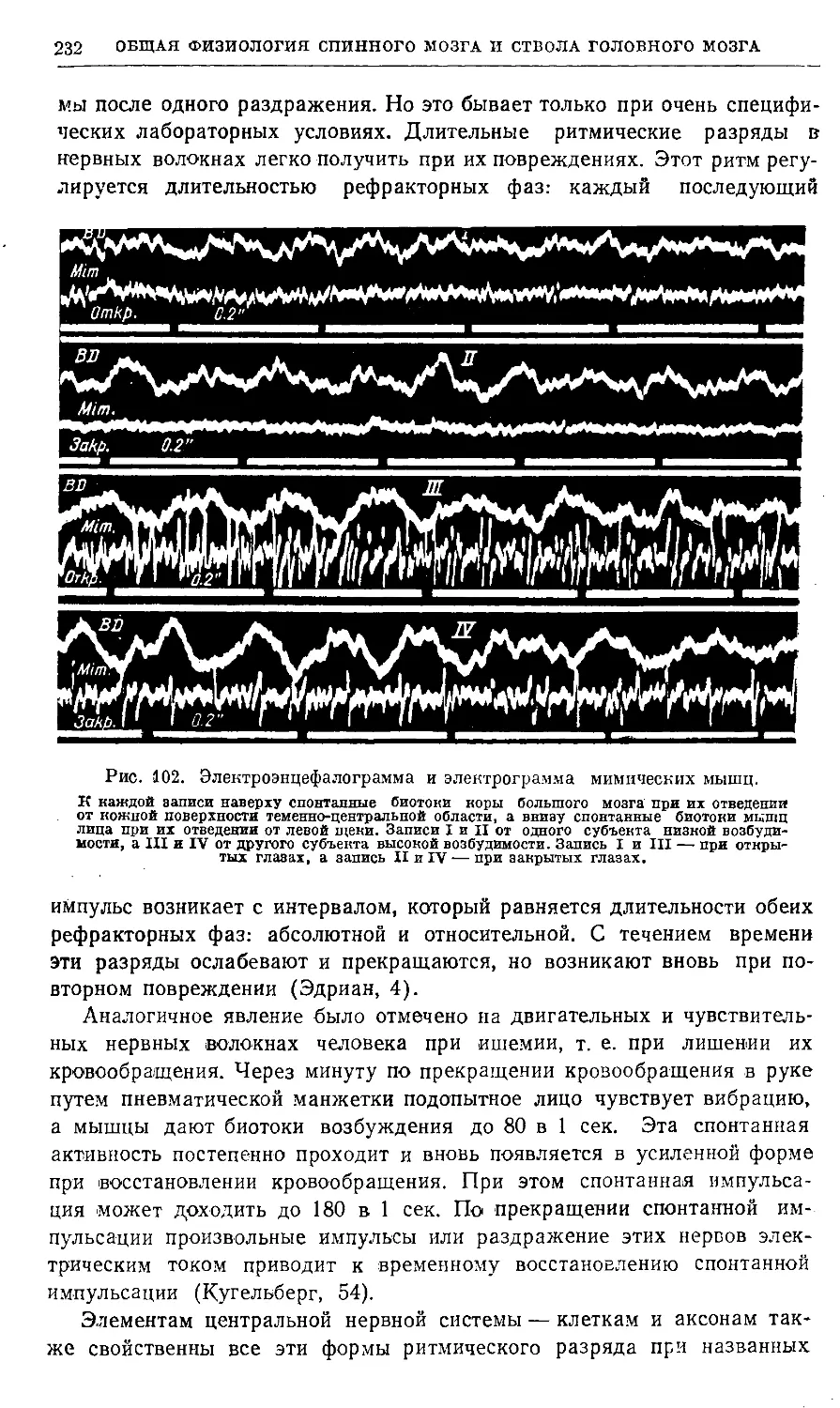
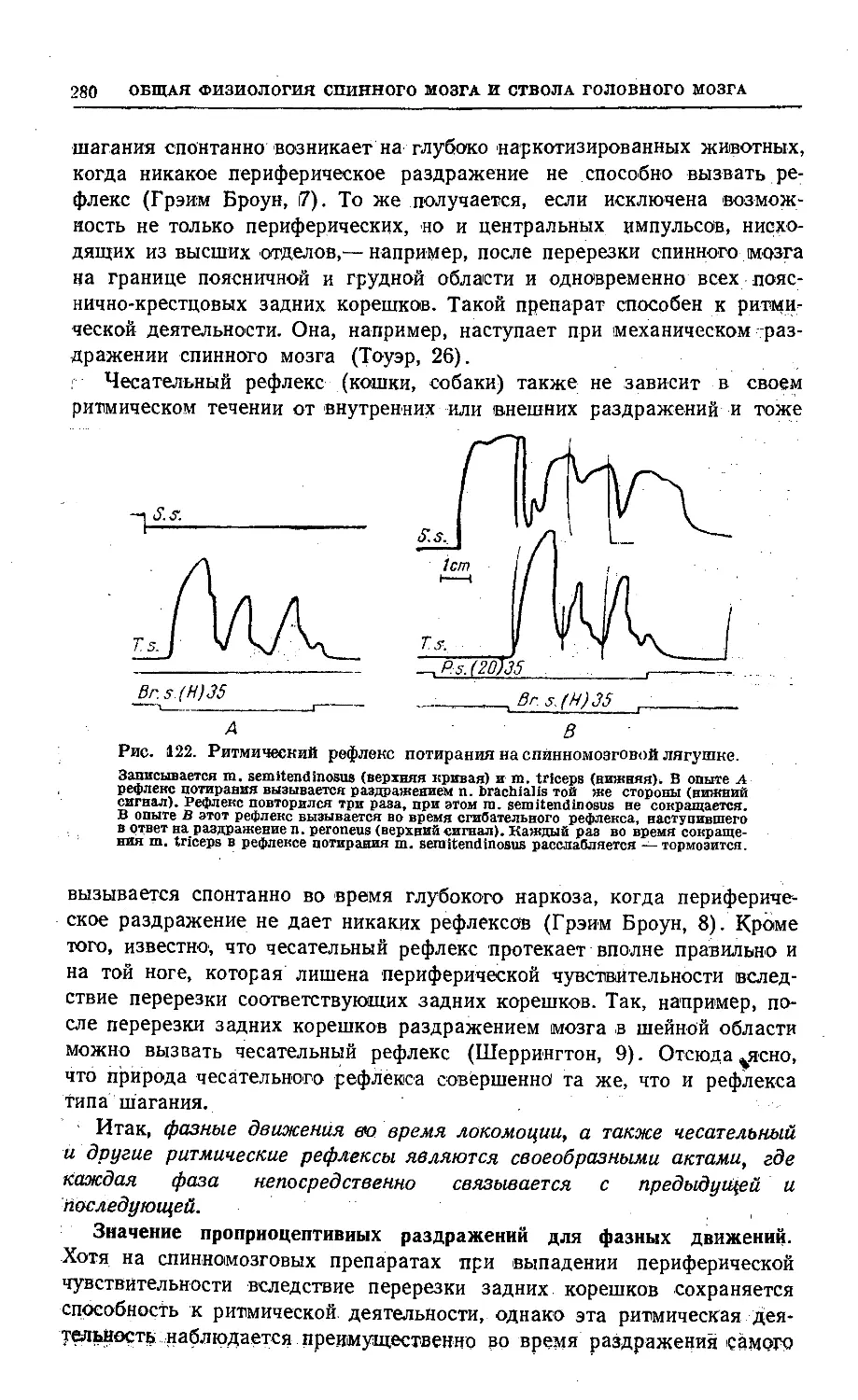
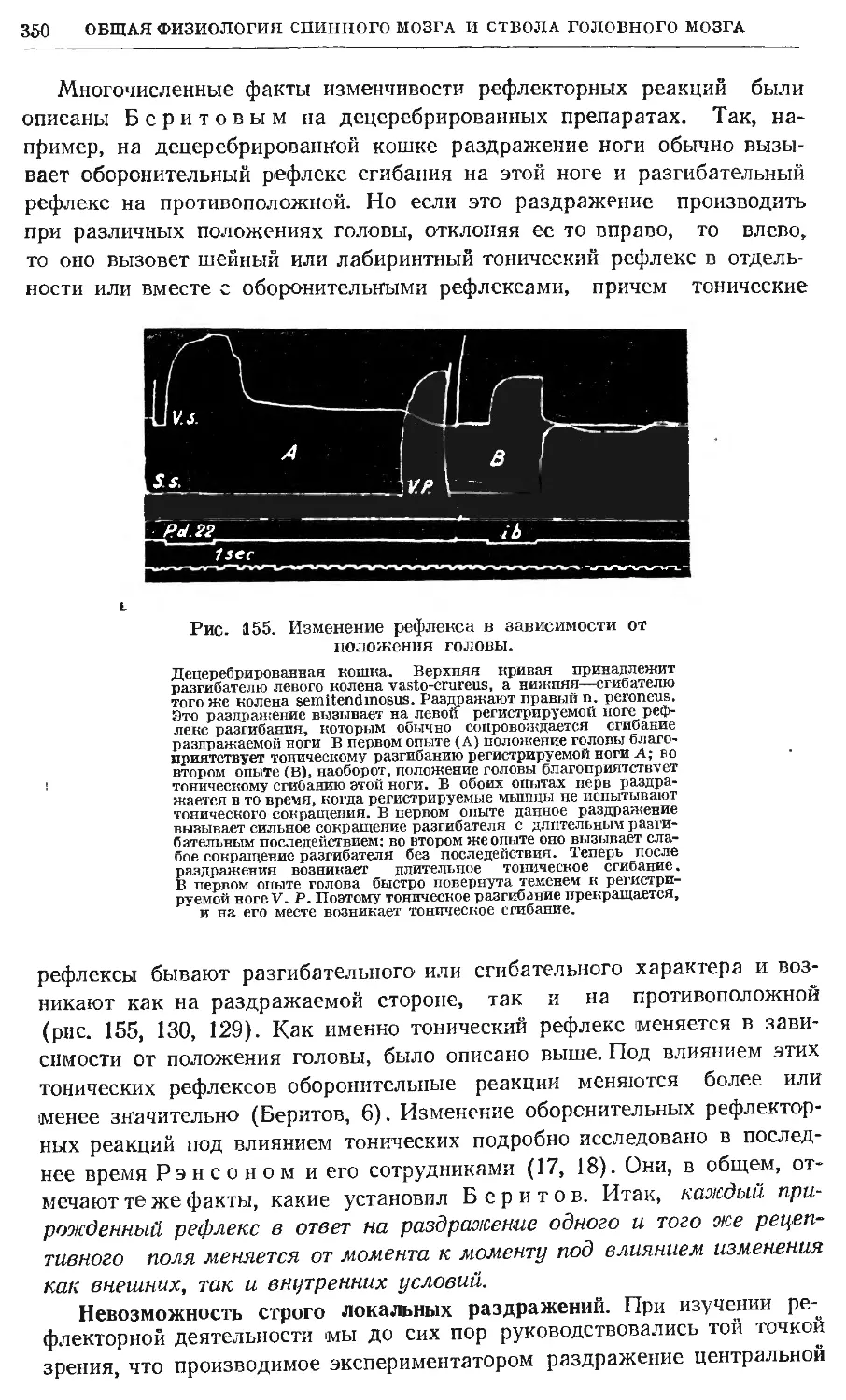
/
























































































































































































































































































































































































































































































































































































































































Автор: Беритов И.С.
Теги: медицина физиология практическая медицина физиология человека мышцы издательство академия наук ссср
Год: 1948




Текст
п
ч
АКАДЕМИЯ НАУК СОЮЗА CG
Академик И. С. БЕРЕТОВ /^
ОБЩАЯ ФИЗИОЛОГИЯ
МЫШЕЧНОЙ И НЕРВНОЙ
СИСТЕМ
ТЕОРЕТИЧЕСКОЕ И ПРАКТИЧЕСКОЕ
РУКОВОДСТВО
ТОМ II
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
Второе, переработанное издание
'Аі
! .
. *
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
Москва 1948 Ленинград
Печатается по постановлению
Редапцибнпо-издательского совета
Академии Наук СССР
редактор издательства Ю. Д. Смирнов
Технический редактор Е. Я. Симкина
РИСО АН СССР М 2664. Издат. № 1003а
Тип. заказ № 3486
Под. к печ. 16/Ш 1948 г. Формат бум. 70X108' lf
Печ. л. 40. Уч.-издат. 52. Тираж 5000*
Цена в переплете 45 руб.
2-я тип. Издательства Академии Наук СССР
Москва, Шубинский пер., д. 1Q
ОГЛАВЛЕНИЕ
Часть пятая
ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА
ГОЛОВНОГО МОЗГА
Глава I. Общее понятие о строении центральной нервной системы позвоночных
животных 9
Глава II. Общее понятие о центральной нервной деятельности 26
Практикум 38
Глава III. Рецептивное поле рефлекса и его значение 41
Практикум 51
Глава IV. Анатомическая локализация координирующих аппаратов 53
Практикум 64
Глава V. Иррадиация рефлекторной деятельности и ее происхождение , . . t. 67
Практикум 80
Глава VI. Основные процессы центральной координации 82
Практикум 104
Глава VII. О природе центрального торможения 108
Практикум ... 151
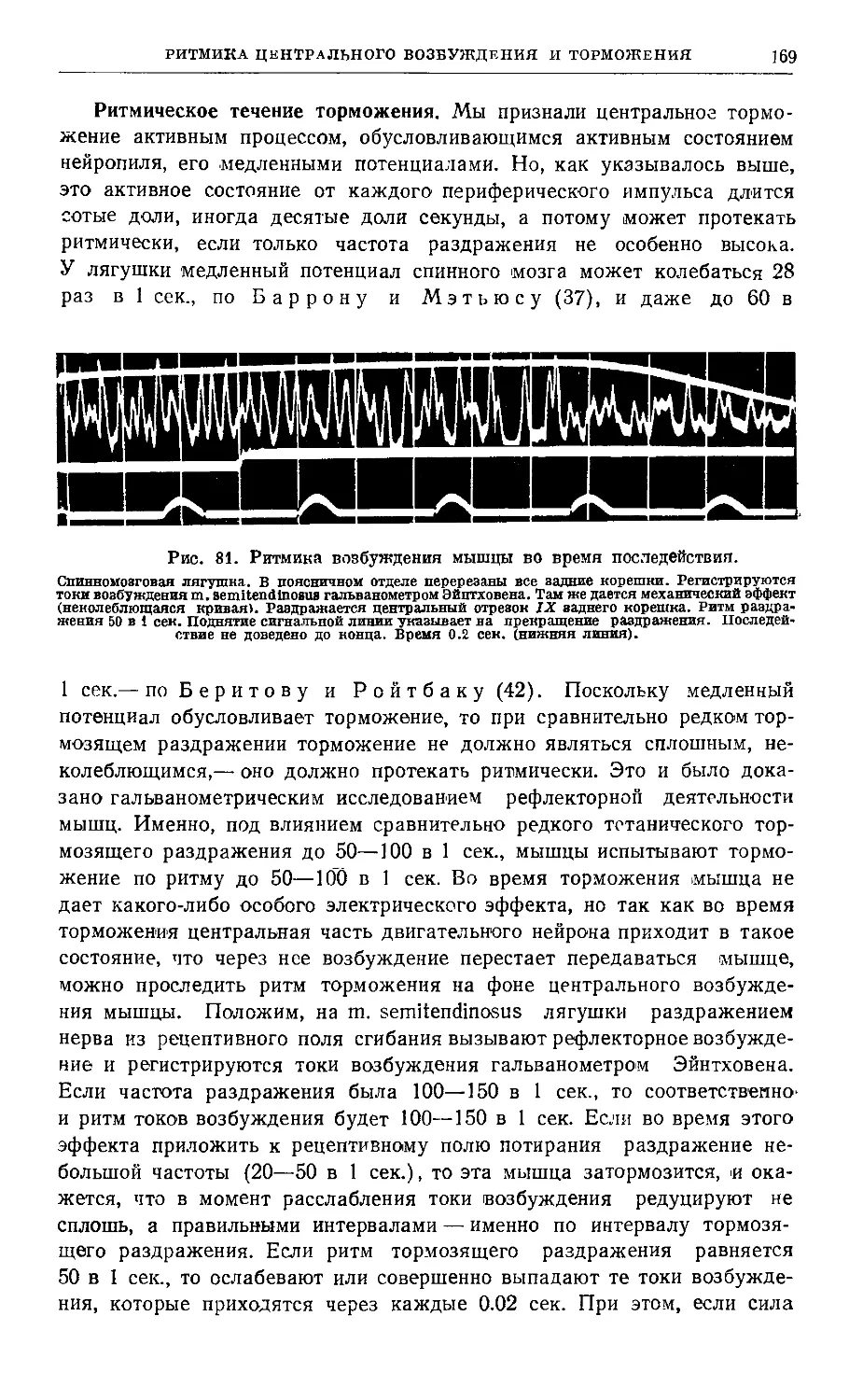
Глава VIII. Ритмика центрального возбуждения и торможения ....... 355
Практикум 3 72
Глава IX. Распространение возбуждения и торможения в нервной системе . . 175
Практикум 190
Глава X. Явление облегчения в центральной нервной системе 192
Практикум 214
Глава XI. Утомление 217
Практикум • 229
Глава XII. Спонтанная ритмическая деятельность центральной нервной
системы 231
Практикум .... • 249
Глава XIII. Шок центральной нервной системы 252
Практикум 259
Глава XIV. Проприоцептивные рефлексы 261
Практикум 276
Глава XV. Ритмические рефлексы 278
Практикум 295
Глава XVI. Тонические рефлексы 301
Практикум 321
Глава XVII. Рефлексы положения и статокинетические рефлексы 325
Практикум 331
Глава XVIII. Локомоция 333
Практикум 341
Глава XIX. Ориентировочные рефлексы 344
Практикум 347
Глава XX. Динамика рефлекторной деятельности 348
Практикум 365
Глава XXI. Координация рефлекторной деятельности у беспозвоночных животных 368
Практикум 387
Часть шестая
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
Глава I. Общие сведения о коре больших полушарий 393
Глава II. Воспринимающая функция коры большого мозга 408
Тлава III. Ответная функция коры большого мозга 423
Практикум 458
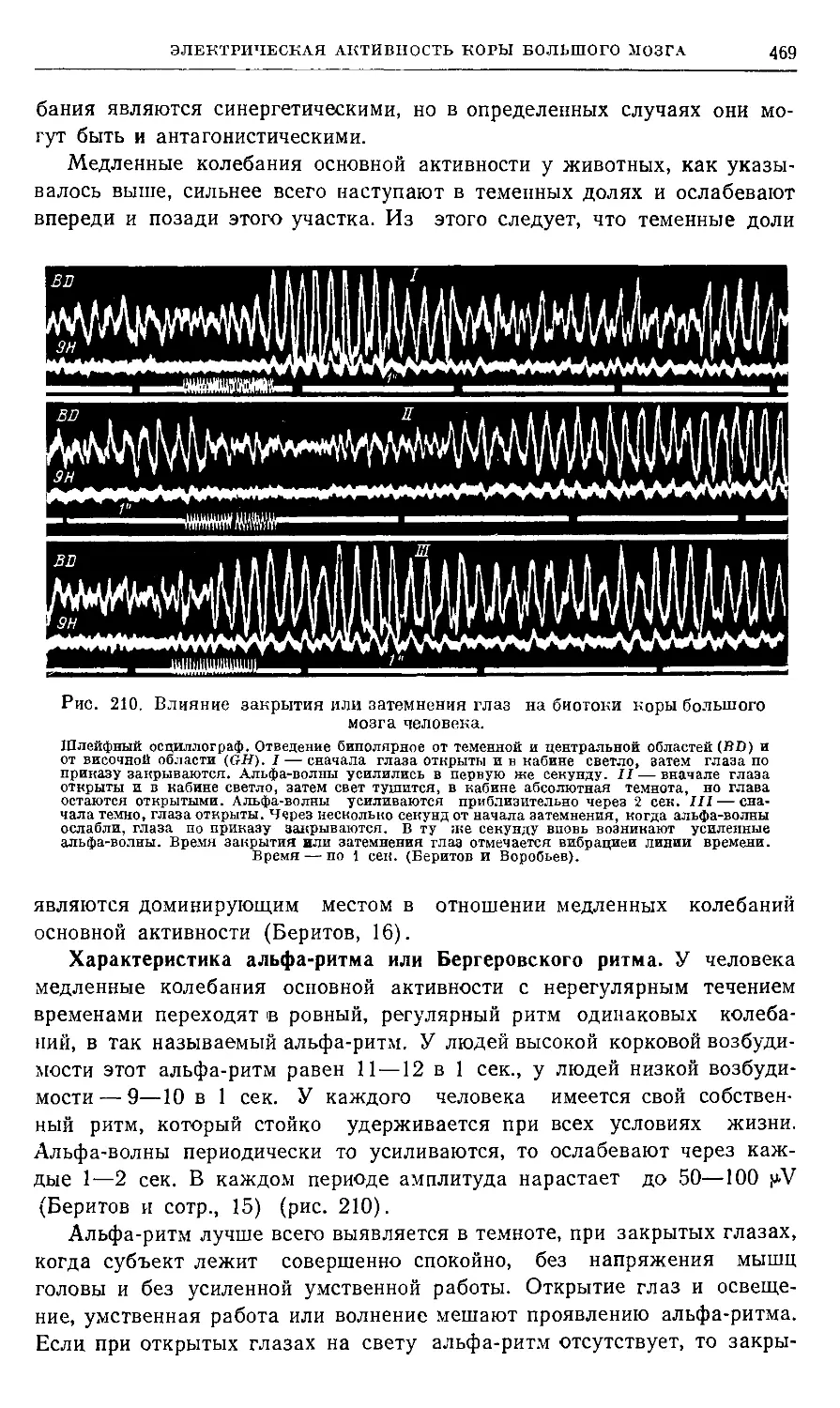
'Глава IV- Электрическая активность коры большого мозга 463
П рактикум 493
Глава V. Образование индивидуально приобретенных рефлексов 499
Глава VI. Типы индивидуальных рефлексов 518
Практикум 528
Глава VII. Закон сопряженной иррадиации возбуждения 536
Глава VIII. Происхождение индивидуальных рефлексов 547
Глава IX. Основные анатомические элементы индивидуальных рефлексов . . 578
Глава X. Угасание и восстановление индивидуальных рефлексов 588
Глава XI. Изменчивость индивидуальных рефлексов под влиянием внешних
раздражений 599
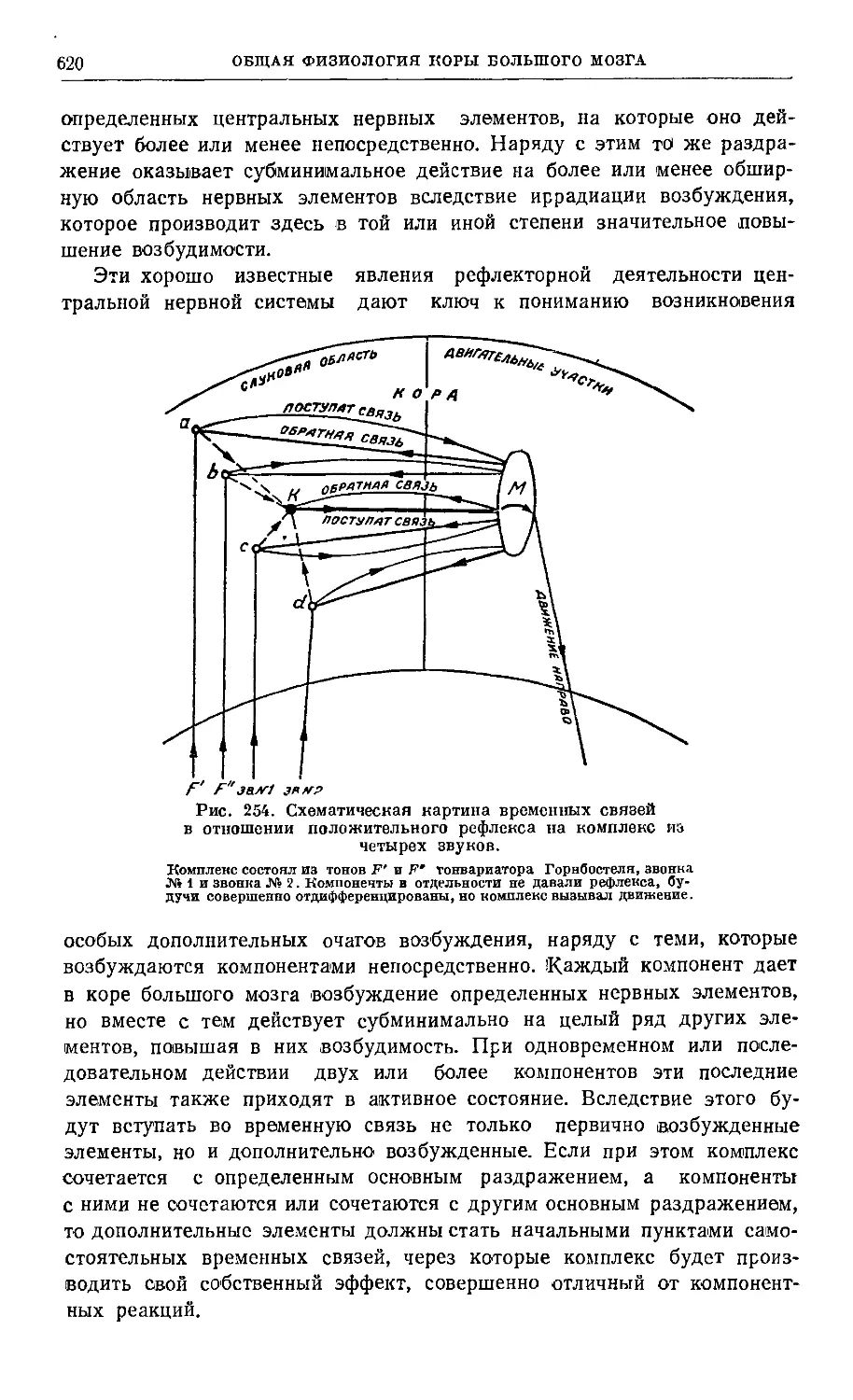
Глава XII. Объединяющая (интегральная) деятельность коры большого мозга 613
Практикум 625
Именной указатель 628
Предметный указатель 636
Часть пятая
ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО
МОЗГА
И СТВОЛА ГОЛОВНОГО ДОЗГА
Глава 1
ОБЩЕЕ ПОНЯТИЕ О СТРОЕНИИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ
СИСТЕМЫ ПОЗВОНОЧНЫХ ЖИВОТНЫХ
Общий тип строения центральной нервной системы. Центральная
нервная система представляет собой очень сложную систему клеточных
скоплений-центров, или нервных ядер, соединенных между собой
пучками нервных волокон, нервными путями. Каждый рецептор связан с
эффекторами — рабочими органами, во-первых, коротким путем через
тот отдел мозга, куда вступает чувствительное волокно от данного
рецептора, и, во-вторых, более длинным путем — через другие отделы
мозга. Предполагается, что координированные реакции организма
всецело обусловливаются деятельностью нервных центров, но в настоящее
время многие выдающиеся гистологи и неврологи не разделяют этого
представления. По их мнению, дифференцированные и
локализированные реакции обусловливаются, известными нервными центрами и
нервными путями. Общие же .реакции, а также объединение локальных
реакций разных органов в одно целое осуществляются, согласно этой новой
концепции, через нейропиль.
Согласно фактическим данным и представлениям Джадсона
Херрика (1), нейропиль является базой развития всех новых
нервных функций в животном царстве. Поэтому в ходе филогенетического
развития животного мира роль и значение нейропиля в
жизнедеятельности организма не ослабевают, а, наоборот, возрастают. Правда,
одновременно развиваются и дифференцированные нервные центры и пути,
но эти элементы не должны больше играть роли в создании новых форм
функциональной деятельности, новых форм поведения. Эта роль всецело
принадлежит нейропилю.
По новейшим исследованиям Лоренте де Но (10), внутри
каждого нервного ядра позвоночных нервные клетки связаны между собой.
Вследствие этого в каждой паре клеток имеется обоюдосторонняя связь:
аксон одной клетки синаптически связан с телом другой клетки. Точно
такая же обоюдосторонняя связь существует как между двумя
соседними, так и между двумя дальними ядрами, а также между ядрами и
нейропилем. При этом каждая группа клеток или каждая клетка
10 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
юбоюдосторонне связана с другими клетками нервными путями разной
сложности, т. е. с включением разного количества нейронов.
Ло ренте де Но (И) установил два типа нервных связей, или
два закона нервных отношений: тип множественной цепочки, по
которому связываются два конечных нейрона через различное число
промежуточных нейронов, и тип замкнутой цепочки, или нейронного круга,
тле промежуточные или ассоциативные нейроны связаны друг с другом,
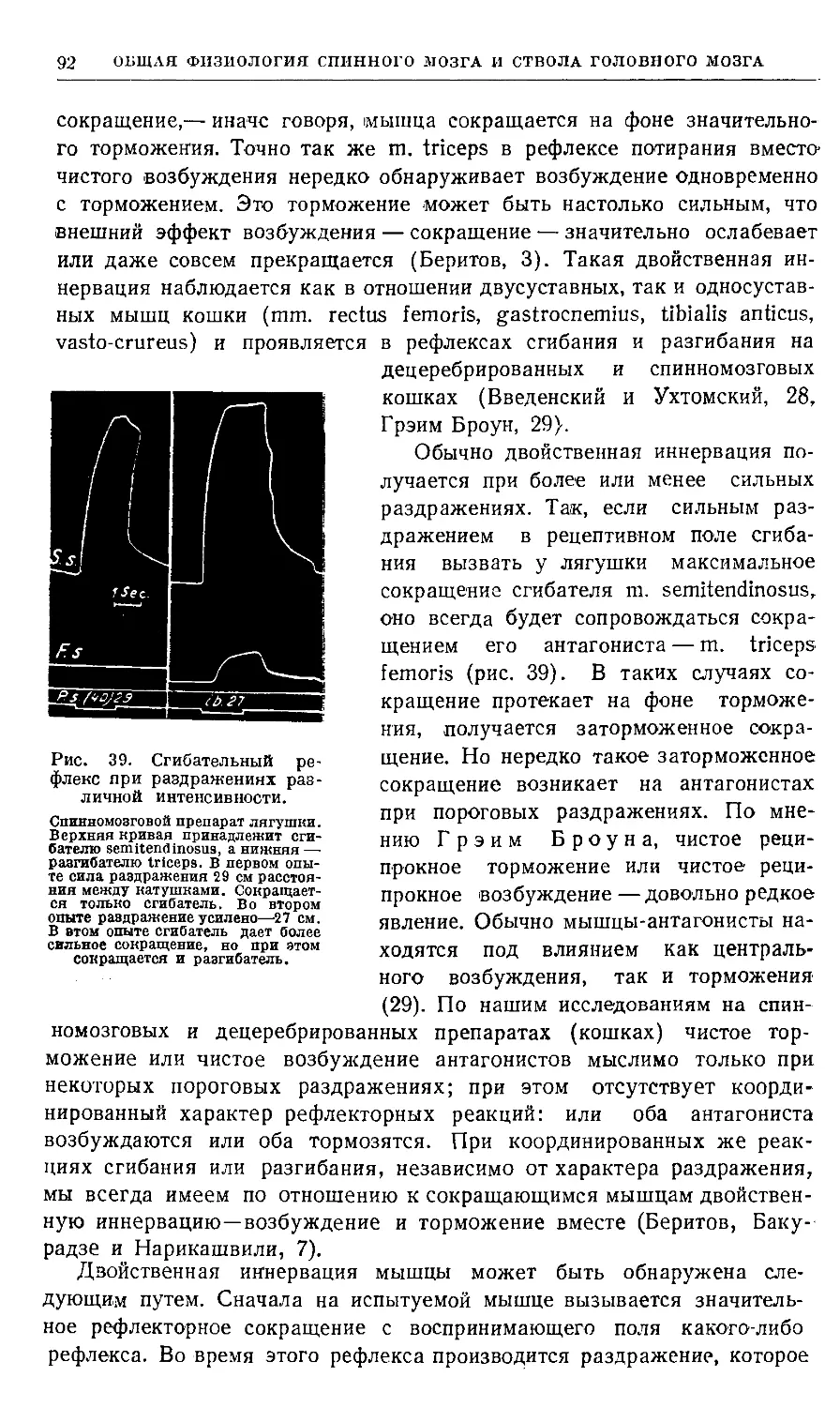
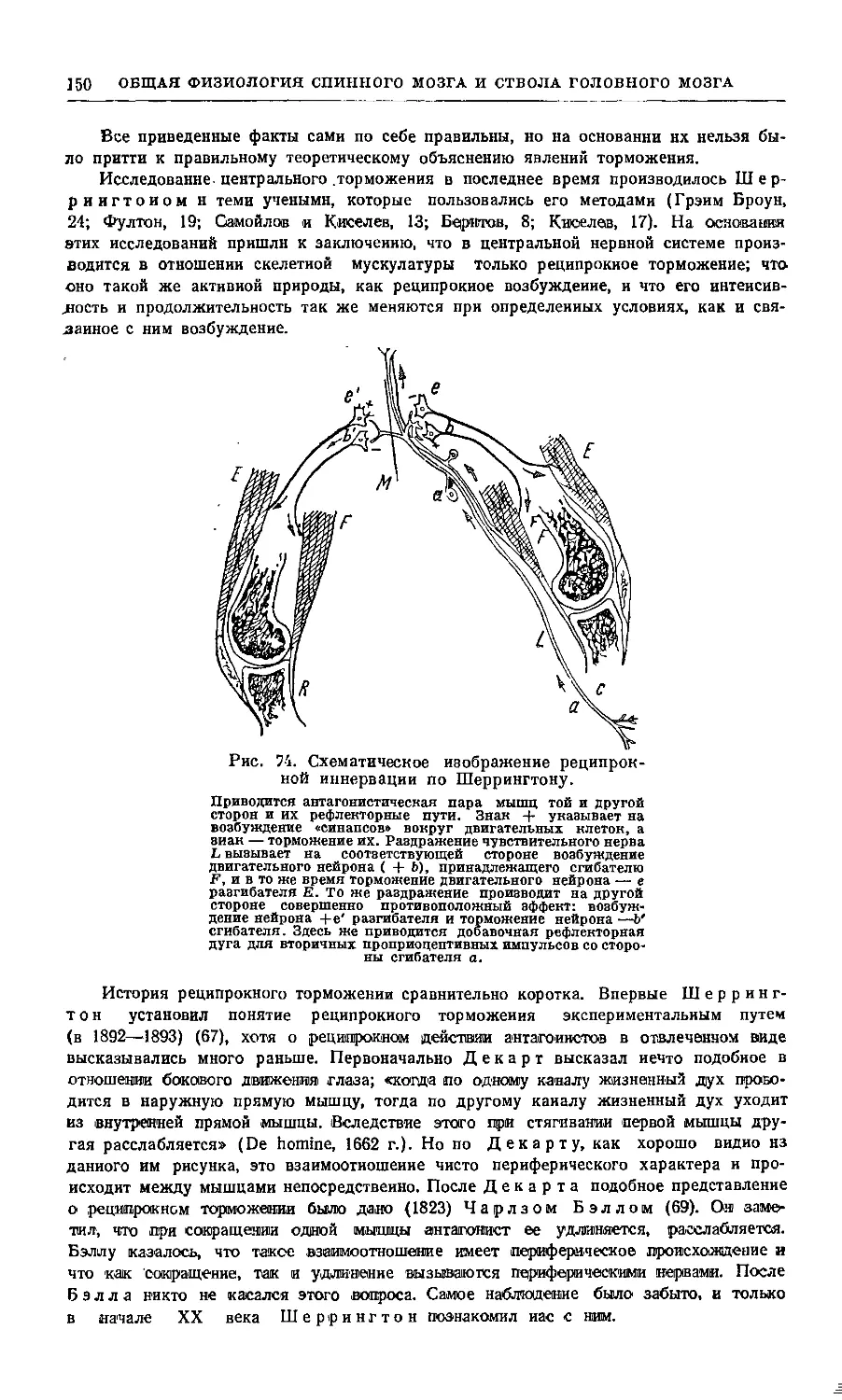
образуя замкнутые круги (рис. 1).
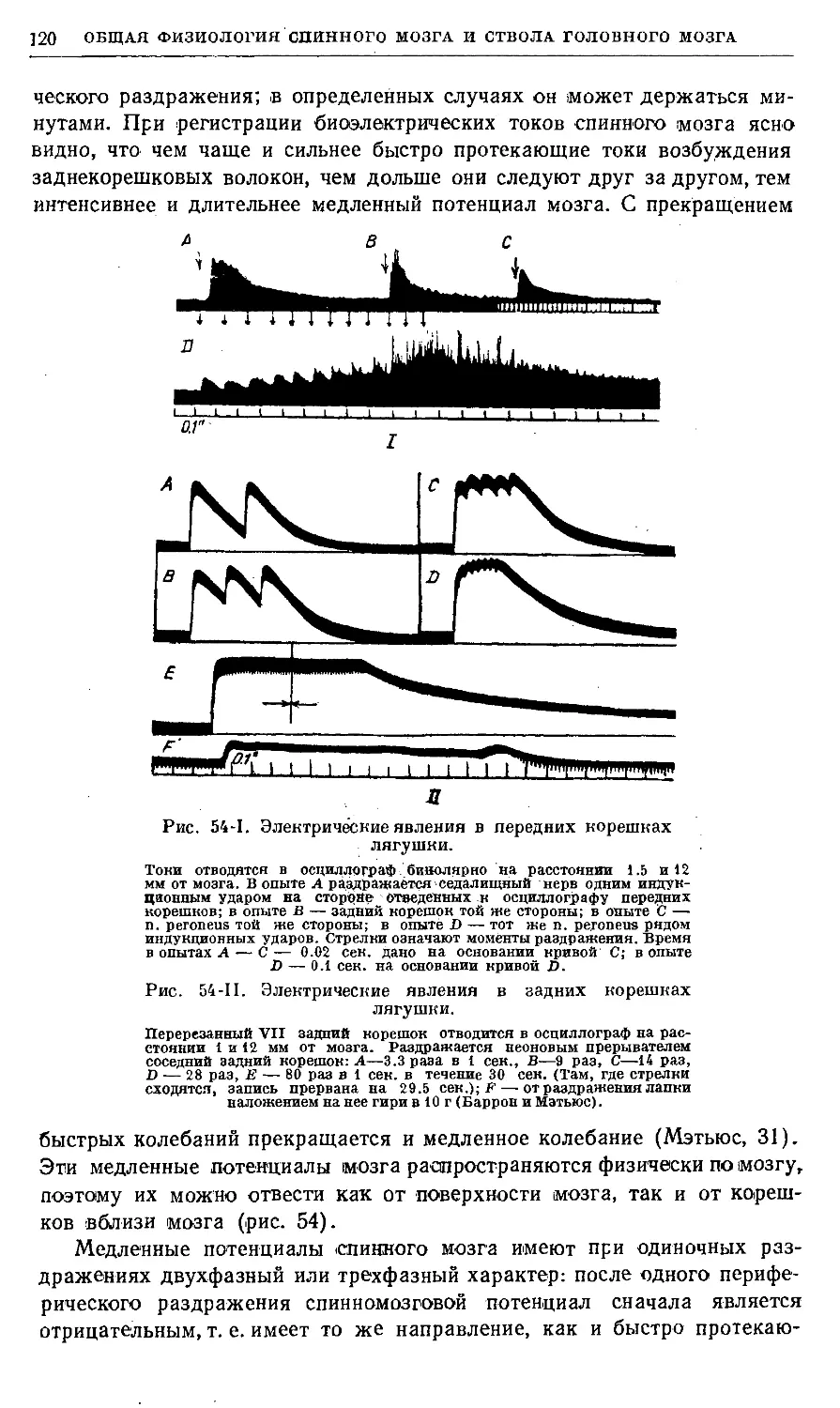
Рис. 1. Схематический рисунок нервных связей.
X — тип множественной цепочки, где афферентный нейрон а/ связан
с эфферентным е как непосредственно, так и через один, два и три
промежуточных нейрона: гі, г2, гЗ; В — тип замкнутой цепочки или
нервного круга, где афферентный нейрон связан с эфферентным
нейроном через промежуточные круги разной сложности: гі—гЗ—гі,
ti—г2—гі, гі—гЗ—г2—г!;С—тип множественной цепочки, где
афферентный нейрон а/ связан с двигательной клеткой М. При этом аксоны
афферентного нейрона и промежуточных нейронов оканчиваются на
двигательной клетке во многих точках как конечными, так и боковыми
синапсами (по Лоренте де Но).
Строение серого вещества спинного мозга. В сером веществе
спинного мозга центральные нервные элементы образуют множество
объединений. Каждое такое объединение имеет определенное
функциональное назначение, а потому их можно назвать физиологическими
единицами центральной нервной системы. В каждой
такой единице нершше клетки связаны между собой по типу как
множественной, так и замкнутой цепочки. В каждой множественной
цепочке возбуждение идет только в одном направлении — от афферентного
чувствительного нейрона к двигательному. В замкнутой же нервной
цепочке возбуждение может распространяться в обоих направлениях:
как от данной клетки по ее аксону к другой клетке, так и наоборот —
по аксону последней клетки к первой. Но, конечно, в каждом элементе
замкнутой цепочки, благодаря синаптическим связям, возбуждение
распространяется только в одном направлении. Каждый такой
объединенный нервный комплекс снабжен своей системой афферентных волокон,
с помощью которых он приходит в действие, и системой эфферентных
О СТРОЕНИИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ ПОЗВОНОЧНЫХ ЖИВОТНЫХ И
волокон, через которые разряды импульсов передаются из этого
комплекса другим аналогичным комплексам или же на периферию, ра(х>
^им органам.
В сером веществе спинного мозга наряду с нервными цепочками
существует нейропиль. Он был описан еще Ретциусом (2) (рис. 2>
как широкопетлистое нейрофиб-
риллярное образование,
заполняющее все свободное от клеток
пространство серого вещества
мозга. Отдельные фибрилли ней-
ропиля местами располагаются
правильными пучками.
Последующие исследования
установили, что все нервные клетки, ден-
дриты и голые осевые цилиндры
вступают в многочисленные
связи с нейрофибриллями нейропиля
(Краузе, 31 и др.) (рис. 3).
Особенно тщательное
исследование нейропиля было
произведено в последнее время X е р р и-
ком (5) и Лоренте де Но
(11). Они находят, что в
центральной нервной системе, и в
частности в спинном мозгу, в
межклеточных пространствах
(кроме глиальных клеток,
кровеносных сосудов и миэлиновых
волокон) существуют два сплете-
яия: одно — из дендритных разветвлений, а другое — из оезмиэлиновых
волоконец. Оба эти сплетения образуют бесчисленные синаптические
связи. Каждый дендрит, пересекая сплетение волоконец, вступает в
связи с множеством волоконец разного происхождения, а каждое
волоконце, пересекая дендритное сплетение, оканчивается на многочисленных
дендритах разных клеток посредством фибриллярных утолщений —
синаптических луковиц. В переднем и боковом рогах дендрятное
сплетение образуется из дендритов двигательных и промежуточных клеток.
Следовательно, одно и то же нервное волоконце вступает в связь как с
клетками двигательных и промежуточных нейронов, так и с их дендри-
тами. Этот нейропиль существенно отличается от нейропиля низших
•беспозвоночных животных, где нервные отростки образуют синцитий, в
•котором отростки одной нервной клетки сливаются с отростками других
клеток (см. т. I, рис. 161, 162 и 163).
В спинном мозгу выделяется нейропиль substantia gelatinosa
Rolando. Эта часть заднего рога состоит из маленьких клеток с сильно
ветвящимися дендритами, которые образуют густое сплетение, а аксоны
Рис. 1. А — нейропиль в спинном
мозгу, образующий войлок между ганг-
лиозными клетками (Ретциус).
12 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
небольшой длины уходят в соседние столбы, откуда их коллатерали
поступают обратно в то же образование Сюда, в substantia gelatinosa,
вступают дендриты других клеточных элементов спинного мозга и
коллатерали заднекорешковых волокон. На примере substantia gelatinosa
ясно видно, что в образовании нейропиля принимают участие не только
дендриты и аксоны клеток, участвующих в проведении возбуждения от
чувствительных нейронов к двигательным, но и особого рода промежу-
Рис. 3. Передний рог в спинном мозгу кролика.
А — В и С — средняя группа клеток, невидимому,
промежуточных; D—медиальная группа двигагельных клеток
Е—вентральная группа двигательных клеток, F—коллатерали
вентральных столбов, 2, 3 и 4 — пучки двигательных нервных
волокон, 1 — возвратные коллатерали двигательных волокон;
К —дорзальная боковая группа двигательных клеток;
L—коллатерали боковых столбов, М — вентральная боковая группа
двигательных клеток Дендритные отростки всех клеток
образуют тончайшую протонлазматическую сеть, в которой
принимают участие и коллатерали нейритов (Краузе).
точные 'нейроны со всеми своими дендритаіми и аксонными разветвле-
ниши. Substantia gelatinosa тянется по всему спинному мозгу в
области задних рогов и заходит в продолговатый мозг (рис. 8). На рис. &
показано это нервное образование в продолговатом мозгу.
Нейрофибриллярные сплетения были найдены также на поверхности
спинного мозга Сюда входит, прежде всего, перимедуллярный нейро-
фибриллярный слой, который образуется, главным образом, из дендри-
тов двигательных и промежуточных клеток и коллатералей
заднекорешковых волоко'н (рис 4). Но здесь же под pia mater находятся
клеточные образования, дендриты которых тоже образуют густук) сеть.
С этим субпиальным нейропилем вступают в связь дендриты от двига-
О СТРОЕНИИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ ПОЗВОНОЧНЫХ ЖИВОТНЫХ J3
тельных и промежуточных нейронов и коллатерали заднекорешковых
волокон. Последние оканчиваются здесь синапсами (Сильвер, 33). Пе-
Рис. 4. Перимедуллярное сплетение спинного
мозга лягушки.
Оно занимает поверхностный слой мозга. Туда вступают
дендриты как от двигательных клеток И, так и от
промежуточных нейронов J; К—комиссуральные клетки, аксоны которых
переходят на другую половину мозга (Р. Кахаль).
римедуллярное сплетение и субпиальный клеточный слой сильнее всего
развиты у рыб и амфибий (Кахаль, 12, 13; Немилов, 4 и др.). У птиц
Рис. 5. Субпиальный нервный слой в спинном мозгу
котенка.
1 — субпиальные клетки; 2—сплетение, образованное дендри-
тами нервных клеток; 3 — начальный отдел дендритов.
Окрашено метиленовой синью (Немилов).
имеется только, остаток перимедуллярного сплетения (Кёлликер, 6 и др.),
У млекопитающих перимедуллярное сплетение имеется только в
14 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
эмбриональной стадии: дендриты из серого вещества достигают
поверхности мозга, но у взрослых они сильно ветвятся, идут в разных
направлениях, переходят на другую сторону через переднюю комиссу-
ру серого вещества, но на поверхности мозга не выходят (Кахаль, 12,
13). Субпиальный слой имеется и у млекопитающих в довольно
развитом виде (Немилов, 4) (рис. 5).
На рис. 6 дается схематическое изображение строения нервных
цепочек и нейропиля спинного мозга.
Рис. 6. Схема нейронно-нейропильного строения спинного мозга позвоночных.
PR — задний корешок; AR — передний корешок; А — клетки афферентных чувствительных
нейронов в спинномозговом ганглии; У — клетки интраспинальных промежуточных нейронов; М —
клетка двигательного нейрона; С — главная коллатераль афферентного нейрона, который
участвует в прямой передаче возбуждения на промежуточные и двигательные нейроны; і — аксон
промежуточного нейрона, который участвует в передаче импульсов на двигательную клетку; N —
нервная клетка, не участвующая в прямей передаче возбуждения от чувствительного нейрона
к двигательному; К — клетка комиссурального нейрона, участвующего в передаче возбуждения
к двигательному нейрону противоположной половины мозга; S — клетка субпиального слоя;
Р— перимедуллярное сплетение. Разветвления дендритов всех означенных клеток образуют ней-
ропиль, в котором принимают участие и нейритные веточки афферентных и промежуточных
нейронов; d — дендриты клеток серого вещества, разветвляющиеся в перимедуллярном слое; dc —
дендриты, переходящие из одной половины мозга в другую; а — аксон субпиальной клетки,
разветвляющийся в нейропиле серого вещества; k — аксон комиссуральной клетки; с — веточки
афферентного нейрона, разветвляющиеся в нейропиле серого вещества мозга; С— веточки того
же нейрона в перимедуллярном слое; с" — веточка афферентного нейрона, переходящая через
заднюю комиссуру на другую сторону; г — возвратные коллатерали двигательных нейронов.
На оисунке можно проследить множественную цепочку, например АСМ, АСігМ, а также замкнутую-
цепочку, или нервный круг, например NnliN (Рисунок составлен по данным Р. Кахаля, Лено-
шека, Догеля, Лоренте де Bot Немилова).
Строение продолговатого мозга. Из нервных образований
головного мозга нужно выделить давно известное сетевидное образование
(formatio reticularis), которое является мощным нейропильным
комплексом. Оно начинается еще в спинном мозгу, затем, расширяясь,
заполняет большую часть продолговатого мозга и постепенно исчезает в
О СТРОЕНИИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ ПОЗВОНОЧНЫХ ЖИВОТНЫХ J&
среднем мозгу. Сетевидное образование занимает в каждой
симметричной половине мозга центральное положение. Оно со всех сторон
окружено нервными ядрами и проводящими путями. Это образование
включает клеточные элементы, которые местами собираются в ядра. Ден-
дриты и аксоны одних клеточных элементов не выходят за пределы
сетевидного образования; другие вступают в многочисленные синапти-
ческие связи со всеми окружающими нейронными образованиями.
Некоторые из нервных ядер прямо погружаются в это образование и
таким образом вступают с ним в связь (рис. 7).
Рис. 7. Поперечный разрез продолговатого мозга на
уровне нижней оливы.
Сетевидное образование и его отношение к ядрам и проводящим
путям в продолговатом мозгу. DO — верхняя добавочная олива;
DT—путь от ядра Дейтерса к спинному мозгу; Fie—дорзальный
мозжечковый путь к спинному мозгу; Got? — вентральный спин-
номозжечковый путь; Неі — путь от оливы к спинному мозгу;
ю — ниншяя олива; NB — ядро Бурдаха; NBI — ядро
Блуменау; Nf8 — ядро лицевого нерва; NG — ядро Голла;
Nhy — ядро подглоточного нерва; Nvd — ядро блуждающего
нерва; NR — ядро Роландо; N12 — п. hypoglossus; PD —
предорзальный пучок; PL— fasciculus longitudinalis posterior;
Py — пирамиды; Ret— сетевидное образование; J?st—tractus
rubrospinalis; Spt — tractus spinothalamicus; TRD—нисходящий
путь тройничного нерва; Ven — четвертый желудочек; VO —
вентральная добавочная олива (из Тилнея).
Не только сетевидное образование, но и ядра продолговатого мозга
богаты густым нервным сплетением. Так, например, на рис. 8
изображена substantia gelatinosa Rolando, которая, как говорилось, тянется
по всему спинному імозгу и имеется также в продолговатом мозгу; на
рис. 9 показано строение верхней оливы, которая считается важным
центром продолговатого мозга. Как видно на обоих рисунках, отростки
клеток образуют густую сеть, с которой вступают в связь
многочисленные разветвления входящих сюда коллатералей чувствительных волокон.
Наиболее детальное микроскопическое исследование ядер
продолговатого мозга в последнее время было дано Херриком на низших
позвоночных, аЛоренте де Но — на млекопитающих. На рис. 10
изображен нейропиль продолговатого мозга по Лорентеде Но.
По Херрику (5), клетки formatio reticularis связываются своими
аксонами положительно со всеми двигательными центрами продолгова-
16 общая физиология спинного мозга и ствола головного мозгд
того и среднего мозга. С другой стороны, около этих клеток
оканчиваются коллатералями многочисленные чувствительные волокна,
вступающие в продолговатый и средний мозг. На основании этих
анатомических данных можно утверждать, что formatio reticularis является важ*
нейшим ассоциативным,
объединяющим органом. Такого же
мнения придерживается выдающийся
невролог Ариёнс Капперс
(7). Он считает это образование
важнейшей координирующей
системой. Она расчоложена между
чувствительной и двигательной
системами всей стволовой части
головного мозга и воспринимает
раздражения из весьма
разнообразных источников, а также
служит для объединения
деятельности как близлежащих, так и
дальних двигательных центров.
Рис. 11 схематически
изображает строение того отдела
продолговатого мозга, через
который, согласно представлениям
Рис. 8. Нейропиль продолговатого мозга Лоренте де Но,
Осуществи — substantia gelatmosa Rolando; С, С — клетки олартга пргЪттрігтппняст тт^атр
плетевидного образования и их связи с eubst. gel,; ВЛЙС1^* рецілеліирил» деніелъ
nV — кодлатщ^^тв^таитіош^водокоя трои- НОСТЬ При раЗДраЖвНИИ ТрОЙНИЧ-
ного нерва.
Строение промежуточного мозга. В промежуточном мозгу X е р-
рик (1) находит нейропнль во всех ядрах и проводящих путях, но,
кроме того, нейропиль здесь образует несколько отдельных полей; у
амфибий эти поля находятся в коленчатом теле, в таламической
области и на передней боковой поверхности (area laieralis tegmenti). У
высших позвоночных животных он также находит эти поля, являющиеся
ассоциативными полями вроде formatio reticularis. Они содержат
клеточные элементы, и тут оканчиваются многочисленные коллатерали от
чувствительных путей разного происхождения (рис. 12).
Таламическая область промежуточного мозга низших позвоночных
животных содержит очень мало дифференцированных нейронных
образований в виде нервных центров. В ней господствует нейропиль, в
котором встречаются восходящие чувствительные пути и нисходящие пути
обонятельной системы. У низших позвоночных таламическая область
является наименее развитой и наименее специализированной частью
мозга. У стоящих же выше животных, в связи с появлением локальных
рефлексов, эта часть мозга сильно развивается и становится главным
центром координации и интеграции этих локальных реакций. С
дальнейшим развитием мозга возникают мощные чувствительные пути,
О СТРОЕНИИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ ПОЗВОНОЧНЫХ ЖИВОТНЫХ 17
идущие от таламической области в кору большого мозга. Это
находится в связи с развитием аналитической способности. Вообще, по
данным X е р р и к а, восходящие чувствительные пути, соединяющие
рецепторы с корой большого мозга, обладают наиболее
совершенным развитием в смысле дифференциации нейронных путей и центров.
Головной мозг амфибий. Особенно подробно изучен головной мозг
у амфибий. Основные сведения о нейропиле были получены Херри-
ком именно на этом материале.
Рис 9. Нейропиль верхней оливы кошки.
Скомбинирован по рисункам Р. Кахаля. Сюда входят главные слуховые
нервные волокна, а уходящие волокна направляются в кору большого
мозга, к мозжечку, к ядру n abducens (Эдингер).
В головном мозгу, кроме хорошо известных дифференцированных
нервных центров и нервных путей, которые служат для более или
менее изолированных рефлекторных реакций, а также для характерных
движений организма в актах поведения, имеется
недифференцированная нервная масса, которая, можно сказать, заполняет весь головной мозг,
проникая во все нервные центры и проводящие пути. Эта
недифференцированная масса нервных элементов в своем развитии прошла разные
стадии, вплоть до образования настоящих нейронов с синаптическими
окончаниями и імякотными оболочками. Одни клеточные элементы ней-
ропиля не совсем развиты: аксоны их похожи на дендриты, они идуг
неправильно, варикозны, с колючками. Другие клеточные элементы
нейропиля хорошо развиты, с пучком дендритов на одном конце и
тонким аксоном на другом. Некоторые аксоны покрыты миэлиновой
оболочкой,— они относятся к далеко идущим нервным путям. Другие
аксоны — немиэлиновые; они или образуют компактные пути, или диф-
фузно ветвятся. Тончайшие разветвления аксонов образуют синаптиче-
ские связи путем контактов на клеточных телах и дендритах нейронов
2 И. С. Беритов 'р111 &]<(* іч,ітт , ¦ ¦¦¦
- - -' -
18 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
в сером веществе или менее интимные контакты на дендритах в белом
івеществе. Этот же нейропиль густым войлоком проникает во все
нервные ядра и проводящие пути. Нейропиль в общем является
связывающей средой или, как говорит Херрик, синаптическим полем,
Рис 10 Поперечный разрез nucleus posterior и tuberculum acusticum у
взрослой кошки.
Зарисован с одного гистологического среза. Рисунок дает представление о протоплазма-
тическом сплетении дендритов. В нем дана только *'«—1/в всего количества окрашенных
клеток. N. г.—nucleus interstitialis, N. h. (а, Ь, с)—nucleus posterior с его тремя отделами,
N. — nucleus lateralis, Та —tuberculum acusticum с четырьмя основными слоями (J, 2,
3, 4). Здесь был применен такой метод окрашивания (метод Сох), который окрашивает
только протоплазматические дендритные отростки Поэтому аксоны не даны здесь. Приведены
только волокна n cocnleans[C Р ] (Лоренте де Но)
где встречаются нейронные элементы разного происхождения и каждый
из поступающих сюда аксонов образует синаптические контакты с
клетками и дендритами находящихся здесь нейронов (рис. 12).
О функциональном назначении нейропиля. Нейропиль отдельных
частей мозга развит различено и связан с разными комплексами
нервных центров и нервных путей. Вполне естественно предположение, что
и функциональное назначение разных отделов нейропиля не одинаково.
Херрик, рассматривая назначение того или другого нейропильного
комплекса с точки зрения анатомических отношений к
дифференцированным нервным центрам и проводящим путям, делает определенные
заключения относительно функций каждого данного нейропильного
комплекса.
На основании анализа анатомических соотношений Херрик делает
следующее заключение о физиологическом назначении нейропиля:
мозговой нейропиль является единственным лабильным и пластичным
образованием. Он служит специально для того, чтобы периферические
возбуждения могли широко влиять на прирожденные реакции в смысле
О СТРОЕНИИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ ПОЗВОНОЧНЫХ ЖИВОТНЫХ ]9
усиления или угнетения; он является материальной базой для памяти
и дрессировки; выполняет общие объединяющие функции и
видоизменяет прирожденные формы поведения для наилучшего приспособления
к внешней среде.
Другой видный невролог — Ал лен (35), много работавший над
изучением нейропиля formatio reticularis позвоночных, над выясне-
Рис. 11. Диаграмма связей нейронов тройничной
системы в продолговатом мозгу.
G. G. — ganglion Gasseri; A—афферентное волокно тройничного
нерва; s. п.—чувствительное ядро; г. tr. —часть сетевидного
образования (formatio reticularis), входящая в тройничную
систему; s. (г. п. — nucleus supratrigeminalis; т. п. —
двигательное ядро тройничного нерва; V. т.—двигательное волокно.
Чувствительное волокно оканчивается синаптически во всех
этих ядрах и в сетевидном образовании как на клетках, так и на
дендритах, участвуя в образовании нейропиля. Аксоны
находящихся здесь клеток также оканчиваются как на клетках, так и
на дендритах. Так, например, клетка В связана через
разветвление аксона с клетками С, D и Nt а также с дендритами.
Клетки F9 /, G, Е с короткими аксонами служат исключительно
для внутриядерных связей. Клетки В, С, D и N служат
главным образом для межядерных связей. Нервные волокна и
клетки образуют замкнутые круги разной сложности. Например,
клетки В и F участвуют в самом простом кругу, который не
выходит из данного чувствительного ядра, но в то же время они
входят в другие нервные круги с участием сетевидного
образования (В, С, F, В), сетевидного образования и двигательного
ядра (В, JV, G, С, F, В), сетевидного образования nucleus
supratrigeminalis и двигательного ядра (В, D, iV, G, С, F, В).
Стрелки указывают направление возбуждения в нервных кругах
(составлено автором но Лоренте де ДЗД.
нием его афферентных и эфферентных связей, предполагает, что сете-
видное образование содержит главные симпатические и
парасимпатические центры для регуляции движений внутренних органов, в частности
оно регулирует сердечный и дыхательный ритмы, кровяное давление, а
также ведает регуляцией тонуса скелетной мускулатуры.
Концепции Херрика и Аллена о функциональном значении
нейропиля слишком общи и не содержат рассмотрения конкретного
физиологического или поведенческого материала. Между тем
анатомические данные о нейропиле настолько значительны, что смело могут
быть использованы для объяснения основных явлений прирожденной
<"" ¦(:
20 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
и индивидуально приобретенной деятельности центральной нервной
системы, но это уже скорее дело соответствующих специалистов.
Мы придаем особенно большое значение следующим положениям в
учении о нейропиле позвоночных животных.
1. Нейропиль позвоночных состоит из густого сплетения дендритных
и аксонных разветвлений со включением клеточных элементов. Аксон-
ные разветвления обладают разной
степенью развития, вплоть до
сформированных волокон с миэлиновы-
ми оболочками и синаптическими
окончаниями. Эти аксоны местами
располагаются пучками, идущими
на более или менее далекое
расстояние.
2. Нейропиль заполняет почти
все части мозга, свободные от
нервных ядер и нервных путей;
элементы нейропиля проникают также в
/{ ? эти последние и вступают с ними в
\ " тесную связь.
3. Нейропиль — не однородная
масса. Он делится на отдельные
поля, из которых каждое имеет
присущие лишь ему особенности и
связано с определенными нервными
центрами и путями.
4. Нейропиль — нервная масса,
постоянно растущая и
развивающаяся. Это субстрат,
объединяющий все нервные центры и пути в
течение всей эмбриональной и
постэмбриональной жизни.
5. Каждый афферентный
чувствительный нейрон находится в связи как с определенными
промежуточными и двигательными нейронами, так и с определенной нейропиль-
ной массой.
6. Нейропиль снабжен более богатой капиллярной системой, чем все
другие элементы нервной системы.
Из этих анатомических положений можно вывести с очень большой
вероятностью следующие физиологические заключения:
а) Нейропиль является одним из наиболее активных нервных
субстратов.
б) Он приходит в активное состояние, подобно нервным клеткам,
под влиянием периферических импульсов.
в) Он должен приходить в активное состояние также под влиянием
промежуточных нейронов.
Рис. 12. Диаграмма нервных связей
в thalamus opt. амфибии Necturus.
Поперечный разрез. Три нейрона из
чувствительного ноля. Их клетки лежат глубоко в
сером веществе перивентрикулярного слоя. Их
дендриты распространяются широко через
белое вещество, где они вступают в синаптиче-
ские связи с окончаниями или коллатералями
различных чувствительных путей и с общим
диффузным нейропилем. Их аксоны идут или
в tractua thalamo-peduncularis—для
производства таламических рефлексов, или
направляются к большим полушариям. Вся дорзальная
часть thalamus opt. представляет «диффузное
ядро», клетки которого не собраны в
отдельные ядра и не имеют своих специфических
связей. На рисунке представлены два отдела
этого диффузного ядра: снаружи коленчатый
нейропиль (genie, neur.) и внутри
промежуточный нейропиль (interm. neur.) (Херрик).
О СТРОЕНИИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ ПОЗВОНОЧНЫХ ЖИВОТНЫХ 21
г) Он должен находиться в активном состоянии все то время, пока
связанные с ним нервные круги находятся в состоянии возбуждения.
д) Возбуждение или вообще активное состояние «ейропильной
массы одного поля должно влиять как на ближайшие двигательные
нейроны, так и на ближайшие промежуточные нейроны и афферентные
волокна.
Конечно, эти положения сами по себе еще ничего не говорят о
функциональных особенностях нейропиля, в отличие от нервных
центров и путей. Между прочим, они не позволяют заключить, что
объединяющая деятельность является функцией исключительно нейропиля.
Эта функция должна принадлежать всей центральной нервной системе,
в частности и нервным ядрам.
Связь периферической нервной системы с центральной нервной
системой. У позвоночных животных чувствительные волокна от
рецепторов поступают в спинной мозг и в стволовую часть головного мозга,
за исключением п. olfactorius, который связывается непосредственно с
передним мозгом, с большими полушариями. Чувствительные,
афферентные волокна головных рецепторов,— как, например, зрительных,
слуховых, обонятельных и вкусовых,— вступают в мозг в виде
самостоятельных нервных стволов. Чувствительные волокна мышечных и
сухожильных рецепторов, интероцепторов внутренних органов*, а также
рецепторов кожи доходят до межпозвоночных ганглиев, по большей
части смешиваясь в нервных стволах с эфферентными волокнами как
соматического, так и вегетативного происхождения.
Начиная же с межпозвоночных ганглиев они разделяются:
афферентные волокна вступают в мозг в составе задних корешков, а эффе*
рентные выходят из мозга — в составе передних корешков. Это правило
корешковых функций было установлено в начале прошлого столетия
анатомом Беллом (1811—1825) (14) и физиологом Мажанди
(1822) (15) и потому получило название законаБелла-Мажанди.
Это распределение эфферентных и афферентных волокон между
корешками, по мнению імногих авторов, не является абсолютным. Вскоре
после установления закона сам Мажанди высказывал
предположение, что задние корешки, возможно, содержат и двигательные волокна.
Исследованиями видных гистологов, как, например, Леношека (16),
Кахаля (1і7), доказано существование центробежных волокон в
задних корешках. Некоторые из этих волокон как будто начинаются даже
в передних рогах от клеток, подобных двигательным клеткам,
отдающим свОи аксоны в передние корешки (Леношек, 16).
Физиологическими опытами также было показано присутствие таких волокон в
задних корешках. Так, например, раздражение периферических отрезков
перерезанных задних корешков у собак вызывает расширение
кровеносных сосудов, повышение температуры конечности (Штрикер, 19).
У лягушки раздражение периферических отрезков задних корешков
вызывает сокращение пищевода, желудка, всего кишечника, мочевого
пузыря (Штейнах, 18).
22 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
В последнее время Кюре с сотрудниками (25) усиленно
пропагандируют мысль о наличии в задних корешках эфферентных волокон,
заведующих движением сосудов, трофикой и секрецией. Они считают
эти волокна преганглионарными волокнами парасимпатической системы.
Гистологические исследования нервной системы у низших
позвоночных (ланцетные, круглоротые) доказывают, что для внутренних органов
эфферентные волокна проходят через дорзальные корешки (Гатчек, 29;
Ариёнс Кдпперс, 28). Существование двигательных волокон скелетной
мускулатуры в задних корешках также доказывается некоторыми
авторами. Однако это наблюдается не всегда и не на всех мышцах и
потому считается аномалией (Вана, 32).
Новейшие исследования дали иное понимание всех этих
гистологических и физиологических фактов. Прежде всего было обнаружено, что
центробежные волокна в задних корешках опять-таки являются
чувствительными. Именно, заднекорешковые чувствительные волокна, войдя
в спинной імозг, могут дать ответвление, которое через другой задний
корешок выходит наружу (Баррон и Мэтьюс, 20). Далее, наличие
двигательного эффекта в мышцах внутренних органов при раздражении
задних корешков также не указывает на существование двигательных
эфферентных волокон в задних корешках. В настоящее время известно,
что чувствительные волокна, начинающиеся в рецепторах внутренних
органов, отдают веточки, которые оканчиваются в гладкой мускулатуре
(Лаврентьев, 21). Таким образом, заднекорешковые чувствительные
волокна не только должны проводить возбуждение от рецепторов в
мозг, но и через указанные веточки производят сокращение. Мы потом
увидим, что заднекорешковые волокна, по существу говоря, проводят
импульсы возбуждения не только от рецепторов в мозг, но и
наоборот,— от мозга на периферию.
Имеются также факты, говорящие за то, что в передних корешках
проходят афферентные, чувствительные волокна. Прежде всего, со
времени Клод Бернара (22) известно, что раздражение
неповрежденных передних корешков вызывает ощущение боли и даже
соответствующую ей голосовую реакцию, подобно тому, как это происходит
при раздражении заднего корешка. Далее, известно, что у человека
болевые ощущения во внутренних органах, а также в конечностях при
сильном сдавливании тканей имеют место при полной деафферентации,
т. е. после перерезки всех задних корешков, имеющих отношение к
раздражаемому органу (Леман, 23). Гистологическое исследование также
как будто указывает на существование центростремительных волокон
в передних корешках. Так, Шеррингтон (24) нашел, что после
перерезки переднего корешка перерождаются некоторые волокна в
центральном отрезке корешка. Они как будто отходят от задних
корешков отдельным пучком и присоединяются к переднему корешку. Но
признается существование и таких афферентных волокон, которые
отходят не от задних корешков.
О СТРОЕНИИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ ПОЗВОНОЧНЫХ ЖИВОТНЫХ 23
Согласно новейшим исследованиям, гистологическое перерождение нервных
волокон наблюдается не только в той части, которая отделяется от клетки, но в
незначительной степени и на стороне клетки. Изменяются как аксоны, так и сами клетки,
в особенности в том случае, если перерезка произведена близко к клеткам (Гейст, 26,
Гинзей, 27). Это обстоятельство, конечно, могло иметь место в опытах некоторых
авторов, которые находили перерожденные волокна в центральном отрезке передних
корешков или в периферическом отрезке задних корешков.
Ощущение боли, вызываемое раздражением передних корешков, следует отнести
за счет возбуждения рецепторов мышц при их сокращении, вызванном раздражением
передних корешков На это указывает, во-п?рзых, факт, изіэестигый еще Клод
Б е ір и а о у, что если раздражать периферический отрезок, то ощущение боли и
голосовая реакция получаются и тсл-е перерезки передних корешков, раздражение же
центрального отрезка переднего корешка этих реакция не вызывает.
Наблюдение Лемана на людях также не представляет неопровержимого
доказательства проведения возбуждения от рецепторов давления и боли через передние
корешки Так,по Ферстеру и сотр (30), ощущение боли в мышцах заднее
конечности наблюдается и в тем случае, если шинное мозг полностью перерезан в грудной
области. Значит, афференгная система болевой чувствительности частью проходят
экстрамедуллярно, через пограничный симпатический столб Следовательно, во всех
тех случаях, когда в данном органе, несмотря на перерезку соответствующих
задних корешков, налицо было ощущение боли, оно могло передаваться к спинному мозгу
через симпатический пограничный столб и задние корешки впереди лежащих
сегментов мозга
Экспериментальное исследование на кошках показывает, что при
раздражении центральных отрезков перерезанных переднил
корешков можно повлиять на деятельность спинного мозга. Это влияние
заключается в некотором угнетении центральной деятельности (Беритов
и Бакурадзе, 34). Точно так же удается повлиять на мозг угнетающим
образом помимсл задних корешков (после перерезки их) раздражением
внутренних полостных органов (мочевой пузырь, прямая кишка,
желудок и т. д) путем растяжения или раздражением соответствующих
чувствительных нервов электрическим током. Эти раздражения угнетают
деятельность спинного мозга. Было установлено, что данное
угнетающее действие осуществляется путем прохождения периферических
импульсов частью через передние корешки, частью помимо них по
нервным сплетениям артериальных сосудов (Беритов и Бакурадзе, 34).
Итак, имеется прямое доказательство проникновения некоторых
чувствительных волокон в спинной мозг помимо задних корешков.
Связь нейронов внутри центральной нервной системы. Каждый
афферентный, т. е. чувствительный, нейрон сейчас же после вступления
в спинной мозг делится на две веточки одна веточка идет в каудаль-
ном направлении, проходя в белом веществе мозга (задние и
боковые столбы) несколько сегментов; другая веточка идет в
краниальном направлении (задние столбы). Длина восходящих веточек
разнообразна, ибо они прерываются на разных уровнях спинного імозга;
некоторые из них достигают продолговатого мозга. Обе веточки отдают
многочисленные боковые ответвления — коллатерали, заходящие в
серое вещество мозга. Здесь они заканчиваются при так называемых
вставочных, или промежуточных, клетках substantia gelati-
24 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
nosa Rolando и столбах Кларка, а также в передних рогах около
двигательных клеток. Особенно много их заканчивается в нейропильной
массе спинного мозга, т. е. ів дендритном сплетении. Коллатерали
афферентных волокон одной симметричной половины мозга переходят
в небольшом количестве и на другую половину по задней комиссуре.
Здесь они заканчиваются в сером веществе, не заходя в передний рог.
Кроме того, связь между обеими половинами осуществляется путем
особых вставочных нейронов, известных под названием ком и с
сура л ь н ы х. Клетки этих нейронов лежат в средней части серого
вещества, а аксоны их переходят по передней комиссуре на другую сторону
и заканчиваются в переднем рогу в- области двигательных клеток. По
передней комиссуре переходят также из одной половины мозга в
другую дендриты двигательных клеток (Кахаль, 12) (рис. 6).
Связь данного афферентного нейрона с двигательными клетками
наиболее интимна © пределах того сегмента, где он вступает в імозг.
Интимность связи, повидимому, прежде всего обусловливается тем, что
здесь от каудальной и краниальной веточек данного афферентного
нейрона отходит наибольшее количество коллатералей. Те же веточки на
дальнейших участках отдают меньше коллатералей (Лапинский, 8).
Каждый афферентный нейрон находится в связи с большим
количеством промежуточных нейронов, а аксоны этих последних стекаются к
ограниченному количеству двигательных нейронов, но любой
афферентный нейрон с любого рефлекторного поля может подействовать через
более или менее сложную цепочку нейронов на данный двигательный
нейрон. Вот это сведение всех нервных путей к одному «общему
конечному пути» и называется принципом к онве р ге н ции (Шерринг-
тон, 9).
Описанное анатомическое устройство! мозга определяет в
значительной мере характер и взаимодействие рефлекторных реакций, как это
будет видно ниже.
Литература
1. Н еггіск С. I., J. Сотр. Neur. Psych., 59, 93, 1934; 59, 233, 1934.
2. R e t z i u s G., Proc. Roy. Soc, London, B. 80, 414, 1908.
3. T i e g s O. W., Austr. Exp. Biol. Med. Sci., 4, 193, 1927; 3, 45, 69, 161, 1936.
4. Немилов А., Гистологическое строение дорзальных корешков и белого
вещества спинного мозга, СПб., 1913.
5. Н еггіск С. I., J. Сотр. Neur. Psych., 58, 481, 1933; Sci., 78, 439, 1933.
6. К о е 1 1 і к е г, Ztschr. math.-nat. К1. К. Akad. Wiss., Wien, 1901; Anat. Anz.,
21, 1902; Ztschr. Wiss. Zool., 72, 1902.
7. Ariens Kappers C. U., Die vergleichende Anat. d. Nervensystems d. Wir-
beltiere u. d. Menschen, 1, 200—206, 272—596, 1920.
8. Lapinsky M., Arch. Psych., 42, 869, 1907.
9. Шеррингтон и сотр., Рефлекторная деятельность спинного мозга., Биомед-
гиз, 1935.
10. Lorente d е N 6, Arch. Neur. Psych., 90, 245, 1933; J. Neur., Psych., 75, 385,
1933; 46, 113, 1934.
11. L о г e n t e d e N 6, J. Neurophysiol., 1, 195, 1938.
О СТРОЕНИИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ ПОЗВОНОЧНЫХ ЖИВОТНЫХ 25-
12. С а ] al R., Arch. Anat. u. Physiol., стр. 319, 1893.
13. Gajal R., Anat. Anz., 5, 85, 111, 1890.
14. Bell Ch., Idea of a new anatomy of the Brain, London, 1811; An exposition of the
natural system of the nerves of the human body, Philadelphia, 1825.
15. Magendie F., A summary of physiology, Trans, from the French by John
Revere, Baltimore, 1822.
16. Lenhossek M., Der feinere Bau des Nervensy stems, Berlin, 1895.
17. Gajal R„ Anat. Anz., 5, 613, 1890.
18. Stein ach S.,Pfl. Arch., 60, 593, 1895; 71, 523, 1898.
19. Strieker Abh. K. Akad. Wiss., Wien, 74, III Abt., 173, 1876.
20. В а г г о n D. H. a. M a 11 h e w s В. Н., J. Physiol., 92, 276, 1938.
21. Лаврентьев Б. И., Личное сообщение в 1941 г.
22. Клод Б е р н а р, Лекции по экспериментальной патологии. Русск. пер., М. —
Л., стр. 146—162, 1937.
23. Lehmann W., Z. ges. exp. Med., 12, 33, 1921.
24. Sherrington Ch., J. Physiol.» 17, 211, 1894.
25. К u г ё К. с сотр., Quart. J. Exp. Physiol., 18, 333, 1928; 20, 51, 1930; 21, 1, 103,
1931; Ztschr. Zellforsch. u. mikr. Anat., 92, 1935.
26. Geist F. В., Arch. Neur. Psych., 29, 88, 1933.
27. Hinseyl. С., Krupp M. A. a. W. T. L h о m о n, J. Сотр. Neur., 67, 205r
1937.
28. A r i ё n s К a p p e г s C. U., The comparative anatomy of the nervous system etc.r
v. I, pp. 276—280, 1936.
29. H a t s с h e k, Verb. anat. Ges. a. d. 5. Versamml. in Wien 1892. Цит. noLehmann'y.
30. F о e r s t e r O., A 11 e n b u r g e r H. u. К г о 11, W. F. Z. ges. Neur., Psych.,
121, 139, 1929.
31. К r a u s е. Цит. по Philippson'y: L'automatie et la centralisation dans le systeme
nerveux des animaux., Bruxelles, 1905.
32. Wan a J., Pfl. Arch., 71, 555, 1898.
33. Silver, J. Сотр. Neurol., 71, 1, 1942.
34. Беритов И. иБакурадзе А., Физиол. журн. СССР, 30, 45, 1941;
Тр. Ин-та физиол. им. Бериташвили, 5, 143, 1943.
35. Allen W. F., J. of the Washington Acad, of Sc, 22, N. 16, 17, 1932.
Глава IT
ОБЩЕЕ ПОНЯТИЕ О ЦЕНТРАЛЬНОЙ НЕРВНОЙ
ДЕЯТЕЛЬНОСТИ
Показатели деятельности центральной нервной системы и ее
возбудители. Деятельность центральной нервной системы не может быть
в полной мере изучена непосредственным наблюдением процессов,
протекающих в мозгу. Правда, центральная деятельность сопровождается
биоэлектрическими токами возбуждения и теплопродукцией, которые
•могут быть приблизительно изучены, но центральная нервная система
представляет собой чрезвычайно сложное образование: состоит из
множества разнообразных групп нервных клеток, разнообразных пучков
нервных волокон и разнообразных нейропильных полей. К тому же все
эти нервные образования весьма сложно переплетаются между собой.
Поэтому электрический и тепловой эффекты центральной нервной
системы являются настолько сложными по своему происхождению, что до
последнего времени было очень трудно правильно определить, какой
именно части мозга или какому комплексу нервных элементов следует
приписать эти эффекты. В настоящее время электрофизиологическая
методика настолько' шагнула вперед, что стало возможным более или
менее детальное изучение деятельности центральной нервной системы
на основании ее электрической активности, но в основном центральная
деятельность исследовалась и исследуется при помощи вызываемых ею
внешних, периферических эффектов. К таким внешним эффектам
относятся сокращение мышц, секреция желез, изменение давления крови в
сосудах и т. д.
Центральную нервную деятельность, с одной стороны, вызывают
раздражения органов чувств, обычно реагирующих на воздействие
внешней среды, так называемые внешние раздражения, например,
световое раздражение сетчатки глаза, звуковое раздражение кортиева
органа, механическое или температурное раздражение кожной
поверхности, химическое раздражение слизистой оболочки ротовой полости, а
также пищеварительного канала и т. д. С другой стороны, центральную
нервную деятельность вызывают раздражения, происходящие внутри
организма, т.е. внутренние раздражения. К таким раздра-
ОБЩЕЕ ПОНЯТИЕ О ЦЕНТРАЛЬНОЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ 27
жениям относится механическое раздражение рецепторов в мышцах
и в сухожилиях от их растяжения, в суставах — от изменения их
положения, в кровеносных сосудах — от их расширения под давлением
крови и т. д.
Понятие о рефлексе. Внешний эффект деятельности центральной
нервной системы, наступающий непосредственно в ответ на
раздражение тоге или другого рецептора, принято называть рефлексом,
рефлекторной реакцией. Понятие о рефлекторном движении как о
непроизвольной реакции впервые было дано философом Декартом (1662)
(1), но термин «рефлекс» за
этим понятием утвердился
позднее, в конце XVIII в.
(Прохаска, 10, Унцер, 13 и др.,
см. подробно у Фиринга, 11).
Слово «рефлекс» означает
«отражение», и оно в известном
смысле хорошо изображает
суть центрального эффекта:
подобно лучам света,
отраженным от зеркала, импульсы
возбуждения, притекающие в мозг
с периферии, отражаются
обратно на периферию, вызывая
движение или секрецию.
Прежде рефлексом
назывался только прирожденный,
наследственный акт. В
настоящее время рефлексом
называют внешние эффекты
центральной деятельности, как
прирожденные, так и
индивидуально приобретенные.
Характер рефлекторной
реакции зависит прежде всего
от того рецептора, который
раздражается, а затем от силы и продолжительности самого
раздражения. Так, если ущипнуть правую заднюю лапку обезглавленной
лягушки, т. е. раздражать кожные рецепторы на этой лапке, то лапка
определенным образом притянется к телу (рис. 13). Если ту же лапку
раздражать не щипком или другого рода повреждающим
раздражением, а легким давлением на вентральную сторону ступни, тогда
наступает рефлекс разгибания: сначала разгибаются пальцы
раздражаемой лапки, а затем и вся конечность.
Характер рефлекса меняется в зависимости и от места раздражения.
Так, если раздражать кожу на правом боку щипком или другим
повреждающим раздражением, то правая задняя лапка опять-таки притянется
Рис. 13. Оборонительный рефлекс сгибания
на спинномозговой лягушке.
А — препарат висит без раздражения; В — то же, но
с раздражением правой задней ноги. Раздраженная
нога согнута во всех суставах. Другая задняя нога
заметно разогнута. Кроме того, правая передняя нога
несколько отклонена назад, а левая передняя
вытянута вперед. Туловище немного изогнуто вправо.
28 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА II СТВОЛА ГОЛОВНОГО МОЗГА
в
к телу, но совершенно иначе, чем при таком же раздражении самой
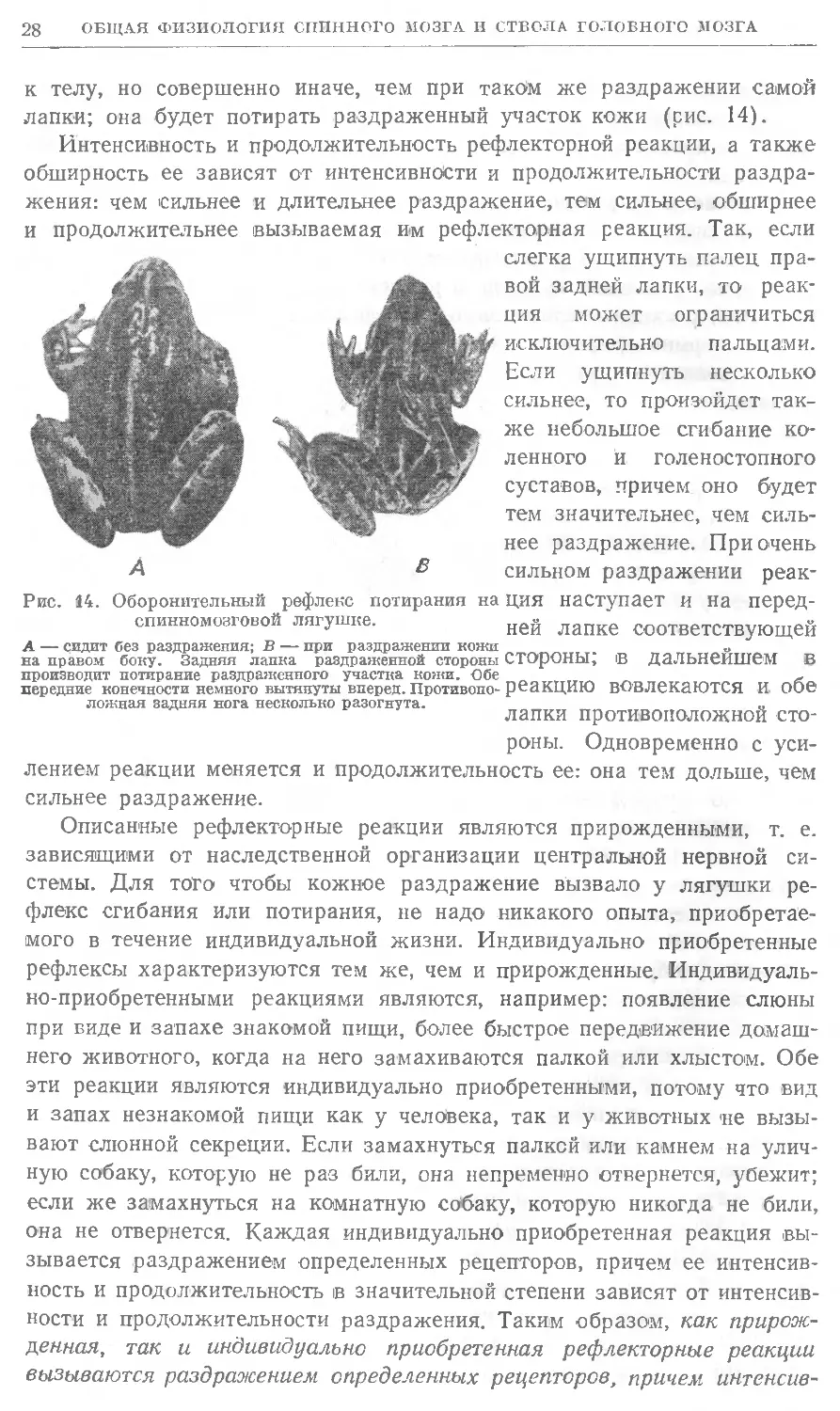
лапки; она будет потирать раздраженный участок кожи (рис. 14).
Интенсивность и продолжительность рефлекторной реакции, а также
обширность ее зависят от интенсивности и продолжительности
раздражения: чем сильнее и длительнее раздражение, тем сильнее, обширнее
и продолжительнее вызываемая им рефлекторная реакция. Так, если
слегка ущипнуть палец
правой задней лапки, то
реакция может ограничиться
исключительно пальцами.
Если ущипнуть несколько
сильнее, то произойдет
также небольшое сгибание
коленного и голеностопного
суставов, причем оно будет
тем значительнее, чем
сильнее раздражение. При очень
сильном раздражении реак-
Рис. 14. Оборонительный рефлекс потирания на ция наступает и на перед-
спинномозговой лягушке. ней лапке соответствующей
А — сидит без раздражения; В — при раздражении кожи ^-^„.т. тт т «° ~
на правом боку. Задняя лапка раздраженной стороны СТОроНЫ, В Дальнейшем іВ
производит потирание раздраженного участка кожи. Обе _-„«.„„.-. ~ ^ *г~
передние конечности немного вытянуты вперед. Противопо- реакцию вовлекаются И 006
ложная задняя нога несколько разогнута. ' „ _ „
лапки противоположной
стороны. Одновременно с
усилением реакции меняется и продолжительность ее: она тем дольше, чем
сильнее раздражение.
Описанные рефлекторные реакции являются прирожденными, т. е.
зависящими от наследственной организации центральной нервной
системы. Для тФго чтобы кожное раздражение вызвало у лягушки
рефлекс сгибания или потирания, не надо никакого опыта, приобретае-
імого в течение индивидуальной жизни. Индивидуально приобретенные
рефлексы характеризуются тем же, чем и прирожденные.
Индивидуально-приобретенными реакциями являются, например: появление слюны
при виде и запахе знакомой пищи, более быстрое передвижение
домашнего животного, когда на него замахиваются палкой или хлыстом. Обе
эти реакции являются индивидуально приобретенными, потому что вид
и запах незнакомой пищи как у человека, так и у животных не
вызывают слюнной секреции. Если замахнуться палкой или камнем на
уличную собаку, которую не раз били, она непременно отвернется, убежит;
если же замахнуться на комнатную собаку, которую никогда не били,
она не отвернется. Каждая индивидуально приобретенная реакция
вызывается раздражением определенных рецепторов, причем ее
интенсивность и продолжительность в значительной степени зависят от
интенсивности и продолжительности раздражения. Таким образом, как
прирожденная, так и индивидуально приобретенная рефлекторные реакции
вызываются раздражением определенных рецепторов, причем интенсив-
ОБЩЕЕ ПОНЯТИЕ О ЦЕНТРАЛЬНОЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ 29
ность и длительность их зависят в значительной мере от интенсивности
и продолжительности данного раздражения.
Характеристика рефлекторной деятельности центральной нервной
системы. Прирожденная рефлекторная деятельность центральной
нервной системы выражается в координировании реакций, протекающих в
том или ином органе, и, кроме того, в установлении определенного
взаимоотношения между этими органами. Если у обезглавленной
лягушки ущипнуть заднюю лапку, то, как было указано выше, эта лапка
притянется к телу, а другая лапка разогнется. Движение как одной, так
и другой лапки является координированным актом. Оба эти движения
обусловливаются сокращением определенной группы мышц и
расслаблением другой группы. Между движениями той и другой лапки
существует определенное соотношение: когда на одной (согнутой) лапке
сокращаются мышцы-сгибатели, тогда на другой (разогнутой) лапке
сокращаются мышцы-разгибатели. Это координированное участие в
рефлексе мышц конечностей зависит непосредственно от деятельности
спинного мозга, которая нормально наступает под влиянием импульсов,
притекающих со стороны кожных рецепторов конечности. Точно так же
движения лапок лягушки, наступающие от раздражения кожи на боку,
предполагают координированное участие определенных мышц. Эта
рефлекторная реакция заключается в притягивании одной задней лапки
к раздраженному участку и в потираний его пальцами; при этом другая
задняя лапка вытягивается, а все тело сгибается в сторону
раздражения. Эта сложная реакция координирована в отношении участия как
разных органов, так и разных мышц в каждом органе. Координация'
производится спинным мозгом. Итак, центральная нервная система
путем рефлекторной деятельности координирует реакции отдельных
органов и устанавливает между ними определенное взаимоотношение.
Центральная нервная система, осуществляя вышеописанную
координацию, достигает жизненно важных целей: она защищает тот или
другой орган от внешних раздражений, регулирует нормальную
деятельность каждого органа в организме, приспосабливает деятельность
разных органов одного к другому. Так, при повреждающем раздражении
ноги последняя притягивается к телу, удаляется от раздражителя и
таким образом избегает дальнейшего раздражения. При вкладывании
сухой пищи в ротовую полость выделяется большое количество жидкой
слюны и происходит увлажнение пищи, что необходимо для
проглатывания ее. Нормальная работа сердца, нормальное дыхание,
передвижение пищи в пищеварительном канале, выделение пищеварительных
соков и т. д.— все эти регулируется рефлекторной нервной деятельностью.
'Каждый из этих органов связан с центральной нервной системой как
чувствительными, так и двигательными или секреторными нервными
путями, и через эти нервные пути центральная нервная система реагирует
по принципу рефлекса: на известные раздражения она производит
определенного рода координированные реакции. Деятельность каждого
органа тесно связана с деятельностью остальных, поэтому нарушение нор-*
30 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
мальной деятельности одного органа влечет за собой нарушение ее и
в других органах. Центральная нервная система рефлекторной
деятельностью устраняет эти нарушения и устанавливает нормальное
действие— иногда очень сложньши путями. Так, например, при повышении
кровяного давления состояние многих органов ухудшается, и прежде
всего ухудшается состояние самого сердца, так как от него требуется
большая работа, чем в норме. В корне аорты и в sftius caroticus
имеются определенные рецепторы, которые механически раздражаются
вследствие повышения давления крови. Отсюда возбуждение передается
по чувствительному нерву в центральную нервную систему. Вслед за
этим чисто рефлекторно наступает расширение всего кровеносного
русла, что вызывает падение кровяного давления до нормы.
Так как рефлекторные реакции служат определенным потребностям
организма, их принято называть целесообразными реакциями. Прежде
многие крупные физиологи думали, что целесообразность рефлекса
обусловливается психической деятельностью мозга. По их мнению, и
спинной мозг обладает способностью к психической деятельности
(Пфлюгер, 14; Лучиани, 15). Но в настоящее время никто не разделяет
этого мнения. Теперь психика, субъективные переживания признаются
основной функцией передней части головного мозга. Целесообразность
же рефлекторной деятельности спинного мозга объясняется тем, что в
спинном мозгу существуют специальные нервные механизмы для
координирования движений каждого органа и системы органов. Эти
механизмы возникли и развились филогенетически, путем естественного
отбора.
Индивидуально-приобретенные реакции, так же как и
прирожденные, служат для защиты органов от внешних раздражений, для
регулирования нормальной деятельности органов, для приспособления
одного органа к другим. Слюна, выделяющаяся при виде знакомой пищи,
служит для нормального пищеварения; убегание собаки при виде
поднятого камня является защитной реакцией от удара и т. д.
Понятие о спонтанной или автоматической деятельности.
Центральная нервная система приходит в активное состояние не только под
влиянием нервных импульсов, притекающих от внешних и внутренних
рецепторов. Она приходит в активное состояние также от воздействия
разного рода специфических (гормонов) и неспецифических
(метаболитов, электролитов) веществ, возникающих внутри организма в
результате обмена веществ в самих нервных центрах и других тканях, в
особенности в органах внутренней секреции. Этот вид нервной деятельности
принято называть спонтанным или автоматическим (Бери-
тов, 19). Спонтанная активность свойственна всем отделам головного
мозга, начиная от коры больших полушарий до продолговатого мозга;
она свойственна в незначительной степени и спинному мозгу. Некото-
.рые отделы мозга — кора большого мозга, кора мозжечка, пpoмeжyтoq-
ный мозг — все время находятся в состоянии усиленной активности.
В определенных случаях спонтанная активность того или другого
ОБЩЕЕ ПОНЯТИЕ О ЦЕНТРАЛЬНОЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ 37
отдела проявляется в определенных внешних реакциях. Спонтанными
являются, например, дыхательные движения, многие движения
новорожденного животного, все так называемые инстинктивные действия
взрослых животных, все движения эмбриона в момент формирования нервно-
мышечной системы (Беритов, 20).
Спонтанная активность центральной нервной системы играет
большую роль в жизнедеятельности организма еще в том отношении, что
она держит нервные центры в состоянии готовности реагировать на
внешние раздражения. Импульсы, возникающие спонтанно в одном
центре, распространяются в некоторой степени на другие центры и на
те рабочие органы, с которыми данный центр связан нервными путями,,
вследствие чего во всех этих образованиях также поддерживается
состояние готовности к реагированию (Беритов, 20).
Понятие о поведении. Деятельность центральной нервной системы
в ответ на воздействие внешней среды проявляется не только в вид©
рефлекторных реакций. Организм отвечает еще особого рода реакциями,
которые не носят рефлекторного характера, а именно — актами
поведения. Поведением называется такая целостная реакция организма*
которая направлена на его непосредственное функциональное
приспособление к внешней среде или же на такое изменение внешней среды,,
которое приспособило бы ее к потребностям организма. Если мы
ущипнем заднюю лапку нормальной лягушки, она сейчас же прыгнет
и убежит в каком-либо направлении, обходя при этом встречающиеся
на пути препятствия. Эту реакцию нельзя считать рефлекторной
реакцией на произведенное раздражение. Возможно, что в самом начале
реакции, в момент первого прыжка, раздраженная лапка впервые
притянулась к телу рефлекторно, т. е. получился рефлекс сгибания лапки.
Но это сгибание не имеет существенного значения в указанной сложной
реакции. Лягушка убежала от раздражителя при своеобразном участии
всех конечностей и всего тела. Эта реакция не является результатом
одного только данного раздражения,— такой же рефлекс убегания мы
получили бы при раздражении кожи на спине. Само направление и
обход препятствий не стоят ни в какой связи с произведенным нами
раздражением. Главную роль здесь, безусловно, играет воздействие
всей среды, прежде всего производимые ею зрительные раздражения.
Наконец, характерное течение этой защитной реакции не зависит от
характера и интенсивности раздражения, лягушка убежала бы таким
же образом не только при любом повреждающем раздражении, но и
при других, даже неповреждающих раздражениях, например при
замахивании на нее рукой. Эта целостная реакция лягушки выражает
приспособление ее к данной внешней среде: она убегает от той среды, где
она получила повреждение или значительное раздражение. Поэтому
данная реакция является поведением, именно защитным поведением, а
не рефлексом. Или, например, при виде знакомой пищи собака тянется
за пищей, бежит к ней, если она лежит далеко от нее; подойдя к пище,
наклоняет голову и сгибает передние ноги, затем захватывает ее с по-
32 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
мощью передних лап или прямо ртом и, наконец, глотает; при этом во
рту выделяется слюна, в желудке — желудочный сок и другие
пищеварительные соки. Вся эта сложная реакция, правда, наступает вслед за
тем, как собака увидела пищу, но не обусловливается этим всецело.
Она обусловливается воздействием всей внешней среды и направляется
к одной цели — принятию и усвоению пищи; таким образом, вся эта
сложная реакция представляет собой пищевой акт поведения.
Рефлекс и поведение. Из наших определений следует, что рефлекс
¦есть проявление центральной нервной деятельности на одном или
многих органах, а поведение — проявление той же центральной нервной
деятельности, но © виде сложной системы движений, направленных на
приспособление всего организма к внешней среде. Однако и эта сложная
целостная реакция, как мы уже видели, осуществляется при помощи
отдельных органов, причем действие каждого органа в поведении
протекает подобно рефлексу, а именно, оно вызывается из
определенных отделов центральной нервной системы, является координированным
и наступает при раздражении определенных рецепторов. Поэтому можно
сказать, что рефлекс является элементом поведения. В приведенном
нами примере защитного поведения лягушки последовательные
сгибание и разгибание конечностей являются следствием раздражения
определенных внешних и внутренних рецепторов самих конечностей. Точно
так же стояние на ногах и выпрямленное положение есть рефлекторная
реакция, наступающая при определенном внутреннем раздражении.
Обход препятствия есть также рефлекс, наступающий в ответ на
раздражение передней части тела, кожи на голове и глаз. Но в акте
поведения івсе эти рефлекторные движения объединяются в один
целостный акт, имеющий целью наилучшим образом защитить организм
в данной внешней среде.
В описанном «пищевом поведении» собаки мы имеем также ряд
элементов, протекающих по принципу рефлекса. Вид знакомой пищи
вызывает передвижение к пище, это — индивидуально приобретенный
рефлекс. Одновременно выделяется слюна, что также является
индивидуальным рефлексом на вид пищи. Наклонение головы к пище есть
частью прирожденный, частью индивидуальный рефлекс на запах и вид
пищи. При наклонении головы передние ноги животного несколько
сгибаются, это — прирожденный рефлекс, вызванный раздражением
мышечных рецепторов шеи, и т. д. Объединение всех этих
прирожденных и индивидуально-приобретенных рефлекторных элементов в
целостный акт поведения происходит через центральную нервную систему.
Эта объединяющая деятельность, обусловливающая поведение, является
функцией определенных отделов мозга, а именно: большого мозга для
высших позвоночных — млекопитающих, промежуточного мозга для
низших позвоночных — рыб и амфибий, и обоих отделов вместе — для
птиц и, повидиімому, для рептилий. Таким образом, рефлексы являются
обычными элементами поведения.
Акты поведения высших позвоночных животных образуются не из
ОБЩЕЕ ПОНЯТИЕ О ЦЕНТРАЛЬНОЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ 33
одних только рассмотренных рефлекторных элементов. Индивидуальное
поведение, приобретаемое на основании индивидуального опыта и
направляемое образами или представлением внешней среды, или
трудовое поведение человека, направляемое сознанием определенной цели
поведения, происходит при участии таких элементов, которые не
являются рефлексами. Это именно те элементы поведения, которые
осуществляются психонервными процессами. Правда, и эти элементы
наступают в ответ на внешнее раздражение и подразумевают
определенные материальные процессы в коре большого мозга, как на это
правильно указывали наши выдающиеся физиологи И. М.
Сеченов (2) и И. П. Павлов (3), но по своей внутренней природе они
представляют собой явления другого порядка. Дело в том, что
материальные процессы, протекающие в мозгу без субъективных
переживаний, должны качественно отличаться от тех материальных
процессов, которые вызывают субъективные переживания. Соответственно
и законы центральной нервной деятельности в том и другом случае не
могут быть одними и теми же. Поэтому, если первая форма нервной
деятельности будет называться рефлекторной, то вторая форма, в
настоящее время еще мало изученная, должна представлять нечто
другое (см. подробно в статьях И. Беритова, 4).
В поведенческих актах имеют место также автоматические, или
спонтанные, реакции, возникающие благодаря спонтанной деятельности
нервных центров. Некоторые автоматические реакции,— например,
дыхательные движения,— наступают в течение всей постэмбриональнюй
жизни. Другие возникают на определенных стадиях индивидуального
развития: например, движение эмбрионов и многие инстинктивные
действия. Некоторые автоматические движения возникают повторно
вследствие периодического изменения внутренней среды, как, например,
периодическая деятельность пищеварительного канала.
Все эти спонтанно наступающие реакции входят в поведенческие
акты как их компоненты. В определенных случаях они играют в них
доминирующую роль.
Рефлекс и нервно-мышечная деятельность. Все основные явления
нервно-мышечной возбудимой системы, как возбуждение, рефракторные
фазы, проведение возбуждения, утомление, пессимальный и
оптимальный эффекты, парабиоз и т. д., также свойственны центральной нервной
системе,—вернее, всем ее нервным элементам. При этом все данные
явления протекают в центральной нервной системе по тем же законам,
как и в нервно-мышечной системе, ибо законы деятельности живой
возбудимой системы везде одни и те же. Однако объяснять ими все то,
что происходит в центральной нервной системе, нельзя. Правда,
спинной імозг и другие отделы мозга состоят из нервных клеток и их
отростков, но они не представляют собой сумму этих клеток и отростков,
а являются особыми целостными органами, деятельность которых имеет
свои специфические особенности. Специфическая деятельность спинного
мозга заключается, между прочим, в координации движений конечно-
3 и. С. Беритов
ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
стей. Эта деятельность протекает согласно особым специфическим
закономерностям. Следовательно, деятельность спинного мозга
качественно отличается от нервно-мышечной деятельности. Поэтому
деятельность спинного мозга нельзя сводить к известным законам
нервно-мышечной деятельности и всецело объяснять законами этой
деятельности. Такое же качественное отличие существует между другими
отделами центральной нервной системы и нервно-мышечной системой.
Но, конечно, знание законов нервно-мьцнечной деятельности абсолютно
необходимо для анализа центральных явлений и установления для них
специфических закономерностей.
Рефлекторная деятельность без центральной нервной сиетемы. Только
у позвоночных животных рефлекторная деятельность является
безусловно функцией центральной нервной системы. Существует целый ряд
животных, у которых нет центральной нервной системы (все
кишечнополостные: гидра, -медуза, актиния), и все-таки раздражение вызывает
у них координированную реакцию. Так, например, у актиний имеется
развитая нервная ткань в виде нейропиля; каждый рецептор
связывается с мышцами через этот нейропиль. Обособленных клеточных
групп в виде нервных узлов или центров у них нет. Однако в ответ на
раздражение определенного участка наружной поверхности в
раздраженном участке наступает защитное движение: в этом участке кожная
поверхность отходит от раздражителя (Будденброк, 5).
У иглокожих нет ..строго обособленной дифференцированной
центральной нервной системы; у них есть нервное кольцо около рта и
радиальные ганглии. Повидимому, эти нервные образования играют роль
нервных центров. Если прикоснуться к одному из лучей морской
звезды, то луч изгибается таким образом, что удаляется от
раздражителя. Это настоящий координированный акт, так как он предполагает
сокращение мышцы на одной стороне луча и расслабление на другой.
Характерно, однако, что такой рефлекс получается на луче, даже если
он отрезан от организма,— следовательно, для этого рефлекса нервное
кольцо около рта не имеет особого значения. Итак, у низших
беспозвоночных животных координированная рефлекторная деятельность
происходит без строго обособленной центральной нервной системы,
через нейропиль (Будденброк, 5).
Центральная нервная система впервые обособляется у червей в виде
скопления 'нервных клеток отдельными узлами. Эти узлы больше всего
развиты в передней части тела и находятся в связи с расположенными
здесь наиболее важными рецепторами — зрения, осязания, обоняния и
вкуса, а также и с наиболее важными двигательными органами,
служащими для захвата пищи. Нервные узлы имеются и в других частях
тела и находятся в связи с определенными двигательными органами.
Центральная нервная система "еще более развита у мягкотелых, и
особенно у членистоногих. Однако у всех беспозвоночных наряду с
рефлексами, происходящими с йомощью нервных узлов, имеются й такие
рефлекторные реакции, которые протекают без участия нервных узлов,
ОБЩЕЕ ПОНЯТИЕ О ЦЕНТРАЛЬНОЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ 35
через нейропиль. Например, известно, что у головоногих обрезанные
щупальцы присасываются к предметам и сворачиваются под влиянием
света (Будденброк, 5).
Появление прирожденных рефлексов на разных ступенях онтогенеза.
Каждбе животное рождается с развитыми механизмами для некоторых
рефлексов, обеспечивающих ему существование в определенных
условиях. Но не все прирожденные рефлексы начинают проявляться с
момента рождения. Рождением называется тот момент в развитии
эмбриона, когда последний начинает принимать пищу из внешней среды через
собственный пищеварительный канал. Этот момент наступает на разных
стадиях эмбрионального развития, а значит, на разных стадиях
развития нервно-мышечной системы. В соответствии с этим одни рефлексы
начинают действовать & Момент рождения, другие возникают
впоследствии, когда развитие организма достигает соответствующей ступени.
Щенок, например, обнаруживает способность стоять й ходить спустя
несколько недель после рождения. Нельзя смотреть на эти рефлексы
как на индивидуально-приобретенные. В действительности стояние и
ходьба—чисто прирожденные рефлексы. Они не проявляются & момент
рождения вследствие недоразвития мышечной и нервной системы. Как
гистологическое, так и физиологическое исследование тканей
показывает, что развитие их еще не закончено. Так, например, известно, что
на раздражение лапки новорожденный щенок отвечает общим
сотрясением и писком без определенного защитного движения, т. е. он не
может согнуть или разогнуть лапки; не может ходить или стоять. Что
стояние и ходьба—прирожденный рефлекс, видно также и из того, что
у целого ряда животных только что родившиеся детеныши вполне
способны стоять и ходить. Так, например, только что родившийся
жеребенок, теленок, поросенок и др. стоят и ходят, как взрослые
животные. Наконец, прирождевность рефлекса ходьбы и стояния можно
доказать и на взрослых животных путем удаления передней части
головного мозга. После такой операции все
индивидуально-приобретенные рефлексы исчезают и вновь не образуются, рефлексы же стояния и
ходьбы остаются (Зеленый, 6).
Онтогенез рефлекторной деятельности в связи со структурным
развитием нервной системы. У холоднокровных, например у амблистом,
каждая более или менее локальная рефлекторная реакция, как дыхание,
локомоция, начинает осуществляться на ранней эмбриональной стадии
в связи с обособлением соответствующих нервных путей из первичной
нервной массы, осуществляющей общие, так- называемые «массовые»,
¦реакции. Зти общие реакции обычно возникают в передней части тела
и потом по' осевой мускулатуре распространяются в направлении к
хвосту* В период формирования жабер к этой общей реакции
присоединяется еще движение жаберного аппарата. Еще позднее, с развитием
конечностей, к общей реакции туловища и жабер присоединяются еще
движения конечностей. На этих стадиях развития нельзя вызвать
локальной реакции жабер и конечностей,— на каждое деятельное раздра-
3*
36 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО* МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
жение наступает одна и та же общая реакция с участием жабер и
конечностей. Лишь позднее, в результате дальнейшего развития нервных
путей, ведущих к мышцам жаберного аппарата, создается такая
обособленная двигательная система, которая может вызывать локальную
реакцию дыхания без возбуждения остальной мускулатуры. Гораздо
позднее, уже у взрослых аксолотлей, обособляются также
двигательные системы, которые обеспечивают производства локальных
локомоторных и глотательных движений. Таким образом, у эмбриона
холоднокровных вначале возникает способность к общим диффузным реакциям,
к «массОівыім» реакциям, а уже позже образуются те или другие
локальные рефлекторные реакции. Но между механизмами общих
целостных реакций и локальных рефлексов существует определенная со-
подчиненность: каждая локальная реакция осуществляется благодаря
торможению общей реакции (Когхилл, 12).
Онтогенез рефлекторной деятельности теплокровных (птицы,
млекопитающие) происходит в общем по тому же типу. У них также
локальные рефлекторные реакции возникают после предварительной стадии
общих рефлекторных реакций. Но эта стадия общих реакций
первоначально осуществляется без участия нервіной системы; эмбрион курицы
уже на 5-й день развития начинает раскачиваться всем телом. Это
происходит в тот период, когда в спинном мозгу еще нет развитых
нервных связей. Эта общая реакция осуществляется, повидимому,
непосредственно мышечной системой подобно пульсации сердечной трубки
эмбриона. На 9-й и 10-й дни развития и позднее также наблюдаются
общие движения, но в этот период общие реакции, вероятно,
осуществляются через центральную нервную систему, ибо к этому возрасту в
переднем и заднем рогах спинного мозга уже имеются основные
нервные элементы (Зурабашвили, 18). Кроме того, в этот период развития
наблюдаются такие спонтанные движения, которые являются
характерными компонентами поведенческих актов вылупившегося цыпленка,—
клевательные движения клюва, разгребательные движения ног,
летательные движения крыльев и т. д. (Чичинадзе, 17).
Таким образом, у высших позвоночных изолированные нервные пути
закладываются уже в стадии образования серого и белого вещества в
спинном мозгу. Как показал У инд л (18) на котятах, образование
локализированных координированных реакций конечностей находится в
связи с появлением в нервном сплетении спинного мозга перицеллюляр-.
ных сплетений и конечных луковиц, т. е. синаптических образований.
Но само развитие спинного и головного мозга у млекопитающих
(крыса, человек) не происходит параллельно и в одинаковой степени во
всех частях мозга: передние части мозга развиваются раньше каудаль-
ных. Поэтому рефлекторные реакции возникают на передних
конечностях раньше, чем на задних. Соответственно, у млекопитающих общие
рефлекторные реакции, охватывающие весь организм, наступают
позднее локальных движений конечностей (Уиндл и сотр., 9).
Формирование первоначальных нервных путей происходит под влия-
ОБЩЕЕ ПОНЯТИЕ О ЦЕНТРАЛЬНОЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ 37
нием внутренних факторов. Прежде всего возникают центральные
нервные ядра функциональных систем, а уже потом, на основании их
функционирования, формируются целостные системы со своими
эфферентными и афферентными путями. Эти ядра закладываются и
развиваются первое время независимо от периферии (Анохин, 8; Уиндл и
Бакстер, 9).
Классификация рефлекторных реакций. Рефлекторные реакции
делятся прежде всего на две большие группы: прирожденные и
индивидуально приобретенные рефлексы. Выше мы уже ознакомились с
обеими группами рефлексов. По своему внешнему проявлению те и
другие ничем характерным не отличаются друг от друга. Основное
различие заключается в условиях вызывания рефлексов той и другой
группы и затем в центральных механизмах, управляющих этими рефлексами.
Каждая группа іможет быть подразделена на целый ряд типов, смотря
по их назначению, но каждый тип представлен в обеих группах, как
в прирожденной, так и в индивидуально-приобретенной, так как у
высших позвоночных любой прирожденный рефлекторный механизм может
быть приведен в действие на основании индивидуального опыта.
Главные типы рефлекторных реакций: а) защитные рефлексы, вроде
чесания, поднятия ноги, сотрясения тела, бодания, оскаливания зубов,
лягания, кусания и т. д.; б) рефлексы, участвующие при завладений
пищей: сосание, жевание, глотание, клевание, лакание, подкрадывание,
нападение; в) пищеварительные рефлексы: выделение слюны и
желудочного сока, движение кишечника, выделение желчи; д) рефлексы,
направленные на защиту рода: обнимательные движения, бегание
самца и самки друг за другом, набухание половых органов и др.; е)
ориентировочные рефлексы, направленные на лучшее восприятие
раздражителей: поднятие ушей, поворот глаз и головы, расширение ноздрей
и т. д.; ж) голосовые рефлексы, вроде лаяния, писка, мяукания, пения
и т. д.; эти рефлексы участвуют в разнообразных актах поведения
защиты ках индивида, так и рода; з) тонические рефлексы, которые
защищают организм от падения >в силу собственной тяжести или же
длительно согласуют действие одного двигательного органа с другими:
стояние на ногах, вертикальное или горизонтальное положение шеи,
сгибание передних и разгибание задних ног при опускании головы,
разгибание передних и сгибание задних ног при поднятии головы,
держание хвоста в приподнятом положении; и) рефлексы положения,
которые придают телу нормальное положение: положение головы
теменем вверх, лицом вниз, туловища спиной вверх, животом вниз
и т. д.
Среди всех классов животных одни прирожденные рефлексы
действуют с момента рождения, другие начинают действовать спустя
некоторое время на разных ступенях постэмбрионального развития, и во
всех этих случаях их появление будет зависеть от прирожденной
организации мышечной и нервной систем.
Индивидуальные же рефлексы постепенно возникают в результате
33 ОВЩ-AJFФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
индивидуального опыта в течение всей жизни животного; они появляют>
ея на основе прирожденных рефлексов.
Порядок изучения прирожденной и индивидуально приобретенной
деятельности. Ввиду определенной качественной разницы между при-»
рожденными и индивидуально-приобретенными рефлексами
прирожденная деятельность центральной нервной системы будет рассмотрена
отдельно от индивидуальной. Так как прирожденная рефлекторная
деятельность является основной функцией сдинного мозга и стволовой
части головного мозга, то прирожденная деятельность будет излагаться
в связи с общей физиологией названнш отделов центральной нервной
системы, а индивидуадьнр-приобретенная деятельность будет изложена
после этого — в связи с изложением физиологии большого мозга, одной
из функций которого она является. Поскольку
индивидуально-приобретенная деятельность возникает нц основе прирожденной и пользуется
нервными механизмами последней, изучение прирожденной деятельности,
конечно, должно предшествовать изучению индивидуально
приобретенной.
Тех специальных функций центральной нервной систему, которые
проявляются по отношению к поведению, мы совсем не будем касаться,
так как это не входит в общую физиологию нервной системы.
Означенные функции являются предметом науки о поведении.
ираптипум
Обнажение спинного и головного мозга лягушки. Для физиологических целей
н^жно вскрыть моэг так, чтобы сохранилась его рефлекторная деятельность, поэтому
операция проводится на слегка наркотизированных животных. Лягушку кладут под
стеклянный колпак и рядом с ней кладут вату, смоченную эфиром или хлороформом.
Как только животное перестанет двигаться и отвечать движением на механическое
раздражение кожи, его заворачивают в полотенце так, чтобы была открыта только
голова. Тремя пальцами левой руки—мизинцем, безымянным и средним — держат
тело животного, а большим и указательным пальцами прочно фиксируют его голову
с боков. Правой рукой срезают ножницами кожу сверху, начиная с глаз, до спины»
после чего производят поперечный разрез черепной коробки, между глазами, а затем
продольные разрезы ио бокам черепной коробки. Оторвав вырезанную пластинку
черепной коробки, обнажают головной мозг. Вытекающую при этом кровь удаляют
марлей или ватой.
После этого укрепляют лягушку в специальный станок (рис. 15) и зажимают
между лопастями (ь) станка боковые выступы ее позвоночника, а зажимом а —
передний край головы. Сначала снимают кожу на спине между лопастями, затем
мускулатуру, пока не обнажится позвоночник. После этого острыми, крепкими ножницами
вырезают верхнюю часть позвоночника, начиная или спереди — от границы
продолговатого мозга, или сзади — от последнего позвонка. В последнем случае сначала
отрезают копчиковую кость у ее основания. В обоих случаях одно острие ножниц
вводят в позвоночный канал между мозгом и верхней стенкой позвоночника на
глубину не более 0,5—1 мм Ближайший позвонок перерезают сначала с одного бака, потом
с другого. После этого верхнюю вырезанную часть позвонка отодвигают назад или
вырывают целиком. Затем таким же путем вырезают верхние части следующих
позвонков. После вскрытия позвоночника виден спинной мозг, прикрытый, крупными
кровеносными сосудами и лимфатическим мешком с белым содержимым. Лимфати-
ОБЩЕЕ ПОНЯТИЕ О ЦЕНТРАЛЬНОЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ 39
ческий -мешок также удаляют, после чего обнажается спинной мозг с дорзальной
поверхности. Сверху видны задние корешки Под ними находятся передние корешки,
которые входят в мозг с вентральной стороны. Передние корешки несколько светлее
задних, поэтому их можно и на-глаз отличить друг от друга. На препарате спинного
мозга надо определить плечевой отдел, состоящий из первых трех сегментов.
Сегменты следует считать по корешкам, от II до IV корешка (у взрослых лягушек
первая пара корешков отсутствует, она имеется только в эмбриональном периоде)
Затем отделить грудной отдел — от V до VII
сегмента включительно и, наконец,
поясничный отдел—от VIII до X сегмента
включительно. Крестцовых сегментов два Их
корешки выходят наружу через копчиковую
кость
Операция должна проводиться с большой
осторожностью, чтобы в момент введения
острия ножниц в позвоночный канал не
повредить мозга или задних корешков. К концу
операции или спустя некоторое чремя после
лее наркоз перестает действовать. Животное
может быть использовано для постановки
физиологических опытов, если на раздражение
кожи оно отвечает рефлекторными реакциями
Вскрытие позвоночника у лягушки можно
производить и без наркоза, но в этом случае
операция затрудняется движениями животного
и обильным кровоизлиянием в оперируемых
"участках.
Наблюдение рефлексов на спинномозговой
лягушке. Путем перерезки мозга в области
атланто-затылочного сочленения получают
спинномозговой препарат, на котором все
рефлексы тела и конечностей вызываются через
<мшой мозг- Рис. 15. Станок для*укрепления ля-
1. Оборонительный рефлекс гушки при вскрытии позвоночника
сгибания на задней лапке. Мехами- а — служит для закрепления головы; &—для
ческое равдл>аакение голени и пальцев (щияок "^^SS^^^^t^SS^0^
пинцетом) вызывает рефлекс сгибания:
раздражаемая лапка притягивается к туловищу.
2. Оборонительный рефлекс потирания на задней лапке
Тип первый, раздражение кожи на латеральной поверхности бедра, на боках
туловища и на спине, а также на передних конечностях вызывает рефлекс потирания одного
типа задняя лапка раздражаемой стороны притягивается к раздражаемому участку
кожи и потирает его пальцами. Тип второй: раздражение кожи над ахилловым
сухожилием вызывает вытягивание обеих задних лапок, при этом нераздражаемая лапка
потирает голеностопным суставом раздражаемый участок кожи. Тип третий-
раздражение вокруг анального отверстия вызывает то сгибание, то разгибание
одновременно обеих задних лапок; при этом обе лапки потирают область анального
отверстия голеностопными суставами. Все рефлексы типа потирания имеют тенденцию
повторяться несколько раз после однократного раздражения.
3. Рефлекс разгибания Если надавить на стопы обеих задних лапок
подвешенного спинномозгового препарата лягушки с вентральной стороны, то пальцы
разгибаются; если давление производить сильно, то наступает общее разгибание
конечностей Этот рефлекс — не оборонительный, он имеет отношение к локомоции
Когда лягушка, пытаясь освободиться из сосуда, в котором она находится, ползет
при этом по его стенке, она опирается на задние сильно разогнутые лапки. Это
разгибание должно представлять описанный сейчас рефлекс.
40 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
4. Рефлекс потира'яия на передней лапке. Если положить
спинномозговой препарат лягушки брюхом вверх, то, раздражая кожу на брюхе, можно
наблюдать рефлекс потираний на передних лапках: передняя лапка раздраженной-
стороны будет производить потирание, а другая вытянется вперед, растопырив-
пальцы.
5. Обннмательный половой рефлекс. Берут лягушку-самца, у
которого, в отличие от самки, имеется мозолистый нарост на большом пальце передней
лапки. Весной надавливание на этот нарост, а также на кожу в грудной области
вызывает у спинномозговой лягушки обнимательное движение обеих передних лапок.
Если раздражение производить толстым, с палец человека, предметом, то лягушка
крепко и длительно обнимает этот предмет. Если поднять предмет, животное
повисает на нем и может держаться так в течение нескольких минут. Подобное движение
может быть вызвано давлением на кожу передней лапки или в грудной области в
любое время года. Но этот рефлекс протекает кратковременно и имеет защитный-
характер. Наблюдается он и на самке.
Литература
1. Descartes R.,De homine figuris et latinitate donatus, 1663, Leiden; Traite
de l'homme, 1662.
2. Сеченов И. M., Рефлексы головного мозга, 1863; Избранные труды, М., 1935.
3. Павлов И. П. 20-летний опыт объективного изучения условной нервной
деятельности (поведения) животных, 1923.
4. Б е р и т о в И. С, Исследование индивидуального поведения собаки. 1—10
сообщения, Физиол. журн. СССР, 18—19, 1934—1935.
5. Buddenbrock W., Grundriss d. vergleichen'den Physiologie, Berlin, 1928*
6. Зеленый Г., Собака без полушарий большого мозга. 1-е и 2-е сообщ., Тр.
Об-ва русск. врачей, 80, 50, 471, 1911—1912.
7. Coghill G. Е., Anatomy and the problem of behavior, Cambridge, 1929. Есть
русск. пер.
8. А н о x и н П., Сб. докладов VI Всес. физиол. съезда, стр. 148.
9. Win die W. F.,a. BaxterR. E...J. Сотр. Neurol, 63, 189, 1936, Windl e
a. Fitzgerald. J. E., J. Сотр. Neurol., 67, 493, 1937.
10. Prochaska G-., Dissertation on the functions of the nervous system, 1784.
11. F e а г і n g Fr., Reflex action and study in the history of physiology a.
psychology, 1930.
12. Coghi 1 1 G. E., J. Gen. Psychol., 48, 3, 1936.
13. U n z e г J. A., Prinzip der Psysiologie. Leipzig, 1771, Цит. по Fulton.
Physiology of the nervous system, Univ. Press. Oxford, 1938.
14. P f 1 ii g e г Е., Die sensoriale Funktionen des Ruckenmarks d. Wirbeltiere. Berlin,
1883.
15. Luciani L., Physiologie d. Menschen, Bd. Ill, S. 351^357, 1904,
16. Ч и ч и н а д з ? Н., Тр. Тбилисск. гос. ун-та им. Сталина, 26а, 61,^1944.
17. ЗурабашвилиА,, Тр. Ин-та физиол. им. Бериташвили, 5, 169, 1943.
18. Win die W. P., J. Сотр. Neurol., 50, 479, 1930.
19. Беритов И., Тр. Ин-та физиол им. Бериташвили, 5, 193, 1943.
20. Беритов И., Об основных формах нервной и психонервной деятельности.
Изд. Акад. Наук СССР, 1947.
Глава III
РЕЦЕПТИВНОЕ ПОЛЕ РЕФЛЕКСА И ЕГО ЗНАЧЕНИЕ
Специфичность рецепторов. С каждого рецептора или
чувствительного органа рефлекторная реакция нормально вызывается
раздражениями только одного качества, С глаза, например, рефлекс вызывается
только светом, с уха — только звуком, с органа вкуса — только
находящимся во рту химическим раздражителем с кожи — прикосновением,
повреждением или изменением температуры. Глаз никогда не
раздражается звуками; наоборот, ухо не раздражается светом. Таким
образом, каждое раздражение действует специфически на один из
рецепторов. В некоторых определенных случаях можно возбудить рецептор
и раздражениями другого рода. Так, например, при давлении на
глазное яблоко ощущается сильный свет, во время прилива крови к голове
в ушах слышится шум (раздражение кортиева органа током кровгі4).
Электрическое или иное раздражение зрительного нерва собаки
вызывает поворот глаза и даже всей головы в сторону раздражения и
затем сужение зрачка как бы от сильного света. Электрическим
раздражением рецепторов кожи можно вызвать ощущение прикосновения,
тепла и холода, давления, боли (Беритов и Дзидзишвили, 13; Дзи-
дзишвили, 15).
Значит, неспецифическое раздражение рецептора производит
совершенно такое же ощущение и движение, какое вызывается
специфическим. На это обстоятельство было обращено внимание еще в начале
XIX в. Выдающийся физиолог Мюллер выдвинул «принцип
специфической энергии», предполагая, что внешняя энергия
раздражающего агента при действии на соответствующий чувствующий орган-
превращается в специфическую энергию последнего. Но специфичность
ощущения того или другого рецептора обусловливается, по Мюлле-
р у, не только разной энергией чувствительных нервов, но и теми
частями головного и спинного мозга, куда эти нервы вступают (1).
В настоящее время иикто не признает существования
специфической энергии в рецепторных или чувствительных нервэ-х. Мы знаем,
что непосредственное раздражение кожных или мышечно-суставных
чувствительных нервов вызывает чувство боли и такое же движение
42 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
органов, как раздражение рецепторов кожи и мышц; каждому
знакомо ощущение колотья в пальцах, возникающее при ударе локтем о
что-либо твердое. Это происходит вследствие раздражения того
чувствительного нерва, который оканчивается в рецепторах пальцев.
Отсюда следует, что каждый периферический рецептор представляет
собой лишь воспринимающий аппарат для определенного рода
раздражений; чувствительный же нерв является проводником возбуждений от
этого аппарата в определенные отделы центральной нервной системы.
Качественное своеобразие субъективного переживания и рефлекторной
реакции при раздражении данного рецепторного аппарата
определяется, главным образом, деятельностью центральной нервной системы.
Правда, в настрящее время известно, что характер реакции и
субъективного ощущения зависит также и от самих рецепторов, а именно,
от частоты и интенсивности импульсов, исходящих от рецептора, но все-
таки можно утверждать, что качество реакции определяется в конечном
счете не специфической функциональной деятельностью рецепторов, не
частотой и интенсивностью исходящих от них импульсов, а деятельностью
связанных с рецептором определенных отделов центральной нервной
•системы.
Воспринимающие, или рецептивные, поля кожных рефлексов.
Впоследствии мы будем иметь дело главным образом с теми
рефлекторными движениями, которые вызываются с рецепторов кожи, поэтому
необходимо ознакомиться с этими рецепторами более подробно. В
коже существуют рецепторы, раздражающиеся при действии всякого
рода повреждаюших агентов* укола, сильного давления, прижигания,
электрического тока, химических веществ и т. д. Густая сеть
свободных нервных окончаний в эпидермисе и должна представлять собой
эти рецепторы (рис. 16,1). Кожа рыб содержит только такие
рецепторы, и, вероятно, они служат так же для восприятия легкого
механического раздражения, как прикосновение. У амфибий в
эпидермисе кожи также в изобилии находятся разветвления чувствительных
волоконец. Они и здесь, вероятно, служат для восприятия всякого
рода механических раздражений. Кроме того* в коже находятся
рецепторы, которые раздражаются прикосновением,— например, клетки
Меркеля, тельца Мейснера и др. (рис. 16, II), Такие рецептары
имеются только у высших животных (начиная с амфибий и рептилий)
Они встречаются в коже там, где нет волос на подушках пальцев, на
ладони, на подошве ноги. Но там, где имеются волосы, осязательные
рецепторы располагаются вокруг волос (рис. 16, III). В коже
находятся и такие чувствительные элементы, которые раздражаются
давлением и растяжением; к ним относятся сложные нервные
образования, расположенные под эпидермисом,— тельца Фатер-Паччини
(рис. 16, IV). Эти тельца были обнаружены у человека, у
млекопитающих, а также у птиц. Все перечисленные рецепторы расположены
в коже вперемежку, поэтому с одного и того же кожного участка
РЕЦЕПТИВНОЕ ПОЛЕ РЕФЛЕКСА И ЕГО ЗНАЧЕНИЕ 4&
можно вызвать как оборонительную реакцию в ответ на
повреждающее раздражение, так и реакции, относящиеся к локомоции, тонусу
и т. д. Так, например, если к вентральной стороне ступни задней лап-
¦'
шп ¦¦¦;¦
-¦•:.р,- ¦.;, •
X
• ;. •¦ : ¦;
У
г/і
в И
¦
I—Окончания нервов в эпителии роговицы кролика. Тончайшие волоконца
видны между эпителиальными клетками; они оканчиваются утолщениями
у самой поверхности роговицы. Начинаются эти волоконца из нервного
сплетения, находящегося под эпителием в клетчатке (Кахаль). II— Тельце
Мейснера из кожи пальца человека. Тангенциальный разрез. Хорошо видна
горизонтально расположенная нейрофибриллярная сеть в протоплазме
рецепторных клеток. Снаружи капсула из соединительно-тканных клеток
<Буке). III — Осязательный волос морской свинки. А — наружный слой
соединительной ткани; В — кольцевой синус; С — кавернозная часть
сосудистого слоя; D — нервное кольцо; Е — внутренний слой соединительной
ткани; F — верхнее расширение его; G — афферентное нервное волокно;
Н—свободное ветвление его; I—эпителиальный слой; J—верхний рядтактиль-
Рис. 16. Рецепторы кожи.
ных менисков; К — кольцевой выступ; L — волос (Телло). IV—Тельце
Паччини из кожи человека: а—афферентное волокно; Ъ-—центральное
зернистое вещество; с — капсула. Лежит глубоко под кожей (Кахаль).
кіі спинномозговой лягушки приложить тупой предмет и слегка
придавить, то это вызовет растопыривание пальцев; если же прижать силь-
44 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
нее, то разогнется вся лапка. Если же ущипнуть кожу на той же
вентральной стороне ступни, то вся лапка согнется и притянется к
корпусу. Это известный защитный рефлекс
сгибания (Балиони, 2; Беритов, 3).
Кожа покрывает весь 'организм. Защитная
реакция іможет быть вызвана с любого ее участка, но
реакция, получаемая с одного участка, не совсем
похожа на реакцию, получаемую с другого участка.
Характер защитной реакции сильно меняется »в
зависимости от участка кожи. Существует несколько
типов рефлекса, и каждый тип имеет определенную
воспринимающую поверхность. Так, например, на
задней лапке лягушки можно вызвать следующие
защитные рефлексы: 1) рефлекс сгибания,
выступающий при раздражении кожи на голени ди стопе;
2) рефлекс потирания, вызываемый с наружной
поверхности бедра, с боков туловища, -со спины и
передних лапок; 3) второй тип рефлекса потирания,
вызывающийся с области анального отверстия;
4) третий тип рефлекса потирания, наступающий
при раздражении кожи над ахилловым сухожилием;
5) рефлекс разгибания, вызывающийся
раздражением голени и ступни противоположной задней
лапки. На рис. 17 показаны некоторые из
перечисленных воспринимающих поверхностей. Известный
Рис. 17. Рецептивные
поля рефлексов на
задней лапке спинномоз- м „ ттт
говой лягушки (левая исследователь центральной нервной системы Ш е р-
сторона) и распростра- р и н г т о н назвал воспринимающую поверхность
рефлекса рецептивным
(воспринимающим) полем. Таким образом, для каждого типа
рефлекса существует определенное рецептивное
поле.
Общее свойство рецепторов. Как указывалось
выше, одни рецепторы представляют собой
свободные окончания чувствительных нервных волокон;
нение задних
корешков поясничного
отдела в коше задней
лапки (правая сторона).
На левой лапке: 1 •—
рецептивное поле рефлекса
сгибания; 2, 3 и 4—поля
рефлексов потирания
разных типов; 5 и 6 — поля
разгибания равных
типов. На правой ноге
кружками обозначено рас-
участием специальных клеток, на которых
оканчиваются чувствительные нервные волокна. В тех
пространение первого
поясничного корешка (vjii другие имеют более или менее сложное строение с
от начала спинного мозга г
по Эккеру), крестиками—
второго корешка {IX от
начала),
треугольниками—третьего корешка (X)
и точками -- четвертого случаях, когда рецепторы являются свободными
{Х1)ш
окончаниями чувствительного волокна, адэкватное
раздражение действует непосредственно на эти
окончания. Таковы свободные окончания чувствительных волокон в
эпителии эпидермиса, в сухожилиях, в склере, в соединительно-тканных
прослойках мышечных оболочек всех внутренних органов. Свободные
окончания образуют своеобразные нейрофибриллярные расширения,
петельки, пластинки. В основном они являются механорецепторами,
возбуждаясь при всяких растяжениях и сокращениях органов, а в опреде-
РЕЦЕПТИВНОЕ ПОЛЕ РЕФЛЕКСА И ЕГО ЗНАЧЕНИЕ
45
ленных случаях, видимо, они же дают импульсы, воспринимающиеся
как боль (Лаврентьев, 19).
Когда рецептор имеет сложное строение с участием специальных
клеток, адэкватное раздражение действует на эти специальные клетки.
Таковы все рецепторы давления и прикосновения, все хеморецепторы,—
например органы вкуса, множество рецепторов кровеносных сосудов,
кишечника, почек и других внутренних органов. Процессы, возникающие
в этих специальных клетках под (влиянием раздражения, являются
источником раздражения для окончаний чувствительного волокна
(Лаврентьев, 19).
Физиологическая деятельность каждого рода рецепторов более или
менее одинакова. Каждый рецептор в ответ на адэкватное — т. е. со
ответствующее ему — раздражение отвечает рядом возбуждений. Это
наблюдается в сетчатке глаза при раздражении светом, в кожных
рецепторах — при давлении, при сотрясении одного волоска, в мышечных
рецепторах — при растяжении мышц. Возникающие здесь импульсы
возбуждения по чувствительным нервным волокнам передаются
центральной нервной системе. Чем сильнее раздражение, тем выше ритм
возбуждения рецепторов и тем дольше он держ'ится. Интенсивность
импульсов возбуждения, получаемых от одного рецептора, одна и та
же (Эдриан, 6). Следовательно, деятельность рецепторов протекает
согласно закону возбуждения; каждый раз рецептор возбуждается с
участием всей возбудимой системы. Но, в отличие от нервного волокна,
возбуждение рецептора после однократного раздражения повторяется
много раз друг за другом. Очевидно, после рдноіго раздражения в ре*
цепторах создаются все необходимые физико-химические условия для
того, чтобы после первого изохронного расщепления и последующего
восстановления возбудимой системы вновь многократно происходило
изохронное расщепление и восстановление этой системы без каких-
либо добэ-вочных внешних раздражений.
Продолжительность отдельных импульсов в чувствительных
волокнах от кожных рецепторов различна. В случае возбуждения тактильных
рецепторов лягушки она варьирует между 0.5 и 1.2 о, при болевых
раздражениях она значительно больше; в некоторых случаях болевые
импульсы продолжаются до 10 о (Дзидзишвили, 20).
Многие рецепторы разряжаются импульсами возбуждения при
полном отсутствии внешних раздражений, т. е. спонтанно. Таковы
сетчатка, кортиев орган, статический орган. Зрительный нерв,
например, обнаруживает импульсы возбуждения при абсолютной темноте, а
VIII нерв — при абсолютной тишине, что доказывается путем
отведения биотоков от соответствующих чувствительных нервов. Это
происходит, повидимому, потому, что физико-химические условия в
межуточной (межклеточной) среде и внутри рецептивных клеток не
вполне одинаковы, вследствие чего межуточная среда является
постоянным источником раздражений для чрезвычайно чувствительных
рецептивных клеток (Беритов и Цкипуридзе, 17). Высокой чувствитель-
46 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
ностью рецептивных клеток, собственно говоря, обусловливается то
явление, что в ответ на каждое кратковременное раздражение онш
разряжаются длительным рядом нервных импульсов. Итак, многим
рецепторам свойственна спонтанная ритмическая деятельность.
Интенсивность и ритм нервных импульсов в чувствительных волокнах
при раздражении разных рецепторов дозольно разнообразны. Особенно
редки и слабы импульсы болевых рецеторов (рис. 18). Высший ритм
возбуждения болевых рецепторов очень низок. По данным Эдриана,
при самых сильных болевых раздражениях (раздавливание и
прижигание кожи) высший ритм возбуждения этих волокон не превышает 20 в
1 сек* (7). С эщм свойством рецепторов хорошо согласуется скорость
проведения и вообще физиологические особенности отходящих от них
чувствительных волокон.
Как уже указывалось, скорость проведения возбуждения в
чувствительных волокнах болевых рецепторов много меньше, чем в
чувствительных волокнах тактильных рецепторов. В начале раздражения ритм
возбуждения этих болевых рецепторов медленно нарастает до
максимума; кроме того, импульсы возбуждения продолжают возникать
длительное время и по прекращении раздражения (Эдриан, 10). Это —
характерное отличие болевых рецепторов от рецепторов других видов.
Импульсы чувствительных волокон, идущих от рецепторов
прикосновения, особенно сильны и часты, причем они наиболее часты в самом
начале раздражения этих рецепторов и прекращаются сейчас же по
окончаний раздражения. Так, например, если направить на тактильные1
рецепторы струю воздуха, прерываемую зубчатым колесом 250—300 раз,
то в отходящих от них нервных волокнах іможко наблюдать 250—300
импульгов в 1 сек., т. е. по одному импульсу возбуждения от каждого
дуновения (Каттел и Хоглэнд, 16). Реакции мышечных рецепторов
характеризуются еще более высоким ритмом. При некоторых малых
растяжениях высший ритм возбуждения чувствительных нервов
невысок, ко при больших растяжениях он сильно нарастает» доходя до 290s
в 1 сек. у лягушек и до 500 в 1 сек. у теплокровных (Мэтьюс, 8), и
даже до 1000 в 1 сек. (Ллойд, 18). Толщина мышечных чувствительных
волокон также характерно отличается от кожных: толщина первых
12—20 у, а вторых — 6—12 р (Ллойд, 18).
, Рис. 18 и 19 иллюстрируют ритмическое течение возбуждения
рецепторов. На рис. 18 представлены электротраммы чувствительного
нерва при раздражениях кожных рецепторов лягушки. На рис. 19 дана
картина токов возбуждения чувствительных нервов при раздражении-
одного мышечного рецептора путем растяжения.
Явление адаптации. Во всех рецепторах частота импульсов падает
при длительном раздражении; как говорят, рецептор приспосабливается
к раздражению, адаптируется. Но скорость адаптации не одинакова у
разных рецепторов. Дольше всего разряжаются импульсами
возбуждения мышечные рецепторы—до 10 сек., ів то время как в рецепторах
кожи разряд затухает ц течение 1 сек. Если рецептор приспосабливаете»
РЕЦЕПТИВНОЕ ПОЛЕ РЕФЛЕКСА И ЕГО ЗНАЧЕНИЕ 4Г
медленно, то ритм возбуждения тем выше, чем сильнее раздражение^
Если же рецептор приспосабливается быстро, то меняется главным
образом продолжительность разряда: более сильное раздражение дает
более длительный ряд импульсов возбуждения. Собственно говоря, от
рецепторов глубокой чувствительности, сигнализирующих о положении
В шлейфный осциллограф отводятся импульсы возбуждения дорзальных нервов лягушки,
возникающие при точечном раздражении кожи спины. Отведение в оп. А монополярное, в остальных
опытах биполярное. А, В и С — эффект тактильного раэдражения различных точек Несмотря
на то, что интенсивность раздражения во всех этих случаях одинакова (прикосновение кончиком;
волоса), адаптация наступает в различное время: в случае А появился один импульс, в случае
В — кратковременная группа импульсов, а на С — показано начало импульсации, которая длилась
0.8". D — эффект укола иглой. Показано начало записи, где наряду с быстрыми импульсами
видны медленные болевые импульсы. Время 0.02 сек. (Дзидзишвили).
-
-
тела, непрерывно идут импульсы. Это наиболее медленно
адаптирующиеся рецепторы. Малейшее передвижение конечностей сразу
вызывает сильный разряд импульсов в рецепторах глубокой
чувствительности, который продолжается, пока движется конечность. Когда
двигательный эффект прекращается, поток импульсов значительно
ослабевает (Эдриан, 10).
Явление адаптации подробно было исследовано на мышечных
рецепторах. Оказалось, что при растяжениях мышцы адаптация
происходит тем скорее, чем меньше нагрузка (рис. 28). После прекращения,
растяжения адаптация не исчезает сейчас же, а остается еще некоторое
время. Например, при нагрузке 2—5 г и разгрузке через 5 сек.
адаптация проходит в течение 10 сек. Повторение нагрузки ускоряет
адаптацию, т. е. начальный высокий ритм падает до более низкого уровня
скорее при повторении нагрузки. Состояние адаптации не ставится в
связь с утомлением; утомление рецептора тоже имеет место, но
адаптация в смысле быстрого падения начального ритма не является
следствием утомления рецептора. Адаптация исчезает после растяжения в.
48 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
течение секунд, в то время как утомление преходит много, позднее,
спустя- несколько минут (Бронк, 9; Мэтьюс, 8), *
По данным Эдриана, наибольшей адаптацией обладают тактильные
рецепторы. Степень их адаптации близка к адаптации нервного
волокна. Однако в различных тактильных рецепторах одной и той же
области кожи адаптация может быть выражена в различной степени (Дзи-
дзишвили, 20). На рис. 18 показано, что при слабом тактильном
раздражении кожи лягушки (прикосновение волоском) может возникать
Рис. 19. Биоэлектрические токи возбуждения чувствительного
мышечного нерва при растяжении мышцы на пальце лягушки.
Записывается осциллографом Мэтьюса. Температура 17° С. А — в момент
растяжения тяжестью в 5 г; В — 2 сек. позднее; С — 5 сек. позднее; D — та
же реакция, записанная при большой скорости фотографического аппарата.
На первых трех рисунках время по 0.2 сек., а на D—0.01 сек. Токи
возникают с большой регулярностью и почти одинаковой высоты (Мэттьюс).
импульсация различной продолжительности: при раздражении
некоторых рецепторов возникают всего лишь 1—2 импульса, при
раздражении же других тактильных рецепторов возникает серия импульсов,
постепенно затухающих в течение почти 1 секунды. При давлении на кожу
или же при болевых раздражениях импульсация может продолжаться
несколько секунд.
Основными сведениями о рецепторах мы обязаны прежде всего
Эдриану (6, 10) него сотруднику Мэ ть ю су (8). Они
регистрировали биоэлектрические токи возбуждения чувствительных волокон,
отходящих от рецепторов. В некоторых опытах они регистрировали токи
возбуждения чувствительного нерва при раздражении лишь одного
рецептора, например одного паччиниева тельца или одного мышечного
веретена. Этим путем Э д р и а н изучил тактильные и болевые кожные
рецепторы, а Мэтьюс — проприоцепторы.
Относительно природы явления адаптации сначала предполагали,
что рецептор обладает определенным запасом потенциальной энергии.
РЕЦЕПТИВНОЕ ПОЛЕ РЕФЛЕКСА И ЕГО ЗНАЧЕНИЕ 49
которым он разряжается при раздражении, а затем при отдыхе вновь
восстанавливает его за счет веществ, поступающих из крови
(Мэтьюс, 8). Но в последнее время при ближайшем исследовании ме-
ханорецепторов кожи лягушки было сделано более конкретное
заключение о происхождении адаптации. Механорецепторы кожи лягушки
древовидно разветвляются в коже и оканчиваются частью в эпидермисе
тончайшей сетью голых волоконец, расположенных между
эпителиальными клетками, частью под эпидермисом — отдельными колбочками
(рис. 16, I). Часть волоконец в эпителии раздражается легким
прикосновением, а колбочки под эпидермисом — давлением, повреждением
и химическими агентами. От раздражения прикосновением возникают
быстрые импульсы типа Л, которые очень быстро адаптируются. От
давления же возникают импульсы типа В я С, которые адаптируются
менее быстро. Если соскоблить эпителий и таким образом удалить
лежащие здесь рецепторы, то механическое раздражение производит
только импульсы типа С. Далее было установлено, что при
механическом раздражении кожи ее эпителиальные клетки выделяют калий
(Хоглэнд, 14). В эпителиальных клетках содержится в 16 раз больше
калия, чем в окружающей их лимфе и плазме. Он там присутствует
большей частью в способной к диффузии форме и вымывается оттуда,
если кожу положить на несколько часов в дестиллированную воду.
Калий — единственное вещество, выделяющееся из кожи под влиянием
механического раздражения.
На этом основании было сделано предположение, что при
механическом раздражении кожи эпителиальные клетки выделяют калий,
который действует на нервные волоконца и понижает их возбудимость.
Вследствие этого механическое раздражение быстро перестает
раздражать нервные окончания, т. е. происходит быстрая адаптация.
Восстановление от адаптации получается вследствие диффузии калия и
вымывания его током крови.
Что в данном случае адаптация действительно обусловлена
химическим веществом, видно из следующего. Если после наступления
адаптации промыть кожу нормальным раствором Рингера, то адаптация
быстро проходит. В то же время промывание кожи раствором Рингера,
содержащим в 15 раз больше калия, чем в норме, заметно ускоряет
адаптацию. Все это свидетельствует о том, что адаптация
обусловливается калием. Можно показать, что калий выделяется эпителиальными
клетками, а не самими нервными волоконцами; это видно из того, что
сейчас же после длительного раздражения кожного чувствительного
нерва электрическим током нервные волоконца в эпителии, несмотря
на длительное возбуждение, не оказываются адаптированными: они
отвечают, как обычно, на механическое раздражение кожи. Значит, от
возбуждения нервных волоконец калий не выделяется, адаптация не
наступает. На основании всех этих фактов Хоглэнд (14) имел
право заключить, что быстрая адаптация голых рецепторов в эпидермисе
обусловливается парализующим действием калия, выделяемого из
эпителиальных клеток. Калий, как известно, в малой концентрации по№-
4 И С Беритов
50
ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
шает возбудимость нервной ткани, а в большой дозе, наоборот,
понижает ее. Нужно думать, что та доза калия, которая выделяется из
эпителиальных клеток, достаточна для того, чтобы подействовать на
нерівные волоконца угнетающим образом. 'Менее быструю адаптацию на
давление и повреждение
кожи лягушки Хоглэнд
(14) считает следствием
отдаленности соответственных
рецепторов от эпидермиса.
Он же считает, что
различная инкапсуляция многих
рецепторов имеет
назначение защитить рецептор от
действия калия,
выделяемого окружающими
эпителиальными клетками при их
механическом раздражении.
Значение рецептивного
поля в координации
рефлекса. То обстоятельство, что
каждый рефлекс имеет свое
рецептивное поле, дает
возможность изучить роль
последнего в координации
рефлекса. Можно взять
чувствительный нерв, отходящий от рецептивного поля данного
рефлекса, и раздражать его механически, химически или
электрическим током. Во всех случаях рефлекс вызывается с точно таким
же постоянством и с такой же координацией, как и при раздражении
самого рецептивного поля. Так происходит и в том случае, если
чувствительный нерв перерезан и раздражается его центральный отрезок.
Например, на спинномозговой лягушке раздражение центрального
отрезка п. peroneus, который получает волокна из рецептивного поля
сгибания, дает вполне координированный рефлекс сгибания. Это было
доказано и на других животных. Так, например, легкое раздражение
кожи на шее, на плечах и спин? кошки, собаки, кролика и других
животных вызывает рефлекс чесания (рис. 20). Этот рефлекс состоит в
притягивании задней ноги раздражаемой стороны к раздражаемому
участку и в ритмических движениях ног. В это время другая задняя
нога разгибается, шея и туловище поворачиваются в раздражаемую
сторону. Данный сложный рефлекс вызывается также электрическим
раздражением чувствительных нервов соответствующего рецептивного
поля (Шеррингтон, 5).
Таким образом, рецептивное поле служит только для восприятия
раздражений, в координации же рефлекса оно не участвует.
Рис. 20. Рецептивное поле рефлекса чесания
на собаке (обозначено на рисунке цифрами)
(А) и рефлекторная дуга чесания (В).
dR> BR— чувствительные нейроны от рецептивного
поля чесания;Рс(, РВ — интраспинальные нейроны или
внутримозговые; FC — двигательные нейроны
(Шеррингтон).
РЕЦЕПТИВНОЕ ПОЛЕ РЕФЛЕКСА И ЕГО ЗНАЧЕНИЕ
51
Практикум
1. Опытное определение рецептивных полей рефлексов на задней лапке
спинномозговой лягушки. Раздражают разные участки кожи механическим путем
(пщпок пинцетом, давление) или индукционными ударами и таким путем находят
рецептивные поля известных рефлексов: сгибания, потирания и разгибания на задних
лапках; рефлексы потирания и обнимания — на передних лапках.
2. Наблюдение рефлексов при раздражении чувствительного нерва. На
спинномозговой лягушке обнажают п. peroneus в области голени. Перевязывают нерв и
перерезают его дистально от нитки. Затем подвешивают препарат к штативу и
раздражают центральный конец нерва механически или электрическим током. Рефлекс
потирания можно вызывать раздражением п. cutaneus femoris lateralis, который
получает волокна от наружной стороны бедра, или же раздражением п. brachialis
передней лапки.
3. Осциллографическая регистрация афферентных импульсов при раздражении
рецепторов кожи лягушки. На коже спины'лягушки делается медиальный раэрез. Под
кожей видны длинные нитевидно тянущиеся кожные нервы (nervi cutanei dorsi
mediates). При ближайшем рассмотрении можно заметить, что это нервы различной
толщины. Отцрепаровыівается лосжут кожи вместе с соеданіяющимся с ним нервом.
Вырезанный лоскут расправляется и помещается на кусочке фильтровальной бумаги
смоченной физиологическим раствором. Нерв перекладывается на электроды,
соединенные с осциллографом через ламповый усилитель. Нужно обратить .внимание на
то, чтобы нерв не высыхал; для этой цели рекомендуется поп>узить нерв с
электродами в парафиновую ванночку, наполненную физиологическим раствором. Перед
начатом отведения биотоков нерв извлекается из ванночки. Поверхность кожи
раздражается прикосновением кончика волоса^ давлением щепинкойі, уколом стеклянной
иглой или же каплей 1—2% соляной кислоты. При раздражении волоском различных
то»ек получаются быстрые импульсы, выражающие возбуждение тактильных
рецепторов. При уколе и при воздействии кислотой, наряду с быстрыми импульсами,
наступают медленные потенциалы. Адаптация на продолжительное тактильное
раздражение наступает быстрее, чем на болевые раздражения.
Литература
LMuller J., Handbuch d. Physiologie des Menschen, Bd. II, 249—275,
1840.
2. BaglioniS., Zur Anal. d. Reflexfunktion. Wiesbaden, 1907.
3. Beritoff J., Pfl. Arch., 151, 171, 1913; Учение об основных элементах
центральной координации скелетной мускулатуры, Петроград, 1916.
4. Беритов И., Тр. Петроградского об-ва естеств., 42, 267, 1913.
5. Sherrington С, The integrative action of the nervous system. London, 1915.
6. AdrianE. D., Erg. Physiol., 26, 501, 1928.
7. ЭдрианЕ., Ученые записки Казанск. ун-та, в. 1—2, стр. 3, 1932.
8. MatthewsB. N. С, J. Physiol., 71, 64, 1931; 78, 1, 1933.
9. Br on k D. W., J. Physiol., 67, 270, 1929.
10. Adrian E. D., The mechanism of nervous action, 1932; Русск. пер. Механизм
нервной деятельности. Биомедгиз, 1935.
11. Э д р и а н Э. Д., Физиол. журн. СССР, 19, 405, 1935.
12. С 1 ark D., Н ugh es J. a. Gasser H. S., Am. J. Physiol., 114, 69, 1935.
13. Беритов И. иДвидзишвили Н., Труды биологического сектора
Грузинского филиала АН СССР, 1, 5, 1934.
14. Hoagland Н., Cold Spr. Harb. Sympos. Quant. Biol., 4, 267, 1936.
15. Дзидзишвили H., Сообщения Грузинского филиала АН СССР, 4, 301,1940.
16. С a 11 е 11 Мс. a. Hoagland. Н., J. Physiol., 72, 392, 1931.
52 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
17. Беритов И. и Цкипуридзе Л., Тр. Ин-та физиол. им. Бериташвили,
Б, 215, 1943; Сообщ. Акад. Наук Груз. ССР, б, 103, 1944.
18. Lloyd D., J. Neurophysiol., 6, 294, 4943; 6, 317, 1943.
19. Лаврентьев Б. И., Журн. общ. биол., 4, 232, 1943.
20. Дзидзишвили Н. Н., Рецепторная функция кожи. Тр. Ин-та физиол. им.
Бериташвили, т. 7, 1948.
Глава IV
АНАТОМИЧЕСКАЯ ЛОКАЛИЗАЦИЯ КООРДИНИРУЮЩИХ
АППАРАТОВ
Роль задних и передних корешков в рефлекторной реакции
спинного мозга. Все чувствительные волокна от рецепторов конечностей,
шен и туловища вступают в спинной мозг через задние корешки
спинного мозга. Чувствительные волокна одного корешка начинаются в
определенной кожно-мышечной области, как это, например, показано
для поясничных задних корешков лягушки на рис. 17.
Передние корешки проводят возбуждение из спинного мозга на
периферию. Они производят сокращение імышц и потому называются
двигательными. Несмотря на то, что передние корешки своими
многочисленными дендритами стоят в связи с разными центральными
элементами мозга, а их волокна имеют коллатерали, которые также
заканчиваются в сером веществе мозга, раздражением одного двигательного
корешка нельзя вызвать внешних двигательных реакций через другие
двигательные корешки, т. е. рефлекторных реакций. Последние можно
-вызвать только раздражением задних корешков или задних и боковых
столбов, где проходят чувствительные волокна. На этом основании был
сформулирован закон одностороннего проведения
возбуждения в центральной нервной системе от чувствительных нейронов к
двигательным.
Значение спинномозговых узлов в координации рефлекса. Прежде
чем возбуждение дойдет от рецептивного поля до спинного мозга, оно
должно пройти через спинномозговой нервный узел, где находятся
клетки чувствительных нейронов. Из каждой такой клетки выходит один
нейрит, который в области узла делится на две ветви. Одна ветвь идет
к рецептивному полю и там оканчивается в рецепторе; другая вступает
в спинной мозг, где дает много боковых ветвей, так называемых кол-
латералей. Эти коллатерали оканчиваются здесь же или в головном
мозгу. Следует решить, какую роль играет этот узел в рефлекторной
деятельности.
Опыт Штейнаха с удалением чувствительных клеток из
спинномозгового узла. В спинномозговых узлах лягушки клетки расположены
54 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
группой в одном месте, именно на дорзальной стороне. Штейнаху
удавалось удалить эту часть так, что при этом не повреждались другие
части узла. Эта операция не прекращала рефлекторного действия через
данный узел. В продолжение нескольких дней рефлекс вызывался
раздражением того воспринимающего поля, чувствительные нервы
которого проходили через оперированный узел. Гистологическая картина
после операции была такова: часть клеток вовсе не существовала,
остальная часть была перерождена, дегенерирована; нейриты же этих
клеток более или менее уцелели. Отсюда было сделано заключение,
что импульсы возбуждения от воспринимающего поля могут проходить
в узле без участия клеток, что они імогут с периферической ветви
нейрита перейти непосредственно на центральную.
Если раздражать чувствительный нерв, а его продолжение до и
после узла поочередно отводить в цепь гальванометра или осциллографа,
то ритм, а также и общий ход биоэлектрических токов возбуждения
будут одни и те же в обоих случаях отведения. іКроме того, при
раздражении заднего корешка впереди узла токи возбуждения появляются
в нервном стволе позади узла; следовательно, возбуждение в этом узле
проводится в обоих направлениях точно так же, как и во всяком
периферическом нервном волокне (Беритов, 4).
Характер рефлекса при раздражении чувствительного корешка.
Роль узла в координации рефлекса можно исследовать, если вызывать
рефлекс непосредственным раздражением самого чувствительного
корешка. В некоторых случаях чувствительные волокна от одного
рецептивного поля проходят через строго определенные задние корешки.
Если такой корешок будет перерезан, а его центральный конец будет
раздражаться, то характер и координация вызванного рефлекса будут
такими же, как и при раздражении самого поля. Например, у лягушки
IX и X корешки получают волокна главным образом с рецептивного
поля сгибания.1 Раздражение их вместе или в отдельности вызывает
рефлекс сгибания (Беритов, 1). То же самое может быть показано и в
отношении других рефлексов; например, Шеррингтон (2) показал
это по отношению к рефлексу чесания.
В некоторых случаях, однако, раздражением чувствительного нерва
не удается вызвать такой же эффект, какой получается при
раздражении рецептивного поля. Нервные стволы и задние корешки большей
частью содержат чувствительные волокна с рецептивных полей разных
рефлексов. Поэтому раздражение их влечет за собой появление
нескольких рефлексов в одном и том же двигательном органе. Следова-
1 Спинной мозг лягушки состоит из 10 сегментов (см. рис. 4 первого тома)
I корешок принадлежит шейной части (Мі); II и III корешки — плечевой
части (корешки М? и М3); IV—VI корешки — грудной (Mt—Мб); VII—IX
корешки— поясничной (М7—Мэ) и X корешок — крестцовой (Мі0). По Эккеру, у
эмбриона имеются две пары шейных корешков; из них передняя потом атрофируется.
Поэтому существующая у взрослых первая пара фактически является второй;
сообразно этому нумерация остальных корешков должна быть увеличена на одну единицу.
Мы придерживаемся последней нумерации, согласно Эккеру.
АНАТОМИЧЕСКАЯ ЛОКАЛИЗАЦИЯ КООРДИНИРУЮЩИХ АППАРАТОВ 55
тельно, вызванное раздражением нервных стволов и задних корешков
движение должно быть более сложным, чем от раздражения одного
какого-либо рецептивного поля. Такую картину очень часто можно
видеть на лягушечьем препарате. Чувствительные нервы с рецептивного
поля рефлекса потирания на бедре обыкновенно входят в мозг через
VIII корешСк, но нередко они частично ©ходят и с IX корешком. Через
IX корешок © мозг входят чувствительные волокна и из рецептивного
поля сгибания. Поэтому очень часто раздражение IX корешка не
вызывает исключительно одного какого-либо рефлекса. Наоборот,
последний представляет смешанное движение, которое включает элементы
рефлекса сгибания и потирания: конечность сгибается, а пальцы
производят ритмические движения, характерные для рефлекса потирания (Бе-
ритов, 3).
Таким образом, спинномозговые узлы, а также узлы черепномозго-
&ых нервов не принимают участия в координации рефлекса.
Следовательно, центральная нервная система, г. е. спинной и головной мозг,
представляет тот единственный орган, в котором протекают процессы
координации рефлекса.
Сегментальное расположение координирующих аппаратов.
Существование каждого рефлекса предполагает наличие известного рода
аппаратов координации в спинном или головном мозгу. Еще С п
алла нц а ни (1778) ясно показал, что у жабы защитный рефлекс
передних конечностей можно наблюдать и в тоім случае, если нервы
передних конечностей находятся в связи с одним только III сегментом, т. е.
когда спинной мозг перерезан впереди и позади данного сегмента.
Отсюда видно, что координирующий аппарат защитного рефлекса должен
находиться в III сегменте. Ринберк (6) повторил этот опыт и на
других животных. Он нашел, например, что у собаки для вызова
грудных сегментальных рефлексов вполне достаточно" наличия одного из
грудных сегментов. Затем Данилевский (7), Бабак (8) и другие
обнаружили, что поясничная часть спинного мозга лягушки способна
на производство ритмических рефлексов, как ходьба и прыгание. Все они
ставили опыты с перерезкой спинного мозга между поясничной и
грудной областями. Следовательно, координирующий аппарат ритмических
рефлексов задних конечностей должен находиться в спинном мозгу, в его
поясничном отделе. Такое же заключение было выведено относительно
ритмических рефлексов у разных высших животных.
Для решения вопроса о местоположении координирующих
аппаратов самым лучшим методом должно считаться локальное (местное)
отравление центральной нервной системы стрихнином. Если отравить
маленькой дозой стрихнина центральную нервную систему (смазать
•0.1% раствором поверхность мозга или приложить к ней смоченную в
этом растворе фильтровальную бумагу величиной в 1—2 мм2), то яд
подействует на мозг характерным образом. Если смазать стрихнином
заднюю поверхность спинного мозга лягушки, то это повлечет за собой
необыкновенное усиление рефлекторных реакций, которые вызываются
56 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
раздражением задних корешков отравленной части мозга (Беритов, 9).
Такое локальное отравление IX—X сегментов спинного мозга у лягушки
способствует усилению тех оборонительных и ритмических рефлексов,
которые обычно вызываются раздражением чувствительных корешков
этих сегментов. Отравление же VII—VIII сегментов с задней
поверхности усиливает тот рефлекс потирания, который обыкновенно
вызывается с того рецептивного поля, где начинаются нервные волокна
VII—VIII задних корешков (Дюссер де Барени, 10; Беритов, 9). В то же
Рис. 21 Рефлекторные реаьции hj отравленного
стрихнином и неотравленного участков мозга.
Спинномозговая лягушка. Поясничный отдел мозга обнажен,
и все задние корешки этого отдела перерезаны. В области
IX—X сегментов с левой стороны на заднюю поверхность
приложен стрихнин. Опыт записан в стадии сильного
отравления. Верхняя кривая от m. semitend. sin., нижняя — от triceps
sin. В опыте А раздражается X задний корешок отравленной
стороны, при этом наступает сильный стрихнинный трехфазный
эффект. В опыте В раздражается X задний корешок правой
стороны. В этом случае на той же конечности получается
слабый сгибательный рефлекс. Внизу сигнальная линия
отмечает время раздражения. На ней цифра в скобках обозначает
ритм раздражения в 1 сек., а без скобок-силу раздражения в
сантиметрах расстояния катушек.
самое время все другие рефлексы с других рецептивных полей проте-
ш кают нормально. Так, например, на рис. 21 дается прекрасная
иллюстрация того, как после стрихнинного отравления IX и X сегментов в
одной симметричной половине мозга раздражение X заднего корешка
отравленной стороны производит сильный стрихнинный эффект на
задней ноге той же стороны, а такое же раздражение того же корешка
другой стороны вызывает на той же ноге нормальный эффект без
признаков стрихнинного отравления.
Вышеприведенные факты ясно указывают, что координация каждого
рефлекса происходит как раз в том участке мозга, куда всходят
чувствительные нервы соответствующего рецептивного поля.
Принцип общего конечного пути. Изучение рефлексов показало,
что начальный нейрон каждого рефлекторного пути, т. е. афферентный,
АНАТОМИЧЕСКАЯ ЛОКАЛИЗАЦИЯ КООРДИНИРУЮЩИХ АППАРАТОВ 57
принадлежит одной какой-либо рецептивной точке, в то время как
конечный, эфферентный путь является общим для всех рефлекторных
путей. Он является общим по отношению к рецепторам всего тела. Эта
конвергенция или схождение всякого рода нервных путей
афферентного, чувствительного характера в общем конечном двигательном пути
происходит при помощи промежуточных нейронов, общих для
определенной группы афферентных нейронов (Шеррингтон, 15).
Общие конечные пути однородны по своей функции: все они
проводят возбуждение из центров к рабочему органу, причем они способны
проводить возбуждение разных ритмов, вплоть до предельного ритма,
характерного для двигательных нервов; когда производится два или
больше раздражений в рецептивном поле одного рефлекса или
вызывается два однородных или гармоничных рефлекса, то общий путь
проводит возбуждение одновременно от рефлекторных путей того и
другого рефлекса. Когда же происходит возбуждение рецептивных
полей двух противоположных рефлексов, общий путь проводит
возбуждение не от обоих рефлексов одновременно, а только от одного. В
таких случаях говорят об антагонистических рефлексах.
Значение двигательных нейронов в координации рефлекса.
Каждый сегмент спинного мозга состоит из двоякого рода нейронных
элементов: 1) двигательных нейронов, которые располагаются в передних
рогах мозга и посылают свои нейриты в периферические органы, и
2) промежуточных нейронов, которые находятся в большом количестве
в задних и боковых рогах и нейриты которых распространяются в
области -мозга. Мы должны исследовать, какое участие принимают в
координации рефлекса те и другие нейроны. Действие стрихнина на
рефлекторный аппарат обнаруживается только в случае отравления им
задней поверхности мозга. Если же смазать стрихнином переднюю
поверхность, то он действует на рефлекторный аппарат совсем
по-другому: нарушает координацию рефлекторных движений, а интенсивность
рефлекторных реакций оставляет без изменения. Это явление
наблюдается только в том случае, если действие стрихнина ограничено
передней половиной мозга и не распространяется на заднюю половину
(Беритов, 11). Этот факт доказывает, что двигательные нейроны,
которые располагаются в передней поло&ине мозга, не принимают
прямого участия в координации рефлекса. Следовательно, центральная
координация рефлекторных реакций должна представлять функцию
задней половины спинного мозга, включающей внутримозговые
промежуточные нейроны.
Задняя половина спинного мозга включает, кроме нервных клеток
промежуточных нейронов, еще нейропиль. Есть основание утверждать,
что в координации рефлекторных реакций участвуют не только
промежуточные нейроны, но и этот нейропиль. Ниже будет приведен ряд
фактических доказательств и соображений в пользу этого положения.
Сейчас лишь отметим в общих чертах роль нервных элементов того и
другого рода в координации рефлекторных реакций. Промежуточные
58 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
нейроны мы считаем проводниками возбуждений в одном
определенном направлении — от определенного чувствительного нейрона к
определенным двигательным нейронам, а нейропиль мы считаем носителем
общих функций, производящим общее понижение или повышение
возбудимости в центральной нервной системе. Значит, согласно нейронно
нейропильчой теории, координирующий аппарат каждого рефлекса
состоит, с одной стороны, из определенных промежуточных клеток и их
аксонов, с которыми рецептивное поле данного рефлекса стоит в
наиболее близкой и тесной связи, а с другой — из окружающего эти
клетки и аксоны нейропиля, в котором принимают главное участие денд-
риты как промежуточных, так и двигательных нейронов.
Вышеприведенное положение, что двигательные нейроны сами не участвуют в
¦координации, что они только проводят координированные импульсы, разделяют не
все физиолога. Например, Филиппсон (12), Введенский (13) и Грэим
Б р о у н (14) приписывают координирующее действие клеткам двигательных нейронов.
Филиппсон утверждает это на основании гистологических наблюдений К р а у з е,
который видел, что, с одной стороны, дендриты двигательных клеток и, с другой
стороны, коллатерали их нейритов осуществляют сложную связь между двигательными
клетками как одной стороны мозга, так и обеих симметрических сторон «Таким
образом,— заключает автор,— двигательные клетки получают возбуждение не только с
чувствительных нейронов, но каждый из них получает возбуждение и с остальных
двигательных клеток. В этом заключается главный элемент координации». Но против
такой координирующей роли коллатералей двигательных нейронов говорит тот факт,
что раздражением передних двигательных корешков нельзя вызвать рефлекторные
двигательные реакции и вообще двигательные реакции через другие двигательные
яейроны. Введенский пришел к подобному выводу, работая над отравленным
стрихнином рефлекторным аппаратом Он заметил, что в одной стадии отравления
торможение протекало в двигательном аппарате, а не в промежуточных нейронах.
Означенное торможение вызывалось раздражением большой частоты и силы, как это
происходит в нервно-мышечном препарате. Позже он воспользовался этим
заключением и отнес торможение нормального координирующего аппарата к функциям
двигательного нейрона.
Однако тот факт, что двигательный нейрон впадает в пеосимальное состояние
при некоторой высокой частоте и силе раздражения, ни в коей мере не может
говорить за его координирующую роль. Пеосимальное явление свойственно всем нервным
элементам, проводящим возбуждение по закону возбуждения с рефракторными
фазами. Двигательный нейрон в целом, в частности его клетки подчиняются этому
закону. Известно, что возбуждение двигательного нейрона антидромной волной создает
рефракторное состояние по отношению к импульсам, приходящим нормальным путем
из афферентных или промежуточных нейронов. Отсюда следует, что ів пессимальном
-состоянии двигательного нейрона, наступающем при некоторой высокой частоте
возбуждения, нужно видеть проявление рефракторной фазы, а не спинномозговой
координации.
Существует еще одно соображение относительно места координации. Ш е р р и н г-
тон (15) и некоторые другие английские физиологи придерживаются того мнения,
•что координирующие иннервации возникают вообще не в клетках нейронов, а вокруг
клеток, именно между клетками двигательных нейронов и оканчивающимися здесь
волокнами чувствительных и промежуточных нейронов. Эта синаптическая теория
предполагает существование вокруг клетки,— между нею и находящимися здесь
нервными окончаниями,— особой физической среды. В этой среде, вследствие чисто
физических процессов диффузия, осмотическое давление, электрические процессы и др.,—
нервные импульсы иногда проходят (возбуждение периферического органа), а иногда
АНАТОМИЧЕСКАЯ ЛОКАЛИЗАЦИЯ КООРДИНИРУЮЩИХ АППАРАТОВ 59
не проходят (торможение этого органа). В настоящее время это представление о роли
синаптической физической среды в координации оставлено. Но синапсы как
морфологические образования продолжают привлекать внимание исследователей. В
настоящее время этим синапсам также приписывается координирующая роль, но теперь и
?амо представление о синапгах другое и сама роль их в координации представляется
иначе. Обо всем этом подробно будет говориться ниже. ,
Анатомические основания рефлекторных путей. Согласно
вышеприведенному заключению, простейший и кратчайший рефлекторный
путь или так называемая рефлекторная дуга у позвоночных
животных должна состоять из трех нейронов: 1) афферентных, или
чувствительных, нейронов, которые приносят периферические импульсы от
рецепторов в центральную нервную систему, 2) промежуточных,
которые проводят эти импульсы к двигательным нейронам, и 3)
эфферентных, или двигательных, нейронов, которые передают их далее
периферическому двигательному органу, так называемому эффектору.
На рис. 22 дана схема рефлекторной дуги координированных
рефлексов сгибания, разгибания и рефлекса потирания с точки зрения
нейронной теории. По этой теории вся рефлекторная реакция с
возбуждением одних мышц и торможением других осуществляется путем
проведения в спинном мозгу афферентных импульсов возбуждения
через нервные клетки и их аксоны.
На рис. 23 дана схема рефлекторной дуги тех же рефлексов,
согласно нейронно-нейропильной теории. Здесь коллатерали заднекореш-
ковых волокон, затем клетки и аксоны промежуточных нейронов
служат для проведения афферентных импульсов возбуждения к мышцам,
а нейропиль — для торможения их. На непременное участие
промежуточных нейронов в рефлекторных реакциях указывают как старые, так
и новейшие гистологические исследования. Так, например, после
перерезки задних корешков кошки на клетках промежуточных нейрояов
оказывается гораздо больше дегенерированных окончаний заднекореш-
ковых волокон, чем на клетках двигательных (Гофф, 21).
Кроме того, известно, что каждый чувствительный нейрон, проходя
по задним столбам спинного мозга, отдает свои коллатерали не в одном
сегменте, а в целом ряде сегментов как впереди места вхождения, так
и позади него. Вследствие этого каждое рецептивное поле и каждый
чувствительный нерв имеют возможность влиять почти на всю
центральную нервную систему. Правда, каждый чувствительный нейрон
связан особенно интимно с двигательными нейронами
соответствующего сегмента и легче всего влияет на эти нейроны. Но факты
показывают, что при определенных условиях данные чувствительные
нейроны способны влиять и на двигательные нейроны других сегментов.
Кроме того, промежуточные нейроны связаны не с одним каким-либо
комплексом двигательных нейронов. Они связаны также более или
менее интимно с двигательными нейронами разных отделов мозга.
Известно, например, что дыхательный центо, который лежит в
продолговатом мозгу, при определенных условиях в состоянии возбудить всю
мускулатуру тела. Следовательно, не может быть речи о строго изолиро-
60 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
ванных нервных путях для рефлекторных реакций. Каково бы ни бьыо
раздражение, и где бы оно ни происходило, вызываемое им
возбуждение должно выходить из пределов соответствующей рефлекторной дуги
5 Q 5 и 5
Передняя
jJOnHQ
ЗаОняя
/іапНо
Передняя
J7QnHQ
Рис. 22. Схематическое изображение
рефлекторных дуг у лягушки
согласно нейронной теории.
Изображены: часть спинного мозга, кожа
на лапке, мышцы-антагонисты и нервные
связи между ними. 1 — чувствительный
нейрон от воспринимающего поля задней
конечности; 2, 2' — промежуточные
нейроны спинного мозга, которые производят
координированную иннервацию в рефлексе
сгибания; 3 — двигательный нейрон
разгибателя; 3' — двигательный нейрон
сгибателя. Плюсы на этих нейронах
обозначают возбуждение, минусы — торможение;
стрелка везде — направление процесса.
V — чувствительный нейрон
воспринимающего поля потирания передних
конечностей; 2*, 2*'— координирующие
нейроны рефлекса потирания: возбуждение
и торможение этой дуги не указаны. А,
А' — клетки чувствительных нейронов;
4 — задний рог серого вещества спинного
мозга; 4'—передний рог этого же вещества;
5 — задние столбы пинного мозга;
5'—передние столбы
Рис. 23. Схематическое изображение
рефлекторных путей по нейронно-
нейропильной теории.
Те же афферентные нейроны и двигательные
нейроны, что и на рис. 22, связываются
между собой не только через
промежуточные нейроны, но и через нейропиль,
образуемый девдритами и коллатералями
нейритов всех этих клеток. От задней лапкв
идут два афферентных нейрона: один—для
сгибательного рефлекса (1А), а другой —
для разгибательного (У А'). Когда
возбуждается первый афферентный нейрон, то
через промежуточный нейрон (2), а также
через коллатераль с возбуждается
сгибатель, а разгибатель через нейропиль
тормозится. Когда же возбуждается второй
афферентный нейрон, то через
промежуточный нейрон (2') и коллатераль с'
возбуждается разгибатель, а сгибатель
через нейропиль тормозится. От передней
лапки идет один афферентный нейрон (А"):
он через вставочный (2*) нейрон
возбуждает разгибатель, а через нейропиль
тормозит сгибатель.
АНАТОМИЧЕСКАЯ ЛОКАЛИЗАЦИЯ КООРДИНИРУЮЩИХ АППАРАТОВ 61
и захватывать в большей или меньшей степени остальную нервную
систему.
Обычно представляют себе, что передача возбуждения с афферентного нервного
волокна на промежуточный и двигательный нейроны происходит через клетки этих
нейронов. Но такое представление отнюдь не обязательно Афферентный и другие
нейроны оканчиваются не только на теле клетки, но и на том холмике, где начинается
аксон клетки. Значит, возбуждение может передаться с нейрона на нейрон и без
участия самого тела клетки. А опытами на ганглии мантии одного из головоногих
моллюсков (Loligo vulgaris) было доказано, что на самом деле сигааптическая передача
возбуждения может происходить без участия клеточных тел. Здесь из гигантских
клеток, расположенных на краю ганглия, выходят гигантские нервные волокна к
мышечным ©олокнам. От этих волокон еще в ганглии отходят коротенькие коллатерали,
которые вступают в синаптичесние контакты с окончаниями нервного волокна, идущего
из центральной нервной системы. Если отрезать клеточную часть, то ганглий вое же
продолжает проводить возбуждение от пресинаптического волокна на постсинаптиче-
ское. Это указывает, что ядро, тельца Ниссля, аппарат Гольджи и другие составные
части клетки не являются необходимыми для осуществления передачи возбуждения с
нейрона на нейрон (Юнг, 22).
Если нельзя говорить о строго локализированных рефлекторных ду-
іах, то тем более нельзя признавать существование строго обособленных
координирующих аппаратов в спинном мозгу. При некоторой
нормальной возбудимости центральной нервной системы каждый
чувствительный нерв вызывает координированную реакцию через тот отдел мозга,
куда входит данный нерв непосредственно. Но при некоторой
повышенной возбудимости в данных координирующих аппаратах последние будут
вовлекаться в работу и при раздражении других чувствительных нервов.
Более того, при нормальной возбудимости центральной нервной системы
данный чувствительный нерв вызывает реакцию исключительно через
определенный комплекс промежуточных, координирующих нейронов; при
некоторой повышенной возбудимости двигательных клеток данный нерв
вызывает реакцию и помимо координирующего аппарата, прямо из
двигательных клеток, и притом тем сильнее, чем они анатомически ближе
к іместу вхождения чувствительного нерва (Беритоз, 9), Отсюда
следует, что фактически нет строго обособленного координирующего
центра, который действовал бы исключительно под влиянием определенных
периферических раздражений. Характер внешней реакции на данное
раздражение определяется главным образом функциональным
состоянием всей центральной нервной системы в момент раздражения, а не
просто анатомическими соотношениями между рецептором и центральной
нервной системой.
Анатомо-функциональный механизм рефлекторной координации.
Каждый координирующий аппарат вместе с соответствующими ему
афферентными и эфферентными нейронами образует единую целостную
систему. Каждая такая система обусловливает определенную целостную
реакцию вроде рефлекса потирания или чесания, рефлексов сгибания и
разгибания, рефлекса положения и т. д. В этой целостной системе,
согласно гистологическим данным Лбренте де Но, неовные элементы
образуют нервные цепочки и замкнутые нервные круги разной сложно-
62 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
сти. Эти цепочки и замкнутые круги составляются главным образом
из промежуточных нейронов, но аксонные разветвления этих нейронов
заканчиваются, с одной стороны, на теле двигательных клеток, а с
другой— в нейропиле. Кроме того, каждая такая система связана более
или менее интимно путем аксонных разветвлений и с другими близкими
и отдаленными системами. С этой точки зрения, координирующий
аппарат в отношении одного какого-либо рефлекса представляет собой
сложную систему нервных элементов с включением промежуточных
нейронов, нейропиля и двигательных клеток, со всеми их синаптически-
ми окончаниями. Вся эта система функционирует каждый раз как
единое целое и выполняет каждый раз одну определенную
координированную реакцию.
К аналогичному заключению приходит А « о х и н (20) на основании изучения
рефлекторных реакций в связи с эмбриональным развитием. Он полагает, что для
каждой целостной реакции, как дыхание, локомоция, плавание, существует свой
объединенный комплекс нервных образований с соответствующими ему периферическими
рабочими органами и рецепторной системой. Но 'Анохин полагает, что эта
объединенная система имеет чисто функциональное значение, не приурочивается к строго
определенному анатомическому субстрату,— потому он и называет ее функциональной
системой. В основу объединенной деятельности он кладет наличность определенной
«структуры возбуждения» или определенной «формы возбуждения». Мы думаем, что
никакой «структуры возбуждения» или «формы возбуждения» без соответствующей
анатомической структуры не существует. Та анатомическая структура, которая дается
в настоящее время ЛорентедеНо и выставляется им как физиологическая
единица, является той анатомической базой, которая обусловливает функциональное
единство нервной деятельности и придает рефлекторной реакции целостный характер.
Эта анатомо-функциональная система зарождается в онтогенезе
первоначально в своей центральной части. Потом уже она своей
деятельностью формирует свой эфферентный аппарат. Этим объясняется то
явление, что если трансплантировать у амблистомы почку конечности
на какую-нибудь другую часть организма, почка вырастаете конечность,
в локомоторный орган. Но эта конечность приходит в действие вместе с
тем органом, к которому почка подсаживалась, например, вместе с
глоточной мускулатурой при подсаживании почки конечности к этой части
тела. Отсюда ясно, что пересаженная почка конечности, находясь под
ближайшим влиянием глотательного центрального ядра, вступила в
прочную связь с глотательным ядром, с глотательной системой
(Анохин, 20).
Рефлекторная дуга беспозвоночных животных. В соответствии с
филогенетическим развитием нервной системы рефлекторный путь в
процессе развития животного мира подвергается существенному изменению.
На нижней ступени развития, например у губок, нет обособленных
нервных элементов. Но безусловно эктодерм ал ьные эпителиальные клетки
являются настолько чувствительными к механическому раздражению,
что через них можно возбудить прилегающие к ним мышечные клетки
(рис. 24,1). Мышечная реакция на раздражение ограничивается
раздраженным участком (Паркер, 23).
АНАТОМИЧЕСКАЯ ЛОКАЛИЗАЦИЯ КООРДИНИРУЮЩИХ АППАРАТОВ 63
У низших кишечнополостных (Hydrozoa) имеются типичные нервные
клетки с отростками. Все они одного типа. Своими
недифференцированными отростками они связываются, с одной стороны, друг с другом,
образуя синцитиальный нейропиль, с другой — с мышечными клетками,
' 2
Рис. 24. Схематическое изображение филогенетического
развития нервных связей.
S — чувствительный нейрон; А — промежуточный нейрон; М—
двигательный нейрон; е — эпителиальные клетки; т —
мышечные клетки. Пунктиром обведены границы центральной нервной
системы: 1 — нервная система у губок; 2 — у H\drozoa, 3 —
простейшая форма нервной системы у Scvphozoa и низших червей;
4—высшая форма нервной системы у беспозвоночных—со
включением нейропиля, в котором принимают участие кроме
чувствительного и двигательного нейронов еще промежуточный нейрон.
Отростки их образуют густое сплетение. 5—высшая форма
нервных связей у позвоночных; она включает кроме ней р о пиля еще
более или менее обособленные промежуточные нейроны.
Нейропиль у беспозвоночных представлен как сплетение разветвления
нервных отростков, где возбуждение переходит от ветви одного
нейрона на ветви другого, вероятно на местах прикосновения.
Нейропиль же позвоночных изображен как синаптическое
поле, где разветвления аксонов синалтически оканчиваются на
дендритах Схема составлена по новейшим исследованиям
строения нервной системы Заварзина, Боцлера, Гоффа, Зурабашви-
ли и др.
Некоторые из этих нервных клеток посылают отростки на поверхность
эктодермы и тем самым играют роль рецептивных клеток. Значит, у
низших кишечнополостных рефлекторная реакция осуществляется, по
существу говоря, через одну нервную клетку, которая одним отростком
воспринимает раздражение, а другим приводит в действие мышечные
клетки (рис. 24, 2).
У высших кишечнополостных (Scyphozoa) нервная система состоит*
¦64 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
из нервных клеток разного рода. Их недифференцированные на
нейриты и дендриты отростки образуют густое сплетение — нейропиль,
который в одних случаях является совершенно синцитиальным. 'Клеточные
нервные элементы нейропиля с одной стороны связаны с эктодерімаль-
ными чувствительными клетками, а с другой — с мышечными клетками.
Поэтому можно утверждать, что рефлекторная дуга этих животных
состоит из чувствительных и двигательных элементов, связанных между
собой нейропилем (рис. 24, 3)
У червей нервная система усложняется еще больше. Кроме
чувствительных и двигательных клеточных элементов у них существуют хорошо
дифференцированные промежуточные клетки. Они все обладают сильно
разветвленными, хорошо дифференцированными на нейриты и дендриты
отростками. Последние переплетены между собой, образуя нейропиль.
Промежуточные клетки с нейропилем скопляются в отдельных
участках в виде нервных узлов (рис. 24, 4). Вследствие этого рефлекторный
путь является еще болзе усложненным. В нем участвуют все три
нервных элемента вместе с нейропилем. В этом рефлекторном пути
возбуждение всегда распространяется в одном направлении. Этот тип
рефлекторных путей характерен для всех высших беспозвоночных животных.
Для производства координированных рефлекторных реакций клеточные
элементы нервных узлов не являются существенно необходимыми.
Бете экспериментально доказал, что и после отделения клеточных
элементов нервный узел сохраняет способность к рефлекторной
деятельности.
У позвоночных животных нервная система получила дальнейшее
существенное развитие. У них хорошо дифференцированные
чувствительные, промежуточные и двигательные нейроны образуют как
нервные пути и нервные центры, так и нейропиль. Связь этих
дифференцированных нервных образований как между собой, так и с нейропилем
осуществляется преимущественно путем синаптических окончаний
(рис. 24,5).
Практикум
Опыт для выяснения роли спинномозговых узлов в координации рефлекса.
Ъерут спинномозговой препарат лягушки, обнажают спинной мозг в поясничной
области Здесь легко найти VIII—XI задние корешки длиной 1—2 см Каждый из них
перерезают как можно дальше от спинного мозга: центральный отрезок раздражают
индукционным током, а эффект наблюдают на задних конечностях. Когда
раздражается VIII корешок (он же I поясничный), то получается большей частью рефлекс поти-
рания, так как этот корешок получает чувствительные волокна с рецептивного поля
рефлекса потирания (наружная поверхность бедра). Если раздражается IX корешок,
то получается рефлекс сгибания, так как этот корешок включает множество
чувствительных волокон от воспринимающего поля рефлекса сгибания Раздражение X
корешка большей частью вызывает рефлекс сгибания по той же причине, как и IX
корешка, но ему часто предшествует рефлекс разгибания, потому что этот корешок
получает чувствительные волокна также с кожи -вентральной стороны стопы и с обла-
•сти ахиллова сухожилия, раздражение которых вызывает рефлекс разгибания. Раз-
АНАТОМИЧЕСКАЯ ЛОКАЛИЗАЦИЯ КООРДИНИРУЮЩИХ АППАРАТОВ 65
дражение XI корешка вызывает своеобразный рефлекс потираний, который
характерен для раздражения © области копчика и анального отверстия. Рефлекс состоит в
приведении задних конечностей к означенной области для удаления раздражающего
агента голеиюстоиньямм суставами'. Этот корешок очень тонок и выходит из копчиковой
кести, потому его трудно ^препарировать для раздражения.
Наблюдение сегментальных рефлексов. Берут лягушку и отрезают1 ей голову
на уровне атланто-затылочного сочленения. Затем перерезают спинной мозг и
туловище іб грудной области. Передние лайки остаются в связи с плечевым отделом
спинного мозга, а задние — с поясничным.
Спустя 10 мин. после операции механическое раздражение передней лапки
вызовет обычные рефлексы: сгибание и разгибание, движение ©перед и назад;
давление же на грудь произведет одновременное движение обеих передних лапок.
Раздражение задних лапок произведет все типичные рефлексы: сгибание и разгибание,
рефлексы потирания разного типа, а иногда рефлексы прыжка и антагонистические
ритмические движения, как это бывает при ходьбе.
Опыт с локальным стрнхнннным отравлением спинного мозга. Берут
спинномозговую лягушку, укрепляют в особом лягушечьем станке и обнажают шинной мозг в
поясничном отделе. Наблюдают, во-первых, рефлекс потирания в области бедра, где
распространяется VIII задний корешок; вонвторьгх, рефлексы сгибания и разгибания,
^которые вызываются с областей стопы и голени, где оканчиваются IX—X задние
корешки. После этого смачивают мелкие кусочки фиільтрова<лъной бумаги {1 мм2
величиной) в 0,1°/о растворе стрихнина и кладут на X сегмент с дорзальной стороны.
Через несколько минут обнаружится, что рефлексы, вызываемые с области стопы и
голени (как сгибания, так и разгибания), значительно усиливаются; теперь они
вызываются таким слабым раздражением, которое до отравления не давало никакого
эффекта. Кроме того, очень часто развивается способность к ритмическому рефлексу,
поочередному сгибанию и разгибанию задних конечностей. Это фазное движение
протекает или совершенно одинаково на обеих задних конечностях, как при прыганий,
или же разгибание одной конечности сопровождается сгибанием другой, т. е.
движение задних конечностей становится антагонистическим, как это бывает при ходьбе.
Усиления центральной деятельности в других сегментах не заметно. Например,
рефлекс потирания с области бедра, координация которого происходит в VIII сегменте,
остается вполне нормальным: ни его интенсивность, ни порог не изменяются.
Литература
1. Beritoff J., Pfl. Arch., 151, 171, 1913; 198, 604, 1923.
2. Sherrington С, Quart. J. Exp. Physiol., 3, 213, 1910; Erg. Physiol., 4, 796,
1904.
3. Беритов И., Pfl. Arch., 200, 564, 1923; Учение об основных элементах
центральной координации скелетной мускулатуры, Петроград, 1916.
4. Беритов И., Роль спинномозговых узлов в проведении возбуждения. Не
опубликовано.
5. Barron D. H.a.Matthews В. Н. С, J. Physiol., 86, 75 и 104, 1935.
6. Rynberk G. v., Fol. NeurobioL, 5, 761, 1911.
7. Danilewsky W., Pfl. Arch., 78, 194, 1899.
8. BabakE., Pfl., Arch., 92, 134, 1903.
9. Beritoff J., Arch. Anat. u. Physiol., 1912, 296; Pfl. Arch., 151, 171, 1913;
Fol. NeurobioL, 7, 487, 1913.
10. D'u sser de Barenne S. G., Fol. NeurobioL, 5, 42, 1911; 6, 277, 1912.
11. Beritoff J., Fol. NeurobioL, 7, 187, 1913.
12. PhilippsonM., L'autonomie et la centralisation dans le syste-me nerveux des
animaux, Bruxelles, 1905.
13. Введенский Н., Работы физиологической лаборатории Петербургского
ун-та, 1, 1906.
5 и. С. Беритов
€6 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
14. Graham В rown Т., Quart. J. Exp. Physiol., 5, 237, 1922; Erg. Physiol., 13,
279, 1913.
15. Sherrington C, Erg. Physiol., 4, 796, 1904; Int. act. of nerv. syst., 1906.
16. С о g h i 1 1 G. E., Anatomy and the problem of behavior, Cambridge , 1929.
17* Minkowsky M., Schw. Med. Wschr., 52, 29, 30, 1922; Zum gegenwertigen
Stand d. Lehre von d. Reflexen. Zurich—Berlin, 1925.
18. Her rick С J., Proc. Nat. Acad. Sci., 1в, 643, 1930.
19. Lorente de N6., Arch. Neur., Psych., 90, 245, 1933.
20. Анохин П. К., Сб. докл. VI Всесоюзного физиол. съезда, стр. 148.
21. Н of f Е. С, Proc. Roy. Soc, В, 111, 175, 1932.
22. Y о u n g I. Z., J. Physiol., 93, 43, 1938.
23. Parker. G. H., The elementary nervous system, Philadelfia a. London, 1919.
Глава V
ИРРАДИАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ
И ЕЕ ПРОИСХОЖДЕНИЕ
Сложность и целостность рефлекса. Мы видели, что каждый
рефлекс имеет свое рецептивное поле и свой координирующий аппарат.
Но, согласно приведенным анатомическим сведениям, раздражение
одного рецептивного поля не приводит к 'возбуждению одного только
соответствующего ему координирующего аппарата. Такое локальное
действие, которое ограничивалось бы исключительно одной
рефлекторной дугой, нужно считать почти ненормальным явлением. Реакция
центральной нервной системы, вызванная внутренним или внешним
раздражением, более или іменее сложна, так как при нормальных условиях
каждое раздражение вызывает действие в разных отделах центральной
нервной системы. Простого, действительно местного рефлекса не
существует,— рефлекс всегда более или менее сложен. «Простой рефлекс,—
говорит Шерринттон (1),— выгодная, хотя невероятная фикция.
Рефлексы бывают различной степени сложности. Поэтому полезно во
время их анализа выделить из каждого соответствующие компоненты,
которые мы можем рассматривать отдельно и, следовательно,
рассуждать о них, как если бы это были простые рефлексы».
Возьмем для примера рефлекс у собаки в ответ на механическое
раздражение задней лапы. В ответ на это раздражение собака
произведет пространную двигательную реакцию: раздражаемая конечность
сгибается, другая задняя конечность, наоборот, разгибается, голова
поворачивается в сторону раздражения, туловище сгибается вместе
с головой, передние конечности передвигаются в раздражаемую
сторону, движется хвост. Такая картина сложного рефлекса в области
скелетной мускулатуры дана на рис. 25. Если бы раздражение было
сильным, тогда собака приняла бы угрожающее положение и издала бы
характерные звуки в виде рычания, лая. Таким образом, при одном
раздражении приходит в действие целый ряд координирующих
аппаратов как в спинном, так и в продолговатом и в среднем мозгу. Кроме
того, это же раздражение вызывает учащение и углубление дыхания.
5*
<58 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
'Следовательно, возбуждение распространяется и на дыхательные
аппараты, которые находятся в спинном и продолговатом мозгу. Эти
реакции касаются только скелетной мускулатуры. Но одновременно данное
раздражение вызывает изменение в кровообращении: в кровеносных
сосудах некоторых органов давление повышается, а в других падает.
При раздражении вовлекаются в реакцию и внутренние органы,
например, сердце, мочевой пузырь. Деятельность этих органов тормозится.
Таким образом, это же раздражение приводит в действие и
координирующие центры кровеносных сосудов и внутренних органов, которые
находятся в спинном и продолговатом мозгу. Но влияние раздражения
Рис. 25. Положение головы и конечностей кошки при рефлекторном
движении.
<х — показано общее состояние ног, когда головной мозг отделен от спинного
(спинномозговой препарат); Ь—общее состояние, когда удалены только большой мозг и
передняя часть среднего мозга (децеребрированный препарат). Здесь все конечности
¦более разогнуты, чем на рисунке а. Это указывает на экстензорное, т. е. разгибатель-
ное, состояние децеребрированного препарата; с — показано состояние, вызванное
раздражением уха? dK— состояние, вызванное раздражением передней конечности,
ие — то же от раздражения задней конечности. Стрелка везде указывает на
раздражаемое место (ДТеррингтон).
не ограничивается только низшими отделами центральной нервной
системы: оно распространяется также и на промежуточный мозг и
большие полушария мозга. От деятельности этих отделов зависит
образование индивидуальных двигательных рефлексов и ощущение болиг а
самое главное — наступление защитного поведения в виде нападения на
врага или убегания от него. Из этого примера ясно, что центральное
действие, вызванное каким-либо периферическим раздражением, не
ограничивается одним каким-либо отделом центральной нервной
системы, что оно распространяется почти на всю центральную нервную
систему, рачиная со спинного мозга и кончая большими полушариями
головного мозга.
Иррадиация рефлекторной деятельности. Указанный выше
сложный характер рефлекторной реакции обусловливается распространением
рефлекторной деятельности в центральной нервной системе из одного
ее отдела на другие. Это явление распространения рефлекторной
деятельности в центральной нервной системе называется
иррадиацией. Для каждого типа рефлекса существует особый порядок ирра-
ИРРАДИАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ И ЕЕ ПРОИСХОЖДЕНИЕ 69
диации. Например, у обезглавленной лягушки при слабом раздражении
в рецептивном поле сгибания задней лапки движется только
раздражаемая лапка. При некотором усилении раздражения наступает
движение и на передней лапке той же стороны. При дальнейшем усиление
раздражения движение наступает и на противоположной стороне,
однако это движение всегда слабее, чем на стороне раздражения. Значит,
иррадиация возбуждения в спинном мозгу происходит по определенному
типу или по определенному закону, впервые сформулированному в 1853 г.
Пфлюгером (7). Аналогичное заключение было сделано в новейшее
время на основании исследования на спинномозговых кошках.
Раздражение одного заднего корешка в поясничной области производит
двигательные реакции скорее и сильнее на соответствующей стороне, чем на
противоположной, скорее и сильнее из соответствующего сегмента,
чем из других сегментов. При этом в краниальном направлении
реакция распространяется дальше, на более дальние сегменты, чем в каудаль-
ном направлении (Беритов и Бакурадзе, 17)*
Однако означенный тип распространения рефлекторной реакции
наблюдается только в том случае, если раздражается одна из
конечностей. В случае раздражения боковой поверхности хвоста или ушной
раковины рефлекс наступает скорее всего на противоположной сторона,
благодаря чему в первом случае хвост и во втором случае голова
отклоняются в противоположную сторону. При раздражении в области
промежности, вблизи от медиальной линии тела, рефлексы могут
наступать на обеих сторонах тела с одинаковой интенсивностью. Например,
у лягушки при раздражении в области ануса наступает рефлекс поти-
рания с одинаковым участием обеих задних лапок.
Определенный тип иррадиации рефлекторной реакции наблюдается
при более или менее одинаковой возбудимости центральной нервной
системы. Рефлекторная реакция ів ответ на раздражение данного
рецептивного поля может существенно измениться, если возбудимость
координирующих аппаратов какого-либо другого рефлекса значительно
повышена. Например, при стрихнинном отравлении IX—X
спинномозговых сегментов лягушки, когда локально повышается возбудимость в
координирующих аппаратах рефлексов сгибания и разгибания, эти
аппараты легко возбуждаются при малейшем деятельном раздражении
рецептивных полей рефлекса потирания (Беритов, 3). То же самое
случается при повышении возбудимости в каком-либо отделе мозга по
какой-либо другой причине. Например, у децеребрированных кошек
сильно повышается возбудимость тонических центров продолговатого
и среднего мозга. Вследствие этого данные центры легко возбуждаются
слабейшими раздражениями любого рецептивного поля: например, от
прикосновения к коже задних конечностей, от дыхательных движений.
В последнем случае при каждом дыхательном движении усиливается на
данной задней конечности тот тонус — сгибательный или разгибатель-
ный, которому благоприятствует положение головы (Беритов, 24)
(о тонических рефлексах см. ниже). Или, например, вскоре после пере-
"70 ОБЩАЯ ФИЗИОЛОГИИ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
резки задних корешков в поясничной и крестцовой областях спинного
мозга возбудимость соответствующих сегментов повышается в такой
мере, что все мышцы задних конечностей сразу приходят в
рефлекторное сокращение от раздражения центрального отрезка любого заднего
корешка отдельными индукционными ударами незначительной силы
{Беритов, 25). Если после перерезки задних корешков проходит много
дней, то повышение возбудимости в оперированном отделе спинного
мозга достигает такой степени, что на деафферентированной задней
конечности возникают значительные рефлекторные движения сгибания или
разгибания при каждом легком вздрагивании животного или при
каждом дыхательном движении в момент выдоха (Кунстман и Орбели, 26).
Только что приведенные факты свидетельствуют о том, что
иррадиация возбуждения в центральной нервной системе происходит всякий
j)03 в гораздо более широких пределах, чем об этом можно судить по
вызываемым ею внешним эффектам, и что наступление того или другого
внешнего эффекта определяется каждый раз не только характером нерв-
пых связей между раздраженным рецептивным полем и
координирующими аппаратами мозга, но и функциональным состоянием этих
координирующих аппаратов.
Происхождение иррадиации. Одним из основных условий
иррадиации центральной деятельности является следующая анатомическая
особенность центральной нервной системы. Чувствительные волокна
заднего корешка оканчиваются не только в том сегменте, куда входит
корешок. Проходя по центральной нервной системе, они отдают колла-
терали и в других сегментах, как в краниальном, так и в каудальном
направлении, хотя все же больше всего коллатералей отходит в том
сегменте, где заднекорешковое волокно вступает в мозг; в соседних
сегментах оно отдает меньше коллатералей, притом тем меньше, чем
дальше расположен сегмент (Лапинский, 2). При этом в краниальном
направлении заднекорешковые волокна распространяются много
дальше, чем в каудальном (Стопфорд, 18 и др.)- Кроме того, от каждого
заднекорешкового волокна гораздо больше коллатералей оканчивается
в соответствующей половине мозга, чем в противоположной (Ка-
халб," 19). Значительная часть заднскорешковых волокон достигает
головного мозга и там заканчивается. Отсюда следует, что
заднекорешковые волокна могут привести в действие не только координирующие
аппараты соответствующего сегмента, но и аппараты целого ряда
сегментов спинного и головного мозга.
Анатомическим строением спинного мозга хорошо объясняется то
явление, что у спинальных животных рефлекторная реакция на
раздражение какой-либо конечности наступает скорее и сильнее на данной
конечности, чем на других; на соответствующей стороне — скорее и
сильнее, чем на противоположной. Но нет сомнения в том, что чем выше
возбудимость координирующих аппаратов других сегментов и чем
большее количество волокон задних корешков одновременно возбуждено, тем
сильнее вовлекаются в реакции и эти координирующие аппараты.
ИРРАДИАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ И ЕЕ ПРОИСХОЖДЕНИЕ 71
Как уже было упомянуто, раздражение одного рецептивного поля
обыкновенно вызывает рефлексы одного определенного типа из одного
соответствующего координирующего аппарата. Отсюда ясно, что в
координирующих аппаратах соседних или отдаленных сегментов
оканчивается так імало коллатералей от данных возбужденных заднекореш-
ковых волокон, что нормальное их возбуждение посредством этих
коллатералей почти невозможно. Но когда возбудимость координирующих
аппаратов значительно повышена, можно легко вызвать их возбуждение
с рецептивного поля любого рефлекса, как уже указывалось выше.
В иррадиации центральной деятельности принимают также
определенное участие координирующие, промежуточные нейроны. Известно,
что каждый координирующий аппарат вызывает внешнюю реакцию не
только в области раздражения, но почти во всем организме; например,
после локального стрихЦинного отравления определенного
координирующего аппарата ів поясничном отделе мозга усиленные рефлекторные
движения при возбуждении этого аппарата возникают не только на
соответствующей конечности, но и на противоположной задней
конечности; в это же самое время сильные движения наступают и на
передних конечностях. Все эти рефлекторные движения вызываются из
одного и того же первично возбужденного координирующего аппарата
(Беритов, 3).
Следовательно, координирующие нейроны одной задней конечности
имеют связь не только с двигательными нейронами другой задней
конечности, но и с двигательными, нейронами передних конечностей.
Значение, такой связи между координирующими и двигательными
нейронами становится понятным при рассмотрении сложного движения чеса-
тельного рефлекса. Как известно, координирующий аппарат чесатель-
.ного рефлекса находится в грудной и шейной частях спинного імозга.
В этом рефлексе в действие приходит, можно сказать, вся мускулатура:
на стороне раздражения задняя лапка производит чесание;
противоположная задняя сильно разогнута, шея и все туловище отклоняются
в сторону раздражения. Отсюда видно, что возбуждение из
координирующего аппарата распространяется на весь двигательный аппарат
спинного мозга.
В иррадиации нервного возбуждения по центральной нервной
системе принимают значительное участие точно так же вторичные
афферентные пути, т. е. те промежуточные нейроны, которые передают
импульсы возбуждения от заднекорешковых волокон к головному мозгу.
Клетки этих волокон лежат в сером веществе задней половины мозга, в
:zona marginalis, substantia gelatinosa и в заднем рогу. Аксоны этих
клеток частью на своей стороне, частью перейдя на другую сторону,
.поднимаются вверх к головному імозгу по белому веществу спинного
мозга. Они передают импульсы от тактильных, болевых, температурных
рецепторов и попутно отдают коллатерали и к серому веществу
спинного мозга (Ариёнс Капперс, 23 и др.). Следовательно, и через эти
72 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
промежуточные нейроны периферические импульсы возбуждения могут
распространяться по всей центральной нервной системе.
В иррадиации центральной деятельности особенно большую роль
играют высшие отделы мозга: продолговатый, промежуточный и
большой. Здесь расположено множество нервных центров, имеющих
непосредственные связи с внешними рецепторами, с одной стороны, и с
координирующими аппаратами спинного мозга — с другой. Через них
происходит объединение деятельности этих аппаратов в разного рода
сложные нервные акты, носящие характер рефлекса или поведения.
Высшие отделы мозга возбуждаются периферическими импульсами
гораздо легче спинного імозга. Потому эти отделы играют чрезвычайно
важную роль в иррадиации центральной деятельности у нормального
животного или препарата, обладающего названными отделами
головного мозга. Так, например, если взять децеребрированную лягушку,
т. е. лягушку с удаленными большими полушариями, то уже небольшое
механическое раздражение лапки будет производить локомоцию, если
препарат не привязан; если же он привязан, то же самое механическое
раздражение будет вызывать рефлекс освобождения. Оба эти
рефлекторных акта, в которых проявляется иррадиация возбуждения почти во
всей центральной системе, обусловливаются деятельностью нервных
центров продолговатого, среднего и промежуточного мозга (Беритов, 13;
Рэнсон и сотр., 14). Отсюда следует, что иррадиация возбуждения, по
П ф л ю г е р у, не_ может иметь места на целых животных. Она является
законом только для спинномозговых препаратов.
В иррадиации центральной деятельности в виде возбуждения у
нормальных позвоночных животных нейропиль не должен принимать
участия, иначе говоря, дендритное сплетение не должно проводить тех
импульсов возбуждения, которые притекают к ним по аксонам
афферентных или промежуточных нейронов. Эти аксоны оканчиваются на
поверхности дендритов, как и на теле нервных клеток, конечными и
боковыми синапсами. Но есть основание утверждать, что при
возбуждении синапсов в дендритах не возникает распространяющийся процесс
возбуждения.
Таким образом, иррадиация возбуждения в центральной нервной
системе зависит преимущественно от того, что, во-первых, каждое
чувствительное волокно заднего корешка посылает коллатерали в разные
отделы мозга, а не 'только в тот, в который оно входит из
спинномозгового узла; во-вторых, каждый координирующий аппарат связан с
помощью нейритов с двигательными нейронами не только того сегмента
спинного мозга, где лежит этот аппарат, но также и с двигательными
нейронами других его сегментов; в-третьих, каждый вторичный
восходящий путь посылает свои коллатерали почти ко всем сегментам и
отделам мозга, через которые он проходит; в-четвертых, объединяющие
центры головного мозга посылают свои нейриты к координирующим
аппаратам не только первично возбужденных отделов мозга, но и
всевозможных других отделов.
ИРРАДИАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ И ЕЕ ПРОИСХОЖДЕНИЕ 73
Роль проприоцептивных раздражений в иррадиации
рефлекторной деятельности. Еще одно обстоятельство способствует иррадиации
рефлекторной деятельности. Известно, что ни одно активное или пас-
Рис. 26. Схема нервно-мышечного
веретена.
Мышечное волокно; а, о' — перерывы в
волокне, сделанные для экономии места;
с — капсула; 6— подкапсулярное
пространство; TS — спиральное нервное окончание
чувствительного волокна (ns) на
мышечном волокне (проприоцептор); М —
обычное мышечное волокно, лежащее рядом;
пт —¦ двигательное нервное волокно;
Р, Р — двигательные нервные окончания;
t, V — сухожилие (по Речо и Фавру).
fa -
Рис. 27. Схема нервно-сухожильного
веретена.
st — веретенообразное утолщение
сухожильного пучка, с которым связан рецептор; Fm,
Fm' — мышечные волокна; t, Т—сухожилие,
в которое переходят мышечные волокна; С —
капсула вокруг пучка сухожильных нитей,
так называемого сухожильного веретена;
/п — чувствительное нервное волокно; Е —
его древовидное разветвление вокруг
капсулы, т. е. сухожильный проприоцептор
(по Речо).
дивное движение не кончается бесследно: за ним следуют раздражения
рецепторов мышц, сухожилий и суставов действующего органа, т. е.
первичное движение производит, со своей стороны, целый ряд
раздражений в области так называемой глубокой чувствительности. Эти
вторичные раздражения вызываются как сокращением мышц, так и пассивным
74 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
растяжением мышц и сухожилий. При этом раздражаются рецепторы
мышц и сухожилий, которые Шеррингтон называет проприоцепто-
рами. Обычно они располагаются в виде длинных нервных образований
спирально вокруг мышечных клеток (рис. 26) или в виде древовидных
образований между сухожильными волокнами (рис. 27). При сокраще-
Рис. 28. Проприоцептивные импульсы скелетной мышцы лягушки.
Отводится в катодный осциллограф г. superficial n. tibialis при растяжении m.
flexor dig. superficialis brevis различными грузами. А—при растяжении 1 г: после
начального эалпа импульсация в течение первой же секунды быстро
прекращается. В и С—при растяжении 2 г: начало импульсации через 4 секунды; D—E
при растяжении 5 г: начало импульсации через 10 сек. Время везде по 0.02 сек.
(Нарикашвили).
ниях и растяжениях мышц они безусловно должны испытывать
механическое раздражение, вероятно, как от растягивания самого рецептора,
так и от сдавливания со стороны сокращенных или растянутых
мышечных и сухожильных іволокон. Так, например, при пассивном сгибании
сустава будут растягиваться мышцы-разгибатели, а в связи с этим
будут раздражаться находящиеся в них проприоцепторы. При пассивном
разгибании, наоборот, будут раздражаться аналогичным образом
проприоцепторы сгибателей.
При растяжении мышц лягушки проприоцепторы разряжаются целым
рядом нервных импульсов, от 5 до 100 и более в 1 сек. При этом
частота их тем выше, чем больше растяжение. На рис. 28 дается
электрограмма чувствительного нерва, оканчивающегося в одной тоненькой
мышце, содержащей всего 3—4 рецептора. Из этого рисунка видно, что
один рецептор реагирует рядом нервных импульсов даже при самом
ничтожном растяжении силой © 1 г (Эдриан, 10).
Итак, опытным путем было доказано, что при напряжении мышц
ИРРАДИАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ И ЕЕ ПРОИСХОЖДЕНИЕ 75
и сухожилий от растяжения или сокращения раздражаются
находящиеся в них рецепторы. Здесь возникает ряд возбуждений, которые потом
через чувствительные нервы достигают координирующего аппарата.
Даже в момент одиночных сокращений раздражаются проприоцепторы
и по чувствительным нервам проходит целый ряд импульсов
возбуждения (Форбс, 9).
Изучение действия проприоцептивных импульсов на спинной мозг
показывает, что эти импульсы действуют возбуждающе главным
образом в отношении той мышцы, рецептор которой раздражается. Так
действует, например, раздражение проприоцепторов, которое
производится, с одной стороны, растяжением мышцы, а с другой — ее
сокращением (Ллойд, 20). В отношении всех остальных мышц они скорее всего
действуют тормозящим образом (Нарикашвили, 21; Беритов и Баку-
радзе, 17), Но известно, что при некотором повышении возбудимости
центральной нервной системы эти же проприоцептивные импульсы
производят сокращение и других мышц (Гедевани, 22). Итак, каждый раз
с наступлением рефлекторного движения от первичного внешнего
раздражения в мышцах возникает множество вторичных проприоцептивных
раздражений. Возникающие импульсы действуют на центральную
нервную систему и вызывают рефлекторные реакции чер^з те отделы мозга,
на которые эти импульсы действуют непосредственно. От них зависит,
между прочим, усиление первично вызванных рефлекторных реакций, а
также наступление разного рода тонических рефлексов*
Таким образом, в иррадиации рефлекторной деятельности
принимают участие и вторичные проприоцептивные раздражения, вызванные
движением, последовавшим за первичным внешним раздражением.
Рефлекторное последействие. На возникновении вторичных
проприоцептивных раздражений основывается та особенность
рефлекторного действия, что по прекращении первичного раздражения оно
продолжается еще некоторое время. Такая удерживающаяся активность
живых тканей называется последействием, в частности в данном
случае — рефлекторным последействием. Чем выше
возбудимость центральной нервной системы и чем более она способна
к активной деятельности, тем продолжительнее последействие.
Наоборот, усталая и маловозбудимая центральная нервная система может и
совсем не давать последействия. Так, например, под влиянием малых
доз ацетилхолина и эзерина, а также при катодной поляризации (кат-
электротон), когда рефлекторная возбудимость повышена,
рефлекторное последействие усиливается; под влиянием же больших доз
ацетилхолина и при анодной поляризации (анэлектротон), когда рефлекторная
возбудимость понижена, рефлекторное последействие ослабевает (Бонне
и Бремер, 16).
Нетрудно доказать, что явление рефлекторного последействия
большей частью зависит от проприоцептивных раздражений; когда какая-
либо задняя конечность совершенно лишена чувствительности вслед-
76 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
ствие перерезки соответствующих задних корешков, рефлекторное
последействие данной конечности сильно ослабевает, а иногда и совсем
исчезает (Балиони, 5; Шеррингтон, 1). Рефлекс при этом вызывается
раздражением центральных отрезков задних корешков.
Возникновение вторичных проприоцептивных раздражений не
является единственным условием рефлекторного последействия. Последнее
зависит также от того существенного свойства центральной нервной
системы, что каждое раздражение вызывает в ней не один, а множество
нервных импульсов. Об этом явлении мы будем ниже говорить более
Рис. 29. Рефлекс сгибания и его последействие.
Спинномозговой препарат охлажденной лягушки. Перерезаны все задние
корешки поясничного отдела, вследствие чего обе конечности лишены
чувствительности. Записывается сгибатель колена m. semitendinosus (верхняя
кривая) и разгибатель колена triceps (нижняя кривая). Сначала раэдражается
центральный отрезок IX заднего корешка (нижний сигнал); это вызывает
сокращение сгибателя, которое после прекращения раздражения продолжается
еще долго. Через 27 сек. раздражается п. brachialis (верхний сигнал),
который вызывает рефлекс потирания, вследствие чего разгибатель сокращается,
а последействие сгибателя ослабевает. Цифры в скобках на сигналах
указывают на ритм раздражения, а цифры без скобок — на расстояния между
катушками в сантиметрах.
