














































































































































































































































Автор: Вартапетов Л.Г.
Теги: географическая зоология зоогеография фауна зоология животные птицы сибирь изучение птиц западная сибирь
Год: 1984




Л.Г.Вартапетов
ПТИЦЫ
таежных междуречий
ЗАПАДНОЙ
СИБИРИ
АКАДЕМИЯ НАУК СССР
СИБИРСКОЕ ОТДЕЛЕНИЕ
БИОЛОГИЧЕСКИЙ ИНСТИТУТ
Л. Г. ВАРТАПЕТОВ
птицы
таежных междуречий
ЗАПАДНОЙ СИБИРИ
Ответственный редактор
д-р биол. наук Ю. С. Р а в к и н
ИЗДАТЕЛЬСТВО «НАУКА»
СИБИРСКОЕ ОТДЕЛЕНИЕ
Новосибирск* 1984
УДК 591.9(5—013) + 591.526 + 598.2/9
Вартапстов Л. Г. Птицы таежных меж-
дуречий Западной Сибири.— Новосибирск: Наука, 1984.
В монографии рассматриваются внутризональные изме-
нения населения птиц таежных междуречий в сравнении с
опубликованными ранее результатами исследования птиц
долины Оби. Приводятся показатели плотности, видового
разнообразия, биомассы и ярусного распределения птиц,
значения трансформируемой ими энергии и соотношения ос-
новных групп потребляемых кормов, а также доминантный и
фаунистический состав населения. Характеризуется подзо-
нально-ландшафтное размещение видов птиц п описывается
их иерархическая классификационная схема по сходству
распределения. В результате выявлены природные режимы,
обусловливающие неоднородность впутрпзональною разме-
щения видов птиц. Приводятся пространственно-типологи-
ческие схемы сообществ птиц и оценивается иерархия факто-
ров окружающей среды и их сочетании, определяющих про-
странственную изменчивость орнитокомплексов.
Книга рассчитана на специалистов в области зоогео-
графии, орнитологии, охотоведения, биоценологии и охраны
природы.
Табл. 28. Ил. 11. Библиогр. НО.
Лев Гургенович Вартапстов
ПТИЦЫ ТАЕЖНЫХ МЕЖДУРЕЧИИ ЗАПАДНОЙ СИБИРИ
Ответственный редактор
Юрии Соломонович Равкин
Утверждено к печати
Биологическим институтом СО АН СССР
Редакторы издательства С. Л. Розина, С. .4. Садко
Художественный редактор В. If. Шумаков
Художник В. В. Растегаев
Технический редактор С. А. Смородинова
Корректоры С. М. Погудина, В. В. Борисова
ИF> .X" 23458
Сдано в набор 29.07.83. Подписано г. печать 26.03.84. МН-03015. Формат Г-0 у ОО1/,*.
Бумага типографская X; 2. Обыкновенная гарнитура. Высокая печать. Усл. печ. л. 15.
Усл. кр.-огт. 15,2. Уч.-изд. л. 19,5. Тираж 1300 экз. Заказ 730. Цена 3 р. 30 к.
Издательство «Наука», Сибирское отделение. 630099, Новосибирск, 99, Советская, 18.
4-я типография издательства «Наука». 630077, Новосибирск, 77, Станиславского, 25.
2001050000-787
В 042(02)—84 223—84—11
© Издательство «Наука», 1984 г.
ВВЕДЕНИЕ
Население птиц таежных междуречий Западной Сибири к на-
стоящему времени изучено явно недостаточно. Освоению между-
речных территорий препятствует их высокая заболоченность, труд-
нодоступность и плохая проходимость. На долю болот приходится
около половины площади таежной зоны Западной Сибири, а на мно-
гих водоразделах — от 60 до 80%. Однако с началом эксплуатации
нефтяных и газовых месторождений резко увеличилась интенсив-
ность использования природных ресурсов таежных территорий.
Кроме того, обсуждаются проекты переброски части обских вод
к югу. Все это может привести к значительным изменениям таежных
биоценозов на междуречьях Западной Сибири и обусловливает ак-
туальность изучения таежных природных комплексов, в том числе
их животных компонентов.
Численность и распределение птиц служат чутким индикато-
ром пространственно-временных измерений биоценозов. Поэтому
количественные исследования позволят не только зафиксировать
исходное состояние сообществ птиц к началу интенсивного хозяйст-
венного освоения таежных междуречий Западной Сибири, но и су-
дить об их последующих изменениях, связанных с интенсивной транс-
формацией ландшафтов этой территории, и, видимо, в значительной
мере прогнозировать ее последствия. Кроме того, исключительная
равнинность Западной Сибири (в том числе ее таежной зоны) делает
этот регион весьма удобным для изучения широтных изменений жи-
вотного мира [Формозов, 1964; и др.]. На междуречьях, в плакор-
ных местообитаниях зональные и подзональные отличия животного
населения проявляются наиболее четко, в отличие от долин крупных
рек, где преобладают азональные факторы [Максимов, 1974; Чер-
нов, 1975]. К тому же долины западно-сибирских транзитных рек
(Оби и Иртыша), в связи с их гораздо более давней и лучшей освоен-
ностью, представляют собой объект традиционного интереса для эко-
логов и зоогеографов и значительно полнее изучены по сравнению
с междуречьями. Поэтому биогеографические исследования на таеж-
ных междуречьях Западной Сибири — неотложная и важная зада-
ча, решение которой представляет большой теоретический и практи-
ческий (инвентаризационный) интерес.
В целом территория таежной зоны Западной Сибири в фаунис-
тическом отношении изучена сравнительно неплохо. История и ре-
3
зультаты эколого-фаунистических исследований птиц обобщены
Г. X. Иогансеном [1943—1961], А. М. Гынгазовым и С. П. Мило-
видовым [1977]. Специально фауне болот всей Западно-Сибирской
равнины посвящена работа А. А. Назарова [1977].
Следует отметить, что болота, особенно верховые, как весьма
характерные местообитания междуречий вызывают в последнее вре-
мя у зоологов повышенный интерес. Так, отдельные сведения об
орнитофауне и населении птиц болот лесной зоны имеются по неко-
торым районам Польши, Финляндии, Белоруссии и Эстонии [Кума-
ри, 1965; A. Dyrcz, J. Oculewicz, В. Wiatr, 1973; Дучиц, 1962, 1975;
О. Jrvinen, L. Sammalisto, 1976; Козлов, Кузьменко, 1977; Бирю-
ков и др., 1979]. В частности, отмечаются значительная видовая
бедность и низкая плотность птичьего населения верховых болот
по сравнению с низинными болотами и суходолами [Владышевский,
1974], а также высокая степень «перелетности» населения и эфемер-
ности их орнитокомплексов. Основная часть обитающих здесь птиц
покидает верховые болота сразу после окончания гнездования.
Число птиц, зимующих на болотах, тоже невелико по сравнению
с лесными местообитаниями. Кроме того, прослеживается значимое
участие в населении арктических видов птиц при «сокращении дли-
ны пролетного пути» [Кумари, 1965]. Все это связано с экстремаль-
ностью условий существования птиц на болотах. Так, по сравнению
с окружающими их суходольными местообитаниями безморозный
период на верховых болотах Западной Сибири на 30—50 дней коро-
че [Потапова, Жуков, 1972], общая продуктивность и биомасса на-
секомых, служащих основным кормом для птиц, тоже обычно зна-
чительно меньше, чем на суходолах.
Сообщества птиц долины Оби изучены сравнительно полно,
результаты исследований приводятся в статье П. А. Пантелеева
[1972а] и монографии Ю. С. Равкпна (1978). Птичье население при-
енисейской тайги описано в статьях Э. В. Рогачевой [1962, 1965]
и Э. В. Рогачевой, Е. Е. Сыроечковского, О. В. Бурского и др.
[1978]. Работы, посвященные населению птиц таежных междуречий
Западной Сибири, как правило, фрагментарны, и число их невели-
ко. Количественные характеристики населения птиц приводятся
В. В. Раевским [1952, 1982], А. М. Гынгазовым, Н. Г. Шубиным
[1968], П. А. Пантелеевым [19726]. Лишь южно-таежные сообщест-
ва позвоночных, в том числе птиц, обследованы полнее [Равкин,
Лукьянова, 1976].
Таким образом, наименее изучены междуречья северных под-
зон. Наши исследования в определенной степени восполняют этот
пробел. Основное внимание уделено сравнительной количественной
характеристике орнитокомплексов междуречий северной, средней
тайги и подтаежных лесов Западной Сибири. Собранные материалы
позволили проанализировать территориальные изменения населе-
ния птиц таежных междуречий (от северной тайги до подтаежных
лесов) и сравнить его особенности в долине Оби и на междуречьях,
используя опубликованные данные предшествующих исследовате-
лей. По этой причине мы полностью придерживались принципов,
4
методик, объемов и сроков работ, указанных Ю. С. Равкиным,
И. В. Лукьяновой [1976] и Ю. С. Равкиным [1978]. Все использо-
ванные нами результаты этих исследователей по южно-таежным
междуречьям и долине Оби в пределах лесной зоны заимствованы
из перечисленных монографий. Стандартизация методик и подходов
обеспечила полную сравнимость наших данных с материалами наз-
ванных авторов.
Основная цель наших исследований заключалась в выявлении
пространственной неоднородности населения птиц таежных между-
речий Западной Сибири и факторов среды, ее определяющих. Особое
внимание уделено описанию типологической структуры сообществ
птиц, которая хотя и определяется ландшафтно-зональной специ-
фикой территории, но не совпадает полностью с ландшафтно-геогра-
фическим ее делением. На неполное совпадение границ сообществ
и ландшафтов указывала, в частности, Н. В. Тупикова [1969, 1979].
Однако в равнинной тайге Западной Сибири, где изменения в ланд-
шафтах весьма постепенны, наиболее важным представляется не
поиск этих довольно условных границ, а обнаружение основных
тенденций территориальных изменений птичьего населения. В вы-
явлении факторов окружающей среды, определяющих эти измене-
ния, состояла наша следующая задача.
Исходя из нее, за наименьшую единицу рассмотрения принято
население ландшафтного урочища, которое выделяется типологи-
чески. Последующее выявление пространственной структуры ор-
нитокомплексов проводится тоже типологически, так как на коли-
чественный состав населения позвоночных различия в условиях
среды влияют больше, чем пространственная сопряженность сооб-
ществ. На строгую зависимость формирования и структуры сообществ
от современных зонально-климатических условий, в отличие от
фауны, состав которой в гораздо большей степени обусловлен исто-
рическими причинами, указывает, в частности, Ю. И. Чернов [1975].
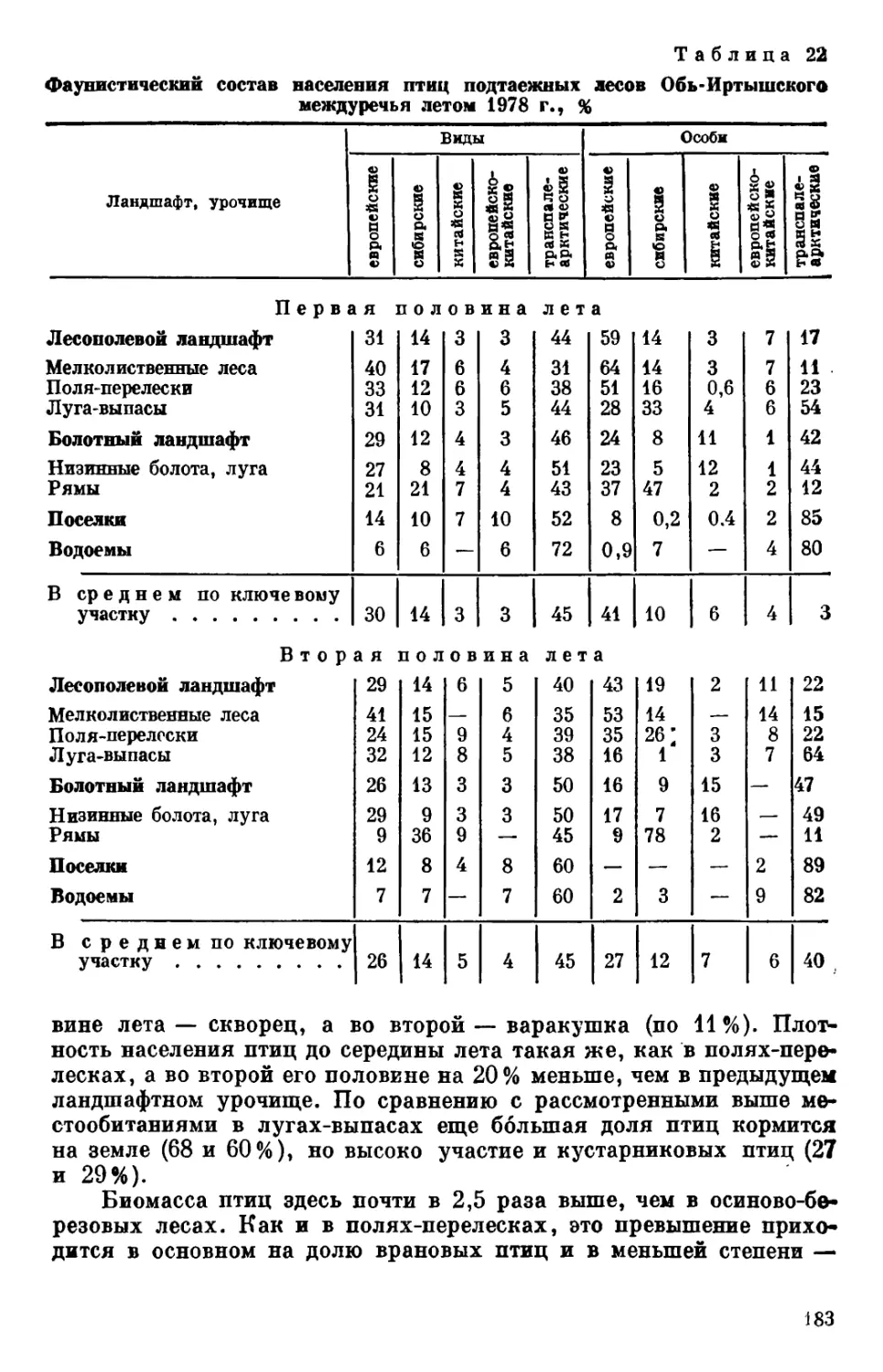
На основании количественных учетов птиц получены показа-
тели видового разнообразия, фаунистического и доминирующего со-
става, плотности населения и его ярусного распределения, биомас-
сы, биоэнергетики и соотношения основных групп потребляемых
кормов (в энергетическом эквиваленте). Материалы позволяют оха-
рактеризовать численность и распределение отдельных видов, а так-
же значимость птиц в трансформации энергии.
Рассматриваются пространственно-типологические структуры
населения птиц и факторы среды, определяющие пространственную
неоднородность их сообществ. В результате анализа этих структур
выявлены наиболее общие факторы, коррелирующие и определяю-
щие территориальные изменения орнитокомплексов. Получена ко-
личественная оценка степени совпадения пространственной неодно-
родности сообществ с различными сочетаниями факторов окружаю*
щей среды (природными режимами), т. е. величина полноты объясне-
ния территориальной изменчивости населения позвоночных выявлен-
ными природными режимами.
Приведенные в монографии количественные характеристики
5
сообществ птиц могут служить основой при разработке охотоустрои-
тельных и природоохранных мероприятий. Возможен пересчет за-
пасов охотничье-промысловых видов с обследованных ключевых
участков на более обширные территории междуречий, в том числе
с использованием картографической или аэрокосмической инфор-
мации о площадях 'природных выделов. Использование аэрокосми-
ческих снимков позволяет проводить достаточно надежные и опера-
тивные оценки ресурсов животного населения [Белов, Вартапетов,
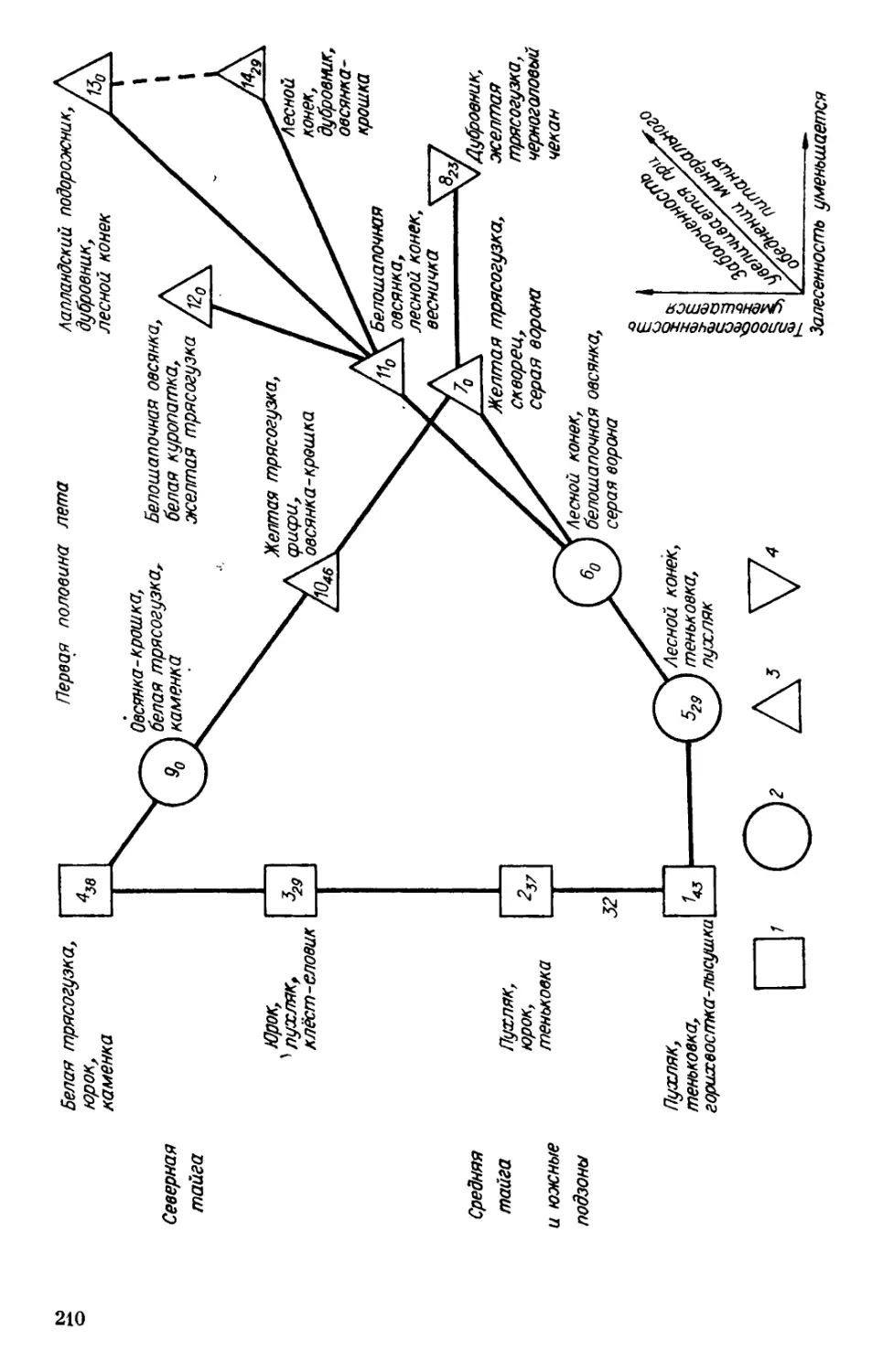
Николаев и др., 1983].
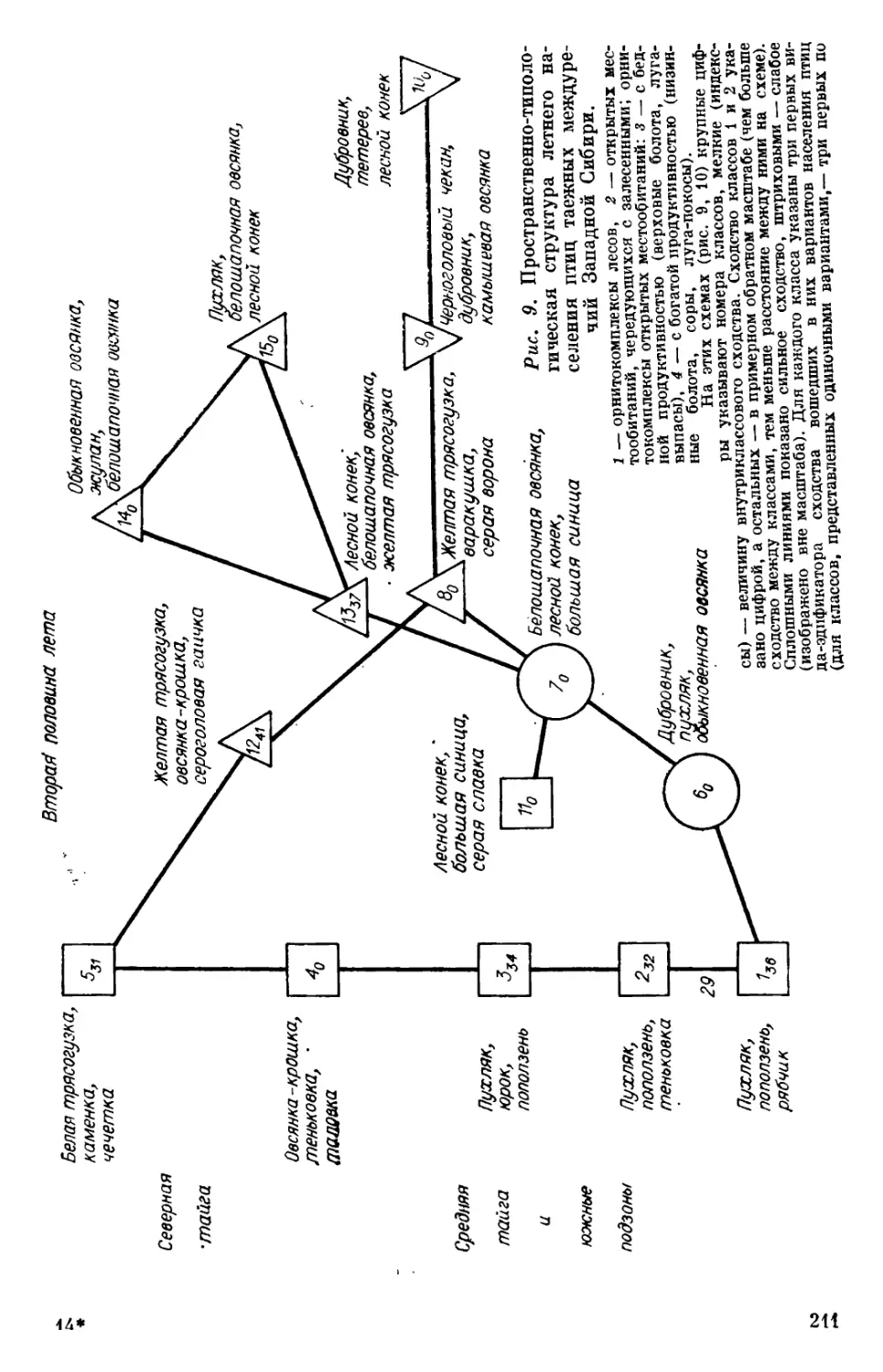
Существенная помощь в сборе, первичной обработке материала
и подготовке рукописи к печати оказана лаборантами и научными
сотрудниками лаборатории зоологического мониторинга Биологиче-
ского института СО АН СССР и студентами московских и новосибир-
ских вузов: С. М. Цыбулиным, Н. С. Степановой, Н. С. Збоевой,
Ю. А. Буйволовым, О. Э. Хатыб-Заде, С. Э. Кругляковым, С. В. Вил-
ковым, М. Г. Вартапетовым, Л. Л. и В. Л. Миловановыми, И. Н. Бо-
гомоловой, И. Ю. Губенко и Н. П. Григорьевой. Большая часть рас-
четов выполнена на БЭСМ-6 Государственного производственного
вычислительного центра СО АН СССР. При этом использованы банк
данных лаборатории зоологического мониторинга БИ СО АН СССР
и набор программ по автоматизированному анализу зоогеографиче-
ских материалов. Математическое обеспечение части расчетов про-
ведено В. И. Шадриной, а большинство использованных алгоритмов
и программ автоматической классификации разработаны В. Л. Ку-
перштохом и В. А. Трофимовым. Постоянная помощь и полезные кон-
сультации на всех этапах исследований и подготовки книги получе-
ны от Ю. С. Равкина. Ряд полезных замечаний по рукописи сделали
А. А. Максимов, А. П. Кузякин, Д. В. Терновский, Э. А. Ирисов,
С. С. Москвитин и Б. С. Юдин. Всем участникам работ и коллегам,
оказавшим помощь в обработке материала и написании книги, автор
выражает искреннюю признательность.
РАЙОНЫ РАБОТ,
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЙ
Большая часть поверхности таежных междуречий Западной Си-
бири представляет собой плоские и слабодренированные пространст-
ва *. Климат всей зоны континентально-циклонический. Северная
часть зоны отличается низкими температурами воздуха зимой и ле-
том, наличием многолетней мерзлоты и сравнительно небольшим ко-
личеством осадков. В южной части летние температуры несколько
выше и на поверхности многолетние мерзлые породы не распростра-
нены. Вся зона избыточно увлажнена.
Широкий спектр лесов почти во всех подзонах включает набор
переходов от темнохвойных до мелколиственных со значительным
участием различных сосновых насаждений. Темнохвойные леса
и их производные, образующие во всех подзонах комплекс урочищ
лесных ландшафтов, занимают наиболее дренированные террито-
рии — склоны речных долин, прирусловые валы и возвышенности
на водоразделах: Древесный ярус в коренных массивах темнохвой-
ной тайги представлен кедром, пихтой, реже елью. В северных под-
зонах для этих лесов характерны разреженный древостой, очень
слабое развитие подлеска и травяного яруса (чаще всего таежное
мелкотравье). На почве густой покров из зеленых мхов и лишайни-
ков. По мере движения на юг густота древесного яруса увеличивает-
ся. В наиболее осветленных участках южно-таежных лесов появля-
ется мощное высокотравье.
В настоящее время в ряде участков темнохвойные леса, исчез-
нувшие в результате пожаров, сплошных и выборочных рубок, по-
вреждений сибирским шелкопрядом и пихтовым усачом, замещены
разновозрастными смешанными и мелколиственными (преимущест-
венно осиново-березовыми) насаждениями. В северных подзонах
в мелколиственных лесах травостой образован в основном вейника-
ми, в южных — типичным широкотравьем. По западинам среди ти-
пичных суходольных темнохвойных и мелколиственных лесов, в до-
линах .^небольших ручьев и речек произрастают полузаболоченные
♦ Здесь приводится лишь краткая характеристика применительно к
районам наших работ, поскольку подробно физико-географические условия
тайги Западной Сибири изложены в специальных публикациях, по которым со-
ставлено данное описание [Крылов, 1961; Западная Сибирь, 1963; Природа и
экономика Привасюганья, 1966; Природные условия освоения междуречья
Обь — Иртыш, 1972].
7
хвойно-лиственные леса. В их древостое доминируют сосна, береза,
реже кедр и пихта. В подлеске встречаются те же кустарники, что
и в суходольных лесах, но более обильны гигрофильные виды. В тра-
вяном покрове господствуют вейник и осоки.
В отдельных районах таежных Обь-Иртышского и Обь-Енисей-
ского междуречий широко распространены сосняки, которые обычно
приурочены к песчаным отложениям надпойменных террас в долинах
крупных притоков. Чаще всего встречаются сосняки-ягельники и
брусничники. Значительные площади занимают гари и вырубки
по соснякам. В северной половине таежных междуречий их восста-
новление происходит за счет сосны, поэтому на таких участках от-
сутствуют смешанные насаждения. К югу увеличивается роль сос-
ново-березовых лесов. Для переходной полосы от суходольных ле-
сов к настоящим болотам характерны рослые рямы. Это угнетен-
ные сосняки с хорошо выраженным древостоем (высота деревьев
10—12 м) и густым кустарничковым ярусом из багульника и Кас-
сандры. На почве густой покров из сфагновых мхов. Осиново-бере-
зовые леса наиболее широко представлены в подзоне подтаежных
лесов. Травяной покров здесь вейниково-осоково-высокотравный.
В настоящее время эти леса в значительной степени сведены и раз-
режены вырубками, распашкой и выпасом скота.
Плоский рельеф, горизонтальное залегание осадочных пород,
слабый врез речных долин, наличие водоупорных горизонтов при
крайне замедленном стоке повсеместно способствуют образованию
больших массивов болот, в ряде случаев преобладающих по площа-
ди. В пределах таежной природно-географической зоны Западной
Сибири выделяют зону верховых сфагновых болот [Романова, 1964].
Для плоских вершин водоразделов, а также отдельных выположен-
ных участков надпойменных террас притоков Оби ландшафт верхо-
вых (олиготрофных) и переходных (мезотрофных) болот особенно
характерен. К северу болота все больше приобретают гетеротроф-
ный (комплексный) характер. Преобладающие по площади верхо-
вые участки чаще чередуются с переходными и низинными. К менее
обводненным частям болотных пространств приурочены сосново-
моховые низкорослые рямы. Это сильно разреженные сосняки с угне-
тенным древостоем и густым кустарничковым покровом из карлико-
вой березки, багульника, подбела и Кассандры. В травяно-кустарнич-
ковом ярусе местами преобладают пушица, осоки и ягодные кустар-
нички. На почве развита мощная подушка из сфагновых мхов, реже
встречаются участки лишайников. В центральных, наиболее обвод-
ненных местах болотных пространств низкорослые рямы сменяются
грядово-мочажинно-озерными комплексами — открытыми сфагно-
выми болотами с мочажинами, озерами и отдельными гривами, по-
росшими угнетенными соснами. По периферии верховых болот и на
месте погребенных речек расположены мезотрофные переходные бо-
лота, как правило, небольшие по площади (мохово-осоковые, кус-
тарничково-моховые и др.).
В притеррасных частях долин таежных рек (притоков Оби) рас-
положены евтрофные (низинные), часто кочкарниковые, залесенные
8
или открытые болота (мохово-осоковые, травяно-моховые). В дре-
весном ярусе залесенных вариантов доминируют береза пушистая
и сосна. Кустарничковый ярус представлен круглолистной и кус-
тарниковой березой, а также ивой сибирской. На почве густой покров
из зеленых и сфагновых мхов. В подзоне подтаежных лесов преобла-
дают низинные осоково-гипновые и тростниково-осоковые болота.
Последние обычно расположены обширными массивами, среди ко-
торых имеются «пятна» сосново-кустарничково-сфагновых рямов.
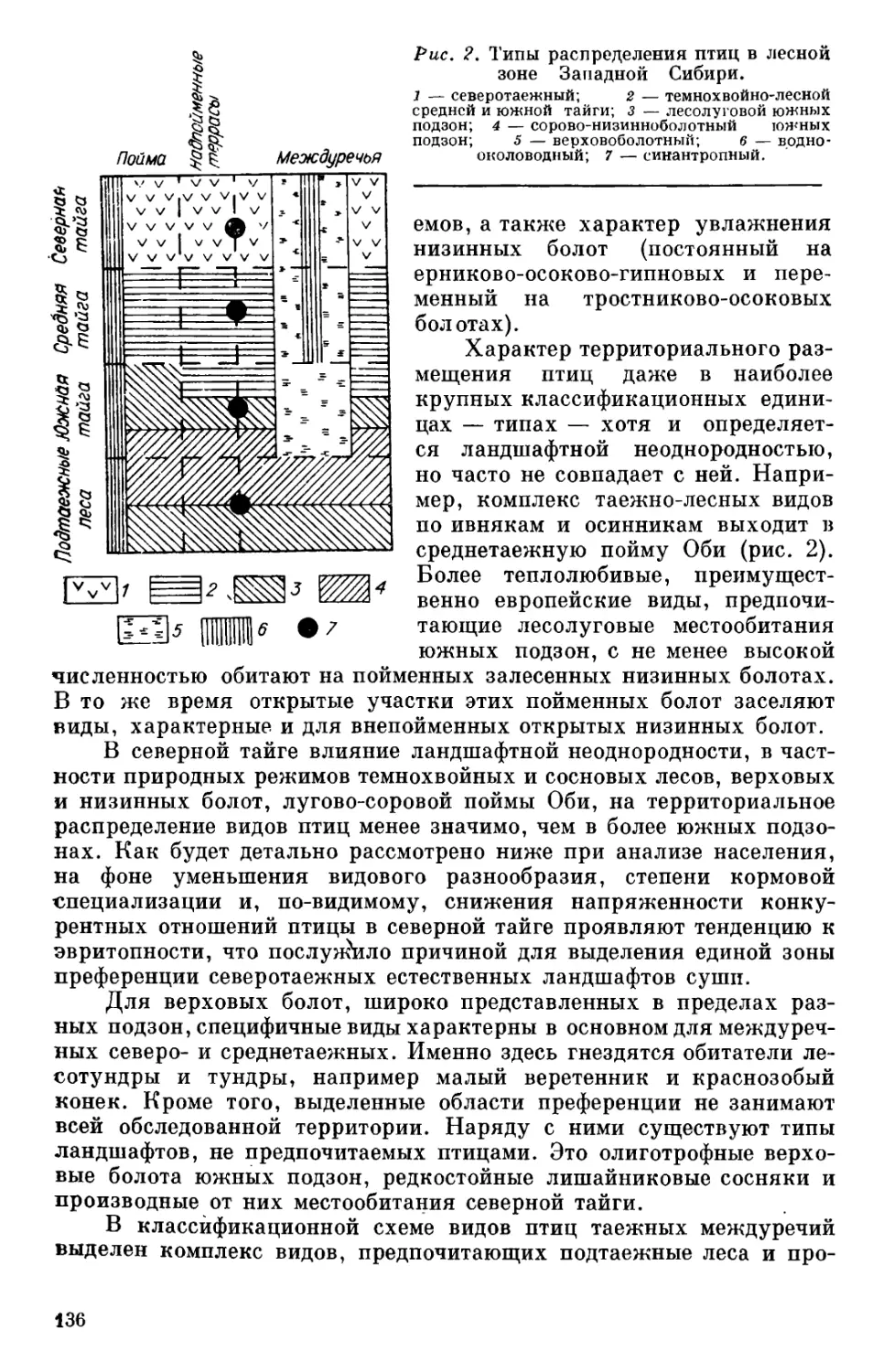
В северной тайге работы проводились в 1976 г. на Обь-Енисей-
ском междуречье в окрестностях пос. Юильск Березовского района
Тюменской области на р. Казым (рис. 1). В более южных подзонах
обследовано Обь-Иртышское междуречье. В средней тайге наблюде-
ния велись в 1977 г. близ пос. Каюково Сургутского района Тюмен-
ской области на р. Бол. Юган, в 450 км к юго-востоку от первого
ключевого участка. По южной тайге использованы данные, собран-
ные в 1967 г. в окрестностях пос. Карагай Бакчарского района Том-
ской области и опубликованные ранее Ю. С. Равкиным и И. В. Лукь-
яновой [1976]. Район работ в подтаежных лесах, обследованный
в 1978 г., располагался в окрестностях пос. Бланка Куйбышевского
района Новосибирской области. Третий и четвертый ключевые участ-
ки находились в общем к юго-востоку от второго (примерно в 600
км), но южно-таежный был одновременно самым восточным. Таким
образом, весь разрез в целом, сос-
тавленный обследованными ключе-
выми участками, пересекает таеж-
ные междуречья Западной Сибири с
юго-востока на северо-запад. Меж-
дуречья мы рассматриваем как
территории, расположенные меж-
ду долинами смежных рек [Сло-
варь..., 1975; Четырехъязычный
словарь терминов..., 1980]. В дан-
ном случае это крупные транзит-
ные реки, пересекающие лесную
зону с юга на север (Обь, Ир-
тыш, Енисей).
Птицы учитывались в север-
ной тайге с 1 июня по 31 августа,
а в более южных подзонах в свя-
зи с фенологическими различиями
наблюдения начинались раньше —
16 мая и продолжались по 31
Рис. 1. Схема района работ.
1 — северная; 2 — средняя; з — южная
тайга; 4 — подтаежные леса; 5 — обсле-
дованные ключевые участки; в — места
проведения работ, материалы которых
использованы для сравнения [Равкин,
Лукьянова, 1976; Равкин, 1978].
ШПШ-?
^4 • 5 6
9
августа. Учеты птиц проводились на постоянных, но не строго фик-
сированных маршрутах. Регистрировались все птицы независимо
от расстояния до них от линии хода наблюдателя. Пересчет на пло-
щадь производился раздельно по группам заметности [Равкин, 1967].
Очень редкие виды птиц, не встреченные на постоянных маршрутах,
учитывались дополнительно при возвращении с учетов и попутно
во время перемещений, не связанных с учетом. На реках подсчет
птиц обычно проводился с моторной лодки. Норма учета составляла
5 км за две недели наблюдений в каждом из выделенных ландшафт-
ных урочищ (на водоемах — не менее 10 км). Результаты учетов по
двухнедельным отрезкам усреднялись за первую и вторую половину
лета (с 16 мая по 15 июля и с 16 июля по 31 августа).
За три полевых сезона на постоянных маршрутах пройдено
965 км. Кроме того, на 576 км птицы подсчитывались с моторной
лодки и на 520 км зарегистрированы птицы, не встреченные на ос-
новных машрутах. Таким образом, суммарная протяженность учет-
ных маршрутов составила 2061 км.
Для расчета биомассы (сырого суммарного веса птиц на 1 км2),
распределения по ярусам (по месту сбора корма) и состава потребля-
емых кормов использованы данные из монографии «Птицы Совет-
ского Союза» (1951—1954). Энергетический поток (энергия сущест-
вования), проходящий через сообщества птиц на 1 км2 в течение су-
ток, рассчитан по В. Р. Дольнику, В. М. Гаврилову и Ч. Кенди [Гав-
рилов, 1977]. При описании численности и распределения видов птиц
и населения в целом используется балльная шкала численности и до-
минирования А. П. Кузякина (1962). Названия птиц даны по
А. И. Иванову (1976), типы фауны приводятся по Б. К. Штегману
(1938), а типы динамики птичьего населения — по Ю. С. Равкину
(1973, 1978).
Сходство сообществ измерялось коэффициентом Жаккара [Jac-
card Р., 1902], модифицированным Р. Л. Наумовым (1964). Для вы-
явления пространственно-типологической структуры сообществ поз-
воночных использовались методы автоматической классификации.
Обычно они предусматривают выделение по матрице коэффициентов
сходства несходных между собой групп максимально похожих объ-
ектов (в нашем случае — вариантов населения). Использованный
нами метод объединяет объекты в одну группу по степени сходства
со всеми остальными анализируемыми вариантами животного насе-
ления [Куперштох и др., 1975; Трофимов, 1978; Трофимов и др.,
1980]*. Полученная структура представляется в виде разделения
по сходству всех вариантов на группы и связей между такими груп-
пами. Это позволяет выявлять основные направления территориаль-
ных изменений сообществ. Далее, исходя из представлений о ланд-
шафтных особенностях обследованной территории, проводится поиск
и отбор факторов среды, в той или иной степени совпадающих с вы-
явленными пространственными изменениями сообществ позвоноч-
ных и составом выделенных групп.
* Описание принципов работы программ заимствовано из указ анных ра-
бот.
10
Для оценки связи пространственной неоднородности населения
и выявленных факторов использован метод факторной классифика-
ции [Трофимов, 1976]. Предлагаемый способ позволяет учесть при
содержательном анализе не одну, а сразу несколько классификаций.
Информация о классифицируемых объектах тоже представляется
в виде матрицы коэффициентов сходства между вариантами населе-
ния. В качестве характеристики «разброса» значений коэффициентов
принят средний квадрат отклонения от среднего по всей матрице
(дисперсия). В результате получается некоторая классификация —
объединение вариантов населения по их максимальному сходству
в незаданное число классов. При этом коэффициенты проб внутри
классов в основном положительны, а между классами — отрица-
тельны.
Далее проводится оценка учтенной этой классификацией дис-
персии коэффициентов сходства. Для этого все коэффициенты вну-
три найденных классов уменьшаются на их среднее, а коэффициенты
сходства между классами увеличиваются на абсолютную величину
их среднего. В результате получается остаточная матрица коэффи-
циентов, дисперсия которой меньше, чем исходной. Разница этих
дисперсий, отнесенная к начальной, составляет долю уменьшения.
В задание для ЭВМ включаются последовательный поиск по оста-
точным матрицам сразу 4—5 классификаций и расчеты по ним. Пер-
вые 2—3 из них обычно хорошо интерпретируются. Для этого зоо-
лог, зная условия среды в местах проведения учетов, экологию ви-
дов и анализируя состав объединенных вариантов, находит для каж-
дого класса фактор среды или их сочетание, общее для всех вошед-
ших в класс проб. Такой фактор, хотя бы предположительно, дол-
жен определять выявленное сходство населения. Если каждому из
классов более или менее четко соответствует какой-либо фактор или
их сочетание, то их набором можно объяснить изменчивость насе-
ления. Множественной мерой связи набора факторов и неоднород-
ности среды служит доля дисперсии, учтенной всеми объясненными
классификациями.
ЧИСЛЕННОСТЬ И РАСПРЕДЕЛЕНИЕ ПТИЦ
ПОВИДОВОЙ ОБЗОР
В этом разделе приводятся данные о ландшафтном распределе-
нии птиц и динамике их численности в летний период, а также опи-
сываются некоторые сезонные явления в жизни птиц. Наши резуль-
таты даются в сравнении с аналогичными материалами по южно-
таежным междуречьям и долине Оби в пределах всей лесной зоны.
Эти сведения заимствованы из публикаций Ю. С. Равкина,
И. В. Лукьяновой [1976] и Ю. С. Равкина [1978]. Кроме того, ис-
11
пользованы данные цо экологии и численности птиц из работ М. П. Та-
рунина [1959] по долине р. Мал. Сосьва (средняя тайга), А. М. Гын-
газова и С. П. Миловидова [1977] в окрестностях Томска (южная
тайга и подтаежные леса), С. С. Москвитина, А. Д. Дубовика,
Б. Я. Горда [1977] по Прикетью (граница южной и средней тайги)
и Э. В. Рогачевой, Е. Е. Сыроечковского, О. В. Бурского и др.
(1978) по среднетаежной долине Енисея. Для упрощения описания
далее в повидовых очерках обычно указываются лишь вышеперечис-
ленные районы, а ссылки на этих авторов не приводятся. В случае
использования других материалов ссылки даются полностью.
Для фоновых видов птиц все повидовые очерки построены, как
правило, по единому плану. Вначале рассматриваются сезонные яв-
ления в жизни птиц и динамика их численности (прилет и пролет,
гнездовой период, сведения о территориальной изменчивости плот-
ности гнездования и экологии размножения птиц, послегнездовые
пространственно-временные изменения численности, включая оцен-
ку послегнездовых кочевок). Затем приводится характеристика осо-
бенностей распределения вида по среднелетним показателям обилия.
Это создает впечатление некоторого повтора со сходными данными
по плотности гнездования. Однако анализ таких показателей не-
обходим в связи с их большей достоверностью (использованы объемы
учетов в 3—7 раз большие, чем для оценки плотности гнездования)
и необходимостью сравнения результатов по видам с различным
временем пребывания на обследованных ключевых участках.
В то же время публикация данных по плотности гнездования оправ-
дывается необходимостью последующих сравнений с материалами
других исследователей, которые часто приводят именно этот пока-
затель. Таким образом, использование в повидовых очерках двух
количественных характеристик (плотности гнездования и средне-
летнего обилия) не повторяет, а взаимно дополняет друг друга. За-
канчиваются повидовые очерки описанием подзональных различий
численности видов на таежных междуречьях и в сравнении с доли-
ной Оби, т. е. по лесной зоне Западной Сибири в целом.
Обилие редких видов в связи со случайностью их встреч чаще
характеризуется показателями, усредненными по большей площади,
чем ландшафтное урочище (ландшафт или ключевой участок в це-
лом), для повышения репрезентативности полученных результатов.
Кроме того, летняя динамика численности этих видов в ряде случаев
не описывается по тем же причинам, а для анализа использованы
лишь средне летние показатели.
За плотность гнездования принято обилие птиц в период ста-
билизации от окончания пролета до начала послегнездовых коче-
вок, а не в крайние сроки начала и конца гнездования. Однако для
куриных птиц этот показатель весьма условен в связи с их малой
заметностью и случайностью встреч в гнездовое время. Плотность
гнездования водоплавающих и редких видов птиц не оценивалась,
что тоже обусловлено невысокой достоверностью имеющихся данных.
При чтении повидовых очерков следует учитывать ряд их осо-
бенностей. В целях наиболее полной сравнимости со сведениями,
12
опубликованными ранее, методы расчета и показатели плотности
гнездования, интенсивности пролета и послегнездовых прикочевок
и откочевок унифицированы с теми, которые использованы Ю. С. Рав-
киным [1973, 1978]. Все сравнения численности видов на между-
речьях и в долине Оби, если это не указывается особо, проводятся
в пределах сходных подзон, что во избежание повторов не оговари-
вается. Показатели обилия приводятся в скобках и соответствуют
числу особей на 1 км2 в ландшафтах суши или на 10 км береговой ли-
нии на водоемах. Средние значения по ключевым участкам и ланд-
шафтам рассчитывались с учетом соотношения площадей, занима-
емых разными урочищами. Описание численности в соответствии
с основной задачей исследования проводится на ландшафтной осно-
ве, однако это не значит, что полученные выводы следует безогово-
рочно распространять на все типологически сходные местообита-
ния лесной зоны Западной Сибири. Напротив, сделанные заключения
относятся преимущественно к территориям обследованных участков
в пределах сроков наших работ.
Описываются только численность и распределение встреченных
нами видов птиц. Виды, зарегистрированные лишь на южно-таеж-
ных междуречьях Ю. С. Равкиным и И. В. Лукьяновой [1976] и в
долине Оби Ю. С. Равкиным [1978], отмечены в алфавитном указа-
теле звездочкой с указанием страниц вышеназванных монографий.
ОТРЯД ГАГАРЫ (GAVHFORMES)
Краснозобая гагара (Gavia stellata Pontopp.)
В северной тайге встречена только в начале июня на крупном
надпойменном озере, окруженном верховыми болотами, и в августе
на небольших озерах среди таких же болот (4 и 2). В средней тайге
отмечена в период гнездования на грядово-мочажинных верховых
болотах и в конце мая и июне (2). С начала июля до середины августа
эти гагары перестают попадаться в учеты в местах гнез-
дования, хотя в конце августа краснозобых гагар вновь виде-
ли на небольших озерках среди грядово-мочажинных верховых
болот (2). В то же время с начала июля эта гагара обычна на круп-
ных надпойменных озерах среди верховых болот (4). На одном из та-
ких озер 1 июля 1977 г. встречены две взрослые краснозобые гагары
с двумя молодыми размером с чирка.
В среднем за лето в северной тайге эта гагара редка на крупных
надпойменных озерах и грядово-мочажинно-озерных комплексах
верховых болот (0,4—0,5). В средней тайге в сходных местообита-
ниях ее в 2—4 раза больше.
Итак, краснозобая гагара четко приурочена к обширным верхо-
вым болотам, встречается только на северо- и среднетаежных между-
речьях. Она совсем не зарегистрирована на междуречьях более юж-
ных подзон и в долине Оби— от северной тайги до подтаежных лесов.
Однако пуховые и лётные птеицы этой гагары наблюдались А. М. Гын-
13
газовым и С. П. Миловидовым [1977] на Оби у г. Стрежевого (сред-
няя тайга). Южный предел гнездового распространения этой гагары
в лесной зоне Западной Сибири те же авторы отмечают по границе
южной и средней тайги.
Чернозобая гагара (Gavia ardica L.)
Встречена на гнездовании в северной тайге в грядово-мочажин-
но-озерных комплексах и низкорослых рямах верховых болот с 15
июня по 15 июля (1 и 0,2). В средней тайге в гнездовое время (с 15
мая по 15 июня) отмечалась на крупных надпойменных озерах (5).
Взрослую гагару с птенцом размером меньше чирка видели в север-
ной тайге 5 августа 1976 г., еще две гагары с птенцом размером с чир-
ка встречены 1 августа 1976 г. на крупном озере, расположенном
в долине р. Глубокий Полуй примерно в 170 км к северу от обследо-
ванного нами ключевого участка. После окончания гнездования
в июле ~ августе часть гагар откочевывает, и в этот период их
становится в 1,5—3 раза меньше, чем на гнездовании.
В среднем за лето в северной тайге чернозобая гагара чаще всего
встречалась в полете или на небольших озерках среди переходных
болот (0,4) и заметно реже — на верховых (0,08). В средней тайге
на верховых болотах ее почти втрое меньше (в среднем 0,07), но в зна-
чительном количестве эта гагара наблюдалась на крупных надпой-
менных озерах (5). В более южных подзонах на междуречьях черно-
зобая гагара не обнаружена, что, видимо, связано с отсутствием круп-
ных рек и озер в обследованных районах.
Итак, основные особенности территориального распределения
чернозобой гагары на таежных междуречьях сходны с таковыми
у предыдущего вида. В долине Обц эта гагара, как и на междуречьях,
обычна на крупных надпойменных озерах и редка на верховых бо-
лотах северных подзон.
По свидетельству А. М. Гынгазова и С. П. Миловидова [1977],
чернозобая гагара гнездится в южной тайге в бассейнах рек Кети
и среднего Васюгана. Южнее, в Обской пойме, зарегистрирована
лишь одна встреча чернозобых гагар в южной тайге в августе
1969 г.
ОТРЯД ПОГАНКИ (PODICIPITIFORMES)
Красношейная поганка {Podiceps auritus L.)
На междуречьях встречена только в средней тайге. В конце
мая стайку этих поганок видели на крупном надпойменном озере
(11). Второй раз поганки отмечены в послегнездовое время (в кон-
це июля) на р. Бол. Юган (1). В долине Оби красношейная поганка
встречалась преимущественно в пойме и реже на крупных надпой-
менных озерах. Как и на среднетаежных междуречьях, эта поганка
повсеместно редка, но на водоемах южных подзон по сравнению с се-
верными ее несколько больше.
44
ОТРЯД ПЛАСТИНЧАТОКЛЮВЫЕ (ANSERIFORMES)
Лебедь- кликун (Cygnus cygnus L.)
В северной тайге с самолета видели двух лебедей-кликунов
31 мая 1976 г. в окрестностях пос. Казым на крупном озере среди
грядово-мочажинно-озерных комплексов верховых болот. В подтаеж-
ных лесах эти лебеди встречены в конце мая и в июне в полете над
лугами-выпасами и рямами (0,02 и 0,3). Таким образом, лебедь-кли-
кун — чрезвычайно редкая птица крайних подзон таежных между-
речий. В долине Оби он наблюдался заметно чаще, но повсеместно
очень или чрезвычайно редок.
Гуси серый и гуменник (Anser anser L.,
A. fabalis L.)
Очевидно, основной пролет гусей проходил до начала наших
наблюдений. Встреченные нами гуси летели молча и на большой
высоте, что затрудняло их видовое определение. В северной тайге
их видели в полете над комплексными верховыми болотами в начале
июня (0,08). В средней тайге они также отмечались на пролете: 16
мая 1977 г. в пяти стаях видели около 200 гусей, но в среднем по тер-
ритории даже во второй половине мая их численность была невели-
ка (0,3). В южной тайге лишь однажды зарегистрирована стая из 16
особей над верховым водораздельным болотом, тоже в середине мая
(0,02). В подтаежных лесах гуси не наблюдались.
Итак, оба вида гусей повсеместно очень или чрезвычайно редки
на междуречьях в пределах собственно таежных подзон (в среднем
по участкам — 0,0009—0,03). В долине Оби в средней и южной тай-
ге их даже несколько меньше, чем на междуречьях (0,0001—0,0005),
а в северной тайге серые гуси встречались чаще (0,1).
Кряква (Anas platyrhynchos L.)
В средней тайге в конце мая прослеживается слабый пролет
кряквы, в подтаежных лесах — средней интенсивности (в среднем
по водоемам — 0,9 и 9). В начале июня кряква обычна в северо-таеж-
ных сосново-озерных ландшафтах и на озерах в поймах крупных
таежных рек (2 и 1). В средней тайге всю первую половину лета кряк-
ва встречалась только на озерах в поймах таежных рек (1). Такая
же численность зарегистрирована на южно-таежных мелких реках.
В подтаежных лесах крякву видели в конце мая в лугах-выпасах,
рямах и на мелких реках (0,2; 2 и 3). В июне кряква попадалась зна-
чительно реже и только на мелких реках (1).
В северной тайге 19 мая 1976 г. найдено гнездо с шестью яйцами
на открытом верховом болоте в 6 м от берега озера, 21 мая 1978 г.
в подтаежных лесах на ряме обнаружено гнездо с кладкой из 10 яиц.
Насиженность яиц 5—7 дней.
После окончания гнездования кряква встречена в северной тай-
ге только в конце июля в низкорослых рямах (2). В средней тайге
15
она обычна в начале августа на грядово-мочажинных верховых бо-
лотах (1), а в конце июля образует заметные скопления на крупных
надпойменных озерах (41). Так, крупное надпойменное озеро Ког-
нен-Тог, соединяясь протокой с р. Бол. Юган и обладая большими
площадями мелководий и илистых отмелей, по-видимому, было осо-
бенно привлекательным для водоплавающих, чаек и куликов в пе-
риод гнездования и особенно послегнездовых кочевок благодаря
своей высокой кормности, что обычно мало характерно для олиго-
трофных водоемов верховых болот. На мелких южно-таежных ре-
ках кряквы в это время тоже заметно больше по сравнению с первой
половиной лета (8), а в подтаежных лесах после окончания гнездо-
вания она совсем не встречалась.
В среднем за лето в северной тайге эта утка редка в сосново-
озерных ландшафтах, низкорослых рямах верховых болот и поймах
крупных таежных рек (0,2—0,4). В средней тайге на верховых бо-
лотах ее примерно столько же, а на крупных надпойменных озерах —
заметно больше (7). В южной тайге кряква встречалась только по
небольшим рекам (2). В подтаежных лесах она крайне редка в лу-
гах-выпасах и обычна в рямах. По берегам мелких рек ее здесь
столько же, сколько в южной тайге.
Таким образом, на таежных междуречьях меньше всего кряквы
зарегистрировано на водоемах северной тайги. К югу от этой подзоны
она отмечена в большем количестве. В долине Оби, особенно в ее
пойме, численность кряквы заметно выше, чем на междуречьях.
Особенно много ее здесь в южной тайге, и меньше — в северных
подзонах и подтаежных лесах. Однако если сравнивать только по
водоемам, прослеживается несколько иной характер распределения.
Самая высокая численность кряквы наблюдается в подтаежной до-
лине Оби, наименьшая — на северотаежных междуречьях и сред-
няя — по всей остальной территории,
Чирок-свистунок {Anas crecca L.)
В северной тайге свистунок обычен в июне в грядово-мочажин-
но-озерных комплексах верховых болот и на старицах крупных рек
(2 и 3). В средней тайге с 15 мая по 15 июня он встречался заметно
чаще, но многочислен не был. Его видели на всех водоемах, откры-
тых верховых болотах, небольших лужах в смешанном лесу (1—5)
и в полете над поселком (0,1). В подтаежных лесах он обычен на мел-
ких речках и лишь иногда наблюдался в полете над лугами-выпаса-
ми (5 и 0,3). Самок с пуховичками видели в средней тайге 17 и 22
июня 1977 г., а 22 августа 1976 г. в северной тайге встречен неле-
тающий молодой чирок-свистунок размером с 2/3 взрослого.
После окончания гнездования обилие чирков-свистунков воз-
росло в конце июня на крупных среднетаежных надпойменных озе-
рах до 130 особей на 10 км береговой линии. На всех остальных об-
следованных водоемах таежных междуречий оно не увеличивалось
либо даже уменьшалось по сравнению с таковым в конце мая и нача-
ле июня. В июле в северной тайге эти чирки совсем не встречались
16
на водоемах. В августе обилие свистунка вновь возрастает почти на
всех водоемах (в северной, средней тайге и подтаежных лесах соот-
ветственно до 17, 43 и 4 особей на 10 км берега). Кроме того, во вто-
рой половине лета чирок-свистунок становится обычным на грядово-
мочажинно-озерных верховых болотах от северной до южной тай-
ги (3—5).
В северной тайге в среднем за лето больше всего этого чирка
на старицах крупных рек (12), заметно меньше в приречных смешан-
ных лесах и на грядово-мочажинно-озерных верховых болотах (по 2),
где он встречался на мелких озерах. Редок на таежных реках (0,2)
и очень редок в сосново-озерных ландшафтах (0,05). В большинстве
местообитаний средней тайги чирка-свистунка несколько меньше,,
чем в северной (в среднем в 1,5 раза), но на водоемах в 4 раза больше.
Особенно много его на крупных надпойменных озерах (70). В юж-
ной тайге свистунок встречался заметно реже (в среднем по терри-
тории участка его здесь в 3 раза меньше, чем в северной тайге).
В подтаежных лесах чирок-свистунок наблюдался только на мелких
реках, но его здесь в 1,5 раза меньше, чем на водоемах северной
тайги.
Таким образом, в среднем по обследованным участкам обилие
чирка-свистунка на таежных междуречьях незначительно убывает
к югу. В сходных местообитаниях долины Оби его лишь немногим
больше. Заметно увеличивается его количество по сравнению с меж-
дуречьями лишь в северотаежной долине с ее широкой, сорово-ост-
ровной поймой. В среднем по водоемам наибольшие показатели ха-
рактерны для северотаежной долины Оби, средние — почти для всей
остальной территории, и минимальная численность этого вида чирка
характерна для южно- и северотаежных междуречий.
Свиязь {Anas penelope L.)
Пролет средней и слабой интенсивности выражен на водоемах
всех подзон в конце мая — начале июня (с севера на юг по подзонам
соответственно 0,5; 3, 5 и 8). Следовательно, интенсивность пролета
в этом направлении возрастает.
В северной тайге в июне свиязь обычна на озерах среди комплекс-
ных верховых и переходных болот (1), редка на озерах, старицах
в поймах крупных рек и очень редка в низкорослых рямах (0,5 и
0,04). В средней тайге в этот период редка на верховых болотах и пой-
менных озерах и заметно чаще наблюдается на крупных надпоймен-
ных озерах (0,6; 0,5 и 6). В пределах обследованных ключевых участ-
ков более южных подзон в начале лета свиязь не встречена.
После окончания гнездования свиязь вновь отмечалась в север-
ной тайге на старицах таежных рек в августе (1). В сходных место-
обитаниях средней тайги встречена в начале июля (3). Наибольшие
скопления свиязи, как и других видов уток, в этой подзоне зарегист-
рированы на крупных надпойменных озерах в конце июля — авгус-
те (73). В южной тайге во второй половине лета свиязь не встречалась,
а в подтаежных лесах появилась на мелких реках только в августе (2).
2 л. Г. Вартапетов
17
В среднем за лето свиязь повсеместно редка в северной тайге,
многочисленна или обычна на среднетаежных озерах, а в остальных
местообитаниях этой подзоны тоже редка. В более южных подзонах
она встречалась только на мелких реках на пролете, поэтому обилие
ее в среднем за лето невелико. Это позволяет считать свиязь редкой
птицей таежных междуречий, предпочитающей обширные озера.
В долине Оби (в среднем по участкам и водоемам) она повсеместно
обычна и встречена в большем числе, чем на междуречьях. Макси-
мальная численность отмечена в пойме Оби, особенно на южно-таеж-
ных сорах, и на крупных озерах среднетаежных междуречий.
Шилохвость (Anas acuta L.)
На водоемах северных подзон во второй половине мая — на-
чале июня отмечен сравнительно интенсивный пролет (11 и 8). В
первой половине лета в северной тайге шилохвость обычна (в лесо-
болотных местообитаниях и поймах крупных притоков Оби). В сред-
ней тайге с 15 мая по 15 июня редка на грядово-мочажинных
верховых болотах (0,8).Самки шилохвости с нелётными молодыми
размером с чирка и больше встречены в северной тайге 7, 15 и 26
июля 1976 г., в средней тайге — 6 августа 1977 г. В подтаежных
лесах самку с восемью пуховичками видели 24 мая 1978 г. на про-
токе, соединяющей два озера в ряму.
После окончания гнездования в северной тайге шилохвость
в среднем по территории встречалась реже, чем в первой половине
лета. Однако она стала многочисленной птицей стариц крупных
таежных рек и грядово-мочажинно-озерных верховых болот (15 и 12),
изредка встречалась в сосново-озерном ландшафте и на крупных
таежных реках. В средней тайге шилохвость становится многочис-
ленной в конце июля на озерах долин крупных рек. Особенно много
ее в августе на крупных надпойменных озерах (15 и 56).
Итак, на таежных междуречьях в среднем за лето шилохвость —
обычная птица северной тайги. В средней тайге ее вчетверо меньше
(в среднем по обследованной территории), но на водоемах вдвое боль-
ше. В южной тайге шилохвость совсем не встречена, и лишь один
раз ее видели в подтаежных лесах. В долине Оби этой утки повсе-
местно значительно больше, чем на междуречьях, особенно в север-
ной тайге. В среднем по обследованным участкам наибольшая ее
численность характерна для северотаежной долины Оби. Значитель-
но меньше ее в долине Оби от средней тайги до подтаежных лесов
и на междуречьях северных подзон. Меньше всего шилохвости за-
регистрировано на междуречьях, южных подзон.
Чир ок-трескунок (Anas qaerquedula L.)
Пролет средней интенсивности идет на водоемах северной тайги
в начале июня, а в остальных подзонах — во второй половине мая
(2—9). В северной тайге в июне трескунок обычен на верховых бо-
лотах и крупных таежных реках (1—2) и редок в их поймах (0,5).
18
В средней тайге в тот же период он встречается преимущественно
на озерах в поймах таежных рек (8) и лишь изредка на самих ре-
ках. В южной тайге обычен в июне на грядово-мочажинно-озерных
верховых болотах и небольших таежных реках (1—2) и совсем не
встречен в подтаежных лесах. Самки с пуховичками наблюдались
22 июня 1977 г. в средней тайге и 1 июля 1976 г. в северной. Здесь
24 июля встречена самка трескунка с девятью оперенными, но не
летающими птенцами.
В послегнездовое время в северной тайге в июле численность
чирка-трескунка возрастает почти в 7 раз, и он становится много-
численным в приречных темнохвойных лесах (76), на небольших
старицах и в заполненных водойг межгривных понижениях. В пер-
вой половине августа трескунок встречен на старицах крупных та-
ежных рек (31), а во второй половине месяца исчезает совсем. В сред-
ней тайге в это время трескунков видели только на старицах таеж-
ных рек в первой половине августа (16). На небольших южно-таеж-
ных реках в тот же период их численность возрастает до 7 особей
на 10 км берега, а на сходных подтаежных реках эти чирки встрече-
ны только в конце июля (1).
В среднем за лето в северной тайге чирок-трескунок предпочи-
тает приречные темнохвойные леса (26), попадаясь здесь на запол-
ненных водой межгривных понижениях. Значительно меньше его
на старицах таежных рек (5) и еще меньше на верховых болотах
и самих реках (0,1—0,5). В средней тайге на водоемах его примерно
столько же, сколько в северной. Больше всего трескунка на стари-
цах таежных рек (9), гораздо ниже его численность на самих реках
(0,01) и крупных надпойменных озерах (0,4). В южной тайге его при-
мерно столько же, сколько в более северных подзонах. Этот чирок
обычен на обширных водораздельных грядово-мочажинно-озерных
верховых болотах и небольших таежных речках (4—6). В подтаеж-
ных лесах ца водоемах его втрое меньше, чем в предыдущих под-
зонах. Здесь трескунок отмечен на небольших реках и низинных бо-
лотах в сочетании с покосными лугами (1 и 0,3).
Итак, на междуречьях примерно одинаковая численность чир-
ка-трескунка зарегистрирована в пределах собственно таежных
подзон и гораздо меньшая — в подтаежных лесах. В долине Оби,
особенно южно-таежной, в среднем по территории его значительно
больше. Однако на водоемах поймы и междуречий зарегистрировано
примерно равное обилие, кроме подтаежных лесов, где в пойме его
заметно больше, чем на междуречьях.
Широконоска (Anas clypeata L.)
В северной тайге нами не отмечена, хотя в долине Оби в преде-
лах этой подзоны летом 1973 г. была обычной и встречалась по всей
пойме. В средней тайге широконоску видели только в июне на круп-
ных надпойменных озерах (3). В южной тайге не встречена, а в под-
таежных лесах зарегистрирована лишь в конце мая — начале июня
2*
19
на западинах, заполненных талыми водами среди обширных низин-
ных тростниково-осоковых болот (1).
Итак, на таежных междуречьях широконоска повсеместно ред-
ка. В долине Оби ее так же мало в пределах средней тайги и значи-
тельно больше в остальных подзонах, особенно в южно-таежной
пойме.
Красноголовый нырок (Aythya ferina L.)
На междуречьях встречен лишь однажды — в подтаежных ле-
сах в полете над лугами, выпасами в конце мая (0,04). В долине Оби
наблюдался в пределах всей лесной зоны (в основном на пойменных
озерах), при этом был редок в северных подзонах и обычен в южных.
К востоку, в среднетаежной долине Енисея, красноголовый нырок
тоже редок, отмечались лишь отдельные встречи. Итак, в тайге За-
падной Сибири красноголовый нырок явно предпочитает пойменные
водоемы Оби в пределах южных подзон.
Хохлатая чернеть (Aythya faligula L.)
В северной тайге в июне многочисленна на водоемах, особенно
на старицах таежных рек (23). В средней тайге в конце мая встре-
чалась в сходных местообитаниях (4). В тот же период ее видели на
небольших речках южных подзон (1—2), а затем не встречали в те-
чение всего лета.
В июле в северной тайге обилие хохлатой чернети в результате
откочевки снижается более чем в 10 раз, и в первой половине авгус-
та она не попадается совсем. К концу этого месяца она вновь появ-
ляется в поймах таежных рек (7). В средней тайге во второй половине
июня чернеть видели на крупных надпойменных озерах и по запа-
динам в приречных смешанных лесах (2—6), реже — на таежных
реках и в их поймах (0,1—0,5). В июле хохлатые чернети концентри-
руются на старицах таежных рек (10) и перестают встречаться на
остальной территории. С 1 по 15 августа они исчезают совсем, одна-
ко в конце месяца появляются на крупных надпойменных озерах
(16).
В среднем за лето в северной тайге хохлатая чернеть предпочи-
тает старицы таежных рек и гораздо реже встречается на самих реках
(9 и 0,3). В средней тайге характер распределения сходен, но здесь
ее на водоемах в 2,5, а в среднем по участку в 5 раз меньше, чем в се-
верной тайге. Еще меньше хохлатой чернети на небольших речках
двух южных подзон, где она встречалась только в мае и в среднем
за лето была редка (0,1—0,3).
В результате следует отметить, что хохлатая чернеть предпочи-
тает поймы таежных рек и крупные надпойменные озера. В долине
р. Кеть (южная и средняя тайга) эта утка тоже предпочитает участки
с хорошо развитой поймой и в устье реки. Поэтому в северных под-
зонах, где нами обследованы крупные таежные реки с хорошо выра-
20
женной поймой, хохлатой чернети заметно больше, чем в южных
подзонах, где учеты проведены на небольших речках.
В долине Оби в среднем по территории хохлатой чернети замет-
но больше, чем на междуречьях, ее численность возрастает к севе-
ру и югу от средней тайги. Если сравнивать отдельно по водоемам,
то наибольшая численность характерна для долины Оби в пределах
подтаежных лесов. В северных подзонах обилие хохлатой чернети
резко снижается и самое низкое отмечено на междуречьях южных
подзон, т. е. однозначных изменений численности в среднем по тер-
ритории и на водоемах не прослеживается.
Морская чернеть (Aythya marila L.)
На таежных междуречьях Западной Сибири нами не встречена,
лишь одна самка морской чернети обнаружена в добыче охотника
в северной тайге 31 мая 1976 г. В долине Оби ее видели также в пре-
делах северной тайги на озерах в июне (1).
Синьга (Melanitta nigra L.)
В северной тайге в июне обычно встречалась на крупных таеж-
ных реках, на старицах и заметно реже — на небольших озерах
среди верховых болот и сосново-озерных ландшафтов (по 2; 0,6 и 0,2).
В средней тайге пролетную синьгу видели в начале июня на неболь-
ших озерках в грядово-мочажинных комплексах верховых болот (1).
В течение июля синьга совсем не наблюдалась, а затем в начале
августа она вновь зарегистрирована в северной тайге на небольших
озерах сосново-озерных ландшафтов и в поймах крупных рек (4 и 0,6).
На крупном надпойменном озере, расположенном среди комплекс-
ных верховых болот, в той же подзоне 5 августа 1976 г. встречены
самка синьги и птенец размером с половину взрослой птицы.
В среднем за лето в северной тайге синьга — редкая утка круп-
ных таежных рек, верховых болот и сосново-озерных ландшафтов
(0,1—0,5). Заметно больше ее только на озерах и старицах в пойме
р. Казым (2). В средней тайге синьги значительно меньше (примерно
в 5 раз). Здесь она встречена лишь на пролете на верховых болотах.
В южных подзонах таежных междуречий синьга нами не зарегистри-
рована. А. М. Гынгазов и С. П. Миловидов [1977] указывают южную
границу её спорадичного гнездования около устья Иртыша (средняя
тайга). В долине Оби синьга встречена только в северной тайге, где
ее заметно меньше, чем на междуречьях.
Итак, в лесной зоне Западной Сибири летом синьга явно предпо-
читает северную тайгу, лишь иногда встречаясь в средней на пролете.
Гоголь (Bacephala clangula L.) 1
В северной тайге в первой половине лета встречен на крупных
реках и верховых болотах (1 и 0,5). В средней тайге с 16 мая до 15
июня стайки гоголей наблюдались на крупных надпойменных озе-
21
pax (16). На таежных реках и старицах их заметно меньше (0,2 и 2).
В южной тайге гоголь редок на небольших таежных речках (0,8).
Самки с выводками из 6—7 птенцов отмечались в средней тайге
16 и 25 июня 1977 г., а в северной — 24, 26 и 29 июля 1976 г. Встре-
ченные 26 июля 1976 г. молодые птенцы были размером со взрослых.
В северной тайге с 15 по 30 июля численность гоголя возрастает
в результате размножения и, по-видимому, прикочевки. На водо-
емах обилие увеличивается почти в 10 раз, а к концу августа — еще
вдвое. В этот период гоголь встречается преимущественно на стари-
цах р. Казым и в небольших заполненных водой межгривных по-
нижениях в приречной темнохвойной тайге (10). На водоемах сред-
ней тайги прослеживается несколько иной ход изменений числен-
ности гоголя. К концу июня обилие возрастает втрое по сравнению
с первой половиной месяца, а затем примерно в 15 раз снижается
в результате откочевки. К концу августа численность этой утки
вновь неуклонно возрастает почти до прежнего максимального
уровня. На небольших таежных реках численность гоголя при не-
высоком ее уровне снижается начиная с конца мая до конца августа
с 1 до 0,3 особей на 10 км береговой линии.
В среднем за лето в северной тайге гоголь многочислен на озе-
рах, старицах крупных таежных рек и небольших залитых водой
западинах в приречных темнохвойных лесах (по 10), обычен в сосно-
во-озерном ландшафте (1) и редок на верховых болотах и крупных
таежных реках (0,1—0,6). В средней тайге на большинстве водо-
емов редок. В среднем по участку его заметно меньше, чем в север-
ной тайге, но на водоемах в 1,6 раза больше за счет скоплений го-
голя на крупных надпойменных озерах (в среднем за лето 22). Го-
раздо реже наблюдался гоголь на южно-таежных речках (0,4) и сов-
сем не встречен в подтаежных лесах.
Следовательно, в северных подзонах на таежных междуречьях
гоголь встречается гораздо чаще, чем в южных. В долине Оби под-
зональные различия его численности не прослеживаются. Гоголя
десь в среднем по территории примерно столько же, сколько на се-
вере- и среднетаежных междуречьях, а на водоемах — несколько
меньше.
Средний крохаль (Mergas serrator L.)
В северной тайге отмечался только до середины июля на гря-
дово-мочажинно-озерных верховых болотах, на крупных таежных
реках и их старицах (0,2—0,3). Нами на междуречьях в более юж-
ных подзонах и Ю. С. Равкиным [1978] в долине Оби этот крохаль
не встречен.
Таким образом, в пределах обследованной нами территории
средний крохаль зарегистрирован и, видимо, гнездился только на
северотаежных междуречьях. Вероятно, он гнездится и южнее,
в средней тайге, так как пуховых птенцов добывали в этой подзоне
на р. Лозьва [Гынгазов, Миловидов, 1977]. В среднетаежной долине
Енисея встречается только на пролете, но, видимо, гнездится по до-
линам его притоков.
22
Большой крохаль (Mergus merganser L.)
В северной тайге наблюдался только в конце июня на крупных
таежных реках и их старицах (0,3 и 1). В средней тайге в тот же пе-
риод обычен на надпойменных озерах и редок в начале июля в низ-
корослых рямах (2 и 0,1). На среднетаежном крупном надпойменном
озере 22 июня 1977 г. встречено три выводка больших крохалей с 2,
4 и 6 пуховыми птенцами.
В южной тайге этого крохаля не видели ни разу и лишь изред-
ка наблюдали в подтаежных лесах в конце июля — начале августа
в полете над низинными болотами и полями-перелесками (0,2 и 0,5).
Итак, большой крохаль повсеместно редок на таежных между-
речьях Западной Сибири. В долине Оби встречался еще реже и толь-
ко на небольших среднетаежных речках. Заметно больше его на Ени-
сее в средней тайге — в среднем за лето 6 птиц на 10 км береговой
линии.
ОТРЯД ХИЩНЫЕ ПТИЦЫ (FALCO NIFORMES)
Скопа (Pandion haliaetus L.)
В северной тайге встречалась постоянно по берегам крупных
таежных рек, их стариц и реже на гарях по соснякам (0,1—0,5).
В средней тайге скопа обычна по берегам крупных озер (1) и очень
редка по берегам таежных рек, на верховых болотах и в сосново-
боровом ландшафте (0,02—0,06). В этой подзоне 15 июня 1977 г.
найдено гнездо скопы на сухой сосне. Вблизи его основания распо-
лагалось гнездо белой трясогузки.
В южных подзонах на междуречьях скопа нами не встречена,
однако зарегистрирована на гнездовании А. М. Гынгазовым и
С. П. Миловидовым (1977) в южной тайге Обь-Ейисейского между-
речья в бассейнах рек Кеть и Чулым. В долине Оби она наблюдалась
на южно-таежных пойменных сорах в начале июня и конце августа
(в среднем за лето по району работ 0,009). В таежной долине Енисея
скопа тоже очень редка и чаще встречалась на его притоках [Сыро-
ечковский, Рогачева, 1980].
Итак, в лесной зоне Западной Сибири скопа повсеместно редка
и предпочитает берега крупных притоков Оби и их стариц в пределах
северной и средней тайги.
Черный коршун (Mildus korschun Gm.)
В северной тайге его видели лишь однажды в начале июня в по-
селках. В средней тайге встречался постоянно, чаще всего в посел-
ках, на крупных таежных реках и их старицах (1 и по 0,4). Реже
коршуна отмечали в сосново-боровом ландшафте и на открытых вер-
ховых болотах (0,06 и 0,005). В южной тайге очень редок в лесном
ландшафте (0,01). В подтаежных лесах все лето черный коршун обы-
чен в поселках и редок в лугах-выпасах (1 и 0,3).
23
Итак, на таежных междуречьях черный коршун предпочитает
поселки и берега водоемов. Его численность неуклонно возрастает
к югу и составляет соответственно от северной тайги к подтаежным
лесам 0,0006; 0,003; 0,007 и 0,02 особи на объединенный 1 км2 ключе-
вого участка. В долине Оби и на междуречьях в пределах северных
подзон показатели обилия сходны, а в южно-таежной и подтаежной
долинах Оби отмечены наибольшие по всей рассматриваемой терри-
тории значения, хотя и здесь он редок.
Орлан-белохвост (flaliaeetus albicilla L.)
В северной тайге на р. Казым самец и самка орлана-белохвоста
были добыты И. Н. Шуховым [1916] 14 мая и 12 июня 1914 г. Нами
он встречен только в средней тайге в конце августа на р. Бол. Юган
(0,3). Кроме того, в первой половине августа молодого орлана-бело-
хвоста видели сидящим на крыше дома в крупном пос. Угут, пример-
но в 30 км к северу от района наших работ. В южной тайге гнездова-
ние зарегистрировано А. М. Гынгазовым и С. П. Миловидовым [1977}
на р. Васюган. В долине Оби встречался гораздо чаще в пределах
всех собственно таежных подзон, предпочитает пойменные водоемы
и северную тайгу, но повсеместно обилие его невелико (очень редок
в северной и южной тайге и чрезвычайно редок — в средней).
Тетеревятник (Accipiter gentilis L.)
В северных подзонах нами не обнаружен, но был добыт охотни-
ком в северной тайге на старице р. Казым 28 августа 1976 г. В юж-
ной тайге редок в лесном ландшафте (0,4). В подтаежных лесах встре-
чен только в конце мая и августа и в среднем за лето редок в лесо-
полевом ландшафте (0,1).
В среднем по обследованным участкам больше всего тетеревят-
ника отмечено на южно-таежных междуречьях и в севере таежной
долине Оби (0,3 и 0,2). Значительно ниже его численность на под-
таежных междуречьях и в долине Оби в пределах срединных подзон
(0,03—0,05), еще ниже — в долине Оби в пределах подтаежных ле-
сов (0,003).
Повсеместно предпочитает темнохвойные, иногда мелколиствен-
ные, и смешанные полузаболоченные леса. С уменьшением залесен-
ности и возрастанием освоенности территорий обилие тетеревятника
снижается.
Перепелятник (Accipiter nisus L.)
В северной тайге на междуречьях не встречен, но в долине Оби
в среднем по району работ редок (0,2). В средней тайге его видели
только в начале июня на вырубках по соснякам и в августе на круп-
ных таежных реках и в поселках (в среднем за лето 0,05 и 0,1).
В южной тайге перепелятник тоже редок в лесном ландшафте (0,8).
24
В подтаежных лесах все лето встречался в поселках и только в конце
мая — в рямах (0,9 и 2).
Итак, на таежных междуречьях перепелятник повсеместно ре-
док, чаще всего он встречался в поселках. В долине Оби его видели
в различных местообитаниях, но здесь его едва ли больше, чем на
междуречьях. В среднем по участкам перепелятник (как и тетере-
вятник) редок на южно-таежных междуречьях и в северотаежной
долине Оби, а на остальной территории — очень или чрезвычайно
редок.
Канюк (Buteo buteo L.)
В северной тайге встречен только в поселках в конце июня и ав-
густа (в среднем за лето — 0,5). В средней тайге в конце мая очень
редок на открытых верховых болотах и редок на крупных таежных
реках (0,03 и 0,3). В южной тайге обычен в лесном ландшафте (2).
В подтаежных осиново-березовых лесах редок в течение всего лета
(0,5) и очень редок в полях-перелесках и рямах (0,09 и 0,01).
В долине Оби канюк встречен лишь в южных подзонах, где он
повсеместно редок. В северных подзонах на Оби он зарегистрирован
У Сургута, Ханты-Мансийска и Березова, т. е. до границы средней
и северной тайги [Гынгазов, Миловидов, 1977]. Таким образом, ка-
нюк предпочитает лесные ландшафты южных подзон. В пределах
этих подзон в среднем по участкам на междуречьях его больше, чем
в долине Оби. Минимальная численность зарегистрирована на се-
веро-и среднетаежных междуречьях.
Большой подорлик (Aguila clanga Pall.)
В северных подзонах нами не зарегистрирован, хотя А. М. Гын-
газовым и С. П. Миловидовым [1977] наблюдался на р. Тым,
а А. Д. Шаронов [1951] считает его гнездящимся до 63° с. ш. Кроме
того, он встречен во второй половине августа 1973 г. в северотаеж-
ной долине Оби на переходном болоте (0,004). В южной тайге боль-
шого подорлика видели в полях-перелесках; в среднем по участку
он чрезвычайно редок. (0,006). В подтаежных лесах встречался пре-
имущественно в открытых лугово-полевых местообитаниях (0,03—
0,09). В долине Оби тоже повсеместно очень редок; в южных подзо-
нах его, видимо, больше, чем в северных и примерно столько же,
сколько на южно- и подтаежных междуречьях.
Полевой лунь (Circus cyaneus L.)
В северной тайге встречен в конце августа в поселках и по бе-
регам крупных таежных рек (0,006 и 0,6). В средней тайге нами не за-
регистрирован, но наблюдался в пойме Оби в начале и конце лета
(0,5). В южной тайге численность этого луня несколько выше, одна-
ко и здесь он повсеместно редок (в среднем по территории — 0,09).
В подтаежных лесах его видели в начале июня и конце августа.
25
В среднем за лето он редок в поселках и мелколиственных лесах
и очень редок в лугах-выпасах (по 0,3 и 0,09).
Итак, полевой лунь, предпочитая открытые местообитания,
повсеместно редок на таежных междуречьях, при этом в южных
подзонах его заметно больше, чем в северной тайге. В долине Оби
зарегистрирована лишь немногим большая численность этого луня.
В результате следует отметить, что полевой лунь отдает предпочте-
ние лесополелуговым местообитаниям южно- и подтаежных между-
речий и пойме Оби в пределах всех лесных подзон.
Степной лунь (Circus macrourus Gm.)
Встречен нами только в подтаежных лесах в полете над низин-
ными болотами в сочетании с покосными лугами в конце мая и пер-
вой половине июня (0,8). В долине Оби Ю. С. Равкиным [1978] не за-
регистрирован.
Луговой лунь (Circus pygargus L.)
Как и степной лунь, отмечен только на подтаежных междуречьях.
В отличие от остальных луней держался здесь в течение всего лета
и встречался несколько чаще. Обычен на низинных болотах в соче-
тании с покосными лугами (1) и редок в остальных лугово-полевых
местообитаниях и по берегам небольших речек (0,1— 0,4).
Пустельга (Cerchneis tinnunculus L.)
В северной и средней тайге нами не встречена. На Енисее в сред-
ней тайге зарегистрирована как редко гнездящаяся птица. В южной
тайге обычна в полях-перелесках и на низинных болотах (5 и 1),
а в среднем по территории редка (0,4). В подтаежных лесах редка
в полях-перелесках, лугах-выпасах и поселках (0,1—0,7) и очень
редка на низинных болотах (0,06). В этой подзоне ее вчетверо мень-
ше, чем в южной тайге. В долине Оби пустельга встречалась тоже
только в южных подзонах, при этом в южной тайге ее больше на меж-
дуречьях, а в подтаежных лесах — в долине Оби, но исходная тен-
денция, по-видимому, сводится к возрастанию численности от между-
речий к долине.
Д е р б н и к (Aesalon columbarius L.)
В северной тайге и подтаежных лесах нами не отмечался.
В средней тайге его изредка видели на грядово-мочажинных вер-
ховых болотах и в приречных смешанных лесах, еще реже (0,4 и
0,1) — по берегам крупных таежных рек (0,04). В южной тайге встре-
чен только на обширных водораздельных верховых болотах (0,2).
Дербник предпочитает открытые участки верховых болот, отмечен
в качестве очень редкой птицы средне-и южно-таежных междуречий.
В долине Оби Ю. С. Равкиным [1978] не зарегистрирован.
26
Кобчик (Erу thro pus vespertinus L.)
В северной тайге чаще всего встречался в сосново-озерном ланд-
шафте и на комплексных верховых болотах (1 и 0,2), реже — в при-
речной темнохвойной тайге и поселках (0,04—0,08). В этой подзоне
7 августа 1976 г. в сосново-озерном ландшафте недалеко от гнезда
встречено два молодых кобчика с кормящими их взрослыми птица-
ми. В средней тайге кобчик нами не зарегистрирован, но в долине
Оби в пределах этой подзоны его наблюдали в пойме в начале ию-
ня (4). В среднем по району работ его здесь примерно столько же,
сколько на северотаежных междуречьях. В южной тайге кобчик от-
мечен повсеместно в открытых и мало залесенных местообитаниях
(0,01—0,3). В подтаежных лесах кобчиков наблюдали только в пер-
вой половине июня в лугах-выпасах и на низинных болотах (0,4—
0,6).
