/






































































































































































Текст
АКАДЕМИЯ НАУК СССР
Н. С. НОВОТЕЛЬНОВА
К. А. ПЫСТИНА
О. Г. ГОЛУБЕВА
Пероноспоровые
грибы—патогены
о
культурных растении
в СССР
АКАДЕМИЯ НАУК СССР
БОТАНИЧЕСКИЙ ИНСТИТУТ им. В. Л. КОМАРОВА
Н. С. НОВОТЕЛЬНОВА,
К. А. ПЫСТИНА. О. Г. ГОЛУБЕВА
Пероноспоровые
грибы—патогены
культурных растений
в СССР
СПРАВОЧНИК ПО ДИАГНОСТИКЕ
И МЕТОДАМ ИССЛЕДОВАНИЯ
8
ЛЕНИНГРАД
«НАУКА»
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
19 7 9
УДК 582(031)632.4 : 281.14(47+57)
Пероноспоровые грибы — патогены культурных растений
в СССР. Справочник по диагностике и методам исследования.
Новотельнова Н. С., Пыстина К. А., Голу-
бева О. Г. Л., «Наука», 1979. 152с.
Книга является диагностическим справочником, в кото-
ром обобщены научные материалы по грибам порядка Рего-
nosporales в аспекте отрицательной роли, свойственной этим
организмам — паразитам растений. Приводятся общие све-
дения, характеризующие эту группу организмов, в частности
положение их в системе грибов, особенности строения на мик-
роскопическом и субмикроскопическом уровнях, КОМПЛ€ЩС
вызываемых ими симптомов поражения растений. Особое
внимание уделено подбору методов, необходимых для изу-
чения структурных вопросов и онтогенеза видов. Для ранней
диагностики болезней приведены наиболее вероятные сроки
поражения растений определенными видами пероноспоровых
грибов, эти виды перечислены по культурам с указанием
признаков болезни, рекомендованы вспомогательные шкалы
учета болезни, сообщены сведения о некоторых особенно
опасных болезнях. Лит. — 227 назв., ил. — 34, табл. — 2.
Ответственный редактор
Н. А. НАУМОВА
40307-583 755 79 3803040000
055(02)-79
© Издательство «Наука»,
1979 г.
ВВЕДЕНИЕ
Рациональное использование и охрана растительного мира —
одна из важнейших современных проблем, к разработке которой
привлечено внимание ботаников многих стран. В задачу охраны
входит защита растений не только от техногенных воздействий,
но и от многих природных факторов, в том числе от патогенных
грибов, особенно сильно поражающих культурные растения в усло-
виях их интенсивного возделывания. Надежное обеспечение на-
шей страны продовольствием и растительным сырьем, получение
максимальных урожаев с каждого гектара требуют осуществле-
ния мероприятий по предотвращению потерь при выращивании
зерна и крупяных культур, свеклы, подсолнечника, хлопка,
картофеля, овощных, плодовых и других ценных растений.
В последние годы многие виды пероноспоровых грибов полу-
чили мировое распространение. Они вызывают опасные «массовые»
болезни культурных растений — сельскохозяйственных, лесных,
лекарственных, декоративных, не менее вредоносные, чем ржав-
чина или головня, и приводят к большим потерям урожаев.
Помимо фитофторы картофеля, известной еще в прошлом веке,
появились болезни, представляющие угрозу для посевов полез-
ных растений. Среди них корневая и прикорневая гниль, вызы-
ваемая питиевыми и фитофторовыми грибами, ложная мучнистая
роса свеклы, подсолнечника, пероноспороз табака, лука, склеро-
спороз ржи и др. Наблюдаются значительное расширение ареалов
одних болезней и вспышки других в регионах, где прежде они
отсутствовали.
Необходимость защиты культурных растений и научного обос-
нования соответствующих мероприятий заставляет исследовате-
лей обратиться к изучению этих заболеваний и их возбудителей
ввиду недостаточности или противоречивости имеющихся по ним
научных материалов. Первостепенное значение для решения этих
практических вопросов имеют исследования по биологии пероно-
споровых грибов в конкретных, местных условиях. Для проведе-
ния углубленных исследований видов, их образа жизни, биологии
и циклов развития, взаимоотношений с питающими растениями и
реакции на воздействие факторов окружающей среды необходимо
владение специальными современными методами работы. Для по-
вышения достоверности результатов и возможности сопоставле-
ния данных отдельных исследований желательна унификация
3
приемов и методов, применяемых при изучении этой группы гри-
бов.
Настоящий справочник содержит информацию о пероноспоро-
вых грибах по результатам оригинальных личных работ авторов,
в частности Н. С. Новотельновой по фитофторовым, пероноспоро-
вым и цистоповым грибам, К. А. Пыстиной по питиевым грибам,
О. Г. Голубевой по анатомическим и цитологическим исследова-
ниям пероноспоровых грибов в тканях высших растений. Авто-
рами использованы также материалы по пероноспоровым грибам
Е. И. Легенькой и Т. В. Енкиной и соответствующая новейшая
отечественная и зарубежная литература.
В книге обобщен материал по методам исследований пероноспо-
ровых грибов, по вызываемым последними болезням растений.
Представлены методы непосредственного изучения самих парази-
тов (гербаризация, изоляция, культивирование, изучение отдель-
ных фаз цикла развития с помощью световой и электронной микро-
скопии), воздействия различных экологических факторов и ин-
фекционного процесса, обусловленного ими (этапов патогенеза),
а также методы учета болезней. Болезни растений, вызываемые
пероноспоровыми грибами, приводятся по культурам в обще-
принятом порядке, и даются краткие ссылки на их местонахожде-
ния по отечественным литературным источникам. В ряде случаев
описания болезней и возбудителей взяты из зарубежных литера*
турных материалов. Это относится, например, к отдельным видам
питиевых, фитофторовых и других грибов, зарегистрированным
во многих странах мира, но не отмеченным еще у нас. Авторы
не ставили задачей дать физиолого-биохимический, иммуноло-
гический и генетический аспекты изучения пероноспоровых гри-
бов, а также прогноз и меры борьбы.
Работа проводилась в лаборатории микологии Отдела низших
растений Ботанического института им. В. Л. Комарова Акаде-
мии наук СССР.
ОБЩИЕ СВЕДЕНИЯ
Пероноспоровые (Peronosporales) — своеобразная группа низ-
ших грибов с особым строением, специфическими свойствами и
особым образом жизни. Большое внимание исследователей обра-
щено на них в связи с их вредоносностью при паразитировании
на высших растениях.
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ
В ранних классических системах порядок Peronosporales отно-
сили к классу Phycomycetes (De Вагу, 1881; Fischer, 1892; Schroe-
ter, 1897; Gaumann, 1926; Fitzpatrick, 1930; Ячевский, 1933;
Курсанов, 1933; Burnett, 1968). Однако позднее, в связи с новыми
данными по топкой морфологической структуре видов, по их
биохимическим особенностям, получили развитие иные взгляды
на систематическое положение порядка. Порядок Peronosporales
рассматривается теперь в составе класса Oomycetes отдела Ооту-
cota царства Mycota (Агх, 1967, 1970) или того же класса Oomycetes,
входящего в подотдел Mastigomycotina отдела Eumycota (Ain-
sworth et al., 1973). По системе Цейпа (Cejp, 1957), порядок Pe-
ronosporales (тоже в классе Oomycetes} отнесен к подотделу Phy-
comycophytina отдела Mycophyta. По Крейзелю (Kreisel, 1969),
оомицеты, в том числе Peronosporales, исключены из Fungi (=Еиту-
cota, — Mycophyta) и отнесены к золотистым водорослям (на основе
сходства структурных признаков и биохимизма видов).
Порядок Peronosporales содержит 4 семейства: Pythiaceae,
Phytophthoraceae, Cystopaceae и Peronosporaceae. Одни виды в своем
развитии довольно тесно связаны с водной средой и почвой и об-
ладают преимущественно сапрофитными свойствами (питиевые
и фитофторовые), другие адаптировались полностью к наземному
существованию и являются весьма вредоносными паразитами
цветковых растений (пероноспоровые, цистоповые).
Сем. Pythiaceae содержит около 100 видов. Это паразиты и
полупаразиты, приуроченные к жизни в воде или в почве на мерт-
вых или живых растениях и животных, с мицелием без гаусториев
и мало дифференцированным спороношением. Основной род —
Pythium. Наиболее известные виды рода Pythium*. Р. debaryanum,
Р. ultimum, Р. aphanidermatum.
Сем. Phytophthoraceae представлено родом Phytophthora, ко-
торый состоит из нескольких десятков видов. Виды обладают
5
сапрофитными свойствами, но способны паразитировать на цвет-
ковых растениях. Они характеризуются мицелием, образующим
иногда гаустории, и мало дифференцированным спороношением.
Широко известны виды Ph. infestans на пасленовых, Ph. cactorum
на розоцветных и других растениях.
Сем. Peronosporaceae содержит 5 родов: Basidiophora, Sclero-
spora, Plasmopara, Bremia и Peronospora, все они облигатные пара-
зиты цветковых растений, характеризуются мицелием с гаусто-
риями, спороносцами древовидной формы. Широко известны
вредоносные виды Sclerospora graminicola на злаках, Plasmopara
viticola на винограде, Р. helianthi на подсолнечнике, Peronospora
tabacina на табаке, Р. schachtii на свекле и др.
Сем. Cystopaceae содержит 1 род Cystopus, в который входит
около 10 видов. Все они облигатные паразиты цветковых расте-
ний, вызывающие уродливости пораженных органов и пустулы
на них. Широко распространены виды С. candidus на крестоцвет-
ных, С. tragopogonis на сложноцветных, С. portulacae на порту-
лаковых.
Таким образом, более примитивные представители порядка
(сем. Pythiaceae, Phytophthoraceae') обладают в той или иной сте-
пени сапрофитными свойствами, тогда как большинство более
высокоорганизованных являются облигатными паразитами
(сем. Peronosporaceae, Cystopaceae\, первые поражают довольно
большой круг растений-хозяев, а вторые чаще приурочены к от-
дельным родам или видам высших растений-хозяев. У некото-
рых видов пероноспоровых грибов установлено наличие специали-
зированных форм и рас. В порядке Peronosporales насчитывается
несколько сотен фитопатогенных видов.
Пероноспоровые грибы характеризуются особыми симптомами
поражения растений и строением органов размножения — веге-
тативного (мицелий), бесполого (спорангиеносцы и зооспорангии,
конидиеносцы и конидии) и полового (ооспоры).
СИМПТОМЫ ПОРАЖЕНИЯ
Симптомы поражения растений, обусловленные пероноспоро-
выми грибами, многообразны и варьируют в зависимости от воз-
будителя. Однако у возбудителей из различных семейств они
довольно четко разграничиваются (см. таблицу).
РАСПРОСТРАНЕНИЕ И ВРЕДОНОСНОСТЬ
Болезни, вызываемые пероноспоровыми грибами, характери-
зуются широким распространением и большой вредоносностью.
Неоднократно изучались пути их возникновения и способы рас-
селения возбудителей (Хохряков и др., 1963а, 19636; Наумов,
1972а, 19726; Горленко, 1975, и др.).
6
Внешние признаки поражения
Семейство Характер поражений растений, пораженные органы Внешний вид гриба на растении
Pythiaceae Загнивание, потемнение, перетяжки на корневой шейке всходов, реже на хорошо сформированных корнях, гниль плодов. Побурение частей рас- тений: корней, клубней, корнеплодов, некротиче- ские пятна на листьях, стеблях, стволах в при- корневой части. Сплетение мицелия в виде войлока на пора- женных частях и почве.
Phytop htho raceae Образование спороноше- ния в виде нежного налета на поверхности субстрата.
Peronosporaceae Задержка роста расте- ний, карликовость; кур- чавость листьев; хлоро- тичные пятна на листьях с угловатыми контурами; у злаков шероховатость листовой поверхности, иногда полное размочали- вание пластинки листа. Нежный или плотный беловатый или серо-фио- летовый налет чаще с ниж- ней стороны листа.
Cystopaceae Уродливости, вздутия, искривления целых расте- ний или их частей. Развитие под эпидерми- сом пустул, раскрываю- щихся и порошащих при созревании.
Широта распространения питиевых грибов обусловлена непри-
хотливостью образа жизни, возможностью существования и со-
хранения в разнообразных условиях, а также многоядностью —
«неразборчивостью» в отношении питающих растений.
Наиболее широко распространенным и вредоносным патоге-
ном является Pythium debaryanum, зарегистрированный во мно-
гих областях СССР, где он поражает растения различных семейств.
Он встречается на овощных, почти повсеместно возделываемых
культурах: огурцах, томатах, капусте, салате, шпинате, на тех-
нических и зерновых культурах: всходах льна, кукурузы. В пар-
никах и теплицах, где выращиваются огурцы, распространен и
Р. ultimum. Р. debaryanum является также одним из частых воз-
будителей корневой гнили в сосновых и еловых питомниках
(рис. 1).
На культурах, возделываемых в открытом грунте, наиболее
часто встречаются следующие виды: Р. hydnosporum, отмеченный
на горохе в Калужской и Ленинградской областях; Р. debary-
anum — на капусте в Ленинградской об л. и Латвийской ССР,
на клевере в Московской и Свердловской областях, на люпине
в Белорусской ССР, на свекле в Калужской, Московской, Ярослав-
ской, Ленинградской областях, на томатах в Ленинградской и
7
Ярославской областях, на льне во Владимирской обл., на всходах
ели и сосны в Ленинградской, Саратовской областях и в Эстон-
ской ССР; Р. irregulare — на огурцах в Ленинградской обл.
(на свекле встречается и в Центрально-черноземной зоне, в Кур-
ской обл.); Р. spinosum — на свекле в Московской обл.; Р. ulti-
тит — на свекле в Московской, Ленинградской, Калининград-
ской областях, на клевере, горохе в Московской обл. (Пыстина,
1972).
Корневые гнили пшеницы в Московской обл., почти на 10%
обусловленные грибами рода Pythium, снижают урожайность
этой культуры в результате выпада растений в предвсходовый
и послевсходовый периоды. В северо-западной зоне снижение
урожайности достигает 5—9%.
Широко распространенным и вредоносным заболеванием яв-
ляется корнеед сеянцев свеклы. Наблюдается повсеместно (за
исключением регионов с сухим и жарким климатом) в западных
районах, включая Ленинградскую обл., Карелии, Центрально-
черноземной зоне, на Украине, в Молдавии, Закавказье. Отме-
чается в Узбекской ССР, хотя в Средней Азии болезнь встречается
реже и менее вредоносна. По экспериментальным данным
Д. Л. Тверского и К. П. Жуковой (1948), Р. debaryanum прояв-
ляет высокую агрессивность по отношению к сеянцам свеклы,
в опытах он вызывал поражение 80% и гибель 50% растений.
Гниль плодов тыквенных, вызванная Р. ultimum, в парниках
встречается довольно часто. Повсеместно она распространена
в Грузии; в некоторых совхозах сильное развитие болезни вы-
зывало до 12% гибели урожая (Асатиани, 1968). В последние
годы в Краснодарском крае отмечается массовое развитие рассад-
ной гнили табака в открытом грунте и в теплицах, вызванное
8
СССР.
Р. perniciosum и Р. aphanidermatum. В отдельные годы гибель
рассады достигала 30% (Богоявленская, Пыстина, 1974).
