/
























































































































































































































































Автор: Баландин С.А. Алексеев Ю.Е. Жмылев П.Ю. Карпухина Е.А.
Теги: морфология растений ботаника биология словарь
ISBN: 5-211-02962-3
Год: 2005




Текст
ЫЛLi; П.Ю. АДE'KuP 115 IO,,
^аРпулин x fc;4-,Баландин e
БИОМОРФОЛОГИЯ
РАСТЕНИЙ
Ил Л IO CT F И ГО В A JI JIЫ Й СЛ О В А РI.
МОСКОВСКИЙ ГОСУДАРСТВЕННЫЙ
УНИВЕРСИТЕТ ИМЕНИ М. В. ЛОМОНОСОВА
БИОЛОГИЧЕСКИЙ ФАКУЛЬТЕТ
ЖМЫЛЕВ П. Ю., АЛЕКСЕЕВ Ю. Е.,
КАРПУХИНА Е. А., БАЛАНДИН С. А.
БИОМОРФОЛОГИЯ
РАСТЕНИЙ
ИЛЛЮСТРИРОВАННЫЙ
СЛОВАРЬ
Учебное пособие
2-е издание,
исправленное и дополненное
МОСКВА • 2005
ББК 28.56
Ж 45
УДК 581.412. (038)
Ответственный редактор:
член-корреспондент РАН, профессор В. Н. Павлов
Рецензенты:
профессор, доктор биологических наук Р. П. Барыкина
профессор, доктор биологических наук Е. И. Курченко
Жмылев П. К)., Алексеев Ю. Е.,
Карпухина Е. А., Баландин С. А.
Ж 45 Биоморфология растении: иллюстрированный словарь.
Учебное пособие. Изд. 2-е, испр. и доп.— М.: 2005.— 256 с., ил.
ISBN 5-211-02962-3
Предлагаемая работа — в 1 орое, исправленное и дополненное, издание
иллюстрированного терминологического словаря по биоморфологии рас-
тений. В отличие от 1-го издания (2002) словарь дополнен основными поня-
тиями эволюционной биоморфологии и биологии развития растений. Он
может служить справочником, позволяющим быстро ориентироваться в
обширном понятийном аппарате этой науки. Словарь содержит около
1300 терминов и понятий. Большое внимание уделено систематизации
близких по смыслу терминов и названий.
Для научных работников, преподавателей и студентов-биологов, свя-
занных с вопросами геоботаники, флористики, экологии и систематики.
ISBN 5-211-02962-3
© П. Ю. Жмылев и др., 2005
© С. А. Баландин, фото на обложке, 2005
Посвящается светлой памяти
основателей отечественной биоморфологии растений,
выдающихся воспитанников кафедры геоботаники
Московского государственного университета,
Ивана Григорьевича и Татьяны Ивановны СЕРЕБРЯКОВЫХ
ВВЕДЕНИЕ
Изучением жизненных форм организмов естествоиспытатели зани-
маются испокон веков. Тем не менее только в последние десятилетия о
биоморфологии (учении о жизненных формах) стали говорить как о са-
мостоятельной науке, сформировавшейся на стыке морфологии, эколо-
гии, систематики, биологии развития и эволюционного учения (Юрцев,
1976; Зозулин, 1976; Серебрякова, 1980; Хохряков, 1981; Правдин, 1986;
Алеев, 1986 и др.).
Термин «жизненная форма» был введен датским ботаником Евгени-
ем Вармингом в конце XIX века применительно к растениям и в настоя-
щее время широко используется не только ботаниками, но и зоологами
как общебиологическое понятие.
Традиционные названия жизненных форм растений (дерево, кустар-
ник, лиана, трава и др.) были заимствованы из разговорного языка. Их
содержание, вероятно, впервые рассмотрено учеником великого Ари-
стотеля Теофрастом, от «Естественной истории растений» которого ве-
дет свое начало тесное переплетение интересов современной биоморфо-
логии, филогенетической систематики растений и палеоботаники. Прак-
тическое применение классификации растений по жизненным формам
(биоморфам), начиная с работ выдающегося
немецкого естествоиспытателя Александра
фон Гумбольдта, получило широкое рас-
пространение в экологии.
Наиболее тесно биоморфология разви-
валась в связи с морфологией растений (осо-
оенно с органографией). Вот почему многие
термины и понятия, используемые при опи-
сании строения жизненных форм, заимство-
ваны из морфологии. В то же время на стыке
интересов морфологии, биологии развития
и физиологии растений сформировался ряд
важных направлений (онтогенетическое и
ритмологическое) и теорий в биоморфоло-
гии (например, теория Н. П. Кренке). И по-
>гому не случайно учение о жизненных
Теофраст
(371-287 гг. до нашей эры)
3
формах до сих пор часто рассматривается как особый раздел морфоло-
гии растений — экологическая морфология.
Определение экологической морфологии было в свое время предло-
жено известными отечественными ботаниками И. Г. Серебряковым и
Т. И. Серебряковой. Они определили ее как науку «о процессах морфоге-
неза растительных организмов и их отдельных органов в зависимости от
факторов внешней среды» (Серебряков, Серебрякова, 1967, с. 1449). Они
же сформулировали и основные методы исследования этой науки:
а) эколого-статистический, б) сравнительный онтоморфологический,
в) экспериментально-онтоморфологический.
Научное направление, основанное И. Г. Серебряковым в результате
синтеза достижений из различных разделов ботаники, в нашей стране яв-
ляется основным в изучении жизненных форм. В процессе его становле-
ния круг решаемых биоморфологами научных и прикладных проблем
постепенно расширялся. Если попытаться выделить среди них наиболее
важные, то к таковым можно отнести: 1) описание жизненных форм и их
классификация, 2) изучение становления жизненных форм в онтогенезе,
3) изучение изменчивости жизненной формы вида в различных экологи-
ческих условиях, 4) выяснение специфики роста и развития (ритма жиз-
ни) разных жизненных форм (одновременно и разных видов) в конкрет-
ных условиях, 5) познание морфогенетических процессов (онтогенеза ор-
ганов), определяемых эндогенными и/или экзогенными факторами,
6) установление коррелятивных связей между биоморфологическими
признаками и физиологическими процессами (интенсивность фотосин-
теза, дыхания и др.), 7) установление адаптивного значения биоморфоло-
гических признаков по отношению к отдельному экологическому факто-
ру или к комплексу факторов почвенной, водной или воздушной среды,
8) физиономическая или фитоценотическая роль биоморф и их спектры
в различных растительных сообществах, а также природных зонах и гор-
ных поясах, 9) эволюционные взаимоотношения жизненных форм.
Значимость перечисленных выше научных проблем, а также возрос-
ший в последнее время интерес к преподаванию биоморфологии на мно-
гих университетских ботанических кафедрах вызывают насущную необ-
ходимость создания иллюстрированного обзора понятий и терминов
этой дисциплины. Настоящее издание — переработанная «версия» на-
шей предыдущей работы (Жмылев и др., 1993, 2002), которая дополнена
основными понятиями эволюционной биоморфологии и биологии раз-
вития растений. Естественно, что при работе над ним авторы использова-
ли как различные словари (Henderson, Henderson, 1953; Jackson, 1953;
Vaczy, 1980; Терминология роста..., 1982; Биологический энциклопедиче-
ский словарь..., 1986; Реймерс, 1991; Меликян, Девятов, 2001, и др.), так
и многочисленные учебные (Ботаника..., 1988; Игнатьева, 1989; Учебно-
полевая..., 1990; Коровина, 1997; Коровкин, 2003, и др.) и научные изда-
ния, в которых комментируются биоморфологические термины и поня-
4
гия (Щербакова, 1960; Серебрякова, 1980; Борисова, 1991; и др.). Особое
внимание уделено работам, вышедшим за последние 10 лет.
Словарь не относится к разряду нормативных изданий. Основная его
цель — представить современную терминологию, используемую в прак-
тике научных исследований по биоморфологии растений. В рамках учеб-
ного пособия выполнить эту задачу в полном объеме чрезвычайно труд-
но, и поэтому проблема составления словника в целом решалась прагма-
тически (например, в него намеренно не включены понятия генетики
развития и биоритмологии). Словарь охватывает только те термины и
понятия, которые имеют более или менее частое употребление при описа-
нии онтогенеза и жизненных форм геммаксилярных растений или отра-
жают существенные с точки зрения авторов явления. Из многочисленных
классификаций учтены только оригинальные, а их варианты не рассмат-
риваются.
При составлении пособия авторы старались при небольшом объеме
достигнуть сочетания лаконизма, удобства пользования и максимально
возможной информативности. В словарь включено около 1300 терминов,
понятий, описаний методов исследования и теорий, сгруппированных в
тематические блоки. Часть из них вынесены в названия статей. Другие —
просто перечислены в словаре с отсылкой к одной из статей, в которой да-
ется объяснение (например: Биоморфа см. форма жизненная).
Принцип расположения терминов и понятий по тематическим бло-
кам обусловил и порядок расположения слов в названиях, состоящих из
двух и более слов. Первым всегда стоит существительное (как правило,
название тематического блока), за которым следует определение (напри-
мер, «Ветвление боковое», а не «Боковое ветвление»). Такой же поря-
док слов принят и в тексте статей, если используемые понятия объясня-
ются в словаре (эти понятия набраны курсивом).
Структура всех статей словаря однотипна. Заголовок статьи включает
название термина (полужирный шрифт), его написание на латинском
языке (в ряде случаев не приводится), справку об этимологии (написание
греческих слов дано латинским алфавитом) и во многих случаях англо-
язычный эквивалент. В тексте статьи понятие, давшее ей название, приво-
дится с прописной буквы в сокращенной форме (например: «По-
бег» — П., «Побег апогеотропный» — П. а.), а термины, разъяснение
которых дается в других статьях, набраны курсивом.
При объяснении терминов авторы стремились отразить всевозмож-
ные точки зрения, не навязывая собственного видения вопроса. Поэтому
во многих статьях наряду с точным (или наиболее распространенным)
значением термина приведены и другие его толкования. В конце статей
указаны автор термина и синонимы. Во многих случаях было трудно ус-
тановить приоритеты в этих вопросах, а при угрожающих размерах си-
нонимики невозможно было избежать некоторой субъективности в вы-
боре предпочтительных названий.
5
Практически ко всем статьям тематических блоков и к ряду статей
небольшого объема предложены литературные источники, в которых
приведены примеры или обсуждается термин, вынесенный в заголовок.
Считаем необходимым отметить, что поскольку «информационный по-
ток» в биоморфологии за последние десятилетия значи гельно вырос, его
тщательный и критический обзор и анализ -— совершенно самостоя-
тельная задача, ждущая своего кропотливого исследователя.
Для уменьшения объема статей в словаре использованы следующие
сокращения: б. м.—~ более или менее; гл. обр.— главным образом; лит.—
литература; наз.— называют, называется; назв.— название; пон.— по-
нятие; раст.— растение; син.— синоним(ы); см.— смотрите; ср.— срав-
ните; терм.— термин; т. о.— таким образом.
Авторы выражают огромную благодарность рецензентам словаря
проф. Р. П. Барыкиной и проф. Е. И. Курченко, а также проф. Н. М. Шс
риной за ценные советы и замечания.
Авторы отчетливо осознают, что настоящее издание не може1 быть
свободно от недостатков и упущений. Все замечания и предложения
по его содержанию с благодарностью будут приняты по адресу: 119899,
Москва, Ленинские горы, МГУ имени М. В. Ломоносова, биологический фа-
культет, кафедра геоботаники — или по электронной почте:
zhmylev@herba. msu. ru
balandin@hcrba. msu. ru
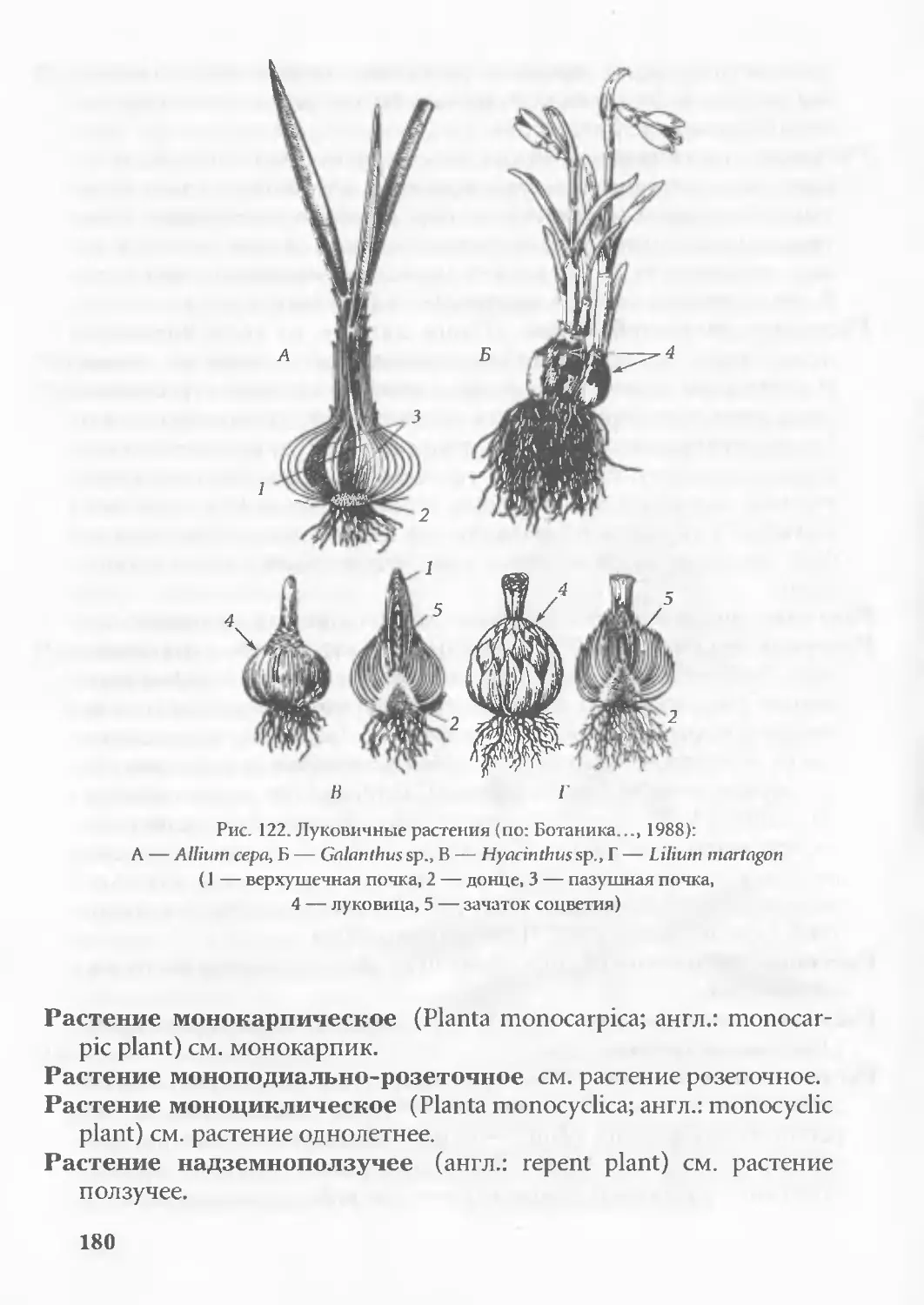
Аббревиация см. анаболия отрицательная.
Абортация верхушки побега — отмирание почки верхушечкой
(почки завершающей) в конце развития побега вегетативного
(Tilia cordata) как проявление возрастных изменений (см. старе-
ние), незаложения цветков (Betula) или из-за ухудшения условий;
А. в. и. предшествует образование отделительного слоя, который
перекрывает поступление в апекс побега питательных веществ;
лит.: Михалевская, 2002.
Автотроф (англ.: autotroph) — организм, способный ассимилиро-
вать СОг и использующий его в качестве главного источника угле-
рода; к А., противопоставляемым гетеротрофам, относятся фото-
трофы (раст., водоросли и бактерии, способные к фотосинтезу) и
хемотрофы (бактерии, окисляющие неорганические вещества).
Адаптация (от лат. adapto •— прилаживаю; англ.: adaptation) — со-
вокупность морфофизиологических, поведенческих (ростовых),
популяционных и других особенностей вида, обеспечивающих
его существование в определенных условиях среды. Нередко А.
рассматривают как процесс изменения организации, в результате
которого снижается уровень смертности из-за внешних условий;
см. модификация.
Акровергенция см. почка.
Акротония (Acrotonia, греч.
akros — верхушка, to-
nes — сила, мощь; англ.:
acrotony) — тип продоль-
ной симметрии побега
(или его прироста), выра-
жающийся в увеличении
длины междоузлий, разме-
ров почек или побегов
ветвления вдоль продоль-
ной оси стебля снизу
вверх (рис. 1); при резкой
Рис. 1. Продольная симметрия побега
(1 — акротония, 2 — мезотония,
3 — базитония)
7
А., например, все междоузлия, кроме последнего, остаются укоро-
ченными; акротонное распределение побегов боковых (ветвление
акротонное) особенно характерно для деревьев; лит.: ботаника...,
1988; Серебряков, 1962; Серебрякова, 1971а.
Акселерация (от лат. acceleratio — ускорение; англ.: acceleration) —
один из основных типов элементарной гетерохронии, при кото-
ром ускорение соматического развития сочетается с неизменной
скоростью полового созревания (ср. неотения). В результате А.
потомок имеет пераморфный облик (достигает половой ;рело-
сти, как и предок, но черты взрослого организма у него появляют-
ся раньше в онтогенезе, рис. 2). В биоморфологии А. часто понима
ют очень широко — как ускорение развития вообще (ср. неоте-
ния); лит.: de Beer, 1958; Юсуфов, 1979; Gould, 1980.
Рис. 2. Изменения цикличности развития и гетерофилии побета в результате
элементарных гетерохроний: 1 — первый год, 2 — второй год, 3 — третий год,
ц — цветок, л в — «взрослый» лист, лю — «ювенильный» лист
Аллелопаразит (Alleloparasitus, от греч. allelus — взаимно, и пара
зитизм) — сапрофит или полусапрофит, с облигатной эндо-
трофной или эктэндотрофной микоризой, получающий питание
из почвы или от других автотрофных раст. посредством своего
микоризного гриба; А.— травянистые многолетники и однолет-
ники, связь которых с грибом рассматривается как симбиотиче-
ский или сбалансированный паразитизм, а при узком понимании
явления паразитизма как мутуализм (Orchidaceae).
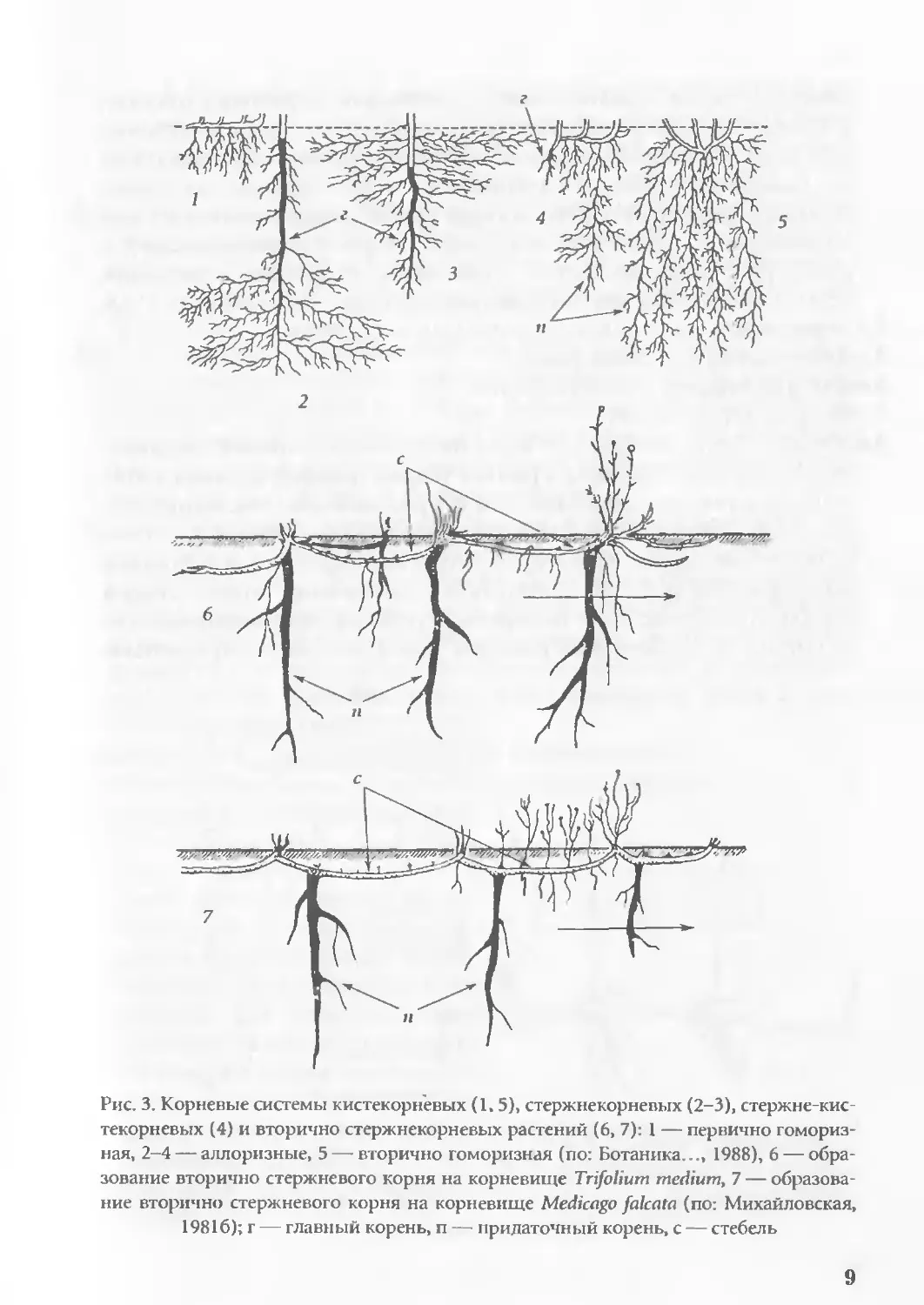
Аллоризия (Allorrhizia, от греч. alios — другой, rhiza—корень;
англ.: allorhizy) — формирование системы корневой раст. из кор-
ней разных по происхождению (рис. 3)— корня главного, i >рня
8
Рис. 3. Корневые системы кистекорневых (1.5), стержнекорневых (2-3), стержне-кис-
текорневых (4) и вторично стержнекорневых растений (6, 7): 1 — первично гомориз-
ная, 2-4 — аллоризные, 5 — вторично гоморизная (по: Ботаника..., 1988), 6 — обра-
зование вторично стержневого корня на корневище Trifolium medium, 7 — образова
ние вторично стержневого корня на корневище Medicago falcate (по: Михайловская,
19816); г — главный корень, п — придаточный корень, с — стебель
9
бокового, корня придаточного (аллоризная корневая система);
в отличие от гоморизофитов, у семенных раст. (аллоризофитов
или раст. биполярных) первый корень зародыша закладывается на
его базальном полюсе и развивается в корень главный, на основе
которого формируется стержневая (раст. стержнекорневые) или
смешанная корневая система (раст. стержне-кистекорневые)', у
некоторых аллоризофитов отмечается вторичная гоморизия',
терм.: Goebel, 1930; лит.: Михайловская, 1981а; Groff, Kaplan, 1988.
Аллоризофит (англ.: allorhizophyte) см. аллоризия.
Амфикотилия см. синкотилия.
Амфисинкотилия см. синкотилия.
Амфифит см. гелофит.
Анаболия (англ.: anaboly) — один из основных способов эволюции
морфологической органов в рамках теории филэмбриогенеза, кото-
рый представляет собой надставку (пролонгацию) последней ста-
дии развития органа, т. е. продолжение развития органа у потом-
ка после той стадии, на которой у предка морфогенез этого органа
заканчивается (рис. 4). В результате добавления новых стадий
морфогенеза происходят изменения строения сформировавшего-
ся органа. А. Н. Северцов выделял также анаболию отрицателъ-
Рис. 4. Схема способов филэмбриогенеза: 1 — удлинение продо оклгельности морфо-
генеза (анаболия, надставка), 2 — сокращение продолжительности морфогенеза
(отрицательная анаболия), 3 — изменение (уклонение) развития на средней стадии
морфогенеза (девиация), 4 — изменение (уклонение) развития на ранней стадии мор-
фогенеза (архаллаксис)
10
ную. В биоморфологии А. часто понимают в широком смысле как
надставку морфогенеза или онтогенеза (ср. гетерохрония); терм.:
Северцов, 1912; син.: надставка, пролонгация, продление разви-
тия; лит.: Северцов, 1945, Тахтаджян, 1964, Северцов, 1981.
Анаболия отрицательная — способ эволюции морфологической ор-
ганов в рамках теории филэмбриогенеза, которым в отличие от
анаболии обозначают выпадение конечной стадии морфогенеза у
потомка (рис. 4) по сравнению с предком (ср. редукция); син.: аб-
бревиация.
Анализ биоморфологический (англ.: life form analysis) — широ-
ко используемое понятие для обозначения процедуры выделения
и описания элементов структуры формы жизненной раст. (см. ме-
тод описания форм жизненных), определения состава жизненных
форм таксона, флоры или фитоценоза и распределения их по эко-
топам (см. А. б. флоры и растительности).
Анализ биоморфологический флоры и растительности — оп-
ределение состава и соотношения жизненных форм (спектра) или
конкретных биоморфологических признаков во флоре или фито-
ценозе с использованием одной или нескольких классификаций
форм жизненных; количественные данные А. б. обычно рассмат-
риваются как индикаторы эколого-ценотических и климатиче-
ских условий; лит.: Raunkiaer, 1934; Серебряков, 1954а; Юрцев,
1976; Полозова, 1981.
Анизокладия (англ.: anisoclady) см. анизофиллия.
Анизокотилия (англ.: anisocotyly) см. гетерокотилия.
Анизоризия (Anisorrhizia, греч.
anisos — неравный, rhiza — ко-
рень) — морфологическое
или/и функциональное разли-
чие корней, расположенных на
разных сторонах одного междо-
узлия (рис. 5); А. характерна, как
правило, для горизонтальных
корневищ, на верхней поверхно-
сти которых корни развиваются
слабее, чем на нижней или бо-
ковых; ср. гетероризия; лит.:
Михайловская, 1981а.
Анизотомия см. ветвление верху-
шечное.
Рис. 5. Столон Ranunculus lingua
с узловыми корнями: 1 — столон,
2 — лист, 3 — придаточные корни
(по: Серебряков, 1952)
11
Рис. 6. Анизофиллия и анизокладия
у Eranthemum pulchellum: 1 — листья,
2 — побеги ветвления (по: Bell,
Bryan, 1998)
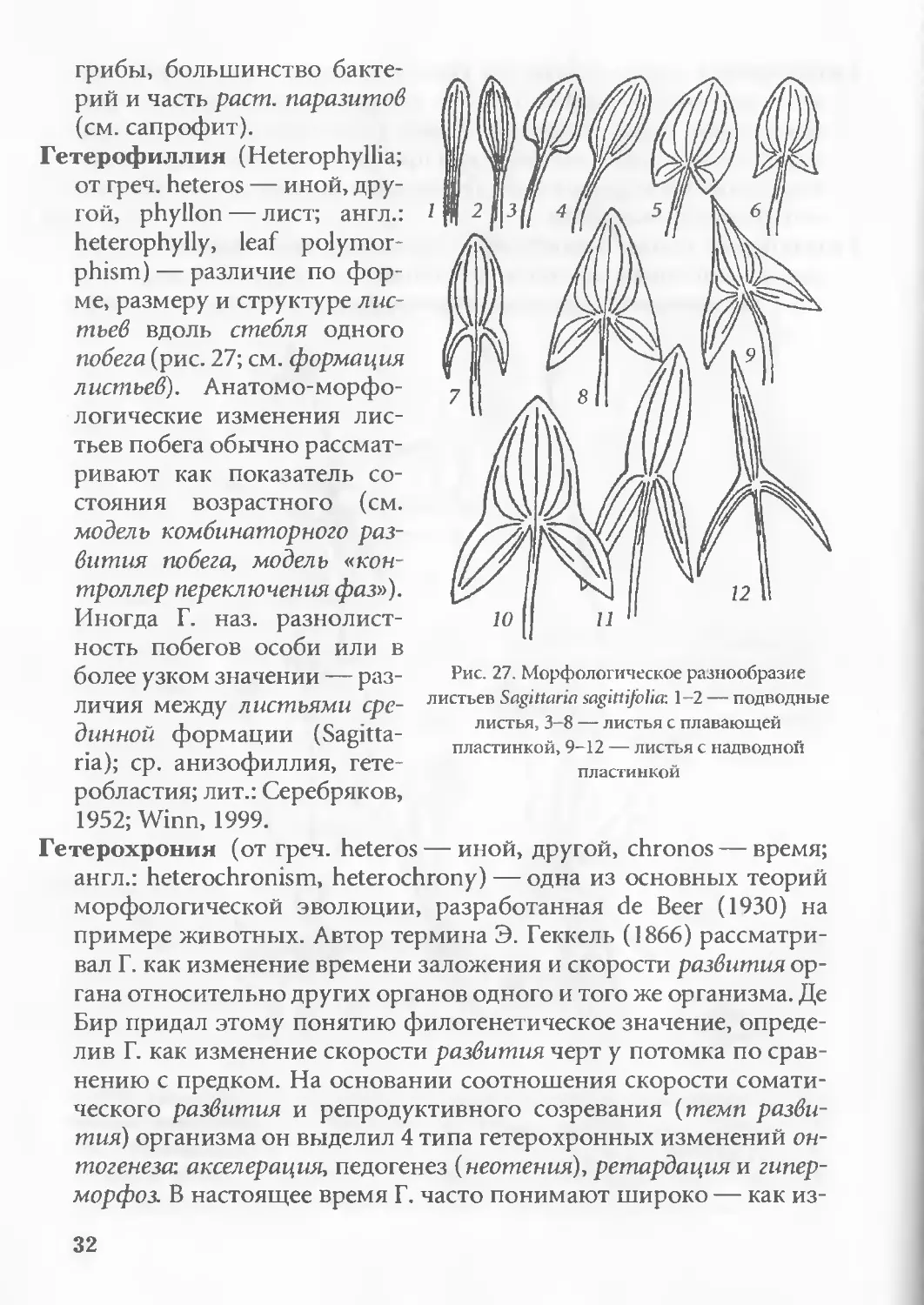
Анизофиллия (Anisophyllia, греч. anisos — неравный, phyllon —
лист; англ.: anisophylly) — форма симметрии побега, при которой
супротивные листья отличаются размером, формой или структу-
рой (нодальная А.; англ.: nodal anisophylly). Выделяют первичную
А., связанную с различиями в заложении примордиев, и вторич-
ную А., как результат ориен-
тации растущего побега. А.
часто сопровождается диф-
ференциацией в развитии
побегов ветвления (анизокла-
дия; англ.: anisoclady), измене-
нием филлотаксиса и асим-
метрией анатомического стро-
ения стебля; в более широком
смысле А. наз. и отличия ли-
стьев на нижней и верхней
стороне побега плагиотропного
(латеральная А.; англ.: lateral
anisophylly; рис. 6); ср. гетеро-
филлия; лит.: Ботаника...,
1988; Dengler, 1999.
Апекс (Apex, от лат. apex — остроконечная верхушка, кончик, вер-
шина; англ.: apex) — верхушка побега {конус нарастания побега) и
корня {А. корня), состоящая из первичной меристемы, обеспечи-
вающей формирование всех частей и первичных тканей (рис. 7);
син.: меристема апикальная, точка роста; лит.: Ботаника..., 1988,
Лотова, 2000; Иванов, 2004.
Апекс корня (англ.: root apex) — верхушка корня, состоящая из
первичной меристемы и защищенная обычно корневым чехли-
ком (рис. 7), который обеспечивает продвижение растущего кор-
ня в субстрате и его геотропическую реакцию (см. тропизм);
у нек-рых корней А. к. функционирует недолго и дифференциру-
ется в корневые волоски (см. корень кластерный); лит.: Михай-
ловская, 1981а; Иванов, 2004.
Апекс побега (англ.: shoot apex) см. конус нарастания побега.
Архаллаксис (от греч. arche — начало, allaxis — изменение; англ.:
archallaxis) — один из основных способов эволюции морфологиче-
ской органов в рамках теории филэмбриогенеза, который пред-
ставляет собой изменение на ранних стадиях развития органа у
потомка по сравнению с предком (рис. 4). В результате изменения
12
Рис. 7. Апекс корня (А), вегетативный апекс побега (Б — элодеи, В — пырея) и фло-
ральный апекс побега (Г — барвинка, Д — календулы): 1—корневой чехлик,
2 — примордий листа, 3 — зачаток листа, 4 — зачаток чашелистика, 5 — примордий
тычинки, 6 — примордий лепестка, 7 — примордий плодолистика, 8 — зачаток цвет-
ка, 9 — зачаток листочка обертки (по: Ботаника. ., 1988)
13
самых ранних стадий морфогенеза происходит резкая и сущест-
венная перестройка всего облика взрослого органа. В биоморфо-
логии к А. относят обычно изменения в развитии зародыша или
числа зачатков на апексе побега:, терм.: Северцов, 1912; лит.: Север-
цов, 1945, Тахтаджян, 1964, Северцов, 1981.
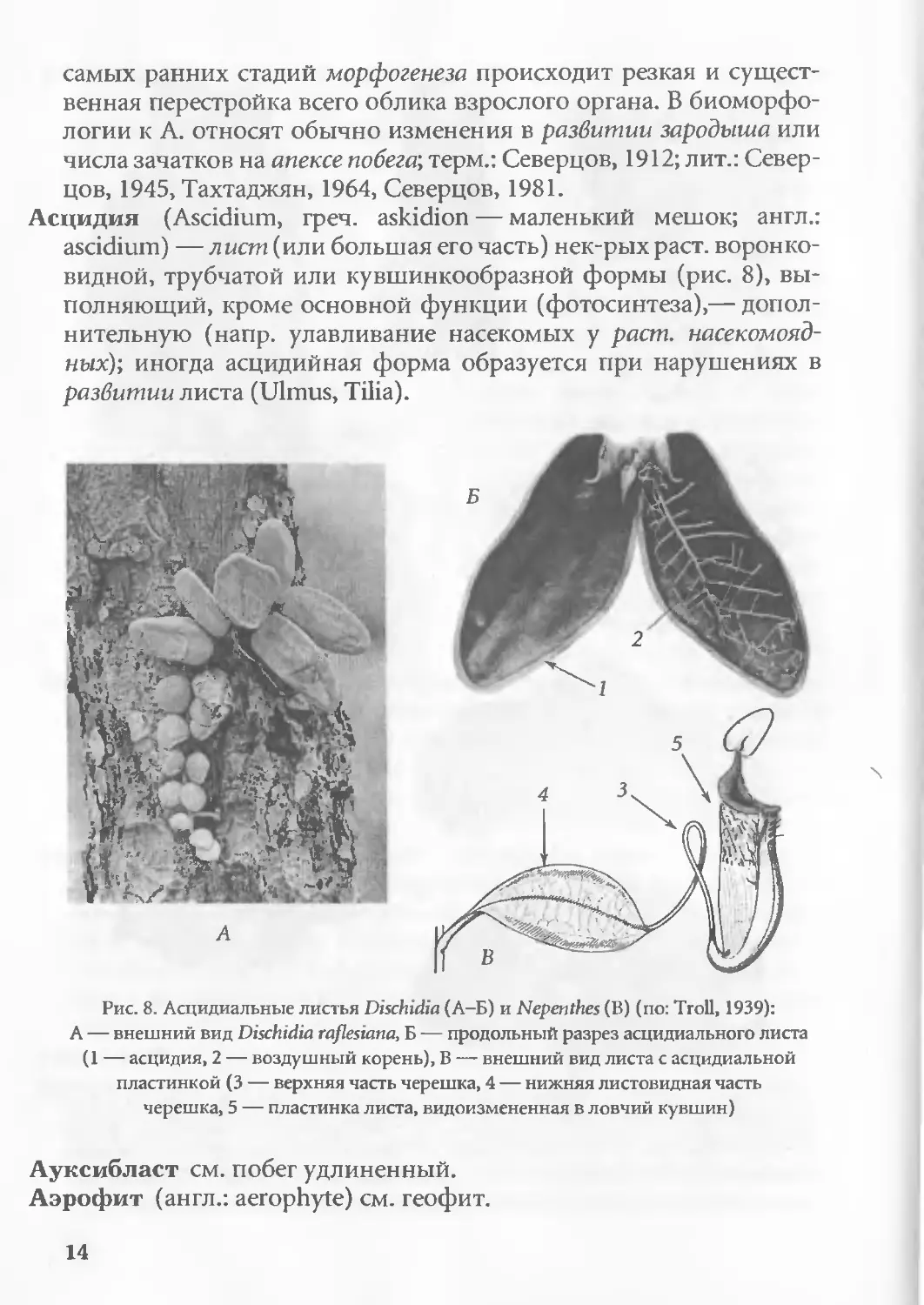
Асцидия (Ascidium, греч. askidion — маленький мешок; англ.:
ascidium) — лист (или большая его часть) нек-рых раст. воронко-
видной, трубчатой или кувшинкообразной формы (рис. 8), вы-
полняющий, кроме основной функции (фотосинтеза),— допол-
нительную (напр. улавливание насекомых у раст. насекомояд-
ных); иногда асцидийная форма образуется при нарушениях в
развитии листа (Ulmus, Tilia).
Рис. 8. Асцидиальные листья Dischidia (А-Б) и Nepenthes (В) (по: Troll, 1939):
А — внешний вид Dischidia raflesiana, Б — продольный разрез асцидиального листа
(1 — асцидия, 2 — воздушный корень), В — внешний вид листа с асцидиальной
пластинкой (3 — верхняя часть черешка, 4 — нижняя листовидная часть
черешка, 5 — пластинка листа, видоизмененная в ловчий кувшин)
Ауксибласт см. побег удлиненный.
Аэрофит (англ.: aerophyte) см. геофит.
14
Базитония (Basitonia, от греч. basis — основание, tones — сила,
мощь; англ.: basitony) — тип продольной симметрии побега (или
его прироста), выражающийся в уменьшении длины междоузлий,
размеров почек или побегов ветвления вдоль продольной оси
стебля снизу вверх (рис. 1); при резкой Б., напр., почки пазушные
закладываются только в нижней части побега; базитонное распре-
деление побегов боковых (ветвление базитонное) характерно для
многих злаков и кустарников (см. кущение); лит.: Ботаника...,
1988; Серебрякова, 1971а.
Биоморфа моноцентрическая — один из типов классификации
форм жизненных по О. В. Смирновой, объединяющий раст., побеги,
корни и почки возобновления взрослых особей которых сосредото-
чены в одном центре (центре разрастания особи)-, гл. обр. раст. ве-
гетативно неподвижные, в конце онтогенеза которых отсутствует
дезинтеграция морфологическая (деревья с нарастанием моноподи-
альным, стержнекорневые монокарпики) или она частичная, позд-
няя, неспециализированная (полукустарники, кустарники, раст.
подушковидные, деревья и стержнекорневые травы); у раст. (плот-
но)рыхлодерновинных отмечается полная неспециализированная
дезинтеграция (злаки, осоки); ср. индивид простой, монофит;
терм.: Смирнова и др. (Ценопопуляции..., 1976); лит.: Ценопопу-
ляции..., 1988.
Биоморфа неявнополицентрическая — один из типов класси-
фикации форм жизненных по О. В. Смирновой, объединяющий
раст., побеги, корни и почки возобновления взрослых особей кото-
рых образуют несколько близко расположенных друг к другу
центров разрастания особи; гл. обр. раст. короткокорневищные
(Asarum europaeum), которые в онтогенезе проходят несколько
фаз развития: первичную моноцентрическую, неявнополицен-
трическую и в ряде случаев — вторичную моноцентрическую; ср.
полифит, индивид сложный; терм.: Смирнова и др. (Ценопопуля-
ции..., 1976); лит.: Ценопопуляции..., 1988.
Биоморфа см. форма жизненная и биоморфология.
Биоморфа явнополицентрическая — один из типов классифика-
ции форм жизненных по О. В. Смирновой, объединяющий раст.,
взрослые особи которых имеют несколько явно выраженных,
отстоящих друг от друга центров разрастания (один из них пер-
вичный); такие участки сосредоточения побегов, корней и почек во-
зобновления могут представлять собой, например, кусты парци-
альные (автономные части индивида сложного); гл. обр. раст. веге-
15
тативно подвижные со специализированной дезинтеграцией
морфологической (размножение вегетативное); ср. полифит;
терм.: Смирнова и др. (Ценопопуляции..., 1976); лит.: Ценопопу-
ляции..., 1988.
Биоморфология (Biomorphologia, от греч. bios — жизнь, morphe —
форма, logos — слово, учение; англ: biomorphology) — учение о
формах жизненных (биоморфах) организмов (в частности, раст.—
фитобиоморфология), их строении, развитии в онтогенезе, рас-
пространении, экологии, биологии (поведении) и эволюции; в
системе биологических наук проблемы Б. находятся на стыке инте-
ресов различных дисциплин: экологии, морфологии, систематики,
биологии развития, физиологии, биогеоценологии, биогеографии
и эволюционного учения; в связи с этим в Б. выделяют несколько
направлений исследования, среди которых: структурная, онтоге-
нетическая, экологическая, географическая, эволюционная Б. и
др.; терм, «биоморфа» введен Б. М. Козо-Полянским (1945) и долго
употреблялся гл. обр. В. Н. Голубевым для обозначения биоло-
го-морфологических особенностей таксонов; им же (Голубев,
1956б), вероятно, впервые использован и термин «биоморфоло-
гия» вместо «экологической морфологии растений» (Голубев,
1965); как синоним учения о жизненных формах терм. Б. впервые
употреблен А. П. Хохряковым (1975а); часто Б. рассматривается
как синоним экологической морфологии; син.: экобиоморфоло-
гия, эпиморфология; лит.: Серебрякова, Серебряков, 1967; Сереб-
рякова, 1972; Юрцев, 1976; Зозулин, 1976; Алеев, 1986; Правдин,
1986; Schmid, 1963; Kull, 1987; Нухимовский, 1997, 2002.
Брахибласт (англ.: brachyblast) см. побег укороченный.
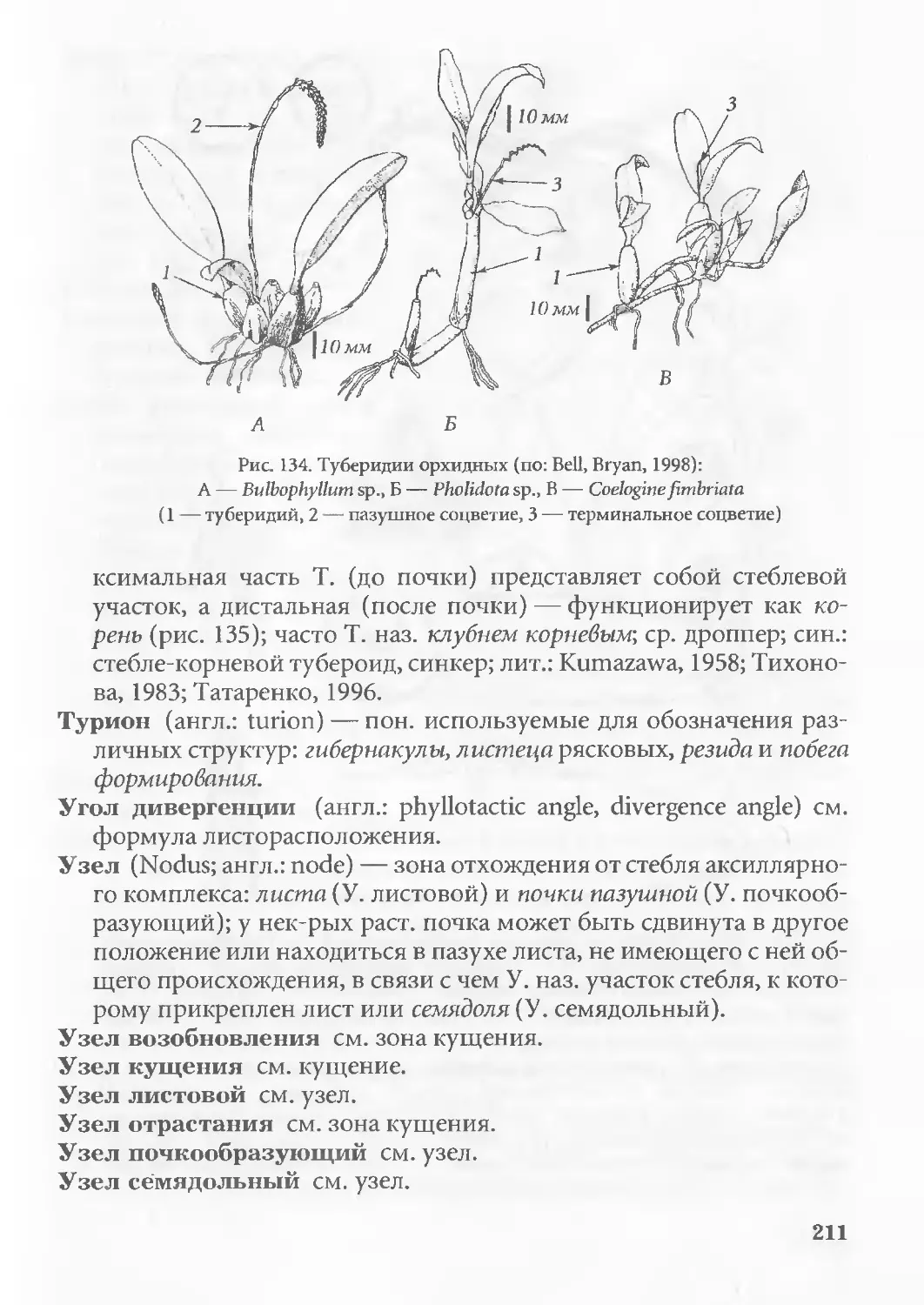
Бульба см. туберидий.
Бульбилла (англ.: bulbil) см. луковичка.
Бульбочка (англ.: tubercle) см. клубенек.
Ветвепад (Cladoptosis, от греч. klados — ветвь, ptosis — падение;
англ.: cladoptosis) — естественное опадение (одно)многолетних
побегов (ветвей) из кроны дерева (Salix fragilis) или кустарника;
обычно рассматривается как один из способов регуляции водного
режима раст. (уменьшение площади испаряющей поверхности);
ср. листопад.
Ветвление (Ramificatio, от лат. ramus — ветвь, сук, ветка, facio —
делать, производить; англ.: branching, ramification) — процесс
роста и формообразования раст., в результате которого возникает
система соединенных между собой в определенном порядке до-
16
черних, однотипных элементов структуры (напр. система побе-
гов, корнейилм сложный лист)-, В. свойственно большинству раст.
и обусловливает увеличение их поверхности соприкосновения с
внешней средой; от характера В. зачастую зависит габитус раст.
(см. поведение раст.); различают два основных типа В.— В. верху-
шечное и В. боковое (рис. 9); часто под В. понимают не процесс,
а результат — взаиморасположение стеблей (корней) относитель-
но друг друга, а их формирование называют нарастанием или во-
зобновлением; лит.: Серебряков, 1952; Грудзинская, 1974; Нухи-
мовский, 1974; Шафранова, 1981; Ботаника..., 1988; Лотова, 2000;
Wilson, 2000; Михалевская, 2002.
Ветвление акромоноподиальное см. нарастание моноподиальное.
Ветвление акросимподиальное см. нарастание симподиальное.
Ветвление акротонное см. акротония.
Ветвление аксиллярное (англ.: axillary branching) см. ветвление
боковое.
Ветвление апикальное (англ.: apical branching) см. ветвление вер-
хушечное.
Ветвление базимоноподиальное см. нарастание моноподиальное.
Ветвление базисимподиальное см. нарастание симподиальное.
Ветвление базитонное см. базитония.
Ветвление боковое (Ramificatio lateralis, от лат. latus — сторона,
бок; англ.: lateral branching) — тип В., при котором точка роста
материнского элемента структуры отделяет один или несколько
зачатков, из которых развиваются дочерние элементы, занимаю-
щие боковое положение (рис. 9); В. б. побеговраст. геммаксилляр-
ных связано с деятельностью почек пазушных (В. пазушное или
аксиллярное) или п. внепазушных (В. внепазушное); при В. б. ма-
теринский элемент растет неопределенно долго, формируя глав-
ную ось (В. моноподиальное; рис. 9) или «точка роста» прекращает
развитие (В. симподиальное, рис. 9), а главная ось образуется из
побегов замещения (см. нарастание).
Ветвление верхушечное (Ramificatio apicalis, от лат. apex — вер-
хушка; англ.: apical branching) — тип В., при котором «точка рос-
та» материнского элемента структуры разделяется на два равных
зачатка (рис. 9), образующих дочерние, равновеликие (дихото-
мия истинная или равновильчитая, изотомия; рис. 9) или нерав-
новеликие (дихотомия неравновильчатая, анизотомия; рис. 9)
элементы, занимающие верхушечное (терминальное) положение
(см. также В. дихоподиальное); при В. в. рост материнского эле-
17
Рис. 9. Основные типы верхушечного (1-3) и бокового (4-7) ветвления: 1 —равно-
вильчатая дихотомия, 2 — неравновильчатая дихотомия, 3 — дихоподиальное,
4 — моноподиальное при супротивном листорасположении, 5 — моноподиальное
при очередном листорасположении, 6 — симподиальное, 7 — ложнодихотомическое
(по: Федоров и др., 1962)
мента прекращается, и главная ось вильчаторазветвленной систе-
мы не формируется; В. в. характерно для некоторых водорослей,
плаунов и, вероятно, папоротников; среди цветковых растений
встречается редко (напр. жилкование лепестков); син.: В. дихото-
мическое, В. апикальное, В. вильчатое.
Ветвление вильчатое см. ветвление верхушечное.
Ветвление вневлагалищное см. ветвление экстравагинальное.
Ветвление внепазушное см. ветвление боковое.
Ветвление внутривлагалищное см. ветвление интравагинальное.
18
Ветвление внутрипочеч-
ное — формирование
почки боковой на побеге в
период его роста внутри-
почечного (рис. 10); часто
такое образование наз.
«почка в почке»; лит.: Ми-
халевская, 2002.
Ветвление дихоподиаль-
ное (Ramificatio dichopo-
dialis, от греч. dicha — от-
дельно, врозь и pous—
нога, здесь — ось) — В.
_ Рис. 10. Внутрипочечное ветвление у дуба:
верхушечное, при кото- , \ п 77
Г у г 1 — катафиллы, 2 — зачатки листьев,
ром размеры дочерних З-боковаяпочкаЩогБотаника...,^)
элементов (напр. стеб-
лей) резко отличны, и больший из них перевершинивает меньший,
создавая видимость продолжения материнского элемента (стеб-
ля); в результате В. д. формируется зигзагообразная главная ось
(дихоподий), внешне напоминающая симподий раст. геммаксил-
лярных (рис. 9); В. д. встречается у нек-рых плаунов, селагинелл,
папоротниковидных и часто неправильно наз. В. симподиалъным.
Ветвление дихотомическое (англ.: dichotomy, dichotomous
branching) см. ветвление верхушечное.
Ветвление интравагинальное (Ramificatio intravaginalis, от лат.
intra — внутри, vagina — влагалище; англ.: intavaginal bran-
ching) — В. боковое преимущественно побегов злаков и осок, в ре-
зультате которого образуются побеги интравагиналъные (рис. 11);
часто В. и. называются кущением или возобновлением интраваги-
нальным; син.: В. внутривлагалищное; лит.: Серебрякова, 1971а.
Ветвление итеративное (Ramificatio iterativa, от лат. itero — вто-
рично делать, повторять; англ.: iterative branching) — беспрерыв-
ное образование за вегетационный период нескольких, следую-
щих друг за другом побегов ветвления, формирующих ось скелет-
ную раст. (рис. 12); В. и. встречается у некоторых однодольных
(Asparagus, Juncus и др.); терм.: Troll, 1964.
Ветвление концентрированное см. кущение.
Ветвление кустовое см. кущение.
Ветвление ложно дихотомическое (Ramificatio pseudodichotomi-
са, от греч. pseudes — ложный, dicha — отдельно, врозь, temnien —
19
Рис. 11. Схема ветвления злаков и осок (по: Смирнов, 1958а):
А — интравагинальное, Б — экстравагинальное, В — смешанное
(1 — побег материнский, 2 — побег интравагинальный,
3 — побег экстравагинальный)
Рис. 12. Итеративное ветвление корневища Carex arenaria
(по: Bell, Bryan, 1998): А — общий вид, Б — схема (1 — побег ветвления,
побег итеративный, 2 — катафилл, 3 — придаточный корень)
20
разрезать) — В. боковое побегов с листорасположением супротив-
ным, в результате которого образуется дихазий (рис. 9); встречается
у некоторых деревьев и кустарников.
Ветвление мезомоноподиальное см. нарастание моноподиальное.
Ветвление мезосимподиальное см. нарастание симподиальное.
Ветвление мезотонное см. мезотония.
Ветвление метельчатое см. ветвление рассеянное.
Ветвление моноподиальное (англ.: monopodial branching) см. на-
растание моноподиальное.
Ветвление пазушное см. ветвление боковое.
Ветвление пучковое (Ramificatio
fascicularis, от лат. fascis — связка,
пучок) — В. боковое побега с чере-
дующимися участками удлинен-
ных и укороченных междоузлий, к
последним из которых приуроче-
ны побеги ветвления (рис. 13); в
результате В. п. создается впечат-
ление «пучкового» распределения
ветвей на материнском стебле
(напр., Eleusine indica, Aeluropus
littoralis); терм.: Jacques-Felix, 1962
(цит. по: Серебрякова, 1971а).
Ветвление рассеянное (Ramificatio
sparsa, от лат. sparsus — разбросан-
ный, рассеянный) — В. боковое по-
бега в области его удлиненных
междоузлий (рис. 13); понятие ис-
Рис. 13. Схема рассеянного (А)
и пучкового ветвления (Б)
пользуется для противопоставле-
ния кущению; у побегов безрозеточ-
ных образование боковых генера-
тивных побегов наз. В. метельчатым.
Ветвление симподиальное (англ.: sympodial branching) см. нарас-
тание симподиальное.
Ветвление смешанное — В. боковое преимущественно побегов зла-
ков и осок, в результате которого образуются два типа побегов: по-
беги экстра- и интравагиналъные (рис. 11); лит.: Серебрякова, 1971а.
Ветвление экстравагинальное (Ramificatio extravaginalis, от лат.
extra — вне, vagina — влагалище; англ.: extravaginal branching) —
В. боковое преимущественно побегов злаков и осок, в результате
21
которого образуются побеги экстравагиналъные (рис. 11); часто
В. э. наз. кущением или возобновлением экстравагинальным; сии:
В. вневлагалищное; лит.: Серебрякова, 1971а.
Ветвь (Ramus; англ.: ramus, branch) — широко используемое поня-
тие для обозначения различных побегов ветвления (напр., ветви
соцветия); в специальном значении В. наз. одревесневшие, мно-
голетние стебли кроныраст. древесных, а для многолетних систем
побеговых раст. травянистых (напр., корневище) используют
назв. «ответвление».
Вивипария (Viviparia, от лат. vivus — живой, pario — рожаю; англ.:
viviparity, vivipary) — прорастание семян до их опадания с мате-
ринского раст. (В. настоящая или живорождение), которое наз.
живородящим раст. (рис. 14); В. свойственна мангровым деревьям
(Rhizophora); обычно В. наз. размножение вегетативное посредст-
вом луковичек или клубеньков, образующихся в области соцветия,
реже—почек выводковых (В. ложная или псевдовивипария;
рис. 15); лит.: Шульц, 1939; Серебряков, 1952; Синнот, 1963; Баты-
гина, Брагина, 2000а.
Рис. 14. Прорастание семени
Rhizophora mangle. 1 — семя,
2 — гипокотиль, 3 — корень
(по: Bell, Bryan, 1998)
Рис. 15. Псевдовивипария
у Lilium sp.: 1 — луковичка
с придаточными корнями
(по: Bell, Bryan, 1998)
22
Вивипария ложная см. вивипария.
Вивипария настоящая см. виви-
пария.
Видоизменение корня см. мета-
морфоз корня.
Видоизменение побега см. мета-
морфоз побега.
Влагалище корневое см. колео-
риза.
Влагалище семядольное (Vagina
cotyledonaris) — базальная часть
семядоли зародыша однодольного
раст. в виде цельного или откры-
того в той или иной степени с вен-
тральной стороны трубчатого об-
разования, охватывающего (за-
ключающего) почечку (рис. 16,17).
Рис. 17. Основные типы прорастания
пальм: А—адмотивно-лигулярное, Б —
ремотивно-лигулярное, В -— ремотив-
но-тубулярное (1 — главный зароды-
шевый корень, 2 — лигула (язычок),
3,4 — первый и второй лист проростка,
5 — семя, 6 — семядольное влагалище,
7 — шейка семядоли (по: Gatin, 1906)
Рис. 16. Надземное (А-Б), подземное (В-Г) и полуподземное (Д) прорастание одно-
дольных: А — лук репчатый, Б — вороний глаз, В — ландыш, Г — финиковая пальма,
Д — традесканция (1—главный корень, 2 — придаточный корень, 3 — семядоля,
4 — влагалище семядоли, 5 ~— гаусторий, 6 — лист, 7 — почка, 8 — чешуевидный
лист, 9 — семенная кожура, 10 — эндосперм) (по: Ботаника..., 1988)
23
1
Рис. 18. Поперечный срез ствола
сосны: 1 — годичные кольца
(по: Федоров и др., 1962)
Возобновление вегетативное (англ.: vegetative renewal р. р.) —
понятие, используемое для обозначения размножения вегетатив-
ного, сезонного возобновления роста раст. или отрастания (англ.:
vegetative renewal), реже — возобновления популяции вследствие
вегетативного размножения.
Возобновление интравагинальное см. ветвление интраваги-
нальное.
Возобновление экстравагинальное см. ветвление экстраваги-
нальное.
Возраст абсолютный — интервал времени от прорастания до мо-
мента наблюдения раст.; у деревьев определяется по годичным
кольцам, или слоям прироста ствола (рис. 18), количеству прирос-
тов годичных, мутовкам, развилкам и др.; у большинства раст.
травянистых— только
путем многолетних на-
блюдений над фиксиро-
ванными особями, в связи
с чем различают В. услов-
ный; В. а. отдельных час-
тей раст. (побега, листа и
др.) определяют во вре-
менных (от возникнове-
ния), пластохронных или
филохронных единицах
(В. собственный); син.:
В. календарный, В. хроно-
логический; лит.: Корча-
гин, 1960 а, б; Работнов,
1960; Стешенко, 1960;
Жуйкова, 1964.
Возраст биологический (англ.: biological age) см. состояние воз-
растное.
Возраст календарный (англ.: calendar age) см. возраст абсолют-
ный.
Возраст общий — В. физиологический побега или системы побегов
(ветвь, ствол, куст парциальный, особь вегетативного происхож-
дения и др.), определяемый как сумма В. абсолютного особи и
В. собственного (абсолютного) побега или системы побегов этой
особи; терм. Кренке, 1940.
Возраст пластохронный см. индекс пластохронный.
24
Возраст собственный см. воз-
раст абсолютный.
Возраст условный — интервал
времени от возникновения са-
мой старой сохранившейся
части раст. до момента наблю-
дения; определяется по остат-
кам надземных побегов, прирос-
ту годичному, числу кустов
парциальных и др. (рис. 19); ве-
личина В. у. зависит от интен-
сивности отмирания много-
летних частей раст. и уменьша-
ется с наступлением старости;
иногда В. у. смешивают с поня-
тием В. абсолютный-, син.: В.
относительный; терм.: Смир-
нова, 1967.
Возраст физиологический
(англ.: physiological age) см. со-
стояние возрастное.
Возраст хронологический см.
возраст абсолютный.
Возрастность см. состояние воз-
растное.
Волчки см. побег формирова-
ния.
Всход (Plantula; англ.: plantlet,
plantling, plantule, seedling) —
пон., обычно рассматриваемое
как синоним терм, «пророс-
ток»; реже проростком наз.
Рис. 19. Пятилетии й участок
корневища Aconitum sp.: 1 — резиды
прошлогодних побегов, 2 — побег
текущего года, 3 — придаточные
корни (по: Федоров и др., 1962)
раст. в период от прорастания до появления первых настоящих
листьев, а В.— раст., несущее семядольные и настоящие листья
(см. Васильченко, 1936; Покровская, 1958).
Габитус (англ.: habit) см. форма жизненная.
Гамокотилия см. синкотилия.
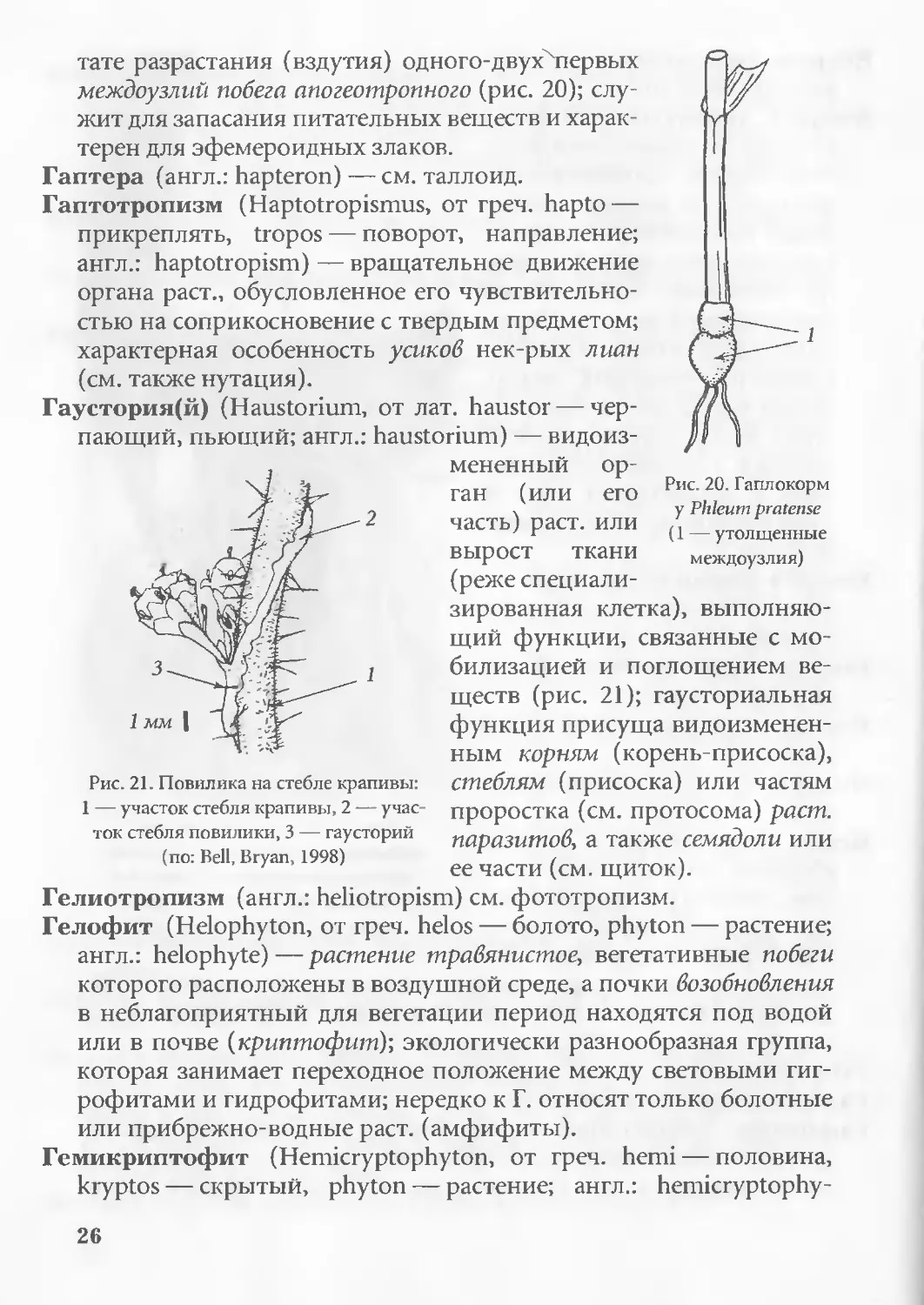
Гаплокорм (Haplocormus, от греч. haploos — одиночный, простой,
kormos — пень, ствол; англ.: haplocorm) — клубневидный участок
стебля нек-рых злаков (Phleum pratense), образующийся в резуль-
25
тате разрастания (вздутия) одного-двуххпервых
междоузлий побега апогеотропного (рис. 20); слу-
жит для запасания питательных веществ и харак-
терен для эфемероидных злаков.
Гаптера (англ.: hapteron) — см. таллоид.
Гаптотропизм (Haptotropismus, от греч. hapto —
прикреплять, tropos — поворот, направление;
англ.: haptotropism) — вращательное движение
органа раст., обусловленное его чувствительно-
стью на соприкосновение с твердым предметом;
характерная особенность усиков нек-рых лиан
(см. также нутация).
Гаустория(й) (Haustorium, от лат. haustor — чер-
пающий, пьющий; англ.: haustorium) — видоиз-
1
Рис. 20. Гаплокорм
у Phleum pratense
(1 — утолщенные
междоузлия)
Рис. 21. Повилика на стебле крапивы:
1 — участок стебля крапивы, 2 — учас-
ток стебля повилики, 3 — гаусторий
(по: Bell, Bryan, 1998)
мененный ор-
ган (или его
часть) раст. или
вырост ткани
(реже специали-
зированная клетка), выполняю-
щий функции, связанные с мо-
билизацией и поглощением ве-
ществ (рис. 21); гаусториальная
функция присуща видоизменен-
ным корням (корень-присоска),
стеблям (присоска) или частям
проростка (см. протосома) раст.
паразитов, а также семядоли или
ее части (см. щиток).
Гелиотропизм (англ.: heliotropism) см. фототропизм.
Гелофит (Helophyton, от греч. helos — болото, phyton — растение;
англ.: helophyte) — растение травянистое, вегетативные побеги
которого расположены в воздушной среде, а почки возобновления
в неблагоприятный для вегетации период находятся под водой
или в почве (криптофит); экологически разнообразная группа,
которая занимает переходное положение между световыми гиг-
рофитами и гидрофитами; нередко к Г. относят только болотные
или прибрежно-водные раст. (амфифиты).
Гемикриптофит (Hemicryptophyton, от греч. hemi — половина,
kryptos — скрытый, phyton — растение; англ.: hemicryptophy-
26
te) — один из типов классификации форм жизненных по К. Раун-
киеру, объединяющий растения травянистые, многолетние, поч-
ки возобновления которых в неблагоприятный для вегетации пе-
риод года расположены на уровне почвы или иногда немного
выше его, защищены листовым опадом и/или снежным покро-
вом (рис. 22); морфологически разнообразная группа, в которой
Рис. 22. Жизненные формы по классификации К. Раункиера (по: Ботаника..., 1988).
На верхнем рис. пунктиром показано положение почек возобновления, на нижнем —
черным цветом зимующие органы, а белым — отмирающие: 1 — фанерофит (а — то-
поль, б — омела), 2 — хамефит (а — брусника, б — черника, в — барвинок), 3 — ге-
микриптофит (а — одуванчик, б — лютик, в — овсяница, г — вербейник), 4 — геофит
(а — ветреница, б — тюльпан), 5 — терофит (а — мак-самосейка, б — семя)
27
различают большое число подтипов по структуре, нарастанию и
консистенции побегов, наличию вегетативной подвижности и др.;
по структуре побега выделяют раст. с побегами розеточными
(Г. розеточный), п. полурозеточными (Г. полурозеточный) и п.
безрозеточными (протогемикриптофит или Г. прямостебель-
ный); Г. распространены весьма широко в луговых, степных и
лесных растительных сообществах внетропических областей;
лит.: Падеревская, 1972.
Гемикриптофит полурозеточный (англ.: half-rosette hemicrypto-
phyte) см. гемикриптофит.
Гемикриптофит прямостебельный (англ.: straight-stalked hemi-
cryptophyte) см. гемикриптофит.
Гемикриптофит розеточный (англ.: rosette hemicryptophyte) см.
гемикриптофит.
Гемифанерофит (англ.: hemiphanerophyte) см. хамефит.
Гемиэпифит (англ.: hemiepiphyte) см. полуэпифит.
Гемиэфемероид (Hemiephemeroidum, от греч. hemi — полу и эфе-
мероид) — зацветающее ранней весной травянистое многолетнее
раст., надземные побеги которого почти полностью или частично
отмирают в летний период; гл. обр. гемикриптофиты, многие из
которых, в отличие от эфемероидов, способны при достаточной
влажности к вторичной вегетации летом (Carex physodes); син.:
полуэфемероид; лит.: Хохряков, 19756.
