о м в книге «Птицы», изданной в серии «Библиотека естество-
знания», изд. Брокгауз-Ефрон, СПб., 1904—1909 гг. Естественно,
оба источника значительно устарели и по своему популярному
характеру лишены желательной точности и полноты материала.
Из иностранных источников ценны: «Птицы» А. X. Эванса
(А. Н. Evans, Birds, 1899), представляющие собой 9-й том в се-
рии «The Cambridge Natural History», London (популярна, но весьма
содержательна), и «Птицы» А. Рейхенова (A. Reichenov, Die
Vogel, Stuttgart, 1913—1914), являющиеся двутомным руковод-
ством по систематической орнитологии, с кратким описанием основ-
ных представителей большей части родов птиц. Основным и капи-
тальным источником остается «Каталог птиц Британского музея»
(«Catalogue of the Birds in the British Museum», London), изданный
в 27 томах в 1874—1895 гг.; здесь дано в сущности описание почти
всех видов птиц земного шара, с главнейшей синонимикой и табли-
цами для определения видов. На основании этого каталога Р. Б. Шар-
пом составлен удобный для справок список видов птиц всего мира,
с указанием распространения (R. В. Sharpe, A Hand-list of the
Genera and Species of Birds, London, 1899—1909), занявший 5 томов.
В настоящее время подобный новый список составляется Дж. Питер-
сом; список доведен пока до 3-го тома, до Psittacifonnes включи-
тельно (J.L. Peters, Check-list of the Birds of the World, Cam-
bridge, Massachusets, vol. I, 1931; vol. II, 1934; vol. Ill, 1937).
Систематический обзор фауны птиц СССР дан С. А. Бутур-
лины м и Г. П. Дементьевы мв книге «Птицы СССР», Мо-
сква, 1934, являющейся определителем и лишенной подробных опи-
саний птиц и специальных экологических данных. Остальные ра-
боты, имеющие в виду СССР в целом, охватывают лишь отдельные
систематические группы, представляя однако и морфологическое
и экологическое описание. Из наиболее важных отметим: М. А. М е н-
з б и р, Птицы России, 2-е изд., вып. 1 (единственный), Москве
1918 (лишь отряды: Colymbijormes, Procellariiformes, Ciconiiformes.
Pelecani/formes и Phoenicopterijormes); В. Л. Б и а н к и, Procellarii
formes и Colymbiformes, в серии «Фауна России», изд. Акад. Наук
1911—1912; Б. К. Ш т е г м а н, Дневные хищные птицы, в cepni
«Фауна СССР», изд. Акад. Наук СССР, 1937.
Из работ, охватывающих значительную часть СССР, важнь
следующие: М. А. М е н з б и р, Птицы России (Европ. часть i
Кавказ; Москва, 1895, 2 тома); его же «Охотничьи и промысло
546
вне птицы Европейской России п Кавказа», Москва, 1901, 2 тома
с атласом; Н. А. X о л о дковский и А. А. Силантьев,
Птицы Европы. Практическая орнитология, с атласом европейских
птиц, СПб., изд. А. Ф. Девриен, 1901, и L. Т а с z а п о w s k i,
Faurte ornithologique de la Siberie orientale, 2 тома, в «Memoires de
I’Acad, de Sc. de St. Petersbourg», VII serie, XXXIX, 1891—1893.
Из крупных иностранных работ важна для познания фауны
СССР сводка Эрн. Хартерта (Ern. Н а г t е г t, Die Vogel der palaark-
tischen Fauna, Berlin, 1903—1922, 3 тома) и дополнение к .ней —
Nachtrag I, 1923 и Erganzungsband, Heft 1—6,.1932—1938, состав-
ленное Штейнбахером и Хартертсм. Эта, в основном систематиче-
ская, работа содержит и определители видов и сжатые сведения по
экологии евразиатских птиц.
Более углубленный обзор филогенетических отношений внутри
класса птиц следует искать в фундаментальных исследованиях
М. Фюрбрингера (М. F u г b г i n g е г): Untersuchungen zur Morpholo-
gic und Systematic der Vogel, zugleich ein Beitrag zur Anatomie der
Stutz- und Bewegungsorgane, Amsterdam, 1888, I и II томы (содержит
детальный обзор систематического значения морфологических при-
знаков, опыт системы и построения родословного дерева), и Zur ver-
gleichende Anatomie des Brust§chulterapparates und der Schulter-
muskeln, напечатанной в «Jenaische Zeitschrift fur Naturwissenschaft»,
t. 29, 1902 (обзор оригинальной системы автора в свете более позд-
них исследований), а также во второй, систематической, части об-
зора птиц Г. Гадова (Н. G a d о w) в «Bronn’s Klassen und Ordnun-
gen des Thierreichs», Bd. VI, Abt. 4; «Птицы» («Vogel»), Leipzig,
1893 (исторический обзор различных классификаций, обзор морфо-
логических черт отдельных групп и их взаимоотношений). Значи-
тельный интерес представляет также книга F.E.Beddard’a, The
Structure and Classification of Birds, London, 1898, по обилию ори-
гинального материала.