Все заболевания культурных растений, вызываемые фито-
фторовыми грибами, крайне вредоносны, но особенно вредоносна
фитофтора картофеля, вызываемая грибом Phytophthora infestans,
которая и в настоящее время несмотря на разработанные меры
борьбы с нею все еще широко распространена во всех странах
мира и причиняет значительный вред урожаю. В годы, благо-
приятные для фитофторы, потери урожая превышают 30% (До-
рожкин, 1959). По данным Н. И. Белозор (1960), фитофтора еже-
годно уничтожает у нас 2—2.5 млн т картофеля и причиняет убытки,
исчисляемые миллионами рублей (рис. 2).
Фитофтороз томатов, вызываемый тем же видом, — одна из
наиболее вредоносных и «массовых» болезней этой культуры.
Иногда от болезни погибает весь урожай томатов, особенно на
плантациях, расположенных вблизи картофельных полей, и
часто после появления болезни на картофеле. Вспышки фито-
фторы томатов могут возникать в любой период вегетации в теп-
лице и в поле.
Фитофтороз земляники, вызываемый Ph. cactorum, особенно
Широко распространен в годы с повышенной влажностью воздуха.
В 1975 г. в Ленинградской обл. он привел к гибели до 25% расте-
ний, а в очагах — до 50% (Андреева, 1977).
Значительные потери фитофтороз причиняет насаждениям
лесных культур на Кавказе, где вид Ph. cinnamomi поражает
сеянцы каштана, пробкового дуба, грецкого ореха (Шишкина,
Цанава, 1973). Он обусловливает так называемую чернильную
болезнь. Кора и древесина корней и стволов при этом чернеют,
9
возникают изъязвления и трещины, из которых вытекает сок,
чернеющий при окислении на поверхности коры, и последняя
становится как бы обугленной или обмазанной мазутом. Пораже-
ние часто приводит к полному усыханию растений (рис. 3,
см. вкл.).
Менее распространены, но очень вредоносны фитофторозы
лекарственных и цветочных растений. В частности, в Приморском
крае отмечёно значительное поражение видом Ph. cactorum жень-
шеня (Бункина, 1963) и в Ленинградской обл. — отдельных
экземпляров лилий.
Не менее вредоносными являются представители семейств
Peronosporaceae и Cystopaceae, а их распространение, несомненно
значительно более широкое, обеспечивается структурными и
биологическими особенностями (обилие пропагативных спор, лег-
кость их отчленения и. рассеивания, способность покоящихся
спор сохраняться и переносить неблагоприятные условия).
К числу наиболее опасных заболеваний винограда (возбуди-
тель — Plasmopara viticola) относится ложная мучнистая роса
(или мильдью), вызывающая преждевременное опадение листьев
и гибель урожая ягод. Заболевание распространено во всех зо-
нах возделывания этой культуры, особенно на Украине, Кав-
казе, в Краснодарском крае, причем виноградники, подвергаю-
щиеся болезни из года в год, полностью выходят из строя через
5—6 лет. По данным Н. П. Олтаржевского, в 1931 г. от болезни
погибло 30% урожая, в 1937 и 1938 гг. в районах Северного Кав-
каза снижение урожая винограда достигало 50—70%. Большие
потери урожая отмечались также в Молдавии и на Нижнем По-
волжье (рис. 4).
Распространение многих видов пероноспоровых грибов носит
Рис. 4. Местонахождения Plasmopara viticola (Berk, et Curt.) Berl. et de T.
на территории СССР.
10
эпифитотийный характер. Гриб Peronospora tabacina происхож-
дением из Австралии в виде панфитотий распространился по Се-
верной и Южной Америке, откуда проник в европейские страны.
Отмечено значительное поражение им растений, в ряде стран
до 90-95%.
В нашей стране экономический ущерб от пероноспороза та-
бака в 1962 г. только по Тернопольской обл. составил свыше
4.5 млн руб. По данным Г. Р. Ибрагимова (1968), пероноспороз
табака был обнаружен в Азербайджане в 1962 г. сначала в парни-
ках, а потом на плантациях и имел весьма ограниченное распро-
странение. Но уже в следующем, 1963 г. 86.1% посадок табака
полностью погибли от пероноспороза. В том же году в Нуха-
Закатальской зоне рассада погибла почти на 100%. Общие по-
тери от пероноспороза по республике превысили 260 тыс. руб.
Автор указывает, что условиями, способствовавшими возникнове-
нию эпифитотий, были частые атмосферные осадки, туманы и
росы, особенно в утренние часы при температуре 15—17°.
Широко распространен и очень вредоносен также возбудитель
ложной мучнистой росы подсолнечника — гриб Plasmopara he-
lianthi. Этот гриб, американского происхождения, в 40-е годы
проник на другие континенты, захватил все страны Европы и
продвинулся в последние годы в Азию. Его распространение но-
сило характер панфитотии. В первые годы появления в Советском
Союзе поражение растений на полях подсолнечника было очень
большим и иногда достигало 70—80%. Позже разработанными
мероприятиями по защите подсолнечника, особенно введением
строгого чередования культур в производственных посевах, по-
тери удалось снизить (Новотельнова, 1966а).
Имеются многочисленные сведения о распространении и вре-
доносности пероноспороза свеклы на Украине. В ряде областей
поражение растений грибом Peronospora schachtii достигало
в 70-х годах 20%, причем гриб вызывает не только выпад расте-
ний. Установлены глубокие изменения, вызванные возбудителем
болезни у растений: нарушение нормальной клеточной струк-
туры; редукция синтеза хлорофилла, сопровождающаяся подав-
лением процесса ассимиляции и фотосинтеза и появлением хлороза;
прекращение роста растений с одновременным усилением про-
цесса дыхания, сопряженного с расходом сахаров, в результате
чего уменьшаются вес ботвы и развитие корнеплодов, снижается
сахаристость последних. Снижаются также и урожай семян, их
всхожесть и энергия прорастания (Кузичева, 1975 и др.). Перо-
носпороз свеклы широко распространен и в других свеклосеющих
районах нашей страны. По данным Е. И. Легенькой (1963), в 1962—
1963 гг. поражение растений в совхозах Курской обл. достигало
25% на маточных и 45% на семенных посевах. Вредоносность
пероноспороза в сильной степени сказывается и на растениях
«фабричной» свеклы: растения отстают в росте от здоровых и те-
ряют более 1/3 веса. В колхозах Краснодарского края паб люда-
И
лось до 20—30% пораженных растений на некоторых полях
в 1962 г.
Пероноспороз огурцов особенно вредоносен в Приморском
крае в связи с особенностями климатических условий. В 1974 г.
в экспериментальном хозяйстве Всесоюзного института растение-
водства поражение растений достигало в июле 55%, а в середине
августа все растения погибли (Плюснина, 1975).
На сильное поражение пероноспорозом гречихи, вызываемое
Peronospora fagopyri, указывает Е. И. Легенькая (1966а), обна-
ружившая в 1964 г. на полях в Белгородской обл. до 63% по-
раженных растений.
Пероноспороз сои, вызываемый грибом Р. manshurica, — вре-
доносное заболевание, наиболее распространенное на Дальнем
Востоке. Экономический ущерб от него очень высок. Болезнь
поражает все органы растений, в том числе бобы и семена. Сни-
жаются урожайность и масличность семян, а также их всхожесть.
По данным 1964—1965 гг., по хозяйствам Дальнего Востока
поражение семян составило 2.98—3.14%.
Пероноспороз, вызываемый Р. pratensis, Р. trifolii-repentis,
Р. trifolii-hybridi, — широко распространенное заболевание ви-
дов клевера в Прибалтике. Однако вредоносность болезни незна-
чительна (Брундза, 1961).
Пероноспороз капусты, вызываемый Р. brassicae, считался
мало вредоносным заболеванием. В Литовской ССР в 1949 г. бо-
лезнь нанесла ощутимый вред культуре ввиду сильного пораже-
ния рассады, достигавшего 40%.
Пероноспороз лука, вызываемый Р. destructor, наносит боль-
шой ущерб культуре. Потери урожая семян в Азербайджане
в 1949 г. были от 46.7 до 94.7% (Абдуллаев, 1951), в Грузии —
39—76% (Шошиашвили, Кирмелашвили, 1950). В Центрально-
черноземной зоне Е. И. Легенькой (1966а) отмечена вспышка
болезни на луке-«репке» в одном из совхозов, где на площади
20 га было поражено 5—56%, причем в ложбинах до 100% расте-
ний.
Значительный ущерб наносит пероноспороз, вызываемый гри-
бом Р. arborescens урожаю семян опийного мака. В Киргизии уро-
жай семян снижается до 70%, а опия-сырца на 4—20% (Зюбин,
1963).
Пероноспороз, вызываемый Р. hyoscyami, оказывает сильное
воздействие на белену. В Западной Сибири, в условиях Мошков-
ского совхоза лекарственных растений, в 1966 г. Т. В. Енкиной
были установлены разные формы проявления болезни и значитель-
ное, до 75%, снижение в полностью пораженных листьях содер-
жания алкалоидов; выпад 20—25% растений на загущенных де-
лянках. Ею отмечено в том же году сильное поражение аптечной
ромашки грибом Р. leptosperma в условиях Новосибирской обл.,
где больные растения составили 50—60%. Болезнь встречается
12
в Литовской, Украинской ССР, в РСФСР — в Ленинградской,
Новосибирской областях.
Белая ржавчина, вызываемая Cystopus candidus, является
одним из наиболее вредоносных заболеваний семенников кресто-
цветных. Это заболевание изучалось Е. И. Легенькой в 1962 г.
в условиях Курской обл. в связи со вспышкой болезни редиса,
поразившей надземные органы растений и вызвавшей искривле-
ние и уродливость стеблей и цветоносов, недоразвитие стручков
и обильные пустулы на них. По данным Легенькой (1966а), в этот
год белая ржавчина семенников редиса причинила значительные
убытки хозяйству одного из колхозов, недобравшего с площади
4.5 га более 10 ц семян редиса и понесшего убыток в размере
1575 руб.
ОСОБЕННОСТИ СТРОЕНИЯ И РАЗВИТИЯ
МИЦЕЛИЙ
Представители порядка Peronospordles имеют несептированный
(неклеточный) мицелий, распространяющийся внутри тканей
растения-хозяина. У грибов различных семейств он имеет свои
особенности. Так, для сем. Pythiaceae характерно, что мицелий
легко выходит из пораженных органов наружу и развивается не
только на их поверхности, но и на почве (например, при болезни
всходов, называемой черной ножкой). Такой экзофитный мицелий
почти не встречается у видов сем. Phytophthoraceae, у которых воз-
душный мицелий может быть обнаружен на разрезанной поверх-
ности органов (клубней, корней), а у грибов остальных двух
семейств воздушный мицелий, как правило, отсутствует и на по-
верхности пораженных частей растений появляются только
органы спороношения, выступающие или через устьица, как это
имеет место у видов сем. Peronosporaceae, или через разорванный
эпидермис — у видов сем. Cystopaceae.
В тканях растений эндофитный мицелий распространяется
большей частью по межклетникам (хотя у представителей Pythi-
aceae и Phytophthoraceae он может пронизывать и клетки), причем
форма его изменяется в зависимости от плотности тканей. В пали-
садной паренхиме листьев, имеющей большие воздушные полости,
он развивается свободно в виде толстых разветвленных гиф (рис. 5,
7) и образует особенно обильные скопления в подустьичных про-
странствах — местах выхода наружу конидиеносцев. При про-
хождении в тканях с очень узкими межклетниками мицелий при-
нимает очертания последних (рис. 5, <2), а иногда под давлением
тканей уплощается и становится древовидным. Мицелий многих
пероноспоровых грибов часто является диффузным и пронизывает
все органы растения-хозяина (корень, стебель, листья, цветки,
плоды).
Мицелий, проходящий по межклетникам, проникает в клетки
растения-хозяина при помощи особых органов питания — га-
усторйев. Последние у разных видов пероноспоровых грибов
различны: пузыревидные (овальные, сферические или булаво-
видные) и нитевидные (короткие, длинные, толстые или тонкие),
простые или разветвленные. Пузыревидные встречаются чаще
у видов родов Plasmopara и Cystopus — Plasmopara nivea (sensu
lato), P. helianthi (рис. 5, 5), а также у Bremia lactucae*, мелкие
у Plasmopara viticola, крупные у Peronospora parasitica (sensu lato);
14
Рис. 5. Особенности мицелия и гаусториев пероноспоровых грибов.
1 — толстые разветвленные межклеточные гифы; 2 — тонкие межклеточные гифы; 3 —
пузыревидные гаустории; 4 — нитевидные гаустории; 5 — нитевидные разветвленные
гаустории.
нитевидные неразветвленные преобладают у видов рода Perono-
spora — Р. viciae (рис. 5, 4), разветвленные — у Р. schachtii
(рис. 5,5). Мицелий и гаустории обычно бесцветные, иногда желто-
ватые (Fraymouth, 1956 и др.).
Цитологические данные
Мицелий пероноспоровых грибов ценоцитный, т. е. не разде-
ленный перегородками и представляющий собой гигантскую много-
ядерную клетку, но иногда в мицелии образуются так называемые
адвентивные септы или псевдосепты. Они возникают при автолити-
ческом разрушении старых частей мицелия, при механических
повреждениях, отделяют мертвые или поврежденные участки.
Оболочка мицелия пероноспоровых грибов состоит из целлю-
лозы и глюканов. Она представляет собой двухфазную систему,
15
Рис. 7. Схематическое строение гаустория.
к — каллозные отложения; ко — клеточная оболочка хозяина; л — ломасома; м — мито-
хондрия; ог — клеточная оболочка гаустория; пг — плазмалемма гаустория; пх — плазма-
лемма хозяина; т — тонопласт; ц — цитоплазма хозяина; эгм — экстрагаусториальный
матрикс; я — ядро.
построенную из аморфного глюканового матрикса и погруженных
в него целлюлозных фибрилл.
В мицелии присутствуют органеллы, свойственные всем эука-
риотическим клеткам: ядра, митохондрии, рибосомы, вакуоли,
эндоплазматический ретикулюм. Для пероноспоровых грибов
характерно наличие в цитоплазме хорошо выраженного аппарата
Гольджи и центриолей. Присутствие этих органелл отличает
клетки пероноспоровых грибов и других фикомицетов от клеток
высших грибов.
В мицелии и гаусториях пероноспоровых грибов часто встре-
чаются плазмалеммосомы (рис. 6, см. вкл.; 7) —лабиринтообразные
структуры, образуемые плазмалеммой на границе цитоплазмы и
клеточной оболочки. О назначении их существуют различные пред-
положения: им приписывают секреторную функцию, участие в об-
разовании клеточной оболочки, функцию поддержания тургора
в клетках.
Митохондрии пероноспоровых грибов разнообразны по форме,
от овальных до нитевидных, и характеризуются трубчатыми кри-
стами, что отличает их от митохондрий других грибов (рис. 6, 7).
Ядра пероноспоровых грибов окружены двойной мембраной,
пронизанной многочисленными порами. По последним данным,
ядра в вегетативном мицелии пероноспоровых грибов диплоидны,
мейоз происходит в гаметангиях, а при кариогамии в оогонии
16
Рис. 3. Усыхание саженцев каштана, вызванное Phytophthora
cinnamomii Rands. (Фот. Н. Цанавы).
Рис. 6. Гаусторий гриба Plasmopara helianthi Novot.
в листе подсолнечника.
диплоидность восстанавливается. Таким образом, в жизненном
цикле пероноспоровых грибов преобладает диплоидная фаза,
гаплоидны только ядра в гаметангиях. Митоз ядер мицелия про-
исходит по внутриядерному типу, т. е. при сохранении ядерной
оболочки. Полюсными структурами, по направлению к которым
происходит расхождение хромосом, служат центриоли. После
расхождения хромосом в ядерной оболочке образуется перетяжка,
и постепенно ядро разделяется на два дочерних ядра.
По внутреннему строению гаустории сходны с межклеточными
гифами. В них присутствуют те же органеллы: митохондрии,
рибосомы, ломасомы, вакуоли, липидные капли. Гаустории
одних представителей пероноспоровых часто безъядерны (виды
Phytophthora, Cystopus), у других в гаусториях имеются одно или
несколько ядер (виды Peronospora).