Генет (англ, genet) см. особь.
Геотропизм (Geotropismus, от греч. geo — земля, tropos — поворот,
направление; англ.: geotropism) — ориентированное движение
органа раст., выражающееся в направлении его роста в ответ на
одностороннее действие силы земного притяжения (см. тро-
пизм); по направлению движения различают рост геотропный,
р. апогеотропныйн р. диагеотропный (рис. 23); терм.: Bucher, 1906;
лит.: Серебряков, 1952; Синнот, 1963.
Геофилия (Geophilia, от греч. geo — земля, phila — любовь, склон-
ность; англ.: geophily) — способность раст. погружать (геофити-
зация) в почву, подстилку и др. субстрат свои многолетние побеги
и/или почки возобновления-, геофитизация травянистых многолет-
ников обычно связана с корнями контрактильными, а кустарни-
ков и кустарничков—с окоренением их полегающих побегов;
Г.— широко распространена среди раст. холодных областей (см.
Голубев, 1956а; Крюкова, 1958; Сенянинова-Корчагина, 1967; Хох-
ряков, Мазуренко, 1985).
28
Рис. 23. Ростовая ориентация побегов Circaea alpina (А)
и Pofygonatum multiflorum (Б) в зависимости от глубины расположения
в почве: 1 — столон, 2 — корневище (по: Rawitscher, 1932)
Геофит (Geophyton, от греч. geo — земля, phyton — растение; англ.:
geophyte) — раст. травянистое, многолетнее, почки возобновле-
ния которого в неблагоприятный для вегетации период года рас-
положены в почве на глубине от 1 до нескольких см (рис. 23); в за-
висимости от характера подземных частей различают Г. корне-
вищные, стеблеклубневые, корнеклубневые (см. клубень),
корнеотпрысковые или корневые, луковичные и др.; шведский
ботаник Ф. Аршуг впервые использовал термин Г. для обозначе-
ния раст. с подземными почками в отличие от аэрофитов (раст. с
надземными почками); позднее К. Раункиер отнес Г. к криптофи-
там; син.: эугеофит; лит.: Падеревская, 1966; Negbi, 1989.
Геофитизация см. геофилия.
Гербоид — терм, введен Л. Е. Гатцук (1973) для обозначения раст.
полудревесных, габитуально сходных с травянистыми поликарпи-
ками и в отличие от полукустарников и полукустарничков харак-
теризующихся удлиненными побегами возобновления и нараста-
нием базисимподиалъным (Linaria alpina, Euphorbia seguieriana);
лит.: Гатцук, 19746.
Гетеробатмия (от греч. heteros — иной, другой, bathmos — степень,
ступень) — неодинаковый уровень эволюционного развития и
специализации органов организма, обусловленный неравномер-
ностью темпа эволюции (мозаичная эволюция) признаков, между
которыми нет тесных функциональных связей; син.: гетеробат-
29
мизм, перекрест специализа-
ции, эволюционная гетеро-
хрония признаков; терм.:
Тахтаджян, 1954; лит.: Тах-
таджян, 1964; Северцов, 1981.
Гетеробластия (Heteroblastia;
от греч. heteros — иной, дру-
гой, blastos — побег, росток,
зародыш; англ.: heteroblas-
ty ) — резкое изменение фор-
мы листьев в онтогенезе
(рис. 24). Г. характерна для
раст. (напр. Acacia), у кото-
рых в результате усложне-
ния (упрощения) первые ли-
стья серии листовой резко от-
личаются от последующих
(развитие гетеробластное).
Изменение формы листьев
в онтогенезе обычно рас-
сматривают как показатель
состояния возрастного (см.
модель комбинаторного раз-
вития побега, модель «кон-
троллер переключения фаз»)',
ср. гомобластия; терм.: Goe-
bel, 1928; лит.: Серебряков,
1952.
Гетерокотилия (Heterocotylia,
от греч. heteros — другой,
иной, kotyle — доля; англ.:
anisocotyly) — различие се-
мядолей проростка по фор-
ме, величине или функции
(рис. 25); напр., в роде
Peperomia одна семядоля вы-
носится на поверхность (фо-
тосинтезирует), другая—ос-
тается в семени (гаусторий);
син.: анизокотилия.
Рис. 24. Гетеробластное (1-2) и гомобластное
(3) развитие проростков Robinia pseudacacia
(1), Caragana arborescens (2) и Casuarina sp.
(3; no: Troll, 1939): c — семядоля, г — гипоко-
тиль, л — листья, к — корень
Рис. 25. Прорастание семян видов р. Peperomia
(по: Серебряков, 1952): А — изокотилия
Р. pellucida, Б — гетерокотилия Р. peruviana
(1 — семядоля, 2 — семя)
30
Гетероризия (Heterorrhizia, от греч. heteros — иной, другой, rhi-
za — корень; англ.: heterorhizy) — морфологическая и/или функ-
циональная неоднородность корней, образующих одну систему
корневую (рис. 26); наиболее ярко проявляется у растений с видо-
измененными корнями (см. метаморфоз корня); ср. анизоризия;
лит.: Михайловская, 1981а.
Гетеротроф (англ.: heterotroph) — организм, использующий в ка-
честве источника углерода экзогенные органические вещества;
к Г., противопоставляемым автотрофам, относятся животные,
Рис. 26. Формирование гетероризии у Ranunculuspcdatus (ориг. рис. Черняковской Е. Ф.):
А — проросток (1—главный корень, 2 — семядоля), Б — ювенильное растение
(3 — придаточный корень), В—Г — виргинильное растение с гетероризией Д — гене-
ративное растение с гетероризией (4 — видоизмененный в клубень придаточный ко-
рень, 5 — отмирающий клубень)
31
Рис. 27. Морфологическое разнообразие
листьев Sagittaria sagittifoha: 1-2 — подводные
листья, 3-8 — листья с плавающей
пластинкой, 9-12 — листья с надводной
пластинкой
грибы, большинство бакте-
рий и часть раст. паразитов
(см. сапрофит).
Гетерофиллия (Heterophyllia;
от греч. heteros — иной, дру-
гой, phyllon — лист; англ.:
heterophylly, leaf polymor-
phism) — различие по фор-
ме, размеру и структуре лис-
тьев вдоль стебля одного
побега (рис. 27; см. формация
листьев). Анатомо-морфо-
логические изменения лис-
тьев побега обычно рассмат-
ривают как показатель со-
стояния возрастного (см.
модель комбинаторного раз-
вития побега, модель «кон-
троллер переключения фаз»).
Иногда Г. наз. разнолист-
ность побегов особи или в
более узком значении — раз-
личия между листьями сре-
динной формации (Sagitta-
ria); ср. анизофиллия, гете-
робластия; лит.: Серебряков,
1952; Winn, 1999.
Гетерохрония (от греч. heteros — иной, другой, chronos — время;
англ.: heterochronism, heterochrony) — одна из основных теорий
морфологической эволюции, разработанная de Beer (1930) на
примере животных. Автор термина Э. Геккель (1866) рассматри-
вал Г. как изменение времени заложения и скорости развития ор-
гана относительно других органов одного и того же организма. Де
Бир придал этому понятию филогенетическое значение, опреде-
лив Г. как изменение скорости развития черт у потомка по срав-
нению с предком. На основании соотношения скорости сомати-
ческого развития и репродуктивного созревания (темп разви-
тия) организма он выделил 4 типа гетерохронных изменений он-
тогенеза: акселерация, педогенез (неотения), ретардациям гипер-
морфоз. В настоящее время Г. часто понимают широко — как из-
32
менения скорости развития особи, органа или ткани (ср.
фил эмбриогенез). Различают Г. элементарные (акселерация, гипер-
мофоз, неотения, прогенез) и Г. комбинированные (акселератив-
ный прогенез, гиперпедоморфоз, неотенический гиперморфоз);
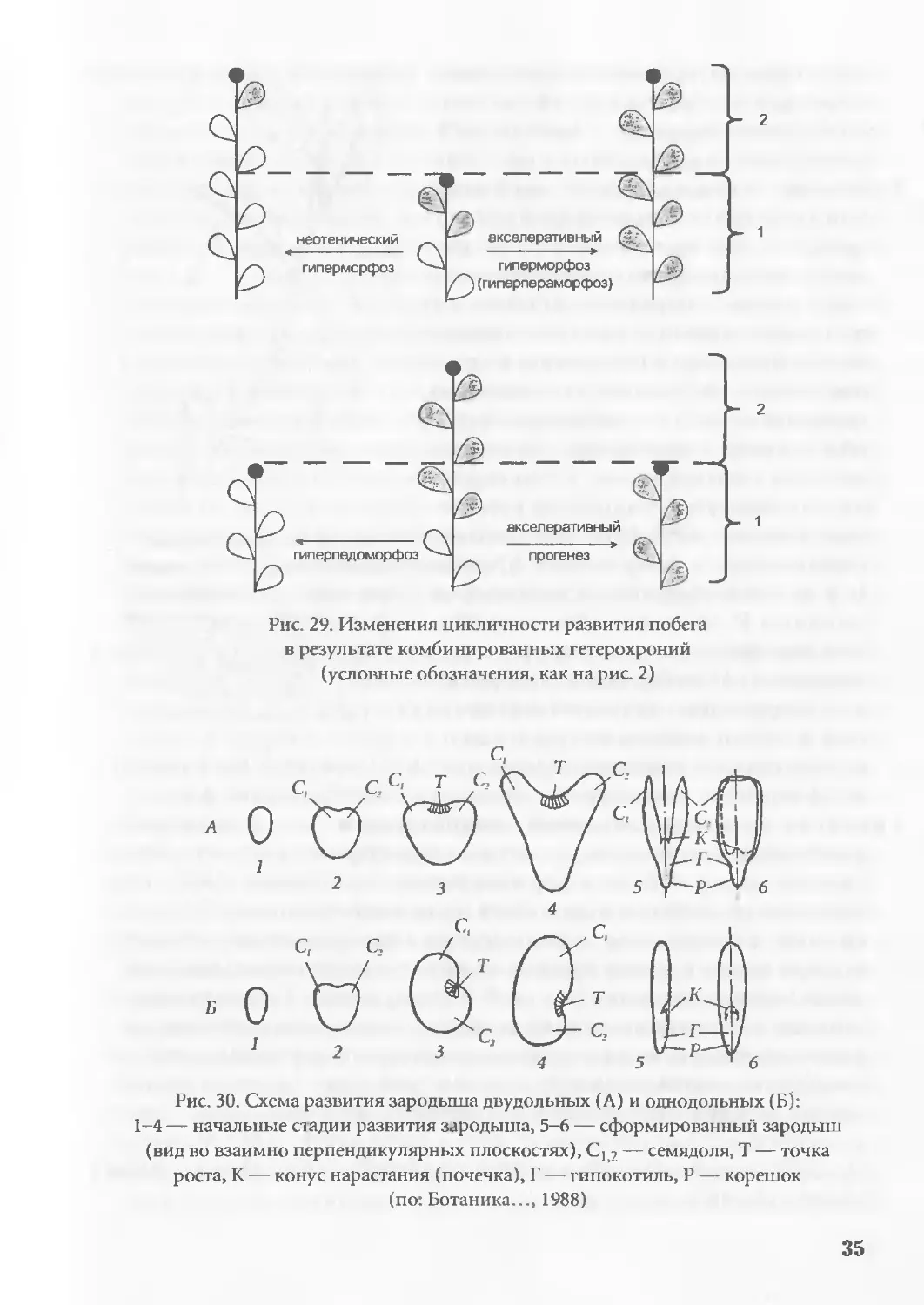
рис. 2, 29; лит.: Арциховский, 1911; de Beer, 1958; Gould, 1980,
Рэфф, Кофмен, 1986; Heterochrony..., 1988; Klingenberg, 1998;
McKinney, 1999; Li, Johnston, 2000; Жмылев, 2004.
Гибернакула (Hibernaculum, от лат. hiberno — зимовать; англ.: hi-
bernaculum, hibernacle, winter bud) — в специальном значении —
турион, т. е. видоизмененные зимующие почки водных раст., со-
держащие большое кол-во зачатков листьев (Utricularia) или/и ви-
доизмененные листья (Hydrocharis); обычно пон. Г. используется
для обозначения различных по происхождению специализиро-
ванных зимующих органов Годновременно органов размножения
вегетативного) некоторых водных раст.: клубнелуковиц рдестов, не
развернувшихся «почек» пузырчаток, реже — зимующих листе-
цов рясок (рис. 28); в англоязычной литературе иногда Г. наз. лю-
Рис. 28. Гибернакулы (по: Sculthorpe, 1967) у Elodea canadensis (1),
Potamogeton pusillus (2), Potamogeton foliosus (3), Potamogeton obtusifolius (4),
Potamogeton vaseyt (5), Potamogeton crispus (6), Myriophyllum verticillatum (7),
Utricularia intermedia (8), Utricularia geminiscapa (9), Utricularia vulgaris (10)
33
бую вегетативную структуру наземных и водных раст., служащую
для переживания неблагоприятных условий (напр. клубень); ср.
почка зимующая; син.: турион; лит.: Sculthorpe, 1967.
Гигантизм (от греч. giganthos — исполин, гигант; англ.: gigantism) —
увеличенные размеры тела потомка (грандизация) в результате ус-
корения его роста, но без изменения синхронизации соматическо-
го и полового созревания (ср. нанизм, гетерохрония). В биоморфо-
логии Г. часто рассматривают как результат гиперморфоза, что со-
ответствует псевдогигантизму; лит.: Цвелев, 1977; Dommergues et
al., 1986; Недолужко, 1997.
Гидрогемикриптофит (англ.: hydrohemicryptophyte) см. гидрофит.
Гидрогеофит (англ.: hydrogeophyte) см. гидрофит.
Гидрокриптофит (англ.: hydrocryptophyte) см. гидрофит.
Гидротерофит (англ.: hydrotherophyte) см. гидрофит.
Гидрофит (Hydrophyton, от греч. hydor — вода, phyton — расте-
ние; англ.: hydrophyte) — водное раст. с плавающими или погру-
женными листьями, почки возобновления которого в неблагопри-
ятный для вегетации период года находятся под водой; морфоло-
гически разнообразная группа криптофитов, среди которых
различают многолетние раст. с почками возобновления на дне во-
доема (гидрогемикриптофиты) или в грунте (гидрогеофиты) и
раст., отмирающие на зиму (гидротерофиты).
Гиперморфоз (от греч. hyper — над, сверх, morphe — вид, форма;
англ.: hypermorphosis) — один из основных типов элементарной
гетерохронии, при котором замедление полового созревания со-
четается с неизменной скоростью соматического развития (ср. ак-
селерация). В результате Г. потомок имеет пераморфный облик,
который формируется в процессе удлинения онтогенеза (рис. 2).
При сочетании Г. с неотенией (неотенический Г.) или с акселера-
цией (акселеративный Г., гиперпераморфоз, рис. 29) облик по-
томка напоминает гигантизм (псевдоУигантизм); лит.: Gould,
1980; Dommergues et al., 1986.
Гипокотиль (Hypocotylus, от греч. hypo — под, вниз, kotyle —
доля; англ.: hypocotyl) — участок стебля проростка, семенного
раст. между узлом семядольным и корешком (рис. 24, 30, 111);
зону перехода Г. в корешок нередко наз. шейкой корневой
(англ.: root collar, peg); син.: колено подсемядольное; лит.: Сереб-
ряков, 1952.
Гипоподий (Hypopodium, от греч. hypo — под, вниз, pous — нога,
здесь — ось; англ.: hypopodium) — первое междоузлие побега бо-
34
Рис. 29. Изменения цикличности развития побега
в результате комбинированных гетерохроний
(условные обозначения, как на рис. 2)
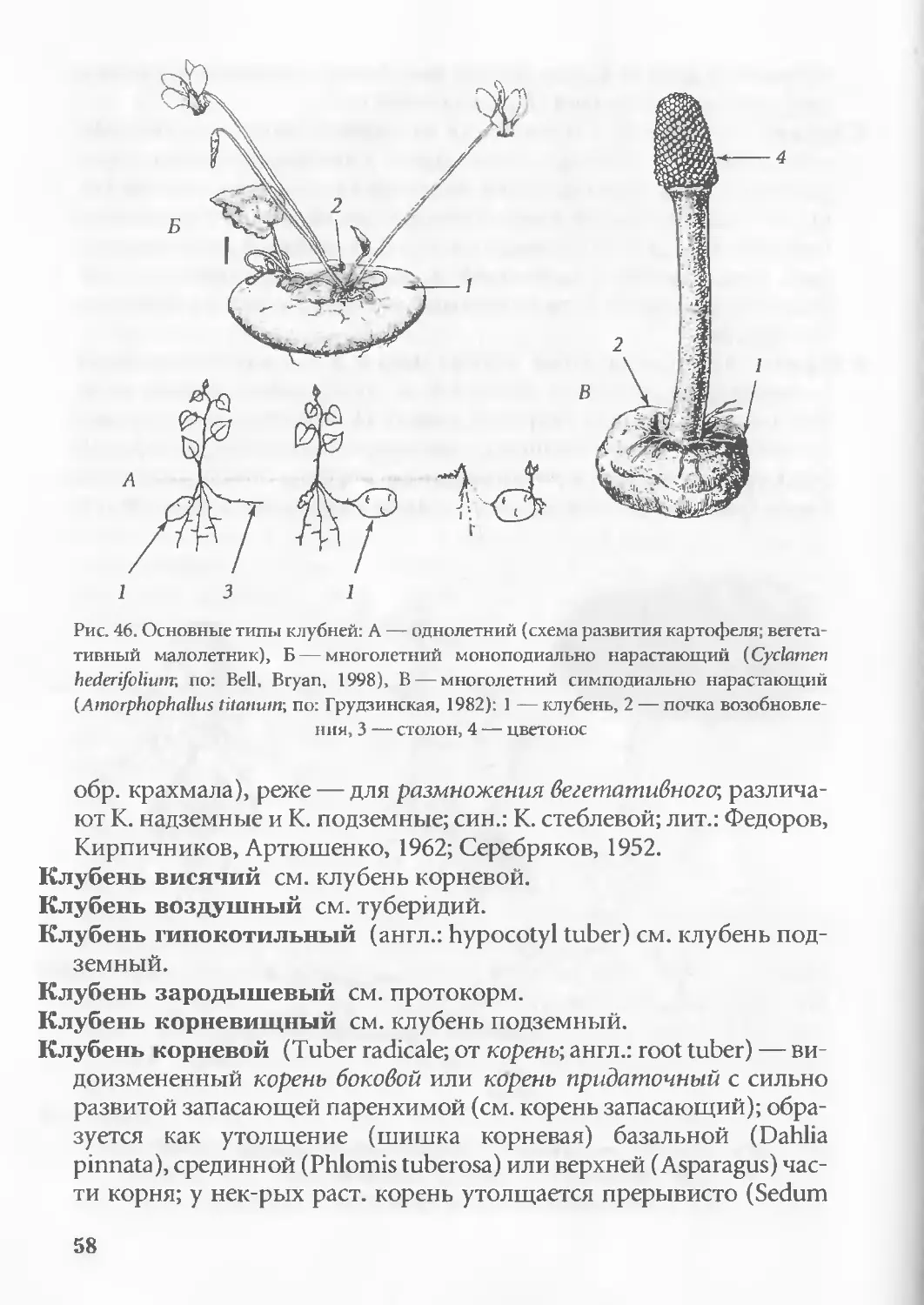
Рис. 30. Схема развития зародыша двудольных (А) и однодольных (Б):
1-4— начальные стадии развития зародыша, 5-6 — сформированный зародыш
(вид во взаимно перпендикулярных плоскостях), С1>2 — семядоля, Т — точка
роста, К — конус нарастания (почечка), Г — гипокотиль, Р — корешок
(по: Ботаника..., 1988)
35
Рис. 31. Типы междоузлий
бокового побега: 1 — материн-
ский побег, 2 — боковой побег,
3 — гипоподий, 4 — мезоподий,
5 — эпиподий, 6 — предлист,
7 — переходный лист,
8 — развитый лист
(по: Федоров и др., 1962)
кового (рис. 31); у некоторых растений
из-за сильного разрастания Г. особен-
но заметен: стрелка у Plantago, фла-
гелла у Saxifraga flagellaris (рис. 130).
Гипотеза аккумуляции мутаций
(англ.: mutation accumulation theory of
aging) — одна из гипотез, рассматри-
вающих старение как неотъемлемую
черту любого организма (побочный
продукт эволюции), которая прояв-
ляется благодаря увеличению в тече-
ние жизни различных повреждений
(включая соматические мутации) или
обусловлена снижением функцио-
нальной деятельности, в частности
нарушением корне-листовой корреля-
ции (Казарян, 1969, 1990) или падения
омоложения (см. теория Н. П. Кренке).
Г. а. м. была предложена английским
зоологом Р. Medawar (1946) для объ-
яснения причин естественной смерти
животных. У листостебельных расте-
ний проявление соматических мута-
ций в жизни особи зачастую не соот-
ветствует представлениям Медавара; лит.: Стрелер, 1964; Lynch et
al., 1999; Orive, 2001; Gavrilov, Gavrilova, 2002; Goldsmith, 2004.
Гипотеза антагонистической плейотропии (англ.: antagonistic
pleiotropy theory of aging) — одна из популярных в геронтологии
гипотез, которая была сформулирована G. Williams (1957) для
объяснения старения и эволюции длительности жизни. Согласно
ей одни те же гены (см. гены старения, гены долголетия) проявля-
ют противоположный эффект на приспособленность (выживае-
мость) молодых и взрослых особей в популяции. Такая антагони-
стическая плейотропия генов должна обусловливать отрицатель-
ные корреляции между уровнем смертности взрослых особей и
скоростью старения, между темпом развития и длительностью
жизни, между интенсивностью отмирания молодых особей и про-
должительностью онтогенеза-, лит.: Charlesworth, 2000; Stearns et
al., 2000; Gavrilov, Gavrilova, 2002; Goldsmith, 2004; Abrams, 2004;
Mitteldorf, 2004.
36
Гипотеза восстановления сомы (англ.: disposable soma theory of
aging) — одна из гипотез, рассматривающих старение как неотъ-
емлемую черту организма. Предложена Т. Kirkwood (1977), кото-
рый предположил, что старение связано не только с накоплением
нарушений на молекулярном и клеточном уровне (см. гипотеза
аккумуляции мутации), но и с формирующимися у организма ог-
раничениями на их исправление (восстановление сомы). Соглас-
но Г. в. с. каждый организм исходно имеет лимитированное «ко-
личество энергии», которая расходуется в процессе жизни на ре-
продукцию и формирование вегетативных (соматических) черт
(продолжительность жизни особи зависит от силы и направления
отбора на оптимизацию модели распределения ресурсов между
генеративными и вегетативными органами). Г. в. с. тесно сопря-
жена с «принципом аллокации» и хорошо объясняет эволюцион-
ные взаимоотношения монокарпиков и поликарпиков. Однако не
всегда продолжительность жизни раст. можно интерпретировать
как проявление конкуренции органов за ресурсы (см. старение
негативное, гипотеза смерти от истощения); ср. гипотеза анта-
гонистической плейотропии; лит.: Злобин, 2000; Hautekeete et al.,
2001; Kirkwood, 2002; Obeso, 2002.
Гипотеза программированной смерти (англ.: programmed death
theory of aging) — одна из гипотез рассматривающих старение
как генетически детерминированную черту высших организмов,
которая сформировалась в процессе их эволюции (см. гены ста-
рения, гены долголетия). Г. п. с. сформулирована немецким био-
логом A. Weismann (1882), который предположил, что запрограм-
мированная смерть (длительность жизни) взрослых особей обес-
печивает успешное развитие потомства благодаря освобождению
пространства и перераспределения доступных ресурсов в популя-
ции. По мнению А. Вейсмана (1914) механизм предопределенной
смерти может заключаться в исходной ограниченности числа по-
следовательных делений соматических клеток. Многие геронто-
логи скептически относятся к Г. п. с. и название «программиро-
ванная смерть» обычно используют для обозначения причины
естественного отмирания клеток (апоптоз), тканей или детерми-
нированных органов, редко — особей, в частности монокарпиков;
лит.: Greenberg, 1996; Ванюшин, 2001; Thomas, 2002; Goldsmith,
2004.
Гипотеза смерти от истощения (нем. Erschopfungstod) — част-
ный случай гипотезы восстановления сомы, предложенный
37
E. Hildenbrand (1882) и Н. Molisch (1938) для объяснения монокар-
пического старения. Согласно Г. с. и. старение и продолжитель-
ность жизни монокарпиков обусловлены расходованием всех пи-
тательных веществ на созревание плодов и семян. Известны две
точки зрения о механизме монокарпического старения. Первая
предполагает, что от развивающихся генеративных органов по-
ступает гормональный сигнал (гормон старения или смерти), ко-
торый инициирует старение вегетативных органов (коррелятив-
ное старение). Вторая рассматривает старение в связи с прису-
щим раст. механизмом повторного перемещения питательных
веществ из отмирающих органов в участки роста (в частности из
листьев в формирующиеся генеративные органы). Не у всех раст.
старение и естественная смерть (продолжительность жизни) со-
ответствуют представлениям этой гипотезы; лит.: Gan, Amasino,
1997; Nooden et al., 2001; Thomas, 2002; Fisher et al., 2002; Hautekeete
et al., 2002; Rossato et al., 2002.
Голопаразит (Holoparasitus, от греч. golos — цельный и parasitus —
паразитизм; англ.: holoparasite) — часто используемое пон. для
обозначения бесхлорофильных раст. паразитов, существующих
целиком за счет автотрофного хозяина; при более широком по-
нимании явления паразитизма к Г. относят и бесхлорофильные
облигатно-микотрофные раст. (аллелопаразиты), которые часто
наз. сапрофитами.
Голопаразитизм (англ.: holoparasitism) см. паразитизм.
Гомобластия (Homoblastia, от греч. homos — равный, одинаковый,
blastos — побег, росток, зародыш; англ.: homoblasty) —- постепен-
ное изменение формы листьев в онтогенезе раст. (рис. 24); харак-
терно для раст., у которых первые листья серии листовой незначи-
тельно отличаются от последующих (развитие гомобластное);
терм.: Goebel, 1928; ср. гетеробластия; лит.: Серебряков, 1952.
Гоморизия (Homorrhizia, от греч. homos — одинаковый, rhiza —
корень; англ.: homorhizy) — формирование системы корневой
раст. из корней одинаковых по происхождению — корней кладо-
генных (рис. 3); у высших споровых (гоморизофитов или раст.
униполярных) первый корень зародыша закладывается обычно
сбоку как придаточный корень побега (первичная Г.), и вся корне-
вая система (первично гоморизная) формируется только из кор-
ней придаточных (кладогенных); Г. семенных раст. (раст. кисте-
корневых) связана с недоразвитием корешка (вторичная Г.) или
ранним отмиранием корня главного в результате чего формирует-
38
ся вторично гоморизная корневая система (Роасеае, Сурегасеае,
Liliaceae и др.); терм.: Goebel, 1930; лит.: Михайловская, 1981а;
Groff, Kaplan, 1988.
Гоморизия вторичная (англ.: secondary homorhizy) см. гоморизия.
Гоморизия первичная (англ.: primary homorhizy) см. гоморизия.
Гоморизофит (англ.: homorhizophyte) см. гоморизия.
Грандизация см. гигантизм.
Грибокорень см. микориза.
Группа экологическая (раст.) — группа раст., одинаково реаги-
рующих на действие какого-либо одного фактора среды (осве-
щенность, влажность, температура и т. п.); к сходным условиям
раст. могут приспосабливаться по-разному (анатомо-морфоло-
гические, физиологические, цитологические и др. приспособле-
ния), поэтому одна Г. э. может объединять разные биоморфы; ср.
форма жизненная, экобиоморфа.
Движение растения (англ.: plant movement) — перемещение особи
или ее части в пространстве; различают Д. р. активное и пассивное
(напр. диссеминация); в связи с прикрепленным образом жизни
многоклеточных раст. к активным Д. р. обычно относят: течение
цитоплазмы, перемещения органоидов клетки или отдельных ор-
ганов {тропизм, настия, нутация); реже активноеД. р. рассматри-
вают как результат освоения пространства в результате размноже-
ния вегетативного (напр. движения клонов фалангой или веером;
соответственно англ.: phalanx growth form, guerilla growth form)
или разрастания особи (перемещения меристем апикальных в
пространстве по траектории маятника, спирали или прямой ли-
нии); лит.: Полевой, Саламатова, 1985.
Двулетник замещающий — малолетник вегетативной с одно-
двухлетней луковицей (Tulipa), клубнелуковицей или клубнем
(Valeriana), ежегодное возоб-
новление роста которого обес-
печивается замещением от-
мирающей луковицы (клубне-
луковицы, клубня) дочерней
(рис. 32); терм.: Ворошилов,
1959.
Девиация (от лат. deviatio — от-
клонение; англ.: deviation) —
один из основных способов эво-
люции морфологической орга-
Рис. 32. Схема развития растения
с двухлетней луковицей
39
нов в рамках теории филэмбриогенеза, который представляет со-
бой изменения на средних стадиях развития органа у потомка по
сравнению с предком (рис. 4). В результате изменения пути мор-
фогенеза происходит усложнение или упрощение сформировав-
шегося органа. В биоморфологии Д. часто понимают в широком
смысле как изменения в развитии органа или онтогенеза (ср. ге-
терохрония)', терм.: Muller, 1864; лит.: Северцов, 1945, Тахтаджян,
1964, Северцов, 1981.
Дегенерация см. редукция.
Дедифференцировка (ация) — см. дифференцировка.
Дезинтегратор (англ.: desintegrator, splitter) см. дезинтеграция мор-
фологическая.
Дезинтеграция морфологическая (англ.: fragmentation) — пон.,
используемое для обозначения различных процессов, обусловли-
вающих размножение вегетативное (сарментация, нормальная
партикуляция, клонирование) и сенильный распад особи-, разли-
чают Д. частичную (автономизация побегов или системы побегов в
результате укоренения) и полную (размножение вегетативное и
партикуляция), Д. неспециализированную (у раст. вегетатив-
но-неподвижных) и специализированную (у раст. вегетативно-
подвижных), Д. раннюю и позднюю (см. партикуляция сениль-
ная); в англоязычной литературе раст., у которых быстро проис-
ходит полная Д. м., наз. дезинтеграторами (англ.: desintegrators,
splitters); при этом раст., у которых связь между раметами сохра-
няется менее 1 года, наз. рестриктивными интеграторами (restric-
tive intergators), а раст., у которых связь между раметами сохраня-
ется более 1 года, наз. экстенсивными интеграторами (extensive
integrators); см. Ценопопуляции..., 1976, 1988.
Дерево (Arbor; англ.: tree) —раст. многолетнее с отчетливо выра-
женной главной осью скелетной — стволом, сохраняющимся до
конца жизни, и кроной, образованной боковыми ветвями и побега-
ми (Д. кронообразующее) или пучком крупных листьев (Д. розе-
точное), как у древовидных однодольных (Palmae), папоротников,
нек-рых двудольных (Senecio); Д. различаются по форме и размеру
ствола (см. фанерофит), кроны и корней (Д. с корнями досковидны-
ми, к. ходульными, пневматофорами и т. д.; рис. 53); Д. наиболее
разнообразны и многочисленны во влажных тропиках; среди кро-
нообразующих выделяют Д. с кронами регулярными или правиль-
ными (пирамидальными, овальными и т. п.) и нерегулярными,
или неправильными (рис. 33), не имеющими четких геометриче-
40
Рис. 33. Форма крон деревьев: 1 — конусовидные, 2 — яйцевидные,
3 — шаровидная (а) и полушаровидная (б), 4 — неправильные,
5 — зонтиковидная 6 — розеточные деревья
(по: Федоров и др., 1962)
41
Рис. 34. Формы одноствольных (а), многоствольных (б),
куртинообразующих (в) и стелющихся (г) деревьев
ских очертаний (шатровидные, раскидистые), а по степени под-
вижности вегетативной и внешнему облику — Д. куртинообра-
зующие, Д. одноствольные и Д. многоствольные (рис. 34); Д. со
стелющимися по земле скелетными осями (стволиками) наз.
стланцами (см. раст. стелющееся), а с ежегодно отмирающими
верхними участками побегов — полудеревом (см. раст. полудре-
весное); лит.: Серебряков, 1962; Чистякова, 1986; Мазуренко, Хох-
ряков, 1991; Halle, Oldeman, Tomlinson, 1978; Watson, 1981.
Дерево бутылочное (англ.: bottle tree) см. ствол.
Дерево длинноксилоризомное (англ.: rhizomatous tree) см. дерево
куртинообразующее.
Дерево кронообразующее (англ.: branching tree) см. дерево.
Дерево ксерогенное см. раст. полудревесное.
Дерево куртинообразующее — Д. многоствольное с отстоящими
друг от друга стволами (рис. 34), которые развиваются из почек на
ксилоризомах (Д. длинноксилоризомное), корнях (см. раст. корне-
отпрысковое) или стволах и ветвях кроны после их полегания
(Tilia cordata); терм.: Чистякова, 1978.
Дерево кустовидное см. дерево многоствольное.
Дерево лептокаульное (англ.: leptocaulis tree) — невысокое дерево
с относительно тонким твердодревесныц стволом, несущим
сильно разветвленную крону из удлиненных многолетних ветвей
(Betula); ср. Д. пахикаульное; терм.: Corner, 1949; лит.: Серебря-
ков, 1962; Гатцук, 1976.
Дерево многоствольное (англ.: multiple-stemmed tree, multicaulis
tree, pleiocormic tree) — Д. с двумя-тремя (Д. немногоствольное)
или большим количеством стволов во взрослом состоянии, кото-
42
рые образуются в результате ветвления базисимподиального или
раннего пробуждения почек спящих в основании ствола (рис. 34)
из-за задержки роста или отмирания главной оси скелетной
(Acer); кустовидная форма роста некоторых Д. одноствольных
рассматривается как реакция на неблагоприятные условия место-
обитания (затенение, засоление и др.); син.: Д. кустовидное,
Д. плейокормное.
Дерево немногоствольное см. дерево многоствольное.
Дерево одноствольное (англ.: monocormic tree)— Д. с единствен-
ным в течение всего онтогенеза стволом (рис. 34), формирую-
щимся в результате нарастания моноподиалъного (Picea) или
н. симподиального (Quercus); у некоторых Д. о. из почек спящих
при спиливании или отмирании ствола могут образовываться
вторичные или порослевые стволы (Д. порослеобразующее).
Дерево пахикаульное (англ.: pachycaulis tree) — невысокое дерево
с неветвящимся или слабо разветвленным (с небольшой кроной)
мягкодревесным, массивным, иногда обратноконической формы
стволом и крупными листьями (Carica); Д. п. преимущественно
распространены в (семи)аридных областях тропического пояса
(ср. Д. лептокаульное); терм.: Corner, 1949; лит.: Серебряков, 1962;
Гатцук, 1976.
Дерево плейокормное (англ.: pleiocormic tree) см. дерево много-
ствольное.
Дерево порослеобразующее см. дерево многоствольное.
Дерево розеточное (англ.: rossette-tree) см. дерево.
Дерево стелющееся см. растение стелющееся.
Дерево-подушка см. растение подушковидное.
Дерновина (Caespes — густое переплетение, клубок корней, стеб-
лей и т. п.; англ.: caespitose, cespitose, clump, bunch, tuft, tussock) —
совокупность многочисленных более или менее плотно располо-
женных друг к другу разновозрастных побегов кущения раст. и их
остатков (пеньков), основания которых образуют многолетнюю
часть раст. (рис. 115), расположенную на поверхности почвы и
разрастающуюся по периферии в ширину (раст. дерновинноё)
или по всей поверхности в высоту, перерастая в кочку (раст. коч-
кообразующеё); в более узком значении Д. наз. совокупность побе-
гов интравагинальных (Д. настоящая), а при ветвлении экстрава-
гинальном используют пон. «куст» (Д. ложная); лит.: Артамонова,
1968; Борисова, Попова, 1971; Персикова, 1959в).
Дерновина ложная см. дерновина.
43
Дерновина настоящая см. дерновина.
Детерминация (от лат. determinatio — ограничение, определение;
англ.: determination) — процесс приобретения клеткой или тка-
нью способности к развитию по одному из возможных направле-
ний (сужение проспективных потенций клетки). В биоморфологии
пон. Д. использует в широком смысле не только как приобрете-
ние клеткой, тканью, органом или особью состояния готовности к
развитию по определенному пути (Д. развития), но и как ограни-
ченный рост побега, корня или листа (см. рост детерминирован-
ный). Различают автономную Д. (за счет внутренних импульсов)
и индуцированную Д. (в результате индукции).
Детки (Bulbil; от луковица; англ.: daughter bulb, bulbet, multiplier) —
дочерние луковицы (клубнелуковицы; англ.: cormel, cormlet), раз-
вивающиеся из пазушных почек материнской луковицы (клубне-
луковицы); обеспечивают размножение вегетативное раст. луко-
вичного или раст. клубнелуковичного (рис. 48); если луковица
однолетняя, Д. отделяются в год своего образования, если много-
летняя— остаются связанными с ней (зубки) до разрушения
кроющих чешуй.
Диаспора (англ.: diaspore) — зачаток раст., естественно отделяю-
щийся от материнской особи и служащий для размножения и рас-
селения; различают Д. вегетативные (клубень, луковица, луковичка
и др.) и генеративные (семя, плод и др.); син.: диссеминула, про-
пагула, зачаток расселения; лит.: Брагина, Батыгина, 2000.
Диссеминула (англ.: disseminule) см. диаспора.
Дифференциация см. дифференцировка.
Дифференцировка (англ.: differentiation) — возникновение после
индукции и детерминации морфофункциональных различий ме-
жду однородными клетками и тканями в морфогенезе и онтогене-
зе (часто в аналогичном смысле используют назв. дифференциа-
ция). В результате Д. формируются специализированные клетки,
ткани и органы. У раст. Д. происходит не только в период форми-
рования зародыша и почки (рост внутрипочечный), но и в процес-
се дальнейшего развития органов (см. метаморфоз), а специали-
зированные клетки могут переходить снова к пролиферации (де-
дифференцировка), например, при регенерации; лит.: Синнот,
1963; Чельцова, 1980; Рост..., 1981; Ростовцева, 1984; Нейфах, Ло-
зовская, 1984; Корочкин, 2002; Иванов, 2004.
Дихотомия истинная (англ.: true dichotomy, isotomy) см. ветвле-
ние верхушечное.
44
Рис. 35. Апикальный контроль развития побегов
ветвления: 1 — апикальное доминирование в теку-
щем сезоне» 2 — снятие апикального доминирования
на второй год, 3 — отсутствие апикального домини-
рования, 4 — граница между годичными прироста-
ми (по: Bell, Bryan, 1998)
Дихотомия ложная (англ.: pseudodichotomy, false dichotomy) см.
нарастание симподиальное.
Дихотомия неравновильчатая (англ.: anisotomy) см. ветвление
верхушечное.
Дихотомия равновильчатая (англ.: isotomy) см. ветвление верху-
шечное.
Доминирование апикальное (англ.: apical dominance) — одна из
форм корреляций ростовых, проявляющаяся в подавлении или
торможении роста почек пазушных апикальной меристемой рас-
тущего побега (рис. 35). В узком смысле различают Д. а. (ингиби-
рование почек, из которых развиваются побеги силлептпические) и
апикальный контроль (ингибирование почек, из которых разви-
ваются побеги пролептические). Д. а. выражено в различной степе-
ни у разных видов: от слабого (напр. побег силлептческии) до пол-
ного подавления (не
заложение почек). Ме-
ханизм Д. а. обычно
объясняется трофиче-
скими или гормональ-
ными корреляциями;
нередко развитие поч-
ки зависит от влияния
листа, в пазухе которо-
го она расположена
(листовое доминиро-
вание); син.: корреля-
тивное ингибирова-
ние; лит.: Синнот, 1963;
Рытова, 1976; Чельцо-
ва, 1980; Ростовцева,
1984; Cline, 1994, 1997;
Wilson, 2000; Shimizu-
Sato, Mori, 2001.
Доминирование листовое см. доминирование апикальное.
Донце (Lecus, от греч. lexus — дно, ложе; англ.: bulb stem, fundus) —
стебель луковицы, обычно дисковидной формы из-за недоразви-
тия его междоузлий (рис. 66).
Дроппер (англ.: dropper — столоновидный) — структура, образо-
ванная в результате вытягивания основания зачатка листа низово-
го и несущая на конце луковицу замещающую (Erythronium);
45
предполагается, что у некото-
рых орхидных (Herminium) раз-
виваются аналогичные структу-
ры, которые несут клубень поч-
ко-корневой и служат для раз-
множения вегетативного (ср. ту-
бероид).
Дуга укороченных междоуз-
лий — участок поворота корне-
вища гипогеогенного вверх (в ре-
зультате изменения направле-
ния его роста}, переходный от
его горизонтальной части (пла-
гиокорма) с междоузлиями удли-
ненными к вертикальной, назем-
ной (ортокорму); обычно соот-
ветствует зоне кущения (рис. 36);
син.: плагиоподий; терм.: Смир-
нов, 1958b.
Единица морфогенеза (англ.:
unit of morphogenesis) — уча-
сток побега вегетативного, об-
разующийся за один период не-
прерывной активности мери-
стемы апикальной (рис. 37);
рассматривается как единица
структуры, морфологические
границы которой могут совпа-
дать с циклом листовым, побе-
гом элементарным (и. монорит-
мичным), побегом ивановым или
приростом деревьев и кустарни-
ков, который образуется за
один период видимого роста;
терм.: Halle, Martin, 1968; лит.:
Halle, 1986.
Единица роста (англ.: growth
unit) см. цикл роста.
Емкость почки — переменная ве-
личина, определяемая как число
Рис. 36. Схема кущения (по: Учебно-поле-
вая..., 1990): 1 —гшагиокорм с удлинен-
ными междоузлиями (корневище), 2 —
дуга укороченных междоузлий (зона ку-
щения или возобновления), 3—-листья,
4 — ортокорм (надземная часть побега),
5 — побеги ветвления (кущения или во-
зобновления )
Рис. 37. Схема ритмичного роста побегов
Hevea brasilensis: А — кривая роста (1 —
длина побега, t — время, 1 — прирост, ог-
раниченный рубцами почечных чешуи,
2 — период непрерывного роста), Б — еди-
ница морфогенеза на участке побега с тре-
мя приростами, В — выделенная единица
морфогенеза с побегами ветвления (3) и
пазушными цветоносами (по: Halle, 1986)
46
листовых зачатков {фитомеров), заключенных в почке от последне-
го видимого примордия до вполне сформировавшегося зачатка
листа; потенциальная готовность почки к раскрыванию связана с
накоплением в ней определенного числа (кванта роста) фитоме-
ров (максимальная емкость), которое рассматривается в качестве
единицы ритма развития побега; терм.: Серебрякова, 1959; лит.: Се-
ребрякова, 1971а.
Живорождение см. вивипария.
Зародыш (Embryo; англ.: emb-
ryo) — зачаток раст. в семе-
ни, развивающийся из зиго-
ты (при половом воспроиз-
ведении) или из других
клеток, принадлежащих за-
родышевому мешку, нуцел-
лусу или интегументу (при
апомиксисе); в зрелых семе-
нах 3., как правило, диффе-
ренцирован на корешок, ги-
Рис. 39. Редуцированные зародыши
Utricularia vulgaris (A), Harvey a coccinea (Б),
Stiga gesnerioides (В), Orobanche hederae (Г),
Aeginetia indica (Д): 1 — апикальный полюс,
2 — базальный полюс зародыша, 3 — заро-
дыш, 4 — запасающая зона, 5 — меристе-
матическая зона, 6 — семенная кожура,
7 — эндосперм (по: Терехин, 1996)
Рис. 38. Схема строения зародыша
злаков: 1 — щиток, 2 — почечка,
3 — колеоптиль, 4 — эпибласт, 5 —
колеориза, 6 — лигула, 7 — заро-
дышевый корень (по: Цвелев, 1976)
47
покотиль, семядоли и почечку (рис. 30). По содержанию в тканях
3. хлорофилла выделяют лейкоэмбриофиты и хлороэмбриофиты.
У нек-рых раст. (Роасеае) 3. имеет своеобразное строение (рис. 38),
у других слабо дифференцирован (недоразвитый 3.) и доразвива-
ется в семени после диссеминации (Ranunculaceae, Papaveraceae)
или представлен только группой однородных клеток (рудимен-
тарный или редуцированный 3.) и развивается в процессе прорас-
тания семени (рис. 39), образуя протосому; лит.: Серебряков, 1952;
Яковлев, Жукова, 1973; Ботаника..., 1988; Терехин, 1996.
Зародыш-проросток см. протокорм.
Зачаток расселения см. диаспора.
Зона возобновления (Zona innovan-
di, от лат. innovatio — возобновле-
ние; англ.: innovation zone) — ниж-
ний участок побега, часто с укоро-
ченными междоузлиями, несущий
почки возобновления; у злаков наз.
зоной кущения (рис. 40); часто к 3. в.
относят и зону резервную.
Зона кущения (Zona fruticandi, от
кущение; англ.: tillering zone) —
нижняя приземная или подземная
часть стебля злаков, состоящая из
укороченных междоузлий, несу-
щих почки возобновления (рис. 36,
40); иногда 3. к. наз. зоной возобнов-
ления или обозначают этим терми-
ном совокупность побегов ветвле-
ния, образовавшихся в результате
кущения; син.: узел возобновления,
узел кущения, узел отрастания;
терм.: Соловьева, 1958; лит.: Сереб-
рякова, 1971а.
Зона обогащения (Zona amplifican-
di, от лат. amplifico — расширять,
увеличивать, приумножать; англ.:
amplification zone) — участок побе-
га, из почек пазушных которого раз-
виваются побеги обогащения; как
правило, 3. о. соответствует фло-
Рис. 40. Морфофизиологические зо-
ны генеративного побега: 1 — зона
возобновления с пазушными почка-
ми, 2 — зона торможения, 3 — зона
обогащения (образование побегов
обогащения соцветия), а — базаль-
ный участок побега без пазушных по-
чек, б — базальный участок побега с
пазушными почками, в — общее чис-
ло метамеров побега
48
ральному участку побега вегетативно генеративного (рис. 40); син.:
зона флоральная.
Зона префлоральная см. зона торможения.
Зона резервная (Zona subsidiaria, от лат. subsidium — резерв) —
участок побега, несущий почки спящие и/или п. зимующие, разви-
тие которых обеспечивает возобновление роста раст. после нару-
шения (отмирания, отчуждения) части его системы побегов (от-
растание) или периода покоя (ср. зона возобновления); у раст.
травянистых часто соответствует понятию резид; терм.: Т. Н. Су-
воровой (1959) введен для обозначения верхней части зоны куще-
ния (см. кущение постгенеративное).
Зона торможения (Zona inhibitionis; от inhibeo — задерживать;
англ.: inhibition zone) — участок побега, расположенный между
зоной возобновления и зоной обогащения и характеризующийся от-
суствием или недоразвитием почек пазушных (рис. 40); син.: зона
префлоральная.
Зона флоральная см. зона обогащения.
Зубки см. детки.
Изменчивость биоморфологическая — внутривидовая измен-
чивости формы жизненной, морфогенеза или онтогенеза (см. поли-
вариантность развития), обусловленная экзогенными (см. моди-
фикация) или эндогенными факторами (И. генотипическая). Ре-
зультаты изучения И. б. обычно рассматривают как основание
для определения направленности эволюции биоморфологической.
В широком смысле к И. б. относят и изменения формы жизненной
в онтогенезе; лит.: Гатцук, 1968; Покровская, 1976; Жукова, 1995.
Изотомия (англ.: isotomy) см. ветвление верхушечное.
Инволюция см. редукция.
Ингибирование коррелятивное см. доминирование апикальное.
Индекс пластохронный (англ, plastochron index) — показатель
пластохронного возраста побега:
PJ.= n +
log L„ - log L„+1
где n — порядковый номер примордия, едва превышающего
10 мм дл., a L — длина и-го листа в мм; методика определения
И. п. основана на допущении постоянного равенства пластохро-
нов в течение всего онтогенеза побега; терм.: Erickson, Michelini,
1957.
Индивид(уум) (англ.: individual) см. особь.
49
Индивид(уум) множественный см. индивид сложный.
Индивид(уум) простой — особь, у которой морфологическая це-
лостность преобладает над автономностью ее частей и отсутству-
ет ясно выраженная морфологическая дезинтеграция (многие де-
ревья, растения стержнекорневые, однолетники)-, в практике И. п.
выделяется по морфологической (физической) непрерывности;
терм.: Левин, 1961.
Индивид(уум) сложный — особь, у которой автономность ее час-
тей преобладает над морфологической целостностью (хотя бы во
взрослом состоянии) и явно выражена морфологическая дезин-
теграция (напр., раст. длиннокорневищное)-, в практике И. с. выде-
ляется по морфологической (физической) связи отстоящих друг
от друга условных особей (англ.: clonal fragment), осуществляе-
мой побегами диагеотропными (корневища, столоны и т. п.); терм.:
Левин, 1961.
Индукция (от лат. induction — побуждение, наведение; англ.: in-
duction) — взаимодействие между частями развивающегося эм-
бриона (эмбриональная И.), в процессе которого одна из них (ин-
дуктор) определяет направление развития другой (детерминация
ткани-мишени). В биоморфологии пон. И. используется в широ-
ком смысле как влияние внешних или внутренних факторов на
клетки, ткани или органы раст., которое приводит к детермина-
ции их развития. В связи с этим выделяют развитие автономное и
развитие индуцированное.
Интегратор рестриктивный (англ.: restrictive intergator) см. де-
зинтеграция морфологическая.
Интегратор экстенсивный (англ.: extensive integrator) см. дезин-
теграция морфологическая.
Интенсификация (от лат. intension — напряжение, усиление, fa-
cio — делаю) — назв. используют для обозначения ускорения
морфогенеза, темпа развития, смены осей скелетных или сокра-
щения продолжительности онтогенеза потомка по сравнению с
предком. И. часто рассматривают как один из основных способов
или принципов эволюции биоморфологической; лит.: Серебряков,
1962; Хохряков, 1975, 1976, 1981; Серебрякова, 1983.
Камптотропизм (Camptotropismus; от греч. campter — поворот, tro-
pos — направление; англ.: camptotropism) — ориентированное
движение органов раст., выражающееся в изменении направления
роста (изгибы) в ответ на одностороннее воздействие различных
факторов среды (см. тропизм); терм.: Bucher, 1906.
50
Канализованность (канализированность) эволюции биомор-
фологической см. направленность эволюции биоморфологи-
ческой.
Карликовость см. нанизм.
Катафилл (Cataphyllum, от греч. kata — приставка со значением
направления вниз, phyllon — лист; англ, cataphyll, scale leaf) —
видоизмененный лист низовой (или его часть) с редуцированной
пластинкой; обычно выполняет функцию специализированной
почечной чешуи (рис. 63); у раст. древесных нередко развиваются
в конце роста видимого, и представляют переходные формы меж-
ду формациями листьев срединных двух приростов одного побега.
Категория форм жизненных — форма жизненная индивида на от-
дельной стадии его онтогенеза (цикла жизненного), которая вы-
полняет различные функции, отличные от функций жизненных
форм других стадий, и занимает свое место в экономике природы
в связи с присущей ей экологией, биологией, приспособлениями
и нормой поведения; пон. К. ф. ж. отражает смену жизненных
форм в онтогенезе индивида, отличие их друг от друга не только
внешне или биологически, но и по выполняемым функциям; у се-
менных раст. выделяют 4 категории: основная (листостебельная),
гаметоносная (пыльца), расселительная (диаспоры) и закрепи-
тельная (проростки); каждая из них выполняет главную (энерге-
тическое и вещественное обеспечение онтогенеза) и побочную
(переживание неблагоприятных условий) функции; терм.: Хох-
ряков, 1981.
Каудекс (Caudex — ствол, бревно; англ.: caudex р. р.) — система по-
бегов многолетнего раст. стержнекорневого, образованная ниж-
ними, многолетними, часто одревесневающими, укороченными
участками побегов (см. резид), несущими почки возобновления и
соединенными с корнем главным (рис. 41); К. характерен для (по-
лупустынных и высокогорных полукустарничков (Rosaceae,
Compositae, Chenopodiaceae) и многолетних монокарпиков (Um-
belliferae); по степени разветвленности различают (рис. 42) про-
стой (одноглавый) и ветвистый (многоглавый) К. (см. каудикул),
а по положению почек возобновления относительно уровня поч-
вы — К. надземный (Cousinia oopoda) и К. погруженный (Poly-
gonum coriarium); часто К. наз. ветвящимся или неветвящимся
корневищем; в зарубежной литературе терм. К. используется в
широком смысле: как стебель раст. древесного или толстое, одре-
весневающее корневище укороченное раст. травянистых; син.:
51
Рис. 41. Схема развития каудекса (по: Фе-
доров и др., 1962): А — проросток, Б —
первый год, В — второй год, Г — третий
год, Д — многолетнее растение (1 — вер-
хушечная почка, 2 — семядоля, 3 — ги-
покотиль, 4 — главный корень, 5 — бо-
ковой корень, 6 — надземная часть побе-
га, 7 — резид, 8 — почка возобновления,
9 — стеблевая часть каудекса)
Рис. 42. Основные формы каудекса (по
Федоров и др., 1962): А — многоглавый,
подземный, Б — одноглавый, надзем-
ный (1—однолетняя часть побега, 2-—
подземная часть каудекса, 3 — надзем-
ная часть каудекса, 4 — зона гипокотиля,
5 — каудикул, 6 — главный корень, 7 —
боковой корень)
стеблекорень; лит.: Федоров, Кирпичников, Артюшенко, 1962;
Нухимовский, 1969а,б.
Каудикул (Caudiculus; от каудекс) — ответвление каудекса (рис. 42);
реже К. наз. облиственный многолетний стебель раст. подушко-
видного.
Каулифлория (Caulifloria; от лат. caulis — стебель, ствол, flos — цве-
ток; англ.: cauliflory) — ра заитиеиз почек спящихцветков или соцве-
тий (побег дополнения) непосредственно на стволе (рис. 43) и на тол-
стых ветвях кроны (рамифлория); встречается преимущественно у
тропических деревьев (Theobroma cacao — см. фото на обложке).
Каулифолия (Caulifolia; от лат. caulis — стебель, ствол, folium —
лист) — развитие из почки спящей малолистного побега укорочен-
52
Рис. 43. Каулифлория у Parmentiera cerifera (А; по: Bell, Bryan, 1998)
и каулифолия у Cercidiphyllum japonicum (Б; по: Федоров и др., 1962):
1 — лист, 2 — плод, 3 — ствол, 4 — цветок)
него {побег дополнения) непосредственно на стволе (рис. 43);
встречается преимущественно у раст. тропиков и субтропиков
(Araucaria); иногда К. наз. каулифилией.
Квант роста — широко используемое пон. для обозначения цикла
листового, емкости почки зрелой, единицы измерения линейного
размера или варьирования размера органа раст.; терм.: Thompson,
1948; лит.: Шмидт, 1968; Серебрякова, 19696; 1971а; Михалевская,
2002.
Кладодий (Cladodium; от греч. klados — ветвь; англ.: cladophyll) —
сплющенный, листообразный, стебель нек-рых раст. аридных об-
' ластей (например, Colletia cruciata), выполняющий функции лис-
та (рис. 44); иногда К. наз. филлокладием; син.: кладофилл.
Кладофилл (англ.: cladophyll) см. кладодий.
Классификация подвижности вегетативной по А. П. Хохря-
кову (1975а) — «шкала интенсификации вегетативной подвиж-
ности высш, раст.», основанная на различной выраженности у
раст. размножения вегетативного; различают простые и партику-
53
Рис. 44. Листообразные побеги у Asparagus densi-
florus (A), Rhipsalidopsis rosea (Б) и Ruscus hypo-
glossum (В): 1 — кладодий, 2 — колючки листо-
вого происхождения, 3 — филлокладий, 4 —
чешуевидный лист (по: Bell, Bryan, 1998)
10 мм
лирующие монофиты, по-
лифиты и раст., рассеиваю-
щиеся вегетативно (см. ви-
випария ложная).
Классификация форм жиз-
ненных (англ.: life forms
classification) — распределе-
ние конкретного множества
форм жизненных (таксона,
фитоценоза, флоры и т. п.)
по определенной системе
иерархически соподчинен-
ных классификационных
категорий; в искусственные
группы объединяются орга-
низмы, сходные по габиту-
су (дерево, кустарники т. д.),
а не по родству (вид, род
и т. д.). Различия между ва-
риантами К. ф. ж. связаны
гл. обр. с выбором наиболее
и наименее значимых при-
знаков биоморфологических,
который часто определяет-
ся задачами конкретного
исследования; в этой связи различают К. ф. ж. биологические (К. ф.
ж. по Раункиеру), морфологические или эколого-физиономические
(К. ф. ж. по Серебрякову), филогенетические (К. ф. ж. таксона), фи-
тоценотические (К. ф. ж. по Зозулину); реже К. ф. ж. строят по осо-
бенностям онтогенеза (К. ф. ж. по Смирновой); для анализа всего
комплекса признаков жизненной формы используются различные
системы эколого-биологических признаков (К. ф. ж. по Голубеву);
лит.: Серебряков, 1962; Серебрякова, 1972; Хохряков, 1979, 1981;
Нухимовский, I960, 1997; Кирпотин, 1999.
Классификация форм жизненных по В. Н. Голубеву (1972) —
«линейная система жизненных форм», разработанная для целей
фитоценологии и флористики на основе принципа политомии:
перечисления совокупности упорядоченных элементарных эко-
лого-биологических признаков раст. в линейной последователь-
ности; классификация форм жизненных заменяется классифика-
54
цией признаков разного значения (морфологических, экологиче-
ских, физиологических, биологических и др.), представленной в
матричной форме: в левой части таблицы располагается список
видов, а в вертикальных столбцах (правой части) отмечается при-
сутствие-отсутствие конкретного признака.
Классификация форм жизненных по Г. М. Зозулину (1961,
1968) — разработанная для целей фитоценологии система жиз-
ненных форм на основе различий раст. по удержанию площади
обитания; типы форм жизненных (раст. вагативное, раст. инси-
дентивное, раст. ирруптивное, раст. реддитивное, раст. рестатв-
ноё) выделяются по способности к восстановлению особи после
уничтожения по какой-либо причине ее надземной части (возоб-
новление вегетативное), подтипы — по длительности сохранения
листьев (раст. вечнозеленое, вечнохвойнозеленое, листопадное,
веткопадное, хвоепадное, безлистное, периодическое), а груп-
пы — по способу удержания площади обитания (разрастание и
размножение вегетативное).
Классификация форм жизненных по Г. Н. Высоцкому (1915) —
первая классификация форм жизненных степных раст., основанная
на различиях в способах размножения вегетативного и выражен-
ности расселения вегетативного, выделены раст. вегетативно-не-
подвижные (осевые и раст. дернованные), раст. ползучие (раст. кор-
невищные, корнеотпрысковые, вегетативные малолетники), мало-
летники (двулетники, однолетники) и ксилофоры (деревья,
кустарники и т. п.).
Классификация форм жизненных по И. Г. Серебрякову
(1962) — наиболее детально разработанная К. ф. ж. раст., основан-
ная на различиях в длительности жизни раст. (для раст. древесных)
и осей скелетных их побеговых систем (для раст. травянистых);
наивысшие ранги классификации (отделы, типы) выделены по
анатомической структуре надземных стеблей и длительности
их жизни (деревья, кустарники, поликарпики, монокарпики и т. п.),
а подчиненные (классы, группы) — по более частным признакам
(направление роста, способ возобновления вегетативного и раз-
множения вегетативного и др.); позднее неоднократно видоизме-
нялась и дорабатывалась многими ботаниками в связи с конкрет-
ной задачей их исследования.
Классификация форм жизненных по К. Раункиеру (С. Ra-
unkiaer) — одна из популярных систем форм жизненных высших
раст., основанная на различиях в положении почек возобновления
55
или верхушек побегов относительно поверхности почвы в тече-
ние неблагоприятного для вегетации времени года (холодного
или сухого); Раункиер выделил 5 основных типов (рис 22): фане-
рофит (Р), хамефит (Ch), гемикриптофит (Н), криптофит (К),
терофит (Th), в каждом из которых позднее им самим и другими
ботаниками установлено большое число подтипов по различным
признакам (ритм развития листвы, особенности размножения ве-
гетативного, характер почечных покровов и др.); различные ва-
рианты классификации широко используются в геоботанике и
флористике для характеристики воздействия среды (гл. обр. кли-
мата) на проявление адаптации раст. (см. спектр биологический,
с. нормальный); нередко в раункиеровские группы вкладывается
иной, расширенный или специальный смысл; лит.: Raunkiaer,
1934; Серебряков, 1962; Серебрякова, 1972.
Классификация форм жизненных по Л. И. Казакевичу
(1922) — классификация форм жизненных, основанная на различ-
ной способности травянистых многолетников к размножению ве-
гетативному, выделены основные группы: раст. стержнекорне-
вые, раст. дерновые, раст. луковичные и клубнелуковичные, раст.
корневищные (стелющиеся и укореняющиеся), раст. корнеот-
прысковые.
Классификация форм жизненных по О. В. Смирновой с соав-
торами (Ценопопуляции..., 1976) — «классификация биоморф
по особенностями хода онтогенеза», разработанная для анализа
численности и пространственной структуры ценопопуляций
раст.; по характеру пространственного размещения побегов, кор-
ней и почек возобновления в онтогенезе особи выделено 3 типа био-
морф (б. моноцентрическая, б. явнополицентрическая и б. неявно-
полицентрическая), которые по степени дезинтеграции морфоло-
гической и последовательности фаз морфогенеза подразделяются
на несколько вариантов; лит.: Ценопопуляции..., 1988.
Клон (Cion, от греч. klon — отпрыск, веточка; англ.: clone) — сово-
купность особей, произошедших от одного раст. (материнской
особи) в результате размножения вегетативного (рис. 114); лит :
Злобин, 1961; Ефейкин, 1970; Касинов, 1972; Андреева, 1987; Це-
нопопуляции..., 1988.
Клубенек (Tuberculum; от клубень-, англ.: bulbil, bulbel) — видоиз-
мененная почка пазушная в области соцветия с утолщенным мя-
систым стеблем (рис. 67); обеспечивает размножение вегетатив-
ное (см. вивипария) нек-рых раст. травянистых (Polygonum
56
viviparum); иногда К. наз. почкой выводковой, луковичкой, клубнем
. надземным, бульбочкой (Dentaria bulbifera).
Клубенек корневой (Tuberculum; от клубень; англ.: root tubercle,
root nodule, root swelling) — небольшое утолщение на корне, обра-
зовавшееся как пролиферация внутренних клеток первичной ко-
ры на проникновение инфекционных нитей азотфиксирующих
бактерий (рис. 45); предполагают, что К. к. является видоизменен-
ным корнем боковым; развитие К. к. особенно характерно для сем.
Fabaceae (ср. корень кораллоидный); иногда К. к. наз. клубень поч-
ко-корневой.
Клубень (Tuber; англ.: tuber, swollen stem р. р.) — видоизмененный
(метаморфоз) мясистый стебель б. м. шаровидной формы из-за
сильного разрастания (вздутия) одного (К. одночленный) или не-
скольких (К. многочленный) междоузлий, несущих катафиллы
(ср. клубнелуковица) и почки пазушные, которые иногда наз. «глаз-
ками» (рис. 46); К. служит для запасания питательных веществ (гл.
Рис. 45. Клубеньки на корнях Alnus glutinosa (А — большой клубенек,
Б — клубеньки на тонких корнях), Acacia pravissima (В), Vicia faba (Г):
1 — боковой корень, 2 — клубенек (по: Bell, Bryan, 1998)
57
Рис. 46. Основные типы клубней: А — однолетний (схема развития картофеля; вегета-
тивный малолетник), Б — многолетний моноподиально нарастающий (Cyclamen
hederifoliuiir, по: Bell, Bryan, 1998), В — многолетний симподиально нарастающий
(Amorphophallus titanium по: Грудзинская, 1982): 1 — клубень, 2 — почка возобновле-
ния, 3 — столон, 4 — цветонос
обр. крахмала), реже — для размножения вегетативного; различа-
ют К. надземные и К. подземные; син.: К. стеблевой; лит.: Федоров,
Кирпичников, Артюшенко, 1962; Серебряков, 1952.
Клубень висячий см. клубень корневой.
Клубень воздушный см. туберйдий.
Клубень гипокотильный (англ.: hypocotyl tuber) см. клубень под-
земный.
Клубень зародышевый см. протокорм.
Клубень корневищный см. клубень подземный.
Клубень корневой (Tuber radicale; от корень; англ.: root tuber) — ви-
доизмененный корень боковой или корень придаточный с сильно
развитой запасающей паренхимой (см. корень запасающий); обра-
зуется как утолщение (шишка корневая) базальной (Dahlia
pinnata), срединной (Phlomis tuberosa) или верхней (Asparagus) час-
ти корня; у нек-рых раст. корень утолщается прерывисто (Sedum
58
telephium) или полностью пре-
вращается в клубень (рис. 47);
син.: корнеклубень, клубень ви-
сячий.
Клубень почко-корневой — видо-
измененная почка пазушная, ось
которой формируется в результа-
те срастания и утолщения укоро-
ченного стеблям корня придаточ-
ного (рис. 67); служит для размно-
жения вегетативного нек-рых
раст. (Ficaria verna); см. дроппер; рис. 47. Форма корневых клубней: 1 —
лит.: Metcalfe, 1938; Барыкина, Гу- Lathyrus tuberosus, 2 — Phlomis tuberosa,
Ленкова, 1990; Барыкина, 1995. 3-4 — Sedum telephium (корневые шиш-
Клубень стеблевой (англ.: Stem ки^’5 HemerocalKsfulva (по: Серебря-
tuber) см. клубень. ков’1952)
Клубнелуковица (Bulbotuber; от луковица и клубень; англ.: corm,
bulbotuber) — видоизмененный подземный стебель (метамор-
фоз), разросшиеся одно или несколько укороченных междоузлий
которого образуют клубневидное, мясистое однолетнее образо-
вание, покрытое сухими, пленчатыми чешуями или/и остатками
оснований отмерших листьев (ср. луковица, клубень); служит
для накопления питатель-
ных веществ и ежегодного
возобновления раст. клуб-
нелуковичного (рис. 48);
встречается у однодольных
и преимущественно среди
Iridaceae и Liliaceae; К.
внешне напоминающую
луковицу, иногда наз. лу-
ковицей кожистой (Colchi-
cum autumnale); лит.: Ро-
дионенко, 1961; Федоров,
Кирпичников, Артюшен-
ко, 1962; Артюшенко, 1970.
Клубнелуковица замещаю-
щая (англ.: replacement
(corm) bulbotuber) — К., об-
разующаяся из почки возоб-
Рис. 48. Строение клубнелуковицы Crocus sp.
(по: Федоров и др., 1962): А — схема. Б — клуб-
нелуковица с детками (1 — стебель моноподи-
ально нарастающей клубнелуковицы, 2 — че-
шуи, 3 — зачатки листьев, 4 — зачаток цветка,
5 — почка возобновления, 6 — придаточные
корни, 7 — пазушные почки, 8 — детки)
59
новления и замещающая отмирающую материнскую К.; обеспечи-
вает ежегодное возобновление раст. клубнелуковичного.
Клубнепочка см. клубенек.
Колено надсемядольное (англ.: epicotyl) см. эпикотиль.
Колено подсемядольное (англ.: hypocotyl, hypocotyl hook) см. ги-
покотиль.
Колеоптиль (Coleoptilis; от i реч. koleos — ножны, футляр и перыш-
ка, англ.: coleoptile) — листоподобное влагалищное образование,
прикрывающее почечку зародыша злаков и имеющее вид конусо-
видного замкнутого колпачка; служит приспособлением, проби-
вающим острым кончиком слой почвы или другие препятствия,
и защищает почечку от механических повреждений (рис. 38);
морфологическая природа К. оценивается по-разному: 1) как вла-
галищный лист зародыша, которому предшествуют щиток и эпи-
бласт; 2) как вырост семядоли, другая часть которой представлена
щитком; 3) как предлист почечки, занимающей боковое положе-
ние; в более широком смысле К. наз. первый лист проростка од-
нодольных; син.: лист влагалищный, колеофилл; лит.: Цвелев,
1976; Лодкина, 1988.
Колеориза (Coleorrhiza; от греч. koleos — ножны, футляр и корень;
англ.: coleorhiza) — пленчатое влагалище вокруг корешка зароды-
ша злаков и цикадовых; выполняет защитную функцию (рис. 38);
морфологическая природа К. оценивается различно: 1) как ниж-
няя часть гипокотиля; 2) как видоизмененный корешок; 3) как на-
ружная часть корневого чехлика; син.: влагалище корневое; лит.:
Цвелев, 1976; Лодкина, 1988.
Колеофилл см. колеоптиль.
Колония клональная — пон., используемое для обозначения ин-
дивидов сложных, у которых каждая относительно автономная сис-
тема побегов образуется из вегетативных зачатков и проходит
собственный онтогенез, аналогичный онтогенезу раст. материн-
ского (индивидума простого); в практике К. к. выделяется по мор-
фологической (физической) связи условных особей, осуществ-
ляемой корнями (см. раст. корнеотпрысковое); терм.: Muller, 1951.
Кольчатки см. побег укороченный.