О строении тела птиц довольно подробные сведения можно полу-
чить из книг: В. Мар шаль, Строение тела птиц (СПб., 1898;
под названием «Анатомия птиц» в том же общедоступном изло-
жении издана вторично в 1902 г.); Автократов, Анатомия
птиц, Москва, 1926 (применительно к целям сельскохозяйственного
птицеводства), и из учебников: В. М. Ш и м к е в и ч, Курс сравни-
тельной анатомии позвоночных животных, 2-е изд., 1922, и И. И.
Шмальгаузен, Основы сравнительной анатомии позвоноч-
ных животных, 3-е изд., 1938. За более подробным изложением мате-
риала приходится обращаться к иностранным сводкам: анатомиче-
ской части сводки «Птицы» Н. Gado w’a и S е 1 е п к’и (в «Bronn’s
Klassen und Ordnungen des Tierreichs», Bd. 6, Abt. 4, 1871—1893);
вышеупомянутым работам M. Фюрбрингера, сводке F. Groeb-
b е 1 s’a («Der Vogel. Bau und Funktion, Lebenserscheinung, Ein-
passung», Berlin, 1932—1937, Bd. I: «Atmungswelt und Nahrungs-
welt»; Bd. II, «For tpflanzungs welt»); обзору птиц E. Strese-
mann ’ а (в «Kukenthal-Krumbach’s Handbuch der Zoologie», Berlin,
1927—1934, Bd. VII, Zweite Halfte, Lief erung 1—8), а также к сравни-
тельно-анатомическим руководствам: J. I h 1 е, P. van К a m p e n,
« 547
H. Nierstrasz. u. J. Versluys, Vergleichende Anato-
mie der Wirbeltiere, Berlin, 1927, и более полной: L. В о 1 к,
Е. Go р per t, Е. К а 11 i u s, u. W. L и b о s с h, Handbuch
der vergleichenden Anatomie der Wirbeltiere, Bd. I—VI, Berlin,
1932—1937.
В этих же источниках можно найти списки частной литературы
по этим вопросам.
О взаимоотношении класса птиц с ближайшими систематическими
группами см. сравнительно-анатомическую литературу: Фюрбрин-
гера, Гадова и др. (см. выше). Взгляды на происхождение птиц
удобно сведены (с указанием литературы) Lambrech t’oM в
Handbuch der Palaeornithologie, Berlin, 1933. Интересный обзор
родственных связей птиц представляет книга G. Heilmann,
Origin of Birds, London, 1926.
По экологии птиц наилучшим обзором являются соответствующие
главы в вышеупомянутых сводках Штреземанна и Грёббельса, снаб-
женных библиографическими указаниями. Специальной сводочной
работы по этому разделу орнитологии мы не имеем, хотя существует
множество прекрасных исследований. Из более обзорных работ
(к тому же содержащих ссылки на литературу) следует указать:
G. М. А 11 е n, Birds and their Attributes, Boston, 1928; M. H i 1 z-
h e i m e r, Vogel — в книге Hilzlreimer und H a e m p e 1, Bio-
logie der Wirbeltiere, Berlin, 1913; W. P. Pycraft, A History of
Birds, London, 1910; J. A. Thomson, The Biology of Birds, Lon-
don, 1923; J. Henderson, The Practical Value of Birds, New
York, 1927; Fr. F inn, Bird Behaviour, Psychical and Physiological.
London, 1919; H. Friedemann, The Cowbirds, Springfield, 1929;
F. H. Herrick, Home Life of Wild Birds, New York, 1905; H. E.
Howard, Territory in- Bird-life, London, 1921; H. Ho.ward,
An Introduction to the Study of Bird Behaviour, Cambridge, 1929;
J. Huxley, Bird-watching and Bird-behaviour, London, 1930;
W. P. Pycraft, The Courtship of Animals, London, 1913; R. A 1-
± u m, Der Vogel und sein Leben, Munster, 1898; S. Ch. Kendeigh,
The Role of Environment in the Life of Birds, «Ecological Mono-
graphs», vol. 4, 1934; Nicholson, How Birds Live, London,
1927; W. Rowan, The Riddle of Migration, Baltimore, 1931;
A. L. Thomson, Problem of Bird Migration, London, 1926;
H. W a c h s, Die Wanderungen der Vogel, «Ergebnisse der Biologic»,
Bd. I, Berlin, 1926, и нек. др.