подробно. Что рефлекторное последействие зависит не только от
вторичных раздражений, ясно видно на препаратах с сильно возбужденной
центральной нервной системой, например на отравленной стрихнином
лягушке. Уничтожение периферической чувствительности у такого
препарата, достигаемое перерезкой задних корешков, вовсе не влечет за
собой потери способности к последействию; последнее может
продолжаться очень долго — несколько секунд и даже минут (рис. 29) (Бе-
ритов, 4).
На мышцах с высокими пластическими свойствами рефлекторное
последействие сокращения сливается с пластическим укорочением,
наступающим в мышце вследствие сокращения. На таких мышцах трудно
решить без экспериментального анализа, какая именно доля
последействия обязана своим происхождением рефлекторному последействию
сокращения, но опытным путем можно легко выявить эту долю. Если
во время рефлекторного последействия произвести такое раздражение,
которое должнго вызвать противоположный рефлекс, то существующее
ИРРАДИАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ И ЕЕ ПРОИСХОЖДЕНИЕ 77
рефлекторное последействие устраняется, тормозится. Если, несмотря
на это, мышца все еще остается до некоторой степени укороченной, то
это уже будет представлять собой пластическое укорочение мышцы.
Это, например, хорошо видно на рис. 29, где последействие
рефлекторного сокращения сгибателя m. semitendkiosus лягушки только частично
устраняется под влиянием рефлекса потирания. Остаточное укорочение
¦сгибателя является пластическим по своему происхождению.
Градация рефлекторной реакции. Интенсивность рефлекторной
реакции меняется очень сильно в зависимости от изменения силы
раздражения: она тем больше, чем сильнее раздражение. Это зависит от
т PdfWO) f г-
Рис. 30. Градация интенсивности и последействия рефлекса.
Спинномозговой препарат лягушки. Региструется m. semitendinosus. Раздражается
п. peroneus. Частота раздражения — 100 в 1 сек. Цифры на кривых означают силу
раздражения в сантиметрах расстояния между индукционными катушками. С
усилением раздражения увеличивается как интенсивность, так и последействие рефлекса.
Опусканием сигнала отмечается продолжительность раздражения.
целого ряда условий. Прежде івсего имеет значение то явление, что
каждый чувствительный нейрон анатомически связан преимущественно
с определенной частью промежуточных нейронов координирующего
аппарата и двигательных нейронов (Купер, Денни-Броун и Шерринг-
тон, 12). Очевидно, чем сильнее будет раздражение, тем больше
рецепторов и чувствительных волокон возбудится, а соответственно, тем
больше координирующих и двигательных нейронов будет вовлечено в
действие, и потому тем интенсивнее будет рефлекторная деятельность.
Кроме того, важное значение имеет нарастание ритма возбуждения
рецепторов по мере усиления раздражения. Так, например, ритм
возбуждения одного мышечного рецептора меняется у лягушки от 5 до 100
в 1 сек., а у теплокровных — до 150 в 1 сек. и выше, причем не только
ритм, но и продолжительность возбуждения рецептора зависит от силы
раздражения (Эдриан, 10).
Принимая во внимание все эти обстоятельства, можно варьировать
силу раздражения таким образом, что с каждым усилением будет
понемногу увеличиваться как число возбужденных чувствительных
нервных волокон, так и ритм распространяющихся по ним нервных
импульсов. Увеличение того и другого фактора поведет к усилению
рефлекторной деятельности благодаря увеличению числа возбужденных
двигательных нейронов. По этой причине интенсивность и продолжительность
ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
рефлекторной деятельности можно градуировать в очень больших
пределах (рис. 30).
Чем толще нерв и чем ниже возбудимость координирующих
аппаратов, тем тоньше градуируется рефлекторный двигательный эффект. Но
каково бы ни было функциональное состояние центральной нервной"
системы, сокращение легко достигает той максимальной интенсивности,
выше которой оно не будет подниматься, несмотря на дальнейшее
усиление раздражения. Наоборот, как мы увидим ниже, с дальнейшим
более или менее значительным усилением раздражения интенсивность,
сокращения будет уменьшаться благодаря влиянию общего торможения.
Фракционирование и конвергенция рефлекторных нервных путей.
Характерно, что при максимальном рефлекторном сокращении
напряжение мышцы значительно меньше напряжения, развиваемого при
максимальном сокращении от раздражения двигательного нерва.; При
этом разные афферентные нервы одного и того же рецептивного поля
производят рефлекторное сокращение с разной степенью максимального
напряжения. Например, по Шеррингтону (27) m. tibialis anticus
кошки при раздражении п. obturatorius superficialis развивает
напряжение в 165 г, что составляет 6.5% максимального напряжения от тета-
нического раздражения двигательного нерва; при раздражении п. saphe-
nus externus — напряжение в 800 г, что составляет 32%; при
раздражении п. plantaris externus— 1240 г, т. е. 50%; а при раздражении кожно-
мышечной ветви п. peroneus—1700 г, т. е. 69%. Кроме того, известно,
что раздражение одного и того же чувствительного нерва вовлекает
разные мышцы в различной степени. Например, при раздражении
п. popliteus in. tensor fasciae femoris развивает 18% того максимального
напряжения, которое можно в ней вызвать раздражением
двигательного нерва, a m. semitendinosus — 80% максимального напряжения. Это
фракционирование максимальных рефлекторных эффектов
обусловливается прежде всего тем, что под влиянием раздражения афферентных
волокон в активное состояние приходят не все двигательные нейроны
данной мышцы, а только часть их, с которой данный афферентный нерв
находится в наиболее интимной анатомической или функциональной
связи (Шеррингтон, 27).
При одновременном раздражении двух афферентных нервов из
одного рецептивного поля наступает рефлекторное сокращение той или
другой мышцы с большим напряжением, чем при раздельном
раздражении каждого из этих нервов, но оно значительно меньше суммы
напряжений, получаемых с того и другого нерва в отдельности. Отсюда
следует, что на значительной части двигательных нейронов данной мышцы
сходятся волокна обоих упомянутых афферентных нервов и лишь
некоторая часть двигательных нейронов связана с каким-либо одним из этих
нервов. Такая конвергенция нервных путей в рефлекторной реакции
наблюдается не только при раздражении афферентных нервов, берущих
начало в пределах ближайших участков рецептивного поля, но и при
ИРРАДИАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ И ЕЕ ПРОИСХОЖДЕНИЕ 79
взаимодействии сравнительно удаленных один от другого нервов, как,
например, п. saphenus internus и п. sartorius (Шеррингтон, 27). Однако
эта конвергенция или перекрытие нервных путей в мозгу не
обязательна. Она имеет место только при нормальной рефлекторной
возбудимости, и притом под влиянием сравнительно сильных раздражений. Если
возбудимость мозга почему-либо снижена, например вследствие
утомления1 или шока, или же если применяется незначительное, околопорого-
рое раздражение афферентных нервов, то рефлекторное сокращение в
ответ на раздражение каждого нерва сопровождается небольшим
напряжением. В таких случаях при одновременном раздражении двух
афферентных нервов получается рефлекторное сокращение, напряжение
которого будет не меньше, а больше суммы напряжений, получаемых с'
каждого нерва в отдельности. Это происходит от того, что при
раздражении двух нервов вовлекаются в реакцию двигательные нейроны, не.
вовлекавшиеся при раздражении одного нерва. Это вовлечение
обусловлено повышением возбудимости в мозгу от иррадиации возбуждения.
Градация, фракционирование и конвергенция рефлекторной
деятельности являются важнейшими функциями координирующих аппаратов
центральной нервной системы. На н*их основывается соответствие
рефлекторной реакции раздражаемому рецептивному полю, силе раздра--
жения и функциональному состоянию центральной нервной системы.
Градация и иррадиация рефлекторной деятельности у
беспозвоночных животных,. У низших беспозвоночных, лишенных центральной
нервной системы, как кишечнополостные и иглокожие, градация и
иррадиация рефлекторной деятельности в очень большой степени зависят от
силы раздражения; и у них чем сильнее раздражение, тем пространнее^
вызываемый эффект. Но для данных животных характерной
особенностью является трудность вызова реакции во всем организме, трудность.
иррадиации возбуждения. У гидры, актинии, морской звезды даже при
сильном раздражении может получиться локальный рефлекторный
эффект. Это явление следует поставить в связь с тем, что в нервной
сети названных беспозвоночных возбуждение распространяется с
декрементом, и чем сильнее начальное возбуждение, вызванное
раздражением, тем дальше оно распространяется (Будденброк, 11).
У высших беспозвоночных имеются нервные центры, из которых
головной узел является наиболее развитым. Этот узел связан с целым
рядом очень важных головных рецепторов: зрения, осязания, обоняния,
благодаря которым он приобрел особое значение в реакциях животного.
Этот же узел связан со всеми другими узлами и автономными
нервными сплетениями каждого двигательного органа. Через эту узловую
систему возбуждение иррадиирует, как у позвоночных, легко и без
декремента. При любой силе раздражения, если движется головная
часть тела, будет двигаться и остальная; если же головная часть
останавливается, то остановится и остальная часть.
80 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
История вопроса об иррадиации. Что возбуждение одного периферического
органа может действовать на всю центральную нервную систему, было давно
замелено как физиологами, так и психологами; например, психолог Б э н (6) в 1863 г. с
полной ясностью изложил принцип! центральной иррадиации. Со сложностью
рефлекса, происходящей вследствие иррадиации, были знакомы уже первые
исследователи центральной нервной системы. Так, например, Пфлюгер {7) привел
соответствующие факты еще в 1853 г. Что же касается положения, отрицающего простейшие
и совершенно локальные рефлекторные реакции, то это—дело новое. Оно впервые
было выставлено Шерринг тоном (1) (і1906) и основывается на физиологических
фактах. Можно привести также мнение современного нам невролога X е р ір и к а
(8), который руководствуется главным образом результатами изучения
соединительных путей между разными отделами центральной нервной системы. На основании
взаимоотношений соединительных нервных путей он утверждает, что каждый
отдельный центр может занимать физиологически господствующее положение по отношению
к какой-либо одной чувствительной системе. Но так как каждый отдельный центр
имеет связь с другими центрами, то деятельность одного центра должна вызвать
деятельность и других центров.
Практикум
Наблюдение иррадиации рефлекторной деятельности. Спинномозговой препарат
лягушки. 1) Если раздражение рецептивного поля сгибания задней конечности яв-
ляется слабым, го оно вызовет движение только раздражаемой конечности. С
усилением раздражения приходит в движение и передняя конечность той же стороны; если
раздражение усилить дальше, рефлекторное действие появится на другой стороне.
2) При раздражении спины (поле потирания) рефлекторная реакция появится сначала
ніа спине, с усилением раздражения приходят в движение задняя и передняя
конечности раздражаемой стороны, а затем обе конечности противоположной стороны.
Градация интенсивности и последействие рефлекса. Спинномозговой препарат
лягушки. Запись m. semitendinosus на кимографе. Для раздражения берут п. peroneus
той же конечности и тетанически раздражают его индукционными ударами. Сначала
находят порог и записывают пороговый эффект. После этого сближают катушки на
1—2 см и опять записывают эффект. Затем снова сближают катушки и снова записывают
аффект и т. д. С усилением раздражения увеличивается как высота сокращения, так
и его последействие. После каждого опыта необходимо дать отдых препарату по
.крайней мере на 2 мин.
Исследование происхождения рефлекторного последействия. Спинномозговой
препарат лягушки (охлажденной или зимней). Обнажают спинной мозг сзади в
поясничном отделе, перевязывают X или IX задний корешок ниточкой как можно дальше от
мозга и перерезают позади ниточки. После того перерезают остальные корешки;
раздражая перевязанный корешок, вызывают рефлекс огибания. Записывается
рефлекторное сокращение m. semitenditnosus на кимографе. Этот опыт обнаружит наличие
рефлекторного последействия и после перерезки корешков.
Какую роль играют проприоцептивные раздражения в рефлекторном
последействии, будет ясно видно, если записать эффект m. semitendinosus до перерезки
корешков, раздражая тот же IX или X корешок, и если затем сравнить этот эффект с
тем, который получится после перерезки.
Литература
ft. Sherrington С, Integrative action of the nervous system, 1906.
2. Lapinsky M., Arch. Psych., 42, 869, 1907.
3. BeritoffJ., Тр. Петерб. об-ва естеств., 41, в. 2, 1910; там же, 42, в. 1, 1913;
РИ. Arch., 151, 171, 1913.
ИРРАДИАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ И ЕЕ ПРОИСХОЖДЕНИЕ
4. Reritoff J., Иав. СПб. биол. лаб., 12, № 2—3, 1912; Arch. Anat. Physiol.,
1913, стр. 1; Zschr. Biol., 64, 161, 1914.
5. В a g 1 i о n i S., Arch. Anat., Physiol. Suppl. 1900, стр. 213; Zur Anal. d. Reflex*
funktion, 1907.
6. Б э h А., Психология, Москва, 1902.
7. PfiugerE. Die sensorischen Funktionen d. Ruckenmarks d. Wirbeltiere, Berlin,
1853.
8. Herrick J., Animal Behavior, 3, 1913.
9. Forbes A., Campbell a. Williams, Am. J. Physiol., 60, 283, 1924.
10. Adrian E., Erg. Physiol., 26, 501, 1928.
11. В uddenbrock W., Hand. Norm. Path. Physiol., 9, 805, 1929.
12. Cooper S., Denny — Brown D. E. a. Sherrington C, Proc. Roy.
Soc, 100, 448, 1926.
13. Беритов И., Элементы центральной координации скелетной мускулатуры,
Петроград, 1916; Журн. эксп. биол. мед., 22, 106 и 117, 1928-
14. Н і n s е у J. С, R a n s о n S. W. а. М с N a t t i n R., Arch. Neur. Psych., 23,
1, 1930.
15. М a 11 h е w s В. Н. С, J. Physiol., 78, 1, 1933; 71, 64, 1931; 72, 153, 1931.
16. BonnetV. а. В г e m e r F., J. Physiol., 90, 45 P, 1937.
17. Беритов И. иБакурадзе А., Физиол. журн. СССР, 27, 337, 1939.
18. S t о р f о г d J. S. В., Sensation and the sensory pathway, 1930.
19. С a j al S. R., Arch. Anat. u. Physiol., 1893, стр. 19.
20. Lloyd D., J. Neurophysiol., 6, 294 и 317, 1943.
21. НарикашвилиС, Юбилейный сборник, посвященный проф. Бериташвили.
1936, стр. 413.
22. Гедевани'Д., Физиол. журн. СССР, 26, 369, 1939.
23. А г і ё n s KappersC. U., The comparative anatomy of the nervous system of
vertebrates includ. man, v. I. 1936.
24. Beritoff J., Pfl. Arch., 202, 265, 1924.
25. Беритов И., Изв. Акад. Наук, 1915, стр. 649; Quart. J. Exp. Physiol., 9, 199,
1915. ' *
26. *K у не т м а н К. И. и О р б ел и Л. А., Изв. Ин-та им. Лесгафта, 9, 187, 1924.
27. Sherrington С. S. и сотр., Reflex activity of the spinal cord, Oxford, 1932,
Русск. пер., Биомедгиз, 1935.
6 и. С. Беритов
Глава VI
ОСНОВНЫЕ ПРОЦЕССЫ ЦЕНТРАЛЬНОЙ КООРДИНАЦИИ
Существуют две степени координации рефлекторных реакций. Есть
координация, которую обнаруживает рефлекс, когда он наступает
только на одном органе или, вернее, когда мы рассматриваем рефлекс
по отношению к одному органу. Но есть еще другая координация,
более сложная, когда рефлекс наступает на многих органах,— вернее,
когда мы рассматриваем наступившую рефлекторную реакцию в целом.
В первом случае координация касается участия мышц в движениях
данного органа, а во втором случае — соотношения и взаимодействия
всех работающих органов.
Принцип координации одного двигательного органа- Каждое
координированное движение конечности или какой-либо части тела
предполагает сокращение одной группы мышц и отсутствие
сокращения в другой, антагонистической группе. Так, например, во время
общего рефлекторного сгибания задней лапы все сгибатели — ram. semiten-
dinosus, sartorius, tibialis, peroneus и др.— сокращаются, а w\
антагонисты, т. е. разгибатели, m. quadriceps (собаки), т. triceps (лягушки),
т. gastrocnemius и др. совсем не сокращаются или сокращаются очень
слабо (рис. 31).
Сокращение каждой группы імышц вызывается деятельностью
координирующего аппарата, откуда возбуждающая иннервация достигает
мышц через двигательный нерв. Но и в отношении группы
недействующих імышц в координирующем аппарате происходит определенный
активный процесс, вследствие чего эти мышцы остаются
несокращенными. Этот процесс затрудняет или же делает невозможным центральное
возбуждение соответствующих мышц из других координирующих
аппаратов. Если бы эти мышцы были рефлекторно сокращены, то в момент
возбуждающей иннервации другой группы мышц сокращение их
уменьшилось бы или даже совершенно прекратилось (рис. 31), Центральный
процесс, лежащий в основе такого задерживающего действия, принято
называть процессом торможения. Таким образом, каждое
координированное движение предполагает возбуждение в одной группе
ОСНОВНЫЕ ПРОЦЕССЫ ЦЕНТРАЛЬНОЙ КООРДИНАЦИИ 83
мышц и торможение в другой, антагонистической группе (Шерринг-
тон, 1). Этот принцип координированной иннервации был установлен
Шеррингтоном. Он же дал ему название реципрокной, т. е.
взаимно сочетанной иннервации. Во время координированного
движения возбуждающая иннервация одной группы мышц так связана с
тормозящей иннервацией другой, что каждый раз центральное
возбуждение одной группы сопровождается
центральным торможением другой.
Шеррингтон и его
сотрудники всесторонне изучили реии-
прокные отношения антагонистов
на млекопитающих. Они, между
прочим, нашли, что продолжитель
ность реципрокного возбуждения
совпадает с продолжительностью
сочетанного торможения и, кроме
того, с течением времени
интенсивность одного процесса меняется з
такой же мере, как интенсивность
другого.
Функциональное деление мышц
конечности на антагонистические
группы. В каждом рефлекторном
движении так или иначе участвуют
все мышцы конечностей, но
характер их участия в каждом движении
разнообразен. В общем можно
разбить всю массу мышц конечности на
две группы: группу сгибателей
суставов и группу разгибателей.
В одних случаях одна группа
возбуждается, а другая тормозится, в
других случаях происходит
обратное. Для каждой группы,
безусловно, существует объединяющий их
нервный центр, ибо возбуждение
мышц каждой группы начинается и
кончается более или менее одновременно; точно так же и торможение
мышц каждой группы происходит более или менее одновременно. Это
объединение осуществляется координирующим аппаратом. Однако
некоторую роль в этом объединении должны играть и проприоцепторы,
ибо известно, что раздражение проприоцелторов при сокращении
флексоров вызывает вторичное рефлекторное сокращение тех ж? мышц
флексоров и одновременно торможение остальных мышц, т. е.
экстензоров. Следовательно, мышцы конечности сами участвуют рефлектор-
Рис. 31. Реципрокная иннервация
антагонистических мышц коленного
сустава.
Децеребрированный препарат кошки.
Верхняя кривая принадлежит сгибателю
колена правой стороны m. semitendinosus,
средняя — односуставному разгибателю
колена vasto-crureus, нижняя-—двусустав-
ному разгибателю rectus femoris.
Раздражается то п. peroneus той же стороны
(верхний сигнал), то противоположный
(нижний сигнал). В первом юпыте (А) сначала
раздражается п. peroneus
противоположной, т. е. левой, стороны, что вызывает
разгибательный рефлекс: оба разгибателя
сокращаются, сгибатель же не
сокращается. Во время этого разгнбательного
рефлекса раздражается на короткое время п.
peroneus той же стороны. Это дает
сокращение сгибателя и в то же время
расслабление разгибателей. Во втором опыте (В)
раздражается только правый п. peroneus;
в ответ на это сокращается сгибатель, а
разгибатели остаются в покое. На
сигнальных линиях цифры в скобках обозначают
ритм раздражений, а цифры после них —
расстояние между катушками в
сантиметрах. Время показано в секундах.
6*
<84 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
ным путем в осуществлении одновременной деятельности флексоров с
¦флексорами и экстензоров с экстензорами.
Односуставные мышцы. При общих сгибательных и разгибательных
.движениях все главные мышцы конечности в отношении реципрокных
шннерваций распадаются на две группы: группу мышц, сокращающихся
при рефлекторных движениях сгибаний, и группу мышцг
сокращающихся при разгибательных движениях. И к той и к другой группе
принадлежат, между прочим, односуставные мышцы. Распределение этих
жышц по группам всецело определяется анатомическим положением.
Если положение данной односуставной мышцы таково, что при
сокращении она сгибает обслуживаемый ею сустав, то она испытывает
возбуждающую иннервацию в сгибательных рефлексах и тормозящую в
разгибательных. Обратное отношение будет наблюдаться у тех одно-
'Суставных мышц, которые занимают такое анатомическое положение,
что при сокращении их получается разгибание обслуживаемого сустава.
Так, например, m. vasto-crureus (головка т. quadriceps), односуставный
разгибатель колена, сокращается в разгибательных рефлексах и
тормозится в сгибательных; m. brachialis является сгибателем локтевого
сустава; эта мышца сокращается в сгибательных рефлексах передней
іноги и тормозится в разгибательных. Таким образом, деление одно-
суставных мышц на группы сгибателей и разгибателей происходит по
очень простому, чисто анатомическому принципу.
Двусуставные мышцы. Большинство наиболее важных мышц
обслуживает не один сустав, а два. Только немногие из нш имеют
одинаковое анатомическое положение при обслуживаемых ими суставах.
Такой мышцей является, например, m. sartorius, при сокращении
сгибающая коленный и тазобедренный суставы. Ее роль в рефлекторных
движениях конечности вполне определяется анатомическим
положением: она сокращается в сгибательных движениях и тормозится ів
фазгибательных. Анатомическое положение остальных двусуставных
мышц таково, что каждая из них действует на обслуживаемые суставы
ф Противоположном направлении: сгибает один из них и разгибает
другой. Это обстоятельство, однако, совершенно не мешает
функциональному делению их на группы сгибателей и разгибателей, так как
одни двусуставные мышцы сокращаются главным образом при
координированных движениях сгибательного характера и тормозятся при
разгибательных движениях, другие же мышцы, наоборот, сокращаются
при разгибательных движениях и тормозятся при сгибательных.
Например, m. semitendinosus по своему анатомическому положению
сгибает коленный сустав и разгибает тазобедренный. Эта мышца
сокращается преимущественно при сгибательных движениях, а при
разгибательных тормозится. М. gastrocnemius по своему положению разгибает
голеностопный сустав и сгибает коленный; эта мышца сокращается
преимущественно в разгибательных движениях, а в сгибательных
тормозится.
ОСНОВНЫЕ ПРОЦЕССЫ ЦЕНТРАЛЬНОЙ КООРДИНАЦИИ 8?
Для наглядного представления механического действия одно- и двусустаяных
мышц дается рис. 32, где изображено расположение мышц задней конечности кошки
по Шеррннгтону.
На этом рисунке представлены следующие мышцы: а) односуставные сгибатели
тазобедренного сустава (N— m. pectineus, S — т. sartorius латеральная часть»
F,F' — tensor fasciae longus el brevis, M—gluteus minimus); b) однооустав'ные
разгибатели тазобедренного сустава (3—adductor minor, 4 — adductor major, 5 — m.
semimembranosus, 6—m. biceps femorisant.); с) односуставные разгибатели коленного
сустава (1 — m. crureus, 9 — m. vastus medialis); d) односуставные сгибатели
голеностопного сустава (Л — m. tibialis anticus, P — m. peroneus); e) односуставной разги*-
батель голеностопного сустава (8 — m. soleus); f) двусуставные мышцы» сгибающие-
Рис. 32. Расположение мышц задней ноги кошки.
/ — с латеральной стороны; II—с медиальной.
колено и разгибающие тазобедренный сустав (В — m. biceps femoris posterior,
G—т. gracilis, T — m. semitendinosus); g) двусуставной сгибатель колена и тазо*
бедренного сустава (S — m. sartorius: медиальная часть); і) двусуставной сгибатель
колена и разгибатель голеностопного сустава (7, 7Ь — m. gastrocnemius) и, наконец
h) трехсуставная мышца (L — extensor digitorum longus), которая разгибает колено,
сгибает голеностопный сустав и разгибает пальцы.
Основной принцип деятельности двусуставных мышц. На
основании анатомического положения двусуставных мышц при том и другом
суставе можно было думать, что они могут сокращаться при всех
рефлекторных движениях. Фактически же, как указывалось выше,
некоторые из них при одних движениях сокращаются, а при других,
наоборот, тормозятся. Значит, руководствуясь только анатомическим
положением, нельзя было решить по отношению к данным мышцам, какую
функциональную роль они исполняют в рефлекторных движениях. Это
можно решить лишь путем физиологического исследования.
Сначала французский ученый Дюшен (39) (1885), затем Ш е р-
рингтон (38) (1910) изучали физиологическую роль некоторых
86 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
мышц во время движений. Заключения их в большинстве случаев
вполне совпадают, в некоторых же случаях, наоборот, сильно
противоречат одно другому. Например, в отношении m. semitendinosus оба
автора утверждали, что эта мышца сокращается при общих сгибатель-
ных движениях. Относительно m. rectus femoris Д ю ш е н утверждал,
что данная мышца сокращается в разгибательных движениях подобно
односуставньш разгибателям (т. vastus interims et medius), III e p-
рингтон же находил, что она сокращается при сгибательных
движениях подобно m. semitendinosus. В настоящее время вопрос о
функциональной роли двусуставных мышц разрешен в том смысле, что они
сокращаются при разгибательных или сгибательных движениях всей
конечности, смотря по характеру анатомического положения их дисталь-
ного конца. Если мышца при сокращении производит разгибание
дистального сустава (m. rectus femoris, т. gastrocnemius, caput longum
tricipitis brachii), тогда она сокращается при общих разгибательных
движениях конечности и тормозится при общих сгибательных. Если же
мышца производит сгибание дистального сустава (т. semitendinosus,
т. biceps brachii), тогда она сокращается при общих сгибательных
движениях и тормозится при общих разгибательных (Беритов, 40).
Итак, принадлежность імышц к антагонистам или синергистам
определяется положением их дистального конца. При этом хотя каждая
мышца и оказывает совершенно противоположное действие на
проксимальный сустав, но это вовсе не мешает выполнению того общего
движения, в котором участвует дистальный конец. Наоборот, при общем
однородном движении конечности двусуставная мышца растягивается,
и это растяжение усиливает действие, производимое дистальиым концом.
Приведенное здесь общее правило о роли двусуставных мышц
вполне применимо к мышцам лягушки. Здесь m. triceps femoris и
т. gastrocnemius, производящие разгибание дистального и сгибание
проксимального суставов, сокращаются преимущественно при
разгибательных движениях, a m. semitendinosus и т. tibialis anticus,
производящие сгибание дистального сустава и разгибание проксимального,
сокращаются главным образом при общих сгибательных движениях. На
рис. 33 дается группировка імышц задней конечности кошки в момент
сгибания и разгибания (по Шеррингтону).
Фиксирующая роль двусуставных мышц по отношению к
проксимальному составу. Можно утверждать, что все двусуставные
и многосуставные мышцы оказывают важное влияние и на
проксимальные суставы, а именно они фиксируют их при тех резких
движениях, которые производятся односуставными мышцами. Так, например,
в тазобедренном суставе существует целый ряд односуставных разги-
баіелей (они соединяют таз с trochanter major femoris). Во время
сокращения эти мышцы тянут бедро вверх и назад. При быстрых
движениях, как прыжок, такое действие может причинить вывих, в
особенности івследствие инерции, если только это действие не умеряется про-
ОСНОВНЫЕ ПРОЦЕССЫ ЦЕНТРАЛЬНОЙ КООРДИНАЦИИ 87
тивоположньш действием таких двусуставных мышц, как m. rectus
femoris, действующих на тазобедренный сустав как сгибатель. Во время
общего разгибательного движения ноги односуставные сгибатели
тазобедренного сустава не сокращаются, а двусуставная m. rectus femoris
сокращается, вследствие чего при общем разгибании ноги эта мышца
активно умеряет быстрое движение, производимое разгибателями
тазобедренного сустава. Следовательно, rn. rectus femoris и подобные ей
двусуставные мышцы играют роль фиксаторов в отношении
проксимальных суставов при движениях, производимых односуставньши раз-
Рис. 33. Расположение мышц задней ноги кошки.
А — вадняя конечность -кошки в момент сгибания; В — то же
в момент разгибания. На А даны те мышцы, которые
сокращаются при сгибании, а на В — мышцы, сокращающиеся при
разгибании. Обозначения те же, что и на рис. 32 (Шеррингтоя).
гибателями. То же самое следует сказать в отношении m. semitendino-
sus и подобных ей імышц: они также являются фиксаторами
проксимальных суставов при быстрых движениях, производимых
односуставньши сгибателями.
Активная природа торможения: Подобно центральному
возбуждению центральное торможение является активным процессом,—
следовательно, во время обоих этих процессов происходит определенная
затрата энергии в возбудимой системе. Что возбуждение элементов
центральной нервной системы представляет собой, по примеру
нервного волокна, активное состояние живой системы, не требует
доказательств, активная же природа торіможения до последнего времени
подвергалась сомнению. Одним из лучших доказательств активной природы
торможения является ослабление тормозящего действия при
длительном раздражении какого-либо чувствительного нерва (Форбс, 2). Если
во время длительного торможения данной группы імышц производить
раздражение какого-нибудь другого чувствительного нерва, вызывающее
сокращение этой группы, то с течением времени это сокращение будет
все усиливаться, несмотря на непрекращающееся тормозящее
раздражение (рис. 34). Значит, рефлекторное торможение при длительном его
88 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
протекании ослабевает, подобно тому как это бывает с возбуждением.
Существует и другое доказательство активной природы торможения.
Как было показано, стрихнинное отравление повышает рефлекторную
деятельность: если отравить
стрихнином один какой-либо сегмент
спинного мозга с задней
поверхности, где лежат координирующие
аппараты определенных рефлексов, то
немедленно происходит усиление
данных рефлексов. При этом от
такого локального стрихникного
отравления координация не
расстраивается, т. е. в какой мере
усиливается реципрокная возбуждающая
иннервация одних мышц, в такой
же мере усиливается тормозящая
иннервация других (Беритов, 3;
Дюссерде Баренн, 4). Если бы
центральное торможение было лишь
слабым возбуждением, как
полагают Ферворн (5) и его
последователи, или же состоянием покоя,
то стрихнин должен был бы
уничтожить реципрокное торможение. Так
как от этого яда возбудимость и
процесс возбуждения сильно
повышаются в координирующем
аппарате, то не может быть и речи о том,
что в отравленном аппарате одни
элементы испытывают слабое
возбуждение или даже отдыхают,
тогда как соседние дают
сильнейшее возбуждение. Нормально
рефлекс потирания у лягушки чаще
всего заканчивается
кратковременным последействием (рис. 35);
после же отравления последействие
этого рефлекса чрезвычайно
усиливается и удлиняется. Несмотря на
это, реципрокная иннервация не
нарушается как во время
раздражения, так я после него — во время последействия. С какой
интенсивностью и как долго сокращается m. triceps, точно на такое же время и с
такой же интенсивностью тормозится ее антагонист — m. semitendinosus
(рис. 36).
Рис. 34. Явление центрального
утомления на лумбальном препарате
кошки.
Регистрируются mm. quadriceps (Q. d. и
Qs) и semitendinosus (S. d и s. s.) обеих
сторон. Длительное тетаническое
раздражение большой силы прикладывается к
VIII ваднему корешку левой стороны
(VIII s) на фоне рефлекторных
вздрагиваний, вызываемых раздражением V
заднего корешка левой стороны (Vs.) и VI
заднего корешка правой стороны (VI d). с
ослаблением сокращения сгибателя
раздражаемой стороны ослабевает торможение
мышц противоположной стороны. На
разгибателе соответствующей стороны
наступает заторможенное сокращение, причем
и на этой мышце, параллельно с
ослаблением сокращения, ослабевает
торможение. На сигнальных линиях цифры
означают силу раздражения в сантиметрах
расстояния между индукционными
катушками (Беритов и Бакурадзе).
ОСНОВНЫЕ ПРОЦЕССЫ ЦЕНТРАЛЬНОЙ КООРДИНАЦИИ 89"
Этими опытами подтверждается наше положение, что процесс
торможения есть процесс sui generis и что стрихнин его не извращает.
Принцип координации нескольких одновременно действующих
органов. Принцип реципрокной иннервации не ограничивается только
иннервацией мышц-антагонистов. Если у кошки раздражать задний
корешок, в который входят чувствительные нервные волокна от хвоста,
то в движение приходит
только хвост, — он
сгибается в сторону
раздражаемого корешка, но в
это же самое время
задняя конечность, вернее,
вся ее мускулатура (как
сгибатели, так и
разгибатели) тормозится.
Аналогичное явление ^имеет
место и при раздражении
IV, V и VI задних
поясничных корешков. В
составе этих корешков
проходят чувствительные
волокна от промежности,
половых органов и оаз-
гибателей бедра, при
тетаническом
раздражении которых вызывается
разгибательный рефлекс.
Обычно раздражение
названных поясничных
корешков производит антагонистическую реакцию только на мышцах
коленного и тазобедренного суставов; мышцы же голеностопного
сустава все тормозятся. То же происходит при изолированных движениях
определенных частей хвоста. Когда приходит в движение основание
хвоста, кончик его остается неподвижным, и наоборот, при движении
кончика хвоста основание его неподвижно. Опытным путем было
доказано, что неподвижная часть испытывает торможение,— следовательно,
когда из координирующего аппарата хвоста поступают двигательные
импульсы в мышцы какой-нибудь части хвоста, мышцы остальных
частей его подвергаются торможению (Беритов, 10), Таким образом,
раздражение определенных чувствительных нервов вызывает в одной
части двигательного органа двигательную реакцию, а в других
частях— одновременное торможение всех мышц.
Далее, опытами на децеребрнрованных животных, т. е. на животных,
лишенных большого мозга (лягушки, щенки), было доказано, что
раздражение головы (сдавливание, прикосновение) вызывает поднятие
и отклонение ее назад и одновременно торможение всей остальной
Рис. 35. Реципрокная иннервация в
рефлексе потирания.
Спинномозговая лягушка. Верхняя кривая от
сгибателя m. semitendinosus, нижняя — от разгибателя
triceps fem. Раздражением п. peroneus вызывается
сгибательный рефлекс. При этом сильно сокращается
сгибатель. Во время этого рефлекса производится
раздражение п. brachialis. Это вызывает сокращение
разгибателя и торможение сгибателя. По
прекращении последнего раздражения разгибатель сейчас же
расслабевает, и вновь начинается сокращение
сгибателя. На сигнальных линиях обычные обозначения.
90 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
мускулатуры тела (рис. 37). То же самое происходит и у нормального
животного (Беритов, 11). Во время акта еды или питья при глотании
Рис. 36. Реципрокная иннервация в рефлексе потирания после
отравления стрихнином координирующего аппарата.
Спинномозговая лягушка. Стрихнином отравлен VIII сегмент. Верхняя
кривая от semitendinosus, нижняя — от triceps. Длительно раздражается п.
superficialis из поля сгибания (нижний сигнал). На фоне раздражения п.
superficial is раздражается на короткое время п. cut. femoralis later, из
поля потирания. Этот нерв вызывает эффект через отравленный сегмент,
деятельность которого чрезвычайно повышена стрихнином. Существующее
сокращение m, semitendinosus расслабевает на все время сокращения т.
triceps в рефлексе потирания. Сокращение т. semitendinosus появляется вновь
в момент ослабления сокращения triceps. С какой постепенностью
расслабевает последействие m. triceps, с такой же усиливается сокращение m.
semitendinosus. Последействие длится около 30 сек. На сигнальных линиях обычные
обозначения.
происходит торможение всей скелетной мускулатуры (рис. 38), то же
бывает и при дефекации (Мельтцер, 12; Ухтомский, 13). Значит, наря-
Anas 14
14
14
Рис. 37. Влияние сдавливания головы на рефлекс потирания.
Бульбарный препарат лягушки (перерезка на границе среднего
и продолговатого мозга).
Верхняя кривая — от m. triceps, нижняя—от m. semitendinosus. Вызывается
рефлекс потирания электрическим раздражением кожи около анального
отверстия. В опытах А и С это раздражение происходит при свободной голове,
а в опыте Б—во время сдавливания головы. В последнем опыте рефлекс
потирания не наступает. На сигнальной линии цифра означает силу электрического
раздражения в сантиметрах расстояния индукционных катушек.
ду с более или менее обширным движением одной части тела
происходит торможение всех остальных частей его (Беритов, И).
ОСНОВНЫЕ ПРОЦЕССЫ ЦЕНТРАЛЬНОЙ КООРДИНАЦИИ 91
• Итак, когда в результате деятельности координирующего аппарата
одного какого-либо рефлекса возбуждается группа мышц одного
органа и тем определяются характер и интенсивность двигательного
эффекта, остальные мышцы того же органа, а также все мышцы соседних
органов оказываются более или менее заторможенными. Это
торможение всех остальных мышц выгодно обеспечивает координированную
деятельность данного органа:
его целесообразная
деятельность не будет нарушаться
рефлекторной деятельностью
других органов, вызванной из
других координирующих
аппаратов (Беритов, 10).
Одновременное
возбуждение и торможение мышцы
приреципрокной иннервации.
Чистая реципрокная
иннервация антагонистических мышц
обычно наблюдается только
при определенных условиях,
и именно—когда определенные
части центральной нервной
системы удалены, все
афферентные пути, за исключением
одного, перерезаны, и
раздражается небольшой
чувствительный нервный ствол,
состоящий из более или менее
однородных волокон ст одного рецептивного поля.
Обычно же даже в искусственных лабораторных условиях одна и
та же мышца,— вернее, один и тот же «конечный общий путь»
испытывает одновременно и возбуждение и торможение. Действие
противоположных влияний в общем конечном пути при двигательных клетках
обычно суммируется алгебраически. Объясняется эта двойственность
иннервации тем, что каждая двигательная клетка в каждый данный
момент испытывает как возбуждающие, так и тормозящие влияния из
самых разных отделов мозга. По мысли Шеррингтона, нормальное
выполнение каждого естественного двигательного акта основано на
сотрудничестве между взаимодействующими процессами возбуждения
и торможения (32).
На нормальных и неповрежденных животных двойственная
иннервация мышц должна быть неизбежным явлением. Но и на препаратах
животных эта двойственная иннервация — самое обычное явление. Так,
на спинномозговой лягушке во время рефлекса потирания случается,
что m. semitendinosus вместо чистого торможения дает заторможенное
Рис. 38. Влияние еды на защитный рефлекс
от электрического раздражения левой
передней ноги. Щенок без коры больших
полушарий.
Верхняя кривая от правой передней ноги,
нижняя — от левой передней. Раздражается левая
нога в опыте А во время лакания молока, а в
опыте В — без него. Во время лакания
фарадическое раздражение ноги в 4 см дает едва
заметную реакцию, а при 3 см наступает значительная
реакция, но только на раздражаемой ноге. В
опыте В раздражение в 4 см дает очень сильную
реакцию, и притом на обеих ногах.
92 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
сокращение,— иначе говоря, мышца сокращается на фоне
значительного торможения. Точно так же m. triceps в рефлексе потирания вместе
чистого возбуждения нередко обнаруживает возбуждение одновременно
с торможением. Это торможение может быть настолько сильным, что
внешний эффект возбуждения — сокращение — значительно ослабевает
или даже совсем прекращается (Беритов, 3). Такая двойственная
иннервация наблюдается как в отношении двусуставных, так и односустав-
ных мышц кошки (mm. rectus femoris, gastrocnemius, tibialis anticus,
vasto-crureus) и проявляется в рефлексах сгибания и разгибания на
децеребрированных и спинномозговых
кошках (Введенский и Ухтомский, 28?
Грэим Броун, 29).
Обычно двойственная иннервация
получается при более или менее сильных
раздражениях. Так, если сильным
раздражением в рецептивном поле
сгибания вызвать у лягушки максимальное
сокращение сгибателя m. semitendinosusr
оно всегда будет сопровождаться
сокращением его антагониста — m. triceps
femoris (рис. 39). В таких случаях
сокращение протекает на фоне
торможения, получается заторможенное
сокращение. Но нередко такое заторможенное
сокращение возникает на антагонистах
при пороговых раздражениях. По
мнению Грэим Броун а, чистое реци-
прокное торможение или чистое реци-
прокное возбуждение — довольно редкое
явление. Обычно мышцы-антагонисты
находятся под влиянием как
центрального возбуждения, так и торможения
(29). По нашим исследованиям на
спинномозговых и децеребрированных препаратах (кошках) чистое
торможение или чистое возбуждение антагонистов мыслимо только при
некоторых пороговых раздражениях; при этом отсутствует
координированный характер рефлекторных реакций: или оба антагониста
возбуждаются или оба тормозятся. При координированных же
реакциях сгибания или разгибания, независимо от характера раздражения,
мы всегда имеем по отношению к сокращающимся мышцам
двойственную иннервацию—возбуждение и торможение вместе (Беритов, Баку-
радзе и Нарикашвили, 7).
Двойственная иннервация мышцы может быть обнаружена
следующим путем. Сначала на испытуемой мышце вызывается
значительное рефлекторное сокращение с воспринимающего поля какого-либо
рефлекса. Во время этого рефлекса производится раздражение, которое
Рис. 39. Сгибательный
рефлекс при раздражениях
различной интенсивности.
Спинномозговой препарат лягушки.
Верхняя кривая принадлежит
сгибателю semitendinosus, а нижняя —
разгибателю triceps. В первом
опыте сила раздражения 29 см
расстояния между катушками.
Сокращается только сгибатель. Во втором
опыте раздражение усилено—27 см.
В этом опыте сгибатель дает более
сильное сокращение, но при этом
сокращается и разгибатель.
ОСНОВНЫЕ ПРОЦЕССЫ ЦЕНТРАЛЬНОЙ КООРДИНАЦИИ 93
дает на данной мышце двойственную иннервацию. Если последнее
раздражение в отдельности давало небольшое сокращение, то во время
комбинации с первым имеющееся значительное сокращение ослабевает
до величины сокращения, получающегося при двойственной иннервации.
Означенное ослабление, несомненно, вызывается торможением
двойственной иннервации (рис. 40). Существует и другого рода
доказательство. Обычно, если сильное раздражение вызывает двойственную
иннервацию со слабым сокращением, то часто по прекращении раздражения
Рис. 40. Одновременное возбуждение и торможение мышц.
Спинномозговой препарат лягушки. Верхняя кривая принадлежит сгибателю
колена m. senntendmosus, нижняя — разгибателю колена m. triceps. Раздражается
п. orachiahs для вызова рефлекса потирания (верхний сигнал) и IX задний корешок
для вызова сгибательного рефлекса (нижний сигнал). Первый опыт (А) представляет
рефлекс потирания, а четвертый опыт (D) — сгибательный рефлекс. В последнем
опыте разгибатель также дает сокращение, которое сразу усиливается по
прекращении раздражения. Во втором и третьем опытах (В и С) происходит комбинация
обоих рефлексов. В обоих случаях значительно уменьшается как сокращение
сгибателя, что указывает на торможение его под влиянием рефлекса потирания, так и
сокращение разгибателя, что указывает на торможение его от рефлекса сгибания.
Время — в секундах.
¦сокращение усиливается (рис. 40 D). Усиление сокращения указывает
>на то, что во время раздражения ослабленное сокращение было обуслов-
.лено торможением, что по прекращении раздражения торможение
прекращается раньше возбуждения и потому мышца имеет возможность
сокращаться сильнее, чем раньше. Это усиление сокращения после
раздражения впервые было отмечено Сеченовым (1869) (17). Он
получал его, применяя сильное раздражение, но его можно наблюдать и
при пороговых раздражениях (Беритов, 27; Грэим Броун, 29).
Двойственная иннервация является неизбежным условием для
осуществления некоторых рефлексов. Таков, например, рефлекс чесания.
В этом рефлексе каждая мышца производит клоническое сокращение,
вроде зубчатого тетануса. Ритмические сокращения, выражающие
чесание, протекают на фоне некоторого постоянного сокращения (рис. 85).
Вследствие этого фонового сокращения нога б продолжение всего
•акта чесания притянута к телу. Было обнаружено, что в период
расслабления между отдельными зубцами сокращения мышца испытывает
94 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
торможение, а именно, при комбинации рефлекса чесания со сгибатель-
ным рефлексом каждый раз во время этого расслабления ослабевает и
сокращение сгибателя, производимое в сгибательном рефлексе (Шер-
рингтон, 1).
Происхождение двойственной иннервации. Явление одновременного
возбуждения и торможения двигательных нейронов зависит от
многих причин. Так, например, в одних случаях двойственная
иннервация наступает оттого, что, кроме первично возбужденного
координирующего центра, благодаря центральной иррадиации возбуждения,
действуют и другие координирующие аппараты. Так, например,
обнаружено, что в рефлексе потирания сокращение сгибателя колена m. semiten-
dinosus происходит вследствие возбуждения координирующего аппарата
сгибания благодаря иррадиации возбуждения (Беритов, 30). Затем
было выяснено, что двойственная иннервация в рефлексах сгибания и
разгибания на децеребрированных кошках зависит от соучастия
тонических рефлексов опять-таки вследствие центральной иррадиации
возбуждения (Беритов, 31). Эти тонические рефлексы будут описаны, ниже.
Двойственная иннервация в рефлексе чесания, вероятно, представляет
природную функцию координирующего аппарата чесательного
рефлекса. За это говорит следующее наблюдение Шерриигтона: чеса-
тельный рефлекс вызывается даже в том случае, если раздражение
является минимальным и если вследствие перерезки задних корешков
устранена возможность возбуждения других координирующих
аппаратов вторичными раздражениями (32).
Двойственная иннервация получается очень легко и постоянно при
раздражении нервных стволов, получающих чувствительные нервные
волокна из разных рецептивных полей. Например, это имеет место при
раздражении седалищного, нерва, п. peroneus, задних корешков и т. п.
Благодаря разной возбудимости нервных волокон, что связано с их
толщиной, при некоторых слабых раздражениях может получиться
активное состояние одного координирующего аппарата с чистым
возбуждением или торможением данной мышцы; но при сильных раздражениях,
вовлекающих в действие все волокна, придут в активное состояние
разнообразные координирующие аппараты, вследствие чего
определённые мышцы одновременно будут испытывать и возбуждение и
торможение.
Существует еще одна причина двойственной иннервации. Так как
возбуждение может распространяться в спинном мозгу и без участия
координирующего аппарата, путем передачи возбуждения от
чувствительного нейрона прямо на двигательный, то при повышении
возбудимости в спинном мозгу двигательные нейроны антагонистических мышц
должны получать как реципрокную иннервацию из соответствующего
координирующего аппарата, так и одновременную возбуждающую
иннервацию прямо от чувствительных волокон. Вследствие этого мышца,
заторможенная под влиянием реципрокных иннервации из координи-
ОСНОВНЫЕ ПРОЦЕССЫ ЦЕНТРАЛЬНОЙ КООРДИНАЦИИ 95
рующего аппарата, должна испытывать также и возбуждение через
коллатерали афферентного нейрона, что внешне выразится в
незначительном сокращении на фоне торможения.
_М ? Т У Г V Т—1
0.02"
мШШШШшМЕШШШ
Рис. 41-1. Биоэлектрические токи возбуждения m. semiten-
dinosus при двойственной иннервации.
Спинномозговая лягушка. Фарадическим раздражением п. cut. fem. lat.
из рецептивного поля потирания вызывается двойственная иннервация;
ритм раздражения дан над кривыми: каждое опускание—размыкатель-
ный удар. А— начало раздражения; В—продолжение его. Вначале
нет токов возбуждения, затем они возникают после каждого
индукционного удара группами. Каждой группе предшествует пауза или
ослабление токов возбуждения (ж). С — сначала идет фарадическое
раздражение п. peroneus из рецептивного поля рефлекса сгибания; затем
присоединяется то же раздражение п. cut. fem. lat. От каждого удара
последнего раздражения устанавливается первым долгом небольшая
пауза (х). Сплошная кривая от механического эффекта. Время — по
0.2 сек.
>bU*Aj^bip^jlb^Ju\*ktyJ,
QM2n
Рис. 41-П. Биотоки возбуждения п. cruralis лягушки при двойственной
иннервации.
Лумбальный препарат. Все мыщцы денервированы. Отводится двигательный нерв разгибателя
колена п. cruralis в катодный осциллограф. Раздражением п. peroneus соотв. стороны вызывается
сгибательный рефлекс. А— начало раздражения, В— период усиленной деятельности; в ответ
на каждый удар раздражения сначала наступает пауза торможения длительностью 15—25?, а затем
эа ней следует длительный разряд импульсов возбуждения.
Шеррингтон и его сотрудники, анализируя явление
двойственной иннервации, приходят к тому мнению, что во время этой
иннервации разные двигательные элементы данной мышцы нннервируются по-
разному, что торможение одних элементов идет рука об руку с
возбуждением других. Однако это положение, которое действительно
96 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
может иметь место в определенных случаях, не исключает другого
положения, когда один и тот же двигательный элемент будет
возбуждаться и тормозиться одновременно. Мы, например, наблюдали в
электрограмме заторможенной імышцы с двойственной иннервацией
поочередную смену периодов торможения и возбуждения. Для
иллюстрации приведем рис. 41-1, где ясно видно, что на каждый
индукционный удар тетанического раздражения п. cutaneus femoralis
lateralis на m. semitendinosus сначала возникает пауза торможения, а петом
группа импульсов возбуждения. Наблюдение показывает, что в этих
условиях опыта чем чаще раздражение, тем короче периоды
возбуждения, а при некоторой высокой частоте (свыше 50 раз в 1 сек.) эти
периоды возбуждения отсутствуют. Но период, возбуждения проявляется по
прекращении раздражения, как изображено на рис. 40 D. Мы думаем,
что в означенных случаях в паузе торможения двигательные нейроны
долучают импульсы возбуждения со стороны промежуточных нейронов,
только они не могут проявиться на периферии вследствие устранения
,их под влиянием торможения.
Аналогичное явление можно наблюдать на разгибателе в сгаба-
¦тельном рефлексе. Для иллюстрации приводится на рис. 41-II
электрический эффект двигательного нерва разгибателя колена (п. cruralis
лягушки) при тетаническом раздражении п. peroneus, который получает
•чувствительные волокна из рецептивного поля сгибания. После того
как установился значительный электрический эффект, через 10—14 а
после каждого удара раздражения биотоки возбуждения
двигательных волокон исчезают. Пауза торможения длится 15—25 о. Скрытый
^период торможения почти соответствует времени проведения нервного
-импульса по афферентному и эфферентному нервным путям. Очевидно,
в данном случае торможение развивается в спинном мозгу сейчас же
ііосле прихода периферических импульсов; последующее возбуждение
проявляется с опозданием на 15—25 о, т. е. при скрытом периоде в
55—35 о.
Общее торможение. Выше мы показали, что рецнпрокную
иннервацию в чистой форме можно наблюдать только при совершенно
.специальных лабораторных условиях. Обычно же возбуждение любой
мышцы сопровождается ее торможением,— иначе говоря, всякий
двигательный эффект мышцы по существу является результатом
алгебраической суммации центрального возбуждения и торможения. Наличие
торможения можно выявить специальными опытами в каждом
конкретном случае. Даже если мышца дает (максимальное сокращение, нельзя
утверждать, что в центральной нервной системе по отношению к ней
протекает только возбуждение. Если раздражение, продолжать до
того момента, когда сокращение ослабеет в силу утомления, или
.если, раздражение ослабить в такой імере, чтобы рефлекторное
сокращение понизилось, то в этих случаях легко можно обнаружить, что
.сокращение .содровещдается торможением. Лучше всего это дости-
ОСНОВНЫЕ ПРОЦЕССЫ ЦЕНТРАЛЬНОЙ КООРДИНАЦИИ 97
гается путем вызывания такого сокращения на фоне более или менее
значительных рефлекторных вздрагиваний, какие обычно появляются,
•если раздражать чувствительный нерв отдельными индукционными
ударами через каждые 1—2 сек. Эти вздрагивания ослабевают или
даже устраняются целиком во время ослабленного тетанического
соглашения (Беритов и Бакурадзе, 26) (рис. 34).
Торможение на сокращенной мышце усиливается с усилением или
учащением раздражения; одновременно усиливается и торможение
Рис. 42. Усиление общего тормошения с усилением раздражения.
Лумбальный препарат кошки. Во время рефлекторных вздрагиваний на
разгибателе (верхняя кривая) от раздражения V заднего корешка правой стороны
производится тетаническое раздражение VII заднего корешка той же
стороны. В опытах А и В с усилением раздражения усиливается как
сокращение сгибателя (нижняя кривая), так и торможение разгибателя. В опытах
С и D при дальнейшем усилении тетанического раздражения торможение
разгибателя усиливается еще больше. Сокращение сгибателя, наоборот,
ослабевает, что свидетельствует о торможении сгибателя под влиянием
усиленного общего торможения (Беритов и Бакурадзе).
других, несокращенных мышц. Пока центральное торможение срав
нительно слабо и вследствие этого центральное возбуждение
превалирует над ним, преимущественно возбуждаются двигательные нейроны
данной мышцы, и торможение не выявляется. Но когда, в связи
с усилением раздражения, центральное торможекие значительно
повышается, оно начинает так сильно влиять на двигательные нейроны,
что возбуждение многих из них становится невозможным, а поэтому
сокращение мышцы ослабевает (рис. 42) (Беритов и Бакурадзе, 26).
Вышеприведенные факты дают нам право заключить, что когда
в каком-либо участке центральной нервной системы возникают
импульсы возбуждения, посылаемые какой-либо группе мышц и
вызывающие определенное движение на данном органе, то в этом же участке
' И. С. Беритов
98 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
возникает процесс торможения не только в отношении мышц,
являющихся антагонистами данной возбужденной группы, но и в отношении всех
без исключения мышц соседних органов, равно как и в отношении
возбужденной группы мышц. Таким образом, центральное торможение
всегда является общим, охватывающим более или менее все
двигательные нейроны активного участка мозга.
Рис. 43. Влияние раздражения кожи холодом на двигательную
реакцию.
Нормальный кролик. Нижняя кривая—движение правой задней конечности,
раздражаемой фарадическим током; средняя — движение левой задней
конечности; верхняя — дыхательные экскурсии: поднятие — выдыхание, опу- -
екание —- вдыхание. Поднятием верхней сигнальной линии обозначается
раздражение правой задней конечности фарадическим током (цифры под этой
линией указывают на силу раздражения в сантиметрах расстояния между
катушками). Поднятие нижней сигнальной линии — охлаждение спины (3°).
В самом низу — время в секундах. В опытах А, В, D и Е наступают
рефлекторные движения конечностей, а дыхательные движения усиливаются.
В опыте С во время охлаждения спины до 3°С рефлекторные движения
конечностей в ответ на то же электрическое раздражение отсутствуют. Кроме того,
ослабевают дыхательные движения (Дзидзишвили).
Отсюда следует, что реципрокное торможение, по Шерринг-
тону, является лишь частью общего торможения по отношению к
антагонистической группе мышц, что реципрокная иннервация мышц, по
существу говоря, является фикцией, ибо торможение наступает в
отношении как несокращенных, так и сокращенных мышц.
Общее торможение при отсутствии рефлекторных движений. Выше
мы рассмотрели такие случаи общего торможения, когда оно приходится
во время рефлекторного движения в одном каком-либо органе. Но
известны и такие случаи, когда общее торможение происходит без каких-
либо признаков возбуждения двигательных органов. Так, например,
известно, что сильное электрическое раздражение чувствительного
нерва, как кожного, так и мышечного, а также сильное механическое
раздражение кожи производят вслед за более или менее
значительной начальной двигательной реакцией общее понижение рефлектор-
ОСНОВНЫЕ ПРОЦЕССЫ ЦЕНТРАЛЬНОЙ КООРДИНАЦИИ
99
ной деятельности. По нашим наблюдениям, это бывает при
сдавливании кожи на голове: после начального движения головы вся
мускулатура организма испытывает общее торможение (Беритов, 14). То же
бывает при сдавливании кожи на ногах, на спине или при
раздражении соответствующих чувствительных нервов (Данилевский, 15;
Беритов и Гогава, 16, Бакурадзе, 36). Аналогичное явление наблю-
Рис. 44. Общее торможение при
слабом химическом раздражении
спинного мозга.
Лумбальный препарат кошки.
Раздражением V и VIII задних корешков пояснично-
крестцовой пбтасти вызываются
рефлекторные вздрагивания на сгибателе (т.
semitendinosus) правой стороны и на
сгибателе и разгибателе (m. quadriceps)
левой. В это время прикладывается ацетил-
холин в растворе 1 : 20 000 локально к
левой дорзо-латеральной поверхности
IV—V сегментов. Рефлекторные
вздрагивания на всех трех мышцах ослабевают
(Беритов и Бакурадзе).
Рис. 45. Общее торможение при
сильном химическом раздражении
спинного мозга.
Та же лумбальная кошка, что и на рис. 44.
Раздражением V заднего корешка правой
и левой сторон вызываются рефлекторные
вздрагивания на трех мышцах. В это время
прикладывается ацетилхолин в растворе
1 : 5000 локально к правой
дорзо-латеральной поверхности III и IV сегментов.
Сначала на всех мышцах рефлекторные
вздрагивания ослабевают, а затем
происходит их усиление, сильнее на
соответствующей правой стороне, чем на левой. Кроме
того, на правом сгибателе наступает
длительное тетаническое сокращение (Беритов
и Бакурадзе).
дается при раздражении смешанных нервов: вслед за начальной '
двигательной реакцией наступает общее угнетение рефлекторной
деятельности (Сеченов, 17; Введенский, 18).
Общее торможение может наблюдаться и без каких-либо
начальных или сопутствующих движений. Это бывает, например, при
раздражении одного чувствительного волокна (Като, 6), при подпороговых
раздражениях чувствительных нервов и задних корешков как
фарадическим током, так и отдельными индукционными ударами (Беритов
и сотр., 7, 24; Гедевани, 25), при раздражениях Холодовых и
тактильных рецепторов на кролике (рис. 43) (Дзидзишвили, 24). То же
7*
J00 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
самое наблюдается при непосредственном раздражении обнаженного
спинного мозга химическими раздражителями, как ацетилхолин, КО
в слабой концентрации (рис. 44 и 45), слабым фарадическим током
или слабым механическим раздражением, как, например,
прикосновением очень мягкой кисточкой (Беритов и сотр., 8, 9, 19, 37). Более силь-
Рис. 46. Общее торможение при
растяжении мочевого пузыря на
торако-лумбальном препарате
кошки.
Те же мышцы, что и па рис. 45.
Мочевой пузырь растягивается посредством
вставленного в него резинового
мешочка; последний соединен с аппаратом
Рива-Роччи и может быть растянут
вдуванием воздуха. На сигнальной линии
У е. цифры означают давление в
миллиметрах ртутного столОа. Растяжение
производится во время рефлекторных
вздрагиваний, вызываемых на обеих
сторонах раздражением заднего столба
в грудной области (Th. s.) и V заднего
поясничного корешка (V d).
Торможение наблюдается уже при растяжении
под давлением в 10 мм; с увеличением
давления до 20, 40 и 100 мм общее
торможение усиливается.
Рис. 47. Общее торможение при
раздражении п. pelvicus.
Опыты на торако-лумбальном препарате
кошки. Регистрация тех же мышц, что и
на рис. 46. В опыте С тетаническое
раздражение п. pelvicus. sin. (P. s.),
производящееся отдельно, совсем не вызывает
сокращений мышц.В опытах А и В это
раздражение производится во время
рефлекторных вздрагиваний, вызываемых
раздражением п. saphenus dex. (Sa. d.) и п.
saphenus sin. (Sa. s.) одиночными
индукционными ударами. Во время сочетания
эти вздрагивания ослабевают.
ное раздражение мозга з этих случаях вызывает возбуждение одних
мышц и торможение других (рис. 45). Такое же общее торможение
получается при раздражении внутренних органов и
вегетативных чувствительных нервов. В этом случае двигательные эффекты
наблюдаются на скелетных мышцах очень редко, исключительно при
высокой возбудимости препарата. Очень часто с усилением
раздражения только усиливается торможение скелетных мышц. Так, например,
на рис. 46 раздражение рецепторов мочевого пузыря путем его
растяжения вызывает общее торможение мышц живота и задних конеч-
иостей при силе давления от 10 до 100 мм ртутного столба, а на
ОСНОВНЫЕ ПРОЦЕССЫ ЦЕНТРАЛЬНОЙ КООРДИНАЦИИ JQ]
рис. 47 раздражение п. pelvicus производит общее торможение при
силе раздражения в 20 и 37 см расстояния катушек, не вызывая при
этом никакого двигательного эффекта (Беритов и Бакурадзе, 34).
Общее торможение наблюдается точно так же при раздражении
чувствительных нервов и самого мозга отдельными индукционными
ударами. Торможение может продолжаться после такого раздражения
Рис. 48, Центральное торможение от одиночных периферических
импульсов.
Лумбальный препарат кошки. Раздражаются два задних корешка VII и VIII левой
стороны. В опыте А раздражение VIII корешка предшествует раздражению Vi7 на
1 сек., и от этого эффект второго раздражения сильно тормозится. В опытах В> С
и D раздражение VII предшествует раздражению VIII на 2, 3 и б сек., и во всех
случаях эффект от второго раздражения оказывается заторможенным на обеих мыщ- и
цах (Беритов и Бакурадзе).
несколько секунд. Так, например, на рис. 48 торможение удерживается
после раздражения заднего корешка около 5 сек. (Беритов и
Бакурадзе, 35, Гедевани, 25). "
Явление общего торможения наблюдалось нами даже в таких
случаях, когда раздражался небольшой участок кожи в 1—2 мм, когда,
следовательно, фактически приходило в возбуждение всего несколько
чувствительных волокон (Беритов и Гогава, 16). Однако
интенсивность торможения в полной мере зависит от величины раздражаемого
участка кожи или от количества возбужденных чувствительных
волокон и от частоты периферических импульсов. Общее торможение
мозга бывает тем интенсивнее, чем больше чувствительных волоков?
возбуждается и чем выше ритм их возбуждения.
При длительных раздражениях общее торможение длится
некоторое время, иногда измеряемое минутами, потом проходит с некоторой
постепенностью. Если же раздражение прекращается раньше этого
времени, то одновременно прекращается и торможение. Так, на
рис. 49 длительно раздражается фарадическим током спинной мозг
кошки (задний столб в грудной области) во время рефлекторных
вздрагиваний, вызываемых раздражением заднего корешка. В самом
]Щ ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
начале, сочетания возникает небольшое сокращение брюшной мышцы.
Одновременно наступает общее торможение, продолжающееся 8 мин.,
но по прекращении раздражения оно тоже быстро прекращается:
рефлекторные вздрагивания восстанавливаются в течение 10—15 сек.
Торможение обычно длится значительно дольше, чем возбуждение.
Если возбуждение сопровождается торможением, то при длительных
раздражениях возбуждение
кончается значительно
раньше торможения. С
прекращением раздражения
торможение тоже прекращается
через несколько секунд.
чрезвычайно характерно то,
что оно проявляется в
ответ на раздражение
чувствительного нерва или
рецептора заметно раньше
возбуждения двигательных
нейронов, раньше двигательной
реакции, вызываемой тем же
раздражением. Так,
например, Киселев (20)
наблюдал, что в
перекрестном разгибательном
рефлексе децеребрированной
кошки торможение возникает
много быстрее, чем возбуждение. При гомолатеральном сгибательном
рефлексе оно наступает даже на 1.5—3 о раньше, чем возбуждение. Геде-
вани (21) показал то же самое на лягушке. Здесь в некоторых
случаях перекрестная реакция начинается общим торможением более чем
на 10<j раньше наступления двигательного рефлекса (рис. SO). Кроме
того Фултон (22) нашел, что торможение разгибателя колена от
раздражения мышечных чувствительных нервов сгибателей на децере-
брированных кошках наступает на несколько сигм раньше, чем
наиболее быстро протекающий коленно-сухожильный разгибательный рефлекс.
Итак, рефлекторная деятельность центральной нервной системы
в ответ на каждое деятельное раздражение начинается общим
торможением, и каждая двигательная реакция центрального происхождения
протекает на фоне общего торможения.
Биологическое значение общего торможения заключается,
во-первых, в охране организма от ненужной траты энергии рабочими
органами при разного рода слабых раздражениях; во-вторых, в
локализации импульсов возбуждения в определенных нервных кругах и
цепочках, вследствие чего на данное раздражение наступает определенная
внешняя реакция, и, в-третьих, в невозможности возбуждения других
амии—имиишміиі. ц^.-——^^^..,.,. ...iM |f іі'иі
Рис. 49. Длительное общее торможение при
тетаническом раздражении заднего столба
спинного мозга.
Торако-лумбальный препарат кошки. На рисунке даны
кривые сверху вниз: quadriceps dex. (Q. d.) semitendi-
nosus dex (S. d.), rectus abdominis (R. a.), semitendinosus
sin (S.s.) и quadriceps sin. (Q.S.). Раздражается левый
задний столб V сегмента грудной области (Th. V s. dor.)
во время рефлекторных вздрагиваний, вызываемых на
мышцах задней конечности раздражением VII заднего
корешка правой стороны отдельными индукционными
ударами (VII d). Первый раз тормозящее раздражение
продолжалось около 20 сек., второй раз—около 1 мин.,
а третий раз — около 6 мин. В последнем случае
кимограф останавливался 5 раз по 1 мин. Рефлекторные
вздрагивания почти целиком тормозятся. Они
восстанавливаются по прекращении тормозящего раздражения
в течение 10—15 сек. После отдыха через 2 и 10 мил.
рефлекторные вздрагивания усиливаются.
ОСНОВНЫЕ ПРОЦЕССЫ ЦЕНТРАЛЬНОЙ КООРДИНАЦИИ ЮЗ
нервных кругов и цепочек под влиянием каких-либо менее значительных
внешних и внутренних раздражений; благодаря этому не нарушается
точность господствующей двигательной реакции. Следовательно, общее
торможение является основным
фактором локализации процессов
возбуждения и координации
движений. Вместе с этим общее
торможение обусловливает действительную
целостность рефлекторной реакции,
ибо наряду с возбуждением
определенных нервных комплексов
происходит торможение всей остальной
части мозга.
Некоординированная
деятельность центральной нервной системы
вследствие повышения
возбудимости. Вполне нормальные
координированные рефлекторные реакции
получаются только при некоторой
нормальной возбудимости
центральной нервной системы. Если
возбудимость ее почему-либо сильно
повысилась,— например, вследствие
повреждения центральной нервной
системы или вследствие стрихнин-
ного отравления,— периферическое раздражение перестает вызывать
координированные реакции. Такое слабое раздражение, которое в
норме не давало никакой центральной реакции, теперь может вызвать
сокращение почти всей мускулатуры тела. Так, например, после
вскрытия и перерезки у кошки спинного імозга в поясничной области
возбудимость мозга так повышается под влиянием всякого рода
субминимальных раздражений, связанных с оперативным вмешательством, что
спустя 2—3 часа раздражение задних корешков отдельными
индукционными ударами производит общее сокращение всех мышц:
антагонисты сокращаются синергично, как в том случае, если бы
непосредственно раздражался их двигательный нерв. Это сокращение наступает
прежде всего и сильнее всего на мышцах, получающих двигательные
нервы от того самого сегмента мозга, куда входит раздраженный
задний корешок. Так, например, на рис. 46, 47 и 48 отдельные
индукционные удары прикладывались к корешкам, чувствительному нерву
н спинному мозгу и всякий раз вызывали одновременное
вздрагивание мышц соответствующей стороны. Такие синергичные эффекты
наступают и от пороговых тетанических раздражений. Если же взять
более сильное фарадическое раздражение, то реакция протекает по
принципу реципрокной иннервации; именно, если раздраженный
задний корешок получает нервные волокна преимущественно от рецеп-
Рис. 50. Торможение предыдущего
двигательного эффекта последующим
раздражением.
Кривые сокращения m. semitendinosus
епинальной лягушки. Раздражается п.рего-
neus той и другой сторон отдельными
индукционными ударами, а—момент
раздражения п. peroneus соответствующей
стороны: / — сокращение, вызванное этим
ударом в отдельности; Ь — момент
раздражения п. peroneus противоположной
стороны: 3 — сокращение, вызванное этим
ударом отдельно; 2 — сокращение при
нанесении обоих раздражений. Несмотря
на раздражение противоположной стороны
перед началом сокращения,
приблизительно спустя 15 а после раздражения
соответствующей стороны, последнее
сокращение оказывается заторможенным
(Гедевани).
104 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
тивного поля разгибания (у кошки такими корешками являются IV,
V и VI задние корешки), то сильное раздражение производит
типичный разгибательный эффект с сокращением разгибателей и
торможением сгибателей. Часто и при сильном раздражении нельзя получить
чистого реципрокн'ого эффекта: заторможенная мышца тоже дает
небольшое сокращение, как это показано на рис. 51 (Беритов, 14).
Из этих наблюдений ясно видно, что при некоторой повышенной
возбудимости возбуждение любого чувствительного нейрона должно
передаться на двигательные нейроны не только через
координирующие аппараты, но и помимо них. Очевидно, при нормальной
возбудимости центральной нервной системы периферические импульсы не
в состоянии возбудить двигательные нейроны помимо промежуточных
координирующих аппаратов, они могут повлиять на них только через
координирующие аппараты. Но при повышенной возбудимости слабое
периферическое раздражение в состоянии возбудить двигательные
нейроны даже без одновременного (возбуждения координирующих
аппэ-ратов. При сильном же раздражении будут возбуждаться как
двигательные, так и промежуточные нейроны.
Вследствие этого рецилрокное торможение, производимое
координирующими аппаратами, будет маскировать эффект непосредственного
возбуждения двигательных нейронов со стороны чувствительных
волокон.
Практикум
Анатомическое определение функций двуеуставных мышц. Берут заднюю
конечность лягушки и снимают с нее кожу. Потом находят места прикрепления главных
двуеуставных мышц и исследуют, как действует данная мышца на соответствующий
дясталъный « проксимальный суставы. Для этого одним пинцетом захватывают кость,
чтобы фиксировать препарат, а другим пинцетом потягивают за мышцу. Пот таком
пстягивании соответствующие суставы приходят в действие* сустав или разгибается,
или огибается. Если исследуют мышцы бедра, конечность удерживают за бецреяную
кость. Для рсследования мышц голени пинцетам захватывают берцовую кость. Таким
способом исследуют mm. sartorius, triceps femoris, semitendinosus, semimembranosus,
gastrocnemius, tibialis anticus, peroneus и др. Некоторые мышцы производят как сгл-
бание, так и разгибание одного и того же сустаюац © зависимости от того, какой угол
образует сустав в момент потягивания. Например, m. ileofibularis разгибает тазобед-
ренный состав е том случае, если угол, составленный последним, больше 90°, и
сгибает, если этот угол меньше 90° (Ломбард и Аббот, 41). Tibialis anticus сгибает
голеностопный сустав, если зтот последний уже согнут до 90°; если же сустав разогнут,
то он не сгибается, столько незначительно уклоняется в сторону.
Наблюдение реципрокной иннервации антагонистов. Берут спинномозговой препарат
лягушки и обнажают сгибатель колена m. sernitendmosus и разгибатель m. triceps.
К перерезанному сухожилию каждой мышцы привязывают нитку для соединения
с миографом. Затем обнажают п. peroneus из поля рефлекса сгибания и п. cutaneus
femoris lateralis или п. brachialis из поля потираний. Эти нервы не переоезз-ются,
а только высвобождаются из окружающей ткани настолько, чтобы они стали
доступны для электрического раздражения. Раздражение п. peroneus дает такой эффект,
как показано на рис. 39; если раздражение слабое, сокращается только m.
sernitendmosus, при сильном раздражении небольшое сокращение наступает и на m. triceps.
ОСНОВНЫЕ ПРОЦЕССЫ ЦЕНТРАЛЬНОЙ КООРДИНАЦИИ
105
Раздражение п. cutaneus femoris lateralis производит сильное сокращение гл. triceps
при отсутствии или ори слабом сокращении m. sermtendinosus. Чтобы удостовериться^
что при возбуждении одной мышцы другая тормозится, прибегают к такому опыту:
сначала вызывают описательный рефлекс, причем записывается сокращение m.
semitendinosus; затем присоединяют рефлекс лотирания, ори котором эта мышца не
сокращается. Во время сочетания обоих рефлексов
данная мышца расслабляется (рис. 35). Это является
показателем того, что в рефлексе потираний
двигательный нейрон т. semitendinosus испытывает
активное торможение.
Усиление реципрокных эффектов под влиянием
стрихнинного отравления. На спинномозгозом
препарате лягушки обнажают плечевую часть спинного
мозга с задней поверхности. Потом препарируют ^еп)
п. brachials на передней лапке и п. peroneus на
задней. Кроме того, выделяют антагонисты колена:
mm. semitendinosus и triceps и соединяют их с
миографами. Обычно раздражение передней лапки
вызывает на задней лапке рефлекс потирания. Задняя ^й[ои)
лайка притягивается к передней для удаления
раздражителя. В этом рефлексе m. triceps сокращается*
a m. semitendinosus тормозится. Опыт с
отравлением заключается в следующем. Определяют то
минимальное раздражение для п. brachialis, которое Рис. 51. Одновременное рефл?к-
вызьгвает на антагонистической паре ясный р?ци- тоРное возбуждение антагони-
прокный эффект. Это достигается путем комбинации' « - * „
• v Лумбальный препарат кошки. Верх-
раздражения п. brachialis с раздражением п. регопе- няя кривая от разгибателя колена
us. Потом прикладывают стрихнин в растворе 0.04- <?' ^ші&і^^^в^Х^
0.5% к плечевой части мозга, как это было пека- peroneus той же стороны, сначала
отдельными индукционными ударами че-
зано выше. Через каждые 3—5 мин. повторяют рез каждую секунду, а затем фаради-
«„ ,«« „T,rt™*« « и ли: г г» in лл ческим током. В первом случае обе
раздражение п. brachialis. Спустя 10—20 мин. ока- мышцы отвечают сокращениями, т. е.
жется, что в отравленном координирующем агаха- Рефлекс протекает без координации,
r r rj а во втором наступает антагонистиче-
рате чрезвычайно усиливаются как возбуждающие, ский эффект: сокращение сгибателя и
торможение разгибателя с небольшим
так и тормозящие процессы по отношению к дан- вздрагиванием вначале. Перед
сигнальной паре антагонистических мышц. Усиление тер- ной линией цифры без скобок обозна-
г г чают силу раздражения в сантиметрах
можания видно из того, что под влиянием реф- индукционной катушки, а в скобках
., ». частоту его при тетаиическом раздра-
лекса потирания сокращение m. semitendinosus в жении. Внизу время в секундах.
слибательном рефлексе затормозится много
интенсивнее, чем до отравления, и что раздражение п. brachialis отдельными
индукционными ударами, которое раньше не давало никаких движений на задней ноге, теперь
будет производить сильные реціипрокные реакции.
Наблюдение координации нескольких одновременно действующих органов. Берут
спинномозговую лягушку и наблюдают рефлекторные реакции на раздражение
передних и задних лапок. При слабом раздражении пальцев или ступни задней лапки*
(прикосновение, легкое сдавливание) наступает сгибание одновременно во всех
суставах. На других лапках не будет движений.
При несколько более сильном сдавливании пальцев лягушка сначала разгибает
лапку, как бы отталкивает от нее раздражитель, а затем сгибает ее. При еще более
сильном раздражении она сначала разгибает обе задние лапки, а затем обе они
сгибаются. В это время движутся и передние лапки, сначала вперед, а затем
отводятся обратно и принимают положение стояния.
При слабом раздражении пальцев передней лапки, данная лапка сгибается.
При более сильном она отводится назад. В это время задняя лапка соответствующей
стороны сначала разгибается, а затем притягивается для потирания передней.
При раздражении в области ануса происходит потирание голеностопными су*
106 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
стчвами: одновременное огибание и разгибание. Если препарат хорошо возбудим,
то в момент разгибания задних лапок обе передние разгибаются и отводятся назад:
происходит прыжок.
Наблюдение общего торможения. Берут нормальную лягушку. Наблюдают
рефлекторные и поведенческие реакции дри раздражении кожи, а затем крепко
перетягивают бинтом голову или переднюю лапку. После этого раздражение кожи совсем
не дает поведенческих реакций, исчезает также рефлекс стояния и переворачивания.
Прикосновение, легкое сдавливание не дают рефлекторных реакций. При раздражениях
пальцев или около ануса рефлексы потираяия и прыжка не вызываются. Вызываются
только рефлексы сгибания при сильных раздражениях кожи. После испытания
раздражения снимают бинт. Все рефлексы и поведенческие реакции сейчас же
восстанавливаются.
Общее торможение можно наблюдать и на спинномозговых препаратах, Крепкое
перетягивание передней конечности приводит к исчезновению рефлексов потираний,
отталкивания и прыжка. Не вызываются реакции на поглаживание н легкое
сдавливание пальцев. Только сильное сдавливание или щипание дает рефлекс сгибания.
Можно заметить, что на стороне перевязки сильнее заторможены рефлекторные
реакции, чем на другой. Сейчас же после снятия бинта рефлекторная деятельность
восстанавливается.
Наблюдение двойственной иннервации. Этот опыт должен ироизвддиггьоя подобно
опыту с реципрокной иннервацией. Записывают на спинномозговой лягушке
антагонистическую пару: m. semitendinosus и т. triceps. Опыт ведут в том же порядке, как
на рис. 40. Берут п. superficialis, раздражение которого дает рефлекс
преимущественно разгибательного характера, и п. peroneus, который дает, наоборот, преимущественно
сгибание. Можно сочетать раздражение п. cutaneus femoris lateralis с раздражением
п. peroneus. Во всех случаях сначала производится такое раздражение одного нерва,
которое дает сильный сгибательный рефлекс с значительным сокращением
сгибателя. Затем во время этого рефлекса прикладывается раздражение к другому нерву,
которое дает на том же сгибателе заторможенное сокращение. В период сочетания
существующее значительное сокращение сгибателя ослабевает до величины
заторможенного сокращения.
Литература
1. Sherrington С. S., Int. action of nervous system, 1906.
2. Forbes A., Quart. J. Exp. Physiol., 5, 149, 1912.
3. В е г i t о f f J., Fol. Neurobiol., 7, 187, 1913; Pfl. Arch., 151, 171, 1913.
4. Dusser de Barenne, Fol. Neurobiol., 5, 42, 1911; 6, 277, 1912.
5. Verworn M., Zschr. Allg. Physiol., 15, 413, 1913.
6. К a t о О., The microphysiology of nerve, 1934.
7. Беритов И., Бакурадз?А. и Нарикашвили С, Тр. Физиол.
ин-та им. Бериташвили, 3, 147, 168 и 173, 1937,
8. Беритов И. иБакурадзеА., Физиол. журн. СССР, 28, 18, 4940.
9. Беритов И. иБакурадз? А. Тр. Ин-та физиол. им. Бериташвили,
5, 1, 1943.
10. Beritoff J., Pfl. Arch., 198, 604, 1923.
11. Беритов И., Журн. эксп. биол. мед., 22, 106 и 114, 1928.
12. MeltzerJ. Das Schluckzentrum, seine Irradiation etc. Inaug. Dissert., Berlin,
1882.
13. Ухтомский А., Тр. Физиол. лаб. Петерб. ун-та, 5, 1911.
14. Б е р ит о в И., Журн. эксп. биол. мед., 11, № 30, 1929; Zschr. Biol., 89, 77,
1929.
15. Данилевский В., Pfl. Arch., 24, 489, 1881.
16. Беритов И. иГогаваМ., Тр. Физиол. ин-та Тбил. ун-та, 3, 265, 1937.
17. Сеченов И. М. Об электрическом и химическом раздражении
чувствительных нервов лягушки. СПб., 1868.
ОСНОВНЫЕ ПРОЦЕССЫ ЦЕНТРАЛЬНОЙ КООРДИНАЦИИ 107
18. We den sky N.. Fol. Neurobiol., 6, 551, 1912.
19. БеритовИ. иБакурадзе A., Acta Medica URSS, 2, 371, 1939.
^30. Киселев M., Казанск. мед. журн., № 4—5, 389, 1931.
21. Г e д e в а н и Д., Тр. Физиол. ин-та им. Бериташвили, 3, 479, 1937.
22. F ul t on J. F. Muscular contraction, etc., 318—320, 1926.
23. ДзидзишвилиН., Физиол. журн. СССР, 26, 354, 1939; Тр. Ин-та физиол.
им. Бериташвили, 4, 108, 1941.
24. БеритовИ. и Баку радзе А., Физиол. журн. СССР, 27, 337, 939.
25. Г е д е в а н и Д., Физиол. журн. СССР, 26, 374, 1939.
26. Б е р и т о в И., Тр. Физиол. ин-та им. Бериташвили, 4, 1, 1941; Бери-
тов И. и Бакурадз?А., ДАН СССР, 26, № 9, 956 и 961, 1941.
27. БеритовИ., Тр. Петерб. об-ва естеств., 42, 267, 1913.
28. Введенский Н. и Ухтомский Ам Раб. физиол. лаб. Петерб. ун-та,
8, 145, 1908.
29. Graham Brown., Quart. J. Exp. Physiol., 4, 331, 1911; 5, 237, 1912.
30. Б e p и t о в И., Изв. Петерб. биол. лаб., 12, 1912.
31. Be ri toff J.s Quart. J. Exp. Physiol., 9, 199, 1915.
32. Sherrington С S. a. SowtonM. E.,Proc. Roy. Soc, 83, 435, 19П.
33. Creed, Denny Brown, Eccles, Liddell a. Sherrington,
Reflex activity of the spinal cord, 1932.
34. Беритов И. иБакурадзе А., Тр. Ин-та физиол. им. Бериташвили, 5,
143, 1943.
35. Беритов И. иБакурадзе А., ДАН СССР, 26, 956 и 961, 1940.
36. Б а к у р а д з е А., Тр. Ин-та физиол. им. Бериташвили, 5, 107, 1943.
37. Беритов И. и Бакурадз? А., Тр. Ин-та физиол. им. Бериташвили,
5, 1, 1943.
38. Sherrington С. S., J. Physiol., 40, 28, 1910.
39. Duchenne G., Physiologie d. Bewegungen. Нем. пер. с франц., 1885.
40. В eritof f J., Pfl. Arch., 194, 365, 1922; Учение об основных элементах
центральной координации скелетной мускулатуры, 1916.
41. Lombard W. Р. и. А Ь b о 11 F. M., Am. J.Physiol., 20, 1007.
*
Глава VII
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ
Гипотеза о пессимальном характере центрального торможения.
Многие авторы рассматривают торможение как результат
интерференции частых импульсов, т. е. как пессиімальное состояние Введенско-
г а Возможность центрального торможения по принципу пессимального
состояния хорошо была доказана самим Введенским на основании
собственных исследований на стрихнинизированных препаратах (1). Он
наблюдал на таких препаратах ослабление рефлекторной реакции под
влиянием более частых и сильных раздражений. Но едва ли явление
пессимального характера имеет іместо при нормальных рефлекторных
или произвольных реакциях, ибо, как показал Эдриан (2), на
изолированных нервных волокнах в рефлекторных реакциях івысший ритм
возбуждения не превоісходит 100—150 в 1 сек. Особенно редкий ритм
получается при тонических сокращениях, где он составляет около 7—20
в 1 сек. (Денни Броун, 3). Между тем известно, что как раз эти
тонические рефлексы тормозятся легче всего даже от раздражения
чувствительного нерва отдельными индукционными ударами.
В последнее время ряд авторов высказывается за такое
происхождение торможения, которое очень близко стоит к пессимальной природе.
Именно, Лоренте де Но (4,5) и Гассер (7,72) полагают, что
нормальная возбуждающая деятельность мозга протекает при пороговой
возбудимости нейронов. Когда же почему-либо возбудимость их ниже
нормы, т. е. когда создается субнормальность, как они говорят, тогда
наступает торможение. А эта субнормальность іможет возникнуть или
вследствие учащения импульсов, притекающих к данному нейрону через
одни и те же синапсы, или вследствие уменьшения частоты импульсов,
притекающих через разные синапсы, когда интервал между импульсами
становится больше периода эффективной сумімации этих импульсов.
Последняя субнормальность получается в том случае, когда возбуждение
двигательных нейронов происходит в результате суммации. Лоренте
д е Н о выдвигает очень сложное представление о картине торможения
вообще и реципрокного торможения в частности. Субнормальность от
учащения импульсов в понимании Лоренте де Но — это еще не пес-
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ
109
симальное явление Введенского, наблюдаемое в нервно-мышечном
препарате при учащении раздражения. Она івыражается только в
блокировании клеток для «слабых» импульсов.
В пользу этого гипотетического представления о центральной
деятельности не имеется прямых доказательств, и оно противоречит многим
фактам. Например, оно не может объяснить явление общего торможения,
когда самое слабое подпороговое раздражение производит торможение
всех мышц, когда оно наступает от одиночных индукционных ударов и
в течение нескольких секунд препятствует наступлению двигательных
эффектов от последующих ударов. Затем, если бы торможение
возникало вследствие субнормальности, создаваемой редкими импульсами, то
повышение возбудимости в центральной нервной системе должно было
бы устранить торможение. Фактически этого не бывает. Как при
локальном стрихнинном отравлении спинного мозга, так и при его
локальном ацетилхолиновом отравлении наряду с повышением возбуждающей
деятельности происходит усиление тормозящей деятельности.
Сам Введенский, получив ослабление рефлекторной реакции под
влиянием более частых и сильных раздражений при общем стрихнинном
отравлении лягушки, считал этот пессимальный эффект не следствием
рефракторности, а проявлением парабиоза, т. е. возникновения р
двигательном центре стойкого неколеблющегося возбуждения под
влиянием сильных и частых импульсов со* стороны чувствующей половины
рефлекторного аппарата,— точнее, со стороны чувствительного нерва и
межцентральных связей (1). К такому же заключению пришел он
вместе с Ухтомским в рабоіе, выполненной на кошках: реципоокное
торможение антагониста разгибателя при сильном сокращении флексора
он считал проявлением парабиоза, вызванного в двигательном центре
разгибателей под влиянием импульсов со стороны сильно действующего
флексорного центра (79). Впоследствии наряду с парабиотическим
торможением Ухтомский (120), Васильев (121) и др. ученики
Введенского допускали и другую форму торможения более
экономной природы. Но парабиотическое объяснение центрального
торможения вообще не может быть оправдано, ибо, как мы указывали выше,
центральное торможение может быть вызвано при таких
периферических раздражениях, которые совершенно не возбуждают ни одного
двигательного центра, причем это торможение охватывает как менее
лабильные центры флексоров, так и более лабильные экстензоров, как
при наличии рефлекторного тонуса, так и при его отсутствии. Да и реци-
прокное торможение одной группы мышц наблюдается не только при
сильном возбуждающем действии центра другой антагонистической
группы мышц, но и при любом возбуждающем действии последнего
центра.
Итак, нормальное центральное торможение спинного мозга, как оно
проявляется в реципрокном или общем торможении, не может быть
уподоблено ни пессималъному состоянию нервно-мышечного аппарата,
ни парабиотическому состоянию нерва.
]]0 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Гипотеза о существовании особых тормозящих нейронов наравне с
возбуждающими. Существование особых тормозящих афферентных
или промежуточных нейронов допускалось многими авторами. Так,
например, по 'Ка то, одни чувствительные волокна производят только
возбуждение, а другие — только торможение.
По нашим первоначальным представлениям, реципрокное торможение являлось-
функцией особых промежуточных нейронов. Предполагалось, что в каждом
координирующем аппарате определенная часть нейронов служит для целей торможения
определенной группы мышц. Основанием для этой гипотезы служил тот факт, что
при стршгинном отравлении задней половины спинного мозга одновременно с
усилением и удлинением реципрокного возбуждения одной группы мышц происходит
усиление и удлинение реципрокного торможения другой группы. Отличительные
особенности возбуждающих и тормозящих нейронов должны были зависеть, по нашему
мнению, от функции конечных разветвлений у дв'игателкных клеток. Мы думали, что
как по возбуждающим, так и по тормозящим нейронам идут обычные импульсы
возбуждения, но через одни конечные разветвления эти импульсы производят
возбуждение двигательной клетки, а через другие — препятствую і возбуждению их, г. е
тормозят их (8).
Если для понимания общего торможения исходить из концепции, что
оно является результатом тормозящего действия определенных
афферентных или промежуточных нейронов на двигательные нейроны, то
тогда пришлось бы допустить, что для осуществления этой функции
каждая из 10 тысяч двигательных клеток, которые имеются в спинном
мозгу лягушки средней величины (Бирге, 9), получает коллатерали от
всех почти афферентных и промежуточных нейронов. Число задне-
корешковых волокон у средней лягушки достигает тоже около1 10 тысяч
(Бирге, 9). Следовательно, нужно было предположить, что у каждой
двигательной клетки лягушки оканчивается свыше 10 тысяч нервных
волокон. У крупных животных этих окончаний должно быть еще
больше. Фактически же около одной двигательной клетки спинного імозга
кошки было найдено только около 100 нейрофибриллярных конечных
утолщений, принадлежащих нервным окончаниям (Шеррингтон, 10)
Отсюда мы должны заключить, что упомянутое предположение о
тормозящей функции определенных нейронов не согласуется с известными
нам анатомическими данными.
Предположение о тормозящих нейронах не может быть оправдано
точно так же с точки зрения современных данных физиологии. В
настоящее время известно, что с любого нерва, с любого участка кожи
можно получить как торможение, так и возбуждение. Также и при
раздражении поверхности мозга можно получить и торможение и
возбуждение как порознь, так и вместе. Заключение (Като (11), что
раздражением мышечных волокон у лягушки 'можно получить только*
торможение, при поверке оказалось неправильным: возбуждается та
мышца, рецепторы которой раздражаются, остальные же мышцы
тормозятся (Нарикашвили, 12). К а то наблюдал именно эти последние
мышцы, потому и получал только торможение. На препаратам с очень
высокой возбудимостью раздражение мышечных чувствительных нервов
может дать рефлекторные реакции и на других мышцах (Гедевани, 18).
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ
пт
Итак, ни с физиологической, ни с анатомической точки зрения нельзя,
допустить, что существуют специальные промежуточные тормозящие и
возбуждающие нейроны или тормозящие и возбуждающие афферентные-
пути в отношении центральной деятельности.
Гипотеза о химической природе торможения. По мнению некоторых
авторов, реципрокное торможение является результатом секреции
какого-то специфического тормозящего вещества. Предполагается, что это
вещество выделяется нервными окончаниями в синапсах около
двигательных нервных клеток (Самойлов и Киселев, 13; Шеррингтон, 14).
Оно действует на клетку и делает ее недоступной для возбуждения со
стороны других возбуждающих нейронов. По этой концепции и передача
возбуждения от одного нейрона к другому происходит путем секреции
особого возбуждающего вещества. Следовательно, в синапсах или в
теле двигательных клеток происходит взаимодействие возбуждающего и
тормозящего вещества, и, очевидно, внешний эффект будет зависеть от
того, какое вещество окажет преобладающее действие. Таким
тормозящим веществом можно было считать ацетилхолин, который, как
известно, в определенных условиях производит торможение рефлекторных
реакций (Швейтцер и Райт, 15; Бабский и сотр., 16). Но против
гуморального происхождения центрального торможения говорит прежде
всего очень малый скрытый период торможения. Как уже указывалось
выше, в определенных случаях оно проявляется даже на несколько сигм
раньше связанного с ним реципрокного возбуждения (Гедевани, 18;
Фултон, 19, Киселев, 17).
Против гуморального) происхождения торможения говорит также тот
факт, что при активном торможении рефлекторного сокращения
сгибателя может происходить выпадение только одного-двух импульсов
возбуждения в ответ на одно тормозящее раздражение, т. е. на один
индукционный удар в области рецептивного поля разгибания (Беритов>
8) (см. ниже о ритмическом течении торможения). Это быстрое
протекание и малый скрытый период торможения и послужили главным
основанием тому, что впоследствии Шеррингтон и Экклс отказались
от химической природы торможения (20).
Новейшие опыты с локальным отравлением спинного мозга ацетил-
холином точно так же не подтверждают химической гипотезы
торможения. Именно на хорошо возбудимых лумбальных препаратах кошки
(перерезка спинного мозга в области последних сегментов грудного
отдела), приготовленных под эфирным наркозом, локальное приложение
ацетилхолина к спинному мозгу в концентрации 1 : 500—1 : 20 000
производит как возбуждение, так и торможение, подобно тому как это
получается при раздражении задних корешков. Именно в случае
приложения ацетилхолина к дорзо-латеральной поверхности пояснично-крест-
цовых сегментов (VII—VIII) одновременно с рефлекторным
сокращением сгибателя происходит рефлекторное торможение разгибателя.
Аналогичное действие оказывает такое же локальное отравление
другими химическими веществами, как, например, хлористым калием, ни-
112 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
котином. Правда, при приложении ацетилхолина в более слабой
концентрации можно наблюдать торможение всех мышц, т. е. общее
торможение. Но опять-таки подобным же образом действуют и другие активные
вещества, как КО и никотин; так же действует и слабое механическое и
электрическое раздражение самого спинного мозга (Беритов и Бакурад-
зе, 21). Здесь выступает такое же явление, какое получается при
слабом электрическом раздражении задних корешков и вообще
чувствительных нервов (Беритов и Бакурадзе, 38).
На препаратах лягушки ацетилхолин очень слабо действует на
обнаженный спинной и головной імозг, даже в таких больших
концентрациях, как 1 : 1000; во многих случаях он совсем не влияет на
рефлекторную деятельность, а когда и влияет, Td это влияние
проявляется в небольшом понижении рефлекторной деятельности (Беритов и
Нивинская, 86). У лягушки центральная нервная система так же мало
чувствительна к ацетилхолину, как и центральная нервная система
кошки при очень плохом функциональном состоянии.
На основании этих наблюдений можно утверждать, что ацетилхолин
является просто химическим раздражителем центральных нервных
элементов, подобно многим другим химическим веществам, и что его
раздражающее действие ничем, по существу, не отличается от
механического и электрического раздражения или от воздействия заднекорешко-
вых импульсов. Во всех этих случаях мы имеем общее торможение без
движений при малых силах раздражения и общее торможение с
двигательной реакцией при сравнительно сильных раздражениях.
Гипотеза о тормозящей функции нейропиля. Выше мы приводили
факты, указывающие на то, что торможение всегда носит общий
характер, что оно может быть вызвано и без возникновения двигательных
реакций. Обычно после прекращения раздражения торможение тоже
быстро прекращается уже в течение нескольких секунд. Это
наблюдается как при тетаническом раздражении чувствительных нервов или
задних корешков, так и при их раздражении отдельными индукционными
ударами. В то же время при длительном тетаническШ раздражении
торможение может длиться гораздо дольше возбуждения: оно
продолжается минутами, после того как двигательные реакции уже полностью
исчезли (Беритов и Бакурадзе, 38). На основании этих и других
наблюдений імы пришли к заключению, что торможение в спинном мозгу
осуществляется не через нервные цепочки и круги імозга, передающие
импульсы возбуждения от чувствительных нервов к двигательным
клеткам, а через другой субстрат спинного мозга, а именно—через нейро-
пиль. Мы нашли, что когда под влиянием заднекорешковых импульсов
или импульсов, исходящих из других источников, нейропиль приходит в
активное состояние, то этоі, в свою очередь, обусловливает торможение
близлежащих нервных клеток и аксонов промежуточных и
двигательных нейронов (Беритов, 38).
Хорошим доказательством этого положения может служить, между
прочим, тот факт, что слабые концентрации таких активных веществ,
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ ИЗ
как ацетилхолин, КС1 и никотин, а. также слабое механическое или
электрическое раздражение, приложенное в одном сегменте к дорзо-
латеральной поверхности мозга, производит общее торможение, иногда
очень значительное, простирающееся на ряд сегментов в обеих
половинах мозга, хотя при этом нет двигательных эффектов, нет и признаков
возбуждения промежуточных и двигательных нейронов, нет также
признаков раздражения задвекорешковых волокон, проходящих в дорзо-
латеральных столбах мозга. Характерно, что при некотором ухудшении
функционального состояния проводящей системы мозга лучше выступает
это изолированное торможение, протекающее без возбуждения
двигательных элементов мозга (Беритов. и Бакурадзе, 21). Мы думаем, что
это общее торможение обусловлено активированием нейропиля
непосредственно внешним раздражением. Для того чтобы получить
двигательные реакции, требуются боілее сильная концентрация активных
веществ и более сильное механическое или электрическое раздражение.
Очевидно, это объясняется тем, что для возбуждения клеток
промежуточных и двигательных нейронов или нервных волокон нужно более
сильное внешнее воздействие.
Выше мы указывали, что общее торможение без двигательных
реакций наблюдается и при раздражении чувствительных нервов. Это
бывает лучше всего выражено на препаратах с низкой рефлекторной
возбудимостью, при малой силе раздражения, и указывает на возможность
такого состояния мозга, при котором его клеточные элементы менее
чувствительны к периферическим импульсам, чем нейропиль. Такая
'возможность прямо вытекает из данных гистологии. Как указывалось
выше, нейропиль всегда1 обладает лучшим капиллярным
кровообращением, чем клеточные элементы. Поэтому последствия наркоза или
ослабленного в силу операции кровообращения должны пройти в нейропиле
скорее, чем в клетках. іК тому же массивные клеточные тела всегда
должны с большим трудом освобождаться от последствий наркоза и
больше страдать от ослабленного кровообращения, чем дендритные
разветвления и конечные волоконца с синапсами.
Общее торможение спинного мозга 'без двигательных реакций
вызывается также растяжением внутренних полостных органов (мочевой
пузырь, прямая кишка, желудок и т. д.), а также раздражением
соответствующих нервов (truncus sympathicus, п. pelvicus, п. hypogastricus)
фарадическим током в условиях перерезки всех задних корешков
грудного, поясничного и крестцового отделов спинного мозга. В этих случаях
общее торможение изучается, в отношении рефлекторных реакций,
вызывающихся раздражением центральных отрезков перерезанных задних
корешков. Возбуждаемые при раздражении (внутренних полостных
органов афферентные чувствительные волокна вступают в мозг частью
через передние корешки, частью через периартериальное сплетение
позвоночных артерий. Так как те же раздражения при целости задних
корешков производят не только общее торможение, но и рефлекторные
двигательные реакции, было заключено, что чувствительные волокна,
8 И. С. Беритов
114 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
вступающие в спинной мозг от внутренних органов через задние
корешки, активируют при посредстве своих коллатералей и клетки
промежуточных и двигательных нейронов спинного мозга и нейропиль. Те же
чувствительные волокна, которые вступают в мозг через другие пути,
повидимому активируют своими коллатералями исключительно
нейропиль (Беритов и Бакурадзе, 82).
Тормозящее действие проявляется не только под влиянием
периферических импульсов. Оно возникает и под влиянием импульсов из про>-
межуточных нейронов. Как известно, когда координирующий аппарат
какого-либо рефлекса приходит в активное состояние, интенсивность и
продолжительность возбуждения одной группы мышц более или менее
совпадают с интенсивностью и продолжительностью торможения другой
группы мышц, относящихся к тому же суставу (Шеррингтон, 22).
Далее, известно, что при стрихнинном отравлении промежуточных
нейронов, наравне с усилением возбуждения одних мышц, происходит
усиление торможения других мышц тех же суставов (Беритов, 23). Это
указывает на то, что промежуточные нейроны, входящие в координирующие
аппараты рефлексов, принимают определенное участие © вызове
торможения.
Известно также, что промежуточные нейроны, передающие
импульсы возбуждения от кожных и проприоцептивных рецепторов к
головному мозгу, тоже вызывают торможение, но не вызывают
двигательных реакций путем прямого влияния на двигательные нейроны. Это
было установлено на кошках путем непосредственного раздражения
вторичных восходящих путей, проходящих в боковых столбах спинного
мозга. Именно раздражение данных путей производит общее
торможение рефлекторных реакций, вызываемых через задние корешки. Эти
пути берут начало в клетках промежуточных нейронов, у которых
оканчиваются коллатерали первичных тактильных, проприоцептивных и
температурных волокон (Беритов и Бакурадзе, 85).
На основании приведенных физиологических фактов, а также
анатомических отношений нейропиля к промежуточным нейронам, можно
заключить, что нейропиль приходит в активное состояние не только
прямо через афферентные нейроны, но и вторичным путем — через
промежуточные нейроны. Когда промежуточные нейроны приходят в
активное состояние от афферентных импульсов, то они, с одной стороны,
действуют на определенные двигательные нейроны, вызывая их
возбуждение, и передают импульсы возбуждения лежащим выше отделам
мозга, а с другой — они активируют нейропиль, производя через него
торможение других двигательных нейронов. Вследствие э^гого, чем
сильнее и длительнее активное состояние промежуточных нейронов, тем
сильнее и длительнее будет как распространение возбуждения по
нервным путям мозга, так и распространение торможения через нейропиль.
Как известно, аксоны двигательных нейронов отдают коллатерали в
спинном мозгу еще до выхода из серого вещества. Эти коллатерали
возвращаются обратно и оканчиваются в сером веществе спинного
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ
115
мозга. Существование коллатералей у двигательных волокон
доказывается главным образом у высших позвоночных и у человека. Их
находили такие выдающиеся гистологи,как Гольджи (80), Кахаль (41),
Л е н о ш е к (69), Кёлликер (42), К р а у з е (40). Но только
Гольджи, Леношек, Краузе находиликоллатералипочти у
каждого двигательного аксона и считали их постоянным явлением; другие
Рис. 52-1. Возвратные коллатерали аксонов двигательных клеток.
Спинной мозг 30-см человеческого зародыша: а, Ь, с — двигательные клетки,
ах — аксоны, г — возвратные коллатерали« двигательных аксонов, d—комиссу-
ральные клетки (Леношек).
авторы наблюдали их лишь у некоторых волокон, и притом не зсегда.
Эти возвратные коллатерали в некоторых случаях (у кроликов.) сильно
ветвятся и образуют нейрофибриллярную сеть таім же, в области
передних рогов. В заднюю половину мозга они не идут (рис. 52 — I и II)
(Леношек, 70). Отсюда следует, что раздражение двигательных корешков
должно влиять на состояние вентральной половины спинного мозга.
Между тем известно, что раздражением передних корешков нельзя вызвать
двигательные реакции из спинного мозга. Значит, эти возвратные
коллатерали не связаны с двигательными же клетками и вообще с клеточными
элементами мозга. Но оказывается, что раздражением одного переднего
корешка можно затормозить рефлекторные вздрагивания, вызываемые
раздражением задних корешков через другие передние корешки. Это
торможение, вызываемое так называемой антидромной волной вюзбуж-
8*
1J6 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
дения, очень слабо, но все же имеет место. Оно простирается на
несколько сегментов и даже захватывает обе половины спинного мозга
(рис. 53) (Беритов; и Бакурадзе, 71). Таким образом, антидромная волна
возбуждения не производит двигательных реакций, но вызывает общее
торможение. Отсюда следует, что возвратные коллатерали от
двигательных аксонов заканчиваются на дендритах передних рогов, т. е. в
нейропиле в области двигательных клеток. На этом основании
можно утверждать, что основная
функция возвратных (Коллатералей
двигательных нейронов заключается в
дополнительном усилении общего торможения
двигательных клеток и их аксонов путем
активирования нейропиля.
Характерно, что раздражением
переднего корешка рефлекторные
двигательные реакции через данный
раздражаемый передний корешок тормозятся в
течение нескольких долей секунды после
одного одиночного раздражения, и
много дольше — (после тетанического
раздражения. Это торможение проявляется
на тех двигательных нейронах, которые
раздражаются после импульсов
возбуждения соответствующих двигательных
клеток (Гассер, 72; Лоренте де Но, 75).
Но на соседних двигательных нейронах
торможение наступает сразу, без
предварительного возбуждения, почти без
скрытого периода, к тому самому
моменту, когда антидромные импульсы
достигают серого вещества мозга. Если к
тому же моменту приходят туда же
импульсы возбуждения, вызванные
раздражением чувствительных нервных
волокон, то эти импульсы не дают внешнего двигательного эффекта или же
дают ослабленный эффект (Ревшоу, 84). Это торможение, конечно,
должно быть тоже связано с активированием нейропиля через
возвратные коллатерали передн?корешковых двигательных волокон, и поэтому
оно может наступить сразу, с приходом антидромных импульсов
возбуждения в нейропиль.
Наконец, общее торможение получается при раздражении
нисходящих пирамидных и экстрапирамидных путей, передающих импульсы
возбуждения от головного мозга к промежуточным и двигательным
нейронам спинного мозга. Известно, что волокна пирамидных путей
оканчиваются в сером веществе задней половины мозга, а волокна
вентро-латеральных столбов, начинающиеся в вестибулярных ядрах, р
Рис. 52-И. Возвратные
коллатерали аксонов двигательных
клеток.
Спинной мозг новорожденного
кролика: Л —аксон двигательной клетки (ах)
и сильно разветвленный возвратный
коллатераль (г), в —тонкое сплетение
из разветвленных возвратных
коллатералей двигательных волокон (а) на
наружной Гранине переднего рога
(Леношек).
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ ]]J
сетевидном образовании продолговатого мозга и в мозжечке, оканчи-
іваются в передней половине мозга, в области переднего рога. Но в
обоих случаях очень мало нисходящих волокон оканчивается прямо
у двигательных клеток,
большинство их заканчивается на
элементах промежуточных нейронов
как на их клетках, так и на ден-
дритах (Ллойд, 90). Совершенно
естественно, что при локальном
раздражении нисходящих путей
получается как двигательная
реакция, так и общее
торможение и что торможение наступает
даже при более слабых
раздражениях, чем двигательная
реакция (Беритов и Бакурадзе, 85).
На более или менее
нормальных препаратах активирование
нейропиля обычно должно
происходить под одновременным
влиянием импульсов из заднеко-
решковых волокон,
промежуточных нейронов и возвратных кол-
латералей двигательных аксонов,
а также нисходящих
пирамидных и экстрапирамидных путей.
Но, понятно, активирование
нейропиля должно происходить под
влиянием импульсов из
промежуточных нейронов намного сильнее
и длительнее, чем из других
образований, ибо, если только
периферические импульсы или
ямпульсы, исходящие из высших
отделов імозга, возбудили
нервные цепочки и круги спинного
мозга, то в дальнейшем
активирование нейропиля будет
протекать, главным образом, под
влиянием этих промежуточных
образований. Интенсивность и
длительность активирования
будут зависеть от того, как долго будут импульсы возбуждения пробегать
в нервных цепочках и как много нервных кругов будет действовать
одновременно. То, что после одного периферического импульса, івызвав-
шего рефлекторное вздрагивание, торможение может продолжаться
Ш>иШи,,1Ш„ШЧ,ш;„ншнш^1и,Ш.1Ш,иши
Рис. 53. Общее торможение от
раздражения переднего корешка.
Лумбальный препарат кошки. Все задние
корешки позади перерезанного участка спинного
мозга, — иными словами, все поясничные и
крестцовые корешки перерезаны. В опытах
Л и В раздражением VII левого заднего
корешка вызываются рефлекторные
вздрагивания m. quadriceps sin. (Q. s.) и т. semiten-
dinosus sin. (S. s.) соответствующей стороны.
В это время производится тетаническое
раздражение VII переднего корешка левой
стороны; корешок этот не перерезан, а потому
производит сокращение т. semitendinosus
соответствующей стороны, и одновременно
тормозятся рефлекторные вздрагивания на т.
quadriceps той же стороны. В опыте С
рефлекторные вздрагивания вызываются на другой
стороне раздражением VII заднего корешка
правой стороны. В это время повторяется
тетаническое раздражение того же переднего
корешка. На обеих мышцах сокращения
заметно ослабевают (Беритов и Бакурадзе).
118 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
несколько секунд, свидетельствует о том, что спинномозговые нервные
цепочки и круги могут приходить в активное состояние и от одного
периферического импульса.
Итак, одна из основных функций нейропиля спинного мозга, пови-
димому, заключается в общем понижении возбудимости нейронных
элементов. Когда под влиянием периферических нервных импульсов
приходят в возбуждение определенные нейронные элементы, тогда через
нейропилъ близлежащие нейронные элементы испытывают понижение
возбудимости. Вследствие этого становится возможным более или менее
изолированное распространение возбуждения от определенных
афферентных нейронов к определенным двигательным нейронам для
производства координированной реакции. Таким образом, нейропиль играет
существенную роль в координации рефлекторных реакций. Можно ска^
зать, что координирующий аппарат каждого рефлекса включает
определенный комплекс промежуточных нейронов вместе с окружающим их
нейропилем.
Общее торможение в связи с эмбриональным развитием
нейропиля. Общее торможение проявляется на самой ранней стадии
эмбрионального развития, когда периферическое раздражение начинает
вызывать общие рефлекторные реакции, охватывающие весь организм. Оно
развивается с некоторой постепенностью. Так, по данным Когхил-
ла (76), у амблистомы торможение наступает при тактильном
раздражении крнечности; первоначально торможение ограничивается
только раздражаемой конечностью: весь организм, кроме раздражаемой
конечности, приходит в деятельное состояние. В следующей стадии
развития, в ответ на тактильное раздражение возникает движение и
раздражаемой конечности. Торможение вообще не выявляется. В третьей
стадии, наоборот, при тактильном раздражении наступает торможение
во всем организме, как в целом. В четвертой стадии тактильное
раздражение вызывает изолированное движение раздражаемой конечности,
как определенный локальный рефлекс при торможении всего
остального организма. Отсюда следует, что общее торможение является
таким же первичным эффектом, как и возбуждение, что оно усиливается
постепенно с развитием рефлекторной деятельности. Когхилл
находит, что в индивидуализации или локализации движений общее
торможение играет решающую роль. По мысли Когхилл а, локальные
рефлексы становятся -возможными благодаря общему торможению всего
остального организма. Такого же -мнения придерживается Гонза-
лец (77), который находит, что процесс, посредством которого
достигается индивидуализация и спецификация некоторых рефлексов, скорее
всего обязан именно этой общей тормозной деятельности, посредством
которой сильно и широко задерживаются все первичные или основные
движения, являющиеся общими целостными реакциями. Отсюда следует,
что целостные формы реакции не прекращаются с развитием организма
и даже не теряют господства над вторичными локальными рефлексамиу
но только временами они устраняются общим торможением.
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ Ц9
Очень важно отметить, что онтогенетическое развитие нейропиля
идет параллельно с развитием общего торможения. Так, например, в
первые дни после рождения котенка, в стадии господства общих
целостных реакций, нейропиль—сплетение нервных отростков в спинном
мозгу—состоит главным образом из тонких волокон, идущих в
поперечном и косом направлениях. Синапсы или перицеллюлярные сплетения
не видны. Постепенно это нервное сплетение становится гуще и
особенно сильно разрастается на третьей и четвертой неделе после
рождения, в тот период, когда появляются синапсы и перицеллюлярные
сплетения, когда возникают локальные реакции при общем торможении
всего остального организма. В это время нейропиль не только
становится гуще, но, кроме того, в нем появляется множество волокон,
идущих вверх и вниз вдоль мозга (Уиндл, 78). Эти наблюдения ясно!
указывают, что локальные реакции связаны не только с обособлением
нервных путей посредством развития синаптических связей, но и с
развитием нейропиля, основного субстрата нервной системы в эмбриональной
стадии развития.
Тормозящая функция нейропиля проявляется у зародыша не только
в ответ на раздражение, но также и спонтанно. Так, было замечено* у
головастиков лягушки, что в период формирования тонических центров
в средних отделах мозга, когда эти центры не дают никаких
двигательных реакций ни в ответ на раздражение, ни спонтанно', средние
отделы мозга оказывают тормозящее действие на двигательные
реакции, вызываемые тактильным раздражением, и на спонтанные
движения из спинного мозга. Это выявляется у головастика значительным
усилением двигательной активности спинного мозга после отделения
средних отделов мозга, при перерезке продолговатого мозга позади
уровня ушных пузырей (Гинг-Хзи Ванг и Це-Вей Лю, 95).
Аналогичное тормозящее действие средних отделов мозга было обнаружено
у зародышей овцы в отношении дыхательных движений (Баррон и Барк-
рофт, 96). Отсюда следует, что нейропиль проявляет свою тормозящую
функцию в данном сегменте мозга спонтанно еще раньше развития в
нем рефлекторных дуг для производства локальных двигательных
реакций, т. е. эмбрионально развивающийся нейропиль способен к
спонтанной деятельности подобно двигательным нервным ядрам и другим ядрам
зародыша, производящим движения еще раньше установления
рефлекторных дуг полностью, раньше образования связей с чувствительными
нейронами.
Медленные потенциалы спинного мозга и их нейропильное
происхождение. При каждом активном состоянии спинного мозга, наряду с
быстро протекающими электрическими колебаниями, наблюдаются
медленные колебания (Бек, 28; Умрат, 29; Гассер и Грэйм, 30 и др.).
Медленные колебания электрического потенциала наступают при всякой
деятельности мозга, как спонтанной, так и при раздражении
чувствительных нервов и кожных рецепторов. Длительность и интенсивность
медленного потенциала зависят бт частоты и длительности черифери-
120 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
ческого раздражения; в определенных случаях он іможет держаться
минутами. При регистрации биоэлектрических токов спинного мозга ясно
видно, что чем чаще и сильнее быстро протекающие токи возбуждения
заднекорешковых волокон, чем дольше они следуют друг за другом, тем
интенсивнее и длительнее медленный потенциал мозга. С прекращением
Рис. 54-1. Электрические явления в передних корешках
лягушки.
Токи отводятся в осциллограф бияолярно на расстоянии і.5 и 12
мм от мозга. В опыте А раздражается седалищный нерв одним
индукционным ударом на стороне отведенных-к осциллографу передних
корешков; в опыте В — задний корешок той же стороны; в опыте С —
п. peroneus той же стороны; в опыте D — тот же п. peronetis рядом
индукционных ударов. Стрелки означают моменты раздражения. Время
в опытах А — С -— 0.02 сек. дано на основании кривой С; в опыте
D — 0.1 сек. на основании кривой D.
Рис. 54-П. Электрические явления в задних корешках
лягушки.
Перерезанный VII задний корешок отводится в осциллограф на
расстоянии 1 и 12 мм от мозга. Раздражается неоновым прерывателем
соседний задний корешок: А—3.3 раза в 1 сек., В—9 раз, С—14 раз,
D — 28 раз, Е — 80 раз в 1 сек. в течение 30 сек. (Там, где стрелки
сходятся, запись прервана на 29.5 сек.); F — от раздражения лапки
наложением на нее гири в 10 г (Баррон и Мэтьюс).
быстрых колебаний прекращается и медленное колебание (Мэтьюс, 31),
Эти медленные потенциалы мозга распространяются физически по мозгу,
поэтому их можно отвести как от поверхности мозга, так и от
корешков вблизи мозга (рис. 54).
Медленные потенциалы -спинного мозга имеют при одиночных
раздражениях двухфазный или трехфазный характер: после одного
периферического раздражения спинномозговой потенциал сначала является
отрицательным, т. е. имеет то же направление, как и быстро протекаю-
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ
12*
щие потенциалы заднекорешкового импульса, потом он становится
положительным. После положительной фазы направление потенциала
вновь может измениться, и он вновь становится отрицательным.. Первая
фаза всегда — намного сильнее второй, а вторая — намного сильнее
третьей. Точно так же не одинакова их продолжительность. Первая
фаза продолжается в спинном мозгу кошки и обезьяны в течение 20—
25 о, а вторая в несколько раз дольше— 100 о и больше (Эрлангер и
Гассер, 33; Гассер и Грэим, 30, Беритов и сотр , 104 и др ) (рис. 55).
Рис. 55. Биотоки спинного мозга кошки
Л>м5альный препарат Все передние корешки перерезаны Раздражается n
tibialis одним ударом релаксационного раздражителя при 4V Порог 0 oV Биотоки
отводятся биполярно в шлеифныи осциллограф Л осциллограмма от задней
поверхности VII поясничного сегмента, В—из глубины (в 1 мм) VII сегмента С от
VI заднего корешьа (не перерезан) на расстоянии 5 мм от мозга D от переднего
корешка на расстоянии 15 мм от мозга Время внизу по 0 02 сеь На всех
осциллограммах первое быстрое колебание — артефакт от раздражения
Следующий за ним через па>з\" около 1 5 з первый быстрый потенциал—от
заднекорешкового импульса За ним уже следуют быстрые и медленные биотоки
центрального происхождения Медленный потенциал двухфазный и продолжается свыше
80 о (Беритов, Бакурадче и Роитбаь)
У лягушки продолжительность фаз больше первая фаза длится 30—40 о»
вторая 140—150 о, а третья, если она заметна, 250—300 о (Беритов и
Войтбак, 98) (рис. 56), но в определенных случаях ясно выраженніле
вторая и третья фазы медленного потенциала могут отсутствовать.
Обычно положительный потенциал отсутствует у теплокровных
непосредственно после перерезки мозга в результате шока Он
восстанавливается постепенно после операции (Юз, Мак Куч и Стюарт, 88) У
лягушек положительный потенциал бывает значительно редуцирован при
плохом функциональном состоянии мозга (Беритов и Роитбак, 98).
122 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Интенсивность отводимых от спинного мозга медленных потенциалов
варьирует в широких пределах в зависимости от условий отведения.
Если спинной мозг лягушки открыт только с дорзальной поверхности,
то при отличном функциональном состоянии мозга (медленные
потенциалы, отводимые от задних корешков на расстояние 1—2 мм, достигают
нескольких сотен f*V. Слабее они отводятся от передних корешков и
еще слабее — от поверхности мозга. Если же спинной мозг вынут из
позвоночника и содержится при сравнительно низкой температуре,
тогда при хорошем функциональном состоянии амплитуда медленных
потенциалов, отводимых от задних корешков, значительно больше —
достигает 1—4 mV (Экклс, 107; Беритов и Ройтбак, 108), Очевидно, это
происходит оттого, что условия ветвления и шунтирования токов
снаружи мозга значительно лучше, когда мозг лежит в позвоночнике, чем
когда он изолирован.
Медленные потенциалы, возникающие внутри спинного імозга,
распространяются й выходят наружу, главным образом электротоническим
путем, по нервным волокнам, и только отчасти путем простого
проведения. И тот и другой путь распространения сопряжен с большим
декрементом. Для установления приблизительной интенсивности ' биотоков
внутри мозга можно поступить таким образом: вставить игольчатые
электроды в серое вещество IX—VIII сегмента спинного мозга
лягушки и пропустить постоянный ток определенного напряжения;
распространяющиеся электротонически от этих электродов токи отводить через
IX передний корешок на расстоянии 2 мм1 от мозга. При этом ^окажется,
что в условиях обнажения мозга только с дорзальной поверхности
отводимый от переднего корешка ток в 130 раз слабее пропускаемого
через мозг. Отсюда можно заключить, что если от IX переднего корешка
отводятся биотоки мозга с амплитудой 150 j*Vf то внутри мозга
источник этого биотока должен иметь 19.5 mV.
Медленные потенциалы, отводимые из спинного мозга, в
определенных случаях сопровождаются быстрыми колебаниями электрического
потенциала (рис. 56-А). Эти потенциалы, безусловно, возникают при
возбуждении промежуточных нейронов. Они значительно интенсивнее при
отведении из глубины мозга, чем от поверхности (Герш, 89; Беритов и
Ройтбак, 98).
Медленный потенциал спинного мозга можно изучить путем
отведения центрального отрезка одного из задних корешков. Если отводящие
электроды расположить близко к імозгу, в нескольких миллиметрах от
него, то каждый раз в момент рефлекторной деятельности отводятся
медленные потенциалы, обычно без сопровождения быстро
протекающими токами (рис. 54-11, 56-В); но при отличном функциональном
состоянии быстрые мозговые потенциалы отводятся и через задний
корешок (Баррон и Мэтьюс, 34; Беритов и сотр., 104; Бонне и Бремер, 32)
{рис. 55).
Точно так же медленные токи спинного мозга удается
зарегистрировать при отведении от передних корешков близко от мозга. Обычно они
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ ]23
сопровождаются быстрыми колебаниями аксонных токов двигательных
волокон (Баррон и Мэтыос, 34; Беритов и Ройтбак, 98) (рис. 54-1,
55-D, 56-С).
При повторных редких раздражениях чувствительного нерва
лягушки медленные потенциалы отводятся как от спинного мозга, так и от
задних корешков в ответ на каждое раздражение. Но если интервал
Рис. 56. Биотоки спинного мозга лягушки при одиночных раздражениях.
Лумбальные препараты. Все передние корешки перерезаны. Из задних корешков перере-
ваны все кроме IX и X или одного IX на одной стороне. На этой стороне раздражается
п. peroneus одним электрическим ударом (релаксационный прерыватель). Регистрация
катодным осциллографом. А — отводится игольчатым электродом из глубины IX
сегмента; В — от перерезанного X заднего корешка на расстоянии 1—2 мм от мозга;
С — от IX переднего корешка на расстоянии 1—2 мм от мозга. Первое быстрое колебание
от биотока вовбужденця ааднекорешковых волокон; за ним следуют мозговые потенциалы.
Масштаб вольтажа: 50[aV—3 мм (Беритов и Ройтбак).
раздражений короче медленного потенциала, то после первого эффекта
все последующие раздражения производят более слабые медленные
потенциалы. При этом ослабевает как первая, так и вторая фаза
медленного потенциала (57-А и В).
При хорошем функциональном состоянии препарата электрический
эффект не ограничивается этими медленными потенциалами, регулярно
наступающими вслед за заднекорешковым импульсом. После первой
фазы регулярного медленного потенциала наступает несколько
медленных колебаний разной продолжительности и амплитуды. Вообще число
их достигает 150 в 1 сек. При отведении от поверхности мозга эти
медленные колебания очень редко осложняются быстрыми колебаниями
(рис. 58-В). При отведении же из глубины мозга или через задние
корешки на фоне медленных потенциалов наступают очень часто мелкие
колебания, выражающие аксонные биотоки возбуждения с
продолжительностью около 2 о (рис. 58-jD). Эти быстрые потенциалы относятся
к биотокам возбуждения промежуточных нейронов, из которых
составляются нервные цепочки и круги мозга (Беритов и Ройтбак, 98).
При частых раздражениях чувствительного нерва лягушки
медленный потенциал наступает в сильной форме в самом начале
раздражения, а затем он представлен в виде очень растянутого колебания
124 общая физиология спинного мозг* и ствол* головного мозга
небольшой амплитуды или же в виде ряда небольших, медленных
колебаний. На фоне медленных потенциалов располагаются быстрые аксон-
ные потенциалы. При отведении биотоков от мозга непосредственно
или от задних корешков они выражают возбуждение главным образом
его нервных кругов и цепочек; при отведении же передних корешков
Рис. 57. Биотоки спинною мозга лягушки при редких повторных
раздражениях.
Лумбальные препараты при таких же условиях отведения, как на рис. 56.
Л-—при отведении от боковой поверхности IX сегмента; В—от X заднего
корешка; С — от IX переднего корешка; D —от IX переднего корешка на
другом препарате. Во всех случаях отведение происходит на стороне
раздражения. Везде первое быстрое колебание от биотока возбуждения заднекореш-
ковых волокон Масштаб вольтажа тот же, что на рис. 56 (Беритов и Ройтбак).
Рис. 58. Биотоки спинного мозга лягушьи при его усиленной рефлекторной
деятельности.
Лумбальные препараты при таком же отведении, как на рис. 56. Л и В —¦ при отведении
боковой поверхности IX сегмента в начале раздражения п. peroneus и в период усиленной
рефлекторной деятельности; С и D — при отведении неперерезанного заднего корешка на
расстоянии 1—2 мм от мозга в начале раздражения п. peroneus и в период усиленной
деятельности. Во всех опытах первое быстрое колебание—артефакт от тока раздражения. Спустя
некоторый скрытый период времени след\ет биоток возбуждения заднекорешковых
волокон; вслед за ним сейчас же начинаются мозговые потенциалы Масштаб вольтажа тот же,
что на рис 56 (Берптов и Ройтбак).
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ
125
быстрые биотоки всегда значительно сильнее, и они выражают,
главным образом, возбуждение переднекорешковых волокон (рис. 59).
При адэкватном раздражении кожи точно так же возникают в
спинном мозгу медленные потенциалы наряду с быстрыми. Медленные
потенциалы обычно предшествуют быстрым, а при некоторых слабых ра-ідра-
0.02" I
йіі##4Ф
Рис. 59. Биотоки спинного мозга лягушки при частых раздражениях.
Те же препараты, что и на рис. 57, и те же условия отведения. Только частота
раздражения 125 в сек. Л — из глубины IX сегмента; В — от X заднего корешка;
С — от IX переднего корешка. Масштаб вольтажа тот же, что на рис. 56 (Беритов и
Ройтбак).
жениях возникают по преимуществу. Они хорошо отводятся как от
задних, так и от передних корешков. При отведении задних корешков
медленные потенциалы проявляются в чистой форме. Лишь при отличном
функциональном состоянии оніи усложняются быстрыми потенциалами
небольшой амплитуды. При всех условиях раздражения кожи и
отведения спинного мозга медленные потенциалы имеют колеблющийся
Рис. 60. Электрические потенциалы спинного мозга лягушки. Лумбальный
препарат.
Все передние корешки перерезаны, а из задних —все, кроме IX и X справа. Раздражается кожа
путем сдавливания пальцев правой задней лапки. В опыте А отводится IX передний корешок
правой стороны, а в опытах В и С — центральный отрезок УШ заднего корешка правой стороны.
Запись В в начале рефлекторной деятельности, а С — в момент >силенной рефлекторной
деятельности.
]26 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
характер, подобно тому как это бывает при раздражениях
чувствительных нервов (Беритов и Ройтбак, 108) (рис. 60).
Итак, каждый раз при рефлекторной деятельности спинного мозга в
нем возникают медленные электрические потенциалы. Они длятся все
время его деятельности, причем интенсивность их периодически то
усиливается, то ослабевает. Происхождение медленных потенциалов мозга
было предметом изучения многих физиологов. Прежде всего было
установлено, что медленные потенциалы спинного мозга не могут быть
уподоблены медленным колебаниям электрического потенциала в нервных
волокнах, которые наступают вслед за быстрым биотоком
возбуждения. Эти медленные токи обладают очень малой электродвигательной
силой; они в несколько раз слабее быстро протекающих токов
возбуждения. Это, например, хорошо видно в опытах Марраци и Лорен-
те де Но, где медленные токи нервных волокон в 10 раз слабее
быстрых (97), В спинном мозгу медленные токи могут достигать
большей интенсивности, почти такой же, как и быстрые колебания, и даже
превосходить их значительно (рис. 55, 56, 57 и 58).
Кроме того, известно, что при одновременном раздражении двух
соседних задних корешков или двух веточек одного заднего корешка
медленный потенциал спинного мозга не является сумімой потенциалов,
получаемых от каждого корешка в отдельности, а значительно ниже этой
суммы. Это также указывает, что медленные потенциалы мозга
происходят не от суммирования низковольтных медленных токов нервных
волокон (Гассер и Грэим, 30; Юз, Мак Куч и Стюарт, 88). На этом
основании авторы относят медленные колебания электрического
потенциала не к нервным волокнам, а к синаптическим окончаниям
чувствительных и промежуточных нейронов на нервных клетках імозга или ко
всему серому веществу мозга, или к дендритной части его, или, наконец,
к перимедуллярному сплетению (Умрат, 29; Юз и Гассер, 6; Гассер и
Грэим, 30; Эрлангер и Гассер, 33; Баррон и Мэтьюс, 34). Мы же
полагаем, что эти медленные электрические колебания возникают при
активировании нейропиля мозга.
Как известно, в нервных волокнах под влиянием подпороговых
раздражений возникают локальные процессы. При одиночных
раздражениях они короче возбуждения. Электрические потенциалы, которыми
сопровождают^ эти процессы, сообразно длятся столько же времени
В йерве лягуніки, например, продолжительность локальных
потенциалов около (У 80 о (Катц, 99). Эти локальные процессы от места
раздражения распространяются по нерву с большим декрементом — на
несколько миллиметров. Аналогичные локальные процессы должны
возникать в нервных элементах мозга, в его клетках и дендритах под
влиянием импульсов, приходящих через синапсы. Судя по тому, что при
одиночных раздражениях чувствительного нерва самый короткий
медленный потенциал мозга лягушки достигает 10 а, можно думать, что
локальный процесс под одним синапсом длится по нескольку о, т. е.
значительно дольше, чем в нервном волокне.
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ
127
Но активация дендритов и клеток должна происходить через скрытые
периоды разной продолжительности, ибо через тонкие голые
разветвления аксонов импульсы возбуждения будут достигать дендритов и
клеток тем позднее, чем длиннее и тоньше аксонные веточки.
Соответственно будет происходить не только пространственная суммация локальных
процессов, но и временная их суммация. Это обусловит длительное
течение медленного потенциала после одного заднекорешкового
импульса.
Мы утверждаем, что в происхождении длительных потенциалов
должны участвовать не только дендриты, но и клеточные образования
двигательных и промежуточных нейронов: медленные потенциалы мозга
должны быть также обусловлены теми локальными изменениями
потенциала внутри клеток, какие возникают под влиянием импульсов из
синапсов. Правда, как указывает Грундфест (83), клеточная масса
симпатических ганглиев дает такие же быстрые токи возбуждения, как
волокна В и С, и по всем своим особенностям она не отличается от
нервного ствола; клеточная же масса центральной нервной системы, при
синхронном возбуждении производит быстрые токи, характерные для
волокон Л. Однако не может быть сомнения в том, что нервные клетки
могут дать длительные локальные потенциалы. При изучении биотоков
передних корешков мы пришли к заключению, что медленные
компоненты этих биотоков выражают, главным образом, локальные потенциалы
двигательных клеток, откуда берут свое начало переднекорешковые
волокна.
Биотоки возбуждения синапсов и голых окончаний должны
являться очень низковольтными процессами, так как эти образования очень
тонки, как симпатические волокна. Поэтому эти биотоки едва ли
принимают существенное участие в образовании медленных потенциалов.
Их удельный вес в этом отношении должен быть во много раз меньше
локальных потенциалов дендритно-клеточной массы, ибо каждый
локальный процесс будет наступать не только в том участке дендрита или
клетки, к которому прикасается синапс, но и на некотором расстоянии
от него.
В новейшее время Экклс (107) иБремер (113), изучая
медленные потенциалы передних и задних корешков спинного мозга,
склоняются относить их происхождение всецело к внутриклеточным локальным
процессам, которые, по их мнению, вызываются «коротким
деполяризующим действием некоего синаптического передатчика». Экклс
называет эти потенциалы с и н а п т и ч е с к и м и, но мы не находим этот
термин вполне удачным. Раз эти потенциалы выражают состояние
клеток, а не синапсов, незачем называть их синаптическими. Кроме того,
нельзя согласиться сЭкклсоми Бремеромв том, что эти
потенциалы возникают только в клетках. Еще большее значение должны иметь
локальные процессы в дендритах, на которых оканчивается наибольшее
количество синапсов.
Изучение медленных потенциалов спинного мозга лягушки показало,
128 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
что первая, отрицательная фаза выражает локальные процессы,
возникающие в клетках и дендритах, главным образом под влиянием задне-
корешковых импульсов. Она лишь удлиняется под влиянием импульсов
из промежуточных нейронов. Вторая же и третья фазы, по нашему
мнению, выражают результат взаимодействия, т. е. алгебраической сум-
мации локальных потенциалов, возникающих в обширных областях
мозга вследствие возбуждения нервных цепочек и нервных кругов
мозга.
аіи*т4>4>
Ч**"***»**^* j ',4/«yi**%N*iMft' ^*-Ч^«м^-Н^И^ N«w*p--w,V««4WW'| "Ч^^"***.
^Sl*;
IP \
200 uV
u.or
Рис. 61. Биотоки спинного мозга лягушки.
Изолированный спинной мозг. Все передние корешки перерезаны. Отводится в
катодный осциллограф IX передний корешок слева на расстоянии 1—2 мм от мозга.
Раздражается п. peroneus sin. релаксационным раздражителем. А — опыт при
пороговой силе раздражения 0.5V; В —при силе раздражения 1 V.
Можно утверждать, что в каждом участке нейропиля локальный
потенциал является однофазным. Это действительно можно видеть,
например, в тех случаях, если он наступает исключительно под влиянием зад-
некорешковых импульсов, главным образом в одном определенном
сегменте. Это наблюдается на лягушечьем препарате при пороговых
раздражениях чувствительного нерва п. peroneus и отведении
соответствующего IX переднего корешка, как, например, на рис. 57-/). То же самое
наблюдается на лягушке при раздражении п. brachialis и отведении X
переднего корешка, когда в грудном и поясничном отделах вовлекаются,
главным образом, двигательные нейроны IX—X сегментов (рис. 62-Л).
Если же рефлекторная реакция разыгрывается более или менее
значительно и промежуточные нейроны многих сегментов принимают в ней
участие, тогда от любого корешка будет отводиться алгебраически
суммированный потенциал: отрицательный — от ближайшего сегмента и
положительный — от дальнейшего. Вследствие этого отводимый биоток
может быть двухфазным или трехфазным. Очень часто включение в
реакцию промежуточных нейронов не просто усиливает медленный
потенциал, вызванный непосредственным действием заднекорешковых
импульсов, а производит новую отрицательную фазу медленного
потенциала (рис. 61).
Для уяснения происхождения двухфазного или трехфазного
медленного потенциала дается рис. 62. На рисунке представлена схема
взаимодействия медленных потенциалов, возникающих внутри мозга.
Средняя кривая сплошной линией дает обычную картину трехфазного
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ 129
медленного потенциала от первично возбужденного участка. Кривые
пунктиром изображают составляющие этого медленного потенциала:
верхняя дает медленный потенциал отводимого участка, а нижняя — в
противоположном направлении, от соседних участков. На нижней
кривой потенциал начинает нарастать несколько позднее и достигает
максимума несколько позднее, чем на верхней кривой; кроме того, она
несколько ниже, чем первая. Все это вполне соответствует
действительности: в соседних участках локальные процессы возникают несколько
позднее и несколько слабее, чем в первично возбужденном участке, а
Рис. 62. А — однофазный медленный потенциал спинного мозга при локальном
активировании двигательных клеток поясничной области от раздражения п. bra-
chialis; В — схема разложения трехфазного медленного потенциала. Объяснение
см. в тексте.
появление локальных процессов в соседних участках должно менять
направление отводимого биотока (Форбс и Морисон, 117; Берятов и
сотр., 104, 118).
Подобный медленный потенциал, но значительно более слабый,
возникает в спинном імозгу при раздражении передних корешков. При
одиночных раздражениях он продолжается около 100 о, а при тетаническом
раздражении — несколько десятых секунды (Гассер, 71; Лоренте де Но,
74). Длительность этого медленного потенциала совершенно такого же
рода, как при раздражении задних корешков, поэтому можно
утверждать, что медленный потенциал при раздражении передних корешков
представляет собой суммацию локальных потенциалов, возникающих
в дендритной массе, во-1-ых, под влиянием импульсов через возвратные
коллатерали двигательных волокон и, во-2-ых, под влиянием
импульсов афферентных волокон, проходящих через передние корешки.
Итак, источником медленных потенциалов, отводимых от спинного
мозга, являются локальные процессы, возникающие в дендритной
массе нейропиля, а также в клетках спинного мозга, под влиянием
импульсов — через заднекорешковые волокна, от промежуточных нейро-
9 И. С. Беритов
130 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
нов и от воздействия импульсов через коллатерали переднекорешковых
волокон.
Об отсутствии способности возбуждаться у дендритов. Мы признали
медленные потенциалы спинного мозга за выражение локальных
процессов, главным образом, дендритной массы. Они возникают под влиянием
импульсов возбуждения со стороны многочисленных синапсов,
принадлежащих окончаниям заднекорешковых волокон и промежуточных
нейронов. Можно было думать, что дендриты, подобно нервным волокнам,
не только производят локальные процессы, но и возбуждаются и
проводят возбуждение. Это воззрение одно время являлось господствующим.
Предполагалось, что дендриты собирают импульсы возбуждения и
проводят их к клеткам, а аксон выводит эти импульсы наружу. Но в
настоящее время известен целый ряд фактов, говорящих против
проводящей функции дендритов. Прежде всего, тончайшие голые волоконца
дендритов, должны обладать активным процессом настолько слабой
интенсивности, сравнительно с обычным нервным волокном, что едва ли
этот процесс в состоянии распространиться до клетки при помощи
вызываемого при этом биотока. Скорее всего процесс, вызываемый в ден-
дритах, всегда является локальным. На это указывает также
многочисленность нервных окончаний на дендритах: нервные синапсы, вроде
тоненьких луковиц, покрывают почти всю поверхность дендритов по
всей длине (Гофф, 39; Зурабашвили, 91). Если бы дендриты
.возбуждались и проводили возбуждение, как нервное волокно, все это количество
нервных окончаний было бы совершенно излишне; более того, оно
мешало бы проведению возбуждения, так как активный процесс,
возникший под одним синапсом, помешал бы проведению возбуждения из
соседних участков.
Наконец, из наблюдений над другими отделами мозга известно, что
дендриты не проводят активного процесса. Так, например, при
локальном электрическом раздражении коры головного мозга активное
состояние верхнего слоя, состоящего, главным образом, из сплетения
дендритов пирамидных клеток, не распространяется на эти клетки (Эдриан, 43).
Итак, дендриты не возбуждаются и не проводят возбуоюдения
подобно нервному волокну. Активное состояние, вызванное 0 области
синапсов под влиянием синаптического тока возбуждения,
локализируется тут же или распространяется дальше с очень большим декрементом,
т. е. является локальным процессом. Область активного ?ейропиля
всецело определяется областью действующих на него нервных импульсов
через заднекорешковые волокнаг аксонные раз&етвления
промежуточных нейронов и возвратные коллатерали двигательных нейронов.
Электротоническая природа нейропильного торможения.
Рассмотрение множества фактов в отношении медленных потенциалов и общего
торможения спинного мозга привело нас к заключению, что медленный
потенциал нейропиля является причиной общего торможения.
Медленные потенциалы порождают токи, которые распространяются по
межуточной жидкости, а также по нервным волокнам электротоническим
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ 131
путем, проникают в близлежащие клетки и аксоны и тем самым
должны оказывать на них анэлектротоническое действие.
Как указывалось выше, интенсивность медленных потенциалов
спинного мозга значительна. Она измеряется сотнями микровольт при
отведении их от корешков. Внутри мозга же она измеряется десятками
милливольт. Например, при определенных условиях опыта, когда от
переднего корешка на расстоянии 2 мм от мозга отводится биоток в 150 (uV,
внутри мозга отвечающий ему источник электричества должен быть
около 20 mV. Потенциал такой величины совершенно достаточен для
анэлектротонического понижения возбудимости нервных элементов. При
пропускании через IX—VIII сегменты постоянного тока 20 mV мы
получали заметное угнетение биотоков возбуждения рефлекторного
происхождения, продолжающееся приблизительно в течение 20 о после
включения тока. Очевидно, то же самое имеет место при
возникновении биотоков такой интенсивности внутри мозга. Медленные
потенциалы, как указывалось выше, периодически то усиливаются, то
ослабевают. Это должно особенно благоприятствовать угнетающему действию
медленных потенциалов на спинной мозг.
Как известно, постоянный ток, в особенности пульсирующий,
оказывает на мозг сильное угнетающее действие; он производит такое
угнетение центральной деятельности, что у животного исчезают рефлексы,
останавливается дыхание и т. д. При соответствующей дозировке и
интенсивности пульсирующего постоянного тока можно поддержать
глубокий наркоз много десятков минут при нормальном дыхании. По
прекращении действия пульсирующего тока животное быстро приходит в
нормальное бодрствующее состояние (Ледюк, 110; Чаговец, 111; Фро-
стиг и сотр., 112; Могендович, 119 и др.)- Нужно думать, что медленные
потенциалы, возникающие внутри мозга, оказывают на нервные
элементы мозга такое же угнетающее действие, как искусственное пропускание
пульсирующего постоянного тока через мозг.
При одновременном изучении спинномозговых потенциалов и
рефлекторного торможения сокращения мышц в определенных случаях
удается наблюдать, что торможение длится почти столько же времени,
сколько и положительный мозговой потенциал (Юз, Мак Куч и Стюарт, 88;
Бонне и Бремер, 115). При повторных раздражениях заднего корешка
или чувствительного нерва (кошки, лягушки) отдельными
электрическими ударами второй удар дает ослабленный рефлекс, если он приходится
во время медленного потенциала. При этом характерно, что ослабевает
также мозговой потенциал на повторное раздражение,— именно,
ослабевает отрицательный медленный потенциал. Это хорошо видно на
рис. 57-А и В, где при отведении от мозга или от заднего корешка после
электрического эффекта' на первый удар раздражения оказываются
ослабленными все последующие эффекты. При этом ослабевает как медлен*
ный потенциал, так и предшествующий ему быстрый потенциал. Это
означает, что под влиянием медленного потенциала первого эффекта
тормозятся не только клетки и дендриты, где возникают локальные
9*
132 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА II СТВОЛА ГОЛОВНОГО МОЗГА
процессы, но и сами заднекорешковые волокна и их коллатерали,
производящие начальный биоток возбуждения (Беритов и сотр., 100, 103, 104,
106).
Тормозящее действие медленных потенциалов при повторных
раздражениях не одинаково проявляется в отношении быстрых и медлен-
Рис. 63. Электрические потенциалы спинного мозга кошки при тетаническом
раздражении.
Лумбальный препарат. Все передние корешки перерезаны. Раздражается п. peroneus 15
раз в 1 сек. Регистрация биотоков шлейфным осциллографом. Отводятся поясничные
корешки слева биполярно через усилители с балансированным входом. А — осциллограмма от
VIII переднего корешка; В — от V переднего корешка; они регистрируются одновременно;
при этом сила релаксационного раздражения IV; С — от VII переднего корешка и D — от
VII заднего корешка регистрируются также одновременно при силе раздражения 2V. В
осциллограмме А а указывает на быстрые потенциалы двигательных нейронов, вызываемые
непосредственно под влиянием заднекорешковых импульсов; b — на быстрые потенциалы,
вызываемые посредством промежуточных нейронов (Беритов, Бакурадзе и Ройтбак).
¦
ных потенциалов, отводимых'от передних и задних корешков. Легче
всего угнетаются быстрые потенциалы, возникающие в. передних
корешках под влиянием заднекорешковых импульсов непосредственно, т. е. по
двухнейронной дуге. Они могут выпасть целиком, в то время как
быстрые потенциалы, возникающие в передних корешках под влиянием
импульсов из промежуточных нейронов, только ослабевают более или
менее значительно. Это хорошо видно на рис. 63, осциллограмма А. При
определенной силе раздражения или, вернее, при достаточно большой
силе медленного потенциала, возникающего от первого удара
раздражения, целиком угнетаются в переднем корешке все быстрые
потенциалы того и другого происхождения, которые должны были
наступать от второго удара. Одновременно в передних корешках угнетаются
почти полностью и медленные потенциалы, как это дано на рис. 63,
осциллограмма С. Но в это время от задних корешков медленные
потенциалы могут отводиться, хотя и в заметно ослабленной форме
(тот же рис., осциллограмма <D).
Торможение переднекорешковых эффектов продолжается в течение
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ
Ш
всего времени действия тетанизирующего раздражения, даже в том
случае, если сами передние корешки не дают медленных потенциалов, или
дают их в очень слабой форме (осцилл. С, рис. 63). Отсюда следует,
что это торможение происходит не благодаря медленным потенциалам
двигательных клеток или вообще переднего рога, а благодаря
медленным потенциалам главным образом нейропиля задней половины
мозга, которые всегда наступают в более интенсивной и более постоян-
ной форме, чем в переднем рогу.
Тормозящее действие медленного потенциала вытекает также из
следующего факта: во время длительного положительного
электрического последействия, вызываемого раздражением переднего корешка,
раздражение задних корешков дает только слабое возбуждение данного
переднего корешка (Гассер, 71, Лоренте де Но, 74). Как уже
указывалось, раздражением передних корешков активируются не только
двигательные клетки, но и нейропиль мозга. Активирование нейропиля
происходит через возвратные коллатерали двигательных волокон и через
афферентные волокна, проникающие _в спинной мозг из вегетативных
органов по передним корешкам. При этом возникает медленный
потенциал, который производит анэлектротоническое угнетение ближайших
двигательных клеток.
Такое же тормозящее действие обнаруживается при определенных
условиях и во время длительного отрицательного потенциала.
Например, при отведении от дорзальных столбов и раздражении заднего
корешка отдельными ударами этот медленный потенциал от одного
раздражения длится 20—30 о и ему предшествует быстрый аксонный ток.
Если второе раздражение происходит через 10 о после первого
раздражения, то отвечающий ему быстрый аксонный ток от раздраженных
заднекорешковых волокон может наступить без изменения;
последующий же медленный отрицательный потенциал вовсе отсутствует (Герш,
89; Беритов и сотр., 100, 106). Но в других случаях, под влиянием
длительного отрицательного компонента, тормозится точно так же и задне-
корешковый эффект. Это, например, хорошо видно на рис. 57 А и С, где
при ритме около 15 в 1 сек. после первого заднекорешкового эффекта
все последующие заднекорешковые эффекты являются ослабленными.
Приведенные факты указывают, что в основе центрального торможения
лежит медленный потенциал в целом, а не только его положительное
колебание, как это полагали некоторые авторы.
Хорошо проявляется тормозящее действие медленных потенциалов
при комбинации раздражения двух чувствительных нервов. Это можно
выявить путем отведения как передних, так и задних корешков.
Например, на рис. 64 дается электрический эффект X переднего корешка
спинного мозга лягушки при комбинации раздражения п. brachialis и
п. peroneus. Раздражение п. brachialis в отдельности дает однофазные
медленные потенциалы значительной амплитуды и длительности, а
раздражение п. peroneus производит после начального заднекорешкового
биотока возбуждения трехфазный медленный потенциал. Оба эти био-
J34 ОЕЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
тока ослабевают на фоне медленных потенциалов, вызываемых от
раздражения п. brachialis.
Если во время спонтанной электрической активности двигательных
клеток, связанной с их механическим раздражением при отведении
биотоков из глубины мозга, производить тормозящее раздражение
чувствительного нерва, тогда удается наблюдать угнетение этих биотоков
наряду с появлением медленных потенциалов (Ллойд, 102). Как это,
например, хорошо видно на рис. 65, угнетение длится во время всего мед-
WQfj V
(J.0P"
В
4A*U;>w^V^v\
*w-.
Рис. 64. Электрический эффект спинного мезга лягушкилю время торможения.
Спинномозговой препарат. Все передние корешки в поясничной области перерезаны. N. brachialis
отпрепарирован по всей длине до plexus brachialis. Отводится X передний корешок слева.
Раздражаются оба нерва релаксационными стимуляторами: п. brachialis—при 2V, а п. peroneus—при IV.
В опыте А дается электрический эффект от раздражения п. brachialis, а в опыте В комбинация
этого раздражения с раздражением п. peroneus.
ленного потенциала; при этом угнетаются большие быстрые потенциалы
от возбуждения двигательных клеток. Спонтанно наступающие быстрые
потенциалы небольшой амплитуды, наоборот, учащаются. Эти быстрые
потенциалы, видимо, возникают от возбуждения нервных кругов, а
потому учащаются под влиянием периферических импульсов. Повидимому,
периферическое раздражение усиленно активирует нейропиль через
нервные круги, и потому соответствующие им биотоки возбуждения
наступают наряду с медленным потенциалом нейропиля.
Что вообще анэлектротоническим действием постоянного тока
можно вызвать угнетение нервных элементов мозга, в частности
двигательных клеток, хорошо можно продемонстрировать пропусканием его через
спинной мозг. Если анод поместить на поверхность одного из
поясничных сегментов мозга, а катод — на соответствующем двигательном
корешке, то в этом случае пропускание небольшого тока, который сам по
себе не дает двигательного эффекта, производит понижение
возбудимости двигательных клеток данного корешка, ибо ток будет входить через
мозг в двигательные клетки, а через их аксоны выходить наружу на
катоде (Баррон и Мэтьюс, 34). При этом будут тормозиться все
рефлекторные реакции, вызываемые через данный корешок в ответ на
раздражение чувствительных нервов.
Если пропускать постоянный ток через весь спинной мозг, приложив
оба электрода по длине мозга, то при любом направлении его
возбудимость центральных ¦ элементов может понижаться. Постоянный ток,
распространяясь от анода к катоду через межуточную жидкость, будет
проникать в клетки и аксоны и тем производить в них анэлектрониче-
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ 135
ское понижение возбудимости. Это замечено было еще Ушинским
(87) в лаборатории Сеченова; он пропускал через переднюю часть
спинного мозга постоянный ток около 1 вольта и наблюдал понижение
рефлекторной деятельности даже без ее предшествующего повышения.
Если медленный потенциал мозга в состоянии вызвать анэлектрото-
ническое понижение возбудимости в тех аксонных и клеточных
элементах, куда этот ток входит, он должен вызвать и повышение
возбудимости, и даже возбуждение, в тех нервных
элементах, где он выходит. И
действительно, при раздражении одного заднего
поясничного корешка кошки и отведении
другого заднего корешка в осциллограф
удается наблюдать длительный потенциал
с разрядом нервных импульсов в период
его нарастания (Баррон и Мэтьюс, 35
Беритов, Бакурадзе и Ройтбак, 104) (рис.
55-С и 66). Очевидно, нейропильный ток,
выходя из заднекорешковых волокон через
перехваты Ранвье, раздражает их и тем
самым производит импульсы возбуждения,
которые уже затем распространяются
антидромно через задний корешок на
периферию. Эти антидромные импульсы по
чувствительным нервам распространяются на
периферию подобно обычным нервным
импульсам и, повидимому, играют
определенную роль в обмене веществ рецепторов и в
регулировании просвета кровеносных
сосудов (Теннис, 76). Точно так же медленный
потенциал двигательных клеток, вызывая
токи, которые распространяются по двигательной клетке, может вызвать
возбуждение аксона. Этим объясняется появление аксонных токов на
фоне медленного потенциала (рис. 57-D) (Беритов и Ройтбак, 98, 106).
Многие известные факты торможения получают объяснение с точки
зрения электротонической природы торможения. Так, например, выше
было отмечено, что торможение двигательных нейронов в спинном мозгу
начинается всегда раньше связанного с ним реципрокного возбуждения
двигательных нейронов антагониста и вообще раньше возбуждения.
Это хорошо согласуется с тем фактом, что при одиночных раздражениях
медленные потенциалы обычно возникают на много сигм раньше
быстрых аксонных потенциалов серого вещества мозга (рис. 56 и 57).
Это же явление дает возможность понять многие факты двойственной
иннервации. Ввиду того, что в ответ на каждое периферическое
раздражение медленный потенциал начинается раньше возникновения
импульсов возбуждения в нервных кругах и двигательных нейронах, пауза
торможения должна предшествовать возбуждению. При отведении от перед-
Рис. 65. Биотоки переднего
рога поясничной-. области
кошки.
А — спонтанная активность из
больших и малых быстрых
потенциалов; В — угнетение
спонтанной активности одиночным
раздражением плечевого
сплетения: от раздражения наступает
двухфазный медленный
потенциал; при этом большие
быстрые потенциалы выпадают, а
малые учащаются (Ллойд).
]36 01ИЦА.Я ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГ4.
них корешков вблизи мозга нередко удается наблюдать, что если
рефлекторная деятельность не очень интенсивна, то вслед за каждым
раздражением в момент нарастания медленного потенциала быстрые
аксонные биотоки пропадают или ослабевают. Это, например, видно на
рис. 57-С. Следовательно, природа торможения такова, что при
возбуждении любого чувствительного нерва или даже любого чувствительного
волокна в центральной нервной системе торможение должно иметь
место наряду с возбуждением.
Выше мы указывали
также, что при всякого рода
слабых раздражениях спинного
мозга, непосредственно или
через задние корешки,
происходит общее торможение без
двигательной реакции.
Изучение же электрических
потенциалов показывает, что при
слабых раздражениях
медленные потенциалы возникают в
мозгу без возбуждения
двигательных нейронов. Этот факт
также согласуется с нашим
утверждением, что общее
торможение обусловлено анэлек-
тротоническим действием
медленных потенциалов.
Выше было отмечено, что
степень торможения в
ближних и отдаленных участках
мозга не одинакова. Если раздражается чувствительный нерв задней
конечности, то общее торможение сильнее на соответствующей стороне,
чем на противоположной, на задних ногах сильнее, чем на передних
(Нарикашвили, 12; Беритов и Гогава, 36; Като, 11). Точно так же при
раздражении задних корешков тормозящее действие проявляется в
ближайших сегментах сильнее, чем в отдаленных. Так, например, при
раздражении V заднего корешка торможение проявляется сильнее по
отношению к рефлекторным реакциям из VI сегмента, чем к реакциям из
VIII и IX сегментов. В последнем сегменте оно может даже
отсутствовать (Беритов и Бакурадзе, 38). Оказывается, что медленные
потенциалы, возникающие при раздражении одного заднего корешка, отводятся
сильнее всего от ближайшего соседнего заднего корешка. От дальнейших
корешков они отводятся тем слабее, чем дальше эти корешки отстоят.
Более слабы потенциалы и от задних корешков противоположной
стороны (Баррон и Мэтьюс, 34; Беритов и сотр., 98, 100). Словом,
распространение медленных потенциалов — такого же рода, как и
распространение общего торможения, вызванного раздражением заднего корешка.
і"л /ч Юв л
Рис. 66 Нейропильный ток вместе
с разрядом нервных импульсов.
Децеребрированная кошка. Один задний корешок
раздражается одиночным размынательным
индукционным ударом, другой задний корешок отводится
к осциллографу. В опытах А и В отводится задний
корешок на расстояние 40 мм от мозга, в опытах С и
D — на расстояние 2 мм от мозга. В опыте А
температура мозга 38°С, в остальных мозг был охлажден
до 36°С. Медленное колебание после охлаждения
усиливается, но и оно хорошо регистрируется
только при отведении на близком расстоянии от мозга.
Внизу время — по 0.01 сек. (Баррон и Мэтьюс).
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ
137
Из всего вышеизложенного видно, что все известные факты общего
торможения совершенно аналогичны фактам, наблюдаемым при
действии постоянного тока. Поэтому в высшей степени вероятно* что
медленные потенциалы, возникающие в активной части мозгового н^йропи-
ля. обусловливают все указанные выше явления общего торможения.
Итак, можно утверждать, что как только периферический импульс
возбуждения достигает спинного мозга, нейропиль тотчас же приходит
в активное состояние; вследствие этого тотчас же, под влиянием
возникших здесь локальных потенциалов, в клетках ближайших
промежуточных и двигательных нейронов, а также в аксонах создается
анэлектротоническое состояние, пониоюающее их возбудимость. Это
анэлектротоническое влияние должно быть тем сильнее, чем ближе
находятся клетки и аксоны к активной части нейропиля.
Гипотеза торможения с точки зрения субнормальности после
возбуждения. Как известно, в нервных волокнах позвоночных животных
после возбуждения наблюдается сначала стадия повышенной, а затем
пониженной, субнормальной возбудимости. Это изменение возбудимости
сочетается с изменением электрического потенциала: ©о время
повышенной возбудимости наблюдается отрицательный электрический
потенциал, а во время пониженной — положительный. Амплитуда этих
потенциалов очень мала — после одного импульса в положительной
фазе она достигает 10 ,uV в А волокнах, а продолжительность их
измеряется несколькими десятками о. При тетанических раздражениях как
амплитуда, так и продолжительность их значительно нарастают (Гассер,
114; Эрлангер и Гассер, 33). Эти электрические последействия
особенно значительны в нервах беспозвоночных.
Гассер (114), Бремер (115а) и другие авторы, основываясь на этих
наблюдениях, предполагают объяснить явления центрального
торможения с точки зрения субнормальности после возбуждения. Они полагают,
что в промежуточных нейронах имеет также место фаза
субнормальности после возбуждения. При некоторой последовательности
периферической импульсации по одним и тем же или по разным чувствительным
волокнам может получиться в отношении некоторых нейронов такое
состояние субнормальности, которое будет тормозить рефлексогенное
действие периферических импульсов.
Характерно, что по наблюдениям Бремера и Бонне (115)
тормозящее действие одних импульсов на последующие может оказаться
в том случае, если первый импульс был подпороговый и даже если
такой подпороговый импульс предшествует на несколько сигм.
Предполагается, что подпороговое раздражение возбуждает также
промежуточные нейроны и что длительное тормозящее действие обусловлено
суммацией субнормальности в промежуточных нейронах. Авторы
приводят ряд доказательств этой гипотезы, например, такое: стрихнин и
другие возбуждающие агенты усиливают и удлиняют торможение. Это, мол,
происходит благодаря увеличению числа активированных нейронов, в
частности — промежуточных нейронов, производящих торможение.
J 38 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Особенно длительный период торможения после одиночного
импульса наблюдается при отравлении вероналом. Угнетение продолжается до
5 сек. Одновременно было замечено удлинение субнормальной
негативной фазы, т. е. периода падения отрицательного потенциала.
Эта гипотеза имеет некоторое сходство с представленной выше
гипотезой нейропильного торможения: в обоих случаях тормозящее действие
сопряжено с медленным потенциалом. Но наряду с этим имеется
существенная разница: гипотеза Гассера предполагает, что медленные
потенциалы возникают в клетках и аксонах промежуточных нейронов в
результате возбуждения. Если бы это было так, то медленные
потенциалы не должны были бы возникать при отсутствии рефлекторной
деятельности, например, после асфиксии или глубокого наркоза, а между
тем они наступают и при этих условиях. Кроме того, как указывалось
выше, медленные потенциалы появляются в мотонейронах без
возбуждения их, или даже медленный потенциал предшествует возбуждению,
проявляющемуся быстрыми аксонными токами. Эти факты с
очевидностью говорят против происхождения медленных потенциалов мозга
подобно низковольтным электрическим последействиям нервного волокна,
а значит против торможения в результате возникновения
субнормальной фазы после возбуждения.
Гипотеза о тормозящей функции нейропиля стволовой части
головного мозга. Хорошо известно, что непосредственным искусственным
раздражением средних отделов імозга можно вызвать, в первую
очередь, общее торможение скелетной мускулатуры. Это явление впервые
было описано Сеченовым (44), а потому носит название
сеченовского) торможения. В определенных случаях к общему
торможению примешиваются фазные и тонические реакции (Болотов, 45;
Гоциридзе, 46). Кат о (11) показал, что у лягушек раздражением
самого переднего отдела промежуточного мозга (lamina terminalis) одной
стороны вызывается общее торможение мышц на противоположной
стороне, причем торможение наступает без каких-либо двигательных
реакций. К а т о же определил, что перекрест тормозящего пути
происходит в продолговатом мозгу, где вообще происходит перекрест
нисходящих путей.
В спинном мозгу эти пути проходят по боковой поверхности мозга,
ибо их раздражение также производит торможение, но только на
соответствующей стороне.
В приведенном выше случае К а т о раздражал мозг тонкими
биполярными микроэлектродами, которые, будучи приложены к мышце,
возбуждали всего несколько мышечных волокон. Эти опыты прямо
показывают, что не может быть и речи о том, что общее торможение из
стволовой части мозга происходит через известные нисходящие
-нервные пучки, ибо нельзя себе представить, что один и тот же небольшой
пучок нервных волокон может вызвать общее торможение всей
мускулатуры одной половины тела, затормозить все 5 тысяч двигательных
нейронов этой половины.
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ
139
Далее, Кат о нашел, что у лягушки только при раздражении
переднего отдела промежуточного мозга получалось общее торможение
при любой силе раздражения. При раздражении же других отделов
мозга — среднего, продолговатого и спинного — торможение получалось
при сравнительно слабых раздражениях (25 см расстояния между
катушками), очень сильное же раздражение (12 см) вызывало
двигательные реакции. Это наблюдалось не только при точечном раздражении
боковой поверхности, но и при раздражении всего спинного мозга
обычными биполярными электродами.
Аналогичные явления общего торможения наблюдаются и у высших
позвоночных при раздражении дна IV желудочка и других отделов
мозга. Так, например, Рожанский (47) обнаружил, что у кошки
при уколах в дно IV желудочка торможение захватывает скелетную
дыхательную и сосудодвигательную системы, при уколе же в область
варолиева моста через заднее четверохолмие торможение захватывает
в основном двигательную систему; слабее всего оно проявляется у этого
животного при уколах в область зрительных бугров. Укол, безусловно,
производит деформацию и частичное разрушение мозга, поэтому его
следует рассматривать как длительное механическое раздражение,
нанесенное в область сетевидного образования. Естественно, что
наступившее торможение проходит очень медленно — в течение 3—4 дней; при
более обширных повреждениях оно прекращается еще позднее. То же
наблюдал Рожанский с сотрудниками у собак, затем у голубей и
других птиц.
В опытах над холоднокровными торможение легче всего
получается при раздражении зрительных чертогов, а в опытах над
теплокровными— при раздражении дна IV желудочка. Это как раз те участки
головного мозга, где нейропильные образования являются наиболее
мощными. Уже на этом основании можно предположить, что и это
общее торможение является функцией нейроттиля.
Точно так же описаны многочисленные случаи общего торможения
при естественном возбуждении стволовой части мозга. 'Как известно,
акт глотания, координирующий аппарат которого находится в
продолговатом мозгу, сопровождается общим торможением всех
двигательных элементов, не участвующих в акте еды. Мельтцер (48)
показал, что при глотании сердечный ритм учащается благодаря
снятию тонического возбуждения с ядра блуждающего нерва,
дыхание становится реже, кровяное давление падает и т. д.
Ухтомский (49) доказал тормозящее 'влияние глотания на скелетную
мускулатуру конечностей. По всей вероятности, аналогичное явление
имеет место и при акте рвоты.
Б ери то в (25) обнаружил, что сдавливание кожи на голове
производит наряду с небольшим движением головы общее торможение
остальной мускулатуры (рис. 37). Это торможение безусловно
вызывается через тройничный нерв при участии среднего и продолговатого
мозга. И действительно, непосредственным раздражением тройничного
140 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Oph.tt.
SO
/"
,^ПГОТ\1\А1\1\АЛ1Ч\Ліи\іиичГ«\Л1вдЛППЛЛПЛ^
сы
&</.
нерва можно вызвать общее торможение, захватывающее почти весь
рефлекторный аппарат, так как при этом тормозятся дыхательные
движения, рефлекторные движения
* мышц передней и задней
конечностей (рис. 67) (Нарикашвили,
94).
Общее торможение
наблюдается также при раздражении
животных светом и звуком. Так,
если в опытах на лягушке звук
или свет воздействовали за 0.1 —
0.5 сек. до механического или
электрического раздражения
кожи, они повышали рефлекторную
реакцию на последнее
раздражение; если же действие звука или
света прекращалось еще
раньше— за 0.5—1 сек.,— они
тормозили ее. Сами по себе ни свет, ни
звук не давали внешних реакций
(Иеркс, 122).
В новейшее время при
изучении на лягушке действия звука
и света на рефлекторную
реакцию от кожного раздражения
было установлено, что звук
усиливает рефлекторные реакции, а
свет, наоборот, угнетает эти
реакции (Иоханес, 52; Бебуришви-
аи, 53).
Аналогичные опыты ставились
на теплокровных животных. Ока-
Рис. 67. Явление общего торможения при залось, что у совершенно нор-
тетаническом раздражении ветвей троинич- J r r
ного нерва на децеребрированной лягушке, мальных кроликов свет и звук
В опыте Л испытывается влияние подпорогового благоприятствуют оборОНИТСЛЬ-
раздраженияп. ophtalmicus (Oph. d.) (с левой
стороны) на дыхательные движения. Они угне- ЦЫМ рефлексам На Электрическое
таются во время раздражения. В опыте В
записываются эффекты на мышцах передней и задней раздражение НОГИ. Но На КрОЛИ-
конечностей. Верхняя кривая— от т. согасо-
radialis dex. (С. г. d.), m. anconeus dex. (A. d.), ках, ОТраВЛеННЫХ ХЛОралОЗОИ,
m. semitendinosus dex. (S. d.) и т. triceps dex.
(Г. d.). Околопороговое раздражение г. maxillo КОГДа КОра ООЛЬШИХ ПОЛУШарИИ,
— mandibular! sin. (M-ms). На всех мышцах,
кроме semitendinosus, наступает сначала тормо- ПОВИДИМОМу, Перестает функЦИО-
жение, которое потом сменяется заторможен-
ным сокращением. По прекращении раздражения НИрОВЗТЬ, те же звук И СВет ТОр-
это сокращение сразу усиливается. На m. semi- r
tendinosus во время раздражения наступает тор- МОЗЯТ оборОНИТелЬИЫ? ОеАлекСЫ
можение, а после раздражения оно сменяется г г т
сокращением (Нарикашвили). (ДЗИДЗИШВИЛИ, 54) (рис. 68).
Итак, активирование
стволовой части головного мозга производит общее торможение наряду с
двигательными реакциями или без них. При этом тормозятся всякого рода
I
M-ms. 3S
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ 141
реакции, как автоматические, вроде дыхательных движений, так и
вызываемые раздражением кожи или чувствительных нервов.
Общее торможение, вызываемое раздражением стволовой части
головного імозга, мы связываем прежде всего с деятельностью нейро-
пиля этой части мозга. Как указывалось выше, мало
дифференцированное сетевидное образование стволовой части головного мозга
состоит в значительной мере из нейропиля. Он существует и во всех
дифференцированных ядрах стволовой части «мозга и на его поверх-
Рис. 68. Тормозящее влияние звука на рефлекторные
реакции.
Кролик после хлоралозного отравления. Верхняя кривая — запись
дыхательных движений; вторая сверху — движения головы; третья —
движения правой задней ноги; нижняя — левой задней ноги.
Раздражается левая задняя нога отдельными индукц. ударами (верхний
сигнал). В опытах Л и С раздражение ноги производится изолированно,
а в опытах В и D — в комбинации со звуком (сильный высокий тон
звукогенератора); при этом в опыте В электрическое раздражение
происходит во время звука и дает ослабленный эффект как на ногах,
так и на дыхании и голове, а в опыте D — через 0.4 сек. после звука и
производит усиленный эффект на раздраженной ноге и ослабление на
всех остальных регистрируемых органах. Внизу — время в секундах
(Дзидзишвили).
ности. Здесь так же, как и в спинном мозгу, нервные круги, из
которых составляются нервные центры и проводящие пути, тесно связаны
с нейропилем. Поэтому в стволовой части мозга каждый раз, наряду
с возбуждением определенных нервных центров и проводящих путей,
т. е. нервных кругов, должно возникнуть активное состояние и в
определенном участке нейропиля. Но, повидимому, ів определенных
случаях нейропиль приходит в активное состояние и при таких
(внешних воздействиях, которые не вызывают двигательной реакции, как,
например, при действии света, при раздражении кожи на голове, а
также при слабом раздражении кожно-мышечиых чувствительных
нервов.
Безусловно, нейропиль спинного мозга играет также значительную
142 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
роль в производстве общего торможения при двигательных реакциях,
вызываемых из стволовой части мозга. Этот нейропиль должен
приходить в активное состояние под действием всякого рода нисходящих
импульсов возбуждения, ибо, как указывалось выше, все нисходящие
нервные пути в значительной мере оканчиваются в нейропиле спинного
мозга. Поэтому и наблюдается общее торможение при их
непосредственном раздражении.
Происхождение тормозящего действия нейропиля стволовой части
головного мозга. Относительно нейропиля спинного мозга мы пришли к
Рис. 69. Ритмический разряд продолговатого мозга Cyprinus
carassius.
А — медленные колебания при одном отведении, когда один электрод лежит на
продолговатом мозгу, на его расширенном месте (вагальный холм), а другой,
заземленный электрод (применяется усилитель!) — на спинном мозгу, как
показывает изображение Ах\ В -—¦ медленные колебания при другом
отведении, когда заземленный конец лежит на среднем мозгу, как показывает Ві.
Извращение направления тока свидетельствует о том, что заштрихованные
участки (вагальный холм) являются электроотрицательными; С и D—разряды
кратковременных импульсов на фоне медленного колебания (Эдриан и Буй-
тендайк).
заключению, что активное состояние его оказывает то или другое
действие на нервные клетки и проводящие пути вследствие поляризующего
влияния возникающих в нейропиле медленных потенциалов.
Аналогичное заключение было сделано нами насчет нейропиля стволовой части
мозга. 'Мы можем даже указать, какие именно медленные потенциалы
должны выражать активное состояние нейропиля, а следовательно,
какие именно токи оказывают -на нейронные элементы электротоническое
действие. Эдриан и Буйтендайк (55) открыли, что совершенно
изолированный головной мозг золотой рыбки (Cyprinus carassius)
спонтанно, через каждые 1—3 сек., разряжается очень медленной волной
электрического тока. Эти волны возникают в продолговатом мозгу и
стоят в связи с работой дыхательного центра. Продолжительность
каждого' колебания равна 0.25—0.5 сек. На фоне этого медленного
колебания авторы наблюдали разряды быстро протекающих токов
возбуждения, характерных для нервных волокон (рис. 69). Эдриан (56)
наблюдал аналогичные медленные колебания электрического потенциала
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ
143
в нервных узлах насекоімых, у которых подобные потенциалы возникают
тоже в связи с дыханием.
Медленные потенциалы (возникают в ядрах стволовой части
головного мозга и в сетевидном образовании продолговатого мозга при
раздражениях чувствительных нервов и рецепторов головы. Если отводить
чувствительный путь вблизи ядра тройничного нерва, тогда при
раздражении какой-либоі ветви тройничного нерва сначала наступают быстрые
колебания потенциала, отвечающие возбуждению чувствительных
волокон, а затем, через 1—2 о — медленные колебания, в том же
направлю ЗА
tSJmsec.
і і t і і і і і і і і і * і
Рис 70 Биотоки возбуждения продолговатого мозга
кошки при раздражении одной ветви тройничного нерва
отдельными электрическими ударами.
Регистрация биотоков катодным осциллографом: 1 — при
отведении чувствительного пути; 2 —¦ при отведении ядра
тройничного нерва, ЗА — при отведении чувствительного пути
при одном раздражении, а ЗВ—3F при двух раздражениях при
интервалах 3, 8, 15, 81 и 145а. Стрелки везде указывают на
момент раздражения (Мак Кинлей и Мэгон).
лении, отвечающие активному состоянию ядра нерва (рис. 70, 1, ЗА)
(Мак Кинлей и Мэгон, 92). Последнее колебание продолжается
2.5—17 о, но вслед за ним наступает более длительное медленное
колебание в противоположном направлении. При отведении от самого
ядра тройничного нерва или от сетевидного образования недалеко от
ядра, первое — быстрое колебание наступает в очень слабой форме,
а второе — медленное, наоборот, в усиленной форме, ноі
продолжительность его та же (рис. 70,2). Такие медленные колебания отводятся и
от других ядер ствола мозга.
Медленные потенциалы возникают в стволовой части головы и при
раздражениях кожи и чувствительных нервов конечностей. У лягушки
они бывают особенно значительны при отведении от зрительных бугров
(рис. 71). В некоторых случаях они достигают 500 j*V и больше. Они
протекают волнами разной длительности и амплитуды, как в спинном
мозгу. Когда раздражается чувствительный нерв при ритме 10—50
в сек. каждый удар производит свой эффект: сначала начальный
быстрый потенциал от возбужденных афферентных путей, а потом
двухфазный или трехфазный медленный потенциал (рис. 71-А). В определенных
случаях наступают более или менее длительные разряды быстрых по-
144 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
тенциалов на фоне медленных, подобно тому как это наблюдалось в
спинном мозгу лягушки. При этом означенный электрический эффект
наступает значительно сильнее при раздражении противоположных
конечностей (Ройтбак, 109).
При более или менее хорошем функциональном состоянии
продолговатого мозга отводимые от него медленные потенциалы покрываются
рядом быстрых. Быстрые потенциалы хорошо отводятся, когда
отводящий электрод находится в глубине или на поверхности самого
активированного ядра. Они значительно слабее или совершенно отсутствуют,
Гис. ~Л. Электрический эффект зрительных Оугров лягушки.
Головной мозг обнажен со всех сторон. Конечности денервироваиы, кроме одной. Огвідятся
зрительный бугор справа с поверхностн. В опыте А раздражается п. peroneus
релаксационным стимулятором при силе 2V, а в опыте В производится щипание пальцев задней
лапки слева (Ройтбак).
когда отводящий электрод находится подальше от этого ядра, например
на поверхности мозга. Так, например, было в опытах Термана при
отведении от ядер Бурдаха и Галля (nucleus ouneatus) и раздражении
чувствительных нервов передней конечности (рис. 72).
Медленные потенциалы ствола головного мозга в ответ на
раздражение чувствительного нерва отдельными электрическими ударами
имеют двухфазный характер. Общая продолжительность — до 160 о, но
первая отрицательная фаза всегда очень короткая — около 5 о; однако она
более высокой амплитуды, чем вторая, положительная фаза. Быстрые
потенциалы возникают главным образом на фоне первой, отрицательной
фазы и в начале второй положительной (рис. 72-А). При этом
направление фаз медленного потенциала извращается при погружении
электродов (Терман, 101). Совершенно такая же картина наблюдается в
спинном мозгу при одиночных раздражениях чувствительных нервов.
При аноксемии, вследствие недостаточной работы сердца, медленные
потенциалы и сопровождающие их быстрые потенциалы исчезают
первыми. Начальные быстрые потенциалы удерживаются еще некоторое
время. При постепенном развитии асфиксии можно наблюдать
исчезновение положительной фазы при сохранении отрицательной (Терман, 101)
(рис.72).
Во время медленного потенциала повторное раздражение того же
нерва дает более слабый медленный потенциал. Например, при
раздражении тройничного нерва и отведении от тройничного ядра второе
раздражение не дает эффекта в.продолговатом мозгу через 10—20а, а за-
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ J45
тем в течение 160 о дает его в ослабленной форме (рис.70). То же самое
наблюдалось при отведении зрительных бугров и раздражении
чувствительных нервов конечностей (рис. 71). Отсюда следует, что в тот самый
период времени, пока ядро дает медленное колебание потенциала,
тормозится повторное возбуждение ядра периферическими импульсами.
Возможность путем локального анэлектротонического воздействия
постоянного тока на продолговатый мозг вызвать торможение
рефлекторных реакций, возникающих при
раздражении как головных нервов, так и спиналь-
ных чувствительных нервов, можно
демонстрировать, пропуская подпороговый
гальванический ток через продолговатый мозг,
причем один полюс прикладывается к
мозгу, а другой, диффузный, помещается в
ротовой полости или под нижней челюстью.
Оказывается, что пропускание слабого
тока— 15—20 микроампер — совершенно
достаточно, чтобы вызвать торможение всех
рефлекторных реакций, дуга которых
проходит через продолговатый мозг. При этом
направление тока не имеет существенного
значения. Как при аноде, так и при катоде
на поверхности мозга угнетаются
дыхательные движения и рефлекторные реакции
освободительного характера, вызываемые
через продолговатый імозг раздражением
кожи конечности. Разница только в том,
что при аноде это угнетение наблюдается
с самого начала, а при катоде — спустя
несколько секунд (Нарикашвили, 94)
(рис. 73). Очевидно, электрический ток при
любом направлении распространяется в
продолговатом мозгу преимущественно
через межуточную жидкость, а уже из этой
жидкости он проникает в клетки и аксоны,
производя тем самым в них анэлектротони-
ческсе понижение возбудимости
(Нарикашвили, 94). Эти опыты хорошо имитируют
не только электротоническую природу
торможения рефлекторной реакции, но и
влияние локальных медленных биотоков на
центры, находящиеся в продолговатом мозгу.
Однако при раздражении головных нервов не исключается
возможность тормозящего влияния вверх и вниз по центральной нервной
системе, но это явление едва ли будет результатом распространения
биоэлектрических токов кейропиля из одного отдела на следующие. Из
Ю и, с. Беритов
Рис. 72. Биотоки nucleus
cuneatus кошки.
Раздражается срединный норв
одним электрическим
ударом. Отводится
микроэлектродом поверхность мозга
на уровне ядра.
1 — при нормальном
кровообращении, 2 и 3 — во время двух
последующих стадий асфиксии,
наступившей вследствие
расстройства кровообращения. Первое
небольшое колебание — артефакт
от раздражающего
электрического удара. Первое быстрое
колебание в электрическом эффекте от
биотока возбуждения
раздраженных чувствительных волокон (Тер-
ман).
все нервные пути и на
146" ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО НОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
наблюдений Термана (101) и др. видно, что биоэлектрические ток»_
в стволовой части улавливаются не дальше нескольких миллиметров
от места их -возникновения. Мы думаем, что распространение
торможения или облегчения не
обусловливается ни распространением медленных
биоэлектрических токов нейропиля, ни
непосредственным распространением актив*
ного состояния нейропиля вдоль по ней-
ропилю центральной нервной системы.
Каждый раз деятельность какого*
либо нервного ядра или нервного
комплекса предполагает, с одной стороны,
возбуждение определенных нервных
кругов, а с другой — активное состояние,
соответствующего нейропиля. Но
поскольку нервные круги данного ядра
головного мозга связаны длинными
аксонами с другими ядрами головного к
спинного мозга, то тем самым сни
связаны и с нейропилем других ядер.
Поэтому в связи с распространением
возбуждения по нервным путям должен
активироваться нейропиль и в других
ядрах; значит, одновременно с
распространением возбуждения по нервным
путям будет происходить распространение
торможения через активируемый пр»
этом нейропиль.
Начальные быстрые потенциалы,
возникающие каждый раз при раздражении
чувствительных нервов, выражают
биотоки возбуждения раздражаемых
чувствительных волокон. Медленные
потенциалы наступают сейчас же вслед за
быстрыми, без паузы, как в спинном
мозгу, или, вернее, эта пауза равняется
длительности биотока возбуждения
чувствительных волокон.
Э д р и а н и другие авторы
приписывают медленные потенциалы,
возникающие при дыхании, дендритной части дыхательных центров.'При
раздражении чувствительных нервов конечности наблюдаемые в
продолговатом мозгу длительные потенциалы Терман объясняет так: он
приписывает происхождение длительной положительной фазы сетевидному
образованию, т. е. нейропилю, а первую отрицательную фазу — самому
ядру Бурдаха и Голля. В последнее время О'Лири и Бишоп (116)
uW-irf^^
у
анор
а
,і*
«¦нипптііііііміііпш»ітиіінитициі«тінмі
-j_VL_Luu
"¦L-*- Ж
L <"*°9
1 -J'
иппнгнипппмтнпиппимн
Рис. 73. Тормозящее влияние
постоянного тока на деятельность
продолговатого мозга децеребри-
рованной лягушки.
Регистрируются дыхательные движения
(А) и мышцы задних конечностей В, с
и D: semitendinosus dex. и sin. (S. d.
и S. е.), triceps dex. и sin. (Г. d., Г. s)
Точечный электрод (анод) лежит в
ромбовидной ямке продолговатого мозга,
диффузный—в ротовой полости. В
опыте А испытывается действие подпорого-
вого постоянного тока на дыхательные
движения. Последние угнетаются
целиком. В опыте С испытывается действие
того же тока на освободительные
движения, вызываемые раздражением
кожи задней лапки. Они также
угнетаются полностью. В опытах В и Л —
освободительные движения от того же
раздражения лапки. На сигнальной
линии цифры означают силу
раздражения в сантиметрах расстояния между
индукционными катушками (Нарика-
швили).
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ
147
наблюдали медленные потенциалы при отведении из глубины
зрительного бугра утки и гуся. Они относят их к дендритному слою мозга. Мы
же находим, что как в этих, так и в других случаях возбуждения
продолговатого мозга медленные потенциалы выражают локальные
биологические процессы, возникающие в клетках и дендритах
продолговатого мозга. По общему признанию, при дыхании активируется сетевидное
образование в средней части продолговатого мозга. В других
отправлениях продолговатого мозга и вообще стволовой части мозга также
участвует сетевидное образование. Каждый раз при активном
состоянии этого нейропильного образования, а также других подобных
структур мозга должны возникать медленные потенциалы, конечно, наряду
с быстрыми аксонными токами нервных кругов, которые также
активируются под влиянием периферических импульсов.
Совершенно естественно, что при отведении биотоков из глубины
ядра, т. е. от основной массы нервных кругов, медленные потенциалы
нейропиля будут отводиться наряду с быстрыми и будут выражать-
результат алгебраической суммации локальных потенциа-лов
отводимого участка и близлежащих участков. Первая отрицательная фаза его
будет происходить от преобладающего значения локальных процессов
отводимого участка, а последующая положительная его фаза — от
преобладающего значения соседних участков. Но если отведение
производится вдали от возбужденных нервных кругов,— например, от
поверхности мозга,— тогда будут отводиться медленные потенциалы,
как более интенсивные, в чистом виде или в сопровождении
быстрых потенциалов очень небольшой амплитуды благодаря
шунтированию их через межуточную жидкость.
Медленные потенциалы, возникающие в продолговатом мозгу и ff
средних отделах головного мозга, имеют настолько большую
амплитуду, что, распространяясь через межуточную жидкость, они должны
действовать анэлектротонически на ближайшие нервные круги и
нервные пути и тем самым вызывать общее торможение всех
рефлексов, происходящих при посредстве средних отделов головного мозга и
продолговатого мозга.
Итак, в то время, когда в продолговатом мозгу и вообще в стволе
головного мозга процесс возбуждения, быстро распространяющийся
по определенным нервным кругам и проводящим путям, производит ту
или другую внешнюю реакцию,— в это время активированный нейро-
пиль через производимые им медленные потенциалы вызывает путем
электротонической поляризации общее торможение близлежащих
нервных кругов и проводящих путей.
Тормозящее действие средних отделов мозга на рефлексы,
вызываемые .раздражением спинальных чувствительных нервов. Торможение,,
возникающее в средних отделах мозга, проявляется в отношении дви*
гательных реакций не только головы, но и передних и задних
конечностей. Так, мы выше указывали, что при сдавливании кожи на голове
или при раздражении чувствительных нервов головы тормозятся
10*
348 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
рефлекторные реакции сгибания, вызываемые раздражением
чувствительных нервов задних конечностей.
Как же происходит это торможение? У нормальных животных или
у животных, лишенных больших полушарий, раздражение конечностей
вызывает рефлекторную реакцию не только через спинной, но и через
головной мозг и даже преимущественно через головной, особенно при
пороговых и слабых раздражениях.
Поэтому можно предположить, что активное состояние нейропиля
средних отделов мозга оказывает тормозящее действие на
рефлекторную деятельность путем анэлектротонического действия на нервные
элементы головного мозга, входящие в длинные нервные пути, и на
центры, находящиеся в продолговатом мозгу.
Итак, промежуточные нейроны средних отделов мозга при
своем возбуждении активируют по пути распространения своих нейритов
нейропилькые поля как среднего и продолговатого, так и спинного
мозга. Вследствие этого должно происходить торможение всех
двигательных нейронов, лежащих в этих отделах мозга.
История вопроса о торможении. Вопрос о торможении был поставлен после
того, как братья Веб ер (1845) открыли тормозящее действие блуждающего нерва
на деятельность сердца. В отношении центральной нервной системы вопрос о
торможении был .впервые выдвинут в 1863 г. Сеченовым (44). Он утверждал на основании
экспериментов, что в головном мозгу существуют особые тормозящие центры,
которые угнетают рефлекторную деятельность. Он перерезал мозг у лягушки в области
зрительных чертогов и раздражал поперечный разрез мозга индукционным током или
же хлористым натрием. От этих раздражений рефлекторные движения задних лапок
угнетались очень сильно. Отсюда он сделал заключение о существовании в головном
мозгу тормозящих механизмов. Впоследствии Сеченов, на основании новых
исследований (1869), утверждал, что для приведения в деятельность тормозящего
механизма требуется более сильное раздражение, чем для вызывания рефлекторного
сокращения (64).
В последнее время ряд ученых поддерживает мнение о существовании
специальных тормозящих центров в промежуточном мозгу (Орбели с сотрудниками, 58;
Като с сотрудниками, 11). Они исходят из такого же рода наблюдений, как и
Сече н о в: раздражая промежуточный мозг лягушки, они наблюдали торможения
рефлекторных реакций на задних конечностях. По их мнению, здесь заложены
центры общего торможения для всего организма. Но специальными исследованиями было
показано, что раздражения зрительных чертогов и среднего мозга могут тормозить
рефлекторные защитные 'реакции на задних лапках потому, что: во-первых, здесь
возбуждаются координирующие аппараты тонических рефлексов, преимущественно
разгибательных (Болотов, 45); во-вторых, здесь же должны приходить в активное
состояние координирующие аппараты тех рефлексов, которые в ответ на механическое
раздражение головы проиаводят общее торможение задней части тела при движениях
передней (Беритов, 25). Наконец, нейропильное образование, функция которого
заключается в торможении, развито у лягушки сильнее в промежуточном мозгу и вообще в
.средних отделах мозга, чем в спинном мозгу.
В последнее время Орбели доказывает, что наблюдавшееся Сеченовым
общее торможение является функцией лежащих здесь центров вегетативной нервной
системы (59). Сотрудница Орбели Тонких (60) произвела опыт перерезки всех
без исключения rami communicantes симпатической системы, и после этого
сеченовского торможения не получилось. Однако это наблюдение не оправдалось при
проверке указанных опытов нашим сотрудником Гоцеридзе (46), а также в олытак
О ПРИРОДЕ ЦЕНТРАЛЬНОГО. ТОРМОЖЕНИЯ
149
К а т о с сотрудниками. Сеченовское торможение хорошо получается и после перареаки
всех rami communicantes. В опытах К а т о весь мозг был вынут из позвоночника и
оставался в связи с задними конечностями только посредством церебросшиалышх
нервов. У К а то раздражение промежуточного мозга давало общее торможение
задней конечности без единого предварительного движения (11).
В последнее время Тонких удалось получить угнетение рефлексов на задних
лапках при раздражении зрительных чертогов после перерезки спинного мозга на
границе III—IV сегмента, сохранив симпатические связи со спинным мозгом (60).
Из этих опытов, как бы с очевидностью, вытекает заключение, что угнетающее
влияние головного мозга на спинной осуществляется именно через симпатическую
систему. Однако, как показал наш сотрудник Гедевани, раздражение п. sympathicus,
вызывая секрецию кожных желез, может определенным образом влиять на скрытый
период рефлекторной реакции, вызываемой кислотным раздражением по методу
Тюрка. Раздражение п. sympathicus, например, может удлинить скрытый период
вследствие обильного выделения из кожных желез секрета, богатого муцином и
щелочами. Этот секрет, обволакивая кожную поверхность, защищает ее от раздражения
кислотой (61).
Итак, нет фактов, говорящих за существование специальных тормозящих центроа
в зрительных чертогах, которые бы через посредство симпатической системы
производили угнетение всей центральной нервной системы.
Вопрос о центральном торможении, поставленный Сеченовым, привлек
внимание выдающихся физиологов. Вслед за Сеченовым этим вопросом занялся
Гольц (62) и еіго ученик Фрейсберг (63). Они стали на ту точку зрения, что
каждый рефлекторный центр, который производит определенное действие в ответ на
данное раздражение, может утратить эту способность, т. е. затормозиться, если он
одновременно активируется под влиянием других раздражений. По предположению
Фрейсберга, «раздражение каждого чувствительного нерва может производить
торможение в каждой точке спинного мозга». Это мнение совершенно противоречит
высказанному Сеченовым предположению, но и оно впоследствии не оправдалось.
Последующие исследователи Бубно© и Гайденгайн (65) (1882)
утверждали, что возбуждение и торможение находятся в полной зависимости от
функционального состояния центральной нервной системы. Если она находится в состоянии покоя,
то слабое раздражение вызывает в ней деятельность; если же она в активном
состоянии, то такое же раздражение производит ее угнетение, т. е. «каждый раз1
существующее состояние уничтожается и заменяется противоположным». Но такое
объяснение центрального торможения вскоре было оставлено.
Введенский подверг детальному исследованию вопрос о происхождении
торможения. Этот автор (11) пришел к заключению, что центральное торможение
обусловлено усилением и учащением раздражений, т. е. является результатом
усиления и учащения импульсов возбуждения. Значит, по Введенскому, центральное
торможение по существу своему представляет пеешмальное состояние центральной
нервной системы. По более поздним воззрениям Введенского (1903),
центральное торможение выражает пар абиотическое состояние, которое создается в нервных
клетках центральной нервной системы вследствие чрезмерного усиления и учащения
импульсов. Это гипотетическое воззрение Введенского не нашло последователей
сче его лаборатории.
Особого взгляда придерживались Ф ерівори (66) и его последователь Фре-
лих (67). Эти авторы утверждали, что торможение представляет собой слабое
возбуждение. Если сильная деятельность периферического органа почему-либо
угнетена то, по их представлениям, это нужно приписать см^не сильной деятельности
на слабую вследствие утомления. Было приведено немало фактов, иллюстрирующих
данное предположение> между прочим и то, что при стрихнинном отравлении всего
спинного мозга реципрокное торможение исчезает. Согласно мнению названных
авторов, это объясняется тем, что в условиях стрихнинного отравления слабое
возбуждение становится невозможным.
J50 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Все приведенные факты сами по себе правильны, но на основании их нельзя
было притти к правильному теоретическому объяснению явлений торможения.
Исследование- центрального .торможения в последнее время производилось Ш е р-
рингтоном и теми учеными, которые пользовались его методами (Грэим Броун,
24; Фултон, 19; Самойлов и Киселев, 13; Беритов, 8; Киселев, 17). На основания
этих исследований пришли к заключению, что в центральной нервной системе
производится в отношении скелетной мускулатуры только реципрокное торможение; что
оно такой же активной природы, как реципрокное возбуждение, и что его
интенсивность и продолжительность так же меняются при определенных условиях, как и
связанное с ним возбуждение.
Рис. 74. Схематическое изображение реципрок-
ной иннервации по Шеррингтону.
Приводится антагонистическая пара мышц той и другой
сторон и их рефлекторные пути. Знак + указывает на
возбуждение «синапсов» вокруг двигательных клеток, а
знак — торможение их. Раздражение чувствительного нерва
L вызывает на соответствующей стороне возбуждение
двигательного нейрона ( + Ь), принадлежащего сгибателю
F, и в то же время торможение двигательного нейрона — е
разгибателя К. То же раздражение производит на другой
стороне совершенно противоположный эффект:
возбуждение нейрона +е' разгибателя и торможение нейрона—Ь'
сгибателя. Здесь же приводится добавочная рефлекторная
дуга для вторичных проприоцептивных импульсов со
стороны сгибателя а.
История реципрокного торможения сравнительно коротка. Впервые III е р р и н г-
т о н установил понятие реципрокного торможения экспериментальным путем
(в 1892—1893) (67), хотя о рецвдірокяом действии антагонистов в отвлеченном виде
высказывались много раньше. Первоначально Декарт высказал нечто подобное в
отношении бокового движения глаза; «когда по одному каналу жизненный дух
проводится в наружную прямую мышцу, тогда по другому каналу жизненный дух уходит
из внутренней прямой мышцы. Вследствие этого при стягивании первой мышцы
другая расслабляется» (De homine, 1662 г.). Но по Декарту, как хорошо видно из
данного им рисунка, это взаимоотношение чисто периферического характера и
происходит между мышцами непосредственно. После Декарта подобное представление
о рецинрокнсм торможении было дано (1823) Чарлзом Бэллом (69). Оя
заметил, что при сокращении одной мышцы антагонист ее удлиняется, расслабляется.
Бэллу казалось, что такое взаимоотношение имеет периферическое происхождение и
что как ¦сокращение, так и удлинение вызываются периферическими нервами. После
Б э л л а никто не касался этого вопроса. Самое наблюдение было забыто, и только
в (на-чале XX века Шеррингтон познакомил нас с ним.
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ
¦ 151
Точка зрения Шеррингтона о реципрокной иннервации была подробно
рассмотрена, но не лишне привести здесь данный им схематический рисунок этой
иннервации {рис. 74). Рисунок этот очень ясш передает теоретическое представление
автора, согласно которому координированная иннервация возникает в той физической
среде, которая существует вокруг двигательных клеток, между этими клетками и
окончаниями других нейронов.
Практикум
Быстрые и медленные потенциалы спинного мозга лягушки. Берут слшзно-
мозговой препарат лягушки и вскрывают поясничную область. Обнажают мозг с
задней и боковой поверхностей и перерезают мозг дополнительно в грудной области.
Затем перерезают все передние корешки, начиная от VII до XI на обеих сторонах и
все задние, кроме IX и X одной стороны.
На этой же стороне обнажаются п. peroneus или п. ischiadicus и берется их
центральный отрезок для раздражения.
Регистрация биотоков производится катодным или шлейфным осциллографом с
помощью однотактного усилителя низких частот. Сеточный электрод в виде
тоненькой нити, смоченной физиологическим раствором, или серебряной иглы,
прикладывается на боковую поверхность IX сегмента на стороне раздражения. Заземленный
электрод подводится к серебряной пластинке 5 X Ю см, на которой лежит препарат.
Поверхность мозга в области отводящего электрода должна быть свободна от
жидкости, чтобы избегнуть ветвления биотоков по этой жидкости.
Раздражается центральный отрезок чувствительных нервов возможно
кратковременными конденсаторными разрядами или индукционными ударами. При этих
условиях растекающийся по нерву электрический ток будет замыкаться через
заземленную пластинку и не будет попадать в усилитель тока через сеточный электрод, или
же будет попадать с такой малой интенсивностью, что не только не помешает
наблюдению биотоков мозга, наоборот, являясь отметкой момента раздражения, послужит
для определения скрытого периода электрического эффекта спинного мозга.
При означенном отведении будут регистрироваться при редких раздражениях до
50 в 1 сек. в ответ на каждый удар сначала быстрое колебание от заднекорешкового
«биотока возбуждения, а затем медленный потенциал от активирования нейропиля.
Аналогичные биотоки будут отводиться от центрального отрезка перерезанного VIII
или X заднего корешка раздраженной стороны. Отведение производится от корешка
на расстоянии 1—2 мм от мозга.
Биотоки мозга отводятся и от передних корешков, если отводящий электрод
приложен достаточно близко к мозгу — на расстоянии 1—2 мм от него. В этом случае
электрический эффект мозга осложняется рядом быстрых колебаний, связанных е
возбуждением переднекорешковых волокон и появляющихся на фоне медленного
потенциала.
Быстрые и медленные потенциалы средних отделов мозга лягушки. Берется кура-
ризованная лягушка. Открывается головной мозг. Сеточный электрод усилителя
прикладывается к одному полушарию среднего мозга —к зрительному бугру, а
заземленный электрод соединяется с серебряной пластинкой, на которой лежит препарат.
Зрительный бугор все время разряжается медленными и быстрыми биотоками, даже
s абсолютной темноте. Освещение глаза производит значительное усиление
биотоков,— гори этом усиливаются как быстрые, так и медленные потенциалы. Тачое же
усиление быстрых и медленных биотоков наблюдается по прекращении света.
При отсутствии кураре можно пользоваться отрезанной головой лягушки, но в
этих условиях нужно вести наблюдение биотоков мозга, когда нет движений глаз.
Литература
1. Введенский Н., Раб. фиэиол. лаб. Петерб. ун-та, 1, 1906.
2. Adrian Е„ Erg. Physiol., 26, 501, 1928.
3. Denny-Brown D., Proc. Roy. Soc. В., 104, 252, 1929.
152 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
4. Lorente deNo, Юбилейный сборник, посвященный проф. И. Бериташвили.
Изд. Груз. фил. АН СССР, 1936.
5. Lorente de N6, J. Neurophysiol., 1, 207, 1938.
6. Hughes Т. a. G a s s e r H. S., Am. J. Physiol., 108, 245, 1934; 108, 307, 1934.
7. GasserH. S., Vol. jubil. publie en l'honneur du prof. J. Demoor. Liege, 212,
1937.
8. Беритов И. Учение об основных элементах центральной координации
скелетной мускулатуры. Пг., 1916.
9. Birge Е., Arch. Anat. u. Physiol., 434, 1882.
10. Cread, Denny-Brown, Eccles, Lidell a. Sherrington,
Reflex Activity of the Spinal ord. Oxford, 1932. Есть русск. пер.: Рефлекторная
деятельность спинного мозга, 1935.
11. К at о G.t The microphysiology of nerve, 1934.
12. H a p и к а ш в и л и С, Юбилейный сборник, посвященный проф. И.
Бериташвили, 413, 1936.
13. Samoiloff А. и. К і s s е 1 е f f M., Pfl. Arch., 215, 698, 1927.
14. Sherrington С. S., Proc. Roy. Soc., В. 97, 519, 1926.
15. S с h w e i t z e г A. a. W r i g h t S., J. Physiol., 89, 165, 1937.
16. Б а бс кий E. В., Доклад VI Всес. физиол. съезда. 189; Учен. зап. каф.
физиол. Моск. пед. ин-та, 1938.
17. Киселев М., Казанск. мед. журн., № 4—5, 389, 1931.
18. Гедевани Д., Доклад на XV Междунар. физиол. конгр., Ленинград, 1935;
Тр. Физиол. ин-та им. Бериташвили, 3, 479, 1937.
19. F u 11 о n J. F., Muscular contraction etc., 318—320, 1926.
20. Eccl es J. С. a. Sherringt on С. S.,Proc. Roy. Soc.,В., 109, 91, 1931.
21. Беритов И. иБакурадзеА., Acta Medica USSR, 2, 271, 1939; Физиол.
журн. СССР, 28, 2 и 18, 1940.
22. Sherrington С. S., Int. act. nerv. syst., 1906.
23. В e г i t о f f J., Fol. Neurobiol., 7, 187, 1913; Pfl. Arch., 157, 171, 1913.
24. Graham Brown. Erg. Physiol., 13, 279, 1913.
25. Б e p и t о в И., Журн. эксп. биол. мед., 8, 106 и 114, 1928
26. Беритов И., Журн. эксп. биол. мед., 11, 1929; Zschr. Biol., 89, 77, 1929.
27. Barron D. Н. а. М a 11 h е w s В. Н. С, J. Physiol., 85, 73, 104, 1935.
28. Beck A., Pfl. Arch., 155, 461, 1914.
29. U m г a t h Ch. u. U m г a t h K., Pfl. Arch., 234, 582, 1934.
30. G a s s e г H. S. a. G г a h a m H. Т., Am. J. Physiol., 103, 203, 1933.
31. Matthews B. H.C.,Proc. Roy. Soc, B. 123, 416, 1937.
32. В о n n e t V. et В r e m e r F., G. R. S. Biol., 128, 806 и 812, 1938.
33. E г 1 a n g e г J. a. GasserH. S., Electr. signs of nervous activity, 1937.
34. В а г г о n D. H. a. M a 11 h e w s B. H. C, J. Physiol., 92, 276, 1938.
35. В а г г о n D. H. a. M a 11 h e w s B. H. C., J. Physiol., 94, 27 P, 1939.
36. Беритов И. иГогава М.,Тр. Физиол. ин-та им. Бериташвили, 3, 265, 1937.
37. Беритов И., Бакурадзе А. и Нарикашвили С, Тр. Физиол.
ин-та им. Бериташвили 3, 1937.
38. Беритов И. иБакурадзеА., Физиол. журн. СССР, 27, 387, 1939.
39. Н of f Е. С., Proc. Roy. Soc, В. Ill, 175, 1932.
40. Philippson M.t Lrautonomie et la centralisation dans le systeme nerveux des
animaux. Bruxelles, 1905.
41. Cajal S. R.,Anat. Anz., 5, 579, 1890.
42. К о 1 1 i k e r A., Z. Wiss. Zool., 51, 1, 1890.
43. Adrian E. D., J. Physiol., 88, 127, 1936.
44. Setschenow J., Physiologische Studien uber d. Hemmungsmechanismen
f. d. Reflextatigkeit d. Ruckenmarks im Gehirn d. Frosches. Berlin, 1863; то же на
русск. яз. Соч., т. I, стр. 18, 1907; Физиология нервных центров, СПб.,189К
45. Болотов В. П., Русск. физиол. журн., 2, 38, 1919.
46. Г о ц и р и д з е А., Журн. эксп. мед , 2, 7, 1929.
О ПРИРОДЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ ] 53
47. Р о ж а не к ий Н. А., Физиол. журн. СССР, 19, 289, 1935; Тез. сообщ. XV
Междунар. конгр. физиологов, 1935.
48. М е 11 z е г J., Das Schluckzentrum, seine Irradiation etc. Inaug. Diss. Berlin, 1882.
49. Ухтомский A.,Tp. Физиол. лаб. Петерб. ун-та, 5, 1911.
50. Bowditsch a. Warren., J. Physiol., 9, 1899.
51. Hofbauer L.,Pfl. Arch. 68, 546, 1897.
52. Johannes Th.,Pfl. Arch.,224, 372, 1930.
53. Бе буришвили H.,Tp. Физиол. ин-та им. Бериташвили, 3, 345, 1937
54. Дэидзишвили Н.,Физиол. журн. СССР, 26, 354, 1939.
55. A d г і а п Е. D. а. В и у t e n d i j k F. J., J. Physiol., 71, 121, 1931.
56. Adrian E., J. Physiol., 72, 132, 1931.
57. Коган Л. О., О применении электро-энцефалографии в исследовании
подкорковой области, 1936.
58. О р б е л и Л., Лекции по физиологии нервной системы. Огиз, 1934.
59. О р б е л и Л., Вегетативная нервная система. БМЭ. М., т. 4, стр. 548, 1928.
60. Тонких А.,Русск. физиол. журн., 10, 86, 1927.
61. Гедевани Д., Доклад на V Всес. съезде физиологов в Москве, 1934.
62. G о 11 z. F.,Beitrage z. Lehre d. Funktion d. Nervencentren d. Frosch., Berlin,
1869.
63. Freusberg A.,Pfl. Arch., 9, 358, 1879.
64. Сеченов И. M., Об электрическом и химическом раздражении
чувствительных нервов лягушки. СПб, 1868.
65. Bubnoff u. Heidenhain., Pfl. Arch., 26, 137, 1881.
66. Verworn M.,Z. AUg. Physiol., 15, 413, 1913.
67. Frohlich F.f Z. AUg. Physiol., 9, 55, 1909.
68. Sherrington С S.,J. Physiol., 13, 621, 1892.
69. Bell Ch., Philos. Trans. Roy. Soc, стр. 289, 1823.
70. Lenhossek M., Der innere Bau d. Nervensystems, Berlin, 1895.
71. БеритовИ. иБакурадзеА., Физиол. журн. СССР, 30, 45, 1941.
72. Gasser Н. S.,J. Neurophysiol., 2, 361, 1939.
73. Е с с 1 е s S. С. а. Р г і t с h a r d J. J., J. Physiol., 89, 43 P, 1937.
74. Bishop G. H.,J. Cell. Сотр. Physiol., 8, 465, 1936.
75. L о r e n t e d e N 6 R., J. Neurophysiol., 2, 402, 1939.
76a. T о e n n i e s J. F., J. Neurophysiol., 1, 378, 1938; 2, 515, 1939.
76b. Когхилл Г. E., Проблемы неврологии и психиатрии. Сб., посвященный
проф. Геймановичу, стр. 21, 1939.
77. Gonzalez A. W., J. Сотр. Neur., 55, 395, 1932; Proc. Soc.Exp. Biol. Med., 21,
111, 1933.
78. W i n d 1 e W. F., J. Сотр. Neur., 50, 479, 1930.
79. ВведенскийН. E. и Ухтомский А. А. Тр. СПб. об-ва естеств., 39,
в. 2, 1908; В в e д e не к и й. Соч., т. IV, стр. 90, 1938.
80. Goldgi CRecherches sur l'histologie des centres nerveux. Arch. Ital. Biol.,
v. Ill et IV, 1883. Цит. по Lenhossek'y (см. № 70).
81. H einbecker F.,Am. J. Physiol., 103, 104, 1933.
82. Беритов и БакурадзеА., Тр. Ин-та физиол. им. Бериташвили, 5,
143, 1943.
83. Grundfest H.,Ann. Rev. Physiol., 2, 213, 1940.
84. R e n s h a w В., J. Neurophysiol., 4, 167, 1941.
85. Беритов И. и Бакурадзе А., Тр. Ин-та физиол. им. Бериташвили,.
5, 1, 1943.
86. Беритов И. и Нивинская О., Тр. Физиол. ин-та им. Бериташвили,
4, 39, 1941
87. Use hi n sky, Pfl. Arch., 37, 17, 1885.
88. S t e w a r t W. В., H u g h e s J. a. M с С о u с h G. P., J. Neurophysiol. 3,
139, 146 и 151, 1940.
154 общая физиология спинного мозга и ствола головного мозга
«9. Hursh J. В., J. NeurophysioL, 3, 166, 1940.
90. Lloyd D. Р. С, Physiol. Rev., 24, 4, 1944.
91. 3 у p а б а ш в и л и А. Д., Сообщ. Акад. Наук Груз. ССР, 5, 647, 1944.
92. М с К і п 1 е у W. А. а. М a g о и п Н. W., Am. J. Physiol., 137, 217, 1942.
93. М agounH. W. aMc Kinley W. A., Am. J.Physiol., 137, 408, 1942.
*94. H арикашвили С. П.,Тр. Ин-та физиол. им. Бериташвили, 3, 463, 1937.
95. Ging-HsiWanga. Tse-Wei Lu. J. NeurophysioL, 7, 151, 1944.
96. Barron D. H.,Biol. Rev., 16, 1, 1941.
97. Marrazzi A. S. a. L о г e n t e de N6 R., J. NeurophysioL, 7, 84, 1944.
98. Беритов И. иРойтбакА., Физиол. жур. СССР, 33, 29 49 и 15?, 1947.
99. Katz В., Ргос. Roy. Soc, В., 124, 247, 1937. .
100. Беритов И., Бакурадз? А. и Ройтбак А., Физиол. журн. СССР, 33, 787, 1947.
101. Therm an Р. О., J. NeurophysioL, 4, 153,1941.
402. Lloyd D. R. С, J. NeurophysioL, 6, 435, 1942.
103. Беритов И., Бюл. эксп. биол. и мед., 22, вып. 6, стр. 3, 1946.
104. Беритов И., Бакурадзе А. и Ройтбак А., О биоэлектрических
потенциалах спинного мозга кошки. Тр. Ин-та физиол. им. Бериташвили, т. 7,
1947.
105. Bonnet V. u. Bremer F.,J. Physiol., 90, 45 P, 1937; Arch. Internat. de
Physiol., vol.211 fasc. 2, p. 153, 1942.
106. Беритов И. и Ройтбак А., О биоэлектрических потенциалах спинного
мозга во время торможения. 1946—1947. Труды физиол. ин-та им.
Бериташвили., 7, 1947.
107. Eccles J. С, J. NeurophysioL, 9, 87, 1946; 9, 139, 1946.
108. Беритов И. и Ройтбак А., Электрические потенциалы спинного мозга
при адэкватных раздражениях кожи. 1946. (Не опубликовано).
109. Ройтбак А., Об электрических потенциалах головного мозга при
раздражениях кожных нервов и рецепторов. 1947. (Не опубликовано).
110. Leduc S., Compt. rend. Acad- de Paris, 135, 194, 878, 1902.
111. TschagovetzW., Pflug Arch., 146, 567, 1912.
112. Frostig J. P. van Harreveld A., Resnik S., Tyler D. B. a. Wiers-
ma C.'A. G., Arch. Neurol, a. Psychiatr, 51, 232, 1946.
113. Bremer F., Bull.de Г Acad. Roy. de med. de Belgique, 9, 148, 1944.
114. Gasser H. SM Юбил. сб., поев. проф. И. Бериташвили, изд. Груз. фил. АН
СССР, 1936.
115. Bremer F. et Bonnet V., Arch. Intern, de Physiol., vo?. LII, fasc. 2, 153,
1942.
116. ОЪеагу J. L. a. Bishop G. H„ J. Cell а. Сотр. Physiol., 22, 73, 1943.
117. Forbes A. a. Mori son B. R., J. NeurophysioL, 2, 112, 1939.
118. Беритов И. и Гедеванишвили Д. Тр. Ин-та физиол. им. Бериташвили,
6, 279, 1945.
119. Могендович М. Р., Сборы, работ сект, флзиол. нер. сист. Гос. Ин-т по
изуч. мозга им. Бехтерева, стр. 45, 1932.
120. Ухтомский А. А., Русск. физиол. журн., 6, 37, 1923.
121. Васильев. Л. Л., Новое в рефлексол. и физиол. н. с, 1, 1925.
122. Jerkes R., РП.'Arch., 107, 207, 1905, J. Сотр. Neur., 14, 1904.
Глава VIII
РИТМИКА ЦЕНТРАЛЬНОГО ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ
Ритмическая природа центрального возбуждения. Возбуждение
в центральной нервной системе протекает ритмически. Это прежде
всего видно из того, что в определенных условиях ритм раздражения
чувствительного нерва точно передается через мозг скелетным
мышцам. Например, установлено, что в сгибательном рефлексе лягушки
мышцы-сгибатели дают биоэлектрические токи возбуждения по ритму
раздражения до 200 в 1 сек., а в том же рефлексе у кошки — до 300 в
J сек. (Беритов, 13; Гофман, 11) (рис. 75). Очевидно, такая точная
передача ритма возбуждения через мозг имеет место только потому,
что нервные цепочки, через которые происходит передача нервных
импульсов в мозгу от чувствительных волокон к двигательным
нейронам, реагируют на периферическое раздражение кратковременными
импульсами, подобно нервному волокну.
Новейшие осциллографические исследования показывают, что от
спинного мозга отводятся наряду с медленными биотоками быстрые
колебания высокой частоты, причем продолжительность их у кошки —
около 1 о, а у лягушки — около 2 о. Эти быстрые биотоки мозга
протекают на фоне медленных (рис. 56-А и 57-А). Они, очевидно,
выражают биотоки возбуждения, во-первых, афферентных чувствительных
волокон, а во-вторых, промежуточных нервных клеток и нервных
волокон, из которых составляются нервные круги или нервные цепочки мозга.
На рис. 56-А и 57-А первые быстрые колебания перед медленным
потенциалом происходят от афферентных волокон, а последующие мелкие
быстрые колебания на фоне медленных — от нервных кругов. При
торможении или утомлении деятельности нервных кругов эти
последние колебания выпадают, а первые большие колебания от
афферентных волокон остаются без изменения (Герш, 14; Ллойд, 15; Беритоів
и Ройтбак, 44, 42).
Биотоки возбуждения афферентных волокон возникают в мозгу
при всяких периферических раздражениях, когда импульсы
возбуждения проникают в мозг через задние корешки. Если же раздражать
¦спинной мозг непосредственно, тогда от него будут отводиться глав-
J56 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
ным образом биотоки возбуждения мозга в виде медленных
потенциалов дендритной массы и быстрых потенциалов от аксонов и клеток
промежуточных и двигательных нейронов. Так, например, на рис. 76
при уколе в серое вещество мозга и при отведении биотоков из
глубины прокола наблюдаются медленные потенциалы разной
длительности и амплитуды, а на фоне их располагаются беспрерывным рядом
быстрые колебания небольшой амплитуды в ритме до 300 в 1 сек.
Рис. 75. Мышечный ритм возбуждения при рефлекторном возбуждении
в сгибательном рефлексе и его зависимость от частоты и силы
раздражения.
Децеребрированная кошка. Токи возбуждения m. senutendmosus, записанные
струнным гальванометром Эйнтховена. Раздражается индукционными ударами п.
peroneus. Вместе с электрическим эффектом записывается механический эффект
(неколеблющаяся кривая). В верхней электрограмче ритм раздражения 15 в і сек.,
сила его около пороговой—30 см расстояния между катушками. Токи возбуждения
следуют точно по ритму раздражения В средней электрограмме ритм раздражения тот
же, но сила раздражения больше — 2 5 см Здесь ритм возбуждения выше, каждый
индукционный удар вызывает по 3—4 импульса, вначале — один сильный,
потом — несколько слабых. В нижней электрограмме ритм раздражения 200 в
1 сек., сила— 15 см (при пороге 20 см). Здесь ритм возбуждения точно следует
за раздражением. Внизу электрограммы сигнальная линия отмечает начало
раздражения. Время—-0.2 сек.
Этот электрический эффект может длиться десятки секунд вследствие
механического раздражения, вызываемого деформацией серого веще-
ства мозга от прокола.
Биотоки возбуждения нервных элементов серого вещества мозга
хорошо отводятся из глубины мозга игольчатым электродом, а также
через задние корешки, коллатерали которых оканчиваются в сером
веществе (рис. 76, 58-С, D). В последнем случае быстрые биотоки
мозга выносятся наружу электротоническим путем.
Если же отведение производится от поверхности мозга, они обычно
отсутствуют. От імозга отводятся более сильные медленные биотоки
РИТМИКА ЦЕНТРАЛЬНОГО ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ 157
кейропиля, а быстрые биотоки возбуждения нервных волокон полностью
шунтируются межуточной жидкостью мозга (рис. 77).
'.з^гзеякгк-
; „** Jv.* <*«*¦$¦¦
%*-^* %,'
250у V
¦v'*^^/*«#'V,~'*'
к!
*Ц^& v V*v**\' I;^^m«w*^>
Рис. 76. Электрическая активность спинного мозга.
Изолированный спинной мозг лягушки. Регистрация биотоков катодным осциллографом.