В среднем по ключевым участкам больше всего этого сокола в
северной тайге (0,3) и значительно меньше в южных подзонах (0,05 и
0,03). В долине Оби зарегистрирована сходная численность по срав-
нению с междуречьями, хотя в северотаежной долине он не отмечен.
При этом к югу лесной зоны кобчик встречался несколько реже.
В отличие от пустельги он заходит дальше на север.
Чеглок (Hypolriorchis subbuteo L.)
В северной тайге изредка встречался на верховых болотах (0,1),
в сосново-озерном ландшафте и по берегам крупных таежных рек
(0,01 и 0,05). В средней отмечен тоже на верховых болотах (0,3),
в сосновых и смешанных лесах, по берегам таежных рек (0,1—0,4)
и вблизи крупных озер (0,08). В этой подзоне 15 июня 1977 г. на опуш-
ке сосняка вблизи верхового болота встречена пара чеглоков, кото-
рые волновались недалеко от гнезда. В южной тайге редок в лесных
местообитаниях в сочетании с полями и обычен на низинных боло-
тах (0,2 и 5). В подтаежных лесах чеглок редок в лесополевом ланд-
шафте (0,2). В среднем по ключевым участкам таежных междуречий
этого сокола больше в срединных таежных подзонах (0,2 и 0,4) и
меньше в крайних (0,06 и 0,1). Сходная тенденция прослеживается
в долине Оби, но чеглока здесь несколько меньше, чем на между-
речьях.
Кречет (Falco gyrfalco L.)
Встречен лишь однажды в конце августа в полете над северо-
таежным поселком (0,02). В более южных подзонах на междуречьях
и в долине Оби кречета ни разу не видели. В период пролета и ко-
чевок встречается в разных местах лесной зоны Западной Сибири
(Тюмень, Тобольский район, Томск), а гнездится севернее, в лесо-
тундре [Гынгазов, Миловидов, 1977]. В среднетаежной долине Ени-
сея встречен дважды в окрестностях пос. Мирное — 11 и 20 мая
1978 г.
27
Сапсан (Falco peregrinus Tunst.)
В северной тайге встречен дважды в августе — в сосново-
озерном ландшафте и на переходном болоте (по 0,3). На последнем,
видимо, гнездился, так как в начале августа 1976 г. видели двух мо-
лодых и двух взрослых птиц недалеко от гнезда. В средней тайге
сапсан тоже редок, встречался в поселках и на грядово-мочажин-
ных верховых болотах (0,4 и 0,1). В южной тайге отмечен также на
обширном грядово-мочажинно-озерном верховом болоте. В подта-
ежных лесах на междуречьях сапсана не видели, но в долине Оби
он обычен на надпойменных низинных болотах (2).
Итак, сапсан повсеместно предпочитает открытые болота. В пре-
делах собственно таежных подзон в среднем по ключевым участкам
на междуречьях отмечено сходное его обилие (0,02—0,03). В долине
Оби сапсан зарегистрирован только в южной тайге и подтаежных
лесах (0,001 и 0,1).
ОТРЯД КУРИНЫЕ (GALLIFORMES)
Перепел (Coturnix coturnix L.)
В. северной тайге не зарегистрирован. В средней тайге перепела
слышали только в начале июня в низкорослых рямах верховых
болот (0,3). Первая встреча с ним в этой подзоне была 2 июня 1977 г.
В южной тайге перепел появился в последней декаде мая 1967 г.,
а в подтаежных лесах — 3 июня 1978 г.
На гнездовании (в июне) в южной тайге перепел многочислен
на низинных болотах (14) и редок на верховых болотах и в полях-
перелесках (0,2—0,8). В подтаежных лесах в этот период он предпо-
читает луга-выпасы и низинные болота, перемежающиеся с сенокос-
ными лугами (3 и 2), и в значительно меньшем числе встречается в
полях в сочетании с перелесками (0,5).
После окончания гнездования (первая половина июля) в южных
подзонах обилие перепела возрастает. Далее в южной тайге числен-
ность его неуклонно снижается. В конце июля перепела встреча-
ются только на небольших полях, окруженных лесами, и к середи-
не августа исчезают совсем. В подтаежных лесах в конце июля и на-
чале августа они попадались в учетах заметно реже. В конце ав-
густа перепелов становится заметно больше, птицы в значитель-
ном числе концентрируются на низинных болотах в сочетании с
покосными лугами (24).
В среднем за лето перепел очень редок на верховых болотах сред-
ней тайги (0,02). В южной тайге, где его заметно больше, он предпо-
читает поля-перелески и низинные открытые болота (13 и 5), зна-
чительно меньше его на верховых и переходных болотах (в среднем —
0,06). В подтаежных лесах перепел обычен на низинных болотах в
сочетании с сенокосными лугами и в лугово-полевых местообитаниях
(1—6). В среднем по территории его втрое больше, чем в южной тайге.
28
Итак, обилие перепела на таежных междуречьях неуклонно воз-
растает к югу. В долине Оби он встречен лишь в пределах южных
подзон, где значительную площадь занимают лесополевые ландшаф-
ты. Численность перепела в них примерно такая же, как и на под-
таежных междуречьях.
Белая куропатка {Lagopus lagopus L.)
В северной тайге обычна на гнездовании по окраинам сосняков-
ягельников и низкорослых рямов верховых болот (в первой полови-
не июня 2—4). В средней тайге с середины мая до середины июня
ее видели только на верховых болотах (4). В сходных местообита-
ниях южной тайги в этот период не обнаружена. После окончания
гнездования в северной тайге белая куропатка не встречена. В сред-
ней тайге ее становится гораздо больше в конце июля, а затем оби-
лие заметно снижается. В южной тайге белая куропатка наблюда-
ется преимущественно на открытых участках верховых болот в кон-
це июня — начале июля, уже после появления молодых, и в этот
период она многочисленна (14). В подтаежных лесах нами не заре-
гистрирована.
В среднем за лето в северной тайге белая куропатка редка в
ягельниковых сосняках и по окраинам верховых болот в низкорос-
лых рямах (0,4—0,5). В средней тайге ее в среднем в 4 раза больше.
Здесь белая куропатка отдает предпочтение грядово-мочажинным
верховым болотам, в меньшем количестве встречаясь в рямах (1—3).
В южной тайге характер распределения и численность в среднем по
территории сходны, хотя в предпочитаемых местообитаниях ее вдвое
больше, чем в средней тайге. На подтаежных междуречьях, как и в
долине Оби, не встречена.
Итак, в пределах таежных междуречий белая куропатка четко
приурочена к верховым болотам собственно таежных подзон. При
этом в северной тайге ее встречено меньше, чем в средней и южной.
В долине Оби в пределах северных подзон ее вчетверо больше, а в
южной тайге почти вдвое меньше по сравнению с междуречьями.
В южно-таежной долине Оби эта куропатка наблюдается в после-
гнездовое время в полях и пойменных лугах. Верховые болота здесь
не обследованы, что, видимо, несколько занижает ее численность.
Глухарь {Tetrao urogallus L.)
В северной тайге на гнездовании в июне обычен в сосново-боро-
вом и сосново-озерном ландшафте (1 и 2). В средней тайге с середины
мая до конца июня глухарь отмечен в приречных смешанных лесах
(3) и на вырубках по соснякам (0,2). В южной тайге он обычен в лес-
ных местообитаниях (3). В подтаежных лесах на гнездовании не об-
наружен.
В северной тайге в послегнездовое время (с 15 июля по 30 ав-
густа) численность глухаря возрастает вдвое. В августе он в значи-
тельном числе начинает встречаться по окраинам переходных болот
29
(10). Еще больше глухаря отмечено в этой подзоне в конце июля
1976 г. на крупнобугристых болотах в 180 км к северу от места наших
работ, в долине р. Глубокий Полуй (15). При урожае голубики по
окраинам этих болот можно было видеть одновременно по два вы-
водка глухарей. В средней тайге обилие глухаря в среднем увеличи-
валось в 2 раза во второй половине лета по сравнению с таковым
на гнездовании. В южной тайге после середины лета глухарь пере-
стает встречаться совсем. В то же время в более северных подзонах,
где преобладают ягельниковые и брусничниковые сосняки, интен-
сивность размножения близка к показателям, приводимым М. И. Со-
рокиной (1974). В подзоне подтаежных лесов глухарь впервые отмечен
нами в учетах в августе в рямах (9). Самок с выводками из пяти
и трех молодых крупнее рябчика наблюдали в северной тайге 12 и
25 июля 1976 г. В средней тайге выводок из пяти пуховичков видели
23 июня 1977 г. В подтаежных лесах выводок пз пяти молодых глу-
харей обнаружен 4 августа 1978 г.
В среднем за лето в северной тайге глухарь чаще всего встре-
чается на переходных болотах, в низкорослых рямах и на гарях
по соснякам (2—3) и заметно реже — в сосновых и темнохвойных
лесах (0,5). В средней тайге он отмечен только в лесных местообита-
ниях (1—3), а в среднем по территории его примерно столько же,
сколько и в северной тайге (0,9). В южно-таежных темнохвойных и
мелколиственных лесах глухаря вдвое меньше, чем в сосново-боро-
вом ландшафте предыдущей подзоны, а в среднем по участку его поч-
ти столько же (0,7). В подтаежных лесах в среднем по району работ
глухаря вчетверо меньше, чем в более северных подзонах.
Итак, на междуречьях глухаря заметно больше в пределах соб-
ственно таежных подзон, особенно в северной и средней тайге, чем
в подтаежных лесах. В долине Оби сходная численность глухаря по
сравнению с междуречьями отмечена в средней тайге и подтаежных
лесах; в северной тайге его заметно меньше. Глухарь совсем не встре-
чен в южной тайге, где в районе работ преобладают лесополевые
местообитания, которых он избегает. По-видимому, в долине Оби
исходная тенденция изменений искажена усиленным антропогенным
влиянием (перепромыслом в северной тайге в связи с близостью круп-
ного поселка и отсутствием пригодных хвойных лесов в южной тайге,
из-за высокой степени сельскохозяйственной освоенности обследо-
ванной нами территории).
Тетерев (Lyrurus tetrix L.)
На междуречьях северной тайги нами не встречен, но зареги-
стрирован в низкорослых рямах северотаежной долины Оби (в сред-
нем за лето — 1). В средней тайге в гнездовое время обнаружен толь-
ко в рямах по окраинам верховых болот (2—3). В южной тайге пред-
почитает осиново-березовые леса и открытые низинные болота (19 и
6), реже встречается по периферии верховых болот. В подтаежных
лесах обычен в лесополевых местообитаниях и на низинных болотах
30
(1—2) и редок в рямах (0,7). В этой подзоне 15 июня 1978 г. в осино-
во-березовом лесу встречена самка с пятью птенцами 1—2-дневного
возраста.
После окончания гнездования в средней тайге обилие и широта
распределения тетерева по местообитаниям возрастают. В августе
при неплохом урожае брусники он начинает встречаться в сосняках
и на грядово-мочажинных верховых болотах (5 и 4). В южной тай-
ге с середины июля до конца августа тетерев в значительном числе
концентрируется на низинных болотах при высоком урожае клюквы
(146). На подтаежных низинных болотах обилие тетерева во второй
половине лета по сравнению с первой возрастает с 1 до 3 особей/км2.
В среднем за лето в средней тайге тетерев предпочитает сосняки
и грядово-мочажинные комплексы верховых болот (2), по крайней
мере в годы урожая брусники. В заметно меньшем количестве он
встречается в рямах по окраинам болот. В южной тайге тетерева
значительно больше, особенно на открытых низинных болотах, в мел-
колиственных лесах и в низкорослых рямах (65, 11 и 9). В подтаеж-
ных лесах его заметно меньше, чем в южной тайге, но несколько
больше, чем в средней. Как и в предыдущей подзоне, здесь он пред-
почитает лесополевые ландшафты и низинные болота (2). В меньшем
числе тетерев встречается в рямах и осиново-березовых лесах (0,3—
0,6).
В итоге можно констатировать, что на междуречьях тетерева
значительно больше в пределах южных подзон, чем в средней тай-
ге. На надпойменных террасах долины Оби тетерев так же немного-
числен, как и на среднетаежных междуречьях. По-видимому, ис-
ходная тенденция — увеличение численности к югу — здесь иска-
жается в результате интенсивной распашки и перепромысла в юж-
ных подзонах, что выравнивает показатели в южной тайге и подтаеж-
ных лесах по сравнению с северными подзонами долины Оби.
Рябчик (Tetrastes bonasia L.)
В гнездовое время (июнь) в северной тайге встречался в при-
речных темнохвойных лесах (3). В сходных темнохвойно-мелколист-
венных лесах средней тайги с 15 мая до 15 июня он тоже обычен (4).
В южной тайге в тот же период наблюдался гораздо шире, предпо-
читая темнохвойные суходольные и смешанные по л у заболоченные
леса. В меньшем количестве рябчик держался в остальных лесных
местообитаниях, в том числе в рослых рямах и на шелкопрядниках,
гарях, вырубках (в среднем по лесному ландшафту — 11). Гнездо
рябчика с пятью яйцами найдено в средней тайге в приречном сме-
шанном лесу под стволом упавшего дерева 17 мая 1977 г.
Выводок из пяти молодых хорошо летающих рябчиков встречен
в северной тайге 26 августа 1976 г. В средней тайге пуховичков
рябчика мы видели 5 и 7 июля 1977 г., а И. К. Приходько [1969]
2 июля 1965 г. наблюдал самку с пятью молодыми рябчиками разме-
ром со скворца и 30 июля того же года — молодых, достигших раз-
мера взрослой особи.
31
В послегнездовое время в северной тайге рябчик не встречался
с начала июля до середины августа, но в конце этого месяца его
обилие возросло в 3 раза по сравнению с периодом гнездования.
В средней тайге показатели обилия колеблются еще больше. Рябчика
не видели на маршрутах с 1 июня до 15 июля, затем зарегистрирован
в среднем за вторую половину лета в количестве в 4 раза большем,
чем на гнездовании. В южной тайге показатели обилия неуклонно
возрастают с начала июля до конца августа и в среднем в 6 раз
превышают таковые во время гнездования. При этом в послегнез-
довый период обилие рябчика по двухнедельным отрезкам изменя-
ется в 3—4 раза. Такие отличия мало достоверны и связаны с
большей случайностью встреч рябчиков на учетных маршрутах
из-за объединения выводков и их кормовых перемещений. Таким
образом, па таежных междуречьях Западной Сибири летние изме-
нения численности рябчиков в основном определяются результатом
их размножения, тогда как в Северо-Восточном Алтае они в зна-
чительной степени связаны с вертикальными перемещениями от-
гнездившихся птиц [Равкин, 1973].
В среднем за лето в северных подзонах рябчик явно приурочен
к приречным темнохвойным и темнохвойно-мелколиственным лесам
{2 и 20). В предпочитаемых местообитаниях южной тайги его в 3 ра-
за больше, чем в средней. Встречается он гораздо шире во всех лес-
ных, лесополевых ландшафтных урочищах и рослых рямах, пред-
почитая елово-кедро-пихтовые и осиново-березовые леса (59—60).
Рябчик отмечался даже по окраинам низинных болот (9). Столь вы-
сокая численность, по-видимому, бывает не часто. Так, В. И. Те-
лепнев (1974) в итоге многолетних наблюдений считал сезон 1967 г.
в южной тайге «урожайным» на рябчика.
Итак, на междуречьях, по крайней мере в годы наших наблюде-
ний, обилие рябчика в пределах собственно таежных подзон неуклон-
но снижалось к северу. В долине Оби в пределах северной тайги ряб-
чик не зарегистрирован. В надпойменных местообитаниях остальных
подзон и на среднетаежных междуречьях его обилие примерно оди-
наково.
ОТРЯД ЖУРАВЛЕОБРАЗНЫЕ (GRUIFORMES)
Коростель (Crex crex L.)
Нами в северной тайге не обнаружен, а в долине Оби его лишь
однажды слышали на низинном пойменном болоте в июле 1973 г.
В средней тайге Обь-Иртышского междуречья чрезвычайно редок—
только один раз 23 июня 1977 г. встречен на открытом низинном
болоте, учеты на котором не проводились. Первый крик коростеля
в подтаежных лесах слышали 29 мая 1978 г. В долине Васюгана
(средняя и южная тайга) появляется, по данным II. К. Приходько
(1969), в начале июня.
В южной тайге в период гнездования (июнь) коростель обычен
на низинных болотах и в полях-перелесках (6 п 4). В подтаежных
32
лесах предпочитает низинные болота в сочетании с лугами-покосами
и луга, используемые под выпас (2 и 4), изредка наблюдается в мел-
колиственных лесах и полях-перелесках (по 0,5). С середины июля
до конца лета коростель встречается заметно реже и только на ни-
зинных болотах южных подзон. В августе его здесь примерно в Обра-
за больше, чем на гнездовании.
В среднем за лето распределение коростеля сходно с таковым
на гнездовании. Он повсеместно предпочитает низинные болота, луга
и в меньшей степени лесополевые местообитания южно-таежных
и подтаежных междуречий. В долине Оби в пределах южных подзон
коростель встречается заметно чаще по сравнению с междуречьями,
предпочитает пойменные луга, особенно южно-таежные, интенсивно
заливаемые в половодье. В долине Оби в северных подзонах отмече-
на наименьшая численность коростеля.
Погоныш (Porzana porzana L.)
В северной тайге на междуречьях нами не обнаружен, но от-
мечался в долине Оби как обычная птица низинных пойменных бо-
лот (июнь — июль 1—2). В средней тайге зарегистрирован на неболь-
ших низинных болотах среди приречных смешанных лесов в первой
половине июня. Первая встреча здесь датируется 31 мая 1977 г.
В южной тайге совсем не встречен на междуречьях и обычен в откры-
тых местообитаниях поймы Оби. В подтаежных лесах на междуречь-
ях его слышали лишь в конце мая на тростниково-осоковых низин-
ных болотах (0,6), а на низинных болотах в пойме Оби погоныш обы-
чен в течение всего лета (3—8).
Итак, погоныш явно предпочитает низинные болота в пойме Оби,
избегает таежных междуречий, встречаясь здесь в небольшом коли-
честве лишь на предгнездовых кочевках.
Серый журавль (Grus grus L.)
Весенний пролет в средней тайге Зауралья отмечен В. В. Раев-
ским [1982] в разные годы с 26 апреля по 11 мая. Прилет наблюдал
И. К. Приходько [1969] в долине р. Васюган в начале мая 1965 г.
В северной тайге в течение всего лета серый журавль встречался
преимущественно по берегам озер, в сосново-озерном ландшафте и
на верховых болотах (в среднем 0,3—0,8). Изредка летящих журавлей
наблюдали в сосново-боровом ландшафте (0,05); В средней тайге они
отмечались нами гораздо реже — только во второй половине августа
10 птиц встречены в полете над поселком. В южной тайге в среднем
по территории их примерно столько же, сколько и в северной. Чаще
всего серых журавлей видели здесь в полях и на низинных болотах
(0,9 и 4). В подтаежных лесах серых журавлей встречалось заметно
больше, чем в предыдущих подзонах.
На гнездовании (июнь) в подзоне подтаежных лесов журавли при-
урочены к тростниково-осоковым болотам и заболоченным участкам
в осиново-березовых лесах (0,7 и 0,2). В июле обилие журав-
ле Г. Вартапетов
33
лей в результате размножения заметно возрастает, они начинают
встречаться шире, в том числе в полях и лугах, используемых под
выпас. Затем обилие вновь снижается в начале августа и возрастает
лишь в конце этого месяца в связи с пролетом слабой интенсивности
(0,8). Пара взрослых птиц с молодым обнаружена 13 июля 1978 г.
Стайки из 5—17 кормящихся и отдыхающих журавлей начали встре-
чаться нам в покосных лугах и в полях с 7—11 августа 1978 г.
Итак, на таежных междуречьях серых журавлей заметно боль-
ше в подтаежных лесах по сравнению с более северными подзонами,
что связано с увеличением здесь площади предпочитаемых ими
местообитаний (низинные болота в сочетании с лугами и полями).
В долине Оби, как и на междуречьях, отмечается тенденция к воз-
растанию обилия серого журавля к северу и югу от средней тайги,
при этом здесь его обычно меньше, чем на междуречьях.
ОТРЯД РЖАНКООБРАЗНЫЕ (СНА RADRII FORMES)
Галстучник (Charadrius hiaticula L.)
В северной тайге нами не зарегистрирован. В пойме Оби в пре-
делах этой подзоны был редок в июне и начале августа 1973 г. В сред-
ней тайге многочислен в августе на илистых отмелях крупного над-
пойменного озера (13). Пара взрослых галстучников добыта здесь
нами 14 августа 1977 г. В. Е. Стрелковым [1976] галстучник заре-
гистрирован в бассейне р. Тым на осеннем пролете с 17 по 28 августа
1971 г. Он считает этого кулика для средней тайги регулярно ле-
тающим, но не размножающимся видом. На междуречьях более
южных подзон галстучник не наблюдался. В долине Оби в пределах
средней и южной тайги многочислен в пойменных лугах на весеннем
пролете во второй половине мая.
В подтаежной части долины Оби галстучник совсем не встречен,
потому что массовый пролет его проходит до начала наших наблюде-
ний. Та же причина низкой его численности и в северной тайге,
поскольку наблюдения начаты здесь на две недели позже, чем в
более южных подзонах. Итак, галстучника следует считать редкой
пролетной птицей таежных междуречий и обычной или многочис-
ленной на пролете в долине Оби.
Малый зуек (Charadrius dubius Scop.)
В северной тайге в начале июня редок по берегам крупных таеж-
ных рек (0,4). С середины июня обычен в поселках (9). Пара зуйков
гнездилась здесь по краю песчаной взлетно-посадочной полосы,
примыкающей к поселку. В более южных подзонах междуречий этот
зуек нами не встречен, но отмечался в небольшом числе в бассейнах
рек Тым и Кеть [Стрелков, 1976]. В долине Оби его наблюдали во
всех подзонах, кроме северной тайги. Чаще всего он встречался по
34
песчаным отмелям. В средней тайге по среднелетним показателям
он очень редок. В более южных подзонах малого зуйка заметно боль-
ше (редок или обычен).
Чибис (Vanellus vanellus L.)
Прилет зарегистрирован И. К. Приходько [1969] в долине р. Ва-
сюган 8—10 мая 1965 г. В северной тайге на междуречьях чибис на-
ми не обнаружен. В долине Оби летом 1973 г. был обычен на надпой-
менных переходных болотах (2) и многочислен в пойменных лугах
и на низинных болотах (по 10). В средней тайге чибис обычен на гнез-
довании (вторая половина мая — июнь) на верховых болотах (2—6)
и редок по берегам водоемов (0,3). Гнездо с четырьмя яйцами найде-
но здесь 21 мая 1977 г. Гнездование чибиса на верховых сфагновых
болотах в Прикетье отмечают также С. С. Москвитин и др. [1977].
В. Е. Стрелков [1974] для среднетаежных верховых болот по р. Тым
приводит плотность гнездования, сходную с нашими показателями
(3—6 особей/км2).
В июле обилие чибиса в средней тайге заметно снижается (с 2 до
0,6 в среднем по территории), а в августе он полностью исчезает.
По-видимому, в это время происходит откочевка чибисов в долину
Оби. В южной тайге чибисов видели только в конце июля на неболь-
ших полях среди осиново-березовых лесов (10), но в среднем за лето
его здесь меньше, чем в средней тайге. В подтаежных лесах в конце
мая — июне наблюдался пролет средней интенсивности (2). В то же
время чибис обычен на гнездовании на низинных болотах в сочета-
нии с покосными лугами и в лугах, используемых под выпас (4 и 9).
Лишь изредка в этот период чибисов встречали в полете над полями-
перелесками (0,08). С середины июня до конца июля численность
чибиса неуклонно уменьшается, а в августе, как и в предыдущих
подзонах, он полностью исчезает.
В среднем за лето чибис редок в средней тайге по берегам водое-
мов и в низкорослых рямах верховых болот (0,08 и 0,5). На грядово-
мочажинных верховых болотах его заметно больше (3). В южной тай-
ге он встречается только в полях-перелесках (1). В подтаежных
лесах чибиса заметно больше по сравнению с более северными подзо-
нами. Здесь он обычен в лугах-выпасах и на низинных болотах в со-
четании с покосными лугами (2—4) и очень редок в лесополевых
местообитаниях (0,08).
Итак, на таежных междуречьях обилие чибиса снижается от
подтаежных лесов к средней тайге в соответствии с уменьшением
площади предпочитаемых им влажных луговых местообитаний. При
отсутствии последних он гнездится на участках открытых верховых
болот, приближающихся по характеру минерального питания кпе-
реходным. В долине Оби, где чибис предпочитает пойменные луга,
его численность заметно выше по сравнению с междуречьями и при-
мерно одинакова в пределах всех подзон.
3*
35
Черныш (Tringa ochropus L.)
В северных подзонах и подтаежных лесах в конце мая — начале
июня наблюдался пролет средней интенсивности (2—3). Только в
этот период пролетных чернышей видели на грядово-мочажинно-
озерных северотаежных верховых болотах (6). В южной тайге в
среднем по территории пролет не выражен, но в шелкопрядниках в
конце мая чернышей встречали в значительном числе на придорож-
ных лужах (26).
На гнездовании (15 июня — 15 июля) в северной тайге черныш
приурочен к берегам таежных рек и приречной темнохвойной тайге
(0,4 и 11). В последней встречался по межгривным понижениям, за-
полненным талыми водами. В средней тайге в гнездовой период
(июнь) встречался гораздо шире, предпочитая берега пойменных озер
и стариц (14). Обычен черныш на заболоченных участках и вблизи
залитых водой западин в сосновых и темнохвойно-мелколиственных
приречных лесах (2—4). Реже чернышей видели по берегам крупных
таежных рек, в поселках и в полете над вырубками по соснякам
(0,3—0,8). Три пуховых птенца обнаружены 23 июня 1977 г. на бе-
регу р. Бол. Юган. В южной тайге черныш часто встречался в лесном
ландшафте (2—7). В подтаежных лесах в гнездовое время он редок
на низинных болотах и по окраинам поселков (по 0,5) и очень редок
в лугах-выпасах.
В послегнездовое время с конца июня до середины августа чис-
ленность черныша повсеместно снижается в 3—10 раз в результате
откочевки. В собственно таежных подзонах он перестает встречаться
с 15 августа, а в подтаежных лесах чернышей видели до конца лета.
По среднелетнпм показателям в северной тайге черныш предпо-
читает приречные темнохвойные леса (8), в значительно меньшем
числе встречается в сосняках и на грядово-мочажинных комплексах
верховых болот (по 2). Редок по берегам таежных рек и на гарях по
соснякам (0,3 и 0,1) и очень редок в поселках (0,02).
В средней тайге в лесных местообитаниях черныша заметно
меньше, а на водоемах гораздо больше, чем в предыдущей подзоне.
Он многочислен на старицах таежных рек (10), обычен в сосняках-
брусничниках и приречных смешанных лесах (1 и 2), редок в осталь-
ных местообитаниях (0,1—0,4), однако совсем не встречен на верхо-
вых болотах. В южной тайге черныша лишь немногим меньше, чем
в средней (в среднем по лесному ландшафту — 1), а в подтаежных
лесах — значительно меньше, чем в более северных подзонах. Он
обычен по берегам небольших речек, в том числе в поселках на при-
дорожных лужах и в лугах-выпасах (3—4). В остальных местооби-
таниях, кроме распаханных участков, редок (0,1—0,4).
Основная особенность подзональных различий численности чер-
ныша сводится к ее уменьшению к северу и югу от средней тайги,
но прослеживается это лишь в виде самой общей тенденции, так как
однозначных различий в среднем по участкам и на водоемах не отме-
чается. Сходные изменения на фоне более высокой численности, осо-
бенно в средней тайге, характерны и для долины Оби.
36
Ф и ф и (Tringa glareola L.)
В конце мая — первой половине июня в северных подзонах
идет пролет средней интенсивности (7 и 3). В подтаежных лесах на
пролете фифи редок в лесополевом и лугово-болотном ландшафтах
(0,2 и 0,3). На гнездовании (15 июня — 15 июля) в северной тайге
предпочитает переходные и верховые болота, а также озера и ста-
рицы в поймах крупных притоков Оби (13—20). В значительно мень-
шем числе встречается в приречных темнохвойных лесах и сосняках
с их производными (2—7). В поселках и по берегам крупных таеж-
ных рек редок (0,5 и 0,1). В средней тайге на гнездовании (июнь)
фифи значительно меньше, чем в северной. Он чаще всего обычен на
верховых болотах и илистых отмелях по берегам надпойменных
озер (2—4). Фифи редок по берегам озер и стариц в поймах таежных
рек (0,5).
В южной тайге по сравнению с более северными подзонами в
гнездовое время фифи еще меньше. Он очень редок в лесном ланд-
шафте (0,02) и обычен на переходных и грядово-мочажинно-озер-
ных водораздельных верховых болотах (2—7). В подтаежных ле-
сах в гнездовое время фифи нами не встречен, хотя в долине Оби
отмечался нередко. Здесь на гнездовании он предпочитает поймен-
ные низинные болота (6—11). Гнездо фифи с четырьмя яйцами най-
дено в северной тайге в низкорослом ряме 10 июня 1976 г., а 19 июля
того же года на небольшом низинном болоте в приречной темнохвой-
ной тайге встречен хорошо летающий слеток.
После окончания гнездования значительная часть популяций
фифи, видимо, откочевывает за пределы таежных междуречий, в том
числе в долину Оби. Обилие его в конце июля — начале августа
снижается в 2—3 раза по сравнению с плотностью гнездования,
а с 15 августа фифи совсем перестает встречаться в собственно таеж-
ных подзонах. В то же время в срединных подзонах долины Оби
происходит непропорционально большое увеличение его численно-
сти (в 5—25 раз по сравнению с плотностью гнездования). В под-
таежных лесах фифи зарегистрирован на послегнездовых кочевках
с 15 июля до конца лета повсеместно, кроме осиново-березовых ле-
сов. При этом обычен в лугах-выпасах, поселках и на низинных бо-
лотах (1—2) и редок в остальных местообитаниях.
В среднем за лето в северной тайге фифи многочислен на верхо-
вых болотах (10—12), обычен в сосново-боровом, сосново-озерном
ландшафтах и на озерах, старицах в поймах таежных рек (1—6).
Редок в небольших таежных поселках и по отмелям Казыма (0,4 и
0,04). В средней тайге фифи значительно меньше, чем в северной.
Он обычен на верховых болотах и по берегам стариц и озер в долинах
таежных рек (1—2). В остальных местообитаниях редок (0,1—0.3)
и совсем не встречен на берегах таежных рек. В южной тайге фифи
еще меньше. Его изредка видели на верховых болотах и в лесном
ландшафте (0,4 и 0,03). В подтаежных лесах его лишь немногим боль-
ше, чем в южной тайге, и значительно меньше, чем в северных под-
зонах. Фифи здесь редок по берегам небольших речек, в лугово-
37
болотных местообитаниях и на окраинах поселков (0,4—0,9) и очень
редок в полях-перелесках (0,01).
Итак, на междуречьях в пределах собственно таежных подзон
фифи явно предпочитает верховые болота и озера в поймах таежных
рек. Обилие его снижается от северной тайги к южной и вновь не-
сколько возрастает в подтаежных лесах. В долине Оби наблюдается
обратная тенденция широтных изменений его численности. В се-
верной тайге и подтаежных лесах его здесь примерно столько же,
сколько на междуречьях, а в срединных подзонах заметно больше,
особенно в лугах и на озерах поймы Оби.
Большой улит (Tringa nebularia Gunn.)
Пролет средней интенсивности выражен только в средней тайге
(1). В северной тайге большой улит появляется 8—13 мая [Венгеров,
1973]. В долине Енисея в пределах этой подзоны первые встречи от-
мечены 4—9 мая, а основной пролет в 1978 г. проходил в 3-й декаде
мая. В южной тайге у с. Карым на Оби и на р. Конца его впервые ви-
дели 7—10 мая [Гынгазов, Миловидов, 1977].
В северной тайге на гнездовании в июне большой улит предпо-
читает комплексные верховые и переходные болота (5—8). В мень-
шем числе наблюдается на вырубках и гарях по соснякам (1 и 2),
в приречных темнохвойных лесах и по берегам озер в поймах таеж-
ных рек (0,6 и 0,5). В средней тайге в тот же период в наибольшем
числе встречается на илистых отмелях надпойменных озер (9), обы-
чен на верховых болотах (2—4) и в лесных местообитаниях (1).
В более южных подзонах большой улит в гнездовое время нами на
междуречьях и в долине Оби не зарегистрирован, хотя, видимо,
гнездится [Стрелков, 1973].
После окончания гнездования в июне в северных подзонах идет
откочевка больших улитов, и обилие их снижается в 3—6 раз по
сравнению с плотностью гнездования. В августе эти кулики пол-
ностью исчезают из лесных ландшафтов междуречий и с верховых
болот, хотя нередки по берегам водоемов, особенно среднетаежных
стариц притоков Оби и надпойменных озер (8 и 16). Видимо, боль-
шая часть популяции в северных подзонах в этот период смещается
в долину Оби, где обилие улита резко возрастает в августе (в среднем
в 10—12 раз по сравнению с плотностью гнездования). В южной
тайге эти улиты лишь изредка встречались на послегнездовых ко-
чевках с середины июля по 15 августа (в среднем по обследованной
территории — 0,01 и 0,002).
В среднем за лето в северной тайге большой улит обычен на
комплексных верховых болотах (3—4). Значительно реже, хотя и
равномерно, отмечался он во всех остальных местообитаниях (0,2—
0,5). В средней тайге большого улита примерно столько же. Он
обычен по берегам озер, на верховых болотах и на вырубках по сос-
някам (1—4) и заметно реже встречался на остальной территории.
В более южных подзонах в среднем за лето этот кулик чрезвычайно
редок.
38
Итак, на междуречьях в северных подзонах численность боль-
шого улита значительно выше, чем в южных. В севере- и средне*
таежной долине Оби его еще больше, соответственно в 2 и 4 раза по
сравнению с междуречьями сходных подзон. К югу от средней тайги
в долине Оби численность большого улита падает.
Травник (Tringa totanus L.)
Встречался только на пред- и послегнездовых кочевках. Травни-
ков видели на берегу надпойменного озера в средней тайге в начале
августа (4) и в подтаежных лесах на придорожной луже в лугах-
выпасах в конце мая (2). В долине Оби этот кулик зарегистрирован
лишь однажды в среднетаежных пойменных лугах. Это позволяет
считать травника чрезвычайно редким в пределах обследованных
участков.
Перевозчик (Actitis hypoleucos L.)
Пролет средней интенсивности отмечен на небольших реках
в южной тайге и высокой интенсивности — в подтаежных лесах
(4 и 43).
На гнездовании (июнь) в северной тайге перевозчик обычен по
песчаным отмелям таежных рек (4) и реже встречается в темно хвой-
ных лесах по их берегам на заполненных водой западинах и на ста-
рицах крупных таежных рек (0,5 и 1). В средней тайге в гнездовой
период (15 мая — 15 июня) обычен на таежных реках и крупных
надпойменных озерах (3—6) и реже наблюдается в смешанных тем-
нохвойно-мелколиственных лесах (0,5). В южных подзонах в этот
же период перевозчиков видели только по берегам небольших речек
(2—3). Гнездо перевозчика с четырьмя насиженными яйцами найде-
но 9 июня 1977 г. в средней тайге на берегу р. Бол. Юган.
После окончания гнездования с конца июня до середины июля
обилие перевозчика повсеместно возрастает в 2—6 раз. На неболь-
ших речках это увеличение, являясь результатом размножения,
не столь велико, тогда как в среднем течении крупных притоков
Оби происходит прикочевка средней и высокой интенсивности (соот-
ветственно 5 и 24 в северной и средней тайге). Во второй половине ле-
та численность перевозчика неуклонно снижается, и к середине ав-
густа по берегам крупных таежных рек его втрое меньше, чем на
гнездовании, а на небольших речках он исчезает совсем.
В среднем за лето в северной тайге перевозчик предпочитает
песчаные отмели крупных таежных рек (6) и в небольшом числе
встречается в темнохвойной тайге по их берегам и на старицах
(1 и 0,5). Сходный характер его распределения прослеживается и в
средней тайге, но здесь на реках численность перевозчика вдвое боль-
ше, чем в предыдущей подзоне. На южнотаежных небольших реках
этого кулика соответственно в 2 и 4 раза меньше, чем в северных под-
зонах, а на сходных реках в подтаежных лесах примерно столько
же, сколько в северной и средней тайге.
39
Итак, перевозчик четко приурочен к берегам рек. Его числен-
ность на междуречьях снижается к югу и северу от средней тайги.
При этом в среднем течении притоков Оби перевозчика больше, чем
в их устьях. В среднем по участкам в долине Оби, и особенно в пойме,
он встречается гораздо чаще, особенно в южной тайге.
Мородунка (Xenus cinereus Giild.)
В северной тайге встречалась с начала лета до середины июля.
В это время обычна на небольших озерках в низкорослых рямах,
сосново-озерном ландшафте и в поймах крупных таежных рек (1—3).
Заметно реже мородунку видели в лесных местообитаниях и по бе-
регам таежных рек (0,3—0,7). В средней тайге ее значительно мень-
ше. Крик самца слышали в этой подзоне только 14 июля 1977 г. в
пойме р. Бол. Юган. Гнездо с четырьмя яйцами найдено В. Е. Стрел-
ковым (1976) в пределах средней тайги на р. Тым 8 июня 1967 г. В юж-
ных подзонах мородунка нами не обнаружена, хотя гнездится в юж-
ной тайге на реках Васюган и Парабель [Гынгазов. Миловидов,
1977].
Таким образом, мородунка встречена нами на таежных между-
речьях только в северных подзонах, но в северной тайге ее гораздо
больше, чем в средней. В долине Оби, особенно в пойме, численность
мородунки выше, чем на междуречьях. Здесь она встречается во
всех подзонах, при этом обилие ее снижается к северо-западу и юго-
востоку от средней тайги.
Круглоносый плавунчик (Phalaropus lobatus L.)
В северной тайге на р. Казым самка круглоносого плавунчика
была добыта И. Н. Шуховым [1916] 1 июня 1914 г., однако мы этого
куличка здесь не видели. На таежных междуречьях нами встречен
лишь однажды в средней тайге на весеннем пролете во второй поло-
вине мая на грядово-мочажинном верховом болоте (2). По свидетель-
ству А. М. Гынгазова, С. П. Миловидова [1977]. в таежной зоне на-
блюдался на многих притоках Оби на пролете, в частности, добывался
на р. Тым в начале июня и конце августа [Стрелков, 1973; 1976].
В долине Оби круглоносый плавунчик обычен в северной тайге в
пойме Оби на пролете и на надпойменных переходных болотах
(встречался до середины июля). В остальных подзонах в пределах
долины Оби его значительно меньше (отмечен лишь на пролете в
конце мая — начале июня), но больше, чем на таежных междуречьях.
Турухтан (Philomachus pugnax L.)
В северной тайге нами не встречен, но в добыче охотника 31 мая
1976 г. был один самец турухтана. В средней тайге турухтанов ви-
дели лишь на пролете в конце мая по берегам таежных рек (0,4) и в
первой половине августа на илистых отмелях надпойменных озер
(2). В южных подзонах нами не обнаружен, хотя отмечался здесь на
40
крупных таежных реках, в том числе на гнездовании [Москвитин
и др., 1977]. В долине Оби турухтана значительно больше (преиму-
щественно на пролете), численность его заметно снижается к югу от
северной тайги.
Кулик-воробей (Calidris minutus Leisl.)
На междуречьях встречен нами всего один раз — в начале июля
1976 г. в северной тайге на переходном болоте (2). В бассейне Оби
в пределах средней тайги отмечался во время миграций В. Е. Стрел-
ковым [1973, 1976]. На р. Кеть в пределах южной тайги, стайки
куликов-воробьев держались в течение всего лета. В долине Оби
этот кулик обнаружен на весеннем пролете в наибольшем числе в
северной и средней тайге и в значительно меньшем — в южной.
При этом в северных подзонах в долине Оби, особенно в пойме,
встречался гораздо чаще, чем на междуречьях.
Белохвостый песочник (Calidris temminckii Leisl.)
В северной тайге зарегистрирован на пролете в начале июня на
крупных таежных реках, старицах и небольших озерках в низко-
рослых рямах (0,9, 1 и 2). В средней тайге держался с 15 мая до
середины июня 1977 г. В наибольшем количестве этого песочника
видели на илистых отмелях надпойменных озер, где он в первой по-
ловине июня многочислен (40) и встречался стайками до 20 особей.
Лишь изредка попадался он на озерках грядово-мочажинных вер-
ховых болот и на таежных реках (0,1 и 0,3). С середины июня до кон-
ца июля белохвостые песочники не встречались и только в августе
вновь отмечены на надпойменных озерах (20).
На междуречьях южных подзон белохвостых песочников мы
ни разу не видели, хотя они обычны в некоторых районах южной
тайги. Например, на р. Кеть держались в течение всего лета. В бас-
сейне средней Оби В. Е. Стрелков [1973] отмечал пролет этого песоч-
ника широким фронтом, особенно осенью. Тем не менее в долине Оби
численность белохвостого песочника значительно выше, чем на
междуречьях, особенно в северной тайге, где повсеместно он много-
числен на предгнездовом и летне-осеннем пролете.
Чернозобик (Calidris alpina L.)
На междуречьях встречен лишь в средней тайге на грядово-
мочажинных верховых болотах в начале августа (2). В бассейне
средней Оби отмечен В. Е. Стрелковым [1973] только на пролете,
В долине Оби весьма многочислен на предгнездовом пролете во вто-
рой половине мая — начале июня в северной и средней тайге и обы-
чен в южной. Таким образом, интенсивность пролета чернозобика
в долине Оби значительно выше, чем на междуречьях.
41
Дупель (Callinago media Lath.)
В северной тайге в августе обычен на переходных и грядово-
мочажинно-озерных верховых болотах (8 и 1). Молодая птица добыта
здесь 8 августа 1976 г. Кроме того, самец дупеля был добыт в долине
р. Казым И. Н. Шуховым [1916] 28 мая 1914 г., а первое токование
тот же автор слышал в середине мая. В средней тайге дупелей видели
лишь изредка во второй половине лета. В конце июля они встречены
в поселках, а в течение августа — по берегам озер в поймах таеж-
ных рек (7 и 2). В южных подзонах таежных междуречий дупель на-
ми не зарегистрирован, хотя отмечен А. М. Гынгазовым и С. П. Ми-
ловидовым [1977] здесь на гнездовании, в частности по рекам Васю-
ган и Шегарка.
Итак, в среднем по обследованным нами участкам дупель повсе-
местно редок. На Обь-Енисейском и Обь-Иртышском междуречьях
он, по-видимому, предпочитает-долины крупных притоков и избе-
гает водоразделов. В долине Оби дупеля повсеместно значительно
больше, чем на междуречьях. Особенно высокая численность заре-
гистрирована в пойме в пределах срединных подзон.
Бекас (Gallinago gallinago L.)
Прилет зарегистрирован С. С. Москвитиным и др. [1977] на
р. Кеть в начале мая. В. В. Раевский [1982] начало токования отме-
чал в средней тайге Зауралья с 23 апреля по 16 мая. Нами в конце
мая — начале июня повсеместно прослежен весенний пролет не-
большой интенсивности. В отличие от дупеля на маршрутах бекас
встречался преимущественно в первой половине лета, в период ин-
тенсивного токования.
На гнездовании в северной тайге (15 июня — 15 июля) обычен
в приречных темнохвойных лесах (преимущественно на небольших
низинных болотах среди этих лесов), на переходных болотах и по
берегам стариц крупных таежных рек (по 2). Значительно реже бе-
касов встречали в низкорослых рямах по окраинам верховых болот
(0,5). В средней тайге в гнездовый период в июне обычен по берегам
крупных надпойменных озер и на вырубках по соснякам, где держал-
ся на небольших участках низинных болот (по 1), редок на залесен-
ной части верховых болот и в приречных смешанных темнохвойно-
мелколиственных лесах (0,2—0,5). В южной тайге и подтаежных ле-
сах четко приурочен к обширным открытым низинным болотам (18 и
4), а в последней из подзон встречался еще и на кочкарниковых и лу-
говых участках среди осиново-березовых лесов (3).
Гнездо с четырьмя яйцами найдено В. Е. Стрелковым [1976] в
средней тайге по р. Тым 26 июня 1971 г., а молодые летающие птицы
добывались с 27 июля. После окончания гнездования бекас встречен
в северной тайге на переходных болотах (11). В южной тайге он в
значительном числе концентрировался на открытых низинных боло-
тах, где обилие его в начале августа почти втрое превышало плот-
ность гнездования. После откочевки во второй половине августа бе-
42
касов здесь было в 6 раз меньше. В остальных подзонах они откоче-
вывали, вероятно, сразу после окончания гнездования, и их не ви-
дели со второй половины июня до конца лета.
Территориальное распределение бекаса по среднелетним пока-
зателям весьма сходно с таковым по плотности гнездования. На
междуречьях численность его меньше всего в средней тайге и зна-
чительно выше в остальных подзонах. В долине Оби бекаса заметно
больше, чем на междуречьях. Здесь он тоже предпочитает низинные
болота как в пойме, так и вне ее. Тенденция подзональных различий
численности бекаса та же, что и на междуречьях.
Лесной дупель (Gallinago megala Swinh.)
Наблюдался на"пролете в конце мая и августа в южной тайге
в лесном ландшафте (2 и 0,6). В подтаежных осиново-березовых ле-
сах лесного дупеля слышали 13 июня 1978 г. (0,3). В восточной части
Западной Сибири в южной тайге и подтаежных лесах отмечен на
гнездовании на реках Кеть и Шегарка [Гынгазов, Миловидов, 1977].
В долине Оби лесного дупеля заметно больше, чем на междуречьях,
особенно в пределах южных подзон. Повсеместно избегает он сомкну-
тых хвойных лесов и верховых болот, отдавая предпочтение лесо-
полевым и лугов о-лесным местообитаниям.
Большой кроншнеп (Numenius arquata L.)
Пролет невысокой интенсивности отмечен в северной тайге
в начале июня и в подтаежных лесах в конце мая (2 и 0,1). На гнездо-
вании в северной тайге (15 июня — 15 июля) обычен на комплекс-
ных верховых болотах (2—3) и изредка встречался в полете над вы-
рубками по соснякам (0,02). В средней тайге наблюдался только в
июне в низкорослых рямах верховых болот (0,4). В южной тайге в
первой половине июня обычен на открытых низинных болотах и очень
редок в лесном ландшафте (2 и 0,05). В подтаежных лесах на гнездо-
вании нами не зарегистрирован, хотя отмечен в этой подзоне гнездя-
щимся к северо-востоку от места наших работ на р. Шегарка [Гын-
газов, Миловидов, 1977].
В средней тайге в бассейне р. Тым 9—11 июня 1971 г. найдены
гнезда с тремя-четырьмя яйцами, И июня зарегистрировано вылуп-
ление птенцов, а стаи молодых летающих птиц видели с 23 июля
[Стрелков, 1976]. В южной тайге Прикетья кладка найдена 6 июня
1974 г., а выводки из четырех особей встречались 27—29 июня 1968 г.
В период послегнездовых кочевок в августе в северной тайге
стайки больших кроншнепов (до 17 особей) постоянно наблюдались
в сосново-озерном ландшафте (в среднем за месяц — 9), а в осталь-
ных местообитаниях они редки. В среднем по территории в северной
тайге в это время прослеживалась прикочевка невысокой интенсив-
ности (3). В средней тайге в течение июля большой кроншнеп не
встречался, а в начале августа он отмечен на илистых отмелях над-
пойменных озер (8), В южной тайге в послегнездовое время большой
43
кроншнеп по-прежнему приурочен к открытым низинным болотам и
становится здесь многочисленным (во второй половине июля — 27),
а затем в августе исчезает. В подтаежных лесах после двухмесячно-
го перерыва он вновь появляется в начале августа. В это время его
видели в полете над поселком (2).
В среднем за лето в северной тайге большой кроншнеп обычен в
сосново-озерном ландшафте и на верховых болотах (1—3), реже
наблюдается -на переходных (0,5) и лишь иногда этих кроншнепов
видели в полете над сосняками-ягельниками и вырубками по ним
(0,01—0,02). В средней тайге они отмечались только в низкорослых
рямах и по берегам надпойменных озер (0,1 и 2), а в среднем по участ-
ку их гораздо меньше, чем в северной тайге. В южной тайге большой
кроншнеп явно предпочитает открытые низинные болота (5), лишь
изредка встречаясь на верховых болотах и в лесных местообитаниях
(0,07 и 0,1). В среднем по территории в южной тайге его несколько
меньше, чем в северной, но значительно больше, чем в средней.
В подтаежных лесах большой кроншнеп прослежен в полете над лу-
гами-выпасами и поселками (0,03 и 0,6). В среднем его здесь пример-
но столько же, сколько в средней тайге, и значительно меньше, чем
в остальных подзонах.
Итак, на междуречьях численность большого кроншнепа выше
всего в северной и в южной тайге. В последней он явно предпочитает
низинные болота. На северотаежных комплексных (гетеротрофных)
болотах большой кроншнеп обычен как на низинных, так и на пере-
межающихся с ними верховых участках. В долине Оби этот кроншнеп
предпочитает обширные низинные и переходные болота. Здесь, как
и на междуречьях, максимальная численность кроншнепа зареги-
стрирована в северной и южной тайге. В пределах одних и тех же
подзон сходная численность в долине Оби и на междуречьях отмече-
на только в северной тайге; в более южных подзонах в долине его
несколько больше.
Средний кроншнеп (Numenius phaeopus L.)
В северной тайге встречен только в начале июня, по-видимо-
му на пролете, в сосново-озерном ландшафте и на комплексных
верховых болотах (0,2 и 0,8). Затем в течение месяца (с середины
июня по 15 июля) его не видели, и вновь отмечен этот кроншнеп в
грядово-мочажинно-озерной части верховых болот в конце июля (1).
В средней тайге в период гнездования (июнь) встречался чаще
всего на грядово-мочажинных верховых болотах (8) и заметно реже —
в периферийных рямах (1—4). В то же время средних кроншнепов
видели по берегам надпойменных озер (2). Гнезд среднего кроншне-
па не находили, но постоянно в одних и тех же местах наблюдали
несколько пар птиц, взлетающих и кружащихся либо садящихся
на деревья с тревожным криком. В 170 км к северу от обследованного
нами ключевого участка на р. Глубокий Полуй 1 августа 1976 г»
встречена стайка средних кроншнепов из двух взрослых и семи мо-
лодых птиц.
44
После окончания гнездования обилие среднего кроншнепа в
результате откочевки сокращается в июле в 10 раз по сравнению
с плотностью гнездования, а в августе он исчезает совсем. В южной
тайге этого кроншнепа не видели, а в подтаежных лесах он встречен
лишь однажды на предгнездовом пролете в конце мая на низинных
болотах в сочетании с лугами (0,6).
В среднем за лето на междуречьях больше всего среднего кронш-
непа отмечено в средней тайге, особенно на верховых болотах, и зна-
чительно меньше в северной тайге. На междуречьях южной тайги
и в долине Оби в пределах лесной зоны летом Ю. С. Равкин [1978] не
встречал этого кроншнепа, но в долине Оби в период миграций он
отмечен у сел Нарым и Батурине [Стрелков, 1973; Гынгазов, Мило-
видов, 1977].
Таким образом, складывается впечатление, что средний крон-
шнеп, большая часть ареала которого лежит в лесотундре, проника-
ет в тайгу по междуречьям. Он гнездится преимущественно по во-
дораздельным верховым болотам северной половины лесной зоны и
экологически викарирует с большим кроншнепом, явно предпочи-
тающим южные подзоны и долину Оби.
Малый веретенник (Limosa lapponica L.)
Зарегистрирован в средней тайге на гнездовании (июнь) пре-
имущественно в открытых центральных частях верховых болот
и в меньшем количестве — в низкорослых рямах (5 и 2). Волнующие-
ся пары малого веретенника с конца мая до начала июля встреча-
лись на постоянных местах (там же, где средние кроншнепы). В июле
малые веретенники почти полностью откочевывают с мест гнездова-
ния, в этот период на верховых болотах их в 20 раз меньше, чем в
июне. В августе они совсем исчезают с верховых болот, но часто
встречаются по берегам надпойменных озер (в среднем за месяц —
11). Взрослая самка и молодая птица добыты соответственно 26 июня
и 28 августа 1977 г. Нами на междуречьях в остальных подзонах и
Ю. С. Равкиным [1978] в долине Оби малый веретенник не заре-
гистрирован, хотя его находили летом, преимущественно в сред-
ней тайге: И. К. Приходько [1969] нар.Васюган, А. М. Гынгазов,
С. С. Москвитин [1965] у Березова, Ханты-Мансийска, на реках Кон-
да и Тым. Последние также считают малого веретенника гнездящим-
ся в лесной зоне Западной Сибири.
Большой веретенник (Limosa limosa L.)
На междуречьях наблюдался нами только в июне в лугах-выпа-
сах и на открытых низинных болотах подтаежных лесов на пролете
(по 2). Имеются указания на гнездование большого веретенника в
пределах лесной зоны на реках Васюган, Тым. Вах и у с. Нарым на
Оби [Гынгазов, Миловидов, 1977].
В долине Оби большой веретенник гнездился и держался в те-
чение всего лета, отдавая предпочтение разнообразным лугово-
45
болотным местообитаниям как в пойме, так и вне ее. По среднелет-
ним показателям здесь его в южной тайге примерно столько же,
сколько на подтаежных междуречьях, а в северных подзонах гораз-
до больше, в основном за счет высокой интенсивности пролета.
В более южных подзонах массовый пролет, по-видимому, проходит до
начала наших наблюдений: например, в южно-таежной долине Оби
С. С. Москвитин (1973) отмечал его 12—14 мая.
Распределение большого и малого веретенников в северной и
средней тайге аналогично таковому большого и среднего кроншне-
пов. На гнездовании Limosa limosa L. явно предпочитает долину
Оби. По долинам крупных притоков оба вида, вероятно, обитают
совместно, а водораздельные болота занимает уже только L. lappo-
nica. Летом 1979 г. нами были обследованы обширные верховые
болота на водоразделе рек Ларь-Еган и Васюган. Здесь гнездились
в значительном числе только малый веретенник и средний кроншнеп,
тогда как в среднем течении р. Ларь-Еган (в 40 км от устья) их уже
полностью замещают L. limosa и Л', arquata.
Сиз а я чайка (Larus сапиз L.)
Пролет средней интенсивности отмечен в средней тайге в начале
июня (5). В период гнездования в северной тайге сизая чайка обычна
(в июне) на переходных болотах и грядово-мочажинных верховых
болотах, а также по берегам рек в пределах поселков (2—8). Заметно
реже встречается по берегам крупных таежных рек вне поселков и
стариц, в сосново-озерном ландшафте и низкорослых рямах (0,1—
0,5). В средней тайге с начала наблюдений (15 мая) до конца июня
эта чайка многочисленна на верховых болотах и по берегам круп-
ных надпойменных озер (11—23)*. В значительно меньшем числе она
встречалась по берегам крупных таежных рек и на их старицах
(0,4 и 1), в рослых рямах по периферии верховых болот, на вырубках
по соснякам (по 0,8) и в поселках на берегах рек (0,2). В подтаежных
лесах сизая чайка обычна в этот период в рямах (7) и иногда отме-
чалась вблизи поселков по берегам рек (0,1) и в лугово-полевых мес-
тообитаниях (0,04). В средней тайге по р. Тым первые летающие вы-
водки встречены 12 июля 1967 г. и 13 июля 1971 г. [Стрелков,
1976].
После окончания гнездования в июле обилие сизой чайки повсе-
местно снижается в 2—6 раз, и в августе она покидает таежные
междуречья. Только на крупных среднетаежных озерах с обширны-
ми илистыми отмелями и мелководьями она в значительном числе
держится до конца лета (в августе — 49).
В среднем за лето в северной тайге сизая чайка обычна в откры-
той части комплексных верховых болот и в поселках по берегам рек
♦ Возможно, что обилие сизой чайки на среднетаежных междуречьях
завышено из-за близости гнездовой колонии сизых чаек и речных крачек к
учетному маршруту на верховом болоте. Обычно такие колонии не характерны
для верховых болот.
46
(1—4) и редка в сосново-озерных ландшафтах и на водоемах (0,3—
0,5). В средней тайге ее втрое больше. Она многочисленна на круп-
ных надпойменных озерах (30), обычна на верховых болотах и стари-
цах крупных таежных рек (1—8) и редка на остальной территории
(0,2—0,4). На междуречьях южной тайги сизая чайка не встречена,
а в подтаежных лесах ее лишь немногим меньше, чем в северной тай-
ге. Она обычна на болотах и по берегам рек (1—2), редка в лугово-
полевых местообитаниях (0,2—0,3) и очень редка в поселках (0,03).
Следовательно, на междуречьях больше всего сизой чайки в
средней тайге и меньше в остальных подзонах. В долине Оби в преде-
лах лесной зоны отмечена сходная численность сизой чайки с неко-
торым уменьшением в средней тайге. При этом в долине ее пример-
но столько же, сколько на среднетаежных междуречьях.
Серебристая чайка (Larus argentatus Pontopp.)
В северной тайге на гнездовании (июнь) обычна в сосново-озер-
ном ландшафте, на комплексных верховых болотах, в поселках и на
старицах таежных рек (1—3). На самих реках редка (0,5). Двух мо-
лодых птиц размером со взрослых видели 23 июля 1976 г. на берегу
р. Казым. В послегнездовый период обилие серебристой чайки в
среднем по участку в июле снижается в 3 раза, а в августе — в 5 раз
по сравнению с плотностью гнездования. На водоемах изменений
численности, имеющих выраженную направленность, не наблюда-
лось. В средней тайге серебристая чайка встречена только в конце
мая на пролете по берегам крупного надпойменного озера (26).
В среднем за лето на водоемах ее столько же, сколько в северной
тайге. В южных подзонах междуречий и долины Оби она не заре-
гистрирована.
Итак, на Обь-Иртышском и Обь-Енисейском междуречьях се-
ребристая чайка чаще всего встречалась в северной тайге и заметно
реже — в средней. В долине Оби прослеживаются сходные подзо-
нальные отличия, но численность здесь заметно меньше, чем на меж-
дуречьях.
Озерная чайка (Larus ridibundus L.)
На Обь-Енисейском и Обь-Иртышском междуречьях нами встре-
чена лишь однажды — в средней тайге на старице крупной таеж-
ной реки в первой половине августа (2). Отмечалась и ранее летом на
притоках Оби в пределах средней тайги, например в среднем тече-
нии р. Тым, А. М. Гынгазовым и С. П. Миловидовым [1977]. В доли-
не Оби озерная чайка в наибольшем количестве наблюдалась в пре-
делах северной тайги и заметно реже — в более южных подзонах.
В долине Оби в среднем по участкам и на водоемах ее повсеместно
больше, чем на среднетаежных междуречьях.
47
Малая чайка (Lams minutus Pall.)
На междуречьях, как и озерная чайка, встречена нами всего
один раз — в средней тайге в первой половине августа на крупном
надпойменном озере (12). Кроме того, отмечена в средней тайге
В. Е. Стрелковым [1973] в среднем течении р. Тым. В северной тай-
ге зарегистрирована Ю. И. Гордеевым [1963] на р. Казым, а в юж-
ной— А. М. Гынгазовым и С. П. Миловидовым [1977] на реках
Васюган и Чулым. В долине Оби малая чайка встречалась в преде-
лах всех собственно таежных подзон и значительно чаще, чем на
междуречьях.
Речная крачка (Sterna hirundo L.)
Пролет невысокой интенсивности отмечался в северных подзо-
нах в конце мая — начале июня (0,4—1). В северной тайге на круп-
ных таежных реках и их старицах эта крачка держалась только
во время пролета в первой половине июня (2—5), а затем не встреча-
лась до конца лета.
На гнездовании в северной тайге (с 15 июня по 15 июля) обычна
вблизи небольших озер на верховых болотах (3—6) и заметно реже
наблюдалась на переходных болотах и в поселках по берегам прито-
ков Оби (0,05 и 0,5). В средней тайге на гнездовании в тот же период
многочисленна на крупных надпойменных озерах (10) и обычна вбли-
зи небольших озер в низкорослых рямах (2). Реже речных крачек
видели на грядово-мочажинных комплексах верховых болот, в по-
селках и на крупных притоках Оби (0,1—0,3). В южной тайге реч-
ная крачка не зарегистрирована, хотя отмечалась здесь на гнездо-
вании по крупным притокам Оби [Стрелков, 1973]. В подзоне
подтаежных лесов в гнездовой период (июнь) обычна на небольших
речках (1), реже видели крачек, пролетавших над лугами и низин-
ными болотами (0,2).
После окончания гнездования речная крачка откочевывает за
пределы северотаежных междуречий, в основном в долину Оби,
где ее обилие в конце августа возрастает в 5—10 раз по сравнению
с плотностью гнездования. В средней тайге на междуречьях к концу
июня обилие этой крачки снижается втрое по сравнению с плотно-
стью гнездования, и к началу августа она перестает встречаться на
большей части территории. В этот период речные крачки концентри-
руются по берегам пойменных и особенно крупных надпойменных
озер. На последних их в первой половине августа в 5 раз больше,
чем во время гнездования, а во второй половине августа речная крач-
ка исчезает совсем. В подтаежных лесах эта крачка к середине июля
почти полностью откочевывает за пределы обследованной террито-
рии, и лишь однажды (во второй половине июля) она встретилась в
рямах (0,2). Таким образом, и в более южных подзонах речная крач-
ка откочевывает с таежных междуречий в июле, но на крупных вы-
сококормных озерах она задерживается до середины августа.
В среднем за лето в северной тайге речная крачка обычна на
комплексных верховых болотах и старицах крупных таежных рек
48
(1—4) и заметно реже встречается на самих реках и переходных бо-
лотах (0,02 и 0,4). В средней тайге в местообитаниях суши ее гораздо
меньше, а на водоемах гораздо больше, чем в северной. В южной
тайге речная крачка нами не зарегистрирована, а в подтаежных ле-
сах этой крачки меньше, чем в северных подзонах, и она повсемест-
но редка (0,3) или очень редка (0,03—0,06).
Итак, на таежных междуречьях речная крачка встречена повсе-
местно, кроме южной тайги. Предпочитает озера различных типов и
в меньшей степени крупные притоки Оби. В долине Оби, особенно в
ее пойме„ в пределах собственно таежных подзон численность этой
крачки значительно выше, а в подтаежных лесах примерно такая
же, как на междуречьях.
Полярная крачка (Sterna paradisaea Pontopp.)
Появилась 25 июня 1976 г. в северной тайге и 22 июня 1977 г.
в средней. На гнездовании в северной тайге в конце июня — первой
половине июля обычна вблизи небольших озер на комплексных
верховых болотах и старицах крупных таежных рек (2—4). Изредка
встречалась на самих реках, в том числе в поселках, по их берегам
(0,8 и 0,05). В средней тайге зарегистрирована в тот же период на
таежных реках и их старицах (0,05 и 7). После окончания гнездова-
ния полярная крачка исчезает в северной тайге в конце июля. В сред-
ней тайге держится до начала августа, в значительном количестве
встречаясь в этот период на крупных притоках Оби.
В среднем за лето в северной тайге полярная крачка обычна на
старицах крупных таежных рек и верховых болотах (по 1), реже
встречается в поселках и на реках (0,02 и 0,4). В средней тайге наб-
людалась только на водоемах, причем в несколько большем числе,
чем в северной (2). В подтаежных лесах лишь однажды (22 июня
1978 г.) пару полярных крачек видели на небольшой речке (верховья
р. Омь), в среднем за лето ее здесь значительно меньше, чем в север-
ных подзонах междуречий (0,3).
Итак, на таежных междуречьях полярная крачка обычна в се-
верных подзонах, где предпочитает старицы крупных притоков Оби,
и лишь изредка встречается в подтаежных лесах. В долине Оби в
пределах лесной зоны Ю. С. Равкиным [1978] не зарегистрирована,
хотя отмечалась на гнездовании у Сургута А. Д. Шароновым [1951].
ОТРЯД ГОЛУБЕОБРАЗНЫЕ (СО LUMBI FORMES)
Сизый голубь (Columba livia L.)
В подтаежных лесах весьма многочислен в поселках, где обилии
его возрастает с 97 особей/км2 в начале июня до 273 в конце июля,
а затем несколько снижается в августе (до 193). В лугово-полевых
местообитаниях в среднем за лето обычен (2—4). В долине Оби отме-
чен только в подтаежных поселках, где его значительно меньше,
4 Л. Г. Вартапетов
49
чем на междуречьях. В более северных подзонах долины Оби и меж-
дуречий нами ни разу не встречен, хотя, по свидетельству А. М. Гын-
газова и С. П. Миловидова [1977], на север таежной зоны Западной
Сибири распространяется до р. Вах и низовий Оби, но встречается там
весьма спорадично и, по-видимому, только в городах и крупных по-
селках.
Клинтух (Columba oenas L.)
В южной тайге встречался только до середины июля на открытых
низинных болотах и в мелколиственных лесах, в том числе переме-
жающихся с полями (0,3—0,5). В подтаежных лесах отмечен в тот же
период в мелколиственных лесах, полях с небольшими участками та-
ких же лесов и на низинных болотах в сочетании с покосными лугами
(2—3). Лишь изредка клинтухов видели в лугах-выпасах (0,02). В на-
чале августа его обилие в среднем по территории сокращается в
20 раз по сравнению с первой половиной лета, а в конце августа он
почти полностью исчезает с подтаежных междуречий.
По среднелетним показателям, клинтух на междуречьях явно
предпочитает лесополевые местообитания, луга и низинные болота
южных подзон. При этом в подтаежных лесах его примерно в 10 раз
больше, чем в южной тайге. В долине Оби, где клинтух встречается
также преимущественно в лесополевых ландшафтах южных подзон,
его примерно столько же, сколько и на подтаежных междуречьях.
Большая горлица (Streptopelia orientalia Lath.)
В северных подзонах таежных междуречий нами не встречена.
В южной тайге многочисленна на гнездовании во второй половине
июня в полях-перелесках (18) и редка в остальных лесных местооби-
таниях (0,4—0,8). В подтаежных лесах в гнездовое время в первой
половине июня обычна в полях-перелесках и по окраинам рямов (1
и 6). После окончания гнездования численность большой горлицы в
южных подзонах возрастает в июле — августе в 2—3 раза, а в под-
таежных лесах с середины августа она перестает встречаться на учет-
ных маршрутах.
В среднем за лето в южной тайге большая горлица чаще всего
встречалась в полях-перелесках (13). Она обычна в лесных местооби-
таниях (2—5), за исключением заболоченных и елово-кедровых ле-
сов, в которых редка (0,2—0,6). В подтаежных лесах ее в среднем по
территории в 5 раз меньше. Чаще всего горлицу видели в полях-
перелесках и рямах (1—2), реже — по берегам рек и на низинных
болотах (0,1 и 0,3) и совсем редко — в остальных местообитаниях
(по 0,01).
Итак, в пределах таежных междуречий большая горлица заре-
гистрирована нами только в южных подзонах. В северных подзонах
она отмечалась в бассейне р. Вах и верховьях р. Таз [Гынгазов, Ми-
ловидов, 1977]. В средней енисейской тайге большой горлицы на
гнездовании примерно столько же, сколько отмечено нами на Обь-
50
Иртышском междуречье в пределах южных подзон (1—5 особей/км2).
В долине Оби от средней тайги до подтаежных лесов численность
большой горлицы одинакова и сходна с таковой на междуречьях в
пределах южных подзон. В северотаежной долине Оби она встрече-
на только в июне, поэтому среднелетние показатели заметно ниже.
ОТРЯД КУКУШКООБРАЗНЫЕ (С UCULIFORMES)
Кукушка (Cuculus canorus L.)
Прилет и первое кукование отмечены нами в подтаежных лесах
15—17 мая 1978 г., а А. М. Гынгазовым и С. П. Миловидовым
11977] — в южной тайге в окрестностях Томска в среднем 16 мая.
В средней тайге в первый раз кукушку слышали 22 мая 1977 г. Про-
лет невысокой интенсивности выражен только в северной тайге (2).
В этой подзоне на р. Казым первое кукование зарегистрировано
И. Н. Шуховым [1916] 23 мая 1914 г*
На гнездовании в северной тайге (16 июня — 15 июля) кукушка
чаще всего встречалась в приречных темнохвойных лесах, на вы-
рубках по соснякам и в низкорослых рямах по окраинам комплекс-
ных верховых болот (3—4) и несколько реже — в центральных
грядово-мочажинно-озерных частях верховых болот и поселках (1 и
2). Наименьшее обилие кукушки зарегистрировано в сосняках-
ягельниках и на гарях по ним, в сосново-озерном ландшафте и на
переходных болотах (0,5—0,9). В средней тайге в гнездовое время
(июнь) кукушка обычна па вырубках по соснякам и в рямах (1—4)
и редка в остальных местообитаниях (0,1—0,6). В южной тайге на
гнездовании в первой половине июля она многочисленна в мелко-
лиственных лесах (15), обычна на открытых низинных болотах (2)
и редка в остальных местообитаниях, кроме угнетенных хвойных ле-
сов, где не встречалась совсем. В подтаежных лесах в период гнездо-
вания (16 июня — 15 июля) кукушка обычна во всех лесолуговых
местообитаниях (1—2) и редка в рямах (0,4). От окончания отклады-
вания яиц до середины августа обилие кукушки повсеместно сни-
жалось в 2—12 раз. В качестве воспитателя птенцов кукушки заре-
гистрирована овсянка-крошка в северной тайге, которая 4 августа
1976 г. выкармливала слетков.
В среднем за лето в северной тайге кукушка предпочитает низко-
рослые рямы по окраинам верховых болот, темнохвойную тайгу и
частичные вырубки по соснякам (2—5). Во всех остальных место-
обитаниях редка (0,4—0,7). В средней тайге ее втрое меньше. Чаще
всего она встречается на вырубках по соснякам, в рямах верховых
болот, а также в поселках (по 1) и реже — в сосняках и центральных
преимущественно открытых частях верховых болот (0,2—0,4).
В южной тайге в среднем по территории кукушки столько же, сколь-
ко в северной. Больше всего ее в суходольных мелколиственных и
полузаболоченных хвойнолиственных лесах, в полях-перелесках и
на открытых низинных болотах (2—3). Значительно реже кукушка
4*
61
встречалась на верховых болотах и в темнохвойно-таежных лесах
(0,01 и 0,5). В подтаежных лесах кукушки лишь немногим меньше,
чем в северной и южной тайге. Она довольно равномерно распределе-
на в лесополелуговых местообитаниях (0,8—1), несколько меньше
кукушки отмечено в рямах (0,4).
Итак, в северных подзонах на междуречьях кукушка предпочи-
тает вырубки среди лесов и рямы по окраинам верховых болот,
а в южных подзонах — осветленные мелколиственные и смешанные
леса и лесополевые местообитания. В долине Оби по сравнению со
сходными подзонами междуречий кукушки в 4—10 раз больше. В до-
лине она концентрируется в пойменных ивняках, а к северу от южной
тайги все чаще встречается в надпойменных местообитаниях, осо-
бенно в хвойно-лиственных лесах и рямах по окраинам верховых
болот. Повсеместно ее численность выше в северной и южной тайге
по сравнению со средней тайгой и подтаежными лесами.
Глухая кукушка (Cuculus saturatus Biyth.)
Первые встречи в северной тайге зарегистрированы 5 июня
1976 г., в средней — 14 мая 1977 г. В южной тайге у Томска первое
кукование датируется А. М. Гынгазовым и С. П. Миловидовым
11977] 22 мая 1972 г.
На гнездовании в северной тайге во второй половине июня глу-
хая кукушка встречалась почти исключительно в приречной темно-
хвойной тайге и лишь изредка — на гарях по соснякам (7 и 0,4)»
В средней тайге в июне редка в приречных смешанных лесах и на
вырубках по соснякам (0,7 и 0,2). В южной тайге во второй половине
июня повсеместно наблюдалась в лесном ландшафте (0,1—0,8), пред-
почитая темнохвойные и полузаболоченные смешанные леса, гораздо
реже ее слышали по окраинам верховых и низинных болот (0,1—0,2).
В подзоне подтаежных лесов глухая кукушка в июне обнаружена
лишь в крупных массивах осиново-березовых лесов (0,7). После
окончания гнездования в середине лета глухая кукушка встречалась
заметно реже (в 3—17 раз по сравнению с плотностью гнездования),
и в августе ее почти нигде не отмечали, кроме южной тайги, где глу-
хая кукушка изредка попадалась в учетах до конца лета.
По среднелетним показателям, в северной тайге глухая кукуш-
ка четко приурочена к приречным темнохвойным лесам (2). В средней
тайге предпочитает сходные смешанные леса (0,2), но в среднем по
территории ее здесь почти вдвое меньше, чем в предыдущей подзоне.
В южной тайге численность глухой кукушки в 7 раз выше, чем в се-
верной. Чаще всего она встречается в мелколиственных, смешанных
полузаболоченных и темнохвойных лесах, не избегая шелкопряд-
ников и полей-перелесков, и в значительно меньшем числе отмечена
в угнетенных лесах (елово-кедровой тайге и рослых рямах) и но
окраинам болот. В подтаежных лесах глухой кукушки примерно
столько же, сколько в средней тайге, а прослежена она только в
обширных осиново-березовых лесах.
52
Итак, на таежных междуречьях глухая кукушка предпочитает
темнохвойно-мелколиственные суходольные леса нормальной пол-
ноты, реже — полузаболоченные. Максимальное обилие этой кукуш-
ки отмечено в южной тайге, к северу и югу от которой оно заметно
снижается. В долине Оби этой кукушки в 2—6 раз меньше, чем на
междуречьях, а характер подзональных различий численности ос-
тается прежним. По сравнению с предыдущим видом численность
глухой кукушки заметно ниже на большей части обследованной тер-
ритории, особенно в пойме Оби. Исключение составляют южно-
таежные междуречья, где преобладают предпочитаемые глухой ку-
кушкой темнохвойные и производные от них леса. Здесь численность
обоих видов примерно одинакова. Кроме того, обилие глухой ку-
кушки обычно возрастает от пойм к междуречьям, а обыкновен-
ной — в обратном направлении.
ОТРЯД СОВЫ (STRIGIFORMES)
Ястребиная сова (Surnia ulula L.)
На междуречьях зарегистрирована только в северной тайге
в начале августа на гарях по соснякам (2), что позволяет считать ее
чрезвычайно редкой (в среднем по участку — 0,01). А. М. Гынгазов
и С. П. Миловидов [1977] находили кладки и выводки этой совы го-
раздо южнее, в окрестностях городов Стрежевого и Томска, по рекам
Шегарка и Чулым. В долине Оби в пределах северных подзон ястре-
биная сова встречалась чаще, чем на междуречьях.
Воробьиный сыч (Glaucidium passerinum L.)
Зарегистрирован на междуречьях только в средней тайге в при-
речных смешанных лесах (в среднем за лето — 5). Видимо, этот по-
казатель сильно завышен, так как при учете 8 июля 1977 г. был встре-
чен выводок из пяти сычей. Один из них слетел с добытым бурунду-
ком в лапах. В среднем по обследованной территории среднетаежных
междуречий редок (0,5) и не найден в долине Оби. По-видимому,
предпочитает крупные лесные массивы, отмечался, в частности,
на среднетаежном Обь-Енисейском междуречье в бассейне р. Тым
[Гынгазов, Миловидов, 1977] и зимой у пос. Ягодного в бассейне
р. Кеть.
Бородатая неясыть (Strix nebulosa J. R- Forst.)
В северной тайге изредка встречалась в приречных темно-
хвойных лесах, на переходных болотах и по берегам крупных таеж-
ных рек (0,5, 0,1 и 0,05). К югу от северной тайги на междуречьях
найдена в бассейнах рек Кеть и Шегарка, в окрестностях Томска и
с. Большой Кемчуг [Гынгазов, Миловидов, 1977; Кисленко, Наумов,
1972]. В долине Оби редка в пределах средней тайги в лесном и пой-
менном ландшафтах.
S3
Болотная сова (Asio jlammeus Pontopp.)
В северных подзонах таежных междуречий нами не зарегистри-
рована. В южной тайге обычна на низинных болотах (1) и редка в лес-
ном ландшафте (0,7). В подтаежных лесах встречалась гораздо ре-
же — только в первой половине июня на обширном низинном болоте
в сочетании с покосными лугами (1). В долине Оби отмечалась так же
редко, как и на междуречьях, но в северных подзонах несколько
чаще, чем в южных.
ОТРЯД КОЗОДОЕОБРАЗНЫЕ (CAP RIMULGI FORMES)
Козодой (Caprimulgus europaeus L.)
В средней тайге встречался до конца июня, преимущественно
на вырубках по соснякам, в приречных смешанных лесах и поселках
(5, 0,1 и 1). В южной тайге численность козодоя лишь немногим
выше, чем в средней, — он держался со второй половины мая до се-
редины августа преимущественно в лесополевых местообитаниях
(2) и в меньшем количестве в темнохвойно-мелколиственных лесах
и низкорослых рямах верховых болот (0,1—0,2). В подтаежных лесах
обилие козодоя примерно такое же, как и в предыдущей подзоне.
В июне и первой половине июля он отмечен в мелколиственных лесах
и рямах (0,4 и 2).
В среднем за лето козодой чаще всего встречался в разреженных
лесных местообитаниях южных подзон и в меньшем количестве —
в средней тайге. В долине Оби зарегистрирован в средней тайге и
в подтаежных лесах примерно в равном количестве в пределах обоих
ключевых участков и в несколько большем, чем на междуречьях.
ОТРЯД СТРИЖЕОБРАЗНЫЕ (APODIFORMES)
Черный стриж (Apus apus L.)
В северной тайге появился 2 июля 1976 г., в средней на месяц
раньше — 3 июня 1977 г., в южной — 7 июня 1967 г., а в подтаеж-
ных лесах первых стрижей видели в конце мая 1978 г. В средней тай-
ге 15 июня 1977 г. наблюдали спаривание черных стрижей в воздухе.
На гнездовании в средней тайге в июне стрижи чаще всего встре-
чались в сосняках-брусничниках, где располагалась их гнездовая
колония, и по берегам крупных надпойменных озер (6 и 9). В зна-
чительно меньшем числе их видели на вырубках по соснякам и
верховых болотах (0,2—0,6). Гнездо черного стрижа найдено в сред-
ней тайге 15 июня 1977 г. в дупле сосны, в 7 м от земли.
После окончания гнездования в июле численность этого стрижа
в среднем возрастает в 2—3 раза по сравнению с плотностью гнездова-
ния. В особенно большом количестве он начинает встречаться на вы-
54
рубках по соснякам и в открытой грядово-мочажинной части верхо-
вых болот (27 и 10). В августе численность черного стрижа сни-
жается вчетверо по сравнению с июлем, а с 19 августа 1977 г, он пе-
рестал встречаться совсем. В остальных подзонах оценить плотность
гнездования и внутрисезонные изменения численности черного стри-
жа не удалось в связи с его малочисленностью и случайностью встреч.
В северной тайге стрижей видели с начала июня до конца августа,
в южной — в июне и июле, а в подтаежных лесах — только в конце
мая и начале июня.
В среднем за лето в северной тайге черные стрижи чаще всего
встречались над старицами крупных таежных рек и темнохвойной
тайгой по их берегам (1 и 0,4) и заметно реже — над самими реками,
сосняками-ягельниками и поселками (0,01—0,05). В средней тайге
черного стрижа гораздо больше. Он многочислен на вырубках по
соснякам (10), обычен в сосняках-брусничниках, над верховыми бо-
лотами и надпойменными озерами (1—5) и редок в остальных место-
обитаниях (0,1—0,4). В южной тайге обилие этого стрижа лишь
немногим выше, чем в северной, но заметно ниже, чем в средней.
Встречается он по всей территории, но повсеместно очень редок
(в среднем — 0,09). Еще меньше черного стрижа в подтаежных лесах,
где он чрезвычайно редок (0,005).
Итак, на междуречьях черный стриж чаще всего встречается в
высокоствольных сосняках и вблизи них. Направленных подзональ-
ных изменений численности не прослеживается. В долине Оби в юж-
ных подзонах его примерно столько же, сколько на междуречьях,
а в средней тайге значительно меньше.
ОТРЯД ДЯТЛООБРАЗНЫЕ (PICIFORMES)
Вертишейка (Jynx torquilla L.)
Пролет средней напряженности (2) выражен только в подтаеж-
ных лесах в конце мая. В северной тайге встречалась лишь в первой
половине июня в сосново-озерном ландшафте, в низкорослых рямах
и в поселках (1—4). В средней тайге вертишеек видели в конце мая в
поселках (0,С) и во второй половине августа на вырубках по сосня-
кам. В южной тайге вертишейка тоже встречалась довольно редко,
только в первой половине июля ее слышали в полях-перелесках (0,7).
В подтаежных лесах вертишейки гораздо больше, чем в остальных
подзонах, и держалась она все лето. На гнездовании с начала июня
до середины июля она концентрировалась в мелколиственных лесах
и по окраинам низинных болот (0,7 и 0,3). В конце июля ее числен-
ность возросла почти втрое по сравнению с плотностью гнездования,
затем снизилась к началу августа, и с середины этого месяца верти-
шейка перестала встречаться совсем.
Итак, предпочитая разреженные и чередующиеся с открытыми
участками лесные местообитания, вертишейка в небольшом числе
отмечена в подтаежных лесах, гораздо меньше ее в остальных под-
55
зонах междуречий. В долине Оби в пределах южных подзон ее при-
мерно столько же, сколько на подтаежных междуречьях, а к северу
численность также резко убывает.
Черный дятел (Dryocopus martius L.)
В крайних таежных подзонах в период гнездования не отмечен.
В средней тайге в июне обычен в небольших таежных поселках (1)
и редок в сосново-боровом ландшафте (0,2—0,5). В южной тайге в то
же время обычен в лесном ландшафте (1), где предпочитает осиново-
березовые леса (2). В период послегнездовых кочевок черный дятел
начал встречаться в северной тайге (в начале июля — 0,4), но затем
его численность неуклонно снижалась до конца августа. В средней
тайге в течение июля черных дятлов не видели, а в августе они вновь
начинают встречаться примерно в том же количестве, что и на гнез-
довании. В южной тайге в июле происходит снижение обилия почти
в 8 раз по сравнению со второй половиной июня. В подтаежных лесах
желна появляется только в начале августа, а к концу этого месяца
ее численность снижается в 3,5 раза. По-видимому, в июле происхо-
дит откочевка большей части гнездящихся птиц, в том числе из юж-
ных подзон в северные, а в августе они смещаются к югу и вновь
начинают встречаться в местах гнездования.
В среднем за лето в северной тайге черный дятел редок в сосня-
ках, производных от них местообитаниях и поселках (0,1—0,5).
В средней тайге встречается несколько чаще в сосново-боровом
ландшафте и поселках (0,6 и 0,8). В южной тайге его примерно в 5 раз
больше, чем в северной и средней, и держится он преимущественно
в темнохвойных и осиново-березовых лесах. В подтаежных лесах
черного дятла примерно столько же, сколько в северной тайге,
встречается он в осиново-березовых лесах, полях-перелесках (0,2—
0,3) и вблизи поселков (0,06).
Следовательно, черный дятел повсеместно предпочитает сомкну-
тые высокоствольные леса, особенно осиново-березовые, и в меньшей
степени — темнохвойные и сосновые. Численность его на междуречь-
ях снижается к северу от южной тайги и в подтаежных лесах. В до-
лине Оби этого дятла повсеместно меньше, чем на междуречьях.
Большой пестрый дятел (Dendrocopus major L.)
На гнездовании в северной тайге до середины июля предпочитал
гари по соснякам, имеющим наибольшее количество сухостойных
дуплистых деревьев (7). В несколько меньшем числе отмечался на
частичных вырубках по соснякам и в приречной темнохвойной тайге
(4 и 3), и еще меньше этого дятла зарегистрировано в остальных
местообитаниях, кроме верховых болот (1—2). В средней тайге на
гнездовании (середина мая — 15 июня) часто встречался в приреч-
ных смешанных лесах, на вырубках по соснякам, в рослых рямах и
поселках (2—4), и лишь изредка — в низкорослых рямах верховых
56
болот (0,5). В южной тайге в тот же период многочислен в шелкопряд-
никах, на гарях и обычен в остальных лесных местообитаниях.
В подтаежных лесах на гнездовании обнаружен в осиново-березовых
лесах и полях-перелесках (1 и 0,2). Птенцов, кричащих в гнездах,
слышали и встречали взрослых птиц с кормом: в северной тайге с
25 июня по 16 июля 1976 г., в средней тайге с 11 по 17 июня 1977 г.