На русском языке можно указать более крупные работы по пе-
релету птиц: М. А. Мензбир, Миграции птиц, Москва, 1934;
Ч: Дик сон, Перелет птиц, СПб., 1895; Д у н к е р, Перелет птиц,
СПб., 1907 (последние две работы сильно устарели) и обзорную ста-
тью А. Н. Йромптова, Перелет птиц и его экспериментальное
изучение («Успехи экспериментальной биологии», т. VII, вып. 2,
1928). Полезный обзор работ по экологии птиц, произведенных
в СССР за последнее двадцатилетие (1917—1937), был сделан А. Н.
Формозовым в «Зоологическом журнале», т. XVI, вып. 5,
1937 (имеется список литературы).
Помимо специальных статей, экологические наблюдения в обилии
543
рассеяны в фаунистической литературе и общих сводках, частью- от-
меченных выше.
О географическом распространении птиц ценный материал можно
получить из следующих книг: Dahl, Oecologische Tiergeographie,
Berlin, ч. I — 1921, ч. II — 1923; Al. Wallace, The Geographical
Distribution of Animals, London, 2 тома, 1876 (значительно устарели);
A. Reichenow, Die Begrenzung zoogeographischer Re^ionen vomi
ornithologischen Standpunkt, «Zoologisches Jahrbuch», Abt. fur Syste-
matic, Bd. 3, Jena, 1888; R. S ha r p, On the Zoo-geographical Areas
of the World Illustrating the Distribution of Birds, Natural Science,
London, 1898, VII; ии'з учебников: В. Г. Г е п т н е р, Общая зоо-
география, Москва, 1936, и И. И. Пузанов, Зоогеография,
Москва, 1937. Из обзоров фауны СССР ценны: М. А. М е н з б и р,
Орнитологическая география Европейской России, «Ученые зап.
Моск, унив.», вып. 2, 1882 г.; Б. К. Ш т е г м а н, Основы орнито-
географического деления Палеарктики, в серии «Фауна СССР»;
«Птицы», вып. 2, 1938.
Библиография орнитологической литературы осуществляется ря-
дом журналов и указателей. Наиболее удобен для пользования еже-
годник зоологической литературы: «Zoological Record, International
Catalogue of Scientific Literature», London, издаваемый c 1864 r.
Каждый том содержит перечисление статей и книг, напечатанных
за год, сгруппированных по, классам и алфавиту авторов; затем
статьи группируются по семействам, разделам орнитологии и от-
дельным проблемам. Кроме того, можно пользоваться следующими
библиографиями: С. G. G i е b е 1, Thesaurus ornithologiae. Reper-
torium der. gesamten ornithologischen Literatur, 3 Bande, Leipzig,
1872—1877; J. T. Zimmer, Catalogue of the Edward E. Ayer Orni-
thological Library. Field Museum of Natural History. Publication
239, Chicago, 1928; C. A. Wood, An Introduction to the Litera-
ture of Vertebrate Zoology based chiefly on the Titles in the Blacker
Library of Zoology... and other Libraries of McGill University,
Montreal, London, 1931; J. V. G a r u s u. Wilh. Engelmann,
Bibliotheca zoologica, Leipzig, 1861, 2 тома, литература за 1846—
1860 гг. «Птицы», т. 2, стр. 1112—1260; G а г u s und Field,
Bibliographia zoologica, Leipzig, 1896—1912; J. V. G а г и s, Bib-
liographia zoologica, Turici, c 1890 г.; B. R e ns c h, Tiergeographie,
литература c 1908 no 1930 гг., «Geographisches Jahrbuch», Bd. 45,
1931; Wilh. Engelmann, Bibliotheca historico-naturalis, Leip-
zig, 1846, литература c 1700 до 1846 гг., «Птицы», стр. 385—419;
О. Tas ch е n b er g, Bibliotheca Zoologica, Leipzig, литература
c 1861 no 1900 гг. «Птицы», стр. 3649—4364, вып. 12, 13 и 14.