В месте проникновения гаустория в клетку растения-хозяина
происходит разрушение клеточной оболочки хозяина за счет
ферментативного и отчасти механического воздействия. Плазма-
лемма клетки растения-хозяина при внедрении гаустория не раз-
рушается, она лишь впячивается внутрь клетки, обволакивая
гаусторий (рис. 6, 7). Плазмалемма хозяина, окружающая гаусто-
рий, не прилегает плотно к оболочке гаустория, а отстоит от нее
на некотором расстоянии. Пространство между оболочкой гаусто-
рия и плазмалеммой хозяина заполнено электроноплотным веще-
ством, которое в литературе часто называют экстрагаусториаль-
ным матриксом. Ширина матрикса может варьировать в зависи-
мости от вида патогена и хозяина, физиологического состояния их
клеток и даже от способа фиксации материала (Carling et al.,
1977). Экстрагаусториальный матрикс играет, по-видимому, су-
щественную роль в установлении взаимоотношений между пара-
зитом и хозяином, но конкретная функция его пока не установ-
лена.
Снаружи от экстрагаусториального матрикса может существо-
вать еще одна оболочка, которая, по всей вероятности, является
продуктом выделения клетки хозяина (рис. 6, 7). Она может окру-
жать весь гаусторий или располагаться только вокруг его основа-
ния в виде воротничка. Гистохимические исследования показали,
что электронопрозрачный материал, из которого она состоит,
является каллозой — веществом полисахаридной природы (Da-
vison, 1968). Степень развития этой оболочки зависит от воспри-
имчивости растения-хозяина к данному патогену (Hohl, Stos-
sel, 1976). У устойчивых хозяев она очень хорошо развита и окру-
жает весь гаусторий, изолируя его от цитоплазмы пораженной
клетки, у восприимчивых форм она может вообще отсутствовать
(Голубева, 1978).
2 Н. С. Новотельнова и др.
17
РЕПРОДУКТИВНЫЕ ОРГАНЫ
Бесполое спороношение
В течение вегетационного периода на поверхности пораженных
пероноспоровыми грибами листьев растений, чаще всего на ниж-
ней стороне, появляются органы спороношения гриба: спорангие-
носцы и споры. Спорангиеносцы большей частью бесцветные,
иногда слегка желтовато-бурые. Морфология спорангиеносцев
у отдельных родов различна, причем особенности их строения
зависят от степени эволюции родов (рис. 8).
Так, у грибов рода Pythium отсутствуют выраженные споран-
гиеносцы и зооспорангии образуются на концах или на протяже-
нии обычных вегетативных гиф. У видов рода Phytophthora име-
ются обособленные спорангиеносцы; они выступают из устьиц
или возникают на воздушном мицелии, обладают ветвлением
простым или симподиальным, иногда концы ветвей, несущих
споры, после отчленения последних способны к дальнейшему
росту, на местах отчленения конидий на ветвях остаются неболь-
шие вздутия, по которым можно судить о количестве образовав-
шихся спор (у Ph. infestans). У представителей сем. Cystopaceae
спорангиеносцы неразветвленные, короткие, булавовидные, ха-
рактерным является образование их под эпидермисом в виде сплош-
ного слоя, они выходят одиночно или группами из устьиц
на поверхности листьев (см. рис. 34).
Вполне дифференцированные спорангиеносцы отличают пред-
ставителей сем. Peronosporaceae (рис. 9). Более просто они устроены
у родов Basidiophora, Sclerospora. Строение спорангиеносцев и их
Рис. 8. Особенности строения спорангиеносцев в семействах Pythiaceae (а)
и Phytophthoraceae (б).
18
Рис. 9. Особенности строения спорангиеносцев у родов сем. Peronosporaceae.
1 — Sclerospora\ 2 — Basidiophora; 3 — Plasmopara', 4 — Peronospora', 5 — Bremia.
конечных ветвей используют (наряду с биологическими особен-
ностями) в качестве систематического признака для разграниче-
ния родов.
У рода Basidiophora спорангиеносцы булавовидные, неразвет-
вленные; зооспорангии возникают на вершине на особых выступах.
Род Sclerospora характеризуется спорангиеносцами короткими,
на вершине обычно беспорядочно разветвленными. Для рода
Plasmopara характерно многократное древовидное ветвление под
прямым углом, с 3—5 затупленными на вершине отрогами на
конечных ветвях, несущих зооспорангии. У рода Peronospora
с подродами Peronospora и Pseudoperonospora (различающимися
по биологическим особенностям) спорангиеносцы с типичным
Дихотомическим ветвлением и конечными заостренными прямыми
или согнутыми отрогами, расположенными под прямым или острым
Углом, а у рода Bremia — тоже дихотомически разветвленные, но
2*
19
Рис. 10. Типы прорастания зооспорангиев (конидий) у пероноспоровых
грибов.
1, 2, 3 — Sclerospora, Basidiophora, Plasmopara\ 4, 5 — Phytophthora; 6 — Peronospora
Bremia,
с концами, имеющими воронковидную (дланевидную) форму с 2—
8 отрогами.
Споры (конидии, зооспорангии) у пероноспоровых грибов ша-
ровидные, яйцевидные, лимоновидные, бочонковидные с бугор-
ком на вершине или без последнего, крупные, примерно 15—
30 мкм и более, бесцветные, реже желтоватые, возникающие оди-
ночно на кончиках отрогов (исключение составляет сем. С у stop а-
сеае, для которого характерно образование зооспорангиев цепоч-
ками). У низших представителей споры (конидии, зооспорангии)
возникают непосредственно на ветвях мицелия (сем. Pythiaceae),
у более высокоорганизованных — на ветвях спорангиеносцев
20
(сем. Phytophthoraceae, Peronosporaceae). По способу прорастания
их делят на зооспорангии и конидии. Для первых характерным
является формирование при прорастании подвижных клеток —
зооспор (рис. 10, 1—4)*, вторые обычно прорастают с образованием
гиф (рис. 10, 6).
У видов, относящихся к родам Sclerospora, Basidiophora, Plas-
mopara и Cystopus и подроду Pseudoperonospora, при наличии ка-
пельно-жидкой влаги прорастание осуществляется зооспорами,
тогда как споры грибов из родов Peronospora и Bremia, прорастая,
образуют мицелиальные ростки. У видов Pythium или Phyto-
phthora характер прорастания зависит от возраста спор и от условий
среды. У видов Phytophthora (рис. 10, 4) обнаружена четкая зави-
симость способа прорастания от температуры: при более низких
температурах (5—15°) образуются чаще зооспоры (косвенное
прорастание), а при более высоких (20—30°) — ростковые трубки
(прямое прорастание). Перед прорастанием происходит разделе-
ние содержимого зооспорангия на отдельные участки, которые
затем выталкиваются наружу в виде оформленных зооспор через
открывающийся на вершине зооспорангия бугорок и активно
движутся в водной среде. У некоторых видов рода Pythium со-
держимое зооспорангия выступает в виде пузыря с тонкой оболоч-
кой через канал бокового выроста, после чего в пузыре форми-
руются зооспоры. Зооспоры пероноспоровых представляют собой
голые комочки протоплазмы довольно крупных размеров (при-
мерно 7—12 мкм) бобовидной формы с 2 тонкими и длинными плаз-
матическими нитями — жгутиками, отходящими у зооспор от
боковой выемки; один из жгутиков снабжен ресничками. Движе-
ние зооспор продолжается в зависимости от условий от несколь-
ких минут до нескольких часов; при отсутствии питающего расте-
ния зооспоры относительно быстро лизируются, а при наличии
соответствующего субстрата тело зооспоры выделяет оболочку,
т. е. она инцистируется, после чего происходит ее прорастание
с образованием мицелиальной трубки, при помощи которой гриб
внедряется в растение обычно через устьице или корневой волосок
и осуществляет заражение.
Цитологические данные
Оболочка спорангия состоит из 2 слоев, в плотной цитоплазме
располагаются митохондрии, ядра, аппарат Гольджи и много-
численные вакуоли. В зрелом зооспорангии обычно присутствуют
2 типа вакуолей: вакуоли, содержащие жировые включения, и
так называемые разделительные вакуоли. Разделительные ва-
куоли, вероятно, являются результатом деятельности аппарата
Гольджи и, по мнению некоторых авторов (Elsner et al., 1967;
Hohl, Hamamoto, 1967), играют важную роль в образовании
зооспор. Как сообщают Хол и Хамамото, образование зооспор
у Phytophthora nicotianae начинается с разделения многоядерной
цитоплазмы на отдельные участки посредством перегруппировки
21
Рис. И. Схематическое строение жгути-
ков (поперечное сечение).
1 —периферические сдвоенные фибриллы; 2 — цен-
тральные фибриллы; 3 — плазмалемма жгутика.
и слияния первоначально равно-
мерно распределившихся в цито-
плазме разделительных вакуолей.
По наблюдениям Элснера с соав-
торами, жгутики будущих зооспор
первоначально заключены в особые
жгутиковые вакуоли. При образова-
нии зооспор происходит слияние
жгутиковых вакуолей с разделитель-
ными и соединение мембран этой системы с плазмалеммой. Таким
образом происходит обособление зооспор и освобождение жгу-
тиков.
Зооспоры пероноспоровых грибов — двужгутиковые клетки
бобовидной формы с передним хлестательным и задним перистым
жгутиком. Перистый жгутик более короткий и несет на себе рес-
нички — мастигонемы — тончайшие нити, наблюдать которые
удается только при помощи электронного микроскопа. Основой
жгутика является аксонема — комплекс трубочек, расположен-
ных параллельно, и в определенном порядке. Электронно-микро-
скопические исследования показали, что аксонема пероноспоровых
грибов, так же как и всех эукариотических организмов, построена
по типу (9-(-2), т. е. состоит из 9 двойных трубочек, расположен-
ных по периферии, и 2 центральных трубочек (рис. 11). Аксонема
погружена в цитоплазму жгутика и окружена мембраной, про-
должающейся в плазмалемму зооспоры.
В основании жгутика в цитоплазме зооспоры располагается
базальное тело, или блефаропласт. — цилиндрическое образова-
ние, каркас которого составлен 9 парами двойных трубочек (в от-
личие от аксонемы центральные трубочки здесь отсутствуют).
Источником образования жгутиков в процессе онтогенеза зооспоры
является центриоль, которая разрастается и превращается в бле-
фаропласт. Блефаропласт образует вырост за пределы тела зоо-
споры, который дает начало жгутику. При движении зооспоры
оба жгутика совершают волнообразные движения, располагаясь
при этом под прямым углом друг к другу. Зооспоры движутся
с очень большой скоростью. При инцистировании зооспоры жгу-
тики втягиваются внутрь, плазмалемма жгутика при этом посте-
пенно сливается с плазмалеммой клетки. Процесс дифферен-
циации жгутиков описан в ряде работ (Попова, Каратыгин,
1976 и др.).
22
Половое спороношение
В определенный период жизненного цикла пероноспорового
гриба происходит образование особых покоящихся спор — ооспор,
служащих для переживания неблагоприятных условий: низких
(или высоких) температур, засухи, изменений в питании и т. п.
Ооспоры остаются жизнеспособными в течение одного года,
а иногда и более продолжительного периода времени.
Образованию ооспоры обычно предшествует половой процесс,
который в общей форме заключается в копуляции — слиянии сфор-
мировавшихся на мицелии половых клеток: оогония и антеридия
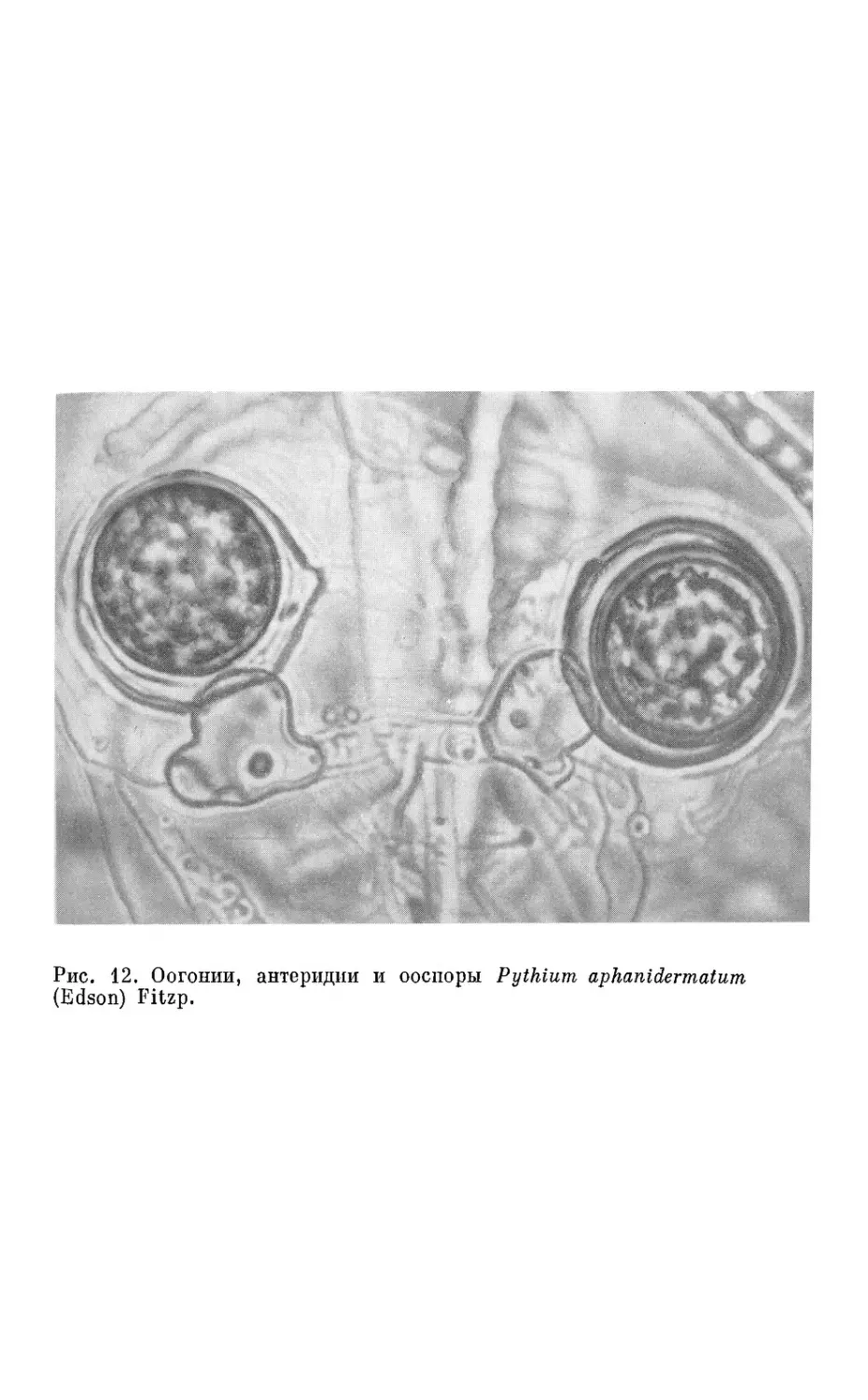
(рис. 12, см. вкл.) — и дальнейшем ядерном процессе оплодотворе-
ния. Оогоний представляет собой женскую половую клетку округ-
лой формы и крупных размеров, антеридий— мужскую половую
клетку булавовидной, пузыревидной или неправильной формы.
В результате процесса оплодотворения образуется ооспора. По-
ловые клетки и ооспоры закладываются всегда в межклетных
пространствах тканей пораженных растений.
У некоторых пероноспоровых грибов женские и мужские поло-
вые клетки образуются на одном и том же мицелии, который в этом
случае является гомоталличным (явление, аналогичное однодом-
ности у высших растений), у других — на разных мицелиях —
гетероталличных (т. е. разнодомных). Ооспоры образуются чаще
у грибов с гомоталличным мицелием. Впрочем, и здесь половой
процесс зависит от сочетания многих факторов, воздействующих
на растение и на гриб.