А— отведение игольчатым электродом из глубины IX сегмента слева через несколько
секунд после проко ла; В-спонтанная активность мозга при отведении дорзо-латеральной
поверхности IX сегмента справа; С—отведение из глубины IX сегмента слева; раздражается
п. peroneus sin. релаксационным стимулятором при 4 v. (Беритов и Ройтбак)
Закон возбуждения и рефракторная фаза- Ритмическое течение
биотоков возбуждения само по себе свидетельствует о том, что
возбудимая система нервных элементов центральной нервной системы
отвечает на раздражение согласно закону возбуждения. Доказать
это можно путем обнаружения в центральной нервной системе
рефракторных фаз — абсолютной и относительной. Но в центральной
нервной системе можно было бы доказать существование рефракторной
фазы вполне убедительно только в том случае, если бы вся
центральная нервная система могла притти в возбуждение одновременно. При
нормальной возбудимости центральной нервной системы это
совершенно недостижимо. Только при стрихнинном отравлении, когда
возбудимость центральной нервной системы очень сильно повышается, вся
центральная нервная система может сразу возбудиться; при таком
условии легко проследить и рефракторные фазы в связи с возбуждением.
Как впервые показал Введенский (1), центральная нервная
система, отравленная стрихнином, уподобляется по своей деятельности
нервно-мышечному препарату. Именно интенсивность деятельности
ее меняется в зависимости от частоты и силы раздражения точно
так же, как и на нервно-мышечном препарате; раздражение
чувствительного нерва индукционными ударами большой частоты и большой
интенсивности вызывает на мышцах через центральную нервную
систему слабый, пессимальный эффект, раздражение же малой силы
и малой частоты — оптимальный эффект. Что наблюдаемый в данных
условиях пессимальный эффект зависит от пессимального состояния
центральной нервной системы, видно из того, что в это время нервы
и мышцы не находятся в пессимальном состоянии, так как прямое
раздражение их производит максимальный внешний эффект. Такое
изменение внешнего эффекта не зависит также от пессимального со-
]58 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
стояния чувствительного нерва. Это видно из того, что пессимальный
эффект может вызываться, когда раздражение одного чувствительного
нерва, вызывающее оптимальный эффект, комбинируется с таким же
раздражением другого чувствительного нерва. Очевидно, в этом случае
пессимальное состояние создается в
центральной нервной системе, на
которую действуют оба нерва независимо
один от другого (Введенский, 1).
Таким образом, интенсивность
деятельности отравленной стрихнином
центральной нервной системы — ее
пессимальное и оптимальное состояние —
зависит, по примеру нервно-мышечного
препарата, от частоты центральных
импульсов, вызванных раздражением;
если в центральной нервной системе
каждый последующий импульс
наступает после рефракторной фазы, тогда
налицо оптимальный эффект; если же
импульсы возникают во время
рефракторной фазы, тогда наступает
пессимальный эффект. Это заключение
принадлежит не Введенскому. Он
рассматривал пессимальное состояние
центральной нервной системы как парабиотиче-
ское явление, т. е. как своеобразное
неколеблющееся состояние возбуждения.
Роль рефракторной фазы в этом
явлении была установлена Тидеманом
(2) и Ф р ё л ихо м (3).
Шеррингтон (5) пробовал установить
рефракторную фазу центральной нервной
системы при ее нормальном состоянии на децеребри-
рованных собаках. В одних опытах он раздражал
чувствительный нерв двумя последовательными
индукционными ударами. Оказалось, что второе
раздражение переставало давать эффект только
при уменьшении интервала ниже величины
рефракторного периода для нервных волокон.
В других опытах он раздражал один за другим
два чувствительных нерва: раздражение второго
нерва вызывало эффект при любом интервале.
Эти опыты как бы говорят против применимости закона возбуждения к
центральной нервной системе. Однако на осноівашга их нельзя отрицать возникновения
рефракторной фазы в центральной нервной системе,так какие соблюдено основное
условие для такого рода опытов, а именно: первое раздражение не вызывает
возбуждения во всей центральной нервной системе. Прівда, в первом ряде 'опытов при
интервале около 1 с второе раздражение не давало эффекта, но это следует приписать
рефракторному состоянию самого^ чувствительного нерва.
Рис. 77. Биотоки VII
поясничного сегмента кошки.
Отводятся микроэлектродом в
катодный осциллограф.
Раздражается плечевое сплетение
одним электрическим ударом. А —
при отведении от дорзальной
поверхности VII сегмента
Остальные кривые при введении
электрода в глубь мозга; цифры
справа указывают на глубину в мм
от дорзальной поверхности. На
дорзальной поверхности и на
глубине до і .5 мм медленные
потенциалы без быстрых, на
глубине 2.2—3.0 мм медленные
потенциалы сопровождаются
быстрыми. Кроме того на глубине
2.2—3.5 мм направление
медленных потенциалов
противоположное сравнительно с
отведением па глубине 0—2.5 мм.
(Ллойд)
РИТМИКА ЦЕНТРАЛЬНОГО ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ 159
Для выяснения этого же вопроса методикой Щ ер р и нгтон а воспользовались
Эдрнан и Ольмстед (6). Они раздражали у кошки чувствительный нерв из.
рецептивного поля сгибания двумя быстро следующими один за другим
индукционными ударами. Эффект сгибания записывался на кимографе. Суммарный эффект
получался в том случае, если интервал между обоими удаір?ми был не меньше 0.0019 сек.
Но отсюда нельзя еще заключить» что этот интервал, как раз и выражает абсолютную-
рефракторную фазу координирующего аппарата. Как и в опыте Шеррингтона,
это время должно выражать (Продолжительность абсолютной рефракториоЯ фазы чую*
ствятельного нерва.
Длительность рефракторных фаз центральных нервных элементов
измерялась непосредственно в отношении двигательных нейронов. Так,
в отношении двигательных клеток глазных мышц кролика, прямым
раздражением ядра глазодвигательного нерва двумя последующими
электрическими ударами, Л о р е н т е де Но установил, что длительность
абсолютной рефракторной фазы не превышает 0.5 о (40). Эта величина
вполне соответствует абсолютной рефракторной фазе, установленной
для нервных волокон группы А у теплокровных. Рефракторная фаза
двигательных клеток определялась также иным способом: один элек*
трический удар наносился на двигательный корешок и тем
производилось возбуждение двигательных клеток спинного мозга антидромным
путем, а другой удар наносился на соответствующий задний корешок
и вызывал разряд импульсов. Двигательные клетки реагировали на эти
разряды через 2.5 о, после антидромного раздражения. За этим
следовал период пониженной возбудимости, длившейся 10—75 о (Экклс, 41).
Но эти величины фаз абсолютной и относительной невозбудимости
двигательных клеток при антидромном возбуждении едва ли следует
считать (выражением рефракторных фаз. Как мы уже указывали выше,
благодаря активированию нейропиля через возвратные коллатерали
возникают длительные электрические потенциалы, которые затем ан-
электротонически действуют на двигательные клетки. Поэтому, когда
после антидромного возбуждения двигательных клеток пытаются
раздражением заднекорешковых волокон определить состояние
возбудимости, то фактически определяют состояние торможения.
Однако есть основание утверждать, что возбуждение аксона может
происходить не только при возбуждении клетки, но и при
возникновении в ней локальных биологических процессов. Так, например, это
можно наблюдать в двигательных нейронах спинного мозга лягушки.
При редких периферических раздражениях небольшой силы в передних
корешках наступает, в ответ на каждое раздражение, сначала
медленный потенциал, который выражает, главным образом, локальные
биологические процессы двигательных клеток, а затем, на фоне их, возникают
быстрые аксонные биотоки возбуждения переднекорешковых волокон
(рис. 57-D). Обычно при умеренных силах раздражения первые залпы
периферических импульсов дают только медленные потенциалы,
постепенно нарастающие от залпа к залпу. Биотоки возбуждения
двигательных волокон возникают, когда локальный потенциал достигает доста-
160 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
точно большой интенсивности. Так как в момент этих биотоков
медленный потенциал не меняется или даже усиливается, мы заключаем, что
в данном случае возбуждение аксонов двигательных клеток возникает
вследствие воздействия клеточных локальных потенциалов на аксоны.
Это происходит в силу замыкания внутриклеточного потенциала через
аксон и межуточную жидкость. Образующийся при этом интенсивный
электрический ток, выходя из аксона наружу, оказывает на аксон
катэлектротоническое действие, вызывая в нем повышение возбудимости
и многократное возбуждение. Но, как указывалось выше, двигательные
клетки могут также возбудиться, т. е. реагировать сразу всем
содержанием возбудимой системы. При этом в них возникает кратковременный
потенциал высокой амплитуды, который и является источником
раздражения соответствующих аксонов. В таких случаях двигательные клетки
не дают медленных потенциалов, а если они уже имелись, то в момент
возбуждения потенциалы исчезают (рис. 57-С).
Итак, нервные цепочки и нервные круги спинного мозга со всеми
своими, аксонами и клетками возбуждаются и проводят возбуждение, по
известному закону возбуждения. Но нервные клетки при некоторых суб-
минимальных воздействиях отвечают локальными процессами.
Соответствующие этим локальным процессам локальные потенциалы при
известной интенсивности являются совершенно достаточными для возбуждения
аксонов.
Значение клеточных локальных процессов в возбуждении двигательных нейронов
впервые было выяснено Шерриягтоном. Он и его сотрудники предположили, что
периферические импульсы непосредственно или через промежуточные нейроны
вызывают в двигательных клетках «местное состояние центрального возбуждения», и
через его посредство обусловливают возникновение быстро рашростраяятощихоя
нервных импульсов. Ш ерраигтон и его сотрудники называют «центральным
состоянием возбуждения» то, что мы называем просто локальным процессом. Шерринг-
т о н локализирует это состояние в двигательных клетках и полагает, что оно
выражается в частичкой деполяризации нервных клеток и тем самым становится
благоприятным условием для возшкнов?ния здесь разряда нервных импульсов. Предполагается,
что при повторении периферических импульсов «местное состояние центрального
возбуждения» или состояние деполяризации усиливается, а при достижении известной
интенсивности клетки разряжаются рядом нервных импульсов, иаправляйощикся по
аксонам к мышцам.
Лоренте де Но (35) предполагает, что «центральное состояние возбуіждения>
обусловлено бомбардировкой двигательных клеток импульсами из нервных мругов
и цепочек. Когда эти импульсы притекают с частотой свыше 2000 в 1 сек., эта
бомбардировка уподобляется действию постоянного тока «ли фарадизации очень высокой
частоты. °
Объяснение Шеррингтоиа и его сотрудников роли местных процессов в
двигательной клетке существенно отличается от данного нами выше. По нашему мнению,
разросшийся местный процесс через соответствующий ему локальный потенциал
сам становится источником многократного возбуждения аксона, а не производит в
клетке же ряд нервных импульсов. Если вообще двигательная клетка возбуждается,
как это должно быть при достаточно сильном воздействии на нее со стороны
синапсов или под влиянием антидромных импульсов, то тогда она должна разрядиться
одним импульсом, одним биотоком возбуждения, а не рядом их. Известно, что это так
и бывает при антидромном возбуждении клетки.
РИТМИКА ЦЕНТРАЛЬНОГО ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ 161
Кроме того, мы находим, что термин «местное состояние центрального
возбуждения» не является точным обозначением того состояния, которое создается в нервных
клетках под 'влиянием притекающих импульсов. Тот процесс, который мы обычно
называем возбуждением, не является локальным и не градуируется в отношении интен-
сивиости. Он возникает всегда с максимальной интенсивностью и тотчас же
распространяется с максимальной интенсивностью. Вследствие этого мы считаем
недопустимым употреблять термин «возбуждение» для* обозначения описанного івъпле местного
процесса. Мы думаем, что этот процесс является местным: усилением 'основного
биологического процесса, которое івіыражается, между прочим, в повышении
возбудимости, в облегчении возникновения и проведения возбуждения.
Ритм рефлекторного возбуждения центральных элементов спинного
мозга и мышцы. Ритм центрального возбуждения чрезвычайно
изменчив. Он зависит, в первую очередь, от функционального состояния
спинного мозга и затем еще от силы и частоты периферического
раздражения,— вернее, от числа и состава возбужденных афферентных волокон
и от частоты возбуждения этих волокон. Ввиду небольшой
продолжительности клеточных и аксонных биотоков возбуждения мозга — у
лягушки около 2 о, а у теплокровных около 1 о —< высший ритм
возбуждения центральных элементов у лягушки должен быть не ниже 500 в
1 сек., а у теплокровных не ниже 1000 в 1 сек. Этот высокий ритм
хорошо выступает при отведении из глубины мозга или через задние
корешки, как это показано на рис. 58-D и 76 в отношении спинного
мозга лягушки и на рис. 77 в отношении спинного мозга кошки. Но,
конечно, если принять во внимание, что нервные клетки и аксоны мозга
возбуждаются не одновременно, то регистрируемый ритм быстрых
колебаний может быть значительно выше.
Ритм центральной иннервации лучше всего изучить путем отведения
биотоков от двигательных нервов- или от передних корешков. При
рефлекторном возбуждении мозга передние корешки показывают высокий
ритм — у лягушки до 500 в 1 сек. (рис. 57 и 59). Они возникают
группами или сплошным рядом, в зависимости от характера раздражения и
функционального состояния препарата. При сравнительно редких
раздражениях до 50 в 1 сек. можно уловить ритм раздражения в
группировке биотоков возбуждения двигательных волокон (рис. 57, 41-И). Но при
более высоких частотах раздражения ритм биотоков возбуждения
двигательного нерва или передних корешков ни в какой мере не
соответствует ритму раздражения (рис. 59-С). Однако, как указывалось выше,
у лягушки мышечные биотоки рефлекторного возбуждения в
определенных случаях могут соответствовать даже ритму раздражения в 200 в
I сек. Это, очевидно, объясняется тем, что в передних корешках мы
имеем собрание двигательных волокон разнообразных мышц, а также
волокон разной проводимости, и потому в них проявляется сложная
алгебраическая суммация биотоков возбуждения этих волокон. Она,
конечно, маскирует в значительной імере характер центральной импуль-
сации к отдельным мышцам или к отдельным пучкам мышечных волокон.
II И. С. Беритов
162 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Двигательная клетка возбуждается не одним импульсом из одного
синапса одного афферентного волокна, а под воздействием многих таких
синапсов как от афферентных волокон, так и от промежуточных
нейронов. Кроме того, аксон двигательной клетки может дать группу
импульсов от воздействия локальных потенциалов, возникающих'при
субминимальном действии этих синапсов. Отсюда следует, что по
биотокам двигательных волокон нельзя точно установить ритм
возбуждения двигательных клеток, а тем более тех нервных элементов, которые
лежат более центрально, т. е. нервных цепочек и нервных кругов. Ритм
возбуждения последних может быть установлен путем отведения
биотоков их из глубины мозга, как это дано на рис. 76 или 77. При
таком методе изучения стало известно, что даже при точечном
отведении микроэлектродами около 50 микронов в диаметре ритм
возбуждения центральных элементов спинного мозга теплокровных достигает
1000 в 1 сек., а у зимней лягушки — 300 в 1 сек. Не может быть
сомнения в том, что эти импульсы возбуждения возникают не в одном
нервном элементе, а в целом ряде их в участке отведения. Понятно,
что суммарный высший ритм возбуждения центральных элементов
должен быть очень высок, ибо суммируются ритмы возбуждения
множества центральных элементов. В соответствии с этим к двигательным
клеткам передних рогов могут притекать из нервных кругов и цепочек
мозга много тысяч импульсов возбуждения в 1 сек.
До последнего времени процесс возбуждения центральной нервной
системы изучался по таким внешним эффектам, как биоэлектрические
токи (возбуждения нерва и мышцы, возникающие в результате
рефлекторной деятельности. Многие авторы обнаружили, что мышца во время
рефлекторного или произвольного сокращения дает различный ритм
возбуждения; она может дать и свойственный ей высший, предельный
ритм возбуждения. Следовательно, ритм рефлекторного возбуждения
мышцы у лягушки может достигать 150—250 в 1 сек., а у
теплокровных — свыше 300 в 1 сек. Основной вопрос заключается в том, в какой
мере этот мышечный ритм соответствует ритму возбуждения
центральной нервной системы. По мнению некоторых ученых, ритм мышечного
возбуждения при рефлекторном сокращении представляет собой
собственный ритм возбуждения мышцы, как это получается на нервно-
мышечном препарате при раздражениях индукционными ударами очень
большой частоты или постоянным током большой интенсивности
(Введенский, 7; Бьюкенен, 8; Гартен, 9). Значит, по мнению этих авторов,
ритм рефлекторных центральных импульсов всегда значительно выше
собственного ритма мышцы. Другие ученые, наоборот, утверждают, что
ритм мышечного возбуждения вполне соответствует ритму центрального
возбуждения (Пипер, 10; Гофман, 11; Диттлер и Гартен, 12).
В этом отношении особенно поучительны наблюдения Диттлера
и Гартена. Эти авторы произвели сравнительное исследование
биоэлектрических токов возбуждения -ц. phrenicus и диафрагмы (собаки,
кролика) во время вдоха и обнаружили, что мышца и нервы имеют при
РИТМИКА ЦЕНТРАЛЬНОГО ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ }63
этом совершенно одинаковый ритм возбуждения. При изменении
температуры тела ритм нерва и мышцы меняется, и притом одинаково. Так,
при температуре 38° их ритм равен ПО—140 в 1 сек.; при температуре
в 28° тот и другой ритм уменьшается до 50 в 1 сек. Это изменение
ритма, несомненно, зависит от центральной нервной системы, так как
подобное же охлаждение одного нерва совершенно не влияло на ритм
возбуждения диафрагмы при вдохе.
Но существует и третье мнение, которое наилучшим образом
учитывает весь фактический материал и вместе с тем объединяет оба
приведенные выше мнения, а именно: во всех тех случаях, когда ритм
центрального возбуждения мышцы меньше собственного ее ритма, он
должен вполне соответствовать ритму центральной иннервации. Наоборот^
когда центральный ритм превышает собственный, тогда мышечный ритм
должен быть собственным, г. е. меньше ритма центральной
иннервации (Беритов, 13).
Это положение подтверждается изучением токов возбуждения
мышцы при рефлекторных сокращениях, вызванных раздражением
чувствительных нервов фарадическим током.
Впервые Гофман (11) показал при помощи 'струнного
гальванометра, что рефлекторный ритм мышечного возбуждения лягушки
зависит от частоты раздражения чувствительного нерва. Рефлекторный ритм
возбуждения мышцы может вполне следовать за ритмом раздражения.
Путем ближайшего исследования было обнаружено, что в сгибательном
оборонительном рефлексе лягушки на мышцах-сгибателях ритм
возбуждения меняется от 20 до 200 в 1 сек., а у собаки — до 300 в 1 сек.
и что величина ритма в значительной степени зависит не только от
частоты, но и от силы раздражения. Так, например, если не очень
сильное раздражение имеет ритм 30 в 1 сек., то возбуждение імышцы
может иметь ритм точно 30 в 1 сек.; при учащении же раздражения
соответственно возрастает и ритм возбуждения (Беритов, 13) (рис, 75).
Однако осциллографическое исследование биотоков передних до-
решков или мышечного нерва показывает, что из спинного мозга
импульсы возбуждения никогда не притекают точно в ритме
раздражения, ибо в ответ на один удар раздражения спинной мозг реагирует
более или менее длинным рядом импульсов. Эта группа импульсов
может быть очень короткой и состоять из нескольких быстрых
потенциалов, как на рис. 78-В, но она может быть более или менее длительной,
как на рис. 57-С или 78-А и С. Следовательно, когда мышечные биотоки
возбуждения протекают в ритме раздражения чувствительного
нерва, то один такой биоток возбуждения должен соответствовать целой
группе нервных импульсов.
При исследовании биотоков возбуждения отдельных нервных и
мышечных волокон во время их произвольной или автоматической
иннервации оказалось, что ритм возбуждения их очень невелик. Э д р и а н и
Бронк (23) отводили токи возбуждения от нескольких нервных во-
11*
164 общая физиология спинного мозга и ствола головного мозга
локон п. phrenicus кошки через усилитель в капиллярный электрометр
и обнаружили здесь при каждом вдохе от 30 до 112 колебаний в 1 сек.
Аналогичные результаты они получили при отведении небольшого
пучка двигательных нервных волокон конечности во время их
рефлекторной иннервации: здесь ритм (возбуждения доходит до 150 в 1 сек. Такой
ритм может свободно передаться мышцам. Те же авторы произвели
сравнительное исследование токов возбуждения нервных и мышечных
волокон задней ноги кошки. При некоторых слабых сокращениях
частота биотоков возбуждения в мышечных волокнах меняется точно так же,
как в неррных волокнах. Так было при сгибательном рефлексе, при
перекрестном разгибательном рефлексе, при экстензорном тонусе. В общем
оказалось, что максимальный ритм центральной иннервации отдельных
мышечных волокон имеет частоту совершенно того же порядка, какая
нужна для (максимального сокращения при непрямом раздражении. Так,
например, ритм нормальной центральной иннервации m. quadriceps
кошки достигает 90 в 1 сек., a m. soleus — 35 в 1 сек. (Купер и Экклс, 30).
Это, несомненно, говорит об экономном течении центральной
деятельности.
В мышцах человека при точечном отведении ритм возбуждения
произвольной иннервации тоже невелик — от 5 до 10 в 1 сек. С усилением
иннервации этот ритм растет, но только при максимальной иннервации
он достигает 50 в 1 сек., т. е. той величины, при которой мышечные
волокна могут развить полный тетанус (Линдслей, 32).
Точно такие же результаты были получены Ваххольдером (24)
и Рихтером (25) на людях во время произвольной иннервации.
Максимальный ритм возбуждения, какой они получили при сильнейших
сокращениях, был 160—250 в 1 сек. Ритм (возбуждения одной
двигательной единицы при произвольной иннервации человека оказывается
особенно мал. Он меняется в связи с усилением сокращения от 3 до
50 в 1 сек., чаще всего — от 10 до 30 в 1 сек. Характерно, что при
утомлении во время сильного и длительного произвольного сокращения
интенсивность импульсов возбуждения уменьшается без изменения
ритма (Линдслей, 32). Очевидно, и у человека при произвольной
иннервации ритм возбуждения мышечных волокон точно передает ритм
центрального возбуждения.
Если другие авторы при отведении всего нервного ствола или всей
мышцы получали во время произвольной иннервации очень высокий
ритм возбуждения, например 300—500 в 1 сек., то это легко
объясняется тем, что возбуждение разных пучков нервных волокон
происходит в разное время и гальванометр или осциллограф регистрируют токи
возбуждения всех этих пучков.
Ритмический разряд импульсов возбуждения от одиночного
раздражения. Когда частота раздражения чувствительного нерва не
превышает 50 в 1 сек., то обычно каждый удар раздражения вызывает не один, а
целый ряд импульсов возбуждения. Количество этих импульсов зависит
РИТМИКА ЦЕНТРАЛЬНОГО ВОЗБУЖДЕНИЯ И ТОРМОШЕНИЯ ]65
от функционального состояния координирующего аппарата. Чем выше
возбудимость координирующего аппарата и вообще способность его к:
деятельности, тем больше количество импульсов возбуждения. В
каждой группе импульсов мышцы первые импульсы обычно интенсивнее
последующих (рис. 75). Это явление часто наблюдается также в
двигательном нерве, но здесь оно не обязательно (рис. 78). Таким
образом, нормально центральная нервная система отвечает на каждое раз-
Рис. 78. Биотоки рефлекторного возбуждения двигательных волокон у лягушки.
Лумбальные препараты. Все мышцы денервированы. Раздражается п. peroneus. Л — от переднего
корешка при ритме раздражения около 15 в 1 сек.; в — от мышечного нерва сгибателей колена
при ритме раздражения око$іо 60 в 1 сек. В этом опыте» в ответ на каждый удар раздражения, сперва
наступает после небольшого однофазного артефакта двуфазное колебание от биотона возбуждения
раздражающихся чувствительных волокон, электротонически распространяющегося от
седалищного нерва на его мышечную веточку к сгибателям колена. Затем» через скрытый период около 20 а,
наступают рефлекторные биотоки возбуждения отводимых двигательных волокон: С —
рефлекторные биотоки возбуждения того же мышечного нерва на другом препарате при ритме раздражения
около 45 в 1 сек.; они наступают группами, но эти группы не следуют ритму раздражения.
дражение рядом импульсов возбуждения. При всех случаях повышения
возбудимости в центральной нервной системе разряд импульсов в ответ
на одиночное раздражение удлиняется. Так бывает, например, после
поляризации спинного мозга катодом или после отравления его
стрихнином, эзерином, никотином (Бонне и Бремер, 33 и др.).
Множественный разряд импульсов в нервных центрах не зависит
существенно от каких-либо дополнительных вторичных раздражений,
например проприоцептивных, ибо известно, что перерезка всех задних
корешков или всех передних, т. е. устранение возможности
дополнительных вторичных раздражений, не устраняет этой характерной
реакции. Тот факт, что локальное отравление одного сегмента стрихнином в
сильной мере благоприятствует последействию рефлекса, вызываемого
через данный сегмент при отсутствии периферических импульсов
вторичного происхождения, является особенно сильным аргументом в
пользу данного положения.
Ритмический разряд центральной нервной системы в ответ на
периферический залп импульсов является одним из обычных проявлений
функциональной деятельности живой системы. Мы уже указывали
выше, что мышечные и нервные волокна в определенных условиях
разряжаются рядом имілульсов в ответ на одно кратковременное
раздражение. Нервным клеткам это явление свойственно в значительно большей
166 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
гллллллллл
в
-^дц^-
степени, чем нервному и мышечному волокнам. Как указывалось выше,
после каждого периферического импульса возбуждения в нервных
клетках возникают локальные процессы, которые длятся сотые или даже
десятые доли секунды. Под влиянием вызываемых ими локальных
потенциалов аксоны
разряжаются длительным рядом
импульсов.
Но длительное течение
ритмического разряда в
центральной нервной системе в
ответ на один периферический
импульс безусловно является
особенностью мозга как
своеобразного структурного
образования. Именно, ритмический
разряд, продолжающийся в
течение многих секунд и даже
минут, должен быть
обусловлен деятельностью нервных
цепочек и кругов. Каждый;
периферический импульс
сначала производит возбуждение
двигательных клеток
непосредственно, помимо нервных
цепочек и кругов, ибо, как
известно, афферентные волокна
посылают коллатерали прямо
к двигательным клеткам. Но
одновременно периферический
импульс передается нервным
кругам и нервным цепочкам
разной сложности и приводит
их в активное состояние.
Нервные круги и цепочки
связаны также с двигательными
нейронами, а потому
импульсы возбуждения будут
достигать двигательных клеток все
время, пока эти круги и
цепочки находятся в
возбуждении. Скрытый период
возбуждения нервной клетки, по Л о-
ренте де Но, составляет около 6 6 а, и, следовательно, если данная
цепочка включает п нейронов, то возбуждение, распространяясь в
цепочке, достигнет двигательной клетки многократно через разные
промежутки от 0.6 о до 0 6 тіс Если круг содержит п нейронов, то через каж-
Рис. 79. Токи возбуждения
глазодвигательного аппарата кролика.
Регистрация токов возбуждения катодным
осциллографом от моага и внутренней прямой мышцы. В опыте
А мо8г отводится микроэлектродом в области
глазодвигательного ядра (ос. п. на рис. 80): первое
колебание в — от петли индукционного удара, второе /
отвечает импульсу нервных волокон — fasciculus long.-
post. на рис. 80, а третье т — двигательному
ядру. Четвертое двухфазное колебание М—от прямой
мышцы глава. В опытах В и о отводится
продолговатый мозг в области form, reticul. (Med. на рис. 80).
В опыте В после одного раздражения от мозга
отводится длинный ряд токов возбуждения, а на мышце
М всего один двуфазный ток возбуждения. В опыте
О от двух раздражений наступает длинный ряд более
сильных токов возбуждения, от жгмттттрт —• всего два
тока возбуждения, причем второй из них во много
pas сильнее первого М. Моменты раздражения
обозначены стрелками. Время — в сигмах. В опытах D
и Е отводятся п. trachlearis при том же раздражении
fasclc. long. post. Первое колебание —от петли тока /,
второе большое колебание — от возбуждения
непосредственно через нервный пучок fasclo. long.
post., а ряд следующих—через длинный путь с
участием промежуточных нейронов. Мелкие колебания
до к после раздражения выражают тоническую им-
пульсацию глазных мышц (Лоренте де Но).
РИТМИКА ЦЕНТРАЛЬНОГО ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ 167
дые 0.6 X па двигательные клетки получат из него по одному импульсу,
тюка не произойдет утомление какого-либо звена в данном нервном
кругу (Лоренте де Но, 35; Рэнсон и Гинзи, 34).
Прохождение периферического импульса к двигательным клеткам
как через промежуточные нейроны, так и без их участия, а также
ритмический разряд нервных кругов в двигательные клетки хорошо
удается уловить на глазодвигательном препарате кролика. Этот
препарат представляет собой fasciculus longitudinalis posterior, который,
между прочим, заканчивается синапсами у двигательных клеток
глазных мышц как непосредственно, так и через fcrmatio reticularis
среднего імозга. Лоренте де Но раздражал fasciculus индукционными
ударами, а глазодвигательное ядро, formatio reticularis или п.
trochlears отводил в осциллограф. При отведении двигательного ядра он
лолучал в определенных случаях два тока возбуждения, из которых
первый соответствовал возбуждению волокон fasciculus, а другой —
возбуждению двигательных клеток (рис. 79). При отведении formatio
reticularis наблюдался длинный разряд импульсов, которые не
вызывали соответствующего разряда в двигательном нерве, ибо последний
отвечал токами возбуждения непосредственно в ответ на нервные
импульсы из fasciculus (79, В и С). Но в других случаях при том же
отведении ЛорентедеНо наблюдал токи возбуждения, вызываемые в
двигательном нейроне как непосредственно импульсами возбуждения
волокон fasciculus, так и ритмическими разрядами из formatio
reticularis (рис. 79, D и Е) (Лоренте де Но, 36).
Здесь же мы приводим рис. 80 по Лоренте де Но, который
дает ясную картину путей прохождения нервных импульсов в
продолговатом мозгу от раздражаемого участка мозга до п. trochlearis.
Ритм возбуждения во время рефлекторного последействия.
Описанное выше многократное возбуждение является главным условием того,
что рефлекторная реакция не кончается вслед за прекращением
раздражения. Как мы уже знаем, после раздражения рефлекс
удерживается более или менее значительное время, иногда в течение нескольких
секунд и даже іминут. Во время этого последействия центральная
нервная система посылает на периферию целый ряд импульсов
возбуждения. Судя по токам возбуждения двигательного нерва и мышцы, ритм
и интенсивность возбуждающих импульсов во время последействия
вначале столь же высоки, как и во время раздражения, затем они
постепенно падают (рис. 81, 57-G).
В условиях устранения вторичных проприоцептивных раздражений
путем перерезки задних корешков последействие рефлекса может
протекать довольно долго — несколько секунд и даже десятков секунд. Это
указывает на то, что длительный ряд возбуждающих импульсов в
последействии может наступить и от одного первичного раздражения, т. е
является результатом длительной самостоятельной деятельности
нервных кругов. На рис. 81 последействие записано как раз в этих уело
виях (Беритов, 18).
168 общая физиология спинного мозга и ствола головного мозгд
Лабильность нервных центров. Судя по ритму рефлекторного
возбуждения двигательного нерва или мышцы, лабильность нервных
центров должна быть несколько ниже, чем двигательных нервов. Это видна
прежде всего из того, что при естественном рефлекторном возбужде-
Рис. 80. Диаграмма нервных путей, связывающих промежуточные
клетки между собой и с глазными мышцами.
У. — п. vestibularis, 1—6 — клетки в первичном вестибулярном ядре;
7—9—клетки formatio reticularis продолговатого Мозга Med и моста Р; 10—12—клетки formatio
reticularis среднего мозга М. о.; Ос. п. — ядро глазодвигательного нерва,* /. I. р. —
lasciculus longitudinalis posterior — пути промежуточных нейронов; Fly Ft и Col. —
положение раздражающих электродов. Диаграмма показывает, какими путями
нервные импульсы должны распространиться от раздражающих электродов до
глазодвигательных ядер. Например, от Ft нервный импульс может дойти до ядра по
короткому пути /. I. р., но он может достигнуть его и длинным путем через средний
мозг т. Ь. с участием клеток 11 и ю. При отведении formatio reticularis
продолговатого мозга импульсы возбуждения будут доходить весьма разнообразными путями
как из первичного вестибулярного центра, так и из formatio reticularis среднего
мозга (Лоренте де Но).
нии небольшого пучка нервных или мышечных волокон самый высокий
ритм намного уступает высшему предельному ритму нервных волокон*
Это видно также из того, что при раздражении чувствительного нерва
прерывистым током возбуждение передается через спинной мозг
двигательному нерву по ритму раздражения у лягушки до 200 в 1 сек., а
у кошки — до 300 в 1 сек., т. е. почти вдвое *ниже высшего предельного
ритма нервного возбуждения.
Кроме того, для нервных центров спинного мозга характерна очень
быстрая изменчивость лабильности в связи с деятельностью. В самом
начале деятельности она выражается в повышении лабильности,
которое тем выше и длительнее, чем слабее раздражение. При более или
менее длительном и сильном раздражении лабильность, наоборот,
быстро падает. В отношении скорости повышения и падения лабильности
в связи с деятельностью нервные центры значительно превосходят
скелетные мышцы.
Это явление изменения лабильности нервных центров должно быть
связано с образованием определенных метаболитов в нервных центрах
в связи с возбуждением. В случае малой концентрации они вызывают
в возбудимой системе повышение возбудимости и лабильности, в случае
же большой концентрации — понижение той и другой. Такого же
мнения держатся в настоящее время многие физиологи (Ухтомский, 38;
Бабский, 39).
РИТМИКА ЦЕНТРАЛЬНОГО ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ 169
Ритмическое течение торможения. Мы признали центральное
торможение активным процессом, обусловливающимся активным состоянием
нейропиля, его .медленными потенциалами. Но, как указывалось выше,
это активное состояние от каждого периферического импульса длится
сотые доли, иногда десятые доли секунды, а потому может протекать
ритмически, если только частота раздражения не особенно высока.
У лягушки медленный потенциал спинного мозга может колебаться 28
раз в 1 сек., по Баррону и Мэтьюсу (37), и даже до 60 в
Рис. 81. Ритмика возбуждения мышцы во время последействия.
Спинномозговая лягушка. В поясничном отделе перерезаны все задние корешки. Регистрируются
токи возбуждения m. semitendinoeus гальванометром Эйнтховена. Там же дается механический эффект
(неколеблющаяся кривая). Раздражается центральный отрезок JX заднего корешка. Ритм
раздражения 50 в 1 сек. Поднятие сигнальной линии указывает на прекращение раздражения.
Последействие не доведено до конца. Время 0.2 сек. (нижняя линия).
1 сек.— по Беритову и Ройтбаку (42). Поскольку медленный
потенциал обусловливает торможение, то при сравнительно редком
тормозящем раздражении торможение не должно являться сплошным,
неколеблющимся,— оно должно протекать ритмически. Это и было
доказано гальванометрическим исследованием рефлекторной деятельности
мышц. Именно, под влиянием сравнительно редкого тотанического
тормозящего раздражения до 50—100 в 1 сек., мышцы испытывают
торможение по ритму до 50—100 в 1 сек. Во время торможения мышца не
дает какого-либо особого электрического эффекта, но так как во время
торможения центральная часть двигательного нейрона приходит в такое
состояние, что через нее возбуждение перестает передаваться мышце,
можно проследить ритм торможения на фоне центрального
возбуждения мышцы. Положим, на m. semitendinasus лягушки раздражением
нерва из рецептивного поля сгибания вызывают рефлекторное
возбуждение и регистрируются токи возбуждения гальванометром Эйнтховена.
Если частота раздражения была 100—150 в 1 сек., то соответственно*
и ритм токов возбуждения будет 100—150 в 1 сек. Если во время этого
эффекта приложить к рецептивному полю потирания раздражение
небольшой частоты (20—50 в 1 сек.), то эта мышца затормозится, и
окажется, что в момент расслабления токи возбуждения редуцируют не
сплошь, а правильными интервалами — именно по интервалу
тормозящего раздражения. Если ритм тормозящего раздражения равняется
50 в 1 сек., то ослабевают или совершенно выпадают те токи
возбуждения, которые приходятся через каждые 0.02 сек. При этом, если сила
170 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
тормозящего раздражения невелика, то через этот интервал меняется
только один ток возбуждения; если же она значительна, то выпадает и
ослабевает несколько токов возбуждения (Беритов, 19) (рис. 82).
Аналогичные опыты были произведены на кошках. Здесь, во время
рефлекторного возбуждения в ритме 200—250 в 1 сек., нерв, с
которого вызывается торможение, раздражается при малом ритме до 100
в 1 сек. В результате, на каждый удар тормозящего раздражения
выпадал то один, то два и более токов возбуждения. Это означает, что
продолжительность одного колебания торможения может быть менее 4 о
(Беритов, 19). Эти фактические данные о ритмическом выпадении токов
Рис. 82. Ритмическое течение торможения.
Спинномозговая лягушка. Записываются токи возбуждения m. semitendinosus (гальванометр Эйнтхо-
вена). Они вызываются раздражением п. peroneus 100 раз в і сек. (рефлекс сгибания) и тормозятся
раздражением п. brachials 30 раз в 1 сек. (рефлекс потирания). Первый сигнал под электрограммои
опусканием обозначает время тормозящего раздражения, а колеблющаяся линия над электрограммои
дает моменты замыкания и размыкания (опускание кривой) первичной катушки при ?том же
раздражении. Во время тормозящего раздражения происходит выпадение одного или двух токов
возбуждения через каждую */>о сек. На электрограмме иксы обозначают выпадение одного тока
возбуждения. Где яет иксов, там выпадало или ослабевало по два тока возбуждения. Время — по 2.2 сек.
возбуждения мышцы, под влиянием торможения были подтверждены
после Беритова целым рядом других авторов (Эдриан, 22; Фултон и
Лидделл, 28; Самойлов и Киселев, 26).
При длительных тетанических раздражениях чувствительных нервов
или корешков торможение является сплошным. Но интенсивность
торможения, конечно, колеблется в связи с изменчивостью активного
состояния нейропиля. Это хорошо выявляется при комбинации тетаниче-
<ского торімозящего раздражения одного чувствительного корешка с
раздражением другого корешка отдельными индукционными ударами.
В момент комбинации одни рефлекторные вздрагивания ослабевают с
разной интенсивностью, а другие даже не меняются (рис. 83).
Разумеется, в центральной нервной системе каждое колебание
торможения двигательных нейронов одной мышцы реципрокно связано не с
оцним импульсом возбуждения антагониста. Каждому периоду
торможения агониста, должно быть, соответствует целая лавина импульсов
возбуждения двигательным нейронам антагониста. Они поступают к ним
со всех нервных кругов и цепочек мозга, возбужденных
периферическим раздражением.
Итак, центральное торможение, подобно возбуждению, протекает
ритмически. Но в координированных рефлекторных реакциях каждому
периоду торможения двигательных нейронов агониста реципрокно соог-
РИТМИКА ЦЕНТРАЛЬНОГО ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ 171
ветствует целая лавина импульсов при двигательных нейронах
антагониста, а не один импульс возбуждения.
Рис. 83. Периодическое изменение интенсивности
торможения при тетаническом тормозящем раздражении.
Лумбальный препарат кошки. Регистрируется mm. semttendi-
nosus (S. d„ S. е.) и quadriceps (Q. d., Q. S.) обеих сторон.
Раздражением VI заднего корешка левой стороны одиночными
индукционными ударами вызываются рефлекторные
вздрагивания этих мышц. Во время нанесения одиночных ударов
производится тетавическое раздражение У эаднего корешка
девой стороны. На фоне вызванного им общего торможения
одни рефлекторные вздрагивания на левой стороне выпадают,
другие в большей или меньшей степени ослабевают, третьи
остаются без изменения или даже усиливаются (Беритов и
Бакурадзе).
Местное состояние центрального торможения. В последнее время Шерринг-
тон и его сотрудники развивают относительно центрального тормозящего действия
такую же гипотезу, как и для «местного состояния центрального
(возбуждения». Они полагают, что центральное торможение возникает в двигательных
клетках под влиянием последовательного ряда импульсов со стороны проме*
жуточных нейронов в виде длительного постоянного состояния торможения. Это
состояние является также местным и способным к суммации при повторных
раздражениях. Оно противоположно в качественном отношении местному состоянию
центрального возбуждения, а потому при одновременном вызывании того и другого
состояния происходит взаимная инактивация. Иначе говоря, «местное состояние
центрального торможения», какого бы высокого уровня оно ни достигло, никак не
проявляется на мышцах; оно лишь препятствует возникновению в двигательных клетках
обычных импульсов возбуждения. Характерно, что это «местное состояние централь-
«ого торможения» не захватывает ни в какой мере даже возбудимой системы
двигательных клеток. Известно, например, что антидромное возбуждение двигательного
нейрона не влияет на это состояние, в то время как оно уничтожает «местное
состояние центрального возбуждения». Поэтому предполагается, что здесь все дело в
изменениях синапсов или поверхностных мембран клетки.
Однако Шеррингтон и сотрудники отмечают также наличие очень коротких
]72 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
периодов торможения, если оно исследуется на фоне рефлекторных возбуждений от
частого тетанического раздражения — как раз при тех условиях, которые были у нас
при исследовании ритмики торможения. Быстрое следование положительных залпов
друг за другом,— указывают авторы,— сокращает, швидимому, продолжительность
тормозного процесса, вызываемого одиночным центростремительным тормозным залпом
(31) Мы думаем, что наличие «длительного торможения» в виде длительного
изменения поверхностных мембран или какого-либо другого изменения в синапсах ил»
промежуточных нейронах не противоречит нашему представлению о ритмической
природе торможения с наименьшей продолжительностью для тормозящего периода в
несколько сигм.
Фактические данные Шеррингтона и сотрудников о центральном
торможении по существу также не противоречат тем представлениям, которые мы развиваем
здесь о природе центрального торможения. Так, например, то наблюдение, что
антидромное возбуждение двигательных клеток не влияет на торможение, хорошо
согласуется с тем нашим положением, что торможение двигательных клеток обусловливается
электротоническим действием нейропильного тока, а это действие, прежде всего,
должно охватывать поверхностные мембраны клеточных тел.
Далее, мы также допускаем длительное состояние центрального торможения пр«г
двигательных клетках. Но мы полагаем, во-первых, что это длительное состояние
возникает через нейропиль, а не непосредственно через промежуточные или афферентные
нейроны; во-вторых, это длительное состояние выражается в понижении
возбудимости,— вернее, в повышении порогов раздражения двигательных клеток по отношению
к биоэлектрическим токам возбуждения оканчивающихся при них промежуточных или
афферентных нейронов.
Практикум
Наблюдение токов возбуждения мышцы при сгибательном рефлексе при помощи-
струнного гальванометра. Спиннсхмозговой препарат лягушки. Обнажен п. peroneus »
отпрепарирован наполовину m. semitendinosus с діисгатгьнюй стороны. На эту часть
мышцы наівязаны на расстоянии 1 см одна от другой омоченные в физиологическом
растворе ватные нити* Эти нити, служащие проводниками биоэлектрического тока,
закрепляются другими концами на платиновых электродах. Последние соединены со
струйным гальванометром Эйнтховена. N. peroneus или кожу на пальцах раздражают
индукционными ударами размой частоты и разной силы и наблюдают на-глаз частоту
колебаний струны на экране. Чтобы мышца не передвигалась и местоположение
отводящих ток нитей не менялось, а тем самым не вызывалось смещение струны от
изменения чисто физических условий отведения, сухожильный конец мышцы соединяют
с изометрическим миографом. Если расположить препарат перед экраном, то на
экране можно следить и за движением миографа. При раздражении нерва отдельным-
размыкательным индукционным ударом также можно вызвать возбуждение мышцы*
Если препарат достаточно чувствителен, то в ответ на один удар мышца будет
реагировать хорошо заметным на-глаз рядом токов возбуждения.
Изменение токов возбуждения мышцы лягушки при центральном торможении-»
Спинномозговая лягушка. Раздражают п peroneus частыми индукционными ударами
100—150 в 1 сек. На m. semitendinosus наблюдают токи возбуждения. Для того чтобы
вызвать торможение, берут п. cutaneus femoralis lateralis или п. brachialis. Его
раздражают в редком ритме 10—25 в 1 сек. Это раздражение сочетается с раздражением*
п. peroneus. Если тормозящее раздражение небольшой силы и производит частичное
ослабление сокращения мышцы, то токи возбуждения при этом окажутся более
редкими, интенсивность же их в определенных случаях может даже увеличиться. Это
бывает в том случае, если без торможения ритм возбуждения был очень высок»
В этом случае увеличение токов возбуждения обусловлено выпадением в мышце
некоторых волн возбуждения, вследствие чего устраняется влияние рефракторной
фазы от предыдущих волн на последующие волны возбуждения Это изменение токов
РИТМИКА ЦЕНТРАЛЬНОГО ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ 173
возбуждения трудно наблюдать на-глаз, а потому колебания струны от токов
возбуждения нужно фотографировать.
Сравнительное наблюдение биотоков рефлекторного возбуждения мышц и нерва
лягушки на катодном или шлейфном осциллографе. Регистрация мышечных биотоков
производится с помощью усилителя переменных токов с однотактным входом.
Сеточный электрод прикладывается к мышце, а заземленный подводится к серебряной
пластинке, на которой лежит препарат. Сеточный электрод может быть игольчатой
серебряной проволокой, которая вкалывается в безнервный участок мышцы. Лучше
всего регистрировать m. sartorius или т. semitendinosus, вытянув мышцу
изометрическим миографом. В этих опытах необходимо, чтобы остальные мышцы, кроме
регистрируемой, были -^ценервированы. Раздражается центральный отрезок
чувствительного нерва (п. peroneus) индукционными ударами или релаксационным
прерывателем.
Регистрация биотоков возбуждения двигательного нерва мышцы производится
после наблюдения мышечных 'биотоков. Перерезается мышечный нерв, и его
центральный отрезок берется на сеточный электрод. Раздражается тот же нерв. При
регистрации биотоков нерва приходится применять значительно большее усиление, чем
для мышцы.
Для точной характеристики эффектов нужно заснять на пленке эти биотоки
мышцы и нерва.
Литература
4. Введенский Н.,Работы физиол. лаб. Петерб. ун-та, 1, 1906.
2. Т ied em an n A.,Z. Allg. Physiol., 10, 183, 1910.
3. Frohlich F.,Z. Allg. Physiol., 9, 55, 1909.
4. Satake J. Z. Allg. Physiol., 14, 79, 1912.
5. Sherrington a. Sowton.,J. Physiol., 49, 331, 1914.
6. Adrian а. О lms ted., J. Physiol., 66, 426, 1921.
7. Введенский Н.,Телеф. исслед. над электрическими явлениями в
мышечных и нервных аппаратах, Петербург, 1884.
8. В и с h a n a n F, J. Physiol., 37, 1908; Quart. J. Exp. Physiol., 1, 211, 1908.
9. Garten S.tZ. Biol., 52, 534, 1909.
10. Piper H, R.,Electrophysiologic menschlicher Muskeln, Berlin, 1912.
41. Hoffmann P.,Arch. Anat. Physiol., 1909, стр. 341.
12. Dittler u. Garten, Z. Biol., 58, 420, 1911.
13. Beritoff J.,Z. Biol., 62, 125, 1913.
14. Hursh J. В., J. Neurophysiol., 3, 166, 1940.
45. Lloyd D. P. С., J. Neurophysiol., 6, 293, 1943.
16. Veszi J..Z. Allg. Physiol., 13, 245, 1915.
47. Hoffmann P.,Arch. Anat. Physiol., стр. 233, 1910.
18. Beritoff J..Z. Biol., 64, 161, 1914.
19. В e г i t о f f J., Z. Biol., 64, 175, 289 и 307, 1914; 80, 171, 1924.
20. Beritoff J.,Arch. Anat. Physiol., 1, 1913.
21. Beritoff J.,Z. Biol., 64, 285, 1914.
22. Adrian E. D., Brain, 47, 399, 1924.
23. A dr i an E. а. В г о nk D.,J. Physiol., 66, 81, 1928; 67, 119. 1929.
24. Wachholder K.,Erg. Physiol., 26, 598, 1928.
25. Richter E. P.,Quart. J. Exp. Physiol., 18, 55, 1927.
26. SamojloffA. u. Kisseleff D.,Pfl. Arch., 215, 698, 1927.
27. Eccles J. С a. Sherrington C. S.,Proc. Roy. Soc. В., 109, 91, 1931.
28. Ful t on J. E. a. Liddel 1 E. G. T.,Proc. Roy. Soc. В., 98, 214, 1925.
29. Киселев М.,Казанск. мед. журн., № 4—5, 1931.
174 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
30. С о о р е г S. a. Eccles J. С, J. Physiol., 69, 377, 1930.
31 «Creed, Denny-Brown, Eccles, Liddell a. Sherrington,,
Reflex activity of the spinal cord, 1932. Есть русск. пер.: Рефлекторная
деятельность спинного мозга, 1935.
32. L і n d s 1 е у D. В., Am. J. Physiol., 114, 90, 1935-
33. Bonnet V. а. В г e m e г F., J. Physiol., SO, 45 P, 1937.
34. R a n s о n S. W. a. Hinsey J. C.,Am. J. Physiol., 94, 471, 1930.
35. Lor en te de N6«J. Neurophysiol., 1, 207, 1938.
36. Lo "ente de N 6, J. Neurophysiol., 2, 402, 1939.
37. В a r io n D. H. a. Matthews B. H. C., J. Physiol., 92, 276, 1938.
38. У x t о m с к и й А. А., Физиол. журн. СССР, 25, 767, 1938.
39. Бабский С. В., Учен. зап. каф. физиол. Моск. пед. ин-та, 1938.
40. Lorente de N6, Am. J. Physiol., Ill, 272 и 283, 1935; 112, 595, 1935.
41. Eccles J. C, Proc. Roy. Soc. В., 107, 557, 1931.
42. Беритов И. и Ройтбак А., Физиол. журн.^СССР, 33, 29, 49 и 157,
1947.
43. Heinbecker P., Am. J. Physiol., 117, 686, 1936.
44. Беритов И., Бюлл. экспер. биол. и мед., 22, вып. 6, 1946.
Глава IX
РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ
В НЕРВНОЙ СИСТЕМЕ
Общее понятие о времени рефлекторной реакции. Временем
рефлекторной реакции называется время от момента раздражения до
начала рефлекторной реакции. Это время иначе называют скрытым
периодом рефлекторной реакции. Оно зависит от скорости проведения
возбуждения и от длины нервных элементов, участвующих в этом
проведении, начиная от рецептора и кончая эффектором; таким
образом, время рефлекторной реакции представляет суммарное врем»,
потребное на возбуждение рецептора, на проведение возбуждения в
чувствительном нейроне, на передачу возбуждения от чувствительного
нейрона на промежуточные и на проведение в этих нейронах, затем на1
передачу возбуждения от этих нейронов на двигательные и на
распространение его по последним и, наконец, на передачу возбуждения от
двигательных нервных окончаний на мышцу, на ее возбуждение и
распространение его по мышечному волокну.
Скорость проведения возбуждения в длинных нервных путях
центральной нервной системы, в частности в спинном мозгу, примерно
такова же как в периферических нервных стволах. В чувствительных
путях, проводящих импульсы возбуждения от мышечных рецепторов,
она является наибольшей. Так, у теплокровных животных в нервных
волокнах, образующих tractus reticulo-cerebellaris и проводящих
афферентные импульсы от мышечных рецепторов, скорость достигает !40 м
в 1 сек. В дорзальных столбах кожной чувствительности скорость —
около 70 м в сек. (Грундфест и Кемпбелл, 49). В волокнах
пирамидных путей скорость разная — от 65 до 18 м в 1 сек. и еще ниже
(Ллойд, 50). Эти скорости проведения в длинных путях зависят, как и-
в нервных стволах, прежде всего от диаметра нервных волокон: они
больше всего в наиболее толстых волокнах. На нервных волокнах
центральной нервной системы нет перехватов Ранвье, но зато у них
имеются многочисленные перехваты в участках отхождения коллате-
ралей, а коллатерали отходят от волокон почти на всем их
протяжении. Весьма возможно, что в миэлиновых волокнах центральной
нервной системы возбуждение распространяется с помощью этих участков -
176 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
импульсы возбуждения перескакивают от одного такого участка к
другому, как в периферических нервных волокнах от одного перехвата
Ранвье к другому.
Передача возбуждения от афферентных нейронов на двигательные
может происходить не только через тот отдел или сегмент мозга, куда
входят возбужденные нервные волокна, но и через другие отделы
мозга, прежде всего через средние отделы и большие полушария
головного мозга, с которыми афферентные нервные волокна связаны
непосредственно или через особые промежуточные нейроны. Значит, для
каждой рефлекторной реакции существует несколько нервных путей:
один путь — наиболее короткий —¦ через тот сегмент, в который
вступают раздраженные афферентные волокна, и затем длинные пути
*через головной мозг. На нормальном животном или на децеребриро-
'Ванных препаратах оба пути могут приходить в действие при каждом
рефлекторном акте. Поэтому скрытый период рефлекторной реакции
носит двойственный характер: сначала рефлекс наступает по
короткому, а потом по длинному пути. Конечно, на спинальных препаратах
мы имеем дело только с короткими путями. Поэтому скрытый период
реакции по коротким путям удобнее всего изучать именно на таких
препаратах.
В скрытом периоде рефлекторной реакции нас больше всего
интересует то время, которое приходится на центральную нервную систему.
Оно устанавливается путем вычитания из всего скрытого периода
реакции того времени, которое нужно для проведения возбуждения от
.рецепторов до мозга и от мозга до эффектора. Вычисленное таким
путем время передачи возбуждения в центральной нервной системе
называется истинным, или редуцированным, временем реакции.
Распространение возбуждения по коротким путям в спинном мозгу.
Распространение возбуждения в спинном мозгу обычно изучается на
спинальных препаратах в отношении сгибательно-оборонительного
рефлекса и перекрестного разгибательного. Впервые оно было изучено
Гельмгольцем на лягушках (1). Он установил, что
продолжительность рефлекторной реакции много больше того времени, какое
нужно для распространения возбуждения по нервному стволу.
Гельмгольц нашел, что в спинном мозгу лягушки для передачи
возбуждения от задних корешков на передние нужно 0.03—0.1 сек.
В таком случае скорость проведения в опинном мозгу не может
превосходить 1—2.5 м, в то время как в нерве скорость проведения
составляет около 25 м.
По исследованиям Вундта (5), возбуждение проходит через
спинной мозг лягушки в 0.008—0.015У сек. Это время оказывалось
значительно большим при передаче возбуждения от одной симметричной
стороны на другую (Розекталь, 4; Бьюкенен, 2).
Скрытый период рефлекторной реакции меняется в зависимости от
силы раздражения. Он уменьшается с усилением раздражения. Он
меняется также в зависимости от температуры: повышение
температуры уменьшает его, а понижение удлиняет. Кроме того, он меняется в
РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ В НЕРВНОЙ СИСТЕМЕ 177
зависимости от утомления: чем больше утомлена центральная
нервная система, тем он больше. Наконец, скрытый период зависит от рода
животных. У теплокровных он много короче, чем у холоднокровных.
По новым исследованиям, наименьшее время рефлекторных
реакций по коротким путям может быть настолько мало, что на спинной
мозг приходится всего несколько сигм. Д ж о л л и (8) нашел, что во
многих случаях у африканской лягушки передача возбуждения с одной
половины мозга на другую происходит без всякой задержки. По Ф о р-
бсу и Греггу (7), & сгибательном рефлексе децеребрированной
кошки на спинной мозг приходится 3—5 о. АпоЭкклсу и Шер-
рингтону (12) на распространение возбуждения в спинном мозгу
теплокровных животных приходится еще меньший промежуток
времени— 2.7 о, который при повторных раздражениях редуцируется до 0.5 о
и менее. У других авторов это время в среднем еще меньше— 1.4 о
(Лис и Эйнарсон, 13). По некоторым опытам запаздывание
передачи возбуждения на клетку глазодвигательных ядер кролика равно
0.5—0.8 о, но при особо благоприятных условиях, когда волна
возбуждения приходит к двигательной нервной клетке в период ее
повышенной возбудимости от непосредственного подпорогового
раздражения индукционными ударами,— скрытый период укорачивается до
0,22 о (Лоренте де Но, 11). Далее, Лоренте де Но показал, что и в
промежуточных клетках скрытый период возбуждения длится не
больше этого времени. Он раздражал на дне IV желудочка (у
кролика) нервные пути, идущие к двигательным ядрам глазных мышц.
Отводя к гальванометру эти мышцы или п. trochlearis, он получал при
одном индукционном ударе два разряда импульсов: один — со
скрытым периодом 0.5—0.8 о, и другой — наступавший не раньше чем
через 1 о. Автор показал, что второй разряд проходит через
промежуточные нейроны, а первый — без их участия, прямо из двигательных
нейронов. Значит, в скрытом периоде второго разряда около 0.5 о
приходится на клетки промежуточных нейронов (рис. 79 и 80).
Специальные исследования, проведенные при усовершенствованной
методике на кошках, показали, что распространение возбуждения в
центральной нервной системе во всех ее отделах, от спинного мозга до
коры включительно, происходит в весьма различные промежутки
времени, но в известных случаях почти без всякой задержки (рис. 84).
Отсюда было сделано заключение, что в спинном мозгу переход
возбуждения от нейрона к нейрону, т. е. от нервных окончаний одного
нейрона на тело клетки другого нейрона, может происходить тем же путем,
что и передача возбуждения от возбужденного участка нервного волокна
к невозбужденному, именно при помощи биоэлектрического тока
возбуждения. В нервных окончаниях, представляющих многочисленные
разветвления с конечными утолщениями, богатыми нейрофибриллярным
веществом, биоэлектрический ток может быть настолько силен, что
будет в состоянии возбудить ту клетку, которую окружают эти окончания
(Леонтович, 20; Беритов, 9).
12 И С. Берптов
178 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Между прочим, установленный Лоренте де Но факт, что на
двигательной клетке может происходить суммирование нервного импульса,
проходящего через синапсы, с подпороговым действием индукцион*
ного удара, в результате чего клетки возбуждаются, является одним из
Хороших прямых доказательств передачи возбуждения с нервного окон-
Рис. 84. Скрытый период электрического эффекта мышцы при
рефлекторной иннервации.
Дедеребрированпая кошка. Струнный гальванометр Эйнтховена. Опыты
после слабого общего отравления стрихнином. Раздражается п.
tibialis d. A — электрограмма индукционного удара. По этому удару
определяется, на сколько времени поднятие электромагнитного сигнала
(указано стрелкой) отстает от начала размыкательного удара. Это
нужно для точного намерения скрытого периода электрического
эффекта; В — электрограмма токов возбуждения сгибателей бедра на
раздражаемой стороне в сгибательном рефлексе; О — электрограмма
возбуждения разгибателей бедра другой ноги в перекрестном разгиба-
тельном рефлексе. Внизу время в 0.01 сек. На электрограмме В токи
возбуждения следуют со скрытым периодом 9.4 о по короткому
рефлекторному пути, а на электрограмме С — сначала очень слабый
эффект (а) по короткому пути со скрытым периодом около 10 о, а
потом большой эффект (Ь) со скрытым периодом в 20.2 а через длинный
рефлекторный путь.
чания на нервную клетку посредством биоэлектрического тока
возбуждения (ом. подробно Экклс, 28).
При нормальной, как и при низкой возбудимости спинномозгового
препарата, для вызова рефлекторной реакции недостаточно раздражать
афферентный нейрон одним индукционным ударом или вообще
одиночным раздражением. Реакция наступает только после ряда раздражений.
Чем чаще и сильнее раздражение, тем скорее она наступает. 'Конечно,
большое значение имеет и функциональное состояние препарата. Время-
реакции тем короче, чем выше возбудимость соответствующего
рефлекторного аппарата. Особенно длительным является время реакции,
когда раздражаются рецепторы. Это зависит от того, что при этом обычно
захватывается небольшое их количество, которое разряжается рядом
импульсов с нарастающей частотой. Первые импульсы, поступающие в
спинной мозг по нескольким чувствительным волокнам, не в состоянии
вызвать рефлекс. Его вызывают последующие и притом более частые
импульсы. Конечно, чем слабее раздражение, тем позднее наступает
рефлекс (Франсуа Франк, 3). В одном опыте Шеррингтона вре-
Распространение возбуждения й торможения в нервной системе 179
мя реакции чесательного рефлекса собаки на слабое раздражение
рецептивного поля равнялось 2.6 сек., а на сильное — 1 сек. (6)
(рис. 85).
Факт зависимости возбуждения спинного мозга и вообще
координирующих аппаратов от числа и ритма периферических импульсов
побудил Л а ті и к а приписать центральной
нервной системе, подобно вегетативной
нервной системе, особую, так называемую
«итеративную» возбудимость. Но, как мы
увидим ниже, «итеративность» центральной
нервной системы основывается на суммации
подпороговых влияний, оказываемых
биотоками возбуждения отдельных синапсов на
нервную клетку. Поэтому мы не находим
целесообразным придерживаться этого
термина Л а п и к а в отношении центральной
нервной системы, подобно тому как мы
отказались применить его в отношении
вегетативной нервной системы.
Время реакции не одинаково для
разных рефлексов. У лягушки сгибательный
рефлекс вызывается тетаническим
раздражением рецептивного поля или
соответствующего чувствительного нерва после более
короткого скрытого периода, чем рефлекс
потирания. Это различие наблюдается,
несмотря на то, что длина рефлекторной дуги
в обоих случаях приблизительно одинакова.
Очевидно, это различие во времени реакции
обусловливается функциональными
особенностями соответствующих координирующих
аппаратов.
Известно также, что у кошки, в ответ
на раздражение мышечных чувствительных
нервов, рефлекс наступает с меньшим
скрытым периодом, чем на раздражение кожных
нервов. Это происходит от того, что, во-первых, чувствительные нервы
от мышечных рецепторов содержат более толстые нервные волокна
(20—12 микронов), чем кожные нервы, а во-вторых, как это мы увидим
ниже, рефлексы от мышечных рецепторов осуществляются через двух-
нейронную дугу путем передачи возбуждения от чувствительного
нейрона прямо на двигательные, а в рефлексах от кожи — через
многонейронную дугу посредством промежуточных нейронов (Ллойд, 50).
На продолжительность времени реакции влияет также длина
нервного пути в пределах спинного мозга. Так, например, на задней ноге
рефлекторная реакция наступает при раздражении передней с большим
скрытым периодом, чем при раздражении задней же ноги
12*
Рис. 85. Продолжительность
скрытого периода рефлекса
чесания в зависимости от
раздражения
Записывается движение всей
задней ноги. Рецептивное поле
чесания раздражают фарадическим
током. Первая ордината на обеих
кривых отмечает начало
раздражения, т вторая ордината — конец
его. В верхнем опыте раздражение
было сильнее, чем в нижнем. В
соответствии с этим в первом случае
скрытый период короче (I сек.), во
втором—длиннее (2.6 сек.),
интенсивность же рефлекса и его
последствие в первом случае больше,
чем во втором. Время — 0.2 сек.
(Шеррингтон).
180 общ\я физиология спинного мозга и ствола головного мозга
Распространение возбуждения по длинным нервным путям через
головной мозг. У нормальных позвоночных животных периферические
раздражения вызывают через головной мозг реакции, носящие
характер поведения. В этом случае спинной мозг со своими координирующими
и двигательными нейронами приходит в активное состояние через
длинные рефлекторные пути, проходящие через головной мозг. Даже у
лягушки при слабых периферических раздражениях конечности защитная
реакция убегания наступает без каких-либо локальных реакций из
спинного мозга. Эго явление было отмечено еще Сеченовым; он же
объяснил его как результат более высокой возбудимости рефлекторных
центров головного мозга (14). У высших позвоночных животных
проведение возбуждения по коротким нервным дугам более затруднено, чем
у лягушек, а потому роль длинных дуг через головной мозг еще более
значительна в рефлекторных реакциях животного. Можно утверждать
вместе с Шеррингтоном и его сотрудниками, что «у более высоко
стоящих животных супраспинальные факторы координации захватили
ту часть функций, которая у более низко стоящих животных падает,
главным образом, на долю спинного мозга» (15). Вследствие этого у
нормальных и децеребрированных кошек, собак, обезьян
периферическое раздражение должно вызывать рефлекторную реакцию по
преимуществу через головной мозг, а значит, и с большим скрытым периодом.
Это и наблюдается на нормальных и децеребрированных кошках в
условиях эксперимента. Раздражение чувствительных нервов вызывает
в мышцах электрическую реакцию с длительным скрытым периодом.
Иногда этому сильному эффекту предшествует небольшой эффект,
слабое активное состояние мышц, происходящее при участии
коротких рефлекторных дуг. Эти двойные и одиночные эффекты наблюдались
и при общем стрихнинном отравлении животного после значительного
повышения возбудимости в центральной нервной системе (рис. 84).
Время реакции второго сильного эффекта для передних ног много
меньше (в среднем 16.3 о), чем для задних (в среднем 23.1 о). Интервал
между первым и вторым эффектом и в первом случае также был короче
(в среднем 8.3 о), чем во втором (в среднем 12 о). Двойные эффекты
отсутствовали при раздражениях коры мозга и на спинномозговых
препаратах, но наблюдались на децеребрированных препаратах (Беритов, 9).
Второй импульс с длинным скрытым периодом мы считаем
результатом возбуждения длинных нервных путей через головной мозг, в то
время как первый импульс мы признаем прошедшим через короткий
нервный путь на уровне того отдела спинного мозга, куда вступают
возбужденные чувствительные волокна. На децеребрированных препаратах
первый импульс нередко наступал через такой длинный скрытый
период, какой точно соответствовал длине нервного пути через головной
мозг. Это получалось чаще всего в перекрестных рефлексах после
стрихнинного отравления. В таких случаях, очевидно, эффект наступал
на противоположной стороне прямо из средних отделов мозга, а именно —
из тонических центров. Как известно, перекрестный разгибательный
рефлекс на децеребрированных препаратах носит тонический характер.
РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ В НЕРВНОЙ СИСТЕМЕ 181
Электрический эффект — ряд мелких электрических колебаний,
вызванных на разгибателе противоположной стороны,—также носит
характер тонического рефлекс». Очевидно, разница скрытых периодов
первого и второго импульсов приходится на разницу длины короткого и
длинного нервных путей. Измерив длину нервных путей, мы высчитали,
что в определенных случаях возбуждение проводится в длинных
нервных путях через головной імозг со скоростью 70—80 >м в 1 сек., как
в нервных волокнах, т. е. без задержки. В других же случаях это
проведение происходит с небольшим замедлением в несколько сигм, с
падением общей скорости распространения возбуждения до 36 м в 1 сек.
Это замедление должно происходить- благодаря тому, что в
нисходящем пути каждый участвующий нейрон, в особенности двигательный,
возбуждается под влиянием ряда импульсов, притекающих к нему
последовательно через одни и те же или разные нервные окончания.
На основании приведенных фактов мы предполагаем, что наиболее
быстрая передача возбуждения в длинных нервных путях головного и
спинного мозга от окончания одного нейрона на тело клетки другого
может происходить тем же путем, что и в нервном волокне, т. е. при
помощи биоэлектрических токов возбуждения.
Происхождение удлиненного скрытого периода рефлекторных
реакций. До самого последнего времени общее мнение физиологов было
таково, что замедление в проведении возбуждения в центральной
нервной системе происходит при передаче возбуждения от одного нейрона
к другому в синаптических областях. Некоторые авторы (Шеррингтон
и др.) предполагали, что в этих областях возбуждение
распространяется не по тому принципу, как в нервном волокне. Возбуждение,
дошедшее до нервных окончаний, производит в синапсах определенные
физико-химические изменения, которые, в свою очередь, раздражающе
действуют на нервную клетку. На эти процессы, по их мнению,
требуется больше времени, чем на проведение возбуждения в нервных
волокнах. Но, как указывалось выше, по'новейшим исследованиям,
возбуждение может передаваться с одного нейрона на другой почти без всякой
задержки. Следовательно, задержка не обусловливается
специфическими особенностями синаптического механизма. Минимальная задержка
в пределах 0.22—0.5 о, наблюдающаяся при точных измерениях,
должно быть, обусловливается тем, что конечное ветвление нервного
волокна, образующее синапсы, является голым, чрезвычайно тонким
образованием, а потому должно проводить возбуждение с очень большим
замедлением.
Как указывалось выше, даже при нормальном функциональном
состоянии центральной нервной системы могут иметь место
рефлекторные реакции с более мли менее длительным скрытым периодом. Но
происхождение этого явления уже иное. Центральные нервные клетки
как промежуточных координирующих, так и двигательных нейронов не
всегда возбуждаются одиночными нервными импульсами. Можно
предположить, что разность потенциалов, возникающая при возбуждении
сина/птических окончаний, не всегда достаточна для возбуждения нерв-
|52 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
ной клетки, несмотря на то, что каждое нервное волокно
заканчивается на одной и той же клетке целым комплексом синапсов, которые в
общем должны дать при возбуждении более высокую разность
электрических потенциалов, чем нервное волокно. Так, например, рефлекс
потирания на нормальной лягушке нельзя вызвать раздражением
чувствительного нерва от соответствующего рецептивного поля отдельными
индукционными ударами. Даже сгибательный рефлекс не всегда
удается вызвать с нерва такими ударами, следовательно, одиночные
периферические импульсы возбуждения не в состоянии возбудить
координирующие аппараты этих рефлексов. Значит, при обычной возбудимости
центральной нервной системы каждая нервная клетка приходит в
активное состояние только после воздействия ряда импульсов со стороны
нервных окончаний. И, конечно, чем ниже возбудимость нервных
клеток, тем больше должен быть ряд действующих на них импульсов,
чтобы вызвать в них возбуждение.
При определенных условиях легко удается вызвать рефлекторную
реакцию раздражением чувствительного нерва отдельными
индукционными ударами. Это случается при большой силе индукционного удара
и при высокой возбудимости центральной нервной системы. Это,
например, наблюдается после операции в результате раздражающего
действия со стороны перерезанных нервов или при стрихнинном
отравлении. Но и в этих случаях скрытый период, рефлекторной реакции
очень часто оказывается удлиненным.
Происхождение удлиненного скрытого периода рефлекторной
реакции при одиночных раздражениях зависит, прежде всего, от того, что
возбуждение проводится в чувствительных волокнах с самой разной
скоростью: например, у теплокровных — от 10 до 100 м в 1 сек. Кроме
того, задн?корешковые волокна спинного імозга, в связи с ответвлением
коллатералей, становятся тоньше. Сообразно должно происходить
падение скорости проведения возбуждения, как это установлено для
двигательных волокон нервных стволов (Эрлангер и Гассер, 27).
Следовательно, даже при раздражении нервного ствола одним
индукционным ударом центральная нервная система получит
периферические импульсы через разные промежутки времени, т. е. залп
последовательных импульсов. В таком случае возбуждение промежуточных
нейронов может быть обусловлено действием ряда периферических импульсов.
В клетках центральной нервной системы происходит не только
временная, но и пространственная суммация. Периферические
импульсы приходят к одной и той же клетке через разные промежутки
времени, и притом к разным участкам. Это последнее обстоятельство,
повидимому, является необходимым условием для возбуждения клетки.
Первые импульсы через соответствующие синапсы производят только
местный процесс, подобно тому как бывает в нервном волокне при под-
пороговых раздражениях. Этот локальный процесс должен
ограничиваться очень небольшим участком, ибо он распространяется с очень
большим декрементом. Но благодаря близкому расположению синапсов
РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ В НЕРВНОЙ СИСТЕМЕ 18&
разных нервных волокон такой локальный процесс, возникший под
действием импульса через один синапс, облегчает реагирование клетки на
импульсы через другие ближайшие синапсы и тем самым обусловливает
пространственную суммацию (Лоренте де Но, 35) (подробно о
суммации см. в следующей главе).
В результате временной и пространственной суммации локальных
процессов происходит возбуждение клетки, которое затем
распространяется и на аксон. Но локальный потенциал, возникающий в клетке,
как указывалось выше, может достигнуть такой интенсивности, что
производит многократное возбуждение аксона. Это проявляется
совершенно ясно, когда изучают электрический эффект в глубине мозга или
переднего корешка вблизи мозга. Обычно вслед за каждым заднеко-
решковым биотоком возбуждения наступает медленный потенциал,
выражающий собой местный клеточный процесс. В некоторых опытах
на лягушке от начала заднекорешкового биотока, возникающего
внутри мозга, до начала медленного потенциала проходит всего 2 о, т. е.
локальный процесс возникает почти без задержки. Быстрые
потенциалы возбуждения двигательных волокон появляются значительно
позднее — через 10—20 о и больше. Они располагаются на
нисходящем колене первого медленного потенциала или последующих
медленных потенциалов. Лишь в условиях высокой возбудимости
мозга или при сильных раздражениях импульсы из заднекорешковых!
волокон или из промежуточных нейронов оказываются в состоянии
возбудить клетку без предварительного медленного потенциала. Из
этих фактов следует, что задержка в передаче импульсов возбуоюде-
ния от нейрона к нейрону происходит каждый раз, когда возбуждение
аксона клетки осуществляется не непосредственным воздействием
синаптических биотоков возбуждения на клетку, а вследствие
медленного потенциала, возникающего в клетке в результате временной
и пространственной суммации локальных процессов.
Биологическое значение того явления, что рецепторы всегда
отвечают рядом импульсов возбуждения и что нервные цепочки обладают
тем же свойством, заключается именно в том, что нормально
промежуточные и двигательные нервные клетки могут быть возбуждены
лишь серией периферических импульсов. Каждая нервная клетка в
мозгу должна подвергнуться одновременному или разновременному
воздействию множества синаптических импульсов, бомбардировке ими,
чтобы она сама или ее аксон возбудились и тем вызвали
периферический эффект. Этим и объясняется обычно наблюдаемое длительное
запаздывание в проведении возбуждения в спинном мозгу.
Гипотеза о гуморальной передаче возбуждения. Положение о
передаче возбуждения с нервного окончания на тело клетки с помощью
биотоков возбуждения разделяют не все физиологи. В настоящее
время некоторые физиологи держатся того мнения, что эта передача
осуществляется химическим путем. Предполагается, что возбужденные
нервные окончания выделяют какое-то активное вещество, которое
потом действует раздражающе на тело клетки. В основе этой гипо-
184 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
тезы химической, или гуморальной, передачи лежат разнообразные
наблюдения,— прежде всего то, что в центральной нервной системе
образуется вещество, обладающее свойством повышать возбудимость.
Такое вещество было впервые обнаружено Габерландоім (16) в
экстрактах разных отделов мозга. В последнее время очень точные
опыты были поставлены в этом направлении в лаборатории Б ы-
кова (38). Раздражали центральный конец блуждающего и
симпатического нервов кошки и собирали кровь, вытекающую в это время
из мозга. Затем эту кровь вводили обратно в мозг через сонную
артерию. Это вызывало такое же изменение дыхания, как и раздражение
названных нервов.
Аналогичный опыт был поставлен с п. depressor. Кровь, собранная
при раздражении этого нерва, вызывала такое же падение кровяного
давления, как и раздражение нерва.
Аналогичные опыты были произведены сотрудниками Быкова —
Горшковой и іКурциныім (39) на перфузируемой рыбьей
голове. Перфузионная жидкость, оттекающая от головы при
раздражении центрального конца блуждающего нерва, действует на сердце
лягушки, задерживая его деятельность. Перфузат же, собранный во
время раздражения тройничного нерва, производит чисто
симпатический эффект. Эти же перфузаты характерным образом влияют на
кровяное давление и на движение тонкой и толстой кишок
(Быков, 37, 88).
Затем было исследовано действие веществ, образующихся в
головном мозгу, на возбудимость самого головного мозга. С этой целью
устраивалось перекрестное кровообращение у двух собак. Головной
мозг одной собаки раздражался, а у другой в это время
исследовалась хронаксия коры большого мозга. Последняя укорачивалась через
10—25 мин. после началаі раздражения мозга. Тот же результат
получался, если у первой собаки были перерезаны спинной мозг и
оба блуждающих нерва. Отсюда ясно вытекает, что в мозгу собаки
при возбуждении образуются какие-то вещества, которые повышают
возбудимость мозга. Так как уменьшение хронаксии в коре мозга
наблюдается при инъекции ацетилхолина и не наблюдается при
инъекции адреналина и питуитрина, авторы заключают, что в
головном мозгу при его деятельности образуются ацетилхолиноподобные
вещества (Бабский с сотрудниками, 34). Они же обнаружили, что
кровь, вытекающая из головного мозга при его судорожной
деятельности, повышает возбудимость спинного мозга (18).
Аналогичные исследования производились в лабораториях Р а-
зенкова и Штерн. Разенков с сотрудниками (23)
установил выделение активных веществ мозгом собаки и лягушки не только
при искусственном раздражении, но и при естественном возбуждении
раздражением чувствительных нервов. Они установили также сходство
этих веществ с ацетилхолином. Однако в этих опытах активное
вещество мозга вызывало не только повышение функциональной
деятельности сердца и сосудистой системы, но и понижение ее (Блинова, 22).
РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ В НЕРВНОЙ СИСТЕМЕ 185
Штерн и ее сотрудники (21) находят, что активные вещества,
образующиеся в головном мозгу при его деятельности, так
называемые метаболиты, распространяются не только через кровь, но
и через спинномозговую жидкость. Эти активные вещества оказывали
на возбудимые ткани (сердце, спинной мозг, меланофоры) иногда
однородное, иногда разнородное действие. Деятельность мозга
вызывалась как его искусственным раздражением (электрический ток,
гистамин, пептон), так и естественным — «болевыми» раздражениями
чувствительных нервов и эмоциями. В образовании этих веществ
большую роль приписывают гипофизу, а в поступлении этих веществ в
кровь — гемато-энцефал'ическому барьеру (Цейтлин, 24; Данилов, 25).
Одни авторы склонны рассматривать приведенные выше
результаты как доказательство химической передачи возбуждения,
предполагая, что активные вещества вроде ацетилхолина выделяются
нервными окончаниями — синапсами. По их мнению, эти вещества, с
одной стороны, действуют на нервные клетки, вызывая в них
возбуждение, а с другой, распространяясь через кровь, производят
функциональные сдвиги в разных отделах мозга (Быков, 37; Кэнкон, 26; Разен-
ков, 23). Другие авторы доказывают, что эти активные вещества
возникают во всей нервной системе как продукты обмена, как
метаболиты; они оказываются активными, но никакого специфического
значения им не придается. Правильнее всего допустить, что всякого
рода активные продукты обмена веществ центральной нервной
системы, так называемые гуморы нервных элементов, играют лишь
некоторую роль в изменении общего функционального состояния
центральных и периферических нервных образований.
В связи с возникновением идеи о химической передаче возбуждения были
исследованы экстракты мозга в отношении содержания в них активных веществ. В
опытах, произведенных в этом направлении, авторы действительно находили в мозгу
ацетилхолшюподобное вещество (Лёви, 3J2; Майн, Тенненбаум и Квостел, 33). После
этого было установлено большое влияние одновалентных и двувалентных катионов,
а также эфира, хлороформа и других веществ на образование в мозгу ацетилхолина
(Стедман, 42). Так, например, ионы калия повышают образование и выход
ацетилхолина из живых нервных клеток, а именно, от 1.5 до 17 у на 1 г веса бычьей
мозговой ткани. Ионы же кальция, наоборот, тормозят образование и выход
ацетилхолина (Манн и сотр.; 33). Эти опыты также приводят к заключению о роли
ацетилхолина в нервной деятельности.
В то же время исследование содержания ацетилхолина в чувствительных нервах
показало, что в этих нервах его содержится чрезвычайно мало или он совершенно
отсутствует. Так, по опытам Ч а н г а и его сотрудников (44), чувствительные иервы
собаки содержат в 50 раз меньше ацетилхолина, а лягушки — в б раз меньше, чем
двигательные нервы тех же животных. Также по опытам Лиссака и Пастора
(45) ацетшіхоли'Н содержится в чувствительных не>рва<х собаки и кошки (п. opticus и
п. saphenus) в значительно меньшем количестве, чем в двигательных нервах. Л ё в и
(33) совсем не нашел его в зрительном нерве и заднекорешковых волокнах быка.
На этом основании нужно отрицать возможность химической передачи возбуждения
с этих нервов в центральной нервной системе. Если в афферентных волокнах нет
ацетилхолина или его очень мало, трудно допустить возможность выделения его их
окончаниями в количестве, возбуждающем нервные элементы мозга.
186 общая физиология спинного мозга и ствола головного мозга
Кроме того, известно, что образование ацетилхолина в эмбриональной ткани
цыпленка достигает максимума уже на 3-й день инкубации, когда еще нет развитой
нервной системы и нет способности к рефлекторным движениям. Это указывает на
то, что ацетилхолин — продукт метаболизма и не связан со специфической
деятельностью центральной нераной системы (Цинг Йяиг-Куо, 40).
Наконец, было исследовано действие ацетилхолигна, как наиболее вероятного
передатчика возбуждения в центральной нервной системе, путем искусственного
введения его в мозг. У одних авторов при малых концентрациях он производит сначала
повышение, затем понижение рефлекторной возбудимости (Бабский и сотр., 34), у
других — только торможение рефлекторной деятельности (Швейтцер и Райт, 41).
Внутривенное или внутриартериальное (мозговые артерии) введение ацетил>о-
лина и эзерина усиливало дыхательные движения без нарушения их координации;
введение же атропина оказывало на эти движения противоположное действие (Ге-
зел и сотр., 47, 48). При локальном приложении ацетилхолина ко дгау IV желудочка
кровяное давление понижалось, а дыхание усиливалось {Кэннон, 26; Гезел и сотр.,
47). На этом основании одіви авторы заключают о гуморальной передаче возбуіжд?-
ния (Кэннон и сотр.; Гезел и сотр ), а другие считают, что ацетилхолин вызывает
только определенные функциональные едшпи (Бабский и сотр., 34).
Гипотеза о химической, или гуморальной, передаче возбуждения в
центральной нервной системе совершенно не считается с
общеизвестными фактами общей физиологии. Так, например, мы выше
указывали, что в спинном и головном мозгу возбуждение распространяется
очень быстро, лишь с небольшой задержкой, а то и без задержки,
как в периферических нервных волокнах; далее, ритм раздражения
чувствительного нерва свободно может передаваться через спинной
мозг на скелетную мышцу с частотой до 150—350 в 1 сек. Все эти
факты таковы, что их совершенно нельзя согласовать с гипотезой о
возникновении возбуждения в центральной нервной системе путем
выделения каких-либо активных химических веществ.
Одним из важных фактов, говорящих против химической передачи
возбуждения, нужно считать отношение центральной нервной системы
к ацетилхолину — предполагаемому передатчику возбуждения. Это
вещество, будучи локально приложено в растворе от 1 : 100 000 до 1 : 500 к
спинному или головному мозгу лягушки, не действует раздражающим
образом, не вызывает рефлекторных двигательных реакций. Оно только
при некоторых малых концентрациях повышает возбудимость, а при
больших концентрациях, как 1 : 500 — 1 : 2000, наоборот, понижает ее
(Беритов и Нивинская, 46). Действие ацетилхолина на спинной імозг
кошки более интенсивно при тех же концентрациях: оно не только
меняет возбудимость, но, сверх того, производит рефлекторные
реакции. Именно, на препаратах, приготовленных под эфирным наркозом,
ацетилхолин в малых концентрациях вызывает общее торможение, а
в больших концентрациях — сначала кратковременное торможение, а
потом обычную реципрокную реакцию возбуждения одних мышц и
торможения других. Одновременно значительно повышается
рефлекторная деятельность отравленного участка мозга (рис. 44 и 45). Но
это действие ацетилхолина не специфично. Аналогичные изменения
наблюдаются при приложении других активных веществ, как КС1 и
никотин, а также при механическом раздражении мозга, как прове-
РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ В НЕРВНОЙ СИСТЕМЕ 187
дение мягкой кисточкой по его поверхности (рис. 46). Слабое
воздействие производит общее торможение, а сильное — торможение и
возбуждение вместе (Беритов и Бакурадзе, 43).
Рис. 86. Явление адаптации спинного мозга к ацетилхолину.
Лумбальный препарат кошки. Регистрируются m. semitendinosus (S. d,,S* е.) и quadriceps (Q. d.»
Q. s.) с обеих сторон. Раздражением правого V заднего корешка одиночными индукционными
ударами вызываются рефлекторные вздрагивания мышц правой стороны» а таким же раздражением
левого VII эаднего корешка — левой. Ацетилхолин в растворе 1 : 2000 прикладывается два раза
с интервалом в 80 сек. к VI сегменту с правой боковой поверхности. Продолжительность
прикладывания отмечена на нижней сигнальной линии. Первый раз ацетилхолин вызвал повышение
рефлекторных вздрагиваний обеих мышц правой стороны, а второй раз не произвел никакого эффекта.
Сам по себе ацетилхолин не дает рефлекторных движений, ибо препарат был приготовлен под хлор-
алозовым наркозом (Беритов и Бакурадзе).
Далее, ацетилхолин, действуя на мозг, не может вызывать
длительно продолжающиеся эффекты: как торможение, так и
возбуждение продолжаются в течение десятка секунд, редко до 1 мин; Затем
он перестает активно действовать,— иначе говоря, спинной мозг
быстро адаптируется к нему. В это время раздражение задних
корешков и какое-либо другое химическое раздражение мозга вызывают
рефлекторные реакции совершенно без изменения (рис. 86). Аналогич-
188 общая физиология спинного мозга и ствола головного мозга
ное действие оказывают никотин и КС1. Оба они не способны
вызывать длительный рефлекторный эффект, но спинной мозг адаптируется
к ним в разной степени: на очень короткое время — к КС1 и на очень
долгое — к никотину.
Наконец, в тех случаях, когда операция производилась под хлор-
алозным наркозом, ни ацетилхолин, ни другие химические
раздражители, как К61 и никотин, при локальном приложении к мозгу не
вызывают рефлекторных реакций. Чувствительность препаратов сама по
себе высока. Они дают спонтанные вздрагивания, а также сильные и
длительные рефлекторные реакции при раздражении задних
корешков. Эти химические вещества только меняют рефлекторную
возбудимость к импульсам чувствительных нервов: в малых концентрациях
они ее понижаюг, а в больших — сначала понижают, а потом
повышают. Отсутствие двигательных эффектов на ацетилхолин, в то время
как периферические импульсы производят сильные и длительные
эффекты, далее, обязательность общей тормозящей фазы в самом
начале действия ацетилхолина, наконец, быстрая адаптация мозга к
химическим раздражителям — лучше всего свидетельствуют о том, что
периферические нервные импульсы производят в центральной нервной
системе координированные процессы возбуждения и торможения без
посредничества ацетилхолина или какого-либо другого химического
вещества (Беритов и Бакурадзе, 43).
Распространение торможения в центральной нервной системе. Мы
предположили выше, что центральное торможение представляет собой
функцию нейропиля и что оно заключается в анэлектротонич?ском
действии медленных токов нейропиля на промежуточные и
двигательные элементы. Но так как это электротоническое действие происходит
сейчас же, как только начинается активное состояние нейропиля, то
торможение двигательных клеток іможет наступить еще раньше,
чем они разрядятся собственными импульсами под влиянием
синоптической импульсации. Этим в значительной мере должно объясняться
то явление, что рефлекторное возбуждение, вызванное одиночным
раздражением одного рецептивного поля, может быть заторможено
таким же раздражением другого поля, даже в том случае, если
последнее происходит спустя несколько сигм после первого,
возбуждающего раздражения ('Киселев, 29; Фултон,, 30). Но если
опережение значительно больше — достигает 10 о, оно должно зависеть от
другого обстоятельства (Гедевани, 31). Как указывалось выше, при
раздражении мышечно-сухожильных, тактильных и Холодовых
рецепторов, а следовательно, и при раздражении соответствующих им
чувствительных волокон, спинной мозг реагирует преимущественно общим
торможением. Общее торможение без двигательных реакций можно
легко получить при слабых раздражениях почти любого
чувствительного нерва. Все эти факты указывают на то, что общее торможение в
очень интенсивной форме легко получается при возбуждении толстых
нервных волокон с высокой возбудимостью. А эти волокна, как
известно, проводят возбуждение со скоростью, в несколько раз пре-
РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ В НЕРВНОЙ СИСТЕМЕ Js9
восходящей скорость проведения тех тонких волокон, которые
проводят возбуждение от болевых рецепторов и обусловливают
оборонительные рефлекторные реакции. Отсюда следует, что при
периферических раздражениях, вызывающих рефлекторные двигательные реакции,
активное состояние нейропиля под влиянием толстых афферентных
волокон может наступать в спинном мозгу намного раньше, чем
возбуждение двигательных клеток по тонким волокнам и, соответственно,
общее торможение возникает в спинном мозгу гораздо раньше
координированной иннервации, соответствующей оборонительной реакции.
Однако есть фактическое основание для утверждения, что через
любое нервное чувствительное волокно торможение может возникать
в спинном мозгу раньше возбуждения. Это утверждение вытекает из
установленного положения, что медленные локальные процессы,
возникающие в клеточной и дендритной массе под влиянием афферентных
импульсов, производят торможение с момента их возникновения.
А как известно, медленные потенциалы возникают тотчас же после
воздействия заднекорешковых импульсов через синапсы на клетки и
на дендриты; их скрытый период, если считать от заднекорешкового
импульса, редко равняется 1 о и больше. Возбуждение же клетки или
аксона, когда оно происходит под влиянием медленных потенциалов
клетки, наступает через 10—20 о после заднекорешкового импульса.
Отсюда ясно, что торможение может предшествовать возбуждению на
10—20 о даже при их возникновении под влиянием возбуждения
одного афферентного нейрона.
Распространение активного состояния в самом нейропиле, путем
перехода локального процесса от одного элемента нейропиля к
другому, очевидно, происходит с очень сильным декрементом, ибо, как
указывалось выше, тормозящее действие, равно как длительное
электрическое колебание того или другого активного участка нейропиля,
тем сильнее, чем ближе он расположен к первично активированному
участку. Однако в определенных случаях торможение может
проявиться и в участках мозга, отдаленных от первично возбужденного
участка нейропиля. Например, при раздражении передней конечности
или ротовой полости тормозятся двигательные элементы задней
конечности. Это явление может обусловливаться не распространением
торможения непосредственно по нейропилю, а активированием отдаленных
участков нейропиля коллатералями возбужденных чувствительных и
промежуточных нейронов.
Точно также общее торможение, наблюдаемое при раздражении
головных рецепторов или вообще при возбуждении головного мозга,
происходит не благодаря распространению торможения по
нейропилю всего мозга или нейропильного биотока по всему мозгу. Это
общее торможение наступает оттого, что в головном мозгу приходит
в активное состояние нейропиль. Это тормозит передаточные станции
и нисходящие пути и тем самым прекращает рефлекторные реакции,
вызываемые через головной мозг.
]§Q ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТЬОЛА ГОЛОВНОГО МОЗГА
Итак, действие торможения более или менее локально и
ограничивается тем районом моЗга, где оно возникло первично под влиянием
приходящих импульсов возбуждения. Если, все же, торможение
является общим, охватывающим почти всю центральную систему, то связано
это с тем, что торможение наступает также во всех тех участках мозга,
куда приходят импульсы возбуждения по проводящим путям.
ТГраппгипум
Наблюдение скрытого периода рефлекторной реакции. Спинномозговая лягушка
Берут сгибатель m. semiltendinosus для записи «а кимографе и п. peroneus —. для
вызова сгибатального рефлекса. Для обозначения момента раздражения употребляют
двуплечий ключ Гельмгольца. Одно плечо его имеет такое же назначение, как ключ
Дюбуа Рэймона, когда он вводится в цепь индукцжшной катушки Второе плечо
введено в особую гальваническую ц?пь, в которую включен электромагнитный сигнал
П?ро сигнала пишет по той же вертикальной линии, как миограф мышцы. До
раздражения это плечо открыто — значит, данная цепь разомкнута, а первое плечо
закрыто — значит, вторичная катушка замкнута на себя. Если закрыть плечо, к которому
присоединен сигнал, последний сделает отметку на кимографе; в то же время
откроется другое плечо и произойдет раздражение нерва. Таким образом, начало
раздражения обозначится на кимографе Время от начала раздражения до начала
рефлекторного сокращения составляет скрытый период рефлекторной реакции. Отсюда
нужно вычесть время, необходимое на распространение возбуждения по нервам и
для возбуждения мышцы. Скрытый период сокращения мышцы можно узнать неио
средственно из опыта: на том же препарате перерезать поясничное сплетение и затем
записать эффект раздражения периферического отрезка таким же образом, как
рефлекторный эффект.
Изменчивость скрытого периода рефлекторной реакции. Записывают на
кимографе рефлекторное сокращение при разных силах раздражения, при разных
температурах, до утомления и после него. При малой силе (раздражения, при утомлении,
при охлаждении скрытый период будет велик; наоборот, при большой силе, при
согревании « после отдыха он будет мал.
Литератур а
!. Helmholtz Н., Benchte uber d. z. Bekannttnach-ing geeign. Verh. K- Preuss.
Akad. Wiss. Berlin, стр. 328, 1845.
2. Buchannan Fh, Quart. J. Exp. Physiol., 1, 1, 1908
3. Franco ls-Fr an k, Le?ons sur les fonctions motrices du cerveau, Pans, 1887.
4. Rosenthal J., Mschr. K- Preuss. Akad. Wiss., Berlin, 1873, стр. 104
5. Wundt W. Untersuchungen z. Mechanik d. Nerven u. Nervenzentren, 2. Abt.,
Stuttgart, 1876.
6. Sherrington C, Int. act. nerv. syst., стр. 20, 1906.
7. Forbes A. a. Gregg A., Am. J. Physiol., 37, 159, 1915.
8. Jolly W. A., Proc Roy. Soc. В., 92, 31, 1920.
9. Bent of f J., Am. J. Physiol , 90, 281, 1929; Тр. Физиол. ин-та им. Бериташвили,
3, 87, 1937.
10. Bent off J., Z Biol , 64, 286, 1914.
11. Lorente de No, Am J. Physiol., Ill, 272, 1935.
12. Eccles J. С a. Sherrington С S.t Proc. Roy. Soc. В., 107, 511, 1931.
13. Leese Ch. E. a. E in arson L., Am. J. Physiol., 109,296, 1934.
14. Сеченов И* М, Физиология нервных центров, 1891.
РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ В НЕРВНОЙ СИСТЕМЕ 191
15. Шеррингтон и сотр. Рефлекторная деятельность спинного мозга, 1935,
16. Haberlandt L., Munch. Med. Wschr., 1929, №30, стр. 1290; Pfl. Arch., 223,
171, 1929; 224, 297, 1930; 226, 781, 1931.
17. Быков К. Мм Доклад на XV Междунар. физиол. конгр., 1935. Тез. сообщ.,стр. 52.
18. Бабский Е. Б. и сотр., Доклад на XV Междунар. физиол. конгр., 1935. Тез.
сообщ., стр. 18.
19. К э н н о н Б. Б., Доклад на XV Междунар. физиол. конгр., 1935.
20. Леонтович А., Биол. журн., 2, 259, 1933.
21. Штерн Л. и сотр., 66. докладов VI Всес. физиол. съезда, стр. 15.
21а. Р о с и н Я. А., Сб. докладов VI Всес. фивиол. съезда, стр. 200.
22. Б л и н о в а А., Сб. докладов VI Всес. физиол. съезда, стр. 206.
23. Р а з е н к о в И. П., Сб. докладов VI Весе, физиол. съезда, стр. 10.
24. Ц?йтлинС. М,, Сб. докладов VI Всес. физиол. съезда, стр. 216.
25. Данилов А. А., Сб. докладов VI Всес. физиол. съезда, стр. 257.
26. Cannon W. В. a. Rosenblueth A., Auton. nerve-effector system, 1937.
27. Erlanger a. Qasser, Electr. signs of nerv. activity, 1937.
28. E с с 1 e s J. C.t Erg. Physiol., 38, 339, 1936.
29. Киселев M., Казанск. мед. вестн., № 4—5, стр. 389, 1931.
30. F u 11 о n J. F., Muse, contract, a. refl. control of movement, стр. 218—320, 1927.
31. Гед?вани Д., Тр. Физиол. ин-та им. Бериташвили, 3, 479, 1937.
32. L о ? w і О. и. Н е 11 а и е г Н., Pfl. Arch., 240, 769, 1938.
33. М a n n P.J.C.Tennenbaum М. a. Q u a s t е 1 J. H., Bioch. J., 32, 243,
1938; 33, 822, 1939.
34. Б а б с к и й Е. Б., Сб. докладов VI Всес. физиол. съезда, стр. 189; Учен. зап.
каф. физиол. Моск. пед. ин-та, 1938.
35. L о г е n t e d е N 6 R., J. Neurophysiol., 2, 402, 1939.
36. L 1 о у d D. Р. С, J. Neurophysiol., 6, 293, 1943.
37. Б ы к о в К. М., Сб. докладов VI Всес. физиол. съезда, стр. 186, 1937.
38. Б ы к о в К. М., Опыты исследований нервно-гуморальной связи, III, 3, 1937.
39. Горшкова С. йКурцин И., Опыт исследования нервно-гуморальных
связей, III, 61, 1937.
40. Zing Yang-Kuo, J. Neurophysiol., 2, 488, 1939.
41. S с h w e i t z e г A. a. W r i g h t S., J. Physiol., 89, 165. 1937.
42. S t e d m a n E. u. Stedman El., Bioch , J., 33, 811, 1939.
43. Беритов И. иБакурадзеА., Acta Medica USSR, 2, 371, 1939; Физиол.
журн. СССР, 28, 3, 1940.
44. Chang H. С, Hhieh W. M., Lee L. Y., L i a T. H., Lim R. K. S.,
Chin. J. Physiol., 14, 19, 1939.
45. Lissak K. u. Pastor J.,Pfl. Arch., 224, 124, 1940.
46. Беритов И. и Нивинская О., Тр. Ин-та физиол. им. Бериташвили,
4, 39, 1947,
47. G e s е 11 R. a. Hansen Е. Т., Am. J. Physiol., 139, 371, 1943.
48. G e s e 11 R., H a n s e n E. T. a. W о r z n i a k J. J., Am. J. Physiol., 138,
776, 1943.
49. Grundfest H. а. С a m p b e 11 В., J. Neurophysiol., 5, 275, 1942.
50. L 1 о у d D. P. C, J. Neurophysiol., 4, 525, 1941.
51. La pic que L., L'excitabilite iterative. Paris, 1936.
52. Беритов И., Бюлл. эксп. биол. и мед., 22, вып. 6, 1946.
Глава X
ЯВЛЕНИЕ ОБЛЕГЧЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ
Общее понятие. Всякое эффективное раздражение, вызывающее
рефлекторную реакцию, производит повышение возбудимости в
центральной нервной системе. При повторных раздражениях возбудимость
повышается особенно сильно. О повышении возбудимости в рефлекторном
аппарате мы заключаем на основании понижения порогов раздражения
соответствующего рецептивного поля или соответствующего
чувствительного нерва, а также на основании усиления периферических
двигательных реакций или электрических реакций самого мозга при
повторении того же раздражения.
Повышение возбудимости в центральной нервной системе при ее
рефлекторной деятельности от одного залпа периферических импульсов
длится различное время, в пределах от нескольких десятых секунды до
нескольких секунд. При тетаническом раздражении и вообще под
влиянием ряда периферических импульсов оно может длиться гораздо
больше— в течение многих минут. Это длительное повышение возбудимости
в центральной нервной системе нельзя рассматривать как свойственную
всякой возбудимой системе супернормальную возбудимость,
наступающую вслед за рефракторными фазами. Такая фаза повышенной
возбудимости имеется и в центральных элементах. Так, например, Лоренте
де Но (11) наблюдал в двигательных клетках кролика тотчас же после
рефракторных фаз кратковременное повышение возбудимости,
длительностью в 7—20 о; в редких случаях оно продолжалось до 80 о.
Вероятно и в клетках промежуточных нейронов продолжительность
супернормальной фазы представляет величину того же порядка. Длительное же
повышение возбудимости, измеряемое секундами и минутами,
безусловно имеет иное происхождение. Этот длительный период повышенной
возбудимости в центральной нервной системе английский физиолог Грэим
Б р о у н (15) назвал явлением облегчения (the phenomenon of
facilitation). Он имел в виду то обстоятельство, что в этот период как
возникновение, так и проведение возбуждения в значительной мере
облегчены Мы воспользуемся этим термином для обозначения указанного
длительного повышения возбудимости в отличие от кратковременной
ЯВЛЕНИЕ ОБЛЕГЧЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ 193
супернормальной фазы, наблюдающейся вслед за рефракторными
фазами.
Явления облегчения после однократного периферического
раздражения. Явление облегчения в центральном рефлекторном аппарате
после одного залгпа периферических импульсов длится от десятых долей
секунды до нескольких секунд. Оно не охватывает данного
рефлекторного аппарата целиком. Оно обнаруживается главным образом по
отношению к определенным мышцам, участвующим в данной
рефлекторной реакции, и стритом, прежде всего, в отношении периферических
иміпульсов из того же рецептивного поля. В определенных случаях
облегчение от раздражений рецептивного поля одного рефлекса можно
наблюдать и в отношении периферических импульсов другого поля с
антагонистическим рефлексом. Но это бывает в тех случаях, когда
почему-либо раздражение этих полей легко дает рефлекторные
сокращения на антагонистических мышцах (Гедевани, 31).
В то же самое время ло отношению к тем мышцам, которые не
сокращаются от данного периферического импульса, центральный
рефлекторный аппарат развивает только торможение. Оно ослабляет
двигательный эффект на данной мышце в ответ на любое другое
раздражение, т. е. на раздражение любого другого рецептивного поля.
Мы думаем, что в самом начале повышение возбудимости, в связи
с возбуждением, происходит в проводящих элементах мозга — в
нервных цепочках и, в особенности, в нервных кругах, которые проводят
периферические импульсы к двигательным клеткам, а также в самих
двигательных клетках.
Нужно думать, что, пока эти нервные цепочки и круги, составленные,
главным образом, из промежуточных нейронов, находятся в активном
состоянии благодаря циркулирующему в них возбуждению, как сами
промежуточные нейроны, так и стоящие с ними в связи двигательные
клетки находятся в состоянии облегчения. Наблюдаемое же
одновременно длительное торможение обусловливается активным состоянием
нейропиля, возникающим благодаря бомбардировке импульсами из
активных нервных кругов и цепочек.
При ближайшем исследовании изменения возбудимости от одного
периферического импульса можно обнаружить длительное состояние
облегчения наравне с кратковременной фазой супернормальной
возбудимости. На спинальной лягушке вслед за рефракторными фазами через
8—10 о после момента раздражения возбудимость повышается; затем
она понижается и через 60—75 о вновь повышается. За этим периодом
повышенной возбудимости опять следует фаза пониженной возбудимости
(Бернштейн, 17) (рис. 87). Эти изменения возбудимости
устанавливаются путем второго раздражения, которое прикладывается к тому же
чувствительному нерву через разные промежутки времени после первого.
Второй период повышенной возбудимости и период последующей
депрессии сильно зависят от интенсивности первого раздражения: они тем
длительнее, чем интенсивнее раздражение.
13 и. с. Беритов
j 94 общая физиология спинного мозга и ствола головного мозга
Путем анализа эффектов изменения возбудимости от одиночных
раздражений было сделано заключение, что первый период облегчения об-
о мое*
Рис. 87* Четыре типа кривых рефлекторной суммации у лягушки.
Раздражается п. ischiadicus короткими разрядами конденсатора. Регистрируется
сокращение m. triceps. Кривая / (со светлыми кружками) изображает суммацию
в случае, когда оба раздражения являются подпороговыми. Ордината — высота
изотонических сокращений, абсцисса — интервал между отдельными ударами в
сигмах. Во всех остальных щ кривых ордината означает высоту изометрического
сокращения в процентах максимального одиночного вздрагивания, абсцисса же —
интервал между ударами в сигмах. Кривая // (с черными кружками) — суммация
от двух максимальных ударов. Кривая III (со светлыми квадратиками)—первый удар
рефлекторно-подпороговый, второй — субмаксимальный. Первая часть кривой
совпадает с кривой II. Кривая JV (с черными квадратиками) — каждый удар рефлек-
торно-пороговый. Начало кривой совпадает с кривой II. На всех кривых высота
сокращения в ответ на второе раздражение, произведенное в отдельности, отмечена
соответствующей горизонтальной линией (Бернштейн).
11 си
11
11
7/
//
Рис. 88. Повышение возбудимости в ответ на раздражение.
Спинномозговая собака. Записывается движение задней ноги. Раздражают рецептивное
поле сгибания фарадическим током. Первый и второй раз раздражение при і 1 см
расстояния между катушками. Это вызывает очень слабые движения. Третий раз
раздражение сильное — 6 см, оно производит сильное сгибание. После этого повторное
слабое раздражение (і 1 см) дает более сильный эффект, чем раньше. Последнее слабое
раздражение производится спустя 30 сек. после сильного эффекта и все-таки дает
в три раза более сильный рефлекс, чем вначале.Время — в секундах.
условливается главным образом повышением возбудимости
двигательных клеток, которое происходит путем передачи импульсов от
афферентных нейронов на двигательные коротким путем, минуя
промежуточные нейроны. Второй период облегчения вызывается более поздним по-
ЯВЛЕНИЕ ОБЛЕГЧЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ 195
еышением возбудимости в тех же двигательных нейронах посредством
запоздалых импульсов, проходящих через промежуточные нейроны.
Что касается периодов депрессии, то можно предположить, что
первый короткий период обусловлен активированием іГейрогшля под
непосредственным влиянием афферентных нейронов,
а второй длительный период — активированием
того же нейропиля посредством иімпульсации из
промежуточных нейронов.
Повышение возбудимости при повторных
периферических импульсах. При тетаничееких
раздражениях чувствительного нерва, в
результате более или менее длительной деятельности,
возбудимость повышается на более значительное
время (рис. 88). На препаратах лягушки, кошки
и собаки повышенная возбудимость может
наблюдаться в течение многих минут после более
или іменее длительного и интенсивного
активного состояния. Специальными опытами было
обнаружено, что эта характерная особенность
центральной нервной системы является свойством
координирующих аппаратов. Установлено, что
длительность состояния повышенной
возбудимости зависит от функционального состояния, а
также от интенсивности деятельности
координирующего аппарата. Чем сильнее вызванная
раздражением деятельность этого аппарата, тем
дольше удерживается в нем повышенная
возбудимость. В последнее время это было
подтверждено прямыми опытами в отношении нервного
механизма глазных мышц. Было доказано, что
явление длительного облегчения рефлекса в
результате повышения возбудимости зависит от
промежуточного нервного механизма, а не от
двигательных нейронов (Лоренте де Но, 11).
Повышение возбудимости двигательных
нейронов при тетаническом раздражении
наблюдается только во время раздражения,— значит, в
течение всего времени, пока двигательные
клетки получают периферические импульсы.
Промежуточные-же нейроны, или, вернее, нервные
цепочки и нервные круги, составленные из этих нейронов, при том же
раздражении отвечают длительным облегчением, продолжающимся в
течение многих секунд после раздражения. Так, если во время тетаниче-
ского раздражения VIII пояснично-крестцового заднего корешка
раздражать соседние корешки той же стороны отдельными
индукционными ударами, то на сгибателе рефлекторные вздрагивания окажутся
усиленными как во время тетанического раздражения, так и в тече-
Рис. 89. Явление
облегчающего последействия
в промежуточных
нейронах.
Лумбальный препарат
кошки. Раздражаются задние
корешки. Регистрируются
мышцы-антагонисты колена
на обеих сторонах. Б опыте
А раздражением правого VII
заднего корешка вызываются
рефлекторные вздрагивания
как на правой стороне, так и
на левом сгибателе. Б это-
время испытывается тетани-
ческое раздражение левого
VIII заднего корешка; по
прекращении
тетанического эффекта отдельные
рефлекторные вздрагивания не из»
меняются. В опыте В
раздражением левого V заднего
корешка вызываются
отдельные вздрагивания на том же
сгибателе. Вновь
производится раздражение левого VIII
заднего корешка, но теперь,
оно приводит к усилению
последующих рефлекторных
вздрагиваний (Беритов и
Бакурадзе).
196 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
ние многих секунд после него (рис. 89-В). В отношении же
перекрестных рефлекторных вздрагиваний от раздражения задних корешков
противоположной стороны тетаническое раздражение проявляет
облегчающее действие только во время самого тетанического
раздражения (рис. 90).
Как мы уже указывали выше, гомолатеральные координированные
реакции наступают на данной конечности через промежуточные нейроны
соответствующей стороны. Значит, на
стороне раздражения возбуждаются
одновременно промежуточные и двигательные
нейроны. Перекрестные реакции вызываются
на данной конечности без участия
промежуточных нейронов соответствующей
стороны, ибо передаточные комиссуральные
нейроны заканчиваются при двигательных
нейронах. Отсюда можно заключить, что
кратковременное повышение рефлекторных
вздрагиваний под влиянием перекрестной
двигательной реакции обусловливается
повышением возбудимости в двигательных
клетках. Очевидно, га отношении
повышения возбудимости, в связи с возбуждением,
двигательные клетки не отличаются по
существу от двигательных нервных волокон.
Поэтому нужно думать, что длительное
повышение рефлекторных вздрагиваний в го-
молатеральных реакциях не должно
обусловливаться повышением возбудимости в
клетках двигательных нейронов.
Большая длительность облегчения в
спинном мозгу безусловно связана с
деятельностью промежуточных
нейронов,—вернее, нервных цепочек и замкнутых кругов,
составляющихся из промежуточных
нейронов. Под влиянием периферических
импульсов эти круги долгое время находятся в
активном состоянии благодаря
многократному пробеганию в них импульсов возбуждения. В каждом нервном
круге циркуляция возбуждения продолжается и после прекращения
периферических импульсов, пока не утомится какое-либо звено этого
круга. Следовательно, в гомолатеральных рефлекторных реакциях
клетки промежуточных нейронов некоторое время после
периферических импульсов должны находиться под влиянием циркулирующих в
нервных кругах импульсов и тем испытывать длительное повышение
возбудимости. Правда, двигательные клетки связаны с
промежуточными клетками путем множественной цепочки, а потому при возбуждении
Рис. 90. Явление облегчения
в двигательных нейронах.
ЛумПальный препарат кпптки.
Раздражаются задние корешки.
Регистрируются мышцы-антагонисты
колена. Раздражением правого У
заднего корешка вызываются
рефлекторные вздрагивания только
мышц соответствующей стороны.
Но они возникают и па левой
стороне, если данное раздражение
сочетать со слабым тетаническим
раздражением (і5—14 см) левого V
корешка (Беритов и Бакурадзе).
ЯВЛЕНИЕ ОБЛЕГЧЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ J97
нервных кругов неизбежно должны посылаться импульсы из последних
к двигательным клеткам. Но с момента прекращения раздражения,—
вернее, с момента прекращения непосредственной импульсации из
афферентных нейронов, двигательные клетки могут перестать
возбуждаться. Есть основание предполагать, что возбуждение в
двигательных клетках нормально возникает в результате суммации местных
процессов, возникающих под влиянием периферических импульсов и
Рис. 91. Облегчение после торможения.
Лумбальный препарат кошки. Вверху кривая m. quadriceps dex.,
внизу — т. quadriceps sin. В опыте А раздражается тетанически
сначала п. tibialis dex. (F. d.), а затем, через 2 сек.—коленосги-
бательный нерв левой стороны (Я. S.). В опыте В раздражается
один коленосгибательный нерв. Из сравнения этих опытов ясно
видно, что после раздражения п. tibialis на разгибателе
соответствующей стороны торможение сменяется состоянием
облегчения, вследствие чего перекрестный разгибательный
рефлекс от коленосгибательного нерва увеличивается очень сильно
(Беритов, Бакурадзе и Нарикашвили).
тех импульсов, которые притекают из промежуточных нейронов.
Вследствие этого сейчас же по прекращении раздражения
двигательные клетки могут перестать' (возбуждаться, в то время как
промежуточные продолжают возбуждаться еще некоторое время благодаря
возбуждению, циркулирующему в нервных кругах (см. также Лоренте де
Но, 29).
Повышением возбудимости промежуточных нейронов от
циркулирующих импульсов, вероятно, объясняется явление лестницы, которое
наблюдается в рефлекторной реакции при повторном периферическом
раздражении. Нужно допустить, что каждый периферический импульс
из заднего корешка не только действует на двигательные клетки
непосредственно, но, кроме того, активирует нервные круги. Это повышает
возбудимость в промежуточных клетках и тем создает в них наилучшие
условия для суммации.
Явление облегчения после торможения. По прекращении
периферического тормозящего раздражения центральное торможение двигатель-
198 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
ных элементов не только тотчас же прекращается, но очень часто даже
заменяется противоположным состоянием — повышением возбудимости.
Это проявляется в облегчении возникновения и проведения возбуждения
в двигательных элементах, которые перед тем были заторможены.
Состояние облегчения легко обнаружить, если после тормозящего
раздражения ів течение нескольких секунд испытывать раздражение
таких чувствительных нервов, которые вызывают возбуждение данных
двигательных элементов. В ответ на такое
раздражение наступает рефлекторное
сокращение, более сильное, чем обычно (рис. 91)
(Грэим Броун, 15). Состояние облегчения
после торможения получается в том случае,
если состояние торможения проходит быстро
по прекращении раздражения. Если же оно
прекращается с большой постепенностью, то
облегчение отсутствует. Состояние облегчения
быстро нарастает сразу по прекращении
раздражения; затем оно медленно снижается1 в
течение нескольких секунд. При этом не
имеет значения, происходит ли торможение в
чистом виде, или же оно сопровождается
возбуждением нервных кругов. В последнем
случае, по прекращении двойственного действия,
если только оно проходит быстро,
торможение сменяется значительным облегчением (Бе-
ритав, Бакурадзе и Нарикашвили, 16). Так,
например, на рис. 93 видно, что сгибатель-
левой стороны после заторможенного тетани-
ческого сокращения дает усиленные
рефлекторные вздрагивания.
Явление облегчения после торможения
наблюдается не только после прекращения
раздражения чувствительных соматических
нервов, но по прекращении всякого рода
раздражений, производящих торможение; например
после электрического раздражения спинного мозга, после
раздражения внутренних органов и вегетативных чувствительных нервов (Бе-
ритов и Бакурадзе, 36, 37) и (рис. 47, 92 и 93).
Повышение возбудимости после торможения наблюдается всего в
течение нескольких секунд. Этим оно отличается от повышения
возбудимости, вызываемого в нейронных кругах распространением процесса
возбуждения. Как указывалось выше, повышение возбудимости в
последнем случае продолжается значительно дольше.
Особенно регулярно явление облегчения получается после
раздражения некоторых кожных рецепторов, которые производят только
общее торможение. Так, например, было обнаружено, что в случаях раз-
Рис. 92. Облегчение после
торможения при
раздражении внутренних органов.
Лумбальный препарат кошки.
Раздражаются рецепторы
прямой кишки расстягиванием
(сигнальная линия Re) Регистри-
руютсяразгибатели(?. d., Q.S.)
и сгибатели (S. d., S. s.) колена
той и другой сторон. В опыте
В растягивание прямой кишки
происходит в отдельности, а в
опытах А и С—во время
рефлекторных вздрагиваний,
вызываемых раздражением задних
корешков VI поясничного
сегмента (Vis. и VI d). Ясно видно
сильное облегчение после
сильного растяжения.
ЯВЛЕНИЕ ОБЛЕГЧЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ 199
дражения рецепторов давления и Холодовых рецепторов, когда не
происходят двигательные реакции во время самого раздражения, реакция
наступает по прекращении раздражения. На фоне наступающего
облегчения вызывается сокращение типа «отдачи» под влиянием разного
рода побочных раздражений (Дзидзишвили, 26). Явление облегчения
легко получается после механического раздражения поверхности спинного
мозга, причем во время самого раздражения не происходит никаких
двигательных реакций (Беритов и Бакурадзе, 25).
Рис. 93. Облегчение после торможения при
раздражении переднего столба спинного мозга.
Лумбальный препарат кошки. Регистрируются те же мышцы,
что и на рис. 92. Фарадическое раздражение передних столбов
III поясничного сегмента на правой стороне производит
торможение рефлекторных вздрагиваний, вызываемых
раздражением задних корешков (VII d. и VI s.). При сравнительно
сильных раздражениях мозга после торможения происходит
облегчение рефлекторных вздрагиваний. При этом облегчение
наблюдается на правой стороне, где торможение было очень
сильное.
Явление облегчения после торможения сл\жит одним из серьезных
доводов в пользу гипотезы об электротонической природе центрального
торможения. Согласно этой гипотезе, торможение обусловливается
анэлектротоническим действием длительных колебаний электрического
потенциала активного нейропиля на эти элементы. Как известно, на
нерве быстрое прекращение анэлектротона влечет за собой
кратковременное повышение возбудимости (рис. 94).
Аналогичное явление, повидимому, имеет место в центральной
нервной системе. Прекращение активного состояния нейропиля
связано с прекращением анэлектротонического действия на двигательные
элементы, а это, в свою очередь, приводит к кратковременному
повышению в них возбудимости.
Это повышение возбудимости, по всей вероятности, заключается в
изменении проницаемости полупроницаемых перепонок в клетках
двигательных нейронов, именно в их разрыхлении, а также в освобождении
и передвижении одновалентных ионов внутри клеток. Благодаря этим
изменениям облегчается передача импульсов возбуждения от
чувствительных и промежуточных нейронов на двигательные нейроны, вероят-
200 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
но в силу облегчения возбуждающего действия биоэлектрических токов
возбуждения синаптических окончаний.
Итак, если по прекращении тормозящего раздражения торможение
кончается быстро, то наступает кратковременное состояние облегчения
рефлекторной деятельности заторможенных нервных элементов. Это
явление облегчения после торможения, видимо, связано с прекращением
анэлектротонического действия нейропильного биотока на эти элементы.
Рис. 94. Повышение возбудимости нерва в области
анода после выключения постоянного тока.
Нервно-мышечный препарат лягушки (седалищный нерв-
икроножная мышца). Слабый постоянный ток (0.14 У)
пропускается через нерв. В области анода производится
пороговое тетаническое раздражение. В каждом опыте оно
испытывается до и после постоянного тока. На первой
сигнальной линии цифры означают силу раздражения в
сантиметрах индукционной катушки.
В опыте А тетаническое раздражение производится сейчас
же после выключения постоянного тока, — оно дает
чрезвычайно сильный эффект; в опыте В — спустя 7 сек. после
" быстрого выключения; здесь также сильный эффект, хотя
слабее, чем в предыдущем опыте. В опыте С постоянный ток
выключается с большой постепенностью в течение 7 сек.,
и сейчас же после этого производится тетаническое
раздражение; оно не дает большого эффекта. Средний сигнал
поднятием отмечает начало убывания постоянного тока,
а полное выключение его отмечается опусканием нижнего
сигнала (Цкипуридзе).
Явление суммации от субминимальных раздражений. Возбудимость
координирующего аппарата повышается и под влиянием недеятельных
субминимальных раздражений. Это видно из того, что если
субминимальные раздражения повторяются быстро одно за другим, они
производят рефлекс. Явление суммации в отношении центральной нервной
системы известно с давних пор. Понятно, что мы будем иметь дело с
явлением суммации в центральной нервной системе в том случае, если
каждое раздражение вызывает возбуждение в раздражаемом рецепторе
или чувствительном нерве. Периферические импульсы возбуждения от
одного раздражения могут быть недостаточны для приведения
координирующего аппарата в действие: для того чтобы он мог притти в
деятельное состояние, необходимо, чтобы раздражения следовали быстро
одно за другим и координирующий аппарат вследствие этого
испытывал воздействие следующих одно за другим периферических
возбуждений (Тарханов, 38).
ЯВЛЕНИЕ ОБЛЕГЧЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ 201
Центральная нервная система обладает способностью к суммации в
большей мере, чем нерв и мышца. Этот факт был отмечен еще
Сеченовым (1868). Он заметил, что некоторые рефлексы совершенно не
вызываются при раздражении чувствительного нерва однократными
раздражениями, как, например, отдельными индукционными ударами,
возбуждающими нерв и мышцу (13). Рефлекс потирания на лягушке не
может быть вызван однократным раздражением нерва или кожи
соответствующего рецептивного поля. Только быстрое повторение
раздражения вызывает этот рефлекс. Но при ненормально повышенной
возбудимости координирующего
аппарата данного рефлекса,—
например, под влиянием стрих-
яинного отравления,— можно
наблюдать рефлекс потирания
и от одного индукционного
удара (Беритов, 3).
Некоторые рефлексы вызываются
отдельными индукционньими
ударами: например, рефлекс
сгибания. Но и здесь порог тета-
нического раздражения
значительно ниже порога
раздражения отдельными ударами.
Кроме того, рефлекс сгибания
может совсем не вызываться
однократным раздражением,
если почему-либо
возбудимость соответствующего
координирующего аппарата
значительно понижена. Именно по
этой причине некоторые
авторы прежде утверждали, что центральная нервная система вообще не
возбуждается однократными раздражениями. Собственно говоря, при
возникновении рефлекторной реакции нормальным путем она никогда
не является результатом одного периферического импульса. Как
указывалось выше, рецепторы разряжаются в ответ на самое минимальное
и кратковременное раздражение рядом или залпом импульсов.
Следовательно, всякий раз координирующий аппарат получает от рецепторов
ряд последовательных импульсов.
Явление суммации наблюдается в центральной нервной системе не
только при раздражениях одного и того же чувствительного нерва или
одного и того же рецептора. Оно имеет место и при раздражении двух
чувствительных нервов или двух участков кожи одного и того же
рецептивного поля. Если приложить подпороговое раздражение к обоим
участкам кожи или к обоим нервам одновременно или быстро одно-
вслед за другим, то рефлекс наступает. Это, например, легко наблюдать
f L
^ЛЛЛЛЛЛЛЛЛЛЛЛJ¦^^Л^VlЛЛJ*lЛЛЛJVlЛЛJVUl/UlЛЛЛJ~^
Sol ' » г-
"I й?л \ г-
Рис. 95. Явление суммации в центральной
нервной системе.
Децеребрированный препарат собаки.
Записывается сгибатель тазобедренного сустава.
Раздражают в поле рефлекса чесания два участка на
расстоянии 8 см один от другого. Раздражение
обоих участков субминимальное и в отдельности
не дает рефлекса (первый опыт). При
одновременном же раздражении получается сильный
чесательный рефлекс. Моменты раздражения
одного (S о.) и другого участка (S 3) отмечаются
опусканием сигнальной линии. Время — в
секундах (Шеррингтон).
202 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
при раздражении двух участков в рецептивном поле рефлексов
сгибания или почесывания (рис. 95). Явление суммации объясняется тем, что
периферические импульсы с одного участка рецептивного поля не в
состоянии возбудить нервные круги спинного мозга, но вызывают в них,
или, точнее, в промежуточных нейронах, повышение возбудимости.
Поэтому когда периферические импульсы приходят к ним
одновременно из двух участков, этого оказывается достаточно для их
возбуждения.
Явление, когда раздражение одного рецептивного поля благоприят-
сівует рефлексу другого рецептивного поля, или же раздражение одной
части данного рецептивного поля благоприятствует вызову эффекта с
другой его части, Шеррингтон назвал «непосредственной
спинномозговой индукцией» (immediate spinal induction) (4). Это же явление
раньше было известно под названием «проторения» (Bahnung), что
означает подготовку пути: одно раздражение как бы расчищает путь
для другого.
Из приведенных фактов следует, что суммация в центральной
нервной системе, как в симілатическом ганглии, бывает двоякой:
пространственной и временной. Пространственная суммация
вытекает из следующих фактов: 1) сравнительно слабое раздражение
чувствительного нерва из рецептивного поля рефлекса сгибания одним
индукционным ударом не производит сгибательного рефлекса, но при
значительном усилении раздражения рефлекс наступает и от одного
удара; 2) если отдельное раздражение того или другого
чувствительного нерва или одного рецептивного поля не дает рефлекса, то
одновременное раздражение двух или более нервов производит его.
Очевидно, пространственная суммация основывается на том, что при
некоторой более или менее низкой возбудимости центральных элементов,
прежде всего промежуточных нейронов, возбуждение небольшого числа
нервных волокон обусловливает возбуждение небольшой группы
синапсов, оканчивающихся на теле клеток Возникающие при этом
биотоки возбуждения вызывают в соответствующих участках клетки
локальные процессы частичного изохронного расщепления возбудимой
системы, без активирования всей клетки. Но когда с усилением
раздражения или, вернее, с распространением раздражения на большее
количество афферентных волокон, происходит значительное увеличение
числа активных синапсов на теле клеток, тогда суммарного
раздражающего действия биотоков возбуждения этих синапсов оказывается
достаточно для возбуждения клеток и при одном периферическом
импульсе.
Временная суммация вытекает из того факта, что когда более или
менее слабое раздражение чувствительного нерва какого-либо
рецептивного поля одним электрическим ударом не дает рефлекса, то оно
производит его при повторении, и притом тем скорее, чем чаще это
лодпороговое раздражение.
Интервал суммации. Первое раздражение оказывает благоприят-
ЯВЛЕНИЕ ОБЛЕГЧЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ 203
ствующее влияние на эффект второго раздражения в течение
определенного интервала времени. На препаратах теплокровных животных
суммация не происходит при интервале менее 4—б <з, что соответствует
продолжительности рефракторных фаз. Суммация наблюдается при
больших интервалах. Если два последовательных раздражения падают
не на один нерв, а на два разных чувствительных нерва из одного
рецептивного поля, суммация происходит при любом интервале, но она не
наблюдается, когда интервал превосходит 10—20 о (Шеррингтон, 12).
На спинальных препаратах холоднокровных животных повышение
возбудимости после субминимального раздражения протекает в два
периода, как это наблюдается и после деятельного раздражения. Так,
например, удлиняя постепенно интервал между двумя субминимальными
одиночными раздражениями (электрические удары), можно наблюдать
две вершины суммации: одна— через 8—10 о, а другая — через 50—
70 ст(Бремер, 18; Бернштейн, 17). На рис. 87 приведены разные случаи
рефлекторной суммации: когда первое раздражение является подпоро-
говым, максимальным или пороговым. Во всех этих случаях
наблюдаются две вершины суммации, причем вторая суммация начинается
приблизительно через один и тот же промежуток времени после первого
раздражения. Предполагается, что эти суммации обусловливаются
повышением возбудимости при двигательных клетках: первый период
возникает под влиянием афферентных импульсов коротким путем, а второй
период — сложным путем, через промежуточные нейроны, т. е. через
нервные цепочки и нервные круги.
Наибольший 'промежуток времени, при котором происходит сумм а-
иия и наступает двигательный эффект, зависит от силы раздражения.
Чем слабее раздражение, тем этот интервал меньше. Но это изменение
интервала зависит от функционального состояния центральной нервной
системы, а потому кривая суммации, отображающая изменения
интервала суммации в зависимости от силы раздражения, определенным
образом характеризует состояние рефлекторного аппарата.
Интервал суммации на разных препаратах одного и того же
животного не одинаков. Так, на спинномозговой лягушке суммация
наблюдается при более значительных силах раздражения, чем на препарате
без больших полушарий. Так, по Ш р и в е р у, при наличии головного
мозга интервал суммации в 1 сек. получается при силе раздражения в
1.5 V, а на спинальных препаратах интервал суммации в 0.25 сек.
достигается при силе раздражения 3 V, а при интервале 0.33 сек.
понадобилось раздражение в 12 V (рис. 96-/). Отсюда следует, что высшие
отделы центральной нервной системы — головной мозг — в большей мере
благоприятствуют суммации, чем низшие — спинной мозг. Это означает,
что функциональное изменение нервных элементов под влиянием
периферических импульсов длится в головном мозгу значительно дольше,
чем в спинном.
С изменением функционального состояния меняется и кривая сум-
204 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
А
Интервал в сек
1
Л
охлаждение
мации. Например, при охлаждении мозга кривая суммации уплощается,
т. е. минимальный вольтаж уменьшается, а предельный интервал
увеличивается. Так, например, в одном эксперименте на спинальной
лягушке при раздражении
чувствительного нерва
разрядами конденсатора
минимальный вольтаж при
комнатной температуре
равнялся 2 V, а частота
раздражения была 12 в 1 сек.; он
повышался до 4.5 V, когда
частота равнялась 4 в 1 сек.»
и становился бесконечно
большим при частоте 3 в
1 сек. Когда же мозг был
охлажден, минимальный
вольтаж при частоте 12 в
1 сек. понизился до 1.5 V,
а при частоте 4 в 1 сек.— до
2V (Шривер, 20). Эта
закономерность изменения
суммации в зависимости от
функционального состояния
называется з а к о н о м
суммации (Шривер, 20;
Лапик, 22). Суммация
зависит также от числа
возбужденных периферических
волокон. Она наступает тем
легче, чем больше
возбужденных чувствительных
волокон. Например, при
раздражении седалищного
нерва суммация в
отношении сгибательного рефлекса
получается легче, т. е. при
меньшем минимальном
вольтаже и большем
предельном интервале, чем при
раздражении п. peroneus
или п. tibialis.
Своеобразное изменение суммации
получается, если у лягушки
охладить не весь мозг, а
только средние отделы головного мозга. Как показывает рис. 96-И,
в этом случае кривая суммации в отношении сгибательного рефлекса
11 ;
12 8 6
1 1
I
2
Интервал в сен.
Рис. 96. I. Влияние высших отделов
мозга на суммацию.
А — лягушка беа больших полушарий; В —
спинномозговая лягушка. Раздражается седалищный
нерв ритмическими разрядами конденсатора. Число
раздражений постоянно — 12, но меняется
интервал—от >/ц сек. до 1 сек. Регистрируется
перекрестный сгибательный рефлекс. На сшгаальном
препарате суммация получается при более значительных
силах раздражения, чем на препарате без полушарий
(Шривер).
II. Влияние охлаждения средних отделов
мозга лягушки на суммацию.
Определяется зависимость между интервалом и
силой раздражения. Условия раздражения — как
в предыдущем случае. Сплошная кривая — слабое
охлаждение; прерывистая — сильное. При одних
и тех же интервалах раздражения во время
охлаждения потребная сила раздражения больше, чем до
и после охлаждения (Шривер).
ЯВЛЕНИЕ ОБЛЕГЧЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ 205
после охлаждения круто поднимается вверх. Это ухудшение суммации
после охлаждения обусловливается выпадением субординационных
влияний со стороны головного мозга (Шривер, 20).
Явление нарастания рефлекторной деятельности. При нормальном
состоянии рефлекторного аілпарата некоторые рефлексы сразу не
вызываются с максимальной интенсивностью. Они нарастают постепенно,
в 10—20 раз медленнее, чем обычное тетаническое сокращение от
раздражения двигательного нерва. Соответственно, производимое в этих
рефлексах возбуждение и торможение постепенно нарастают. Это
постепенное установление оптимальной деятельности нужно считать
характерным признаком, например, рефлекса потирания на лягушке
(Беритов, 10) и перекрестного разгибательного рефлекса на децереб-
рированных и спинальных препаратах кошки (Шеррингтон, 9). Шер-
рингтон называет это явление «вовлечением» (recruitment),
подразумевая, что оно обусловлено постепенным увеличением числа
возбужденных двигательных волокон. Оборонительный рефлекс сгибания на
лягушке и на других животных принадлежит к противоположному
типу. Он обычно устанавливается, подобно периферическому тетанусу,
почти сразу, без постепенного нарастания. Характерно, что и токи
возбуждения разгибателя в рефлексе потирания начинаются с очень
небольшой высоты и затем постепенно нарастают, в то время как в
рефлексе сгибания токи возбуждения обычно с самого начала значительны
(Беритов, 10). При некоторых пороговых раздражениях и рефлекс
сгибания может начаться с большой постепенностью. На рис. 97 в опыте
Л показан эффект разгибателя колена в рефлексе потирания, где токи
возбуждения вместе с механическим эффектом нарастают постепенно,
а в опыте В дается эффект сгибателя в рефлексе сгибания; здесь токи
возбуждения начинаются с больших колебаний, причем механический
эффект хотя и дает нарастание, но оно продолжается всего % сек.
Постепенное нарастание рефлекторной деятельности при всех
силах раздражения должно явиться особенностью соответствующего
координирующего аппарата, а не двигательных нейронов, ни тем
более самой мышцы. Это видно прежде всего из того, что в рефлексе
потирания подобное нарастание наблюдается и на сгибателях-
m. sartorius и т. iliofibularis, которые участвуют в нем подобно
разгибателю m. triceps (Беритов); на кошках же разгибатель колена,
m. vastocruralis, может дать моментальное сокращение в некоторых
разгибательных рефлексах (Шеррингтон, 9). Нужно думать, что
координирующие аппараты определенных рефлексов находятся в таком
взаимоотношении с чувствительными заднекорешковыіми волокнами,
что под влиянием периферических импульсов они приходят в действие
с большой постепенностью, ввиду постепенного вовлечения 'все
большего и большего количества координирующих нейронов. В соответствии
с этим количество возбужденных двигательных нейронов
увеличивается также постепенно, чем и объясняется медленное нарастание
периферического эффекта.
206 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Итак, для определенных рефлексов с участием промежуточных
нейронов очень характерно постепенное нарастание центральной
деятельности, обусловленное постепенным вовлечением промежуточных„
а вслед за ними и двигательных нейронов в активное состояние.
Некоторые рефлексы производятся без участия промежуточных
нейронов, путем передачи возбуждения от чувствительных к двигательным,
как это имеет место в рефлексе разгибания при раздражении
сухожильных рецепторов. Эти рефлексы наступают без нарастания. То же
может случиться при искусственном раздражении чувствительных нер-
Рис. 97. Л — нарастающий рефлекторный эффект на m. triceps спинномозговой
лягушки.
Раздражается п. cut. fern. Iat. из рецептивного поля потирания. Частота раздражения 100 в 1 сек-
Сила — 25 см расстояния между катушками. Колеблющаяся кривая от токов возбуждения в общем
следует ритму раздражения, что указывает на большую силу последнего. Сплошная кривая .—
от механического эффекта. Время 0.2 сек.
В—ненарастающий рефлекторный эффект на m. semitendinosus. Раздражается п. peroneus из
рецептивного поля сгибания. Частота раздражения — 100 в 1 сек. Сила — 30 см расстояния между
катушками. Токи возбуждения следуют по ритму раздражения в самом начале кривой, а потом
отдельными участками. Сплошная кривая от механического эффекта. Отрезок С является
продолжением В через 2 сек. Внизу — время 0.2 сек.
вов мышцы и вообще нервных стволов, содержащих сухожильные и
мышечные чувствительные волокна. Но такое быстрое нарастание
рефлекторных сокращений, как это бывает при раздражении
чувствительных нервов, совершенно не характерно для нормальных рефлекторных
реакций, вызываемых при участии промежуточных нейронов.
Изменение электрических потенциалов спинного мозга при
облегчении. Изучение электрической активности спинного мозга при повторных
раздражениях чувствительных нервов дает возможность проследить,
как меняются электрические потенциалы мозга в период облегчения.
Лучше всего для этой цели брать раздражение небольшой частоты,
10—30 в 1 сек., и небольшой интенсивности, а отводить потенциалы от
переднего корешка вблизи мозга. Опыты на спинном мозгу лягушки
при отличном функциональном состоянии препарата показывают, что
первый удар раздражения обычно вызывает после начального задне-
корешкового биотока возбуждения один медленный потенциал, который
возникает от локальных процессов в дендритно-клеточной массе перед-
ЯВЛЕНИЕ ОБЛЕГЧЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ 207
него рога непосредственно под влиянием заднекорешковых импульсов.
При нескольких последующих ударах медленный потенциал нарастает,
а иногда усложняется появлением нового медленного потенциала,
который наступает через 10—20 а после заднекорешкового импульса. Этот
второй медленный потенциал возникает в том же переднем рогу под
влиянием импульсов из промежуточных нейронов. Одновременно появ-
Рис. 98—/. Электрическая активность спинного мозга лягушки при облегчении.
Сшгаальный препарат. Все передние корешки перерезаны. Отведение от IX переднего
корешка слева на расстоянии 2 мм от мозга. А—раздражается п. peroneus sin.
релаксационным стимулятором при 0,4 V; В — тоже, при силе 0,8 V.
ляются быстрые потенциалы от возбуждения переднекорешковых
волокон (рис. 98-1). В случае наличия двух медленных потенциалов
возникают две группы быстрых импульсов: одна на фоне первого медленного
потенциала, другая на фоне второго. Первая группа, должно быть,
получается непосредственно от воздействия заднекорешковых импульсов,
а вторая — от воздействия импульсов из промежуточных нейронов.
Затем эти быстрые потенциалы сильно нарастают и учащаются.
Медленные потенциалы, наоборот, ослабевают, но появляются все время с
разной продолжительностью и разной амплитудой. Быстрые потенциалы
располагаются на их фоне. При сравнительно большой силе
раздражения медленные потенциалы того и другого происхождения возникают в
самом начале раздражения (рис. 61). В таких случаях быстрые
разряды появляются на фоне того и другого потенциала уже со второго
удара (рис. 61 и 98-1).
При изучении биотоков мозга путем отведения их через
центральный отрезок заднего корешка или от поверхности мозга, можно
проследить главным образом изменение медленных потенциалов. Они
выражают локальные процессы дендритно-клеточной массы задней
половины мозга. Как указывалось выше, первый потенциал значительно
сильнее последующих вследствие тормозящего действия первого
медленного потенциала на близлежащие нервные элементы. Последующие
медленные потенциалы при частоте раздражения 10—30 в 1 сек.
постепенно нарастают (рис. 57-А и В). При отличном функциональном
состоянии препарата электрический эффект усложняется появлением
дополнительных медленных потенциалов (рис. 58). Регулярные медленные
потенциалы после заднекорешкового импульса обусловлены локальными
процессами, возникающими под влиянием заднекорешковых импульсов,
20S ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА. И СТВОЛ \ ГОЛОВНОГ07МОЗГЛ
а дополнительные медленные потенциалы — от локальных процессов
под влиянием импульсов из промежуточных нейронов. В этих случаях
от задних корешков отводятся также быстрые потенциалы, которые
выражают биотоки возбуждения клеток и аксонов промежуточных
нейронов (рис. 58-С и D).
Итак, при повторных раздражениях, облегчающая деятельность
спинного мозга выявляется сначала в отношении локальных процессов,
а затем б* отношении импульсов возбуждения, причем и тот и другой
эффект развивается под влиянием заднекорешковых импульсов раньше,
чем под влиянием импульсов из промежуточных нейронов.
Происхождение суммации в центральной нервной системе.
Возбуждение промежуточных нейронов спинного мозга нормально происходит
не от действия одного периферического импульса через один синапс,
а в результате возбуждения целого комплекса синапсов,
принадлежащих, вероятнее всего, разным афферентным и промежуточным
нейронам. Отсюда следует, что как активирование координирующего
аппарата повторными подпороговыми раздражениями или двумя
одновременными подпороговыми раздражениями, так и явление нарастания
рефлекторной реакции и явление лестницы обусловливаются одними и
теми же причинами: когда возбуждение одного чувствительного
волокна или целого пучка этих волокон производит активирование
определенных промежуточных и двигательных нервных клеток, в других
промежуточных и двигательных клетках оно производит определенные
локальные биологические процессы, повидимому, в смысле частичного
перевода гетерохронного состояния возбудимой системы в изохронное.
Это изменение основного биологического процесса обусловливает
определенное физико-химическое изменение в данных клетках, приводящее
к повышению в них возбудимости. Благодаря этому при повторном
раздражении данного нерва или при одновременном раздражении двух
нервов и эти клетки должны разрядиться импульсами возбуждения.
Нужно предположить, что во всех этих случаях возбуждение
чувствительных нервов влияет на клетки промежуточных и двигательных
нейронов главным образом токами возбуждения своих нервных
окончаний, своих синаптических образований. Эти токи производят
изменение концентрации ионов между клеткой и синапсами и в самих
клетках, как это обычно бывает в возбудимых тканях при пропускании
электрического тока. Кроме того, синаптический ток возбуждения
вызывает в клетке локальный биологический процесс, а именно — переход
части возбудимой системы из гетерохронного состояния в изохронное.
Это явление также должно благоприятствовать физико-химическому
изменению клетки. В настоящее время есть основание утверждать, что
это изменение зависит от того, что при пропускании тока через
возбудимую систему нервных клеток внутри клеток повышается рН, т. е.
происходит изменение реакции содержимого клетки в -сторону ущела-
чивания. Это обстоятельство, по данным Б е т е и других, должно
вызвать повышение возбудимости. Данное предположение хорошо согла-
ЯВЛЕНИЕ ОБЛЕГЧЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ 209
суется и с новейшими наблюдениями на нервных волокнах. Именно,
Л е м а н (28) установил, что повышение рН в нервных волокнах
сопровождается повышением возбудимости, а понижение рН, -наоборот,
понижением возбудимости. Возможно, при быстром следовании
периферических ижіульсов или при увеличении притока одновременных
импульсов к данной клетке концентрация водородных ионов в ней
достигает той предельной зеличины, при которой происходит
возбуждение клетки.
Шеррингтон и его сотрудники предполагают, что явление суммации в
центральной нервной системе под влиянием периферических импульсов происходит в
двигательных клетках. Это согласуется с тем их представлением, что координация
центральных процессов является івсецело функцией сигаадсов при двигательных
клетках. Далее, Шеррингтон и его сотрудники полагают, что местное изменение в
центральной нервной системе, лежащее в основе суммации, является тем
«центральным состоянием возбуждения» в двигательных клетках, о котором уже говорилось
выше. При повторных периферических импульсах субминимального характера
«центральное состояние возбуждения» постепенно усиливается и, наконец, достигает того
уровня, когда оно становится причиной разряда обычных нервных импульсов. Чем
чаще раздражение, тем скорее достигается этот уровень.
Введенский также изучал вопрос о суммации в центральной нервной
системе. Он тоже локализовал явление суммации в двигательной клетке и смотрел на
него как на «пробуждение к жизни ряда слабых (остаточных) возбуждений под
влиянием новой, (пришлой волны 'возбуждения» (32). Эти объяснения явлений
суммации, данные Шеррингтоном и Введенским, собственно говоря, сходны с
развивавшимся нами выше. Основное расхождение касается терминологии. То, что
мы называем усилением основного биологического процесса, разумея под этим
местное и преходящее изменение, лежащее в основе местного повышения возбудимости,
Шеррингтон называет «центральным состоянием возбуждения», а
Введенский — «слабым остаточным возбуждением». Кроме того, мы находим, что явление
суммации может происходить не только в двигательных, но и в промежуточных
нейронах, в последних даже в более эффективной форме, чем в двигательных.
Итеративная возбудимость и гетерохронизм центральной нервной системы. Как
указывалось выше, Л а п и к причисляет клеточные элементы мозга, специально
промежуточные клетки, к итеративным системам, возбуждающимся только в результате
многократного воздействия периферических импульсов. По Л а п и к у, это
происходит благодаря тому, что хронаксия промежуточных нервных клеток значительно
длительнее хронаксин афферентных нервных волокон, что существует определенный
гетерохронизм между этими нервными образованиями. Следовательно, повторная
импульсация со стороны афферентных волокон необходима,-,по Л анику, для того,
чтобы повлиять на функциональное состояние промежуточных клеток и тем укоротить
их хронаксию в такой мере, чтобы гетерохронизм сменился изохронизмом (35).
Однако нет никаких оснований утверждать, что клетки промежуточных нейронов
епинного мозга не могут возбудиться от одного периферического импульса через одну
определенную группу синапсов только потому, что хронаксия этих клеток
значительно больше хронаксии афферентных волокон. Все дело в том, что, как указывалось
выше, при нормальном функциональном состоянии промежуточных клеток
раздражающее действие биотоков возбуждения определенной группы синапсов, сходящихся
от возбужденных афферентных волокон на теле этих клеток, может оказаться подпо-
роговым. В таких случаях для возбуждения промежуточных клеток требуется
многократное действие биотоков возбуждения от одних и тех же активных синапсов,
потому что это приводит к повышению возбудимости данных клеток. Но данное явление
имеет место и в отношении таких наиболее возбудимых образований с очень короткой
хронаксией, как нервные волокна группы А. В этих волокнах при слабом подпорого-
бом раздражении возбуждение возникает не сразу при первом же раздражении, а
Н и. С. Беритов
2)0 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
после многократного раздражения. В обоих случаях мы имеем дело с временной сум-
мацней подпороговых воздействий.
Против особой итеративной возбудимости нервных клеток мозга говорит с
очевидностью н факт пространственной суммацин. В то время как при слабых
раздражениях, при возбуждении небольшого количества афферентных волокон, требуете»
многократное повторение раздражения, при сильных раздражениях, когда
возбуждается большое количество волокон, рефлекс наступает от одного раздражения, от
одного индукционного удара.
Облегчающее и тормозящее действие головного мозга на спинной.
Головной мозг оказывает определенное влияние на степень возбудимости
спинного мозга. Высшие отделы головного мозга, в особенности кора
большого мозга, должны иметь особенно большое значение, ибо они,
как мы увидим ниже, все время находятся в активном состоянии,
выражающемся в биотоках разной частоты и интенсивности. Это
активное состояние наблюдается не только при бодрствующем состоянии,
но и во время сна и наркоза, хотя и значительно слабее. Спонтанное
активное состояние каждого отдела головного мозга передается всем
его отделам, а также спинному мозгу. Эдриан и Моруци (30) в
своих исследованиях показали путем осциллографической записи
биотоков коры большого мозга и нисходящих корковых путей в
продолговатом мозгу, что не только в бодрствующем состоянии, но и во время
наркотического сна нисходящие, пирамидные пути разряжаются
импульсами более или менее синхронно с двигательными участками коры. Эта
деятельность нисходящих корковых ^іутей не всегда производит
периферическую реакцию. Движение конечности, например, происходит
только при некоторой высокой частоте и интенсивности разряда
импульсов. Характерно, что всякое периферическое раздражение рецепторов
или нерва, приводящее к усиленной активации коры мозга, усиливает
также корковую импульсацию в отношении спинного мозга и стволовой
части головного діозга (Эдриан и Моруци, 30). При низкой частоте
и интенсивности этой импульсации спинной мозг не возбуждается, но
определенным образом реагирует на такую импульсацию, вследствие
чего возбудимость в нем повышается. Это создает благоприятные
условия для активирования спинного мозга под влиянием всякого рода
внешних и внутренних раздражений.
Итак, стволовая часть головного мозга и спинной мозг все время
находятся под влиянием корковой импульсации, вследствие чего они
пребывают все время в состоянии некоторой повышенной возбудимости,
в состоянии облегчения.
Большое значение лмеют также те средние отделы, откуда
регулируется тонус скелетной мускулатуры; а именно — средний мозг с его
красными ядрами. Было показано, что разрушение или охлаждение
среднего мозга меняет кривую суммации спинного мозга: она круто
поднимается вследствие сильного укорочения интервала суммации
(Шривер, 20) (рис. 96). Аналогичным образом действует торможение
тонических рефлексов, производимое сжиманием головы позади
челюстей (Лапик, 22).
ЯВЛЕНИЕ ОБЛЕГЧЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ 211
Так же действуют наркотики; например, хлорал, бромистые соли,
которые понижают івозбудимость в средних отделах мозга. Наоборот,
такое вещество, как пикротоксин, которое вызывает повышение
возбудимости и судорожные сокращения, уплощают кривую суммации
спинного імозга (Шривер и Першман, 21). Из всех этих фактов следует, что
тонические центры, находясь все время под влиянием проприоцептив-
ных, )Экстероцептивных и интероцептивньах раздражений, посылают в
спинной мозг импульсы, поддерживая тем самым возбудимость на
некотором высоком уровне, т. е. производя в ней общее облегчение. Если
почему-либо действие тонических центров прекращается от охлаждения,
торможения или наркотиков, спинной мозг перестает получать из них
импульсы и возбудимость его понижается. Если же тоническая
деятельность усиливается (согревание, пикротоксин), возбудимость спинного
мозга повышается. Характерно, что указанные изменения возбудимости
или кривой суммации в определенных случаях происходят параллельно
с изменением субординационной хронаксии периферических
двигательных нервов. Когда благодаря понижению активности среднего мозга
возбудимость спинного мозга понижалась и кривая суммации круто
поднималась вверх, тогда хронаксия удлинялась, т. е. субординация
выпадала и хронаксия становилась конституционной.
В некоторых случаях происходила диссоциация этих явлений: субординационная
хронаксия менялась без изменения возбудимости «шинного мозга, или, наоборот,
субординация не менялась, а менялась только возбудимость спинного мозга. Так как
спинномозговая суммация происходит главным образом в промежуточных нейронах,
а субординация осуществляется через двигательные нейроны, то Л а п и к считает
возможным предположить, что высшие отделы мозга оказывают определенное
субординационное влияние как на промежуточные, так и на двигательные нейроны в
смысле укорочения хронаксии. Нормально возбудимость промежуточных нейронов низка
и характеризуется длинной хронаксией. Благодаря этому между чувствительными
элементами и этими промежуточными нейронами существует большой гетерохронизм.
Импульсы среднего мозга, действуя на промежуточные нейроны, укорачивают их
хронаксию,—я тем (самым уменьшается гетерохронизм; в результате облегчается
переход возбуждения от чувствительных нервов через промежуточные на
двигательные. Так как субординация двигательных нервов может происходить независимо от
изменения закона суммации рефлекторного аппарата, то это дает право
предположить, что средний мозг действует также непосредственно на двигательные нейроны.
По Л а п и к у, передача импульсов возбуждения от чувствительных волокон на
двигательные происходит только после установления изохронизма между клетками
мозга и этими волокнами. Но еще никто не измерял возбудимость двигательных
клеток хіроткошетріич?сіки, и это положение Л а п и к а является чисто гипотетическим.
Высшие отделы головного мозга влияют на возбудимость низших
отделов головного мозга и на спинной мозг не только изменением своей
спонтанной импульсацни. Они влияют в ту или другую сторону и при
каждом активном состоянии под влиянием периферических раздражений.
Еще Бубнов и Гайдентайн (1881) показали, что на морфини-
знрованных собаках тактильное раздражение кожи в определенных
случаях влияет облегчающим образом на рефлекторные реакции
передних конечностей, вызываемые раздражением коры мозга; в других слу-
14*
212 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО ЛОЗГА
чаях, наоборот, то же раздражение тормозит двигательный эффект от
коркового раздражения. На норхмальных животных прессорчое и холо-
довое раздражение также может как облегчить оборонительный
рефлекс, вызываемый раздражением конечности, так и затормозить его.
Торможение происходит обычно во время тактильного и температурного
раздражения, а облегчение — после него (Дзидзишвили, 26). Также
действует раздражение головных рецепторов. Например, у лягушки
зрительное или слуховое раздражение само по себе не дает
двигательных рефлексов на конечностях, но благоприятствует оборонительным
двигательным рефлексам, если зрительное или слуховое раздражение
Рис. 98 — II. Явление торможения и облегчения
от воздействия света у лягушки.
Регистрируется движение аадней конечности.
Рефлекторное сгибание ее вызывается электрическим раздражением
кожи размыкательными ударами (Р). В опытах 1 я 3 раз-
мыкательный удар в отдельности вызывает значительное
поднятие лапки; в опыте 2 тот же удар в сопровождении
света — зажигания электрической лампы — производит
сильно ослабленную реакцию. В опыте 4 тот же удар
в момент потухания света дает максимальное поднятие
лапки (Бебуриіпвили).
производится раньше раздражения рецептивных полей этих рефлексов.
Рефлекс сгибания выражен сильнее при раздражении соответствующего
рецептивного поля, если ему предшествует зрительное или слуховое
раздражение (Мерцбахер, 6; Иеркс, 7; Бебуриіпвили, 34), но, с другой
стороны, свет тормозит двигательную реакцию, если оборонительный
рефлекс вызывается во время действия света (Иоханес, 33;
Бебуриіпвили, 34) (рис. 98-П). У нормальных кроликов звук и свет
благоприятствуют оборонительным рефлексам, но у кроликов,
отравленных хлоралозой, с пониженной корковой деятельностью, те же
звук и свет понижают рефлекторную деятельность; прекращение их
ведет, наоборот, к облегчению оборонительных рефлексов
(Дзидзишвили, 26).
Очень интересную иллюстрацию облегчающего действия высших
отделов мозга на спинной дал Демидов (8) из лаборатории И. П.
Павлова. У собаки, лишенной больших полушарий, рефлекторная
деятельность очень сильно повышается вследствие повышения- возбудимости
центральной нервной системы. Демидов заметил на таких собаках,
ЯВЛЕНИЕ ОБЛЕГЧЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ 213
что звуковое раздражение сильно- благоприятствует чесательному
рефлексу, вызываемому в ответ на раздражение соответствующего
рецептивного поля (на плечах, ушах, шее). Опыт ставился так: сначала
производилось слабое механическое раздражение в поле чесания, в ответ
на это рефлекс чесания или совсем не вызывался, или вызывался в
очень слабой форме. Во время этого раздражения производилось
звучание какого-нибудь музыкального инструмента. В отдельности звучание
не давало чесательного рефлекса и вообще никакого движения на
ногах, но в комбинации со слабым раздражением рецептивного поля
чесания оно усиливало существующий рефлекс чесания или вызывало его
с самого начала, если рефлекса не было.
Все ,эти опыты единогласно свидетельствуют о том, что головной
мозг оказывает на рефлекторную деятельность спинного мозга усак
облегчающее, так и тормозящее действие, и что это действие
определенным образом зависит от внешних раздражений. При этом то или
другое внешнее раздражение может обусловить подобное действие
головоного мозга даже в том случае, если оно само по себе не вызывает
внешних двигательных реакций.
Происхождение облегчающего и тормозящего действия высших
отделов мозга на низшие. Облегчающее действие внешних раздражений
безусловно должно зависеть в определенных случаях от
распространения нервных импульсов из головного мозга в спинной. От этого в
проводящих путях и нервных кругах спинного мозга возбудимость
повышается так же, как при раздражении соответствующего рецептивного
поля или чувствительного нерва. Облегчающее действие коры большого
мозга и тонических центров, повидимому, объясняется иррадиацией
исходящих от них импульсов по спинному мозгу. Но едва ли можно
утверждать, что облегчающее действие головного мозга на спинной мозг
происходит всецело этим путем. Звук и свет обычно не вызывают
двигательных реакций ни у лягушки, ни у кролика; они не дают реакций на
конечностях. Несмотря на это, они действуют на спинной мозг и
меняют рефлекторные реакции на конечностях. Отсюда следует, что те
нервные комплексы головного мозга, которые воспринимают звуковыя
и световые раздражения, не должны быть соединены
дифференцированными нервными путями с двигательными нейронами конечностей. Тем
не менее, звук и свет в определенных случаях то повышают, то
понижают их возбудимость. Очевидно, механизм облегчающего действия
головного мозга на спинной мозг в таких случаях совершенно иной.
Нужно думать, что в облегчающем <и. тормозящем действиях
головного мозга определенную роль играет также чисто электротоиическое
распространение биотоков нейропиля средних отделов мозга на нервные
пути и нервные круги головного мозга. Звук и свет, возбуждая
соответствующие нервные круги средних отделов мозга, активируют .также
нейропиль этих отделов мозга. Этим самым такие раздражения произ-
іводят через нейропильный ток торможение рефлекторных реакций;
вызываемых через средние отделы мозга: например, дыхательных движе-
214 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
пий, освободительных движений, которые наступают на привязанных
животных при любом повреждающем раздражении (Бебуришвили, 34;
Дзидзишвили, 26). Этот же нейропильный ток в момент своего
исчезновения, в связи с прекращением раздражения, должен оказывать
противоположное действие на рефлекторные пути средних отделов мозга, т. е.
должен облегчать наступление рефлекторных реакций.
Однако нейропильный ток средних отделов мозга должен действовать
не только на нервные круги средних отделов мозга, но и на нисходящие
нервные пути и нервные круги, соединяющие головной мозг со спинным.
Нейропильный ток, входя в аксоны и клетки средних отделов мозга,
должен производить в них анэлектротоническое понижение
возбудимости. Этот же ток, распространяясь по осевым цилиндрам нисходящих
путей и нервных кругов электротонически, должен вызывать в них
повышение возбудимости в связи с выхождением их наружу через
перехваты Ранвье и нервные окончания — синапсы, т. е. здесь должно
происходить известное пери?лектротоническое действие. Вследствие этого
наряду с угнетением рефлекторных реакций, дроисходящих через
средние отделы мозга, должно получаться облегчение реакций,
происходящих через нижние отделы, .через спинной мозг. Этим, вероятно, и
объясняется, что очень часто звук и свет вместо угнетения рефлекторных
реакций производят их усиление. Повидимому, это является
результатом облегчения рефлекторной деятельности ів спинном мозгу под
влиянием периэлектротонического действия головного нейропильного тока
на спинальные нервные пути.
Итак, в основе тормозящего и облегчающего действия высших
отделов мозга на низшие лежит, с одной стороны, ослабление и усиление
той постоянной импульсации, которой подвергаются двигательные
клетки низших отделов со стороны высших отделов, а с другой —
электротоническое действие нейропильного тока средних отделов мозга на
нервные круги этих же отделов мозга и на те проводящие пути, которые
соединяют их с низшими отделами мозга.
Практикум
Наблюдение повышения возбудимости в ответ на деятельное раздражение.
Спинномозговая лягушка. Рефлекс сгибания. Записывается m. semitendinosus.
Раздражается п. peroneus индукционными токами частотой 30—50 ударов в 1 сек. Опыт
начинается спустя 10—15 мин. после операции. Сначала находят порог раздражения и
записывают на кимографе пороговый эффект. Затем раздражают сильными токами я
записывают полученный рефлекторный эффект. После этого, не останавливая
кимографа, снова отыскивают пороговое раздражение: оно окажется более слабым, чем
было до сильного раздражения.
Наблюдение суммации. На том же препарате скачала находят порог
раздражения п. peroneus для сгибательного рефлекса в отношении отдельных индукционных
ударов, а эатем в отношении титанического раздражения. В следующий раз берут
m. triceps femoris и п. cutaneus femoralis lateralis или п. brachialis и повторяют тот
же опыт для рефлекса потираіния. Отдельные удары совсем не будут давать
рефлекса потгаранюя. Его будет вызывать тетаническое раздражение, (притом при значительно
большем сближения каггуішек, чем сгабательный рефлекс. В этих опытах надо соблю-
ЯВЛЕНИЕ ОБЛЕГЧЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ 215
дать однородность физических условий. Размыкание и замыкание переичиой цепи для
получения одиночных ударов производятся через тот же прерыватель тока, который
*слуіжит длія тетаіниічешоігю раздражения.
Повышение возбудимости вследствие иррадиации возбуждения. Спинномозговая
лягушка. Записывается гл. semitendinosus. Раздражают из поля рефлекса сгибания
для чувствительных нервов: п. peroneus и п. superficialis. Сначала находят порог
тетаническсго раздражения для одного из нервов. Потом сильно раздражают тетали*
чески другой нерв и после этого снова находят порог раздражения первого нерва.
При этом произойдет понижение порогового раздражения. N. peroneus входит
главным образом в IX сегмент, п. superficialis — в X сегмент. Поэтому повышение
возбудимости будет происходить вследствие иррадиации возбуждения.
Можно взять для раздражения п. superficialis и п. cutaneus femoralis lateralis;
этот последний нерв отдает нервные волокш VIII сегменту. В данном опыте сначала
испытывают раздражение п cutaneus femoralis lateralis; обычно оно не дает
сокращения m. semitendinosus, а если и дает его, то ів очень слабо 3 форме. Потом сильно
раздражают п. superficialis, что вызывает повышение возбудимости в X сегменте»
Вскоре после этого вновь лробуют раздражать п. cutaneus femoralis lateralis. В ответ
на это m. semitendinosus дает значительное сокращение. Такой эффект должен
вызываться из X сегмента под влиянием возбуждения, иіррадиирующего из VIII сегмента.
Как известно, раздражение п. cutaneus femoralis lateralis вызывает очень
интенсивное торможение m. semitendinosus, и если в данном случае после раздражения
п. superficial^ раздражение п. cutaneus femoralis lateralis дает сильное сокращение,
то причина должна заключаться только в возбуждении X сегмента вследствие
иррадиации возбуждения.
Влияние света и звука на рефлекторные реакции лягушки. Нормальную ляпушку
привязывают к дощечке с помощью куска материи, имеющей вырезы для
конечностей. Задние конечности свободно висят в воздухе. Размах их движений можно
измерить по миллиметровой шкале или записать на кимографе, соединив лапку с
миографом. Путем раздражения кожи конечности отдельными индукционными ударами
.вызываются освободительные движения. Затем это раздражение испытыаается во
время звучания электрического звонка и освещения электрической лампой от
автомобильного фонаря, укрепленного на расстоянии 5—6 см от глаз. Прослеживаются
изменения оборонительных реакций на электрическое раздражение лапки во время
света и звука.
Затем то же раздражение лапки применяется сейчас же вслед за прекращением
света и звука. Устанавливается, как меняются освободительные рефлекторные
реакции во время света и звука и вскоре после них.
Литер ату р а
1. Forbes A., J. Physiol , 31, 102, 1912.
2. Wedensky N., Fol. Neurobiol., 6, 591, 1912.
3. В e г i t о f f J., Arch.Anat. Physiol., 1912,5стр. 266; Tp СПб. об-ва естеств., 42,
в. 4, стр. 245, 1913.
4. Sherrington С. I., Int. Act. Nerv. Syst, 1906.
5.?B ubnoff u. Heidenhain Pfl. Arch., 20, 137, 1881.
6. Merzbacher, Harward Psych. Stud., 1, 1903.
7. Yerkes R., Pfl. Arch., 107, 1905.
8. ДемидовВ., Условные (слюнные) рефлексы у собаки без передних половин
обоих полушарий, СПб. Дисс. из лаб. Павлова, 1909.
9. Sherrington С, Ргос. Roy. Soc., В, 95, 407, 1923; Ргос. Roy. Soc., В.,
97, 488, 1925.
10. В е г і t о f f J., Z. Biol., 64, 161, 176, 285 и 298, Ш4.
11. Lorente de N6. Юбилейный сборник проф. И. Бериташвили. Изд. Груз,
отд. АН СССР, 1936.
12. Cread R. S.t Denny-Brown DM Eccles I. C, Liddell G.
216 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
a. Sherrington С. S , Reflex activity of the spinal cord. 1932. Есть русск
пер. Биомедгиз, 1935.
13. СеченовМ. И., Об электрическом и химическом раздражении чувствующих
спинномозговых нервов лягушки, СПб., 1868.
14. В ег it off J., Pfl. Arch., 202, 265, 1924.
15. Graham Brown Т., Quart. J. Exp. Physiol., 0, 81, 101 117 и 131, 1915..
16. Б e p и t о в И., Бакурадзе А. и НарикашвилиС, Тр. Физиол.
ин-та им. Бериташвили, 3, 173, Тбилиси, 1937.
17. Bernstein S., Am. J. Physiol., 120, 798, 1937.
18. Bremer Ft, С R. S. Biol., 103, 509, 1930; С R. S. Biol., 104, 819, 1930.
19. Lorento de N6, R.( Am. J. Physiol.» Ill, 2,2a, 289, 1935; 112, 595, 193=*.
20. Schriever H., Z. Biol., 04, 1933; 95, 105,1934; Erg. Physiol., 38, 877, 3936.
21. Schriever H. a. Perschmann, Pfl. Arch., 236, 447, 1935.
22. L a p i с q u e L., Cold Spr. Harb. Symp. Quant. Biol., 4, 98, 1936.
23. ГедеваниД., Тр. Физиол. ин-та им. Бериташвили, 3, 479, 1937.
24. Беритов И. и Бакурадзе А., Физиол. журн. СССР, 27, 387, 1939.
25. Беритов И. иБакурадзе А., Физиол. журн. СССР, 28, 18, 1940.
26. Д з и д з и ш в и л и Н., Физиол. журн. СССР, 26, 354, 1939; Сообщ. Груз. фил.
АН СССР, 1, 217, 1940.
27. Ц к и я у р и д з е Л., Тр. Физиол. ин-та им. Бериташвили, 3, 331, 1937.
28. LehmannJ.E., Am. J. Physiol., 118, 600, 1937.
29. L о r e n t e d e N 6., J. Neurophysiol., 2, 402, 1937.
30. Adrian E. D. a. Moruzzi, J. Physiol., 97, 153, 1939.
31. Гедевани Д., Физиол. журн. СССР, 26, 384, 1939; 28, 18, 1940.
32. В в е д е н с к и й Н. Е., Соч., т. IV, стр. 47, 1938.
33. Johannes Th., Pfl. Arch., 224, 372, 1930.
34. Бебуришвили H., Тр. Физиол. ин-та им. Бериташвили, 3, 345, 1937.
35. Lapicque L., С R. Acad. ScL, Paris, 155, 70, 1912; Cold. Spr. Harb. Symp.,
Quant. Biol., 4, 98, 1936.
36. Беритов И. и Бакурадзе А., Тр. Йн-та физиол. им. Бериташвили, 5,.
I, 1943.
37. Беритов И. иБакурадзе А., Тр. Ин-та физиол. им. Бериташвили. 5,
125, 1943.
38. Т а р х а н о в И., Известия Имп. Анад. Наук, 1870, т. 7, стр. 717.
Г и а в а XI
УТОМЛЕНИЕ
Общее понятие. Центральная нервная система, как и всякая живая
ткакь, требует для своей деятельности питательных веществ и
кислорода. Если она не обладает всем этим в достаточной мере, она более или
менее быстро утомляется. Ф е р в о р н, которому мы обязаны первыми
исследованиями в этом направлении, различает двоякого рода
утомление: утомление в собственном смысле слова (Ermudung), обусловленное
главным образом накоплением продуктов обмена веществ, и истощение
(Erschopfung), вызываемое недостатком кислорода (1). Ф е р в о р н
пришел к этому заключению на основании следующих опытов.
Отравленные стрихнином лягушки после первоначальных судорожных
припадков приходят в такое состояние, что дрипадки эти становятся все
слабее и слабее и наступают все реже и реже. Наконец, после каждого
припадка центральная нервная система лягушки лишается возбудимости
на более или менее значительное время. Периферическое раздражение
в этот момент эффекта не дает. Эта фаза абсолютной невозбудимости
после каждой новой судороги все увеличивается во времени, и, наконец,
происходит окончательная потеря возбудимости. Такое же изменение
рефлекторной деятельности наблюдается и в том случае, если перед
отравлением заменить кровь физиологическим раствором, который был
осзобожден от кислорода путем кипячения. Если после стадии судорог
в период полной невозбудимости переменить физиологический раствор
на новую порцию бескислородного физиологического раствора, то до
некоторой степени рефлекторная возбудимость восстанавливается на
небольшое время. Значит, в данном случае абсолютное утомление
центральной нервной системы было вызвано отравляющим действием
продуктов обмена веществ. Если в описанном опыте произвести
промывание мозга физиологическим раствором, насыщенным кислородом, то
происходит более значительное восстановление рефлекторной
возбудимости. Отсюда совершенно ясно вытекает, что в жизнедеятельности
центральной нервной системы кислород играет существенную роль; чем
меньше в ней запас кислорода, тем скорее падает ее возбудимость, тем
скорее происходит ее утомление.
218 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Согласно более поздним наблюдениям Б ал ион и (2), спинной мозг
лягушки, вынутый из позвоночного канала и лишенный
кровообращения, может сохранить возбудимость в течение 1—2 дней, если его
держать в атмосфере чистого кислорода. На воздухе такой препарат
сохраняет возбудимость только в течение 2 часов при комнатной
температуре (18—20°).,Чем ниже температура, тем дольше сохраняется
возбудимость.
Из продуктов обмена веществ, влияющих на центральную нервную
систему в смысле утомления, известную роль должна играть
углекислота. Повидимому, еще более существенное значение должны иметь не-
аолные продукты сгорания. В упомянутых опытах Б алию ни, падение
возбудимости в изолированном спинном мозгу, должно быть,
происходило не только из-за отсутствия кислорода, а главным образом
вследствие накопления ядовито действующих продуктов обмена веществ,
окисляющихся при наличии кислорода до углекислоты (Винтерштейн, 3).
Обмен веществ в мозгу. По приблизительным подсчетам Винтер-
штейна, 1 г спинного мозга в течение 1 часа потребляет 250—
350 імм3 кислорода (в атмосфере чистого кислорода). По исследованиям
Босса (4), потребление кислорода еще значительнее, причем оно не
одинаково в головном и спинном імозгу; головной мозг потребляет
кислорода на 13—20% больше, чем спинной. Мозг теплокровного
животного потребляет больше кислорода, чем мозг холоднокровного: 1 г
головного мозга теплокровного потребляет в 1 час 815 мм3 кислорода, а
холоднокровного — 682 мм3. Если головной и спинной мозг не будут
обеспечены соответствующим количеством кислорода, то при работе они
неминуемо будут утомляться.
Во время нормальной рефлекторной деятельности, вызываемой
раздражением кожи или чувствительного нерва, не удалось наблюдать
повышения газообмена в спинном мозгу: отдача углекислоты и
потребление кислорода происходят во время рефлекторной деятельности
приблизительно с той же скоростью, как и в отсутствие этой деятельности
(Винтерштейн, 9). Только при стрихнинном отравлении было отмечено
небольшое повышение газообмена. Заметно повышенный газообмен
наблюдается при непосредственном электрическом раздражении спинного
мозга (Винтерштейн, 9).
Точно так же в головном мозгу собаки при нормальном
кровообращении не ^удалось обнаружить увеличения потребления кислорода и
выделения углекислоты в связи с работой. Лондон и сотрудники
установили это по опытам на ангиостомированных собаках, у которых
определялись количества кислорода и углекислоты в притекающей и
оттекающей крови (13). ч
Оказывается, что при рефлекторной деятельности спинного мозга
усиленно потребляется глюкоза. В спокойном состоянии спинной мозг
лягушки потребляет в 1 час в среднем 0.37 мг глюкозы на 1 г мозга.
Раздражение лапки путем щипания вызывает повышение потребления
УТОМЛЕНИЕ
219
сахара ;на 100% и больше. Это повышение потребления сахара не
сопровождается повышенным потреблением кислорода. При этом сахар не
расщепляется до молочной кислоты (Винтерштейн и Френкель-Конрат,
10). Иногда ^наблюдается ничтожное увеличение содержания молочной
кислоты (Лондон и сотр., 13). Но это может получиться и от усиления
тока крови, что обычно имеет место при усиленной деятельности мозга.
По новейшим исследованиям, не отрицается образование молочной
кислоты из глюкозы и окисление ее в мозгу, но предполагается, что это
окисление происходит не непосредственно, как в мышцах, а сложным
путем — через предварительное образование.пировиноградной кислоты
(К'востел, 20).
Для жизнедеятельности центральной нервной системы имеет
существенное значение также присутствие ионов натрия. Так, например,
Балиони, изучая влияние разных растворов на изолированный
спинной мозг, нашел, что изотоничный раствор хлористого натрия нельзя
заменить изотоничным раствором какого-либо другого вещества
(глюкозы, глицерина, хлористого калия, хлористого лития).
Физиологический раствор можно заменить только изотоничными растворами
натриевых же солей (двууглекислым натрием, азотнокислым натрием).
Спинной імозг теряет способность к рефлекторной деятельности в изотонич-
яом растворе глюкозы спустя 2—5 часов (в атмосфере кислорода).
Бели затем перенести препарат в физиологический раствор, то
возбудимость его вновь возвращается. Важное значение имеет соотношение
калия и кальция в жизнедеятельности мозга. Известно, например, что
при возбуждении мозга во время эпилептических припадков или при
длительном раздражении мозга электрическим током происходит
задержка калия и выделение кальция. В периоде же угнетения,
наблюдаемом вслед за возбуждением, происходит задержка кальция и
выделение калия (Кассиль, 14).
В центральной нервной системе, конечно, происходит также распад
белка и потребление его. При этом образуется, между прочим, аммиак.
Раздражение, повидимому, не усиливает белкового обмена. Наркоз же
сильно влияет на белковый обмен, снижая отдачу аммиака
(Винтерштейн и Гиршберг, 25). Выход из мозга аминосодержащих веществ
при состоянии возбуждения был отмечен в оттекающей из .мозга крови
также в условиях нормального обмена веществ (Рубель, Фрид и іК'ис-
ллнекий, 21).
В результате обмена веществ возникает, между прочим, ацетилхо-
лин. Он, повидимому, усиленно образуется в нервных элементах. В
мозгу лягушки ацетилхолин находится в количестве 4—5 у на 1 г мозга.
Из этого количества 20% растворяется в воде, а 80% в ней
нерастворимо, но растворяется в алкоголе. Эта часть должна быть соединена с
липоидом, по всей вероятности, при участии белка (Лёви, 15).
Предполагается, что ацетилхолин в мозгу синтезируется из холина и двууксус-
ной кислоты (Стедман и Стедман, 16).
220 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Участие центральных элементов в рефлекторном утомлении.
Впервые Ферворн (1) показал, что в утомлении рефлекторного аппарата
эфферентная часть дуги не участвует. Отравленная стрихнином
лягушка после ряда судорог совершенно теряет способность к рефлекторной
деятельности, но если раздражать переднюю поверхность мозга в
грудной области, тг наступают движения задних лапок. Этот эффект должен-
вызываться раздражением передних и боковых столбов, волокна
которых оканчиваются около клеток двигательных нейронов (Балиони, 5).
Это ясно указывает на отсутствие утомления в эфферентной части
рефлекторной дуги, т. е. в двигательных нейронах.
Вообще из опытов на стрихнинизированных препараіах лягушки
ясно следует, что в результате длительной судорожной деятельности
утомление координирующего рефлекторного аппарата, т. е.
промежуточных нейронов, наступает гораздо раньше, чем утомление
двигательных нейронов (Введенский, 22). Аналогичное заключение нужно
сделать на основании опытов на нормальных препаратах. Так, если
раздражать чувствительный нерв из поля сгибания,— например п. peroneus,—
тетаническими токами умеренной силы до полного прекращения сгиба-
тельного рефлекса и сейчас же вслед за этим раздражать другой
чувствительный нерв тогЪ же поля,— например п. tibialis,— то последнее
раздражение вызовет не ослабленный рефлекторный эффект, а,
наоборот, усиленный. Но и при сильном тетаническом раздражении первого
нерва, продолжаемом до наступления полного утомления, раздражение
второго нерва произведет эффект,— только он будет несколько
ослаблен (Ли и Эверинхэм, 18; Форбс, 19). Так получается даже тогда,,
когда второе раздражение производится во время первого, в период
полного прекращения рефлекса (Шеррингтон, 6). Отсюда очевидно,
что во время утомления рефлекторного аппарата в результате
длительного раздражения одного участка рецептивного поля двигатаяьные
нейроны сохраняют способность почти к максимальной деятельности.
В новейшее время это подтвердил Лоренте де Но (11),
который показал неутомляемость двигательного нейрона глазных мышц при
непосредственном раздражении 5 раз в 1 сек. Но если раздражение в-
том же ритме производилось через пр'омежуточный механизм,
утомление непременно наступало и проявлялось в виде ослабления сокращения
глазных мышц.
Итак, утомление рефлекторного аппарата обусловливается падением
возбудимости и ослаблением деятельности в промежуточных нейронах,
т. е. в тех центральных элементах, где протекают главные
координирующие процессы.
Приведенный выше опыт с раздражением двух нервов из поля
одного рефлекса имеет большое значение еще и в другом отношении.
Утомление рефлекса в результате раздражения одного чувствительного
нерва или одного участка данного рецептивного поля не обусловливается
утомлением всего координирующего аппарата данного рефлекса. При
некоторых умеренных раздражениях данного чувствительного нерва
УТОМЛЕНИЕ
221
утомляется только определенная часть этого аппарата, наиболее тесно
с ним связанная. Остальная часть совсем не утомляется,— наоборот, ее
возбудимость и реакционная способность повышаются. Только при
сильных раздражениях данного чувствительного нерва утомляется не только
тесно связанная с ним часть координирующего аппарата, но и
остальная его часть. При этом последняя утомляется в меньшей степени.
Из этих наблюдений следует, что при координированной центральной
деятельности фракционированию подвергаются первично не
двигательные, а промежуточные нейроны, из которых составляются
координирующие аппараты. Как из этих физиологических наблюдений, так и из
гистологических исследований видно, что определенные рецепторы, или
чувствительные волокна, связаны преимущественно с определенным
комплексом промежуточных нейронов, входящих в координирующий
¦аппарат данного рефлекса. 'А комплекс промежуточных нейронов, пови-
димому, связан преимущественно с определенной группой двигательных
нейронов. Благодаря этому во время каждой координированной
иннервации в активном состоянии должна находиться определенная часть
координирующего аппарата с определенной группой двигательных
нейронов. Несмотря на это конвергенция рефлекторных путей происходит
не только на двигательных клетках, но в некоторой-мере еще раньше,
на промежуточных нейронах координирующего аппарата. Этим и
должно объясняться то явление, что утомление рефлекторного аппарата
раздражением одного участка рецептивного поля, одного его
чувствительного нерва определенным образом влияет на рефлекс, вызываемый
со стороны другого участка или другого нерва того же рецептивного
поля, сначала в смысле его повышения, а потом в смысле понижения.
Итак, при длительном раздражении одного участка рецептивного
поля или одного чувствительного нерва утомляется прежде всего одна
определенная часть координирующего аппарата, наиболее тесно
связанная с раздраженными рецепторами или чувствительными волокнами.
Изменение электрической активности спинного мозга цри его
утомлении. При длительных раздражениях чувствительных нервов или
самого опийного мозга, электрическая активность его в период утомления
меняется характерным образом. После начальной стадии облегчения,
когда электрический эффект постепенно нарастает, усиленный эффект
удерживается некоторое время — более или менее длительно, в
зависимости от функционального состояния животного. Затем электрический
эффект постепенно ослабевает. При отведении от передних корешков
быстрые потенциалы всегда исчезают раньше медленных, а из
медленных исчезают раньше те, которые зависят от промежуточных нейронов,
т. е. нерегулярные потенциалы разной амплитуды и длительности.
Позднее всего исчезают регулярные медленные потенциалы, наступающие
вслед за заднекорешковым биотоком возбуждения. Эти биотоки
возбуждения ослабевают последними, и притом при более частых и длительных
раздражениях (рис. 99)
222 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Точно так же, при отведении от задних корешков или от
поверхности мозга, дополнительно возникающие медленные потенциалы
исчезают при утомлении раньше регулярных медленных потенциалов. Если
при этом наблюдались и быстрые потенциалы, то они исчезают вместе
с нерегулярными медленными потенциалами (Беритов и сотр., 27).
Рис. 99. Изменение электрических потенциалов спинного мозга
при утомлении.
Спинальный препарат лягушки. Все передние корешки перерезаны.
Отводится IX передний корешок слева на расстоянии 2 мм от мозга. Раздражается
п. peroneus sin. релаксационным стимулятором при 0.5 V (порог 0.05 V).
А—начало раздражения; В—через 1 сек.; С—через 5 сек.; D—через 30 сек.;
?"—через 3 мин.
Итак, судя по изменениям электрической активности при
продолжительной деятельности спинного мозга, прежде всего утомляются
промежуточные нейроны, значительно позднее — двигательные; при этом и те
и другие нервные элементы первоначально утомляются в отношении
возбуждения, а затем, значительно позднее,— в отношении
локальных процессов. В момент исчезновения медленных потенциалов задне-
корешковые элементы внутри спинного мозга (конечные разветвления
заднекорешковых волокон с их синапсами), которым принадлежит
начальный быстрый потенциал, обнаруживают лишь небольшое
утомление или даже не показывают его вовсе.
УТОМЛЕНИЕ
223
Структурные изменения центральной нервной системы при
длительной деятельности. При длительной деятельности ослабление последней
может происходить в результате определенных структурных изменений
синаптичесшх образований. При исследовании изменений,
происходящих в растущем конусе перерезанного нервного волокна, а также в
Рис. 100. Явление утомления на лумбальном препарате кошки.
Продолжение опыта, представленного на рис. 34. Два раза повторяется длительное тетаниче-
ское раздражение левого VIII заднего корешка на фоне рефлекторных вздрагиваний всех
мышц. После падения тетанического сокращения на сгибателе и разгибателе
соответствующей стороны торможение ослабевает, но не исчезает: после того как сокращение мышц
сошло на-нет, торможение еще долго удерживается на всех мышцах (Беритови»Бакурадзе).
кожных окончаниях чувствительного нерва в результате химического,
температурного, механического или иного раздражения было
обнаружено, что в случае сильного и длительного раздражения происходит
сморщивание, укорочение самого окончания (Спейдель, 17).
Аналогичное явление может иметь место при длительном возбуждении синапсов:
в результате сморщивания последних может увеличиться пространство,
отделяющее синапсы от тела клетки, к которому они прилегают. Таким
образом, ухудшаются условия передачи возбуждения с синапсов на
тело клетки.
Роль торможения в ослаблении двигательного эффекта при
длительном раздражении. Когда во время длительного раздражения
двигательный эффект начинает ослабевать и, наконец, исчезает, это еще не
выражает перехода от активного состояния центральной нервной
системы « покою. Обычно в это время центральная нервная система обнару-
^24 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
живает состояние торможения. В это время все рефлекторные реакции,
вызываемые на утомленной конечности, оказываются ослабленными.
Кроме того, очень часто тотчас же по прекращении раздражения
наступает облегченное состояние: все двигательные реакции на этой
конечности оказываются усиленными.
Впервые явление общего торможения в момент ослабления
рефлекторного сокращения мышц при длительном раздражении было
отмечено Сеченовым (12). Но Сеченов наблюдал это явление только
в случае применения сильного раздражения. Затем Введенский
(22) описал его на стрихнинизированных препаратах лягушки. В
первых стадиях отравления, когда одиночное эффективное раздражение
вызывает сильные общие вздрагивания и судороги, тетаническое
раздражение любого чувствительного нерва частотой 20—100 в 1 сек.
производит более или -менее' быстро протекающий тетанус, сменяющийся
длительным общим торможением. В момент расслабления мышц
никакое другое раздражение .чувствительных нервов не способно вызвать
двигательный эффект. В определенных случаях по прекращении
раздражения вновь возникает сокращение,— очевидно, в результате
облегчения, наступающего вслед за торможением.
Но и на нормальном препарате легко установить, что очень часто
ослабление рефлекторного сокращения во время длительного
раздражения сопровождается общим торможением. Так, например, на лумбаль-
ных препаратах кошки в момент ослабления рефлекторного
сокращения, вызываемого длительным раздражением одного заднего корешка,
одиночные рефлекторные вздрагивания, вызываемые с другого заднего
корешка, ослабевают, а по прекращении длительного раздражения,
наоборот, усиливаются (рис. 83).
Частота тетанического раздражения имеет определенное значение
для быстроты расслабления: при некоторых высоких частотах
сокращение ослабевает ввиду развития сильного торможения. Это наблюдается
на препаратах как лягушки (Гогава, 23), так и кошки (Беритов и Ба-
курадзе, 24). Это указывает, что процесс торможения усиливается не
только с усилением раздражения, но и с учащением его. Однако в
определенных случаях быстрое ослабление двигательного эффекта
происходит при всякой силе и частоте раздражения. На лумбальных
кошках, например, это наблюдается в отношении мышц колейа при
раздражении IX—X задних корешков, через которые вступают в мозг
чувствительные волокна из хвоста и промежности. При всякой частоте
раздражения следует после начального сокращения общее торможение на
мышцах колена той и другой конечности. Это наблюдение указывает,
что здесь мы имеем не ослабление сокращения благодаря
рефракторной фазе, т. е. не пессимальный эффект в нервах или центрах, а
настоящее торможение.
Характерно, что когда при длительном тетаническом раздражении
рефлекторное сокращение ослабевает, то наблюдаемое при этом тормо-
УТОМЛЕНИЕ
22
жение имеет колеблющийся характер. Если в это время вызывать
рефлекторные вздрагивания раздражением какого-либо другого заднего
корешка,— вздрагивания то ослабевают, то нет, а то даже усиливаются
(рис. 83,100). Такую изменчивость угнетения мы объясняем колебанием
интенсивности торможения. Если бы ослабление двигательного эффекта
было обусловлено утомлением, то такая изменчивость угнетения не
могла бы иметь место.
Это торможение при длительном раздражении не только
сопровождает двигательные эффекты, но происходит и по прекращении их.
При этом оно может длиться в чистом виде в течение многих десятков
секунд, хотя с течением времени и ослабевает, как это хорошо видно на
рис. 100. Значит, при длительном активном состоянии центральной
нервной системы ослабление тормозящего действия наступает значительно
позднее, чем прекращается возбуждающее действие.
Это, очевидно, стоит в связи с тем обстоятельством, что нервные
элементы утомляются в отношении локальных процессов, которые
своими потенциалами производят торможение, значительно позднее, чем в
отношении возбуждения.
Выше было указано, что при комбинации тормозящего раздражения
с возбуждающим тормозящее действие на двигательные нейроны
проявляется тем сильнее, чем слабее возбуждение последних. Отсюда
следует, что при длительном раздражении все условия благоприятствуют
проявлению общего торможения.
На основании всех этих фактов нужно заключить, что ослабление
центральной спинномозговой деятельности должно существенно
зависеть от общего центрального торможения. Так как последнее возникает
при каждом активировании центральной нервной системы и длится
дольше центрального возбуждения, то его угнетающее действие
на двигательные нейроны доляшо иметь место и при длительной
утомляющей деятельности. Отсюда следует, что ослабление центрального
двигательного эффекта представляет собой более сложное и
качественно иного рода явление, чем ослабление двигательного эффекта нервно-
мышечного препарата. Можно даже утверждать, что первоначальное
ослабление центрального двигательного эффекта^ по существу говоря,
обусловливается главным образом не утомлением каких-либо нервных
элементов, а центральным торможением.
Об утомлении центральной нервной системы в отношении
торможения. Каждый раз при длительном тетаническом раздражении
чувствительных нервов торможение с течением времени постепенно ослабевает.
Это хорошо видно, между прочим, на рис. 100: одиночные вздрагивания,
вызываемые через те же двигательные нейроны раздражением других
нервов, восстанавливаются во время тетанического раздражения. Это
ослабление торможения совпадает с ослаблением сокращений в
определенной группе мышц. Регистрация биотоков спинного мозга показывает,
что при длительных раздражениях быстрые колебания потенциала от
промежуточных нейронов нервных кругов ослабевают и исчезают парал-
1о И. С. Беритов
226 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
лельно с теми медленными потенциалами, которые возникают
одновременно с ними. Отсюда становится совершенно вероятным, что
ослабление центрального торможения при рецшрокных реакциях обусловлено
утомлением промежуточных нейронов, ибо это последнее обстоятельство,
fB свою очередь, приводит к ослаблению активного состояния нейропиля.
Если тетавическое раздражение вызывало заторможенное сокращение,
то с течением раздражения торможение проходит, но от этого
сокращение не восстанавливается, ибо как торможение, так и возбуждение
двигательных клеток вызывалось через промежуточные нейроны.
Итак, при длительных периферических раздражениях ослабление
центрального торможения, если оно вызывается наряду с возбуждением
двигательных клеток, обусловливается главным образом утомлением
промежуточных элементов: вследствие этого ослабевает как
возбуждающее действие их прямо на двигательные клетки, так и тормозящее
действие их через нейропилъ.
Как указывалось выше, общее торможение может быть вызвано и
таким раздражением кожной поверхности или чувствительных нервов,
которое не дает двигательных рефлекторных реакций. Это дает право
утверждать, что общее торможение может быть вызвано и .без
активного участия промежуточных нейронов. Исследование биотоков
спинного мозга показывает, что и в случаях отсутствия возбуждения
промежуточных элементов медленные потенциалы наступают при
раздражениях чувствительных нервов; это наблюдается при слабых
раздражениях на хорошо функционирующем препарате и при всех силах
раздражения вскоре после исчезновения рефлекторной деятельности. При
редких раздражениях, до 60 в 1 сек., медленные потенциалы следуют вслед
за каждым заднекорешковым импульсом непосредственно. Если
раздражение продолжается несколько десятков секунд или несколько минут,
эти медленные потенциалы ослабевают и, наконец, исчезают полностью.
Эги медленные потенциалы возникают © дендритах и клетках нейропи-
ля спинного мозга под влиянием заднекорешковых импульсов. Эти
потенциалы довольно значительны. При отведении их от задних корешков
они достигают нескольких сот микровольт. Поэтому они не могут не
оказать анэлектротонического действия на нервные элементы мозга и
тем не обусловливать торможение. Последнее можно утверждать, между
прочим, и на том основании, что после электрического эффекта от
первого раздражения все последующие эффекты на повторные
раздражения оказываются ослабленными, если они попадают во время
медленного потенциала. Ослабевают не только медленные потенциалы, но и
биотоки возбуждения заднекорешковых волокон или их коллатералей в
опинном мозгу (Беритов и Ройтбак, 26).
Отсюда следует, что в отсутствии двигательных реакций центральное
торможение ослабевает при длительных периферических раздраоісениях
вследствие утомления дендритной и клеточной массы нейропиля в
отношении локальных процессов, вызываемых заднекорешковыми
импульсами.
УТОМЛЕНИЕ
227
Развитие рефлекторного утомления в зависимости от интенсивности
раздражения. Утомление координирующего аппарата рефлекса
происходит тем скорее, чем слабее раздражение. Слабое раздражение
перестает вызывать рефлекс очень быстро, в течение нескольких секунд;
сильное раздражение производит сильный и долго длящийся рефлекс
(Шеррингтон, 6) (рис. 101). Это парадоксальное явление зависит,
вероятно, от следующих причин: слабое раздражение приводит в действие
небольшое число промежуточных нейронов, а сильное раздражение —
тем больше нейронов, чем оно сильнее; в последнем случае эти нейроны
приходят в действие не все сразу, а последовательно — один за другим.
Основное условие для этого явления должно заключаться в том, что
координирующие промежуточные нейроны, или, вернее, нервные круги
и цепочки, получают не одинаковое количество коллатералей от каждого
чувствительного нейрона соответствующего рецептивного поля. Одни
нервные цепочки и круги получают их больше, другие меньше.
Вследствие этого при раздражении одной труппы чувствительных нейронов
прежде всего будут приходить в действие те нервные круги и цепочки,
которые получают от них наибольшее количество коллатералей, а
затем уже другие круги и цепочки, по мере повышения в них
возбудимости под влиянием того же раздражения. Чем меньше коллатералей
оканчивается на данных нервных кругах и цепочках, тем позднее они
придут в активное состояние, так как тем позднее возбудимость
промежуточных нейронов достигнет такой степени, что они смогут
возбудиться.
Во время деятельности возбудимость промежуточных нейронов
падает быстро, гораздо быстрее, чем в мышце или нерве. Чем сильнее
падает возбудимость этих нейронов, тем сильнее должно быть
применяемое раздражение, т. е. тем больше ^должно быть число
возбужденных чувствительных волокон для вызова возбуждения. Отсюда понятно,
что каждый деятельный координирующий нейрон тем іскорее должен
перестать возбуждаться, чем слабее раздражение, т. е. чем меньше
возбужденных, действующих на него «чувствительных волокон.
Наконец, в более быстром прекращении двигательного эффекта при
слабом тетаническом раздражении существенную роль, должно быть,
играет общее «торможение. Как уже говорилось выше, общее
торможение проявляется сильнее при слабом возбуждающем действии
периферических импульсов на двигательные клетки; кроме того, торможение
наступает еще раньше возбуждения двигательных клеток. Поэтому
быстрое ослабление двигательного эффекта при слабых раздражениях,
должно быть, обусловлено главным образом тормозящим действием ней-
ропиля на двигательные клетки. Когда же, в связи с усилением
раздражения, возбуждающее действие на двигательные клетки со стороны
афферентных и промежуточных нейронов нарастает, тогда должно
ослабевать тормозящее действие нейропиля на эти двигательные клетки.
Итак, во время сильного раздражения рефлекторная деятельность
является более продолжительной, чем во время слабого, так как, во-
]5*
228 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
первых, чем сильнее раздражение, тем дольше должно продолжаться
активное состояние промежуточных нейронов, несмотря на падение
возбудимости в силу утомления; во-вторых, угнетающее действие общего
торможения на двигательные клетки при сильном возбуждающем
действии со стороны чувствительных и промежуточных нейронов должно
проявляться слабее, чем при слабом их действии.
Повышение возбудимости в центральной нервной системе под
влиянием утомляющего раздражения. Как бы сильно ни было
раздражение, активное состояние координирующего аппарата все же ослабевает
и, наконец, совсем прекращается, если раздражение длится достаточно
Рис. 101. Повышение возбудимости во время раздражения,
вызвавшего утомление.
Спинномозговой препарат собаки. Записываются движения задней
ноги. Ногу раздражают четыре раза; сначала раздражают стопу
слабым тетаническим раздражением (расстояние между катушками
10 см), затем голень—более сильным раздражением (9 см), а вскоре после
него-опять стопу тем же слабым раздражением дважды. За все время
этих опытов кимограф не останавливается. Во всех опытах раздражение
продолжается до полного прекращения вызываемого сгибательного
рефлекса. Как высота, так п длительность рефлекса во втором случае
значительно больше, чем во всех остальных, а высота и длительность
рефлекса в третьем и четвертом случаях больше, чем в первом. Период
раздражения обозначается поднятием сигнальной линии. Время — в
секундах.
долго. Но этим не кончается действие раздражения на центральную
нервную систему,— оно продолжает действовать на ее возбудимость.
Мы уже указывали выше, что длительное утомляющее возбуждение
производит общее повышение возбудимости в центральной нервной
системе. Так, например, известно, что длительное раздражение одного
участка рецептивного поля, наравне с утомлением рефлекса, производит
повышение возбудимости по отношению к другому раздражению того
же поля. Это показано на рис. 101, полученном на спинномозговой
собаке. На кимографе записывается сгибательный рефлекс задней ноги
(движение всей ноги). Ногу раздражают в двух местах: на голени и на
стопе. Раздражение голени слабое; эффект, вызванный им, быстро
прекращается, утомление развивается в 2—3 сек. (запись 1, 3, 4). Раздра-
УТОМЛЕНИЕ
229
жение стопы сильнее; эффект от него длится 20 сек. (запись 2). После
этого раздражения эффект с голени возрастает, хотя эффект со стопы
был доведен до утомления. Этот повышенный эффект наблюдается и
спустя 15 сек.
Согласно приведенной выше концепции, явление общего повышения
возбудимости в центральной нервной системе, в результате
длительного утомляющего раздражения, не может быть объяснено только
повышением возбудимости в тех нейронных элементах, у которых
оканчиваются раздраженные чувствительные нейроны. Правда, эти
афферентные нейроны должны действовать на ряд нейронных элементов
центральной нервной системы субминимально. Это должно происходить,
прежде всего, оо отношению к тем элементам, с которыми
чувствительные волокна анатомически связаны в недостаточной мере интимно
(малое количество коллатералей, недостаточное развитие нервных
окончаний у этих коллатералей). Однако едва ли можно объяснить этим
обстоятельством общее повышение возбудимости в центральной нервной
системе. Оно может быть обусловлено в некоторой мере активированием
нейропиля, его электротоническим и периэлектротоническим действием
на нейронные элементы.
Если признать нейропильный биоток причиной торможения, т. е.
понижения возбудимости нервных клеток и нервных волокон, вследствие
его анэлектротонического действия на эти нервные элементы, то
безусловно следует тогда признать и возможность повышения
возбудимости в нервных клетках, а также в других нервных элементах: во-первых,
вследствие периэлектротонического действия нейропильного биотока на
отдаленные нервные элементы и, во-вторых, в момент прекращения
анэлектротонического действия на ближайшие нервные элементы.
Практикум
1. Наблюдение зависимости утомления координирующего аппарата от
интенсивности раздражения. Сіплгшюмозговая лягушка. Раздражают п. peroneus, а
сокращение m. semitendinosus записывают на кимографе. Сначала записывают сокращение
при длительном пороговом раздражении. Затем интенсивность раздражения
увеличивается, и снова записывают при его длительном применении. Чем сильнее будет
раздражение, тем дольше ирвдется его продолжать для полного ослабления
рефлекса.
2. Наблюдение повышения возбудимости одновременно с утомлением рефлекса.
На спинномозговом препарате лягушки раздражение одного из нервов рецептивного
поля сгибания, например, п. peroneus, до полного утомления рефлекса, повышает
возбудимость в отношении других нервов, например, п. superficiahs или tibialis. Это
повышение возбудимости есть результат иррадиации возбуждения.
3. Наблюдение частичного утомления координирующего аппарата.
Спинномозговая лягушка. Регистрируется на кимографе т. semitendinosus. Раздражают п.
peroneus и п. tibialis. Для каждого нерва своя цепь для раздражения фарадическим
током. Сначала записывают пробные максимальные эффекты путем раздражения того
и другого нерва. Затем длительно, до абсолютного утомления, раздражают п.
peroneus. Сначала длительно раздражают его при одной силе, затем, когда двигательный
эффект значительно упадет, увеличивают силу раздражения на несколько сантимет-
230 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
ров и вновь раздражают длительно. Во время полного прекращения эффекта
пробуют раздражать другой нерв тем же максимальным раздражением, какое
применялось до утомления. Оно произведет эффект, но только более слабый, чем до
утомления.
Иногда п. tibialis дает очень слабый огибательный рефлекс, притом в самом
начале раздражения, но потом он заменяется разгибательным. Это происходит
вследствие преобладающего действия тех нервных волокон, которые идут с рецептивного
поля разгибательных рефлексов. В этом случае нужно переменить препарат.
Литература
і. Verworn M.f Arch. Anat. Physiol., Supplbd., 1900, 152; Die Biogenhypothese,
Jena, 1903.
2. В a g 1 i о n i S., Z. a 11 g. Physiol., 9, 1, 1909.
3. Winterstein H., Arch. Anat. Physiol., Supplbd. 1900, 177.
4. Boss E., Biol., 78, 161, 1923; Ztbl. Physiol., 21, 869, 1908; Z. allg. Physiol.,
в, 315, 1907.
5. В a g 1 i о n i S., Zur Analyse d. Reflexfunktion etc., 1907.
6. Sherrington C, Int. Act. Nerv. Syst., 1906.
7. В e г i t о f f J.t E г g. Physiol., 23, 33, 1924.
8;Wedensky N.. Fol. Neurobiol., 6, 591, 1912.
9. WintersteinH., Pfl. Arch., 224, 749, 1930; Bioch. Z., 232, 196, 1931; Natur-
wiss., 247, 1931.
10. Winterstein H. u. Frankel-Conrat, Bioch. Z., 247, 178, 1932.
11. L о rente de No, Юбилейный сборник проф. И. Бериташвили. Изд. Груз,
отд. АН СССР, 1936.
12. Сеченов И. М., Об электрическом и химическом раздражении
чувствительных спинномозговых нервов лягушки, 1868.
13. London Е., Kotschnef N., Dubinsky a. Katzwa A., Pfl.
Arch., 232, 160, 1933.
14. К а с с и л ь Г. Н., Сб. докладов VI Всес. Физиол. съезда, стр. 454.
15. Loewi О., Pfl. Arch., 239, 430, 1937.
16. Stedman E. a. Stedman EI., Bioch. J., 31, 817, 1938.
17. S p e i d e 1 С. С, Cold Spr. Harb. Symp. Quant. Biol., 4, 13, 1936.
18. Lee a. E v e r i n g h a m, Am. J. Physiol., 24, 384, 1909.
19. F о г b e s A., Am. J. Physiol., 31, 102, 1912.
20. Q u a 8 t ё 1 J. H., Physiol. Rev., 19, 435, 1939.
21. P у б e л ь В. М., Фрид А. И. иКислинский А. Н., Физиол. журн.
СССР, 27, 58. 1939.
22. В в е д е н с к и й Н. Е., Соч., т. 4, 5, 1938.
23. Г о г а в а М., Тр. инст. Физиол. им. Бериташвили, 5, 161,1943.
24,- Беритов И. и БакурадзеА., ДАН СССР, 26, 261, 965, 1940.
25. W і n t е г s t е і п Н. и. Н і г s с h b е г g Е., Bioch. Z., 166, 138, 1925.
26. Б е р и т о в И. и Р о й т б а к А., Об электрических потенциалах спинного
мозга лягушки. Труды Ин-та физиол. им. Бериташвили, 7, 1947.
27. Беритов И. и Р о й т б а к А., Об изменениях электрических потенциалов
спинного мозга при длительных раздражениях. Печатается в сборнике,
посвященном памяти Н. Е- Введенского, 1948.
Глава XII
СПОНТАННАЯ РИТМИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ
ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Общее понятие. Центральная нервная система обнаруживает
ритмическую деятельность не только рефлекторно, т. е. в ответ на
раздражение внешних или внутренних рецепторов,— она проявляет
ритмическую деятельность все время более или менее интенсивно независимо
от этих раздражений. Эта так называемая спонтанная деятельность
свойственна в большей или меньшей степени всем отделам головного
мозга: коре большого мозга, промежуточному мозгу, мозжечку,
среднему мозгу, продолговатому мозгу. При нормальном состоянии дыхания
и кровообращения, при нормальном обмене веществ, каждый отдел
головного мозга, или, вернее, каждое ядро его, производит разряды
имиульсов.
Спинной мозг обладает свойством спонтанной ритмической
деятельности в меньшей мере, но и он обнаруживает ее в условиях полной
деафферентации, т. е. в условиях отсутствия периферических импульсов
после перерезки задних корешков (Дюссер де Баренн, 1; Руч, 2). На
лягушке редко удается наблюдать в спинном мозгу спонтанную
электрическую активность — как при отведении биотоков от поверхности мозга,
так и из глубины его, от серого вещества (Беритов и Ройтбак, 53)
(рис. і76). Она часто отсутствует в спинном мозгу лягушки при наличии
ее во всех отделах головного мозга. Высшие же отделы мозга
показывают ритмическую деятельность даже во время глубокого сна или
наркоза, когда только очень сильные периферические раздражения
вызывают рефлекторные реакции.
Спонтанная деятельность в значительной мере свойственна многим
рецепторам, как об этом уже указывалось выше. Периферические
же нервные стволы, а также периферические ганглии у млекопитающих,
и среди них симпатические ганглии, не обладают способностью к
спонтанной ритмической деятельности (Проссер, 3). Но в определенных
условиях и они могут давать ритмические разряды автоматически, без
повторных внешних раздражений. Мы уже выше познакомились с
такими ритмическими разрядами в нервных и мышечных волокнах в виде
.многократного повторного изохронного расщепления возбудимой систе-
232 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
мы после одного раздражения. Но это бывает только при очень
специфических лабораторных условиях. Длительные ритмические разряды в
нервных волокнах легко получить при их повреждениях. Этот ритм
регулируется длительностью рефракторных фаз: каждый последующий
Рис. 102. Электроэнцефалограмма и электрограмма мимических мышц.
К каждой записи наверху спонтанные биотоки коры большого мозга при их отведении
от кожной поверхности теменно-центральной области, а внизу спонтанные биотоки мышц
лица при их отведении от левой щеки. Записи I и II от одного субъекта низкой
возбудимости, а III и IV от другого субъекта высокой возбудимости. Запись I и III — при
открытых глазах, а запись II и IV — при закрытых глазах.
импульс возникает с интервалом, который равняется длительности обеих
рефракторных фаз: абсолютной и относительной. С течением времени
эти разряды ослабевают и прекращаются, но возникают вновь при
повторном повреждении (Эдриан, 4).
Аналогичное явление было отмечено на двигательных и
чувствительных нервных волокнах человека при ишемии, т. е. при лишении их
кровообращения. Через минуту по прекращении кровообращения в руке
путем пневматической манжетки подопытное лицо чувствует вибрацию,
а мышцы дают биотоки возбуждения до 80 в 1 сек. Эта спонтанная
активность постепенно проходит и вновь появляется в усиленной форме
при восстановлении кровообращения. При этом спонтанная импульса-
ция может доходить до 180 в 1 сек. По прекращении спонтанной
импульс ации произвольные импульсы или раздражение этих нервов
электрическим током приводит к временному восстановлению спонтанной
им-пульсации (Кутельберг, 54).
Элементам центральной нервной системы — клеткам и аксонам так*
же свойственны все эти формы ритмического разряда при названных
СПОНТАНН АЯ РИТМИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ ЦЕНТР. НЕРВН. СИСТЕМЫ 233-
специальных условиях. Но центральная нервная система
характеризуется способностью к постоянной ритмической деятельности при самых
нормальных условиях.
Рис. 103. Биотоки в разных отделах мозга лягушки при их спонтанной
ритмической деятельности.
Регистрация производилась катодным осциллографом с помощью однотактного
усилителя переменного тока. Усиление 1 200 000 раз. Головной мозг обнажен сверху.
Лягушка обездвижена к\раризацией. Биотоки отводятся к сетке точечным
электродом— заостренной хлорированной серебряной проволокой. Диффузный заземленный
электрод — большая серебряная пластиньа находится под животом, А —биотоки
lobus olfactonus, В—биотоьи большого мозга, С—биотоки того же отдела на другом
препарате, D — среднего мозга сейчас же после приложения электрода, Е — то же
спустя некоторое время,/7—продолговатого мозга в области вхождения VIII нерва;
G—то же на другом препарате. В последнем случае производилось звуковое
раздражение—звук тонвариатора Штерна 1160 колебаний в 1 сек. (белая линия сверху);
Я — биотоки мозжечка; J — спинного мозга на границе с продолговатым. Время—
0.02 сек.
Спонтанная деятельность стволовой части головного мозга~
Спонтанная деятельность промежуточного, среднего и
продолговатого мозга в основном носит двоякий характер: спонтанная
деятельность одних центров проявляется на периферии в виде фазных
движений рабочих органов. Такова, например, периодическая
деятельность дыхательного центра в продолговатом мозгу, периодическая
деятельность мигательных центров в средних отделах головного мозга,,
постоянная ритмическая деятельность координирующих центров імими-
234 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
веских мышц. Что дыхательные и мигательные движения возникают
¦автоматически независимо от внешних раздражений, можно было 'бы
^предположить на основании собственного опыта. Но оказывается, что
мышцы лица, создающие мимику, характерное выражение лица, также
шее время возбуждены в большей или меньшей степени. Чем выше
возбудимость субъекта, тем сильнее их спонтанное возбуждение (рис. 102)
*(Бериташвили, Гедеванишвили и Воробьев, 5). В этом отношении
мимические мышцы лица характерно отличаются от скелетных мышц
конечностей.
Спонтанная деятельность мышц головы может быть произвольно
дли рефлекторно значительно усилена или ослаблена. Например, при
закрывании глаз электрическая активность мимических мышц во
многих случаях значительно ослабевает (рис. 102). Возможно, что
афферентная импульсация из раздражающихся при закрывании глаз
рецепторов глазных мышц и глазных яблок тормозит средние отделы мозга, а
.значит, и исходящую из этих отделов эфферентную импульсацию по
отношению к мышцам лица.
Спонтанная деятельность ствола мозга, не проявляющаяся во
внешних движениях. Спонтанная деятельность центральной нервной
системы очень часто не проявляется на периферии. Очевидно, при этом
двигательные нейроны не вовлекаются в действие. Она всецело
ограничивается промежуточными нейронами. Такая автоматическая
деятельность нервных центров выражается в возникновении электрических
потенциалов двоякого рода: медленные колебания потенциала с частотой
'От одного до нескольких десятков в секунду, и быстрые колебания с
частотой до нескольких сот в секунду.
Спонтанная деятельность каждого отдела головного мозга имеет
<гвои особенности. Она различается по характеру течения быстрых
потенциалов, именно—по высоте отдельных колебаний и по их ритму,
а также по длительности и течению медленных потенциалов и по
присутствию или отсутствию их (Шпигель 6; Доу, 7; Либет и Жерард, 8;
Беритов и Цкипуридзе, 9). Это хорошо видно на рис. 103, где
приводится автоматическая ритмическая деятельность разных отделов
головного мозга лягушки. Эти особенности, очевидно, обусловливаются
структурными особенностями нервной ткани разных отделов мозга. Отсюда
-следует, что по характеру и течению медленных и быстрых потенциалов,
продуцируемых разными отделами головного мозга или разными
участками одного отдела, можно заключить о характере автоматической
•деятельности серого вещества соответствующих отделов мозга.
Однако не всякий участок головного мозга обнаруживает
способность к спонтанной электрической активности. При точечном отведении
того или другого участка мозга лягушки легко установить, что
спонтанная электрическая активность со всеми своими характерными быстрыми
и медленными волнами наблюдается только в тех участках, где серое
вещество находится в непосредственной близости к отводящему
электроду: напр. у лягушки — при отведении зрительной покрышки среднего
СПОНТАННАЯ РИТМИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ ЦЕНТ. НЕРВ. СИСТЕМЫ 235
мозга, дорзальной поверхности верхнего отдела продолговатого мозга,
корковой поверхности мозжечка. Если же поверхностно лежит более
или менее толстый слой белого вещества, то в этих участках
спонтанная электрическая активность не наблюдается или наблюдается в очень
слабой форме; она значительно усиливается, если отводящий биотоки
Рис. 104. Спонтанные электрические потенциалы коры
мозжечка кошки.
Монополярное отведение, катодный осциллограф. А — осциллограмма при
отведении от передней части (lobus anterior); В—от полушария мозжечка (crus I іоЪиз
ansiformis); С—от передней части мозжечка после децеребрации (Цкипуридзе),
игольчатый электрод вставить глубоко в мозг до соприкосновения с
серым веществом. Так, например, спонтанные биотоки не отводятся от
вентральной поверхности среднего и продолговатого мозга лягушки или
при отведении от белого вещества большого мозга кошки (Беритов и
сотр., 9, 28).
Все это свидетельствует о том, что спонтанная электрическая актив,
ность есть функция серого вещества головного мозга.
Электрические потенциалы, отводимые от головного мозга, точно
отображают главным образом электрическую активность серого
вещества в отводимом участке мозга, а не всего мозга или всего отдела
мозга, где этот участок находится. Благодаря этому характер
электрических колебаний может изменяться при малейшем передвижении
отводящих электродов вдоль по поверхности мозга. Так, например,
электрическая активность продолговатого мозга лягушки в области ядер
слухового нерва значительно сильнее, чем на 1 мм выше и ниже
этого участка; или, например, в середине той или другой половины
мозжечка она значительно сильнее, чем на ее краях (Беритов и сотр., 9).
Спонтанная активность мозжечка занимает особое положение среди
других отделов мозга. У высших позвоночных (млекопитающие и пти-
236 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
цы) она выражается в трех видах электрической активности: быстрые
потенциалы высокого ритма до 400 в 1 сек.; медленные потенциалы в
ритме около 40 в 1 сек. и медленные потенциалы в ритме 5—10 в 1 сек.
Последние потенциалы зависят от нервных связей с большими
полушариями. После перерезки их,— например, при децеребрации,—
медленные потенциалы исчезают (рис. 104-С). Они легко исчезают и при
разного рода необычных раздражениях рецепторов, например, при
раздражении шумами (Бакурадзе и Цкипуридзе, 46; Тен Кате и Виггерс, 47).
Быстрые потенциалы и медленные потенциалы высокого ритма
наблюдаются у всех позвоночных (Цкипуридзе, 51).
Спонтанная деятельность некоторых ядер ствола мозга имеет
периодический характер: она периодически значительно усиливается.
Это характерно не только для дыхательного центра продолговатого
імозга, но и для некоторых других центров. Так, например, у кошки
некоторые ядра промежуточного мозга (nucl. medialis centralis, nucl. me-
dialis dorsalis, nucl. medialis lateralis), расположенные медиально от
восходящей афферентной системы, дают правильно периодически разряды
больших волн по 5—10 в 1 сек. через каждые 3—5 сек. Разряды
большею частью наступают в этих ядрах одновременно. Они совпадают во
времени с подобными вспышками в коре большого мозга, но н? зависят
от коры, ибо наступают и после ее удаления (Морисон, Филней и Ло-
троп, 33).
Спонтанная активность нервных узлов беспозвоночных. Нервные узлы
беспозвоночных также обладают способностью к спонтанной активности.
Это было показано в отношении нервной цепочки гусеницы (Эдриан, 36) >
изолированной нервной цепочки водяного жука (Эдриан, 37), в
отношении изолированной нервной цепочки дождевого червя (Моор и Брэ-
дуей, 38), в отношении зрительного ганглия рака (Проссер, 3), в
отношении брюшной цепочки медицинской пиявки (Беритов, 40), в
отношении изолированных ганглиозных скоплений сердечного нерва Limulus
(Хейнбекер, 41). При этом было установлено, что спонтанная
деятельность нервных узлов сопряжена с мышечной активностью* с усилением
¦спонтанной активности в нервной цепочке происходит усиление
мышечных сокращений в соответствующих сомитах (Моор и Брэдуей, 38;
Беритов, 40).
Таким образом, можно утверждать, что и у беспозвоночных
животных усиление спонтанной активности в определенных нервных узлах
обусловливает усиление мышечной деятельности соответствующих
двигательных органов.
Влияние периферических раздражений на спонтанную
деятельность. На спонтанную деятельность данного центра
преимущественно! оказывает влияние раздражение того рецептора, который
связан с ним непосредственно. Так, у лягушки на спонтанную
ритмическую деятельность среднего мозга, где первично оканчивается
зрительный нерв, влияет раздражение глаза светом (освещение или
затемнение глаза) или движение предметов в поле зрения лягушки.
СПОНТАННАЯ РИТМИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ ЦЕНТР. НЕРВ. СИСТЕМЫ 237
Эта влияние является перекрестным: правый глаз действует на левый
холмик среднего мозга. На ритмическую деятельность передней части
продолговатого мозга, где первично заканчивается слуховой нерв,
сильно влияют звуки и вибрации (рис. 103).