♦ Для литературы, выходящей на русском языке: Л. П. Саба-
неев, Указатель книг и статей охотничьего и зоологического со-
держания, 1883. Литература до конца 70-х гг. «Птицы», стр. 319—
479; «Указатель русской литературы по математике и естественным
наукам», изд. Киевского общества естествоиспытателей, выходил
с 1872 по 1891 гг.; F. Th. К б р р е n, Bibliotheca Zoologica Rossica,
Literatur liber die Tierwelt Gesammthrussland bis zum Jahre 1885
incl. (Вышли только два тома—I и II, СПб., 1905); А. Богданов,
549
Материалы для истории научной и прикладной деятельности в Рос-
сии по зоологии и сопредельным с ней отраслям знания (1850—1888),
«Труды Зоологического отделения Общества любителей .естество-
знания, антропологии и этнографии», тт. I—IV, Москва, 1888—1892;
Г. А. Кожевников, Отчет о литературе по позвоночным
(1885—1889); там же, т. VIII, Москва, 1893; Библиография рус-
ской зоологической литературы за 1931 г. Приложение к тому XII
«Зоологического журнала», Москва, 1933; в дальнейшем списки
помещаются в самом журнале. С 1933 по 1937 гг., издавался в Москве
на русском языке небольшой «Реферативный биологический журнал»,
для обзора иностранной литературы.
Большинство специально-орнитологических журналов, издавае-
мых сейчас в количестве не менее 40 названий, посвящают часть
своих страниц реферированию новой литературы по птицам. У нас
с 1910 по 1917 гг. издавался в Москве «Орнитологический вестник»,’
и с 1926 по 1929 гг. в Томске издавался «Журнал Сибирского орни-
тологического общества «Uragus».
Можно рекомендовать следить за следующими журналами: «The
Ibis», издаваемый Британским орнитологическим союзом в Лон-
доне (с 1859 г.); «Journal fur Ornithologie», издаваемый Германским
орнитологическим обществом в Берлине (с 1853 г.) (последние годы
рефераты в журнале отсутствуют и публикуются в небольшом жур-
нале «Ornithologische Monatsberichte»); .«Alauda», издаваемый Фран-
цузским орнитологическим обществом в Париже; «Айк», издаваемый
Американские орнитологическим союзом в Филадельфии (с 1876 г.).
В своей основе эти журналы посвящены разработке систематики,
экологии и географического распространения птиц, и работы мор
фологического и физиологического характера находят себе место
преимущественно в других специальных изданиях. Следить за ними
можно по рефератам обзорных журналов, из которых лучшими яв-
ляются: «Berichte fur Wissenschaftliche Biologie» и «Zoologischer
Bericht», издаваемым в Берлине.
Орнитологические научные общества являются широко распро-
страненной формой общения и организации людей, интересующихся
этой областью знания и стремящихся работать по ее развитию. Они
существуют почти во всех странах. У нас в СССР имеется Орнито-
логическое общество имени акад. М. А. Мензбира, с центром в Мо-
скве, и Сибирское орнитологическое общество с центром в Томске.
Международными объединениями ученых, помимо зоологических
конгрессов, созывалось уже девять специальных орнитологических
конгрессов, проходивших: в 1884 г. в Вене, в 1891 г. в Будапеште,
в 1900 г. в Париже, в 1905 г. в Лондоне, в 1919 г. в Берлине, в 1926 г.
в Копенгагене, в 1930 г. в Амстердаме, в 1934 г. в Оксфорде и в
1938 г. в Руане. Каждый конгресс в известной мере подводит
итог работам в области орнитологии и хорошо отражает современ-
ные ему направления и методы работы исследователей.
ОГЛАВЛЕНИЕ
Стр.