Колючка (Spina; англ.: spine, thorn, prickle) — острый, твердый, как
правило, одревесневший вырост (рис. 49), образующийся в ре-
зультате метаморфоза побега (англ.: stem spine; Crataegus), листа
(англ.: leaf spine; Cactaceae) или корня придаточного (англ.: root
spine; корень-колючка нек-рых лиановидных пальм); часто К. наз.
60
Рис. 49. Колючки барбариса (1-2), груши (3) и боярышника (4)
(по: Ботаника..., 1988): 1 — побег с колючками листового происхождения,
2 — переход от листа к колючке, 3-4 — колючки побегового
происхождения (п — пазушный побег)
также шипы (англ.: emergence, prickle), представляющие собой
выросты эпидермиса (Rosa).
Комплекс аксиллярный см. узел.
Контрактильность (Contractilitas; от лат. cum — вместе, trachere —
тянуть, тащить; англ.: contractily, root contraction) — процесс со-
кращения корней, благодаря которому основания побегов раст. с
почками возобновления втягиваются в почву; К. широко распро-
странена среди травянистых многолетников (см. геофилия) и, как
правило, обнаруживается по поперечной морщинистости корня
(рис. 50); укорочение корней связано с определенной их частью и
зависит гл. обр. от изменения формы клеток первичной коры,
происходящего в результате роста или потери влаги и запасных
веществ тканями корня; лит.: Грушвицкий, 1952; Михайловская,
1981а.
Контроль апикальный см. доминирование апикальное.
Конус нарастания побега (англ.: shoot axon hill, shoot apex) — вер-
хушка побега (апекс побега), состоящая из первичной меристемы,
61
Рис. 50. Контрактильные корни Menyanthes trifoliala
1 — побег, 2 — поперечная морщинистость на контрактильных корнях
(по: Федоров и др., 1962)
62
обеспечивающей образование всех его частей и первичных тканей;
размеры и форма К. н. п. различны у разных раст. (рис. 7) и изменя-
ются в течение пластохрона (рис. 102), большого и малого циклов
развития и в зависимости от внешних условий; наиболее резкие
изменения связаны с переходом побега в генеративную фазу разви-
тия (образование цветков); обычно К. н. п. рассматривают как
часть целого раст., которая с одной стороны зависит от влияния
других органов, а с другой — как самоопределяющийся и домини-
рующий центр развития (см. доминирование апикальное) контро-
лирует рост раст. и структуру взрослых побегов (листорасположе-
ние, длину междоузлии)-, иногда К. н. п. наз. только верхнюю часть
апекса (точка роста побега), расположенную выше зоны образова-
ния примордиев; лит.: Серебря-
ков, 1963; Синнот, 1963; Чельцо-
ва, 1980; Ростовцева, 1984.
Корень (Radix; англ.: root) — осе-
вой орган раст. с радиальной сим-
метрией, неопределенно долго
нарастающий в длину благодаря
деятельности его меристемы апи-
кальной, которая никогда не об-
разует листовых зачатков и
обычно защищена корневым
чехликом; К. ветвится эндогенно,
служит в основном для прикреп-
ления раст. к субстрату, поглоще-
ния воды, питательных веществ
и синтеза различных физиологи-
чески активных соединений (фи-
тогормонов); у многих раст. вы-
полняет добавочные функции
(см. метаморфоз корня); по по-
ложению в пространстве выделя-
ют К. вертикальный, К. горизон-
тальный и К. косой, а по месту
их происхождения — К. главный,
К. боковой и К. придаточный
(рис. 51); сочетание различных
корней определяет тип и форму
системы корневой раст., корни в
Рис. 51. Основные типы корней растения:
1 — побег, 2 — корень главный, 3 — ко-
рень придаточный, 4 — корень скелет-
ный, 5 — корень ростовой, 6 — корень
сосущий, 7 — корень боковой
63
которой могут отличаться длительностью существования (К. одно-
летний и К. многолетний), темпом роста (К ростовой) и функцио-
нальной ролью {К. скелетный, К. сосущий, К. контрактильный,
К. цепляющийся, К. опорный, К. размножения и др.); лит.: Серебря-
ков, 1952; Федоров, Кирпичников, Артюшенко, 1962; Михайлов-
ская, 1981; Plants roots..., 1991;Таршис, 1975, 2003.
Корень адвентивный (англ.: adventitious root) см. корень прида-
точный.
Корень аксиллярный (англ.: axile root) см. корень кладогенный.
Корень ассимилирующий (Radix assimilans; англ.: green root,
assimilative root) — К., содержащий в значительном количестве
хлорофилл и выполняющий функцию ассимиляции; у некоторых
раст. (Podostemonaceae; Lithops) из-за редукции зеленых листьев
фотосинтез полностью осуществляется К. а.
Корень боковой (Radix lateralis; от лат. latus — бок, сторона; англ.:
lateral root, branch root) — К. любого порядка ветвления (рис. 51);
часто обозначается как К. вторичный (К. б. на К. главном), К. тре-
тичный (К. б. на К. вторичном) и т. д., или как К. первого, второго
и т. д. порядка ветвления; в более узком значении К., закладываю-
щийся эндогенно вблизи от апекса К. главного или К. материн-
ского (ср. К. придаточный); син.: К. ризогенный, К. корнеродный.
Корень вентиляционный см. пневматофор.
Корень вертикальный (Radix verticalis; от лат. vertex — вершина,
верх; англ.: vertical root) — К., расположенный по отношению к
поверхности почвы вертикально; терм., обычно используется для
обозначения К. растущего отвесно вниз (положительный геотро-
пизм); син.: К. отвесный.
Корень видоизмененный см. метаморфоз корня.
Корень воздушный (Radix аегеа; от лат. аег — воздух; англ.: aerial
root, crampon) — К. придаточный, развивающийся в воздушной
среде (рис. 52); часто выполняет дополнительные функции (см.
К. опорный, К. цепляющийся); у многих эпифитов дождевых
тропических лесов (Orchidaceae, Bromeliaceae) К. в. запасают вла-
гу, поглощая ее из воздуха с помощью специализированной тка-
ни — веламена (англ.: velamen); син.: К. надземный.
Корень временный см. корень однолетний.
Корень вторично-стержневой см. растение кистекорневое.
Корень вторичный см. корень боковой.
Корень втягивающий (англ.: contractile root) см. корень контрак-
тильный.
64
Рис. 52. Воздушные корни Ficus pumila (А) и Pandanus nobilis (Б):
1 — цепляющиеся корни на побеге корнелазящей лианы, 2 — корни подпорки
в основании ствола (по: Bell, Bryan, 1998)
Корень гипокотильный (англ.: hypocotyl root) см. корень кладо-
генный.
Корень главный (Radix primaria; от лат. primus — первый; англ.:
primary root, tap root, main root) — К., формирующийся из кореш-
ка зародыша семени; иногда наз. К. первичным; у большинства
двудольных раст. К. г. развивается сильнее, чем корни боковые и
поэтому наз. К. стержневым (рис. 51); у однодольных К. г. обычно
недолговечен, и система корневая формируется К. придаточными^
син.: К. основной.
Корень главный зародышевый (англ.: main seminal root, primary
root, embryonic root) см. корешок.
Корень горизонтальный (Radix horizontalis; от греч. horizon — го-
ризонт; англ.: horizontal root) — К., расположенный параллельно
поверхности почвы (рис. 53); обычно достигает большой длины
(у Populus tremula до 30 м) и обеспечивает устойчивость раст.,
напр. на подвижных субстратах (псаммофиты); К. г., располага-
ясь близко от поверхности почвы, нередко оголяется ветром и
образует отпрыски корневые (см. К. размножения).
Корень детерминированный (англ.: determinate root) — корень с
ограниченным ростом (см. рост детерминированный) в длину в
результате дифференциации меристемы апикальной (см. К. кла-
стерный); назв. используется для противопоставления корням с
длительным ростом (К. недетерминированный).
65
Рис. 53. Основные типы скелетных корней деревьев: 1 — горизонтальные,
2 — якорные, 3 — досковидные, 4 — ходульные, 5 — столбовидные,
6 — утолщенное основание ствола (по: Jenik, 1978)
Корень досковидный (Radix tabuliformis; от лат. tabula — стол,
доска, forma — форма; англ.: tabular (plank-buttress) root) —
обычно К. боковой, развивающийся горизонтально близ поверх-
ности почвы и обладающий несимметричным вторичным утол-
щением, в результате которого верхняя (надземная) часть К.
разрастается в ширину интенсивнее нижней (подземной); К. д.
обеспечивают устойчивость деревьев с крупными кронами, спо-
собствуют усилению аэрации подземных частей раст. и являются
характерной особенностью ряда тропических раст. (рис. 53).
Корень дыхательный (англ.: breathing (aerating) root) см. пневма-
тофор.
Корень запасающий (Radix proventicia; от лат. pro — приставка со
значением «для», venio — расти, вырастать; англ.: storage root) — К.
утолщенный, слабо ветвящийся с сильно развитой запасающей па-
66
ренхимой, расположенной в первичной коре, древесине или серд-
цевине; по форме и происхождению различают К. клубневидные
или шишковатые (Sedum telephium), К. конусовидные, или кони-
ческие (Daucus sativus), К. шаровидные (Raphanus sativus), К. редь-
ковидные или репчатые (Beta vulgaris); в любом К. есть некоторое
количество запасных веществ, но К. з. обычно наз. мясистый К. с
сильно развитой паренхимой (см. клубень корневой, корнеплод).
Корень зародышевый (англ.: embryo root) см. зародыш.
Корень кладогенный (Radix cladogena; от греч. klados — ветвь, по-
бег, genesis — происхождение, потомство; англ.: cladogenous ro-
ot) — К. придаточный, развивающийся эндогенно на разных
участках стебля, в связи с чем различают К. пазушный (К. аксил-
лярный), К. узловой (К. надузловой и К. подузловой), К. междо-
узлиевый, К. гипокотильный; син.: К. стеблеродный.
Корень кластерный (англ, cluster root) — короткий (2-4 мм дл.)
корень боковой с ограниченным ростом в длину (К. детерминиро-
ванный) и волосками корневыми, расположенными на апексе;
обычно К. к. многочисленны и образуют эллипсовидные «щетки»
(кластеры) на поверхности корней; характерны для представите-
лей сем. Fabaceae, Myricaceae, Betulaceae (Alnus), Elaeagnaceae, Ca-
suarinaceae, Proteaceae, Moraceae, которые растут на бедных фос-
фатами местообитаниях; предполагается, что К. к. служат для
усиления поглощения (прежде всего фосфатов)в отсутствии ми-
коризы-, син.: протеоидный корень; лит.: Skene, 1998, 2000.
Корень клубневидный см. корень запасающий.
Корень коммуникационный см. корень скелетный.
Корень конический см. корень запасающий.
Корень контрактильный (Radix contractilis; от лат. cum — вместе,
trachere — тянуть, тащить; англ.: contractile root) — К. вертикаль-
ный, обычно длинный и утолщенный, с резко выраженной спо-
собностью к сокращению (контрактильности) в продольном на-
правлении, в результате чего изменяется положение почек возоб-
новления по отношению к уровню почвы; у многих раст.
стержнекорневых следы контрактильности заметны по наличию
поперечных складок («морщин») поверхностных тканей в базаль-
ной части К. (рис. 50); не только специализированный К. к., но и
любой растущий К. вызывает напряжение в основании побега и
втягивает его в глубь почвы (см. геофилия); син.: К. втягиваю-
щий, К. сокращающийся; лит.: Скрипчинский, Максименко, 1950;
Михайловская, 1981а.
67
Корень коралловидный (англ.: coraloid root) см. микориза.
Корень кораллоидный (англ.: coralloid root р. р.) — сильно раз-
ветвленный корень боковой с отрицательным геотропизмом,
имеющий вид коралловидного образования; предполагается, что
ограниченный рост и коралловидная форма обусловлена про-
никновением синезеленых водорослей (Nostoc, Anabaena), фик-
сирующих азот (ср. клубенек корневой); К. к. характерны для ци-
кадовых (Cycas, Stangeria, Encephalartos и др.); в зарубежной лите-
ратуре назв. coralloid root часто используют для обозначения К.
коралловидного; лит.: Grobbelaar et al., 1986.
Корень корнеродный см. корень боковой.
Корень косой (Radix obliqua; от лат. obliquus — косой) — К., рас-
положенный наклонно по отношению к поверхности почвы.
Корень материнский — назв. используется для обозначения К. лю-
бого порядка ветвления, который несет К. боковой и является по от-
ношению к нему основным.
Корень междоузлиевый (англ.: internode root) см. корень кладо-
генный.
Корень многолетний (англ.: perennial root) см. корень.
Корень надземный см. корень воздушный.
Корень надузловой см. корень кладогенный.
Корень недетерминированный (англ.: indeterminate root) см. ко-
рень детерминированный.
Корень однолетний (Radix annua; от лат. annus — год; англ.:
annua] root) — К., живущий один год; обычно тонкие, слабо вет-
вящиеся корни, выполняющие функцию всасывания; у многих
пустынных раст. К. о развиваются только во влажные периоды
года; син.: К. эфемерный, К. временный; лит.: Шалыт, 1960.
Корень опорный (англ.: supporting root, stilt root, prop root, brace
root) — назв. используется для обозначения К., образующихся на
надземной части стебля, достигающих земли и часто одревесне-
вающих; К. о. служат дополнительной опорой для раст. (рис. 53);
утолщенные К. о., поддерживающие обширную крону дерева наз.
К. столбовидными (К.-подпорка; англ.: columnar root) или колон-
новидными «стволами» (Ficus benghalensis); у нек-рых тропиче-
ских деревьев (Pandanaceae) К. о. развиваются в нижней части
ствола и достигают земли в наклонном направлении (К. ходуль-
ные; англ.: stilt root, prop root).
Корень основной см. корень главный.
Корень отвесный см. корень вертикальный.
68
Корень пазушный см. корень кладогенный.
Корень партикулирующий — назв. используется для обозначения
гл. обр. К. главного, отмирание тканей центральной части которого
в продольном направлении обусловливает его частичное или пол-
ное расщепление (партикуляцию); син.: К. расщепляющийся.
Корень первичный см. корешок.
Корень питающий см. корень сосущий.
Корень поглощающий см. корень сосущий.
Корень подпорка (англ.: columnar root) см. корень опорный.
Корень подузловой см. корень кладогенный.
Корень придаточный (Radix adventicia; от лат. ad — приставка в
значении дополнительности, venio — приходить, возникать; англ.:
additional root, adventitious root) — К., образующийся эндогенно на
стебле (К. кладогенный; рис. 51) или на листе (Begonia, Saintpaulia);
К. п. широко распространены среди сосудистых раст. и имеют
большое значение в их размножении вегетативном; у нек-рых раст.
выполняют особые функции (см. К. ассимилирующй, К. опорный,
К. цепляющийся и др.); иногда к К. п. относят и К. ризогенные, кото-
рые закладываются вдали от меристема апикальной К. главного или
К. материнского, что затрудняет разграничение между К. п. и К. бо-
ковыми; син.: К. адвентивный; лит.: Barlow, 1986.
Корень протеоидный (англ.: proteoid root) см. корень кластерный.
Корень размножения (Radix propagationis, от лат. propagare —
размножать, разводить, распространять) — К. обычно горизон-
тальный, на котором образуются отпрыски корневые (рис. 99,119;
см. раст. корнеотпрысковое); терм.: Казакевич, 1922; лит.: Васи-
левская, 1957а; Барыкина, 1958; Кондратьева-Мельвиль, 1959;
Peterson, 1975.
Корень расщепляющийся см. корень партикулирующий.
Корень редьковидный см. корень запасающий.
Корень ризогенный (англ.: rhizogenic root) см. корень боковой.
Корень ростовой — назв. используется для обозначения быстро
растущих К., обычно слабо ветвящихся, недолговечных или со
временем одревесневающих (К. скелетный) и обеспечивающих
расширение границ ризосферы раст. (рис. 51).
Корень скелетный — назв. используется для обозначения много-
летних, обычно одревесневающих К., отличающихся крупными
размерами и несущих К. сосущие и К. ростовые (рис. 51); К. с. явля-
ются основой для формирования системы корневой раст.; иногда
К. с. наз. К. коммуникационными.
69
Корень сокращающийся см. корень контрактильный.
Корень сосущий (англ.: feeding root) — назв. используется для
обозначения обычно тонких интенсивно ветвящихся К., отми-
рающих в конце вегетационного сезона (К. однолетний) или на
второй год после образования и обеспечивающих увеличение об-
щей поглощающей поверхности системы корневой раст.; син.:
К. питающий, К. поглощающий.
Корень стеблеродный (англ.: shoot-borne root) см. корень кладо-
генный.
Корень стержневой (англ.: tap root, primary root) см. корень
главный.
Корень столбовидный (ангд.: columnar root) см. корень опорный.
Корень узловой (англ.: nodal root) см. корень кладогенный.
Корень ходульный (англ.: stilt root, prop root) см. корень опорный.
Корень цепляющийся (англ.: clasping root) — назв. используется
для обозначения К. кладогенных, обеспечивающих прикрепление
нек-рых лиан к опоре (Hedera helix), в связи с чем последние наз.
лианами корнелазающими (рис. 52); иногда К. ц. неправильно наз.
корнями-присосками; син.: К.-прицепка.
Корень эфемерный см. корень однолетний.
Корень якорный (англ.: strut root, anchor root) — назв. использует-
ся для обозначения К. главного или К. бокового древесных раст.,
глубоко уходящего в почву и препятствующего их выворачива-
нию при механических нагрузках (рис. 53).
Корень-колючка (англ.: spine root) см. колючка.
Корень-присоска см. гаусторий.
Корень-прицепка (англ.: holdfast root) см. корень цепляющийся.
Корешок (Radicula, уменьшительное от корень; англ.: radicle, embry-
onic root) — зародышевый главный корень; закладывается на
поздних стадиях развития зародыша и развивается как базальное
продолжение гипокотиля проростка (рис. 30); у нек-рых орхид-
ных, водных и паразитных раст. из-за недоразвития или отсутст-
вия К. образуются зародышевые корни придаточные; син.: корень
первичный, К. главный зародышевый.
Кормус см. растение листостебельное.
Корневище (Rhizoma, от греч. riza — корень; англ.: rhizome) — ви-
доизмененный многолетний подземный побег или система побе-
гов с укороченными междоузлиями и чешуевидными листьями
(катафиллами) или остатками отмерших листьев; обычно пон. К.
используют для обозначения многолетнего, подземного или над-
70
земного, ползучего побега или системы побегов, которая нарас-
тает симподиалъно (К. симподиальное, К. определенное) или мо-
ноподиально(К. моноподиальное или К. неопределенное) и посте-
пенно отмирает с базальной стороны (ср. каудекс); К. служит для
отложения запасных веществ, размножения вегетативного (см.
сарментация) и ежегодного возобновления, роста травянистых
многолетников, кустарничков и кустарников (см. ксилоризом);
по длине междоузлий различают К. укороченное и К. удлиненное,
а по происхождению — К. эпигеогенное, К. гипогеогенное и К. сме-
шанное (рис. 54); терм.: впервые употреблен Эрхартом (1787) для
папоротников (Holtum, 1929); син.: ризом; лит.: Серебряков, 1952;
Федоров, Кирпичников, Артюшенко, 1962; Серебряков, Серебря-
кова, 1965; Stevens, 1966.
Корневище вторично-подземное см. корневище эпигеогенное.
Корневище гипогеогенное (Rhizoma hypogeogeneum, от греч. hy-
po — под, внизу, geo — земля, genos — род, происхождение) —
К., стебель которого несет катафиллы и развивается из почки,
расположенной на подземном побеге раст. (К. подземного про-
исхождения); К. г. обычно нарастает симподиалъно, на боль-
шем своем протяжении диагеотропно или плагиотропно (см.
К. удлиненное) и обусловливает развитие клона раст. длиннокор-
невищного (ср. К. эпигеогенное); реже К. г. укороченное (Orobus
vernus) и нарастает моноподиалъно (рис. 54); терм.: Серебряков,
Серебрякова, 1965; син.: К. первично-подземное, ризом гипогео-
генный.
Корневище гипоэпигеогенное см. корневище смешанное.
Корневище деревянистое (англ.: ligneous rhizome) см. ксилоризом.
Корневище лептоморфное (Rhizoma leptomorphum, от греч. 1ер-
tos — тонкий, morphe — вид, форма; англ.: leptomorph rhizo-
me) — К. гипогеогенноес нарастанием моноподиалъными междоуз-
лиями удлиненными (К. удлиненное), из почек пазушных которого
развиваются одиночные надземные побеги апогеотропные
(рис. 55; ср. К. пахиморфное); терм.: McGlure (1966) ввел для опи-
сания К. бамбуков.
Корневище ложноползучее (Rhizoma pseudorepens, от греч. pseu-
dos — ложь, лат. repo — ползти) — побег ползучий плотнодерно-
винных злаков (Deschampsia cespitosa), развивающийся вначале
апогеотропно (см. побег интравагинальный), но позднее — диа-
геотропно, полегая из-за удлинения гипоподиян всех междоузлий-,
терм.: Hackel, 1882; лит.: Серебрякова, 1971а.
71
Корневище укороченное (Rhizoma abbreviatum, от лат. ab — при-
ставка, здесь со значением преувеличения, brevis — короткий;
англ.: pachycaul rhizome, short rhizome) — К., стебель которого об-
разован междоузлиями укороченными (рис. 54); характерно для
раст. короткокорневищных иногда К. у. с длинным стеблем наз. К.
удлиненным (Nuphar lutea); лит.: Голубев, 1958; Любарский, 1973.
Корневище эпигеогенно-гипогеогенное см. корневище сме-
шанное.
Корневище эпигеогенное (Rhizoma epigeogenum, от греч. epi —
на, geo —- земля, genos — род, происхождение) — К., стебель ко-
торого несет остатки отмерших листьев, развивается из почки,
расположенной на надземном побеге (ср. К. гипогеогенное), и за-
нимает подземное положение в результате котрактилъности
или/и засыпания опадом (рис. 54; К. вторично-подземное); сте-
бель К. э. обычно образован междоузлиями укороченными и на-
растает симподиалъно, реже К. э. удлиненное и нарастает моно-
подиалъно; терм. Серебряков, Серебрякова, 1965.
Корнедонце (Rhizolecus; от корень и донце) — подземная часть од-
нолетников вегетативных из рода Eremurus, которая представля-
ет собой крайне укороченное корневище (внешне напоминает дон-
це луковицы), покрытое волокнистыми или пленчатыми (как у
клубнелуковицы) остатками оснований листьев и несущее по пе-
риферии радиально расходящиеся, утолщенные корни запасаю-
щие, терм.: Хохряков, 1965.
Корнеклубень см. клубень корневой.
Корнеотпрысковость (Soboliferous, от лат. soboles — поросль, от-
прыск, побег, fero — носить) — образование отпрысков корневых
(рис. 99, 119); среди раст. корнеотпрысковых различают К. обли-
гатную (Convolvulus arvensis) и К. факультативную, связанную с
повреждением корня раст., обычно не образующего корневые от-
прыски (Trifolium montanum); К. широко распространена в при-
роде и относится к одному из способов размножения вегетатив-
ного; лит.: Михайловская, 1981а.
Корнеплод (англ.: storage root)— назв., используемое в обиходе для
обозначения сочного, питательного корнестеблевого образова-
ния двулетнего культурного раст.; в формировании К. принима-
ют участие в разном соотношении корень главный и разрастаю-
щаяся нижняя часть стебля (гипокотиль): у моркови утолщается
гл. обр. корень, у репы — нижняя часть стебля (рис. 56); нередко
К. рассматривают как метаморфоз корня, обусловленный отложе-
74
Рис. 56. Корнеплоды моркови (1), репы (2)
и свеклы (3-4): пунктиром показана граница
стебля и корня (по: Ботаника..1988)
нием запасных ве-
ществ (см. корень за-
пасающий); лит.: Фе-
доров, Кирпичников,
Артюшенко, 1962;
Ботаника..., 1988.
Корреляция ростовая
(англ.: growth correla-
tion) — зависимость
роста и развития од-
них органов (частей)
индивидуума от дру-
гих. Одним из прояв-
лений К. р. является
доминирование апи-
кальное; лит.: Син-
нот, 1963.
Корреляция функциональная (англ.: correlation) — согласован-
ное развитие органов растения посредством метаболитов (тро-
фические К.) или фитогормонов (гормональная К.), которые обу-
славливают пространственную организацию индивидуума. Взаи-
мосвязь органов раст. в онтогенезе наиболее ярко проявляется в
корреляциях ростовых, полярности и симметрии; лит.: Синнот,
1963; Полевой, 1981; Шампанья, 1983; Батыгин, 1986; Полевой, Са-
ламатова, 1991.
Кочан (англ.: head) — видоизмененная почка двулетника (капуста),
выполняющая функции запасания влаги и питательных веществ,
которые используются для развития генеративных органов на
втором году жизни раст. (рис. 67)
Кочка (Grumulus, от лат. grumus — куча, холм; англ.: tussock) —
высокая, плотная дерновина полусферической или цилиндриче-
ской формы, почки возобновления которой находятся на несколь-
ко см или дм выше уровня почвы; нижняя часть К. (тело или пье-
дестал К.) образована частично или полностью отмершими ос-
татками побегов (корневищ) и пронизана многочисленными
корнями придаточными, а верхняя (крона К.) — плотно располо-
женными друг к другу побегами кущения (Eriophorum vaginatum);
размеры и плотность К. зависят от возраста и условий место-
обитания раст. кочкообразующего, рост апогеотропный побегов
кущения которого обусловливает ежегодное нарастание дерно-
75
вины вверх у моло-
дых и образование К.
у взрослых особей
(рис. 57); лит.: Кур-
кин, 1954.
Криптокотилия (англ.:
cryptocotyly) см. про-
растание подземное.
Криптофит (Crypto-
phyton, от греч. kryp-
tos — скрытый, спря-
танный, phyton —
растение; англ.: cryp-
tophyte) — один из
основных типов клас-
сификации форм жиз-
ненных по К. Раункие-
ру, объединяющий раст. травянистые (многолетники), почки во-
зобновления которых в неблагоприятный для вегетации период
года расположены на некоторой глубине в почве (геофит) или
под водой (гелофиты, гидрофит); рис. 22.
Крона (Сота, от греч. коте — волосы на голове; англ.: crown) —
совокупность ветвей и побегов (система побеговая) раст. древес-
ного, образующихся в результате многократного ветвления ство-
ла (стволика) и несущих листья, цветки и плоды; строение и
внешний облик К. зависят от эндогенных (тип ветвления, разли-
чия в направлении и скорости роста ветвей и побегов и др.) и эк-
зогенных факторов (напр., освещенность) и изменяются с возрас-
том раст.; один из методов анализа ее структуры — выявление
моделей архитектурных; при описании формы К. (рис. 33) ее об-
лик обычно сопоставляется с геометрическими телами (К. кону-
совидная, шаровидная и т. п.), предметами (К. зонтиковидная,
вееровидная и т. п.) или индивидуальными признаками (К. пла-
кучая); терм. К. обычно используется для обозначения совокуп-
ности ветвей ствола дерева кронообразующего; в более широком
значении К. наз. и розетки листьев раст. древовидных (деревьевро-
зеточных), и совокупность ветвей стволиков кустарника, кустар-
ничка и т. п.; лит.: Серебряков, 1962; Федоров, Кирпичников, Ар-
тюшенко, 1962; Fisher, 1986.
Крона кочки см. кочка.
76
Ксеноиаразит (Xenoparasitus, от греч. xenos — чужой и парази-
тизм) — раст., для которого характерен паразитизм на высших
раст. (гл. обр. на цветковых и голосеменных), редко на водорос-
лях (ср. аллелопаразит); К.— травянистые многолетники и одно-
летники, редко кустарники и кустарнички (Loranthaceae), фа-
культативно или облигатно паразитирующие на корнях, корневи-
щах или надземных побегах, К. питаются за счет гаустория и
развиваются в тканях (Rafflesia) или вне тканей (Rhinanthus) раст.
хозяина, в связи с чем различают эндо- (паразит стеблевой) и эк-
топаразитов (паразит корневой); лит.: Терехин, 1977.
Ксилоподий см. резид.
Ксилоризом(а) (Xylorrhizoma, от греч. xylo — древесина и корневи-
ще) — многолетнее, одревесневающее, корневище гипогеогенное с
междоузлиями удлиненными и диагеотропное на большем своем
протяжении (Salix polaris); иногда К. наз. также и погребенные,
полегающие ветви (см. корневище эпигеогенное) кустарников и
кустарничков (Salix reticulata); терм.: Дервиз-Соколова, 1966; син.:
корневище деревянистое (одревесневающее).
Куст (англ.: bush, shrub,
clump, tuft, bunch) —
широко используе-
мое пои. для обозна-
чения не только сис-
темы побегов кустар-
ника, полукустарни-
ка или кустарничка
(англ.: shrub), но и
дерновины злаков и
осок (англ.: clump,
tuft, bunch), поросли
деревьев и б. м. плот-
но стоящих побегов
нек-рых двудольных
трав; по аналогии с
классификацией по-
Рис. 58. Основные типы систем побегов у кустарников и
кустарничков (по: Серебряков, 1962): А — первичный
куст, Б — парциальный куст, В — замещающий куст
(1 — главная ось, 2 — боковые оси, 3 — главный ко-
рень, 4 — ксилоризом, 5 — придаточные корни)
бегов различают К.
первичный, К. заме-
щающий, К. парци-
альный и К. материн-
ский (рис. 58).
77
Куст замещающий — система побегов многолетних, формирую-
щаяся в результате ветвления побега замещения (побега формиро-
вания) кустарников и кустарничков (рис. 58); терм.: Серебряков,
Чернышева, 1955.
Куст материнский см. куст.
Куст парциальный — система побегов многолетних кустарника
или кустарничка, обладающая вторичной системой корневой (ср.
К. первичный) и формирующаяся на ксилоризоме, соединяющем
ее (К. субпарциальный или субпарциаль) с К. материнским или
К. первичным (рис. 58); часто К. п. наз. дочернюю особь (парциаль)
в клоне раст. длиннокорневищного; терм.: Серебряков, Доманская,
Родман (1954) ввели для обозначения структурных элементов
(пучков побегов, сегментов дерновины, групп побегов), обособив-
шихся в результате отмирания старых участков куста; лит.: Се-
ребряков, 1962.
Куст первичный — система побегов многолетних, обладающая пер-
вичной системой корневой и формирующаяся в результате ветвле-
ния побега первичного кустарника или кустарничка (рис. 58); терм.:
Серебряков, Чернышева, 1955; лит.: Серебряков, 1962.
Куст субпарциальный см. куст парциальный.
Кустарник (Frutex; англ.: frutex, shrub) — многолетнее раст. дере-
вянистое более (60) 80 см высотой, у которого главный ствол б. м.
выделяется только в начале жизни, а затем отмирает или теряется
среди равных ему осей скелетных (стволиков), образующихся в
результате кущения приземного/надземного (К. аэроксильный)
или подземного (К. геоксильный или настоящий); взрослое раст.
(рис. 59) всегда имеет много надземных скелетных осей (стволи-
ков), последовательно сменяющихся в онтогенезе К. (см. цикл
основной) и связанных друг с другом своими базальными участ-
ками (см. ксилоподий); по форме роста различают К. прямостоя-
чие (Salix саргеа), полупростратные, или гемипростратные (см.
раст. стелющиеся) и К. стелющиеся (шпалерные) или стланики
(Alnaster), а по способности к размножению вегетативному— К. с
ксилоризомами (Salix) и отпрысками корневыми (Prunus spinosa);
К. с ежегодно отмирающими верхними участками побегов наз.
полукустарниками (см. раст. полудревесное); лит.: Серебряков,
1962; Голубев, 1973; Гатцук и др., 1974; Мазуренко, Хохряков,
1977; Мазуренко, 1980, 1986; Дервиз-Соколова, 1982; Лучник,
1988.
Кустарник аэроксильный см. кустарник.
78
Рис. 59. Строение геоксильного (А—Б — орешник) и аэроксильного
(В — сирень, Г — каргана) кустарника (1 — замещающий стволик, 2 — ксилоподий,
3 — отмирающий стволик); по: Ботаника..., 1988
Кустарник гемипростратный см. растение стелющееся.
Кустарник геоксильный см. кустарник.
Кустарник ксерогенный см. растение полудревесное.
Кустарник настоящий см. кустарник.
Кустарник стелющийся см. растение стелющееся.
Кустарник шпалерный см. растение стелющееся.
Кустарничек (Fruticulus; от лат. frutex — кустарник; англ.: fruticu-
lose, dwarf shrub, small shrub) — низкорослый от 5-7 до 50-60, ре-
же до 80 см высотой кустарник (рис. 60); аналогично классифика-
ции кустарников выделяют К. аэроксильные (Calluna vulgaris), К.
геоксильные (Beckea linifolia), К. шпалерные или стланички
(Dryas), К. полупростратные или гемипростратные и т. п.; К. рас-
пространены гл. обр. в высокогорьях и тундрах, где часто приоб-
ретают подушковидную форму роста; К. с ежегодно отмирающи-
ми верхними участками побегов наз. полукустарничками (см.
раст. полудревесное); лит.: Серебряков, 1962; Голубев, 1968а, 1973;
Дервиз-Соколова, 1982; Мазуренко, 1982,1986; Шутов, 2000.
Кустарничек гемипростратный см. растение стелющееся.
Кустарничек ксерогенный см. растение полудревесное.
Кустарничек стелющийся см. растение стелющееся.
Кустарничек шпалерный см. растение стелющееся.
79
Рис. 60. Строение кустарничков (по: Ботаника..., 1988):
А-Г — развитие длиннокорневищного кустарничка (черника),
Д — схема клона черники, Е — андромеда (1 — ксилоризом,
2 — стволики, 3 — парциальный куст, 4 — ксилоподий)
80
Кущение (Fruticatio, от лат. frutico — пускать ветви, давать побеги,
куститься; англ.: tillering, branching р. р.) — интенсивное ветвле-
ние побега в области его нижних (приземных или подземных),
укороченных междоузлий (см. зона кущения) с образованием во-
круг него компактной группы («куста») из нескольких побегов бо-
ковых, наиболее широко распространено у злаков и осок (см. дер-
новина); иногда К. рассматривают как ветвление базитонное одно-
летников или еще в более узком смысле — злаков в первый год их
жизни (фаза кущения); в самом широком значении К.— образо-
вание «куста» из побегов ветвления; син.: ветвление концентриро-
ванное, ветвление кустовое; лит.: Смелов, 1958; Серебрякова,
1962, 1969а; Нухимовский, 1974.
Кущение вторичное см. кущение постгенеративное.
Кущение интравагинальное см. ветвление интравагинальное.
Кущение первичное см. кущение прегенеративное.
Кущение постгенеративное (Fruticatio postgenerativa, от лат. post —
после, generare — рождать, производить) — ветвление побега злака
после отмирания его однолетней флоральной части (соломины);
син.: К. вторичное; терм.: Серебрякова, 1971а.
Кущение прегенеративное (Fruticatio praegenerativa, от лат. ргае —
впереди; generare — рождать, производить) — ветвление побега
злака до начала вытягивания первых длинных междоузлий его со-
ломины; син.: К. первичное; терм.: Серебрякова, 1971а.
Кущение экстравагинальное см. ветвление экстравагинальное.
Лейкоэмбриофит(ы) — назв. предложено М. С. Яковлевым и
Г. Я. Жуковой (1973) для обозначения раст., ткани зародыша кото-
рых не содержат хлорофилла (бесцветный зародыш); напр. пред-
ставители Boraginaceae, Caprif oliaceae, Ranunculaceae, Saxif raga-
ceae; ср. хлороэмбрифит.
Лиана (Liana, от лат. ligo — связывать; англ.: liana, liane, climber, vi-
ne) — раст. лазящее с длинными побегами, не способными само-
стоятельно сохранять вертикальное положение и использующи-
ми в качестве опоры другие раст., скалы, постройки и т. п.; стебли
большинства Л. растут быстро в длину и незначительно в шири-
ну, имеют сильно вытянутые междоузлия и значительную протя-
женность зоны роста; проводящие пучки в стебле изолированы
друг от друга паренхимой, что обеспечивает его гибкость и проч-
ность; среди Л. есть раст. деревянистые и раст. травянистые, од-
нолетники и многолетники, эпифиты и раст. паразиты; по спо-
собу освоения опоры различают (рис. 61): Л. опирающаяся, Л. кор-
81
Рис. 61. Способы освоения опоры лианами: 1 — цепляющиеся корни Hedera helix (по:
Федоров и др., 1962), 2 — закручивающиеся черешки Clematis jacktnanii, 3 — обвива-
ние опоры усиком Vitis atnurensis, 4 — вьющийся стебель Aristolochia manshurerisis,
5 -— прикрепление к опоре с помощью шипообразных чешуи почек Celastrus flagellaris
(по: Головач, 1973)
82
нелазящая, Л. вьющаяся, Л. усиконосная; некоторые Л. обладают
несколькими способами лазания (Celastrus flagellaris); иногда к Л.
относят только лазящие раст. древесные; ок. 4/s всех Л. сосредото-
чено в тропиках; лит.: Дарвин, 1941; Серебряков, 1962; Головач,
1973; Dawson, 1986.
Лиана вьющаяся (Liana volubilis, от лат. volvo — вращать, кру-
тить; англ.: voluble liana; twining climber) — раст. лазящее, побеги
которого в результате нутации обвивают опору по винтовой ли-
нии (рис. 61); среди всех Л. это наиболее обширная группа, к ко-
торой относятся виды родов Aristolochia, Schizandra, Humulus;
большинство Л. в. завиваются против часовой стрелки, реже осо-
би одного вида обвивают опору в разных направлениях (Solanum
dulcamara); син.: раст. вьющееся.
Лиана корнелазящая (Liana radicibus, от корень и лат. cibus — еда,
пища, корм; англ.: root climber) — раст. лазящее, закрепляющееся
на опоре с помощью корней цепляющихся (рис. 61), которые раз-
виваются на стебле обычно со стороны, обращенной к опоре и
проникают в ее неровности (Hedera helix); специфические выде-
ления корней некоторых Л. к. способствуют освоению ими также
опор с гладкой поверхностью (Ficus pumila).
Лиана опирающаяся (Liana suffulta, от лат. sub(suf) — приставка
со значением «под», fulcio — подпирать; англ.: scrambler, hook
climber) — раст., не имеющее специальных приспособлений для
лазания (корней цепляющихся, вьющихся стеблей, усиков), длин-
ные побеги которого взбираются на опору, полегая (опираясь) на
нее; большей частью кустарники, реже раст. травянистые
(scrambler), стебли которых удерживаются на опоре с помощью
жестких волосков, шипиков, колючек (hook climber; рис. 61) или
черешков листьев и побегов ветвления, отстоящих часто под пря-
мым углом или загнутых книзу (виды р. Galium); к Л. о. относятся
ротанговые пальмы и ряд роз; некоторые кустарниковые Л. о. наз.
раст. ветвелазящими.
Лиана усиконосная (Liana cirrosa, от лат. cirrus — усик; англ.: ten-
dril climber) — раст. лазящее, закрепляющееся на опоре с помо-
щью чувствительных на прикосновение твердых предметов уси-
ков стеблевого или листового происхождения (см. гаптотро-
пизм); большая группа Л., к которой относятся виды родов Vitis,
Passiflora, Bryonia и др. (рис. 61); часто к Л. у. относят раст. листо-
лазящие (рис. 61).
Лигнификация см. одревеснение.
83
Линия эволюции форм жизненных восходящая см. направлен-
ность эволюции биоморфологической.
Линия эволюции форм жизненных нисходящая см. направ-
ленность эволюции биоморфологической.
Лист (Folium; англ.: leaf) — боковой придаток стебля, характери-
зующийся ростом детерминированным (редко р. недетерминиро-
ванным) и выполняющий функции фотосинтеза, транспирации и
газообмена; закладывается в результате деления клеток в опреде-
ленном участке периферической зоны апекса побега (см. примор-
дий), местоположение которого контролируется типом листорас-
положения раст.; взрослый Л. состоит из б. м. ясно различимых
частей (рис. 62): пластинки и основания Л. (Л. сидячий), которые
часто соединены черешком (Л. черешковый); различают Л. с не-
сколькими (Л. сложный) и с одной пластинкой (Л. простой), фор-
ма которой изменяется в связи с гетерофилией, положением Л. на
стебле (см. формация листьев), возрастом (см. серия листовая) и
видом раст. (см. метаморфоз Л.); по длительности жизни Л. (см.
листопад) выделяют раст. вечнозеленые, раст. зимнезеленые и
раст. листопадные, лит.: Серебряков, 1952; Федоров, Кирпични-
ков, Артюшенко, 1956; Ботаника..., 1988; Лотова, 2000.
Лист верховой (Folium superius, от лат. super — верхний) — Л.,
расположенный в верхней части побега или его прироста и по сво-
ей форме и размерам заметно отличающийся от Л. срединного;
Рис. 62. Основные части листа (по: Ботаника..., 1962):
А — черешковый лист, Б — сидячий лист, В — лист с прилистниками,
Г — лист с влагалищем, Д — сложный лист (1 — пластинка, 2 — основание,
3 — влагалище» 4 — прилистники, 5 — черешок, 6 — пазушная почка,
7 — интеркалярная меристема)
84
Рис. 63. Листовые серии и формации листьев (по: Ботаника..., 1988): 1-4 — листовая
серия у проростка земляники, 5-9 — форма верховых листьев у земляники, 10 — сре-
динные листья и верховой (а — покрывало) лист у белокрыльника, 11-14 — листовая
серия проростка гороха, 15-20 — листовая серия проростка ясеня, 21 — формации
листьев у ландыша (б — верховые, в —низовые или катафиллы, г — срединные
листья), 22 — низовые листья шиповника, 23 — почечные чешуи и срединный лист
орешника, 24 — низовые листья чемерицы (катафиллы)
85
обычно Л. в. наз. Л., расположенный около цветка или соцветия
(гипсофилл; англ.: hypsophyll): прицветник, прицветничек и т. п.
(рис. 63); см. формация листьев.
Лист влагалищный см. колеоптиль.
Лист детерминированный (англ.: determinate leaf) см. рост детер-
минированный.
Лист зародышевый см. семядоля.
Лист недетерминированный (англ.: indeterminate leaf) см. рост
недетерминированный.
Лист низовой (Folium basale, от лат. basis — основание, нижняя
часть) — Л., расположенный в основании побега (Л. нижний) или
его прироста и отличающийся небольшими размерами и редуци-
рованной листовой пластинкой (рис. 63); часто имеет вид чешуи
или пленки, лишенной зеленой окраски, и представляет собой ви-
доизмененное основание Л. (катафилл); у раст. травянистых Л.
н. иногда наз. прикорневым, см. формация листьев.
Лист первичный (англ.: initial leaf) см. примордий; иногда Л. п.
наз. первый настоящий Л., образующийся при прорастании семе-
ни вслед за семядолей(ями).
Лист поддерживающий см. лист подпирающий.
Лист подпирающий (Folium fulciens, от лат. fulcio — подпирать,
поддерживать; англ.: subtending leaf) -— Л., в пазухе которого на-
ходится почка пазушная; иногда Л. п. наз. Л. кроющим; син.: Л.
поддерживающий.
Лист прикорневой см. лист низовой.
Лист примордиальный см. примордий.
Лист семядольный см. семядоля.
Лист срединный (Folium caulinum, от лат. caulis — стебель; англ.:
foliage leaf) — Л., расположенный в средней части побега или его
прироста и отличающийся максимально развитой листовой пла-
стинкой (рис. 63; номофилл или настоящий ассимиляционный
лист); Л. с. часто наз. Л. стеблевым; см. формация листьев.
Лист стеблевой см. лист срединный.
Листец (Frons; англ.: frond) — назв., используемое для обозначения
вегетативного «тела» рясковых, не дифференцированного на сте-
бель и лист и внешне напоминающего слоевище низших раст.
(рис. 64); часто в том же смысле используют назв.: побег, лист, фил-
локладий, вайя, щиток, пластинка, листоветвь, фронд, турион.
Листоветвь см. листец.
Листоветка см. филлокладий.
86
6
Рис. 64. Строение расгения (А) и листеца (Б) многокоренника обыкновенного (по:
Hegelmaier, 1868): Л(1 — материнский листец, Л] — дочерний листец первой и второй
(Л2) генерации, 1 — кармашек, 2 — ряды вытянутых клеток о г узла к ножке, 3 — нож-
ка, 4 — придаточный корень, 5 — узел, 6 — жилки, 7 — проксимальный («стебле-
вой») участок, 8 — дистальный («листовой») участок
Листопад (Defoliatio, от лат. de — приставка со значением отделе-
ние, лишение и лист; англ.: def oliation, leaf fall, leaf abscis-
sion) — опадение листьев в связи с их старением; обычно подго-
тавливается цитологическими и биохимическими изменениями
клеток у основания черешка (зона отделения) и характерно для
деревьев и кустарников, редко — трав (Urtica); листья могут опа-
дать одновременно (раст. листопадное) в определенный период
года (сезонный Л.) или постепенно в течение длительного време-
ни (см. раст. вечнозеленое); в сезонном климате Л. рассматривает-
ся как приспособление к уменьшению транспирации в неблаго-
приятных условиях; иногда причиной Л. являются различные по-
вреждения; лит.: Серебряков, 1951; Одишария, 1974.
Листорасположение (Phyllotaxis; от греч. phyllon — лист, taxis —
расположение; англ.: phyllotaxis, phyllotaxy, leaf arrangement) —
порядок размещения листьев на стебле, отражающий радиаль-
ную симметрию в структуре побега-, определяется порядком зало-
жения примордиев на конусе нарастания побега и характеризуется
формулой Л.; различают Л. очередное, Л. супротивное и Л. мутов-
чатое (рис. 65), для описания и интерпретации которых исполь-
зуют различные понятия и гипотезы (см. теория К. Шимпера и
А. Брауна, теория К. Уордлоу, теория Л. Плантефоля); нередко
Л. на стебле может отличаться от Л. в почке, изменяться в онтоге-
незе или под воздействием внешних факторов; син.: филлотаксис;
лит.: Житков, 1983, 1988; Croizat, 1971; Schwabe, 1984; Kunz, 2001.
87
Листорасположение двурядно-
супротивное (англ.: opposite
decussate leaf arrangement) см.
листорасположение супротив-
ное.
Листорасположение ложно-му-
товчатое (англ.: pseudowhorled
leaf arrangement, false whorl leaf
arrangement) см. листорасполо-
Рис. 65. Листорасположение:
А — очередное (спиральное),
Б — супротивное, В — накрест
супротивное, Г — мутовчатое
жение мутовчатое.
Листорасположение мутовча-
тое (Phyllotaxis verticillata, от
лат. verticillus — мутовка; англ.:
(по: Лотова, 2000)
whorled leaf arrangement, verti-
cillate leaf arrangement) — Л.,
при котором от каждого узла побега отходит три и более листа
(рис. 65); возникает при б. м. одновременном закладывании на
одном уровне конуса нарастания нескольких примордиев, обра
зуюгцих общий узел; примордии следующей мутовки обычно за-
кладываются над промежутками между примордиями предыду-
щей мутовки; у некоторых раст. (Peperonia) Л. м. образуется в ре-
зультате недоразвития междоузлий при Л. очередном или Л.
супротивном (т. наз. ложномутовчатое Л.); часто Л. супротивное
рассматривается как частный случай Л. м.
Листорасположение накрест-супротивное (англ.: opposite de-
cussate leaf arrangement) см. листорасположение супротивное.
Листорасположение очередное (Phyllotaxis alterna, от лат. alter
nus — попеременный; англ.: alternate leaf arrangement) — Л., при
котором от каждого узла побега отходит один лист (рис. 65); воз-
никает при закладывании примордиев по одному последовательно
на разных уровнях и сторонах конуса нарастания, гак что основа-
ния примордиев одного цикла листового полностью покрывают
окружность стебля; в зависимости от угла дивергенции различают
Л. двурядное или двурядно-очередное (Iris; Роасеае), трехрядное
(Carex) и т. д. (соотвественно англ.: distichous, tristichous leaf
arrangement); син.: Л. спиральное, Л. рассеянное.
Листорасположение перекрестно-супротивное см. листораспо-
ложение супротивное.
Листорасположение рассеянное см. листорасположение оче-
редное.
88
Листорасположение спиральное см. листорасположение оче-
редное.
Листорасположение супротивное (Phyllotaxis opposita, от лат.
орропеге — противопоставлять, ставить против; англ.: opposite
leaf arrangement) — Л., при котором от каждого узла побега отхо-
дят два листа, расположенных супротивно (рис. 65); соседние па-
ры листьев могут быть расположены строго друг над другом
(Л. двурядно-супротивное) или б. м. под прямым углом относи-
тельно друг друга (Л. перекрестно- и накрест-супротивное); часто
Л. с. рассматривают как частный случай Л. мутовчатого-, лит.: Бо-
таника..., 1988; Лотова, 2000.
Листосложение (англ.: ptyxis) см. почка.
Листосмыкание (vernation, prefoliation) см. почка.
Ложнодву(семя)дольность см. псевдодикотилия.
Ложноодно(семя)дольность см. псевдомонокотилия.
Луковица (Bulbus; англ.: bulb) — видоизмененная почка (метамор-
фоз) или видоизмененный подземный стебель дисковидной фор-
мы (донце) из-за сильно укороченных междоузлий, несущий плот-
но расположенные друг к другу сочные чешуи, служащие для на-
копления воды и питательных веществ (ср. клубнелуковица);
встречается у однодольных и преимущественно среди Liliaceae и
Amaryllidaceae (см. раст. луковичное); Л. может быть образована
чешуями разного происхождения: видоизмененными листьями
низовыми (Tulipa), основаниями листьев (Allium ursinum) или, ча-
ще всего, сочетанием тех и других (Galanthus nivalis); в некоторых
Л. выделяют чешую влагалищную и покровную; Л. различают
(рис. 66) по форме, количеству чешуй, длительности жизни
(Л. однолетняя, Л. многолетняя), строению и расположению че-
шуй (Л. полутуникатная, Л. туникатная, Л. черепитчатая), раз-
витию почек пазушных (Л. простая, Л. сложная)-, лит.: Серебряков,
1952; Родионенко, 1961; Федоров и др., 1962; Артюшенко, 1970;
Байтулин, Рахимбаев, Каменецкая, 1986; Баранова, 1999, 2000.
Луковица воздушная см. луковичка.
Луковица замещающая (Bulbus succedaneus, от лат. succedo — за-
мещать, сменять; англ.: replacement bulb) — Л., образующаяся из
почки возобновления и замещающая отмирающую материнскую
Л. (рис. 32, 66); обеспечивает ежегодное возобновление раст. с Л.
однолетней (Tulipa).
Луковица имбрикатная (англ.: imbricated bulb) см. луковица чере-
питчатая.
89
Луковица кожистая см. клубнелуковица.
Луковица концентрическая см. луковица туникатная.
Луковица ложная — назв. используется для обозначения утол-
щенной за счет плотного расположения оснований ассимили-
рующих листьев подземной части стебля некоторых корневищ-
ных луков; в отличие от настоящей Л. такое образование не имеет
сочных, запасающих чешуи (Allium fistulosum).
Луковица многолетняя (Bulbus perennis, от лат. per— приставка
со значением усиления действия, направленного через что-либо,
annus — год; англ.: perennial bulb) — Л., длительность жизни ко-
торой превышает один год (Narcissus); терм, используется для
противопоставления Л. однолетней (рис. 66); у Л. м. ежегодно
происходит полное или частичное обновление чешуй, при этом
отмирает и нижняя часть донца; реже (Ungernia) освобождающая-
ся от чешуй часть донца отмирает не сразу, в результате чего об-
разуется короткое корневище (раст. корневищно-луковичное).
Луковица однолетняя (Bulbus annus, от лат. annus — год; англ.:
annual bulb) — Л., длительность жизни которой не превышает
одного года (рис. 66); у раст. (Tulipa) с Л. о. ежегодно в конце веге-
тационного периода из почки возобновления образуется Л. заме-
щающая.
Луковица пленчатая см. луковица туникатная.
Луковица полутуникатная (Bulbus semitunicatus, от лат. semi —
полу, пол, tunica — оболочка, корка) — Л. с широкими чешуями,
каждая из которых не полностью охватывает (покрывает) сле-
дующую за ней; ср. Л. туникатная.
Луковица простая (Bulbus simplex) — Л., донце которой не несет
сильно разросшихся почек пазушных (деток); терм, используется
для противопоставления Л. сложной.
Луковица сложная (Bulbus compositus) — Л., донце которой несет
сильно разросшиеся пазушные почки (детки), одинакового раз-
мера с основной (материнской) Л. (Allium sativum).
Луковица туникатная (Bulbus tunicatus, от лат. tunica — оболоч-
ка, корка; англ.: tunicated bulb) —Л. с широкими чешуями, каж-
дая из которых, срастаясь (соприкасаясь) своими краями, полно-
стью охватывает (покрывает) расположенную под ней (Allium се-
ра); Л. концентрическая, Л. пленчатая.
Луковица черепитчатая (Bulbus imbricatus, от лат. imbrex — че-
репица; англ.: imbricated bulb) — Л., узкие чешуи которой не при-
крывают рядом расположенные чешуи, а только краями соприка-
91
«е
о
й?
Рис. 66. Основные типы луковиц (по: Федоров и др., 1962): А — многолетняя моноподиально нарастающая, Б — много-
летняя симподиально нарастающая, В — однолетняя (1 — донце, 1 а — отмершая часть донца, 2 — чешуи, 3 — отмершие
чешуи, 4 — цветонос, 5 — цветоносы прошлых лет, 6 — зачатки листьев, 7 — зачаток цветка, 8 — почка возобновления,
9 — придаточные корни, 10 — замещающая луковица, 1998-2000 — годичные приросты донца)
саются с ними (Lilium kesselringianum); син.: Л. чешуйчатая, Л.
имбрикатная.
Луковица чешуйчатая см. луковица черепитчатая.
Луковичка (Bulbillus, от луковица-, англ.: bulbil, bulbel, brood-
bud) — почка пазушная в области соцветия, с утолщенными, мя-
систыми листочками (рис. 15,67); обеспечивает размножение веге-
тативное (см. вивипария) некоторых раст. травянистых (Fes-
tuca vivipara); иногда Л. наз. клубеньком или почкой выводковой;
син.: луковичка выводковая, луковица воздушная, бульбилла;
лит.: Баранова, 2000.
КН
Рис. 67. Видоизменения почки (по: Ботаника..1988): 1 — пазушная почка, 2 — луко-
вичка, 3 — клубенек, 4 — клубень почко-корневой, 5-7 — развитие кочана капус-
ты (а — проросток, 6 — сформированный кочан, в -— верхушечная почка после
перезимовки), к — придаточный корень, кн — конус нарастания, оп — ось побега,
с — семядоли)
92
Луковичка выводковая см. луковичка.
Макробласт (англ.: macroblast) см. побег удлиненный.
Малолетник — пон. обычно используется для обозначения моно-
карпиков, длительность жизни которых не превышает одного
(раст. однолетнее) или двух лет (раст. двулетнее). В более широ-
ком смысле к М. относят монокарпики и поликарпики (олигокарпи-
ки) с продолжительностью онтогенеза до 5-15 лет. Разнообразная
группа преимущественно растений травянистых (редко растения
дерявянистые), которые отличаются формой жизненной, выражен-
ностью старения (см. старение монокарпическое, старение нега-
тивное) и размножением. В последнем случае выделяют М. генера-
тивные, которые размножаются только семенами (англ.: short-lived
plants), и малолетники вегетативные, лит.: Марков, 1990.
Малолетник вегетативный—раст. травянистое, с размножением
вегетативным и однолетними (однолетник вегетативный; англ.:
pseudoannual) или малолетними (2—3 года) дочерними особями
(рис. 114); в более широком смысле к М. в. относят не только раст.
с малолетними раметами, образующимися в результате специа-
лизированного (Trientalis) или неспециализированного (Sax fra-
ga) вегетативного размножения, но и раст. кистекорневые с не-
долговечными резидами (Ranunculus) и двулетники замещающие-
ся-, терм.: Высоцкий, 1915; лит.: Любарский, 1961.
Малолетник генеративный см. малолетник.
Мегафанерофит (англ/, megaphanerophyte) см. фанерофит.
Междоузлие (Internodium, от лат. inter — между, nodus — узел;
англ.: internode) — участок стебля между двумя узлами побега; по
соотношению длины (Д) и ширины (Ш) различают М. укорочен-
ное (Д < Ш; англ.: short internode) и М. удлиненное (Д > Ш; англ/,
long internode), а по положению на побеге — гипоподий, мезопо-
дий, эпиподий (рис. 31) и М. субфлоральное, которое у нек-рых
раст. наз. стрелкой.
Междоузлие удлиненное (англ.: long internode, elongated interno-
de) см. междоузлие.
Междоузлие укороченное (англ.: short internode) см. междоузлие.
Мезант — мало используемое пон. для обозначения раст., цветущих
в середине периода своей вегетации; терм.: Алехин, 1925; ср. про-
ант, метант.
Мезокотиль (Mesocotylus, от греч. mesos — середина и kotyle — до-
ля; англ.: mesocotyle) — участок стебля проростка злаков, распо-
ложенный между щитком и колеоптилем и образующийся в ре-
93
зультате растягивания (роста интеркалярного) узла семядольного
при прорастании семени; обеспечивает продвижение почечки к
поверхности почвы; нек-рые рассматривают образование М. как
результат конгенитального срастания шейки семядоли с гипоко-
тилем; иногда М. обозначают среднюю часть семядоли однодоль-
ных (шейку семядольную) или межсемядольное междоузлие, фор-
мирующееся у нек-рых двудольных (Aeginetia indica); терм.: Cela-
kovsky, 1897; лит.: Лодкина, 1988.
Мезоподий (Mesopodium, от греч. mesos — средний, промежуточ-
ный, pous — нога, здесь ось; англ.: mesopodium) — междоузлие по-
бега бокового, ограниченное узлами предлистьев (рис. 31).
Мезотония (Mesotonia, от греч. mesos — средний, tonos — сила,
мощь; англ.: mesotony) — тип продольной симметрии побега
(или его прироста), выражающийся в изменении длины междоуз-
лий, размеров почек или побегов ветвления таким образом, что
наиболее крупные из них приурочены к средней части стебля
(рис. 1); мезотонное распределение побегов боковых (ветвление ме-
зотонное) встречается у нек-рых травянистых многолетников;
лит.: Серебрякова, 1971а.
Мезофанерофит (англ.: mesophanerophyte) см. фанерофит.
Меристема (Meristema, от греч. meristos — делимый; англ.: meris-
tem) — образовательная ткань раст., долго сохраняющая способ-
ность к делению и образованию новых клеток; по положению
различают М. апикальную, М. интеркалярную и М. латеральную
(прокамбий, камбий, перицикл, феллоген); лит.: Ботаника...,
1988; Лотова, 2000.
Меристема апикальная (англ.: apical meristem) см. апекс.
Меристема интеркалярная (Meristema intercalare, от лат. inter-
calo — вводить, вставлять, добавлять; англ.: intercalary meristem) —
меристематический участок, отделившийся от М. апикальной и от-
граниченный от нее дифференцированными тканями; обычно рас-
полагается в основании растущего листа, у основания междоузлия
и функционирует непродолжительное время.
Метамер (Metamerus, от греч. meta — между, после, через, meros —
часть, доля; англ.: metamere) — участок побега, включающий узел
с листом (листьями), почкой пазушной и нижележащее междоуз-
лие (элементарный М.; рис. 97); у нек-рых раст. (Роасеае) почка
располагается на нижнем конце междоузлия (почка внепазуш-
ная); в более широком смысле М. наз. любые повторяющиеся эле-
менты структуры раст. (см. метамерность): участок побега, огра-
94
ниченный одним циклом листовым, побег генеративный (см. мо-
дуль), систему побегов, куст парциальный и т. п.: син.: фитомер;
лит.: Серебряков, 1952; Житков, 1976; Шафранова, 1980; Барыки-
на, Буленкова, 1983; Цвелев, 2000.
Метамер элементарный см. метамер.
Метамерность (англ.: metamerism) — морфологическое отображе-
ние ритмичности роста и формообразования раст., проявляю-
щееся в повторяемости элементов структуры вдоль оси побега
(метамеров элементарных), а в более широком значении и в сис-
теме побегов (метамеры разных уровней от побега до куста парци-
ального)', ср. модулярность; лит.: Шафранова, 1980; White, 1984.
Метаморфоз корня (от греч. metamorphosis — превращение;
англ.: root (modification) metamorphosis) — изменение внешнего
облика и внутреннего строения корня, вызванное сменой выпол-
няемых им функций: симбиоз (см. клубенек корневой, корень ко-
раллоидный), отложение запасных веществ (см. К. запасающий),
защита (см. колючка), питание эктопаразита (см. гаустория); ино-
гда М. к. наз. также видоизменения К., связанные с выполнением
им дополнительных функций: фотосинтеза (см. К. ассимилирую-
щий), газообмена (см. пневматофор, К. воздушный, К. досковид-
ный), дополнительной опоры (см. К. опорный, К. цепляющийся),
микоризы и др.; лит.: Федоров, Кирпичников, Артюшенко, 1962;
Михайловская, 1981а; Ботаника..., 1988; Лотова, 2000.
Метаморфоз листа (от греч. metamorphosis — превращение; англ.:
leaf (modification) metamorphosis) — изменение внешнего облика
и внутреннего строения листа, вызванное сменой выполняемых
им функций (см. усики, колючки, катафиллы); иногда М. л. наз.
гетерофилию, видоизменения, обусловленные развитием гете-
робластным или выполнением дополнительной функции (напр.
асцидии некоторых раст. насекомоядных); лит.: Лотова, 2000.
Метаморфоз побега (от греч. metamorphosis — превращение; англ.:
shoot (modification) metamorphosis) — изменение внешнего обли-
ка и внутреннего строения побега, вызванное сменой выполняе-
мых им функций (см. колючка, усики, кладодий, филлокладий
и др.); М. п. может происходить на разных этапах его онтогенеза
(см. онтогенез побега), в связи с этим выделяют метаморфоз почки;
часто М. п. наз. видоизменения побега, связанные с выполнением
дополнительных функций (см. столон, корневище, клубень, клуб-
нелуковица, луковица и др.); лит.: Серебрякова, 1984; Ботаника...,
1988; Барыкина, Буленкова, 1987, 1990; Лотова, 2000.
95
Метаморфоз почки (англ.: bud (modification) metamorphosis) —
изменение внешнего облика и внутреннего строения почки, вы-
званное сменой выполняемых функций (см. кочан, луковица);
часто видоизмененные почки являются органами размножения ве-
гетативного-, при этом по их строению различают луковички, клу-
беньки и клубни почко-корневые (рис. 67); лит.: Ботаника..., 1988;
Барыкина, Туленкова, 1987, 1990.
Метант — мало используемое пон. для обозначения раст., цветущих
в самом конце своей вегетации (Allium); терм.: Алехин, 1925; ср.
проант, мезант.
Метод морфогенетических рядов — наиболее широко распро-
страненный подход реконструкции эволюции биоморфологиче-
ской, предполагающий сравнительный анализ развития форм
жизненных современных родственных видов (Серебряков, Сереб-
рякова, 1969). Проецирование результатов сравнительного ана-
лиза на филогенетические построения позволяет сформулиро-
вать кодекс признаков, реконструировать габитус предка и опре-
делить основные направления эволюции. Для объективизации
предположений обычно используют дополнительные данные по
географии, экологии и хромосомным числам современных ви-
дов; лит.: Серебряков, 1964; Голубев, 1965; Серебряков, Серебря-
кова, 1969; Серебрякова, 1972; Гатцук и др., 1974; Барыкина, 1979;
Савиных, 1979.
Метод описания архитектурных моделей (англ.: method of ar-
chitectural models description, architectural analysis) — разработан-
ный на примере тропических деревьев метод описания моделей
архитектурныхраст. древесных; М. о. а. м. предполагает изучение
деятельности конуса нарастания побега при формировании сис-
темы побегов в онтогенезе раст.; рост и форма побеговой системы
анализируется с использованием трех архитектурных элементов:
единица морфогенеза, модуль и модель архитектурная; архитек-
турные модели выделяют на основании нескольких признаков:
длительность функционирования и специализация меристем
апикальных (рост детерминированный или рост недетерминиро-
ванный), характер ветвления и нарастания (нарастаниемоноподи-
альное и н. симподиальное), направление роста (рост ортотроп-
ный или р. плагиотропный) и ритмичность роста; для раст. тра-
вянистых сходный подход предполагает описание моделей
побегообразования; лит.: Halle, Oldeman, Tomlinson, 1978; Антоно-
ва, Азова, 1999.
96
Метод описания форм жизненных по Е. С. Смирновой (1990) —
разработанный для практических целей метод выделения «морфо-
логических типов» орхидных на основе морфологического кода,
формула которого включает основные параметры формы роста
(наличие корневища, клубня, клубнелуковицы, направление роста
побега и характер его облиствления) и системы побегов (ветвление и
специализация побегов); различия форм жизненных генеративных
раст. определяются по таблице, в левой части которой располага-
ются виды, а в правой — числовые обозначения морфологическо-
го кода и состава серии листовой (число членов в формации листьев
и длина междоузлия).
Метод описания форм жизненных по И. Г. Серебрякову
(1964а) — широко используемый метод, основанный на изуче-
нии морфогенеза вегетативных органов на протяжении онтогене-
за раст. при непосредственном наблюдении за развитием модель-
ных особей, выращенных из семян, или на основании реконст-
рукции по ряду разновозрастных особей; различия форм
жизненных определяются у взрослых особей по структуре осей
скелетных (раст. древесные, раст. полудревесные, раст. травяни-
стые), длительности жизни (многолетники, однолетники), харак-
теру системы корневой, структуры побега и др.
Метод описания форм жизненных по Л. Е. Гатцук (1974а, б) —
один из возможных методов анализа внешнего строения форм
жизненных, основанный на выделении соподчиненных единиц
структуры (метамеров последовательно возрастающего порядка)
в побеговой системе раст. геммаксиллярных: метамера, побега эле-
ментарного, побега моноподиального (побега скелетного и побега
обогащения), системы побега скелетного, системы побеговмонопо-
диальных (оси скелетной главной, о. с. закрепления и возобновле-
ния), куста первичного, куста замещения и куста парциального;
различия между жизненными формами определяются по типу
метамерных образований, создающих системы побегов раст., ха-
рактеру их «сочленения» и положению последнего относительно
почвы или другого субстрата.
Метод описания форм жизненных по М. Т. Мазуренко и
А. П. Хохрякову (1977) — один из методов анализа строения и
развития форм жизненных (преимущественно раст. деревяни-
стых), основанный на выделении структур, выполняющих глав-
ную роль в формировании габитуса (рис. 68): побега (побег пер-
вичный, п. ветвления, п. формирования, п. дополнения и др.) и сис-
97
Рис. 68. Основные типы побегов и их систем в строение деревянистого
растения: 1 — первичный побег, 2 — побег ветвления, 3 — побег дополнения,
4 — побег формирования, 5 — система первичного побега, 6 — система
побега дополнения, 7 — система побега формирования
98
Рис. 69. Коралловидные
корни сосны (по: Сереб-
ряков, 1952)
темы побега (СПП, СПВ, СПФ, СПД); при этом онтогенез раст.
рассматривается как чередование циклов развития соподчинен-
ных систем побегов, среди которых выделяют основной цикл; лит.:
Мазуренко, 1986.
Микориза (Mycorrhiza, от греч. mykes — гриб,
rhiza — корень; англ.: mycor(r)hiza) — сим-
биотическая система мицелия гриба и корня
раст.; различают два основных типа: экто-
микоризу, когда гриб покрывает кончик
корня плотным чехлом (гифальной манти-
ей), и эндомикоризу, когда гриб внедряется
внутрь клеток паренхимы; эктомикоризные
корни короткие (ср. корень кластерный),
разбухшие, разветвленные и иногда имеют
вид коралловидного образования (рис. 69;
корень коралловидный); син.: грибокорень.
Микоризом(а) (Mycorrhizoma, от греч. mykes — гриб и корневище;
англ.: mycorhizome) — метаморфизированный в результате про-
никновения гиф микоризного гриба подземный стебель гл. обр.
сапротрофных орхидных; М.
развивается из апикальной
зоны протокорма в виде мя-
систого, нередко разветвлен-
ного образования; из мери-
стематических центров (или
центра) последнего образу-
ются надземные побеги или
последующие членики М.
(рис. 70); иногда М. наз. кор-
невище папоротников с эн-
дотрофной микоризой.
Микрофанерофит (англ.: microphanerophyte) см. фанерофит.
Миниатюризация см. нанизм.
Многовариантность развития см. поливариантность развития.
Многолетник см. растение многолетнее.
Многосемядольность см. поликотилия.
Модель архитектурная (англ.: architectural model) — видимое вы-
ражение генетической программы развития растения, проявляю-
щееся во взаимном расположении модулей (побегов) в пределах
общей «конструкции» взрослого раст.; выделенные 24 М. а. (на-
Рис. 70. Формирование микоризома
Orchis ustulata (по: Burgeff, 1936)
99
званы по имени выдающихся ботаников) можно объединить в
3 основных группы: модель модулярная (М. a. Chamberlain, М. а
Holtum, М. a. Koriba, М. a. Leeuwenberg, М. a. Prevost, и М. a. Tom-
linson), м. частично модулярная (см. модель модулярная) и м. не-
модулярная; терм.: Halle, Oldeman, 1970; лит.: Halle, Oldeman,
Tomlinson, 1978; Kull, 1987.