Содержимое ооспоры заключено в многослойную оболочку,
состоящую из внутреннего тонкого и светлоокрашенного слоя, так
Рис. 13. Прорастание ооспоры Рис. 14. Прорастающая ооспора
с образованием зародышевого с образованием ростковой трубки
зооспорангия. с зародышевым зооспорангием
на вершине.
23
называемого эндоспория, и наружного, большей частью толстого
И" окрашенного в золотисто-желтые или коричнево-бурые тона,
называемого экзоспорием. Поверхность ооспор у различных пред-
ставителей разная: гладкая, сетчатая, складчатая, бугорчатая
или щетинистая, что используется в качестве систематического
признака для распознавания отдельных видов. Величина ооспор
18—60 мкм в диам. Она различна у видов одного рода грибов.
Например, у довольно близких видов Plasmopara отмечаются и
крупные ооспоры (Р. asterea), и небольшие (Р. helianthi).
Ооспоры некоторых пероноспоровых грибов прорастают
вскоре после образования (рис. 13), но в большинстве случаев
требуют продолжительного периода покоя, причем под влиянием
неблагоприятных факторов среды: замерзания и оттаивания, вы-
мокания и высушивания — их жизнедеятельность не подавляется,
а активизируется. В весенний период ооспоры прорастают и
обусловливают первичное заражение растений в поле. Процесс
прорастания ооспор прослежен только у незначительного числа
видов. Так, например, известно, что ооспоры Р. viticola при про-
растании образуют ростковую трубку, на вершине которой воз-
никает крупный «зародышевый» зооспорангий с большим коли-
чеством зооспор (рис. 14); у Cystopus candidus зооспорангий
выпячивается непосредственно из ооспоры (рис. 13). Для мно-
гих видов пероноспоровых грибов установлено, что ооспоры обра-
зуют только ростки. Обычно ооспоры являются источником ин-
фекции растений.
Цитологические данные
Оогонии и антеридии начинают развиваться как терминальные
или интеркалярные вздутия гиф, которые отделяются перегород-
кой от остальной части гифы. После отделения оогония пере-
городкой его оболочка утолщается и содержимое дифференци-
руется на два участка: центральный гомогенный — ооплазму и
периферический вакуолизированный — периплазму. Вначале
ядра распределены по всему оогонию, а затем они мигрируют
в периплазму. В ооплазме остается только одно ядро, которое при
оплодотворении сливается с мужским ядром, проникающим
в ооплазму через оплодотворяющую трубку (полинодий) антери-
дия. Признаком происшедшего процесса оплодотворения явля-
ется образование в оогонии жировых капель.
Электронно-микроскопические исследования, которые начали
проводиться в последнее время, раскрыли характер цитологиче-
ских изменений, происходящих в оогонии в процессе его превра-
щения в ооспору (МсКееп, 1975). В молодом оогонии содержатся
многочисленные ядра, митохондрии, диктиосомы, цитоплазмати-
ческие пузырьки с содержимым разной плотности, хорошо разви-
тый эндоплазматический ретикулюм. В процессе созревания
ооспоры большая часть ядер и митохондрий дегенерируют, часть
24
Рис. 12. Оогонии, антеридии и ооспоры Ру thium aphanidermatum
(Edson) Fitzp.
Рис. 18. Оогонии с шиповатой оболочкой у Pythium acanthicum Drechs.
цитоплазматических пузырьков мигрирует к центру оогония и,
сливаясь, образует одну крупную центральную глобулу, состоя-
щую из запасных веществ. Цитоплазматические пузырьки другой
разновидности переходят на периферию оогония, где сливаются
с образованием многочисленных липидных капель. Цистерны
эндоплазматического ретикулюма располагаются вдоль плазма-
леммы оогония, где принимают участие, по-видимому, в секреции
оболочки ооспоры, которая образуется изнутри под оболочкой
оогония. Периплазма становится все более и более вакуолизиро-
ванной, вероятно вследствие расходования веществ периплазмы
на построение оболочки ооспоры. В результате ядро зиготы и
остатки цитоплазмы оказываются заключенными в небольшом
пространстве между запасающей глобулой и липидными каплями.
Образовавшаяся оболочка ооспоры состоит из 3 слоев: первичного
и вторичного эндоспория и толстого складчатого экзоспория.
Структура и состояние ооспоры — обезвоженность цито-
плазмы, обилие запасных питательных веществ, толстая обо-
лочка — обусловливают способность ооспоры к длительному пере-
живанию неблагоприятных условий.
ЦИКЛ РАЗВИТИЯ
Цикл развития каждого пероноспорового гриба имеет свои
характерные особенности, которые трудно вместить в единую
схему. В общих чертах в нем различают следующие фазы раз-
вития.
В ранневесенний период прорастают покоящиеся ооспоры.
Первичное заражение растений осуществляется внедрением в рас-
тения (через корневые волоски, устьица) ростковых трубок зооспор,
вышедших в окружающую среду — почву в условиях насыщения
ее водой, или непосредственно ростком ооспоры. Внутри тканей
растения-хозяина развивается мицелий.
В течение весенне-летнего периода на поверхности поражен-
ных органов развивается спороношение — спорангиеносцы с зоо-
спорангиями (или конидиями). Последние прорастают с обра-
зованием зооспор (или непосредственно мицелием) и инфицируют
здоровые растения; это может повториться неоднократно в тече-
ние лета.
В осенний период (иногда летом и даже весной) внутри тканей
питающего растения закладываются оогонии и антеридии и как
результат полового процесса развиваются ооспоры.
Ооспоры зимуют и при этом сохраняют жизнеспособность.
Прорастанием ооспоры весной начинается новый цикл развития
пероноспорового гриба.
Известны и более сложные формы цикла развития. Так, неко-
торые представители обладают способностью зимовать не только
при помощи ооспор, но и в виде мицелия в клубнях, корнях или
семенах; тогда весеннее возобновление болезни обусловлено
25
РиС. 15. Цикл развития гриба Plasmopara viticola (Berk, et Curt.) Berl. et de
1 — прорастание ооспоры и образование зооспор; 2 — внедрение ростка зооспоры в ткань
растения-хозяина; 3 — внутритканное развитие гриба с образованием мицелия и гаусто-
риев; 4—6 — первая и последующие генерации спор; 7 — переход к зимующей стадии
развития, образование оогониев, антеридиев, ооспор; 8 — ооспоры в растительных
остатках.
другим источником, в частности развитием мицелия. У некоторых
видов имеются ооспоры и многолетний мицелий, сохраняющийся
из года в год в корнях или других органах многолетних высших
растений (рис. 15).
Развитие пероноспоровых грибов зависит от условий внешней
среды; это легко можно заметить при наблюдениях за появлением
болезни в поле. Решающее значение имеют высокая влажность
воздуха, обильные осадки и ночные росы наряду с относительно
низкой температурой; эти условия особенно благоприятны для
заражения растений и массового проявления болезней, вызывае-
мых грибами, образующими зооспоры.
Прорастание спор этих грибов целиком зависит от наличия
капельно-жидкой влаги, которая необходима для выхода зооспор
26
и для прорастания конидий; однако последние иногда способны
прорастать при высокой влажности воздуха в отсутствие воды.
Чаще всего прорастание имеет место в капельках дождя или росы.
При заражении подземных частей растений большую роль играет
влажность воздуха и почвы. Развитию болезней способствуют
сорняки, которые быстро образуют плотный покров, затрудняю-
щий аэрацию почвы и поддерживающий высокую влажность.
Фактором, лимитирующим развитие болезней и возбудителей,
является также температура. При оптимальной температуре про-
растание происходит быстро — в течение 15—30 мин, тогда как
при неблагоприятной — медленно или вовсе не происходит.
Длительность движения зооспор также зависит от температуры.
Наиболее благоприятна для образования, прорастания и движе-
ния зооспор температура в пределах 15—18°, а для некоторых
видов и более низкая.
В соответствии с этим пониженная температура является' осо-
бенно благоприятной для заражения растений. Однако развитию
большинства болезней более способствует повышенная темпера-
тура: инкубационный период в этих случаях короче, чем при
низкой температуре (см. с. 65).
МЕТОДЫ ИССЛЕДОВАНИЯ
При изучении пероноспоровых грибов используются как ста-
рые классические, так и более современные методы микологиче-
ских и фитопатологических исследований. Ниже приводятся
некоторые из них.
ГЕРБАРИЗАЦИЯ
Сбор видов пероноспоровых грибов на образцах пораженных
растений и их сохранение необходимы для постановки правиль-
ного диагноза и документации. Обследованию и сбору подлежат
растения с признаками, характерными для поражения пероно-
споровыми грибами. При взятии образцов не следует ограничи-
ваться сбором только пораженных частей (листьев, клубней и т. п.),
в ряде случаев важно брать больное растение целиком. Обсле-
дование и сбор образцов растений с паразитирующими на них
пероноспоровыми грибами производят начиная с ранневесеннего
и кончая позднеосенним периодом. Большинство изучаемых бо-
лезней обнаруживается весной, так как питиевые грибы, многие
ложно-мучнисторосяные и цистоповые поражают растения в пе-
риод всходов. Поскольку не все виды пероноспоровых способны
к образованию ооспор в весенний период, собирать образцы сле-
дует также и осенью. Развитие гриба и течение болезни необ-
ходимо проследить начиная с момента всходов и до уборки уро-
жая. Следует набрать больше материала, что позволит отобрать
лучшие образцы для определения вида, а следовательно, и при-
чин болезни.
Собранный материал в зависимости от формы заболевания
и участка поражения фиксируется разными способами (Хохря-
ков, 1936). Травянистые растения с налетами, пятнистостями
и пустулами высушивают в ботанической папке или пресс-сетке.
Засушенные образцы перекладывают в конверты из плотной бу-
маги, к которым заполняют этикетку. Оставшиеся образцы после
определения вида гриба — возбудителя болезни оставляют на
сохранение в микологических гербариях, где располагают в си-
стематическом порядке. Образцы хранятся в сухом прохладном
помещении, каждый вид уложен в специальные папки из плотной
бумаги для предотвращения поломки. При просмотре образцов
папки не переворачивают, а перекладывают одну на другую, сверху
вниз, а затем снизу вверх. Корни, корневища, клубни, участки
28
стволов, стеблей с признаками изменения окраски, с уродливо-
стями, выростами, ранами, язвами фиксируются в различных
жидкостях: спирте (70%), формалине (1—2 части продажного
формалина на 40 частей воды), растворе поваренной соли (10%).
Чтобы объект сохранил зеленую окраску, его кипятят 1—2 мин
в 1%-ном растворе медного купороса, а затем промывают в воде
и помещают в одну из вышеперечисленных жидкостей. Для образ-
цов, зафиксированных в жидкостях, этикетки пишут простым
карандашом на пергаментной или другой, несильно размокающей
бумаге и закладывают внутрь банок, а на банки наклеивают та-
кие же этикетки. Пробки заливают парафином.
Составленная таким образом коллекция служит наглядным
справочным материалом в случае затруднений при установлении
точного диагноза болезней растений, а также отражает видовой
состав грибов, свойственных данному району, области.
ИЗМЕРЕНИЕ, ОБРАБОТКА ДАННЫХ
При изучении морфологических особенностей и изменчивости
видов производят измерение отдельных органов изучаемых гри-
бов, особенно спор, что необходимо для таксономических работ.
Для измерения объектов пользуются окулярным микрометром
с произвольно разделенной шкалой. Определение величины деле-
ния производят с помощью объективного микрометра, на котором
1 мм точно разделен на 100 частей. Совместив направления объек-
тивного и окулярного микрометров в поле зрения микроскопа
при разных комбинациях объективов и окуляров, отмечают
число делений объективного микрометра, точно покрываемых
делениями окулярного микрометра. Помножив первое число на
100, делят его на второе и получают величину 1 деления окуляр-
ного микрометра, выражая его в микронах. Сделав такой расчет
для всех возможных оптических комбинаций микроскопа, состав-
ляют таблицы для облегчения пересчета при измерении объектов
в последующей работе.
Для анализа материала исследования отдельные данные груп-
пируются в ряды или таблицы. Если материал достаточно одноро-
ден, а вариация внутри его чисто количественная, то вычисляют
среднюю арифметическую
X Xi
* = ~7Г>
где х — средняя арифметическая, х. — значение любой варианты,
2 — знак суммирования, п — количество суммируемых единиц.
Средняя арифметическая позволяет заменить индивидуальные
значения признаков некоторой средней величиной. Она указывает
значение признака, наиболее характерного для данной совокуп-
ности, но не показывает всего разнообразия отдельных данных.
29
Отчасти степень этого разнообразия можно установить, опреде-
лив вариационный размах признака, т. е. разницу между макси-
мальным и минимальным значениями определенного ряда призна-
ков. Но такой способ влечет за собой ошибки, так как крайние
значения вариационного ряда могут меняться в зависимости от
количества вариант и, кроме того, при одних и тех же пределах
вариации распределение вариант может быть различным. По-
этому для выявления различий между отдельными данными (ва-
риантами) надо их сравнить друг с другом или с какой-либо по-
стоянной величиной. В качестве последней можно взять среднюю
арифметическую х и вычислить среднее квадратиче-
ское отклонение, иначе — стандартное от-
клонение
1/ 2 (^ — ^)2
с = Г •
Например: при измерении длины конидий у Peronospora para-
sitica получены следующие данные: 23.4, 21.0, 17.8, 21.6, 23.8,
22.0, 17.3, 17.6, 19.3, 24.2 мкм. Средняя арифметическая будет
2+ 23.4+21.0+17.8+21.6+23.8+22.04-17.34-17.6+19.3+24.2
Исходя из средней арифметической вычисляется стандартное
отклонение
1/2 (^ — *)2
Q=V =
!/(23.4 — 20.8)2 + (21.0 — 20.8)2 + . . . + (24.2 — 20.8)2 п п
= у 9 —2.7.
Значит, наиболее характерное значение длины конидий для
Р. parasitica равно 20.8 + 2.7 мкм.
Часто при анализе опыта, при изучении какого-либо процесса
средняя арифметическая недостаточно точно характеризует мате-
риал, тогда пользуются средней геометрической, вычисляют коэф-
фициент вариации, средние ошибки и т. д. Более подробно этот
вопрос освещен в книгах П. Ф. Рокицкого (1967) и Е. С. Смир-
нова (1969).
ЦИТОЛОГИЧЕСКОЕ ИЗУЧЕНИЕ ЖИВОГО
И ФИКСИРОВАННОГО МАТЕРИАЛА
Микроскопическое изучение клеток пероноспоровых и других
грибов затруднено вследствие того, что эти клетки отличаются
малыми размерами, в их содержимом, прозрачном и бесцветном,
слабо выявляются структурные элементы. При обычном микро-
скопировании удается наблюдать только очертания оболочек,
перегородки между клетками, вакуоли и мелкие включения ци-
30
топлазмы. Более подробное изучение клеток требует примене-
ния фазово-контрастной или темнопольной микроскопии. Разра-
ботаны также методы прижизненной окраски клеток некоторыми
красителями.
Наблюдения в темном поле
В темнопольной микроскопии вместо обычного конденсора
применяется параболоид- или кардиоид-конденсор, освещающий
объект сбоку. Прямые лучи в объектив не попадают, и фон оказы-
вается темным. Темное поле позволяет вести наблюдения за изме-
нением степени дисперсности коллоидов протопласта, выявить
явление коагуляции, наступающее при отмирании клеток. Цито-
плазма живых клеток оптически пуста, черного цвета. Включения
протоплазмы, находящиеся в непрерывном броуновском движении,
и оболочки клеток ярко светятся отраженным светом. При отми-
рании клеток наблюдается прекращение движения частиц, про-
топлазма становится оптически видимой, начинает светиться.
Метод наблюдения в темном поле особенно пригоден для изуче-
ния грибов в культуре, так как при изучении срезов тканей, пора-
женных грибами, сильно светятся целлюлозные оболочки и вклю-
чения клеток растения-хозяина. Темнопольные наблюдения де-
лают возможным распознавание живых и мертвых клеток без
проращивания спор и без применения красителей.