ОТ АВТОРА................................................... 3
ОБЩАЯ ХАРАКТЕРИСТИКА КЛАССА ПТИЦ............................ 5
МОРФОЛОГИЯ ................................................. 9
1. Общие покровы.......................................... —
2. Скелет................................................ 37
3. Мышечная система . . . .•............................. 71
4. Органы дыхания и голосовой аппарат.................... 89
5. Кровь и кровообращение............................. 105
6. Пищеварительная система и пищеварение................ 117
7. Органы выделения и размножения ...................... 130
8. Нервная система и органы чувств .................... 140
ЭКОЛОГИЯ ................................................ 166
1. Обитаемая среда и основные способы передвижения........ —
2. Обзор приспособлений к характеру местообитания и способу добыва-
ния пищи............................................... 194
3. Размножение и индивидуальное развитие................ 210
4. Климатические факторы и вызываемые ими явления в жизни птиц . 252
5. Приспособления, связанные с стремлением к самосохранению . . . 292
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ............................ 303
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ КЛАССА ПТИЦ...................... 333
КРАТКИЙ СИСТЕМАТИЧЕСКИЙ ОБЗОР КЛАССА ПТИЦ........... 349
Подкласс I: Saururae. — Ящерохвостые птицы . . ........... —
Отряд Archaeoptepygiformes............................. —
Подкласс II: Ornithurae. —Веерохвостые птицы ................................... —
Надотряд 1: Odonthognathae.—Зубочелюстные. 350
Отряд 1: Hesperornithiformes.— Гесперорнисы................................. —
Сем. 1: Hesperornithidae. —
Сем. 2: Enaliornithidae............... 352
Сем. Ichthyornithidae.......................................... —
Надотряд 2: Palaeognathae. —Древненебные птицы......................... —
Отряд 1: Struthioni formes. — Страусы............................. 354
Сем. Struthionidae. — Страусы.........'........................ —
Отряд 2: Rheiformes. — Нанду...................................... 356
Сем. Rheidae. — Нанду ....................................... 357
Отряд 3: Casuariiformes. — Казуарообразные........................ 358
'Сем. 1: Casuariidae.—Казуары...................'................ 359
Сем. 2: Dromaedidae. —Эму....................................... 360
Отряд 4: Aepyornithiformes. —Мадагаскарские страусы............. 361
Сем. Aepyornithidae........................................... —
Отряд 5: Apterygiformes. — Бескрылые............................ 362
Сем. Apterygidae. — Киви...................................... —
• Отряд 6: Dinornithiformes. —Моа................................. 364
Сем. Dinornithidae. — Моа...................................... —
Отряд 7: Tiriamiformes. —Скрытохвосты..................•.......... 365
Сем. Tinamidae.—Скрытохвосты.................................. —
Надотряд 3: Neognathae. — Йовонебные птицы........................... 366
551
Отряд 1: Sphenlsclformes. — Пингвины............................... Збб
Сем. Spheniscidae. — Пингвины................................. 367
Отряд 2: Procellariiformes. —Трубконосые........................... 371
Сем. Procellariidae. — Трубконосые......................... ........
Отряд 3: Colymbiformes.— Гагарообразные......................... 374
Подотряд 1: Colymbi. —Гагары.......................................... 375
Сем. Colymbidae.*—Гагары........................................ —
Подотряд 2: Podicipedes. —Поганки....................................... —
Отряд 4: ^elecaniformes. —Веслоногие.................................. 377
Подотряд 1; Phaethontes. —Фаэтоны........................................ 378
Сем. Phaethontidae. — Фаэтоны.................................... 379
Сем. 1: Pelecanidae. — Пеликаны.................................. —
Сем. 2: Cyphornithidae........................................ . 380
Сем. 3: Pelagornithidae..................................... . —
Сем. 4: Sulidae. —Олуши ........................................ 381
Сем. 5: Phalacrocoracidae.—Бакланы.......................... . . 382
Сем. 6: Anhingidae.—Змеешейки . •............................... 383
Подотряд 3: Fregathae. — Фрегаты . . . ......................., . . . 384
Сем. Fregathidae. — Фрегаты —
Подотряд 4: Odontopteryges......................................... 385
Сем. Odontopterygidae........................................... —
Отряд 5: Ciconiiformes. —Листообразные............................ —
Подотряд 1: Ardeae. — Цапли........................................ 386
Сем. 1: Ardeidae. — Цапли . . ..............*.................... —
Сем. 2: Cochleariidae . :....................................... —
Подотряд 2: Scopides. — Молотоглавы.................................. 387
Сем. Scopidae. — Молотоглавы.................................... —
Подотряд 3: Balaenicipetes. —Китоглавы . . . .....................388
Сем. Balaenicipetidae. — Китоглавы.............................. —
Подотряд 4: Ciconiae. —Аисты........................................ 389
Сем. Ciconiidae.—Аисты. ........................................ —
Подотряд 5: Threskiornithides. — Ибисы.............................. 391
Сем. Threskiornithidae. — Ибисы .... ... —
Отряд 6; Phoenicopteriformes. —Фламинго........................... 392
Сем. Pboenicopteridae. — Фламинго............................ —
Отряд 7: Anseriformes. — Гусеобразные............................. 394
Подотряд 1: Anhimae. — Паламедеи . .................................... —
Сем. Anhimidae. — Паламедеи.................. ч.............. —
Подотряд 2: Anseres. —Гусиные ....................................*. 396
Сем. 1: Mergidae. —Крохали...................................... 397
Сем. 2: Anatidae. — Утки.......................................... —
Сем. 3: Anseridae.—Гуси........................................... —
Сем. 4: Cygnidae. —Лебеди. . . .............................. —
Отряд*8: Falconiformes. —Дневные хищные птицы......................400
Подотряд 1: Cathartae. — Американские грифы.......................... 401
Сем. 1: Cathartidae . •........................................ 402
Сем. 2: Terathornithidae......... . . <......................... —
Подотряд 2: Sagittarii. —Птицы секретари............................... —
Сем. Sagittariidae. — Птицы секретари............... ...... . —
Подотряд 3: Accipitres. — Нормальные хищные шицы .................... 403
Сем. 1: Accipitridae. —Ястребиные................................. —
Сем. 2: Falconidae. — Соколиные................................. 404
Отряд 9: Galliformes. — Курообразные ............................. 405
Подотряд Г/Galli. — Куры..................................................