и в подтаежных лесах 13 июня 1978 г. Трех слетков большого пестро-
го дятла видели в средней тайге 5 июля 1977 г.
В послегнездовое время (во второй половине июля) в северной
тайге до 90 % популяции большого пестрого дятла откочевывает за
пределы обследованной территории, а в августе они вновь появляют-
ся в значительном числе. Обилие его в это время превышает гнездо-
вое в 8—9 раз. В более южных подзонах в начале июля численность
большого пестрого дятла возрастает в результате размножения и
прикочевки средней интенсивности (2—5), но к концу лета примерно
то же число особей откочевывает (2—6). По-видимому, во второй по-
ловине лета часть дятлов откочевывает к югу, так как их численность
последовательно возрастает в южно-таежных приобских борах в
конце июля и в Северо-Восточшш Алтае в первой половине августа
ТРавкин, 1973]. Однако не исключены кочевки обратной направ-
ленности, определяющие столь резкое возрастание численности
дятлов в августе в северной тайге.
В среднем за лето в северной тайге большой пестрый дятел
предпочитает вырубки, гари по соснякам и сосново-озерный ланд-
шафт (6—8), в меньшем числе встречаясь в коренных лесных форма-
циях: в приречной темнохвойной тайге и лишайниковых сосняках
(2—4). Еще меньше его на комплексных верховых болотах (0,1 —
0,6), где он держался преимущественно по небольшим лесным гривам
и участкам низкорослых рямов. В средней тайге этого дятла пример-
но столько же, сколько в северной. Предпочитает вырубки по сосня-
кам и приречные темнохвойно-мелколиственные леса с примесью
сосны (6—8). На остальной территории, за исключением открытой
части верховых болот, обычен (2—4). В южной тайге его почти втрое
больше, чем в предыдущих подзонах. Он многочислен на шелкопряд-
никах и гарях, во влажных и заболоченных лесах (10—27). В ос-
тальных местообитаниях лесного ландшафта обычен (3—8) и лишь
изредка появляется на окраинах открытых низинных болот, избегая
обширных верховых. В подтаежных лесах большого пестрого дятла
в 3 и 8 раз меньше, чем соответственно в северных подзонах и южной
тайге. Он предпочитает осиново-березовые леса (3). На послегнездо-
вых кочевках встречается в рямах и в лугово-полевых местообита-
ниях (1 и 0,4).
Таким образом, следует отметить, что численность большого
пестрого дятла выше в лесных и производных от них местообитаниях
с большим числом дуплистых и сухостойных деревьев, а в кустар-
ничковых и лишайниковых сосняках северных подзон она невелика.
В среднем по территории на междуречьях его больше всего в южной
тайге, меньше в северных подзонах и особенно в подтаежных лесах.
При сравнении с долиной Оби следует отметить, что наибольшая
57
численность этого дятла характерна для южно-таежных междуре-
чий и подтаежных надпойменных террас, включая южно-таежные
приобские сосняки, расположенные на последних. Наименьшая чис-
ленность зарегистрирована в северотаежной пойме Оби и на подта-
ежных междуречьях. На остальной территории отмечены средние
показатели.
Белоспинный дятел (Dendrocopus leucot os Bechst.)
В северной тайге не зарегистрирован. В средней наблюдался в
рослых рямах по окраинам верховых болот во второй половине
июня (1). В южной тайге его встречали все лето. На гнездовании в
конце мая — июне обычен в хвойно-лиственных полузаболоченных,
осиново-березовых лесах и на шелкопрядниках в сочетании с гаря-
ми и вырубками (4—7). На подтаежных междуречьях нами не на-
блюдался, но в долине Оби в пределах той же подзоны часто встречал-
ся в пойменных ивняках, в том числе на гнездовании.
В среднем за лето на междуречьях чрезвычайно редок в средней
тайге и обычен в южной (0,003 и 1). Таким образом, белоспинный дя-
тел предпочитает осветленные мелколиственные и полузаболоченные
хвойно-лиственные леса, вырубки и шелкопрядники, а также осин-
ники и ивняки в поймах крупных рек. Больше всего этого дятла
встречено в подтаежной долине Оби и на южно-таежных междуречьях
и значительно меньше — в средней тайге.
Малый пестрый дятел {Dendrocopos minor L.)
На междуречьях северной тайги нами не наблюдался. Однако
в долине Оби летом 1973 г. был обычен в пойменных ивняках и редок
в надпойменных смешанных лесах. В средней зарегистрирован во
второй половине августа на вырубках по соснякам и в поселках
(2 и 0,2). В южной тайге держался все лето. На гнездовании (июнь)
встречен в елово-кедровых лесах (1). Во второй половине июля ста-
новится обычным в разных лесных местообитаниях, и численность
его в среднем по территории заметно возрастает, а позже, в августе,
до 70% популяции откочевывает из района работ. В подтаежных
лесах этого дятла видели лишь однажды — вне учета, в ивняках
поймы небольшой речки (верховья р. Омь). В среднем за лето на
междуречьях малый пестрый дятел редок в южной и очень редок в
средней тайге и подтаежных лесах. В долине Оби, где этот дятел
предпочитает пойменные ивняки и осинники, его заметно больше,
особенно в северных подзонах.
Трехпалый дятел (Picoides tridactylus L.)
На гнездовании в северной тайге (до середины июля) обычен
в сосняках-ягельниках и низкорослых рямах (1—3) и редок в сосно-
во-озерном ландшафте (0,3). В средней тайге обычен на вырубках
по соснякам (3). В южной тайге в тот же период обычно встречается
58
в смешанных и мелколиственных лесах (1—2) и реже — в рослых
рямах (0,5). В послегнездовое время его численность в северных под-
зонах в конце июля — начале августа возрастает вдвое. Во второй
половине августа обилие этого дятла вновь снижается в 3—4 раза
по сравнению с плотностью гнездования. В южной тайге после-
гнездового увеличения плотности не прослеживается, напротив,
трехпалые дятлы не встречались с начала июля до середины августа.
Во второй половине августа они начинают попадаться вновь на
послегнездовых кочевках, но в меньшем числе, чем на гнездовании.
В среднем за лето в северной тайге трехпалый дятел чаще всего
встречен в сосновых и темнохвойных лесах и в сосново-озерных
ландшафтах (1—2) и несколько реже — в остальных местообитани-
ях, кроме открытых комплексных верховых болот и поселков (0,4—
0,9). В средней тайге его примерно в 10 раз меньше, здесь он зареги-
стрирован в сосняках-брусничниках и на вырубках по ним (0,4 и 2).
В южной тайге трехпалого дятла почти вдвое меньше, чем в северной,
но в 6 раз больше, чем в средней. Он отдает предпочтение темнохвой.-:
ным и мелколиственным лесам и редок в остальных местообитаниях
лесного ландшафта.
Итак, на междуречьях, где он концентрируется по соснякам,
трехпалого дятла больше всего в северной тайге, меньше в южной и
особенно в средней. В долине Оби, где этот дятел предпочитает
надпойменные хвойно-лиственные леса и пойменные ивняки, про-
слеживается обратный характер подзональных различий численно-
сти (больше всего в средней тайге, меньше в южной и северной).
В целом для обследованных участков наибольшая численность
трехпалою дятла характерна для среднетаежной долины Оби и се-
веротаежных междуречий и меньшая — для остальной территории.
В подзоне подтаежных лесов в долине Оби, как и на междуречьях,
этот дятел нами и Ю. С. Равкиным [19781 не зарегистрирован, хотя
спорадично встречается на юге до северной лесостепи и ленточных
боров Приобья [Гынгазов, Миловидов, 1977].
ОТРЯД ВОРОБЬИНЫЕ (PASSERIFORMES)
СЕМЕЙСТВО ЖАВОРОНКИ (ALAUDIDAE)
Полевой жаворонок (Alauda arvensis L.)
На междуречьях зарегистрирован только в подтаежных лесах.
В конце мая прослеживается пролет средней интенсивности (2).
На гнездовании в первой половине июня обычен в лугово-полевых
местообитаниях (1—2). В послегнездовое время численность полево-
го жаворонка сначала возрастает примерно в 4 раза, а затем в ре-
зультате откочевки снижается во второй половине лета до уровня
плотности гнездования. В июле и августе он перестает встречаться
в полях, лугах-выпасах и концентрируется на участках сенокосных
лугов, перемежающихся с низинными болотами. По среднелетним
59
показателям, явно предпочитает покосные луга в сочетании с ни-
зинными болотами (5). В значительно меньшем числе отмечается
в полях-перелесках и лугах-выпасах (0,1—0,6).
В долине Оби от подтаежных лесов до средней тайги его пример-
но вдвое меньше, чем на подтаежных междуречьях. Наблюдается в
долине преимущественно в подтаежных полях-перелесках, проникая
к северу по пойменным лугам.
Рогатый жаворонок (Eremoppila alpestris L.)
Наблюдался только на предгнездовом пролете. В северной тай-
ге в начале июня многочислен в поселках (127). Стайки рогатых жа-
воронков в это время постоянно встречались здесь на краях песчаной
взлетно-посадочной полосы. И. Н. Шуховым [1916] стайки в 40—50
особей наблюдались 22 мая 1914 г. в долине р. Казым. В средней тай-
ге их видели в конце мая на грядово-мочажинных верховых болотах
(0,5). В южных подзонах нами не зарегистрирован. В долине Оби
кроме северной тайги встречался на весеннем пролете чаще, чем на
междуречьях, и преимущественно в пойменных лугах.
СЕМЕЙСТВО ЛАСТОЧКИ (HI KU NDINIDAE)
Береговая ласточка (Riparia riparia L.)
Появилась в поселках, расположенных по берегам рек, в север-
ной тайге 7 июня 1976 г., в средней — 27 мая 1977 г. В средней тайге
Зауралья на р. Мал. Сосьва прилет зарегистрирован М. П. Таруни-
ным [1959] 2 июня 1931 г., у Томска появляется в среднем 18 мая.
В июне на гнездовании в северной тайге наиболее многочислен-
на в поселках вблизи береговых обрывов, в которых располагались
ее колонии (52). Сравнительно много береговушек встречалось по
берегам крупных таежных рек и на открытых участках верховых
болот (12—17) и гораздо меньше — в приречных темнохвойных ле-
сах, сосняках-ягельниках и на гарях по ним (2—3). В средней тайге
в гнездовое время тоже предпочитает поселки (454). В значительно
меньшем числе встречается в сосняках-брусничниках и на открытых
частях верховых болот (15—16) и в еще меньшем — в низкорослых
рямах, в приречных смешанных лесах и по берегам рек (1—6). Мини-
мальное обилие зарегистрировано на вырубках по соснякам и рослым
рямам (0,2 и 0,4). В Прикетье, на границах южной и средней тайги,
яйца находили 6 июня 1962 г., вылупление птенцов отмечено 6 июля
1968 г., а 17 июля в той же колонии были птенцы, готовые к вылету.
В июле, после окончания гнездования, в северной тайге в по-
селках и на крупных притоках Оби береговой ласточки становится
в 2—3 раза больше. К началу августа в результате откочевки ее
обилие в среднем по району работ снижается почти в 15 раз по срав-
нению с гнездовым периодом. Держится она только в поселках и
после 15 августа перестает встречаться совсем. В средней тайге про-
60
слеживаются сходные послегнездовые изменения численности этой
ласточки. В начале июля в поселках ее становится вдвое больше,
чем на гнездовании, а к середине августа большинство береговушек
улетает — последних видели 24 августа 1977 г.
В среднем за лето в северной тайге береговая ласточка предпочи-
тает поселки и берега крупных притоков Оби (68 и 24), заметно реже
встречается на верховых болотах и в сосново-озерном ландшафте
(2—7), а самое низкое обилие зарегистрировано в темнохвойных и
сосновых лесах (по 0,5). В средней тайге ее втрое больше (в среднем
по обследованному участку), береговая ласточка здесь явно отдает
предпочтение поселкам (366). В остальных местообитаниях обычна
или редка. На междуречьях, где в южных подзонах в районах наших
работ отсутствовали пригодные для ее гнездования обрывы, эта лас-
точка не встречена.
Таким образом, следует отметить, что береговая ласточка рас-
пространена на междуречьях весьма неравномерно, преимуществен-
но по крупным притокам Оби, достигая особенно высокой численно-
сти в поселках вблизи высоких отвесных берегов, пригодных для ее
гнездования. В долине Оби в пределах северных подзон ее примерно
столько же, сколько на междуречьях, а в южных — несколько
больше. При этом ее численность в небольших поселках по берегам
крупных притоков Оби обычно выше, чем в крупных населенных
пунктах долины, удаленных от гнездовых колоний.
Деревенская ласточка (Hirundo rustica L.)
К началу наших наблюдений в средней тайге (14 мая 1977 г.)
деревенские ласточки уже прилетели в крупный пос. У гут и были
здесь многочисленны. В южной тайге первые из них появились 29 мая
1967 г., а массовый прилет их в подтаежных лесах отмечен 20 мая
1978 г.
На гнездовании в северных подзонах деревенская ласточка не
зарегистрирована. Однако гнездилась в крупных среднетаежных по-
селках, которые нами не обследованы. В мелких населенных пунк-
тах, где проводились наблюдения, не встречалась. На южно-таежных
междуречьях учеты в поселках не проводились. В гнездовое время
(в июне) деревенская ласточка в южной тайге лишь изредка попада-
лась в лесополевых местообитаниях и на низинных болотах (0,1—
0,6). Однако в южно-таежных приобских селах она весьма многочис-
ленна (371). В подтаежных лесах гнездится в поселках (437). В не-
большом числе встречается за их пределами по берегам речек и в
лугах, используемых под выпас (8 и 14). В Прикетье, на границе
средней и южной тайги, гнездо с шестью яйцами найдено 3 июня
1961 г., а в июле повсеместно в гнездах находили птенцов. В южной
тайге по р. Чулым кладки в пять и шесть яиц найдены 15 июня и
6 июля, а голых птенцов видели 17 июля 1969 г. [Гынгазов, Мило-
видов, 1977].
В послегнездовой период в южной тайге с 15 июля по 15 ав-
густа начинает встречаться значительно чаще. Становится много-
61
численной в полях-перелесках (14—18) и обычной в лесном ланд-
шафте (в среднем — 2). После окончания гнездования в первой по-
ловине июля в подтаежных поселках обилие деревенских ласточек
возрастает после размножения в 1,8 раза. Затем к концу июля чис-
ленность снижается на 20% по сравнению с предыдущим периодом в
результате откочевки части популяции. В начале августа происхо-
дит интенсивная прикочевка, которая увеличивает обилие почти
вдвое. К концу месяца значительная часть популяции покидает эти
поселки, и происходит трехкратное снижение численности. В тот же
период в долине Оби отмечено еще более резкое увеличение числен-
ности деревенских ласточек в конце июля в южно-таежных, а в кон-
це августа в подтаежных селах. Столь резкие изменения численности
этих ласточек в послегнездовой период, несомненно, вызваны их ин-
тенсивной откочевкой к югу и с междуречий в долину Оби.
В среднем за лето в южной тайге деревенская ласточка обычна
в полях-перелесках (5), а на остальной территории, кроме верховых
болот, редка. В подтаежных поселках зарегистрирована наиболее
высокая ее численность (571), гораздо реже она встречается в приле-
жащих к поселкам лугах-выпасах (И) и еще реже — на низинных
болотах и по берегам рек (3 и 8).
Итак, на междуречьях деревенская ласточка явно предпочитает
подтаежные поселки. В долине Оби в пределах южных подзон в сред-
нем ее примерно столько же, сколько на подтаежных междуречьях.
К северу отмечена только до средней тайги и в незначительном коли-
честве. Она отдает предпочтение крупным населенным пунктам, обыч-
но окруженным сельскохозяйственными землями, и реже встреча-
ется в мелких таежных поселках.
СЕМЕЙСТВО ТРЯСОГУЗКОВЫЕ (MOTAC1LLIDAE)
Желтая трясогузка (Motacilla flava L.)
В средней тайге появилась 19 мая 1977 г., в остальные подзоны
прилетела до начала наших наблюдений. В северной тайге на р. Ка-
зым прилет зарегистрирован И. Н. Шуховым [1916] 27 мая 1914 г.
В средней тайге на р. Мал. Сосьва появляется 19 мая, а в южной
в окрестностях Томска — с 27 апреля по 20 мая [Гынгазов, Милови-
дов, 1977]. Пролет средней и высокой интенсивности во второй поло-
вине мая выражен повсеместно, кроме северной тайги (5—11). Осо-
бенно большие скопления пролетных трясогузок в этот период на-
блюдали на южно-таежных открытых низинных болотах (161).
На гнездовании в северной тайге (июнь) многочисленна на вер-
ховых и переходных болотах (60—150) и обычна в сосново-озерном
ландшафте (9). В средней тайге больше всего желтой трясогузки
гнездится на грядово-мочажинных верховых болотах (26) и значи-
тельно меньше — в рямах и приречных темнохвойно-мелколиствен-
ных лесах (1—2). В южной тайге гнездится только на грядово-моча-
62
жинных обширных водораздельных верховых болотах (16). На гнез-
довании в подтаежных лесах предпочитает суходольные луга, ис-
пользуемые под выпас, и низинные тростниково-осоковые болота в со-
четании с сенокосными лугами (24—63). В гораздо меньшем количе-
стве встречается в поселках и по берегам небольших рек (0,3 и 4).
Гнезда желтой трясогузки с пятью и шестью яйцами находили 11 и
13 июня 1976 г. в северной тайге, но уже 1 июня видели самца, вол-
нующегося с кормом. В той же подзоне 13 июля 1976 г. видели сам-
ку с кормом, а 16 июля — слетков с кормящими их взрослыми пти-
цами. В средней тайге 9 июня 1977 г. наблюдали постройку гнезда.,
Сразу после окончания гнездования (в июле) в северной тайге
начинается откочевка, в результате которой плотность популяции
снижается к началу августа более чем на 70% по сравнению с таковой
на гнездовании. На фоне общего снижения численности в июле про-
исходит ее рост в 1,3—4 раза в сосново-озерных ландшафтах, низко-
рослых рямах и на переходных болотах в результате концентрации
здесь молодых птиц. В августе идет пролет, не увеличивающий чис-
ленности, так как он менее интенсивен по сравнению с откочевкой
местных птиц. Однако в этот период трясогузки начинают встречать-
ся гораздо шире, в том числе в сосняках-ягельниках, на вырубках и
гарях по ним. В августе нередко видели стайки желтых трясогузок
по 30—40 птиц. Резкое увеличение численности в первой половине^
августа происходит только в поселках (325), где постоянно кормятся
смешанные стайки желтых и белых трясогузок. В средней тайге поч-
ти все отгнездившиеся желтые трясогузки откочевывают в июле,
а в августе прослеживается их пролет средней интенсивности (6)~
В этот период они встречаются гораздо шире, чем на гнездовании,
и в особенно большем числе в поселках с середины до конца авгус-
та (38). В южных подзонах значительное количество желтых трясо-
гузок тоже прикочевывает после окончания гнездования. В августе
идет пролет (4—7), и концентрация трясогузок в полях и населенных
пунктах возрастает (41—60).
В среднем за лето желтая трясогузка в северной тайге многочис-
ленна на комплексных верховых болотах и в сосново-озерном ланд-
шафте (14—95), а в сосново-боровом ландшафте обычна (5). В сред-
ней тайге желтой трясогузки почти в 7 раз меньше, чем в северной.
Предпочитает эта трясогузка грядово-мочажинные верховые болота.
В меньшем числе встречается в низкорослых рямах, по берегам
водоемов и в поселках (2—8). В сосново-боровом ландшафте редка
(0,3). В южной тайге ее почти столько же, сколько в средней, но в 8
раз меньше, чем в северной. По среднелетним показателям, предпо-
читает поля-перелески и низинные болота (12—24), обычна на вер-
ховых болотах и в лесном ландшафте (2—5). В подтаежных лесах
желтой трясогузки втрое меньше, чем в северной тайге, но в 2 раза
больше, чем в средней и южной. Чаще всего встречается она в сухо-
дольных лугах, используемых под выпас (63), довольно многочис-
ленна на низинных болотах, чередующихся с покосными лугами»
и в поселках (13—16), обычна в полях-перелесках и по берегам рек
(по 2) и редка в остальных местообитаниях (0,3—0,9).
63
В результате следует отметить значительно более высокую
численность желтой трясогузки в крайних подзонах таежных между-
речий (особенно в северной тайге) по сравнению со срединными.
При сравнении с долиной Оби выявляется, что больше всего этой
трясогузки на северотаежных междуречьях и в долине Оби без под-
таежных лесов. В последних отмечен несколько меньший уровень
численности. Минимальное обилие вида характерно для междуречий
срединных подзон. При этом в южных подзонах и среднетаежной
долине Оби желтая трясогузка предпочитает низинные болота и
луга, особенно пойменные. С продвижением к северу все большее
предпочтение отдает открытым грядово-мочажпнным верховым и
переходным, а в северной тайге — гетеротрофным болотам.
Желтоголовая трясогузка
(Motacilla citreola Pall.)
Пролет отмечался в средней тайге (устье р. Кеть) 6 июля
1966 г. В южной тайге желтоголовая трясогузка встречена только во
второй половине мая на предгнездовом пролете в мелколиственных
лесах. В подтаежных лесах на гнездовании в первой половине июня
многочисленна на низинных болотах и в лугах-выпасах по берегам
речек (12—19). В конце июня обилие ее в результате размножения
возрастает в среднем вдвое и молодые желтоголовые трясогузки кон-
центрируются на низинных тростнпково-осоковых болотах (42).
Затем эта трясогузка перестает встречаться в течение месяца, и толь-
ко в конце августа, на послегнездовых кочевках, в небольшом числе
отмечается вновь на тех же болотах и в полях (4—7). В среднем за ле-
то она явно предпочитает низинные болота (15). В небольшом числе
встречается в лугах-выпасах и по берегам рек (2 и 3) и в еще мень-
шем — в полях-перелесках и поселках (0,6 и 0,3).
Итак, желтоголовая трясогузка явно предпочитает подтаежные
низинные болота. Встречается в отличие от желтой трясогузки по
наиболее увлажненным участкам, значительно уступая ей по чис-
ленности на суходольных лугах. В долине Оби максимальная чис-
ленность желтоголовой трясогузки зарегистрирована тоже в преде-
лах подтаежных лесов. В более северных подзонах ее значительно
меньше (повсеместно редка) и примерно столько же на южно-таежных
междуречьях.
Белая трясогузка (Motacilla alba L.)
Пролет заканчивается в северной тайге в начале июня, а в ос-
тальных подзонах — во второй половине мая. С севера на юг интен-
сивность его по подзонам равна соответственно 10, И, 9 п 5 особей
на объединенный 1 км2 ключевого участка.
На гнездовании во второй половине июня в северной тайге
больше всего белых трясогузок встречено в поселках (172). В зна-
чительном числе они держались в сосняках-ягельниках и на вы-
рубках по ним, в сосново-озерном ландшафте, на верховых болотах
64
и старицах крупных таежных рек (11—47). Заметно меньше этих
трясогузок зарегистрировано по берегам самих рек, в приречной
темнохвойной тайге и на гарях по соснякам (3—4). В средней тайге
с 1 по 15 июля белая трясогузка многочисленна в поселках и по бе-
регам крупных надпойменных озер (33 и 12), обычна на грядово-
мочажинных верховых болотах, в сосняках-брусничниках и на вы-
рубках по ним (3—7) и редка по берегам крупных таежных рек (0,6).
В южной тайге обычна в полях-перелесках (9). В подзоне подтаежных
лесов отмечена в поселках и по берегам небольших речек (4 и 1).
Гнезда белых трясогузок с пятью — семью яйцами находили в се-
верной тайге 19 и 26 июня 1976 г., а в средней — 27 мая, 30 июня и
1 июля 1977 г.
В послегнездовое время в северной тайге в результате размноже-
ния и прикочевок извне обилие белой трясогузки возрастает в тече-
ние июля почти в 4 раза, а затем в августе снижается вдвое. Особен-
но много белых трясогузок встречалось в общих стайках с желтыми
в начале августа в поселках (454). В средней тайге послегнездовая
прикочевка прослеживается в конце июня и в начале августа (3—5
в среднем по участку). Больше всего белой трясогузки отмечено в
июле в поселках, а в августе — по илистым отмелям крупных над-
пойменных озер (154 и 193). В южной тайге белые трясогузки в после-
гнездовое время (июль — август) лишь изредка встречались в полях-
перелесках (0,1—3). В подтаежных лесах с середины июня до конца
июля численность их также невелика (0,2—0,4), но к концу августа
она возрастает почти в 10 раз в результате прикочевки. В период
кочевок эта трясогузка в наибольшем числе отмечалась в лугах, ис-
пользуемых под выпас (3). Первые встречи молодых вместе с выкарм-
ливающими их взрослыми птицами датируются в средней тайге
22—24 июня 1977 г., а в северной — 5 июля 1976 г.
В среднем за лето в северной тайге белая трясогузка предпочи-
тает поселки (296). Многочисленна она в сосново-боровом, сосново-
озерном ландшафтах и по берегам таежных рек (10—63) и обычна на
комплексных верховых болотах и на отмелях по старицам таежных
рек (2—8). В средней тайге ее в 5 раз меньше. Больше всего этой тря-
согузки встречено в поселках и на отмелях крупных надпойменных
озер (81 и 70), гораздо меньше — на вырубках по соснякам и на гря-
дово-мочажинных верховых болотах (по И) и еще меньше — в осталь-
ных местообитаниях (3—6). Минимальное обилие зарегистрировано
в приречных смешанных лесах и рослых рямах верховых болот
(по 0,4). В южной тайге белая трясогузка в течение всего лета дер-
жалась в полях-перелесках (26). В подтаежных лесах ее примерно
столько же, сколько и в южной тайге. Белая трясогузка предпочи-
тает поселки, луга, используемые под выпас, и берега речек (7—13).
В меньшем количестве встречается в полях-перелесках и на низин-
ных болотах (4 и 0,3), но избегает рямов верховых болот.
Подводя итог, следует отметить, что белая трясогузка повсемест-
но предпочитает поселки и открытые пойменные местообитания, но
в северной тайге распространена наиболее широко: в значительном
числе встречается в редкостойных сосняках и сосново-озерных ланд-
5 Л, Г. Вартапетов
65
шафтах. Обилие этой трясогузки на таежных междуречьях резко
уменьшается к югу. Сходная тенденция прослеживается и в долине
Оби, но здесь снижение с северо-запада на юго-восток гораздо более
плавное. Поэтому в северной тайге в долине и на междуречьях отме-
чена наиболее высокая и примерно равная численность белой трясо-
гузки. В южнее расположенных подзонах в долине ее значительно
больше, чем на междуречьях.
Горная трясогузка
(Motacilla cinerea Tunst.)
В средней тайге горную трясогузку отмечали с конца июня до
начала августа по берегам крупных рек (0,4) и в конце августа на
илистых отмелях надпойменных озер (2). Кроме того, однажды она
встречена в подтаежных осиново-березовых лесах во второй полови-
не августа (2) на придорожных лужах. В долине Оби лишь изредка
наблюдалась в осиново-березовых южно-таежных лесах и поселках
средней тайги. Таким образом, горная трясогузка не зарегистри-
рована нами и Ю. С. Равкиным [1978] в северной тайге и в качестве
редкой птицы отмечалась в более южных подзонах. Однако
А. М. Гынгазов и С. П. Миловидов [1977] указывают на ее распро-
странение в Западной Сибири к северу до лесотундры.
Степной конек (Anthus richardi Vieill.)
Встречен только на подтаежных междуречьях. В первый раз
стайку степных коньков видели 24 мая 1977 г. В начале июня идет
довольно интенсивный пролет (14). На гнездовании (15—30 июня)
обычен на сенокосных лугах, чередующихся с низинными болотами,
и лугах-выпасах (6—7). В послегнездовое время (в июле) его числен-
ность во время кочевок в 4—5 раз выше, чем на гнездовании. Вторая
волна кочевок прослеживается в конце августа. В среднем за лето
больше всего конька зарегистрировано на участках лугов, череду-
ющихся с низинными болотами (21). В полях-перелесках и лугах-
выпасах обычен (1—9), в поселках и по берегам небольших рек ре-
док (0,1—0,3).
Лесной конек (Anthus trivialis L.)
Во второй половине мая в южных подзонах выражен пролет
средней интенсивности (в южной тайге — 5, в подтаежных ле-
сах — 9).
На гнездовании в северной тайге не обнаружен. В средней тай-
ге (первая половина июня) предпочитает рямы, в меньшей степени —
открытые грядово-мочажинные верховые болота (21—41). В сосново-
боровом ландшафте обычен (1—2). В южной тайге предпочитает
переходные и низинные болота (38 и 37), в меньшем числе встречает-
ся в шелкопрядниках и низкорослых рямах (16 и 15). В лесных место-
обитаниях обычен (1—9) и только в темнохвойной тайге редок (0,9).
66
В подтаежных лесах его больше всего в полях в сочетании с пере-
лесками (34), значительно меньше на низинных болотах, в рямах и
осиново-березовых лесах (12—18) и еще меньше в лугах, используе-
мых под выпас (8). Гнезда лесного конька находили в Прикетье (на
границе средней и южной тайги) с 21 мая по 15 июня, в большинстве
из них было по пять яиц. В средней тайге 16 июня 1977 г. мы наблю-
дали постройку гнезда, 21 июня встречены молодые птицы (три вы-
водка, в одном из которых было четыре слетка), а 24 июня видели
лесного конька, волнующегося с кормом у гнезда.
После окончания гнездования увеличение обилия лесных конь-
ков в средней тайге невелико. Оно делается заметным только в конце
июля. Особенно много их в этот период держится в рямах и поселках
(86 и 190). В августе в результате откочевки численность повсеместно
снижается и становится почти в 1,5 раза ниже, чем на гнездовании.
В южной тайге в результате размножения уже в начале июля обилие
лесных коньков возрастает вдвое по сравнению с периодом гнездова-
ния. К концу июля около 25% популяции откочевывает, а в августе
прослеживается прикочевка лесных коньков с севера (10). Особенно
много встречено их во второй половине августа на гарях и в шелко-
прядниках (160), низкорослых рямах, на низинных и переходных
болотах (63—93). В подтаежных лесах прослеживается примерно
такой же ход численности лесного конька, как и в южной тайге. Од-
нако максимальное его обилие отмечено не в августе на послегнездо-
вых кочевках, а в начале июля, после вылета молодых. Тем не менее
в этот период часть отгнездившихся птиц исчезает сразу после окон-
чания гнездования, поэтому фактическое увеличение на 20% меньше
расчетного. Итак, повсеместно прослеживается два периода увеличе-
ния численности лесных коньков: в конце июня — начале июля и в
конце июля — начале августа. Наряду с послегнездовыми кочевка-
ми они, видимо, в некоторой степени определяются двумя волнами
вылета лесных коньков, описанными С. С. Москвитиным [1974].
По среднелетним показателям, в средней тайге лесной конек
предпочитает поселки (76), в меньшем числе встречается на верховых
болотах и вырубках по соснякам (16—46). Значительно меньше его
в сосновых и приречных темнохвойно-мелколиственных лесах (2 и 7).
В южной тайге лесного конька примерно столько же, сколько в сред-
ней. Многочислен он на низинных, переходных болотах и в низко-
рослых рямах, а также в шелкопрядниках, мелколиственных лесах и
полях-перелесках (19—57). В остальных местообитаниях обычен
(3—7). В подтаежных лесах численность лесного конька примерно
такая же, как и в предыдущих подзонах. Больше всего его в осиново-
березовых лесах и полях среди них (34—47). В лугово-болотных мес-
тообитаниях обилие этого конька значительно ниже (7—10) и самое
низкое в поселках (0,9).
Итак, лесной конек предпочитает разреженные лесные и лесо-
полевые местообитания. В значительном числе встречается по окраи-
нам внепойменных болот, особенно в рямах и пойменных лесах.
В сомкнутых темнохвойных и сосновых лесах, а также в открытых
местообитаниях его обилие снижается. На междуречьях от средней
5*
67
таиги до подтаежных лесов отмечена сходная численность лесного
конька. В долине Оби обилие его неуклонно снижается к северу,
и в южных подзонах оно немного больше, а в средней тайге несколь-
ко меньше, чем на междуречьях. На северотаежных междуречьях
лесной конек не зарегистрирован, а по долине Оби проникает к севе-
ру гораздо дальше, по крайней мере до северной тайги.
Зеленый конек (Anthas hodgsoni Richm.)
В северной тайге песню в первый раз слышали 3 июня 1976 г.
В более южных подзонах прилетает раньше; так, в средней тайге
впервые встречен нами 19 мая 1977 г., а в южной по Чулыму отмечен
А. М. Гынгазовым и С. П. Миловидовым [1977] 11 мая 1969 г. Во вто-
рой половине мая в средней и южной тайге идет пролет (5).
На гнездовании в северной тайге (вторая половина июня) много-
числен на переходных и грядово-мочажинно-озерных верховых бо-
лотах (13—16), но держится преимущественно по их окраинам и на
участках открытых болот и рямов в сосново-боровом ландшафте,
где его численность еще выше — до 58 особей/км2 [Вартапетов,
1980]. Зеленый конек обычен на вырубках и гарях по соснякам,
в сосново-озерном ландшафте и низкорослых рямах (1—8) и редок в
сосняках-ягельниках (0,3). В средней тайге в тот же период предпо-
читает рослые рямы, окраины небольших таежных поселков и при-
речные смешанные леса (5—7). В сосняках-брусничниках его не-
сколько меньше (3) и еще меньше на верховых болотах (0,4). В южной
тайге в наибольшем числе этот конек встречен на гнездовании в рос-
лых рямах и елово-кедровой тайге (32—34). Значительно меньше
зеленых коньков видели в мелколиственных и полузаболоченных
смешанных лесах (11—14). В подтаежных лесах в гнездовое время не
наблюдался. Гнезда зеленых коньков в Прикетье (на границе средней
и южной тайги) с четырьмя и пятью яйцами находили 2 июня 1971 г,
и 24 июня 1963 г. Молодые коньки встречены нами в северной тайге
7 и 24 июня 1976 г.
После окончания гнездования в северной тайге обилие зеленых
коньков уменьшилось в июле вдвое по сравнению с плотностью
гнездования и не изменялось (в среднем по участку) до конца лета.
В августе эти коньки перестают встречаться на открытых верховых
и переходных болотах и в сосняках, но в значительном числе отме-
чены на гарях, в низкорослых рямах и в поселках (12—23). В сред-
ней тайге прослеживается сходный характер послегнездовых пере-
распределений и изменений численности, но зарегистрирован некото-
рый ее спад в начале августа и подъем в конце этого месяца, вызван-
ный, по-видимому, началом осенних кочевок. В южной тайге в конце
июля свыше 90% популяции коньков откочевывает, хотя некоторая
часть птиц, конечно, гибнет. К концу августа численность зеленого
конька в результате прикочевки резко возрастает и становится
почти на 70% выше, чем на гнездовании. В подтаежных лесах этот
конек встречен лишь однажды — в начале августа в полях-пере-
лесках (1).
68
В среднем за лето в северной тайге зеленый конек предпочитает
гари по соснякам (10), где держится преимущественно по небольшим
участкам рямов. В значительном числе встречается в приречной
темнохвойной тайге, по окраинам комплексных верховых болот и
небольших таежных поселков (6—8). Значительно меньше его на
грядово-мочажинно-озерных верховых болотах, вырубках по сосня-
кам и в сосново-озерном ландшафте (2—3) и крайне мало в сосняках-
ягельниках (0,05). В средней тайге зеленого конька примерно столь-
ко же, сколько в северной. Держится он преимущественно в при-
речных темнохвойно-мелколиственных лесах (6), особенно во время
кочевок. В остальных местообитаниях обычен (2—3) и только на вы-
рубках по соснякам редок (0,1). В южной тайге его в среднем втрое
больше, чем в предыдущих подзонах. Повсеместно многочислен зеле-
ный конек в лесном ландшафте (11—26) и только в темнохвойной
тайге обычен (2).
В результате следует отметить, что на таежных междуречьях
самая высокая численность зеленого конька зарегистрирована в
южной тайге. К северу и особенно к югу от нее численность снижается.
При сравнении с долиной Оби заметно, что в среднем по участкам
зеленого конька больше всего на южно-таежных междуречьях.
Здесь преобладают производные от темнохвойных, в том числе угне-
тенные и часто заболоченные леса, предпочитаемые этим коньком.
Значительно меньше зеленого конька насчитывалось в долине Оби
в пределах всех подзон и на северо- и среднетаежных междуречьях,
где превалируют хорошо дренируемые и редкостойные леса. Мини-
мальное обилие зарегистрировано на подтаежных междуречьях с наи-
более разреженными и сухими осиново-березовыми лесами.
Итак, в лесной зоне Западной Сибири обилие зеленого конька
наиболее резко снижается на подтаежных междуречьях и в пойме
Оби в пределах всех подзон. Именно здесь по численности преоблада-
ет лесной конек. Весьма характерно распределение этих двух конь-
ков в северной тайге Западной Сибири. Лесной конек значительно
более многочислен в северотаежной долине Оби на надпойменных
верховых и переходных болотах. Однако с продвижением на восток,
на Обь-Енисейском междуречье, лесной конек перестает встречаться
совсем и в сходных местообитаниях его замещает зеленый. Та же осо-
бенность отмечена для полян и вырубок в лиственничниках приени-
сейской северной тайги [Рогачева, 1961]. Еще севернее, в северота-
ежных редколесьях, лесной конек не встречен даже в долине Оби,
и его полностью замещает здесь зеленый конек [Вартапетов и др.,
1980]. На наш взгляд, описанные выше особенности распределения
этих двух близких видов позволяют внести некоторую детализацию
в имеющиеся представления об их экологическом викариате [Матюш-
кин, Кулешова, 1972; Равкин, 1978; Гынгазов, Миловидов, 1977].
В основном на территории таежной зоны Западной Сибири она сво-
дится к следующему. В северной тайге Обь-Енисейского междуречья
и долины Енисея зеленый конек полностью замещает лесного, в том
числе в предпочитаемых последним местообитаниях. На остальной
части рассматриваемой территории оба вида встречаются совместно,
69
при этом обычно по численности преобладает лесной конек. Особен-
но заметно его преобладание в «нетаежных» лесах (в пойме Оби,
сосняках и мелколиственных- лесах южных подзон).
Краснозобый конек (Anthus cervina Pall.)
В конце мая — первой половине июня в северных подзонах
отмечен пролет средней интенсивности (2—5). На гнездовании в
северной тайге (первая половина июля) многочислен на переходных
болотах и обычен на грядово-мочажинно-озерных верховых болотах
(12 и 4). К концу июля его численность уменьшилась вдвое, а в пер-
вой половине августа он перестал встречаться совсем. В конце
августа краснозобый конек появился вновь на послегнездовом про-
лете. В этот период в особенно большом количестве он отмечен на пе-
реходных болотах, в поселках и приречной темнохвойной тайге
(19—29). Значительно меньше краснозобых коньков видели на вер-
ховых болотах (6—7). В это время коньки, в основном молодые, часто
встречались стайками по 10—15 особей. В средней тайге они держа-
лись только во второй половине мая — первой половине июня на
грядово-мочажинных участках верховых болот (25). Часто наблюда-
лись стайками по 12—15 птиц, особенно в конце мая.
В среднем за лето в северной тайге краснозобый конек предпочи-
тает переходные болота (8), в меньшем числе встречается на верхо-
вых болотах, в темнохвойной тайге и поселках (2—4). В средней тай-
ге его обилие примерно в 2 раза ниже, чем в северной, при этом
краснозобый конек обычен на грядово-мочажинных верховых боло-
тах (6). В долине его примерно столько же в средней тайге и значи-
тельно больше в северной. В последней зарегистрирована макси-
мально высокая численность на надпойменных переходных, поймен-
ных низинных болотах и в лугах-сорах.
Таким образом, краснозобый конек встречен нами только в
пределах северных подзон. Наиболее высокая численность его за-
регистрирована в северотаежной долине Оби и значительно мень-
шая — на остальной территории.
СЕМЕЙСТВО СОРОКОПУТЫ (LANIIDAE)
Жулан (Lanius collurio L.)
Нами в северной тайге не зарегистрирован, хотя его отмечали
на р. Казым и в долине Оби [Johansen, 1944—1961; Равкин, 1978].
В средней тайге жуланов видели только в июле на верховых боло-
тах (3). В южной тайге на гнездовании (15 июня — 15 июля) много-
числен на шелкопрядниках и гарях (18) и обычен в полях-перелес-
ках, на низинных и верховых болотах (2—3). В подзоне подтаежных
лесов в тот же период держался в осиново-березовых лесах (2).
Гнезда жулана с пятью и шестью яйцами находили в окрестностях
70
Томска 12 и 17 июля 1977 г. Слетки в разные годы встречались с 2
по 14 августа.
После окончания гнездования в южной тайге обилие жулана
возрастает в конце июля в 4 раза, а затем в течение месяца до 95%
популяции откочевывает за пределы района работ. В подтаежных
лесах послегнездовое увеличение численности еще большее, в основ-
ном за счет прикочевки средней интенсивности (8), но после 15 ав-
густа жуланы не встречались. В конце июля они наблюдались чаще,
чем на гнездовании, преимущественно в полях-перелесках и мелко-
лиственных лесах (31 и 14) и в меньшем числе — на низинных боло-
тах (3).
В среднем за лето в средней тайге жулан редок на верховых
болотах (0,9). В южной тайге его значительно больше. Предпочитает
шелкопрядники и гари (23). В остальных местообитаниях он обычен
(1—8). При этом жулан избегает хвойных и смешанных лесов. В под-
таежных лесах его примерно столько же, сколько в южной тайге.
Предпочитает мелколиственные леса, нередко чередующиеся с поля-
ми (4—5), в небольшом числе встречается на низинных болотах (0,4).
Таким образом, на междуречьях жулана значительно больше в пре-
делах южных подзон, чем в средней тайге. В долине Оби по сравне-
нию с междуречьями его примерно столько же в пределах южных
подзон, а в северных подзонах значительно меньше, чем в южных.
Повсеместно предпочитает разреженные мелколиственные леса,
лесополевые местообитания и окраины болот.
Большой сорокопут (Lanius excubitor L.)
В северной тайге встречался с начала июля до середины ав-
густа на комплексных верховых болотах (в среднем за этот период 3).
В низкорослом ряме на берегу р. Казым 9 июля 1976 г. видели четы-
рех молодых птиц. В средней тайге этот сорокопут отмечен во второй
половине июля на верховых болотах (3). В южной он зарегистриро-
ван на низинных болотах с середины июня до конца июля (12).
В подтаежных лесах большого сорокопута видели лишь однажды, в
лугах-выпасах в конце июня (4).
Итак, мы этого сорокопута повсеместно встречали на летних ко-
чевках и лишь один раз видели молодых птиц. По среднелетним пока-
зателям большой сорокопут, как и жулан, повсеместно тяготеет к
открытым и частично залесенным местообитаниям (в основном к бо-
лотам). При этом в северной тайге численность его выше, чем в более
южных подзонах, что едва ли достоверно и, скорее, связано с ред-
костью и случайностью встреч большого сорокопута. В долине Оби
обилие его обычно выше, чем на междуречьях, а в южных подзонах
выше, чем в северных.
71
СЕМЕЙСТВО СВИРИСТЕЛЕВЫЕ (BOMBYCILLIDAE)
Свиристель (Bombycilla garrulus L.)
В северной тайге пролет заканчивается в начале июня, в сред-
ней — во второй половине мая. Напряженность его в северных под-
зонах невелика (1—2). В пределах южных подзон свиристели в этот
период не встречены. Во время гнездования (вторая половина июня)
в северной тайге свиристели обычны в приречной темнохвойной тай-
ге, на гарях по соснякам, на грядово-мочажинных верховых болотах
и в поселках (3—6). На частичных вырубках по соснякам и низко-
рослых рямах редки (по 0,3). В средней тайге на гнездовании сви-
ристель предпочитает вырубки по соснякам (13). Обычен в рослых
рямах и сосновых лесах (1—3) и редок в поселках (0,8). В средней
тайге по р. Тым массовое гнездостроение и откладывание яиц отме-
чено с 29 мая по 8 июня 1966 г. [Москвитин, 1965].
После окончания гнездования в июле численность свиристелей
не возрастает, так как часть отгнездившихся птиц, по-видимому,
вскоре откочевывает. Послегнездовая прикочевка прослеживается
в северной тайге во второй половине августа (2). Птицы в этот период
концентрируются в приречной темнохвойной тайге и на гарях по
соснякам (16—34). В средней тайге уже в июле заметна прикочевка,
в результате которой птиц становится в 4 раза больше, чем на гнездо-
вании. Особенно много птиц держалось в рослых рямах (44) во время
урожая голубики и морошки. Однако в течение месяца свиристели
откочевывают и после 15 августа перестают встречаться совсем.
В южных подзонах они отмечены только на послегнездовых кочев-
ках. Так, в южной тайге в значительном числе держались в шелко-
прядниках во второй половине августа (44). В первой половине ав-
густа их видели в подтаежных рямах (1).
В среднем за лето в северной тайге свиристель предпочитает при-
речные темнохвойные леса и гари по соснякам (7—9) и в меньшем
числе встречается в сосняках и по периферии верховых болот (1—2).
В остальных местообитаниях редок (0,1—0,6). В средней тайге его
вдвое меньше. Держится преимущественно на вырубках по соснякам
и в рослых рямах (6—8) и в меньшем числе — в сосновых и при-
речных смешанных лесах (1—3). В остальных местообитаниях, кро-
ме грядово-мочажинных участков верховых болот, редок (0,1—0,4).
В результате следует отметить, что свиристель отдает предпочте-
ние смешанным лесам, рямам, вырубкам и гарям по соснякам. Его
обилие снижается с севера таежных междуречий, где он гнездится
регулярно, к югу, где встречен лишь на послегнездовых кочевках.
В долине Оби и на междуречьях в пределах одних и тех же подзон
отмечена сходная численность свиристеля.
72
СЕМЕЙСТВО ДРОЗДОВЫЕ (TURDIDAE)
С о л о в е й - к р а с и о ш е йк a {Calliope calliope Pall,)
Прилет в южной тайге в окрестностях Томска зарегистрирован
с 24 мая по 5 июня 1962 и 1964 гг. [Москвитин, 1972]. В 1967 г. крас-
н ошейка появилась на южно-таежных междуречьях 23 мая, а в сред-
нетаежной долине Оби — 1 июня 1970 г.
В средней тайге нами при учетах птиц не встречена, лишь од-
нажды во второй половине июня 1977 г. слышали далеко поющего
самца на опушке приречного смешанного леса вблизи поселка. В
средней тайге Зауралья этот соловей отмечен В. В. Раевским (1982)
в качестве редкой птицы. В южной тайге на гнездовании обычен в
мелколиственных лесах, в том числе в сочетании с полями (4). Хвой-
ных лесов избегает. В подтаежных осиново-березовых лесах встре-
чен только в первой половине июня (0,3).
Итак, по среднелетним показателям обилие красношейки на
междуречьях резко снижается к северу и югу от южной тайги. В до-
лине Оби в пределах подтаежных лесов красношейки несколько
больше, чем на южно-таежных междуречьях, а с продвижением на
север численность этого соловья убывает, хотя здесь его заметно
больше, чем в тех же подзонах на междуречьях. Наиболее высокая
численность характерна для пойменных лугов-ивняков и залесен-
ных низинных болот и несколько меньшая — в пределах южных
подзон для осиново-березовых лесов, в том числе в сочетании с
полями.
Варакушка {Cyanosylvia svecica L.)
В конце мая — начале июня повсеместно прослеживается про-
лет, наиболее интенсивный в подтаежных лесах (7) и более слабый
в южной и северной тайге (1—2).
В северной тайге на гнездовании (15 июня—15 июля) обычна
в низкорослых рямах. В южной тайге в гнездовое время не зарегист-
рирована, хотя на пролете обычна в полях-перелесках и на низин-
ных болотах (6—8). В подтаежных лесах на гнездовании (июнь)
многочисленна в полях-перелесках и лугах-выпасах, а также по
окраинам поселков (12—17). Обычна в осиново-березовых лесах и
на низинных болотах в сочетании с покосными лугами (4—9). Гнез-
да варакушки с пятью и семью яйцами находили в окрестностях
Томска 3 июня 1939 г. и 30 мая 1970 г., а в южной тайге по р. Васю-
гану с шестью яйцами — 6 июня 1965 г. [Гынгазов, Миловидов,
1977]. Те же авторы в северной тайге по р. Таз с 18 июля отмечают
лётных птенцов. В Прикетье, на границе средней и южной тайги,
оперившиеся птенцы наблюдались в гнездах 3 и 8 августа 1968 г»
Нами в северной тайге слеток варакушки встречен 12 августа 1976 г.
В северной тайге отгнездившиеся птицы откочевывают к сере-
дине июля (в течение месяца после этого не встречаются). Появля-
ются варакушки вновь на осенних кочевках с середины августа на
73
окраинах поселков и в низкорослых рямах (8 и 2). На междуречьях
южной тайги в послегнездовое время, как и на гнездовании, не от-
мечены. В подтаежных лесах в результате размножения обилие ва-
ракушки возрастает вдвое во второй половине июля, а затем к кон-
цу августа до 75% птиц откочевывают, хотя часть их, несомненно,
гибнет. В этот период варакушка перестает встречаться на низин-
ных болотах по окраинам поселков и концентрируется на полях (в
основном, кукурузных) и лугах, используемых под выпас.
В среднем за лето в северной тайге варакушка обычна в приреч-
ных темнохвойных лесах, низкорослых рямах и поселках (1—2),
редка в сосняках-ягельниках и на грядово-мочажинных верховых
болотах (0,2—0,5). При этом в последних и в сосново-боровом ланд-
шафте встречена лишь на пролете. В средней тайге не зарегистри-
рована. В южной тайге ее вдвое меньше, чем в северной. Обычна
она в полях, перелесках и на низинных болотах (по 2) и редка в мел-
колиственных лесах (0,1). В этой подзоне на междуречьях ее видели
только на весеннем пролете. В подтаежных лесах варакушки гораз-
до больше, чем в предыдущих подзонах. В течение всего лета она
держится преимущественно в лугово-полевых местообитаниях (17—
20), где предпочитает кустарники в перелесках и на заболоченных
западинах. Значительно меньше ее в осиново-березовых лесах, на
низинных болотах и в поселках (3—6).
Подводя итоги, следует отметить, что излюбленными местооби-
таниями варакушки на междуречьях являются лесополевые урочи-
ща южных подзон, а в северной тайге — залесенные болота. Наи-
большая численность зарегистрирована в подтаежных лесах, замет-
но меньшая — в северной тайге и минимальная — в южной. В до-
Лине Оби прослеживается сходный характер подзональных разли-
чий численности варакушки, но здесь ее повсеместно в 2—10 раз
больше, чем на междуречьях. Наиболее предпочитаемые местооби-
тания варакушки — залесенные пойменные низинные болота, в
меньшей степени — остальные пойменные и подтаежные внепоймен-
ные лесополевые местообитания.
Синехвостка (Tarsiger cyanurus Pall.)
В северной тайге первую песню слышали 8 июля 1976 г. До се-
редины июля синехвостка обычна в сосняках-ягельниках и на вы-
рубках по ним (1—2). В сосняках 2 июля 1976 г. наблюдали самца
и самку, волнующихся с кормом. В конце августа синехвостка поя-
вилась вновь на послегнездовых кочевках в приречных темнохвой-
ных лесах (8). В средней тайге ее видели лишь однажды — во вто-
рой половине июля в приречном смешанном лесу (2). В более южных
подзонах междуречий не наблюдалась. В долине Оби она зарегист-
рирована только в южно-таежных сосняках в июне и конце июля
(в среднем за лето — 3).
Итак, на междуречьях и в долине Оби синехвостка встречается
в качестве редкой птицы лишь там, где имеются сосняки, и в смеж-
74
ных с ними темнохвойных и смешанных лесах. Однако в южной тай-
ге Прииртышья летом 1968 г. синехвостка была обычной птицей
темнохвойных и смешанных лесов.
Горихвостка-лысушка (Phoenicurus phoenicurus L.)
В конце мая — начале июня повсеместно прослеживается окон-
чание пролета, наиболее интенсивного в средней тайге (7) и выра-
женного слабее в остальных подзонах (1—3).
На гнездовании в северной тайге (июнь) горихвостка-лысушка
в наибольшем числе встречена в поселках и на гарях по соснякам
(8—9). Несколько меньше ее в сосняках-ягельниках, на вырубках
по ним и в сосново-озерном ландшафте (4—6). В средней тайге в тот
же период многочисленна в сосняках-брусничниках и на вырубках
по этим лесам (по 14). В приречных темнохвойно-мелколиственных
лесах обычна (6) и редка в рослых рямах по периферии верховых бо-
лот (0,5). В южной тайге в первой половине июля предпочитает шел-
копрядники (78). Несколько меньше ее в рослых рямах и полуза-
болоченных смешанных лесах (40—54) и еще меньше в темнохвой-
ных и мелколиственных лесах, в том числе в сочетании с небольши-
ми полями (10—28). В подтаежных лесах на гнездовании предпочи-
тает поля-перелески (12). Заметно реже встречается в осиново-бе-
резовых лесах и рямах (2—3) и совсем редко — в лугах, используе-
мых под выпас (0,5). Гнезда этой горихвостки с восемью яйцами най-
дены в южной тайге у с. Бакчар 17 июня 1977 г., а 17 июня 1965 г.
и 27 июня 1976 г. в долине р. Васюган в гнезде были обнаружены
птенцы. В средней тайге на р. Тым птенцы в гнездах найдены
30 июня 1935 г. и 2 июля 1936 г. [Гынгазов, Миловидов, 1977]. В се-
верной тайге слетки отмечены нами 21—23 июля 1976 г., а 22 июля и
5 августа того же года мы видели молодых птиц размером со
взрослых.
В послегнездовое время (в июле — августе) в результате отко-
чевки и гибели численность горихвостки-лысушки повсеместно сни-
жается (на 80—90% в конце августа по сравнению с плотностью гнез-
дования). Только в южной тайге это уменьшение происходит рань-
ше, уже в конце июля, а во второй половине августа оно компенси-
руется прикочевкой горихвосток из северных подзон. В результате
их численность становится такой же, как на гнездовании.
В среднем за лето в северной тайге горихвостка-лысушка чаще
всего встречалась в поселках и сосново-озерном ландшафте (по 7)»
В темнохвойных и сосновых лесах с их производными ее несколько
меньше (2—5) и еще меньше в низкорослых рямах по периферии
верховых болот (0,2). В сходных местообитаниях средней тайги чис-
ленность лысушки в среднем почти в 2 раза выше. Она предпочитаес
смешанные приречные, сосновые леса и вырубки по ним (8—9). В по-
селках и рослых рямах ее заметно меньше (2—4). В южной тайге
обилие этой горихвостки соответственно в 3 и 6 раз выше, чем в пре-
дыдущих подзонах. Она концентрируется в разреженных, освет-
ленных лесах и шелкопрядниках (25—33). В меньшем числе встре-
75
чается в полях-перелесках и темнохвойной тайге (11—15) и еще
меньше ее по окраинам открытых низинных болот и в низкорослых
рямах (1—2). В подтаежных лесах горихвостки-лысушки примерно
столько же, сколько в северной тайге, и в 5 раз меньше, чем в южной.
Держится она преимущественно в осиново-березовых лесах, в том
числе в сочетании с полями (5—6). Значительно меньше ее в рослых
рямах и лугах-выпасах (0,6 и 1).
В результате следует отметить, что горихвостка-лысушка пред-
почитает осветленные смешанные и светлохвойные леса. Обилие
ее снижается в сомкнутых темнохвойных и сильно разреженных мел-
колиственных насаждениях. Наиболее высокая численность отмечает-
ся на междуречьях южной тайги, а к северу и югу она заметно сни-
жается. В лесных ландшафтах долины Оби этой горихвостки в южной
тайге почти столько же, а в остальных подзонах больше, чем на ме-
ждуречьях. Кроме того, в долине тоже прослеживается некоторое
снижение численности в северной тайге по сравнению с остальными
подзонами (в сходных местообитаниях). В среднем по обследованным
участкам наиболее высокое обилие горихвостки-лысушки характер-
но для южно-таежных междуречий и подтаежной долины Оби, вклю-
чая приобские сосняки, и заметно меньше — для всей остальной
территории.
Луговой чекан (Saxicola rubetra L.)
Этот чекан отмечен нами примерно на 300 км севернее указан-
ной ранее границы ареала у г. Сургута [Иванов, 1976]. Прилет в
северной тайге зарегистрирован 20 июня 1976 г., в средней тайге в
долине Оби первая встреча датирована 28 мая 1972 г., а на между-
речьях он появился лишь 19 июня 1977 г. В южных подзонах приле-
тает до 15 мая. Пролет средней интенсивности зарегистрирован в
подтаежных лесах в конце мая (8).
В северной тайге на гнездовании (вторая половина июля) обы-
чен на переходных болотах (1), а в средней — в низкорослых рямах
(4). В южной тайге встречается гораздо чаще, преимущественно на
низинных болотах (46), и в меньшем числе — на верховых (4—8).
В подтаежных лесах в первой половине июня отдает предпочтение
низинным болотам, чередующимся с покосными лугами (9), и в мень-
шем числе отмечается в лугах-выпасах и полях-перелесках (4 и 1).
Гнезда с семью яйцами находили в подтаежных лесах на р. Шегар-
ка 4 июня 1955 г., с пятью голыми птенцами — 24 июня 1966 г. в
южной тайге по р. Васюган [Гынгазов, Миловидов, 1977]. В после-
гнездовое время (в июле) численность лугового чекана несколько
возрастает в результате размножения, но одновременно часть от-
гнездившихся птиц откочевывает. G 1 по 15 августа эти чеканы не
встречены совсем. Только в подтаежных лесах вслед за откочевкой
местных отгнездившихся птиц во второй половине июня отмечена
прикочевка (8). В июле следует еще более интенсивная прикочевка,
по-видимому из северных подзон (12). В августе обилие снижается
вновь и становится вчетверо меньше, чем на гнездовании.
76
В среднем за лето в северной тайге луговой чекан редок на
комплексных верховых болотах и в поселках (0,4—0,5). В средней
тайге его лишь немногим больше (в среднем по району работ). Здесь
этот чекан обычен на вырубках по соснякам (1) и редок в самих сос-
няках и низкорослых рямах верховых болот (по 0,5). В южной тай-
ге его примерно в 10 раз больше, чем в средней. Предпочитает он
открытые низинные и переходные болота (24—48). В меньшем чис-
ле встречается на верховых болотах и в полях-перелесках (2—3).
В подтаежных лесах лугового чекана больше, чем в остальных под-
зонах (соответственно в 30 и 2,5 раза по сравнению с северными под-
зонами п южной тайгой). Держится он преимущественно на покос-
ных лугах в сочетании с низинными болотами (11) и в меньшем чис-
ле — в остальных лесолуговых местообитаниях (3—6).
Следовательно, луговой чекан предпочитает открытые лугово-
болотные местообитания, на междуречьях его обилие резко снижа-
ется с юга на север. В долине Оби в северной тайге не зарегистриро-
ван, а в остальных подзонах его численность несколько возрастает
к югу, особенно в сходных местообитаниях. В среднем по участкам
больше всего лугового чекана отмечено на подтаежных междуречь-
ях. Средняя численность характерна для долины Оби и междуречий
южной тайги и минимальная — для северных подзон.
Черноголовый чекан (Saxicola torquata L.)
В северной тайге прилет отмечен 10 июня 1976 г., в средней —
27 мая 1977 г., в подтаежных лесах черноголовый чекан появляется
до середины мая. Во второй половине мая — начале июня идет про-
лет, наиболее интенсивный в южных подзонах (10—12).
На гнездовании в северной тайге во второй половине июня встре-
чен на низкорослых рямах и переходных болотах, расположенных по
периферии верховых (14 и 10). В средней тайге на гнездовании не
зарегистрирован. В южной держится преимущественно на откры-
тых низинных и переходных болотах (20—26) и в гораздо меньшем
количестве — в низкорослых рямах верховых болот (3). В подта-
ежных лесах чекан многочислен на низинных болотах и в лугах,
используемых под выпас (68 и 20). Гнездо с шестью яйцами найдено
в подтаежных лесах 15 июня 1978 г. В Прикетье (на границе сред-
ней и южной тайги) свежая кладка отмечена 17 июня 1969 г.; в дру-
гом гнезде с шестью яйцами 24 июня 1967 г. наблюдалось вылупле-
ние птенцов. В 1968 г. основная масса птенцов покинула гнезда к
3 июля.
После окончания гнездования (15—30 июля) в северной и юж-
ной тайге большая часть отгнездившихся птиц откочевывает. Во вто-
рой половине августа прослеживается прикочевка черноголовых че-
канов, более заметная в южной тайге (0,4 и 4). В подтаежных лесах
чеканы держатся в течение месяца, их численность в июле возраста-
ет вдвое по сравнению с плотностью гнездования, а в августе свыше
90% популяции откочевывает.
У7
В среднем за лето в северной тайге черноголовый чекан обычен
на верховых болотах (2—4). В сходных местообитаниях средней
тайги редок (0,5 — встречен только на пролете). В южной тайге
его вдвое больше, чем в северной. Он предпочитает открытые низин-
ные болота (29); в небольшом числе встречается в полях-перелесках
и на верховых болотах (по 1). В сходных местообитаниях подтаеж-
ных лесов его в 2,5 раза больше, чем в южной тайге. Здесь черного-
ловый чекан предпочитает низинные болота (70). В меньшем числе
встречается в полях-перелесках и суходольных лугах, используе-
мых под выпас (3—17).
Итак, на междуречьях черноголовый чекан явно отдает пред-
почтение низинным болотам. Его численность резко возрастает к
югу, и на подтаежных междуречьях она наиболее высока. В долине
Оби черноголовый чекан тоже предпочитает открытые низинные бо-
лота. В меньшем числе встречается в пойменных лугах. В среднем
по участкам обилие его здесь несколько ниже, чем на подтаежных
междуречьях, и примерно такое же, как в южно-таежных не долин-
ных местообитаниях. Наименьшая численность черноголового че-
кана характерна для междуречий северных подзон.
Каменка (Oenanthe oenanthe L.)
В северной тайге в начале июня идет пролет средней интенсив-
ности (3). В остальных подзонах в конце мая напряженность проле-
та невелика (0,5—0,9). На гнездовании в северной тайге (15 июня —
15 июля) предпочитает поселки (138). В сосново-озерном ландшафте
(по сухим гривам с сосновым редколесьем) на частичных вырубках,
гарях по соснякам и на переходных болотах многочисленна (10—
23). Обычна в сосняках-ягельниках и по берегам стариц таежных
рек (1—6). В низкорослых рямах редка (0,6). Избегает каменка тем-
нохвойные леса и грядово-мочажинно-озерные верховые болота.
В южнее расположенных подзонах на гнездовании нами не заре-
гистрирована, хотя отмечалась ранее в средней тайге — по р. Мал.
Сосьва, в южной — у г. Тобольска и в подтаежных лесах — на,
р. Шегарка [Гынгазов, Миловидов, 1977]. Гнезда каменки с восемью
яйцами находили в северной тайге с 21 по 24 июня 1976 г. В то же
лето слетков встречали 15 .и 23 июля, а 5 августа видели молодых
птиц размером со взрослых. В средней тайге молодые птицы наблю-
дались на вырубках по соснякам уже 12 июля 1977 г.
В послегнездовое время (с 16 по 31 июля) обилие каменки воз-
растает в 3,5 раза в результате размножения. В августе в результате
откочевки за пределы обследованного участка ее становится втрое
меньше, чем в предыдущий период. В средней тайге она встречена
на весеннем пролете в конце мая в поселках и на грядово-мочажин-
ных верховых болотах (13 и 3). Вновь появились каменки в первой
половине июля на послегнездовых кочевках и на вырубках по сос-
някам (6). В южной тайге их видели только во второй половине мая
в частично открытых местообитаниях лесного ландшафта и на вер-
ховых болотах (2—7). В те же сроки каменка зарегистрирована в
78
подтаежных лесах на низинных болотах и в поселках (по 2). Кроме
того, она наблюдалась в последних в послегнездовое время (в конце
июля — 8).
По среднелетним показателям, в северной тайге каменка пред-
почитает поселки (114), в значительном числе встречается в сосняках
и производных от них местообитаниях (14—38) и реже — на
комплексных верховых болотах (1—3). В более южных подзонах на-
блюдается гораздо реже, только на весеннем пролете и летних кочев-
ках, в основном в поселках и лишь изредка в остальных местооби-
таниях (в среднем по участкам по 0,1).
Итак, наиболее высокая численность каменки отмечена на се-
веротаежных междуречьях, где она приурочена к лишайниковым
соснякам и поселкам. В более южных подзонах междуречий, а в
долине Оби — от северной тайги до подтаежных лесов обилие ка-
менки значительно ниже. Здесь она встречается в основном на про-
лете и кочевках. В небольшом количестве гнездится по окраинам до-
линных северотаежных поселков.
Чернозобый дрозд (Turdus atrogularis Jar.)*
Первая встреча в северной тайге приходится на 11 июня 1976 г.,
в средней — на 22 мая 1977 г. В южных подзонах во второй полови-
не мая отмечался пролет средней интенсивности (1—2), после чего
чернозобый дрозд не наблюдался в течение всего лета. В этот период
в южной тайге он встречен в лесном ландшафте и по окраинам вер-
ховых болот (2—9). В подтаежной подзоне в то же время чернозобых
дроздов видели в полях-перелесках, лугах-выпасах и осиново-бере-
зовых лесах (2—8).
На гнездовании в северной тайге (15 июня — 15 июля) предпо-
читает приречную темнохвойную тайгу (25) и в меньшем количестве
встречен на гарях по соснякам (2). В средней тайге в июне обычен
в приречных темнохвойно-мелколиственных лесах и сосняках-брус-
ничниках (1—2). После окончания гнездования в северных подзо-
нах прослеживается откочевка, особенно в начале августа (1—4).
Однако во второй половине августа обилие возрастает в среднем на
2—6 особей на объединенный 1км2 ключевых участков. По соотно-
шению этих показателей можно предположить, что часть отгнездив-
шихся птиц в данный период смещается на междуречьях к югу.
В среднем за лето в северной тайге чернозобый дрозд предпочи-
тает приречную темнохвойную тайгу (15), обычен в поселках й на
гарях по соснякам (1—2) и очень редок в остальных местообитаниях
(0,01—0,05). Избегает верховые болота и сосново-озерные ландшаф-
ты. В средней тайге его несколько больше (за счет интенсивной при-
кочевки в конце августа). Чаще всего этот дрозд встречается в при-
речных смешанных лесах и поселках (4—5) и заметно реже — в сос-
няках-брусничниках, на вырубках по ним и, во время урожая го-
♦ Вслед за Л. С. Степаняном (19/8) считается нами самостоятельным ви-
дом.
79
лубики, в низкорослых рямах (1—2). В южных подзонах, где заре-
гистрирован только на кочевках, его заметно меньше (0,3—1 в раз-
ных лесных местообитаниях).
Следовательно, чернозобый дрозд чаще встречается в приречных
лесах, при этом в северных подзонах на междуречьях обилие его вы-
ше, чем в южных. В долине Оби он чаще наблюдался в различных
лесных местообитаниях, хотя не избегал и пойменных лугов, ивня-
ков и болот, особенно в период урожая ягод. При этом в средней
тайге его примерно столько же, сколько на междуречьях северных
подзон, а в северной тайге в долине значительно больше (за счет
значительной концентрации его здесь на послегнездовых кочевках
в августе).
а Рябинник (Tardus pilaris L.)
На междуречьях северных подзон нами не зарегистрирован,
хотя в долине Оби распространен до северной границы лесной зоны.
На междуречьях южной тайги на гнездовании тоже не отмечен. На-
чинает встречаться здесь в конце июня на послегнездовых кочевках,
интенсивность которых наиболее велика во второй половине июля
и августа (8 и И). В среднем за лето больше всего рябинников заре-
гистрировано на низинных болотах и шелкопрядниках в результате
их концентрации здесь в период урожая ягод (30—38). В остальных
местообитаниях он обычен или редок. На междуречьях подтаежных
лесов рябинника значительно меньше — встречается только на
пред- и послегнездовых кочевках в конце мая и июле. В среднем за
лето он обычен в осиново-березовых лесах и полях-перелесках (1—3),
редок в лугах-выпасах (0,3).
Таким образом, в лесной зоне Западной Сибири рябинник оп-
ределенно предпочитает разреженные, осветленные и островные
леса долины Оби, как это отмечалось и ранее для южной тайги [Рав-
кин, Лукьянова, 1976]. Особенно высокая численность рябинника
характерна для долины Оби в подтаежных лесах. Средние показа-
тели отмечены в долине в пределах более северных подзон и на юж-
но-таежных междуречьях (на последних встречен только во второй
половине лета). Наконец, минимальные значения обилия рябинни-
ка зарегистрированы на подтаежных междуречьях, где его видели
лишь во время кочевок.
Белобровик (Tardus iliacus L.)
На междуречьях в период гнездования в северной тайге (июнь)
предпочитает приречную темнохвойную тайгу (4). В небольшом ко-
личестве встречается на гарях по соснякам и в поселках (0,3—0,5).
В средней тайге (16 мая — 15 июня) обычен в приречных смешанных
лесах и на вырубках по соснякам (3—4). Редок в рослых рямах по
периферии верховых болот (0,5). В южной тайге (июнь) обычен в
темнохвойных лесах (4). На междуречьях подтаежных лесов на
гнездовании не зарегистрирован. Белобровиков здесь видели однаж-
80
ды во второй половине мая в полях-перелесках (0,2). В Прикетье
на границе средней и южной тайги 23 мая 1962 г. и *31 мая 1971 г.
в гнездах было по 5 яиц. Вылупление птенцов в 1963 г. началось
16 июня. Слетки пойманы 8 июня 1963 г. и 13 июля 1968 г. Нами в
северной тайге слеток встречен 4 июля 1976 г.
В среднем за лето в северной тайге белобровик обычен в при-
речных смешанных лесах и на гарях по соснякам (2—4), редок в
поселках (0,2). В средней тайге его несколько больше. Здесь этот
дрозд предпочитает приречные смешанные леса и примыкающие к
ним участки вырубок по соснякам (по 2). На рямах верховых болот
он редок (0,1—0,5). В южной тайге белобровика заметно меньше,
чем в северных подзонах, — он предпочитает темнохвойные леса (1)
и лишь изредка встречается в мелколиственных (0,03). Еще меньше
белобровиков в подтаежных лесах, где их видели только на пред-
гнездовых кочевках.
Таким образом, на Обь-Енисейском и Обь-Иртышском между-
речьях белобровик чаще всего встречается в смешанных лесах по
долинам крупных притоков. Его численность снижается к северу
и особенно к югу от средней тайги. В долине Оби в пределах север-
ных подзон обилие белобровика несколько больше, чем в южных,
и повсеместно значительно выше, чем на междуречьях. Для этого
дрозда, так же как и для рябинника, разреженные, островные сме-
шанные и мелколиственные леса долины Оби, в том числе пойменные,
оказываются гораздо более привлекательными по сравнению с круп-
ными массивами монотонных, в основном сосновых, темнохвойных
и смешанных лесов на междуречьях.
Певчий дрозд {Tardus philomelos Brehm.)
В северной тайге в первый раз песню слышали 6 июня 1976 г.
южнее певчий дрозд появлялся до 15 мая. В средней тайге на Мал.
Сосьве прилет зарегистрирован 9 мая. На междуречьях северной
тайги встречался только в первой половине июня. В этот период
редок в приречной темнохвойной тайге и сосново-озерном ландшаф-
те (0,2—0,4). На гнездовании в средней тайге (15 мая — 15 июня)
предпочитает приречные смешанные леса (7) и в небольшом коли-
честве держится в сосняках-брусничниках (1). В южной тайге встре-
чается гораздо чаще. В темнохвойных и мелколиственных лесах
многочислен (14), обычен в остальных местообитаниях лесного ланд-
шафта (2—8) и только в елово-кедровой тайге редок (0,2). В под-
таежных лесах нами не зарегистрирован. В Прикетье (граница юж-
ной и средней тайги) гнезда с шестью яйцами находили 14 и 18 мая
1969 г., а птенцов видели только 21 июня.
В послегнездовое время (с 15 июня по 15 июля) в средней тайге
обилие певчего дрозда упало в 4 раза по сравнению с периодом гнез-
дования, а во второй половине лета он исчез полностью. В южной
тайге, наоборот, прослеживается интенсивная прикочевка и к кон-
цу июля этих дроздов становится в 13 раз больше, чем на гнездова-
нии, после чего в августе большая их часть откочевывает.
б л. Г. Вартапетов
81
В среднем за лето в северной тайге певчий дрозд очень редок в
приречной темнохвойной тайге и в сосново-озерном ландшафте (0,04
и 0,05). В средней тайге его несколько больше. Он предпочитает
приречные смешанные леса (2), в меньшем числе встречается в сос-
няках-брусничниках и на вырубках по ним (0,05 и 0,3). В южной
тайге численность певчего дрозда значительно выше, чем в северных
подзонах. В осиново-березоЪых лесах и шелкопрядниках он много-
числен (10—37), обычен в остальных местообитаниях лесного ланд-
шафта (4—8) и редок в рослых рямах (0,7).
Следовательно, на междуречьях певчий дрозд чаще всего зани-
мает мелколиственные, смешанные и темнохвойные леса. Его чис-
ленность резко уменьшается к северу от южной тайги. В долине
Оби в пределах северных подзон он так же малочислен, как и на ме-
ждуречьях, и держится в сходных местообитаниях. В долине Оби
в пределах южной тайги площадь предпочитаемых им ландшафтных
урочищ невелика, и здесь обилие этого дрозда значительно ниже,
чем на южно-таежных междуречьях. В подтаежных лесах зарегист-
рирована средняя численность певчего дрозда, здесь его меньше,
чем на южно-таежных междуречьях, но больше, чем на всей осталь-
ной рассматриваемой территории.
Дрозд-деряба (Turdas viscivorus L.)
В северной тайге нами не зарегистрирован, хотя наблюдался
в Приказымье И. Н. Шуховым (1915). В средней тайге обычен на
гнездовании (июнь) в сосняках-брусничниках и на вырубках по
ним (2—4). В южной тайге в гнездовой период (15 мая — 15 июня)
встречен лишь однажды, в конце мая в полях-перелесках (2). В под-
таежных лесах на гнездовании не зарегистрирован, хотя отмечался
здесь в начале июня на предгнездовых кочевках в осиново-березо-
вых лесах (0,3). В послегнездовое время в июле происходит падение
численности дерябы в срединных подзонах или его полное исчезно-
вение. Однако в конце июля — августе прослеживается прикочевка
этих дроздов (1—2). Дерябы начинают встречаться значительно ши-
ре, в том числе в темнохвойных и смешанных лесах, в рямах верхо-
вых болот и по окраинам низинных, а также на шелкопрядниках и
гарях в период урожая ягод.
По среднелетним показателям, в средней тайге этот дрозд пред-
почитает сосняки и вырубки по ним (по 2), в меньшем числе наблю-
дается на рямах верховых болот (1) и в приречных смешанных ле-
сах (0,1). В южной тайге обилие его лишь немногим ниже. Чаще все-
го деряба отмечается в местообитаниях, где открытые участки чере-
дуются с залесенными (2). В лесных местообитаниях повсеместно
редок (0,4—0,7). В подзоне подтаежных лесов, где его значительно
меньше, очень редок в осиново-березовых лесах (0,04).
Итак, на междуречьях наибольшая численность дерябы отме-
чена там, где имеются высокоствольные сосняки, хотя в период пос-
легнездовых кочевок он не избегает и остальных залесенных и час-
тично открытых местообитаний. В долине Оби встречен лишь в под-
82
таежных лесах, где его несколько меньше, чем на южно- и средне-
таежных междуречьях. К северу от подтаежных лесов Ю. С. Рав-
киным (1978) в долине не зарегистрирован, однако к северу по Оби
в сосновых борах отмечался до г. Сургута [Гордеев, 1965].
СЕМЕЙСТВО СЛАВКОВЫЕ (SYLVIIDAE)
Обыкновенный и пятнистый сверчки
(Locustella naevia Bodd., L. lanceolata Temm.)
По песне эти виды сверчков плохо различимы, поэтому они
учитывались вместе. На большей части обследованной территории
ареалы их перекрываются, однако обыкновенного сверчка стано-
вится больше к западу и по южной периферии лесной зоны Западной
Сибири, а пятнистого — к востоку, и, кроме того, он значительно
дальше проникает к северу. В северной и средней тайге Приобья
Ю. С. Равкиным (1978) добывались только пятнистые сверчки. В
подтаежных лесах долины Оби, по сведениям того же автора, из
15 добытых сверчков лишь один был обыкновенным. В 400 км к за-
паду, на междуречьях этой же подзоны, все три отстрелянных нами
сверчка были обыкновенными. Первую песню этих сверчков слыша-
ли в подтаежных лесах 24 мая 1978 г., в южной тайге — 5 июня
1967 г., а в средней — лишь 20 июня 1977 г.
На гнездовании в средней тайге (15 июня — 15 июля) пятни-
стый сверчок держался по окраинам поселков (2). В южной тайге
обыкновенного и пятнистого сверчков значительно больше. Пред-
почитают они открытые низинные болота и шелкопрядники в сочета-
нии с гарями и вырубками (35—57). Значительно ниже обилие этих
сверчков в мелколиственных и смешанных полузаболоченных ле-
сах, а также в полях-перелесках (2—4). Редки они на переходных
болотах (0,4). Р. Л. Наумов и Г. С. Кисленко (1965) приводят для
приенисейской южной тайги вдвое большую плотность гнездования
по сравнению с аналогичными показателями в наиболее предпочи-
таемых местообитаниях той же подзоны Приобья. Гнездо с шестью
яйцами найдено этими авторами 29 июня 1961 г., а первые молодые
птицы в разные годы встречены 23—28 июля. В подзоне подтаежных
лесов на гнездовании сверчки предпочитают низинные болота (28) и,
кроме того, встречаются в осиново-березовых лесах и лугах-выпа-
сах (8 и 12).
В послегнездовое время они полностью исчезают в средней тай-
ге в конце июля. В более южных подзонах наблюдаются до конца
лета, но в августе их становится в 3—10 раз меньше, чем на гнездо-
вании, так как часть популяции откочевывает за пределы обсле-
дованных районов.
В среднем за лето в средней тайге обыкновенный и пятнистый
сверчки — редкие птицы небольших таежных поселков (0,7). В юж-
ной тайге их значительно больше, особенно на открытых низинных
болотах (37). Сравнительно часто встречаются они на шелкопряд-
б*
83
никах, вырубках и гарях (12) и реже в полузаболоченных смешан-
ных, мелколиственных лесах и на переходных болотах (2, 0,7 и 0,1).
В подзоне подтаежных лесов этих сверчков в среднем по участку
вдвое больше, чем в южной тайге. Они многочисленны на низинных
болотах (14), обычны в березово-осиновых лесах и лугах-выпасах
(4—6), редки в полях-перелесках (0,3).
В результате можно отметить, что численность обыкновенного
и пятнистого сверчков на междуречьях возрастает к югу в соответст-
вии с увеличением площади предпочитаемых низинных болот. Од-
нако на самих низинных болотах в южной тайге их больше, чем в
подтаежных лесах. Сходная тенденция изменения их обилия харак-
терна и для долины Оби, но здесь эти сверчки значительно дальше
(вплоть до северной тайги) и в большем числе проникают к северу,
чем на междуречьях. В долине Оби пятнистый и обыкновенный
сверчки в наибольшем количестве встречены тоже на низинных бо-
лотах, особенно пойменных, а также в лугах, ивняках поймы Оби.
Барсучок (Acrocephalus schoenobaenus L.)
В северной тайге первая и единственная встреча двух рядом
поющих самцов отмечена на низинном болоте 11 июня 1976 г. В сред-
ней тайге аналогичная встреча произошла 2 июня 1977 г., и барсуч-
ки здесь тоже, по-видимому, не гнездились. В подтаежных лесах
первую песню слышали 31 мая 1978 г., а в июне идет пролет бар-
сучков, наиболее интенсивный в первой половине месяца (18).
На гнездовании в подтаежных лесах (первая половина июля)
барсучок обычен на низинных болотах и в лугах, используемых под
выпас (4—9). К концу июля почти все отгнездившиеся птицы отко-
чевывают за пределы района работ, а в начале августа в результате
прикочевки их становится почти столько же, сколько на гнездовании.
По среднелетним показателям обилия, барсучок на междуречьях
предпочитает подтаежные низинные тростниково-осоковые болота,
совсем не зарегистрирован в южной тайге и в качестве редкой птицы
отмечен в более северных подзонах, где наблюдения проводились
на более крупных притоках Оби. В долине Оби барсучок повсемест-
но отдает предпочтение пойменным низинным болотам и его гораздо
больше, чем на междуречьях. Наиболее высокая численность ха-
рактерна для долины в пределах южных подзон, несколько мень-
шая — в северных подзонах и на подтаежных междуречьях. Мини-
мальное обилие отмечено на междуречьях северных подзон, где бар-
сучок держится лишь в поймах крупных притоков Оби.
Садовая камышевка (Acrocephalus dumetorum Blyth.)
В северной тайге садовая камышевка не встречена. В средней
тайге первую песню слышали 7 июня 1977 г., в южной — 5 июня
1967 г., а в подтаежных лесах массовый прилет отмечен 31 мая 1978 г.
В первой половине июня идет пригнездовой пролет, наиболее интен-
84
сивный в южной тайге (32) и выраженный слабее в остальных под-
зонах (1—3).
На гнездовании в средней тайге (июль) садовая камышевка в
качестве обычной птицы держится в поселках (3). В южной тайге
(первая половина июля) в наибольшем числе гнездится в шелкопряд-
никах, по вырубкам и гарям (31). Несколько меньше ее в полях-
перелесках и темнохвойной тайге (16—20). В подтаежных лесах с
15 июня по 15 июля зарегистрирована в лугах-выпасах (3) и осиново-
березовых лесах (18). Полные кладки из 4—6 яиц найдены в окрест-
ностях Томска с 12 июня по 10 июля, а вылет птенцов отмечался
2—15 июля. В Прикетье с 13 по 30 июля 1968 г. наблюдались взрос-
лые камышевки, выкармливающие птенцов в гнездах.
В средней тайге после окончания гнездования (с 1 по 15 августа)
садовые камышевки перестают встречаться, но в небольшом числе
наблюдаются вновь в конце месяца на послегнездовых кочевках.
В южной тайге интенсивная прикочевка тоже отмечена в конце ав-
густа (10). В этот период садовых камышевок становится в 6 раз
больше, чем на гнездовании. В подтаежных лесах уже в конце июля
около 80% отгнездившихся птиц откочевывает (хотя часть их, не-
сомненно, гибнет), а к середине августа они исчезают совсем.
В среднем за лето в средней тайге эта камышевка обычна в по-
селках (1) и редка в низкорослых рямах (0,5 — встречена только
на пролете). В южной тайге ее значительно больше. Здесь садовая
камышевка многочисленна в мелколиственных лесах, полях-пере-
лесках и шелкопрядниках (25—29). В темнохвойных и полузаболо-
ченных смешанных лесах обычна (7—8) и редка в елово-кедровой
тайге (0,7). В подтаежных лесах ее в среднем втрое, а в предпочитае-
мых местообитаниях вдвое меньше, чем в южной тайге. В наиболь-
шем числе она встречается в осиново-березовых лесах (13), а на ос-
тальной территории редка.
В результате следует констатировать наиболее высокую чис-
ленность садовой камышевки на междуречьях южной тайги. Зна-
чительно ниже ее обилие в подтаежных лесах и особенно в средней
тайге. В лесной зоне Западной Сибири в целом садовая камышевка
явно предпочитает суходольные местообитания с развитым кустар-
никовым ярусом, где открытые участки чередуются с залесенными
(шелкопрядники, поля-перелески, лугово-лесные участки высоких
пойм), а также в большом числе заселяет мелколиственные леса,
особенно разреженные, осветленные и островные, в том числе пой-
менные. В долине Оби подзональные отличия ее численности анало-
гичны, но показатели в 2—10 раз выше, чем на междуречьях.
Пере смешка (Hippolais icterina Vieill.)
На междуречьях зарегистрирована только в подтаежных лесах.
В первый раз ее слышали 13 июня 1978 г., хотя, возможно, она поя-
вилась раньше. На гнездовании (15 июня — 15 июля) многочислен-
на в березово-осиновых лесах (22). В послегнездовое время в конце
июля, видимо в результате откочевки, ее становится в 10 раз мень-
85
ше, чем на гнездовании, а в августе она исчезает совсем. В долине
Оби отмечена в пределах южных подзон в надпойменных мелколист-
венных и пойменных лесах. Однако ее здесь значительно меньше,
чем на подтаежных междуречьях.