При раздражении чувствительных нервов или участков кожи на
конечностях изменение спонтанной активности можно обнаружить во всех
отделах стволовой части головного імозга. Опыты на лягушке показали,
что раздражение кожных нервов сильнее всего активирует средние
¦отделы мозга, где сходятся все афферентные пути (Ройтбак, 35) (рис. 71).
Спонтанная активность коры мозжечка не меняется заметным
образом под влиянием внешних раздражений (Бакурадзе и Цкипуридзе, 48).
Только тактильные раздражения кожи в области распространения
тройничного нерва оказывают некоторое возбуждающее влияние на
латеральный участок мозжечка лягушки (Беритов и Цкипуридзе, 9).
У высших позвоночных спонтанная активность коры мозжечка заметно
меняется под влиянием раздражения коры большого мозга (Доу, 49;
Уолкер, 50 и др.). Однако даже резкие изменения потенциалов коры
большого мозга не всегда сопровождаются изменениями спонтанной
активности мозжечка (Бакурадзе и Цкипуридзе, 48). ha спонтанную
деятельность плечевого отдела спинного мозга действует раздражение
кожных и мышечных рецепторов передней конечности; при этом на
правую половину спинного мозга значительно сильнее действует
раздражение кожных и мышечных рецепторов правой передней
конечности, чем левой передней. Во всех этих случаях влияние
периферических импульсов выражается в усилении и учащении существующей
автоматической ритмики. Усиление ритмической деятельности
центральной системы может сопровождаться наступлением движений, но это не
обязательно. Звуки и вибрации повышают ритмическую деятельность
продолговатого мозга ^ очень сильно, не вызывая внешних движений.
Свет также усиливает электрическую активность среднего мозга без
вызова внешних движений. Легкое кожное раздражение —
прикосновение или небольшое пассивное растяжение мышц может вызвать
усиление ритмической деятельности спинного мозга без рефлекторных
реакций (Беритов и Цкипуридзе, 9).
Происхождение спонтанной деятельности ствола мозга.
Автоматическая деятельность центральной нервной системы в значительной
мере зависит от спонтанной деятельности рецепторов. Это былоі
доказано в отношении автоматической деятельности покрышки среднего
імозга и дорзального и вентрального ядер VIII нерва у лягушки. Их
автоматическая деятельность значительно ослабевает после удаления
рецепторов или перерезки соответствующих чувствительных нервов
(Беритов и Цкипуридзе, 9, 10).
Однако спонтанная ритмическая деятельность названных и
других отделов головного мозга наблюдается и при отсутствии
периферических влияний. Так, например, на лягушке она наблюдается после
полной кураризации, когда животное лежит без всяких движений, т. е.
23S ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
при отсутствии проприоцептивных раздражений. Ритмическая деятель*
ность головного мозга сохраняется и после удаления у курарйзованной
лягушки всех внутренних органов, т. е. когда исключено также
действие всех интероцептивных раздражений, или даже в том случае,,
¦если у такой лягушки отрезать голову и на ней изучить электрическую
активность головного імозга (Беритов и Цкипуридзе, 9, 10).
Спонтанная деятельность сохраняется в головном мозгу лягушки и
тогда, когда он вынут из черепа; сохраняется даже в небольшом
изолированном кусочке імозга. Например, если отделить небольшой
кусочек мозга весом около 0.1 імг от lobus olfactorius, то и в нем будет
наблюдаться автоматическая ритмическая деятельность (Либет и Же-
рард, 8; Беритов и сотр., 9). На основании этих наблюдений следует
заключить, что автоматическая ритмическая деятельность
головного імозга обусловливается ритмическим самовозбуждением
промежуточных нейронов мозга. Самовозбуждение характерно для живой
возбудимой системы вообще, как об этом уже говорилось выше, но,
несомненно, оно свойственно промежуточным нейронам в большей мере,
чем всем другим нервным элементам. Иначе говоря, изохронное
расщепление значительной части возбудимой системы нервной клетки,
дающее начало распространяющемуся импульсу возбуждения в аксоне,,
возникает в клетке под влиянием тех изменчивых физикохимических
условий, какие могут иметь место внутри и снаружи нервных клеток.
Импульсы возбуждения, распространяясь по аксонам, активируют и
нервные круги и нейропиль. Возбуждение нервных кругов проявляется
в быстрых колебаниях электрического потенциала высокого ритма, а
активное состояние нейропиля — в медленных колебаниях низкого
ритма.
Особенности ритмической деятельности центральной нервной
системы, несомненно, определяются, во-первых, физиологическими
особенностями нервных кругов, а во-вторых, структурой нервных кругов,—
прежде всего нервными связями между разными нервными ядрами. Но
само возникновение спонтанных импульсов возбуждения в нервных
элементах мозга зависит, конечно, от раздражающего) действия физико-
химических условий межуточной жидкости мозга. Например,
значительная роль ионов Na доказана специальными опытами: лягушки,
помещенные на несколько дней или на неделю в дистиллированной
воде, теряют 30—50% своих хлоридов. Одновременно сильно
редуцируется спонтанная электрическая активность головного мозга. Если
такой лягушке ввести в спинной лимфатический мешок небольшое
количество NaCl, спонтанная электрическая активность восстанавливается (Пик
и Миллер, 34). Вообще известно, что повышение ионов Na, К, или рН, а
также уменьшение Са в головном мозгу лягушки благоприятствуют
быстрым волнам, т. е. спонтанные электрические потенциалы становятся
более частыми и высокими; уменьшение же Na, понижение рН,
увеличение Са и Mg, наоборот, благоприятствуют .медленным потенциалам,
т. е. спонтанные потенциалы становятся более длительными и низкими
СПОНТАННАЯ РИТМИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ ЦЕНТ. НЕРВ. СИСТЕМЫ 239*
(Либет и Жерард, 8). 'Аналогичные факты известны в отношении кош*
ки. Например, небольшое повышение содержания калия в крови на:
20—50% вызывает, наряду с повышением рефлекторной деятельности,
усиление электрической активности в среднем мозгу. Повышение же
содержания кальция понижает и то и другое (Дубнер и Жерард, 29).
Возможно, что угнетающее и облегчающее действие ионов
обусловливается влиянием этих ионов на окислительные процессы мозга.
Существуют прямые наблюдения, указывающие на существенную роль
окислительных процессов в происхождении спонтанной электрической
активности. Именно, установлено, что в нервных узлах беспозвоночных
спонтанное ритмическое возбуждение клеточных элементов
обусловливается окислительными процессами. Чем интенсивнее эти
окислительные процессы, тем интенсивнее происходят разряды импульсов. Если
поместить изолированный нервный узел в атмосфере азота, то через
несколько минут спонтанная деятельность ослабевает и затем совершенно*
прекращается. В воздухе спонтанная деятельность восстанавливается.
Так же действуют цианиды и углекислота, которые тормозят действие
дыхательных энзимов и тем ослабляют окислительные процессы (Прос-
оер и Бюел, 11). Спонтанная ритмическая деятельность в головном*
мозгу ухудшается, когда препарат отмирает: ритм биотоков
уменьшается, амплитуда их падает. Но характерно, что она продолжается еще
некоторое время после того, как рефлексы перестают вызываться. Очень.
характерно отношение мозжечка лягушки к обмену веществ. Весной и
летом мозжечок дает сильную спонтанную активность до 400 импульсов
в 1 сек. В период зимней спячки она сходит на-нет, но ее можно
искусственно возродить длительным согреванием лягушки (Цкипупид-
зе, 43). Эти факты указывают, что спонтанная электрическая активность
центральной нервной системы находится в определенной зависимости от
ее функционального состояния,— иначе говоря, от интенсивности
основного обмена веществ. Вероятно, чем лучше условия для восстановления
возбудимой системы ганглиозных клеток после возбуждения, т. е., чем
интенсивнее обмен веществ внутри ганглиозных клеток, тем чаще
происходит «самопроизвольное» возбуждение нервных кругов, тем
интенсивнее каждое такое возбуждение и, сообразно, тем сильнее спонтанная
электрическая активность.
Периодическая деятельность нервных кругов и происхождение
быстрых и медленных биотоков спонтанной электрической активности.
Электрическая активность мозга, как уже указывалось, проявляется в
двоякого рода биотоках: быстрых и (медленных. Быстрые биотоки, по
общему мнению, возникают в результате распространения импульсов
возбуждения по нервным кругам. Для каждого комплекса нервных
кругов характерна периодичность в возникновении нервных импульсов.
Иначе говоря, в каждом нервном кругу после возникновения ряда
нервных импульсов наступает кратковременный покой, после чего вновь
получается разряд. Это хорошо видно на примере дыхательного центра
и координирующих аппаратов фазных движений (Гольст, 30 и др.)> а
^240 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
также при деятельности любого отдела или центра головного мозга.
Так, например, при освещении глаза лягушки или при движении
предмета в поле зрения в среднем мозгу импульсы возбуждения
возникают или усиливаются периодически (10). То же наблюдается в
продолговатом мозгу при определенных условиях. Это особенно
характерно для спонтанной ритмической деятельности большого и
промежуточного мозга, а также среднего мозга и обонятельных долей (9). Такая
способность к периодической ритмической деятельности безусловно
стоит в связи с особенностями структуры мозга. Каждый нервный
комплекс состоит из нервных кругов разной сложности; каждый круг — из
нервных волокон, нервных клеток и окончаний нервных волокон на теле
нервных клеток, иначе говоря синапсов. Установлено, что нерв*-
ные волокна? трудно утомляются, они могут работать в течение многих
часов без утомления. Значительно легче утомляются нервные клетки и
возможно нервные окончания, синапсы. Сообразно, возбуждение мо- ;
жет вращаться по нервному кругу до тех пор, пока не произойдет утом- \
ление этих образований. А утомление их непременно произойдет, и
притом очень быстро, ибо возбуждение вращается в нервном кругу
непрерывно. Чем меньше крут, тем чаще будут возбуждаться соответ- ;
ствующие нервные клетки и поэтому тем раньше произойдет их
утомление. Вследствие этого прекратится распространение возбуждения
в данном кругу. Такие нервные круги перейдут в состояние покоя и
вновь возбудятся только тогда, когда пройдет утомление нервных
клеток.
В каждом периоде деятельности нервных кругов последние
разряжаются рядом быстрых волн самой разной амплитуды — от нескольких
микровольт до 100 микровольт. Длительность этих волн также очень
изменчива — от нескольких тысячных до нескольких сотых секунды.
Обычная длительность одного биотока возбуждения в аксонах, в
нервных волокнах, а, возможно, также в нервной клетке или синаптических
окончаниях — значительно меньше. У теплокровных животных она
меньше одной тысячной секунды. Но при активном состоянии нервных
кругов происходит слияние быстро следующих друг за другом биотоков
возбуждения, возникающих в разных составных частях этих кругов.
Вследствие этого получается, что биотоки нервных кругов, во-первых,
более длительны, чем в нервных волокнах, а во-вторых, они весьма
разной длительности и амплитуды. Сказанное касается всех отделов
мозга — и в частности коры большого мозга, ее быстрых волн,
именуемых бета-волнами.
Но, конечно, если отведение биотоков производится от небольшого
пучка волокон белого вещества под корой или в других отделах мозга,
то при их возбуждении получаются не упомянутые выше медленные и
быстрые колебания электрического потенциала, а обычные аксонные
биотоки возбуждения, очень высокого ритма, почти до 1000 ів 1 сек.,
как свойственно нервным стволам теплокровных животных. Это хорошо
видно из исследований Эдриана и Моруци (31).
СПОНТАННАЯ РИТМИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ ЦЕНТР. НЕРВ. СИСТЕМЫ 241
Нервные импульсы, периодически возникающие в нервных кругах,
•обусловливают периодическое активирование нейропиля в ближайших
участках мозга. А так как в каждом участке нейропиля биотоки
протекают более длительно, чем биотоки возбуждения нервных кругов, то
обычно, ввиду совместного отведения нервных кругов и нейропиля
быстрые токи проявляются главным образом на восходящем колене
длительного биотока. Это было выявлено при изучении электрической
активности среднего мозга лягушки (10), затем при эзерино-ацетилхо-
линовом отравлении коры большого мозга кошки (Беритов и
сотрудники, 32).
Итак, каждый нервный комплекс, содержащий замкнутые нервные
круги, периодически приходит в активное состояние спонтанно или под
влиянием внешних импульсов, производя каждый раз последовательный
ряд импульсов после некоторого периода покоя, наступающего вслед"
ствие утомления клеток. Электрическая активность периодически
возбуждающегося нервного комплекса выражается, с одной стороны, в
группировке -.быстрых колебаний аксонного происхождения, а с
друзой — в ритмическом течении медленных колебаний нейропильного
происхождения.
Облегчающее и тормозящее действие спонтанной активности.
Спонтанная активность головного мозга, в особенности коры большого мозга,
-оказывает определенное влияние на степень возбудимости спинного
мозга. Это происходит путем распространения импульсов возбуждения
зіо нисходящим путям из спонтанно возбужденных центров. Так,
например, Эдриан и Моруци (52) показали, путем одновременного
изучения биотоков коры мозга и пирамидных .путей в продолговатом мозгу,
что не только в бодрствующем состоянии, но и во время
наркотического сна нисходящие пирамидные пути разряжаются импульсами
возбуждения более или менее синхронно с двигательными участками
коры. Так же должны действовать и другие спонтанно активирующиеся
дентры головного мозга. При низкой частоте и интенсивности эта
спонтанная импульсация не возбуждает спинного мозга, но в значительной
степени повышает в нем возбудимость, создает состояние облегчения.
Благодаря этому спинномозговые центры оказываются более
чувствительными к раздражению разного рода периферических рецепторов.
Этим объясняется известный факт уменьшения хронаксии — так
называемой субординационной хронаксии,— под влиянием центральной
нервной системы. Спонтанная активность мозжечка, наоборот, оказывает
тормозящее действие на рефлекторную деятельность стволовой части
головного мозга и на спинной мозг. Всякое усиление спонтанной
активности,— например, от непосредственного раздражения мозжечка
электрическим током,— сопровождается ослаблением тонуса скелетных
імышц (рис. 137). Обратно действует ослабление спонтанной активности,
наступающее, например, от локального отравления новокаином,
стрихнином. Это ведет к усилению тонической деятельности на гомолате-
ральной стороне тела (Доу, 7).
^6 я. С. Берчтов
242 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГ V
Итак, спонтанная деятельность нервных центров головного мозга'
играет существенную роль не только в производстве определенных'
внешних реакций, но и в общих изменениях возбудимости нервных
центров в смысле как общего облегчения, так и общего торможения.
Ритмическая деятельность дыхательного центра. Деятельность
дыхательного центра проявляется на периферии ,в виде дыхательных
движений. Он также все время находится под влиянием периферических
раздражений и в большой мере*
зависит от них в своей деятельно*
сти. Одни периферические
раздражения усиливают
деятельность дыхательного центра,
другие тормозят ее (рис. 105).
Неона сохраняется и при
исключении внешних раздражений,
подобно тому как координирующий
аппарат фазных рефлексов,
находящихся в спинном мозгу, также-
может развивать ритмическую
деятельность при полном
исключении периферических импульсов-
(Гезел, Аткинсон и Броун, 12).
Установлено, что у взрослых.
животных автоматическая
деятельность дыхательного центра
обусловливается раздражающим
действием химического состава
крови. Она наблюдается в
экспериментальных условиях после-
двусторонней перерезки ваго-сим-
патических пучков и депрессоров;
и одновременного удаления всей:
области каротидных синусов, т. е.
после удаления всех механо- и
хеморецепторов, принимающих
участие в регуляции
дыхательных движений (Винтерштейн, 59):.
Ритмическая деятельность
дыхательного центра наблюдается №
у зародышей. Она наступает как
а ответ на раздражение, так и
спонтанно, т. е. помимо каких-
либо внешних раздражений (Баркрофт и Баррон, 13). В этом случае
координирующий аппарат дыхательмых движений приходит в активное
состояние совершенно спонтанно, т. е. под влиянием нормального
химического состава крови, как сердечная мышца. Известно, что у эм-
Рис. 105. Запись дыхательных движений
ротовой полости лягушки.
А — большие дыхательные движения полностью
прекращаются во время раздражения п.
trigeminus; В — большие дыхательные движения при
зрительном раздражении заменяются малыми
дыхательными движениями; О — малые
дыхательные движения при постукивании по столу,
вызывающем вибрацию стола, превращаются в
большие (Ройтбак).
СПОНТАННАЯ РИТМИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ ЦЕНТ. НЕРВ. СИСТЕМЫ
бриона двусторонняя перерезка блуждающего нерва не вызывает
типичных изменений в ритме и амплитуде дыхательных движений
(Аршавский, 56, Крючкова, 57).
і Путем изучения биоэлектрических токов изолированного ствола
мозга золотой рыбки Эдриан и Буйтендик (14) установили,
что в клеточных элементах дыхательного центра спонтанно возникают
длительные колебания потенциала по дыхательному ритму, которые
сопровождаются рядом быстрых колебаний (рис. 69). Впоследствии
Эдриан (15) изучил ритмическую деятельность дыхательных нервных
элементов изолированной нервной цепочки водяного жука ( Dytiscus
marginalis ) и тоже наблюдал медленные колебания потенциала в
ритме дыхания и быстрые колебания на фоне медленных.
На основании современных исследований автоматической
ритмической деятельности дыхательного центра можно дать следующее
объяснение. Координация дыхательных движений, главным образом их
ритмичность, обусловливается деятельностью определенной/части сетевид-
ного образования продолговатого мозга (formatio reticularis). Оно
приходит в активное состояние, во-первых, под постоянным влиянием
раздражающего действия химического состава крови и, во-вторых, под.
влиянием периферических импульсов, временами притекающих от
разных рецепторов организма. Нервный импульс, возникающий спонтанно
в сетевидном образовании, распространяется по его нервным кругам и
тем обусловливает, с одной стороны, разряд двигательных импульсов
через nn. phrenici и nn. costales, а с другой — активирование дендритного
сплетения. Распространение возбуждения пел нервным кругам
производит быстро протекающие колебания электрического потенциала, а
активное состояние дендритного сплетения — медленные колебания. Эти
медленные токи, достигнув определенной амплитуды, оказывают на
нервные круги анэлектротоническое действие и тем понижают их
возбудимость, т. е. производят торможение. : Вследствие этого нервные
круги перестают проводить возбуждение, от чего прекращается как
двигательная иннервация дыхательных мышц, так и более или менее
значительное активное состояние нейропиля, а следовательно, и его
анэлектротоническое действие на нервные круги. Последние вновь
начинают действовать, под влиянием ©нутренней среды и разного рода
периферических импульсов. Как только начинается дыхательное
движение, сейчас же присоединяется сильнейшая афферентная импульсация
со стороны дыхательных органов, в особенности через пп. phrenici, co-
stales, vagi и trigemini. Чувствительные волокна этих нервов должны
возбуждаться каждый раз во время дыхательных движений. Эта
афферентная импульсация усиливает деятельность и нервных кругов и
дендритной части сетевидного образования, в результате чего вновь
наступает торможение нервных кругов и вновь прекращаются дыхательные
движения и т. д.
Искусственное раздражение чувствительных нервов, имеющих
отношение к органам дыхания, сильно влияет на дыхание, как в смысле
16*
ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
усиления, так и угнетения. Так например, раздражение п. trigeminus
тормозит дыхательные движения (рис. 105). Раздражение посторонних
чувствительных нервов, например, нервов конечностей, также очень
сильно влияет на дыхание. Очевидно, импульсы возбуждения этих
нервов, действуя на продолговатый мозг, активируют не только нервные
круги, но и нейропиль дыхательных центров и вообще сетевидное обра-
зовани? продолговатого мозга. Поэтому это раздражение также влияет
на дыхательные, центры как в смысле усиления дыхательных движений,
так и ослабления или полного угнетения их. Чаще всего
периферическое раздражение производит торможение дыхательных движений. Это,
очевидно, стоит ів связи с тем, что дыхательные центры находятся в
обширном нейропильном образовании, которое легко активируется
периферическими раздражениями.
У высших позвоночных центры вдоха и выдоха существуют раздела
но, но в близком соседстве друг с другом. Это доказано прямыми
опытами. Например, у кошки центром вдоха является центральное
сетевидное образование непосредственно над нижней оливой. Локальное
раздражение его вызывает вдох, длящийся все время, пока
производится раздражение. Дорзальное сетевидное образование, лежащее
впереди краниального конца вдыхательного образования, служит для
выдоха (Рэнсон и сотр., 16, 17). В этом случае теоретическое толкование
дыхательной автоматии остается тем же. Но, кроме того, придется
допустить, что нервные круги вдыхательного сетевидного образования
довольно тесно связаны с нейропилем выдыхательного центра и,
наоборот, нервные круги последнего центра тесно связаны с нейропилем
вдыхательного. Поэтому, когда нервные круги центра вдоха находятся
в возбужденном состоянии, усиленно активируется нейропиль центра
выдоха и сообразно тормозятся нервные круги последнего. Обратное
будет происходить при возбуждении нервных кругов центра выдоха.
Очередность в деятельности парных центров осуществляется, повиди-
мому, потому, что период спонтанного возбуждения нервных кругов
каждого центра проходит быстро вследствие утомления и одновременно
снимается торможение с другого центра. Быстрое прекращение
торможения, как известно, сопровождается облегчением. Благодаря этому "-в
момент прекращения торможения нервные круги другого'центра легко
подвергаются возбуждению под влиянием непрекращающихся
периферических импульсов и постоянно действующих активных веществ
внутренней среды.
Спонтанная деятельность центральной нервной системы
зародыша. Для растущего организма в эмбриональном периоде
чрезвычайно характерна беспрерывная спонтанная деятельность
нервно-мышечных элементов и, в особенности, центральной нервной системы. Эта
деятельность начинается периодическим движением сердечной трубки;
затем происходит периодическое движение амниона и всего тела, а-в
более поздней стадии точно так же периодически наступают локальные
движения отдельных органов (Монталчини и Визинтини, 20; Куо, 21;
СПОНТАННАЯ РИТМИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ ЦЕНТ. НЕРВ. СИСТЕМЫ
Уиндл и Орр, 22 и 23; Страус и Веддел, 24). По наблюдениям Чичи-
надзе (18), при развитии цыпленка сердечная трубка начинает
пульсировать уже в конце 2-го дня инкубации. На 5-й день развития, когда
амнион замыкается и эмбрион опускается в етб полость, появляются
спонтанные движения в ©иде медленных ритмических раскачиваний
всего тела вместе с амнионом приблизительно 14—16 раз в 1 мин. с
амплитудой 2—2.5 мм. На 6-й день прибавляются ритмические
движения головы и хвостовой части; эти части двигаются иногда
одновременно—сходятся, расходятся,— а иногда изолированно: например, голова
отклоняется то вверх, то вниз, то вбок. На 7—8-й день количество
карательных движений увеличивается до 20—24 в 1 мин., и они
совершаются то в горизонтальном, то в вертикальном направлении. К этому
времени усиливаются и учащаются также движения головы и хвостовой
части. Характерно, что совершенно такие же движения наступают при
уколе в спину. В этот же период появляются движения конечностей.
Паузы видимого покоя длятся от 11 до 25 сек.
На 8—9-й день, в связи с развитием конечностей и головы,
намечаются новые формы движений: клевательное движение клюва,
размахивание крыльев, изолированное разгибание одной задней лапки и т. п*
Значит, уже на 8—9-й день появляются также и спонтанные движения»
которые являются характерными элементами поведенческих актов у
взрослых животных.
В последующие дни развития зародыша цыпленка число локальных
движений растет. Голова движется о разные стороны 4—6 раз в імин.;
стопа и пальцы движутся отдельно; коленный сустав то сгибается, то
разгибается, причем разгибается быстрым толчком, а сгибается
медленно, и т. п. Но что характерно,— одно движение, начавшись
совершенно спонтанно, ведет к ряду других движений, благодаря тому что
первично движущийся орган наталкивается на другой орган и
раздражает его. Так, например, крыло сначала ударяет бедро и затем, в
ответ на это раздражение, бедро движется медленно, а пальцы быстро
расходятся, производя разгребательные движения. Если при движении
головы клюв коснется голени, то в ответ на это данная конечность
движется то назад, то вперед, то вниз. Если при сталкивании пальцы
касаются челюсти, то клюв раскрывается и голова быстро отклоняется
назад.
На 14—15-й день заканчивается развитие рефлекторного механизма,
и у зародыша можно наблюдать почти все элементы поведенческих
актов только что вылупившегося цыпленка. Начиная с 15-го дня
спонтанные движения постепенно затихают. Общие движения наступают
реже, а на 17—18-й день совсем исчезают. Локальные движения
продолжаются, но наступают реже; периоды покоя удлиняются, доходя на
19-й день до 6 мин. Часто возникают энергичные движения,
направленные как бы на высвобождение из скорлупы. Это, собственно говоря,
подлинный поведенческий акт, специфичный для цыпленка в скорлупе.
Они начинаются толчками клюва, ножки или крыла; задняя лапка ка-
246 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
сается головы и «яйцевого зуба» и затем быетро и сильно
отталкивается назад.
Наблюдая за развитием, эмбриона цыпленка, мы видим прежде
всего спонтанные сокращения мышц: начиная со 2—3-го дня инкубации
сокращается сердечная трубка, а на 5-й день движется все тело. Но
характерно, что сокращение как сердечной трубки, так и всего тела не
является обусловленным деятельностью центральной нервной системы.
Развитие структуры спинномозгового рефлекторного аппарата
происходит намного позднее (Уиндл, 25, Кахаль, 27). По наблюдениям
Зурабашвили, спинной мозг 4—6-дневных цыплят находится на
начальной стадии своего развития: весь спинной-мозг содержит только
униполярные и 'биполярные клетки при полном отсутствии
межклеточной сети нервных отросткбв; дендриты очень коротки и без ветвлений,
а аксоны некоторых клеток прослеживаются довольно далеко,—
например, аксоны нервных клеток принимают участие в образовании
переднего! корешка. Ясно видно, что на 4—6-й день развития между
нервными элементами спинного мозга нет нервных связей. Это же
установил много раньше Кахаль (27). Следовательно, все
спонтанные движения зародыша цыпленка в этот период развития, а также
двигательная реакция на укол спины, о чем говорилось выше,
осуществлялись мышечной тканью без участия нервных элементов.
Очевидно, в этот период развития не только сердечная трубка,
но и вся мышечная система обладают способностью к спонтанной
деятельности. В 9-дневном возрасте спинной мозг показывает
существенное изменение. В спинном мозгу зародыша четко выступают ядра
(группы клеток) в переднем и заднем роге, дендриты ветвятся и
покрываются шипиками, появляются волоконца в межклеточном
пространстве и в коммиссурах, дендриты и аксоны развиты лучше и выходят за
пределы серого вещества, устанавливая связь ме'жду разными
участками. Эта межнейронная связь осуществляется подрастанием аксонов
к клеткам и дендритам и образованием на них синапсов (Лаврентьев,
26). Как раз в этот период начинают проявляться локальные движения
с элементами чисто поведенческого характера. Начиная с 13-го дня все
ясней и ясней вырисовывается межклеточная сеть — нейропиль — в
результате дальнейшего роста и разветвления дендритов и, значит, все
теснее и многообразнее становятся связи между клеточными
элементами заднего и переднего рогов. Сообразно с этим производимые
эмбрионом движения становятся все сложнее и совершеннее.
Итак, в период формирования нервных элементов спинного мозга —
роста аксонов и дендритов — и установления связей между ними —
возбудимость их так высока, а происходящий в это время обмен веществ,
повидимому, так интенсивен, что создается естественное условие для
спонтанного возбуждения этих нервных элементов. Это спонтанное
возбуждение обусловливает деятельность рефлекторных центров и
рефлекторных путей, что в свою очередь в чрезвычайной мере должно cnocdtf-
ствовать быстрейшему развитию этих же центров и нервных путей.
СПОНТАННАЯ РИТМИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ ЦЕНТ. НЕРВ. СИСТЕМЫ 247
Спонтанная активность в постэмбриональном периоде развития.
В период постэмбрионального развития животных и человека также
проявляется спонтанная активность ряда органов или
преимущественная деятельность их под влиянием определенных внешних раздражений.
Так, например, цыпленок после вылупления клюет положительно всей
реагирует клеванием на разного рода раздражения: зрительные,
слуховые, тактильные (Попов, 44). Новорожденный младенец производит
ротовую реакцию сосания и искания пищи не только в ответ на
прикосновение к губам, но и спонтанно; в случае голода одновременно с общей
ре-акцией (общее движение и крик) он производит также движения
сосания и искания груди (поворот головы с открытым ротиком) при
отсутствии в это время соответствующего внешнего раздражения (Бехтерев
и Щелованов, 45).
Игры молодых животных, а также человека в детском возрасте,
равно как повышенная подвижность животных и человека в раннем
возрасте, являются выражением продолжающегося после рождения
функционального и структурного развития организма. Некоторые
нервные центры!, некоторые рабочие механизмы продолжают развиваться
и после рождения. Вследствие этого возбудимость их повышена и это
облегчает вовлечение их в реакцию как спонтанно, так и под влиянием
внешних и внутренних раздражений. Эти движения у молодых
животных известны как игры. Игры свойственны всем млекопитающим; они
выявляются тогда, когда развитие нервной и мышечной систем уже
подходит к концу, когда в достаточной мере сформированы
рефлекторные механизмы рабочих движений рта и конечностей. Но к этому
времени и кора большого мозга достигает значительного онтогенетического
развития. Поэтому определенные движения конечностей и головы,
наступающие более или менее спонтанно, -быстро индивидуализируются.
Эти движения быстро складываются в серию связанных между собой
индивидуальных реакций, представляющих типичную для данного вида
животных игру. Вследствие этого только первоначально основные
двигательные компоненты игры наступают в результате непосредственного
возбуждения или самовозбуждения соответствующих нервных центров.
Впоследствии же они могут возникать у каждого молодого животного
индивидуальным путем от одного видаі играющего животного или от
вида своего партнера в игре, или даже только от внешней обстановки, где
они играли. Мне, например, приходилось наблюдать, что если несколько
раз во время игры котенка с мячиком пустить метроном, то потом один
звук метронома вызывает движения игры и при отсутствии мячика.
Итак, начальный период постэмбрионального развития, подобно
эмбриональному периоду, характеризуется повышением возбудимости
определенных нервных центров. Вследствие этого и в данный период
онтогенетического развития животных определенным нервным центрам
свойственна спонтанная двигательная активность, а также повышенное
реагирование на внешние или внутренние раздражения.
Спонтанная деятельность центральной нервной системы в инстинк-
248 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
тивных действиях. Существует целый класс поведенческих актов,
которые в основном зависят от спонтанной активности центральной
нервной системы. Это так называемые инстинктивные действия. Известный
специалист по изучению поведения животных Вл. Вагнер (42)
утверждал, что в определенных случаях так называемого инстинктивного
действия поведение животных определяют не внешние стимулы, а
какое-то внутреннее состояние, которое побуждает животное к
определенному целесообразному действию. Это утверждение сейчас находит свое
научное обоснование в спонтанной активности нервных центров.
Лучше всего изучать инстинктивные действия у высших беспозвоночных,
ибо у них инстинктивные действия очень часто совершенно не зависят
от воздействия внешнего імира; они всецело обусловливаются
внутренними изменениями. Так, например, пчела, из так называемых
одиночных, делает кокон (ячейку) и наполняет его медом для прокорма своей
будущей личинки. Если проколоть иголкой кокон снизу, то приносимый
пчелой мед будет вытекать и кокон останется пустым. Однако, пчела
слетает за медом столько раз, сколько нужно для того, чтобы
наполнить целый кокон. Отсутствие меда в ячейке после прокола не
стимулирует дальнейшей его доставки. Она запечатывает пустой кокон, как
будто бы он был полон.
Этого рода факты хорошо уясняют природу типичного
инстинктивного действия: при наступлении того или другого инстинктивного
действия существенно важно не внешнее раздражение, а очередное
повышение возбудимости и обусловленное этим усиление спонтанной
активности в определенных нервных комплексах центральной нервной
системы. Усиление спонтанной активности может происходить или
вследствие наследственных структурных изменений в нервных центрах, в
связи с морфогенезом, или вследствие наследственных изменений в
других тканях, которые продуктами своего обмена веществ чрезвычайно
благоприятствуют возбуждению данных центров, или же вследствие
периодической автоматической деятельности нервных центров,
происходящей под влиянием активных веществ (электролитов, метаболитов)
внутренней межуточной жидкости.
То, что нервные узлы беспозвоночных животных, подобно головному
мозгу позвоночных, обладают способностью к спонтанной активности,
уже показано выше. При этом было установлено, что малейшее
изменение концентрации электролитов — одновалентных и двувалентных
ионов — играет существенную роль в повышении и понижении этой
активности (Моор и Брэдуэй, 38).
Очевидно, в данном ряде инстинктивных действий порядок их
наступления определяется порядком наследственных изменений во
внутренней среде организма. Этот порядок, должно быть, установился в ходе
филогенетического развития в результате естественного отбора.
¦ ¦'У позвоночных животных инстинктивные элементы также широко
представлены в актах поведения. Такова, например, постройка гнезда у
птиц, вывод птенцов и их кормление; постройка норок полевыми мы-
СПОНТАННАЯ РИТМИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ ЦЕНТ. НЕРВ. СИСТЕМЫ 249=
шами и собирание запаса зерен и т. д. Изучение этих инстинктивных
действий приводит также к заключению, что как наступление
определенных инстинктивных действий, так и смена их во времени
обусловливаются, главным образом, наследственно меняющимися внутренними
факторами. Так, например, из наблюдений над гнездованием и
насиживанием у птиц следует заключить следующее. Во-первых, и гнездование
и насиживание обусловливаются состоянием повышенной возбудимости
определенных нервных центров под влиянием внутренних факторов, в
данном случае—под влиянием гормонов половой железы; во-вторых,
эти центры разряжаются импульсами под влияниам определенного рода
внешних раздражений, а также, повидимому, спонтанно; эти импульсы
обусловливают, с одной стороны, возбуждение определенных рабочих
органов, а с другой — торможение всех остальных, как это вообще
имеет место при каждом активировании координирующих аппаратов
центральной нервной системы; в-третьих, с окончанием выделения
гормонов со стороны половой железы сопряжено прекращение насиживания.
Однако, инстинктивные действия позвоночных животных или даже
высших беспозвоночных не обязательно должны протекать всецело
спонтанно прирожденным путем. Лишь первое инстинктивное действие
после рождения или вылупления из яйца вызывается прирожденным
путем. Если же инстинктивное действие повторяется и животное способно
к образованию индивидуально-приобретенных рефлексов, тогда
инстинктивное действие более или менее индивидуализируется, ибо оно
усложняется или видоизменяется на основании индивидуального опыта,
под влияниам той среды, в которой оно протекает. Более того, у
высших животных и у человека инстинктивное действие может измениться
под влиянием психонервной деятельности в такой мере, что у
животных примет характер индивидуального поведения, а у человека —
характер сознательного поведения (Беритов, 48).
Практикум
Опыты над дыхательными движениями лягушки. Автоматическую деятельность
дыхательного центра можно изучить путем записи движений дна ротовой
полости лягушки. Нормальную лягушку осторожно привязывают бинтами за 4
конечности к доске, в которой прорезаны два отверстия для передних конечностей и одно
отверстие для регистрации движений дна ротовой полости. Голова может свободно
двигаться по отношению к туловищу. Регистрация производится так же, как
регистрация сердечных сокращений. Сердечный пинцет прикрепляется к коже дна ротовой
полости. Нитку от пинцета привязывают к легкому миографу, который записывает
дыхательные движения на кимографе.
При постановке опыта необходимо оградиться от посторонних раздражений, от
движения предметов и людей, от манипуляции экспериментатора в поле зрения
лягушки, а также от вибрационных раздражений, как сотрясение стола от ходьбы.
Время от времени будут наступать общие освободительные движения, но это не
помешает наблюдениям. При продолжительных опытах над-о увлажнять кожу лягушки.
Ритмические движения дна ротовой полости двоякого типа: одни движения
большой амплитуды, совпадающие с движением, носовых отверстий, дыхательной щели и
брюшных стенок и связанные с вентиляцией легких; с ними чередуются движения
:250 общая физиология спинного мозга щствола головного^мозга
малой амплитуды, во время которых остальные части дыхательного аппарата
бездействуют. Этими малыми движениями осуществляется веапчаляция ротовой полости;
слизистая оболочка этой полости обладает дыхательной функцией.
Влияние раздражений на автоматическую деятельность дыхательного центра.
Раздражение кожи и 'Чувствительных нервов на- голове, растяжение мышц [вызывают
торможение (рис. 105-Л). Так же действует свет, хотя и слабее (рис. 105-В).
Слабое околопороговое раздражение п. peroneus усиливает дыхательные
движения. Так же действуют вибрации от постукивания тго столу (рис. 105-С).
Исследование спонтанных биотоков возбуждения в разных отделах головного
мозга лягушки. Перерезается опийной мозг на границе с головным мозгом или
берется изолированно голова лягушки. Еще лучше взять кураризованную лягушку.
Открывается весь головной мозг сверху; голова прикрепляется прочно к пробковой
пластинке булавками. В качестве отводящ'?го электрода берут заостренную
хлорированную серебряную иглу. Острие иглы прикладывается к отводимому участку
головного мозга. Другой отводящий электрод в виде серебряной пластинки лежит под
препаратом. Биотоки регистрируются катодным осциллографом.
Если усилитель однотактный, то игольчатый электрод соединяется с сеткой, а
пластинка с землей. Если же усилитель двутактный, с балансированным входом, тогда
-игольчатый электрод соединяется с любой входной клеммой усилителя.
Наблюдают на экране спонтанные биотоки различных отделов головного мозга,
прикладывая игольчатый электрод к поверхности обонятельных долей больших полу-
лгхарий, промежуточного мозга, среднего мозга, мозжечка и продолговатого мозга.
Затем наблюдается изменение этих биотоков под влиянием периферических
раздражений, а именно — усиление биотоков среднего мозга при раздражении сетчатки
светом или передвижением предмета в поле зрения лягушки; усиление биотоков
продолговатого мозга в области вхождения VIII нерва или на расстоянии 1—2 мм от
него при звуках и вибрациях: человеческий разговор, свисток, вибрации от проведения
палочкой или кисточкой по пробковой пластинке, от приложения общего конца
-Движущегося камертона к той же пластинке.
Лите атура
.1. Dusser de Barenne a. Breve e, J. Physiol., 61, 81, 1928.
2. R u с h Т. С, Am. J.Physiol., 114, 457, 1936.
3. ProsserCL, Gold Spr. Harb. Symp. Quant. Biol., 4, 339, 1936.
4. Adrian EM Proc. Roy. Soc, В., 106, 596, 1930.
5. Б?ритов И., Гедеванишвили Д., Воробьев А., Тр. Ин-та
физиол. им. Бериташвили, 5, 358, 1943.
6. S р і е g е 1 Е. A., Am. J. Physiol., 118, 569, 1937.
7. D о w R. S., J. Physiol., 04, 67, 1938.
8. L i b e t B. a. G e r a r d R. W., J. Neurophysiol., 2, 153, 1939. ^
9. Беритов И. и ЦкипуридзеЛ., Сообщ. Акад. Наук Груз. ССР, 2, № 9,
1941; 3, № 1, 1942; 5, № 1, 1944.
10. Беритов И. и Цкипуридзе Л., Тр. Ин-та физиол. им. Бериташвили,
5, 215, 1943.
11. Р г о s s е г С. L. а. В и е h 1 С. С, J. Cell. Сотр. Physiol., 14, 287, 1939.
12. G e s е 11 R., Atkinson А. N. а. В г о w n R. С, Am. J. Physiol., 128,
629, 1940.
13. Barcroft J. a. Barron D. H., Физиол. журн. СССР, 22, 278, 1937.
14. Adrian E. D. a. Buytendijk F., J. Physiol., 71, 9, 1931.
15. Adrian E. D., J. Physiol., 72, 132, 1931; Механизм нервной деятельности,
пер. Биомедгиз, 1935.
16. Pitts R. F., Magoun H. W., a. Rans on S. W., Am. J. Physiol., 126,
673, 1939.
17. Pitts R. F., J. Сотр. Neurol., 72, 605, 1940.
СПОНТАННАЯ РИТМИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ ЦЕНТ. НЕРВ. СИСТЕМЫ 251
18. Чичинадзе Н., Тр. Тбил. гос. ун-та им. Сталина, 26-а, 61, 1944.
119. ЗурабашвилиА. Д., Тр. Ин-та физиол. им. Бериташвили, б, 169> 1943;
Сообщ. АН Груз. ССР, 4, 477, 1943.
'.20. Montalcini R. L. eVisantini F., Boll. d. Soc. Ital. d. Biol. Sperim.,
13, 976, 1938.
21. Kuo J., J. Exp. Zoology, 61, 395, 1932.
22. Windle W. F. a. Orr D. W., J. Сотр. Neurol., 50, 287, 1934.
23. Orr D. W. a. W i n d 1 e W. F., J. Сотр. Neurol., 60, 271, 1934.
24. S t г a u s W. a. W e d d e 1 G., J. NeurophysioL, 3, 358, 1940.
25. Windle W. F.,J. Сотр. Neurol. 50, 479, 1930.
26. Л а в p e h t ь e в В. И., Арх. биол. наук, 48, 194, 1938.
27. Ramon и. С а ] а 1. S., Anat. Anz., 5, 609, 631, 1890.
28. Б е р и т о в И., Брегадзе А. и Цкипуридзе Л., Сообщ. АН Груз,
ССР, 3, № 2, 1942.
29. Dub пег Н. Н. a. Gerard R. W., J. NeurophysioL, 2, 142, 1939.
30. Hoist E., E. Pfl. Arch., 237, 655, 1938.
31. Adrian E. D. a. M о r u z z i, J. Physiol., 97, 155, 1939.
32. Беритов И., Брегадзе А. и Цкипуридзе Л., Тр. Ин-та физиол.
им. Бериташвили, 5, 241, 1943.
33. Moris on R. S.,Filney К. H. a. Lothrop G. N., J.
NeurophysioL, 6, 243, 1943.
-34. Pick E. P. and M i 11 e r M. M., J. NeurophysioL, 8, 47, 1945.
35. P о й т б а к А., Об электрических потенциалах головного мозга лягушки при
раздражениях нервов и рецепторов. 1947. Не опубликовано.
36. Adrian Е. D., J. Physiol., 22, 132, 1931.
37. A d г і а п Е. D., J. Physiol., 70, р. XXXIV, 1930.
38. М о о г е A. R. and В г a d w а у W. Е., J. Cell. а. Сотр. Physiol. 25, 181, 1945.
39. Bernard G., J. NeurophysioL, 5, 32, 1942.
40. Б e p и т о в И. С, Тр. Тбилисск. гос. ун-та им. Сталина, 27-а, 29, 1945.
41. Heinbecker P., Amer. J. Physiol., 117, 686, 1936.
42. В а г н е р В. А. Этюды по сравнительной психологии. Вып 1, 2, 3 и 4. Л. 1925.
43. Беритов И. и Цкипуридзе Л. Сообщ. АН' Груз. ССР, 6, 719, 1945.
44. Попов] Н., Сиб. Арх. тер. и клин, медйц., 2, 232, Томск, 1927.
45. Бехтерев В. иЩелованов, Сборы. Новое в рефлексологии и физиологии
нервной системы, т. 1, 125, 1925.
46. БакурадзеА. иЦкипуридзеЛ. Тр. Инст. Физиол. им. Бериташвили,
7, 1947.
47. Т е п - С a t е J. a. W і g g е г s К., Arch. Neerland. Physiol., 2S, 423, 1942.
48. Бакурадзе А. иЦкипуридзеЛ. Тр. Инст. Физиол. им. Бериташвили,
7, 1947.
49. Dow R. S., J. NeurophysioL, 2, 543, 1939.
50. Wo Ike r, A. E., J. NeurophysioL, 1, 16, 1938.
51. Цкипуридзе Л., Докл. на научной сессии Тбил. гос. универс. им. Сталина,
1947.
52. AdrianE. D. a.Moruzzi G., J. Physiol., 97, 153, 1939.
53. Беритов И. иРойтбак А.,Тр. Ин-та физиол. им. Бериташвили, т. 7,
1947 г.
54. Kugelberg E., Brain, 69, 310, 1946.
55. Winter stein H., Pfl. Arch., 138, 159, 1911;-—Arch. int. pharmacodyn., 73,
302, 1946.
56. A p ш а в с к и й Н. А., Физиол. журн. СССР, 29, 417, 517, 1940;, 32, 495, 1946.
57. К рю чко в а А. П., Физиол. журн. СССР, 24, 523, 1938.
58. Беритов И., Об основных формах нервной и психонёрвной; деятельности.
Изд. АН СССР, 1947.
Глава XIII
ШОК ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Общее понятие. У позвоночных животных перерезка толстого
чувствительного нерва и, ів особенности, самого мозга производит ів
центральной нервной системе характерное изменение, подобного которому
не знает ни мышечная, ни периферическая нервная система. Именно,
центральная нервная система парализуется на некоторое время. При этом
рефлекторная деятельностыможет ослабнуть до такой степени, что даже
сильнейшими раздражениями нельзя вызвать оборонительных
движений. Абсолютная недеятельность йотом заменяется глубокой
депрессией, и это явление проходит с большой постепенностью в течение
часов, дней и даже месяцев. Такое состояние возникает вообще в
результате всякого рода чрезмерных раздражений центральной нервной
системы, — например, от сотрясения. Подобная депрессия рефлекторной
деятельности известна под названием шока, что, означает удар,
сотрясение. Это название было дано английским ученым Маршал
Холлом (1).
Явление шока обнаруживается у различных позвоночных животных
в различной степени. Чем выше ступень, которую занимает животное на
зоологической лестнице, тем сильнее и длительнее бывает у него
состояние шока. У человека и у высших обезьян после перерезки спинного
мозга конечности на долгое время, а иногда и навсегда, утрачивают
способность к рефлекторной деятельности. Восстановление
рефлекторной деятельности, если и случается, то никогда не бывает полным, даже
в отношении наиболее устойчивых оборонительных рефлексов.
У кошек и собак рефлекторная деятельность ослабевает после
перерезки спинного мозга в меньшей мере и в значительной мере
восстанавливается спустя несколько дней или недель. Хорошо
восстанавливается оборонительный сгибательный рефлекс, хуже—рефлекс
чесания, еще хуже — сухожильный рефлекс и перекрестный экстензорный
рефлекс. У кроликов шок выявляется еще слабее. У них, например,
сухожильный рефлекс исчезает всего на 10—15 мин. (Шеррингтон, 2).
У амфибий при этих условиях явление шока продолжается очень
недолго.- ют одной до нескольких десятков минут, а иной раз вообще не
ШОК ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ 253
поддается учету. У птиц шок протекает сильнее, чем у амфибий, но
много слабее, чіем у млекопитающих (Шеррингтон, 2).
Состояние шока при перерезке мозга проявляется не в одинаковой
степени во всех элементах спинного мозга. Путем изучения биотоков
спинного мозга было установлено, что у собак и кошек состоянию
шоковой депрессии после перерезки спинного мозга главным образам
подвергаются промежуточные нейроны, причем сильнее перекрестные ком-
миссуральные нейроны для перекрестных рефлексов, чем гомолатераль-
ные — для гомолатеральных рефлексов. У обезьян шоковая депрессия
промежуточных нейронов выступает еще сильнее, но, кроме того,
заметной депрессии подвергаются и двигательные нейроны (Юз, Коуч
и Стюарт, 14).
После перерезки мозга угнетенное состояние нервных элементов
выступает гораздо сильнее в каудальном направлении, чем в краниальном.
Кроме того, в каждом данном отделе мозга угнетенное состояние
зависит от его близости к разрезу мозга. Например, если мозг перерезан в
шейном отделе спинного мозга, то состояние шока проявляется на
передних конечностях сильнее, чем на задних. У низших млекопитающих
и, вероятно, у івсех низших позвоночных состояние шока наблюдается
главным образом в отношении скелетной мускулатуры. Из
вегетативных органов, повидимому, шоку подвергается только сосудистая
система. Сосудодвигательные рефлексы, вызываемые раздражением
афферентных нервов, в первое время после наступления шока отсутствуют.
Они восстанавливаются одновременно с восстановлением рефлексов
на скелетной мускулатуре (Шеррингтон 2, 6). Но у высших
позвоночных, а также и у человека, шок в -одинаковой мере захватывает как
соматическую, так и вегетативную систему. Так, у обезьян после
перерезки спинного мозга наступает паралич кишечника, мочевого пузыря;
кожа становится сухой; пилоэрекция и сосудодвитательные рефлексы
отсутствуют (Закс и Фултон, 12). У человека при сильных травмах
мозга кожа становится холодной и сухой, пульс учащается, температура
понижается, кровяное давление падает (Гринкер, 13). Характерно, что
эти шоковые явления в области вегетативной системы проходят с
большой постепенностью в течение многих дней, значительно позднее, чем
нарушения в области соматических рефлексов (Закс и Фултон, 12).
Шок и наркоз. Явление шока, наступающее при повреждениях
периферических чувствительных нервов, имеет место только в том
случае, если эти повреждения происходили при нормальной возбудимости
центральной нервной системы. Они устраняются совсем, если
повреждение происходит при глубоком наркозе центральной нервной системы,
вызываемом с помощью хлороформа, эфира и других наркотиков.
При пониженной возбудимости центральной нервной системы шок
хотя и имеет место, но в значительно более слабой степени; он тем
слабее, чем ниже была возбудимость. Этим и объясняется, почему все
операции на высших позвоночных животных производятся под наркозом.
Точно также, если при операциях на человеке прибегают к наркозу
254 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
или к (местной анэстезии, то это делают не только для того, чтобы
пациент не чувствовал боли, но главным образом, для того, чтобы
избегнуть или ослабить явления шока.
При перерезках мозга шок наблюдается и в том случае, если
перерезка произведена в условиях 'глубокого наркоза. После прекращения
действия наркотика рефлекторная деятельность как соматическая, так.
и вегетативная, оказывается пониженной на более или менее
продолжительное время.
Происхождение шока. О природе шока впервые высказался Гольц
(10). Он признал шок явлением торможения, которое вызывается
травмой. По его мнению, при травме не только непосредственное
повреждение, но и развивающийся затем воспалительный процесс или.
процесс дегенерации и регенерации, действуют как раздражение,
которое вызывает торможение и тем самым понижает рефлекторную
деятельность не только в ближайших, но и в отдаленных участках
центральной нервной системы.
Однако Шерри-нгтон (2), особенно тщательно изучивший это-
явление, разнообразными опытами показал, что шок центральной
нервной системы не представляет собой торможения.
1. Если бы шок был торможением, он обнаруживался бы в
краниальном направлении с такой же силой, как и в каудальном.
2. После перерезки шейнЪго отдела мозга давление крови падает^
вазомоторные рефлексы исчезают; спустя несколько дней давление
крови возвращается к норме. Если после этого снова перерезать мозг
нижепрежнего іместа, то давление крови падает значительно слабее, и
притом на очень короткое время. Если же шок был бы торможением,
вызванным травмой, то во втором случае падение давления должно было»
бы быть таким же сильным, как и в первом.
3. Судя іпо характеру изменения рефлекторных движений во время
шока, последний скорее похож на утомление, чем на торможение. Так„
например, рефлекс чесания то вызывается, то не вызывается, а когда
вызывается, протекает нерегулярно; электрическое раздражение кожи
его не вызывает, а механическое вызывает. Все это наблюдается при»
утомлении чесательного рефлекса.
Таким образом, по мнению Шеррингтона, во время шока
состояние рефлекторного аппарата очень похоже на его утомление.
Судя по реакциям двигательных органов, шок возникает как бы в-
результате затруднений для проведения возбуждения в рефлекторных
дугах; иначе говоря, он как бы происходит от ослабления
межнейронных связей в рефлекторной дуге, между прочим, благодаря
затруднению передачи возбуждения от чувствительных нейронов к двигательным.
Затруднения в лередаче возбуждений, согласно первоначальному
мнению Шеррингтона, наступают вследствие перерезки каких-то
нисходящих путей. Эти пути, повидимому, начинаются не в больших
полушариях, ибо удаление большого імозга не ведет к глубокой депрессии,
характерной для шока; она имеет место после перерезки позади варолие-
ШОК ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
255
ва моста. Что в происхождении явления шока действительно играет роль
прекращение каких-то центральных импульсов, об этом свидетельствует
ослабление рефлекторной деятельности спинного мозга при устранении,
влияния головного м/озга без какого бы то ни было раздражения
нервной системы. Этого можно достигнуть методом Тренделенбурга
(3) — охлаждением спинного мозга до исчезновения в нем
проводимости. Если охладить локально какой-либо сегмент спинного мозга и
таким образом прекратить в этом участке проведение возбуждения, та
деятельность находящейся позади него части спинного мозга
ослабевает: затрудняется вызов рефлекторных реакций на задних ногах,
кровяное давление падает. Явления, развивающиеся в этом случае, ничем
не отличаются от происходящих при перерезке спинного мозга.
Охлаждение не вызывает никаких движений животного, так как оно
происходит постепенно. Охлажденный участок нервной системы находится в
состоянии парабиоза. По прекращении охлаждения проводимость
постепенно возвращается к норме и рефлекторная деятельность в заднем
отделе вполне восстанавливается. Тренделенбург заключает, что в
нормальных условиях возбудимость спинного мозга зависит от влияния
возбуждений, иррадирующих из головного мозга. Когда же вследствие
Охлаждения это влияние прекращается, возбудимость в спинном мозгу
падает, а потому ослабевает и рефлекторная деятельность. И
действительно, в настоящее время хорошо известно, что нервные центры
головного мозга — дыхательный, тонические, сосудодвигательные
испытывают спонтанную активность и потому играют существенную роль в
рефлекторной возбудимости спинного мозга. Так, например, известно,
что во время дыхательных движений рефлекторная деятельность
усиливается. Усиливаются все онинальные рефлексы: сухожильные, чеса-
тельный, тонические и т. д. Чем сильнее деятельность дыхательного
центра, тем сильнее его положительное влияние на рефлексы (;Киш\
Блэир и Гаррей, 9 и др.). Отсюда следует, что дыхательный центр
определенным образом связан нейронными путями с двигательными
нейронами спинного мозга и через эти связи распространяет на них свое
возбуждающее влияние.
Выше было подробно рассмотрено облегчающее действие спонтанной
активности головного мозга на спинной мозг, а также вообще
облегчающее действие мозга на двигательные нервы и на мускулатуру. На
этом основывается субординирующее действие мозга, т. е. уменьшение
хронаксии двигательного нерва и скелетной мускулатуры, Мы так же
указывали на зависимость этого облегчающего и субординирующего
действия от периферических раздражений, ибо периферическими
раздражениями обусловливается интенсивность деятельности локомоторных,
тонических, дыхательных, сосудодвигательных центров стволовой части
головного мозга.
Но активное облегчающее действие высших отделов мозга
проявляется не только в виде облегчения спинномозговых рефлексов, айв
другой чрезвычайно важной форме. Именно, мы выше указали, что нор-
256 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
мально у высших позвоночных периферическое возбуждение не
вызывает рефлекторной реакции непосредственно из спинного мозга.
Сначала оно достигает высших центров, а уже потом возвращается к
координирующим аппаратам спинного мозга. Обычно прямой путь к
координирующим аппаратам бездействует. Наши опыты на кошках над
проведением возбуждения в центральной нервной системе показали, что не
только при низкой возбудимости спинного мозга, но и во время
повышения возбудимости от неконвульсивных доз стрихнина
периферический рефлекторный эффект от раздражения рецепторов и
чувствительных нервов конечности получается, главным образом, через длинные
пути головного мозга (7).
На основании этих данных нужно предположить, что одним из
главных факторов явления шока у высших позвоночных при перерезках
мозга является разрыв длинных путей, нисходящих из головного мозга,
/по которым происходит постоянная импульсация к спинному мозгу.
В происхождении шока существенную роль играет еще другой
фактор. Перерезка мозга производит длительное механическое
раздражение его. Раздражаются при этом как нейропиль, так и восходящие и
нисходящие нервные пути. Эти пути связаны с нейропилем всего
спинного мозга, вследствие чего последний также приходит в длительное
активное состояние. Чем сильнее деформация мозга при перерезке, тем
сильнее и длительнее раздражение нисходящих путей и самого нейропи-
ля, тем сильнее и длительнее нейропильное угнетение. Далее, при
любой перерезке мозга некоторое кратковременное угнетение
рефлекторной деятельности всегда наблюдается »и в отношении передней части
мозга, т. е. в краниальном направлении. Это видно из опытов Гольца,
а также Шеррингтона. Это кратковременное угнетение совершенно
такого же рода как и при перерезке нервного ствола, причем оно
повторяется при каждом повторении перерезки мозга. Эти факты говорят
за то, что явление шока обязано своим происхождением также и
центральному угнетению тормозящего характера, которое должно
происходить каждый раз при перерезке мозга вследствие активирования ней-
ропиля. ,
Характерно, что продолжительность шока зависит от способа
перерезки. Когда перерезка производится острым инструментом, шок
проявляется слабее и длится более короткое время, чем при грубом
разрушении определенного участка мозга (Гольц, 10). Далее, известно, что у
высших позвоночных механическое повреждение средних отделов мозга,
¦в особенности продолговатого, производит длительное угнетение
рефлекторной деятельности (Рожанский, 8). То обстоятельство, что у
высших позвоночных послеоперационное угнетение тем сильнее и
длительнее, чем больше развито нейропильное образование в области
повреждения и чем сильнее было повреждение, свидетельствует также о
правильности того предположения, что это угнетение обусловлено
тормозящим действием нейропиля на нисходящие пути и нервные ядра.
ШОК ЦЕНТРАЛЬНОЙ'НЕРВНОЙ СИСТЕМЫ 257
Вызванное операцией перерезки раздражение перерезанного отдела
мозга с течением времени проходит. Вследствие этого ослабевает
активация нейропиля, и потому нервные пути освобождаются от угнетающего
влияния со стороны раздраженного нейропиля, и возбудимость их
должна возвратиться к норме. Но обычно возбудимость не остается в
норме, спустя несколько дней она становится значительно выше нормы.
Это, повидимому, объясняется тем, что перерезанные афферентные и
нисходящие нервные пути с течением времени дегенерируют. В
поврежденных волокнах в период дегенерации спонтанно возникают
импульсы возбуждения, которые, действуя на двигательные и
промежуточные клетки, при которых они оканчиваются, повышают в них
возбудимость.
Выше мы указывали, что если после восстановления рефлекторной
деятельности вновь перерезать спинной мозг несколько ниже, чем в
первый раз, то это вызовет незначительный шок, который продолжится
более короткое время. Это объясняется тем, что основные пути, идущие
из головного мозга, уже были перерезаны и дегенерировали,—
следовательно, вторичная перерезка могла вызвать раздражение или
выключить только тот пучек нервных волокон, который начинается в
небольшом участке спинного мозга ниже уровня первой перерезки. Отсюда
следует, что вызванное второй перерезкой небольшое и
кратковременное угнетение спинномозговых рефлексов может быть объяснено
незначительным активированием нейропиля от раздражения
небольшого пучка нервных волокон, начинающегося ниже уровня первой пере-
резки.
Явление шока имеет место не только при перерезке мозга или
толстых нервных стволов, но и при сильном и длительном раздражении
чувствительных нервов и кожных рецепторов, а также при сотрясении
мозга. Далее, известно, что этот так называемый травматический щок
случается тогда, когда повреждение или раздражение мозга и
периферических нервных стволов происходит в бодрствующем состоянии
животного. Эти факты указывают на то, что чрезвычайно усиленное
активное состояние центральной нервной системы играет существенную роль
в происхождении травматического шока. Как уже указывалось выше,
усиленное активное состояние должно вызвать понижение
рефлекторной деятельности прежде всего вследствие истощения координирующих
аппаратов в результате их чрезвычайно интенсивной деятельности, а
потом вследствие угнетения их, а также двигательных клеток под
влиянием усиленного активного состояния нейропиля, обусловленного
длительной импульсацией со стороны раздражаемых периферических
нервов или нервных центров. Но в случаях травматического шока, когда
он вызывается сильным и длительным раздражением нервов или
мозга, в происхождении шокового угнетения рефлекторной деятельности
наиболее существенное значение должен иметь другой фактор.
Работами Л. Ш те р н и ее сотрудников (И) в последнее время было
установлено, что во время шокового угнетения, вызванного у кошки или
.17 и. С. Беритов
25S ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
собаки сильным и длительным раздражением чувствительных нервов
или кожных рецепторов, меняется проницаемость гемато-энцефаличе-
ского барьера (она, например, повышается в отношении
кристаллоидов) , меняется химический состав спинномозговой жидкости
(например, коэффициент К/Са уменьшается вследствие увеличения
содержания кальция и уменьшения калия). В результате этих изменений
меняются и физиологические свойства спинномозговой жидкости.
Спинномозговая жидкость, взятая у собаки в состоянии такого шока,
угнетает, рефлекторную деятельность, лягушки, замедляет и даже
останавливает изолированное сердце лягушки. Все эти изменения в
спинномозговой жидкости отсутствуют при раздражении чувствительных.нервов
на наркотизированных животных. На этом основании предполагается,
что под влиянием сильных и длительных раздражений чувствительных
нервов в химическом составе спинномозговой жидкости происходят
определенные сдвиги благодаря усиленной активации вегетативных
центров, в частности гипофиза. Эти сдвиги, в свою очередь действуя
на центральную нервную систему, обусловливают понижение
соматической и вегетативной рефлекторной деятельности.
В определенных случаях эти химические факторы, безусловно,
должны играть доминирующую роль в происхождении шокового угнетения.
Это лучше всего видно из того, что совершенно аналогичное шоковое
угнетение можно вызвать искусственным изменением химического
состава спинномозговой жидкости путем введения некоторых химических
веществ,— например, пептона, гистамина, солей кальция и т. д.,— в
общее русло крови или непосредственно в спинномозговую жидкость
(токсический шок).
Поскольку происхождение травматического шока, подобно
токсическому, связано с определенными изменениями химического состава
спинномозговой жидкости, то естественно ожидать, что всякое
изменение его в противоположном направлении должно повысить возбудимость
центральной нервной системы и тем самым вывести ее из состояния
шока. Это достигается введением в спинномозговую жидкость
фосфорнокислого калия, который повышает коэффициент К/С а до нормы (И).
В боевой обстановке тяжело-е шоковое состояние может наступить и
при небольшом ранении, т. е. от незначительной, на-глаз, травмы.
В таких случаях в происхождении шока играет существенную роль не
ранение, а то психофизиологическое состояние, в котором находится
боец в момент боя. Сильнейшие внешние раздражения, а также
чрезмерные эмоционально-психические возбуждения обусловливают
сильнейшую активацию симпато-адреналовой системы, вследствие чего
кровь перегружается адреналином, что, в свою очередь, приводит к
тяжелому расстройству кровообращения с недостаточным
кровоснабжением и аноксическими явлениями, между прочим, в мозгу и мышцах.
При таком состоянии организма любая травма может вызвать тяжелое
шоковое состояние (Цкиманаури, 15).
ШОК ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ «;59
Итак, шок по своему происхождению представляет собой сложное
явление, которое в различных случаях зависит от разных факторов в
разной степени. Я?ление шока при перерезках мозга должно
обусловливаться: во-первых, разрывом нисходящих нервных путей, по которым
осуществляется вся рефлекторная деятельность головного мозга; во-
вторых, тормозящим дейст&ием поврежденного отдела мозга на эти
пути и близлежащие ядра; в-третьих, утомлением координирующих
аппаратов в случае их длительной и интенсивной деятельности. При
периферических раздражениях явление шока должно зависеть, во-первых, от
утомления координирующих аппаратов и, во-вторых, от тормозящего
действия активного нейропиля на нейронные элементы спинного мозга.
Но в тех случаях, когда мы имеем дело с длительным и сильным
возбуждением центральной нервной системы, связанным с травмой или
с чрезмерным эмоционально-психическим состоянием, тогда шоковое
состояние обусловливается, главным образом, не непосредственным
действием импульса на центральную нервную систему, а изменением
химического состава крови и спинномозговой жидкости, угнетающе дей-
ітвующим на центральную нервную систему.
Практикум
Наблюдение шока на лягушке. Берут лягушку, перерезают спинной мозг под
продолговатым и сейчас же находят пороги раздражения для сгибательного рефлекса
задней или передней лапки и рефлекса потирания. Раздражается кожа
индукционными ударами в рецептивном поле сгибания. Определяют пороги несколько раз один
вслед за другим на одном и том же участке кожи через промежутки в 2—3 мин.
Пороговая сила последующих раздражений будет меньше, чем первоначального.
Состояние шока в спинном мозгу лягушки держится значительно дольше, если
грубо повредить продолговатый моэг,— например, разрушить его тупым предметом.
Это повреждение вызывает длительное судорожное сокращение всех мышц. Сейчас
же после прекращения судорог рефлекторная деятельность оказывается сильно
пониженной: рефлекс сгибания вызывается в сильно ослабленной форме, а рефлекс
потирания отсутствует. В течение нескольких минут это состояние проходит:
рефлекторная деятельность восстанавливается. Это состояние шока, повидимому,
обусловливается главным образом утомлением клеточных или синаптических элементов
спинного мозга.
Состояние шока у лягушки можно вызвать также, подбрасывая ее до потолка и
давая ей упасть на пол. Нужно подбросить ее так 2—4 раза подряд. После каждого
нового подбрасывания рефлекторная деятельность ухудшается. После 3-го или 4-го
падения рефлекторная деятельность исчезает полностью. Сейчас же после падения
лягушка лежит без движения, не дышит, не реагирует на раздражения кожи. Но
уже в течение первой минуты рефлекторная деятельность начинает
восстанавливаться: на болевое раздражение лапка сгибается в колене, но рефлекс потирания
отсутствует, лягушка не передвигается, не переворачивается со спины на живот. Еще
позднее вызывается рефлекс потирания, лягушка прыгает при щипке,
переворачивается со спины на живот, но спонтанная локомоция отсутствует. Она
восстанавливается позднее всего.
\1*
260 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Литература
1. Marshall Hall, Synopsis of the Diastaltic Nervous System. London, 1853.
2. Sherrington C.,' Int. Act. Nerv. Syst., 1906.
3. Tr en del enburg. P., Pfl. Arch., 133, 305, 1910; 135,460, 1910,137,515, 1911,
Zbl. Physiol., 1914.
4. Pike G. H.,Am. J. Physiol., 24, 124, 1909; 30. 436, 1912; Quart J. Exp.
Physiol., 7, 1, 1913.
5. D a n i 1 e w s к у V., Pfl. Arch., 78, 194, 1899.
6. Шеррингтон и сотр. Рефлекторная деятельность спинного мозга, 4935.
7. Беритов И., Тр. Физиол. ин-та им. Бериташвили, 3, 87, 1936.
8. Рожанский Н.А., Физиол. журн. СССР, 19, 289, 1935.
9. King С. Е., Blair Е. A., G а г г е у W. Е., Am. J. Physiol., 97, 329, 1931
10. G о 1 t z Fr. u. E w a 1 d J. R., Pfl. Arch., 63, 362, 1896.
11. Штерн Л., Шок. Тр. конф. по проблеме шока, стр. 10, 1937; см. также Сб ,
посвящ. проф. Когану-Ясному, 1940.
12. Sachs A. L. a. F u 11 о n J. F., J. Neurophysiol., 3, 258, 1940
13. G rinker R. R., Neurology, 786, 1937.
14. Hughes J., Mc Couch G. P., a. S t e w a r t W. В., J. Neurophysiol.,
3, 139, 146 и 151, 1940.
15. ЦкиманауриГ. A f Сборн. тр. Инст. гигиены труда и проф-забол.
Тбилиси, 1947; '
Глава XIV
ПРОПРИОЦЕПТИВНЫЕ РЕФЛЕКСЫ
Общее понятие. Проприоцеитивными рефлексами
называются такие реакции центральной нервной системы, которые наступают в
ответ на раздражение проприоцепторов, т. е. рецепторов мышц,
сухожилий и суставов. Раздражение их нормально происходит во время
самого движения конечности. Происходящее при этом сокращение одних
мышц и пассивное растяжение антагонистов являются причиной
механического раздражения рецепторов.
Свойства мышечных рецепторов и роль их в рефлекторной
деятельности были подробно исследованы Эдрианом и его сотрудником
Мэтью сом (20). Они изучили их как у холоднокровных (лягушек),
так и у теплокровных животных (кошек, морских свинок) и пришли к
более или менее одинаковым результатам. Значительное количество
рецепторов, представленных в мышце в виде спиральных образований в
мышечных веретенах, возбуждается при растяжении (рис. 26—27).
Чувствительный нерв, связанный с одним таким рецептором, дает при
каждом растяжении серию импульсов, ритм которых, сначала высокий,
постепенно падает (рис. 28). Раздражение этих рецепторов получается
уже при 1—2 г нагрузки. Но чем больше растягивающая сила, тем
выше начальный ритм. При некоторых больших нагрузках он может
достигнуть у лягушек 290 в 1 сек., а у млекопитающих 500 в 1 сек. Во
всех случаях импульсы, с прекращением растяжения, исчезают.
Полагают, что при растяжениях возбуждаются толстые афферентные
волокна, оканчивающиеся в мышечном веретене кольцеобразно-спиральными
окончаниями Руффини (рис. 106-6).
Мышечные рецепторы возбуждаются также при сокращениях, но
только в том случае, когда сокращение довольно сильно. Так,
например, это можно наблюдать при сильном тетаническом раздражении
двигательного нерва (рис. 107 Л и С). Чувствительный нерв мышцы дает
при этом довольно редкий ритм возбуждения. Предполагают, что при
этом возбуждаются особые рецепторы мышечных веретен, которые
связаны с тонкими нервными волокнами (рис. 106-d), Они, повидимому,
262 общая физиология спинного мозга и ствола головного мозга
.
№11
іііёіі
II
Рис. 106. Схематическое изображение концевых органон в мымі
це млекопитающих (Денни-Броун).
От каждого отдельного мышечного волокна идет сухожильное волокно
к апоневрозу. На рисунке изображены 4 двигательных нервных волокна
<а), иннервирующих 23 мышечных волокна. В действительности гораздо более
значительное число мышечных волокон иннервируется каждым отдель іым
двигательным нервным волокном; мышечные волокна, ипнервируемые одпой
нервной клеткой, собраны в группы. Схематически изображенное
мышечное веретено состоит из 3 тонких мышечных волокон, которые иннер-
вируются при посредстве двигательных конечных пластинок 3 двигательными
нервными волокнами (предполагается, что эти волокна берут начало в трех
разных двигательных нервных клетках).
Кольцеобразно-спиральное нервное окончание, находящееся в связи с
толстым афферентным нервным волокном 6, охватывает тонкие, так
называемые интрафузальные мышечные волокна на уровне наибольшего диаметра
веретена. Мышечные волокна также имеют нервные окончания второго
афферентного волокна d. В более сложных мышечных веретенах, содержащих
большое число интрафузальных волокон, имеется гораздо более
значительное количество этих окончаний, а также и окончаний двигательных
нервных волокон. Сухожильный аппарат Гольджи распространяется на
небольшое количество волокон. Соответствующие им мышечные волокна ипнер-
вируются различными двигательными нервными клетками; д —
соответственный афферентный нерв. Афферентные волокна (с) направляются к стенкам
артериол (е) и к окружающей их соединительной ткани. Они переплетаются
с симпатическим сплетением (sp.).
проприоцшітивные рефлексы
263
раздражаются (механически вследствие сокращения мышечных волокон
мышечного веретена. Эти волокна, а также соответствующие им
двигательные волокна обладают низкой возбудимостью. Этим и объясняется
необходимость сильных раздражений.
Когда двигательный нерв раздражается субмаксимально, тогда
мышечные проприоцепторы не возбуждаются. Более того, если они уже
были возбуждены в силу растяжения и чувствительные волокна давали
А
W
8
ПИіііМ
П1111П111
Ж-Н4
Рис. 107. Электрическая реакция мышечных рецепторов
от сокращения m peruueus longus кошки.
Соответствующий мышечный нерв отводится к осциллографу
Мэтьюса. Сплошная кривая — напряжение мышцы, быстро
колеблющаяся и ток л действия нерва. Л и В—при начальном
напряжении в 25 г; С — при начальном напряжении в 900 г.
В опыте А раздражение максимальное, в опыте В —
субмаксимальное, а в опыте С — супрамаксимальное. Электрический
аффект до сокращения мышц—от начального растяжения мышцы,
В опыте А большие токи от раздражения нерва непосредственно,
а мелкие от раздражения рецептора. До сокращения ритм этих
токов 40, а во время него около 300 в 1 сек. После
расслабления постепенно устанавливается прежний ритм. В опыте
В при субмаксимальном раздражении во время сокращении
одни только большие токи от раздражения нерва
непосредственно. Токи возбуждепия от раздражения рецепторов спвгрм
отсутствуют. Они появляются лишь в момент расслаб лени і
мышцы. В опыте С при сунрамаксимальном раздражении оольшие
токи — цт раздражения нерва, а малые — от возбуждения
рецептора. Они наступают почти вдвое чаще, чем во время
расслабления: около 140 в 1 сек. Шэтыос).
токи возбуждения, то эти токи при таком сокращении исчезают.
Происходит это оттого, что при сокращении перестает действовать
механическое раздражение от растяжения. Это отрицательное
действие сокращения на разряды мышечных веретен ссобенно'ясно
сказывается при одиночных сокращениях, что видно на рис. 108.
Возбуждение в связи с растяжением возникает и в рецепторах
сухожилия (рис. 106-g). Но для этого требуется более сильное растяжение,
чём для рецепторов мышцы, а именно 20—200 г и больше. При
сокращениях мышцы они также возбуждаются, и притом тем чаще, чем
сильнее начальное растяжение (рис. 109).
Как уже указывалось выше, основное назначение проприоцептивных
реакций заключается в усилении движений, наступающих рёфлекторно
в ответ на внешнее раздражение. Наряду с этим они имеют и Другое
назніачейие.
264 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Проприоцептивны? рефлексы, омотря по тому, где лежат
раздражаемые рецепторы, классифицируются на сухожильные рефлексы,
вызываемые раздражением
миотатические рефлексы,
торов.
сухожильных рецепторов, и мышечные или
вызываемые раздражением мышечных рецеп-
Сухожильные
рефлексы. С давних пор
известно, что механическое
которое
ударом по
вызывает
сокраще-
раздражение,
производится
сухожилию,
рефлекторное
ние той мышцы, которой
принадлежит
раздражаемое сухожилие.
Например, при ударе по
ахиллову сухожилию
сокращается икроножная
мышца, причем стопа
разгибается; при ударе по
коленному сухожилию
выше или ниже
надколенника сокращается
четырехглавая мышца,
причем колено разгибается.
Несмотря на явно разги-
бательное движение, это
разгибание не является
вполне
координированным: при этом не
сокращаются, например,
другие разгибатели
конечности. Кроме того, такое
сокращение не
сопровождается реципрокным тор
можением сгибателей (Шеррингтон, 2). Этот сухожильный рефлекс
обязан своим происхождением раздражению проприоцепторов как
самого сухожилия, так и мышцы, ибо при ударе по сухожилию
происходит сотрясение всей мышцы (Гофман, 25).
Сухожильный рефлекс характеризуется тем, что протекает в виде
одиночного сокращения; мышца получает из центральной нервной
системы один пульс возбуждения, как это видно из рис. ПО и 111
(Гофман, 4; Дитлер, 5; Ллойд, 14). В обычном координированном рефлексе
сокращение мышцы всегда протекает тетанически: оно вызывается рядом
быстро ^следующих один за другим центральных нервных импульсов.
Это бывает даже в том случае, если раздражение рецептивного поля
Рис. 108. Влияние одиночного сокращения на
биоэлектрические токи возбуждения чувствительных
волокон от мышечных рецепторов.
Мышцы на пальце лягушки всего с одним мышечным
веретеном. Биоэлектрические токи (быстро колеблющаяся
кривая) чувствительных волокон возникают от растяжения
мышечных рецепторов. В опытах А — D — изотоническое
состояние: А — в самом начале нагрузки тяжестью в 2 г;
В — 1 сек. после нагрузки; С — 5 сек. после нагрузки и
D — 15 сек. после нагрузки; Е — изометрическое
состояние — 2 сек. после растяжения. В каждом опыте
вызывается одиночное сокращение раздражением двигательного нерва
(сплошная кривая). Во время него токи возбуждения
исчезают. Оки вновь возникают в учащенном виде в момент
расслабления. Время указано в опыте D (Мэтьюс).
ПРОИРИОЦЕИТИВНЫЕ РЕФЛЕКСЫ
265
вызывается отдельным индукционным ударом (Гофман, 6; Беритов, 7).
Скрытый период сухожильного рефлекса всегда очень короток и
очень мало меняется от опыта к опыту. При регистрации одним и тем
же методом скрытого периода этого рефлекса и еще какого-либо
другого рефлекса координированного типа непременно окажется, что при
г—| 1 ( 1 1 , 1 , . . »
Рис. 109. Электрическая реакция сухожилььшо
рецептора m. soleus от растяжения и сокращения.
В опыте J начальное напряжение — 100 г, а в опыте II—40 г.
В первом случае возникают токи возбуждения с ритмом около
20 в і сек. Во втором случае они отсутствуют. В обоих случаях
во время одиночного сокращения, вызванного раздражением
мышечного нерва, возникают токи возбуждения высокого ритма.
Они располагаются попарно, ибо соответствуют двум
сухожильным рецепторам. Здесь же даны механограммы. С расслаблением
сокращения токи возбуждения от сокращения исчезают
(наверху время по 0.05 сек.) (Мэтьюс).
Рис. 110. Биоэлектрический ток возбуждения разгибателя колена
четырехглавой мышцы человека после удара молоточком но коленному
сухожилию,
Верхняя кривая обозначает время, по одному колебанию в 1/ш сек. Под ней дается
электрограмма мышцы, полученная посредством струнного гальванометра; первая фаза
показывает появление биоэлектрического тока под проксимальным отводящим электродом,
а вторая -— под дистальным электродом. В самом низу кривая обозначает момент
раздражения; поднятие линии соответствует удару молотка (Гофман).
сухожильном рефлексе на прохождение возбуждения в мозгу тратится
меньше времени, чем при любом другом рефлексе (Гофман, 4; Иендрас-
сик, 3). У кошек на это тратится не более 0.8 о (Ллойд, 14). На этом
ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
основании было сделано предположение, что нервный путь
сухожильного рефлекса состоит из дву^с нейронов, из чувствительного и
двигательного. Повидимому, рецепторы сухожилий через чувствительные нейроны
находятся в особой интимной связи
с двигательными нейронами
соответствующих мышц. Благодаря
этому при их раздражении
периферические импульсы свободно
передаются к двигательным нейронам.
Значит, эти проприоцептивные
рефлексы не носят характера обычной
координированной реакции. Они не
предполагают участия
промежуточных нейронов спинного и головного
мозга. Таким образом, простейший
рефлекторный путь, как он
выявляется в сухожильных рефлексах у
позвоночных животных, должен
состоять из двух нейронов:
чувствительного и двигательного.
Положение о простейшем
составе нервного пути из двух нейроном
для осуществления рефлекса
хорошо согласуется с гистологическими
данными. Известно, что коллатерали
чувствительных нейронов
оканчиваются не только в задних и
боковых рогах серого вещества мозга, т. е. у промежуточных нейронов, но
и в передних рогах, на клетках двигательных нейронов (Кёлликер,
Кахаль, Леношек, Эдингер, 1; Гофф, 23; Капперс, 22а). Это
гистологическое наблюдение и послужило Введенскому, Шерринг-
тону и другим .ученым поводом для того, чтобы признать достаточным
для любой рефлекторной реакции наличие двух нейронов в составе
рефлекторного пути. На основании определенных фактов мы пришли к
заключению, что все координированные, рефлекторные реакции должны
протекать с участием промежуточных нейронов и нейропиля.
Современные гистологические данные также согласуются с этим утверждением,
так как чувствительные волокна, идущие с рецептивных полей;
оканчиваются главным образом в задних и боковых рогах. Только в
сухожильных и миотатических рефлексах передача возбуждения от чувстви
тельного нейрона на двигательный происходит без участия
промежуточных нейронов, но одновременно обязательно должен вовлекаться в
реакцию и нейропиль.
При ударе молоточком по сухожилию через кожу или
непосредственно происходит не только рефлекторное сокращение соответствующей
-мышцы, но и торможение ее. Это торможение проявляется после возбуж-
Рис. 111. Миотатический рефлекс от
ш. gastrocnemius кошки.
Л — механическая кривая растяжения
мышцы; В — проприоцептивные импульсы
возбуждения задних корешков от этого
раздражения; С—двигательные импульсы
передних корешков в ответ на данные
проприоцептивные импульсы (Ллойд).
ЫРОИРИОЦЕПТИВНЫК РЕФЛЕКСЫ 2Ь7
дения и продолжается около 0.1 сек. (Гофман, 4, 6). Если мышца
находилась в тоническом сокращении и давала токи возбуждения, то при
сухожильном рефлексе эти токи на некоторое время исчезают, что
наступает после одного сильного биотока возбуждения, отвечающего су-
Рис. 112. Сухожильный рефлекс мышцы теплокровного
животного во время тонического возбуждения.
Наверху механограмма: первое колебание кривой а—от сотрясения
ударом молотка, а второе о — от сокращения. Внизу токи
возбуждения мышцы. Во время сокращения сейчас же после сильного тока
возбуждения, отвечающего сухожильному рефлексу, токи тонического
возбуждения выпадают (Денни-Броун).
хожильному рефлексу (рис. 112). Эта фаза торможения настолько
значительна, что служит препятствием для наступления последующего
сухожильного рефлекса вскоре после первого. Явление это подвергалось
специальному исследованию. Было установлено, что период торможения
в общем продолжается все время, пока продолжается сокращение. На
этом основании было сделано заключение, что, в связи с сухожильным
рефлексом, прекращается раздражение некоторых проприоцепторов,
заключенных в мышечных веретенах и обусловливающих тоническое
рефлекторное сокращение. В момент сокращения мышцы эти рецепторы
перестают растягиваться, а следовательно, перестают и раздражаться
(Фултон, 27; Мэтьюс, 20). Одновременно наступает торможение. От
него ослабевают токи возбуждения мышцы при произвольном
сокращении. Торможение не ограничивается раздражаемой мышцей. Оно
распространяется и на другие мышцы.
Кроме того, известно, что после фазы торможения наступает фаза
облегчения с усилением токов возбуждения произвольного сокращения
(Гофман, 25). Все это указывает на то, что в периоде ослабления
рефлекторной деятельности при вызове сухожильного рефлекса мы имеем
подлинное центральное торможение. Это центральное торможение
безусловно вызывается активированием нейропиля как под
непосредственным влиянием периферических импульсов от сухожильных рецепторов,
так и через промежуточные нейроны. При этом нужно иметь в виду,
что удар молоточка по сухожилию происходит через кожу и
вследствие этого одновременно раздражаются и кожные рецепторы, а, как
268 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
мере
ных
того,
точка
сение
раздражением
рецепторов,
при ударе
происходит
мышцы. Это
Рис. 113. Влияние реципрокного торможения на
сухожильный рефлекс.
Децеребрированный препарат кошки; записывается
четырехглавая мышца. Через каждые 2—3 сек. происходит
механическое раздражение коленного сухожилия, причем
разгибатель колена в ответ на каждое раздражение дает
одиночное сокращение. В это время происходит слабое
фарадическое раздражение чувствительного нерва той
же ноги (поднятие сигнальной линии). Это вызывает
расслабление существующего тонуса и выпадение
сухожильных рефлексов. После раздражения тонус и
сухожильные рефлексы восстанавливаются постепенно.
Нижняя линия отмечает время в секундах (Шеррингтон)
указывалось выше, раздражение кожи давлением вызывает общее
торможение даже в том случае, если оно не дает двигательных реакций.
Поэтому торможение при сухожильных рефлексах должно быть
обусловлено в некоторой
кож-
Кроме
моло-
сотря-
также
должно производить
раздражение мышечных
рецепторов, и это раздражение,
как імы увидим ниже, также
вызывает общее
торможение. Итак, торможение при
сухожильных рефлексах,
наступающих при ударе
молоточка, имеет сложное
происхождение. Оно должно
быть обусловлено
активированием промежуточных
нейронов и нейропиля под
влиянием периферических импульсов со стороны сухожильных,
мышечных и кожных рецепторов.
Сухожильный рефлекс чрезвычайно неустойчив. Легче всего он
угнетается центральным реципрокным торможением,— например, под
влиянием сгибательного рефлекса (рис. 113). Он не вызывается даже
непосредственно после произвольного сокращения. Его угнетает всякого
рода рефлекторная деятельность (глотание, кашель), а также
умственная работа: например, чтение, решение арифметической задачи
(Ломбард, 12; Боудич, 13).
Понижение и повышение возбудимости спинного імозга очень
отзываются на этом рефлексе. Поэтому во врачебной практике пользуются
сухожильным рефлексом для установления ненормальной рефлекторной
возбудимости центральной нервной, системы.
Миотатические рефлексы. Так называется рефлекторное сокращение
мышцы разгибателя, вызываемое раздражением мышечных рецепторов
при растяжении той же мышцы. Миотатические рефлексы были
изучены наиболее подробно Ше ррин г тоном на децеребрированных
животных. Он-то и дал рефлексу на растяжение это название. Миотатиче-
ский рефлекс имеет много общего с сухожильным рефлексом: обычно
он наступает только на той мышце, рецепторы которой раздражаются.
Но у тонизированных препаратов он может проявиться и на других
мышцах (Самойлов и Киселев, 18). Скрытый период миотатического-
рефлекса обычно очень мал, как и сухожильных рефлексов (рис. 116}
(Асаяма, 10; Шеррингтон, 11'; Ллойд, 14). Подобно сухожильным реф-
ПРОПРИОЦЕПТШШЫЕ РЕФЛЕКСЫ 269
лексам и эти сродные им мышечные рефлексы очень легко угнетаются
как во время рецилрокного торможения, так и во время разного рода
рефлекторной и умственной деятельности. Ввиду этого сходства
сухожильного рефлекса с мышечным рефлексом Гофман объединил оба
рефлекса под одним названием собственных рефлексов мышцы.
кг
3.S
2
1.5
1
5
О
Рис. 114. Миотатический рефлекс m. quadriceps
на децеребрированнон кошке.
Мышца соединена с изометрическим миографом. Кривая Т представляет
растяжение мышцы в миллиметрах; кривая М — напряжение, которое
нарастает до 3.5 кг; кривая Р — напряжение, которое получается при
том же растяжении после перерезки мышечного нерва, т. е. в
отсутствии миотатического рефлекса: при растяжении напряжение
увеличивается, но значительно меньше. Абсцисса дает время в секундах.
С течением времени рефлекс ослабевает. Ордината дает напряжение в
килограммах (Шеррингтон).
Миотатические мышечные рефлексы удерживаются все то время,
пока продолжается раздражение, но последействие их очень невелико; по
прекращении растяжения рефлекс сейчас же прекращается. Рефлекс
вызывается как быстрым, так и постепенным растяжением. Он
вызывается даже при очень небольшом растяжении мышцы — всего на 0.8°/о
нормальной длины (рис. 114). Развиваемое при этом напряжение тем
сильнее, чем больше растяжение, но даже при наиболее сильных
растяжениях оно не достигает величины, достигаемой при раздражениях
двигательного нерва. Это указывает на то, что в миотатических рефлексах
двигательные нервные волокна возбуждаются не все одновременно
(Шеррингтон).
При сильных растяжениях ритм возбуждения мышцы высок — он
может достигать 200—250 в 1 сек. Это происходит вследствие
неодновременного поступления нервных импульсов через разные нервные
волокна, ибо при отведении ограниченного участка мышцы или при
уменьшении числа участвующих двигательных нейронов до 1—2 число
импульсов при растяжении мышцы вариирует в пределах от 7 до 25
в 1 с?к. При увеличении натяжения імышцы происходит учащение им-
М
eJL
270 ОБЩАЯ ФИЗИОЛОГИЯ. (ЛШіШОіО МОЗГА Ы СТВОЛА ГОЛОВНОГО МОЗГА
пульсов, но в общем оно не превосходит 30—40 в і сек. (Шерриштоіі
и сотр., 17).
При пассивном растяжении тонической мышцы-разгибателя у де-
церебрированной кошки существующие
биоэлектрические токи возбуждения этой мышцы
сначала сильно нарастают в течение 0.52—
1 сек., потом, еще во время растяжения,
ослабевают (Самойлов и Киселев, 18).
Особенно сильное нарастание как токов
возбуждения, так и напряжения в этой мышце
происходит при одновременном пассивном
вытяжении обоих разгибателей — ипсилатерально-
го и контралатерального m. quadriceps (Кисе-
лев, 19).
Отравление стрихнином не влияет на
миотатические рефлексы. Этот рефлекс не
усиливается после стрихнинного отравления. Он
вызывается даже в том случае, когда перестают
вызываться все остальные рефлексы в
результате истощения спинного мозга в связи с
длительными судорогами.
Миотатические рефлексы не имеют
самостоятельного значения в поведении животного,
но играют вспомогательную роль при
производстве всех других рефлекторных реакций,—
например, в статическом рефлексе, в
тонических рефлексах конечностей и т. д. Известно,
что после перерезки чувствительных нервов
мышцы или после впрыскивания новокаина в
мышцу тоническое сокращение последней в
значительной мере ослабевает.
Все вышеприведенные факты указывают
на то, что нервный путь разгибательных мио-
татических рефлексов не содержит в спинном
мозгу координирующих нейронов, что,
следовательно, возбуждение с чувствительных
нейронов прямо переходит на двигательные.
Миотатические рефлексы на разгибателе,
подобно сухожильным рефлексам,
сопровождаются общим торможением. Оно настолько
значительно, что при этом ослабевают или
целиком устраняются рефлекторные вздрагивания, вызываемые на
других мышцах раздражением чувствительных нервов или задних
корешков отдельными индукционными ударами. Так, например, при
растяжении одной из мышц колена происходит усиление рефлекторных
вздрагиваний растягиваемой мышцы, и одновременно они тормозятся на всех
Рис. 115. Общее
торможение от раздражения про-
приицепторов растяжением
мышцы.
Лумбальныіі препарат кошки.
Раздражением правого V
заднего корешка вызываются
рефлекторные взппагивания всех
мышц. Растпгичается
разгибатель m. rectus femoris (Я. /. d.)
(одна из головок четырехглавой
мышцы) грузом в 100 г. Во
время растяжения
рефлекторные вздрагивания ослабевают
как на разгибатете со'>ти"'"Т^\т.
кмцеп стороны т. V3.st.iis cxtermis
(V. •*.), так и на сгибателе т.
st-iiiiten ninosus (S. s.) и
разгибателе m. guadriecps (Q. s.)
противоположной стороны. На
самом rectus femoris рефлектпрпьн*
взіпагивакия усиливаются
(Беретов и Бакурадзе).
ііроприоцептивные рефлексы 27 Г
других мышцах как данной конечности, так и другой одноименной
(рис. 115).
При кратковременных растяжениях мышцы последняя дает такое же
одиночное сокращение, как при сухожильном рефлексе. Такая же
реакция наблюдается при раздражении чувствительного нерва мышцы одним
индукционным ударом (рис. 116). Но в случае кратковременного
растяжения в рецепторах мышцы возникает ряд
импульсов, причем двигательный нейрон
реагирует только на один из первых про-
приоцептивных импульсов. Остальные не
дают эффекта (Ллойд, 14). Это, должно
быть, происходит под влиянием
торможения, вызываемого в это самое время через
нейропиль теми же проприоцептивными
импульсами. Между прочим, на рис. 116
ясно видно, что после биотока возбуждения
двигательного корешка в ответ на
раздражение мышечного нерва заметно ослабевает
первый биоток возбуждения от одновремен-
но раздражаемого кожного нерва.
Итак, импульсы возбуждения от
чувствительных нервов разгибателя действуют
исключительно на двигательные нейроны
данной мышцы и притом без участия
промежуточных нейронов. Одновременно эти
импульсы активируют нейропиль и тем.
самым вызывают торможение.
Так как миотатические рефлексы на
разгибателях задних конечностей
наблюдаются и на лумбальных препаратах, то
ясно, что центральный механизм этих
рефлексов лежит в поясничном отделе
спинного мозга (Денни-Броун и Лиддел, 15).
Правда, на децеребрированных препаратах эти рефлексы наблюдаются
много лучше, но это объясняется тем, что на них разгибатели
находятся в состоянии повышенного тонического возбуждения, что
благоприятствует выявлению миотатических рефлексов на разгибателях.
Миотатические рефлексы координированного характера. Не только
растяжение разгибателей, но и растяжение сгибателей вызывает
рефлекторную реакцию. В отличие от разгибательных миотатических
рефлексов эта реакция менее постоянна и протекает слабей и быстрей;
кроме того, она имеет координированный характер: одновременно с
сокращением растягиваемой мышцы-сгибателя происходит сопряженное
торможение, расслабление антагониста-разгибателя. Если разгибатель
находится в тоническом сокращении, он при этом расслабляется — и
тем значительнее, чем сильнее растяжение и чем большее число сгиба-
Рис. 116. Биотоки переднего
ьо решка при раздражении
мышечного и кожного нервов
спинальной кошки.
Отводится 1 сакральный передний
корешок. А — при раздражении
мышечного нерва (п. Kaslrocnerriius)
одним электрическим ударом. В —
при раздражении кожного норна
(i.suralis) одним ударом. С — при
одновременном раздражении обоих
нервов одним ударом (Ллойд).
272 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
телей растягивается (рис. 117). Аналогичное торможение
получается при раздражении чувствительных нервов сгибателей (Шер-
рингтон, 11). И эти миотатические рефлексы наблюдаются на лумбаль-
ных препаратах. Следовательно и здесь координирующие аппараты
лежат в поясничном отделе мозга (Денни-Броун и Лиддел, 15).
Растяжение сгибателей, вызывающее реципрокное торможение
разгибателя, также сопровождается общим торможением,
распространяющимся на все мышцы как раздражаемой конечности, так и другой одно-
I I і У [ L 1 -.—L,
сен 1" 2» зн *"
Рис 117. Миотатический рефлекс на ю. quadriceps
децеребрированной кошки и его торможение от
растяжения сгибателя ш. biceps.
Г — кривая растяжения; М — кривая напряжения до
перерезки двигательного-нерва сгибателя; Р— тоже после
перерезки. В момент,, указанный стрелкой, происходит растяжение
сгибателя. Кривая М немедленно падает до уровня кривой Р.
На абсциссе время в секундах, на ординате напряжение
в килограммах (Шеррингтон).
именной. Так, при пассивном растяжении сгибателя калена івызывается
торможение не только разгибателя колена, но также и
антагонистической пары на другой конечности.
Не только пассивное растяжение, но и активное сокращение
сгибателей производит раздражение рецепторов мышцы, и вследствие этого
наступает не только усиленное реципрокное торможение разгибателей,
но и общее торможение. При этом на противоположной ноге
тормозятся и разгибатели и сгибатели (рис. 118).
Отсюда следует, что чувствительные нервы сгибателей передают
возбуждение двигательным нейронам через промежуточные нейроны и
одновременно активируют нейропиль не только непосредственно, но и
через те же промежуточные нейроны.
Торможение, производимое при сокращении сгибателя, должно
иметь, между прочим, подсобное значение в производстве нормального
шагательного рефлекса. В сгибательной фазе разгибатель
растягивается, но рефлекторное сокращение на нем отсутствует — не только
вследствие реципрокного торможения под влиянием сгибательной фазы
самого шагательного рефлекса, но н вследствие торможения под влия-
ЛРОПРИОЦЕПТИВНЫК РЕФЛЕКСЫ
273
аием сокращения сгибателей. По окончании сгибательной фазы
торможение заменяется облегчением, и так как разгибатель остается еще
некоторое время растянутым, то создается условие для его
рефлекторного сокращения, которое способствует наступлению последующей
разгибательной фазы.
При совершенно нормальных условиях .рефлекторной деятельности и
сухожильные, и миотатические рефлексы ограничиваются сокращением
одной мышцы, рецепторы которой
раздражаются. Но в случае па-
галогического или искусственно
вызванного повышения
возбудимости в центральной нервной
системе, при подергивании или
растяжении мышцы сокращение
наступает и на других мышцах
той и другой стороны. Так,
например, сухожильный рефлекс
одной стороны сочетается с
разгибанием другой конечности.
Этот разгибательный рефлекс
носит название филиппсонов-
ского разгибания, по
имени Филиппсона, впервые
описавшего перекрестный
разгибательный рефлекс (28). В
определенных случаях наступает
сокращение всех скелетных мышц.
Так, например, при растяжении
m. quadriceps наблюдается
поворот туловища в
противоположную от растягиваемой мышцы
сторону, разгибание передней
конечности той же стороны,
сгибание другой передней конечности
и поворот хвоста (Киселев, 22).
Рефлекторный пластический
тонус. Мышцы-экстензоры во
время децеребрационной
ригидности обнаруживают высокую
степень пластичности: они
сохраняют любое приданное им
положение. Если сгибать колено де-
церебрированной кошки и таким
образом растягивать
четырехглавую мышцу, то сначала в ней будет нарастать напряжение, которое
будет оказывать сопротивление растяжению. Но когда разгибатель
18 II. ( Веритов
Рпс. И8. Общее торможение ""от
раздражения проприоцепторов в
результате соьращения.
На лумбальном препарате кошки
раздражением периферического отрезка левого VIII
переднего корешка вызывается тетаничеекое
сокращение левого сгибателя. Во время этого
сокращения тормозятся рефлекторные
вздрагивания, вызываемые раздражением правого
VIII заднего корешка. Оба индуктория без
сердечника.
При пороговых раздражениях переднего
корешка (16 см) торможение проявляется на
разгибателе соответствующей стороны (Q. S.)
и видимо также на самом левом сгибателе
(S. s.). При сильном раздражении (13 см)
торможение наступает и на мышцах
противоположной стороны. На сгибателе (S. d.)
сильные рефлекторные вздрагивания ослабевают, а
на разгибателе (Q. d) слабые вздрагивания
выпадают (Беритов и Бакурадзе).
274 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
растягивается на 7 мм,. Напряжение сразу прекращается и конечность
застывает в приданном ей положении. Это называется реакцией
удлинения. Если же разогнуть колено и тем самым пассивно
укоротить разгибатель, рефлекторное напряжение последнего ослабевает
и мышца застывает во вновь приданном ей положении. Это называется
реакцией укорочения (Шеррингтон, 17) (рис. 119). Реакцию
Рис. 119. Рефлекс «укорочения» и «удлинения» на децеребри-
рованной кошке.
Регистрируется движение коленного сустава, на который действует
изолированная односуставная головка m. quadriceps vastocrurbus. s, s, s, s —¦
рефлекс «укорочения», вызванный пассивным разгибанием колена, а 1, 1, I, 1—
рефлекс «удлинения», вызванный пассивным сгибанием колена, т. е.
растяжением мышцы. 60°, И0° и 92° обозначают углы сгибания в коленном
суставе (Шеррингтон).
удлинения Шеррингтон объясняет так. Он полагает, что при некотором
сильном растяжении разгибателя происходит раздражение особых
мышечных рецепторов, вызывающее рефлекторное торможение
растягиваемого разгибателя и сокращение противоположного. Выпадение
центральных импульсов к разгибателю и ведет к тому, что эта
мышца сохраняет длину, которой она достигла к моменту торможения
(Шеррингтон и сотр., 17). Но Самойлов и Киселев показали, что
здесь торможение растянутого разгибателя зависит не от растяжения.
Они установили, что пассивное растяжение всегда вызывает миотатиче-
ский рефлекс, распространяющийся не только на разгибатель
противоположной стороны, но и на мышцы туловища хвоста. Торможение же
наступает по прекращении растяжения в связи с пассивным
укорочением растянутых мышц. В этот момент происходит раздражение
особого рода рецепторов, вызывающее торможение миотатического рефлекса.
Это происходит моментально в самом начале укорочения, а потому
мышца может застыть в растянутом положении. Укороченная мышца
застывает в одном положении, ибо она не может также удлиниться
вследствие отсутствия рефлекса растяжения. Пластичность імышцы, по
мнению Самойлова и Киселева (18), осуществляется путем
борьбы этих двух противоположных реакций.
ПРОПРИОЦЕЛТИВНЫЬ РЕФЛЕКСЫ 27о
Миотатические рефлексы на лягушке. Приведенные выше
результаты были получены в опытах на теплокровных животных. На
холоднокровных миотатические рефлексы мало изучены. Известно, что
растяжение разгибателя производит возбуждение разгибателей той же
конечности и общее торможение всех остальных мышц как
соответствующей, так и противоположной стороны, хотя и в более слабой степени.
Нечто аналогичное происходит и при растяжении сгибателей.
Растяжение сгибателя благоприятствует сокращению сгибателей той же ко-
Рис. 120. Мпотатичесыіе рефлексы на спинномозговой ляглшкс.
.Записывается m. semitendmosus. Фарадическим раздражением и. реюпеиь
(нижний сигнал) вызывается рефлекторное сокращение m. semitendmosus
В опыте А во время этого рефлекса производится одно растяжение m. tncep&
(0» J. два раза т. gastrocnemius (д). Каждый раз сокращение т. semitendm jsub
расслабевает и восстанавливается после растяжения. В опыте В вместо
растяжения происходит электрическое раздражение центрального отрезка
мышечного нерва m triceps (верхний сигнал). Сокращение m semitendmosus
расслабевает и восстанавливается после раздражения. Цифры > называют расскш
ние ме*ду индукционными катушками в сантиметрах (Нарикашвили).
нечности и вызывает торможение разгибателей. О возбуждении
растянутых мышц легче всего судить по биоэлектрическим токам
возбуждения. Торможение же других мышц легко можно установить на
основании комбинации рефлекторного сокращения их с растяжением
антагонистов (Нарикашвили, 21) (рис. 120). Те же результаты получаются
при раздражении чувствительных мышечных нервов. При
электрическом раздражении чувствительного нерва одной мышцы антагонисты
тормозятся. Однако бывают и такие случаи, когда раздражение
мышечных рецепторов или мышечных нервов производит общее торможение
как антагонистов, так и синергистов. Специальными опытами К а то и
его сотрудники (9) доказали, что те чувствительные мышечные
волокна, которые производят общее торможение на лягушках, начинаются в
мышечных веретенах от рецепторов Руфини. К а то предполагает, что
чувствительные волокна мышечных веретен, с одной стороны, идут к
головному імозгу и тем обусловливают мышечное чувство, а с другой —
они своими коллатералями соединены с двигательными клетками
передних рогов и производят их торможение. Мы же думаем, что афферент-
27о ОБЩАЯ ФИЗИ0Л0ГИЯ"СПИНН0ГОТМОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
ные восходящие пути из проприоцептивных волокон служат для
передачи импульсов возбуждения к головному мозгу, а коллатерали
находятся в связи с нейропилем и через него производят общее
торможение.
Практикум
Наблюдение сухожильного рефлекса. Подопытное лицо кладет одно колено на
другое. Слегка ударяют перкуссионным молотком или каким-либо тупым предметом
по колейному сухожилию ниже надколенной косточки. В ответ на это колено
разгибается, что указывает на сокращение четырехглавой мышцы. Путем ощупывания
нетрудно убедиться, что упомянутая мышца действительно сокращается, а все
остальные мышцы находятся в покое.
Наблюдение миотатического рефлекса на лягушке. Берется спинномозговая лія-
гушка и записывается на кимографе гл. semitendinosus. Раздражением п. peroneus
вызывается рефлекторное сокращение. Раздражение должно быть пороговым или
умеренной силы. Если во время этого сокращения растянуть трехглавую или
икроножную мышцу той же ноги, сокращенный сгибатель сейчас же расслабевает.
Эффект получается и при очень слабых растяжениях. Расслабление сгибателя
получается и при растяжении разгибателей другой ноги, но значительно слабее.
Наблюдение миотатических рефлексов на кошке. Животное усыпляют эфиром
под стеклянным колпаком, а затем переносят на операционный стол, надевают голо-
водержатель и привязывают спиной вниз. Быстро производят трахеотомию и вводят
специальную трахеальную У-образную трубку общим отверстием в трахею. Из двух
других отростков этой трубки один опускают в склянку, на дне которой находится
вата с эфиром, а другой оставляют открытым для поступления чистого воздуха и
надевают резиновую трубку, которая может быть сужена зажимом до любой
величины. Этим регулируется поступление чистого воздуха в легкие, т. е. степень нарко*
за. Наркоз должен быть настолько глубок, чтобы животное во время операции не
реагировало движениями. Дыхание и пульс во время операции должны быть
хороши. В случае остановки дыхания трубка вынимается из склянки и дыхание
восстанавливается ритмическим сжатием и отпусканием грудной клетки.
С наступлением глубокого наркоза перевязывают сонные артерии, перерезают на
одной стороне п. obturatorius при выходе из foramen obturatorium и седалищный нерв
около колена; на центральный конец п. peroneus или п. tibialis навязывают нитку.
Затем высвобождают на этой ноге сухожильный конец m. semitendinosus, привязывают
нитку и перерезают его пониже нитки. Так же поступают с m. quadriceps. После
этого животное поворачивают брюхом вниз, обнажают спинной мозг в области
последних грудных сегментов и перерезают его. Кровоизлияние останавливают
наложением тампонов поверх раны. Укрепляют животное в этом положении. Оперированную
ногу фиксируют за бедро выше колена и за голень повыше голеностопного сустава.
Спустя чае приступают к опытам. Соединяют сначала одну, потом другую мышцу
с изометрическим миографом. Малейшее растяжение мышцы вызывает увеличение
налряжекш в ией. Растяжение одной мышцы отзывается на напряжении другой мыш-.
цы: последнее ослабевает.
Раздражением п. peroneus отдельными индукционными ударами вызывают на
мышце отдельные рефлекторные вздрагивания. На фоне этих вздрагиваний
производится растяжение одной из мышц. В результате растяжения регистрируемой мышцы
рефлекторные вздрагивания на ней усилятся, а при растяжении другой они, наоборот,
ослабнут.
Описанная операция и способ фиксации не обеспечивают вполне точной запцси
мышечных сокращений, так как кроме регистрируемых мышц будут сокращаться и
другие мышцы той и другой конечности, а кроме того, будет двигаться передняя
часть тела. Вследствие этого регистрируемая мышца будет испытывать более или
ПРОПРИОЦЕПТИВНЫЕ РЕФЛЕКСЫ
277
менее значительные механические сдвиги при движениях конечностей животного.
Однако описанный, сильно упрощенный способ вполне достаточен для наблюдения
проприоцептивных рефлексов. Он может быть применен без предварительного
обучения. Но если поставить себе целью избежать означенных помех для точной
регистрации мышечных сокращений, то нужно применить более сложную операцию и сложную
методику, которые нужно предварительно изучить. Эта методика будет описана
ниже, в главе XVI.
Литература
1. EdingerL., Vorlesungen iiber d. Bau d. nervosen Zentralorgane, Bd. I, Leipzig,
1911.
2. Sherrington C, Int. Act. Nerv. Syst., 1906; Proc. Roy, Soc., 52, 556, 1892.
3. Jendrassik E., Arch Klin. Med., 52, 569, 1894.
4. Hoffmann P., Untersuchungen iiber d. Eigenreflexe (Sehnenreflexe)
men^chlicher Muskeln, Berlin, 1922.
5. D i 11 1 e r R u. G u n t e г., Pfl. Arch., 155, 251, 1914,
6. Hoffmann P., Arch. Anat. Physiol., стр. 233, 1910.
7. В e г i t о f f J., Z. В і о 1., 62, 125, 1913.
8. S herring t on C.,Q. J. Exp. Physiol., 2,109,1909.
9. KatoG., The Mierophysiology of Nerve, 1934.
10. A s а у a m a Ch., Q. J. Exp. Physiol., 9, 265, 1915.
11. L i d d e 1 1 E. G. T. a.SherringtonC, Proc. Roy. SocM В., 96, 212, 1924;
97, 267, 1925.
12. Lombard W. P., Am J. Psychol. Oct. 1887.
13. Bowditch u. Warren., J. Physiol., 11, 25, 1890.
14. Lloyd D. P. C, J. Neurophysiol., 6, 294, 317, 1943.
15. D e n n y-B г о w n D. E. a. L i d d e 1 1 E. G. Т., J. Physiol., 63, 144, 1927.
16. С о о p e г S. a. Creed R.S..J. Physiol., 64, 199, 1927.
17. Шеррингтон и сотр., Рефлекторная деятельность спинного мозга, 1935.
18. Sam о ] 1 о f f А. и. К isselef f M., Pfl. Arch., 218, 268, 1927; 220, 424, 1928.
19. Kisseleff M., Pfl. Arch., 225, 357, 1930.
20. M a t t h e w s B. H. C, J. Physiol., 78, 1, 1933; 71, 64, 1931; 72, 153, 1931.
21. Нарикашвили. Юбилейный сборник, поев. проф. И. Бериташвили, стр.
413, 1936.
22. Киселев М. А., Сб. докладов VI Всес. фивиол. съезда, стр. 156.
22 а. А г і ё n s Kappers G. A., The comparative anatomie of the nervous system
etc. 1936.
23. H о f f E. C, Proc. Roy. Soc, В., Ill, 175, 1932.
24. Winterstein H. u. H irschberg E., Bioch. Z., 156, 138, 1925.
25. Hoffmann P., Erg. Physiol., 36, 15, 1934.
26. Беритов И. иБакурадз? А., Об явлении общего торможения при
раздражении мышечных рецепторов. Не опубликовано.
27. Fulton J. F. a. I. Pi-Sun пег., Am. J. Physiol., 83, 554, 1928.
28. P h i 1 i p p s о n M., L'autonomie et la centralisation dans le syste-me nerveux
des animaux, Bruxelles, 1905.
Глава XV
РИТМИЧЕСКИЕ РЕФЛЕКСЫ
Понятие о ритмических рефлексах. Некоторые оборонительные
рефлексы, например рефлексы потираний у лягушки и чесания у
млекопитающих, а также движения конечностей іво время ходьбы, прыгания
и т. д., протекают ритмически. Движущаяся ко-нечность поочередно то
разгибается, то сгибается. При однородных движениях трех главных
суставов конечности распределение мышц на группы возбужденных и
заторможенных происходит по обычному «принципу координированного
движения. Так, например, во время ходьбы в фазе разгибания
возбуждаются все односуставные разгибатели и те из двусуставных мышц,
которые по своему анатомическому положению способны разогнуть
обслуживаемый ими дистальный сустав; тормозятся же односуставные
сгибатели и те из двусуставных, которые *могут сгибать дистальный сустав
(Шеррингтон, 1). Если движение ноги не одинаково во всех суставах,
то распределение імышц является иным. Так, например, в рефлексе
потирания лягушки тазобедренный сустав постоянно согнут, а коленный
тб сгибается, то разгибается. В соответствии с этим, участие
сгибателей колена m. semitendinosus и т. sartorius не одинаково: обе мышцы
двусуставные, но только m. semitendinosus по своему анатомическому
положению сгибает коленный сустав, а тазобедренный разгибает,
т. sartorius же сгибает оба сустава. В рефлексе потирания в момент
разгибания колена m. sartorius сокращается, a m. semitendinosus
тормозится, а в момент сгибания первая мышца перестает сокращаться, а
вторая перестает тормозиться. Очевидно-, m. sartorius в момент
разгибания колена исполняет, во-первых, активную роль—своим сокращением
он сгибает тазобедренный сустав,— во-вторых, он исполняет роль
фиксатора в отношении коленного сустава. М. semitendinosus не
сокращается ни при одном из движений колена, ибо это сокращение
произвело бы двигательный эффект, совершенно противоположный рефлексу
потирания (Беритов, 2). На рис. 121 дается иллюстрация реципрокных
иннервации в ритмическом рефлексе типа шагания. Фазные сокращения
антагонистов протекают на фоне некоторого постоянного сокращения.
На сгибателе это происходит потому, что раздражение нерва произвел-
РИТМИЧЕСКИЕ РЕФЛЕКСЫ
279
дит, наравне с фазным рефлексом, еще оборонительный рефлекс
сгибания. Возбуждающая иннервация этого рефлекса препятствует полному
расслаблению под влиянием фазного торможения. Разгибатель в момент
сгибания также не расслабляется полностью. Это происходит оттого,
что эта мышца во время фазных движений испытывает тоническое
сокращение. В данном опыте ритмический эффект на разгибателе
продолжается и по прекращении раздражения.
Рефлекс потирания у лягушки, подобно чесательному рефлексу у
собаки, по существу является ритмическим рефлексом. Но здесь
характерно, что при ритмическом течении потирания сгибатель колена
m. semitendinosus производит
ритмическое торможение без последующего
возбуждения после каждой фазы
торможения. Каждый раз во время
сокращения разгибателя колена (m.
triceps) сгибатель его тормозится; когда
же разгибатель расслабляется,
сгибатель не сокращается (рис. 122).
Чередование сгибания и
разгибания. Ниже мы подробно ознакомимся
с одним свойством защитных
рефлексов: защитный рефлекс существенно
меняется в зависимости от положения
двигательного органа. Так, если нога
пассивно или активно разогнута, то
раздражение вызывает ее сгибание;
наоборот, если нога согнута, то на
раздражение она ответит разгибанием.
Причина такого изменения рефлекса
заключается в том, что каждому
положению ноги соответствует
раздражение определенных проприоцепторов
мышц, сухожилий и суставов
(Магнус, 4). Эти раздражения действуют на
спинной мозг, предрасполагая его к
производству противоположного движения. Вследствие этого
последующее ©нешнее раздражение вызовет движение, противоположное
существующему положению. Но по существу возникновение и исчезновение
противоположных двигательных фаз ритмического рефлекса типа
шагания не зависят от характера первичного раздражения или от вторичных
проприоцептивных раздражений мышц, сухожилий и суставов. Одна
фаза сменяется другой и в том случае, если совершенно исключена
возможность вторичных раздражений. Так, например, после перерезки
всех задних корешков данной конечности последняя не утрачивает
способности к ритмическому рефлексу шагания (Сеченов, 21; Грэим Бро-
уц, 5; Беритов, 6). Или, например, известно, что ритмический рефлекс
Рис. 121. Ритмический рефлекс типа
шагания.
Децеребрированная кошка. Верхняя
кривая принадлежит triceps bi achii,
разгибателю локтевого сустава, а нижняя —
сгибателю этого сустава, biceps bracbii.
Раздражаются п. radiahs. На обеих
мышцах эффект протекает ритмически: когда
одна сокращается, другая расслабляется
Or). Первые три антагонистические фазы
обозначены цифрами 7, 2 и 3. Момент
раздражения обозначается оп> еканием
сигнальной чинш*. Время - - в секун га\.
280 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
шагания спонтанно возникает на глубоко наркотизированных животных,
когда никакое периферическое раздражение не способно вызвать
рефлекс (Грэим Броун, 7). То же получается, если исключена
возможность не только периферических, но и центральных импульсов,
нисходящих из высших отделов,— например, после перерезки спинного мозга
на границе поясничной и грудной области и одновременно всех лояс-
нично-крестцовых задних корешков. Такой препарат способен к
ритмической деятельности. Она, например, наступает при механическом
раздражении спинного мозга (Тоуэр, 26).
Чесательный рефлекс (кошки, собаки) также не зависит в своем
ритмическом течении от внутренних или внешних раздражений и тоже
Рис. 122. Ритмический рефлекс потирания на спинномозговой лягушке.
Записывается т. semitendinous (верхняя кривая) и т. triceps (нижняя). В опыте Л
рефлекс потирания вызывается раздражением п. brachialis той же стороны (нижний
сигнал). Рефлекс повторился три раза, при этом m. semitendinosus не сокращается.
В опыте В этот рефлекс вызывается во время сгибательного рефлекса, наступившего
в ответ на раздражение п. peroneus (верхний сигнал). Каждый раз во время
сокращения m. triceps в рефлексе потирания m. semitendinosus расслабляется —тормозится.
вызывается спонтанно во время глубокого наркоза, когда
периферическое раздражение не дает никаких рефлексов (Грэим Броун, 8). Кроме
того, известно, что чесательный рефлекс протекает вполне правильно и
на той ноге, которая лишена периферической чувствительности
вследствие перерезки соответствующих задних корешков. Так, например,
после перерезки задних корешков раздражением мозга в шейной области
можно вызвать чесательный рефлекс (Шеррингтон, 9). Отсюда ^ясно,
что природа чесательного рефлекса совершенно та же, что и рефлекса
типа шагания.
Итак, фазные движения &о время локомоцищ а также чесательный
и другие ритмические рефлексы являются своеобразными актамщ где
каждая фаза непосредственно связывается с предыдущей и
последующей.
Значение проприоцептивных раздражений для фазных движений.
Хотя на спинномозговых препаратах при выпадении периферической
чувствительности вследствие перерезки задних корешков сохраняется
способность к ритмической деятельности, однако эта ритмическая дея-
уельйостьнаблюдаетсяпреимущественно ро время раздражения самого
РИТМИЧЕСКИЕ РЕФЛЕКСЫ
281
спинного мозга или же какого-либо чувствительного нерва. После
прекращения раздражения эти движения ограничиваются всего
несколькими фазами: например, на спинномозговой лягушке — двумя. Но если
периферическая чувствительность сохранена, то эта ритмическая
деятельность может продолжаться долгое время и после прекращения
раздражения. Это наблюдается как на лягушке (Беритов, 6), так и на
теплокровных животных (Грэим Броун, 8). Ясно, что наличие
периферической чувствительности, посредством .вторичных проприоцептивных
раздражений, благоприятствует протеканию ритмических рефлексов.
Роль проприоцепторов лучше всего проявляется в фазных
движениях, наступающих при раздражении коры большого мозга. Эти
фазные движения всецело зависят от проприоцептивных раздражений.
Конечности, лишенные проприоцепторов, вследствие перерезки задних
корешков, производят только длительное сгибание. Можно предположить;
что проприоцептивные импульсы от сокращенных или растянутых во
время движений мышц, передаваясь в центральную нервную систему,
обусловливают периодическое возбуждение и торможение мышц
(Смите, Меттлер и Куллер, 27).
Особенно легко вызывается ритмический рефлекс, когда препарат
(епинальная кошка, собака) висит в воздухе, т. е. при выключении
кожных раздражений. Но стоит только задержать одну лапу в одном
положении или наложить на кожу конечности или на палец небольшой
зажим и тем самым произвести кожное раздражение, которое вызовет
общее торможение, как ритмический рефлекс сразу прекращается.
После снятия зажима ритмическое шагание сейчас же начинается вновь
(Шеррингтон, 15).
Как известно, растяжение мышцы обусловливает также
раздражение проприоцепторов, которое возбуждает растянутую мышцу и
тормозит все остальные. Отсюда можно заключить, что периодическая смена
фаз сгибания и разгибания обусловливается раздражением
проприоцепторов растянутых мышц, которое в каждой фазе производит
возбуждение растянутых мышц и общее торможение всех остальных мышц и тем
самым приводит к смене данной фазы на противоположную.
Координирующий аппарат ритмических рефлексов. Ритмический
рефлекс, шагания на задних конечностях наблюдается не только на
едцнальных, но и на лумбальных препаратах. Следовательно!,
координирующий аппарат лежит в поясничной области спинного мозга. На спи-
нцльных препаратах он возбуждается при раздражении дорзальной
части поперечного разреза боковых столбов. Стрихнинное отравление в
дозах, не вызывающих судорог, значительно благоприятствует этому
рефлексу (Шеррингтон, 15). Очевидно, эти нервные пути боковых стол-
бор, оканчиваются у промежуточных нейронов, образующих
координирующие аппараты шагательного рефлекса.
Координирующий аппарат ритмических рефлексов должен
существовать независимо от оборонительных и тонических рефлексов. Об этом
мы.^ключае^ на основании взаимоотношений этих рефлексов. Раздра-
282 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
жением чувствительного нерва ноги можно вызвать как один, так и
другой рефлекс. На препаратах, склонных к ритмическому рефлексу
шагания, слабое одностороннее раздражение производит шагательный
рефлекс на раздражаемой конечности. Более сильное раздражение
начинает вызывать также сгибательный оборонительный рефлекс: сильные
ритмические движения наступают на фоне оборонительного сгибатель-
ного рефлекса, а еще более сильное раздражение вызывает обычный
оборонительный рефлекс (рис. 123). Но что при этом имеет место и
-
Рис. 123. Ритмический рефлекс шагания
при разных силах раздражения.
Децеребрированная кошка. Верхняя кривая от разгибателя
локтевого сустава triceps Ьг., нижняя от сгибателя того ж?
сустава—biceps. Раздражается п. radial is фарадическим током.
В опыте А раздражение слабое в условных единицах Кронеке-
ра—10 ед., в опыте В раздражение в 4 раза сильнее — 40 ед.,
а в опыте С в 40 раз сильнее — 400 ед. Слабое раздражение
производит ритмический рефлекс шагания, среднее —
ритмический рефлекс на фоне сгибательного рефлекса, а сильное —
чистый сгибательный рефлекс. Время — в секундах.
ритмическая иннервация шагания, видно из того, что с ослаблением
сгибательного рефлекса, в силу утомления координирующего аппарата
сгибания, начинает проявляться ритмическая иннервация шагания,
координирующий аппарат которой труднее поддается утомлению (рис. 124).
Известно, что шагательный рефлекс на препаратах кошки и собаки
может продолжаться долгое время, в течение многих минут с точно
определенным ритмом (Шеррингтон, 11).
У лягушки координирующий аппарат Шагательного рефлекса задних
конечностей лежит во II и III поясничных сегментах (IX и X сегменты
по Эккеру); это было установлено путем локального стрихніинного
отравления соответствующих спинномозговых сегментов с задней
поверхности. Обычно на спинальных или лумбальных препаратах
раздражение задней конечности производит ритмический рефлекс шагания из
РИТМИЧЕСКИЕ РЕФЛЕКСЫ
283
двух-трех движений и при том только при сильных раздражениях.
После же указанного локального отравления ритмический рефлекс
вызывается одним прикосновением к коже задней лапки в области
распространения IX и X задних корешков, причем он составляется из
15—20 фаз сгибания и разгибания. В большинстве случаев эти
антагонистические движения относились к шагательному типу и
начинаются не флексией раздражаемой конечности, как на нормальном
препарате, а экстензией. В это
время другая задняя нога
сгибается, но при одновре-
ног, а иногда и одной
получается длительный
ритмический рефлекс типа прыгания
или плавания (Беритов, 6).
Многофазный
ритмический рефлекс у лягушки
получается и при стрихнинном
отравлении IX и X
сегментов в одной симметрической
половине, а именно — путем
отравления с боковой
поверхности. В этом случае
ритмический рефлекс всегда
шагательного типа и
протекает всегда дольше и
активнее на конечности
отравленной стороны, чем на
противоположной. При этом
ритмический рефлекс
вызывается каждый раз только при раздражении отравленной стороны, а
раздражение неотравленной стороны вызывало обычно локальные реакции
сгибания, потирания или разгибания. При раздражении этой конечности
может также получиться ритмический рефлекс, но он протекает
совершенно так же, как и при непосредственном раздражении отравленной
стороны, из чего следует, что эта ритмическая реакция обусловлена
деятельностью отравленной половины (Беритов, 6).
При отравлении VIII сегмента, т. е. первого поясничного, не
получается усиления ритмического рефлекса типа шагания. В этом случае
усиливается рефлекс потирания, вызываемый раздражением
рецептивного поля потирания на бедре, т. е. из области распространения VIII
заднего корешка. Далее было установлено, что отравление тех же IX
и X сегментов с передней стороны не ведет к усилению ритмического
рефлекса. Наконец, было обнаружено, что, если периферическая
чувствительность устранена перерезкой IX и X задних корешков, то после
отравления соответствующих сегментов с задней поверхности можно
Рис. 124. Ритмический рефлекс при сильном
раздражении чувствительного нерва.
Тот же децеребрированный препарат, что и на рис. 123.
Те же мышиы. Сильное раздражение п. radiahs—1000
ед. Кронекера, Первый раз во время раздражения
наступает сгибательный рефлекс, а после раздражения--
ритмический. Второй раз во время длительного
раздражения сгибательный рефлекс сменяется ритмическим, а
іретий раз о самого начала возникает ритмический
рефлекс.
284 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
вызвать ритмический рефлекс в несколько фаз раздражением IX или
X заднего корешка. Раздражением же других корешков и
чувствительных нервов из рецептивного поля потирания будут вызываться на
задних ногах совершенно нормальные рефлексы без всяких признаков
стрихнинного отравления (рис. 21).
' Отсюда нами было сделано заключение, что у лягушки координация
ритмических рефлексов типа шагания, прыгания и плавания на задних
конечностях происходит в IX и X сегментах, а именно — в их задней
Рис. 125.Ритмический рефлекс при одновременном раздражении
двух симметричных нервов.
Децеребрированная кошка. Верхняя кривая — от разгибателя правого
колена, а нижняя—от разгибателя левого колена. Под ними верхний сигнал
отмечает раздражение п. peroneus правой стороны, а нижний— того же нерва
левой стороны. Каждый раз при двустороннем раздражении наступает на
обеих ногах ритмический рефлекс шагательного типа, во время сокращения
разгибателя одной стороны происходит торможение другой. При
одностороннем же раздражении длительное торможение одной стороны совпадает
с длительным сокращением другой Наверху время — по 0.2" (Шеррингтон)
половине, причем эти координирующие аппараты лежат симметрично в
каждой половине данных сегментов и каждая сторона управляет,
главным образом, ритмическим движением соответствующей конечности, но
в некоторой мере она вовлекает в ритмическую деятельность и другую
конечность.
На препаратах без большого мозга координирующие аппараты
ритмических рефлексов приводятся .в активное состояние раздражением
рецепторов конечностей. Если раздражением этой конечности вызывается
также какой-либо нефазный рефлекс, как оборонительный рефлекс
сгибания или тонический рефлекс разгибания, тогда фазный рефлекс
уступает место нефазному вследствие господства нефазной реципрокной
иннервации в «общем конечном пути». Но если каким-либо образом
устранить действие этой нефазной иннервации,— путем ли утомления
соответствующего координирующего аппарата, путем ли вызова другой
нефазной иннервации противоположного характера.— то ритмический
рефлекс получит возможность проявить себя в полной мере. Так,
например, на рис. 124 сильное периферическое раздражение вызывает
оборонительный сгибательный рефлекс, а при длительном раздражении,
с утомлением соответствующего координирующего аппарата,
оборонительный рефлекс сменяется ритмическим. На рис, 125 видно, что раз-
РИТМИЧЕСКИЕ РЕФЛЕКСЫ
285
дражение чувствительного нерва одной стороны вызывает сильное тор-
можение разгибателя на соответствующей стороне и сильное
сокращение разгибателя на другой; при одновременном же раздражении
чувствительных нервов на обеих сторонах получается ритмический
рефлекс обеих мышц. Очевидно, в результате алгебраической сумма-
Рис. 126. Сокращение отдачи на
децеребрированной кошке.
Регистрируются разгибатель локтевого
с> става (верхняя кривая) и сгибатель
(нижняя кривая). Раздражается
фарадическим током лучевой нерв той же стороны
во время экстензорного тонуса, В момент
сгибательного рефлекса тонус расслабевает
и вновь проявляется в усиленной форме
после раздражения.
Рис. 127. Сокращение
отдачи на децеребрированной
кошке при отсутствии
экстензорного тонуса.
Тот же препарат и те же
мышцы. Раздражается лучевой нерв.
После сгибательного рефлекса
по прекращении раздражения
наступает сильное тоническое
сокращение.
ции нефазных торможения и возбуждения общий конечный путь
освобождается от их нефазного действия и координирующий аппарат
ритмического рефлекса получает возможность проявить свою
ритмическую деятельность на мышцах.
Явление отдачи и его происхождение. При изучении
антагонистических иннервации обращают особое внимание на одно чрезвычайно
распространенное явление. Если раздражение вызывает разгибательный
рефлекс, то по прекращении раздражения он очень часто сменяется
286 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА Й С?ВОЛА ГОЛОВНОГО'МОЗГА
сгибательным, и, наоборот, если при раздражении рефлекс был сгиба-
тельного характера, то по прекращении раздражения наступает разги-
бательный рефлекс. В обоих случаях на заторможенной мышце после
раздражения наступает сокращение. Это сокращение после торможения
было подробно изучено Шеррингтоном на децеребрированных
кошках © отношении омены сгибательного оборонительного рефлекса
тоническим разгибанием. В этом случае смена торможения
сокращением происходит на разгибателях. Шеррингтон дал этому
сокращению название сокращение в силу отдачи (rebound
contraction), а тот физиологический закон, по которому это явление
возникает, назвал последовательной спинномозговой
индукцией (successive spinal induction) (11), Это название должно
выражать то общее свойство спинного мозга, что в каждом
заторможенном центре по прекращении торможения возникает повышенная
деятельность, как бы вследствие освобождения энергии, накопленной
во время торможения.
Сокращение, в силу отдачи, на разгибателях децеребрированной
кошки наблюдается как в случае наличия экстензорного тонуса на
раздражаемой, конечности^ так и при его отсутствии. В первом случае
тонус ослабевает во время сокращения сгибателя и вновь появляется
после него в усиленной форме в івиде отдачи; а во втором случае
заторможенная мышца во время раздражения не меняет своей длины
и дает сокращение в силу отдачи после раздражения. В обоих случаях
сокращение в силу отдачи носит длительный тонический характер
(рис. 126 и 127).
Явление отдачи было впоследствии открыто на сгибателях как
децеребрированных, так и спинальных животных. В этом случае
перекрестный разгибательный рефлекс сменялся сгибательным тотчас же
по прекращении раздражения: на сгибателе после торможения
наступало сокращение. Явление отдачи подробно изучено учеником Ш е р -
рингтона Грэиім Броуном, ноионв общем .придерживался
того толкования, какое было первоначально дано Шеррингтоном.
Грэим Броун обратил внимание также на то обстоятельство,что
когда заторможенная мышца дает сокращение в силу отдачи,
антагонист, сокращавшийся во время раздражения, теперь быстро
расслабляется. Здесь получается торможение после сокращения, как бы отдача
в виде торможения. Это явление, понятно, связано с тем, что мы здесь
фактически имеем смену одного рефлекса другим, одних реципрокных
иннервации*—другими, противоположными (7, 12).
Позднее явление отдачи было изучено на разных препаратах кошки,
кролика и лягушки {Беритов, 17). Было обнаружено, что на задних
конечностях препаратов кролика, находящихся в сгибательном тонусе,
легко можно наблюдать смену оборонительного рефлекса разгибания
тоническим сгибанием. То же самое было установлено на лягушках, у
которых задние конечности обладают сгибательным тонусом. У этих
Животных задние конечности на более или менее значительное повреж«
РИТМИЧЕСКИЕ РЕФЛЕКСЫ 287
дающее раздражение отвечают разгибанием, после чего наступает
тоническое сгибание.
Явление отдачи легко наблюдать на лягушке при раздражении
чувствительного корешка или нерва, получающего волокна от рецептив-
в
Рис 128. Общее торможение и рефлеьс потираний на
спинномозговой лягушке.
Раздражается VIII задний корешок, получающий чувствительные
волокна с рецептивного поля потирания на бедре Верхняя кривая—
от сгибателя колена, а нижняя — от разгибателя.
О п ы т А. Первое более слабое раздражение (35 см) производит общее
горчожение во время раздражения и однократный рефлекс потирания
после раздражения, второе раздражение (30 см) производит сначала
общее торможение, а затем трехкратный рефлекс потирания.
ОпытВ. На том же препарате раздражение VIII корешка
производится во время сгибательного рефлекса, вызываемого раздражением
п. peroneus. Во время раздражения сокращение расслабевает, что
указывает на торможение сгибателя одновременно с торможением
разгибателя.
ных полей двух рефлексов, сгибания и разгибания, напр., при
раздражении IX и X задних корешков (рис. 40). Далее явление отдачи было
установлено после общего торможения. Оно наступает по прекращении
раздражения в виде реципрокной иннервации определенного типа. Так,
например, очень часто при раздражении рецептивного поля потирания
288 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА Й СТВОЛА ГОЛОВНОГО МОЗГА
или его чувствительных нервов сначала наступает общее тормржение, а
затем рефлекс потирания. /¦;.,,:г . :; г: г. ¦'"¦;. .-.:v "У- 'т
Если раздражение прекратить до начала рефлекса, то последний
наступит в виде отдачи, т. е. по прекращении раздражения (рас. 128).
Явление отдачи на скелетных мышцах наблюдается и при
раздражении спинного мозга, а также рецепторов внутренних органов и вегета-
на рис. 92 оно наступило на
сгибателях колена после
торможения, вызванного
растяжением прямой кишки
(Беригов и Бакурадзе, 28,
29).
Далее было установлено,
что на децеребрированных
кошках явление отдачи
обязано своим происхождением
тоническому состоянию
препарата. Когда препарат
расположен к разгибательному
тонусу, тогда имеет место
явление отдачи на
разгибателях. Когда же, наоборот,
он расположен к сгибатель-
ному тонусу, тогда
наблюдается явление отдачи на
сгибателях.
Очень ясно выступает
зависимость отдачи от
тонического состояния препарата
при изучении этого явления
в связи с шейными и лаби-
положениях головы,
благоприятных для экстензорного тонуса данной конечности, и вообще при
наличии разгибательного тонуса, явление отдачи наблюдается на
разгибателях. При положениях головы, благоприятных для сгибательного
тонуса, и вообще при наличии сгибательного тонуса, то же явление
наблюдается на сгибателях (рис. 129, 130) (см. подробно о значении
положения головы в главе XVI).
Путем анализа этого рода явлений імы пришли к заключению, что
на децеребрированных животных смена торможения возбуждением, в
связи с прекращением раздражения чувствительных нервов,
обусловлена тем, что, наряду с возбуждением спинномозговых координирующих
аппаратов сгибания и разгибания, приходят в активное состояние
координирующие аппараты тонических рефлексов в продолговатом мозгу.
Вследствие этого мышцы конечности получают одновременно сильные,
быстропротекающие иннервации из спинальных координирующих аппа-
тивных чувствительных нервов. Например,
Рис. 129. Явление отдачи и его зависимость
от тонических рефлексов.
Децеребрированная кошка со сгибатель ной тонической
ригидностью. Верхняя кривая — от сгибателя локтевого
сустава, нижняя — от разгибателя того же сустава.
Раздражается п. brachial is другой стороны. В опыте А
вызывается перекрестный разгиОательный рефлекс при
положении головы, благоприятном тоническому
сгибанию, а в опыте В — при положении, не благоприятном
этому тонусу. В первом случае наступает после
раздражения длительная отдача на сгибателе.
ринтными тоническими рефлексами. При
РИТМИЧЕСКИЕ РЕФЛЕКСЫ 289
ратов и слабые длительно протекающие из топических. В случае
противоположного характера этих иннервации во время раздражения, пока
действуют спинальные аппараты, тоническая иннервация не может
проявить себя, но после раздражения происходит быстрое прекращение
спинальных иннервации, и тогда продолжающаяся тоническая
иннервация начинает влиять на общий конечный путь и вызызать
соответствующую реакцию. Таким образом, возбуждение данной мышцы после тор-
можения на децеребрированных препаратах есть ?е что иное, как
Рис. 13и. Явление отдачи и его зависимость от тонических
рефлексов.
Децеребрированная кошка с разгибательной тонической ригидностью.
Верхняя кривая — от triceps brM разгибателя локтевого сустава; нижняя — от
biceps Ьг., сгибателя того же сустава. Раздражается с той же стороны п. radla-
lis. Наступает оборонительный сгибательный рефлекс. В опыте Л положение
головы благоприятно для разгибательного рефлекса данной ноги, а в опыте В,
наоборот,— для сгибательного тонуса. В первом случае разгибатель дает
отдачу после торможения, во втором случае не дает.
проявление тонического рефлекса, вызванного раздражением, но
заторможенного оборонительным рефлексом во время раздраоісения (Бери-
тов, 10).
Анализ многообразных примеров отдачи на спинномозговых
препаратах лягушки показал нам, что она в определенных случаях
обусловливается возбуждением двух разнородных оборонительных рефлексов —
сгибания и разгибания. Благодаря длительному тоническому
последействию сгибательного рефлекса и сильной, но кратковременной разгиба-
тельной иннервации, во время раздражения наступает разгибательный
рефлекс, а после раздражения— сгибательный.
Все вышеприведенные фактические данные указывают на то, что для
вызова сокращения в силу отдачи необходимо наличие посторонних
центральных импульсов возбуждения, направленных к тем двигатель-
19 и. С. Беритов
4
ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА.
ным нейронам, которые перед тем были заторможены периферическим
раздражением. Если почему-либо нет центральных импульсов
возбуждения одновременно с торможением двигательных нейронов сокращение
отдачи после торможения отсутствует. Так, например, обычно бывает
при раздражении в рецептивном поле потирания лягушки. В рефлексе
потирания на спинальных лягушках m. semitendinosus тормозится, и
это торможение сгибателя не сменяется сокращением. После каждого
торможения мышца приходит в покойное состояние. Так бывает при
торможении любой интенсивности и продолжительности. Сокращения
появляются на m semitendinosus после торможения, если одновременно
раздражался нерв из рецептивного поля сгибания (рис. 122 и 128).
Облегчение после торможения, о котором говорилось выше, играет
определенную роль в происхождении сокращения отдачи. Именно от
него зависят те сложные и быстро развивающиеся сокращения отдачи,
которые наступают после торможения в связи с прекращением
раздражения. Во всяком случае опыты показывают, что в фазе облегчения
центральные импульсы возбуждения производят гораздо более сильные
двигательные эффекты, чем в ее отсутствии (рис. 91). Можно
утверждать, что при наличии значительного тонического сокращения
сокращение отдачи возникает главным образом благодаря фазе облегчения на
основании тех двигательных импульсов, которые обусловливают тониче
ское сокращение (Беритов, Бакурадзе и Нарикашвили, 25).
Этой же фазе облегчения обязано своим происхождением
наступление сокращения отдачи при отсутствии всякого тонического сокращения
и при наличии раздражения чувствительных волокон только из одного
рецептивного поля. Так, например, мы привели выше случги явления
отдачи при раздражении такого чувствительного нерва, который
содержит афферентные волокна от одного рецептивного поля, именно —
рефлекса потирания. Очень часто во время раздражения получается общее
торможение, а после раздражения в виде отдачи возникает характерный
для раздражаемого нерва рефлекс потирания (рис. 128). Анализ этого
явления привел нас к убеждению, что здесь отдача возникает
вследствие раздражения одних и тех же волокон и происходит при участии
одного и того же координирующего аппарата Мы думаем, что
афферентные волокна рецептивного поля в определенных случаях
производят такое сильное общее торможение двигательных нейронов через
нейропиль, что импульсы возбуждения, притекающие к ним через
промежуточные нейроны, не в состоянии вызвать их возбуждение. С
прекращением раздражения общее торможение через нейропиль снимается,
и сейчас же, вследствие прекращения анэлектротонического состояния,
вызвавшего угнетение возбудимости двигательных нейронов, наступает
обратное функциональное изменение — повышение возбудимости тех же
нейронов. По прекращении раздражения промежуточные нейроны
продолжают еще некоторое время действовать при посредстве нервных
кругов и от этого сейчас же наступает рефлекс потирания в усиленной
форме. Если раздражение продолжается долго, деятельность промежу-
РИТМИЧЕСКИЕ РЕФЛЕКСЫ
291
точных нейронов может усилиться в такой мере, что двигательные
нейроны начинают реагировать возбуждением, наступает рефлекс'
потирания. Но наблюдаемое при этом сокращение носит явно
заторможенный характер, ибо если раздражение в этот момент прекратить,
сокращение сейчас же усиливается. Очевидно, то общее торможение/
которое имеет место до наступления сокращения, продолжается и во
время него.
Итак, в определенных случаях сокращение в силу отдачи может
иметь место и при участей одного координирующего аппарата, если
только общее торможение, наступающее во время раздражения,
является более или менее значительным; вследствие этого во время
раздражения оно препятствует наступлению рефлекса, а после раздражения
фазой «облегчения», наоборот, благоприятствует ему.
Историческая справка об явлении отдачи. Явление отдачи после торможения было
впервые изучено Сеченовым. Он вызывал ее при раздражении седалищного
нерва лягушки. При некоторых средних и больших силах раздражения начальное
рефлекторное движение сменяется покоем. Эта фаза покоя была им признана
проявлением торможения специальных центров, ибо (вслед за прекращением раздражения
в этой фазе (покоя обычно начиналась новая двигательная фаза. Сеченов дал
этому іяівлеіниію такое объяснение: во время раздражения происходит возбуждение
двух антагонистически действующих рефлекторных механизмов, нз которых один
дает рефлекторное движение, а другой угнетает его. При более слабых
раздражениях верх берет двигательный, .а яри сильных — угаетателыгый рефлекторный
механизм. Но в то время как происходит задержка двигательных импульсов,
возбуждаемый орган продолжает заряжаться энергией и в течение остановки, длящейся
минуты; к концу раздражении энергии накопляется столько, что по прекращении
тормозящих влияний она (переливается, так оказать, через край, выражаясь усиленным рядом
«разрядов» (21).
В настоящее время вышеприведенная гипотеза Шеррингтона о
происхождении отдачи оставлена самим автором. Ое, как и другие азторы, раэзвшает
объяснение, по существу не отличающееся от Сеченовского (Фултон, 13;
Рэнсон, 14). Они полагают, что отдача обусловлена тем, что нервные стволы
содержат смешанные чувствительные волокна с различной центральной реакцией.
Раздражение одной группы волокон рефлекторно вызывает на периферии
сокращение, а раздражение другой — угнетение. Вследствие этого периферический эффект
раздражения двойствен: когда возбуждающий эффект протекает дольше
тормозящего, то по прекращении раздражения может проявиться сокращение в виде отдачи;
значит, чистое торможение, не сопровождающееся длительным возбуждением, не
может дать отдачи в виде возбуждения. Шеррингтон находит причину отдачи
исключительно в одновременном возбуждении нервов из разных рецептивных полей и
не приписывает ей какой-либо роли в нормальной координации (15). Это толкование,
безусловно, вполне применимо для определенных случаев, когда действительно
раздражается нерв, заключающий в себе чувствительные волокна из рецептивных полей
двух рефлексов, как X задний корешок и п. tibialis лягушки (Беритов, 17). Но оно
совершенно не применимо для понимания той отдачи, которая обычно получается
на децеребрированных препаратах. Последняя всецело зависит от деятельносги
лабиринтных и шейных тонических центров, которые возбуждаются при раздражении
любого чувствительного нерва.
Рэнсон со своими сотрудниками опубликовал в последнее время серию
исследований рефлекторной деятельности спинальных и децеребрированных кошек; авторы
пришли в общем к такому же пониманию этого явления, какое было развито нами
выше. Именно, они заключают, что периферическое раздражение производит двойной
19*
292 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
эффект в экстензорных двигательных нейронах. В течение раздражения
преобладает тормозящий эффект. Он препятствует сокращению мышцы разгибателя во время
раздражения и тормозит существующий в ней тонус. Но по прекращении
раздражения тормозящий эффект убывает быстрее, чем возбуждающий, что
обусловливает отдачу возбуждения и вызывает сокращение (Рэнсон и Гинзей, 14) Эта
формула совершенно одинакова с той, которая дается нами. Но на вопрос, откуда
берется длительно протекающий возбуждающий эффект разгибателя, упомянутые
авторы отвечают иначе Они выставляют гипотезу о длительном течении возбуждения
в разгибательных нейронах, разгибательном спинномозговом полуцентре Мы же, на
основании эксперимента, пришли к заключению, что на децеребрирозанных
препаратах этот эффект исходит из тонических разгибательных центров шейных и
лабиринтных тонических рефлексов
Гипотеза Рэнсона не может быть применена уже потому, что явления отдачи
возбуждения бывают и на сгибателях. Так, например, случается, что периферическое
раздражение вовлекает в действие сгибательные тонические центры (6). При этом
следует уіказаггь, что сокращение сгибателей и разгибателей в (виде отдачи
даже на сшдаальных и лумбальных препаратах должно быть обусловлено участием
тонического сгибательного и разгабателыного центров; известна, например, что лум-
бальный препарат кошки может стоять на задних ногах и вообще раэшбать их
тонически в течение многих минут (Беритов, 16). Следовательно, во время раэдраіжения
рецептивного поля сгибания при вовлечении спинальных тонических центров
разгибатели должны давать сокращение в виде отдачи.
О происхождении смены антагонистических иннервации в
ритмических рефлексах. Происхождение чередования противоположных фаз
до сих пор еще не выяснено окончательно. По этому вопросу
существует несколько предположений. Так, например, в первое время Ш е р-
р и н г т о н объяснял смену противоположных фаз явлением отдачи.
Предполагалось, что каждый раз фаза торможения «индуцирует» фазу
возбуждения. Но, как уже указывалось выше, Шеррингтон
Отказался от этого гипотетического объяснения явления отдачи.
В наиболее законченном виде гипотеза о деятельности центрального
механизма ритмических движений разработана Грэим Броуноім
(12). По его представлению вышеупомянутые механизмы состоят из
двух антагонистических центров (полуцентры), которые приводят©
действие антагонистические мышцы. Деятельность одного полуцентра
вызывает длительное разгибание и одновременно ослабление деятельности
сгибательного полуцентра. Второй же полуцентр дает длительное
сгибание и одновременное ослабление деятельности разгибательчого
полуцентра. Следовательно, по представлению Грэим Броуна, во
время ритмического движения превалирует деятельность то одного
полуцентра, то другого. Причинами же смены деятельности
полуцентров Грэим Броун считает утомление и отдачу возбуждения.
Когда вследствие какой-либо причины нарушается равновесие в
деятельности обоих полуцентров, деятельность одного из них
становится господствующей. Если это сгибательный полуцентр, то он
вызовет сгибание и одновременно специальными тормозящими
путями произведет торможение деятельности второго разгибательного
полуцентра. Так будет продолжаться до того времени, пока не наступит
утомление сгибательного полуцентра. Вследствие утомления
деятельность этого центра ослабевает и уменьшается его тормозящее действие
РИТМИЧЕСКИЕ РЕФЛЕКСЫ
293
на разгибательный полуцентр. Это обусловит восстановление
равновесия между обоими центрами. Но когда разгибательный полуцентр
освободится от тормозящих влияний, в нем разовьется усиленная
деятельность. Происходит своего рода отдача возбуждения: заторможенный
разгибательный полуцентр развивает усиленную деятельность.
Деятельность же сгибательного полуцентра, наоборот, тормозится под влиянием
пришедшего в усиленное деятельное состояние разгибательного
полуцентра. Это будет продолжаться до тех пор, пока не наступит
утомление разгибательного полуцентра. В связи с этим утомлением произойдет
отдача возбуждения в сгибательном центре, и тогда деятельность
сгибательного полуцентра опять займет господствующее положение.
Чередование деятельности полуцентров будет продолжаться до тех пор,
пока длится раздражение и пока у центров не иссякнет способность к
деятельности. По представлению автора, полуцентры состоят из клеток
двигательных нейронов. Следовательно, каждая клетка одного центра
посылает отростки к клеткам другого, противоположного центра. С
помощью этих отростков и осуществляется торможение одного центра
другим.
Эта гипотеза в настоящее время не может быть признана
удовлетворительной. Во-первых, она основывается на гипотезе отдачи
возбуждения, т. е. последовательной индукции, с которой мы уже
ознакомились и которую мы отвергли. Во-вторых, происхождение
координирующих процессов она приписывает исключительно двигательным нейронам,
что также совершенно неприемлемо.
На основании современных представлений о центральных процессах
возбуждения и торможения можно дать следующую гипотезу о
происхождении смены фаз в ритмических рефлексах: можно предположить,
что центральный механизм фазного рефлекса в спинном мозгу устроен
так, что' когда его нервные цепочки и нервные круги приходят в
активное состояние и производят возбуждение двигательных нейронов
определенной группы мышц, в это самое время они активируют нейропиль
и через медленные потенциалы тормозят по преимуществу
двигательные нейроны антагонистической группы мышц. Вероятно в этой реци-
прокной связи некоторую роль играют и возвратные коллатерали
двигательных нейронов. Как известно, при антидромном возбуждении
двигательного корешка тормозятся соседние двигательные нейроны из других
корешков. То же самое должно наблюдаться при рефлекторном их
/возбуждении. Известно, что антидромные импульсы тоже вызывают
медленные потенциалы, которые могут производить торможение.
Смена фаз в ритмическом рефлексе, например, сгибания на
разгибание, вероятно обусловливается прежде всего тем, что период
деятельности первично возбужденных нервных кругов в центральном
аппарате фазного рефлекса проходит быстрб вследствие быстрого утомления
промежуточных элементов. Сообразно, быстро ослабевает и активное
состояние нейропиля, благодаря чему анэлектротоническое действие его
медленных потенциалов на антагонистическую группу нейронов, т. е. на
294 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
группу нейронов разгибателей, сменяется на катэлектротоническое
действие, т. е. на облегчение. Это является благоприятным условием для
возбуждения данной антагонистической группы под влиянием всякой
периферической импульсации и даже под влиянием импульсов
последействия со стороны утомленных промежуточных элементов.
Возбуждение антагонистической группы, конечно, может сопровождаться
торможением первой группы нейронов, т. е. сгибателей, через возвратные
коллатерали, идущие от двигательных нейронов разгибателей.
При наличии нормальной периферической чувствительности в этой
смене двигательных фаз, конечно, большую роль должны играть про-
приоцептивные импульсы. Как известно, при сокращении каждой группы
(мышц исходящие от них проприоцептивные импульсы возбуждающе
:действуют на данную (возбужденную группу двигательных нейронов и
"тормозят все остальные нейроны, но по прекращении движения они
быстро ослабевают. Это должно вызывать состояние облегчения з
заторможенных нейронах и создать благоприятные условия для смены
существующей фазы движения противоположной.
Характеристика и происхождение ритмической деятельности
плавников рыб. Ритмическая деятельность плавников у рыб при локомоции
-характерно отличается от аналогичной деятельности конечностей у
позвоночных животных. Она совершенно не меняется у данной пары
плавников после перерезки всех соответствующих задних корешков. Так,
например, у карпа (Тіпса) перерезаются все задние корешки, кроме I и
II пары, которые относились к передним плавникам. Такая рыба плавает
нормально с участием всего тела. Движения возникают от раздражения
головных рецепторов (Гольст, 24). Отсюда следует, что у рыб
ритмическая деятельность плавников во время локомоции по существу імало
зависит от периферических раздражений. В ответ на раздражение кожи
около того1 или другого плавника могут наступить сильные
рефлекторные реакции на данное раздражение, но без изменения самого ритма
движения данного плавника и всех остальных. По исследованиям
Голье та, для каждого плавника существует свой автоматический
центр, и каждый такой центр может работать независимо от других
своим собственным ритмом. Но обычно один из этих ритмических
центров является доминирующим: он определенным образом влияет на
ритмическую деятельность других плавников, соподчиняя их работу
¦своей путем повышения или понижения определенной фазы в их ритме.
Это обусловливается главным образом путем воздействия ритмической
деятельности одного автоматического центра на другие автоматические
центры. В этой координации двигательные клетки не играют никакой
роли. Значит, данная координация целиком представляет функцию
специальных промежуточных нейронов. При этом деятельность таких
центров более или менее автономна, ибо когда раздражаются кожные
рецепторы, периферические импульсы могут достигнуть определенных
промежуточных нейронов и затем двигательных клеток, минуя ритмические
центры.
Гольст полагает, что каждый плавниковый координирующий центр
РИТМИЧЕСКИЕ РЕФЛЕКСЫ
2^5
выполняет свою координирующую функцию не путем посылки
двигательных импульсов то к одной группе двигательных клеток, то к
другой, а тем, что он реципрокно меняет их чувствительность к
периферическим или центральным влияниям; иначе говоря, когда данный центр
действует на одну группу двигательных клеток облегчающим образом,
на другую — антагонистическую группу он влияет тормозящим образом.
Таким образом, у рыб координирующий центр плавательных движений
определяет главным образом частоту ритмических движений, амплитуда
же этих движений зависит от всей суммы периферических импульсов,
притекающих к двигательным клеткам в данный момент (Гольст, 24).
Эта форма координации ритмических движений рыб отличается от
ритмической деятельности наземных позвоночных животных, у которых
движения всех конечностей во время ритмического рефлекса всегда
взаимно координированы и никогда не обнаруживают самостоятельных,
независимых ритмов. Поэтому Гольст, исследовавший плавательную
ритмику рыб, называет наблюдаемую при этом координацию
относительной. Эта относительная координация является первоначальной,
примитивной формой координации, очень похожей на координацию
сердечных движений. Абсолютная форма координации высших позвоночных
животных, очевидно, должна была развиться в результате перехода от
водного образа жизни к наземному.
Практикум
Наблюдение ритмического рефлекса. Рефлекс попирания на спинномозговой
лягушке. Раздражается уксусной или серной кислотой (10—20%) рецептивное поле
потирания путем прикладывания к коже смоченных в кислоте кусочков
фильтровальной бумаги (1—2 мм). В ответ на это задняя нога раздражаемой стороны
притягивается к корпусу и несколько раз подряд потирает раздражаемый участок кожи.
Для прекращения рефлекса нужно удалить бумажку и несколько раз обмыть кожу
водой.
Шагательный рефлекс на спинномозговой лягушке. Обнажается спинной мозг в
поясничной области. Затем он отравляется с дорзальной стороны в области IX—X
сегментов путем прикладывания к нему кусочков фильтровальной бумаги, смоченных
в 0 4—0 5% растворе солянокислого стрихнина. Отравление должно быть точно
локальным. Обнаженное место мозга освобождают лигнином от тканевой жидкости и
крови. Рекомендуется удалить все внутренности, чтобы «обескровить» препарат.
Раздражение задних ног до отравления дает однофазные движения, а после
отравления через 10—20 мин. те же раздражения вызывают фазные движения но типу
шагания или ітрыгания. Число фаз может дойти до 20.
Эти опыты лучше всего удаются на зимних лягушках при низкой комнатной
температуре, ибо в таких условиях обескровленные препараты лягушки могут жить
часами.
Значение периферической чувствительности в ритмических рефлексах. Опыт со
стрихнинным отравлением ставится на таких спинномозговых препаратах, у которых
все задние корешки в поясничной области перерезаны. После отравления
электрическое или механическое раздражение перерезанных задних корешков вызывает
несколько фазных движений за время раздражения. Если раздражение
кратковременно, наступят только две фазы с разгибательной в начале. Эта смена фаз может
происходить как во время раздражения, так и после него.
2% ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Наблюдение «сокращения отдачи» на спинальных лягушках. Перерезают мозг
и выделяют m. semitendinosus и т. triceps наполовину; соединяют их с миографами
для записи сокращений на кимографе. Раздражение производится при помощи ключа
Гельм гольца и момент раздражения отмечается на кимографе. Перо
электромагнитного сигнала должно находиться на одном уровне с миографами. Электроды
прикладывают к коже на вентраданую поверхность ступни соответствующей лапки.
Здесь находится рецептивное поле рефлексов слибан>ия и разгибания. При
умеренных силах раздражения можно наблюдать во время него быстрое разгибание
<* сокращением т. triceps и торможением т. semitendinosus; после раздражения
наступает сгибание: сокращается заторможенная мышца. Смена разгибания сгибанием
С =*-
Рис. 131. Аппарат для искусственного дыхания.
Здесь же дан кран Кронекера в разрезе. N — неподвижная медная трубка, которая имеет сбоку
отверстие О и перегородку в просвете как раз в этом месте L; М — подвижная муфта, которая
изнутри имеет выемку в одном месте. Если эта выемка приходится над отверстием, как показано на
рисунке, воздух может проходить через трубку. Если же муфту повернуть так, чтобы выемка отошла
от отверстия, тогда воздух перестанет проходить. Остальные объяснения даны в тексте.
может произойти еще во время раздражения. Во время более сильных раздражений
может наступить сгибание; после раздражения сгибание на время сменяется
разгибанием, причем сокращается заторможенная мышца m. triceps и тормозится сонращен-
Hdfl m. semitendinosus.
Наблюдение сокращения отдачи на децеребрированных кошках. Кошку
помещают под стеклянный колпак и кладут туда смоченную эфиром или хлороформом
вату. Вдыхание этих веществ вызывает наркоз. Глубина наркоза определяется по
дыханию: сначала дыхание становится неравномерным, иногда останавливается; если
дыхание прекращается надолго, следует только встряхнуть кошку вместе с
колпаком, чтобы оно восстановилось. В дальнейшем дыхание становится ровным и
глубоким. Во время обеих стадий животное находится в значительном экстензорном тонусе,
который нужно приписать возбуждению экстешорных центров наркотическим
веществом. Наконец, дыхание становится поверхностным. В этот момент наркоз является
наиболее глубоким; рефлекторный тонус исчезает, и конечности совершенно расслаб-
РИТМИЧЕСКИЕ РЕФЛЕКСЫ
297
ляются; самое сильное раздражение последних не дает рефлекторного движения. Как
только дыхание становится поверхностным, животное переносят на операционный стол
и привязывают животом вверх.
Операция начинается с трахеотомии: перерезают кожу на шее продольно
посередине, обнажают трахею и пересекают до половины. В периферический конец трахеи
вставляют трахеальную канюлю. Эта канюля соединяется с аппаратом для
искусственного дыхания, назначению которого—сохранить глубокий нааркоз во время всей
операции путем введения наркотического (вещества в легкие. Аппарат для искусственного
дыхания показам на рис. 131: небольшой мех периодически приводится в движение
рукой или электрическим мотором. Воздух из моха через резиновую трубку поступает
в У-обршвую трубку, каждая ветвь которой соединяется с краном Кронекера.
Рис 132. А — положение головы во время
децеребрации и уровень перерезки мозга (VV).
t —средний зубец tentorium cerebelH; h — hyoideum; I,
2 и-3— первый, второй и третий шейные позвонки. Ход
позвоночных артерий показан сбоку у первого позвонка
стрелкой. Когда хотят приготовить спинномозговой
препарат, обрезают спереди остистый отросток второго
позвонка и затем в этом месте крепко завязывают вокруг
позвоночника толстую нитку. Положение этой лигатуры
показано на рисунке штриховыми линиями. Лигатура
придавливает позвоночные артерии. В — первые
три позвонка сверху. Ясно видно
положение лигатуры для сдавления позвоночных артерий
(Шеррингтон).
В этом кране просвет шириной <в 0.5 см может быть уменьшен до нуля.
Один такой кран соединен с вульфовой склянкой, в которой находится хлороформ или
?фир или же смесь хлороформа с эфиром. Воздух проходит через слой наркотика и
вместе с парами последнего через трубку присоединяется к току чистого воздуха,
идущего из другого крана Кронекера. Воздух в смеси с парами наркотика вгоняется
в легкие через трахеальную канюлю. Регулируя просвет кранов Кронекера, можно
по желанию изменять количество наркотического вещества, направляющегося в легкие.
Так, например, уменьшением просвета в первом кране уменьшают его поступление.
Для того чтобы регулировать количество поступающего в легкие воздуха, трахеаль-
ная канюля имеет боковое отверстие, через которое весь лишний воздух выходит
наружу.
После трахеотомии производят ваготомию, т. е. перерезку обоих блуждающих
нервов. Эта операция производится во избежание остановки сердца вследствие
возбуждения указанных нервов в момент перерезки головного мозга. Затем
перевязывают обе сонные артерии во избежание больших кровотечений при вскрытии черепа и
перерезке мозга.
298 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
После операции на шее животное подвергается децеребрации. Для этой цели его
переворачивают спиной вверх. Прежде чем начать эту операцию, нужно
удостовериться, что животное находится в глубоком наркозе: сильное сдавливание лапы,
хвоста, ушей не должно давать движений; прикосновение к роговице глаза не
должно вызывать закрытия век. Операция производится так. Разрезают кожу вдоль
головы посередине и удаляют височные мышцы с обеих сторон. Затем просверливают
череп трепаном в области височной кости и через полученное отверстие костными
щипцами снимают верх черепной коробки, который прикрывает большие полушария.
С обнаженного мозга снимают dura mater и, наконец, тупым шпаделем перерезают
весь мозг впереди tentorium в области среднего мозга (рис. 132). Весь передний,
отрезанный отдел мозга вынимают нз черепной коробки и туда вкладывают немного ваты.
Во избежание больших кровотечений при вскрытии черепной коробки и при перерезке
мозга нужно временно прекратить движение крови в позвоночных артериях: ассистент
придавливает их, сжимая шею большим и указательным пальцем позади 1 шейного
позвонка (рис. 132-В). Позвоночные артерии при переходе от II позвонка к I лежат
поверхностно по бокам тела позвонка, и в этом месте их можно очень легко сжать.
После децеребрации сжатие артерий прекращается, так же как и наркоз. Чтобы
прекратить кровотечение из мозга, поднимают голову вверх на 10—15 мин. В течение
всей операций необходимо следить за глубиной наркоза: он должен быть настолько
сильным, чтобы животное все время было недвижимо, в противном случае развивается
длительный и сильный шок, а самое производство операции очень затрудняется
сильным и продолжительным кровотечением.
После операции препарат оставляют в покое, пока не прекратится кровотечение
и пройдет наркоз. Спустя несколько минут после децеребрации развивается
ригидность, т. е. экстензорный тонус; ригидность наступает сильнее на передних ногах, чем
на задних. Одновременно возвращается способность к оборонительным рефлексам.
Временами наблюдают сгибательный и сухожильный рефлексы, рефлекс мигания на
прикосновение к роговице глаза.
После децеребрации теплорегуляция в организме прекращается: животное
остывает и от этого рефлекторная деятельность исчезает. Чтобы сохранить эту
деятельность, нужно искусственно согревать животное. Для этой цели в операционном столе
имеется большая выемка (30X15 см), покрытая жестью. Под жестью помещены
электрические лампочки, согревающие через нее препарат. К'роме того, препарат
солравается сверху электрической лампочкой, или электрической медицинской
грелкой, или же покрывается колпаком, который внутри имеет обычную
электроспираль для соір?вашя. На рис. 133 показан такой согревательный операіциовныя стол
и колпак для согревания. Однако при согревании нужно соблюдать большую
осторожность, чтобы не перегреть животное.
Рефлекторные реакции, и в частности сокращение отдачи, можно наблюдать
через час после операции. Освобождают конечность у животного и раздражают,
сдавливая пальцем, то заднюю, то переднюю ногу. Если раздражение слабое, то
наступает сгибание раздраженной ноги и разгибание противоположной одноименной
ноги. Если раздражение сильное, то после раздражения нерва нога дает усиленное
разгибание.
Миографическим путем сокращение отдачи проще всего регистрируется таким
образом. Еще до децеребрации на одной задней конечности перерезывается п. obtu-
ratorius, все мышечные нервы, идущие к сгибателям колена. Они отходят от
седалищного нерва в области тазобедренного сочленения. Затем перерезают п. tibialis и
п. peroneus в области коленного сочленения. Из п. cruralis перерезаются все веточки,
за исключением тех, которые идут к m. quadriceps. Перерезаются также все мелкие
односуставные мышцы вокруг trochanter major бедра, а также m. psoas magnus и
т. psoas minor. На коленное сухожилие навязывают нить для соединения с
миографом. Затем перерезают сухожилие ниже надколенника и освобождают все
сухожилие от окружающей ткани.
На другой задней ноге перерезается седалищный нерв. Можно перерезать и
РИТМИЧЕСКИЕ РЕФЛЕКСЫ
299
п. obturatorius, а также нерв сгибателей колена и п. cruralis. Децеребрация
происходит после этой операции.
Животное укладывают в брюшном положении и оперированную ногу фиксируют в
горизонтальном положении винтами. Один винт проходит через головку бедра в
области trochanter major, а другой — через другой конец бедра. Свободный конец обоих
винтов укрепляется в зажимах. Нить от сухожилия четырехглавой мышцы соединяется
с миографом. На п. tibialis или п. peroneus накладывают электроды Шеррингтона.
Такие же электроды накладывают на седалищный нерв другой стороны.
Рис. 133. Операционный стол с принадлежностями для согревания
животного сверху и снизу и для фиксации его.
Л—намордник; В~винтодершатель с винтом; С—колпак для согревания сверху с
занавесочкой; D—привод электрического тока для колпака; Е—вилка для согревания стола.
Прочие объяснения даны в тексте.
Опыт 1. Слабое раздражение нерва регистрируемой ноги вызывает расслабление
разгибателя. Если конечности имели тонус, то при раздражении тонус на
раздражаемой стороне ослабевает, тормозится. Очень часто слабое раздражение вместо
расслабления производит усиление тонического сокращения; при некотором усилении
раздражения сначала происходит кратковременное усиление тонуса, а потом
расслабление.
Опыт 2. Сильное раздражение производит полное торможение тонуса, а после
прекращения раздражения наступает на раздражаемой стороне силіьное
тоническое разгибание — явление отдачи. Если вновь раздражать во время сокращения
отдачи, разгибатель расслабится и после раздражения вновь даст сильное тоническое
сокращение. Если мышца без тонуса, то во время раздражения внешнего эффекта не
получится, а после раздражения может наступить тоническое сокращение отдачи.
Опыт 3. Если повернуть голову теменем к раздраженной ноге и так раздражать
нерв, то после раздражения сокращение отдачи или не наступит, или наступит, но в
более слабой форме.
Опыт 4. Если раздражать седалищный нерв другой стороны, то на
регистрируемой конечности наступает тонический, разгибательный рефлекс. При
одновременном раздражении той и другой стороны может получиться ритмическое сокращение.
Если состояние препарата хорошее, можно наблюдать ритмический рефлекс и
под влиянием одного раздражения.
Опыт 5. Раздражается нерв соответствующей стороны с такой силой, при коте-
300 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
рой вызывается торможение, но не получается сокращение отдачи. По прекращении
раздражения в течение нескольких секунд пробуется раздражение нерва
противоположной стороны. Эта проба будет приходиться на фазу облегчения и поэтому будет
давать более сильный перекрестный рефлекс ш разгибателе, чем обычаю.
Литература
1. Sherrington С, J-, Physiol., 40, 28, 1910.
2. Beritoff J., Arch. Anat. Physiol., стр. 296, 1912.
3. Graham Brown, Erg. Physiol., 13, 279, 113; Quart. J. Exp. Physiol., 4,
331, 1911.
4. Magnus R., Pfl. Arch., 130, 219, 1909; 134, 545—584, 1910.
5. Graham Brown, Quart. J. Exp. Physiol., 5, 237, 1912; Proc. Roy. Soc, B^
84, 308, 1911.
6. В e r i t о f f J., Pfl. Arch., 151, 171, 1913; Тр. Петерб. об-ва естеств., 42, 267,
1913.
7. Graham Brown, Proc. Roy. Soc, 81 B, , 278, 1912; J., Physiol., 48, 18, 1914;
Erg. Physiol., 13., 279, 1913.
8. Graham Brown, Proc. Roy. Soc, B. 84, 555, 1912.
9. Sherrington C, Quart. J. Exp. Physiol., 3, 213, 1910.
10. Beritoff J.v Quart. J. Exp. Physiol., 9, 199, 1915; Изв. Акад. Наук, стр. 64S
и 853, 1915.
11. S h е г г і n g t о п С. S-, Int. Act. Nerv. Syst., 206—212, 1906.
12. Graham Brown, Erg. Physiol., 15, 691 и 723, 1916.
13. F u 11 о n J. E., Muscular contraction a. the reflex control of movement, London,
1926.
14. R a n s о n S. W. a. H i n s ? у J. C, Arch. Neur. Psych., 20, 247, 1931.
15. Sherrington C. S. и сотр., Reflex activity of the spinal cord, Oxford, 1932.
16. Б e p и t о в И., Изв. Акад. Наук, стр. 1117, 1915.
17. Б е р и т о в И., Pfl. Arch. 201, 445, 1923; 200, 562, 1923; Учение об основных
элементах центральной координации скелетной мускулатуры, СПб., 1916.
18. A dr і an Е. D. а. В и у t e n d i j k F. J. J., J. Physiol., 71, 9, 1931.
19. Adrian Е. D., J. Physiol., 72, 132, 1931; Механизм нервной деятельности,
пер., Биомедгиз, 1935.
20. Setschenow I., Pfl. Arch. 25, 281, 1881.
21. С е ч е н о в И. М., Об электрическом и химическом раздражении
чувствительных спинномозговых нервов лягушки, 1868.
22. С е ч е н о в И. М., Физиология нервных центров, 1891.
23. S h е г г і n g t о п С. S., Proc. Roy. Soc. В., 86, 219, 1913.
24. Hoist E., Pfl. Arch., 235, 345, 1935; 237, 356, 655, 1936.
25. Б е р и т о в И., Бакурадзе А. и Нарикашвили С, Тр. Ин-та
физиол., 3, 147, 1937.
26. Tower S. S.f Arch. Сотр. Neurol., 67, 109, 1937.
27. Smits G. G., M e 111 e г F. a. Culler E., J. Neurophys., 3, 182, 1940.
28. Беритов И. и Бакурадзе А., Тр, Ин-та физиол. им. Бериташвили,
5, 1, 1943.
29. Беритов И. и Бакурадзе А., Тр. Ин-та физиол. им. Бериташвили,
5, 125, 1943.
Глава XVI
ТОНИЧЕСКИЕ РЕФЛЕКСЫ
Общее понятие. Тоническим рефлексом называют такой
прирожденный акт, который обеспечивает тому или другому органу
определенное устойчивое положение в организме, — например, выпрямление ноіг
при стоянии, вертикальное держание головы, выпрямленное положение
лозвоночника.
Тонические рефлексы могут продолжаться целыми часами, иногда
целый день и дольше, не вызывая утомления.
Тоническая деятельность центральной нервной системы имеет особое
назначение. Во-первых, она защищает организм или части организма
от падения под влиянием собственной тяжести и, во-вторых, длительно
удерживает двигательные органы в одном положении. Тоническое
движение никогда, ни у одного вида животных не зависит от больших
полушарий, от психонервных процессов. Животное стоит и держит
голову прямо и после удаления больших полушарий. Животное может
стоять на ногах даже после удаления всего головного мозга,— значит,
стояние может осуществляться благодаря деятельности спинного мозга.
Рецептивное поле тонического рефлекса. Тонические рефлексы
характерно отличаются от оборонительных рефлексов тем, что
вызываются главным образом под влиянием внутренних раздражений.
Однако существуют и такие, которые вызываются внешними раздражениями,
только в этом случае раздражение не является повреждающим, т. е.
болевым, как это бывает при защитных рефлексах. Во всяком случае,
каждый тонический рефлекс имеет собственное рецептивное поле
(Магнус, 1).
Координация тонических рефлексов. Координация всех тонических
рефлексов происходит по тому же принципу, что и координация
защитных рефлексов. Значит, в каждом тоническом рефлексе одна группа
мышц сокращается, а другая, антагонистическая, тормозится. Состав
мышц той и другой групп совершенно такой же, как в оборонительных
движениях; например, при тоническом разгибании ноги сокращается
та же группа разгибателей, что и при оборонительном разгибании,
группа же сгибателей, наоборот, тормозится (Беритов, 2).
302 общая физиология спинного мозга и ствола головного мозга
Координирующие аппараты тонических рефлексов лежат в
центральной нервной системе, как раз в тех отделах ее, куда входят
чувствительные нейроны от рецептивного поля данного рефлекса (Магнус, 3).
Рефлекторная дуга тонического рефлекса должна состоять по крайней
мере из трех нейронов: чувствительного,
промежуточного, или координирующего, и
двигательного.
Сгибательный тонический рефлекс на
спинномозговой лягушке. Этот рефлекс
заключается в одновременном сгибании обеих задних
конечностей. Он особенно интенсивен и
длителен на зимних или охлажденных лягушках
(Бидерман, 4). Как нормальная,
неповрежденная, так и спинномозговая лягушка
обычно держит задние конечности в тоническом
сгибании; лягушка принимает сидячее
положение, которое прекращается только во
время оборонительных и локомоторных движений.
Тоническое сгибание зависит от
особенностей координирующего аппарата; на каждое
короткое раздражение координирующий
аппарат отвечает длительно протекающей
деятельностью; кроме того, длительности реакции
благоприятствуют вторичные раздражения,
возникающие в мышцах, сухожилиях,
суставах и в самой коже вслед за первичным
движением. Обычно лягушка держит обе лапки
согнутыми. Это состояние прекращается после
перерезки нервов, когда мышцы лишаются
центральных влияний. Тоническое сгибание
задних конечностей проявляется значительно
слабее, когда лягушка висит в воздухе: лапки
опускаются вниз, но в главных суставах
заметно сгибание. Если на одной стороне
перерезать поясничное сплетение, тоническое
сгибание в главных суставах соответствующей
лапки пропадает и вследствие этого лапка
опускается больше (Бронджист, 5) (рис.134).
Рецептивное поле этого тонического рефлекса
то же, что и защитного сгибательного
рефлекса: оно охватывает кожную поверхность главным образом стопы и
голени. При сидячем положении механическое и химическое раздражение
кожи в этом поле, должно быть, является основньш условием
тонического сгибания. Химическое раздражение происходит путем выделения
кислой секреции из кожных желез, которое, видимо, особенно усиливается
при рефлекторных реакциях (Гедевани, 51), При висячем положении
¦іМ****Ж«*-**і«тт*~,<.іШ. ¦¦ - •¦. -. .«»-.•;
Рис. 134. Сгибательный
тонический рефлекс по Бронд-
жисту.
Спинномозговая лягушка
подвешена за голову на штативе.
На правой стороне перерезано
седалищное сплетение. Правая
задняя нога опущена вниз
больше левой вследствие
прекращения сгибательного тонуса;
неповрежденная же нога немного
согнута под влиянием этого
тонуса во всех главных суставах.
ТОНИЧЕСКИЕ РЕФЛЕКСЫ
313
механический фактор раздражения кожи отсутствует, но химический
остается, и он вместе с ироприоцептивными раздражениями сгибателей
должен главным образом обусловливать тоническое сгибание лапок.
Раздражение, произведенное вне рецептивного поля сгибания,—
например, в рецептивном поле потирания,— не благоприятствует этому
рефлексу: наоборот, оно действует на него тормозяще (Беритов, 6).
В этом тоническом сгибании отношение между мышцами антагонистами
такое же, как и в защитных рефлексах: одна группа мышц длительно
Рис 135. Тонический сгибательный рефлекс.
Децеребрир о ванный кролик. Первая кривая от
разгибателя m. Vasto-crureus левой стороны. Вторая — от
сгибателя semitendmosus правой ноги, а третья — от
разгибателя m. Vasto-crureus этой же ноги. Раздражают
седалищный нерв правой стороны. После раздражения
кимограф останавливался четыре раза по 1 мин.
Усиленное тоническое сгибание правой ноги, вызванное
раздражением, удерживается при постепенном ослаблении
более 4 мин.
сокращается, другая длительно тормозится (Беритов, 2). Вследствие
тонического сгибания лягушка принимает сидячее положение: она сидит
с плотно поджатыми задними лапками, опираясь на передние
конечности и на таз. Такое положение принимает как цельная лягушка, так и
спинномозговой препарат ее после каждого защитного движения.
Следовательно, вышеописанный рефлекс представляет тонический рефлекс
сидения.
Координация тонического рефлекса сгибания задних ног происходит
в поясничном отделе спинного мозга. Это видно из того, что рефлекс
сохраняется и в том случае, если поясничный отдел мозга отделен от
остального спинного мозга.
Сгибательный тонический рефлекс на кролике. Совершенно такой
же тонический рефлекс наблюдается и на кроликах. И здесь он имеет
такое же назначение, как у лягушки: с помощью его животное
принимает сидячее положение. На децереброванных препаратах этот
рефлекс может быть вызван слабым раздражением кожной поверхности
ноги или ее чувствительного нерва; сильное же раздражение сперва
304 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
вызывает оборонительные движения Этот тонус протекает длительно —
до нескольких минут после каждого раздражения (Беритов, 7) (рис. 135).
Децеребрационная ригидность. Так называется разгибательный
тонус, наступающий у млекопитающих животных в первое время после
удаления больших полушарий и промежуточного мозга, т. е. после
децеребрадии. Подробное экспериментальное исследование этого
явления принадлежит Шер,рингтону (8). Он изучал его на децеребриро-
ванных кошках, собаках и обезьянах. Обычно вместе с большими
полушариями и промежуточным мозгом удаляют также передние холмы
четверохолмия. Эта операция производится во время глубокого
хлороформного или эфирного
наркоза. После операции, как
только наркоз начинает
проходить, на ногах и во всем
теле развивается
децеребрационная ригидность: прежде
всего сокращаются все те
мышцы, которые должны
поддерживать организм и
его органы от падения в
силу тяжести. Антагонисты их
одновременно испытывают
торможение. В результате,
все конечности разгибаются,
голова и шея поднимаются
вверх, туда же отгибается
хвост и выгибается спинной
хребет (рис. 136). Во время
сильного ригидного
состояния сокращаются точно та*
же сгибатели, притом тем
сильнее, чем больше
ригидность. Несмотря на это, раз-
гибательное положение
сохраняется, вследствие того,
что механическое действие
разгибателей на суставы сильнее, чем сгибателей (Вахгольдер, 37).
Во время децеребрационной ригидности оборонительный рефлекс
сгибания и чесательный рефлекс наступают с меньшей интенсивностью.
Это происходит оттого, что мышцы-сгибатели конечности испытывают
сильное тоническое торможение. Децеребрационная ригидность хорошо
выражена в любом положении препарата: в спинном, боковом и т. д.
Происхождение децеребрационной ригидности. Если во время
децеребрационной ригидности перерезать спинной мозг в грудной области,
задние ноги тотчас же лишатся ригидности, а на передних, наоборот,
ригидность усилится. Это указывает, что децеребрационная ригидность
Рис. 136. Децеребрирэванная кошка в
состоянии экстензорной ригидности.
Децеребрация произведена по методу анемии.
Перевязаны сонные артерии и базальная артерия
в области варолиева моста. Прекращение
кровообращения в передней части мозга равносильно
децеребрации. После перевязки развивается экс-
тензорный тонус, как после децеребрации. Рис. А
показывает животное в состоянии экстензорной
ригидности. Рис. В—то же самое при
одновременном выключении мозжечка (Поллок и Девис).
ТОНИЧЕСКИЕ РЕФЛЕКСЫ
305
обусловливается деятельностью головного мозга. Прежде всего следует"
отметить, что децеребірационный экстензорный тонус той или иной
конечности, в частности, зависит и от проприоцептлвных раздражений
самой конечности. Было доказано, что при выпадении мышечной
чувствительности вследствие инъекции кокаина в мышцу-разгибатель
экстензорный тонус конечности ослабевает и даже совсем исчезает
(Магнус и де Клейн, 10; Шеррингтон, 11). Известно также, что после
прекращения экстензорного тонуса, когда мышцы вполне расслабились,
тонус их можно восстановить, применяя пассивные движения, т. е.
производя раздражение проприоцепторов (Баиет и Пенфильд, 12). Но
нельзя приписывать децеребрационный тонус конечности исключительно
проприоцептивным раздражениям,— он может быть вызван на
конечности даже в том случае, когда чувствительные нервы ее перерезаны
(Магнус и де Клейн, 10). Этот тонус по своему составу очень сложен; он
включает многочисленные лабиринтные и шейные тонические рефлексы,
которые возникают вследствие возбуждения тонических
координирующих аппаратов под влиянием раздражений, производимых мозговым
разрезом, и других раздражений, связанных с операцией на голове
(Беритов, 22).
Существенную роль играет сильное раздражение красных ядер.
Известно, что перерезка мозга позади красных ядер, т. е. разрыв пирамидных
путей и удаление красных ядер, не приводит к развитию достаточно
сильной децеребрлционной ригидности. Для этого требуется
повреждение красных ядер, перераздражение их, как это, например, случается
при перерезке мозга через передние холмы, где лежат эти ядра (Кюре
и сотр., 20; Тоуэр и Петере, 39). Очевидно, децеребрационная
ригидность обусловливается в первую очередь теми нервными импульсами,
которые периодически возникают в перерезанных или поврежденных
нервных центрах и нервных волокнах.
По данным Эдриана (36), чувствительные волокна
млекопитающих бывают особенно активны после перерезки. Возникающие в них
после перерезки разряды импульсов убывают в течение нескольких іминут,
но вновь возобновляются при малейшем изменении в окружающих
условиях, например, при смачивании, струе теплого или прохладного
воздуха, сотрясении. В болевых нервных волокнах, в случае их
повреждения, импульсы небольшой частоты продолжаются часами.
В условиях децеребрации происходит перерезка громадного количества
чувствительных нервных волокон как в связи с перерезкой мозгового
ствола, так и вследствие вскрытия черепа, операции на шее и т. д.
Очевидно, все эти нервы дают разряды импульсов, действующих на
близлежащие тонические центры и тем обусловливающих их в зысшей
степени активное состояние. С течением времени эти нервные разряды от
повреждения ослабевают, и, очевидно, вследствие этого ослабевает и
ригидное состояние животного. Децеребрационная ригидность
получается также при анемии передней части большого мозга, которая вы-
20
И. С. Беритов
306 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
зывается путем одновременной перевязки сонных и базальной артерий
(Поллок и Дэвис, 27). Но и в этом случае причина ригидности должна
быть та же, что и при перерезке мозга. Удушение передней части
головного мозга в связи с анемией должно быть источником
значительных раздражений для нисходящих нервных путей, оканчивающихся в
тонических центрах.
Интересно отметить, что разгибательный децеребрационный тонус
не возникает или выражен слабо, если разрез мозга произведен впереди
красных ядер, в области зрительных чертогов.
Таким образом, нормальная функциональная деятельность красного
ядра мешает развитию децеребрационного разгибательного тонуса (Ра-
демакер и Магнус, 30). Повидимому, красные ядра играют
существенную роль в распределении тонуса в организме и возбудимости в спи-
нальных центрах.
Отношение мозжечка к тонусу. Тоническая деятельность средних
отделов головного мозга регулируется из мозжечка. Это доказывается
новейшими опытами с удалением мозжечка и с электрическим
раздражением коры мозжечка. Впервые Дюссер де Баренн (40)
показал, что удаление мозжечка у собак ведет к усилению экстен-
зорного тонуса конечностей и туловища. Затем это было
подтверждено другими авторами в отношении собак и других животных (Бремер
и Лей (41); Кунстман и Орбели (42); Попов (43) и др.). При этом
установлено, что этот усиленный тонус не является результатом
раздражения вследствие операции, ибо повышенный экстензорный тонус
сохранялся в продолжение 10 лет (Столанд и сотр., 44). Отсюда было
сделано заключение, что мозжечок оказывает на тонус скелетных мышц
постоянное тормозящее действие.
Опыты с раздражением мозжечка электрическим током выявили
характер тормозящего действия на тонус скелетных мышц. Сначала
Шеррингтон (46), затем Бремер (47) и др. показали, что
электрическое раздражение мозжечка производит торможение децере-
брационной ригидности, причем чем лучше функциональное состояние
мозжечка, тем значительнее тормозящее действие (Миллер и Бран-
тинг, 47). Это тормозящее действие обнаруживается лучше всего при
раздражении передней части мозжечка. Оно вовсе не имеет места при
раздражении полушарий (Бремер, 46).
Удаление мозжечка не препятствует вызову тонических рефлексов
(Беритов и Магнус, 48). Наоборот, оно усиливает дец?ребрационную
ригидность и благоприятствует наступлению шейных и лабиринтных
тонических рефлексов (Поллок и Девис, 27) (рис. 136).
Своеобразно действует в отношении тонуса мышц отравление коры
мозжечка стрихнином. Стрихнинное отравление одного полушария
мозжечка вызывает усиление тонуса скелетных імышц на
соответствующей стороне тела (Миллер, 19 и др.). Это явление объясняется
тем, что стрихнин не усиливает спонтанную активность в коре
мозжечка, а, наоборот, ослабляет ее, что, конечно, в свою очередь, ослаб-
ИОНИЧЕСКИЕ РЕФЛЕКСЬі
307
ляет постоянное тормозящее действие коры мозжечка на тонус мышц
(Доу, 50). Наоборот, раздражение коры мозжечка электрическим
током производит усиление быстрых биоэлектрических потенциалов
спонтанной активности и параллельно вызывает усиленное
торможение тонуса мышц (Доу, 50) (рис. 137). Отсюда было сделано заклю-
Рис. 137. Влияние спонтанной активности мозжечка на тонус скелетной
мускулатуры.
Наверху осциллограмма от передней доли мозжечка, внизу—механограмма (изометрическое
сокращение) мышцы-разгибателя (т. plantaris). Раздражается передняя доля мозжечка электрическим
током. А—до раздражения; J?—сейчас же после раздражения: при этом электрическая активность
значительно повышена, а напряжение сильно подавлено—механограмма опустилась ниже
записывающей бумаги; С — через 7 сек. электрическая активность мозжечка сильно ослабла, а
напряжение мышцы поднялось высоко; Z>—через 60 сек. в период стабилизации электрической активности
и тонуса (Доу).
•
чение, что быстрые электрические потенциалы мозжечка как раз
соответствуют тем импульсам возбуждения, которые производят
торможение тонуса мышц. Нужно думать, что это торможение
осуществляется путем усиленной активации нейропиля тонических центров
импульсами возбуждения, идущими из мозжечка. При этом здесь
возникают медленные потенциалы, производящие анэлектротонич?ское
угнетение нервных кругов тонических центров.
Итак, мозжечок в норме выполняет тормозящую функцию по
отношению к тонусу скелетной мускулатуры. Эта функция, повидимому,
осуществляется путем распространения импульсов возбуждения из
мозжечка к нейропилю тонических центров. Все те симптомы, которые
наблюдаются у безмозжечковых животных и у человека — атаксия,
дисметрия, асинергия и др.— хорошо объясняются выпадением или
ослаблением этой функции.
Сгибательный тонус децеребрированноіго препарата. На децеребри-
рованных препаратах, кроме описанного разгибательного тонуса, можно
наблюдать в определенных условиях также сгибательный тонус,
развивающийся вслед за разгибательным тонусом после его ослабления
и исчезновения. Сгибательный тонус представляет рефлекторный акт, по
своему внешнему проявлению совершенно противоположный разгиба-
тельному тонусу (Беритов, 2). Он также складывается из шейных и
лабиринтных сгибательных тонических рефлексов и вызывается
раздражением поперечного разреза мозга. Сделанные в последнее время на
хронически оперированных собаках наблюдения показывают, что экстен-
зорный тонус через несколько дней непременно сменяется тонусом фле-
20*
303 ОЁЩ\Я ФИЗИОЛОГИЯ СПИЙНОГО^МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГ4
ксорным. В дальнейшем флексорный тонус остается навсегда, экстен-
зорный же появляется только временами, в связи с определенным
положением головы (Бацет и Пенфильд, 12).
Экстензорная и флексорная ригидность спинномозгового препарата.
На кошках и собаках спустя полчаса или более после перерезки
под продолговатым мозгом и после прохождения наркоза развивается
экстензорная ригидность. Она проявляется на задних ногах гораздо
п5
Рис. 138. Различные положения го лавы при
вращении животного вокруг битемпоральной оси.
При положении 360° (0) и 180° экстензорный тонус
минимальный. При положениях в пределах 65—100°, а
также при положении от 295 до 255° обычно наблюдается
усиленный экстензорный тонус (Радемакер иГоугерверф).
сильнее, чем на передних, и выражается в активном сопротивлении при
пассивном сгибании. Она бывает настолько значительна, что животное
может стоять на выпрямленных ногах (Филиппсон, 34).
Эта экстензорная ригидность, несомненно, зависит от раздражений,
идущих от поперечного разреза мозга; в случае отсутствия ригидности
ее можно вызвать механическим раздражением разреза мозга,
например, прикосновением. Развитию ригидности благоприятствуют
периферические раздражения определенных рецептивных полей,
именно — кожи на хвосте, между бедрами, на брюхе. Эти раздражения
могут вызвать на задних ногах экстензорную ригидность при любом
положении спинномозгового препарата. Экстензорная ригидность
наблюдается и на лумбальных препаратах. Очевидно, экстензорная ригидность
задних ног спинномозгового препарата всецело обусловливается
деятельностью разгибательных координирующих центров поясничной части
спинного мозга; эти центры возбуждаются в данном случае раздраже-
ТОНИЧЕСКИЕ РЕФЛЕКСЫ
309
ниями из поперечного разреза спинного мозга, а нормально —
раздражениями кожи на хвосте, между бедрами и на брюхе.
На спинномозговых препаратах также наблюдается флексарная
ригидность. Она обнаруживается на задних ногах, когда препарат
лежит брюхом на столе, с отведенными в стороны бедрами. При спинном
или боковом положении, а также когда препарат удерживается в
воздухе, флекссрная ригидность отсутствует. Она зависит исключительно
от пролриоцептивных и экстероцептивных (т. е. кожных) раздражений
Рис. 139. Лабиринтные тоническиегрефлексы децеребриро-
ванной кошки при разных положениях головы.
А — при нулевом положении, максимально неблагоприятном для экс-
тензорного тонуса; В — при максимально благоприятном положении
для этого тонуса; голова обращена вверх от 'Гнулевого положения на
J 85° (Радемакер и Гоугерверф).
задних конечностей в рецептивном поле огибания. Характерно, что
пассивное движение передней части животного определенным образом
влияет на эту ригидность; поднятие ее усиливает ригидность, а
опускание, наоборот, ослабляет; вращение передней части животного вокруг
продольной оси вызывает усиление флексорной ригидности на одной
задней ноге, а именно на той стороне, к которой поворачивается спина,
а на другой задней лапе — ослабление ее. На этой стороне флексорная
ригидность может даже замениться экстензорной (Дюссер де Баренн
и Косков, 35).
Лабиринтные тонические рефлексы. Так называются своеобразные
тонические ірефлексы, вызывающиеся при изменениях головы в
пространстве. Эти рефлексы подробно были изучены Магнусом и де
Клейном (1, 10, 13) на децеребрированных препаратах кошки, собаки и
кролика. Они установили следующее: когда положение головы животного
таково, что нижняя челюсть обращена вниз и плоскость ротовой щели
составляет с горизонтальной плоскостью угол около 45° (на рис. 138 это
положение обозначено нулем), тогда описанный выше экстензорный
тонус децеребрированного препарата является наименьшим, иногда
даже заменяется флексорным тонусом (рис. 139 Л). Если поворачивать
животное, исходя из указанного положения, вокруг битемпоральной оси
310 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
в ту или другую сторону, экстензорный тонус нарастает. По позднейшим
исследованиям учеников Магнуса, максимальный экстензорный тонус
при вращении головы в пространстве наступает два раза, т. е.
существует два положения, благоприятствующие экстензорному тонусу: при
положении головы 100—125° и 295—255° (рис. 139 В и 140 В).
Существуют также два положения для іминимального экстензорного тонуса.
Одно из этих положений было обнаружено еще Магнусом: это то
Рис. 140. Лабиринтные тонические рефлексы децереб-
рированной кошки при разных положениях головы.
А и О — при нулевом положении; В и D — при положении
головы в 285°, максимально благоприятном для экстензорного
тонуса. При С и D пробуется степень экстензорного тонуса на
пассивное сгибание (Радемакер и Гоугерверф).
положение, когда ротовая щель опущена вниз и составляет с
горизонтальной плоскостью угол в 45°. Если это положение с минимальным
разгибательным тонусом обозначить нулем, то другое такое положение
наступает при угле около 180°. Как івидно на рис. 139 и 140, при
минимальных положениях передние ноги расслаблены. При максимальном
положении (В), когда голова повернута от нулевого положения на 85°,
на конечностях наступает сильная экстензорная ригидность. Выраженная
ригидность наблюдается и в том случае, когда голова повернута на 285°
от того же нулевого положения. На рис. 140 дается снимок
пассивного сгибания и разгибания конечностей при различных
положениях головы. В минимальном положении передние ноги свободно
подвергаются пассивному сгибанию (С), в максимальном же они не могут
быть согнуты (D).
Наличие двух минимальных и максимальных положений головы
было установлено не только путем изучения напряжения мышц, но и
путем регистрации биоэлектрических токов при тоническом возбужде-
ТОНИЧЕСКИЕ РЕФЛЕКСЫ
311
л тт
.,*>:
- „„„.„у
*Ч-?4, v V С «
нии разгибателей и сгибателей передней конечности. Наименьшие токи
получаются при нулевом положении и при 180°. Они сильно нарастают
при поворачивании головы от 65° до 110—125°, а затем при поворачивании
от 295° до 235°. При этом токи возбуждения появляются и нарастают
не только на разгибателе, но и
на сгибателе, однако на
последнем всегда значительно слабее
(Радемакер и Гоугерверф, 31).
На рис. 141 показаны токи
возбуждения, возникающие при
вращении децеребрированной кошки
от 0 до 180°. Аналогичная
картина получается при вращении от
180° до 360°.
Лабиринтные изменения
тонуса конечностей наступают также
при вращении головы вокруг
фронто-окципитальной оси.
Когда голова опущена вниз или
повернута вверх и1 притом
сохраняет точно медиальное
положение, это соответствует
минимальному положению для экстензор-
ного тонуса. Если же
поворачивать голову вокруг
фронто-окципитальной оси влево или вправо
от медиального положения, то
получается усиление экстензор-
ного тонуса децеребрированного
препарата. Это бывает при
поворачивании в пределах от 80° до
110°. Следовательно, когда
животное лежит на боку, то это
создает условия, благоприятные
для экстензорного тонуса
(Радемакер и Гоугерверф, 31).
Лабиринтные рефлексы
относятся к тоническим, так как
продолжаются часами — в течение
всего времени, пока удерживается данное положение головы. Их
называют лабиринтными, так как они вызываются раздражением
лабиринтных рецепторов. Известно, что после разрушения лабиринтов
или перерезки VIII пары нервов эти рефлексы исчезают (Магнус и де-
Клейн, 13).
Координация лабиринтных рефлексов происходит в продолговатом
Рис. 141. Биоэлектрические токи
разгибателя m. tricepsJ(BepXHHH кривая) и
сгибателя т. biceps br. (нижняя кривая).
1 — при нулевом положении головы; 2 — при
положении 25°; з — при положении 45°; 4 — при
положении 75е (во всех этих положениях биотоки
очень слабые); 5 —при положении 85° токи заметно
сильнее; 6-—при положении 105° токи еще
сильнее; 7 — при положении 125° токи чуть слабее;
8 — при положении 145° токи совсем слабые; 9~
10 —при положении 165 и 180° токов нет вовсе
(Радемакер и Гоугерверф).
312 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Мозгу, именно там, где главным образом оканчивается п. vestibularis,
т. е. в ядре Дейтерса, находящемся в передней части продолговатого
мозга. Поэтому лабиринтные рефлексы не исчезают при удалении
всего головного мозга, вплоть до продолговатого, т. е. включая варолиев
мост и мозжечок (Магнус, 3).
Лабиринтные тонические рефлексы на шее и туловище. Под
влиянием лабиринтов находится, кроме конечностей, мускулатура шеи и
туловища. Если положение головы в пространстве таково, что оно
обусловливает разгибание конечностей, тогда, в соответствии с этим,
сокращаются все мышцы туловища и шеи, удерживающие их от падения
вследствие тяжести, т. е. мышцы, поднимающие шею и туловище.
Поэтому голова и хвост отгибаются назад к спине, спиіна горбится.
Наоборюгг, при том положении головы, которое вызывает на
конечностях минимальный разгибательный или сгибатсльный тонус,
сокращение мышц, поднимающих голову, ослабевает и даже совсем
прекращается. Зато сокращаются антагонистические мышцы, вследствие чего
голова пригибается, хвост опускается (Магнус и де Клейн). Каждый
лабиринт совершенно одинаково воздействует на тоническое состояние
обеих сторон. По разрушении одного лабиринта изменение положения
головы в пространстве вызывает на ногах такие же рефлексы, как
и при целости обоих лабиринтов. Однако тоническое действие
лабиринтов на шейную мускулатуру не является билатеральным, двусторонним:
каждый лабиринт влияет только на мускулатуру противоположной
стороны шеи. Это видно из того, что по разрушении правого лабиринта на
противоположной левой стороне шеи мышечный тонус выпадает.
Поэтому, если голова не фиксирована, она поворачивается направо, где
мышечный тонус остался без изменения. Координация этих рефлексов
происходит там же, где и координация лабиринтных рефлексов
конечностей, т. е. в ядре Дейтерса, недалеко от того места, где оканчивается
п. vestibularis (Магнус, 1).
Тонические рефлексы со стороны лабиринтов на глазные мышцы.
Изменение в положении лабиринтов в пространстве действует также
и на г'лазные мышцы. Когда изменяется нормальное положение головы,
происходит раздражение определенных лабиринтных рецепторов,
которое вследствие сокращения соответствующих глазных імышц вызывает
т&ксе изменение в положении глазного яблока, что сетчатка мак бы
стремится все время остаться в одной и той же плоскости. Но этот
лабиринтный тонический рефлекс никогда не достигает полностью этой
цели,— положение сетчатки в пространстве все-таки оказывается
сильно измененным. Например, если животному, держащему голову
в нормальном, горизонтальном, положении, нагнуть ее вниз так, чтобы
головаі по битемпоральной оси повернулась на 60°, тогда глаза
повернутся кверху только на 38°. Следовательно, сетчатка сдвигается в
отношении горизонта на 22°, т. е. полной компенсации не происходит. Если
голову, находящуюся в нормальном, горизонтальном, положении,
повернуть мордой вверх на 45°, так чтобы левый глаз оказался наверху, то
ТОНИЧЕСКИЕ РЕФЛЕКСЫ
313
этот последний повернется в своей орбите книзу, а правый, наоборот,
повернется кверху. Но и в этом случае достигается только частичная
компенсация, так как плоскость зрения сдвигается от горизонта на 25°
Было доказано, что описанные главные движения действительно
лабиринтного происхождения, так как после двустороннего разрушения
лабиринтов эти тонические рефлексы на глазных мышцах исчезают.
Понятно, что для получения тонических рефлексов на глазных
мышцах необходимо сохранить целость глазодвигательных ядер
в среднем мезгу. Для этих опытов нужно брать такой децеребрирован-
ный препарат, у которого головной мозг перерезан на передней границе
среднего мозга (Магнус и сотр., 14, 15, 16, 24).
У человека и у тех животных, у которых глаза расположены в
фронтальной плоскости и поле зрения одного глаза в значительной мере
перекрывает поле зрения другого, тонический рефлекс на глазных
мышцах очень незначителен. Эти рефлексы великолепно вызываются у
животных с боковым расположением глаз, где поле зрения одного глаза
не покрывает полу зрения другого. Таковы кролик и морские свинки,
на которых собственно и изучены эти рефлексы.
Иннервация глазных імышц в этих тонических рефлексах является
рецилрокной. При самых разных положениях животного в пространстве
верхняя и нижняя прямые мышцы почти всегда реагируют реципрокно.
То же самое следует сказать и относительно верхней и нижней косых
мышц; в редких случаях наблюдается исключение из этого правила
(Дюссер де Баренн и де Клейн, 32). Это положение оспаривает Л о-
ренте де Но (33). Он утверждает, что глазные движения не
обусловлены реципрокной иннервацией глазных мышц, но Дюссер
де Баренн и де Клейн находят, что соответствующее
наблюдение ЛорентедеНо является результатом несовершенной методики.
Сведения о лабиринтных рецепторах и способе их раздражения.
Внутреннее ухо состоит из двух частей: 1) улиткообразного
слухового аппарата, где помещается кортиев орган, и 2) статического
органа, который состоит из преддверия и трех полукружных каналов
(рис. 142). Лабиринтные рефлексы вызываются как раз из статического
аргана через преддвериый нерв (п. vestibularis). В статическом органе
существуют особые рецепторы, или чувствительные органы, maculae
utriculi et sacculi в преддверии и cristae в ампулах полукружных
каналов. Устройство этих органов одно и то же: складка-утолщение
перепончатой оболочки, на которой расположены клетки ресничного
эпителия. В макулах на реснички эпителия опираются кристаллы углекислого
кальция, так называемые отолиты. В кристах же реснички
свободно мерцают. Здесь также есть отолиты; они склеены между собой
клейким веществом и в большом количестве окружают реснички крист.
Вся совокупность отолитов и ресничек называется купулой (cupula).
Раздражение чувствительных клеток происходит следующим
образом. Когда меняется положение головы, меняется и положение купул
в силу тяжести. Купулы наклоняются в ту или другую сторону, и соот-
314 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
вепственно раздражается та или другая группа клеток. Очевидно, Піри
каіжідом положении головы будут раздражаться определенные
рецепторы, что, в свою очередь, влияет на деятельность координирующего
аппарата определенного тонического рефлекса. Раздражение соответ-
Sacc
pndol
Лтр апі —V-
R sup і
В cochlear is
Рис. 142. Схематическое изображение лабиринта и его
иннервация.
Центральное место рисунка занимает utriculus, на дне которого
находятся отолиты, изображенные черным контуром. Слева к utriculua
присоединяются ампулы горизонтального и переднего вертикального
полукружного канала, а справа, — ампула заднего канала. Кристы с купу-
лами в этих ампулах показаны схематически. Посередине utriculus в
него входит общее колено (cms communae) обоих вертикальных
каналов. Под utriculus находится sacculus. Их соединяет узкое колено
ductus endolymphaticus. Отолит расположен на медиальной стенке
sacculus. Он состоит из основной части (редкий штрих) и дорзальной
(частый штрих). Sacculus соединен узким каналом (ductus reuniens) с
улиткой. Иннервация рецепторов: проксимальный корешок VIII нерва,
пройдя gangl. proximaie Scarpae 2, заканчивается в виде ramus superior
в чувствительных элементах ампул горизонтального и переднего
вертикального полукружного каналов, затем в макуле utriculus; кроме
того, он дает боковую ветвь 4 к дорзальной стороне чувствительного
эпителия sacculus. Дистальная ветвь VIII нерва представлена ramus
inferior (черн.), который, пройдя через gangl. distale Scarpae 2,
заканчивается в основной части макулы sacculus и дает ветвь к ампуле заднего
вертикального канала. Кроме того, маленькой веточкой 5 он
присоединяется к дистальному корешку слухового нерва (п. cochlearis) (Бурлет).
ствующих рецепторов продолжается все время, пока сохраняется
существующее положение головы. Сообразно с этим и тонический
рефліекс удерживается на все это время.
Функции отолитных аппаратов. Благодаря исследованиям Маг-
н у с а и его учеников известно, что в возникновении лабиринтных
рефлексов шеи, туловища и конечностей наибольшее значение имеет
раздражение отолитами воспринимающих органов utriculus, т. е. макул.
Эти рефлексы исчезают только после повреждения utriculus. Что
касается компенсаторных движений глаз, то они зависят от раздражения
макул sacculus. Магнусом было доказано, что эти макулы раздра-
ТОНИЧЕСКИЕ РЕФЛЕКСЫ
315
жаются как при давлении сверху, так и при повисании на них отолитов.
Наибольшее раздражение получается в том случае, когда отолиты
находятся е горизонтальном положении и висят на макулах; минимум же
раздражения получается, когда в том же горизонтальном положении
отолиты лежат на макулах.
Такой же модус раздражения принят Магнусом и для макул
utriculus. Но и здесь необходимо отметить следующее: когда в saccu-
lus существующее положение должно обусловливать минимум
раздражения, чувствительный орган находится в состоянии покоя. То же
происходит и в utriculus в отношении компенсаторных движений глаз,
но по отношению к шее, туловищу и конечностям положение
минимального раздражения не означает отсутствия возбужідейия. Даже
и при таком положении отолитов макулы раздражаются, вследствие
чего развивается тоническое сокращение на шее, туловище и
конечностях; это тоническое сокращение распространяется на сгибательные
мышцы конечностей и мышцы, поднимающие шею и туловище (17)
Экспериментальное исследование отолитных функций — дело
позднейшего времени. За 1919—1924 гг. из лаборатории Магнуса вышел
ряд работ по этому вопросу. До того существовало много гипотез
о деятельности отолитов; между прочим» считалось, что тонические
рефлексы конечностей вызываются раздражением рецепторов ампул,
т. е. деятельностью купул. Эта гипотеза была опровергнута прямыми
опытами: купулы не играют никакой роли в вызове тонических
рефлексов на конечностях.
Шейные тонические рефлексы. Шейными тоническими рефлексами
называются рефлексы, вызываемые при изменениях положения головы
по отношению к туловищу. Эти рефлексы также подробно были
изучены Магнусом и де Клейном на децеребрированных животных.
Они заключаются в следующем. Когда шея вращается вокруг
медиальной оси, наступает тонический (рефлекс одного типа: на лицевой
стороне, куда обращена морда животного, передняя и задняя ноги
тонически разгибаются, а на другой, теменной стороне, существующий
экстензорный тонус передней и задней ног ослабевает или совсем
прекращается; в определенных условиях экстензорный тонус заменяется
флексорныім. Этот рефлекс носит название шейного
тонического рефлекса вращения.
Когда шея наклоняется на правый бок, правая передняя и правая
задняя ноги тонически разгибаются, левые же тонически сгибаются,
т. е. рефлекс протекает так же, как и при вращении головы; этот
рефлекс носит название шейного тонического рефлекса
наклонения. Если шея отогнута назад, в сторону спины, на 90°
от медиальной линии, то вызывается другой рефлекс: обе передние
ноги усиленно разгибаются, а на обеих задних существующий разгиба-
тельный тонус ослабевает или совсем прекращается, часто заменяясь
сгибательным тонусом. При наклонении шеи вниз к груди на 90° от
медиальной линии наступает совершенно противоположный рефлекс:
316 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
передние ноги лишаются разгибательного тонуса и иногда принимают
сгибательный, а задние ноги усиленно разгибаются. Этот рефлекс
называется шейным тоническим рефлексом поднятия и
Опускания головы. Его можно наблюдать на кошке, собаке,
обезьяне и морской свинке. На кролике же при поднятии и отгибании
шеи назад к спине получается разгибание всех конечностей, при
наклонении же шеи вниз к груди все конечности сгибаются. Существует
еще один своеобразный шейный тонический рефлекс, который можно
вызвать надавливая на остистый отросток VII шейного позвонка (ver-
Рис. 143. Шейные тонические рефлексы на поднятие и опускание шеи.
А — нормальная кошка. Когда кошке сверху показывают мясо, передние ноги у нее разгибаются,
а задние несколько сгибаются. В — нормальная кошка. Кошке показывают мясо снизу. Вследствие
опускания шеи вниз передние ноги согнуты, а задние разогнуты. С — кошка, лишенная шейных
рецепторов. Перерезаны шейные задние корешки I, II, III. При показывании мяса кошка поднимает
голову кверху, однако передние ноги не разгибаются (Магнус и Левеи).
tebra promiinens): этот рефлекс заключается в сгибании всех
конечностей. Магнус назвал его рефлексом vertebra prominens.
Описанные выше рефлексы возникают вследствие раздражения рецепторов
мышц, сухожилий и суставов шеи и продолжаются до тех пор, пока
удерживается данное положение шеи. Все они, за исключением
рефлекса vertebra prominens, возникают при раздражениях в области
распространения первых трех шейных задних корешков. После перерезки
означенных корешков все шейные рефлексы, кроме последнего, уже не
вызываются (Магнус и Левен, 18, 16) (рис. 143).
Влияние изменения положения головы по отношению к туловищу
не ограничивается одним только шейным тоническим рефлексом.
Одновременно изменяется положение головы в пространстве и потому,
наряду с шейным тоническим рефлексом, наступает лабиринтный
рефлекс. Если, например, животное лежит на правом боку, то<
вращение шеи в левую сторону мордой вверх вызовет, во-первых, шейный
рефлекс на вращение, т. е. разгибание обеих левых ног и сгибание
обеих правых, и, во-вторых, лабиринтный рефлекс с разгибанием всех
ног (рис. 144).
Координация шейных рефлексов происходит в первых трех
сегментах шейного отдела мозга, т. е. там, куда входят чувствительные
волокна от рецептивных полей этих рефлексов. Это видно из того, что
тоНріческие рефлексы 3-17:
после удаления всего головного мозга указанная [рефлекторная
деятельность все же не исчезает (Магнус, 3).
Тонические рефлексы с шеи на глазные мышцы. Если голову
кролика зафиксировать в медиальном положении, а туловище
постепенно выводить из этоіго положения, то станет изменяться и
положение глаз. Это явление впервые обнаружил Б а р а н и (26). Он же за-
г sec
Рис. 144. Шейные и лабиринтные тонические рефлексы.
Децеребрированная кошка. Записывается пара
антагонистических мышц правой передней ноги: сгибатель
локтевого сустава m. biceps Ьг. (верхняя кривая) и разгибатель
его m. triceps Ьг. (нижняя кривая). Препарат укреплен
на левом боку, поэтому ротовая щель находится в
вертикальной плоскости. Когда голова поворачивается
нижней челюстью вверх (FP), разгибатель резко сокращается.
Этот рефлекс представляет комбинацию шейного и
лабиринтного разгибательных тонусов, так как при данном
вращении шеи положение головы таково, что оно
благоприятствует лабиринтному экстензорному тонусу. Когда
голова поворачивается в противоположном направлении
(нижняя челюсть обращена вниз — SP), тоническое
сокращение разгибателя ослабевает наиболее сильно. Это
тоже результат двоякого рода рефлексов: 1) сгибатель-
ного шейного рефлекса на вращение теменем направо и
2) сгибательного лабиринтного рефлекса соответственно
нулевому положению головы. При среднем положении
головы (ротовая щель в вертикальной плоскости—MP)
разгибатель показывает некоторое тоническое
сокращение чисто лабиринтного происхождения, так как в это
время положение головы в отношении туловища является
нейтральным. Время — в секундах.
ключил, что эти глазные рефлексы — шейного происхождения. Однако
его заключение не вытекало само собой из собранного им фактического
материала. Этот вопрос был основательно изучен де Клейном (16)
опять-таки в лаборатории Магнуса. По исследованиям де Клей-
н а, каждому положению головы в отношении туловища соответствует
определенное положение глазного яблока.
Если фиксировать голову кролика в медиальном положении, то
передвижением туловища, т. е. вращением шеи в ту или другую
сторону при неподвижном положении головы, можно вызвать следующие
рефлексы:
1. Если согнуть туловище в горизонтальной плоскости (по, дорзо-
вентральной оси) в какую-либо сторону, например вправо, то оба
глаза поворачиваются влево: справа сокращается m. ,reofcus internus,
слева — т. rectus externus.
2. Если вращать туловище по фронтальной оси, то при поднятии
спины вверх глазное яблоко поворачивается вниз вследствие
сокращения m. obliquus inferior. При опускании же туловища вниз (приближе-
318 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
нием к челюсти) оба глаза поворачиваются вверх вследствие
сокращения обоих mm. obliqui superiores.
3. Если повернуть туловище по сагиттальной оси спиной вверх,
глаз повернется по вертикали вниз. Если же животное окажется
грудью вверх, то и глаз повернется по вертикали вверх.
Движения глаз не обладают при этом большой амплитудой,
необходимо значительно повернуть туловище, для того чтобы получить
заметное изменение их положения.
Рис. 145. Шейный тонический рефлекс на 9-летнем ребенке.
На рис. А лицо ребенка повернуто налево. В соответствии с этим на
левой стороне верхняя и нижняя конечности разогнуты во всех главных
суставах, а на правой стороне, куда обращено темя, все главные
суставы согнуты. На рис. В голова повернута лицом вправо. Здесь правые
верхняя и нижняя конечности разогнуты, а левые согнуты (Магнус).
Лучше всего эти рефлексы наблюдать на препаратах с обоими
разрушенными лабиринтами. В этом случае вращение головы в ту или
другую сторону вызовет движение глаз, если только туловище
фиксировано.
Тонические рефлекторные движения глаз, так же как и остальные
нормальные глазные движения, являются реципрокными; например,
когда сокращается m. rectus interims, то m. rectus externus
расслабляется, тормозится. Рецептивное поле этих рефлексов то же, что и для
шейных тонических рефлексов на конечностях; следовательно, оно состоит
из рецепторов шейных мышц, суставов и сухожилий, чувствительные
нервы которых входят в спинной мозг через первые три шейные задние
корешка. Координирующий же аппарат, подобно другим шейным
тоническим рефлексам, должен находиться в первых трех шейных
сегментах:
тонические рефлексы
319
Шейные и лабиринтные тонические рефлексы на нормальных
животных и на человеке. Постановка соответствующих опытов на
нормальных животных очень затруднительна. Животное оказывает активное
сопротивление пассивному изменению положения головы: нужно івзять
ее руками и употребить некоторое усилие, чтобы ее повернуть. На
хорошо прирученных животных, как, например, на кроликах,
производство таких опытов все же возможно. Можно поставить эти опыты
также на котятах, на детях-идиотах (Магнус и сотр., 18, 21). Для
примера на рис. 145 мы приводим шейный тонический рефлекс на
вращение головы на ребенке 9 лет, у которого большой мозг с рождения
был лишен своих функций. Но и на каждом взрослом и совершенно
нормальном животном можно обнаружить наличие шейных тонических
рефлексов во время их обычных поведенческих актов. Например, если
кошке или собаке показать пищу сверху, они поднимают голову вверх
и в то же время сильно разгибают передние ноги, а задние сгибают
(рис. 143 Л). Это шейный тонический рефлекс на поднятие шеи. Если
же показать пищу снизу, они опускают голову, одновременно сгибают
передние ноги и несколько разгибают задние (рис. 143 В). Это
тонический рефлекс на опускание шеи. Когда кошка ползает под шкафом,
у нее нет возможности стоять на выпрямленных ногах: она
передвигается с согнутыми ногами. Это сгибание есть тонический рефлекс
с vertebra prominens, возникший вследствие трения остистого отростка
этого позвонка о дно шкафа. На спящем человеке также легко
наблюдать шейный тонический рефлекс. Например, если у спящего ребенка
голова наклонена вправо, правая рука будет разогнута, а левая
согнута.