Модель архитектурная Attims (англ.: Attims model, названа в честь
Y. Attims) — модель дерева одноствольного, кронообразующего, по-
ликарпического с пазушными соцветиями на ветвях кроны; фор-
мируется аналогично М. a. Rauh, отличаясь постоянным (нерит-
мичным) ростом ствола и ветвей кроны (рис. 71); представители
сем. Avicenniaceae (виды р. Avicennia), Betulaceae (Alnus incana),
Euphorbiaceae (Phyllanthus polygonoides), Rhizophoraceae (виды
p. Rhizophora) и др., среди которых много деревьев с корнями опор-
ными; лит.: Halle, Oldeman, Tomlinson, 1978.
Рис. 71. Архитектурная модель Attims (по: Halle et al., 1978)
Модель архитектурная Aubreville (англ.: Aubreville’s model, на-
звана в честь A. Aubreville) — модель дерева одноствольного, кро-
нообразующего, поликарпического с пазушными соцветиями на
ветвях кроны; формируется аналогично М. a. Fagerlind, отличаясь
ростом недетерминированным побегов ветвления (рис. 72); в ос-
новном представители сем. Combretaceae, Euphorbiaceae и
Sapotaceae; лит.: Halle, Oldeman, Tomlinson, 1978.
100
Рис. 72. Архитектурная модель Aubrcville (по: Halle et al., 1978)
Модель архитектурная Chamberlain (англ.: Chamberlain’s model,
названа в честь С. J. Chamberlain) — модель дерева одноствольно-
го, розеточного, поликарпического с терминальными соцветиями;
формируется в результате ветвления акросимподиального и обра-
зования одинаковых побегов замещения с ростом ортотропным
(рис. 73); древовидные однодольные (Agavaceae, Агасеае и др.),
двудольные (Apocynaceae, Araliaceae, Bignoniaceae и др.); лит.:
Halle, Oldeman, Tomlinson, 1978.
101
Модель архитектурная Champagnat (англ.: Champagnat’s model,
названа в честь Р. Champagnat) — модель дерева одноствольного,
кронообразующего, поликарпического; формируется аналогично
М. a. Mangenot, но изменение ориентации побега замещения обу-
словлено не ростом, а собственной массой (рис. 74). Представите-
ли сем. Boraginaceae (виды р. Cordia), Caprifoliaceae (Sambucus
nigra), Rosaceae (Rosa canina, Rubus idaeus) и др.; лит.: Halle,
Oldeman, Tomlinson, 1978.
Рис. 74. Архитектурная модель Champagnat (no: Halle et al., 1978)
Модель архитектурная Cook (англ.: Cook’s model, названа в честь
О. F. Cook) — модель дерева одноствольного, кронообразующего,
поликарпического с пазушными или терминальными соцветиями
на ветвях кроны; формируется в результате непрерывного нарас-
тания моноподиального ствола и постоянного опадения нижних
ветвей кроны (рис. 75); некоторые двудольные из сем. Euph-
orbiaceae (Phyllanthus urinaria), Moraceae (Castilla elastica), Rubia-
ceae (Canthium glabriflorum) и др.; лит.: Halle, Oldeman, Tomlinson,
1978.
Рис. 75. Архитектурная модель Cook (no: Halle et al., 1978)
102
Модель архитектурная Corner (англ.: Corner’s model, названа в
честь Е. J. Н. Corner) — модель дерева одноствольного, розеточно-
го, поликарпического с пазушными соцветиями; формируется в
результате нарастания моноподиалъного и роста недетерминиро-
ванного (рис. 76); неветвящиеся (исключая соцветие) растения
древовидные из сем. Palmae (Cocos nucifera), Compositae (виды
p. Espeletia), Euphorbiaceae (виды p. Euphorbia), Sterculiaceae (виды
p. Cola) и др.; среди растений травянистых аналогично развива-
ются некоторые монокарпики; лит.: Halle, Oldeman, Tomlinson,
1978.
Рис. 76. Архитектурная модель Corner:
1 — рост постоянный, 2 — рост ритмичный
(по: Halle et al., 1978)
103
Модель архитектурная Fagerlind (англ.: Fagerlind’s model, названа
в честь F. Fagerlind) — модель дерева одноствольного, кронообра-
зующего, поликарпического с терминальными соцветиями на вет-
вях кроны', формируется в результате нарастания моноподиального
ствола и симподиального ветвей кроны, которые из-за перевершини-
вания занимают плагиотропное положение (рис. 77); в отличие от
М. a. Petit нарастание ствола характеризуется ритмичным ростом-,
только двудольные из сем. Bombacaceae, Magnoliaceae, Rosaceae,
Rubiaceae и др.; лит.: Halle, Oldeman, Tomlinson, 1978.
Рис. 77. Архитектурная модель Fagerlind (по: Halle et al., 1978)
Модель архитектурная Holtum (англ.: Hokum’s model, названа в
честь R. Е. Holtum) — модель дерева одноствольного, розеточного,
монокарпического с терминальным соцветием; характеризуется
нарастанием моноподиальным и ростом детерминированным (рис.
78); неветвягциеся (исключая соцветие) растения древовидные из
сем. Palmae (Corypha elata), Boraginaceae (виды p. Echium), Compo-
sitae (Wilkesia gymnoxiphium), Crassulaceae (виды p. Aeonium),
Lobeliaceae (виды p. Lobelia) и др.; среди растений травянистых
аналогично развиваются многие монокарпики; терм.: Halle, Olde-
man, 1970; лит.: Halle, Oldeman, Tomlinson, 1978.
Рис. 78. Архитектурная модель Holtum (no: Halle et al., 1978)
104
Модель архитектурная Koriba (англ.: Koriba’s model, названа в
честь Kwan Koriba) — модель дерева одноствольного, кронообра-
зующего, поликарпического с терминальными соцветиями; фор-
мируется в результате ветвления акросимподиального и образова-
ния одинаковых побегов клиноапогеотропных (рис. 79); в отличие
от М. a. Leeuwenbergодин из побегов ветвления позднее становится
сильнее, переходит к росту апогеотропному и продолжает нарас-
тание ствола или ветвей {побег замещения); только двудольные из
сем. Apocynaceae, Bignoniaceae, Euphorbiaceae и др.; лит.: Halle,
Oldeman, Tomlinson, 1978.
Рис. 79. Архитектурная модель Koriba (по: Halle et al., 1978)
Модель архитектурная Leeuwenberg (англ.: Leeuwenberg’s mo-
del, названа в честь A. J. М. Leeuwenberg) — модель дерева одно-
ствольного, кронообразующего, поликарпического с терминальны-
ми соцветиями; формируется в результате ветвления акросимпо-
диального и образования одинаковых побегов замещения с ростом
ортотропным (рис. 80); в отличие от М. a. Chamberlain после цве-
тения каждый побег образует более одного побега ветвления; в ос-
новном двудольные (Apocynaceae, Araliaceae, Compositae, Eupho-
rbiaceae и др.), реже однодольные (Agavaceae, Pandanaceae и др.)
растения древовидные, травянистые и древесные; терм.: Halle,
Oldeman, 1970; лит.: Halle, Oldeman, Tomlinson, 1978.
Рис. 80. Архитектурная модель Leeuwenberg (по: Halle et al., 1978)
105
Модель архитектурная Mangenot (англ.: Mangenot’s model, назва-
на в честь G. Mangenot) — модель дерева одноствольного, кронооб-
разующего, поликарпического-, формируется в результате нараста-
ния мезосимподиального ствола и изменения направления роста
побега замещения с ортотропного на плагиотропное (рис. 81); пред-
ставители сем. Ericaceae (Vaccinium corymbosum), Melastomataceae
(виды p. Mouriri), Thymelaeaceae (виды p. Dicranolepsis) и др.; лит.:
Halle, Oldeman, Tomlinson, 1978.
Модель архитектурная Massart (англ.: Massart’s model, названа в
честь J. Massart) — модель дерева одноствольного, кронообразую-
щего, поликарпического с этажированным (симметричным) рас-
положением ветвей кроны, которые несут как правило пазушные
соцветия; формируется в результате нарастания моноподиального
ствола и побегов ветвления, которые характеризуются ростом пла-
гиотропным (рис. 82); отличается от М. a. Roux ритмичным рос-
том ствола; голосеменные (Araucaria columnaris, Abies alba, Taxus
baccata и др.) и двудольные из сем. Ebenaceae, Myristicaceae,
Rhizophoraceae и др.; лит.: Halle, Oldeman, Tomlinson, 1978.
Рис. 82. Архитектурная модель Massart (по: Halle et al., 1978)
106
Модель архитектурная McClure (англ.: McClure’s model, названа
в честь F. A. McClure) — модель дерева гбуртинообразующего, мо-
нокарпическокого или поликарпическокого; формируется в резуль-
тате ветвления базисимподиального и образования двух типов по-
бегов (рис. 83); оси скелетные (стволики) развиваются как побеги
анизотропные с плагиотропной подземной частью и ортотроп-
ной надземной, несущей олиствленные и генеративныве побеги
боковые; в основном древовидные однодольные и растения травя-
нистые (Costaceae, Bambusoideae, Liliaceae, Smilacaceae и др.); лит.:
Halle, Oldeman, Tomlinson, 1978.
Модель архитектурная Nozeran (англ.: Nozeran’s model, названа
в честь R. Nozeran) — модель дерева одноствольного, кронообра-
зующего, поликарпического с терминальными соцветиями; фор-
мируется аналогично М. a. Prevost, отличаясь ростом плагиотроп-
ным побегов, образующих крону (рис. 84); в основном представи-
тели сем. Euphorbiaceae и Sterculiaceae; лит.: Halle, Oldeman,
Tomlinson, 1978.
Рис. 84. Архитектурная модель Nozeran (по: Halle et al., 1978)
107
Модель архитектурная Petit (англ.: Petit’s model, названа в честь
Е. Petit) — модель дерева одноствольного, кронообразующего, поли-
карпического с терминальными соцветиями на ветвях кроны;
формируется в результате нарастания моноподиальног о ствола и
симподиального ветвей кроны, которые из-за роста плагиотропно-
го побегов ветвления ориентированы горизонтально (рис. 85); в
отличии от М. a. Fagerlind ствол нарастает непрерывно; в основ-
ном двудольные из сем. Malvaceae и Rubiaceae, редко однодоль-
ные (Pandanaceae); лит.: Halle, Oldeman, Tomlinson, 1978.
Рис. 85. Архитектурная модель Petit (по: Halle et al., 1978)
Модель архитектурная Prevost (англ.: Prevost’s model, названа в
честь М. F. Prevost) — модель дерева одноствольного, кронообра-
зующего, поликарпического с терминальными соцветиями; фор-
мируется в результате ветвления акросимподиального и образова-
ния 2 типов побегов ветвления (рис. 86): силлептических и пролеп-
тических; силлептические побеги развиваются из самых верхних
почек и образуют ветви кроны, которые впоследствии занимают
горизонтальное положение; пролептические побеги развиваются
из почек расположенных ниже, растут ортотропно и формируют
ствол дерева (побеги замещения); в основном представители сем.
Apocynaceae и Boraginaceae; лит.: Halle, Oldeman, Tomlinson, 1978.
Рис. 86. Архитектурная модель Prevost (по: Halle et al., 1978)
108
Модель архитектурная Rauh (англ.: Rauh’s model, названа в честь
W. Rauh) — модель дерева одноствольного, кронообразующего, по-
ликарпического с пазушными соцветиями на ветвях кроны; фор-
мируется в результате ритмичного нарастания моноподиального
ствола и ортотропных ветвей кроны (рис. 87); обычная модель
среди тропических и бореальных деревьев (виды р. Acer, Arcto-
carpus, Fraxinus, Hevea, Musanga, Pinus, Quercus, и др.); лит.: Halle,
Oldeman, Tomlinson, 1978.
Рис. 87. Архитектурная модель Rauh (no: Halle et ah, 1978)
Модель архитектурная Roux (англ.: Roux’s model, названа в честь
J. Roux) — модель дерева одноствольного, кронообразующего, по-
ликарпического с пазушными соцветиями на ветвях кроны; фор-
мируется в результате нарастания моноподиального ствола и побе-
гов ветвления, которые характеризуются ростом плагиотропным
(рис. 88); отличается от М. a. Massart непрерывным ростом ствола;
в основном двудольные из сем. Annonaceae (Nylopia discreta),
Euphorbiaceae (Phyllanthus koghiensis), Rubiaceae (Coff ea arabica)
и др.; лит.: Halle, Oldeman, Tomlinson, 1978.
Рис. 88. Архитектурная модель Roux (no: Halle et al., 1978)
109
Модель архитектурная Scarrone (англ.: Scarrone’s model, названа
в честь F. Scarrone) — модель дерева одноствольного, кронообра-
зующего, поликарпического с терминальными соцветиями на вет-
вях кроны; формируется в результате ритмичного нарастания мо-
ноподиального ствола и ветвления симподиального ветвей кроны,
которые характеризуются ростом ортотропным (рис. 89); в ос-
новном двудольные из сем. Apocynaceae (Cerberiopsis comptonii),
Compositae (виды р. Bidens), Ericaceae (виды р. Rhododendron) и
др., редко однодольные из сем. Pandanaceae (виды р. Pandanus);
лит.: Halle, Oldeman, Tomlinson, 1978.
Рис. 89. Архитектурная модель Scarrone (по: Halle et al., 1978)
Модель архитектурная Schoute (англ.: Schoute’s model, названа в
честь J. С. Schoute) — модель дерева многоствольного или курти-
нообразующего, розеточного или кронообразующего, поликарпиче-
ского с пазушными соцветиями; формируется в результате ветв-
ления дихотомического и образования одинаковых (стволиков)
побегов ортотропных или плагиотропных (рис. 90); раст. древо-
видные в основном из сем. Palmae. (виды р. Hyphaene); лит.: Halle,
Oldeman, Tomlinson, 1978.
Рис. 90. Архитектурная модель Schoute (по: Halle et al., 1978)
НО
Модель архитектурная Stone (англ.: Stone’s model, названа в честь
В. С. Stone) — модель дерева одноствольного, кронообразующего, по-
ликарпического с терминальными соцветиями на ветвях кроны,
формируется аналогично М. a. Scarrone, отличаясь непрервным на-
растанием ствола (рис. 91); в основном представители сем. Рап-
danaceae (виды р. Pandanus) и Compositae (Mikania cordata); лит.:
Halle, Oldeman, Tomlinson, 1978.
Модель архитектурная Tomlinson (англ.: Tomlinson’s model, на-
звана в честь Р. В. Tomlinson) — модель дерева многоствольного
или куртинообразующего, розеточного, поликарпического с терми-
нальными или пазушными соцветиями; формируется в результа-
те нарастания базисимподиального и образования одинаковых по-
бегов (стволиков) из почек, расположенных под землей или на
уровне почвы (рис. 92); раст. древовидные, часто с корневищами
удлиненными из сем. Palmae (Rhapis excelsa), Euphorbiaceae (виды
p. Euphorbia), Lobeliaceae и др.; среди травянистых аналогично
развиваются многие растения длиннокорневищные и столонообра-
зующие (Сурегасегае, Роасеае и др); терм.: Halle, Oldeman, 1970;
лит.: Halle, Oldeman, Tomlinson, 1978.
Модель архитектурная Troll (англ.: Troll’s model, названа в честь
W. Troll) — модель дерева одноствольного, кронообразующего, по-
ликарпического; формируется в результате нарастания моноподи-
ального или мезосимподиального ствола и роста плагиотропного
побегов ветвления или замещения, базальный участок которых по-
сле становится вертикальным (рис. 93); представители сем. Eup-
horbiaceae (виды р. Phyllanthus), Fagaceae (виды р. Fagus), Legu-
minosae (виды р. Bauhinia, Albizzia), Tiliaceae (Tilia platyphyllos),
Ulmaceae (виды p. Ulmus); лит.: Halle, Oldeman, Tomlinson, 1978.
Ill
Рис. 92. Архитектурная модель Tomlinson: 1 — рост детерминированный,
2 — рост недетерминированный (по: Halle et al., 1978)
Рис. 93. Архитектурная модель Troll: I — нарастание моноподиальное,
2 — нарастание симподиальное (по: Halle et al., 1978)
112
Рис. 94. Схематическое изображение модели
«комбинаторного развития» (по: Poethig,
1990): А — виргинильное растение, Б — ге-
неративное растение, 1 — экспрессия юве-
нильных черт, 2 — экспрессия взрослых ве-
гетативных (виргинильных) черт, 3 — экс-
прессия генеративных черт, 4 — цветок
Модель комбинаторного раз-
вития побега (англ, combina-
torial model of shoot develop-
ment) — одна из теоретических
моделей развития побега гене-
ративного (однолетника), ко-
торая была предложена
R. S. Poethig (1990) для объясне-
ния возрастных изменений (см.
состояние возрастное), прояв-
ляющихся как гетеробластия,
гетерофилия, темп развития и
топофизис. Согласно М. к. р.
разные фазы морфогенеза или
онтогенеза регулируются неза-
висимыми перекрывающими-
ся программами (рис. 94), что
обусловливает различную ско-
рость полового созревания и
соматического развития расте-
ния (ср. модель «контроллер пе-
реключения фаз»)-, лит.: Tefler,
1998; Diggle, 1999; Ork-
wiszewski, Poethig, 2000.
Модель «контроллер переключения фаз» (англ, controller of pha-
se switching) — одна из теоретических моделей развития монокар-
пика, которая была предложена G. W. Haughn et al. (1995) для объ-
яснения различий индивидуального развития мутантов Arabidopsis
thaliana. Согласно М. к. п. ф. изменения состояния возрастного рас-
тения контролируется уровнем активности гипотетического веще-
ства (контроллера), так что время перехода от одной фазы разви-
тия особи (побега генеративного) к последующей (в частности темп
развития) определяется скоростью изменения критического уров-
ня активности контроллера (рис. 95). М. к. п. ф. отражает традици-
онную точку зрения на развитие растения — каждая фаза онтоге-
неза является необходимым шагом для последующих возрастных
изменений (Синнот, 1963; Куперман, 1977); ср. модель комбина-
торного развития побега; лит.: Koornneef et al., 1998; Diggle, 1999.
Модель модулярная (англ.: model with modular growth, modular
model) — группа моделей архитектурных (модель архитектурная
113
Рис, 95. Схематическое изображение действия «контроллера переключения фаз» раз-
вития Arabidopsis thaliana (по: Haughn et al., 1995): A —развитие дикого типа при не-
прерывном освещении, Б — развитие дикого типа или поздно цветущего мутанта на
коротком дне, В — развитие райо цветущего мутанта на коротком дне, 0 — переход от
розеточной фазы к формированию цветоноса, 1 — переход к генеративному разви-
тию (образование боковых генеративных побегов), 2 — переход к генеративному раз-
витию главного побега, (---) — пунктирной линией показано изменение уровня
«контроллера»
Chamberlain, м. a. Holtum, м. a. Koriba, м. a. Leeuwenberg, м. a. Prevost,
и м. a. Tomlinson), конструкция которых образована одним или
несколькими модулями (все побеги характеризуются ростом де-
терминированным)', у некоторых деревьев ветви кроны или ствол
нарастают моноподиально (часть побегов с детерминированным
ростом) в связи с чем выделяют модели частично модулярные
{модель архитектурная Nozeran, м. a. Fagerlind, м. a. Scarrone);
терм.: Halle et all, 1978; лит.: Prevost, 1978.
Модель немодулярная (англ.: nonmodular model) — группа моде-
лей архитектурных, в конструкции которых нет модулей, и систе-
ма побегов {нарастание моноподиалъное) формируется на основе
только единиц морфогенеза (напр. модель архитектурная Rauh).
Модель побегообразования — наследственно закрепленный тип
формирования системы побеговой, характерный для того или
иного вида раст. и определяемый в первую очередь особенностя-
ми функционирования меристем {м. верхушечной и м. интерка-
лярной) и типом ветвления', по способу нарастания {н. моноподи-
алъное, н. симподиалъное) осей скелетных {корневища, каудекса) и
выраженности розеточности побегов у травянистых многолетни-
ков умеренной зоны различают 4 М. п. (рис. 96): М. п. моноподи-
алъная (розеточная и длиннопобеговая) и М. п. симподиальная
(полурозеточная и длиннопобеговая); лит.: Серебрякова, 1977,
1981, 1987.
114
Рис. 96. Основные модели побегообразования травянистых многолетников:
1 — полурозеточная симподиальная, 2 — длиннопобеговая симподиальная,
3 — розеточная моноподиальная, 4 — длиннопобеговая моноподиальная
(по: Серебрякова, 1981)
Модель побегообразования моноподиальная — обобщенная схе-
ма строения побегового «тела» травянистых многолетников с нарас-
танием моноподиалъным; система побеговая таких раст. построена
из повторяющихся элементов структуры двух типов: побега от-
крытого (многолетний полициклический побег, формирующий
ось скелетную) и побега закрытого (однолетний моноциклический
побег, несущий цветки); по структуре открытого побега различают
М. п. моноподиальную розеточную (рис. 96; побеги розеточные) и
длиннопобеговую (рис. 96; побеги удлиненные); первая встречается
среди раст. кистекорневых и раст. стержнекорневых, побеговая
система которых представлена корневищем или каудексом; вто-
рая— среди раст. ползучих; терм.: Серебрякова, 1977,1981.
Модель побегообразования симподиальная — обобщенная схе-
ма строения побегового «тела» травянистых многолетников с на-
растанием симподиалъным; система побеговая таких раст. построе-
на из повторяющихся элементов структуры одного типа — мно-
голетних участков (резидов) побегов закрытых (монокарпических
поли(ди)циклических побегов); по структуре закрытого побега
115
различают М. п. симподиальную полурозеточную (рис. 96; побег
полурозеточный) и длиннопобеговую (рис. 96; побег удлиненный)',
терм.: Серебрякова, 1977, 1981.
Модель ростовых единиц (англ.: growth unit model) см. модель
строения побега фитонная (фитомерная).
Модель строения побега (англ.: models of shoot construction) —
метод интерпретации расчлененности побега на структурные эле-
менты {лист, междоузлие, узел, почка пазушная); выделяют 6 ком-
плементарных моделей (рис. 97), отличающихся взглядами на
происхождение структурных элементов и формирование из них
побега {М. с. п. классическая, М. с. п. «фертильный лист», М. с. п.
«лист-кожица», М. с. п. фитонная, М. с. п. метамерная, М. с. п.
континуальная); лит.: Rutishauser, Sattler, 1985; Гатцук, 1995; Куз-
нецова, 1995.
Модель строения побега «лист-кожица» (англ.: leaf-skin model
of shoot construction) — модель, предполагающая, что основны-
ми элементами структуры побега являются лист и внутренняя
часть стебля; при этом кора стебля формируется в результате уд-
линения листовых следов, которые как «кожа» окружают цент-
ральную часть стебля (рис. 97); основана на представлениях о
единстве листа и периферии стебля; лит.: Rutishauser, Sattler, 1985;
Гатцук, 1995.
Рис. 97. Модели строения побега сосудистых растений
(по Rutishauser, Sattler, 1985): 1 — классическая модель, 2 — фертильный лист,
3 — лист-кожица, 4 — фитонная, 5 — метамерная (а — апекс побега,
1-4 — последовательно формирующиеся: б^ — эпибластемы,
Bj-д — листья, Г[_4 — фитоны, д!4 — метамеры)
116
Модель строения побега «фертильный лист» (англ.: fertile-leaf
model of shoot construction) — модель, предполагающая, что ос-
новным элементом структуры побега является лист «несущий»
почку (фертильный лист); основана на представлении о единстве
происхождения листа и почки пазушной {почки эпифилъной), ко-
торые возникают в результате бифуркации эпибластемы (при-
мордия); т. о. побег представляет собой сумму раздвоенных эпи-
бластем (рис. 97); лит.: Rutishauser, Sattler, 1985; Гатцук, 1995.
Модель строения побега классическая (англ.: classical «stem-and-
leaf» model of shoot construction) — популярная модель, предпола-
гающая, что побег состоит из двух основных элементов (листа и
стебля), между которыми отсутствуют переходы (рис. 97); основа-
на на представлении о независимом происхождении листа и стеб-
ля; почки в данной модели не рассматриваются; лит.: Rutishauser,
Sattler, 1985; Гатцук, 1995.
Модель строения побега континуальная (англ.: continuum model
of shoot construction) — модель, предполагающая единство проис-
хождения всех элементов структуры побега (лист, узел, междоузлие,
почка пазушная); согласно этой модели любой побег образуется в
результате деятельности одной и той же меристемы апикальной
(дифференциации одного конуса нарастания на зачатки стебля,
листьев и пазушных почек) и представляет собой единое целое;
лит.: Sattler, 1974; Rutishauser, Sattler, 1985; Гатцук, 1995.
Модель строения побега метамерная (англ.: metameric model of
shoot construction) — модель, предполагающая, что основным
элементом структуры побега является метамер, т. е. лист (розет-
ка листьев) и междоузлие, расположенное под его узлом (рис. 97);
син.: модель ростовых единиц; лит.: Rutishauser, Sattler, 1985; Гат-
цук, 1995.
Модель строения побега фитонная (англ.: phytonic model of sho-
ot construction) — модель, предполагающая, что основным эле-
ментом структуры побега является фитон, т. е. лист (розетка ли-
стьев) и участок стебля, расположенный под его узлом (рис. 97);
при этом стеблевая зона одного фитона представляет сегмент
(полоску) стебля протяженностью в два или более междоузлий
при листорасположении очередном (напр. длина фитона при лис-
торасположении 2/5 составляет 5 междоузлий); основана на пред-
ставлении о единстве листа, его листового следа и прилегающих
тканей; син.: модель ростовых единиц; лит.: Rutishauser, Sattler,
1985; Гатцук, 1995.
117
Модель частично модулярная (англ.: partially modular model) —
см. модель модулярная.
Модификация (от лат. modus — мера, вид, facio — делаю; англ.:
modification) — ненаследственные изменения признаков (фено-
типа) организма, возникающие под влиянием изменившихся ус-
ловий внешней среды. Длительные изменения морфологических
признаков (адаптивные М.) в результате эписелективной или
эпигенетической эволюции могут стать наследственными; лит.:
Хохряков, Мазуренко, 1983; Кирпичников, 1983; Северцов и др.,
1993.
Модуль (англ.: module) — простая морфогенетическая побеговая
единица структуры, характеризующаяся ростом детерминиро-
ванным (ср. побег) благодаря формированию терминального
цветка или соцветия, усика (Landolphia dulcis), колючки (Carissa
macrocarpa) или паренхиматизации меристемы апикальной
(Apocynaceae); в более широком смысле М. наз. повторяющиеся
элементы структуры раст. разного уровня: лист с почкой пазуш-
ной, метамер, систему побегов и т. п. (ср. метамерность); терм.:
Prevost, 1967.
Модулярность (англ.: modularity) — повторение в модели архитек-
турной раст. одинаковых элементов структуры — модулей; боль-
шинство раст. являются мультимодулярными; к немодулярным
относятся, например, монокарпические Lobelia; в более широком
смысле М. рассматривают как синоним пон. метамерность; лит.:
Prevost, 1978; Halle, 1986.
Модус эволюции морфологической см. способ эволюции мор-
фологической.
Монокарпик (Monocarpic; от греч. monos — один, единственный,
karpos — плод; англ.: monocarp, monocarpic plant, semelparous
plant) — раст., которое после первого цветения (раст. гапаксант-
ное) и плодоношения полностью отмирает (Libanotis intermedia)
из-за расходования всех питательных веществ (см. гипотеза смер-
ти от истощения). В зависимости от продолжительности жизни
различают однолетники (М. моноциклические; англ.: annual),
двулетники (М. дициклические; англ.: biennial) и многолетники
(М. полициклические; англ.: pluriennial), а среди последних —
раст., живущие 3-5 лет (М. плейоциклические); терм.: Candolle de,
1827; син.: раст. монокарпическое, раст. семельпароузное; лит.:
Janzen, 1976; Ojala, 1988; Young, 1990; Krumbiegel, 1998, 1999;
Hautekeete et al„ 2001; Metcalf et al., 2003.
118
Моноподий (Monopodium, от греч. monos — один, pous — нога,
здесь — ось; англ.: monopodium) — многолетний осевой орган
раст. (ствол, ветвь, стебель, корень, корневище, побег), образован-
ный в результате деятельности одной и той же меристемы апикаль-
ной (рис. 98); при ветвлении М. каждый дочерний элемент также
Рис. 98. Схема моноподиального (А-Б) и симподиального (В-Г) нарастания (по: Бота-
ника. .., 1988): А — последовательное образование побегов у ели (1-4 — порядки ветв-
ления), Б—12-летнее дерево (5 — моноподий), В — последовательное образование
побегов у купены (6 — вегетативные побеги, 7 — цветоносные побеги, 8 — резид),
Г — взрослое растение (9 — симподий)
119
М., а образующаяся система наз. моноподиальной (напр. система
побегов моноподиальная); см. также нарастание моноподиальное.
Монофит (Monophyton, от греч. monos — один, единственный, phy-
ton — растение) — раст., не способное к естественному размноже-
нию вегетативному и на протяжении всего онтогенеза представ-
ленное одной особью (простой М.); к М. относятся гл. обр. раст. ве-
гетативно-неподвижные (деревья, кустарники аэроксильные, раст.
стержнекорневые, однолетники и т. п.); нек-рые из них (раст. по-
душковидные, кустарнички шпалерные) в результате партикуляции
сенильной образуют компактные группы из недолго живущих до-
черних особей (партикулирующий М.); терм.: Хохряков, 1975а.
Морфогенез (Morphogenesis, от греч. morphe — вид, форма, gene-
sis — происхождение, возникновение; англ.: morphogeny) — про-
цесс формообразования (заложение, рост и развитие) у раст.
структур различного уровня: органов (органогенез), тканей (гис-
тогенез) и клеток (цитогенез или клеточная дифференцировка); в
биоморфологии пон. М. используется для обозначения процесса
развития побегов (онтогенез побега) или их систем, корней, листь-
ев, реже—формы жизненной (ср. онтогенез); лит.: Ботаника...,
1988; Лотова, 2000.
Морфология растений (Morphologia, от греч. morphe — форма, lo-
gos — слово, учение; англ.: plant morphology, phytomorphology) —
учение о форме, строении и происхождении отдельных структур
раст.; в широком смысле М. р. изучает формы на всех уровнях от
особи до клеточных органоидов; выделяют специальные (карполо-
гия, палинология и др.) и общие разделы М. р.; к последним отно-
сятся теоретическая М. р. (рассматривает общие проблемы проис-
хождения структур), описательная М. р. или органография (клас-
сификация разнообразия форм), экологическая М. р. (изучение
изменчивости формы под воздействием внешних факторов) и др.;
в более узком значении М. р. изучает формы только макроструктур
(лист, побег, цветок и т. п.); в этом случае из нее выделяют как само-
стоятельные науки анатомию, эмбриологию, цитологию раст. и
др.; в последние годы из экологической М. р. выделился раздел о
формах жизненных раст. (биоморфология).
Надставка см. анаболия.
Нанизм (от греч. nanos — карлик; англ.: nanism, dwarfism) —
уменьшенные размеры тела потомка (миниатюризация) в ре-
зультате замедления его роста, но без изменения синхронизации
соматического и полового созревания (ср. гигантизм, гетерохро-
120
Александр Гумбольдт
(1769-1859)
выдающийся немецкий естествоиспытатель,
один из основателей биоморфологии растений
Иван Григорьевич Серебряков
(1914-1969)
выдающийся отечественный ботаник, морфолог,
основатель советской школы биоморфологии растений,
автор первых учебников (монографий) «учения о жизненных
формах», в которых сформулированы основные методы
и направления биоморфологических исследований
Татьяна Ивановна Серебрякова
(1922-1986)
выдающийся отечественный ботаник, морфолог,
внесший значительный вклад в становление отечественной
школы биоморфологии растений, в развитие
биоморфологического анализа таксонов, теоретической
и эволюционной биоморфологии
Виталий Николаевич Голубев
(р. в 1926 г.)
известный отечественный ботаник,
внесший существенный вклад в развитие
биоморфологического анализа растительного
покрова, классификации жизненных
форм и фитофенологии
Борис Александрович Юрцев
(1932-2004)
выдающийся отечественный ботаник, флорист,
систематик и ботанико-географ, внесший существенный
вклад в развитие теоретической биоморфологии
растений
Андрей Павлович Хохряков
(1933-1998)
выдающийся отечественный ботаник, флорист,
систематик, внесший значительный вклад в развитие
биоморфологического анализа таксонов и эволюционной
биоморфологии растений
ния). В биоморфологии Н. часто рассматривают как результат
прогенеза, что соответствует псевдонанизму, или как реакцию
раст. на неблагоприятные условия среды, которая проявляется в
уменьшении общих размеров особи (миниатюризация); син.:
карликовость; лит. Dommergues et al., 1986.
Нанофанерофит (англ.: nanophanerophyte) см. фанерофит.
Направленность эволюции биоморфологической — осуществ-
ление одного или нескольких из реально возникавших или теоре-
тически возможных направлений изменения формы жизненной
(онтогенеза, морфогенеза) исходного таксона в результате дейст-
вия факторов эволюции на онтогенетическом и популяционном
уровне. Обычно выделяют нисходящую (редукционную) и вос-
ходящую линию эволюции форм жизненных (соответственно от
деревьев к травам или наоборот); син.: канализованность (кана-
лизированность) эволюции биоморфологической; лит.: Серебря-
ков, 1962; Серебрякова, 1972, 1973; Хохряков, 1975а, 1981; Гатцук,
1976; Северцов, 1990; Недолужко, 1997; Воронцов, 1999; Цвелев,
2005.
Нарастание (Accrescentia; англ.: growth) — увеличение размеров
раст. или его части (напр., побега); обычно используется для ха-
рактеристики роста оси скелетной многолетнего раст.; различают
Н. моноподиальноеи Н. симподиальное (рис. 98); иногда пон. Н. ис-
пользуют в том же смысле, что и пон. ветвление.
Нарастание моноподиальное (Accrescentia monopodialis, от греч.
monos — один, pous — нога, здесь — ось; англ.: monopodial
growth) — рост раст., ось скелетная которого формируется (на-
растает) в результате неопределенно продолжительной деятель-
ности одной и той же меристемы апикальной (рис. 98) и представ-
ляет собой побег вегетативный, состоящий из нескольких прирос-
тов (моноподий); образование побегов боковых у таких раст.
обычно наз. ветвлением моноподиальным, которое может иметь
характер акротонии, базитонии или мезотонии (соответственно,
ветвление акро-, бази- и мезомоноподиальное); Н. м. характерно
для хвойных деревьев (Picea) и нек-рых раст. травянистых
(Geum); терм.: Troll, 1935.
Нарастание симподиальное (Accrescentia sympodialis, от греч.
syn — вместе, pous — нога, здесь — ось; англ.: sympodial growth) —
рост раст., ось скелетная которого формируется (нарастает) в ре-
зультате перевершинивания (рис. 98) и представляет собой состав-
ную ось (симподий), состоящую из многолетних частей нескольких
121
побегов замещения (напр.: ствол, ветвь, корневище); в зависимости
от числа замещающих побегов различают монохазий, дихазий
(ложная дихотомия), плейохазий, а от их положения на побеге ма-
теринском— Н. акро-, мезо- и базисимподиальное (см. акротония,
мезотония, базитония); образование побегов боковых при Н. с.
обычно наз. ветвлениемсимподиальным (ветвлением акро-, мезо-,
базисимподиальным); терм.: Troll, 1935.
Неотения (от греч. neos — незрелый, юный, teino — растягиваю,
удлиняю; англ.: neoteinia, neoteny) — один из основных типов
элементарной гетерохронии, при котором замедление соматиче-
ского развития сочетается с неизменной скоростью полового со-
зревания (ср. акселерация). В результате Н. потомок имеет педо-
морфный облик (педоморфоз) — половозрелый организм с юно-
шескими чертами (рис. 2). В биоморфологии Н. часто называют
ускорение полового созревания (ср. прогенез), а результат — обо-
значают как нанизм; син.: ретардация, фетализация; терм.:
Э. М. Кольман (1884); лит.: Арциховский, 1911; Тахтаджян, 1964;
Васильченко, 1965; Gould, 1980; Серебрякова, 1983.
Номофилл см. лист срединный.
Нутация (Nutatio, от лат. nuto — качаться, колебаться; англ.: nuta-
tion) — вращательное движение растущего органа раст., обуслов-
ленное смещением по кругу местного ускорения роста клеток в
зоне растяжения; свойственна верхушкам побегов раст., но наибо-
лее ярко проявляется у вьющихся усиков (гаптотропизм) и стеб-
лей лиан; к Н. нередко относят колебательные движения прекра-
тивших рост листьев и прилистников, связанные с изменением
тургора в клетках листовых сочленений; терм.: Ю. Сакс, 1870;
лит.: Полевой, Саламатова, 1985.
Однолетник (англ.: annual) см. растение однолетнее.
Однолетник вегетативный (англ.: pseudoannual) см. малолетник
вегетативный.
Однолетник озимый (англ.: winter-annual) см. растение однолетнее.
Однолетник яровой (англ.: summer-annual) см. растение одно-
летнее.
Одревеснение (англ.: lignification) — процесс изменения клеточ-
ных оболочек в результате заполнения их лигнином (лигнифика-
ция), который начинается после окончания роста клеток и сопро-
вождает их старение. В результате О. формируются специализи-
рованные ткани с мертвыми клетками, имеющими твердые,
прочные стенки (напр., механические ткани, сосуды). В широком
122
смысле О. раст. наз. и опробковение клеток (появление в оболоч-
ке суберина), различая растения деревянистые и растения травя-
нистые; лит.: Ботаника..., 1988; Лотова, 2000.
Олигокарпик (англ.: oligocarpic) см. поликарпик.
Олигомеризация (от греч. oligos — немногочисленный, незначи-
тельный, meros — часть; англ.: oligomerization) — один из резуль-
татов функциональной дифференциации и редукции в эволюции
организмов, проявляющийся как уменьшение числа гомологич-
ных органов или структур организма (ср. полимеризация); терм.:
Догель, 1936, 1954; лит.: Тахтаджян, 1964; Заморский, 1971, 1980;
Хохряков, 1975, 1981; Мейен, 1977; Северцов, 1981; Терехин, 1991.
Омоложение (англ.: rejuvenescence) — процесс усиления жизнедея-
тельности организма, связанный с интенсификацией синтеза бел-
ков и нуклеиновых кислот, активацией роста и клеточных деле-
ний, возникновением и накоплением эмбриональных тканей и
общим усилением физиологических функций; проявляется у
раст. в образовании новых меристем, обусловливающих форми-
рование новых побегов, корней, обновление системы побеговой и
системы корневой, а при размножении вегетативном (напр. псев-
довивипарии) — длительное существование клона; лит.: Кренке,
1940; Дубровицкая, 1961; Сабинин, 1963; Ефейкин, 1966; Биология
развития..., 1975.
Онтобиоморфа см. фаза онтогенеза.
Онтогенез (Ontogenesis; от греч. on — существо, genesis — проис-
хождение, возникновение; англ.: ontogeny) — генетически обу-
словленная последовательность этапов развития особи от ее заро-
ждения в результате полового (полный О.) или вегетативного
размножения (сокращенный О.) до ее естественного отмирания
(см. старение) или преждевременной смерти (сокращенный или
обрывающийся О.). В биоморфологии изучение индивидуального
развития раст. обычно не включает этапы формирования зароды-
ша (эмбриогенез, этап эмбриональный), а охватывает только пе-
риод онтоморфогенеза. В ходе О. происходит изменение состоя-
ния возрастного раст., на основании чего выделяют периоды онто-
генеза или фазы онтогенеза (см. модель комбинаторного развития
побега, модель «контроллер переключения фаз»). В более широком
значении пон. О. используют для описания полного цикла разви-
тия клона (синонтогенез, О. клона или сложный О.); син.: индиви-
дуальное развитие, простой О., простой или большой жизненный
цикл раст., большой цикл развития раст.; лит.: Левин, 1963, 1964,
123
1966; Гупало, Скрипчинский, 1971; Серебрякова, I97I6; Хохряков,
1973; Скрипчинский, 1977; Жукова, 1983; Батыгин, 1986.
Онтогенез клона см. онтогенез.
Онтогенез обрывающийся см. онтогенез.
Онтогенез побега (полный) — жизнь побега от момента его зало-
жения до отмирания всех его частей; в О. п. различают 3 основных
этапа, периода или фазы развития: внутрипочечную или эмбрио-
нальную (см. рост внутрипочечный), внепочечную или постэм-
бриональную (см. цикл жизненный малый) и фазу вторичной
деятельности побега-, лит.: Серебряков, 1952, 1959а; Скрипчин-
ский и др., 1970; Серебрякова, 1971а.
Онтогенез полный см. онтогенез.
Онтогенез простой см. онтогенез.
Онтогенез сложный см. онтогенез.
Онтогенез сокращенный см. онтогенез.
Онтоморфогенез (Ontomorphogenesis — от греч. on — существо,
morphe — вид, форма, genesis — происхождение, возникнове-
ние) — генетически обусловленная последовательность этапов
развития особи от прорастания до ее естественного отмирания
(см. старение) или преждевременной смерти (ср. онтогенез)-, ср.
онтогенез.
Орган размножения вегетативного (англ.: vegetative multiplica-
tion organs, organs (morphological structures) of vegetative repro-
duction) — пон., используемое для обозначения б. м. видоизме-
ненных побегов (корневище, столон, флагелла, клубень, луковица
и т. п.) и почек (клубенек, луковичка, почка выводковая и т. п.), при
отделении которых от раст. образуются дочерние особи (см. раз-
множение вегетативное); лит.: Серебряков, 1952.
Органогенез (англ.: organogeny, organogenesis) см. морфогенез.
Ортокорм см. дуга укороченных междоузлий.
Ортостиха (Orthostichy, от греч. orthos — прямой, правильный, sti-
chos — ряд, линия; англ.: orthostichy) — условная вертикальная
линия, соединяющая основания листьев, расположенных строго
друг над другом на стебле (рис. 133); при уменьшении угла дивер-
генции и приближении его значения к 137,5° (зачатки листьев в
почке) 0. выделяются условно или вообще не обнаруживаются.
Особь (Individuum, от лат. in — не, divide — разделять, делить;
англ.: individual, bion) — живой организм, образующийся в ре-
зультате размножения полового (генет; англ.: genet) или размно-
жения вегетативного (рамет; англ.: ramet), взаимосвязь элементов
124
которого обусловлена его морфологической и физиологической
целостностью (неделимостью); относительная автономность по-
бегов, выражающаяся в потенциальной способности давать само-
стоятельные О., обусловливает наличие у раст. некоторых коло-
ниальных черт; в связи с этим различают индивид простой, и.
сложный и колонию клональную, син.: индивид(уум); лит.: Козо-
Полянский, 1937; Чайлахян, 1956; Левин, 1961, 1963; Сенянинова-
Корчагина, 1967.
Особь материнская (англ.: maternal (parent) individual) см. клон.
Ось скелетная — мощный боковой стебель, возникший в резуль-
тате кущения из приземных или подземных почек спящих мате-
ринского побега^ в отличие от верхних боковых ветвей кроны О. с.
укореняется и формирует структурную основу кустарника гео-
ксильного; обычно используют в более широком смысле называя
О. с. многолетние системы побегов, составляющие основу формы
жизненной раст., терм.: Серебряков, Доманская, Родман, 1954.
Отбег см. столон.
Отпрыск корневой (Soboles; англ.: root shoot, ratoon, root-borne
shoot) — побег надземный, образующийся из почки придаточной,
расположенной на корне (рис. 99); служит для размножения веге-
тативного гл. обр. двудольных многолетних раст. стержнекор-
невых (Cirsium, Sonchus), которые наз. раст. корнеотпрысковыми
(см. сарментация); О. к. образуются на приповерхностных корнях
горизонтальных или в базальной части корня главного (см. корне-
отпрысковость); син.: поросль корневая.
Пазуха листа (Axilla; англ.: axil) — угол, образованный листом
подпирающим и участком стебля, расположенным выше его узла.
Паразит корневой (англ.: root parasit) см. ксенопаразит.
Паразит стеблевой (англ.: branch parasit) см. ксенопаразит.
Паразитизм (Parasitismus, от греч. para — возле, мимо, вне; sites —
хлеб, пища; англ.: parasitism) — форма взаимоотношений двух
растений, принадлежащих к разным видам, при которой одно из
них (паразит) использует другое (хозяина) в качестве источника
пищи и осуществляет посредством него регуляцию своих отноше-
ний с окружающей средой (антагонистический или паразитный
симбиоз); по степени зависимости раст.-паразита от раст.-хозяи-
на различают облигатный и факультативный П., а по способности
раст.-паразита к фотосинтезу выделяют голопаразитов (голопара-
зитизм; англ.: holoparasitism) и полупаразитов (полупаразитизм;
hemiparasitism); реже встречается сверхпаразитизм — поселение
125
3
Рис. 99. Образование корневых отпрысков у Cerasus fruticosa: А — схема расположения
побегов (1 — боковой корень, 2 — главный корень, 3 — главный побег, 4 — гипоко-
тильный побег, 5 — корневой отпрыск, 6-—побег ветвления), Б — внешний вид
корневого отпрыска, В — придаточные почки на боковом корне (7-8 — продольный
разрез, 9-10 — внешний вид)
126
раст. паразита (Viscum moniliforme) на другом раст. паразите
(Viscum orientale); часто П. рассматривают более широко и относят
к раст. паразитам не только ксенопаразитов, но и аллелопаразитов
(облигатно микотрофные раст.), различая среди них сапрофиты и
полусапрофиты; лит.: Терехин, 1977; Kuijt, 1969.
Парастиха (Parastichy, от греч. para — возле, мимо, вне, stichos —
ряд, линия; англ.: parastichy) — условная линия, которая соединя-
ет косые ряды тесно примыкающих друг к другу боковых придат-
ков побега (листьев) и спирально закручивается от центра к пери-
ферии вокруг оси с разной степенью наклона (рис. 133); пон. ис-
пользуется при описании листорасположения в почке, на конусе
нарастания или укороченном побеге; обычно выделяются П.,
идущие в противоположных направлениях (против и по часовой
стрелке), и по количественному отношению между ними опреде-
ляют формулу листорасположения.
Партикула (Particula — частица, кусочек) — часть раст. стержне-
корневого, образовавшаяся в результате его полной партикуляции
и способная к существованию как самостоятельная особь непро-
должительное время; в более широком смысле П. отождествляют
с особью дочерней, образовавшейся в результате размножения ве-
гетативного, наз. ее партикуляном, парциалью или особью пар-
циальной; терм.: Высоцкий, 1915.
Партикулян см. партикула.
Партикуляция (Particulatio, от партикула) — продольное расщеп-
ление каудекса и системы корневой в результате некроза тканей
(естественная или некротическая П.), происходящего при старе-
нии раст. стержнекорневого (рис. 100); различают полную и час-
тичную П.; у нек-рых раст. П., происходящая в результате обо-
собления проводящих пучков каудекса, обуславливает формиро-
вание жизнеспособных дочерних особей (рис. 100; размножение
вегетативное); нередко пон. П. рассматривают в более широком
смысле, включая сюда распад раст. дернованных (напр. злаки) и
раст. длиннокорневищных (см. парциаляция); в связи с этим вы-
деляют П. нормальную, П. сенильную и П. травматическую; терм.:
Высоцкий, 1915; лит.: Шалыт, 1965; Нухимовский, 19736; Ценопо-
пуляции..., 1976, 1988; Барыкина и др., 1976; Барыкина, 2000.
Партикуляция нормальная — размножение вегетативное спе-
циализированными, неразносимыми органами раст.: столон, кор-
невищем т. п. (рис. 100); терм.: Работнов, 1969.
Партикуляция сенильная — распад раст. при старении с образо-
127
Рис. 100. Партикуляция у Aconitum orientate (A-В; по: Барыкина и др., 1976) и Salsola
dendroides (Г; по: Рахманина, 1962): А — каудекс взрослого растения (1 — щелевидные
углубления), Б — схема поперечного среза стеблевого участка каудекса с обособляе-
мыми проводящими пучками (2 — наружная перидерма, 3 — внутренняя перидерма,
4 — вторичная флоэма, 5 — камбий, 6 — вторичная ксилема, 7 — некротический уча-
сток), В — корневой партикул, Г — развитие сенильной п артикуляции (8 — зона от-
мирания, 9 — расщепление каудекса)
ванием самостоятельных недолгоживущих особей (рис. 100; см.
также период сенильный); терм.: Работнов, 1969.
Партикуляция травматическая — образование дочерних особей в
результате механического отчленения от раст. (животными или
человеком) неспециализированных органов вегетативного раз-
множения-, терм.: Работнов, 1969.
Парциаль см. партикула.
Парциаляция — терм, предложен Корчагиным (19606) для обозна-
чения распада раст. длиннокорневищного в отличие от партикуля-
ции раст. стержнекорневого.
Пасынки см. побег обогащения.
128
Педоморфоз (от греч. paidos — дитя, morphe — форма; англ.: ре-
domorphosis, paedomorphosis) — один из способов эволюцион-
ных изменений облика организма, посредством которого выпада-
ет взрослая стадии развития {онтогенез потомка заканчивается
стадией, которая соответствует личиночной стадии предка). В на-
стоящее время под П. понимают морфологический результат
прогенеза и неотении. В биоморфологии к педоморфозу часто от-
носят нанизм; терм.: Garstang, 1922; лит.: Gould, 1980.
Пенек см. резид.
Перевершинивание (Metacoryphismus) — перерастание одного
дочернего элемента (напр. стебля) и его отклонение в сторону
другим, большим по размеру дочерним элементом при ветвле-
нии дихоподиальном (рис. 9); часто П. наз. рост побегов формирова-
ния, образующих симподий (рис. 98); лит.: Серебряков, 1952.
Перекати-поле (Pervolventes, от лат. per — приставка со значением
усиления, volvo — катить, вращать) — многолетнее травянистое
раст. стержнекорневое со специализированным механизмом дис-
семинации: переносом семян на соцветии, которое в результате
интенсивного ветвления и переориентации его веточек приобре-
тает шаровидную форму и перекатывается ветром по земле после
отламывания (перегнивания) стебля у основания; П. п. часто рас-
сматривается как особая форма жизненная, которая более широ-
ком смысле объединяет некоторые однолетники, раст. кистекор-
невыеи раст. луковичные; лит.: Алехин, 1939.
Период виргинильный (Periodus virginalis; от лат. virgo — девст-
венный, целомудренный; англ.: virginile (period) stage) — период
жизни раст. от прорастания диаспоры до первого цветения (см.
темп развития) или формирования луковичек и клубеньков вме-
сто цветков {псевдовивипария). В течение П. в. морфологический
облик раст. существенно меняется, что связано с изменением его
состояния возрастного (рис. 101); в связи с этим выделяют возрас-
тные состояния: проросток, раст. ювенильное, раст. имматурное,
раст. виргинилъное (обозначается соответственно: pl, j, im, v); ино-
гда П. в. наз. П. ювенильным; син.: П. прегенеративный, П. девст-
венный; терм.: Работнов, 1945; лит.: Ценопопуляции..., 1976.
Период возрастной см. период онтогенеза.
Период вторичной деятельности побега см. фаза вторичной
деятельности побега.
Период генеративный (Periodus generativa, от лат. generare — рож-
дать, производить; англ.: generative (period) stage) — период жизни
129
раст. от первого до последнего формирования генеративных орга-
нов или их видоизменений, напр., при псевдовивипарии (ср. раз-
витие генеративное); у поликарпиков по количеству генеративных
побегов, соотношению процессов роста и отмирания обычно выде-
ляют (рис. 101) молодые (ранние), средневозрастные (зрелые), ста-
рые (поздние, стареющие) генеративные раст. (обозначаются соот-
ветственно: gp g2, g3); многие раст. по различным причинам цветут
в П. г. не каждый год (временно не цветущие); син.: П. плодоноше-
ния, П. половой зрелости, П. дифинитивный, П. жизни взрослой,
П. репродуктивный; лит.: Ценопопуляции..., 1976.
Период девственный см. период виргинильный.
Период дефинитивный см. период генеративный.
Период жизни взрослой см. период генеративный.
Период латентный (Periodus latens, скрытый, невидимый; англ.:
latent (period) stage) — период жизни, в течение которого раст. на-
ходится в состоянии покоя первичного в виде семени, плода или
иного зачатка (обозначается индексом se); терм.: Работнов, 1945.
Период онтогенеза (англ.: ontogeny (period) stage) — промежуток
времени индивидуального развития особи, охватывающий опре-
деленный, законченный этап возрастных изменений ее морфоло-
гического строения (рис. 101); обычно различают П. эмбриональ-
ный, П. латентный, П. виргинильный, П. генеративный, П. сениль-
ный-, П. о. не всегда соответствует фазе онтогенеза; син.: П.
возрастной; лит.: Ценопопуляции..., 1976, 1988.
Период плодоношения см. период генеративный.
Период половой жизни см. период генеративный.
Период постгенеративный (англ.: postgenerative (period) stage) см.
период сенильный.
Период прегенеративный (англ.: pregenerative (period) stage) см.
период виргинильный.
Период развития побега эмбриональный см. рост внутрипочеч-
ный.
Период репродуктивный см. период генеративный.
Период сенильный (Periodus senilis; от лат. senex — старый, стар-
ческий; англ.: senile (period) stage) — период жизни раст. от по-
следнего цветения (или образования диаспор при псевдовивипа-
рии) до полного отмирания (рис. 101); П. с. не выражен в онтоге-
незе монокарпиков; у пол икарпиков по количеству отмерших час-
тей, наличию ювенильных черт (формы листьев, характеру побе-
гов и др.) различают раст. субсенильные (старые вегетативные) и
131
130
Рис. 101. Периоды онтогенеза и возрастные состояния длиннокорневищного (А), дерновинного (Б)
и стержнекорневого (В} растения (по: Ботаника..., 1988)
раст. сенильные (обозначаются соответственно ss, s); в популяци-
ях некоторых поликарпиковсенильные и субсенильные особи мо-
гут отсутствовать (см. старение); син.: П. старческий, П. постгене-
ративный; терм.: Работнов, 1945; лит.: Ценопопуляции..., 1976.
Период старческий см. период сенильный.
Период эмбриональный (Periodus embryonalis, от греч. embry-
оп — зародыш; англ.: embryonal (period) stage) — период раз-
вития зародыша раст. от момента его возникновения до отде-
ления от материнской особи; иногда в П. э. включают П. ла-
тентный.
Период ювенильный (juvenile (period) stage) см. период вирги-
нильный.
Перышко (Plumula, от лат. pluma — перышко, пух) — зачаток пер-
вого листа проростка злака.
Петилоид (англ.: petiloide) см. филломорфа.
Плагиокорм см. дуга укороченных междоузлий.
Плагиоподий см. дуга укороченных междоузлий.
Пластохрон (Plastochronus, от греч. plastos — вылепленный, сфор-
мированный, chronos — время; англ.: plastochrone) — временная
единица ритма роста внутрипочечного побега, определяемая как
интервал времени между
заложением двух последо-
вательных примордиев (или
двух пар примордиев при
листорасположении супро-
тивном; рис. 102); в более
широком смысле — ин-
тервал времени между на-
чалом двух последователь-
но повторяющихся собы-
тий, таких как закладка
примордия, достижение
определенной стадии раз-
вития листа (ср. филло-
хрон) и т. п.; иногда П. ис-
пользуют для определения
онтогенетического возрас-
та побега или его частей
(см. индекс пластохрон-
ный); терм.: Askenasy, 1880.
Рис. 102. Изменения формы конуса нарастания
побега в течение двух пластохронов: I—V — пер-
вый пластохрон, V-IX —- второй пластохрон,
АВ — граница конуса нарастания, С, D — пазу-
хи примордиев, Bj, в2—верхушка примордия
(по: Серебрякова, 1963)
132
Плейокотилия (англ.: pleico-
tyledony) см. поликотилия.
Плеть см. столон.
Плодушки см. побег укоро-
ченный.
Плюмула (англ.: plumule) см.
почечка.
Пневматофор (Pneumato-
phorum, от греч. pneyma —
дыхание, воздух, phoros —
несущий; англ.: pneumato-
phore, pneumor(r)hizae, bre-
athing root) — корень, рас-
тущий вверх (отрицатель-
ный геотропизм) от корне-
вища или подземного кор-
ня раст., и в своей верхней
части, расположенной над
поверхностью почвы, име-
ющий чечевички (рис.
103); обеспечивает подзем-
ные органы раст. кислоро-
дом (К. вентиляционный,
К. дыхательный); развива-
ется у раст., обитающих на
заболоченных почвах (Ta-
xodium distichum) и в при-
ливной полосе морских по-
бережий (Avicennia).
Побег (Cormus; англ.: shoot,
sprout р. р., tiller s. 1.,
spray) — один из основных
органов высших раст.,
представляющий собой не-
разветвленный стебель, не-
сущий листья, почки и/или
другие придатки и обра-
зующийся в результате дея-
тельности одной и той же
меристемы апикальной
Рис. 103. Прямые (1, 5 — по: Федоров и др.,
1962) и коленчато изогнутые (2—4 — по: Bell,
Bryan, 1998) дыхательные корни Sonneratia sp.
(1, 5), Mitragina ciliata (2), Ganystylus sp. (3),
Symphonia gabonensis (4)
133
(рис. 104); снизу П. ог-
раничен шейкой кор-
невой (П. главный) или
своим основанием (17.
ветвления), сверху —
цветком (П. закры-
тый), усиком (Landol-
phia dulcis), колючкой
(Carissa macrocarpa),
рубцом (абортация
верхушки П.) или поч-
кой, недоразвитой в
Рис. 104. Схема развития побега (по: Учебно-поле
вая..., 1990): А — внутрипочечная стадия, Б — види-
СВЯЗИ С паренхимати- МЫЙ рост, В — сформированный побег с одним при-
ЗациеЙ апИКаЛЬНОЙ ростом (элементарным побегом), Г — сформирован-
мрпигтрмы СП пт- ный побег с двумя приростами (элементарными
1*1 V- > 1 V 1 С 1*1 1Э1 (11. v J.
„х п побегами), 1 — граница приростов
крытый); часто II. наз. r r г
стебель, несущий бо-
ковые придатки, участок стебля, образующийся к моменту наблю-
дения или за один цикл видимого роста (ср. прирост); П.— уни-
версальный структурный элемент метамерного строения раст.,
ритмичность роста которого обусловливает симметрию продоль-
ную в его морфологическом строении; для интерпретации строе-
ния П. используют модели строения побега-, лит.: Серебряков, 1952;
Нухимовский, 1971; Гатцук, 1974а,б; Тихонов, 1977; Sattler, 1974;
Михалевекая, 2002.
Побег автономный (Cormus autonomus, от греч. autos — сам, по-
mos — закон) — П. ветвления, несущий корни кладогенные (П.
антагонист); при партикуляции или клонировании раст. укорене-
ние побегов может обусловливать размножение вегетативное;
терм.: С. П. Смелов (1947) ввел для обозначения одного из перио-
дов развития П. злаков (ср. П. сосунок).
Побег адвентивный (Cormus adventivus, от лат. advenio — прихо-
дить, прибывать; англ.: adventitious shoot) — П., развивающийся
из почки придаточной; различают П. внепазушный (см. почка
внепазушная) и отпрыск корневой (см. раст. корнеотпрысковое);
син.: П. придаточный.
Побег акрофильный (Cormus acrophilus, от греч. асго — верхний,
phyllon — лист) — П. розеткообразущий, стебель которого несет
надземную «розетку» листьев (см. дерево розеточное); терм.:
Смирнова, 1970.
134
Рис, 105. Направления роста побегов:
1 — апогеотропное (ортотропное),
2 — анизотропное, 3 — геотропное,
4 — диагеотропное (плагиотропное),
5 — изотропное, 6 — клиноапогеотропное
(ортотропное), 7 — клиногеотропное
(плагиотропное)
Побег анизотропный (Cormus
anisotropus, от греч. anisos —
неравный, trope — поворот,
направление) — П., изменя-
ющий направление роста в
процессе своего развития под
влиянием (эндо)экзогенных
факторов (рис. 105), напр.
столоны Adoxa moschatellina;
ср. П. изотропный; син.: П. ге-
теротропный; лит.: Серебря-
ков, 1952.
Побег анизофильный (Cormus
anisophyllus, от греч. anisos —
неравный, phyllon — лист;
англ.: anisophyllous shoot) —
П. с неодинаковыми листьями
на разных сторонах оси или, в
более узком значении, на од-
ном и том же узле при листо-
расположении супротивном
или л. мутовчатом (рис. 6; ср.
П. изофильный, П. гетерофильный).
Побег антагонист см. побег автономный.
Побег апогеотропный (Cormus apogeotropus, от греч. аро — при-
ставка, здесь в значении отрицания, обратности, geo — земля,
trope — поворот, направление; англ.: apogeotropic shoot) — пря-
мостоячий П., вертикальный рост которого обусловлен отрица-
тельным геотропизмом (рис. 105); часто обозначается как П. ор-
тотропный} терм.: Hackel, 1882; син.: П. вертикальный, П. прямо-
стоячий.
Побег афильный (Cormus aphyllus, от греч. а — в сложных словах
приставка не-, без-, phyllon — лист; англ.: aphyllous shoot) — в
специальном значении — быстрорастущий, одревесневающий
П. апогеотропный бамбуков, несущий на большем своем протя-
жении катафиллы и только на верхушке и на побегах ветвле-
ния — листья ассимилирующие (Molisch, 1926; цит. по Серебряко-
ва, 1971а); в обычном смысле — стебель без листообразных при-
датков (напр., усы) или только с катафиллами (напр. корневище),
или с опавшими листьями (П. безлистный).
135
Побег безлистный см. побег афильный.
Побег безрозеточный (Cormus erosulans, от лат. е — приставка со
значением отсутствия чего-либо, rosa — розетка; англ.: erosulate
shoot) — П„ в онтогенезе которого нет фазы «розетки» (рис. 106),
а в морфологической структуре — участка с укороченными меж-
доузлиями и ассимилирующими листьями, образующими розет-
ку (ср. П. удлиненный); терм. Воронов, 1952; лит.: Серебряков,
1952; Голубев, 1965; Серебрякова, 1971а.
Побег бесплодный см. побег вегетативный.
Побег боковой (Cormus lateralis, от лат. latus — сторона, бок; англ.:
laterate shoot, sucker) — П., развивающийся из почки пазушной
(рис. 31; П. пазушный); в более широком смысле — побег ветвле-
ния бокового (ср. П. адвентивный).
Побег брахиморфный (Cormus brachymorphus, от греч.
brachys — короткий, morphe — вид, форма) — терм, заимство-
ван В. Н. Голубевым (1965) из антропологии для обозначения П.
укороченного у раст. древесного.
Побег вегетативно-генеративный (Cormus mixtus — побег сме-
шанный) — П„ стебель которого несет листья и/или катафиллы,
цветок или цветки, собранные в простое соцветие.
Побег вегетативный (Cormus vegetativus, от лат. vegetus — пол-
ный сил, деятельный; англ.: vegetative shoot) — П., стебель кото-
рого несет только листья и/или катафиллы без генеративных ор-
ганов; часто П. в. наз. вегетативный участок (прирост) П. вегета-
тивно-генеративного; син.: П. бесплодный, П. стерильный.
Побег вертикальный см. побег апогеотропный.
Побег ветвления — наз., используемое для обозначения П. любого
порядка ветвления верхушечного или ветвления бокового (П. пер-
вого, второго и т. д. порядка); в зависимости от функционального
значения различают П. возобновления, П. дополнения, П. замеще-
ния, П. кущения, П. обогащения и др.; син.: П. дочерний, П. произ-
водный.
Побег вневлагалищный см. побег экстравагинальный.
Побег внекустовой см. побег диагеотропный.
Побег внепазушный см. побег адвентивный.
Побег внутривлагалищный см. побег интравагинальный.
Побег внутрикустовой см. побег интравагинальный.
Побег водяной (англ.: water sprout) см. побег формирования.
Побег возобновления (Innovatio — возобновление; англ.: innova-
tion shoot) — П. ветвления, развивающийся из почки зимующей и
136
Рис. 106. Структурно-морфологические типы Побегов: 1-3 — укороченный (1,3 — ро-
зеточный), 4-6, 9, 10 — удлиненный (безрозеточный), 7, 8, 11 — полурозеточный (по:
Учебно-полевая..., 1990), А — удлиненные и укороченные побеги березы, Б — розе-
точный побег подорожника с пазушными цветоносами (В — продольный разрез;
к — корневище, п — придаточный корень), Г — полурозеточный побег живучки
(Д — схема строения; р — розетка, с- столон, ц — цветонос (по: Ботаника..., 1988)
137
обеспечивающий возобновление роста оси скелетной травянисто-
го (в более широком смысле и деревянистого) раст. после периода
покоя (рис. 107); нередко П. в. наз. прирост годичный (см. П. го-
дичный); ср. П. замещения.
Побег восходящий см. побег анизотропный.
Побег генеративный (Cormus generativus, от лат. generare — рож-
дать, производить; англ.: generative shoot) — П. ветвления с реду-
цированными листьями (брактеями, чешуями) или безлистный,
несущий один или несколько цветков, собранных в простое со-
цветие; всегда П. однолетний; к П. г. относятся: П. обогащения
сложного соцветия, безлистный цветонос; в более широком
смысле — П„ несущий генеративные органы; часто П. г. наз. гене-
ративный прирост П. вегетативно-генеративного; син.: П. репро-
дуктивный, П. фертильный, П. цветоносный.
Побег гетерогенный (Cormus heterogeneus, от греч., heteros — дру-
гой, genos — род, происхождение) — П. с неоднородной морфо-
логической структурой; наиболее ярко гетерогенность П. прояв-
Рис. 107. Функционально-ритмологические типы побегов: 1 — побег возобновления,
2 — почка возобновления, 3 — почечное кольцо, 4 — побег обогащения, 5 — прирост
(иванов побег, элементарный побег), 6 — побег формирования (по: Серебряков, 1952;
Ботаника..., 1988)
138
ляется, когда он завершается цветком или безлистным цветоно-
сом; в более узком значении (Мазуренко, Хохряков, 1977) — П. с
чередующимися укороченными и удлиненными приростами.
Побег гетеротропный см. побег анизотропный.
Побег гетерофильный (Cormus heterophyllus, от греч. heteros —
другой, pnyllon — лист; англ.: heterophyllous shoot) — П. с раз-
личными листьями, форма, размер и структура которых изменя-
ется от основания стебля к его верхушке (см. формация листьев);
в более узком значении — П. с неодинаковыми листьями средин-
ной формации; к П. г. относятся побеги нек-рых водных раст.
(напр. стрелолист); ср. П. анизофильный.
Побег гипокотильный (Cormus hypocotyleus, от греч. hypo — ни-
же, под, kotyle — чашеобразная структура, здесь — семядоля;
англ.: hypocotyl shoot) — П. ветвления, формирующийся из поч-
ки, расположенной на гипокотиле.
Побег главный (Cormus primarius, от лат. primus — первый; англ.:
primary shoot) — первый по происхождению П„ развитие кото-
рого начинается с момента формирования проростка и заканчи-
вается прекращением меристематической деятельности его кону-
са нарастания; П. г. снизу ограничен шейкой корневой; син.:
П. первичный, П. первый.
Побег годичный (Cormus annotinus, от лат. annus — год; англ.:
annual shoot, current twig) — П. данного года вегетации, цикл раз-
вития которого длится один вегетационный период (ср. П. одно-
летний); часто П. г. наз. годичный прирост.
Побег гомогенный (Cormus homogeneus, от греч. homos — одина-
ковый, genos — род, происхождение) — в узком смысле (Мазу-
ренко, Хохряков, 1977) — П. сложный, все приросты которого уд-
линенные или укороченные; в более широком значении — П. ве-
гетативный с однородной морфологической структурой (ср. П.
гетерогенный); пон. «гомогенность» можно использовать для оп-
ределения однородности П. по любому признаку его морфологи-
ческой структуры, однако, как правило, гомогенность П. относи-
тельна, что связано с его сложным метамерным строением.
Побег гомотропный см. побег изотропный.
Побег горизонтальный см. побег диагеотропный.
Побег двулетний (Cormus biennis, от лат. bis — два раза, дважды,
annus — год; англ.: biennial shoot) — в широком смысле любой
П., живущий в течение двух лет; в специальном значении исполь-
зуется как синоним П. бициклического.
139
Побег диагеотропный (Cormus diageotropus, от греч. dia — при-
ставка со значением через, поперек, geo — земля, trope — пово-
рот, направление; англ.: diageotropic shoot) — П., растущий гори-
зонтально (рис. 105); часто П. д. наз. длинный, горизонтальный
участок П. анизотропного (корневище)-, терм.: Hackel, 1882; син.:
П. внекустовой, П. горизонтальный; лит.: Серебряков,. 1952.
Побег диатропный см. побег изотропный.
Побег дициклический (Cormus dicyclicus, от греч. di — два, kyk-
los — круг) — П. генеративный или П. вегетативно-генератив-
ный, живущий два года от развертывания почки до плодоноше-
ния и отмирания или перехода в фазу вторичной деятельности
(напр. резид); рис. 108; ср. П. двулетний; терм.: Warming, 1883.
Рис. 108. Цикличность развития побегов: А — внутрипочечная стадия развития побе-
га, Б — моноциклический побег, В—дициклический побег, Г — трициклический
(полициклический) побег (1-3 — годичные приросты, 4 — лист, 5 — пазушная почка,
6 — цветок)
Побег долихоморфный (Cormus dolichomorphus, от греч. doli-
chos — длинный, morphe — форма) — терм, заимствован В. Н. Го-
лубевым (1965) из антропологии для обозначения П. удлиненного у
раст. древесного.