Сем. 1: Cracidae.—Краксовые..................................... 406
Сем. 2: Gallinuloididae .......................................... “
Сем. 3: Megapodiidae.—Большеногие куры ......................*• 407
Сем. 4: Phasianidae. —Фазановые .. ............................... —
Подотряд 2: Opisthocomi. —Гоацины . ........................... 409
Сем. Opisthocomidae. — Гоацины................................. —
Б62
Отряд 10: Turniciformes. — Трехперстки ................... . , / 410
Сем. 1: Pedionomidae......................................... 411
Сем. 2: Turnicidae.—Трехперстки................................ —
Отряд И: Gruiformes. —Журавлеобразные......................... 412
Подотряд 1: Mesoenades. —Мадагаскарские пастушки................. 413
Сем. Mesoenatidae............................................ —
Подотряд 2: Ralli. — Пастушки........................................ —
Сем. Rallidae. — Пастушки.................................. 414
Подотряд 3: Heliornithes. —Лапчатоноги............................. 415
Сем. Heliornithidae............................ Г............ —
Подотряд 4: Rhinocheti. —Кагу...................................... 416
Сем. Rhinichetidae........................................... —
Подотряд 5: Eurypygae. —Солнечные цапли.......................... 417
Сем. Eurypygidae. — Солнечные цапли.......................... —
Подотряд 6: Cariamae. —Серимы . . ............................. 418
Сем. 1: Cariamidae. —Серимы..................................... —
Сем. 2: Hermosiornidae........................................ 419
Сем. 1: Phororhacidae............................................ —
Сем. 2: Brontornithidae.......................................... —
Подотряд 8: Diatrymae..............................’.................. —
Сем. Diatrymidae............................................ 420
Подотряд 9: Psopniae— Трубачи......................................... —
См. Psophiidae................................................... —
Подотряд 10: Grues. —Журавли.......................................... —
Сем. 1: Aramidae. —Арамы...................................... 421
Сем. 2: Gruidae. — Настоящие журавли......................... 422
Подотряд И: Glides. —Дрофы........................................ 423
Сем. Otidae. — Дрофы......................................... —
Отряд 12: Charadriiformes. —Ржанкообразные .................... 424
Подотряд 1: Jacanae. —Яканы....................................... 425
Сем. Jacanidae. —Яканы..................................... 426
Сем. Thi nocoridае.......................................... —>
Подотряд 3: Limicolae. —Кулики........................................ —
Сем. 1: Burhinidae.—Авдотки.................................... 427
Сем. 2: Dromadidae. — Рачьи ржанки............................. 428
Сем. 3: Glareolidae. —Тиркушки................................... —
Сем. 4: Chionididae. —Белые ржанки............................. 429
Сем. 5: Chapadriidae. — Ржанковые................................ —
Подотряд 4: Lari. — Чайки........................................... 431
Сем. 1: Laridae. —Настоящие чайки.............................. 432
Сем. 2: Stercorariidae. — Поморники............................ 433
Сем. 3: Sternidae.—Крачки........................................ —
Сем. 4: Rhynchopidae. — Водорезы................................. —
Подотряд 5: Alcae. — Чистики........................................ 434
Сем. Alcidae. — Чистики...................................... —
Отряд 13: Соlumbiformes. —Голубеобразные........................ 436
. Подотряд 1: Pterocletes. — Рябки .'................................. —
Сем. Syrrhaptidae. —Саджи и рябки . .’..................... 437
Подотряд 2: Columbae. —Голуби....................................... 438
Сем. 2: Columbidae. —Настоящие голуби............................. —
Отряд 14: Cuculiformes. — Кукушки................................. 441
Подотряд 1: Musophagi. —Бананоеды....................................... —
Сем. Musophagidae. — Бананоеды...................’........... 442
Подотряд 2: Cuculi. —Кукушки............................................ —
Сем. Cuculidae. — Кукушки.................................... 443