На здоровых взрослых людях не удается наблюдать шейные и
лабиринтные тонические рефлексы в связи с пассивным или активным
изменением положения головы. Но в последнее время было обнаружено
изменение возбудимости и напряжения мышц под влиянием положения
головы. Так, например, наклонение головы вперед вызывает
уменьшение мышечного напряжения, а отклонение назад — его повышение.
Характерное изменение претерпевает диафрагма человека: при повороте
головы повышается тонус диафрагмы с той стороны, в которую
поворачивается голова. То же наблюдается у децеребрированных животных
(Крестовников и сотр., 38).
Основное условие для вызова шейных и лабиринтных тонических
рефлексов. Магнус и его ученики утверждали, что шейные и
лабиринтные рефлекоы вызываются непосредственно раздражением
соответствующих рецепторов шеи и лабиринтов, но это обычно происходит
только при сильных раздражениях рецепторов — при быстром
вращении или же при искусственном раздражении этих рецепторов, например,
при раздражении рецепторов лабиринта горячей водой. Если же
изменение положения головы производить очень осторожно, чтобы не
причинить животному значительных раздражений, соответствующий
тонический рефлекс не вызывается. Он наступает только в том случае, если
320 ОБЩАЯ ФИЗИОЛОГИЯ СіТИЙНОГО МОЗГА И СТВОЛА ГОЛОВНОГО ЗГОЗГА
экспериментатор крепко сжимает голову животного при совершении
пассивного движения шеи, вследствие чего получается раздражение шеи
или головы, а также в случае, если экспериментатор производит
механическое раздражение какого-либо участка кожи на теле животного:
щипанием хвоста или ушей, пассивным сгибанием и разгибанием ноги,
раздражением чувствительного нерва. Иногда бывает достаточно
провести рукой ло шерсти животного, а часто произвести и совсем
незначительные внутренние и внешние раздражения, чтобы вызвать
тонический рефлекс,
соответствующий данному положению
головы (рис. 146). Отсюда
следует, что раздражение
шейных и лабиринтных
рецепторов только благоприятствует
деятельности определенных
координирующих аппаратов в
смысле повышения их
возбудимости; что же касается вы
зова этой деятельности,
приведения координирующих
аппаратов в активное состояние,
то это зависит от разного рода
побочных раздражений.
Значит, периферические
импульсы, действующие на
координирующие аппараты со стороны
рецепторов шейных и
лабиринтных рефлексов, носят
преимущественно
субминимальный характер. Эти импульсы
не вызывают рефлекса, т. е. возбуждения координирующего аппарата;
они только повышают его возбудимость; вследствие этого каждому
положению головы должно соответствовать повышение возбудимости
определенного координирующего аппарата. Если в это время
происходит какое-либо другое более или менее значительное раздражение, то
вызванное им возбуждение, иррадиируя по центральной нервной
системе, достигает также тонического центра с повышенной
возбудимостью и приводит его в активное состояние (Беритов, 2, 28).
Продолжительность тонического рефлекса, вызванного побочным
раздражением при данном положении головы, зависит от
продолжительности этого раздражения. Тонический рефлекс всегда имеет
последействие, которое тем продолжительнее, чем сильнее побочное
раздражение. Понятно, что если это раздражение слабо и кратковременно,
тонический рефлекс продолжается недолго, несмотря на положение
головы, благоприятное для данного рефлекса. Отсюда следует, что
характерное течение тонических рефлексов, между прочим, обусловли-
Рис. 146. Тонические рефлексы в ответ на
внешнее раздражение.
Децеребрированная кошка. Животное лежит на
правом Ооку. Верхняя кривая принадлежит разгибателю
колена vasto-crureus, нижняя — сгибателю колена
semitendinosus. В опыте А положение головы
благоприятствует тоническому сгибанию левой ноги; в
опыте В—-тоническому разгибанию. В обоих опытах
раздражается ушная раковина. В первом опыте
раздражение вызывает сгибание, а во втором
разгибание. То и другое положение головы устанавливалось
путем вращения шеи без вызова тонического рефлекса.
ТОНИЧЕСКИЕ РЕФЛЕКСЫ
321
•вается продолжительностью и интенсивностью побочных раздражений.
Для вызова тонических ірефлексов на децеребрированных препаратах
обычно бывает достаточно одного более или менее быстрого
насильственного изменения положения головы. Тонический рефлекс наступает
непосредственно вслед за этим изменением. Происходит эта не просто
вследствие раздражения рецепторов шеи и лабиринтов, а потому чтоде-
церебрированные препараты имеют большую рану на шее и на голове,
являющуюся источником интенсивных раздражений. Сам экстензорный
тонус децеребрированного препарата зависит от того, что
координирующие аппараты! тонических рефлексов беспрерывно возбуждаются
раздражениями, исходящими из раневых поверхностей. Если на
децеребрированных препаратах господствует сначала экстензорный тонус, а
затем флексорный, то это потому, что сначала берет верх деятельность
экстензорных координирующих аппаратов, а затем уже флексорных.
Следовательно, во время экстензорного децеребрационного тонуса
изменение положения головы вызывает лабиринтные и шейные тонические
рефлексы непосредственно в силу того, что каждое новое положение
головы повышает возбудимость в определенных координирующих
аппаратах и тем благоприятствует возникновению усиленной деятельности
их под влиянием раневых раздражений.
Практикум
Наблюдение шейных и тонических рефлексов на децеребрированных кошках.
Методика децеребрации была описана в предыдущей главе. Опыты с тоническими
рефлексами производятся через 1—2 часа после операции, когда произойдет более или
менее значительное восстановление рефлекторной деятельности. При этих опытах
препарат совершенно освобождают от веревок, которыми он был привязан к
операционному столу.
Опыты над шейными тоническими рефлексами, а) Животное лежит на спине в
медиальном положении. Вращают голову около медиальной оси — один раз мордой
направо, другой раз налево. При этом наступают шейные тонические рефлексы на
вращение шеи.
б) Животное находится в том же положении. Поворачивают голову около
вертикальной оси к туловищу сначала в одну сторону и наблюдают состояние конечностей,
затем в другую сторону. При этом наступают шейные тонические рефлексы на
наклонение.
в) Животное находится в том же положении. Сгибают голову вниз вентрально,
наблюдают состояние конечностей; затем отклоняют назад дорзально. При этом
наступают шейные тонические рефлексы на вентральное и дорзальное сгибание головы,
т. е. на ее опускание и поднятие.
г) Следующий раз повторяют те же опыты при боковом положении животного.
Результаты будут во многих случаях те же, но может быть и значительная разница.
При указанных (изменениях положения головы по отношению к туловищу ме*
няется также положение головы в пространстве. Это, в свою очередь, вызывает
лабиринтные тонические рефлексы. Следовательно, шейные тонические рефлексы обычно
наступают совместно с лабиринтными. А в связи с разным исходным положением
животного одно и то же вращение, поворачствакие или дорзальное и вентральное сгвба-
ние головы, дает разные изменения положения головы в пространстве. Это, конечно,
влияет на характер наступающих тонических реакций. Так, при спинном положении
21 И. С. Беритов
ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
вентральное и дорзальное сгибание головы изменяет ее положение в пространстве.
Следовательно, в этом случае должен получиться не только шейный тонический
рефлекс, но и лабиринтный. При дорзальном сгибании, когда ротовая щель опущена
вниз и образует с горизонтальной плоскостью угол около 45° (на рис. 138 это будет
при 270°) наступает значительный лабиринтный разгибательный рефлекс.
Такой же лабиринтный экстензорный тонус будет иметь место при сильном
вентральном сгибании, когда плоскость ротовой щели образует с горизонтальной
плоскостью угол около 135° (по рис. 138 это будет при 90—100е). Когда же вентральное
сгибание головы невелико, а именно ротовая щель образует над горизонтальной
плоскостью угол 45° (рис. 138), этот экстензорный тонус должен отсутствовать.
Опыты над лабиринтными тоническими рефлексами. Лабиринтные тонические
рефлексы можно наблюдать при вращении всего животного вокруг битемпоральной
Рис. 147. Тонические рефлексы на децеребрированной лягушке после перерезки
VIII нерва правой стороны.
А—типичное положение, которое принимает лягушка: голова наклонена теменем на правую
сторону; это положение вызвано выпадением тонуса в шейных мышцах левой стороны после перерезки
правого VIII нерва. Кроме того, на правой стороне передняя и задняя конечности согнуты, а на
левой разогнуты. Это—тонический шейный рефлекс, возникший вследствие наклонения головы
теменем направо. В—та. же лягушка после слабого механического раздражения. На левой стороне
разгибательный тонус еще более усилился. С—голова приведена в медиальное положение. Вместе
с этим и конечности приходят в симметричное положение.
или фронто-окципитальной оси. Для этой цели животное прикрепляют к доске так,
чтобы оно не падало на бок при вращениях, зат-ем вращают доску вместе с животным
вокруг битемпоральной или фронто-окципитальной оси.
Роль посторонних раздражений в вызове лабиринтных и шейных тонических
рефлексов. Шейный тонический рефлекс наступает сразу, тотчас же после изменения
положения головы, только в том случае, если это изменение производится быстро, почти
порывисто. Если же изменять положение головы медленно и с большой осторожг
ностью, чтобы при этом не производить более или менее значительных побочных
раздражений, то тонический рефлекс не наступит или будет выражен очень слабо. В этих
случаях пассивное сгибание и разгибание конечностей или раздражение кожи
произведут значительный тонический рефлекс, отвечающий данному положению головы.
Наблюдение шейных и лабиринтных тонических рефлексов на децеребрирован-
ных лягушках. Вскрывают ножницами черепную коробку и перерезают мозг на уровне
среднего. Всю переднюю обрезанную часть мозга удаляют и в черепную коробку
вкладывают вату. После этой операции сбоку продолговатого мозга тонким и
заостренным ланцетом перерезают с одной стороны VIII нерв. При этом перерезается
и п. vestibularis как составная часть VIII нерва. От этого шейные мышцы
противоположной стороны теряют тонус, и голова лягушки поворачивается теменем в сторону
перерезанного нерва. Одновременно обе конечности этой стороны, в особенности
передняя, тонически сгибаются, а обе лапки на другой стороне разгибаются. Тоническая
асимметрия конечностей представляет собой шейный тонический рефлекс от поворота
головы. Это видно из того, что если насильственно придать голове медиальное
положение, то тоническая асимметрия исчезает (де Клейн, 23) (рис. 147).
ТОНИЧЕСКИЕ РЕФЛЕКСЫ
323
На этом препарате можно убедиться в том, что изменения положения головы
недостаточно для того, чтобы вызвать тонический рефлекс: если голове придать вполне
медиальное положение и этим самым уничтожить существующий асимметрический
тонус на конечностях, а потом осторожно повернуть голову к поврежденной стороне»
то конечности продолжают оставаться в симметрическом положении. Но стоит к этому
присоединить раздражение какой-нибудь конечности, туловища или головы, как
конечности, вследствие возникновения шейного тонического рефлекса, тотчас же примут
асимметрическое расположение.
Ли тература
1. Magnus R., Munch. Med. Wschr., 1912; 681. Korperstellung, Berlin, 1924.
2. В e г i ? о f f J., J. Physiol. 49, 147, 1915; Изв. Росс. Акад. Наук, стр. 649, 1915.
3. Magnus R., Pfl. Arch , 159, 224, 1914.
4. Biedermann W., Pfl. Arch., 80, 420, 1900.
5. Brondgeest, Arch. Anat. Physiol., стр. 704, 1860.
6. Beritoff J., Arch. Anat. Physiol., стр. 1,1913, Изв. СПб. биол.лаб., 12, 29,1912.
7. Беритов И., Учение об основных элементах центральной координации
скелетной мускулатуры. Пгд., стр. 97, 1916
8. Sherrington С, J. Physiol., 22, 319, 1897.
9. Beritoff J., Quart J. Exp. Physiol., 9, 199, 1915; Изв. Росс. Акад. Наук,
стр. 854, 1915.
10. Magnus R. u. de Kleyn A., Pfl. Arch., 145, 455, 1912.
11. Sh er ringt on C, J. Physiol., 47, 196, 1913.
12. В a z e 11 a. P e n f i e 1 d, Brain, 45, 185, 1922.
13. M a g n u s u. de Kleyn, Pfl. Arch , 158, 178, 1913.
14. V a n der H о e v e u. de К 1 e у n, Pfl. Arch., 169, 241, 1917; 178, 179, 1920.
15. Magnus u. de К 1 e у n, PfJ. Arch., 169, 241, 1917; 178, 179, 1920.
16. De Kleyn A.f Pfl. Arch., 186, 82,1921.
17. M a g n u s u. de К 1 e у n A., Pfl. Arch., 18*5, 6 и 61, 1921; 194, 407, 1922.
18. Magnus u. Storm van Leeuwen, Pfl. Arch., 159, 157, 1914.
19. Magnus u. de Kleyn A., Pfl. Arch., 147, 403, 1912.
20. Magnus u. de Kleyn A., Pfl. Arch., 154, 163, 1913.
21. Magnus u. de К 1 e у n A., Munch, med. Wschr., стр. 2566, 1913, Pfl. Arch.,
160, 429, 1915.
22. В e г i t о f f J., Pfl. Arch., 199, 248, 1923; Изв. Росс. Акад. Наук, стр. 1117, 1915.
23. De К 1 е у ii A., Pfl. Arch., 159, 218, 1914
24 Rademaker G., Klin. Wschr., 1, 40, 1923; Magnus u. Rademaker,
Schw. Neur. Psych., 13, 408, 1913.
25. R a d e m a k e г G. Die Bedeutung der roten Kerne und des ubrigen Mitelhirns
fur Muskeltonus, Korperstellung und Labyrinthreflexe. Berlin, 1926.
26. Bar an у R., Z. Physiol., 20, 298, 1917.
27. Pollock L. u. Davis L., J. Сотр. Neurol , 50, 377, 1930.
28. Б e p и t о в И., Мед. биол. журн., 4, в. 3—4, стр. 106, 1928; Acta oto-laryng,
12, 468, 1928.
29. KureK.,Shinosaki, Nagano u. Imagawa, Z. Ges. Exp. Med., 45,
246, 1925.
30. Magnus R. u. Rademaker G., Arch. Neur. Psych., IB, 408, 1923.
31. Rademaker G. et Hoogerwerf S., Arch. Neerl. Physiol., 16, 305.
32. D u s s e r d e В а г e n n e J. G. a. de К 1 e у n A., Acta oto-laryng., 16, 97,
1931.
33. Lorentede N6, Acta oto-laryng, 9, 163, 1926.
34. Philippson M. L'autonomie et la centralisation dans le systeme nerveux
des animaux. Bruxelles, 1905.
35. Dusser de Barenne a. Koskoff I., Am. J. Physiol., 107, 441, 1934.
21*
324 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
36. Adri an Е. D. The mechanism of nervous system, 1932, Механизм нервной
деятельности, Биомедгиз, 1935.
37. W а с h h о 1 d е г К., PfL Arch., 221, 66, 1928.
38. Крестовников А. Н. и сотр., Сб. докладов VI Всес физиол. съезда, стр. 337.
39. ThauerG. u. Peters G., PfL Arch., 242,54,1939.
40. Dusser de Barenne J. G., Alexander a. Marburg, Handb. d.
Neurol, des Ohres,l, 589, 1923.
41. В rem or F. etLey R., Bull. de. l'Acad. Roy, d. Med. d. Belgique, 60, 1927.
42. Кунстман К. и ОрбелиЛ., Физиол. Журн. СССР, 15, 549, 1932.
43. Попов Н. Ф., Сборн. «Высшая нервная деятельность», 1, 93, 1929.
44. Stoland О. О., Greer В. Н. a. Blood R., Amer. J. Physiol., 119, 410,
1937.
45. Sherington С. S., Shafer's Text book of physiology, 2, 884, 1900.
46. Bremer F., Arch. Internal. Physiol, 19, 189, 1922.
47. Miller F. R. а. В ran ting F. G., Brain, 45, 104, 1922.
48. Beritoff J. u. Magnus R., PfL Arch., 178, 124, 1914.
49. Miller F. R., Physiol. Rev., 6, 124, 1926.
50. Dow R. S., J. Physiol., 90, 67, 1938.
51. Гедевани Д., Физиол. журн. СССР, 19, 1042, 1935.
Глава XVII
РЕФЛЕКСЫ ПОЛОЖЕНИЯ И СТАТОКИНЕТИЧЕСКИЕ
РЕФЛЕКСЫ
Общее понятие. Нормальное животное обычно находится в точно
определенном положении: ногами, животом и мордой вниз, спиной и
теменем вверх. Таково нормальное положение каждого животного. Если
животное по какой-либо причине
вышло из этого положения, оно
сейчас же возвращается к нему
рефлекторным путем. Эти
рефлексы впервые были
исследованы в лаборатории Магнуса
(1914—1924) и были названы
им рефлексами
положения [1]. Нормальное положение
является исходным для каждого
акта поведения. Как видно из
опытов Магнуса и его
сотрудников, сохранение нормального
положения является сложным
рефлекторным актом. В нем уча- „ п лЛО т> *
г ^ r J Рис. 118. Рефлексы положения на голове
СТВует ряд рефлексов, ВО Глав- кролик без большого мозга. Животное удержи-
HVfn пппи нгпярт пагЪпоігп ТЭ1.ТОТ-Т вается в вертикальном положении головой вниз,
nytu риль шраеі реірлекц БЫЗЫ- л —лабиринты целы. Голова находится в нор-
рярмый ич гтятнііагіглгл лпгяио мальном положении: она повернута теменем вверх.
°йШМИ "^ чаіичеі шу орі ана. в __ дабиринты разрушены. Голова опущена
Координирующий аппарат ЭТОГО теменем вниз (Магнус).
рефлекса лежит в среднем мозгу.
У высших позвоночных после децеребрации рефлексы положения
исчезают, если только разрез прошел позади четверохолмия.
Рефлексы положения имеют тонический характер, так как
продолжаются все время, пока существует ненормальное положение
животного. В течение этого времени координирующие центры посылают
к определенной группе мышц импульсы небольшой частоты, 10—25
в 1 сек.
326 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Рефлексы положения .из лабиринтов на голове. Если взять
нормальное животное (кошку, собаку, кролика) и завязать ему глаза или
животное, у которого удален большой мозг, и придать телу животного
ненормальное положение в пространстве: подвесить головой вниз, или
поднять в вертикальном положении головой вверх, или держать тело в
горизонтальном положении животом вверх,— то во всех этих случаях
голова будет удерживаться в одном и том же, совершенно нормальном
положении: теменем вверх и мордой вниз (рис. 148-Л и 149-Л).
Рис. 149. Рефлексы положения на голове.
Кролик без большого мозга. Животное удерживается спиной вверх.
А —лабиринты целы. Рефлексы положения на голове нормальны.
В — оба лабиринта разрушены. Нормальное положение головы
отсутствует: голова повисла теменем вниз (Магнус).
После разрушения обоих лабиринтов этот рефлекс совершенно
исчезает. При ненормальном положении тела положение головы будет
также ненормальным (рис. 148-В и 149-В). Это указывает на то, что при
ненормальном положении животного в пространстве нормальное
положение головы устанавливается рефлекторно, вследствие раздражения
лабиринтных рецепторов.
Опытным путем было установлено, что рецептивным полем рефлекса
положения головы является sacculus со своими рецепторами: их
максимальное раздражение получается, когда отолиты повисают на імакулах;
наоборот, минимум раздражения получается, когда отолиты опираются
на макулы. Такие рефлексы положения головы исчезают после
перерезки позади среднего мозга и, наоборот, продолжают обнаруживаться
у животных, у которых перерезка произведена впереди среднего мозга;
поэтому считают, что координирующий аппарат этих рефлексов должен
находиться ів среднем мозгу (Магнус, 1; Дюссер де Баренн, 2).
Рефлексы положения, вызываемые с туловища на голове. Как
было указано выше, после разрушения лабиринтов животное теряет
нормальное положение головы, если его держать в воздухе. Но если
такой препарат уложить на стол в боковом положении, голова сейчас
же примет нормальное положение. Этот рефлекс,1 очевидно, вызывается
асимметрическим раздражением кожи. Когда животное лежит на
правом боку, у него кожа раздражается только с этой же стороны.
Значение асимметрического раздражения кожи на одном боку для восста-
ГЕФЛЕКСЫ ПОЛОЖЕНИЯ И СТАТОКИНЕТИЧЕСКИЕ РЕФЛЕКСЫ 327
новления нормального положения головы видно из того, что если сверху
положить на левый бок доску и чуть придавить ее, голова сейчас же
утрачивает нормальное положение; стоит только отнять доску, как
голова возвратится опять в нормальное положение. Очевидно, рецепторы
этого рефлекса лежат в коже туловища, координирующий же аппарат,
по исследованиям Магнуса, находится в среднем імозгу. По
удалении его означенный рефлекс исчезает.
Рефлексы положения, вызываемые с шеи на мышцах туловища.
Если животное лежит на боку и голова приходит в нормальное
положение, то вслед за этим последует выпрямление сначала передней
части туловища, а потом задней. Этот рефлекс не зависит от лабиринтов,
он наблюдается и на безлабиринтных препаратах. Здесь мы имеем дело
с такого рода рефлексом: при боковом положении головы от
асимметрического раздражения рецепторов кожи на туловище голова
выпрямляется, приходит в нормальное положение. При этом происходит
поворот шеи, а это раздражает шейные проприоцепторы; от их раздражения
и зависит наступление рефлекса положения на передней части
туловища. Последнее же движение, в свою очередь, раздражая
проприоцепторы передней части туловища, вызывает выпрямление задней его части
(Магнус, 1).
Рефлексы положения, вызываемые с туловища на мышцах
туловища. Нормальное положение туловища зависит не только от того, что
голова находится в нормальном положении. Если уложить кролика на
бок и фиксировать голову в этом же бо-ковом положении так, чтобы она
не могла притти в нормальное положение, то и тогда туловище может
принять нормальное положение, т. е. перевернуться спиной вверх. Этот
рефлекс вызывается особенно легко, если животное лежит на
шероховатой поверхности. Он обусловливается асимметрическим раздражением
кожи туловища: если сверху на другой бок положить доску, слегка
надавить на нее и тем самым достигнуть симметрического" раздражения
кожи, туловище останется в боковом положении. Этот рефлекс также
обусловливается деятельностью среднего! мозга (Магнус, 1).
Рефлекс положения, вызываемый с туловища на мышцах тела,
имеется у лягушки. Если положить децеребрированную лягушку спиной
вниз, она сейчас же перевернется на брюхо. Этот рефлекс не зависит
от лабиринтов, так как лягушка переворачивается и после их
удаления. Координирующий аппарат этого рефлекса лежит ів продолговатом
мозгу, так как он вызывается после перерезки мозга по верхней
границе продолговатого мозга и не вызывается на спинномозговых
препаратах. Рецептивным же полем являются кожа и проприоцепторы мышц
на спинной стороне тела (Беритов, 11),
Итак, голова и туловище приходят в нормальное положение
при посредстве двоякого рода механизмов: положение головы зависит
от действия лабиринтных и туловищных рефлексов положения, а
нормальное положение туловища — от действия шейных и туловищных
рефлексов положения. '
328 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Рефлексы положения, вызываемые с глаз на голове. Нормальное
положение головы зависит я от глаз. Кошка, собака или обезьяна с
разрушенными лабиринтами держат голову в нормальном положении,
если они могут видеть. В каком бы положении ни находилось тело,
голова примет нормальное положение: теменем вверх и мордой вниз.
Доказать, что этот рефлекс положения зависит от глаз, очень просто:
если безлабиринтным животным завязать глаза, голова теряет
нормальное положение.
Данный рефлекс положения имеется не у всех животных,— его нет,
например, у кроликов и морских свинок. Если у какого-либо из
названных животных разрушить лабиринты и затем держать его в воздухе,
то голова у него повиснет, как у мертвого, теменем вниз, даже в том
случае, если глаза открыты. Рефлекс положения с глаз вызывается
только на голове. Выпрямление головы уже вторичным путем вызывает
шейные и туловищные рефлексы положения. Этот рефлекс положения
исчезает по удалении большого мозга (д? Клейн и Магнус, 4).
Значение красных ядер для рефлексов положения.
Координирующие аппараты рефлексов положения, как уже отмечалось, лежат в
среднем мозгу. Опытным путем было доказано, что находящиеся в
среднем мозгу красные ядра являются центральным органом
координации рефлексов положения. Если перерезать мозг позади красных ядер
или нисходящие пути tractus rubrospinalis, идущие из красных ядер, то
рефлексы положения исчезнут; если же перерезка мозга производится
впереди красных ядер, то все эти рефлексы сохранятся. (Мозжечок не
необходим для рефлексов положения: удаление его не ведет к
уничтожению рефлексов положения с лабиринтов или от асимметричного
раздражения туловища. Но, повидимому, мозжечок необходим для
рефлексов положения от рецепторов туловища на туловище же. Были случаи
исчезновения этого рефлекса у кроликов после удаления мозжечка.
Предполагается, что в отношении этого рефлекса положения
восходящий чувствительный путь проходит через імозжечок. Характерно, что
координация этого тонического рефлекса не происходит в красных
ядрах, он продолжает вызываться и после перерезки нервных путей
(tractus rubrospinalis) позади перекреста Фореля. Повидимому,
координирующий центр туловищных тонических рефлексов лежит позади
красных ядер, где-то поблизости от них (Магнус, 8, 9; Радемакер,
8, 7). Как уже указывалось, этот рефлекс на лягушках вызывается и
после удаления всего среднего мозга вместе с мозжечком, т. е.
после перерезки мозга нг\ верхней границе продолговатого (Беригов, 11).
Статокинетическіие рефлексы. Рецепторы лабиринтов
раздражаются не только от изменения положения в пространстве, но и при
передвижении с нарастающей скоростью,— все равно, какое будет движение —
прямолинейное или вращательное. В ответ на такое раздражение
наступают своеобразные рефлексы в области головы и конечностей.
Рефлексы, вызывающиеся ускоренным вращательным движением,
называются рефлексами вращения, а вызывающиеся ускоренным пря-
РЕФЛЕКСЫ ПОЛОЖЕНИЯ И СТАТО КИНЕТИЧЕСКИЕ РЕФЛЕКСЫ 329
молинейным движением,— называются рефлексами
прогрессивного движения. Эти рефлексы также были открыты и
подробно изучены в лаборатории Магнуса. Все статокинетические
рефлексы являются тоническими, так как продолжаются в течение
всего времени ускоренного движения.
Рефлексы вращения. Если посадить животное на вращающийся диск
по направлению радиуса головой к окружности, то при вращении диска
направо голова поворачивается налево. По прекращении вращения
диска голова движется в противоположном направлении, т. е. в
направлении вращения. Иногда во время вращения и вскоре после него
возникает так называемый нистагм, т. е. голова начинает
поворачиваться то в одну, то в другую сторону. Эти статокинетические рефлексы
одинаково хорошо наблюдаются на кошках, кроликах, голубях,
щенках, а также на лягушках.
Когда животное (кролик, собака) вращается на диске, то
одновременно с головой поворачиваются и глаза в сторону, противоположную
вращению диска. По окончании вращения глаза производят обратное
движение, т. е. в направлении вращения.
Часто наблюдается нистагм глаз как во время вращения, так
и после него. Этот статокинетический рефлекс наблюдается в любом
положении животного, будет ли оно находиться в сидячем или боковом
положении или же будет повешено в воздухе головой вниз. ВС всех
случаях, если животное вращается вокруг своей вертикальной оси, глаза
поворачиваются в горизонтальной плоскости в направлении,
противоположном движению.
Эти рефлекторные движения головы и глаз имеют то значение, что
своим обратным направлением они пытаются удержать то зрительное
поле, какое было до вращения.
Рефлекс прогрессивного движения на мышцах головы и
конечностей. Если усадить какое-либо домашнее животное на доску и быстро
поднимать и опускать ее, то животное производит следующие
характерные движения: при поднятии оно опускает вниз голову, сгибает ноги,
приседает, пригибается; при спускании же вызываются обратные
движения: голова поднимается вверх, ноги разгибаются, животное встает,
приподнимается. Эти явления можно хорошо наблюдать на животных,
если их поднимать и опускать в лифте. При остановке лифта (при
движении вверх или вниз) реакция животного меняется на диаметрально
противоположную. Например, когда прекращается поднятие,
животное поднимает голову, ноги у него разгибаются, особенно передние. Это
движение конечностей, оказывается, не зависит от одновременных
движений головы и шеи. То же явление наблюдается, если фиксировать
голову и после этого поднимать и опускать животное. Не зависят эти
реакции и от глаз: они наблюдаются и при завязанных глазах. Все эти
рефлексы исчезают после разрушения лабиринтов,— значит, их
рецептивное поле лежит в статическом органе (де 'Клейн и Магнус). Эти
рефлексы можно хорошо наблюдать и на лягушке.
330 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Приготовление к прыжку. Если животное держать в вертикальном
положении головой вниз и затем быстро опустить, передние ноги
сейчас же разогнутся, как будто животное собирается прыгнуть. Конеч
ности разгибаются, чтобы принять на себя всю тяжесть тела в момент
соприкосновения с полом. Эта
реакция исчезает после
разрушения лабиринтов, значит,
рецептивное поле данного
рефлекса также лежит в
лабиринте.
"Как видно из работ
лаборатории Магнуса, реакции
на вращение и прогрессивное
движение не зависят от
раздражения рецепторов sacculus
и utriculus. Это заключают из
того факта, что после
удаления отолитов рефлексы
стояния и положения исчезают
полностью, в то время как
рефлекторные реакции на
вращение и прогрессивное
движение остаются. Так как
последние исчезают только после
удаления лабиринтов, то, пови-
димому, они вызываются
раздражением рецепторов,
лежащих в ампулах. Однако
известно, что рецепторы utriculus
также должны играть роль в
вызывании рефлекторных
реакций на прогрессивное
движение. Так, например, если
голова находится в нормальном
медиальном положении,
отолиты utriculus располагаются
горизонтально и давят сверху
на імакулы. При быстром
опускании животного вниз
отолиты слабее давят на імакулы
и этим могут вызвать
разгибание передних ног, как при
отклонении головы назад. При остановке движения вниз отолиты будут
сильнее давить на імакулы и тем самым прекратят усиленное
разгибание передних ног.
Координирующий аппарат рефлекторных реакций на вращение и про-
Рис. 150. Кинематографический снимок
переворачивания животного в воздухе при
падении спиной вниз.
Ясно видно, что сначала поворачивается в нормальное
положение голова (•?, 4, б), а потом вращается
туловище и происходит разгибание передних ног
(Магнус).
I
РЕФЛЕКСЫ ПОЛОЖЕНИЯ И СТАТОКИНЕТИЧЕСКИЕ РЕФЛЕКСЫ 331
грессивное движение лежит в продолговатом мозгу, в области
вхождения VIII нерва. Но на кошках эти реакции не вызываются, если мозг
перерезан позади четверохолмия (Радемакер, і7), а на лягушках они
получаются хорошо после удаления большого и промежуточного мозга
(Беритов).
Вращение животного при падении. Нормальное, а также
лишенное большого мозга животное (кошки, собаки, обезьяны, лягушки), при
падении спиной вниз с некоторой высоты переворачивается в воздухе
раньше, чем упадет на землю. Животное падает брюхом вниз при
выпрямленных ногах (рис. 150). При этом рефлексе поворачивается и
приходит в нормальное положение сначала голова: это обычный
рефлекс положения с лабиринтов. Поворот головы вызывает
раздражение рецепторов шеи, что, в свою очередь, вызывает рефлекс
положения с шеи на переднюю часть туловища. Раздражение рецепторов
последней вызывает выпрямление задней части туловища. Благодаря
указанным рефлексам животное винтообразно вращается в воздухе,
причем передние ноги разгибаются. Это — рефлекс приготовления к
прыжку, который вызывается также с лабиринтов (Магнус, 6).
Практикум
Наблюдение рефлексов положения на кролике. Предварительно удаляют большой
мозг. Для этого сначала дают наркоз и производят трахеотомию и перевязку артерий,
как описано в предыдущей главе. Потом вскрывают большой мозг и удаляют его.
Наркоз прекращают тотчас же после этого. Спустя 10—30 минут кролик принимает
свое обычное сидячее положение. Он сидит на задних ногах, передние разогнуты,
голова поднята и находится в медиальном положении.
Опыты: 1. Если такой препарат взять в руки и менять его положение в
пространстве, голова всегда будет принимать нормальное положение: теменем вверх,
челюстями вниз. Это будет лабиринтный рефлекс положения на голове.
2. Если животное положить на бок, то сначала примет нормальное положение
голова, а потом туловище. Это будет рефлекс положения с туловища на голове.
3. Если при боковом положении животного голову удерживать в боковом
положении и не дать ей принять нормальное положение, то туловище все же его примет.
Это будет туловищный рефлекс положения на туловище.
4. Если препарат держать в воздухе спиной вверх и быстро опускать вниз в
вертикальном направлении, то передние ноги сейчас же будут сильно разгибаться. Это
будет рефлекс подготовки к прыжку, вызванный поступательным движением.
5. Если положить животное на доску и быстро поднимать ее и опускать, то
получим статокинегический .рефлекс на прямолинейное (поступательное движение.
6. Если посадить животное на доску и вращать его влево или вправо, то получим
статохинетический рефлекс на вращательное движение.
Все перечисленные рефлексы исчезнут, если перерезать ствол мозга позади
среднего мозга.
Рефлексы положения на лягушке. На лягушке также легко наблюдать ряд
рефлексов положения и статокинетических рефлексов. Для этого можно брать как
нормальное, так и лишенное большого мозга животное.
1. Если лягушку положить на спину, она сейчас же перевернется. Этот рефлекс
положения не исчезает после удаления среднего мозга и даже после перерезки
продолговатого мозга выше вхождения VIII нерва.
332 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Что этот рефлекс ве зависит от лабиринтов, видно из того, что оя наступает и
после удаления лабиринтов или перерезки VIII нерва с обеих сторон.
2. Посадить лягушку без больших полушарий на дощечку и вращать ее в
горизонтальной плоскости; при вращении вправо голова, а иногда и вся передняя часть
тела, поворачивается влево, при вращении влево лягушка повернется вправо.
3. Поднимать и опускать дощечку с лягушкой вверх и вниз. При поднимании
голова опустится, а передние ноги согнутся; обратное получится при опускании. Эти
статокинетические рефлексы получаются и при медленных движениях дощечки.
Аналогичные опыты можно ставить на совершенно нормальных кроликах, на
щенках, а также на птицах (голубь).
Литература
I.Magnus R., Pfl. Arch., 163, 405, 1916; 174, 134, 1916; 193, 326, 1922; Когрег-
stellung, Berlin, 1924.
2. Dusser de Barenne u. Magnus., Pfl. Arch., 180, 75, 1920.
3. De К 1 e у n A. u. M a g n u s, Pfl. Arch., 194, 407, 1922; 186, 6, 61, 1921.
4. De К 1 e у n A. u. M a g n u s, Pfl. Arch., 180, 291, 1920.
5. De К 1 e у n A. u.Magnus, Pfl. Arch., 186, 39, 1921.
6. M agnus R., Arch. Neerl. Physiol., 7, 218, 1922.
7. R a d e m a k e r G. G. J., Klin. Wschr., 1, 404, 1923.
8. Magnus R. u. Rademaker, Schw. Neur. Psych., 13, 408, 1913.
9. M agnus R., Dtsch, med. Wschr., стр. 501, 1923.
10. R a d ? m a k e г G., Die Bedeutung der roten Kerne und des ubrigen Mittelhirns
fur Muskeltonus, Korperstellung und Labyrinthreflexe, Berlin, 1926.
11. Beritoff J., Acta oto-laryng., 12, 492, 1928.
12. С о о p e г S. a. E с с 1 e s J. C, J. Physiol., 69, 377, 1930.
43. Wachholder K., Verhl. Dtsch. Orthopad. Ges., 33. Kongr. Giessen, 1938.
Глава XVIII
ЛОКОМОЦИЯ
Общее понятие. Одной из функций центральной нервной системы
является локомоция. Локомоция есть врожденный, чисто рефлекторный
акт перемещения. Известно, что* локомоция 'может протекать
непроизвольно, бессознательно. Каждый из нас может ходить во время
увлекательной беседы, не обращая на ло<комоцию никакого внимания.
Объективно может быть доказано, что локомоция животных не нуждается
в индивидуальном опыте. После удаления больших полушарий собака
и другие животные не теряют способности к передвижению. Кролики
и морские свинки сохраняют способность к локомоции даже после
удаления головного імозга, включая и средний, т. е. после децеребрации.
Более того, в настоящее время известно, что даже обезглавленные
животные обладают этой способностью. Так, например, обезглавленная
курица іможет еще перемещаться в пространстве, обезглавленные утки
еще плавают в воде; даже обезглавленные собаки и кошки сохраняют
некоторые элементы локомоции.
Только у человека локомоция не является всецело прирожденным
актом. Человек научается ходить. Долго болеющие дети разучиваются
ходить.
Локомоция представляет очень сложный акт. Она складывается иэ
четырех элементов: из фазных движений конечностей, статического
тонуса, рефлекса положения и рефлекса равновесия. К локомоции
относятся: шагание, бегание, прыгание, плавание и т. д. Для всех этих
движений перечисленные элементы, безусловно, необходимы.
Фазные движения. Как известно, во время локомоции движение
конечностей складывается из двух главных фаз: сгибания и разгибания.
В каждой локомоции эти фазы протекают характерным образом. Кроме
того, в каждой локомоции характерными являются также
взаимоотношения между фазами той и другой ноги. Для иллюстрации здесь
приводятся фазные движения человека при шагании.
I фаза (рис. 151, правая нога, R 10—20). Правая нога опирается
на пол и принимает на себя тяжесть всего тела. Вначале она немного
согнута во всех трех главных суставах, т. е. в тазобедренном, колен-
334 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
ном и голеностопном, затем все более и более разгибается. При этом
она из вытянутого вперед положения принимает вертикальное
(14—17 Ег), затем оттягивается назад. В это время туловище
переносится вперед.
Левая нога в начале этой фазы разогнута целиком и отклонена
назад, касаясь пола большим пальцем. Затем она начинает сгибаться во
всех суставах и переносится вперед. В тот момент, когда правая
принимает вертикальное положение, левая находится на одном уровне с
правой. После этого левая начинает разгибаться в коленном суставе
Рис. 151. Ходьба.
На переднем плане нарисованы правые ноги, на заднем — левые. Ноги, находящиеся в сгибательной
фазе, поперечно исчерчены (Sr— 1 — 9 и Sr— 21), белыми ногами изображены в периоде опоры,
т. е. в разгибательной фазе (10—20), В момент Ег 7 и Ег 17 ноги отходят от земли, сначала пятой,
потом пальцами, Аг и Аг указывают моменты опоры всей стопой левой и правой ноги (О. Фишер)
и переносится вперед. Когда разгибание правой достигает максимума,
левая вытягивается вперед максимально, до прикосновения к полу
сначала подошвой, а затем пальцами. Она переставляется на такое же
расстояние вперед от правой ноги, на какое была отставлена назад.
В этот момент обе ноги прикасаются к полу.
II фаза (правая нога Sr I-R 10). Выше была описана одна фаза
движений для правой ноги, а другая —для левой. По окончании этих
движений происходит смена фаз: на правой ноге начинается то самое
движение, которое раньше было на левой, а на левой, наоборот,
повторяется движение правой. При этой омене фаз происходит
перемещение центра тяжести с правой ноги на левую. Это перемещение
происходит в последний момент первой фазы и следует за разгибательным
движением голеностопного сустава правой ноги (братья Вебер, 1;
Марей, 2; Браун и Фишер, 3; Фишер, 4; Бернштейн, 15).
Назначение сгибательной фазы заключается в переносе собственной
тяжести согнутой ноги, а разгибательной фазы — в переносе тяжести
всего остального тела, а также центра тяжести с одной ноги на другую.
Как видно из описания и рисунка, на каждой ноге одна фаза
является тіо преимуществу сгибательной, а другая — разгибательной, но
локомоция
335
по существу в каждой фазе имеется элемент как сгибания, так и
разгибания. Координация імышц в той и другой фазе происходит по
известному уже принципу; в фазе сгибания сокращаются и тормозятся те
же односуставные и двусуставные імышцы, как и в оборонительном
рефлексе сгибания. Координация ж? разгибательной фазы происходит,
как в оборонительном рефлексе разгибания (Шеррингтон, 5; Беритов, 6).
Как указывалось выше, координирующие аппараты фазных движений
лежат в спинном мозгу: для задних ног—в поясничном отделе, а для
передних — ів плечевом.
Мы рассмотрели фазное движение ног при шагании человека, но в
общем такое же фазное движение мы наблюдаем при ходьбе
четвероногих животных. Характер фазных движений каждой ноги при других
типах локомоции несколько отличается от описанных выше. Фазы эти
меняются также в отношении продолжительности.
Очень сильно вариирует соотношение фаз всех ног между собой.
Во время ходьбы четвероногих животных две ноги по диагонали
сгибаются, а две другие — разгибаются. Так ходят кошка, собака, лошадь
и др. Но ходьба может происходить иначе: передняя и задняя ноги
одной стороны одновременно сгибаются, в то время как на другой
стороне обе ноги разгибаются (верблюд, иноходь лошади). Есть и такие
животные, у которых обе передние или задние ноги движутся
одинаково. Так, например, у лягушки во время тірыгания все ноги
одновременно тб сгибаются, то разгибаются; у нее же во время плавания
все ноги сгибаются и разгибаются одновременно. Некоторые животные
во время ходьбы передними ногами шагают, а задними — прыгают,
например кролики. Словом, соотношение фазных движений ног
чрезвычайно разнообразно. Но каково бы ни было соотношение фазных
движений во время локомоции, всегда фазные движения каждой ноги
протекают по одному и тому же типу, и, следовательно, координация их
также должна происходить одним и тем же путем. На рис. 152 дана
иллюстрация реципрокной иннервации во время фазных движений.
Статический тонус опоры, или стояние при локомоции. Все
животные, обладающие конечностями, передвигаются на более или менее
выпрямленных ногах. Во время локомс-пии все конечности, туловище и
шея находятся в разгибательном тонусе, благодаря чему организм
защищен от падения в силу собственной тяжести. Этот статический тонус
представляет собой второй существенный элемент локомоции. Во время
локомоции конечности и другие органы непрерывно находятся в
статическом тонусе.
В моменты фазного сгибания статический экстензорный тонус не
выпадает целиком: он только ослабевает. Степень экстензоркого тонуса
меняется в зависимости от быстроты локомоции. Чем быстрее следуют
одно за другим фазные движения, тем сильнее экстензорный тонус на
конечностях, шее и хвосте. Эта зависимость выступает очень рельефно
при сравнении тонуса шагающей лошади со скачущей. Последняя
держит конечности более выпрямленными, шею и хвост более
приподнятыми по сравнению с шагающей лошадью.
336 общая физиология спинного мсзга и ствола головного мозга
Разгибательный тонический рефлекс всегда сопровождает ло-комо-
щію. Он возникает одновременно с фазными движениями, если только
не существовал уже до того (рис. 152). Если лежащей собаке
незаметно ущипнуть ногу или хвост, она сейчас же становится на ноги и
передвигается. Это происходит автоматически, чисто рефлекторным пу-
Рис. 152. Работа антаюниетов во время обычной
для кролика локомоции.
Децеребрированный препарат. Записываются разгибатели
колена той и другой ноги-m. vastocrureus и сгибатель колена
одной ноги-m. semitendinosus.
Во время раздражения седалищного нерва правой стороны
происходит ряд одновременных сокращений обоих разгибателей
с постепенным нарастанием на них тонуса Во время аі их
сокращений сгибатель тормозится, существующий на нем сіибатель-
ный тонус выпадает. Каждый раз сейчас же после сокращепия
разгибателей происходит сокращение сгибателя Сначала, когда
прыгание производилось быстро, сокращение сгибателл было
слабое и кратковременное, затем оно стало более сильным
и длительным Это указывает, что животное после каждого
прыжка переходит в сидячее положение.
тем. Если вырезать у собаки большой мозг, как раз тот отдел мозга,
который является седалищем индивидуальных рефлексов, то и тогда,
при указанном механическом раздражении, собака встает на ноги и
передвигается.
Рецептивное поле статического рефлекса опоры конечностей
находится в коже на пальцах с вентральной стороны и затем в мышцах и
суставах пальцев. Давление на подушечки пальцев или дорзальная
флексия пальцев (отгибание вверх) вызывает разгибание всей
раздражаемой ноги. На другие ноги рефлекс не простирается. С прекращением
раздражения рефлекс прекращается. Этот рефлекс легко наблюдать как
на нормальных животных, так и на спинальных и лумбальных
препаратах (Магнус, 10; Шён, 11).
Большое значение в поддержании статического тонуса и вообще
выпрямленного состояния имеет «миотатический рефлекс растяжения».
При стоянии все мышцы, противодействующие силе тяжести, более или
локомоция
337
менее растянуты. Как только тот или другой орган поддастся
усиленному действию силы тяжести, сейчас же произойдет новое растяжение
этих імышц, вследствие чего в них наступит миотатический рефлекс
сокращения и выпрямленное положение органа будет удержано (Шер-
рингтон, 13).
Статический рефлекс разгибания характерно отличается от всех
других видов разгибания: в нем сокращаются все мышцы конечности —
как разгибатели, так и сгибатели. Это обусловливает прочную
фиксацию суставов, что, безусловно, необходимо для локомоции. При этом
на передней ноге сокращаются как все двусуставные, так и все одно-
суставные мышцы, на задней же ноге сокращаются из сгибателей
только те, которые служат для фиксации (Блэк Ричард, 12).
Вентральная флексия пальцев (пригибание вниз), а также
голеностопного сустава на задней ноге вызывает противоположный эффект:
разгибательный статический тонус исчезает и наступает обычный сги-
бательный тонус во всех суставах данной конечности с сокращением
сгибателей и торможением разгибателей. Этот рефлекс является
исключительно проприоцептивным, т. е. вызывается раздражением рецепторов
мышц и сухожилий пальцев (Шён, 11, Магнус, 10).
Оба статических рефлекса играют важную роль при локомоции.
Разгибательный рефлекс представляет положительную опорную фазу,
которая превращает конечность, состоящую из подвижных частей, в
несгибающийся опорный аппарат. Сгибательный же рефлекс
представляет отрицательную фазу: она как бы удаляет задвижку, которая при
стоянии держит ногу фиксированной, и тем придает ей подвижность
(Шён, 11).
В координации акта стояния участвует почти вся центральная
нервная система, так как в самом стоянии организма участвует вся
скелетная мускулатура. Но статический рефлекс каждой конечности, а также
фазные движения ее координируются в том отделе мозга, куда входят
чувствительные нервы данной конечности. Так, например, при стоянии
координация в отношении задних ног происходит в поясничной части
спинного мозга, так как и после перерезки спинного мозга в грудной
области задняя часть тела не лишается статического тонуса. Она не
только дает характерные эффекты на дорзальную и вентральную
флексию пальцев, но и фазные движения; тем самым она может
осуществить и настоящую локомоцию. Это, например, бывает у кошки. Если
после перерезки мозга в грудной области раздражать хвост при
брюшном положении іживотного, то задние ноги производят фазные
движения и одновременно тонически разгибаются. Вся задняя часть тела
поднимается, и если ее удержать от падения, она передвинется. По
прекращении раздражения фазные движения тотчас же прекращаются,
разгибательный же тонус постепенно ослабевает, задняя часть тела
постепенно опускается. Отсюда следует, что координирующий аппарат,
производящий характерную иннервацию задних ног животного при
статических рефлексах, лежит в поясничном отделе спинного мозга
(Беритов, 6).
22 и. С. Беритов
33S ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Рефлексы положения во время локомоции. Как указано выше,
ненормальное положение головы или туловища вызывает рефлекторным
путам выпрямление головы, т. е. приведение ее в нормальное
положение. Известно, что это происходит от асимметрического раздражения
лабиринтов и туловища. Выпрямление головы влечет за собой сгибание
шеи и раздражение шейных рецепторов, что, в свою очередь, вызывает
вначале выпрямление передней части туловища, а затем, вследствие
этого, и выпрямление задней его части. Так как нормальная локоімо-
ция, да и локомоция вообще, возможна
только при нормальном положении животного,
т. е. теменем и спиной вверх, животом и
мордой вниз, то понятно, что этот рефлекс
положения должен играть существенную роль в
процессе локомоции. Как известно, при
разрушенных лабиринтах и завязанных глазах
голова животного теряет способность
принимать нормальное положение, если только туло-
Рис. 153. Точки опоры вер- вище испытывает симметричное раздражение.
Отсюда видно,, что если бы рефлексы
положения отсутствовали, то падение животного
во время локомоции было бы частым
явлением, а само производство локомоции после
падения было бы невозможно, так как
животное было бы лишено возможности принять
нормальное положение.
Во время локомоции центр тяжести
постоянно перемещается то в одну, то в другую сторону. Если при этом
животное не падает, то это происходит благодаря рефлексам,
выражающимся в противоположных движениях. Эти активные движения
называются рефлексами равновесия.
В теле человека при свободном стоянии на сдвинутых ногах
вертикальная линия, опущенная из центра тяжести, проходит через подошву
правой стопы на 1 см от внутреннего края стопы и на 15,5 ом от
переднего ее края (Б'аслер, 14) (рис. 153).
При разного рода движениях центр тяжести перемещается то в
одну, то в другую сторону, сообразно с чем меняется и положение
вертикальной линии, опущенной из центра тяжести. Когда человек садится
на корточки, центр тяжести перемещается так, что опущенная из центра
тяжести вертикальная линия проходит между его ногами (рис. 153);
когда он наклоняется вперед, центр тяжести тоже перемещается
вперед; когда он отгибается назад, центр тяжести уходит назад; при
наклонении набок центр тяжести переходит на бок. Во всех случаях
передвижения центра тяжести человек не падает, так как вертикальная
линия, опущенная из центра тяжести, не выходит из области опоры
обеих ног вследствие своевременного возникновения противоположных
движений рефлекторного характера. Следовательно, одну из функций
тикальной линии из центра
тяжести при стоянии на
ногах.
Точка—при свободном стоянии.
Квадрат—при держании в
руках длинного шеста, одинаково
уравновешенного.
Треугольник—при сидении на корточках
(Баслер).
Рефлекс равновесия.
локомоция 339
центральной нервной системы человека составляет недопущение
выхода проекции центра тяжести за пределы линии, очерчивающей
область опоры нижних конечностей. У четвероногих животных
центральная нервная система защищает животное от падения тем, что не
допускает выхода вертикальной линии центра тяжести из области, лежащей
между точками опоры всех четырех конечностей.
Все компенсаторные движения, защищающие организм от падения,
представляют собой чисто прирожденные рефлексы. Они всегда
зависят от периферического раздражения. (Каждый знает из личного опыта,
что при поскальзывании равновесие восстанавливается сразу,
совершенно независимо от наших желаний или воли. Когда человек дремлет
сидя, можно наблюдать следующее явление: туловище и голова
постепенно наклоняются вперед; когда наклонение достигает определенного
угла и вертикальной линии центра тяжести грозит выход из области
оперы, то сразу наступает быстрое выпрямление. Затем снова следует
медленное наклонение, а затем снова быстрое выпрямление. Это,
конечно, происходит без сознания, без участия воли. Во время дремоты
возбудимость центральной нервной системы находится еще на такой
высоте, что реагирует на раздражение проприоцепторов спины
рефлексом выпрямления.
Рефлекс равновесия имеет большое значение для локомоции. Без
этого рефлекса движение животных было бы чрезвычайно ограничено
и в высокой степени опасно. Так, например, каждый раз, когда
кончается разгибательная фаза данной ноги (у человека, птицы) или пары ног
(у собаки, верблюда) происходит их отталкивание от опоры, для того
чтобы перебросить приходящийся на них центр тяжести на другую ногу
или на другую пару ног. Для этого, конечно, все животное должно
наклоняться в сторону последних и если бы каждый раз одновременно не
происходило компенсаторного движения противоположного характера,
животное падало бы на этот бок, тем более что в данный момент при
быстрых передвижениях ноги, на которые переносится центр тяжести,
еще не опираются на землю.
Компенсаторное движение выпрямления вызывается раздражениями
проприоцепторов. Так, например, боковое наклонение в одну сторону
производит растяжение мышц на другой стороне, что вызывает
раздражение находящихся в них проприоцепторов. На нормальных животных,
а также на децеребрированных препаратах можно заметить и другого
рода компенсаторные движения. Когда передняя часть тела
наклоняется набок, соответствующая передняя нога тотчас же вытягивается в
ту же сторону и вперед, чем устраняется возможность падения. Этот
рефлекс, очевидно, вызывается проприоцептивными раздражениями
туловища и шеи (Беритов, 8). У человека также бывают компенсаторные
движения: когда от толчка тело наклоняется влево, наряду с
рефлексом выпрямления правая рука вытягивается вправо. Когда тело
наклоняется назад от толчка спереди, обе руки вытягиваются вперед. Эти
движения, конечно, благоприятствуют удержанию центра тяжести в
пределах опоры ног.
22*
ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Пока еще не выяснено, где происходит координация рефлексов
^равновесия. Возможно, что координация этих рефлексов, как и вообще
всех рефлексов, происходит там, куда входят чувствительные нервы их
рецептивных полей, т. е. в данном случае в спинном мозгу. Однако не
подлежит сомнению, что для возникновения рефлексов равновесия
необходим головной мозг, а именно: для высших млекопитающих (собаки,
кошки)—задние две трети зрительных чертогов, а для кролика —
передние две трети варолиева моста. После удаления этих частей мозга
способность к рефлексам равновесия исчезает, несмотря на то, что
тонические рефлексы и фазные движения сохраняются (Лаутон, 9).
Направление локомоции. При локомоции животное следует как
по прямолинейному направлению вперед, так и по всякому другому
направлению. Направление движений определяется начальным
положением головы: животное движется в том направлении, куда обращена
его голова; когда голова находится точно в медиальном полоіжении,
животное направляется прямо вперед; когда голова, под влиянием
какого-либо раздражения, повернута вправо, тогда животное во время
локомоции поворачивает тоже вправо. По Магнусу, это явление
обусловливается тем, что при повороте головы вправо наступает шейный
проприоцептивный рефлекс на передних ногах: правая передняя нога
усиленно выпрямляется, и одновременно на левой передней разгибатель-
ный тонус ослабевает, в силу чего центр тяжести переходит на правую
йогу. Это ведет к тому, что в последующей локомоции движение
открывается левой ногой, которая является свободной. В результате
получается поворот всего тела вправо, и, значит, локомоция протекает в
направлении направо'. Отсюда следует, что направление в локомоции
обусловливается главным образом действием шейного тонического
рефлекса, а потому и животное, лишенное большого мозга, может
передвигаться в том или другом направлении, смотря по положению головы.
Взаимоотношение между статическим тонусом и фазными
движениями. Мы рассмотрели элементы локомоции и пришли к заключению,
что как фазная, так и статическая рефлекторная деятельность
вызывается внутренними и внешними периферическими раздражениями, что
координирующие аппараты обоих рефлексов для каждого органа
лежат в одном и том же отделе мозга, куда входят чувствительные нервы
данного движущегося органа. Из этого следует, что деятельность
одного элемента локомоции немыслима без деятельности другого. Когда
раздражается нога, это должно вызвать как фазный, так и статический
рефлекс. Если первоначально начнется деятельность одного из этих
элементов, то непременно за ней последует действие и другого
элемента вследствие вторичных раздражений. Следовательно, условия
деятельности координирующих аппаратов тонического и фазного
рефлексов таковы, что наступление одного рефлекса должно неизбежно
вызвать другой. Если, например, сперва был вызван статический рефлекс,
то путем вторичных проприоцептивных раздражений, которые
производятся первичным тоническим движением, будет вызван и фазный
рефлекс. Если же сначала возник фазный рефлекс, то вторичные раздра-
локомоция
341
жения сейчас же вызовут статический рефлекс. Таким образом, явление
локомоций — сочетанное наступление фазного рефлекса и экстензорно-
го тонуса — представляет собой результат взаимодействия одного
рефлекса с другим путем вторичных раздражений.
Регуляция локомоций. Как было указано выше, координация
разных элементов локомоций происходит в разных отделах імозга. Фазные
движения конечностей координируются в поясничном и плечевом
отделах мозга. Рефлекс стояния координируется также в этих отделах.
Рефлексы положения во время локомоций координируются в средних
отделах мозга, здесь же осуществляется рефлекс равновесия.
Очевидно, для выполнения нормальной локомоций существует
специальный регулирующий центр. У аімфибий, птиц и млекопитающих
регулирующий локомоцию центральный нервный аппарат лежит в
промежуточном мозгу. Это заключение вытекает, прежде всего, из того
факта, что животные (собаки, кошки, кролики), лишенные больших
полушарий, производят локомоцию совершенно нормально (Гольц, 16;
Дюссер д? Баренн, 17 и др.). Локомоция протекает нормально и при
удалении большого мозга, включая весь neostriatum и почти весь
paleostriatum (Дрезель, 18; Магнус, 19). Исчезновение нормальной
локомоций связано с удалением промежуточного мозга. Центральный
аппарат, регулирующий локомоцию, занимает определенное место в
промежуточном імозгу, именно — в подбугорном отделе (subthalamus),
дорзально от сосковидного тела. Установлено, что локальным
электрическим раздражением этого участка регулярно вызываются равноімер-
ные шагательные и бегательные движения (Уоллер, 20). При перерезке
у кошки ствола імозга впереди этого участка нормальная локомоция
сохраняется, если же перерезка происходила позади него, она исчезает
(Гинеей, Рэнсон и Мак Нэттин, 21).
Итак, главная регуляция локомоторных движений, выражающаяся
с одной стороны, в объединении отдельных элементов локомоций в акт
передвижения, а с другой стороны, в установлении порядка отдельных
фаз в локомоций, осуществляется в подбугорном отделе
промежуточного мозга. [
Практикум
Наблюдение локомоций на препаратах лягушки. 1. Препарат без больших
полушарий. Ножницами вырезают черепную крышку и заостренным скальпелем
перерезают мозг позади больших полушарий. После этого лягушка сохраняет нормальную
локомоцию. 2. Децеребрированный препарат. Вместе с большим мозгом удаляется и
промежуточный. В этом случае способность локомоций сохраняется. 3. Обезглавленный
или спинномозговой препарат. Вырезается весь головной мозг; перерезка происходит
на уровне calamus scriptorius. Этот препарат лишен способности к локомоций.
На каждом из этих препаратов можно поставить следующие опыты: а) спустя
10—15 мин. после операции щиплют кожу на лапке и смотрят, переместится препарат
или не переместится; б) обращают внимание на передние ноги во время локомоций-
если они при движении выпрямлены и животное на них опирается, то ясно, что
локомоция полная, статический рефлекс существует; г) берут за одну переднюю лапку и
пытаются поднять препарат вверх и опрокинуть его на спину если препарат солротив-
342 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
ляется таким целесообразным движением, как вытягивание вперед другой передней
ноги или поворачивание всего тела, нужно считать, что он владеет способностью к
рефлексу равновесия.
Наблюдение локомоции на децеребрированных кроликах и голубях. Дец-еребра-
ция кролика производится таким же способом, как описано выше в отношении кошки.
При децеребрации голубей трахеотомию можно не делать; наркоз их производится
путем прикладывания к носовым отверстиям ваты, пропитанной хлороформом или
эфиром, но и в этом случае необходимо перевязать сонные артерии. Децеребрирован-
ные кролики и голуби обладают способностью к локомоции: стоят на ногах,
перемещаются на выпрямленных ногах. На этих препаратах хорошо наблюдать рефлексы
равновесия. Необходимо только, чтобы не весь средний мозг был удален, так как при
его полном удалении рефлекс равновесия исчезает.
На этих же препаратах легко наблюдать зависимость направления локомоции от
положения головы. Если кролик держит голову хоть немного повернутой направо, он
будет кружиться в правом направлении.
Локомоторное движение на спинномозговых кроликах или кошках. Наркоз и
операция на шее (трахеотомия, перевязка сонных артерий и перерезка блуждающих
нервов) производятся, как при децеребрации. Затем переворачивают животное спиной
вверх. Разрезают кожу на затылке по средней линяй до Ш-го шейного позвонка и
вокруг шеи на уровне атланто-затылочного сочленения. Верхний остистый отросток
П-го шейного позвонка освобождают от мышц, а передний край его до тела позвонка
обрезают костными щипцами. С помощью кривой и тупой иглы накладывается
крепкая лигатура вокруг позвонка по его переднему краю (рис. 132-В). Предварительно
на этом уровне перерезают дорзальный слой мышц для удобства накладывания
лигатуры и перевязывают яремные вены. Этой лигатурой достигается перевязка
позвоночных артерий. Здесь же накладывается вторая лигатура, охватывающая всю шею,
за исключением кожи и трахеи. Далее, следует отрезание головы по атланто-затылоч-
ному сочленению. Отрезав голову, поднимают шею вверх, чтобы приостановить
кровотечение из мозга. Необходимо, конечно, чтобы к этому времени уже производилось
искусственное дыхание и животное находилось в глубоком наркозе. Если после
операции прекратить наркоз, то рефлекторная деятельность на давление задней конечности
проявится через 10—20 мин. Обезглавленный препарат следует согревать искусственно.
Описанный способ обезглавливания принадлежит Шеррингтону. Он имеет
ту невыгодную сторону, что мозговая рана остается открытой и, несмотря на
прикрывание ватой, ее трудно уберечь от охлаждения. Поэтому лучше удовлетвориться
перерезкой мозга без обезглавливания. Для этой цели после операции на шее
обнажают membrana obturatoria, прикрывающую атланто-затылочное сочленение, и
перерезают ее по средней линии; края разреза отодвигают в сторону до обнажения мозга,
который и перерезают острым скальпелем приблизительно на 1—2 мм выше уровня
первой пары шейных корешков. Чтобы избежать повреждения позвоночных артерий,
последние нужно перевязать здесь же с обеих сторон.
На спинномозговых препаратах наблюдают: рефлексы сгибания при сдавливании
пальцев, рефлекс разгибания при сдавливании кожи в промежности, коленосухожиль-
ный рефлекс при ударе тупым краем скальпеля на сухожилие четырехглавой мышцы,
рефлекс чесания при раздражении кожи на плечах, на шее; наблюдают также
ритмический рефлекс локомоторного характера: препарат поддерживают за спинной хребет
и длительно раздражают кончик хвоста. В ответ на это обе задние ноги выпрямляются
и производят ритмическое движение по типу шагания.
Наблюдение статического рефлекса опоры. Лучше всего иаблюдать этот рефлекс
на нормальных кошках и щенках или же на животных, лишенных большого мозга.
Животное удерживают в воздухе, взяв его за живот. Ноги свободно висят. Если
надавить на пальцы какой-либо ноги снизу с вентральной стороны, то данная нога
будет разгибаться. Вся конечность станет твердой наощупь. При последующей
вентральной флексии пальцев будет наблюдаться обратное явление: разгибание пройдет,
а вместо него наступит обычный сгибательный рефлекс.
локомоция
343
Литература
1. Weber Ed. u. Wil., Mechanik der menschlichen Gewerkzeuge, 1836.
3. M а г e y, La machine animale, Paris, 1873; Le mouvement, Paris, 1884; C. R. Acad.
Sci. 1882—1898.
3. В r a u n u. Fischer, Abh. Math.-nat. KL, K. Sachs. Ges. Wiss. Leipzig, 21,
№. 4, 1895.
¦4. Fischer, Тот же журн.,25, № 1, 1899; там же, 26, № 3, 1900 и 28, № 5 и 6, 1903.
5. Sherrington С, J. Physiol., 40, 28, 1910.
¦6. Beritoff J., Pfl. Arch., 199, 248, 1923; Изв. Росс. Акад. Наук, 1915, стр. 1117
7. Magnus R. u. v. Leeuwen St. Pfl. Arch., 159, 157, 1914.
8. Беритов И., Учение об основных элементах центральной координации
скелетной мускулатуры, Пг., 1916, стр. 100.
9. L aught on N. В., Am. J. Physiol., 70, 358, 1924.
40. Magnus R., Lancet, 1925, стр. 551—556, 585—588.
11. Schoen R., Pfl. Arch., 214, 21 u. 48, 1926.
«2. Blake Prichard, Pfl. Arch,, 214, 148, 1926.
13. Шеррингтон и его сотр., Рефлекторная деятельность спинного мозга, 1935.
14. Basler A., Pfl. Arch., 221, 708, 1929.
15. Бернштейн Н. А., Попова Т. и Могилянская 3, Техника
изучения движений, М., 1934.
46. Goetz F., Pfl. Arch., 51, 570, 1892.
17. Dusser de BarenneJ. G., Arch, neerl. de Physiol., 4, 31, 1920.
18. D r e s e 1 K., Klin. Wschr., 3, 2231, 1924.
19. Magnus R., Korperstellung, Berlin, 1924.
20. Waller W. H., J. Neurophysiol., 3, 300, 1940.
21. Hinsey I. C, Ranson S. W. a. Nat tin R. F., Arch. Neurol. Psych.
Chicago, 23, 1, 1930.
Глава XIX
ОРИЕНТИРОВОЧНЫЕ РЕФЛЕКСЫ
Общее понятие. Если незаметно прикоснуться к животному сзади
или сбоку, то оно повернет голову в сторону раздражения или даже
повернется в эту сторону всем туловищем. Это не оборонительный
рефлекс, так как раздражение не является повреждающим и само животное
не производит агрессивных движений. Услыхав необычный звук,
животное поворачивает голову в ту сторону, откуда он слышен,
поднимает уши и направляет их на звук. Если животное видит необычный
предмет, оно направляет глаза и голову на этот предмет и даже
подходит к нему. Когда доходит неожиданный запах, у животного
расширяются ноздри, дыхание становится глубоким. Когда в рот животного
попадает незнакомая пища, оно производит характерное движение
губами и языком, направленное на лучшее соприкосновение пищи с
ротовой полостью. Назначение всех перечисленных реакций одно и то же:
наилучшее восприятие необычных раздражений, наилучшее
ознакомление с их качествами. Поэтому такие рефлексы называются
ориентировочными. У высших позвоночных животных, начиная с рептилий,
все акты поведения на внешнее раздражение начинаются с
ориентировочной реакции.
Происхождение ориентировочных реакций. Ориентировочные
рефлексы животных и человека протекают обычно совершенно непроизвольно.
Когда человек при неожиданном шуме оборачивается назад, это,
безусловно, происходит помимо его воли. Если в поле зрения сбоку
движется какой-либо предмет, сейчас же голова и глаза поворачиваются
в соответствующую сторону для лучшего восприятия движущегося
предмета. Это тоже происходит непроизвольно, бессознательно. В обоих
случаях субъект не чувствует желания производить указанные движе-
рия. Это не заученные, привычные движения. Например, у
новорожденных животных с развитой нервной системой самая тонкая
ориентировочная реакция обнаруживается сейчас же после рождения:
жеребенок, теленок, поросенок и др. в первый же день рождения обнаруживают
в совершенстве все ориентировочные реакции.
ОРИЕНТИРОВОЧНЫЕ РЕФЛЕКСЫ
345
Итак, ориентировочные реакции являются прирожденными,
наследственными рефлексами.
Нервный механизм ориентировочных рефлексов. Почти все
ориентировочные рефлексы протекают через кору больших полушарий.
Конечно, это происходит у животных, имеющих развитую кору больших
полушарий, а именно — у млекопитающих. Известно, что у высших
позвоночных— у собаки, кошки — по удалении больших полушарий
совершенно исчезают ориентировочные рефлексы на зрительные и
обонятельные раздражения, а на слуховые раздражения они вызываются в очень
слабой форме: слабые звуки совсем не вызывают ориентировочных
рефлексов; довольно сильные звуки, вроде свиста, звучания дудки,
хлопанья в ладоши, вызывают движение головы и встряхивание ушей.
Что касается вкусовых и кожно-механических раздражений, они, по-
видимому, могут производить ориентировочные реакции и помимо
больших полушарий (Гольц, 2; Зеленый, 3).
Ориентировочные рефлексы на сильный звук, на кожное
раздражение вызываются даже у децеребрированных животных, например, у
кошки (Форбс, 1), но у нормальных млекопитающих все эти
раздражения дают ориентировочные реакции, главным образом через кору
большого мозга.
У низших позвоночных — рыб, амфибий, которые не обладают
развитой корой мозга, типичные ориентировочные рефлексы отсутствуют.
У птиц, которые уже обладают развитой корой, ориентировочные
реакции могут вызываться и помимо больших полушарий (Шрадер, 4).
Координация ориентировочных рефлексов происходит в том отделе
мозга, где оканчиваются чувствительные нервы от соответствующих
чувствительных органов. Например, координация ориентировочных
движений на звуки происходит в среднем мозгу, где оканчивается п. cochle-
aris, передающий возбуждение из кортиева органа. Кора больших
полушарий совсем не принимает участия в координации ориентировочных
движений, она является лишь одной из передаточных станций.
Периферический импульс от рецептора, с одной стороны, прямо вступает в
координирующий аппарат, а с другой,— минуя этот аппарат, доходит
до коры. Отсюда уже через особые корковые участки, периферический
импульс переходит в координирующий аппарат. Таким образом,
ориентировочный рефлекс обладает двоякого рода рефлекторной дугой:
короткой, лежащей всецело за пределами больших полушарий и
состоящей из трех нейронов — чувствительного, координирующего и
двигательного,— и длинной, проходящей через большие полушария. Эта
длинная дуга должна состоять не менее чем из 6 нейронов: 1)
чувствительного — прерывающегося где-нибудь в стволовой части головного імозга
еще до больших полушарий; 2) промежуточного — связывающего этот
отдел мозга с корой больших полушарий; 3) коркового — связывающего
промежуточный нейрон с расположенными в коре же двигательными
участками; 4) коркового нисходящего пирамидного — начинающегося в
двигательных точках и заканчивающегося в координирующем аппарате;
346 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
5) координирующего промежуточного и 6) двигательного. Известно, что
слабый периферический импульс не вызывает ориентировочной реакции
через короткую дугу, в то время как через длинную он вызывает
сильную реакцию. Это явление, повидимому, объясняется тем, что кора :
больших полушарий играет роль усилителя периферических импульсов,
ибо возбуждение иррадиирует в коре мозга и тем самым вовлекает в
работу значительную ее часть.
Отличительная особенность ориентировочных рефлексов.
Отличительной чертой ориентировочных рефлексов является их быстрое
ослабление и исчезновение при быстром повторении одного и того же
обычного раздражения. Спустя некоторое время после исчезновения
ориентировочный рефлекс можно снова вызвать, повторяя то же
раздражение, но на этот раз рефлекс исчезает еще скорее. После этого
интервал, необходимый для восстановления рефлекса, еще более
удлиняется и в некоторых случаях может растянуться на несколько часов
или даже дней. Так бывает и в тех случаях, когда необычное
раздражение не оказало заметного действия на организм животного, т. е. когда
вслед за ним не возникло ни защитного, ни пищевого поведения или
поведения для сохранения рода.
Каждый врожденный рефлекс защитного типа тоже ослабевает при
повторных раздражениях и может даже совсем исчезнуть, но защитный
рефлекс быстро восстанавливается; достаточно небольшого отдыха,
чтобы вызвать его снова с большой силой. Как бы часто ни повторять этот
рефлекс, он исчезает вследствие утомления только на очень короткий
срок.
Ориентировочный рефлекс на повторное раздражение исчезает
значительно быстрее и на более длительный период времени, причем
период отдыха для восстановления бывает тем продолжительнее, чем
чаще данный рефлекс повторяется (Беритов, 5; Попов, 6).
Если же необычное раздражение вызвало наряду с ориентировочным
рефлексом какое-либо полезное для организма действие в смысле
оборонительного, пищевого или родового движения, то при повторных
раздражениях ориентировочный рефлекс может даже усилиться; в таких
случаях даже слабые раздражения могут вызвать сильную
ориентировочную реакцию.
Указанные особенности ориентировочных рефлексов характерны и
для всех тех реакций, которые вызываются у высших позвоночных через
кору больших полушарий. Таковы все индивидуально приобретенные
рефлексы.
Природа изменения ориентировочных рефлексов в связи с их
повторением, повидимому, та же, что и угасания индивидуально
приобретенных рефлексов; поэтому происхождение означенного изменения
ориентировочных рефлексов будет рассмотрено одновременно с угасанием
индивидуальных рефлексов.
ОРИЕНТИРОВОЧНЫЕ РЕФЛЕКСЫ
347
Практипум
Наблюдение ориентировочных рефлексов на голубях. Голубь помещен в клетку.
Клетка закрыта со всех сторон материей, но спереди имеется отверстие, достаточное
для наблюдения за движениями голубя. Когда голубь находится в спокойном
состоянии, дают звук какого-нибудь инструмента (свисток, звонок, метроном). В ответ
на раздражение голубь вытянет голову, посмотрит по сторонам. Может случиться, что
он сделает общее движение и даже переменит место, если только раздражение было
достаточно сильно. Когда голубь успокоится, снова производят тот же звук. Голубь
опять будет двигать головой, сделает общее движение. Если повторять звуковое
раздражение, (реакция ослабевает и, в конце концов, совсем исчезает. Если в это время
дать звучать другому инструменту, ориентировочная реакция опять появится,
причем ее можно вызывать повторно несколько раз. Если дать небольшой отдых
(5—10 мин.), ориентировочный рефлекс на то же звучание восстановится, но при
повторениях он будет ослабевать быстрее по сравнению с первым опытом..
Аналогичные опыты можно поставить на кролике.
Наблюдение ориентировочных реакций на кролике, лишенном большого мозга.
Удаляют большие полушария вместе с промежуточным мозгом. Мозговой ствол
перерезают впереди четверохолмия, но так, чтобы задние холмы не были
повреждены. Спустя 2 часа после операции кролик в ответ на любой шум поднимает уши и
приподнимает голову. На сильный шум кролик может даже подняться на ноги и
побежать.
Литература
і. Forbes a. Sherrington, Am. J. Physo1. 15, 36", 1914.
2. G о 1 t z F., PfJ. Arch., 51, 570, 1892 и другие работы в том же журнале.
3. Зеленый Г., Труды Об-ва русск. врачей в Петербурге, 1911.
'., Schrader М. J., Pf]. Arch., 41, 75, 1887.
5. Be гі toff J., Pr'L Arch., 213, 370, 1926.
6 Попов H., Русск. физиол. журн., З, 10, 1921.
Глава XX
ДИНАМИКА РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ
Общее понятие изменчивости рефлексов. Непостоянство и
изменчивость центральной реакции — явление общее. Каждый прирожденный
рефлекс подвержен изменениям. Уже первые исследователи рефлексов
отметили их изменчивость [Кюршнер (1841), (1); Пфлюгер (1853), (2);
Сеченов (1863), (3)]. Сеченов, например, наблюдал, что при
раздражении задней ноги в тот момент, когда она разогнута, ответная реакция
является оборонительным сгибательным рефлексом; если раздражение
происходит в момент сгибания той же ноги, то ответная реакция
представляет оборонительный разгибательный рефлекс.
Таким образом, на одно и то же внешнее раздражение наступает
двоякого рода оборонительный рефлекс: сгибательный и
разгибательный, и оба раза он представляет собой целесообразное удаление от
раздражителя.
Аналогичная изменчивость рефлекторной деятельности была
обнаружена целым рядом физиологов и была доказана по отношению почти
к любому рефлексу. Так, перекрестный разгибательный рефлекс легко
может замениться сгибательным. Это бывает, например, если вызывать
перекрестный рефлекс ударом молотка по коленному сухожилию одной
ноги. Другая нога будет разгибаться в том случае, если в момент
раздражения она была согнута; наоборот, она будет сгибаться, если
предварительно была разогнута (Магнус, 4). Изменяется также рефлекс по-
тирания, вызываемый на лягушке раздражением кожи около анального
отверстия. Если задние лапки были разогнуты, потирание начинается их
сгибанием; если они были согнуты, тогда оно начинается их
разгибанием (Бикель, 10).
Очень легко наблюдать изменчивость рефлекса на хвосте кошки.
Если кошка лежит на краю стола на левом боку и хвост свисает вниз,
то раздражение кончика хвоста вызывает поднятие его вверх, т. е.
отклонение вправо. Если же уложить кошку на правый бок и раздражать
хвост, то и тогда он поднимется вверх, но в этом случае отклонение
произойдет влево (Магнус, 4), (рис. 154).
ДИНАМИКА РЕФЛЕКТОРНОЙ ДЕ ТГЗТЬНОЗГД
349
Условия изменчивости рефлексов. Явление
изменчивости рефлексов подробно было исследовано
Магнусом (1910—1913). Отмеченные
Сеченовыми другими изменения рефлексов, по
исследованиям Магнуса, зависят от внутренних
раздражений. Именно, каждое пассивное движение
сопровождается раздражением проприоцепторов
мышц, сухожилий и суставов, которое производит
на центральную нервную систему определенное
влияние, вызывая в ней функциональное изменение
«eine Schaltung», по выражению Магнуса,
вследствие чего центральная нервная система
устанавливается на вызов противоположного движения.
Изменение рефлекса іможет зависеть и от
внешних раздражений. На обезглавленных кошках,
собаках, кроликах и т. д. раздражение шеи, плеч и
спины производит рефлекс чесания: задняя нога
раздраженной стороны притягивается к
раздраженному участку и производит ритмическое
движение чесания. Г е р г е н (5), ученик известного
физиолога Гольца, заметил, что если собака,
лишенная больших полушарий, лежит, например, на
правом боку, то при настойчивом и усиленном
раздражении рецептивного поля чесания с правой
стороны чесательный рефлекс может наступить, но
чесание будет производиться не правой задней
ногой, а левой. Это изменение рефлекса также было
исследовано Магнусом. Оказалось, что
основным условием этой изменчивости является
раздражение кожи на боку туловища при лежании. Если
смазыванием кокаином уничтожить здесь
чувствительность, тогда то же раздражение при том же
положении животного будет давать чесательный
рефлекс по обычному типу, т. е. на правой задней
НОГе (4) . ка лежит на левом боку
у края стола. Хвост ви-
Большую роль в изменчивости рефлекторных ре- SF чеРез кРаи вниз-
г г у г л г~ Раздражение кончика
акций играют раздражения в области вегетатив- хвоста вызывает
поданных органов, так называемые интероцептивные
раздражения. Так, например, іКоштоянц (31)
показал, что у рыб плавательный пузырь, а у
аксолотля гомолог его — легочная полость через свои
рецепторы играют важную роль в координации
движений и в тонусе мускулатуры. Выключение
этих рецепторов приводит к дисритмии и дискоординации движений и к
выпадению тонуса туловищной мускулатуры.
Рис. 154. Изменение
рефлекса на хвосте
в зависимости от его
положения.
Лумбальный препарат
децеребрированной
кошки, т. е. такой аппарат,
на котором после деце-
ребрирования била
произведена перерезка
спинного мозга на уровне XII
грудного сегмента. Кош-
тие хвоста вверх, т. е.
отклонение его вправо
(по кинематогр афическо-
му снимку Магнуса).
При другом боковом
положении на такое же
раздражение хвост
отклонился бы налево.
350 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Многочисленные факты изменчивости рефлекторных реакций были
описаны Беритовым на децеребрированных препаратах. Так,
например, на децеребрированаой кошке раздражение ноги обычно
вызывает оборонительный рефлекс сгибания на этой ноге и разгибательный
рефлекс на противоположной. Но если это раздражение производить
при различных положениях головы, отклоняя ее то вправо, то влево,
то оно вызовет шейный или лабиринтный тонический рефлекс в
отдельности или вместе с оборонительными рефлексами, причем тонические
Рис. 155. Изменение рефлекса в зависимости от
положения головы.
Децеребрированная кошка. Верхняя кривая принадлежит
разгибателю левого колена vasto-crureus, а нижняя—сгибателю
того же колена semitendinosus. Раздражают правый п. peroneus.
Это раздражение вызывает на левой регистрируемой ноге
рефлекс разгибания, которым обычно сопровождается сгибание
раздражаемой ноги В первом опыте (А) положение головы
благоприятствует тоническому разгибанию регистрируемой ноги А; во
втором опыте (В), наоборот, положение головы благоприятств\ет
тоническому сгибанию этой ноги. В обоих опытах нерв
раздражается в то время, когда регистрируемые мышцы не испытывают
тонического сокращения. В первом опыте данное раздражение
вызывает сильное сокращение разгибателя с длительным разги-
бательным последействием; во втором же опыте оно вызывает
слабое сокращение разгибателя без последействия. Теперь после
раздражения возникает длительное тоническое сгибание.
В первом опыте голова быстро повернута теменем к
регистрируемой ноге У. Р. Поэтому тоническое разгибание прекращается,
и на его месте возникает тоническое сгибание.
рефлексы бывают разгибательного или сгибательиого характера и
возникают как на раздражаемой стороне, так и на противоположной
(рис. 155, 130, 129). Как именно тонический рефлекс меняется в
зависимости от положения головы, было описано выше. Под влиянием этих
тонических рефлексов оборонительные реакции меняются более или
менее значительно (Беритов, 6). Изменение оборонительных
рефлекторных реакций под влиянием тонических подробно исследовано в
последнее время Рэ неоном и его сотрудниками (17, 18). Они, в общем,
отмечают те же факты, какие установил Б е р и т о в. Итак, каждый
прирожденный рефлекс в ответ на раздражение одного и того же
рецептивного поля меняется от момента к моменту под влиянием изменения
как внешних, так и внутренних условий.
Невозможность строго локальных раздражений. При изучении ре-^
флекторной деятельности мы до сих пор руководствовались той точкой
зрения, что производимое экспериментатором раздражение центральной
ДИНАМИКА РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ
351
нервной системы является локальным, так как оно происходит в
определенной части периферического чувствительного органа. Фактически
эта точка зрения неправильна. Мы не можем даже представить таких
идеальных условий, при которых животное испытывало бы только одно
раздражение. Представим себе, что собака или кошка спокойно лежит
на правом боку, и для большей убедительности допустим, что это
животное лишено больших полушарий, т. е. что нервная система его
лишена способности к восприятию целого ряда раздражений: животное не
видит, не сЛышит, не обоняет и т. д. Можно ли сказать о таком
животном, что центральная нервная система его пребывает в абсолютном
покое? Отнюдь нет,— и при этом условии животное получает множество
раздражений. Приведем некоторые из них:
а) Если нога согнута, то разгибатели ее пассивно растянуты, и
следовательно, рецепторы их подвергаются раздражению. Как показал
Шеррингтон (7), пассивное растяжение разгибателей одной ноги
производит миотатический рефлекс на тех же мышцах, а иногда и на
других разгибателях.
б) Животное прикасается к полу правым боком. Как указывалось
выше, такое прикосновение раздражает кожу и этим самым изменяет
функциональное состояние центральной нервной системы; это устраняет
рефлекс чесания на правой задней ноге.
в) Если голова наклонена вниз, то происходит раздражение
определенных полукружных каналов, а это, как известно, благоприятствует
общему тоническому сгибанию ног.
г) Если в это же время шея повернута набок, тогда раздражаются
рецепторы шеи, что благоприятствует тоническому разгибанию ног на
той стороне, куда обращена морда, и сгибанию их на другой стороне.
Если даже в идеальном случае спокойного состояния животного
центральная нервная система получает целый ряд раздражений, то
совершенно естественно ожидать, что она будет подвергаться несравненно
более многочисленным раздражениям, когда животное находится в
беспокойном состоянии или передвигается, в особенности если
центральная нервная система не повреждена и животное видит, слышит, обоняет
и т. д.
Но, как известно, рефлекторная реакция центральной нервной
системы на данное раздражение зависит но только от происходящих в
настоящее время раздражений, но и от предшествующих. Так, например,
как указывалось выше, у собаки вскоре после более или менее сильного
раздражения конечности, вызвавшего оборонительную реакцию, слабые
кожные раздражения той же конечности производят более
значительную реакцию. Более того, звук, вообще не вызывающий двигательной
реакции на ногах, может ее вызвать, если ему предшествует
значительный двигательный рефлекс в ответ на раздражение конечности.
Таким образом, когда исследователь производит одно избранное им
раздражение, он, безусловно, присоединяет его к целому ряду других,
уже существующих, а также предшествующих. Мы видели, как велико
352 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
значение всей 'суммы раздражений для течения рефлекса в одном
определенном направлении, как мало нужно іменять во внешних условиях,
чтобы рефлекс существенно изменился в ответ на определенное
раздражение. Итак, когда исследователь производит какое-либо раздражение,
он не получит такого локального эффекта, который соответствовал бы
только ранному раздражению. Эффект будет более или менее сложным,
соответствующим целому ряду раздражений, вследствие активирования
разнородных координирующих центров (Беритов, 8). На этом
основании «можно утверждать, что деятельность центральной нервной системы
в ответ на данное раздражение всегда является более или менее
сложной и притом целостной, ибо она представляет собой объединенную
реакцию в ответ на всю сумму предшествующих и притекающих в
данный момент афферентных импульсов. Ввиду того что вся система
активных нервных центров, участвующих в рефлекторной реакции на
данное раздражение, меняется от момента к моменту в зависимости от их
функционального состояния,— можно называть эту систему
функциональной, как предлагает Анохин (40). Понятие функциональной
системы предполагает, во-первых, что рефлекторная реакция
нормального животного на данное раздражение всегда является результатом
сложной и целостной деятельности всего мозга, и, во-вторых, что эта
реакция постоянно меняется от момента к моменту благодаря
беспрерывным изменениям в функциональном состоянии центральной нервной
системы, возникающим под влиянием многих предшествующих и
одновременных внешних и внутренних раздражений.
Происхождение изменчивости рефлекса. Главный вопрос
заключается в выяснении причины изменения рефлексов от побочных
раздражений. Выше было показано, что раздражение вызывает не только
возбуждение, но и повышение возбудимости как в тех промежуточных и
двигательных нейронах, с которыми первично связаны возбужденные
чувствительные нейроны, так и во всех нервных путях центральной
нервной системы, по которым возбуждение передается из одного отдела
мозга в другие. Кроме того, нами выше показано, что возбужденные
чувствительные волокна активируют нейропиль как в том отделе мозга,
куда они вступают первично, так и по всему пути их распространения.
Этим обусловливается общее торможение всех остальных нервных
путей и нервных центров. Все указанные явления играют существенную
роль в изменении рефлексов. Когда в каком-либо отделе центральной
нервной системы от определенного раздражения возбудимость
повышается, то рефлекс может вызываться из этого отдела центральной
нервной системы под влиянием раздражений как того же рецептивного
поля, так и других полей. Когда же, наоборот, в каком-либо отделе
мозга возбудимость значительно понижается, рефлекс не вызывается из
этого отдела не только при раздражении посторонних рецептивных полей,
но и соответствующего рецептивного поля.
Близкое соседство координирующих аппаратов имеет также большое
ДИНАМИКА РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ 353
значение в изменениях рефлекса. Известны факты, которые указывают
на то, что влияние одной рефлекторной реакции на другую тем сильнее,
чем ближе расположены координирующие аппараты этих рефлексов
(Беритов, ?). Приведем результаты анализа нескольких характерных
случаев изменчивости рефлекса.
1. Как было показано, координирующий аппарат рефлекса потирания
у лягушки заложен в грудных и в I поясничном сегментах, а
координирующий аппарат сгибания и разгибания задних ног — во II и III
поясничных сегментах. Рефлекс потирания обычно нельзя вызвать
раздражением рецептивного поля сгибания, но если грудные сегменты
отравить локально с задней поверхности стрихнином и тем поднять
возбудимость координирующего аппарата рефлекса потирания, то можно
вызвать этот рефлекс и с поля рефлекса сгибания (Беритов, 9).
2. Как было отмечено выше, на децеребрированном препарате
сначала развивается общая тоническая экстензия, а затем общая
тоническая флексия. Известно, что и тот и другой тонус зависят от
возбуждения соответствующих координирующих аппаратов, происходящего под
влиянием раздражений, вызванных разрезом мозга и вообще операцией
на голове. Одновременно с возбуждением в этих аппаратах происходит
повышение возбудимости. Этим нужно объяснить то характерное
явление, что на децеребрированном препарате каждое раздражение
действует на координирующие аппараты определенных тонических
рефлексов. Так, например, слабое раздражение задней ноги в рецептивном
поле сгибания обычно вызывает оборонительный рефлекс сгибания, а во
время экстензорного тонуса, вследствие повышения возбудимости
координирующих аппаратов последнего, око усиливает существующую
тоническую экстензию; наоборот, во время флексорного тонуса действие
того же раздражения только усиливает тоническую флексию. Если
периферическое раздражение более или :менее сильно, то оно вызывает
оборонительный сгибательный рефлекс вместе с тем или другим
тоническим рефлексом (Беритов, 6).
3. Как было сказано, пассивное сгибание ноги благоприятствует
разгибательному рефлексу, а пассивное разгибание — сгибательному.
Это изменение происходит от того, что при пассивном сгибании
растягиваются разгибатели и производимое при этом раздражение их
рецепторов действует на центральную нервную систему. Действие
выражается прежде всего в возбуждении и повышении возбудимости
двигательных нейронов разгибателей, но в некоторой малой степени, в виде
субминимального раздражения, оно должно действовать и на
координирующий аппарат разгибания, повышая в нем возбудимость. Поэтому
последующее периферическое раздражение ноги произведет эффект
прежде всего из данного координирующего аппарата. При пассивном
разгибании будут растягиваться сгибатели, и раздражение
рецепторов сгибателей будет действовать на центральную нервную систему,
вызывая повышение возбудимости в координирующем аппарате сгибания
-3 и. С. Беритов
354 общая физиология спинного мозга и стіюла головного мозга
и в двигательных нейронах сгибателей. Вследствие этого последующее
периферическое раздражение вызовет прежде всего рефлекс сгибания.
4. Изменение всех тонических рефлексов в зависимости от
положения головы основывается также на повышении возбудимости в
определенных координирующих аппаратах. Когда положение головы меняется
осторожно, это не вызывает рефлекса, но этого должно быть достаточно
Рис. 156. Изменчивость рефлекторных реакций в зависимости от
положения головы.
Децеребрированная кошка. Наверху кривая разгибателя локтевого сустава m. triceps
Ьг., внизу сгибатель его — biceps Ьг. Раздражается лучевой нерв. Сила раздражения
пороговая. В опыте Л и С положение головы благоприятствует экстензорному тонусу,
а в опыте В — флексорному. В последнем случае наступает ритмический рефлекс с
антагонистическим участием обеих мышц, в первом и третьем случаях — сильное
тоническое разгибание с кратковременным сгиоанием вначале.
для повышения возбудимости в тех координирующих аппаратах, на
которые действуют раздраженные рецепторы растянутых мышц шеи и
статического органа. Только по этой причине всякое периферическое
раздражение производит вполне определенный тонический рефлекс, как
раз тот самый, который соответствует данному положению головы
(рис. 155).
5. В определенных случаях децеребрированные препараты кошки в
отсутствии децеребрационной экстензорной ригидности склонны давать
при кожных раздражениях конечностей ритмический рефлекс типа
шагания. На таких препаратах при положениях головы, благоприятных
экстензорному тонусу данной конечности, периферическое раздражение
производит тоническое сокращение, а при положении головы,
благоприятном для флексорного тонуса, то *:ке самое периферическое
раздражение производит на задних ногах ритмический рефлекс шагания
ДИНАМИКА РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ 355
(рис. 156). Очевидно, это происходит вследствие того, что возбудимость
в спинномозговых центрах ритмического рефлекса шагания сильно
повышена под влиянием раневого раздражения центральной нервной
системы. Поэтому они легко (вовлекаются в реакцию. Когда двигательные
нейроны не захвачены реципрокной иннервацией сильного экстензорно-
го тонуса, периферическое раздражение активирует их через центры
ритмического рефлекса.
6. Отмеченное выше устранение рефлекса чесания на правом боку
децеребрированной собаки, в случае лежания ее на этом боку,
объясняется следующим образом. Раздражение кожи на боку от давления
производит общее торможение, особенно сильное в. правой половине
спинного мозга. Вследствие этого раздражение рецептивного поля
рефлекса чесания на правой стороне не вызовет на ней этого рефлекса. Но
если применить сравнительно сильное раздражение, рефлекс чесания
может наступить на другой стороне благодаря иррадиации (возбуждения
на координирующий аппарат чесательного рефлекса противоположной
половины тела.
Итак, изменение рефлекторной деятельности в ответ на определенное
раздражение зависит главным образом от того, что одновременно с
координирующим аппаратом, на который действует раздраженное
рецептивное поле, возбуждаются и другие координирующие аппараты,
возбудимость которых сравнительно повышена под влиянием каких-
либо внутренних или внешних раздражений.
Это правило совозбуждения касается только нормальной
центральной нервной системы, которая вследствие различных раздражений,
испытываемых ею, несомненно должна иметь в одних отделах или в
одних нервных комплексах большую возбудимость, чем в других.
Шеррингтон и его сотрудники несколько иначе объясняют изменение
рефлекторных реакций на децеребрированных препаратах в зависимости от положения
головы, а также от общего тонического состояния конечностей. Они полагают, что
изменение обусловливается соответствующим функциональным изменением
двигательных элементов. Так, например, при лабиринтных разгибательных рефлексах или при
положении головы, благоприятном для этих рефлексов, двигательные элементы
экстензоров находятся в состоянии «подпорогового» возбуждения. Так упомянутые авторы
называют в данном случае состояние, которое мы называем состоянием повышенной
возбудимости. Поэтому периферическое раздражение приводит эти элементы в
активное состояние по преимуществу. Нельзя отрицать, что при активном тоническом
рефлексе двигательные элементы определенных мышц находятся в состоянии
повышенной возбудимости, но роль их в извращениях рефлексов должна быть
второстепенной. Периферическое раздражение прежде всего действует на координирующие
аппараты, а мы знаем, как легко они вовлекаются в сферу реакции иррадиирующим
возбуждением, если только в них в некоторой мере повышена возбудимость под
влиянием активных или субминимальнык периферических раздражений.
Изменчивость рефлекторных реакций в зависимости от характера
самого раздражения. Выставленное выше положение о роли побочных
раздражений в изменчивости рефлекторных реакций является
основным по своему значению в нормальной деятельности центральной нерв-
23*
355 ОБЩАЯ ФИЗИОЛОГИЯ СШШНОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
ной системы. Но в определенных условиях на характер рефлекторной
реакции влияет также характер самого раздражения, его качественные
особенности, его сила и длительность.
Обычно одна и та же кожная поверхность служит рецептивным
полем разного рода рефлексов. Это обусловливается тем, что на небольшом
участке кожной поверхности имеются
разного рода рецепторы, которые
возбуждаются при качественно разных воздействиях.
Так, например, при легком давлении на
внутреннюю поверхность стопы лягушки
наступает быстро протекающий разгиба-
тельный рефлекс, длящийся не более 1 сек.
Продолжительность его не зависит от силы
или продолжительности давления. Он
представляет собой не что иное, как рефлекс
прыжка (Балиони, 11; Беритов, 9). Обычно
после рефлекса прыжка наступает
тоническое сгибание, которое выражает переход
животного в сидячее положение. Это
тоническое сгибание наступает, повидимому,
под івлиянием самого экстензорного толчка
от производимых им кожных и проприо-
цептивных раздражений. Такой же рефлекс
вызывается с вентральной стороны стопы
при слабом раздражении фарадическим
током (рис. 157) или же при раздражении
чувствительного нерва п. superficialis, от-
кожи (Беритов, 12, 9). Аналогичный экстен-
зорный рефлекс от давления на вентральную поверхность ступни
наблюдается на спинальных собаках (Шеррингтон, 13).
При сильном раздражении кожи на внутренней стороне лапки,
электрическом или імеханическом, рефлекс прыжка вначале отсутствует.
Рефлекс начинается оборонительным сгибанием, а уже потом это
сгибание прерывается на один момент рефлексом прыжка. Часто смена
реакции происходит после раздражения. Очевидно, это изменение ре
флекторных реакций обусловливается тем, что при слабом
механическом или электрическом раздражении возбуждается преимущественно
координирующий аппарат рефлекса прыжка, а при сильном
раздражении возбуждается также координирующий аппарат сгибательного
рефлекса. В этом случае получается комбинация того и другого
рефлексов. Так, на рис. 158 от сильного раздражения кожи над ахилловым
сухожилием разгибательный рефлекс прыжка наступает после
раздражения; в это время сгибательный рефлекс прерывается на короткий
момент. Но на этом же препарате при более слабом раздражении
рефлекс прыжка наступает в самом начале, причем смена прыжка на
сгибание происходит еще во время раздражения.
Рис. 157. Рефлекс прыжка и
последующее тоническое
сгибание.
Спинномозговая лягушка. Верхняя
кривая от m. triceps femoris.
нижняя т. semitendinosus.
Раздражают кожу над ахилловым
сухожилием фарадическим током
(частота— 100, сила— 10 см
расстояния между катушками). Рефлекс
прыжка сменяется тоническим
сгибанием.
ходящего от этого участка
ДИНАМИКА РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ 357
При раздражении чувствительных нервных стволов, получающих
нервные волокна от разных рецептивных полей, изменения рефлектор-
Рис. 158. Рефлекс прыжка после оборонительного
сгибания.
Спинномозговая лягушка. Верхняя кривая от m. semitendinosus,
нижняя — от triceps fem. Раздражается кожа над ахилловым
сухожилием. Частота индукционных ударов — 20 в сек , а сила—¦
10 см расстояния между катушками. Во время раздражения
наступает сгибательный оборонительный рефлекс; после
раздражения происходит сначала разгибание на один короткий
момент, а затем тоническое сгибание.
ных реакций наблюдаются очень часто в зависимости от силы и
продолжительности раздражения. Как известно, чувствительные волокна
по своей возбудимости и скорости проведения распадаются на
несколько групп. Имеются указания
на то, что они начинаются
в разного рода рецепторах.
Следовательно, при
некоторых малых силах
раздражения будет возбуждаться
одна надболее возбудимая
группа1 волокон — и
соответственно рефлекторная
реакция будет одного типа.
При некоторой умеренной
силе будет возбуждаться и
другая группа волокон
умеренной возбудимости —
соответственно, конечно, будет
меняться и рефлекторная
реакция. С усилением
раздражения она может извратиться один или два раза (Грэим Броуч, 15;
Рис. 159. Рефлеко сгибания и разгибания при
различных силах раздражения.
Спинномозговая лягушка. Верхняя кривая от triceps
fem. нижняя—отш. semitendinosus. Раздражается IX
задний корешок (частота—50 в f сек ). Опыты А и О
при слабых раздражениях (30 см). Опыты В и D при
сильных раздражениях (23 см), а опыт Е при
умеренной силе. Внизу—время в секундах.
35R общая физиология спинного мозга и ствола головного мозга
Беритов, 14, 12, 9). Это положение иллюстрирует рис. 159: слабое
раздражение вызывает значительное сгибание, сильное раздражение —
сильное разгибание, а среднее раздражение дает слабый,
неопределенный эффект. Очевидно, при слабом раздражении пришел в активное
состояние только координирующий аппарат сгибательного рефлекса, при
сильном — работали координирующие аппараты сгибательного и разги-
бательного рефлексов, но рещшрокная иннервация разгибания была
Рис. 160. Смена рефлекса потираний рефлексом сгибания при повторном
раздражении кожи над ахилловым сухожилием.
Спинномозговая лягушка. Верхняя кривая m. triceps fern., нижняя — т. semitendinosus.
Сила раздражения все время одна — 11.5 см. В опыте А раздражение повторяется 7 раз:
по 10 сек. раздражения и по 10 сек. отдыха. Вначале сокращается только разгибатель,
а седьмой раз только сгибатель. В опыте В. произведенном после 2-минутного отдыха,
сокращается один разгибатель. Зубцы на кривой сокращения указывают на повторные движения
потираний. Внизу время: 1 зубец — 2 сек.
сильнее сгибания; при умеренной же силе раздражения оба аппарата
давали реципрокную иннервацию более или менее одинаковой
интенсивности, а потому периферический эффект того и другого аппарата
оказался взаимно заторможенным.
Извращения рефлекторных реакций наблюдается точно так же в
связи с продолжением или повторением раздражения. Так, например,
при длительном раздражении в рецептивном поле сгибания сгибатель-
ный рефлекс может смениться рефлексом разгибания (Шеррингтон, 13,
Введенский и Ухтомский, 41). Рефлекс потирания, вызываемый
раздражением кожи над ахилловым сухожилием, может смениться
рефлексом сгибания, как это показано, например, на рис. 160 (Беритов, 7, 14).
Это изменение рефлекса обусловливается разной утомляемостью
координирующих аппаратов конкурирующих рефлексов. Очевидно, с
самого начала возбуждаются координирующие аппараты обоих рефлексов,
но так как реципрокная иннервация из аппарата рефлекса потирания
оказывается сильнее сгибательной, то вначале верх берет первая.
С повторением опыта координирующий аппарат потирания утомляется.
Это утомление наступает раньше, чем в координирующем аппарате
сгибания, а потому, когда потирание перестает вызываться, наступает
рефлекс сгибания. После длительного перерыва опять получается рефлекс
потирания вследствие отдыха координирующего аппарата потирания.
Такое извращение рефлекторных реакций в связи с усилением и
продолжением раздражения чувствительного нерва со смешанным содер-
ДИНЛЛГИК\ РРФЛЕКТОРНОИ ДЕЯТЕЛЬНОСТИ 359
жанием волокон представляет собой обычное явление. Шеррингтон
даже кладет именно это обстоятельство в основу всех ненормальных
реакций или, вернее, изменчивости рефлекторных реакций, но, как мы
\же подробно рассмотрели выше, основным условием изменчивости этих
реакций является внутреннее функциональное состояние центральной
нервной системы, а не состав нерва и характер действующего на него
раздражения. Это обстоятельство играет определенную роль в чисто
лабораторных условиях, когда деятельность рефлекторного аппарата
вызывается раздражением нервных стволов. В нормальной
жизнедеятельности организма изменчивость рефлекторных реакций меньше всего
зависит от одновременного возбуждения одним и тем же раздражением
двух рецептивных полей или отходящих от них нервных волокон.
Итак, динамика деятельности центральной нервной системы может
быть изображена в немногих словах так. В нормальном и
бодрствующем состоянии животное не дает простых и локальных
совершенно неизменных рефлексов. Центральная реакция всегда более или
менее сложна и изменчива. Это зависит от того, что животное в
бодрствующем состоянии каждый миг получает целый ряд внутренних и
внешних раздражений. Каждое раздражение повышает возбудимость
прежде всего и больше всего в тех нервных кругах и нервных
цепочках, на которые оно действует сильнее всего, а потом, более умеренно,
и в других нервных кругах, по которым возбуждение иррадиирует.
Одновременно каждое раздражение, действуя на нейропиль, производит
общее торможение ближайших нервных кругов и двигательных
нейронов. Повышенная возбудимость удерживается на более или менее
продолжительное время и после раздражения. Результатом всего этого яв--
ляется тот факт, что действие каждого нового раздражения не
ограничивается одним центральным аппаратом. Возбуждение, иррадиируя по
центральной нервной системе, вызывает деятельность во всех тех
нервных комплексах, где возбудимость быля достаточно повышена, поэтому
рефлекторный эффект всегда возникает в результате соучастия целого
ряда центральных нервных образований. Координированный,
направленный характер данного рефлекторного эффекта происходит от того, что,
с одной стороны, реципрокные иннервации одних координирующих
аппаратов согласованно сочетаются с иннервациями других; а с другой
стороны, когда эти реципрокные иннервации имеют противоположный
характер, внешний эффект одних, более слабых иннервации
уничтожается, т. е. тормозится, под влиянием других более сильных иннервации.
Об изменении рефлекторной деятельности вследствие
перекрещивания нервов. Характерные изменения наблюдаются в центральной
деятельности высших позвоночных животных при перекрещивании нервов
(Флуранс, 30; Бете, 16; Кеннеди, 28; Осборн и Кильвингтон, 29;
Анохин, 27). Методика заключается в перерезке двух нервов и сшивании
центрального конца одного нерва с периферическим концом другого как
той же конечности, так и другой, с ней одноименной. После врастания
первого нерва в рабочий орган второго паралич последнего проходит, но
360 общая физиология спинного мозга и ствола головного мозга
он вступает в такие отношения с центральной нервной системой, каких
у него раньше не было.
Первое такое сшивание было произведено Флурансом (1842)
на курах. Он сшил центральный отрэзок нерва опускателей и
сгибателей крыла с периферическим концом нерва подчимателей и
разгибателей того же крыла. Так как центральный отрезок нерва сохраняет
способность к росту, то сгибательный нерв врастает в мышцы-разгибатели,
а разгибательный нерв — в мышцы-сгибатели. Таким образом,
кардинально было изменено анатомическое отношение координирующего
аппарата к мышцам. Несмотря на это, спустя несколько месяцев, вполне
восстановилась нормальная функция крыла: при полете оно
действовало как нормальное крыло. Очевидно, координирующие аппараты крыла
перестроились,— они как бы потеряли свою прирожденную форму
деятельности и восприняли новую форму.
Происхождение изменчивости рефлекторной деятельности в
зависимости от оперативного изменения нервных связей стало предметом
специального исследования в самое последнее время; многочисленные
анастомозы нервов произвели для этой цели Анохин иАсратян. Так,
например, Анохин установил анастомоз імежду блуждающим нервом и
plexus brachialis. Центральный конец блуждающего нерва был сшит с
периферическим концом плечевого сплетения. После восстановления
двигательной и чувствительной функции передней конечности
раздражение последней (почесывание, разминание) первоначально вызывает
рефлекторным путем явления кашля и рвоты, характерные для
раздражения чувствительных волокон блуждающего нерва. Но спустя
некоторое время, через 10—12 месяцев, те же раздражения передней
конечности перестают давать эти реакции; вместо них они вызывают
характерные для данной конечности локальные двигательные реакции. При
болевом раздражении от уколов ,в это время еще вызывалось рвотное
движение, но впоследствии и оно исчезало. Такое раздражение
вызывало оборонительную реакцию ноги и взвизгивание, как у нормальной
собаки.
Таким образом, первоначально раздражение рецепторов передней
конечности давало совершенно необычную для этих рецепторов
реакцию. Очевидно, рефлекторная реакция г* ответ на раздражение передней
конечности изменилась по существу благодаря тому, что в переднюю
конечность вросли чувствительные нервы, которые передают
периферические импульсы тем координирующим аппаратам продолговатого мозга*
которые производят кашлевые и рвотные движения. Впоследствии
установилась нормальная рефлекторная реакция на раздражение конечности
в смысле оборонительного сгибания. Анохин предположил, что в этом
случае центры продолговатого мозга кардинально изменили свою
форму деятельности: на те периферические импульсы, нга которые они
раньше отвечали рвотными движениями, они теперь стали отвечать сгиба-
тельным рефлексом. Произошло не просто кратковременное
функциональное изменение нервных центров, а основательная перестройка их
ДИНАМИКА РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ 361
путем установления новых нервных связей в отношении органа, нервы
которого были перекрещены. Анохин полагает, что после того как
произошло врастание чувствительных и двигательных нервных волокон
блуждающего нерва в мышцы передней конечности, каждый раз при
движении конечности от возбуждения двигательных волокон
блуждающего нерва раздражаются проприоцелторы мышц, сухожилий и
суставов передней ноги. От этих рецепторов возбуждение дередается по
чувствительным волокнам блуждающего нерва в ядро последнего. Это
приводит к взаимодействию между определенными ipynnavn двигательных
и чувствительных нейронов в ядре блуждающего нерва, всчедствие чего
между ними устанавливаются новые синаптические отношения, новые
нервные связи, подобно тому как это имеет место, по данным К о г-
х и л л а (30), при образовании рефлекторных дуг в эмбриональном
периоде развития нервной системы амблистомы. Таким образом, по
Анохину, ядро блуждающего нерва,— вернее, продолговатый мозг,—
так перестраивает свою работу, что вполне заменяет работу
координирующих центров, образовавшихся эмбрионально в плечевом отделе
спинного мозга и до момента перекреста нервов, управлявших
координацией движений передней ноги.
Однако из опытов Асратяна (32) ясно видно, что во всех
опытах Анохина, а также других авторов, работавших над птицами и
млекопитающими, все вышеописанные явления восстановления
нормальной рефлекторной деятельности после перекрещивания нервов
обусловливаются деятельностью коры больших полушарий. Именно,
А с р а т я н установил, что если после восстановления нормальной
функции, нарушенной перекрестным сшиванием блуждающего и
медиального нервов, удалить большие полушария, то нормальное
функционирование исчезает: раздражение кожи конечности (почесывание,
разминание) теперь вновь производит явления кашля и рвоты,
характерные для раздражения блуждающего нерва.
Асратян же показал, что разного рода компенсации, какие
получаются после ампутации конечностей, половинной перерезки спинного
мозга, разрушения лабиринтов, совершенно исчезают после удаления
больших полушарий и совсем не наступают, если полушария были
предварительно удалены.
На основании этих данных можно утверждать, что у высших
позвоночных все известные компенсации рефлекторных нарушений являются
функцией коры больших полушарий или, точнее,
индивидуально-приобретенными рефлексами.
Аналогичные опыты, проведенные под руководством Асратяна
на низших позвоночных, как птицы и амфибии, свидетельствуют о *
том, что у них компенсация некоторых рефлекторных нарушений может
происходить и без участия большого мозга. В отношении птиц (голуби)
показано, что рефлекторные нарушения, вызванные односторонней
половинной перерезкой спинного мозга, восстанавливаются почти в полной
мере при участии большого мозга, хотя в известной степени восстанов-
362 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
ление может происходить и без его участия (Карамян, 33). У амфибий
(лягушки) рефлекторные нарушения, вызванные половинной перерезкой
спиннгго мозга, компенсируются вообще без участия не только
большого мозга, но и промежуточного (Вишневецкая, 32). Отсюда следует, что
у низших позвоночных животных компенсация некоторых рефлекторных
нарушений или перестройка рефлекторной деятельности
обусловливается не только большим мозгом, но и нижележащими отделами головного
мозга.
Перестройка рефлекторной деятельности изучалась также в связи с
трансплантацией сухожильного конца одной мышцы к месту
прикрепления другой, антагонистической мышцы, а также в связи с врастанием
-периферического конца нерва одной мышцы в антагонистическую
мышцу. Трансплантация сухожильных концов производилась на главных
мышцах кошки, собаки и обезьяны. Вскоре после срастания сухожилий
наблюдалось полное функциональное приспособление почти без
предварительной стадии тренировки (Марина, 34; Ольмстед, Моргути, Ианаги-
сана, 35). Такое же быстрое приспособление функций наблюдалось в
отношении мышц задней конечности кошки: часть разгибателя
четырехглавой мышцы коленного сустава переставлялась в такое место, что
при сокращении она должна была производить сгибание, а не
разгибание. После сживления эта часть восприняла сгибательную
функцию: сокращалась в рефлексе сгибания (Анохин, 27). Особенно много
случаев таких перестроек известно на людях: любая синергетическая
группа мышц может быть диссоциирована путем трансплантации
соответствующих сухожильных концов, т. е. путем превращения в
антагонистическую (Шерб, 36). Но у амфибий не наблюдается
аналогичной перестройки при такой же трансплантации сухожильных концов
(Вейсс, 37); не наблюдается она и у низших млекопитающих, как
крысы (Сперри, 38). На этом основании нужно заключить, что способность
перестраивать основные формы координированных движений
конечности путем тренировки развивается постепенно от низших позвоночных
к высшим (Сперри, 38). Это заключение хорошо согласуется,
во-первых, с данным выше выводом — что перестройка в основном
происходит при участии коры большого мозга путем образования
индивидуально приобретенных реакций, а во-вторых, с установленным нами
положением, что способность к образованию автоматизированной
индивидуальной рефлекторной реакции почти отсутствует у амфибий и
что она значительно ниже у низших млекопитающих, например,
кроликов, чем у высших — например собак (Беритов, 39).
Итак, наряду с изменчивостью рефлекторной деятельности на основе
функциональных изменений координирующих аппаратов, непрестанно
происходящих под влиянием внешних и внутренних раздражений,
известна еще такая изменчивость рефлекторной деятельности, которая
основывается на возникновении новых нервных связей с периферическим
органом, а также новых морфологических соотношений в центральной
нервной системе.
ДИНАМИКА РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ
363
Гипотеза о пластичности центральной нервной системы. Явления изменчивости
рефлекторной деятельности вследствие функционального изменения центральной
нервной системы, а также вследствие возникновения новых структурных соотношений в
ней дали повод к отрицанию локализации центров и существования определенных
нервных путей для каждого рефлекса. Именно Бете и Фишер (16) полагают, что
организм обладает способностью приспосабливаться к внешним и внутренним
изменениям через центральную нервную систему и тем самым всякий раз проявляет
целесообразную работу отдельных органов. Эту способность Бете называет
пластичностью и полагает, что в явлениях пластичности, т. е. в новых установках функций,
ответственны главным образом центральные нервные аппараты, но определенным
образом участвуют также рецепторы и эффекторы. При создании этой теории авторы
основываются на известных фактах. Они, между прочим, приводят ряд фактов замены
фуькции одной комичности другой на человеке, на четвероногих животных,
насекомых, раках, многоножках, морских звездах. У всех этих животных удаление одной
или нескольких конечностей не устраняет локомоции; она осуществляется путем
остальных конечностей, но при этом роль каждой из них изменена сравнительно с
нормой. Другой ряд фактов касается изменений локомоторной деятельности в связи с
фиксацией одной или нескольких конечностей. Приводятся также отмеченные выше
опыты с перекрещиванием нервных стволов и пересадкой сухожильных концов одной
мышцы к месту прикрепления других мышц.
Авторы отказываются дать объяснение своей новой теории, они считают это
преждевременным. Но на основании собранного материала Бете со своим
сотрудником находят, что классическая теория центров Шеррингтона, Магнуса и др.
требует в строгом смысле слова «предустановления» всего того, что может
происходить в центральной нервной системе. Факты же, по мнению Бете, все более и
более приводят нас к признанию той теории, что на базе известной предварительной
стабилизации все время происходит индивидуальная эволюция, в процессе которой в
каждый момент могут открыться новые возможности (16).
Приводимый авторами фактический материал касается не только рефлекторных
реакций, но и актов поведения. Если иметь в виду только факты изменчивости
рефлекторных реакций, то нужно сказать, что они нами уже объяснены или могут быть
объяснены на основании функционального изменения центральной нервной системы
в виде повышения или понижения возбудимости то в одних, то в других нервных
образованиях под влиянием разного рода внешних и внутренних раздражений, а также, в
определенных случаях, на основании образования новых связей. В изменчивости
рефлекторных реакций, соответственно изменениям внешней и внутренней среды,
безусловно проявляется пластичность, приспосабливаемость центральной нервной системы,
но в этих явлениях нет ничего загадочного, противоречащего теории локализации
центров и проводящих путей. Нужно только признать, согласно новейшим
морфологическим и физиологическим данным, что не только анатомическая локализация
нервных центров и проводящих путей определяет характер центральной деятельности в
ответ на данное раздражение, но также их функциональное состояние, которое
создается в них изменчивым действием внешней и внутренней среды.
Мы не можем входить в рассмотрение природы изменения актов поведения
животных и человека, так как акты поведения не являются предметом данного руководства.
Но, конечно, приспосабливаемость нормального животного и человека к внешней
среде, как она проявляется в актах поведения, имеет в своей основе закономерности,
отличные от тех, которые излагались нами выше по поводу изменчивости
рефлекторных реакций.
Теория доминанты. В последнее время было предложено несколько теорий для
понимания динамики центральной деятельности. Одна из этих теорий принадлежит
А. Ухтомскому. Формула ее такова: если в каком-нибудь центре протекает
достаточно стойкое возбуждение, то оно становится фактором для деятельности остальных
центров; возбужденный центр притягизает возбуждение из всех других источников и,
кроме того, тормозит деятельное состояние остальных центров, которое должно было
364 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛ \ ГОЛОВНОГО МОЗГ\
возникнуть в ответ на соответствующие периферические раздражения Ввиду того, что
это стойкое возбуждение является господствующим фактором для деятельности вс^й
центральной нервной системы, принцип, на котором зиждется эта деятельность,
Ухтомский назвал принципом доминанты (1) Учение о доминанте включаег в
себя два гипотетических момента первый — тормозящее влияние из очага стойкого
возбуждения одного центра на другие, второй — притягивание возбуждения из
других центров и усиление вследствие этого собственной деятельности доминантною
центра
Основанием для .первого момента доминанты Ухтомский считает, между
прочим, тот факт, что во время актов глотания и дефекации задние конечности испы
тывают торможение (20) Но, как было указано выше, это явление обусловлено тем,
что в координирующих аппаратах глотания и дефекации вместе с возбуждением по
отношению к одной группе двигательных нейронов, обслуживающих органы глотания
и дефекации, возникает общее торможение влияющее на всю остальную группу дви
гательных нейронов
Основанием для второго момента Ухтомский считает то наблюдение, что во
время рефлексов глотания и дефекации раздражение коры больших полушарий, обычно
вызывающее движения конечностей, теперь уже их не вызывает, а если и вызывает
то в очень слабой степени, но зато усиливает существующую реакцию —рефлексы
глотания и дефекации. Согласно теории доминанты, это явление истолковывается
следующим образом возбуждение, возникшее в больших полушариях — в двигатель
ной области задних конечностей, изменяет свой обычный путь к задним конечностям
и направляется вместо этого к центрам глотания и дефекации, усиливая их деятель
ность Мы объясняем это явление так, как было указано выше если какой нлбудъ
координирующий аппарат приходит в деятельное состояние, то в нем происходчт
повышение возбудимости, которое иногда может сохраниться в течение долгого времени
Например в координирующем аппарате глотания эта фаза повышенной возбуди
мости растягивается до 20 сек (Исаяма, 21) Понятно, что возбудимость в
координирующих аппаратах глотания и дефекации может возрасти настолько, что под вли
янием возбуждений, иррадиирующих из двигательных точек задних конечностей по
экстрапирамидным путям, эти аппараты придут в активное состояние, или же уси
лится существующее в них активное состояние Если одновременно с этим не вызы
вается соответствующий рефлекс на задних конечностях, то происходит это только
потому, что во время усиленного глотания и дефекации двигательные нейроны задних
конечностей испытывают усиленное торможение из тех же координирующих аппаратов
глотания и дефекации
Для доказательства второго момента также приводят установленный мной факт
что на спинномозговой лягушке, при отравлении стрихнином координирующего аппа
рата рефлекса потирания, этот рефлекс можно вызывать и из поля сгибательного реф-
jeKca, как если бы вследствие повышения возбудимости координирующий аппарат
потирания стал бы притягивать возбуждение из координирующего аппарата сгибания
Но одновременно с обнаружением этого факга мы подробно изучили и причины его
происхождения Было доказано, что при раздражении рецептивного поля сгибания в
деятельное состояние приходит как координирующий аппарат потирания, возбудимость
которого была повышена, так и нормальный координирующий аппарат сгибания Когда
обе иннервации сталкиваются в общем конечном пути при двигательных нейронах,
эффект реципрокной иннервации из аппарата сгибания, как более слабый, легко
тормозится под влиянием сильной противоположной иннервации из координирующего
аппарата потирания (Беритов, 22)
Последователи теории доминанты из лаборатории Ухтомского (Виноградов
26, Ветюков и др ) приводят и другие наблюдения с целью ее подтверждения, но все
они могут быть хорошо объяснены, как и вышеприведенные, помимо теории доминан
ты (Беритов, 23)
Впоследствии Ухтомский (24) связал теорию доминанты с теорией парабиоза
Введенского Он выдвинул положение, что — «доминанта е^ть преддверие пара-
ДИНАМИКА РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ 365
<5ноза». Прекращение доминанты ставилось в зависимость от усиления
концентрируемого или накапливаемого в доминантном очаге возбуждения. Это усиление
возбуждения создает в доминантном очаге торможение, пессимальный эффект,
согласно теории Введенского о парабиозе нервно-мышечного препарата. Значит,
прекращение доминантного процесса обусловливается наступлением полного парабиоза
в доминантном очаге возбуждения. Это предполагает, что в центрах, наряду с реци-
*
прокным торможением, существует парабиотическое торможение. По этому поводу
был также произведен ряд исследований (Конради, 25 и др.), но мы не считаем
нужным входить в их разбор для выяснения правильности этого нового положения,
поскольку мы находим, что основные явления центральной деятельности можно
объяснить убедительнее и без гипотетических допущений теории доминанты.
Практикум
1. Наблюдение изменчивости рефлексов на лягушке. Опыт Сеченова. 1.
Спинномозговой препарат лягушки. Изменение оборонительного рефлекса задней
конечности при раздражении ее в рецептивном поле сгибания в зависимости от
положения ноги. Когда задние конечности притянуты к телу, сильное раздражение
пальцев вызывает разгибание ног; когда же они разогнуты, то же раздражение
производит их сгибание.
2. Опыт Бикеля. Изменение рефлекса потирания, вызызаемого раздражением
кожи около анального отверстия. Если обе ноги согнуты, раздражение прежде всего
вызывает разгибание. Если они разогнуты, то же раздражение сначала производит их
сгибание. Если одна нога согнута, а другая разогнута, тогда от раздражения первая
разгибается, а вторая сгибается (10).
3. Наблюдение изменчивости рефлексов на кошках по Магнусу. Этот отіьгт лучше
всего поставить на лумбальных препаратах, т. е. на животных, у которых спинной
мозг перерезан на границе грудной и поясничной областей. Спустя 1—2 часа
наблюдаются рефлексы. Очень хорошо получается изменчивость рефлекторного движения
хвоста в зависимости от его положения. Если хвост отклонен вправо, тогда
раздражение его кончика — прикосновение, сдавливание и т. д.— вызовет его движение
налево; движение является обратным при том же раздражении, если хвост был
отклонен влево. Если же хвост наклонен вниз, тогда то же раздражение вызывает его
поднятие.
4. Наблюдение извращения рефлекса вследствие искусственного повышения
возбудимости в координирующем аппарате другого рефлекса. Спинномозговой препарат
лягушки. Обнажают поясничный отдел мозга с задней стороны и перерезают первый
задний корешок (VIII по счету). Лигнином удаляют жидкость с поверхности мозга и
затем отравляют стрихнином первый поясничный сегмент, прикладывая кусочки
фильтровальной бумаги, смоченные в 0,1% растворе стрихнина, к задней поверхности
мозга. Спустя несколько минут сильно повышается чувствительность рефлекторных
реакций потирания, вызываемых из отравленного сегмента путем раздражения
перерезанного заднего корешка. В то же время раздражение задней ноги в рецептивном
поле сгибания производит такой сгибательный эффект, в котором ясно заметны
элементы рефлекса потирания: нога сгибается во всех суставах, как в рефлексе
сгибания, но при этом пальцы задней ноги производят ритмическое движение потирания.
Движения потирания обусловлены активированием отравленного сегмента
иррадиацией возбуждения, вызванного раздражением рецептивного поля сгибания. При
этом не происходит возбуждения отравленного сегмента вторичным раздражением
рецептивного поля потирания, ибо это поле лишено чувствительности благодаря
перерезке VIII заднего корешка.
5. Измечеьие рефлекса при усилении и удлинении раздражения. Спинномозговая
лягушка. Записывается гл. triceps femoris и m. semitendinosus. Раздражается
п. tibialis или его веточка, п. superficial. Эти нервы получают чувствительные во-
366 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
локна от кожи на пальцах вентральной стороны ступни и над ахилловым
сухожилием. Поэтому при одной силе может получиться рефлекс разгибания: сократится
т. triceps, а при другой — рефлекс огибания: сократится гл. semitendinosus.
Если данное раздражение дает сгибание, то при некотором удлинении или
повторении раздражения сгибание сменяется разгибанием. Если же раздражение
вначале дает разгибание, то при удлинении или повторении раздражения оно может
смениться сгибанием.
Вместо чувствительного нерва можно раздражать электрическим током кожу
над ахилловым сухожилием и на чувствительных препаратах получить отсюда как
сгибание, так и разгибание, а также смену их в связи с усилением или удлинением
раздражения.
Литература
1. Kurschner G., Добавление к книге Mar&chall Hall, Marburg, 1840.
2. Pfliiger Ed., Die sensor. Function d. Ruckenmarks d. Wirbeltiere, Berlin, 1853.
3. Сеченов П., Мед. вестн. СПб, 1863; Избранные труды, стр. 167, 1935.
4. MagnusR., РП. Arch., 130, 219, 1909; Ы4У 545 и 5»*, 1910.
5. GergenR., РП. Arch., 14, 340, 1876.
6. Beritoff J., Quart. J. Exp. Physiol., 9, 199, 1915; Изв. Росс. АкадЛіаук,
стр. 649 и 853, 1915.
7. Sherrington и сотр., Reflex activity etc., 1932; русск. пер.; Рефлекторная
деятельность спинного мозга, 1935.
8. Beritoff J., Erg. Physiol., 20, 408, 1922.
9. Beritoff J.,Pfl. Arch., 151, 171, 1913; Тр. Петерб. об-ва естеств., 41, в. 2, 1910.
10. В і с k e 1 A., Arch. Anat. Physiol., 1900, стр. 485..
11. В a g 1 і о n i S., Archivio de fisiologia, 1, 575, 1904; Zur Analysed. Reflexfunk-
tion. Wiesbaden, 1907.
12. Беритов И., Учение об основных элементах центральной координации
скелетной мускулатуры. Пг., 1916.
13. Sherri n gton С. S., Int. Act. Nerv. Syst., 1906.
14. Beritoff J.; Pfl. Arch., 200, 562, 1923; 201, 445, 1923.
15. Graham Brown, Quart. J. Exp. Physiol., 4, 331, 1911; 5, 237, 1913 и другие
работы.
16. В e t h e Al. u. Fischer E., Die Anpassungsfahigkeit (Plastizitat) d. Nerven-
systems, Bethe's Handb. norm. path. Physiol., 15, H. 2, стр. 1045, 1931.
17. R a n s о n S. W., Der Nervenarzt, 4, jg. 1931. H. 4.
18. Ranson S. W. a. H i n s e у J. C, Arch. Neur. Psych., 26, 247, 1931.
19. Ухтомский А., Русск. физиол. журн., 6, 33, 1923; Новое в рефлексологии
и физиологии нервной системы, 1, 60, 1925; 2, 3, 1926.
20. Ухтомский А., Тр. об-ва естеств. в СПб., 41, в. 2, 1911.
21. I say a ma S., Z., Biol., 82, 338, 1925.
22. Б е р и т о в И.. Труды Об-ва естеств. в СПб., 41, в. 2, стр. 245, 1911.
23. Б е р и т о в И., Мед.-биол. журн., 3-й год, в. I, стр. 82, 1927.
24. Ухтомский А., Парабиоз и доминанта. Сб. «Парабиоз», 1927.
25. К о н р а д и Г. П.. Сб. раб. физиол. лаб. Ленингр. ун-та, стр. 118, 1930.
26. В и н о г р а д о в М., Русск. физиол. журн., 6, 47, 1923; Новое в рефлексологии
и физиологии нервной системы, 1, 67, 1925.
27. Анохин П. и сотр., Проблема центра и периферии в физиологии нервной
деятельности. 1935.
28. KennedyB., Phi]. Trans. Roy. Soc. London, 194, 1901: Proc. Roy. Soc. London,
В., 87, 331, 1913.
29. Osborn W. A. a. Kilving ton В., Brain, 33, 260, 1911.
30. Flourens M. P.. Ann. Sci. Natur., 13, 113, 1828.
31. Коштоянц X. С, Сб. докладов VI Всес. физиол. съезда, 1937, стр. 268.
ДИНАМИКА РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ 367
32. Л с р а т я н Э., Сб. докладов VI. Всес. физиол. съезда, стр. 158.
33. К а р а м я н А. И., Бюлл. эксп. биол. и мед ., 7, 481, 1939.
34. Marina A., Dtsch. Ъ. Nervenheilk. 44, 138, 1912.
35. Olmsted J., Morgutti M. a. Yanagisana R., Am. J. Physiol.>
116, 245, 1936.
36. Sherb R., Schweiz. Med. Wschr. 68, 354, 1938.
37. Weiss P., Biol. Rev., 11, 494, 1936.
38. S perry R. W.. J. Сотр. Neurol., 73, 379, 1940.
39. Б?ритовй., Вестн. АН СССР, № 10, 84, 1939.
40. Анохин П. К., Невропатология и психиатрия, 9, 31, 1940.
41. Введенский Н. Е. и Ухтомский А. А, Тр, СПб.. Общества естество-
испыт , 39, вып. 2, 1908; Собр. соч., Н. Е. Введенского, 4. 90, 1938.
Глава XXI
КООРДИНАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ
У БЕСПОЗВОНОЧНЫХ ЖИВОТНЫХ
Принципы координации рефлекторных реакций у низших
беспозвоночных. У тех многоклеточных беспозвоночных, которые лишены
центральной нервной системы, рефлекторная реакция определяется просто
диффузным проведением возбуждения >в нейропиле, проникающем во
все ткани животного. Нервные отростки клеток, образующие нервную
сеть, не дифференцированы на нейриты и дендриты. Так как в таких
нервных элементах возбуждение распространяется с декрементом, то
реакция животного по существу зависит от силы раздражения. Она тем
сильнее и обширнее, чем больше сила раздражения (гидра, медуза,
актиния, (морская звезда). Декрементное проведение возбуждения у
малоподвижных животных является единственной формой рефлекторной
координации. Вследствие этого действие кожного раздражения
простирается на прилегающие группы мышц и этим самым данный участок
организма устраняется от раздражителя. Так, например, у гидры, при
слабом механическом раздражении одного щупальца, наступает
сокращение только одного этого щупальца; при несколько более сильном
раздражении сокращаются все щупальцы, при еще более
сильном—сокращается все тело (Паркер, 13; Иордан, 1 и др.). Характерно, что внутри
нейропиля возбуждение распространяется во все стороны одинаково,
поэтому можно сделать глубокие надрезы в нейропильном организме
или органе, не прекращая распространения возбуждения от одного
участка ткани ко всем другим. Но у некоторых низших беспозвоночных
существуют такие органы, где возбуждение распространяется по нейро-
пилю только в одном направлении, т. е. где нейропиль является более
или менее поляризованным, как и в центральных нервных узлах
высших животных. Таковы, например, шупальца актинии (Паркер, 13).
По принципу декрементного проведения возбуждения
координируются движения и таких низших форм беспозвоночных, у которых уже
имеется дифференцированная центральная нервная система, как,
например, улитка, плоские черви. У этих животных йервная система в целом
КООРДИНАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ У БЕСПОЗВОНОЧНЫХ 369
реагирует как нейропиль, несмотря на существование
дифференцированных нервных центров и нервных путей.
Но у низших высоко подвижных беспозвоночных имеется также
нервная система с бездекрементным проведением, благодаря чему в
реакцию может вовлекаться все тело животного. Она образует длинные
нервные пучки внутри диффузного нейропиля. Эти пучки более
возбудимы и проводят возбуждение в одном определенном направлении, и
притом без декремента. Через эту систему осуществляется быстрая
реакция всего тела. Такая система имеется, например, у медузы.
Благодаря ей в каждой части зонтика іможет возникнуть спонтанное
сокращение, и каждый раз это сокращение распространяется на весь зонтик.
Обычно сокращение сначала возникает на краю зонтика; здесь
находится наиболее развитая бездекрементная система. Но и по удалении
краевой части зонтик продолжает сокращаться целиком, хотя и в
более медленном ритме. Значит, бездекрементная система существует
во всем зонтике. Такая система, повидимому, имеется и у актиний
{Aiptasia annulata), ибо при механическом раздражении одного
щупальца может наступить мгновенное сокращение всего тела.
Итак, уже у низших беспозвоночных — у кишечнополостных — наряду
с нейропилем с декрементным проведением возбуждения имеется более
развитая нервная система с бездекрементным проведением возбуждения.
Эта система осуществляет целостные реакции всего животного в ответ
на каждое деятельное раздражение.
Нейропиль низших беспозвоночных с его декрементным и
бездекрементным проведением возбуждения осуществляет также более сложные
координированные движения. Например, у медузы, когда добыча
улавливается краевым щупальцем, оно сокращается и поднимает добычу
вверх, направляя ее к центру, где находится хобот. Одновременно
хобот изгибается в сторону добычи,—т. е. к тому самому краевому
участку, где находится добыча,— приходит в соприкосновение с добычей и
поглощает ее. Вся эта сложная реакция производится нейропилем. Это
доказывается тем, что если обвести разрезом один краевой участок ней-
ропильной субумбреллы, то его раздражение опять вызывает движение
хобота; однако последний более не направляется к раздраженному
краевому участку. Нормально от каждого краевого участка возбуждение
достигает хобота через нейропиль сильнее всего по прямому
радиальному пути; значит, мускулатура хобота получает импульсы к сокращению
сильнее всего по этому радиусу, и этим определяется его движение к
раздражаемому участку. После перерезки этого прямого пути
возбуждение достигает хобота через нейропиль по другим обходным путям, и
потому хобот направляется к другим участкам (Романее, 17).
Нейропиль осуществляет также плавательные движения медузы.
У медузы нет антагонистической мускулатуры, имеется только одна
мускулатура в субумбрелле, которая, сокращаясь, уменьшает внутренний
объем колокола. Ей противостоит эластический студень, который при
сокращении мышцы претерпевает эластическую деформацию. После
24 и. С. Беритов
370 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
сокращения мышца расслабевает, а эластический студень своим
напряжением восстанавливает форму колокола. При каждом сокращении
Шда выбрасывается под давлением наружу из колокола в одном
направлении, и вследствие этого животное передвигается в противоположном
направлении. Локомоторные движения при плавании повторяются одно
за другим в очень правильном ритме и всегда охватывают сразу всю
мускулатуру. Они возникают под влиянием раздражения краевых
рецепторов, но всецело регулируются деятельностью нейропиля. В период
сокращения нейропиль находится целиком в активном состоянии
возбуждения; отсюда поступают в імышцу импульсы возбуждения. Во время
расслабления и паузы нейропиль находится в активном состоянии
торможения. В этот период никакие раздражения краевых рецепторов не
способны вызвать сокращение мышцы. Многие авторы это состояние
невозбудимости относят к рефракторной фазе нейропиля (Буденброк, 3
и др.)- Но вероятнее, что в этом случае мы имеем в нейропиле период
невозбудимости тормозного характера. Нужно думать, что нейропиль
у медузы совершенно заменяет нервнь'й узел высших беспозвоночных.
Подобно этому нервному узлу нейропиль обусловливает смену фазы воз-
буждения на торможение, независимо от периферических влияний.
Природа смены фазы возбуждения на фазу торможения, и наоборот,
неизвестна, но по аналогии со сменой фаз у высших животных можно
считать ее проявлением анэлектротонического действия медленных
токов, которые возникают в нейропиле с декрементным проведением.
Импульсы возбуждения для локомоторных движений медузы возникают
нормально в краевых рецепторах, откуда они по бездекрементной
системе достигают мышц. Но одновременно эти импульсы активируют
нейропиль. Возникающие здесь медленные токи, распространяясь по
межуточной жидкости, могут действовать анэлектротонически на краевые
рецепторы и бездекрементную систему. Вследствие этого возбудимость
в них должна понизиться и должны прекратиться как их спонтанное
возбуждение, так и возбуждение под влиянием внешней среды. Это
приводит к прекращению сокращения мышц или к паузе, с одной
стороны, а с другой — к прекращению активного состояния нейропиля, а
значит, и к исчезновению медленных токов. Это последнее
обстоятельство является основным фактором для последующей двигательной фазы.
Вследствие исчезновения нейропильного тока возбудимость в краевых
рецепторах и бездекрементной проводящей системе восстановится: они
вновь возбудятся спонтанно или под влиянием внешней среды. Вновь
сократится мускулатура субумбреллы и вновь произойдет
активирование нейропиля с последующим торможением краевых рецепторов и
проводящей системы и т. д.
Происхождение нейропильного тока должно быть связано с теми
физико-химическими процессами, которые протекают в нервных
волокнах низших беспозвоночных во время возбуждения, а эти процессы, как
уже указывалось выше, сопровождаются очень сильными и
длительными токами. Например, в коммиссуральном нерве беззубки от одиночного
КООРДИНАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ У БЕСПОЗВОНОЧНЫХ 371
раздражения возникает многофазное изменение потенциала: после
начального сравнительно быстрого отрицательного колебания
продолжительностью около 1 сек. наступает длительное положительное
колебание, продолжающееся многие секунды. При тетанических
раздражениях происходит тоже отрицательное отклонение. По прекращении
раздражения развивается сильно выраженное последействие, состоящее из
двух фаз — короткой отрицательной и очень длительной положительной
фазы, продолжающейся в течение многих минут (Кан, 18). Безусловно
такие длительные электрические последействия должны иметь место
везде в нейропиле при его активном состоянии, поэтому совершенно
естественно допустить, что у медузы угнетение нервной деятельности
после каждого периода возбуждения обусловливается анэлектротониче-
ским действием медленного тока нейропиля на краевые рецепторы и
бездекрементную проводящую систему.
У некоторых низших беспозвоночных существует и другой принцип
координации, который регулирует периодичность фазных движений.
Этот принцип заключается в том, что при сокращении одних мышц
происходит растяжение других, и это растяжение становится основным
условием для их сокращения. Так, например, ритмическое сгибание
луча офиуры то в одну, то в другую сторону обусловливается тем, что
каждый раз сокращение возникает на той стороне, где мышцы
растянуты. Импульсы возбуждения по нервной сети распространяются к тем
мышцам, которые растянуты (Юкскюль, 11). Аналогичное явление
наблюдается на хвосте, кошки, как мы это видели выше, но у кошки это
обусловливается повышением возбудимости в соответствующих
координирующих аппаратах путем раздражения проприоцепторов растянутых
мышц. В луче офиуры едва ли можно предположить существование
подобного механизма. Здесь мы имеем чисто периферическое явление.
Растяжение, повидимому, повышает возбудимость растянутых мышц, и
благодаря этому распространяющиеся через нейропиль импульсы
возбуждения вызывают сокращение преимущественно в растянутых
мышцах.
Подобный механизм координации существует также у низших
червей. Так, например, известно, что у немертин при раздражении головы
сначала возникает сокращение продольных мышц на голове и
происходит утолщение головного участка тела; затем, постепенно переходя от
участка к участку, волна сокращения, наконец, достигает
противоположного конца. Такое же распространение сокращения имеет место при
раздражении хвоста. В этом случае волна сокращения распространяется
в обратном направлении. Здесь она простирается тем дальше, чем
сильнее раздражение (Буденброк, 3). В последнем случае мы, повидимому,
имеем координацию, обусловленную декрементным проведением
возбуждения, но в первом случае, когда двигательная реакция свободно
распространяется от головного конца к хвосту, едва ли все цело
объясняется бездекрементным проведением возбуждения. Здесь должно
играть большую роль то обстоятельство, что сокращение каждого участ-
24"
372 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
ка ведет к растяжению ближайшего соседнего участка; в растянутом
участке должны раздражаться находящиеся здесь кожно-мышечные
рецепторы. На этом основании можно предположить, что
распространение двигательной реакции в хвостовом направлении происходит по
так называемому принципу цепного рефлекса: нервные импульсы
рецепторов, возникающие в каждом растянутом участке, производят
через соответствующий участок нейропиля рефлекторное сокращение
соответствующих сегментов.
Принципы координации движений у высших червей. Высшие
беспозвоночные, как кольчатые черви, моллюски, членистоногие, обладают
дифференцированной центральной нервной системой: каждый
двигательный орган имеет свой нервный узел, работающий более или менее
автономно. Но, кроме того, существует более развитый узел в головной
части. Развился он вследствие его прямой связи с главными
рецепторами зрения, обоняния и благодаря обслуживанию ротового отверстия с
его сложными органами захвата пищи. Этот головной узел находится в
связи со всеми остальными нервными узлами, и через эти связи он
іможет распространять свое влияние на все двигательные органы (Буден-
брок, 3; Ганстрём, 4).
Из множества типов передвижения и рефлекторных реакций мы
рассмотрим лишь несколько наиболее характерных: перистальтические
движения по земле и в воде у червей, передвижение с помощью ноги
улитки и хватательные движения у членистоногих.
Тело кольчатых червей состоит из большого количества
однообразных сегментов. КаждЬій сегмент снабжен своим нервным центром в
виде парного ганглиозного узла, чоторый более или іменее самостоятельно
управляет движением данного сегмента. В узлах, во-первых, лежат
двигательные клетки, аксоны которых оканчиваются в мышцах;
во-вторых, клетки межцентральных нейронов, аксоны которых оканчиваются
в нервных узлах соседнего сегмента; в-третьих, здесь оканчиваются
аксоны межцентральных нейронов, а также чувствительных нейронов из
кожно-мышечного мешка соответствующего сегмента. Наконец, здесь
имеется нейропиль, образуемый из разветвлений дендритов и аксонов
всех упомянутых нервных элементов. Кроме того, в нервном узле
дождевого червя проходят три очень толстых нервных волокна, которые
тянутся по всей брюшной цепочке животного; клетки этих волокон, по-
видимому, лежат в переднем узле цепочки. Наряду с организованной
центральной нервной системой у всех червей существует еще
периферический нейропиль, нервное сплетение которого проникает во все
органы, и в частности, в мышечную систему.
Дождевой червь лишен конечностей и передвигается при помощи
мышц кожно-мышечного мешка, в котором различают внешние
кольцевые и внутренние продольные 'мышцы. Эти мышцы действуют
антагонистически: когда сокращаются кольцевые мышцы сегмента, продольные
находятся в расслаблении; при этом сегмент суживается и удлиняется.
Когда же сокращаются продольные імышцы сегмента, кольцевые рас-
КООРДИНАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ У БЕСПОЗВОНОЧНЫХ 373
слабевают; при этом сегмент утолщается и укорачивается (Гогава, 26)
(рис, 161). При передвижении волна сокращения начинается с
головного конца и охватывает поочередно все сегменты до; заднего концаг
причем в каждом сегменте сначала наступает удлинение, а затем
укорочение. При этом животное передвигается вперед, ибо хитинки, распо-
Рис. 161. Антагонистическая иннервация мышц серединных
сегментов дождевого червя.
Верхняя кривая от кольцевой мускулатуры; нижняя—от продольной
мускулатуры. А—во время спонтанной деятельности; В- при
электрическом раздражении нервной цепочки. Верхний сигнал отмечает
момент раздражения. Хорошо выраженный антагонизм в обоих
опытах (Гогава).
ложенные на брюшной стороне по бокам сегмента, направлены своими
острыми концами назад и так упираются в землю, что при укорочениях
и удлинениях тело животного может передвигаться только вперед
(Иордан, 1).
Это распространение волны сокращения не обусловливается всецело
деятельностью головного ганглия. Здесь мы имеем случай так
называемого цепного рефлекса: при укорочении головных сегментов следующие
за ними ближайшие сегменты растягиваются. При этом раздражаются
рецепторы кожно-мышечного мешка. Исходящие из них импульсы
поступают в соответствующие нервные узлы, вызывая через
межцентральные нейроны сокращение кольцевых мышц данных сегментов, т. е.
удлинение, благодаря чему эти сегменты передвигаются вперед, следуя за
передними укороченными сегментами. После удлинения наступает
сокращение продольных мышц. Вероятно, это происходит в результате
раздражения некоторых рецепторов в связи с активным удлинением
сегментов. За этим следует пассивное растяжение следующих
сегментов, которое сейчас же приводит к их активному удлинению и
передвижению вперед. Таким путем волны сокращения с передвижением
достигают хвостового конца.
Наряду с вышеописанной локомоцией, в которой каждый сегмент
371 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
участвует по типу цепного рефлекса, у дождевого червя имеется еще
общая рефлекторная реакция, охватывающая сразу все животное. Это,
например, бывает при более или менее значительном раздражении
переднего конца тела: ©се животное, в результате сокращения
продольных мышц, укорачивается сразу во всех сегментах, вследствие чего оно
быстро отодвигается назад — защитная реакция. Эта реакция
осуществляется гигантскими нервными волокнами. В них возбуждение
распространяется быстро и без декремента — со скоростью 150 см в 1 сек.
Они отдают коллатерали во всех узлах и, вероятно, через них
производится сокращение імышц во всех сегментах (Херрик, 12).
Другой тип передвижения имеет место у пиявок. Они ходят и
плавают. При хождении важную роль играют присоски. Прикрепление
передней присоски ведет к сокращению продольных мышц и
расслаблению кольцевых, причем задний конец переносится вперед и, затем,
вследствие более сильного сокращения брюшных продольных мышц, все
тело изгибается, так что задняя присоска оказывается рядом с передней,
и движение заканчивается присасыванием задней присоски. Сейчас же
передняя присоска освобождается и начинается сокращение кольцевых
и расслабление продольных мышц, благодаря чему животное
вытягивается головой вперед. Как одно, так и другое движение начинается
с головы и постепенно распространяется назад, до задней присоски.
В этой двигательной реакции сегментальные рефлексы по принципу
цепного рефлекса участвуют в очень слабой степени. По нашим
наблюдениям, импульсы, исходящие из головного узла, безусловно играют
существенную роль в распространении волны сокращения. Это видно из
того, что после перерезки нервной цепочки посредине передняя часть
совершает локомоцию значительно сильнее задней части и даже
передвигается без ее участия: последняя пассивно тащится за передней.
После же перерезки мускулатуры без повреждения нервной цепочки
задняя часть вполне следует за передней, ритмически удлиняясь и
укорачиваясь, подобно передней части.
Удлинение и укорочение пиявки начинаются с головного конца,
причем для удлинения периферические импульсы возникают в рецепторах
передней присоски, а для укорочения — в рецепторах задней присоски.
В обоих случаях импульсы сначала достигают головного ганглия, а
отсюда уже вызывается соответствующее движение передних сегментов.
Хождение прекращается после отрезания головы. Животное более
не передвигается, но сможет производить при раздражении как
удлинение, так и укорочение всего тела. Удлинение происходит при более или
менее значительном раздражении кожи, вслед за которым всегда
следует укорочение. Но укорочение может наступить с самого начала
после раздражения. Это бывает при слабых раздражениях кожи.
Плавательные движения происходят таким образом: кольцевые
мышцы расслаблены; дорзовентральные мышцы сокращены, вследствие
чего тело становится плоским и принимает вертикальное положение.
При плавании фазную деятельность испытывают только продольные
КООРДИНАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ У БЕСПОЗВОНОЧНЫХ 375
мышцы. При этом спинные и брюшные продольные мышцы выступают
¦как антагонисты. Предполагают, что сокращение распространяется по
принципу цепного рефлекса. Положим, что волна сокращения возникает
в спинных мышцах передних сегментов. При этом брюшные імышцы
растянутся, что приведет к раздражению соответствующих рецепторов,
Рис. 162. Антагонистическая и синэргическая деятельность продольной
и кольцевой мышц пиявки.
Вверху—спинная продольная мышца от передней половины тела беэ головы, внизу—
кольцевая мышца от одного переднего сегмента. Эта часть пиявки так
отпрепарирована и фиксирована, что движение продольной мышцы не влияет механически на
кольцевую. Пороговое раздражение нервной цепочки—14.5 см расстояния
индукционной катушки — производит сокращение продольной мышцы и расслабление кольцевой
(опыты В и С). При сильном раздражении (8 см) сокращаются обе мышцы, но после
раздражения обе мышцы расслабевают ниже того уровня, который был до
раздражения (опыт А) (Беритов и Гогава).
а это, в свою очередь, приведет к рефлекторному сокращению брюшных
мышц ближайших сегментов. В результате, в данных сегментах
растягиваются спинные мышцы; от этого растяжения рефлекторное
сокращение наступает в спинных мышцах следующих за ними сегментов и
таким путем довольно быстро волнообразное движение достигает
заднего конца. Передача возбуждения от одного сегмента к другим
происходит по нервной цепочке, а потому если удалить один сегмент без
перерезки нервной цепочки, обе части тела плавают, как целое
животное. Для плавательных движений головной мозг не является
необходимым. Животные свободно плавают в воде и после отрезания как
переднего, так и заднего конца.
Координация кольцевых и продольных мышц при активном удлине-
нении и укорочении тела обусловлена антагонистической работой этих
мышц: когда продольные мышцы сокращаются, кольцевые удлиняются,
и наоборот. Это хорошо выявляется путем миографической записи
376 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
одной части тела. Путем электрического раздражения той же части тела
или другой, с которой первая имеет нервную связь, или же
изолированной нервной цепочки, можно вызвать удлинение регистрируемой части:
в это время наряду с удлинением продольных мышц происходит
сокращение кольцевых. То же бывает при спонтанных движениях (Бери-
тов, 16, 23) (рис. 162, 163).
Во всех случаях рефлекторной деятельности расслабление той или
другой мышцы пиявки является активным процессом,— оно обусловли-
Рис. 163. Ритмическая антагонистическая реакция мышцы пиявки.
Такой же препарат другой пиявки, как на рис. 162. Производится длительное,
тетаническое раздражение нервной цепочки в течение 7 мин. Во время раздражения
продольная мышца расслабевает, а кольцевая очень слабо сокращается. После
раздражения наступает ритмическая антагонистическая реакция мышц (Беритов и
Гогава).
вается торможением. Оно как будто реципрокно: при наличности
сокращения одной мышцы другая, антагонистическая мышца тормозится. Но
в действительности торможение наблюдается на обеих мышцах,
одновременно как при слабых, так и при сильных раздражениях (рис. 162,
163).
При миографической записи антагонистических мышц пиявки и
электрическом раздражении одной части тела, т. е. рецепторов или
нервной цепочки, обнаруживается, что торможение наступает на них
уже при пороговых раздражениях и усиливается с усилением
раздражения. Это происходит несмотря на то, что с усилением раздражения
усиливается также сокращение данной мышцы. Торможение хорошо
изучить на той мышце, которая находится в рефлекторном тоническом
сокращении. При пороговых и околопороговых раздражениях
торможение проявляется в виде расслабления тонического сокращения, причем
оно быстро проходит вслед за прекращением раздражения (рис. 162 В).
При сильных же раздражениях торможение хорошо проявляется на
антагонистических мышцах в виде расслабления тонуса вслед за
быстрым прекращением сокращения, вызванного раздражением
(рис. 162 Л). В этих случаях торможение продолжается и после раз-
КООРДИНАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ У БЕСПОЗВОНОЧНЫХ 377'
дражеиия от нескольких секунд до нескольких десятков секунд. Это-
видно из того, что пороговое электрическое раздражение той или
другой части тела, до того вызывавшее значительное сокращение данной
мышцы, в это время или совсем не дает ее сокращения или дает
ослабленное сокращение (Беритов и Гогава, 16). Но здесь мы имеем не
смену возбуждения торможением. Последнее наступает еще во время
раздражения в присутствии сокращения. Это хорошо проявляется в случае-
комбинации с сильным тоническим сокращением. Последнее ослабевает
Рис. 164. Возбуждение и торможение продольной спинной мышцы
пиявки из задней половины тела.
Нервная цепочка удалена. Один конец мышцы раздражается тетанически, а через
всю мышцу пропускаются одиночные индукционные удары. В опыте А раздражением
мышцы одиночным размыкательным индукционным ударом вызывается длительное
„ сокращение (нижний сигнал). В опыте В производится тетаническое раздражение,
которое вызывает небольшой тетанический эффект (верхний сигнал). Опыт С
начинается с тетанического раздражения. Во время него два одиночных индукционных
удара не дают эффекта, но отвечающий им длительный эффект наступает по
прекращении тетанического раздражения. Повторное тетаническое раздражение производит
ослабление сокращения. Внизу время — 2 сек. (Беритов и Гогава).
в самом начале раздражения, еще до наступления самого сокращения .
(рис. 163 Л).
Однако одновременное возбуждение и торможение в рефлекторных
реакциях пиявки наблюдается не только при сильных раздражениях, но
и при всех деятельных раздражениях, если только пробовать их при
отсутствии тонуса, когда мышца значительно расслаблена. В таких
случаях возбуждение проявляется в виде сокращения мышцы, которое с
усилением раздражения тоже усиливается. О наличии торможения
свидетельствует в данном случае прежде всего усиление сокращения по
прекращении раздражения, т. е. явление отдачи (Беритов, 23).
Итак, на пиявках при рефлекторной деятельности возбуждение
мышц всегда сопровождается торможением, но в определенных случаях,,
когда на одной мышце доминирует возбуждение, на антагонисте,
наоборот, доминирует торможение. Этим обусловливается возможность
антагонистических реципрокных реакций на мышцах пиявки.
378 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
Реципрокное торможение, проявляющееся при антагонистических
реакциях кольцевых и продольных мышц пиявки, безусловно возникает
в центральной нервной системе, в нервных узлах брюшной цепочки. Это
доказывается, между прочим, тем, что при удалении нервной цепочки
антагонистические реакции более не наступают. Однако кожно-мышеч-
ный мешок после удаления нервной цепочки не совсем лишен
рефлекторной деятельности, и в этом случае продольная мышца дает
тоническое сокращение как спонтанно, так и в ответ на раздражение кожных
Рис. 165. Биотоки возбуждения кожно-мышечного мешка 'пиявки при наличии
нервной цепочки (Л и В) и после удаления ее (С—F).
Отводятся биполярно в катодный осциллограф небольшой участок с межполюсным
расстоянием около 8 мм. А — при тоническом сокращении с участием нервной цепочки
отдельные колебания достигают 200 p.V. На один момент наступает спонтанно сильное
сокращение. В этот момент отдельные колебания достигают 500 ;j.V; В—спонтанное расслабление
тонуса при наличии нервной цепочки; электрические колебания сходят на-нет; С —
тоническое сокращение в отсутствии нервной цепочки, в этот момент длина кожно-мышечного
мешка была 18 мм; D — в момент ослабления тонуса того же препарата — длина того же
кожно-мышечного мешка увеличивается до 24 мм; Е — препарат без нервной цепочки;
реакция на раздражение мышечных рецепторов растягиванием; F—тот же препарат при
раздражении кожных рецепторов прикосновением кисточки. В обоих последних случаях
в начале значительные эффекты, которые быстро затухают в течение нескольких секунд.
рецепторов. Это хорошо выявляется при регистрации как
механического, так и электрического эффекта мышцы. Так, например, на рис. 164
записано тоническое сокращение препарата пиявки без брюшной
цепочки после электрического раздражения кожных рецепторов, а на рис. 165
показан электрический эффект от такого же препарата как при
спонтанном сокращении (С), так и при спонтанном расслаблении (D), затем
дается электрическая реакция от раздражения мышечных рецепторов
путем растяжения кожно-мышечного мешка (Е), а также кожных
рецепторов путем прикосновения на один момент мягкой кисточкой (F).
КООРДИНАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ У БЕСПОЗВОНОЧНЫХ 379
Однако следует отметить, что
тоническое сокращение кожно-мышечно-
го мешка при наличии нервной
цепочки развивает гораздо большее
напряжение. Наблюдаемая при этом
электрическая реакция неизмеримо
сильнее. Это хорошо видно на рисунке 165,
где дается случай спонтанного
тонического сокращения и расслабления
при участии центральной нервной
системы. В этом случае отдельные
колебания в период тонуса достигают
200 yV и выше (Л), в то время как
в отсутствии нервной цепочки при
тонусе отдельные колебания не
превосходят 15 jj-V (СиО).
На препаратах без нервной
цепочки при раздражении кожных
рецепторов тоже іможно наблюдать
торможение. Если мышца была тонически
сокращена, она расслабевает во время
или после раздражения кожных
рецепторов (рис. 164 С), но и при
отсутствии тонуса раздражение этих
рецепторов также может вызвать
торможение. Оно выявится после
раздражения в виде рефлекса «отдачи»
(рис. 164 С). Сокращение получается
при таких же силах раздражения,
как на препаратах с нервной
цепочкой. Торможение получается на них
ари сравнительно сильных тетаниче-
ских раздражениях или при очень
сильных индукционных ударах. При
этом как возбуждение, так и
торможение бывает общим, простирается
одновременно на продольные и
кольцевые мышцы данного сегмента (Бе-
ритов и Гогава, 16). Отсюда следует,
что в сегментах пиявки рефлекторная
реакция осуществляется двумя
механизмами: центральным механизмом с
участием нервных узлов брюшной
нервной цепочки — путем передачи
возбуждения от окончаний
центростремительных чувствительных нейронов
Рис. 166. Схема нервных путей для
центрального и периферического
сегментальных рефлексов у Hirudo
medicinal is.
Схема составлена на основании
анатомических данных Ливанова и нашего
физиологического исследования. В — кожные
рецепторы; 5 — чувствительные волокна
от этих рецепторов; Р — мышечный
рецептор; S'—нервное волокно этого
рецептора; G — нервный узел в брюшной
цепочке; N, N'—двигательные клетки
двоякого типа; М — поперечный разрез
продольной мускулатуры; О — неврилемма
для всего пучка волокон, внутри которого
может происходить передача импульсов от
чувствительных волокон к двигательным:
К — К' — линия перерезки. После такой
перерезки раздражение кожного рецептора
будет производить двигательный эффект
исключительно путем передачи импульсов
возбуждения от чувствительных волокон
к двигательным в общем для них нервном
стволе.
380 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
к клеткам центробежных двигательных нейронов, и периферическим
механизмом путем передачи возбуждения от сравнительно толстых
чувствительных волокон к двигательным волокнам в общем для них
нервном стволе (рис. 166). В обоих случаях передача возбуждения, повиди-
мому, происходит посредством биотоков возбуждения. Эксперименты
Джаспера и Монни? (25) доказали возможность перехода
возбуждения в общем нервном стволе для беспозвоночных. Можно
считать, что смешанный нерв у беспозвоночных животных является таким
же передаточным органом, как и нервный узел (см. схематический
рис. 166, где R — S — N — п — М представляет длинный путь для
центрального рефлекса через ганглий, a R — S — п — М — короткий
путь для периферического рефлекса через общий нервный ствол).
Итак, каждое периферическое раздражение — возбуждение кожного
или мышечного рецептора — может вызвать рефлекторную двигательную
реакцию, во-первых, локально, в раздраженном сегменте, путем
передачи возбуждения к двигательным нервным волокнам при посредстве
смешанного нервного ствола и, во-вторых, в целом ряде сегментов
или во всем животном — путем распространения возбуждения по
брюшной нервной цепочке.
Периферическому рефлексу ів значительной степени обязано своим
происхождением тоническое сокращение, а также длительное
последействие тетанического сокращения и длительное сокращение в ответ на
раздражение одним индукционным ударом. Эти длительные
рефлекторные сокращения осуществляются благодаря возбуждению рецептивных
клеток мышцы под влиянием механического и электрического
воздействия возбужденных мышечных волокон и передаче возбуждения через
их аксоны к двигательным волокнам в общем для них нервном стволе.
Одним из основных центральных процессов координации у пиявки
является центральное торможение, протекающее в нервных узлах. При
помощи него осуществляется координированное выключение одних
двигательных нейронов во время возбуждения других. Нужно думать, что
это центральное торможение является функцией нейропиля,
занимающего центральное место в каждом нервном узле. Каждый раз, когда
возбуждается группа двигательных нейронов,— например, иннервирую-
щих кольцевую мускулатуру,— активируется соответствующий им
участок нейропиля. Здесь возникают медленные биотоки, как в цервном
узле насекомых по Эдриану (24). Медленные биотоки тормозят
другую группу нейронов, иннервирующих продольную мускулатуру. Это
торможение осуществляется, по нашему мнению, анэлектротоническим
действием медленных биотоков активированного нейропиля на
невозбужденную группу двигательных нейронов.
Другим основным процессом координации у пиявки является
периферическое торможение, осуществляемое путем особых центробежных
нейронов, связывающих нервные узлы с мускулатурой. Каждый раз,
когда возбуждаются определенные центробежные волокла,
вызывающие сокращение одной группы мышц, одновременно возбуждаются и
КООРДИНАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ У БЕСПОЗВОНОЧНЫХ 381
другие центробежные волокна, которые — в случае, если они не будут
угнетены в нервном узле нейропильным биотоком,— тормозят
сокращение другой группы мышц периферическим путем, подобно тому, как это
установлено у ракообразных. Последнее делает невозможным
нарушение центральной координации вследствие активирования
периферического рефлекторного механизма.
Торможение, наблюдаемое на кожно-мышечном мешке без нервной
цепочки, вызывается раздражением тормозящих центробежных волокон
путем передачи возбуждения от кожных чувствительных волокон
в общем для них стволе. Благодаря сравнительно низкой возбудимости
они возбуждаются при несколько более сильном раздражающем
воздействии, чем центробежные возбуждающие волокна.
Природа периферического торможения, вероятно, гуморальная, как
торможение сердца у позвоночных животных. С этим хорошо
согласуется факт длительного тормозящего последействия.
Координация локомоторных движений у низших моллюсков. Из
разнообразных движений моллюсков мы рассмотрим локомоцию улитки
(Helix). Локомоция улитки совершается с помощью ноги, состоящей,
в основном, из одной мышцы. При движении на поверхности ноги
возникают волны сокращения, поочередно проходящие в направлении от
заднего конца к переднему. Одновременно на всю ногу приходится до
10—12 волн. Каждая волна проходит всю ногу в течение приблизительно
30 сек. со скоростью 0.1—0.2 см в 1 сек. Каждая сокращенная часть
отделяется от земли и передвигается вперед; таким образом происходит
передвижение вперед всей ноги вместе с телом.
Распространение волны сокращения объясняется следующим
образом. Впереди и позади волны сокращения імышца растягивается. Это
растяжение происходит благодаря выжиманию кровяной жидкости из
сокращенного участка в соседние расслабленные участки. Растяжение
повышает возбудимость. Вследствие этого в следующей фазе
сокращение наступает в растянутых участках. Импульсы для сокращения
поступают из нервной системы.
У улитки имеется центральный ганглий в головной части и еще ряд
ганглиев: специально для ноги — педальный ганглий, для внутренних
органов — висцеральные ганглии, плевральный ганглий. Все ганглии
соединены с церебральным комиссурами. Кроме того, имеется нервная
сеть — нейропиль. У Helix локомоторные движения зависят
исключительно от педального ганглия: после его экстирпации эти движения
исчезают. У Umax и Aplisia нога сохраняет способность к
передвижению и после удаления ганглия: повидимому здесь фазные движения
обусловливаются импульсами как из ганглия, так и из нервной сети.
Нейропиль ноги активируется раздражением кожных рецепторов ноги и
активируясь производит ее сокращение. Но педальный ганглий
определенным образом регулирует его активность. Это видно из того, что если
удалить педальный ганглий, то под влиянием активного состояния ней-
ропиля мышца ноги сокращается очень сильно и до наступления смерти
382 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
остается в состоянии максимального сокращения. Но если, по
возможности, уберечь ногу от всевозможных периферических раздражений, то
сокращение развивается только спустя несколько часов. Отсюда
следует, что педальный ганглий каким-то образом прекращает действие
импульсов, приходящих от рецепторов в нейропиль.
Головной ганглий не играет роли в координации передвижения, ибо
оно происходит почти нормально и после экстирпации головного ганглия,
но он определенным образом регулирует эти движения: тормозит их,
приводя в состояние покоя; затем, под влиянием головных рецепторов,
придает им то одно, то другое направление. Это видно, между прочим,
из того, что после удаления головного ганглия или его наркотизации
кокаином движения ноги становятся непрерывными и однообразными,
тонус мышц повышается; кроме того, эти мышцы по отношению к
искусственному раздражению становятся более возбудимыми, чем до
удаления головного ганглия.
Нейропиль улитки и вообще моллюсков играет роль центрального
органа — через него осуществляются разного рода локальные рефлексы.
В этом нейропиле возбуждение распространяется во все стороны, и
притом с декрементом, поэтому происходящие через него рефлексы тем
интенсивнее и обширнее, чем сильнее раздражение.
Нейропиль улитки богат клеточными скоплениями. Местами он имеет
определенное строение — в виде веревочной лестницы. Этим, вероятно,
и обусловливается то, что в определенных случаях нейропиль
производит сложные координированные движения, как волнообразные
локомоторные движения ноги улитки. Нейропиль имеется у всех
моллюсков,— в частности, у высших головоногих моллюсков, у которых он
также обусловливает локальные рефлекторные реакции. Так, например,
на щупальцах головоногих моллюсков присоски работают рефлекторно
через свой собственный нейропиль: при раздражении их внутренней
поверхности присоски присасываются, даже тогда, когда они совершенно
изолированы (Буденброк, 3; Иордан, 1; Балиони, 15).
Координация хватательных движений у членистоногих. Наиболее
высокоорганизованные из беспозвоночных — членистоногие —- в связи с
их передвижением с помощью членистых конечностей, с очень сложным
устройством ротового аппарата и хватательных органов — производят,
кроме локомоции, множество локальных движений конечностей.
Соответственно этому они обладают наиболее развитой центральной нервной
системой и более сложными механизмами нервной координации. В
основе координированных движений конечностей лежит антагонистическая
иннервация мышц. Она наиболее изучена в отношении клешни рака при
ее хватательных движениях, а потому будет изложена здесь подробно.
Исследованиями целого ряда физиологов на нервно-мышечном
препарате клешни рака было установлено, что мышцы клешни снабжаются
двоякого рода нервными волокнами: одни вызывают в них процесс
возбуждения, другие — торможение. Если перерезать только тормозящие
волокна, то соответствующая мышца при всех рефлекторных воздей-
КООРДИНАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ У БЕСПОЗВОНОЧНЫХ 333
rt-h
ствиях только сокращается; если же, наоборот, перерезать
возбуждающий нерв, то при рефлекторном воздействии эта мышца совсем не
сокращается,— все время остается расслабленной. В нормальных же
условиях эта мышца отвечает ритмической
реакцией, т. е. то сокращается, то расслабляется
(Гофман, 6).
Гистологическое исследование мышц
ракообразных показывает, что одни мышцы, как,
например, размыкатель (абдуктор) клешни речного
рака, снабжаются двумя нервными волокнами,
из которых одно, более толстое, является
возбуждающим, а другое, более тонкое,
тормозящим (рис. 167), (Мангольд, 8; Гарревельд, 21),
а другие мышцы снабжаются тремя нервными
волокнами. Так, например, сгибатель проподита
и разгибатель карпоподита имеют тройную
иннервацию.) Из этих волокон одно нервное
волокно, наиболее толстое, производит быстрое
сокращение, другое, несколько более тонкое,—
медленное сокращение, а третье, чрезвычайно
тонкое,— торможение (Гарревельд и Вирсма, 20,
22). Все они образуют густое сплетение вокруг
мышечного волокна, из которого выходят
троякого рода веточки, оканчивающиеся в каждом
мышечном волокне. Вследствие этого одно и то
же мышечное волокно может дать как
медленное, так и быстрое сокращение, причем и то и
другое могут тормозиться тормозящим
волокном. У многих морских ракообразных каждое
мышечное волокно получает по 4—5 нервных
волокон разной толщины. Физиология подобной
иннервации еще не изучена.
Иннервация клешни речного рака хорошо
исследована. Установлено, что при рефлекторных
движениях через одни нервные волокна мышцы
сокращаются, а через другие — тормозятся. Сле-
довательноТ антагонистическая реципрокная
иннервация клешни рака имеет не центральное
происхождение, а периферическое. Если отрезать клешню и
раздражать на ней нервы, то изолированная клешня отвечает
координированной реакцией: на слабое раздражение она размыкается, а на
сильное замыкается (рис. 168). Было установлено, что эти
координированные реакции зависят от разных порогов тормозящего и
возбуждающего нервных волокон. При слабых раздражениях приходит в активное
состояние возбуждающее волокно размыкательной мышцы и
тормозящее— замыкательной; при сильных раздражениях приходит в действие
Рис. 167. Схема
иннервации мышц клешни
речного рака.
1 — неподвижная ветвь
клешни — проподит, 2 — запи-
рательная (приводящая)
мышца, з—тормозящее
нервное волокно открывающей
(отводящей) мышцы
(пунктиром), 4 — возбуждающее
нервное волокно запиратель-
ной мышцы, 5—тормозящее
нервное волокно запиратель-
ной мышцы (пунктиром), 6—
возбуждающее нервное
волокно открывающей мышцы,
7 — ишиоподит, 8 —¦ меро-
подит, 9 — карпоподит, ю—
открывающая мышца, 11 —
подвижная часть клешни —
дактплоподит (по Иордану).
334 общая физиология спинного мозга и ствола головного мозга
¦возбуждающее нервное волокно замыкателя и тормозящее
размыкателя. То же происходит и при рефлекторном возбуждении клешни через
нервные узлы в результате раздражения рецепторов. При слабых
периферических раздражениях клешня раскрывается вследствие активного
состояния двигательного волокна размыкателя и тормозящего —
замыкателя. При сильных раздражениях клешня закрывается вследствие
активного состояния двигательного волокна замыкателя и
тормозящего — размыкателя (Бидерман, 8; Гофман, 6).
В последнее время Иордан (1) и его сотрудники (Сегаар, 2 и др.),
в результате исследования той же клешни рака, подтвердили данные
Бидерманаи Гофмана в отношении изолированной клешни рака.
Кроме того, оказалось, что при раздражении головных узлов на клешне
-получается картина реакции, обратная имеющей место при перифериче-
fn V'.* fstf 2'ога
1і1Пі11ч1П1Ичіни1,чііч11111м.1.і1чім,..11м1111.111ім1чі i ... ,
Рис. 168. Реакция клешни Cancer pagurus на сильное и слабое
электрическое раздражение.
Цифры означают расстояние индукционных катушек в см. При сильных раздражениях
О—4.0 см клешня закрывается, и притом тем слабее, чем слабее раздражение. При слабых
раздражениях 4.5—5 см клешня открывается. Каждое раздражение производится 2 раза
(из Буденброка).
*ском раздражении. Так, при периферическом раздражении клешня
размыкается или размыкатель ее испытывает усиление тонуса через
определенные изолированные нервные волокна, а при раздражении головного
ганглия тот же размыкатель через те же волокна тормозится.
Раздражение клешни через другие нервные волокна вызывает возбуждение
замыкателя; при центральном же раздражении через эти волокна
сначала также происходит возбуждение замыкателя, но при продолжении
раздражения — его торможение и т. д. (рис. 169). Отсюда авторы
заключают, что в клешне рака и в других органах беспозвоночных
мышцы-антагонисты лишены специфических тормозящих или возбуждающих
нервных волокон; каждое нервное волокно может быть как
возбуждающим, так и тормозящим, и центральное раздражение, т. е. раздражение
головного ганглия, действует на нервные волокна клешни рака
противоположно их непосредственному раздражению.
Это заключение, отрицающее существование специфических
возбужденных или тормозящих нервов, нельзя считать вполне обоснованным.
В данных опытах авторы вызывали сокращение или торможение мышц
клешни рака рефлекторным путем, а не прямым раздражением
двигательных и тормозящих нервных волокон. Вследствие этого все взаимо-
координация рефлекторной деятельности у беспозвоночных 385
действие периферического и центрального раздражения должно было
осуществляться при участии нервных узлов брюшной цепочки и
головного мозга, Повидимому, при периферическом раздражении самой
клешни на ней вызывается через брюшную цепочку один рефлекс, а при
раздражении головного ганглия — другой, который -может быть
противоположен первому и потому способен устранить рефлекс, вызванный
через брюшную цепочку.
Природа процесса торможения в изолированной клешне рака еще
не выяснена. Предполагают, что явление торможения мышц
членистоногих животных имеет гуморальное происхождение, подобно
торможению некоторых вегетативных органов позвоночных животных (Пак-
тин, 19).
Регуляция нервной деятельности у беспозвоночных животных.
Иордан (1, 9) и его сотрудники утверждают, что координирующая
деятельность центральной нервной системы беспозвоночных протекает
совершенно иначе, чем у позвоночных. 'Иорданом было создано
целое учение об этой деятельности, которое в общих чертах
заключается в следующем. Ганглии, нервные центры, действуют на
протекание рефлексов или на удержание тонуса не посредством отдельных
импульсов. Электрическое раздражение ганглиев, например, не влияет
на рефлексы. Нервные центры обладают способностью приходить в
особое длительно протекающее активное состояние, не выявляемое в
ритмических импульсах. Состояние активности, в отличие от быстро про
текающего процесса возбуждения, называется стационарным
(Сегаар, 2). Под этим состоянием активности разумеется повышенная
подвижность частиц возбудимой системы нервных клеток с высоким
обменом веществ. Состояние активности каким-то неизвестным образом
влияет на обмен веществ и на деятельность других нервных узлов и
периферических органов. Чем выше состояние активности в нервных
центрах, тем интенсивнее протекает обмен веществ па периферии, тем
возбудимее животное, тем живее становятся спонтанные движения.
Наоборот, чем ниже состояние активности в центрах, тем ниже
деятельность периферических органов, тем более заглушены двигательные
реакции на периферии. Авторы думают, что высший головной узел,
находясь в состоянии покоя, действует на низшие узлы, как незаряженный
проводник на связанный с ним заряженный, а при активном состоянии
высшего узла — наоборот. Словом, каждый раз энергия центров как бы
стремится к выравниванию. Отсюда эта гипотеза была названа
гипотезой выравнивания энергии (Иордан).
В основу этого учения легли многочисленные факты. Если, например,
у аплизии удалить головной узел, то происходит понижение порогов
периферического раздражения и наступают беспрерывные
незаторможенные движения. А если удалить ножной узел, то нога съеживается,
твердеет, т. е. наступает максимальный тонус. Согласно учению
Иордана это происходит оттого, что в норме головной узел нахо-
25 И. С. Веритов
38b ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА
дится в некоторой низкой степени активного состояния. Вследствие
этого энергия низших центров переходит в головной центр, и соответ
ственно периферические двигательные органы находятся в состоянии
низкой возбудимости и малой подвижности. Удаление высшего центра, по
мнению Иордана, действует на низшие растормаживающим образом,
так как энергия последних не будет более уходить к высшему центру
Это проявится в усилении спонтанных движений. Ножной узел в норме
также находится в состоянии покоя. Поэтому из периферической
нервной сети энергия переходит в ножной узел и от этого сам двигательный
орган находится в состоянии некоторого пониженного тонуса, т. е.
торможения. После удаления ножного узла тормозящее влияние на
двигательный орган снимается, так как энергия периферической сети
перестает распространяться к ножному узлу. Вследствие этого нога
впадает в тонус. Далее, если в головном узле усилить активное состояние
действием хлористого натрия, то животное делается более подвижным;
если же этот узел смазать кокаином и тем понизить его активное
состояние, то животное перестает двигаться. Предполагается, что в
первом случае из головного узла энергия переходит, между прочим, в
ножной узел, а отсюда в нервную сеть, и потому нога становится более
подвижной. В случае же кокаинизации головного узла, наоборот,
энергия из ножного узла переходит в головной, а из нервной сети в ножной»
вследствие чего нога становится неподвижной.
Аналогичная картина получается при смазывании ножного ганглия
хлористым натрием и кокаином: в первом случае тонус ноги
усиливается, а во втором ослабевает. Согласно учению, все эти влияния на
мускулатуру йоги осуществляются из нервных узлов не непосредственно,
а через периферическую нервную сеть ноги. Когда, например, в ножном
узле активное состояние почему-либо нарастает или падает, в
периферической нервной сети ноги создаются соответственно более или менее
благоприятные условия для тонической деятельности.
Однако С е г а а р (2), ученик Иордана, работая на раках,
сделал наблюдение, в корне противоречащее учению Иордана — а
именно, он наблюдал, что при некотором активном состоянии подглоточного
узла, вызванном действием хлористого натрия, происходит понижение
активного состояния клешневого узла. Обратную картину автор
наблюдал при понижении активного состояния подглоточного узла под
влиянием кокаина. Эта форма нервной регуляции,— говорит автор,— до сих
пор не была известна у беспозвоночных, но была хорошо известна
у высших позвоночных. Эта новая форма названа им «реципрокной
координацией». Головной узел действовал так же, как подглоточный.
Автор указывает, что это взаимодействие происходит не путем
импульсов, а каким-то неизвестным способом. Обычные, известные, по
Иордану, отношения автор обнаружил и при изучении действия
клешневого узла на клешню.
Эти противоречивые фактические наблюдения в значительной мере
лишают гипотетическое учение Иордана убедительности
КООРДИНАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ У ВЕСПОЗВОДОЧНЫХіГ 387
^лсктротоническая роль нейропиля в регуляции рефлекторной
деятельности у беспозвоночных. Описанные Иорданом и Сегааром
явления центральной координации у беспозвоночных, мне кажется,
чрезвычайно напоминают явления, наблюдаемые у позвоночных животных и
приписываемые нами действию нейропиля. У беспозвоночных нейропиль
является основным структурным образованием. Активное состояние его
должно сопровождаться длительным изменением электрического
потенциала. Это подтверждается отдельными осциллографическими опытами.
Гак, например, Э д р и а н (5) показал, что изолированный нервный узел
насекомых все время периодически работает в ритме дыхания. Эта
работа проявляется, с одной стороны, в разрядах быстропротекающих
токов возбуждения во время каждого дыхательного движения, а с дру*
гой — в возникновении одной длительной волны за все время этого
разряда. Длительное изменение потенциала Э д р и а н* приписывает
дендритной области ганглия, т. е. нейропилю, ибо, собственно говоря,
нейропиль образуется главным образом из разветвлений дендритов.
Можно предположить, что длительное изменение потенциала,
обусловленное активным состоянием нейропиля, производит указанное выше
электротоническое понижение возбудимости в близлежащих нервных
узлах и периэлектротоническое повышение в удаленных нервных узлах.
С этой гипотезой хорошо согласуются разного рода факты, приводимые
Иорданом. Например, по удалении головного узла у аплизии
подвижность ноги повышается, ибо этим устраняется постоянное анэлек-
тротоническое периодическое действие длительного потенциала
головного мозга на ножной узел. Усиление активного состояния головного узла
также обусловливает высокую подвижность ноги, ибо в этом случае
либо проявляется периэлектротоническое повышение возбудимости в
нервном узле ноги, либо же эта подвижность обусловливается
усиленным действием нервных разрядов.
По С е г а а р у, такое же усиление активного состояния головного
узла рака вызывает значительное понижение активного состояния клеш-
невого узла, что можно поставить в связь с усиленным анэлектротони-
ческим действием нейропиля головного мозга ь на клешневой узел.
Таким образом, мы думаем, что позвоночные животные унаследовали
от своих беспозвоночных предков как структуру нейропиля, так и его
основные функции торможения и облегчения.
Ир актикум
I. Наблюдение над медицинской пиявкой. Сначала наблюдают локомоцию,
плавательные движения ш рефлекторные реакции на механическое раздражение кожи но
нормальной пиявке. Затем наблюдают сегментальные рефлексы после отрезания
головы и хвоста, т. е. после удаления головного и хвостового нервных узлов. Можно
применить также локальное фзірадическое раздражение кожи.
Когда оба конца препарата зафиксированы,— например, если он приколот к
пробковой пластинке, тогда при слабом локальном раздражении сначала наступает
укорочение сегментов в области раздражения, а потом происходит удлинение, коте-
25*
ЗЧЧ ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВНОГО МОЗГА'
рое постепенно распространяется вдоль всего препарата. При сильном раздражении
сразу укорачивается весь препарат.
Если один конец укрепить булавкой, то можно наблюдать, как другой свободный
конец то вытягивается путем удлинения сегментов, то притягивается обратно путем
их укорочения, так же как это делает нормальная пиявка, когда она присасывается
задней присоской.
Когда же концы препарата не зафиксированы, тогда наблюдаются и другого
рода рефлекторные реакции: например, при слабом раздражении на спинной стороне
переднего конца этот конец вытягивается, спина выпячивается, а задний конец
изгибается в брюшную сторону. При сильном раздражении сразу укорачивается весь
препарат.
2. Наблюдение антагонистической деятельности продольных и кольцевых мыши
на медицинской пиявке. На передней или задней части серединных сегментов пиявки
(после отрезания головы и хвоста) выделяется брюшная цепочка. Выделяется она
вместе с прилегающей тканью, чтобы не повредить цепочки. Другая часть препарата
пиявки служит для кимографической регистрации движений. Для миографического
изучения продольной мускулатуры поступают так: один конец препарата со стороны
нерва фиксируется, а на другой конец навязывается нитка для соединения с
миографом.
Укорочение препарата свидетельствует о сокращении продольной мускулатуры,
а удлинение — о торможении этого сокращения. При удлинении сокращается
кольцевая мускулатура; но это сокращение не давало бы эффекта удлинения, если бы
одновременно не происходило торможение продольной мускулатуры.
На таком препарате наблюдают спонтанное тоническое укорочение и удлинение,
а затем влияние тетанического электрического раздражения нервной цепочки, а
также механического или электрического раздражения кожи (кожных рецепторов). При
наличии тонуса слабое раздражение кожи или нервной цепочки производит
расслабление тонуса, а при отсутствии тонуса слабое раздражение производит
небольшое сокращение. Сильное раздражение нервной цепочки или кожного покрова
производит всегда сокращение. Оно будет тем выше, чем сальнее раздражение.
По прекращении раздражений сокращение быстро кончается, а одновременно
ослабевает и тонус. Если тонуса не было, то нередко после раздражения сокращение,
усиливается — наступает сокращение «отдачи». Эти факты демонстрируют наличие
торможения и (Во время возбуждения при сильных раздражениях.
3. Наблюдение рефлекторного возбуждения и торможения мышц у пиявки после
удаления центральной нервной системы. Взять серединные сегменты и вырезать из
них брюшную цепочку, затем один конец зафиксировать, а другой соединить с
миографом. Можно наблюдать небольшое тоническое укорочение продольной
мускулатуры. Механическое, а также электрическое раздражение кожи производит
сокращение. Сокращаются те сегменты, где происходит раздражение. В этом
проявляется передача возбуждения от кожных рецепторов через кольцевой нерв на
двигательные нервы.
Сильное тетаническое раздражение производит сильное сокращение продольной
мускулатуры, которое расслабевает сейчас же по прекращении раздражения. При
этом ослабевает и тонус, если он был до этого. В этом выявляется периферическое
торможение.
4. Изучение координации продольной и кольцевой мускулатуры у пиявки. Из
одной половины выделяют нервную цепочку и удаляют безнервные сегменты. Здесь
на стороне выделенного нерва высвобождаются несколько сегментов кожно-мышеч
ного мешка путем его круговой перерезки без повреждения нервной цепочки. С
одной стороны кольцо фиксируется булавкой, а с другой стороны оно соединяется
ниткой с миографом. При сужении кольца миограф будет подниматься. Другая
неповрежденная половина фиксируется в области разреза, а свободный конец соединяется с
миографом для записи продольной мускулатуіры. Нужно это сделать так, чтобы
исключить чисто механическое взаимодействие между кольцом сегментов и
неповрежденной половинкой.
КООРДИНАЦИЯ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ У БЕСПОЗВОНОЧНЫХ 389
Таким путем регистрируется, во-первых, спонтанная антагонистическая
деятельность кольцевой и продольной мускулатуры, во-вторых, рефлекторные реакции воз
буждения и торможения при раздражениях нервной цепочки.
Когда продольная и кольцевая мышцы находятся в состоянии тонического
сокращения, тогда слабое тетаническое раздражение нервной цепочки вызывает
расслабление тонуса на обеих мышцах. Если обе мышцы совершенно расслаблены, тогда слабое
раздражение вызывает сокращение на обеих мышцах. Если одна из мышц тонически
сокращена, то, при слабом раздражении, сокращенная расслабнет, з расслабленная
сократится.
ііііп.иі.иітті.,1„1„і.і.чін,міі ...»....,......»...ч.»......,..ч..,і..............,.!...........
Рис. 169. Взаимодействие рефлексов на клешне рака (Cancer pagurus).
Поднятие рычага миографа указывает на замыкание клешни, опускание—на
открытие. При х начинается сильное тетаническое раздражение нервов клешни, которое
вызывает замыкание клешни. Это раздражение продолжается все время. Временами
(С) присоединяется кратковременное раздражение головного ганглия, которое
, производит размыкание клешни — торможение замыкателя. При О прекращается
мозговое раздражение (по Иордану).
Если раздражение цепочки сильное, то сокращаются обе мышцы, и сокращение
тем сильнее, чем больше сила раздражения* Но по прекращении раздражения обе
мышцы быстро расслабевают, а при этом расслабевает и существующий до
раздражения тонус.
5. Наблюдение антагонистической иннервации на клешне рака. Отрезанная
клешня фиксируется в штативе таким образом, чтобы только подвижная ее часть (дактило-
подит) оставалась свободной. Она соединяется ниткой с миографом. В два
последующие членика (мероподит и карпоподит) вкалываются булавочные электроды так,
чтобы нервный ствол оказался между ними (рис. 167). Электроды соединяются с
индукториумом. Сначала определяют силу тетанического раздражения, при которой
производится закрывание клешни. Этот эффект возникает в результате активирования
возбуждающего нервного волокна замыкателя и тормозящего волокна размыкателя.
Затем раздвигают катушки и находят порог, при котором клешня активно
раскрывается. При этом в активное состояние приходят возбуждающие волокна размыкателя
в тормозящие волокна замыкателя. Раздражение при промежуточных расстояниях
катушки будет вызывать то закрывание, то открывание клешни.
6. Взаимодействие периферического и центрального раздражения в отношении
клешни рака. Фиксируется голова — прудь животного и одна клешня. Для
периферического раздражения булавочные электроды вставляются в мероподит и в карпопо
дит регистрируемой клешни, а для центрального раздражения другая пара
электродов втыкается в голову через панцырь справа и слева у основания глаз. Достаточно
<Звдьное тетаническое раздражение клешни производит ?е замыкание, а достаточно
сильное раздражение головы, т. е. головного ганглия, вызывает судорожное
выпрямление туловища и конечностей и открывание клешни, Если это раздражение
производить во время периферического раздражения клешни, вызывающего замыкание, то
каждый раз при центральном раздражении замыкание клешни прекратится (рис. 169)
390 ОБЩАЯ ФИЗИОЛОГИЯ СПИННОГО МОЗГА И СТВОЛА ГОЛОВЙОГО МОЗГА
•4итррятурн
1. Jordan Н., AUgemeine vergleichende Physiologie d. Tiere, Berlin u. Leipzig,
1929; Verb, Dtsch. Zool. Ges. 32. Jg., 195, 1928
2. Segaar I., Z. Vergl. Physiol., 10, 120, 1930,
3. BuddenbrockW., Vergleichende Anatomie d. Nervensystema d, Wirbelloseu
Handb. norm. Path. Physiol., 9, 805, 1929.
4 HanstromB., Vergleichende Anatomie d. Nerven systems d. Wirbellosen Tierc»
Berlin, 1928.
5. Adri an E. D„ J. Physiol., 72, 132, 1931.
6. Hoffmann P., Z. Biol., 63, 4119 1934.
7. M a n g о 1 d E., Z, AUg. Physiol., 5, 135, 1905
8. Bidermann W., Sitzb. Akad. Wiss. Wien, 96, 3, 5, 8, 1887.
9. Иордан Г., Физиол. журн. СССР, 19, 141, 1935.
10. И о р д а н Г., Практикум сравнительной физиологии. Госиздат, стр. 192, 1934.
11. U е х k u 11 J., Z. Biol., 46, 1905.
12. Н е г гі с к С. J., Neurobiological foundation of animal behavior, USA^ 1924
13. P а г ке г G. H., The elementary nervous system. USA, 1912.
14. U e x к ii 11 J., Umwelt und Innerwelt der Tiere. Berlin, 1909.
15. Baglioni S., Winterstein's Handb. d. Physiol., Bd. 4, 23, 1913.
16 Беритов И. иГогава M., Тр. Тбилисск. гос. ун-та им. Сталина, 27а
№ 1, 1945.
1/ Romanes J., J. Internat. Sci. Ser. 1893. Цит. но Buddenbrock'y.
18. К a h И. Л., Сб. докладов VI Всесоюз. физиол. съезда, стр. 82, 1937.
19. Р a n ti n С. F. A., J. Exp. Biol., 13, 159, 1936.
20. Van H ar re ve 1 d A. a. Wiersma С. A., J. Exp. Biol., 14, 448, 1937.
21. Van H a r r e v e 1 d A., J. Сотр. Neurol.,70, 267, 1939; J. Exp. Biol., 16, 398, 1939
22. Van HarreveldA. and Wier sm a C. A., J. Physiol., 88. 78, 1936
23. Беритов И., Тр. Тбилисск. гос. ун-та, 27а, 29, 1945.
24. Adrian Е. D., J. Physiol., 72, 132, 1931.
25. Jasper Н. Н. a. Monnier. J. Cellur. Сотр. Physiol., 11, 259, 1938.
26. Гогава М., О координации сегментальных рефлекторных реакций у дожде
вого червя 1947 (не напечатано)
Часть шестая
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ
БОЛЬШОГО МОЗГА
Глава 1
ОБЩИЕ СВЕДЕНИЯ О КОРЕ БОЛЫПИХ^ПОЛУШАРИЙ
Главные функции большого мозга. Функцией большого мозга
является, во-первых, восприятие тех раздражений, которые действуют на
органы чувств На расстоянии. Так, раздражение органов зрения, слуха
и обоняния происходит на расстоянии, и от каждого «дистантного
рецептора» раздражение воспринимается корой большого імозга
непосредственно. Во-вторых, большому мозгу принадлежит главная роль в
производстве ориентировочных реакций на всевозможные раздражения.
В-третьих, он является усилителем центральных возбуждений: слабые
периферические раздражения, не способные вызвать периферический
эффект через спинной мозг или стволовую часть головного мозга,
производят его в сильной форме через кору больших полушарий.
В-четвертых, он является главным нервным субстратом для производства
прирожденных и индивидуально приобретенных актов поведения, а также
индивидуально автоматизированных двигательных и секреторных
реакций. Наконец, в-пятых, наиболее сложной функцией коры большого
мозга является психо-нервная деятельность, благодаря которой
возникают все ощущения, представления и суждения.
У высших позвоночных удаление коры большого мозга уничтожает
все эти функции полностью или ослабляет их в значительной степени
У низших позвоночных кора больших полушарий развита слабее и
означенные функции не сосредоточиваются в коре полностью. Так, у
рыб удаление большого мозга не влияет на двигательную способность
и на их отношение к внешней среде (Штейнер, 1). Такие рыбы по
своему поведению не отличаются заметно от нормальных: они различают
съедобное вещество от несъедобного, ищут и принимают пищу; поведе-
те их в отношении других рыб такое же, как у нормальных.
Удаление больших полушарий у амфибий приводит к тому же
результату. Лягушка без полушарий передвигается, ловит мух; зимой
впадает в спячку и зарывается в землю, весной пробуждается. Такая
лягушка не наталкивается на препятствия, обходит их, что указывает на
394
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
сохранение зрения (Шрадер, 2). Она утрачивает означенные рефлек
торные и поведенческие реакции только в том случае, если одновремен
но с большими полушариями удален и промежуточный мозг.
Рептилии и птицы с удалением большого мозга только частично
утрачивают упомянутые функции. Голубь без большого мозга сначала
теряет способность произвольно принимать пищу, но потом эта
способность восстанавливается. Он не теряет зрения полностью, поэтому при
полетах не наталкивается на препятствия. Днем он хоДиіг, а ночью стоит
неподвижно и спит. Но в то же время он не узнает голоса птенцов, не
пугается кошки; самка такого голубя не узнает самца и т. д.
Если вместе с корой вырезать подкорковые узлы, полосатое тело,
тогда голубь долгое время остается совершенно беспомощным и даже
не клюет пищи. Впоследствии он несколько оправляется и начинает
клевать, но не способен выводить птенцов (Роджерс, 3; Шрадер, 2).
Голуби без больших полушарий обнаруживают способность к
индивидуальным актам поведения с целью защиты и питания. Они научаются,
например, подходить на пищевой сигнал к кормушке и клевать зерно
(Беритов, 11).
У (Млекопитающих удаление больших полушарий, как уже
указывалось, «окончательно устраняет все описанные выше функции. Сначала
Гольц (4), затем другие (Ротман, 17; Зеленый, 5) экстирпировалн
всю кору головного мозга у собаки и оставляли ее жить более года.
В течение этого времени ни одна из упомянутых функций не
восстанавливалась. Такая собака большей частью спит, обычно просыпаясь лишь
для отправления естественных потребностей или же под влиянием
голода. Проснувшись, она встает и медленно ходит или же стоит на месте,
упершись головой в стену. Встречающихся на ее пути предметов она
не обходит. Не очень тихие звуковые раздражения (свист, дудка,
хлопанье ладонями) (вызывают явственную реакцию, а именно — движение
юловы и встряхивание ушей. Но под влиянием этих звуков собака не
меняет своего поведения. Сама она за еду не принимается; пищу нужно
класть ей в рот, только тогда она ее проглатывает (Зеленый, 5). Она
геряет все заученные движения или индивидуально приобретенные
рефлексы и не приобретает их вновь. Зеленый пытался образовать
у такой собаки индивидуальный слюнной рефлекс на звуковое
раздражение: тон С2 духового камертона, при первых испытаниях вызывавший
двигательную реакцию, сочетался с прирожденным слюнным рефлексом
от 0.25% раствора соляной кислоты. Сочетание было произведено
324 раза, но звук камертона не приобрел свойства индивидуального
раздражителя,— он не вызывал слюнного рефлекса. Характерно, что
если у собаки сохранен небольшой остаток коры, то способность к
индивидуальным рефлексам у нее не уничтожается. Так, у одной собаки
остался небольшой участок коры над полосатым телом и в одной из
височных долей на основании мозга и у нее удалось после очень долгой
работы образовать индивидуальный оборонительный рефлекс на звук.
ОБЩИЕ СВЕДЕНИЯ О КОРЕ БОЛЬШИХ ПОЛУШАРИЕ 395
ло индивидуального слюнного рефлекса образовать не удалось
(Зеленый и Полтырев, 15).
Было произведено также удаление коры большого мозга у обезьян
V них после этой операции нервная регуляция страдает еще в большей
Рис. 170. Слон нервных клеток и'нервных пучков в коре большого
мозга человека.
Схематическое изображение наиболее типичной'для коры шестислойной
архитектоники. С левой стороны даны клеточные слои: / — зональный или моле
кулярный слой; // — внешний зернистый; ///—пирамидный; IV—внутренний
зернистый; V — ганглионарный или внутренний пирамидный; VI —
полиморфный. С правой стороны даны расположения нервных пучков в тех же
слоях (по Бродмачу и Фохту).
і
мере, чем у других -'животных и, кроме того, в значительной степени
нарушается жизнеспособность. Обезьяны без большого мозга могли
жить максимум 26 дней, в то время как собаки без него живут годами
Такие об&зьяны могут стоять на ногах, сидеть, двигать головой,
схватывать предметы рукой, но локомоция у них расстроена очень сильно.
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
Кдода они спят, шум их будит. Вообще звуки действуют на них,
вызывая ориентировочные реакции (Карплюс и Крейдль, 19; Магнус, 9).
Аналогичные наблюдения были проведены на людях, родившихся
без коры большого мозга. Один такой ребенок был под наблюдением
37г года. Большой мозг его представлял тонкостенную кисту, не
содержавшую миэлиновых нервных элементов. Палеоэнцефалон и другие
отделы мозга не были повреждены. Все те нервные пути, которые
начинаются р коре, отсутствовали. Ребенок с момента
рождения умел сосать молоко. Все время спал
Голод его не будил; для кормления приходилось
его будить. Первоначально все конечности были
неподвижны. На четвертой неделе начались
судорожные движения: нижние конечности были
судорожно разогнуты, а верхние согнуты.
Пальцы были сильно согнуты. Руками он не мог
держать предметы. Кроме молока, никакой другой
пищи проглотить не мог, даже если ее клали
ему в рот. Он ничего не видел, но свет вызывал
сужение зрачка. Сильные звуки вызывали
только вздрагивание. Ребенок не чувствовал
повреждающих раздражений кожи, но реагировал на
поглаживание головы и на давление на нее-
если, например, он плакал, то при таких
раздражениях головы затихал. Это — обычный
рефлекс, который наблюдается у животных после
удаления большого мозга. Ребенок не улыбался.
Мимика у него полностью отсутствовала. Он не
менял положения и первоначально совсем не
издавал звуков. Впоследствии обнаружил
способность к плачу. Итак, ребенок без большого
мозга обнаруживал еще меньше активности, чем
обезьяна боз большого імозга, и особенно імало —
в сравнении с собакой Гольца (Эдингер и
Фишер, 10).
Строение коры большого мозга. іКора
больших полушарий содержит различного
характера нейроны. Их клетки располагаются в коре
определенными слоями. У высших позвоночных имеется шесть строго
определенных слоев. Из них два слоя — второй и четвертый,— так
называемые внешний и внутренний зернистые слои, включают массу
совершенно недифференцированных эмбриональных клеток. Первый, так
называемый зональный, слой состоит главным образом из сплетений
дендрит ов. Нервных клеток в нем очень мало. Он образуется главным
образом из разветвлений дендритов от пирамидных клеток и аксонов от
нервных клеток афферентных путей и еще от некоторых непирамидных
клеток коры мозга.
39ф
Рис. 171. Кора большо
го мозга мыши.
Л—наружный слой из
сплетения клеточных отростков;
В~слой мчтіых пирамидных
клеток; С — слой средней
величины пирамидных
клеток; D—слой больших
пирамидных клеток; Е—с юй
яйцевидных или полиморфных
телец; ^—белое вещество. От
всех клеток наиболее
толстый дендрит поднимается к
поверхности мозга и
принимает участие в образовании
наружного сплетения
(Р.-Кахаль).
ОБЩИЕ СВЕДЕНИЯ О КОРЕ БОЛЬШИХ ПОЛУШАРИЙ 3$7
Третий, пятый и шестой слои наиболее богаты клеточными
элементами (рис. 170). В коре большого мозга насчитывают до 15 млрд.
нервных клеток, из них 6 млрд. приходятся на долю маленьких. Величина
клеток колеблется очень широко: от 8—9 до 150 микронов. Основная
Рис. 172. Диаграмма составных частей коры большого мол а.
С левого края — клеточная картина париетальной коры взрослой мыши, окрашенной по Нисслю.
В середине рисупка тела и дендриты представленных в коре типов клеток. Аксоны их не зарисованы
1 — пирамиды II слоя; 2 и 3 — пирамиды III слоя; 4, 5, 6 — звездчатые пирамиды; 7, 8, 9 — длиь
ные, глубоко лежащие пирамиды; 10— короткие пирамиды; 11— средние пирамиды; 12, лЗ —
короткие пирамиды VI слоя; 14, 15, 16 — веретенообразные клетки; 17 — глубоко лежащие
звездчатые клетки. С правого края приведены афферентные волокна: a, b — специализированные тала-
мические волокна; с, d—неспециализированные афферентные волокна; е, /— ассоциативные волокна.
Все корковые клетки располагаются регулярно на более или менее близком расстоянии друг от
друга, так что дендриты их образуют очень густое сплетение, синаптически сочлененное с нейро-
фибрнллярным сплетением, которое образуется афферентными и ассоциативными волокнами.
Дендриты клеток одного типа, как, например, 5 и 10, не выходят из соответствующего слоя.
Другие- клетки, например 14, отдают дендриты ко всем слоям. У большинства клеток дендриты
сконцентрированы в определенных слоях (Лоренте де Но).
масса клеток имеет форму пирамид, и потому они называются
пирамидными. Все они направлены вершинами к поверхности мозга.
Нейриты их вступают в подкорковое вещество мозга, а из дендритов
один, наиболее толстый, начинающийся от вершины клеток, идет к
наружной поверхности мозга и здесь разветвляется в наиболее
поверхностно лежащем зональном слое (рис. 171).
Пирамидные клетки впервые появляются в коре головного мозга у
рептилий. У них намечается кора, разделенная на слои. В наиболее
развитой части, bulbus olfactorius, имеется три слоя. Пирамидные клетки
отдают верхушечные дендриты к поверхности коры, где из них
образуется густое сплетение. У птиц строение коры в общем такое же, как и у
рептилий. Наибольшее развитие пирамидная система получает только
398 ОБШАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
у млекопитающих. У них число пирамидных клеток сильно возрастает
Одновременно они сильно дифференцируются по своим формам я
взаимным связям (Ариёнс Капперс, 18).
Кора содержит также множество пучков нервных волокон, которые.
с одной стороны, вступают сюда из других отделов мозга и таким
образом объединяют периферические органы чувств с корой, а с другой,—
представляют нейриты корковых нейронов, которые объединяют между
собой различные корковые участки, а также связывают кору с другими
Рис. 173. Типы цитоархитектонического строения коры больших
полушарий человека.
Столбик 1—тип двигательной области; 2—фронтальный тип; 3—париетальный
тип; 4 — полярный тип; 5 — гранулярная, зернистая кора (Экономо).
подкорковыми отделами центральной нервной системы. Через эти во
локва корковая деятельность влияет на другие отделы мозга. На
рис. 172 дается схематическое соотношение цитоархитектоники коры.
типов клеток с их дендритами, афферентных и ассоциативных нервных
волокон поЛоренте де Но (20).
Строение коры, так называемая, архитектоника, неодинаково в раз
ных областях коры большого мозга. Ширина всего коркового слоя на
фронтальном разрезе и каждого слоя клеток, количество слоев,
клеточный состав, т. е. величина, форма, расположение клеток, в каждом
участке коры чрезвычайно вариируют (рис. 173). Эти вариации
занимают определенные, ясно отграниченные территории. В большом мозг\
млекопитающих еще Мейнерт насчитывал 11 хорошо отграниченных
больших корковых полей. На основании гистологического и
физиологического исследований каждое такое поле делится еще на несколько
ОБЩИЕ СВЕДЕНИЯ О КОРЕ БОЛЬШИХ ПОЛУШАРИЙ 3<Нг
самостоятельных участков (Бродман, 14; Фохт, 13). Значительная
диффередоировка коры признается уже у высших млекопитающих и
особенно релика у человека. По данным Ц, Фохт и О. Фохт, імозго-
«ная^повер^ность человека может быть разделена на 200 полей.
Приводимая дифференциация в строении коры в общем совпадает
l данными физиологии и клинического материала о локализации
функций в коре мозга. Особенно показательны исследования неврологов
Ц. Фохт и О Фохт с электрическим раздражением коры обезьяны
ВЕДРО ^
\1*Г(ЩЕНЬ + ja +tfOH{4rtffr
*БЕД+РО }
(За *
Ш
Г поэаон. т " іосдтго і
+ хвосхлнус +позвоночник 1тпсгеяпа$Ц.
% голень ХА *+++ ± + іпозвоічочні
>* БЕДРО Л*Ьлппдт«л іяппАтил*
6а*
+ ДОПЛТНА
+
+ ПЛЕЧО
+
+ ПРЕДПЛЕЧЬЕ \
ЗАПЯСТЬЕ
ЛОПА+ТМа +
+ +
ПРЕДПЛЕЧЬЕМ
¦ +
ЗАПЯ+СТЬЕ
і + г
ГЛАЗд
КНАРУЖИ
ГгЛАЗОі
ННАРЖ1
+ + ЗАПЯСТЬЕ J ¦
+ ВЕРХН0Н?Чн\РАЗГПАЛьАраЗГ ^ПО/ГЬ
+ \СГНБ. іОЛЬЦ? *
Г9с
+ глаза
хщ
fC язык
сгив*пал.
*Д*?|
/
7V ?ЬЦ$х
bjorscrbfr^
вмЕЗте "
<*<?
,нмжн
лицЯы.^
* 6aa ЧЕЛН*
X ^.т
№б\бЬа\ЯЗь1Н / Л
* * -% J челюсть ним***
глаза
ннаружи
і уши »•
, глаза 4
ікнаРшні
+76J ??уз}
+j* глаза t
ЩЛРХЖА
*~? я*
я
/So.
глаза
ННАРХЖИ
\ДдИЖ?НИЕШЗ
глазя
Рис. 134. Деление коры церкопитека (обезьяны) на отдельные ноля на основании
физиологического и гистологического исследования.
Кору раздражают электрическим током. Там же указывается, какие именно части тела реагировали
на раздражение. Цифры обозначают нумерацию отдельных полей по гистологическим исследованиям
(Ц. и О. Фохт).
церкопитека. Они обнаружили полное соответствие добытых этим путем
физиологических результатов с гистологически установленной
архитектоникой мозга животных. На основании своих исследований Ц. Фохт
и О. Фохт составили предположительную карту полей для
человеческого мозга, которая оказалась очень сходной с той, которая была
составлена невропатологом Фёрстером для человеческого мозга на
основании раздражения коры при операциях (рис. 174).
Микроскопическая структура коры высших позвоночных в новейшее
время наиболее детально была изучена Лоренте де Но (20). Он
изучил ее у разных (млекопитающих, до обезьяны включительно.
Подобно Рамон'-и-Кахалю, он находит, что клетки всех слоев коры
мозга у млекопитающих образуют своими сильными ветвящимися ден-
дритами густой войлок (рис. 174). Многообразный клеточный состав
наружных двух слоев коры мозга совсем не дает эфферентных нисхо
400
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
s.a.
дящих аксонов. Аксоны клеток коротки и оканчиваются частью в том
же слое, где лежат клетки, частью в соседних слоях, выше или ниже
этого слоя. Иначе говоря, все клетки первых двух слоев являются
ассоциативными. Только в нижних четырех слоях значительное количество
клеток отдает эфферентные, выходящие из коры волокна, но и здесь
не мало клеток с короткими
осевыми цилиндрами,
осуществляющими связь
элементов этого слоя между
собой и с вышележащими
слоями (рис. 175). Очень
характерно существование
так называемых
«возвратных» аксонов и коллатера-
лей. Осевые цилиндры
некоторых клеток сначала
идут вниз, а потом
поворачивают обратно и
заканчиваются в вышележащих
слоях. Во многих случаях
поворачивает обратно не весь
аксон, а его коллатерали
(рис. 177). Все эти аксоны
и их коллатерали
оканчиваются многочисленными
утолщениями, т. е. синапсами в
том или другом слое коры
на поверхности тел клеток
и их дендритов. На клетках
и дендритах эти синапсы
располагаются сотнями,
иногда беспрерывным слоем
(рис. 176).
Афферентные волокна, приходящие в кору из нижних отделов мозга,
заканчиваются преимущественно в третьем и четвертом слоях, лишь
немногие из них заходят еще в первый, зональный слой. Остальным
слоям они редко отдают коллатерали (рис. 172). Лоренте де Н о (20)
находит, что эти афферентные волокна своими окончаниями приходят
в множественный синаптический контакт с находящимися здесь дендри-
тами и телами клеток. Отсюда следует, что пирамидные клетки нижних
слоев могут получать периферические импульсы главным образом в своей
дендритной части, распространяющейся во втором и третьем слоях.
Но ввиду многочисленных связей нижних пирамид с ассоциативными
клетками второго и третьего слоев, пирамиды должны получать
импульсы от афферентных волокон также и через эти клетки, и притом
довольно многочисленные и в интенсивной форме.
Рис. 175. Связь афферентного волокна с
клетками и дендритным разветвлением коры.
I, 2, 3—пирамидные клетки, 4 — ассоциативная клетка.
Аксоны пирамидных клеток уходят ез коры, а аксон
ассоциативной клетки связан с пирамидными клетками 2
и ч и отчасти с дендритным сплетением, а/ —
афферентное волокно, сильно ветвясь, заканчивается синаптически
несколькими синапсами на пирамидной клетке / и на
ассоциативной клетке и множеством синапсов в дендритном
сплетении. Стрелки указывают на направление импульсов
возбуждения в аксонах. На данном рисунке афферентное
волокно передает импульсы возбуждения непосредственно
только одной пирамидной клетке, двум же остальным—
через ассоциативную клетку (Лоренте де Но).
ОБЩИЕ СВЕДЕНИЯ О КОРЕ'БОЛЫПИХ ПОЛУШАРИИ 401
Синапсы
коротких
аксонов
Синапсы
Офф€>/>
Залома*
Синапсы
«ъ р. аксонов
Синапсы
/ ассои; путей
) Синапсы аксонов
\пениетых нлетон
Синапсы
Из данного краткого описания видно, что в коре большого мозга,
гак же как и в других отделах мозга, нервные элементы образуют
замкнутые круги разной сложности. Каждый такой круг имеет свою
группу афферентных и
эфферентных волокон (рис. 177) и,
по предположению Л о р е н т е
д е Н о, представляет
самостоятельную физиологическую
единицу (20). В этой системе
возбуждение может
распространяться во всех
направлениях,— как от афферентного
волокна к эфферентному, так и
наоборот,— хотя в каждой
частичке ее импульсы
возбуждения могут итти только в одном
направлении. Все нервные
круги окружены густым войлоком
нервных отростков, отходящих
от участвующих в нервных
кругах клеток. Но в этом ней-
ропиле принимают участие
также многочисленные нервные
клетки с короткими аксонами
и сильно разветвляющимися
дендритами (рис. 175). Ней-
ронно-нейропильная структура
коры мозга не похожа на
другие формы этой "структуры в
других отделах центральной
нервной системы; она более
развита, более высоко
организована и, очевидно, предназначена для выполнения более сложных
функций, протекающих в коре большого мозга. Джадсон Херрик
детально изучил нейропиль головного мозга у низших позвоночных, а
именно — у амфибий и рыб. Он нашел, что главная масса большого
мозга представляет собой нейропиль, но чем выше животное, тем
последний более дифференцирован. На рис. 178 дается нейропиль в той
примитивной форме, как он представлен у низших позвоночных, у
амблистомы. Херрик полагает, что локализированные нервные
центры служат для высоко специализированных функций, для их большей
эффективности во всех стереотипных или автоматизированных формах
поведения. Нейропильная же масса считается основным субстратом для
объединения локализированных аппаратов при высших формах нервной
деятельности, как восприятие, представление, творчество. Вся
динамичность корковой деятельности связывается с нейропильным субстратом
26 и. С. Беритов
Синапсы
Рис. 176. Пирамидная клетка с двумя
дендритами у основания и с
верхушечным дендритом.
На дендрита* и на теле клетки оканчивается
множество синапсов от разнообразных ней-
роноь (Лоренте де Но).
402 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
коры мозга. Предполагается, однако, что с филогенетическим
развитием нервной ткани и этот нейропиль меняет свои свойства: в коре
большого мозга высших млекопитающих и человека он становится высоко
Рис. 177. Диаграмма нервных кругов в коре большого мозга млекопитающих
Ру. J и Ру. 2 — пирамидные клетки третьего слоя; В. с.— корзиночные клетки; Str.—
звездчатые клетки. Шероховатые отростки — дендриты, а гладкие — аксоны (a). A. f. — афферентные
волокна, приходящие в кору из низших отделов мозга или из других отделов коры. Как
аксоны корковых клеток, так и афферентные, ветвясь, заканчиваются синапсами как на клетках,
так и на дендритах. Из клеток и аксонов образуются замкнутые нервные круги разной
сложности. Эти круги обведены штриховыми линиями вдоль аксонов. Как дендриты, так и си-
вантические окончания представ чены на них в сильно редуцированном виде. Стрелки
указывают направление распространения возбуждения в кругах (составлен автором по Ло-
ренте де Но).
специализированным аппаратом интеллектуальной работы. Таково
(мнение выдающегося американского невролога Джадсона Херрика-
Мы же думаем, что нейропиль сам по себе не может обладать высшими
психическими качествами, как это полагает X е р р и к. Этими
свойствами должна обладать целиком кора большого мозга с участием нейро-
пиля и нервных кругов, из которых складывается кора мозга.
Нисходящие корковые пути. Нервные волокна, нисходящие из коры
большого мозга, проходят сначала через белое подкорковое вещество
лучистого венца. Они передают корковые импульсы всем нижним
отделам центральной нервной системы. Часть этих волокон берет начало в
гигантских пирамидных клетках Б е т ц а, которые лежат в пятом
корковом слое двигательной области (см. поле 4 на рис. 174). Они
направляются в спинной мозг и называются пирамидными путями.
Эти пути проходят через ножки большого мозга, варолиев мост и
продолговатый мозг. В последнем ояи частично перекрещиваются. Одна
ОБЩИЕ СВЕДЕНИЯ О КОРЕ БОЛЬШИХ ПОЛУШАРИЙ 403
часть волокон переходит на другую сторону, а потом через боковые
столбы оканчивается в спинном мозгу. Другая часть остается на той же
стороне и передними столбами вступает в спинной мсзг. Эти
пирамидные пути перерождаются, если удалить кору (рис. 179).
Рис. 178. Нейропиль мозга амблистомы по Херрику.
Горизонтальный разрез через серое вещество дорзальной части
переднего обонятельного ядра. Метод Голь лжи. Увел. 100 раз. А —
окрашен только нейропиль. Клеточные тела не окрашены. Они
выступают кружками. Тонкие аксоны, образующие этот нейропиль,
идут от клеток переднего ядра, от обонятельной луковицы и,
вероятно, от primorriium piriforme (р. ріг.) и primordium hippocampi
(p. hip.); v. 1 —боковой желудочек. В- приводится связь обычного
нейрона с нейропилем (Херрик).
Наряду с пирамидными пучками, достигающими спинного мозга без
перерыва, существуют другие корковые пучки, прерывающиеся в
головном мозгу. Почти все ядра головного мозга получают корковые
волокна того же характера, как пирамидные пучки (рис. 179 — р", р')>
например, зрительные бугры, красные ядра, ядра продолговатого мозга
и варолиева моста. Эти пути называются экстрапирамидными.
Они начинаются во всех двигательных участках коры, а именно, в по-
26*
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
лях 1, 2, 3, 4, 5, б, 8 и 22, приведенных на рис. 174. По существу
между этими пучками и пирамидными нет разницы,— все они прерываются
около координирующих аппаратов и двигательных нейронов.
Исследованиями Монакова (6) и Ариёйс Капперса (7) установлено,
что корковые центробежные волокна прерываются около тех же
элементов мозга, где оканчиваются коллатерали чувствительных заднекореш-
ковых волокон. Так, в каждом
сегменте спинного мозга волокна
пирамидных пучков, подобно заднеко-
решковым волокнам, заканчиваются
в передних и боковых рогах,
вероятно на одних и тех же интра-
спинальных нейронах, из которых
составляются координирующие
аппараты спинномозговых рефлексов,
а также в нейропиле и на
двигательных клетках. На чисто
физиологическом основании подобное
заключение было сделано М у н-
ком (8). Именно, по его
наблюдениям, между рефлекторными
координирующими механизмами и
определенной частью двигательной
сферы противоположного полушария
существует тесная связь, какой нет
между той же двигательной сферой
и другими рефлекторными
центрами.