Итак, пересмешка встречается преимущественно в подзоне оси-
ново-березовых лесов и проникает в южную тайгу по разреженным,
преимущественно мелколиственным, лесам в долинах крупных рек,
при этом ее численность снижается к северу.
Ястребиная славка (Sylvia nisoria Bechts.)
На междуречьях встречена нами лишь однажды в подтаежных
осиново-березовых лесах в начале июня (2). В долине Оби в преде-
лах той же подзоны зарегистрирована тоже всего одна встреча с
этой славкой, когда из выводка была добыта взрослая самка [Сто-
палов и др., 1973].
* Садовая славка (Sylvia borin Bodd.)
На междуречьях в северной тайге песню садовой славки слы-
шали всего один раз — в приречной темнохвойной тайге 23 июня
1976 г. В средней тайге прилет и первая песня зарегистрированы
3 июня 1977 г., в южной тайге— 25 мая 1967 г., в подтаежных ле-
сах — 29 мая 1978 г.
На гнездовании в средней тайге (15 июня — 15 июля) садовая
славка держится в приречных темнохвойно-мелколиственных лесах
(12), предпочитая их прибрежные участки с преобладанием в древо-
стое березы с подлеском из черемухи. В южной тайге в тот же период
концентрируется в смешанных, мелколиственных лесах (по 9) и
полях-перелесках (12). Избегает хвойные леса и болота. В подзоне
подтаежных лесов тоже предпочитает осиново-березовые леса (3),
в меньшем количестве встречается в полях-перелесках (10) и в лу-
гово-болотных местообитаниях (2).
После окончания гнездования численность садовой славки в
результате откочевки и отчасти недоучета повсеместно снижается в
3—5 раз, а в средней тайге она исчезает совсем. В начале августа
эта славка начинает встречаться вновь на послегнездовых кочевках,
и ее становится примерно столько же, сколько на гнездовании.
Только в подтаежных лесах такое увеличение не прослеживается,
здесь обилие ее неуклонно снижается с конца июня до начала ав-
густа. Во второй половине августа садовая славка перестает встре-
чаться либо обилие ее становится минимальным.
В среднем за лето в средней тайге садовая славка обычна в при-
речных смешанных лесах и небольших поселках среди этих лесов
(4—6). В южной тайге в среднем по участку ее примерно столько же,
а в предпочитаемых местообитаниях несколько больше. Держится
она в смешанных, мелколиственных лесах (2 и 4), в том числе в со-
четании с полями (10). В подтаежных лесах садовой славки вдвое
больше, чем в предыдущих подзонах. Она предпочитает осиново-
86
березовые леса (10), обычна в полях-перелесках и лугах-выпасах
на месте лесов (3—7) и редка по окраинам низинных болот (0,6).
Следовательно, на междуречьях обилие садовой славки выше
всего в мелколиственных и смешанных лесах. К северу от подтаеж-
ных лесов оно снижается. В долине Оби ее либо столько же, либо не-
многим больше, чем на междуречьях. При этом максимальная чис-
ленность характерна для южно-таежных мелколиственных лесов
как в пойме, так и вне ее. В северных подзонах долины Оби садовой
славки тоже заметно меньше, чем в южных, но в долине эта славка
распространена к северу значительно дальше, чем на междуречьях.
Серая славка {Sylvia communis Lath.)
На междуречьях в северных подзонах нами не зарегистрирова-
на, хотя отмечалась как редкая птица на гнездовании на границе
южной и средней тайги в Прикетье и в средней тайге Зауралья на
р. Мал. Сосьва. В южной тайге первая встреча датирована 24 мая
1967 г., а в подтаежных лесах — 30 мая 1978 г. Предгнездовой про-
лет (в июне) отмечен только в подтаежных лесах (9).
На гнездовании в южной тайге (вторая половина июня) мно-
гочисленна в полях-перелесках (52) и обычна на низинных болотах
(9). В подтаежных лесах в период гнездования (15 июня — 15 июля)
встречалась заметно чаще. Предпочитает мелколиственные леса и
поля-перелески (25—39), в меньшем числе отмечается в лугово-болот-
ных местообитаниях (13—14). Гнезда с кладками в пять свежих
яиц зарегистрированы в окрестностях Томска в разные годы с 16
по*28 июня, а массовое появление слетков наблюдалось с 29 июня
по 15 июля.
В послегнездовое время (в июле) в южной тайге почти все от-
гнездившиеся птицы откочевывают. В начале августа идут кочевки
средней интенсивности (7), в результате которых обилие серой слав-
ки становится почти в 2 раза больше, чем во время гнездования.
В подтаежных лесах к концу июля после вылета молодых птиц чис-
ленность возрастает вдвое, а затем, к концу августа, Почти все пти-
цы откочевывают за пределы района работ, по-видимому смещаясь
в долину Оби, где их численность в этот период резко возрастает.
В среднем за лето в южной тайге серая славка многочисленна
в полях-перелесках (20), обычна в остальных местообитаниях лес-
ного ландшафта и на низинных болотах (2—3) и редка в темнохвой-
ной тайге (0,6). В подтаежных лесах ее в среднем в 6 раз больше,
чем в южной тайге, однако в сходных ландшафтных урочищах обеих
подзон (поля-перелески) обилие этой славки одинаково. Предпочи-
тает она мелколиственные леса (33), но встречается и в лугово-бо-
лотных местообитаниях (8—12).
В результате следует отметить, что серая славка предпочитает
разреженные мелколиственные леса и лесополевые местообитания.
Она распространена на междуречьях только в пределах южных
подзон. Обилие этой славки снижается к северу. В пределах под-
87
таежных лесов в долине Оби и на междуречьях численность серой
славки одинакова. Однако в долине южной тайги ее больше, чем
на междуречьях, что обусловлено большим числом пригодных для
этой славки местообитаний.
Славка-завирушка {Sylvia curruca L.)
В северной тайге первую песню слышали 8 июня 1976 г., в сред-
ней — 19 мая 1977 г. В более южных подзонах славка-завирушка
появилась до начала наблюдений, а с середины мая по 15 июня идет
пролет (6—16).
На гнездовании в северной тайге в июне славка-завирушка пред-
почитает приречную темнохвойную тайгу (6), изредка встречается
в сосняках (0,4). В средней тайге ее заметно больше. В тот же период
эта славка многочисленна в поселках и приречных смешанных ле-
сах (10—12). В южной тайге (вторая половина июня) обычна в мел-
колиственных и полузаболоченных смешанных лесах (3—4). В под-
зоне подтаежных лесов (июнь) обычна в полях-перелесках, осиново-
березовых лесах (4—6) и лишь изредка наблюдается по окраинам
низинных болот (0,5). Полные кладки из пяти-шести яиц находили
вблизи Томска в разные годы с 30 мая по 5 июня. На р. Мал. Сось-
ва гнездо с шестью яйцами обнаружено 1 июня. В Прикетье пять
гнезд с пятью-шестью яйцами найдены в первой половине июня.
Молодые птицы группами до пяти особей встречены нами 8 июля
1976 г. в северной тайге и 24 июня 1977 г. в средней.
В послегнездовое время (в конце июля) в северной тайге в ре-
зультате кочевок молодых птиц обилие славки-завирушки возраста-
ет почти в 10 раз по сравнению с плотностью гнездования, тогда как
в августе она перестает встречаться совсем. В средней тайге после-
гнездового увеличения не прослеживается, а в августе численность
сокращается почти в 10 раз. Однако в этот период кочующие славки-
завирушки в значительном числе отмечены в поселках (25—38).
В южной тайге прослеживаются две волны послегнездовой прико-
чевки: в начале июля и в конце августа (5 и И). В это время славка-
завирушка становится многочисленной в шелкопрядниках (24 и
51), хотя на гнездовании здесь не зарегистрирована. В подтаежных
лесах она исчезает сразу после окончания гнездования в начале
июля.
В среднем за лето в северной тайге славка-завирушка предпо-
читает приречную темнохвойную тайгу (14), изредка встречаясь в
сосняках-ягельниках и по окраинам переходных болот (по 0,2).
В средней тайге ее вдвое больше. Держится она в небольших таеж-
ных поселках (20), в основном на кочевках, и в приречных смешан-
ных лесах (6). В южной тайге славки-завирушки почти втрое больше,
чем в средней, но в местообитаниях, предпочитаемых этой славкой,
обилие ее одинаково. Многочисленна она в мелколиственных лесах,
шелкопрядниках, на гарях и вырубках (11—20), обычна в остальг
ных местообитаниях лесного ландшафта (4—8) и редка в рослых ря-
мах (0,6). В подтаежных лесах славки-завирушки примерно столь-
88
ко же, сколько в средней тайге, и в 3 раза меньше, чем в южной.
Здесь она предпочитает мелколиственные леса (5), в меньшем числе
встречается в полях-перелесках (2) и лугово-болотных местообита-
ниях (0,1—0,7).
Таким образом, славка-завирушка чаще всего наблюдается в
мелколиственных и смешанных приречных лесах. На междуречьях
ее численность максимальна в южной тайге и снижается в осталь-
ных подзонах. Однако в наиболее излюбленных местообитаниях
южной и средней тайги отмечено равное количество этой славки.
В долине Оби она предпочитает пойменные ивняки, осинники, зале-
сенные низинные болота, надпойменные осиново-березовые и разре-
женные северотаежные темнохвойные леса. Обилие славки-зави-
рушки в долине Оби гораздо выше, чем на междуречьях, причем
отмечен противоположный характер отличий, т. е. в крайних подзо-
нах ее больше, чем в срединных.
Весничка (Phylloscopus trochilus L.)
Повсеместно, кроме подтаежных лесов, во второй половине
мая — начале июня идет пролет средней интенсивности (2—5). В
средней тайге на р. Мал. Сосьва прилет зарегистрирован 21 мая,
нами в той же подзоне весничка отмечена 17 мая 1977 г., но, видимо,
прилетела она раньше.
На гнездовании в северной тайге (вторая половина июня) боль-
ше всего веснички было в низкорослых рямах и приречной темно-
хвойной тайге (10—12). Несколько меньше ее на гарях по соснякам
и в сосново-озерном ландшафте (по 8). Еще меньше в сосняках-ягель-
никах, на комплексных верховых болотах и в поселках (2—5).
В средней тайге, по-видимому, она не гнездилась, хотя с 15 мая до
середины июня зарегистрирована на весеннем пролете в приречных
темнохвойно-мелколиственных лесах, на вырубках по соснякам
(2—4), в поселках и сосняках-брусничниках (по 0,5). Позднее (с
середины июня по 15 июля) не встречалась. В южной тайге на гнез-
довании во второй половине июня отмечена в мелколиственных ле-
сах и полях-перелесках (2—5). В подтаежных лесах в тот же период
обилие ее гораздо выше. Весничка многочисленна здесь в осиново-
березовых лесах, полях-перелесках и на низинных болотах (21—44).
На последних держалась в основном по окраинам в кустарниках и
в небольших березовых колках среди этих болот. В лугах, исполь-
зуемых под выпас, где имеются куртины кустарников и перелески,
и рямах обычна (7—9).
В послегнездовое время в июле — начале августа во всех под-
зонах около 85—100% местных птиц откочевывает, хотя часть их,
несомненно, гибнет. В конце лета прослеживается прикочевка, бо-
лее интенсивная в начале августа в южных подзонах, а во второй по-
ловине этого месяца — в северных. Напряженность кочевок в край-
них подзонах выше, чем в срединных (соответственно 8, 12 и по 2).
В среднем за лето в северной тайге весничка предпочитает сос-
ново-озерный ландшафт и частичные вырубки по соснякам (13—
89
16). Несколько меньше ее в приречных темнохвойных лесах и ря-
мах верховых болот (8 и 10) и еще меньше в сосняках, на гарях по
ним и на открытых участках комплексных верховых болот (3—5).
В средней тайге численность ее в 6 раз ниже. Обычна весничка в
приречных смешанных лесах и поселках (2—4) и редка в сосняках,
на вырубках по ним и в рослых рямах (0,5—0,6). В южной тайге
ее вдвое меньше, чем в северной, но втрое больше, чем в средней.
Держится весничка в смешанных полузаболоченных, мелколиствен-
ных лесах и полях-перелесках (1—6). В подтаежных лесах обилие
веснички значительно выше, чем в предыдущих подзонах. Особен-
но много ее в мелколиственных лесах и полях-перелесках (24 и 25),
втрое меньше в лугово-болотных местообитаниях и совсем мало в
поселках (0,3).
Итак, на междуречьях весничка предпочитает мелколиственные
леса либо местообитания, в которых открытые участки чередуются с
залесенными. При этом в крайних подзонах веснички значительно
больше, чем в срединных. В долине Оби она с наиболее высокой чис-
ленностью заселяет мелколиственные леса в южных подзонах, а в
северных — пойменные ивняки. В подтаежных лесах и особенно в
северной тайге обилие веснички в долине выше, чем на междуречь-
ях, в основном за счет интенсивной послегнездовой прикочевки.
Теньковка (Phylloscopus collybita Vieill.)
Во второй половине мая — начале июня идет пролет, наиболее
интенсивный в южной тайге (24) и слабее выраженный в остальных
подзонах (3—9). На гнездовании в северной тайге (15 июня — 15 ию-
ля) держится преимущественно в приречных темнохвойных лесах
(32) и реже попадается на гарях, вырубках по соснякам и в низко-
рослых рямах (0,5—1). В средней тайге в тот же период многочис-
ленна в приречных смешанных лесах и поселках (14—24). В послед-
них концентрируется по опушкам леса и на отдельно стоящих высо-
ких деревьях. На вырубках по соснякам обычна (2). В южной тайга
больше всего этой пеночки в темнохвойных и полузаболоченных сме-
шанных лесах (53 и 46) и заметно меньше в остальных лесных место-
обитаниях (12—28). В подтаежных лесах предпочитает поля-пере-
лески и осиново-березовые леса (15 и 26). В меньшем числе встреча-
ется в лугах-выпасах и рямах (5 и 8). В разные годы кладки из пя-
ти-шести яиц находили в окрестностях Томска с 8 июня по 10 июля,
а оперившихся птенцов встречали 10—22 июня. Молодые хорошо
летающие птицы наблюдались нами 24 июля 1976 г. в северной
тайге.
В послегнездовое время (во второй половине июля) двукратное
увеличение численности прослеживается только в подтаежных ле-
сах. На остальной территории теньковка после вылета молодых,
по-видимому, сразу же откочевывает. В августе в срединных подзо-
нах идет прикочевка средней интенсивности (5—7). В крайних под-
зонах, напротив, в августе обилие снижается, так как часть птиц,
90
вероятно, перемещается в долину Оби, где в этот период идет интен-
сивная прикочевка теньковок.
В среднем за лето в северной тайге теньковка многочисленна
в приречных темнохвойных лесах (36), обычна в сосняках и рямах
(1—2) и редка в остальных местообитаниях (0,5—0,8). В средней
тайге ее вдвое больше, хотя максимальные показатели обилия та-
кие же, как в северной тайге. Теньковка здесь предпочитает приреч-
ные смешанные леса и небольшие поселки среди этих лесов (22 и
46). В меньшем числе встречается она в сосняках, вырубках по ним
(4 и 1) и в рослых рямах (0,1). В южной тайге теньковки соответст-
венно в 9 и 4 раза больше, чем в северной и средней. Больше всего
ее встречено в мелколиственных лесах (46). Кроме того, она много-
численна во всех остальных местообитаниях лесного ландшафта
(12—39) и редка на открытых низинных болотах (0,4). В подтаежных
лесах обилие теньковки несколько выше, чем в северных подзонах,
но почти втрое ниже, чем в южной тайге. Многочисленна она в оси-
ново-березовых лесах и полях-перелесках (19 и 15) и обычна в луго-
во-болотных местообитаниях (2—4).
Следовательно, на междуречьях теньковка предпочитает сме-
шанные, темнохвойные и мелколиственные леса. Численность ее
максимальна в южной тайге, затем она снижается к югу и особенно
к северу. В долине Оби теньковка предпочитает в общем те же лес-
ные местообитания, что и на междуречьях, и, кроме того, пойменные
ивняки, особенно подтаежные, расположенные в сравнительно высо-
кой и сухой пойме Оби. Численность теньковки и здесь убывает к
северу, при этом в пределах срединных подзон ее несколько меньше,
а в крайних подзонах значительно больше, чем на междуречьях.
Такой характер отличий, так же как и у веснички, по-видимому,
связан с позднелетней откочевкой части птиц в периферийных под-
зонах с междуречий в долину Оби.
Зарничка (Phylloscopus inornatus Blyth.)
Нами на междуречьях северной тайги и подтаежных лесов не
зарегистрирована. В долине Оби наблюдалась в пределах северной
тайги летОхМ 1973 г. в качестве обычной птицы в полузаболоченных
лесах и на залесенных низинных болотах (2—3). В остальных место-
обитаниях редка. В средней тайге нами отмечена в конце мая и на-
чале июля в приречных темнохвойно-мелколиственных лесах (в
среднем за лето редка — 0,5). В южной тайге ее видели в конце мая—
июне и на осенних кочевках во второй половине августа. В среднем
за лето численность зарнички здесь почти в 20 раз выше, чем в сред-
ней тайге. В наибольшем числе она встречена в мелколиственных и
темнохвойных лесах (5 и 3). Заметно меньше ее в елово-кедровых и
полузаболоченных смешанных лесах (1 и 0,4).
Итак, наибольшая численность зарнички отмечена на южно-
таежных междуречьях, значительно меньше ее на междуречьях
средней тайги и в долине Оби.
91
Т а л о в к a (Phylloscopus borealis Blas.)
В северных подзонах первую песню слышали 6 июня 1976 и
1977 гг. В южной тайге в первый раз встречена 5 июня 1967 г. В
Прикетье, на границе южной и средней тайги, появилась 11 июня
1963 г. В июне идет пролет средней интенсивности в северной и юж-
ной тайге (по 2) и слабой — в средней тайге (0,2).
На гнездовании (первая половина июля) на междуречьях се-
верной тайги таловка держится почти исключительно в приречных
темнохвойных лесах (18), в небольшом числе отмечена на переход-
ных болотах и в сосново-озерном ландшафте (1 и 0,2). В средней тай-
ге в июле этих пеночек не видели. В южной тайге в тот же период они
встречены в шелкопрядниках и рослых рямах (3 и 2). В подтаежных
лесах в течение всего лета таловка не наблюдалась. Кладку из шести
яиц находили в приенисейской средней тайге 29 июня 1978 г., птен-
цы вылетели из гнезда 20 июля. В северной тайге слетки таловки
отмечены нами 24 июля 1976 г.
После вылета молодых птиц (15 июля — 15 августа) численность
таловки возрастает в 2—4 раза. В северной тайге одновременно с
появлением молодых птиц идет прикочевка этих пеночек, наиболее
интенсивная в конце июля — начале августа (2). К концу августа
в северной тайге численность снижается и становится почти такой
же, как на гнездовании. Часть популяции, по-видимому, смещается
к югу, в среднюю и южную тайгу, так как здесь в тот же период оби-
лие таловки возрастает примерно на 2 особи на объединенный 1 км2
ключевого участка.
В среднем за лето в северной тайге Обь-Енисейского междуречья
таловка предпочитает темнохвойные леса вдоль притоков Оби (28),
в меньшем числе встречается на гарях и в сосново-озерном ландшаф-
те (по 2) и редка по окраинам верховых болот (0,2—0,6). Вырубок и
открытых участков верховых болот избегает. В средней тайге ее почти
в 4 раза меньше, чем в северной. Обычна она в приречных смешан-
ных лесах (3) и редка на вырубках по соснякам (0,2). В южной тайге
таловки примерно столько же, сколько в средней, и втрое меньше,
чем в северной. Обычная птица елово-кедровых и осиново-березовых
лесов (по 2), она в остальных урочищах лесного ландшафта редка
(0,6-0,9).
Таким образом, на междуречьях таловка чаще встречалась в
темнохвойных и смешанных лесах по долинам притоков Оби. Боль-
ше всего ее отмечено в северной тайге и значительно меньше в сред-
ней и южной. В долине Оби таловка предпочитает примерно те же
местообитания. Кроме того, в северной тайге эта пеночка многочис-
ленна в пойменных ивняках и на надпойменных залесенных низин-
ных болотах, а в более южных подзонах преимущественно наблю-
дается в надпойменных осиново-березовых лесах. В долине Оби за-
регистрированы сходные численность и характер ее подзональных
различий по сравнению с междуречьями. Только в северной тайге
в долине Оби таловки значительно больше, чем на междуречьях,
за счет интенсивной послегнездовой прикочевки.
92
F Зеленая пеночка (Phylloscopus trochiloides Sund.)
Появилась в подтаежных лесах 27 мая 1977 г., в южной тайге
в конце мая — начале июня 1967 г. В средней тайге прилет в 1977 г.
зарегистрирован нами 2 июня, а в северной — 14 июня 1976 г. В под-
таежных лесах в конце мая идет пролет (1). На гнездовании (июнь)
в северной тайге зеленая пеночка обычна в приречных темнохвой-
ных лесах (4). В сходных среднетаежных смешанных лесах тоже
обычна (5). В южной тайге в тот же период многочисленна в лесах
различных типов (14—28) и обычна в елово-кедровой тайге и рослых
рямах (5—6). Гнезда с шестью яйцами находили в окрестностях
Томска в разные годы 8—12 июня, а слетков видели 15—20 июля
1974 г.
В послегнездовое время в июле зеленая пеночка перестает встре-
чаться совсем. Только в средней тайге она наблюдалась постоянно в
течение всего лета. При этом в июле ее примерно столько же, а в ав-
густе вдвое меньше, чем на гнездовании. В остальных подзонах зе-
леная пеночка появляется вновь в августе на послегнездовых ко-
чевках, однако в это время ее обычно гораздо меньше по сравнению
с гнездовым периодом.
В среднем за лето в северных подзонах зеленая пеночка обычна
в приречных темнохвойных и смешанных лесах (2—3) и редка в
среднетаежных сосняках-брусничниках (0,4). В южной тайге (в
пределах сходных местообитаний) ее примерно в 3 раза больше.
Она предпочитает высокоствольные темнохвойные и осиново-бере-
зовые леса (по 8), в меньшем числе встречается в угнетенных влаж-
ных и заболоченных лесах (1—4). Еще меньше ее на шелкопрядни-
ках и в полях-перелесках (0,6 и 0,4). В подтаежных лесах обилие
этой пеночки значительно ниже, чем в южной тайге. Здесь она от-
мечена лишь на кочевках в осиново-березовых лесах и перелесках
среди полей и лугов (0,4—0,7).
Следовательно, на междуречьях зеленая пеночка предпочитает
темнохвойные и мелколиственные леса. Ее численность снижается
к северу и югу от южной тайги. В долине Оби, где эта пеночка тоже
отдает предпочтение суходольным лесным урочищам, отмечен сход-
ный уровень численности, и только в южной тайге, где в пределах
района работ мало заселяемых зеленой пеночкой местообитаний,
ее заметно меньше, чем на междуречьях.
СЕМЕЙСТВО МУХОЛОВКИ (MUSCICAPIDAE)
Серая мухоловка (Muscicapa striata Pall.)
Прилет вблизи Томска наблюдался в разные годы 12—21 мая,
в средней тайге на р. Мал. Сосьва — 15 мая 1931 г., нами в подтаеж-
ных лесах отмечен 27 мая 1978 г.
В северной тайге серая мухоловка не встречена. В средней на
гнездовании (первая половина июня) обычна в приречных смешан-
93
ных лесах и сосняках-брусничниках (3 и 4). В южной тайге обычна
в елово-кедровых и мелколиственных лесах (6 и 7) и шелкопрядни-
ках (10). В подтаежных лесах в тот же период держалась в осиново-
березовых лесах (12). Гнезда с четырьмя яйцами находили в конце
мая 1968 г. в Прикетье. В окрестностях Томска в разные годы клад-
ки с пятью яйцами наблюдали с 5 по 21 июня. В средней тайге по
р. Тым кладка из пяти сильно насиженных яиц найдена 9 июня
1967 г. [Гынгазов, Миловидов, 1977]. Те же авторы отмечали слет-
ков в южной и средней тайге с 20 июля по 5 августа.
Во второй половине июля в средней тайге численность серых
мухоловок возрастает почти втрое по сравнению с плотностью гнез-
дования. Эти мухоловки становятся многочисленными в сосняках-
брусничниках и обычными на вырубках по ним (12 и 4). В августе
они перестают встречаться. В южной тайге уже в конце июля обилие
серой мухоловки возрастает примерно так же, как в предыдущей
подзоне, после чего около 90% популяции откочевывает. С середины
июля по 15 августа идет прикочевка средней интенсивности (6),
в результате которой птиц становится вдвое больше, чем на гнездо-
вании. В подтаежных лесах изменение численности определяется
в основном размножением местной популяции. Обилие возрастает
до середины июля, когда серых мухоловок становится втрое больше,
чем на гнездовании, а затем с 15 июля до середины августа идет ин-
тенсивная откочевка птиц (32), и в конце августа они перестают
встречаться совсем.
В среднем за лето в средней тайге серая мухоловка обычна в сос-
няках-брусничниках (3), редка на вырубках по ним и в приречных
смешанных лесах (0,5 и 0,4). В южной тайге ее примерно в 5 раз
больше. Предпочитает она мелколиственные леса (10), в меньшем
числе встречается в темнохвойной тайге и шелкопрядниках (2—6) и
лишь изредка — в заболоченных лесах (0,6—0,7). В подтаежных
лесах обилие этой мухоловки в среднем по району работ лишь не-
многим ниже, а в предпочитаемых местообитаниях в 1,6 раз выше,
чем в южной тайге. Держится она в осиново-березовых лесах и по-
лях-перелесках на их месте (16 и 2).
Следовательно, на междуречьях серая мухоловка чаще всего
встречается в суходольных мелколиственных, темнохвойных и сос-
новых лесах. При этом в южных подзонах ее значительно больше,
чем в средней тайге. В долине Оби численность серой мухоловки не-
сколько выше, и только в южной тайге, где мала доля пригодных
местообитаний, она ниже по сравнению с междуречьями. Обилие
этой мухоловки в долине монотонно убывает от подтаежных лесов до
северной тайги, наиболее высокая ее численность характерна для
пойменных лесов южных подзон.
Мухоловка-пеструшка (Ficedula hypoleuca Pall.)
В подтаежных лесах первая встреча приходится на 20 мая
1978 г., в средней тайге по Мал. Сосьве — на 12 мая 1931 г. и
16 мая 1932 г., а в Прикетье — на 8 мая 1964 г. Во второй половине
94
мая — начале июня в южных подзонах идет пролет средней интен-
сивности (1—7).
В северных подзонах мухоловка-пеструшка не зарегистрирова-
на. Вюжной тайге на гнездовании (вторая половина июня) предпо-
читает темнохвойную тайгу (19), в меньшем числе встречаясь в ос-
тальных местообитаниях (3—7). В подтаежных лесах отмечена с
15 мая до середины июня. В этот период обычна в лугах, используе-
мых под выпас, где держится по небольшим березовым колкам и в
осиново-березовых лесах (2—5). В конце июня — июле не наблюда-
лась, что, видимо, связано со снижением ее заметности в конце гнез-
дового периода. Отмечена вновь в южных подзонах на послегнездо-
вых кочевках в конце июля — первой половине августа (1—2).
По среднелетним показателям этой мухоловки на междуречьях
больше всего в южно-таежных лесах и заметно меньше — в подтаеж-
ных (8 и 2). В долине Оби ее численность гораздо выше, особенное
пойменных ивняках.
СЕМЕЙСТВО ОПОЛОВНИКИ (AEGITHALIDAE)
Ополовник (Aegithalos caudatus L.)
В северной тайге на междуречьях нами не зарегистрирован.
На гнездовании отмечен только в южных подзонах. В южной тайге
во второй половине мая обычен в мелколиственных и смешанных
полузаболоченных лесах, полях-перелесках и шелкопрядниках (4—
9). В подтаежных лесах в тот же период встречен в осиново-березо-
вых лесах и полях-перелесках на их месте (6 и 3). Массовое появле-
ние слетков в южной тайге по рекам Кеть и Васюган отмечено во вто-
рой половине июня [Гынгазов, Миловидов, 19771.
В послегнездовое время (в конце июня) ополовник появился
на кочевках в средней тайге. Возможно, он встречался раньше и
гнездился, но не попадал в учеты в связй с малочисленностью и пло-
хой заметностью во время гнездования. По крайней мере, 22 июня
1977 г. в пойменном ивняке на р. Бол. Юган видели двух слетков,
одного из которых кормила взрослая птица. До конца лета ополов-
ник держался в приречных темнохвойно-мелколиственных лесах
(в среднем с середины июня до конца августа — 6). Обилие его не-
уклонно возрастало с начала июля до конца августа. В среднетаеж-
ной долине Енисея тоже происходили кочевки ополовников, наибо-
лее выраженные в середине мая и сентябре 1978 г. В южной тайге
наблюдались два периода прикочевки — в начале июля и конце ав-
густа (29 и 22). В подтаежных лесах увеличение обилия в конце ию-
ня (в 8 раз по сравнению с плотностью гнездования) в равной степе-
ни определялось результатом размножения и прикочевкой. С начала
июля к августу в результате откочевки численность неуклонно сни-
жалась (почти в 5 раз по сравнению с предыдущим периодом). Воз-
можно, в это время ополовник смещается в долину Оби, по крайней
мере в пределах последней зарегистрирована интенсивная прикочев-
95
ка, в результате которой обилие ополовника в подтаежных лесах в
июле — августе резко возрастает (на 55 особей на объединённый
1 км2 ключевого участка);
В среднем за лето в среднетаежных приречных смешанных ле-
сах ополовник обычен (4). В южной тайге в лесном ландшафте его в
5 раз больше. Предпочитает он осиново-березовые леса (31). Не-
сколько меньше ополовника в темнохвойных лесах и шелкопрядни-
ках (18 и 25). Обычен он в угнетенных, влажных и заболоченных ле-
сах и полях-перелесках (2—4). В подтаежных лесах ополовника поч-
ти вдвое меньше, чем в южной тайге. Он держится в осиново-березо-
вых лесах и полях-перелесках на их месте (20 и 4).
Следовательно, на междуречьях максимальная численность
ополовника отмечена в березово-осиновых лесах в южной тайге, не-
сколько меньшая — в подзоне подтаежных лесов, а минимальная
характерна для средней тайги, где его видели лишь на кочевках.
В долине Оби в наибольшем числе ополовник встречен в пойменных
ивняках. По сравнению с междуречьями его здесь значительно боль-
ше в подтаежных лесах и меньше в южной тайге, а в средней тайге
зарегистрирована сходная численность. Однако описанные разли-
чия среднелетних показателей определяются интенсивностью после-
гнездовых кочевок и, видимо, могут отличаться по годам. В южных
подзонах ополовника больше, чем в средней тайге, а в северной тай-
ге он не зарегистрирован.
СЕМЕЙСТВО СИНИЦЫ (PARIDAE)
Пухляк (Paras montanus Bald.)
Во второй половине мая идут предгнездовые кочевки, наиболее
выраженные в южной тайге (10) и незначительные в средней и под-
таежных лесах (1—2).
На гнездовании (июнь) в северной тайге пухляк встречался толь-
ко в приречных темнохвойных лесах (10). В средней тайге он много-
числен в сходных темнохвойно-мелколиственных лесах (24) и обычен
в поселках и сосняках (1 и 6). В южной тайге в тот же период его
больше, чем в предыдущих подзонах. Многочислен во всех урочи-
щах лесного ландшафта (34—55) и только в полях-перелесках обы-
чен (6). В подтаежных лесах пухляка в 7 раз меньше, чем в южной
тайге. Держится во время гнездования в осиново-березовых лесах,
полях-перелесках и рямах (12, 2 и 5). Постройка дупел наблюдалась
в окрестностях Томска в разные годы с 10 мая по 7 июня; 12 июня
1964 г. в двух гнездах было семь и восемь свежих яиц, а слетков ви-
дели с середины июля. В северной тайге мы встретили молодых птиц
1 июля 1976 г.
В послегнездовое время (в июле) численность пухляка повсе-
местно возрастает в 4—6 раз, что несколько превышает результатив-
ность размножения и определяется, видимо, кочевками молодых
птиц. Вторая волна прикочевок идет в августе, наибольшая интен-
96
сивность ее отмечена в южной тайге (190), слабее — в средней (15)
и минимальная — в крайних подзонах (1—2). Во время кочевок стай-
ки пухляков начинают встречаться на верховых и низинных боло-
тах, кроме северотаежных. В приенисейской средней тайге тоже
прослеживаются две волны кочевок пухляка: одна в третьей декаде
июля, направленная преимущественно на север, другая — в конце
августа, когда птицы перемещались в основном на юг.
В среднем за лето в северной тайге пухляк многочислен в при-
речных темнохвойных лесах (22), обычен на гарях по соснякам (2)
и редок в поселках (0,4). Избегает сосняков и комплексных верхо-
вых болот. В средней тайге пухляка почти в 12 раз больше, и встре-
чается он гораздо шире. Многочислен в лесных местообитаниях,
включая вырубки по соснякам (28—54), а на остальной территории
обычен (2—8), В южной тайге его в 4 раза больше, чем в средней.
Максимальные показатели обилия характерны для темнохвойных,
мелколиственных, полузаболоченных лесов и шелкопрядников
(102—289) и более низкие — для остальных местообитаний лесного
ландшафта (47—86). Самая низкая численность пухляка зарегист-
рирована на верховых и низинных болотах, где его видели на после-
гнездовых кочевках (1—4). В подтаежных лесах этой синицы соот-
ветственно в 2 и 8 раз меньше, чем в средней и южной тайге, но в 6
раз больше, чем в северной. Предпочитает она осиново-березовые ле-
са (26), довольно много ее в полях-перелесках и рямах (18 и 14),
в небольшом числе встречена в лугово-болотных местообитаниях
(по 1).
В результате следует отметить, что на междуречьях пухляк
занимает леса различного типа, проникая в лесополевые местообита-
ния и по рямам на окраины болот. Наибольшее его обилие харак-
терно для южной тайги, к северу и югу от которой показатели сни-
жаются, что особенно проявляется в северной тайге. В долине Оби
эта тенденция внутризональных изменений численности прослежи-
вается в меньшей степени, так как в южной тайге, где на обследован-
ном участке леса занимают небольшую площадь, пухляка в среднем
по территории меньше, чем в остальных подзонах. В среднем по об-
следованным участкам лесной зоны Западной Сибири наиболее вы-
сокое обилие пухляка отмечено на южно-таежных междуречьях,
минимальное — на северотаежных междуречьях, а средние показа-
тели характерны для всей остальной территории.
Сероголовая гаичка (Parus cinctus Bodd.)
Встречена только в северных подзонах. На гнездовании в се-
верной тайге (июнь) держалась в темнохвойных, сосновых лесах,
на гарях, вырубках по ним, в сосново-озерном ландшафте и посел-
ках (2—5). В средней тайге в это время нами не зарегистрирована,
однако отмечена гнездящейся на р. Мал. Сосьва, где 15 июля
1939 г. из гнезда был взят птенец. В северной тайге молодых серого-
ловых гаичек в одной стайке со взрослыми видели 22 июля 1976 г.
7 Л. Г. Вартапетов
97
В послегнездовое время (первая половина июля) в северной
тайге обилие возрастает втрое. Особенно резко оно увеличивается
за счет прикочевки с середины июля по 15 августа (И). В этот пе-
риод стайки гаичек начинают встречаться на комплексных верховых
болотах (3—4). В средней тайге сероголовые гаички появляются
только в июле — августе на послегнездовых кочевках. В это время
они обычны в рослых рямах и на вырубках по соснякам (1—3). По-
видимому, с середины июля начинаются широкие кочевки сероголо-
вых гаичек, в результате которых часть популяций, отгнездившихся
севернее, смещается к югу.
В среднем за лето в северной тайге сероголовая гаичка много-
численна в сосново-озерном ландшафте, темнохвойных и сосновых
лесах (10—16) и обычна во всех остальных местообитаниях (1—6).
В средней тайге ее в 100 раз меньше, чем в северной. Здесь она встре-
чена лишь на кочевках в рослых рямах и на вырубках по соснякам
(1—2). В среднем по району работ на северотаежных междуречьях
этой гаички в 4 раза больше, чем пухляка. Однако последний пре-
обладает по обилию в приречных темнохвойных лесах с наиболее
высокими кормными и защитными свойствами, тогда как в остальных
местообитаниях встречена почти исключительно сероголовая гаич-
ка. К югу от северной тайги преобладает по численности или встре-
чается только пухляк. Так, в средней тайге, по нашим данным, пух-
ляка в среднем за лето в 18 раз больше, чем сероголовой гаички (в
тех местообитаниях, где они встречались вместе). По среднегодовым
показателям, приводимым В. В. Раевским [1982], соотношение чис-
ленности этих гаичек в средней тайге Зауралья составляет 3 :17 в
пользу пухляка, хотя, по свидетельству того же автора, оно может
сильно различаться по годам.
Итак, на таежных междуречьях сероголовая гаичка встречена
преимущественно в сосново-боровом ландшафте северной тайги и
лишь изредка попадается в средней тайге, В долине Оби она заре-
гистрирована только в пределах северной тайги. При этом в сходных
местообитаниях ее примерно столько же, сколько на междуречьях,
а в среднем по участкам значительно меньше.
Московка (Parus ater L.)
В северной тайге встречена только в первой половине июня в
приречных темнохвойных лесах (8). На гнездовании в июне в средней
тайге обычна в сходных смешанных лесах (4). В южной тайге (пер-
вая половина июня) многочисленна в те^нохвойных и полузаболо-
ченных смешанных лесах (11—17). В подтаежных лесах в течение
всего лета не зарегистрирована. Кладки из семи яиц находили в
окрестностях Томска 8 июня, в средней тайге по р. Тым — 14 июня,
где было четыре и восемь яиц; пять оперившихся птенцов видели в
южной тайге 18 июня 1971 г. [Гынгазов, Миловидов, 1977].
В послегнездовое время во второй половине июля обилие этой
синицы в средней тайге возрастает в 4 раза по сравнению с плот-
98
ностью гнездования. Затем в первой половине августа московка пе-
рестает встречаться и появляется вновь на кочевках лишь во второй
его половине. В этот период она распространяется гораздо шире,
чем на гнездовании: держится в рослых рямах, приречных смешан-
ных лесах и на вырубках по соснякам (0,8, 4 и 10). В южной тайге
в то же время идут кочевки, максимально интенсивные во второй
половине июля (26), а затем к концу лета более половины популяции
откочевывает за пределы обследованной территории.
В среднем за лето в северной тайге московка обычна в приреч-
ных темнохвойных лесах (2). В сходных местообитаниях средней
тайги ее втрое больше. Обычна она в приречных смешанных лесах,
на вырубках по соснякам (6 и 2) и редка в рослых рямах (0, 2). В
лесных местообитаниях южной тайги этой синицы соответственно в
10 и 3,5 раза больше, чем в северной и средней* Особенно много ее
в темнохвойной тайге вследствие интенсивной прикочевки в июле —
августе (в среднем за лето — 118). Она многочисленна в остальных
лесных местообитаниях и шелкопрядниках (14—22), обычна в ело-
во-кедровой тайге (2) и редка в рослых рямах (0, 1).
Таким образом, московка предпочитает высокоствольные леса,
особенно темнохвойные. На междуречьях больше всего ее отмечено в
южной тайге, при этом к северу численность неуклонно снижается.
В долине Оби она встречена в средней тайге и подтаежных лесах
там, где имеются высокоствольные леса нормальной полноты. В сред-
нем по участкам ее здесь примерно столько же, сколько на
среднетаежных междуречьях.
Большая синица (Parus major L.)
На междуречьях северной тайги не встречена. В средней ее лишь
однажды видели в поселках в первой половине июля (4). В южной
тайге отмечена только на кочевках во второй половине мая и авгус-
та в полях-перелесках и мелколиственных лесах (в среднем по лес-
ному ландшафту за эти периоды — 1иЗ). В подтаежных лесах на
гнездовании (15 мая — 15 июня) обычна в перелесках среди полей и
лугов, используемых под выпас, а также в осиново-березовых лесах
(2—6). Стайки молодых кочующих птиц начали встречаться здесь
после 1 июля 1978 г.
В послегнездовой период в подтаежных лесах обилие большой
синицы в первой половине июля резко возрастает в результате разм-
ножения местных популяций и прикочевки высокой интенсивности
(20). С середины июля до конца августа в подтаежных лесах идет от-
кочевка, вследствие которой обилие снижается в 4 раза. Во время
кочевок в июле — августе эта синица начинает встречаться в посел-
ках (в среднем за этот период — 9). По-видимому, большая часть
отгнездившихся птиц в этот период смещается из таежной зоны к
югу в лесостепь [Равкин, 1978].
По среднелетним показателям, большая синица в средней тайге
редка в поселках (0,5). Значительно больше ее в южной тайге, где
она обычна в полях-перелесках (4) и редка в березово-осиновых ле-
7*
99
сах (4 и 0,3). В подтаежных лесах обилие большой синицы в 25 раз
выше, чем в южной тайге. Предпочитает она осиново-березовые леса
(28), в значительном числе встречается в полях-перелесках (10) и
вдвое меньше ее в березовых колках среди лугов-выпасов и в посел-
ках (4 и 5).
Следовательно, на междуречьях больше всего этой синицы в
подтаежных березово-осиновых лесах, к северу от которых числен-
ность ее резко снижается. В долине Оби ее повсеместно больше (пред-
почитает пойменные леса и поселки). Распространена к северу го-
раздо дальше (до северной тайги), а уменьшение обилия в том же
направлении значительно более плавное, чем на междуречьях.
Князек (Paras су anus Pall.)
Встречен нами только в подтаежных лесах. На гнездовании
(15 мая — 15 июня) отмечен в перелесках среди полей и лугов, ис-
пользуемых под выпас, а также в осиново-березовых лесах (11, 4 и
2). По-видимому, одновременно с гнездованием в начале июня идут
кочевки средней интенсивности (3). В послегнездовое время
(15 июня — 15 июля) обилие снижается втрое по сравнению с плот-
ностью гнездования, что связано с откочевкой большей части от-
гнездившихся птиц. Во второй половине июля идет волна после-
гнездовых кочевок (6), после чего к концу лета численность вновь
сокращается до прежнего уровня. В среднем за лето больше всего
князька отмечено в перелесках среди полей и в осиново-березовых
лесах (8 и 5) и значительно меньше в лугах-выпасах (1), где он дер-
жался в небольших березовых колках.
Итак, на междуречьях князек встречен нами только в подтаеж-
ных лесах, хотя отмечался и севернее, в южной тайге на гнездова-
нии по долине Васюгана [Гынгазов, Миловидов, 1977]. В долине
Оби эта синица наблюдалась в пойменных ивняках. Ее здесь значи-
тельно меньше, чем на подтаежных междуречьях, но распростране-
на она дальше — к северу, по крайней мере, до южной тайги, при
этом численность князька снижается в том же направлении.
СЕМЕЙСТВО [ПОПОЛЗНИ (SITTIDAE)
Поползень (Sitta europaea L.)
На гнездовании в северной тайге (июнь) обычен в приречных
темнохвойных лесах, на гарях, вырубках по соснякам (2—4) и ре-
док в сосново-озерном ландшафте (0,5). В средней тайге в тот же пе-
риод предпочитает приречные темнохвойные леса (12), в меньшем
числе встречается в поселках и на вырубках по соснякам (4 и 0,2).
В южной тайге (15 мая — 15 июня) многочислен в темнохвойных и
смешанных лесах (19—24), обычен в мелколиственных (8) и редок в
шелкопрядниках (0,5). Гнезда поползня находили в окрестностях
Томска в разные годы с 9 по 25 мая, а вылет птенцов отмечен с 14 по
100
18 июня 1970—1972 гг. В Прикетье гнездо с тремя голыми птенцами
найдено 6 июня 1963 г. В средней тайге 17 июня 1977 г. мы видели
поползня, волнующегося с кормом. В северной тайге 3 июля 1976 г.
встречен слеток, а 6 августа видели молодых птиц, по размеру не
отличавшихся от взрослых.
В послегнездовое время (первая половина июля) обилие по-
ползня возрастает в северных подзонах вдвое в результате размно-
жения. В южной тайге в тот же период прослеживается 6-кратное
увеличение численности в связи с интенсивной прикочевкой (17).
В северной тайге прикочевка птиц идет в основном в конце июля (2).
Затем численность снижается и к концу августа становится вдвое
меньше, чем на гнездовании. По-видимому, большая часть птиц сме-
щается к югу. В результате в августе в средней и южной тайге идет
прикочевка высокой интенсивности (соответственно 10 и 57). В пе-
риод кочевок поползень начинает встречаться заметно шире, в том
числе на среднетаежных верховых болотах, а в южной тайге в пе-
релесках среди полей и рослых рямах. В долине Енисея в пределах
средней тайги тоже отмечены интенсивные кочевки поползня к севе-
ру в начале августа и к югу в конце этого месяца.
В среднем за лето в северной тайге поползень многочислен в
приречных темнохвойных лесах (14), обычен на вырубках, гарях
по соснякам (1 и 2) и редок в остальных ландшафтных урочищах
(0,1—0,6). В лесных ландшафтах средней тайги его почти в 5 раз
больше. Предпочитает поползень приречные смешанные леса и в
меньшем числе встречается на вырубках по соснякам (21 и 11). Еще
меньше поползня отмечено в поселках и сосняках (4 и 6), а мини-
мальное его обилие характерно для верховых болот (1). На послед-
них встречался лишь на послегнездовых кочевках в августе. В лес-
ных ландшафтах южной тайги поползня в 10 и 3 раза больше, чем в се-
верной и средней тайге. Предпочитает он темнохвойную тайгу (90),
в значительном числе встречается в остальных местообитаниях лес-
ного ландшафта (14—49) и в небольшом — в полях-перелесках. В от-
личие от более северных подзон, в южной тайге совершенно избе-
гает болот. На подтаежных междуречьях в течение всего лета не за-
регистрирован, хотя в долине Оби в пределах той же подзоны, как
правило, обычен, а в сосняках даже многочислен за счет интенсивной
прикочевки птиц в конце августа.
В результате следует отметить, что поползень повсеместно пред-
почитает темнохвойные и смешанные (темнохвойно-мелколиствен-
ные) леса. На междуречьях его численность возрастает от северной
тайги к южной. В долине Оби, где поползень в значительном числе
встречается в пойменных ивняках, его обычно больше, чем на между-
речьях. Однако в южной тайге, где в пределах обследованного
участка отсутствуют предпочитаемые им темнохвойные леса, обилие
поползня значительно ниже.
101
СЕМЕЙСТВО ПИЩУХОВЫЕ (CERTHIIDAE)
Пищуха (Certhia familiaris L.)
В северной тайге нами не зарегистрирована. В средней обычна
на послегнездовых кочевках в июле — августе в приречных темно-
хвойно-мелколиственных лесах (в среднем за этот период — 6).
В южной тайге встречалась несколько чаще, хотя по среднелетшш
показателям ее здесь примерно столько же, сколько в средней тайге.
При этом пищуха обычна в смешанных полузаболоченных и темно-
хвойных лесах (8 и 4). В подтаежных лесах ее значительно меньше,
чем в предыдущих подзонах. Зарегистрирована она в осиново-бере-
зовых лесах (в среднем за лето — 0,3).
По среднелетним показателям, максимальное обилие пищухи
на междуречьях отмечено в средней и южной тайге (в темнохвойных
и смешанных лесах) и значительно меньше ее в подтаежных осиново-
березовых лесах. В долине Оби, там, где имеются предпочитаемые
ею высокоствольные темнохвойные леса (в средней тайге), пищухи
примерно столько же, сколько на междуречьях, а в остальных под-
зонах она не зарегистрирована.
СЕМЕЙСТВО ОВСЯНКОВЫЕ (EMBERIZIDAE)
Белошапочная овсянка (Emberiza leucocephalos Gm.)
В северной тайге на междуречьях летом 1976 г. и в долине Оби
в 1973 г. не зарегистрирована, хотя добыта И. Н. Шуховым [1915]
в среднем течении р. Казым 27 мая 1914 г. Окончание пролета в
средней тайге и подтаежных лесах отмечено во второй половине мая
(3-4).
В средней тайге на гнездовании (июнь) держится на верховых
болотах, где обилие возрастает от периферии к центру с увеличением
площади открытых участков (в рослых рямах — 1, в низкорослых —
6 п в грядово-мочажинных комплексах — 10). Изредка встречалась
в сосняках-брусничниках (0,1). В южной тайге ее заметно больше.
В первой половине июня концентрируется в перелесках среди полей
(40), в меньшем числе наблюдается в мелколиственных лесах и на
верховых болотах (4—7). В подтаежных лесах в июне ее примерно
столько же, сколько в южной тайге. Предпочитает поля в сочетании
с перелесками и рямы, расположенные среди низинных болот (26 и
37). Изредка встречается в открытых лугово-болотных местообита-
ниях (0,5). Гнезда с двумя и шестью яйцами находили в окрестностях
Томска 30 июня 1963 г. и 2 июля 1972 г. В южной тайге по р. Васю-
ган гнездо с шестью яйцами найдено 7 июня 1965 г. и у с. Бакчар
14 июня 1971 г. с тремя свежими яйцами. Здесь же 17 июня 1971 г.
встречены слетки [Гынгазов, Миловидов, 1977].
После вылета молодых в средней тайге до 95 % местной популя-
ции откочевывает в начале июля. Затем овсянки появляются вновь в
102
значительном числе с середины июля, при этом их численность воз-
растает до конца августа втрое по сравнению с гнездовым периодом.
Во время кочевок во второй половине лета белошапочная овсянка
начинает встречаться в большем числе местообитаний, чем на гнездо-
вании, например появляется в приречных смешаннах лесах и посел-
ках (3 и 8). В южных подзонах после окончания размножения ее оби-
лие во второй половине июня — начале июля возрастает вдвое. Од-
новременно с вылетом молодых в южной тайге идет интенсивная
прикочевка (13). Затем во второй половине июля численность этой
овсянки в подтаежных лесах и южной тайге снижается и становится
даже немного меньше, чем на гнездовании. Следующая волна коче-
вок равной интенсивности в обеих подзонах (по 10) идет в первой по-
ловине августа в подтаежных лесах и в конце этого месяца — в юж-
ной тайге.
В среднем за лето в средней тайге белошапочная овсянка пред-
почитает грядово-мочажинные верховые болота (10), в меньшем чис-
ле встречается в низкорослых рямах и поселках (6 и 8). В последних
ее видели лишь во время кочевок. Еще меньше этой овсянки в рос-
лых рямах и приречных смешанных лесах (2 и 3), а минимальное
обилие зарегистрировано в сосняках-брусничниках (0,5). В южной
тайге численность белошапочной овсянки в 2,5 раза выше, чем в
средней. Многочисленна она на верховых болотах, в шелкопрядни-
ках и полях-перелесках (14—70), обычна в рослых рямах и осиново-
березовых лесах (1 и 3). В подтаежных лесах белошапочной овсянки
примерно столько же, сколько в южной тайге. Предпочитает она
здесь рямы и поля-перелескй (24 и 39), меньше ее на низинных боло-
тах (2). Изредка встречается в осиново-березовых лесах, лугах-вы-
пасах и поселках (0,1—0,8).
Следовательно, на таежных междуречьях белошапочная ов-
сянка отдает предпочтение лесополевым местообитаниям и верховым
болотам. При этом в южных подзонах ее больше, чем в средней тай-
ге. В северной тайге эта овсянка не зарегистрирована. В пределах
более южных подзон белошапочной овсянки в долине Оби примерно
столько же, сколько на междуречьях, и только в южной тайге зна-
чительно больше. В последней подзоне лесополевые надпойменные
террасы долины Оби гораздо более привлекательны для белошапоч-
ной овсянки, чем таежные междуречья.
Овсянка-крошка (Emberiza pusilia Pall.)
На подтаежных междуречьях эти овсянки появились 24 мая
1978 г. В средней тайге пролетные стайки (около 10 особей) начали
встречаться после 27 мая 1977 г. Предгнездовой пролет идет в конце
мая — первой половине июня, наиболее интенсивен он в северной
тайге (15) и слабее выражен в подтаежных лесах и средней тайге
(1-4).
На гнездовании (вторая половина июня) в северной тайге ов-
сянка-крошка многочисленна в сосняках-ягельниках, на гарях по
103
ним, в сосново-озерных ландшафтах и на верховых болотах (10—
31). Обычна на переходных болотах и в приречной темнохвойной тай-
ге (5 и 8). Совсем не встречена она в поселках и на частичных выруб-
ках по соснякам. В средней тайге в тот же период эта овсянка обычна
на вырубках по соснякам и в рямах (4—6), а в южной тайге ее лишь
изредка встречали в низкорослых рямах (0,3). В подтаежных лесах
на гнездовании не зарегистрирована. В северной тайге гнездо с клад-
кой из шести яиц найдено 24 июня 1976 г. В то же лею первые слет-
ки встречены 8 июля, а массовый вылет молодых птиц отмечен
24 июля. Овсянка-крошка, выкармливающая птенца кукушки, встре-
чена 4 августа 1976 г.
После вылета молодых в северной тайге численность этой овсянкп
возрастает вдвое (к концу июля). Затем идет интенсивная откочевка
(20), в результате которой обилие в начале августа становится мень-
шим, чем на гнездовании. Во второй половине августа наблюдается
прикочевка птиц, по-видимому, более северных популяций (7). В
средней тайге послегнездовое увеличение численности более чем в
5 раз превышало плотность гнездования, так как вылет молодых птиц
шел одновременно с прикочевкой этих овсянок. Вторая волна прико-
чевки так же, как и в предыдущей подзоне, прослеживается в сред-
ней и южной тайге во второй половине августа. Во время кочевок
овсянка-крошка начинает встречаться там, где ее не было на гнездо-
вании (в поселках, приречных смешанных лесах, полях-перелесках).
Итак, повсеместно прослеживаются две волны послегнездовых коче-
вок: одна — в конце июля, при вылете молодых птиц, и другая —
в конце августа, в результате которой часть отгнездившихся попу-
ляций смещается к югу.
В среднем за лето в северной тайге овсянка-крошка почти пов-
семестно многочисленна. Больше всего ее встречено в низкорослых
рямах, приречной темнохвойной тайге и в сосново-озерных ландшаф-
тах (33—40), т. е. там, где имеется хорошо развитый ярус угнетен-
ных сосен или кустарников. Заметно меньше ее в редкостойных сос-
няках, на вырубках, харях и открытых верховых болотах (10—26).
Минимальное обилие отмечено в поселках и на частичных вырубках
по соснякам (по 2). В средней тайге этой овсянки почти в 4,5 раза
меньше. Здесь она повсеместно обычна (2—6) и лишь в рослых ря-
мах многочисленна (14). В южной тайге численность овсянки-крошки
в 31 раз ниже, чем в северной, ее встречали лишь в полях-перелесках
(на послегнездовых кочевках) и в низкорослых рямах (16 и 0,04).
В подтаежных лесах этой овсянки еще меньше (в 7 раз по сравнению
с южной тайгой). Здесь ее видели только на предгнездовом пролете.
По среднелетним показателям обычна в поселках (2) и редка в луго-
во-болотном ландшафте (0,3).
В результате следует отметить, что на междуречьях северной
тайги овсянка-крошка — один из наиболее многочисленных и широ-
ко распространенных видов птиц. В пределах остальных подзон ее
обилие резко убывает к югу. В долине Оби она зарегистрирована
только в северных подзонах, при этом в северной тайге ее значитель-
но больше, а в средней — несколько меньше, чем на междуречьях»
104
Таким образом, по рассматриваемой территории в целом численность
овсянки-крошки снижается в ряду: северотаежная долина Оби —
северотаежные междуречья — более южные подзоны.
Овсянка-ремез (Ember iza rustica Pall.)
В конце мая — начале июня в северных подзонах идет пролет,
наиболее интенсивный в средней тайге (4) и слабее выраженный в
северной (0,4). В гнездовой период на междуречьях северной тайги
(15 июня — 15 июля) держится в приречных темнохвойных лесах
(10). В средней тайге (июнь) встречена в сходных смешанных лесах
и сосняках (8 и 1). В среднетаежной долине Енисея овсянки-ремеза
примерно столько же, сколько ее отмечено нами на Обь-Иртышском
междуречье в пределах той же подзоны (2—3 особи на объединенный
1 км2 ключевого участка). В южной тайге (середина мая — конец
июля) она предпочитает смешанные полузаболоченные леса (12),
встречается в перелесках среди полей (4). На междуречьях подтаеж-
ных лесов в течение всего лета не зарегистрирована. Гнезда с пятью
яйцами найдены в окрестностях Томска 5 и 24 июня 1974 г., а нелет-
ный птенец пойман 26 июля 1959 г. В среднетаежной долине Енисея
гнездо с четырьмя яйцами найдено 4 июня 1974 г., а молодые птицы
размером со взрослых встречались здесь с последних чисел июля.
После окончания гнездования (в июле) увеличение численности
не прослеживается. К началу августа в средней тайге большая часть
птиц откочевывает, а в северной и южной тайге они перестают встре-
чаться совсем. В середине августа эти овсянки вновь появляются на
кочевках. Их численность в северной тайге становится примерно та-
кой же, как на гнездовании, а в средней и южной в 3—6 раз боль-
шей.
В среднем за лето в северной тайге овсянка-ремез обычна в при-
речных темнохвойных лесах (8). В сходных смешанных лесах в сред-
ней тайге ее лишь немногим больше (10). Реже встречается она в сос-
няках (1), на вырубках по ним и в поселках (0,1 и 0,5). В южной тай-
ге ее примерно столько же, сколько в средней. Предпочитает она
смешанные полузаболоченные леса (8), в меньшем числе зарегистри-
рована в мелколиственных лесах и полях-перелесках (по 4).
Таким образом, овсянка-ремез на таежных междуречьях чаще
всего встречена в смешанных полузаболоченных и приречных тем-
нохвойных лесах. Обилие этой овсянки в средней и южной тайге
несколько выше, чем в северной. В долине Оби в пределах северной
тайги ее в среднем примерно столько же, сколько на междуречьях;
особенно много ее в полузаболоченных смешанных лесах, а в более
южных подзонах, где на обследованных участках такие леса отсутст-
вуют, численность овсянки-ремеза значительно ниже.
Дубровник (Emberiza aureola Pall.)
В подтаежных лесах прилет зарегистрирован 21 мая 1978 г.,
а после 27 мая начали встречаться большие стаи пролетных птиц,
в основном самцов (до нескольких сотен особей). В южной тайге в
105
1967 г. дубровник появился 19 мая, но в массе — только 1 июня»
В средней тайге в 1977 г. прилет зарегистрирован 2 июня, а в север-
ной — 10 июня 1976 г.
В гнездовой период па междуречьях северной тайги дубровник
концентрируется по периферии комплексных верховых болот — в
низкорослых рямах и на переходных болотах (вторая половина ию-
ня — 8 и 11). Немного меньше его на грядово-мочажинно-озерных
верховых болотах (2). В средней тайге в тот же период держится на
верховых болотах, при этом на открытых участках его больше, чем
на залесенных (5 и 2). В южной тайге в первой половине июля оби-
лие дубровника значительно выше, особенно на открытых низинных
болотах (265). Реже встречался он в полях-перелесках и на верхо-
вых болотах (1—4). В подтаежных лесах (первая половина июня)
зарегистрирован в лугах, используемых под выпас, и на низинных
болотах в сочетании с покосными лугами (13 и 25). Гнездо с четырь-
мя яйцами найдено в этой подзоне 26 июня 1978 г.; 15 июля 1976 г.
в северной тайге видели волнующуюся самку дубровника с кормом.
После окончания гнездования (в июле) в северной тайге числен-
ность дубровника возрастает в 2 раза. В августе после откочевки
численность снижается и становится такой же, как на гнездовании.
В средней тайге в первой половине июня одновременно с вылетом мо-
лодых идет прикочевка птиц средней интенсивности (5). В конце ию-
ля все дубровники улетают с мест гнездования (верховые болота),
но продолжают встречаться в поселках, а после 15 августа исчезают
совсем. В южной тайге в августе идет интенсивная откочевка (12),
в результате чего к концу лета дубровников становится почти вдвое
меньше, чем на гнездовании. Одновременно с массовым отлетом часть
их, видимо более дальние мигранты, в августе задерживаются в от-
дельных местообитаниях. Например, в полях-перелесках в это вре-
мя насчитывается до 130 особей/км2, что в 32 раза превышает плот-
ность их гнездования. В подтаежных лесах в первой половине июля
откочевывает свыше 30% местной популяции. В конце июля идет
прикочевка высокой интенсивности (28). В августе еще большая
часть дубровников откочевывает (41), и к концу лета их становится
в 5 раз меньше, чем на гнездовании. Итак, с середины июля до сере-
дины августа большая часть дубровников на междуречьях смещает- \
ся к югу, а затем, видимо, откочевывает за пределы обследованной
территории.
В среднем за лето в северной тайге дубровник многочислен на
переходных, обычен на верховых болотах (10 и 3) и редок в сосново-
озерных ландшафтах (0, 2). В средней тайге его примерно столько
же; он обычен на верховых болотах и в поселках (2—4), причем в
последних встречен лишь на послегнездовых кочевках. В южной
тайге дубровника почти в 8 раз больше, чем в северных подзонах.
Он предпочитает низинные болота (205), в меньшем числе встре-
чаясь на верховых болотах (кроме грядово-мочажинно-озерных) и
в лесополевых местообитаниях (10—12). Обычен в смешанных полу-
заболоченных лесах, шелкопрядниках и на переходных болотах
(1—4) и редок в мелколиственных лесах (0,6). В подтаежных лесах
106
его на 20% больше, чем в южной тайге, и в 9 раз больше, чем в се-
верных подзонах. Держится преимущественно на низинных боло-
тах в сочетании с покосными лугами (38). В меньшем числе встре-
чается в остальных местообитаниях (2—8).
Итак, дубровник на междуречьях южных подзон предпочитает
открытые низинные болота, а в северных подзонах, где такие болота
отсутствуют, держится на верховых. В южных подзонах его гораздо
больше, чем в северных. Еще выше численность этой овсянки в доли-
не Оби, при этом в предпочитаемых открытых пойменных местооби-
таниях и на низинных болотах южно-таежных междуречий отмечена
сходная и самая высокая численность. Уменьшение обилия дубров-
ника к северу в долине выражено гораздо слабее, чем на междуре-
чьях, и в основном при переходе от средней к северной тайге.
Камышевая овсянка (Emberiza schoeniclus L.)
На междуречьях северной тайги нами не встречена, однако ле-
том 1973 г. зарегистрирована в качестве многочисленной птицы в
северотаежной долине Оби, особенно в ее пойме. На междуречьях
средней тайги всего один раз (22 июня 1977 г.) видели двух самцов
Камышевой овсянки в кустах на берегу надпойменного озера. В юж-
ной тайге тоже встречалась редко — только на весеннем пролете во
второй половине мая (9). В подтаежных лесах ее значительно больше
чем в предыдущих подзонах. На гнездовании камышевая овсянка
обычна на открытых низинных тростниково-осоковых болотах (8).
В послегнездовое время (вторая половина июля) идет интенсивная
прикочевка этих овсянок (25), но уже в начале августа их обилие
становится таким же, как на гнездовании, а к середине августа
камышевые овсянки перестают встречаться в учетах.
В среднем за лето камышевая овсянка редка в средней и южной
тайге. В подтаежных лесах ее в 12 раз больше, чем в предыдущих
подзонах. Она многочисленна на низинных болотах (14) и редка в
перелесках среди полей (0,3).
Таким образом, на междуречьях больше всего Камышевой ов-
сянки в подтаежных лесах на тростниково-осоковых низинных бо-
лотах и значительно меньше в срединных подзонах. В долине Оби
она тоже предпочитает низинные болота и пойменные луга. Макси-
мальное обилие зарегистрировано в северной тайге, оно неуклонно
снижается к юго-востоку, т. е. в обратном направлении по сравне-
нию с междуречьями. Поэтому от северной до южной тайги Камыше-
вой овсянки здесь гораздо больше, а в подтаежных лесах значитель-
но меньше, чем на междуречьях.
Лапландский подорожник (Calcarius lapponicus L.)
Встречен только на пролете. В южной тайге подорожников часто
видели во второй половине мая — начале июня в низкорослых ря-
мах (220), гораздо реже — на переходных и низинных болотах (34 и
3). В подтаежных лесах редок в конце мая в рямах (0,1). В долине
Оби не зарегистрирован, видимо, пролетал до начала наблюдений.
107
СЕМЕЙСТВО ВЬЮНКОВЫЕ (FRING1LLIDAE)
Зяблик (Fringilla coelebs L.)
Во второй половине мая — начале июня повсеместно идет окон-
чание пролета (1—4). На междуречьях северной тайги пение зябли-
ка слышали только в первой половине июня. В этот период он обы-
чен в приречной темнохвойной тайге и редок в сосняках-ягельни-
ках (2 и 0,8).
На гнездовании в средней тайге (первая половика июня) обычен
в приречных смешанных лесах (5). В южной тайге в тот же период
предпочитает мелколиственные леса (6), в меньшем числе встречает-
ся в заболоченных сосновых и смешанных лесах (1—2). На гнездо-
вании (июнь) в подтаежных лесах зяблика заметно больше. Он мно-
гочислен в осиново-березовых лесах (12) и обычен в лугово-полевых
местообитаниях (1—9), где держится в колках.
Постройка гнезда отмечена в средней тайге на сосновой гриве
среди верхового болота 21 мая 1977 г. В той же подзоне на р. Ларь-
Еган разновозрастные птенцы найдены 23—27 июня 1976 г., а в юж-
ной тайге на р. Парабель — 9—14 июня 1970 г. [Гынгазов, Мило-
видов, 1977].
После вылета молодых обилие зяблика в середине июля обычно
возрастает втрое по сравнению с плотностью гнездования. К середине
августа в средней тайге около 90% птиц откочевывают, и их стано-
вится в 2,5 раза меньше, чем на гнездовании. Прикочевка птиц более
северных популяций последовательно прослеживается сначала в
южной тайге во второй половине июля, а затем в подтаежных лесах
в середине августа (по 6). К концу лета в южной тайге их становится
втрое меньше, а в подтаежных лесах вдвое больше, чем на гнездо-
вании.
В среднем за лето в северной тайге зяблик — редкая птица
сосновых и темнохвойных лесов (0,2—0,4). В сходных местообита-
ниях средней тайги его в 15 раз больше. Он обычен в приречных
смешанных лесах (6), редок в сосняках-брусничниках и на вырубках
по ним (0,6 и 0,1). В южной тайге обилие зяблика в 1,5 раза выше,
чем в средней. Предпочитает он мелколиственные леса (7), в мень-
шем количестве встречается в остальных местообитаниях лесного
ландшафта. В подтаежных лесах его соответственно в 4 и 3 раза
больше, чем в средней и южной тайге. Здесь зяблик тоже концент-
рируется в осиново-березовых лесах (25), значительно меньше его в
полях-перелесках и лугах, используемых под выпас (1—5,'«
В результате следует отметить, что на междуречьях зяблик пред-
почитает мелколиственные и смешанные леса, при этом его обилие
неуклонно снижается к северу. В долине Оби зяблик тоже чаще все-
го встречался в мелколиственных лесах как в пойме, так и вне ее.
Здесь зарегистрированы сходная численность и такой же характер
ее внутризональных изменений, как и на междуречьях.
108
Юрок (Fringilla montifringilla L.)
Во второй половине мая — первой половине июня повсеместно
идет пролет высокой интенсивности в таежных подзонах (15—21)
и значительно более слабый в подтаежных лесах (2).
На гнездовании (15 июня — 15 июля) в северной тайге юрок
предпочитает приречные темнохвойные леса (57). Вчетверо ниже его
численность в сосняках-ягельниках и на гарях по ним (И—16)«
Почти в 10 раз меньше юрка по сравнению с максимальной плот-
ностью гнездования в сосново-озерных ландшафтах и низкорослых
рямах по окраинам комплексных верховых болот (4—8). Минималь-
ное обилие отмечено на частичных вырубках по соснякам (0,4)<
В средней тайге в июне больше всего юрков держится в приречных
смешанных лесах (22), несколько меньше — в сосняках-бруснични-
ках и на вырубках по ним (14 и 18). Значительно меньше их отмече-
но в рямах верховых болот (2—4), а минимальное обилие зарегист-
рировано в грядово-мочажинных комплексах тех же болот и посел-
ках (0,2—0,5). В южной тайге (вторая половина июня) наиболее
часто юрок встречался в мелколиственных лесах и на шелкопрядни-
ках (23 и 25), в меньшем числе — в остальных урочищах лесного
ландшафта и в низкорослых рямах (10—19), а также на переходных
болотах (3). Юрок избегает лишь открытых болот. В подтаежных
лесах он гнездится в меньшем количестве, чем в предыдущих под-
зонах, в основном в осиново-березовых лесах и лишь иногда в ря-
мах (в первой половине июня — 12 и 1). Гнезда находили в окрест-
ностях Томска с шестью яйцами 26 мая и с пятью голыми птенцами
12 июня 1974 г. Слетки встречены нами в средней тайге после 21 ию-
ля 1977 г.
После окончания гнездования численность юрка обычно возрас-
тает в 2—2,5 раза. В средней тайге увеличения не прослеживается,
видимо из-за откочевки, которая продолжается до середины авгус-
та, когда юрков становится вдвое меньше, чем на гнездовании. В ос-
тальных подзонах обилие тоже снижается к середине этого месяца
(откочевывает 75—100% популяции). Во второй половине августа
к югу от северной тайги идет прикочевка, наиболее заметная в сре-
динных подзонах (6—10) и незначительная в подтаежных лесах
(0,6).
В среднем за лето на междуречьях северной тайги юрок предпо-
читает темнохвойные и меньше — сосновые леса (55 и 22). Он много-
числен на гарях по соснякам, в низкорослых рямах и сосново-озер-
ных ландшафтах (10—13) и обычен на вырубках и открытых верхо-
вых болотах (6 и 1). В средней тайге его на 20% меньше в среднем по
району работ, а в предпочитаемых местообитаниях почти в 2 раза
меньше. Здесь юрок многочислен в сосново-боровом ландшафте (20—
29), обычен на верховых болотах и в поселках. В южной тайге юрка
почти столько же, сколько в северной, но в предпочитаемых место-
обитаниях в 1,6 раза меньше. Повсеместно многочислен он в лесном
ландшафте (10—35) и обычен на болотах (1—2), кроме грядово-мо-
чажинных верховых. В подтаежных лесах обилие юрка в 4 раза ни-
109
же, чем в южной тайге. В наибольшем числе он зарегистрирован в
осиново-березовых лесах (11), значительно меньше его в полях-
перелесках, рямах и поселках (1—2).
Следовательно, на междуречьях юрок широко распространен в
лесах различного типа, отдавая предпочтение разреженным и освет-
ленным лесным местообитаниям. Его минимальная численность отме-
чена на подтаежных междуречьях и существенно большая — в пре-
делах более северных подзон. В предпочитаемых местообитаниях
прослеживается тенденция увеличения обилия юрка к северу. В до-
лине Оби он предпочитает сходные местообитания и, кроме того,
пойменные ивняки. В долине Оби юрка примерно столько же, сколь-
ко в таежных подзонах междуречий, и только в северной тайге го-
раздо больше вследствие интенсивной послегнездовой прикочевки.
Чиж (Spinus spinus L.)
В северной тайге на междуречьях нами не зарегистрирован,
хотя отмечался Ю. И. Гордеевым [1963] в среднем течении р. Казым
8 июля 1962 г. В средней тайге стайки чижей начали встречаться во
второй половине июня. В этот период чиж многочислен в сосняках
(14), обычен в приречных смешанных лесах и редок в поселках (3 и
0,5). В первой половине июля не встречался и вновь появился на ко-
чевках во второй половине этого месяца на вырубках по соснякам и
в приречных смешанных лесах (1 и 0,2). В южной тайге на гнездова-
нии (первая половина июня) чиж отмечен в елово-кедровой тайге и
смешанных полузаболоченных лесах. В конце июня одновременно с
вылетом молодых идет интенсивная прикочевка чижей. В результа-
те их становится в среднем по району в 15 раз больше, чем на гнез-
довании. Во второй половине лета большая часть чижей откочевы-
вает, и с середины августа они перестают встречаться совсем. В под-
таежных лесах на междуречьях чиж не обнаружен.
По среднелетним показателям, больше всего чижа зарегистриро-
вано в южной тайге, т. е. там, где имеются наиболее крупные масси-
вы лесов, в том числе темнохвойных. Значительно меньшая числен-
ность отмечена на среднетаежных междуречьях и в долине Оби в
пределах южных подзон. В целом в южных подзонах чижа больше,
чем в северных, но встречается он в основном там, где имеются хвой-
ные леса.
Щегол (Carduelis carduelis L.)
В северных подзонах на междуречьях нами не зарегистрирован.
В южной тайге на гнездовании (15 мая — 15 июня) щегол редок в
березово-осиновых лесах и полях-перелесках (0,2 и 0,4). В подтаеж-
ных лесах в тот же период держится в лугах, используемых под вы-
пас (4). После окончания гнездования щеглы перестают встречаться
на учетных маршрутах в южной тайге с середины июня до середины
Июля, а в подтаежных лесах с 15 июля по 15 августа. В августе они
110
вновь появляются на кочевках, примерно в толМ же количестве, что
и на гнездовании.
В среднем за лето на междуречьях щегол предпочитает лугово-
полевые местообитания. При этом в подтаежных лесах его лишь
немногим больше, чем в южной тайге. Численность щегла в долине
Оби в пределах южных подзон, где он часто встречался в разрежен-
ных осиново-березовых лесах, значительно выше, чем на между-
речьях.
Чечетка (Acanthis flammea L.)
Пролет высокой интенсивности идет в северной тайге в первой
половине июня (14). На гнездовании (15 июня — 15 июля) в той же
подзоне чечетка встречена повсеместно, кроме центральных (грядо-
во-мочажинно-озерных) частей верховых болот. Больше всего ее
держится в приречных темнохвойных лесах (18) и несколько мень-
ше на гарях по соснякам, в сосново-озерных ландшафтах и поселках
(6—8). Еще меньше чечетки в сосняках-ягельниках и на вырубках
по ним (по 2), а минимальное обилие зарегистрировано по перифе-
рии комплексных верховых болот в низкорослых рямах и на пере-
ходных болотах (0,4 и 0,5). В средней тайге численность чечетки зна-
чительно ниже. В первой половине лета она редка в сосново-боровом
ландшафте и в поселках (по 0,4) и очень редка на верховых болотах
(0,03). В северной тайге 10 июня видели пару чечеток, которые бес-
покоились, как около гнезда, но само гнездо не найдено.
В послегнездовое время (с 15 июля по 15 августа) в северной
тайге обилие чечетки возрастает сначала после вылета молодых,
а затем в связи с интенсивной прикочевкой птиц более северных
популяций. Особенно много чечетки во время кочевок держалось в
конце июля — начале августа на гарях по соснякам (145). С середи-
ны августа свыше 95% птиц откочевывало, видимо к югу и в долину
Оби. По крайней мере, с 15 июля по 15 августа в северотаежной до-
лине Оби обилие чечетки резко возросло (в 26—28 раз по сравнению
с плотностью гнездования). Послегнездовая кочевка идет и в средней
тайге в начале августа. В южной тайге чечетка встречена лишь на
весеннем пролете и в конце лета в полете над мелколиственными ле-
сами и открытыми низинными болотами (0,1 и 3). В подтаежных ле-
сах в течение всего лета не зарегистрирована.
В среднвхМ за лето на междуречьях северной тайги чечетка пред-
почитает гари по соснякам (58). Она многочисленна в темнохвойных,
сосновых лесах и на вырубках по ним (10—20), обычна во всех ос-
тальных местообитаниях (1—6). В средней тайге ее в 50 раз меньше.
Чаще всего она встречается в сосняках, приречных смешанных ле-
сах и поселках на их месте (0,1—0,5) и реже в остальных местообита-
ниях (0,2—0,05). Еще ниже численность чечетки в южной тайге.
Итак, чечетка в некоторой степени тяготеет к разреженным лес-
ным местообитаниям. Ее обилие выше всего на междуречьях в се-
верной тайге и резко снижается в более южных подзонах. В долине
Оби в пределах северной тайги ее значительно больше (в результа-
111
те интенсивной послегнездовой прикочевки), а в более южных под-
зонах примерно столько же, сколько на междуречьях. Таким обра-
зом, в среднем по обследованным участкам лесной зоны Западной
Сибири чечетка многочисленна в северной тайге и обычна или редка
в более южных подзонах.
Чечевица {Carpodacas erythrinus Pall.)
Прилет приходится повсеместно на последнюю декаду мая. Так,
в северной тайге чечевица прилетела до 1 июня 1976 г., в средней
появилась 28 мая 1977 г., в южной — 21 мая 1967 г. и в подтаеж-
ных лесах — 20 мая 1977 г. Пролет средней интенсивности идет в
подтаежных лесах с конца мая до середины июня (5). В северной
тайге встречена только на кочевках в поселках в начале июня и ав-
густа (4 и 2).
На гнездовании (июнь) в средней тайге обычна в поселках и
сосняках-брусничниках (9 и 3) и редка на вырубках по соснякам
(0,5). В южной тайге больше всего чечевицы отмечено в елово-кедро-
вой тайге с небольшими участками гарей и в полях-перелесках (вто-
рая половина июня — 12 и 16). Значительно меньше ее в хвойно-
лиственных полузаболоченных лесах и шелкопрядниках (3 и 6).
В подтаежных лесах в тот же период предпочитает луга, используе-
мые под выпас, и осиново-березовые леса (4 и 5). В меньшем числе
встречается на низинных болотах, в полях-перелесках и поселках
(1—2). В послегнездовое время (в июле) показатели обилия стано-
вятся меньшими по сравнению с периодом гнездования — видимо,
часть птиц откочевывает.
В средней тайге чечевица перестает встречаться с середины
июля, а в подтаежных лесах исчезает на месяц позднее. В южной
тайге держится до конца августа, но в это время ее в 4 раза меньше,
чем на гнездовании. Следовательно, к середине августа 90—100%
отгнездившихся птиц откочевывает за пределы обследованных нами
районов.
В среднем за лето на междуречьях северной тайги чечевица —
редкая птица поселков (0,8). В средней тайге ее заметно больше.
Здесь она тоже предпочитает поселки и прилежащие к ним сосняки
(4 и 1). Значительно меньше ее на вырубках по соснякам (0,1).
В южной тайге обилие чечевицы в среднем по ключевому участку
почти в 5 раз больше, чем в предыдущей подзоне. Предпочитает она
поля-перелески (9), в меньшем числе встречаясь в лесных местооби-
таниях (1—3) и на низинных болотах (0,4). В подтаежных лесах че-
чевицы в среднем втрое больше, чем в южной тайге. В наибольшем
числе она держится в разреженных осиново-березовых лесах (6),
несколько меньше ее в остальных местообитаниях (1—2) и особенно
мало в рямах (0,01), где ее видели лишь на пролете.
Таким образом, на таежных междуречьях чечевица предпочи-
тает осиново-березовые леса южных подзон, в том числе в сочетании
с полями. В северных подзонах встречается преимущественно по вы-
рубкам и в поселках. Ее обилие резко снижается с юга на север.
112
Сходное уменьшение прослеживается в долине Оби, но здесь чече-
вицы значительно больше, чем на междуречьях, особенно в высо-
ких поймах и лесополевых ландшафтах надпойменных террас.
ЩУР (Pinicola enucleator L.)
На междуречьях встречался нам только в северной тайге. С на-
чала июня до конца июля обычен в сосняках-ягельниках и на выруб-
ках по ним (9 и 1). В течение двух месяцев численность щура была
постоянной, а в августе он перестал встречаться совсем. В долине
Оби зарегистрирован тоже только в пределах северной тайги, но ви-
дели его реже, чем на междуречьях. В средней тайге нами не заре-
гистрирован, однако отмечался на гнездовании в средней тайге Зау-
ралья М. П. Таруниным [1959] в долине р. Мал. Сосьва, а также
В. В. Раевским [1982] на территории бывшего Кондо-Сосьвинского
заповедника.
Клест-еловик (Loxia curvirostra L.)
Массовое гнездование, по-видимому, проходит до начала наших
наблюдений, по крайней мере, изменение обилия клеста-еловика в
течение лета весьма беспорядочно. Однако повсеместно преобладает
откочевка этих клестов, и их численность в результате снижается в
15—100 раз со второй половины мая к концу августа. На между-
речьях в среднем за лето в северной тайге клест-еловик предпочитает
вырубки по соснякам (16), в меньшем числе встречаясь в остальных
лесных местообитаниях и поселках (2—8). Сходный характер распре-
деления прослеживается и в средней тайге, но здесь еловиков держа-
лось почти вдвое меньше. В южной тайге обилие примерно в 1,5 раза
выше, чем в северной. Чаще всего этот клест встречался в мелко-
лиственных лесах (11) и реже в остальных местообитаниях (1—6).
В подтаежных лесах нами не зарегистрирован, хотя отмечался в пре-
делах этой подзоны в долине Оби в основном в мелколиственных и
сосновых лесах (2 и 0,9).
Таким образом, однозначных изменений численности клеста-
еловика на междуречьях не прослеживается. В среднем по районам
работ и лесным ландшафтам самое высокое его обилие отмечено в
южной тайге, т. е. там, где имеются большие площади лесных масси-
вов. Однако в наиболее предпочитаемых местообитаниях в северных
подзонах численность его выше, чем в южной тайге. В долине Оби в
средней тайге, где имеются предпочитаемые еловиком темнохвойные
леса, его даже больше, чем на междуречьях, а в южных подзонах
значительно меньше.
Белокрылый клест (Loxia leucoptera Gm.)
Для этого клеста тоже характерны резкие изменения числен-
ности в течение лета, хотя происходили они иначе, чем у еловика.
В северной тайге обилие возрастало с начала июня до конца июля с
в Г, Т. Вартапстов
113
0,3 до 9 особей на объединенный 1 км2 района работ, а затем оно
вновь снижалось в августе (до 0,6). В средней тайге наиболее интен-
сивные кочевки происходили с середины июня до конца июля.
Стайки белокрылых клестов появились после 15 июня, и до середины
июля численность была довольно высокой (5), а затем к середине
августа большинство птиц откочевало. В южной тайге и в подтаежных
лесах этот клест нами не зарегистрирован.
В среднем за лето в северной тайге белокрылый клест предпочи-
тает приречные темнохвойные леса (20). В меньшем количестве встре-
чается в сосняках, рямах и поселках (1—5). В средней тайге обычен
в приречных смешанных и сосновых лесах (2—4), очень редок на вы-
рубках и в низкорослых рямах (0,09 и 0,06).
Итак, отдавая предпочтение темнохвойным и смешанным лесам,
белокрылый клест встречается на междуречьях только в пределах
северной и средней тайги. В долине Оби в этих же подзонах обилие
его несколько выше, поскольку здесь большую площадь занимают
предпочитаемые им темнохвойные леса. Весьма характерно соотно-
шение численности белокрылого клеста и еловика. В северной тай-
ге или встречался только белокрылый клест (в долине Оби), или его
было значительно больше, чем еловика (па междуречьях). В средней
тайге численность обоих видов примерно одинакова, а дальше к
югу летом распространен только еловик, а белокрылый клест не за-
регистрирован.
Снегирь (Pyrrhula pyrrhula L.)
Встречен нами на междуречьях только в средней и южной тай-
ге. В последней во второй половине мая идет пролет невысокой ин-
тенсивности (3). На гнездовании (первая половина июня) в средней
тайге обычен в приречных смешанных лесах (4). В южной тайге
(июнь) предпочитает мелколиственные, смешанные полузаболочен-
ные и темнохвойные леса (15—23). В меньшем числе наблюдается в
остальных местообитаниях лесного ландшафта.
В послегнездовое время снегири начинают встречаться в боль-
шем количестве местообитаний — появляются на вырубках по сос-
някам и в поселках (по 4). Однако в первой половине июля в средней
тайге часть птиц, видимо, откочевывает, и в среднем по району ра-
бот численность остается до конца лета примерно такой же, как во
время гнездования. В южной тайге в июле обилие в результате
размножения возрастает вдвое, а затем в августе большая часть птиц
откочевывает.
В среднем за лето в средней тайге снегирь предпочитает приреч-
ные смешанные леса (4). В меньшем числе встречается он на выруб-
ках по соснякам и в поселках (по 0,5). В южной тайге в лесном
ландшафте снегиря в 6 раз больше. Он многочислен в темнохвойных
мелколиственных лесах и на шелкопрядниках (10—29) и обычен в
остальных местообитаниях лесного ландшафта (1—8).
Таким образом, снегирь отмечен нами только в срединных под-
зонах таежных междуречий, т. е. там, где имеются предпочитаемые
114
им леса нормальной полноты, в основном темнохвойные. При этом
в южной тайге обилие его значительно выше, чем в средней. В доли-
не Оби в среднем по районам работ его лишь немногим больше,
чем на среднетаежных междуречьях, а в северной тайге значительно
меньше. Однако в лесных ландшафтах долины Оби, там, где есть
предпочитаемые снегирем темнохвойные леса, его численность столь
же высока, как и на южно-таежных междуречьях.