Отряд 15: Psittaciformes, — Попугаи............................... 444
Сем. Psittacidae. — Попугаи.................................. 445
Отряд 16: Strigiformes. —Совы..................................... 448
Сем. 1: Tytonidae.—Сипухи......................................... —
х/*35 Орнитология—1209 653
Сем. 2: Strigidae. — Нормальные совы......................... 448
Отряд 17: Capri mu !gi formes. — Коаодоеобразные............... 450
Сем. Г. Steatornithidae.—Жиряки................................ —
Сем. 3: Podargidae. — Совиные козодои.......................... —
Сем. 4: Nyctioiidae.—Исполинские козодои..................... 452
Сем. 5: Caprimulgidae. — Настоящие козодои..................... —
Отряд 18: Micropodiformes.—Стрижеобразные.................: . . . 454
Подотряд 1: Micropodii. —Стрижи................................... 455
Сем. Micropodidae. — Стрижи.................................. —
Подотряд 2: Trochili. — Колибри................................... 456
Сем. Trochilidae.— Колибри.................................. —
Отряд 19: Coliiformes. — Птицы-мыши (мышевки)................... 458
Сем. Coliidae. — Птипы-мыши.................................. —
Отряд 20: Trogoniformes. —Трогоны............................... 459
Сем. Trogonidae. — Трогоны................................. 460
Отряд 21: Coraciiformes. —Ракшеобразные......................... 461
Подотряд 1: Coraciae. —Ракши...................................... 462
Сем. 1: Leptosomatidae.—Кирумбо................................ —
Сем. 2: Brachypteraciidae. —Земляные ракши................... 463
Сем. 3: Coracildae. —Древесные ракши........................... —
Подотряд 2: Halcyones. —Зимородки................................... —
Сем. Alcedinidae. — Зимородки.............................. 464
Подотряд 3: Meropes. —Щурки...................................... 465
Сем. Meropidae. — Щурки..................................... —
Подотряд 4: Momoti. —Момоты....................................... 466
Сем. Momotidae.—Момоты .....................•............... —
Подотряд 5: Todi. —Тоди (плоскоклювы) ............................. —
Сем. Totidae. — Тоди (плоскоклювы)......................... 467
Подотряд 6: Upupae. — Удоды......................................... —
Сем. Г. Lpupidae. — Настоящие удоды. . . .........- . . . . —
Сем. 2: Phoeniculidae.—Древесные удоды. .'................... 468
Подотряд 7: Bucerotes. — Птицы-носороги............................. —
Сем. Bucerotidae. — Птицы-носороги........................ 469
Отряд 22: Piciformes. —Дятлообразные........................... 470
Подотряд 1: Galbulae. —Якамары ................................... 471
Сем. 1: Galbulidae. — Настоящие якамары.......... —
Сем. 2: Bucconidae.—Пуховки (ленивки)........................ 472
Подотряд 2: Rhamphasti. —Перцеяды (туканы)........................ 473
Сем. Rhamphastidae. — Перцеяды.............................. —
Подотряд 3: Capitonides........................................... 474
Сем. 1: Indicatoridae. —Медоуказчики . —
Сем. 2: Capitonidae. —Бородастики............................ 475
Подотряд 4: Pici. —Дятлы.......................................... 476
Сем. 1: Jyngidae. —Вертишейки................................. —
Сем. 2: Picidae. —Дятлы........................................ —
Отряд 23: Passeriformes. — Воробьинообразные................... 478
Подотряд 1: Eurylaemi. —Рогоклювы................................. 480
Сем. Eurylaemidae. — Рогоклювы.............................. —
Подотряд 2: Clamatores. — Кричащие воробьиные.................. 481
Сем. 1: Acanthisittidae . .................................. —
Сем. 2: Philepittidae....................................... —
Сем. 3: Pittidae. — Питты................................. 482
Сем. 4: Tyrannidae.—Тираны.................................. —
Сем. 5: Pipridae. —Мананины . ........................... 483
Сем. 6: Cotingidae.—Котинги................................. —
Сем. 7: Phytotomidae.—Траворезы........................... 484
Сем. 9: Pteroptochidae........................................ —
Сем. 10: Formicariidae.—Муравьеловки........................ 485
Сем. И: Dendrocolaptidae. — Древолазы........................