Однако это нельзя понимать в
том смысле, что после разрыва
какого-нибудь пирамидного пучка
определенный двигательный орган
должен потерять способность к
корковым реакциям. 'Каждый
координирующий аппарат действует н? на
один только двигат ельный орган,
например, не на одну, а на
несколько конечностей. Поэтому любой
двигательный орган может приходить в действие не только через
наиболее 'прямой пирамидный путь, но и через ряд других пирамидных
путей, оканчивающихся при разных координирующих аппаратах. Кроме
того, известно, что двигательные органы приходят в действие также
через экстрапирамидные пучки, которые прерываются в стволовой части
мозга, например, >в красных ядрах. Через нервные волокна, образующие
нисходящие пути из данных ядер, производятся такие же движения, как
и через прямые корковые пучки. Следовательно, перерезка прямых кор-
404
Рис. 179. Схема пирамидного пучка.
га — передний корешок; рі — боковой
пирамидный пучок; ра — передний пирамидный
пучок; col — коллатерали; р", р' — акстра-
пирамидные пучки, идущие к ядрам
черепных нервов; р — пирамидный пучок внутри
мозговых полушарий и в мозговом стволе;
XI, XII, X, IX, VI, VII, V, III,
IV—корешки и ядра соответствующих черепных
нервов; сі— внутренняя капсула (Бехтерев).
і
ОБЩИЕ СВЕДЕНИЯ О КОРЕ БОЛЬШИХ ПОЛУШАРИЙ
405
ковых пучков, оканчивающихся в спинном мозгу, не устраняет
способности к «произвольным» и вообще корковым движениям.
Прямые пирамидные пути особенно сильно развиты у человека и
обезьян. Это зависит от степени развития коры больших полушарий.
Рис. 180. Монаковский к
другие нисходящие пучки
th — зрительные бугры; ct —
нервные пучки» идущие из коры
к зрительным буграм; пг—нрас-
ные ядра; tr — пучки,
соединяющие зрительные бугры с
красными ядрами; /и—монаковский
пучок; trt — пучок, идущий из
зрительных бугров к nucleus
reticularis; nrt—nucleus
reticularis; tho — пучок зрительных
бугров к продолговатому мозгу;
га — передние корешки
(Бехтерев).
Рис. 181. Схема восходящих
корковых путей.
гр — задние корешки; /г — волокна
боковых столбов, которые
присоединяются к петле Рейля; / — волокна
боковых столбов, которые проходят к
зрительным буграм по внутренней стороне
петли; rlXs, rVs — чувствительные
волокна IX и V головных нервов; nIXsu
nVs — ядра тех же нервов;
fsc—восходящие волокна от IX и V* головных
нервов; nd — nucl. den Latum; cca —
передняя ножка мозжечка; пг — красные
ядра; th — зрительные бугры; сі —
nucleus lentifonnis; cts— корковые
пучки из зрительных бугров (Бехтерев).
У птиц прямые пирамидные пути развиты меньше, чем у
млекопитающих. У низших позвоночных сильнее развиты непрямые пути,
прерывающиеся в красных ядрах. Среди млекопитающих у человека и обезь
ян начинающиеся в красных ядрах нисходящие пути, именуемые м о-
наковскими, развиты значительно слабее, чем у собаки и кошки
идти у других низших (млекопитающих (рис. 180). Если на одной стороне
перерезать ножку мозга, а на другой — боковой столб в спинном мозгу,
где проходит монаковский пучок, то на стороне с перерезанным
боковым столбом способность к «произвольным» движениям ослабеваем
40b ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
больше, чем при перерезке только ножки мозга. Этим доказывается
участие монаковского пучка в «произвольных» движениях. Но не один
он участвует в «произвольных» движениях: после означенной двойной
операции животное еще сохраняет способность к этим движениям.
Кроме того, путем искусственного раздражения коры можно вызвать
двигательный эффект. Отсюда очевидно, что корковое движение может быть
вызвано на периферии через любой корковый путь — как через
пирамидный, непосредственно оканчивающийся в спинном мозгу, так и через
экстрапирамидные, прерывающиеся в ядрах головного мозга.
Чувствительные или восходящие корковые пути. Все заднекореш-
ковые волокна и вообще все волокна, идущие от рецепторов, имеют
связь с корой больших полушарий, но ни одно из них не достигает
большого мозга без перерыва. Каждое волокно прерывается где-либо
ів спинном мозгу или в ядрах головного мозга, а отсюда начинаются
новые нейроны, направляющиеся к коре. Но и эти нейроны не
достигают мозга: они в большинстве случаев кончаются в зрительных буграх
(рис. 181). Некоторые нейроны второго порядка кончаются в
четверохолмии; например, нейроны слухового и зрительного нервного пути
(рис. 183, 185). Спинномозговые чувствительные пути все проходят
сначала через петлю Рейля, а затем через зрительные бугры. Таким
образом, восходящие корковые пути должны прерываться минимум
дважды, а потому состоять по крайней мере из грех нейронов.
Большая часть чувствительных восходящих путей перекрещивается,
однако существуют и неперекрещивающиеся пучки, например,
некоторые кожно-мышечные и зрительные. Совсем не перекрещивается
обонятельный путь.
Перекрещивание обычно происходит в области второго нейрона
(рис. 181). Первый, т. е. заднекорешковый, нейрон, а также третий,
кончающийся в коре, не перекрещиваются. Так как часть заднекореш-
ковых волокон кончается в спинном мозгу, а часть в продолговатом, то
и перекрещивание происходит частью в спинном, частью в
продолговатом імозгу.
Литература
1. Steiner, Die Funktionen d. Zentralnervensystems u. ihre Phylogenese. Braun
schweig, 1888.
2. Schrader, Pfl. Arch., 41, 75, 1887; 44, 175, 1888.
3. Roggers F., J. Сотр. Neurol, 35, No 1, 1922.
4. GoltzF., Pfl. Arch., 13, 1, 1879; 14, 412, 1877; 20, 1, 1879; 2t>, 1. 1881; 28, 579,
1882; 34, 450, 1884; 51, 570, 1892.
5. ЗеленыйГ., Тр. Об-ва русск. врачей в СПб., 1911; Revue med., № 2, 191, 1929.
6. MonakowC. v., Z. Psych. Physiol. Sinn., 54, 1909; Localisation im Grosshirn
Wiesbaden, 1924.
7. Ariens Kappers, Fol. NeurobioL, 1, 373, 1908; 2, 261, 1908; Brain, 44.
125, 1921.
8. MunkH., Sitzber. Berl. Akad. Wiss., 1893t 759.
9. M a g n u s R., Pfl. Arch., 193, 396, 1922.
t0 EdingeTU. Fischer, Pfl. Arch., 152, 535, 1912
ОБЩИЕ СВЕДЕНИЯ О КОРЕ БОЛЬШИХ ПОЛУШАРИЙ 407
11. Beritoff J., РП. Arch., 213, 370, 1926.
12. Minkowsky M., Zum gegenwartigen Stand d. Lehre von den Reflexen, 1925;
см. также кбил. сб., иосвящ. 70-летию акад. Павлова, стр. 261, 1925.
13. V о g t С. и, О., Naturwiss., 14 Jg., Н. 50/51; J. Pt-ych. Neur., Bd. 25, Erganzunga
heft, 1918.
34. Brodmann K.f Vergleichende Lokalisationslehre d. Grosshirnrinde. Leipzig,
1925.
15. Z e 1 i о n у G. P. u. PoltireffS. S., Berichte d. Gesellschaft russischer
Physiol. Nam. J. Setchenow, И. 3, 1929 (на нем. яз.).
16. H e г гі с k С. J. , J. Сотр. Neurol., 69, 93 и 239, 1934
17. Rothmann. Arch. Auat. Physiol., 1908, стр. 103.
18. Ariens Kappers, The comparative anatomy of the nervous system etc., 1936.
19. Karplus I. P. a. Kreidl A., Arch. Anat. u. Phy.iol. 1914, стр. 155.
^0. Lorentede N6 R., J. Psych. Neurol., 46, 381, 1933; ib. 46, 113. 1934.
Глава II
ВОСПРИНИМАЮЩАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА
Воспринимающие области в коре большого мозга. Человек
и высшие позвоночные животные отвечают на раздражения не
только деятельностью спинного мозга или стволовой части головного
мозга, но также и деятельностью коры больших полушарий.
Центральная нервная система устроена так, что возбуждение каждого
рецептора неизбежно оказывает свое влияние и на кору больших
полушарий.
Но как указывалось выше, существуют рецепторы, возбуждение
которых действует преимущественно на кору. К таким рецепторам
относятся органы зрения и обоняния, и в меньшей степени — орган
слуха.
В соответствии с шестью рецептивными органами в коре
больших полушарий существует шесть воспринимающих областей
(Мунк, 2; Феррье, 1). Шестым рецептивным органом являются
рецепторы мышц, сухожилий и суставов. Для восприятия
возбуждений от этих рецепторов в коре существует особая воспринимающая
поверхность.
Воспринимающая функция занимает в коре большого мозга
определенные территории только у млекопитающих. У низших
млекопитающих,— например, у кролика или кошки,— каждая функция
занимает значительную часть мозговой поверхности, перекрывая одна
другую, но в каждой воспринимающей территории имеется одна
оптимальная область. При удалении этой области данная функция
страдает очень сильно, но полностью не исчезает. У высших животных
каждая из воспринимающих функций все более и более
сосредоточивается в соответственной оптимальной области. Вследствие этого
значительная часть мозговой коры освобождается от воспринимающих
функций. У высших обезьян почти у3 всей коры, а у человека % всей
коры не играют существенной роли в отношении воспринимающей
функции, но, по клиническим наблюдениям, даже у человека в некоторой
очень малой степени воспринимающая функция сохраняется во всех
отделах коры мозга (Монаков, 5; Гольдштейн, 26)
ВОСПРИНИМАЮЩАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА 409
Воспринимающая область для зрения. Большой мозг лягушки,
повидимому, совсем не участвует в зрении. Лягушка без больших
полушарий <в отношении зрения не отличается от нормальной (Шрадер,
39). У птиц, в частности у голубей, большой мозг безусловно владеет
зрительной функцией, ибо с удалением коры происходят некоторые
расстройства в области зрения: поведение птицы свидетельствует об
отсутствии узнавания знакомой среды. Но голубь с удаленной корой
избегает препятствий и при полетах хорошо различает предметы, на
которые садится. Значит, подкорковые
центры и промежуточный мозг у птиц
так же владеют зрительной функцией,
как и у амфибий. У рептилий,
повидимому, зрительная функция
представлена в імозгу, как у птиц
(Шрадер, 39).
Кора большого мозга
млекопитающих (кошки, собаки) обладает
зрительной функцией в полной мере. На
основании опытов школы И. П. П а в-
л о в а можно утверждать, что для
высшего анализа и синтеза зрительных
раздражений служит определенная
область в затылочной части коры, а
именно — в ее верхней, задней ч
медиальной поверхностях (рис. 182).
По исследованиям М у н к а на
собаках и других высших позвоночных,
а также на человеке, зрительная область одного полушария связана с
сетчаткой обоих глаз. Внутренняя, носовая часть сетчатки связывается
с корой противоположного полушария. Наружная, височная часть
сетчатки связывается с полушарием той же стороны (рис. 183). Это
перекрещивание происходит в хиазме зрительных нервов. От центральной
макулярной части сетчатки волокна идут в соответствующее и
противоположное полушарие (Ринг, 38). У низших млекопитающих, а также у
птиц, рыб и амфибий имеется полное перекрещивание зрительных
волокон. Поэтому у этих животных сетчатка одного глаза связывается толь
ко с одним противоположным полушарием.
У обезьян, а также у человека зрительная область помещается в
верхнем отделе затылочной доли (рис. 184). Одностороннее
разрушение ее вызывает частичную слепоту обоих глаз. Согласно
исследованиям Минковского (6), у животных существует более или
менее выраженное территориальное соответствие между сетчаткой и
корой мозга. Так, он находит, что передняя часть зрительной
воспринимающей области связывается главным образом с нижней частью
сетчатки, но в то же время каждый ее участок имеет множество свя
зей со всей воспринимающей областью. К аналогичному заключению
Рис. 182. Топография зрительной (А)
и слуховой воспринимающей
области (В) у собаки.
Эти воспринимающие области обозначены
точками. Чем гуще точки, тем больше в
соответствующем участке воспринимающая
способность (Лу чиани).
410 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
пришли и относительно представительства сетчатки в коре
человеческого мозга (Геншен, 7).
На зоологической лестнице позвоночных зрительная область впер-
Рис. 183. Зрительные восходящие пути.
V — оптическая система глаз; CV—зрительная область коры больших
полушарий; ОС — подкорковые зрительные пути; fa—волокна,
переходящие с четверохолмия в fasciculus praedorsalis; s—волокна из
хиазмы, идущие ко дну третьего желудочка. Остальные обозначения см. на
самом рисунке (Бехтерев).
вые развивается у птиц. Она занимает поверхность полушарий
впереди и сверху мозговых ножек и прикрывает мозговой желудочек.
Если у новорожденных животных вырезать глазные яблоки, то
зрительный нерв и его продолжение в мозгу перерождаются;
перерождаются также corpus geniculatum lateralis, pulvinar и передние холмы
четверохолмия; в зрительной же области затылочных долей заме-
ВОСПРИНИМАЮЩАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА 411
чается остановка развития. По экстирпации зрительной области у
молодых животных перерождаются corpus geniculatum lateralis, puU
vinar и передние холмы; несколько позднее дегенерирует зрительный
нерв (Монаков, 5)
Рис. 184 Воспринимающие области коры большого мозга
человека.
На рисунке дана кожно-мышечная, слуховая, зрительная и вкусовая
воспринимающие области. Они занимают сравнительно небольшие
территории. Конгао-мышечная область представлена вместе с
двигательными участками, которые располагаются по преимуществу впереди ро
ландовой борозды (Эдингер).
Воспринимающая поверхность слуха. Воспринимающая поверхность
слуха находится у собак и кошек впереди и позади сильвиевой борозды
(рис. 182-В). До последнего времени основным методом исследования
анатомических границ слуховой функции, как и других
воспринимающих функций, являлось удаление определенных частей коры мозга.
Этим путем было установлено, что слуховая функция осуществляется
главным образом позади сильвиевой борозды в височных долях. Но
данный метод не является точным, ибо собаки и кошки, лишенные всей
коры, реагируют на звуки рефлексами. Даже децеребрированная кошка
проявляет акустические рефлексы (Форбс и Шеррингтон, 29). На
кошках после удаления коры Дюссер де Баренн наблюдал
способность локализации направления звуков. Спустя б—7 недель после
полного удаления коры кошка шла по направлению к звуку, в
особенности когда была голодная (Дюссер де Баренн, 28).
В настоящее время пределы воспринимающих областей в коре
больших полушарий устанавливаются электрографическим путем.
Раздражается рецептор и исследуется изменение электрического потенциала в
том или другом участке коры мозга. Этим путем было показано, что у
кошки и собаки воспринимающая функция слуха осуществляется в
4J2 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
области верхней части gyrus Sylvii, средней, а также передней и
задней части g>rus ectosylvius (Корнмюллер, 31; Бремер и Доу, 32; Тун-
тури, 36),
При этом восприятие звуков разной частоты производится разными
участками коры, и эти участки располагаются сообразно с высотой зву
ков (Ларионов, 18). Путем изучения корковых потенциалов при
Рис. 185 Схема восходящих чувствительных путей слухового нерва.
Даны важнейшие пути Улитковый, первичный нейрон оканчивается сейчас же при вхожде
нии в мозг в nucleus ventrahs и несколько выше tuberculum acusticum Отсюда начинается
второй нейрон, который оканчивается или в верхней оливе соответствующей стороны, или же
в той же оливе другой стороны, или же через etna acustica в заднем двухолмии Из олив
третий нейрон заканчивается или в ваднем двухолмии или в nucleus laqueus Из последнего
ядра начинается четвертый нейрон к коленчатому телу Из двухолмия и коленчатого тела
четвертый и пятый по счету нейроны поступают в кору мозга. Названия ядер и путей даны
на рисунке (Эдингер)
разных звуках было установлено, что в средней части gyrus ectosylvius
звуки более высокой частоты воспринимаются передними ее участками,
а низкие — задними; причем последовательные октавы располагаются
друг за другом вдоль коры через более или менее одинаковые
интервалы. Кроме этой области звуки воспринимаются также в вентральных
частях gyrus ectosylvius,— именно, низкие звуки — в переднем колене
этой извилины, а высокие —в заднем. От каждого уха звуки
воспринимаются в обоих полушариях, но более сильно —в противоположном
полушарии (Тунтури, 36).
В высшей степени вероятно, что означенная область коры мозга —
gyrus ectosylvius — имеет отношение главным образом к восприятию
звуковых раздражений. Что же касается дифференциации и синтеза
их, то эти функции должны осуществляться главным образом в височной
области, как это было показано работами школы Павлова и др
ВОСПРИНИМАЮЩАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА 413
Разрушение одного уха вызывает у новорожденных животных
недоразвитие слуховой области (Мунк, 2).
У человека слуховая область намечается в середине верхней
височной извилины и в так называемой гешлевской извилине, которая
начинается в глубине сильвиевой борозды и направляется к той же
височной извилине (Геншен, 7). На рис. 185 показан весь восходящий
путь от улитки до коры мозга.
Обонятельная воспринимающая область. Обонятельная область
занимает комплекс area pyrifarmis и gyrus hippocampi. Эта корковая
область представляет продолжение lobus olfactorius. На зоологической
лестнице первоначально эта область намечается среди рептилий (Эдин-
гер, 4), но по работам Дюссер де Баренна (28) известно, что
обонятельные реакции сохраняются у кошек после удаления всего нео-
кортекса. Оперированные таким образом животные находили пищу по
запаху.
Первоначально топографическое положение обонятельной
воспринимающей области было установлено на основании экстирпации мозга
у животных и клинических наблюдений на людях. В настоящее время
оно было подтверждено на основании электрографического
исследования коры мозга, а именно, было установлено, что у кроликов и собак
при раздражении обонятельного рецептора пахучими веществами
возникают биоэлектрические токи возбуждения в виде усиленных
ритмических колебаний потенциала коры в области lobus pyriformis и gyrus
hippocampus (Бун-Иши Хасама, 27; Аллея, 47; Эдриан, 50).
Воспринимающая область для вкуса. Вкусовая область у
млекопитающих лежит в пределах сильвиевой борозды между слуховой и
кожно-мышечной бороздами (рис. 186). Частью она распространяется
и на эти области. По мнению Бехтерева, она должна находиться
гам, где лежат двигательные точки глотания и жевания.
На основании изучения военных повреждений последнего времени,
а также по опытам на обезьянах была уточнена корковая локализация
вкусовой чувствительности. Она находится в кожно-мышечной
воспринимающей области лица (см. ниже) и перекрывает участок общей
чувствительности языка в близком соседстве с центрами жевания и
глотания (Бёрнштейн, 49 и др.). Значит, новейшими наблюдениями
подтверждается первоначальное положение Бехтерева.
Воспринимающая область кожно-мышечных раздражений. Эти
области лежат в передней части больших полушарий: у кошек и собак —
вокруг крестовидной борозды (sulcus cruciatus); у человека и обезьян
они занимают центральные извилины по обеим сторонам роландовой
борозды.
У собак и кошек воспринимающая область кожных раздражений в
отдельности простирается за пределы крестовидной области в gyrus
coronarius й в передние части gyrus marginalis, suprasylvius и ectosylvius
(Дюссер де Баренн, 20) (рис. 186). У человека и у обезьян она лежит по
преимуществу в задней центральной извилине. У обезьян она безусловно
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
шире, простираясь вперед до sulcus arcuatus и назад — до сильвиевой
борозды (Дюссер де Баренн, 25) (рис. 187, 184). Воспринимающая
область імышечно-сухожильно-суставных раздражений у собаки и
кошки находится по преимуществу в сигмовидной извилине; у человека
и у обезьян она помещается главным образом в передней центральной
извилине. Обычно эта область объединяется с кожной областью под
одним названием кожно-мышечной воспринимающей области.
Несомненно, что и та и другая области являются самостоятельными, хотя в
значительной степени и перекрывают одна другую (Красногорский 11;
Монаков, 5; Флексиг, 9; Редлих, 10).
Восходящие пути кожно-мышечных рецепторов перекрещиваются
приблизительно на том же уровне мозгового ствола, где вступают в
мозг, значит, они достигают коры
противоположного полушария (рис, 181).
Некоторая часть восходящих
чувствительных путей, несомненно, не пере
крещивается и вступает в кору
соответствующего пол>шария. Это видно
из физиологических наблюдений: с
удалением одного полушария
передача в кору кожно-мышечных
возбуждений с той же стороны тела не
уничтожается.
У обезьян и, вероятно, у человека
корковая проекция тактильных
рецепторов от разных участков кожи распо
лагается в задней центральной
извилине противоположного полушария
приблизительно в том же порядке,
как эти кожные участки располага
ются на теле. На самом верху задней
извилины проецируется пятка, а за
тем сверху вниз идут проекции
остальной стопы, голени, бедра, туловища; затем направление проекций
меняется: после туловища — затылок, за ним шея, плечи, предплечие,
кисть, пальцы; в самом низу опять ряд сверху вниз: щека, лицо, глаза,
нос, губы, рот, язык, гортань. Это расположение указывает, что
корковая проекция происходит согласно сегментальным дерматомам, с
некоторыми извращениями в отношении шейного отдела (Вулси, Маршал и
Бард, 48)
Характеристика воспринимающих областей. Главная функция
указанных воспринимающих областей заключается не просто в восприятии
периферических раздражений, а в анализе и синтезе этих раздражений
в установлении разницы между данными раздражениями, в разложении
каждого внешнего воздействия на компоненты и затем в связывании их
одного с другим в одно целое и каждого компонента или всего внешнего
414
Рис. 186. Топография вкусовой
(С) и кожно-мышечной (D)
воспринимающих областей у
собаки.
Точки обозначают соответствующие
поверхности. Большими точками
обозначены участки лкмга,
обладающие наибольшей воспринимающей
функцией (Лучиани).
ВОСПРИНИМАЮЩАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА 415
воздействия в целом с какой-нибудь целесообразной деятельностью
организма. Эта характеристика воспринимающих областей
принадлежит И. П. Павлову: она является результатом работ, выполненных
под его руководством многочисленными учениками. В соответствии с
этой основной аналитической функцией воспринимающие области были
названы просто анализаторами зрения, слуха, обоняния, вкуса, кожи
(осязание, температура) и мышц (двигательный анализатор). В
лаборатории Павлова анализатором называется вся центрипетальная,
Рис. 187. Кожно-мышечная воспринимающая область
у обезьяны (Macacous).
Установлено путем стрихнинного отравления. Точечными
линиями указаны границы чувствительных зон задней конечности,
передней конечности и головы, прерывистой линией
ограничивается двигательная зона, заднюю границу этой зоны составляет
роландова борозда {FR), между зоной передних и задних
конечностей узкой полоской идет зона туловища, F, S —сильвиева
борозда; S. іип — ыіісиь lun itus,.? ore.—sulcus arcuatus, F. %p
f is sura luLerpauetalis (Дюссер де Б аренд).
афферентная часть рефлекторной дуги — от рецептора до коры
включительно. Так, например, зрительным анализатором называется весь
нервный аппарат, начиная от сетчатки глаза — до зрительной
воспринимающей области включительно,— значит, весь тот нервный аппарат,
который изображен на рис. 183. Но так как главная функция этого
аппарата — аналитическая деятельность — зависит от коры большого
мозга, то название анализатора обычно применяют к корковой части
всего центрипетального аппарата, т. е. к воспринимающей области
коры. Этот термин Павлов ввел в то время, когда им признавалась
только аналитическая функция коры мозга. Позднее (1923)
Павлов (12) приписал коре мозга, кроме того, и синтетическую функцию
как на этом много раньше настаивал выдающийся русский невролог
Бехтерев (16). Поэтому в настоящее время термин «анализатор»
больше не подходит для обозначения воспринимающей области коры
мозга.
Локализация воспринимающих функций. В пределах каждой
воспринимающей области существует локализация определенных функций.
4)6 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
Периферическое раздражение каждого рода воспринимается
отдельным участком коры большого мозга. Во всяком случае, это установлено
в наиболее изученных областях зрения, слуха и кожно-мышечных
раздражений. Относительно зрительной области, например, известно,
что центральное зрение, соответствующее центральной ямке (желтому
пятну) сетчатки, локализируется в fissura calcarina. Здесь же многие
авторы локализируют цветоощущение. О слуховой области известно,
что корковое представительство различных звуков — разное (Мунк, 2).
Ларионов (18) из лаборатории Бехтерева пытался доказать
Рис. 188. Поверхность коры больших полушарий человека в области роландовой
борозды.
А — соответствующими пиниями ограничены двигательные области: языка, рта, лица, пальцев,
руки, плеча, туловища и ноги; В — теми же линиями ограничены воспринимающие области для
тех же органов. Движение данного органа и восцриятие раздражений из данного органа
производится в пределах всей соответствующей корковой области (Пенфильд и Болдрей).
это специальными исследованиями. Путем частичной экстирпации коры
височной области собаки он обнаружил, что корковые участки,
воспринимающие музыкальные звуки, располагаются в порядке их
естественного положения в отношении высоты. Значит, более близкие по высоте
звуки обладают более близким корковым представительством. То же
самое известно и относительно кожно-мышечной области. При
отравлении определенных участков этой области стрихнином получается
повышение периферической кожной чувствительности в определенной
кожной поверхности. Например, при отравлении стрихнином того участка
мозга, где лежат двигательные точки правой передней ноги на правой
ВОСПРИНИМАЮЩАЯ ФУНКЦИЯ КОРЫ БОЛЫПОГА МОЗГА 417
передней ноге наблюдается повышение кожной чувствительности на
прикосновение, а также чувствительности глубоко лежащих тканей на
давление. Другая передняя нога не обнаруживает этих изменений (Бери-
10B, 19; Дюссер де Баренн, 20). Далее, путем электрического
раздражения коры головного мозга на людях было показано, что при
электрическом раздражении задней центральной извилины подопытное лицо
ощущает раздражение в виде прикосновения, царапания и щипания в
определенных участках тела, на руке или ноге, в зависимости от
участка раздражения (Кушинг, 21; Фолькенбург, 22).
У каждого индивида существуют в коре свои особые точки для
вызывания означенных ощущений в том или другом участке кожи и
мышечного чувства движения. Эти точки расположены по обе стороны
роландовой борозды в постцентральной и прецентральной извилинах.
Они не стандартны: не только у разных людей они лежат в разных
местах, но и у одного и того же индивида меняют с течением времени свое
место (Пенфильд и Болдрей, 33). При этом они не выходят из
определенных границ данных извилин, как видно на рис. 188,
составленном Пенфильдом и Болдреем. Эти авторы обнажали мозг
у больных людей в целях хирургического удаления заболевших частей
мезга и применяли электрическое раздражение коры для установления
очага заболевания. Раздражение производилось без наркоза, и
больные сами активно участвовали в установлении ответных реакций на
раздражение. Таким путем Пенфильд и Болдрей обнаружили
наличие точек для разного рода кожных ощущений, как щипание, зуд,
холод, тепло, чувство движения без движения, прилива крови,
утолщения или вздутия в отношении языка, а также побуждение к движению.
Но они не обнаружили точек, соответствующих типичной сильной боли.
Только несколько раз указывалось на ощущение боли в конечности.
На этом основании авторы находят, что боль мало представлена или
даже совсем не имеет представительства в коре мозга. Все ощущения
в громадном большинстве случаев локализировались на
противоположной стороне, лишь в отношении лица, глаза и языка ощущения
получались билатерально. Изолированно на гомолатеральной стороне
ощущения не наступали.
Однако в настоящее время многие авторы не разделяют
представления о точечной территориальной локализации воспринимающих
функций. Прежде всего следует отметить, что у собак удаление
какой-либо означенной выше воспринимающей поверхности не ведет к
полному исчезновению соответствующей воспринимающей функции.
Так, например, после удаления височной доли слуховая функция не
исчезает целиком и навсегда: индивидуальные рефлексы на звук все
еще образуются, и даже сохраняется способность к элементарной
дифференциации звуков (Павлов, 41; Калишер, 42). Весьма возможно, что
отдельные элементы коры, воспринимающие звук, рассеянные по всей
коре, обладают тем меньшей аналитической и синтетической
способностью, чем дальше они отстоят от височной доли, являющейся ядром
27 и. С. Беритов
418
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
слуховой воспринимающей области (Павлов, 41). Более того, у
низших млекопитающих, как кролик, белые крысы, восприятие
периферических раздражений не должно являться основной функцией коры.
Удаление коры мозга, так называемого неопаллиума, т. е. височных,
затылочных, а также теменных и лобных долей, не ведет ни к глухоте,
ни к слепоте. Кролики реагируют на малейшие шумы,
дифференцируют свет от темноты, видят предметы, обходят препятствия (Тен Кате
и Герк, 44).
У кошек удаление коры мозга неокортекса без повреждения
полосатого тела не ведет к слепоте или глухоте: они слышат звуки и
реагируют на них ориентировочной реакцией, отличают свет от темноты,
обходят препятствия. Могут передвигаться по направлению звука или
лучей света (Тен Кате, 45; Гирндт и Лемпке, 46). В отношении
локализации воспринимающих функций, очевидно, кошки занимают
промежуточное положение между кроликами и собаками.
Птицы и рептилии безусловно лишены локализации
воспринимающих функций в коре мозга. Доказано, что у птиц удаление неопаллиума
ке сопровождается потерей слуха или зрения (Беритов, 34; Баянду-
ров, 43). Следовательно, кора переднего мозга у птиц не является
главным носителем воспринимающих функций и не содероюит в себе даже
признаков дифференциации на отдельные воспринимающие области.
То же, з общем, можно сказать относительно низших млекопитающих.
У высших млекопитающих воспринимающая функция головного мозга
всецело переходит в кору — неокортекс. Одновременно с переходом
происходит дифференциация коры в отношении этой функции. У собак
воспринимающие области широко перекрывают друг друга, у обезьян
этого нет: у них эти области строго разграничены. У человека же почти
2/з коры свободны от воспринимающей функции.
Внешнее воздействие среды всегда так сложно, а действие каждого
компонента настолько обширно вследствие корковой иррадиации
возбуждения, что едва ли можно говорить о локализации влияния внешней
среды или какого-либо ее элемента на один определенный участок коры
мозга. На это указывает, повидимому, и то, что электрическим
раздражением участков коры мозга никогда не удается вызвать такое
ощущение или представление, какое обычно получается под влиянием
воздействия внешней среды. Например, при раздражении зрительной области
получаются просто световые ощущения в виде мигания, света или
сверкания, которые проецируются в пространстве. Но при этом нет никакого
видения предмета и внешней среды (Краузе, 23; Лёвенштейн и Бор-
хардт, 24). Итак, территориальная локализация воспринимающих
функций в коре большого мозга главным образом касается качества внешних
раздражений, но не самых ощущений от внешних предметов и явлений,
лежащих в основе представления об этой среде.
Методы изучения территориальной дифференциации корковой
воспринимающей способности. Эту дифференциацию экспериментально
изучали следующим образом: обнажали большой мозг и вырезали в
ВОСПРИНИМАЮЩАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА 419
коре одного или обоих полушарий определенные области. Операция
производилась в асептических условиях во избежание инфекции. После
такой операции животные могут жить по нескольку лет, поэтому вполне
возможно определить, какая функция преимущественно пострадала под
влиянием данной операции. Таким путем было установлено, что если у
собаки вырезать затылочные доли в обоих полушариях, то она
слепнет. Она теряет способность ориентироваться в пространстве при
помощи зрения: наталкивается на препятствия, не узнает знакомой
среды и т. д. Если вырезать какую-либо воспринимающую область
только в одном полушарии, собака теряет способность к восприятию
возбуждений от соответствующего рецептора противоположной
стороны. Так, например, если у собаки вырезать слуховую область в
правом полушарии, слух страдает на левое ухо. Если вырезается такая
воспринимающая область, восходящие пути которой перекрещиваются
не целиком, а частично, то соответствующая функция частично теряется
в отношении обоих рецепторных органов. Так происходит при
вырезании зрительной области в одном полушарии.
Если воспринимающие области вырезаются в одном или в обоих
полушариях не целиком, а частично, то соответствующие функции
исчезают не полностью, а только более или менее значительно
ослабевают. Чем больше вырезаемый участок коры, тем сильнее страдает
соответствующая функция. Затем имеет значение, какой именно
участок вырезается из данной области. Каждая область имеет небольшое
центральное место с большой воспринимающей способностью и
обширную периферию со слабой воспринимающей функцией (рис. 186).
Конечно, при удалении центрального места аналитическая функция
данной воспринимающей области страдает больше, чем при удалении
периферических участков. Вообще при обширном разрушении
воспринимающей области выпадает более сложная аналитико-синтетическая
деятельность, а при частичном разрушении ее — менее сложная. Так,
например, при небольшом частичном разрушении в зрительной области
устраняется способность различать предметы по их форме, но
сохраняется способность отличать сильный свет от слабого, движущийся
предмет от недвижущегося. Если разрушенная часть более обширна, то
исчезает аналитическая способность в отношении движения. Различие
же света от темноты исчезает при полном удалении зрительных
областей (Топоров, 13; Кудрин, 14).
Более новым способом для изучения территориального
разграничения воспринимающих областей является легкое стрихнинное
отравление коры большого мозга. Это отравление вызывает повышение
восприимчивости периферических раздражений. Пока этим методом
изучали воспринимающую область кожных и проприоцептивных
раздражений. Приемы изучения таковы: обнажают кору мозга и
определенный небольшой участок ее смазывают очень слабым раствором
стрихнина; затем животное ставят на пол и наблюдают его реакции на
кожное и проприоцептивное раздражение. Таким образом были исследо-
27*
420
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
ваны Дюссер де Варенном (20, 25) кошки и обезьяны.
Результаты уже сообщались выше.
Возбудимость воспринимающих областей. Еще недавно считалось
бесспорным фактом, что обширный отдел коры больших полушарий,
где сосредоточена воспринимающая функция, невозбудим для
непосредственного раздражения, так как не дает двигательных реакций.
Обычно при раздражении коры внешний эффект получался только
с определенных участков, поэтому и возбудимость приписывалась
только этим так называемым двигательным или секреторным
участкам. В последнее время это мало вероятное утверждение было
отвергнуто. Прежде всего, как мы указывали выше, у людей
электрическое раздражение воспринимающей области дает субъективные
ощущения. Кроме того, Грэим Броун (15) доказал на
человекоподобных обезьянах, что раздражением задней центральной извилины,
которая относится к кожной воспринимающей области и раньше считалась
невозбудимой, можно вызвать двигательный эффект, если
предварительно раздражать двигательный участок в передней центральной
извилине. Так, например, если здесь раздражать двигательный участок
сгибания пальцев, то раздражением задней центральной извилины
вызывается двигательный эффект, а именно — сгибание же пальцев.
Эти факты с очевидностью указывают, что воспринимающая область
коры обладает возбудимостью, как и всякое другое нервное
образование. Нужно думать, что между воспринимающей областью и
двигательными участками не существует строго определенных прирожденных
связей. Поэтому двигательная реакция в ответ на раздражение
рецепторов тотчас же после рождения животного должна зависеть,
во-первых, от возбудимости двигательных точек и, во-вторых, от их близости
к возбужденному воспринимающему участку. Мы знаем, что
ориентировочная реакция головы и глаз вызывается при любом необычном
раздражении глаза. То же следует сказать по поводу всех других
корковых движений в первое время после рождения. Вызванное необычным
раздражением возбуждение иррадиирует по всей коре и приводит в
активное состояние прежде всего все наиболее возбудимые и наиболее
близкие к возбужденной воспринимающей поверхности двигательные
участки. То же должно было происходить в опыте Грэим Броуна
на обезьянах. Возбуждение, вызванное в воспринимающей области,
иррадиировало по коре и вызывало эффект из раздраженного
двигательного участка вследствие повышения в нем возбудимости. Правда,
воспринимающая область взрослых животных должна владеть массой
определенных нервных связей с двигательными участками, которые
возникли на основании индивидуального опыта. Но эти связи не должны
функционировать сейчас же после операции. Вообще в условиях
операции воспринимающая кора не может сохранить нормальную
возбудимость; она неизбежно должна сильно пострадать: кора лишена
оболочек и подверглась кровоизлияниям, охлаждена; кроме того, она
более или менее механически повреждена и испытывает еще влияние
ВОСПРИНИМАЮЩАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА 421
наркоза. Поэтому вскоре после операции раздражение
воспринимающей области или совсем не вызовет возбуждения, или вызовет настолько
слабое, что оно не сможет подействовать на двигательные участки и
произвести отсюда периферический эффект.
Возбуждение воспринимающих областей. Выше мы показали, что
воспринимающие участки коры большого мозга у человека и животных
возбуждаются как под влиянием периферических импульсов,
приходящих туда по афферентной системе от рецепторов, так и под влиянием
непосредственного раздражения. Процесс возбуждения в коре мозга,
как и в других частях центральной нервной системы, в настоящее
время изучается путем исследования возникающих при этом биотоков
возбуждения. Электрической активности коры мы посвящаем
специальную главу, где будут подробно рассмотрены все формы корковой
электрической активности и их происхождение. Мы здесь укажем
только, что в воспринимающих областях коры большого мозга, как в
любом другом отделе центральной нервной системы, мы имеем как
быстрые колебания электрического потенциала, в ритме 100—200 в
1 сек. и выше, выражающие возбуждение нервных кругов, так и
медленные колебания, сопутствующие активированию нейропиля, в
ритме 4—40 в 1 сек. Эти формы электрической ативности возникают
в ненаркотизированной коре при нормальном функциональном
состоянии независимо от внешних раздражений, т. е. спонтанно. На глубоко
наркотизированных животных спонтанная ритмическая активность в
коре мозга отсутствует, но появляется при непосредственном
раздражении коры или при сильных раздражениях рецепторов. При этих
условиях электрическая активность дает колебания, характерные как
для возбуждения нервных кругов, в виде быстрых колебаний аксониого
характера, так и для активирования нейропиля в виде медленных
колебаний (Беритов, Брегадзе и Цкипуридзе, 34; Беритов и Гедева-
нишвили, 35).
Литература
1. Ferrier D., Vorlesungen ub. Hirnlokalisation, Leipzig u. Wien, 1892.
2. MunkH., Ueber die Funktionen der Grosshirnrinde, 2. Aufl. Berlin, 1890; Ueber
die Funktionen \on Him u. Ruckenmark. Berlin, 1909.
3. Bechterew W., Die Funktionen d. Nervencentra, Dtsch. Ausgabe, 1.
4. Edinger L., Vorlesungen uber Bau d. nervosen Zentralorgane. Leipzig, 1908—
1911.
5. MonakowC. v., Die lokalisation im GrossMrn. Wiesbaden, 1914, S. 343.
6. Minkowski M, Pfl. Arch., 141, 171, 1911.
7. HenschenS. E, Klinische u. anatomiscbe Beitrage z. Pathologic d. Gehirns.
Upsala, 1890—1903.
8. BecbterewW., Arch. Anat Phvsiol. Supplbd., 1900, S. 145.
9. F 1 e с h s i g P., Neurol. Zbl., IB, 606, 1894; 14, 1118, 1895; 17, 977, 1898; 18, 1060,
1899; 22, 202, 1903; Gehirn u. Seele, Leipzig, 1896; D. Lokalisation d. geistigen
Vorgange. Leipzig, 1896.
10. Redlich, W. Klin. Wschr., Nr. 2, 24—30, 1893.
И. КрасногорскийН., О процессе задержки и о локализации кожного и
двигательного анализатора etc. Дисс, СПб. 1911.
12 ПавловИ, 20-летнпй опыт объективного изучения высшей нервной
деятельности. Л., 1923.
13. Топоров Н, Условные рефлексы с глаза при удалении затылочных долей
больших полушарий собаки. Дисс, 1908,
422
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
14. Кудрин А., Условные рефлексы при удалении задних половин больших
полушарий. Дисс, 1910.
15. Graham Brown, J. Physiol., 48, 1914.
16. Бехтерев В., Общие основы рефлексологии человека, стр. 157, 1923.
17. Павлов И., 20-летний опыт объективного изучения высшей нервной
деятельности, 3-е изд., стр. 358, 1925.
18. Ларионов В., О корковых центрах слуха. Дисс, 1898.
19. Беритов И., Русск. физиол. журн., т. 1, 1917.
20. Dusser d e Barenne J. G., Q. J. Exp. Physiol., 9, 355, 1916.
21. G us hi n g H., Brain, 32, 44, 1909.
22. Volkenburg, Z. ges. Neur. Psych., 24, 294, 1914.
23. Krause, Klin. Wschr. Nr. 8, 1263.
24. Lowenstein u. Borchardt, Dtsch. Nervenheilk., 58, 264,
25. Dusser de Barenne, Proc. Roy. Soc, B, 96, 272, 1924; Dtsch. Z.
Nervenheilk., 83, 273, 1924.
26. Goldstein K., Die Lokalisation in der Grosshirnrinde, Handb. r.orm. path.
Physiol., 10, 1927.
27. В u n-I с h i H a s a m a. Pfl. Arch., 234, 748, 1934.
28. Dusser de Barenne J. G., Arch. Neur. Psych., 30, 884, 1933.
29. Forbes A. a. Sherrington C. S., J. Physiol., 35, 367, 1914.
30. В u n-I с hi H a s a m a., Pfl. Arch., 236, 36, 1935.
31. KornmiillerA. Е., Die bioelektrischen Erscheinungen d. Hirnrindfelder etc.
Leipzig, 1937.
32. Bremer F. a. DowR. S., J. Neurophysiol., 2, 308, 1939.
33. P e n f i e 1 d S. H. a. Boldrey., Brain, 60, 389, 1938.
34. Беритов И., Брег адзе А. иЦкипуридзеЛ., Сообщ. АН Груз. ССР,
3, 169, 1942; Тр. Ин-та физиол. им. Бериташвили, 5, 240, 1943.
35. Беритов И. иГедеванишвилиД., Тр. Ин-та физиол. им. Бериташвили,
6, 279, 1945.
36. Tunturi A. R., Am. J. Physiol., 141, 397, 1944.
37. В е г і t о f f J., Pfl. Arch., 213, 370, 1926.
38. Ring R., Gehirn und Auge. Miinchen, 1923.
39. Schrader., Pfl. Arch., 41, 75, 1887; 44, 175, 1889.
40. Adrian E., J. Physiol., 87, 83P, 1936; 89, IP, 1437.
41. П а в л о б И., Лекции о работе больших полушарий, 1927.
42. KalischerO., Sitzber. Preuss. Akad. Wiss. Berlin, VIII—IX, стр. 204, 1907;
Berl. Physiol. Ges. 6 Febr., 1914.
43. Б а я н д у p о в Б. И., Тр. Томск, мед. ин-та б, 1937.
44. Теп С ate u. A. W. v. Н er k, Arc л. Nearl. Physiol., 18, 1, 1933.
45. Ten Cate, Arch. Neerl. Physiol., 19, 191, 1934.
46. Girndt O. u. Lerapke H., PfJ. Arch., 239, 544, 1937.
47. Allen W. F.f Am. Jour. Physiol., 139, 553, 1943.
48. Woolsey C. N., Marshall, W. H. a. Bard P., J. Neurophysiol., 6,287, 1943.
49. Born stein W., Yale J. Biol. Med., 12, 719, 1940; ibid. 13, 113, 1941.
50. Adrian E. D.5 J. Physiol., 100, 459, 1942.
Глава III
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА
Общее понятие. Каждое движение млекопитающих животных и
человека, именуемое заученным или привычным, вызывается через
воспринимающие области в коре больших полушарий. Если собаку
(научить подавать переднюю лапу в ответ на какой-нибудь звук, то это
движение несомненно будет вызываться через слуховую
воспринимающую область. После удаления слуховой области заученное движение
исчезает. Если такое же движение было выработано на показывание
хлыста, то оно исчезает после удаления зрительной области. Одним
словом, каждое индивидуально приобретенное движение осуществляется
при обязательном участии какой-либо воспринимающей области.
Мы знаем, что кора больших полушарий участвует также в
прирожденных движениях. Так, нормально все ориентировочные реакции
производятся через кору. Известен еще целый ряд корковых
изолированных движений, являющихся прирожденными рефлексами. К таким
движениям относятся кожные рефлексы, например подошвенный, который
состоит в сгибании большого пальца ноги и вызывается при
механическом раздражении подошвы. Все эти корковые движения предполагают
участие воспринимающих областей коры больших полушарий.
В осуществлении коркового движения участвуют не только
воспринимающие области коры, но еще и другие участки. В каждой
воспринимающей области существуют особо дифференцированные участки,
через которые производятся все без исключения корковые реакции на
периферии. Возбуждение из любой воспринимающей области коры не
может распространиться на подкорковые отделы центральной нервной
системы помимо этих дифференцированных участков. Они тем
характерно и отличаются от остальной коры, что после вскрытия черепа и
обнажения коры большого мозга искусственное раздражение их дает
определенный внешний эффект, двигательный или секреторный. Поэтому
принято называть эти ответные участки двигательными или
секреторными участками коры. Таких участков в коре много
и каждый служит для вызова определенного рода движений или
секреций. Ответные участки каждой воспринимающей области, повиди-
424
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
мому, предназначены для производства тех двигательных или
секреторных актов, которые нормально наступают в ответ на раздражения
соответствующего периферического рецептора. Ответные двигательные
и секреторные участки в коре мозга, вероятно, являются узловыми
точками, где связываются друг с другом разные гармонично
действующие корковые нервные круги.
Распределение двигательных участков в коре больших полушарий.
В зрительной воспринимающей области существуют такие
двигательные участки, откуда искусственное раздражение вызывает
движение глаз и головы, сужение зрачка. Когда раздражается такой участок
в левом полушарии, голова и глаза поворачиваются направо, как будто
бы глаза раздражались с правой стороны светом (рис. 174).
Рис. 189. Головной мозг разных животных.
/ — мозг собаки сверху; S — крестовидная борозда; F — сильвиева борозда,* О — обонятельная
луковица; 1 — двигательный участок для мышц шеи и затылка; 2 — для разгибания и отведения
передних ног; 3 — для сгибания и приведения передних ног; 4 — для движения задних ног; 5 —
для движения лица; 6 — для бокового движения хвоста; 7 — для оттягивания назад передней ноги;
8 — для поднятия плеча и перестановки ноги вперед (шагание); 9 — для закрытия век. II — вид
мозга собаки сбоку. Те же обозначения. Кроме того, а, а обозначает двигательный участок для
поднятия углов рта; Ъ — для раскрытия рта и движения языка; с, с — platisma; d — раскрытие
глаз. III— большой мозг кролика. W — головной мозг голубя. V — головной мозг лягушки.
VI—головной мозг рыбы (Caprinus carpio). На этих рисунках везде О означает обонятельную
луковицу; 1 — большой мозг; 2 — зрительный бугор среднего мозга; 3 — мозжечок; 4 —
продолговатый мозг (Ландуа).
В слуховой области имеются такие участки, раздражение
которых вызывает движение ушей, а также глаз и головы. И здесь
раздражение их в левом полушарии производит движение вправо, как будто
правое ухо раздражается звуком (рис. 189, 174).
Во вкусовой области и вблизи от нее помещаются участки,
откуда вызываются жевание, глотание, движения губ и языка, а также
секреция слюны и желудочного сока (рис. 189, 174).
В обонятельной области также существуют двигательные
участки, откуда вызывается движение ноздрей, например сужение их, как
это бывает при сильном неприятном запахе. Отсюда же вызываются
разные пищевые реакции, как жевание, глотание, выделение слюны,
движение головы вперед и назад, движение языка, а также резкие
вдыхательные движения и расширение зрачка (Риоч и Бреннер, 95).
Особенно обширны и многочисленны двигательные участки в к о
ясно-мышечной воспринимающей области. Здесь имеются особые
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА 425
участки для движения передних конечностей и особые для задних;
существуют также отдельные участки для головы, шеи, туловища,
хвоста. У высших позвоночных, как кошка и собака, эти участки
расположены по обе стороны крестовидной борозды (рис. 189). У человека
и у обезьян они расположены главным образом впереди роландовой
борозды, т. е. в передней центральной извилине (рис. 174). Но
некоторое количество их имеется и в задней центральной извитщне (рис. 188)
(Пенфильд и Болдрей, 68).
С развитием класса млекопитающих количество и дифференциация
двигательных участков растут. Дифференциация идет так далеко, что
каждое движение любой конечности имеет свой особый участок. Так,
например, можно найти такие точки, откуда вызывается сгибание
данной ноги по преимуществу, а также такие, которые дают разгибание ее
(Шеррингтон и Геринг, 8) (рис. 174). Особенно многочисленны и тонко
дифференцированы двигательные участки у высших обезьян и у
человека. При раздражениях передней центральной извилины было
получено у обезьян около 400 разных движений (Лейтон и Шеррингтон, 49).
С этой зоны могут вызываться изолированные движения не только
отдельных конечностей, но и отдельных суставов и даже отдельных мышц
(Ц. Фохт и О. Фохт, 32). На этом основывается возможность
производства тонких рабочих движений человека.
У человека электрическим раздражением коры можно вызвать
такие тонко дифференцированные движения, которые являются частями
трудового процесса, т. е. привычными движениями, образовавшимися в
результате упражнения (Бергер, 46). Это указывает на то, что в
двигательной /области коры мозга, в результате упражнения, образуется
множество разнообразных нервных комбинаций, которые служат для
вызова трудовых движений.
Двигательные точки кожно-мышечной области не имеют
постоянного анатомического положения; они располагаются у разных людей в
разных местах и с течением времени меняют положение даже у одного
и того же человека. Но представительство каждого двигательного
органа н? выходит из определенных границ. Эти границы для главных
органов довольно хорошо совпадают с точками их ощущений. Эти границы
показаны на рис. 188. Пенфильд и Болдрей, на основании
наблюдений которых составлен этот рисунок, обнаружили, что чем
больше пользуется человек тем или другим двигательным органом, тем
чаще встречаются соответствующие двигательные точки в коре
большого імозга. В этом отношении на первом месте стоит рука,—¦ вернее,
кисть,— затем губы и локтевой сустав. В отношении чувствований
также больше всего точек інаходят для руки, а затем для губ и языка.
Итак, в каждой воспринимающей области существуют участки,
искусственное раздражение которых производит совершенно такое же
движение, какое обычно вызывается нормальным возбуждением этой
воспринимающей области в ответ на раздражение соответствующего
рецептора.
426
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
Характеристика двигательных реакций, наступающих по
пирамидным и экстрапирамидным путям. По новейшим исследованиям Ф ё р~
стера (59), на людях, движения, вызванные с коры по пирамидным
и экстрапирамидным путям, характерно отличаются друг от друга.
Осуществляемые через пирамидные пути корковые реакции являются
более или менее изолированными реакциями одной группы мышц или
одной только мышцы, или даже части одной мышцы. В то же время
через экстрапирамидные пути наступают сложные движения, целиком
распространяющиеся на один двигательный орган или на систему
органов, как, например, поворачивание глаз, головы и туловища в одну
сторону с разгибанием конечностей на этой стороне. Аналогичные
наблюдения были сделаны на высших позвоночных — на обезьянах,
собаках и кошках (Фултон, 60).
Пирамидный путь, как известно, начинается в участке 4, который у
человека лежит вдоль роландовой борозды, а у животных впереди
центральной борозды. Следовательно, означенные изолированные
реакции прежде всего получаются при раздражении этого участка. Однако
изолированные реакции могут быть получены и при раздражении
участка 6. Но и в этом случае они фактически вызываются из участка 4
вследствие перехода возбуждения из участка 6 © участок 4. В случае
разрушения участка 4 или неглубокого разреза коры между этими
участками, раздражение 6 не дает изолированных реакций.
Экстрапирамидный путь начинается в участках 1, 2, 5, 6, 8, 22, и
соответственно этому сложные движения вызываются со всех этих
точек. Даже при раздражении участка 4, если оно интенсивно и длится
долго, двигательный эффект постепенно усложняется путем включения
© реакцию экстрапирамидных путей (Фултон, 60). Обычно при
некоторых слабых раздражениях определенного двигательного участка
движение наступает на одном из органов противоположной стороны. При
сильных же раздражениях одновременно приходят в движение и
другие конечности. Так, например,'если данная передняя нога сгибается,
то другая передняя нога может разогнуться. Из задних ног, наоборот,
соответствующая первой разгибается, а противоположная сгибается.
Одним словом, движения ног представляют такую картину, какая
бывает при шагании (Ухтомский, 33) (рис. 190). Если раздражать
одновременно две смежные точки, из которых одна дает сгибание, а
другая— разгибание данной ноги, то в ответ может наступить фазное
движение всех ног по типу шагания. Можно отыскать и такую точку,
откуда будет вызываться одновременное разгибание передних или
задних ног. Словом, и в этом случае раздражением двигательных
участков в кож'но-мышечной .воспринимающей области мы получаем такие
движения, которые характерны для раздражения соответствующих
рецепторов кожной поверхности.
Моторная зона характеризуется еще и тем, что с участков ее
краткими отдельными раздражениями можно вызвать двигательные
эффекты в виде одиночных сокращений определенной группы мышц. С пре-
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА
427
моторной зоны эти эффекты не получаются. При сильных
раздражениях премоторной зоны можно получить эффект, но только при
повторных раздражениях по 2 раза в 1 сек. и чаще, причем «премоторный
эффект» наступает после большого скрытого периода и по мере
повторения распространяется на все большее и большее число мышц. Кроме
того, эти сокращения проходят с большим замедлением.
Способность премоторной зоны к двигательным реакциям настолько
связана с моторной, что обе зоны нужно рассматривать как
функционально единую систему (Уисс и Обрадор, 61).
Ответная функция коры в отношении вегетативных органов.
Двигательная область коры большого мозга, а также премоторная
зона включают и такие двигательные точки, которые вызывают реакции на
органах, иннервируемых вегетативной нервной системой. Так,
например, раздражением коры в участках 4 и 6 можно вызвать повышение
или понижение кровяного давления и перераспределение крови в
организме. Оно вызывает также возбуждение или торможение мигательной
перепонки (у кошки). Эти реакции совершенно таковы, как и при
раздражении афферентных нервов, например седалищного (Ветохин,
63; Грин и Гофф, 64; Морисон и Риоч, 65). После повреждения у
обезьян участка 4 и 6 в коре мозга одного полушария устраняется
теплорегуляция в определенных участках кожи противоположных
конечностей (Пинкстон и Риоч, 67).
Однако ответная функция коры в отношении вегетативных органов
не ограничивается двигательной и премоторной зонами. Вегетативные
реакции вызываются и из других двигательных участков. Известно,
что некоторые вегетативные реакции, например, сердечно-сосудистые,
вызываются и вне двигательных участков: например, из височно-темен-
ной области (Бехтерев и іМиславский, 98; Гофф и Грин, 97). Обширные
вегетативные реакции вызываются из затылочной области, где Геде-
в а н и (96) обнаружил у кошек кортикальный центр агрессивной
реакции.
Историческая справка. Дштатшьные участки в коре больших полушарий
впервые былл обнаружены Фричем иГитцигом (1870) (2). Они установили т
собаках существование в крестовидной области коры таких участков, электрическое
раздражение которых дает сокращение мышц на конечностях и на других частях
тела. После них Феррье (3) обнаружил, что вызванные из коры двигательные
эффекты являются определенными, вполне координированными актами (1873—1875).
Лучиани и Тамбурина (4) определили положение возбудимых областей
специально в отношении каждого движения (1878). Они же впервые установили, что
эти области не совсем одинаковы у разных индивидов или даже в разных
полушариях одного и того же животного. Соответствующие опыты на обезьянах впервые
проделал Феррье (1877), а затем Бифор и Горслей (1887) (5), наконец, на
орангутанах Шеір рн нгтон и Грюнбаум (6) установили, что у высших обезьян
дьигательные точки существуют только в передней центральной извилине и в глубине
центральной, т. е. роланцовой, борозды, что они отсутствуют в задней центральной
извилине. На людях соответствующие экспериментальные исследования производили
Феррье, Біифор и Горслей (1890), Бехтерев (1899) (7) и др. Опыты
ставились случайно во время хирургических операций на голове и было установлено
полное сходство большого мозга человека и высших обезьян в отношении локализа-
428
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
цим двигательных участков. Однако, на осно.в,аішш' опытов всех гтои'М-енновааиых
исследователей, известно, что двигательные участки не ограничиваются описанной
областью, а встречаются и вне этих пределов в разных долях больших полушарий.
Филогенетическое р'азвитіие ответной функции коры мозга. Кора
большого мозга холоднокровных животных, повидимому, совершенно
лишена двигательной функции, в том виде, как это изложено выше.
У лягушек ее, бесспорно, нет, ибо у них нет типичной коры (Херрик,.
56). У рептилий имеется настоящая кора большого мозга, и она,
повидимому, не лишена двигательной функции. Но электрическое
раздражение коры у черепах и ящериц дает двигательные эффекты лишь при
таких силах раздражения, что они могли быть вызваны
распространением тока на подкорковые центры (Б и к е л ь, 75; Д ж о н с т о н, 76; Бре-
мер и сотр., 79). Более определенные результаты получаются при
электрическом раздражении коры большого мозга у аллигаторов.
Двигательные реакции наступают только с определенных участков. Они не
вызываются с близлежащих полосатых тел, а после наркоза перестают
вызываться (Бэгли и Рихтер, 77). Эта разница между черепахой и
аллигатором іможет быть объяснена тем, что у аллигатора кора мозга
развита немного больше.
У птиц многие исследователи также наблюдали двигательные
реакции при раздражении коры імозга (Бикель, 75; Роджерс, 78). Но
первые исследователи наблюдали их лишь при очень сильных
электрических раздражениях, и можно' было предполагать, что эти эффекты
вызывались раздражением подкорковых узлов (ectostriatum).
Наиболее убедительные корковые реакции получались в опытах К а л и-
шера (1), изучившего электрическую возбудимость разных птиц, в
частности попугаев. Он нашел в передней части полушарий
определенные точки для движения языка, челюстей и для издания голоса, затем
несколько позади — другие точки для движения ног и крыльев, а в
затылочной части — для движения глаз. В новейшее время подлинные
двигательные корковые реакции наблюдали у птиц (у голубей) Б р е-
мер с сотрудниками (79). Они вызывали их при довольно слабом
электрическом раздражении дорзолатеральной поверхности мозга, а
также затылочных долей. Эти реакции заключались главным образом
в движениях головы и глаз, причем вызываемые движения были такого
же характера, как и спонтанные. Кокаинизация поверхности мозга, а
также эфирный наркоз устраняли электрическую возбудимость. На этом
основании нужно считать, что неудачные результаты прежних авторов
объясняются тем, что они работали на сильно наркотизированных
птицах.
Двигательная функция свойственна коре большого мозга всех
млекопитающих, но развита она у них неодинаково. Прежде всего это видно
из того, что количество обособленных двигательных участков и
вызываемых отсюда разнообразных движений у кошек и собак меньше, чем
у обезьян. У первых эти движения носят характер простого сгибания
и разгибания разных суставов конечности с приведением и отведением
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА
429
бедра, пронацией и супинацией лапы; затем возникают движения глаз,
лицевых мышц и хвоста. У обезьян корковые движения более тонки и
разнообразны, в особенности в дистальных суставах; у них можно
получить отдельные движения пальцев. Имеются хорошо развитые
участки для движения лицевых 'мышц, языка и гортани. Кроме того эти
участки более обособлены друг от друга, чем у кошек и собак. Так,
например, участки для мускулатуры лица, задней и передней
конечностей лежат отдельно.
А К
Crt dex
sin t3
dex IS
sm 13
Рис. 190* Антагонистические эффекты
электрического раздражения кортикальных участков
одинаковой функции в правом и левом
полушариях.
Кошка. Записываются сгибатель (FI) и разгибатель (Ext)
колена на той и другой задних ног. В опыте А и С
раздражается двигательный участок задней ноги в правом
полушарии, а в опыте В и D — в левом. В обоих случаях
сильнее сокращается сгибатель противоположной стороны
и слабее разгибатель одноименной стороны. Антагонисты
не сокращаются, — наоборот, они несколько удлиняются,
что указывает на торможение (Ухтомский).
У человека мы видим дальнейшее развитие в этом направлении.
С развитием двигательной функции в коре імозга роль ее вообще в
двигательных реакциях животного становится все более и более
значительной. Это хорошо видно из опытов с экстирпацией двигательных
участков. У собак и кошек при удалении двигательных участков в
области крестовидной борозды наблюдается только небольшой недостаток
мышечного тонуса на противоположной стороне, но этот недостаток
быстро исчезает.
У обезьян после аналогичной операции в области роландовой
борозды наблюдается на противоположной стороне сильное расстройство
430
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
движений, которое с течением времени проходит только частично.
Спустя еще 2—3 дня наблюдается слабый мышечный тонус и подавление
сухожильных рефлексов. Все эти изменения потом проходят. Сильно
расстраиваются движения пальцев. Только через неделю они в
некоторой мере восстанавливаются, но
расстройство тонких движений ди-
стальных суставов остается на
долгое время.
У антропоидов, как гиббоны и
шимпанзе, удаление двигательной
области ведет к еще более
сильным и длительным расстройствам.
У них наблюдается настоящий
паралич, отсутствие произвольных
движений, который проходит в
области бедра через 2—4 дня, в
области колена — через 4 дня, в
области голеностопного сустава —
через 24 дня, а в области
пальцев — через 40—60 дней.
На основании этих данных
следует заключить, что двигательная
функция коры большого мозга
развивалась постепенно по
филогенетической лестнице позвоночных
животных, причем по мере развития
она приобретала все более и более
доминирующую роль в
двигательных реакциях животного.
Онтогенез ответной функции
коры мозга. У высших позворючных —
кошек и собак — кора] мозга
возбудима уже в первые сутки после
рождения (Бари, 69; Михайлов,
70). В первые дни двигательная
реакция в" ответ на раздражение
интенсивность реципрокного торможения, - *, « . .
вызываемого из коры. Это корковое раз- КОрЫ В ООЛаСТИ SUICUS CrUCiatUS НО-
дражение влияет тормозящим образом так- «
же на тоническое сокращение разгибателя СИТ координированный характер
левой стороны (Ухтомский), .,
приведения и отведения, сгибания
и разгибания всей ноги. А начиная
с 7-го дня, двигательная реакция может ограничиться изолированным
движением отдельных суставов. При этом двигательные эффекты
наступают на передних ногах регулярнее, чем на задних. Значит, в
отношении задних конечностей наблюдается некоторое отставание в
развитии соответствующих корковых точек (Клебанова и Лейбсон, 71).
Онтогенетическое развитие каждой двигательной функции, как ука-
U. sin 20
Crt d!l
ib.
Sec
ib.
Рис. 191. Выявление реципрокного
торможения при корковой
иннервации.
Те же мышцы, что и на рис. 190. Сначала
раздражается одна кора (правое
полушарие), а затем кора в комбинации с
раздражением левого чувствительного нерва
п. ulnaris sin. (верхн. стнал). Последнее
раздражение дает одновременно
сокращение сгибателя и разгибателя на
противоположной правой стороне и
расслабление на разгибателе соответствующей
стороны. Под влиянием коркового раздражения
правый сгибатель расслабляется
одновременно с усилением сокращения правого
разгибателя. В этом проявляется большая
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА 431
зывалось выше, происходит в определенных анатомических границах.
Но если у молодых животных разрушить всю двигательную зону
конечностей, тогда у них двигательная функция развивается в
ближайших уцелевших областях коры. Так, например, если у обезьян в
детском возрасте удалить всю двигательную область (поля 4 и 6),
двигательная возбудимость обнаруживается через некоторое время в
окружающей коре, а именно, в переднем крае центральной извилины, в
области 6а и 66 в большей степени, чем это бывает у нормальных
обезьян или после удаления тех же участков в позднем возрасте.
Движения, вызываемые с необычной зоны, не так совершенны и требуют
более сильных раздражений, чем это бывает при раздражении обычной
двигательной зоны. Отсюда следует, что в молодом возрасте может
происходить некоторая реорганизация немоторной области,
находящейся в соседстве с моторной, если последняя была удалена (Кеннард и
Мак Куллох, 92).
Общая характеристика корковых движений. Корковое движение,
прирожденное или индивидуально приобретенное, в отношении
координации ничем не отличается от прирожденного двигательного рефлекса.
Если при виде хлыста собака становится на задние ноги и
сгибает.передние, то она выполняет такие же координированные акты сгибания и
разгибания, какими являются прирожденные спинномозговые рефлексы
сгибания и разгибания. Или, например, если на звуковое раздражение
нормальный кролик двигает ушами и поднимает голову, то и здесь мы
имеем дело с таким же координированным актом, как при подобном
же раздражении у кроликов, лишенных коры большого мозга.
Очевидно, координация корковых и некорковых движений должна быть одна
и та же и должна происходить по одному и тому же принципу.
Опытами Геринга и Шеррингтона (8), Ухтомского (33)
и др., было доказано, что в корковых движениях сгибания и
разгибания, вызываемых непосредственно раздражением коры,
мышцы-антагонисты испытывают такую же реципрокную иннервацию, как в
аналогичных рефлекторных движениях. Например, при разгибаниях,
вызванных раздражением коры, разгибатели сокращаются, а сгибатели
тормозятся (рис. 190, 191). То же наблюдается при произвольных
движениях сгибания и разгибания. Так, Бете и Каст (9) воспользовались
больными, у которых была ампутирована рука по методу Зауербру-
х а — специально для ношения искусственной руки. Этот метод
заключается в том, что после ампутации руки в локтевом суставе, в
сухожилиях главных мышц сгибателя (m. biceps brachii) и разгибателя (m.
triceps brachii) делается щель, которая внутри выстилается кожей. В щель
вставляются слоновые косточки, служащие для связывания сухожилия
с протезом (рис. 192). Бетеи Каст связывали эти сухожилия с
миографами и производили кимографическую запись произвольных
движений. Когда больным предлагали сгибать руку, то сгибатель сокращался,
а разгибатель тормозился. Обратное наблюдалось, когда больной
пытался разогнуть руку (рис. 193). Аналогичные данные были полу-
432
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
*
чены Ваххольдером (57) при исследовании у человека токов
действия антагонистических мышц при волевом сокращении. Таким
образом было подтверждено, что при всяких корковых движениях
сгибания и разгибания мышцы испытывают реципрокную иннервацию.
Взаимодействие между корковыми реакциями, вызываемыми
непосредственным раздражением коры и разного рода спинальными и
другими реакциями, совершенно такое же, как и между отдельными не-
Рис. 192. Ампутированная рука по способу Зауербруха.
Наверху слоновая косточка проходит через сухожилие сгибателя
in. biceps br., от нее с двух сторон идут нитки к миограф}'. Внизу
видна такая же косточка от разгибателя m. triceps br. Вверху рука
дана в покойном состоянии, а внизу — при сильном сокращении
сгибателя. На верхнем рисунке косточка сгибателя приходится ч\ ть
впереди косточки разгибателя, а ниже она отодвигается назад на
несколько сантиметров. Об этом можно судить по сантиметровой шкале,
поіМ еще иной между руками. Этим отодвиганием назад и осуществляется
сгибание пальцев искусственной руки (Бете и Каст).
корковыми реакциями. Так, например, рефлекторные сокращения мышц
конечности, вызванные раздражением чувствительных нервов,
ослабевают во время реципрокного коркового торможения (рис. 191).
Наоборот, сами корковые реакции сильно тормозятся под влиянием таких
некорковых реакций, как глотание и дефекация (Ухтомский, 33).
Отсюда ясно, что по внешнему проявлению нельзя судить о том,
каково данное движение: корковое или некорковое. Однако, если взять
не одно движение в отдельности, а все поведение животного за
известный промежуток времени, то можно путем анализа определить
происхождение данного движения. Так, например, корковое движение
может протекать с такой последовательностью, которая не встречается
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА 433
среди прирожденных рефлексов. Впрочем, существуют и такие
движения, которые вызываются исключительно из коры. Таково, например,
движение большого пальца у человека и обезьян и противопоставление
его другим пальцам. После удаления коры это движение исчезает.
Следовательно, данное движение всегда является корковым.
Раздражением коры мозга вызывается не только сгибательное или
разгибательное движение конечностей, но и более или менее
длительные фазные движения их. Фазные движения не имеют в коре мозга
собственного представительства.
Возникновение их всецело обусловливается
действием вторичных проприоцептивных
раздражений, ибо они исчезают после
деафферентации конечностей, после
перерезки соответствующих задних
корешков. Координация корковых фазных
движений происходит в спинном мозгу, и,
очевидно, производится теми же
морфологическими образованиями, как и
фазные движения спинальных животных
(Ф. Меттлер и Ц. Меттлер, 83).
Локализация координации корковых
движений. Все корковые движения
координируются не в коре, а в спинном
мозгу и в стволовой части головного мозга.
Если удалить кору в определенном
месте и потом раздражать подкорковое
вещество, то вызывается такое же
координированное движение, какое было
характерно для удаленного участка коры. Так, например, если удалить
двигательный участок передних ног в одном полушарии и затем
раздражать лежащее под ним нервное вещество или соответственную
часть во внутренней капсуле, то это также вызовет сгибание одной
передней ноги и разгибание другой (Геринг и Шеррингтон, 8).
Очевидно, координирующие аппараты корковых и некорковых движений
должны быть одни и те же, а роль двигательных участков должна быть
такой же, как роль рецептивного поля однородного прирожденного
рефлекса. Значит, двигательные участки в коре больших полушарий
являются передаточными станциями для корковых возбуждений, они
передают эти возбуждения к определенным координирующим
аппаратам. Эта передача происходит или прямо по пирамидным путям, без
перерыва, или по экстрапирамидным путям посредством ядер головного
.мозга, где оканчиваются нейриты корковых пирамидных клеток и
начинаются новые нисходящие пути.
Координация корковых вегетативных реакций. Подобно корковым
двигательным реакциям координация вегетативных реакций
происходит не в коре, а под корой, в вегетативных центрах головного и спин-
28 и. С. Беритов
Рис. 193. Произвольная
иннервация мышц руки,
ампутированной по Зауербруху.
Наверху кривая разгибателя m.
triceps Ът., а внизу — сгибателя т.
biceps Ьг. Поочередно происходит
то разгибание, то сгибание руки.
Рука разгибается а сгибается по
команде: «сгибать» и «разгибать».Все
время сохраняются правильные
реципрокные отношения (Бете и
Каст).
434
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
ного імозга. В головном мозгу эти центры находятся, главным образом,
в hypothalamus и продолговатом мозгуг но есть указания, что они
находятся также в thalamus и corpus striatum. В промежуточном мозгу
и полосатом теле происходит интеграция наиболее сложных
вегетативных реакций, которые включают разные периферические механизмы
как вегетативной, так и соматической системы: так происходит,
например, регуляция теплопродукции. Вегетативные центры продолговатого
мозга ведают менее сложными функциями: поддержанием кровяного
давления на одном уровне, секрецией слюны и других
пищеварительных соков и т. д. Спинной мозг выполняет более простые вегетативные
функции: например, рефлекторное расширение кровяного русла
конечностей.
Кора производит ©сякого рода сдвиги в вегетативных органах через
означенные вегетативные центры, но при этом выполняет специальную
функцию: она согласует работу вегетативных органов с работой
соматических. Этим объясняется, что активация каждого двигательного
участка сопровождается определенными вегетативными сдвигами,
благоприятствующими выполнению двигательной функции. Так, например,
известно, что раздражение участков 4 и 6 может вызвать перемещение
крови из внутренних органов в імышцы. Эта реакция происходит даже
в том случае, еош мышцы почему-либо не сокращаются: например,
вследствие кураризации (Грин и Гофф, 97). Несомненно, когда
корковые импульсы приводят мышцы в сокращение, они одновременно
перегоняют определенное количество крови из внутренних органов в
мышцы.
Итак, кора больших полушарий целесообразно объединяет работу
соматических и вегетативных центров, находящихся вне коры, и тем
обусловливает рациональное обслуживание рабочих органов разного
рода внутренними функциями.
Изменчивость корковых реакций. Двигательные реакции,
вызываемые раздражением одного и того же двигательного участка,
меняются независимо от силы и характера раздражения, т. е. в связи с
функциональным состоянием животного. Непостоянство корковых
двигательных реакций было отмечено еще первыми исследователями этих
реакций (Броун и Шеррингтон, 84 и др.), но только в последнее время
оно стало предметом тщательного исследования. Так, Кларк и Уорд
(85) заметили, что на кошках неанестезированных и нефиксированных
раздражение одной и той же точки коры путем хронически
вживленных электродов вызывало разные двигательные реакции, вв зависимости
от положения животного. Затем Уорд (86) подробно исследовал это и
установил, что корковые двигательные реакции конечностей меняются
от положения головы и конечностей, подобно тому как меняются спи-
нальные рефлексы при тех же условиях. Так, если передняя нога была
разогнута, то при раздражении соответствующего двигательного
участка коры она сгибается; если, наоборот, нога была согнута, тогда она
разгибается. Точно так же влияет изменение положения головы. На*
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА 435
пример, если голова была повернута направо, то правая передняя нога
отвечала на корковое раздражение соответствующей двигательной
точки разгибанием и аддукцией; если голова была повернута налево,
тогда на той же правой конечности наступало сгибание с аддукцией.
Менялся также скрытый период корковых реакций при тех же
условиях. Все эти изменения были
обусловлены раздражением проприоцепторов, ибо
они в значительной мере устранялись после
перерезки соответствующих задних
корешков, после деафферентации конечности.
Очевидно, двигательный участок той или
другой конечности через пирамидные и
экстрапирамидные пути влияет не на один
определенный координирующий аппарат
или на одну определенную группу
двигательных нейроновt а на все
координирующие аппараты этой конечности^ вызывая
внешнюю реакцию из того аппарата^ где
возбудимость была повышена под влиянием
проприоцептивных раздражений.
Следовательно, в определенных случаях изіменчи- Рис. 194. Произвольные, дви-
вость корковых двигательных реакций ооі- жения мышц руки, ампутиро-
ваной по методу Зауербруха.
ределяется теми же закономерностями тт m
1 А Наверху кривая разгибателя m. tn-
рефлекТОрНОЙ Деятельности, ЧТО И ИЗМеНЧИ- ceps Ьг., внизу —сгибателя т. Ьі-
1 ceps Ьг. При В—одновременное сокт;
ВОСТЬ СПИНальНЫХ рефлеКСОВ. ращение антагонистов; при V0—изо-
г т лированное сокращение разгибате-,
Синергетическая ИННерваЦИЯ аНТагОНИ- ля; н* нем существующее
сокращение усиливается, а на сгибателе оно -
СТОВ при КОрКОВОЙ ДеятеЛЬНОСТИ. РеЦИПООК- расслабляется; при Я0—изолирован-
Г ное сокращение сгибателя, разгиба-
НаЯ ИННерваЦИЯ аНТаГОНИСТОВ ЯВЛЯетСЯ Не тель в это время расслабляется.
Внизу — время в секундах (Бете).
единственным принципом координированной .
деятельности. Эта иннервация была
установлена на основании изучения рефлекторных движений сгибания
и разгибания конечностей, но мы уже видим, что среди рефлекторных
реакций существует одна сложная реакция, н? являющаяся реципрок-
но координированной. Это — статический рефлекс опоры, в котором
сокращаются все без исключения мышцы конечности. При раздражение
коры можно также получить не только реципрокную иннервацию
антагонистов, но и одновременное сокращение или одновременное их
расслабление (Риоч и Розенблют, 53). Точно так же, на основании про->
извольных реакций на человеке можно сказать, что любая пара імышц
или антагонистических мышечных групп двигательных органов может
испытывать как реципрокную антагонистическую, так и синергетиче-
скую корковую иннервацию. Рука человека, например, при
произвольном сгибании и разгибании получает реципрокную иннервацию, но при
поднятии и опускании, при сжатии и раскрывании кулака, и вообще
во время фиксации суставов, часть антагонистов иннервируется синерт
гетично: они одновременно сокращаются, одновременно тормозятся
28*
436
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
(Бете и К,2СТ, 9). Бете и Каст впервые записали эти синергичные
координированные реакции миографическим путем у тех лиц, рука
которых была ампутирована по способу Зауербруха. Когда эти лица
пытались поднять и опустить руку, антагонисты — сгибатель и
разгибатель локтевого сустава,— то сокращались, то тормозились
одновременно (рис. 194).
Эта же синергетическая^ деятельность была обнаружена во время
произвольной фиксации суставов при сжатии кулака, путем изучения
биоэлектрических токов антагонистических мышц. Например, при
сжатии кулака разгибатель локтевого сустава m. triceps и сгибатель его
^ *WVwv»
\fVf**%^
^Y^V4MWW*
Рис. 195. Биоэлектрические токи возбуждения при сжатии и раскрытии
кулака.
Верхняя электрограмма от triceps br., нижняя — от biceps Ьг. Каждый раз при
сжатии обе мышцы дают токи возбуждения. Посередине между ними прямая линия
относится к локтевому суставу: последний совершенно не двигается. Наверху—время
в секундах (Вагнер).
m. biceps дают токи возбуждения, которые тем сильнее, чем сильнее
сжатие. При раскрытии же кулака токи возбуждения на обеих мышцах
исчезают (Вагнер, 34; Вахгольдер, 45) (рис. 195).
Разумеется, в названных движениях поднятия и опускания руки,
сжатия и раскрытия кулака синергетическая деятельность не
простирается на все мышцы. Так, например, при сложении руки в кулак и
при ее раскрытии антагонисты локтевого сустава сокращаются синер-
гетиЧ'НО, в то время как мышцы пальцев работают реципрокно.
Назначение синёргетического сокращения антагонистов всегда одно и то же.
Это — фиксация сустава или суставов в одном положении. Так,
например, когда рука поднимает тяжесть, она должна находиться в одном
каком-либо положении. Фиксация руки в одном положении
осуществляется путем синергетической иннервации мышц локтевого сустава;
при этом она сопровождается тем более сильным сокращением
антагонистов, чем большая тяжесть поднимается. Без фиксации того
или другого сустава путем одновременного сокращения антагонистов
немыслимо рабочее движение руки, так как удерживание в руке какой-
либо тяжести, особенно перенос ее с места на іместо, не может
происходить без такой фиксации. Итак, функция антагонистов заключается
не только в производстве движений руки в одном определенном
направлении, которое совершается благодаря реципрокной иннервации, но и
в фиксации руставов при рабочих движениях путем синёргетического
сокращения антагонистов. .
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА
437
Бете и Каст обнаружили у тех же оперированных людей
возможность произвольного сокращения одной мышцы. В это время
антагонист не будет тормозиться, или будет заторможен только в очень
малой степени, или же, наоборот, немного сокращен. Рефлекторным
путем изолированное сокращение одной мышцы может быть вызвано
только раздражением проприоцепторов в виде сухожильно^мышечных
рефлексов, причем эти рефлексы происходят без участия
координирующих аппаратов. Кроме того, как уже указывалось выше, как у высших
животных, так и у человека при слабом электрическом раздражении
коры в участке 4, где начинается пирамидный путь, получается
изолированная реакция определенной мышцы или даже части ее (Фёрстер,
59; Фултон, 60).
На этом основании можно утверждать, что произвольное синергети-
ческое сокращение антагонистов или изолированное сокращение одной
мышцы происходит без участия координирующих механизмов спинного
мозга. Эти эффекты должны вызываться через те корковые нервные
пути, которые оканчиваются прямо при клетках двигательных нейронов.
Следует отметить, что по новейшим анатомическим исследованиям,
расположение нисходящих корковых путей и заднекорешковых волокон
почти одинаково. По исследованиям Ариёнс Капперса (10), у
всех низших млекопитающих (однопроходных, сумчатых, грызунов,
копытных и рукокрылых) обе проводящие системы проходят в задних
столбах. Только у высших млекопитающих (хищных и приматов)
расположение нисходящих корковых путей несколько иное вследствие
вторичных изменений.
Анатомические связи между двигательными участками коры
и координирующими механизмами. Эти связи всегда являются
прирожденными. Известно, что многие животные (лошадь, корова, морская
свинка) рождаются с развитой корой и развитыми анатомическими
связями между корой и координирующими аппаратами. Эти животные
с самого рождения дают все ориентировочные движения и локомоцию.
У них искусственное раздражение коры вызывает ' соответствующий
двигательный эффект с самого рождения (Тарханов, 28; Бехтерев, 14).
У других животных (собака, кошка) кора и ее связи заканчивают свое
развитие после рождения. Искусственное раздражение ее в первые дни
после рождения не вызывает определенных корковых движений, их
можно наблюдать на таких животных после того, как закончилось
органическое развитие коры и ее анатомических связей. Впрочем, кора
их не лишена -некоторой возбудимости даже в первые дни после
рождения: в ответ на искусственное раздражение ее можно наблюдать
общие неопределенные движения конечностей. Отсюда следует, что у
этих животных одна часть анатомических связей между корой и
другими отделами мозга формируется в эмбриональный период, другая же
часть образуется впоследствии, в период постэмбриональной жизни.
Ни та, ни другая часть этих нервных связей не представляет
неизменных и ничем не заменимых нервных путей для осуществления опре-
43?
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО ,МОЗГА
деленных корковых реакций. Опыты показывают, что каждый
координирующий аппарат связан прежде всего с одним определенным
двигательным участком коры, но, как было упомянуто выше, он может быть
приведен в действие с других двигательных участков. Для иллюстрации
этого явления приведем здесь один очень поучительный факт. О р б е-
л и удалял у собаки всю верхнюю половину большого мозга с
включением всех двигательных участков конечностей. Несмотря на это,
собака сохранила способность к локомоции на обонятельные и
звуковые раздражения. Локомоция, несомненно, вызывалась через
двигательные участки уцелевших воспринимающих областей и представляла
собой наиболее развитую форму ориентировочных движений. На
физиологическом конгрессе в Базеле (1889) Гольц демонстрировал собаку,
у которой было вырезано левое полушарие, и, следовательно, правые
конечности «были лишены соответствующих корковых двигательных
участков. Несмотря на это, собака производила заученные движения
правой ногой. Геринг (13) сделал подобное же наблюдение на
обезьянах. После удаления левого полушария правая рука производила
заученные движения. Прямыми опытами было доказано, что
действительно в этих случаях одно полушарие імозга начинает выполнять
двигательные функции по отношению к обеим половинам тела. Так,
О с б о р н и Кильвингтон (35) показали, что если у собаки
центральный конец одного нервного ствола плечевого сплетения левой
стороны срастить с периферическим отрезком того же нерва правой
стороны, то через 10 (месяцев после операции раздражение правого
двигательного участка вызывает движение правой лапы, хотя нормально оно
вызывало реакцию только на левой; в то же время в левом полушарии
соответствующий двигательный участок оказался недеятельным. Таким
образом, ясно, что в этом случае в правом полушарии развилась новая
функция по отношению к правой ноге. Опыты Франца (50) и Л е-
шли (51) дали аналогичные результаты. Тренировкой они
образовывали у животных (кошки, обезьяны) определенные навыки,— например,
приучали вынимать корм из закрытого ящика,— а потом разрушали
лобные доли в обоих полушариях. После операции приобретенные
навыки временно исчезали, но путем новой тренировки вновь развивались,
только на этот раз была необходима более длительная тренировка.
Эти факты с очевидностью подтверждают то основное положение,
что каждая нога сохраняет способность к произвольной локомоции и
заученным движениям даже в том случае, если удаляется корковый
участок, откуда начинается корковый путь к соответствующим
координирующим аппаратам. Повидимому это происходит вследствие
приспособления других существующих двигательных участков и непрямых
корковых путей для проведения возбуждения от оставшегося отдела
большого мозга к тем же координирующим аппаратам.
Возбудимость двигательных участков. Возбудимость двигательных
участков на прямое раздражение очевидна, так как непосредственное
раздражение их производит внешний эффект. Для того чтобы вызвать
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГ\
439
двигательный эффект с коры, -нужно приложить очень сильное
раздражение, сильнее, чем требуется для мышцы. Объясняется это прежде
всего тем, что двигательными элементами являются не поверхностные
слои коры мозга, а глубоко лежащие — четвертый и пятый. В
двигательной области четвертый и пятый слои соответствуют пятому и
шестому слоям обычной архитектоники вследствие исчезновения
внутреннего зернистого слоя. Здесь начинается большая часть пирамидных
нервных волокон. Но и физиологические опыты последнего времени
определенным образом указывают на это. ~Д ю с с е р де Баренн (36)
особым аппаратом нагревал в прецентральной извилине обезьяны (Ма-
caccus rhesus) верхние слои определенных двигательных участков коры
до умерщвления, т. е. вызывал термокоагуляцию верхних слоев; потом
производил электрическое раздражение. Оказалось, что умерщвление
верхних трех слоев н? влияет на пороги раздражения этих участков в
отношении двигательных эффектов. Точно так же совершенно не
меняется характер вызываемых двигательных реакций. Отсюда следует,
что двигательные реакции на электрическое раздражение коры
наступают путем возбуждения тех больших и гигантских пирамидных кле-
току которые имеются в четвертом и пятом слоях.
Возбудимость коры очень сильно меняется в зависимости от разного
рода внешних воздействий. Если эти воздействия таковы, что ухудшают
обмен веществ (понижение температуры, сильное электрическое
раздражение, хлороформ, эфир и т. д.), то возбудимость коры понижается.
Когда внешние агенты улучшают обмен веществ (повышение
температуры, отдых), возбудимость повышается. Особенно сильно повышается
она под (влиянием стрихнинного отравления. Если к обнаженному
двигательному участку коры приложить кусочки фильтровальной бумаги,
смоченные в 0.1% растворе стрихнина, то спустя несколько минут
замечается сильное падение порогов раздражения, т. е. происходит
большое повышение возбудимости в этом участке. Одновременно сильно
возрастают двигательные реакции в ответ на раздражение
отравленного участка. Кроме того, замечается спонтанное наступление
правильного ряда движений с реципрокной иннервацией. Эти движения,
очевидно, вызываются под влиянием всякого рода слабых внешних и
внутренних раздражений вследствие повышения возбудимости
отравленной коры. Для стрихнинного отравления коры характерно, что
данные движения стрихнинного происхождения усиливаются при
раздражении периферических рецепторов как противоположной, так и
соответствующей сторон, а также коры того и другого большого полушария.
Это, конечно, происходит в силу иррадиации возбуждения по всей
центральной нервной системе и вовлечения отравленного стрихнином
двигательного участка в сферу реакции вследствие повышения в нем
возбудимости (Беритов, 19) (рис. 196, 197).
Возбудимость двигательного участка меняется очень сильно в ответ
и на деятельное и на недеятельное раздражение (Бубнов и Гейден-
гайн, 25). На нормальном препарате одиночное раздражение коры эф-
440
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
фекта не дает: такое раздражение всегда будет субминимальным.
Значит, эффект тетанического раздражения всегда является результатом
суіммации, т. е. повышения возбудимости, но повышение возбудимости
особенно сильно и длительно после деятельного раздражения (Грэим
Броун, 37) (рис. 198). После возбуждения коры естественным путем —
от раздражения рецепторов повышенная возбудимость может удержи-
мЩш&Жк\кк v tUwf ьніММк Ш
Cor. s. /4cm
^7і%ігИіиЬ^^
Рис. 196. Стрихнинные подергивания левой
передней ноги кошки при локальном
отравлении соответствующего
двигательного участка в правом полушарии.
Записываются сгибатели (biceps br. et bracn. s.
верхняя кривая) и разгибатель (triceps br.
нижняя кривая) левой передней ноги. Ряд спонтанных
сокращений сгибателя при покое его на
разгибателя. Раздражение в неотравленной зоне
передних ног другого полушария (поднятие
сигнала) производит значительное усиление и
учащение спонтанных сокращений сгибателя.
Рис. 197. Стрихнинный эффект на
правой передней ноге кошки при
локальном отравлении
соответствующего двигательного участка в
правом полушарии.
Верхняя кривая от сгибателя правой
передней ноги, а нижняя — от разгибателя
ее. Спонтанные слабые сокращения на
разгибателе; на сгибателе они отсутствуют.
При раздражении лучевого нерва правой
стороны на разгибателе происходит
усиление вздрагиваний, а на сгибателе
наступает значительное тетаническое
сокращение, которое прерывается в момент
каждого вздрагивания. Это свидетельствует
о реципрокном торможении сгибателя в
момент каждого сокращения разгибателя.
ваться не только секунды, но минуты и целые часы. Повышенная
возбудимость имеет существенное значение в отношении изменчивости
двигательных эффектов. Так, Грэим Броун (18) показал, что если
сначала раздражать двигательную точку сгибания, а вскоре после
этого точку разгибания, то при втором раздражении можно получить
сгибание вместо разгибания. Это явление объясняется тем, что в уча-
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА
441
стке сгибания возбудимость после раздражения повышается до такой
степени, что слабое раздражение разгибательной точки путем
иррадиации из нее возбуждения прежде всего вызывает внешнюю реакцию из
сгибательной точки.
Из приведенных фактов относительно корковой возбудимости ясно
следует, что данный двигательный участок в коре мозга может быть
Рис. 198. Явление суммации и облегчения при раздражении коры.
Обезьяна Macaccus rhesus. Верхняя кривая от сгибателя плеча m. supinator
longus, а нижняя — от разгибателя triceps br. Раздражается двигательный
участок в противоположном полушарии. Всего 6 тетанических раздражении
продолжительностью около 1 сек. с интервалами около 4 сек. Первое
раздражение было субминимально: оно не дало эффекта, второе дало ничтожный
эффект — слабое сокращение на сгибателе, а все последующие раздражения
произвели прогрессивно нарастающие эффекты. Внизу — время в секундах
(Грэим Броун).
возбужден любым периферическим раздражением в силу иррадиации
возбуждения, если только возбудимость двигательного участка по ка-
кой-либо причине более или менее повышена.
Возбуждение двигательных участков. Возбуждение двигательных
участков вызывается всеми раздражениями, которые вообще действуют
на нервно-мышечную ткань, например, механическим, как
прикосновение, давление; электрическим, как постоянный и индукционный ток;
химическим, как креатин и т. д. Нормально эти участки возбуждаются,
во-первых, спонтанно, вследствие самовозбуждения и, во-вторых, при
раздражении периферических рецепторов. Это, между прочим, видно
из опытов со стрихнинныод отравлением определенных двигательных
участков. Характерный стрихнинный эффект вызывается из
отравленного участка іне только его непосредственным раздражением, но и
разного рода периферическими раздражениями, а также и спонтанно (Бе-
ритов, 19).
442
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
Процессы возбуждения в коре большого мозга до последнего
времени изучались по тем биоэлектрическим токам возбуждения, какие
вызываются отсюда іна мышцах. Этим путем было установлено, что
возбуждающий импульс в коре большого мозга продолжается такое же
короткое время, как и в центральной нервной системе вообще.
Корковое возбуждение, безусловно, также сопровождается абсолютной
рефракторной фазой и следует одно за другим ритмически. Это хорошо
видно из таких опытов, где кора раздражается быстрым рядом
индукционных ударов, а корковый эффект регистрируется от мышцы в виде
электрических токов возбуждения. Эти токи меняются в зависимости
от частоты и силы раздражения коры совершенно так же, как было
установлено для спинномозговых рефлексов (Гофман, 15; Купер и
Денни-Броун, 41). Это, конечно, было бы невозможно, если бы
возбуждение коры »не протекало ритмически или если бы кора мозга не
была способна к высокому ритму возбуждения.
Ритмическое течение возбуждения в двигательных участках
подтверждается и при изучении электрических явлений непосредственно в
них самих. Раздражение двигательных участков или их спонтанное
возбуждение всегда производит ритмическое колебание потенциала
очень изменчивой частоты, до 100—200 в 1 сек. и выше. Эти
электрические явления должны выражать деятельность как раз тех нервных
кругов, которые при этом возбуждаются в области двигательных участков.
Возбуждение нервных кругов сочетается с активированием
соответствующего нейропильного участка.
Наряду с быстрыми колебаниями потенциала в двигательных
участках возникают медленные колебания разной длительности, которые
должны представлять собой результат суммации множества биотоков
разновременно реагирующих дендритных участков коркового нейропиля
(Беритов и сотр., 90).
При одновременном изучении биотоков двигательной зоны коры и
движений конечностей у животных оказалось, что возникающие в коре
медленные биотоки в ритме 20—40 в 1 сек. находят свое выражение
в ритме клонических подергиваний соответствующей конечности. При
параллельной же регистрации биотоков двигательного участка коры и
соответствующей мышцы было установлено, что означенные
электрические колебания © коре в ритме 20—40 в 1 сек. более или менее
совпадают с группировкой быстрых колебаний соответствующей мышцы.
Именно, каждому медленному колебанию потенциала в коре отвечает
одна группа быстрых колебаний в мышце (Эдриан, 74) (рис. 199).
Вследствие этого ритмическое течение биотоков в мышце принимает
двоякий характер: с одной стороны, мышечные биотоки в высоком
ритме 150—200 в 1 сек., а с другой,— их группировки в ритме 20—40 в
1 сек. Ритм группировок мышечных биотоков отвечает ритму
медленных колебаний в коре нейропильного происхождения, а ритм самих
мышечных биотоков 150—200 в 1 сек., должно быть, обусловливается
возбуждением пирамидных клеток импульсами возбуждения нервных кру-
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА 443
гов. Как подробно будет изложено в следующей главе, нервные круги
в коре большого мозга возбуждаются не беспрерывно, а п?риодически-
Это обусловливает как периодическое активирование нейропиля в ритме
20—40 в 1 сек., так и периодические разряды импульсов, которые через
пирамидные и экстрапирамидные пути производят периодическое
возбуждение скелетных мышц.
Рис. 199. Биотоки возбуждения коры и мышцы после
электрического раздражения двигательного участка передней конечности
обезьяны.
А — электрограммы: X — при отведении в осциллограф ближайшего
участка; У—несколько более удаленного участка, Z — при отведении мышц
передней конечности; В — электрограммы в таком же опыте: X ¦— при
отведении ближайшего участка, У — при отведении отдаленного двигательного
участка задней конечности, Z—-от мышц передней конечности (Эдриан).
При отведении корковых биотоков человека от кожной поверхности
черепа одновременно с биотоками возбуждения скелетных мышц
удается также наблюдать как группировки мышечных биотоков, так и
соответствие этих группировок с ритмом медленных колебаний в коре
мозга. Ритм их чаще всего 10—30 в 1 сек. Но> конечно, ввиду того
что всякая мышца получает корковые импульсы из одного
соответствующего двигательного участка коры, а отводятся в осциллограф
потенциалы громадной ее области, т. е. множества двигательных и
недвигательных участков, большею частью не бывает полного соответствия
между отдельными группировками мышечных биотоков и медленными
колебаниями коры. ,
Аналогичные результаты получали Эдриан и'Моруци (80) при
одновременном изучении биотоков двигательных участков коры и
нисходящих пирамидных путей ів продолговатом мозгу животного. Между
этими токами существует большая согласованность, но и в этом
случае очень часто одной волне коркового потенциала соответствует
несколько быстропротекающих импульсов в пирамидных путях. Иной раз
после воздействия на кору некоторых ядов, как стрихнин, пирамидные
пути разряжаются в очень высоком ритме — до 500—1000 в 1 сек., но
небольшими группами: по 10—80 колебаний.
Приведенные выше наблюдения указывают, что двигательные участки
коры большого мозга, или, точнее, нервные круги их, разряжаются
импульсами в таком высоком ритме, какой вообще свойствен нервному
444
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
волокну. Но ввиду периодической деятельности нервных кругов, как
активирование нейропиля коры мозга, так и возбуждение пирамидных
и экстрапирамидных путей происходит периодически в ритме 10—40 в
1 сек.
Многие авторы, изучающие произвольную иннервацию скелетных
мышц струнным гальванометром, наблюдали, что ритм 40—50 в 1 сек.
доминирует над другими более редкими и частыми ритмами. Они
считают этот ритм за основной ритм .возбуждения центральных элементов.
Более высокий ритм объясняют разновременной деятельностью
центральных элементов (Эдриан и Бронк, 44; Правдич-Неминский, 38;
Рихтер, 43 и др.)- Предполагалось, что возникновение импульсоів
возбуждения в значительном числе центральных элементов совпадает во
времени и этим самым обусловливается низкий ритм 40—50 в 1 сек.;
некоторое же число их возбуждается в разное время и ведет к
возникновению імелких іволн высокого ритма (Эдриан). И действительно, при
отведении к гальванометру небольшого пучка мышечных волокон,
отвечающих одной двигательной единице, иннервируемой одним
двигательным нервным волокном, ритм возбуждения меняется всего от 5 до 50 в
1 сек. (Ваххольдер, 45; Рихтер, 43; Линдслей, 58). При самых слабых
сокращениях двигательной единицы этот ритм составляет 5—10 в 1 сек.
С усилением произвольного сокращения ритм возбуждения повышается,
но не превышает 50 в 1 сек. Наблюдаемое при этом усиление
сокращения ©сей мышцы зависит не только от учащения возбуждений
двигательной единицы, но в значительной мере и от увеличения числа
активных двигательных единиц (Линдслей, 58). Очевидно, более
высокий ритм, 150—250 в 1 сек., регистрируемый с больших участков
-мышцы, обусловливается разновременным течением возбуждения во
многих двигательных единицах. Отсюда следует, что нормальный ритм
возбуждения корковых двигательных элементов при произвольной
иннервации очень низок: 5—50 в 1 сек.
Этот низкий ритм, конечно, нельзя считать выражением лабильности
корковых двигательных элементов, ибо, как указывалось выше,
корковые элементы могут отвечать и более высоким ритмом возбуждения,,
вдвое-втрое превосходящим нормальный. Очевидно, низкий ритм, до
50 в 1 сек., является наиболее благоприятным ритмом возбуждения
корковых двигательных элементов,— таким ритмом, при котором они
длительно могут работать без утомления. И действительно, Линдслей
(58) показал, что при длительных произвольных сокращениях, до 15—
30 мин., нормальный ритм корковой иннервации не меняется,— значит,
не происходит утомления двигательных элементов коры. Меняется
только амплитуда токов возбуждения мышцы, что должно быть, зависит
от утомления менее лабильных координирующих элементов спинного
мозга или самой мышцы.
Проведение возбуждения через кору большого мозга. До
последнего времени среди физиологов господствует мнение, что в коре мозга
возбуждение распространяется с очень малой скоростью, задерживаясь
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА
445
особенно долго при переходе с одного нейрона на другой. Так,
например, по очень старым наблюдениям Франка (16), на возбуждение
коры мозга и на переход его к подкорковым нервным путям требуется
до 0.02 сек. По новейшим исследованиям Бергера и др., это время
также очень велико. По Бергеру (46), скорость распространения
возбуждения в коре и вообще в центральной нервной системе человека
равняется 5—7 м в 1 сек. Путем специального исследования нами было
обнаружено, что замедление в распространении возбуждения не является
свойством коры мозга. Здесь возбуждение может распространяться
почти с такой же скоростью, как и в нервных стволах. При некоторых
условиях опыта задержка длится 0.002—0.004 сек. (Б ер и то в, 40).
Но это время, может быть, тратится, главным образом, на
распространение возбуждения по тонким разветвлениям нервных волокон коры
мозга. і
Возбуждение в коре мозга, как и вообще в центральной нервной
системе, распространяется исключительно по нервным путям,
состоящим из аксонов. Дендриты не возбуждаются и не проводят
возбуждения. Это видно лучше всего из опытов Эдриана (74); раздражая
поверхностный слой коры в двигательной области, он вызывал в нем
активное состояние без вызова периферических движений. Как
известно, в позерхностном слое коры заканчиваются верхушечные
дендриты пирамидных клеток из глубоких слоев коры. Очевидно, если бы
дендриты возбуждались подобно аксану, то при раздражении
поверхностного слоя должны были бы возбуждаться через верхушечные
дендриты также и пирамидные клетки, а значит, и пирамидные пути, и тем
самым они вызывали бы движение. За невозможность распространения
возбуждения по деядритам, а значит, по нейропилю, говорит также тот
факт, что при раздражении одного двигательного участка
электрическим током поверхностный медленный ток отводится только на
расстоянии нескольких миллиметров от раздражаемой точки. Весьма
возможно, что он фактически отводится только от тех участков коры, которые
раздражаются непосредственно петлями тока.
Явление облегчения и угнетения в коре большого мозга. Явление
облегчения в коре больших полушарий подробно было изучено в
последнее время Дюссер де Варенном (72). Он изучал облегчение
в коре большого мозга обезьян, раздражая двигательные участки коры
подряд через разные интервалы и наблюдая движения конечностей. Он
установил, что раздражением двигательного участка коры создается
облегчение не только по отношению к последующему раздражению
данного участка, но и по отношению к последующему раздражению
соседних функционально объединенных участков коры. Облегчение
удерживается в течение нескольких секунд, иногда довольно долго, до 20 сек.
Но когда оно проходит, функциональное состояние только на один
момент возвращается к норме: потом оно переходит в новое состояние,
проявляющееся в ослаблении двигательных реакций в ответ на
последующее раздражение того же или соседних участков коры (рис. 200).
446 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
Это состояние ослабленной корковой деятельности Дюссер де
Баре н н называет угасанием, но гораздо правильнее было бы называть
его угнетением, ибо фактически мы имеем здесь временное угнетение
активной деятельности перед тем возбужденного коркового участка.
Кроме того, термин «угасание» давно уже был введен школой
И. П. Павлова для обозначения другого рода явлений из
индивидуально приобретенной корковой деятельности.
л сЛ_аА
/і
г
Т TV1 ГГ1МЧП |'tJl Ч11ЦПЦЧ1 ТТГТТ1 'ГП
?jl
а а а
LJ Lf-
г м ч 11111 ц і і і и 1111 и і и 11 г, і ч 11 і 11 111 и м і і і иг
Рис. 200. Явление облегчения и угнетения корковых
двигательных реакций обезьяны Масасса mulatto,.
В опыте А раздражается один и тот же двигательный участок тремя
одинаковыми раздражениями, но через разные интервалы. Второе
раздражение через 14 сек. дало ослабленный эффект, третье—через I1/» сек.,
наоборот, — усиленный, и притом с сильно уменьшенным скрытым
периодом. ВГопыте В применялось второе раздражение черезД1/, сек., и
опять-таки получился сильно облегченный эффект. В опыте С второе
раздражение воздействовало через 4 сек.; оно дало смешанный эффект:
амплитуда уменьшилась, как при угнетении, а скрытый период
укоротился, как при облегчении. В опыте D два разнородных раздражения
а и Ь воздействуют на один и тот же двигательный участок. Раздражение
а облегчает последующий ответ на а, но это же раздражение
угнетает последующий ответ на о при том же интервале. В конце дается
ответ на Ъ в отдельности (Дюссер де Баренн).
Период угнетения вслед за облегчением продолжается много
дольше облегчения: 5—10 мин. и больше. Продолжительность и
интенсивность облегчения и угнетения определенным образом зависят от
характера как предшествующего, так и последующего раздражений.
Аналогичные результаты были получены Гедеванишвили (88)
при раздражении двигательных участков у кошки. Но если раздраже-.
ние производить в немоторной зоне, которая сама по себе
двигательных эффектов не дает, тогда оно угнетает двигательный эффект,
вызываемый раздражением двигательных участков коры, с самого начала
без предшествующей фазы облегчения. Это, например, хорошо видно
на рис. 201, где раздражение двигательной зоны не дает эффекта во
время раздражения немоторной, теменной доли.
Кроме того, Беритов и Гедеванишвили (81), показали,
что раздражение коры мозга облегчает и угнетает точно так же
двигательные реакции, вызываемые раздражением чувствительных нервов.
Как известно, на нормальном животном, при нормальной его
возбудимости, пороговое раздражение чувствительных нервов производит дви-
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА \\7
гательный эффект преимущественно через кору большого мезга. Эти
рефлекторные эффекты облегчаются, если их вызывать во время
раздражения соответствующего двигательного участка в противоположном
полушарии или вскоре после него. Наоборот, рефлекторные реакции
тормозятся, если их вызывать по
прекращении коркового раздражения, т. е.
после фазы облегчения. Торможение
рефлекторных реакций может получиться и
во время раздражения соответствующего
двигательного участка, если только
последнее — подпороговое и поэтому не
дает двигательного эффекта или если
этот эффект прекратился в связи с уіч>
млением раздражаемого двигательного
участка (рис. 202).
При раздражении немоторной зоны в
теменных и затылочных долях імозга
тормозятся также рефлекторные
реакции, вызываемые раздражением
чувствительных нервов. Торможение
наступает с самого начала раздражения и
может удержаться по прекращении его
в течение нескольких минут (рис. 203).
Большая длительность тормозящего
последействия является характерной
особенностью коры большого мозга.
Спинномозговое торможение, как известно,
обычно прекращается в течение
нескольких секунд после прекращения
раздражения.
При раздражении коры мозга в одном
участке торможение простирается на всю
кору, но, повидимому, сильнее на соответствующее полушарие, чем на
другое. Иначе говоря, тормозятся рефлекторные реакции, вызываемые
раздражением как гомолатерального двигательного участка, так и кон-
тралатерального, как противоположного нерва, так и гомолатерального.
Отсюда следует, что наблюдаемое при корковом раздражении
торможение является общим.
Угнетение двигательных корковых реакций наблюдается и при
раздражении чувствительных нервов. Оно наступает через некоторый
промежуток времени после раздражения, даже в том случае если
раздражение чувствительного нерва подпороговое. При этом было
установлено, что и это обусловлено торможением, осуществляющимся
в коре и простирающимся на оба полушария, т. е. общим (Гедева-
нишвили, 88).
Было установлено, что в период угнетения корковой деятельности
Рис. 201. Явления общего
торможения в норе большого мозга
кошки.
Регистрируются разгибатели и сги*
батели голеностопного сустава:
gastrocnemius dex. и sin. (G. d.t G.s.),
tibialis anticus dex. и sin. (т. d.yT. s.).
В опытах А ж С раздражается
двигательный участок в правом полушарии
(сигнальная линия Сот. d.); при этом
наступает сокращение всех мышц.
В опыте В это же раздражение
произошло во время раздражения
немоторной зоны противоположного
полушария (Cor. s.) и оно вызвало лишь
очень слабый двигательный эффект на
левом сгибателе (Беритов и Гедевани).
418
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
ослабевает и спонтанная электрическая активность коры больших
полушарий. Это угнетение также является общим, ибо простирается на оба
полушария. Оно наступает как при раздражении коры в моторной и
немоторной зоне, так и при раздражении чувствительных нервов (Гедева-
Рис. 202 Явление облегчения и угнетения в коре большого мозга кошки.
Раздражается тетанически через биполярные электроды двигательный участок правой задней
конечности в коре левого полушария (нижний сигнал) и п. saphenus (верхний сигнал). Регистрируются
сокращения m. tibialis (нижняя кривая) и т. gastrocnemius (верхняя кривая) правой стороны.
В опыте А раздражается одна кора, эффект наступает при 9 см расстояния между катушками. В опыте
В происходит комбинация раздражения п. saphenus с подпороговым раздражением коры—10
см расстояния между катушками, а, / — п. saphenus раздражается в отдельности, d — п. saphenus
и кора раздражаются одновременно — рефлекторный эффект значительно усилен, Ъ — п. saphenus
раздражается сейчас же после коры — рефлекс значительно эаторможен; с, е —-п. saphenus
раздражается через 25—30 сек., после раздражения коры — рефлекс опять заторможен, но слабее (Бе-
ритов и Гедевани).
дишвили, 89). Этим путем было доказано, что данное торможение
действительно осуществляется в коре большого мозга.
Происхождение облегчения и торможения в коре мозга. Явление
облегчения и торможения находится в определенной связи" с
медленными изменениями электрического потенциала. Как указывалось выше,
в каждом участке коры мозга после каждого активного состояния,
вызванного непосредственно раздражением или распространением
импульсов возбуждения из других участков коры, проявляется медленное
колебание электрического потенциала. Сначала оно имеет
отрицательный знак, а затем положительный. Эти медленные колебания
потенциала имеют такой же характер, какой свойствен и спинномозговым
потенциалам, но только здесь они значительно более длительны и
интенсивны. Путем сопоставления медленных колебаний с периодами
облегчения и угнетения, Дюссер де Баренн (73) установил, что
период облегчения совпадает с отрицательным медленным колебанием,
а период угнетения — с положительным. Чем сильнее и активнее
состояние данного участка, тем длительнее и сильнее выступают сначала
облегчение и медленное отрицательное колебание, а затем угнетение
и медленное положительное колебание (рис. 204). Между прочим,
хорошо известно, что период облегчения зависит от возбуждающего
последействия раздражения: облегчение проявляется тем сильнее и
длительнее, чем сильнее и длительнее возбуждающее последействие.
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА 449
Характерно, что быстрые колебания спонтанного характера
ослабевают или исчезают в период медленного положительного колебания
и вновь появляются в конце его или, вернее, в конце периода
угнетения. Отсюда, следует, что данный активный участок коры становится
недеятельным в период положительного колебания не только в
отношении последующих раздражений коры, -
но и в отношении спонтанно
возникающих корковых импульсов.
С точки зрения нейропильно-ней-
ронной теории строения мозга,
активное состояние в коре мозга должно
выражаться, во-первых, в
возбуждении замкнутых кругов, во-вторых, в
активировании нейропиля. Быстро
протекающие разряды, как они даны
на рис. 199, безусловно выражают
токи возбуждения нервных кругов.
Длительное же отрицательное
последействие, во время которого наблюдается
облегчение, должно быть обусловлено
физико-химическими изменениями
клеточных элементов, входящих в
нервные круги прежде всего пирамидных
клеток. Нужно думать, что в связи с
возбуждением и локальными
процессами в клеточных телах, с одной
стороны, происходит избыточное
образование ионов, а с другой,—
разрыхление протоплазматических мембран.
Это было бы совершенно достаточно
для возникновения медленного
отрицательного колебания, а также
состояния облегчения, ибо означенное
физико-химическое изменение должно
улучшать условия проведения возбуждения по соответствующим
нервным кругам.
Последующий период угнетения сопровождается, как указывалось
выше, медленным положительным колебанием коркового потенциала.
В это время отсутствуют или ослабевают токи возбуждения спонтанного
происхождения. Они появляются и достигают нормы приблизительно к
концу положительного колебания. Отсюда следует, что фаза угнетения
корковой деятельности происходит от анэлектротонического действия
положительного колебания на нервные круги. Длительные
положительные колебания значительной продолжительности, обязательно
наступающие при каждом активировании (мозга, повидимому, являются, как уже
указывалось выше, выражением активного состояния нейропиля. На
29 и. С. Беритов
Рис. 203. Общее торможение в коре
большого мозга кошки.
Регистрируются те же мышцы. В опыте А
раздражение п. saphenus dex. (S. d.)
вызывает сильные сокращения на сгибателях и
небольшое сокращение в правом
разгибателе. В опыте В то же раздражение нерва
происходит во время раздражения немотор-
пои зоны в теменной доле того же
полушария. Теперь оно дает ничтожный
двигательный эффект. В опыте С через 2 мин.
повторяется раздражение нерва, и опять
оно дает сильно ослабленный двигательный
эффект. В опыте D через 4 мин. оно
вызывает обычный двигательный эффект
(Беритов и Гедевани).
450 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
этом основании можно предположить, что и угнетение корковой
деятельности в вышеприведенном случае является функцией нейропиля.
Итак, в коре больших полушарий возбуждение нервных кругов
обусловливает облегчение данных кругов к последующей деятельности в
результате повышения в них возбудимости; активирование же
ближайшего коркового нейропиля производит угнетение этих нервных кругов
вследствие анэлектротонического действия на них нейропильного тока.
С этим положением о происхождении коркового облегчения и
угнетения хорошо согласуются наши наблюдения — что при подпороговом
раздражении коры мозга, когда нет двигательных реакций, когда,
значит, не происходит возбуждения пирамидных клеток, облегчение
отсутствует после раздражения, а иногда и во время него, в то время как
угнетение проявляется хорошо как после раздражения, так и во время
него. Как известно, по Э д р и а н у, при подпороговом раздражении
поверхностного слоя коры мозга наблюдаются медленные колебания
потенциала в раздражаемой области. Этими медленными колебаниями
потенциала, вероятно, и обусловливается означенное угнетение
рефлекторных реакций и во время подпорогового коркового раздражения и
после него.
Выше мы говорили, что при наличии двигательных реакций в ответ
на раздражение коры сначала проявляется облегчение, а потом
угнетение. Этот факт, конечно, не указывает на то, что в периоде облегчения
нейропиль не находится в активном состоянии. Он испытывает это
состояние и даже, несомненно, более сильное, чем спустя некоторое
время после раздражения. Только пока нервные круги находятся в
состоянии возбуждения, сопровождаемого повышением возбудимости,
нейропильный ток не в состоянии произвести в них анэлектротониче-
ское понижение возбудимости. Что вообще нейропиль находится в
активном состоянии и в периоде двигательных реакций, видно из
смешанных эффектов, когда предыдущее раздражение влияет на ответ
последующего как облегчающим, так и угнетающим образом.
Например, скрытый период укорачивается, а амплитуда уменьшается. По
всей вероятности, уменьшение скрытого периода происходит от того,
что второе раздражение производит возбуждение тех нервных кругов,
где возбудимость была особенно повышена под влиянием предыдущего
раздражения, а уменьшение амплитуды обусловливается тем, что часть
нервных кругов или их звеньев, возбужденных первым раздражением,
вскоре после окончания возбуждения подвергается угнетению под
влиянием существующего длительного нейропильного тока.
Мы также указывали выше, что облегчение в коре происходит во
время коркового раздражения и некоторое время после него, угнетение
же может длиться после раздражения до 10 мин., т. е. в течение
многих минут после того, как первично возбужденные нервные круги
пришли в состояние покоя. Нужно думать, что нейропиль продолжает
активироваться, благодаря существованию в нем специальных коротких
замкнутых кругов, с участием звездчатых клеток с короткими аксонами
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА 451
(см. на рис. 177 нервный круг х — х, состоящий из двух звездчатых
клеток и их аксонов). Эти круги приходят в активное состояние
благодаря своим связям с первично возбужденными сложными нервными
кругами, а также благодаря прямой связи с отдаленными участками
коры большого мозга и с низкими отделами мозга. Повидимому, когда
сложные круги перестают работать, короткие, собственные нервные
круги нейропиля продолжают еще действовать и тем обусловливают
длительное активирование нейропиля, а значит, и длительное,
медленное колебание потенциала.
Зысота
}ижений
$&
Тони
^озс/уждения
Медл 1/~
налегания
Рн
Раздр
у
Нислот*іост ь
2'
д'
4'
5'
Рис. 204. Диаграмма соотношения между облегчением и угнетением
двигательных корковых реакций обезьяны Масасса mulatto, (А),
изменениями токов возбуждения (В), медленными колебаниями потенциала (С) и
рН (D) в раздражаемом участке коры.
Раздражение производится в ритме 60 в 1 сек. в течение 5 сек. (Дюссер де Баренн).
Нужно думать, что корковый нейропиль проявляет свою тормозящую
функцию не только в двигательных участках коры, но и вообще во всех
участках коры большого мозга, поскольку медленные колебания
потенциала наблюдаются везде, во всех участках коры. Это хорошо видно по
многим наблюдениям. Так, например, Бишоп и его сотрудники (91)
наблюдали, что в зрительной области во время первых двух
медленных колебаний потенциала, наступающих после быстрых колебаний от
раздражения зрительного нерва, повторное зрительное раздражение не
производит соответствующих быстрых колебаний. Кроме того, во время
этих медленных колебаний исчезают спонтанные колебания. Обычно
после периода угнетения спонтанные колебания усиливаются, а
повторное раздражение дает свой эффект. Очевидно, первое и второе
медленные колебания потенциала, выражающие активное состояние
нейропиля, анэлектротонически действуют на нейронные круги и угнетают их.
Последующая фаза облегчения, повидимому, возникает вследствие
поляризации, произведенной первыми двумя фазами медленного коле-
29*
452
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
бания. Прекращение нейропильного медленного колебания действует на
эти же круги подобно размыканию анэлектротона, производя в них
повышение возбудимости, т. е. облегчение.
С концепцией о тормозящей функции нейропиля хорошо согласуется
также следующий факт. У бодрствующего животного (кошка с
хроническими отводящими электродами, вставленными через
черепную коробку) почти вся кора реагирует на каждое раздражение,
только характер этих изменений несколько иной в соответствующей
корковой области по сравнению с остальными частями коры. Так,
например, при звуке учащаются и усиливаются электрические
колебания, в слуховой области, и в то же время происходит ослабление их в
двигательной, кожно-мышечной и зрительной областях (Рейнбергер и
Джаспер, 93). Можно .предположить, что усиление электрических
колебаний в слуховой области обусловливается усиленной деятельностью
нервных кругов этой области, а выпадение быстрых колебаний в других
областях обусловливается частью распространением нейропильного
тока из слуховой области, частью преимущественным активированием
нейропиля в других областях через ассоциационные нейроны из
слуховой области.
Производимое раздражением коры торможение рефлекторных
реакций в определенных случаях зависит, между прочим, от деятельности
подкорковых центров ствола головного мозга, где оканчиваются
экстрапирамидные нисходящие пути. Например, Тоуэр (87) показала,
что раздражением извилин в области крестовидной борозды (кошка)
можно получить торможение тонического сгибания и разгибания
конечностей в условиях перерезки пирамидных путей. Такое же торможение
тонических реакций можно получить при некотором подкорковом
раздражении нисходящих экстрапирамидных путей, причем как в случае
раздражения коры, так и нисходящих экстрапирамидных путей
торможение не простирается на фазные движения конечностей. Отсюда ясно
следует, что корковое раздражение в состоянии вызвать торможение
точно так же и в тонических центрах ствола головного мозга через
нисходящие экстрапирамидные пути, оканчивающиеся в тонических
центрах.
Производимое раздражением коры мозга торможение фазных
движений конечностей и других частей тела также может происходить не
только в коре, но и в подкорковых центрах. Это хорошо видно из наблюдений
той же Тоуэр (87). По ее опытам, фазные движения, вроде
дыхания и шагания, тормозятся при раздражении не только определенных
участков коры, но и соответствующих экстрапирамидных путей,
например, в области внутренней капсулы; при этом тонические реакции не
тормозятся. Опыты ясно показывают, что ов торможении фазных
движений должны участвовать как кора, так и подкорковые центры
промежуточного и среднего мозга, откуда вызываются фазные движения.
История вопроса о тормозящей функции коры большого мозга. Явление
угнетения, наблюдаемое в коре мозга, мы признали проявлением торможения, обусловлен-
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА 453
ного активным состоянием нейропиля. Это торможение носит такой же общий
характер, как и торможение в других отделах мозга, т. е. активное состояние данного
участка нейропиля оказывает торможение на все близлежащие нервные круги, на все
их клеточные и аксонные элементы. С распространением активного состояния в ней-
рспиле коры мозга распространяется и торможение. Значит, общее торможение
свойственно коре, как и другим отделам мозга. Раньше мы утверждали, что кора
большого мозга не знает тормозящих процессов, но мы тогда же имели в виду прежде*
всего то реципрокное торможение, которое обусловливает координацию движений.
А это торможение действительно происходит не в коре. Известно, что раздражением
двигательной зоны в области крестовидной борозды (собака, кошка) и в области
ргландовой борозды (обезьяны) вызывается антагонистическая иннервация мышц
конечностей. Раздражением двигательной зоны, в особенности ее премоторной зоны,
можно вызвать также общее торможение антагонистов (Риоч и Розенблют, 53).
С этих же участков вызывается и общее возбуждение антагонистов. Но во всех этих
случаях мы имеем дело с торможением, складывающимся не в коре, ибо такие же
явления торможения получаются при раздражении подкоркового вещества (Геринг и
Шеррингтон, 8; Морисон и Риоч, 65). Реципрокное торможение как основной элемент
координации протекает исключительно в координирующих механизмах вне коры в
шинном и головном моэгу. Шеррингтон (54) считает, что «центральное торможение
(имеется в виду реципрокное — И. Б.)у возникающее где-либо кроме точки,
расположенной в непосредственной близости от двигательной нервной клетки (спинного
мозга), ио имеющимся экспериментальным данным, почти неизвестно».
При своем отрицательном отношении к тормозящей функции коры большого
мозга мы имели в виду еще господствующее среди физиологов мнение о тормозящем
действии коры мозга на рефлекторную деятельность низших отделов. И эта
тормозящая функция большого мозга не отвечает действительности, не вытекает прямо из
фактов. Из всего того, что мы знаем о действии коры мозга на низшие отделы,
следует заключить скорее всего обратное: о постоянном облегчающем действии большого
мозга и вообще высших отделов на низшие, как об этом уже говорилось выше.
Господствующее ныне представление о тормозящей функции коры больших
полушарий в отношении низших отделов мозга опирается прежде всего на последствия
экстирпации этой коры. Именно, со времени Гольца (1884) (31) известно, что
после удаления коры, в особенности всей передней половины больших полушарий,
неизменно происходит повышение рефлекторных функций животного. На этом
основании Г о л ь ц утверждал, что в лобных долях лежат тормозящие центры и что
повышение возбудимое! и в этих случаях объясняется их удалением. В последнее время
такое же заключение было сделано Калишером (20). Однако из- других
исследований стало известно, что и удаление задней половины больших полушарий ведет к
послеоперационному повышению возбудимости .(Фано и Либертини, 24). На этом основании
тормозящая функция приписывалась всей коре. Новейшие исследования из
лаборатории Павлова также подтвердили ф?кт порышения рефлекторной возбудимости
после экстирпации коры. Авторы этих исследований рассматривают это каік
результат ослабления тормозящих процессов, будто бы всегда протекающих в коре наравне
с возбуждающими процессами (Орбели, 12; Эльяссон, 21; Демидов, 22; Кураев,
23 и др.).
Однако все эти заключения не вытекают из наблюдаемых фактов. Наблюдаемое
при корковых операциях повышение рефлекторной возбудимости должно быть
объяснено совершенно иначе. Из общей физиологии центральной нервной системы
известно, что всякое поранение последней является источником частью деятельных, но
еще в большей мере недеятельных субмнимальных раздражений и что эти
раздражения всегда ведут к повышению возбудимости всей нервной системы. Заживление
мозговой раны есть процесс длительный, и, повидимому, рана никогда не
излечивается вполне, так как громадное большинство оперированных животных все-таки
погибает преждевременно. Отсюда ясно, что и повышение возбудимости центральной
нервной системы должно продолжаться все то время, пока животное живо.
454
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
Заслуживает особого внимания, что даже такой сторонник тормозящих
механизмов в головном мозгу, как Сеченой, не находит возможным толкозать явление
повышения рефлекторной деятельности после обезглавливания как результат
устранения тормозящих механизмов. Он также считает его результатом слабых раздражений
поперечного разреза мозга, главным образом его нисходящих нервных путей:
во-первых, в силу самого разреза, а во-вторых, под влиянием крови и других неизвестных
агентов (47). Сеченову хорошо было известно, что слабые химические и
электрические раздражения поперечного разреза опийного мозга повышают рефлекторную
деятельность последнего.
Повышение возбудимости под влиянием вышеозначенных раздражений
нисходящих корковых путей должно быть особенно сильно в координирующих и
двигательных нейронах спинного мозга после удаления передней половины больших
полушарий, так как при этом перерождаются начальные пункты пирамидных путей, которые
оканчиваются при этих нейронах. Этим объясняется, что разрушение передней
половины одного полушария производит повышение рефлекторной возбудимости главным
образом на противоположной стороне тела, т. е. там, где оканчиваются поврежденные
нисходящие пути (Демидов, 22).
Другое основание для предположения тормозящих процессов в коре авторы
видят в явлениях угнетения рефлекторной деятельности под влиянием коркового
раздражения. Первое исследование в этом направлении принадлежит Бубнову и Гей-
денгайну (25). В период повышенной возбудимости эти авторы наблюдали на одной
мышце передней ноги собаки, отравленной морфином, что сильные раздражения самой
ноги или соответствующего коркового двигательного участка вызывают значительное
сокращение мышцы с длительным последействием. В момент же последействия
слабые раздражения той же ноги или коры вызывают более или менее значительное
ослабление существующего рефлекторного последействия. Отсюда они заключили, что
«раздражение производит в клетках, находящихся в покое, возбуждение, а в
деятельных— торможение. Вследствие этого каждый раз существующее состояние клетки
прекращается и превращается в противоположное состояние». Авторы при этом
имели в виду центральную нервную систему — и кору больших полушарий в частности.
Между тем они не имели права делать такое обобщение. Они сами указывали, что
данное угнетение тонического последействия наблюдалось и после удаления
существующего двигательного участка. Значит, и то явление угнетения, которое
наблюдалось при раздражениях коры, могло локализоваться исключительно в подкорковых
центрах.
Что в действительности это так, видно из рефлекторного последействия,
наблюдаемого ш морфини&иірованных собаках, несомненно тонического по своей природе
и исходящего из некорковых отделов головного мозга; оно подобно тонусу,
наступающему в связи с перерезкой среднего мозга. В настоящее время точно установлено, что
рефлекторный тонус при дацеребрация и тонус при наркотизации имеют совершенно
одинаковое происхождение: сильное возбуждение подкорковых тонических центров
{Ротфельд, 26). Затем работами последнего времени было установлено, что
тоническое сокращение мышц на децеребрированных животных, как в опытах Бубнова и
Г е й д е н г а й н а, легко подвергается не только усилению, но и ослаблению под
влиянием периферического раздражения. Это зависит, с одной стороны, от силы
раздражения, а с другой — еще в большей мере от степени возбудимости
координирующих механизмов. Так, например, если положение головы благоприятствует раэгиба-
тельному тонусу данной .ноги, то слабое раздражение в рецептивном поле сгибания
произведет его усиление, а сильное — его ослабление. Усиление будет обусловлено
возбуждением тонических центров разгибания, а ослабление—-возбуждением
оборонительного рефлекса 'Огибания (Беритов, 127).
Итак, фактических оснований для утверждения о тормозящем действии коры
больших полушарий на низшие отделы центральной нервной системы не имеется.
Правда, мы говорили, что нейропиль коры мозга производит торможение, но под
влиянием этого торможения ни в коей мере не может произойти угнетение рефлек-
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
455
торной деятельности низших отделов мозга. Когда какой-либо участок нейропиля в
коре мозга ^находится в активном состоянии, он может затормозить
электротоническим действием своего тока только те нервные клетки и аксоны, которые лежат в
пределах активированного нейропиля или в близком соседстве с ним. Но, конечно,
поскольку спонтанная деятельность коры большого мозга оказывает облегчающее
действие на низшие отделы мозга, то при всех обстоятельствах, угнетающих эту
активность более или менее значительно, должно получиться некоторое понижение
возбудимости в стволе головного мозга и в спинном мозгу во всех тех нервных
комплексах, где оканчиваются пирамидные и экстрапирамидные пути. Вследствие этого будет
происходить ослабление рефлекторной деятельности в данных комплексах. Но это
не может считаться результатом тормозящего действия коры большого мозга на
низшие отделы центральной нервной системы.
Корковая эпилепсия. Если двигательный участок коры
раздражается очень сильно или длительно, то получается длительное
судорожное сокращение скелетных мышц всех двигательных органов,
продолжающееся от нескольких секунд до многих минут. Это называется
эпилептическим припадком. Если раздражается двигательный участок в
правом полушарии, то судорога начинается в мышцах противоположной
стороны тела, а именно — в том двигательном органе, корковый участок
которого подвергался раздражению. Затем судорога распространяется
из этого участка по одноименной стороне тела, откуда переходит на
противоположную. Судорожное сокращение каждой мышцы сначала
является клоническим, затем тоническим и в конце снова клоническим.
Во второй раз эпилептический припадок может быть вызван с того
же коркового участка при более слабом и коротком раздражении. Если
раздражение коры было не очень сильно и возбудимость была
понижена, то судорожное состояние может ограничиться одной группой
мышц или одной ногой, участок которой подвергался раздражению
(Фрич и Гитциг, 2; Гитциг, 29; Ферье, 3).
У человека корковая эпилепсия развивается при заболевании коры:
при раздражениях и вообще травмах мозга, когда имеет место
воспаление, нагноение, дегенерация. Но могут быть эпилептические припадки
и при совершенном отсутствии травмы мозга. Это бывает в случае
значительного повышения возбудимости в коре большого мозга, как,
например, при усиленной продукции некоторых активных веществ в
железах внутренней секреции. Но во всех случаях раздражение коры
является моментом, безусловно необходимым для вызова этой
эпилепсии, а потому ее называют корковой эпилепсией (Джексон, 94).
Эпилептический припадок может случиться и при раздражении других
отделов головного мозга, но этот эпилептический припадок
характерно отличается от коркового: он наступает и протекает
одновременно во всем теле.
Если удалить какие-либо двигательные участки в коре, то в
соответствующих группах мышц не наступит эпилептической судороги. Так,
например, если вырезать двигательный участок одной ноги, то на данной
ноге корковые судороги исчезают, а на другой ноге и других частях
тела протекают в обычном порядке. Это указывает, что распространение
456
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
судорожных сокращений от одной части тела на другие обусловливается
распространением сильных возбуждений от одного двигательного
участка на другие (Лучиани, 30; Бубнов и Гейденгайн, 25).
Типичные эпилептические судороги отсутствуют также в условиях
деафферентации конечностей. Если какая-либо конечность лишена
периферической чувствительности путем перерезки задних корешков, то
при судорожном припадке от раздражения коры эта конечность не
участвует в судорогах: она не производит фазных движений локомоторного
типа или клонических сокращений мышц. На ней наступает только
тоническая фаза: конечность приходит в положение флексии и
остается в нем во время судорожного припадка. Очевидно, афферентные
проприоцептивные импульсы конечности, возникающие в результате
движений, являются существенным условием для развития на ней
обычной эпилептической судороги (Меттлеры, 83, Кряжев, 82).
Во время эпилептического припадка антагонистическая деятельность
мышц не исчезает: она хорошо выступает в любой период этого
припадка. Правда, обычно обе антагонистические группы пребывают в
сокращении, но каждый раз усиление сокращения в одной группе мышц
сопровождается ослаблением в другой. Повидимому, во время
эпилептического припадка возбуждение, перебегая в коре от одного
двигательного участка к другому, одновременно приводит в активное состояние
целый ряд координирующих аппаратов, производящих как фазные, так
и тонические движения данной ноги. В результате, к каждой паре
антагонистических мышц центральная нервная система должна
посылать как возбуждающие, так и тормозящие иннервации. В зависимости
от преобладания тех или других иннервации данная группа мышц
сокращается то больше, то меньше (Беритов, 17).
При раздражении воспринимающей области коры также наступают
эпилептические припадки. Особенно хорошо изучены те из них, которые
вызываются замораживанием маленьких участков. Вызываемые этим
путем судороги развиваются спустя некоторое время после
замораживания, и притом с большой постепенностью, начиная с какого-либо
органа, группы мышц или даже с одной мышцы. Они принимают форму
настоящего эпилептического припадка только через 1—5 часов после
начальных вздрагиваний. Эти припадки не зависят от соучастия
двигательных участков коры. Они возникают от непосредственного влияния
замороженного участка на подкорковые центры фазных и тонических
движений при условии легкого наркоза, или анемии, или одновременного
раздраженного состояния какого-либо чувствительного нерва
(Сперанский, 48), т. е. при условии ненормального повышения возбудимости в
подкорковых центрах под влиянием одновременного и длительного
раздражающего действия с разных сторон. Эпилептический припадок,
развивающийся при сильных раздражениях двигательных участков коры,
безусловно также производится с участием подкорковых центров фазных
и тонических движений. Судорожная деятельность должна наступить
в них сейчас же после приложения раздражения к коре, очевидно вви-
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА.
457
ду могучего воздействия двигательной зоны коры на подкорковые
центры.
Происхождение корковой эпилепсии. В тех случаях, когда
эпилептические припадки наступают вследствие раздражения или вообще
травмы коры мозга, совершенно естественно приписать их наступление
усиленному возбуждению нервных кругов под влиянием
раздражающего или повреждающего агента. Можно вызвать их на любом животном
при непосредственном электрическом раздражении двигательных
участков, если применять раздражение достаточно большой силы. Но
известно, что при довольно высокой возбудимости коры судороги наступают
уже при пороговых силах раздражения, а при особенно высокой
возбудимости они наступают спонтанно, т. е. помимо внешних
раздражений.
Далее известно, что при незначительной возбудимости коры
судороги не распространяются на весь организм — они могут ограничиться
одной конечностью (Беритов и сотр., 89, 90), а при довольно высокой
возбудимости судороги будут наступать и распространяться на
скелетную мускулатуру уже при пороговых раздражениях (Беритов, 19). Эти
факты с очевидностью указывают, что основным условием для
наступления эпилептических судорог в одних случаях является более чли
менее сильное и непосредственное раздражение коры, а в других
случаях—особое ее физиологическое состояние.
Сильное раздражение коры приводит большое количество нервных
кругов в более или іменее синхронное возбуждение, производя при этом
сильные электрические разряды, вследствие чего непременно будут
раздражаться близлежащие невозбужденные нервные круги. Это будет
происходить как путем иррадиации усиленных импульсов возбуждения
из первично возбужденных нервных кругов, так и с помощью
раздражающего действия возникающих здесь сильных электрических
разрядов. Известно, что если после перерезки спинного мозга отравить
животное стрихнином, то активное состояние задней части спинного мозга
может легко передаться через перерезанный участок к передней части
мозга с помощью судорожных биотоков заднего отдела спинного мозга
(Бремер, 102). Такое же явление может происходить в коре большого
мозга. Понятно, чем выше возбудимость коры, тем скорее и легче
осуществится распространение судорожной активности в коре мозга как
одним, так и другим способом.
Но когда в коре большого мозга возбудимость чрезвычайно
повышена под влиянием таких активных веществ, как стрихнин, эзерин и
ацетилхолин, и вследствие этого эпилептические судороги наступают
чрезвычайно легко, как при малейшем внешнем раздражении, так и
сами собой — спонтанно,— тогда можно утверждать, что существенным
условием для наступления судорог является общее повышение
возбудимости в коре больших полушарий. Оно, как об этом подробно будет
изложено ниже, сопровождается усилением спонтанной электрической
активности корковых элементов, а именно, появлением длинных серий
458 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
судорожных разрядов помимо внешних раздражений (см. подробно о
-судорожных разрядах в следующей главе). Вот это спонтанно
возникающее судорожное состояние может являться причиной
эпилептических судорог. Как усиленная импульсация из нервных кругов,
возбужденных спонтанно более или менее интенсивно, так и распространение
сильных электрических разрядов из того же нервного комплекса могут
<:тать возбудителями пирамидных клеток двигательной области. Во
многих случаях так называемой генуинной эпилепсии мы
должны иметь дело именно с такого рода происхождением эпилептических
судорог (Беритов и сотр., 100).
Если принять во внимание, что спонтанная активность коры
большого мозга обусловливается в значительной мере спонтанной деятель-
ностыо длинных нервных кругов, начинающихся в ядрах
промежуточного мозга (см. следующую главу), то станет совершенно понятно, что
судорожное состояние коры мозга в определенных случаях может
возникнуть от усиленной спонтанной активности промежуточного мозга.
Экспериментальным путем это было доказано Дюссер де
Варенном. Путем раздражения ядер промежуточного мозга или путем
отравления их стрихнином он вызывал судорожное возбуждение в коре.
При раздражении же или стрихнинизации коры он наблюдал усиление
активности в промежуточном мозгу (101).
Из всего вышесказанного следует, что в основе корковых
эпилептических припадков лежит усиленная активность нервных кругов коры
большого мозга, которая происходит или под влиянием сильного
непосредственного раздражения коры, или же благодаря чрезвычайному
повышению возбудимости. При последнем условии эпилептический
припадок может наступить под влиянием пороговых раздражений, а также
помимо них и вообще помимо раздражений вследствие чрезвычайного
повышения спонтанной активности.
Практикум
Наблюдение корковых двигательных участков на кошках или собаках.
Трахеотомия. Глубокий наркоз поддерживается хлороформом или эфиром путем
искусственного дыхания, как это уже описано выше. Сначала освобождают черепную крышку
от кожи и прилегающих мышц. Потом трепаном делают отверстие в области
височной кости и расширяют его, выламывая верхнюю часть черепной крышки. Ножницами
снимают твердую оболочку, и этим обнажают большие полушария в лобной и
теменной долях. В лобной доле находится крестовидная борозда. Раздражение впереди и
позади этой борозды (индукционкьвми ударами п|ри 14—10 см расстояния катушек
вызывает различные движения в зависимости от того, какой участок раздражается
(рис. 1891. Раздражение производится платиновыми электродами.
Наблюдение облегчения и торможения в коре большого мозга. 1. Находят
двигательный участок для передней или задней конечности. Повторяют пороговое тетани-
¦ческое раздражение через промежутки в 10—20 сек. Само раздражение продолжается
каждый раз ровно 2 сек. Двигательный эффект на конечности будет нарастать, что
укажет на наступление облегчения в раздражаемом участке коры мозга.
2. Затем берут околопороговое раздражение, дающее значительный эффект на
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА
459
конечностях, и повторяют его через интервалы в 1—2 мин. Двигатеіьный эффект
будет ослабевать. В этом выражается торможение, проявляющееся после периода
облегчения.
3. Если в ответ на электрическое раздражение коры наступает сильное общее
движение судорожного характера, тогда период торможения продолжается после него
до 10 мин. и дольше: пороговое раздражение этого же или другого двигательного
участка не будет давать эффектов в течение этого времени.
4. Такое же длительное угнетение корковых эффектов получается после
спонтанных судорожных движений.
5. Производят тетаническое раздражение двигательного участка коры мозга во
время и после тетанического раздражения какого-либо воспринимающего участка в
теменной или затылочной доле того же полушария (немоторная зона). Двигательный
участок раздражается пороговой силой, которая дает более или менее постоянный
двигательный эффект. Воспринимающий участок раздражается приблизительно такой
же силой. Но лучше здесь раздражение усилить на 1—2 см расстояния катушек. Клк
во время раздражения воспринимающего участка, так и в течение нескольких минут
после него раздражение двигательного участка будет вызывать ослабленный эффект
Наблюдение отравления стрихнином корковых двигательных участков. На
участок, откуда вызывается сгибание противоположной передней «оги, накладывают
кусочки фильтровальной бумаги (1—2 мм), смоченные в 0.1% растворе стрихнина
Предварительно снимают жидкость с данного участка лигнином, чтобы стрихнин не
мог распространиться на другие участки. Спустя 2—5 мин. передняя нога
противоположной стороны начинает давать сгибательные вздрагивания через правильные
короткие интервалы 1—4 раза в 1 сек. Амплитуда движений сначала в течение 5—10 мин.
ірастет, потом уменьшается. Стрихяияный эффект кончается через 20—30 мині. Такие
спонтанные движения происходят также на шее и на другой передней ноге, только
движение эгой ноги носит разгибательныч характер и происходит одновременно ее
сгибанием первой. В то же время путем пороговых раздражений ус гака вливается, что
зозбудимость отравлеыного участка значительно повышается. Повышение возбудимости
замечается и в симметричном участке другого полушария. Вызванный отсюда
пороговый эффект совершенно таков же, как если бы раздражался отравленный участок
Это указывает на то, что данный эффект вызывается из отравленного участка путем
иррадиации возбуждения из неотравленного.
Наблюдение эпилептического припадка. На том же препарате после описанных
опытов раздражают сильными индукционными ударами какой-нибудь двигательный
участок. Если этот участок находится в правом полушарии и ведает сгибанием левой
передней ноги, то сначала судорожное подергивание начнется на левой передней
ноте, затем на левой стороне туловища, шеи и лица, и, наконец, начнет подергиваться
задняя нога на левой стороне. После этого судороги распространятся на правую
сторону: на заднюю ногу, потом на переднюю, на мышцы шеи и лица. В этот момент
на левой стороне судороги или несколько ослабевают, или продолжаются с прежней
интенсивностью.
Наблюдение биоэлектрических токов возбуждения мышцы человека при
произвольной иннервации. При наблюдении биотоков на катодном или шлейфовом
осциллографе отведение мышечных биотоков происходит пластинчатыми электродами. Они
прикладываются на плече над сгибателями при межполюсном расстоянии в 5 см.
Достаточно самого небольшого напряжения, чтобы осциллограф обнаружил сильные
мышечные биотоки. Обычно мышечные биотоки обнаруживаются и при отсутствии
какого-либо произвольного сокращения, т. е. от непроизвольного рефлекторного
сокращения мышц.
При отведении к струнному гальванометру без предварительного усиления лучше
всего его производить игольчатыми электродами, которые вставляются в мышцу
через кожу. Электродами служат булавки в б мм длиной, снабженные плоской
головкой. Головка должна быть покрыта непроводником тока, напр. эмалью. Булавки,
предварительно продезинфицированные, вкалываются до головок и бинтом гак укреп-
460
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
ляются на плече, чтобы при движениях его они не выпадали. Подопытное лицо берет
тяжесть, которую то поднимает, то опускает, сгибая локтевой сустав. Каждый ряз
при поднятии, когда происходит работа, а также при опускании, когда приходится
удерживать тяжесть от падения, гальванометр показывает быстрый ряд токов
возбуждения. Чем больше будет тяжесть, тем сильнее будут эти токи возбуждения.
Литература
I. Kalischer О., Sitzber. К. Preuss. Akad. Wiss., 1900, стр. 722—4, 428; Abh.
К. Preuss. Akad. Wiss., 1905, стр. 1.
2. F r i t s с h G. u. HitzigE., Arch. Anat. Physiol., 1870, стр. 300; 1873, 397;
Arch. Psych. 3, 231, 1872.
3. Ferrier D. Vorlesungen iiber d. Hirnlokal. Dtsch. von Weiss. Leipzig, 1882.
4. Luciani L, u. Tamburini. Sulla funzione del cervello. Regio-Emil'o, 1878.
5. В e e v о г С. E. a. H о г s 1 e у V., Philos. Trans. Roy. Soc, 181, 49, 125, 1891;
185, 39, 1895; J. Physiol., 23, 10, 1899.
6. Grunbaum A. S. F. a. Sherrington C. S., Proc. Roy. Soc, 69, 206, 1902;
72, 152, 1904.
7. Bechterew W., Arch. Anat. Physiol., Suppl., 1899, стр. 543.
8. HeringE. u. Sherrington С. S, Pfl., Arch. 68, 222, 1897; Erg. Physiol. lr
Abt. 2, стр. 503, 1902.
9. BetheA. u. KastH., Pfl. Arch., 198, 604, 1922.
10. Ariens Kappers, Fol. Neurobiol., 1, 509, 1908.
II. Munk H., Sitzber. Berl. Akad. Wiss., 1893, стр. 759.
12. О p б e л и Л., Тр. об-ва русских врачей. Март — май, 1908.
13. HeringE., Pfl. Arch., 70, 1898.
14. Bechterew W., Neurol. Cb]., 17, 148, 1898; Physiologie d. motorischen Hirn-
rindenregion, 1886.
15. H о f f m a n n P., Arch. Anat. Physiol., Suppl., 1910, стр. 286.
16. F r a n с k Fr., Lemons sur les fonctions motrices du cerveau. Paris, 1887.
17. Беритов И., О корковой эпилепсии. 1917. Не напечатано.
18. Graham Brown, Erg. Physiol., 13, 1913.
19. Беритов И., Русск. физиол. журн., 1, 1917.
20. Kalischer О., Berl. physiol. Ges., 6, Februar, 1914.
21. Эльяссон М., Исследование слуховой способности собаки в нормальных
условиях и при частичном двустороннем удалении коркового центра слуха.
Дисс, 1908.
22. Демидов В., Условные рефлексы у собаки без передней половины обоих
полушарий. Дисс, 1909.
23. К у р а е в С., Исследование собак с нарушенными передними долями больших
полушарий в позднейший послеоперационный период. Дисс, 1912.
24. Fa no G. u. Libertini G., Arch. Hal. Biol., 24, 438—446, 1895.
25. В u b n о f f V. u. R. H e i d e n h a i n, Pfl. Arch., 26, 137, 1881.
26. Rothfeld J., Pfl. Arch., 148, 564, 1912.
27. В e r i t о f f J., Q. J. Exp. Physiol., 9, 199, 1915; Изв. Росс. Акад. Нзук, 1915,
стр. 649 и 853.
28. Тарханов И., О психомоторных центрах и их развитии у людей и
животных, СПб., 1879.
29. Hitzig Е., Hughlings Jackson u. motorische Rindencentren im Lichte physi-
ologischer Forschung. Berlin, 1901.
30. Luciani L, Physiologie des Menschen. Jena, 3, стр. 584, 1907.
31. Goltz F., Pfl. Arch., 42, 450, 1884.
32. V о g t C. u. V о g t O., Naturwiss., 14. Jg., H. 50—51; Jahrb. Physiol. Neur., Bd.
25, Ergenzungsheft, 1919.
33. Ухтомский А., О зависимости кортикальных двигательных эффектов от
побочных центральных влияний. Юрьев, 1911.
ОТВЕТНАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА
461
34. Wagner R., Z. Biol., 83, 120, 1925.
35. OsbornW. а. К і 1 v і n g t о п В., Brain, 33, 260, 1911.
36. Dusser de Barenne, Pfl. Arch., 233, 529, 1933.
37. Graham Brown, Quart. J.Exp. Physiol., 9, pp. 81, 101, 117, 131, 1915.
38. Правдич -Неминский В., Pfl. Arch., 209, 364; 210, 223, 1925.
39. Fischer M., Pfl. Arch., 233, 738, 1934.
40. Б e p и t о в И., О скорости проведения возбуждения в коре больших полушарий;
Не напечатано.
41. С о о р е г S. a. D. D е п п у-В г о w п, Ргос. Roy. Soc. В. 102, 222, 1927.
42. Dusser de Barenne a. Breve e, J. Physiol., 61, 81, 1926.
43. Richter C. P., Quart. J. Exp. Physiol., 18, 55, 1927.
44. A d r i a n E. a. BronkD., J. Physiol., 67, 119, 1929.
45. Wachholder R., Erg. Physiol., 26, 568, 1928.
46. Berger H., Arch. Psych. Nervenkr., 77, 321, 1926.
47. Сеченов И. иПашутин В., Neue Yersuche am Him u. Ruckenmark d. Fro-
sches, Berlin, 1865.
48. Сперанский А. Д., Эпилептический приступ. Л. 1932.
49 LeytonA. S. F. a. Sherrington С. S.s Quart. J. Exp. Physiol., 11,
No. 2, 1917.
50. F i a n z S. I., Arch. Psychol., No. 2, 1, 1907.
51. L a s h 3 e у N. S., Arch. Neur. Psych., 12, 249, 1924.
2. Анохин П. и сотр. Проблема центра и периферии в физиологии нервной системы,
1935.
53. R і о с h Mc D. a. R о s e"n b 1 u e t h A., Am. J. Physiol., 113, 663, 1935.
54. Шеррингтон и сотр. Рефлекторная деятельность спинного мозга, 1935.
55. Dusser de Barenne J. G., Arch. Neur. Psych., 30, 884, 1933.
56. H e r r i с k С J u d s о n, The amphibian forebrain. VII—X communications,
J. Сотр. Neurol., 58, 481, 1933; 58, 737, 1933; 59, 93, 1934.
57. Wachholder KM Pfl. Arch., 214, 1926.
58. Li n d sley D. В., Am. J. Physiol., 114, 90, 1935.
59. ForsterO., Lancet, 2, 309, 1931.
60. Fulton J. F., Medicine, 15, 297, 1936.
61. Wy ss O. A. M. а. О bra dor S., Am. J. Physiol., 120, 42, 1937.
62. В о у о п + о n E. P. a. Mines M., Am. J. Physiol., 106, 175, 1933.
63. Ветохин И. Сб. докладов VI Всес. физиол. съезда, 251.
64. G г е е п Н. D. а. Н о f f Е. С, Am. J. Physiol., 118, 641, 1937.
65. М о г і s о n R. S. а. М с R і о с h D., Am. J. Physiol., 220, 257, 1937.
66. W у s s О. А. М., Verh. freien Verein schweiz. Physiol. Juni, стр. 26, 1937.
67. P i n k s t о n J. О. a. M с R i о с h D., Am. J. Physiol., 121, 49, 1938.
68. P e n f i e 1 d W. а. В о 1 d г е у Е., Brain, 60, 389, 1937.
69. Бари А., О возбудимости мозговой коры новорожденных животных. Дисс, 1898.
70. Michailov S., Pfl. Arch., 133, 45, 1910.
71. Клебанова Е. А. иЛейбсонЛ. Г., Физиол. жур. СССР, 25, 418, 1938.
72. Dusser deBarenneJ. G., Am. J. Physiol., 116, 39, 1936; Dusser de
Barenne a. McCullochW. S., Am. J. Physiol., 118, 510, 1937.
73. Dusser de Barenne J. G. a. McCullochW. S.f J. Neurophysiol.,
2, 356, 1939.
74. Adrian E. D., J. Physiol., 88, 127, 1936.
75. Bickel A., Arch. Physiol. Lpz., 1901, стр. 52; Pfl. Arch., 72. 190, 1898.
76. J о h n s t о n J. В., Compar. Neurol., 25, 393, 1915; 26, 475, 1916.
77. В a g 1 e у С. a. L a n g w о r t h у О. R., Arch. Neurol. Psych. Chicago. 16, 154.
1926; В a g 1 e у a. R i с h t e г С. Р., ib. 11, 257, 1924.
78. Rogers F. Т., J. Compar. Neuiol., 35, 61, 1922.
79. В r e m e г E., D o> R. 8. a. M о r u z z i G., J. Neurophysiol., 2, 478, 1939.
SO. AdrianE. D. a. Moruzzi G., J. Physiol., 97, 153, 1939.
31. БеритовИ. иГедеваниД., Сообщ. АН Груз. ССР, 2, 157. 1941.
462 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
82. Кряжев Я. Физиол. журн. СССР, 30, 169, 1941.
S3. М е t t 1 е г F. А. а. М е 11 1 е г С. С, J. Neurophysiol., 3, 536, 1940.
84. Graham Brown Т. a. Sherrington С. S., Proc. Soc, 85 В, 250, 1912.
85. С 1 а г к S. L. a. W a r d I. W., Arch. Neur. Psych. Chicago, 38, 927, 1937.
86. Ward J., WM J. Neurophysiol., 1, 463, 1938.
87. Tower S. S., Brain, 69, 408, 1936.
-88. Гедеванишвили Д., Тр. Ин-та физиол. им. Бериташвили, б, 39, 1943.
89. Гедеванишвили Д. Тр. Ин-та физиол. им. Бериташвили, б, 263, 1943.
90. БеритовИ. иГедеванишвилиД., Тр. Ин-та физиол. им. Бериташвили,
6, 279, 1945.
91. Ві sho р G. Н. a. O'L е а г у J., Am. J. Physiol., 117, 292, 1936, Bart 1е у П.,
O'Leary J- a. Bishop G. H., Am. J. Physiol., 120, 604, 1937.
92. К e n n a r d M. A. a. M с С u 11 о с h W. S., J. Neurophysiol., 6, 181, 1943.
93. Rheinberger M. B. a. Jasper H. H., Amer. J. Physiol. 119, 186, 1937.
94. Taylor J., Selected Writings of John Hughlings Jackson. London, 1932.
95. Rioch D. Mc. a. Brenner С D., J. Сотр. Neurol., 68, 491, 1938.
S6. Гед?вани Д. M., Кардиоваскулярные'сдвиги при раздражении
кортикального центра агрессивной реакции. 1945. Печатается в Тр. Ин-та физиол. им.
Бериташвили, т. 7, 1947.
97. Green N. D. а. Но if Е. С, Amer. J. Physiol., 118, 641, 1937; ibid., 117,
411, 1936.
98. Bechterew W. a. Mislawski N., Neurol. Zbl., 5, 193, 1886.
99. Беритов И., Брегадзе А. и Цкипуридзе Л., Тр. Ин-та. физиол.
им. Бериташвили, 5, 240, 1943.
100. Беритов И., Бакурадзе А. и Дзидзишвили. Н., Тр. Ин-та
физиол. им. Бериташвили, 5, 283, 1943.
101. Dusser de Barenne J. G. a. Mc. Culloch W. S., Am. J. Physiol.,
127, 620, 1939; J. Neurophysiol., 1, 127, 364, 1938.
102. Bremer F., Proc. Soc. Exp. Biol. Med., 46, 627, 1941.
Глава IV
ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШОГО МОЗГА
Общее понятие. Кора большого мозга, подобно другим отделам^
головного мозга, в течение всей своей жизненной деятельности
продуцирует электричество, источником которого является активное состояние-
нервных элементов. На животных корковая электрическая активность,
изучается путем отведения биотоков непосредственно от обнаженного-
мозга. На людях ее изучают путем отведения корковых биотоков от
кожной поверхности черепа, куда они проникают путем физического,
распространения через оболочки мозга, черепную кость и кожу.
Потенциалы, продуцируемые в коре, не велики. Они измеряются при
отведении от поверхности мозга несколькими десятками микровольт
(миллионных вольта). Поэтому детальное изучение корковой электрической
активности как при отведении от кожной поверхности черепа, так и
непосредственно от поверхности коры стало возможным только
благодаря развитию техники усиления электрического тока при помощи
электронных ламп и применению безинертного регистрирующего аппарата^
как катодный осциллограф,-или аппарата с очень небольшой инерцией,
как шлейфный осциллограф.
Биотоки коры мозга, как и других отделов головного мозга, состоят
из медленных колебаний потенциала чрезвычайно изменчивой
продолжительности от 0.2 до 0.05 сек. у кошек (Беритов и сотр., 11) (рис. 205)>.
от 0.1 до 0.03 сек. у обезьян (Беритов и Гедеванишвили, 23) (рис. 207).
Подобные потенциалы возникают также в коре большого мозга
человека (Беритов и сотр., 16) (рис. 208). Эти нерегулярные медленные
потенциалы свойственны всем отделам коры большого мозга. Наряду с
медленными колебаниями потенциала в коре большого мозга всегда
наступают быстрые колебания потенциала также с очень меняющейся
продолжительностью. При отведении от обнаженной коры у кошек и
обезьян она меняется от 0.012 до 0.003 сек. (рис. 205 D и 212). По
амплитуде они значительно ниже медленных колебаний. Быстрые
колебания потенциала наступают на фоне медленных. Они наступают
также во всех отделах коры большого мозга, и их принято называть
бета-волнами. Эти медленные нерегулярные потенциалы и быстрые
464
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
колебания на фоне их характеризуют спонтанную деятельность всех
отделов коры большого мозга при их нормальном функционировании,
и потому можно считать эту электрическую активность основной
электрической активностью коры.
Рис. 205. Электрическая активность коры большого мозга кошки.
Катодный осциллограф. Биполярное отведение обнаженной коры с межполюсным
расстоянием 3 мм. А—от лобной доли; В — от теменной; С — от затылочной. Во всех случаях сначала
лается период сравнительного затишья, а затем период усиленной деятельности; D — от
теменной доли при более быстром вращении фотографического аппарата (Беритов, Брегадзе и Цкипу-
ридзе).
У человека в определенных отделах коры мозга и при
определенном физиологическом состоянии временами устанавливается
правильный ритм медленных колебаний с продолжительностью около 0.1 сек.
Этот регулярный ритм медленных колебаний впервые был отмечен
Бергером (3), а потому некоторые авторы называют его Бергеров-
ским ритмом. Большинство авторов называют их альфа-волнами. Альфа-
волны у человека наступают, как правило, в затылочных и теменных
долях и отсутствуют в лобных долях. В височных и центральных долях
у нормальных людей они наступают не всегда и не с такой
интенсивностью, как в теменных и затылочных (рис. 209).
Регулярные медленные колебания наблюдаются и в коре большого
мозга животных. Они обычно очень быстро и сильно изменяются как в
отношении длительности, так и в отношении ритма. Наиболее ровным
ЭЛЕКТРИЧЕСКАЯ \KJHRHOCTb КОРЫ БОЛЬШОГО МОЗГА 465
ритмом они наступают во время более или менее глубокого наркоза,
причем амплитуда их, а также и ритм, тем меньше, чем глубже наркоз.
В бодрствующем состоянии в отсутствии наркоза медленные
потенциалы также могут протекать совершенно ровным ритмом. Это, например,
было установлено для жевательной двигательной зоны кролика в
лобной области, где наступают медленные потенциалы двоякого рода:
Рис 206 Электрокортикогра уіма жевательной зоны зрительной области кролика
Отведение происходит биполярно через вживленные электроды в шлеифный осциллограф. А — от
ггеа striata и В — от жевательной зоны в лобной области одного животного, с — от жевательной
-зоны другого животного. Все осциллограммы при совершенно спокойном состоянии животного.
Время по 0 02 сеь (Гедевани).
редким ритмом по 3—5 колебаний в 1 сек. и высоким ритмом по
50—65 колебаний в 1 сек. Последние потенциалы протекают очень
ровным ритмом, располагаясь на фоне первых — более медленных; причем
они усиливаются периодически, а именно, каждый раз во время
восходящего колена медленного потенциала редкого ритма (Гедевани, 72)
(рис. 206). Описанные быстрые и медленные колебания
электрического потенциала наступают помимо внешних раздражений, сами
собой, и потому можно считать их автоматическими, спонтанными. Вся-
кое внешнее раздражение, влияющее на кору большого мозга, влияет
и на спонтанную электрическую активность в смысле усиления или
ослабления ее.
Корковые биотоки, отводимые от одного участка коры или от одного
участка кожной поверхности, выражают те физиологические процессы,
которые протекают в отводимом участке коры, а не во всей коре. Они
возникают в корковых слоях, а не под корой. От данного участка коры
будут отводиться биотоки соседних участков, вследствие чисто
физического распространения, только в том случае, если они очень большой
интенсивности; притом эти биотоки будут отводиться чрезвычайно
ослабленными (Рубин, 62; Беритов, Брегадзе и Цкипуридзе, 11; Кеннард
и Нимс, 69).
Общий характер спонтанных биотоков коры мозга не во всех ее
участках одинаков. Разные архитектонические поля производят и
биотоки разного характера (Корнмюллер, 43). Более того, сравнительные
исследования спонтанных биотоков коры мозга на разных животных
(собака, кролик, крыса, морская свинка) показали определенную связь
между морфологическими особенностями коры мозга и ее спонтанными
биотоками (Саркисов, 41).
чо
° С И. Беритов
466 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
При отведении от коры большого мозга непосредственно, и в
особенности от кожной поверхности черепа, к биотокам коры
примешиваются биотоки возбуждения мышц головы. Эти токи могут быть
настолько сильны, что вследствие физического распространения будут
в некоторой степени маскировать корковые биотоки. Это особенно
часто бывает при отведении мышечных участков черепа,— например,
Рис. 207. Электрическая активность коры большого мозга обезьяны (Macaccus rhesus).
Шлейфный осциллограф. Биотоки регистрируются от двух участков. Биполярное отведение через
двутактные усилители с емкостной связью. А — от симметричных участков центральных долей
справа (Са) и слева (С$); В — от симметричных участков затылочных долей справа (Ad) и слева
(.4s); С — от левой центральной доли около височной линии (C's) и около -медиальной (Cs); D —-от
правой затылочной доли {Ad) и от правой лобной доли (Dd) в период затишья; Е — те же участки,
что и в опыте D, но в период усиленной деятельности. Время—везде по 0.02 сек. (Беритов и Гедева-
нишвили).
при отведении затылочной области, где лежит затылочная мышца
(см., например, рис. 209-/), от передней половины лобной области, где
находится лобная мышца, и от височной области, где лежит височная
мышца (рис. 209-V). Так как мышечные биотоки всегда высокой
частоты и часть их всегда большой амплитуды в виде кратковременных
толчков или игл, то их легко можно отличить от корковых биотоков.
Можно их избегнуть, если отводить корковые биотоки при спокойном
лежачем положении от безмышечных участков черепа; у человека — это
теменные и центральные области.
ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 467
Характеристика медленных колебаний основной активности. Как
указывалось выше, медленные потенциалы основной активности
наступают во всех отделах, но интенсивность их не одинакова. У кошек они
сильнее всего в теменных долях, несколько слабее в лобных и еще
слабее в затылочных долях (Беритов и сотр., 12) (рис. 205). Такая же
разница у обезьян (макаки): в затылочных и лобных долях — слабее,
Рис. 20Я Основная электрическая активность коры большого мочга
человека.
Репістрацип биотоков нпейфным осциллографом. Поперечное биополпрное отведение
о г ьошной поверхности черепа с правоп стороны, как показано на риг. -ічі.
Межполюсное расстояние 5—b см. АтА' — биотоки от затылочной области; ВтВ' ¦— от теменной
области, СтС — от задней і раницы постцрнтралыюи извилины; JJmD' — от передней
границы предентральной извилины, EmF/ —от лобной области. Все осцитюграмчы
заенпгы при открытых глазах на свету. Шшзу время — по 0.02 сек.
чем в теменных и центральных долях (Беритов и Гедеванишвили, 23)
(рис. 207). Медленные колебания потенциала периодически значительно
усиливаются. Это наблюдается как у кошек, так и у обезьян. Вспышки
наступают во всех участках коры одновременно, но с разной
интенсивностью и продолжительностью, через интервалы от нескольких секунд
до нескольких минут (Беритов и сотр. 11, 23; Морисон, Финлей и Лот-
роп, 67) (рис. 205 и 207). В период усиленной деятельности
медленные колебания значительно уравниваются в отношении и амплитуды и
продолжительности, причем с повышением амплитуды они становятся
более регулярными. Такое уравнивание медленных колебаний у кошек
проявляется значительно слабее, чем у обезьян.
У человека при отведении корковых биотоков от кожной поверхности
черепа основная электрическая активность выступает слабо;
соответствующие медленные потенциалы очень низкой амплитуды, чаще всего
10—20 jaV (рис. 208). Они, конечно, сильно ослаблены вследствие по-
30"
468
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
бочного замыкания корковых биотоков через тканевую жидкость (Бе-
ритов, 16).
При одновременной регистрации основной электрической активности
от двух участков мозга оказывается, что медленные потенциалы часто
Рис. 209. Альфа-ритм коры большого мозга человека.
Регистрация шлейфным осциллографом. Отведение биотоков продольное биполярное с
правой стороны. Положение электродов, как на рис. 221. АВ— биотоки от
затылочной области; ВС — от теменной; С—от центральной области (постцентральной и пре-
центральной извилин); DE — от задней половины лобной области; GH — от височной
области. Все осциллограммы — при закрытых глазах. Осциллограммы I, //, III и IV —
от того же субъекта, как рис. 208, а У — от другого. Везде хорошо выраженные альфа-
волны, кроме лобной доли, где они совершенно отсутствуют.
совпадают во времени, а иногда и по интенсивности. Эта синхронность
лучше всего проявляется при отведении симметричных участков темен-
но-центральной области. Она выступает лучше у человека, чем у
животных; лучше при отведении больших участков, чем малых; лучше при
отведении симметричных участков обоих полушарий, чем от
несимметричных участков одного полушария. Чаще всего эти синхронные коле-
ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 469
бания являются синергетическими, но в определенных случаях они
могут быть и антагонистическими.
Медленные колебания основной активности у животных, как
указывалось выше, сильнее всего наступают в теменных долях и ослабевают
впереди и позади этого участка. Из этого следует, что теменные доли
Рис. 210. Влияние закрытия или затемнения глаз на биотоки коры большого
мозга человека.
Шлейфный осциллограф. Отведение биполярное от теменной и центральной областей (BD) и
от височной области (GH). J — сначала глаза открыты и в кабине светло, затем глаза по
приказу закрываются. Альфа-волны усилились в первую же секунду. II — вначале глаза
открыты и в кабине светло, затем свет тушится, в кабине абсолютная темнота, но глаза
остаются открытыми. Альфа-волны усиливаются приблизительно через 2 сек. III —
сначала темно, глаза открыты. Ччрез несколько секунд от начала затемнения, когда альфа-волны
ослабли, глаза по приказу закрываются. В ту же секунду вновь вознинают усиленные
альфа-волны. Время закрытия или затемнения глаз отмечается вибрацией линии времени.
Время —по 1 сек. (Беритов и Воробьев).
являются доминирующим местом в отношении медленных колебаний
основной активности (Беритов, 16).
Характеристика альфа-ритма или Бергеровского ритма. У человека
медленные колебания основной активности с нерегулярным течением
временами переходят ів ровный, регулярный ритм одинаковых
колебаний, в так называемый альфа-ритм. У людей высокой корковой
возбудимости этот альфа-ритм равен 11 —12 в 1 сек., у людей низкой
возбудимости— 9—10 в 1 сек. У каждого человека имеется свой
собственный ритм, который стойко удерживается при всех условиях жизни.
Альфа-волны периодически то усиливаются, то ослабевают через
каждые 1—2 сек. В каждом периоде амплитуда нарастает до 50—100 j*V
(Беритов и сотр., 15) (рис. 210).
Альфа-ритм лучше всего выявляется в темноте, при закрытых глазах,
когда субъект лежит совершенно спокойно, без напряжения мышц
головы и без усиленной умственной работы. Открытие глаз и
освещение, умственная работа или волнение мешают проявлению альфа-ритма.
Если при открытых глазах на свету альфа-ритм отсутствует, то закры-
470 ОБЩДЛ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
вание глаз приводит к его появлению со скрытым периодом около
7з—7г сек., т. е. существующий до этого основной ритм переходит в
альфа-риты. Если же альфа-ритм был до закрывания глаз, то при
закрывании их он значительно усиливается (рис. 210). Так же действует
темнота. Если потушить свет при открытых глазах, альфа-волны
усиливаются, или если их не было до темноты, то появляются. Только в
этом случае скрытый период длиннее—больше 1—2 сек. (Беритов и
Воробьев, 18) (рис. 210-//).
Рис. 211. Биотоки коры большого мозга человека.
Регистрация шленфным осциллографом. Продольное биполярное отведение корковых биотоков
от кожной поверхности черепа. Глаза закрыты. I — биотоки от симметричных участков
затылочных областей, II— то же от теменных, III — то же от задней половины лобных долей.
В разных участках одного полушария альфа-волны наступают в
большинстве случаев одновременно и одновременно же усиливаются и
ослабевают (рис. 210). Но так бывает не всегда. Временами
синхронность нарушается и альфа-волны наступают разновременно. Это
происходит потому, что альфа-ритм сравниваемых участков разнится на
одну или половину волны. Лучше всего синхронность проявляется при
отведении симметричных участков, но моментами они также расходятся
или из синергетических становятся антагонистическими (Беритов, 16)
(рис. 211).
Регулярный альфа-ритм 9—12 в 1 сек. бывает только у человека, у
животного его нет. Как указывалось выше, у животных (кошка,
обезьяна) медленные колебания временами значительно нарастают и при этом
уравниваются. Но эти периодические вспышки не обладают
определенным ритмом; последний быстро меняется в больших пределах;
например, у обезьян (макаки) от 10 до 35 в 1 сек., чаще всего около 20 в
1 сек. Такие медленные колебания не усиливаются при закрытии глаз.
При значительном понижении возбудимости в коре,— например, под
влиянием барбитуратов,— медленные потенциалы могут иметь
ровный ритм около 10 в 1 сек. Но это не естественный альфа-ритм чело-
ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 471
века, наблюдающийся при бодрствующем состоянии, а ритм,
наступающий вследствие глубокого наркоза (Беритов и Гедеванишвили, 23).
Альфа-волны, достигая большой амплитуды, двуфазны; при
небольших амплитудах они однофазны; причем эта одна фаза может быть как
отрицательной, так и положительной. При малых амплитудах альфа-
волны нередко сливаются, т. е. не успевает предшествующая волна
достигнуть абсциссы, как начинается следующая волна. Слившиеся
альфа-волны мы называем «эта-волнами», в отличие от всех других
нормальных волн, уже обозначенных предыдущими буквами греческого
алфавита (Беритов, Бакурадзе и Дзидзишвили, 15).
Центральным местом возникновения альфа-волн является в
полушариях теменная область на границе с затылочной. Здесь они возникают
сильнее всего и регулярнее всего. Впереди и позади этой области альфа-
волны постепенно ослабевают (Эдриан и Иамагива, 32; Беритов, 16)
(рис. 209). Некоторые авторы признают также существование
второго центра для альфа-волн в лобных долях (Рубин, 62; Лумис и
сотр., 9), но это не оправдалось исследованиями других авторов
(Беритов и сотр., 15, 16).
Теменная область вместе с area praeoccipitalis является главнейшим
ассоциативным центром. Здесь сходятся проекции основных
рецепторов: зрения, слуха, кожной и мышечной чувствительности. От
постцентральной извилины отходит отросток в теменную область вдоль sulcus
interparietalis. Он пересекает всю теменную область и в затылочной
доле соединяется со зрительной областью. Этим достигается очень
тесная связь между воспринимающими областями зрительной и кожно-
мышечной чувствительности. Кроме того означенная теменно-затылоч-
ная область! тесно связана с thalamus, с его nucleus lateralis. Здесь же
имеются хорошо развитые межкорковые пути tractus centro-parietalis,
которые связывают теменную область с двигательными корковыми
центрами. Очевидно, теменная область является важнейшим
структурным образованием, в котором осуществляется значительная часть
нервных связей, и потому она, вероятно, является основным нервным
субстратом для возникновения альфа-ритма.
Характеристика быстрых колебаний. Продолжительность быстрых
потенциалов коры большого мозга, так называемые «бета-волны», при
отведении непосредственно от поверхности коры, как уже указывалось
выше, очень разнообразна. В общем, у кошек можно было насчитать
до 120 в 1 сек. (рис. 205-D); у обезьян при отведении от поверхности
коры удается зарегистрировать до 200—300 в 1 сек. (рис. 212); у
человека при отведении от кожной поверхности в безмышечных участках
ритм бета-волн бывает от 80 до 200—250 в 1 сек. Но такая высокая
частота быстрых колебаний может быть наблюдаема при достаточно
большой скорости фотографирования, выше 50 см в 1 сек.
Амплитуда бета-волн обычно небольшая — 10—20 микровольт, но
может быть и больше. Особенно сильными бета-волны бывают при
непосредственном отведении от коры, особенно если электроды глубоко
472
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
проникают в кору. В таких случаях волны могут усилиться до 60
микровольт. Особенно слабыми они бывают при отведении от кожной
поверхности черепа. Вообще отведение через кость и кожу ослабляет
биотоки в 2—3 раза (Беритов и сотр., 11, 17, 23).
Бета-волны двух участков обычно не синхронны, но если они
достигают значительной амплитуды, особенно при отведении двух близких
участков одного полушария или двух симметричных участков, то многие
Рис. 212. Спонтанная электрическая активность коры большого мозга
обезьяны (Macaccus rhesus).
Шлейфный осциллограф. Заснято при большой скорости. А— отводятся большие
симметричные участки обнаженной коры с межполюсным расстоянием около 20 мм, один
электрод находится в затылочной области, а другой — в центральной. Верхняя
осциллограмма (ACd) справа, нижняя {ACs) слева. В — отводятся маленькие симметричные
участки от центральных долей справа (Cd) и слева (Cs). Межполюсное расстояние — по
3 мм (Беритов и Гедеванишвили).
из них бывают синхронны. Особенно часты синхронные волны при
отведении больших участков при большом межполюсном расстоянии.
Бета-волны усиливаются и ослабевают вместе с медленными
колебаниями. При периодическом усилении медленных колебаний
усиливаются иногда и быстрые бета-волны (Беритов и Гедеванишвили, 23)
(рис. 207 D и Е).
В литературе обычно указывается, что ритм бета-волн колеблется в пределах
17—45 в 1 сек. (Джаспер и Гоуке, 63; Лемер, 64; Шварц и Керр, 65 и др.). Редко кто
наблюдал до 90 в 1 сек. и выше (Рорахер, 58; Бергер, 3; Шпильберг, 57). Низкий ритм
бета-волн у многих авторов объясняется тем, что регистрация корковых биотоков
производилась у них при очень низкой чувствительности регистрирующего аппарата
и, кроме того, при малой скорости зааіиои, кіак 2—3 ем >в 1 сек. При этих условиях
малые быстрые колебания совсем не регистрировались, а большие, если они следовали
довольно быстро друг за другом, должны были слиться в одно длительное
колебание, которое и регистрируется как одна бета-волна. В этих случаях их трудно
отличить от медленных колебаний основной активности. Непременным условием для
выявления действительного бета-ритма является большая чувствительность
регистрирующего аппарата и достаточно большая скорость движения фотографического
аппарата— до 50 см в 1 сек. Другой 'причиной к установлению низкого ритма для бета-
ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 473-
волн является применение фильтров дитя устранения мышечных токоз высокого ритма,
а затем механическая регистрация биотоков путем специальных чершмочишущих
приборов. При этих условиях, конечно, не могут быть зарегистрированы быстрые биотоки
коры, и потому высший ритм бета-волн является сильно уменьшенным
Происхождение быстрых колебаний потенциала коры большого
мозга. Как быстрые, так и медленные потенциалы в коре большого
мозга могут возникать, при отсутствии внешних и внутренних
раздражений, в результате самовозбуждения нервных кругов самой коры.
Но, конечно, самовозбуждение одного круга может повлечь за собой
возбуждение целого ряда других нервных кругов оолее или менее
отдаленной области коры, с которой он находится в структурной связи.
Существование ассоциативных волокон обеспечивает тесное
взаимодействие разных участков одного полушария, существование коммиссураль-
ных волокон — переход возбуждения от одного полушария на другое,
а наличие кортико-таламических и таламо-кортикальных связей
обусловливает тесное взаимодействие между корой большого мозга и
промежуточным мозгом. Отсюда понятно, что нормальная спонтанная
активность ів каждом участке коры большого мозга обусловливается
спонтанной деятельностью всей коры и промежуточного мозга. Вместе с тем
доказано, что у животных удаление значительной области коры
существенно не меняет характера электрической активности оставшейся
коры (Беритов и сотр., 11). Точно так же доказано, что перерезка
ствола мозга позади промежуточного существенно не влияет на
эту активность (Бр?мер, 25; Демпсей и Морисон, 24). Только при
глубоком наркозе такая перерезка может повлечь исчезновение корковой
активности (Леви и Гаммон, 60; Форбс и Морисон, 61). Но корковая
активность в значительной мере меняется при бодрствующем состоянии
коры после перерезки проводящих путей между'промежуточным мозгом
и корой. Она в общем ослабевает, но периодические взрывы
сохраняются и даже усиливаются. Отсюда следует, что спонтанная
электрическая активность в смысле периодического усиления всецело зависит от
коры, от ее само&озбужбения, а в остальном — от совместного
возбуждения коры и промежуточного мозга (Демпсей и Морисон, 24; Дюссер
це Баренн, 50; Морисон и Демпсей, 26).
Характерно, что между промежуточным мозгом и корой существует
такое же взаимодействие, как между симметричными участками обоих
полушарий: спонтанные медленные колебания определенных участков
промежуточного мозга и коры являются © значительной мере
синхронными (Демпсей и Морисон, 24; Дюссер де Баренн, 50).
Быстрые колебания выражают собой биотоки возбуждения от пучка
нервных волокон или от нервных кругов, возбуждающихся в коре более
или менее одновременно. Когда эти круги возбуждаются совершенно
одновременно, тогда, конечно, продолжительность соответствующих
быстрых колебаний будет наиболее короткая, как продолжительность
одного аксонного тока возбуждения, т. е. менее 1 о. Но так как в массе
нервных волокон абсолютной синхронности возбуждения не может быть,
474
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
то продолжительность биотоков возбуждения нервных пучков должна
быть всегда больше,— тем больше, чем больше разнообразия между
чувствующими нервными кругами, прежде всего в отношении длины
нервных кругов и их функционального состояния.
Синхронность бета-волн от двух разных участков коры
обусловливается івозбуждением тех длинных нервных кругов, которые являются
общими для данных отводимых участков. В случае механического
раздражения коры, при котором в определенных нервных кругах наступает
усиленная деятельность, усиленные 'быстрые колебания отводятся как
от раздраженных, так и от соседних нераздраженных участков. Эти
колебания являются целиком синхронными и в большинстве случаев
антагонистическими. Это обусловливается, по нашему мнению, тем, что
от ближайших соседних участков отводятся корковые биотоки, которые
возникают в раздраженном участке и распространяются отсюда чисто
физически.
Происхождение медленных колебаний основной активности.
Медленные колебания потенциала коры большого мозга, как уже указывалось
в предыдущей главе, отображают активное состояние нейропиля. Ней-
ропиль богато представлен >во всех слоях коры большого мозга;
особенно сильно он развит в верхних двух слоях (см. рис. 171 и 172).
Этот нейрипиль активируется при спонтанной активности коры,
во-первых, афферентной системой, которая спонтанно возбуждается в
передаточных станциях афферентной системы, и, во-вторых, спонтанно
возбуждающимися нервными кругами. Мы считаем, что медленные потенциалы
получаются не путем слияния быстрых колебаний бета-волн, а в
результате слияния местных процессов, возникающих в дендритной массе
нейропиля, а также в клеточных элементах коры, т. е. тех длительных
процессов, которые возникают локально в дендритах и теле клеток в
области синапсов под влиянием импульсов из афферентной системы и
нервных кругов. При этом сливается очень большое количество местных
процессов (благодаря образованию каждым аксоном множества
синапсов на теле клеток и дендритах), поэтому интенсивность суммарного
электрического потенциала от активирования дендритной массы обычно
значительно превосходит интенсивность токов возбуждения нервных
кругов. Этим объясняется, что при отведении от поверхности коры
мозга и, тем более, от кожной поверхности черепа медленные биотоки
могут выступать без осложнения их быстрыми биотоками.
Существуют факты, указывающие на то, что медленные колебания
потенциала в коре мозга в основном происходят от активирования
нейропиля. Прежде всего за это говорят опыты Э д р и а н а (39) с
электрическим раздражением разных слоев коры мозга и отведением в
осциллограф ближайших участков мозга на наркотизированных
животных (.кролик, кошка, обезьяна). При раздражении самого
поверхностного слоя коры обычно наступают монофазные токи с
продолжительностью около 100 о. Эти токи ограничиваются соседними к
раздражаемой области участками (рис. 213); они исчезают после тепловой коагу-
ЭЛККТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 475
ляции верхних слоев. Автор предполагает, ч*то они возникают от
активирования горизонтальных клеток Кахаля или поверхностного разветвления
верхушечных дендритов, приходящих сюда от пирамидных клеток. Это
активное состояние распространяется в верхнем слое с большим декре
ментом, всего на несколько миллиметров, и является главным образом
результатом деятельности тех нервных элементов, на которые распро-
Рис 213 Биотоки двигательного участка і оры больших потушарий
и соответствующей мышцы } обезьяны
Раздражается двигательный участок руьи Частота раздражения 18 раз в 1 сек.
Нижняя электрограмма — от двиі ательного участка верхняя — от мышц руки В
опыте А — электрические волны в виде длительных отрицательных колебании от
поверхностного слоя коры мозга при отсутствии биотоков мышц руки В опыте В
электрические колебания наступают в коре одновременно с электрическими
колебаниями в мышцах руки В этом опыте корковые биотоки отличаются от первого опыта
тем, что здесь за отрицательным колебанием следует небольшое поло*ительное Это
указывает на участие глубоких слоев коры больших полушарии (Эдриан)
страняется прямое действие электрического раздражения. Это активное
состояние может не сопровождаться двигательными эффектами, даже
если раздражается верхний слой в двигательной области коры мозга
(рис. 213 А). Движения возникают при сильном раздражении, когда
раздражающий ток проникает глубоко в третий-четвертый слои, где
лежат пирамидные клетки, дающие начало пирамидным путям (рис. 213-в).
Эти наблюдения прямо указывают на то, что при раздражении
поверхностного слоя мы имеем дело не с возбуждением нервных кругов, а с
активированием какого-то другого образования. Основным образованием
в верхних слоях коры является нейропиль — сплетение разветвлении
верхушечных дендритов пирамидных клеток глубоких слоев и дендритов
горизонтальных клеток верхнего слоя. Отсюда следует, что медленные
колебания потенциала, возникающие в верхних слоях коры при
непосредственном электрическом раздражении, представляют собой нейро-
пильный ток от активирования поверхностного нейропиля коры мозга.
При поверхностном отведении коры нейропильный ток отводится
вместе с аксонными токами нервных кругов. Обычно в каждом
электрическом эффекте коры, наступающем в ответ на короткое
раздражение какого-либо рецецтора, быстрые колебания предшествуют
медленным. 'Анализ этих явлений приводит авторов к утверждению, что
медленные потенциалы возникают самостоятельно, в результате
суммирования более медленных процессов, чем аксонные биотоки возбуждения.
Например, авторы находят, что при световом раздражении медленные
потенциалы возникают во втором и первом слоях коры, где господ
ствует нейропиль с его дендритной імассой, а быстрые токи—в
средних слоях (Маршалл, Тальборт и Адес, 73). Но при изолированном
отведении от определенных нервных слоев микроэлектродами удается
отводить медленные и быстрые потенциалы независимо друг от друга. Так,
476 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
Форбс и сотрудники (51) показали, что при раздражении определенных
проводящих путей, идущих к аммониевому рогу, в последнем возникают
медленные потенциалы продолжительностью 130—160 о без какого-либо
осложнения их быстрыми. И когда случаются быстрые потенциалы, они
протекают на фоне медленных без признаков слияния с последними.
При этом было установлено, что медленные потенциалы возникают в
слое пирамидных клеток и их дендритных разветвлений, где, по Ло-
ренте де Но (53), синаптически оканчиваются афферентные пути.
Медленные потенциалы могут быть положительными и
отрицательными, однофазными и двуфазными (рис. 214-С). Все это зависит от
места отведения. Точно так же можно отвести быстрые потенциалы
отдельно от медленных, если, например, однополюсный микроэлектрод
вставить в аммониев рог до слоя пирамидных клеток (stratum pyrami-
dale) (рис. 214, А—В).
Итак, можно утверждать, что как в рассмотренных, так и во всех
других случаях медленные колебания потенциала в коре большого
мозга возникают в активированном нейропиле коры, что эти колебания
могут получиться в чистой форме, независимо от быстрых аксонных
токо&, когда нейропиль отводится изолированно от нервных кругов.
Происхождение синхронизации медленных колебаний. Основная
электрическая активность в каждом участке коры большого мозга
определяется, во-первых, его функциональным состоянием и степенью
самовозбуждения и, во-вторых, воздействием притекающих к нему со
стороны нервных импульсов. Поэтому два разных участка всегда будут
производить медленные колебания разного ритма и разной
интенсивности. Но обычно некоторое количество медленных колебаний является
синхронным.
Синхронность медленных биотоков в тех или других участках коры
мозга зависит, в первую очередь, от функционального состояния
доминирующей (теменной) области и от ее ассоциативных связей со
всеми остальными участками коры, Чем сильнее активное состояние
доминирующей области и чем многочисленнее ее связи с каким-либо
другим участком, тем сильнее взаимодействие между ними и тем
больше должно быть у них синхронных медленных колебаний.
Синхронность медленных колебаний в разных участках коры
обусловливается прежде всего общностью нервных кругов. Участки коры,
объединенные ассоциативными и коммиссуральными волокнами,
безусловно будут активироваться более или менее одновременно благодаря
распространению импульсов возбуждения из одного участка в другие по
этим волокнам. Соответственно с этим и нейропильные медленные токи,
возникающие в этих участках под влиянием распространяющихся
импульсов, будут протекать более или менее синхронно.
іКогда одновременно отводятся доминирующая теменная область и
какой-либо соседний участок впереди или позади нее, синхронные
медленные колебания могут являться антагонистическими. Это
объясняется тем, что в доминирующей области сильно активирован нейропиль
ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 477
глубоко лежащих слоев, во-первых, вследствие самовозбуждения
нервных кругов коры, а во-вторых, под влиянием афферентных импульсов,
притекающих в третий и четвертый слои коры по1 нервным кругаім из
100 ^милласен
Рис. 214. Биотоки слоя пирамидных клеток g. hippocampus.
Запись катодным осциллографом. Токи отводятся
микроэлектродом. Опыт А — спонтанные биотоки при быстром вращении
съемочного аппарата, внизу время—500 колебаний в 1 сек. Опыт В—при
медленном вращений. Здесь даны группы быстро потухающих
колебаний, видимо от небольшой группы клеток, возбуждаемых
циркулирующими по нервным кругам импульсами. Опыт С
—медленные волны g. hippocampus при раздражении одним
электрическим ударом area entorhinalis. Токи отводятся однополюсным
микроэлектродом: в опыте 1 — от поверхности коры, в опыте 9 —
от вентрального края верхней части аммониева рога на глубине
3 мм. Все остальные — на разных глубинах между ними (Реншоу,
Форбс и Морисон).
промежуточного мозга. В соседних же областях будут активироваться
главным образом поверхностно лежащие слои вследствие иррадиации
импульсов из доминирующей области, ибо ассоциативные волокна
оканчиваются преимущественно в верхних слоях коры. Как известно,
направление биотоков, отведенных от поверхности коры мозга,
противоположно, если они возникают то в наружных, то so внутренних слоях
коры (Форбс и Морисон, 61; Беритов и сотр., 13).
478
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
Медленные колебания могут стать антагонистическими при
отведении разных участков впереди и позади доминирующей области также
и потому, что активная масса нервных кругов и нейропиля должна
уменьшаться во все стороны, начиная от центра доминирующей
области. Сообразно с этим, если отводятся корковые биотоки впереди и
позади доминирующей области и притом параллельно медиальной линии, то
непременно медленные колебания окажутся в значительной мере
антагонистическими, ибо в одной паре электродов, позади доминирующей
области, задний полюс будет активен менее переднего, а в другой паре
электродов, лежащей впереди ее, он будет активен больше переднего.
Происхождение альфа-волн. Альфа-волны, наблюдаемые у человека,
безусловно являются видоизменением периодически усиливающихся
медленных колебаний основной активности. Доминирующая область для
альфа-волн — та же самая теменная область, как и для этих медленных
колебаний; альфа-волны являются синхронными, и притом синергети-
ческими или антагонистическими, как медленные потенциалы во время
основной активности при тех же условиях отведения; альфа-волны
также периодически усиливаются и ослабевают, как это характерно для
медленных потенциалов основной активности у животных. Поэтому
можно утверждать, что альфа-волны человека филогенетически
развились в связи с развитием мозга из медленных колебаний основной
активности, а именно — из тех периодически наступающих усиленных
разрядов, которые являются характерными для коры животных. Нужно
думать, что возникновение альфа-ритма обусловлено таким развитием
структуры коры и ее связей с подкорковыми образованиями, что оно в
значительной мере улучшило в теменных долях синхронность работы
нервных кругов.
Альфа-волны возникают в коре большого мозга. Но, вероятно, эти
чолны у человека обусловливаются связанной деятельностью коры и
промежуточного мозга, т. е. возникают в результате самовозбуждения
нервных кругов, объединяющих кору и промежуточный мозг. Когда
нервные круги возбуждаются со стороны, под влиянием вообще
периферических импульсов, тогда, наоборот, альфа-ритм расстраивается или
совершенно исчезает. Так же действуют усиленная умственная
деятельность и утомительная физическая работа (Бергер, 3; Воробьев и Дзидзи-
швили, 19). Очевидно, когда кортико-таламические нервные круги,,
участвующие в возникновении альфа-волн, возбуждаются не путем само-
возбуждения, а насильственно, под влиянием притекающих импульсов^
обычная синхронная деятельность нервных кругов нарушается и альфа-
ритм исчезает.
1 Альфа-волны, протекающие по ритму 9—12 в 1 сек., не
исчерпывают всю спонтанную деятельность затылочно-теменной
области. Частотный анализ биотоков этой области, произведенный
Ливановым (42), Слепяном (75) и др., показывает, что во
время альфа-ритма наряду с альфа-волнами наступают медленные
колебания других частот. Так, С л е п я н обнаружил другие частоты в виде
ЭЛЕКТРИЧЕСКАЯ АКТИіШОСіЬ КОРЫ БОЛЬШОГО ЛГОЗГА 479
гармоник, (с отношением 1 : 2 : 3), как, напр., 9.2 Hz : 18.4 Hz : 27.6 Hz,
и субгармоник, как 7г, Уз, частоты 4.6 Hz, 3.1 Hz, а также в виде
таких некратных колебаний, как 2/3, 4/s, 5/г- Отсюда следует, что во
время альфа-ритма в коре большого мозга активируются нервные круги
разной длины, причем наибольшее количество их приходится на круги
определенной длины, которые обусловливают альфа-ритм.
Но в патологических случаях и даже у нормальных людей иногда
доминирует не альфа-ритм, а другие ритмы, обычно
обнаруживаемые частотным анализом (Беритов, 76; Кэиз и Буци, 78). Отсюда
следует, что в определенные моменты в затылочно-теменной области
доминирует деятельность не тех нервных кругов, которые обусловливают
альфа-ритм, а других, с другим ритмом медленных колебаний.
Происхождение альфа-волн и вообще медленных колебаний корковой
электрической активности некоторые авторы объясняют биохимическими процессами,
протекающими в ганглиозных клетках. Благодаря таким процессам корковые клетки усваигатот
вещества, необходимые для их деятельности (Рорахер, 3). Подтверждением этого
взгляда считается тот факт, что при повышении температуры тела под влиянием
высокочастотных токов, затем при внутреннем введении тироксина, когда обмен
веществ в мозгу должен повыситься, частота альфа-волн повышается (Гогленд, 27;
Рубин, Коген и Гогленд, 28). Не может быть сомнения <э том, что повышение обмена
веществ или повышение температуры тела должно способствовать повышению а іь-
фа-ритма. Но это обстоятельство ни в коем случае не объясняет самого
происхождения альфа-волн, их регулярного течения и большой синхронизации в разных участка<
коры.
Другие авторы рассматривают альфа-волны как результат суммации быстры\
колебаьий — бета-волн, выражающих возбуждение нервных клеток и нервных воло
кон (Эдриан и Мэтьюс, 4; Бартлей и Бишоп, 55). Но против такого понимания,
прежде всего, говорят опыты с отведением от коры медленных и быстрых токов
раздельно друг от друга (Реншоу, Форбс и Морисон, 51, 61). Против этого говорит
также тот факт, что бета-волны наступают на фоне медленных колебаний с такой же
интенсивностью, как и при отсутствии их (Беритов 12; Беритса и сотр., 17).
Происхождение второй фазы у альфа- и бета-волн. Альфа и бета-
волны очень часто имеют синусоидальный характер, т. е. они состоят из
двух противоположных фаз, как бы полуволн одинаковой амплитуды,
но в большинстве случаев они не носят характера типичной синусоиды.
У альфа-волн, например, при их малой амплитуде вторая фаза
выражена значительно слабее первой или даже отсутствует, затем, когда
альфа-волны большой амплитуды, то переход первой фазы
альфа-волны ко второй фазе очень часто происходит с большей быстротой, чем
переход от второй фазы одной альфа-волны к первой фазе
последующей волны; наконец, от второй фазы одной альфа-волны до первой
фазы другой иногда включается небольшая пауза, во время которой
кривая удерживается около абсциссы. При этом все означенные
особенности проявляются без всякой периодичности. Поэтому імне
кажется, что альфа-волны, по существу говоря, протекают не
синусоидально, что вторая фаза осциллограммы альфа-волны имеет
самостоятельное биологическое происхождение. То же самое следует сказать насчет
бета-волн (Беритов и сотр., 15).
Результаты частотного анализа медленных колебаний во время
альфа-ритма, а именно — наличие субгармоник и некратных колебаний,
480
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
также свидетельствует о том, что представляемые последними
биологические процессы в коре мозга не носят характера простых
синусоидальных колебаний. То же самое было обнаружено методом накладывания
.альфа-волн: форма главных колебаний, определяющих характерный
вид энцефалограмм, т. е. форма альфа-волн, совершенно не
соответствует синусоиде (Слепян, 75).
Происхождение первой и второй фазы альфа- и бета-волн должно
быть именно такой природы, как это вообще известно для двуфазных
колебаний потенциала в нервной системе, т. е. эти фазы отвечают двум
самостоятельным процессам, возникающим в разное время в разных
>частках нервной системы. Можно, например, утверждать, что первая
фаза альфа-волн обусловливается преимущественным активированием
средних слоев коры большого мозга, происходящим под влиянием
импульсов, притекающих сюда из таламуса по длинным нервным кругам.
Возбуждение этих нервных кругов производит группу бета-волн;
одновременно активируется дендритная масса в тех же слоях коры,
вследствие чего возникает первая фаза альфа-волны. Вторая же фаза альфа-
волны получается от активирования нервных кругов и дендритной
массы наружных слоев коры, происходящего несколько позднее, благодаря
передаче импульсов возбуждения из средних слоев к наружным
посредством ассоциативных нейронов с короткими аксонами. Так
обусловливается новый разряд бета-волн и вторая фаза альфа-волн (Бери-
тов, 12).
Когда возбудимость коры понижена, вследствие чего переход
возбуждения из средних слоев к наружным задерживается, тогда выпадает
вторая фаза альфа-волны. Благодаря этому происходит, между прочим,
удлинение первой фазы, и может случиться, что новая альфа-волна
наступит раньше, чем кончится предыдущая альфа-волна,— вернее, ее
первая фаза. Вследствие этого происходит слияние их, образование
эта-волн.
Происхождение благоприятствующего влияния закрытия и1
затемнения глаз на альфа-ритм. Как указывалось выше, закрытие и
затемнение глаз благоприятствуют альфа-ритму. Было установлено, что это
вызывается не всецело лростым ограничением световых раздражений.
Известно, что закрытие глаз благоприятствует альфа-ритму и при
абсолютной темноте (см. рис. 210-///). Очень часто закрытие или
затемнение глаз благоприятствует альфа-ритму не все время закрытия, а
только в самом начале, в течение нескольких секунд. Наконец, при
многократном повторении закрытия глаз или затемнения друг за другом,
благоприятствующее влияние их н*а альфа-ритм ослабевает и даже
может исчезнуть. Все эти факты свидетельствуют о том, что закрытие
глаз благоприятствует альфа-ритму не только благодаря ограничению
световых раздражений. Путем анализа этого явления мы пришли к
заключению, что закрытие глаз благоприятствует альфа-ритму не только
прирожденным путем, благодаря понижению корковой возбудимости
ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 481
вследствие устранения световых раздражений, но и индивидуально-
рефлекторным путем, благодаря тому, что происходящие при закрытии
глаз раздражения проприоцепторов глазных мышц и век и экстеро-
рецепторов оболочки глазных яблок являются индивидуальным
сигналом к усиленному синхронному возбуждению нервных кругов теменно-
затылочной области (Беритов и Воробьев, 18).
Так как закрытие глаз с затемнением происходит всегда перед
сном, когда субъект спокойно лежит в постели, при отсутствии
слуховых, зрительных и других раздражений, то закрытие глаз и
прекращение света предшествуют усилению альфа-ритма, наступающему в связи
с прекращением внешних раздражений. Отсюда совершенно естественно,
что прекращение света и раздражение рецепторов глазных мышц и
глазного яблока становятся индивидуальным сигналом к альфа-ритму,
т. е. на все эти раздражения устанавливаются временные связи со
всеми теми нервными комплексами в коре мозга, которые обусловливают
своей деятельностью альфа-ритм.
Что вообще внешние раздражения могут стать индивидуальным
сигналом к альфа-ритму, видно из того, что можно образовать
индивидуальный рефлекс альфа-ритма на постороннее раздражение,—
например, на звук. Путем многократного сочетания звука с затемнением,
которое приводит к альфа-ритму, звук в отдельности также начинает
вызывать альфа-ритм (Беритов и Воробьев, 18). Очевидно, временные
связи на звук установились со всеми нервными комплексами в коре
мозга, обусловливающими возникновение альфа-ріитма.
Точно так же открытие глаз препятствует альфа-ритму не только
прирожденным путем, благодаря возбуждающему действию освещения
глаз на затылочную область коры мозга, но и
индивидуально-рефлекторным путем, благодаря тому что происходящие при открытии глаз
раздражения рецепторов глазных мышц и век, а также оболочки
глазных яблок являются индивидуальным сигналом к асинхронному
возбуждению нервных кругов теменно-затылочной области, т. е. к
расстройству или исчезновению альфа-ритма. Этот индивидуальный рефлекс
выработался безусловно на основе совпадения открытия глаз с
освещением их, производящим расстройство или исчезновение альфа-ритма
вследствие нарушения синхронной деятельности нервных кругов,
устанавливаемой в результате самовозбуждения нервных кругов коры и
промежуточного мозга. Что раздражения, происходящие при открытии
глаз, действуют подобно свету независимо от освещения глаз, видно из
того, что открытие глаз ослабляет или устраняет альфа-ритм даже в
абсолютной темноте. А что проприоцептивные и экстероцептивные
раздражения, происходящие при открытии глаз, должны действовать
индивидуальным путем, об этом можно заключить из того, что при
многократном сочетании освещения глаза со звуком последний становится
индивидуальным сигналом к прекращению альфа-ритма (Джаспер и
Шагес, 74).
31 И. С. Беритов
482 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
Онтогенез спонтанной электрической активности коры большого
мозга. Онтогенетическое развитие спонтанной электрической
активности было прослежено в отношении медленных колебаний у обезьян и
человека. У обезьян сейчас же после рождения наблюдаются редкие
длительные колебания частотой 2—3 в 1 сек. очень низкой амплитуды.
С этого момента до конца б месяцев происходит прогрессивное
нарастание амплитуды и частоты медленных колебаний. За этот период
времени частота медленных колебаний достигает 7—8 в 1 сек. В
дальнейшем частота еще более увеличивается и в конце 2-го года
достигает 10—12 в 1 сек. У взрослых обезьян наблюдается такая же частота
и еще большая (Кеннард и Нимс, 69; Беритов и Гедеванишвили, 23).
У человека спонтанные медленные колебания впервые появляются
в возрасте от 27г до 6 месяцев Вначале они наступают моментами,
затем становятся постоянными, правильно ритмичными. Ритм
колебаний сперва низкий: 4—5 в 1 сек.; длина волн большая- 60—175 <з. С
возрастом длина волн уменьшается, а ритм нарастает. Нормальный
альфа-ритм взрослого человека 9—12 в 1 сек. достигается в возрасте от
10 до 12 лет. Появление спонтанных биотоков связывают с развитием
функций коры. Они появляются, когда ребенок начинает узнавать и
хватать предметы (Бергер, 3; Линдсли, 56; см. подробно у Шпиль-
берг, 57).
Влияние периферических и корковых раздражений на спонтанную
электрическую активность. Всякое периферическое раздражение
действует на кору большого мозга, усиливая или ослабляя ее, прежде
всего в том участке, куда поступают периферические импульсы со
стороны рецепторов. Периферические импульсы влияют главным образом
на ту активность, которая возникает в соответствующих
воспринимающих полях коры, в проекционных областях. Этот эффект заключается п
усилении электрических колебаний в определенной области коры, с
которой связаны возбужденные афферентные пути: например, в
затылочной области под влиянием зрительных раздражений или в центральной
области, под влиянием тактильных раздражений и болевого (укол)
раздражения противоположной конечности (Бремер и Доу, 44; Экторс, 40;
Саркисов, 41)
Но в определенных условиях за этим локальным эффектом следует
более или іменее длительный ряд медленных колебаний с очень
регулярным ритмом, который не локален и может проявиться в разных
участках коры. Этот эффект повидимому связан с активированием
ассоциативных связей (Демпсей, Морисон Р. и Морисои Б, 7).
Всякое периферическое раздражение производит также
торможение спонтанной электрической активности. Оно проявляется в
соответствующей воспринимающей области после периода повышенной
электрической активности, а в остальных участках коры — в большей или
меньшей степени с самого начала. Аналогичное изменение наступает
при искусственном раздражении коры большого мозга Раздражение
любого участка коры, моторного или немоторного, производит значи-
ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 483
тельное усиление электрической активности в раздраженном участке и
одновременно общее торможение электрической активности в обширных
областях коры (Гедеванишвили, 14) (рис. 215). Угнетение корковой
электрической активности сопровождается угнетением двигательных
бес, Ж В ШШ
Рис. 215. Биотоки коры большого мозга кошки.
Наркоз барбитуратом. Биполярное отведение коры через отверстия с межполюсным
расстоянием 3 мм. Шлейфный осциллограф. А — верхняя кривая — электрокортикограмма левой
затылочной области, средняя = левой двигательной области, нижняя — мехаяо
грамма m. tibialis dex. Раздражается двигательная зона в левом полушарии (И.5 см
РК) — 2 раза. Раздражение вызывает угнетение спонтанной электрической активности в
двигательной зоне и сокращение мышцы. На фоне этого угнетения второе раздражение
вызвало ослабленный двигательный эффект и еще большее угнетение спонтанной активности.
В — верхняя кривая — электрокортикограмма левой затылочной области, средняя —
правой двигательной зоны, нижняя — механограмма m. tibialis dex. Раздражается
двигательная зона в правом полушарии (12.7 см РК). Раздражение вызывает угнетение спонтанной
корковой активности как в раздраженном участке, так и в левой затылочной области; значит
угнетение наступило по всей коре. На мышце сокращение наступало после
раздражения— явление «отдачи» (Гедеванишвили).
»
эффектов, вызываемых раздражением чувствительных нервов или
двигательных участков коры. Это свидетельствует о том, что ослабление
корковой электрической активности при раздражении коры или
чувствительных нервов является следствием понижения корковой
возбудимости или, иначе говоря, общего торможения.
Изменение электрической активности во время наркоза и сна.
Было установлено, что с углублением наркоза электрическая
активность ослабевает, спонтанные колебания наступают периодически
небольшими группами: с углублением наркоза интервалы между ними
нарастают, а сами группы укорачиваются. Частота медленных колебаний
при разном наркозе разная, но она не падает заметно с углублением
наркоза. 'Меняется при углублении наркоза главным образом ампли-
31*
484
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
туда колебаний: она падает вплоть до исчезновения медленных
колебаний (Бичер и 'Мак Доно, 8). В это время всякое периферическое
раздражение приводит к электрической активности строго определенных
участков коры, куда входит возбужденная афферентная система.
Активность прекращается вслед за прекращением раздражения. Здесь
мы имеем дело как бы с пробуждением мозга в одном олределенном
участке. Это наблюдается у обезьян в зрительной области при
освещении глаза, в слуховой области при звуках, в задней центральной
извилине при щипании пальцев и т. д. (Эдриан, 49). При раздражении
чувствительного нерва, например п. saphenus, в ритме до 30 в 1 сек.
в ответ на каждый удар в кожно-мышечной воспринимающей области
(средняя часть позади сигмоидной борозды) наступает небольшой
комплекс колебаний из 2—3 фаз с последующим длительным
отрицательным последствием. Весь комплекс продолжается 140 о. В соседних
участках эти колебания быстро исчезают (Бартлей и Хейнбекер, 35).
Точно так же при электрическом раздражении зрительного нерва
« зрительной области коры мозга возникают сложные комплексы
изменений потенциала, состоящие из быстрых двуфазных и (медленного
трехфазного колебания с общей продолжительностью около 250 о При
этом оказалось, что медленные трехфазного колебания отводятся со
средних слоев коры, где оканчиваются зрительные восходящие пути
/Бишоп и сотр., 37, 38).
Во время нормального сна у животных (кошки) основная
электрическая активность коры больших полушарий снижается: электрические
потенциалы высокого ритма от 50 до 70 в 1 сек. временами выпадают.
Взамен их возникают длительные медленные потенциалы 2—3 в 1 сек.
Иногда быстрые потенциалы высокого ритма располагаются на фоне
длительных. Пробуждение животного вызывает исчезновение
длительных потенциалов и появление потенциалов высокого роста (Рейнбергер
и Джаспер, 79; Цкипуридзе, 80) (рис. 216). Эти длительные медленные
потенциалы впервые возникают или лучше всего выступают в
зрительной области (Кларк и Уорд, 81), а также в моторной (Цкипуридзе, 80).
То же самое наблюдалось на изолированном головном мозгу кошки. Во
время сна корковые потенциалы наступали с частотой 2—4 в 1 сек.
{Бремер, 82). Все эти наблюдения на животных производились с
помощью хронических (вживленных) электродов.
Во время сна у человека электрическая активность коры большого
мозга меняется очень своеобразно. Альфа-волны в затылочных и
теменных долях первоначально в период дремотного состояния несколько
усиливаются; с наступлением сна, наоборот, они становятся ниже
амплитудой и наступают более короткими группами. Промежутки
затишья постепенно увеличиваются. Наконец альфа-ритм затухает.
Нерегулярные медленные колебания основной электрической
активности начинают усиливаться еще в дремотном состоянии. С
наступлением сна они еще более усиливаются, особенно в центрально-лобных
ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШОГО МОЗГ 4 485
областях. Моментами эти медленные колебания наступают в
нерегулярном ритме 14, 16, 18, 22 в 1 сек. (рис. 217).
Иногда альфа-ритм затухает без одновременного усиления
основной электрической активности. При этом не регистрируются ни
медленные колебания потенциала, ни быстрые бета-волны. Период затишья
длится то короткое время, то долгое, и после него основная
электрическая активность івсегда усиливается.
Рис. 216. Олектрокортикограмма кошки.
А—во время сна, В—сейчас же после пробуждения. Отведение от area striata
вживленными электродами (Цкипуридзе).
Во время глубокого сна появляются медленные дельта-подобные
волны. Они наступают то одиночками, то группами, и притом более или
менее синхронно как в симметричных, так и в несимметричных
участках коры. На фоне их располагаются более кратковременные
колебания основной электрической активности (рис. 217). Но синхронизм
дельта-подобных колебаний, подобно регулярным колебаниям
основной активности во время сна, выражен слабее, чем у медленных волн
основной активности или у альфа-волн в бодрствующем состоянии
'(Бакурадзе и Нарикашвили, 10; Дэвис и сотр., 9, 47 и др.).
Характерно, что в периоде глубокого сна, перед пробуждением в-
определенных областях коры часто возникают сильные двуфазные
колебания, так называемые, судорожные разряды (Бакурадзе и
Нарикашвили, 10). Видимо это всегда бывает при судорожных
вздрагиваниях спящего.
Слабые световые и звуковые раздражения, не вызывающие
пробуждения, в большинстве случаев не влияют заметно на электрическую
картину мозга. Лишь длительные дельта-подобные волны основной
активности под влиянием звука (шума) в некоторых случаях
изменяются: они ослабевают и часто затухают, т. е. во время сна шум
действует на дельта-подобные волны, как на альфа-волны в бодрствующем
состоянии (Бакурадзе и Нарикашвили, 10).
Усиление потенциалов основной электрической активности с
наступлением сна следует рассматривать как результат улучшения условий
синхронизации в деятельности собственных нервных кругов коры. Это
обусловливается выпадением обычной афферентной импульсации
через промежуточный мозг, ибо во время сна прежде всего угнетается
деятельность промежуточного імозга, как передаточной станции для
4S6
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
периферических импульсов (Рожанский, 77). Благодаря устранению
периферических влияний нервные круги коры, активируемые в
результате самовозбуждения, так влияют друг на друга по ассоциативным
и коммиссуральным связям, что развивают более синхронную работу,
чем это бывает в бодрствующем состоянии.
Рис. 217. Электрическая активность коры большого мозга человека во время сна.
Запись шлейфным осциллографом. В каждой записи верхняя кривая от затылочно-теменных долей,
а нижняя — от центральпо-лобных. А — перед сном; до закрытия глаз слабые нерегулярные
колебания основной электрической активности; с закрытием глаз (зак.) появляется альфа-ритм:
сильно—в теменно-затылочной области, слабо — в центрально-лобной. В — дремотное состояние через
?0 мин. после разрешения спать; в обоих участках основная активность усилена, моментами ровный
ритм 18—22 в 1 сек.; альфа-волны ослаблены. С — через 30 мин. от начала засыпания; спит, на зов
не реагирует. Основная электрическая активность еще больше усилена, особенно в лобной области,
местами ровный ритм 14—16—22 в 1 сек.; альфа-волны отсутствуют. П — через 1 ч. 45 м. храпит.
Альфа-волны отсутствуют; основная электрическая активность еще более усилена. Временами
наступают дельта-подобные волны длительностью в 0.25—0.4 сек., на которых наложены более
слабые потенциалы короткой продолжительности в 0.1—0.05 сек. Эти волны значительно сильнее
в лобной области, чем в затылочяо-теменной (Бакурадзе и Нарикашвили).
В некоторых участках коры імозга синхронизация деятельности
нервных кругов настолько повышается, что электрический разряд
уподобляется судорожному. Очевидно, в случае возникновения такого
разряда в двигательных областях коры мозга наблюдается судорожное
вздрагивание спящего.
Первоначальное усиление альфа-волн в самом начале дремотного
состояния объясняется закрытием и затемнением глаз, которым всегда
начинается дремотное состояние. Главную роль играет происходящее
при этом устранение световых и других периферических раздражений.
Оно способствует лучшей синхронизации деятельности длинных
нервных кругов, производящих альфа-волны. Последующее ослабление и
исчезновение альфа-волн должно зависеть от значительного угнетения
ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 487
деятельности промежуточного мозга. При этом перестают работать
длинные нервные круги между корой и промежуточным мозгом,
обусловливающие альфа-волны.
Явление затишья после исчезновения альфа-ритма, по-видимому,
происходит главным образом потому, что в результате полного
выпадения периферических влияний через промежуточный мозг корковая
возбудимость заметно понижается. Это обусловливает такое снижение
электрической активности, что она не может быть уловлена при
отведении от кожной поверхности черепа. Затишье ни в коем случае не
оеначает, что во время сна кора перестает быть активной. Мы знаем
ло наблюдениям на обезьянах, что даже аіри глубоком сне, когда
раздражение кожи не производит рефлекторных реакций, обнаженная
кора продолжает быть спонтанно активной. Наблюдается только
заметное ослабление спонтанной активности (Беритов и Гедеванишви-
ли, 23).
Электрическая активность и функциональное состояние коры
большого мозга. Когда функциональное состояние коры улучшается и
ее возбудимость повышается, тогда спонтанная электрическая
активность усиливается: повышается амплитуда медленных колебаний
основной активности, повышается и амплитуда быстрых колебаний
(бета). Та же картина наблюдается при повышении возбудимости от
других причин, например, при отравлении эзерином коры у животных
(Беритов, Брегадзе и Цкипуридзе, 13). То же наблюдается у человека
при гипервентиляции, при абсцессах мозга. У человека наряду с
усилением колебаний основной активности усиливаются также альфа-волны,
но при этом происходит уменьшение той области, где эти последние
наступают. Альфа-волны начинают ограничиваться теменной областью,
доминирующей областью для альфа-волн. Ритм альфа-волн при этом
меняется очень мало: он повышается, но не более чем на 1—2 цикла,
он не становится выше, чем 12—13 колебаний в 1 сек. Когда
возбудимость повышается особенно сильно, как это иногда наблюдается при
абсцессах в коре, альфа-волны совсем исчезают (Беритов и сотр.,
21,29).
Когда же функциональное состояние коры большого мозга
животных ухудшается, корковая возбудимость падает, тогда спонтанная
электрическая активность ослабевает. Ослабевают медленные волны
основной активности, а также бета-волны. Так бывает, например, при
охлаждении коры, при отравлении кокаином (Беритов и сотр., 19), при
искусственной задержке кровообращения в мозгу путем перевязки
сосудов (Правдич-Неминский, 5; Андреев и Шпильберг, 57). Так, в
коре большого мозга кошки после остановки кровообращения сначала
исчезают быстрые волны, а потом через 20 сек. исчезает вся
электрическая активность, причем в коре она исчезает раньше, чем в других
отделах головного мозга (Джаспер, 66). То же бывает при
искусственной задержке дыхания (Корнмюллер, 43).
488 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
У человека тоже наблюдается ослабление корковой электрической
активности при ухудшении функционального состояния, но тут при
некотором малом понижении возбудимости происходит усиление альфа-
волн; при этом расширяется область их наступления; они начинают
наступать с большой регулярностью в височных и центральных долях.
Бели же возбудимость понижается еще в большей степени, тогда
медленные колебания основной активности и бета-волны понижаются еще
сильнее, но теперь ослабевают и альфа-волны; последние часто
сливаются, образуя эта-волны, и постепенно ограничиваются т?менно-заты-
лочной областью. Ритм меняется мало; он уменьшается на 1—2 цикла,
но не становится меньше 9—8 в 1 сек. При особенно сильном
понижении возбудимости альфа-волны совсем исчезают, или сохраняются их
следы в теменных областях. Так бывает, например, после повреждения
головного мозга воздушной ударной волной (при взрывах), а также
при огнестрельных черепных ранениях в отсутствии гнойного
воспаления (Беритов и сотр., 22, 29).
Нередко, одновременно с ослаблением альфа-волн в теменно-заты-
лочных областях, происходит заметное .усиление медленных волн
основной активности; особенно сильно это проявляется в центральных
долях и в задней половине лобных, как это наблюдается во время
нормального сна. Так бывает часто при черепных ранениях (Беритов, 29).
Характерно меняется отношение закрытия и затемнения глаз к
электрической активности коры. Как при сильном повышении
корковой электрической активности, так и при сильном понижении ее,
закрытие и затемнение глаз у человека перестают благоприятствовать
альфа-ритму или благоприятствуют очень слабо, и притом с большим
скрытым периодом. Так, например, на рис. 218 приводится случай
пониженной электрической активности: альфа-волны совершенно
отсутствуют; их не вызывает и закрытие глаз. Это указывает на то, что
функциональное состояние коры значительно изменено и что
автоматическая, индивидуально-приобретенная деятельность в отношении
альфа-волн понижена очень сильно. Было установлено, что в такой
момент у человека страдает память, наблюдается некоторая
психическая заторможенность (Беритов и сотр., 30).