Дубонос (Coccothraustes coccothraustes L.)
Встречен только в средней и южной тайге. В средней тайге его
видели в основном на предгнездовых кочевках в конце мая — пер-
вой половине июля. В этот период дубонос повсеместно редок в сос-
ново-боровом ландшафте (0,3) и очень редок в рослых рямах (0,01).
В поселках держался до конца июня, возможно гнездился, и в сред-
нем за время своего пребывания обычен (7).
В южной тайге его наблюдали все лето. На гнездовании (первая
половина июня) обычен в мелколиственных лесах и полях-перелес-
ках (2—7). В послегнездовое время (во второй половине июня) оби-
лие дубоноса в результате размножения возрастает в 2,5 раза, после
чего в июле до 90% популяции откочевывает. В августе идет прико-
чевка (8). Особенно много дубоноса начинает встречаться в это вре-
мя в шелкопрядниках и темнохвойных лесах (42 и 6).
В среднем за лето в средней тайге дубонос обычен в поселках
(3), повсеместно редок в сосново-боровом ландшафте (0,3) и очень ре-
док в рослых рямах по окраинам верховых болот (0,01). В южной
тайге его значительно больше. Предпочитает он осиново-березовые
леса и шелкопрядники (7 и 16), в меньшем числе встречаясь в осталь-
ных местообитаниях лесного ландшафта.
В итоге складывается впечатление о предпочтении дубоносом
разреженных лесов, вырубок и шелкопрядников. В долине Оби в
пределах подтаежных лесов дубоноса так же много, как и на южно-
таежных междуречьях. Его обилие в долине снижается к северо-
западу, особенно резко в северной тайге.
СЕМЕЙСТВО ТКАЧИКОВЫЕ (PLOCEIDAE)
Домовый воробей (Passer domesticus L.)
На междуречьях повсеместно встречен нами только в поселках
В северной тайге, судя по показателям обилия, прослеживается два
периода размножения: в первой половине июня и во второй полови-
не июля. Численность в это время держится на довольно сходном
уровне (49—64 особей/км2). Возможно, в конце мая — начале июня
гнездятся оседлые птицы, а в июле — прилетевшие в начале лета
из более крупных и южных поселков в долине Оби. По свидетельст-
ву местных жителей, раньше вся популяция домовых воробьев была
перелетной, и только в последние годы часть птиц стала оставаться
В*
115
на зиму. По-видимому, в северной тайге Обь-Енисейского между-
речья еще идет процесс заселения небольших таежных поселков до-
мовым воробьем, так как дальше к востоку в пос. Нумто в июле
1974 г. этот воробей не встречен. В средней тайге на гнездовании
его почти в 1,5 раза больше, чем в северной (80). В южной тайге в
поселках учеты не проводились. В подтаежных лесах домового во-
робья на гнездовании (вторая половина мая) в 12 раз больше, чем в
средней тайге (955).
’ В послегнездовое время численность домового воробья возрас-
тала в 2,6—3,3 раза, т. е. увеличение, видимо, определялось резуль-
татом размножения местных птиц. В северной тайге прослеживалось
два периода такого увеличения: в конце июня и начале августа. За-
тем большая часть домовых воробьев откочевывает, и в конце августа
их становится меньше, чем на гнездовании. В более южных подзо-
нах прослеживается лишь один период увеличения численности в
связи с вылетом молодых. В средней тайге он приходится на конец
июля, после чего часть домовых воробьев откочевывает, хотя в конце
августа их еще вдвое больше, чем на гнездовании. В подтаежных
лесах максимальная численность отмечена в начале июля, затем
часть птиц откочевывает, и в конце августа их становится примерно
столько же, сколько на гнездовании. Возможно, часть домовых во-
робьев во второй половине лета смещается в крупные южно- и под-
таежные села, в том числе в долинах крупных рек. По крайней мере,
обилие этих воробьев здесь последовательно возрастает в 3—4 раза
по сравнению с предыдущим периодом в южной тайге в первой поло-
вине августа и в конце этого месяца в подтаежных лесах.
Таким образом, на междуречьях в небольших таежных посел-
ках северных подзон численность домового воробья значительно
ниже, чем в подтаежных селах. В более крупных поселках долины
Оби его примерно столько же в пределах подтаежных лесов и зна-
чительно больше по сравнению с междуречьями в более северных
подзонах.
Полевой воробей (Passer montanus L.)
В северотаежных поселках полевые воробьи появились 19 июня
1976 г. Плотность гнездования (конец июня — первая половина
июля) составляла здесь 20 особей/км2. В этих поселках 8 июля виде-
ли полевого воробья, который волновался около гнезда и залетал с
кормом под конек крыши. Во второй половине июля после вылета
молодых обилие возросло в 2,5 раза по сравнению с гнездовым пери-
одом. Затем в начале августа воробьев становится в 5 раз больше,
чем на гнездовании, в результате интенсивной прикочевки (92), кото-
рая во второй половине августа сменяется еще более значительной
откочевкой птиц (120).
На междуречьях средней тайги полевой воробей нами не заре-
гистрирован. В южной тайге отмечен в полях-перелесках в начале
и середине лета (2—23). В подтаежных лесах на гнездовании (вто-
рая половина мая) его численность максимальна, после чего она не-
116
уклонно снижается и в конце июля составляет лишь 1G особей/км2.
В конце августа этих воробьев становится в 2,5 раза больше, чем в
предыдущий период, но меньше, чем на гнездовании. Таким образом,
летом на подтаежных междуречьях преобладает откочевка полевых
воробьев. По-видимому, птицы из более северных подзон смещаются
к югу — в лесостепь и в крупные подтаежные поселки в долине Оби.
По крайней мере, в последних их численность со второй половины
мая к первой половине августа возрастает в 17 раз. В среднем за лето
полевой воробей многочислен в северотаежных поселках (42). В юж-
но-таежных полях-перелесках его намного меньше (5), но в среднем
по участку несколько больше, чем в предыдущей подзоне. Однако в
южно-таежном Прииртышье, где учеты в поселках проводились, по-
левого воробья значительно больше, чем на междуречьях северных
подзон. В подтаежных лесах его численность гораздо выше, чем в
предыдущих подзонах, но несколько ниже, чем в южно-таежном
Прииртышье. Держится он преимущественно в поселках (50), в мень-
шем числе встречается в окрестных полях-перелесках и лугах, ис-
пользуемых под выпас (3 и 0,07).
Итак, численность полевого воробья на таежных междуречьях
несколько возрастает к югу, хотя в северо- и подтаежных поселках их
отмечено почти равное количество. В долине Оби этого воробья зна-
чительно больше там, где имеются сельскохозяйственные земли, т. е.
в южных подзонах, а в северной тайге примерно столько же, сколько
на междуречьях.
СЕМЕЙСТВО СКВОРЦЫ (STURNIDAE)
Скворец (Sturnus vulgaris L.)
На междуречьях северной тайги скворца мы не встречали, одна-
ко в небольшом числе он отмечался в этой подзоне в поселках доли-
ны Оби. В средней тайге на учетах не зарегистрирован, хотя скворцы,
видимо, гнездились в пос. Угут, более крупном, чем тот, в котором
проводились наши наблюдения. В южной тайге, где на междуречьях
поселки не обследованы, отмечен только на весенних кочевках во
второй половине мая в полях-перелесках (9). В подтаежных лесах на
гнездовании (вторая половина мая) предпочитает поселки (330), в не-
большом числе встречается в осиново-березовых лесах и лугово-бо-
лотных местообитаниях (23—27) и в еще меньшем — в полях-пере-
лесках (6). В конце мая 1978 г. птенцы еще находились в гнездах, но
вскоре молодые вылетели, и уже в начале июня большая часть птиц
откочевала за пределы района работ. В результате в первой половине
июня их стало втрое меньше, чем на гнездовании. Откочевка про-
должалась до начала июля, когда численность скворцов упала в 12
раз по сравнению с гнездованием, а с середины июля они не встреча-
лись совсем.
По среднелетним показателям в районах работ, скворец чрезвы-
чайно редок в южной тайге и обычен в подтаежных лесах. В послед-
117
них его больше всего отмечено в поселках и лугах-выпасах вблизи
них (114 и 20). Значительно меньше скворца в осиново-березовых
лесах, полях-перелесках и на низинных болотах (3—5).
В долине Оби, кроме поселков, скворцы в большом числе дер-
жались в открытой луговой пойме и лесо по левых надпойменных
ландшафтах южных подзон, где их обилие гораздо выше, чем на
междуречьях. В долине скворец распространен к северу значительно
дальше, чем на междуречьях, при этом в северных подзонах его чис-
ленность резко снижается по сравнению с южными.
СЕМЕЙСТВО ИВОЛГИ (ORIOLIDAE)
Иволга (Oriolus oriolus L.)
В средней тайге иволгу в первый раз слышали 24 мая 1977 г.
В подтаежных лесах прилет отмечен 27 мая 1978 г., а в южной тай-
ге — 3 июня 1977 г. В Прикетье появилась 4 июня 1963 г. На гнездо-
вании (вторая половина июня) в южной тайге встречена в смешанных
полузаболоченных и мелколиственных лесах (1 и 3). В подтаежных
лесах держится преимущественно в перелесках среди полей (первая
половина июня — 6), в меньшем числе отмечена в небольших колках
на низинных болотах и лугах (2) и изредка наблюдается в осиново-
березовых лесах (0,4).
В послегнездовое время (в июле — первой половине августа)
в южной тайге иволга перестала встречаться совсем и отмечена
вновь только на кочевках в конце августа (3). В подтаежных лесах
численность ее неуклонно снижалась с середины июня, а во второй
половине августа все птицы покинули район работ.
По среднелетним показателям, на междуречьях в пределах юж-
ных подзон, где иволга предпочитает мелколиственные леса, отмече-
на ее примерно одинаковая численность. В долине Оби ее больше,
особенно в пойменных лесах, при этом численность возрастает от
средней тайги к подтаежным лесам.
СЕМЕЙСТВО ВРАНОВЫЕ (CORVIDAE)
Сойка (Garrulus glandarius L.)
В северной тайге на междуречьях встречена лишь однажды —
во второй половине июня на окраине верхового болота (0,03). В сред-
ней тайге нами не зарегистрирована, хотя отмечалась ранее в долине
Оби в пределах той же подзоны в июне 1972 г. в темнохвойных и сме-
шанных лесах (4 и 8). На гнездовании (вторая половина мая — пер-
вая половина июня) наблюдалась только в южной тайге. В этот пе-
риод чаще всего встречалась в темнохвойных лесах (8) и значительно
реже — в мелколиственных лесах, шелкопрядниках (по 1) и в полях-
перелесках (0,1). Во второй половине июня ее не встречали, а в нача-
118
ле июля идет интенсивная прикочевка (25), после которой числен-
ность опять снижается во второй половине лета примерно в 6 раз.
В подзоне подтаежных лесов сойка зарегистрирована только во
второй половине июля в осиново-березовых лесах (4).
По средне летним показателям больше всего сойки отмечено на
южно-таежных междуречьях и несколько меньше — в подтаежной
долине Оби (5 и 1 особь/км2 ключевого участка). На остальной терри-
тории она встречалась нерегулярно и в значительно меньшем числе*
В лесах южных подзон обилие ее выше, чем в северных, но из-за
редкости встреч довольно сложно судить о предпочитаемости сойкой
тех или иных ландшафтных урочищ*
К у к ш a [[Perisoreus infaustus L.)
В северных подзонах окончание гнездования приходится на пер-
вую половину июня. В северной тайге в этот период кукша много-
численна в сосняках-ягельниках (14) и обычна в приречных темно-
хвойных лесах и низкорослых рямах (2—4), в средней — встречена
только в сосняках-брусничниках (3). В южной тайге во второй поло-
вине мая она в наибольшем числе держалась в темнохвойных (23) и
в значительно меньшем — в березово-осиновых и полузаболоченных
смешанных лесах (4 и 7). В подзоне подтаежных лесов в течение всего
лета нами на междуречьях не зарегистрирована, а в долине Оби —
обычная птица сосняков (в среднем за лето — 2).
После окончания гнездования (вторая половина июня) в север-
ной тайге обилие кукши снижается в 8 раз по сравнению с началом
июня, а в более южных подзонах она перестает встречаться совсем.
В июле — августе появляется вновь на послегнездовых кочевках,
но численность в это время обычно ниже, чем в период гнездования*
Только в средней тайге обилие возрастает в результате прикочевки,
наиболее интенсивной в конце августа (8).
В среднем за лето в северной тайге кукша предпочитает сосново-
озерные ландшафты, темнохвойные и сосновые леса и вырубки по
ним (1—4). Реже встречается на гарях по соснякам и на верховых
болотах (0,2—0,5). В средней тайге ее примерно столько же, сколько
в северной. Предпочитает она сосняки-брусничники (9). В меньшем
числе встречается на вырубках по ним и в смешанных приречных
лесах (0,1 и 1). Еще меньше кукши отмечено на верховых болотах
(0,04), где она появляется только на послегнездовых кочевках. В юж-
ной тайге в среднем по району работ ее втрое меньше, чем в предыду-
щих подзонах, хотя в предпочитаемых местообитаниях почти столько
же. Больше всего кукши зарегистрировано в темнохвойной тайге(7)
и значительно меньше — во всех остальных урочищах лесного ланд-
шафта (0,1—1).
Таким образом, кукша повсеместно предпочитает хвойные леса,
особенно сосняки. В северных подзонах таежных междуречий чис-
ленность ее значительно выше, чем в южной тайге. В долине Оби, там,
где в пределах обследованных участков нет сосняков, кукши повсе-
местно меньше, чем на междуречьях.
119
Сорока (Pica pica L.)
В северной тайге на междуречьях нами не встречена, однако ле-
том 1973 г. была многочисленна в долине Оби в поселках и окружа-
ющих их лесах (в среднем за лето — 14—36 особей/км2).
В период гнездования (вторая половина мая) в средней тайге
сорока держалась в поселках (63), хотя гнезд здесь не находили,
п лишь изредка видели ее по берегам крупных таежных рек (0,3)«
В южной тайге отмечена в осиново-березовых лесах и полях-перелес-
ках (0,9 и 2). В подтаежных лесах ее значительно больше. Много-
численна сорока в лугово-полевых местообитаниях и поселках (13—
28), обычна по берегам небольших рек, в рямах и осиново-березовых
лесах (1—7). В этой подзоне 9 июня 1978 г. на ивовом кусте на высоте
2,5 м от земли найдено гнездо с семью голыми птенцами.
В послегнездовое время численность сороки в среднем не воз-
растает, только в поселках в июне — начале июля ее становится в
в 1,2—3,5 раза больше, чем в гнездовой период. Во второй половине
лета обилие сороки обычно снижается по сравнению с периодом
гнездования (там, где она держится постоянно). По-видимому, в это
время на таежных междуречьях сороки откочевывают из местооби-
таний, прилежащих к поселкам, где они гнездились, и часть их сме-
щается к югу, в лесостепь и в долину Оби.
В среднем за лето в средней тайге сорока многочисленна в
поселках (42), обычна в окружающих их приречных смешанных
лесах (1), редка в сосняках-брусничниках и очень редка по берегам
таежных рек (0,1 и 0,04). В южной тайге ее примерно столько же.
Предпочитает она поля-перелески, в меньшем числе встречается в
березово-осиновых лесах (6 и 0,3). В подтаежных лесах численность
сороки в 10 раз выше. Больше всего ее отмечено в поселках (49),
значительно меньше — в лугово-полевых местообитаниях (13—15)
и совсем мало — в болотном ландшафте и по берегам небольших рек
(0,9 и 3).
Итак, максимальное обилие сороки зарегистрировано на между-
речьях в поселках и прилежащих к ним лугово-полевых местообита-
ниях. При этом в подтаежных лесах ее значительно больше, чем в бо-
лее северных подзонах. В долине Оби зарегистрирована не менее
высокая численность, чем на подтаежных междуречьях. Особенно
много сороки в пойме в пределах южных подзон (иногда даже боль-
ше, чем в поселках).
Кедровка (Nucifraga caryocatactes L.)
На гнездовании (первая половина июня) в северной тайге обычна
в приречных темнохвойных лесах, на гарях по соснякам (1—5)
редка в сосняках-ягельниках (0,8). В средней тайге (вторая полови-
на мая — первая половина июня) обычна в небольших таежных по-
селках и рослых рямах (1 и 4) и редка в сосново-боровом ландшафте
(0,3—0,4). В южной тайге (вторая половина мая) в наибольшем числе
встречалась в елово-кедровых лесах (17). Значительно меньше кед-
120
ровки держалось в темнохвойных и мелколиственных лесах (по 3),
а минимальное ее обилие зарегистрировано в полях-перелесках и
шелкопрядниках (0,2—0,4). В подтаежных лесах в первой половине
лета кедровка нами не встречена. В средней тайге (на р. Мал. Сосьва)
постройка гнезд отмечена 8 апреля 1935 г., а 2 июня найдено гнездо
с оперившимися птенцами. В северной тайге слетки кедровки встре-
чены нами 1 и 14 июня 1976 г.
В послегнездовое время во второй половине июня — июле оби-
лие возрастает в 3—5 раз по сравнению с таковым во время окончания
гнездования. В северной тайге в конце лета идет прикочевка кедро-
вок (2), а в более южных подзонах — откочевка примерно такой же
интенсивности. Во время кочевок (в первой половине августа) кед-
ровка появляется в подтаежных осиново-березовых лесах (0,4).
Поскольку ни один год наших наблюдений на междуречьях не
приходился на урожай кедра, направленных массовых перемещений
птиц наблюдать не удавалось. Стайки кедровок (до восьми особей)
начали встречаться в северной тайге после 23 июля.
В среднем за лето в северной тайге кедровка предпочитает при-
речные темнохвойные леса и частичные вырубки по соснякам (4 и 3).
Несколько меньше ее в остальных лесных местообитаниях и посёлках
(1—2) и значительно меньше — на комплексных верховых болотах
(0,2—0,6), где кедровки держались преимущественно на участках
низкорослых рямов. В средней тайге ее примерно столько же, сколь-
ко в северной. Встречалась она в основном в сосновых лесах (2—3).
Год наших наблюдений был неурожайным для кедра, и кедровок
нередко видели с сосновыми шишками. Значительно меньше их оби-
тало в смешанных приречных лесах и на верховых болотах (0,3—
0,5). В южной тайге обилие кедровки в среднем в 2 раза больше, чем в
северных подзонах. Держится она преимущественно в темнохвойных,
смешанных и мелколиственных лесах и шелкопрядниках (4—7),
в значительно меньшем числе наблюдается в рослых рямах и полях-
перелесках (по 0,3). В подтаежных лесах ее значительно меньше, чем
в предыдущих подзонах. Встречена она лишь в осиново-березовых
лесах (0,01).
Таким образом, следует отметить, что численность кедровки
выше там, где имеются темнохвойные леса, хотя большинство лет
наших наблюдений и Ю. С. Равкина в долине Оби характеризова-
лись неурожаем кедра. В тех районах, где нет темнохвойных лесов
или их площадь очень мала (подтаежные междуречья и долина Оби
в пределах южных подзон), обилие кедровки значительно ниже да-
же в урожайные для кедра годы.
Галка (Corvus monedula L.)
В северной тайге на междуречьях нами не зарегистрирована,
однако летом 1973 г. в долине Оби была редкой и чрезвычайно редкой
в поселках и прилежащих к ним лесах (0,2 и 0,005). В средней тайге
галку видели только во второй половине мая в поселках (2). В южной
тайге она изредка встречалась с середины мая до конца июня и в
121
первой половине августа в осиново-березовых лесах (в среднем за
лето — 0,06). В подзоне подтаежных лесов в период гнездования
(вторая половина мая) галка многочисленна в поселках и осиново-
березовых лесах (71 и 11), обычна в лугах, используемых под выпас
(7), где часто встречалась в стаях с другими видами врановых, и очень
редка в полях-перелесках (0,08).
В подтаежных лесах во второй половине июня в среднем по рай-
ону работ обилие галки возрастает вдвое. Особенно много ее становит-
ся в полях-перелесках и поселках (46 и 13). В июле численность рез-
ко снижается (почти в 8 раз), а в августе галка почти совсем перестает
встречаться.
По среднелетним показателям, в целом по районам работ галка
чрезвычайно и очень редка в средней и южной тайге. В подтаежных
лесах ее значительно больше. Предпочитает она поселки (26), в мень-
шем числе встречается в лесополевом ландшафте (2—7).
В долине Оби предпочитает сходные местообитания, особенно
в пределах южных подзон. Следовательно, в лесной зоне Западной
Сибири численность галки на подтаежных междуречьях и в южных
подзонах долины Оби значительно выше, чем в более северных под-
зонах.
Грач (Corvus frugilegus L.)
На междуречьях северной тайги лишь однажды (в первой поло-
вине июня) видели грача в поселке (4). В средней тайге он нами не
зарегистрирован, но в долине Оби в пределах этой подзоны его регу-
лярно встречали до конца июня в пойме Оби, в поселках и на зале-
жах вблизи них (1—14). В южной тайге грача видели только во вто-
рой половине мая в полях-перелесках (0,4).
В подтаежных лесах грача больше, чем в предыдущих подзонах,
хотя наблюдался он тоже преимущественно во второй половине мая.
В этот период он в наибольшем числе держался в лугах, используе-
мых под выпас (11), в смешанных стаях с серыми воронами и галками.
Значительно меньше грачей встречено в то же время в поселках и на
низинных болотах, чередующихся с покосными лугами (1 и 0,09).
В полях-перелесках грачей видели во второй половине мая и июня
(0,3 и 0,07). В пределах обследованных нами районов на гнездовании
не зарегистрирован. Однако гнездовая колония около 30 гнезд най-
дена на границе подтаежных лесов и лесостепи недалеко от пос. Чу-
маково Новосибирской области, примерно в 50 км к юго-западу от
обследованного нами подтаежного ключевого участка.
По среднелетним показателям, грач на междуречьях повсеместно
очень или чрезвычайно редок. В долине Оби его несколько больше
(редок или очень редок). Численность его тоже возрастает к югу.
Серая ворона (Corvus cornix L.)
На междуречьях в период гнездования в северной тайге (пер-
вая половина июня) серая ворона чаще всего встречалась в низко-
рослых рямах и поселках (8 и 9), хотя в последних не гнездилась. Зна-
122
чительно меньше ее держалось в сосново-озерных ландшафтах, по
берегам водоемов и на открытых участках верховых болот (0,8—2)
и еще меньше — в сосново-боровом ландшафте (0,2). В средней тайге
во второй половине мая максимальное обилие отмечено в поселках
(8), значительно меньше ее на верховых болотах и по берегам водо-
емов (0,1—0,2). В то же время в южной тайге предпочитает низинные
болота (1). В меньшем числе встречается на верховых болотах и в
лесном ландшафте. В подтаежных лесах во время гнездования самое
высокое обилие зарегистрировано в поселках (27). Втрое меньше се-
рой вороны в лугово-полевых местообитаниях (7—12) и более чем в
10 раз меньше в осиново-березовых лесах и на низинных болотах (2—
3). Минимальное обилие зарегистрировано в рямах (0,03).
В северной тайге 2 июня 1976 г. видели ворону с кормом в клю-
ве, а 6 июня на сосне найдено гнездо с двумя птенцами. Молодые
птицы в средней тайге встречены 3 июля 1977 г.
В послегнездовое время (во второй половине июня — первой
половине июля) обилие повсеместно возрастает в 2—5 раз. В августе
в таежных подзонах идет прикочевка ворон (2—3), и в то же время
наблюдается образование стай (в северотаежных поселках до 40 осо-
бей). В подтаежных лесах в результате прикочевки средней интенсив-
ности (5) обилие возрастает уже в первой половине июля, а затем оно
еще более интенсивно снижается к середине августа, когда ворон
становится вдвое меньше, чем на гнездовании. По-видимому, во вто-
рой половине лета часть местных ворон смещается с таежных между-
речий к югу — в лесостепь и в крупные поселки долины Оби.
В среднем за лето в северной тайге серая ворона многочисленна
в поселках (32), обычна на верховых болотах и в сосново-озерном
ландшафте (1—6) и редка по берегам водоемов и в сосново-боровом
ландшафте. В средней тайге ее примерно в 2 раза меньше. Предпочи-
тает поселки, верховые болота и берега крупных надпойменных
озер (1—4). В меньшем числе встречается в остальных местообита-
ниях (0,005—0,4). В южной тайге серой вороны почти столько же,
сколько в средней. Обычна она в лесном ландшафте и на низинных
болотах (по 1) и очень редка на верховых (0,004). В подтаежных лесах
ее гораздо больше, чем во всех предыдущих подзонах. Предпочитает
она поселки (46), в несколько меньшем числе встречается в лугово-
полевых местообитаниях (18—29) и в еще меньшем — в осиново-
березовых лесах, лугово-болотном ландшафте и по берегам водоемов
(0,3-4).
Таким образом, на междуречьях максимальная численность
серой вороны отмечена в поселках и подтаежных лугово-полевых
местообитаниях. В крайних подзонах ее больше, чем в срединных,
т. е. численность возрастает с уменьшением залесенности и «таеж-
ности» обследованных участков. В долине Оби, преимущественно в
пойме, обилие серой вороны, как правило, выше, чем на междуречь-
ях, и только в подтаежных лесах — ниже. В целом по лесной зоне
Западной Сибири общий характер распределения серой вороны при-
мерно такой же, как у сороки. Максимальное обилие зарегистриро-
вано в более открытых и освоенных человеком местообитаниях —
123
в долине Оби, особенно в пойме и на подтаежных междуречьях; оно
снижается на междуречьях более северных подзон, слабее освоенных
и сильнее залесенных.
Черная ворона (Corvus corone L.)
Встречена нами только в средней тайге во второй половине
мая в полете над поселками и берегами крупных таежных рек (0,6 и
0,3). В долине Оби Ю. С. Равкиным [1978] отмечена в подтаежных
лесах. Встречается в Прикетье (в южной и средней тайге Обь-Енисей-
ского междуречья) и в верховьях р. Кеть уже более многочисленна,
чем серая ворона.
Ворон (Corvux corax L.)
В таежных подзонах держится в течение всего лета. В северной
тайге численность ворона неуклонно возрастала (видимо, шла при-
кочевка птиц) и в конце августа стала в 20 раз выше, чем в начале
июня. В средней и южной тайге наибольшее обилие зарегистрировано
в июле, после вылета молодых. В этот период ворона было в 10 раз
больше, чем в начале, и в 3—4 раза больше, чем в конце лета. Моло-
дые впервые встречены в северной тайге 19 июня 1976 г., а в средней
22 июня 1977 г. В подтаежных лесах ворона видели только во второй
половине мая в рямах (2).
В среднем за лето в северной тайге ворон встречался по берегам
крупных таежных рек, в том числе в поселках и приречных темно-
хвойных лесах, а также на гарях и в сосново-озерных ландшафтах
(0,1—0,4). Реже его видели в сосняках и на верховых болотах (0,002
и 0,01). На междуречьях средней тайги ворона примерно столько же,
сколько в северной. Держится он в сосново-боровом ландшафте и по
берегам крупных таежных рек (0,1— 0,2). Очень редок в рямах вер-
ховых болот (0,04). В южной тайге ворон тоже распространен повсе-
местно и примерно в равном количестве встречается в лесном ланд-
шафте и на верховых болотах (0,09 и 0,07), в значительно мень-
шем — на низинных болотах (0,001). В подтаежных лесах числен-
ность его заметно ниже, чем в предыдущих подзонах, встречен он
только в рослых рямах, (0,03).
Следовательно, на подтаежных междуречьях ворона гораздо
меньше, чем в более северных подзонах. В долине Оби в южных под-
зонах обилие его ниже, а в северных выше, чем на междуречьях, осо-
бенно северной тайги. Высокая численность ворона, как и других
видов врановых, в обследованном северотаежном участке долины
Оби, по-видимому, весьма локальна, так как их привлекали отходы
находящейся недалеко зверофермы. Для интенсивно трансформиро-
ванных в результате хозяйственной деятельности территорий (под-
таежные междуречья и южные подзоны долины Оби) характерна зна-
чительно более низкая численность ворона, чем для менее освоенных
южно-таежных междуречий и северных подзон лесной зоны Западной
Сибири.
124
ОБЩИЕ ОСОБЕННОСТИ РАСПРЕДЕЛЕНИЯ ВИДОВ
Для выявления основных особенностей территориального рас-
пределения птиц междуречий и долины Оби в пределах лесной зоны
использованы алгоритм и программа факторной классификации, раз-
работанные В. А. Трофимовым [Трофимов, 1976; Трофимов, Равкин,
1980]. Как указывалось ранее [Равкин, Шадрина, 1980; Равкин,
1983], предложенный метод особенно удобен при анализе размещения
птиц по обширной территории, при континуальном распределении
видов и значительном их числе (в нашем случае 212 видов птиц
в первой половине лета и 193 во второй). Для суждения о мере сход-
ства распределений видов вычислялся нецентрированный коэффи-
циент линейной корреляции. Этот показатель значительно более
экономичен для расчетов на ЭВМ по сравнению с применявшимся
ранее Ю. С. Равкиным и В. И. Шадриной [1980] коэффициентом ран-
говой корреляции Кендэла [Кендэл, 1975].
Использованная программа разбивает анализируемую сово-
купность на незаданное число групп видов со сходным характером
распределения. Далее крупные классы делятся на более мелкие с по-
мощью той же программы. Одновременно с этим при интерпретации
выделившиеся мелкие классы объединяются в более крупные в слу-
чае значительного перекрывания списков урочищ, в которых виды,
вошедшие в тот или иной класс, имеют одновременно максимальную
численность. Поскольку выделение таких урочищ вручную весьма
затруднительно, в связи с уникальностью распределения и громозд-
костью списков видов и местообитаний, В. Л. Куперштохом разрабо-
тана специальная программа «Территориальный анализ классов».
Последняя позволяет выделить набор урочищ, предпочитаемых той или
иной группой видов со сходным характером территориального рас-
пределения.
Полученное разделение встреченных видов на группы служит
лишь основой, на которой проводится последующее упорядочение
их количества и состава. Целый ряд видов, встреченных в не свой-
ственных им местообитаниях, перенесен при классификации в соот-
ветствующие им группы. Например, пролетные кулики (круглоно-
сый плавунчик, чернозобик, хрустан), которых видели в полете над
лесными местообитаниями или отдыхающими на берегах небольших
озер среди верховых болот, включены в одну группу с остальными
куликами и водоплавающими птицами. Черныш, в наибольшем числе
встречавшийся в южно-таежных шелкопрядниках, по придорожным
западинам, наполненным водой, но не связанный с самими шелко-
прядниками, тоже перенесен в группу околоводных птиц. Подобные
перестановки осуществлялись во всех случаях, когда вследствие
редкости и случайности встреч отдельные виды птиц зарегистриро-
ваны в явно не свойственных им местообитаниях.
Таким образом, объем выделенных групп и дробность классифи-
кации определяются, с одной стороны, результатами разделения
данной выборки с помощью ЭВМ, а с другой — предметными сообра-
жениями орнитолога-интерпретатора, основанными на знании терри-
125
ториального размещения видов и их экологии. Так, для таежных
междуречий и всей лесной зоны Западной Сибири достаточной оказа-
лась трехступенчатая (тип — класс — подкласс) классификацион-
ная схема, приведенная ниже. Однако следует учитывать, что по-
добные схемы справедливы лишь для конкретных территорий, взя-
тых в указанные периоды при данном масштабе рассмотрения (в ран-
ге ландшафтного урочища). Очевидно, что одни и те же виды в соот-
ветствии с их сезонным перераспределением и отличиями в общем
характере пространственного размещения на разных схемах могут
относиться к разным группам.
В первой половине лета целесообразно выделить 6 типов пре-
ференции (предпочитаемости) птицами местообитаний таежных меж-
дуречий Западной Сибири.
1. Птицы, предпочитающие северотаежные и отчасти средне-
таежные леса (бородатая неясыть, овсянка-крошка, юрок)*:
1.1 —приречные темнохвойные и смешан-
ные леса северных подзон (свиристель, черно-
зобый дрозд, белобровик, таловка, овсянка-ремез, чечетка,
белокрылый клест);
1.2 — северотаежные лишайниковые сос-
няки и вырубки по ним (синехвостка, сероголовая
гаичка, щур).
2. Птицы, предпочитающие южно-таежные лесные ландшафты
(перепелятник, воробьиный сыч, глухарь, козодой, кукушка, трех-
палый дятел, рябинник, пищуха, клест-еловик, ворон):
2.1 —темнохвойные и производные от них
лесные местообитания (глухая кукушка, белоспин-
ный дятел, зеленый конек, кедровка):
2.1.1 — темнохвойную тайгу (длиннохвостая неясыть, желто-
головый королек, московка, кукша),
2.1.2 — мелколиственные, смешанные и темнохвойные леса
(рябчик, большой пестрый дятел, горихвостка-лысушка, пев-
чий дрозд, славка-завирушка, садовая камышевка, теньков-
ка, корольковая и зеленая пеночки, мухоловка-пеструшка,
ополовник, пухляк, поползень, чиж, снегирь),
2.1.3 — мелколиственные леса (тетерев, лесной дупель, чер-
ный и малый пестрый дятлы, соловьи (свистун и синий)**,
певчий сверчок, дубонос, сойка);
2.2 — шелкопрядники (жулан, деряба, толстоклювая
пеночка, обыкновенная овсянка);
2.3 — елово-кедровую тайгу и рослые рямы
(зарничка, мухоловки — малая и мугимаки);
2.4 — южно-таежные пол я-п ерелески (осоед, ка-
нюк, большой подорлик, полевой лунь, пустельга, перепел,корос-
* Для типов и классов распределения птиц указаны виды с неопределен-
ным характером распределения, которые не удается отнести к более мелким
классификационным подразделениям.
♦♦ Эти соловьи встречались в приручьевых смешанных лесах среди мел-
колиственных.
126
тель, большая горлица, мохноногий сыч, удод, соловей-красно-
шейка, таежный сверчок, белошапочная овсянка).
3. Птицы, предпочитающие подтаежные леса и по низинным бо-
лотам, полям и осиново-березовым лесам проникающие в южную
тайгу (кобчик, чеглок, погоныш, болотная сова, обыкновенный и
пятнистый сверчки):
3.1 — осиново-березовые леса южных под-
зон и поля на их месте (лесной конек, садовая и се-
рая славки, серая мухоловка, чечевица):
3.1.1 — разреженные осиново-березовые подтаежные леса (те-
теревятник, пересмешка, ястребиная славка, большая си-
ница, зяблик),
3.1.2 — подтаежные поля-перелески и разреженные осиново-
березовые леса (клинтух, вертишейка, варакушка, весничка,
князек, иволга);
3.2 — южно-таежные осоково-гипновые ни-
зинные болота (бекас, большой кроншнеп, желтая
трясогузка, большой сорокопут, луговой чекан, дубровник);
3.3 — подтаежные тростниково-осоковые
низинные болота в сочетании с покосны-
ми лугами (степной и луговой луни, серый журавль, боль-
шой веретенник, полевой жаворонок, желтоголовая трясогузка,
степной конек, черноголовый чекан, барсучок, камышевая
овсянка).
4. Птицы, предпочитающие йерховые болота (белая куропатка):
4.1 —-северных подзон (кулик-воробей, малый веретенник,
средний кроншнеп, краснозобый конек, на пролете гуси —
серый и гуменник и круглоносый плавунчик);
4.2 — южной тайги (дербник, сапсан, на пролете —
лапландский подорожник).
5. Птицы, предпочитающие поселки и прилежащие к ним луга,
используемые под выпас:
5.1 — поселки северной тайги (белая трясогузка,
каменка, на пролете — рогатый жаворонок);
5.2 — поселки средней тайги и южных под-
зон (черный коршун, сизый голубь, деревенская ласточка,
домовый и полевой воробьи, скворец, сорока, галка, черная
ворона);
5.3 — луга, используемые под выпас (чибис,
щегол, грач, серая ворона, травник на пролете).
6. Птицы, предпочитающие водоемы и их берега (краснозобая и
чернозобая гагары, красношейная поганка, лебедь-кликун, кряква,
чирки свистунок и трескунок, свиязь, широконоска, гоголь, боль-
шой крохаль, скопа, малый зуек, хрустан, черныш, большой улит,
перевозчик, турухтан, белохвостый песочник, сизая чайка, речная
крачка, черный стриж, береговая ласточка, горная трясогуз-
ка):
6.1 — северотаежные во д"о е м ы и окружаю-
щие их комплексные верховые болота
127
(шилохвость, хохлатая чернеть, синьга, луток, средний крохаль,
фифи, мородунка, серебристая чайка, полярная крачка).
Во второй половине лета схема содержит также шесть типов
распределения птиц.
1. Птицы, предпочитающие северотаежные леса и среднетаежные
сосняки:
1.1 —приречную темнохвойную тайгу (бо-
родатая неясыть, трехпалый дятел, свиристель, синехвостка,
чернозобый дрозд, белобровик, таловка, сероголовая гаичка,
овсянка-крошка, юрок, ворон, белокрылый клест);
1.2 — л и ш а й н и к о в ы е сосняки, вырубки и
гари по ним (ястребиная сова, чечетка, щур, клест-
еловик);
1.3 — среднетаежные сосняки-бруснич-
ники (глухарь, кукша).
2. Птицы, предпочитающие южно-таежные лесные ландшафты
(тетеревятник, кукушка, мухоловка-пеструшка, рябинник):
2.1 — темнохвойные и производные от них
лесные и лесополевые местообитания
(мухоловка-мугимаки, чиж):
2.1.1 — темнохвойную тайгу (черный дятел, корольковая
пеночка, желтоголовый королек, московка, поползень, пи-
щуха),
2.1.2 — мелколиственные леса (перепелятник, канюк, глухая
кукушка, соловьи (свистун и красношейка), певчий дрозд,
зарничка, серая мухоловка, ополовник, пухляк, овсянка-
ремез, зяблик, иволга);
2.2 — шелкопрядники (осоед, большой пестрый дятел,
жулан, дубонос);
2.3 — лесополевые местообитания (славка-за-
вирушка, •Теньковка):
2.3.1 — темнохвойные и мелколиственные леса, поля на их
месте и шелкопрядники (рябчик, белоспинный дятел, лесной
и зеленый коньки, горихвостка-лысушка, садовая камышев-
ка, снегирь, сойка, кедровка),
2.3.2 — поля-перелески (пустельга, перепел, чибис, лесной
дупель, большая горлица, козодой, удод, малый пестрый
дятел, деряба, садовая славка, обыкновенная и белошапоч-
ная овсянки, щегол, чечевица).
3. Птицы, предпочитающие лесополевые и лугово-болотные
местообитания подтаежных лесов и по низинным болотам проникаю-
щие в южную тайгу:
3.1 —поля-перелески, луга, используе-
мые под выпас, и разреженные осиново-
березовые леса (клинтух, вертишейка, варакушка,
пересмешка, серая славка, весничка, зеленая пеночка, большая
синица, князек);
3.2 — южно-таежные низинные осоково-
гипновые болота (полевой лунь, кобчик, чеглок, тете-
128
рев, серый журавль, бекас, большой кроншнеп, болотная сова,
луговой чекан, обыкновенный и пятнистый сверчки, дубровник);
3.3 — подтаежные низинные тростниково-
осоковые болота в сочетании с покосны-
ми лугами (большой подорлик, луговой лунь, коростель,
полевой жаворонок, желтоголовая трясогузка, степной конек,
черноголовый чекан, барсучок, камышевая овсянка).
4. Птицы, предпочитающие грядово-мочажинно-озерные вер-
ховые болота (дербник, сапсан, белая куропатка):
4.1 — северной тайги (кречет, фифи, дупель, средний
кроншнеп, краснозобый конек, большой сорокопут).
5. Птицы, предпочитающие поселки (полевой воробей, серая
ворона):
5.1 — северной тайги (желтая и белая трясогузки,
каменка);
5.2 -средней тайги и южных подзон (черный
коршун, сизый голубь, деревенская ласточка, домовый воро-
бей, сорока, галка).
6. Птицы, предпочитающие водоемы и их берега (красношейная
поганка, чирок-трескунок, синьга, большой крохаль, орлан-бело-
хвост, чернозобик, серебристая и озерная чайка, черный стриж):
6.1 — берега таежных рек и их поймы (скопа,
малый зуек, черныш, перевозчик, полярная крачка, береговая
ласточка);
6.2 — крупные надпойменные озера (красно-
зобая и чернозобая гагары, кряква, чирок-свистунок, свиязь,
шилохвость, хохлатая чернеть, гоголь, галстучник, малый
веретенник, травник, большой улит, турухтан, белохвостый
песочник, сизая и малая чайки, речная крачка, горная трясо-
гузка).
Таким образом, приведенные классификационные схемы весьма
сходны для первой и второй половины лета. Постоянным в течение
всего сезона остается состав наиболее крупных классификационных
единиц — типов, тогда как списки классов и особенно подклассов
могут меняться в соответствии с послегнездовым перераспределением
птиц, а для редких видов — из-за случайности их встреч. Число не-
делимых подразделений примерно одинаково в той и другой половине
лета (19 и 17). Во второй половине лета выпадают три группы птиц,
предпочитающих наиболее олиготрофные южно-таежные верховые
болота, подтаежные разреженные осиново-березовые леса, сухие и
монотонные участки которых малопривлекательны для птиц во время
послегнездовых кочевок, а также луга-выпасы, особенно страдающие
во второй половине лета от пастбищной дигрессии. Большая часть
птиц после окончания гнездования либо перемещается с этих место-
обитаний в соседние, либо полностью покидает территорию таежных
междуречий и, видимо, смещается в долину Оби.
Фаунистический состав выделенных типов распределения птиц
сводится к следующему. Первый тип (птицы, предпочитающие север-
ную и частично среднюю тайгу) невелик и однороден. В него входят
9 Л. Г. Вартапето!
129
13 и 18 видов, 100 и 94% которых сибирские *. Второй тип (птицы,,
предпочитающие южно-таежные леса) наиболее крупный (62 и 54 ви-
да) и разнородный. Больше всего в нем представителей сибирского
типа фауны (42 и 30%) при высоком участии европейских видов
(25 и 30%). Сравнительно много транспалеарктов (19 и 25%) и зна-
чительно меньше европейско-китайских и китайских видов, на долю
которых приходится 13 и 16%.
Третий тип (птицы, предпочитающие подтаежные леса) занимает
второе место по разнообразию после предыдущего (37 и 28 мдов).
Для него характерно равное участие европейских видов и транспале-
арктов (42 и 39, 39 и 43%). Остальное приходится на европейско-
китайские, китайские, монгольские, монголо-тибетские и тибетские
виды (по 3—7%).
Следующий, четвертый, тип (птицы, предпочитающие верховые
болота) — наименьший по видовому разнообразию (11 и 8 видов),
но обладающий весьма характерным фаунистическим составом. Здесь
преобладают арктические виды (55 и 33%) за счет пролетных и
гнездящихся тундровых птиц. Сравнительно велико участие транс-
палеарктов (36 и 33%), значительно меньше европейских и сибир-
ских видов (9—16%).
Самая большая • доля транспалеарктов в следующем, пятом,
типе (птицы, предпочитающие поселки) — 67 и 75%, а остальное
приходится на европейские, европейско-китайские, арктические
и средиземноморские виды.
Последний, шестой, тип (птицы, предпочитающие водоемы)
тоже характеризуется преобладанием транспалеарктов (42 и 53%).
Несколько меньше сибирских и арктических видов (30 и 16, 15
и 21 %) и значительно меньше европейских и голарктических (3—6%).
Таким образом, при значительной гетерогенности орнитофауны
Западной Сибири [Johansen, 1943—1961; Формозов, 1964; Гынгазов,
Миловидов, 1977; и др.] представители различных фаунистических
типов на таежных междуречьях Западной 'Сибири имеют явно выра-
женные зоны преференции. Сибирские или таежные виды предпочи-
тают лесные, особенно темнохвойные, местообитания от северной до
южной тайги. Европейские виды занимают в основном разреженные
осиново-березовые леса южной периферии лесной зоны, поля-пере-
лески и в меньшей степени — низинные болота. Транспалеаркты
отдают предпочтение поселкам и водоемам и отчасти открытым боло-
там различных типов. Арктические виды чаще встречаются на вер-
ховых болотах и в меньшей степени — на водоемах. Доля остальных
типов фауны невелика, и распределяются они следующим образом.
Европейско-китайских видов больше всего в темнохвойных и произ-
водных от них южно-таежных лесах. Представители остальных, бо-
лее южных, фаунистических комплексов (китайского, тибетского,
монгольского, монголо-тибетского) превалируют среди видов, пред-
почитающих подзону подтаежных лесов, особенно низинные болота,
* Здесь и далее цифры приводятся соответственно для первой и второй
половины лета.
13°
чередующиеся с участками остепненных разнотравно- и галофитно-
злаковых лугов.
На основе классификационных схем можно выделить природные
режимы (сочетания факторов среды), определяющие размещение
птиц, зарегистрированных нами на таежных междуречьях Западной
Сибири. Наиболее значимы следующие природные режимы: 1) северо-
и среднетаежных темнохвойных и сосновых лесов; 2) темнохвойных
и производных от них лесов южной тайги, включая мелколиственные
леса, шелкопрядники и небольшие поля; 3) подзон подтаежных ле-
сов, южно-таежных низинных болот и северотаежных гетеротроф-
ных болот; 4) верховых грядово-мочажинно-озерных болот; 5) водо-
емов и их берегов; 6) поселков и прилежащих к ним лугов, исполь-
зуемых под выпасы. Менее значимы природные режимы: лесов раз-
личного породного состава и полноты древостоев, низинных и вер-
ховых болот разной подзональной принадлежности, поселков север-
ной тайги и более южных подзон, водоемов и верховых болот север-
ной тайги.
Некоторая условность и перекрывание выделенных зон префе-
ренции связаны со значительной трансгрессией распределения от-
дельных видов. В отличие от схем с заранее заданной (жесткой)
рубрикацией предложенные списки природных режимов, по-види-
мому, лучше отражают особенности эколого-географического рас-
пределения видов на обширных территориях.
Объединение собранных нами материалов и опубликованных
ранее аналогичных данных по долине Оби позволило проанализи-
ровать особенности территориального распределения видов птиц в
пределах всей лесной зоны Западной Сибири в первой половине лета
и привести их в виде следующей схемы.
1. Птицы, предпочитающие северную тайгу (ястребиная сова,
бородатая неясыть, овсянка-крошка):
1.1 — лесные ландшафты (свиристель, юрок):
1.1.1 — темнохвойные леса в долине Оби (сероголовая гаичка,
чечетка, ворон),
1.1.2 — приречные темнохвойные и смешанные леса долин
притоков Оби (чернозобый дрозд, таловка, овсянка-ремез,
белокрылый клест),
1.1.3 — междуречные сосняки-ягельники и вырубки по ним
(синехвостка, щур);
1.2 — б о л отн о-л у гово-сор овую пойму Оби,
надпойменные переходные и низинные бо-
лота (чирок-свистунок, шилохвость, фифи, круглоносый пла-
вунчик, турухтан, гаршнеп, краснозобый конек, камышевая
овсянка).
2. Птицы, предпочитающие междуречные южно-таежные лес-
ные ландшафты и проникающие в среднюю тайгу по надпойменным
темнохвойным и смешанным лесам и пойменным ивнякам (тетере-
вятник, перепелятник, глухарь, вяхирь, воробьиный и мохноногий
сычи, козодой, белоспинный и трехпалый дятлы, зеленый конек,
серая мухоловка, клест-еловик, дубонос):
9*
131
2.1 — леса южной и средней тайги:
2.1.1 — темнохвойные и производные от них (рябчик, длинно-
хвостая неясыть, корольковая и зеленая пеночки, желтоголо-
вый королек, пухляк, московка, поползень, пищуха, чиж,
снегирь, кукша, кедровка),
2.1.2 — осиново-березовые леса южно-таежных междуречий и
ивняки, осинники среднетаежной поймы Оби (глухая кукуш-
ка, большой пестрый дятел, горихвостка-лысушка, пев-
чий дрозд, садовая камышевка, славка-завирушка, теньковка,
ополовник);
2.2 — южно-таежные местообитания:
2.2.1 — мелколиственные леса (тетерев, черный дятел, со-
ловьи — свистун и синий, сойка),
2.2.2 — рослые рямы и смешанные полузаболоченные леса
(мухоловки — малая и мугимаки),
2.2.3 — шелкопрядники (жулан, деряба, толстоклювая пе-
ночка),
2.2.4 — поля-перелески (осоед, канюк, большой подорлик,
полевой лунь, пустельга, перепел, большая горлица, удод,
таежный сверчок, белошапочная овсянка).
3. Птицы, предпочитающие внепойменные лесополевые и пой-
менные лугово-лесные ландшафты южных подзон (весничка):
3.1 — лесополевые внепойменные ландшаф-
т ы (клинтух, вальдшнеп, лесной конек, рябинник, пересмешка,
ястребиная и серая славки, большая синица, обыкновенная
овсянка, зяблик, иволга);
3.2 — пойменные ивняки, осинники, луга и
залесенные низинные болота (кобчик, лесной
дупель, кукушка, вертишейка, малый пестрый дятел, соловей,
соловей-красношейка, варакушка, белобровик, садовая славка,
бурая пеночка, зарничка, мухоловка-пеструшка, князек, урагус,
чечевица).
4. Птицы, предпочитающие луга, открытые низинные болота
и пойменные соры южных подзон (большой кроншнеп, большой со-
рокопут):
4.1 — пойменные луга, соры (выпь, коростель, жел-
тая трясогузка);
4.2 — подтаежные долинные низинные ер-
никово-осоково-гипновые болота (сапсан,
погоныш, серый журавль, бекас, болотная сова, желтоголовая
трясогузка, луговой и черноголовый чеканы, певчий, обыкно-
венный и пятнистый сверчки, вертлявая камышевка, барсучок,
бормотушка, дубровник, на пролете — полярная овсянка);
4.3 — подтаежные междуречные низинные
тростниково-осоковые болота в сочета-
нии с покосными лугами (степной и луговой луни,
полевой жаворонок, степной конек).
5. Птицы, предпочитающие верховые преимущественно гряДо-
во-мочажинно-озерные болота междуречий (дербник, чеглок, белая
132
куропатка, большой улит, на пролете — лапландский подорожник):
5.1 — северо- и среднетаежных междуре-
чий (средний кроншнеп, малый веретенник).
6. Птицы, предпочитающие поселки (черный коршун, сизый го-
лубь, деревенская ласточка, горная и белая трясогузки, каменка,
щегол, коноплянка, домовый и полевой воробьи, скворец, сорока*
галка, грач, черная и серая вороны, близ берегов Оби и ее притоков
береговая ласточка).
7. Птицы, предпочитающие водоемы (гуси — серый и гумен-
ник, серая утка, морская чернеть, белоглазый нырок, малый зуек,
перевозчик, мородунка, белохвостый песочник, сизая чайка, чер-
ный стриж):
7 Л — пойменные:
7.1.1 — северотаежные озера и протоки (кулик-сорока, ще-
голь, на пролете — камнешарка),
7.1.2 — среднетаежные озера и окрестные луга (в основном
на пролете — турпан, тулес, золотистая и бурокрылая ржан-
ки, галстучник, хрустан, чибис, черныш, травник, кулик-
воробей, чернозобик, дупель, большой веретенник, рогатый
жаворонок),
7.1.3 — южно-таежные озера, соры, песчаные отмели Оби (чер-
ношейная и красношейная поганки, лебедь-кликун, свиязь,
чирок-трескунок, широконоска, красноносый и красноголовый
нырки, луток, лысуха, поручейник, озерная и малая чайки,
черная, светлокрылая, малая и речная крачки),
7.1.4 — берега Оби в пределах подтаежных лесов (шилоклюв-
ка, зимородок);
7.2 — внепойменные (преимущественно
крупные озера):
7.2.1 — северных подзон (краснозобая и чернозобая гагары,
скопа, синьга, средний и большой крохали, полярная крачка,
серебристая чайка),
7.2.2 — подтаежных лесов (серошейная поганка, чомга, кряк-
ва, хохлатая чернеть, гоголь).
Во второй половине лета схема также содержит 7 типов распре-
деления птиц.
1. Птицы, предпочитающие северную тайгу (ястребиная сова,
бородатая неясыть, кречет):
1.1 —надпойменные лесные ландшафты и
пойменные ивняки:
1.1.1 — надпойменные полузаболоченные смешанные леса
(щур, овсянка-ремез),
1.1.2 — надпойменные суходольные темнохвойные и смешан-
ные леса и пойменные ивняки (кукушка, зеленый конек, чер-
нозобый дрозд, весничка, таловка, сероголовая гаичка, ов-
сянка-крошка, юрок, чечетка, белокрылый клест, ворон);
1.2 —п о й м у Оби (фифи):
1.2.1 — луга, соры, ивняки (кулик-сорока, турухтан, боль-
шой веретенник, озерная чайка, камышевая овсянка),
133
1.2.2 — открытые низинные болота (гаршнеп, краснозобый
конек).
2. Птицы, предпочитающие темнохвойно-лесные ландшафты
средней и южной тайги (тетеревятник, перепелятник, зарянка, ма-
лая мухоловка);
2,1__средней тайги (трехпалый дятел, глухарь, клест-
еловик, длиннохвостая неясыть, белоспинный дятел, свиристель,
кукша, кедровка);
2.2 — южной тайги:
2.2.1 — темнохвойные и мелколиственные леса (черный дятел,
рябчик, московка, корольковая пеночка, желтоголовый коро-
лек, мухоловка-мугимаки, пухляк, поползень, пищуха, чиж,
снегирь),
2.2.2 — мелколиственные леса (канюк, глухая кукушка, соло-
вьи — свистун и красношейка, певчий дрозд, зарничка),
2.2.3 — сосняки (седой дятел, синехвостка),
2.2.4 — шелкопрядники (большой пестрый дятел, дубонос,
сойка),
2.2.5 — поля-перелески (осоед, пустельга, перепел, деряба,
белошапочная овсянка).
3. Птицы, предпочитающие внепойменные лесополевые и пой-
менные лугово-лесные ландшафты южных подзон (кобчик, полевой
жаворонок):
3.1 — внепойменные осиново-березовые ле-
с а (зеленая пеночка, жулан, пересмешка, ястребиная славка);
3.2 —п ойменные ивняки, луга, осинники и
зал.е сенные низинные болота (чибис, лесной
дупель, клинтух, большая горлица, филин, козодой, вертишей-
ка, малый пестрый дятел, лесной конек, большой сорокопут,
соловей, горихвостка-лысушка, рябинник, белобровик, вара-
кушка, садовая камышевка, славки — садовая, серая и зави-
рушка, теньковка, бурая пеночка, мухоловки — серая и пест-
рушка, ополовник, большая синица, князек, обыкновенная
овсянка, зяблик, урагус, чечевица, иволга).
4. Птицы, предпочитающие открытые низинные болота южных
подзон (дупель, погоныш, болотная сова):
4.1 —южно-таежные ерниково-осоково-
гипновые (полевой лунь, чеглок, тетерев, серый журавль,
бекас, большой кроншнеп, луговой и черноголовый чеканы,
обыкновенный и пятнистый сверчки, дубровник);
4.2 —п одтаежные ерниково-осоково-гип-
новые и тростниково-осоковые в соче-
тании с лугами (луговой лунь, коростель, желтоголо-
вая трясогузка, степной конек, вертлявая камышевка, бормо-
чу шка).
5. Птицы, предпочитающие верховые болота междуречий (дерб-
ншц сапсан, белая куропатка, чернозобик на послегнездовых ко-
чевках, средний кроншнеп).
6. Птицы, предпочитающие поселки и прилежащие к ним мес-
тообитания (рогатый жаворонок на пролете, береговая ласточка близ
134
берегов Оби и ее притоков, горная трясогузка, коноплянка, черная
ворона):
6.1 — пойменные и северотаежные поселки
(каменка, желтая и белая трясогузки, скворец);
6.2 — крупные поселки и прилежащие к ниц
ивняки, луга и смешанные леса (сорока, серая
ворона);
6.3 — поселки ’южных подзон (сизый голубь, дере-
венская ласточка, щегол, домовый и полевой воробьи, галка»
грач).
7. Птицы, предпочитающие водоемы (лебедь-кликун, серый
гусь, большой крохаль, малый зуек, черныш, черный стриж, зимо-
родок):
7.1 — пойменные:
7.1.1 — северотаежные протоки Оби (орлан-белохвост, ще-
голь, серебристая чайка, на пролете — гуменник),
7.1.2 — среднетаежные пойменные водоемы (перевозчик, мо-
родунка),
7.1.3 — южно-таежные соры и берега Оби (черный аист, чирок-
трескунок, широконоска, красноголовый нырок, луток, боль-
шой подорлик, болотный лунь, поручейник, малая чайка,
светлокрылая, черная и малая крачки, колючехвост, певчий
сверчок, барсучок)*;
7.2 — внепойменные:
7.2.1 — преимущественно крупные озера высокой кормности
на северо- и среднетаежных междуречьях (чернозобая и крас-
нозобая гагары, чирок-свистунок, свиязь, шилохвость, синьга,
гоголь, скопа, галстучник, большой улит, травник, белохвос-
тый песочник, малый веретенник, сизая чайка, речная и по-
лярная крачки),
7.2.2 — подтаежные надпойменные озера (красношейная по-
ганка, чомга, кряква, хохлатая чернеть, черный коршун).
Приведенные выше классификационные схемы содержат по 7
типов распределения птиц. Для краткости изложения мы. будем назы-
вать их по'порядку расположения в классификационной схеме:
северотаежный, темнохвойно-лесной южной и средней тайги, лесо-
луговой южных подзон, сорово-низинноболотный южных подзон,
верхово-болотный, водно-околоводный, синантропный. Названия
этих типов соответствуют наиболее значимым природным режимам
(сочетаниям факторов среды), определяющим пространственную
неоднородность внутризонального распределения видов птиц. Менее
значимыми оказались следующие факторы и их сочетания: состав и
полнота древостоев, заболоченность, подзональный режим средней
тайги и южных подзон, влияние вырубок, распашки, пожаров и унич-
тожения лесов сибирским шелкопрядом, пойменный режим, тип водо-
♦ Для южно-таежных пойменных соров характерны интенсивное затопле-
ние в половодье и спад воды в середине лета, что определяет сочетание насе-
ляющих их обитателей водоемов и болотно-кустарниковых птиц.
135
Рис, 2, Типы распределения птиц в лесной
зоне Западной Сибири.
7 — северотаежный; 2 — темнохвойно-лесной
средней и южной тайги; <3 — лесолуговой южных
подзон; 4 — сорово-низинноболотный южных
подзон; 5 — верховоболотный; в — водно-
околоводный; 7 — синантропный.
емов, а также характер увлажнения
низинных болот (постоянный на
ерниково-осоково-гипновых и пере-
менный на тростниково-осоковых
болотах).
Характер территориального раз-
мещения птиц даже в наиболее
крупных классификационных едини-
цах — типах — хотя и определяет-
ся ландшафтной неоднородностью,
но часто не совпадает с ней. Напри-
мер, комплекс таежно-лесных видов
по ивнякам и осинникам выходит в
среднетаежную пойму Оби (рис. 2).
Более теплолюбивые, преимущест-
венно европейские виды, предпочи-
тающие лесолуговые местообитания
южных подзон, с не менее высокой
численностью обитают на пойменных залесенных низинных болотах.
В то же время открытые участки этих пойменных болот заселяют
виды, характерные и для внепойменных открытых низинных болот.
В северной тайге влияние ландшафтной неоднородности, в част-
ности природных режимов темнохвойных и сосновых лесов, верховых
и низинных болот, лугово-соровой поймы Оби, на территориальное
распределение видов птиц менее значимо, чем в более южных подзо-
нах. Как будет детально рассмотрено ниже при анализе населения,
на фоне уменьшения видового разнообразия, степени кормовой
специализации и, по-видимому, снижения напряженности конку-
рентных отношений птицы в северной тайге проявляют тенденцию к
эвритопности, что послуя?ило причиной для выделения единой зоны
преференции северотаежных естественных ландшафтов суши.
Для верховых болот, широко представленных в пределах раз-
ных подзон, специфичные виды характерны в основном для междуреч-
ных северо- и среднетаежных. Именно здесь гнездятся обитатели ле-
сотундры и тундры, например малый веретенник и краснозобый
конек. Кроме того, выделенные области преференции не занимают
всей обследованной территории. Наряду с ними существуют типы
ландшафтов, не предпочитаемых птицами. Это олиготрофные верхо-
вые болота южных подзон, редкостойные лишайниковые сосняки и
производные от них местообитания северной тайги.
В классификационной схеме видов птиц таежных междуречий
выделен комплекс видов, предпочитающих подтаежные леса и про-
136
пикающих по низинным болотам в южную тайгу. В аналогичной схе-
ме для всей лесной зоны он заменен двумя отдельными комплексами::
лесолуговым и сорово-низинноболотным южных подзон, в соответ-
ствии с появлением в выборке пойменных лесов в сочетании с луга-
ми и большей представительностью внепойменных лесополевых
ландшафтов. Таким образом, три комплекса видов со сходным харак-
тером распределения — темнохвойно-лесной, лесолуговой и сорово-
низинноболотный — занимают соответственно три зоны преференции
с различной степенью залесенности. Первая из них соответствует
наиболее залесенным местообитаниям, вторая представлена разре-
женными лесами, залесенными болотами и ландшафтными урочища-
ми, где зелесенные участки чередуются с открытыми, а третья за-
нимает открытые местообитания. Сходное деление по залесенности
предложено при аналогичной классификации для южной тайги
Западной и Средней Сибири [Равкин, 1983].
Птицы, предпочитающие водоемы, являются в значительной сте-
пени сборной группой, которая распадается на две подгруппы:
птицы внепойменных и пойменных водоемов. Первая представлена н
основном водоплавающими и околоводным видами, предпочитаю-
щими озера среди верховых болот северо- и среднетаежных междуре-
чий. Значительная концентрация птиц прослеживается во второй
половине лета на озерах, которые обладают высокой кормностью
и обширной площадью мелководий, что в целом малохарактерно для
олиготрофных водоемов, расположенных среди верховых болот.
Наряду с прикочевкой местных птиц (чирок-свистунок, свиязь,
шилохвость, гоголь) появляются виды, гнездящиеся по северной пе-
риферии лесной зоны и, видимо, в лесотундре (галстучник, турухтан,
белохвостый песочник, синьга). Вторая группа представлена вида-
ми, чаще всего широко распространенными, но достигающими высо-
кой численности на пойменных водоемах и прилежащих лугах.
Кроме того, в пойму проникают представители всех остальных комп-
лексов, за исключением птиц верховых болот.
Таким образом, следует констатировать значительную гетеро-
генность видового состава птиц в пойме Оби и отсутствие единого,
«азонального» комплекса пойменных видов. Одновременно с этим
существует определенная «зональность» в распространении выделен-
ных нами комплексов в пойме Оби, которая сводится к следующему.
Пойменно-водный комплекс видов распространен в пределах всех
подзон. Лесолуговой и сорово-низинноболотный типы занимают
подтаежные леса и южную тайгу, проникая на внепойменные терри-
тории по открытым низинным болотам и лесополевым ландшафтам.
Для среднетаежной поймы характерен преимущественно таежно-
лесной комплекс видов южной и средней тайги, распространенных
по ивнякам, осинникам и тополевникам. Для лугов и водоемов,
особенно во время весеннего пролета, в средней тайге характерны
арктические виды птиц. По-видимому, в остальных подзонах их
пролет проходил до начала наших наблюдений, так как в средней
тайге учеты птиц фенологически начаты наиболее рано. Наконец,
для лугов, соров и ивняков северотаежной поймы характерен северо-
137
‘Таежный комплекс видов, общий с внепойменными территориями.
Таким образом, в пойме Оби в пределах лесной зоны с севера на юг
происходит смена комплексов видов со сходным характером распре-
деления, не совпадающая с изменением подзональных природных
режимов и с аналогичной сменой видовых комплексов на внепоймен-
ных территориях.
В течение летнего сезона состав выделенных типов претерпевает
определенные изменения, которые сводятся к следующему. Во вто-
рой половине лета количество видов, относящихся к темнохвойно-
лесному типу распределения, уменьшается с 54 до 40, а их доля от
общего числа видов птиц снижается с 26 до 21%. Одновременно с
этим участие видов, составляющих лесолуговой комплекс, возрастает
с 14 до 20%. Это вызвано послегнездовой перекочевкой целого ряда
видов птиц, например теньковки, славки-завирушки, горихвостки-
высушки, ополовника, мухоловки-пеструшки, из темнохвойно-
таежных лесов междуречий в лесолуговую пойму Оби в пределах юж-
ных подзон.
Так же как и для отдельно взятых таежных междуречий, для
лесной зоны в целом характерно снижение доли видов, предпочита-
ющих верховые болота (с 7 до 2 %), что определяется их послегнездо-
вой откочевкой. Доля видов остальных комплексов относительно
стабильна в течение лета и составляет для северотаежного комплекса
10 и 13%, сорово-низинноболотного 12 и 10%, водно-околоводного
24 и 25% и синантропного 8 и 10%.
Основные особенности пространственных изменений фаунистиче-
ского состава выделенных комплексов видов со сходным распределе-
нием в лесной зоне Западной Сибири примерно те же, что и на та-
ежных междуречьях, и могут быть представлены в виде приведен-
ной ниже схемы. Отличия сводятся к возрастанию в долине по срав-
нению с междуречьями доли европейских видов за счет преоблада-
ния последних в лесолуговых местообитаниях поймы Оби в преде-
лах южных подзон и к увеличению участия арктических видов, в том
числе пролетных, среди птиц, предпочитающих северную тайгу.
Тип распределения -
Северотаежный
Темнохвойно-лесной южной и
средней тайги
Лесолуговой южных подзон
Сорово-низинноболотный юж-
ных подзон
Верховоболотный
Водно-околоводный
Синантропный
Фаунистический состав
А рктосибирский
Европейско-китайско-сибирский
Сибирско-европейский при высо-
ком участии транспалеарктов
Преобладание транспалеарктов
при высоком участий арктических
и сибирских видов
Явное преобладание транспале-
арктов при участии европейских
и европейско-китайских видов
ПРОСТРАНСТВЕННАЯ НЕОДНОРОДНОСТЬ
НАСЕЛЕНИЯ ПТИЦ
СЕВЕРНАЯ ТАЙГА
’ СОСНОВО-БОРОВОЙ ЛАНДШАФТ
В сосняках- ягельник ах в течение всего лета по
обилию преобладают (табл. 1) белая трясогузка (18 и 11%) и юрок
(17 и 12%)*. Кроме того, во второй половине лета доминируют че-
четка и каменка (19 и 10%). Плотность населения птиц составляет
169 и 136 особей/км2 (табл. 2).
Подавляющее большинство птиц кормится на земле и в кронах
(табл. 3). Биомасса и величина трансформируемой ими энергии в
сосняках равны 7 и 8 кг/км2 и 4 тыс. ккал/сут-км2. В первой поло-
вине лета по биомассе доминируют серебристая чайка и белая тря-
согузка (14 и 10%), а во второй половине — глухарь (40%). Энер-
гетические потребности птиц в основном удовлетворяются за счет
беспозвоночных (80 и 78%), гораздо меньше в рационе птиц доля
семян и сочных плодов (17 и 16%), роль вегетативных частей расте-
ний и позвоночных незначительна (табл. 4).
Видовое разнообразие в сосняках довольно велико в первой
половине лета и резко уменьшается во второй (всего встречено 35
и 22 вида, из них фоновых 25 и 17). По числу особей преобладают
представители сибирского типа фауны (52 и 59%), на втором месте —
транспалеаркты (32 и 35%) и на третьем — представители европей-
ского типа (16 и 5%, табл. 5). Таким образом, если не принимать
во внимание транспалеарктов, фаунистический состав населения ев-
ропейско-сибирский.
В северотаежных соСняках Онежского полуострова во второй
половине лета 1969 и 1970 гг. В. Т. Бутьев и Е. С. Гусаков [1976]
насчитывали в 1,2 раза больше птиц, чем мы в сосняках на р. Ка-
зым. В прионежских Босняках доминировали пухляк* еловик и
весничка.
На частичных вырубках по соснякам до-
минирует белая трясогузка (47 и 20%), кроме того, в первой поло-
вине лета преобладает клест-еловик (10%), а во второй — те же
виды, что и в сосняках, и весничка (17%). Плотность населения птиц
в первой половине лета чуть меньше, чем в сосняках, а во второй
половине — в 1,5 раза больше.
Ярусное распределение птиц близко к таковому в сосняках*
но еще выше доля птиц, держащихся на земле, и соответственно ни-
же участие кронников, что связано здесь с большим количеством от-
крытых участков. Биомасса и количество трансформируемой энер-
♦ Здесь и далее, если не оговаривается особо, цифры приводятся соответ»
ственно для первой и второй половины лета.
139
Таблица 1
Население птиц с основ о-борового ландшафта северной тайги Обь-Енисейского
междуречья летом 1976 г., особей/км2
Вид В среднем по ланд- шафту Сосняки- ягельники Частичные вырубки Гари по соснякам Приречная темнохвойная тайга
1 2 3 4 5 6
п ервая половина лета
Всего 199 169 | 1154 1 145 347
Юрок 34 28 9 18 70
Белая трясогузка 27 31 72 12 13
Чечетка 15 14 2 14 20
Овсянка-крошка 15 15 2 19 13
Каменка 12 15 10 14 —
Клест-еловик 9 9 16 6 12
Теньковка 7 2 1 1 28
Горихвостка-лысушка 7 9 5 8 —
Щур 7 9 2 — —
Черныш 5 5 — 0,2 8
Пухляк 4 — — 3 19
Кукша 4 5 0,5 — 3
Весничка 4 3 — 4 8
Белокрылый клест 4 2 0,2 3 13
Сероголовая гаичка 4 4 5 7 2
Таловка 3 0,2 — 3 16
Свиристель 3 2 0,2 3 7
Фифи 3 2 3 3 8
Чернозобый дрозд 3 — — 1 18
Чирок-трескунок 3 — — — 19
Поползень 2 — 2 3 8
Кукушка 2 1 4 0,7 5
Большой пестрый дятел 2 1 6 7 3
Трехпалый дятел 2 3 — — —
Овсянка-ремез 2 —- — — 12
Славка-завирушка 1 0,3 — — 5
Кедровка 1 0,4 5 2 4
Зеленый конек 1 0,1 3 4 3
Варакушка 1 1 — — 2
Синехвостка 1 2 2 — —
Ласточка береговая 1 1 — 2 1
Белобровик 1 — — 0,5 6
Большой улит 0,8 0,7 1 1 1
Глухая кукушка 0,6 — — 0,1 3
Глухарь 0,6 — — 5 1
Бекас 0,6 0,3 — 0,07 2
Мородунка 0,6 0,7 — — 0,5
Московка 0,5 — — — 3
Зеленая пеночка 0,5 — — — 3
Серая ворона 0,5 0,6 3 0,1 0,007
Белая куропатка 0,5 0,7 — —• —
Хохлатая чернеть 0,5 — — —' 3
Серебристая чайка 0,5 0,7 — —- —•
Рябчик 0,4 — — — 2
Зяблик 0,3 0,3 — — 0,7
140
Продолжение табл. 1
1 2 3 4 5 6
Мухоловка-пеструшка 0,2 1
Перевозчик 0,05 — — — 0,3
Полярная крачка 0,05 -— — — 0,3
Ворон 0,02 0,01 — — 0,1
Певчий дрозд 0,01 — - — 0,07
Скопа 0,002 — — 0,02 —
Большой кроншнеп 0,0004 — 0,02 — —
В т о р а я по ловина лета
Всего 216 492 | 1 136 197 | 272
Чечетка 28 26 32 103 1
Сероголовая гаичка 23 24 6 2 31
Юрок 19 16 2 8 40
Белая трясогузка 19 15 39 32 25
Овсянка-крошка 17 5 3' 32 58
Каменка 13 14 37 22
Теньковка 10 3 0,7 — 44
Желтая трясогузка 9 7 8 1 18
Большой пестрый дятел 7 8 6 5 2
Таловка 7 — — 1 40
Белокрылый клест 7 3 4 2 26
Чирок-трескунок 6 — — — 32
Пухляк 5 — — — 25
Весничка 5 3 33 6 8
Славка-завирушка 4 — — — 22
Поползень 4 0,7 — 1 19
Кедровка 3 3 0.7 0,2 . 3
Зеленый конек 3 — — 16 9
Свиристель 3 о,1 — 11 11
Чернозобый дрозд 3 0,02 0,1 9 12
Гоголь 3 — — — 19
Г орихвостка-лыс ушка 2 1 4 1 4
Кукша 2 2 2 1 4
Клест-еловик 1 — 17 11 —
Черныш 1 — — — 7
Трехпалый дятел 1 . 0,7 0,7 1 2
Краснозобый конек 1 / — — — 6
Щур 1 2 — — —
Белобровик 0,9 — — 4 3
Глухарь 0,8 1 — 1 —
Серая ворона 0,7 1 0,6 0,003 —
Рябчик 0,5 — — — 3
Чирок-свистунок 0,5 — — — 3
Синехвостка 0,5 — — — 3
Овсянка-ремез 0,5 — — — 3
Фифи 0,5 — 0,3 — 3
Перевозчик 0,4 — — — 2
Черный дятел 0,2 0,3 0,2 0,2 —
Зеленая пеночка 0,2 — — — 0,9
Бородатая неясыть 0,2 — — — 1
Черный стриж 0,1 0,02 — — 0,7
Ворон о,1 — 0,02 0,2 0,7
Серый журавль 0,7 0,07 о,1 — 0,1
141
Окончание табл. 1
1 2 3 4 5 6
Скопа 0,05 — — — 0,3
Сова ястребиная 0,05 — — 0,7 —
Глухая кукушка 0,02 — — 0.2 —
Кукушка 0,02 — 0,1 0,2 —
Кобчик 0,02 — — , — 0,1
Большой кроншнеп 0,02 0.03 — — —
Ласточка береговая 0,01 — — — 0,07
Серебристая чайка 0,005 — 0,2 — 0,01
Полярная крачка 0,002 — — — 0,01
гии такие же, как в сосняках-ягельниках. Доминанты по биомассе
те же, что и по обилию, кроме того, в первой половине лета — се-
рая ворона и кедровка (22 и 12%). В состав потребляемых кормов
входят лишь беспозвоночные, семена и сочные плоды, а соотношение
их близко к таковому в сосняках. Видовое разнообразие птиц до
середины лета в 1,5 раза меньше, а в конце лета такое же, как в сос-
няках. Фаунистический состав населения здесь также европейско-
сибирский, но с еще более высоким участием транспалеарктов, ко-
торые во второй половине лета преобладают над представителями
всех типов фауны по числу видов и особей.
Таблица 2
Плотность и разнообразие летнего населения птиц северной тайги Обь-Еннсей-
ского междуречья летом 1976 г.
Ландшафт, урочище Плотность населения Отмечено видов
всего фоновых
1 1 II I 1 II 1 1 II
Половина лета
Сосново-боровой ландшафт 199 212 32 30 25 21
Сосняки-ягельники 169 136 34 25 22 17
Частичные вырубки 154 197 23 24 19 13
Гари по соснякам 145 272 30 28 23 21
Приречная темнохвойная тайга 347 492 42 42 35 33
Сосново-озерный ландшафт 152 317 37 33 21 20
Комплексные верховые болота 248 118 38 28 28 16
Низкорослые рямы Грядово-мочажинно-озерные комп- 262 135 46 27 34 15
лексы 228 115 39 30 26 19
Переходные болота 276 113 30 28 23 13
Поселки 624 1019 37 32 23 21
Пойменные озера, старицы 81 85 30 14 18 10
Крупные таежные реки (р. Казым) 44 52 23 19 6 6
В среднем по ключевому
участку 210 199 35 30 25 19
142
Таблица 3
Ярусное распределение птиц в северной тайге Обь-Енисейского междуречья
летом 197 b г., %
<з> и св и св 6 я
Ландшафт, урочище ев 5 1 X ь «а - На воде В воздухе
К га и Вч
Первая половина лета
Сосново-боровой ландшафт 47 8 39 3 2 0,5
Сосняки-ягельники 52 6 38 2 0,4 0,6
Частичные вырубки 66 4 25 5 — —
Гари по соснякам 48 6 38 7 0,01 1
Приречная темнохвойная тайга 37 12 41 3 6 0,3
Сосново-озерный ландшафт 76 7 12 2 3 0,01
Комплексные верховые болота 76 9 5 0,3 7 3
Низкорослые рямы Грядово-мочажинно-озерные ком- 69 15 9 0,8 4 2
плексы 73 7 3 — 10 6
Переходные болота 84 10 3 0,4* 3 —
Поселки 80 2 4 0,5 0,5 14
Пойменные озера, старицы 44 — 1 — 47 . 7
Крупные таежные реки (р. Казым) 34 — — — 14 52
В среднем по ключевому
участку 65 9 18 1 5 2
Вторая половина лета
Сосново-боровой ландшафт 38 15 37 6 5 0,07
Сосняки-ягельники 39 13 40 7 — 0,01
Частичные вырубки 46 19 31 4 0,1 —
Гари по соснякам 45 3 50 3 — —
Приречная темнохвойная тайга 35 19 30 5 И 0,2
Сосново-озерный ландшафт 73 8 11 6 1 0,06
Комплексные верховые болота 70 15 6 0,5 9 0.2
Низкорослые рямы Грядово-мочажинно-озерные ком- 79 8 10 0,7 0,7 0,5
плексы 60 llf 4 0,1 17 0,1
Переходные болота 80 14 4 1 0,3 0,2
Поселки 91 1 2 0,3 0,8 5
Пойменные озера, старицы 13 — — — 84 3
Крупные таежные реки (р. Казым) 43 — — — 8 48
В среднем по ключевому
участку 58 12 20 . 4 ; 4 0,2
На гарях по соснякам все лето преобладают овсян-
ка-крошка (13 и 12%) и чечетка (10 и 38%). В первой половине лета
в число доминантов входят также юрок и каменка (12 и 10%), а во
второй — белая трясогузка (12%). Таким образом, доминирующий
143
Таблица 4
Биомасса и биоэнергетика населения птиц северной тайги Обь-Енисейского меж-
дуречья летом 1976 г.
Ландшафт, урочище Биомасса, кг/км2 Трансформируемая энергия
Всего, тыс. ккал/сут-км2 В том числе (%) за счет
беспозво- ночных семян, плодов вегетатив- ных час- тей расте- ний позвоноч- ных
Первая половина лета
Сосново-боровой ландшафт 11 6 80 12 7 1
Сосняки-ягельники 7 4 80 17 1 2
Частичные вырубки 7 4 92 8 — —
Гари по соснякам 20 5 73 7 19 —
Приречная темнохвойная тайга 25 11 80 7 13 0,2
Сосново-озерный ландшафт 23 6 72 7 10 И
Комплексные верховые болота 18 8 82 2 5 И
Низкорослые рямы Грядово-мочажинно-озерные комп- 19 9 89 1 5 5
лексы 19 8 76 3 5 16
Переходные болота 15 8 89 0,4 5 6
Поселки 22 15 89 9 — 2
Пойменные озера, старицы 40 9 63 7 10 20
Крупные таежные реки (р. Казым) 6 2 66 9 5 21
В среднем по ключевому
участку 16 7 79 6 7 8
Вторая половина лета
Сосново-боровой ландшафт 14 6 78 11 9 2
Сосняки-ягельники 8 4 78 16 6 0,6
Частичные вырубки 6 4 78 21 — 1
Гари по соснякам 10 6 67 29 3 2
Приречная темнохвойная тайга 42 17 80 4 13 2
Сосново-озерный ландшафт 29 10 87 3 6 4
Комплексные верховые болота 18 5 75 0,6 19 5
Низкорослые рямы Грядово-мочажинно-озерные комп- 7 4 93 2 2 2
лексы 21 6 71 0,2 21 8
Переходные болота 23 5 73 0,8 24 2
Поселки 56 29 84 13 — 3
Пойменные озера, старицы 44 И 66 2 30 2
Крупные таежные реки (р. Казым) 7 2 74 — 3 2?
В среднем по ключевому
участку 19 7 80 5 и 4
144
Таблица 5
Фаунистический состав населения птиц северной тайги Обь-Енисейского меж-
дуречья летом 1976 г., %
Виды Особи
Ландшафт, урочище европейские сибирские арктические транспалеар- ктические европейские сибирские арктические транспале- арктические
Первая половина лета
Сосново-боровой ландшафт 14 43 4 31 10 57 0,3 31
Сосняки-ягел ьники 18 41 3 32 9 55 0,4 35
Частичные вырубки 9 52 — 35 4 30 — 64
Гари по соснякам 10 50 — 37 9 59 — 29
Приречная темнохвойная тайга 14 45 2 31 12 63 о,1 22
Сосново-озерный ландшафт И 41 — 41 9 37 — 50
Комплексные верховые болота И 35 9 38 3 20 2 68
Низкорослые рямы 7 37 9 41 7 34 2 51
Г рядово-мочажинно-озерные комплексы 10 36 5 38 2 19 2 72
Переходные болота 10 30 10 40 3 11 2 74
Поселки 8 24 5 51 3 3 7 87
Пойменные озера, старицы — 33 10 53 — 22 4 72
Крупные таежные^реки (р. Ка- зым) — 30 9 57 — 7 2 90
В среднем по ключевому
участку 10 35 7 38 7 36 1 52
Вторая Сосново-боровой ландшафт : п о 12 • лов 46 И Н £ 4 1 ле 33 т а 10 59 0,5 30
Сосняки-ягельники 16 52 — 32 ' 5 59 — 35
Частичные вырубки 17 42 — 38 19 32 — 48
Гари по соснякам И 61 — 25 3 69 — 23
Приречная темнохвойная тайга 14 43 5 31 16 57 1 24
Сосново-озерный ландшафт 9 45 — 39 9 26 — 65
Комплексные верховые болота 9 34 7 43 5 34 5 47
Низкорослые рямы 7 33 4 44 2 35 1 53
Грядово-мочажинно-озерные комплексы 10 27 10 43 7 32 5 48
Переходные болота И 39 7 36 5 38 9 40
Поселки 16 22 6 47 0,9 1 0,7 96
Пойменные озера, старицы 7 29 7 57 2 43 0,4 54
Крупные таежные реки (р. Ка- зым) 16 16 5 58 2 3 0,6 91
В среднем по ключевому
участку 11 39 4 39 8 41 1 47
14Г>
Ю л. Г. Вартапетов
состав сходен с таковым в предыдущих местообитаниях со сно во -бо-
рового ландшафта. Исключение составляла овсянка-крошка, ко-
торая встречалась здесь в западинах с низкорослыми рямами. В ос-
тальных лесных местообитаниях такие западины отсутствуют.
Плотность населения птиц до середины июля почти такая же,
как и на вырубках, а во второй половине лета она возрастает в 1,4
раза. Распределение птиц по ярусам аналогично таковому в сосня-
ках-ягельниках, но во второй половине лета несколько больше до-
ля кронников.
Биомасса птиц и количество трансформируемой ими энергии
соответственно в 2 и 1,5 раза превышают эти показатели в сосняках.
По биомассе все лето доминирует глухарь (79 и 30%) и во второй по-
ловине лета — чечетка (13%). Долевое участие основных групп по-
требляемых птицами кормов почти такое же, как и в предыдущих
урочищах этого ландшафта, но в первой половине лета в рационе не-
сколько выше содержание вегетативных частей растений, а во вто-
рой — семян и сочных плодов. Видовое разнообразие на гарях чуть
меньше, чем в сосняках, но больше, чем на частичных вырубках.
Фаунистический состав населения европейско-сибирский.
В приречной темнохвойной тайге по оби-
лию преобладают в первой половине лета юрок, а во второй — се-
роголовая гаичка и овсянка-крошка (20, 18 и 12%). Плотность на-
селения птиц почти втрое больше, чем в сосняках. Распределение
по ярусам сходно с таковым в остальных лесных урочищах, но до-
ля птиц, держащихся на воде, выше за счет особей тех видов водо-
плавающих, которые встречены по западинам и небольшим стари-
цам р. Казым среди темнохвойной тайги.
Биомасса и количество трансформируемой энергии также боль-
ше, чем в ранее рассмотренных местообитаниях, и соответственно
в 4 и 3,5 раза выше, чем в сосняках. Все лето по биомассе домини-
руют чирок-трескунок (по 31%), глухарь (в первой половине лета —
13%) и гоголь (во второй половине — 33%). По сравнению с други-
ми лесными местообитаниями в рационе птиц темнохвойной тайги
несколько выше участие вегетативных частей растений.
Видовое разнообразие, как и предыдущие показатели, здесь
выше, чем в ранее рассмотренных ландшафтных урочищах. Фау-
нистический состав населения европейско-сибирский. Доля транс-
палеарктов здесь также сравнительно велика, но ниже, чем в дру-
гих местообитаниях.