Прижизненное окрашивание
Некоторые водорастворимые красители обладают способностью
проникать через клеточные оболочки и избирательно окрашивать
содержимое клеток, не вызывая при этом гибели последних.
Это нейтральный красный, метиленовый синий, нильский голу-
бой, толуидиновый синий, бисмарк коричневый, цианин, эозин
и др. Раствор красителя берется в слабой концентрации, от 1:1000
до 1:500 000. Время окрашивания исчисляется несколькими минута-
ми и более точно подбирается для каждого объекта в отдельности.
Существует способ комбинированной окраски нейтральным
красным и толуидиновым синим, предложенный С. В. Дмитрие-
вой и 3. Э. Беккер (1962), позволяющий наблюдать возрастные
изменения в клетках мицелия в условиях культуры. При окраске
нейтральным красным в вакуолях клеток грибов образуются
волютиновые гранулы, состоящие, как полагают, из полифосфа-
тов и РНК. Различным возрастным состояниям клеток соответ-
ствует различная концентрация этих веществ в вакуолях и в ци-
топлазме. Методика окраски следующая.
Мицелий окрашивается прижизненно 0.02%-ным раствором
нейтрального красного с образованием в вакуолях гранул крас-
ного цвета. Затем мицелий фиксируется 70%-ным спиртом. После
вымывания красителя остаются слабо преломляющие свет обра-
зования. Затем тот же мицелий окрашивается 0.04%-ным раство-
31
ром (водным) толуидинового синего в течение 1—2 мин. Толуиди-
новый синий красит РНК ортохроматически в синий цвет, а поли-
фосфаты метахроматически — в пурпурный цвет. В вакуолях
молодого мицелия окраска, выявляющая полифосфаты, сильно
положительная, но отрицательная в отношении РНК. Окраска
более старого мицелия (48-часовая культура) дает такой же ре-
зультат, но при этом четко выявляются включения РНК вокруг
тонопласта. Резко меняется картина в 72-часовом мицелии: толу-
идиновый синий дает очень слабую окраску гранул на полифос-
фаты и интенсивную на РНК. В старом вакуолизированном ми-
целии (96-часовая культура) гранул полифосфата нет, но четко
видны гранулы РНК.
Способы фиксации материала
для световой микроскопии
Спирт с формалином
(части)
Спирт 70%-ный — 94
Формалин — 6
Срок действия состава не ограничен, и последующей промывки
не требуется. Этот фиксатор используется, если требуется выявле-
ние только мицелия в тканях без тонких деталей.
Спирт с уксусной кислотой
(части)
Спирт абсолютный — 2
Уксусная кислота крепкая — 1
Уксусная кислота действует как хороший ядерный фиксатор.
Срок действия фиксатора — 24 ч, для нежных объектов —
меньше. После фиксации материал промывается и сохраняется
в 70 %-ном спирте.
Жидкость К а р н у а (части)
Абсолютный спирт — 6
Хлороформ — 3
Уксусная кислота ледяная — 1
Срок действия фиксатора также ограничивается несколькими
часами. Достоинство этого состава — способность быстро прони-
кать в растительные ткани.
Фиксатор Навашина (части)
Хромовая кислота (1%-ная) — 10
Формалин (40%-ный от продажного) — 4
Уксусная кислота ледяная — 1
После фиксации материал тщательно промывается в воде и со-
храняется в 70 %-ном спирте. Фиксатор Навашина употребляется
для фиксации хромосом.
32
Жидкость Флемминга (части)
Хромовая кислота (1%-ная) — 25
Уксусная кислота ледяная — 10
Осмиевая кислота (2%-ная) — 10
Широко применяется в ботанических и зоологических иссле-
дованиях, особенно при цитологических исследованиях, связан-
ных с изучением ядра.
Действие уксусной кислоты, которая является ядерным фикса-
тором, дополняется действием осмиевой кислоты — быстродей-
ствующего протоплазматического фиксатора. Хромовая кислота
действует в качестве протравы, способствуя поглощению краси-
телей тканями. Следствием присутствия в фиксаторе осмиевой
кислоты является почернение объекта, причем оно тем интенсив-
нее, чем больше материал содержит жиров. Устранение темной
окраски достигается помещением материала в слабый раствор
перекиси водорода.
Фиксатор Аллена
Хромовая кислота — 1 г
Уксусная кислота — 1 мл
Мочевина — 0.5 г
Вода — 100 мл
Предложен для фиксации мицелия ржавчинных грибов в тка-
нях растений, но, по-видимому, может быть применим и для фик-
сации пероноспоровых грибов.
Люминесцентная микроскопия
Для выявления мицелия грибов в тканях растений и для наблю-
дения происходящих в нем функциональных изменений может
применяться люминесцентная микроскопия.
Для немногих видов грибов характерна собственная люминес-
ценция, т. е. способность их клеток светиться под воздействием
ультрафиолетовых лучей. Но в большинстве случаев для изуче-
ния клеток грибов с помощью люминесцентного , микроскопа тре-
буется специальная окраска — флуорохромирование. Она осно-
вана на том, что обработанные флуорохромом ткань растения-
хозяина и мицелий паразитического гриба светятся по-разному.
Кроме того, характер свечения мицелия меняется в зависимости
от его онтогенетического состояния и физиологических особен-
ностей. Так, например, живые клетки грибов, окрашенные акри-
диновым оранжевым, при определенном значении pH раствора
светятся зеленым светом, а отмершие — красным (Strugger, 1943).
Таким образом, флуорохромирование дает возможность определить
состояние мицелия в тканях растения-хозяина.
Для изучения грибов в тканях растений использовались раз-
личные красители: эозин, эритрозин, акридиновый оранжевый,
аурамин и др. Л. И. Пшедецкая и Н. П. Черепанова (1961а),
испробовав различные флуорохромы для зучения Мицелия
3 Н. С. Новотельнова и др. 33
Phytophthora injestans в листьях картофеля, пришли к выводу,
что наилучшие результаты дают аурамин и акридиновый оран-
жевый.
Окрашивать флуорохромами можно как живой, так и фикси-
рованный материал. Фиксирующей жидкостью служит смесь
спирта с формалином: 40%-ного формалина — 6 частей, 70%-ного
спирта — 94 части. Другой способ фиксации — паром в течение
30—60 мин. Краситель берется в очень слабой концентрации
(1 : 1000), время окраски — 10—20 мин.
Метод люминесцентной микроскопии с успехом используется
для определения жизнеспособности спор фитопатогенных грибов,
в том числе и пероноспоровых. А. К. Вялых и В. П. Ланецкий
(1974) окрашивали зооспорангии грибов из рода Cystopus акриди-
новым оранжевым и наблюдали различное по цвету свечение у про-
росших, непроросших и мертвых зооспорангиев, что позволило
разработать способ быстрой оценки качества спорового материала.
Метод флуоресценции дает возможность получить сведения о жиз-
неспособности спорангиев популяции за 10—20 мин.
Для наблюдения флуоресценции клеток используются люми-
несцентные микроскопы МЛ-1, МЛ-3.
Методы окраски мицелия грибов
Быстрый способ окраски хлопчатобумажным синим. Приме-
няется для дифференциации мицелия в тканях растений. Объект
фиксируется нагреванием на предметном стекле в капле лактофе-
нола (10—50%-ный раствор фенола в молочной кислоте), затем
помещается в каплю 1%-ного водного или молочнокислого рас-
твора красителя на 1 мин и после этого промывается в воде.
Гифы гриба приобретают синюю окраску.
Окраска иодом. Применяется для изучения строения клеточных
оболочек. Раньше применяли для окраски жгутиков зооспор.
Готовится раствор иода в иодистом калии или глицерине. Капля
раствора добавляется к препарату, приготовленному на воде.
Окрашивание ведется в течение 10—15 мин, после чего препарат
промывается в воде.
Окраска сафранином и светлым зеленым. Препараты окраши-
ваются в 1%-ном спиртовом растворе сафранина в течение 24 ч,
проходят через серию спиртов, обезвоживаются в абсолютном
спирте. Затем производится окрашивание препаратов в насыщенном
спиртовом растворе светлого зеленого (Lichtgriin) в течение
30 мин, после чего — проводка по восходящей серии спиртов,
просветление в ксилоле и заключение в канадский бальзам.
Этот и последующие способы окраски пригодны для препаратов,
полученных при заключении материала в парафин, и приготовле-
ния микротомных срезов по общепринятой методике. В резуль-
тате гифы гриба окрашиваются в зеленый цвет, ядра — в красный
цвет, а клетки растения-хозяина приобретают розовую окраску.
34
Окраска сафранином, анилиновым синим и пикриновой кисло-
той. Срезы фиксированного материала помещаются в 1%-ный
раствор сафранина, после чего их окрашивают в смеси анилинового
синего и пикриновой кислоты (25 мл насыщенного водного раствора
ацилинового синего и 100 мл такого же раствора пикриновой
кислоты). Препараты промываются в дистиллированной воде,
в нескольких сменах 96%-ного спирта и в абсолютном спирте,
просветляются в гвоздичном масле и заключаются в канадский
бальзам.
Метод Стоутсона. Срезы, освобожденные от парафина, прово-
дят по нисходящей серии спиртов до воды и помещают на 1 ч
в краситель следующего состава: тионин — 0.1г, 5%-ный раствор
фенола в дистиллированной воде — 100 мл, после чего следуют
проводка по спиртам восходящей серии до абсолютного и окраши-
вание в насыщенном растворе оранжа Ж в абсолютном спирте
с одновременной дифференциацией предыдущей окраски, про-
ходящей в течение 0.5—1 мин и контролируемой под микроскопом.
В результате гифы гриба окрашиваются в фиолетовый цвет, клет-
чатка — в желтый или зеленый, ядра — в голубой.
Метод Колпин—Равна. Материал фиксируется в жидкости
Флемминга. В качестве красителя используется концентрирован-
ный раствор орсеина ВВ в 3%-ной уксусной кислоте. После про-
мывания в воде следует окрашивание концентрированным раство-
ром анилинового синего в уксусной кислоте, затем дифференциа-
ция в 90 %-ном спирте, обезвоживание и заключение в канадский
бальзам. Гифы гриба окрашиваются в синий цвет, целлюлозные
оболочки клеток растения — в голубой, ядра и протоплазма —
в красный.
Тройная окраска по Флеммингу (метод Аллена). Метод разра-
ботан для окрашивания мицелия ржавчинных грибов, но может
быть применен и к пероноспоровым. Фиксирующей жидкостью
служит фиксатор Аллена. Для окрашивания готовят раствор
сафранина в абсолютном спирте (50 г спирта и 0.5 г красителя).
Через 4 дня к раствору прибавляют 10 мл воды. Окрашивают
в течение 5—12 ч, промывают в воде и окрашивают в 2 %-ном вод-
ном растворе генцианвиолета. Затем опять промывают в воде
и окрашивают 1%-ным раствором оранжа Ж в воде или гвоздич-
ном масле в течение 1—3 мин, после чего следует промывание
в воде или гвоздичном масле, обезвоживание и заключение в баль-
зам. Хроматин в гифах гриба окрашивается в красный цвет,
протоплазма — в оранжевый, клетчатка — в фиолетовый.
Заключение материала в парафин
Цитологические исследования, как правило, предполагают
получение тонких микротомных срезов изучаемого материала.
При цитологическом изучении грибов в ряде случаев этого не
требуется. Такие структуры, как гифы, сами по себе достаточно
3*
35
тонки, поэтому тотальные препараты гиф оказываются пригод-
ными для изучения строения составляющих их клеток. Однако
при изучении мицелия и гаусториев, располагающихся внутри
тканей растений, первым этапом работы является приготовление
срезов.
При повседневной микроскопии можно удовольствоваться
срезами «от руки» и изготовлением временных препаратов, однако
для анатомических исследований срезы делаются с помощью за-
мораживающего микротома. Готовятся постоянные препараты,
требующие предварительной, довольно длительной подготовки
материала — обезвоживания и заключения в парафин или дру-
гую среду. Фиксатор должен быть в избытке, кусочки объекта
малыми.
Пропитывание объекта парафином осуществляется после пред-
варительного полного обезвоживания материала. Зафиксирован-
ный материал проводится через серию спиртов возрастающей
концентрации, обезвоживание завершается в двух сменах абсо-
лютного спирта. Дальнейшая подготовка объектов заключается
в замене пропитывающего их спирта «промежуточной жидкостью»,
т. е. средством, растворяющим парафин и хорошо смешивающимся
со спиртом. Чаще всего такими жидкостями служат ксилол или
хлороформ. Для проводки цспользуются абсолютный спирт и
химически чистые, обезвоженные ксилол или хлороформ.
Ниже приводится общая схема подготовки материала для за-
ключения в парафин. Удобнее всего делать проводку в пеницил-
линовых бутылочках или бюксах, в которых сменяются одна за
другой все требующиеся жидкости.
Водный фиксатор
Спиртовой
фиксатор
I
30%-ный
50%-ный
70%-ный
80%-ный
96%-ный
100%-ный (1)
100%-ный (2)
По 1 ч в каждом спирте
ф ( 3 : 1 1
Смесь спирта и хлороформа < 1 : 1 > По 2 ч в каждой смеси
| 1 : 3 J
Хлороформ-2 } По 2“3 4 в каждой смене
После выдерживания в хлороформе-2 материал помещается
в бюкс с новой порцией чистого хлороформа, туда же кладется
небольшое количество парафиновой стружки. Бюкс ставят на
несколько минут в термостат с температурой 40° для растворения
парафина. После этого в бюкс осторожно заливается расплавлен-
ный парафин (толщина слоя залитого парафина — не менее 0.5 см),
бюкс с закрытой крышкой снова помещают в термостат, выдер-
живают 4—6 ч и переносят в термостат с температурой 60°, затем
36
через 2 ч снимают крышку с бюкса и оставляют его в термостате
до полного испарения хлороформа. Степень испарения может быть
проверена на вкус — исчезновение сладкого вкуса говорит о пол-
ном испарении хлороформа.
В некоторых случаях вместо ксилола или хлороформа в каче-
стве растворителя парафина используют кедровое масло. Этот
способ заключения в парафин применяют при заливке нежных
объектов, таких как гифы грибов, полученные в культуре, по-
скольку хлороформ и ксилол часто вызывают их сжатие.
Способ быстрой заливки в парафин через кедровое масло опи-
сывается в книге А. А. Прозиной (1960 : 46): «Кедровое масло
наливается в цилиндр с притертой пробкой на одну треть его
высоты. Сверху на масло наливается абсолютный спирт, а затем
в него помещаются кусочки материала. Кусочки, пропитываясь
сначала смесью спирта с маслом, а затем маслом, постепенно опу-
скаются на дно цилиндра, где их оставляют лежать некоторое
время. После этого спирт осторожно сливают и заменяют смесью
из кедрового масла пополам с парафином, оставляют стоять пол-
часа и затем переносят материал в чистый парафин, который через
некоторое время заменяют порцией чистого парафина. Таким
образом можно через 3 часа получить пропитанный парафином
материал, вполне готовый к резке».
Заливка в парафин очень мелких объектов, таких как споры
грибов, часто вызывает трудности при проводке, так как при
перемене растворов 'объекты могут просто потеряться. Чтобы
избежать этого, материал помещают в пробирку, наливают не-
обходимый реактив, а перед сливанием помещают пробирку
в ручную центрифугу. При центрифугировании материал оседает
на дно, жидкость осторожно сливают и заменяют следующей.
Методика фиксации материала
для электронной микроскопии
Наиболее распространенный способ — фиксация глутаровым
альдегидом с постфиксацией четырехокисью осмия. Этот способ
с успехом применяется для изучения грибов, относящихся к раз-
ным систематическим группам.
За день до фиксации готовятся следующие растворы.
1. Фосфатный буфер, pH 7.4. Для получения 100 мл буфера
смешиваются 20 мл 0.1 М раствора КН2РО4 и 80 мл 0.1 М раствора
Na2HPO4. pH раствора зависит от качества исходных реактивов,
поэтому после смешения растворов необходимо измерить pH по-
лученного буфера и если величина pH не соответствует требуемой,
то добиться сдвига pH в нужную сторону добавлением того или
другого раствора.