Побег дополнения — П. ветвления, развивающийся из почки спя-
щей и выполняющий функцию обрастания оси скелетной кустар-
ника или кустарничка:, к П. д. также относятся колючки побегово-
го происхождения и укороченные П. генеративные (каулифло-
рия); развитие П. д. обычно усиливается в конце онтогенеза или
при механических повреждениях раст.; ср. П. формирования.
140
Побег дочерний см. побег ветвления.
Побег жировой (жирующий) (англ.: nourishing shoot) см. побег
формирования.
Побег закрепления — наз. используется для обозначения орто-
тропного, укорененного участка П. анизотропного, образующего-
ся при размножении вегетативном (корневище, столон).
Побег закрытый см. побег, побег монокарпический.
Побег замещения (Cormus succedaneus, от лат. succedo — замещать,
сменять; англ.: replasement shoot) — П. ветвления, перерастающий
П. материнский и обеспечивающий дальнейшее нарастание оси
скелетной древесного раст., у травянистых раст. такой П. обычно
наз. побегом возобновления; терм.: Серебряков, Чернышева, 1955.
Побег Иванов (Cormus iohanni, англ.: lammas-shoot) — второй за
вегетационный период прирост П. древесного раст., примерно
совпадающий по времени с древним восточнославянским празд-
ником «Иванов день» (рис. 107); в Англии и Сев. Америке П. и.
наз. Ламмасовы побеги (праздник святого Ламмаса), в Герма-
нии — побеги Иоганна; часто П. и. неправильно наз. П. силлепти-
ческий; Серебряков, 1950, 1952; Соколов, Артюшенко, 1957; Груд-
зинская, 1960, 1964; Михалевская, 2002.
Побег изотропный (Cormus isotropus, от греч. isos — равный, оди-
наковый, trope — поворот, направление; англ.: isotropic shoot) —
П., направление роста которого не изменяется в течение всего его
цикла развития (ср. П. анизотропный) и обусловлено односто-
ронним воздействием факторов среды (см. тропизм); П. и. может
быть ориентирован вдоль градиента раздражителя (П. ортотроп-
ный), под прямым углом к нему (П. диатропный) или под любым
другим углом (П. плагиотропный); обычно отмечают геотропизм
П. и., различая П. диагеотропный, П. апогеотропный и П. геотроп
ный (рис. 105); син.: П. гомотропный; лит.: Серебряков, 1952.
Побег изофильный (Cormus isophyllus, от греч. isos — равный, оди-
наковый, phyllon — лист; англ.: isophyllous shoot) — П. с одинако-
выми листьями на разных сторонах оси; ср. П. анизофильный.
Побег интравагинальный (Cormus intravaginalis, от лат. intra —
внутри, vagina — влагалище, обертка; англ.: intravaginal shoot) —
П. ветвления, растущий апогеотропно (см. П. апогеотропный),
реже — анизогеотропно (корневищеложно-ползучее), внутри вла-
галища своего кроющего листа (рис. 11); трудности установления
внутривлагалищного роста П. злаков и осок связаны со скоро-
стью разрушения кроющего листа, в связи с чем различают П.
141
Рис. 54. Основные типы корневищ: А — укороченное гипогеогенное (Bambusa vulgaris), Б — укороченное эпигеогенное (Nym-
phaea alba), В — удлиненное гипогеогенное (Eleocharis palustris), Г — смешанное (Oxalis acetosella), Д — смешанное (Majanthemum
bifolium) 1 — гипогеогенное удлиненное корневище, 2 — катафиллы, 3 — почка возобновления, 4 — придаточные корни,
5 — листовые рубцы, 6 — эпигеогенное укороченное корневище (по: Федоров и др., 1962).
Корневище моноподиальное
(англ.: monopodial rhizome)
см. корневище.
Корневище неопределенное
см. корневище.
Корневище определенное см.
корневище.
Корневище пахиморфное
(Rhizoma pachymorphum, от
греч. paxus — толстый, mor-
phe— вид, форма; англ.: ра-
chymorph rhizome) — К. гипо-
геогенное с нарастанием сим-
подиальным и междоузлиями
укороченными (К. укорочен-
ное), из почек пазушных кото-
рого в результате кущения об-
разуются компактные группы
надземных побегов апогео-
тропных (рис. 55; ср. К. лепто-
морфное); терм. McClure
(1966) ввел для описания К.
Рис. 55. Основные типы корневищ бамбуков
(по: McClure, 1966): А — пахиморфное, Б —
лептоморфное (1 — надземный побег, 2 —
пазушная почка, 3 — рубец от катафилла,
4 — удлиненное междоузлие, 5 — укорочен-
ное междоузлие)
бамбуков; лит.: Серебрякова, 1971а.
Корневище первично-подземное см. корневище гипогеогенное.
Корневище ползучее (англ.: creeping rhizome) см. корневище уд-
линенное.
Корневище симподиальное (англ.: sympodial rhizome) см. корне-
вище.
Корневище смешанное (Rhizoma amphigeogenum, от греч. amphi —
двоякий, geo — земля, genos — род, происхождение) — К., пред-
ставляющее собой систему подземных побегов, часть из которых
надземного (К. эпигеогенноё), другие — подземного происхожде-
ния (К. гипогеогенное)-, рис. 54; терм.: Серебряков, Серебрякова,
1965; син.: К. гипоэпигеогенное, К. эпигеогенно-гипогеогенное.
Корневище удлиненное (Rhizoma elongatum, от греч. е — при-
ставка, здесь со значением усиления, longus — длинный; англ.:
elongated rhizome, leptocaul rhizome, long rhizome p. p.) — К., сте-
бель которого образован междоузлиями удлиненными (рис. 54);
характерно для раст. длиннокорневшцных, К. у. раст. деревянис-
тых наз. ксилоризом; син.: К. ползучее; лит.: Любарский, 1973.
73
Евгений Варминг
(1841-1924)
выдающийся датский ботаник, основатель учения
о жизненных формах растений
Кристиан Раункиер
(1860-1938)
известный датский ботаник,
создатель одной из самых популярных
классификаций жизненных форм растений
позднеинтравагинальный — развивающийся после отмирания
кроющего листа; син.: П. внутривлагалищный, П. внутрикусто-
вой; лит.: Серебрякова, 1971а.
Побег итеративный (Cormus iterativus, от лат. itero — вторично
делать, повторять; англ.: iterative shoot) — П. ветвления, разви-
вающийся пролептически (П. пролептический) в результате ветв-
ления итеративного (рис. 12); терм.: Troll, 1964.
Побег клиноапогеотропный (Cormus clinoapogeotropus, от греч.
klino — наклонять, аро — приставка, здесь со значением отрица-
ния, обратности, geo — земля, trope — поворот, направление) —
П., растущий наклонно вверх (рис. 105); частный случай П. пла-
гиотропного или П. ортотропного; син.: П. косоапогеотропный.
Побег клиногеотропный (Cormus clinogeotropus, от греч. klino —
наклонять, geo — земля, trope — поворот, направление) — П. рас-
тущий наклонно вниз (рис. 105); частный случай П. плагиотропно-
га, син.: П. косогеотропный.
Побег косоапогеотропный см. побег клиноапогеотропный.
Побег косогеотропный см. побег клиногеотропный.
Побег косо-экстравагинальный (Cormus plagio-extravaginalis, от
греч. plagios — косой, боковой, поперечный, лат. extra — вне,
vagina — влагалище) — П. клиноапогеотропный, разрывающий
кроющий лист своим основанием, а не верхушкой (см. П. экстра-
вагинальный); син: П. клино-экстравагинальный; терм.: Артамо-
нова, 1963; лит.: Серебрякова, 1971а.
Побег крупноквантовый — наз. введено Т. И. Серебряковой
(19696) для обозначения П., развивающегося из почки, содержа-
щей несколько циклов листовых (см. емкость почки); ср. П. мел-
коквантовый.
Побег кустящийся (Cormus ramificans, от лат. ramus — ветвь) —
П. апогеотропный злаков, интенсивно ветвящийся в своей базаль-
ной части (в области укороченных междоузлий); терм.: Смелов,
1966.
Побег кущения (Cormus fruticationis, от лат. frutex — куст; англ.:
tiller) — П. ветвления злаков, развивающийся из почки, располо-
женной в зоне кущения П. материнского; в более узком значе-
нии — П., образующийся в фазу кущения раст. виргинилънога,
син.: П. отрастания.
Побег материнский (Cormus maternus, от лат. mater — мать) — П.
главный или П. любого порядка ветвления, который дал боковое
ответвление и в силу чего является по отношению к этому П. бо-
142
ковому{П. дочернему) побегом основным (рис. 31); син.: П. основ-
ной, П. несущий, П. производящий.
Побег мелкоквантовый — наз. введено Т. И. Серебряковой (19696)
для обозначения П„ развивающегося из почки, содержащей толь-
ко один цикл листовой (см. емкость почки); ср. П. крупнокванто-
вый.
Побег многолетний (Cormus perennis, от лат. per — приставка со
значением действия, направленного через; annus — год; англ.:
perennial shoot) — в широком смысле — любой П., живущий не-
сколько лет; в специальном значении наз. используется как сино-
ним П. полициклического.
Побег монокарпический — П., цикл жизненный малый которого
заканчивается образованием из его меристемы апикальной цвет-
ка или соцветия {побег закрытый); после плодоношения П. м. от-
мирает полностью или частично, но тогда его нижняя часть {ре-
Зид, подиум), на которой располагаются почки возобновления, вхо-
дит в состав многолетней системы побеговой {каудекса, корневища
и т. п.); по длительности развития различают П. моноцикличе-
ский, П. дициклический, П. трициклический, П. полициклический
(рис. 108); обычно П. м. наз. побеги возобновления раст. травяни-
стых, лит.: Серебряков, 1952; Серебрякова, 1971а; Гатцук, 19746.
Побег моноподиальный (Cormus monopodialis, от греч. monos —
один, единый, podos — нога) — в специальном значении — П.,
рост которого никогда не завершается образованием генератив-
ных органов (П. открытый); часто в его границы включают па-
зушные соцветия (см. система побегов) и тогда наз. П. поликар-
пическим; в широком смысле наз. П. м. полностью совпадает с
понятием «побег» (П. одноосный); терм.; Василевская, 19576.
Побег моноритмичный см. побег элементарный.
Побег моноциклический (Cormus monocyclicus, от греч. monos —
один, kyklos — круг) — П. генеративный или П. вегетативно-ге-
неративный, цикл развития которого от развертывания почки, до
плодоношения и отмирания длится один год (рис. 108); П. м. не
всегда является П. однолетним, т. к. его базальная часть может
входить в состав оси скелетной (см. резид); терм.: Warming, 1883;
лит.: Серебряков, 1952.
Побег надземный (Cormus epigaeus, от греч. epi — на, над, сверх,
geo — земля; англ.: epigeal shoot, above-ground shoot) — наз.
обычно используется как противопоставление П. подземному
(англ.: underground shoot) и для обозначения П. ортотропного в
143
отличие от П. наземно-ползучего (см. П. размножения); реже —
П. ортотропный наз. П. наземным.
Побег несущий см. побег материнский.
Побег обогащения (Cormus amplificandi, от лат. amplif ico — расши-
рять, увеличивать, приумножать) •— П. однолетний, развиваю-
щийся как П. пролептический или П. силлешпический обычно в
верхней части стеблям обеспечивающий увеличение (обогащение)
общей фотосинтезирующей или репродуктивной поверхности
растений (рис. 107); П. о. культурных растений наз. «пасынками».
Побег однолетний (Cormus annotinus, от лат. annus — год) — П„
полный онтогенез которого длится не более одного года; обычно
развивается как П. пролептический или П. силлептический; к П. о.
относятся П. обогащения, столон, П. генеративный и др. (ср. П.
моноциклический); син.: П. эфемерный.
Побег одноосный см. побег моноподиальный.
Побег ортотропный (Cormus orthotropus, от греч. orthos — пря-
мой, trope — поворот, направление; англ.: orthotropic shoot) —
частный случай П. изотропного; обычно П. о. наз. П. надземный,
вертикальное положение которого обусловлено отрицательным
геотропизмом (рис. 105; П. апогеотропный и П. клиноапогетроп-
ныи); лит.: Серебряков, 1952.
Побег основной см. побег материнский.
Побег открытый см. побег, побег моноподиальный.
Побег пазушный см. побег боковой.
Побег пассивно-экстравагинальный см. побег экстравагиналь-
ный.
Побег первичный см. побег главный.
Побег первый см. побег главный.
Побег плагиотропный (Cormus plagiotropus, от греч. plagios — ко-
сой, боковой, поперечный, trope — поворот, направление; англ.:
plagiotropic shoot) — частный случай П. изотропного; обычно
П. п. наз. побег, обладающий б. м. продольным геотропизмом
(рис. 105), включая П. диагеотропный, П. клиноапогеотропный и
П. клиногеотропный; лит.: Серебряков, 1952; Сабинин, 1963; Син-
нот, 1963.
Побег плодущий (Cormus fertilis, от лат. fero — производить, рож-
дать; англ.: spur) в плодоводстве — укороченный П. вегетатив-
но-генеративный.
Побег подземный (англ.: underground shoot) см. побег надземный.
Побег позднеинтравагинальный см. побег интравагинальный.
144
Побег ползучий (англ.: repent shoot) — неспециализированный,
удлиненный, укореняющийся побег, стелющийся по поверхно-
сти почвы и несущий только формацию срединных листьев, реже
и низовых (ср. столон); у многих раст. ползучих П. п. могут вре-
менно изменять направление роста: приподнимаясь (Veronica
officinalis) или зарываясь в субстрат (Glechoma); в более широком
смысле к П. п. относят и специализированные побеги размноже-
ния; лит.: Серебрякова, 1981.
Побег поликарпический см. побег моноподиальный.
Побег полициклический (Cormus polycyclicus, от греч. polys —
много, kyklos — круг) — П. вегетативно-генеративный, цикл
развития которого от развертывания почки до плодоношения и
отмирания или перехода в фазу вторичной деятельности длится
много лет (рис. 108); не каждый П. многолетний является П. п.;
терм.: Warming, 1883; лит.: Серебряков, 1952.
Побег полурозеточный (Cormus semirosularis, от лат. semi — напо-
ловину, rosa — розетка; semirosette shoot) — П. вегетативно-гене-
ративный с сильно сближенными ассимилирующими листьями в
основании (розеточная часть), за которым следует префлоральный
участок с удлиненными междоузлиями и ассимилирующими ли-
стьями часто меньшего размера (рис. 106); частный случай П. ро-
зеткообразующего; лит.: Серебряков, 1952; Голубев, 1965; Серебря-
кова, 1971а.
Побег посредник — наз. введено Г. М. Денисовой (1960) для обо-
значения П. апогеотропного или П. клиноапогеотропного, входя-
щего в состав куста парциального длиннокорневищного злака и
от подземной части которого отходит п. диагеотропный.
Побег придаточный см. побег адвентивный.
Побег продолжения см. побег замещения.
Побег производный см. побег ветвления.
Побег производящий см. побег материнский.
Побег пролептический (Cormus prolepticus, от греч. prolepis — уп-
реждение; англ.: proleptic shoot) — П. ветвления, возникающий из
незимующей почки, но находившейся некоторое время в состоя-
нии покоя; к П. п. относится П. итеративный; иногда П. п. непра-
вильно наз. побегом Ивановым; терм.: Spath, 1912; лит.: Грудзин-
ская, 1960.
Побег прямостоячий см. побег апогеотропный.
Побег разветвленный см. система побегов.
Побег раздвинутоузловой см. побег удлиненный.
145
Побег размножения (Cormus propagationis, от лат. propagare —
размножать, разводить, распространять; англ.: propagation sho-
ot) — специализированный П., обеспечивающий размножение ве-
гетативное раст.; к П. р. относятся столон, усы (англ.: runner),
корневище, которые часто наз. побегами наземно- или подземно-
ползучими или, реже — П. убегающими.
Побег разрастания (Cormus accrescentiae, от лат. accrescere — воз-
растать, увеличиваться) — П. удлиненный, обеспечивающий веге-
тативное разрастание раст. в горизонтальной плоскости; син.: П.
расселения.
Побег расселения см. побег разрастания.
Побег репродуктивный см. побег генеративный.
Побег ритма см. побег элементарный.
Побег розеткообразующий — назв. введено Т. И. Серебряковой
(1967) для обозначения П., который в цикле своего развития име-
ет фазу «розетки», а в структуре — участок с укороченными меж-
доузлиями и ассимилирующими листьями, образующими розет-
ку; по степени облиственности префлоралъного участка различа-
ют П. полурозеточный и собственно П. розеточный (рис. 106);
лит.: Серебрякова, 1971а.
Побег розеточный (Cormus rosulans, от лат. rosa — розетка; англ.:
rosette shoot) — П. укороченный, стебель которого несет более
трех ассимилирующих листьев, образующих так наз. «розетку»
(рис. 106); у травянистых раст. «розетка» как правило, приземная, у
деревянистых — надземная (П. акрофильныи); если П. вегетатив-
но-генеративный, то его префлоральный участок представлен
стрелкой; лит.: Серебряков, 1952; Голубев, 1965; Серебрякова, 1971а.
Побег сближенноузловой см. побег укороченный.
Побег силлептический (Cormus syllepticus, англ.: sylleptic sho-
ot) — П. ветвления, развивающийся из почки без периода покоя и
одновременно с продолжающимся ростом его П. материнского;
иногда П. с. неправильно наз. побегом Ивановым; терм.: Spath, 1912;
лит.: Грудзинская, 1960.
Побег скелетный (Cormus sceletalis, от греч. sceletus — скелет) —
побег раст. многолетнего, в структуре которого есть многолетний
участок (у раст. травянистых — резид); представляет собой ос-
нову формирования оси скелетной раст.; терм.: Гатцук, 1974а.
Побег скрытогенеративный (Cormus cryptogenerativus, от греч.
kryptos — скрытый, лат. generate — рождать, производить) —
у злаков П. вегетативно-генеративный, верхний (флоральный)
146
участок которого не развивается в силу экзо(эндо)генных при-
чин; терм.: Смелов, 1947.
Побег скрытый см. почка спящая.
Побег слепой (Cormus caecus — от лат. caecus — слепой) — как
правило, однолетний, удлиненный П. вегетативный злаков; лит.:
Серебрякова, 1971а.
Побег смешанный см. побег вегетативно-генеративный.
Побег сосунок—фаза развития П. злаков, характеризующаяся от-
сутствием корней придаточных; терм.: Смелов, 1947.
Побег специализированный см. побег размножения.
Побег стерильный см. побег вегетативный.
Побег трициклический (Cormus tricyclicus, от греч. tria — три, kyk-
los — круг) — П., цикл развития которого от развертывания почки
до плодоношения и отмирания или перехода в фазу вторичной
деятельности длится три года (рис. 108); терм.: Warming, 1883.
Побег убегающий см. побег размножения.
Побег удлиненный (Cormus elongatus, англ.: elongated shoot, long
shoot) — П. с расставленными узлами, длина междоузлий которо-
го превышает их ширину (рис. 106); П. у. раст. древесныхназ. аук-
сибластом (макробласт); лит.: Серебряков, 1952; Голубев, 1965;
Паутов, 1987
Побег укороченный (Cormus abbreviatus, англ.: short shoot,) — П. с
сильно сближенными узлами и короткими междоузлиями, длина
которых значительно меньше их ширины (рис. 106); П. у. раст.
древесного наз. брахибласт (плодушки, кольчатки), а длинный П.
у. иногда — П. сближенноузловым; лит.: Серебряков, 1952; Голу-
бев, 1965; Паутов, 1987.
Побег фертильный см. побег генеративный.
Побег фитофильный (Cormus phytophilus) — П., в цикле разви-
тия которого есть фаза облиственного стебля.
Побег формирования (Cormus formativus) — П., развивающийся
из почки спящей и служащий основой для формирования главной
оси скелетной (стволика) кустарников^ кустарничков (рис. 68,107);
от других побегов ветвления отличается крупными размерами и
усиленным ростом; терм.: предложен М. Т. Мазуренко и А. П. Хох-
ряковым (1977) для обозначения побегов, именуемых турион,
П. возобновления, поросль стеблевая (англ.: stump sprout), П. замеще-
ния, П. ростовой (англ.: leading shoot), П. порослевой (англ.: sobole),
П. нарастания, П. сильный, П. обновления, П. водяной (англ: water
sprout), П. жирующий (англ.: nourishing shoot), волчок; П. ф. выпол-
147
няет гл. обр. функцию омоложения побеговой системы и завоева-
ния пространства раст. {ксилоризом)-, лит.: Мазуренко, 1986.
Побег цветоносный см. побег генеративный.
Побег экстравагинальный (Cormus extravaginalis, англ.: extrava-
ginal shoot) — П. диагеотропный или П. анизотропный, проры-
вающий верхушкой влагалище своего кроющего листа (рис. 11;
П. вневлагалищный); в некоторых случаях П. э. развивается после
разрушения своего кроющего листа (П. пассивно-экстраваги-
нальный); лит.: Серебрякова, 1971а.
Побег элементарный — наз. используется для обозначения при-
роста побега, образующегося за один цикл видимого роста
(рис. 107; напр., побег иванов)-, терм.: Грудзинская, 1960; син.:
П. моноритмичный, П. ритма.
Побег эпикотильный (Cormus epicotyleus, англ.: epicotyl shoot) —
П. ветвления, развивающийся из почки, расположенной на эпико-
тиле.
Побег эфемерный см. побег однолетний.
Побегообразование (англ.: shoot formation, sprout formation) —
широко используемое назв. для обозначения различных процес-
сов, обусловливающих формирование системы побеговой: ветв-
ления, нарастания, возобновления вегетативного, цикла жизненно-
го малого.
Поведение растения (англ.: plant behavior) — способность раст.
реагировать на воздействия экзогенных и эндогенных факторов;
в отличие от животных проявляется прежде всего в ростовых ре-
акциях (ростовое поведение): ритм и направление роста побега и
корня, ветвление, движение раст. (см. расселение вегетативное),
обновление адсорбирующей поверхности (поддержание жизни
особи)-, к П. р. относят также различные видовые адаптации, обес-
печивающие размножение и расселение, физиологические реак-
ции на изменение внешней среды и взаимодействия с окружаю-
щими организмами в биогеоценозе; син.: образ жизни, стратегия
роста раст.; лит.: Arber, 1950; Серебрякова, 1980; Silverstown,
Gordon, 1989; Bell, Bryan, 1998.
Подвижность вегетативная см. расселение вегетативное.
Подиум см. резид.
Подушка воздушная см. растение подушковидное.
Подушка дерновинная см. растение подушковидное.
Подушка плотная (англ.: hard (polster) cushion) см. растение по-
душковидное.
148
Подушка радиально-(полу)шаровидная (англ.: radial-hemisphe-
ric (polster) cushion) см. растение подушковидное.
Подушка радиально-плоская (англ.: radial-flat (polster) cushion,
flat-topped cushion) см. растение подушковидное.
Подушка розеточная (англ.: rosette (polster) cushion) см. растение
подушковидное.
Подушка рыхлая (англ.: lax (polster) cushion) см. растение подуш-
ковидное.
Подушка-сообщество см. растение подушковидное.
Покой абсолютный (англ.: absolute dormancy) см. покой растения.
Покой биологический (англ.: biological dormancy) см. покой рас-
тения.
Покой вынужденный (англ.: induced dormancy) см. покой рас-
тения.
Покой глубокий (англ.: true dormancy) см. покой растения.
Покой органический (англ.: organic dormancy) см. покой растения.
Покой первичный см. покой растения.
Покой растения (англ.: dormancy (rest, quiescence) of plant) — фи-
зиологическое состояние семян (П. первичный), почек, вегетатив-
ных органов (корневища, луковицы и т. п.) или всего раст., во время
которого прекращается видимый рост, но сохраняются скрытые
процессы структурообразования (напр. рост внутрипочечный)',
рассматривается как приспособление для переживания неблаго-
приятный внешних условий в определенные периоды онтогенеза
или сезоны года; различают несколько типов П., среди которых два
основных: П. глубокий (П. органический, абсолютный или биоло-
гический), вызванный внутренними причинами (ингибиторами
роста) и П. вынужденный, обусловленный факторами внешней
среды; лит.: Серебряков, 19596, 1966; Ворошилов, 1960; Сабинин,
1963; Тюрина, 1979; Шампанья, 1983.
Поливариантность онтогенеза см. поливариантность развития.
Поливариантность развития — реализация вариантов общей
программы морфогенеза и онтогенеза (П. онтогенеза, П. индиви-
дуального развития) в различных условиях среды, отражающих
изменчивость биоморфологическую. Выделяют несколько типов
(морфологическая, ритмологическая и др.), которые обычно рас-
сматривают как модификации или как проявление мобилизаци-
онного резерва внутривидовой изменчивости; син.: многовари-
антность развития; лит.: Жукова, 1995; Жукова, Глотов, 2001.
Поликарпик (Polycarpic; от греч. polys — многий, karpos — плод;
149
англ.: polycarpic) — раст., неоднократно цветущее и плодонося-
щее в течение своего онтогенеза (ср. монокарпик); большинство
многолетников— П„ среди которых иногда выделяют раст. с не-
продолжительным периодом генеративным (олигокарпик, раст.
олигокарпическое, см. также малолетник). Разнообразная группа
растений травянистых и растений деревянистых, которые отли-
чаются формой жизненной, продолжительностью жизни (см. ма-
лолетник) и старением (см. старение негативное)', терм.: Candolle,
1827; син.: раст. поликарпическое, раст. итеропароузное.
Поликотилия (Polycotylia, от греч. polys — много, kotyle — доля;
англ.: polycotyledony) — наличие у зародыша и проростка более
двух семядолей-, как обычное явление встречается у голосеменных
(до 10-15 семядолей и более); у покрытосеменных увеличение
числа семядолей — следствие тератологических отклонений (рас-
щепления); син.: многосемядольность, плейокотилия.
Полимеризация (от греч. polys— многочисленный, обширный,
meros — часть; англ.: polymerization) — один из способов морфо-
логической эволюции организмов, благодаря которому увеличи-
вается число гомологичных органов или структур (ср. олигомери-
зация). Различают первичную П. (возникновение у потомка но-
вых полимерных структур) и вторичную П. (увеличение числа
органов, существующих у предка); терм.: Догель, 1929,1954; лит.:
Тахтаджян, 1964; Хохряков, 1974, 1975, 1981; Мейен, 1977; Замор-
ский, 1971, 1980; Северцов, 1981; Терехин, 1991.
Полифит (Polyphyton; от греч. polys — много, phyton — расте-
ние) — раст., способное к образованию клона в результате естест-
венного, специализированного (расползающийся П.) или неспе-
циализированного (партикулирующий и куртинообразующий
П.) размножения вегетативного; к П. относятся гл. обр. раст. веге-
тативно-подвижные и нек-рые раст. короткокорневищные, раст.
кистекорневые, лианы, раст. луковичные, раст. дерновинныеи др.;
терм.: Хохряков, 1975а.
Полудерево см. растение полудревесное.
Полукустарник (англ.: subshrub, half-shrub, semishrub, semi-
(suf)frutex) см. растение полудревесное.
Полукустарничек (англ.: dwarf (subshrub) semischrub) см. растение
полудревесное.
Полупаразит (Hemiparasitus, греч. hemi — полу, parasitus — пара-
зит; англ.: hemiparasite, semi-parasite, partial parasite) — часто ис-
пользуемое пон. для обозначения раст. паразитов, способных к
150
фотосинтезу (зеленые паразиты); П.— раст. с облигатным
(Viscum) и факультативным (Euphrasia) паразитизмом; при более
широком понимании явления паразитизма к П. относят и обли-
гатно-микотрофные раст. (аллелопаразиты) с зелеными листья-
ми, которые часто наз. полусапрофитами (Pyrolaceae, Burmannia-
сеае и др.); лит.: Терехин, 1965, 1977; Бейлин, 1968.
Полупаразитизм (англ.: hemiparasitism) см. паразитизм.
Полусапрофиты (англ.: semi-saprophyte, hemisaprophyte) см. полу-
паразит.
Полустланец см. растение стелющееся.
Полустланик см. растение стелющееся.
Полустланичек см. растение стелющееся.
Полуэпифит (Hemiepiphyton, от греч. hemi — полу- и эпифит;
англ.: hemiepiphyte) — разнообразная группа раст. травянистых
и раст. деревянистых, поселяющихся на других раст. (эпифиты),
но вначале (вторичные эпифиты) или во второй половине онто-
генеза (первичные эпифиты), связанные корнями с почвой; син.:
семиэпифит, гемиэпифит.
Полуэфемероид см. гемиэфемероид.
Полярность (Polaritas, от греч. polos — полюс; англ.: polarity) —
свойственная организмам специфическая ориентация процессов
и структур в пространстве, приводящая к возникновению морфо-
физиологических различий на противоположных концах или
сторонах клеток, тканей, органов или организма в целом; наибо-
лее ярко выражена у высших раст. и обнаруживается уже на ран-
них стадиях онтогенеза в виде оси с апикальным (почечка) и ба-
зальным (корешок) полюсами (биполярные растения), вокруг ко-
торой позднее симметрично возникают боковые образования:
побеги, листья, почки, и т. п. (см. рост биполярный); у споровых
раст. корешок закладывается сбоку и ось полярности формирует-
ся позднее (униполярные растения; см. рост униполярный); осно-
ванная на морфофизиологических взаимоотношениях противо-
положных полюсов, П. определяет физиологические и структур-
ные градиенты, разнокачественность метамеров, способность к
восстановлению утраченных частей (регенерации) и др.; лит.: Мо-
лотковский, 1961; Синнот, 1963; Сабинин, 1963.
Поросль корневая (англ.: root shoot) см. отпрыск корневой.
Поросль стеблевая (англ.: soboles) см. побег формирования.
Почечка (Plumula, от лат. pluma — перышко; англ.: plumule) —
почка верхушечная зародыша и проростка, содержащая зачаток
151
эпикотиля и примордии первых листьев; П. зародыша сильнее
развита у раст. с прорастанием подземным, чем с прорастанием
надземным; син.: плюмула.
Почка (Gemma; от лат. geno — рожать, (по)рождать, производить
на свет; англ.: bud, gemma) — неразвернувшийся побег или при-
рост, зачаточный стебель которого покрыт катафиллами (П. за-
крытая) и/или разновозрастными зачатками листьев (П- голая),
возникающими в акропетальной последовательности и тесно
примыкающими друг к другу (рис. 109), защищая конус нараста-
ния побега (П. вегетативная) или зачаток соцветия (П. генера-
тивная); возвышение и загибание наружных (нижних) молодых
листьев над апексом связано с их неравномерным ростом (акро-
вергенция листовых зачатков). Взаиморасположение листьев и
цветковых покровов в П. наз. почкосложением (почкосмыкани-
ем) или листосмыканием, а состояние пластинки (свернутое,
плоское и т. п.) каждого листа в П.— листосложением. Топогра-
фически различают П. боковую и П. верхушечную, функциональ-
но — П. возобновления и П. спящую, а по времени развития побе-
га— П. начальную, П. промежуточную и П. завершающую. П.—
ростовая единица, отражающая особенности раннего периода
развития побега (см. рост внутрипочечный); формирования и
строения П. во многом предопределяют морфологический облик
взрослого побега; лит.: Серебряков, 1952; Шилова, 1967, 1988; Ры-
това, 1972; Серебрякова, 1971а, 1983а; Падеревская, 1984; Шорина,
Михалевская, 2000.
Рис. 109. Строение закрытых (А-Б) и голых (В-Г) почек (по: Ботаника..., 1988): А — ве-
гетативная почка дуба, Б — генеративно-вегетативная почка бузины, В — зимующая
почка калины-гордовины, Г — продольный разрез верхушечной почки злака (1 — за-
чатки листьев, 2 — зачатки цветков, 3 — почечные чешуи, 4 — пазушные почки)
152
Почка адвентивная (англ.: adventitious bud) см. почка прида-
точная.
Почка аксиллярная (англ.: axillary bud) см. почка пазушная.
Почка бисериальная (англ.: biserial bud) см. почка добавочная.
Почка боковая (Gemma lateralis, от лат. latus — бок, сторона; англ.:
lateral bud) — П., расположенная на боковой стороне стебля (П.
стеблевая), в пазухе (П. пазушная) или вне пазухи листа (П. прида-
точная, П. внепазушная); развитие П. б. обусловливает ветвле-
ние раст.
Почка в почке см. ветвление внутрипочечное.
Почка вегетативная (Gemma vegetativa, от лат. vegetare — ожив-
лять; англ.: vegetative bud) — П. зрелая, содержащая зачатки толь-
ко органов вегетативных (рис. 109): листьев, стебля и, нередко,
почек пазушных (см. ветвление внутрипочечное); иногда П. в. не-
правильно наз. почкой листовой (см. побег вегетативный); син.:
п. ростовая.
Почка верхушечная (Gemma apicalis, от лат. apex — вершина;
англ.: apical bud) — П., занимающая терминальное положение на
стебле (П. стеблевая); обеспечивает нарастание побега в длину и
образование его новых фитомеров; иногда П. в. наз. почкой тер-
минальной, или П. конечной.
Почка влагалищная см. почка пазушная.
Почка внепазушная (Gemma extraaxillaris, от лат. extra — за, вне,
выше, axilla — подмышка; англ.: extra-axillary bud) — IL, распо-
ложенная на боковой стороне стебля (П. стеблевая) вне пазухи
листа; внепазушное положение почки может быть первичным
(и. придаточная) или вторичным, в результате перемещения
(конкаулесценции) почки, изначально заложенной в пазухе лис-
та, вверх по стеблю или на лист.
Почка внешняя см. почка пазушная.
Почка внутренняя см. почка пазушная.
Почка возобновления (Gemma innovationis; от лат. innovare — во-
зобновление; innovation (renewal, perennating) bud) — любая IL,
которая после некоторого периода покоя формирует новый побег
(П. боковая) или прирост (П. верхушечная). Обычно П. в. называ-
ют почку (рис. 107), период относительного покоя которой связан
с наступлением зимнего (П. зимующая) или другого (П. покоя-
щаяся) неблагоприятного для вегетации времени года (ср. почка
спящая). По степени сформированности побега в почке выделяют
три группы растений; лит.: Серебряков, 1952; Вихирева-Василько-
153
ва, 1962; Гамцемлидзе, 1979; Кузнецов, 1985; Шорина, Михалев-
ская, 2000.
Почка выводковая — терм, используется для обозначения П. эпи-
фильной, специализированной как орган размножения вегетатив-
ного некоторых папоротников (Asplenium viviparum) и цветковых
раст. (Bryophyllum); П. в. образует небольшой побеге корешками
придаточными, который опадает с раст. и дает начало новой особи
(см. эпифиллия); иногда П. в. наз. луковичку, клубенек или буль-
бочку (см. вивипария); обычно понятие П. в. применяют преиму-
щественно к низшим раст.; лит.: Батыгина, Брагина, 20006.
Почка генеративная (Gemma generativa, от лат. generare — рож-
дать, производить; англ.: flower bud) — П. зрелая, содержащая за-
чатки структур только генеративной сферы раст.: цветка (тогда П.
г. наз. бутоном) или соцветия и цветоноса; иногда П. г. наз. П.
цветочной, репродуктивной или неправильно — П. плодовой
(англ.: fruit bud).
Почка генеративно-вегетативная (Gemma generativo-vegetativa,
от лат. generare — рождать, производить, vegetare — оживлять;
англ.: reproductive (mixed) bud) — П. зрелая, содержащая зачатки
органов вегетативных и генеративных: листьев, стебля, цветка или
соцветия (рис. 109); иногда П. г.-в. наз. П. смешанной (mixed bud).
Почка голая (Gemma nuda, от лат. nudus — голый; англ.: naked
bud) — П„ не имеющая специализированных покровов (ката-
филлов) или не закрытая другими, замкнутыми, защитными об-
разованиями: колеоптилем, предлистом (рис. 109); иногда П. г.
наз. П. открытой; в буквальном смысле «открытых» П. нет, т. к.
молодые зачатки любой П. обычно прикрыты более взрослыми
листьями и, располагаясь в пазухе листа, б. м. защищены его ос-
нованием; следует различать П., исходно лишенные защитных
покровов, и П. с опавшими почечными чешуями (G. deperulata);
первые — характерная черта многих деревьев влажных тропиков
и субтропиков; у раст. древесных умеренных широт такие П.
встречаются как исключение (Frangula); терм.: Mirbel, 1815.
Почка добавочная (Gemma accessoria, от лат. accedere — подхо-
дить, присоединяться, прибавляться; англ.: accessory bud, super-
nummerary bud) — П., расположенная сбоку (П. коллатеральная)
или сверху (П. сериальная) от основной (англ.: main bud), обычно
более крупной П. пазушной и т. о. являющаяся добавочной (П. по-
бочной) по отношению к ней; формирование П. д. связано с дея-
тельностью пазушной меристемы и происходит в центробежной
154
(П. коллатеральные), нисходящей или восходящей последова-
тельности (П. сериальные); сериальное (вертикальное) располо-
жение почек может быть одно- (Lonicera) и двурядным (Acacia)
или уни- и бисериальным; П. коллатеральные встречаются у раст.
(гл. обр. однодольных) с широким основанием листьев (Allium
sativum); лит.: Серебряков, 1952.
Почка дормитивная см. почка спящая.
Почка дремлющая см. почка спящая.
Почка завершающая — почка верхушечная, из которой развивает-
ся генеративный прирост или последний прирост побега вегета-
тивного (ср. почка начальная, почка промежуточная)-, син.: почка
финальная; лит.: Шорина, Михалевская, 2000.
Почка закрытая (Gemma tecta, от лат. tectum — крыша; англ, bud
with scales) — П., имеющая специальный защитный покров (рис.
109); иногда различают П., снабженную катафиллами (G. tecta), и
П., скрытую под замкнутым защитным образованием, колеопти-
лем или предлистом (G. abscondita; Роасеае); ср. П. голая; лит.: Па-
деревская, 1958.
Почка зародышевая (Plumula, уменьшительное от лат. pluma —
перышко; англ.: plumule) — почечка зародыша семени, располо-
женная между семядолями (двудольные) или смещенная семядо-
лей в боковое положение (однодольные); нередко наз. плюмулой
и объединяют с пон. эпикотиль.
Почка зимующая (Gemma hibernans, от лат. hiberno — зимовать;
англ.: winter bud) — наз используется для обозначения разных об-
разований: П. возобновления наземных раст. и специализированно-
го зимующего органа нек-рых водных раст. (см. гибернакула).
Почка зрелая (Gemma matura, от лат. maturus — зрелый, спелый,
взрослый) — П., содержащая предельное (максимальное) число
зачатков (см. емкость почки); терм.: С. П. Смелов (1937) ввел для
обозначения П„ которая при отделении от раст. способна укоре-
ниться; лит.: Серебрякова, 1971а.
Почка инициальная см. почка начальная.
Почка интермедиальная см. почка промежуточная.
Почка коллатеральная (англ.: collateral bud) см. почка добавочная.
Почка конечная см. почка верхушечная.
Почка корневая (англ.: root bud) см. почка придаточная.
Почка листовая (англ.: leaf bud) — наз. иногда используют для
обозначения П. вегетативной (Gemma folirera) или П. эпифиль-
ной (G. foliaris).
155
Почка наружная см. почка пазушная.
Почка начальная — почка боковая, из которой начинает разви-
ваться побег ветвления или его первый прирост (ср. почка проме-
жуточная, почка завершающая); син. почка инициальная; лит.:
Шорина, Михалевская, 2000.
Почка незрелая см. емкость почки.
Почка неприкрытая см. почка пазушная.
Почка основная (англ.: main bud) см. почка пазушная.
Почка открытая см. почка голая.
Почка пазушная (Gemma axillaris, от лат. axilla — подмышечная
ямка; англ.: axillary bud) — П., расположенная в пазухе листа или
катафилла (почка начальная); нередко наз. П. аксиллярной; рас-
положение П. и. относительно друг друга подчиняется в основ-
ном характеру листорасположения; иногда в пазухе листа закла-
дывается не одна, а несколько почек обычно меньшего размера
(см. П. добавочная), чем П. основная; к П. п. относятся так наз. П.
прикрытые или П. погруженные (П. внутренние), которые в от-
личие от обычных П. п. (П. неприкрытые, П. внешние, П. наруж-
ные или П. свободные) скрыты (П. скрытая) листовым рубцом,
прилистниками, листовой подушкой, расширенным основанием
черешка (П. черешковая) или влагалищем (П. влагалищная).
Почка плодовая (англ.: fruit bud) см. почка генеративная.
Почка побочная см. почка добавочная.
Почка погруженная см. почка пазушная.
Почка покоящаяся (англ.: dormant (resting) bud) см. почка спящая.
Почка покрытая см. почка закрытая.
Почка превентивная см. почка спящая.
Почка преформированная (от лат. praeformo — заранее образую,
предобразую; англ.: preformated bud) — почка возобновления, ко-
торая содержит все зачатки будущего побега генеративного (П. ге-
неративно-вегетативная, П. генеративная). Halle et al. (1978) ис-
пользовали название «преформация» в более широком смысле,
как развитие побега за один цикл видимого роста, заложение за-
чатков которого соответствует единице морфогенеза. Противопо-
ложный случай представляет «неоформация» — заложение до-
полнительных зачатков во время внепочечной фазы развития по-
бега и их раскрывание в том же году; лит.: Шорина, Михалевская,
2000; Михалевская, 2002.
Почка придаточная (Gemma adventicia, от лат. ad — приставка в
значении дополнительности, venio — приходить, становиться,
156
возникать; англ.: adventitious bud) — П. боковая (см. почка началь-
ная), закладывающаяся вне прямой связи с меристемой апикаль-
ной, а в результате дедифференциации постоянных тканей. П. п.
могут закладываться на корне (П. корневая), листе (П. эпифильная;
см. эпифиллия) или стебле (П. внепазушная). П. п. развиваясь на
корнях (см. отпрыск корневой) или листьях (Bryophyllum), обеспе-
чивают размножение вегетативное, а при повреждениях на стеб-
ле — восстановление утраченных частей; син.: П. адвентивная.
Почка прикрытая см. почка пазушная.
Почка промежуточная — почка верхушечная, из которой разви-
вается второй или последующие приросты (кроме последнего)
побега полициклического или побега многолетнего (ср. почка на-
чальная, почка завершающая); син.: почка интермедиальная; лит.:
Шорина, Михалевская, 2000.
Почка резервная см. почка спящая.
Почка репродуктивная см. почка генеративная.
Почка ростовая см. почка вегетативная.
Почка свободная см. почка пазушная.
Почка сериальная (англ.: serial (superposed) bud) см. почка доба-
вочная.
Почка скрытая см. почка пазушная.
Почка смешанная (англ.: mixed bud) см. почка генеративно-веге-
тативная.
Почка спящая (Gemma dormiens, от лат. dormio — спать; англ.:
dormant (resting) bud) — П. пазушная или П. придаточная, нахо-
дящаяся в состоянии относительного покоя (П. покоящаяся) на
протяжении нескольких лет (рис. 110); в одних случаях П. с. дрем-
лет (П. дремлющая) в течение всей жизни раст., ежегодно нарас-
тая примерно на толщину годичного кольца древесины и образуя
скрытую веточку (Побег скрытый), в других — пробуждается по-
сле повреждения ствола (ветви) или естественного старения сис-
темы побегов (П. превентивная), образуя побег формирования
(П. резервная); П. с. характерны для лиственных деревьев, кустар-
ников, кустарничков и некоторых многолетних трав; син.: П. дор-
митивная; лит.: Любименко, 1900; Серебряков, 1952; Ляшенко,
1958; Падеревская, 1957; Михалевская, 2002.
Почка стеблевая см. почка боковая.
Почка терминальная см. почка верхушечная.
Почка финальная см. почка завершающая.
Почка цветочная (англ.: flower bud) см. почка генеративная.
157
Рис. 110. Спящие почки (по: Ботаника..., 1988):
1 — нарастание спящей почки, 2,4, 5 — ветвление спящей почки,
3 — скопление спящих почек в основании ствола
(а — скрытый побег, б — пазушная почка)
Почка черешковая см. почка пазушная.
Почка эпифильная см. почка придаточная.
Почкосложение (англ.: ptyxis) см. почка.
Почкосмыкание (англ.: vernation, prefoliation) см. почка.
Правило кратных отношений — подмеченная А. В. Гурским
(1948) закономерность кратного соотношения между числом ли-
стьев (цикловметамеров или квантов) на брахибластах и ауксиб-
ластах одного и того же раст.; лит.: Шмидт, 1968.
Правило П. А. Смирнова (1958) — «правило центробежного раз-
вития вневлагалищных подземных диагеотропных побегов» —
развитие корневища гипогеогенного только в противоположную
сторону от побега материнского-, закономерность, свойственная
всем раст. длиннокорневищным.
Правило уподобления — подмеченная И. Г. Серебряковым (19646)
закономерность изменения ритма развития и побегообразования у
раст.: при естественном или искусственном расселении раст. их
ритм сезонного развития изменяется в сторону феноритмотипа,
свойственного большинству аборигенных видов раст.; учет П. у.
позволяет моделировать изменения ритма развития у широко рас-
пространенных видов при интродукции; у нек-рых раст. при пере-
носе в новые условия ритм сезонного развития не меняется.
158
Предлист (Prophyllum, от греч. pro — перед, раньше, вместо и phyl-
lum — лист; англ.: prophyllum) — редуцированный первый лист
(однодольные) или первые два листа (двудольные) побега боково-
го (рис. 31); нередко П. видоизменен в колючку или усик; в облас-
ти соцветия П. наз. прицветничком; син.: профилл, профиллум.
Предстебель см. прокаулом.
Признаки биоморфологические (англ.: biomorphological (sign)
characters) — широко используемое пон. для обозначения качест-
венных (реже и количественных) признаков разного уровня, от-
ражающих особенности конкретной формы жизненной (элементы
ее структуры) или ее отдельных структур (побега, системы побегов
и т. п.); обычно к П. б. относят признаки вегетативного тела раст.,
доступные при изучении биоморфы снаружи, без использования
анатомических и других более тонких методов (П. эпиморфоло-
гические); П. репродуктивной сферы (цветки, плоды, семена
и т. п.), которые существенно не влияют на общий габитус, обыч-
но не включаются в характеристику жизненной формы; в нек-
рых случаях (для раст. травянистых) может учитываться облик
соцветия или в назв. биоморфы — образ жизни раст. (эпифиты,
раст. паразиты); при более широком понимании явления «жиз-
ненная форма» к П. б. также относят признаки, характеризующие
экологию раст., способы его опыления и диссеминации (перека-
ти-поле) и др.
Признаки эпиморфологические см. признаки биоморфологиче-
ские.
Примордий (Primordium, от лат. primus — первый, ordior — начи-
нать; англ.: primordium) —- нерасчлененный зачаток листа, в виде
бугорка или валика на конусе нарастания, побега, состоящий из
б. м. однородных меристематических клеток (лист примордиаль-
ный или первичный); П. наз. также зачаток побега или зачатки
частей цветка и др. (рис. 7).
Принцип(ы) эволюции биоморфологической — главные функ-
циональные и структурные взаимодействия органов (структур)
в онтогенезе, которые предоставляют основу для эволюции био-
морфологической посредством разных способов (см. способ
эволюции морфологической). В эволюционной биоморфологии
не всегда различают способы и принципы эволюции, относя к
последним интенсификацию, полимеризацию, олигомеризацию,
гетеробатмию и рекапитуляцию; лит.: Северцов, 1990; Терехин,
1991.
159
Прирост (англ.: accretion, current twig) — широко используемое
пон. для обозначения увеличения размеров разных частей раст. на
неопределенную или б. м. определенную величину за счет актив-
ности меристематических тканей; часто годичный прирост побега
наз. побегом годичным (П. элементарный), а возобновление рос-
та побегов во второй половине лета — приростом вторичным
или побегом Ивановым (рис. 107).
Прирост истинный см. резид.
Проант (англ.: hysteranthous plant) — наз. введено А. А. Алехиным
(1925) для обозначения раст., цветущих до или в самом начале
распускания листьев (Corylus).
Пригенез (от греч. pro — вперед, genesis — происхождение; англ.:
progenesis) — один из основных типов элементарной гетерохро-
нии, при котором ускорение полового созревания сочетается с не-
изменной скоростью соматического развития (рис. 2). В результа-
те П. потомок имеет педоморфный облик (педоморфоз), форми-
рующийся в течение сокращенного онтогенеза. При сочетании П.
с акселерацией (рис. 29) облик потомка напоминает нанизм (псев-
донанизм). В биоморфологии П. часто называют неотенией или
аббревиацией; терм.: Giard (1887); лит.: Gould, 1980; Серебрякова,
1983; Жмылев, 2004.
Прокаулом (Procauloma, от греч. pro — до, перед, kaulos — стебель;
англ.: procaulome) — недифференцированный нитевидный про-
росток грушанковых, развивающийся униполярно из базальной
области зародыша (рис. 113); в естественных условиях существует
только при контакте с подходящим грибным мицелием; часто П.
наз. протосомой или предстеблем; терм.: Velenovsky, 1892, 1905.
Пролонгация см. анаболия.
Пропагула (англ.: propagule) см. диаспора.
Прорастание (Germinatio, от лат. germino — производить на свет;
англ.: germination) — процесс перехода диаспоры из состояния по-
коя к активной деятельности, в результате которой образуется
проросток или раст. ювенильное (напр. при псевдовивипарии); раз-
личают П. надземное а П. подземное (рис. 16,111); лит.: Трофимов,
1963; Васильченко, 1936; Серебряков, 1952; Грушевицкий, 1963;
Mayer, Poljakoff-Mayber, 1975; Артюшенко, 1990; Tillich, 1995.
Прорастание адмотивное (Germinatio admotiva, от лат. admo-
veo — приближать; англ.: admotive germination) — форма П. под-
земного, при котором проросток остается в непосредственной
близости к семени (ср. П. ремотивное); при П. а. апикальная часть
160
7
Рис. 111. Надземное (А, Б) и подземное (А, В) прорастание двудольных:
А — начало прорастания, 1 — главный корень 2 — боковые корни, 3 — гипокотиль,
4 — придаточные корни, 5 — семядоля, 6 — чешуевидный лист, 7 -—эпикотиль
(по: Ботаника..., 1988)
семядоли остается в семени и выполняет гаусториальную функ-
цию, а ее базальная часть образует короткое влагалище семядоль-
ное с язычком (лигулой) в верхней части (прорастание адмотив-
но-лигулярное; рис. 17); характерно для некоторых однодольных;
терм.: Martins, 1823-1850.
Прорастание гипогеальное (англ.: hypogeal germination) см. про-
растание подземное.
Прорастание гипокотилярное (Germinatio hypocotylaris, от гипо-
котиль-, англ.: hypocotylar germination) — форма 17. надземного,
при котором разрастающийся гипокотиль выносит семядоли на
поверхность земли (рис. 111); ср. П. котиледонарное.
Прорастание котиледонарное (Germinatio cotyledonaris, от семя-
доля) — форма П. надземного, при котором гипокотиль не разрас-
тается, а семядоли выносятся на поверхность земли посредством
удлинения их черешков (Clematis, Anemone); терм.: Барыкина,
Чубатова, 1981.
Прорастание криптокотилярное (англ.: cryptocotylar germinati-
on) см. прорастание подземное.
161
Прорастание надземное (Germinatio epigaea, от греч. epi — на,
над, сверх, geo — земля; англ.: epigeal germination) — П. семени,
при котором вытягивающийся гипокотиль (П. гипокотилярное)
или семядольные черешки (П. котиледонарное) выносят семядоли
на поверхность почвы (рис. 16,111; ср. П. подземное); чаще встре-
чается у двудольных; при П. н. семядоли освобождаются от по-
кровов (явносемядольность или фанерокотилия), принимают б.
м. листовидную форму и фотосинтезируют; у нек-рых раст.
(Amygdalus) они едва приподнимаются над уровнем почвы и ос-
таются бледными и мясистыми (П. полунадземное; англ.: mixed
germination); син.: П. фанерокотилярное, П. эпигеальное.
Прорастание подземное (Germinatio hypogaea, от греч. hypo — под,
внизу, geo — земля; англ.: hypogeal germination) — П. семени, при
котором гипокотиль недоразвивается, семядоли остаются частично
или полностью в семени, а почечка зародыша выносится на по-
верхность земли вытягивающимся мезокотилем или эпикотилем
(рис. 16, 111; ср. П. надземное); чаще встречается у однодольных;
при П. п. семядоли обычно не зеленеют, остаются скрытыми в семе-
ни (скрытосемядольность или криптокотилия) и выполняют гау-
сториальную или запасающую функцию; различают П. адмотив-
ное и П. ремотивное, син.: П. гипогеальное, П. криптокотилярное.
Прорастание полунадземное (англ.: mixed germination) см. про-
растание надземное.
Прорастание ремотивное (Germinatio remotiva, от лат. removeo —
отодвигать, удалять; англ.: remotive germination) — форма П. под-
земного, при котором проросток отодвигается от семени, иногда на
значительное расстояние, благодаря вытягиванию шейки семя-
дольной (рис. 17); П. р. характерно для пальм, но встречается и у
других однодольных; различают П. ремотивно-лигулярное и П. ре-
мотивно-тубулярное, терм.: Martius, 1823-1850.
Прорастание ремотивно-лигулярное (Germinatio remotivo-ligu-
laris; англ.: remotive-ligulate germination) — форма П. ремотивного
нек-рых пальм, при котором первый настоящий лист проростка
выходит наружу через разрыв влагалища семядольного, образую-
щего язычок (лигулу); рис. 17; син.: П. ремотивно-язычковое, П.
язычковое.
Прорастание ремотивно-трубчатое (англ.: remotive-tubular
germination) см. прорастание ремотивно-тубулярное.
Прорастание ремотивно-тубулярное (Germinatio remotivo-tubu-
laris; англ.: remotive-tubular germination) — форма П. ремотивного
162
некоторых пальм, при котором первый настоящий лист пророст-
ка выходит наружу через верхушечное отверстие «трубки», обра-
зованной влагал ищем семядольным (рис. 17); син.: П. трубчатое, П.
ремотивно-трубчатое.
Прорастание ремотивно-язычковое см. прорастание ремотив-
но-лигулярное.
Прорастание трубчатое см. прорастание ремотивно-тубулярное.
Прорастание фанерокотилярное (англ.: phanerocotylar germinati-
on) см. прорастание надземное.
Прорастание эпигеальное (англ.: epigeal germination) см. прорас-
тание надземное.
Прорастание язычковое см. прорастание ремотивно-лигулярное.
Проросток (Plantula; англ.: plantlet, plantling, plantule, seedling) —
по-разному трактуемое пон.; в самом широком смысле использу-
ется для обозначения раст., в строении которого отчетливо выра-
жены структуры зародыша: семядоли, корешок, гипокотиль (дву-
дольные), колеоптиль, колеориза, мезокотиль (однодольные); в
более узком значении — раст. в период от прорастания до обра-
зования эпикотиля, первого настоящего листа (ср. всход); П. не-
которых раст. с недоразвитым или редуцированным зародышем
наз. протосомой-, лит.: Васильченко, 1960; Рысина, 1973.
Протогемикриптофит (англ.: protohemicryptophyte) см. гемикрип-
тофит.
Протокорм (Protocormus от
греч. protos — первый, kor-
mos — пень, ствол; англ.:
protocorm) — проросток
орхидных в виде б. м. ок-
руглого подземного обра-
зования, состоящего из од-
нородных паренхимных
клеток; развивается из ме-
таморфизированного кор-
невого полюса зародыша
в естественных условиях
обычно после проникнове-
ния в семя гиф микоризно-
го гриба (некоторые бази-
диомицеты); на последней
стадии развития П. выделя-
0,1 мм
А Б
Рис. 112. Проросток Listera cot data А — первая
стадия развития, Б — схема строения (1 — апекс
корня, 2 — апекс побега, 3 — ризодерма, 4 —
клетки коры, содержащие гифы гриба, 5 —
крахмалоносный слой, 6 — проводящий пу-
чок); по: Виноградова, 1996
163
ются две зоны: базальная и апикальная; в апикальной зоне эндо-
генно дифференцируется апекс надземного побега (или микоризо-
ма) и корня (рис. 112); П. иногда наз. протосомой, проэмбрио
(англ.: proembryo), зародышем-проростком или зародышевым
клубнем; терм.: Bernard, 1909; лит.: Андронова, Батыгина, Василье-
ва, 2000.
Протосома (Protosoma, от греч. protos — первый, первоначальный
и soma — тело) — общее название проростков (прокаулом, про-
токорм и др.) высокоспециализированных паразитных цветко-
вых раст.; П.— недифференцированное вегетативное тело (сома),
развивающееся из корневого или стеблевого полюса зародыша
и выполняющее функции поиска и освоения растения-хозяина
или грибного симбионта, реже — размножения вегетативного
(рис. 113); терм.: Терехин, Никитичева, 1968; лит.: Терехин, 1977.
Рис. 113. Прорастание Lathraea squamaria (А—Г), Orobanche pallidiflora (Д-Ж; по: Тере-
хин, Никитичева, 1981), Pyrola rotundifolia (3, Л), Orthilia secunda (И) и Maneses uniflora
(К; по: Терехин, 1977): 1 — апекс побега, 2 — гаусторий, 3 — клубенек, 4 — корень
растения хозяина 5 — протосома, 6 — семя, 7 — апикальный полюс зародыша,
8 — базальный полюс зародыша, 9 — эндосперм
Профилл(ум) (англ.: prophyll(um) см. предлист.
Проэмбрио (англ.: proembryo) см. протокорм.
Псевдобульба (англ.: pseudobulb) см. туберидий.
164
Псевдовивипария (англ.: pseudovivipary) см. вивипария.
Псевдогигантизм (англ.: pseudogigantism) см. гигантизм.
Псевдодикотилия (Pseudodicotylia, от греч. pseudes — ложный,
di — два, kotyle — доля; англ.: pseudodicotyly) — наличие второй
семядольной доли в результате расщепления (схизокотилия, ши-
зокотилия; англ.: schizocotyly) единственной семядоли однодоль-
ного раст.; син.: ложнодву(семя)дольность.
Псевдомонокотилия (Pseudomonocotylia, от греч. pseudes — лож-
ный, monos — один, kotyle — доля; англ.: pseudomonocotyly) —
отсутствие одной из двух семядолей у двудольного раст. в резуль-
тате редукции или синкотилии (Corydalis, Ranunculus); иногда
пон. П. применяют и в случае амфикотилии; син.: ложноодно(се-
мя)дольность.
Псевдонанизм (англ.: pseudonanism) см. нанизм.
Развитие (англ.: development) — процесс взаимосвязанных коли-
чественных (рост) и качественных (дифференцировка) преобра-
зований клеток, тканей, органов и особи, связанных с их морфоге-
незом или онтогенезом. Рост и дифференцировка в онтогенезе
растения осуществляется в результате регуляции развития, детер-
минации и индукции развития. Предложены две модели Р.: модель
комбинаторного развития, модель «контроллера переключения
фаз»; лит.: Сабинин, 1963; Синнот, 1963; Леопольд, 1968; Биоло-
гия..., 1975; Полевой, 1981; Уоринг, Филипс, 1984; Батыгин, 1986;
Полевой, Саламатова, 1991; Лутова и др., 2000; Корочкин, 2002;
Oborny, 2003.
Развитие автономное — детерминация развития и дифференци-
ровка под влиянием возрастных изменений или других внутрен-
них факторов (напр., цветение нейтральных видов); ср. развитие
индуцированное; лит.: Чайлахян, 1975.
Развитие вегетативное см. развитие генеративное.
Развитие генеративное — процесс заложения и развития репро-
дуктивных органов (цветков, семян, плодов). Часто Р. г. противо-
поставляют вегетативному развитию (соматическое развитие),
в процессе которого раст. приобретает способность к цветению
(детерминация) или в более широком смысле и к размножению ве-
гетативному (репродуктивное развитие); лит.: Гупало, Скрип-
чинский, 1971.
Развитие гетеробластное см. гетеробластия.
Развитие гомобластное см. гомобластия.
Развитие индивидуальное см. онтогенез.
165
Развитие индуцированное — детерминация развития и диффе-
ренцировка, происходящая в результате индукции определенны-
ми экзогенными факторами (индукторы). Длина дня, температу-
ра и др. факторы внешней среды воспринимаются как индукторы
после достижения компетенции (способности воспринимать ин-
дуцирующее воздействие и специфически реагировать на него) и
до начала детерминированного развития (напр., после фотопе-
риодической индукции взрослых раст. их дальнейшее развитие
не нуждается в оптимальной для цветения длине дня); лит.: Чай-
лахян,1975.
Развитие репродуктивное см. развитие генеративное.
Развитие сезонное — развитие раст., в течение года связанное с
периодическими изменениями погоды и эндогенным ритмом
(биологические часы). В течение года раст. претерпевает законо-
мерные морфологические изменения (смена фенофаз), отражаю-
щие ритмичность роста (сезонный ритм развития) в определен-
ных условиях среды. По характеру Р. с. выделяют несколько фено-
ритмотипов.
Развитие соматическое см. развитие генеративное.
Размножение (Reproductio, от лат. ге — приставка со значением
повторности, produce — производить, рождать; англ.: reproduc-
tion, multiplication, propagation) — присущее всем организмам
свойство воспроизведения себе подобных, обеспечивающее не-
прерывность и преемственность жизни; обычно выделяют три
основных формы: Р. бесполое, Р. вегетативное и Р. половое-, часто
Р. рассматривают как частный случай воспроизведения, в резуль-
тате которого происходит увеличение численности особей одного
раст.; лит.: Левин, 1961; Ботаника..., 1988; Лотова, 2000.
Размножение бесполое (англ.: asexual reproduction) — в узком
значении пон. используется для обозначения различных спосо-
бов Р. путем образования неполовых клеток (агамная цитогония,
шизогония); в более широком смысле к Р. б. относят и Р. вегета-
тивное.
Размножение вегетативное (англ.: vegetative (multiplication, pro-
pagation) reproduction, clonal reproduction) — увеличение числен-
ности особей одного раст. и образование клона (клональный рост;
англ.: clonal growth) в результате искусственного или естественно-
го отделения (см. сарментация) от материнской особи (рис. 114)
вегетативных диаспор (специализированное Р. в.) или жизнеспо-
собных частей ее системы побеговой (неспециализированное Р. в.,
166
Рис. 114. Образование клона в результате вегетативного размножения у растений с
надземными столонами (1, 2), короткими корневищами (3, 4), клубнелуковицами
(5-8), клубнями (9) и луковицами (10): р — раметы; (по: Bell, Bryan, 1998)
дезинтеграция морфологическая)-, часто к Р. в. относят партикуля-
цию и нек-рые формы возобновления вегетативного (поросль пне-
вая, двулетник замещающий и др.); классификация способов Р. в.
обычно основаны на морфологических различиях органов Р. в.
(столон, корневище, луковица, клубень, клубнелуковица, псевдови-
випария и др.); лит.: Серебряков, 1952; Любарский, 1967; Барыки-
на, 2000; Шорина, 2000а.
Размножение генеративное см. размножение половое.
Размножение половое (англ.: sexual (seed) reproduction) — пон.
используется для обозначения различных способов Р., при кото-
рых новый организм развивается обычно из зиготы, образую-
167
щейся в результате слияния разнополых клеток (гамет); син.:
Р. генеративное.
Рамет (англ.: ramet) см. особь.
Рамифлория (англ.: ramiflory) см. каулифлория.
Расселение вегетативное (англ.: vegetative (spreading) expan-
sion) — в узком значении пон. используется для обозначения
быстрого роста диагеотропного или роста плагиотропного удли-
ненных побеговраст. вегетативно-подвижных (специализирован-
ная вегетативная подвижность); в более широком смысле Р. в.
можно рассматривать и как расширение площади обитания особи
(см. поведение раст.) в результате кущения, полегания побегов
и т. п. (неспециализированная вегетативная подвижность); неред-
ко Р. в. рассматривают как синоним размножения вегетативного.
Растение акрофилльное см. растение розеточное.
Растение безрозеточное (Planta erosulans, от лат. е — приставка
со значением отсутствия чего-либо; rosa — розетка; англ.: erosu-
late plant) — одна из форм жизненных, объединяющая раст. тра-
вянистые с удлиненными междоузлиями надземных побегов (см.
побег безрозеточный) и ассимилирующими листьями, не обра-
зующими т. н. розетку; терм.: Воронов, 1952; лит.: Голубев, 1965;
Серебрякова, 1971а.
Растение бескорневое (англ.: rootless plant) — раст. талломное
или реже раст. листостебельное, у которых корни не закладыва-
ются или исчезают в начале онтогенеза, а функцию всасывания
осуществляет тело раст. или ризоиды; среди листостебельных
раст. к Р. б. относятся многие растения-паразиты, а также ряд
эпифитов (Tillandsia) и водных раст. (Utricularia).
Растение бесстебельное см. растение розеточное.
Растение биполярное (англ.: bipolar plant) см. полярность и рост
биполярный.
Растение вагативное (Planta vagativa, от лат. vagari — кочевать,
блуждать) — один из типов классификации форм жизненных по
Г. М. Зозулину, объединяющий однолетники и двулетники-, среди
Р. в. выделяются (группы) раст. плавающие и прикрепляющиеся,
а среди последних (подгруппы) — раст. озимые, односезонные и
эфемеры; терм.: Г. М. Зозулин, 1961.
Растение вегетативно-неподвижное —часто используемое назв.
для обозначения раст., не имеющих специализированных побегов
размножения, и для их противопоставления раст. вегетативно-
168
подвижным-, в более широком смысле к Р. в.-н. иногда относят и
раст., не имеющие побегов разрастания.
Растение вегетативно-подвижное — раст., образующее побеги
удлиненные (корневищаудлиненные, столоны, побеги ползучие, усы,
флагеллы), которые в результате быстрого роста диагеотропного
или роста плагиотропного и укоренения, обусловливают расселе-
ние вегетативное, а при отмирании — образование клона (см. по-
лифит); Р. в.-п. нередко наз. растениями наземно- или подземно-
ползучими; терм.: Высоцкий, 1915.
Растение ветвелазящее см. лиана опирающаяся.
Растение вечнозеленое (Planta sempervirens, от лат. semper — все-
гда, постоянно, vireo — быть зеленым; англ.: evergreen (semper-
virent) plant) — раст. многолетнее, каждый лист которого живет
более одного года; преимущественно раст. тропиков и субтропи-
ков, в умеренных широтах — нек-рые деревья (Picea), кустарнички
(Ericaceae), редко травы (см. раст. зимнезеленое); в более широком
смысле Р. в. наз. также и раст., у которых листья живут недолго, но
опадают постепенно, после развертывания новой генерации листь-
ев (гл. образом деревья влажных тропических лесов с круглогодич-
ной облиствленностью кроны).
Растение взрослое вегетативное (англ.: adult (mature) vegetative
plant) см. растение виргинильное.
Растение виргинильное (Planta virginalis, от лат. virgo — девст-
венный, целомудренный; англ.: virginile plant) — особь вегетатив-
ная, в морфологическом облике которой появляются основные
черты (листья, побеги, система корневая и т. д.), характерные для
взрослого раст. (обозначается индексом V); у раст. с продолжи-
тельным периодом виргинильным нередко выделяют молодые и
взрослые вегетативные раст. (см. период виргинильный); часто к
Р. в. относят и раст. имматурные или объединяют под этим на-
званием все молодые и взрослые (англ.: adult (mature) vegetative
plant) вегетативные (не цветущие) особи; син.: раст. молодое или
взрослое вегетативное; терм.: Работнов, 1945; лит.: Ценопопуля-
ции... 1976, 1988.
Растение вьющееся (англ.: voluble (twining) plant) см. лиана вью-
щаяся.
Растение гапаксантное (англ.: hapaxanthous plant) см. монокарпик.
Растение геммаксиллярное (Planta gemmaxiliaris, от лат. gem-
ma — почка и axilla — пазуха, подмышечная ямка) — раст. с оп-
ределенной последовательностью заложения листа и почки, в ре-
169
зультате которой последняя занимает пазушное положение {поч-
ка пазушная}-, к Р. г. относятся покрытосеменные и часть голо-
семенных (исключая семенные папоротники, саговники и боль-
шинство беннеттитовых), у которых пазушная почка и ее крою-
щий лист представляют собой единый морфо-биологический
комплекс аксиллярный, элементы которого развиваются под непо-
средственным и определяющим влиянием друг друга; терм,: Гат-
цук, 1974а.
Растение генеративное см. период генеративный.
Растение гетеробластное (англ.: heteroblastic plant) см. гетеро-
бластия.
Растение гипогеогенно-длиннокорневищное (англ.: (long-)rhi-
zome plant) см. растение длиннокорневищное.
Растение гипогеогенно-короткокорневищное (англ.: (short-)rhi-
zome plant р. р.) см. растение короткокорневищное.
Растение гомобластное (англ.: homoblastic plant) см. гомобластия.
Растение двулетнее (англ.: biennial plant) см. растение монокар-
пическое.
Растение деревянистое (англ.: woody (ligneous) plant) — пон., ис-
пользуемое для обозначения раст. с многолетними деревянисты-
ми побегами и почками возобновления, расположенными над зем-
лей; к Р. д., противопос-
тавляемым раст. травя-
нистым, относятся раст.
древесные и раст. полу-
древесные (рис. 128).