В. Т. Бутьев и Е. С. Гусаков [1976] с конца июля до начала
сентября 1969 и 1970 гг. в долинных ельниках Онежского полуост-
рова насчитывали 167 особей/км2. В качестве доминантов отмечены
пухляк, юрок и чечетка. Нами в темнохвойных лесах на р. Казым
в августе 1976 г. зарегистрирована в 1,7 раза большая плотность
населения птиц, а доминировали пухляк, овсянка-крошка, талов-
ка и теньковка.
В среднем по ландшафту доминируют юрок и
белая трясогузка (в первой половине лета 17 и 14%), а во второй
половине их сменяют чечетка и сероголовая гаичка (13 и 11%).
146
Плотность населения птиц в течение всего лета составляет около
200 особей/км2. Большинство птиц в первой половине лета держится
на земле, чуть меньше их в кронах и значительно меньше — в кус-
тах. Во второй половине лета доля наземных птиц ниже, а кустар-
никовых и водных — выше. Биомасса птиц в сосново-боровом ланд-
шафте равна 11 и 14 кг/км2, а количество трансформируемой ими
энергии около 6 тыс. ккал/сут-км2. По биомассе преобладают глу-
харь (16 и 18%), чирок-трескунок (12 и 17%), и, кроме того, во вто-
рой половине лета — гоголь (18%). Энергетические потребности
птиц удовлетворяются в основном за счет беспозвоночных, но выше
доля вегетативных и генеративных частей растений. Количество
встреченных видов птиц (в пересчете в среднем на одно урочище)
составляет 32 и 30, а фоновых — 25 и 21. Фаунистический состав
населения европейско-сибирский при значительном участии транс-
палеарктов.
СОСНОВО-ОЗЕРНЫЙ ЛАНДШАФТ
Доминируют по обилию овсянка-крошка (22 и 10%), белая
трясогузка (19 и 31%) и каменка (13 и 17%, табл. 6). Плотность на-
селения птиц в первой половине лета такая же, как и на вырубках,
а во второй — в 1,7 раза выше. Подавляющее большинство птиц
держится на земле и значительно меньше — в кронах. Такой ха-
рактер ярусного распределения птиц в некоторой степени соответст-
вует комплексному характеру сосново-озерного ландшафта, состоя-
щего из лесных (сосновые гривы) и болотных (низкорослые рямы
в понижениях) участков, чередующихся с озерами. Биомасса птиц
составляет 23 и 29 кг/га, а количество трансформируемой ими энер-
гии — 6 и 10 ккал/сут-га, что выше сходных показателей для сос-
ново-борового ландшафта, но ниже таковых в темнохвойной тайге.
Все лето по биомассе доминируют глухарь (27 и 31%) и серая воро-
на (11 и 12%), и лишь в первой половине — серый журавль и сереб-
ристая чайка (24 и 12%). По сравнению с населением лесных место-
обитаний, в сосново-озерном ландшафте среди потребляемых пти-
цами кормов ниже доля семян и сочных плодов, но довольно велико
участие позвоночных. Видовое разнообразие здесь несколько выше,
чем в сосново-боровом ландшафте, но ниже, чем в темнохвойной тай-
ге. Всего встречено 37 и 33 вида птиц, из них 21 и 20 фоновых. По
долевому участию в населении на первом месте стоят транспалеарк-
ты (по числу особей 50 и 65%), на втором — сибирские виды (37 и
26%) и значительно меньше доля европейских видов (9% по числу
особей).
КОМПЛЕКСНЫЕ ВЕРХОВЫЕ БОЛОТА
В низкорослых рямах в течение всего лета по чис-
ленности доминируют желтая трясогузка (24 и 38%) и овсянка-
крошка (19 и 22%). Плотность населения птиц в первой половине
10*
147
Таблица 6
Летнее население птиц сосново-озерного ландшафта северной тайги Обь-Ени-
сейского междуречья в 1976 г., особей/км2
Вид Плотность Вид Плотность
Пер в а я по ловина лета
Всего ... 152 II 1 1
Овсянка-крошка 33 Серый журавль 1
Белая трясогузка 29 Белокрылый клест 1
Каменка 20 Кряква 0,7
Юрок 10 Кукушка 0,7
Весничка 8 Бекас 0,7
Желтая трясогузка 7 Таловка 0,4
Горихвостка-лысушка 5 Поползень 0,3
Серая ворона 5 Большой улит 0,3
Чечетка 5 Вертишейка 0,3
Фифи 5 Свиристель 0,3
Зеленый конек 4 Дубровник 0,3
Сероголовая гаичка 3 Мородунка 0,3
Большой пестрый дятел 2 Трехпалый дятел 0,3
Глухарь 2 Кукша 0,2
Кобчик 2 Певчий дрозд 0,1
Серебристая чайка 2 Синьга 0,1
Теньковка 1 Средний кроншнеп 0,07
Сизая чайка 1 Чеглок 0,02
Шилохвость 1
В т о рая по ловина лета
Всего ... 317 | 1
Белая трясогузка 97 Гоголь 2
Каменка 55 Белокрылый клест 2
Овсянка-крошка 33 Поползень 1
Желтая трясогузка 22 Кукушка 0,7
Весничка 18 Зеленый конек 0,7
Сероголовая гаичка 16 Кобчик 0,7
Большой пестрый дятел 15 Серебристая чайка 0,7
Юрок 9 Фифи 0,6
Горихвостка-лысушка 9 Серый журавль 0,5
Серая ворона 7 Ворон 0,3
Чечетка 7 Шилохвость 0,2
Большой кроншнеп 6 Чернозобая гагара 0,2
Трехпалый дятел 3 Сапсан 0,2
Таловка 3 Синьга 0,2
Глухарь 3 Чирок-свистунок 0,1
Кедровка 2 Большой улит 0,07
Кукша 2
лета в 1,6 раза выше, а во второй такая же, как в сосняках. Боль-
шая часть птиц держится на земле, кустарники и кроны привлекают
их значительно меньше. Биомасса и величина трансформируемой
птицами энергии до середины июля более чем вдвое выше, а во вто-
рой половине лета такие же, как в сосняках. В течение всего лета
148
Таблица 7
Население птиц комплексных верховых болот северной тайги Обь-Енисейского
междуречья летом 1976 г., особей/км?
Вид В среднем по ландшафту Низкорослые рямы Грядово-моча- жинно-озер- ные комплек- сы Переходные болота
1 2 3 4 5
Пер вая половина лета
Всего ... 246 | 262 228 | 276
Желтая трясогузка 105 62 93 161
Овсянка-крошка 26 49 24 14
Фифи 21 20 22 21
Береговая ласточка 8 5 14 —
Весничка 7 17 3 8
Зеленый конек 7 6 5 10
Дубровник 7 6 4 14
Белая трясогузка 6 10 6 3
Большой улит 6 8 5 5
Черноголовый чекан 6 7 7 5
Обыкновенная крачка 5 3 8 0,4
Юрок 4 14 2 0,7
Чечетка 4 4 5 2
Сизая чайка 3 0,07 5 2
Краснозобый конек 3 2 3 5
Черныш 2 — 4
Кукушка 2 6 1 1
Шилохвость 2 3 2 3
Свиристель 2 2 1 3
Каменка 2 3 — 6
Большой кроншнеп 2 3 2 1
Серебристая чайка 2 2 2 2
Полярная крачка 2 1 2 1
Серая ворона 1 4 0,5 1
Варакушка 1 5 0,3
Бекас 1 0,7 .—. 3
Мородунка 0,9 2 1
Большой сорокопут 0,6 3 —
Луговой чекан 0,6 1 0,3
Чирок-трескунок 0,6 0,2 1
Сероголовая гаичка 0,6 3 —
Свиязь 0,6 0,03 0,7 0,7
Теньковка 0,5 2 0,1
Чирок-свистунок 0,5 — 1 —
Кукша 0,4 1 0,3 —
Чернозобая гагара 0,4 о,1 0,7 —
Синьга 0,4 0,2 0,7 —
Черный дятел 0,3 — — 1
Таловка 0,3 0,3 — 1
Кобчик 0,3 0,3 0,3 0,3
Большой пестрый дятел 0,2 1 —
Трехпалый дятел 0,2 1 —
Вертишейка 0,2 1 —
Кулик-воробей 0,2 — — 0,7
Белая куропатка 0,2 1 —
149
Продолжение табл. 7
1 2 3 4 5
Гоголь 0,2 1 0,07
Средний кроншнеп 0,2 1 — —
Кедровка 0,1 0,3 0,07 —
Белохвостый песочник 0,1 0,7 — —
Длинноносый крохаль 0,1 — 0,2 —
Славка-завирушка 0.08 — — 0,3
Горихвостка-лысушка 0,06 0,3 — —
Гусь серый 0.02 0,1 — —
Сойка 0,01 — 0,02 —
Ворон 0,01 — 0,02
Вторая поло вина лет а
Всего . . . 122 135 115 115
Желтая трясогузка 36 52 33 29
Овсянка-крошка 21 30 13 29
Шилохвость 6 — 12
Сероголовая гаичка 6 4 7 6
Большой сорокопут 5 4 7 1
Дубровник 5 0,7 6 7
Краснозобый конек 5 2 4 10
Юрок 3 7 2 1
Серая ворона 3 4 3 1
Весничка 3 3 3 3
Зеленый конек 3 11 0,7 2
Бекас 3 — 11
Дупель 3 — 5 0,7
Ч ирок-свистунок 2 — 3 —
Глухарь 2 — — 6
Фифи , 2 0,07 3 0,7
Белая трясогузка 1 0,7 2 0,3
Каменка 1 2 2
Серебристая чайка 1 0,2 2 —
Кукушка 0,8 4 — —
Большой улит 0,7 0,7 1 0,07
Чечетка 0,6 1 0,7 0,1
Луговой чекан 0,6 —. — 2
Белокрылый клест 0,6 2 0,1 0,7
Полярная крачка 0,6 — 1 0,3
Кедровка 0,5 1 0,3 0,5
Сизая чайка 0,5 — 1 .—.
Большой пестрый дятел 0,4 0,3 0,2 0.9
Свиристель 0.4 2 — —
Черноголовый чекан 0,4 — 0,7 —
Средний кроншнеп 0,4 — 0,7 —
Краснозобая гагара 0,4 — 0,7 —
Чеглок 0,2 0,7 0,2 —
Трехпалый дятел 0,1 0,7 —
Кряква 0,1 0,7
Варакушка 0,1 0,7 — —
150
Окончание табл. 7
1 2 3 1 1 4 5
Большой кроншнеп од — 0,2 —
'Таловка 0.08 — — 0,3
Кукша 0,06 0,2 — 0,07
Поползень 0,06 — — 0,2
Сапсан 0,06 — — 0,2
Бородатая неясыть 0,06 — — 0,2
Серый журавль 0.02 — — 0,07
Ворон 0,007 0,01 0,01 —
по биомассе доминирует серая ворона (10 и 31%). В первой полови-
не лета в число преобладающих видов входят также серебристая
чайка и шилохвость (14 и 13%), а во второй — кряква и желтая тря-
согузка (14 и 13%). Среди потребляемых птицами кормов больше
всего беспозвоночных, значительно меньшая часть приходится на
вегетативные части растений и позвоночных.
В низкорослых рямах отмечено в первой половине лета в 1,5 ра-
за больше видов птиц, чем в лесных местообитаниях (в основном за
счет пролетных), а во второй — чуть меньше, чем в лесах. Преобла-
дают транспалеаркты, а без их учета большая часть особей прихо-
дится на представителей сибирского типа фауны (34 и 35%), значи-
тельно ниже доля европейских видов (7 и 2%), т. е. состав населения
европейско-сибирский, но со значительно более высоким участием
транспалеарктов, чем в лесных сообществах птиц.
В низкорослых рямах на Елогуе (примерно в 100 км от Енисея,
на границе северной и средней тайги) в первой половине июля 1958 г.
Э. В. Рогачева [1962] насчитывала 423 особи/км2 девяти видов, сре-
ди которых преобладали пухляк и овсянка-крошка. Пересчет про-
веден нами по средней дальности обнаружения [Вартапетов и др.,
1980]. В низкорослых рямах на р. Казым птиц в 1,6 раза меньше.
В этот период здесь отмечено 23 вида птиц, преобладали овсянка-
крошка и желтая трясогузка.
В грядово-мочажинно -озерных комплек-
сах два первых доминанта по обилию те же, что и в низкорослых
рямах (табл. 7). Кроме того, в число преобладающих видов в первой
половине лета входит фифи (11%), а во второй половине —шило-
хвость (10%). Плотность населения птиц здесь в 1,2 раза ниже, чем
в предыдущем местообитании. Ярусное распределение в целом сход-
но с таковым в низкорослых рямах, но несколько большее число
птиц держится на воде и меньшее — в кустарниках и кронах де-
ревьев.
Биомасса птиц в первой половине лета такая же, а во второй —
втрое выше, чем в низкорослых рямах. Все лето доминирует сереб-
ристая чайка (по 14%), в первой половине — сизая чайка (12%),
а во второй — шилохвость (49%). Количество трансформируемой
энергии сходно с таковым в предыдущем местообитании. Соотноше-
ние основных групп потребляемых кормов почти такое же, как в
рямах (выше участие позвоночных и зелени). В грядово-мочажинно-
151
озерных комплексах встречено 39 и 30 видов птиц, из них фоно-
вых — 26 и 19. По числу особей преобладают транспалеаркты. До-
ля их здесь даже несколько выше, чем в предыдущем местообита-
нии, тогда как представителей сибирского типа фауны ниже.
На переходных болотах, территориально смеж-
ных с верховыми, в течение всего лета по обилию доминирует желтая
трясогузка (58 и 26%), и только во второй половине — овсянка-
крошка (26%). Плотность населения птиц в первой половине лета
несколько выше, а во второй такая же, как и в грядово-мочажинно-
озерной части верховых болот. Распределение по ярусам близко
к таковому в других урочищах верховых болот, но доля птиц, дер-
жавшихся на земле, еще выше, что соответствует наибольшей сте-
пени открытости этого местообитания.
Биомасса птиц в первой половине лета немного ниже, а во вто-
рой — чуть выше, чем в грядово-мочажинно-озерных комплексах
(15 и 23 кг/км2). До середины июля по биомассе, как и в предыду-
щих местообитаниях, преобладают серебристая чайка и шилохвость
(19 и 17%), а также желтая трясогузка (18%), а во второй полови-,
не лета — глухарь (81%). Показатели трансформируемой энергии
те же, что и в грядово-мочажинно-озерных комплексах.
Соотношение основных групп потребляемых кормов примерно
такое же, как и в остальных местообитаниях верховых болот. Ви-
довое разнообразие птиц на переходных болотах несколько мень-
ше, чем в вышеуказанных урочищах, особенно в первой половине
лета. Здесь также преобладают транспалеаркты. Если их не прини-
мать во внимание, то состав населения можно считать европейско-
сибирским.
В среднем по ландшафту в течение всего лета в
число доминантав по численности входят желтая трясогузка (43 и
30%) и овсянка-крошка (10 и 18%). Плотность населения птиц здесь
в первой половине лета в 1,2 раза выше, а во второй — в 1,8 раза
ниже, чем в сосново-боровом ландшафте. Ярусное распределение
птиц в разных местообитаниях сходно. Подавляющее большинство
птиц держится на земле, значительно меньшая часть населения
приходится на кустарниковых птиц, водоплавающих и кронников.
Биомасса птиц на комплексных верховых болотах в среднем
в 1,4 раза выше, чем в лесных местообитаниях. В течение всего лета
доминирует шилохвость (И и 29%), кроме того, в первой полови-
не — серебристая чайка, а во второй — глухарь (15 и 28%). Вели-
чина трансформируемой энергии тоже немного больше, чем в сосно-
во-боровом ландшафте. Среди потребляемых птицами кормов пре-
обладают беспозвоночные, значительно ниже доля вегетативных
частей растений и позвоночных и минимальная — семян и сочных
плодов. Видовое разнообразие, как и плотность населения птиц,
в первой половине лета немного больше, а во второй — чуть мень-
ше, чем в сосново-боровом ландшафте. По числу видов и особей пре-
обладают транспалеаркты. Без их учета фаунистический состав на-
селения можно считать европейско-сибирским.
152
ПОСЕЛКИ
Доминируют широко распространенные в северной тайге виды:
белая трясогузка (30 и 39%), каменка (в первой половине лета —
21%) и желтая трясогузка (во второй половине лета — 19%), а так-
Таблица 8
Летнее население птиц поселков северной тайги Обь-Енисейского междуречья
в 1976 г., особей/км2
Вид Плотность Вид Плотность
Первая половина лета
Всего . . . 624 1
Белая трясогузка 190 Чечевица 1
Каменка 129 Грач 1
Ласточка береговая 88 Серебристая чайка 1
Домовый воробей 85 Сер о головая гаичка 1
Рогатый жаворонок 43 Кедровка 0,7
Желтая трясогузка 14 Фифи 0,7
Полевой воробей 13 Канюк 0,3
Горихвостка-лысушка 10 Варакушка 0,3
Малый зуек 9 Большой улит 0,3
Чечетка 8 Вертишейка 0,3
Серая ворона 7 Черный коршун 0,3
Весничка 6 Белобровик 0,3
Клест-еловик 5 Зеленый конек 0,2
Большой пестрый дятел 2 Кобчик 0,1
Сизая чайка 2 Черный стриж 0,07
Чернозобый дрозд 2 Обыкновенная крачка 0,04
Поползень 1 Полярная крачка 0,04
Кукушка 1 Черныш 0,03
Свиристель 1
Вторая половина лета
Всего ... 1019 J
Белая трясогузка 402 Серебристая чайка 2
Желтая трясогузка 196 Кедровка 2
Каменка 99 Чечетка 2
Домовый воробей 78 Чернозобый дрозд 1
Полевой воробей 70 Сероголовая гаичка 1
Серая ворона 56 Пухляк 6,7
Ласточка береговая 49 Канюк 0,7
Зеленый конек 15 Чечевица 0,7
Белокрылый клест 10 Луговой чекан 0,7
Краснозобый конек 7 Ворон 0,4
Сизая чайка 6 Малый зуек 0,3
Овсянка-крошка 5 Свиристель 0,1
Горихвостка-лысушка 4 Кобчик 0,07
Весничка 4 Серый журавль 0,01
Большой пестрый дятел 3 Кречет 0,007
Варакушка 3 Лунь полевой 0,002
153
же синантропный вид — домовый воробей и, кроме того, береговая
ласточка, связанная с обрывами р. Казым (в первой половине лета
по 14%, см. табл. 8). В поселках отмечено максимальное в северной
тайге суммарное обилие птиц (624 и 1019). Распределение по ярусам
здесь весьма специфично: максимальное число птиц держалось на
земле, а следующей по обилию была группа воздухореев.
Показатели биомассы и биоэнергетики в поселках соответствен-
но в 2—3 раза выше, чем в естественных ландшафтах. По биомассе
преобладают белая трясогузка (19 и 16%) и серая ворона (16 и 50%)
и только в первой половине лета — каменка и домовый воробей
(15 и 12 %). Соотношение основных групп кормов, за счет которых
удовлетворяются энергетические потребности птиц, и видовое раз-
нообразие примерно такие же, как и в окружающих поселки лесных
местообитаниях. По количеству видов и особенно особей в поселках
абсолютно преобладают транспалеаркты (87 и 96% особей).
ВОДОЕМЫ
На озерах, расположенных в пойме р. Ка-
зым, в первой половине лета по обилию доминируют хохлатая
чернеть, белая трясогузка и фифи (20, 16 и 15%, табл. 9), а во вто-
рой половине — чирки: свистунок и трескунок (26 и 12%), го-
голь и шилохвость (24 и 18%). Плотность населения птиц составля-
ет 81 и 85 особей на 10 км береговой линии. Более половины птиц
кормится в воде, меньшее количество — на земле, около уреза воды,
и еще меньше — в воздухе.
Биомасса птиц составляет 40 и 44 кг на 10 км береговой линии.
В первой половине лета по биомассе преобладают хохлатая чернеть
(29%), шилохвость, серый журавль и синьга, а во второй — те же
виды, что и по обилию. Населением птиц на озерах трансформиру-
ется 9 и 11 тыс. ккал/сут на 10 км берега. Энергетические потреб-
ности удовлетворяются главным образом за счет беспозвоночных
(63 и 66%) при сравнительно большом участии вегетативных частей
растений и позвоночных. Всего здесь встречено 30 и 14 видов птиц,
из них 18 и 10 — фоновые. Преобладают транспалеаркты при зна-
чительном участии сибирских видов.
На крупных таежных реках (среднее течение
р. Казым) доминируют береговая ласточка (52 и 48%), белая трясо-
гузка (14 и 27%) и перевозчик (в первой половине лета — 16%).
Здесь насчитывается вдвое меньше птиц, чем на озерах. Ярусное
распределение отличается значительной долей воздухореев (52 и
48%), меньше птиц держится на земле по берегу (34 и 43%) и срав-
нительно мало — на воде (14 и 8%).
Биомасса и количество трансформируемой энергии соответствен-
но в 7 и 5 раз меньше, чем на озерах. Преобладают по биомассе синь-
га и гоголь (в первой половине лета — 35 и 11%), а также серебри-
стая чайка (во второй половине лета — 41%). Энергетические по-
154
Таблица 9
Летнее население птиц водоемов северной тайги Обь-Енисейского междуречья
в 1976 г., особей/км?
Вид Пойменные озера, ста- рицы Крупные таежные реки (р. Казым) Вид Пойменные озера, стари- цы Крупные таежные реки (р. Казым)
п е р в а я поло вина лета
Всего . . . 81 44 1 1 1
Хохлатая чернеть 16 0,3 Краснозобая гагара 1 —
Белая трясогузка 13 6 Чернозобый дрозд 0,7 0,2
Фифи 12 0,07 Скопа 0,7 0,1
Шилохвость 7 0,3 Длиннохвостый крохаль 0,7 0,2
Ласточка береговая 6 23 Большой улит 0,7 —
Мородунка 3 0,3 Каменка 0,7 —
Синьга 2 1 Овсянка-крошка 0,7 —
Обыкновенная крачка 2 0,7 Луток 0,7 —
Серебристая чайка 2 0,4 Чирок-трескунок 0,3 1
Полярная крачка 2 0,6 Белохвостый песочник 0,3 0.3
Кедровка 2 — Большой крохаль 0,3- 0,1
Чирок-свистунок 2 — Кряква 0,3 —
Перевозчик 1 7 Свиязь 0,3 —
Серая ворона 1 0,5 - Гоголь — 1
Сизая чайка 1 0.3 Черныш — 0,4
Серый журавль 1 — Малый зуек — 0,1
Бекас 1 — Бородатая неясыть — 0,1
Вторая поло вина лета
Всего . . . | 85 52 j 1
Чирок-свистунок 22 0,3 Полярная крачка 0,3 0,3
Гоголь 20 0,3 Скопа 0,3 0,4
Шилохвость 15 0,5 Перевозчик — 5
Чирок-трескунок 10 — Серебристая чайка — 2
Желтая трясогузка 6 1 Серая ворона — 1
Белая трясогузка 4 14 Большой улит — 0,6
Хохлатая чернеть 2 0,3 Ворон — 0,5
Черный стриж 2 0,1 Сизая чайка — 0,3
Фифи 1 — Черныш — 0,2
Синьга 1 — Полевой лунь — 0,2
Ласточка береговая 0,7 25 Чеглок — 0,1
Свиязь 0,7 —
требности птиц удовлетворяются преимущественно за счет живот-
ных кормов, доля вегетативных частей растений в рационе птиц
значительно ниже, чем на озерах.
Видовое разнообразие птиц на р. Казым несколько меньше,
чем на озерах, а число фоновых видов вдвое меньше. Подавляющее
большинство птиц приходится на транспалеарктов.
155
*
* *
Таким образом, в северной тайге в состав доминантов по обилию
чаще всего входят в сосново-боровом ландшафте юрок и белая тря-
согузка, а на комплексных верховых болотах — желтая трясогузка
и овсянка-крошка. Заметно реже в лесах преобладают чечетка,
клест-еловик и каменка, и только по второй половине лета — серо-
головая гаичка и весничка. На болотах лишь изредка доминируют
фифи и шилохвость. Характерный доминант поселков — домовый
воробей. На водоемах преобладают перевозчик, чирки — свистунок и
трескунок, гоголь и шилохвость.
Максимальное суммарное обилие птиц отмечено в населенных
пунктах (624 и 1019). Вдвое меньше птиц зарегистрировано в приреч-
ной темнохвойной тайге. В первой половине лета на комплексных
верховых болотах птиц в 2,5, а в сосняках и их производных в 4 ра-
за меньше, чем в поселках. После середины лета за счет интенсивной
откочевки птиц из болотных местообитаний суммарное обилие в них
уже вдвое ниже, чем в сосняках.
В населении северной тайги преобладают птицы, кормящиеся
на земле, лишь на водоемах во второй половине лета на первое место
выходят водные птицы. В лесных местообитаниях доля кронников
несколько ниже по сравнению с наземными птицами и значительно
меньше кустарниковых птиц. На водоемах и в поселках сравнитель-
но велико участие воздухореев.
Показатели биомассы и биоэнергетики изменяются почти так
же, как и плотность населения, но уменьшение биомассы птиц на
верховых болотах во второй половине лета не прослеживается по
причине значительного и стабильного участия в населении крупных
видов (глухарь, шилохвость, серебристая чайка и серая ворона).
В населении лесных местообитаний по биомассе преобладают как
воробьиные (те же, что и по численности), так и крупные неворобьи-
ные птицы (глухарь, чирок-трескунок и гоголь). В поселках по био-
массе доминируют серая ворона и белая трясогузка; на водоемах —
чаще всего утки (шилохвость и гоголь), кроме них в первой полови-
не лета — хохлатая чернеть, синьга и серый журавль, а во вто-
рой — чирок-свистунок и серебристая чайка. Энергетические по-
требности птиц удовлетворяются в основном за счет беспозвоночных.
Кроме того, в лесах заметную роль играет потребление семян и соч-
ных плодов, а на водоемах — вегетативных частей растений и по-
звоночных.
Видовое разнообразие птиц изменяется в том же направлении,
что и плотность населения (от 42 видов в темнохвойной тайге до 28
на переходных болотах), за исключением поселков, где оно сравни-
тельно невелико. Фаунистический состав населения птиц северной
тайги в целом европейско-сибирский, но в сосново-озерном ландшаф-
те, на верховых болотах и особенно в поселках и на водоемах пре-
обладают транспалеаркты.
156
Летом 1973 г. в сходных местообитаниях северотаежной долины
Оби состав доминантов был примерно такой же, какой отмечен нами
на междуречьях. Лишь сероголовая гаичка и каменка преобладали
только на последних. В долине Оби по сравнению с междуречьями
отмечены в 2,4—8 раз большие показатели плотности населения и
в 3—3,5 раза большие значения биомассы и биоэнергетики птиц.
По видовому разнообразию превышение не столь велико; в долине
Оби, как правило, встречалось в 1,1—1,4 раза больше видов птиц,
чем в сходных ландшафтах междуречий. Различия в фаунистиче-
ском составе населения обычно незначительны. На междуречьях по
сравнению с долиной возрастает доля транспалеарктов и несколько
снижается участие европейских видов.
В северной половине подзоны в редкостойных елово-листвен-
ничных лесах в окрестностях бывшей фактории Пос-Полуй в июле
1976 г. на междуречьях нами насчитывалось в 2,4 раза меныпе-птиц,
чем в долине Оби [Вартапетов и др., 1980]. В этих лесах в долине
Оби преобладали овсянка-крошка, таловка, юрок и сероголовая га-
ичка, а на междуречьях — только первые два вида и, кроме того,
желтая трясогузка. Видовое разнообразие населения птиц на
междуречьях также было в 1,3 раза ниже, чем в долине
Оби.
СРЕДНЯЯ ТАЙГА
СОСНОВО-БОРОВОЙ ЛАНДШАФТ
В сосняках-брусничниках в течение всего лета
доминируют юрок (20 и 10%) и пухляк (13 и 29%), и только в первой
половине лета — горихвостка-лысушка (11%, см. табл. 10). Плот-
ность населения птиц здесь составляет 140 и 133 особи/км2 (табл. 11).
Примерно половина птиц кормится в кронах деревьев и несколько
меньше — на земле (табл. 12). Значительно участие в населении
воздухореев (в основном за счет черного стрижа).
Биомасса птиц составляет 5 и 26 кг/км2 (табл. 13). Столь резкое
ее увеличение в конце лета связано со значительной прикочевкой
глухаря и тетерева в период созревания брусники. Доминируют по
биомассе в первой половине лета юрок и кедровка (13 и 11%), а во
второй — глухарь и тетерев (72 и 11%). Энергетические траты птиц
составляют по периодам Зи 6 тыс. ккал/сут-км2. Большая часть энер-
гетических потребностей удовлетворяется за счет беспозвоночных
при увеличении во второй половине лета доли вегетативных частей
растений.
Всего здесь встречено 38 и 31 вид птиц, из которых 21 и 20 фо-
новые. Фаунистический состав населения европейско-сибирский
(табл. 14). По данным П. А. Пантелеева [1972&], пересчитанным
Ю. С. Равкиным на 1 км2, в сосняках по Конде на границе средней и
южной тайги в июне 1959 г. насчитывалось птиц на 30% меньше,
157
Таблица Ю
Население птиц сосново-борового ландшафта средней тайги Обь-Иртышского
междуречья летом 1977 г., особей/км2
Вид В среднем по ландшафту Сосняки-брус- ничники Приречные смешанные леса Вырубки по соснякам
1 2 3 4 5
Первая половина лета
Всего ... 188 | 140 | 230 | 152
Юрок 27 28 27 26
Пухляк 25 18 32 9
Теньковка 16 — 30 2
Г орихвостка-лысушка 13 16 10 И
Поползень 8 0,2 15 0,1
Большой пестрый дятел 7 4 8 13
Овсянка-ремез 6 2 10 0,2
Зяблик 6 0,5 10 0,2
Славка-завирушка 6 — 11
Зеленый конек 5 3 8 0,2
Свиристель 4 6 2 12
Клест-еловик 4 7 0,3 24
Береговая ласточка 4 8 2 0,08
Белокрылый клест 4 7 3 0,08
Садовая славка 4 — 7
Овсянка-крошка 3 6 — 4
Черныш 3 2 4 0,2
Белая трясогузка 3 6 — 5
Московка 3 —. 5
Воробьиный сыч 3 — 5
Черный стриж 2 5 — 4
Певчий дрозд 2 0,6 4 0,1
Кедровка 2 3 1 4
Зеленая пеночка 2 4
Белобровик 2 — 4 4
Чиж 2 4 0,8
Чернозобый дрозд 2 1 2 4
Снегирь 2 — • 3 1
Весничка 1 0,2 2 1
Серая мухоловка 1 2 0,8
Глухарь 1 — 2 0,1
Бекас 1 — 2 0,8
Рябчик 1 — 2
Пищуха 1 — 2
Кукша 0,9 0,9 1 0,2
Деряба 0,9 2 — 1
Лесной конек 0,8 0.8 0,2 7
Чечевица 0,8 2 — 0,2
Черный дятел 0,7 0,5 1 0.1
Фифи 0,6 0,6 0,6 0,2
Кукушка 0,6 0,8 0,3 2
Луговой чекан 0,5 1 2
Зарничка 0,5 1
Перевозчик 0,5 1
Желтая трясогузка 0,5 — 1 —
Барсучок 0,4 0,8 .
Погоныш 0,4 0,8
Ополовник 0,4 — 0.8 —
158
Продолжение табл. 10
1 2 3 1 4 5
Большой улит 0,3 0,5 0,1 1
Чирок-свистунок 0,3 — 0,5 —
Хохлатая чернеть 0,3 — 0,5 —
Козодой 0.3 -— 0,08 4
Гоголь 0,3 — 0,5 —
Глухая кукушка 0,2 — 0,4 0,1
Дубонос 0.3 0,6 0,02 0,05
Таловка 0,1 — 0,2 0,5
Черный коршун 0,1 — 0,2 0,08
Трехпалый дятел 0,1 — — 2
Каменка 0,1 -— — 2
Чеглок 0,08 0,2 —
Чечетка 0,07 0,08 0,05 0,2
Сероголовая гаичка 0,06 — — 1
Ворон 0,05 0,02 0,05 0,2
Скопа 0,05 — — 0,8
Гуменник 0,05 — 0,1 —
Сизая чайка 0,04 — — 0,6
Белошапочная овсянка 0,02 0,05 — —
Серая ворона 0,01 0,005 0,02 0,01
Перепелятник 0,01 — — 0,2
Речная крачка 0,005 0,01 — 0,01
Вторая половина лета
Всего ... 221 133 292 203
Пухляк 58 38 75 51
Рябчик 20 — 38 —
Поползень 20 И 27 22
Юрок 15 13 14 32
Лесной конек 10 3 14 25
Теньковка 10 7 13 —
Береговая ласточка 9 * 3 15 —
Кукша 8 17 1 0,07
Овсянка-крошка 7 5 8 4
Овсянка-ремез 5 — 9 —
Чернозобый дрозд 5 1 8 1
Горихвостка-лысушка 4 1 6 9
Белошапочная овсянка 4 1 6 —
Московка 4 — 7 3
Ополовник 4 — 7 —
Зеленый конек 3 3 4 —
Черный стриж 3 5 — 17
Весничка 3 1 5 —
Белая трясогузка 3 3 0,8 17
Большой пестрый дятел 3 0,9 4 4
Снегирь 3 — 6 —
Таловка 3 — 5 —
Пищуха 3 — 5 —
Серая мухоловка 2 4 — 1
Глухарь 2 6 — —
Садовая славка 2 — 4 —
Зеленая пеночка 1 0,7 2 —
Сорока 1 0,2 2 —
159
Окончание табл. 10
1 2 3 4 5
Тетерев 1 3 —
Кедровка 0,9 2 — 2
Зяблик 0,8 0,7 1 —
Деряба 0,8 1 0,2 4
Чечетка 0,7 0,9 0,7 0,02
Черный дятел 0.5 — 0,9 0,07
Трехпалый дятел 0,5 0,7 — 0,3
Славка-завирушка 0,5 — 1 —
Клест-еловик 0,5 0,1 0,7 0.7
Синехвостка 0,4 — 0,7
Серая ворона 0,3 0,2 0,4 —
Ворон 0,2 — 0,3
Белобровик 0.2 — 0,3 —
Сероголовая гаичка 0,2 -— — 3
Свиристель 0,1 0,1 — 1
Чеглок 0,1 — 0,2
Дербник 0,1 0,2 —
Большой улит 0,06 — — 1
Чиж 0,06 — 0,07 0,3
Белокрылый клест 0,05 о,1 — 0,1
Луговой чекан 0,04 — — 0,7
Малый пестрый дятел 0,04 — — 0,7
Кукушка 0,03 0,07 — 0,07
Желтая трясогузка 0.006 — — 0,1
Вертишейка 0.004 — — 0,07
Таблица 11
Плотность и разнообразие летнего населения птиц средней тайги Обь-Иргыш-
ского междуречья летом 1977 г.
Ландшафт, урочище Плотность населения Отмечено видов
всего фоновых
1 1 и I II 1 1 11
Половина лета
Сосново-боровой ландшафт 188 221 47 32 26 21
Сосняки-брусничники 140 133 38 31 21 20
Вырубки по соснякам 152 203 49 29 24 18
Приречные смешанные леса 230 292 55 38 33 26
Верховые болота 133 90 36 26 20 13
Рослые рямы 90 117 31 23 14 13
Низкорослые рямы Грядово-мочажинные комплексы 119 171 90 86 39 26 29 22 23 16
верховых болот 38 И
Поселки 806 918 47 37 27 24
Надпойменные озера 168 552 25 26 21 22
Пойменные озера, старицы 67 73 18 17 14 и
Крупные таежные реки (р. Бол. Юган) И 16 28 17 2 4
В среднем по ключевому
участку 165 161 42 30 23 18
160
Таблица 12
Ярусное распределение птиц в средней тайге Обь-Иртышского междуречья ле-
том 1977 г., %
Ландшафт, урочище На земле в ьу- : стах В кро- нах На ство- лах На воде 1 В воздухе
Первая Сосново-боровой ландшафт п 0 J 30 I о в И I 15 та ле 42 т а 9 0,5 4
Соспяки-брусничники 29 4 54 3 0,01 9
Вырубки по соснякам 30 4 49 10 1 5
Приречные смешанные леса 31 20 36 И 0,7 0,9
Верховые болота 71 8 3 0,8 12 5
Рослые рямы 62 6 25 6 1 0,5
Низкорослые рямы 70 9 3 0,9 12 5
Грядово-мочажинные комплексы 74 5 0,7 — 13 7
Поселки 32 7 2 1 0,1 58
Надпойменные озера 18 1 — — 79 2
Пойменные озера, старицы 58 — — — 42 —
Крупные таежные реки (р. Бол. Юган) 49 — 0,1 — 12 39
В среднем по ключевому
участку 46 12 26 5 5 6
Вторе Сосново-боровой ландшафт 1 я по 34 ЛОВЕ 9 I н а л 39 : е т а 12 6
Сосняки-брусничники 32 7 46 10 .— 6
Вырубки по соснякам 34 1 42 15 — 8
Приречные смешанные леса 35 10 37 13 — 5
Верховые болота 76 2 8 4 4 5
Рослые рямы 75 0,8 18 4 — 1
Низкорослые рямы 79 2 9 4 0,01 5
Грядово-мочажинные комплексы 71 2 4 3 14 6
Поселки 56 И 4 0,6 — 29
Надпойменные озера 36 — — — 64 0,3
Пойменные озера, старицы 48 — — — 49 3
Крупные таежные реки (р. Бол. Юган) 80 — 0,4 — 15 5
В среднем по ключевому
участку 46 7 30 9 1 6
чем встретили мы в тот же период 1977 г. в юганских сосняках. В
кондинских сосняках преобладали юрок и горихвостка-лысушка,
а на р. Бол. Юган — они же и береговая ласточка. Высокое обилие
последней связано с местной спецификой территории — близостью
гнездовой колонии на береговом обрыве р. Бол. Юган и высокой ак-
тивностью ласточек в период выкармливания птенцов. Видовое раз-
нообразие птиц также было сходным. Всего встречено 41 и 33 вида,
из них 25 и 22 фоновые. Таким образом, сравниваемые показатели
11 Л. Г. Вартапетов
161
Таблица 13
Биомасса и биоэнергетика населения птиц средней тайги Обь-Иртышского
междуречья летом 1977 г.
Ландшафт, урочище Биомасса, кг/км* Трансформируемая анергия
Всего, тыс. ккал/еут-км® В том числе (%) за счет
беспозво- ночные семян, плодов вегета- тивных частей растений позвоноч- ных
Первая половина лета
Сосново-боровой ландшафт 10 5 87 5 6 2
Сосняки-брусничники 5 3 90 10 — 0,6
Вырубки по соснякам 9 5 85 9 0,4 6
Приречные смешанные леса 15 6 86 3 9 2
Верховые болота 16 6 76 1 5 17
Рослые рямы 6 3 94 2 3 2
Низкорослые рямы 13 5 77 2 4 18
Грядово-мочажинные комплексы 25 8 75 0,4 8 17
Поселки 32 20 90 9 0,2 1
Надпойменные озера 82 20 54 0,9 17 29
Пойменные озера, старицы 16 5 60 — 33 6
Крупные таежные реки (р. Бол. Юган) 1 0,5 70 — И 20
В среднем по ключевому 13 |
участку 1 6 81 3 6 10
В т о р i Сосново-боровой ландшафт 1 Я П 0 22 • л О В I 8 I н а л 74 [ е т а 4 22 0,4
Сосняки-брусничники 26 6 74 6 20 —
Вырубки по соснякам 6 5 99 0,3 —
Приречные смешанные леса 21 10 73 3 23 0,7
Верховые болота 8 3 78 9 9 4
Рослые рямы 4 3 91 3 6 0,2
Низкорослые рямы 5 3 91 7 — 2
Грядово-мочажинные комплексы 15 5 63 12 18 7
Поселки 31 22 72 26 — 2
Надпойменные озера 226 59 48 — 43 9
Пойменные озера, старицы 19 6 64 0,3 28 9
Крупные таежные реки (р. Бол. Юган) 2 0,8 75 — 4 21
В среднем по ключевому
участку 15 6 76 7 16 2
населения птиц среднетаежных сосняков весьма близки, несмотря
на их удаленность друг от друга (450 км но широте) и разные годы
наблюдений.
На вырубках по соснякам постоянно доминирует
юрок (17 и 16%), кроме него в первой половине лета — клест-ело-
вик (16%), а во второй — пухляк, лесной конек и поползень (25,
162
Таблица 14
Фаунистический состав населения птиц средней тайги Обь-Иртышского между-
речья летом 1977 г., %
Ландшафт, урочище Виды Особи
европейские сибирские арктические транспале- арктические европейские сибирские арктические транспале- арктические
Первая половина лета
Сосново-боровой ландшафт 23 36 — 33 29 50 — 14
Сосняки-брусничники 26 37 — 26 20 52 — 20
Вырубки по соснякам 22 41 — 31 21 62 — 17
Приречные смешанные леса 24 38 — 29 33 48 — И
Верховые болота 15 27 12 42 31 18 7 38
Рослые рямы 19 32 3 35 43 34 0,2 14
Низкорослые рямы 13 26 5 51 40 18 3 34
Грядово-мочажпнные комплексы 8 29 18 39 18 16 14 46
Поселки 17 30 — 40 6 4 — 81
Надпойменные озера 4 24 8 56 2 23 7 63
Пойменные озера, старицы Крупные таежные реки (р. Бол. — 22 6 72 — И 6 83
Юган) 4 18 11 64 2 5 2 91
В среднем по ключевому 17 | 31 1 1 36
участку 1 7 36 29 1 3 25
Втора я половина лета
Сосново-боровой ландшафт 23 43 — 26 16 69 — 19
Сосняки-брусничники 26 42 — 23 17 72 — 8
Вырубки по соснякам 21 48 — 24 28 60 — И
Приречные смешанные леса 24 42 — 21 15 68 — 11
Верховые болота 9 37 9 39 42 29 0,6 25
Рослые рямы 17 43 9 26 54 39 2 4
Низкорослые рямы 12 38 — 42 47 29 — 19
Грядово-мочажинные комплексы 7 34 7 52 30 27 2 42
Поселки 24 30 — 41 28 6 — 63
Надпойменные озера — 19 19 62 — 31 6 63
Пойменные озера, старицы Крупные таежные реки (р. Бол. 6 18 6 65 1 14 1 83
Юган) 6 6 6 76 1 12 4 83
В среднем по ключевому 56 0,2 16
участку 18 34 5 35 24
12 и 11%). Плотность населения птиц до середины лета такая же,
как в сосняках, а во второй половине — на 50% выше. Ярусное
распределение птиц мало отличается от такового в сосняках.
Биомасса и энергетические затраты птиц в первой половине ле-
та в 1,8 и 1,6 раза больше, чем в сосняках, а во второй — в 4 и 1,2 pa-
ll*
163
за меньше. До середины лета по биомассе доминируют скопа *,
большой пестрый дятел и клест-еловик (15 и по 12%), а во второй
его половине — те же виды, что и по обилию, лишь поползня сменя-
ет черный стриж. Сравнительно высокая численность последнего
объясняется близостью его гнездовой колонии в сосняках. Энерге-
тические потребности птиц удовлетворяются преимущественно за
счет беспозвоночных. Видовое разнообразие такое же, как в сосня-
ках. Фаунистический состав населения европейско-сибирский.
В приречных смешанных лесах, как и в от-
дельных лесных местообитаниях, постоянно доминирует пухляк
(14 и 26%), кроме того, в первой половине лета — теньковка и юрок
(13 и 12%), а во второй — рябчик (13%). Здесь насчитывается вдвое
больше птиц, чем в сосняках. Распределение птиц по ярусам сходно
с таковым в предыдущих урочищах сосново-борового ландшафта,
по доля птиц, кормящихся в кустарниках, значительно выше, что
соответствует характеру ярусной структуры этих лесов.
Показатели биомассы и трансформируемой энергии в смешан-
ных лесах также в среднем в 1,5 и 2 раза выше, чем в остальных лес-
ных местообитаниях. В первой половине лета по биомассе домини-
рует глухарь, а во второй — рябчик (42 и 70%). Соотношение ос-
новных групп потребляемых кормов почти такое же, как в сосняках,
лишь немного больше доля вегетативных частей растений и меньше
процент участия семян и сочных плодов. Видовое разнообразие птиц
в смешанных лесах также в 1,5 раза выше, чем в сосняках. Фаунис-
тический состав населения европейско-сибирский.
П. А. Пантелеев [1972а] в июне 1959 г. в сходных смешанных
лесах по р. Конде (на границе южной и средней тайги) насчитывал
птиц на 26% меньше, чем отмечено нами в сходных лесах по р. Бол.
Юган. В лесах по р. Конда доминировал чиж, а по р. Бол. Юган —
пухляк и теньковка. Близко видовое разнообразие в сравниваемых
лесах (39 и 41 вид). Таким образом, хотя население этих среднета-
ежных лесов и сходно по ряду показателей, оно различается больше,
чем орнитокомплексы рассмотренных выше сосняков.
В среднем по ландшафту постоянно доминирует
пухляк (13 и 26%), и в первой половине лета — юрок (14%). Здесь
насчитывается по периодам 188 и 221 особь/км2. Наибольшее коли-
чество птиц кормится в кронах и на земле, заметно меньше — в
кустах и на стволах. Биомасса птиц составляет 10 и 22 кг/км2, а ве-
личина трансформируемой энергии — 5 и 8 ккал/сут«км2. По био-
массе доминируют глухарь (32 и 35 %) и рябчик (во второй половине
лета — 35%). Затраты энергии удовлетворяются в основном за счет
беспозвоночных при заметном участии семян, сочных плодов и веге-
тативных частей растений (особенно во второй половине лета). Всего
в сосново-боровом ландшафте встречено 47 и 32 вида птиц, из них
26 и 21 фоновые. Фаунистический состав населения европейско-си-
бирский.
* Скопа гнездилась сравнительно недалеко от учетного маршрута и поэто-
му постоянно регистрировалась нами, что, несомненно, завышает ее численность.
<64
ВЕРХОВЫЕ БОЛОТА
В рослых рямах доминируют лесной конек (36 и 52%),
овсянка-крошка (11 и 15%), и после середины июля — пухляк
(12%, табл, 15). Здесь насчитывается в 1,3 раза меньше птиц, чем
в сосняках. Большая часть их кормится на земле, но сравнительно
велика и доля кронников. Биомасса птиц и трансформируемая ими
энергия в первой половине лета примерно такие же, а во второй зна-
чительно меньше, чем в сосняках. По биомассе, так же как и по оби-
лию, преобладают лесной конек (13 и 32%) и, кроме того, в первой
половине лета — гуменник и свиристель (13 и 12%), а во второй —
белая куропатка. Среди потребляемых кормов преобладают беспоз-
воночные, а во второй половине лета несколько увеличивается роль
семян, сочных плодов и вегетативных частей растений. В среднем
здесь встречено в 1,3 раза меньше видов птиц, чем в сосняках. Фау-
нистический состав населения в отличие от сосняков-брусничников,
где доминируют сибирские виды, сибирско-европейский при сущест-
венном участии транспалеарктов.
В низкорослых рямах преобладают по обилию лес-
ной конек (38 и 42%), и в первой половине лета — сизая чайка (10%).
Обилие последней, возможно, несколько завышено из-за близости
к учетному маршруту гнездовой колонии этой чайки, хотя в целом на
обследованной территории такие колонии единичны. В первой по-
ловине лета в низкорослых рямах отмечено птиц на 30 % больше,
чем в рослых рямах, а во второй — на столько же меньше. По срав-
нению с предыдущим урочищем, в низкорослых рямах зарегистри-
ровано больше птиц, держащихся на воде, земле и в воздухе, и со-
ответственно меньше — в кронах деревьев.
Биомасса птиц в низкорослых рямах в 1,8 раза выше, чем в рос-
лых, в основном за счет чаек и гагар, которые встречаются здесь на
небольших озерах. В первой половине лета по биомассе доминирует
сизая чайка (42%), а во второй — серая ворона и лесной конек (33
и 19%). Величина трансформируемой энергии в первой половине ле-
та в 1,6 раза больше, а во второй — такая же, как и в рослых рямах.
Состав потребляемых птицами кормов близок к таковому в предыду-
щем урочище, лишь необычно велика доля позвоночных (18 и 2%),
в основном за счет рыбы, поедаемой на рямовых озерах сизыми чай-
ками, гагарами, речными крачками. Общее число встреченных и фо-
новых видов в 1,1—1,6 раза больше, чем в рослых рямах. Фаунисти-
ческий состав населения здесь сибирско-европейский при значи-
тельном участии транспалеарктов.
На грядово-мочажинных комплексах в
число доминантов тоже постоянно входит лесной конек (16 и 27 %),
кроме того, в первой половине лета — белая трясогузка (13% X
а во второй — желтая трясогузка и белошапочная овсянка (24 и
10%). Плотность населения птиц до середины лета в 1,4 раза боль-
ше, а во второй половине лета такая же, как и в низкорослых рямах*
Ярусное распределение птиц сходно с таковым в низкорослых ря-
мах, лишь немного ниже доля кронников и выше участие в населе-
165
Таблица 15
Население птиц верховых болот средней тайги Обь-Иртышского междуречья
летом 1977 г., особей/км?
Вжд В среднем по ландшафту Рослые рямы Низкорослые рямы Грядово-мо- чажинно-озер- ные комплек- сы
1 2 3 4 5
Первая половина лета
Всего ... 136 90 119 171
Лесной конек 38 32 45 27
Сизая чайка 13 0,6 12 16
Желтая трясогузка 10 1 8 16
Белая трясогузка 9 — 3 22
Дубровник 7 4 7 7
Овсянка-крошка 7 10 . 7 5
Береговая ласточка 6 0,4 5 9
Белошапочная овсянка 6 0,6 5 10
Краснозобый конек 4 — — 12
Большой улит 3 1 2 6
Белая куропатка 3 0,2 2 6
Средний кроншнеп 3 0,5 2 4
Чибис 3 — 1 6
Юрок 3 4 3 2
Фифи 3 2 3 3
Зеленый конек 2 4 0,6 3
Малый веретенник 2 — 1 3
Жулан 2 — 2 1
Черный стриж 1 0,08 0,2 3
Свиристель 1 12 0,6 —
Речная крачка 1 — 1 2
Большой пестрый дятел 0,9 5 1 —
Пухляк 0,6 — 1 —
Кукушка 0,6 2 0,7 0,05
Луговой чекан 0,6 — 1 —
Тетерев 0,6 0,5 1 —
Чирок-свистунок 0,6 -— 0,5 1
Садовая камышевка 0,6 —. 1 —
Краснозобая гагара 0,6 — —> 2
Серая ворона 0,5 0,8 0,5 0,3
Горихвостка-лысушка 0,3 5 — —
Кедровка 0,3 2 0,2 0,05
Каменка 0,3 — — 0,8
Черноголовый чекан 0,3 — — 1
Круглоносый плавунчик 0,2 — — 0,5
Певчий дрозд 0,2 —• .— 0,5
Чеглок 0,1 — 0,2 __
Бекас 0,1 0,1 0,2
Скопа 0,1 —. 0,2 —
Большой кроншнеп 0,1 — 0,2 —
Шилохвость 0,1 •— — 0,4
Гуменник 0,08 0,2 — 0,2
Кукша 0,08 0,2 0,2
Свиязь 0,07 0,1 — 0,2
166
Продолжение табл. 15
1 2 3 4 5
Черный дятел 0,06 — 0,1 —
Весничка 0,06 1 —— —
Белокрылый клест 0,06 — 0,1 —
Синьга 0,06 — 0,2
Перепел 0,05 —— 0,08 —
Ворон 0,04 0,05 0,06 —
Чечетка 0,03 —- 0,055
Рогатый жаворонок 0,03 —. — 0,1
Чернозобая гагара 0,02 —- — 0,05
Большой крохаль 0,01 0,02 —
Белоспинный дятел 0,01 0,2 — —
Теньковка 0,01 0,2 — —
Белобровик 0,01 0,2 — —
Белохвостый песочник 0,006 —. — 0,02
Канюк 0,003 — — 0,008
Дубонос 0,001 0,02 — —
Вторая половина лета
Всего ... 89 | 1 117 1 1 90 1 | 86
Лесной конек 35 61 38 23
Желтая трясогузка 9 0,4 3 21
Белошапочная овсянка 7 3 6 9
Пухляк 6 14 7 3
Овсянка-крошка 5 18 6 —-
Белая трясогузка 3 0,7 4 0,7
Зеленый конек 2 — 4 —.
Черный стриж 2 0,7 2 3
Береговая ласточка 2 1 2 2
Серая ворона 2 0,02 3 2
Поползень 2 3 0,9 3
Большой пестрый дятел 2 2 3 —
Шилохвость 2 — 6
Юрок 1 1 2 —
Деряба 1 2 2 —•
Тетерев 1 0,07 0,3 3
Чернозобый дрозд 1 — 2 —
Чирок-свистунок 1 «— —• 4
Большой сорокопут 0,8 — 1 0,7
Кедровка 0,6 1 0,06 0,5
Белобровик 0,6 — 1 —
Чеглок 0,5 — 0,7 0,2
Дубровник 0,4 — 0,7 —
Фифи 0,3 —— 0,4 <К2
Свиристель 0,3 4 0,1
Большой улит 0,2 0,03 — 0,7
Речная крачка 0,2 —. — 0,7
Жулан 0,2 — —- 0,7
Краснозобая гагара 0,2 — 0,7
Чернозобик 0,2 — — 0,7
Кряква 0,1 —- ——• 0,3
Чернозобая гагара 0,1 — 0,01 0,4
Чибис 0,1 •— — 0,3
Белая куропатка 0,1 2 — —
Сероголовая гаичка о,1 2 — —
167
Окончание табл. 15
1 2 1 1 3 4 5
Средний кроншнеп 0,06 — — 0,2
Сапсан 0,06 —- — 0,2
Клест-еловик 0,04 0,7 — —
Кукушка 0,04 — 0,07 —
Чечетка 0,03 — — 0,1
Свиязь 0,03 — — о,1
Московка 0,02 0,3 — —
Белокрылый клест 0,01 — 0,02 —
Ворон 0,004 0,07 — —
Черный коршун 0,003 — — 0,01
Малый веретенник 0,002 0,03 — -
нии водных птиц, что соответствует значительной открытости и боль-
шему числу водоемов в этих центральных частях верховых болот.
Биомасса птиц на грядово-мочажинных комплексах в 2 и 3 ра-
за выше, чем в низкорослых рямах. Доминируют в первой половине
лета сизая чайка, белая куропатка и краснозобая гагара (29, 15 и
14%), а во второй половине — шилохвость и тетерев (33 и 19%).
Энергетические затраты птиц в 1,6 раза выше, чем в низкорослых
рямах. Состав кормов сходен с таковым в предыдущем местообита-
нии, а доля позвоночных даже выше, чем в низкорослых рямах (так-
же за счет потребления на озерах рыбы гагарами и чайками).
Видовое разнообразие птиц в грядово-мочажинных комплексах
примерно такое же, как и в низкорослых рямах. Почти половина
суммарного обилия птиц приходится на транспалеарктов. Если не
принимать их во внимание, то по числу особей преобладают европей-
ские виды, а по количеству видов — сибирские. Заметно участие
в населении арктических видов, особенно в начале лета за счет про-
лета краснозобого конька.
В среднем по ландшафту постоянно доминирует
лесной конек (29 и 39%), в первой половине лета — сизая чайка,
а во второй ее сменяет желтая трясогузка (по 10%). Плотность на-
селения птиц по периодам в 1,4 и 2,5 раза ниже, чем в сосново-боро-
вом ландшафте. Подавляющее большинство птиц кормится на зем-
ле, заметно участие в населении водоплавающих.
Биомасса в первой половине лета в 1,6 раза больше, а во вто-
рой — почти в 3 раза меньше, чем в сосново-боровом ландшафте.
До середины лета по биомассе доминируют сизая чайка и белая ку-
ропатка (35 и 12%), а позднее — шилохвость, серая ворона, тете-
рев и лесной конек (20, 16, 14 и 10%). Энергетические потребности
птиц по периодам в 1,2 раза вышей в 2,7 раза ниже, чем в лесных ме-
стообитаниях, и удовлетворяются в основном за счет беспозвоноч-
ных при сравнительно высоком участии позвоночных животных и
небольшой доле растительных кормов.
На верховых болотах в пересчете на одно урочище встречено
36 видов птиц в первой половине лета и 26 видов во второй, в том
числе фоновых соответственно 20 и 13, Преобладают по числу осо-
168
бей европейские виды, а по количеству видов — сибирские при вы-
сокой доле транспалеарктов. В первой половине лета заметно учас-
тие арктических видов (в основном за счет пролетных). По сравне-
нию с сосново-боровым ландшафтом, здесь заметно ниже участие в
населении таежных (сибирских) видов и выше доля европейских
видов, особенно транспалеарктов.
ПОСЕЛКИ
В течение всего лета доминируют береговая ласточка (58 и 29%)
и домовый воробей (12 и 22%), а во второй половине лета, кроме
того, лесной конек и белая трясогузка (16 и 10%, табл. 16). В по-
селках насчитывается соответственно в 4 и 8 раз больше птиц, чем
в лесах и на верховых болотах. В распределении по ярусам харак-
терна высокая доля наземных птиц и воздухореев при заметном
участии птиц, кормящихся в кустарниках.
Показатели биомассы и биоэнергетики соответственно в 2 и 4 ра-
за выше, чем в естественных ландшафтах. По биомассе постоянно
доминируют сорока (38 и 22%) и береговая ласточка (22 и 13%),
а во второй половине лета — домовый воробей и лесной конек (21
и 11%). Энергетические потребности удовлетворяются в основном
за счет беспозвоночных при заметном участии сочных плодов и семян.
Видовое разнообразие птиц в поселках в первой половине лета такое
же, как в сосново-боровом ландшафте, а во второй — даже немного
выше, что связано со значительной концентрацией здесь птиц из
окрестных лесных местообитаний во время послегнездовых кочевок*
По числу особей и видов преобладают транспалеаркты при заметном
участии представителей европейского и сибирского типов фауны*
ВОДОЕМЫ
На надпойменных озерах все лето доминирует чирок-свистунок
(16 и 18%), кроме того, в первой половине — речная крачка и си-
зая чайка (14 и 13%), а во второй — белая трясогузка, свиязь и
шилохвость (24, 13 и 10%, табл. 17). Плотность населения птиц по
периодам составляет 168 и 552 особи на 10 км береговой линии. В
среднем за лето здесь держалось в 1,7 раза больше птиц, чем на озе-
рах в среднетаежной пойме Оби в 1972 г. [Равкин, 1978]. По-види-
мому, столь высокая концентрация птиц связана с тем, что озеро,
на котором проводились учеты, обладало высокой кормностью и зна-
чительной площадью мелководий и илистых отмелей, что привле-
кало большое количество уток, чаек и куликов, особенно в после-
гнездовой период. Обычно такие озера не характерны для верховых
болот, среди которых они расположены. Здесь отмечено даже в 1,3
и 2 раза больше водоплавающих, чем на южно-таежных озерах в
поймах Иртыша и Оби летом 1968 и 1970 гг. [Вартапетов и др.,
1972; Равкин и др., 1972]. Распределение птиц по ярусам характер-
169
Таблица 16
Население птиц поселков средней тайги Обь-Иртышского междуречья летом
1977 г., особей/км*
Вид Плотность 1 Вид Плотность
Пер Всего ... | в а я поло 806 | вина лета 1 1
Береговая ласточка 466 Садовая камышевка 1
Домовый воробей 100 Пятнистый сверчок 1
Белая трясогузка 72 Большая синица 1
Сорока 53 Черный коршун 0,9
Теньковка 22 Черный дятел 0,8
Славка-завирушка 18 Сизая чайка 0,7
Клест-еловик 10 Кедровка 0,6
Чечевица 8 Чечетка 0,5
Дубонос 6 Галка 0,5
Серая ворона 6 Кукушка 0,4
Овсянка-крошка 4 Вертишейка 0,2
Лесной конек 4 Черная ворона 0,2
Пухляк 4 Гуменник 0,2
Поползень 4 Чиж 0,1
Большой пестрый дятел 4 Юрок 0,2
Зеленый конек 3 Черныш 0,2
Каменка 3 Фифи 0,2
Го рихвостка-л ысу шка 2 Свиристель 0,2
Белошапочная овсянка 2 Весничка 0,2
Чернозобый дрозд 2 Белокрылый клест 0,2
Овсянка-ремез 1 Речная крачка 0,1
Снегирь 1 Ч и рок-свистунок 0,05
Желтая трясогуска 1 1 Черный стриж 0,02
Козодой 1
В т о рая поло вина лета
Всего ... | 918 |
Береговая ласточка 266 Кедровка 2
Домовый воробей 203 Кукушка 2
Лесной конек 147 Дупель 2
Белая трясогузка 90 Черный коршун 1
Теньковка 71 Садовая камышевка * 1
Сорока 30 Черный дятел 0,7
Славка-завирушка 21 Черныш 0,7
Белошапочная овсянка 13 Большой пестрый дятел 0,7
Пухляк И Дербник 0,7
Садовая славка 8 Большой сорокопут 0,7
Юрок 7 Сапсан 0,7
Чернозобый дрозд 7 Клест-еловик 0,5
Горихвостка-лысушка 6 Чечетка 0,3
Желтая трясогузка 6 Черный стриж 0,2
Овсянка-крошка 5 Серый журавль 0,2
Поползень 4 Свиристель 0,07
Весничка 3 Перепелятник 0,07
Серая ворона з j Малый пестрый дятел 0,07
Дубровник з 1
170
Таблица 17
Летнее население птиц водоемов средней тайги Обь-Иртышского междуречья
1977 г., особей/10 км береговой линии
Вид Надпойменные озера Пойменные озера Крупные таеж- ные реки
1 2 3 4
Первая половина лета
Всего . . . | 168 67 И
Чирок-свистунок 38 2 о,3
Речная крачка 23 — 0,2
Сизая чайка 22 2 0,3
Гоголь 16 1 0,08
Белохвостый песочник 10 — 0,08
Белая трясогузка 9 3 0,5
Чернозобая гагара 6 — —
Шилохвость 6 — —-
Серебристая чайка 6 — —
Большой улит 5 2 од
Черный стриж 4 — —
Свиязь 4 4 0,2
Красношейная поганка 3 — —
Фифи 2 4 —
Серая ворона 2 0,8 0,08
Хохлатая чернеть 2 1 0,05
Скопа 2 0,2 0,08
Широконоска 2 — —•
Камышевая овсянка 2 — —•
Средний кроншнеп 1 — ОД
Краснозобая гагара 1 — —
Чирок-трескунок 0,8 13 0,02
Бекас 0,5 1 —
Чибис 0,5 — 0,08
Большой крохаль 0,5 — —
Перевозчик •—. 16 3
Черныш —— И 0,4
Полярная крачка — 4 0,02
Кряква 1 —-
Мородунка —. 0,5 —
Черный коршун —— 0,2 0,08
Береговая ласточка —— — 4
Ворон — .— 0,2
Турухтан — — ОД
Желтая трясогузка — — од
Горная трясогузка •— •—• ОД
Чеглок •— — 0,08
Сорока — — 0,08
Канюк — — 0,08
Черная ворона — — 0,08
Кедровка — — 0,02
Вторая половина лета
Всего ... 552 73 16
Белая трясогузка 131 10 5
Чирок-свистунок 103 16 1
171
Окончание табл. 17
1 2 3 4
Свиязь 73
Шилохвость 56 5 —
Сизая чайка 39 — 0,2
Гоголь 27 0,3 —
Речная крачка 26 4 0,3
Желтая трясогузка 15 — 3
Кряква 14 — —
Белохвостый песочник 13 — —
Большой улит И 5 0,07
Галстучник 9 — —
Малый веретенник 7 — —
Хохлатая чернеть 5 3 .—-
Чернозобая гагара 4 — —
Малая чайка 4 — —-
Большой кроншнеп 3 — - —
Краснозобая гагара 3 — —
Фифи 2 — —
Серая ворона 2 — 0,4
Береговая ласточка 1 2 0,7
Травник 1 — —
Черныш 0,7 9 —
Чеглок 0,7 — 0,07
Горная трясогузка 0,7 — 0,3
Турухтан 0,7 — •—
Перевозчик — 10 3
Чирок-трескунок — 5 .—
Полярная крачка — 1 0,9
Черный коршун — 0,7 0,8
Скопа — 0,7 —
Дупель — 0,7 —
Озерная чайка — 0,7 —
Красношейная поганка — 0,3 .—•
Ворон — — 0,2
Перепелятник — — 0,1
Орлан-белохвост — — 0,1
Дербник — — 0,07
ное для водоемов. Большинство птиц кормится на воде, при замет-
ном участии птиц, встречающихся на земле по берегу, особенно за
счет многочисленных стаек трясогузок и пролетных куликов, кор-
мящихся на илистых отмелях.
Биомасса птиц составляет по периодам 82 и 226 кг на 10 км бе-
реговой линии. По биомассе так же, как и по обилию, в течение все-
го лета доминирует чирок-свистунок (по 16%) и кроме него в первой
половине — чернозобая гагара, гоголь, сизая и серебристая чайки
(17, 14, 12 и 10%), а во второй — свиязь и шилохвость (24 и 21%).
Количество трансформированной энергии составляет по периодам
20 и 59тыс. ккал/сут-10км береговой линии. Энергетические потреб-
ности птиц удовлетворяются за счет беспозвоночных и вегетативных
частей растений при высоком участии позвоночных (в основном за
счет рыбы, поедаемой гагарами, чайками и крачками). На надпой-
менных озерах встречено 25 и 26 видов птиц, из них 21 и 22 фоно-
172
вых, В населении птиц преобладают транспалеаркты и сибирские
виды при заметном участии арктических.
На пойменных озерах и старицах в течение
всего лета доминируют перевозчик (24 и 14%), черныш (16 и 12%)
и чирки: в первой половине лета трескунок, во второй — свисту-
нок, а также белая трясогузка (19, 22 и 14%). В среднем за лето
здесь насчитывается в 5 раз меньше птиц, чем на надпойменных
озерах. Большинство птиц держится на земле по береговой линии
и немного меньше на воде. Биомасса птиц и трансформируемая ими
энергия на пойменных озерах соответственно в 8,6 и 7,2 раза ниже,
чем на надпойменных. По биомассе доминируют чирок-трескунок
(32 и 10%), свиязь (в первой половине лета — 19%), чирок-свисту-
нок, шилохвость и хохлатая чернеть (во второй половине — 29, 22
и 11%). Состав потребляемых кормов почти такой же, а видовое раз-
нообразие птиц в 1,5 раза ниже, чем на надпойменных озерах. Пре-
обладают транспалеаркты при заметном участии сибирских видов.
На крупных таежных реках (р. Бол. Юган) по-
стоянно доминирует перевозчик (27 и 12%), кроме него в первой
половине лета — береговая ласточка (36%), а во второй — белая
и желтая трясогузки (31 и 19%). В среднем за лето на реке насчи-
тывается в 5 раз меньше птиц, чем на пойменных озерах и старицах.
Для ярусного распределения характерны высокая доля птиц, дер-
жащихся по береговым отмелям, и значительное число воздухореев.
Биомасса птиц и количество потребляемой ими энергии также в
8—9 раз меньше, чем на пойменных озерах. По биомассе в первой
половине лета доминируют ворон и свиязь (14 и 11%), а во второй —
черный коршун, орлан-белохвост и чирок-свистунок (30, 18 и 16%).
Энергетические потребности птиц удовлетворяются почти полно-
стью за счет животных кормов, в том числе при сравнительно высо-
ком участии позвоночных.
Видовое разнообразие птиц довольно высокое (28 и 17 видов
по периодам), но фоновые из них лишь 2 и 4, т. е. большинство ви-
дов редкие. В населении птиц, как и на других водоемах, преобла-
дают транспалеаркты.
*
* *
Итак, в средней тайге в лесных местообитаниях чаще всего до-
минирует пухляк, а на верховых болотах — лесной конек. Гораздо
реже в число доминантов в сосново-боровом ландшафте входит юрок,
только в первой половине лета — теньковка, горихвостка-лысушка
и клест-еловик, а во второй — рябчик, лесной конек и поползень.
На верховых болотах лишь изредка преобладает овсянка-крошка,
до середины лета — свиристель, белая трясогузка и сизая чайка,
а во второй половине — желтая трясогузка и белошапочная овсян-
ка. Характерные доминанты поселков — береговая ласточка и до-
мовый воробей, а водоемов — чирок-свистунок, перевозчик (только
173
в первой половине лета — сизая чайка и речная крачка, во вто-
рой — белая трясогузка и свиязь). По сравнению с населением се-
верной тайги среди доминантов не встречены чечетка, сероголовая
гаичка, каменка, весничка, зато впервые как доминанты отмечены
пухляк, лесной конек, поползень и белошапочная овсянка.
Наибольшая плотность населения птиц зарегистрирована, как
и в предыдущей подзоне, в поселках (806 и 918). В приречных сме-
шанных лесах и в сосняках с их производными птиц соответственно
в 3,3 и 5,5 раза меньше, и минимальное их количество отмечено на
верховых болотах (133 и 90). Таким образом, в смешанных приреч-
ных лесах и на верховых болотах средней тайги птиц в 1,6 раза мень-
ше, чем в сходных местообитаниях северной тайги, но в среднем по
сосново-боровым ландшафтам обеих подзон их примерно равное
количество.
Ярусное распределение птиц в средней тайге близко к таковому
в северной, лишь в лесах на первое место по обилию выходят крон-
ники и увеличивается доля птиц, держащихся на стволах деревьев,
что соответствует большей сомкнутости древостоев этой подзоны.
Участие в населении наземных птиц соответственно снижается. По-
казатели биомассы и количество трансформируемой птицами энер-
гии изменяются так же, как плотность населения птиц. При этом в
естественных ландшафтах суши они, как правило ниже, а в посел-
ках и на водоемах выше, чем в северной тайге.
В лесных местообитаниях по биомассе чаще всего преобладают
глухарь и юрок, и только во второй половине лета — рябчик. На
верховых болотах до середины лета обычно доминируют сизая чай-
ка и белая куропатка, а во второй половине — лесной конек, шило-
хвость, серая ворона и тетерев. В поселках преобладают сорока и
береговая ласточка, и только во второй половине лета — домовый
воробей и лесной конек, а на водоемах — чирок-свистунок и, кроме
того, в первой половине лета — чернозобая гагара, гоголь и сизая
чайка, а во второй — шилохвость и свиязь. Из числа видов, преоб-
ладающих по биомассе в северной тайге, в средней не встречены в
качестве доминантов чечетка, каменка, желтая трясогузка, сереб-
ристая чайка, зато впервые отмечены как преобладающие виды ряб-
чик, тетерев, пухляк, лесной конек, сизая чайка, свиязь и чернозобая
гагара.
Состав потребляемых птицами кормов сходен с таковым в пре-
дыдущей подзоне при несколько меньшем участии в рационе семян
и сочных плодов и большем — позвоночных (в основном за счет ры-
боядных видов). Максимальное видовое разнообразие отмечено в
приречных смешанных лесах и поселках (37—55 видов). Несколько
ниже оно в сосняках и на вырубках (29—49) и минимально на вер-
ховых болотах (23—39) и водоемах (17—28). Таким образом, на во-
доемах и верховых болотах средней тайги встречено примерно та-
кое же количество видов птиц, как и в сходных ландшафтах север-
ной тайги, а в среднетаежном сосново-боровом ландшафте и посел-
ках их разнообразие в 1,5 раза выше, чем в аналогичных северота-
ежных местообитаниях.
174
Фаунистический состав лесных сообществ птиц европейско-си-
бирский. На верховых болотах доля сибирских и европейских ви-
дов заметно меньше. На этих болотах, а особенно в поселках и на
водоемах, преобладают транспалеаркты. По сравнению с аналогич-
ными показателями для птичьего населения северной тайги в сред-
ней возрастает доля европейских видов и уменьшается участие си-
бирских видов и транспалеарктов.
В сходных местообитаниях среднетаежной долины Оби летом.
1972 г. чаще всего преобладали те же виды, что и на междуречьях.
Только в долине как доминанты отмечены белобровик, серая мухо-
ловка и белая куропатка, а только на междуречьях — теньковка,
еловик, овсянка-крошка и сизая чайка, но эти различия, скорее
всего, случайны. Показатели плотности населения, биомассы и био-
энергетики птиц в аналогичных урочищах долины лишь в 1,5—2 ра-
8а выше, чем на междуречьях, т. е. увеличение их по направлению
от междуречий к долине Оби гораздо слабее, чем в северной тайге^
Это связано главным образом с высокой численностью птиц за счет
большей интенсивности послегнездовых кочевок и осеннего пролета
в северотаежной части долины Оби по сравнению с более южными
подзонами [Равкин, 1978] и с междуречьями северной тайги.
В сходных местообитаниях долины и междуречий зарегистри-
ровано примерно равное видовое разнообразие птиц, и лишь в ни-
зовьях таежных рек (Ларь-Еган) в первой половине лета встречена
почти вдвое больше видов птиц, чем в их среднем течении (р. Бол.
Юган). Это связано в долине Оби с близостью Ларьеганского сора
и широкой поймы с высоким видовым разнообразием населения птиц.
Резких отличий в фаунистическом составе населения не наблюда-
ется, лишь доля представителей китайского типа фауны в сходных
местообитаниях увеличивается в долине по сравнению с между-
речьями.
ЮЖНАЯ ТАЙГА
Характеристика населения птиц этой подзоны опубликована
ранее Ю. С. Равкиным и И. В. Лукьяновой [1976], поэтому здесь
приводятся лишь основные различия между населением южной тай-
ги и более северных подзон.
В лесном ландшафте южной тайги главным доминантом стано-
вится пухляк, а юрок в отличие от средней и северной тайги почти
ни в одном из ландшафтных урочищ не входит в число преобладаю-
щих видов. В темнохвойных лесах южной тайги в качестве доминан-
та впервые отмечена московка. На верховых болотах здесь в отличие
от сходных средне- и северотаежных ландшафтов ни разу в качестве
доминантов не встречены овсянка-крошка, желтая трясогузка п
сизая чайка. Зато впервые в число преобладающих видов входят
лапландский подорожник (на пролете), жулан и обыкновенная ов-
сянка. Луговой чекан и дубровник характерны для обширных южно-
17У
таежных низинных болот, которые реже встречаются в более север-
ных подзонах и нами не обследованы.
В лесах южной тайги насчитывалось втрое больше птиц, чем
в более северных подзонах. В некоторой степени это связано с пре-
обладанием в северной и средней тайге сосняков, а в южной — бо-
лее продуктивных и многоярусных смешанных и мелколиственных
лесов. При сравнении только темнохвойных и смешанных лесов сле-
дует отметить, что в южной тайге суммарное обилие птиц соответст-
венно в 1,8 и 2,9 раза выше, чем в северной и средней. На верховых
болотах прослеживаются обратные соотношения. Так, в южной тай-
ге насчитывается в 1,7 и 1,1 раза меньше птиц, чем в северной и
средней, что обусловлено большей олиготрофностью верховых бо-
лот южной тайги в отличие от таких же болот в более северных под-
зонах.
В ярусном распределении птиц в отличие от северной тайги в
лесах выше доля кронников и ниже — наземных видов. По сравне-
нию со средней тайгой лишь немного возрастает доля птиц, кормя-
щихся в кронах во второй половине лета. Распределение птиц по
ярусам на верховых болотах южной тайги и более северных подзон
довольно сходно. Биомасса птиц и количество трансформируемой
ими энергии в южной тайге отличаются от более северных подзон
так же, как и плотность населения птиц. В среднем по подзоне здесь
отмечены в 2,3 и 3 раза большие показатели биомассы, чем в север-
ной и средней тайге. Величина трансформируемой энергии соответ-
ственно в 2,6 и 3 раза выше, чем в более северных подзонах. Среди
доминантов по биомассе в южной тайге впервые встречается кряква
(на верховых болотах). В соотношении потребляемых кормов не про-
слеживается значительных отличий по сравнению с более северными
подзонами, лишь несколько ниже потребление беспозвоночных и
соответственно выше — семян, сочных плодов и вегетативных час-
тей растений. Видовое разнообразие, так же, как и плотность насе-
ления птиц, в лесных ландшафтах несколько выше, чем в северной
и средней тайге, а на верховых болотах — заметно ниже. В фаунис-
тическом составе населения птиц южной тайги по сравнению со
средней и особенно с северной возрастает участие представителей
европейского и китайского типа фауны и снижается участие сибир-
ских видов и особенно транспалеарктов.
ПОДТАЕЖНЫЕ ЛЕСА
ЛЕСОПОЛЕВОЙ ЛАНДШАФТ
В осиново березовых лесах в первой половине
лета ни один из видов не входит в число доминантов. В этот период
самыми многочисленными были лесной конек, серая славка и вес-
ничка (по 9%, табл. 18). Во второй половине лета кроме первых двух
видов (15 и 12%), доминировали большая синица, пухляк и зяблик
176
Таблица 18
Население птиц лесополевого ландшафта подтаежных лесов Обь-Иртышского
междуречья летом 1978 г., особей/км?
Вид В среднем по ландшафту Мелколист- венные леса Поля-перелес- кж Луга-выпасы
1 2 3 4 5
Первая половина лета
Всего ... 387 436 313 327
Лесной конек 40 37 51 10
Весничка 32 40 23 12
Серая славка 32 40 23 11
Теньковка ’ 24 30 16 4
Зяблик 19 28 7 2
Пухляк 19 28 6 1
Большая синица 18 29 3 2
Ополовник 16 24 4 —
Серая мухоловка 13 22 0,8 —
Белошапочная овсянка 13 2 34 0,8
Садовая славка 13 15 11 1
Садовая камышевка 13 21 0,2 2
Серая ворона 11 0,5 26 32
Юрок И 17 3 —.
Пересмешка 8 14 — —
Горихвостка-лысушка 8 8 10 1
Скворец 8 6 6 35
Варакушка 8 4 13 14
Сорока 8 2 16 16
Чечевица 7 И 1 2
Галка 6 3 13 3
Желтая трясогузка 6 2 2 72
Славка-завирушка 6 8 3 ’ 0,2
Сверчки — обыкновенный и
пятнистый 6 9 —. 7
Князек 5 4 7 2
Луговой чекан 4 4 3 5
Дубровник 3 3 1 11
Белая трясогузка 3 2 4 2
Большой пестрый дятел 2 4 0,2 —
Клинтух 2 2 3 0,02
Рябинник 2 2 3
Иволга 2 1 4 0,5
Кукушка 2 2 1 2
Полевой воробей 1 — 4 —
Мухоловка-пеструшка 1 2 — 1
Жулан 1 2 —. .—
Веретенник 1 0,8 2 —
Черноголовый чекан 1 — — 19
Чернозобый дрозд 1 0,5 2 2
Тетерев 0,8 0,8 1 —
Сизый голубь 0,8 —. 2 1
Коростель 0,7 0,7 0,2 3
Бекас 0,7 1 —- 1
Чибис 0,5 — 0,06 8
Зеленая пеночка 0,5 0,8 0,02
12 л. Г. Вартапвтов
177
Продолжение табл. 18
1 1 2 3 4 5
Ласточка деревенская 0,4 — — 7
Перепел 0,4 .— 0,5 4
Серым журавль 0,4 0,6 ОД 0,3
Щегол 0,4 — 0,8 2
Степной конек 0,3 . — 0,02 5
Полевой лунь 0,3 0,5 — 0,2
Черныш 0,3 — — 5
Пищуха 0,3 0,5 — —
Славка ястребиная 0,3 0,5 ——• —
Глухая кукушка 0,2 0,4 — —
Грач 0,2 — 0,09 3
Сизая чайка од — 0,5 0,3
Желтоголовая трясогузка 0,2 — — 3
Барсучок 0,2 — — 3
Козодой 0,2 0,3 — —
Камышевая овсянка 0,2 — 0,5 —
Полевой жаворонок 0,1 — 0,2 1
Канюк 0,1 од 0,2 —
Пустельга 0,09 — 0,2 0,4
Большая горлица 0,07 — 0,2 —
Луговой лунь 0,07 — 0,2 —
Большой кроншнеп 0,06 — — 1
Чеглок 0,06 — — 1
Перевозчик 0,06 — — 1
Большой веретенник 0,06 — 1
Большой сорокопут 0,06 — — 1
Тетеревятник 0,06 0,1 — •—
Соловей красношейка 0,05 0,08 — —
Деряба 0,05 0,08 — •—
Лесной дупель 0,05 0,08 — ——
Фифи 0,03 — — 0,5
Травник 0,03 — — 0,5
•Большой подорлик 0,02 — 0,05 —
Белобровик 0,02 — 0,05 —
Черный стриж 0,02 0,02 — 0,05
Кобчик 0,01 — — 0,2
Кряква 0,01 — — 0,2
Чирок-свистунок 0,01 — — 0,2
Крачка речная 0,005 — — 0,08
Лебедь кликун 0,002 — — 0,03
Свиязь 0,001 — — 0,01
Красноголовый нырок 0,001 — — 0,01
Вторая половина лета
Всего • • • | 244 197 321 257
Лесной конек 33 30 42 12
Пухляк 25 23 33 1
Большая синица 23 27 18 7
Серая славка 20 24 16 4
Белошапочная овсянка 16 — 47 0,7
Зяблик 13 21 1 0,7
Весничка 12 3 28 7
Ополовник И 15 5 —
Варакушка И 2 22 2$
178
Продолжение табл. 18
1 1 2 3 4 5
Жулан 8 7 11 —
Князек 8 7 10 —
Теньковка 7 4 12 0,2
Луговой чекан 5 7 2 7
Серая мухоловка 5 7 3 —
Серая ворона 4 0,03 8 26
Желтая трясогузка 4 —-> 2 50
Сорока 4 — 8 12
Черноголовый чекан 3 —— 7 15
Дубровник 2 — 6 4
Садовая славка 2 2 2 4
Степной конек 2 — 3 14
Клинтух 2 — 5 2
Белая трясогузка 2 —’ 3 13
Камышевая овсянка 2 3 —— —
Большой пестрый дятел 1 2 0,7 1
Сизый голубь 1 — 3 7
Юрок 1 2 0,2 —
Мухоловка-пеструшка 1 2 — -—
Рябинник 1 — 3 0,7
Полевой воробей 1 — 3 0,2
Большая горлица 1 — 3 0,03
Деревенская ласточка 1 — — 16
Серый журавль Сверчки — обыкновенный и 0,9 1 0,8 0,3
пятнистый 0,9 1 0,7 0,7
Тетерев 0,9 0,3 2 —
Чечевица 0,8 — 2 2
Горихвостка-лысушка 0,8 0,7 1 —
Галка 0,8 — 0,4 10
Перепел 0,7 — 2 0,07
Канюк 0,6 1 — —
Сойка 0,6 1 — —
Черный дятел 0,6 0,7 0,4 —
Зеленая пеночка 0,5 — 1 2
Иволга 0,5 0,3 0,7 0,7
Пересмешка 0,4 0,7 — •—
Чеглок 0,4 0,7 — —
Горная трясогузка 0,4 0,7 —— «—
Пустельга 0,4 — 1 . 1
Желтоголовая трясогузка 0,3 — 1 1—
Щегол 0,2 — — 3
Черныш 0,2 0,3 -— -—
Фифи 0,1 —— 0,03 2
Большой подорлик 0,1 0,2 — 0,2
Вертишейка 0,1 0,2 — —
Тетеревятник о,1 — 0,3 —
Зеленый конек 0,1 —- 0,3 —
Большой крохаль 0,07 —• 0,2 —
Черный коршун 0,04 — — 0,7
Луговой лунь 0,04 — — 0,7
Глухая кукушка 0,04 0,07 — —
Кедровка 0,04 — 0,1 —
Барсучок 0,01 — — 0,2
Большой улит 0,001 — — 0,02
12*
179
Таблица 19
Плотность и разнообразие населения птиц подтаежных лесов Обь-Иртышского
междуречья летом 1978 г.
Ландшафт, урочище Плотность населения Отмечено видов
всего фоновых
I II 1 1 1 п I П
Половина лета
Лесополевой ландшафт 387 244 55 40 37 27
Мелколиственные леса 428 197 52 34 34 22
Поля-перелески 310 321 52 46 32 34
Луга-выпасы 326 257 61 40 44 25
Болотный ландшафт 318 200 40 22 23 15
Низинные болота, луга 347 222 52 34 33 24
Рямы 106 42 28 И 14 6
Поселки Водоемы (особей/10 км берего- 2626 2679 29 25 19 16
вой линии) 57 34 18 15 14 9
В среднем по ключевому
участку 375 246 46 32 29 21
(14, 12 и 11 %). Плотность населения птиц составляет 428 особей/км2
в первой половине лета и резко уменьшается во второй (197, табл. 19).