2. Фосфатный буфер, pH 8.0. Приготавливается путем смеше-
ния 0.1 М растворов тех же солей в следующем соотношении:
0.5 мл КН2РО4 и 89.5 мл Na2HPO4.
37
3. 3%-ный раствор глутарового альдегида на фосфатном бу-
фере, pH 7.4. Глутаровый альдегид выпускается обычно в виде
25%-ного раствора. Для получения 3%-ного раствора на 1 мл
25%-ного раствора добавляется 7.3 мл фосфатного буфера. По-
мутнение раствора при добавлении буфера говорит о недостаточ-
ной чистоте исходного раствора альдегида и о непригодности его
для фиксации.
4. 2%-ный раствор четырехокиси осмия. За 2—3 дня до фик-
сации готовится 4%-ный раствор четырехокиси осмия. Он служит
исходным раствором для приготовления забуференного 2%-ного
раствора. Ампулу с кристаллами реактива, завернутую в бумагу,
разбивают и кристаллы четырехокиси вместе с осколками ампулы
высыпают в темную бутыль с притертой пробкой, в которую затем
добавляют нужное для получения 4%-ного раствора количество
дистиллированной воды. Все операции проводятся под тягой
с большой осторожностью, так как четырехокись осмия очень
ядовита.
Полученный 4%-ный раствор хранится в холодильнике в бу-
тыли, завернутой в темную бумагу, под стеклянным колпаком.
2%-ный раствор готовится путем смешения 4 мл 4%-ного раствора
четырехокиси осмия, 1 мл фосфатного буфера (pH 8.0) и 2 мл
дистиллированной воды.
5. Абсолютный спирт. Для получения абсолютного спирта
в 100%-ный спирт засыпается окись кальция за 2—3 дня до ис-
пользования спирта. Существует и другой способ, используемый
в световой микроскопии: в 96%-ный спирт помещается прокален-
ный безводный медный купорос (на 1 л спирта 200 г купороса);
купорос завертывается в бумагу так, чтобы получились две гильзы,
по 100 г каждая; дня через три-четыре гильзы нужно заменить
новыми. Всего желательно сменить купорос в банке со спиртом
раза три в течение 10 дней.
6. Абсолютный ацетон. Обезвоживание ацетона осуществляется
с помощью засыпки в него прокаленного хлористого кальция.
7. Серия спиртов: 30, 50, 70, 85, 90, 96%-ного.
В растворы глутарового альдегида, четырехокиси осмия и
в буфер, оставшийся после приготовления растворов, добавляется
сахароза до 2—2.5%-ного раствора.
Фиксируемый материал разрезается на кусочки размером
примерно 2x6 мм и помещается в пробирки с 3%-ным раствором
глутарового альдегида на фосфатном буфере. Время фиксации
1.5—2 ч. Для ускорения проникновения фиксатора в ткани про-
бирки помещаются в колбу, из которой откачивается воздух
с помощью ручного насоса Камовского или водоструйного насоса.
Откачка воздуха прекращается, когда кусочки ткани опустятся
на дно пробирки и из них перестанут выделяться пузырьки воз-
духа.
После фиксации материал промывается в трех сменах буфера,
по 20 мин в каждой, и помещается для постфиксации в 2%-ный
38
раствор четырехокиси' осмия на фосфатном буфере (pH 8.0) на
3—4 ч. Затем начинается обезвоживание материала в серии спир-
тов восходящей концентрации: 30, 50, 70 %-ном. В каждом спирте
материал выдерживается по 10—15 мин, а в 70%-ном спирте
остается на ночь.
На следующий день зафиксированные объекты помещаются
в 1.5%-ный раствор уранилацетата в 70%-ном спирте на 2 ч,
после чего продолжается обезвоживание их в 85, 90, 96 %-ном
спиртах, в двух сменах абсолютного спирта и в смеси (1 : 1)
спирта с ацетоном. В каждом спирте материал выдерживается
по 10—15 мин. Завершается обезвоживание в двух сменах абсо-
лютного ацетона, по 20 мин в каждой.
Следующий этап подготовки материала для изучения его под
электронным микроскопом,— заключение его в эпоксидную смолу
Эпон илиАралдит.
Смола Аралдит включает в себя три компонента, которые соеди-
няются вместе непосредственно перед использованием в следую-
щем соотношении (мл):
Аралдит СУ212 — 10
Уплотнитель НУ — 10
Катализатор БУ064 — 0.4
Компоненты смол сливаются вместе и тщательно перемеши-
ваются. Лучше всего пользоваться для перемешивания магнитной
мешалкой. Если же перемешивание ведется вручную, то делать
это следует с осторожностью, плавными движениями, не допуская
образования в смоле мелких пузырьков воздуха.
Перед заключением материала в смолу проводится постепенное
пропитывание его смолой. Для этого готовятся смеси смолы и
ацетона с возрастающей концентрацией смолы, в которых мате-
риал выдерживается определенное время.
Смола (в частях по объему) Ацетон Время выдерживания в смеси (мин)
0.5 9.5 15
1 9 30
2 8 30
3 7 50
4 6 50
5 5 Остается на ночь
В последней смеси материал выдерживается всю ночь в про-
бирках с закрытыми пробками. Утром на следующий день пробки
из пробирок вынимаются для испарения ацетона. Спустя 3—4 ч
начинается заливка объектов в смолу. Кусочки материала поме-
щаются в заранее подготовленные пронумерованные и наполнен-
ные смолой желатиновые капсулы. Капсулы ставят в эксикатор
с хлористым кальцием на двое суток. После этого они переносятся
39
в термостат с температурой 40° на сутки, затем — в термостат
с температурой 60°, где остаются на двое суток. При 60* проис-
ходят полимеризация и затвердевание смолы.
Материал, заключенный в смолу, может храниться неограни-
ченно долго. С блоков, полученных в результате заливки, приго-
тавливаются ультратонкие срезы (5-10-2 мкм толщ.) с помощью
специального прибора — ультрамикротома. После контрастиро-
вания в растворах цитрата свинца и уранилацетата срезы про-
сматриваются и фотографируются в электронном микроскопе.
Для более подробного ознакомления с методом подготовки
материала при изучении в электронном микроскопе можно реко-
мендовать книгу Б. Уикли «Электронная микроскопия для на-
чинающих» (1975).
Оборудование, инструменты и посуда,
необходимые для цитологического изучения материала
Иглы препаровальные
Скальпель
Лезвия от безопасных бритв
Пинцеты разных размеров
Предметные и покровные стекла
Часовые стекла
Пробирки разного диаметра и длины
Пробки корковые
Колбы с широким дном
Цилиндры мерные, различных объе-
мов
Чашки Петри
Кристаллизатор для промывки пре-
паратов
Воронки стеклянные разного размера
Пенициллиновые бутылочки
Стеклянные цилиндры с притертыми
пробками для проводки препаратов
Алюминиевые бюксы для заливки
в парафин
Склянки разного размера для реак-
тивов и красителей
Капельницы
Баночки со стеклянными палочками
для канадского бальзама
Пипетки
Банки с широким горлом и притер-
тыми крышками для промывания
и хранения предметных стекол
Фарфоровые стакан для варки пара-
фина, ступка и пестик
Химические стаканы разного раз-
мера
Водоструйный насос или насос Ка-
мовского
Электрическая плитка
Электрический столик с температу-
рой нагрева не выше 50е' для рас-
правления парафиновых срезов
Эксикатор
Термостат
Змеевик для расплавления парафи-
новых срезов и для расплавления
парафина
Спиртовка
Марля
Фильтровальная бумага
Ножницы
Асбестовая сетка
Огнеупорные чашки для прокали-
вания хлористого кальция и мед-
ного купороса
Коробки с ячейками для хранения
постоянных препаратов
Кисточки мягкие для расплавления
парафиновых срезов
Лабораторные весы
Биологический микроскоп МБИ
Микротом салазочный или рота-
ционный
ИЗОЛИРОВАНИЕ И КУЛЬТИВИРОВАНИЕ
PYTHIACEAE И PHYTOPHTHORACEAE
Литиевые грибы можно выделять из природных мест обитания
с помощью приманок. В качестве последних используют стериль-
ные зерна конопли, муравьиные коконы, кусочки вареного корня
40
канатника. Приманки помещают в бюксы с исследуемым мате-
риалом, залитым стерильной водой. Для ограничения развития
сапрофитных бактерий эту воду слегка подкисляют. Когда на
приманке разовьется гриб, кончики гиф переносят в другие бюксы
со стерильной водой или на твердую среду. Пересев производят
однократно или несколько раз.
Для выделения питиевых грибов из почвы применялись разно-
образные методы. Н. А. Наумов (1937) рекомендует небольшие
комочки почвы (0.5—1.0 г), взятой под пораженными растениями,
помещать в широкую и плоскую стеклянную посуду, воды при-
ливать столько, чтобы над почвой был слой ее высотой в 1 см.
На поверхность раскладывают приманки, а воду подкисляют.
Через 24 ч приманки переносят в чистую воду, в которой остав-
ляют до появления роста и спороношения изолируемых грибов.
Таким методом выделяли Pythium debaryanum, Р. intermedium,
Р. monospermum, Р. rostratum, Р. vexans.
Многие виды Pythium можно с успехом выделять методом пря-
мого посева на искусственную агаровую или желатиновую среду,
на которой они легко культивируются. Мередит (Meredith, 1940)
изолировал виды Pythium рассеиванием измельченной почвы на
поверхность водного агара в чашки Петри. Мак-Лауглин (McLaug-
lin, 1947) предложил помещать измельченную почву под слой
водного агара. Уоркап (Warcup, 1950) применял смесь исследуе-
мой почвы (около 10 мг) в чашках Петри с теплой агаровой средой,
селективной для видов Pythium. Из выросших колоний при после-
дующих пересевах отбирались изоляты, относящиеся к этому
роду. Ранде и Доп (Rands, Dopp, 1934) исследуемую почву поме-
щали в вегетационные сосуды, куда высаживали восприимчивые
растения, а из последних в дальнейшем выделяли патогенные
виды. Шринивасан (Srinivasan, 1958) несколько видоизменил эту
методику для выделения видов грибов, патогенных на сахарном
тростнике. Кусочки стебля сахарного тростника раскладывают
на поверхность почвы в вегетационном сосуде, которую обильно
смачивают водой, и затем покрывают перевернутым сосудом
меньшего размера. Опыт проводят в темноте. Через 2—3 недели
стебли вынимают, отмывают стерильной водой и помещают в за-
крытый стерильный стакан или чашку Петри. Развившийся
через 2—3 суток ватообразный мицелий Pythium переносят на
питательную среду. Особенно эффективен метод Баниасхеми
(Banihashemi, 1970). Из прикорневой зоны больных растений
берется почва в количестве 40—60 г, помещается в стерильный
химический стакан и заливается стерильной водой на 1 см выше
уровня почвы. В этот же стакан на одну треть погружается све-
жий здоровый огурец, вымытый и продезинфицированный, с по-
верхности этиловым спиртом. Через 4—7 суток на огурце появ-
ляются признаки развития гриба (инкубация при 20°). Огурец
моют, поверхность дезинфицируют спиртом и помещают в пустой
стерильный стакан, где инкубируют при 20° до проявления роста
41.
мицелия. Из кусочков этого мицелия или зараженной ткани огурца,
взятых выше «водной линии», после перенесения их на питатель-
ную среду развивается чистая культура гриба. Хайн и Луна
(Hine, Luna, 1963) разработали методику для выделения из почвы
Р. aphanidermatum. Ломтики клубня картофеля толщиной 3 мм
после дезинфекции в течение 1 ч в водном растворе антибиотика
помещают в чашки Петри на анализируемую почву и инкубируют
12—15 ч при 31°. Затем ломтики извлекают, промывают стериль-
ной водой и раскладывают на голодный агар с антибиотиком.
Через сутки при 20° на агаре разрастается мицелий гриба.
К. А. Пыстина (1973) для изучения почвенных питиевых
грибов использовала капиллярные приспособления Перфильева—
Габэ. Капилляры, закрепленные на стеклянной пластинке, за-
полняются агаризированной вытяжкой из семян конопли и поме-
щаются в почву на глубину около 5 см. Через 2—3 суток пластинки
вынимают, очищают поверхность от приставших частиц и содер-
жимое капилляров просматривают под микроскопом. Автор
находит эту методику очень удобной, простой и быстрой по выпол-
нению, оговаривая, что она непригодна для дальнейшего выделе-
ния изолятов гриба в чистую культуру из-за развития сапрофит-
ных микроорганизмов.
Вообще основная трудность выделения видов Pythium в чи-
стую культуру заключается в необходимости освобождения их от
бактерий и других грибов, обычно развивающихся совместно
с питиевыми грибами. Поэтому обязательна обработка поражен-
ных грибом частей растений (чаще всего корневой системы) анти-
биотиками. После тщательной промывки в обычной и стерильной
воде больные корни рекомендуется помещать на 5 мин в 0.1 %-ный
раствор гипохлорита натрия. При этом следует иметь в виду,
что отдельные виды Pythium чувствительны к гипохлориту натрия
и могут погибнуть вместе с почвенными сапрофитами, в связи
с чем Гаррисон (Harrison, 1955) предложил применять ауреоми-
цин, а ранее Шультц (Schultz, 1942) — роданистый натрий.
В качестве дезинфектора используется и0.1 %-ныйраствор сулемы.
Из обработанных антисептиками и промытых водой корней выре-
зают небольшие кусочки на границе пораженной и здоровой ча-
стей, помещают в бюксы со стерильной водой. Воду, мутнеющую
из-за развития бактерий, меняют несколько раз. Через сутки
кусочки корня отжимают стерильной фильтровальной бумагой
и помещают на слегка подсушенные пластинки агара в чашки
Петри. Еще через сутки из появившейся колонии гриба отсеивают
кончики гиф, повторяя отвивку несколько раз, чтобы добиться
свободной от бактерий культуры гриба.
В случае неудачи получения чистой культуры гриба Мередит
и другие исследователи (Meredith, 1940; Sleeth, 1945) предлагают
следующую технику ш^сева гриба. В чашки Петри наливают ага-
ровую среду до толщины 5 мм. Застывшая среда разрезается на
4—5 секторов. Кусочки зараженного растения или частицы почвы
42
помещаются в центр каждого сектора. Затем сектор с инокулюмом
переносят в другие чашки Петри, причем агаровую пластинку
переворачивают верхней поверхностью вниз, чтобы инокулюм
прилип к агаровой пластинке, а края блока — к дну чашки.
Если не переворачивать агаровую пластинку, то следует накрыть
ее другой агаровой пластинкой таким образом, чтобы инокулюм
оказался между ними. Гифы гриба прорастают сквозь агар и,
освободившись от бактерий, достигают его поверхности. Иногда
при перевертывании агар растрескивается, и тогда культуру не
избавить от загрязнения. Ввиду этого Махачек (Machacek,
1934) предложил колонию гриба в чашке Петри покрыть стериль-
ным стеклянным диском 20 мм в диам.; диск благодаря плотному
прилеганию к агару препятствует распространению бактерий.
Д. Л. Тверской (1955) указывает, что воздушный мицелий неко-
торых видов Pythium способен разрастаться по стеклу чашки
Петри и при перенесении его на агаровую среду дает чистую куль-
туру. По Шмитхеннеру (Schmitthenner, 1964), освободиться от
бактерий в культуре можно путем добавления в среду родани-
стого натрия, поскольку чувствительность к нему бактерий в 20 раз
выше, чем грибов. Для этой же цели используют фунгицид ПХНБ,
однако он не влияет на рост грибов рода Fusarium, развивающихся
при корневых гнилях совместно с Pythium.