Растение дерновинное
(Planta caespitosa; англ.:
c(a)espitose (turf-forming)
plant, bunchgrass, tussock
grass, см. также дернови-
на} —широко используе-
мое назв. для обозначе-
ния одной из форм жиз-
ненных раст. кистекорне-
вых, надземные побеги ко-
торых образуют дернови-
ну (рис. 115); обычно к
Р. д. относят только мно-
голетники из сем. Роасеае
Рис. 115. Дерновинные травянистые растения:
А — Cobresia royleana, Б — Linosyris villosa
(по: Серебряков» 1964а)
170
и Сурегасеае, почки возобновления у которых расположены ниже
уровня почвы (Stipa), на поверхности почвы (раст. собственно дер-
новинные) или над ней (раст. кочкообразущие); плотность дерно-
вины зависит от характера кущения, в связи с чем различают Р.
плотнодерновинные или плотнокустовне (Nardus stricta) и Р. рых-
лодерновинные или рыхлокустовые (Calamagrostis arundinacea); в
более широком смысле к Р. д. относят и другие раст. короткокорне-
вищные; лит.: Персикова, 1959а,б; Рытова, 1987; Серебрякова, 1971а.
Растение дициклическое (англ.: d cyclic plant) см. растение одно-
летнее.
Растение длиннокорневищное (Planta longe rhizomatosa; англ.:
long rhizome plant) — раст. кистекорневое, многолетнее, травяни-
стое или деревянистое (см. ксилоризом) с подземным корневищем
удлиненным (рис. 116), соединяющим надземные кусты парци-
альные или побеги (колония клональная); Р. д. приурочены к
хорошо аэрируемым, достаточно влажным почвам; по происхож-
дению корневища различают раст. эпигеогенно- (Asarum) и гипо-
геогенно-длиннокорневищные (Mercurialis); в более широком
значении к Р. д. относят и некоторые раст. стержнекорневые
(раст. длиннокорневищно-стержнекорневые) с отмирающим во
взрослом состоянии корнем главным и многоглавым каудексом,
отдельные «ветви» которого имеют гипогеогенное происхожде-
ние (Galium); лит.: Любарский, 1965, 1973, 1967; Bell, Tomlison,
1980.
Растение длительновегетирующее (Planta longe vegetans) — раст.
травянистое, облиствленное в течение всего или большей части
года; к Р. д. относятся раст. вечнозеленые (Asarum) и раст. лет-
не-зимнезеленые (Geum).
Растение древесное (Planta arborea; от дерево; англ.: arboreal
(woody) plant) — один из основных типов форм жизненных, выде-
ляемых в сезонном климате по эколого-морфологическому при-
знаку: наличие многолетних надземных осей скелетных (побегов) с
почками возобновления (фанерофиты, хамефиты); среди Р. д. раз-
личают деревья, кустарники и кустарнички (рис. 128); часто Р. д.
отличают от Р. травянистых по анатомическим признакам стеб-
ля и говорят о травянистой и деревянистой консистенции побе-
гов, имея в виду количественные отношения одревесневших (см.
одревеснение) и неодревесневших тканей; последнее позволяет
провести 6. м. четкую границу между деревянистыми и травяни-
стыми фанерофитами бессезонного климата; лит.: Гатцук, 1976.
171
Рис. 116. Длиннокорневищные травянистые растения:
1 — Agropyrum repens, 2 — Veronica longifolia, 3 — Convallaria majalis,
4 — Paris quadrifolia, 5 — Carex pilosa (по: Ботаника..., 1988)
172
Растение древовидное (англ.: tree-like (dendroid, dendritic, arbo-
roid) plant) см. дерево.
Растение живородящее (англ.: viviparous plant) см. вивипария.
Растение зимнезеленое (Planta hieme frondescens, от лат. hiems —
холодное полугодие, зима, frondeo — быть покрытым листьями;
англ.: winter-green (chimochlorous) plant) — в широком смысле
травянистое раст., зимующее с зелеными листьями; различают Р.
летне-зимнезеленое и собственно зимнезеленое (см. Р. вечнозеле-
ное); в более узком смысле Р. з. наз. зимующие эфемеры и некото-
рые эфемероиды (Роа bulbosa) с перерывом вегетации в летнее
время (англ.: summer-dormant plant); лит.: Лапшина, 1928; Воро-
нов, 1952; Серебрякова, 1976.
Растение имматурное (Planta immature, от лат. im — приставка со
значением отрицания, maturus — зрелый, взрослый; англ.: imma-
ture plant) — особь вегетативная, в морфологическом облике ко-
торой сочетаются признаки раст. ювенильного и раст. виргиниль-
нога сохранение отдельных элементов первичной структуры {ко-
рень, побег) и появление взрослых черт (смена типа нарастания,
начало ветвления, образование побегов диагеотропных и т. д.) или
формирование листьев, системы побегов, системы корневой пере-
ходного (полувзрослого) типа (обозначается индексом im); см.
период виргинильный; син.: раст. прематурное, раст. полувзрос-
лое; терм.: Уранов, 1960; лит.: Ценопопуляции... 1976, 1988.
Растение инсидентивное (Planta insidens, от лат. insido — садить-
ся) — один из типов классификации форм жизненных по Г. М. Зо-
зулину, объединяющий раст., особи которых не занимают отдель-
ной площади обитания (группы: эпифиты, сапрофиты, парази-
ты)-, терм.: Зозулин, 1968.
Растение ирруптивное (Planta irruptiva, от лат. irrumpo — втор-
гаться, вламываться) — один из типов классификации форм жиз-
ненных по Г. М. Зозулину, объединяющий раст. многолетние, побеги
и корни у которых обеспечивают разрастание и размножение веге-
тативное (вторжение и захват площади обитания других раст.);
среди Р. и. выделяются группы расползающихся (Р. короткокорне-
вищное, Р. рыхлодерновинное, Р. корнеотпрысковое), перебрасываю-
щихся (Р. длиннокорневищноё) и разбрасывающихся раст. (Р. сто-
лонообразующее, Р. длиннокорневищное, Р. корнеотпрысковое);
терм.: Зозулин, 1961.
Растение итеропаро(у)зное (англ.: iteroparous plant) см. поли-
карп ик.
173
I
Рис. 117. Клубнелуковичное растение
(Crocussp.): 1 —продольный разрез,
2,3 — клубнелуковицы с детками
Растение карликовое (англ.: dwarf plant) см. карликовость.
Растение кистекорневое (Planta radicibus fibrillosis praedita, англ.:
plant with fibrillose (racemose) root system) — раст., корень главный
которого не развивается или отмирает в первые годы жизни, а сис-
тема корневая во взрослом состоянии образована только корнями
кладогенными (рис. 127); различают корневые системы мочкова-
тую (злаки и другие однодольные), бахромчатую и кистевидную
(двудольные соответственно с горизонтальным и вертикальным
корневищем)-, нередко к Р. к. относят раст., главный корень кото-
рых долго не отмирает, но не выделяется ни толщиной, ни дли-
ной среди кладогенных корней (Р. стержне-кистекорневое); в не-
которых случаях в результате вторичного утолщения одного из
них может образовываться ложный или т. н. вторично-стержне-
вой корень, который внешне напоминает главный корень стерж-
некорневых раст. (корневая система вторично-стержнекорневая);
лит.: Михайловская, 1981а; Голубев, 1962.
Растение клубне луковичное (Plan-
ta bulbotuberifera; от клубнелукови-
ца:, англ.: bulbotuberiferous plant) —
форма жизненная кистекорневых
травянистых раст., подземная часть
которых видоизменена в клубнелу-
ковицу (рис. 117); в зависимости от
положения на клубнелуковице поч-
ки возобновления различают Р. к. с
нарастанием моноподиалъным
(Crocus) и симподиальным (Со1-
chicum); Р. к.— геофиты, гл. обр.
эфемероиды аридных областей;
обычно их относят к многолетни-
кам, хотя в большинстве случаев
ежегодно материнская клубнелуко-
вица отмирает, замещаясь дочер-
ней (малолетник вегетативный);
у нек-рых (Gladiolus) образуется не-
сколько дочерних клубнелуковиц
(размножение вегетативное), реже
клубнелуковицы соединены корне-
вищем (Tritonia); лит.: Андреева,
1979; Артюшенко, 1970. (по: Ботаника..., 1988)
174
Рис. 118. Растения с клубнями корневого (1-4) и стеблевого (5-10) происхождения
(по: Bell, Bryan, 1998): кк — корневой клубень, пк — придаточный корень, ок — отми-
рающий клубень, ск — стеблевой клубень, пп — пазушная почка, п — побег, чл — че-
шуевидный лист, рчл — рубец от чешуевидного листа (1 — Chlorophytum comosutn,
2 — Dahlia sp., 3— Dioscorea sp., 4 — Kedrostris africana, 5 — Helianthus tuberosus,
6—Cissus tuberosa, 7 — Solarium tuberosum, 8 — Sinningia speciosa, 9 — Ballota nigra,
10 — Anredera gracilis)
175
Растение клубнеобразующее (англ.: tuberous plant) — сборная
группа, объединяющая раст. травянистые с надземными клубня-
ми (туберидиями и луковичками), с клубнями подземными на стеб-
ле (Р. стеблеклубневое), на корнях (Р. корнеклубневое), на столо-
нах (Р. столонноклубневое) и корневищах (Р. корневищноклубне-
вое), а в более широком смысле — раст. с клубнелуковицами и
выводковыми почками (рис. 118); лит.: Голубев, I960; Коровкин,
1983, 1984; Маркаров и др., 2001; Седельникова, 2002.
Растение корневищноклубневое см. растение клубнеобразующее.
Растение корневищнолуковичное см. луковица многолетняя.
Растение корнеклубневое (англ.: plant with root tuber) см. расте-
ние клубнеобразующее.
Растение корнеотпрысковое (Planta sobolifera, от лат. soboles —
побег, поросль, отпрыск, fere — носить; англ.: soboliferous (root
sucker, burr) plant) — P., образующее отпрыски корневые (рис. 119);
раст. древесные и травянистые, среди которых много сорных (см.
также корнеотпрысковость); лит.: Высоцкий, 1926; Голубев, Голу-
бева, 1964; Кондратьева-Мельвиль, 1957; Котт, 1956.
Рис. 119. Развитие корневых отпрысков Rubus idacus:
1 — корень, 2 — корневой отпрыск (по: Bell, Bryan, 1998)
176
Растение коротковегетирующее (Planta breviter vegetans) — раст.
травянистое, вегетирующее только в благоприятное для развития
время года: весной и осенью (Phlomis puberula), весной и летом (см.
гемиэфемероиды) или летом и осенью (Р. летнезеленое).
Растение короткокорневищное (Planta breviter rhizomatosa; англ.:
short rhizome plant) — P. кистекорневое, многолетнее, травянистое
с хорошо выраженным подземным, мясистым корневищем укоро-
ченным, постоянно нарастающим с апикальной стороны и отми-
рающим с базальной части (рис. 120); в зависимости от происхож-
дения корневища разичают Р. эпигеогенно- (Betonica officinalis) и
гипогеогенно-короткокорневищные (Anemone nemorosa), Р. к. с
нарастанием моноподиальным (Butomus umbellatus) и симподиаль-
ным; в более широком смысле к Р. к. относят Р. дерновинные и Р.
(собственно) кистекорневые, со слабо выраженным корневищем
(из-за быстрого отмирания с базальной стороны) и мочковатой
системой корневой (Ranunculus); Голубев, 1958; Любарский, 1973.
Рис. 120. Короткокорневищные травянистые растения (по: Ботаника..., 1988):
1 — Viola mirabilis, 2 — Fragaria vesca, 3 — Geutn male, 4 — Iris germanica
(а-д — приросты корневища)
177
Рис. 121. Летне-зимнезеленые травянистые расте-
ния: А —- Geurn urbanum (сентябрь), Б — Succisa
praeinorsa (конец октября), 1—листья весенне-
летней генерации, 2 — зимующие листья летне-
осенней генерации, 3 — перезимовавшие листья,
прошлогодней генерации, 4 — верхушечная почка
(по: Серебряков, 1952)
Растение кочкообразующее (англ.: tussock-forming plant) — час-
то используемое назв. для обозначения одной из форм жизненных
раст. кистекорневых (травянистых многолетников преимущест-
венно из сем. Роасеае и Сурегасеае), рост апогеотропный побегов
кущения которых обусловливает ежегодное нарастание дерновины
вверх и образование кочки; в результате почки возобновления у та-
ких раст. располагаются над поверхностью почвы, а отмирающие
части побегов формируют «тело» кочки (рис. 57), которое ис-
пользуется раст. в качестве «субстрата» (Carex caespitosa); лит.:
Куркин, 1954.
Растение лазящее (Planta scandens; англ.: climber) см. лиана.
Растение летнезеленое (Planta aestate frondescens, от лат. aestas —
теплое время года, лето, frondeo — быть покрытым листьями;
англ.: summer-green plant) — раст. облиствленное в летний и
безлистное в зимний пе-
риод года; к Р. л. отно-
сятся деревья, кустарни-
ки (Р. листопадные) и
раст. травянистые гл.
обр. сезонного климата.
Растение летне-зимнезе-
леное — раст. травяни-
стое с несколькими гене-
рациями листьев в бес-
снежный период года, из
которых листья послед-
ней генерации (осенней)
сохраняются под снегом
(см. раст. зимнезеленое)
до весны следующего го-
да (рис. 121); иногда раз-
личают облигатные и фа-
культативные Р. л.-з.;
лит.: Серебрякова, 1976.
Растение листолазящее
(англ.: leaf climber) —ли-
ана, закрепляющаяся на
опоре с помощью чувст-
вительных на прикосно-
вение твердого предмета
178
(см. гаптотропизм) черешков растущих листьев (англ.: twining
leaf petiole climber) или удлиненных верхушек листовых пласти-
нок (Gloriosa superba); рис. 61.
Растение листопадное (Planta decidua; англ.: deciduous plant) —
раст., сбрасывающее б.’ м. одновременно все листья в конце веге-
тации {листопад сезонный); гл. обр. деревья и кустарники, реже
травы, находящиеся в безлистном состоянии от неск. дней (влаж-
ные тропические леса) до 8-9 месяцев (умеренный пояс); ср.
Р. вечнозеленое; лит.: Одишария, 1974; Серебряков, 1951.
Растение листостебельное (Planta cormosa, от греч. kormos —
пень, ствол; англ.: cormophyta, cormobiont) — один из типов
структурной организации раст., характеризующийся расчлене-
нием тела раст. (кормуса; англ.: cormus) на вегетативные органы
{лист, стебель, корень); к Р. л. относятся плауновидные, хвоще-
видные, папоротниковидные, голосеменные и цветковые раст.;
нек-рые из них в ходе эволюции утратили корни {растения бес-
корневые) или листостебельность (см. листец, талоид, филломор-
фа); ср. растение талломное; син.: кормобионт (англ.: cormo-
biont).
Растение ложноподушковидное см. растение подушковидное.
Растение луковичное (Planta bulbifera; от лат. bulbus — луковица;
англ.: bulb(ace)ous (bulbiferous, bulb bearing) plant) — форма жиз-
ненная кистекорневых травянистых растений, подземная часть
стебля которых видоизменена в луковицу (рис. 122); в зависимо-
сти от положения на луковице почки возобновления различают Р.
л. с нарастанием моноподиальным (Galanthus L.) и симподиальным
(Hyacinthus L.); Р. л.— геофиты, гл. обр. эфемероиды аридных об-
ластей; некоторые из них (Galanthus nivalis L.) обладают способ-
ностью к саморегуляции глубины залегания луковицы; лит.: Ар-
тюшенко, 1970; Баранова, 1986, 1999; Бочанцева, 1956; Головкин,
1965; Седельникова, 2002; Черемушкина, 2004.
Растение малолетнее (англ.: short-lived plant, short cycle plant) см.
малолетник.
Растение микотрофное (Planta mycotrophica; англ.: mycotrophic
plant) см. сапрофит.
Растение многолетнее (Planta perennis; от лат. per — приставка со
значением действия направления — через, annus — год; англ.:
perennial (long-lived) plant) — пон., используемое обычно для
обозначения раст., живущих более двух, реже — 5-10 лет (ср. ма-
лолетник); различают Р. монокарпические и Р. поликарпические.
179
Рис. 122. Луковичные растения (по: Ботаника..., 1988):
А — Allium сера, Б — Galanthus sp., В — Hyacinthussp., Г — Lilium martcigon
(1 — верхушечная почка, 2 — донце, 3 — пазушная почка,
4 — луковица, 5 — зачаток соцветия)
Растение монокарпическое (Planta monocarpica; англ.: monocar-
pic plant) см. монокарпик.
Растение моноподиально-розеточное см. растение розеточное.
Растение моноциклическое (Planta monocyclica; англ.: monocyclic
plant) см. растение однолетнее.
Растение надземноползучее (англ.: repent plant) см. растение
ползучее.
180
Рис. 123. Однолетние растения:
А — Phleum panicularum,
Б — Matricaria lamellata
(по: Серебряков, 1964)
Растение надземностолонное (англ.: flagellant plant, plant with
runners) см. столон.
Растение наземноползучее (англ.: repent plant) см. растение веге-
тативно-подвижное.
Растение насекомоядное (Planta insectivora; от лат. insecta — насе-
комые и voro — пожирать; англ.: insectivorous plant) — автотроф-
ное раст., способное улавливать насекомых и частично перевари-
вать их с помощью протеолитических ферментов и органических
кислот; гл. обр. тропические раст., листья которых видоизменены
в ловушки (асцидии), покрыты железистыми волосками, выде-
ляющими липкое вещество (Aldrovanda), активно захлопываются
(Dionaea) или сворачиваются (Drosera) в результате тургорных
настий; син.: раст. плотоядное, раст. хищное; лит.: Дарвин, 1949;
Lloyd, 1942; Juniper, Robins, Joel, 1989.
Растение однолетнее (Planta annua;
от лат. annus — год; англ.: annual
plant) — раст., онтогенез которого
длится не более одного года
(рис. 123): обычно от нескольких не-
дель (эфемеры) до нескольких меся-
цев; гл. обр. раст. пустынь и полу-
пустынь, в умеренном поясе — сор-
няки (Viola arvensis), полупаразиты
(Euphrasia) и зерновые культуры; в
зависимости от ритма сезонного
развития различают Р. о. яровые
(Р. моноциклические) или летне-
осенние и Р. о. озимые (Р. дицикли-
ческие) или осенне-весенние (см.
малолетник); син.: однолетник; лит.:
Марков, 1990; Маркова, 1973; Нечае-
ва, Василевская, Антонова, 1969;
Krumbiegel, 1998.
Растение олигокарпическое (англ.:
oligocarpic plant) см. поликарпик.
Растение-паразит (Planta parasitica;
англ.: parasite plant) — широко ис-
пользуемое назв. для раст. с различ-
ным уровнем (голопаразит, полупа-
разит) и формой паразитизма (ксе-
181
нопаразит и аллелопаразит); лит.: Бейлин, 1968; Терехин, 1977;
Kuijt, 1969; Dawson, 1986.
Растение плейоциклическое (англ.: pleiocyclic plant) см. моно-
карпик.
Растение плотнодерновинное см. растение дерновинное.
Растение плотнокустовое см. растение дерновинное.
Растение плотоядное (англ.: carnivorous plant) см. растение насе-
комоядное.
Растение подземно-ползучее (англ.: rhizome plant) см. растение
ползучее.
Растение подземностолонное (англ.: diageic plant, plant with under
ground stolones) см. столон.
Растение подушковидное (Planta pulviniformis; от лат. pulvinus —
подушка, forma — форма, вид; англ.: cushion (polster) plant) —
раст. многолетнее, стержнекорневое, травянистое или деревяни-
стое (кустарник, кустарничек), реже полудревесное (полукустар-
ник, полукустарничек), обычно вечнозеленое с облигатной (Р. п.
настоящее или P.-подушка) или факультативной (Р. ложнопо-
душковидное) подушковидной формой роста, характеризую-
щейся плотным, «этажированным» расположением побегов с не-
значительным ежегодным приростом и часто ветвлением акро-
тонным, в результате чего формируется надземная, однородная и
ровная (в виде «подушки») сферическая или б. м. плоская поверх-
ность побеговой системы раст., внутренний объем которой за-
полнен в той или иной степени торфяной массой (из выветри-
вающихся листьев) и корнями придаточными (рис. 124); Р. п.—
хамефиты, распространенные гл. обр. в высокогорьях и тундрах;
по форме поверхности различают радиально-шаровидные и ра-
диально-плоские подушки, а по компактности и наличию запол-
няющего материала — плотные, рыхлые и воздушные (незапол-
ненные) подушки; в более широком смысле к Р. п. относят также
невысокие деревья (деревья подушковидные) и травянистые кис-
текорневые раст. с плотным расположением побегов возобновле-
ния: раст. дернованные (подушки дерновинные) и раст. розеточ-
ные (подушки розеточные или подушки-сообщества); лит.: Rauh,
1939,1940; Борисова, 1962; Зайцева, 1949; Культиасов, 1962а,б; Се-
ребряков, 1962; Стешенко, 1973; Удалова, 1978; Волков, 2002,2003.
Растение ползучее (англ.: creeping (crawling, repent) plant, creeper,
crawler) — раст. кистекорневое, многолетнее, травянистое, только
с побёгами ползучими (Lysimachia nummularia); в более широком
182
3
3
Рис. 124. Растения-подушки (по: Ботаника, 1988):
А — радиально-шаровидная подушка, Б — радиально-плоская подушка,
В — Azorelki sp. (1 — главный корень, 2 — придаточный корень,
3 — система акротонно ветвящихся побегов)
значении к Р. п. или надземноползучим относят и Р. надземносто-
лонные (рис. 126), а раст. с подземными столонами и корневищами
удлиненными выделяют как Р. подземноползучие (ср. Р. стелющееся);
лит.: Голубев, 1961; Савиных, 1986; Серебрякова, 1981.
Растение поликарпическое (Planta polycarpica; англ.: polycarpic
plant) см. поликарпик.
Растение полициклическое (Planta polycyclica; англ.: polycyclic
plant) см. монокарпик.
Растение полудревесное — пон., используемое для обозначения
разных форм жизненных разных раст. деревянистых (деревьев, кус-
тарников, кустарничков), у которых ежегодно часть побегов опа-
дает или отмирают верхние участки надземных побегов (рис. 125,
128); распространены гл. обр. в аридных областях; по общему об-
лику, высоте раст., степени отмирания и др. признакам выделяют
плохо отграниченные друг от друга группы Р. п.: полудеревья,
или деревья ксерогенные (Haloxylon aphyllum), полукустарники
(ксерогенные кустарнички), или примитивные кустарнички
183
Рис. 125. Полукустарнички Artemisia santolina (1) и Bupleurum exaltatum (2):
черным цветом показаны многолетние части растений (по: Ботаника..1988)
(Salsola arbuscula), и настоящие (ксерогенные кустарники) или ис-
тинные кустарнички (Atriplex сапа); см. также гербоид; лит.: Бес-
палова, 1965; Гатцук, 1973; Голубев, 19686, 1969, 1973; Мулкиджа-
нян, 1965; Банковская, 1957; Стешенко, 1955, 1956.
Растение полупаравит (Planta hemiparasitica; англ.: hemiparasite
(semi-parasite, partial parasite) plant) см. полупаразит.
Растение полурозеточное (Planta subrosulans; от лат. sub — при-
ставка co значением некоторой степени; rosa — розетка; англ.:
semirosette plant) — одна из форм жизненных, объединяющая
раст. травянистые, нижние междоузлия надземных побегов кото-
рых (см. побег полурозеточный) сильно укорочены, и ассимили-
рующие листья образуют т. н. розетку, а верхние междоузлия
(цветоноса) — вдпянуты, вследствие чего листья расставлены и
не образуют розетку; часто Р. п. наз. и нарастающие моноподиаль-
но травы (раст. моноподильно-розеточные), у которых побеги ске-
летные — побеги розеточные, а побеги генеративные — побеги без-
розеточные (Alchemilla); при более широком понимании розеточ-
184
ности к Р. п. относят и раст. безрозеточные, у которых в развитии
их побегов полициклических есть фаза розетки (см. побег розетко-
образующий); лит.: Голубев, 1965; Серебрякова, 1971а.
Растение полутравянистое — многолетник с однолетними одре-
весневающими надземными побегами и многолетними деревяни-
стыми подземными побегами (Sophora, Karelinia caspica); терм.:
Прозоровский, 1936.
Растение реддитивное (Planta redditiva, от лат. reddo — отдавать
назад) — один из типов классификации форм жизненных по
Г. М. Зозулину, объединяющий раст. многолетние, практически
не имеющие почек возобновления и почек спящих и в результате
при уничтожении их надземной части «уступающие» площадь
обитания другим раст.; гл. обр. тропические раст., среди которых
выделяются (группы) Р. вечнозеленые и Р. листопадные; терм.: Зо-
зулин, 1961.
Растение рестативное (Planta restativa, от лат. resto — оставаться,
сохраняться) — один из типов классификации форм жизненных по
Г. М. Зозулину, объединяющий раст. многолетние, возобновляю-
щиеся в случае уничтожения их наземных частей за счет деятель-
ности почек спящих и почек возобновления («сопротивляются за-
хвату площади другими раст.»); среди Р. р. выделяются группы
удерживающихся (Р. стержнекорневое, Р. кистекорневое, Р. клуб-
необразующее, Р. луковичное), продвигающихся (Р. короткокорне-
вищное) и нарастающих (Р. плотнодерновинное, Р. рыхлодерновин-
ное); терм.: Зозулин, 1961.
Растение розеточное (Planta rosularis; от лат. гоза — розетка;
англ.: rosette plant) — одна из форм жизненных, объединяющая
раст. с сильно укороченными междоузлиями надземных побегов,
ассимилирующими листьями, образующими т. н. розетку (см.
побег розеточный), и безлистными цветоносами (Plantago); гл.
обр. травянистые многолетники (раст. бесстебельные), розеточ-
ная структура побегов которых обычно рассматривается как
адаптация к различным неблагоприятным условиям среды (низ-
кая температура, недостаток влаги, сильная освещенность); раст.
древовидные, крона которых образована розетками листьев, наз.
раст. акрофилльными; нередко к Р. р. относят нарастащие моно-
подиально травы (раст. моноподиально-розеточные), у которых
побеги скелетные розеточные, а побеги генеративные безрозеточ-
ные (Alchemilla); лит.: Голубев, 1965; Серебрякова, 1971а.
Растение рыхлодерновинное см. растение дерновинное.
185
Растение рыхлокустовое см. растение дерновинное.
Растение семельпаро(у)зное (англ.: semelparous plant) см. моно-
карпик.
Растение сенильное см. период сенильный.
Растение стеблеклубневое (англ.: plant with stem tuber) см. расте-
ние клубнеобразующее.
Растение стелющееся (англ.: prostrate (trailer, trailing) plant) —
P. деревянистое с полегающими, укореняющимися осями скелет-
ными и из-за более интенсивного их роста превышающими по дли-
не недолговечные прямостоящие боковые ветви (рис. 126); распро-
5гм
Рис. 126. Стелющиеся (Б) и ползучие (В-Г) растения:
А—Б — Juniperus turkestanica (А — древовидная форма, Б — стланец; по: Серебряков,
1964а), В — Potentilla anserina, Г — Lysimachia nummularia (по: Ботаника..., 1988)
186
странены гл. обр. в высокогорьях, субарктических и субантаркти-
ческих широтах; облигатные и факультативные стелющиеся
деревья (Pinus montana) наз. стланцами (реже — стланиками), кус-
тарники (Salix) — стланиками или шпалерными кустарниками, а
кустарнички (Ericaceae) — стланичками или шпалерными кустар-
ничками; у нек-рых Р. с. скелетные оси полегают не полностью или
часть из них остается прямостоячими, в связи с чем выделяют по-
лустланцы, полустланики или гемипростратные кустарники, пот
лустланички или гемипростратные кустарнички; ср. Р. ползучее;
лит.: Дервиз-Соколова, 1967; Колищук, 1968а,б; Мазуренко, 1978;
Серебряков, 1962.
Растение стержне-кистекорневое — Р., система корневая кото-
рого во взрослом состоянии образована корнем главным и корня-
ми кладогеными; стержне-кистекорневая (смешанная) система яв-
ляется переходной между собственно стержнекорневой и кисте-
корневой, в силу чего не имеет четких границ (рис. 127); обычно к
Р. с.-к. относят раст., главный корень которых поздно отмирает
и/или явно выделяется среди массы корней кладогенных (ср. Р.
стержнекорневое); лит.: Голубев, 1962; Михайловская, 1981а.
Рис. 127. Корневая система Plantago media (А),
Р. lanceolata (Б) и Р. major (Б): 1 — главный корень,
2 — придаточный корень (по: Голубев, 1965)
187
Растение стержнекорневое (Planta radice palari (verticali) praedita;
англ.: plant with tap root, taproot plant)— раст., система корневая
которого во взрослом состоянии образована корнем главным и его
ответвлениями (корни боковые; рис. 127); часто к Р. с. относят
раст., у которых в небольшом или значительном числе развива-
ются и корни кладогенные, но при этом главный корень сильно
развит и хорошо заметен среди остальных корней (ср. Р. стерж-
не-кистекорневое); лит.:Голубев, 1962; Игнатьева, 1965; Михай-
ловская, 1981а.
Растение столонно-клубневое см. растение клубнеобразующее.
Растение столонообразующее (Planta stolonifera; англ.: stolonate
(stoloniferous) plant) см. столон.
Растение субсенильное см. период сенильный.
Растение суккулентное (Planta succulenta; англ.: succulent plant)
см. суккуленты.
Растение талломное (Tallobiont, tallophyt, от греч. tallos — молодая
ветка, росток, побег; англ.: tallophyte, tallogen plant) — один из ти-
пов структурной организации раст., характеризующийся отсутст-
вием расчленения тела раст. (таллома или слоевища; англ.: tallus)
на вегетативные органы (лист, стебель, корень); к Р. т. относятся
водоросли, слизевики, грибы, лишайники, нек-рые моховидные и
заростки высших споровых (первично талломные); среди семен-
ных растений нек-рые утратили листостебельность и характеризу-
ются вторично талломным типом организации тела (см. листец,
таллоид, филломорфа); ср. растение листостебельное.
Растение травянистое (Planta herbacea; от лат. herba — трава, зе-
лень; англ.: herbaceous plant, herb) — один из основных типов
форм жизненных, выделяемый в сезонном климате по эколого-
морфологическому признаку — отсутствию прямостоячих над-
земных или надводных живых стеблей, переживающих неблаго-
приятный сезон года (условия внутри «кочек» раст. дерновинных,
в частности некоторых злаков и осок, принимаются за сходные с
подземными); Р. т.— терофиты, гемикриптофиты и криптофи-
ты, почки возобновления которых располагаются на корневищах,
каудексах, клубнях, луковицах или столонах (рис. 128); часто Р. т.
выделяют по анатомическим признакам (слабое одревеснение, рас-
сеченная или сильно паренхиматизированная стела, отсутствие
перидермы и др.), которые позволяют провести относительную
границу между деревянистыми (напр., дерево кронообразующеё) и
травянистыми фанерофитами (напр., дерево розеточное) только в
188
150
Рис 128. Соотношение многолетних и отмирающих частей у деревянистых (A-В), по-
лудревесных (Г) и травянистых (Д) растений: А — дерево, Б — кустарник. В — кус-
тарничек, Г — полукустарник, Д — травянистый многолетник (многолетние части
показаны черным, отмершие — двойным пунктиром, отмирающие — одинарным
пунктиром, цифрами — возраст отдельных элементов) (по: Ботаника..., 1988)
бессезонном климате; лит.: Гатцук, 1976; Гатцук и др., 1974; Сереб-
рякова, 1973; Хохряков, 1976; Elten, 1991.
Растение униполярное (англ.: unipolar plant) см. полярность и рост
униполярный.
Растение хищное (англ.: carnivorous plant) см. растение насекомо-
ядное.
Растение эпигеогенно-длиннокорневищное см. растение длин-
нокорневищное.
Растение эпигеогенно-короткокорневищное (англ.: short-rhizo-
me plant р. р.) см. растение короткокорневищное.
Растение ювенильное (Planta juveniles; от лат. juvenis — молодой,
юный; англ.: juvenile plant) — особь вегетативная, не имеющая
связи с семенем и отличающаяся несформированностью морфо-
логических признаков взрослого раст.: листья иной формы и рас-
положения, отсутствие ветвления или другой тип нарастания, со-
хранение зародышевых структур (первого корня, побега) и др.
189
(обозначается индексом j); у нек-рых раст. семядоли сохраняются
долго (Quercus), у других — возможно усложнение системы кор-
невой (см. период виргинильный); часто к Р. ю. относят и про-
ростки или реже под этим названием объединяют все молодые
особи неспособные к восприятию индукции цветения (ювениль-
ный этап — от прорастания до появления способности к цвете-
нию); син.: Р. юношеское; терм.: Богдановская-Гиенэф (1926) ис-
пользовала для обозначения периода виргинильного; лит.: Ценопо-
пуляции... 1976, 1988.
Растение-подушка (англ.: cushion (polster) plant) см. растение по-
душковидное.
Регенерация (Regeneratio, от лат. ге — приставка, означающая по-
вторность действия, genero — производить, создавать; англ.: re-
generation) — широко используемое назв. для обозначения раз-
личных процессов воссоздания целостной особи: от восстановле-
ния нарушенных тканей (реституция) или отчужденных органов
(репродукция) до сезонного возобновления роста раст. (Р. типа
репродукции) и образования целого организма из его части при
размножении вегетативном (иногда наз. реституцией); в специ-
альном значении под Р. понимают процесс восстановления на-
сильственно отторженных частей раст. (при искусственном раз-
множении раст. черенками, частями корневищ и т. п.); ср. реите-
рация; лит.: Кренке, 1950; Дубровицкая, 1961; Юсуфов, 1982.
Регуляция развития — направленное изменение скорости и ха-
рактера (индукция) развития, обусловленное внешними и внут-
ренними факторами. Предполагают, что основным принципом
организменного уровня Р. р. раст. является взаимодействия оча-
гов меристемы (доминирующих центров), которые проявляются
в корреляциях, обеспечивающих целостность индивидуума; лит.:
Синнот, 1963; Полевой, 1981; Батыгин, 1986.
Редукция (от лат. reductio — возвращение, отодвигание назад;
англ.: reduction) — один из основных способов эволюции морфоло-
гической органов в рамках теории филэмбриогенеза, который
представляет собой недоразвитие (рудиментация) или полное ис-
чезновение органа (афанизия) у потомка по сравнению с пред-
ком. В широком смысле редукцией обозначают не только измене-
ние органов в эволюции (инволюция), но и упрощение структу-
ры органов или тканей в онтогенезе (дегенерация); ср. анаболия
отрицательная; лит.: Северцов, 1945, Северцов, 1981, Терехин,
1991; Кузнецова, 1998.
190
Резид (Residuum, от лат. residuus — оставшийся, сохранившийся) —
базальный, многолетний участок побега монокарпического много-
летних травянистых раст.; как правило, несет почки возобновления
и морфологически соответствует зоне возобновленияили зонекуще-
ния (рис. 19,41,98); для нек-рыхраст. деревянистыхв том же смыс-
ле используются терм, ксилоподий, «турион»; син.: прирост ис-
тинный, пенек или подиум; терм.: Нухимовский, 1969 а,б, 1997.
Реитерация (англ.: reiteration) — развитие из почек спящих побегов
формирования, на основе которых формируется сходная с мате-
ринским деревом модель архитектурная (повторение архитектур-
ной модели); пробуждение спящих почек может быть связано с
условиями произрастания и механическими нарушениями, в свя-
зи с чем различают Р. адаптивную и травматическую (ср. регене-
рация, рост квантированный); лит.: Oldeman, 1974.
Рекапитуляция (от лат. recapitulatio — повторение; англ.: recapitu-
lation) — повторение в эмбриогенезе потомка признаков взрос-
лого предка. Эта закономерность послужила основой для «биоге-
нетического закона» Э. Геккеля (1866) — «онтогенез есть быстрое
и краткое повторение филогенеза». Согласно ему Р. являются ре-
зультатом эволюции онтогенеза посредством изменения послед-
них стадий развития. Позднее было показано, что Р. проявляются
только в эволюции органов (см. филэмбриогенез) при анаболиях
или благодаря акселерации и гиперморфоза. Принцип Р. помогает
реконструировать направления морфологической эволюции жи-
вотных при отсутствии палеонтологических материалов. В био-
морфологии такой подход используют в более широком смысле
(на первых стадиях онтоморфогенеза проявляются палеоморф-
ные черты развития). Однако специфика развития и организации
раст. резко ограничивает использование принципа Р.; лит.: Ко-
зо-Полянский, 1937; Голубев, 1959; Северцов, 1970; Мирзоян,
1974; Цвелев, 1977; Недолужко, 1996а, б, 1997.
Ретардация (англ.: retardation) см. неотения.
Ризокорм(ус) (англ.: rhizocormus) см. побег автономный.
Ризология (от греч. riza — корень, logos — слово, учение) — наука
о развитии корней и корневых систем, их строении, функцио-
нальной специализации и эволюции. В более широком смысле Р.
рассматривают как дисциплину, изучающую структуру подзем-
ных органов растений, включая систему корневую и подземные
стебли (корневище, каудекс, столон, клубень, луковица и др.); лит.:
Серебряков, 1952; Таршис, 1975, 2003; Рысин, Рысина, 1987.
191
Ризом (Rhizoma; англ.: rhizome) см. корневище.
Ризом гипогеогенный см. корневище гипогеогенное.
Ризом эпигеогенный см. корневище эпигеогенное.
Ризостиха (англ.: rhizostichy) см. ризотаксис.
Ризотаксис (Rhizotaxis, от греч. rhizoma — корень, taxis — располо-
жение; англ.: rhizotaxis) — порядок размещения корней боковых,
отражающий симметрию первичного строения корня материнско-
го; корни боковые обычно закладываются напротив участков
протоксилемы и после прорастания через ткани первичной коры
формируют правильные продольные ряды (ризостихи); в зависи-
мости от числа участков протоксилемы количество ризостих раз-
лично: 2-5 (у двудольных) и более (у однодольных); лит.: Михай-
ловская, 1981а.
Ритм развития сезонный см. развитие сезонное.
Ритм роста (англ.: growth rhythm) см. рост.
Ритмичность роста (англ.: growth rhythm) — чередование процес-
сов (ритмов или циклов) интенсивного и замедленного роста
(вплоть до полной остановки), обусловленное внешними и внут-
ренними факторами, которые рассматриваются как индукторы и
триггеры наследственно закрепленных морфогенетических про-
цессов; различают эндогенный (сохраняющийся при постоянных
внешних условиях) и экзогенный ритм (обусловленный клима-
тическими факторами); лит.: Сабинин, 1963; Грудзинская, 1964;
Серебряков, 1966; Юрцев, 1976; Михалевская, 2002.
Рост (Crescentia, от лат. cresco — расти, вырастать; англ.: growth) —
необратимое увеличение размеров и массы раст., связанное с но-
вообразованием элементов его структуры: клеток, тканей, орга-
нов; часто Р. понимают только как увеличение размеров и массы
раст.; общий характер Р. раст. выражается в виде сигмоидной кри-
вой (кривая Р. большая), параметры которой зависят от эндоген-
ных и экзогенных факторов (см. поведение раст.); основные фазы
Р. (деление клеток, их растяжение и дифференциация) происхо-
дят в меристеме, деятельность которой обусловливает открытый
и периодически возобновляющийся Р. раст. (см. ритмичность Р.);
различают Р. апикальный и Р. интеркалярный; лит.: Сабинин,
1963; Синнот, 1963; Серебряков, 1966; Зотин, 1974; Иванов, 1974;
Рост..., 1981; Кефели, 1984.
Рост апикальный (Crescentia apicalis, от лат. apex — вершина;
англ.: apical growth) — основной тип неограниченного Р. стебля л
корня в длину, связанный с меристематической деятельностью
192
конуса нарастания побега и корня; обусловливает ориентацию ор-
ганов раст. в пространстве (см. тропизм, настии); син.: Р. верху-
шечный.
Рост апогеотропный (Crescentia apogeotropus, от греч. аро — при-
ставка, здесь со значением обратности, geo — земля, tropos — по-
ворот, направление; англ.: apogeotropic growth) — Р. апикальный
побега или реже корня (напр. пневматофоры), направленный вер-
тикально вверх (отрицательный геотропизм); у нек-рых раст.
стебель «растет» вниз, что связано напр. с недостаточной механи-
ческой прочностью для его поддержания в вертикальном положе-
нии (раст. ампельное); обычно Р. а. побега наз. ростом ортотроп-
ным (рис. 105).
Рост биполярный (англ.: bipolar growth) —развитие ргсс. из заро-
дыша, на апикальном полюсе которого находится почка зароды-
шевая, а на базальном — корешок; Р. б. в основном характерен для
аллоризофитов; ср. рост униполярный.
Рост верхушечный см. рост апикальный.
Рост видимый (побега) — пон. используемое для обозначения
увеличения размеров побега; рассматривается как фаза развития
побега (см. онтогенез побега, цикл жизненный малый), следую-
щая за ростом внутрипочечным (от момента раскрывания почки
до прекращения увеличения линейных размеров); син.: рост вне-
почечный; лит.: Серебряков, 1952; Михалевская, 2002.
Рост внепочечный см. рост видимый.
Рост внутрипочечный (побега) — фаза (период, этап) развития
побега от момента его заложения (или заложения фитомеров его
очередного прироста) до развертывания почки; характеризуется
временными (пластохрон) или структурными единицами роста
(цикл листовой, емкость почки, квант роста); син.: фаза (период,
этап) эмбрионального развития побега; лит.: Серебряков, 1952;
Шилова, 1988; Михалевская, 2002.
Рост вставочный см. рост интеркалярный.
Рост геотропный (Crescentia geotropus, от греч. geo — земля, tro-
pos — поворот, направление; англ.: geotropic growth) — Р. апи-
кальный корня или реже побега (напр. раст. ампельное), направ-
ленный вертикально вниз (положительный геотропизм; рис. 105);
у нек-рых раст. корни растут кверху (напр. пневматофоры), что
обусловлено часто доминированием факторов хемотропизма.
Рост детерминированный (англ.: determinate growth) — непродол-
жительный рост осевых или боковых элементов растения, обу-
193
словленный их быстрым созреванием (рост контролируемый воз-
растом); обычно используется для характеристики роста листа
(лист детерминированный; англ.: determinate leaf) или реже корня
(см. корень детерминированный), которые быстро достигают пре-
дельных размеров; для побегов пон. Р. д. отражает относительно
быстрое формирование терминального соцветия (цветка), усика,
колючки или естественное отмирание меристемы апикальной
(см. цикл жизненный малый); ср. рост недетерминированный.
Рост диагеотропный (Crescentia diageotropus, от греч. dia — при-
ставка со значением поперек, geo — земля, tropos — поворот, на-
правление; англ.: diageotropic growth) — Р. апикальный побега или
корня, направленный горизонтально (поперечный тропизм); к
диагеотропно растущим органам раст. относятся побеги и корни
размножения (рис. 105); обычно Р. д. побега наз. ростом плагио-
тропным (см. побег диагеотропный).
Рост интеркалярный (Crescentia intercalaris; от лат. intercalarius —
вставной, добавочный; англ.: intercalary growth) — тип Р. стебля и
листа в длину, связанный с деятельностью меристемы интерка-
лярной; наиболее ярко выражен у однодольных; длительный Р. и.
побегов злаков наз. Р. телескопическим или суставчатым; син.:
Р. вставочный; лит.: Серебрякова, 1971а.
Рост квантированный — Р. раст., обусловленный меристематиче-
ской деятельностью конуса нарастания и морфометрически про-
являющийся в форме правила кратных отношений; в более
широком смысле к Р. к. (Р. модулярному) относят сериальное по-
вторение в структуре раст. элементов различного уровня от ме-
тамера до моделей архитектурных (см. реитерация); лит.:
Шмидт, 1968; Серебрякова, 1971а.
Рост клональный (англ.: clonal growth) см. размножение вегета-
тивное.
Рост модулярный (англ.: modular growth) см. рост квантиро-
ванный.
Рост недетерминированный (англ.: indeterminate growth) — про-
должительный рост осевых или боковых элементов растения (по-
бег, лист, корень); обычно используется для характеристики роста
побега, развитие которого продолжается в течение нескольких лет
за счет деятельности меристемы апикальной, которая долго (см. по-
бег полициклический) или вообще (см. побег открытый) не обра-
зует терминального соцветия (цветка); у нек-тых раст. листья рас-
тут в течение нескольких лет (напр. Guarea rhopalocarpa) и также
194
рассматриваются как органы с Р. н. (недетерминированный лист;
англ.: indeterminate leaf); ср. рост детерминированный.
Рост ортотропный (Crescentia orthotropus; англ.: orthotropic growth)
см. рост апогеотропный.
Рост плагиотропный (Crescentia plagiotropus; англ.: plagiotropic
growth) см. рост диагеотропный.
Рост суставчатый см. рост интеркалярный.
Рост телескопический см. рост интеркалярный.
Рост униполярный (англ.: unipolar growth) — развитие раст. из за-
родыша, при котором побег главный формируется как продолже-
ние апикального полюса зародыша, а первый корень является бо-
ковым органом (корешок закладывается сбоку, как корень прида
точный побега); Р. у. характерен для первичных гоморизофитов;
часто пон. Р. у. используют для характеристики развития раст.,
у которых корешок недоразвивается или быстро отмирает (нек-
рые вторичные гоморизофиты); ср. рост биполярный.
Рост усиления — закономерное увеличение диаметра побега в на-
чале его развития в области укороченных междоузлий; наиболее
выражен у однодольных и морфологически проявляется, напр.,
у злаков в обратноконической форме нижней части их побегов;
терм.: Troll, 1937-1939, цит. по: Серебрякова, 1971а.
Рудиментация см. редукция.
Сапрофит (Saprophyton, от греч. sapros — гнилой, phyton — расте-
ние; англ.: saprophyte) — гетеротрофный организм, использую-
щий для питания органические соединения мертвых организмов;
среди покрытосеменных С. традиционно наз. лишенные хлоро-
филла микотрофные раст. (англ.: mycotrophic plant) из сем.
Gentianaceae, Orchidaceae, Burmanniaceae, Thismiaceae, Triurida-
ceae, Pyrolaceae и др. (ср. голопаразит); для большинства из них
установлено, что все питание они получают из почвы или от дру-
гих автотрофных раст. только через посредство своего микориз-
ного гриба и относятся не к сапротрофам, а к раст.-паразитам
(аллелопаразиты); лит.: Терехин, 1965, 1977.
Сарментация (от лат. sarmentum — лоза, отпрыск, ус, плеть) — об-
разование побега (отпрыска) из почки верхушечной или почки
боковой столона, корневища или корня (отпрыск корневой), ко-
торый после укоренения отделяется от материнского растения
(см. размножение вегетативное) и дает начало дочерней особи
(рамета); лит.: Барыкина, 2000.
Сверхпаразитизм см. паразитизм.
195
Связник семядоли см. шейка семядольная.
Семиэпифит (англ.: hemiepiphyte) см. полуэпифит.
Семя (Semen; англ.: seed, semen) — орган размножения и расселения
(см. диаспора) цветковых и голосеменных раст., образующийся в
результате оплодотворения или у некоторых раст. спонтанно
(агамоспермия, апомиксис); в типе состоит из покровов (семен-
ной кожуры), зародыша и запасающей ткани (эндосперма, перис-
перма). Выделяют С. безбелковые (без эндосперма) и белковые
(с эндоспермом). С.— форма существования особы на первых ста-
диях ее онтогенеза (см. период латентный), характеризующаяся
различными специфическими адаптивными морфологическими
особенностями и физиологическими механизмами расселения
(анемохория, зоохория и т. п.), покоя и прорастания (см. катего-
рия форм жизненных); лит.: Артюшенко, 1990; Терехин, 1996;
Forbis et al., 2002.
Семядоля (Cotyledon, от греч. kotyle — доля; англ.: cotyledon, seed
(leaf) lobe, seminal leaf) — первый листовой орган раст., возни-
кающий в семени на апикальном полюсе зародыша до начала
формирования почечки-, по форме, анатомическому строению и
функции часто резко отличается от настоящих листьев-, при про-
растании надземном нередко зеленеет и фотосинтезирует, а при
прорастании подземном выполняет запасающую (Quercus) или
гаусториальную функцию (см. щиток); исключая крайние случаи
(см. поликотилия, псевдомоно(ди)котилия), одна С. характерна
для проростков однодольных, две — двудольных; зародыш мно-
гих раст.-паразитовне имеет С. (Orobanchaceae); син.: лист семя-
дольный, Л. зародышевый; лит.: Серебряков, 1952.
Серия листовая (англ.: leaf series) — ряд листьев, последовательно
образующихся при прорастании раст. (рис. 63); изменения фор-
мы листа в С. л. связаны с переходом раст. из ювенильного к
взрослому состоянию возрастному (см. гетеро-, гомобластия).
Симметрия (Symmetria, от греч. symmetria — соразмерность; англ.:
symmetry) — закономерность в строении и форме раст. или его
отдельных органов, проявляющаяся в неизменности строения
или/и формы относительно геометрической оси; тело раст. или
его орган, как правило, имеет продольную ось, через которую
можно провести много, две, одну или ни одной плоскости сим-
метрии; в связи с этим различают радиально-симметричное (ко-
рень), билатерально-симметричное (зародыш), мносимметричное
(лист) и ассимметричное строение; в более широком смысле к
196
С. относят и повторяемость структуры вдоль оси (продольная
или поступательная С.; см. метамерность).
Симподий (Sympodium, от греч. syn — вместе, pous — нога, здесь —-
ось; англ.: sympodium) — многолетний осевой орган раст. (ствол,
ветвь, корневище), образованный в результате деятельности мери-
стем апикальных нескольких дочерних элементов, последователь-
но сменяющих друг друга по принципу «перевершинивания»
(рис. 98); при ветвлении С. каждый его дочерний элемент также С.,
а образующаяся система наз. симподиальной (напр. система побе-
гов симподиальная); см. также нарастание симподиальное.
Синкер (англ.: sinker) см. тубероид.
Синкотилия (Syncotylia, от греч. syn — вместе, kotyle — доля;
англ.: syncotyly) — слияние двух семядолей на ранних фазах эм-
бриогенеза; в первоначальном значении — более или менее пол-
ное срастание семядолей своими краями; различают срастание се-
мядолей краями (амфикотилия, амфисинкотилия) с образовани-
ем в той или иной степени бокаловидной структуры и полное или
частичное срастание смежными сторонами (гамокотилия).
Синонтогенез см. онтогенез.
Система корневая (Systema radicale; англ.: root system) — совокуп-
ность всех корней особи, развивающаяся в течение ее онтогенеза
и достигающая полного формирования у взрослого раст. (рис. 3);
в образовании С. к. участвуют корни разного происхождения (см.
корень главный, корень боковой, корень придаточный), в связи с
чем выделяют раст. стержнекорневое (С. к. стержневая), раст. кис-
текорневое (С. к. кистевидная или мочковатая) и раст. стержне-
кистекорневое (С. к. смешанная); по глубине проникновения кор-
ней в субстрат различают К. с. глубинную (длина корней 2-10 и бо-
лее м; англ.: deep root system), поверхностную (длина корней 0,2-
0,5 м; англ.: shallow root system) и универсальную (с глубинными и
поверхностными корнями), а по степени развития — К. с. экстен-
сивную (корни охватывают объем почвы большого диаметра) и
интенсивную (корни густо пронизывают объем почвы небольшо-
го диаметра); иногда в С. к. раст. выделяют физиологически непре-
рывные подсистемы (комплекс корней обособленный) — сово-
купность корней и их ответвлений, связанных только с одним побе-
гом или формирующейся на его основе системой побегов-, см. также
аллоризия, гоморизия; лит.: Казакевич, 1928; Сабинин, 1949; Голу-
бев, 1962; Красильников, 1970; Михайловская, 1981а; Рысин, Рыси-
на, 1987; Plants roots..., 1991; Барыкина, 1999а; Таршис, 1975, 2003.
197
Система корневая аллоризная см. аллоризия.
Система корневая вторично-гоморизная см. гоморизия.
Система корневая вторично-стержневая см. растение кистекор-
невое.
Система корневая гоморизная см. гоморизия.
Система корневая кистевидная см. растение кистекорневое.
Система корневая мочковатая см. растение кистекорневое.
Система корневая первично-гоморизная см. гоморизия.
Система корневая смешанная см. растение стержне-кистекор-
невое.
Система корневая стержневая см. растение стержнекорневое.
Система побега (англ.: shoot system) — совокупность побега мате-
ринского и развивающихся из его почек боковых побегов дочерних; в
зависимости от функциональной значимости материнского по-
бега различают системы: побега ветвления (СПБ), побега первично-
го (СПП), побега дополнения (СПД), побега формирования (СПФ),
побега скелетного, побега годичного и др.; лит.: Серебряков, 1952;
Гатцук, 1974а; Мазуренко, Хохряков, 1977.
Система побегов моноподиальная см. моноподий.
Система побегов симподиальная см. симподий.
Система побегов см. система побеговая.
Система побеговая (англ.: shoot system) — широко используемое
назв. для обозначения единиц структуры разного уровня: от сис-
темы побега до системы побегов индивида сложного, отдельные
части которой (напр., кусты парциальные) связаны между собой
корневищами удлиненными или корнями размножения; строение С.
п. определяется структурой входящих в нее побегов, типом и ин-
тенсивностью их ветвления, направлением роста, обусловливаю-
щим ориентацию побегов, и др.; один из способов описания
строения С. п.— выявление модели архитектурной или модели
побегообразования; лит.: Нухимовский, 1971; Смирнова, 1980; Гат-
цук, 1974а.
Система форм жизненных линейная см. классификация форм
жизненных по В. Н. Голубеву.
Скрытосемядольность см. прорастание подземное.
Слоевище (англ.: thallus) см. растение талломное.
Состояние возрастное (англ.: age state) — физиолого-биохимиче-
ское состояние особи, отражающее этап ее онтогенеза в момент на-
блюдения. С. в. нежестко скореллировано с возрастом абсолютным
и определяется по морфологическим признакам, предположи-
те
тельно обусловленными возрастными (онтогенетическими) изме-
нениями (см. старение, модель комбинаторного развития побега,
модель «контроллера переключения фаз»)-, син.: возраст физиологи-
ческий, возраст биологический, возрастность, состояние онтогене-
тическое; лит.: Кренке, 1940; Ценопопуляции... 1976, 1988.
Соцветие (Inflorescentia, от лат. inflorescere — начинать, цвести, за-
цветать; англ.: inflorescence) — в самом общем смысле С. наз. со-
вокупность цветков, расположенных на верхушке (С. терминаль-
ное) или в средней
части (С. интеркаляр-
ное) побега (главной
оси). По развитию
прицветных листьев
различают С. фрон-
дозное (облиствлен-
ное), брактеозное и
голое (эбрактеозное).
По степени разветв
ленности выделяют С.
простое (ветвление не
превышает двух по-
рядков) и сложное
(ветвление более двух
порядков с образова-
нием парциальных,
элементарных или ча-
стных С.). По типу на-
растания и порядку
развития цветков вы-
деляю! С. цимозное
(закрытое, определен-
ное) и рацемозное
(ботрическое, откры-
тое, неопределенное).
Рис. 129. Число синфлоресценций и их положение в
побеговых системах однолетников (А) и многолетни-
ков с симподиальным (Б) или моноподиальным (В)
нарастанием: 1 — листья, 2 — отмершие листья, 3 —
брактеи, 4 — семядоли, 5 — конгу р объединенного
соцветия (синфлоресценции), 6 — контур главного
соцветия и ко-флоресценции, 7 — одиночные цветки,
8 — верхушечная почка, 9 — пазушная почка возоб-
новления (по: Учебно-полевая..., 1990)
Первое характеризу-
ется нарастанием сим-
подиальным, базипе-
тальным развитием
цветков и центробеж-
ным их раскрывани-
199
ем, а второе — нарастанием моноподиалъным, акропетальным
развитием цветков и центростремительным их раскрыванием.
Поскольку С. представляет видоизмененную часть побеговой
системы растения, которая б. м. отграничена от вегетативной сфе-
ры и обычно полностью или почти полностью отмирает после
цветения и плодоношения, то флоральный участок побега во-
зобновления (или прироста) нередко рассматривают как струк-
турно-функциональную (цветоносную) зону, именуемую син-
флоресценцией (англ, synflorescence) или объединенным С.
(рис. 129). При таком подходе С. обозначают часть системы побе-
га, состоящей из флорального участка побега возобновления (сте-
бель первого порядка побега закрытого или открытого) и отходя-
щих от него побегов генеративных (побегов обогащения), которые
развиваются силлептически (.побег силлептический) или пролеп-
тически (побег пролептический) и в свою очередь способны вет-
виться (см. зона обогащения). В результате синфлоресценция мо-
жет объединять С. разных иерархических уровней: главное С.
(терминальное С. побега возобновления) и С. паракладиев или
ко-флоресценции (англ, co-florescence), которые представляют
собой терминальные С. побегов обогащения (рис. 129); лит.: Бота-
ника..., 1988; Кузнецова и др., 1992; Лотова, 2000.
СПВ, СПД, СПП, СПФ — см. система побега.
Спектр биологический (англ.: biological spectrum) — соотношение
форм жизненных или выделенных по каким-либо признакам био-
морфологических групп растений конкретной территории, флоры,
типа растительности и т. п„ выраженное в процентах; впервые ста-
тический метод при изучении закономерностей распределения
жизненных форм в зависимости от климата применил К. Раункиер,
который соотношение 10 групп жизненных форм (суккуленты
стеблевые — S, эпифиты — Е, мега- и мезофанерофиты — ММ,
микрофанерофиты — М, нанофанерофиты — N, хамефиты —Ch,
гемикриптофиты — Н, геофиты — G, гело- и гидрофиты — НН,
терофиты — Th) наз. С. б. или спектром фитоклиматическим,
а эталонный спектр — спектром нормальным; иногда С. б. наз. С.
биоэкологическим, С. жизненных форм или С. экологическим.
Спектр биоэкологический см. спектр биологический.
Спектр глобальный см. спектр нормальный.
Спектр жизненных форм см. спектр биологический.
Спектр нормальный — эталонный С. используемый для анализа
С. биологического и отражающий процентное соотношение форм
200
жизненных всей Земли; впервые построен К. Раункиером для
400 видов, произвольно выбранных из «Index Kewensis» (фанеро-
фитов 43 %, хамефитов 9 %, гемикриптофитов 27 %, геофитов
4 %, гидрофитов 1 %, терофитов 13 %, эпифитов3 %); считается,
что отклонения от С. н. отражают воздействие среды (гл. обр.
климата) на характер адаптации раст.; иногда С. н. наз. спектром
глобальным.
Спираль филотаксиса генетическая см. теория К. Шимпера и
А. Брауна.
Способ (модус) эволюции морфологической — процесс или ре-
зультат эволюционных изменения развития тканей, органов,
формы жизненной или онтогенеза. В рамках теорий филэмбриоге-
неза и гетерохронии выделяют несколько способов на основании
сравнения строения и развития потомка с предком. В биоморфо-
логии выделение С. э. м. обычно связано с методом морфогенети-
ческих рядов. См. также эмбрионизация, полимеризация-, лит.: Се-
верцов, 1945; Тахтаджян, 1964; Северцов, 1981; Серебрякова, 1983;
Терехин, 1991; Барыкина, 19996; Жмылев, 2004.
Старение (англ.: senescence, aging) — эндогенно и экзогенно обу-
словленный процесс возрастных изменений организма, прояв-
ляющийся в ослаблении его жизнедеятельности и, в конечном
итоге, в естественном отмирании (см. период сенильный). С. раст.
протекает гетерохронно (с различной скоростью в разных клет-
ках, тканях, органах) и гетеротропно (неодинаково в разных
структурах). У многоклеточных раст. С. отдельных клеток, тканей
и органов не ведет к С. всей особи (или клона) сразу, т. к. в течение
онтогенеза одновременно с отмиранием отдельных структур
(напр. апоптоз, листопад, веткопад и т. п.) идет их новообразова-
ние (омоложение). В результате С. поликарпиковили клоня зачас-
тую проявляется в постепенном преобладании отмирания над но-
вообразованием (напр., вырождение клона). Механизм С. не из-
вестен и многочисленные гипотезы объясняют только отдельные
стороны этого явления (гипотеза программированной смерти, ги-
потеза аккумуляции мутаций, гипотеза антагонистической
плейотропии, гипотеза восстановления сомы, гипотеза смерти от
истощения). По выраженности С. выделяют организмы с посте-
пенным (у многолетних поликарпиков), лавинообразным (у мо-
нокарпиков) и незначительным старением (старение негатив-
ное)-, лит.: Williams, 1957; Стрелер, 1964; Казарян, 1969; Ефейкин,
1970; Зотин, 1974; Биология развития..., 1975;Zelawski, 1985; Юсу-
201
фов, 1988; Gan, Amasino, 1997; Юсуфов, 1992; Silverstown et al.,
2001; Thomas, 2002; Thomas et al., 2000, 2003; Gavrilov, Gavrilova,
2002; Kirkwood, 2002; Goldsmith, 2004.
Старение коррелятивное см. гипотеза смерти от истощения.
Старение монокарпическое см. гипотеза смерти от истощения.
Старение негативное (англ, negative senescence) — название ис-
пользуется для организмов у которых: 1) уровень смертности в
популяции не увеличивается с возрастом, 2) интенсивность раз-
множения возрастает или не уменьшается с возрастом, 3) не обна-
руживается морфолого-физиологического проявления старения
или снижения устойчивости особи с возрастом. Среди растений С.
н. обнаружено у некоторых многолетних (напр., Pinus longaeva) и
малолетних поликарпиков (напр., Plantago lanceolata). Среди моно-
карпиков смерть без старения возможна только в случае экзоген-
ного контроля продолжительности жизни однолетника (напр.,
Ricinus communis в сезонном климате); лит.: Finch, Austad, 2001;
banner, Connor, 2001; Roach, 2001; Silverstown et al., 2001; Vaupel et
al„ 2004.
Ствол (Trupcus; англ.: trunk) — основной многолетний стебель де-
рева (ось скелетная основная или лидерная), видоизмененный в
типе в результате вторичного утолщения; развивается из почечки
зародыша и нарастает моноподиалъно (Picea) или симподиально
(Quercus) прямо вверх в течение нескольких десятков лет; сохра-
няется в течение всей жизни дерева, ветвится в той или иной сте-
пени (см. крона) и служит опорой для кроны, транспорта раство-
ров (сокодвижение) и запасания влаги и питательных веществ; у
настоящих деревьев ежегодно утолщается за счет действия кам-
бия; у раст. древовидных С. не имеет вторичного утолщения (Ра1-
тае), представляет собой полый стебель (С. ложный; англ.: pse-
udostem), образованный влагалищами крупных листьев (Zingibe-
rales) или утолщается за счет особой меристемы (Dracaena);
форма и предельная высота С. зависят от видовой принадлежно-
сти раст. и условий его местообитания; обычно С. имеет конусо-
видную форму (С. сбежистый), но у некоторых раст. (Brachy-
chiton) вздутый (бочонковидный) из-за сильного развития водо-
запасающей паренхимы (дерево бутылочное), у других раст.
полегает (стланец) или вьется (лиана древовидная), а у ряда полу-
эпифитов (первичных эпифитов) образуется из спускающихся к
земле переплетенных корней (С. корневой); часть С. мелких де-
ревьев или стланцев наз. стволиками.
202
Стволик (Trunculus; от ствол) — широко используемое назв. для
обозначения основных осей скелетных различных форм жизнен-
ных с одревесневающими побегами или их основаниями (рис. 59,
60): ствола мелких деревьев (С. главный), наиболее утолщенных
многолетних стеблей, образующих куст (составные скелетные
оси) кустарников, кустарничков, полукустарников, полукустар-
ничков и др. (С. скелетный или замещающий).
Стебель (Caulis; «стебель» от древнерусск. стебль, стъбль, стьбло;
англ.: stem, caulis) — осевая часть побега, состоящая из узлов и меж-
доузлий и образующаяся за счет роста верхушечного и роста интер-
калярного; выполняет гл. обр. опорную (механическую) и проводя-
щую функции, а иногда — запасающую, защитную и др. (см. ме-
таморфоз побега); часто С. наз. ось скелетную деревянистого раст.
(ствол, ветвь)-, лит.: Федоров, Кирпичников, Артюшенко, 1962.
Стеблекорень см. каудекс.
Стланец см. растение стелющееся.
Стланик см. растение стелющееся.
Стланичек см. растение стелющееся.
Столон (Stolo; корневой побег; англ.: stolon) — видоизмененный,
одно(двух)летний побег ползучий или участок побега с длинными,
тонкими междоузлиями и чешуевидными (катафиллы) или асси-
милирующими листьями нижней формации (плеть); обеспечи-
вает размножение вегетативное (см. сарментация) и расселение
(рис. 130); различают С. подземные и С. надземные, последние
иногда наз. усами, флагеллами, плетьми или отбегами; в связи с
этим среди раст. столонообразущих выделяют раст. подземносто-
лонные (англ.: diageic plant; Trientalis europaea) и раст. надземно-
столонные (Fragaria); лит.: Серебряков, 1952; Серебрякова, 1981.
Столон надземный (англ.: runner, flagellum) см. столон.
Столон подземный (англ.: stolon) см. столон.
Стрелка (англ.: scape) см. междоузлие.
Структура побега (англ.: shoot structure) — часто используемое
понятие для обозначения морфологической или анатомической
неоднородности в строении побега-, обычно отмечают изменения
метамеров от основания побега к его верхушке по одно- или мно-
говершинной кривой, форма которой отражает процесс развития
побега в его онтогенезе (напр., изменения длин междоузлий по ти-
пу акро-, бази- или мезотонии); иногда различают морфофизио-
логические зоны побега генеративного (рис. 40; по Troll, 1964): зо-
на кущения или возобновления, зона торможения (префлоральный
203
Рис. 130. Образование надземных столонов у Saxifraga flagellaris (1; по: Серебряков:
1952), Fragaria vesca (2 — общий вид растения, 3 — верхушка столона) и Rubus saxatilis
(4): г — столоновидный гипоподий, р — дочерняя розетка, л — чешуевидный лист
(по: Ботаника..., 1988)
204
участок), зона обогащения (сборное соцветие) и зона резервная;
при этом С. и. может быть записана в виде формулы структуры
побега; лит.: Серебрякова, 19696; 1971а.
Суккулент (Succulentis, от лат. sucus — сок, влага, сочность; англ.: su-
cculent) — раст. многолетнеес сочными, мясистыми л истъями или
стеблями, в водоносной паренхиме которого запасается большое
количество воды; С.— ксерофиты, распространенные гл. обр. в
пустынях Америки и Южн. Африки, среди которых различают С.
листовые (Crassula) и стеблевые (Cactaceae); нередко С. рассматри-
вают как особую форму жизненную раст. древовидных (Euphor-
biaceae) и раст. травянистых (Sedum); син.: раст. суккулентное.
Сформированность побега в почке — понятие используется для
определения состава зачатков побега генеративного в зрелой почке
возобновления (см. почка зрелая). Выделяют 3 группы растений:
1) растения, у которых почка возобновления содержит зачатки всех
элементов побега, включая зачатки соцветия и цветков (см. почка
преформированная, почка генеративная, почка вегетативно-гене-
ративная; 2) растения, у которых почка возобновления содержит за-
чатки только вегетативной сферы побега, а соцветие и цветки за-
кладываются после ее раскрывания (см. почка вегетативная);
3) растения, у которых почка возобновления содержит зачатки
только части вегетативной сферы побега и остальные элементы
(листья, цветки) закладываются после ее раскрывания (см. почка
вегетативная). У многих растений в начале вегетационного сезона
раскрывается несколько почек возобновления, степень сформиро-
ванное™ которых может отличаться в зависимости от положения
в системе побеговой; лит. Серебряков, 1947, 1952; Борисова, Малы-
шева, 1993; Шорина, Михалевская, 2000; Карпухина и др., 2002.
Схизокотилия (англ.: schizocotyly) см. псевдокотилия.
Таллоид (англ.: talloid) — один из основных элементов структуры
вегетативного тела представителей пор. Podostemales, имеющий
плоско листовидную или лентовидную форму и выполняющий
функции фотосинтеза и разрастания особи (рис. 131); развивается
из базального полюса зародыша (гипокотиля) и прикрепляется к
субстрату с помощью гаптер и ризовидных волосков; на верхней
поверхности или по краям Т. эндогенно закладываются почки, из
которых развиваются вегетативные и репродуктивные побеги; у
нек-рых раст. апекс Т. прикрыт псевдочехликом, в связи, с чем
предполагают, что Т. имеет корневую природу (рис. 132); син.:
таллус, слоевище; лит.: Cusset, Cusset, 1988,1989; Rutishauser, 1991.
205
Рис. 131. Схема формирование таллоида у Indotristicha ramosissima (А) и Dalzellia
ceylanica (Б): 1 — гипокотиль, 2 — семядоля, 3 — апекс, 4 — листья, 5 — таллоид,
6 — псевдочехлик, 7 — гаптера, 8 — ризовидные волоски, 9 — инициаль, 10 — почка,
11 — вегетативные побеги, 12 — генеративный побег, 13 — эпикотиледонарный лист,
14 — листочки (по: Cusset G-, Cosset, 1988)
Таллом (англ.: thallus) см. растение талломное.
Тело кочки см. кочка.