Почти половина птиц кормится в кустарниковом ярусе и заметно
меньше — в кронах деревьев и на земле (табл. 20).
Биомасса птиц в березово-осиновых лесах составляет 14 и
11 кг/км2, а величина трансформируемой ими энергии — 10 и
5 тыс. ккал/сут-км2 (табл. 21). В течение всего лета по биомассе до-
минирует серый журавль (23 и 50%). Энергетические потребности
населения птиц удовлетворяются в основном за счет беспозвоночных
при значительно меньшем участии семян и сочных плодов (7 и 6%)
и позвоночных животных (3 и 10%). Необычно высокая доля пот-
ребления позвоночных приходится на серого журавля, который не*
редко встречался по луговинам и на небольших кочкарниковых ни-
зинных болотах среди осиново-березовых лесов.
Видовое разнообразие довольно велико в первой половине лета
и значительно ниже во второй (всего встречено по периодам 52 и
34 вида птиц, из них 34 и 22 фоновые). По числу особей преобладают
европейские виды (64 и 53 %) при значительно меньшем участии си-
бирских (по 14%) и транспалеарктов (11 и 15%, табл. 22). На долю
европейско-китайских видов приходится еще меньшая часть насе-
ления (7 и 14%). Итак, фаунистический состав населения осиново-
березовых лесов довольно разнороден. Его можно считать сибирско-
европейским при значительном участии европейско-китайских ви-
дов и транспалеарктов.
В полях, чередующихся с перелесками,
отмечено два постоянных доминанта — лесной конек (16 и 13%)
м белошаиочная овсянка (11 и 15%). Во второй половине лета к
<80
Таблица 20
Ярусное распределение птиц в подтаежных лесах Обь-Иртышского междуречья
летом 1978 г., %
Ландшафт, урочище На земле В кустах В кронах На ство- лах На воде в воздухе
Первая половин а лета
Лесополевой ландшафт 37 41 21 0,7 од 0,2
Мелколиственные леса 26 46 27 1 —. 0,07
Поля-перелески 58 30 12 — 0,2 —
Луга-выпасы 68 27 2 — 0,2 2
Болотный ландшафт 43 53 2 — 1 0,6
Низинные болота, луга 42 55 1 — 0,9 0,6
Рямы 59 21 12 — 6 2
Поселки 82 0,8 од — — 17
Водоемы 58 — — — 30 12
В среднем по ключевому
участку 42 44 И 0,4 0,5 2
Вторая половин а лета
Лесополевой ландшафт 36 38 25 0,8 — 0,6
Мелколиственные леса 18 43 36 1 — 0,4
Поля-перелески 43 41 16 0,3 — —
Л у га-выпасы 60 29 5 0,4 — 6
Болотный ландшафт 31 64 3 0,3 — 1
Низинные болота, луга 30 67 1 — — 1
Рямы 41 2 50 7 0,2 —•
Поселки 73 0,3 0,2 — — 27
Водоемы 57 — — — 13 30
В среднем по ключевому
участку 42 44 И 0,4 0,5 2
ним добавляется пухляк (10%). Плотность населения птиц до се-
редины июля в 1,4 раза ниже, чем в мелколиственных лесах, а во
второй половине лета — в 1,6 раза выше. Большая часть птиц кор-
мится на земле, особенно в первой половине лета (58 и 43%), и в
кустарниках (30 и 41%). Значительно меньшая доля населения при-
ходится на кронников (12 и 16%).
Биомасса птиц и величина трансформируемой ими энергии в
полях-перелесках соответственно в 2 и 1,5 раза выше, чем в осиново-
березовых лесах. По биомассе все лето преобладает серая ворона
(45 и 18%), только в первой половине — сорока, только во второй —
серый журавль (12 и 20%). Энергетические потребности птиц в пер-
вой половине лета почти полностью удовлетворяются за счет беспоз-
воночных при незначительном участии семян и сочных плодов (94
181
Таблица 21
Биомасса и биоэнергетика населения птиц подтаежных лесов Обь-Иртышского
междуречья летом 1978 г.
Ландшафт, уро- чище Биомасса, кг/км2 Трансформируемая энергия
Всего, тыс. ккал/сут-км2 В том числе (%) за счет
беспозво- ночных семян, плодов вегетатив- ных ча- стей рас- тений позвоноч- ных
Первая половина лета
Лесополевой ландшафт 21 11 92 6 — 2
Мелколиственные леса 14 10 90 7 — 3
Поля-перелески 29 13 94 5 —> 0.9
Луга-выпасы 35 15 95 3 —• 2
Болотный ландшафт 24 10 77 13 0,7 8
Низинные болота, луга 26 11 78 14 0,3 8
Рямы 10 4 77 9 6 9
Поселки 150 93 61 38 — 0,3
Водоемы 15 4 62 0,3 33 5
В среднем по ключевому 24 | 83 |
участку 11 12 | 0,2 4
Вторая половина лета
Лесополевой ландшафт 16 7 76 17 — 6
Мелколиственные леса 11 5 84 6 — 10
Поля-перелески 22 10 68 28 — 4
Луга-выпасы 28 11 87 10 — 3
Болотный ландшафт 14 6 77 14 2 7
Низинные болота, луга 15 7 78 15 — 7
Рямы 11 2 59 6 35 0,06
Поселки 176 99 33 67 — 0,3
Водоемы 5 2 77 1 18 4
В среднем по ключевому участку 17 8 71 22 0,5 6
и 5%). Во второй половине доля беспозвоночных в рационе птиц
снижается, тогда как доля, семян и сочных плодов возрастает (68
и 28%).
Видовое разнообразие до середины лета такое же, как и в оси-
ново-березовых лесах, а во второй половине — в 1,4 раза выше.
Фаунистический состав населения птиц в полях-перелесках сибир-
ско-европейский при более высоком по сравнению с мелколиствен-
ными лесами участии транспалеарктов (23 и 22% по числу особей).
В лугах-выпасах все лето доминируют желтая тря-
согузка (22 и 19%) и серая ворона (по 10%), а также в первой поло-
182
Таблица 22
Фаунистический состав населения птиц подтаежных лесов Обь-Иртышского
междуречья летом 1978 г., %
Виды Особи
о о 0) о s а> 2g g 3
Ландшафт, урочище о 5 К К 58 fl § S о 3 « к За
g а. о fl 55 о и о о. о « i>y 55
о р< ё ь ag SSi о а S <о ев ь ag
п о S S к && 4) S § &S1
Первая половина лета
Лесополевой ландшафт 31 14 3 3 44 59 14 3 7 17
Мелколиственные леса 40 17 6 4 31 64 14 3 7 И
Поля-перелески 33 12 6 6 38 51 16 0,6 6 23
Луга-выпасы 31 10 3 5 44 28 33 4 6 54
Болотный ландшафт 29 12 4 3 46 24 8 11 1 42
Низинные болота, луга 27 8 4 4 51 23 5 12 1 44
Рямы 21 21 7 4 43 37 47 2 2 12
Поселки 14 10 7 10 52 8 0,2 0.4 2 85
Водоемы 6 6 — 6 72 0,9 7 — 4 80
В ср е д н е м по ключе вому
участку 30 14 3 3 45 41 10 1 6 4 3
Вторая половина лета
Лесополевой ландшафт 29 14 6 5 40 43 19 2 11 : 22
Мелколиственные леса 41 15 — 6 35 53 14 — 14 15
Поля-перелески 24 15 9 4 39 35 26’ 3 8 22
Луга-выпасы 32 12 8 5 38 16 1 3 7 64
Болотный ландшафт 26 13 3 3 50 16 9 15 — 47
Низинные болота, луга 29 9 3 3 50 17 7 16 — 49
Рямы 9 36 9 — 45 9 78 2 — И
Поселки 12 8 4 8 60 — — — 2 89
Водоемы 7 7 — 7 60 2 3 — 9 82
В среднем по ключевому
участку 26 14 5 4 45 27 12 7 6 40
вине лета — скворец, а во второй — варакушка (по 11%). Плот-
ность населения птиц до середины лета такая же, как в полях-пере-
лесках, а во второй его половине на 20% меньше, чем в предыдущем
ландшафтном урочище. По сравнению с рассмотренными выше ме-
стообитаниями в лугах-выпасах еще большая доля птиц кормится
на земле (68 и 60%), но высоко участие и кустарниковых птиц (27
и 29%).
Биомасса птиц здесь почти в 2,5 раза выше, чем в осиново-бе-
резовых лесах. Как и в полях-перелесках, это превышение прихо-
дится в основном на долю врановых птиц и в меньшей степени —
183
на голубей. Все лето по биомассе доминирует серая ворона (46 и
47%), и только в первой его половине — сорока (10%). Интенсив-
ность энергетического потока, проходящего через популяции птиц,
почти такая же, как в полях-перелесках. Соотношения основных
групп потребляемых кормов (в энергетическом эквиваленте) в этих
местообитаниях также близки между собой, лишь потребление се-
мян и сочных плодов (преимущественно зерна) после середины лета
в полях-перелесках значительно выше, чем в лугах-выпасах.
Видовое разнообразие здесь в среднем за лето наиболее высокое
(всего встречено по сезонам 61 и 40 видов птиц, из них 44 и 25 фоно-
вых). По числу особей в лугах-выпасах преобладают транспалеарк-
ты, а без их учета фаунистический состав населения можно считать
европейским (28 и 16%) при значительном участии европейско-ки-
тайских (6 и 7%), а во второй половине лета — монгольских видов
(5%, приходящихся на степного конька).
В среднем по ландшафту в течение всего лета до-
минирует лесной конек (10 и 14%), и во второй половине лета в ка-
честве содоминанта зарегистрирован пухляк (10%). Плотность на-
селения составляет 387 птиц на объединенный 1 км2 в первой поло-
вине лета и в 1,5 раза снижается во второй. Наибольшее и примерно
равное число птиц кормится на земле и в кустах. Доля кронников
в суммарном обилии птиц заметно ниже по сравнению с двумя пре-
дыдущими группами.
Биомасса птиц по периодам соответственно равна 24 и 14 кг/км2.
Доминируют серый журавль и серая ворона (11 и 31, 27 и 14%).
Интенсивность потока энергии, трансформируемого населением птиц,
составляет 11 и 7 тыс. ккал/сут-км2. Энергетические потребности
птиц более чем на 90% удовлетворяются за счет беспозвоночных
при невысоком участии в питании птиц семян, сочных плодов и поз-
воночных. В среднем на одно урочище в лесополевом ландшафте
встречено 55 и 37 видов птиц, в том числе 40 и 27 фоновые. Фаунис-
тический состав населения сибирско-европейский при высоком уча-
стии транспалеарктов, особенно по количеству видов.
БОЛОТНЫЙ ЛАНДШАФТ
На низинных болотах, чередующихся с лугами,
в течение всего лета доминируют черноголовый чекан (по 24%) и
дубровник (12 и 16%). После середины июля в число преобладающих
видов входят также камышевая овсянка и степной конек (12 и 11 %).
Суммарное обилие птиц в первой половине лета на 20% меньше,
чем в осиново-березовых лесах, а во второй — такое же (табл. 23).
В отличие от местообитаний лесополевого ландшафта на низинных
болотах подавляющее большинство птиц кормится в кустарниках
(55 и G7 %) и на земле (42 и 30%).
Биомасса птиц примерно в 1,5 раза выше, чем в осиново-бере-
зовых лесах (за счет большего количества крупных видов), а интен-
сивность энергетического потока, проходящего через популяции
134
Таблица 23
Население птиц болотного ландшафта подтаежных лесов Обь-Иртышского меж-
дуречья летом 1978 г., особей/км^
Вид В среднем по ландшафту Низинные боло- та Рямы
1 2 3 4
Первая половина лета
Всего | 319 | 348 | 106
Черноголовый чекан 72 82
Дубровник 35 40 2
Желтая трясогузка 24 27 0,5
Желтоголовая трясогузка 22 25 —
Барсучок 18 20 —
Степной конек 17 19 -—
Сверчки-обыкновенный и пятни- стый 13 15
Луговой чекан 12 14 «—
Лесной конек 11 10 15
Весничка 10 10 12
Серая славка 10 11 —
Скворец 8 9 —
Белошапочная овсянка 7 3 37
Варакушка 7 8 —
Полевой жаворонок 5 6 —
Сорока 5 5 2
Камышевая овсянка 4 4 .—
Перепел 4 4 —
Сизый голубь 4 4 —
Чибис 3 3 .—
Бекас 3 3 —
Сизая чайка 2 2 4
Кукушка 2 2 0,4
Серая ворона 2 2 од
Чечевица 2 2 0,02
Серый журавль 2 2 0,008
Деревенская ласточка 2 2 -—
Клинтух 2 2 -—
Коростель 2 2 —
Теньковка 1 — 8
Пухляк 1 — 8
Тетерев 0,9 1 0,5
Славка-завирушка 0,9 1 —
Садовая славка 0,9 1 —
Большой веретенник 0,9 1 —
Луговой лунь 0,9 1 •—
Черныш 0,7 0,8 0,1
Овсянка-крошка 0,6 0,5 1
Юрок 0,5 4
Большая горлица 0,5 — 4
Широконоска 0,4 0,5 —
Садовая камышевка 0,4 0,5 —
Каменка 0,4 0,5 —
Иволга 0,4 0,5 —
Чирок-трескунок 0,4 0,5 —
Степной лунь 0,4 0,4 —
185
Продолжение табл. 23
1 2 1 1 3 4
Кряква 0,2 — 2
Козодой 0,2 — 2
Горихвостка-лысушка 0,2 — 2
Болотная сова 0,2 0,2 —
Вертишейка 0,2 0,2 .—
Погоныш 0,2 0,2
Средний кроншнеп 0,2 0,2 —
Кобчик 0,09 0,1 —
Речная крачка 0,09 0,1 —
Пустельга 0,09 0,1 .—
Фифи 0,07 0,05 0,2
Перепелятник 0,06 0,5
Ворон 0,06 0,5
Лебедь кликун 0,02 — 0,2
Белая трясогузка 0,02 0,02 —
Грач 0,02 0,02 —>
Канюк 0,002 — 0,02
Лапландский подорожник 0,002 — 0,02
Свиязь 0,002 — 0,02
Втор ая половин а лета
Всего 200 222 42
Черноголовый чекан 48 54
Дубровник 31 35 1
Камышевая овсянка 24 27
Степной конек 21 24
Серая славка Сверчки обыкновенный и пят- 11 13 —
нистый 11 12 .
Перепел 8 9
Теньковка 8 9
Луговой чекан 6 7 ,
Пухляк 4 2 22
Тетерев 3 3
Деревенская ласточка 3 3
Лесной конек 2 2 4
Фифи 2 2 0,7
Желтоголовая трясогузка 2 2
Полевой жаворонок 2 2
Коростель 2 2
Барсучок 2 2
Желтая трясогузка 2 2 .
Белошапочная овсянка 2 1 7
Вертишейка 0,9 1
Серый журавль 0,9 1
Жулан 0,9 1
Луговой лунь 0,9 1
Серая ворона 0,9 0,9 0,7
Весничка 0,8 0,9
Сорока 0,7 0,8 0,7 0,7
Кукушка 0,7 0,3
Большая горлица 0,6
Белая трясогузка 0,6 0,7 —
186
Окончание табл. 23
* 1 2 1 3 4
Большой пестрый дятел 0,4 — 3
Глухарь 0,4 — 3
Большой подорлик 0,2 0,2 —
Клинтух 0,09 0.1 —
Чибис 0,06 0,07 —•
Большой крохаль 0,06 0,07 —
Свиристель 0,04 — 0,3
Речная крачка 0,008 — 0,07
птиц, в сравниваемых местообитаниях примерно равна. Все лето
по биомассе доминирует серый журавль (42 и 36%), и только во вто-
рой его половине — тетерев (19%). По энергетической значимости
в рационе птиц, так же как и в лесополевом ландшафте, преоблада-
ют беспозвоночные при сравнительно высоком участии семян и соч-
ных плодов (в среднем — 76 и 14%).
На низинных болотах зарегистрировано примерно такое же чис-
ло встреченных и фоновых видов птиц, как и в мелколиственных
лесах. По числу особей преобладают транспалеаркты, а без их уче-
та фаунистический состав населения китайско-европейский.
В рямах, расположенных островами среди низинных бо-
лот, в течение всего лета преобладает белошапочная овсянка (35 и
17%), кроме нее в первой половине лета доминируют лесной конек
и весничка (14 и 11 %), а во второй — пухляк (52%). Здесь держится
почти вчетверо меньше птиц, чем на низинных болотах. Распреде-
ление птиц по ярусам в первой половине л$т? близко к таковому
в полях-перелесках, а во второй — резко возрастает участие кронни-
ков, на долю которых приходится около половины суммарного оби-
лия птиц.
Биомасса п величина трансформируемой птицами энергии здесь
соответственно в 2 и 3 раза ниже, чем на низинных болотах. В пер-
вой половине лета наибольшая доля суммарной биомассы прихо-
дится на крякву, серебристую чайку, лебедя-кликуна и белошапоч-
ную овсянку (25, 18, 14 и 11 %), а во второй половине — на глуха-
ря (87%). Соотношение основных групп потребляемых кормов до
середины лета сходно с таковым на низинных болотах, а позже рез-
ко возрастает потребление вегетативных частей растений.
Видовое разнообразие птиц по периодам в рямах в 2 и 3 раза
меньше, чем на низинных болотах и в осиново-березовых лесах*
В отличие от всех предыдущих местообитаний в рямах преобладают
представители сибирского (таежного) типа фауны, а фаунистический
состав населения европейско-сибирский.
В среднем по ландшафту на протяжении всего
лета доминируют черноголовый чекан и дубровник (по 24, 12 и 16%).
Во второй половине лета в число доминантов входят также камыше-
вая овсянка и степной конек (12 и 10%). Последний встречался по
участкам покосных лугов в сочетании с солонцами. По среднелетним
показателям, в болотном ландшафте держится в 1,2 раза меньше
187 ,
птиц, чем в лесополевом. Биомасса и величина трансформируемой
энергии в сравниваемых ландшафтах довольно сходны.
Все лето по биомассе преобладает серый журавль (39 и 32%),
и, кроме того, во второй половине лета — тетерев и глухарь (17 и
11%). В первой половине лета среди потребляемых кормов доля
беспозвоночных несколько ниже, чем в лесополевом ландшафте,
при более высоком участии семян, сочных плодов и позвоночных.
После середины июля долевое участие кормов в их энергетическом
эквиваленте в обоих ландшафтах сходно. Видовое разнообразие и
число фоновых видов птиц в болотном ландшафте в 1,4—1,8 раза
ниже, чем в лесополевом. Примерно половина встреченных видов и
суммарного обилия птиц приходится на транспалеарктов. На вто-
ром месте представители европейского типа фауны, которых, как и
сибирских видов, здесь заметно меньше, чем в предыдущем ланд-
шафте. Значительно возрастает по сравнению с лесополевыми место-
обитаниями доля европейско-китайского и особенно китайского фа-
унистического типа вместе взятых, которые выходят на третье ме-
сто по долевому участию в населении птиц. Таким образом, если
не принимать во внимание транспалеарктов, фаунистический состав
населения по числу видов сибирско-европейский, а по количеству
особей китайско-европейский.
ПОСЕЛКИ
В населенных пунктах в течение всего лета по обилию преоб-
ладают домовый воробей (62 и 56%) и деревенская ласточка (17 и
27%). В среднем за лето здесь держится в 9 раз больше птиц, чем
в естественных ландшафтах суши (табл. 24). Больше всего птиц кор-
мится на земле (82 и 73%), а остальная часть населения приходится
в основном на воздухореев (17 и 27%).
Показатели биомассы и величины трансформируемой энергии
в поселках в среднем в 8 и 11 раз выше по сравнению с предыдущими
ландшафтами подтаежных лесов. В течение всего лета по биомассе
преобладают домовый воробей (33 и 27%), сизый голубь (25 и 38%)
и серая ворона (12 и 16%), и только в первой половине лета — скво-
рец (10%). По энергетической значимости в рационе птиц в первой
половине лета превалируют беспозвоночные, а на втором месте —
семена и плоды (61 и 38%). После середины лета соотношение этих
групп кормов меняется в пользу семян и сочных плодов (33 и 67%)
преимущественно за счет зерноядных видов (сизый голубь). Видовое
разнообразие птиц в населенных пунктах почти вдвое ниже, чем
в лесополевом ландшафте. По числу особей в населении абсолютно
преобладают транспалеаркты (85 и 89%).
ВОДОЕМЫ
На мелких реках (верхнее течение р. Омь) по обилию
все лето доминирует деревенская ласточка (12 и 29%), только в пер-
вой половине лета — перевозчик, а во второй — черныш (25 и 15%),
188
Таблица 24
Летнее население птиц поселков подтаежных лесов Обь-Иртышского между-
речья в 1978 г., особей/км1
Вид Плотность Вид ПЛОТНОСТЬ
Первая половина лета
Всего . . . | 2626 || 1 1
Домовый воробей 1616 Юрок 2
Деревенская ласточка 458 Перепелятник 1
Скворец 200 Черный коршун 1
Сизый голубь 126 Садовая камышевка 1
Полевой воробей 68 Желтая трясогузка 0,6
Сорока 50 Каменка 0,5
Серая ворона 36 Перевозчик 0,5
Галка 23 Лесной конек 0,5
Варакушка 10 Желтоголовая трясогузка 0,5
Белая трясогузка 9 Пустельга 0,2
Дубровник 6 Щегол 0,2
Черныш 6 Грач 0,2
Чечевица 4 Белошапочная овсянка 0,2
Овсянка-крошка 4 Сизая чайка 0,05
Большая синица 2
Вторая половина лета
Всего ... 2679 ||
Домовый воробей 1506 фифи 1
Деревенская ласточка 722 Черныш 1
Сизый голубь 220 Черный коршун 1
Серая ворона 58 Весничка 0,7
Сорока 48 Перепелятник 0,7
Желтая трясогузка 30 Полевой лунь 0,7
Галка 29 Степной конек 0,7
Полевой воробей 27 Черный дятел 0,1
Белая трясогузка 18 Большой улит 0,07
Большая синица 9 Большой кроншнеп 0,07
Каменка 3 Чирок-свистунок 0,03
Дубровник 2 Горлица большая 0,03
Лесной конек 1
Суммарное обилие птиц по периодам составляет 57 и 34 особи на 10 км
береговой линии (табл. 25). Более половины птиц кормится по бере-
гу около уреза воды (58 и 57%), значительно меньше —• в воде (30
и 13%) и в воздухе (12 и 30%).
Биомасса птиц и величина трансформируемой ими энергии со-
ставляют по периодам 15 и 5 кг и 4 и 2 тыс. ккал/сут-10 км берего-
вой линии. В течение всего лета по биомассе преобладают свиязь
(20 и 14%)» чирок-свистунок (12 и 20%) и серая ворона (17 и 29%).
Кроме того, в число доминантов по биомассе в первой половине лета
входит кряква, а во второй — сорока (26 и 13%). Энергетические
189
Таблица 26
Летнее население птиц мелких рек подтаежных лесов Обь-Пртышского между*
речья в 1978 г., особей /10 км береговой линии
Вид Плотность 1 вид Плотность
Первая пол овина лета
Всего ... 57 S
Перевозчик 14 I Сизая чайка 2
Деревенская ласточка 7 1 Чирок-трескунок 2
Серая ворона 5 Белая трясогузка 1
Чирок-свистунок 5 Черныш 1
Желтая трясогузка 4 Большой веретенник 1
Желтоголовая трясогузка 4 Речная крачка 0,5
Свиязь 4 Луговой лунь 0,5
Кряква 3 Полярная крачка 0,5
Сорока 2 Большая горлица 0,2
Вторая половина лета
Всего ... 34 | 1 1
Деревенская ласточка ю I Свиязь 1
Черныш 5 1 Желтоголовая трясогузка 0.7
Серая ворона з В Луговой лунь 0,7
Чирок-свистунок з Чирок-трескунок 0,3
Сорока 3 1 Желтая трясогузка 0,3
Перевозчик з 1 Степной конек 0,3
Фйфи 2 1 Сизый голубь 0,3
Белая трясогузка 1 D
потребности птиц удовлетворяются главным образом за счет беспоз-
воночных (62 и 77 %) при сравнительно высоком участии вегетативных
частей растений (33 и 18%). Видовое разнообразие птиц здесь даже
ниже, чем в поселках. За эту и другую половину лета встречено соот-
ветственно 18 и 15 видов птиц, из них 14 и 9 фоновые. По числу осо-
бей преобладают транспалеаркты (80 и 62%).
♦
♦ *
Таким образом, в подзоне подтаежных лесов чаще всего доми-
нирует по обилию лесной конек в осиново-березовых лесах, полях-
перелесках и рямах. Для лесных местообитаний в качестве доминан-
тов весьма характерны серая славка и во второй половине лета —
большая синица и пухляк. Наиболее специфичен состав доминантов
на низинных болотах (черноголовый чекан и дубровник) и в посел-
ках (домовый воробей и деревенская ласточка). Последняя также
входит в число преобладающих видов на водоемах вместе с перевоз-
чиком и чернышом. Доминирующие виды подтаежных лесов и южной
190
тайги довольно сходны по составу, но значительно отличаются от на-
бора доминантов средней и особенно северной тайги. По сравнению
с более северными подзонами в подтаежных лесах возрастает участие
лесного конька и белошапочной овсянки в птичьем населении, а доля
и широта доминирования пухляка, наоборот, резко снижаются. В от-
личие от южной тайги здесь ни разу не встречены в качестве доми
пирующих видов московка, теньковка и поползень, зато впервые
отмечены как преобладающие виды серая славка, черноголовый че-
кан, степной конек и серая ворона.
Максимальная плотность населения отмечена в поселках (в сред-
нем за лето примерно 2,6 тыс. особей/км2). Соответственно в 8 и 9 раз
меньше птиц держится в лесополевом ландшафте и на низинных бо-
лотах и в 34 раза меньше в рямах. В среднем по обследованной терри-
тории суммарное обилие птиц в подтаежных лесах в 1,5 и 2 раза выше,
чем в северной и средней тайге, и в 4 раза ниже, чем в южной. При-
мерно такой же характер отличий прослеживается в большинстве
сходных ландшафтов и урочищ подтаежных лесов и остальных под-
зон. Только на верховых болотах подтаежных лесов птиц даже не-
много меньше, чем в южной тайге.
Распределение птиц по ярусам в целом соответствует соотно-
шению последних в разных местообитаниях подтаежных лесов. По
сравнению с более северными подзонами значительно возрастает
удельный вес кустарниковых птиц. Наибольшая биомасса птиц
зарегистрирована в поселках (в среднем за лето — 161 кг/км2). В 5—
6 раз меньшие показатели характерны для лесолугополевых место-
обитаний и низинных болот и в 12—16 раз меньшие для осиново-бере-
зовых лесов и рямов. Для птичьего населения подтаежных лесов
и более северных подзон характерны примерно такие же различия
по биомассе, как и по плотности населения. Среди доминантов по
биомассе в подтаежных лесах наиболее широко распространены серая
ворона (в лугово-полевых местообитаниях) и серый журавль (на ни-
зинных болотах и в мелколиственных лесах). Кроме того, в поселках
в качестве преобладающих видов по биомассе чаще всего отмечались
домовый воробей и сизый голубь, а на водоемах — свиязь и чирок-
свистунок. По сравнению с более северными подзонами в подтаеж-
ных лесах из списка доминантов выпадают белая куропатка, бекас,
рябчик, зато впервые встречен сизый голубь.
Территориальные различия в интенсивности энергетического
потока, проходящего через популяции птиц, в подтаежных лесах
примерно такие же, как в отношении плотности их населения. Боль-
шая часть энергетических потребностей птиц удовлетворяется за
счет беспозвоночных при незначительном участии остальных групп
кормов. Только на водоемах и в рямах возрастает потребление веге-
тативных частей растений, а в поселках — семян и сочных плодов,
особенно во второй половине лета за счет зерноядных видов.
Видовое разнообразие птиц выше всего в лесополевом ландшаф-
те (55 и 40 видов), почти такое же на низинных болотах и вдвое
ниже в поселках и рямах. В пересчете в среднем на одно урочище
видовое разнообразие птиц в подтаежных лесах примерно такое же,
191
как в средней и южной тайге, но в 1,2 раза больше, чем в северной.
Фаунистический состав населения лесополевых местообитаний обыч-
но сибирско-европейский. В открытых местообитаниях (на низин-
ных болотах и в лугах) преобладают транспалеаркты, если их не при-
нимать во внимание, то состав населения можно считать китайско-
европейским или европейским. Для рослых рямов характерен евро-
пейско-сибирский фаунистический состав, а в поселках и на водо-
емах абсолютно преобладают транспалеаркты. По сравнению с более
северными подзонами в подтаежных лесах резко сокращается учас-
тие представителей сибирского типа фауны, но несколько возрастает
доля европейских, китайских видов и транспалеарктов (последних
по сравнению только со срединными таежными подзонами).
В сходных местообитаниях подтаежных лесов в пределах долины
Оби в 1970 г. чаще всего доминировали те же виды, что и на между-
речьях. Только в долине Оби в качестве доминантов встречался ря-
бинник, а на междуречьях — черноголовый чекан и белошапочная
овсянка. Показатели плотности населения, биомассы и биоэнергетики
птиц в сходных урочищах долины Оби в 1,6 — 3 раза выше по сравне-
нию с междуречьями. Видовое разнообразие населения в первой
половине лета на междуречьях примерно такое же, как и в долине
Оби, а во второй — в 1,4 раза меньше. В подтаежных лесах на между-
речьях несколько возрастает участие транспалеарктов и соответствен-
но снижается доля сибирских видов птиц.
ВНУТРИЗОНАЛЬНЫЕ ИЗМЕНЕНИЯ
НАСЕЛЕНИЯ ПТИЦ
ВРЕМЕННАЯ ДИНАМИКА ЛЕТНЕГО НАСЕЛЕНИЯ ПТИЦ
СЕЗОННЫЕ ЯВЛЕНИЯ В ЖИЗНИ ПТИЦ
В связи с пред- и послегнездовыми перемещениями птиц их на-
селение в течение летнего периода постоянно меняется. Количество
мигрантов оценивалось по числу птиц, отдыхающих и кормящихся
на обследованной территории [Равкин, 1973]. Птицы, пролетающие
транзитом, пересчитывались на 1 км2 с поправкой на скорость пере-
мещения, поэтому их доля в населении невелика. Приведенные в пре-
дыдущем разделе показатели числа мигрирующих и гнездящихся
птиц для отдельных видов суммировались для населения в целом в
среднем по территориям ключевых участков. Подобные расчеты
проведены по 120 видам, преимущественно фоновым, из 174, встре-
ченных нами на таежных междуречьях. Они позволяют отличать
перераспределения птиц из одних урочищ и ландшафтов в другие
внутри обследованных площадей от прикочевки извне или откочевки
за пределы ключевых участков,
192
Интенсивность весеннего пролета оценивалась по превышению»
показателей обилия птиц в конце весны и начале лета над плот-
ностью их гнездования. Поэтому напряженность весеннего пролета
характеризуется не только транзитным перемещением птиц, но и их
местным предгнездовым перераспределением. Поскольку начало
наших наблюдений, как правило, совпадало с окончанием пролета,
местные перемещения птиц, по-видимому, преобладали над тран-
зитными.
Послегнездовое уменьшение численности принималось за отко-
чевку, т. е. не оценивалась величина гибели молодых птиц. Кроме
того, послегнездовая откочевка птиц часто бывает завуалирована
их прикочевкой, которая в ряде случаев происходит одновременно
с вылетом молодых.
Несмотря на указанные допущения, полученные результаты
достаточны для выявления географических различий сезонных ас-
пектов орнитокомплексов и оценки влияния мигрантов на состав
и плотность летнего населения птиц лесной зоны Западной Сибири*
Предгнездовой пролет. Наиболее интенсивный про-
лет птиц, как и начало их гнездования, повсеместно проходил до на-
чала наших наблюдений. В северной тайге количество пролетных
птиц в первой половине июня значительно больше, чем во второй,—
93 и 9 особей/км2 ключевого участка (рис. 3, а). В начале июня среди
мигрантов преобладают овсянка-крошка, юрок, чечетка и белая тря-
согузка (по 11—16%)*, а в конце этого месяца — каменка и гори-
хвостка-лысушка (33 и 22%).
В средней тайге зарегистрирована примерно такая же интенсив-
ность пролета, что и в северной, но проходит он на две недели рань-
ше. Среди пролетных птиц здесь тоже преобладают юрок и белая тря-
согузка в конце мая (22 и 12%), с середины мая — теньковка и в пер-
вой половине июня — овсянка-крошка (по 17%).
В южной тайге интенсивность пролета втрое больше, чем в пре-
дыдущих подзонах. Во второй половине мая идет пролет преимущест-
венно юрка и теньковки (20 и 32%). Суммарное количество мигран-
тов на объединенный 1 км2 ключевого участка примерно такое же,
как в средней тайге. В начале июня напряженность пролета возраста-
ет вдвое, в основном за счет пролета лапландского подорожника
(59%), а в конце месяца становится минимальной. В июне среди про-
летных птиц сравнительно велико участие садовой камышевки (19%)*
Завершают предгнездовой пролет в конце июня иволга и таловка
и в начале июля — певчий сверчок.
В подтаежных лесах пролетных птиц примерно в 2,5 раза боль-
ше, чем в северных подзонах, но на 20% меньше, чем в южной тайге
(за весь период пролета). В конце мая мигрантов в подтаежных ле-
сах даже больше, чем в южной тайге, а к концу июля число их сни-
жается по сравнению с предыдущим периодом более чем в 5 раз. Сре-
♦ Здесь и далее в скобках указывается суммарное обилие птиц на объеди-
ненный 1 км2 ключевого участка, а для доминантов — доля в процентах от
этого показателя.
13 л, г. Вартапетов
193
Рис. 3. Динамика летних миграций и плот-
ность гнездования птиц таежных междуречий
Западной Сибири.
Интенсивность, особей/км2: а — предгнездового про-
лета, б — гнездования, в — послегнездовой откочев-
ки, г — послегнездовой прикочевки; 1 — северная,
2 — средняя, 3 — южная тайга, 4 — подтаежные
леса.
Рис. 4. Видовое разнообразие гнез-
дящихся и мигрирующих птиц в лес-
ной зоне Западной Сибири.
а — предгнездовой пролет, б — гнездо-
вание, в — послегнездовая откочевка, г —
послегнездовая прикочевка; 1 — Обь-
Енпсейское и Обь-Иртышское междуречья,
2 — долина Оби.
ди пролетных птиц во второй половине мая преобладает черноголо-
вый чекан (9%), в течение всего июня — барсучок (19%), кроме того,
в первой половине этого месяца — дубровник и степной конек (21
и 14%), а во второй — обыкновенный сверчок и серая славка (35
и 40%).
Итак, наибольшая интенсивность пролета в летний период за-
регистрирована в конце мая, и только в южной тайге максимальное
194
количество пролетных птиц наблюдалось в первой половине июня
(за счет лапландского подорожника). Завершение пролета обычно
приходится на конец июня, лишь в средней тайге, по-видимому
в связи с ранней и теплой весной 1977 г., он закончился к середине
июня. В собственно таежных подзонах на пролете преобладают лесные
и лесотундровые виды птиц, а в подтаежных лесах — птицы преиму-
щественно открытых лугово-кустарниково-болотных местообитаний*
В долине Оби напряженность пролета значительно выше, особенна
в северных подзонах. Так, если в подтаежной и южно-таежной доли-
не Оби мигрантов всего в 1,4 и 2,2 раза больше, чем на междуречьях
тех же подзон, то в средней и северной тайге отмечено соответствен-
но 12- и 20-кратное превышение. Таким образом, с юга на север лес-
ной зоны Западной Сибири пролет птиц в долине Оби становится
более интенсивным по сравнению с междуречьями. Возможно, эта
связано с малой экологической привлекательностью обследованных
нами районов северо- и среднетаежных междуречий, где преобладают
верховые болота и лишайниково-кустарничковые сосняки. Крома
того, в южных подзонах большая часть птиц пролетает, по-видимо-
му, до начала наших наблюдений, тогда как в северных во второй
половине мая — начале июня пролет идет еще весьма интенсивно*
При этом большинство пролетных лесотундровых и тундровых птиц
отдают предпочтение долине по сравнению с междуречьями. Видовое
разнообразие мигрантов в долине, как правило, выше, чем на меж-
дуречьях, и только в подтаежных лесах наблюдается обратное соот-
ношение (рис. 4, а).
Гнездование. У многих видов гнездовой период начинает-
ся до начала наших наблюдений. В северной тайге в первой поло-
вине июня, а в остальных подзонах во второй половине мая показа-
тели суммарного обилия гнездящихся птиц уже близки к максималь-
ным за весь период (см. рис. 3, б). Так, в северной тайге количества
гнездящихся птиц почти не изменяется во второй половине июня
по сравнению с первой (159 и 152 особей/км2), а в начале июля умень-
шается втрое по сравнению с предыдущим периодом. В июне среди
гнездящихся видов преобладают желтая трясогузка и овсянка-крош-
ка (30 и 10 %), в первой половине июля — юрок, фифи и каменка
(22, 15 и 13%), а во второй еще продолжают гнездиться краснозобый
конек, щур, полевой и домовый воробьи.
В средней тайге плотность гнездования в 1,4 раза ниже, чем
в северной. Максимальное число гнездящихся птиц отмечено в пер-
вой половине июня, после чего оно несколько уменьшается во вто-
рой половине июня и особенно резко падает в первой половине июля
(117, 93 и 8). Среди гнездящихся птиц с середины мая до конца июня
преобладают лесной конек и юрок (18 и 10%), а в первой половине
июля — теньковка и садовая славка (50 и 38%).
В южной тайге плотность гнездования соответственно в 1,6
и 2,2 раза выше, чем в северной и средней. Суммарное обилие гнез-
дящихся птиц возрастает со 181 до 231 особи на объединенный 1 км2
со второй половины мая к первой половине июня, а затем к концу
июня становится таким же, как в конце мая, В июле число гнездя-
13 ♦
195
щихся птиц последовательно снижается в первой и второй половине
месяца (62 и 5). С 16 мая по 30 июня на гнездовании преобладают
пухляк, а с 16 мая по 15 июля — теньковка (14 и 12%). Только в пер-
вой половине июля среди гнездящихся птиц доминирует дубровник
и во второй — певчий сверчок (21 и 80%).
В подтаежных лесах плотность гнездования еще выше, чем в пре-
дыдущих подзонах (соответственно в 1,8; 2,5 и 1,1 раза). Наиболь-
шее число гнездящихся птиц, так же как в средней и южной тайге,
зарегистрировано в первой половине июня (239). Затем этот пока-
затель последовательно снижается во второй половине июня и пер-
вой половине июля соответственно в 1,2 и 3,8 раза. Во второй поло-
вине мая преобладает скворец (И %), с середины мая до конца июня —
черноголовый чекан и весничка (14 и 13%), с середины июня до се-
редины июля — серая славка, и только в первой половине июля до-
минируют обыкновенный сверчок и теньковка (по 16%).
Таким образом, на таежных междуречьях Западной Сибири наи-
большее суммарное обилие гнездящихся птиц зарегистрировано
в первой половине июня, а окончание гнездования обычно проис-
ходит в первой половине июля. В северной тайге в связи с фенологи-
ческими различиями гнездование запаздывает по сравнению с более
южными подзонами примерно на две недели. Плотность гнездова-
ния уменьшается к северу в соответствии со снижением теплообес-
печенности и продуктивности биоценозов, но вновь возрастает
в северной тайге по сравнению со средней за счет высокой численности
лесотундровых птиц, в том числе на комплексных верховых болотах.
Для долины Оби характерны сходные изменения плотности гнездо-
вания со сменой широты местности, однако показатели здесь в 1,7—
3 раза выше, чем па междуречьях; особенно велико это превышение
в северной тайге. Видовое разнообразие гнездящихся птиц как в до-
лине, так и на междуречьях снижается к северу от южной тайги
и в подтаежных лесах (см. рис. 4, б). Последнее, по-видимому, обус-
ловлено небольшим набором местообитаний в этой подзоне (преобла-
данием массивов осиново-березовых лесов в сочетании с полями и
низинных болот).
Послегнездовая откочевка начинается одно-
временно с гнездованием значительного числа птиц (см. рис. 3, в).
В северной тайге количество откочевывающих неуклонно возрастает
со второй половины июня до первой половины августа (с 17 до 143),
а затем к концу августа снижается вдвое по сравнению с предыдущим
периодом. В конце июня откочевывают рано гнездящиеся птицы,
преимущественно кукша и клест-еловик. В начале июля в откочевку
вовлекается большее число особей и видов птиц. Преобладают среди
откочевывающих видов весничка, зеленый конек, черноголовый че-
кан и береговая ласточка (27, 23, 16 и 11%). Кроме того, с начала
июня до конца августа идет откочевка наиболее массового вида се-
веротаежных междуречий — желтой трясогузки (11 %). По видимому,
часть желтых трясогузок откочевывает сразу после окончания гнездо-
вания, а затем их сменяют птицы, отгнездовавшиеся севернее —
в северотаежных редколесьях и лесотундре, что определяет столь
196
длительную откочевку этого вида. В начале августа число птиц,
покидающих северотаежные междуречья, еще увеличивается, среди
них преобладают белая трясогузка, юрок, овсянка-крошка и камен-
ка (21, 15, 14 и 13%), а во второй половине августа к ним присоеди-
няются чечетка и сероголовая гаичка (32 и 11%).
В средней тайге откочевывающих птиц лишь на 5% больше, чем
в северной, но откочевка начинается значительно раньше — уже
в первой половине июня (9). Ее интенсивность возрастает до сере-
дины июля, а затем снижается к концу августа (соответственно 112
и 41). Начинает откочевку в первой половине июня клест-еловик.
Во второй половине июня более половины откочевывающих птиц со-
ставляют лесные коньки при высоком участии снегиря и певчего
дрозда (54, 18 и 14%). В июле среди откочевывающих птиц преобла-
дает юрок, а в августе — опять лесной конек (11 и 22%)*, кроме
того, в первой половине августа — овсянка-крошка и во второй —
желтая трясогузка (13 и 27%).
В южной тайге интенсивность откочевки изменялась примерно
так же, как в средней, но наибольшие ее значения в первой половине
июля почти втрое выше, чем в предыдущей подзоне. Количество от-
кочевывающих птиц здесь в 1,7 раза больше, чем в средней тайге.
Наиболее рано, в середине июня, начинают откочевку клест-еловик,
поползень и желтоголовый королек (39, 22 и 20%), а к концу этого
месяца среди откочевывающих птиц уже преобладают мухоловка-
пеструшка и певчий дрозд (33 и 20%). В первой половине июля их
сменяют пухляк и юрок, а во второй — теньковка и горихвостка-
лысушка (34, 13, 28 и 14%). С начала до середины августа среди
птиц, покидающих южно-таежные междуречья, преобладает снегирь,
а в конце этого месяца — дубровник (29 и 69%).
В подтаежных лесах количество откочевывающих птиц соответст-
венно в 2,5; 2,4 и 1,4 раза больше, чем в предыдущих подзонах. Мак-
симальная интенсивность откочевки, как и в северной тайге, наблю-
дается в первой половине августа. В первой половине июля среди
птиц, покидающих подтаежные леса, доминируют скворец и юрок
(68 и 22%), во второй половине месяца их сменяют князек и лесной
конек (39 и 33%), а в первой половине июля — весничка и садовая
славка (37 и 10%). В первой половине августа преобладают черно-
головый чекан, дубровник и камышевая овсянка (21, 14 и 11%),
во второй — деревенская ласточка (12%).
Итак, общее количество откочевывающих птиц снижается с юга
на север таежных междуречий в соответствии с запаздыванием фе-
нологического развития природы. В долине Оби прослеживается
сходная тенденция. Доля птиц, покидающих таежные междуречья
после окончания гнездования, от числа местных (тех, что гнездились
и вывелись) значительно выше, чем в долине Оби, где в летний пе-
риод преобладает иммиграция. Видовое разнообразие откочевываю-
♦ Вторая волна откочевки лесного конька в августе, как указывалось в
повидовом обзоре, по-видимому, связана с одновременным вылетом молодых
второго цикла размножения и пролетом птиц более северных популяций.
197
щих птиц повсеместно снижается к северу и югу от средней тайги
(см. рис. 4, в). Увеличение числа встреченных видов в последней
связано с одновременными кочевками таежных видов и птиц, гнез-
дящихся севернее, в северотаежных редколесьях и лесотундре.
При этом на междуречьях число откочевывающих видов повсеместно
больше, чем в долине Оби.
Послегнездовая прикочевка обычно начинает-
ся через две недели после начала откочевки местных птиц и одновре
менно с окончанием гнездования (см. рис. 3, г). Ее интенсивность
в северной тайге в конце июня — начале июля невелика, но резко
возрастает в конце июля (10 и 65), а затем в августе снижается при-
мерно в 1,5 раза. Наиболее рано (с 15 июня по 15 июля) идет при-
кочевка клестов (еловика и белокрылого). Во второй половине июля
среди прикочевывающих птиц преобладают белая трясогузка, ка-
менка и весничка (25, 17 и 11%), с середины июля по 15 августа —
сероголовая гаичка, а только в первой половине этого месяца —
чечетка и большой пестрый дятел (И, 34 и 22%). Во второй половине
августа их сменяют овсянка-крошка, весничка и краснозобый конек
(19, 16 и 14%).
В средней тайге общее количество прикочевавших птиц в 1,6
раза больше, чем в северной. Интенсивность прикочевки неуклонно
возрастает с середины июня до конца августа (с 22 до 92). Первыми
(в конце июня) прикочевывают большой пестрый дятел и белокры-
лый клест *. В первой половине июля доминант послегнездового
пролета — дубровник, а во второй — лесной конек (по 23, 29 и 34%).
В первой половине августа среди прикочевывающих птиц преобла-
дают желтая трясогузка, пухляк и поползень (24 и по 16%), а в кон-
це месяца — рябчик и юрок (28 и 11%).
В южной тайге прослеживается такой же ход прикочевки птиц,
как и в средней, но суммарное количество мигрантов здесь в 4 раза
выше. Особенно возрастает интенсивность послегнездового пролета
в начале июля и конце августа. Наиболее рано (в конце июня) при-
кочевывает кедровка. В июле и августе среди прикочевывающих птиц
преобладают пухляк, только в июле — московка (25 и 15%), и,
кроме того, в первой половине июля — ополовник, поползень и
сойка, а во второй — певчий дрозд (21, 16, 12, 29%).
В подтаежных лесах прикочевывает в 3,4 раза меньше птиц, чем
в южной тайге, но в 1,8 и 1,2 раза больше, чем в северных подзонах.
Временные изменения количества прикочевывающих птиц примерно
такие же, как в северной тайге, т. е. максимальная интенсивность
послегнездового пролета приходится на вторую половину июля.
Первыми прикочевывают галка и ополовник (во второй половине
июня). В первой половине июля доминанты послегнездовых коче-
вок — большая синица, пухляк и лесной конек, а во второй — камы-
шевая овсянка и дубровник (29, 15, 13, 36 и 31%). В начале августа
♦ В тех случаях, когда число прикочевывающих видов невелико (до 3),
они перечисляются, но доля каждого из них не оценивается, поскольку все эти
виды являются доминирующими.
198
среди прикочевывающих птиц преобладают весничка, лесной конек,
белошапочная овсянка и деревенская ласточка (19, 17 и по 16%),
а в конце месяца — степной конек, перепел и зяблик (34, 25 и 14%).
Итак, максимальная интенсивность послегнездовой прикочевки
(как и весенне-летнего пролета) зарегистрирована в южной тайге,
она уменьшается к северу и в подтаежных лесах в соответствии со
снижением разнообразия местообитаний и их экологической привле-
кательности для мигрирующих птиц. В срединных, самых «таеж-
ных», подзонах идет последовательная прикочевка преимущественно
лесных видов, и максимальные ее значения зарегистрированы в кон-
це августа. В периферийных подзонах прикочевывают в основном
птицы разреженных лесов, полуоткрытых и открытых местообита-
ний. Наибольшая интенсивность послегнездового пролета здесь за-
регистрирована на месяц раньше, чем в срединных подзонах,— во
второй половине июля.
Суммарное обилие прикочевывающих птиц в долине Оби зна-
чительно выше, чем на междуречьях (в 3—15 раз). Особенно велико
это превышение в северотаежной долине Оби, где, видимо, одно-
временно идут две волны прикочевки: из лесотундровых и северо-
таежных редколесий в южную часть северной тайги и с междуречий
в долину. Видовое разнообразие птиц, прикочевывающих после
окончания гнездования, на междуречьях и в долине Оби изменя-
лось в целом примерно так же, как их суммарное обилие, но макси-
мальное количество видов характерно для подтаежной долины
Оби (см. *рис. 4, г).
Суммарная доля птиц, мигрирующих на таежных междуречьях,
от среднелетнего их обилия по сравнению с долиной Оби невелика.
Она составляет по 22% в периферийных подзонах и возрастает в
средней и особенно в южной тайге до 28 и 37%. Аналогичный пока-
затель для долины Оби 48—55%. На основании этого следует счи-
тать, что летние изменения плотности птичьего населения на таеж-
ных междуречьях определяются главным образом их гнездованием,
результатом размножения и откочевкой. Влияние послегнездовой
прикочевки существенно только в южной тайге. В долине Оби влия-
ние миграций на состав и плотность летнего населения птиц значи-
тельно сильнее, чем на междуречьях.
ПОДЗОНАЛЬНО-ЛАНДШАФТНЫЕ ОСОБЕННОСТИ
ВНУТРИЗОНАЛЪНОЙ ДИНАМИКИ ПЛОТНОСТИ
НАСЕЛЕНИЯ ПТИЦ
Существенной особенностью динамики населения птиц таеж-
ных междуречий является сходный характер изменений птичьего
населения в пределах аналогичных ландшафтов всех подзон [Вар-
тапетов, 1979в]. Так, в лесных ландшафтах преобладает равновес-
ный тип динамики. Увеличение численности, определяемое весенним
пролетом птиц, как правило, невелико и лишь в северной тайге вы-
ражено сильнее (рис. 5, а—г). В лесах этой подзоны в первой поло-
вине июня возрастает обилие юрка, чечетки, овсянки-крошки, бе-
199
Май Июнь Июль Август
Рис. 5. Временная динамика лет-
него населения птиц таежных
междуречий Западной Сибири.
а — в среднем по лесным ландшафтам,
б— в среднем по ключевым участкам,
в — в темнохвойных лесах, г — на вер-
ховых болотах, д -— в поселках,
е — на водоемах; 1 — северная, 2 —
средняя, 3 — южная тайга, 4 — под-
таежные леса.
лой трясогузки и каменки,
часть популяций которых
гнездится севернее, в редко-
лесьях и лесотундре, и вновь
появляется в северной тайге
после окончания гнездова-
ния (во второй половине
июля).
В подзонах, располо-
женных к югу от северной
тайги, пролет большей части
птиц, видимо, идет до на-
чала наших наблюдений.
Послегнездовое увеличение суммарного обилия птиц здесь проис-
ходит на две недели раньше, чем в северной тайге, что так же, как
и большая интенсивность пролета в последней, связано с фенологи-
ческими различиями северной тайги и более южных подзон. В сред-
нетаежных лесах возрастание обилия птиц в послегнездовой период
невелико (в 1,7 раза по сравнению с гнездовым периодом), что, ви-
200
димо, обусловлено откочевкой местных популяций сразу после
скончания гнездования к югу и в долину Оби. В конце августа вновь
прослеживается увеличение плотности населения птиц, связанное
с их прикочевкой с севера таежных междуречий Западной Сибири
и с местным перераспределением. В этот период здесь резко возраста-
ет обилие юрка, овсянок (крошки и ремеза), кукши, поползня, пух-
ляка и чернозобого дрозда.
В лесных местообитаниях южной тайги послегнездовое увели-
чение обилия несколько выше, чем в предыдущей подзоне, но также
не пропорционально интенсивности размножения. Очевидно, и
здесь часть популяций птиц сразу после окончания гнездования от-
кочевывает, хотя их доля от суммарного обилия меньше, чем в более
северных подзонах. Так же, как и в предыдущей подзоне, в конце
августа здесь происходит увеличение плотности населения птиц
за счет прикочевки юрка, королька, пухляка, поползня и зяблика.
А. А. Ананин и С. С. Москвитин [1978] летом 1976 г. в окрестностях
с. Бакчар, в 50 км к западу от места наших работ в южной тайге,
также отмечали два пика трехкратного увеличения численности
птиц — в конце июля за счет массовых перекочевок молодняка и
в конце августа с первой волной миграции птиц с севера.
В районах наших работ в северной и средней тайге преобладают
сосняки, а в южной — мелколиственные леса, поэтому, чтобы иск-
лючить влияние состава лесообразующих пород на динамику птичь-
его населения, проведено сравнение изменений суммарного обилия
птиц в темнохвойных лесах трех таежных подзон. В результате вы-
явился такой же характер временных изменений птичьего населения
этих лесов, что и по лесным ландшафтам в целом. Отличия сводятся
к тому, что в северотаежных темнохвойных лесах во второй половине
июля происходит значительно более резкое увеличение плотности
населения птиц, чем в среднем по сосново-боровому ландшафту
(в 3,5 раза по сравнению со второй половиной июня). Это, видимо,
обусловлено лучшими кормовыми и защитными свойствами темно-
хвойных лесов и, следовательно, их большей привлекательностью
для птиц по сравнению с сосняками. Так, во второй половине июля
в северотаежных темнохвойных лесах резко возрастает обилие юрка,
пеночек (таловки и теньковки), овсянки-крошки, зеленого конька
и сероголовой гаички.
В подтаежных (березово-осиновых) лесах послегнездовое уве-
личение плотности птичьего населения минимально по сравнению
со сходными показателями населения лесных ландшафтов севернее
расположенных подзон. Численность птиц здесь после окончания
гнездования возрастает всего в 1,5 раза по сравнению с гнездовым
периодом. Далее с середины июля и до конца августа наблюдается
неуклонное и значительное (почти 5-кратное) уменьшение плотности
птичьего населения. Откочевывает большая часть популяций гнездив-
шихся в подтаежных лесах лесного конька, большой синицы, вес-
нички, чечевицы, юрка, серой мухоловки, садовой камышевки, серой
славки и других видов птиц. Послегнездовая прикочевка птиц в под-
таежных лесах не прослеживается. Непривлекательность осиново-
201
березовых лесов этой подзоны для послегнездовых мигрантов, по-
видимому, определяется некоторым дефицитом увлажнения в июле
августе по сравнению с лесными местообитаниями более северных
подзон.
На таежных реках также преобладает равновесный тип дина-
мики населения птиц, за исключением северотаежных (р. Казым),
где плотность населения изменялась по иммиграционному типу
(см. рис. 5, д, е). Она определялась обилием береговой ласточки, ко-
торое увеличивалось вплоть до окончания вылета молодых во вто-
рой половине июля, а затем резко уменьшалось в августе.
На верховых болотах таежных междуречий плотность птичьега
населения изменяется по эмиграционному типу (см. рис. 5, г). Мак-
симальная численность птиц здесь наблюдается в начале лета в юж-
ной тайге за счет пролета лапландского подорожника, в средней —
белошапочной овсянки, белой трясогузки, краснозобого конька,
а в северной — овсянки-крошки и фифи. Далее обилие птиц не-
уклонно снижается — наиболее резко в северной тайге и более плав-
но — в южных подзонах. Снижение суммарного обилия птиц во вто-
рой половине лета связано с тем, что большая часть их популяций
сразу после окончания гнездования покидает эти болота (желтая
трясогузка, фифи, овсянка-крошка, белошапочная овсянка, лесной
конек).
В таежных поселках прослеживается тип динамики населения
птиц, который можно считать иммиграционно-эмиграционным, т. е.
суммарное обилие значительно увеличивается от начала к середине
лета, а в августе вновь уменьшается. Большинство птиц, гнездящих-
ся в населенных пунктах, в значительном числе задерживаются здесь
в послегнездовой период (домовый и полевой воробьи, сорока, белая
трясогузка, деревенская и береговая ласточки). Последняя гнездится
по обрывистым берегам рек, на которых часто расположены таежные
поселки. Высокая концентрация птиц в населенных пунктах проис-
ходит также за счет кормящихся и отдыхающих здесь птиц сосед-
них ландшафтов (желтая трясогузка, лесной конек, большая синица,
юрок и чернозобый дрозд).
Итак, во временных изменениях населения птиц таежных меж-
дуречий Западной Сибири преобладают процессы динамического рав-
новесия и эмиграции, и только в поселках наблюдается иммиграция
птиц. Большая часть популяций гнездящихся птиц после оконча-
ния гнездования покидает пределы таежных междуречий и, видимо,
смещается в долину Оби, вследствие чего в среднем по территории
междуречий преобладает динамически равновесный тип динамики на-
селения, а в долинах крупных рек — У-образный.
ТЕРРИТОРИАЛЬНЫЕ ИЗМЕНЕНИЯ
ДОМИНИРУЮЩЕГО И ФАУНИСТИЧЕСКОГО СОСТАВА
И ОБОБЩЕННЫХ ПОКАЗАТЕЛЕЙ НАСЕЛЕНИЯ ПТИЦ
Наибольшие отличия в составе доминирующих видов характер-
ны для птичьего населения крайних таежных подзон. Только в се-
верной тайге в качестве преобладающих видов зарегистрированы
202
сероголовая гаичка, чечетка и каменка [Вартапетов, 19796]. По срав-
нению с более южными подзонами здесь отмечены большее количест-
во доминантов и значительные изменения их состава в течение лета.
На аналогичную особенность указывал П. П. Второв [1976], изу-
чавший экосистемы высокогорий, которые так же, как и северная
тайга, характеризуются низкой стабильностью окружающей
среды.
Списки видов птиц, преобладающих в трех расположенных
южнее подзонах, более сходны. При этом в средней тайге специфич-
ные доминанты не зарегистрированы. В основном здесь доминируют
виды, преобладающие и в южной тайге, и в меньшей степени — севе-
ротаежные. В качестве доминантов, характерных только для южной
тайги и подтаежных лесов, отмечены дубровник, луговой и черно-
головый чеканы, распространение которых на междуречьях связано
с обширными низинными болотами, отсутствующими в более север-
ных подзонах. Специфичные доминанты подтаежных лесов — се-
рая славка, большая синица и степной конек.
Роль наиболее многочисленных доминантов в населении птиц,
как правило, постепенно изменяется по направлению с севера на юг.
Так, доля юрка в сообществах птиц лесных местообитаний к югу
уменьшается, а пухляка, поползня и большого пестрого дятла —
увеличивается от северной тайги к южной. На верховых болотах
в том же направлении снижается участие в птичьем населении жел-
той трясогузки и овсянки-крошки и возрастает доля белошапочной
овсянки.
Многие исследователи отмечали, что плотность и разнообразие
населения птиц в пределах лесной зоны к северу убывают [Гладков,
1958; Новиков, 1960; Дубинин, Торопанова, 1960; Ходашова, 1966;
Чернов. Ходашова, 1966; Пузаченко, 1967; Бутьев, Рубинштейн,
1979; MacArthur R., 1965; Margalef R., 1968], в частности в связи
с изменением подзонального климата. Для птичьего населения та-
ежных междуречий Западной Сибири эта особенность прослеживает-
ся лишь в виде самой общей тенденции, имеющей значительное число
отклонений. Показатели плотности и видового разнообразия умень-
шаются как к северу от южной тайги, что совпадает со снижением
теплообеспеченности ландшафтов, так и к югу, при переходе к под-
таежным лесам, что также связано со снижением продуктивности
биоценозов вследствие некоторого дефицита увлажнения, интенсив-
ной распашки, перевыпаса скота и сенокошения (рис. 6). Сказывают-
ся и фенологические различия более северных подзон и подтаежных
лесов, где большая часть мигрантов пролетает, видимо, до начала
наших наблюдений, а степень послегнездовой откочевки птиц наи-
более велика. Все это уменьшает среднелетнее суммарное их обилие
до уровня, более низкого, чем в южной тайге, тогда как плотность
гнездования птиц здесь даже несколько выше по сравнению с по-
следней.
В средней тайге (в среднем по обследованным территориям и в
темнохвойных лесах) наблюдается уменьшение показателей разно-
образия и плотности населения птиц по сравнению с соседними таеж-
203
Видовое
Особей/13 км
береговой
линии
Видов на одно
урочище
Рис. 6. Плотность и видовое разно-
образие населения птиц таежных
междуречий Западной Сибири.
А — в среднем по лесным ландшафтам;
J5 — темнохвойные леса; В — верховые
болота; Г — поселки; Д — в среднем по
ключевым участкам; Е — водоемы.
1 — северная, 2 — средняя, з — южная
тайга, 4 — подтаежные леса. Для видо-
вого разнообразия: 5 — фоновых видов;
б — всего видов. I — первая полови-
на, II — вторая половина лета.
Биомасса Биоэнергетика
У/
7 Йр /-Х ЕШЗ
J в!
I 1 I Г I I I I I 1 1 I
40 0 40 30 0 30
кг/км2 тыс.ккал/сут-км2
ЮОЮ 5 0 5
кг/10км береговой тыс.ккал/сутбОкм
линии береговой линии
Рис. 7. Биомасса и биоэнергетика
населения птиц таежных междуре-
чий Западной Сибири.
Ус л. обозн. см. на рмс. 6.
ними подзонами. Сходная осо-
бенность отмечается в долинах
Оби и Енисея [Равкин, 1978;
Анзигитова и др., 1979]. В не-
которой степени это связано с
увеличением численности птиц
лесотундры и редколесий (гипоарктические и бореально-гипоаркти-
ческие виды) на севере таежных междуречий [Кищинский, 1977*
1979]. С юга на север происходит снижение обилия многих лесных
видов птиц, которое не компенсируется возрастанием численности
птиц редколесий и лесотундры, вследствие чего уменьшение плотно-
сти птичьего населения от южной к северной тайге сохраняется как
общая тенденция. В Северном Заангарье также отмечается значи-
тельное (почти двухкратное) снижение суммарного обилия птиц при
переходе от южной к средней тайге [Гибет, Артамошин, 1977].
Целый ряд исследователей отмечают зависимость плотности
и разнообразия населения птиц от мозаичности и комплексности
204
местообитаний [MacArthur R., MacArthur J., 1961; Кулешова, 1968;
Roth R., 1976; и др.]. По-видимому, мозаичное чередование лесных
и болотных биотопов в северной тайге также служит одной из при-
чин увеличения суммарных показателей населения птиц в отлична
от более монотонных территорий средней и южной тайги, где чере-
дуются крупные массивы болот и высокобонитетных лесов.
Кроме того, большая плотность населения птиц в северной тайге
по сравнению со средней обусловлена более высокой интенсивностью
летних кочевок в первой. Однако в северной тайге напряженность
летних кочевок многих видов птиц резко различается в разные годы
[Леонович, 1976]. Тем не менее Ю. С. Равкин [1978] в 1973 г. также
отмечал большую интенсивность летних миграций в северотаежной
долине Оби по сравнению с более южными подзонами.
На верховых болотах прослеживается обратный ход изменений
суммарных значений по сравнению с суходольными ландшафтами,
т. е. показатели плотности и разнообразия населения птиц к северу
увеличиваются. Кроме уже отмеченных причин возрастание числен-
ности птиц здесь может быть вызвано увеличением к северу богатства
минерального питания за счет гетеротрофности, особенно в северной
тайге, где преобладающие по площади массивы верховых болот че-
редуются с переходными и низинными. Особенно высокая плотность
населения птиц характерна для тех территорий болот, где неболь-
шие открытые участки мозаично чередуются с залесенными [Варта-
петов, 1980].
Сходное возрастание плотности и разнообразия населения птищ
с юга на север наблюдается на торфяных болотах Финляндии и до-
лины Оби [Jarvinen О., Sammalisto L., 1976; Равкин, 1978]. Фин-
ские орнитологи считают, что такое увеличение объясняется также
оптимальным режимом увлажнения северных торфяных болот, спо-
собствующим развитию водных беспозвоночных, и ростом на север-
ных болотах роли кустарникового и кустарничкового покрова, до-
полнительно привлекающего птиц. Кроме того, в северной тайге уве-
личивается количество суходольных территорий, пригодных для
гнездования птиц, обитающих на верховых болотах (редколесья
и берега водоемов). На таежных реках видовое разнообразие и осо-
бенно плотность птичьего населения возрастают в крайних таежных
подзонах по сравнению со срединными.
Общее количество встреченных видов с севера на юг по подзонам
составляет 101, 123, 144 и 111, т. е. в подтаежных лесах этот пока-
затель заметно снижается по сравнению с южной тайгой, что связано
с меньшим числом обследованных местообитаний.
Число встреченных видов в пересчете в среднем на одно урочи-
ще в северной тайге составляет 35, в средней — 42, в южной — 44
и в подтаежных лесах — 46, а фоновых — соответственно 25, 23,
28 и 29. Таким образом, количество встреченных видов с юга на се-
вер таежных междуречий Западной Сибири уменьшается в большей
степени, чем фоновых, т. е. широта распределения последних по ланд-
шафтным урочищам относительно возрастает. Как отмечалось ранее,
использование большого числа местообитаний увеличивает стабиль-
205*
яость популяций в неблагоприятных условиях [Den Boer Р., 1968;
Levins, R., 1969], что имеет важное значение при снижении тепло-
обеспеченности ландшафтов с юга на север таежных междуречий За-
падной Сибири. Увеличение широты распределения видов птиц к се-
веру, по-видимому, связано и с возрастанием сходства природных
условий разных местообитаний в том же направлении, в частности
с выравниванием общей продуктивности лесов и верховых болот.
Показатели биомассы и биоэнергетики изменяются, как прави-
ло, так же, как и плотность населения птиц (рис. 7). Лишь на южно-
таежных реках отмечено превышение показателей по сравнению
с более северными подзонами, что определяется большим обилием
здесь уток (в основном кряквы). По биомассе из неворобьиных в лес-
ных ландшафтах чаще всего доминируют куриные, на верховых бо-
лотах — чайки, утки и реже врановые. Последние также преобла-
дают по биомассе в лугополевых местообитаниях подтаежных лесов
и в поселках всех подзон. На водоемах по биомассе чаще всего доми-
нируют утки и реже чайки, а из воробьиных обычно преобладают
наиболее многочисленные доминанты по обилию.
Территориальные различия суммарных показателей сообществ
птиц по профилю: междуречья — надпойменные террасы — пойма
Оби представлены на рис. 8 на примере изменений плотности их на-
селения в первой половине лета. Как отмечалось и ранее, суммарное
Рис. 8. Внутризональные изменения плотности населения
птиц в лесной зоне Западной Сибири в первой половине
лета.
А — междуречья; Б — надпойменные террасы; В — пойма Оби.
1 — верховые болота*, 2 — лесные и лесополевые ландшафты;
3 — пойменные лесолуговые и лугово-соровые ландшафты; 4 —
низинные болота.
206
обилие птиц неуклонно возрастает от междуречий через надпоймен-
ные террасы к пойме. При этом максимальное количество птиц за-
регистрировано на пойменных низинных болотах, а минимальное —
на верховых болотах междуречий. Только в южной тайге прослежи-
вается уменьшение плотности населения птиц в лесных местообита-
ниях на надпойменных террасах по сравнению с междуречьями. Эта
определяется высокой степенью распашки лесополевых надпоймен-
ных террас в отличие от менее освоенных междуречий.
Внепойменные низинные болота по сравнению с пойменными
и лесными ландшафтами характеризуются меньшей плотностью птичь-
его населения, но большей, чем на верховых болотах. На последних
происходит значительное обеднение сообществ птиц, по сравнению
с лесными и пойменными ландшафтами, особенно на юге таежных
междуречий. Это обусловливается бедным минеральным питанием
их фитоценозов, ухудшением здесь ландшафтного климата и, следо-
вательно, снижением общей продуктивности биоценозов. С продви-
жением к северу продуктивность биоценозов последних снижается,
а на верховых болотах, по-видимому, несколько возрастает. Кроме
вышеуказанных причин подобное изменение продуктивности может
определяться значительным уменьшением мощности торфяной за-
лежи в условиях северной тайги. Ослабевает влияние торфа, изоли-
рующего растительность от минерализованных грунтовых вод, и,
следовательно, возрастает богатство минерального питания фито-
ценозов верховых болот.
Такое сближение природных условий с юга на север таежных
междуречий, как отмечалось выше, ведет к увеличению широты рас-
пространения и выравниванию территориального распределения
отдельных видов птиц, а значит, нивелирует различия суммарных
показателей сообществ. В северной тайге на комплексных (гетеро-
трофных) верховых болотах птиц даже несколько больше, чем в лес-
ном ландшафте, в котором преобладают по площади одноярусные
лишайниковые сосняки.
Та же тенденция прослеживается с продвижением в более се-
верные подзоны и зоны. Так, А. А. Кищинский [1976] отмечает воз-
растание уровня биомассы птиц на фоне снижения зоомассы, общей
биомассы и биологической продуктивности сообществ при переходе
от бореальных экосистем к зональным субарктическим.
Еще севернее, в тундровой зоне, сообщества животных на боло-
тах в ряде случаев характеризуются более высокими суммарными
показателями, в том числе разнообразием, по сравнению с зональ-
ными местообитаниями [Чернов, 1978]. При этом для тундр харак-
терно особенно равномерное распределение видов. Это, по мнению
того же автора, является следствием обеднения фаунистического
состава и уменьшения возможностей экологического викариата,
что в ряде случаев снижает напряженность межвидовых конкурент-
ных отношений. Сходная тенденция проявляется, видимо, и с юга
на север в лесной зоне.
Основные территориальные изменения остальных обобщенных
показателей сообществ птиц (видового разнообразия, биомассы и био-
207
энергетики) сходны с описанными для плотности населения в первой
половине лета, поэтому их диаграммы не приводятся. Отличия носят
частный характер и сводятся к следующему. Во второй половине лета
становится значительно меньше птиц на верховых болотах между-
речий, зато резко возрастает их численность в северотаежной долине
и подтаежной лесолуговой пойме Оби. Максимальная биомасса ха-
рактерна для северотаежной поймы Оби в течение всего лета и для
южно-таежной — во второй половине лета в связи с концентрацией
уток на сорах. Показатели биомассы птиц на верховых болотах в ряде
случаев непропорционально велики, по сравнению с суммарным оби-
лием птиц, что связано со встречами водоплавающих птиц на фоне
низкой плотности населения.
В распределении птиц таежных междуречий по ярусам одно-
значные изменения прослеживаются нечасто. В лесных местообита-
ниях от северной к южной тайге уменьшается доля наземных птиц
в населении и возрастает участие кронников, что соответствует уве-
личению сомкнутости древостоев в том же направлении. В подтаеж-
ных лесах по сравнению с остальными подзонами возрастает доля
наземных и кустарниковых птиц. Виды, кормящиеся на стволах де-
ревьев, отсутствуют на верховых болотах в южной тайге, но начи-
нают встречаться в северной и средней, хотя участие их в населении
птиц незначительно. Соотношение основных групп кормов, за счет
которых удовлетворяются энергетические потребности птиц, в ана-
логичных ландшафтах таежных подзон сходно, а наблюдаемые раз-
личия невелики.
Фаунистический состав населения птиц лесных ландшафтов
(по числу особей) чаще всего европейско-сибирский и только в под-
таежных лесах сибирско-европейский. При этом участие сибирских
видов в птичьем населении к югу неуклонно уменьшается, а евро-
пейских увеличивается. На верховых болотах северных подзон, а
также на лугах и низинных болотах и особенно в поселках и на водо-
емах преобладают транспалеаркты. Участие их в птичьем населении
естественных ландшафтов суши возрастает к северу и к югу от юж-
ной тайги. На верховых болотах значимо участие в птичьем населе-
нии арктических видов, преимущественно за счет пролетных и реже —
гнездящихся птиц.
Такая гетерогенность фаунистического состава орнитокомплек-
сов верховых болот, особенно междуречных грядово-мочажинно-
юзерных, связана с наличием открытых пространств, большим чис-
лом водоемов и, видимо, некоторой их экологической близостью
к тундровым территориям [Вартапетов, 1979а, б]. По-видимому,
наряду с хорошо известным расселением более южных форм к се-
веру по долинам крупных рек, в таежной зоне Западной Сибири се-
верные виды проникают к югу по водоразделам, т. е. преимущест-
венно по верховым болотам. Для западно-сибирской тундры и лесо-
тундры сходная особенность отмечена, в частности, В. И. Телеги-
ным [1963].
Заметные различия фаунистического состава в долине Оби и на
междуречьях прослеживаются нечасто. Обычно в пойме (кроме се-
208
веротаежной) европейских и китайских видов больше, чем на над-
пойменных террасах. На междуречьях, особенно на северотаежных
верховых болотах, выше участие в птичьем населении транспале-
арктов по сравнению с остальными территориями.
ПРОСТРАНСТВЕННО-ТИПОЛОГИЧЕСКАЯ СТРУКТУРА
НАСЕЛЕНИЯ ПТИЦ
Пространственно-типологическая структура населения птиц
таежных междуречий Западной Сибири выявлена с помощью авто-
матической классификации [Трофимов, 1978; Трофимов и др., 1980]
и представлена в виде схем на рис. 9. Для первой половины лета
наиболее удачным признано разделение при коэффициенте агре-
гации 0,4 *.
На схеме вертикальный ряд (классы 1—4) отражает изменения
лесных сообществ птиц, направленные от вариантов населения ко-
ренных и производных формаций южной тайги (темнохвойная тай-
га, мелколиственные и смешанные полузаболоченные леса) к сооб-
ществам среднетаежных приречных смешанных лесов и угнетенных
лесов южной тайги (елово-кедровая тайга и рослые рямы). Сходство
между первыми двумя классами составляет 32 единицы, т. е. срав-
нительно велико. Далее изменения сходства птичьего населения на-
правлены к среднетаежным соснякам и вырубкам и северотаежным
приречным темнохвойным лесам. Вертикальный ряд заканчивается
орнитокомплексами разреженных северотаежных сосняков-ягель-
ников и производных от них местообитаний. Сходство лесных орни-
токомплексов средней тайги и более южных подзон определяется
участием в них пухляка. В наиболее богатых южно-таежных сооб-
ществах (темнохвойных, смешанных и мелколиственных лесов), ко-
торые относятся к 1-му классу, сходство эдифицируют, кроме того,
горихвостка-лысушка и теньковка. С обеднением орнитокомплексов
в угнетенных южно-таежных и среднетаежных лесах, особенно сос-
няках (классы 2 и 3), все большим становится участие юрка. В раз-
реженных северотаежных ягельниковых сосняках, на гарях и выруб-
ках по ним (4-й класс) сходство определяется тоже присутствиехМ
юрка и кроме него белой трясогузки и каменки.
Таким образом, изменения птичьего населения в вертикальном
ряду определяются широтным снижением теплообеспеченности и свя-
занным с ним уменьшением продуктивности лесных биоценозов.
* Варьируя коэффициент агрегации, можно получить структуры разной
степени «сжатия». Варианты населения птиц поселков имеют крайне слабое
сходство со всей остальной выборкой, поэтому они исключены из расчета при
выявлении пространственно-типологической структуры. Тем не менее степень
антропогенного влияния — весьма сильный, но локально проявляющийся фак-
тор, определяющий пространственную организацию птичьего населения лишь
в 3 вариантах из 33. Поэтому, когда оценивалась связь неоднородности насе-
ления птиц с проявлением факторов среды, сведения о сообществах птиц по-
селков были включены в расчеты.
14 Л. Г. Вартапетов
209
Северная
тайга
Средняя
тайга
и южные
подзоны
Белая трясогузка,
юрок,
каменка
Юрок,
5 пухляк,
клёст-еловик
Пухляк,
юрок,
теньковка
Пухляк,
теньковка,
горихвостка-лысушка
Первая половина
лета
32
Белошапочная овсянка
белая куропатка,
желтая трясогузка
Овсянка-крошка,
белая трясогузка,
каменка
Желтая трясогузка,
срисри,
овсянка-крошка
4зв
Лапландский подорожник,
дубровник,
лесной конек
130
&
Лесной конек,
теньковка,
пухляк
Белошапочная
овсянка,
лесной конек,
весничка 1
Лесной
конек,
дубровник,
овсянка-
крошка
Лесной конек,
белошапочная овсянка,
серая ворона
Желтая трясогузка,
скворец,
серая ворона
^23/
VДубровник,
желтая
трясогузка,
черноголовый
чекан
h
И
Залесенность уменьшается
♦ 7k
Белая трясогузка,
каменка,
чечетка
Вторая! половина лета
Обыкновенная овсянка,
Северная
•тайга
Средняя
тайга
и
южные
подзоны
Овсянка -кротка,
теньковка, •
таловка
Пухляк,
юрок,
поползень
Пухляк,
поползень,
теньковка
Пухляк,
поползень,
рябчик
$34
?32
29
Чв
Лесной конек,'
большая синица,
серая славка
• желтая трясогузка
&о \Желтая трясогузка,
* варакушка,
серая ворона
Белошапочная овсянка,
лесной конек,
большая синица
лесной конек
Черноголовый чекан,
дубровник,
камыш евая овсянка
Дубровник,
пухляк,
обыкновенная овсянка
Рис. 9. Пространственно-типоло-
гическая структура летнего на-
селения птиц таежных междуре-
чий Западной Сибири.
2 __ орнитокомплексы лесов, 2 — открытых мес-
тообитаний, чередующихся с залесенными; орни-
токомплексы открытых местообитаний: з — с бед-
ной продуктивностью (верховые болота, луга-
выпасы), 4 — с богатой продуктивностью (низин-
ные болота, соры, луга-покосы).
На этих схемах (рис. 9, 10) крупные циф-
ры указывают номера классов, мелкие (индекс-
_____величину внутриклассового сходства. Сходство классов 1 и 2 ука-
зано цифрой, а остальных — в примерном обратном масштабе (чем больше
сходство между классами, тем меньше расстояние между ними на схеме).
Сппптиыми линиями показано сильное сходство, штриховыми — слабое
(ичобоажГно вн” масштаба). Для каждого класса указаны три первых ви-
п1-эпнФика?ора сходства вошедших в них вариантов населения птиц
(для классов, представленных одиночными вариантами, три нерв х по
обилию вида). Осп координат примерно соответствуют основным направлениям территори-
альных изменений сходства сообществ птиц и коррелирующих с ними факторов среды.
В первой половине лета в 1-й класс входят варианты населения 15, 17, 19, 20; во 2-й —
И, 16, 21; в 3-й — 4, 9, 10; в 4-й — 1, 2, 3; в 5-й — 18, 26; в 6-й — 27; в 7-й — 28; в 8-й —
22, 29; в 9-й — 5* в 10-й — 6, 7, 8; в 11-й — 30; в 12-й — 23; в 13-й — 24; в 14-й — 12,
13, 14, 25.
Во второй половине лета в 1-й класс входят варианты населения 15, 16, 17, 19, 20; во
2-й—11 2Г в 3-й—9, 10; в 4-й—4; в 5-й—1, 2, 3, 5;в 6-й—18; в 7-й — 27: в 8-й — 28;
в 9-й ’29' в 10-й — 22: в 11-й — 26; в 12-й — 6, 7; 8; в 13-й — 12, 13, 14, 24, 25; в 14-й —
23, в 15-й — 30.
Население ландшафтов и урочищ
Северная тайга
С о с н о в о - б о р о в о й ландшафт (1 — сосняки-ягельники, 2 — вырубки, 3 — га-
ри, 4 — приречная темнохвойная тайга); сосново-озерный ландшафт’(5); комплек-
сные верховые болота (в — низкорослые рямы, 7 — грядово-мочажинно-озерные комплек-
сы, 8 — переходные болота).
Средняя тайга
Сосново-боровой ландшафт (.9 — сосняки-брусничники, 10 — вывубкп но
соснякам, 11 — приречные смешанные леса); верховые болота (72 — рослые рямы, 13 —
низкорослые рямы, и — грядово-мочажинные комплексы).
Южная тайга
Лесной ландшафт (15 — темнохвойная тайга, 16 — слово-кедровая тайга, 17— мел-
колиственные леса, 18 — поля-перелески, 19 — шелкоирядникп, 20 — смешанные полуза-
болоченные леса, 21 — рослые рямы); низинные болота (22); верховые бо-
лота (23 — грядово-мочажпнно-озерные комплексы, 24 — низкорослые рямы, 25 — пе-
реходные болота).
4 Подтаежные леса
Лесополевой ландшафт (26 — мелколиственные леса, 27 — поля-перелески, 28 —
луга-выпасы); низинные болота (29 — низинные болота, луга, 30 — рямы).
При этом выделенные классы вариантов орнитокомплексов не со-
впадают полностью с подзональным делением таежных междуречий
Западной Сибири. Обедненные сообщества более южных подзон,
как правило, объединяются с наиболее богатыми из расположенных
севернее. Отчасти это обусловлено постепенностью изменений птичь-
его населения и взаимопроникновением фитоценозов соседних под-
зон [Ильина и др., 1976], хотя в некоторой степени демонстрирует
и собственную изменчивость населения птиц, которая не совпадает
с ландшафтно-зональной неоднородностью территории.
Следующий ряд отражает тенденцию изменений орнптокомплек-
сов южной тайги и подтаежных лесов, связанную с увеличением за-
болоченности, снижением залесенностп и продуктивности биоцено-
зов (классы 1, 5—8). Уменьшение последней определяется созданием
монокультурных агроценозов в полях-перелесках южных подзон
и перевыпасом скота и сенокошением при переходе к последователь-
но расположенным в рассматриваемом ряду сообществам лугов-
выпасов и низинных болот в сочетании с покосными лугами. Весьма
характерно объединение в одну группу орнитокомплексов южно-
таежных полей-перелесков и подтаежных мелколиственных лесов
(5-й класс). Оба местообитания представляют собой осиново-березо-
вые леса, которые в значительной степени трансформированы хо-
зяйственной деятельностью человека: в первом случае в резуль-
тате распашки небольших полей внутри мелколиственных лесов,
а во втором — вследствие длительных, но частичных вырубок.
Сходство орнитокомплексов этих местообитаний определяется при-
сутствием в них лесного конька, теньковки и пухляка. Следующий,
212
6-й класс представлен сообщество .! птиц более обширных подтаежных
полей-перелесков, в которых преобладают лесной конек, белошапоч-
ная овсянка и серая ворона. Далее в ряду последовательно распо-
ложены орнитокомплексы подтаежных лугов-выпасов (7-й класс)
и низинных болот (8-й класс). Эдификатор сходства для обоих клас-
сов — желтая трясогузка, только для первого свойственны еще скво-
рец и серая ворона, а для второго — дубровник и черноголовый
чекан.
Следующий, северотаежный ряд изменений соединяется с пре-
дыдущим и в некоторой степени аналогичен ему (классы 4, 9, 10).
Только влияние антропогенной трансформации местообитаний здесь
не прослеживается. Этот ряд начинается от северотаежных лишай-
никовых сосняков и далее, через последовательно расположенные
сообщества птиц сосново-озерного ландшафта и комплексных верхо-
вых болот, соединяется с классом, представленным населением под-
таежных лугов-выпасов и принадлежащим к предыдущему ряду.
Сходство населения птиц сосново-озерного ландшафта и комплекс-
ных верховых болот определяется присутствием овсянки-крошки,
кроме того, для первого так же, как для сосняков, характерны белая
трясогузка и каменка, а для последних — желтая трясогузка. Не-
сколько неожиданное сходство северотаежных комплексных верхо-
вых болот и подтаежных лугов определяется высокой численностью
желтой трясогузки на фоне обеднения их орнитокомплексов, вызван-
ное в первом случае суровостью подзонального климата, а во вто-
ром — интенсивной пастбищной дигрессией.
Последний ряд изменений связан с обеднением минерального
питания и снижением продуктивности биоценозов при переходе от
полей-перелесков к верховым болотам. Индивидуальность орнпто-
комплексов последних (на фонё пониженной плотности населения
и видового разнообразия) значительно выше, чем всех ранее рас-
смотренных сообществ птиц. Поэтому каждый вариант населения
обычно образует отдельную группу, и лишь сообщества среднетаеж-
ных верховых болот были объединены в единый класс с довольно вы-
соким внутренним сходством, но слабо связанный с остальной выбор-
кой (соответственно 29 и 15 единиц).
Сходство верховых болот средней тайги и южных подзон обыч-
но эдифицируют лесной конек, белошапочная овсянка, дубровник.
Для южно-таежных водораздельных грядово-мочажинных болот
в качестве преобладающего вида характерна белая куропатка, а в
подтаежных рямах — весничка. Кроме того, в ряде случаев пре-
обладают пролетные птицы (в южно-таежных рямах — лапландский
подорожник, на среднетаежных верховых болотах — овсянка-
крошка).
Для второй половины лета наилучший вариант пространствен-
но-типологической структуры выявлен при коэффициенте агрегации
0,2. Полученная схема (см. рис. 9) весьма сходна с таковой для пер-
вой половины лета, поэтому здесь и далее подробных описаний прост-
ранственно-типологических структур сообществ птиц во второй по-
ловине лета не приводится. Указываются только основные отличия
213
по сравнению с аналогичными схемами для первой половины лета.