По литературным данным, питиевые грибы довольно стойки
по отношению к антибиотикам, поэтому последние используют
в качестве ингибиторов бактерий. Для освобождения культуры от
сопутствующих микроорганизмов рекомендуют разнообразные
соединения: хлортетрациклин и стрептомицин в водном агаре
(0.5 мг сухого аптечного препарата на 50 мл среды), смесь агри-
мицин (15% стрептомицина и 15% окситетрациклина). Хайн
(Hine, 1962) сообщает о росте Р. ultimuiri и Р. debaryanum на сре-
дах, содержащих 100 ppm сульфата стрептомицина и 100 ppm пи-
марицина. Некоторые препараты не влияют на рост мицелия
гриба, но подавляют прорастание его спор. Выбор антибиотика
зависит от вида исследуемого питиевого гриба. По данным Ваар-
тайи (Vaartaja, 1960), на рост Р. debaryanum не влияет добавление
в среду стрептомицина, однако рост гриба ослабевает в присут-
ствии хлортетрациклина, неомицина и новобиоцина. Следует
использовать несколько антибиотиков в комплексе.
При выращивании питиевых грибов в культуре среда должна
быть нейтральной или слабо кислой. Для освобождения от бакте-
рий величина pH может быть доведена до 4. Оптимальная темпе-
ратура для роста питиевых грибов 20—25°. Для отдельных видов
оптимум значительно выше: так, например, для Р. aphaniderma-
turn — 34°.
Для успешного изолирования фитофторовых грибов рекомен-
дуется, как правило, производить посев немедленно после взятия
образцов; дезинфицировать с поверхности пораженные органы
растений и отделять для посева небольшие участки в зоне, грани-
43
чащей со здоровыми тканями; закладывать испытуемые образцы
в свежие (незрелые) неповрежденные плоды и одновременно на
специальные среды; наблюдения над ростом и развитием изолятов
проводить возможно дольше, поскольку некоторые виды Phyto-
phthora образуют вегетативные и репродуктивные структуры лишь
после длительного культивирования. Так, Томпкинс и Такер
(Tompkins, Tucker, 1937) наблюдали образование спороношений
у видов Phytophthora лишь спустя 4 мес после начала наблюдений.
Общие указания по методам сбора, транспортировки и приемам
изучения фитофторовых грибов даются Блэкуелл (Blackwell,
1944).
Для выделения видов Phytophthora из почвы наиболее широкое
применение имеет приманочный метод (или метод «ловушек»).
Ph. cinnamomi изолируется непосредственно из почвы питомников
древесных культур с использованием яблок в качестве дифферен-
цирующего субстрата. Почвенные образцы составляют из 24 от-
дельных проб, смачивают дистиллированной водой и в виде не-
больших комочков помещают внутрь яблока в 2 углубления на
каждом. При комнатной температуре заметное развитие гриба
наблюдается через 5—10 дней.
Для выделения Ph. cinnamomi Ньюгук (Newhook, 1959) уда-
ляет сначала верхний слой почвы, примерно до глубины 5 см,
и пробы отбирает на глубине 10 см. Исследуемая почва хорошо
перемешивается и увлажняется. В яблоках, обеззараженных с по-
верхности спиртом, вырезают сверлом отверстия глубиной 5—7 см,
которые заполняют влажной почвой и прикрывают; яблоки инку-
бируют в течение 2—3 недель. Технику приманок или посева
гриба непосредственно на питательный субстрат применяют Клем-
мер и Накано (Klemmer, Nakano, 1962): частицы зараженной
грибом почвы авторы переносят на среды с низким содержанием
сахара (для исключения развития бактерий) или с антибиотиком
пимарицином (100 ppm), который задерживает развитие сапро-
фитных грибов на двое и более суток. При изолировании Ph. cin-
namomi из садовой почвы, взятой под грушевыми деревьями (Came-
ron, 1962а), проводится посев почвенных частиц на 2%-ный мо-
лочный агар или в мякоть яблок с последующим перенесением
кусочка ткани плода с растущим на ней грибом на картофельно-
декстрозный агар (последний готовится с добавлением 100 г ва-
реных яблок на 1 л среды).
Мунро (Munro, 1945) предлагает изолирование Ph. cinnamomi
из корней цинерарий производить следующим образом: корни
с розоватой окраской (ранняя стадия поражения) отделять от
растения и разрезать на небольшие кусочки: последние дезинфи-
цировать с поверхности 0.1%-ным раствором сулемы и помещать
на картофельный агар. Рост обнаруживается на 2—3-й день.
Гриб выделяется из корней быстро и легко.
Метод выделения Ph. cinnamomi из коры пораженных де-
ревьев, предлагаемый А. К. Шишкиной (1946), включает предва-
44
(ригельную дезинфекцию коры спиртом, фламбирование, асепти-
ческое удаление поверхностного слоя. Кусочки пораженной коры,
вырезанные в виде небольших блоков в местах, пограничных со
здоровой частью, помещаются в пробирки с сусло-агаром. Основ-
ное условие, необходимое для успешного изолирования, — взятие
образцов лишь с вегетирующих растений и немедленная обра-
ботка их in situ. Гриб культивируется на картофельно-декстроз-
ной, кукурузной, глицериновой средах, навозной вытяжке с ага-
ром, древесине дуба с добавлением пивного сусла, разбавлен-
ного водой (1 : 1), в стерильной дождевой воде.
Изолирование из почвы Ph. cactorum — возбудителя гнили
стволов деревьев плодовых и других культур — производится
следующим образом (Schwinn, 1960). Наколотые с поверхности
яблоки закапывают на 3/4 в почву непосредственно в природных
условиях и для защиты от грызунов прикрывают сверху проволоч-
ной сеткой. Через 6 суток гриб достаточно хорошо заметен по
пятнам гнили на плодах. Кусочки тканей из этих участков пере-
носят на бобовый агар для получения чистой культуры. Вероят-
ность выделения гриба из почвы повышается с устранением влия-
ния погодных условий. С этой целью почвенные образцы, взятые
из верхнего слоя почвы (до 20 см), помещают в ящики 24x24 см
и в каждый закладывают по 4 «ловчих» яблока. Ящики содержат
в камерах искусственного климата при 24° и 90—100% относи-
тельной влажности воздуха, опрыскивают ежедневно дождевой
водой при помощи распылителя, равномерно и сильно увлажняю-
щего почву. Через 5—6 дней на яблоках появляются бурые уча-
стки, свидетельствующие о присутствии Ph. cactorum. Однако
в этих условиях быстрое гниение яблок обусловливает появление
сопутствующих сапрофитных микроорганизмов, развитие кото-
рых можно предотвратить, используя ненаколотые яблоки, но
с более длительной экспозицией их в почве (до 8—10 дней).
Приманки в виде кусочков коры стволов и ветвей деревьев дают
менее успешные результаты. Автор рекомендует для большей
вероятности обнаружения гриба отбирать образцы почвы на раз-
ных участках питомника, составлять их из 10—12 проб, хранить
образцы в прохладном и защищенном от высыхания месте и ис-
следовать не позже 8 недель после отбора.
При изучении потенциала инокулюма в почве, зараженной
Ph. nicotianae, и выделении гриба (Nusbaum et al., 1952) в ка-
честве теста применяют титрованную естественно зараженную
почву, добавленную в предварительно прогретый паром песок
с посаженными в него восприимчивыми растениями. Для изуче-
ния возможности развития в почве Ph. infestans с дальнейшим его
изолированием закладывают в почву кусочки здоровых клубней
картофеля, на которые гриб переходит из почвы (Наумова, 1939;
Бордукова, 1941). Аналогичный метод применяется и другими
исследователями (Lacey, 1965) в работах с разными видами
Phytophthora (Banihashemi, 1970). В опытах Н. А. Наумовой
45
(1939) c Ph. infestans почва с растертыми перезимовавшими
листьями больных растений вызвала заражение стеблей кар-
тофеля.
Изолирование гриба из картофельных клубней, пораженных
Ph. erythroseptica, легко удается (Pethybridge, 1913) при соблюде-
нии мер предосторожности от загрязнения посторонними микро-
организмами. Рекомендуются выбор клубней, не слишком сильно
пораженных гнилью, обработка их поверхности щеткой в про-
точной воде, а затем обязательная дезинфекция сулемой, форма-
лином (обычные концентрации) или спиртом. После подсушивания
из разделенного пополам клубня вырезают кусочки пораженной
ткани на границе со здоровой, возможно дальше от поверхности,
и переносят в чашки Петри на твердую питательную среду.
Аткинсон (Atkinson, 1965) выделяет виды Phytophthora из
древесных растений, обрабатывая пораженные корни сначала
70%-ным спиртом, а затем просушивая их при помощи стерильной
бумаги. Небольшие отрезки корней закладываются затем в чашки
Петри на подкисленный водный агар или в мякоть незрелого
яблока. Тот же автор получает изоляты гриба из почвы методом
«приманочного хозяина». Он помещает сеянцы или кусочки кор-
ней в естественно зараженную почву, взятую из питомника.
Изолирование Ph. fragariae из корней земляники успешно
осуществляется погружением отрезанных частей корня с изме-
ненной окраской в водный или овсяный агар и инкубированием
при 15° (Hickman, Goode, 1953).
Для выделения Ph. syringae Клебан (Klebahn, 1909) исполь-
зует непосредственный посев пораженных частей побегов расте-
ний сирени на агаровые среды, приготовленные на отварах сливо-
вом, морковном или из листьев сирени, а также посев на стериль-
ные ломтики моркови, картофеля, хлеба. Фрецци (Frezzi, 1950)
применяет 1 %-ный картофельно-декстрозный агар. Сьюел
и Уилсон (Sewell, Wilson, 1963) рекомендуют для выделения
Ph. syringae из коры плодовых деревьев такой способ. Вырезан-
ный из пораженной коры блок размером 5 мм3 длительно обра-
батывается водопроводной водой (24—48 ч) и инкубируется при
лабораторных условиях в неглубоком слое дистиллированной
воды в течение 12 ч. Развивающийся мицелий закладывается
в плоды яблок, откуда затем, после образования бурых пятен,
выделяется чистая культура гриба. Тот же вид Ph. syringae с тру-
дом изолируется без сопутствующих бактерий из пораженных
листьев фенхеля, а дезинфекция спиртом или сулемой, так же как
и подкисление среды, часто подавляют рост гриба (Noviello,
Snyder, 1962). Рекомендуется следующая техника выделения
гриба. Пораженные участки листьев, пограничные со здоровыми,
промывают в проточной воде 24 ч и трижды в сменяющейся сте-
рильной дистиллированной воде; затем разрезанные на небольшие
кусочки листья помещают на агар Чапека, кукурузный агар или
агар из фенхеля (листья фенхеля — 200 г, глюкоза — 10 г, ди-
46
стиллированная вода — 1л, агар — 2% с добавлением 20 ppm
пенициллина и 20 ppm стрептомицина) и культивируют при 21°
следует строго соблюдать температурные условия, так как
ph. syringae не растет при температуре выше 25°). Через 4—5 дней
производится пересев кончиков гиф на искусственные среды
или внутрь плодов яблок как дополнительная мера для избавле-
ния от бактерий.
Томпкинс и Такер (Tompkins, Tucker, 1937) при изолировании
Ph. capsici и Ph. drechsleri из плодов дыни переносят асептически
небольшие кусочки тканей из краевой зоны пораженного участка
плода на сусло-агар в чашки Петри. Через 48 ч культивирования
при комнатной температуре гриб, взятый на периферии выросшей
колонии, пересевается в пробирки.
Выделение Ph. cryptogea из стеблей томатов и других растений
(Pethybridge, Lafferty, 1919) осуществляется относительно легко
помещением в асептических условиях небольших кусочков стеб-
лей, дезинфицированных сулемой (1 : 1000, 1 мин) и промытых
стерилизованной водой, в чашки Петри, на питательные среды
(картофельно-глюкозный агар и др.) с последующим внесением
мицелия в пробирки.
Культивирование видов Phytophthora осуществляется в основ-
ном на агаровых средах, причем большинством исследователей
предпочтение отдается картофельно-декстрозной агаровой среде.
Непременное выращивание видов на этой среде — одно из стан-
дартных условий, необходимых для идентификации полученных
данных. Частое применение в работах по культивированию фито-
фторовых грибов находят и другие агаровые среды: овсяная,
фасолевая, гороховая, кукурузная среды, приготовленные на
декоктах из плодов, корней и листьев растений, а также стерили-
зованные и нестерилизованные асептически приготовленные
естественные субстраты.
Правильный выбор питательных сред для изолирования и куль-
тивирования видов Pythium и Phytophthora — основное условие
для экспериментальной работы с этими грибами (Самуцевич,
1931; Наумов, 1937; Новотельнова, Пыстина, 1978, и др ).
При посеве на искусственные селективные среды рекоменду-
ется применять ряд испытанных в экспериментах антибиотиков.
Определенные концентрации (обычно 50—100 ppm) пенициллина,
пирамицина, нистатина, бацитрацина, неомицина, полимиксина,
ванкомицина, к которым мало чувствительны виды Phytophthora,
тормозят развитие почвенных бактерий. Изолированные штаммы
в виде чистых культур должны содержаться в условиях, сохра-
няющих гриб жизнеспособным длительное время (Пшедецкая,
Черепанова, 19616). Хорошие результаты дает хранение гриба
под слоем масла.
Широко применяется дифференциация видов по стойкости
к температурному фактору и к ядам. Вследствие того что виды
Phytophthora обладают различной термоустойчивостью, последняя
47
определяется при их идентификации, для каждого изолята уста-
навливаются критические температурные точки роста гриба на
питательных средах. При определении ядоустойчивости к пита-
тельной среде добавляются определенные концентрации малахито-
вой зелени (1 : 4-106, 1 : 8-106).
Для идентификации видов Phytophthora необходимо изучение
морфологии репродуктивных органов. Образование последних
в природных и искусственных условиях у многих видов отсутствует
или затруднено по тем или иным причинам. В связи с этим пред-
ложены различные методы, способствующие выявлению органов
бесполого и полового размножения.
Отдельные виды Phytophthora, не спороносящие в обычных ус-
ловиях культивирования, переходят к бесполому спороношению
при перенесении гриба с высокопитательной твердой среды на
жидкую, «голодную» — в дистиллированную воду, растворы ми-
неральных веществ, на естественные субстраты.
Образования обилия зооспорангиев у Ph. nicotianae var.
parasitica добиваются выращиванием гриба на субстратах из
люцерны, а также батата (Klotz, Wolfe, 1960; Wong, Varchese,
1966).
С целью образования зооспорангиев у Ph. drechsleri мицелий
культуры трехнедельного возраста с овсяного агара переносят
в раствор Петри, в котором зооспорангии развиваются через 48 ч,
или в нестерильную почвенную взвесь, где образование зооспо-
рангиев происходит раньше (через 24 ч) и в большем количестве
(Tompkins et al., 1936). Формирование зооспорангиев у Ph. erythro-
septica стимулируется помещением мицелия в воду и водные
экстракты соответствующих растений (Pethybridge, 1913). Обиль-
ные зооспорангии у Ph. citrophthora могут быть получены в ре-
зультате заливки чашек Петри с культурой гриба (на картофельно-
глюкозной агаровой среде) водой или раствором KNO3 (0.01 мл)
или путем переноса мицелия с твердой среды в воду или раствор
KNO3 (0.01 мл) при экспозиции 28°; зооспорангии образуются
через 1—2 дня.
Для получения спороношения у гриба Ph. cinnamomi предла-
гается ряд способов: перенос гриба с твердой среды на жидкую
(Pethybridge, Lafferty, 1919), с горохового отвара в дистиллиро-
ванную воду (Leonian, 1934), в почвенные экстракты, которые
стимулируют образование зооспорангиев (Mehrlich, 1935). Зоо-
спорангии гриба образуются на мицелии на агаровых дисках,
помещенных в чашки Петри с почвенным экстрактом. Применя-
ется также культивирование гриба: в нестерильной воде с бро-
шенными в нее семенами конопли, в нестерильной воде из пруда,
разведенной дистиллированной водой, в стерильном растворе
Петри, в нестерильном почвенном экстракте, при контаминации
с бактериями рода Pseudomonas (Manning, Crossan, 1966). Мунро
(Munro, 1945) предлагает способы индуцирования спороношения,
включающие помимо использования обычных твердых сред и сте-
48
рилизованной воды среду с высоким содержанием азота: карто-
фельный агар с добавлением 1%-ного KNO3 (pH 5.7) и экстракт
болотной почвы (pH 4.6). Примерно те же условия для получения
ооспор у Ph. ctnnamomi рекомендуются и другими авторами.