Тело растения (англ.: plant body) — часто используемое пон. для
обозначения совокупности вегетативных органов особи {системы
побегов и системы корневой)', форма Т. р. обусловлена типом
структурной организации, симметрией, полярностью и ростом;
син.: вегетативное тело.
206
Рис. 132. Внешний вид Podostemum ceratophyllum (по: Philbrick, Novelo, 2004):
А — внешний вид растения; Б — генеративный побег; 1 — корень;
2 — побег; 3 — цветок; 4 — лист
Темп развития — скорость полового созревания организма. Для
растений пон. введено Т. А. Работновым (1950) и Урановым
(1960) как время наступления периода генеративного (продолжи-
тельность периода виргинильного). Как правило, с T. р. сопряжена
продолжительность жизни особи; лит.: Серебряков, 1952; Райкова,
1965; Василевская, Борисовская, 1968; Серебрякова, 19716; Harper,
White, 1974; Bender et al., 2000; Silverstown et al., 2001.
Теория см. гипотеза.
Теория возрастной цикличности см. теория Н. П. Кренке.
207
Теория К. Уордлоу (Wardlow,
1947) — популярная теория
«физиологических полей», ос-
нованная на эксперименталь-
ном методе; согласно Т. каж-
дый примордий и дистальная
часть апекса представляют со-
бой центры роста и окружены
физиологическими полями
определенного диаметра, в
пределах которых ингибиру-
ется возникновение других
ростовых центров (примор-
диев); образование новых
примордиев связано с освобо-
ждением места (расхождение
ингибирующих полей в ре-
зультате роста апекса); рис.
133; лит.: Серебрякова, 1963;
Синнот, 1963; Schwabe, 1984.
Теория К. Шимпера и
А. Брауна (Braun, 1835) — од-
на из популярных теорий, ос-
нованная на математическом
методе анализа филлотаксиса;
согласно Т. правильное рас-
пределение листьев на стебле
определяется одной спиралью
(основная генетическая спи-
раль филлотаксиса), соеди-
няющей листья последова-
тельно их возниковению на
апексе побега (рис. 133); сосед-
ние листья разделены углом ди-
вергенции, который выражает-
ся в долях от 360° и считается
константным для определен-
ной фазы развития раст.; для
характеристики филлотаксиса
используется формула листо-
Рис. 133. Теории листорасположения (по:
Ботаника..., 1988): А — одной генетической
спирали (1-12 — порядок возникновения
листьев, 14 — генетическая спираль); Б —
множественных геликоид (1-5—левоза-
крученные и I—VIII — правозакрученные па-
растихи); В — физиологических полей (1 —
поля торможения, 2 — зачатки листьев)
208
расположения; лит.: Серебрякова, 1963; Синнот, 1963; Schwabe,
1984.
Теория Л. Плантефоля (Plantefol, 1946-1947) — популярная Т.
«множественных листовых геликоид», основанная на экспери-
ментальном методе; согласно Т. примордии располагаются на не-
скольких листовых спиралях (геликоидах), которые соответству-
ют группам контактных парастих (рис. 133); предполагается, что
на верхнем конце каждой геликоиды находится листородный
центр, который перемещается акропетально по апексу и образует
новые примордии; характер расположения листьев определяется
наклоном и количеством геликоид, синхронной (листорасполо-
жение мутовчатое) или асинхронной (листорасположение очеред-
ное) деятельностью их листородных центров; лит.: Серебрякова,
1963; Синнот, 1963; Schwabe, 1984.
Теория Н. П. Крейкс (1940) — «теория циклического старения и
омоложения раст. в онтогенезе» (теория возрастной циклично-
сти), согласно которой развитие раст. и его частей определяется
взаимоотношением двух противоположных процесов старения и
омоложения-, на развитии любой части раст., имеющей собствен-
ный возраст, сказывается и возраст организма в целом (см. воз-
раст общий); т. о. онтогенетическое старение особи представляет
собой процесс неравномерного прогрессивного падения омоло-
жения последовательно возникающих «дочерних меристем» по-
бега и корня; на основе этой теории для селекции и растениеводст-
ва разработаны рекомендации по определению скороспелости и
скороплодности раст. в конкретных условиях культуры; лит.: Гу-
пало, 1954; Сабинин, 1963.
Теория циклического старения и омоложения см. теория
Н. П. Кренке.
Терофит (Therophyton, от греч. theros — лето, phyton — растение;
англ.: therophyte) — один из типов в классификации форм жизнен-
ных по К. Раункиеру (рис. 22), объединяющий раст., переживаю-
щие неблагопрятный для вегетации период года в виде семян (од-
нолетник, эутерофит); преимущественно однолетние травы, ха-
рактерные для пустынь, полупустынь и южных степей Сев.
полушария (Brassicaceae), а также сорняки полей лесной зоны.
Тип биологический см. классификация форм жизненных по К. Ра-
ункиеру.
Тип структурной организации растения — пон. используется
для описания степени дифференциации тела растения на органы;
209
различают 2 основных типа: растения листостебельные (Согшо-
phyta) и растения талломные (Thallophyta).
Торчок — угнетенная особь дерева на первых стадиях его онтогене-
за-, в неблагоприятных условиях у многих деревьев рост главной
оси скелетной задерживается из-за периодического отмирания по-
бегов замещения, в результате либо боковые ветви догоняют или
обгоняют ее по высоте, формируя б. м. зонтиковидную крону, ли-
бо трогаются в рост почки спящие (Т. с кустовидной формой рос-
та, или многоствольный); длительность жизни Т. различна у раз-
ных видов и в разных условиях; терм.: Корнаковский, 1882; лит.:
Белостоков, 1983; Чистякова, 1986.
Точка роста побега см. конус нарастания побега.
Точка роста см. апекс.
Тропизм (Tropismus, от греч. tropos — поворот, направление; англ.:
tropism) — ориентированное движение органа раст., выражаю-
щееся в направлении роста или его изменении (изгибы) в ответ на
одностороннее воздействие различных факторов среды (рис. 23):
гравитации (геотропизм), света (фототропизм), химических ве-
ществ (хемотропизм) и др.; обусловлен (не)равномерным ростом
клеток на разных сторонах побега, корня и т. п. из-за (асиммет-
ричного распределения ауксинов и ингибиторов роста; в зависи-
мости от реакции на раздражение различают положительный, от-
рицательный и поперечный Т.; органы, располагающиеся вдоль
градиента раздражителя наз. ортотропными, под прямым углом к
нему — диатропными, а под любым другим углом — плагио-
тропными (см. рост); иногда Т. понимают в более широком смыс-
ле (включая нутации и некоторые настические движения); лит.:
Синнот, 1963; Полевой, Саламатова, 1985.
Туберидий (Tuberidium, от лат. tuber — клубень; англ.: tuberidi-
um) — клубень надземный орхидных (гл. обр. эпифитов), образую-
щийся в результате разрастания одного (клубень одночленный)
или нескольких (клубень многочленный), часто верхних, междоуз-
лий побега бокового с ассимилирующими листьями и/или цветоно-
сом на верхушке (рис. 134); часто наз. псевдобульбой, клубнем воз-
душным, бульбой; лит.: Тихонова, 1981; Смирнова, 1990.
Тубероид (англ.: tuberoid) — одно(двух)летний подземный столо-
новидный орган некоторых орхидных, несущий почку возобновле-
ния и покрытый волосками, напоминающими корневые волоски;
развивается из нижних почек однолетнего побега монокарпическо-
го в горизонтальном направлении за счет роста апикального; про-
210
Рис. 134. Туберидии орхидных (по: Bell, Bryan, 1998):
А — Bulbophyllum sp., Б — Pholidota sp., В — Coelogitie fimbriata
(1 — туберидий, 2 — пазушное соцветие, 3 — терминальное соцветие)
ксимальная часть Т. (до почки) представляет собой стеблевой
участок, а дистальная (после почки) — функционирует как ко-
рень (рис. 135); часто Т. наз. клубнем корневым; ср. дроппер; син.:
стебле-корневой тубероид, синкер; лит.: Kumazawa, 1958; Тихоно-
ва, 1983; Татаренко, 1996.
Турион (англ.: turion) — пон. используемые для обозначения раз-
личных структур: гибернакулы, листеца рясковых, резида и побега
формирования.
Угол дивергенции (англ.: phyllotactic angle, divergence angle) см.
формула листорасположения.
Узел (Nodus; англ.: node) — зона отхождения от стебля аксиллярно-
го комплекса: листа (У. листовой) и почки пазушной (У. почкооб-
разующий); у нек-рых раст. почка может быть сдвинута в другое
положение или находиться в пазухе листа, не имеющего с ней об-
щего происхождения, в связи с чем У. наз. участок стебля, к кото-
рому прикреплен лист или семядоля (У. семядольный).
Узел возобновления см. зона кущения.
Узел кущения см. кущение.
Узел листовой см. узел.
Узел отрастания см. зона кущения.
Узел почкообразующий см. узел.
Узел семядольный см. узел.
211
1
Рис. 135. Строение и развитие тубероида Perularia ussuriensis (1-12) и Platanthera minor
(13-15) (по: Kumazawa, 1958): 1 13 — подземная часть растения летом (а — тубероид),
2—3 — схема поперечного строения тубероида в проксимальной (2) и средней (3) час-
ти, 4-6 — стадии роста тубероида, 7 — почка на тубероиде (лето), 8-12 — схема про-
дольного разреза тубероида на разных этапах его развития, 14 — проксимальный уча-
сток тубероида, 15 — схема строения стебля (6 — проксимальный участок старого ту-
бероида, в — почка, г — центральный цилиндр (стеблевого происхождения),
д — ткань подобная корневому чехлику, е — сектор стеблевого происхождения,
Ж] — ж3 — последовательные чешуи и зачатки листьев, з — ризодерма, и — прошло-
годний тубероид, к - прида точный корень, л — стела корневого происхождения,
м — надземный побег, х — апекс корня)
212
Усик (Cirrus; англ.: tend-
ril) — видоизмененный
лист (или его часть)
или побег боковой, слу-
жащий для закрепле-
ния на опоре нек-рых
раст. (см. лиана);
рис. 136.
Усы см. флагелла.
Учение о формах жиз-
ненных см. биомор-
фология.
Фаза вторичной дея-
тельности побега —
последний перед старе-
нием этап (период) он-
тогенеза побега много-
летнего, в течение кото-
рого он видоизменяет-
ся (метаморфоз побега)
и выполняет функцию
запасания питательных
веществ (корневище, ре-
зид) и коммуникации
(ствол, стволик).
Фаза кущения см. ку-
Рис. 136. Усики листового (13) и побегового (4-5)
происхождения (по: Ботаника..., 1988): 1 — Vicia
sepium, 2 — Lathyrus aphaca, 3 — Cucurbita pepo,
4-5 — Vitis vinifera (a-6 — видоизменения соцветия в
усик, 5 — схема симподиального нарастания стебля)
щение.
Фаза онтогенеза—определенный момент в ходе индивидуально-
го развития особи, улавливаемый по внешнему изменению ее
формы жизненной (напр., смена системы побегов, типа системы
корневой и т. д.); выделение Ф. о. (онтобиоморф) основано на раз-
личиях жизненной формы одного раст. (деревья, кустарники, кус-
тарнички, реже многолетние травы) в разные моменты его онто-
генеза и не всегда совпадает с периодом онтогенеза-, син.: этап или
фаза морфогенеза.
Фаза развития побега эмбриональная см. рост внутрипочечный.
Фаза фенологическая см. фенофаза.
Фанерокотилия (англ.: phanerocotyly) см. прорастание надземное.
Фанерофит (Phanerophyton, от греч. phaneros — видимый, откры-
тый, явный, phyton — растение; англ.: phanerophyte) — один из
213
типов в классификации форм жизненных по К. Раункиеру, объеди-
няющий растения, почки возобновления которых расположены вы-
соко над землей в течение всего года (рис. 22); к Ф. относят деревья,
кустарники, деревянистые лианы (Ф. лазящие), эпифиты и стебле-
вые суккуленты (Ф. суккулентные); по произвольно установлен-
ному интервалу высот различают мегафанерофиты (выше 30 м),
мезофанерофиты (8-30 м), микрофанерофиты (2-8 м) и нанофа-
нерофиты (0,3—2 м); в каждом из этих подтипов выделяют группы
по ритму развития листвы, характеру почечных покровов, конси-
стенции стебля (Ф. деревянистые и Ф. травянистые) и т. п.
Фанерофит деревянистый (англ.: ligneous phanerophyte) см. фа-
нерофит.
Фанерофит травянистый (англ.: herbaceous phanerophyte) см. фа-
нерофит.
Фенология (от греч. phainomena — явления, logos — слово, учение;
англ.: phenology) — наука о сезонных явлениях живой природы,
сроках и причинах их наступления. Фенологические исследова-
ния раст. (фитофенология) включают изучение развития сезонно-
го популяций, особей или побегов в зависимости от погодных явле-
ний и микроклимата; терм.: Р. Ремюр (1735); лит.: Шнелле, 1961;
Зайцев, 1978, 1981; Шульц, 1981; Похилько, 1993; Елагин, 1994;
Минин, 2000; Жмылев и др„ 2002, 2003.
Феноритмотип (англ.: phenological groups) — фенологическая
группа, объединяющая раст. со сходной длительностью, сроками
начала и завершения вегетации и сменой основных фенофаз; раз-
личают: 1) по длительности вегетации раст. длителъновегети-
рующие, раст. коротковегетирующие и эфемеры; 2) по времени
цветения—ранне- (средне-, поздне-)весенние, ранне-(средне-,
поздне-)летние и осенние; 3) по соотношению фаз цветениям ве-
гетации — проанты, мезанты, метанты; 4) по характеру перези-
мовывания — раст. зимнезеленые и раст. летнезеленые, а по дли-
тельности вегетации — раст. вечнозеленые и раст. листопадные и
др.; одно и то же раст. в разных географических районах может
относиться к разным Ф. (см. правило уподобления); терм.: Бори-
сова, 1965; лит.: Серебряков, 19546, 19646; Серебрякова, 1976.
Фенофаза (англ.: phenophase, phenological phase) — этап развития
сезонного раст., определяемый по внешним изменениям (раскры-
вание почки, облиствление, цветение, диссеминация, листопад и
др.). Обычно выделяют основные фенологические фазы (вегета-
ция, бутонизация, цветение, плодоношение, отмирание, покой),
214
которые подразделяются на подфазы (напр., начало и конец) и
обозначаются буквами, цифрами или особыми значками.
Фетализация см. неотения.
Филлодий (Phyllodium, от греч. phyllon — лист, eidos — вид; англ.:
phyllode) — уплощенный в виде листовой пластинки черешок лис-
та', выполняет функции фотосинтеза вместо редуцированной
пластинки листа и свойственен гл. обр. раст. аридных областей
(Acacia melanoxylon).
Филлокладий (Phyllocladium, от греч. phyllon — лист, klados —
ветвь; англ.: phylloclade) — листообразный стебель нек-рых раст.
аридных областей (рис. 44), рано
прекращающий рост и выполняю-
щий функции листа (Ruscus; ср.
кладодий); син.: листоветка.
Филломорфа (англ.: phyllomorph) —
основной элемент структуры веге-
тативного тела представителей сем.
Gesneriaceae, имеющий листовид-
ную форму и выполняющий функ-
ции побега (рис. 137); состоит из
пластинки, нарастающей за счет ба-
зальной меристемы, и петилоида
(черешка), на котором находятся
меристематические зоны (выемоч-
ная и отделенная меристема); Ф.
развивается из семядоли (первичная
или семядольная Ф.) или отделен-
ной меристемы (вторичная, тре-
тичная и т. д. Ф.) и неопределенно
долго нарастает в длину за счет ба-
зальной и выемочной меристем
(рост интеркалярный); терм.: Jong,
Burtt, 1975; лит.: Halle, Delmotte,
1973; Серебрякова, 19836; Tsukaya,
1987.
Филлотаксис (англ.: phyllotaxis,
phyllotaxy) см. листорасположение.
Филлохрон (Phyllochronus, от греч.
phyllon — лист, chronos — время;
англ.: phyllochrone) — период рос-
Рис. L37. Схема развития филломор-
фы Streptocarpus fanniniae (по: Jong,
Burtt, 1975): А-Д — развитие семя-
дольной филломорфы (1, 2 — семя-
доли, 3 — гипокотиль, 4 — корень,
5 — вторичная филломорфа, 6 — пе-
тилоид, 7 — базальная меристема,
8 — выемочная меристема, 9 — отде-
ленная меристема)
215
та одного листа, или его части, не совпадающий с ростом после-
дующего или предшествующего ему другого листа побега-, по
шкале Ф., отражающей сроки окончания роста пластинок после-
довательных листьев, определяют частоту развертывания листьев
на побеге; последняя обычно устанавливается по числу листьев,
развернувшихся за определенный срок, или по интервалу време-
ни между появлением двух последовательных листьев; терм.:
Bunting, 1965, цит. по: Рытова, 1976.
Филэмбриогенез (от греч. phylon — род, племя, embryon — заро-
дыш, genesis — происхождение) — одна из основных теорий
морфологической эволюции, разработанная А. Н. Северцовым
(1910, 1945) на примере животных. В первоначальном вариан-
те— теория эволюционных изменений органов (ср. гетерохро-
ния), которая позднее, в том числе и в биоморфологии, стала ис-
пользоваться в более широком смысле (для описания эволюци-
онных изменений морфогенеза и онтогенеза). Согласно Ф.
изменения строения взрослого организма зависят не только от
степени перестройки морфогенеза, но и от времени их возникно-
вения. На основании этого выделяют 12 способов эволюции морфо-
логической, среди которых наиболее известны анаболия, девиация,
архаллаксис и редукция (рис. 4); терм.: Северцов (1910); лит.: Тах-
таджян, 1950,1964, Северцов, 1970, 1981; Серебрякова, 1983, Тере-
хин, 1991, Жмылев, 2004.
Фитобиоморфология см. биоморфология.
Фитомер (англ.: phytomer) см. метамер.
Флагелла (Flagellum, от лат. flagrum — кнут, бич, плеть; англ.:
flagellum) — надземный, тонкий, ползучий, безлистный стебель,
образующийся в результате вытягивания первого междоузлия (ги-
поподия) побега ветвления (Saxifraga flagellaris; рис. 130); обеспе-
чивает размножение вегетативное и расселение, Ф. часто наз. сто-
лоном надземным или плетью; син.: усы.
Форма жизненная (Biomorpha, от греч. bios — жизнь, morpha —
форма, образ; англ.: biomorph, life form, growth form, biological
form, dormancy form) — общий облик (габитус) раст., обуслов-
ленный своеобразием его системы надземных и подземных орга-
нов вегетативных (см. признак биоморфологический), форми-
рующихся в онтогенезе в результате роста и развития раст. в оп-
ределенных условиях среды; в разных условиях один и тот же вид
может иметь разные Ф. ж., но несколько видов, сходных по эколо-
го-ценотическим параметрам в результате конвергенции могут
216
иметь одну Ф. ж.; в связи с этим Ф. ж. часто наз. единицу экологи-
ческой классификации раст.; в отличие от групп экологических Ф.
ж. отражает приспособленность габитуально сходных раст. ко
всему комплексу факторов внешней среды (ср. экобиоморфа,
экоморфа); габитус каждого раст. изменяется в онтогенезе, поэто-
му в классификации форм жизненных под Ф. ж. понимают сово-
купность взрослых особей, а для смен жизненных форм в онтоге-
незе индивида используют названия фаза морфогенеза или период
онтогенеза, реже — категория форм жизненных, которые отража-
ют их габитуальное своеобразие и различие в образе жизни; часто
вместо терм. Ф. ж. используются близкие пон. (тип биологиче-
ский, форма роста) или в связи с различиями в подходах и целях
исследования в пон. Ф. ж. вкладывается дополнительный смысл
(см. ценобиоморфа, онтобиоморфа, фенобиоморфа, экобиомор-
фа); син.: биоморфа, эпиморфа; терм.: Warming, 1884; лит.: Се-
ребряков, 1962; Серебрякова, 1972, 1980; Шафранова, 1990; Нухи-
мовский, 1973а, 1997; Борисова, 1991; Шорина, 20006.
Форма роста (англ.: growth form) — габитус раст., выделяемый по
структуре побегов (напр., раст. с побегами удлиненными) или на-
правлению их роста; в последнем случае различают Ф. р. раст.
прямостоячего, раст. полегающего, раст. лазящего, раст. ползуче-
го и т. п.; часто используется как «нейтральный термин» для отра-
жения своеобразия жизненной формы (напр. Ф. р. раст. длинно-
корневищного называется геофильной, а раст. подушковидно-
го— подушковидной и т. п.) или экологически обусловленных
форм вида, отличающихся габитуальными признаками, а в более
широком значении — как синоним терм, форма жизненная; терм.:
DuRietz, 1931.
Формация листьев (англ.: leaf formation) — группа (категория)
листьев, выделяемых по морфологическому сходству и топогра-
фическому положению на стебле; различают Ф. л. низовых, сре-
динных и верховых, границы между которыми не всегда отчетли-
во выражены (рис. 63); морфолого-анатомические различия меж-
ду листьями низовыми, листьями срединными и листьями
верховыми обусловливают гетерофиллию побега и связаны с изме-
нениями ритма листообразовательной деятельности его конуса
нарастания; иногда закономерное изменение формы и размера
листьев вдоль стебля наз. метаморфозом листа.
Формула листорасположения (англ.: phyllotaxis (formula) frac-
tion) — угловое расстояние между медианами листьев (угол дивер-
217
генции), выраженное дробным отношением, в котором числитель
обозначает число оборотов по стеблю генетической спирали фил-
лотаксиса между двумя листьями, расположенными один под дру-
гим на ортостихе; а знаменатель — кол-во листьев в цикле листо-
вом (рис. 133); используется для характеристики листорасположе-
ния взрослого побега (см. теория К. Шимпера и А. Брауна).
Формула структуры побега — способ краткой характеристики
развития и структуры побега генеративного, при котором различ-
ные параметры представлены комбинацией чисел (а, б, Bj_n, г);
Ф. с. п. впервые предложена Ю. Е. Алексеевым (1996) как комби-
нация 4 параметров: а) длительность цикла жизненного малого; б)
число метамеров базального участка побега, не имеющих почек
пазушных (рис. 40); В|_Г1) порядковые номера узлов зоны возобнов-
ления (рис. 40); г) общее число метамеров побега (рис. 40).
Фототропизм (Phototropismus, от греч. photos — свет, tropos — по-
ворот, направление; англ.: phototropism) — ориентированное
движение органа раст., выражающееся в направлении его роста в
ответ на одностороннее освещение (см. тропизм); часто наз. ге-
лиотропизмом; корни обычно обнаруживают отрицательный Ф.;
стебли — положительный, а листья—поперечный Ф.; предпо-
лагают, что в рецептации света принимают участие каротиноиды
и флавины; лит.: Синнот, 1963.
Фототрофы см. автотрофы.
Фронд см. листец.
Хамефит (Chamaephyton, от греч. chamai — наземный, phyton —
раст.; англ.: chamaephyte) — один из основных типов в классифи-
кации форм жизненных по К. Раункиеру, объединяющий раст., поч-
ки возобновления которых в неблагоприятное для вегетации время
года расположены невысоко (до 20-30 см) над поверхностью поч-
вы и обычно защищены почечными чешуями и/или снежным по-
кровом (рис. 22); морфологически разнородная группа, в которой
различит полукустарники (гемифанерофиты или X. полукустар-
никовые), раст. подушковидные (X. подушковидные), X. сукку-
лентные (суккуленты листовые), X. активные (раст. с побегами диа-
геотропными; напр. Lysimachia nummularia), X. пассивные (раст.
с полегающими побегами; напр. Stellaria holostea) и др.
Хамефит активный см. хамефит.
Хамефит пассивный см. хамефит.
Хлороэмбриофит(ы) — назв. предложено М. С. Яковлевым и
Г. Я. Жуковой (1973) для обозначения раст., у которых ткани за-
218
родыша содержат хлорофилла (зеленый зародыш); напр. предста-
вители Convolvulaceae, Dipsacaceae, Tiliaceae; ср. лейкоэмбриофит.
Цветение (англ.: flowering, flowerage, blossom, bloom) — пон. ис-
пользуют для обозначения: 1) периода жизни цветка от его раскры-
вания до увядания; 2) этапа развития сезонного (фенофаза); 3) пе-
риода онтогенеза монокарпика (период генеративный), а в более
широком смысле — периода жизни раст. от заложения в почке за-
чатков цветка до засыхания околоцветника или тычинок (ср. раз-
витие генеративное); лит.: Аксенова и др., 1973; Бернье и др., 1985.
Ценобиоморфа — терм, введен Б. А. Юрцевым (Петровский, Мат-
веева, 1968) для обозначения биоморф, сходных по ценотическим
свойствам.
Цикл (ритм) роста — совокупность процессов, образующих за-
конченный период роста; длительность Ц. р. измеряется в раз-
личных временных (см. пластохрон, филлохрон, циклохрон), ре-
же — структурных единицах роста (цикл листовой, единица мор-
фогенеза, квант роста, метамер, модуль и др.).
Цикл жизненный (англ.: life cycle) — совокупность всех последо-
вательных фаз развития раст. от одного исходного состояния (зи-
готы при размножении половом, диаспоры при размножении веге-
тативном) до его повторения (до формирования зрелой особи,
способной дать начало следующему поколению — зиготе или ди-
аспоре); различают одно-, дву- и многолетние Ц. ж.; иногда пон.
Ц. ж. используют в том же смысле, что и цикл жизненный большой;
син.: Ц. ж. сложный, цикл развития, цикл морфогенеза; лит.: Ле-
вин, 1959, 1963, 1964, 1966; Хохряков, 1981; Жукова, 1983.
Цикл жизненный большой см. онтогенез.
Цикл жизненный малый — внеп очечная фаза развития побега
(см. рост внепочечный) от прорастания (побег главный) или раз-
вертывания почки начальной (побег ветвления) до цветения и пло-
доношения; по длительности этой фазы и надземной жизни гл.
обр. у раст. травянистых различают побеги моноциклические, по-
беги олигоциклические (ди-, трициклические) и побеги полицикли-
ческие; в зависимости от условий среды и возрастного состояния
растения длительность Ц. ж. м. может изменяться; часто под Ц. ж.
м. понимают полный онтогенез побега; терм.: Смелов, 1937; син.:
цикл развития малый; лит.: Серебряков, 1948, 1949, 1952, 1959а;
Серебрякова, 1971а.
Цикл листовой — совокупность листьев, расположенных на спи-
рали филлотаксиса генетической, не считая последнего листа, на-
219
ходящегося на одной ортостихе с первым; обычно листья одного
Ц. л. образуются друг за другом непрерывно, с связи с чем Ц. л.
рассматривают как структурную единицу роста внутрипочечного-,
син.: цикл листовых зачатков, цикл метамеров.
Цикл листовых зачатков см. цикл листовой.
Цикл метамеров см. цикл листовой.
Цикл основной (ОЦ) — период развития ситемы побега формиро-
вания (стволика) — основной единицы структуры системы побе-
говой (полу)кустарника или (полу)кустарничка; различают три
этапа Ц. о.: рост (образование побега формирования из почки спя-
щей), стабилизация (формирование системы этого побега) и от-
мирание (отмирание побегов его кроны); нижняя часть побега
формирования не отмирает, а входит в состав ксилоподия; из по-
чек последнего образуются новые побеги формирования, разви-
вающиеся сходным образом (последовательная смена стволиков
куста)-, терм.: Лучник, 1960; лит.: Мазуренко, Хохряков, 1977.
Цикл развития — широко используемое назв. для обозначения
периода развития метамеров разного уровня: побега (Ц. р. малый)
системы побега (напр., цикл основной), особи (Ц. р. большой) и др.
Цикл развития большой см. онтогенез.
Цикл развития малый см. цикл жизненный малый.
Циклохрон (от греч. kyklos — круг, chronos — время) — времен-
ная единица ритма роста видимого, определяемая как интервал
времени для образования одного побега элементарного; ср. пла-
стохрон; терм.: Грудзинская, 1993.
Шейка корневая (англ.: root collar, peg) см. гипокотиль.
Шейка семядольная (Collum cotyledonare; англ.: cotyledonary pe-
tiole) — утонченная средняя часть семядоли зародыша и проростка
некоторых однодольных, связывающая базальную (влагалище
семядольное) и апикальную (гаусторий) части семядоли; иногда
наз. мезокотилем; син.: связник семядоли, средняя часть семя-
доли.
Шишка корневая см. клубень корневой.
Щиток (Scutellum, от лат. scutra — плоское блюдо, чаша; англ.: scu-
tellum) — видоизмененная в щитовидный гаусторий дистальная
часть семядоли зародыша злаков; одной своей стороной прилегает
' к эндосперму, другой — б. м. охватывает остальную часть заро-
дыша (рис. 38); иногда Щ. рассматривают как семядолю или видо-
измененную ось зародыша; терм.: Gaertner, 1788-1807; лит.: Лод-
кина, 1988.
220
Эволюция биоморфологическая (англ.: life-form, growth form,
habit form evolution) — процесс изменения формы жизненной
(Э. форм жизненных) и онтогенеза (Э. онтогенеза), связанный с
приспособлением растений к окружающей среде (адаптация).
В связи с отсутствием или неполнотой палеонтологических дан-
ных для реконструкции Э. б. используют различные подходы (ме-
тод морфогенетических рядов, рекапитуляция), а для решения
проблемы направленности эволюции (см. направленность эволю-
ции биоморфологической) обычно привлекают различные косвен-
ные свидетельства, требующие специальной аргументации (из-
менчивость биоморфологическая, поливариантность развития,
полиплоидия и др.). Э. б. происходит на основе принципов эволю-
ции морфологической и посредством различных способов эволюции
морфологической; син.: эволюция соматическая; лит.: Серебряков,
1955; Цвелев, 1970, 2005; Серебряков, Серебрякова, 1972; Хохря-
ков, 1973, 1975, 1981; Гатцук и др„ 1974; Юсуфов, 1976; Голубев,
1981; Василевская и др., 1983; Лодкина, 1983; Терехин, 1991; Недо-
лужко, 1997; Барыкина, 19996.
Эволюция онтогенеза см. эволюция биоморфологическая.
Эволюция соматическая см. эволюция биоморфологическая.
Эволюция форм жизненных см. эволюция биоморфологическая.
Экобиоморфа (Oecobiomorpha, от греч. oecos — дом и форма жиз-
ненная) — «совокупность видов (иногда и внутривидовых таксо-
нов), имеющих сходные форму роста, биологические ритмы,
а также эколого-физиологические, в том числе приспособитель-
ные и средообразовательные особенности»; при выделении Э.
учитывается форма жизненная и характер местообитания (напр.,
травы корневищные мезофильные); часто Э. рассматривают как
«типовые адаптационные организменные системы, существую-
щие в определенных условиях среды» (Лавренко, Свешникова,
1965) и выделяют по морфологическим, биологическим, анато-
мическим, экологическим, физиолого-биохимическим призна-
кам; реже Э. наз. экологически обусловленные формы вида, отли-
чающиеся габитуальными признаками, а в самом широком смыс-
ле — синонимизируют с понятием «форма жизненная»; терм.:
Быков, 1962, 1983; лит.: Борисова, 1991.
Экобиоморфология см. биоморфология.
Экоморфа (Oecomorpha, от греч. oecos — дом, morpha — форма,
образ) — группа видов, выделяемых по их сходному отношению
к фитоценозу (ценоморфы: лесные, степные и т. п.), климату (фа~
221
нерафиты, хамефиты, гемикриптофиты, криптофиты, терофи-
ты), освещению (гелиоморфы), температуре (термоморфы),
почвенному плодородию (трофоморфы) и увлажнению (гигро-
морфы); ср. экобиоморфа; часто под Э. понимают экологически
обусловленные формы вида, отличающиеся габитуальными при-
знаками, а в более широком смысле — форму жизненную раст.;
терм.: Бельгард, 1950.
Эктопаразит (англ.: ectoparasite) см. ксенопаразит.
Элемент структуры биоморфы см. признаки биоморфологи-
ческие.
Эмбрионизация (от греч. embrion — зародыш; англ.: embryoniza-
tion) — один из способов эволюции онтогенеза, посредством
которого часть постэмбриональных стадий развития предка
включены в эмбриогенез потомка. В биоморфологии Э. обычно
понимают широко, включая педоморфоз, возникновение почки,
увеличение емкости почки, образование почки преформированной
(Э. вегетативная) и живорождение; терм.: Шмальгаузен (1940) как
«эмбрионализация», А. А. Захваткин (1948) как «эмбрионизация
развития»; лит.: Северцов, 1981; Хохряков, 1975, 1981.
Эндопаразит (англ.: endoparasite) см. ксенопаразит.
Эпибласт (Epiblastus, от греч. epi — на, над, сверх, blastos — росток,
зародыш, побег; англ.: epiblast) — складкообразный вырост, рас-
положенный напротив щитка с наружной стороны почечки заро-
дыша многих злаков; рассматривают как рудимент второй семядо-
ли, лопасть щитка, колеоризы или гипокотиля; терм.: Ришар, 1809,
цит. по: Серебряков, 1941.
Эпикотиль (Epicotylus, от греч. epi — на, над, сверх, при, kotyle — до-
ля; англ.: epicotyl, embryonic shoot) — междоузлие проростка между
узлом семядольным и узлом первого настоящего листа (рис. 111);
у нек-рых раст. (Fragaria) Э. и последующие междоузлия недораз-
виваются (проростки розеточного типа); часто Э. наз. почечку или
плюмулу, син.: колено надсемядольное; лит.: Серебряков, 1952.
Эпиморфа см. форма жизненная.
Эпиморфология см. биоморфология.
Эпиподий (Epipodium, от греч. epi — на, над, сверх, pous — нога,
здесь — ось; англ.: epipodium) — междоузлие побега бокового, ог-
раниченное снизу узлом предлиста, а сверху — узлом первого на-
стоящего л иста (рис. 31).
Эпифилл (Epiphyllum, от греч. epi — на, phyllum — лист; англ.: epi-
phyll) — раст., поселяющееся на листьях других раст.; водоросли,
222
мхи, редко — цветковые раст., распространенные гл. обр. во
влажных тропиках и субтропиках (ср. эпифит).
Эпифиллия (англ.: epiphylly) — образование структур (листа, поч-
ки выводковой, побега, соцветия) на листе (фертильном) в результа-
те онтогенетических перемещений (при закладке и формирования
органов) или гетеротопии (по: Zimmerman, 1959), т. е. изменения
положения меристемы (листа, почки выводковой, побега, соцве-
тия) благодаря ремеристематизации тканей фертильного листа
или закладки почки адвентивной на его примордии (рис. 138);
у нек-рых растений Э. формируется при повреждении листьев кле-
щами; лит.: Dickinson, 1978.
Рис. 138. Расположение эпифильных структур (по: Dickinson, 1978)
Эпифит (Epiphyton, от греч. epi — на, phyton — растение; англ.:
epiphyte) — растение, поселяющееся на другом раст. (гл. обр. на
стволах и ветвях деревьев) и в течение всего онтогенеза связанное
с ним, но не являющееся раст.-паразитом, а получающее питат.
вещества из окружающей среды (голоэпифит или облигатный
Э.); разнообразная группа гл. обр. раст. травянистых, встречаю-
щихся преимущественно во влажных тропических лесах; разли-
чают Э. с незначительной специализацией побегов, Э. с приспо-
соблениями для накапливания мелкозема (гнездовые Э.) или во-
ды (цистерновые Э.); в более широком смысле к Э. относят
полуэпифиты и эпифиллы; лит.: Федоров, 1950; Dawson, 1986;
Benzing, 1987; Kress, 1989; Luttge, 1989; Smith, 1989.
Эпифит гнездовой (англ.: nest epiphyte) см. эпифит.
Эпифит облигатный (англ.: true (obligatory) epiphyte) см. эпифит.
Эпифит цистерновый (англ.: tank epiphyte) см. эпифит.
Этап вторичной деятельности побега см. фаза вторичной дея-
тельности побега.
Этап морфогенеза см. фаза онтогенеза.
Этап развития побега эмбриональный см. рост внутрипочечный.
Этап эмбриональный см. период эмбриональный.
223
Этап ювенильный см. растение ювенильное.
Эутерофит (англ.: eutherophyte) см. терофит.
Эфемер (Ephemera; англ.: ephemer) — травянистое, однолетнее раст.
(см. терофит), развивающееся во влажное время года, весной (яро-
вые Э.) или осенью (озимые или зимующие Э.) и отмирающее в
период летней засухи; раст. пустынь, полупустынь и отчасти сте-
пей, обычно раноцветущие и быстро завершающие цикл своего
развития; лит.: Коровин, 1935; Волкова, 1958; Проскоряков, 1950.
Эфемер зимующий см. эфемер.
Эфемер озимый см. эфемер.
Эфемер яровой см. эфемер.
Эфемероид (Ephemeroidum) — травянистое многолетнее раст. с
отмирающими в летний период надземными побегами; гл. обр.
раноцветущие геофиты аридных областей (Tulipa) и широколи-
ственных лесов (Corydalis); терм.: Коровин, 1935; лит.: Хохряков,
19756, Барыкина, Потапова, Степанова, 1986.
Явносемядольность см. прорастание надземное.
ЛИТЕРАТУРА
Аксенова Н. П„ Баврина Т. В., Константинова Т. Н. Цветение и его фотопериоди-
ческая регуляция.— М.: Наука, 1973.— 294 с.
Алеев Ю. Г. Экоморфология.— Киев: Наук, думка, 1986.— 424 с.
Алексеев Ю. Е. Осоки (морфология, биология, онтогенез, эволюция).— М.: Ар-
гус, 1996 — 251 с.
Алехин В. В. Фитосоциология (учение о растительных сообществах) и ее послед-
ние успехи у нас и на Западе // Методика геоботанических исследований.—
Л.; М.: 1925 — С. 7-75.
Алехин В. В. Некоторые соображения относительно жизненной формы «перека-
ти-поле» // Президенту АН СССР акад. В. Л. Комарову к 70-летию со дня ро-
ждения и 45-летию научной деятельности.— Л., 1939.— С. 97-107.
Андреева И. И. Морфогенез некоторых южноафриканских клубнелуковичных
растений из семейства Iridaceae И Бюл. Главного Бот. сада АН СССР.—
1979,— Вып. III.— С. 31-39.
Андреева И. И. Старение клонов и продуктивность гладиолуса гибридного // Ин-
тенсификация возделывания полевых культур и морфологические основы
устойчивости растений.— М., 1987.— С. 63-70.
Андронова Е. В., Батыгина Т. Б., Васильева В. Е. Протокорм // Эмбриология цвет-
ковых растений. Терминология и концепции. Т. 3. Системы репродукции.—
СПб.: Мир и семья, 2000.— С. 329-329.
Антонова И. С., Азова О. В Архитектурные модели кроны древесных растений //
Ботан. журн — 1999,— Т. 84, № 3,— С. 10-32.
Артамонова И. К. Некоторые особенности вегетативного возобновления Роа
longifolia Trin. И Научн. док. высш, школы. Биол. науки.— 1963, № 3.—
С. 119-124.
Артамонова И. К. Формирование дерновины луговика извилистого (Deschampsia
flexuosa (L.) Trin.) в условиях высокогорий Кавказа И Бюлл. Моск. Об-ва ис-
пытат. природы. Отд. биол.— 1968.— Т. 73, вып. 5.— С. 89-97.
Артюшенко 3. Т. Амариллисовые СССР. Морфология, систематика и использо-
вание.— Л.: Наука, 1970.— 180 с.
Артюшенко 3. Т. Атлас по описательной морфологии высших растений. Се-
мя.— Л.: Наука, 1990.— 204 с.
АрциховскийВ.М. О педогенезе у растений // Изв. СПб. бот. сада.— 1911.— Т. 11,
вып. 1.— С. 1-6.
Байтулин И. О., Рахимбаев И. Р, Каменецкая И. И. Интродукция и морфогенез
дикорастущих луков Казахстана.— Алма-Ата: Наука, 1986.— 156 с.
Баранова М. В. Структура, классификация и направления эволюционных преоб-
225
разований луковичных растений // Ботан. журн.— 1986.— Т. 71, № 10.—
С.1308-1320.
Баранова М. В. Луковичные растения семейства Лилейных (география, биомор-
фологический анализ, выращивание).— СПб.: Наука, 1999.— 229 с.
Баранова М. В. Луковица. Луковичка И Эмбриология цветковых растений. Тер-
минология и концепции. Т. 3. Системы репродукции.— СПб.: Мир и семья,
2000 — С. 321-329.
Барыкина Р. П. Особенности образования корневых отпрысков у белой акации
(Robinia pseudoacacia L.). И Бюл. Моск, об-ва испытат. природы. Отд. биол.—
1958.— Т. 63, вып. 4 — С. 57-71.
Барыкина Р. П. Чистяк весенний / Биологическая флора Московской области.
Вып. 10.— М.: Изд-во МГУ; Изд-во Аргус, 1995.— С. 75-82.
Барыкина Р. П. Полиморфизм корней и корневых систем в семействе Ranuncu-
laceae И Моск, об-ва испытат. природы. Отд. биол.— 1999а.— Т. 104,
Вып. 3.— С. 26-31.
Барыкина Р. П. О некоторых модусах преобразования онто-, органно- и гистоге-
неза в соматической эволюции семейства Ranunculaceae И Бюл. Моск, об-ва
испытат. природы. Отд. биол.— 19996.— Т. 104, Вып. I.— С. 49-53.
Барыкина Р. П. Сарментация. Партикуляция И Эмбриология цветковых расте-
ний. Терминология и концепции. Т. 3. Системы репродукции.— СПб.: Мир
и семья, 2000.— С. 302-310.
Барыкина Р. П., Гуланян Т. А., Чубатова Н. В. Морфолого-анатомическое иссле-
дование некоторых представителей рода Aconitum L. секции Lycoctonum DC. в
онтогенезе // Бюл. Моск, об-ва испытат. природы. Отд. биол.— 1976.— Т. 81,
вып. 1.— С. 99-116.
Барыкина Р. П., Гуленкова М. А. Элементарный метамер побега цветкового расте-
ния//Бюл. Моск, об-ва испытат. природы. Отд. биол.— 1983.— Т. 84,
Вып. 4.—С. 114-124.
Барыкина Р. П„ Гуленкова М. А. Метаморфоз надземного побега, его адаптивное
значение И 5 Всес. шк. по теор. морф. раст. 15-18 сент., 1987.— Львов,
1987.— С. 24-29.
Барыкина Р. П., Гуленкова М. А. Метаморфоз и его значение в жизни растений И
Бюл. Моск, об-ва испытат. природы. Отд. биол.— 1990.— Т. 95, вып. 5.—
С. 103-111.
Барыкина Р. П, Потапова Н. Ф., Степанов Б. П. Морфогенез побегов некоторых
лесных эфемероидов рода Anemone L. // Науч. докл. высш, школы. Биол. нау-
ки.— 1986.— № 7 — С. 86-92.
Барыкина Р. П., Чубатова Н. В. О типах прорастания и первых этапах онтогенеза
в роде Clematis L. // Жизненные формы: структура, спектры и эволюция.—
М.: Наука, 1981 —С. 111-140.
Батыгин Н. Ф. Онтогенез высших растений.— М.: Агропромиздат, 1986.-101 с.
Батыгина Т. Б., Брагина Е. А. Вивипария // Эмбриология цветковых растений.
Терминология и концепции. Т. 3. Системы репродукции.— СПб.: Мир и се-
мья, 2000а.— С. 39-62.
Батыгина Т. Б., Брагина Е. А. Выводковая почка И Эмбриология цветковых рас-
226
тений. Терминология и концепции. Т. 3. Системы репродукции.— СПб.:
Мир и семья, 20006.— С. 315-321.
Бейлин И. Г Цветковые полупаразиты и паразиты.— М.: Наука, 1968.— 117 с.
Белостоков Г. П. Системы стеблесложения подроста древесных растений // Бюл.
Моск, об-ва испытат. природы. Отд. биол.— 1983.— Т. 88, вып. 1.—
С. 87-104.
БельгардА. Л. Лесная растительность на юго-востока УССР.— Киев: Изд-во Ки-
евск. гос. ун-та, 1950.— 263 с.
Бернье Ж., Кине Ж.-М., СаксР. Физиология цветения. Т. 1-3.— М.: Агропромиз-
дат, 1985.
Беспалова 3. Г. О жизненной форме «полукустарничек» // Проблемы современ.
ботаники. Т. 2.— М.; Л.: Наука, 1965.— С. 65-69.
Биологический энциклопедический словарь.— М.: Советская энциклопедия,
1986 — 831 с.
Биология развития растений.— М.: Наука, 1975.— 229 с.
Богдановская-Гиенеф И. Д. К вопросу о семенном возобновлении в луговых со-
обществах // Зап. Лен. с.-х. Ин-та — 1926.— Т. 3.— С. 216-253.
Борисова И. В., Попова Т. А. Возрастные этапы формирования дерновины степ-
ных злаков // Ботан. журн.— 1971.— Т. 56, № 5.— С. 619-629.
Борисова И. В. Биолого-морфологическая характеристика травянистых подуш-
ковидных растений Северного Кавказа И Проблемы ботаники. Т. 6.—
1962 — С. 336-345.
Борисова И. В. О понятиях «биоморфа», «экобиоморфа» и «архитектурная мо-
дель» // Ботан. журн.— 1991.— Т. 76, № 10.— С. 1360-1367.
Борисова И. В. Ритмы сезонного развития степных растений и зональных типов
степной растительности Центрального Казахстана И Труды Ботан. ин-та
им. В. Л. Комарова. Сер. 3. Геоботаника, вып. 17. Биология и экология расте-
ний целинных районов Казахстана.— М.— Л.: Наука, 1965.— С. 64-99.
Борисова И. В., Малышева Г. С. Биолого-морфологические признаки растений,
определяющие время их зацветания И Бот. жур. 1993.— Т. 78, № 7.—
С. 63-71.
Ботаника: Морфология и анатомия растений: Учеб, пособие для студентов пед.
Ин-тов по биол. и хим. спец. / А. Е. Васильев, Н. С. Воронин, А. Г. Еленевский и
др. 2-е изд., перераб.— М.: Просвещение, 1988.— 480 с.
Бочанцева 3. П. Онтогенез тюльпанов И Труды ботан. сада АН УзССР.— 1956.—
Вып. 5.— С. 71-108.
Брагина Е. А., Батыгина Т. Б. Диаспора И Эмбриология цветковых растений.
Терминология и концепции. Т. 3. Системы репродукции.— СПб.: Мир и се-
мья, 2000 — С. 420-428.
Быков Б. А. Доминанты растительного покрова Советского Союза. Т. 2.— Алма-
Ата: Изд-во АН Каз. ССР, 1962.— 436 с.
Быков Б. А. Экологический словарь.— Алма-Ата: Наука, 1983.— 216 с.
Ванюшин Б. Ф. Апоптоз у растений // Успехи биол. хим.— 2001.— Т. 41.—
С. 3-38.
Василевская В. К. Анатомия образования почек на корнях некоторых древесных
227
растений // Вести. Ленинград, гос. ун-та Сер. биол., вып. 1.— 1957а.—
№3 —С. 5-21.
Василевская В. К. Особенности ветвления хлопчатника предельного и непредель-
ного типа // Труды ин-та земледелия АН Туркм. ССР. Т. 1.— Ашхабад,
19576 — С. 1-24.
Василевская В. К., Борисовская Г. М. Влияние листьев и верхушечного генератив-
ного побега на пазушные почки Cosmos bipinnatus Cav. И Вотан, журн.—
1968.— Т. 53, N 9 — С. 1189-1195.
Василевская В. К., Борисовская Г. М„ Баранов М. П. Некоторые проблемы морфо-
логической эволюции вегетативных органов высших растений// Вести. ЛГУ.
Виол — 1983 — № 15, вып. 3,— С. 36-47.
Васильченко И. Т. Всходы деревьев и кустарников.— М., Л.: Изд-во АН СССР,
I960.— ЗО2с.
Васильченко И. Т. Неотенические изменения у растений.— М., Л.: Наука, 1965.—
83 с.
Васильченко И. Т. О значении морфологии прорастания семян для системати-
ки высших растений и истории их происхождения // Тр. Вотан, ин-та АН
СССР, 1936.— Сер. I. Флора и систематика высших растений, вып. 3.—
С. 7-66.
Вейсман А О жизни и смерти// Смерть и бессмертие / Новые идеи в биологии.
Вып. 1.— СПб., 1914.— С. 1-66.
Виноградова Т. Н. Ранние стадии развития Тайника сердцевидного в природных
условиях И Бюл. Моск, об-ва испытат. природы. Отд. биол.— 1996.— Т. 101,
вып. 4.— С. 82-91.
Вихирева-Василькова В. В. О почках возобновления арктических растений И Во-
тан. жури.— 1962.— Т. 47, № 8.— С. 1141-1151.
Волков И В. Морфологическое строение подушковидных растений юго-восточ-
ного Алтая // Вотан, журн.— 2002.— Т. 87, № 8.— С. 76-83.
Волков И. В. Подушковидные растения юго-восточного Алтая.— Томск: ТГПУ,
2003 — 199 с.
Волкова Е. Н. Особенности развития пустынных эфемеров в условиях Москвы //
Труды ип-та ботаники АН Туркм. ССР. Т. 4.— 1958.— С. 97-126.
Воронов А. Г. О перезимовывании травянистых растений И Вотан, журн.—
1952 — Т. 37, № 62 — С. 173-180.
Воронцов Н. Н. Развитие эволюционных идей в биологии.— М.: УНЦ ДО МГУ,
Прогресс-Традиция, АФБ, 1999.— 640 с.
Ворошилов В. Н. Лекарственная валериана.— М.: Изд-во АН СССР, 1959.— 159 с.
Ворошилов В. Н. Ритм развития у растений — М.: Изд-во АН СССР, 1960.— 135 с.
Высоцкий Г. Н. Ергеня. Культурно-фитологический очерк И Труды по приклад,
бот., генет. и селекц.— 1915.— Т. 5.— С. 1113-1464.
Высоцкий Г. Н. О некоторых формах корнеотпрыскового возобновления // Тру-
ды сигьско-господ. боташки.— 1926.— Т. 1, вып. 2.
Гамцемлидзе 3. Г. О почках возобновления субнивальных растений Казбеги
(Центральный Кавказ) // Изв. АН Груз. ССР, Сер. биол.— 1979.— Т. 5,
№ 4,— С. 342-350.
228
Гатцук Л. Е. О внешней структуре «полудревесных растений» и возможных
принципах их разграничения // Тез. докл. 5-го делегат, съезда ВБО.— Киев,
1973 —С. 118-119.
Гатцук Л. Е. Геммаксиллярные растения и система соподчиненных единиц по-
бегового тела // Бюл. Моск, об-ва испытат. природы. Отд. биол.— 1974а.—
Т. 79, вып. 1 — С. 100-113.
Гатцук Л. Е. К методам описания и определения жизненных форм в сезонном
климате // Бюл. Моск, об-ва испытат. природы. Отд. биол.— 19746.— Т. 79,
вып. 3.— С. 84-100.
Гатцук Л. Е. Содержание понятия «травы» и проблема их эволюционного поло-
жения И Проблемы экологической морфологии растений.— М.: Наука,
1976 —С. 55-130.
Гатцук Л. Е. Комплементарные модели побега и их синтез И Ботан. журн.—
1995 — Т. 80, № 6 — С. 1-4.
Гатцук Л. Е. Морфогенез копеечника кустарникового (Hedysarum fruticosum
Pall.) при переменном уровне песчаного субстрата и предполагаемый облик
его предка И Вопросы морфогенеза цветковых растений и строения их попу-
ляций.— М.: Наука, 1968.— с. 52-81.
Гатцук Л. Е.,Дервиз-Соколова Т. Г, Иванова И. В., Шафранова Л. М. Пути перехо-
да от кустарниковых форм к травянистым в некоторых таксонах покрытосе-
менных И Проблемы филогении высших растений.— М.: Наука, 1974.—
С. 16-37.
Головач А. Г. Лианы, их биология и использование.— Л.: Наука, 1973.— 259 с.
Головкин Б. Н. О продолжительности жизни луковичных растений И Ботан.
журн.— 1965.— Т. 50, №11.— С. 1642-1645.
Голубев В. Н. О биологическом значении геофилии у травянистых растений //
Ботан. журн.— 1956а.— Т. 41, № 2.— С. 236-242.
Голубев В. И. К онтогенезу корневищ кистекорневых растений И Ботан. журн.—
19566 — Т. 41, № 2 — С. 248-253.
Голубев В. Н. О короткокорневищных растениях И Бюл. Моск, об-ва испытат.
природы. Отд. биол.— 1958.— Т. 63, вып. 3.— С. 97-103.
Голубев В. Н. О некоторых особенностях морфогенеза жизненных форм травя-
нистых растений лесолуговой зоны в связи с их эволюцией// Ботан. журн.—
1959 — Т. 44, № 12.— С. 1704-1716.
Голубев В. Н. К морфолого-генетической характеристике клубневых растений И
Бюл. Моск, об-ва испытат. природы. Отд. биол.— 1960.— Т. 65, вып. 5.—
С. 81-92.
Голубев В. Н. К морфолого-генетической характеристике ползучих растений //
Бюл. Моск, об-ва испытат. природы. Отд. биол.— 1961.— Т. 66, вып. 4.—
С. 53-66.
Голубев В. Н. Основы биоморфологии травянистых растений Центральной лесо-
степи // Труды Центрально-Чернозем. гос. зап. им. В. В. Алехина, 1962.—
Вып. 7.— 510 с.
Голубев В. Н. Эколого-биологические особенности травянистых растений и рас-
тительных сообществ лесостепи.— М.: Наука, 1965.— 286 с.
229
Голубев В. Н. О морфогенезе шпалерных вегетативно-подвижных кустарников
Крымской Яйлы // Биол. науки.— 1968а.— № 9.— С. 58-63.
Голубев В. Н. О морфогенезе моноподиальных полукустарничков Крымской
Яйлы // Бюл. Моск, об-ва испытат. природы. Отд. биол.— 19686.— Т. 73,
вып. 4.— С. 63-71.
Голубев В. Н. О морфогенезе симподиальных полукустарничков Крымской
Яйлы // Укр. ботан. журн.— 1969.— № 2.— С. 37-43.
Голубев В. Н. Принцип построения и содержание линейной системы жизненных
форм покрытосеменных растений И Бюл. Моск, об-ва испытат. природы.
Отд. биол.— 1972 — Т. 77, вып. 6 — С. 72-80.
Голубев В. Н. К определению понятий жизненных форм полукустарничков, по-
лукустарников, кустарничков и кустарников // Бюл. Никитск. Ботан. сада.—
1973,— Вып. I,— С. 9-12.
Голубев В. Н. Проблемы эволюции жизненных форм и филогения растений//
Проблемы эволюционной морфологии и биохимии в систематике и филоге-
нии растений.— Киев: Наук. Думка, 1981.— С. 3-29.
Голубев В. Н., Голубева И. В. О факультативной корнеотпрысковости клевера гор-
ного в условиях луговой степи И Ботан. журн.— 1964.— Т. 49, № И —
С. 1624-1628.
Грудзинская И. А. О периодичности развития (побегообразовании у дуба) // Ито-
ги и перспективы исследований развития растений.— Л.: Изд-во АН СССР,
1959 — С. 82-102.
Грудзинская И. А. Летнее побегообразование у древесных растений и его класси-
фикация // Ботан. журн.— 1960.— Т. 45, № 7.— С. 968-978.
Грудзинская И. А. Некоторые итоги изучения онтогенеза побегов дуба (Quercus
roburL.). И Ботан. журн.— 1964.— Т 49, № 3.— С. 321-337.
Грудзинская И. А. Об основных типах ветвления (критические заметки) И Ботан.
журн — 1974 — Т. 59, № 4 — С. 572-577.
Грудзинская И. А. Семейство аронниковые (Агасеае) / Жизнь растений. Т. 6.— М.:
Просвещение, 1982 — С. 466-493.
Грудзинская И. А. Циклохрон — новая единица биологического времени// Докл.
РАН — 1993.— Т. 329, № 3.— С. 802.
Грушвицкий И. В. «Втягивающие корни» — важная биологическая особенность
женьшеня (Ранах ginseng С. А. М.) И Ботан. журн.— 1952.— Т. 40, № 5.—
С. 682-685.
Грушвицкий И. В. Подземное прорастание и функции семядолей И Ботан.
журн,— 1963 — Т. 48, № 6 — С. 909-915.
Гупало П. И. Пути дальнейшего развития теории онтогенеза растений (К вопро-
су о теории возрастной цикличности Н. П. Кренке) // Успехи совр. биол.—
1954,—Т. 38, вып. 1.—С. 111-121.
Гупало П. И. Физиология старения и омоложения высших растений И Биол. раз-
вития растений.— М.: Изд-во Наука, 1975.— С. 198-213.
Гупало П. И., Скрипчинский В. В. Физиология индивидуального развития расте-
ний.— М.: Изд-во Колос, 1971.— 224 с.
Гурский А. В. Закономерности роста древесных растений // Сообщ. Тадж. фил.
АН СССР.— 1948,— Т. 4.— С. 19-22.
230
Дарвин Ч. Лазящие растения. Соч. Т. 8.— М.; Л.: Изд-во АН СССР, 1941.— 541 с.
Дарвин Ч. Насекомоядные растения. Соч. Т. 7.— М.; Л.: Изд-во АН СССР,
1949.— 650 с.
Денисова Г. М. Некоторые особенности побегообразования у Роа alpina L. И Во-
тан. журн.— 1960.— Т. 45, № 10.— С. 1552-1554.
Дервиз-Соколова Т. Г. Анатомо-морфологическое строение Salix polaris Wahlb. и
S. phlebophylla Anderss. // Бюл. Моск, об-ва испытат. природы. Отд. биол.—
1966 — Т. 71, вып. 2.— С. 28-38.
Дервиз-Соколова Т. Г. О стланиковом характере роста деревянистых растений на
примере Salix viminalis L. // Науч. докл. высш. шк. Биол. науки.— 1967.—
№ 11 — С. 64-71.
Дервиз-Соколова Т. Г. Жизненные формы ив северо-востока СССР. // Ботан.
журн.— 1982.— Т. 67, № 7 — С. 975-982.
Догель В. А. Олигомеризация гомологичных органов как один из главных путей
эволюции животных.— Л., 1954.
Дубровицкая Н. И. Регенерация и возрастная изменчивость растений.— М.:
Изд-во АН СССР, 1961.— 230 с.
Елагин И. Н. Времена года в лесах России.— Новосибирск: Наука, 1994.— 270 с.
Ефейкин А. К. Меристема и проблема омоложения растений И Докл. Ереван,
симпоз. по онтогенезу высш. раст.— Ереван, 1966.— С. 195-202.
Ефейкин А. К. Природа образовательной ткани растений и вопрос о старении и
вырождении побегов И Онтогенез высших растений (Сообщ. Ереванск. сим-
поз.) — Ереван: Изд. Акад, наук Армян. ССР, 1970.— С. 225-231.
Житков В. С. Величина основания листа как критерий для классификации форм
филлотаксиса и характера метамерности побега цветковых растений И
Журн. общей биол.— 1983.— Т. 44, № 6.— С. 802-822.
Житков В. С. О метамере побега в связи с разными формами филлотаксиса. И
Матер. 5-го Моск, совещания по филогении растений (декабрь 1976 г.), М.:
Наука, 1976 — С. 53-56.
Житков В. С. Способ листорасположения как отражение организации побега И
Актуальные вопросы ботаники в СССР: Тезисы докл. VIII делегат, съезда
ВБО.— Алма-Ата: Наука, 1988.— С. 331.
Жмылев П. Ю. Эволюция жизненных форм растений: суждения и предположе-
ния// Журн. общ. биол.— 2004.— Т. 65, № 3.— С. 232-249.
Жмылев П. Ю., Алексеев Ю. Е., Карпухина Е. А. Основные термины и понятия
биоморфологии растений.— М.: Изд-во МГУ, 1993.— 149 с.
Жмылев П. Ю„ ЖмылеваА. 11, Карпухина Е. А., Титовец А. В. Возможные причи-
ны изменения сезонного развития растений в связи с потеплением климата //
Вест. РУДН. Сер. Экология и безопасность жизнедеятельности.— 2003.—
№ 3 (9).— С. 98-103.
Жмылев П. Ю., Карпухина Е. А., Жмылева А. П. Изменения ритма сезонного раз-
вития растений в связи с глобальным потеплением климата // Актуальные
проблемы экологии и природопользования. Вып. 3. Сб. науч, трудов.— М.:
РУДН., 20026,— С. 41-46.
Жуйкова И. В. Особенности роста и определение возраста некоторых растений
231
Хибин // Проблемы Севера (Природа). Вып. 8.— М., Л.: Наука, 1964.—
С. 118-129.
Жукова Л. А. Онтогенез и циклы воспроизведения растений. // Журн. общ. био-
логии.— 1983,— Т. 44, № 3 — С. 361-374.
Жукова Л. А. Популяционная жизнь луговых растений.— Йошкар-Ола: РИИК
«Ланар», 1995.— 224.
Жукова Л. А., Глотов Н. В. Морфологическая поливаринтность онтогенеза в
природных популяциях растений // Онтогенез — 2001.— Т. 32, № 6.—
С. 455-461.
Зайцев Г. Н. Фенология травянистых многолетников.— М.: Наука, 1978.— 149 с.
Зайцев Г. Н. Фенология древесных растений.— М.: Наука, 1981.— 120 с.
Зайцева М. Г. О развитии формы растений-подушек И Сообщ. Таджик, фил.
АН СССР — 1949 — Вып. 17,— С. 16-20.
Заморский А. Д. Полимеризация организма // Журн. общ. биол.— 1971.— Т. 32,
№ 3 — С. 332-340
Заморский А. Д. О концепции полимеризации и олигомеризации в морфоло-
гии// Арх. анатом., гистол., эмбриол.— 1980.— Т. 78, № 6.— С. 5-18.
Злобин Ю. А. К познанию строения клонов V actinium myrtillus L. // Ботан.
журн,— 1961.— Т. 43, № 3 — С. 414-419.
Злобин Ю. А. Репродуктивное усилие / Эмбриология цветковых растений. Тер-
минология и концепции. Т. 3. Системы репродукции.— СПб: Мир и семья,
2000 — С. 247-251.
Зозулин Г. М. Система жизненных форм высших растений И Ботан. журн.—
1961— Т. 46, № 1,— С. 3-20.
Зозулин Г. М. Схема основных направлений и путей эволюции жизненных форм
семенных растений // Ботан. журн.— 1968.— Т. 53, № 2.— С. 223-233.
Зозулин Г. М. Аспекты учения о жизненных формах растений в биосферном пла-
не // Проблемы экологической морфологии растений.— М.: Наука, 1976.—
С. 45-54.
Зотин А. И. Термодинамический подход к проблемам развития, роста и старе-
ния.— М.: Наука, 1974.— 184 с.
Иванов В. Б. Клеточные основы роста растений.— М.: Наука, 1974.— 222 с.
Иванов В. Б. Меристема как самоорганизующаяся система: поддержание и огра-
ничение пролиферации клеток И Физиол. раст.— 2004.— Т. 51, № 6.—
С. 926-941.
Иванов В. Б. Меристема как самоорганизующаяся система: поддержание и огра-
ничение пролиферации клеток И Физиол. раст.— 2004.— Т. 21, № 6.—
С. 926-941.
Игнатьева И. П. О жизненном цикле стержнекорневых и кистекорневых травя-
нистых поликарпиков И Ботан. журн.— 1965.— Т. 50, № 7.— С. 903-916.
Игнатьева И. И. Онтогенетический морфогенез вегетативных органов травяни-
стых растений (Учебное пособие).— М.: Изд-во МСХА, 1989.— 62 с.
Казакевич Л. И. Материалы к биологии растений Юго-Востока России. I. Глав-
нейшие типы вегетативного возобновления и размножения травянистых
многолетников И Известия Саратовск. с.-х. опытн. станции.— 1922.— Т. 3,
вып. 3, 4.— С. 99-117.
232
Казакевич Л. И. Некоторые типы корневых систем растений Нижнего Поволжья
// Дневн. Всес. съезда ботан. в Ленинграде в январе 1928.— Л., 1928.—
С. 231-232.
Казарян В. О. Старение высших растений.— М.: Наука, 1969.— 313 с.
Казарян В. О. Физиологические аспекты эволюции от древесных к травам.— Л.:
Наука, 1990.— 348 с.
Карпухина Е. А., ЛамыкинаД. Е., Ж мыл ев П. Ю., Жмылева А. П. Время цветения и
структурные адаптации растений / Актуальные проблемы экологии и при-
родопользования (вып. 3).— М.: Изд-во РУДН, 2002.— С. 54-59.
Косинов В. Б. Понятие поколения и проблема изменчивости особей одного поко-
ления у организмов, размножающихся почкованием И Онтогенез.— 1972.—
Т. 3, № 4 — С. 360-370.
Кефели Е. И. Рост растений: учебное пособие.— М.: Колос, 1984.— 175 с.
Кирпичников В. С. Модификационная изменчивость и ее роль в эволюции // Раз-
витие эволюционной теории в СССР.— Л.: Наука, 1983.— С. 155-164.
Кирпотин С. Н. О целесообразности использования физиономического подхода
и принципов теории симметрии при выделении и изучении жизненных
форм растений И Krylovia — 1999.— Т. 1, № 1.— С. 15-25.
Козо-Полянский Б. М. Основной биогенетический закон с ботанической точки
зрения.— Воронеж, 1937.— 253 с.
Козо-Полянский Б. М. Случаи превращения биоморф культурных растений и их
значение//Труды Воронеж гос. ун-та.— 1945.— Т. 13, №1.— С. 46 50.
Колищук В. Г. До классифп<ацп життэвих форм сланских деревних рослин И Укр.
ботан. журн.— 1968а.— Т. 25, № 3.— С. 59-66.
Колищук В. Г. О морфологической эволюции от деревьев к травам в ряду стелю-
щихся форм растений // Ботан. журн.— 19686.— Т. 58, № 8.— С. 1029-1041.
Кондратъева-Мельвилъ Е. А. Образование корневых отпрысков у некоторых тра-
вянистых двудольных И Вести. Ленингр. гос. ун-та.— Сер. биол.— 1957.— №
3, вып. I.— С. 22-37.
Кондратъева-Мелъвилъ Е. А. Об образовании почек на корнях Rubus idaeus L. //
Бот. Журн,— 1959 — Т. 44, № 5 — С. 651-657.
Коровин Е. П. Эфемеры в растительном покрове восточной Бетпак-Дала // Труды
Среднеазиат. гос. ун-та. Сер. VIII-Ь. Ботаника, вып. 24. / Результаты Бетпак-Да-
линской экспедиции Среднеазиат. гос. ун-та,— 1935.— Вып. 7.— С. 3-15.
Коровина О. Н. Иллюстрированное руководство по морфологии цветковых рас-
тений / Под ред. М. Г. Агаева — СПб.: Изд. ВИР, 1997.— 154 с.
Коровкин О. А. Морфогенез вегетативных органов Helianthus tuberosus L. при вы-
ращивании растений из семян // Известия Тимирязев, с.-х. акад.— 1983.—
Вып. I.— С. 48-56.
Коровкин О. А. Морфогенез вегетативных органов Solanum tuberosum L. при вы-
ращивании растений из клубней // Известия Тимирязев, с.-х. акад.— 1984.—
Вып. 4.— С. 27-34.
Коровкин О. А. Основные термины и понятия морфологии высших растений:
Учебное пособие — 2-е перераб. и доп. изд.— М.: Изд-во МСХА, 2003.—
100 с.
233
Корочкин Л. И. Биология индивидуального развития (генетический аспект).—
М.: МГУ, 2002 — 264 с.
Корчагин А. А. Определение возраста деревьев умеренных широт // Полевая гео-
ботаника. Т. 2.— М., Л.: Изд-во АН СССР, 1960а.— С. 209-240.
Корчагин А. А. Определение возраста и длительности жизни кустарников И По-
левая геоботаника. Т. 2 — М., Л.: Изд-во АН СССР, 19606.— С. 241-248.
Котт С. А. Вегетативное размножение травянистых многолетников при помо-
щи нитевидных корней И Бюл. Моск, об-ва испытат. природы. Отд. биол.—
1956,— Т. 61, вып. 3,— С. 92-94.
Крамер П. Д., Козловский Т. Т. Физиология древесных растений.— М.: Лесн.
пром., 1983.— 463 с.
Красильников П. К. Классификация корневых систем деревьев и кустарников И
Лесоведение.— 1970.— № 3.— С. 35-44.
КренкеН. П. Регенерация растений.— М„ Л.: Изд-во АН СССР, 1950.— 674 с.
КренкеН. П. Теория циклического старения и омоложения растений и практиче
ское ее применение.— М.: Сельхозгиз, 1940.— 135 с.
Крюкова Л. Н. К вопросу о геофилии растений // Ботан. журн.— 1958.— Т. 43,
№ 3 — С. 425- 128
Кузнецов Г. В. Морфогенез почек возобновления и ритм развития бобовых юго-
восточного Алтая // Бюл. Моск, об-ва испытат. природы. Отд. биол.—
1985,— Т. 90, № 4,— С. 89-96.
Кузнецова Т. В. К истории развития представлений о плане строения побега сосу-
дистых растений и о месте филлома в нем // Ботан. журн.— 1995.— Т. 80,
№7 —С. 1-11.
Кузнецова Т. В. Редукционные явления в области соцветия: сущность и роль ре-
дукции в эволюции модульных объектов // Журн. общ. биол.— 1998.—
Т. 59, № 1— С. 74-103.