Характерно, что исчезновение альфа-ритма и снижение памяти у
человека наблюдается при повреждении теменной области в одном
полушарии. Очевидно, теменные области обоих полушарий так тесно
связаны коммиссуральными путями, что повреждение теменной области
в одном полушарии ведет к нарушению функциональной деятельности
теменных областей в обоих полушариях.
Спонтанная электрическая активность патологического характера.
Электрическая активность коры большого мозга может меняться в
смысле ослабления или усиления, без нарушения основного характера
нормальной электрической активности. В этом случае мы имеем дело
с улучшением или ухудшением нормального физиологического состоя-
ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШОГО МОЗГА
489
ния коры. Но спонтанная электрическая активность коры мозга может
принять совершенно особый характер. В самой легкой форме это будет
заключаться в появлении более сильных двуфазных разрядов —
быстрого колебания потенциала вначале и последующего медленного
колебания в противоположном направлении. Обе фазы вместе могут
Рис. 218. Пониженная электрическая активность коры большого мозга
человека-эпилептика.
Регистрация корковых биотоков путем отведения от кожной поверхности черепа в шлейфный
осциллограф. А — электроэнцефалограммы от затылочной области; В — ЭЭГ от теменной области. В обоих
случаях глаза закрыты. Альфа-волны отсутствуют, но хорошо выступает основная электрическая
активность с медленными и быстрыми колебаниями потенциала.
продолжаться 0.05—0.2 сек. После каждого такого разряда
спонтанная активность ослабевает или исчезает на разный срок, измеряемый
десятыми долями секунды. Эти ненормальные разряды могут
наступить изолированно в одном участке как спонтанно, так и в ответ на
внешние раздражения. Так бывает, например, у животных при легком
локальном отравлении коры большого мозга ацетилхолином или
стрихнином (Дюссер де Баренн и сотр., 48; Беритов и сотр., 13, 23)
(рис. 219). Так как в случае отравления двигательной зоны эти
двуфазные разряды сопровождаются судорожным подергиванием
соответствующей конечности, то их принято называть судорожными
(Корн'мюллер, 43). Но фактически они бывают в любом участке коры
при достаточно высокой возбудимости, и когда они возникают
изолированно ів немоторной зоне, то не сопровождаются судорожным
сокращением мышц.
При более или менее значительном локальном стрихнинном или
ацетилхолиновом отравлении судорожные электрические разряды
наступают длительными сериями. Высокая возбудимость отравленного
участка является основным условием для наступления этих разрядов.
Конечно, при этом условии они возникают как спонтанно, в результате
самовозбуждения нервных кругов, так и под влиянием
периферических импульсов. Но при сильных периферических раздражениях или
490 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
при сильном раздражении самой коры судорожные разряды наступают и
при нормальной или даже пониженной корковой возбудимости (Гедева-
нишвили, Г4). При сильном раздражении судорожные разряды
появляются длительными сериями в правильном ритме от 4 до 20 в 1 сек.
Так бывает и спонтанно, если возбудимость была достаточно высокая
(Корнмюллер, 43, Беритов и сотр., 13, 23), а также при эпилепсии или
при наличии инородного тела в коре или под корой при черепных
ранениях (Беритов и сотр., 21, 22, 29).
Рис. 219. Одиночные судорожные разряды коры большого мозга обезьяны
Macaccus rhesus.
Локальное отравление ацетилхолином в области отводящих электродов. Начальный
период отравления. Верхняя кривая (Гл.) при биполярном отведении из нижних
слоев коры мозга; нижняя кривая (Пов.)—то же от поверхности мозга на том же уровне.
Межполюсное расстояние в обоих случаях 5 мм. В середине записи сильные
судорожные разряды наступили одновременно в глубоких и верхних слоях коры. На краях
и а и и си небольшие судорожные разряды возникают раздельно то в глубоких, то в
поверхностных слоях. Внизу время — по 0.02 сек. (Беритов и Гедевани).
Характерно, что когда очаг раздражения находится в коре,
судорожные разряды являются локальными. Так бывает, например, при
электрическом раздражении небольшого участка коры и при локальном
отравлении небольшого участка коры такими активными веществами,
как ацетилхолин и стрихнин. Было установлено, что при электрическом
раздражении или локальном отравлении судорожные разряды не
наблюдаются даже на расстоянии нескольких миллиметров от первично
возбуждаемого участка. Так, например, при отравлении двигательной
зоны передней ноги эти разряды не отводятся от двигательных зон
задней ноги. Они отсутствуют даже в симметричных участках другого
полушария (Дюссер де Баренн, 48; Беритов и сотр., 13, 23). Более
того, в начальной стадии отравления ацетилхолином судорожные
разряды імогут возникать изолированно то в поверхностных, то в глубоких
слоях коры отравленного участка (рис. 219).
В случае отравления двух участков коры мозга в каждом из них
могут наступить судорожные разряды своим собственным ритмом, без
заметного взаимодействия. Но во всех случаях, если судорожные
разряды наступают в одном участке, ів соседних участках того же
полушария, а также в симметричном участке другого полушария,
происходит торможение спонтанной электрической активности как быстрых,
так и медленных колебаний.
Судорожные разряды, вызванные фарадическим раздражением
коры, можно поддержать на долгое время раздражением определенных
ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 491
участков коры отдельными электрическими ударами и отдельными
эфферентными импульсами в ритме 1-—4 в Г сек. (Розенблют и Кэннон,
70). Это происходит благодаря повышению возбудимости в
раздраженном участке. Этот участок первым долгом вовлекается в ответ на
новое раздражение коры отдельными ударами. Было установлено,
что передача судорожной деятельности от одного полушария к другому
происходит по длинным комиссуральным нервным путям, а
распространение их в пределах одного полушария — по коротким нервным
связям: от раздраженных клеток — к соседним и так далее, все дальше
и дальше от раздраженного участка (Розенблют, Бонд и Кэннон, 71).
Судорожные разряды в некоторых случаях наступают во всей
коре, и притом совершенно одновременно, но интенсивность разрядов
может быть в одном полушарии больше, чем в другом, или даже они
могут наступить в одном полушарии при отсутствии их ів другом. Это,
например, может случиться при общем повышении возбудимости в
коре, но при наличии очага раздражения вне коры в одной половине
ствола мозга. Усиленные импульсы, исходящие из этого очага, цости
гают коры более или менее одновременно, но действие их будет
сильнее в одном полушарии, чем в другом. Благодаря этому судорожные
разряды будут проявляться в одном полушарии сильнее, чем в другом
(рис. 220).
Происхождение судорожных разрядов объясняется усиленным
возбуждением большого комплекса нервных кругов сначала в
раздраженном или отравленном участке, а затем и в соседних участках. Первая
фаза каждого разряда зависит от возбуждения первично возбужденных
нервных кругов (отравленного участка в случае самовозбуждения,
¦раздраженного участка в случае раздражения), а вторая фаза — в
противоположном направлении: от возбуждения ближайших нервных кр}-
гов в соседних участках. Усиленная иррадиация импульсов
возбуждения из нервных кругов будет производить усиленное активирование
нейропиля как в первичном возбужденном, так и в ближайших
участках. Активирование нейропиля в последних участках будет
начинаться и кончаться позднее, чем в первично возбужденном, поэтому при
отведении первично возбужденного участка нейропильный ток сначала
должен протекать в одном направлении, а потом в другом. По этой
причине, когда судорожные разряды возникают поодиночке, вслед за
второй быстрой фазой от возбуждения нервных кругов ближайших
участков наступает медленная фаза того же направления от
активирования соответствующего им нейропильного субстрата.
Угнетение спонтанных биотоков в соседних участках во время
судорожных разрядов, должно быть, зависит от того же медленного биотока,
возникающего при активировании нейропиля. Этот ток, распространяясь
физически из судорожно-возбужденных участков, должен действовать
анэлектротонически на соседние участки и тем самым вызвать в них
угнетение спонтанной активности.
492
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
При сильном понижении корковой возбудимости спонтанная электри*
ческая активность также существенно меняется: в этом случае
возникают одиночками или сериями медленные колебания большей продол-
жительности, чем обычные медленные колебания основной электрической
активности, или альфа-волны, так называемые дельта-волны с
продолжительностью от 0.2 до 0.5 сек. и больше. Это бывает при разных забо-
Рис. 220. Судорожные биотоки коры большого мозга у эпилептика.
Биотоки отводятся от симметричных участков затылочной области. Один пластинчатый
электрод (сетка) лежит на затылке, а другой (земля) на мочке уха. Глаза сначала
вакрыты, затем открываются. Верхняя запись от правого полушария дает более
сильные разряды, и притом более часто (С), чем от левого полушария. Когда судорожный
разряд справа наступает сравнительно сильно, тогда он проявляется и слева, и притом
совершенно синхронно. Между судорожными разрядами синхронные дельта-волны.
Альфа-волны отсутствуют. Бета-волны сильно ослаблены. Такая же картина
электрической активности наблюдалась у этого эпилептика во всех других областях коры
(Беритов, Бакурадзе и Дзидзишвили).
леваниях мозга. Если возбудимость понижена во всей коре, тогда мы
имеем дельта-волны во всей коре. Так бывает при повреждении
головного мозга взрывной волной (Беритов и сотр., 30). Когда же
возбудимость понижена локально в одном участке, тогда и дельта-волны
возникают локально ів этом участке; кругом этого участка ів нормальных
участках электрическая активность может быть более или менее
нормальна. Так бывает при локальном повреждении мозга при черепных
ранениях, при опухолях мозга и т. п. (Вальтер, 31; Беритов и сотр., 22, 29).
Дельта-волны бывают у нормального человека во время глубокого
сна (Блэк и Жерард, 33; Бакурадзе и Нарикашвили, 10). Дельта-волны
часто возникают после ряда судорожных волн в связи с истощением
коры большого мозга. Это, например, хорошо видно на рис. 220, где
дельта-волны наступают в ритме 4—5 в 1 сек. В том случае, когда
дельта-волны наступают в разных отделах коры большого мозга, они чаще
всего являются вполне синхронными. В некоторых случаях
дельта-волны как будто образовались путем слияния альфа-волн или медленных
колебаний основной активности; так бывает, например, вовремя сна. Но
обычно при патологическом состоянии коры большого мозга
дельта-волны возникают самостоятельно (Беритов и сотр., 20).
Дельта-волны всецело обусловливаются сильным понижением
возбудимости и действием какого-либо раздражающего агента в самой коре
или под корой. Когда дельта-волны являются синхронными, возбуждаю-
ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 493
щие их импульсы, должно быть, возникают не в коре, а где-либо иод
корой,— например, в промежуточном мозгу, где очаг раздражения может
захватить обширные пути, идущие к коре мозга.
Историческая справка о биоэлектрических токах коры большого мозга.
Биоэлектрические токи были открыты в коре мозга в 1876 г. К это ном. Оиі установил, что
токи возбуждения коры могут быть отведены от всей ее поверхности. Если какой-
либо участок мозга приходит в деятельное состояние, он становится
электроотрицательным по отношению к другим недеятельным участкам. При освещения глаза,
например, отрицательной становится затылочная область; жевание и движение головы
ьызываіот электроотрицательность в лобных долях. Почти одновременно с Кэтоном
этот вопрос разрабатывал В. Я. Данилевский и пришел к аналогичным
результатам.
Первая попытка регистрации биоэлектрических токов коры больших полушарий
была осуществлена Прявдич-Неминским с помощью струнного гальванометра
(1925). Он установил, что при отведении от моторной и оптической коры животных
получаются спонтанные колебания потенциала двоякого рода: медленные волны в
низком ритме, 11—16 в 1 сек.— волны первого порядка, на которые наслаивались волны
меньшей амплитуды и меньшей продолжительности, и притом более высокого ритма,
20—32 в 1 сек.— волны второго порядка. При раздражении седалищного нерва
характер электрических колебаний несколько изменялся.
На основании своих наблюдений Правдич-Неминский заключил, что кора
большого мозга обладает специфической способностью к ритмической деятельности.
Ьто было первое обоснованное заявление об автоматической ритмической
деятельности коры мозга. Аналогичные результаты были получены Фишером (6). Он
отводил в струнный гальванометр обнаженную кору у разных животных (кролики, кошки,
собаки, обезьяны). У всех он наблюдал спонтанные редкие колебания потенциала при
самом тщательном исключении раздражения органов чувств. Они не были одинаковы
при отведении от разных участков. Периферическое раздражение влияло на них.
Так, освещение одного глаза вызывало в area striata противоположной стороны
характерные токи возбуждения и в начале освещения и по окончании его. То же
наблюдалось от воздействия разных звуков, а также от механического раздражения кожи
при отведении определенных участков коры мозга (Фишер, 6; Бун-Ичи Хасама, 1, 2).
Точно такие же спонтанные биоэлектрические токи двоякого ритма наблюдались в
коре переднего мозга у птиц (Бремер и сотр., 46). Существенный перелом получился
в этой исследовательской работе, когда стали применять осциллографы. Первое
подробное осциллографическое исследование биотоков коры большого мозга было
произведено Эдрианом иМэтьюсом (4) при отведении маленьких участков коры
у животных. Эти авторы не только зарегистрировали биотоки мозговой коры, но и
подробно изучили их происхождение. Например, они установили, что кора большого
мозга обнаруживает биоэлектрические колебания все время, даже при глубоком
наркозе, когда периферические раздражения не дают рефлекторных реакций.
Биоэлектрические явления в коре большого мозга человека были обнаружены
Бергером в 1929 г. (3). Он наблюдал у людей регулярные электрические волны,
когда глаза испытуемых были закрыты и внимание ничем не было занято. Полученные
при этом кривые были названы им электроэнцефалограммами. Бергер
отметил на электроэнцефалограмме человека двоякого рода волны: одни — большие
медленные, с правильным ритмом около 10 в 1 сек., так называемые альфа-волны,
и другие, более мелкие и частые, так называемые бета-иолны.
Практикум
Регистрация биотоков коры большого мозга человека. Корковые биотоки у
человека обычно отводятся от кожной поверхности черепа. Лучше всего отводить бипо
494
ОБЩАЯ ФИЗИОЛОГИЯ КОГЫ БОЛЬШОГО МОЗГА
лярно с межполюсным расстоянием в 5—8 см. Серебряные пластинчатые электроды в
64 мм2, обтянутые ватой, смоченные в і0% растворе хлористого натрия с примесью-
небольшого количества щелочи, прикладываются к определенным участкам черепа.
Кожа с волосами в этих участках предварительно протирается смесью спирта и
эфира для очистки от жира.
М. front.
N. supraorb.
N. auric-temp.
М. masset.
Рис. 221. Расположение мышц и нервов на голове и
электродов для продольного биполярного отведения.
Точки обозначают местоположение отводящих электродов.
Они расположены на расстоянии 3 см от медиальной линии.
Точка А — на задней границе затылочной области на linea
nuchae; В — на передней границе затылочной области; С — на
передней границе теменной области; D — на передней границе
прецентральной извилины; Е — на середине лобной области;
F — на передней границе лобной области; Я и G — в височной
области. Между соседними точками —5—6 см. При наложении
электродов на точки А и В отводится затылочная область, от
В ж С — теменная, С и D — центральная область
(постцентральная и прецентральная извилины), D и Е —задняя
половина лобной области, Е и F — передняя половина лобной
области; от Я и G — височная область.
N. auric.
Отведение может происходить как продольно, параллельно к медиальной линия»
так и поперечно. При продольном отведении электроды можно расположить на
расстоянии 3 см от медиальной линии с межполюсным расстоянием по 5—6 см. Как
показано на рис. 221, при таком продольном отведении от точек А и В отводится
затылочная область, от В и С — теменная, от С и D — центральная (прецентральная и
постцентральная извилины), от D и Е — задняя половина лобной области и от Е и F —
передняя іполовин-а лобной. Кроме того отводится височная область несколько ©ыше
слухового отверстия от G и Я. Для поперечного отведения электроды располагаются
иначе: один ряд электродов располагается на расстоянии 1 см от медиальной линии,
а другой — на 1—2 см выше височной линии. Точки поперечного отведения и их
отношение к коре большого мозга даны на рис. 222.
При отведении затылочной и височной областей, а также передней половины
лобной области вместе с корковыми биотоками будут отводиться и мышечные токи от
подлежащих мышц. Поэтому нельзя получить корковые биотоки в чистом виде из
этих участков- мышечные биотски высокого ритма обязательно будут примешиваться
ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 493
к корковым биотокам. Для наблюдения корковых биотоков в чистом виде лучше-
всего отводить продольно около медиальной линии из теменно-центральной области..
Подопытные лица находятся в экранированной железом кабине, для
предохранения главным, образом от переменного городского тока. Кабина заземлена.
Подопытное лицо лежит на спине на
диване. Ему дается инструкция
совершенно расслабить мускулатуру»
лежать покойно в самом удобном
для него положении и, по
возможности, ни о чем не думать.
Внутри кабины находятся
предварительные усилители со
своими аккумуляторами и
анодными батареями. Снаружи кабины
находятся конечные усилители и
регистрирующий прибор; катодный
или шлейфный осциллограф. За
поведением испытуемого во
время опыта наблюдение ведется
через щель в стенке кабины.
Кабина освещается
электрической лампой в 25 ватт, которая
находится над крышкой кабинки^
и свет проникает внутрь через
щель, покрытую металлической
сеткой. Так как некоторая доля
городского тока все же может
проникнуть внутрь кабины, то
необходимо, чтобы в кабине все
провода, соединяющие
аккумуляторы и анодные батареи, а также отводящие электроды с усилителями были
заэкранированы.
Подопытное лицо лежит в освещенной кабинке с открытыми глазами и
закрывает и открывает глаза по приказу, спокойно, без напряжения мышц на лице.
Для обследования характера корковой электрической активности от каждого
субъекта поочередно отводятся все области, как показано на рис. 221 и 222. После
визуального наблюдения осциллограмма каждой области фотографируется. Для съемки,
медленных волн при достаточно малой скорости вращения фотографического
аппарата, например 3—4 см в 1 сек. Для съемки быстрых бета-волн необходимо брать,
большую скорость = 12—24 см в 1 сек. Каждый раз регистрация происходит при
открытых и закрытых глазах. При медленной съемке это делается без остановки
аппарата. Через 3—4 сек. после начала съемки дается приказ закрыть глаза; спустя
4—5 сек. дается новый приказ открыть глаза, и через 3—4 сек. после этого съемка.
кончается.
У людей высокой возбудимости при отведении мышечных участков вместе с
корковыми биотоками непременно будут отводиться и мышечные биотоки. Путем
инструкции расслабить мышцы лица: не хмуриться, не зажимать челюстей, не улыбаться,—
можно добиться такого значительного ослабления этих токов, что они не помешают
наблюдению корковых биотоков. Иногда мышечные участки дают большие быстрые
колебания продолжительностью около 0.005—0.01 сек. с довольно ровным ритмом
10—20 в 1 сек. Эти пикообразные колебания происходят от рефлекторного
возбуждения какого-либо мышечного пучка, лежащего под одним из электродов. Стоит только
переместить электрод на 1—2 см, чтобы они исчезли.
Иногда обнаруживаются чрезвычайно быстрые иглообразные колебания элек?
Рис. 222. Расположение электродов при
поперечном отведении и отношении их к долям-
большого мозга.
Am — area striata; Ar—на границе аг. praeoccipitalis;
и аг. occipito-temporalis; Вт — аг. praeoccipitalis;
В — на границе аг. angularis и аг. tempor. sup.;
Cm—на гравице аг. parietalis и ar. praeparietalis^
С—на границе ar. ar. supramarojinalis, tempor. sup.;
u temp, transversapost.; Dm — ar. praecentralis gi-
gantopyramidalis; D'—нижний край ar. postcentralis;
Em — на границе ar. front intermedia u ar. front,
granularis; E'—ar. operculars,- Fm— ar. praefrontalis;
F', — ar. triangularis. Между соответствующими
электродами около 5 см. Кроме того, два электрода G иг
Я на височной области.
496
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
трического тока,— быстрее, чем 0.001 сек. Они наступают залпами совершенно
одинаковой высоты и исчезают при передвижении электродов на несколько миллиметров.
Вероятно, они возникают от возбуждения кожных чувствительных нервов.
При отведении лобной области обычно вместе с биотоками отводятся и биотоки
глазных мышц и век, если они сокращаются. Поэтому при съемке этих областей
нельзя двигать глазами и мигать.
Регистрация биотоков коры большого мозга кошки. Под эфирным наркозом
обнажается одно полушарие большого мозга: лобная область с сигмовидной бороздой,
теменная и затылочная области. Твердую мозговую оболочку лучше оставить, тогда
кора меньше повреждается. В лобную кость вставляется маленький стерженек с
винтовой нарезкой и на нем устанавливаются обычные платиновые электроды. Концы
электродов с межполюсным расстоянием 5—8 мм прикладываются к коре мозга
поочередно то в одной, то в другой области для установления характера спонтанной
электрической активности.
Регистрация корковых биотоков происходит катодным или шлейфным
осциллографом через усилитель переменного тока. Усиление должно быть в миллион раз
и выше, ибо отводимые биотоки измеряются несколькими десятками микровольт.
Биотоки будут двоякого рода: медленные—в ритме 15—30 в 1 сек. и быстрые — от 100
до 120 в 1 сек. и выше.
При отведении корковых биотоков в затылочной области испытывается
раздражение глаз светом. В ответ на раздражение спонтанная активность усилится; заметно
действует и звуковое раздражение. При этих опытах наркоз должен быть очень
слаб. Если отводится не зрительная область коры, тогда то же зрительное
раздражение угнетает спонтанную электрическую активность.
При отведении сигмовидной извилины в двигательной зоне левого полушария
для правой передней конечности раздражение чувствительного нерва или кожи на
этой конечности вызовет усиленные колебания потенциала в отводимой двигательной
зоне.
Для того чтобы проследить влияние черепного покрова мозга кожи и кости на
интенсивность отводимых корковых биотоков, следует отводить кору то
непосредственно, то через черепную кость и кожу.
Наблюдение судорожных электрических разрядов на кошке. Сначала наблюдают
путем катодного или шлейфного осциллографа обычную картину из быстрых и
медленных колебаний спонтанной электрической активности, затем отравляют
стрихнином отведенный участок коры путем приложения кусочков фильтровальной бумаги,
намоченных в 1% растворе стрихнша. Через 1—2 мин. спонтаншя электрическая
активность временами исчезает и заменяется серией судоіроіжных разрядов. Если
отравление производится в двигательной зоне какой-либо конечности, то эти судорожные
разряды будут совпадать с подергиваниями данной конечности. При отведении других
неотравленных участков судорожные разряды или отсутствуют, или значительно
ослабевают. Обычно они наблюдаются в симметричных участках другого полушария
или в нескольких миллиметрах от отравленного участка того же полушария.
Такие же более или менее локальные судорожные разряды можно вызвать в коре
сильным электрическим раздражением. В области сигмовидной извилины эти разряды
сопровождаются судорожным движением конечностей. В теменно-затылочной области
они наступают без двигательных судорог.
Литература
1. Bun-Ichi Hasama, Pfl. Arch., 234, 748, 1934.
2. Bun-Ichi Hasama, Pfl. Arch., 236, 36, 4935.
3. В e г g e г Н., Arch. Psych, u. Nervenkr., 87, 527, 1929; 94, 16, 1931; 97, 6, 1932.
4. A d r i a n E. D. a. M a 11 h e w s B. H. C., J. Physiol., 81, 440, 1934.
5. Prawdicz-Neminski W.W., Pfl. Arch., 209, 364, 1925; 210, 223, 1925.
6. Fischer M., Pfl. Arch., 233, 738, 1934; 230, 167, 1932.
ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 497
?. Dempsey Е. W., Morison R. S. a. Morison В. R., Am. J. Physiol.,
131, 718, 1940.
8. BeceherH. К. а. М с DonoughF. К., J. Neurophysiol., 2, 289, 1939.
9. L о о m i s A. L., H а г v ? у Е. N. a. Hobart G. A., J. Neurophysiol., I,
414, 1938.
40. БакурадзеА. иНарикашвили С, Тр. Ин-та физиол. им.
Бериташвили, 6, 377, 1945.
11. Бериташвили И., Брегадзе А. и Цкипуридзе Л., Сообщ. Акад.
Наук Груз. ССР, 3, № 2, 1942.
12. Беритов И., Тр. Ин-та физиологии им. Бериташвили, б, 193, 1943.
13. Беритов И., Брегадзе А. иЦкипуридзе Л., Тр. Ин-та физиол.
им. Бериташвили, б, 241, 1943.
14. Гедеванишвили Д., Тр. Ин-та физиол. им. Бериташвили, б, 268, 1943
45. Беритов И., БакурадзеА. иДзидзишвили Н., Тр. Ин-та физиол.
им. Бериташвили, б, 283, 1943.
16. Беритов И., Тр. Ин-та физиол. им. Бериташвили, б, 327, 1943.
17. Беритов И., Гедеванишвили Д. и Воробьев А., Тр. Ин-та
физиол. им. Бериташвили, б, 358, 1943.
18. Беритов И. и Воробьев А., Тр. Ин-та физиол. им. Бериташвили, б,
369, 1943.
19. Воробьев А. иДзидзишвили Н., Тр. Ин-та физиол. им. Бериташвили,
б, 387, 1943.
20. Беритов И., БакурадзеА. иДзидзишвили Н., Тр. Ин-та физиол.
им. Бериташвили, б, 405, 1943.
21. Беритов И., Бакурадзе Н. иДзидзишвили Н., Тр. Ин-та физиол.
им. Бериташвили, 5, 473, 1943.
22. Беритов И., БакурадзеА. иДзидзишвили Н., Тр. Ин-та физиол.
им. Бериташвили, б, 495, 1943.
23. Беритов И. и Гедеванишвили Д., Тр. Ин-та физиол. им.
Бериташвили, 6, 273, 1945.
24. Dempsey Е. W. a. Morison R. S., Am. J. Physiol., 138, 283, 1943.
25. Bremer Fr., C. R. Foe. Biol., 127, 383, 1938; 118, 1235, 1935.
26. M о r i s о n R. S. a. D e m p s e у E. W., Am. J. Physiol., 138, 297, 1943.
27. H о a g 1 a n d H., Am. J. Physiol., 116, 604, 1936.
28. R u b i n M. А., С о h e n L. H. a. H о a g 1 a n d H., Endocrinol., 21, 536, 1937.
29. Беритов И., Тр. Ин-та физиол. им. Бериташвили, 6, 319, 1945.
30. Беритов И., Ананьев Б. и Воробьев А., Юбил. сборник,
посвященный проф. Узнадзе, 1945, стр. 1.
31. Walter W. G., Lancet, Nr. 2, 305, 1936.
32. A d г і a n E. D. a. Yamagiwa К., Brain, 68, 323, 1935.
33. BlakeH.a. Gerard R. W., Am. J. Physiol., 119, 692, 1937.
34. Rhei nbe rge r M. B. a. Jasper H. H., Am. J. Physiol., 119, 186, 1937.
35. В а г t h 1 e у S. H. a. H e i n b e с k e r P., Am. J. Physiol., 121, 21, 1938.
36. Pen fie Id W. a. Boldrey E., Brain, 60, 389, 1937.
37. В ar thle у S. H., O1 Lear у J. a. Bishop G. H., Am. J. Physiol., 120, 604,
1937.
38. BishopG.H.a. O'Leary J., Am. J. Physiol., 117, 292, 1936.
39. A d r i a n E., J. Physiol., 83, 127, 1936; Физиол. журн. СССР, 24, 6, 1938
40. Ее tors L., Arch internal, physiol., 45, 267, 1936.
41. С a p к и с о в С. А., Тр. Ин-та мозга, вып. 3—4, 443, 1938.
42. ЛивановМ. Н., Тр. Ин-та мозга, вып. 3—4, 487, 1938.
43. Kopnmuller А. Е., Die bioelectrischen Erscheinungen d. Hirnnndfelder etc.
Leipzig, 1937.
44. В r e m e r F. a. D о w R. S., J. Neurophysiol., 2, 308, 1939.
45. S pie ge 1 E. A., Am. J. Physiol., 118, 569, 1937.
46. В r e m e r F., DowR. S.a. MoruniG., J, Neurophysio' , 2, 473, 1939
32 и. О. Беритов
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
47. D a vi s Нм D a vi s F., L о о m i s A., Harvey E. a. Hobart (j., J.
Neurophysiol., 2, 500, 1939.
48. Dusser de В a r e n n e J. G. a. M с С u 1 loch W. S., Am. J. Physiol., 114r
692, 1936; Z. Ges. Neurol, u. Psychol., 162, 817, 1938.
49. Adrian E. D., Proc. Roy. ttoo.f B, 126, 433, 1939.
50. Dusser de Barenne, J. Neurophysiol., 1, 69, 1935.
51. Renshaw В., F о r b e s A. a. Morison B. R., J. Neurophysiol., 3, 74, 1940.
52. Ухтомский А. А, Шестое совещание no физиологическим проблемам АК
СССР. Тез. докл., стр. 47, 1940.
53. Lorentede N 6.5 J. Psychol. Neurol., 46, 113, 1934.
54. Beritoff J., Pfl. Arch. 213, 370, 1926.
55. В a r t 1 e у S. a.BishopG., Am. J. Physiol., 103, 159 и 173, 1933.
56. Lindsley D., Sci., 84, 354, 1936.
57. Шпильберг П. И., Усп. совр. биол., 12, 290, 1940; Физиол. журн. СССР.
27, 195, 1940.
58. Rohracher Н., Zschr. Psychol., 136, 308, 1935.
59. A dri an E. a. Moruzzi G., J. Physiol., 97, 155, 1939.
60. Lew у F. H. a. G a m m о n G. D., J. Neurophysiol., 3, 383, 1940.
6,1. ForbesA. a. MorisonB. R., J. Neurophysiol., 2, 112, 1939.
62. Rubin M. A., J. Neurophysiol., 1, 313, 1938.
63. Jasper H. H. a. HawkeW. A„ Arch. Neurol. Psych., 39, 885, 1938.
64. Lemere Fr., Brain, 60, 118, 1937.
65. Schwartz H. G. а. К e r r A. S., Arch. Neurol. Psych., 43, 547, 1940.
66. J a s p e г Н. H., Annual review of physiol., 3, 377, 1941.
67. MorisonR.S. FinleyK, H. a. LothropG. N., J. Neuiophysiol., 6, 254„
1943.
68. Kennard M. A., J. Neurophysiol., 6, 233, 1943.
69. К e n n a r d M. A. a. NimsL. F., J. Neurophysiol. 5, 325, 1942.
70. R о s e n b 1 u e t h А. а. С a n n о n W. В., Am. J. Physiol., 135, 690. 1942.
71. RosenbluethA., Bond D. a. Cannon W., Am. J. Physiol., 137, 681.
1942.
72. Гедавани Д., Об электрических потенциалах в жевательной зоне. 1947
Готовится к печати.
73. Marshall W. Н., Talbot S. A. a. AdesH. W., J. Neurophysiol., 6, 1,
1943.
74. Jasper H. a. Shagass Ch., J. Exp. Psychol., 28, 373, 1941.
75. С л e п я и Л. В., Тр. Ин-та физиол. им Бериташвили, 6, 403, 1945.
76. Б е р и т о в И., Тр. Ин-та физиол. им. Бериташвили, 6, 425, 1945.
77. Рожанский Н. А., Физиол. журн. СССР, 19, 289, 1935.
78. Case J. а. Вису Р. С, J. Neurophysiol., 1, 245, 1938.
79. Rheinberger М. В. a. Jasper Н. Н., Am. J. Physiol., 119, 186, 1937
80. Цкипуридзе Л. Р. Биоэлектрическая деятельность коры больших полу-
шапий и коры мозжечка во время сна. 1947. Готовится к печати.
81. Clark S. L. a. Ward J. W., J. Neurophy^ol., 8, 99, 1945.
82. Bremer F., Bollet. d. Soc, Ital d. Biol. Speriment., 13, 271, 1938.
Глава V
ОБРАЗОВАНИЕ ИНДИВИДУАЛЬНО ПРИОБРЕТЕННЫХ
РЕФЛЕКСОВ
Общие сведения об индивидуальных рефлексах. Одну из главных
функций большого мозга составляет индивидуально приобретенная
рефлекторная деятельность. Результатом ее являются всякого рода
привычные или заученные движения и привычные секреции. Человек или
животное реагирует такими заученными действиями на определенные
раздражения. Лошадь ускоряет шаг, когда замечает над собой хлыст,
но валяющийся на земле хлыст не вызывает такой реакции. У человека
при виде знакомой пищи выделяется слюна. Ни одно, ни другое
действие не находится в прямой прирожденной зависимости от данных
раздражений. Вид поднятого хлыста ускоряет бег лошади потому, что
когда-то за таким видом последовал удар по телу, что вызвало
усиленную локомоцию,— лишь после этого вид поднятого хлыста над
головой лошади связался с усиленной локомоцией. Вид знакомой пищи
вызывает секрецию слюны потому, что когда-то подобная пища пойала
в рот и вызвала секрецию,— лишь после этого вид ее связался с
выделением слюны. Эти явления хорошо известны в обиходной жизни, но
их лабораторное экспериментальное исследование — сравнительно новое
/ело. В связи с этим исследованием была поставлена главная
проблема физиологии большого мозга: выяснение тех закономерностей,
согласно которым протекают физиологические процессы в коре
большого мозга.
Систематическое выяснение физиологических процессов путем
исследования индивидуально приобретенных рефлексов было начато в
России под руководством выдающегося физиолога И, П. Павлова.
Первый труд принадлежит его ученику Толочинову (1902)
(1). Затем последовал ряд других работ самого Павлова (2) и его
многочисленных учеников. За 30 лет Павловым было создано
обширное и весьма оригинальное учение о сложной нервной
деятельности в коре большого мозга. В этом же направлении работал другой
выдающийся русский ученый — В. Бехтерев (5). Он тоже руководил
работами многочисленных учеников для выяснения этой чрезвычайно
важной проблемы.
32
500 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
В лабораториях Павлова индивидуально приобретенную
деятельность коры большого імозга изучали на собаках с помощью
индивидуально приобретенных слюнных рефлексов. Эти рефлексы образовывались
путем сочетания посторонних раздражителей с раздражением ротовой
полости съедобным или отвергаемым веществом,— например, раство-
Рис. 223. Метод Павлова для миографической записи
слюнного рефлекса.
Сверху слева изображено определение количества слюны,
вытекающей из выведенного наружу протока слюнных желез.
Сверху справа даны 3 опыта с кривыми выделения слюны. В каждом
опыте первая линия принадлежит кривой рефлекса, где каждая
вертикальная линия равняется одной капле слюны, другая
линия указывает на момент раздражения; третья линия — на
время в секундах. Первый опыт А — дает секрецию слюны во
время еды пищевого вещества; второй В — при виде пищи;
третий с — при освещении экрана, что служило
индивидуальным раздражителем. Внизу изображено сложное устройство для
автоматической записи слюнного рефлекса на кимографе с
помощью капсул Марея е, с'. Перед собакой находится экран для
дачи индивидуального светового сигнала. На полке миска Ь для
пищи (по Павлову).
ром соляной кислоты. В случае съедобного вещества авторы имеют
дело с пищевым поіведением, а в случае несъедобного — с защитным
поведением. Авторы, однако, изучали не все поведение, а только
секрецию слюны. Для этой цели протоки больших слюнных желез —
подчелюстных и околоушных — путем операции выводились наружу, на
щеку. Выделяемая наружу слюна собиралась в пробирки через
приклеенные к щеке стеклянные воронки (рис. 223) или же отсчитывалась
в кубических сантиметрах по стеклянной трубке, которая связывалась
ОБРАЗОВАНИЕ ИНДИВИДУАЛЬНО ПРИОБРЕТЕННЫХ РЕФЛЕКСОВ 501
герметически с отверстием протока слюнных желез, как это показано
на рис. 232. Иногда велась графическая запись выделяемых капель
слюны (рис. 223).
В лаборатории Бехтерева индивидуально приобретенная
рефлекторная деятельность изучалась при помощи двигательных рефлексов
как на животных, так и на человеке. Индивидуальные двигательные
рефлексы образовывались путем сочетания посторонних раздражений с
сильным электрическим раздражением кожи на конечностях; это
раздражение вызывает поведение освобождения из станка и вообще из
данной обстановки опыта. Но и в этом случае авторы изучали не все
поседение, а только один элемент его — движение одной только
раздражаемой конечности. Изучение шло 'миографическим путем:
движение раздражаемой конечности записывалось на кимографе. Иногда,
наряду с этим, записывалось и дыхание.
При изучении индивидуально приобретенных рефлексов животное
изолируется в отдельную комнату, с тем, чтобы во время опыта на
него не действовали посторонние раздражители помимо воли
экспериментатора. Животное ставят в станок и укрепляют в нем так, что оно
не может свободно двигаться. Экспериментатор находится снаружи и
наблюдает не за всем поведением животного, а только за тем или
другим компонентом поведения. В случае пищевого поведения он
наблюдает главным образом за выделением слюны, а в случае
защитного поведения освобождения — главным образом за движением
конечностей.
В последнее время индивидуальная деятельность изучается и в ряде
других лабораторий как у нас, так и за границей. В общем, в этих
лабораториях работа ведется по методу Павлова или Бехтерева.
Существенное изменение в (методику внесено Беритовым. Он
производит запись движений не только раздражаемой, но также какой-
либо другой конечности и головы. Это дает возможность следить за
общим состоянием животного и за его ориентировочными движениями
(рис. 233).
Направление Павлова и Бехтерева. В лабораториях Павлова и
Бехтерева собрали большой фактический материал относительно
внешних условий происхождения, развития и изменения индивидуально
приобретенных реакций. Наряду с этим, с самого начала они задались
целью выяснить физиологическую природу наблюдаемых явлений с
течки зрения нервных процессов, причем основные теоретические воззрения
относительно центральной нервной деятельности вырабатывались
главным образом Павловым. Мы отмечаем у Павлова два периода
в развитии теоретических представлений на основе полученных
фактических результатов. В первый период как сам Павлов, так и его
ученики стремились объяснить явления индивидуальной деятельности, исходя
из общеизвестных законов прирожденной нервной деятельности. Этот
начальный период был короток: продолжался от 1902 до 1907 г. С самого
начала он не удовлетворял Павлова. Это было еще! до появления
502 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
основательных и обширных работ Шеррингтона, Грэим Броу-
йз, Магнуса и др. Естественно, что Павлов не находил тогда з
общей физиологии центральной нервной системы таких положений, на
которые он мог бы опираться при анализе крайне сложных явлений
индивидуальной рефлекторной деятельности, поэтому в дальнейшем
этот первоначальный способ объяснения явлений был постепенно
оставлен, и с 1907 г. устанавливается уже новое направление и для
объяснения каждого сложного явления создается специальное гипотетическое
представление о нервных процессах.
Основные гипотезы были созданы Павловым в период от 1907
до 1910 г. Едва ли можно отрицать, что в то время толькс/ с помощью
этих гипотез можно было объяснять и систематизировать быстро
накапливающиеся факты. Но этот второй период продолжался и в то
время, когда состояние общей физиологии центральной нервной системы
в значительной степени изменилось. В течение последних 30 лет со
времени появления известного труда Шеррингтона: «The
integrative action of the nervous system» (1906) развитие общей
физиологии центральной нервной системы шло быстрым темпом. К
настоящему времени оно достигло такой высоты, что нельзя не
руководствоваться данными общей физиологии центральной нервной системы как
при объяснении сложных явлений индивидуальной деятельности, так и
при установлении новых законов специально для этой деятельности. Но
школа Павлова, как и школа Бехтерева, которая в отношении
теории в общем следует за первой, еще и теперь продолжает
развивать далее те же гипотезы, которые были созданы в 1907—1910 гг.
Правда, школой Павлова воспринимаются некоторые достижения
общей физиологии центральной нервной системы, но все то
гипотетическое, что было создано в прежние годы, господствует и сейчас почти
без изменения.
Направление Беритова. Первая попытка к установлению
основных закономерностей индивидуально приобретенной деятельности на
основе общепризнанных законов нервной деятельности была
произведена Беритовыім (4). Он специально изучал каждое характерное
явление индивидуальной деятельности, анализируя его с точки зрения
общеизвестных законов прирожденной нервной деятельности, и в то
же время стремился выявить специфические качественные особенности
индивидуальной нервной деятельности. Этим путем он установил
основные специфические закономерности данной деятельности и затем
предпринял систематическое изложение всего учения об индивидуальной
деятельности с точки зрения этих закономерностей. Важнейшие
фактические результаты были собраны им главным образом в 1916—1919 гг.
Первое сообщение Беритова об индивидуальной деятельности
поязилось в 1920 г. на грузинском языке (6) и в 1922 г. на русском
языке (7) затем одновременно в 1924 г. на английском и немецком
языках (4). Систематическое же изложение всего учения об
индивидуальной деятельности было дано в 1927 г. и в 1932 г. (16, 17).
ОБРАЗОВАНИЕ ИНДИВИДУАЛЬНО ПРИОБРЕТЕННЫХ РЕФЛ&КСОВ ?03
Терминология индивидуальных рефлексов. Индивидуально
приобретенный рефлекс, образуемый на почве прирожденного рефлекса, в
школе Павлова называется условным. Это название было дано
Павловым еще в первом исследовании об индивидуальных слюнных
рефлексах в диссертации Толочинова в 1902 г. Авторы дают этому
термину такое толкование. Индивидуально приобретаемые рефлексы в
своем возникновении и развитии зависят от ряда условий; изменение
условий всегда іведет к ослаблению или даже к исчезновению рефлекса.
Значит, существование индивидуального рефлекса всегда зависит от
наличия определенных условий. Индивидуально приобретенные
рефлексы противопоставляются прирожденным рефлексам, которые будто бы
являются стереотипными актами, не зависящими от определенных
условий. Поэтому эти рефлексы были названы безусловными. Но,
как теперь известно, каждый прирожденный рефлекс, раньше
казавшийся неизменным, постоянным актом, может не только меняться, но
также и исчезать или превращаться в полную противоположность в
зависимости от определенных внешних и внутренних условий. Как мы
уже видели, исследованиями Магнуса, Грэим Броуна, Бери-
това и др., не только обнаружены многочисленные случаи
изменчивости рефлексов, но выяснено также происхождение изменчивости
каждого двигательного рефлекса.
Характерно, что чрезвычайную изменчивость как количественную,
так и качественную, отмечает Степанов (18), один из уче'ников
Павлова, и по отношению к слюнным прирожденным рефлексам.
Было найдено, например, что вязкость слкУны, выделяемой собакой на
пищевое вещество, меняется, если за несколько минут до дачи пищи
влиіь ей в рот отвергаемое вещество или произвести сильное звуковое
раздражение наподобие грома. Так же действует снятие животного со
станка, повторение кормления через короткий промежуток времени.
Было доказано, что в этой изменчивости слюнной секреции
индивидуальные слюнные рефлексы не участвуют. Таким образом, на основании
современных данных физиологии центральной нервной системы,
прирожденные рефлексы не я&ляются стереотипными, постоянными
реакциями. При определенных изменениях внешних и внутренних условий
они так же изменчивы, как и индивидуально приобретенные рефлексы.
Исходя из приведенных данных, можно считать, что термины —
«усіовный» применительно к индивидуально приобретенным рефлексам
и «безусловный» применительно \к прирожденным — не отвечают
действительному состоянию наших знаний о рефлексах.
Бехтерев и его школа называют индивидуально приобретенные
рефлексы сочетательными. Этим термином обозначается
происхождение индивидуально приобретенных рефлексов. Он не отражает
наиболее характерной черты обозначаемых им рефлексов. Из общей
физиологии центральной нервной системы известно, что любой,
с виду индифферентный, раздражитель может вызвать тот или другой
рефлекс, если он следует вскоре после такого раздражения, которое
504
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
произвело значительный внешний эффект. Например, после сильного
электрического раздражения одной ноги звучание электрического звонка
может вызвать оборонительное движение этой ноги. Это происходит
по той причине, что под влиянием деятельного раздражения в
определенных нервных центрах возбудимость повышается ів такой мере, что
после этого любое периферическое раздражение путем иррадиации
возбуждения может вызвать деятельность этих центров. Словом, мы
будем иметь в буквальном смысле слова сочетательный двигательный
рефлекс. Сгибательное движение ноги на звонок вскоре после
электрического раздражения н? есть, конечно, индивидуально приобретенный
рефлекс. Аналогичное явление можно наблюдать и на спинальных
препаратах животных. Так, например, раздражение рецептивного поля
потирания сейчас же после рефлекса сгибания дает не потирание, а
сгибание (Уфлянд, 27). На основании этих данных мы находим, что термин
сочетательный для обозначения индивидуально приобретенных
рефлексов не является вполне подходящим.
Мы считаем, что было бы много лучше, если бы индивидуально
приобретенные рефлексы назывались не иначе, как индивидуально
приобретенными или просто индивидуальными. Этим будет обозначено
как происхождение их на основании индивидуального опыта, так
и противопоставление их видовым, прирожденным рефлексам.
Соответственно и раздражение, вызывающее индивидуально приобретенный
рефлекс, следовало бы называть индивидуальным.
Что же касается тога рефлекса, на основе которого образуется
индивидуальный рефлекс, мы предлагаем называть его основным
рефлексом, а то раздражение, которое его вызывает, основным
раздражением. Ниже мы увидим, что таким основным
рефлексом может служить как прирожденный, так и любой более или менее
прочный индивидуальный рефлекс.
Основное условие для образования индивидуальных рефлексов.
Каждый внешний агент, действующий на рецепторы, может быть
превращен в такой раздражитель, который вызовет определенный
секреторный или двигательный акт. Для этого нужно, чтобы этот внешний
агент действовал одновременно с каким-либо секреторным или
двигательным актом, или предшествовал этому акту, или следовал
вскоре после него. При этом образуемый индивидуальный секреторный
или двигательный рефлекс в общем носит такой же характер, как
и основной рефлекс.
Так, школа Павлова установила, что индивидуальное
раздражение вызывает точно такую же секрецию, как и основное раздражение.
Например, если основным раздражением служило несъедобное
вещество, например, 0.1—0.2% раствор соляной кислоты, то и на основное и
на индивидуальное раздражение выделялась жидкая слюна, бедная
органическими веществами. Наоборот, когда основным раздражением
служило пищевое вещество (хлеб или мясо), на оба раздражения
получалась густая слюна, богатая ферментами (Вульфсон, 8; Зальгейм, 9;
ОБРАЗОВАНИЕ ИНДИВИДУАЛЬНО ПРИОБРЕТЕННЫХ РЕФЛЕКСОВ 505
Болдырев, 28). Разница была только количественная: в
индивидуальном рефлексе слюна выделялась всегда в меньшем количестве, чем в
основном.
Двигательный индивидуальный рефлекс протекает точно так же
и в такой же форме, как и основной, на почве которого он
образовался. Так, например, если звук тонвариатора Штерна — 200 колебаний
4^W
Я/7.
200
Fa3 г0° *-*soo 500
"¦"¦'¦' ' 'H..H.I..II1I»») і1(іішіііиііі»..гііііііиншіц1|ніНиіііііііц..нііи.»...і
Рис. 224. Образование индивидуального рефлекса на
правой передней ноге на звук тонвариатора 200
колебаний в 1 сек.
Собака Боб. Верхняя кривая — от движения головы Г, средняя—от движения левой
задней ноги 3. л. и нижняя — от движения иравой передней ноги Я. п. Запись
происходит справа налево, как показывает стрелка как на этом, так и на всех
рисунках по индивидуальным рефлексам.
Опыт 1 дает 10-е сочетание раздражений. Сначала производится индивидуальный звук
(поднятие нижнего сигнала с числом 200), ноторый вызывает сильный рефлекс
сгибания правой передней ноги (нижняя кривая) и еще раньше —ориентировочное
движение (верхняя кривая, где поднятие кривой происходит от опускания головы,
а опускание кривой — от поднятия головы). Во время звука, спустя 10 сек. от
начала его, производится электрическое раздражение ноги, вызывающее второе
поднятие правой передней ноги (опускание верхнего сигнала с цифрой 5, обозна-
чающей число сантиметров расстояния между индукционными катушками). В опыте
2 производится необычный звук тонвариатора в 500 колебаний в 1 сек. Собака
сначала сильно встряхивает голову, затем поднимает переднюю ногу. В опыте 3 звук
500 колебаний повторяется, он дает сравнительно слабое движение передней ноги.
В опыте 4 не было специального раздражения. Рефлекс наступил, вероятно, на шум
кимографа. В опыте 5 производится 11-е сочетание: индивидуальный звук 200
колебаний в отдельности дает сильнейший эффект. В опыте 6 производится необычный
звук органной трубы 512 колебаний в 1 сек. (Fa3), он не дает никакого движения.
Была ориентировочная реакция, но она не была записана, так как перед этим опытом
нить, передающая движение головы, сошла с роликов и защемилась. Нижняя
линия отмечает время в секундах.
в 1 сек. сочетается с рефлекторным сгибанием правой передней ноги,
которое вызывается электрическим раздражением последней, то после
нескольких сочетаний данный звук в отдельности вызовет такое же
сгибание правой передней ноги вместе с теми сопутствующими
движениями, которые обычно производятся электрическим
раздражением: поворот головы в сторону раздражения, общее беспокойство,
визг, углубленное дыхание и т. д. (рис. 224).
Однако внешний индивидуальный эффект обычно определенным
образом меняется с количеством сочетаний. Первоначально после двух-
трех сочетаний индивидуальное раздражение производит
преимущественно ориентировочное движение головы, общее беспокойство и
передвижение, углубленное дыхание. Что же касается поднятия ноги, как
это бывает от электрического раздражения, то оно проявляется позднее,
506 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
после многократного сочетания, в тот же день или в следующие дни
работы. Но и в дальнейшем индивидуальный рефлекс не остается в
этом виде. После нескольких дней работы индивидуальное раздражение
начинает вызывать только сгибание раздражаемой ноги, а все другие
движения перестают вызываться (Протопопов, 13; Израэльсон, 11;
Бехтерев, 12; Беритов, 4, 17) (рис. 226).
Образование индивидуального рефлекса путем обратного сочетания
индивидуального раздражения с основным. Для образования
индивидуального рефлекса обычно прибегают к такому способу сочетания:
сначала начинается индивидуализируемое раздражение, а потом іво
время него или после него производится основное раздражение. Но
рефлекс образуется и при обратном порядке сочетания: если сначала
производится основное раздражение, а затем индивидуальное. Таким
путем были образованы индивидуальные рефлексы как на животных —
собака, голубь (Беритов, 17; Баяндуров, 19),—так и на людях (Шнир-
ман из лаборатории Бехтерева, 15). Но только для образования
рефлекса этим путем требуется несколько больше сочетаний, и
образуемый при этом рефлекс непрочен и не так интенсивен, как при обычном
порядке сочетания. Кроме того, индивидуальный рефлекс в данных
условиях не всегда удается образовать, для этого надо, чтобы основное
раздражение было не очень сильно и чтобы индивидуальное
раздражение производилось не .во время основного, а после него.
Первоначально лаборатория Павлова отрицательно относилась к
возможности образования индивидуальных слюнных рефлексов путем
обратного порядка сочетания. Один из учеников его,
Крестовников (20), не мог образовать этот рефлекс. Но до Крестовникова
другому ученику Павлова, Пименов у (21), удалось образовать
сильный рефлекс и при обратном порядке сочетания. В настоящее
время школа Павлова на основании последних исследований, также
признает возможность образования индивидуальных рефлексов путем
обратного сочетания раздражения (Крепе, 29; Павлова, 30 и др.).
Образование индивидуальных рефлексов высшего порядка. Для
образования индивидуальных рефлексов можно воспользоваться не
только любым -прирожденным рефлексом, но1 и любым уже
существующим индивидуальным рефлексом ("Кашеринова, 22; Зеленый, 24).
Основным условием является большая прочность и интенсивность того
индивидуального рефлекса, который берется в качестве основного
(Чеботарева, 23).
В области двигательных рефлексов такой индивидуальный
рефлекс был образован впервые Из р а эл ьсон о м(11). Эти
индивидуальные рефлексы, образуемые на основе индивидуального же, в
лаборатории Павлова называются вторичными рефлексами. В общей
физиологии вторичными двигательными рефлексами называются
рефлексы, возникающие от раздражения мышечно-суставных рецепторов
в связи с движением конечности. Когда, например, конечность
производит рефлекторное движение от первичного периферическою раз-
ОБРАЗОВАНИЕ ИНДИВИДУАЛЬНО ПРИОБРЕТЕННЫХ РЕФЛЕКСОВ 5?7
доажения, то в мышцах, сухожилиях и суставных поверхностях
происходит раздражение находящихся здесь рецепторов, которые в свою
очередь, могут вызвать рефлекторное движение. Поэтому
индивидуальный рефлекс, образованный на основе индивидуального же
рефлекса, мы предлагаем называть, по примеру Ленца (25),
индивидуальным рефлексом высшего порядка.
Индивидуальные рефлексы на людях. Изучение индивидуальных
рефлексов іна людях производилось тем же методом, как и на
собаках. На них образовывались как пищевые слюнные, так и оборонитель-
Рис 225. Капсула для вывода слюны наружу при работе над слюнными
рефлексами.
I—по Лешли, II — по Ющенко. Подробное описание см.
в тексте.
яые двигательные рефлексы. Человек изолировался от наблюдателя.
Основным раздражением служили кусок пастилы, бисквит и т. п. Они
вызывали выделение слюны в количестве 30—60 капель в 1 мин.
Раствор кислоты оказался неподходящим основным раздражением, так
как он дает очень незначительную слюнную секрецию (Ленд, 25). В
качестве индивидуальных раздражений применяли удары метронома,
звонок и т. д. На основании этих опытов авторы пришли к тем же
результатам, какие были уже получены на собаках (Ющенко, 31; Ленд, 25).
На людях слюнную секрецию изучают при помощи капсулы,
изобретенной американским ученым Лешли. Это маленькая круглая
серебряная чашечка с двойной стенкой (рис. 225). Центр 'чашечки (Л),
а также пространство между стенками (В) соединены с трубками. Во
рту капсула прикладывается к слизистой оболочке так, чтобы центр
ее приходился над отверстием протока околоушной слюнной железы.
Путем разрежения воздуха в пространстве между двойными стенками
капсулы достигают того, что капсула присасывается к слизистой
оболочке. Центральная трубка капсулы (а) резиновой трубкой соединяется
с измерительным прибором. Этим путем может быть отмечено выхож-
дение каждой капли слюны.
Индивидуальные двигательные рефлексы образовывались на людях
путем сочетания с электрическим раздражением руки или ноги,
которое вызывало оборонительное движение. Эти опыты производились
первоначально только учениками Бехтерева (Лукина, 35; Мясищев,
36; Шнирман, 15).
Скорость образования индивидуальных рефлексов. Образование
индивидуальных рефлексов происходит не с одинаковой скоростью.
Искусственные слюнные рефлексы, т. е. такие, которые вызываются ий-
5$S ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
дивидуальными раздражителями, не имеющими прямого отношения
к основному раздражению ротовой полости,— например электрический
звонок,— образуются даже в наиболее совершенных условиях
павловской методики после десятка сочетаний и становятся постоянными
после (многодневной работы. Много быстрее образуются слюнные
рефлексы в ответ на какое-либо свойство основного раздражения, например
на вид и запах пищи, т. е. так называемые натуральные слюнные
рефлексы.
Медленное образование искусственных рефлексов наблюдается лишь
при образовании первых рефлексов. После того как был образован
и укреплен один рефлекс, второй рефлекс путем сочетания с тем же
основным раздражением образуется и укрепляется значительно быстрее
первого, а третий — быстрее второго (Кашеринова, 22; Крыжановский,
37 и др.)- В определенных случаях повторный слюнной индивидуальный
рефлекс может образоваться после двух-трех подкреплений едой и
сделаться значительным и постоянным уже ів первые дни работы (см.
работы в Трудах физиологических лабораторий акад. Павлова за 1926—
1929 гг.).
Большое значение в отношении скорости образования рефлекса
имеет предварительное приучение животного к обстановке опыта. В
новой обстановке животное производит ориентировочные движения,
стремится выйти из данного ему положения. Это обусловливает
повышение возбудимости в центральной нервной системе,— в частности,
в коре большого мозга,— а такие обстоятельства, как мы увидим
ниже, затрудняют образование и укрепление индивидуального рефлекса.
Поэтому, прежде чеім начать образование рефлекса на собаке,
необходимо приучить ее к обстановке опыта в течение нескольких, иногда
даже імногих дней.
До последнего времени было принято думать, что индивидуальный
оборонительный рефлекс на ногах образуется в первый же день работы,
после 2—6 сочетаний (Беритов, 16, 17; Протопопов, 13). Но обычно
о каждом данном случае, еще раньше получения индивидуального
движения на ногах, индивидуальные движения появляются на голове
в виде ориентировочной реакции, а также на голосовом органе в виде
визга или воя, на дыхательном аппарате — в виде учащения и
усиления дыхания. Эти индивидуальные движения, охватывающие почти
весь организм, не только возникают на какое-нибудь
индифферентное раздражение после одного сочетания его с основным
раздражением, но оказываются настолько прочными, что передаются на другой
и на третий день. В этой двигательной реакции мы имеем не
индивидуальный рефлекс, а индивидуальное поведение освобождения
животного из станка. Поведение освобождения из станка наступает у собаки
при каждом болезненном раздражении в станке, и всякое
индифферентное раздражение в виде звука или света, какое производится
в это время, становится индивидуальным сигналом к поведению
освобождения. Этот поведенческий акт возникает в результате психо-
ОБРАЗОВАНИЕ ИНДИВИДУАЛЬНО ПРИОБРЕТЕННЫХ РЕФЛЕКСОВ 509
нервной деятельности, воспроизводящей сложную реакцию
неприятного эмоционального состояния и освобождения из станка
(Беритов, 55, 60).
Нужно многократное сочетание основного электрического
раздражения конечности, производящего сгибание ноги наряду с общим
поведенческим движением освобождения, с индивидуализируемым
индифферентным раздражением, чтобы последнее стало индивидуальным сиг
налом к поднятию ноги или к индивидуальному рефлексу сгибания
ноги. На собаках это достигается в результате одного или нескольких
десятков сочетаний в течение нескольких дней работы. Образование
индивидуального рефлекса на ноге объективно выражается в том, что
он наступает с большим постоянством и с одинаково малым скрытым
периодом. Фактически индивидуальный оборонительный рефлекс
сгибания ноги есть автоматизированный элемент поведенческого акта
освобождения, который наступает ів ответ на индивидуальное
раздражение, без участия представления (Беритов, 55, 60).
У людей оборонительные рефлексы на почве сочетания с
электрическим раздражением пальца образуются в большинстве случаев много
медленнее, чем у собак. У подростков 16—17 лет, по данным Осипо-
в о й, оборонительный индивидуальный рефлекс образуется ів среднем
после 14—16 сочетаний и больше. У детей младшего возраста он
образуется в среднем несколько быстрее. Умственно-отсталые дети
показывали наиболее быстрое образование рефлекса, в среднем, поел?
5 сочетаний. У взрослых людей скорость образования индивидуальных
оборонительных рефлексов чрезвычайно вариирует. Так, в опытах
разных авторов из Института мозга им. В. 'М. Бехтерева они возникали
у одних подопытных лиц после 10, у других после 100 сочетаний в
течение нескольких дней работы; у третьих они не образуются и после
нескольких сотен сочетаний. У некоторых испытуемых
индивидуальный рефлекс вырабатывался уже в первый день работы после
нескольких сочетаний (Лукина, 35; Мясищев, 36; Шнирман, 15 и др.).
Фактические данные, полученные в опытах на людях, заставляют
признать интеллектуальную, сознательную деятельность препятствием
для быстрого образования индивидуальных рефлексов по обычному
методу, применяемому при работе на животных. Специальными опытами
было обнаружено, что у людей индивидуальный двигательный рефлекс
н? проявляется, несмотря на многократные сочетания с основным
раздражением, не вследствие неспособности к индивидуально
приобретенной рефлекторной деятельности, а вследствие присутствия у человека
выісшей формы психонервной деятельности — 'сознания (Беритов и
Дзидзишвили, 38).
У высших позвоночных, например у обезьяны, индивидуальные
рефлексы образуются также очень медленно. Большое значение имеет
одомашнивание обезьяны. В камерной обстановке индивидуальные
рефлексы образуются скорее у одомашненных и сильных обезьян, чем уди-
і&их и слабых. Оборонительные и главным образом ориентировочные
510
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
реакции очень мешают образованию индивидуальных рефлексов у не-
одомашненных обезьян (Каминский, 54).
Скорость образования индивидуальных реакций у животных разных
классов и разного возраста. У низших млекопитающих, например у
кроликов, для образования пищевых и оборонительных
индивидуальных реакций требуется часто по нескольку десятков сочетаний — много
больше, чем у собак. Так же обстоит дело с птицами, например с
голубями, у которых оборонительные индивидуальные реакции образуются
после 40—60 сочетаний и упрочиваются после 200—300 сочетаний (Бе-
ритов, 14; Баяндуров, 19). Пищевые индивидуальные реакции у птиц
образуются заметно быстрее, иногда после десятка сочетаний (Беритов,
14), а то и после 2—3 сочетаний (Баяндуров, 53).
Но в последнем случае едва ли мы имеем дело с настоящим
индивидуальным рефлексом. В таких случаях, несомненно, проявлялось
индивидуальное пищевое поведение, направляемое представлением
определенной внешней среды и местоположения пищи. А такие
поведенческие акты, как известно, у птиц и млекопитающих после одного
первоначального переживания вызываются в течение многих часов и даже
многих дней при возникновении данного представления под влиянием
какого-либо компонента соответствующей внешней среды (Беритов, 55).
Особенно трудно образуются индивидуальные реакции у низших
позвоночных; например, у амфибий требуются десятки и сотни
сочетаний, чтобы образовать индивидуальную реакцию на свет и на
кваканье, издаваемое другой лягушкой; на другие звуки индивидуальные
реакции не образуются (Беритов, 40). Заметно быстрее образуются
они на рыбах (Фролов, 39).
Скорость образования и укрепления индивидуальных реакций не
одинакова на разных стадиях онтогенеза. Были проведены специальные
опыты для выяснения этого вопроса на щенках разного .возраста. На
переднюю лапку щенка навязывались тонкие провода от
индукционного аппарата для вызывания оборонительного рефлекса
электрическим током. Щенок не был привязан. Он свободно передвигался на
столе, окруженном загородкой. Индивидуальным сигналом служили
звуки, главным образом электрический звонок. У щенков 12 дней от
роду индивидуальные оборонительные реакции образовались после 13
сочетаний, -но не упрочились в такой мере, чтобы сохраниться до
следующего дня. У более взрослых щенков, 20—23 дней от роду, реакции
получались уже на четвертом сочетании, но и здесь в течение первого
дня они не упрочились 'в такой мере, чтобы передаться на следующий
день. У еще более взрослого щенка, 30 дней от роду, реакции
образовались быстро и уже в первый день работы укрепились в такой мере,,
что не исчезли и на другой день (Топурия, 41).
У новорожденных животных, у которых развитие нервной системы
является совершенно законченным к моменту рождения, как, например,
у морских свинок, индивидуальные рефлексы, конечно, образуются уже
в первый день после рождения. Весьма возможно, что у них индивиду-
ОБРАЗОВАНИЕ ИНДИВИДУАЛЬНО ПРИОБРЕТЕННЫХ РЕФЛЕКСОВ 51Г
альные рефлексы могут образоваться еще до рождения (Голубева, 56).
Однако следует отметить, что во всех этих случаях изучения
индивидуальных реакций на низших позвоночных, а также на щенках и
морских свинках авторы имеют дело с поведением животного, с его
целостной реакцией приспособления. И метод исследования был такой,
какой применяется для изучения поведения: наблюдение за всем
поведением животного в свободно подвижных условиях. Поэтому данные
выше результаты по существу касаются индивидуального поведения
животных в связи с их филогенезом и онтогенезом. При этом даже не
ставится цель изучить исключительно движение раздражаемой ноги или
определенной ротовой реакции, как это должно быть при изучении
рефлекса. Обычно ведется письменная регистрация всего поведения. Да и
реакция животного в данных условиях работы не дает повода
останавливать наше внимание на движении того или другого органа, так как
животное реагирует на раздражения всем организмом, и притом
длительными изменчивыми движениями всех его органов.
В связи с поведенческим характером индивидуальных реакций стоит,
между прочим, то явление, что означенные реакции могут образоваться
через небольшое число сочетаний, как у взрослых, так и у
новорожденных животных (Беритов, 57). Эти индивидуальные поведенческие
реакции возникают и протекают не по тем закономерностям, как
индивидуальные рефлексы. Но после того как поведенческий акт
повторился много раз и принял автоматический характер, он уподобляется
рефлексу, ибо каждый элемент его начинает протекать по принципу
рефлекса (Беритов и сотр., 55). Поэтому по наблюдениям над
автоматизированными поведенческими актами или даже над каким-либо
элементом его можно изучать индивидуальную рефлекторную деятельность.
Мы, например, пользовались для этой цели главным образом той
начальной реакцией, которая наступала на голове или на ногах вслед за
индивидуальным сигналом.
Рецепторная генерализация и дифференциация индивидуальных
рефлексов. В самом начале образования индивидуального рефлекса на
один определенный раздражитель рефлекс наступает не только от
данного раздражителя, но и от других того же качества, а иногда и
другого качества. Как говорят, рефлекс генерализован. Однако отнюдь не
во всех случаях генерализация делается в такой мере обширной,
чтобы любое раздражение того же качества вызывало индивидуальный
рефлекс (Бурмакин, 32; Купалов, 34). Так, в наших опытах ,во время
выработки индивидуального оборонительного рефлекса ноги у одной
собаки на звук тонвариатора 200 колебаний в 1 сек. в первый же день
работы рефлекс вызывался звуками тонвариатора лишь до 500
колебаний, т. е. звуками менее чем на октаву выше обычного, а из
посторонних звуков иного тембра,— например, органных труб, в это же самое
время рефлекс вызывался звуками от 200 до 320 колебаний в 1 сек.
Из звуков тонвариатора наиболее близкие к обычному звуку
производили наиболее сильные рефлексы. Более отличные звуки тонвариатора.
532 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
например, 600 колебаний в 1 сек., а также звуки органной трубы,
512 колебаний в 1 сек. обычного рефлекса не вызывали.
При специальном исследовании генерализации было обнаружено, что
генерализация индивидуального рефлекса происходит не сразу, а
постепенно (Шевалев, 33; Купалов, 34). Так, при образовании
двигательного рефлекса на звук тонвариатора 300 колебаний в 1 сек. в первый
день работы рефлекс был очень слабо генерализован. Он вызывался
необычными звуками тонвариатора 350 колебаний в 1 сек., но звуки
400—500 колебаний н? давали его. В следующие дни генерализация
индивидуального рефлекса становилась все больше и больше. Так, на
второй день звуки 400—450 колебаний вызывали рефлекс, а звуки
500 колебаний не вызывали; на четвертый день звуки 500—510
вызывали рефлекс, а 550—600 не вызывали. Лишь на пятый день
дальнейшая генерализация приостановилась, даже пошла на убыль: звук
500 колебаний иногда не давал эффекта. В одном случае на одной
собаке генерализация вновь образуемого рефлекса достигла .максимума
уже на второй день. Бывали и такие случаи, когда слабая
генерализация, имевшая место в первый день работы, ничуть не прогрессировала
в следующие дни, а наоборот, еще более ограничивалась (Беритов, 17).
Характерно, что генерализация рефлекса на данный индивидуальный
звук тонвариатора происходила по отношению к звукам ниже и выше
обычного звука в одинаковой мере, как это еще показал Эльясон(42).
С течением времени генерализация постепенно проходит, т. е.
рефлекс дифференцируется (Зеленый, 10; Эльясон, 42; Протопопов, 13).
Так, в одном опыте при образовании рефлекса на звук тонвариатора
200 колебаний в 1 сек. более или менее отдаленные звуки другого
тембра уже в первые дни не вызывали рефлекса. Наиболее же близкие
звуки того же тонвариатора с разницей до полутона (до 208
колебаний) перестали давать рефлексы в течение 2—3 недель. Таким
образом, этот процесс дифференциации рефлекса может продолжаться
целые недели.
Дифференциация рефлекса в значительной мере зависит от
характера собаки. В наших опытах на легко возбудимых собаках тонкая
дифференциация при образовании рефлекса на звук тонвариатора ни
разу не достигалась; звуки, отличные от индивидуального на целый тон,
производили рефлекс даже спустя много недель работы. В то же время
на спокойных собаках такая дифференциация получалась в течение
нескольких дней; так, на одной собаке при образовании рефлекса на звук
тонвариатора 200 колебаний в 1 сек. уже на третий день звук 225
колебаний, отличный от обычного на один тон, не давал рефлекса.
Дифференциация может быть так тонка, что необычные звуки, отличные от
индивидуального всего на 7в тона, уже не будут вызывать рефлекса
(Беляков, 43; Протопопов, 13).
Более или менее обширная генерализация двигательного рефлекса
наблюдается только при образовании первого рефлекса. В случае
образования новых рефлексов путем сочетания новых индивидуальных раз-
ОБРАЗОВАНИЕ ИНДИВИДУАЛЬНО ПРИОБРЕТЕННЫХ РЕФЛЕКСОВ 513
дражений с одним и тем же основным раздражением, например с
электрическим раздражением одного и того же участка кожи какой-либо
ноги, рефлекс образуется очень быстро, и притом только с очень
небольшой генерализацией, а именно по отношению к близким
раздражителям того же качества.
Это явление первоначальной генерализации и последующей
дифференциации индивидуального рефлекса по отношению к раздражениям
рецепторов называется рецепторной генерализацией и
дифференциацией.
Эффекторная генерализация и дифференциация индивидуальных
рефлексов. Выше мы имели в виду факты генерализации и
дифференциации индивидуальных рефлексов по отношению к раздражению
рецепторов.
Но наряду с этим происходит еще другого рода генерализация и
дифференциация, именно, по отношению к внешним движениям. При
образовании первого оборонительного рефлекса путем сочетания
индивидуализируемого раздражения с электрическим раздражением ноги
первоначально индивидуальный двигательный рефлекс более или менее
обширен: движется все животное; оно не только поднимает
раздражаемую ногу, но производит движение всех конечностей, шеи и туловища,
порывисто дышит, визжит. Все движения животного направлены на то,
чтобы освободиться из станка, и поэтому, как уже указывалось выше,
являются чисто поведенческим актом. Но потом, спустя несколько дней
регулярного сочетания, индивидуальный сигнал вызывает не эту
поведенческую реакцию, а более или менее ограниченные движения тела
В конце концов, вся индивидуальная реакция ограничивается
поднятием раздражаемой ноги и небольшим движением головы, т. е.
происходит полная двигательная дифференциация. Генерализация и
дифференциация по отношению к внешней реакции называются эффектор-
нон генерализацией и дифференциацией.
Обширный поведенческий эффект освобождения в начале
образования рефлексов бывает не при всех условиях опыта. Прежде всего его
не бывает на собаках очень низкой возбудимости, или он наступает у
них в очень слабой форме. Затем этот генерализованный эффект
отсутствует при повторном образовании однородных рефлексов. После
образования и дифференцирования двигательного рефлекса на одной ноге
новый рефлекс на той же неге образуется очень быстро без всякой
двигательной, т. е. эффекторной, генерализации.
Вторичная рецепторная и эффекторная генерализация
индивидуальных рефлексов. Любой хорошо дифференцированный рефлекс
может при некоторых определенных условиях подвергнуться новой
генерализации. Так, в конце опытного дня, после 2—3 часов стояния в
станке, собака всегда обнаруживает более или менее сильное
беспокойство. Обычно это выражается в визге, в беспрерывном поворачивании
головы к дверям. Иногда оно проявляется в бурной форме в виде
порывистых движений освобождения. В этот именно период все индиви-
33 И. С. Беритов
514 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
дуальные раздражители начинают вызывать сильные общие движения
освобождения наряду с поднятием ноги. Такая вторичная
генерализация индивидуальной реакции на короткое время была замечена также
после сильных необычных раздражений,— например, после сильного и
неожиданного звука (Беляков, 43). Ее легко можно получить также
при помощи быстро следующих одно за другим электрических
раздражений как с обычным сочетанием — со звуком, так и без него.
Далее, вторичная генерализация может получиться при легком
утомлении (Абуладзе, 44), при стрихнинном отравлении (Никифоров-
ский, 45) и вообще при повышенной нервной возбудимости.
Генерализация и дифференциация индивидуальных рефлексов на
разных ступенях филогенеза и онтогенеза. У низших млекопитающих
(кролик) рецепторная генерализация наблюдается в большей мере, чем
у собаки. У них, например, не удается отдифференцировать от
индивидуального звука близкие необычные звуки (на 7г—2Х12 тона выше или
ниже) (Брегадзе, 46). В этом отношении кролики напоминают птиц.
У голубей и кур рефлекс на звуки дифференцируется,, как и у
кроликов, очень плохо (Беритов, 14; Завадовский и Рохлина, 52), но на
цвета и фигуры — хорошо. Можно образовать пищевой рефлекс на один из
цветов, на одну из фигур, на движение фигуры в одном направлении,
и все остальные цвета, фигуры и направления движения фигур
отдифференцировать (Абуладзе, 47; Баяндуров, 53; Беритов и сотр., 58, 59).
В отношении звуков дифференциация достигается у птиц труднее и
достигнутая дифференциация расстраивается легче, чем у
млекопитающих. У птиц также можно отдифференцировать звуки на целый тон
или на полтона выше обычного индивидуализируемого, но эта
дифференциация непрочна и не всегда достигается. Наоборот, легко
образуется и дифференцируется индивидуальный оборонительный рефлекс на
тактильные раздражения: индивидуальный рефлекс отсутствует уже при
первых пробах раздражения в необычном участке кожи (Баяндуров, 53).
Рецептивная дифференциация индивидуальных рефлексов
происходит не одинаково на разных стадиях онтогенетического развития.
Специальными опытами на щенках разных возрастов было найдено, что v
них оборонительный индивидуальный рефлекс является более
генерализованным, и это состояние удерживается на большее время, чем v
взрослых собак. Следовательно, на щенках дифференциация
происходит более медленным темпом, чем на взрослых собаках. На одном
щенке, с которым работа началась, когда ему было 20 дней,
необычный звук, на 4 тона ниже обычного, производил рефлекс, несмотря на
многократные испытания в течение 16 дней и несмотря на 150
сочетаний обычного звука с основным электрическим раздражением (Топу-
рия, 41).
Весьма характерно для щенков то явление, что обширная
поведенческая реакция самозащиты не так быстро сменяется изолированным
поднятием ноги, как у взрослых. На индивидуальный сигнал щенок
усиленно двигает головой, визжит и часто дышит, дрожит всем телом,
ОБРАЗОВАНИЕ ИНДИВИДУАЛЬНО ПРИОБРЕТЕННЫХ РЕФЛЕКСОВ 515
передвигается с места на место. Такой поведенческий характер
индивидуальной оборонительной реакции бывает у взрослых собак только в
первые дни образования рефлекса, и притом у собак очень
возбудимого типа. Если же начато образование рефлекса на 12-дневных
щенках, то диффузный характер рефлекса удерживается в течение
многих дней работы без ослабления.
На основании того, что было сказано выше относительно
генерализации и дифференциации рефлекса у животных разных возрастов и
разных классов, а также относительно скорости образования и
укрепления у них рефлексов, мы приходим к заключению, что у собаки
развитие способности к индивидуальной рефлекторной деятельности в
онтогенезе повторяет развитие этой деятельности в филогенезе.
Образование индивидуальных рефлексов на сложное раздражение.
Как секреторный, так и двигательный рефлексы могут быть образованы
на разного рода сложные комплексы раздражений. Эти комплексы
могут состоять как из одного рода, так и из разного рода раздражений,
например света и звуков, действующих одновременно или же
последовательно. Во всех случаях развитие рефлекса происходит в одном и том
же виде: первоначально не только комплекс, но и его компоненты дают
рефлекторную реакцию. Чем сильнее физиологическое действие
компонента, тем интенсивнее вызываемый им рефлекс. Впоследствии, после
длительного ряда сочетаний с основным раздражением, компоненты
перестают вызывать рефлекс; последний наступает только на комплекс
(Бабкин, 48; Палладии, 49; Зеленый, 10; Фурсиков, 50; Беритов, 4,17).
Здесь мы имеем своеобразную генерализацию и дифференциацию
индивидуального рефлекса по отношению к компонентам
индивидуального раздражения. Это свойство отдифференцирования целого комплекса
от компонентов у человека выражено в большей мере, чем у собак
(Иванов-Смоленский, 51).
Л и т е і» а т у р а
\ Т о і о t s с Ь і и о v, Contribution a 1'etude de la physiologie et de la psychologie
des glandes salivaires. Forhandlingar vid. Nord. Naturforskareoch Lakaremotet,
1903.
2. ПавловИ., 20-летний опыт объективного изучения высшей нервной дея
тельности (поводения) животных Л., 1923.
3. Бехтерев В., Объективная психология. СПб., 1914—1917; Общие основания
рефлексологии человека, Л., 1923.
4. В е гі t о f f J., Brain., 47, 109, 359, 1924; J. Psych. Neur. 30, 217, 1924; 82, 29,
253, 265, 1925.,
5. Бехтерев В., Обозрение психологии, неврологии и экспериментальной
психологии, 12, 512, 1907.
6. Беритов И., Физиология животных, 1, 1920 (на груз. яз.).
7. Беритов И., Общая физиология мышечной и нервной системы. Тифлис, 1922
S, Вульфсон С, Работа слюнных желез. Дисс, 1898.
9. ЗальгеймА., Работа слюнных желез до и после перерезки п. glossopharyngei
и lingualis. Дисс , 1904.
10. 3 е л е н ы й Г., Материалы к вопросу о реакции собаки на звуковые
раздражения. Дисс, 1907.
33*
516 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
Й1. Израэльсон Ж.? О топической дифференцировке
сочетательно-двигательного рефлекса на кожные и мышечные раздражения. Дисс, 1913.
12. Б е х т е р е в В., Обозрение психиатрии, неврологии и экспериментальной
психологии, стр. 449, 1910.
13, Протопопов В., О сочетательно-двигательной реакции на звуковые
раздражения. Дисс, 1909.
?4. Beritoff J., РЛ. Агсік, 213, 370, 1926.
15. Ш н и р м а н А., Новое в рефлексологии и физиологии нервной системы, 1, 218,
1925.
16. Beritoff J., tjfcer die individuell-erworbene Tdtigkeit des Zentralnerven-
systems, J. Psych. Neur., 33, 113, 1927.
17. Беритов И., Индивидуально приобретенная деятельность центральной
нервной системы. Тифлис, 1932.
48. Степанов Г., Русск. физиол. журн., 3, 114, 1921.
19. Б а я н д у р о в В.. Журн. эксп. биол. и мед., 4, № 10—11, 210, 1926.
20. Крестовников А., Тр. Об-ва русских врачей, 1913.
21. Пименов П., Особая группа условных рефлексов. Дисс, 1907.
22. Кашеринова Н. Материалы к изучению условных слюнных рефлексов
на механическое раздражение кожи у собаки. Дисс, 1908.
23. Чеботарева О. Дальнейшие материалы к физиологии условного торможения.
Дисс, 1912.
24. ЗсленыйГ, Арх. биол. наук, 14, 458, 1909.
25. Л е н ц А., Русск, физиол. журн., 5, 93, 1922.
26. Добротворская, цит. по Бехтереву: Обозрение психиатрии,
неврологии и экспериментальной психологии, 12, 512, 1907.
27. У ф л я н д Ю., РИ. Arch. 207, 87, 1925; Русск. физиол. журн., 8, 132, 1925.
28. Болдырев В., Тр. Об-ва русских врачей, 72, 321, 1904.
29. К р е п с Е., Тр. Физиол. лаб. акад. Павлова, б, 5, 1933.
30. Павлова В., Тр физиол. лаб. акад. Павлова, б, 21, 1933.
31. Ющенко А., Условные рефлексы ребенка. Госиздат, Москва, 1928.
32. Бурмакин В., Процесс обобщения условного звукового рефлекса у собаки
Дисс, 1909.
33. Ш е в а л е в Е., О процессе дифференциации сочетательно-двигательного
рефлекса на тактильные раздражения и т. д. Дисс, 1913.
34. Купа лов П., Арх. биол. наук, 19, 20, 1915.
35. Лукина А., НоЕое в рефлексологии и физиологии нервной системы, 1, 205,
1925.
36. М я с и щ е в В.. Новое в рефлексологии и физиологии нервной системы, 1, 291,
1925.
37. Крыжановскйй И., Условные звуковые рефлексы при удалении
височных областей и т. д. Дисс, 1909.
38. Беритов И., иДзидзишвили Н.,Тр. биол. сект. Груз. фил. АН СССР, 1.
1934.
39. Фролов Ю., РП. Arch., 208, 261, 1925; 220, 339, 1928.
40. Беритов И., Изв. Тифл. ун-та, 10, 225. 1929 (на груз. яз.).
41. Т о п у р и я Ш., Изв. Тифл. ун-та, 10, 225, 1929 (на груз. яз.).
42. Эльясон М., Исследование слуховой способности собаки и т. д. Дисс, 1908.
43. Беляков В., Материалы к физиологии диф|)еренцировки внешних
раздражений. Дисс, 1911.
44. Абуладзе К., Русск. физиол. журн. 9, 169, 1927.
45. Н и к и ф о р о в с к и й Н., Фармакология условных рефлексов как метод их
изучения. Дисс, 1910.
46. Б ре гад зе А., Журн. эксп. биол. и мед., 33, 385, 1929.
47. Абуладзе К., Русск. физиол. журн., 12, 75, 1929.
48. Б а б к и н Б., Тр. Об-ва русских врачей, 77, 197, 1910; 78, 249, 1911; 80, 1913.
49. Палладии А., Тр. Об-ва русск. врачей, 73, 393, 1906.
ОБРАЗОВАНИЕ ИНДИВИДУАЛЬНО ПРИОБРЕТЕННЫХ РЕФЛЕКСОВ 517
50. Ф у р с и к о в Д., Доклад на II Всеросс. патоневр. съезде в Ленинграде.
51. Иванов-Смоленский А., Мед. биол. журн., 2-я ч., в. 3, стр. 84, 1925.
52. Завадовский Б. и Рохлина М., Мед. биол. журн. 3-я ч., в. 3, стр. 83,
1927.
53. Б а я н д у р о в Б. И., Услогные рефлексы у птиц, Тр. Томск, мед. ин-та, 5, 1937
54. К а м и н с к и Й С. Д., Сб. докладов VI Всесоюз. физиол. съезда, стр. 182.
55. Б е р и т о в И. и сотр., 10 сообщений в Физиол. журн. СССР за 1933—1935 гг.
56. Г о л у б е в а Е. А., Арх. биол. наук, 54, 132, 1939.
57. Б е р и т о в И. Проблемы биологии и медицины. Сб., посвящ. Л. С. Штерн. 1935.
стр. 69.
58. Беритов И. и Чичинадзе Н., Тр. биол. сект. Закавк. фил. АН СССР.
1, 105, 1934.
59. Беритов И. и Ахметели М., Бюлл. эксп. биол. и мод., 2, 1С9, 1936.
60. Беритов И., 05 основных формах нервной и психонервной деятельности.
Изд. АН СССР, 1947.
61. О си нов а В., Сб. Новое в рефлекс, и физиол. нервн. системы, т. II, стр. 291
192U
Глава VI
ТИПЫ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ
Совпадающие, или наличные рефлексы. Индивидуальный рефлекс
наступает вскоре после начала индивидуального раздражения, если
основное раздражение следует спустя несколько секунд после начала
этого раздражения. Такие рефлексы называются в лаборатории
Павлова совпадающими или наличными. Скрытый период
слюнной индивидуальной реакции в совпадающих рефлексах
измеряется обычно несколькими секундами. Наблюдаемая при этом
двигательная пищевая реакция — глотание, облизывание — наступает раньше
слюнной реакции. Но всегда пищевой реакции предшествует
ориентировочная реакция головы.
По новейшим опытам, скрытый период слюнной реакции может
продолжаться около 1 сек.
В индивидуальных двигательных рефлексах скрытый период
реакции много короче. Кратчайший скрытый период реакции конечности
составляет около 1 сек. Скрытый же период реакции от самого
электрического раздражения равен 0.15—0.20 сек.,— следовательно, время,
необходимое для распространения возбуждения через кору больших
полушарий, составляет около 0.8 сек. Индивидуальная
ориентировочная реакция головы всегда наступает раньше индивидуального
движения ног. Кратчайший скрытый период ее равен приблизительно 0.55 сек
(Беритов, 1).
По исследованию Быкова и Петровой (30), с применением
более совершенной методики, скрытый период индивидуальной
ориентировочной реакции оказался много меньше. Например, при пищевых
двигательных рефлексах на звонок после 600 сочетаний
ориентировочная реакция головы начиналась минимум через 0.08 сек. Это было в
начале опытного дня. При повторении скрытый период удлинялся до
0.20 сек. При кожно-оборонительном рефлексе на звонок движение
головы начиналось со скрытым периодом в 0.06 сек., при повторении
период удлинялся до 0.09 сек.
На людях скрытый период индивидуальной двигательной реакции
руки был определен Мясищевым (31) в 0.2—0 3 сек., но в некото-
1И11Ы ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ оі9
рых случаях доходил до 0.1 сек. Этот скрытый период обычно больше,
чем рефлекс от электрического раздражения. В некоторых единичных
случаях скрытый период индивидуальной реакции оказывался точно
таким же, как в реакции от электрического раздражения (Беритов и
Дзидзишвили, 32).
Совпадающий двигательный рефлекс может протекать в виде одного
или двух быстропротекающих сгибаний, которые кончаются еще во
время индивидуального раздражения,
лли в виде длительного поднятия ноги,
или в виде длительного ритмического
движения, которое продолжается и после
раздражения. Тот или другой характер
рефлекса зависит от способа применения
основного электрического раздражения.
Если основное раздражение является
однократным и применяется по одному
разу продолжительностью в 1—2 сек., то
индивидуальный рефлекс наступает в
виде одного или нескольких быстро проте-
<ающих сгибательных движений. Это
показано на рис. 224. Если же основное
раздражение применяется многократно
во время каждого индивидуального
раздражения, и притом невзирая на наличие
или отсутствие индивидуального
рефлекса, то этот последний рефлекс начинает
протекать длительно в виде поднятия
ноги (рис. 226) или в виде ритмических
движений ее. На рис. 231 приводится
один такой рефлекс.
Запаздывающие рефлексы. При
определенных условиях образования
индивидуального рефлекса этот рефлекс
начинается не сразу в начале
индивидуального раздражения, а спустя некоторое
время после него, например, спустя 10—60 сек. и позднее от начала
последнего. Такие рефлексы называются в лаборатории Павлова з а-
паздывающими (Васильев, 3). Некоторые авторы из лаборатории
Павлова называют запаздывающие рефлексы, с небольшим
запаздыванием в 5—10 сек., отставленными.
При образовании запаздывающего рефлекса запаздывание пол>-
чается не с самого начала его образования. Первоначально этот
рефлекс является совпадающим (рис. 227), лишь потом скрытый период
постепенно нарастает до срока основного раздражения (Завадский, 4).
Это может быть как при образовании слюнного запаздывающего
рефлекса, так и двигательного оборонительного (Беритов, 5) (рис. 228).
\
Лп.
300
200
21S
Рис. 226. Состояние сильной
дифференциации рефлекса на правой
передней ноге на звук тонвариа-
тора 200 колебаний в 1 сек.
Собака Боб. В опыте J необычный
звук— 215 колебаний в 1 сек., только
на 1/* тона выше обычного, не даеі
рефлекса, не появляется даже
ориентировочное движение. В опыте 2
обычный звук — 200 колебаний вызывает
длительное сгибание правой передней
ноги с поворачиванием головы. В этом
опыте дифференцированный звук—300
колебаний в 1 сек. — применялся во
время индивидуального рефлекса и не
повлиял на него*
520
ОБЩАЯ ФИЗИОЛОГИЯ КЭРЫ БОЛЬШОГО МОЗГА
Фоб
Чес.
"I—*"-
|мии>.і»иПііПііі;іуііммітгпніиіПППіім.-Мі1пПпі .а-і:
По отношению к двигательной реакции впоследствии наступает
еще стадия прерывистых движений: собака во время индивидуального
сигнала то поднимает ногу, то опускает ее (Беритов, 5). На рис. 228
даются три сочетания, где хорошо выявляется эта стадия в первых
двух сочетаниях; в третьем же
сочетании рефлекс является чисто
запаздывающим.
Неактивная фаза запаздывающего
рефлекса чрезвычайно легко
заменяется активной. При разнообразных
условиях рефлекс перестает наступать с
запаздыванием, он становится
совпадающим. Завадский изучил
подробно это явление на слюнных
рефлексах. Оказалось, что всевозможные
необычные раздражители, если они
приходятся во время недеятельной
фазы, устраняют эту фазу. Так же
действует двухдневное голодание. Это
наблюдается также в двигательных
рефлексах. Каждый раз в период
беспокойства от долгого стояния в
станке, а также после сильных электриче-
Рис. 227. Стадия совпадающего ре- пячггпяжений чяттячпыияюший
флекса при образовании запаздываю- ских раздражении запаздывающий
щего эффекта на правой передней рефлекс превращается в совпадающий
ноге на чесание спины. (Беритов, 5).
Собака Мура. 4-й день работы. Рефлекс _
начинается спустп 3—4 сек. от начала че- Последовательные рефлексы. В ОП-
сания (чес.) и продолжается около 6и сек.;
к этому времени производится электриче- ределенНЫХ УСЛОВИЯХ ИНДИВИДуалЬ-
ское раздражение (верхний сигнал). Верх- „ -
пня кривая от головы, средняя — от зад- ныи рефлекс начинается не во вре-
ней левой, нижняя — от правой передней
ноги. ' мя раздражения, а некоторое
время спустя. Так бывает в том случае,
если основное раздражение каждый раз производится не во время
индивидуального раздражения, а через некогоро? время после него
(рис. 229) (Пименов, 6; Гроссман, 7; Беритов, 9). В лаборатории
Павлова такие рефлексы называются следовыми. Полагают, что
в данных условиях рефлекс образуется на след индивидуального
раздражения, на его остаточное возбуждение. Такое толкование факта
следования рефлекса после раздражения не соответствует
действительности (9, 27), поэтому мы предлагаем называть эти рефлексы
последовательными, отмечая этим наступление их после индивидуального
раздражения.
Последовательные рефлексы, подобно запаздывающим, не с самого
начала образуются как таковые. Несмотря на то, что основное
раздражение происходит после индивидуального, первоначально рефлекс
является совпадающим. Впоследствии индивидуальный рефлекс частью
наступает во время раздражения, частью после него. Еще позднее он
ТИПЫ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ
521
наступает главным образом после раздражения (Добровольский, 8;
Беритов, 9). Последовательный рефлекс может развиться также сразу,
т. е. без предварительной стадии совпадающего рефлекса. Это
наблюдалось при определенных условиях на детях (Леонов, 33).
'6 4
ih 4
Г
lb s
¦vTUWr
1_1 Г
MUllltimuiiltiilliniiiiiHiii.iiiiniiimiMHi '1l{'-4'HI'l'mrimtillilHiH.M»i итіітіпщщмі|і»'"Ц"''"Щ'^"'"-''"''''"''"""Д'''"'""'М'М|1ПГ1НМЩ
Рис. 228. Образование заназдывающэго рефлекса на правой
передней ноге на чесание спины.
Собака Мура. 11-й день образования этого рефлекса. В опытах 7 и 2 дается
прерывистый, а в опыте з запаздывающий рефиекс. Кривая относится к
правой передней ноге. Поднятие нижней сигнальной линии обозначает
время чесания, опускание верхнего сигнала — момент электрического
раздражения правой передней ноги.
Последовательные слюнные рефлексы дифференцируются слабо, но
зато генерализуются сильнее совпадающих. При некоторой
настойчивости в подборе наиболее благоприятной методики можно достигнуть
Jr*VfVl
Л
Лп.
<~і Н-
¦v-u-
imtniitiiiiitnrntitniiitntni.Ptiiiiimiiiiti.tiHii,»|fHi||--fiiiiii іііііірмічіііііиііііміііііііііііініииіііп!ііііііі»імшіііііиіпіігиі
Рис. 229. Образование последовательного рефлекса на свет
электрической лампы на левой задней ноге.
Собака Мура. 6-й день работы. В опыте 1 свет произвел последовательный
рефлекс на левой задней ноге (верхний кринан), которая на этот раз
подверглась электрическому раздражению (10 см расстишшя между катушками).
В опыте 2 через 3 мни. после первого, свет вызвал как последовательный,
так и совпадающий рефлексы. В этом опыте свет не сочеталсп с
электрическим раздражением. 3 мин. позднее (опыт а) спет вызвал только
последовательный рефлекс. На сигнальной линии С отмечает время раздражения
светом электрической лампы, а цифра ю — момент раздражения
электрическим током.
значительной дифференциации (Гроссман, 7; Фролов, 10). Но сна
очень непрочна и с течением времени или по другим причинам вновь
исчезает. В опытах над двигательными рефлексами в условиях
образования последовательных рефлексов на той ноге, на которой уже были
образованы другие рефлексы, дифференциация достигалась так же
быстро, как это наблюдается в аналогичных условиях при образовании
других рефлексов. Вообще последовательный рефлекс образуется и
дифференцируется тем скорее, чем короче интервал между концом инди-
,522 ОіэЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА.
видуального раздражения и началом основного (Фролов, 10). При
некоторых больших интервалах (в 1—2 мин.) рефлекс может совсем не
образоваться (Гроссман, 7). Наоборот, если интервал равняется
нескольким секундам, последовательный рефлекс образуется и упрочивается,
как совпадающий (Петрова, 11; Белиц, 12).
Последовательные слюнные рефлексы легко переходят в
совпадающие, подобно тому как это бывает с запаздывающими рефлексами
(Гроссман, 7; Добровольский, 8). Это, например, часто наблюдается
после электрического раздражения (рис. 229). Последовательные
слюнные рефлексы непрочны и легко угасают, т. е. при пробах без сочетания
с основным раздражением могут быть вызваны раза два, не более
(Добровольский, 8).
К последовательным рефлексам следует отнести индивидуальные
рефлексы на прекращение раздражителя. Эти рефлексы были изучены
впервые Зеленым (34). Он вводил собаку в экспериментальную
комнату, когда там стучал метроном. Стук продолжался 5—20 мин., а затем
прекращался. Спустя 5—30 сек. по прекращении стука метронома собаке
вливали в рот 0,5-процентный раствор соляной кислоты. Через 4—10 мин.
метроном опять приводился в действие. После 21 раза совместного
действия кислоты и прекращения стука метронома образовался слюннной
рефлекс на прекращение стука. Подобно обычным последовательным
рефлексам рефлекс на прекращение звука мало устойчив и, кроме того,
мало специфичен. Не только прекращение обычного звука, но и всякого
другого звука вызывает рефлекс слюноотделения.
Отрицательные рефлексы. Если сочетать какой-либо необычный
раздражитель с индивидуальным раздражителем и не подкреплять этой
комбинации основным раздражением, то индивидуальный рефлекс
наступает в ослабленной форме или совсем не наступает. При
последовательном многократном повторении этой комбинации замечается
ослабление отрицательного действия необычного раздражителя на
индивидуальный рефлекс. После некоторого числа комбинаций
необычный раздражитель перестает совсем влиять на рефлекс. Если
продолжать еще дальше сочетание необычного раздражителя с
индивидуальным, то спустя некоторое время он вновь начинает действовать на
рефлекс ослабляющим образом, и наконец, вновь начинает устранять его
полностью (Васильев, 3; Миштовт, 20) (рис. 230). При этом сколько
бы раз ни повторялось сочетание с индивидуальным рефлексом,
отрицательное действие более не ослабевает. Такое явление установлено в
лаборатории Павлова и носит название условного тормо
ж е н и я.
Это название дано явлению на основании той гипотезы, с помощью
которой Павлов его объясняет. Мы же называем его
отрицательным индивидуальным рефлексом, имея в виду факт устранения или
отрицания рефлекса. Индивидуальным рефлексом мы на?ываем его на
том основании, что он обладает такими же свойствами, как и
описанные выше положительные индивидуальные рефлексы.
ТИПЫ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ
528
тг
е
IF
"~с5
200
2Q0
Что мы здесь действительно имеем дело с отрицательными
индивидуальными рефлексами, видно из следующего ряда фактов:
1. Данное раздражение теряет приобретенное отрицательное
действие легко и окончательно, если перестать с ним работать. Это
происходит тем легче, чем меньше
была предыдущая работа с А Ж Л
ним (Чеботарева, 19).
2. При повторении
отрицательного раздражения без
сочетания с положительным
индивидуальным раздражением
отрицательное действие его
исчезает. Аналогичным
образом теряется положительное
действие положительного
индивидуального сигнала при
повторении его без сочетания
с прирожденным рефлексом
(Чеботарева, 19).
3. Угасание отрицательного
рефлекса путем повторения
без сочетания с
положительным индивидуальным
рефлексом не является абсолютным.
После перерыва отрицательное
действие восстанавливается, но
всегда не полностью. Для
полного восстановления необходимо повторное сочетание с положительным
рефлексом (Чеботарева, 19).
4. При выработке отрицательного действия на данное раздражение
первоначально это свойство приобретается не только им, но и другими
необычными раздражениями того же качества, т. е. происходит
генерализация отрицательного действия (Миштовт, 20; Кржышковский, 21).
Лишь позднее, в результате длительной работы, может быть достигнуто
полное дифференцирование данного раздражения в его отрицательном
действии (Николаев, 16; Лепорский, 28).
Мысль о возможности образования отрицательного индивидуального
рефлекса высказывалась и самим Павловым (17) и многими его
сотрудниками, например Фольбортом (15), Николаевым (16).
Они рассматривают раздражение, вызывающее отрицательный рефлекс
как сигнал к торможению. Лишь некоторые авторы пытаются на
основании анализа явлений провести более детальную аналогию между
отрицательным и положительным индивидуальными рефлексами
(Николаев, 16).
Отрицательный рефлекс обычно образуют так: сперва начинается
постороннее раздражение, а спустя несколько секунд после его начала
— гоо 20о
И1.1 U п,пиіш-ипиіппит; і,мтіт.|,ім1ч|М ііпіпмпмі...щР(тт.1>
Рис. 230. Действие отрицательного
индивидуального раздражителя — света электри
ческой лампы на положительный
индивидуальный рефлекс правой передней ноги — на
звук тонвариатора 200 колебаний в 1 сек
Собака Боб. В опыте 1 сначала производится отри
дательное индивидуальное раздражение—свет
электрической лампы (средний сигнал). К нему
присоединяется положительный индивидуальный звук тон-
вариатора 200 колебаний в 1 сек. (нижний сигнал),
и последний не вызывает рефлекса. В опыте 2
производится та же комбинация в обратном порядке.
Лампа зажигается во зремя происходящего рефлекса
на звук. Последний во время комбинации очень
постепенно ослабевает и спустя іО сек. исчезает
полностью. В опыте 3 производится сочетание
индивидуального рефлекса на звук тонвариатора 200
колебаний в і сек. с основным электрическим
раздражением (верхний сигніл с цифрой 6 служит для
обозначения силы раздражения в сантиметрах расстояний
между индукционными катушками). В опыте 4 спустя
4 мин. после опыта 3 дается такая же комбинация.
как в опыте 1. Но на этот раз рефлекс не исчезав)
иод влиянием света, он только становится слабее
524
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
присоединяется положительное; затем оба раздражения действуют
вместе, вместе же и кончаются. При этом образуется, так сказать,
совпадающий отрицательный рефлекс. В этом случае индивидуальный
рефлекс устраняется только при совпадении с отрицательным
раздражением. Но можно образовать и последовательный отрицательный
рефлекс, если положительное индивидуальное раздражение следует
после индифферентного. Фролову (10) удалось образовать таким
путем последовательный отрицательный рефлекс сначала с
интервалом в 0.5 мин., а потом удлинить этот интервал до 1 мин. Здесь
индивидуальный рефлекс устраняется последействием отрицательного
раздражения. Наконец, отрицательный рефлекс удалось образовать при
обратном порядке сочетания, т. е. когда сначала идет положительное
индивидуальное раздражение, а затем к нему присоединяется
постороннее. При этом получается совпадающий отрицательный рефлекс
(Фурсиков, 18).
Отрицательные рефлексы чрезвычайно непостоянны. Всякое
условие, влияющее на усиление положительного индивидуального рефлекса,
благоприятствует ослаблению и даже исчезновению отрицательного
действия на него отрицательного индивидуального рефлекса. Так
действует длительный отдых, предварительное сочетание положительного
индивидуального раздражения с сильным основным раздражением,
беспокойное состояние животного. На рис. 230 свет электрической
лампочки является индивидуальным отрицательным раздражителем (опыты J
и 2). Но тотчас же после сочетания индивидуального положительного
рефлекса с электрическим раздражителем (опыт 5), свет не устраняет
положительного рефлекса в последующей комбинации (опыт 4),
Вторичные проприоцептивные рефлексы. Индивидуальные
двигательные рефлексы образуются не только на раздражение внешних
рецепторов, но и на проприоцептивные раздражения, т. е. раздражения
рецепторов мышц, сухожилий и суставов.
Что вообще проприоцептивные раздражения могут стать
индивидуальными раздражителями, было уже известно в лабораториях
Павлова и Бехтерева: пассивное поднятие ноги превращалось в
индивидуальный сигнал к слюнной секреции или к оборонительному
поднятию ноги (Красногорский, 35; Архангельский, 36; Израэльсон, 37).
При этом было доказано, что здесь важно именно проприоцептчвное
раздражение, а не раздражение кожи или глаза видом согнутой ноги.
В данном случае проприоцептивное раздражение является первичным
индивидуальным раздражением.
Проприоцептивные раздражения становятся индивидуальными также
в том случае, если они возникают вторично под влиянием первичного
оборонительного рефлекторного движения, и это происходит при
образовании каждого индивидуального оборонительного рефлекса, так как
каждый раз с основным раздражением сочетается не только
индивидуальное раздражение, но и те проприоцептивные раздражения, которые
возникают во время первичных рефлекторных движений (Беритов, 27)
ТИПЫ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ
525
Очень демонстративно проявляется индивидуальный
оборонительный рефлекс на проприоцептивное раздражение, если раза два подряд
раздражать ногу электрическим раздражением в момент ее опускания.
В следующий раз тотчас же после опускания собака дает сильную
общую реакцию: визжит, двигает головой и ногами. Эта
оборонительная реакция, очевидно, возникает в результате образования
индивидуального оборонительного рефлекса на проприоцептивные
раздражения, происходящие в момент опускания ноги.
Такого рода вторичные рефлексы образуются особенно быстро при
сочетании индивидуального сигнала с многократным электрическим
раздражением. После нескольких дней работы рефлекс как на
индивидуальный сигнал, так и на электрическое раздражение принимает
характер длительного сгибания: нога после начального сильного
сгибания удерживается в более или менее согнутом положении десятки
секунд. Все это последействие рефлекса является вторичным проприо-
цептивным рефлексом индивидуального происхождения.
Вторичные проприоцептивные рефлексы, конечно, образуются под
влиянием повторных электрических раздражений также при отсутствии
сочетания с индивидуальным раздражением, т. е. без участия
индивидуального рефлекса на какое-либо внешнее раздражение (Беритов, 27).
Вторичные рефлексы, несомненно, не только всецело обусловливают
последействие индивидуального рефлекса, но и влияют на первичное
индивидуальное движение в смысле его усиления.
Ритмические рефлексы. При определенных условиях двигательный
индивидуальный рефлекс протекает ритмически. Нога много раз то
опускается, то поднимается; иногда фазы сгибания и разгибания
следуют одна за другой с довольно правильным ритмом за все время
индивидуального сигнала, но опускание редко бывает полным. Новое
поднятие начинается еще раньше, чем нога соприкоснется с полом.
Часто первые опускания бывают значительнее, чем последующие. От
этого размах движения постепенно уменьшается, и напоследок нога
надолго остается в согнутом положении.
Ритмическое повторение рефлекса наблюдается в случае сочетания
индивидуального раздражения с многократным электрическим
раздражением и представляет собой следствие образования проприоцептивных
рефлексов вторичного происхождения (Беритов, 9). После того как
образовался ритмический рефлекс, он вызывается не только
индивидуальным, но и основным электрическим раздражением. Но это бывает
не всегда. Иногда электрическое раздражение производит поднятие
ноги на все время его действия, тогда как индивидуальный рефлекс
протекает ритмически (рис. 231).
Интероцептивные индивидуальные рефлексы. Подобно проприоцеп-
тивным и экстероцептивным индивидуальным рефлексам образуются
и интероцептивные слюнные и оборонительные рефлексы, т. е. на
раздражение внутренних органов. Так, например, вливанием воды
(200 см3) в желудок через желудочную фистулу многократно вызывал-
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГЛ
ся повышенный диурез, т. е. повышенное мочеотделение. Впоследствии
вливание воды на 1—2 мин. (вода потом выливалась обратно) также
вызывало повышение мочеотделения. Таким образом, орошение стенок
желудка водой превратилось в индивидуальный сигнал к усиленной
деятельности почек (Быков,41, 45). Если кратковременное орошениг
стенок желудка сочетается с электрокожным раздражением или
раздражением полости рта пищей или
кислотой, то сравнительно быстро
образуются индивидуальный оборонительный
или индивидуальный пищевой и
кислотные рефлексы на вливание воды в
желудок (Айрапетьянц, 42).
Эти рефлексы дифференцируются.
Например, можно образовать
индивидуальный рефлекс на орошение стенок
желудка теплой водой при температуре
36°, отдифференцировать орошение водой
при температуре 26° (Айрапетьянц, 42).
Рефлекс времени. Если повторять
основное раздражение — кормление —
в сочетании с индивидуальным
раздражением или без него через строго
определенный промежуток времени,
измеряемый минутами (до 30 мин.), то после
100—200 повторений слюнный рефлекс
наступает сам собой, без какого-либо
сигнала к сроку основного раздражения.
Это явление впервые наблюдалось 3 е-
л е н ы м (29) и было названо рефлексом времени. Зеленый
применял основное раздражение каждые 10 мин. В промежутках
животное нередко івпадает в сонное состояние, но оно не мешает
наступлению рефлексов в срок,— наоборот, благоприятствует своевременному
его наступлению.
Рефлекс наступает только один раз после последнего основного
раздражения, так что начальным моментом для отсчета времени является
эффект, вызываемый основным или соответствующим индивидуальныу
раздражением, если последнее обычно употреблялось в сочетании с
основным (Феокритова, 24; Стукова, 25).
Можно также образовать индивидуальный двигательный рефлекс
времени. В качестве основного рефлекса берется оборонительный
рефлекс сгибания, вызванный электрическим раздражением кожи. Если,
например, он производится через каждые 5 мин., то рефлекс времен;*
будет наступать через 5 мин. после основного раздражения (Бери-
тов, 27).
Рефлекс времени в последнее время был получен также на
черепахе. Именно, Никифоровский (38) образовал оборонительный
326
380
JL
гэо sso зве
Рис. 231. Ритмический рефлекс на
левой задней ноге в ответ на звук
тонвариатора 29 J колебаний в сек.
Собака Белый. 5-й день образования
рефлекса. Генерализованное состояние.
В опытах J и 2 ритмический рефлекс
вызван необычным звуком 386
колебаний в 1 сек., а в опыте 3 — обычным
индивидуальным звуком.
ТИПЫ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 527
рефлекс втягивания головы на промежуток времени 4 мин. Основное
раздражение состояло в механическом раздражении головы.
Рефлекс времени на собаке образуется при условии применения
основного раздражения через один и тот же промежуток. В остальное
время собака испытывает действие одной лишь обычной обстановки.
В эту обычную обстановку входит не только вид комнаты, но и стояние
на ногах, связанное с целым рядом проприоцептивных раздражений,
затем механическое раздражение кожи лямками и т. д. Повидимому,
обычная обстановка является индивидуальным раздражителем для
рефлекса времени. Это видно, например, из того, что если сначала
образовать такой рефлекс в присутствии экспериментатора и затем
изолировать собаку от него в особую комнату, то существующий рефлекс
времени исчезает окончательно, и наступает вновь лишь после новой
долгой работы (Дерябин, 26).
Кроме того, рефлекс времени развивается по типу запаздывающего
рефлекса: сначала индивидуальный рефлекс наступает в различные
сроки после предшествующего основного раздражения, и только
постепенно начало его переносится к сроку основного раздражения; при
зюм, как при запаздывающем рефлексе, сонное состояние в неактивной
фазе благоприятствует наступлению рефлекса точно к сроку. Если
собака не засыпает и вообще находится в беспокойном состоянии, то
рефлекс наступает до срока, иногда много раз, лишь значительно
усиливается к сроку. Возникшее в промежутке беспокойное движение
также заметно усиливается к сроку раздражения (Беритов, 27).
Произвольные индивидуальные рефлексы. Так называются
индивидуально-приобретенные рефлексы, образуемые по методу
индивидуальных рефлексов на произвольные акты. Как уже указывалось выше,
характерный для корковой деятельности человека альфа-ритм может
вызываться и угнетаться индивидуальным путем. Например, если
несколько раз сочетать звук с затемнением глаз, он становится
индивидуальным сигналом к появлению альфа-ритма. Если же звук
сочетается с освещением глаз, тогда он становится индивидуальным сигналом к
исчезновению альфа-ритма. Оказывается, индивидуальным сигналом
к альфа-ритму может явиться такой произвольный акт, как произнесение
слова «блок», если повторять его беззвучно, т. е. без произношения
вслух и в это время освещать глаз. Само по себе это слово никак не
влияет на корковую электрическую активность, но после такого
сочетания слово «блок» угнетает альфа-ритм. У одного субъекта это случилось
после 5 сочетаний, а у другого после 4 сочетаний. Интервал между
сочетаниями определяется субъектом. На основании инструкции он
должен был после беззвучного произнесения слова «блок» зажать
резиновую грушу, с помощью которой зажигалась электрическая лампа.
Временами экспериментатор выключал ток, груша зажималась, но
лампа не зажигалась. Несмотря на отсутствие света при слове «блок»-
альфа-ритм исчезал (Джаспер и Шагас, 43).
523
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
Очевидно, не только внешнее раздражение, но и любой
произвольный умственный процесс может стать индивидуальным сигналом по
отношению к определенному корковому процессу.
К произвольным индивидуальным рефлексам следует относить все
ассоциации, возникающие в результате заучивания, т. е. многократного
повторения определенного ряда восприятий. Это, например, имеет
место при заучивании стихотворения. Специальными опытами над
заучиванием бессмысленных слогов в определенном порядке
устанавливали характер связей между этими слогами. Было, между прочим,
установлено, что каждое воспроизведение одного слога ведет не только к
воспроизведению следующего за ним, но и всех последующих слогов,
перескакивая через промежуточные, только переход одного слога к
следующему за ним непосредственно более постоянен и устойчив, чем
переход к отдаленным слогам. Было также установлено, что при
заучивании бессмысленных слогов в одном порядке воспроизведение одного
слога ведет к воспроизведению не только последующих, но и
предыдущих слогов, однако этот обратный переход менее постоянен, требует
больше повторений, чем обычный переход в поступательном порядке
(Эббингхаус, 44).
Практикум
Методы исследования. Индивидуальные слюнные рефлексы изучаются в школе
Павлова на слюнных железах собаки. Собака помещена в особой камере, куда не
полагается входить во время опыта. Экспериментатор находится >в другой комнате.
Собака стоит в станке, к которому привязана так, что не может ни лежать, ни
ходить, а только может стоять или приседать на задние ноги, если несколько опустить
лямки, которыми она привязана к станку (рис. 232). Обстановка в комнате собаки
всегда одна и та же. Здесь находится кормушка, в которой животному <в
определенное время подается еда, здесь же находятся различные аппараты для
индивидуальных раздражений,— тонвариатор Штерна, метроном, электрически"'! звонок,
электрическая лампа, вертушка для раздражения глаз и т. д. Все эти аппараты, так же как и
кормушка, приводятся в действие из комнаты экспериментатора.
Проток слюнных желез выведен наружу по методу Глинского и приживлен на
щеке или на подбородке животного. Для учета количества выделенной слюны
приклеивается посредством менделеевской замазки маленький стеклянный пузырь,
широким открытым отверстием к месту выхода протока, предварительно выбритому.
Стеклянный пузырь снабжен двумя отростками в виде тонких трубочек. Одна из этих
трубочек посредством каучуковой трубки соединяется с горизонтальной стеклянной
трубкой длиной в 60 см и диаметром 3—4 мм, которая находится в комнате
экспериментатора. Эта трубка наполнена водой. Выделившаяся слюна вытесняет некоторое
количество воздуха из пузыря, а воздух в свою очередь вытесняет соответствующий
объем воды из стеклянной трубки. Другой трубчатый отросток пузыря соединен со
стеклянной банкой, куда пересасывается из пузыря выделившаяся слюна. Таким
образом, экспериментатор в состоянии, наблюдая за трубкой, отсчитать количество
слюны, выделившейся из железы, и затем удалить ее из пузыря (рис. 232). Слюнная
секреция может регистрироваться на кимографе. В этом случае из горизонтальной
трубки жидкость каплями падает на рычаг Мареевской капсулы. Эта капсула
посредством каучуковой трубки связана с другой капсулой, которая отмечает на кимографе
вертикальную черточку от падения каждой капли (рис. 223). Во время опыта
соблюдается тишина. Комната собаки остается закрытой (см. подробно книгу
Подкопа е в а. Методика изучения условных рефлексов. Госиздат, 1926).
ТИПЫ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОЙ
529
Индивидуальный слюнной рефлекс образуется следующим образом. Каждый раз*
во время еды производится одно и то же внешнее раздражение. Это раздражение
после нескольких сочетаний с едой вызывает такую же слюнную реакцию, как еда.
Слюнная секреция бывает не только при еде, но и при любом химическом
раздражении ротовой полости, например, соляной кислотой, перцем и т. д. Она бывает и при
определенном механическом раздражении, как всыпание в рот измельченных
камешков, песку и т. д. Поэтому для образования индивидуального слюнного рефлекса на
данное раздражение нет необходимости сочетать его с едой,— этот рефлекс обра-
Рис. 232. Методика, применяемая в настоящее время в лабораториях Павлова
для изучения слюнных рефлексов.
А — отделепие собаки; в — отделение экспериментатора Собака стоит в станке. К — кормушка?
Р — перегородки, укрывающая кормушку; на правой стороне перегородки вырез, через который
кормушка подается собаке потмгиванием веревки; а - пузырь над слюнпои фистулой; б —
стеклянный приборчик для отсчитывания слюны. Система трубок оос /—для отсчитывания
выделившейся слюны; бнтл-длп удаления слюны; С- стена, отделяющая комнату собаки от
экспериментатора (составлен по рисункам Нодкопаева).
зуется при сочетании его с любым раздражителем ротовой полости, который вызывает
секрецию слюны. Так как секреция слюны значительно меняется как количественно,
так и качественно, в зависимости от качества раздражителя ротовой полости, то
сообразно с этим и индивидуальный слюнной рефлекс меняется как качественно, так и
количественно. Индивидуальный слюнной рефлекс может образовываться на любое
внешнее раздражение, как вызывающее определенную реакцию в организме, —
например электрическое раздражение ноги,— так и вполне недеятельное,
индифферентное, вроде обычного звука или обычного вида предмета. Так как экспериментатор
и собака находятся в разных комнатах, то как кормление собаки, так и
индивидуализируемые раздражения производятся автоматически из комнаты экспериментатора.
Для этого существуют сложные приспособления (см. цитированную выше книгу Под-
копаева).
Индивидуальный оборонительный рефлекс образуется путем сочетания
электрического раздражения кожи на ноге с индивидуализируемым раздражением. Изучение
ведется путем регистрации на кимографе движений ног и головы. В школе
Бехтерева, где впервые применен этот метод, регистрировали движение только одной
ноги, на которой образовывался индивидуальный рефлекс, и кроме того дыхательные
движения. В опытах Беритоэа регистрировались сразу две ноги и голова: одна
нога—на которой образовывается индивидуальный рефлекс, другая нога — для
наблюдения за общим движением животного; голова же служила для наблюдения за
ориентировочной реакцией.
Электроды надеваются на нижнюю часть голени н предплечья или же немного
выше пальцев. На раздражаемом участке шерсть предварительно выбривается.
Раздражение производится из комнаты экспериментатора. Движения ног и головы пере-
34 и. С. Беритов
530- ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
даются к миографам при посредстве ниток через систему блоков. Индивидуальным
раздражением могут служить звуки тонвариатора Штерна, метронома, электрический
звонок, звуки металлического цилиндра, свет электрической лампы, чесание спины,
как это делается и в лаборатории Павлова. Время каждого раздражения
регистрируется на кимографе электромагнитными сигналами. Обстановка опыта схематически
показана на рис 233.
Рис. 233. Схематическое изображение остановки олыга при изучении индиви
дуальных двигательных рефлексов.
В комнате А находится собяка. Она привязана к станку на столе лямками ал. От головы и
от левой и правой передних ног протянуты толстые нити ааа, bbb, ссс через блоки к миографам
гд, ей, мсз в другую комнату В, где стоит кимограф и где экспериментатор производит все
нужные для опыта маялпуляции, Т — тонвариатор, который резиновой трубкой (Я) соединен с
аппаратом для нагнетания воздуха. К — ключ от вторичной цепи индуктория, откуда ток идет
по проводам к электродам для раздражения. На рисунке вид опыта очень упрощен.
Вышеописанная методика, вырабатывавшаяся годами для точной регистрации
слюнных и двигательных реакций, мало годится для практических занятий со
студентами. Для освоения ее студентам понадобится так много времени, что для постановки
основных опытов у них времени не останется. Кроме того, для практических занятий
йужна такая методика, которая могла бы легко быть создана в любой лаборатории
без особенных затрат. Такой методикой является изучение индивидуальных реакций в
свободно подвижных условиях. Она применяется для изучения поведения животных,
но может быть использована и для изучения индивидуальных рефлексов. Для этой
цели сначала путем многократного повторения одного и того же индивидуального
акта поведения в одной и той же обстановке производят автоматизацию
индивидуального поведения. В индивидуальных автоматизированных актах поведения каждый
отрезок поведения является индивидуальным рефлексом, наступающим в ответ на
воздействие внешней среды (Беритов, 39, 40). Обычно для изучения индивидуальных
рефлексов мы пользуемся начальным отрезком поведения, начальным движением
животного, наступающим непосредственно в ответ на индивидуальное раздражение.
Выработка пищевого двигательного рефлекса у голубей. Голубь сидит в клетке.
Вместо клетки можно взять стол, огороженный проволочной сеткой. Чтобы голубь не
улетал, у него обрезают маховые перья на крыльях. В углу стола стоит кормушка —
маленький деревянный ящик с крышкой. Экспериментатор снимает крышку
потягиванием за веревку. Открытую кормушку, куда кладут хлебные крошки или зерно,
можно подвесить на нитках на определенной высоте и в определенный момент ее
опускать. Соответствующая методика для изучения как пищевого, так и защитного
поведения индивидуальных рефлексов у мелких животных, в частности, у голубей,
дана на рис. 234.
Прежде всего голубя приучают к тому, чтобы он подходил к кормушке и клевал
корм, как только кормушку опустят или откроют. На это понадобится несколько дней.
Рис. 234. Методика изучения индивидуальных рефлексов на мелких животных.
Стол огорожен проволочной сеткой. Он разделен картонной перегородкой. Впереди другой стол,
на котором расположены приборы для индивидуального раздражения: метроном, электрический
звонок и тонвариатор Штерна, затем индукторий с аккумулятором для основного электрического
раздражения. Там же стоит соска (с) для основного пищевого раздражения при работе со щенками.
Электроды для накладывания на ногу висят на проволочной стенке (э).
Чтобы избегнуть лишней траты времени на приручение, лучше всего предоставить
студентам уже прирученных голубей. Само это приручение есть образование
индивидуальной пищевой реакции: при виде опускания или открывания кормушки голубь
подходит к ней, даже если он не видит самого корма. Это будет так называемый
натуральный индивидуальный рефлекс. Но для изучения индивидуальных рефлексов
лучше пользоваться искусственными рефлексами.
Искусственный индивидуальный рефлекс образуется таким образом: сначала
производится индивидуализируемый звук или свет (метроном, электрический звонок,
музыкальный тон тонвариатора, зажигание электрической лампочки, причем комнага
затемняется или клетка покрывается материей), и затем через 5—10 сек. от начала
раздражения присоединяется открывание или опускание кормушки с кормом. При
индивидуализируемом сигнале голубь подходит и клюет в течение 10—12 сек.; затем
кормушку закрывают или поднимают. Одновременно прекращается сигнал.
У хорошо прирученных голубей пищевой двигательный рефлекс на
искусственный сигнал развивается уже в первый день работы, после 4—10 комбинаций
индивидуального сигнала с кормлением. У совершенно новых, еще не вполне прирученных
птиц для развития рефлекса может потребоваться несколько дней работы, несколько
десятков сочетаний. Индивидуальный пищевой рефлекс заключается в усиленных
ориентировочных движениях, в передвижении в сторону кормушки, затем в
глотательных движениях, наступающих еще раньше, чем голубь начнет клевать зерно. Этот
слджный. индивидуальный рефлекс в первые дни работы иногда не наступает, но в
следующие дни наступает регулярно и с большой интенсивностью.
• • •• ¦
34*
532
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
Дифференциация рефлекса у голубей. Если пищевой рефлекс был образован на
музыкальный звук, то стук метронома с самого, начала не вызовет пищевого
рефлекса, т. е. голубь не пойдет к кормушке. Точно так же не вызовет его зажигание
электрической лампочки, но музыкальные звуки вызывать его будут. Чтобы эти звуки
также не давали рефлекса, нужно их пробовать повторно, не открывая, конечно, при
этом кормушки и не давая еды. Таким путем легко можно достигнуть того, что звуки,
различающиеся только на несколько тонов, перестанут вызывать пищевую
реакцию.
Если индивидуальным сигналом является определенный звонок, то метроном я
музыкальные звуки с самого начала не дадут эффекта, но другие необычные звонки
будут его вызывать. При повторении, однако, и они перестанут вызывать данный
рефлекс; особенно легко удается эта дифференциация, если необычный звук раздается в
другом месте, с другой стороны клетки.
Образование индивидуальных оборонительных рефлексов у голубей. В качестве
основного раздражения пользуются электрическим раздражением. Фарадический ток
от индуктория Дюбуа-Реймона подводится легким мягким шнуром к ноге голубя.
Концы проводов наматываются ©а бинт на протяжении нескольких миллиметров я
затем этим бинтом обвязывают ногу так, чтобы провода, т. е. электроды, приходились
на оголенной кожной поверхности пятки. Электрическое раздражение вызывает
сильное ориентировочное движение головы, взлет голубя, т. е. движение крыльев,
топтание ногами на месте. Если это раздражение сочетать со звуками или светом, как это
описано выше, то после нескольких или многократных сочетаний эти звуки или свет
<5удут вызывать индивидуальный оборонительный рефлекс, вернее — индивидуальное
поведение. После укрепления этой реакции можно изучить ее дифференциацию, как
в случае пищевой двигательной реакции.
Образование пищевого двигательного и оборонительного рефлексов у кроликов.
Хорошим объектом для изучения индивидуальных реакций является также кролик.
Опыты на нем можно вести совершенно так же, как на голубях, в клетке или на столе,
огороженном проволочной сеткой. У кроликов можно образовать как пищевую, так и
оборонительную реакцию. Основным пищевым рефлексом может служить поедание
капусты; пища подается в кормушке, как голубям. Основной оборонительный рефлекс
вызывается электрическим раздражением ноги. Электроды накладывают на то место
ноги, где шерсть предварительно была выбрита. Оборонительный рефлекс у кролика
выражается в сильном ориентировочном движении, подпрыгивании, сильном ударе
задних ног, писке и т. д.
Кролики, как и голуби, очень пугливы, и прежде чем начать на них образование
индивидуальных реакций, следует хорошенько приучить их к себе, к поеданию пищи
из кормушки как только ее опустят или откроют, и вообще к условиям опыта. Только
после этого приступают к образованию искусственных индивидуальных реакций.
Затем изучают те же явления, что и в опытах на голубях.
Образование индивидуальных рефлексов у щенков. Наилучшим объектом для
изучения индивидуальных реакций по данному здесь упрошенному методу являются
двух-трехмесячные щенки. Они легко привыкают к экспериментатору и вообще к
условиям опыта. Кроме того, индивидуальные реакции у них образуются и
дифференцируются быстрее, чем у голубей и кроликов. У них также можно образовывать как
пищевые, так и оборонительные реакции. В качестве основного пищевого рефлекса
можно взять молоко и подавать его из соски, какую употребляют для грудных детей,
но можно подавать молоко с хлебом и из маленькой чашки, которая стоит в кормушке.
Кормушку открывают потягиванием веревки. Оборонительная реакция вызывается
электрическим раздражением, как у кроликов. Ставятся такие же опыты, как на
кроликах и голубях.
Образование индивидуальных реакций у взрослых собак по методу свободных
движений. Для изучения индивидуальных реакций на взрослых собаках лучше всего
пользоваться методикой, которая служит для изучения у них поведения. Эта методика
настолько проста, что может легко применяться в любой лаборатории. Собаку
приучают лежать в одном месте, лучше всего на определенной лежанке, и есть хлеб из
ТИПЫ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ
533
кормушки» которая стоит где-нибудь в комнате и не должна быть видна собаке: ее
можно заслонить шкафом или ширмой. Кормушка представляет собой ящик с
круглым- отверстием. В определенные моменты под это отверстие автоматически,
притягиванием веревки, передвигается миска с едой (рис. 235), но» конечно, можно взять
любой ящик с крышкой. В ящик кладут миску с пищей, открывают его потягиванием
веревки. Кормушку открывают на 5—10 сек., чтобы собака могла захватить несколько
а
гмшпхцшіш/муя
Ь___/м»»м.;..>щ
Рис. 235. Кормушка для работы на собаках.
Наверху — наружный вид; внизу — в разрезе. Кормугака в
обоих случаях закрыта Потягиванием веревки (а) вращается
ось (о), и чашка (с) становитсп на место дощечки (d) дод
отверстием кормушки (6)
небольших кусков хлеба, и затем закрывают. Приучают также собаку после еды
сейчас же возвращаться обратно на лежанку. Вся эта предварительная работа
производится в течение 3—5 дней.
Для образования индивидуальных пищевых двигательных реакций дают тот или
иной искусственный сигнал и затем подзывают собаку к кормушке. Вместо слова и
подзывания можно пользоваться шумом кормушки, который производится
открыванием и закрыванием крышки. Уже в результате первых кормлений у собак
вырабатываются индивидуальные реакции на шум открывания кормушки: на этот шум собака
срывается с лежанки и бежит к кормушке. После ряда сочетаний этих реакций с
каким-либо посторонним звуком последний становится сигналом к этой пищевой
реакции.
Сигнальные раздражения даются со стола экспериментатора, который *стоит в
нескольких метрах от собаки за ширмой так, чтобы собака не могла видеть пи
экспериментатора, ни его движений В ширме имеется продольная щель, через
которую наблюдают за животным. Сигнальные раздражители могут стоять и рядом с
кормушкой. Для быстроты образования рефлекса это имеет большое значение, так
как они значительно быстрее превращаются в индивидуальные сигналы, когда
находятся у кормушки. В таких случаях раздражители являются как бы натуральными
сигналами, как вид пищи.
Конечно, мы и здесь обращаем главное внимание на начальное движение собаки,
которое наступает непосредственно в ответ на искусственное раздражение.
Порядок регистрации наблюдаемых реакций. Наблюдаемые у всех животных
реакции подробно записываются в тетрадь.
Как видно из примерной записи, в 1-м столбце отмечают время опыта (в котором
часу производится раздражение); во 2-м — какое раздражение производится;
53 t ОБЩАЙ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
I II
>
^ Время
12 ч. 5 м.
Характер
pajrfpaw e гоя
Звонок -f-еда
Ш
Порядковый
номер
раздражения
J (SO)
IV
Продолжи-
тель 'Ость
индивид)аль-
ного раздра-
)* ения
10 сек.
V
Положение
ссба си до
раздраи ечия
Лежит,
повернув
голову
направо
VI
Характер реакции
На 1 сек. поднимает
голову; на 2 сек.
встает и идет к кормушке;
после закрытия
кормушки на 5 сек.
поворачивается от кормушки; не
останавливаясь,
возвращается на лежанку
в 3-м — порядок сочетания; при этом ставят две цифры: одна указывает порядок
сочетания за день работы, а вторая — за весь период работы с данным сочетанием,
в 4-м столбце дается продолжительность индивидуального раздражения; в 5-м —
указывается положение собаки на лежанке в момент раздражения: стоит, сидит, лежит,
куда смотрит и т. д.; в 6-м столбце подробно записывают, какие возникают реакции
и через какой промежуток времени от начала раздражения они начинаются.
Каждый день работа продолжается 1—2 часа, В день можно сделать 10—15
сочетаний. В случае работы над пищевыми реакциями животное с утра не кормят.
Экспериментатор обязан каждый день подводить итоги своим наблюдениям:
отмечать в тетради, сколько раз применялось то или другое раздражение за этот день;
какой получился результат; затем нужно сравнить этот результат с полученными в
предыдущие дни; отметить также все новые моменты в реакциях животного.
Литература
1. В е і- t о і f J., Brain, 47, 109, 1924.
2. В е г і t о f f J.. B-ain, 47, 359, 1924.
3. В а с и л ь e в П., Тр. Об-ва русск. врачей, 73, 389, 1906.
4. Завадский И., Материалы к вопросу о торможении и растормаживании
условных рефлексов. Дисс, 1908
5. Be r i t of f J., P^ych. Neur., 32, 262, 1925.
6. Пименов И., Особая группа условных рефлексов. Дисс, 1907.
7. Гроссман Ф., Материалы к физиологии следовых условных слюнных
рефлексов, Дисс, 1909.
8. Добровольский В., О пищевых следовых рефлексах. Дисс ,. 1911
9. В е ri t о f f J., J. Psych. Neur., 32, 29, 1925.
10. Фролов И., Арх. биол. наук, 24, 102, 1924.
11. Петрова М., Арх. биол. наук, 20, 1, 1914.
12 Белиц М., Об условных следовых рефлексах. Дисс, 1917.
13. Кунстманн К., Известия Научного ин-та им. Лесгафта в Ленинграде, 7,
59, 1923.
14. В е г і t о f f J., J. Psych. Neur., 32, 253, 1925.
15. ФольбортГ., Тр. Об-ва русск. врачей, 80, 245, 1910.
16. Николаев П., К физиологии условного торможения. Дисс, 1910.
17. Павлов И., 20-летний опыт. 3-е изд., 1925.
18. Ф у р с и к о в Д., Тр. Физиол. лаб. акад. Павлова, 1. 2, 1925.
19. ЧеботареваО., Дальнейшие материалы к физиологии условного торможения.
ДисСм 1907.
ТИПЫ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ
535
.20. Миштовт Г., Выработка торможения искусственного условного рефлекса на
слюнные железы. Дисс, 1907.
21. К р ж ы ш к о в с к и й К., К физиологии условного тормоза. Дисс, 1909.
22. Зеленый Г., Русск. физиол. журн., 6, 155, 1923.
23. Бехтерев В., Общие основания рефлексологии человека. 2-е изд., стр. 264.
24. Феокритова Ю., Время как условный возбудитель слюнной железы. Дисс,
1912.
25 Стукова М., Дальнейшие материалы к физиологии времени. Дисс, 1914.
26. Дерябин В., Дальнейшие материалы к физиологии времени как условного
возбудителя слюнных желез. Дисс, 1916.
27. В е г і toff J., Ueter d. indiv.-erworb. Tatigkeit d. ZNS, J. Psych. Neur., 33, 113,
1927. Индивидуально приобретенная деятельность центральной нервной
системы. Тифлис, 1932.
28. Лепорский Н.? Материалы к физиологии условного торможения. Дисс.
т. 1, ч. II.
29. 3 е л е н ы й Г., Материалы к вопросу о реакции собаки на звуковые раэдражения
Дисс, 1907.
30. Бы ков К. иПетрова М., Тр. Физиол. лаб. акад. Павлова, т. II, в* 1* стр; §1,
1927.
51. Мясищев В., Новое в рефлексологии и физиологии нервной системы» 1, 291
1925.
52. БеритовИ. и Дзидзишвили Н., Тр. биол. сект. Груз. отд. АН СССР,
т. 1,ч. II, стр. 1, 1933.
53. Леонов В. PfL Arch., 214, 304, 1926.
"34. Зеленый В., Доклад в Об-ве русск. врачей в Петербурге 20 мая 1907.
35. Красногорский Н., О процессе задержания и локализации кожного и
двигательного анализатора и т. д. Дисс, 1911.
56. Архангельский В., Арх. биол. наук, 22, 5S, 1922.
57. Израольсон Ж., О топической дифференцировке сочетательного рефлекса
etc Дисс из лаб. Бехтерева, 1913.
38. Никифоровский П., Русск. физиол. журн., 12, 483, 1929.
39. БеритовИ. иБрегадзеА., Мед.-биол. журн., 5-й г., в. 4, 83, 1929.
40. Б е р и т о в И., Советск я психоневрология, № 3, 1933.
41. Быков К. М., IV Всес оъезд физиологов. Тезисы и авторефераты докладов,
стр. 41. 1930; Пробл. биол. и мед., Сб., посвящ. J]. Штерн, стр. 73, 1935.
42. АйрапетьянцЭ. Ш., Сб. докладов VI Всес физиол. съезда, стр. 176.
1937
43. Jasper Н. a. Shagass Ch., J. Exper. Psych., 28, 503, 1941.
44. Эббингхаус, Основы психологии. 1912. Пер. с нем.
¦45 Быі.ов К. Мм Кора головного мозга и внутренние органы. Изд. Военно-Морск*
Акат > Кирор 1942
Глава VII
ЗАКОН СОПРЯЖЕННОЙ ИРРАДИАЦИИ ВОЗБУЖДЕНИЯ
Определение закона сопряженной иррадиации возбуждения и его
физиологические основания. При изучении корковых реакций обращает
на себя внимание следующее характерное явление. Каждый раз
вызванный новым внешним раздражением корковый рефлекс, будь он
индивидуальный или прирожденный, представляет очень обширный и
сложный эффект. При повторении этого раздражения эффект
становится все более и более простым и ограниченным. Так, например, более
или менее сильное электрическое раздражение ноги первоначально
вызывает не только рефлекс сгибания данной ноги, но и движения других
ног; животное передвигается взад и вперед в станке, дрожит всем
телом. Оно обнаруживает также усиленную ориентировочную и
голосовую реакции и углубленное дыхание. Очевидно, электрическое
раздражение ноги производит этот обширный эффект в силу самой
прирожденной организации центральной нервной системы, как через кору,
так и помимо нее, как представлено на рис. 236. Но >в общем вся
реакция имеет определенный поведенческий характер: она направлена на
освобождение из станка. При ежедневном многократном повторении
электрического раздражения в одном и том же участке ноги эта
реакция постепенно упрощается и, наконец; в определенных случаях, огра-
ничивается движением раздражаемой ноги и небольшим
ориентировочным движением головы, т. е. происходит полное выпадение
поведенческого акта освобождения и вся реакция на основное раздражение
ограничивается рефлексом сгибания ноги.
Аналогичное изменение претерпевает двигательная реакция на
индивидуальный сигнал. При образовании индивидуального двигательного
рефлекса на ноге первоначально индивидуальный сигнал вызывает не
только рефлекторное сгибание ноги, но и обширную поведенческую
реакцию освобождения. Впоследствии, с упрочением рефлекса на ноге,
поведенческая реакция освобождения постепенно ослабевает и,
наконец, исчезает совсем. Индивидуальный сигнал производит лишь
поднятие ноги и поворот головы без визга и углубленного дыхания.
Такое же упрощение реакции наблюдается при многократном
применении всякого другого раздражения, которое первоначально дает об-
ЗАКОН СОПРЯЖЕННОЙ ИРРАДИАЦИИ ВОЗБУЖДЕНИЯ 537
ширную реакцию. Так, например, у взрослых животных сильное, не»
обычное звуковое раздражение первоначально вызывает всегда обшир*
ную двигательную реакцию ориентировочного характера: быстрое
поднятие головы и поворачивание ее то в одну, то в другую сторону;
вскакивание на ноги, если животное лежало; передвижение в противо-
Рис. 236. Рефлекторные пути прирожденного сгибателъного рефлекса, голосового*
рефлекса и ориентировочного движения шеи у собаки.
Для каждого рефлекса дан короткий нервный путь через подкорковый отдел мозга и длиппый
через кору больших полушарий. ЛФферснтный путь длп всех этих рефлексов один. Он изображен
толстой черной липией в сопровождении штриховой. Начинается он на ноге, где производится
электрическое раздражение ?', и кончается и коре большого мозга, в кожной воспринимающей
области В. Короткий путь сгибательного рефлекса проходит через спинний мозг ///; здесь лежит его
координирующий аппарат Длинный путь атого рефлекса включает еще кожную воспринимающую
область и двигательный участок / в коре же. Для ориентировочною движения шеи короткий путь
проходит через шейный отдел мозга II, где лежит его координирующий аппарат. Дтипнып путь
этого движения включает еще ту же самую кожную область в коре и двигательный уча«ток 2 для
шейного движения. Для юлооового рефлекса нервный путь проходит через продолговатый мозг I;
эдесь лежит координирующий аппарат мышц гортани. Длинный путь его включает ту же
кожную область и двигательный участок з длп гортани. Стрелки в коре и в путях обозначают
направление иррадиации возбуждения.
положном направлении от источника звука. С повторением
раздражения эта реакция значительно упрощается, ограничиваясь поворотом
головы в одну сторону. У новорожденных и молодых животных все
раздражения являются необычными, поэтому у таких животных
ориентировочные реакции протекают более или менее бурно. Итак, можно
утверждать, что каждое новое раздражение производит через кору
больших полушарий на периферии обширный и очень сложный эффект;
если, обычное, часто повторяемое раздражение производит внешний
эффект* то последний всегда ограничивается одним очень простым
движением (Беритов, 1).
?38
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
На основании вышеприведенных и других фактов было сделано
очень важное в теоретическом смысле заключение: каждый раз,
Когда возбуждаемый очаг какого-либо участка коры большого мозга не
связан определенным образом с какими-либо ответными корковыми
аппаратами, возбуждение из него иррадиирует более или менее обширно
по всей коре больших полушарий, приводя в активное состояние
разнообразные двигательные и секреторные аппараты, обусловливая
определенный поведенческий акт. Когда же возбуждаемый очаг связывается
определенными путями с определенными ответными аппаратами, то
возбуждение из одного очага иррадиирует преимущественно через эти
пути. Это связывание определенными путями нужно понимать не в
смысле возникновения новых нервных путей, а в смысле
приспособления уже существующих нервных кругов к преимущественному
проведению возбуждения в определенном направлении. Нужно думать, что
если электрическое раздражение ноги сначала дает обширные и
сложные реакции, а после многократного повторения вызывает только
сгибание одной ноги, то это происходит потому, что первоначально
периферическое возбуждение так или иначе действует на разнообразные
дифференцированные нервные центры и проводящие пути и тем самым
вызывает сложные двигательные реакции поведенческого характера
В связи же с многократными сочетаниями электрического раздражения
с индивидуализируемым, в коре большого мозга развиваются
определенные нервные круги, которые связывают корковые участки,
воспринимающие болевое и индивидуализируемое раздражения, с двигатель
ными участками. Это должны быть как раз те нервные круги, которые
связывают данные корковые участки посредством наилучше
функционирующих элементов. Образование новых дифференцированных путей
следует понимать прежде всего как результат повышения возбудимости
и проводимости в данных кругах. Благодаря этому возбуждение
перестает более или менее значительно распространяться в коре мозга по
другим нервным кругам и, таким образом, перестает активизировать
другие корковые области как двигательные, так и воспринимающие.
Путем дальнейшего анализа этих фактов мы пришли к следующему
заключению о физиологической закономерности, лежащей в основе
описанных явлений и вообще корковой деятельности. Из каждого
возбужденного очага возбуждение иррадиирует во всей коре больших
полушарий, но интенсивность возбуждения в каждом данном нервном пути,
начинающемся от возбужденного очага, зависит не только от степени
возбудимости данного пути, но также и от возбудимости всех
остальных начинающихся от него же путей, а именно, возбуждение иррадиирует
по данному пути тем сильнее, чем выше его возбудимость и чем ниже
возбудимость всех остальных путей. Эту закономерность мы назвали
законом сопряженной иррадиации возбуждения. При
этом мы имеем в виду то обстоятельство, что по смыслу этого закона
усиленная иррадиация возбуждения по наиболее возбудимому нервному
ЗАКОН" СОПРЯЖЕННОЙ ИРРАДИАЦИИ ВОЗБУЖДЕНИЯ 539
кругу совершенно определенным образом сопряжена с ослабленной
иррадиацией его по всем остальным кругам.
С точки зрения закона сопряженной иррадиации возбуждения,
одновременно с повышением возбудимости в одних наиболее активных нерв-
'ных кругах, которые исходят из данного очага, непременно должно
происходить понижение ее во всех других кругах, исходящих из toro же
очага. Повышенная возбудимость поддерживается в каждом нервном
пути, если последний возбуждается регулярно. Чем интенсивнее
деятельность нервных кругов, тем болыпб повышается их возбудимость.
Но если из всех нервных путей, отходящих от данного очага
возбуждения, какой-либо один путь развивается особенно сильно и возбудимость
соответствующего нервного круга становится особенно высокой, то
возбуждение^иррадиирует по этому кругу сильнее, чем по другим; а чем
меньше будет возбуждение иррадиировать по другим кругам, тем
меньше будет поддерживаться в них повышенная возбудимость,—
следовательно, тем больше она будет в них снижаться.
Очень возможно, что кортикальная иррадиация возбуждения всегда
является сопряженной, так как нельзя себе представить, чтобы у
взрослых животных хотя бы одна точка была связана со всеми другими
точками коры посредством совершенно одинаково возбудимых путей.
Только у новорожденных корковая иннервация может быть
несопряженной. Мы говорим «может быть», ибо у новорожденных некоторые
точки коры больших полушарий должны быть соединены между собой
более возбудимыми нервными путями, чем другие. Это должно быть, во-
первых, в силу неодинакового филогенетического развития нервных
путей: одни точки должны быть связаны между собой более интимными
прирожденными путями, чем другие; например, корковый участок,
воспринимающий раздражение кожи одной ноги, должен быть связан
более интиімно с корковым двигательным аппаратом данной ноги, чем
других ног; во-вторых, онтогенетическое развитие разных нервных
элементов коры происходит на разных стадиях эмбриональной и
постэмбриональной жизни (Флексиг, 5). Кроме того, известно, что
возбудимость нервных элементов, очень низкая на первой ступени
эмбрионального развития, постепенно нарастает по мере развития эмбриона, и в
этом развитии, несомненно, одни элементы опережают другие (Болафио
и Артон, 6). Закон сопряженной иррадиации возбуждения вместе с
общеизвестными законами деятельности центральной нервной системы
совершенно достаточен для понимания любого явления из области
индивидуальных рефлексов.
О физико-химических основаниях закона сопряженной иррадиации
возбуждения. Закон сопряженной иррадиации возбуждения был
выведен путем изучения деятельности сложного сплетения нейронов, и
потому нужно предположить, что такое распространение возбуждения
обусловливается свойствами этого сплетения. Закон сопряженной
иррадиации возбуждения формулируется совершенно так же, как закон
540 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
ветвления электрического тока в системе проводников разного
сопротивления. Сила тока в данном проводнике разветвленной системы
обратно пропорциональна сопротивлению данного проводника и прямо
пропорциональна сопротивлению остальных проводников, т. е. она тем
больше, чем меньше сопротивление данного проводника и чем больше
сопротивление всех прочих проводников взятой системы. Это заставляет
искать объяснения закона сопряженной иррадиации возбуждения в
условиях распространения биоэлектрических токов возбуждения
нервных элементов в нервной ткани.
Как уже говорилось раньше, все нервные пути в центральной
нервной системе образуют замкнутые круги, т. е. каждая клеточная группа
не только отдает нервные отростки к другим клеточным группам, но и
получает их от других клеточных групп (Лоренте д,е Но, 13). В этих
нервных кругах нейрофибрилли и нейроплазма осевых цилиндров и
нервных клеток обладают почти в 5 раз меньшим электрическим сопро-
тивлением, чем оболочки нервных волокон и поверхностные протоплаз-
матические перепонки клеток.
Известно, что электропроводность всех живых тканей очень низка.
Самая протоплазма клеток, из которых составляются ткани,
представляет собой относительно хороший проводник, но оболочки, протоплаз-
матические перепонки обладают очень низкой проводимостью или же
почти совершенно лишены электропроводности (Гебер, 3; Като, 11).
Специально о нервных волокнах известно, что их сопротивление в
продольном направлении значительно меньше, чем в поперечном; это в
основном зависит от того, что при пропускании тока в поперечном
направлении нервного ствола электрический ток должен пройти через
большее число полупроницаемых перепонок, чем в продольном направлении.
Установлено, что биоток возбужденного участка является
существенным звеном в распространении процесса возбуждения. Именно
этот ток при своем распространении возбуждает соседние еще не
возбужденные участки. Мы же на основании определенных фактов
полагаем, что ток возбуждения влияет определенным образом не только на
близлежащие невозбужденные части, но и на отдаленные нервные
образования. Это происходит вследствие того, что ток возбуждения
замыкается в нервных кругах. В нервных путях из миэлиновых волокон
проведение электрического тока возбуждения, несомненно, происходит
главным образом через осевые цилиндры, но в коре мозга, где
значительная часть нервных волокон лишена миэлиновых оболочек, ток
возбуждения должен проводиться по нервным кругам, а также в некоторой
мере через омывающую их жидкость (Эдриан, 9). Проведение тока
возбуждения происходит по нервным кругам электротонически, независимо
от распространения возбуждения, и притом даже в том случае, если
возбуждение не распространяется, т. е. не переходит от первично
возбужденного нейрона на другие. Естественно, ток возбуждения буцет про-
ЗАКОН СОПРЯЖЕННОЙ ИРРАДИАЦИИ ВОЗБУЖДЕНИЯ 541
водиться в нервных кругах малого сопротивления с большей силой и,
наоборот, в нервных кругах большего сопротивления с меньшей силой,
и будет производить здесь такие же физиологические изменения, какие
свойственны вообще действию электрического тока.
Конечно, в хорошо развитых нервных кругах, состоящих из цепи
нейронов, ток возбуждения будет проводиться главным образом по
осевым цилиндрам. Лишь в некоторой малой мере он должен
распространиться также через промежуточное вещество нервного сплетения,
именно — через лимфу, кровь и нейропиль. Нужно иметь в виду, что
оболочки представляют поляризующиеся образования, которые
оказывают проходящим токам известное противодействие. Это уменьшает
интенсивность и крутизну нарастания токов, выходящих через оболочки
наружу в промежуточную среду (Гильдемейстер, 10). Отсюда понятно,
что если периферическое возбуждение, придя в воспринимающую
область коры мозга, попадает в хорошо развитый нервный круг, то ток
еозбуждения этого нервного круга будет распространяться, главным
образом, в данном кругу.
Сопротивление цепи нейронов проведению тока возбуждения,
возникшего в какой-нибудь нервной клетке, должно быть обусловлено
длиной нервного пути, поперечником осевых цилиндров, в особенности
же проницаемостью протоплазматических перепонок нервных окончаний,
затем расстоянием между синаптическим окончанием одного нейрона и
телом клетки другого и, наконец, концентрацией ионов в этой
промежуточной среде. Следовательно, чем короче нервный путь, чем толще
осевой цилиндр, чем более проницаемы для ионов протоплазматические
перепонки нервных окончаний, чем тоньше синаптические
промежуточные среды и чем больше концентрация в последних ионов, тем меньше
сопротивление данного нервного пути, а значит, тем с большей силой
распространится здесь ток возбуждения.
Допустим теперь, что возбуждается афферентный нейрон,
оканчивающийся в комплексе нервных кругов коры большого мозга. Ток
возбуждения афферентного нейрона будет распространяться наиболее
сильно вдоль того нервного круга, сопротивление которого меньше всего,
и соответственно он распространится в других нервных кругах
значительно слабее. Как известно, ток возбуждения будет действовать
характерным образом на весь нервный путь, через который он проводится, а
именно, увеличится концентрация ионов (преимущественно
одновалентных) в синапсах и вообще на протоплазматических перепонках
данного круга. Это обусловит, по Геб е ру (3) и Э бб еке (4),
разрыхление протоплазматических перепонок и тем самым будет
благоприятствовать как интенсивности, так и скорости проведения процесса
возбуждения. Но ток возбуждения распространится из данного
нервного элемента в разных нервных путях нервного комплекса не с
одинаковой силой. Соответственно, и изменение в распределении ионов и
проницаемости протоплазматических перепонок в нервных путях малого
сопротивления, т. е. там, где ток возбуждения распространится с боль-
542
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
шей силой, должно происходить сильнее, чем в путях большого
сопротивления. Вследствие этого иррадиация возбуждения в нервном кругу
малого сопротивления будет происходить сильнее, чем в других кругах
и притом тем сильнее, чем меньше будет его сопротивление проведению
электрического тока и чем больше это сопротивление во всех других
кругах.
Ток возбуждения, распространяющийся через нервные круги,
должен действовать и на нейропиль, на дендритную массу, где
заканчивается бесчисленное количество коллатералей из нервных кругов и от
афферентной системы. Дендриты нейропиля, будучи голыми, не должны
участвовать в образовании замкнутых цепей нервных кругов, но раз
коллатерали из нервных кругов и афферентной системы оканчиваются
на дендритах, то безусловно ток возбуждения нервных кругов, электро*
тонически распространяющийся из нервных кругов через
коллатерали, должен действовать и на нейропиль. Весьма возможно, что этот
диффузно распространяемый ток является достаточным для
активирования нейропиля. Иначе говоря, нейропиль может быть настолько
чувствительным, что будет реагировать на действие электротонически
распространяющегося тока возбуждения. С этим хорошо согласуется факт
наступления общего торможения в коре большого мозга при таких
раздражениях рецепторов или коры мозга, которые не дают двигательных
реакций. Во всяком случае ток возбуждения может так изменить
чувствительность нейропиля, что последний легко придет в активное
состояние при последующем распространении возбуждения из первично
возбужденного очага или же под влиянием импульсов, притекающих к
нейропилю через афферентную систему. Итак, каждый раз при
возбуждении данного нервного круга как импульсы возбуждения, так и ток
возбуждения должны активно действовать не только на связанные с ним
другие нервные круги, но и на связанный с ним нейропиль. При этом
данный нервный круг будет действовать на связанный с ним нейропиль
тем сильнее, чем слабее ток возбуждения будет распространяться из
этого круга на другие связанные с ним нервные круги. ,
Итак, можно с большим основанием утверждать, что сопряженный
характер иррадиации возбуждения обусловливается физиологическими
изменениями, возникающими в нервных кругах путем распространения
биоэлектрического тока возбуждения.
От чего же может зависеть разница нервных путей в отношении
физико-химических свойств, которые определяют их различную
электропроводность?
Морфологические основания закона сопряженной иррадиации. Нужно
думать, что повторная деятельность определенных нервных путей
между нервными комплексами большого мозга, воспринимающими
индивидуализируемое и основное раздражение, производит в этих
путях, наряду с определенными физико-химическими изменениями, чисто
анатомические изменения. Прежде всего следует указать на то морфо*
логическое изменение, которое наблюдается в нервных элементах в
ЗАКОН СОПРЯЖЕННОЙ ИРРАДИАЦИИ ВОЗБУЖДЕНИЯ 543
связи с тренировкой животных. Так, например, А г д у р (7) показал на
мышах, что при кратковременной тренировке значительно нарастает
число нервных клеток в ганглиях, а также число толстых нервных
волокон; кроме того, значительно увеличивается нейрофибриллярная
масса ,в нервных элементах. Наоборот, после изолирования спинного
мозга от периферических влияний в результате перерезки всех задних
корешков и нисходящих путей (перерезка мозга на границе грудного
отдела) отмечается утоньшение осевых цилиндров и уменьшение
величин нервных клеток и дендритной массы в спинном мозгу, а также в
периферических нервных стволах и в нервных окончаниях
двигательного нерва (Тоуэр, 15). В этих исследованиях мы находим,
следовательно, подтверждение основного положения относительно
морфологического изменения нервных элементов в связи с их деятельностью.
Морфологическое изменение нервных элементов должно решительно
влиять на сопротивление проведению электрического тока в случае их
утолщения в смысле его уменьшения, а в случае утоныпения — в смысле
его увеличения.
Далее, как утверждает Эббеке (4), разрыхленный в результате
возбуждения нервный участок даже после процесса возбуждения
сохраняет разрыхленное состояние, вследствие чего при повторном
возбуждении разрыхление перепонки становится глубже и держится
дольше, чем в предыдущий раз. Повидимому, последствия
разрыхления в цепи нейронов тем значительнее и глубже, чем чаще и сильнее
возбуждаются эти элементы.
Отсюда мы можем заключить, что в наиболее деятельных нервных
путях как прирожденных, так и индивидуальных рефлексов,
нейрофибриллярная масса и нейроплазма нервных элементов, а также
проницаемость их протоплазматических перепонок должны быть больше,
чем в других, менее деятельных нервных путях. Соответственно,
электрическое сопротивление нервных путей в них должно быть меньше»
чем в других путях.
Кроме того, можно принять, что при усилении деятельности
наступает еще одно характерное морфологическое изменение. А р и ё н с
Капперс (8) установил, что в филогенетическом развитии нервных
путей рост главных дендритов, а также удлинение клеточного тела
происходят в том направлении, откуда притекает к клетке наибольшее
число возбуждений, т. е. навстречу к этим импульсам возбуждения.
Осевые цилиндры, наоборот, вырастают в том самом направлении, в
котором, по словам автора, действует «ток раздражения», но вернее
было бы сказать — в сторону распространения процесса возбуждения.
Последнее явление было отмечено, между прочим, К а х а л е м в
отношении эмбрионального развития нервной системы. Когда аксоны
приближаются к какому-либо нервному ядру, то из этого ядра навстречу
этим аксонам вытягиваются дендриты и тела клеток. В данном случае
растущий аксон выступает как активный, возбужденный нервный
элемент.
544
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
Тот биологический процесс, который лежит в основе этого явления,
Ариёнс Капперс называет нейробиотаксисом (8). Он полагает,
что означенное передвижение дендритов и тела клетки в сторону
наиболее сильно возбужденных нейритов происходит от
гальванотропического действия возбужденных элементов. Аксоны вырастают в сторону
распространения их собственного тока возбуждения, т. е. в
направлении от клетки; дендриты же и тело соседних клеток передвигаются
навстречу этому току. Ввиду этих морфологических изменений, связанных
с деятельностью, мы склонны думать, что когда цепь нейронов
находится в состоянии деятельности, то узенькое промежуточное
пространство на месте перехода от одного нейрона к другому, или синапс в
старом понимании, вследствие взаимного влечения нервных элементов,
согласно закону нейробиотаксиса, становится еще уже. Это уменьшение
промежуточного слоя, безусловно, приведет к повышению
электропроводности и, значит, к улучшению проведения возбуждения в данной
цепи нейронов.
Сам АриёнсКапперс(8) не раз подробно останавливался на
том, что филогенез нервных связей подобен развитию ассоциативных,
т. е. временных, связей в индивидуальной жизни, что закон
нейробиотаксиса аналогичен закону образования ассоциаций,так как
одновременные или последовательные раздражения не только вызывают
определенные ассоциации в нашей психике, но и определенные материальные
изменения в нервной системе.
Итак, нужно думать, что как в прирожденных рефлекторных путях,
так и во временных связях индивидуальных рефлексов синапсы, как
промежуточная среда между нейронами, должны стать тоньше в связи
с усиленным функционированием. Соответственно, электропроводность
упомянутых рефлекторных путей и временных связей должна быть
больше, чем у других, мало функционирующих нервных путей.
Морфологическое развитие нервных элементов, которое
наблюдается в связи с усиленной деятельностью, сопровождается их
функциональным развитием. Рост нейрофибриллярной массы означает
увеличение возбудимой системы, потому что, как известно, возбуждение
распространяется в нервных волокнах через нейрофибрилли. С
увеличением же возбудимой системы повышается возбудимость и скорость
проведения возбуждения.
Итак, те нервные пути, которые морфологически и функционально
наиболее развиты, обладают наиболее высокой электропроводностью.
Вследствие этого из каждого первично возбужденного нервного
комплекса биоэлектрический ток возбуждения с наибольшей силой пойдет
по тем нервным кругам, которые являются и морфологически и
функционально наиболее развитыми. Этим самым данные круги будут
наиболее подготовлены для последующего распространения возбуждения.
Однако биоэлектрический ток не при всех условиях может играть
существенную роль в иррадиации возбуждения. Если, в результате
длительной тренировки, возбужденный нервный комплекс в коре мозга
ЗАКОН СОПРЯЖЕННОЙ ИРРАДИАЦИИ ВОЗБУЖДЕНИЯ 545
связан чрезвычайно хорошо развитыми нервными связями только с
одним нервным аппаратом, то возбуждение будет распространяться
преимущественно по этим связям при всех условиях распространения
биоэлектрического тока возбуждения.
Специфичность закона сопряженной иррадиации возбуждения.
Названный закон был выведен нами на основании изучения
индивидуально приобретенной деятельности коры больших полушарий. Эта
деятельность является одной из форм центральной нервной
деятельности. По существу она представляет собой автоматизированную
деятельность, протекающую на основе особых морфологических изменений
в коре мозга, возникающих в силу многократных повторений одного и
того же акта в более или менее одинаковых внешних и внутренних
условиях. Вследствие этого закон сопряженной иррадиации
возбуждения является характерной специфической закономерностью для
автоматизированной, индивидуально приобретенной деятельности коры
большого мозга.
Закон сопряженной иррадиации возбуждения не должен быть
применяем ко всякой другой форме деятельности коры мозга, например, к
ее психо-нервной деятельности как сознательной, характерной для
человека, так и бессознательной — животных.
Закон сопряженной иррадиации возбуждения не применим и к
центральной деятельности спинного мозга и стволовой части головного
мозга. Правда, и здесь все нервные связи образуются в эмбриональном
периоде развития по принципу временных связей, как это показал
Когхилл (12), и здесь возбуждение распространяется
преимущественно по одним нервным путям, а по другим идет в незначительной степени,
но это не обусловлено сопряженной иррадиацией возбуждения.
Биоэлектрический ток возбуждения, распространяясь в нервной системе с
большой скоростью еще раньше возбуждения, может повлиять на
нервные пути в смысле повышения их функционального состояния. Несмотря
на это, морфологическое состояние нервных путей и нервных связей в
спинном мозгу и стволовой части головного мозга при их нормальном
состоянии настолько стабильно благодаря длительному постоянному
упражнению, что означенное предварительное электротоническое
распространение биоэлектрического тока возбуждения не в состоянии
изменить функциональное состояние этих нервных путей в такой мере,
чтобы оно играло существенную роль в характере распространения
возбуждения в этих отделах центральной нервной системы.
В определенных случаях биоток возбуждения при своем физическом
распространении может даже возбудить клеточные нервные элементы
спинного мозга и других отделов мозга. Это, например, может иметь
место при стрихнинном отравлении клеточных элементов в связи с
сильным повышением возбудимости в этих элементах (Бремер, 16).
Более того, не исключается возможность возбуждения нервных волокон
от раздражения их под влиянием чисто физически распространяющегося
тока возбуждения (Баррон и Мэтьюс, 17; Теннис, 18; Гранит и
35 И. С. Беритов
546
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
сотр., 19). Несмотря на это, физическое распространение биотока
возбуждения не должно играть решающей роли в характере иррадиации
возбуждения вне коры большого мозга.
Литература
I. Беритов И., Новое в рефлексологии и физиологии нервной системы, 2, 192&.
2. Н е г m a n n L., Handbuch d. Physiologie, Bd. II, Abt. 1, стр. 194, 1879.
3. Ho be r R., Klin. Wschr., 4 Jg. N. 28, 1925; Physikalische Ghemie d. Zelle
u. Gewebe, 1924.
4. Ebbecke U., Pfl. Arch., 195, 555, 1922.
5. FlechsigP., Gehirn u. Seele. Leipzig, 1896; Lokalisation d. geistigen Vorgange
Leipzig, 1896.
6. Bolaffio M. u. А г t о n, Arch. Sci. Biol., 5, 457, 1924.
7. Agduhr E., J. Psych. Neur., 25, 463, 1920.
8. Ariens KappersC. U., Die vergleichende Anatomie d. Neroensystems.
Haarlem, Bd. I, 1921; Acta psychiatrie et neurologie, 2, 118, 1927.
9. Эдриан Э. Д., Механизм нервной деятельности. Пер. Биомедгиз, 1935.
10. G і 1 d е m е і s t е г М., Z. Biol., 83, 145, 1925.
II. К ato G., The microphysiology of nerve, 1934.
12. С о g h i 1 1 G. В., Anatomy and the problem of behavior. Есть русс, пер., 1934.
13. Lorente de N6 R., J. Psych. Neur., 45, 381, 1933; 46, 113, 1934.
14. Ariens KappersC. U., The comparative anatomy of the nervous system etc.
v. 1, 1936.
15. Tower S. S., Arch. Compar. Neurol., 67, 109, 1937.
16. Bremer F., Proc. Soc. Exp. Biol. Med., 46, 627, 1941.
17. В arron D. H., J. Neurophysiol., 3, 403, 1940.
18. Toennies J. F., J. Neurophysiol., 1, 378, 1938; 2, 515, 1939.
19. G га nit R., Leksel 1 L. a. Skoglund С R., Brain, 67, 125, 1944.
Глава Till
ПРОИСХОЖДЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ
Понятие о временных связях. Из факта образования нового
рефлекса на какое-либо индифферентное раздражение вследствие
сочетания его с каким-либо рефлекторным действием само собой вытекает,
что при этих условиях между нервным комплексом, воспринимающим
данное раздражение, и деятельностью того или другого рабочего органа
возникает определенная связь. Так как эта связь по существу является
временной, развивается при определенных условиях и держится, пока
продолжают держаться эти условия, то Павлов дал ей название
временной связи. Она противопоставляется постоянной связи,
обусловливающей прирожденные рефлекгорные реакции.
Временные нервные связи, несомненно, возникают в коре большого
мозга между теми нервными комплексами, которые возбуждаются
индивидуализируемым и основным раздражениями. По представлениям
Павлова, а также Бехтерева, эти временные связи имеют
всегда одно направление от воспринимающего очага слабого раздражения к
очагу более сильного раздражения или от очага индивидуализируемого
раздражения к очагу основного раздражения. В противовес этому, по
представлениям Беритова, каждый раз при образовании
индивидуального рефлекса возникают временные связи обоих направлений: как
от очага индивидуального раздражения к очагу основного раздражения,
т. е. к нервным комплексам, возбуждаемым основным раздражением,—
так называемые поступательные временные связ и,— так
и, наоборот, от нервных комплексов, возбуждаемых основным
раздражением, к комплексу, возбуждаемому индивидуальным раздражением,—
так называемые обратные временные связи (1) (рис. 237).
Индивидуальный рефлекс, образуемый при обычном сочетании
раздражений, безусловно, обязан своим происхождением
преимущественной деятельности поступательных временных связей. Однако ©
определенных случаях очень хорошо обнаруживается также наличие действия
обратных связей. Это бывает, если индивидуализируемое раздражение
само по себе вызывает характерный эффект, отличный от вызываемого
основным раздражением. В этом случае внешний эффект соответствует
35 *¦
548 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
то действию поступательной связи, то действию обратной, смотря по
тому, какова степень развития и деятельности той и другой связи. При
определенных условиях можно наблюдать на периферии совместный
эффект от той и другой связи. Это, например, бывает, когда поедание
сахарного песка сочетается с электрическим раздражением ноги. Если
собаке всыпать в рот сахарный песок и во время еды раздражать ее
электрическим током, то в первые дни собака при этом раздражении
перестает есть песок; впоследствии она будет продолжать есть его и
3d* до Злектр.раздрл
Рис. 237. Схематический рисунок временных связей
индивидуального двигательного рефлекса на звук
тонвариатора.
Показаны очаг возбуждения в слуховой области коры мозга от
раздражения звуком, другой очаг в кожной—от электрического
раздражения и еще третий — двигательный участок. Все они
соединены временными связями, как поступательными (более
жирные линии), так и обратными (более тонкие линии). Стрелка
показывают направление иррадиирующего возбуждения по этим
связям.
во время электрического раздражения, а еще позднее будет его есть
только при электрическом раздражении: сахарный песок всыпан в рот, но
собака его не ест до наступления электрического раздражения (Савич, 3).
Так как электрическое раздражение производилось вслед за
всыпанием сахарното песка, то, очевидно, индивидуальный рефлекс поедания
Сахара в ответ на электрическое раздражение кожи представляет
результат сильного развития обратной связи. Всыпание песка н? дает
рефлекса, так как возбуждение из очага, воспринимающего вкусовое
раздражение, устремляется главным образом в двигательные участки
и кожную воспринимающую область, а на аппараты пищевой реакции,
согласно принципу сопряженной иррадиации, оно действует с
субминимальной интенсивностью. Из двигательных участков и кожной
воспринимающей области возбуждение направляется, главным образом, по
путям хорошо развитых обратных связей обратно в очаг вкусового раз-
ПРОИСХОЖДЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ §4$
дражения, а по направлению к двигательным координирующим
аппаратам, согласно тому же принципу, оно идет с субминимальной
интенсивностью. И на этот раз возбуждение вкусового очага
преимущественно направляется в двигательные участки и кожную воспринимающую
область по поступательным связям, а затем снова обратно и т. д. В
результате не получается ни движения, ни секреции. 'Когда
присоединяется электрическое раздражение, действие обратных связей от очага
электрического раздражения значительно усиливается. Это производит
возбуждение очага вкусового раздражения с такси интенсивностью, что
оно вызывает отсюда обычный эффект еды: движение рта и секрецию.
Приведем еще один очень демонстративный случай. Если слабое
механическое раздражение, как почесывание ноги кисточкой, сочетать
с вливанием в рот слабого раствора соляной кислоты, то после ряда
сочетаний это почесывание производит не только секрецию слюны, но и
движение ноги и голосовую реакцию, как будто бы ноги подвергались
сильному повреждающему раздражению (Петрова, 4). До этого
почесывание не давало никаких двигательных реакций. Оно и теперь н?
дает их с других кожных участков. Поэтому двигательная реакция в
ответ на почесывание обычно раздражаемого участка кожи должна
быть приписана деятельности обратных временных связей.
Возбуждение из индивидуального очага распространяется главным образом к
секреторному и двигательному аппаратам пищевой реакции, а отсюда-
оно распространяется в некоторой мере через обратные связи, обратно
к очагу индивидуального раздражения. От этого существующее здесь
возбуждение усиливается в такой степени, как будто бы оно вызывалось
сильным механическим раздражением кожи. При этом характерно, что
двигательная реакция на почесывание тем сильнее, чем слабее секреция
слюны. Очевидно, это получается потому, что распространение
возбуждения из секреторного аппарата происходит согласно принципу
сопряженной иррадиации: чем больше оно уходит в очаг механического
раздражения, тем меньше его идет к подкорковым центрам слюнной
секреции.
Происхождение временных связей. Положим, вырабатывается
индивидуальный сгибательный рефлекс на правой передней ноге на
звуковое раздражение. Для этого звук сочетают с электрическим
раздражением этой ноги. В результате такой комбинации в коре большого
мозга возникают три очага возбуждения; один—ів звуковой
воспринимающей области, второй — в кожной воспринимающей области и
третий — в двигательных участках, которые, возбуждаясь через кожную
область коры, вызывают движение на периферии (рис. 236, 237). В это
время возбудимость «всех этих очагов тоже становится наиболее
высокой. Все эти очаги,— вернее нервные комплексы из определенных
нервных кругов, соединены между собой нервными путями разной
сложности. Эти связи, по всей вероятности,* тзкл^еі принадлежат к типу церв^
ных кругов, но только чрезвычайно сложного состава. Вследствие этого
устанавливаются поступательные связи, ведущие от каждого очага к
550
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
двум остальным очагам, и обратные связи от последних очагов к
первому. При сочетании основного раздражения с индивидуальным,
возбуждение из каждого возбуждаемого очага иррадйирует по всему
полушарию, но сильнее всего действует на два остальных очага, где
возбудимость была более всего повышена. Это воздействие может быть
настолько сильным, что уже при первом сочетании (будет в состоянии
дать определенный внешний эффект. Для примера приведем такой
случай. Если ногу раздражать слабым электрическим током и в то же
©ремя производить сильное звучание, вроде электрического звонка, то
звук значительно усилит движение ноги, вызванное электрическим
током. В этсГ время корковые нервные комплексы, возбуждаемые
электрокожным раздражением и звуком, оказывают друг на друга гальва-
йотропическое действие. Это обусловливает улучшение імежнейронных
связей между данными комплексами. Так как наиболее короткие
нейронные пути, соединяющие эти очаги, возбуждаются сильнее других
путей, то и возбудимость и проводимость этих путей должны повыситься
больше, а синаптическая связь стать значительно интимнее, чем в других
путях. Разумеется, с каждым новым сочетанием возбудимость нервных
комплексов и проводимость связывающих путей повышаются все более
и более, а синаптические связи становятся все интимнее, так как
каждое новое сочетание происходит в то время, когда еще не прошли все
функциональные и структурные последствия от предыдущего сочетания.
Вследствие этого с каждым новым сочетанием возбуждение из очагов
все интенсивнее распространяется по коротким путям. Уже после
нескольких сочетаний влияние, произведенное внешним раздражением в
одном очаге, приводит к возбуждению других очагов. Например,
возбуждение звуковой воспринимающей области приводит к столь сильному
возбуждению кожной области и двигательных участков коры, что на
периферии вызывается движение ноги.
В результате взаимодействия возбужденных корковых очагов,
образованные между ними короткие нервные пути с повышенной
возбудимостью и представляют собой поступательные и обратные временные
связи. По одним временным связям возбуждение проводится в
направлении от очага индивидуального раздражения к ответным
двигательным и секреторным участкам коры, а по другим, наоборот, от
последних участков—к очагу индивидуального раздражения. Все эти пути,
очевидно, возникают из наиболее активно действующих нервных
кругов вследствие их определенного функционального и морфологического
развития, как об этом уже говорилось выше (утолщение осевых
цилиндров и их синалтических окончаний, увеличение тела клеток и их
вытягивание в сторону притекающих импульсов, уменьшение синаптнческих
сред между окончаниями v телом клеток, разрыхление
полупроницаемых мембран, повышение возбудимости и проводимости этих путей).
Скорость образования тех и других связей зависит прежде всего
от степени физиологического действия сочетаемых раздражений.
Экспериментальным путем установлено, что чем выше физиологическое дей-
ПРОИСХОЖДЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ
551
ствие основного раздражения, т. е. чем больше повышается
возбудимость соответствующих корковых центров от этого раздражения, тем
скорее образуется поступательная временная связь при обычном
порядке сочетания. С другой стороны, чем сильнее физиологическое
действие индивидуализируемого раздражения, тем скорее образуется
обратная временная связь.
Степень развития той и другой связи, их прочность, Еозбудимость и
деятельность в каждом рефлексе зависят от подготовленности
взаимодействующих очагов к образованию временных связей. Чем более
будут подготовлены очаги возбуждения основного раздражения, т. е. чем
выше будет их возбудимость, тем скорее разовьется образуемая с ними
поступательная временная связь. Таким же образом, чем больше будет
подготовлен в этом отношении очаг индивидуального раздражения, тем
скорее и сильнее разовьется образуемая с ним обратная связь.
Приведем соответствующие данные для пояснения того и другого положения.
Возьмем сначала положение относительно поступательной временной
связи. При последовательном образовании ряда индивидуальных
двигательных рефлексов путем сочетания с одним и тем же основным
раздражением, первый из них образуется гораздо труднее, чем
последующие. Это происходит вследствие того, что конечный корковый пункт
поступательной временной связи — двигательный аппарат
раздражаемой ноги — подготовляется настолько, что каждая новая
поступательная связь с ним образуется гораздо быстрее, чем в первом рефлексе.
Рассмотрим теперь положение с обратной связью. Когда
индивидуальное раздражение продолжается десятки секунд до присоединения
основного, как это бывает при образовании запаздывающего рефлекса,
развитие очага индивидуального раздражения находится в очень
благоприятных условиях. Образуемая с ним обратная связь имеет
возможность хорошо развиваться. Вследствие этого возбуждение, вызванное
индивидуальным раздражением, через поступательные связи приходит
в двигательный аппарат, а отсюда, распространяясь по принципу
сопряженной иррадиации, переходит преимущественно через развитые
обратные связи в очаг индивидуального раздражения, а на периферию
к рабочему органу распространяется с субминимальной интенсивностью.
Только при утомлении обратных связей, т. е. при определенном физико-
химическом их изменении, возбуждение может распространиться на
периферию с большей интенсивностью и вызвать там заметный эффект.
Следовательно, недеятельная фаза запаздывающих рефлексов обязана
своим происхождением большому развитию обратных связей
вследствие длительного действия индивидуального раздражения (Бери-
тов, 15).
Происхождение 'рецепторной генерализации и дифференциации
индивидуальных рефлексов. Как указывалось выше, явление рецепторной
генерализации имеет место в начале образования индивидуального
рефлекса. Нами было выяснено, что это явление обусловливается
повышением возбудимости в коре, в особенности в тех нервных комплексах,
552
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
которые воспринимают индивидуальное и основное раздражения.
Повышение возбудимости есть результат повторного возбуждения этих
очагов и иррадиации возбуждения из них по всей коре.
Как указывалось выше, в начале образования индивидуального
рефлекса возбуждаемые ©о время сочетаемых раздражений корковые
воспринимающие участки не связаны хорошо развитыми нервными
путями с какими-либо ответными участками. Поэтому нужно думать, что
под влиянием сочетаемых раздражений прежде всего возбуждаются
нервные круги, которые имеются в корковых областях,
воспринимающих индивидуальное и основное раздражения. Соответственно и
возбудимость их должна повыситься прежде всего в этих областях.
Вследствие этого постороннее раздражение,, приходящее в указанные
области, и прежде всего раздражение того же качества, как
индивидуализируемое, будет иррадиировать в них настолько сильно, что окажется
в состоянии активировать здесь те нервные круги, в которых
возбудимость была повышена особенно сильно. Следовательно, 'постороннее
раздражение должно вызвать такой же внешний эффект, как основное
или индивидуальное раздражение. Естественно, чем слабее постороннее
раздражение и чем дальше соответствующий ему воспринимающий очаг
отстоит от очагов индивидуального и основного раздражений, тем
слабее должен быть вызываемый им внешний эффект.
Когда новое индивидуальное раздражение сочетается с тем же
самым основным раздражением, которое уже раньше служило для
образования индивидуальных рефлексов на том же животном,— например,
если сначала был образован индивидуальный двигательный рефлекс на
звук путем сочетания с электрическим раздражением правой передней
ноги, а затем новый рефлекс на какое-либо другое раздражение путем
сочетания его с электрическим раздражением той же ноги, то новый
рефлекс образуется без значительной генерализации, в то время как
при образовании первого рефлекса генерализация была более или
менее значительна. Этот факт служит хорошим подтверждением данного
выше объяснения. Корковые очаги возбуждения от электрического
раздражения тесно связаны при помощи временных связей с очагом
индивидуального раздражения уже сущертвующего рефлекса и также
посредством хорошо тренированных путей с корковым двигательным
аппаратом раздраженной ноги. Поэтому при образовании нового
индивидуального рефлекса возбуждение от основного раздражения
распространяется главным образом по этим хорошо возбудимым нервным
связям и, согласно закону сопряженной иррадиации возбуждения, не
может иррадиировать по другим нервным путям коры в такой мере, чтобы
повышать здесь возбудимость до степени, достаточной для
генерализации рефлекса.
В период генерализации рефлекс может вызываться необычными
раздражениями как через временные связи из очага индивидуального
раздражения, так и помимо них, непосредственно из коркового
двигательного аппарата (Беритов, 15). Было установлено, что более или
ПРОИСХОЖДЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 553-
менее близкие необычные раздражения производят эффект через
временные связи. Это видно, между прочим, из того, что чем ближе
необычное раздражение по своему качеству к обычному индивидуальному,
тем сильнее его рефлекторное действие. Мы считаем в высшей степени
вероятным, что чем ближе стоит необычное раздражение к
индивидуальному, тем интимнее функциональная и анатомическая связи между
соответствующими воспринимающими участками коры мозга. В
соответствии с этим при обычных сочетаниях повышение возбудимости в
каком-либо соседнем участке вокруг очага индивидуального раздражения
происходит тем сильнее, чем ближе находится этот участок или чем
интимнее он связан с очагом индивидуального раздражения. Поэтому
близкие необычные раздражения должны действовать на очаг
индивидуально-то раздражения сильнее и этим вызывать через временные
связи более сильный индивидуальный рефлекс, чем более отдаленные
необычные раздражения.
Далее; если индивидуальный рефлекс вызывать много раз без
сочетания с основным раздражением и этим ослабить индивидуальный
рефлекс, то после этого и необычные раздражения перестают его
вызывать. То же бывает, если, наоборот, повторно вызывать рефлекс
каким-либо близким необычным раздражением до его прекращения.
После этого нельзя вызвать рефлекс в ответ на обычное раздражение,
а также на другие более или менее необычные раздражения. Это
происходит потому, что они вызываются через одни и те же временные
связи, начинающиеся от очага индивидуального раздражения.
Существуют также определенные факты, свидетельствующие о
действии необычных раздражений прямо на двигательные участки, без
участия временных связей, благодаря общей иррадиации возбуждения
в коре больших полушарий. Так, например, если после полной
дифференциации рефлекса на правой передней ноге приложить сильное
электрическое раздражение к какой-либо другой ноге, которое производит
бурную реакцию, то в течение нескольких минут после этого
индивидуальное раздражение будет производить рефлекс на этой ноге, с
корковым двигательным аппаратом которой не было временных связей, а не
на правой передней ноге. Известно также, что слабое раздражение
какой-либо ноги, на которой не было индивидуального рефлекса,
вызывает двигательный эффект не на этой ноге, а на той, где был
образован индивидуальный рефлекс (Беритов, 15) (рис. 238).
Совершенно аналогичный факт обнаружил Затманзон (19) при
изучении индивидуальной деятельности по методу Фурсикова и
Воскресенского. После образования индивидуального
оборонительного рефлекса у собаки на левой задней лапе на стук метронома
он обнажил кору большого мозга и отравил стрихнином двигательные
участки правой передней ноги. После операции, но еще до отравления,
метроном давал обычную слабую сгибательную реакцию на левой
задней ноге; после же отравления он производил сильную двигательную
реакцию на правой передней ноге. Очевидно, это происходило, по-
?54 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
мимо временных связей, в силу общей иррадиации возбуждения п?
нервным кругам и активирования этим возбуждением двигательного
участка правой передней ноги, где возбудимость была чрезвычайно
сильно повышена стрихнином.
Итак, необычные раздражения вызывают индивидуальный рефлекс
путем общей иррадиации возбуждения, с одной стороны, прямым
'действием на корковый двигательный аппарат, помимо временных связей,
а с другой,— через очаг индивиду^
алъного раздражения, который
приводит е действие тот же
двигательный аппарат посредством
временных связей.
Как указывалось выше, реЦеп-
торная генерализация рефлекса
обусловливается повышением
возбудимости в нервных кругах вокруг
очага индивидуального
раздражения, вследствие иррадиации
возбуждения из этого очага, а также из
очагов, возбуждаемых
электрическим раздражением. Но в
некоторый период работы достигается
значительное укрепление
временных связей между очагом
индивидуального и основного
раздражений. Этот момент становится
поворотным пунктом в развитии
индивидуального рефлекса, так как,
согласно закону сопряженной
иррадиации возбуждения, дальнейшее
развитие означенных временных связей должно быть связано с
усилением иррадиации возбуждения по путям временных связей и
одновременно с ослаблением ее по остальным нервным путям. А это должно
привести к понижению возбудимости как вокруг начального пункта
временных связей в воспринимающей области индивидуального
раздражения, так и вокруг воспринимающего очага и двигательных участков,
возбуждаемых основным раздражением.
Вместе с этим, конечно, возбудимость должна понизиться и во всей
остальной коре, так как в связи с укреплением временных связей,
возбуждение, вызываемое индивидуальным раздражением, будет
направляться все больше через временные связи и двигательные участки по
'пирамидным путям к периферическому органу и все м'еньше
распространяться вообще в коре больших полушарий (Беритов, 15).
Кроме того, с дальнейшим укреплением временных связей на звук
в слуховой воспринимающей области прежде всего потеряют
повышенную возбудимость те нейронные комплексы, которые были связаны
г
Зп
8
а
10
—ишпші_і
11М11Г1М11М)Пі)ИИ1Иі!і1П)1П1іНі1МММі1ИіН11»іПі1111И)іИ>
Рис. 239. Извращение оборонительного
рефлекса на электрическое раздражение
правой задней ноги в связи с
существованием хорошо развитых
индивидуальных рефлексов на правой передней
ноге.
Собака Боб. В опыте / несколько раз
производится слабое электрическое раздражение
правой задней ноги (10 см расстояния между
катушками). Оно дает значительное сгибание
на правой передней ноге, а на правой задней
не дает эффекта. В опыте 2 берется более
сильное раздражение (8 см); оно вызывает сначала
небольшое сгибание правой задней с общим
движением, а затем длительное изолированное
сгибание правой передней.
ПРОИСХОЖДЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 555
наименее интимно с очагом индивидуального раздражения. Поэтому,
при образовании рефлексов на музыкальный звук прежде всего шумы
и более отдельные по высоте и по тембру звуки перестают вызывать
сильное возбуждение и вследствие этого перестают производить эффект
как через воспринимающий очаг индивидуального раздражения, так и
-непосредственно через корковые двигательные участки. Затем перестают
вызывать такой эффект точно так же более близкие звуки, потому что
спустя некоторое время и они застают соответствующие нейронные
комплексы воспринимающей области в состоянии малого повышения
возбудимости или даже совсем без повышения. Рефлекторная деятельность
на посторонние раздражения другого качества естественно отсутствует
вследствие одновременного понижения возбудимости во всей остальной
коре.
Итак, рецепторная дифференциация индивидуальна го рефлекса в
значительной ^іере обусловливается понижением возбудимости рокруг
очагов индивидуального и основного раздражений и вообще в коре
большого мозга, благодаря преимущественной иррадиации коркового
возбуждения, вызываемого индивидуальным и осно&ным
раздражениями, по путям временных связей.
Однако не вся дифференцировка рефлекса может быть объяснена
понижением корковой возбудимости, т. е. устранением прямого
действия необычных раздражений на воспринимающие очаги и корковые
двигательные участки данного индивидуального рефлекса, а только
более грубая, наблюдающаяся уже в первые дни работы.
Дифференцировка необычных звуков, наиболее близких к обычному, как
указывалось івыше, является затруднительной и требует многодневной работы.
Необходимо применять их многократно изо дня в день, чтобы они
перестали вызывать рефлекс. Каждый раз под влиянием необычного
раздражения сначала возбуждается определенный корковый очаг,
соответствующий этому раздражению, а затем, путем иррадиации по нервным
кругам, все элементы временных связей индивидуального рефлекса.
При этом ів коре должны разыгрываться нервные процессы такого же
характера, как при образовании отрицательного индивидуального
рефлекса, когда необычное раздражение систематически сочетается со
слабым индивидуальным рефлексом. В последнем случае сначала
возбуждается кора мозга от необычного раздражения, а затем все
корковые элементы индивидуального рефлекса. Вследствие этого данное
необычное раздражение после нескольких сочетаний становится
отрицательным сигналом по отношению к индивидуальному рефлексу. При
тонкой дифференциации рефлекса мы также должны иметь дело с
образованием отрицательного рефлекса, причем необычное раздражение
становится отрицательным сигналом по отношению к индивидуальному
рефлексу. Это явление обусловливается преимущественным развитием
обратных временных связей от всех корковых элементов рефлекторной
дуги индивидуального рефлекса к очагу необычного раздражения.
Вследствие обратных связей возбуждение означенных элементов рас-
556
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
пространяется главным образом к очагу необычного раздражения и
поэтому, согласно закону сопряженной иррадиации, распространяется
на периферию с меньшей интенсивностью (Беритов, 15). С
образованием прочных обратных связей в коре мозга получается чрезвычайна
деятельный нервный * круг из поступательных и обратных временных
связей. Это обусловит непрерывное вращение импульсов возбуждения
по данному кругу как во время раздражения, так и некоторое время
после него. Одновременно этими же импульсами возбуждения будет
все время активироваться соподчиненный нейропиль, т. е. тот его
участок, с которым данный нервный круг связан многочисленными синаті-
тическими окончаниями. Возникший здесь нейропильный ток не может
не оказать анэлектротонического угнетающего .влияния на ближайшие
нервные клетки и аксоны. Сильнее всего нейропильный ток
подействует на те нервные элементы, которые не возбуждены, а такими
элементами должны быть те аксоны пирамидных клеток, которые не
входят в означенный нервный круг, т. е. которые являются
эфферентными, уходящими из конечного двигательного участка в подкорку в
составе пирамидных и экстрапирамидных путей. Значит, каждый раз,
когда обратные связи включаются в работу, возбуждение, циркулирующее
по замкнутому кругу, активирует нейропиль, и этим самым последний
должен также оказывать отрицательное действие на индивидуальный
рефлекс.
Наиболее яркое доказательство образования обратных временных
связей при тонкой дифференциации дается в одной работе
Подкопа е в а из лаборатории Павлова (10). Он образовал пищевой
слюнный рефлекс на механическое раздражение кожи посредством
надавливания тупыми иглами, так называемой кололкой, на голень
левой задней ноги собаки. Это раздражение не болезненно, оно не
вызывает оборонительной реакции. В период дифференциации
необычных кожных участков раздражение последних вызывало двигательную
реакцию, — резкие оборонительные движения частей тела, где
находится дифференцируемый кожный участок. Если, например,
необычное место раздражения находилось на левой передней ноге, то в
момент дифференцируемого раздражения отдергивалась эта нога; иногда
отдергивания ноги совпадали с ритмом раздражения. При
раздражении на плече собака резко отводила плечо; при раздражении на
бедре — отдергивала заднюю ногу и приседала на задние лапы. Во
время этих реакций слюнная реакция отсутствовала. Словом,
получилась такая картина, как будто бы данные необычные кожные участки
раздражались болезненно или повреждающе щипанием или
электрическим током. Очевидно, тактильное, неболезненное механическое
раздражение кожи стало индивидуальным сигналам к оборонительным
реакциям.
Это явление могло получиться только вследствие
преимущественного развития обратных временных связей от корковых элементов
обычного индивидуального рефлекса к корковым очагам, возбуждае-
ПРОИСХОЖДЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 557
мым раздражением необычных, дифференцируемых участков кожи.
При раздражении дифференцируемого участка кожи возбуждение
соответственного коркового участка по поступательным связям вызывает
деятельность в нервных элементах пищевого слюнного рефлекса, а
возбуждение этих элементов иррадиирует обратно через обратные
связи к первичному очагу возбуждения от кожного раздражения. От
этого здесь получается такое усиленное возбуждение, какое бывает при
повреждающем раздражении соответствующего кожного участка. При
этом слюнная реакция отсутствует, так как возбуждение элементов
пищевого рефлекса иррадиирует, главным образом, по обратным
связям и, согласно закону сопряженной иррадиации, распространяется с
субминимальной интенсивностью к слюноотделительным центрам
продолговатого мозга.
Итак, трудность дифференциации наиболее близких к
индивидуальному раздражению {необычных раздражений основывается на том, чіо
эти раздражения вызывают (рефлекс через временные связи, которые
образовались в ответ на индивидуальное раздражение.
Дифференциация этих необычных раздражений зависит прежде всего от
дальнейшего ограничения повышения возбудимости в области очага
индивидуального раздражения, а затем от развития обратных временных
связей между очагами необычных раздражений и корковыми элементами
индивидуального рефлекса и, наконец, от тормозящего действия
активируемого при 'этом нейропиля на ближайшие пирамидные и
экстрапирамидные пути.
Обычно для полной дифференциации необычные раздражения
повторяются систематически до полного угасания вызываемого ими
рефлекса (Розенталь, 11; Соломонов, 12). Но в определенных случаях
полная дифференциация достигается и путем долгого
систематического сочетания индивидуального раздражения с основным, т. е. без
многократных проб необычных раздражений. В таком случае близкие
необычные раздражения могут не производить эффекта при первой
ж? пробе (Снегирев, 14). Очевидно, в последнем случае полная
дифференциация происходит исключительно путем достаточно сильного
понижения возбудимости в нейронных кругах вокруг очагов
индивидуального и основного раздражений в связи с очень сильным
развитием временных связей на индивидуальное раздражение.
Роль психонервной деятельности в явлениях рецепторной генерализации и
дифференциации индивидуальных рефлексов. При изучении индивидуального поведения
высших позвоночных животных (шищ и млекопитающих) было установлено, что
пищевое или оборонительное поведение первоначально осуществляется путем
психонервной деятельности вследствие ©озиикновения у животных психонешного комплекса
представления местоположения пищи или врага с соответствующим эмоциональным
тя&режишыит. Лишь потом, вследствие многократного повторения дачного
индивидуального поведения при одной и той же внешней обстановке, когда индивидуальное
поведение автоматизируется, последнее протекает, в общем, по принципу цепного
рефлекса (Беритов, 38, 39)
К&к указывалось выше, в условиях изучения индивидуальных рефлексов то ме-
558
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
тоду Павлова или Бехтерева, мы также имеем дело с поведенческими актами-
Поэтому возникает вопрос, какую роль играет псстхонервная деятельность в реакциях,,
вгблюдаем-ьгх у животных в процессе образования и дифференциации индивидуальные
рефлексов.
Специальное изучение вопроса о происхождении индивидуальных реакций в-
самом начале их образования в условиях работы по методу Павлова и Бехте-.
рева показало, что первоначально как индивидуальная двигательная реакция, так
и секреторная, возникающие в ответ на экстероцептивные раздражения, должны
наступать благодаря возникновению представления пищи или обстановки болевого
раздражения, а не через временные связи. Так, например, не может быть сомнения в
том, что в начальный период выработки индивидуального оборонительного рефлекса,
буйная реакция животного в ответ на звуковой сигнал, более буйная, чем в ответ
на самое электрическое раздражение, представляет собой индивидуальный акт
поведения, направленный на освобождение из станка и вообще из экспериментальной
камеры, где животное подвергалось болевому раздражению. Это поведение, очевидно,
вызывается представлением обстановки камеры, которое, должно быть,
сопровождается воспроизведением чрезвычайно неприятных ощущений (Беритов, 41).
По опытам на людях мы знаем, что в аналогичных случаях при образовании
индивидуальных оборонительных рефлексов индивидуальные сигналы вызывают
чрезвычайно неприятные ощущения (Беритов и Дзидэишвили, 40). Характерно, что в первое
время животное производит буйную реакцию освобождения вскоре после установки
в станок; оно пытается освободиться также и впоследствии, и притом помимо
индивидуальных сигналов. Очевидно, это происходит в результате (возникновения
соответствующего психонервного комплекса представления и неприятных ощущений, которые
наступают под влиянием обстановки камеры или «самовозбуждения» соответствующих,
иеіихоьервных элементов.
Пожалуй, можно утверждать, что начальный период генерализованного состояния
индивидуальных реакций на экстероцептивные раздражения по существу обязан своим
происхождением исихон?раной деятельности. Лишь вследствие многократного
сочетания индивидуализируемого сигнала с едой или электрическим раздражением
образовываются временные связи, и, сообразно, психонервный процесс отступает на задний
план. Это, в свою очередь, обусловливает дифференциацию рефлекса, т. е. все
другие раздражения, кроме индивидуального, перестают вызывать индивидуальную
реакцию.
Следует отметить, <что в стадии генерализации рефлекса очень часто после
однократной пробы (постороннего дифференцируемого раздражения, без дачи осчовчого
раздражения, собака перестает давать © течение некоторого времени индивидуальный
рефлекс и на обычныч индивидуальный сигнал. Это явление, известное как
тормозящее последействие, точно так же объясняется как результат психонервной
деятельности. Так как при необычном1 раздражении основное раздражение не произвол л тс я„
у собаки возникает психонервный процесс представления отсутствия основного
раздражения (напр., в случае неподкрешгения едой, в виде закрытой кормушки). Очевидно,
это представление сопровождается соответствующим эмоциональным возбуждением, и
благодаря этому нормальная реакция н? наступает в ответ на индивидуальный
сигнал, пока это представление достаточно свежо. Впоследствии после многократного
повторения дифференцируемого раздражения без сопровождения основным
раздражением устанавливаются временные связи с преимущественным развитием обратных, и*
тогда дифференцируемый раздражитель перестает вызывать рефлексы уже благодаря
деятельности этих обратных связей.
Признавая определенную роль психонервной деятельности в генерализации и
дифференциации индивидуальных рефлексов, мы тем самым не нарушаем вышеизложенной
концепции насчет физиологической деятельности коры большого мозга, объясняющей
г?черали?ацию и дифференциацию индивидуальных 'рефлексов. Следует только
допустить, что те сложные физиологические процессы, которые имеют место прч
генерализации и дифференциации первоначально, возникают не прямо от воздействия инди-
ПРОИСХОЖДЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 559*
«дуального сигнала, а через репродуцируемый этим сигналом психонеріэный процесс
таредс гаівления.
Происхождение последовательных рефлексов. При
последовательном сочетании основное раздражение производится через некоторое
время после индивидуального раздражения. В этих случаях
взаимодействие [возбужденных очагов осуществляется таким образом.
Основное раздражение действует на очаги индивидуального раздражения,,
где возбудимость была только что повышена предшествовавшим
индивидуализируемым раздражением. Последнее же раздражение действует
на очаги основного раздражения после того, как повышение
возбудимости, (вызванное предыдущим сочетанием в этих очагах основного
раздражения, в значительной мере прошло. В связи с этим действие
возбуждения из очага индивидуального раздражения на очаги
основного происходит імного слабее, чем в случае совпадения раздражений,
что в значительной мере задерживает образование поступательных
временных связей между ними. Об этом свидетельствует малая прочность
последовательного рефлекса, т. е. быстрая угасаемость его, и затем
невозможность образования рефлекса при сравнительно больших
интервалах между индивидуализируемым и основным раздражениями. Раз-
при последовательном сочетании раздражений временные связи
развиваются слабо, то, согласно закону сопряженной иррадиации
возбуждения, при каждом сочетании возбуждение распространяется в коре
больших полушарий из элементов временных связей в большей мере,
чем при совпадающем раздражении, когда временные связи достигают
значительного развития. Следовательно, при последовательном
сочетании возбудимость всей коры должна повышаться в большей мере, чем
при совпадающем сочетании. В действительности так и бывает, о чем
свидетельствует большая генерализация последовательного рефлекса,
большая трудность и даже невозможность полной его дифференциации,
если только интервал между раздражениями был достаточно велик.
Как указывалось выше, возбуждение, вызванное основным
раздражением, иррадиирует по коре при последовательном сочетании сильнее,
чем при совпадающем. Это должно благоприятствовать образованию
обратных временных связей, так как иррадиированное возбуждение
должно действовать на очаг индивидуального раздражения, где
возбудимость была только что повышена индивидуальным раздражением. Пока
обратная связь не достигла еще значительного развития,
индивидуальное раздражение способно давать совпадающий индивидуальный
рефлекс через поступательные связи. Но как только в значительной мере
разовьются обратные связи, возбуждение из двигательного аппарата
иррадиирует преимущественно через них к очагу индивидуального
раздражения (Беритов, 2, 15). Это, конечно, должно подействовать
отрицательно на периферический эффект. Кроме того, нужно иметь в
виду, что во время возбуждения нервных кругов поступательных и
обратных связей соподчиненный им нейропиль также находится в
усиленном активном состоянии. Благодаря этому нисходящие эффектор-
560
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
-К, t—і г1_/7
1
лые пути должны испытывать анэлектротоническое действие, как это
уже указывалось выше. Круговорот возбуждения прекратится, когда
произойдет утомление обратных или поступательных связей. В связи с
прекращением круговорота возбуждения в нервных кругах ослабнет
активное состояние нейропиля, а значит, и его угнетающее действие на
эфферентные пирамидные пути.
Как было упомянуто, последовательный рефлекс развивается в
некоторых случаях и без предварительной совпадающей стадии.
Следовательно, обратные временные связи с самого же начала
упрочиваются в такой степени, что уже в начале образования рефлекса
препятствуют во время индивидуального раздражения проведению
возбуждения через поступательные связи к периферическому органу. По всей
вероятности, это явление имеет место в том случае, если
индивидуализируемое раздражение производит сильное физиологическое действие
(звучание электрического звонка, удары метронома) и продолжается
более длительное время (30 сек. и более), как это было в опытах
Леонова (20). При таких
условиях возбудимость очага
ЛЛХ индивидуального раздражения
должна сильно повышаться и
этим самым значительно
благоприятствовать образованию
и укреплению обратных
связей.
_, лпгч „ Существование обратных
Рис. 239. Отношение света к своему
последовательному рефлексу на левой ноге. связей прежде всего видно из
Собака Мура. После первого применения света С это ТОГО, ЧТО В ТОТ ПерИОД, КОГДЭ
(Повторяется 3 раза с перерывом в 3, 8 и 11 сек. Каш- 0 «
дай раз свет длится по 10 сек. Последовательного ре- Совпадающий рефлекс Превра-
флекса не было, но позднее, спустя 33 сек., еще раз
повторенный свет вызвал обычный последовательный ЩаеТСЯ В ПОСЛеДОВатеЛЬНЫИ,
рефлекс. Очевидно, при быстром следовании
световых раздражений одного за другим каждое последую- СИЛЬНО Нарастает ОГрИЦатеЛЬ-
ацее раздражение действует отрицательно на
последовательный рефлекс от предыдущего. Верхняя НОе ДеиСТВИб ИНДИВИДуЗЛЬНОГО
кривая — от левой задней ноги, нижняя — от
правой передней. раздражения на
индивидуальный рефлекс. Это
проявляется, между прочим, в том, что последовательный рефлекс не
наступает, если по прекращении раздражения возобновить его, не
дождавшись наступления рефлекса (рис. 239). Отрицательное действие
начинает проявляться еще в период совпадающего рефлекса: в связи с
прекращением раздражения совпадающий рефлекс усиливается. Так было,
например, при образовании последовательного рефлекса на свет
электрической лампы. Этот раздражитель до образования на пего
индивидуального рефлекса не влиял отрицательно на индивидуальный
рефлекс, с момента же образования последовательного рефлекса он
стал обнаруживать сильное отрицательное действие.
Характерно, что периферический эффект через поступательные
временные связи обычно устранялся не только во время индивидуального
раздражения. В опытах Добровольского (21) основное раздра-
UllllllllllllUllllmillllllllinniiimlhiiiliiiinil 1...I, ,, (|Ш|
ПРОИСХОЖДЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ
561
жение следовало спустя 2 мин. после индивидуального, а
индивидуальный последовательный рефлекс начинался через разные более или
менее значительные промежутки времени, причем эти промежутки
удлинялись с укреплением рефлекса. Отсюда следует, что отрицательное
действие индивидуального раздражения на рефлекс происходит не
только во время раздражения, но и некоторое время после него,—
следовательно, прекращение раздражения не является сигналом к
последовательному рефлексу. Нужно думать, что возбуждение, пришедшее по
обратным связям в очаг индивидуального раздражения, отсюда вновь
распространяется к корковым двигательным или секреторным
аппаратам. Из корковых очагов возбуждение вновь направляется ів очаг
индивидуального раздражения и опять обратно к ответным очагам.
Разумеется, этот круговорот возбуждения будет происходить как во
время индивидуального раздражения, так и по прекращении его, до тех
пор, пока не произойдет утомление обратных или поступательных связей.
Быстрота, с которой наступает утомление, зависит от степени развития
этих связей. Чем более они развиты, тем дольше должна
продолжаться их деятельность. Но как только наступит утомление обратных
связей, и в связи с этим ослабнет тормозящее действие активированного
при этом нейролиля, возбуждение из конечных пунктов
поступательных временных связей начнет усиленно иррадиировать по пирамидным
путям и вызывать периферический эффект. Конечно, может случиться,
что при этом также произойдет утомление поступательных связей,
тогда последовательный рефлекс совсем не наступит. Это, например,
получается при быстром повторении индивидуального раздражения, еще
раньше, чем наступит последовательный рефлекс на первое
раздражение: повторное раздражение устраняет последовательный рефлекс на
первое раздражение и не дает своего последовательного рефлекса
(Беритов, 2, 15). Об этом же свидетельствуют наблюдения Гр о с с м а-
н а (16). На одной подопытной собаке ему не удалось образовать
последовательный слюнной рефлекс на чесание, несмотря на 400 сочетаний.
Безусловно, временные связи при этом образовались, так как под
влиянием экстрараздражителей, которые присоединялись к чесанию,
последовательный рефлекс обнаруживался полностью. Значит, у этой собаки
при данных условиях обратные связи развились лучше, чем
поступательные, вследствие чего при круговороте возбуждения поступательные
связи утомлялись раньше обратных. От экстрараздражителей,
производивших ориентировочную реакцию, возбудимость конечных ответных
пунктов поступательных связей повышалась, и от этого становился
возможным вызов периферического эффекта. Гроссману удалось на той
же собаке образовать последовательный рефлекс, после того как
предварительно он образовал совпадающий рефлекс на чесание, т. е. после
того, как предварительно он в значительной мере укрепил
поступательные временные связи.
Итак, при образовании последовательных рефлексов развитие
обратных связей находится в несколько лучших условиях, чем при образо-
36 и. С. Беритов
562 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
вании совпадающих. Поэтому во время индивидуального раздражения
и некоторое время после него возбуждение вращается в нервном кругу
из поступательных и обратных связей, пока не произойдет утомление
обратных связей, вследствие чего возбуждение из конечных пунктов
поступательных связей начнет усиленно иррадиировать по пирамидным
путям. Кроме того, пока данный нервный круг находится в
возбуждении, пирамидные аксоны подвергаются торможению со стороны
активного нейропиля. Таким образом, последовательный рефлекс наступает
после того, как обратные связи утомились и ослабло тормозящее
действие нейропиля на пирамидные аксоны.
Происхождение отрицательных рефлексов. Как указывалось выше,
при систематическим сочетании необычного раздражения с
определенным индивидуальным рефлексом необычное раздражение становится
индивидуальным сигналом — начинает вызывать тот же индивидуальный
рефлекс, т. е. образуется индивидуальный рефлекс высшего порядка.
Так бывает, если основной индивидуальный рефлекс интенсивен. Если
же он слаб, то такое необычное раздражение приобретает свойство
отрицательного действия на этот индивидуальный рефлекс. 'Мы привели
ряд доказательств того, что здесь мы имеем типичный индивидуальный
рефлекс отрицательного типа. Следовательно, при образовании
отрицательного рефлекса должны развиваться как поступательные, так и
обратные (временные связи, а именно: поступательные связи — от очага
отрицательного индивидуального раздражения ко всем корковым
очагам основного положительного рефлекса и обратные — от последних к
первому. Из самого характера явлений следует, что в отрицательном
рефлексе обратные связи развиты сильнее поступательных, и поэтому,
согласно закону сопряженной иррадиации, возбуждение
распространяется из элементов поступательных связей в обратные связи в такой
мере, что оно идет на периферию и достигает координирующих
аппаратов с субминимальной интенсивностью. Деятельность обратных связей
отрицательного рефлекса преобладает над поступательными настолько,
что она способна устранить всякий другой рефлекс, вызываемый через
конечные пункты поступательных связей отрицательного рефлекса. По
обратным связям возбуждение будет иррадиировать не только из
поступательных связей отрицательного рефлекса, но и из поступательных
связей сочетаемого с ним положительного индивидуального рефлекса.
К этому прибавится еще усиленное активирование нейропиля при
возбуждении нервных связей отрицательного рефлекса, а это будет влиять
отрицательно, между прочим, и на двигательный участок, на его
пирамидные клетки и их аксоны, вызывая их угнетение посредством нейро-
пильного тока. Но устранится ли последний рефлекс полностью или
нет — это будет зависеть, конечно, от его интенсивности. Если он
достаточно силе^н,— например, если он перед тем многократно сочетался
с основным раздражением,— отрицательный раздражитель только
ослабит его. Если же он перед тем был ослаблен многократным повторе-
ПРОИСХОЖДЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ
563
нчем без сочетания с основным раздражением, отрицательный
раздражитель устранит его полностью.
Очень характерно, что образование отрицательного рефлекса
происходит тем быстрее, чем сильнее необычное раздражение по сравнению
с комбинируемым положительным раздражением. Фурсиковым (5)
было установлено, что если физиологическое действие отрицательного
раздражения слабее положительного индивидуального раздражения, то
отрицательный рефлекс развивается медленно; кроме того, в этих
случаях он слаб и непостоянен. Напротив, если отрицательный
раздражитель является более сильным агентом сравнительно с положительным,
то отрицательный рефлекс образуется много быстрее; при этом он
бывает гораздо постояннее и сильнее. Фурсиков находит, что, в общем,
для образования отрицательного рефлекса требуется более сильное
индивидуализируемое раздражение, чем для образования
положительного.
Нетрудно объяснить физиологическую природу этих фактов, если
принять во внимание, от чего зависит преимущественное развитие той
и другой временной связи. Как выше было показано, преимущественное
развитие обратных ©ременных связей обусловливается тем, что
физиологическое действие первого комбинируемого раздражения
превосходит действие второго. Если данный внешний агент регулярно
предшествует индивидуальному раздражению и сила его физиологического
действия, в сравнении с индивидуальным раздражением, больше, то
обратные связи, а именно временные связи от нервных элементов,
возбуждаемых положительным индивидуальным раздражителем, к очагам
внешнего агента должны развиться сильнее, чем поступательные связи.
Это обусловит усиленный круговорот возбуждения по нервным кругам
из поступательных и обратных связей во время действия данного
агента и затруднит наступление периферического эффекта.
Одновременно наступает усиленное активирование нейропиля, которое будет
действовать тормозящим образом, между прочим, на конечный пункт
поступательных временных связей, в особенности на его пирамидные
аксоны. Вследствие этого внешний агент становится отрицательным
индивидуальным сигналом. Так возникает отрицательный рефлекс. Если
физиологическое действие внешнего агента относительно слабее, чем
действие положительного раздражения, то обратные связи должны
отставать в своем развитии, и тогда отрицательный рефлекс или
совсем не образуется, или если и образуется, то он очень слаб по
интенсивности (Беритов, 15).
Отрицательный раздражитель своими развитыми обратными связями
оказывает на состояние животного характерное действие. Если оно
находится в беспокойном состоянии — визжит, беспокойно двигает
головой и ногами,— то отрицательный раздражитель устраняет это
состояние: животное успокаивается и даже начинает дремать. Следовательно,
отрицательный раздражитель обнаруживает действие, совершенно
противоположное тому, которое наблюдается при положительном индиви-
36?
564
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
дуальном рефлексе. Отсюда следует, что в отрицательном рефлексе
образуются "хорошо развитые обратные связи со всеми теми
элементами коры, 'которые обычно приходят в деятельное состояние от поло*
жительного индивидуального рефлекса, а именно — с элементами
голосовой и ориентировочной реакции, двигательных реакций конечностей и
дыхания (Беритов, 15).
tb
іЬ
ib
<Ь
200
iliNlimimUnimililliiiililiiuii» .iim^flpfljmnnni.Hnfinmimri.HH.f. u.liru^»t|mim.rlilii-iil..rii.nm4ium»iii.uninii..i.»
Рис. 240. Взаимодействие между положительным и отрицательным
рефлексами у собаки.
Собака Боб. Положительный индивидуальный раздражитель — тон 200
колебаний в 1 сек. (нижний сигнал); отрицательный раздражитель — свет
электрической лампы (средний сигнал). В опыте 1 тон вызывает обычный
положительный рефлекс. В опыте 2 тон дается во время света. В этом случае тон не
дает положительного рефлекса. Он, наоборот, производит отрицательную
реакцию: животное опускает голову (поднятие кривой) и засыпает.
Повторение тона через 15 сек. после светового раздражения также не дает
положительного индивидуального рефлекса. В опыте 3 через 1 мин. после второго
•опыта тот же тон вновь производит только более сильное опускание головы.
Лишь через 2.5 мин. после светового раздражения тон вызвал снова обычный
положительный рефлекс.
Характерная картина реакции получается в том случае, если во
время действия отрицательного раздражителя почему-либо запоздало его
отрицательное действие и как раз в это время был применен
положительный раздражитель: сейчас же после присоединения последнего
животное успокаивается и начинает дремать. Повидимому, с прибавлением
положительного сигнала усиленно начал работать нервный круг из
обратных и положительных временных связей. Одновременно усилилось
активное состояние соподчиненного нейропиля. Все это вместе
обусловило усиленное отрицательное действие на конечный пункт
поступательных связей. Рис. 240, иллюстрирующий это явление, показывает, что
даже повторное применение положительного раздражителя через
минуту после отрицательного дает усиленную отрицательную реакцию и что,
следовательно, повышенная возбудимость в обратных связях
удерживается некоторое время после отрицательного раздражения.
До сих пор 'Мы имели в виду такие отрицательные рефлексы, при
образовании которых внешний агент, действующий как отрицательное
раздражение, предшествует положительному рефлексу. Но імы
указывали, что отрицательный рефлекс образуется также при обратном
порядке сочетания, т. е. когда данный агент действует во время
положительного рефлекса. Фурсиков (17) образовал подобный рефлекс,
ПРОИСХОЖДЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 565
применив в качестве отрицательного раздражения сильный шум,
вызываемый вращением кимографа; для положительного же раздражения у
него был применен стук метронома. В этом случае поступательные связи,
которые вели от корковых элементов, возбуждаемых первым,
положительным раздражением — метрономом, к очагу второго, отрицательного
раздражения — шума, развились сильнее, чем обратные связи из очагов
последнего раздражения к очагам первого, так как второе раздражение
являлось физиологически более сильным. Следовательно, при
образовании отрицательного рефлекса путем обратного порядка сочетания
отрицательное действие обусловливается деятельностью поступательных
связей. Но и теперь отрицательное действие на индивидуальный
рефлекс зависит от усиленного вращения возбуждения по замкнутому
нервному кругу и одновременно от усиленного активирования нейропиля,
ибо оба эти обстоятельства должны производить торможение
эфферентных путей.
О происхождении экспериментального сна. По работам лаборатории
Павлова известно, что во время опыта некоторые собаки при
определенных условиях засыпают, а именно, это наблюдалось во время
отрицательного рефлекса (Анреп, 23; Фурсиков, 5), во время
недеятельной фазы последовательного (Добровольский, 21; Белиц, 24; Леонов, 20)
и запаздывающего рефлексов (Петрова, 4; Безбокая, 6), затем вовремя
действия дифференцированного раздражения с угашенным рефлексом
(Коган, 27; Бирман, 26), а также во время необычного раздражения
при угасании ориентировочного рефлекса (Розенталь, 11; Чечулин, 28).
Некоторые собаки засыпают, стоя в станке в экспериментальной
комнате и без перечисленных выше условий. Так, например, собака
может заснуть в станке уже в первый день постановки ее в станок, значит
еще раньше образования какого-либо искусственного индивидуального
рефлекса и даже еще до применения электрического раздражения.
Другие собаки засыпают впоследствии, более или менее быстро, в
связи с характером работы. Так, например, особенно сонливыми
становятся собаки при работе с механическими (Ерофеева, 29; Петрова, 4;
Рожанский, 22) и температурными раздражителями кожи (Шишло и
Соломонов, 12).
Нас интересует то явление сонливости, которое определенным
образом связано с действием индивидуальных раздражений.
Прежде всего укажем на то, что экспериментальный сон
вызывается действием таких раздражителей, которые, судя по вышеизложенному,
возбуждают хорошо развитые обратные связи, а именно, действием
индивидуальных раздражителей последовательного, запаздывающего и
отрицательного рефлексов. Так же действуют дифференцированные
раздражители, ибо они также обладают хорошо развитыми обратными
связями, и еще индивидуальные раздражители с угасшим рефлексом,
ибо при угасании индивидуального рефлекса, как это подробно будет
выяснено ниже, мы также имеем дело с хорошо развитыми обратными
связями.
566
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
Итак, во всех известных случаях экспериментального сна мы имеем
дело с такими индивидуальными раздражениями, корковые очаги кото-
рых обладают хорошо развитыми обратными связями с корковыми
элементами какого-либо положительного рефлекса.
Теперь остается рассмотреть вопрос, почему именно
индивидуальные раздражители с хорошо развитыми обратными связями вызывают
сонливость. Прежде всего
укажем на несколько
характерных черт сонного состояния.
1. Когда сон еще является
поверхностным, отношение
животного к
дифференцированным звукам и
индивидуальным раздражителям
нормально. Во время дремоты
индивидуальный раздражитель дает
рефлекторную реакцию.
Дифференцированные звуки либо
не дают никакого видимого
эффекта, либо даже
углубляют сон. Кроме того, во
время сна отрицательное действие
дифференцированного
раздражителя на индивидуальный
раздражитель не прекращается.
2. Если собака имеет склонность ко сну, то она при
продолжительном действии дифференцированного раздражителя засыпает, однако
сейчас же после прекращения раздражения она просыпается. Значит,
действие дифференцированного раздражителя продолжается во время
сна (рис. 241).