Подотряд 3; Menurae. —Примитивные воробьиные....................
.654
Сем. 1: Atrichornithidae.................................... 485
Сем. 2: Menuridae. — Птицы-лиры............................. 486
Подотряд 4: Oscines. — Певчие воробьиные.......................... —
Сем. 1: Alaudidae. —Жаворонки............................... 487
Сем. 2: Palaeospizidae...................................... 488
Сем. 3: Hirundinidae. — Ластечки.............................. —
Сем. 4: Campephagidae.— Личинке еды........................... —
Сем. 5: Brachypoaidae. — Коротконогие дрозды................ 489
Сем. 6: Motacillidae.—Трясогузки.............................. —
Сем. 7: Cinclidae.—Оляпки................................... 490
Сем. 8: Troglodytidae.—Крапивники........................... —
Сем. 9: Mimidae.—Пересмешники............................... 491
Сем. 10: Turdidae. — Дрозды..................................... —
Сем. И: Zeledoniidae. —Водяные дрозды........................ 492
Сем. 12: Timaliidae. —Тималии................................... —
Сем. 13: Sylviidae. — Славковые................................. —
Сем. 14: Muscicapidae.—Мухоловки.............................. 493
Сем. 15: Pachycephalidae. —Толстоголовые сорокопуты........... 494
Сем. 16: Prionopidae. —Лесные сорокопуты........................ —
Сем. 17: Vangidae............................................. 495
Сем. 18: Laniidae.—Сорокопуты................................... —
Сем. 19: Artamidae. —Ласточковые сорокопуты................... 496
Сем. 20: Pseudochelidonidae..................................... —
* Сем. 21: Bombycillidae.—Свиристели.............................. —
Сем. 22: Sturnidae.—Скворцы.................................. 497
Сем. 23: Oriolidae.—Иволги.................................. . —
Сем. 24: Dicruridae. —Дронго.................................. 498
Сем. 25: Cracticidae............................................ —
Сем. 26: Corvidae. — Врановые................................... —
Сем. 27: Paradiseidae. —Райские птицы......................... 499
Сем. 28: Paradoxornithidae. — Камышовые синицы................ 500
Сем. 29: Paridae. — Синицы...................................... —
Сем. 30: Sittidae. — Поползни................................. 501
Сем. 31: Hyposittidae,—Мадагаскарские поползни................ 502
Сем. 32: Cerlhiidae. — Питухи.................................. —
Сем. 33: Nectariniidae. — Нектарницы............................ —
Сем. 34: Meliphagidae.—Медососы............................... 503
Сем. 35: Dicaeidae........................................... 504
Сем. 36: Zosteropidae. — Белоглазки............................ —
Сем. 37: Vireonidae.......................................... 505
Сем. 38: Prunellidae. — Завирушки.............................. —
Сем. 40: Thraupidae. —Танагры............................ 506
Сем. 41: Tersiiridae....................................... —
Сем. 42: Plcceidae. —Ткачики............................. 507
Сем. 43: Frirgiliidae. — Вьюрки.......................... 508
Сем. 44: Drepanididae. —Гг вайе кие цветочницы........... 509
Сем. 45: Coerebidae.—Цветочницы.......’................... .—
Сем. 46: Icteridae. —Американские иволги................. 510
ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ ПТИЦ.................................... 511
РУКОВОДЯЩАЯ ЛИТЕРАТУРА........................................ 545
Отв. редактор Е. М. Мастыко. Техн, редактор А. Н. КоварскаЯ.
Корректор М. И. Коровин,
Сдано в набор 1Э/VIII 1939 г. Подписано к печати 8/1 1940 г.
Леноблгорлит № 82 Тираж 1209 экз. Печ. л. 348Л. + 2 вклейки
Авт. л. 41,1. Уч.-авт. л. 42,1. Формат бумаги 60X92. Кол. печ. зн. в 1 печ. л. 48.288.
Цена 21 руб., переплет 1 руб. Заказ № 1209. Бумага Вишерского бумкомбината
Отпечатано в ФЗУ 2-й типографии ОГИЗа РСФСР треста „Полиграфкнига" «Печатный Двор*
им. А. М. Горького. Ленинград. Гатчинская, 26.