Культиасов И. М. Некоторые особенности морфогенеза жизненной формы по-
душковидных растений И Докл. Тимирязев, с.-х. акад.— 1962а.— Вып. 77.—
С. 575-579.
Культиасов И. М. Эколого-морфологические особенности подушковидных эс-
парцетов Средней Азии // Ботан. журн.— 19626.— Т. 47, № 5.— С. 645-657.
Куперман Ф. М. Морфофизиология растений. Морфофизиологический анализ
этапов органогенеза различных жизненных форм покрытосеменных расте-
ний.— М.: Высш, шк., 1977.— 287 с.
Куркин К. А. Некоторые биоэкологические особенности осоки дернистой (Carex
caespitosa L.) // Вести. Моск. гос. ун-та. Сер. физ.-мат. и естеств. наук.—
1954,— № 12,— С. 83-93.
Лавренко Е. М., Свешникова В. М. О синтетическом изучении жизненных форм
на примере степных дерновинных злаков // Журн. общ. биол.— 1965.—
Т. 23, №3.—С. 12-37.
Лапшина Е. И. О перезимовывании высших растений по наблюдениям в окрест-
ностях Петергофа // Труды Петергоф, естеств.-научн. ин-та.— Л., 1928.—
Вып. 5,— С. 36-98.
234
Левин Г. Г. Возрастные изменения у растений (Анализ некоторых понятий и
представлений) И Ботан. журн.— 1966.— Т. 51, № 12.— С. 1774-1795.
Левин Г. Г. Жизненные циклы растений, их связи и эволюция (обзор литерату-
ры) // Ботан. журн.— 1963,— Т. 48, № 7.— С. 1039-1060.
Левин Г. Г. Индивидуальность и жизненные циклы растений // Ботан. журн.—
1964 — Т. 49, № 2 — С. 272-280.
Левин Г. Г. Проблема индивидуальности у растений // Ботан. журн.— 1961.—
Т. 43, № 3.— С. 432-447.
Левин Г. Г. Чередование поколений, цикл развития и онтогенез растений // Бо-
тан. журн.— 1959,— Т. 44, № 7.— С. 943-953.
Левина Р. Е. Многообразие и эволюция форм размножения растений.— М.—
Учпедгиз, 1961.— 71 с.
Леопольд А. Рост и развитие растений.— М.: Мир, 1968.— 494 с.
Лодкина М. М. Черты морфологической эволюции растений, обусловленные
спецификой их онтогенеза И Журн. общ. биол.— 1983.— Т. 44, № 2.—
С. 239-253.
Лодкина М. М. Эволюционные отношения однодольных и двудольных по дан-
ным изучения зародыша и проростков // Ботан. журн.— 1988.— Т. 73,
№ 5 — С. 617-630.
Лотова Л. И. Морфология и анатомия высших растений.— М.: Эдиториал
УРСС, 2000 — 528 с.
Лутова Л. А., Проворов Н. А., Тиходеев О. Н. и др. Генетика развития.— СПб: Нау-
ка, 2000.— 539 с.
Лучник 3. И. Декоративная долговечность кустарников в культуре.— Новоси-
бирск: Наука, 1988.— 102 с.
Лучник 3. И. Обрезка кустарников.— М Изд во с.-х. Лит-ры, 1960.— 94 с.
Любарский Е. Л. К экологической анатомии корневища некоторых длиннокор-
невищных растений-гидрофитов И Ботан. журн.— 1965.— Т. 50, № 1.—
С. 119-123.
Любарский Е. Л. О биоморфологической границе между длиннокорневищны-
ми и короткокорневищными растениями // Экология.— 1973.— № 2.—
С. 94-95.
Любарский Е. Л. Об эволюции вегетативного возобновления и размножения тра-
вянистых поликарпиков И Ботан. журн.— 1961.— Т. 46, № 7.— С. 959-968.
Любарский Е. Л. Экология вегетативного размножения высших растений.— Ка-
зань, 1967.— 180 с.
Любименко В. Н. О спящих почках // Труды С.-Петербург, о-ва естествоиспытат.
Отд. ботан.— 1900.— Т. 30, вып. 3.— С. 195-259.
Ляшенко Н. И. О спящих почках кустарников // Ботан. журн.— 1958.— Т. 43,
№7.—С. 1039-1047.
Мазуренко М. Т. Биоморфологические адаптации растений Крайнего Севера.—
М.: Наука, 1986.— 208 с.
Мазуренко М. Т. Вересковые кустарнички Дальнего Востока (структура и морфо-
генез).— М.: Наука, 1982.— 184 с.
235
Мазуренко М. Т. О жизненных формах стелющихся лесных растений И Ботан.
журн — 1978 — Т. 63, № 4 — С. 593-603.
Мазуренко М. Т. Рододендроны Дальнего Востока (структура и морфогенез).—
М.: Наука, 1980 — 229 с.
Мазуренко М. Т., Хохряков А. П. Структура и морфогенез кустарников.— М.:
Наука, 1977.— 158 с.
Мазуренко М. Т., Хохряков А. П. Классы метамеров деревьев И Журн. общей
биол.— 1991 — Т. 52, № 3,— С. 409-421.
Маркаров А. М, Головко Т. К., Табаленкова Г. Н. Морфофизиология клубнеобра-
зующих растений.— СПб: Наука, 2001.— 208 с.
Марков М. В. Популяционная биология розеточных и полурозеточных малолет-
них растений.— Казань, 1990.— 185 с.
Маркова Л. Е. Изменение онтогенеза у однолетних растений Средней Азии в свя-
зи с условиями водоснабжения И Пробл. биогеоценол. геобот. и ботан.
геогр.— Л.: Наука, 1973.— С. 170-180.
Мейен С. В. Олигомеризация и полимеризация в эволюции древнейших высших
растений // Значение процессов полимеризации и олигомеризации в эволю-
ции— Л.: ЗИН АН СССР, 1977.
Меликян А. П., Девятов А. Г. Основные карпологические термины. Справоч-
ник.— М.: Тов. науч. изд. КМК, 2001.— 47 с.
Минин А. Фенология русской равнины: материалы и обобщения.— М.: Изд-во
ABF/АБФ, 2000 — 160 с.
Мирзоян Э. Н. Развитие учения о рекапитуляции.— М.: Наука, 1974.— 368 с.
Михайловская И. С. Корни и корневые системы растений.— М.: МГПИ
им. В. И. Ленина, 1981а.— 136 с.
Михайловская И. С. Анатомические особенности корневищ некоторых много-
летних трав // Жизненные формы: структура, спектры и эволюция.— М.:
Наука, 19816.—С. 141-160.
Михалевская О. Б. Морфогенез побегов древесных растений. Этапы морфогенеза
и их регуляция.— М.: МПГУ, 2002.— 66 с.
Михалевская О. Б. О внутри— и внепочечной фазах в развитии элементарных по-
бегов древесных растений // Онтогенез — 2002.— Т. 33, № 4.— С. 258-263.
Молотковский Г. X. Полярность развития растений.— Львов: Изд-во Львовск.
ун-та, 1961.— 259 с.
Мулкиджанян Я. И. К классификации жизненной формы «полукустарник» //
Проблемы современ. ботаники. Т. 2.— М., Л.: Наука, 1965.— С. 88-90.
Недолужко В. А. Эволюция жизненных форм покрытосеменных растений с по-
зиций биогенетического закона И Комаровские чтения. Вып. 42.— Владиво-
сток, 1996а — С. 5-29.
Недолужко В. А. Биогенетический закон и эволюция биоморф цветковых расте-
ний И Материалы IX Моск, совещ. по филогении растений.— М., 19966.—
С. 93-95.
Недолужко В. А. Древесные растения: Проблема эволюции жизненных форм.—
Владивосток, 1997.— 120 с.
НейфахА. А., Лозовская Е. Р. Гены и развитие организма.— М„ 1984.
236
Нечаева Н. Т., Василевская В. К., Антонова К. Т. Экологическая классификация
однолетних растений Каракумов И Ботан. журн.— 1969.— Т. 54, № 1.—
С. 1689-1704.
Нухимовский Е. Л. О термине и понятии «каудекс». 2. Современное состояние
вопроса И Вести. Моск. гос. ун-та. сер. Биол.-почвовед.— 1969а.— № 1.—
С. 55-62.
Нухимовский Е. Л. О термине и понятии «каудекс». 3. Многообразие каудексов и
отличия их от других структурных образований // Вести. Моск. гос. ун-та.
Биол.-почвовед.— 19696.— № 2.— С. 71-77.
Нухимовский Е. Л. Осевая и побеговая системы семенных растений. И Известия
Тимирязевск. с.-х. акад.— 1971.— Вып. 1.— С. 54-66.
Нухимовский Е. Л. О понятии «жизненная форма» // Сборник науч, работ Всесо-
юзн. науч.-исслед. ип-та лекарств, раст.— 1973а.— Вып. 5.— С. 222-232.
Нухимовский Е Л. О соотношении понятия «партикуляция» и «вегетативное
размножение» И Бюл. Моск, об-ва испытат. природы— 19736 .— Т. 78,
Вып. 5,— С. 107-120.
Нухимовский Е. Л. Ветвление и кугцение семенных растений И Известия Тимиря-
зевск. с.-х. акад.— 1974.— Вып. 2.— С. 50-62.
Нухимовский Е. Л. Проблемы классификации в биоморфологии семенных расте-
ний. И Успехи совр. биол.— 1980.— Т. 90, № 2.— С. 286-307.
Нухимовский Е. Л. Основы биоморфологии семенных растений. 1. Теория орга-
низации биоморф.— М.: Недра, 1997.— 630 с.
Нухимовский Е. Л. Основы биоморфологии семенных растений. 2. Габитус и
формы роста в организации биоморф.— М.: Оверлей, 2002.— 859 с.
Одишария К. Ю. Продолжительность жизни листьев растений // Труды Сухум,
ботан. сада.— 1974.— Вып. 20.— С. 30-40.
Падеревская М. И. К биологии спящих почек дуба // Учен. зап. Курск, гос. пед.
ин-та. Цикл естеств.-геогр.— 1957.— Вып. 4.— С. 20-38.
Падеревская М. И. К вопросу о значении почечных чешуй // Учен. зап. Курск, гос.
пед. ин-та. Цикл, естеств.-геогр.— 1958.— Вып. 7.— С. 33-42.
Падеревская М. И. Особенности почек возобновления геофитов Стрелецкой сте-
пи // Ботан. журн.— 1966.— Т. 51, Ne 1.— С. 100-104.
Падеревская М. И. Особенности почек возобновления некоторых гемикрипто-
фитов Стрелецкой степи И Ботан. журн.— 1972.— Т. 57, № 9.— С. 1138- 1152.
Падеревская М. И. Сравнительная характеристика структурных особенностей
почек растений типов жизненных форм в условиях среднерусской лесостепи
// Флора и растит, среднерус. лесостепи.— Курск, 1984.— С. 46-74.
Паутов А. А. Строение сформированных удлиненных и укороченных годичных
побегов у древесных двудольных // Ботан. журн.— 1987.— Т. 72, № 12.—
С.1631-1636.
Персикова 3. И. Большой жизненный цикл щучки Deschampsia caespitosa L. И
Учен. зап. Моск. гос. пед. ин-та им. Потемкина.— 1959а.— Т. 100, вып. 5.—
С. 111-149.
Персикова 3. И. Формирование дерновин и жизненный цикл белоуса торчащего
237
(Nardus stricta L.) // Бюл. Моск, об-ва испытат. природы. Отд. биол.—
1959в.— Т. 64, вып. 5 — С. 61 68
Персикова 3. И. Формирование и жизненный цикл некоторых дерновинных зла-
ков// Науч. докл. высш. шк. Биол. науки.— 19596.— № 3.— С. 160-163.
Петровский В. В., Матвеева Н. В. Обсуждение вопроса о зональных типах расти-
тельного покрова на научном семинаре лаборатории растительности Крайне-
го Севера Ботанического института Академии наук СССР И Ботан. журн.—
1963 — Т. 53, № 5 — С. 721-731.
Покровская В. М. Классификация возрастных состояний растений И Вест. Моск,
ун-та,— 1958 — № 4.— С. 199-201.
Покровская Т. М. Онтоморфогенез и жизненная форма как индикатор среды на
примере некоторых клеверов И Проблемы экологической морфологии рас-
тений.— М.: Наука, 1976.— 156-166.
Полевой В. В. Система регуляции у растений И Вест. ЛГУ.— 1981, № 21.—
С. 105-109.
Полевой В. В., Саламатова Т. С. Эволюция способов движения у растений И Эво-
люция функций в растительном мире.— М.: Изд-во ЛГУ, 1985.— С. 188-200.
Полевой В. В., Саламатова Т. С. Физиология роста и развития растений.— Л.:
ЛГУ, 1991.— 238 с
Полозова Т. Г. Жизненные формы сосудистых растений в различных подзонах
Таймырской тундры И Жизненные формы: структура, спектры и эволю-
ция.— М.: Наука, 1981.— С. 265-281.
Похилько А. А. Сезонная динамика растительных сообществ Хибинских гор.—
СПб: Наука, 1993.— 144 с.
Правдин Ф. Н. Учение о жизненных формах как общебиологическая проблема И
Жизненные формы в экологии и систематике растений.— М., 1986.— С. 3-8.
Прозоровский А. В. О биологических типах растений пустыни // Ботан. журн.—
1936 — Т. 21, № 5.— С. 559-563.
Проскоряков Е. И. О происхождении однолетних эфемеров И Труды Узбек, гос.
ун-та. Нов. сер.— 1950.— N° 43.— С. 71-88.
Работнов Т. А. Биологические наблюдения на субальпийских лугах Северного
Кавказа И Ботан. журн.— 1945.— Т. 30, № 4.— С. 167-177
Работнов Т. А. Методы определения возраста и длительности жизни у травяни-
стых растений И Полевая геоботаника. Т. 2.— М., Л.: Изд-во АН СССР,
I960 — С. 249-262.
Работнов Т. А. Некоторые вопросы изучения ценотических популяций //
Бюл. моек, об-ва испытат. природы. Отд. биол.— 1969.— Т. 74, вып. 1.—
С. 141-149.
Работнов Т. А. Фитоценология: Учеб, пособие.— 3-е изд., перераб. и доп.— М.:
Изд-во МГУ,1992.— 352 с.
Райкова И. А. Морфобиологические особенности растений Памира при их куль-
тивировании в Ташкенте / Проблемы современной ботаники. Т. 2.— М.:
Наука, 1965,— С. 97-105.
Рахманина А. Т. Биолого-экологическая характеристика караганных сообществ
238
(из Salsola dendroides Pall.) в Кура-Араксинской низменности И Эколого-гео-
ботанические и агромелиоративные исследования в Куро-Араксинской низ-
менности Закавказья.— М., Л.: Изд-во АН СССР, 1962.— С. 204-297.
Рачковская Е. И. К биологии пустынных полукустарничков И Труды Ботан.
ин-та АН СССР. Сер. 3. Геоботаника, 1957.— С. 3-87.
Реймерс Н. Ф. Популярный биологический словарь.— М.: Наука, 1991.— 544 с.
Родионенко Г. И. Род Ирис — Iris L. (Вопросы морфологии, биологии, эволюции
и систематики).— М.: Изд-во АН СССР, 1961.— 215 с.
Рост растений и дифференцировка.— М.: Наука, 1981.
Ростовцева 3. II. Рост и дифференциация органов растений (апикальное нарас-
тание и органогенез).— М.: Изд-во МГУ, 1984.— 152 с.
Рысин Л. II., Рысина Г. П. Морфоструктура подземных органов лесных травяни-
стых растений.— М.: Наука, 1987.— 208 с.
Рысина Г. П. Ранние этапы онтогенеза лесных травянистых растений Подмоско-
вья.— М.: Наука, 1973.— 215 с.
Рытова Н. Г. Морфогенетические факторы, ограничивающие кущение злаков в
вегетативной фазе (к вопросу о листовом и апикальном доминировании) И
Ботан. журн.— 1976.— Т. 61, № 12 — С. 1670-1679.
Рытова Н. Г. Связь между продолжительностью прорастания почек и строением
побегов у Festuca pratensis Huds. И Ботан. журн.— 1972.— Т. 57, № 11.—
С. 1393-1400.
Рытова Н Г. Формирование боковой почки и строение дерновины у Nardus
stricta L. (Роасеае) И Морфогенез и ритм развития высш. раст.— М., 1987.—
С. 82-87.
Рэфф Р, Кофмен Т. Эмбрионы, гены и эволюция. М: Мир, 1986.— 404 с.
Сабинин Д. А. О значении корневой системы в жизнедеятельности растений.—
М„ Л.: Изд-во АН СССР, 1949.— 47 с.
Сабинин Д. А. Физиология развития растений.— М.: Изд-во АН СССР, 1963.—
194 с.
Савиных Н. П. Побегообразование и взаимоотношения жизненных форм в сек-
ции Veronica рода Veronica L. И Бюл. Моск, об-ва испытат. природы. Отд.
биол.— 1979.— Т. 84, вып. 3.— С. 92-105
Савиных Н. П. К вопросу о происхождении ползучих травянистых растений И
Бюл. Моск, об-ва испытат. природы.— 1986.— Т. 91, вып. 1.— С. 64-71.
Северцов А. Н. Эволюция и эмбриология // Дневник XII съезда русских естество-
испытателей и врачей.— М., 1910.— С. 262-275.
Северцов А. Н. Этюды по теории эволюции. Индивидуальное развитие и эволю-
ция.— Киев, 1912.— 300 с.
Северцов А. Н. Собрание сочинений. Т. 3. Общие вопросы эволюции.— М„ Л.:
АН СССР, 1945,— 528 с.
Северцов А. С. К вопросу об эволюции онтогенеза И Журн. общ. биол.— 1970.—
Т. 31, №2,— С. 222-235.
Северцов А. С. Введение в теорию эволюции.— М.: МГУ, 1981.— 318 с.
Северцов А. С. Направленность эволюции.— М.: МГУ, 1990.— 272 с.
239
Северцов А. С., Креславский А. Г., Черданцев В. Г. Три механизма эволюции // Со-
временные проблемы теории эволюции.— М.: Наука, 1993.— С. 17-42.
Седелъникова Л. Л. Биоморфология геофитов в Западной Сибири.— Новоси-
бирск: Наука, 2002.— 308 с.
Селезнёва В. А. Насекомоядные растения// Природа.— 1955.— Ле 12.— С. 110-111.
Сенянинова-Корчагнна М. В. Геофилия и ее значение в сложении структуры рас-
тительного сообщества (О целостности организма высших растений) //
Учен. зап. Ленинград, гос. ун-та. Сер. геогр. наук.— 1967.— № 327, вып. 19.—
С. 7-96.
Серебряков И. Г. Некоторые данные по истории развития листьев черемухи и ли-
пы // Вести. Моск, ун-та.— 1947.— № 7 — С. 105-117.
Серебряков И. Г. Структура и ритм в жизни цветковых растений. Ч. 1 // Бюл.
Моск, об-ва испытат. природы. Отд. биол.— 1948.— Т. 53, вып. 2.— С. 49-66;
Ч. 2. там же.— 1949.— Т. 54, вып. 2.— С. 47-62.
Серебряков И. Г. Развитие Ивановых побегов у ели И Бюл. Моск, об-ва испытат.
природы. Отд. биол.— 1950.— Т. 55, вып. 6 — С. 68-70.
Серебряков И. Г. О длительности жизни листа и факторах, ее определяющих И
Учен. зап. Моск. гос. пед. ин-та им. Потемкина.— 1951.— Т. 19, каф. ботан.,
вып. 1.— С. 3-18.
Серебряков И. Г. Морфология вегетативных органов высших растений.— М.:
Изд-во «Советская наука», 1952.— 390 с.
Серебряков И. Г. Биолого-морфологический и филогенетический анализ жиз-
ненных форм покрытосеменных // Учен, зап Моск, гос пед. ин-та им.
В. П. Потемкина.— 1954а.— Т. 37, каф. бот., вып. 2.— С. 21-89.
Серебряков И. Г. О методах изучения ритмики сезонного развития растений в
геоботанических стационарах // Докл. совещ. по стационарным геобот. ис-
след —Л., 19546,—С. 145-159.
Серебряков И. Г. Основные направления эволюции жизненных форм у покрыто-
семенных растений И Моск, об-ва испытат. природы. Отд. биол.— 1955.—
Т. 60, Вып. 3 — С. 71-91.
Серебряков И. Г. Типы развития побегов у травянистых многолетников и факто-
ры их формирования И Вопросы биологии растений / Учен, записки Моск,
педаг. ин-та им. В. П Потемкина.— 1959а.— Т. 100, каф. ботан., вып. 5.—
С. 3-38.
Серебряков И. Г. Период покоя у некоторых травянистых и древесных растений
Подмосковья И Вопросы биологии растений / Учен, записки Моск, педаг.
ин-та им. В. П. Потемкина.— 19596 — Т. 100, каф. ботан., вып. 5.— С. 39-51.
Серебряков И. Г. Экологическая морфология растений (жизненные формы по-
крытосеменных и хвойных).— М.: Высш, школа, 1962.— 377 с.
Серебряков И. Г. Жизненные формы высших растений и их изучение И Полевая
геоботаника. Т. 3.— М., Л.: Наука, 1964а.— С. 146-208.
Серебряков И. Г. Сравнительный анализ некоторых признаков ритма сезонного
развития растений различных ботанико-географических зон СССР. И Бюл.
Моск, об-ва испыт. природы. Отд. биол.— 19646.— Т. 69, вып. 5.— С. 62-75.
Серебряков И. Г. Соотношение внутренних и внешних факторов в годичном
240
ритме развития растений (к истории вопроса) И Ботан. журн.— 1966.— Т. 51,
№ 7 — С. 923-938.
Серебряков И. Г., Демонская Н. П., Родман Л. С. О морфогенезе жизненной фор-
мы кустарника на примере орешника // Бюлл. Моск, об-ва испыт. природы.
Отд. биол.— 1954.— Т. 59, вып. 2.— С. 57-70.
Серебряков И. Г., Иорданская Н. Н. О морфогенезе жизненной формы кустарника
на примере бересклета бородавчатого Euonymus verrucosa Scop. И Ботан.
журн,— 1954,— Т. 39, № 5 — С. 768-773.
Серебряков И. Г., Серебрякова Т. И. О двух типах формирования корневищ у тра-
вянистых многолетников // Бюл. Моск, об-ва испыт. природы. Отд. биол.—
1965 —Т 70, вып. 2 —С. 67-81.
Серебряков И. Г., Серебрякова Т. И. Экологическая морфология высших растений
в СССР И Боган, журн,— 1967 — Т. 52, № 10,— С. 1449-1471.
Серебряков И. Г., Серебрякова Т. И. Жизненные формы покрытосеменных и их
эволюция в отдельных систематических группах И Ботан. журн.— 1969.—
Т. 54, № 9,— С. 1321-1326
Серебряков И. Г., Серебрякова Т. И. Некоторые вопросы эволюции жизненных
форм цветковых растений // Ботан. журн.— 1972.— Т. 57, № 5.— С. 417-433.
Серебряков И. Г., Чернышева М. Б. О морфогенезе жизненной формы кустарнич-
ка у черники, брусники и некоторых болотных Ericaceae // Бюл. Моск, об-ва
испыт. природы. Отд. биол.— 1955.— Т. 60, вып. 2.— С. 65-77.
Серебряков К. К. Очерки по истории ботаники. Ч. 1.— М.: Изд-во Наркомпроса,
1941 —245 с.
Серебрякова Т. И. О некоторых морфологических закономерностях побегообра-
зования у овсяницы луговой (Festuca pratensis) И Бюл. Моск, об-ва испыт.
природы. Отд. биол.— 1959.— Т. 64, вып. 3.— С. 89-98.
Серебрякова Т. И. К вопросу о побегообразовании и кущении у злаков (по пово-
ду статей Т. Н. Суворовой) И Ботан. журн.— 1962.— Т. 47, № 3.— С. 427-432.
Серебрякова Т. И. Строение и деятельность верхушки побега И Ботан. журн.—
1963,— Т. 48, № 5.— С. 699-712.
Серебрякова Т. И. Типы побегов и эволюция жизненных форм в семействе зла-
ков И Науч, доклады высш. шк. Биол. науки.— 1967.— № 2.— С. 61-73.
Серебрякова Т. И. Побегообразование и жизненные формы некоторых овсяниц
(Festuca L.) в связи с их эволюцией И Вопросы морфогенеза цветковых расте-
ний и строение их популяций.— М.: Наука, 1968.— С. 7 -51.
Серебрякова Т. И. Ветвление и кущение в семействе Роасеае И Ботан. журн.—
1969а.— Т. 54, № 6,— С. 858-871.
Серебрякова Т. И. О ритмичности в структуре и онтогенезе побегов у злаков //
Бюл. Моск, об-ва испыт. природы. Отд. биол.— 19696.— Т. 74, вып. 4.—
С. 60-71.
Серебрякова Т. И. Морфогенез побегов и эволюция жизненных форм злаков.—
М.: Наука, 1971а.— 360 с.
Серебрякова Т. И. Типы большого жизненного цикла и структура наземных по-
бегов у цветковых растений И Бюл. Моск, об-ва испытат. природы. Отд.
биол.— 19716.— Т. 76, вып. 1.— С. 105-119.
241
Серебрякова Т. И. Учение о жизненных формах растений на современном этапе И
Итоги науки и техники. / Ботаника. Т. 1.— М.: ВИНИТИ, 1972.— С. 84-169.
Серебрякова Т. И. К вопросу об эволюционных взаимоотношениях древесных и
травянистых жизненных форм цветковых растений И Бюл. Моск, об-ва ис-
пытат. природы. Отд. биол.— 1973.— Т. 78, вып. 3.
Серебрякова Т. И. Некоторые итоги ритмологических исследований в разных бо-
танико-географических зонах СССР И Проблемы экологической морфоло-
гии растений.— М.: Наука, 1976.— С. 216-238.
Серебрякова Т. И. Об основных «архитектурных моделях» травянистых много-
летников и модусах их преобразования И Бюл. Моск, об-ва испытат. приро-
ды. Отд. биол.— 1977.— Т. 82, вып. 5.— С. 112-128.
Серебрякова Т. И. Еще раз о понятии «жизненная форма» у растений // Бюл.
Моск, об-ва испытат. природы. Отд. биол.— 1980.— Т. 85, вып. 6.— С. 75-86.
Серебрякова Т. И. Жизненные формы и модели побегообразования наземно-
ползучих многолетних трав И Жизненные формы: структура, спектры и эво-
люция.— М.: Наука, 1981.— С. 161-179.
Серебрякова Т. И. Почка как этап развития побега // Тез. докл. 7-го Делегат, съез-
да ВБО. Донецк, 11-14 мая.— Л., 1983а.— С. 232-233.
Серебрякова Т. И. О некоторых модусах морфологической эволюции цветковых
растений И Журн. общей биол.— 19836.— Т. 44, № 5.— С. 579-593.
Серебрякова Т. И. О некоторых модусах морфологической эволюции цветковых
растений И Журн. общ. биол.— 1983в.^- Т. 44, №Дг— С. 579 593.
Серебрякова Т И. Метаморфоз у растений: онтогенетический и эволюционный
аспекты // Бюл. Моск, об-ва испытат. природы. Отд. биол.— 1984.— Т. 89,
вып. 5,— С. 94-102.
Серебрякова Т. И. О вариантах моделей побегообразования у многолетних трав //
Морфогенез и ритм развития высших растений.— М., 1987.— С. 3-19.
Синнот Э. Морфогенез растений,— М.: Изд-во иностр, литер., 1963.— 600 с.
Скрипчинский В. В. Эволюция онтогенеза растений.— М.: Наука, 1977.— 84 с.
Скрипчинский В. В., Дударь Ю. А., Скрипчинский В. В., Шевченко Г. Т. Методика
изучения и графического изображения морфогенеза монокарпического по-
бега и ритмов сезонного развития травянистых растений И Труды Ставро-
польск. НИИ сельск. хоз-ва. Вып. X / Морфогенез растений. Ч. 2.— Ставро-
поль, 1970.— С. 3-15.
Скрипчинский В. В., Максименко Л. Д. Втягивающие корни и самозарывание лу-
ковиц у пролески сибирской (Scilla sibirica Andr.) И Сборник научно-исследо-
вательских работ студентов Ставроп. с.-х. ин-та.— Черкесск: Ставроп. кн.
Изд-во, 1950,— Вып. I,— С. 116-120.
Словарь ботанических терминов (Под общей ред. Дудки И. А.).— Киев: Наук,
думка, 1984.— 308 с.
Смелов С. П. Вегетативное возобновление злаков И Ботан. журн.— 1937.— Т. 22,
№ 3,— С. 296-325.
Смелов С. П. Основные этапы жизни лугового злака И Ботан. журн.— 1947.—
Т. 32, № 2.— С. 79-89.
242
Смелов С. П. О причинах различий в кущении луговых злаков в первый и после-
дующие годы жизни И Ботан. жури.— 1958.— Т. 43, № 6.— С. 774-780.
Смелов С. П. Теоретические основы луговодства.— М.: Изд-во Колос, 1966.—
367 с.
Смирнов П. А. Флора Приокско-Террасного государственного заповедника: Спи-
сок растений, собранных и зарегистрированных в окрестностях с. Лужки на
р. Оке Московской обл. / Труды Приокско-Террасного государственного за-
поведника. Вып. 2.— М.: 1958а.— 246 с.
Смирнов П. А. О Hierochloe odorata авторов среднерусской флоры // Бюл. Моск,
об-ва испытат. природы. Отд. биол.— 1958b.— Т. 63, вып. 5.— С. 77-82.
Смирнова Е. С. Морфологические типы многолетних цветковых растений
тропиков и субтропиков // Журн. общ. биол.— 1970.— Т. 31, вып. 5.—
С. 578-587.
Смирнова Е. С. Биоморфологические структуры побеговой системы тропиче-
ских и субтропических цветковых растений в природе и оранжерейной куль-
туре И Интродукция тропических и субтропических растений.— М., 1980.—
С. 52-91.
Смирнова Е. С. Морфология побеговых систем орхидных.— М.: Наука, 1990.—
208 с.
Смирнова О. В. Жизненный цикл пролески сибирской (Scilla sibirica Andr.). И На-
учи. докл. высш. шк. Биол. науки.— 1967.— № 9.— С. 76-84.
Соколов С. Я., Артюшенко 3. Т. Ивановы побеги у сосны И Ботан. журн.—
1957,— Т. 42, № 5 — С. 741-745.
Соловьева Н.А К изучению формирования и развития Agropyrum repens (L.) Р. В.
И Докл. АН СССР,— 1958 — Т. 122, № з.
Стешенко А. П. О методах определения возраста и длительности жизни пустын-
ных полукустарничков И Полевая геоботаника. Т. 2.— М., Л.: Изд-во АН
СССР, I960 — С. 263-278.
Стешенко А. П. Образование полукустарничковой формы роста в условиях вы-
сокогорий 11амира И Изв. АН Тадж. ССР. Отд. естеств. наук.— 1955.—
Вып. 12 — С. 143-148.
Стешенко А. П. Определение возраста растений-подушек (Sibbaldia tetrandra
Bunge) в условиях высокогорий Памира И Ботан. журн.— 1973.— Т. 58,
№7 —С. 1004-1011.
Стешенко А. П. Формирование структуры полукустарничков в условиях высо-
когорий Памира И Труды ин-та ботан. АН Тадж. ССР.— 1956.— Т. 50.—
С. 1-160.
Стрелер Б. Время, клетки и старение.— М.: Мир, 1964.— 251 с.
Суворова f. Н. Кущение злаков И Ботан. журн.— 1959.— Т. 44, № 9.— С. 1291—
1298.
Таршис Г. И. Подземные органы многолетних травянистых растений.— Сверд-
ловск: СГПИ, 1975.— 135 с.
Таршис Л. Г. Структурное разнообразие подземных органов высших расте-
ний.— Екатеринбург: УрО РАН, 2003.— 172 с.
243
Татаренко И. В. Орхидные России: жизненные формы, биология, вопросы охра-
ны.— М.: Аргус, 1996.— 207 с.
Тахтаджян А. Л. Теория филэмбриогеиеза А. Н. Северцова и эволюционная
морфология растений И Проблемы ботаники. Т. 1.— М., Л.: АН СССР,
1950.— С. 222-231.
Тахтаджян А. Л. Вопросы эволюционной морфологии растений.— Л.: ЛГУ,
1954 —213 с.
Тахтаджян А. Л. Основы эволюционной морфологии покрытосеменных.— М.,
Л.: Наука, 1964.— 235 с.
Терехин Э. С. О терминах «сапрофит», «полусапрофит» и «полупаразит» (в связи
с характером биотических отношений некоторых покрытосеменных расте-
ний) И Ботан. журн.— 1965.— Т. 50, № 1.— С. 60-69.
Терехин Э. С. Паразитные цветковые растения (Эволюция онтогенеза и образ
жизни).— Л.: Наука, 1977.— 218 с.
Терехин Э. С. Проблемы эволюции онтогенеза семенных растений И Тр. БИН
РАН. Вып. 2 — СПб: БИН, 1991 — 69 с.
Терехин Э. С. Семя и семенное размножение.— СПб: Мир и семья-95,1996.— 377 с.
Терехин Э. С., Никитичева 3. И. Постепенное развитие паразитных Angiospermae.
1. Метаморфоз И Ботан. журн.— 1968.— Т. 53, № 1.— С. 39-57.
Терехин Э. С., Никитичева 3. И. Семейство Orobanchaceae. Онтогенез и филоге-
нез.— Л.: Наука, 1981.— 228 с.
Терминология роста и развития высших растений / М. X. Чайлахян, Р. Г. Бутен-
ко, О. Н. Кулаева, В. Т. Кефели, Н. П. Аксенова.— М.: Наука, 1982.— 96 с.
Тихонов В. И. Побег как высший этап эволюции формы тела растений И Журн.
общей биол.— 1977.— Т. 38, № 6.— С. 841-848.
Тихонова М. Н. К вопросу о терминологии, касающейся специализированных
утолщенных побегов орхидных // Ботан. журн.— 1981.— Т. 66, № 11.—
С. 1628-1630.
Тихонова М. Н. К вопросу о природе подземных органов некоторых представи-
телей трибы Orchideaell Бот. журн.— 1983.— Т. 68, № 5.— С. 648-652.
Трофимов Т. Т. О типах прорастания семян некоторых многолетников. И Ботан.
журн — 1963.— Т. 48, №11,— С. 1583-1597.
Тюрина М. М. Развитие представлений о состоянии покоя у древесных растений
И Физиология раст.— 1979.— Т. 26, № 5.— С. 899-907.
Удалова Р. А. Морфологический анализ некоторых подушковидных кактусов И
Вотан, журн.— 1978.— Т. 63, № 2.— С. 256-263.
Уоринг Ф., Филипс И. Рост растений и дифференцировка.— М.: Мир, 1984.—
572 с.
Уранов А. А. Жизненное состояние вида в растительном сообществе И Бюл.
Моск, об-ва испытат. природы. Отд. биол.— 1960.— Т. 67, вып. 3.— С. 77-92.
Учебно-полевая практика по ботанике: Учеб, пособие для биол. спец, вузов / М.
М. Старостейкова, М. А. Буленкова, Л. М. Шафранова, Н. И. Шорина.— М.:
Высш. шк„ 1990.— 191 с.
Федоров А. А. Древесные эпифиты и фикусы-удушители в тропических лесах
Китая И Ботан. журн.— 1959.— Т. 44, № 10.— С. 1409-1424.
244
Федоров A. A., Кштнчнтшв М )., Артюшенко 3. Т. Атлас по описательной мор-
фологии вы, iiinx р.п к нпп Лист.— М., Л.: Изд-во АН СССР, 1956.— 301 с.
Федоров А. А.. Кнрничникчв М Артюшенко 3. Т. Атлас по описательной мор-
фологии выыппх р.п к пип. ( гсбель и корень.— М., Л.: Изд-во АН СССР,
1962 — 349,.
ХохряковА. II. »peMvpy<i.i и их культура.— М.: Наука,1965.— 126 с.
Хохряков А. II. О,.iiii.ii- ыкономерности эволюции онтогенеза у растений И
Бюл. Моск, он па iuiii.ii.ii природы. Отд. биол.— 1973.— Т. 78, вып. 1.—
С. 59 69.
Хохряков А. II Формы и П1И1.1 полимеризации в эволюции растений И Журн.
общ. биол 1974 I 15, №3.— С. 331-345.
Хохряков А II. l.iKonoMcpiiocni эволюции растений.— Новосибирск: Наука,
1975а — 199,
ХохряковА. ПЛ ома i иче, к ш июлюция однодольных.— М.: Наука, 19756.— 194 с.
Хохряков А. II Циклы рывшия побегов в связи с проблемой «травянистости»//
Проблемы >кол<>1 и'пткои морфологии растений.— М.: Наука, 1976.—
С. 141 155.
ХохряковА. II. 11 рип пн 11 i.i клас. ификации жизненных форм растений И Изв. АН
СССР. Сер. <>по г 1979. № 4,—с. 586-598.
Хохряков А. П >вол1оция бпоморф растений.— М.: Наука, 1981.— 165 с.
Хохряков А. 11., Mil крепко М. Г. Биоморфологическая эволюция как активный
процесс (па примере экстремальных условий высоких широт и высокого-
рий) И Ьиоморфология растений Дальнего Востока.— Владивосток: ДВНЦ
АН СССР, 198 > С. 3 16.
Хохряков А. II Ma lypciiKO М. Т. Геофилия как один из основных путей экологи-
ческой июлюции опоморф растений в Арктике и Субарктических высоко-
горьях И Iioi.ni.жури 1985.— Т. 70, №7.— С. 876-884.
Цвелев Н. Н. О направлениях соматической эволюции покрытосеменных расте-
ний И Бюл. Моск, оо ва испыгат. природы. Отд. биол.— 1970.— Т. 75,
Вып. 2.— С. 76 86.
Цвелев Н. Н. Злаки СССР.— Л.: Наука, 1976.— 788 с.
ЦвелевН. Н. О первичной жизненной форме покрытосеменных растений И Бюл.
Моск, об-ва испытат. природы. Отд. биол.— 1977.— Т. 82, вып. 1.— С. 79-88.
Цвелев Н. Н. Концепция фи гомера и эволюция высших растений // Эмбриоло-
гия цветковых растений. Терминология и концепции. Т. 3. Системы репро-
дукции.— СПб.: Мир и семья, 2000.— С. 350-354.
Цвелев Н. Н. Проблемы теоретической морфологии и эволюции высших расте-
ний.— СПб: Тов. пауч. издан. КМК, 2005.— 407 с.
Ценопопуляции растений (Основные понятия и структура).— М.: Наука,
1976.— 214 с.
Ценопопуляции растений (Очерки популяционной биологии) / Л. Б. Заугольно-
ва, Л. А. Жукова, А. С. Комаров и др.— М.: Наука, 1988.— 184 с.
Чайлахян М. X. Онтогенез и целостность растительного организма И Ботан.
журн,— 1956,— Т. 41, № 4 — С. 487-509.
245
Чайлахян М. X. Автономный и индуцированный механизмы регуляции цвете-
ния растений // Физиол. раст.— 1975.— Т. 22, вып. 6.
ЧепьцоваЛ. П. Рост конусов нарастания побегов в онтогенезе растений.— Ново-
сибирск: Наука, 1980.— 190 с.
Черемушкина В. А. Биология луков Евразии.— Новосибирск: Наука, 2004.—
280 с.
Чистякова А. А. О жизненной форме и вегетативном разрастании липы сердце-
видной //Бюл. Моск, об-ва испытат. природы. Отд. биол.— 1978.— Т. 83,
вып. 2 —С. 129-137.
Чистякова А. А. Жизненные формы деревьев и их эколого-ценотическая обу-
словленность // Жизненные формы в экологии и систематике растений.—
М., 1986,— С. 70-75.
Шалыт М. С. Эфемерные корни и их значение И Труды Первой научной конф,
препод, биол., с.-х. и хим. дисциплин педагог, ин-тов УССР, Симферополь,
27-29 мая 1957 г,— I960.— С. 49-94.
Шалыт М. С. Партикуляция у высших растений И Проблемы современной бота-
ники. Т. 2.— М., Л.: Наука, 1965.— С. 117-121.
Шампанья П. Покой почек, корреляция между органами и морфогенез древес-
ных растений // Физиол. раст.— 1983.— Т. 30, № 3.— С. 587-601.
Шафранова Л. М. О метамерности и метамерах у растений И Журн. общей
биол.— 1980,— Т. 41, № 3.— С. 437 447.
Шафранова Л. М. Ветвление растений: процесс и результат И Жизненные фор-
мы: структура, спектры и эволюция.— М.: Наука, 1981.— С. 179-213.
Шафранова Л. М. Растение как жизненная форма (к вопросу о содержании поня-
тия «растение») И Журн. общей биол.— 1990.— Т. 51, № 1.— С. 72-89.
Шилова Н. В. Приспособление вечнозеленых кустарничков к условиям Севе-
ра.— Л.: Наука, 1967.— 63 с.
Шилова Н. В. Ритмы роста и пути структурной адаптации тундровых расте-
ний.— Л.: Наука, 1988.— 212 с.
Шмальгаузен И. И. Пути и закономерности эволюционного процесса.— М„ Л.:
АН СССР, 1940 — 230 с.
Шмидт В. М. О квантированности роста и органогенеза у растений И Вести. Ле-
нинград. гос. ун-та. Биол.— 1968.— № 3, вып. 1 — С. 99-11.
Шнелле Ф. Фенология растений.— Л.: Гидрометеоиздат.— 1961.— 258 с.
Шорина Н. И. Вегетативное размножение И Эмбриология цветковых растений.
Терминология и концепции. Т. 3. Системы репродукции.— СПб.: Мир и се-
мья, 2000а,— С. 299-302.
Шорина Н. И. Жизненная форма И Эмбриология цветковых растений. Термино-
логия и концепции. Т. 3. Системы репродукции.— СПб.: Мир и семья,
20006 — С. 415-420.
Шорина Н. И., Михалевская О. Б. Почка // Эмбриология цветковых растений.
Терминология и концепции. Т. 3. Системы репродукции.— СПб.: Мир и се-
мья, 2000.— С. 310-315.
Шульц Г. Э. Случай экспериментальной вивипарии у злака И Ботан. журн.—
1939.— Т. 24, № 3 — С. 197-208.
246
Шульц Г. Э. Общая фено ни ин Ч 11 is i I*>81 186 с.
Шутов В. В. Структура динамика и ion ioiihhiciiiic кустарпичкоиых рас тенпи
Кострома: КГ’ГУ, 2()00ю tot) с
Щербакова А. А. История рус • кой tioi лпгкч кон (морфологической) герминоло
гии И Труды ин ia п< inpiiii е< к I । ио шан ня и lexHUKH. 'Г. 32. / I к порпя био
логич. наук. Вып 7. МИ щ ио Л11 < .( t 1’, 1960.— С. 203- 250.
ЮрцевБ. А. Жиснсиныс формы один и > у июных объектов ботаники И 11робле-
мы экологической морфо.........hi | а< кыш. - М.: Наука, 1976 ('. 9 44.
Юсуфов А. Г. 11 н । сч p.i । in и oiiioiiiii i.i у высших растении в ходе >но поции //
Журн. общей опои !•)/(> 1. 1/, № о,--С. 803-814.
Юсуфов 4. Г. Акцелс рации он ки < нс ia у цветковых растений и пущ ее дос i иже
ния//Журн оОщ оной 1'1/4 Г 40, № 2.— С. 165-175.
ЮсуфовА. Г. Мехапп imii р и'-рапип рас 1епий.— Ростов: Рос i он уп i, 1‘>Н’а.
176 с.
ЮсуфовА. Г. Роль прощч сои pei -! играни и в онтогенезе растении // Ж< рн очщеп
биол.— 19820. 1 И. Nuft ( /88 798.
ЮсуфовА. Г.. шдогепная pel уляции <| i а реи и я растений и ее эволюционно' ша
чение// Проб кмы макро шо iiomin. М.: Наука, 1988.
ЮсуфовА. I Биолш ия с lap, пня пи, псовых растений.— Махачкала И i ш Да
гестап. уп та, |9‘> ‘ ’(>1 с
Яковлев М. С., Жум»&1 /. Я. 1Ьжры юс сменные растения с зеленым и бесци, i iii.im
зародышем (хлоро и лс йко >мориофигы).— Л.: Наука, 1973.— 116 с.
Abrams A. Evolutionary biologv moil.ility and lifespan // Nature-2004.— V. 4 ’ I
P. 1048.
Askenasy E. Ubi i einc пене Methode, uin die Vertheilung der Wachsthumsintensltat in
wachsenden I heilen zu bestininu-n // Verh. Naturh.-medic. Ver. Heidelberg.
1880 -V. 2. -S. 7(1 153
BarlowP. IV. Adventioons mots of whole plants: their forms, funcoons and evolution
// New root foi mation in plants and cut tings.— Martinus Nijhoff Publ., Dor-
drecht, 198o P. 67 lit)
Bell A. D., Bryan A. Plant form. An illustrated guide to flowering plant morphol-
ogy.— Oxford, New York. I okyo (Ixford Univ. Press, 1998.— 341 p.
Bell A. D., Tomlinson P B. Adaptive arc hitecture in rhizomatous plants // Bot. J. Linn.
Soc— 1980. — V. 80, N 2- P 125 160.
Bender M. H , Bask tn j M., Baskin ( C. Age of maturity and life span in herbaceous,
polycarpic perennials // Bot. Rev. - 2000.— V. 66. N 3.— P. 311-349.
Bernard N. (.’evolution dans la synibio.se. Les Orchidees et leurs champignons com-
maesaux // Ann. Sci natur. bot biol., veget.— 1909.— Ser. 9.— P. 1-196.
Braun A., Dr. Carl Schimpers. Vorti-ще iiber die Moglichkeit eines wissenschaftlichen
Verstandnisses dcr Blaltstellung// Hora.— 1935.— Bd. 35.
Bucher H. Anatomische Veriinderungcn bel gewaltsamer Krum mung und geotro-
pischer Induktion // lahrb. Wiss. Bot.— 1906.— B. 43, N 1.— S. 271-360.
Burgeff H. Samenkeiinung del Orchideen und Entwicklung ihrer Keimpflanzen.—
Verlag von Gustav Pislier, Jena, 19*16. - 312 s.
247
Benzing D. H. Vascular epiphytism: taxonomic participation and adaptive diversity //
Ann. Missouri Bot. Gard.— 1987.— V. 74, N 1.— P. 183-204.
Candolle Au. P. de. Organographie vegetale. 2 Vols.— Paris, 1827.— Vol. 1, 558 p.,
Vol. 2, 304 p.
Celakovsky L. S. Uber die Homologien des Grasembryos П Bot. Zeitung.— 1897.—
Jg. 55 — S. 141-172.
Charlesworth B. Fisher, Medawar, Hamilton and evolution of aging // Genetics —
2000,—V. 156 —P 927-931.
Cline M. G. Concepts and terminology of apical dominance П Amer. Journ. Bot.—
1997,— V. 84, N 9,— P. 1064-1069.
Cline M. G. The role of hormones in apical dominance. New approaches to an old prob-
lem in plant development // Physiol. Pl.— 1994.— V. 90.— P. 230-237.
Corner E. J. H. The Durian theory of the origin of the modern tree // Ann. Bot.—
1949.— V. 13, N 52 — P. 367-414.
Croizat L. La filotaxie: sus principios, metodos de investigacion у de docencia. Sim-
bolismo // Bol. Acad, cienc. fis., mat. natur.— 1971.— N 93.— P. 53-99.
Cusset G., Cusset C. Etude les Podostemales. 12. Biogeographie evolutive de Tristicha
trifaria (Bory ex Willd.) Spreng. // Bull. Mus. natur Hist. nat. Paris.— 1989.—
Ser. 11, sect. B. Adansonia, N 1.— P. 39-70.
Cusset G., Cusset C. Etude sur les Podostemales. 10. Structures florales et vegetatives des
Tristichaceae // Bull. Mus. natur Hist, nat., Paris.,— 1988.— Ser. 10, sect. В Adan-
sonia, N 2.— P. 179-218.
Dawson J. W. The vines, epiphytes and parasites of New Zealand forests // Tuata-
ra— 1986,— V. 28, N 2.— P. 43-69.
De Beer G. R. Embryos and ancestors.— Clarendon Press, Oxford, 1958.— 198 p.
Dengler N. G. Anisophylly and dorsiventral shoot symmetry // Int. Journ. Pl. Sci.—
1999,— V. 160 — P. 67-80.
Dickinson T. A. Epiphylly in angiosperms 11 Bot. Rev.— 1978.— V. 44, N 2.—
P. 181-232.
Diggle P. K. Heteroblasty and evolution of flowering phenologies// Int. Journ. Pl.
Sci.— 1999 — V. 160. № 6.— P. 123-134.
Dommergues J.-L., DavidB., Marchand D. Les relations ontogenese-phylogenese: appli-
cations paleontologiques // Geobios, 1986.— N 19, fasc. 3.— P. 335-356.
Du Rietz G. E. Life-forms of terrestrial flowering plants.— Uppsala, 1931.— p. 1-95.
Elten G. What is a herb? (with examples from the tropical «Savanna» of Brasil and the
humid temperate zone of Poland) // Veroff. Geobot. Inst. ETH, Stiftung Riibel,
Zurich.— 1991,— B. 106.— P. 288-304.
Erickson R. O., Michelini F. ]. The plastochron index. 11 Amer. J. Bot.— 1957.— V. 44,
N 4 — P. 297-305.
Finch С. E„ AustadS. N. History and prospects: symposium on organisms with slow ag-
ing // Exper. Geront.— 2001.— V. 36.— P. 593-597.
Fisher ]. Branching patterns and angles in trees // On the economy of plant form and
function.— Cambridge Univ. Press, Cambridge, 1986.— P. 493-523.
Fisher M. С. T„ Eissenstat D. M., Lynch J. P. Lack of evidence for programmed root se-
248
nescence in common bean (Phaseolus vulgaris) grown at different levels of phos-
phorus supply // New Phytol.— 2002.— V. 153.— P. 63-71.
Forbis T.A., FloydS. K., de QueirozA. The evolution of embryo size in angiosperms and
other seed plants: implications for the evolution of seed dormancy // Evol.—
2002,— V. 56, N 11,— P. 2112-2125.
Gaertner J. De Fructibus et Seminibus plantarum.— Stuttgart, Leip’ig, 1788-1807.—
384 s.
Gan S„ Amasino R. M. Making sense of senescence // Pl. Physiol.— 1997.— V. 113.—
P. 313-319.
Garstang W. The theory of recapitulation: A critical restatementa of the biogenetic law
// Journ. Linn. Soc. Zool.— 1922.— V. 35.— P. 81-101.
Gatin C. L. Recherches anatomiques et chimiques sur la germination des Palmiers //
Ann. sci. natur. Bot. (Paris).— 1906.— Ser. 9, t. 3.— P. 191-315.
Gavrilov L. A., Gavrilova N. S. Evolutionary theories of aging and longevity. // Sci.
World Journ.— 2002,— V. 2 — P. 339-356.
Goebel K. Organographie der Pflanzen.— Jena, 1928.— 642 s.; 1930.— S. 642-1378.
Goldsmith T. C. The evolution of aging. [Electronic resource],— 2004.— Mode of ac-
cess: http: //www.azinet.com/aging/Aging_Book.pdf.
Gould S. J. Ontogeny and phylogeny.— Belknap Press, Harvard Univ. Press, Cam-
bridge, Massachusetts, London, Englsnd, 1980.— 501 p.
Greenberg/. T. Programmed cell death: A way of life for plants // Proc. Natl. Acad. Sci.
USA.— 1996 — V. 93,— P. 12094-12097.
Grobbelaar N., Hattingh W., Marshall J. The occurrence of coralloid roots on the South
African species of the Cycadales and their ability to fix nitrogen symbiotically //
S.-Afr. Tydskr. Plantk.— 1986.— V. 52, N 5.— P. 467-471.
Groff P. A., Kaplan D. R. The relation of root systems to shoot systems in vascular plants
// Bot. Rev.— 1988.— V. 54, N 4,— P. 387-422.
HackelE. Monographia Festucarum europaearum.— Kassel, Berlin, 1882.— 216 s.
Halle E., Oldeman R. A. A., Tomlinson P. B. Tropical trees and forests: an architectural
analysis.— Berlin: Springer-Verlag, 1978.— 312 p.
Halle F. Modular growth in seed plants// Phil. Trans. R. Soc. Lond.— 1986.— B. 313.—
P. 77-87.
Halle F„ Delmotte A. Croissance et floraison de la Gesneriaceae Africaine Epithema te-
nue С B. Clarke // Adansonia.— 1973,— V. 13 — P. 273-287.
Halle F., Martin L. Etude de la croissance rhythmique chez 1’Hevea (Hevea brasillensis
Mull.-Arg. Euphorbiacees.-Crotonoidees) // Adansonia.— 1968.— V. 8, fasc. 4.—
P. 475-503.
Halle F., Oldeman R. A. A. Essai sur 1’architecture et la dynamique de croissance des
arbres tropicaux.— Paris: Masson, 1970.— 178 p.
Harper J. L., White J. The demography of plants // Ann. Rev. Ecol. Syst.— 1974.—
V. 5,— P. 419-463.
Haughn G. W., Shultz E. A., Martinez-Zapater ]. M. The regulation of flowering in
Arabidopsis thaliana: meristems, morphogenesis and mutants 11 Can. Journ. Bot.—
1995.— V.—73.—P. 959-981.
249
Hautekeete N.-C., Piquot Y., Van Dijk H. Investment in survival and reproduction
along a semelparity-iteroparity gradient in the Beta species complex // Journ. Evol.
Biol.— 2001 — V. 14 — P. 795-804.
Hautekeete N.-C., Piquot Y„ Van Dijk H. Variations in ageing and meristematic activity
in relation to flower-bud and fruit excision in the Beta species complex // New
Phytol.— 2002 — V. 154 — P. 641-650.
Hegelmaier F. Die Lemnaceae — eine monographishe Untersuchungen.— Leipzig,
1868,— 169 s.
Henderson J. F., Henderson W. D. A dictionary of scientific terms (Pronunciation, deri-
vation and definition of terms in biology, botany, zoology, anatomy, cytology, ge-
netics, embryology, physiology).— D. Van Nostrand Company, WC., New-York,
1953 —506 p.
Heterochrony in Evolution. A multidisciplinary approach.— Plenum Press, New York,
London, 1988.
Holm T. The application of the term «rhizome» 11 Rhodora.— 1929.— V. 31, N 1.—
P.6-17.
jackson B. D. A glossary of botanic terms with their derivation audaccent.— Hafner
Publ. CO WC., New-York, 1953.— 481 p.
Janzen D. H. Why bamboos wait so long to flower // Ann. Rev. EcoL Syst.— 1976.—
V. 7 —P. 347-391.
Jenik J. Roots and root systems in tropical trees: morphologic and ecologic aspects I
Tropical trees as living systems.— Cambridge Univ. Press, Cambridge, 1978.—
P. 323-349.
Jones C. S. An essay on juvenility, phase change, and heteroblasty in seed plants. // Int.
Journ. Pl. Sci.— 1999,— V. 160.— P. 105-111.
Jong K, Burtt B. L. The evolution of morphological novelty exemplified in the growth
patterns of some Gesneriaceae IJ New Phytol.— 1975.— V. 75.— P. 297-311.
Juniper B. E„ Robins R. J., Joel D. M. The carnivorous plants.— London etc., Academic
Press, 1989.— 353 p.
Kirkwood T. B. L. Evolution of ageing// Meeh. Ageing Devel.— 2002.— V. 123, Iss.
7.— P. 737-745.
Kirkwood T. B. L. Evolution of aging // Nature.— 1977.— N 270.— P. 301-304.
Klingenberg С. P. Heterochrony and allometry: the analysis of evolutionary change in
ontogeny // Biol. Rev.— 1998.— V. 73.— P. 79-123.
Koornneef M„ Alonso-Blanco C, Peeters A. Genetic control of flowering time in Ara-
bidopsisll Annu Rev. Pl. Physiol.— 1998.— V. 49.— P. 345-370.
Kress W. J. The systematic distribution of vascular epiphytes 11 Vase. Plants Epiphyt.—
Berlin, 1989 — P. 234-261.
Krumbiegel A. Growth form of annual vascular plants in central Europe // Nord. Journ.
Bot — 1998 — V. 18, N 5,— P. 563-575.
Krumbiegel A. Growth form of biennial and pluriennial vascular plants in central Eu-
rope // Nord. Journ. Bot.— 1999.— V. 19, N 2 — P. 217-226.
Kuijt J. The biology of parasitic flowering plants.— Berkley-Los Angeles, Univ. Cali-
fornia Press, 1969.— 246 p.
250
Kull U. Neuere Aspekte der Pflanzenmorphologie // Jahresh. Ges. Naturk. Wurttem-
berg.— 1987.— 142 —S. 5-45.
Kumazawa M. The sinker of Platanthera and Perularia, its morphology and develop-
ment // Phytomorphology.— 1958.— V. 8, N 1-2.— P. 137-145.
Kunz M. Dinamical modes of phyllotaxis// Physica. D.— 2001.— V. 157, N 1-2.—
P. 147-165.
Liittge U. Vascular epiphytes: Setting the scene // Vase. Plants epiphyt.— Berlin,
1989.—P. 1-14.
LannerR. M., Connor К F. Does bristlecone pine senesce? // Exper. Geront.— 2001.—
V. 36 — P. 675-685.
Li P., Johnston M. O. Heterochrony in plant evolutionary studies through the twentieth
century 11 Bot. Rev. — 2000.— V. 66, № 1.— P. 58-88.
Lloyd F. E. The carnivorous plants.— Waltham, Mass., 1942.— 270 p.
Lynch M., Blanchard J., Houle D„ Kibota T„ SchultzS., Vassilieva L., Willis J. Perspective:
spontaneous deleterious mutation // Evol.— 1999.— V. 53, N. 3.— P. 645-663.
Martins K. F. P. Historia naturalis Palmarum. 3 Vols.— Lipsiae, 1823-1850.— Vol. I,
198 p„ Vol. II, III, 350 p.
Mayer A. M„ Poljakoff-Mayber A. The germination of seeds.— London: Pergamon
Press, 1975.— 192 p.
McClure F. A. The bamboos: a fresh perspective.— Cambridge: Harward Univ. Press,
1966 — 347 p.
McKinney M. I,. Heterochrony: Beyond words // Paleobiol.— 1999.— V. 25.—
P. 149-153.
Metcalf J. C., Rose К E., Rees M. Evolutionary demography of monocarpic perennials //
Trends Ecol. Evol.— 2003.— V. 18, N 9.— P. 471-480.
Metcalfe C. R. The morphology and mode of development of axillary tubercules and
root tubers of Ranunculus ficaria 11 Ann. Bot.— 1938.— V. 2, N 5.— P. 145-157.
Mirbel C. F. Elemens de physiologic vegetale et de botanique. 3 Vols.— Paris, 1815.—
924 p.
Mitteldorf J. Ageing selected for its own sake // Evol. Ecol. Res.— 2004.— V. 6.—
P. 1-17.
Molisch H. The longevity of plants.— Sci. Press, Lancaster, Pa., 1938.— 226 p.
Muller С. H. The significance of vegetative reproduction in Quercus 11 Madrono-
1951— V. 11, N 1,— P. 120-137.
Negbi M. Theophrastus on geophytes // Bot. J. Linn. Soc.— 1989.— V. 100, N 1.—
P. 15-43.
Nooden L. D., Penney J. P. Correlative controls of senescence and plant death in Ara-
Lidopsis thaliana (Brassicaceae) // Journ. Exper. Bot.— 2001.— V. 52, N 364.—
P. 2151-2159.
Obeso ]. R. The costs of reproduction in plants// New Phytol.— 2002.— V. 155.—
P. 321-348.
Obomy B. External and internal control on plant development // Bull. SFI — 2003.—
V. 18, N 1,— P. 1-11.
Ojai a A. Eliukiertojen evoluutiosta monivuotisilla kasveilla 11 Luonuon tutkija.—
1988,— V. 92, N 1,— P. 4-9.
251
Oldeman R. A. A. L’architecture de la foret guyanaise // Mem. O. R. S. T. О. M.—
1974 — N 73 — P. 1-204.
Drive M. E. Somatic mutations in organisms with complex life histories П Teor. Pop.
Biol.— 2001.— V. 59, Iss. 3.— P. 235-249.
Orkwiszewski J. A. J., Poethig R. S. Phase identity of the maize leaf is determined after
leaf initiation // Proc. Nat. Acad. Sci. USA.— 2000.— V. 97. Iss. 19.— P. 10631-
10636.
Peterson R. L. The initiation and development of root buds // The development and
function of roots.— Acad. Press, London, 1975.— P. 125-161.
Philbrick C. Th., Novelo A. R. Monograph of Podostemum (Podostemaceae) П System-
atic Botany Monographs, vol. 70.— The American Society of Plants Taxonomists,
2004 — 106 p.
Plantefol L. Fondements d’une theorie phyllotaxique nouvelle: la theorie des helices
multiples // Ann. Sci. nat. Bot.— 1946.— Ser. 11, N 7 — p. 153-229; 1947.—
Ser. 11, N 8.— P. 1-66.
Plants roots: the hidden half.— N. Y„ Basel, Hong Kong: Marsel Denner, Inc., 1991.—
948 p.
Poethig R. S. Phase change and the regulation of shoot morphogenesis in plants // Sci-
ence-1990—V. 250,— P. 923-930.
Prevost M. F. Architecture del quelques Apocynacees ligneuses // Mem. Soc. Bot. Fr.—
1967,— V. 114 — P. 24-36.
Prevost M. F. Modular construction and its distribution in tropical woody plants //
Tropical trees as living systems.— Cambridge Univ. Press, 1978.— P. 223-231.
Rauh W. Uber polsterformigen Wuchs. Ein Beitrag zur Kenntnis der Wuchsformen der
hoheren Pflanzen.— Halle (Saale), 1939.— 244 s.; (Nova acta Leopoldina. N. F.—
Bd. 7, N 49.— S. 267-508.
Rauh W. Die Wuchsformen der Polsterpflanzen. Zweiter Tell // Bot. Archiv, Leip-
zig.— 1940,— Bd. 40, Heft 3,— S. 375-462.
Raunkiaer C. The life form of plants and statistical plant geography.— Oxford,
Clatrendon, 1934.— 632 p.
Rawitscher F. Der Geotropismus der Pflanzen.— Jena, Verlag von Gustav Fischer,
1932,— 420 s.
Roach D. A. Environmental effects on age-dependent mortality: a test with a perennial
plant species under natural and protected conditions 11 Exper. Geront.— 2001.—
V. 36,— P. 687-694.
Rossato L„ MacDuff J. H., Laine P., le Deunff E., Ourry A. Nitrogen storage and
remobuization in Brassica napus L. during the growth cycle: effects of methyl
jasmonate on nitrate uptake, senescence, growth and VSP accumulation // Journ.
Exper. Bot.— 2002.—V. 53, N 371 — P. 1131-1141.
Rutishauser R. The development morphology of Indotristicha ramosissima (Podoste-
maceae, Tristichoided) // Pl. Syst. Evol.— 1991.— V. 178.— P. 195-223.
Rutishauser R., Sattler R. Complementarity and heuristis value of contrasting models in
structural botany. I. General considerations // Bot. Jahrb. Syst.— 1985.— V. 107.—
P. 415-455.
252
Sattler R. A new conception of the shoot of higher plants 11 J. Theor. Biol.— 1974. —
V. 47, N 2,— P. 367-382.
Schmid E. Die Erfassung der Vegetationseinhelt mit floristischen und epimorpho
logischen Analysen 11 Berichte der Deut. Bot. Gessellsh.— 1963.— Bd. 73.—
S. 276-324.
Schwabe W. W. Phyllotaxis // Positional controls in plant development.— Cambridge:
Cambridge Univ. Press, 1984.— P. 403-440.
Sculthorpe C. D. The biology of aquatic vascular plants.— London: Edward Arnold
(Publ.) Ltd., 1967,—610 p.
Shimizu-Sato Mori H. Control of outgrowth and dormancy in axillary buds // Pl.
Physiol —2001. V. 127 —P. 1405-1413.
Silverstown ]., Franco M„ Perez-Ishiwara R. Evolution of senescence in iteroparous pe-
rennial plants П Evol. Ecol. Res.— 2001.— V. 3.— P. 393-412.
Skene K. R. Cluster roots: some ecological considerations // Journ. Ecol.— 1998.—
V. 86,— P. 1060-1064.
Skene K. R. Pattern formation in cluster roots: some developmental and evolutionary
considerations // Ann. Bot.— 2000.— V. 85.— P. 901-908.
Smith J. A. C. Epiphytic Bromeliads // Vase. Plants Epiphyt.— Berlin, 1989.—
P. 109-138.
Spath H. Der Johannistrieb.— Berlin, 1912.— 91 s.
Steams S. C., Ackermann M., Doebeli M., Kaiser M. Experimental evolution of aging,
growth and reproduction in fruitflies 11 PNAS — 2000.— V. 97, № 7.— P. 3309-
3313.
Stevens O. A. Rhizomes, stolons and roots // Castanea.— 1966.— V. 31, N 1.—
P. 140-145.
TelferA., Poethig R. S. HASTY: a gene that regulates the timing of shoot maturation in
Arabidopsis II Development — 1998.— V. 125.— P. 1889-1898.
Thomas H. Ageing in plants // Meehan. Ageing Devel.— 2002.— V. 123.— P. 747-753.
Thomas H., Ougham H. J., Wagstaff C., Stead A. D. Defining senescence and death //
Journ. Exper. Bot.— 2003.— V. 54, N. 385,— P. 1127-1132.
Thomas H., Thomas H. M., Ougham H. Annuality, perenniality and cell death // Journ.
Exper. Bot — 2000.—V. 51, N. 352,— P. 1781-1788.
Thompson A. W. On growth and form.— N. Y.: Cambridge Univ. Press.— 1948.—
1116 р.
Tillich Hans-Jiirgen. Seedlings and systematics in Monocotyledons 11 Monocotyledons:
systematics and evolution.— Kew: Royal. Bot. Gard,— 1995.— P. 303-352.
Troll W. Die Infloreszenzen. Bd. 1.— Jena, 1964.— 615 s.
Troll W. Vergleichende Morphologic der hoheren Pflanzen.— Berlin: Springer,
1935,—235 s.
Troll W. Vergleichende Morphologic der hoheren Pflanzen. Bd. Vegetationsorgane.—
Berlin: Verlag von Gebriider Borntraeger, 1939.— S. 957-2005.
Tsukaya H. Determination of the unequal fate of cotyledons of a one-leaf plant,
Monophyllaea// Develop.— 1987.— V. 124.— P. 1275-1280.
Vaczy C. Dictionar botanic poliglot (latina, romana, engleza, germana, franceza,
maghiara, rusa).— Bucuresti, 1980.— 1017 p.
253
Vaupel J W., Baudisch A., Dolling M., Roach D. A., Gampe J. The case for negative se-
nescence.— [Electronic resource].— 2004.— Mode of access: http: www.demogr.
mpg.de.
Velenovsky J. Uber die Biologie und Morphologic der Gattung Moneses // Rozpravy
Trida, 1892.— 11, Cislo. 39 — S. 147-159.
Velenovsky J. Uber die Keimpflanzen der Pirolaceen // Bull. Internal. Acad. Sci.
Bohem— 1905 — B. 7,— S. 17-21.
Wardlaw C. W. Tensile stress as a factor in fern phyllotaxis // Nature.— 1947.—
V. 160, N 4072 — P. 678.
Warming E. Uber perenne Gewachse // Bot. Centralblatt.— 1884.— B. 18, N 19.—
S. 16-22.
Wanning E. Uber Sprossbau. Uberwinterung, Verjungung 11 Engler’s bot. Jahrb., 1883.
White J. Plant metamerism // Perspectives on plant population ecology.— Sunderland,
Massachusetts, 1984.— P. 15-47.
Williams G. C. Pleiotropy, natural selection and evolution of senescence // Evol.—
1957 —V. 11,—P. 398-411.
Wilson B. F. Apical control of branch growth and angle in woody plants // Amer. Journ.
Bot — 2000.—V. 87, N 5.— P. 601-607.
Winn A. A. The functional significance and fitness consequences of heterophylly // Int.
Journ. Pl. Sci.— 1999,— V. 160.— P. 113-121.
Ibnng T. Evolution of semelparity in Mount Kenya Lobelias // Evol. Ecol.— 1990.—
V.4.N2 —P. 157-171.
Zelawski W. Starzenia organizmu roslinnego. Cz. 11. Zagadnienia szczegolowe // Wiad.
bot — 1985,— V. 29, N 3,— P. 203-212.
Zimmerman W. Die Phylogenie der Pflanzen, 2nd ed.— Stuttgart: Verlag Gustav
Fischer, 1959.
Жмылев П. Ю_, Алексеев Ю. Е.,
Карпухина Е. А., Баландин С. А.
БИОМОРФОЛОГИЯ РАСТЕНИЙ
ИЛЛЮСТРИРОВАННЫЙ СЛОВАРЬ
Учебное пособие
Подписано в печать 12.12.2005 г.
Формат 60x84/16. Печ. л. 16. Печать офсетная.
Бумага офсетная. Тираж 800 экз. Заказ № 558.
ИПП «Гриф и К», г. Тула, ул. Октябрьская, 81-а.