3. Индивидуальное раздражение не в состоянии вызывать
двигательный оборонительный рефлекс без пробуждения. Пробуждение
является необходимым условием. Сначала животное открывает глаза,
поднимает голову, производит общее движение туловищем, и только
потом наступает индивидуальный рефлекс на ноге. Часто случается, что
индивидуальный раздражитель будит, но не дает рефлекса на ногах, и,
по Рожанскому, пробуждение животного есть необходимое условие
для наступления индивидуального двигательного рефлекса. Но он же
наблюдал наступление индивидуального слюноотделения без
пробуждения и без какой-либо одновременной двигательной реакции (22).
4. Механическое или электрическое раздражение кожи может
вызывать рефлекс и во время сна, без пробуждения. Так, например, во
время ( глубокого сна электрическое раздражение вызывает слабый
визг, движение раздражаемой ноги, поворот головы в
раздражаемую сторону, но глаза не открываются, голова не поднимается. С пре-
/7/7.
300
300
¦ мпиіппіініііщ.і.м.м.пмм,.піііііиііпіи.мтщі.ипіі.піпн.нм.мі
Рис. 241. Снотворное действие
дифференцированного раздражителя звука тонвариатора
300 колебаний в 1 сек.
Собака Боб. Звук применялся дважды по полминуты.
Оба раза собака сразу из состояния бодрствования
переходит в сонное (поднятие верхней кривой); затем
каждый раз после звука пробуждается: поднятие
головы (опускание кривой) и небольшое движение
правой передней ноги.
ПРОИСХОЖДЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ
567
кращением раздражения визг сейчас же прекращается, нога опускается,
голова возвращается в медиальное положение (рис. 242). Конечно, все
эти рефлексы значительно ослаблены против того, что наблюдается при
бодрствующем состоянии. В тот самый период, когда электрическое
раздражение не .в состоянии разбудить животное, индивидуальный
раздражитель, звук тонвариатора 200 колебаний в секунду в состоянии
вызвать пробуждение.
5. Все вышеприведенные раздражители, вызывающие засыпание
животного, совсем лишены способности давать ориентировочные
реакции. Можно сказать, что чем более угашена ориентировочная реакция
\\ ^ ] \ і Л
_± 4 4 S S 6 7
—і г— * г- » 1 1 1
И1И1Ш1і1ЩЩЩігптиитпі.іі.ііі»цг "ИНЧШМ" " "' ііінмпіііііінініиі
Рис. 242. Действие электрического раздражения кожи
во время сна.
Собака Боб. В самом "начале работы, еще до образования
индивидуальных рефлексов. На 15-й день после первой установки
в станок. Собака спит: глаза закрыты, задние конечности
расслаблены, собака не опирается на них, а свободно висит на
лямках; голова сильно опущена вниз; храпит. В опыте 1
электрическое раздражение правой передней ноги при 7—4 см
производит небольшое движение без пробуждения. Только голова
немного поднимается (быстрое опускание кривой Г глаза
не открываются, и после раздражения голова вновь
наклоняется вниз. В опыте 2 сильное электрическое раздражение
производится во время сна и после пробуждения. Первый раз
оно пробудило собаку и дало слабое движение, а второй раз
вызвало сильнейшее отдергивание ноги.
на данный раздражитель, тем сильнее его усыпляющее действие.
Электрическое раздражение тоже не пробуждает, если оно не вызвало
ориентировочной реакции.
6. Пробуждающим действием обладают всякого рода необычные
посторонние раздражения, которые не применялись раньше в данной
экспериментальной обстановке и потому способны давать
ориентировочное движение. При повторном применении они перестают его вызывать.
В таком случае они перестают также будить животное. При этом
характерно, что пробуждающим действием обладает начало
необычного раздражения, от которого, собственно говоря, зависит
ориентировочная реакция. Если животное не пробудилось в начале его, то и
продолжение не даст пробуждения. Так же действуют резкие звуки —
звонок и удары по металлическому цилиндру,— которые в
бодрствующем состоянии животного оказывали сильное отрицательное действие
Из этих наблюдений следует, что животное засыпает при таких
раздражениях, которые не способны производить ориентировочную реак-
568 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
цию, а пробуждается при таких раздражениях, которые такую реакцию
вызывают (Беритов, 15).
Это положение дает нам ключ к пониманию снотворного действия
раздражителей, обладающих хорошо развитыми обратными связями.
Эти связи существуют в каждом данном случае со всеми теми
элементами, которые действуют в индивидуальном положительном рефлексе,
т. е. с корковыми двигательными участками освободительных и
ориентировочных движений, а также дыхания, голосовой реакции и
открывания глаз. Обратные связи отклоняют возбуждение иэ всех этих
элементов по закону сопряженной иррадиации возбуждения. Кроме того,
возбуждение обратных связей сопряжено с усилением активного
состояния нейропиля и его тормозящего действия на двигательные
участки. Вследствие этого животное лишается возможности производить
какое-либо движение. С отсутствием ориентировочных и других
движений стоит в связи полное
отсутствие проприоцептивных
раздражений, а это обусловливает
понижение возбудимости в
промежуточном мозгу, а также в коре
больших полушарий, в
особенности в проприоцептивной
воспринимающей области и
двигательных участках, находящихся в
интимной связи с этой областью.
Это в свою очередь
обусловливает ослабление спонтанной
активности в этих отделах
головного мозга, а также понижение
статического или пластического
рефлекторного тонуса,
нормально (в значительной мере
зависящего от проприоцептивных
раздражений, и затем понижение
или даже полное выпадение
рефлексов положения, т. е.
характерное явление сна.
В лаборатории П а ел о в а засыпание собак первоначально
связывалось с ограничением свободы движений по условиям эксперимента
(ноги в лямках, голова привязана). Это заключение было сделано
на основании того наблюдения, что сонливые в станке собаки после
снятия из станка обнаруживали бодрое состояние (Рожанский, 22;
Бирман, 26). Но Бирман показал, что названные выше раздражители
производят на свободе такое же снотворное действие, как и в станке.
Отсюда следует, что ограничение движений просто путем заключения
в станок или путем действия обратных временных сзязей оказывает
одно и то же снотворное действие.
38
и
ШіШіііПМіМПіШіііііііипиіміпіішііщ.піііііі.іііп-іііішпіііміп»,
Рис. 243. Влияние необычных звуков на
сон.
Собака Боб. В опыте 1 применяется один удар по
металлическому цилиндру ц. Это раздражение
применялось ранее только несколько раз. Собака
пробуждается (опускание верхней кривой) и
через довольно большой скрытый период
поднимает правую переднюю ногу (нижняя кривая),
как бы от индивидуального раздражителя.
В опыте 2 применяется звонок, который
систематически в течение многих дней сочетался о
индивидуальным рефлексом для образования
отрицательного рефлекса. Он не будит животное,
только в самом начале голова немного
поднимается.
ПРОИСХОЖДЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 569
Как известно, обстановка, в которой происходит засыпание,
становится индивидуальным возбудителем сна. Это известно каждому по
личному опыту. То же было обнаружено в наблюдениях над собаками.
Если несколько раз усыпить собаку в экспериментальной комнате
действием снотворных раздражений, то затем собака, входя в эту комнату,
очень быстро приходит в сонливое состояние (Павлов и Воскресенский,
18; Бирман, 26). В данном случае опытная обстановка является тем
раздражителем, который обладает развитыми обратными связями с
корковыми элементами индивидуальных рефлексов. Известно, что
экспериментальная комната делается индивидуальным возбудителем всех
тех рефлексов, которые образуются в этой комнате. Легко возбудимые
собаки, как только становятся в станок, визжат, надолго поднимаю!
ту ногу, на которой был образован рефлекс. Очевидно, у этих собак
хорошо развились поступательные временные связи імежду нервными
элементами, воспринимающими обстановку, и теми корковыми
нервными элементами, которые участвуют в индивидуальных рефлексах. Но,
конечно, если в данной обстановке вызывать сон применением
определенных раздражений, устраняющих индивидуальные и ориентировочные
рефлексы, то лучше всего должны развиться обратные временные
связи между нервными элементами, воспринимающими обстановку, и
корковыми двигательными элементами индивидуальной и ориентировочной
реакций.
Итак, наступление сна при индивидуальных раздражениях
происходит & тех случаях, когда имеет место преимущественное развитие
обратных связей. Это обусловливает полное отсутствие проприоцептив-
ных раздражений в связи с выпадением ориентировочных и других
движений. Отсутствие tyce проприоцептивной импульсации приводит к
понижению возбудимости и в связи с этим к ослаблению спонтанной и
рефлекторной деятельности в промежуточном мозгу, а также в коре
большого мозга, т. е. ко сну.
Раз наступление сна в экспериментальной обстановке зависит от
деятельности обратных связей между двигательными участками коры
и очагами возбуждения коры обычной обстановки, то укрепление
поступательных связей между ними должно приводить к исчезновению
сонного состояния. Это и наблюдалось в опытах Петровой (4). Она
вызывала сон у собак ежедневным многократным применением
одного и того же запаздывающего рефлекса, а затем устраняла его
полностью путем образования многочисленных новых совпадающих
рефлексов. Очевидно, первоначально при работе с запаздывающим
рефлексом между двигательными участками коры и очагами возбуждения
от обычной обстановки опыта образовались сильно развитые
обратные связи, что вело ко сну в этой обстановке. С образованием
совпадающих рефлексов сон перестает наступать в обычной
обстановке, так как теперь, наоборот, восприятие обстановки связывается с
двигательными участками коры более развитыми поступательными связями.
570
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
В лаборатории Павлова было замечено, что в определенных
условиях сам экспериментатор становится индивидуальным возбудите-
лем сна. Если, например, систематически приготовлять пищу перед
животным, смотреть ему пристально в глаза и в это самое время не
давать есть, то как процедура приготовления пищи, так и вид
экспериментатора и пристальный его взгляд становятся возбудителями сна
(Бирман, 26). Здесь опять-таки мы имеем последствие усиленного
развития обратных связей. При описанном поведении экспериментатора
животное сначала тянется к пище, т. е. у него наступает ряд обычных
индивидуальных реакций при виде пищи: вытягивание шеи,
передвижение всего тела к пище, чавкание, выделение слюны и т. п. Так как за
этим не следует захвата пищи, то этот сложный индивидуальный
рефлекс на поведение экспериментатора угасает, а угасание его
способствует развитию обратных временных связей. Вследствие деятельности
этих связей все поведение экспериментатора, и, в частности, его
пристальный взгляд, становится возбудителем сна.
Аналогичное явление мы должны иметь при усыплении человека
гипнотизером. Первый раз гипнотизер усыпляет человека при полной
тишине, в полутьме, приказывая ему не двигаться; при этом он
проводит рукой по голове, по лицу этого человека (так называемые пассы).
Отсутствие шума и разговоров, слабое освещение, неподвижность —
все это благоприятствует понижению возбудимости центральной
нервной системы. Раздражение головы путем поглаживания и
сдавливания ведет к небольшому ее движению и одновременно к рефлекторному
обездвижению остальной мускулатуры тела. Как указывалось выше,
специальными опытами на животных (лягушки, собаки) было
установлено, что под влиянием механического раздражения головы возникает
определенный рефлекс с возбуждающей иннервацией по отношению к
мышцам головы и тормозящей иннервацией ко всей остальной
мускулатуре. Вследствие этого торможения выпадают тонические рефлексы и
локомоция, а дыхание и защитные рефлексы на повреждающие
раздражения значительно ослабевают. При некоторой длительности
раздражения это состояние неподвижности переходит в соч. Путем
экспериментального анализа было выяснено, что сонное состояние есть результат
понижения возбудимости центральной нервной системы вследствие
полного выпадения проприоцептивных раздражений во время активного
обездвижения животного и значительного ослабления рефлекторного
действия внешних раздражений в силу означенного торможения (Бери-
тов, 25). Аналогичный рефлекс обездвиживания, несомненно, имеет
место у человека при поглаживании и сдавливании кожи головы, поэтому
и здесь наступлению сна благоприятствует не только отсутствие разного
рода внешних и внутренних раздражений, но и активное обездвижение
общим торможением всей скелетной мускулатуры. Общее торможение
должно в значительной мере ослаблять рефлекторное действие всякого
рода посторонних раздражений, а также действие произвольной
иннервации.
ПРОИСХОЖДЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 571
Опытами на людях было установлено, что общее торможение,
вызываемое сдавливанием кожи на голове, сильно ослабляет разного
рода боли (зубная, раневые, головные) (Цкипуридзе и Чичинадзе, 42). Это
очевидно, происходит вследствие понижения возбудимости в
промежуточном мозгу.
Если гипнотизеру удалось вызвать сон у человека описанным выше
способом, то после этого один вид гипнотизера в той же обстановке
или определенные движения рук (пассы) гипнотизера, а также
определенные слова, вроде «засни^, которыми он сопровождал первое
усыпление, могут оказаться совершенно достаточными для быстрого
усыпления того же пациента. Нужно думать, что это получилось у пациента
от развития обратных связей в коре больших полушарий, а именно от
двигательных участков ориентировочной реакции и других движений
к тем очагам, которые воспринимали всю обстановку, включая
манипуляции гипнотизера. Чем чаще повторяется усыпление пациента одним
и тем же гипнотизером, тем сильнее будет усыпляющее действие того
или другого определенного движения гипнотизера.
Как указывалось выше, пробуждение происходит от таких
раздражений, которые вызывают ориентировочную реакцию. Известно, что
ориентировочная реакция происходит главным образом через
промежуточный мозг и кору большого мозга. В том и другом отделе центральной
нервной системы имеются такие двигательные участки, через которые
вызываются ориентировочные и другие двигательные реакции. Так,
например, в обоих отделах головного мозга имеются двигательные участки
для движения ушей и головы, для движения глаз и головы и т. д.
Первые движения, какие могут быть вызваны периферическим
раздражением как во время бодрствования, так и во время сна, являются
ориентировочными; очевидно, возбуждение иррадиирует раньше всего и
сильнее всего к двигательным участкам ориентировочной реакции.
Характерно, что даже индивидуальные раздражители как в состоянии
бодрствования, так и во время сна сначала производят ориентировочное
движение, а затем уже индивидуальное движение ноги. Разница только
в том, что во время сна индивидуальное движение ноги запаздывает
много больше, чем при бодрствовании. Не может быть сомнения, что
пробуждение, вызываемое индивидуальными раздражителями, как и
необычными раздражениями, связано, в первую очередь, с
возбуждением тех корковых ориентировочных аппаратов, которые лежат в
пределах соответствующей воспринимающей области.
Однако ориентировочный рефлекс наступает в ответ на
индивидуальное раздражение не только вследствие близости коркового
ориентировочного аппарата к воспринимающему очагу данного раздражения. Как
бы глубоко ни было общее понижение корковой возбудимости во время
сна, оно не может быть везде, во всех элементах, одинаково низким.
Возбудимость тех элементов, которые участвуют в хорошо развитых
индивидуальных рефлексах, и во время сна остается выше возбудимости
других элементов. Только поэтому индивидуальные раздражители спо-
572 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
собны будить животное в большей мере, чем разного рода
неиндивидуальные. Нужно полагать, что временные связи, идущие от очага
индивидуального раздражения к корковым двигательным аппаратам
ориентировочной реакции, обладают большей возбудимостью, чем
остальные связи, и поэтому двигательные аппараты ориентировочной
реакции скорее и сильнее приходят в действие от иррадиирующего
возбуждения, чем другие аппараты.
Итак, пробуждение от сна под влиянием индивидуального
раздражения происходит, главным образом, вследствие усиленной иррадиации
возбуждения по временным связям от очага индивидуального
раздражения к корковым двигательным участкам ориентировочной реакции.
Далее, пробуждение, открытие глаз, выпрямление конечностей,
поднятие головы нельзя считать непосредственным результатом
возбуждающего действия внешнего раздражения на кору больших полушарий.
Внешнее раздражение с хорошо развитыми обратными связями также
действует на кору и, очевидно, на двигательные корковые аппараты;
однако такое раздражение не будит. Очевидно, пробуждение возникает
вторично под влиянием наступления ориентировочного движения головы.
С движением головы связан целый ряд таких раздражений, которые,
действуя на средние отделы головного мозга, обусловливают мышечный
тонус животного: раздражение рецепторов шеи от движения ее по
отношению к туловищу, раздражение статического органа —
лабиринтов — от движения головы в пространстве. Вызываемые отсюда
тонические рефлексы простираются на всю скелетную мускулатуру — и, в
частности, на мускулатуру глаз. Действием этих рецепторов
возбудимость промежуточного мозга возвращается к норме; при этом
восстанавливается нормальная спонтанная активность нервных центров
головного мозга, усиливается тонус конечностей и всего тела,
устанавливается нормальное положение животного, производятся
компенсаторные движения глаз.
Итак, ориентировочное движение головы является возбудителем
целого ряда очень сложных тонических рефлексов, поддерживающих
нормальное, выпрямленное положение животного. Но вслед за этими
тоническими движениями возникают проприоцептивные раздражения во
всей мускулатуре. Эти вторичные раздражения, действуя на
центральную нервную систему, в частности на промежуточный мозг и на кору
больших полушарий, повышают их возбудимость, даже могут вызывать
со своей стороны разного рода рефлексы, как локомоцию или даже
оборонительное движение ноги, если только соответствующий корковый
аппарат находился в состоянии повышенной возбудимости. Это мы
видим, например, на рис. 241, где после пробуждения от необычного
раздражения собака подняла правую переднюю ногу, на которой был
образован ряд индивидуальных рефлексов.
Чрезвычайно поучителен тот факт, что во время глубокого сна можно
вызвать электрическим раздражением ноги довольно сильное оборони*
ПРОИСХОЖДЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 573
тельное движение без пробуждения животного. Очевидно, это зависит
от того, что произведенные движением ноги проприоцептивные
раздражения не являются достаточными для таких сложных рефлексов, как
рефлексы положения, которые установили бы нормальное тоническое
состояние животного и подняли бы возбудимость промежуточного мозга
и коры в достаточной мере.
Само собой понятно, что не только прирожденные, но и
индивидуальные рефлексы, как наблюдал Рожанский, могут наступать без
пробуждения, если только при этом ориентировочная реакция будет
отсутствовать. Так обстояло дело в некоторых случаях с индивидуальными
слюнными рефлексами: собака реагировала выделением слюны без
пробуждения при отсутствии ориентировочной реакции. Очевидно это
бывает в таких случаях, когда временные связи с корковым
секреторным аппаратом являются более развитыми, чем с корковыми
двигательными участками ориентировочной и ротовой реакции.
Итак, в какой мере неподвижность животного, и в особенности
отсутствие ориентировочной реакции, является основным условием
наступления сна с его пониженной таламической и корковой
деятельностью, в такой же мере появление движения, в особенности
ориентировочного, является главным условием пробуждения с его характерными
симптомами восстановления нормального тонуса и положения, а также
повышения таламической и корковой возбудимости.
Выше мы рассмотрели происхождение экспериментального сна,
связанного с образованием обратных временных связей. Но известно, что
явление сна не является функцией коры большого мозга: животные,
лишенные коры мозга, продолжают спать, даже еще больше, чем
нормальные животные. Не нарушается также обычная периодичность
сна и бодрствования. Отсюда следует, что обычный сон у животных
является функцией промежуточного мозга (Рожанский, 36 и др.). Но,
конечно, как это видно из вышеизложенного, промежуточный мозг играет
существенную роль и в происхождении экспериментального сна.
Когда индивидуальное раздражение перестает давать ориентировочную
реакцию и вообще двигательные реакции благодаря развитию обратных
временных связей в коре мозга, тогда не только кора, но и все
подкорковые центры головного мозга перестают получать с периферии
проприоцептивные импульсы. Это должно привести к понижению
возбудимости, а также спонтанной и рефлекторной деятельности всех центров
головного мозта и, между прочим, тонических центров скелетной
мускулатуры, дыхательных, сосудодвигательных и других центров,
вследствие чего ослабевает постоянная импульсация координирующих аппа-
^атоа "и. дата^т^ъъъга. ^ей^оноъ станкого мозга со стороны этих
центров. Этим, должно быть, объясняется как ослабление тонуса в
организме, так и выпадение спинальных рефлексов, обычно вызываемых
периферическим раздражением конечностей.
Учение Павлова о происхождении индивидуальных рефлексов. Образование
индивидуального рефлекса. Механизм образования временной связи Павлов представ-
574
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
ляет так: «Если после того, как новое индифферентное раздражение
(подразумевается возбуждение — И. Б.) достигло больших полушарий, в центральной нервной
системе возникает очаг сильного возбуждения, то ранее возникшее индифферентное
раздражение (возбуждение — И. Б.) начинает концентрироваться к этому
возбужденному очагу, известным образом прокладывает к нему путь, а отсюда — к
соответствующему рабочему органу, и тем самым оно становится возбудителем этого органа»
(30). (В этой цитате, как и в последующих цитатах Павлова, очень часто термин
«раздражение» употребляется вместо возбуждения). Из данной цитаты видно, что, по
Павлову, основное условие образования временной связи заключается в том, что
участок коры, находящийся в состоянии сильного возбуждения, притягивает к себе
все существующие возбуждения из других участков.
Хотя это положение и выведено на основании определенного фактического
материала, но оно противоречит данным общей физиологии центральной нервной системы. По-
общей физиологии, возбуждение, возникнув в одном участке коры, должно ирради-
ировать затем во всей коре больших полушарий. Чем сильнее деятельность данного
возбужденного очага, тем сильнее должно быть иррадиирующее из него возбуждение.
Спрашивается: существуют ли в общей физиологии какие-либо несомненные
указания на то, что, наоборот, какой-либо возбужденный участок вследствие его сильного
возбуждения сам становится центром притяжения более слабых возбуждений? —
Ничего подобного физиология центральной нервной системы не знает. Из
современной физиологии совершенно ясно, что любой отдел центральной нервной системы
может функционально так измениться, что легко будет возбуждаться под влиянием
возбуждений, иррадиирующих из друігих участков. Но у всех авторов речь идет не о
притягательной силе возбужденного состояния, а о существовании повышенной
возбудимости в данном отделе центральной нервной системы.
И. П. Павлов до последнего времени признавал при образовании
индивидуального рефлекса возникновение временных связей только в одном направлении —
от более слабого очага возбуждения к более сильному, т. е. от корковых рецептор-
ных центров, возбуждающихся под влиянием внешнего стимула, к корковым центрам
пищевой реакции и затем к подкорковым центрам слюноотделения. В последние годы
своей жизни И. П. Павлов изменил это свое мнение и допускал возникновение
временных связей и в другом, обратном направлении: от корковых центров пищевой
реакции к слабому очагу от индивидуального раздражения. Сейчас вслед за ним
разделяют это мнение и его ученики: Быков, Анохин и др. Однако, «ак заявляет
Быков, вопрос этот все еще не является решенным и требует дополнительных
исследований (37).
Генерализация индивидуального рефлекса* Для объяснения генерализации
Павлов выдвигает хорошо известное правило иррадиации. Раздражение,
возникшее в больших полушариях, непременно разливается, иррадиирует по большим
полушариям. Это, по Павлову, первый закота раздражения. Данное правило применялось
в таком виде. Возбуждение, вызванное раздражением, дойдя до коры и не встретив
очага возбуждения, иррадиирует по своему анализатору, а также за пределы его на
другие анализаторы. Поэтому, когда под влиянием прирожденного рефлекса в
центральной нервной системе появляется сильный очаг возбуждения, последний
притягивает к себе не только возбуждение из очага индивидуального раздражения, но и
все возбуждения со всех точек коры, возбудившихся под влиянием иррадиации
возбуждения. По этой причине временная связь образуется не только между пищевым
центром (так называются центры слюнной секреции и двигательной пищевой реакции
в разных отделах мозга) и очагами индивидуального раздражения, но и между этим
пищевым центром и каждым другим возбужденным участком (31). Таким образом,
факт генерализации рефлекса объясняется образованием многочисленных временных
связей в силу иррадиации возбуждения из очага индивидуального раздражения.
Значит, согласно учению Павлова, каждый необычный раздражитель вызывает
индивидуальный рефлекс через свои собственные временные связи.
Это предположение также не может быть оправдано с точки зрения общей
ПРОИСХОЖДЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ
575
физиологии. Возбуждение распространяется из своего первичного очагэ в течение
всего того времени, пока оно удерживается в данном первичном очаге. Возбуждение
последнего следует за раздражением, а потому вскоре после прекращения
раздражения должно прекратиться не только возбуждение в первичном очаге, но вслед за ним
и его иррадиация по большим полушариям. Кроме того, сама иррадиация
возбуждения в коре должна происходить очень быстро, как это вообще совершается в
центральной нервной системе. В течение очень небольшой доли секунды возбуждение
может захватить всю центральную нервную систему. Продолжительность активного
состояния по прекращении раздражения к каждой возбужденной точке зависит от
функционального состояния последней; в одном центре активное состояние
продолжается несколько больше времени, в другом — несколько меньше, независимо от
того, как оно вызвано: непосредственно с периферии по восходящим путям или в
силу иррадиации возбуждения по коре (Беритов, 32).
Если исходить из таких положений, то станет яснга, что точка зрения П а в л о в а
на генерализацию рефлекса не может быть правильной. Если бы генерализация была
обусловлена образованием многочисленных временных связей, как это было указано
выше, тогда она должна была бы совсем отсутствовать в таких случаях, когда
индивидуальное раздражение прекращается раньше основного на несколько секунд или
даже минут, т. е. при образовании последовательных индивидуальных рефлексов.
Невозможно, чтобы активное состояние коры продолжалось по нескольку секунд и
минут после прекращения индивидуального раздражения. Например, действие звука,
как субъективное, так и объективное, кончается почти в ту же секунду, когда
прекращается звук. Несмотря на это, последовательный рефлекс на звук образуется при
применении его на несколько минут раньше основного раздражения, причем в данных
условиях индивидуальный рефлекс бывает более генерализован, чем совпадающий
рефлекс, как это, например, известно из опытов Пименова (33).
Дифференциация индивидуального рефлекса. Для объяснения этого явления
Павлов пользуется двумя допущениями. Первое: возбуждение обладает свойством
концентрироваться в известное русло, в определенные линии и точки мозга; второе:
вокруг очагов возбуждения в коре больших полушарий развивается процесс
торможения. Об этом торможении заключают из того факта, что если сначала употребить
необычное раздражение, которое во время генерализации давало рефлекс, а потом,
спустя несколько секунд или минут подействовать индивидуальным раздражением, го
последнее может совсем не дать рефлекса или же вызовет его в слабой форме.
Павлов поясняет это так: «Надо дать пройти известному времени, надо дать время
уйти из нервной системы этому задерживающему процессу, чтобы в силу этого
деятельное раздражение снова приобрело силу» (31).
По данным общей физиологии, торможение, подобно возбуждению, вызывается
раздражением. Оно возникает в центральной нервной системе в общем с такой же
скоростью, как и возбуждение. Во время дифференциации рефлекса не видно очага
торможения в коре большого мозга; есть только очаг возбуждения. По Павлову,
" это торможение возникает вокруг очага возбуждения в силу особого закона, по
которому возбуждение само создает вокруг своего очага торможение (Павлов, 34;
Ерофеева, 29).
В последнее время этот закон называют иначе законом сукцессивной
индукции (Фурсиков, 35). Этот термин принадлежит Шеррингтону. Но как
мы уже говорили выше, Шеррингтон пользовался им совершенно в другом
смысле. По его мнению, торможение возникает вслед за возбуждением в тех самых
нервных элементах, где существовало возбуждение. Но фактически нельзя признать
ни закона контрастов, ни сукцессивной индукции. Единственное, что на самом деле
существует в этом смысле — это кратковременное облегчение вслед за быстрым пре
кращением торможения, причем облегчение происходит в тех самых нервных эле*
ментах, которые были заторможены, а не вокруг них. Правда, мы показали выше, что
всякое деятельное раздражение рецепторов и самой коры может оказать тормозящее
действие на определенные нервные пути, но это происходит от общего торможения,
.576
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
которое возникает во всех отделах мозга при их активном состоянии. Это общее
торможение играет определенную роль в дифференциации индивидуального рефлекса.
Но в основе дифференциации лежит не это, а образование обратных временных
связей.
Но если бы даже закон сукцессивной индукции на самом деле существовал, то и
тогда данное объяснение дифференциации не было бы удовлетворительным. Если
торможение — активный процесс, то оно должно было бы существовать вокруг очага
возбуждения, пока этот очаг находится в активном состоянии под влиянием
раздражения; во всяком случае, торможения не могло бы быть спустя несколько часов а дней
после раздражения. Между тем, дифференцированное состояние продолжается не
только во время индивидуального раздражения, но даже спустя много дней после
него. Нам кажется совершенно ясным, что торможение ни в коем случае не можег
быть признано причиной дифференциации индивидуального рефлекса.
Литература
1. Beritoff J., Brain, 47, 109, 1924.
2 В е гі t of f J., J. Psych. Neurol., 32, 29. 1925; 32, 262, 1925.
3 С а в и ч А., Дальнейшие материалы к вопросу о влиянии пищевых рефлексов
друг на друга. Дисс, 1913.
4 Петрова М., К учению об иррадиации возбуждения и торможения. Дисс,
1914
Г.. Ф у р с и к о в Д., Арх. биол. наук, 22, 83, 1922.
6. Безбокая М., Материалы к физиологии условных рефлексов. Дисс, 1913.
7. Красногорский Н., О процессе задерживания и локализации и т д.
Дисс, 1911.
8. В е г і t о f f J., J. Psych. Neurol., 30, 217, 1924.
f 9. Beritoff J., Pfl. Arch., 213, 370, 1926.
10. Подкопаев H., Тр. физиол. лаб. акад. И. Павлова, 1, 187, 1926.
11. Розен та ль И., Арх. биол. наук, 23, 247, 1924.
12. Соломонов О., О тепловых условных и снотворных рефлексах с кожи
собаки. Дисс, 1910.
13 Снегирев Ю., Клинические монографии, январь 1911.
14. Фридман С, Дальнейшие материалы к физиологии дифференцирования
внешних раздражений. Дисс, 1912.
15. В егі t of f J„ Ueber d. individuell-erworbene Tatigkeit d. Z. N. 8., J. Psych.
Neurol., 33, 113, 1927; Индивидуально приобретенная деятельность
центральной нервной системы. Тифлис, 1932.
16. Гроссман Ф., Материалы к физиологии следовых условных слюнных
рефлексов. Дисс, 1910.
17. Фурсиков Д., Тр. физиол. лаб. акад. Павлова, сб. 1, в. 1, стр. 1, 1925.
18. Павлов И., 20-летний опыт, стр. 183, 1923.
19. 3 а л м а н з о н А., Тр. Ин-та высшей нервной деятельности, в. 1, стр. 39, 1929.
20. Леонов В., Pfl. Arch., 214, 304, 1926.
21. Добровольский В., О пищевых следовых рефлексах. Дисс , 1911.
22. Р о ж а н с к и й Н., Материалы к физиологии сна. Дисс, 1913.
23. Анреп Г., Русск. физиол. журн., 1, 1, 1917.
24. Белиц М., О следовых условных рефлексах. Дисс, 1917.
25. Б е р и т о в И., Z. Biol., 89, 59, 1930; Журн. эксп. биол. и мед., 30, 93, 1929.
26. Б и р м а н, В , Экспериментальный сон. Дисс, 1925.
27 Коган Б., Об иррадиации и концентрации угасания торможения в коре
больших полушарий. Дисс, 1914.
28. Чечулин С, Арх. биол. наук, 23, 142, 1923.
29. Ерофеева М, Электрическое раздражение кожи как условный возбудитель
и. т. д. Дисс, 1912.
30. Павлов И., 20-летний опыт, стр. 72, 1923.
31. Павлов И., 20-летний опыт, стр. 114, 1923.
ПРОИСХОЖДЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 577
32. Беритов И.. Русск. физиол. журн., 1, 6, 1917.
33. Пименов П., Особая группа услэвных рефлексов. Дисс, 1907.
34. П а в л о в И.. Лекции о работе больших полушарий головного мозга. Госиздат.
1927.
35. Фурсиков Д., Арх. биол. наук, 23, в. 2, 125, 1923.
36. Р о ж а н с к и й Н. А., Физиол. журн. СССР, 19, 289, 1935; Тезисы сообщ. 15-го
Межд. конгр. физиологов, 1935.
37. Айрапетянц Э. Ш\, Балакшина В. Л., Быков К. М.,
Подкопа е в Н. А, и Р и к к л ь А. В. Курс лекций по физиологии выстой нервной
деятельности. Гос. уч.-пед. изд.- 1941, стр. 92.
38. Беритов И. и Брегадзе А., Медико-биол. журн.. 1929, вып. 4, стр. 83.
39. Беритов И., Тр. биол. сект. Гр. отд. Закфил. АН СССР,т. I, ч. 1,1934» стр. і.
40. Беритов И. и Дзядзишвили Н., Те же труды, т. I, ч. 2, 1934, стр. 75.
41. Беритов И., Об основных формах нервной и психонервной деятельности.
1947 г.
;2. Цкипуридзе Л. н Минина две Н.( Сообщ. АН Г р. ССР, 5, 311 и 439,1944.
37 и. с. Беритов
Глава IX
ОСНОВНЫЕ АНАТОМИЧЕСКИЕ ЭЛЕМЕНТЫ ИНДИВИДУАЛЬНЫХ
РЕФЛЕКСОВ
Восходящая часть. Афферентная часть нервного пути, по которому
периферический импульс, вызванный индивидуальным или основным
раздражением, доходит до коры, до соотв. воспринимающей области, та же
самая, что и у прирожденных корковых рефлексов. Это, между прочим,
видно из того, что то самое раздражение, которое вызывает
ориентировочную реакцию, может быть превращено в возбудитель
индивидуального рефлекса. Восходящая часть нервного пути начинается на
периферии в рецепторах и кончается в коре большого мозга. Эта часть всегда
состоит из нескольких нейронов, т. е. восходящий путь прерывается
несколько раз, пока достигает коры (рис. 181, 183, 185).
При образовании индивидуального рефлекса не имеет
существенного значения — где происходит основное раздражение: в рецепторе,
в проводящих путях спинного мозга или в конечном участке:
непосредственно в коре большого мозга. Известно, что индивидуальный
рефлекс образуется, если основное раздражение прикладывается к
задним и боковым столбам спинного мозга (Луке и Гант, 20). Он
образуется также, если основное раздражение прикладывается к
двигательной зоне и таким образом вызывается движение конечности. При этом
условии рефлекс образуется на конечности даже в том случае, если
она лишена периферической чувствительности путем перерезки задних
корешков, т. е. при невозможности возбуждения коры вторичным путем
от раздражения проприоцепторов (Кряжев, 21).
Корковая часть. Та воспринимающая область коры большого мозга,
где кончается восходящий путь, возбуждаемый индивидуальным
раздражением, представляет начало поступательной временной связи. Этот
очаг, воспринимающий индивидуальное раздражение, прежде всего
связан с областью коры, воспринимающей основное раздражение, в
нашем случае — с кожной областью, которая возбуждается
электрическим раздражением ноги; затем он связан с теми двигательными
участками коры, откуда возбуждение кожной области дает внешний
эффект, т. е. с двигательными участками ноги, с участками голосовой
реакции, ориентировочной реакции и др. (рис. 244). Когда основным
ОСНОВНЫЕ АНАТОМИЧЕСКИЕ ЭЛЕМЕНТЫ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 579
раздражением является пищевое вещество, положенное в рот, очаг
индивидуального раздражения связывается с вкусовой воспринимающей
областью и с теми двигательными и секреторными участками коры,
через которые вызывается корковая реакция, состоящая в выделении
слюны, жевании, глотании и т. д.
Участие проприоцептивных рефлексов. Условия развития
индивидуального рефлекса таковы, что участие коры в индивидуальных
рефлексах не может ограничиться упомянутыми элементами. Как известно,
Рис. "ikk Рефлекторные пути индивидуального сгибательного рефлекса на
ноге и ориентировочного рефлекса на шее на аьук и двигательного рефлекса
ноги на элеьтричесьое раздражение ьоги.
В — проприоцептивпая воспринимающая область коры большого мозга; С — кожная
Область; D — слуховая; Е — кожный участок на йоге с электродами для раздражении;
отсюда начинается восходящий путь к кожной воспринимающей области. Здесь же в
мышцах начинается восходящий путь и' для приприоцептивного рефлекса (непрерывная линия
с штриховой). 1 — поперечный разрез спинного мозга с координирующим аппаратом для
сгибания ноги о. II — поперечны» разрез среднего мозга, через который проходит
слуховой путь к слуховой области и к координирующему аппарату движения шеи. /// —
поперечный разрез шейного отдела мозга с координирующим аппаратом для движения шеи Ь'.
Стрелки везде обозначают направление возбуждении в рефлекторных путях. Зачерненные
участки в воспринимающих областях указывают па воспринимающие очаги возбуждения,
а обведенные — на двиг. тыльные участки: 1 — для сгибания ноги, 2 — для поворачивания
шеи. Двойные линии, соединяющие эти участки, обозначают временные связи. Наиболее
длинный рефлекторный путь индивидуального сгибания на звук начинается в ухе,
проходит черен средний мозг //, через воспринимающие области V, С и Б, а затем через
двигательный участок / спускается в спинной мол- к координирующему аппарату сгибании Ь.
Наиболее же короткий путь проходит в коре воспринимающей области звука /), а отеюдп прямо
К двигательному участку ноги /. Рефлекторный путь проприонептивного сгибательного
рефлекса: наиболее длинный начинается в мышцах ноги а", проходит через те же норковые
воспринимающие обиасти и затем через двигательные участии / спускается к Koop.timupvto-
щ< му аппарату сгибания; наиболее короткий же будет проходить в норе проприоцептииную
воспринимающую область и затем прямо к двигательному участку ноги /. Рефлекторный;
прирожденный путь ориентировочного движения шеи па энчк начинается н ухе, затем
через средний мозг направляется, с одной стороны, к координирующему аппарату шеи ///
(короткий рефлекторный путь), а с другой — через слуховую область и двигатолі.ныіі
участок 2 к тому же координирующему аппарату (длинный рефлекторный путь).
каждое движение конечности или другого органа вызывает целый ряд
вторичных проприоцептивных раздражений в суставах, мышцах,
сухожилиях. Все эти раздражения, как уже указывалось выше, действуют
на кору и производят здесь в определенных частях возбуждение и
повышение возбудимости. В силу этого очаги индивидуального и основного
раздражений вступают во взаимодействие как между собой, так и с
37*
580 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
очагами, которые возбуждаются проприоцептивными раздражениями.
В соответствии с этим временные связи образуются, с одной стороны,
между очагами индивидуального и основного раздражения, а с
другой— между ними и очагами проприоцептивных раздражений. В
результате этого наравне с первичным индивидуальным рефлексом возникает
вторичный проприоцептивный индивидуальный рефлекс в ответ на
вторичное раздражение. Как мы уже указывали, существование
проприоцептивных индивидуальных рефлексов проявляется, между прочим, в
том, что индивидуальный рефлекс не прекращается вместе с
первичным индивидуальным раздражением, но продолжается еще некоторое
время после него, иногда несколько минут.
Итак, в развитии индивидуального рефлекса участвуют следующие
корковые элементы: 1) участок коры, воспринимающий первичное
индивидуальное раздражение; 2) участок коры, воспринимающий основное
раздражение; 3) участки коры, воспринимающие проприоцептивные
раздражения и 4) двигательные или секреторные участки. Возникшие
между ними индивидуальные пути имеют различную сложность, так как
из каждого элемента возбуждение может переходить в другой как
непосредственно, так и через тот или другой или другие элементы (Беритов, 1).
Нас здесь интересуют те индивидуальные пути, которые возникают
между очагами, воспринимающими первичное индивидуальное
раздражение, с одной стороны, и ответными двигательными и секреторными
участками — с другой. Между ними должен существовать один простой
и прямой путь, который непосредственно связывает эти элементы коры.
Этот путь включает одну поступательную и одну обратную связь,
т. е. представляет собой наиболее простой замкнутый нервный круг
Но наряду с этими должны существовать и другие, более сложные пути,
со включением коркового участка, воспринимающего основное
раздражение, или участка, воспринимающего проприоцептивное раздражение;
это будут нервные круги с двумя нервными связями. Очевидно,
существуют также пути с тремя связями, именно те, которые включают оба
только что упомянутых корковых участка (рис. 244). Наравне с ними
могут существовать и другие, еще более сложные пути. Например,
если животное видит произведенное им движение, то в образовании
сложных путей должна участвовать и зрительная область коры; если
индивидуальный раздражитель является сложным, каждая из его
составных частей будет создавать очаг возбуждения в коре и
принимать участие в образовании сложных путей.
Итак, гог или иной индивидуальный рефлекс в действительности
осуществляется не просто при участии каких-либо немногих очагов
возбуждения, имеющих прямое отношение к индивидуальному и основному
раздражению, а представляет собой результат очень сложной
деятельности почти всей коры большого мозга.
Мы все время говорим о тех или иных очагах возбуждения и
временных связях между ними, как об определенных элементах
рефлекторной дуги индивидуального рефлекса. Однако все это нужно пони-
ОСНОВНЫЕ АНАТОМИЧЕСКИЕ ОЛЕМЕНТЫ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 581
мать условно, как результат аналитического изучения сложного
явления, когда мы искусственно разбиваем сложное явление на части и
изучаем какую-либо составную часть отдельно. Фактически никаких
обособленных, совершенно самостоятельно функционирующих
рефлекторных дуг или временных связей, конечно, не существует. В
каждый момент корковой деятельности мы имеем дело с синтетической
работой очень сложного нервного комплекса. Лишь в целях
облегчения изучения этой работы нам приходится подходить аналитически,
выделяя из этого комплекса ту или другую часть и рассматривая ее,
как будто бы она имеет совершенно самостоятельное существование.
Из означенных временных путей наиболее деятельным и
возбудимым должен являться короткий простой путь, так как по условиям опыта
этот путь должен развиваться наиболее сильно. Это вытекает из закона
сопряженной иррадиации возбуждения: данный очаг действует на
другой по простому и короткому пути сильнее, чем по длинным и
сложным путям. Следствием усиленного развития односложных путей
является то, что, согласно закону сопряженной иррадиации,
возбуждение из очага индивидуального раздражения распространяется по этим
путям с большей интенсивностью, чем по всем остальным, более
сложным и менее развитым путям. Это обстоятельство, в свою очередь,
благоприятствует преимущественному развитию коротких путей и в то
же время заглушению более сложных путей. Значит, можно сказать, что
с упрочением индивидуального рефлекса корковая деятельность
происходит главным образом в коротких прямых временных путях. Участие
сложных путей проявляется только в самом начале образования
рефлекса, когда короткие пути еще не выделяются своим усиленным
развитием. Это бывает и впоследствии, если почему-либо повышается
возбудимость сложных путей,— например, после сильного
электрического раздражения. Деятельность сложных путей проявляется в том,
что двигательная реакция на индивидуальное раздражение является
очень сложной и протекает бурно в виде беспокойного движения ног,
головы, туловища и сильного стона, что указывает на участие целого
ряда двигательных участков в коре большого мозга. При участии только
коротких путей двигательная реакция ограничивается тем
двигательным органом, который подвергается электрическому раздражению, и
небольшим ориентировочным движением головы. Итак, при упрочении
индивидуального рефлекса односложные временные пути более раз-
виты и обладают большей возбудимостью, чем многосложные, и поэтому
рефлекс происходит главным образом через эти односложные пути
(Беритов, 1).
Нисходящая часть. От ответных двигательных и секреторных участков
возбуждающие импульсы переходят в низшие отделы центральной
нервной системы. Нисходящие пути, по которым происходит передача
корковых импульсов, не приобретены индивидуальным путем; они
прирожденные, через них происходят все прирожденные корковые реакции.
582 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
Следовательно, между корой и другими отделами центральной нервной
системы не существует временных связей. Корковые импульсы в
индивидуальных рефлексах, очевидно, передаются по пирамидным путям,
которые оканчиваются в координирующих аппаратах, и по тем
экстрапирамидным путям, которые прерываются в ядрах среднего и
продолговатого мозга. Отсюда следует, что координация индивидуальных
рефлексов происходит в тех самых координирующих аппаратах, откуда
вызываются соответствующие прирожденные рефлексы (рис. 244).
Нервный путь развитого индивидуального рефлекса по своей
сложности не особенно отличается от пути коркового прирожденного
рефлекса. Разница заключается в том, что индивидуальный рефлекс
происходит путем возбуждения определенных временных связей между
очагом индивидуального раздражения и определенными ответными
участками; в прирожденных же корковых рефлексах эти ответные аппараты
приходят в активное состояние вследствие общей иррадиации
возбуждения из области, возбужденной внешним раздражением.
О корковых элементах индивидуального слюнного рефлекса.
Впервые Бабкин высказался о корковом составе нервных элементов
индивидуального слюнного рефлекса. Он был того мнения, что временная
связь в индивидуальных слюнных рефлексах устанавливается между
элементами коры, воспринимающими, с одной стороны, индивидуальное
раздражение, а с другой — вкусовое (3). Таким образом, согласно его
мнению, возбуждение из очага, воспринимающего индивидуальное
раздражение, передается сначала по путям временной связи в корковый
вкусовой отдел, а уже отсюда в подкорковый слюнный центр.
Когда впоследствии в школе Павлова создалось определенное
представление о пищевом центре, авторы высказались за образование
временных связей между этим центром и очагом индивидуального
раздражения (Былина, 4 и др.). Под пищевым центром подразумеваются
определенные центральные элементы, которые регулируют отделение
желез пищеварительного канала, в особенности слюнных и желудочных,
а затем деятельность мускулатуры для захвата и проглатывания пищи.
После того как ученик Павлова Тихомиров (23) на основании
экстирпации различных частей коры мозга пришел к отрицанию
слюноотделительного центра в коре мозга, существование которого
доказывалось опытами Бехтерева и Миславского, Павлов и его
сотрудники придерживались того представления, что пищевой центр
в основном лежит под корой и что временные связи образуются между
корой и подкорковыми центрами.
Однако это положение разделялось не всеми в школе Павлова.
Некоторые авторы придерживались схемы Бабкина. Они считали,
что раз мозговой конец ротового анализатора, т. е. вкусовая область,
воспринимающая раздражение ротовой полости, находится в коре мозга,
то временные связи должны образоваться прямо между этим
«мозговым концом ротового анализатора» и корковым очагом индивидуального
раздражения (Дерябин, 5). Сейчас, в самое последнее время, это мне-
ОСНОВНЫЕ АНАТОМИЧЕСКИЕ ЭЛЕМЕНТЫ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 583
ние уже стало господствующим. Школа Павлова признает
существование коркового представительства пищевого центра в виде
слюноотделительных и других секреторных центров и образования временных
связей между корковым очагом индивидуального раздражения и этим
корковым пищевым центром, как ответной или исполнительной
системой (Купалов, 18).
Однако схема Бабкина, которая в последнее время считается в
лаборатории Павлова наиболее вероятной, не может быть признана
полной, и вот почему. В коре больших полушарий в тесной связи с
признанной вкусовой областью, или, по терминологии Павлова,
ротовым анализатором, находятся корковые аппараты отделения слюны
и желудочного сока, а также целый ряд двигательных участков,
имеющих отношение к акту еды: жевания, глотания, движения губ и языка
и т. д. Эти участки, несомненно, представляют сосредоточенные
начальные пункты центробежных путей коры, откуда корковые импульсы
передаются подкорковым сокоотделительным и двигательным центрам
ротовой реакции. Мы думаем, что если признавать существование в
коре особых секреторных и двигательных участков в тесной связи с
вкусовой областью, то нельзя оспаривать возбуждение этих участков в
связи с возбуждением вкусовой области. А потому временные связи
должны образоваться не только с этой областью, но параллельно также
с указанными секреторными и двигательными участками коры.
Несомненно, что в индивидуальных слюнных рефлексах
значительную роль играют и проприоцептивные индивидуальные связи на
вторичные раздражения, связанные с двигательной частью ротовой реакции.
Эти раздражения должны были стать индивидуальными, так как
воспринимающие их корковые участки безусловно связываются со
вкусовой областью и секреторным аппаратом коры. Условия опыта были
более чем благоприятны для образования вторичных индивидуальных
рефлексов. В период сочетания кормление продолжается целую минуту,
а вливание кислоты во время действия индивидуального раздражителя
производится не раз, а два-три раза в минуту. Таким образом,
вторичные раздражения от каждого предыдущего акта еды или от каждой
отрицательной ротовой реакции, вызываемой вливанием кислоты,
сочетаются не только с одновременным, но и с последующим
возбуждением вкусовой области и секреторных участков в коре от последующего
приема пищи или вливания кислоты. Действие этих вторичных
временных связей проявляется в том, что слюнный рефлекс дает значительное
последействие, в особенности если первичное индивидуальное
раздражение нарочно укорачивается, как это бывало в опытах ОрбелииПет-
ро в о й.
Итак, корковая часть дуги слюнного индивидуального рефлекса
должна состоять из целого ряда путей разной сложности, образуемых
между начальным пунктом, воспринимающим первичное индивидуальное
раздражение, и конечным пунктом, представляющим собой секреторный
584 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
и двигательный участки ротовой реакции. Передаточными станциями в
этих временных путях являются, во-первых, вкусовая область и,
во-вторых, проприоцептивная область, которая возбуждается мышечно-сустав-
ными раздражениями головы; но, конечно, передаточными станциями
являются и другие области, воспринимающие многочисленные внешние и
внутренние раздражения одновременно с раздражением ротовой полости,
а также многие двигательные участки, участвующие во всем пищевом
двигательном акте животного.
Об образовании временных связей вне коры мозга. У высших
позвоночных животных и у людей в их постэмбриональной жизни временные
связи возникают на основании индивидуального опыта, безусловно,
только в коре больших полушарий. Это было установлено в лабораториях
Павлова путем экстирпации. Все индивидуально приобретенные
рефлексы исчезают после удаления коры мозга.
Однако Зеленому удалось образовать индивидуальный рефлекс
при таких условиях, что можно считать возможным образование его и
помимо коры мозга. Зеленый (6) совместно с Руковяткиной
образовали индивидуальное сгибание правой передней ноги на звук
после удаления левого полушария и разрушения улитки левой стороны.
Точно так же удалось образовать такой же рефлекс на правой стороне
на механическое раздражение кожи правой ноги после удаления левого
полушария. Ведь возбуждение от рецепторов правой стороны поступает
главным образом в левое полушарие. И если, невзирая на отсутствие
левого полушария, рефлекс все-таки образовывался, то это уже должно
свидетельствовать о возможности образования рефлекса помимо коры
большого мозга. Вскоре после этого Зеленый с Полтыревым (7)
опубликовали новую работу, где они утверждают, что у собаки
образуются индивидуальные двигательные рефлексы на звуки почти при
полном удалении коры большого мозга. Кора была удалена без
повреждения подкорковых узлов и зрительных бугров, из нее не был удален только
небольшой слой над corpus striatum и затем небольшая часть ,внсочной
доли в основании мозга на левой стороне. Собака вела себя так, как это
характерно для собаки без мозговой коры. Такая собака не
ориентируется в пространстве, наталкивается на окружающие предметы, глотает
пищу, вложенную ей в заднюю половину рта; не поднимает опущенную
в холодную воду лапу. И вот на такой собаке удалось образовать
индивидуальный оборонительный рефлекс на охотничий свисток путем
сочетания последнего с электрическим раздражением ноги: на звук собака
обычно поднимала раздражаемую ногу. Рефлекс образовался после
104 сочетаний; он был все время генерализован; другие звуки также
давали рефлекс, но после угасания на необычные звуки обычный свисток
давал его. На раздражители другого качества: запахи, свет,
прикосновение,— образовать индивидуальные рефлексы не удалось.
Специальные опыты, поставленные на кошке, обнаружили, что после
удаления всей коры (neopallium) хорошо образуются и упрочиваются
рефлексы на звуковые раздражения. Кроме того, быстро достигается
ОСНОВНЫЕ АНАТОМИЧЕСКИЕ ЭЛЕМЕНТЫ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 580
дифференциация индивидуальных звуков от отдельных звуков другого
качества,— например, при образовании пищевого двигательного рефлекса
на электрический звонок этот звук может быть отдифференцирован от
метронома, от шумов. Но тонкая дифференциация в отношении
интенсивности и высоты звука не достигается. Например, не удается
отдифференцировать определенный звук свистка Гальтона от других звуков того
же свистка. Способность к образованию индивидуальных рефлексов на
раздражители другого качества исчезала полностью в связи с удалением
коры мозга. Это в особенности касается зрительных индивидуальных
рефлексов. Исчезла не только способность к этим рефлексам, но вообще
исчезло зрение. Кошки без area striata в затылочных долях оказывались
совершенно слепыми (Тен-Кате, 16). Однако кошка, лишенная неокор-
текса без повреждения полосатого тела, сохраняет примитивное зрение;
она не наталкивается на препятствия, отличает свет от темноты, идет по
направлению к свету (Гирндт и Лемпке, 19). У низших
млекопитающих— у кроликов, например,— после удаления всей коры мозга
способность к образованию индивидуальных рефлексов сохраняется
полностью на все раздражители — слуховые, зрительные, обонятельные
и др. Сохраняется в полной мере и способность к дифференциации
раздражителей. В этом отношении не наблюдалось разницы между
оперированными и нормальными кроликами. Носителем зрительной
индивидуальной способности признается следующая система узлов, имеющих прямую
связь с area striata: с. geniculatum, с. quadrigeminum, pulvinar и п.
lateralis thalami (Тен-Кате и Герк, 16). Звуковая же индивидуальная
способность признается функцией другой системы подкорковых узлов.
На основании приведенных сведений нужно думать, что на низшей
ступени филогенеза млекопитающих способность к образованию
автоматических индивидуальных рефлексов связана с нервными центрами
промежуточного и среднего мозга. Эти центры в отношении каждого рода
рецепции образуют взаимно связанные системы, нарушение которых в
одной части ведет к нарушению индивидуальной рефлекторной
способности. По мере филогенетического развития млекопитающих эта
способность постепенно переходит в кору. У кошек она еще хорошо
представлена в подкорковых узлах в отношении звуковых раздражений. У собак
эта способность еще более ограничена, допуская лишь обобщенные
реакции. Нужно думать, что у обезьян и человека эта способность целиком
перешла в кору мозга.
У низших позвоночных индивидуальные рефлексы, повидимому,
образуются помимо больших полушарий. Так, известно, что у рыб
индивидуальные рефлексы образуются и хорошо дифференцируются после
удаления больших полушарий переднего мозга. У них удается
дифференцировать рефлексы после удаления переднего мозга. Так, например,
один из новейших авторов утверждает, что ему удалось у лягушки,
лишенной большого мозга, легко образовать индивидуальный
оборонительный рефлекс на свет электрической лампы (Леутский, 9). Наши же
опыты показали, что этот рефлекс на свет не что иное, как прирожден
?S6
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
ный поведенческий акт, что он даже не меняется в какой-либо мере в
результате индивидуального опыта (Бебуришвили и Чичинадзе, 10).
Бесспорные индивидуальные реакции наблюдаются у костистых рыб
(Фролов, 11; Бул, 12). Фролов, например, образовывал прочные
оборонительные индивидуальные реакции у таких рыб после 20—39
сочетаний, но так как большой мозг рыб не имеет настоящего коркового
образования, то можно утверждать, что у них временные связи
образуются без коры, даже не в большом мозгу, а в промежуточном, который
является у них наиболее развитой частью головного мозга.
Исследования на голубях показали, что удаление коры большого
мозга, с сохранением из большого мозга полосатого тела — meso-
striatum и ectostriatum, не ведет к исчезновению обычной
индивидуальной рефлекторной деятельности (Беритов, 13). Но если удалить весь
большой мозг, тогда индивидуальная рефлекторная деятельность
исчезает полностью (Попов, 14, Баяндуров, 15). Баяндуров пытался
образовать новый индивидуальный рефлекс, производя до 500 сочетаний
в течение многих месяцев после такой операции, но совершенно без
успеха. Отсюда можно заключить, что у птиц индивидуальная
рефлекторная деятельность не является исключительной функцией коры мозга.
Несмотря на высокую степень развития этой деятельности, она повиди-
мому осуществляется преимущественно подкоркой, а именно — частями
полосатого тела. Отсюда следует, что в связи с филогенетическим
развитием головного мозга от рыб к птицам функция индивидуально
приобретенной рефлекторной деятельности переходит из промежуточного мозга
в полосатое тело.
Итак, временные связи, безусловно, образуются, помимо коры
большого мозга, в нижних отделах головного мозга. Но развитие их
происходит много медленнее и не достигает такой степени, как в коре мозга.
Повидимому, развитие временных связей вне коры мозга протекает по
иным закономерностям, чем в коре мозга.
Литература
1. Беритов И., Brain, 47, 198, 1924; Индивидуально приобретенная деятельность
центральной нервной системы, 1932.
2. П а в л о в И., 20-летний опыт. Лекции о работе больших полушарий головного
мозга 1927.
3. Бабкин В., Опыт систематического изучения сложнонервных явлений у
собаки. Дисс, 1904.
4. Былина А., Простое торможение условных рефлексов. Дисс, 1910.
5. Дерябин В.,К физиологии времени как условного возбудителя слюнных желез
Дисс, 1916.
6. ЗеленыйГ., Тр. Ленингр. об-ва естеств., 58, в. 1, стр. 83, 1928.
1. Z е 1 і о п у G. и. Р о 1 t і г е f f S„ Ber. d. Ges. d. russ. Physiol., H. 3, 1929.
8. Schrader M., РЛ. Arch., 44, 175, 1889.
9. Леутский К., Русск. физиол. журн., 12, 235, 1929.
10. Бебуришвили Н. и Чичинадзе Н., Тр. Тифлисск. гос. ун-та, т. II,
127. 1936.
основріые анатомические элементы индивидуальных рефлексов 587
И. Froloff J., РП. Arch., 201, 261, 1926; 220, 339, 1928.
12. Bull Н. О., J. Man. B.ol. Assoc, 15, 485, 1928.
13. В e r i t о t f J., Pil. Arch., 213, 370, 1926.
14. Попов и Черников А. Тр. Ill Всес. съезда физиологов, стр. 300, 1928
15 БаяндуровВ. И., Тр. Томск, мед. ин-та, 5., 1937.
16. Ten CateJ.u. van HerkA W. Arch. Neerland de Physiol. 18, 1—42, 1933
17. Ten CateJ., Arch. N erland. Physio!., 19, 191, 1934.
18. К у n а л о в П. С.: Арх. биол. наук, 54, 5, 1939.
19. Girntit О. u. Lempke H., Pfl. Arch., 290, 544, 1937.
20. LoucksR.B.a.GanttW. Н., J. Сотр. Psychol., 25, 425, 1938
21. Кряжев Б. Я.. Природа, 4, 44, 193S.
22. С и 1 I е г Е. а. Л1 е t t I е г F. А., .1. Сотр. Psychol., 18, 291, 1934.
23. Тихомиров Н. Опыт строго объективн. исслед. функций больших полу
шарий у собаки. Дисс. из лаб. Павлова, 1906.
г—^»и
Глава X
УГАСАНИЕ И ВОССТАНОВЛЕНИЕ ИНДИВИДУАЛЬНЫХ
РЕФЛЕКСОВ
Общее понятие об угасании индивидуальных рефлексов.
Индивидуальный рефлекс, повторяясь один за другим без сочетания с основным
раздражением, ослабевает и затем исчезает, по термину Павлова —
угасает (Бабкин, 1, Перельцвейг, 2). При угасании рефлекса меняется
не только интенсивность, но и скрытый период его: последний удлиняется
(рис. 245). Чем прочнее индивидуальный рефлекс, тем дольше его надо
угашать. Слабые и молодые рефлексы угасают быстрее старых и
сильных.
Последовательные рефлексы, следующие спустя значительное время
после индивидуального, угасают много быстрее совпадающих рефлексов
(Добровольский, 9). Это зависит от малой прочности последовательного
рефлекса. Когда основное пищевое раздражение производится череа
минуту и более после индивидуализируемого, т. е. когда индивидуальный
слюнный рефлекс является не особенно прочным, то он угасает после
одного-двух применений; но если последовательный индивидуальный
рефлекс довольно прочен, как было в наших опытах с оборонительными
рефлексами, когда основное электрическое раздражение следовало
спустя 30 сек. вслед за индивидуализируемым, то рефлекс может быть
повторен до 7 раз.
Скорость угасания зависит в большой мере от состояния возбудимости
коры большого мозга. Угасание происходит тем позднее, чем выше
возбудимость.
Чем это повышение возбудимости вызвано — при этом не имеет
значения: каким-либо сильным посторонним раздражением, рубцом
в мозгу, легким утомлением или иной причиной. Так, в опытах
Башмурина и Мюльберга (11) из лаборатории Зеленого на
оперированных собаках с экстирпацией одного полушария угасание
оборонительного рефлекса произошло после 605 повторений.
Индивидуальный рефлекс угасает также сам собой в результате дли-
УГАСАНИЕ И ВОССТАНОВЛЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ
Ш
тельных перерывов; при
этом чем слабее, чем
моложе рефлекс, тем скорее он
угасает. Так, двигательный
рефлекс, который только что
образовался за 1 день
работы, не всегда
обнаруживается через 1—2 дня, в то
время как хорошо развитой
старый рефлекс сохраняется
месяцами.
Любой индивидуальный
рефлекс может угаснуть и
от разового применения
индивидуального раздражения,
если последнее является
длительным. Так, например,
если продолжать
индивидуальное раздражение долгое
время, то даже наиболее
прочный рефлекс
удерживается только некоторое
время, не более нескольких
минут, и затем постепенно
ослабевает, угасает. Но
такое угасание никогда не
является абсолютным. Если
раздражение прекратить и
вновь его произвести спустя
несколько минут, то рефлекс
наступит с новой силой.
Индивидуальный рефлекс
угасает сам собой, если угасить
какой-либо другой
однородный индивидуальный рефлекс
(Бабкин, 1, Зеленый, 4).
Однородными рефлексами
называют такие рефлексы,
которые образуются путем
сочетания с одним и тем же
основным раздражением: с
одним и тем же пищевым
веществом или электрическим
раздражением одного и того же
участка кожи. Это явление
А
Л.п\
W
.J -т-
200
шшшищшшшщшщшшшщщщщшшшіщщ
и
ib
J~1F ПР Эй*
ішттііішіітшшлшшішішшшшшшщщщшшшшимшишішл
сз
гоо
гоо ггод~
ЩЦШlVAШШШШШШШMШЛЦШШШiШЩ|ЩЩШШШШiШШШШШШШJШШШШU
п
и
гЛ
т.
!Сб
сз
IF
ib
ib
ituuu-.il
iimuuii-.i in ииш.ніи шипи
200
ііі'іпіп-иі||іііііі[іпі
Рис. 245. Угасание рефлекса на правой передней
ноге при быстром повторении индивидуального
раздражения — звука 200 колебаний в 1 сек.
Собака Боб. Каждое новое раздражение начинается
спустя Ю сек. по окончании эффекта от предыдущего
раздражения и продолжается около 10 сек. (нижний
сигнал). Запись А дает начало опыта—первые два эффекта.
На записи В представлен момент окончательного
угасания этого рефлекса. Между А и В выпущена запись из 12
эффектов в течение 10 мин. Здесь 17-е и 18-е звучания не
производят эффектов. Но во время этого звучания
другое индивидуальное раздражение — метроном (М)
(верхний сигнал)—вызывает значительный рефлекс. Метроном
был испробован потом еще три раза в отдельности и
каждый раз давал рефлекс. Эти записи пропущены. Затем
спустя 10 сек. после последнего метрономного рефлекса
было испробовано новое индивидуальное раздражение —
зажигание и потухание электрической лампы г«; оно потом
повторялось еще три раза. На записи С представлен
последний эффект светового раздражения (верхний сигнал)
спустя 5 мин. после записи В. После светового
раздражения звук 200 колебаний снова дает рефлекс. Произошло
восстановление рефлекса, но незначительное, так как при
повторении звука через 20 сек. снова не было рефлекса.
Однако, когда промежуток был удвоен, рефлекс на звук
наступил. Запись D дает конец данной серии опытов
спустя 3.5 мин. после С. Здесь звук в 200 колебаний
производит эффект при больших промежутках 37—48 сек., но не
дает его при 28 сек., а рефлекс на свет обнаруживает
более глубокое угасание: свет не дает рефлекса и при
промежутке в 1.5 мин.
590 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
угасания в лаборатории Павлова называется вторичным
угасанием. Чем прочнее первично угашенный рефлекс, тем сильнее
вторичное угасание другого рефлекса. Но вторичное угасание характерна
отличается от первичного. Через некоторый промежуток времени
вторичное угасание проходит бесследно, в то время как первичное
проходит только частично.
Все случаи угасания индивидуального рефлекса, наступающие
вследствие несочетания с основным раздражением, по своим последствиям
двоякого рода. В одних случаях угасание временное и окончательно
проходит через некоторое короткое время,— для его восстановления не
требуется ничего больше, кроме отдыха. В других случаях раз
достигнутое угасание удерживается потом в большей или меньшей степени
значительное время и не проходит полностью в связи с отдыхом. Этот послед-
ний род угасания характерен для индивидуальной корковой деятельности.
Его не бывает при спинномозговой и вообще при прирожденной
рефлекторной деятельности.
Происхождение непреходящего угасания индивидуального рефлекса.
Этот род угасания индивидуального рефлекса безусловно зависит от
изменения механизма временных связей, а это изменение вообще
характерно отличается от утомления прирожденного рефлекса. В последнем
случае отдых вполне восстанавливает рефлекс, изменение же механизма
временных связей не проходит бесследно. Если повторять рефлекс один
за другим много раз до полного угасания, без сочетания с основным
рефлексом, то после любого отдыха он будет вызываться с меньшей
интенсивностью. Очевидно, повторная деятельность без сочетания с
основным раздражением характерно меняет временные связи. Этот
механизм возник и развился на почве взаимодействия между очагами
индивидуального и основного раздражений во время их сочетания, но
если 'многократно повторяется только индивидуальное раздражение, то
взаимодействия не бывает. Действие происходит главным образом в
одном направлении, от очага индивидуального раздражения. Вот эта
односторонность действия и должна служить причиной расстройства
механизма временных связей (Беритов, 3).
То, что другие корковые элементы не играют существенной роли в
угасании индивидуального рефлекса, может быть доказано
экспериментальным путем. В отношении начальных пунктов временных связей,
г. е. воспринимающих индивидуальные раздражения, это может быть
доказано следующим образом. Такое необычное раздражение, как
звонок, обычно устраняет индивидуальный рефлекс на какой-либо другой
звук: в это время звуковое индивидуальное раздражение не дает
рефлекса. Оказывается, что если сначала звонок продолжается 15—30 мин.
без перерыва, а во время него пробуется индивидуальный звук, то и
тогда звонок действует на индивидуальный рефлекс отрицательным
образом. 'Если бы слуховая область коры утомлялась длительным
раздражением звонка, то не было бы отрицательного действия на индивидуальный
рефлекс. Отсюда ясно, что если при повторном индивидуальном звуке
УГАСАНИЕ II ВОССТАНОВЛЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 59Г
соответствующий рефлекс исчезает и не восстанавливается в связи с
отдыхом, то это не может быть обусловлено утомлением слуховой
области коры.
Конечные пункты временных связей, а именно, ответные
двигательные и секреторные участки коры, обычно также не принимают участия в
непроходящем угасании индивидуальных рефлексов. Это с очевидностью*
обнаруживается из следующего опыта. Если вполне угасить один
индивидуальный рефлекс несколько раз подряд, то вместе с этим не угасают
окончательно и другие однородные рефлексы, т. е. те, которые имеют
одни и те же конечные пункты.
Положим, например, угас рефлекс на данной ноге на звук 200
колебаний в секунду; спустя некоторое время после этого другой
индивидуальный звук — стук метронома—может вызвать на той же ноге
обычный неослабленный рефлекс. Это показано на рис. 245-В. Так как
конечные пункты временных связей в обоих рефлексах одни и те же, то,
очевидно, угасание первого рефлекса не было вызвано утомлением
ответных двигательных участков.
Итак, непреходящее угасание индивидуального рефлекса происходит
от изменений в механизме временных связей. Ни воспринимающие
области, ни ответные двигательные и секреторные участки не играют
существенной роли в угасании рефлекса.
О характере изменения временных связей при непреходящем
угасании. Выше было указано, что развитие временных связей должно быть
обусловлено определенным функциональным и морфологическим
развитием нервных элементов. А именно, прежде всего здесь должно иметь
значение увеличение нейрофибриллярного вещества, затем уменьшение
промежуточных слоев, так называемых синапсов, в цепи нейронов.
Нужно полагать, что в условиях многократного вызова индивидуального
рефлекса без сочетания с основным раздражением, поступательные
временные связи от очага индивидуального раздражения к двигательным
участкам претерпевают обратное морфологическое развитие. Особенно
сильно это должно выразиться в отношении синапсов, так как, по
смыслу закона нейробиотаксиса Ариёнс Каппсрса, уменьшение
пространства между двумя нервными элементами мыслимо только при
одновременном возбуждении того и другого элемента с разных сторон.
В условиях же угасания рефлекса эти элементы возбуждаются только с
одной стороны.
При угасании рефлекса должны изменяться и обратные связи,
т. е. связи от двигательных участков к очагу индивидуального
раздражения, но только не в смысле ослабления, а, наоборот, в смысле
дальнейшего укрепления. Дело в том, что возбудимость конечного пункта
обратных связей — в очаге индивидуального раздражения — должна
повышаться в связи с частым повторением индивидуального раздражения и
с очень трудной утомляемостью коры, воспринимающей индивидуальное
раздражение. Это должно благоприятствовать упрочению обратных
связей в силу непрекращающейся иррадиации возбуждения из двига-
592 ОНЩАН ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
тельных участков и других элементов поступательных временных связей
по существующим обратным связям к очагу индивидуального
раздражения.
Значит, с каждым повторением индивидуального раздражения без
комбинации с основным раздражением обратные связи должны
становиться все сильнее и сильнее сравнительно с поступательными связями.
Благодаря этому при индивидуальном сигнале возбуждение из конечных
двигательных или секреторных пунктов поступательных связей будет
иррадиировать более сильно в обратные связи и сообразно, согласно
закону сопряженной иррадиации, оно должно все меньше
распространяться по нисходящим путям к координирующим аппаратам. С развитием
обратных связей должно усиливаться их влияние на соподчиненный им
нейропиль, поэтому каждый раз при возбуждении обратных связей
должен усиленно активироваться и этот нейропиль. Соответственно должно
усилиться общее торможение, т. е. анэлектроническое действие нейро-
пильного тока на ближайшие нервные круги — ив частности на конечные
эфферентные пункты временных связей. Значит, при угасании мы имеем
такую же картину взаимодействия поступательных и обратных связей,
какая имеет место в недеятельную фазу запаздывающего или
последовательного рефлекса. Таким образом, первичное глубокое угасание
рефлекса, вероятно, является результатом непреходящего ослабления посту-
нательных и усиления обратных связей (Беритов, 3).
Усиленное развитие обратных связей при глубоком угасании
индивидуального рефлекса прежде всего обнаруживается в том, что при
угасании старого, наиболее прочного рефлекса угасают сами собой и все
остальные однородные рефлексы. Они перестают вызывать рефлекс при
любом интервале после пробы первично угашенного рефлекса. Затем об
этом свидетельствует наступление сна у животного при повторном
угасании рефлекса, как это уже говорилось выше.
Означенное изменение временных связей должно наблюдаться только
при многократном угашении рефлекса подряд в течение одного или
нескольких дней. Разовое угасание в течение одного опыта как при
однократном длительном раздражении, так и при повторных раздражениях
короткой продолжительности обычно проходит бесследно для временных
связей. Значит, первичное разовое угасание в течение одного дня не
обусловливается изменениями самого механизма временных связей.
Происхождение преходящего угасания индивидуального рефлекса.
Преходящее угасание индивидуального рефлекса, особенно когда оно
получается при быстром повторении индивидуального раздражения через
10—20 сек. или при однократном длительном применении
индивидуального раздражения, должно быть, происходит тем же самым путем, что и
ослабление спинномозгового рефлекса при тех же условиях
раздражения, а именно, под влиянием утомления и общего торможения.
Доказано, что утомление при преходящем угасании происходит не в тех
нервных комплексах коры, которые воспринимают индивидуальное
раздражение. Выше мы указали, что звуки могут оказывать на кору опре-
УГАСАНИЕ И ВОССТАНОВЛЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 5СЗ
деленное действие в течение многих десятков минут, без заметного
ослабления, в то время как рефлекс на индивидуальный звук может
угаснуть в течение нескольких минут или даже в течение менее 1 минуты.
Опыты свидетельствуют, что преходящее угасание должно наступать
от утомления или временных связей, или конечных пунктов временных
связей, или тех и других вместе. Так, например, если иметь дело с двумя
однородными рефлексами, можно обнаружить, что при угасании одного
наиболее сильного рефлекса заметно ослабевает сравнительно слабый и
молодой рефлекс (Бабкин, 1). Вторичное угасание рефлекса проходит
быстро. Его можно заметить только в том случае, если второй рефлекс
испытывается во время угашенного рефлекса или спустя несколько
секунд после него. Так как для этих рефлексов общими являются
конечные пункты временных связей и затем весь нисходящий нервный путь, а
сами временные связи различны, то, очевидно, угнетающее действие
одного рефлекса на другой происходит или в ответных двигательных и
секреторных участках коры, или же где-либо в подкорковых элементах.
Но мы знаем, что даже после абсолютного угасания индивидуального
рефлекса основное раздражение может вызывать прирожденный рефлекс
без заметного ослабления, значит вся некорковая нисходящая часть
нервного пути с координирующими аппаратами, при угасании
индивидуального рефлекса, видимо, не утомляется. Следовательно, при
угасании одного индивидуального рефлекса быстро проходящее вторичное
угасание другого индивидуального рефлекса, должно быть,
обусловливается ослаблением деятельности исключительно в конечных
корковых элементах временных связей.
Утомление временных связей также наблюдается в определенных
случаях с большой отчетливостью. Например, если угасить рефлекс на
одно раздражение и затем перейти сейчас же к другому, вызывающему
однородный рефлекс, ставя опыты при тех же условиях, то последнее
может вызывать рефлекс подряд много раз. Если затем спустя 10 сек.
после последнего рефлекса возвратиться к первому раздражению, то
угасший рефлекс на первое раздражение может оказаться
восстановленным. Это, например, видно на рис. 245. Значит, оживление угасшего
рефлекса происходит, несмотря на беспрерывную деятельность
конечных пунктов временных связей. Отсюда совершенно ясно, что
первичное угасание произошло* благодаря утомлению поступательных
временных связей первично угашенного рефлекса.
Нужно предположить, что в коре большого мозга, как и в спинном
мозгу, в результате длительного или повторного индивидуального
раздражения происходит не только подлинное утомление определенных
элементов мозга, но и их усиленное угнетение под влиянием общего
торможения, вызываемого через нейропиль коры мозга при этом
раздражении. Очевидно, ослабленные от деятельности нервные круги в коре
мозга сильнее поддаются влиянию общего торможения, а потому
угасание однородных рефлексов в данном случае должно быть обязано своим
38 п. С. Беритов
594 еъ'ЦЧК ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО 310ЗГА
происхождением главным образом совместному действию утомления
и общего торможения временных связей в этих нервных кругах.
При некотором повышении возбудимости в коре мозга, как
указывалось выше, угасание индивидуального рефлекса сильно затрудняется:
оно не наступает даже при повторении его сотни раз без сочетания
с основным раздражением. Это, видимо, обусловливается повышением
возбудимости в корковых нервных кругах — ив частности, в элементах
нервных связей. Благодаря этому они быстро восстанавливаются после
каждого активного состояния и не подвергаются заметному влиянию
со стороны общего торможения. Такое же явление имеет место в
спинном мозгу: при значительном повышении возбудимости рефлекторные
реакции повторяются друг за другом без ослабления значительно
больше времени, и влияние общего торможения проявляется значительно
слабее, чем при некоторой низкой возбудимости спинного1 мозга.
Итак, первичное разовое угасание индивидуального рефлекса при
быстром повторении последнего в течение одного опыта или при одном
длительном действии индивидуального раздражения обусловливается
утомлением и общим торможением, с одной стороны, конечных
пунктов поступательных связей, т. е. двигательных или секреторных
участков коры, а, с другой,— самих поступательных связей.
Восстановление индивидуальных рефлексов. После разового угасания
рефлекса вследствие длительного беспрерывного раздражения или
нескольких повторных раздражений он может восстановиться сам
собой спустя более или менее значительное время после отдыха.
Если же рефлекс угашался систематически несколько раз подряд без
подкрепления его основным раздражением, то для полного
восстановления рефлекса отдых недостаточен. Восстановление может быть
получено применением одного основного раздражения или путем вызова
какого-либо однородного рефлекса, или путем применения какого-либо
постороннего необычного раздражения (Бабкин, 1; Завадский, 6;
Подкопаем, 12). Но это восстановление не будет полным и окончательным.
После него рефлекс удерживается всего только несколько минут, а
затем угасает (Бабкин, 1; Завадский, 6). Это временное восстановление
объясняется тем, что основное раздражение повышает возбудимость
в конечных нервных элементах временных связей; пока повышенная
возбудимость существует, слабое возбуждение, пришедшее через
временные связи, может привести эти элементы в активное состояние. Но,
понятно, после того как возбудимость возвратилась к норме, вновь
должна исчезнуть возможность вызывания индивидуального рефлекса.
Полное восстановление угасшего рефлекса происходит только
путем сочетания индивидуального раздражения с основным, с помощью
которого был образован данный рефлекс (рис. 246). Восстановление
рефлекса при этом условии происходит быстро, но все же не сразу.
Устойчивость рефлекса возрастает с каждым новым сочетанием. Чем
глубже было угасание, тем больше сочетаний требуется для полного
восстановления (Кашеринова, 13). Характерно, что сочетание одного реф-
УГАСАНИЕ II ВОССТАНОВЛЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 595
лекса ведет к восстановлению лишь данного рефлекса, но не других,
также угашенных рефлексов.
Восстановление угасшего рефлекса путем сочетания с основным
раздражением представляет собой процесс, обратный первично.му
угасанию рефлекса. Оно, несомненно, основывается на восстановлении
ослабленных поступательных временных связей, так как более или менее
полное угасание рефлекса, как мы показали, связано главным образом
—л kfLL^lriilrU^L—d
1
UU fL
1
J/7.
mj
? i
300
virui ¦"¦-*" u-r-xnr
"\r
S . M M
200
300
200 200
i и г
200
Рис. 246. Восстановление угашенного индивидуального рефлекса^на правой
передней ноге на звук тонвариатора Штерна 200 колебаний в сек.
Собака Боб. В опыте / дается сначала проба звука 200 колебаний, который эффекта не дает, затем
влияние дифференцированного звука тонвариатора Штерна 300 колебаний на метрономный рефлекс.
Последний был слабее, чем потом в отдельности. В опытах 2, Я и 4 происходит подкрепление звука
200 колебаний. В третьем сочетании звук уже дает рефлекс. Опыт 5 записан спустя 14 мин. после
подкреплений. Здесь звук 200 колебаний повторялся друг за другом четыре раза, один раз во время
звука 300 колебаний. В последнем случае звук 2 00 колебаний не дал рефлекса, в остальных случаях
он дал его.
с некоторыми изменениями, вызываемыми продолжительной
односторонней деятельностью в механизме этих связей. Поэтому можно утверждать,
что восстановление есть процесс, совершенно подобный новообразованию
рефлекса. Иначе говоря, восстановление угасшего рефлекса путем
сочетания с соответствующим основным раздражением обозначает новое
морфологическое и функциональное развитие поступательных временных
связей (Беритов, 3).
Разумеется, при восстановлении поступательных временных связей
угасшего рефлекса дело идет не об устранении в них утомления,
произведенного в период угасания. Как бы сильно ни было утомление, оно
не может продолжаться после угасания целые дни, и притом у
совершенно нормального животного: оно должно проходить в течение нескольких
минут, самое большее — в течение нескольких часов. Это, например,
хорошо известно по работам на спинальных животных.
В случае глубокого угасания старого и прочного рефлекса мы
596 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
отметили усиленное развитие обратных связей параллельно с
ослаблением поступательных. Конечно, после восстановления последних путем
сочетания с основным раздражением деятельность обратных связей
не может сказаться на эффекте в такой мере, как раньше. Вследствие
сильного возбуждения конечных двигательных и секреторных участков
коры и нисходящих эфферентных путей при индивидуальном сигнале
возбуждение не будет всецело иррадиировать в обратные связи, и
активированный при этом нейропиль больше не будет анэлектротониче-
ски угнетать эти эфферентные пути. Поэтому не будет препятствий к
распространению возбуждения из конечных пунктов к рабочему органу
н к наступлению периферического двигательного эффекта (Беритов, 3).
Угасание и восстановление по учению Павлова. Первоначально юекоторые ученики
Павлова рассматривали угасание рефлекса как проявление утомления в
определенных корковых частях рефлекторной дуги. Впервые высказался в этом смысле
Бабкин (1904) (1). Так, разовое угасание рефлекса путем повторения его через
определенный промежуток времени — от 2 до 3 мин.*—автор приписывает утомлению
тех корковых центров, которые воспринимают индивидуальное раздражение. Это
предположение было сделано на том основании, что после угасания рефлекса на одно
индивидуальное раздражение такой же рефлекс не переставал вызываться на другое
индивидуальное раздражение. Автор работал с так называемыми «натуральными
условными рефлексами». Индивидуальным раздражителем в таких случаях являлся
тот или другой внешний признак пищевого вещества: например, вид пищи, запах ее,
звук, сопровождающий пищу перед кормлением (хруст сухаря) и т. д. В этих условиях
при угасании рефлекса на один внешний признак другие внешние признаки не теряли
слюногонного свойства. Но автор приводит и такие факты, на основании которых он
считает возможным участие вкусового центра в угасании рефлекса. Так, он находит,
что, после повторного многократного угасания индивидуального рефлекса и оживления
его одним и тем же вкусовым веществом, без значительного отдыха между двумя
последовательными угасаниями, полное восстановление рефлекса не может быть
достигнуто. Отсюда он сделал предположение об утомлении вкусового центра при
означенном угасании рефлекса.
Дальнейшее развитие взгляда Бабкина было сделано Перельцвейгом
(1907) (2). Этот автор признает лишь два существенных момента в угасании
индивидуального слюнного рефлекса: порчу временных путей и понижение возбудимости
ротового центра, т. е. вкусового по обычной терминологии. Порчей временных путей
он объясняет главным образом первичное угасание индивидуального рефлекса, а
понижением возбудимости ротового центра — вторичное угасание другого однородного
рефлекса. Но при детальном рассмотрении факта вторичного угасания автор не
удовлетворяется этим объяснением. Кроме того, он становится на обычную для школы
Павлова точку зрения о взаимодействии возбужденных точек. Именно он допу
екает, что одновременно с понижением возбудимости ротового центра, в силу
расходования потенциальной энергии на секрецию слюны, происходит повышение
возбудимости в очаге индивидуального раздражения, а это в свою очередь производит
угнетение вкусового центра.
Павлов еще в период работы Бабкина, т. е. в 1904 г., не находил
возможным объяснять явление угасания истощением, т. е. утомлением рефлекторной дуги,
так как, по его словам, дело идет о повторных раздражениях слабой силы (7). Но и
объяснение Перельцвейга не нашло поддержки. Начиная с 1908 г. все авторы
поддерживают взгляд Завадского, что угасание есть исключительно проявление
торможения (Потехин, 14; Горн, 15 и др.). По словам Завадского, условное
возбуждение рефлекса при повторении переходит в торможение рефлекса. Это счи-
УГАСАНИЕ И ВОССТАНОВЛЕНИЕ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 597
тается следствием ненужности такого слюнного рефлекса, который не сочетается со
вкусовым раздражением. Значит, развитие торможения вместо возбуждения
объясняется с точки зрения целесообразности.
Авторы локализируют торможение, производящее угасание, в той корковой
области, которая воспринимает раздражение. Они обосновывают это тем, что если
бы, наоборот, торможение было локализировано во вкусовой воспринимающей области
или, как теперь называют, в пищевом центре, то восстановление вторично угашенного
однородного рефлекса должно было бы происходить одновременно с восстановлением
первично угашенного. В действительности восстановление вторично угашенного
рефлекса всегда происходит раньше восстановления первично угашенных (Горн, 15 и др.).
Временное оживление индивидуального рефлекса без сочетания с основным
раздражением путем одного основного раздражения или иным путем первоначально
объясняли повышением возбудимости в центре слюнных желез. «Мы можем
принять,— говорит Павлов,— что благодаря прямому рефлексу (основное
раздражение.— И. Б,) раздражимость центра слюнных желез повышается, и более слабое
раздражение, действие объекта на расстоянии (вид и запах пищи.— И. ?.), вновь
приобретает способность к действию». Это же объяснение повторяют Бабкин и
Пере льцвейг. Но, начиная с Завадского (1908), который установил точку
зрения на угасание как на внутреннее торможение, все авторы рассматривают оживление
угасшего рефлекса под влиянием изолированного применения основного рефлекса
как расторможение угашенного рефлекса.
Восстановление индивидуального рефлекса путем сочетания с основным ротовым
раздражением первоначально объясняли в школе Павлова как результат
повышения возбудимости в центре слюнных желез. Начиная с Бабкина (1901) до К а ш е-
р и нов ой (1908), это мнение, повидимому, являлось господствующим. Но потом,
когда Завадский (1908) обосновал взгляд на угасание как на внутреннее
торможение, было выдвинуто новое объяснение: что всякое восстановление есть
«расторможение угасательного торможения». Так как «угасательное торможение»
локализируется в анализаторе индивидуального раздражения, то, по мнению
авторов, восстановление должно быть обусловлено прекращением торможения в этом
анализаторе.
На данном этапе развития наших знаний о корковой деятельности мы также
считаем торможение ответственным за угасание индивидуального рефлекса. Но, ©о-пер-
вых, это торможение совершенно такое же общее, нейропильное торможение, какое
бывает в любом другом отделе мозга, а во-вторых, оно не всецело обусловливает
угасание, преходящее или непреходящее, а только играет некоторую роль в его
происхождении.
Литература
1. Бабкин Б. Опыт систематического изучения сложнонервных явлений у
собаки. Дисс, 1904.
2. Перельцвейг И. Материалы к учению об условных рефлексах. Дисс,
1907.
3. BeritoffJ. tJber d. individuell erworbene Tatigkeit d. ZNS, J. Psych. Neurol.
33, 113, 1927; Индивидуально приобретенная деятельность центральной нервной
системы, 1932.
4. Зеленый Г. Материалы к вопросу о реакции собаки на звуковые раздражения.
Дисс, 1907.
5. Ее гі toff J., J. Psych. Neurol., 30, 217, 1924.
6. Завадский И. Материалы к вопросу о тормошении и расторможении
условных рефлексов. Дисс, 1908.
7. Павлов И. 20-летний опыт ..., 1-е изд., стр. 35.
8. Былина П. Простое торможение условных рефлексов. Дисс, 1910.
9. Добровольский В. О пищевых следовых рефлексах. Дисс, 1911-
398 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
10. Абуладзе К. Русск. физпол. журн.. 9, 160, 1927.
11. БашмуринА. иМюльбергВ.. Beitr. d. Ges. d. russ. Physiol, в. 3. 1923.
12. П о д к о п э е в В., Тр. физпол. лаб. акад. Павлова, 2, в. 1, стр. 117, 1927.
33. Кашерииова Н. Материалы к изучению условных слюнных рефлексов.
Дисс, 1911.
14. П о т е х и н С, К физиологии внутреннего торможения условных рефлексов.
Дисс, 1911.
15. Г о р н Э. Материалы к физиологии внутреннего торможения условных рефлексов.
Дисс, 1912.
16 S р е і d е 1 С. С, Cold Spr. Harb. Symp, Quant. Biol., 4, 13, 1936
Глава XI
ИЗМЕНЧИВОСТЬ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ ПОД
ВЛИЯНИЕМ ВНЕШНИХ РАЗДРАЖЕНИЙ
Изменчивость индивидуальных рефлексов под влиянием необычных
раздражений. Постоянство индивидуального рефлекса чрезвычайно
условно. Он очень легко меняется и даже исчезает, если внешняя
обстановка, при которой он возник, изменяется,— например, введением
необычного звука, света или движения. Если такое необычное изменение
вызывает ориентировочные движения, то во время него, а также
некоторое время после него обычный индивидуальный звук не дает
рефлекса (рис. 247). Но каждое необычное раздражение при повторении
теряет способность вызывать ориентировочные реакции: одновременно
оно перестает угнетать индивидуальный рефлекс (Розова, 1).
Происхождение отрицательного действия необычного раздражения.
Необычное раздражение вызывает возбуждение главным образом в
соответствующей воспринимающей области коры большого мозга. Здесь
оно приводит в активное состояние двигательные участки
ориентировочных движений. Возбуждение из этих участков распространяется на
периферию к двигательным органам, но в некоторой мере оно ирради-
ирует и по всей коре больших полушарий. Если необычное раздражение
достаточно сильно, результатом иррадиации возбуждения в коре імозга
іможет явиться возбуждение двигательных участков разнообразных
воспринимающих областей. Это вызовет вздрагивание всего тела и
даже локомоцию, что, в свою очередь, произведет раздражение проприо-
цепторов во всех движущихся органах. Вследствие всех этих
первичных и вторичных раздражений возбудимость повышается как в
первично возбужденных воспринимающих областях, так и в некоторой степени
во всей коре больших полушарий. Разумеется, возбудимость
повышается также и в области временных связей. Вследствие этого возбуждение,
вызванное индивидуальным раздражением, сильно иррадиирует в коре
из всех очагов временных связей и поэтому, согласно закону
сопряженной иррадиации возбуждения, достигает конечных пунктов временных
связей с меньшей интенсивностью, чем обычно.
Постороннее необычное раздражение не только возбуждает разные
600 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
нервные круги в коре мозга, в том числе нервные круги, образующие
временные связи, и тем повышает в них возбудимость, но должно
активировать также богатое нейропильное сплетение дендритов во всех
слоях коры мозга. Выше імы видели, что на совершенно бодрствующих
животных одно зрительное раздражение меняет электрическую
активность не только в зрительной области, но и во всех остальных частях
коры. Изменение активности в остальных частях коры заключается в
ослаблении спонтанной
электрической активности; мы приписали
это анэлектротоническому действию
\ і \ медленных токов активного нейро-
*—— пиля на нервные круги. Нужно
думать, что и в отрицательном
действии необычного раздражения на
индивидуальный рефлекс активное
состояние дендритного нейропиля
играет определенную роль. Именно,
можно предположить, что ирради-
ирующее по нервным кругам возбу-
Рис. 247. Действие необычного посте- ЖДенИе активирует нейропиль, при-
роннего раздражения на индивидуаль- чем в нем возникают
распространяющиеся медленные токи, которые
действовать анэлектрото-
круги.
Пп\
V
ц
гоо
20Q гоо
ШНИПИЩПП iiilimimmi....nmmm„,|m.n иугчЫЩГГШНПШ?
ный рефлекс.
металлического цилиндра — оно прерывает
мин. после
опыта 7, сначала производится
индивидуальное раздражение, которое вызывает обычный Ч6СКОГО раСПрОСТраНСНИЯ
рефлекс. К нему вскоре присоединяетсн не- ^ Twn и ст Tow-
обычный звук металлического цилиндра, что ВОЗОуЖДеНИЯ. 1 ЗКИМИ
производит только ослабление рефлекса.
Собака Боб. В опыте 1 сначала дается
необычное злучание металдичесного цилиндра ц по ДОЛЖНЫ
удару через каждую секунду. В этот момент
собака дремала. На звуки она пробуждается, НИЧеСКИ HQ ЭТИ Же НврВНЫ?
поднимает голову (опускание кривой). К не- ^ ^ »
обычному звучанию на несколько секунд при- UCOOeHHO СИЛЬНО ОНИ ДОЛЖНЫ ДСИ-
соединяется индивидуальный звук 200
колебаний в 1 сек., потом это необычное раздра- СТВОВаТЬ На ТС КЛ?ТКИ И аКСОНЫ, КО'
жение прекращается, а индивидуальное про-
должается еще 17 сек. Но последнее не дает ТОрЫе Не ВХОДЯТ В Замкнутые Кру-
рефлекса — как во время необычного звука, лв
так и ппгтте него Индивидуальное різлражр- ГИ И ПОТОМу Не ИСПЫТЫВЭЮТ КЭТ-
ние по прекращении было возобновлено опять ^ л „ л „ ллл
спустя 5сек.; на этот раз оно вызвало обычный ЭЛекТрОТОНИЧесКОГО ПОВЫШеНИЯ ВОЗ-
рефлекс — поднятие правой передней ноги ^ „л лтл «.,,„„
Во время последействия вновь дается звучание буДИМОСТй В Силу распространения
металлического цилиндра — оно прерывает -¦ %
последействие. В опыте 2, спустя 4 мин. после ВОЗбуЖДвНИЯ, а также ЧИСТО фИЗИ"
биотокоз
нервными
элементами являются прежде всего
аксоны пирамидных клеток, дающие
начало пирамидным и экстрапирамидным путям. Следовательно,
тормозящее действие нейропиля сильнее всего должно проявиться fB
отношении пирамидных аксонов. Вследствие этого возбуждение,
притекающее по временным связям к пирамидным клеткам, не сможет
распространиться на периферию и вызвать там внешний эффект.
Таким образом, отрицательное действие необычного раздражения на
индивидуальный рефлекс происходит от того, что под влиянием
иррадиации возбуждения из соответствующей воспринимающей области и
двигательных \участко& ориентировочной реакции, а также из
воспринимающей области проприоцептивных раздражений возбудимость
повышается во всей коре в сильной степени. Вследствие этого возбуждение
ИЗМЕНЧИВОСТЬ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 601
из времеяшх связей в значительной мере иррадиирует во всей коре и
поэтому достигает конечных двигательных элементов с ослабленной
интенсивностью. Одновременно активируется и нейропиль Так как
аксоны пирамидных клеток должны подвергаться особенно сильно
угнетающему действию нейропильного тока, то весьма вероятно, что
отрицательное дейст&ие необычного раздражения в некоторой мере
обусловлено угнетением проведения возбуждения из пирамидных клеток на
периферию.
Как было указано, отрицательное действие наблюдается только в
тех случаях, если необычное раздражение вызывает ориентировочную
реакцию. Последняя представляет результат деятельности особых
корковых двигательных участков. Деятельность этих участков происходит
под влиянием общей иррадиации возбуждения из очагов необычного
раздражения. Поэтому прекращение ориентировочной реакции должно
служить показателем того, что общая иррадиация прекращается или
очень сильно ослабевает. Кроме того, раз нет ориентировочных
движений, нет и раздражений проприоцепторов. В соответствии с этим
возбудимость нервных кругов не повышается в такой імере, чтобы это
послужило условием для о^цей иррадиации возбуждения в более или
менее значительной степени из элементов временных связей. В то же
время нейропиль коры не активируется в такой степени или в таком
объеме, чтобы вызывать в. аксонах пирамидных клеток значительное
анэлектротоническое понижение возбудимости Значит, необычное
раздражение при его многократном ^повторении потому перестает
действовать отрицательно, что оно более не повышает возбудимости в нервных
кругах и не активирует в достаточной мере нейропиля в коре больших
полушарий.
Отрицательное действие дифференцированного раздражения на
индивидуальный рефлекс. Из необычных раздражений следует
выделить одну группу, которая характеризуется тем, что при повторении
она не теряет способности к отрицательному действию. Сюда
относятся так называемые необычные дифференцированные раздражения,
которые во время генерализации вызывали рефлекс, а потом перестали
его вызывать. Эти дифференцированные раздражения устраняют
индивидуальный рефлекс при любом повторении, несмотря на то, что они
не вызывают ориентировочных движений. Для примера укажем на
рис. 248, где необычный дифференцированный звук, 512 колебаний в
секунду, не производя никаких внешних движений, тем не менее
устраняет индивидуальный рефлекс на звук 200 колебаний в секунду (Эльяс-
сон, 3; Беляков, 4).
Отрицательное действие дифференцированных раздражений не
одинаково. Чем ближе стоит данный дифференцированный раздражитель
к индивидуальному, тем сильнее его отрицательное действие на
индивидуальный рефлекс (Эльяссон, 3; Красногорский, 5).
Происхождение отрицательного действия дифференцированных
раздражений. Как указывалось выше, при дифференциации ближайших
6Q2 ОГ.ЩШ ФіПИОЛО' Hit КОРЫ ЬО.'іЬШОГО ЛЮ.'іГА
раздражений развиваются обратные связи от воспринимающего очага
индивидуального раздражения к очагам необычных раздражений.
Очевидно, отрицательное действие таких дифференцированных
раздражений на индивидуальный рефлекс прежде всего должно зависеть от
деятельности обратных связей. Однако несомненно, что отрицательное
действие дифференцированных раздражений зависит и от следующего
обстоятельства: корковые очаги
дифференцированных раздраже<
ний, производящих
отрицательное действие, находятся очень
близко к очагу индивидуального
раздражения, т. е. к начальному
пункту временных связей. Это не
может не являться причиной
отрицательного действия
дифференцированных раздражений. Нужна
небольшая иррадиация
возбуждения из очагоз этих раздражений,
чтобы, с одной стороны, вовлечь
нервные круги, связанные с
временными связями, а с другой —
активировать нейропиль в
области этих связей. Возбуждение
нервных кругов обусловит
иррадиацию возбуждения из
элементов временных связей, а
активирование нейропиля должно
подействовать угнетающим образом
на конечные двигательные
элементы временных связей, в
особенности на аксоны пирамидных
клеток. Вследствие этого
индивидуальный сигнал не сможет
производить внешний эффект. Эти*1
хорошо объясняется то явление,
что чем ближе необычные дифференцированные раздражения к
индивидуальному раздражению, тем сильнее отрицательное действие их на
индивидуальный рефлекс.
Итак, отрицательное действие посторонних дифференцированных
раздражений зависит от того, что они повышают возбудимость как в
обратных временных связях, так и в нервных кругах, включающих
временные связи данного индивидуального рефлекса, и одновременно
активируют нейропиль в области временных связей. Этим
обусловливается прекращение распространения возбуждения из пирамидных
клеток на периферию.
flmnillllllllllllimiimillllllilllirtMiiiinitt.iiM.iM.i.M.tn.mmu»,,,..,,,»
Рис. 248. Взаимодействие необычного
дифференцированного раздражения и
индивидуального рефлекса на собаке.
Собака Воб. Верхняя кривая — от головы,
средняя—от левой передней ноги, нижняя—от правой
передней ноги. Индивидуальное раздражение —
звук тонвариатора200 колебанийвісек. (нижний
сигнал), необычное раздражение — звук тонва-
риатора 512 колебаний в 1 сек. (средний
сигнал). В опыте 1 сначала происходит
дифференцированное раздражение и оно не вызывает
рефлекса; затем, 3 сек. спустя, — индивидуальное
раздражение, и оно дает движение ноги и
ориентировочную реакцию (опускание верхней кривой); в
опыте 2, спустя несколько минут после
предыдущего опыта вновь дается индивидуальный звук;
в опыте з сначала дается дифференцированный
звук 512 колебаний, затем присоединяется
индивидуальный звук 200 колебаний, который
продолжается по прекращении дифференцированного
звука еще 25 сек. Индивидуальный звук вызвал
в самом начале только поднятие головы, но вслед
за тем собака постепенно опускает ее и начинает
дремать. Затем происходит перерыв
индивидуального звука на И сек. После возобновления
рефлекса снова не наступает. На этот раз
индивидуальный звук продолжается 10 сек., и затем
вновь происходит перерыв на 22 сек. Теперь, при
возобновлении, индивидуальный звук производит
обычный рефлекс, но в более слабой форме, чем
в предыдущем опыте. Между опытами 2 и 3
индивидуальный звук сочетался с электрическим
раздражением. Этот опыт не был записан. Время—
по 2 сек.
ИЗМЕНЧИВОСТЬ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 603
Как дифференцированное раздражение, так и необычное постороннее
раздражение, производящее ориентировочную реакцию, оказывают на
индивидуальный рефлекс более слабое отрицательное действие, когда
они начинаются во ,время существующего рефлекса. Это хороню видно
на рис. 249. Объясняется это тем, что возбудимость временных связей
значительно повышена, когда они находятся в действии. Кроме того,
под влиянием тока возбуждения в нервных кругах и аксонах
пирамидных клеток возникает катэлектротоническое состояние. Вследствие
этого постороннее раздражение должно действовать значительно
дольше и сильнее, чтобы из элементов временных связей возбуждение
U
П.л.
ib ib JS
JTL^ П 1 * —I ' * 1
200
¦ \ _J 1 L_l Т.
ib ib 200
ііііищііііііПіі1пімипц|пАД11'""'ЧИ11'М||""""и""/11"ЧИ1иМ1 """¦""" іиі|іііііі,іпіиііініі.інаіитііпінніііііітіііиііііішщм||""|м
Рис. 249. Действие необычного раздражения (электрического звонка) на
индивидуальный рефлекс правой передней ноги на звук тонвариатора 200 колебаний в 1 сек.
Собака Боб. В опыте 1 индивидуальное раздражение производится после того, как начался звонок,
и оно не дает рефлекса. В опыте 2 индивидуальное раздражение производится сначала, а затем уже
присоединяется звонок. Здесь индивидуальный рефлекс наступает, лишь немного ослабевая при
повторении звонка. В опыте 3 звонок испытывается два раза по 1.5 сек.; каждый раз к нему
присоединяется индивидуальное раздражение, которое продолжается после него еще 10 сек.; несмотря
на это, рефлекса нет. Затем звонок испытывается во время последействия индивидуального рефлекса;
первый раз он только немного ослабляет это последействие, а второй раз прекращает его.
могло сильнее иррадиировать и чтобы нейропиль был в состоянии
произвести анэлектротоническое понижение возбудимости в аксонах
пирамидных клеток.
Об отрицательном последействии посторонних раздражений на
индивидуальный рефлекс. Как необычное постороннее раздражение,
производящее ориентировочную реакцию, так и постороннее
дифференцированное раздражение действуют отрицательно на индивидуальный рефлекс
и в том случае, если указанное раздражение предшествует
индивидуальному. Когда индивидуальный рефлекс слаб и непрочен, а сила
постороннего раздражения значительна, это отрицательное последействие
последнего іможет продолжаться в течение многих минут. Но если
индивидуальный рефлекс ів достаточной імере упрочился, то он не
устраняется в полной мере, даже в тех случаях, когда следует спустя
несколько секунд после постороннего раздражения. Так, например, на
рис. 248 посторонний дифференцированный звук 512 колебаний в 1 сек.
совершенно устранил индивидуальный рефлекс, когда индивидуальное
раздражение началось во івремя него (опыт 3). Но спустя 3 сек. после
дифференцированного раздражения индивидуальное вызвало обычный
рефлекс (опыт 1). Он был только чуть ослаблен по сравнению с тем
рефлексом, который вызывался без предшествующего
дифференцированного раздражения (опыт 2).
Отрицательное последействие посторонних раздражений, повидимому,
604
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
основывается на длительно продолжающемся возбуждении нервных
кругов, которое постепенно затухает. Сообразно с этим, оно должно
обусловить постепенно затухающее активное состояние нейропиля,
которое в свою очередь производит своим током постепенно затухающее
анэлектротоническое понижение возбудимости в элементах временных
связей и в аксонах пирамидных клеток. Отсюда понятно, что пока
индивидуальный рефлекс слаб ©виду слабого развития временных связей,
нейропильное угнетение может оказать отрицательное действие на
данный рефлекс и спустя
значительное время после
прекращения раздражения.
л. С укреплением уже
рефлекса этот интервал
отрицательного действия должен
укоротиться.
Отрицательное
"последействие может быть оч?нь
длительным, если индиви-
п
Чес
(
* і і гоо
іШИ.іііЦШіііШіііііііііііііііііііііМііііііі/.ііііііцііш.'ііііЦіііііі т III ШПіИШИ
Рис. 250. Взаимодействие двух разнородных
рефлексов на собаке.
дуальное раздражение
тянется после сочетания с по-
Мура. Один рефлекс — сгибание левой задней СТОрОННИМ без Перерыва ИЛИ
l звук 200 колебаний в і сек. (нижний сигнал); г
индивидуальное
раздражение может продолжаться
несколько десятков секунд,
не давая рефлекса.
Чем
Собака
ноги на „ _. __ . _ . ,
второй рефлекс—сгибание правой передней ноги на чеса- с небОЛЬШИМ ИбрерЫВОМ, K3K
ние кожи на спине (верхний сигнал). Верхняя кривая
принадлежит голове, средняя — левой задней ноге, ниж- на рИС 249. В ТЭКИХ СЛУЧЭЯХ
няя—правой передней ноге. В опыте 1 сначала вызывает- г
ся рефлекс на левой задней ноге на звук 200 колебаний в
1 сек. К нему присоединяют на короткое время чесание.
Во время комбинации рефлекс на задней ноге исчезает,
и сейчас же возникает рефлекс на правой передней ноге.
Повторное звучание вновь вызывает рефлекс на левой
задней ноге. В опыте 2 сначала производится рефлекс на
чесание: сильное сгибание правой передней ноги. К нему
присоединяют на короткое время звук 200 колебаний в
1 сек., что вызывает исчезновение рефлекса на правой слабее ИНДИЗИДуаЛЬНЫИ 0?-
передней ноге, но вместе с тем рефлекс на левой задней
ноге не возникает. Такое сочетание происходит второй флеКС, Т?М ПОЗДНее ОН НаСТу-
раз: теперь 8вук не дает своего рефлекса и не устраняет
существующего рефлекса на чесание. Третий раз дается пает. При ЭТОМ не ИМ6СТ
один звук, он вызывает обычный эффект. Время — г
в секундах. существенного значения,
сколько времени
продолжалось само сочетание обоих раздражений: 10 сек., і сек. или еще
меньше. Так, в одном опыте отрицательное последействие длилось после
сочетания с необычным 25 сек. в случае сильного рефлекса, 52- сек. в
случае слабого рефлекса и всего 5 сек. в случае одновременного вызоза
обоих рефлексов. На рис. 249 видно, что предварительного сочетания в
1—2 сек. достаточно, чтобы оказать на индивидуальный рефлекс
длительное отрицательное последействие.
Всестороннее изучение описанного явления показало, что большая
длительность отрицательного последействия постороннего раздражения
на индивидуальный рефлекс, если последний сочетается с ним,
обусловливается тем, что отрицательное последействие, вызванное посторонним
раздражением, поддерживается затем самим индивидуальным
раздражением. В период сочетания возбуждение, вызванное индивидуальным
раздражением, распространяется из элементов временных связей пре-
ИЗМЕНЧИВОСТЬ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 605
имущественно в коре мозга в случае необычного постороннего
раздражения н по обратным связям—tB случае дифференцированного раз
дражения. (Кроме того, эти элементы временных связей тормозятся от
анэлектротонического влияния нейропильного тока в особенности
сильно в аксонах пирамидных клеток. Это состояние безусловно должно
поддерживаться в течение некоторого времени и одним
индивидуальным раздражением, ибо пока имеется налицо повышенная
возбудимость нервных кругов, индивидуальное раздражение может возбуждать
их и одновременно через них же активировать
нейроииль. Но достаточно прервать
индивидуальное раздражение на 10—20 сек. и тем
самым понизить возбудимость всей коры и
обратных связей, чтобы возобновление
индивидуального раздражения вызвало обычный
индивидуальный рефлекс (Беритов, 2) (рис.
248).
Изменчивость одного индивидуального
рефлекса под влиянием другого. Когда мы имеем
дело с двумя разнородными рефлексами, т. е.
такими, которые были образованы на почве
разных основных раздражений, то при
комбинации их получается то один рефлекс, то
другой; и ів то же время этот рефлекс протекает
или слабее, или сильнее норма чьного эффекта.
Бывают случаи, когда в момент комбинации
оба рефлекса исчезают или же оба наступают
(рис. 250). Каждый возможный случай может
быть заранее предугадан, если состояние
обоих рефлексов заранее хороши известно.
Например, в том случае когда оба рефлекса
обладают примерно одинаковой
интенсивностью, комбинация их может дать
ослабленный эффект. При комбинации слабых
рефлексов разной прочности менее прочный
ослабевает до исчезновения, а другой, наоборот,
может усилиться по сравнению с нормой (Беритов, 2) (рис. 251).
Изменчивость одного рефлекса под влиянием другого зависит от степени
возбуждения и возбудимости корковых элементов временных связей и
вообще нервных кругов в коре больших полушарий, затем от
прочности временных связей и, наконец, от степени активного состояния
нейропиля. Так, например, отрицательное действие одного рефлекса на
другой имеет место, когда один рефлекс прочнее и сильнее другого*
Чем сильнее будут возбуждаться корковые элементы данного рефлекса,
тем сильнее и длительнее будет общее возбуждение нервных крутое
временных связей, а также всей коры, благодаря возбуждению про-
приоцепторов от сильной двигательной реакции. Это обусловит повыше-
О п
Чес
2О0
ZOQ
ll.llltlll.l «HMIini IIMIMliilt.iim. r,r
Рис 251 Взаимодействие
двух ралюродпых рефлок
сов на собаке.
Собака Мура. Те же рефлексы,
что и на предыдущем рисунке.
Верхняя кривая — головы,
средняя — от задней левой,
нижняя — от правой передней
ноги Сначала чесанием
(индивидуальный раздражитель)
вызывается сгибание правой
передней ноги, во время этого
рефлекса индивидуальный звук 200
колебаний производит
небольшое усиление существующего
рефлекса на чесание
Впоследствии же звук вызывает обычный
рефлекс на левой задней ноге
Время — в секундах Кривые
читать справо налево.
606 ОБЩА.Я ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГ V
ние возбудимости вообще в коре мозга. Благодаря же повышенной
возбудимости должна происходить иррадиация возбуждения из
элементов временных связей, что приведет к ослаблению и даже исчезновению
более слабого рефлекса. Кроме того, чем сильнее и длительнее будет
возбуждение нервных кругов, телг сильнее и длительнее будет
активирование нейропиля. Это также создает условия для ослабления более
слабого рефлекса, ибо
активированный нейропиль подействует
угнетающе на нервные круги, и
в особенности на пирамидные
клетки и аксоны, участвующее
в слабом рефлексе.
Второй случай, когда один
рефлекс благоприятствует
другому, должен зависеть от того, что
в известных случаях
возбуждение, иррадиируя из корковых
нервных кругов одного рефлекса
по всей коре, более всего повлияет
на нервные круги другого
рефлекса, возбудимость которых
была повышена, а это может
повлечь за собой усиление
рефлекса.
Если оба рефлекса одинаковы
по прочности и интенсивности и
потому влияние одного рефлекса
на другой более или менее
одинаково,— может произойти
уменьшение обоих рефлексов или же
оба они останутся без изменения
(рис. 252). В этом явлении нейропиль должен играть определенную
роль. Если под влиянием иррадиации возбуждения из нервных кругор
обоих рефлексов произойдет более или менее значительное усиление
активного состояния нейропиля, то произойдет ослабление рефлексов
вследствие усиленного анэлектрического действия нейропильного тока
на аксоны пирамидных клеток, которые не входят в эти круги. Если же
при взаимодействии обоих рефлексов анэлектротоническое действие
нейропиля не усилится в достаточной мере, тогда оба рефлекса могут
проявиться без изменения.
Когда імы имеем дело с однородными индивидуальными рефлексами,
то при комбинации их большей частью получается эффект обычной
величины. Но в определенных условиях рефлекс может усилиться,
ослабнуть и даже исчезнуть. Так, например, если угасить рефлекс
повторением индивидуального раздражения без сочетания с основным, а затем
Vec.
200
м.шиттмпп) ним нити.і;ппііііініііпіігіііттімінтііініііі11і
Рис. 252. Одновременное наступление двух
разнородных рефлексов.
Собака Мура. Верхняя кривая записывает
движение левой задней ноги, нижняя—-движение правой
передней. Сначала производится звук 200
колебаний в 1 сек., который вызывает индивидуальный
рефлекс —поднятие левой задней ноги; к нему
ярисоединяется чесание спины; оно также
производит рефлекс — поднятие правой передней ноги.
Собака держится в стоячем положении вследствие
того, что опирается на лямкп.
изменчивость индивидуальных. РЕФлыиоь 607
сочетать его с другим индивидуальным раздражениелг, то можно
наблюдать ослабление или даже исчезновение рефлекса на второе раз-
дражение. Если тем же путем ослабить оба рефлекса в отдельности,.
то в момент комбинации может получиться еще значительный рефлекс
(рис. 253).
То явление, что один индивидуальный рефлекс нормальной
интенсивности не действует отрицательно на другой однородный рефлекс,
зависит от того, что при комбинации раздражений, хотя и происходит
некоторое усиление активного
состояния нейропиля, но
одновременно усиливается в
такой же мере возбуждающее
действие нервных кругов того —"*¦¦¦ » ¦ ^ г і lL—-гзд-
и другого рефлекса на общие
для них пирамидные клетки. ^—^р г^
Благодаря этому анэлектрото- ¦¦¦¦¦¦¦>... "¦""¦'¦"""""¦'"'"¦"""¦"¦"""¦"¦"""ініі"11""! ¦ «у
ническое действие нейропиль- рис 253. Действие метрономного индиви-
НОГО тока не в СОСТОЯНИИ влиять дуального рефлекса правой передней ноги
на однородный рефлекс на звук /UU
колена ЭТИ КЛетКИ И ИХ аКСОНЫ В баний в 1 сек
ТаКОЙ Мере, ЧТОбы ОСЛабиТЬ Собака Боб. В опыте 1 вначале производится не-
г обычное звучание электрического звонка К нему
ВЫЗЫВаеМЫЙ Через НИХ ПерИ- присоединяется индивидуальный сигнал — стук
метронома АГ. По прекращении звонка наступает
(ЬерНЧеСКИЙ эффект несколько ослабленный индивидуальный рефлеке
т г тт на метроном. Во время него индивидуальный звук
ОтПИИателЬНОе ЛеЙСТЕИе 2 00 колебаний в 1 сек. вызывает полный рефлекс.
vi^maiwion^ Mv,iri4,iwi в опыте 2 сначала вызывается обычный метроном-
УГашенНОГО рефлекса На ОДНО- ный рефлекс. Во время негоi испытывается дваж-
уісхыл^т^ v г^Ч> *-" м ды индивидуальный звук 200 колебании Первый
рОДНЫЙ рефлекс быЛО уже раз он произвел небольшое усиление существую-
объяснено выше. Было
указано, что кратковременное
ослабление слабого рефлекса сейчас же после угасания сильного однородного
рефлекса зависит от утомления конечных пунктов временных связен.
Положительное же действие одного слабого рефлекса на другой
однородный рефлекс малой интенсивности происходит потому, что
благодаря иррадиации возбуждения возникает усиленная деятельность
нервных кругов и это в свою очередь сильно повышает деятельность
общих для них пирамидных клеток. При этом, конечно, усиливается и
активное состояние нейропиля, но, повидимому, его аиэлектротониче-
ское действие на эти клетки и их аксоны не становится таким
значительным, чтобы вызвать значительное ослабление периферического
эффекта.
Итак, отрицательное и положительное действие одного
индивидуального рефлекса на другой происходит согласно принципам, лежащим
вообще в основе корковой деятельности (Беритов, 2).
Учение Павлова об изменчивости индивидуальных рефлексов под влиянием
внешних раздражений. По Павлову и Бехтереву, всякое отрицательное действие
внешнего раздражения гаа индивидуальный рефлекс обусловлено торможением. В
настоящее вреуя признаются два типа торможения: внешнее и внутреннее.
Внешнему торможению приписывают отрицательное действие посторонних необыч
60S ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
ных раздражителей на индивидуальный рефлекс, затем действие электрического
раздражения ноги или раздражения ротовой полости на индивидуальный рефлекс,
или, наконец, одного пищевого индивидуального рефлекса на другой. Механизм этого
торможения, по словам Павлова, заключается в следующем: если какой-нибудь
другой пункт центральной нервной системы приводится в деятельное состояние
соответствующими внешними или внутренними раздражениями, то это сейчас же ведет к
уменьшению или полному исчезновению возбудимости центра нашего условного
рефлекса, т. е. наш условный рефлекс слабеет или временно совершенно перестает
обнаруживаться (6).
Как мы указывали выше, отрицательное действие, производимое необычными
постоянными раздражениями, стоит в определенной связи с ориентировочной реакцией.
Отсюда следует, что в создании внешнего торможения первенствующую роль играет
возбуждение центров, производящих ориентировочную реакцию. Чем сильнее
ориентировочная реакция, тем сильнее торможение индивидуального рефлекса. То же самое
следует сказать по поводу тормозящего действия пищевого безусловного или
условного раздражителя. Чем больше возбужден пищевой центр путем данного
раздражения ротовой полости, тем сильнее торможение последующего индивидуального
слюнного рефлекса другим ротовым раздражением {Савич, 19). Так же действует
разрушительное электрическое раздражение на слюнной рефлекс (Ерофеева, 20).
Во всех этих случаях, по учению Павлова, происходит одно и то же явление*
сильно возбужденный центр тормозит другие центры и перетягивает к себе
возбуждение из этих центров (6).
Выше в общей физиологии спинного мозга и стволовой части головного мозга, а
также коры большого мозга мы указывали, что периферическое раздражение, с
возбуждением определенной цепи нейронов, производит общее понижение возбудимости,
общее торможение через нейропиль. Но как уже указали выше, характер воздействия
необычного раздражения, вызывающего ориентировочную реакцию на
индивидуальные рефлекеы, такое, что отрицательное действие нельзя признать исключительно
результатом торможения коры, т. е. нейропильного торможения, по нашему пониманию.
Известные факты говорят о неизбежном повышении возбудимости в нервных кругах
коры, ибо в ответ на данное раздражение наступает обширная двигательная реакция,
при этом происходит раздражение многочисленных проприоцепторов, и в частности —
статического органа. Затем известно, что означенные посторонние раздражителя
способствуют генерализации дифференцированного рефлекса, пробуждению спящего
животного, оживлению угасшего рефлекса и т. д. На основании этих фактов можно
утверждать, что отрицательное действие необычного постороннего раздражения,
вызвавшего более или менее значительный двигательный рефлекс, обусловливается не
просто торможением, какое наблюдается в спинном мозгу или в других отделах
мозга, а представляет собой сложное явление, обусловленное как этим торможением,
так и повышением возбудимости в многочисленных нервных кругах коры мозга.
Внутреннему торможению приписывается: угасание рефлекса (Коган, 10),
дифференциация его (Красногорский, 5). запаздывание его (Потехин, 23; Завадский, 24),
недеятельная фаза последовательного рефлекса (Гроссман, 25), отрицательные
рефлексы (Лепорский, 26) и сои (Павлов, 11; Бирмаи, 27). Во всех случаях основное
условие для их наступления одно и то же: когда индивидуальное раздражение
происходит в течение более или менее продолжительного времени, не сопровождаясь
безусловным, оно перестает давать рефлекс вследствие развития внутреннего
торможения. Характерной чертой внутреннего торможения Павлов считает его большую
лабильность, так как все эти явления торможения устраняются оод влиянием каких-
либо лостороиіних раздражений, вызывающих ориентировочную реакцию. Рефлекс
перестает тормозиться или, как говорят в лаборатории Павлова, происходит
расторможение заторможенного рефлекса.
Каждый вид внутреннего торможения наделен свойством иррадиации и
концентрации, подобно возбуждению. Здесь, в процессе внутреннего торможения, это
правило выступает, по словам Павлова, еще отчетливее и неоспоримее, чем в
процессе возбуждения (28). Так, например, утверждают, что при угасании раздражения
ИЗМЕНЧИВОСТЬ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 609
торможение впервые возникает в той области коры, которая воспринимает угасаемое
раздражение; отсюда оно иррадиирует в другие воспринимающие области, а затем
снова концентрируется в исходной области, исчезнув в других местах (Горн, 9).
Кроме того, характерно, что иррадиация внутреннего торможения происходит
не так быстро, как это признается в общей физиологии в отношении рецштрокного или
общего торможения, а настолько медленно, что распространяется в одном полушарии
или небольшой части его в течение нескольких минут, и притом у разных
животных с разной скоростью (Коган, 10; Красногорский, 5 и др.)* Но в школе Павлова
считают, что иррадиация возбуждения также протекает очень медленно — в течение
нескольких минут ш расстояние (нескольких миллиметров.
Однако в самое последнее время выдвигают новое представление, по которому
иррадиация и концентрация не составляют неотъемлемого свойства процесса
торможения. Этим свойством оно обладает только в определенных условиях. Так,
например, Подкопаев (29) находит, что во время тормозного условного раздражения
торможение не иррадирует: процесс внутреннего торможения, возникающий в коре
больших полушарий под влиянием тормозного условного раздражителя, остается
строго концентрированным в очаге своего возникновения в течение всего времени,
пока действует тормозящий раздражитель. Наоборот, по его наблюдениям, в это
время происходит повышение возбудимости вокруг очага процесса торможения. Эта
новая концепция в корне противоречит существующему учению о внутреннем
торможении.
В последнее же время в школе Павлова нашли нужным признать двоякого
рода иррадиацию возбуждения и торможения: статическую и динамическую.
Статической иррадиацией возбуждения называется явление, наблюдаемое во время
генерализации рефлекса. Предполагается, что вокруг очага индивидуального раздражения
возникает поле возбуждения, которое постоянно и в течение более или менее
значительного времени не меняется; благодаря ему каждое необычное раздражение
производит индивидуальный рефлекс (Анреп, 16; Розенталь, 17).
Статическую иррадиацию торможения видят при дифференциации рефлекса.
Предполагается, что івокруіг воспринимающего очага дифференцированного^
раздражения создается поле торможения, вследствие чего всякое необычное раздражение
перестает вызывать рефлекс (Фурсиков, 18).
Динамическая иррадиация торможения признается при так называемом угаса-
тельном торможении, а также при условном торможении, т. е. отрицательном
рефлексе, и в других случаях — когда отрицательное действие раздражителя проходит
быстро.
Природу внутреннего торможения авторы представляют чрезвычайно неясно.
Павлов рассматривает внутреннее торможение как явление истощения нервных
элементов. От продолжительного раздражения истощается данный пункт, и в нем
наступает в связи с истощением недеятельное состояние, или состояние сна (т. е.
внутреннее торможение — И, Б.), Это состояние недеятельности в виде сна,
происшедшее в данной клетке, не остается в ней одной, а распространяется все дальше и
дальше и, наконец, іге только захватывает полушария, но опускается и на низшие
отделы головного мозга. Следовательно, нервные клетки, которые не работали,
переживают состояние, подобное состоянию работавшей, тратившейся клетки (II).
Согласно данной концепции, сонное состояние является результатом
возникновения какого-то химического процесса в определенной точке мозга и распространения
отсюда на всю кору и подкорку. Я не буду говорить о всей невероятности такого
допущения, тем более что сам Павлов не настаивает на этом$ считая такое
распространение темным пунктом в явлении. Но характерно, что Рожанский, один из
учеников Павлова, который изучил явление сна в экспериментальных условиях, в
последнее ©ремя высказывает диаметрально противоположное мнение о локализации
и распространении торможения во время сна. Исходя из данных наступления сонного
состояния при непосредственных раздражениях средних отделов головного мозга,
автор приходит к заключению, что сонное торможение локализируется в подкорке, а
отсюда уже распространяется вверх на кору и вниз на спинной мозг (30). Конечно,
39 и. С. Беритов
610
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
это понимание Рожанского в корне противоречит учению об условных
рефлексах, ибо исходный пункт внутреннего торможения локализируется таким образом не
в коре, а в подкорке.
В возникновении внутреннего торможения при данных условиях Павлов видит
определенную биологическую ценность. Этим торможением постоянно коррегируетс»
и совершенствуется сигнализационная деятельность больших полушарий. Если в
данное время сигнальный условный раздражитель повторно не сопровождается
безусловным деловым раздражением, то он, как убыточный, напрасно вызывающий трату
энергии, на время лишается физиологического действия. Из ряда высказываний
Павлова видно, что по его представлениям торможение не является активным
процессом, требующим траты энергии, подобно возбуждению. За время тормозного
периода, оставаясь свободной от работы, клетка 'восстанавливает свой нормальный
состав. Но в других случаях Павлов высказывается совершенно наоборот, а именно,
что торможение подобно раздражению, т. е. возбуждению; что это есть функция
нервных клеток, представляющая собой определенную фазу физико-химического
процесса, происходящего в этих клетках под влиянием многочисленных раздражений.
Характерно, что безусловное раздражение, фактически более деятельное, по мысли
Павлова, благоприятствует переходу клетки в тормозное состояние в меньшей
мере, чем всякое другое раздражение. Безусловный раздражитель даже задерживает
торможение, вызванное другими раздражениями.
Вся эта концепция внутреннего торможения полна противоречий: то оно —
активный процесс, то оно — абсолютный покой; то оно возникает от всякого более
длительного и частого раздражения, то задерживается в своем развитии таким раздражением,
как безусловный раздражитель.
Ясно, что внутреннее торможение не имеет ничего общего с торможением,
известным в общей физиологии. Согласно нынешнему состоянию общей физиологии,
торможение, как оно проявляется в прирожденных двигательных рефлексах, есть
активный процесс. Подобно возбуждению, торможение быстро наступает и быстро же
проходит. Так же быстро, как возбуждение, даже быстрее его, торможение действует
на двигательные нейроны.
Выше мы также пользовались понятием торможения в смысле общего нейро-
пильного торможения, свойственного, между прочим, и коре мозга при теоретическом
рассмотрении взаимодействия индивидуальных рефлексов. Но, во-первых, понимание
этого торможения совершенно иное, чем внутреннее торможение Павлова, а во-
вторых, оно было признано не специфическим и не характерным для понимания
индивидуальной рефлекторной деятельности.
В одном из последних сообщений Павлов (15) рассматривает возбуждение и
торможение с точки зрения изменения возбудимости в отдельных точках коры. Он
считает возможным представлять торможение как явление особой отрицательной
возбудимости. Если условное раздражение перестает по какой-либо причине
вызывать рефлекс,— например, в результате угасания,— то оно превращается, по
Павлову, в тормозящий акт. В этом случае, по его словам, корковые участки теряют
свою прежнюю положительную возбудимость и приобретают новую, отрицательную
возбудимость. По_ существу это новое воззрение, конечно, совершенно не отличается
от прежнего: отрицательная возбудимость представляет собой только новое
обозначение понятия «состояние торможения».
Для объяснения сложных явлений индивидуальной деятельности школа Павло-
в а создала много других гипотез, о которых мы не считаем нужным здесь
распространяться. Все эти гипотезы иллюстрируются массой фактического материала. Несмотря
на это, подходя к ним критически, нетрудно усмотреть в них искусственность,
глубокие противоречия и полное расхождение с общеизвестными законами нервной
деятельности. 1
J Подробно об этом н вообще об индивидуальной деятельности см. в монографии
Беритова: Индивидуально приобретенная деятельность центральной нерзчой
системы.
ИЗМЕНЧИВОСТЬ ИНДИВИДУАЛЬНЫХ РЕФЛЕКСОВ 611
Практикум
После образования пищевого индивидуального рефлекса по методу свободных
движений на' одном из следующих животных: голубе, кролике, щенке, собаке,—
производится ряд опытов над изменчивостью индивидуальных рефлекеов.
Угасание индивидуального рефлекса. Повторяется индивидуальное ^раздражение
одно за другим через 1—2 мин. без кормления. После того как рефлекс угас,
интервал удлиняется вдвое-трое: в этом случае рефлекс наступит, но вновь угаснет после
нескольких повторений через этот удлиненный интервал. Дальнейшее удлинение
интервала может вновь привести к восстановлению рефлекса. Но, наконец, рефлекс так
угасает, что отдых не ведет к восстановлению. В этом случае для восстановления
надо сочетать индивидуальный сигнал с кормлением. Восстановление произойдет уже
после одного сочетания. Опыт угасания нужно производить в начале работы, как>
только индивидуальный рефлекс станет наступать более или менее регулярно, а потом
после значительного упрочения рефлекса повторить его.
Действие необычных посторонних раздражений. Сначала производится какое-
либо необычное раздражение, которое вызывает усиленную ориентировочную'
реакцию. Например, если индивидуальным сигналом был свет или музыкальный звук, то
в качестве необычного раздражения можно взять электрический звонок, который
может находиться тут же в клетке или рядом с ней. Если во время такого необычного
звука произвести индивидуальное раздражение — индивидуального рефлекса не
будет. Но если данное необычное раздражение повторить много раз и дождаться
значительного ослабления ориентировочной реакции, то индивидуальный пищевой сигнал
произведет во время такого необычного раздражения свой обычный рефлекс. -
Отрицательное действие дифференцированного раздражения. После того как в
период дифференциации необычное раздражение пересгіло вызывать пищевой двягэ-'
тельный рефлекс, пробуют сочетать его с индивидуальным раздражением. В этом
случае индивидуальный пищевой рефлекс не наступает.
Отрицательное последействие необычных посторонних и дифференцированных
раздражений. Сначала дается необычное раздражение, затем пріисоедіиняется
индивидуальное; спустя несколько секунд необычное прекращается, а индивидуальное
продолжается 10—30 сек.; индивидуальный рефлекс будет отсутствовать как во время
необычного раздражения, так и после него, т. е. несмотря на действие индивидуаль--
ного раздражения. Только при сильно упроченных рефлексах пищевой рефлекс
может начаться вскоре после прекращения необычного раздражения.
Затем пробуют то же индивидуальное раздражение по прекращении необычного
раздражения, спустя 20—30 сек. и более. Рефлекс непременно наступит. Если этот
интервал времени будет равняться нескольким секундам, тогда только
индивидуальный рефлекс не наступит, а если и наступит, то с опозданием. Чем моложе или
менее прочен рефлекс, тем больше будет этот период отрицательного последействия.
Аналогичные опыты можно проделать на оборонительных реакциях голубей и щенков,
если у них выработать индивидуальные автоматизированные оборонительные акты
поведения по указанному выше методу свободных движений.
Литература
1. Розова А., О взаимоотношениях различных видов внешнего торможения
условных рефлексов. Дисс, 1914.
2. BeritoffJ., Uber d. individuell erworbene Tatigkeit d. ZNS, J. Psych. Neurol.,
33, 113, 1927; Индивидуально приобретенная деятельность центральной нервной
системы, 1932.
3. Э л ь я с с о н М., Исследование слуховой способности собаки в нормальных
условиях и при частичном двустороннем удалении коркового центра слуха. Дисс.
1908.
39*
612 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
4. Беляков В., Материалы к физиологии дифференцировки внешних
раздражений. Дисс, 1911.
5. КрасногорскийН.,0 процессах задержки и локализации кожного и
двигательного анализатора в коре больших полушарий. Дисс, 1911.
6. Павлов И., 20-летний опыт, стр. 145, 1923.
7. Павлов И., 20-летний опыт, стр. 222, 1923.
8. Павлов И., 20-летний опыт, стр. 229, 1923.
9. Горн Эм Материалы к физиологии внутреннего торможения. Дисо, 1909.
10. Коган Б., Об иррадиации и концентрации угасательного торможения. Дисс,
1914.
11. Павлов И., 20-летний опыт, стр. 222—229, 1923.
12. Петрова М., Арх. биол. наук, 25, 3, 1925.
13. Павлов И., 20-летний опыт, стр. 232, 1923.
14. Фурсиков Д., Арх. биол. наук, 23, 195, 1923.
15. Павлов И., 20-летний опыт, 5-е изд., стр. 339, 1932.
16. Анреп Г., Арх. биол. наук, 20, 262, 1917.
17. Розенталь., Арх. биол. наук, 23, 13, 1923.
18. Фурсиков Д., Арх. биол. наук, 23, 2, 1923.
19. Савич А., Дальнейшие материалы к вопросу о влиянии пищевых рефлексов
друг на друга. Дисс, 1913.
20. Ерофеева М., Электрическое раздражение кожи как условный возбудитель.
Дисс., 1913.
21. Дерябин В., К физиологии внутренних торможений. Дисс, 1914.
22. Schloesser М., Arch. Anat. Physiol., стр. 303, 1880.
23. Поте хин С, К физиологии внутренних торможений. Дисс, 1911.
24. Завадский И., Материалы к вопросу о торможении, Дисс, 1908.
25. Г р о с с м а н Ф., К физиологии следовых условных рефлексов. Дисс, 1910.
26. Лепорский Н., Материалы к физиологии условного тормошения. Дисс,
1911.
27. Б и р м а н В., Экспериментальный сон. Дисс, 1925.
28. Павлов И., 20-летний опыт, стр. 144, 1923.
29. Подкопаев Н., Арх. биол. наук, Юбил. сб. Павлова, стр. 297, 1925.
30. Рожанский Н. Ам Физиол. журн. СССР, 19, 289, 1935. Тезисы сообщений
XV Межд. конгр. физиологов, 1935.
Глава XII
ОБЪЕДИНЯЮЩАЯ (ИНТЕГРАТИВНАЯ) ДЕЯТЕЛЬНОСТЬ КОРЫ
БОЛЬШОГО МОЗГА
Общее понятие. В образовании каждого индивидуального
рефлекса проявляется вполне определенно объединяющая деятельность коры
большого мозга. Если раздражения из внешнего мира происходят
одновременно или быстро одно за другим, они объединяются как >бы в
единое раздражение. Это, імежду прочим, проявляется в том, что
впоследствии каждый компонент объединенного раздражения производит
такой эффект, как будто бы все эти компоненты действовали вместе.
Иначе говоря, компонент перестает выступать как самостоятельный
раздражитель; он замещает тот комплекс, в котором был объединен.
Так, мы выше указывали, что если комбинировать два деятельных
раздражения, — например, электрическое раздражение ноги и
раздражение ротовой полости пищевым веществом,— то после нескольких
комбинаций каждое раздражение в отдельности дает такой же эффект,
как если бы применялись оба раздражения вместе, собака будет
давать как оборонительную реакцию, так и реакцию еды. Только чем
сильнее будет одна реакция, тем слабее будет другая.
Такое же объединение получается при комбинации какого-либо
недеятельного раздражения, не производящего характерного внешнего
эффекта, с каким-либо деятельным раздражением, вызывающим
определенный эффект. Это объединение очень демонстративно
проявляется в следующем факте. Многими авторами при работе с
пищевыми двигательными рефлексами было отмечено, что если регулярно
имеете с прекращением индивидуального сигнала насильственно
прекращать еду, закрывая кормушку или удаляя пищу, то начало и
прекращение акта еды определенным образом связывается
индивидуальным сигналом. Именно, если во время еды прекратить сигнал, животное
перестает есть, а с возобновлением сигнала снова начинает есть. Это
наблюдалось на разных животных (Беритов, 1; Брегадзе, 2).
Очевидно, основное раздражение образует с индивидуальным раздражением
одно целое раздражение.
614
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГ\
Объединяющая деятельность коры большого мозга проявляется не
только в образовании индивидуального рефлекса на какой-либо
простой раздражитель, т. е. в связывании простого раздражения с
основным. Она проявляется еще ів большей мере, когда
индивидуальный рефлекс образуется на какой-либо сложный оаздражитель.
В последнем случае мы имеем, с одной стороны, объединение частей
сложного раздражителя с основным раздражением, а с другой, —
объединение частей сложного раздражителя в одно целое и связывание
этого объединенного целого с основным раздражением. До сих пор
мы имели в виду главным образоім объединение какого-либо простого
раздражителя с основным. Что же касается объединения частей
сложного раздражителя внешнего мира в одно целое и затем связывания
его с основным раздражением, то это объединение, или
интегрирование, осталось вне нашего рассмотрения.
До последнего времени объединяющая деятельность коры большого
мозга очень мало изучалась экспериментально, хотя в лаборатории
Павлова эта проблема была поставлена с самого же начала. В
диссертации Зеленого (1907) объединяющей деятельности уделено
большое внимание. Он образовал индивидуальный слюнной рефлекс
на комплекс звуков и затем изучал взаимоотношение между
данными компонентами и комплексом. Зелен ый установил, что
первоначально и компоненты дают рефлекс, подобно комплексу, но только
слабее комплекса. Если компоненты одинаковы по интенсивности, то
они вызывают одинаковые рефлексы, которые почти вдвое слабее, чем
целый комплекс; если же один компонент сильнее другого, то в
отдельности он дает больший эффект, чем слабый. Разница наблюдается как
в скрытом периоде, так и в величине секреции. Далее, Зеленый
установил, что с течением времени компоненты перестают давать
рефлексы, в то время как комплекс, воздействующий сейчас же после
них, вызывает его без ослабления. Этой стадии полной
дифференциации компонентов предшествует такой период, когда испытание
компонентов отражается на величине комплексного рефтекса: последний
ослабевает. Зеленый подробно изучил отношение компонентных
рефлексов друг к другу и установил, в общем, такие же отношения, как
между однородными рефлексами (3). Но эти фактические результаты
не получили дальнейшего развития, не привели к выяснению
физиологических основ объединяющей деятельности коры большого мозга. Все
последующие работы школы Павлова, а также Бехтерева в
этом направлении касаются, как и работа Зеленого,
констатирования объединяющей деятельности, или, как теперь принято называть
ів этих лабораториях, синтетической способности, наряду с
аналитической (Бабкин, 4; Палладии, 5; Фурсиков, 6; Иванов-Смоленский, 7;
Яковлев, 8; Лукина, 9).
Иванов-Смоленский (7) приходит к выводу об
образовании из комплекса разных раздражителей одного «целого, слитного
раздражения» и отсюда заключает о том, что «в коре больших полу-
[
ОБЪЕДИНЯЮЩАЯ ДЕЯТЕЛЬНОСТЬ КОРЫ БОЛЬШОГО МОЗГ\ 615
шарий происходит процесс интегрирования (сложного синтеза)» (1928).
Но и этот автор ограничивается только одним констатированием факта.
Объединяющая деятельность коры большого мозга изучалась Б е-
ритовым совместно с Брегадзе по методу свободных движений
(10). Собаку приучали лежать на лежанке в одном месте, а по сигналу
она шла к кормушке, которая находилась вне поля ее зрения. Кончив
еду, собака возвращалась на лежанку. После автоматизации движений
изучалось все поведение собаки: ее движение от лежанки к кормушке
во время индивидуального сигнала, ее пребывание у кормушки, а
также обратное движение к лежанке,— наконец, ее поведение в
промежутках между опытами. Но для выяснения объединяющей
деятельности принималось во внимание не все поведение животного, а только
первая рефлекторная реакция в ответ на искусственный сигнал.
Характеристика рефлекторных реакций на комплекс раздражений.
При образовании индивидуального пищевого поведения на комплекс
трех или четырех одновременных звуков, вначале, когда комплекс
начинает вызывать пищевое движение, а именно рефлекторное движение
собаки к кормушке, компоненты также производят это движение,
только с большим скрытым периодом и в более слабой форме. Но
после нескольких десятков сочетаний комплекса с едой компоненты
перестают его вызывать.
Далее, оказалось, что в период дифференциации комплексного
рефлекса комплекс вызывает движение к кормушке только при
совершенно одновременном действии всех компонентов. Если сперва
начинается действие одного компонента, а потом другого и третьего, то
движение не вызывается. Кроме того, оказалось, что компонент,
переставший вызывать пищевое движение, не совсем недеятелен,— он
продолжает действовать, но только иным образом. Именно, он препятствует
наступлению пищевого движения на комплекс. Если сначала пустить
компонент и потом, во время его действия или вскоре после него,
подействовать всем комплексом, то движение не наступает. Или,
например, если выработать новое пищевое движение к кормушке на новый
звук, а затем испытать его во время одного из компонентов
означенного комплекса, то пищевого движения не будет.
Итак, в то время как комплекс звуков оказывает на животное
положительное влияние, вызывая рефлекторное движение к кормушке,
дифференцированные компоненты действуют на него отрицательно,
устраняя рефлекторное движение.
После этих опытов была проведена серия опытов с целью
выработать пищевое движение на компоненты, но так, чтобы іпри этом
комплекс его не вызывал. Так, например, мы взяли два музыкальных звука,
на один из них выработали рефлекторное движение к левой кормушке,
а на другой — аналогичное движение к правой кормушке. Затем мы
добились того, что комплекс этих звуков, их одновременная комбина*
ция, не вызывал ни того, ни другого движения. Собака оставалась
лежать на лежанке. Это было достигнуто в результате многократного
616 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
повторения комбинации звуков без сочетания с едой. При такой
комбинации собака первоначально шла к кормушке, но тогда мы
кормушек не открывали. После многих проб собака перестала ходить на
комбинацию раздражений. В то же время на тот. и другой компонент
в отдельности она шла правильно. Более того, было установлено, что
когда комбинация звуков перестает вызывать движение к кормушке,
она действует на это движение отрицательно. Именно, если вскоре
после комплекса пробовать один компонент, то он не вызывает
движения к кормушке.
Таким образом, опытами была установлена полная возможность
того, чтобы комплекс звуков стал индивидуальным, положительным
сигналом к рефлекторному, ^пищевому движению, а компоненты
действовали отрицательно, т. е. становились отрицательным
индивидуальным сигналом, и, наоборот, чтобы компоненты были индивидуальными
положительными сигналами и в то рюе время комплекс их действовал
отрицательно. Этот фактический материал указывает прежде всего на
то, что целое нельзя считать простой суммой компонентов, что
физиологическое действие комплекса, целого, вместе с его внешним
проявлением может совершенно отличаться от физиологического действия
компонентов и их внешних проявлений.
Аналогичные результаты были получены при образовании
индивидуального рефлекторного движения на сложную световую фигуру и на
комплекс звуковых и световых раздражений (Брегадзе, 11). И в этом
случае, при сочетании с едой только комплекса, компоненты
первоначально давали пищевое движение, а потом переставали его давать,—
пищевое движение вызывалось только комплексом.
В дальнейшем 'было образовано пищевое рефлекторное движение
на два и на три последовательных звуковых раздражений. Собаку
призывали к кормушке через несколько секунд после данного ряда
раздражений. Первоначально пищевое движение наступало еще во
время звуков. Дальнейшее развитие рефлекса шло, как при образовании
обычного последовательного рефлекса. В этот начальный период
каждый звук отдельно также вызывал последовательное пищевое
движение, но после нескольких или многократных испытаний звуки в
отдельности переставали его вызывать. Последнее вызывалось только
целым последовательным комплексом (Беритов, 12).
Исходя из этого фактического материала, можно утверждать, что
при образовании индивидуального рефлекторного движения на
комплекс раздражений первоначально это движение вызывалось не
просто данным комплексом, как единым целым раздражением, а всеми
входящими компонентами самостоятельно. Эти компонентные рефлексы
были тем значительнее и прочнее, чем сильнее было физиологическое
действие компонента. Следовательно, каждый корковый очаг,
воспринимающий компонент, связан особыми временными связями с
элементами коры, производящими движение к кормушке. Эти связи
развиваются тем сильнее, чем сильнее физиологическое действие компонента»
ОБЪЕДИНЯЮЩАЯ ДЕЯТЕЛЬНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 617
как это вообще установлено* для образования рефлекса на одиночный
раздражитель. Очевидно, в этот период работы поступательные связи
развиты лучше обратных.
Для изучения характера временных связей производилось угашение
компонентных рефлексов. Оказалось, что после угасания рефлекса на
один компонент комплексный рефлекс, а также другие компонентные
рефлексы остаются; но угасание всех компонентов подряд и, в
особенности, комбинации их непременно ведет к временному угасанию
рефлекса на комплекс. Угасание же комплекса путем повторения беа
сочетания с едой вело к угасанию рефлекса на компоненты. Отсюда
следует, что в начальный период образования положительного
индивидуального рефлекса на сложный раздражитель каждый компонент
производит рефлекс через свои собственные временные связи, а ком-
плекс вызывает его через временные связи всех входящих в него
компонентов.
Далее было установлено, что в последующий период образования
двигательного рефлекса на сложный раздражитель компоненты
перестают давать рефлекс по той же самой причине, по какой происходит
обычная дифференциация рефлекса вообще по отношению к
необычным раздражителям, т. е. благодаря преимущественному развитию
обратных ©ременных связей. Что это именно так, доказывается
следующими фактами. Если сначала пустить хорошо
отдифференцированный компонент, а затем через 3—5 сек. присоединить к нему другой
компонент или комбинацию компонентов, которые еще дают рефлекс,
то в таком случае рефлекс может не наступить. Таким образом,
дифференцированный компонент действует отрицательно на
недифференцированные компоненты. Дифференцированный компонент устраняет
также другой однородный пищевой двигательный рефлекс. Он
устраняет даже рефлекс на комплекс, а именно — рефлекса не будет, если
сначала пустить сильно дифференцированный компонент, а через
3—5 сек. присоединить к нему все остальные компоненты одновременно.
Очевидно, под влиянием дифференцированного звука обратные
временные связи приходят в состояние повышенной возбудимости. Это
обусловливает усиленное распространение возбуждения но нервному
кругу из корковых элементов поступательных связей через общие для
них двигательные участки коры в обратные связи и в то же время
усиленное активирование соподчиненного нейропиля. Вследствие этого
возбуждение не может распространяться через эфферентные пути в
такой мере, чтобы вызвать внешний эффект.
Отсюда вытекает, что в период дифференциации компонентов
комплексного рефлекса компоненты перестают давать двигательный
рефлекс благодаря преимущественному развитию обратных связей от дви
гательных участков к корковым очагам, воспринимающим компоненты,
к двигательным участкам, и что поэтому полощительный двигательный
рефлекс на действие комплекса не может быть обусловлен временными
связями компонентов.
Последнее заключение следует еще с большей убедительностью из
?18 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
следующего факта: когда компоненты сочетаются с едой, а комплекс
с ней не сочетается, то, в конце концов, устанавливается такое
соотношение, что компоненты вызывают рефлекс, а комплекс его не
вызывает. Судя по характеру работы, в отношении компонентов безусловно
были образованы положительные рефлексы с преимущественным
развитием поступательных связей между соответствующими
воспринимающими очагами и двигательными участками коры, ведущими к
кормушке. На комплекс же был образован несомненно отрицательный
рефлекс. Это было! видно из отрицательного действия на
компонентный рефлекс. Каждый раз после комплекса в течение нескольких
секунд компоненты не давали рефлекса или производили его с большим
опозданием. Кроме того, комплекс производил определенный
двигательный эффект, характерный для отрицательного сигнала, именно:
собака вначале давала быстрое ориентировочное движение, иногда
даже вскакивала, но сейчас же после этого ложилась и низко
опускала голову.
Отрицательный рефлекс, как известно, обусловливается
преимущественным развитием обратных связей. В отрицательном рефлексе на
комплекс обратные временные связи должны были образоваться от
ответных двигательных участков к корковым нервным элементам,
возбужденным комплексом. Но эти последние не могут быть теми
корковыми элементами, которые [воспринимают компоненты непосредственно.
Они связаны с теми же двигательными участками посредством хорошо
развитых поступательных связей. Очевидно, временные связи
комплексного рефлекса начинаются в особам комплексе нервных элементов,
возбуждающихся только совместным действием всех компонентов. С
этими элементами образуются и те поступательные связи, которые
обусловливают комплексный положительный рефлекс при полной
неактивности компонентов, и обратные связи, обусловливающие комплексный
отрицательный рефлекс.
Когда комплекс состоит из последовательных раздражений,
объединяющая деятельность коры осуществляется, с одной стороны, путем
связывания каждого предыдущего компонента с последующим и затем
путем такого развития временных связей между соответствующими
компонентными очагами и ответными двигательными участками, что
только при обычном следовании компонентов получается характерная
для комплекса реакция; с другой стороны, эта объединяющая
деятельность осуществляется путем возникновения особых очагов,
возбуждающихся исключительно действием всего последовательного комплекса и
связывания его с ответными двигательными участками посредством
особых временных связей (Беритов, 12).
Физиологическое основание объединяющей деятельности. Выше
мы видели, что объединяющая деятельность осуществляется не просто
установлением временных связей между теми корковыми элементами,
которые возбуждаются компонентами непосредственно. Наравне с
таким объединением выступает и другого рода объединение, которое как
ОБЪЕДИНЯЮЩАЯ ДЕЯТЕЛЬНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 619
раз имеет существенное значение в создании единого целого из
многочисленных частей одного и того же или разного качества. Это
объединение заключается в возбуждении особых элементов в коре большого
мозга исключительно под совместным действием всех компонентов и
затем в образовании временных связей между ними, с одной стороны,
и компонентными очагами возбуждения и ответными двигательными
участками — с другой.
Как известно, обычно слабое недеятельное раздражение кожи или
нерва в отдельности не дает рефлекса, но при быстром повторении
раздражений одного за другим оно становится деятельным и может
вызвать сильный рефлекторный эффект. Так при нормальной
возбудимости центральной нервной системы большинство рефлексов не
вызывается отдельными индукционными ударами, но возникает при
повторном приложении таких ударов к чувствительному нерву или к коже,
причем этот эффект ни в коем случае не может быть признан
произведением эффекта от разового раздражения, помноженного на число,
указывающее, сколько раз раздражение было повторено. Или,
например, известно', что слабое раздражение одного участка кожи или
одного нерва может не дать рефлекторного эффекта; но если такое слабое
раздражение приложить сразу к двум нервам, или двум участкам
кожи, или вообще к двум рецепторам, может получиться сильный
рефлекс. Такое действие обнаруживается при применении двух
раздражений не только в одном рецептивном поле (Шеррингтон, 13), но и в
разных, как, например, кожа и кортиев орган уха (Демидов, 14).
Ясно, что и в этом случае эффект от двух раздражений не может быть
признан суммой тех действий, какие вызывают эти раздражения в
отдельности.
Но в обоих примерах происхождение эффекта от недеятельных ©
отдельности раздражений обусловливается общим свойством живой
возбудимой ткани: субминимальное раздражение, хотя и не вызывает
возбуждения, но производит повышение возбудимости.
В отношении приведенных случаев дело обстоит так: слабое
периферическое раздражение возбуждает рецептор или чувствительный
нерв. Отсюда возбуждение передается в центральную нервную систему
и действует на более или менее обширный отдел через посредство
коллатералей чувствительных нервных волокон. Но оно не производит
здесь возбуждения — потому ли, что возбудимость центральной
нервной системы была слишком низка, или оттого, что ©'значенное
периферическое влияние не было достаточно сильно. Но как бы то ни было,
действие слабого периферического раздражения не проходит бесследно.
Оно повышает возбудимость центральной нервной системы, и
вследствие этого следующее или одновременно возникшее такое же слабое
раздражение может оказаться достаточным для вызова активного
состояния с наступлением периферического эффекта. Но и в случае
применения такого раздражения, которое с самого начала дает рефлекс,
действие его не ограничивается приведением в активное состояние
620 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
определенных центральных нервных элементов, на которые оно
действует более или менее непосредственно. Наряду с этим то же
раздражение оказывает субминимальное действие на более или менее
обширную область нервных элементов вследствие иррадиации возбуждения,
которое производит здесь в той или иной степени значительное
повышение возбудимости.
Эти хорошо известные явления рефлекторной деятельности
центральной нервной системы дают ключ к пониманию возникновения
F F" з&лгз jft мр
Рис. 254. Схематическая картина временных связей
в отношении положительного рефлекса на комплекс и&
четырех звуков.
Комплекс состоял из тонов F' и F" тонвариатора Горнбостеля, звонка
№ 1 и звонка № 2. Компоненты в отдельности не давали рефлекса,
будучи совершенно отдифференцированы, но комплекс вызывал движение.
особых дополнительных очагов возбуждения, наряду с теми, которые
возбуждаются компонентами непосредственно. Каждый компонент дает
в коре большого мозга возбуждение определенных нервных элементов,
но вместе с тем действует субминимально на целый ряд других
элементов, повышая в них возбудимость. При одновременном или
последовательном действии двух или более компонентов эти последние
элементы также приходят в активное состояние. Вследствие этого
будут вступать во временную связь не только первично (возбужденные
элементы, но и дополнительно возбужденные. Если при этом комплекс
сочетается с определенным основным раздражением, а компоненты
с ними не сочетаются или сочетаются с другим основным раздражением,
то дополнительные элементы должны стать начальными пунктами
самостоятельных временных связей, через которые комплекс будет
производить свой собственный эффект, совершенно отличный от
компонентных реакций.
ОБЪЕДИНЯЮЩАЯ ДЕЯТЕЛЬНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 621
Итак, комплексный раздражитель производит такое
физиологическое действие, которое всегда обширнее суммы физиологического
действия компонентов, вследствие 0озбуждения под влиянием
комплексного раздрао/сения таких нервных элементов, на которые компоненты в
отдельности действуют рубминимально. Это дополнительное
возбуждение новых .нервных элементов придает корковой деятельности объеди-
rf F"o8.NU8.N2
Рис. 255, Схематическая картина временных связей, когда
комплекс из тех же звуков, как на рис. 254, действовал
отрицательно, не вызывая своего положительного рефлекса;
компоненты же в отдельности производили положительный
рефлекс.
пенный, целостный характер угри всяком комплексном раздражении. Но
когда комплексный раздражитель становится совершенно
самостоятельным индивидуальным сигналом, производящим отличную от
компонентов реакцию, тогда имеет место образование особых временных
связей на означенные, рополнительно возбужденные элементы.
Далее, опыты указывают, что физиологическое действие компонента
не меняется по существу при его вхождении в комплекс. Оно только
расширяется в том смысле, что данный компонент в комплексе
вследствие одновременного действия ряда других компонентов 'активирует
такие нервные элементы, на которые он в отдельности действует
субминимально.
Согласно этому положению, при применении комплекса каждый
компонент производит свое специфическое физиологическое действие, а
следовательно, должен возбудить прежде всего свои собственные
временные связи. И если, несмотря на это, наступает один только
комплексный рефлекс, то это должно быть обусловлено действием
временных комплексных связей.
622 ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
На рисунках 254, 255, 256 представлена схематически
предполагаемая картина основных, наиболее деятельных временных связей для
каждого представленного здесь комплексного рефлекса.
На всех приведенных схемах линии ведут от
компонентов-раздражителей в кору к соответствующим воспринимающим элементам
(а, Ь, с, d). Отсюда штриховыми линиями обозначаются нервные связи
с дополнительным комплексным элементом (к). Как воспринимающие
Рис. 256 Схематическая картина временных связей для
отрицательного рефлекса на комплекс звуков A -j- В, причем
компонент Л в отдельности давал рефлекс к правой
кормушке, а компонент В— к левой.
очаги компонентов, так и этот дополнительный участок соединены с
двигательными корковыми участками (М), производящими движение
направо или налево, поступательными и обратными временными
связями. Стрелки указывают направление, а толщина линий — степень их
развития. Даны также нисходящие пути от двигательных участков.
В случае отрицания рефлекса эти пути изображены штрихами.
Переход возбуждения от компонентных поступательных временных связей
в обратные связи к дополнительному очагу также изображен
стрелками.
О локализации объединяющей деятельности. Необходимо
отметить, что все эти временные связи с объединяющим центром из
дополнительно возбужденных нервных элементов должны являться точно
локализированными в коре большого імозга. Но эту локализацию ни в
коем случае нельзя считать точно детерминированной наследственной
организацией коры большого мозга. Участие дополнительных нервных
элементов обусловливается иррадиацией возбуждения из фокальных
воспринимающих участков, а эта иррадиация в коре большого мозга в
каждый данный момент определяется не наследственной организацией
ее, а функциональным состоянием существующих нервных элементов,
ОБЪЕДИНЯЮЩАЯ ДЕЯТЕЛЬНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 623-
которое зависит от многочисленных условий. Это положение, в
частности, было выставлено американским ученым Леш л и (16) на
основании опыта. Он, например, изучал влияние частичного удаления
затылочных долей на тренировку крыс в лабиринте, которая состояла в
создании определенной привычки к различной освещенности. Оказалось, что
способность к образованию этой привычки ничуть не страдает от такой
операции, если только она не превосходит одной трети івсей
затылочной доли. При этом не имеет значения, где именно находился
удаленный участок. Оперированная крыса тренируется так же быстро, как и
нормальная. Наоборот, операция сильно влияет на существующую
тренировку. Она исчезает после операции и восстанавливается путем новой
тренировки, тем позднее, чем больше был удаленный участок мозга.
На основании этих и других подобных опытов можно утверждать, что
образованная привычка всегда точно локализирована в коре больших
полушарий, хотя ее начальное образование не связано с какой-либо
определенной частью мозга,
О нервных элементах объединяющей деятельности. Из работ
школы Павлова хорошо известно, что вся кора мозга собаки
обладает воспринимающей функцией. Так, например, зрительная сливается
со слуховой и даже с кожно-мышечной. По всей вероятности, у собак
и подобных им животных как воспринимающая, так и объединяющая
деятельность коры по отношению к раздражениям одного качества
выполняется соответствующей воспринимающей областью. Возможно,
однако, что центральные, так называемые фокальные, участки
воспринимающей области менее заняты объединяющей деятельностью, что
для этого служит как раз периферическая часть воспринимающей
области. Что же касается объединяющей деятельности по отношению к
разного рода раздражениям,— например, звуку и свету,— то с большой
вероятностью можно утверждать, что эта деятельность осуществляется
периферическими пограничными участками воспринимающих областей.
Не м-ржет быть сомнения, что при двух разнородных раздражениях,
вследствие иррадиации возбуждения, эти пограничные участки скорее
всего и сильнее всего должны подвергаться совместному возбуждению.
Очевидно, и дополнительные очаги возбуждения должны возникать при
этих условиях главным образом в пограничных, периферических частях
воспринимающих областей.
Считается общепризнанным, что у высших животных, как обезьяна,
а также у человека, воспринимающая функция в основном занимает
небольшую территорию коры мозга. Кроме того, у обезьян
воспринимающие территории не только отграничены одна от другой, но одна треть
коры даже почти свободна от таких территорий, т. е. не играет
существенной роли в восприятии периферических раздражений. Полагают,
что у человека две трети всей коры более или менее свободны от
воспринимающей функции. Сначала Флексиг (18), а затем ряд
физиологов и неврологов признали эти участки коры на основании их
гистологического строения и некоторых физиологических данных за
специальные центры, которые служат для связывания, объединения воспри-
€24
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
нимающих областей. Поэтому Ф л е к с и г называет эти центры а с с о-
ци атиівн ыім и. Неврологи считают их анатомическим субстратом
интеллекта Так, выдающийся невропатолог Гольдштейн (17) находит
возможным локализировать чисто качественные и чисто составные
части единого психонервного процесса в соответствующих сенсорных
(воспринимающих) и двигательных областях коры мозга. Что же
касается процесса, который придает сенсорному и двигательному явлению
впервые характер единого психического и который находит свое
выражение в образовании особого переживания предмета, понятия, імысли,
специфического переживания речи, автор признает необходимым для
такого процесса участие в нем всей коры, и прежде всего той области,
которая лежит между двигательными и сенсорными областями. Следует
упомянуть еще современного невролога Эббека (19), который также
указывает, что у человека две трети коры, представляющие более
новое филогенетическое образование, эмбрионально созревающее позднее,
являются высшими ассоциативными центрами. Этот автор признает
существование ассоциативных центров также в каждой воспринимающей
области. Наш ленинградский невролог Блуменау (19) приписывает
сочетательную функцию тем отделам коры, которые лежат вне
воспринимающих областей, т. е. он считает эти отделы тем центральным
механизмом, при помощи которого устанавливаются временные связи. Но,
как мы уже указали выше, временные связи образуются из всех
корковых элементов, в частности из воспринимающих и двигательных
элементов. С этой стороны не должно быть существенной разницы между
последними элементами и теми, из которых состоят так называемые
ассоциативные элементы. Согласно нашим исследованиям, целостное
действие комплексного раздражителя,— иначе говоря, определенной
внешней ситуации,— обусловливается деятельностью тех нервных
элементов, которые возбуждаются одновременным действием всех
компонентов данной ситуации. По нашему мнению, эти дополнительно
возбуждаемые элементы дсуіжны принадлежать главным образом к тем
корковым участкам, которые лежат вне воспринимающих территорий
и признаются неврологами за седалище интеллекта. Так как эти
участки объединяют между собой разнообразные раздражители внешней
среды в одно целое, в одну ситуацию, то справедливо можно было бы
назвать их объединяющими областями.
Фронтальная область коры мозга впереди двигательной зоны, пови-
димому, осуществляет объединяющую работу в отношении
соматических и вегетативных функций. Эти функции не имеют за собой
локализированных проекций. Та и другая функция одинаково захватывают
всю премоторную зону (6 я по Фохту, см. рис. 174). Это заключение
было сделано Фултоном (20) на основании известных данных
относительно раздражения и экстирпации премоторной зоны у обезьян.
Отсюда, например, происходит корковая интеграция жевания и слюнной
секреции, угнетение кишечной моторики при беге и т. д. Характерно,
что в отсутствии моторной зоны после перерождения пирамидных пу-
ОБЪЕДИНЯЮЩАЯ ДЕЯТЕЛЬНОСТЬ КОРЫ БОЛЬШОГО МОЗГА f?5
тей, премоторная зона способна производить корковые рефлексы.
Удаление же премоторной зоны вызывает тяжелые расстройства как в
области соматической, так и в области вегетативной системы,— в
частности, исчезают привычные тонкие движения пальцев. Последнее
обстоятельство ясно свидетельствует об объединяющей функции
премоторной зоны ів отношении индивидуальных рефлексов.
Возбуждение объединяющих областей, по нашему мнению, не
должно сопровождаться субъективным переживанием. Это должно быть
исключительным свойством воспринимающих элементов. Но, разумеется,
посредством объединяющих элементов могут притти в активное
состояние воспринимающие элементы — как путем временных связей, так и
помимо них, путем общей иррадиации возбуждения. В этих случаях
мы будем иметь дело с возникновением представлений с их обычным
субъективным содержанием.
Итак, у низших млекопитающих, как собака, у которых
воспринимающие области, видимо, занимают всю кору, объединяющая
деятельность, по всей вероятности, осуществляется главным образом
периферическими пограничными участками этих областей. У высших же
млекопитающих,— как у обезьян,— а затем у человека, воспринимающие
области занимают не всю кору, а лишь определенные, вполне
отграниченные территории, и у них объединяющая деятельность, по всей
вероятности, является основной функцией той части коры, которая лежит
между воспринимающими территориями.
Практикум
Образование индивидуальной пищевой реакции на комплекс звуков. Эти опыты
производятся тем же методом, как и вообще опыты с образованием индивидуальных
реакций. Для этих опытов годятся любые животные: голуби, кролики, щенки,
взрослые собаки.
Легче всего работать на щенках и взрослых собаках: у них индивидуальные
реакции на сложный раздражитель образуются и дифференцируются быстрее всего
В качестве комплексного раздражителя лучше всего брать музыкальные звуки
фисгармонии или тонвариатора Штерна, но если их нет, можно пользоваться
электрическими звонками. Подбираются два или три таких звонка, которые ясно отличаются
один от другого по тембру и высоте звука.
1. Образование положительной пищевой двигательной реакции на комплекс трех
звуков. Хорошо, если для опыта берутся звуки разной интенсивности. Для этих опытов
нужно брать таких животных, которые уже служили для других опытов с
индивидуальными рефлексами, т. е. хорошо приученных к опытной обстановке. Дается
комплекс звуков, и затем через 2—5 сек. животное привлекается к кормушке при
помощи шума, издаваемого открыванием кормушки. Животное ест пищу 10—15 сек., затем
кормушка закрывается. В тот же момент прекращается и комплекс звуков.
На второй день после образования индивидуальной реакции на комплекс в виде
движения животного к кормушке испытывается по-очереди каждый компонент в
отдельности по одному или по два раза. Животное пойдет к кормушке, но кормушку
открывать не надо. Каждый раз после пробы компонентов производится обычное
сочетание комплекса с едой. Так же поступают впоследствии каждый день, пока компонень
ты не перестанут вызывать пищевую реакцию. Это достигается прежде всего в
отношении слабого компонента. Потом становятся недеятельными и другие.
40 И. С. Бзритов
626-
ОБЩАЯ ФИЗИОЛОГИЯ КОРЫ БОЛЬШОГО МОЗГА
После того как компоненты перестанут действовать, испытывают одновременное
действие двух компонентов; комбинация двух сначала дает пищевую реакцию, а
затем іакже перестает давать ее. Ее дает только комплекс.
После того как произошла дифференциация компонентов и их неполных
комбинаций, допытывается их отрицательное действие на комплекс. Так, например, если
сначала дать один компонент, а затем присоединить все остальные, индивидуальной
реакции не будет. Она не наступит также, если сначала повторить компонент
несколько раз один за другим, а затем, спустя несколько секунд, испытать комплекс.
2. Образование отрицательной индивидуальной реакции на комплекс
одновременных звуков и положительных реакций на компоненты. Берется тот же комплекс
звуков: компоненты его систематически сочетаются с едой, а комплекс не сочетается.
Для этих опытов можно взять то же животное, на котором были произведены лре-
дыдущие опыты. Первоначально комплекс вызывает движение животного к кормушке,
как и компоненты, но через несколько дней работы он перестает давать это
движение. Легко обнаружить, что после этого комплекс оказывает отрицательное дейстзие
па компонентные реакции. Если начать раздражение комплексом, а затем комплекс
прекратить, оставив один из компонентов, то раздражение можно продолжать 10 сек
я оно не произведет пищевой реакции.
Образование последовательной индивидуальной реакции на три следующих одни
за другим звука. Берут три разных звонкіа и заставляют их звонить один за другим
по 5 сек. без промежутков. Пища дается после третьего звонка, спустя 2—3 сек.
В первый день пищевая реакция будет совпадать: она будет наступать во время
второго звонка, еще позднее — во время третьего. Лишь на второй или на третий
день — или еще позднее — реакция станет последовательной, т. е. будет наступать
после звонков.
В эти первые дни следует испытывать и компоненты в отдельности. Они будут
давать сначала совпадающие, а потом последовательные двигательные реакции, но
после нескольких проб они перестанут вызывать эту реакцию. Комплекс же дает ее,
но только яри применении в обычном порядке. Если изменить порядок путем
перестановки звонков или обратного следования их, то обычного движения к кормушке
не будет, или же оно произойдет только при первых пробах.
Литература
і. Beritoff J., Pfl. Arch., 213, 370, 1926.
2. Б p e г а д з e А., Журн. экспер. биол. и мед. № 33, 385, 1929.
3. Зеленый Г., Материалы к вопросу о реакции собак на звуковыераздраже-
ния. Дисс, 1907.
4. Б а б к и н В., Тр. Об-ва "усек, врачей, 77, 197, 1910; 78, 219, 1911; 80, 203, 1912,
5. П а л л а д и н А., Тр. Об-ва русск. врачей, 73, 393, 1906.
6. Фурсиков Д., Доклад на II Всеросс. психоневрол. съезде 8 янв. 1922 г. в
Ленинграде.
7. Иванов-Смоленский А., Мед. биол. журн., 2, в. 3, стр. 84, 1925; Тр.
Физиол. лаб. акад. Павлова, 2, в. 1, стр. 47, 1927.
8. Я к о в л е в В., Тр. физиол. лаб. акад. Павлова, 2, в. 1, стр. 58.
9. Лукина А., Новое в рефлексологии и физиологии нервной системы, 1, 205,
1925.
10. БеритовИ. иБрегадзе, А., Мед биол. журн., 5, в. 3, стр. 131, 1929; б,
в,. 4 1929; 6, в. 4, 1929; 6, в. 1, стр. 104, 1930.
11. Брегадзе А., Мед. биол. журн., 6, в. 6, стр. 483, 1930.
12. Беритов И., Индивидуально приобретенная деятельность центральной
нервной системы, 1932.
43. Sherrington G. S, The integrative action of the nervous system, New York,
1906
ОБЪЕДИНЯЮЩАЯ ДЕЯТЕЛЬНОСТЬ КОРЫ БОЛЬШОГО МОЗГА 627
14. Демидов В., Условные следовые рефлексы у собаки без передних половин
обоих полушарий. Дисс, 1909.
*5. Введенский Н., Fob neurobiol., 6, 591, 1912.
16. L а ч h 1 е у К., Brain mechanism and intelligence. Chicago, 1929. Русск. пер.—
Мозг и интеллект, 1933.
17. Goldstein К., Handb. Norm. Path. Physiol., 10, 600, 4927.
18. F 1 e с h si g., Gehirnu. Seele, 1896; Lokalisation d. geistigen Vorgange. Leipzig,
1896.
19. Блуменау А., Материалы 1-го Всесоюзн. съезда по изучению поведения,
человека, стр. 361, 1930.
20. Фултон И. Ф., Междунар. конгресс физиологов, 1935. Тез. сообщ., стр. 42.
*
ИМЕННО^ УКАЗАТЕЛЬ
А
Абуладзе К. 514, 588
Айрапегьянц Э. 525
Ананьев Б. 488, 492
Андреев Л. 487
Анохин П. 37, 62, 352, 359, 360, 361, 574
Анреп Г. 565, 609
Архангельский В. 521
Аршавский И. 243
Асрагян Э. 360, 361, 362
Ахметели М. 514
Б
Бабкин Б. 515, 582, 588, 589, 594, 596,
597, 614
Бабский Е. 169, 186
Бакурадзе А. 23, 69, 75, 88, 92, 97, 99,
100, 101, 112, ИЗ, 114, 116, 117, 122,
132, 135, 136, 187, 188, 196, 197, 198,
199, 224, 236, 237, 270, 273, 288, 290,
459. 469, 471, 479, 435, 486, 487, 488,
490, 492
Бари А. 429
Башмуран А. 583
Баяндуров Б. 418, 505, 510, 514, 586
Бебуришвили Н. 140, 212, 214, 586
Безбокая М. 565
Белиц М. 522, 565
Беляков В. 514, 602
Бернштейн Н. 334
Бехтерев В. 16, 247, 404, 405, 410, 413,
415, 416, 427, 437, 499, 500, 502, 503,
506, 507, 524, 547, 558, 582, 607, 614
Бирман В. 565, 568, 569, 608
Блуменау А. 624
Болдырев В. 504
Болотов В. 138, 148
Брегадзе А. 235, 241, 421, 464, 465, 473,
477, 487, 489, 490, 514, 613, 616
Бурмакин В. 511
Быков К- 184, 185, 518, 526, 574
Былина А. 582
В
Вагнер В. 248
Васильев Л. 358
Васильев П. 519
Введенский Н. 58, 92, 99, 108, 109, 149,
158, 162, 209, 220, 224, 266, 364, 365,
478
Ветохин И. 427
Веткжов А. 304
Виноградов М. 364
Вишневецкая 362
Воробьев А. 234, 469, 470, 478, 479, 481,
488, 492
Воскресенский Л. 553, 569
Вульсон С. 504
Г
Гедеванишвили (Гедевани) *Д. 75, 99, 101,
102, 103, ПО, 111, 129, 149, 188, 193,
234, 302, 421, 427, 446, 447, 448, 463,
465, 466, 467, 471, 472, 479, 482, 483,
487, 489, 490
Гогава М. 99, 101, 136, 224, 372, 374,
375, 376, 379
Голубева Е. 511
Горн Э. 597, 609
Горшкова С. 185
Гоциридзе А. 138, 148
Гроссман Ф. 520, 521, 522, 561, 608
д
Данилевский В. 55, 99, 493
Данилов А. 185
Демидов В. 212, 453, 454, 619
Дерябин В. 527, 583
ИМЕННОЙ УКАЗАТЕЛЬ
6?9
Дзидзишвили Н. 41, 45, 47, 48, 93, 99,
НО, 199, 212, 214, 458, 469, 471, 479,
487, 488, 490, 492, 509, 519, 558
Добровольский В. 521, 522, 560, 565, 588
Дубинский 218, 219
Е
Ерофеева М. 565, 575, 608
3
Завадский И. 514, 519, 520
Залманзон А. 553
Зальгейм А. 504
Зеленый Г. 35, 394, 395, 506, 512, 515,
522, 526, 584, 588, 589, 614
Зурабашвили А. 36, 130, 246
И
Иванов-Смоленский А. 515, 614
Израэльсон Ж. 505, 506, 524
К
Каменский А. 219
Каминский С. 510
Кан И. Л. 371
Карамян А. 362
Кассиль Г. 219
Катцва А. 218, 219
Кашеринова Н. 506, 507, 594, 597
Киселев М. 102, 111, 150, 170, 188, 268,
270, 273, 274
Клебанова Е. 429
Коган Б. 565, 608, 609
Конради Г. 365
Кочнева Н. 218, 219
Коштоянц X. 349
Красногорский Н. 414, 524, 602, 608, 609
Крепе Е. 506
Крестовников А. 319, 506
Кржышковский К. 523
Крыжановский И. 508
Крючкова А. 243
Кряжев Я. 456, 579
Кудрин А. 415
Кунстман 70, 306
Купалов П 511, 512, 583
Кураев С. 453
Курцин И. 185
Л
Лаврентьев Б. 21, 22, 45, 246
Лапинский М. 70
Ларионов В. 412, 416
Лейбсон Л. 429
Ленд К. 501
Леонов В. 521, 560, 565
Леонтович А. 177
Лепорский Н. 523, 608
Леутский К. 585
Ливанов М. 478
Лондон Е. 218, 219
Лукина А. 507, 509, 614
М
Миславский Н. 427, 582
Михайлов С. 429
Миштовт Г. 522, 523
Могендович М. 131
Могилянская 3. 334
Мюльберг В. 588
Мясищев В. 507, 509, 518
Н
Нарикашвили С. 75, 92, 99, ПО, 136, 140,
145, 197, 198, 275, 290, 485, 486, 492
Немилов А. 13, 14
Нивинская О. 112, 186
Никифоровский Н. 514, 526
Николаев П. 523
О
Орбели Л. 70, 148, 306, 438, 453, 583
Осипова В. 509
II
Павлов И. 33, 212, 409, 412, 415, 417,
418, 446, 454, 499, 500, 501, 502, 503,
504, 506, 512, 522, 523, 524, 547, 556,
558, 565, 568, 569, 573, 574, 575, 582,
583, 588, 590, 597, 607, 608, 609, 610,
614, 623
Павлова В. 506
Палладии А. 515, 614
Перельцвейг И. 588, 596, 597
Петрова М. 518, 522, 549, 565, 569, 583
Пименов П. 506, 520, 575
Подкопаев Н. 528, 556, 594, 609
Полтырев С. 395, 588
Попов Т. 247, 306, 586
Потехин С. 608
Правдич-Неминский В. 444, 487, 493
Протопопов В. 505, 508
630
р
Разенков И. 184, 185
Рожанский Н. 139, 144, 256, 480, 565,
566, 568, 573, 609, 610
Розенталь И. 557, 565, 601
Розова А. 599
Ройтбак А. 121, 122, 123, 124, 126, Г32,
135, 155, 157, 169, 226, 231, 237, 242
Рохлина М. 514
Рубель В. 219
Руковяткина 584
С
Савич А. 548, 608
Самойлов А. 111, 150, 170, 268, 270, 274
Саркисов С. А. 456, 482
Сеченов И. 33, 93, 99, 135, 138, 148, 180,
201, 224, 279, 291, 348, 349, 451
Слепян Л. 478, 479
Снегирев И. 557
Соломонов О. 557, 565
Сперанский А. 456
Степанов Г. 503
Стукова М. 526
Т
Тарханов И. 200, 437
Тихомиров Н. 582
Тологинов И. Ф. 499, 503
Тонких А. 148, 149
Топоров Н. 419
Топурия Ш. 510, 514
У
Уфлянд Ф. 504
Ухтомский А. 90, 92, 109, 358, 426, 429,
431
Ушинский Н. 135, 139, 169, 363, 364, 432
Ф
Феокритова Ю. 526
Фольборт Г. 523
Фрид А. И. 219
Фролов Ю. 510, 521, 522, 524, 586
Фурсиков Д. 515, 553, 563, 564, 565, 575,
609, 614
ц
Цейтлин С. 185
Цкиманаури Г. 258
Цкипуридзе Л. 45, 234, 235, 236, 237,
УКАЗАТЕЛЬ
238, 239, 241, 421, 464, 465. 473, 477.
484, 485, 487, 489, 571
Ч
Чаговец В. 131
Чеботарева О. 506, 523
Чегулин С. 565
Чичинадзе Н. 36, 245, 514, 571, 586
Ш
Шевалев Е. 512
Шишло А. 565
Шпильберг П. 472, 482, 487
Шпирман А. 506, 507, 509
Штерн Л. 184, 185, 257
Щ
Щелованов 247
Э
Эльяссон М. 453, 512, 602
Ю
Ющенко А. 507
Я
Яковлев В. 614
А
Ades Н. W. (Адес) 475
Adrian Е. D. (Эдриан) 45, 46, 47, 48, 74,
77, 108, 130. 142, 146, 156, 164, 170,
210, 232, 236, 240, 241, 243, 261, 305,
380, 387, 413, 442, 443, 444, 445, 450,
471, 474, 475, 479, 484, 493, 540
Agduhr Е. (Агдур) 643
Aktinson A. W. (Актинзон) 242
Allen W. F. (Аллен) 19, 413
Arton (Артон) 539
Asajama Ch, (Асаяма) 268
В
Babak Е. (Бабак) 55
Bagley С. (Багли) 428
Baglioni S. (Бальони) 44, 76, 218. 219,
220, 356, 382
Вагапу R. (Барани) 317
Barcroft J. (Баркрофт) 119, 242
Bard P. (Бард) 414
Barron D. H. (Баррон) 22, 119, 122, 123,
126, 134, 135, 136, 169, 242, 545
Barthley S. (Бартлей) 479
ИМЕННОЙ УКАЗАТЕЛЬ
631
Basler А. (Баслер) 338
Baxter R. Е. (Бакстер) 37
Bazett (Бацет) 305, 308
Beck А. (Бек) 119
Beeher Н. К. (Бигер) 484
Beevor С. Е. (Бифор) 427
Bell Ch. (Белл) 21, 150
Berger H. (Бергер) 425, 445, 464, 472,
478, 482, 493
Bernstein S. (Бернштейн) 193, 203
Bethe А. (Бете) 64, 209, 359, 363, 431,
432, 433, 435, 436, 437
Bickel А. (Бикель) 348, 428
Biedermann W. (Бидерман) 302, 384
Birge Е. (Бирге) ПО
Bishop G. Н. (Бишоп) 146, 451, 479, 484
Bolaffio М. (Болаффио) 539
Blair Е. А. (Блэир) 255
Blake Н. (Блэк) 492
Blake-Prichard (БлэкьПричард) 337
Blood R. (Блуд) 306
Boldrey (Болдрей) 416, 417, 425
Bond D. (Бонд) 491
Bonnet V. (Боннэ) 75, 122, 131, 137, 165
Borchard (Борхард) 418
Bornstein W. (Бернштейн) 413
Boss Е. (Босс) 218
Bowditch (Боудич) 268
Bradway W. Е. (Бродуэй) 236, 248
Branting F. G. (Брантинг) 306
Braun (Браун) 334
Bremer F. (Бремер) 75, 122, 127, 131, 137,
138, 165, 203, 206, 412, 428, 457, 473,
482, 484, 493, 545
Brenner С. D. (Бреннер) 424
Brodmann R. (Бродмал) jju, 399, 495
Brondgeest (Бронджест) 302
Bronk D. W. (Бронк) 48, 164, 444
Brown R. С. (Броун) 242
Bubnoff (Бубнов) 149, 211, 439, 454, 456
Bucy P. С. (Буци) 479
Buchanan F. С. (Бьюкенен) 152, 176
Buddenbrock W. (Будденброк) 34, 35 79
370, 371, 372, 382
Buehl С. С. (Бюел) 239
Bujtendijk F. J. (Буйтендик) 142, 243
Bull H. О. (Бул) 586
Bun-Ichi Hasama (Бун-Иши Хасама) 413
493
С
Cajal R. (Кахал) 13, 14, 21, 24, 43, 70,
115, 246, 266, 399, 543
Campbell В. (Кемпбел) 75, 175
Cannon W. В. (Кэнон) 185, 186
Case J. (Кэиз) 479
Cattell Mck (Каттел) 46
Chang H. С. (Чанг) 185
Clark S. L. (Кларк) 434, 484
Coghill С. Е. (Когхил) 36, 118, 361, 545
Cohen L. H. (Коген) 479
Cooper S. (Купер) 77, 164, 442
Culler E. (Куллер) 281
Cushing H. (Кушинг) 417
D
Davis F. (Дэвис) 485
Davis H. (Дэвис) 485
Davis L. (Дэвис) 306
Dempsey E. W. (Демпеей) 473, 482
Denny-Brown D. E. (Денни-Броун) 77,271.
272, 442.
Descartes R. (Декарт) 27, 150
DMtler R. (Диттлер) 162, 163, 264
Dow R. (Доу) 234, 237, 241, 307» 411, 482,
493
Dresel К- (Дрезель) 341
Dubner H. H. (Дубнер) 239
Duchenne G. S. (Дюшен) 85, 86, 108
Dusser de Barenne S. G- (Дюссер де Бз-
ренн) 56, 88, 231, 306, 309, 313, 227,
228, 229, 341, 413, 415, 417, 420, 439,
445, 446, 448, 451, 458, 473, 489, 490
Е
Ebbecke U. (Эббеке) 541, 543, 624
Ebbinghaus (Эбингхаус) 528
Eccles J. С. (Экклс) 111, 122, 127, 159,
164, 177, 178
Ecker А. (Эккер) 54
Fetors L. (Экторс) 482
Edinger L. (Эдингер) 17, 266, 396, 411,
412, 413
Einarson L. (Эйнарсон) 177
Erlanger J. (Эрлангер) 121, 126, 137, 162
Everingham (Эверинхэм) 220
Ewald J. К. (Эвальд) 254
F
Fano G. (Фано) 453
Fearing Fr. (Фиринг) 27
Ferrier D. (Феррье) 408, 427, 455
Filney К. Н. (Филней) 236, 467
Fischer (Фишер) 334, 359, 363, 396, 493
Fiechsig P. (Флексиг) 414, 539, 623, 624
Flourens M. P. (Флуранс) 359, 360
Foerster О. Н. (Ферсгер) 23, 426, 437
Forbes А. (Форбс) 75, 87, 129, 177, 220,
632
ИМЕННОЙ УКАЗАТЕЛЬ
345, 411, 473, 476, 477, 479
Franck Fr. (Франк) 445
Franco's-Frank (Франсуа-Франк) 178
Frankel-Conrat (Френкель-Конрат) 219
Franz S. (Франц) 438
Freusberg А. (Фрейсберг) 144
Fritsch G. (Фрич) 427, 455
Frohlich F. Z. (Фрелих) 149, 158
Frostig J. P. (Фростиг) 131
Fulton J. F. (Фултон) 102, 111, 150, 170,
188, 253, 267, 291, 426, 437, 624
G
Gammon G. D. (Гаммон) 473
Garrey N. E. (Гаррей) 255
Gasser H. S. (Гассер) 108, 116, 119, 121,
126, 129, 133, 137, 182, 420
Garten S. (Гартен) 162, 163
Geist F. В. (Гейст) 23
Gerard R. W. (Жерард) 234, 238, 239, 492
Gergen R. (Герген) 349
Gessell R. (Гезелл) 186, 242
Gildemeister M. (Гильдемейстер) 541
Ging-Hsi Wang (Гинг-Хзи Ванг) 119
Girndt О. (Гирндт) 418, 585
Goldgi С. (Гольджя) 115
Goldstein К. (Гольдштейн) 408, 624
Goltz F. (Гольтц) 149, 254, 256, 345, 349,
394, 438, 453
Gonsalez A. W. (Гонзал?ц) 118
Graham Brown Т. (Грэим Броун) 58, 92,
150, 192, 198, 279, 280, 281, 286, 292,
357, 434, 440, 441, 501, 503
Graham Н. Т. (Греим) 119, 121, 126
Granit R. (Гранит) 546
Green Н. D. (Грин) 427, 434
Greer В. Н. (Грир) 306
Gregg А. (Григг) 177
Grinker R. R. (Гринкер) 253
Grunbaum А. (Грюнбаум) 427
Grundfest Н. (Грундфест) 127, 175
Н
Haberlandt L. (Габерландт) 185
Hansen Е. Т. (Гаязен) 186
Hanstrom В. (Ганстрем) 372
van Harreveld А. (Гарревельд) 131, 383
Harvey Е. W. (Гарвей) 471, 485
Hatschek (Гатшек) 22
Hawke W. А. (Гоуки) 472
Heidenhain R. (Гейденгайи) 149, 211, 439
454, 456
Heinbecker P. (Хейнбеккер) 236, 481
Heliauer Н. (Гелауэр) 185
Helmholtz Н. (Гельмгольц) 176
Henschen S. Е. (Геншен) 410, 413
Hering E. (Геринг) 425, 431, 433, 438, 452
van Herk A. W. (Герк) 418, 585
Herrick С. Judson (Джадсон Херрик) 9,
11, 15, 16, 17, 18, 19, 20, 80, 374, 401,
402, 403, 428
Hhich W. М. (Хич) 185
Hinsey J. С. (Гинзей) 23, 72, 167, 292,
341, 350
Hirschberg Е. (Гиршберг) 219
Hitzig Е. (Гитциг) 427, 455
Hoagland Н. (Хоглэнд) 46, 49, 50, 479
Hobart G. А. (Гобарт) 471, 485
НбЬег R. (Гебер) 540, 541
Hoff Е. С. (Гофф) 59, 130, 266, 427, 434
Hoffmann P. (Гофман) 155, 162, 163, 264,
265, 267, 269, 383, 384, 442
Hoist Е. Е. (Гольст) 239
Hoist V. (Гольст) 294, 295
Hoogerwerf S. (Гоугерверф) 308, 309, 310,
311
Horsley V. (Горслей)' 427
Hughes J. (Хьюгз) 121, 126, 131, 253
Hursch J. В. (Герш) 112, 133, 155
J
Jackson (Джексон) 455
Jasper Н. Н. (Джаспер) 380, 452, 472, 481,
484, 487, 527
Jendrassik Е. (Иендррассик) 265
Johannes Th. (Иоганнес) 140, 212
Johnston J. (Джонстои) 428
Jolly W. А. (Джоли) 177
Jordan H. (Иордан) 368, 373, 382, 384,
385, 386, 38?, 389
Jsayama S. (Исаяма) 364
Jerkes R. (Иеркс) 140, 212
К
Kalischer О. (Калишер) 417, 428, 453
Karplus J. P. (Карплюс) 396
Kast H. (Каст) 431, 432, 433, 436, 437
Kato G. (Като) 99, 110, 136, 138, 139, 148,
149, 275, 540
Katz В. (Катц) 126
Kennard M. А. (Кеннард) 431, 465, 482,
487
Kennedy В. (Кеннеди) 359
Kerr A. S. (Kepp) 472
Katton (Кэтон) 493
Kilvington В. (Кильвипгтон) 359, 438
King С. Е. (Кинг) 255
ИМЕННОЙ УКАЗАТЕЛЬ
633
De Юеуп А. (Де Клайп) 305, 309, 311,
312, 313, 315, 317, 319
Kolliker А. (Кёлликер) 115
Kornmiiller А. (Корнмюллер) 412, 489, 490
Koskoff J. (Косков) 309
Krause (Краузе) 11, 12, 58, 115, 418
Kreidl А. (Крейдль) 396
Kugelberg Е. (Кугельберг) 232
Kuo J. (Куо) 244
Кигё К. (Кюре) 22
Kurschner G. (Кюршнер) 348
L
Lapicque М. (Лапик) 179, 204, 209, 211
Lapinsky М. (Лапинский) 24
Lashley К. S. (Лешли) 628
Leduc S. (Ледюк) 131
Leese Ch. Е. (Лис) 177
Lehmann W. Z. (Леман) 22, 23, 209
Leksell L. (Лекселл) 546
Lemere Fr. (Лемер) 472
Lempke Н. (Лемпке) 585
Lennossek М. (Леношек) 21, 115, 266
van Leuwen Strom (Левен) 316, 319
Lewy F. H. (Леви) 473
Libertini G. (Либертини) 453
Liddell E. G. (Лидделл) 170, 271, 272
Lindsley D. В. (Линдслей) 444, 482
Lloyd D. (Ллойд) 46, 75," 117, 134, 155,
179, 264, 265, 268 277
Loomis A. L. (Лумис) 471, 285
Lorente de N6 (Лоренте де Но) 9, 10, 11,
15, 16, 18, 19, 61, 62, 108, 116, 126, 129,
133, 159, 160, 166, 167, 177, 178, 183,
192, 195, 197, 220, S13, 397, 398, 399,
400, 401, 402, 476, 540
Loucks R. В. (Луке) 578
Luciani L. (Лучиани) 30, 414, 427, 456
M
Mc Couch G. P. (Мак Куч) 121, 126, 131,
253
Mc Culloch W. S. (Мак Куллох) 431, 469
Mc Dokough F. К. (Мак Доко) 474
Mc Kmley W. А. (Мак Кинлей) 142
Mc Nattin R. 72, 341
Mc Rioch D (Мак Риоч) 424, 427, 435,
453
Magendie F. (Мажанди) 21
Magnus R. (Магнус) 279; "301, 302, 305,
306, 309, 310, 311, 312, 313, 314, 315,
316, 317, 319, 325, 326, 327, 328, 329,
330, 331, 336, 337, 340, 341, 348, 349,
363, 396. 501, 503
Magoun H. W. (Магон) 141
Mangold E. (Мангольд) 383
Mann P. J. С. (Манн) 185
Marey (Марей) 334
Marina А. (Марина) 362
Marrazzi A. S. (Марраци) 126
Marshall Hall (Маршалл Холл) 252
Marshall W. H. (Маршалл) 414, 475
Matthews Br. (Мэтьюс) 46, 48, 49, 120,
122, 123, 126, 134, 135, 136, 169, 261,
264, 267, 479, 493, 545
Meinert (Мейнерт) 398
Meltzer J. (Мельтцер) 90
Merzbacher (Мерцбахер) 212
Mettler С. С. (Мегтлер) 433, 456
Mettler F. А. (Меттлер) 281, 433, 456
Miller F. А. (Миллер) 306
Minkowsky M. (Минковский) 409
Monakow С. (Монаков) 404, 408, 411,414
Monnier A. M. (Монье) 380
Montalcini R. L. (Монтальчини) 244
Moore A. R. (Moop) 236
Morgutti M. (Моргутти) 362
Morison B. R. (Морисон) 129, 452, 473,
477, 479, 482
Morison R. S. (Морисон) 236, 427, 467,
473, 482
Moruzzi G. (Моруци) 210, 240, 241, 443,
493
Miiller J. (Мюллер) 41
Mtiller M. M. (Мюллер) 238
Munk H. (Мунк) 404, 408, 409, 413, 416
JST
Nims L. F. (Нимс) 465, 482
О
Obrador S. (Обрадор) 427
O'Leary J. L. (О'Лири) 146, 484
Olmsted J. (Ольмстед) 159, 362
Orr D. W. (Opp) 245
Osborn W. А. (Осборн) 359, 438
P
Pantin L. (Паятин) 385
Parker G. H. (Паркер) 62, 69, 368
Pastor I. (Пастор) 185
Penfield W. (Пенфильд) 305, 308, 416,
417, 425
Perschmann (Першман) 211
Pfluger E. (Пфлюгер) 30, 72, 80, ai8
Philippson M. (Филиппсон) 58, 273. 308
Pick E. P. (Пик) 238
Pinkston J. О. (Пннкстод) 4g7
Piper H. R. (Пнпер) 162 =
?34
ИМЕННОЙ УКАЗАТЕЛЬ
Pollock L. (Поллок) 306
Prochaska G. (Прохаска) 27
Prosser С. L. (Проссер) 231, 236, 239
Q
-Quastel I. H. (Квостел) 185, 219
В
Rademaker G. (Радемакер) 306, 308, 309,
310, 311, 228
Ranson S. W. (Рэнсон) 72, 167, 244, 291,
292, 341, 350
Hedlich W. (Редлих) 414
Renshaw В. (Реншоу) 116, 479
"Resriik S. (Резник) 131
Retzius G. (Ретцнус) 11
Rhednberger M. В. (Рейнбергер) 452, 484
Kichter С. Р. (Рихтер) 164
Richter E. P. (Рихтер) 428, 444
Ring R. (Ринг) 409
Roggers F. (Роджерс) 384, 428
¦Rohracher H. (Popaxep) 472, 479
Romanes J. (Романее) 369
Rosenblueth А. (Розенблют) 435, 453, 491
Rosenthal J. (Розенталь) 176
Rothfeld I. (Ротфельд) 454
Rothmann (Ротманн) 394
Rubin M. А. (Рубин) 465, 471, 479
Ruch Т. С. (Руч) 231
Rijnberk G. (Ринб?рк) 55
S
Sachs A. L. (Закс) 253
Schagass Ch. (Ularac) 481, 527
Schoen R. (Шен) 336, 337
Schrader M. I. (Шрадер) 394, 409
Schriever H. (Шривер) 203, 204, 205, 210,
211
Schwartz H. G. (Шварц) 472
Schweitzer А. (Швейцер) 111, 186
Segaar J. (Ceraap) 384, 385, 386, 387
Sherb R. (Шерб) 362
Sherrington Ch. (Шеррингтон) 22, 44, 50,
54, 57, 58, 67, 68, 74, 76, 77, 78, 79, 80,
83, 85, 86, 87, 91, 94, 95, 98, 110, 111,
114, .150, 151, 158, 159, 160, 171, 172,
177, 178, 180, 181, 202, 203, 205, 209,
220, 227, 252, 253, 254, 256, 264, 266,
268, 269, 270, 272, 274, 278, 280, 281,
282, 284, 286, 291, 292, 304, 305, 306,
335, 337, 351, 355, 356, 358, 359, 363,
411, 425, 427, 431, 433, 434, 452, 501,
502, 575, 619
Silver I. (Сильвор) 13
Smits C. G. (Смите) 281
Spalanzani (Спаланцани) 55
Speidel С. С. (Спейдель) 223
Sperry R. W. (Сперри) 362
Spiegel E. А. (Шпигель) 234
Stedman E. I. (Стедман) 185, 219
Stedman E. L. (Стедман) 185, 219
Steinach (Штейнах) 21, 54
Steiner (Штейнер) 393
Stewart W. В. (Стюарт) 121, 126, 131,
253
Stoland О. О. (Столанд) 306
Stopford I. S. (Стопфорд) 13, 70
Straus W. (Страус) 245
Strieker К- (Штрикер) 21
T
Talbort S. А. (Тальборт) 475
Tamburini (Тамбурини) 427
Taylor D. В. (Тейлор) 131
Ten-Cate J. (Тен-Кате) 236, 418, 585
Tennenbaum M. (Тенненбаум) 185
Therman O. J. (Терман) 144, 146
Tiedemann А. (Тидеман) 158
Toennies J. F. (Теннис) 135, 545
Tower S. S. (Тоуэр) 280, 452, 543
Trendelenburg P. (Тренделленбург) 255
Tse-Wei-Lu (Це-Вей-Лю) 119
Tunturi A. R. (Тунтури) 412
U
Uexkull J. (Юкскюль) 371
Umrath Ch. (Умрат) 119, 126
Umrath К- (Умрат) 119, 126
Unzer J. (Унцер) 27
V
Van der Hoeve (Ван дер Гофф) 313
Verworn (Ферворн) 88, 149, 217, 220
Visantini F. (Византини) 244
Vogt С. (Фогт) 395, 399, 425
Vogt О. (Фогт) 399, 425
Volkenburg (Фолькенбург) 417
W
Wachholder К. (Вахольдер) 164, 304, 432,
436, 444
Wagner R. (Вагнер) 436
Waller W. H. (Уоллер) 341
Walter W. G. (Вальтер) 492
Wana J. (Вана) 22
Ward J. W. (Уорд) 434, 489
Warren (Варрен) 268
Weber W. (Вебер) 148, 334
ИМЕННОЙ УКАЗАТЕЛЬ
635
Weber Е. (Вебер) 334
Weddel G. (Ведель) 245
Weiss P. (Вейсс) 362
Wiersma С. А. (Вирзма) 131, 383
Wiggers К. (Виггерс) 236
Windle W. F. (Уиндл) 36, 37, 119, 245, 246
Winterstein Н. (Винтерштейн) 218, 219,
242
Wolker А. Е. (Уолкер) 237
Woolsey С. W. (Вулси) 414
Worzniak J. (Ворцниак) 186
Wright S. (Райт) 111, 186
Wundt W. (Вундт) 176
Wyss О. А. (Уисс) 427
Z
Zing-Yang'-Kuo (Цинг-Янг-Куо) 186
Y
Yamagiwa К. (Иамагива) 471
Yanagisana R. (Ианагизана) 362
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
А Г
Автоматическая деятельность 20
Адаптация рецепторов 46, природа ее 48
Альфа-ритм 464, характеристика 469,
происхождение 478, вторая фаза 479,
действие закрытия глаз 480, опыты 493
Антагонисты — понятие 83, принцип
деления односуставных мышц 83, двусу-
ставных мышц 85, еинергетическая
деятельность 435
Антидромное возбуждение 159
Афферентная система 21, в коре 400, 406
Б
Бета-волны 471, происхождение 473
В
Возбуждение в ц. н. с. 155, спонтанное
221, возбуждение при посредстве
медленных потенциалов 159, антидромное
159, стационарное 385, двигательных
участков 440
Возвратные коллатерали двигательных
нейронов 114, роль в торможении 115
Воспринимающие функции коры 408,
зрения 409, слуха 411, обоняния 413,
вкуса 413, кожно-мышечных раздражений
413, характеристика 414, локализация
415, методы изучения 418, возбудимость
420, возбуждение 421
Восстановление индивидуальных
рефлексов 594
Временные связи — понятие 547,
происхождение 549, корковые элементы
двигательных индивидуальных рефлексов
578, слюнного рефлекса 582, вне коры
584
Время рефлекторной реакции 175,
редуцированное 176, происхождение 181, по
коротким путям 176, по длинным 180,
опыты 190
Выравнивание энергии 385
Генерализация индивидуальных
рефлексов— рецепторная 511, эффекторная 513,
вторичная 513, происхождение 551, роль
психонервной деятельности 557
Гигантские нервные волокна 371
Гипноз 570
Гипотеза выравнивания энергии 385
Градация рефлекса 77, у беспозвоночных
79, опыты 80
Гуморальная передача возбуждения 183,
критика 186
д
Двигательные участки в коре — понятие
423, распределение 424, характеристика
426, вегетативных органов 427,
историческая справка 427, филогенез 428,
онтогенез 430, координация 431,
синергетическая деятельность 435, связь с
координирующими аппаратами 437,
возбудимость 438, возбуждение 440,
проведение возбуждения 444, облегчение 445,
угнетение 445, происхождение
облегчения и угнетения 448, опыты 458
Двойственная иннервация 91,
происхождение 94, опыты 106
Декреметное прозедение возбуждения 368
Дельта-волны 492
Дендриты — роль в нейропиле 11,
локальные процессы 126, отсутствие
проводимости 130
Децеребрированный препарат — операция
296, ригидность 304, происхождение 304»
сгибательный тонус 306
Динамика рефлекторной деятельности —
изменчивость 348, невозможность
локальных раздражений 350,
происхождение изменчивости 352, 359
Дифференциация индивидуальных
рефлексов— рецепторная 511, эффекторная 513»
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
637
вторичная 513, происхождение 551, роль
психонервной деятельности 557, опыты
611
Дифференцированное раздражение 511,
отрицательное действие 601, опыты 611
Доминанта 365
3
Замкнутые круги-цепочки 10
Закон Белла-Мажанди 21
Закон возбуждения в рецепторах 45
Закон возбуждения в ц. н. с. 157
Закон локализации координирующих
центров 56
Закон одностороннего проведения
возбуждения 52
Закон сопряженной иррадиации
возбуждения 536, о физико-химических основах
539, морфологические основания 542,
специфичность 542
Закон сукцессивной индукции 575
Закон суммации 204
Запаздывающий индивидуальный рефлекс
519
И
Изменчивость прирожденной рефлекторной
деятельности 348, происхождение
изменчивости 352, характерные случаи
353, тонических рефлексов 354, от
характера раздражения 355, от
перекрещивания нервов 359, опыты 365.
Изменчивость корковых реакций 434,
индивидуальных рефлексов 599, 605
Индивидуальные рефлексы — понятие 499,
направления 501, терминология 503,
образование 504, высшего порядка 506,
«а людях 507, скорость образования 507,
у разных животных 510, у
новорожденных 510, на сложные раздражители 515,
типы 518, происхождение 547, опыты
528, корковые элементы 582
Индивидуальные рефлексы —
анатомические элементы 578, восходящая часть
578, корковая 578, нисходящая 581,
изменчивость 599
Индукция непосредственная
спинномозговая 202, сукцессивная 286
Интероцептивные индивидуальные
рефлексы 525
Иррадиация рефлекторной деятельное гя
68, происхождение 70, роль проприо-
цепторов 73, у беспозвоночных 79,
история вопроса 80, опыты 80
Итеративная возбудимость ц. н. с. 179,
ігетерохронизм ц. и. с. 209, критика 209
К
Клешня рака 383, иннервация 383
Координация — понятие 29, значение
спинномозговых узлов 53, значение
двигательных нейронов 57, роль
промежуточных нейронов 57, анатомо-функциональ-
ный механизм 61, основные процессы
82, одного сустава 83, нескольких
органов 89, опыты 105
Координация у низших беспозвоночных
368, у высших червей 372, у пиявок
374, у низших моллюсков 381, у
членистоногих 382, опыты 387
Координирующие аппараты — локализация
55, связь с корой 437, координация
корковых движений 433, корковых
вегетативных реакций 433
Конвергенция нервных путей 78
Кора большого мозга — функции 393,
строение 396, микроскопическая
структура 399, воспринимающие функции 408,
ответная функция 423, синергетическан
деятельность 435
Корковые пути нисходящие 402,
восходящие 406
Л
Лабильность ц. н. с. 168
Лабиринтные тонические рефлексы на
ногах 309, на шее и туловище 312, на
глазных мышцах 312
Лабиринты — строение 313, способ их
раздражения 313
Локальные процессы в дендритах 126, в
клетке 127, медленные потенциалы 126
Локомоция — понятие 333, фазные
движения 333, статический тонус 335,
рефлекс равновесия 338, направление 340,
опыты 341
Локомоция у беспозвоночных 370. 373,
374
М
Медленные потенциалы спинного мозга
119, от корешков 122, регулярные и
нерегулярные 123, при адекватном
раздражении 125, многофазный характер 120,
однофазный 128, происхождение 136,
роль локальных процессов в дендритах
126, в клетках 127, тормозящее
действие 130, ствола мозга 142, опыты 151,
роль в передаче возбуждения 160
Местное состояние центрального
возбуждения 161
Местное состояние центрального
торможения 171
€38
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Методы свободных движений 530
Миотатические рефлексы 268, у лягушки
275
Мозжечок—спонтанная активность 237,
отношение к тонусу 306
Монаковские пучки 405
Мышцы односуставные 84, двусуставчые
85, принципы их деятельности 84, 85,
фиксирующая роль 86, опыты 109
Н
Нарастание рефлекторной деятельности
205
Нейропиль спинного мозга 9, 11,
назначение 18, тормозящая функция 112,
ствола мозга 16, тормозящая функция 138,
у беспозвоночных 368, роль его в
регуляции рефлекторной деятельности 387
Нейропиль в коре 401, функция по Хер-
рику 401, роль в индивидуальных
рефлексах 599
Некоординированная деятельность ц. и. с.
103
Нервные круги 10, в коре 400
Нервные цепочки 10
О
Облегчение в ц. н. с.— понятие 192, после
однократного раздражения 193, при
повторных раздражениях 195, после
торможения 197, электротоническая
природа 199, электрические потенциалы 206,
опыты 214
Облегчение в коре 445, происхождение 448
Облегчающее действие головного мозга
на спинной 210
Обмен веществ в ц. н. с. 218
Обратные связи 547
Общее торможение — понятие 96, без
двигательных реакций 98, отношение к ре-
ципрокному торможению 98,
биологическое значение 102, опыты 106
Общее торможение в эмбриональном
развитии 118, спонтанное 119, роль
медленных потенциалов 130, электротоническая
природа 130, ствола мозга 138, у
беспозвоночных 376, в коре 447
Объединяющая деятельность коры —
понятие 613, физиологические основания
618, локализация 622, о нервных
элементах 623, опыты 625.
Ориентировочные рефлексы — понятие 344,
происхождение 344, нервный механизм
345, отличительные особенности 346,
значение при пробуждении от сна 571
Основная активность коры 464,
характеристика 467, происхождение 474, опыты»
493
Ответная функция коры 413, в отношении
вегетативных органов 427,
филогенетическое развитие 428
Отолитный аппарат—строение 313,
функция 314
Отрицательный индивидуальный рефлекс
522, происхождение 562
Отрицательное действие посторонних
раздражений 599, дифференцированных 601,
последействие 603, одного рефлекса на
другой 605, опыты 611
Отставленный индивидуальный рефлекс
519
И
Передача возбуждения в ц. н. с. 177, 181,
гуморальная 183, критика 186
Перимедулярное сплетение 13
Пессимальный эффект в ц. н. с. 158
Пирамидные клетки 397, пути 402,
характеристика 426
Плавательные движения медузы 369, у
пиявок 374
Пластический тонус 273
Принципы — конвергенции 24, 78, общего-
конечного пути 56, координации одного
сустава 83, нескольких органов 89
Поведение — понятие 51, и рефлекс 32, и
спонтанная деятельность 33, методы 530
Последовательная спинномозговая
индукция 286
Последовательный индивидуальный
рефлекс 520, происхождение 558
Произвольные индивидуальные рефлексы
527
Проприоцептивные рефлексы — понятие
261, сухожильные 263, миотатические
268, координированные 271, и
пластический тонус 273, у лягушки 275,
опыты 276
Проприоцептивные индивидуальные
рефлексы 524
Проприоцепторы — строение 73, роль в
иррадиации 73, импульсы 74, значение
при фазных движениях 280
Р
Раздражения внешние 26, внутренние 26
Распространение возбуждения в ц. ч. с. по
коротким путям 176, по длинным 180
Распространение торможения 188
Реакция удлинения л укорочения 274, у
беспозвоночных 372
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
639>
Регуляция рефлекторной деятельности у
беспозвоночных 385
Рефлекторная деятельность — понятие 27,
индивидуально-приобретенная 28,
характеристика 29, сложность и целостность
67, 352, иррадиация 63, градация 77,
нарастание 205, динамика 348, без ц. н. с.
378
Рефлекс времени 526
Рефлекс обнимания Ш
Рефлекс — понятие 27, индивидуально-
приобретенный 28, характеристика 29,
назначение 29, и поведение 32, и
нервно-мышечная деятельность 33, без
ц. н. с. 34, онтогенез его 35,
классификация 37, опыты 38
Рефлекс потирания 28, 39, 353
Рефлекс равновесия 333
Рефлекс разгибания 39, изменчивость 353
Рефлекс сгибания 28, 39, изменчивость
353
Рефлекс чесания 50, 201, 355
Рефлекс шагания 354
Рефлексы сухожильные 263, миотатиче-
ские 268, тонические 301
Рефлекторная дуга — понятие 59,
беспозвоночных 62
Рефлекторное последействие 75, опыты
80, ритм возбуждения 167, роль
нервных кругов 166, у беспозвоночных 380
Рефлекторные пути — анатомические
основания 59, отсутствие локализации 59
Рефракторная фаза в ц. н. с. 157,
двигательных клеток 159
Рецептивное поле — понятие 42, значение
в координации 50, опыты 51
Рецепторы — понятие 40, строение 43,
свойства 44, импульсация 46, адаптация
46, опыты 51
Ритмическая деятельность плавников у
рыб 294
Ритмические рефлексы — понятие 278,
чередование фаз 279, значение проприо-
цепторов 280, координация 281,
происхождение смены фаз 291, опыты 295
Ритмические индивидуальные рефлексы
525
Ритмика центрального торможения 169,
опыты 172
С
Сегментальные рефлексы 55, опыты 62, у
беспозвоночных 372
Сетевидное образование 15
Сеченовское торможение 139
Сгибательный рефлекс — оборонительный
28, 29, тонический 303
Синапсы 11
Синаптические потенциалы 127
Синаптическое поле 18
Совпадающие индивидуальные рефлексы
518, происхождение 549
Сокращение отдачи 286, происхождение
288, история вопроса 291
Сочетательные рефлексы 503
Спинной мозг — строение 10, обнажение
его 38
Спинномозговые препараты — операция
242, регидность 308
Спинномозговые узлы — значение в
координации 52, опыт Штейнаха 52, опыты
61
Спонтанная деятельность мозга — понятие
20, 231, рецепторов 46, 231, ствола
мозга 233, без внешних реакций 234,
влияние внешних раздражений 236,
происхождение 237, периодичность 239,
облегчающее и тормозящее действие 241,
дыхательного центра 242, у зародыша-
244, в постэмбриональном периоде 247,
при инстинктивных действиях 248, опь*-
ты 249
Спонтанная деятельность н. с. у
беспозвоночных 236, 378
Спонтанная деятельность в коре 463,
влияние периферических раздражений
482, во время наркоза и сна 483,
патологические изменения 488, фу нкцио -
нальные изменения 487
Статический тонус 335
Стато-кинетические рефлексы вращения
328, прогрессивного движения 329,
приготовления к прыжку 330, при падении
331, опыты 331
Стационарное возбуждение 385
Строение спинного мозга 10,
продолговатого мозга 14, промежуточного мозга-
16, головного мозга амфибий 17, коры
396
Субординационная хронаксия 211
Субпиальный слой 13
Substantia gelatinosa Rolandi — строение
11
Субординационное действие —
облегчающее и тормозящее 210
Судорожные разряды 489, опыты 496
Сукцессивная индукция по Шеррингтону
286, по Павлову 575
Суммация временная и пространственная
182, 202, от субминимальных
раздражений 200, интервал 202, закон 204,
происхождение 208, опыты 214
Сухожильные рефлексы 263
640
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Т
Теория доминанты 365
Тонические рефлексы — понятие 300,
рецептивное поле 301, координация 301,
сгибания у лягушки 302, у кролика 303,
лабиринтные 309, шейные 315, у
нормальных животных и у человека 319,
механизм 319, опыты 321, изменчивость
354
Торможение в коре 445, происхождение
448, история вопроса 452, внешнее 607,
внутреннее 608
Тормозящее действие медленных
потенциалов 130, 142, средних отделов мозга
на спинальные рефлексы 147, 210
Тормозящие нейроны 109, у
беспозвоночных 379, 384
Торможение — понятие 82, реципрокное 83,
активная природа 87, одновременно с
возбуждением 91, общее 96, гипотезы:
пессимальная 108, химическая 111,
субнормальная 137, ритм 169, утомление
222, у беспозвоночных 376,
периферическое 379, опыты 104
У
Угасание индивидуальных рефлексов 588,
происхождение 590, опыты 611
Условные рефлексы 503, учение Павлова
об образовании их 573, об угасании 596,
об изменчивости 607
Утомление торможения 225
Утомление ц. и. с.— понятие 217, роль
центральных элементов 220, и
электрические потенциалы 221, структурные
изменения 223, роль торможения 223,
зависимость от силы раздражения 227,
и повышения возбудимости 228, опыты
229
Ф
Фракционирование нервных путей 78
Функциональная система 352
X
Химическая природа торможения 111
ц
Центральная нервная система — строение
9, единица 10, связь с периферической
н. с. 21, возбудители ее 26, у
беспозвоночных 34, онтогенез 35, обнажение
38.
Центральное состояние возбуждения 160
Центральное торможение — пессимальный
характер 108, тормозящие нейроны 109,
химическая природа 111, роль нейропиля
112, электротоническая природа 130, от
постоянного тока 134, и
субнормальность 137
Цепочки замкнутые 10, множественные 10
Ш
Шейные тонические рефлексы вращения
315, наклонения 315, поднятия и
опускания 316, на глазных мышцах 317,
опыты 322
Шок ц. н. с.— понятие 252, и наркоз 252,
происхождение 254, травматический 257,
опыты 259
Э
Экстрапирамидные пути 403, функция 426
Электрическая активность коры 464,
понятие 464, характеристика медленных
потенциалов 467, альфа-ритм 464, 469,
быстрые потенциалы 471,
происхождение 473, основная активность 464, 467,
474, влияние периферических
раздражений 482, при наркозе и во время сна
483, при функциональных изменениях
487, в патологии 488, историческая
справка 493, опыты 493
Электротоническая природа торможения
130, электротоническое возбуждение
задних корешков 136
Экспериментальный сон 565,
происхождение 568
Опечатки и исправления
Стр.
39
74
101
101
126
166
170
249
303
341
349
3?9
419
419
419
435
440
465
479
483
489
Ыо
Строка
9 св.
Подп. к рис.
28, 2 и 3 сн.
Подп. к рис.
48, 4 св.
Там же, 6 св.
9 сн.
4 сн.
Подп. к рис.
82, 1 сн.
8 сн.
3 сн.
13 св.
Подп. к рнс.
154, 7 св.
2 сн.
19 сн.
17 сн.
17 сн.
4 св.
Подп. к рис.
196, 8 св.
Подп. к рис.
206
19 св.
Подп. к рис.
215, 5 сн.
Поди, к рис.
218, 2 сн.
9 сн.
Напечатано
отделить
начало импульсации через
раздражение VIII корешка
предшествует раздражению
VII
раздражение VII
предшествует раздражению VIII
0.180
6.6
Время—по 2.2 сек.
оградиться
децереброванных
у амфибий
аппарат
374
обширном
частичном
—менее
аддукцией
при покое его на
разгибателя
зоны зрительной
внутреннем
dex.
глаза закрыты
психологии
Должно быть
определить
начало импульсации и через
раздражение VII корешка
предшествует раздражению
VIII
раздражение VIII
предшествует раздражению VII
0.5
0.6
Время—по 0.2 сек.
оградить животное
децеребрнрованных
у рептилий
препарат
172
частичном
обширном
—также менее
абдукцией
при покое разгибателя
зоны и зрительной
внутривенном
sin
глаза сначала оікрыты,
а затем закрыты
психиатрии
И. С Беритов