Ряд лесных орнитокомплексов представлен уже не четырьмя,
а пятью классами, так как население птиц северотаежных приреч-
ных лесов выделяется в самостоятельный 4-й класс (преобладает
таловка). Для населения северотаежных сосняков-ягельников, га-
рей и вырубок по ним и сосново-озерных ландшафтов характерны
те же эдификаторы сходства, что и в первой половине лета, но юрка
сменяет чечетка. В южно-таежных лесах эдификатором сходства,
характерным для второй половины лета, является рябчик. Кроме
того, в этот период ряд широко распространенных видов (юрок, пух-
ляк, поползень) откочевывает из северной тайги в среднюю и южную.
Таким образом, в послегнездовое время возрастает различие между
лесными северотаежными орнитокомплексами и населением более
южных подзон.
Население птиц подтаежных осиново-березовых лесов образует
отдельный класс (11-й), представляющий собой отклонение от ос-
новного ряда. Это связано с откочевкой большей части популяций
гнездившихся здесь птиц. Среди оставшихся преобладают во второй
половине лета лесной конек, большая синица и серая славка. Воз-
растают индивидуальные различия между южно- и подтаежными
низинными болотами, образующими отдельные классы. В качестве
преобладающих видов, характерных для второй половины лета,
в южной тайге отмечен тетерев, а в подтаежных лесах — камышевая
овсянка. На грядово-мочажинно-озерных верховых болотах преоб-
ладает обыкновенная овсянка, а в рямах — пухляк. На северотаеж-
ных комплексных верховых болотах в послегнездовое время в числе
эдификаторов сходства появляется сероголовая гаичка.
Таким образом, основные направления территориальных из-
менений сообществ птиц совпадают с изменением залесенности, под-
зонального климата (широтным снижением теплообеспеченности ланд-
шафтов), состава лесообразующих пород, заболоченности и мине-
рального питания фитоценозов и продуктивности биоценозов. Кроме
того, довольно сильный, но более частный фактор, определяющий
пространственную изменчивость населения птиц,— антропогенное
влияние, проявляющееся через застроенность территории, распашку,
перевыпас и сенокошение.
Объединение наших материалов с опубликованными ранее дан-
ными позволяет выявить пространственно-типологическую структу-
ру населения птиц лесной зоны срединной части Западной Сибири.
Лучшее разбиение в первой половине лета получено при коэффициен-
те агрегирования 0,9. На схеме (рис. 10) лесные орнитокомплексы
представлены в виде двух вертикальных рядов, изменения в которых
определяются снижением теплообеспеченности с возрастанием ши-
роты местности (от подтаежных лесов к северной тайге) и соответст-
венно уменьшением продуктивности лесных фитоценозов.
Первый из этих рядов (классы 1—3) образует население птиц
преимущественно темнохвойных лесов. В 1-й класс входит лишь
один вариант — население южно-таежных междуречных темнохвой-
ных (елово-кедрово-пихтовых) лесов. Характерный эдификатор
214
сходства птичьего населения всех лесов, кроме северотаежных.—
пухляк. В южно-таежных темнохвойных лесах к нему добавляются
московка и теньковка. В следующий, 2-й класс этого ряда входят
варианты населения среднетаежных надпойменных темнохвойных
и смешанных лесов. Характерные эдификаторы сходства — юрок
и зеленый конек. Последний, 3-й класс представлен сообществами
птиц северотаежных надпойменных темнохвойных и смешанных
полузаболоченных лесов. Кроме юрка в качестве преобладающих
видов здесь зарегистрированы овсянка-крошка и чечетка.
Следующий вертикальный ряд, связанный с предыдущим, об-
разуют орнитокомплексы нетемнохвойных лесов (пойменных, мелко-
лиственных, сосновых). Его образуют классы с 4-го по 6-й. Началь-
ный в этом ряду 4-й класс представлен орнитокомплексами смешан-
ных, мелколиственных и сосновых лесов южных подзон и осветленных
лесов средней тайги (пойменных ивняков, осинников, надпойменных
осиново-березовых и приречных темнохвойно-мелколиственных ле-
сов с примесью сосны по долинам притоков Оби). Сходство этих
вариантов населения птиц определяется участием в них теньковки
и горихвостки-лысушки. Следующий в этом ряду 5-й класс представ-
лен сообществами среднетаежных сосняков-брусничников, вырубок
по ним и приречных темнохвойных лесов северной тайги. Эдифика-
торы сходства — юрок, пухляк и клест-еловик. Ряд замыкается 6-м
классом, в который входят орнитокомплексы сосняков-ягельников,
вырубок, гарей по ним и сосново-озерных ландшафтов междуречий
северной тайги. Их сходство эдифицирует белая трясогузка, юрок
и каменка.
Несколько в стороне от этого ряда, но связанные с ним и друг
с другом находятся классы 7 и 16. Они представлены соответственно
населением северотаежных надпойменных смешанных лесов и зале-
сенных болот, пойменных ивняков-лугов. Их сходство, как и во всех
местообитаниях северной тайги, определяется участием в населении
овсянки-крошки, а в лесных сообществах — юрка. Кроме того, на
залесенных надпойменных болотах и в пойме Оби в число эдифика-
торов сходства входит весничка. В смешанных суходольных лесах
(7-й класс) в числе трех первых преобладающих видов зарегистриро-
вана сорока, что связано с близостью поселка и зверофермы,
отходами которой питались врановые.
Горизонтальные ряды рассматриваемой схемы демонстрируют
изменения сообществ птиц, вызванные уменьшением залесенности
при переходе от темнохвойных к производным и пойменным лесам,
полям, чередующимся с перелесками, пойменным лугам, сорам
и открытым низинным болотам. При этом выделяются два свя-
занных между собой верхних ряда, соответствующих изменениям
населения птиц в северной тайге, и два близких и связанных друг
с другом нижних ряда, отражающих сходные территориальные из-
менения в более южных подзонах. Нижний горизонтальный ряд со-
ставляют уже описанные 1-й и 4-й классы, а также классы с 8-го по
10-й. В 8-й класс вошли варианты населения южно-таежных поймен-
ных ивняков и осинников в сочетании с лугами. Их сходство, так же
215
W
O>
$ *
I к
I
Залесенность уменьшается
Северная
тайга
Средняя
тайга
южные
подзоны
Первая
половина лета
Пухляк,
московка,
теньковка
Юрок,
пухляк,
клест-
еловик
Овсянка -
крошка,
юрок,
сорока
Овсянка-
крошка,
юрок,
чечетка
Юрок,
пухляк,
зеленый
конек
$42
Белошапочная овсянка,
белая куропатка,
рюелтая трясогузка
Белая
трясогузка,
юрок,
каменка
Желтая трясогузка,
овсянка-крошка,
qxupu
овсянка,
16
15.
12
13.
Черноголовый чекан,
желтая трясогузка
Дубровник,
рябинник,
теньковка
Пухляк,
теньковка,
горихвостка-
лысушка
Дубровник,
желтая трясогузка,
скворец
Дубровник,
сверчки
певчий,
обыкновенный
и пятнистый
Лапландский
подорожник,
дубровник,
лесной конек
Овсянка -
крошка
юрок,
весничка
Лесной конек
белошапочная
овсянка
серая
славка
Береговая
ласточка
дубровник,
фифи
елтая трясогузка
фифи,
овсянка-крошка
Желтая
трясогузка
овсянка -
крошка,
дубровник
Турухтан,
овсянка-крошка,
краснозобый
конек
17.
Лесной конек,
теньковка
юрок
Лесной конек,
белошапочная
дубровник
Чернозобик,
галстучник,
рогатый жаворонок
Рис. 10. Пространственно-типологическая структура насе-
ления птиц лесной зоны Западной Сибири.
Условные обозначения те же, что и на рис. 9.
В первой половине лета в 1-й класс входят варианты населения
34; во 2-й — 24, 25; в 3-й — 9, 11; в 4-й—20, 26, 30,31, 35—38,-40, 47;
в 5-й — 4, 18, 19; в 6-й — 1—3, 5, 27; в 7-й — 10; в 8-й — 48, 49; в 9-й
41, 50—52, 62; в 10-й — 55, 56, 60; в 11-й —63, 64; в 12-й — 39, 45, 53,
61; в 13-й — 46, 54, 58, 59; в 14-й — 32; в 15-й — 33; в 16-й—
12, 14—16; в 17-й — 17; в 18-й — 13; в 19-й — 6; в 20-й — 7, 8; в 21-й—
21—23, 28, 29, 42, 57; в 22-й — 44; в 23-й — 43.
Во второй половине лета в 1-й класс входят варианты населения
24, 34; во 2-й — 25, 35, 36, 40; в 3-й — 18, 19; в 4-й — 3, 4; в 5-й —
1, 5; в 6-й — 2; в 7-й — 20, 37, 38, 47; в 8-й — 41, 49, 59, 64; в 9-й —
50—52, 63; в 10-й — 26, 30, 31, 45; в 11-й — 39, 58, 61; а 12-й — 46,
48, 62; в 13-й — 33, 56; в 14-й — 9—12, 14, 15; в 15-й — 13, 16, 17;
в 16-й — 32; в 17-й — 6—8; в 18-й — 57; в 19-й — 44; в 20-й — 21,
22, 28, 53, 54, 60; в 21-й—42, 43; в 22-й — 23, 29; в 23-й—27; в 24-й—
55.
Население урочищ и ландшафтов
Северная тайга
Междуречные ландшафты — сосново-боровой (1 — сосняки-
ягельники, 2 — вырубки по соснякам, 3 — гари по соснякам, 4 — при-
речная темнохвойная тайга), сосново-озерный (5), комп-
лексные верховые болота (6 — низкорослые рямы, 7 —
грядово-мочажинно-озерные комплексы, 8 — переходные болота). Над-
пойменные ландшафты — лесной (9 — темнохвойная тайга, 10 —
смешанные суходольные леса, 11 — то же, полузаболоченные), болот-
ный (12 — низкорослые рямы, 13 — переходные болота, 14 — низин-
ные залесенные болота). Пойменные ландшафты — лугово-соровый
(15 — ивняки-луга, 16 — луга-соры), низинные болота (17).
Средняя тайга
Междуречные ландшафты — сосново-боровой (18 — сос-
няки-брусничники, 19 — вырубки по соснякам, 20 — приречный сме-
шанный лес), верховые болота (21 — рослые рямы, 22 — низ-
корослые рямы, 23—грядово-мочажинные комплексы). Надпойменные
ландшафты — лесной (24 — темнохвойная тайга, 25 — смешанные
леса, 26 — березово-осиновые леса, 27 — залежи), верховые бо-
лота (28 — низкорослые рямы, 29 — грядово-мочажинно-озерные
комплексы). Пойменные ландшафты — лесолуговой (зо —
ивняки, 31 — осинники, 32 — луга-покосы, зз — луга-выпасы).
Южная тайга
Междуречные ландшафты — лесной (34 — темнохвойная тайга,
35 — елово-кедровая тайга, 36 — смешанные полузаболоченные леса,
37 — березово-осиновые леса, 38 — шелкопрядники, 39 — поля-перр-
лески, 40 — рослые рямы), низинные открытые болота,
(41). в е р х о в ы е болота (42 — переходные болота, 43 — низко-
рослые рямы, 44 — грядово-мочажпнно-озерные комплексы). Надпой-
Вторая половина лета
Желтая Хелта#
трясогузка, трясогузка,
овсянка-крошка, . лесной конек,
сероголовая гаичка белошапочная
а овсянка
Овсянка крошка,
белая
трясогузка,
каменка
Желтая
трясогузка,
варакушка,
серая ворона
$
Овсянка- крошка,
белая трясогузка,
свиристель
овсянка,
белошапочная
овсянка, жулан
Овсянка - крошка,
чечетка,
юрок
7-----—— —
Белая *--1
трясогузка, Обыкновенная
каменка,
весничка
8
''зо
37
Пухляк,
поползень,
московка
Пухляк,
поползень,
рябчик
Овсянки
крошка и
камышевая,
дубровник
[Дубровник
рябинник,
лесной конек
Дубровник,
скворец,
певчий
сверчок
---------
Чирок- 4—
трескунок,
чирок -
свистунок,
лесной конек
Лесной
конек
пухляк,
белошапоч-
ная овсянка
Пухляк,
лесной
конек
дубровник
Пухляк,
белошапочная
овсянка.
лесной
конек
Пухляк,
юрок,
поползень
Пухляк,
поползень,
большой
пестрый
дятел
Пухляк,
лесной конек,
теньковка
Лесной конек,
желтая трясогузка
белошапочная овсянка
Дубровник,
желтая трясогузка,
турухтан
Садовая
камышевка,
пухляк,
лесной кочек
Дубровник,
желтая трясогузка,
уири
менные ландшафты — лесополевой {45 — березово-осиновые леса, 46 — поля-пере-
лески), сосново-боровой {47 — сосняки). Пойменные ландшафты — лесолуго-
вой (48 — ивняки-луга, 49 — осинники-луга, 50 — луга-выпасы, 51 — луга-покосы,
52 — соры).
Подтаежные леса
Междуречные ландшафты — лссополевой {53 — осиново-березовые леса, 54 — по-
ля-перелески, 55 — луга-выпасы), болотный (56-- низинные болота, луга, 57 —
рямы). Надпойменные ландшафты — лесополевой (58 — осиново-березовые леса,
59 -поля-перелески), низинные болота (60 — открытые, 61 — залесенные). Поймен-
ные ландшафты —лесолуговой {62 — луга-ивняки), низинные болота
(63 — открытые, 64 — залесенные).
как и остальных орнитокомплексов пойм и низинных болот более
южных подзон, определяется присутствием дубровника, и, кроме
того, рябинника и теньковки. Следующий в этом ряду 9-й класс пред-
ставлен сообществами птиц пойменных лугов, соров южных подзон
и южно-таежных междуречных открытых низинных болот. Сходство
эдифицирует желтая трясогузка, а в пойменных вариантах населе-
ния — Скворец. Ряд замыкается 10-м и 11-м классами, представлен-
ными соответственно орнитокомплексами подтаежных внепойменных
открытых низинных болот, лугов-выпасов и пойменных низинных
болот. Сходство пойменных болот определяется присутствием сверч-
ков певчего, обыкновенного и пятнистого, а внепойменных—участием
в их населении черноголового чекана и желтой трясогузки.
Два класса — 14-й и 15-й — имеют слабое сходство с основным
рядом и образуют отклонения от него. Они представлены сообщест-
вами птиц среднетаежных пойменных лугов-выпасов и лугов-поко-
сов. В первом случае отличия от описанного выше ряда связа-
ны с обеднением населения в результате перевыпаса (преобладают
береговая ласточка, дубровник, фифи), а во втором — значительным
участием в населении пролетных тундровых птиц (преобладают чер-
нозобик, галстучник, рогатый жаворонок).
Следующий горизонтальный ряд, связанный с предыдущим,
начинается от 1, 2 и 4-го классов (лесные орнитокомплексы средней
тайги и южных подзон). Далее в нем последовательно расположены
классы 12-й и 13-й, первый из которых представлен вариантами на-
селения осиново-березовых лесов, надпойменных залесенных ни-
зинных болот и небольших полей в сочетании с перелесками в преде-
лах южных подзон, а во второй вошли орнитокомплексы более об-
ширных подтаежных полей, чередующихся с перелесками, и осино-
во-березовых лесов. В качестве эдификатора сходства для обоих
классов зарегистрирован лесной конек. Общность сообществ птиц
мелколиственных лесов и залесенных надпойменных низинных болот
связана с участием в их населении теньковки и юрка, а сходство
орнитокомплексов полей-перелесков определяется присутствием в
них белошапочной овсянки и серой славки.
Два верхних горизонтальных ряда рассматриваемой схемы от-
носятся к северной тайге. Один из них демонстрирует изменения
сообществ птиц в долине Оби от темнохвойных лесов через надпой-
менные залесенные болота и пойменные луга-ивняки к открытым
пойменным низинным болотам (классы 3, 16 и 17). Кроме описанных
218
эдификаторов сходства северотаежного населения птиц на низинных
болотах в число преобладающих видов входят турухтан и краснозо-
бый конек. Верхний горизонтальный ряд, связанный с предыду-
щим, отражает изменение сообществ птиц на северотаежных между-
речьях при переходе от сосняков, гарей и вырубок по ним и сосново-
озерных ландшафтов к комплексным верховым болотам (классы 19 и
20). Характерные эдификаторы сходства орнитокомплексов этих
болот — желтая трясогузка и фифи. Кроме того, сообщества птиц
открытых низинных и верховых болот северной тайги (17-й и 20-й
классы) имеют сходство с населением птиц открытых надпойменных
переходных болот той же подзоны (18-й класс), а те, в свою очередь,
с птицами пойменных лугов южных подзон (9-й класс). Последнее
определяется участием в их орнитокомплексах дубровника и желтой
трясогузки. Таким образом, образуется ряд изменений орнитоком-
плексов открытых местообитаний (классы 9, 17, 18, 20), связанный
с переходом от южно-таежных пойменных лугов и соров к северо-
таежным открытым болотам. Среднетаежные пойменные луга, как
указывалось выше, в первой половине лета имеют высокую инди-
видуальность орнитокомплексов и не входят в этот ряд.
Длинный диагональный ряд изменений, прослеживаемый на
схеме, отражает слабое сходство сообществ птиц олиготрофных
верховых болот средней тайги и южных подзон с остальными вариан-
тами населения. Этот диагональный ряд, в свою очередь, состоит из
двух ветвей. Одна из них демонстрирует изменения сообществ птиц
при переходе от мелколиственных внепойменных и пойменных ле-
сов (классы 12 и 8) к полям-перелескам (13-й класс), когда сни-
жается продуктивность биоценозов при создании монокультурных
агроценозов, и далее — к верховым болотам (21-й класс), когда про-
исходит ухудшение их минерального питания. Другая ветвь изме-
нений населения демонстрирует ухудшение минерального питания
в более «чистом» виде при переходе от темнохвойных лесов к сосно-
вым в пределах средней тайги и далее к залесенным гетеротрофным
северотаежным болотам, а затем к олиготрофным верховым. Послед-
ние представлены 21-м классом, в который вошли варианты населе-
ния всех верховых среднетаежных и подтаежных и переходных
южно-таежных болот. С ним связаны 22-й и 23 классы, образован-
ные сообществами птиц крайне обедненных верховых водораздель-
ных болот южной тайги. Сходство орнитокомплексов верховых бо-
лот эдифицируют лесной конек, белошапочная овсянка, дубровник.
Только на грядово-мочажинно-озерных частях этих болот в южной
тайге в число преобладающих видов входят белая куропатка и жел-
тая трясогузка, а в низкорослых рямах — лапландский подорожник
на пролете.
Для второй половины лета наилучшим признано разделение
всех вариантов населения птиц на 24 класса при коэффициенте агре-
гирования 1 (см. рис. 10). Общая схема пространственно-типологи-
ческой структуры населения птиц в этот период сохраняется, однако
по сравнению с первой половиной лета появляются некоторые от-
личия, которые сводятся к следующему. В результате послегнездо-
219
вых”кочевок птиц нивелируются различия между темнохвойными и
остальными лесными орнитокомплексамп, поэтому изменения в них
отражаются не двумя, а одним рядом (классы 1—5). В связи с тем
же выравнивающим эффектом местных перекочевок птиц возрастает
сходство открытых и залесенных местообитаний, поэтому горизон-
тальные ряды становятся короче. Одновременно схема значительно
растягивается по вертикали, что определяется увеличением разли-
чий между северной тайгой и более южными подзонами. Эта тенден-
ция при объединении материалов по лесной зоне Западной Сибири
прослеживается сильнее, чем на отдельно взятых междуречьях, за
счет послегнездовой прикочевки птиц лесотундровых и северотаеж-
ных редколесий в долину Оби в пределах северной тайги. Кроме то-
го, за счет откочевки значительного числа отгнездившихся птиц
уменьшаются отличия орнитокомплексов олиготрофных верховых
болот. Они, в свою очередь, становятся весьма сходными с населе-
нием наиболее обедненных во второй половине лета мелколиствен-
ных лесов на подтаежных междуречьях. Сходство вариантов насе-
ления птиц низкорослых рямов и мелколиственных лесов, которые
образуют единую группу (20-й класс), возрастает также за счет
кочующих в этих местообитаниях в небольшом числе стаек пухля-
ка. Выделяются крайне обедненные варианты населения подтаеж-
ных лугов-выпасов и среднетаежных залежей (классы 23, 24), кото-
рые слабо связаны лишь с олиготрофными верховыми болотами. Спе-
цифика населения птиц залежей определяется появлением по запа-
динам, наполненным водой, чирков — свистунка и трескунка, ко-
торые входят в число преобладающих видов.
Состав эдификаторов сходства населения птиц во второй поло-
вине лета по сравнению с первой пе претерпевает значительных из-
менений. Однако в полях-перелесках только во второй половине лета
характерны пухляк и садовая камышевка, в северотаежных при-
речных смешанных и темнохвойных лесах — свиристель, а на меж-
дуречных комплексных верховых болотах — сероголовая гаичка.
Таким образом, если пренебречь частными отличиями отдель-
ных классов, пространственно-типологическую структуру населения
птиц лесной зоны Западной Сибири схематично можно представить
на плоскости в виде прямоугольной трапеции и далеко выходящего
за ее пределы диагонального ряда. Последний демонстрирует терри-
ториальные изменения орнитокомплексов при увеличении заболо-
ченности с обеднением минерального питания фитоценозов. Осталь-
ная часть схемы представляется в виде горизонтальных рядов, от-
ражающих уменьшение залесенности в северной тайге и более юж-
ных подзонах, и вертикальных, которые определяются снижением
теплообеспеченности (возрастанием суровости подзонального кли-
мата) в лесных и открытых ландшафтах. Трапециевидная форма
описываемой схемы связана с тем, что в северной тайге различия
между открытыми и залесенными местообитаниями меньше, чем в
€олее южных подзонах, за счет высокой численности широко рас-
пространенных видов птиц северотаежных и лесотундровых редко-
лесий. Кроме того, трапеция состоит из верхней и нижней половин,
220
которые соответствуют сообществам птиц северной тайги и более
южных подзон.
Таким образом, наиболее резкие изменения сходства орнито-
комплексов наблюдаются при переходе от средней к северной тайге,
что определяется уменьшением численности лесных (таежных)
видов и возрастанием обилия лесотундровых птиц. Однако это
справедливо преимущественно для междуречий и надпойменных
террас, тогда как в пойме Оби значительные изменения населения,
как это показано ранее, отмечены при смене южно-таежных орнито-
комплексов среднетаежными [Равкин, 1978]. В фаунистическом
отношении на основе анализа ареалов птиц в Западной
Сибири Г. X. Иогансен (1943—1961), А. М. Гынгазов, С. П. Мило-
видов (1977) и В. В. Раевский (1982) указывают на существование
пределов распространения многих видов с юга на север до 61—62°
с. ш., что определяет большее сходство среднетаежной орнитофауны
с южно- и подтаежной, чем с северотаежной и лесотундровой.
Набор факторов, вызывающих пространственные изменения на-
селения птиц лесной зоны Западной Сибири, сходен с таковым на
междуречьях. Кроме того, прослеживается влияние рельефа (раз-
личия пойм, надпойменных террас и междуречий), которое тесно
связано с залесенностью.
Итак, в территориальных изменениях сообществ птиц преоб-
ладают три основных тренда, которые соответствуют снижению
теплообеспеченности, уменьшению залесенности и увеличению за-
болоченности при обеднении минерального питания. В отличие от
ранее опубликованных аналогичных схем для южной тайги и доли-
ны Оби в пределах лесной зоны [Равкин, Лукьянова, 1976; Равкин,
1978], влияние заболоченности без обеднения минерального питания
(при переходе от лугов к сорам и низинным болотам и от лесов к за-
лесенным низинным- болотам) почти не прослеживается. Это связано
с преобладаниехМ в нашей выборке верховых болот, что соответству-
ет доминированию в лесной зоне Западной Сибири верхового типа
заболачивания. Кроме того, мы не указываем продуктивность сооб-
ществ среди основных маркеров территориальных изменений орни-
токомплексов, как это делалось ранее, поскольку она оказывается
тесно скоррелированной сразу с двумя факторами: с теплообеспе-
ченностью и заболоченностью при обеднении минерального питания.
Представление трех основных тенденций изменений сообществ
в двухмерном пространстве приводит к значительным искажениям
их пространственной структуры. Поэтому макроструктуру терри-
ториальных изменений сообществ птиц лесной зоны Западной Сиби-
ри лучше изображать в виде пирамиды в трехмерном факторном про-
странстве (рис. 11). В связи с большей степенью абстрагирования
основание пирамиды преображается из трапеции в прямоугольник.
В пределах последнего по оси X происходят изменения сообществ
птиц, определяемые уменьшением залесенности от темнохвойных ле-
сов к пойменным лугам, а по оси У направлены изменения орнито-
комплексов, связанные со снижением теплообеспеченности от южных
подзон к северной тайге. От основания пирамиды к ее вершине (по
221
Рис. 11. Макроструктура терри-
ториальных изменений сообществ
птиц лесной зоны Западной Си-
бири (ординационная схема)»
оси Z) идут изменения насе-
ления птиц, которые опреде-
ляются уменьшением продук-
тивности биоценозов, уве-
личением заболоченности
при обеднении минерального
питания от лесных место-
обитаний через поля-перелес-
ки или малопродуктивные
сосняки к верховым боло-
там.
Эта ординационная схе-
ма не имеет принципиаль-
ных отличий от предложен-
ной ранее для лесной зоны
долины Оби [Равкин, 1978].
Различия в трактовках скорее сводятся к преобладанию действия
тех или иных факторов, определяющих основные тренды террито-
риальных изменений сообществ, и к иной ориентации схемы в фак-
торном пространстве. Так, уменьшение продуктивности заменяется
на увеличение заболоченности при обеднении минерального пита-
ния (причины этого объяснялись выше). Слабое проявление заболо-
ченности без обеднения минерального питания позволяет не изобра-
жать его в виде самостоятельного тренда, а показать отличия сооб-
ществ птиц низинных болот и лугов расположением первых над вто-
рыми. В результате одна ось оказывается свободной, и по ней отоб-
ражается снижение теплообеспеченности ландшафтов (возрастание
суровости подзонального климата). Это, в свою очередь, позволяет
заменить трактовку четырех подзональных пирамид, встроенных
друг в друга [Равкин, 1978], на единую «пирамидальную» структу-
ру. Если первая предполагает равную степень подзональных отли-
чий, то последняя в некоторой степени отражает различия между
северной тайгой и более южными подзонами.
ПРОСТРАНСТВЕННАЯ ОРГАНИЗАЦИЯ
[НАСЕЛЕНИЯ ПТИЦ
Рассмотренные в предыдущем разделе пространственно-типо-
логические структуры населения птиц таежных междуречий и лес-
ной зоны срединной части Западной Сибири позволили выявить ос-
новной набор факторов, определяющих территориальные изменения
222
Таблица 26
Сила связи неоднородности факторов среды с пространственной изменчивостью
населения птиц лесной зоны Западной Сибири в первой и второй половине лета
(учтенная дисперсия, %)
Фактор Междуречья Долина Оби и междуречья
I 1 II I. 1 П
Залесенность Подзональное изменение кли- 36 23 31 22
мата 19 25 12 15
Антропогенное влияние Состав лесообразующих по- 15 12 17 14
род 12 6 И 7
Заболоченность 10 8 8 4
Минеральное питание 9 8 7 5
Продуктивность 4 5 7 5
Рельеф — — 4 3
Всего I 71 I 62 I 63 I 55
орнитокомплексов. Набор таких факторов, сила и иерархия их
проявления и степень совпадения сочетаний факторов и террито-
риальной неоднородности сообществ птиц создают представления о
пространственной организации их населения. В табл. 26 приводятся
индивидуальные оценки связи этих факторов с территориальной не-
однородностью сообществ птиц. Факторы частично скоррелированы,
поэтому множественные оценки всегда меньше, чем суммы индиви-
дуальных.
На таежных междуречьях в первой половине лета наиболее
значимым фактором оказалась залесенность местообитаний, на вто-
ром месте — подзональное изменение климата (снижение тепло-
обеспеченности), а на третьем — антропогенное влияние, включая
застроенность, вырубки, распашку, сенокошение и пастбищную
дигрессию. Во второй половине лета эта иерархия воздействия фак-
торов среды на население птиц несколько изменяется. Как указыва-
лось выше, возрастает влияние смены подзонального климата (в ос-
новном за счет отличий северотаежных сообществ птиц) и выходит
на первое место. За счет местных перекочевок менее значимым ста-
новится влияние залесенности и особенно состава лесообразующих
пород, который занимает предпоследнее место. При объединении
материалов по междуречьям и долине Оби расчеты показали пример-
но сходную иерархию проявления факторов среды. Лишь антропо-
генное влияние в первой половине лета выходит на второе место пос-
ле залесенности в результате большей освоенности долины Оби *.
♦ Столь высокая оценка антропогенного влияния связана также с учас-
тием в расчетах вариантов населения птиц подтаежных лесов — наиболее ос-
военной подзоны. При исключении их из выборки, т. е. в пределах собственно
таежных подзон, влияние антропогенной трансформации местообитаний на
территориальную неоднородность сообществ птиц оказывается вдвое меньшим
(8 и 7 % в долине Оби и 7 и 4 % на междуречьях). Особенно снижается этот пока-
затель на междуречьях во второй половине лета в результате прикочевки птиц
из окрестных ландшафтов в небольшие таежные поселки.
223
Таблица 27
Природные режимы, определяющие территориальную неоднородность летнего
населения птиц таежных междуречий Западной Сибири
№ группы Природные режимы Учтенная дисперсия, %
1 2 3
I Первая половина лета 1. Северотаежных лесов и среднетаежных сосняков 2. Комплексных верховых болот северной тайги 3. Среднетаежных смешанных приречных и южно-таеж- ных лесов 4. Полей-перелесков и подтаежных (березово-осиновых) лесов 5. Лугов-выпасов и низинных болот южных подзон 6. Среднетаежных верховых и южно-таежных переход- ных болот 7. Верховых болот южных подзон 8. Низкорослых рямов южной тайги 9. Крупных поселков (подтаежных) 10. Мелких поселков (средне- и северотаежных) 43
II 1. Северной тайги, среднетаежных грядово-мочажин- ных комплексов верховых болот и подтаежных лугов- выпасов 2. Северотаежных поселков и частичных вырубок по соснякам 3. Лесов и производных от них местообитаний (кроме северотаежных сосняков) 4. Подтаежных низинных болот в сочетании с лугами 5. Южно-таежных болот, кроме грядово-мочажинно-озер- ных верховых 6. Средне- и подтаежных рямов и подтаежных полей- перелесков 7. Грядово-мочажинно-озерных верховых болот южной тайги 29
III 1. Местообитаний северной тайги, где открытые участ- ки перемежаются с залесенными (частичные выруб- ки, сосново-озерный ландшафт, низкорослые рямы) 2. Северотаежных открытых комплексных верховых и южно-таежных низинных болот 3. Приречных (смешанных и темнохвойных) лесов се- верной и средней тайги 4. Сосняков, южно-таежных рослых рямов и елово- кедровой тайги 5. Южно-таежных производных лесных местообитаний п подтаежных рямов 6. Подтаежных лесов и переходных болот южной тай- ги 7. Верховых болот южной и средней тайги 10
Всего 82
224
1 Окончание табл. 27
1 1 2 1 3
Вторая половина лета
I 1. Темнохвойных лесов северной тайги 2. Северотаежных сосняков и производных от них мес- тообитаний, включая сосново-озерный ландшафт 3. Комплексных верховых болот северной тайги 4. Среднетаежных сосняков 5. Среднетаежпых смешанных прпречных и южно-таеж- ных лесов 6. Полей-перелесков южной тайги 7. Подтаежных мелколиственных лесов и полей-пере- лесков 8. Низинных болот южной тайги 9. Подтаежных лугов-выпасов 10. Низинных болот в сочетании с лугами в подзоне подтаежных лесов И. Верховых болот южной и средней тайги 12. Южно-таежных грядово-мочажинно-озерных комп- лексов верховых болот 13. Северотаежных (мелких) поселков 14. Среднетаежных (мелких) поселков 15. Подтаежных (крупных) поселков 51
II 1. Комплексных верховых болот, сосново-озерного ландшафта северной тайги и грядово-мочажинных комплексов верховых болот средней тайги 2. Среднетаежных лесов и рямов и угнетенных лесов южно-таежной и подтаежной подзон 3. Лесов и производных от них местообитаний южной тайги 4. Лесополевых и лугово-болотных местообитаний подтаежных лесов 5. Верховых болот южной тайги подтаежных мелколист- венных лесов 17
III 1. Северной тайги и подтаежных лугов-выпасов 2. Угнетенных южно-таежных лесов и среднетаежных сосняков 3. Южно-таежных темнохвойных и мелколиственных лесов нормальной полноты 4. Среднетаежных смешанных лесов и рямов, южно- таежных шелкопрядников и верховых болот и лесо- полевых местообитаний южных подзон 5. Низинных болот 6. Средне- и южно-таежных грядово-мочажинно-озер- ных комплексов верховых болот и подтаежных рямов 14
Всего * . • « • 82
Кроме того, по объединенным материалам прослеживается влияние
рельефа, правда довольно слабое и в значительной степени скорре-
лированное с залесенностыо. В результате следует отметить, что
сила связи территориальной неоднородности сообществ птиц с факто-
рами среды по объединенным данным оказалась ниже, чем на отдель-
15 Л. Г. Вартапетов
225
Таблица 28
Природные режимы, определяющие территориальную неоднородность летнего
населения птиц лесной зоны Западной Сибири
№ группы Природные режимы Учтенная дисперсия, %
1 2 3
I Первая половина лета 1. Северотаежной долины Оби 2. Лишайниково-кустарничковых сосняков, гарей и вы- рубок по ним и сосново-озерных ландшафтов север- ных подзон 3. Междуречных северотаежных комплексных верховых болот и подтаежных лугов-выпасов 4. Южно-таежных междуречных и среднетаежных тем- нохвойных, производных от них и пойменных лесов 5. Среднетаежных надпойменных залежей ♦ 6. Среднетаежных пойменных лугов-покосов ♦ 7. Среднетаежных пойменных лугов-выпасов при вы- сокой пастбищной дигрессии 8. Лесополевых ландшафтов подтаежных лесов и южно- таежной долины Оби и подтаежных надпойменных залесенных низинных болот 9. Южно-таежной поймы Оби и подтаежных пойменных лугов-ивняков и залесенных низинных болот 10. Открытых низинных болот южных подзон И. Верховых среднетаежных и переходных южно-таеж- ных болот 12. Верховых болот междуречий южных подзон 13. Низкорослых рямов южно-таежных междуречий * 14. Мелких (таежных) поселков на междуречьях север- ных подзон 15. Крупных и средних поселков долины Оби и подтаеж- ных лесов 39
II 1. Северотаежных междуречий, пойменных лугов-соров и надпойменных низкорослых рямов 2. Средне- и северотаежных темнохвойных и смешан- ных лесов, северотаежных надпойменных залесен- ных низинных болот и пойменных ивняков-лугов 3. Северотаежных низинных открытых пойменных бо- лот и среднетаежных лугов-покосов 4. Северотаежных вырубок по соснякам 5. Лесов нормальной полноты южных подзон и сред- нетаежных пойменных лесов 6. Угнетенных южно-таежных междуречных и средне- таежных нетемнохвойных лесов, вырубок по сосня- кам и рослых рямов 7. Полей-перелесков, внепойменных залесенных низин- "ныг и верховых болот южных подзон 8. Лугов, соров, открытых низинных болот, обширных полей в сочетании с перелесками южных подзон; северотаежных надпойменных переходных болот и среднетаежных пойменных лугов-выпасов 9. Подтаежных пойменных низинных болот 17
* В населении местообитаний, отмеченных звездочкой, значительную долю состав-
ляли пролетные птицы.
226
П р’о должение табл. 28
1 1 2 3 4 5 6 7 8 9 10 11 3
10. Грядово-мочажинно-озерных верховых болот сред- ней и южной тайги 11. Переходных болот и низкорослых рямов на южно- таежных междуречьях 12. Поселков
III 1. Северотаежных темнохвойных, смешанных и сосно- вых лесов, вырубок, гарей по ним и среднетаежных приречных смешанных лесов 2. Северотаежных надпойменных верховых болот и пойменных лугов-соров, междуречных открытых верховых болот северных подзон, южно-таежных пойменных лугов 3. Рямов и сосново-озерных ландшафтов междуречий северных подзон 4. Внепойменных южно- и среднетаежных лесов 5. Лесополевых ландшафтов и пойменных лесов юж- ных подзон, среднетаежных мелколиственных лесов, пойменных осинников, лугов-выпасов и подтаежных низинных болот и лугов 6. Низинных и переходных болот южных подзон 7. Пойменных открытых низинных болот и соров юж- ных подзон, среднетаежных лугов-покосов 8. Верховых болот (кроме северотаежных) 14
Всего 70
В т о"р а я половина лета
37
1. Северотаежных лишайниковых сосняков и производ-
ных от них местообитаний
2. Северотаежной долины Оби и темнохвойной тайги
по долинам ее притоков
3. Междуречных северотаежных комплексных верховых
болот и подтаежных лугов-выпасов
4. Южно-таежных лесополевых ландшафтов и лесов
средней тайги и южных подзон
5. Среднетаежных залежей и пойменных лугов-выпасов
6. Среднетаежных пойменных лугов-покосов, южно-
таежных низинных болот и пойменных лесолуговых
ландшафтов, подтаежных открытых низинных пой-
менных болот и надпойменных полей-перелесков
7. Подтаежных пойменных лугов-ивняков и залесен-
ных низинных болот
8. Подтаежных междуречных низинных болот в соче-
тании с лугами
9. Верховых болот (за исключением северотаежных и
крайне обедненных), мелколиственных лесов и по-
лей-перелесков
10. Крайне обедненных верховых болот (южно-таежных
грядово-мочажинно-озерных комплексов и подтаеж-
ных рямов)
11. Поселков
15*
227
Продолжение табл. 28
1 2 3
II 1. Сосняков-ягельнпков и вырубок по ним на северо- таежных междуречьях 2. Северотаежных долпнных лесов, низкорослых рямов, поименных ивняков п междуречных гарей по сос- някам 3. Комплексных верховых болот, сосново-озерных ланд- шафтов северотаежных междуречий, открытых пе- реходных и верховых надпойменных болот в преде- лах северных подзон 4. Северотаежных низинных болот и поименных лугов в пределах северных подзон 5. Сосняков-брусничников на среднетаежных между- речьях 6. Южно-таежных лесов, приречных смешанных и тем- нохвойных л?сов по долинам притоков Оби в север- ных подзонах 7. Полей-перелесков п низинных болот южных под- зон, подтаежных осиново-березовых лесов, поймен- ных лугов-ивняков, южно-таежных пойменных ив- няков и осинников в сочетании с лугами 8. Лугов-выпасов и низинных болот на подтаежных междуречьях 9. Пойменных открытых низинных болот и соров в пре- делах южных подзон 10. Пойменных лугов и поселков южной тайги И. Рямов средней тайги и южных подзон, долинных средне- и южно-таежных мелколиственных лесов, среднетаежных вырубок по соснякам и пойменных ивняков, осинников 12. Мелких таежных поселков на северо- и среднетаежных междуречьях 13. Крупных и средних поселков долины Оби и подтаеж- ных лесов 22
III 1. Темнохвойных и сосновых лесов, вырубок и гарей по ним и сосново-озерных ландшафтов на северотаеж- ных междуречьях 2. Северотаежных надпойменных темнохвойных и смешанных лесов и пойменных ивняков-лугов 3. Северотаежных надпойменных болот и пойменных лугов-соров 4. Темнохвойных и смешанных лесов средней п южной тайги 5. Южно-таежных елово-кедровых и угнетенных сосно- вых лесов (рослых рямов) 6. Сосновых и смешанных лесов на среднетаежных междуречьях 7. Осиново-березовых лесов в долине Оби в пределах южных подзон и полей-перелесков на подтаежных междуречьях 14
-228
I Окончание табл. 28
г 1 2 3
8. Среднетаежных лугов-покосов 9. Южно-таежных пойменных лугов-ивняков и осин- ников 10. Низинных болот южных подзон И. Северотаежных комплексных и среднетаежных гря- дово-мочажинно-озерных верховых болот 12. Южно-таежных верховых болот и среднетаежных рямов на междуречьях 13. Поселков в пределах северных подзон 14. Поселков в долине Оби в пределах южных подзон 15. Поселков на подтаежных междуречьях
Всего .... | 73
но взятых междуречьях (множественная оценка — на 7—8?^, а ин-
дивидуальные, кроме антропогенного влияния,— на 1—10%). Осо-
бенно сильно уменьшилось влияние подзональных отличий.
Однако полнота объяснения территориальной неоднородности
населения птиц отдельными факторами среды оказалась недостаточ-
ной. Кроме того, указанные факторы в значительной степени скор-
релированы, а птицы обычно реагируют не на отдельные факторы,
а на их неразложимые сочетания (природные режимы). Поэтому
для оценки степени совпадения природных режимов и простран-
ственной изменчивости сообществ птиц мы использовали экспрес-
сный алгоритм факторной классификации [Трофимов, 1976]. Ре-
зультаты этой классификации, представленные в виде списков при-
родных режимов и степени их совпадения с пространственной не-
однородностью населения птиц таежных междуречий Западной Си-
бири (оценивается величиной учтенной дисперсии), приведены в
табл. 27.
Факторная классификация сообществ птиц по лесной зоне сре-
динной части Западной Сибири показала, что неоднородность насе-
ления лучше всего аппроксимируется природными режимами, пе-
речисленными в табл. 28. Выявленные наборы природных режимов
при множественной аппроксимации определяют около 82% терри-
ториальной неоднородности сообществ птиц на таежных междуречьях
и 70—73% по всей лесной зоне Западной Сибири, что на 11—20%
больше, чем было получено по выделенным градациям тех же факто-
ров (без учета их сочетаний). Зто позволяет считать приведенные
списки природных режимов достаточно полными, а полученное с их
помощью объяснение территориальной изменчивости населения птиц
довольно надежным для последующей пространственной индикации
сообществ птиц на необследованных таежных территориях За-
падно-Сибирской равнины.
229
ЗАКЛЮЧЕНИЕ
Проведенные исследования населения птиц таежных междуре-
чий Западной Сибири позволили объединить полученные материалы
с аналогичными опубликованными данными по долине Оби. В ре-
зультате анализа выявлен общий характер территориальных изме-
нений в сообществах птиц таежных междуречий и лесной зоны
срединной части Западной Сибири, основные особенности которых
сводятся к следующему.
1. Подзонально-ландшафтное размещение видов птиц опреде-
ляется природными режимами: северной тайги, темнохвойно-лесных
ландшафтов срединных подзон, лесолуговых и лесополевых ланд-
шафтов южных подзон, пойменных соров и низинных болот южных
подзон, верховых болот, поселков и прилежащих местообитаний,
водоемов. Эти режимы как комплексы условий окружающей среды
позволяют выделить типы распределения птиц по предпочитаемое™
территорий с действием указанных режимов. Типы подразделяются
на подтипы, что совпадает с различиями в составе и полноте древо-
стоев, с особенностями подзонального режима средней тайги и юж-
ных подзон, влияния вырубок и распашки, пожаров и уничтожения
лесов сибирским шелкопрядом, пойменного режима, типа водоемов и
характера увлажнения низинных болот.
2. В сообществах птиц чаще доминируют широко распростра-
ненные виды, характерные для двух и более подзон. Доминанты,
специфичные лишь для одной подзоны, встречаются реже и в основ-
ном в северной тайге. Доля преобладающих видов в сообществах
птиц неуклонно возрастает от южной тайги к северной, так как чис-
ленность их падает к северу в значительно меньшей степени, чем
второстепенных видов.
3. Максимальная плотность гнездования птиц на таежных меж-
дуречьях зарегистрирована в подтаежных лесах, откуда она неук-
лонно снижается к северу до средней тайги, что совпадает с увели-
чением суровости подзонального климата и уменьшением продуктив-
ности сообществ, а затем несколько возрастает в северной тайге за
счет видов, гнездящихся преимущественно в лесотундровых и севе-
ротаежных редколесьях. Для долины Оби характерны аналогич-
ные широтные изменения плотности гнездования, однако обилие
гнездящихся птиц здесь в 1,7—3 раза больше, чем на междуречьях.
230
4. Число откочевывающих после окончания гнездования птиц
возрастает с севера на юг таежных междуречий в соответствии с бо-
лее ранним фенологическим развитием природы в южных подзонах.
Сходная тенденция прослеживается и в долине Оби, но доля отко-
чевывающих птиц от числа отгнездившихся здесь значительно ниже
и снивелирована иммиграцией птиц.
5. На таежных междуречьях интенсивность пред- и послегнездо-
вого пролета уменьшается к северу от южной тайги и в подтаежных
лесах вследствие снижения разнообразия местообитаний, их эколо-
гической привлекательности для мигрирующих птиц и уменьшения
площади лесных ландшафтов. В долине Оби количество пред- и
послегнездовых мигрантов в 3—20 раз больше, чем на междуречьях,
причем это превышение с юга на север возрастает. Таким образом,
в лесной зоне Западной Сибири с продвижением от подтаежных ле-
сов к северной тайге пролет птиц по долине Оби становится более
интенсивным по сравнению с междуречьями.
6. Доля мигрантов в населении птиц таежных междуречий по
сравнению с долиной Оби невелика, а большая часть отгнездивших-
ся птиц, по-видимому, смещается с междуречий в долину Оби. Это
определяет преобладание на междуречьях равновесного и эмигра-
ционного типов динамики населения птиц, а в долине Оби —
V-образного.
7. Плотность, разнообразие, биомасса населения птиц и величи-
на трансформируемой энергии уменьшаются к северу от южной тай-
ги и в подтаежных лесах, что совпадает со снижением продуктивно-
сти сообществ при увеличении суровости подзонального климата,
а в подтаежных лесах определяется некоторым дефицитом увлажне-
ния, возрастанием интенсивности антропогенного воздействия, мень-
шим набором местообитаний и, видимо, их малой привлекатель-
ностью для пред- и послегнездовых мигрантов. Однако в северной
тайге суммарные показатели сообществ птиц несколько выше, чем
в средней, в связи с обогащением минерального питания фитоцено-
зов комплексных верховых болот, большей напряженностью лет-
них кочевок и высокой численностью гипоарктических и бореально-
гипоарктических видов птиц вблизи оптимумов их ареала.
8. Суммарные показатели населения птиц в пределах сходных
подзон снижаются в направлении от поймы Оби к междуречьям.
При этом наиболее высокие значения характерны для пой-
менных лесолуговых ландшафтов и особенно низинных болот, да-
лее они снижаются в ряду: лесные и лесополевые ландшафты — вне-
пойменные низинные болота — верховые болота надпойменных тер-
рас и междуречий. В лесных ландшафтах с юга на север таежных
междуречий прослеживается тенденция к уменьшению суммарных
показателей орнитокомплексов, а на верховых болотах они, наобо-
рот, возрастают в том же направлении. В результате в северной тай-
ге плотность, разнообразие, биомасса населения птиц и величина
трансформируемой энергии характеризуются более выровненными
показателями на верховых болотах по отношению к лесным ландшаф-
там и на междуречьях в сравнении с поймой Оби.
231
9. Сходство северотаежных сообществ птиц (с учетом численно-
сти видов) по сравнению с более южными подзонами также возраста-
ет за счет высокой численности птиц северотаежных и лесотундро-
вых редколесий и значительного выравнивания территориального
распределения птиц, что в некоторой степени связано с увеличением
сходства природных условий ландшафтов северной тайги. Это при-
водит к значительным различиям между орнитокомплексами север-
ной тайги и более южных подзон, тогда как последние близки между
собой.
10. Фаунистический состав лесных орнитокомплексов чаще все-
го европейско-сибирский и только в подтаежных лесах сибирско-
европейский. При этом доля сибирских видов по направлению к се-
веру возрастает, а европейских снижается. Значительных различий
в фаунистическом составе населения птиц долины Оби и междуречий
не прослеживается, но в первой в ряде случаев больше европейских
и китайских видов, а на последних — транспалеарктов. Верховые
болота, особенно их междуречные грядово-мочажинно-озерные мас-
сивы, характеризуются наибольшей разнородностью фаунистиче-
ского состава, в том числе относительно высоким участием аркти-
ческих видов, что, видимо, связано с большим числом водоемов, на-
личием открытых пространств и их некоторой экологической бли-
зостью к тундровым территориям.
11. Несмотря на значительную гетерогенность населения птиц
лесной зоны Западной Сибири, представители основных слагающих
его типов фауны имеют вполне обособленные, хотя иногда перекры-
вающиеся зоны преференции, которые физиономически сходны с об-
ластями формирования того или иного фаунистического комплекса.
Так, арктические виды занимают преимущественно подзону северной
тайги, обширные верховые болота на междуречьях более южных под-
зон и на пролете — водоемы. Представители сибирского типа фауны
отдают предпочтение таежно-лесным ландшафтам срединных подзон.
Европейские виды приурочены в основном к лесолуговым поймам,
низинным болотам и лесополевым ландшафтам южных подзон.
12. Пространственно-типологическая структура населения птиц
лесной зоны Западной Сибири представима на плоскости в виде тра-
пеции и выходящего за ее пределы диагонального ряда. При этом
горизонтальные ряды орнитокомплексов соответствуют их измене-
ниям, связанным с уменьшением залесенности при переходе от тем-
нохвойных лесов к пойменным лугам и открытым низинным болотам.
Вертикальные ряды демонстрируют территориальную смену сооб-
ществ птиц при уменьшении теплообеспеченности ландшафтов от
южных подзон к северной тайге. Наиболее длинный, диагональный
ряд отражает изменения населения птиц при переходе от всех осталь-
ных местообитаний к олиготрофным верховым болотам. Этот ряд ха-
рактеризуется увеличением заболоченности при обеднении минераль-
ного питания фитоценозов Ориентируя это изображение в трехмер-
ном факторном пространстве, при увеличении степени абстрагиро-
вания можно получить ординационную схему пространственных
изменений населения птиц в виде пирамиды. В этом случае ее осно-
232
ванне отражает изменения сообществ, связанные с трендами умень-
шения залесенности и теплообеспеченности. По третьей оси (от ос-
нования к вершине пирамиды) идут изменения населения птиц,
совпадающие с увеличением заболоченности при обеднении мине-
рального питания.
13. Наиболее значимыми факторами, определяющими простран-
ственную организацию населения птиц таежных междуречий и всей
лесной зоны Западной Сибири, следует считать: залесенность, антро-
погенное влияние (включая застроенность, распашку, вырубку ле-
сов, пастбищную дигрессию, сенокошение и зарегулированность сто-
ка Оби), подзональное изменение климата (широтное снижение теп-
лообеспеченности) и определяемого им подзонального природного
режима в целом, состав лесообразующих пород, заболоченность,
богатство минерального питания фитоценозов и общую продуктив-
ность биоценозов. Влияние рельефа более слабое и прослеживается
лишь по всей лесной зоне срединной части Западной Сибири. Соче-
тания этих факторов, проявляющиеся как природные режимы, опре-
деляют около 82% территориальной изменчивости сообществ птиц
на таежных междуречьях и примерно 72% — в лесной зоне Западной
Сибири. Такая полнота объяснения позволяет считать наборы вы-
явленных факторов достаточно надежными для последующей про-
странственной индикации сообществ на необследованных таежных
территориях Западно-Сибирской равнины.
ЛИТЕРАТУРА
Ананин А. А., Москвигпн С. С. Динамика населения птиц южной части Васю-
ганья.— В кн.: II Всесоюз. конф, ко миграциям птиц. Тез. сообщ. Алма-
Ата, 1978, с. 77-78.
Анзмгитова II. В., Бадаев С. А., Бурский О. В. и др. Некоторые зональные осо-
бенности населения птиц приенисейской тайги. — В кн.: VII Всесоюз.
зоогеогр. конф. Тез. докл. М.: Наука, 1979, с. 92—93.
Белов С. Н., Вартапетов Л. Г., Николаев В. II. и др. Охотничьс-промысловые
ресурсы и особенности животного населения (на примере Среднего При-
обья).— В кн.: Космические исследования природных комплексов Сиби-
ри и Дальнего Востока. Новосибирск: Наука, 1983, с. 135—140.
Бирюков В. П., Козлов В. П., Кузьменко В. Я. Материалы к орнитофауне бо-
лот Белорусского Поозерья.— В кн.: Новые проблемы зоологической
науки и их отражение в вузовском преподавании. Тез. докл. науч. конф,
зоол. пед. ин-та. Ч. 2. Ставрополь, 1979, с. 212.
Бутьев В. Т., Гусаков Е. С. Послегнездовое население птиц тайги Онежского
полуострова.— В кн.: Фауна и экология животных. М., 1976, с. 50—66.
Бутьев В. Т., Рубинштейн Н. А. Структура населения птиц тайги Европейской
территории СССР. — В кн.: Новые проблемы зоологической науки и их
отражение в вузовском преподавании. Тез. докл. науч. конф. зоол. иед.
ин-та. Ч. 2. Ставрополь, 1979, с. 221—223.
Вартапетов Л. Г. Особенности пространственных изменений сообществ позво-
ночных таежных междуречий Западной Сибири.— В кн.: VII Всесоюз.
зоогеогр. конф. Тез. докл. М.: Наука, 1979а, с. 101—104.
Вартапетов Л. Г. Внутризональные отличия населения птиц таежных между-
речий Западной Сибири.— Там же, 19796, с. 99—101.
233
Вартапетов Л. Г. Динамика плотности населения птиц таежных междуречий
Западной Сибири в гнездовой и послегнездовой периоды. — В кн.: Тез.
Всесоюз. конф, молодых ученых «Экология гнездования и методы ее изу-
чения». Самарканд, 1979в, с. 43—44.
Вартапетов Л. Г. Внутриландшафтная структура населения птиц северной тай-
ги Приказымья.— В кн.: Проблемы зоогеографии и истории фауны. Но-
восибирск: Наука, 1980, с. 156—166.
Вартапетов Л. Г., Фомин Б. Н., Равкин Ю. С. Водоплавающие птицы Уват-
ского района Тюменской области.— В кн.: Ресурсы водоплавающих
птиц СССР, их воспроизводство и использование. Вып. 2. М.: Изд-во
МГУ, 1972, с. 42-43.
Вартапетов Л. Г., Цыбулин С. М., Равкин Ю. С. и др. Географическая изменчи-
вость населения птиц северной тайги Западной Сибири, — В кн.: Проб-
лемы зоогеографии и истории фауны. Новосибирск: Наука, 1980, с. 145—
156.
Венгеров М. П. Миграция куликов в пойме нижней Оби. — В кн.: Фауна и эко-
логия куликов. Вып. 2. М.: Изд-во МГУ, 1973, с. 94—95.
Владышевский Д. В. Значение трофического фактора для птиц в различных эко-
логических ситуациях.— В кн.: Экология популяций лесных животных
Сибири. Новосибирск: Наука, 1974. 142 с.
Второв П. Н. Птицы в экосистемах горных стран.— В кн.: Орнитология.
Вып. 12. М.: Изд-во МГУ, 1976, с. 55—60.
Гаврилов В. М. Биоэнергетика миграций.— В кн.: Методы изучения миграции
птиц. Материалы Всесоюз. школы-семинара. М., 1977, с. 7—16.
Тибет Л. А. Артамошин А. С. Птицы тайги Северного Заангарья.— В кн.: Фау-
на и систематика позвоночных Сибири. Новосибирск: Наука, 1977,
с. 286-292.
Гладков Н. А. О географической изменчивости числа видов и численности осо-
бей.— В кн.: Проблемы зоогеографии суши. Львов, 1958, с. 57—63.
Гордеев Ю. И. Некоторые данные по птицам Казыма,— В кн.: Орнитология.
Вып. 6. М.: Изд-во МГУ, 1963, с. 469-470.
Гордеев Ю. И. К орнитофауне восточной части Ханты-Мансийского националь-
ного округа (Тюменская область).— В кн.: Орнитология. Вып. 7. М.:
Изд-во МГУ, 1965, с. 466—467.
Гынгазов А. М., Москвитин С. С. О распространении некоторых птиц в лесной
зоне Западной Сибири.— Там же, с. 71—74.
Гынгазов А. М., Миловидов С. П. Орнитофауна Западно-Сибирской равнины.
Томск: Изд-во ТГУ, 1977. 350 с.
Гынгазов А. М., Шубин Н. Г. О растительности и численности куриных птиц
в Западной Сибири.— В кн.: Ресурсы тетеревиных птиц в СССР. Мате-
риалы совещ. М., 1968, с. 13—15.
Дубинин Н. П., Торопанова Г. А. Некоторые закономерности распространения
птиц лесной зоны.— В кн.: Орнитология. Вып. 3. М.: Изд-во МГУ, 1960,
с. 114-121.
Дучиц В. Н. К изучению орнитофауны верховых и низинных болот Белорусской
ССР.— В кн.: Экология и миграция птиц Прибалтики. Рига, 1962,
с. 317—322.
Дучиц В. Н. Орнитофауна верховых болот заповедника. — В кн.: Березинский
заповедник. Вып. 4. Минск, 1975, с. 184—190.
Западная Сибирь/Под ред. Г. Д. Рихтера. М.: Изд-во АН СССР, 1963. 488 с.
Иванов А. И. Каталог птиц СССР. Л.: Наука, 1976. 276 с.
Ильина И. С., Лапшина Е. И., Махно В. Д. и др. Растительность Западно-Си-
бирской равнины. Карта М 1 : 1 500 000. М.: ГУГК, 1976.
Кендэл М. Д. Ранговые корреляции. М., 1975. 214 с.
Кисленко Г. С., Наумов Р. Л. Некоторые сведения о неясытях Красноярского
края.— В кн.: Орнитология. Вып. 10. М.: Изд-во МГУ, 1972, с. 339—342.
Кищинский А. А. Численность, биомасса птиц и трансформация ими энергии
в субарктических экосистемах.—Экология, № 5, 1976, с. 71—78.
Кищинский А. А. Понятие о гипоарктической и эоарктической авифаунах.—
В кн.: VII Всесоюз. орнитол. конф. Киев: Наукова думка, 1977, с. 65—
234
К и ш ине кий А. А. Пространственные и экологические связи авифауны Северо-
Восточной Азии. Автореф. докт. дис. М., 1979. 47 с.
Козлов В. П., Кузьменко В. П. К орнитофауне верховых болот Белорусского
Поозерья.— В кн.: VII Всесоюз. орнитол. конф. Киев: Наукова думка,
1977, с. 68-69.
Крылов Г. В. Леса Западной Сибири. Новосибирск: Наука. 1961. 252 с.
Кузякин А. П. Зоогеография СССР. — Учен. зап. Моск. обл. пед. ин-та им.
Н. К. Крупской, 1962, т. 52, с. 3—182.
Кулешова Л. В. Анализ структуры птичьего населения в связи с ярусностыо
леса.— В кн.: Орнитология. Вып. 9. М.: Изд-во МГУ, 1968, с. 108—120.
Кумари Э. В. Верховые болота Эстонии как местообитания птиц. — В кн.: Ор-
нитология. Вып. 7. М.: Изд-во МГУ, 1965, с. 36—43.
Куперштох В. Л., Трофимов В. А., Равкин Ю. С. Новый способ выявления про-
странственной макроструктуры животного населения (на примере птиц).—
В кн.: Роль животных в функционировании экосистем. М., 1975, с. 162—
165.
Леонович В. Л. Миграция и гнездовой консерватизм у некоторых видов птиц
со специализированным питанием.— В кн.: Орнитология. Вып. 12. М.:
Изд-во МГУ. 1976, с. 87—94.
Максимов А. А. Структура и динамика биоценозов речных долин. Новосибирск:
Наука, 1974. 260 с.
Матюшкин Е. II., Кулешова Л. В. Пятнистый конек в Среднем Сихотэ-Алине:
опыт анализа структуры ареала.— В кн.: Орнитология. Вып. 10. М.:
Изд-во МГУ. 1972. с. 182-193.
Москвитян С. С. К авифауне лесной зоны Западной Сибири. — Там же, 1972а,
с. 372-373.
Москвитин С. С. О распространении и экологии соловьев в междуречье Оби и
Енисея.— Там же, 19726, с. 173—181.
Москвитин С. С. Заметки по биологии воробьиных Западно-Сибирской равни-
ны.— В кн.: Биология. Т. 4. Томск: Изд-во ТГУ, 1974, с. 9—14.
Москвитин С. С., Дубовик А. Д., Горд Б. Я. Птицы долины р. Кеть. — В кн.:
Фауна и систематика позвоночных Сибири. Новосибирск: Паука, 1977,
с. 245—279.
Назаров А. А. Фауна болот. — В кн.: Научные предпосылки освоения болот
Западной Сибири. М.: Наука, 1977, с. 148—160.
Наумов Р. Д. Птицы в очагах клещевого энцефалита Красноярского края. Ав-
тореф. канд. дис. М.. 1964. 19 с.
Наумов Р. Л., Кисленко Г. С. Гнездование пятнистого и певчего сверчков в Крас-
ноярском крае.— В кн.: Орнитология. Вып. 7. М.: Изд-во МГУ, 1965,
с. 83—86.
Новиков Г. А. Географическая изменчивость плотности населения лесных птиц
в европейской части СССР и в сопредельных странах.— Зоол. журн.,
1960, т. 39, вып. 3, с. 433—447.
Пантелеев П. А. О птичьем населении долины Оби и примыкающих ландшафтов
в Нарымском крае.— В кн.: Орнитология. Вып. 10. М.: Изд-во МГУ,
1972а, с. 161—172.
Пантелеев П. А. Материалы к количественной характеристике авифауны юж-
ной тайги Зауралья.— Там же, 19726, с. 374—377.
Потапова Л. С., Жуков В. М. Местный климат и микроклимат междуречья
Обь — Иртыш.— В кн.: Природные условия освоения междуречья Обь —
Иртыш. М., 1972, с. 94—107.
Природа и экономика Привасюганья. Томск, 1966. 266 с.
Природные условия освоения междуречья Обь — Иртыш. М., 1972. 260 с.
Приходько И. К. Некоторые наблюдения над птицами Васюганья.— В кн.:
Вопросы биологии и агрохимии. Томск: Изд-во ТГУ, 1969, с. 111—119.
Птицы Советского Союза. Т. 1—6. М.: Наука, 1951—1954.
Пузаченко Ю. Г. Географическая изменчивость обилия и структуры населения
птиц лесных биоценозов.— В кн.: Орнитология. Вып. 8. М., 1967, с. 109—
122.
Равкин Ю. С. К методике учета птиц лесных ландшафтов.— В кн.: Природа оча-
гов клещевого энцефалита на Алтае. Новосибирск, 1967, с. 66—75.
235
Равкин 10. С. Птицы Северо-Восточного Алтая. Новосибирск: Наука, 1973.
344 с.
Равкин Ю. С. Птицы лесной зоны Приобья. Новосибирск: Наука, 1978. 287 с.
Равкин Ю. С. Пространственная организация населения птиц лесной зоны. Но-
восибирск: Наука, 1984. 280 с.
Равкин Ю. С., Фомин Б. Н., Вартапетов Л. Г. и др. Учеты водоплавающих птиц
в Томской области.— В кн.: Ресурсы водоплавающих птиц в СССР, их
воспроизводство и использование. Вып. 2. М.: Изд-во МГУ, 1972, с. 44—
45.
Равкин Ю. С., Лукьянова И. В. География позвоночных южной тайги Западной
Сибири. Новосибирск: Наука, 1976. 338 с.
Равкин Ю. С., Шадрина В. И. Факторная классификация птиц по характеру их
пространственного распределения.— Изв.Сиб. отд-ния АН СССР, 1980,
вып. 1, с. 65—69.
Раевский В. В. Опыт количественной характеристики зимней фауны птиц тайги
Западной Сибири.— В кн.: Методы учета численности и географического
распределения наземных позвоночных. М., 1952, с. 130—136.
Раевский В. В. Позвоночные животные Северного Зауралья. М.: Наука, 1982.
146 с.
Рогачева Э. В. Численность и распределение птиц нижнего Елогуя.— В кн.:
Орнитология. Вып. 5. М.: Изд-во МГУ, 1962, с. 118—134.
Рогачева Э. В. Зональные особенности населения птиц енисейской лесотундры
и тайги. — В кн.: Новости орнитологии. Материалы IV Всесоюз. орнитол.
конф. Алма-Ата, 1965, с. 320—322.
Рогачева Э. В., Сыроечковский Е. Е. О влиянии деятельности человека на фау-
ну енисейской тайги.— Учен. зап. Моск. обл. пед. ин-та им. Н. К. Круп-
ской, 1962, т. 59, с. 197—215.
Рогачева Э. В., Сыроечковский Е. Е., Бурский О. В. и др. Птицы среднетаежно-
го Енисея: фауна, ее охрана и рациональное использование.— В кп.: Ох-
рана фауны Крайнего Севера и ее рациональное использование. М., 1978,
с. 30—165.
Романова Е. А. Типы болотных массивов и закономерное их распределение на
территории Западной Сибири.— В кн.: Типы болот СССР и принципы их
классификации. Л., 1974, с. 167—174.
Словарь общегеографических терминов. Т. 2. М.: Прогресс, 1975. 394 с.
Сорокина Л. И. Ландшафтное распределение глухаря в средней тайге Запад-
ной Сибири.— В кн.: Материалы VI Всесоюз. орнитол. конф. Ч. 2. М.,
с. 296-297.
Степанян Л. С. Состав и распределение птиц фауны СССР. М.: Наука, 1978.
392 с.
Стопалов В. С., Вартапетов Л. Г., Бурский О. В. К орнитофауне Западной и
Средней Сибири.— В к,и.: Материалы науч, совещ. зоологов пед. ин-тов.
Владимир, 1973, с. 338—339.
Стрелков В. Е. Кулики бассейна средней Оби,— В кн.: Фауна и экология ку-
ликов. Вып. 2. М., 1973, с. 72—73.
Стрелков В. Е. К экологии размножения куликов и чаек в бассейне Средней
Оби.— В кн.: Материалы VI Всесоюз. орнитол. конф. Ч. 2. М.: Изд-во
МГУ, 1974, с. 141-143.
Стрелков В. Е. Кулики и чайки бассейна реки Тыма.— В кн.: Биология. Томск,
1976, с. 30-40.
Сыроечковский Е. Е., Рогачева Э. В. Животный мир Красноярского края. Крас-
ноярск, 1980. 360 с.
Тарунин М. П. Птицы реки Малой Сосьвы,— Ежегодник Тюмен. обл. краев,
музея, 1959, вып. 1, с. 124—148.
Телегин В. И. Долины северных рек как места концентрации и пути проник-
новения таежных животных на крайний север.— В кп.: Природа поймы
реки Оби и ее хозяйственное освоение. Томск: Изд-во ТГУ, 1963, с. 343—
349.
Телепнев В. Г. К динамике численности тетеревиных птиц в Томской обла-
сти.— В кн.: Материалы VI Всесоюз. орнитол. конф. М., 1974, с. 297—299.
233
Трофимов В. А. Модели и методы качественного факторного анализа матрицы
связи,— В кн.: Проблемы анализа дискретной информации. Ч. II. Ново-
сибирск, 1976, с. 24—36.
Трофимов В. А. Качественный факторный анализ матрицы связей в простран-
стве разбиений со структурой.— В кн.: Модели агрегирования социально-
экономической информации. Новосибирск, 1978, с. 91—106.
Трофимов В. А., Равкин Ю. С. Экспресс-метод оценки связи пространственной
неоднородности животного населения и факторов среды.— В кн.: Коли-
чественные методы в экологии животных. Л., 1980, с. 113—115.
Трофимов В. А., Куперштох В. Л., Равкин Ю. С. К проблеме выявления про-
странственно-типологической структуры сообществ.— В кн.: Проблемы
зоогеографии и истории фауны. Новосибирск, 1980, с. 41—58.
Тупикова Н. В. Зоологическое картографирование. М., 1969. 250 с.
Тупикова Н. В., Комарова Л. В. Принципы и методы зоологического картогра-
фирования. М.: Изд-во МГУ, 1979. 191 с.
Формозов А. II. Равнинность Западной Сибири и связанные с ней особенности
животного мира. — В кн.: Развитие и преобразование географической
среды. М.: Наука, 1964, с. 201—221.
Ходашова К. С. О географических особенностях структуры населения позво-
ночных животных.— В кн.: Зональные особенности населения наземных
животных. М., 1966, с. 7—32.
Чернов Ю. И. Природная зональность и животный мир суши. М.: Наука, 1975.
225 с.
Чернов Ю. II. Структура животного населения Субарктики. М.: Наука, 1978.
167 с.
Чернов Ю. II., Ходашова К. С. Некоторые закономерности изменения структу-
ры животного населения в зональном аспекте.— В кн.: IV Межвуз. зоо-
геогр, конф. Тез. докл. Одесса, 1966. 306 с.
Четырехъязычный энциклопедический словарь терминов по физической гео-
графии. М.: Советская энциклопедия, 1980. 703 с.
Шаронов А. Д. Некоторые результаты изучения фауны птиц в таежной зоне
Западной Сибири.— Докл. АН СССР, 1951, т. 78, № 3. с. 1057—1059.
Штегман Б. К. Основы орнптогеографического деления Палеарктики.— В ки.:
Фауна СССР. Птицы. Т. 1. Вып. 2. М.— Л., 1938. 156 с.
Шухов II. Н. Птицы Обдорского края.— Ежегодник зоол. музея Акад, наук,
1915, т. 20, № 35-39, с. 167-238.
Шухов II. Н. Река Казым и ее обитатели.— Ежегодник Тобол, губерн. музея,
1916, вып. 26, с. 1—56.
Den Boer Р. J. Spreading of risk and stabilisation of animal numbers.— Acta bio-
theoretica, 1968, v. 18, p. 165—194.
Dyrcz A., Okulewicz J., Wiatr B. Ptaci Leczynsko-Wlodawskiego W’okresio lo-
gowym (suwsgliedhieniem liscbnosci ptacow wodnych).— Acta zool, Cra-
cov, 1973, v. 18, N 15, p. 475—528.
Jaccard P. Lois de distribution florale dans la zone alpine. — Bull. Soc. Vaund.
. Sci. Nat., 1902, v. 38, p. 69—130.
Jarvinen O., Sanimalisfo L. Regional trends in the avifauna of Finnish peatland
bogs.—Ann. Zool. Fennici, 1976, N 13, p. 31—43.
Johansen H. Die Vogelfauna Westsibiriens.— J. fur Ornithologie, 1943—1961,
N 1-102.
Levins R. Some demographic and genetic consequenses of environmental hetero-
geneity for biological control.— Entomol. Soc. Amer. Bull., Washington,
1969, v. 15, p. 237-240.
MacArthur R. H. Patterns of species diversity. — Biol. Rev., 1965, N 40, p. 510 —
533.
MacArthur R. H., MacArthur J. W. On bird species diversity. — Ecology, 1961,
v. 42, p. 594-598.
Margalef R. Perspectives in Ecological Theory. Chicago: Univ, of Chicago Press.
1968. Ill p.
Roth R. R. Spatial heterogeneity and bird species diversity.— Ecology, 1976,
v. 57, N 4, p. 773—782.
237
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
РУССКИХ НАЗВАНИЙ ВИДОВ ПТИЦ1
Аист черный 75**
Барсучок 84
Бекас 42
Белобровик 80
Бормотушка 100*
Вальдшнеп 63**
Варакушка 73
Веретенник большой 45
— малый 45
Вертишейка 55
Весничка 89
Воробей домовый 115
— полевой 116
Ворон 124
Ворона серая 122
— черная 124
Выпь 75**
Вяхирь 60*
Гагара краснозобая 13
,— чернозобая 14
Гаичка сероголовая 97
Галка 121
Галстучник 34
Гаршнеп 49*
Глухарь 29
Гоголь 21
Голубь сизый 49
Горихвостка-лысушка 75
Горлица большая 50
Грач 122
Гуменник 15
Гусь серый 15
Дербник 26
Деряба 82
Дрозд певчий 81
— чернозобый 79
Дубонос 115
Дубровник 105
Дупель 42
— лесной 43
Дятел белоспинный 58
— пестрый большой 56
— пестрый малый 58
— седой 83**
Дятел трехпалый 58
— черный 56
Жаворонок полевой 59
— рогатый 60
Жулан 70
— сибирский 81*
Журавль серый 33
Зарничка 91
Зарянка 83*
Зимородок 66**
Зуек малый 34
Зяблик 108
Иволга 118
Каменка 78
Камнешарка 47*
Камышевка вертлявая 9
— садовая 84
Канюк 25
Кедровка 120
Клест белокрылый ИЗ
— еловик ИЗ
Клинтух 50
Князек 100
Кобчик 27
Козодой 54
Колючехвост 87**
Конек зеленый 68
— краснозобый 70
Конек лесной 66
— степной 66
Коноплянка 131*
Королек желтоголовый 110*
Коростель 32
Коршун черный 23
Крачка малая 59*
— полярная 49
— речная 48
— светлокрылая 66**
— черная 66*
Кречет 27
Кроншнеп большой 43
— средний 44
Крохаль большой 23
— средний 22
Кряква 15
Кукушка 51
— глухая 52
Кукша 119
Кулик-воробей 41
— сорока 39*
Куропатка белая 29
Ласточка береговая 60
— деревенская 61
Лебедь-кликун 15
Лунь болотный
— луговой 26
— полевой 25
Лунь степной 26
Луток 25*
Лысуха 52**
1 Указатели русских и латинских названий относятся к повидовому обзо-
ру. Виды, встреченные в долине Оби Ю. С. Равкиным [1978], отмечены одной
звездочкой, а зарегистрированные только в южной тайге Ю. С. Равкиным,
И. В. Лукьяновой [1976] —двумя. Описание их см. на указанных страницах
соответствующих публикаций.
238
Мородунка 40
Московка 98
Мухоловка малая 112*
— мугимаки 121**
— пеструшка 94
— серая 93
— сибирская 120**
Неясыть длиннохвостая 64*
— бородатая 53
Нырок белоглазый 73**
— красноголовый 20
— красноносый 73**
Овсянка белошапочная 102
— камышевая 107
— крошка 103
— обыкновенная 119*
— полярная 126*
— ремез 105
Ополовник 95
Орлан-белохвост 24
Осоед 79**
Пеночка бурая 106*
— зеленая 93
— корольковая ПО**
— толстоклювая Щ**
Перевозчик 39
Перепел 28
Перепелятник 24
Пересмешка 85
Песочник белохвостый 41
Пищуха 102
Плавунчик круглоносый 40
Поганка красношейная 14
— серощекая 19*
— черношейная 68**
Погоныш 33
Подорожник лапландский 107
Подорлик большой 25
Поползень 100
Поручейник 58**
Пустельга 26
Пухляк 96
Ржанка золотистая 36*
— бурокрылая 36*
Рябинник 80
Рябчик 31
Сапсан 28
Сверчок 83
— певчий 95*
— пятнистый 83
— таежный 111**
Свиристель 72
Свиязь 17
Синехвостка 74
Синица большая 99
Синьга 21
Скворец 117
Скопа 23
Славка-завирушка 88
— садовая 86
— серая 87
— ястребиная 86
Снегирь 114
Сова болотная 54
— ястребиная 53
Сойка 118
Соловей 84*
— красношейка 73
— свистун 104**
— синий 85*
Сорока 120
Сорокопут большой 71
Стриж черный 54
Сыч воробьиный 53
— мохноногий 64*
Таловка 92
Теньковка 90
Тетерев 30
Тетеревятник 24
Травник 39
Трясогузка белая 64
— горная 66
— желтая 62
— желтоголовая 64
Тулес 36*
Турпан 23*
Турухтан 40
Удод 82**
Улит большой 38
Урагус 132*
Утка серая 15*
Фифи 37
Хрустан 38*
Чайка малая 48
— озерная 47
— серебристая 47
— сизая 46
Чеглок 27
Чекан луговой 76
— черноголовый 77
Чернеть морская 21
— хохлатая 20
Чернозобик 41
Черныш 36
Чечевица 112
Чечетка 111
Чибис 35
Чиж 110
Чирок-свистунок 16
— трескунок 18
Чомга И*
Шилоклювка 39*
Шилохвость 18
Широконоска 19
Щегол ПО
Щеголь 44*
Щур из
Юрок 109
230
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
ЛАТИНСКИХ НАЗВАНИЙ ВИДОВ ПТИЦ
Acanthis flammea 111 Accipiter gentilis 24 — nisus 24 Acrocephalus dumetorum 84 — paludicola 97* — schoenobaenus 84 Actitis hypoleucos 39 Aegithalos caudatus 95 Aegolius funereus 64* Aesalon columbarius 26 Alauda arvensis 59 Alcedo atthis 66* Anas acuta 18 — clypeata 19 — crecca 16 — penelope 17 — platyrhynchos 15 — querquedula 18 — strepera 15* Anser anser 15 — fabalis 15 Anthus cervina 70 — hodgsoni 68 — richardi 66 — trivialis 66 Apus apus 54 Aquila clanga 25 Arenaria interpres 47* Asio flammeus 54 Aythya ferina 20 — fuligula 20 — marila 21 — nyroca 73** Bombycilla garrulus 72 Botaurus stellaris 75** Bucephala clangula 21 Buteo buteo 25 — pygargus 26 Coccothraustes coccothraustes 115 Columba livia 49 — oenas 50 — palumbus 60* Corvus corax 124 — corone 124 — cornix 122 — frugilegus 122 — monedula 121 Coturnix coturnix 28 Crex crex 32 Cuculus canorus 51 — saturatus 52 Cyanosylvia svecica 73 Cygnus cygnus 15 Dendrocopos leucotos 58 — major 56 — minor 58 Dryocopus martius 56 Emberiza aureola 105 — citrinella 119* — leucocephalos 102 — pallasi 126* Emberiza pusilia 103 — rustica 105 — schoeniclus 107 Eremophila alpestris 60 Erithacus rubecula 83* Erythropus vespertinus 27 Eudromias morinellus 38* Falco gyrfalco 27 — peregrinus 28 Ficedula hypoleuca 94 Fringilla coelebs 108 — montifringilla 109
Calcarius lapponicus 107 Calidris alpina 41 — minutus 41 — temminckii 41 Calliope calliope 73 Cannabina cannabina 131* Caprimulgus europaeus 54 Carduelis carduelis 110 Carpodacus erythrinus 112 Cerchneis tinnunculus 26 Certhia familiaris 102 Charadrius dubius 34 — hiaticula 34 Chlidonias leucoptera 66** Chlidonias nigra 57* Ciconia nigra 75** Circus aeruginosas 77** — cyaneus 25 — macrourus 26 Fulica atra 52** Gallinago gallinago 42 — media 42 — megala 43 * Garrulus glandarius 118 Gavia arctica 14 — stellata 13 Glaucidium passerinum 53 Grus grus 33 Haematopus ostralegus 39* Haliaeetus albicilla 24 Hippolais caligata 100* — icterina 85 Ilirundapus caudacutus 87** Hirundo rustica 61 Hypotriorchis subbuteo 27 Jynx torquilla 55
240
Lagopus lagopus 29
Lanius collurio 70
— cristatus 81*
— excubitor 71
Larus argentatus 47
— canus 46
— minutus 48
— ridibundus 47
Larvivora cyano 85*
Limosa limosa 45
— lapponica 45
Locustella certhiola 95*
— fasciolata 111**
— lanceolata 83
— naevia 83
Loxia leucoplera 113
— curvirostra 113
Luscinia luscinia 84*
Lymmocryptes minimus 49*
Lyrurus tetrix 30
Melanitta fusca 23*
— nigra 21
Mergus albellus 25*
— merganser 23
— serrator 22
Milvus korschun 23
Motacilla alba 64
— cinerea 66
— citreola 64
— flava 62
Muscicapa sibirica 120**
— striata 93
Netta rufina 73**
Nucifraga caryocatactes 120
Numenius arquata 43
— phaeopus 44
Oenanthe oenanthe 78
Oriolus oriolus 118
Pandion haliaetus 23
Parus ater 98
— cinctus 97
— cyanus 100
— major 99
— montanus 96
Passer domesticus 115
— montanus 116
Perisoreus infaustus 119
Pernis apivorus 79**
Phalaropus lobatus 40
Philomachus pugnax 40
Phoenicurus phoenicurus 75
Phylloscopus borealis 92
— collybita 90
Phylloscopus fuscatus 106*
— inornatus 91
— proregulus 110**
— schwarzii 111**
— trochiloides 93
— trochilus 89
Pica pica 120
Picoides tridactylus 58
Picus canus 83**
Pinicola enucleator 113
Pluvialis apricarius 36*
— dominica 36*
Podiceps auritus 14
— cristatus 11*
— griseigena 10*
— nigricollis 68**
Poliomyas mugimaki 121**
Porzana porzana 33
Prunella modularis 125*
Psoudaedon sibilans 104**
Pyrrhula pyrrhula 114
Recurvirostra avosetta 39*
Regulus regulus 110*
Riparia riparia 60
Saxicola rubetra 76
— torquata 77
Scolopax rusticola 63**
Siphia parva 112*
Sitta europaea 100
Spinus spinus 110
Squatarola squatarola 36*
Sterna albifrons 59*
— birundo 48
— paradisea 49
Streptopelia orientalis 50
Strix nebulosa 53
— uralensis 64*
Sturnus vulgaris 117
Surnia ulula 53
Sylvia borin 86
— communis 87
— curruca 88
— nisoria 86
Tarsiger cyanurus 74
Tetrao urogallus 29
Tetrastes bonasia 31
Tringa erythropus 44*
— glareola 37
— nebularia 38
— ochropus 36
— stagnatilis 58**
— totanus 39
Turdus iliacus 80
— philomelos 81
— pilaris 80
— atrogularis 79
— viscivorus 82
Upupa epops 82**
Uragus sibiricus 132*
Vanellus vanellus 35
Xenus cinereus 40
ОГЛАВЛЕНИЕ
Введение................................................................ 3
Районы работ, материалы и методы исследований....................... 7
Численность и распределение птиц...................................... И
Повидовой обзор....................................................... —
Отряд Гагары (Gaviiformes)..................................... 13
Отряд Поганки (Podicipitiformes)............................... 14
Отряд Пластинчатоклювые (Anseriformes)......................... 15
Отряд Хищные птицы (Falconiformes)............................. 23
Отряд Куриные (Galliformes).................................... 28
Отряд Журавлеобразные (Gruiformes)............................. 32
Отряд Ржанкообразные (Charadriiformes).......................... 34
Отряд Голубеобразные (Columbiformes)............................ 49
Отряд Кукушкообразные (Cuculiformes)........................... 51
Отряд Совы (Strigiformes)...................................... 53
Отряд Козодоеобразные (Caprimulgiformes)....................... 54
Отряд Стрижеобразные (Apodiformes).............................. —
Отряд Дятлообразные (Piciformes)............................... 55
Отряд Воробьиные (Passeriformes)............................... 59
Семейство Жаворонки (Alaudidae)................................ —
Семейство Ласточки (Hirundinidae)............................. 60
Семейство Трясогузковые (Motacillidae)........................ 62
Семейство Сорокопуты (Laniidae)............................... 70
Семейство Свиристелевые (Bombycillidae)....................... 72
Семейство Дроздовые (Turdidae)................................ 73
Семейство Славковые (Sylviidae)............................... 83
Семейство Мухоловки (Muscicapidae)............................ 93
Семейство Ополовникп (Aegithalidae)........................... 95
Семейство Синицы (Paridae).................................... 96
Семейство Поползни (Sittidae)................................ 100
Семейство Пищуховые (Certhiidae)............................. 102
Семейство Овсянковые (Emberizidae)............................. —
Семейство Вьюрковые (Eringillidae)........................... 108
Семейство Ткачиковые (Ploceidae)............................. 115
Семейство Скворцы (Sturnidae)................................ 117
Семейство Иволги (Oriolidae)................................. 118
Семейство Врановые (Corvidae).................................. —
Общие особенности распределения видов................................ 125
Пространственная неоднородность населения птиц.......................... 139
Северная тайга....................................................... —
Средняя тайга........................................................ 157
Южная таЙ1а........................................................ 175
Подтаежные леса...................................................... 176
Внутризональныо изменения населения птиц.............................. 192
Временная динамика летнего населения птиц............................ —
Сезонные явления в жизни птиц..................................... —
Подзонально-ландшафтные особенности внутризональиой дина-
мики плотности населения птиц................................. 199
Территориальные изменения доминирующего и фаунистического
состава и обобщенных показателей населения птиц.................. 202
Пространственно-типологическая структура населения птиц............ 209
Пространственная организация населения птиц.......................... 222
Заключение.............................................................. 230
Литература.............................................................. 234
Алфавитный указатель русских названий видов птиц........................ 238
Алфавитный указатель латинских названий видов птиц...................... 241