Зооспорангии образуются в изобилии при 24° и в темноте, и при
продолжительном освещении, и при чередовании освещения и
темноты (Zentmyer, Marshall, 1959). А. К. Шишкина и Н. И. Ца-
нава (1973) индуцирование ооспор получали на стерильных мор-
ковных блоках.
Для образования ооспор у видов Phytophthora предлагаются
различные искусственные агаровые среды, приготовленные на
растительных отварах (овсяная, фасолевая, томатная, рапсовая
и др.)» а также естественные субстраты, антисептированные тем
или иным путем. На них в местах соприкосновения колоний гете-
роталличных штаммов формируются оогонии и ооспоры.
С целью индуцирования образования ооспор у видов Phyto-
phthora широко применяется добавление в среды стеринов; наилуч-
ший эффект достигается при помощи бета-ситостерина. Суспен-
зия бета-ситостерина приготовляется растворением его в количе-
стве 30 мг в 4—10 мл СН2С12 и разведением до 100 мл дистиллиро-
ванной и деионизированной водой. Все среды стерилизуются
автоклавированием в течение 20 мин при 1.5 ат.
Индуцирование прорастания ооспор Ph.infestans (Romero, Erwin,
1969) включает следующие манипуляции. Охлажденные в холо-
дильнике ооспоры смывают с агаровых блоков в стерильную де-
ионизированную воду, фильтруют через керамическое сито и осаж-
дают в цилиндре с водой, после чего их переносят пипеткой в чашку
Купера (с низкой крышкой для непосредственного микроскопи-
рования и предотвращения загрязнения) с 4 мл деионизированной
воды. При 20° прорастание наступает обычно через 8 дней и про-
должается неделю и более.
Выделение моноспоровых культур
с помощью микроманипулятора
Для работы необходимо иметь стеклянную камеру для мани-
пулирования; кольца Ван-Тигема; 3 стерильные чашки Петри;
2 стерильные пипетки; чистые предметные и покровные стекла;
стерильный агар в пробирке; стеклянные иглы для манипулиро-
вания (толщина кончика не должна превышать 2 мкм); эксикатор;
петлю для нанесения культуры; спиртовку, спички, пинцет,,
тушь, перо.
Последовательность операций. На верхнюю поверхность по-
кровных стекол тушью наносят точки. Покровные стекла (с точ-
ками и без последних) стерилизуют над пламенем спиртовки и
Помещают в стерильную чашку Петри на кольца Ван-Тигема,
тушью вниз. В другую стерильную чашку Петри помещают на
Дно простерилизованное над пламенем спиртовки предметное
4 H. С. Новотельнова и др. 4 9-
стекло и на него простерилизованную также фламбированием
камеру для манипулирования. Поверхность всех покровных стекол
покрывают разогретым агаром с помощью пипетки. На покровное
стекло без точек наносят петлей водную суспензию из проросших
спор и дают подсохнуть. Ланолином приклеивают камеру к пред-
метному стеклу и на нее пинцетом кладут агаром вниз покровное
стекло с суспензией спор (ближе к кончику иглы) и чистое, с на-
несенными точками. Камеру устанавливают под объективом
микроскопа, вводят в нее иглу таким образом, чтобы кончик был
в центре поля зрения. Объектив фокусируют на нижнюю поверх-
ность агара, чтобы стали видны споры. Иглу располагают ниже
агара, но так, чтобы была видна ее тень. Подцепленную иглой
спору переносят на покровное стекло (одну спору против каждой
точки). Стекла с нанесенными на них спорами приклеивают лано-
лином к крышке чашки Петри, агаром вниз. Чашку помещают
в эксикатор и выращивают колонии.
Видоизмененная техника выделения монозооспоровых куль-
тур, без соблюдения стерильности при культивировании моно-
споровых изолятов не на искусственных, а на естественном суб-
страте, приводится для Phytophthora infestans М. В. Патрикеевой
(1970), для Peronospora tabacina — В. А. Виноградовым (1977).
ИЗОЛИРОВАНИЕ И КУЛЬТИВИРОВАНИЕ
PERONOSPORACEAE И CYSTOPACEAE
Для многих паразитных грибов разработаны методы выделе-
ния и выращивания, предложены селективные среды. Имеющиеся
методы включают культивирование в стерильных условиях на
искусственных средах, круглогодичное культивирование на жи-
вых растениях в нестерильной обстановке, выращивание на орга-
нах растений и их частях в асептических условиях, применение
метода культуры тканей.
Много попыток было сделано исследователями по инкубиро-
ванию облигатных грибных паразитов в стерильных условиях
на синтетических и природных средах. Испытывались стерили-
зованные органы растений, соки и минеральные питательные рас-
творы с добавлением сахаров, микроэлементов, витаминов,
аминокислот, различных стимуляторов роста, растительных
экстрактов, агара.
Так, Пиот (Pioth, 1957) доказывает возможность культивиро-
вания гриба Plasmopara viticola на искусственных средах in vitro.
Сначала автором было использовано несколько вариантов сред,
содержащих соли, органические кислоты, сахара, аминокислоты
и витамины, но роста гриба на них не было получено. Предпо-
ложив, что неудача зависит от отсутствия в средах «точек роста,
аналогичных устьицам» (Haftpunkte), Пиот поместила в них ку-
сочки ипорки (синтетический материал губчатой структуры типа
пенопласта). На питательной среде с ипоркой при внесении орга-
50
нических кислот — винной, яблочной и особенно щавелевой —
автору удалось обнаружить в культуре образование отдельных
*онидиеносцев Р. viticola (состав среды на 1 л: FeCl3-6H2O —
0.027 г, MgSO4-7H2O - 0.24 г, Ca(NO3)2.4H2O - 0.118 гг
КН2РО4 — 0.682 г, анейрин — 10 мкг, бета-индолилуксусная
кислота — 10 мкг, аспарагин — 1г, винная кислота — 200 мкг,
щавелевая кислота — 100 мкг, яблочная кислота — 100 мкг,
лимонная кислота — 10 мкг, 1 мл раствора смеси микроэлемен-
тов; pH 5.4). Конидиеносцы выступают всегда только в местах
соприкосновения ипорки со стенкой сосуда и никогда не наблю-
дались в середине кусочков ипорки. При отсутствии в среде бета-
индолилуксусной или щавелевой кислот рост гриба не наблю-
дался. Наложение коллодиевых или целлофановых пленок для
уменьшения величины пор ипорки с нанесением на них уколон
тонкими иглами не улучшало результатов опыта. Хороший рост
гриба получен, по словам автора, применением метода Зергеля
(маленькие чашки Петри с питательной жидкой средой, покрытой
фильтровальной бумагой, на которую нанесен инокулюм, поме-
щаются в открытом виде в большие чашки Петри), однако при
этом культуры загрязняются бактериями. Пересевы гриба из.
одной чашки в другую удаются, но также всегда сопровождаются
увеличением загрязнения культуры; добавление раствора анти-
биотика небацетина подавляет рост бактерий, но одновременно-
действует угнетающе на гриб.
Наиболее распространенным и простым способом выращивания
облигатных паразитов является длительное культивирование
их на живых растениях в оранжерее с реинфекцией. Мелхасу
(Melhus, 1911) удалось получить таким путем многие генерации
грибов Cystopus candidus, Peronospora parasitica. Метод широко
применяется в настоящее время в работах по изучению иммуни-
тета сельскохозяйственных растений, при отборе селекционного
материала и т. д.
Имеется ряд исследований, показывающих возможность выра-
щивания грибов сем. Peronosporaceae на отдельных органах ра-
стений и их частях в условиях асептики не только в течение
вегетационного периода, но и круглогодично. Исследования по
культивированию гриба Peronospora brassicae провели Гуттенберг
и Шмоллер (Guttenberg, Schmoller, 1958). Не получив положи-
тельных результатов в работе с искусственными средами, авторы
разработали метод выращивания гриба на корне брюквы: корне-
плоды моют теплой водой, дезинфицируют в течение 1 ч (сулема
0«2°/о-ная и сапонин 0.1%-ный), споласкивают дистиллированной
Водой, очищают от кожуры и разрезают на ломтики, которые по-
мещают затем в чашки Петри (без добавления воды). Для приго-
товления инокулюма, не заспоренного посторонними организ-
мами, производится заражение растений, выращенных в стериль-
ной почве. Перед окончанием инкубационного периода листья
срезают, дезинфицируют теми же химикатами в течение 1 минг
4*
51
споласкивают многократно водой и закладывают во влажные
камеры. При 16—18° через 18 ч образуется обильное спороношение.
Заражение ломтиков брюквы осуществляется при помощи конидий.
Инокулюмом может служить и мицелий. После инокуляции через
4—5 дней на ломтиках наблюдается образование конидиеносцев,
а потом и воздушного мицелия.
Петрлик и Штыс (Petrlik, Stys, 1962) разработали метод куль-
тивирования гриба Р. ( = Pseudoperonospord) humuli — возбуди-
теля пероноспороза хмеля — на черенках хмеля. Последние за-
готавливают на зиму и оставляют в питомнике прикрытыми поч-
вой. По мере надобности их отмывают от приставших почвенных
частиц и хранят на протяжении 10—14 дней в холодном месте при
6—8°. Перед инокуляцией черенки очищают от коры и из них
готовят небольшие блоки с сохранением у подкорковых тканей
сосудистых пучков и сердцевины. Блоки погружают в суспензию
зооспор гриба, после чего помещают на влажную фильтроваль-
ную бумагу в чашки Петри и ставят в термостат. При 20° и 95—
100% влажности через 4—5 дней сначала на подкорковых тканях,
а впоследствии на всей поверхности блока появляются отдельные
группы конидиеносцев, которые постепенно сплетаются в виде
густого налета серого цвета. Возникающие зооспорангии служат
для приготовления суспензии — инокулюма для новых блоков.
Гриб в культуре сохраняет свойства, присущие ему в естествен-
ных условиях, и способен с той же агрессивностью заражать
листья хмеля даже после 6-месячного культивирования в лабо-
раторных условиях при осуществлении повторных пассажей
через каждые 6—8 дней. Таким образом гриб культивируется
в течение всего года.
Имеется несколько работ по культивированию гриба Р. taba-
cina Adam — возбудителя пероноспороза табака. Необходимость
иметь под рукой жизнеспособные споры этого гриба в достаточном
для заражения растений количестве побудило исследователей
Вольфа и Мак-Лина (Wolf, McLean, 1940) разработать метод полу-
чения гриба in vitro. Проростки табака выращиваются в стеклян-
ных сосудах емкостью около 1.5 л, содержащих 500 г стерилизо-
ванной паром почвы. Инокуляция проводится распылением вод-
ной суспензии спор, после чего сосуды с растениями закрываются
завинчивающимися крышками и ставятся в инкубационную ка-
меру, в которой поддерживается температура около 15° и постоян-
ное освещение (25 вт). На инокулированных растениях возникает
спороношение, однако необычные условия выращивания (высокая
влажность и длительное освещение) изменяют нормальный су-
точный ритм продукции конидий и вызывают необычный для гриба
морфогенез.
Работа Изара (Izard, 1960) была предпринята с целью раз-
работки метода культивирования того же гриба в лаборатории,
позволяющего изучать неясные вопросы его биологии, испытывать
фунгициды, уточнять способ их воздействия на гриб, а также изы-
52
скивать устойчивые к патогену виды и сорта растений. Изар вы-
резал из листьев испытуемых растений Nicotiana glutinosa и
ДО. tabaccum var. barley кусочки в виде дисков диаметром 12—
14 мм. Для получения более тонких и просветленных тканей
опытные растения, служащие для изготовления дисков, выра-
щивают при ослабленном освещении. При подготовке к инокуля-
ции диски раскладывают по 15 штук нижней стороной кверху
в чашках Петри на 3 слоя фильтровальной бумаги, пропитанной
питательным солевым раствором (состав, °/0: виннокислый нат-
рий — 0.5, фосфорнокислый калий двухзамещенный — 0.1,
азотнокислый калий — 2, азотнокислый кобальт — 0.006, хло-
ристый калий — 0.03, сернокислый марганец — 0.001, серно-
кислый магний — 0.05), а затем инокулируют обычными спосо-
бами (нанесением нескольких капель дистиллированной воды на
нижнюю поверхность пораженных листьев табака со свежим нале-
том, а отсюда с помощью стеклянной иглы на инокулируемые
диски или снятием сухих спор с листа и непосредственным внесе-
нием их в капли воды на диски). Условия проведения опытов
следующие: температура днем 20°, ночью 10—15°, освещение днев-
ное, чашки устанавливаются на окне,, ориентированном на север.
После 24 или 48 ч инфекционные капли с дисков отсасывают при
помощи тонкой полоски фильтровальной бумаги, после чего диски
переворачивают нижней стороной вниз, к бумаге, насыщенной
питательным раствором. Внутри тканей листа мицелий хорошо
развивается уже на 4-й день культивирования и благодаря про-
зрачности тканей настолько заметен при микроскопировании, что
не требует изготовления срезов и применения окраски. Однако
вскоре диски начинают желтеть и на 7-е сутки некротизируются
и засыхают.
Шеферд, Стюарт и Мандрик (Shepherd et al., 1963) усовершен-
ствовали метод Изара, который при проверке не дал им желае-
мого результата в отношении длительности наблюдения над раз-
витием гриба. Модификация заключается в добавлении к среде
водного раствора кинетина (1 мг/мл). На этой среде неинокули-
рованные диски остаются зелеными и внешне здоровыми в течение
3 недель, тогда как на среде без кинетина желтеют на 5—7-й день.
Инокуляция дисков осуществляется нанесением в каплю воды
в центре диска 2-102 конидий гриба, инкубация — при 20° и
световом режиме, обеспечивающем 16 ч света (флуоресцентная
лампа 800 вт) и 8 ч темноты. На инокулированных дисках через
двое суток появляются некрозы, на 6—7-й день — спороношение;
конидиеносцы возникают только на зеленых частях дисков и не
наблюдаются в зоне некроза. В последующие дни количество
конидиеносцев возрастает. Прибавление к раствору кинетина,
параамидобензойной и никотиновой кислот удваивает образова-
ние конидиеносцев. Авторы использовали этот метод для выделе-
ния моноспоровых клонов гриба Р. tabacina. Каждая выделенная
спора переносилась на диск в каплю среды; клоны успешно пас-
53
сировались 3 раза, однако наблюдалась частичная потеря жизне-
способности гриба, проявляющаяся в редукции числа конидиенос-
цев при каждом последующем пассаже. Для преодоления этого»
явления авторы рекомендуют пассировать первый моноспоровый
изолят через лист растения табака, прикрепляя диск к поверхности
листа при помощи тонкого слоя 1%-ного дифко-агара; мицелий
из диска быстро врастает в лист и на последнем образует обильное
спороношение. Предлагаемая авторами техника дает возможность
изоляции и поддержания клоновых линий патогена и изучения
физиологических рас гриба. Тем же методом проведено исследова-
ние воздействия 148 метаболитов на споруляцию Р. tabacina
(Shepherd, Mandryk, 1964).
Сходный метод предложен Лехоцким (Lechoczky, 1965) для
возбудителя мильдью винограда. Листья винограда промыва-
ются химически чистой водой, просушиваются и из них буравчи-
ком вырезаются диски 18 мм в диам. Они раскладываются по-
10 штук в чашки Петри на поверхность воды, тоже химически
чистой (очищенной в ионообменнике), налитой по 50 мл в каж-
дую чашку, инокуляция проводится нанесением на диски 5 капе-
лек густой суспензии (5-103 зооспорангиев в 1 мл). Опыт прово-
дится при комнатной температуре (19—25°) при рассеянном осве-
щении. Образование оогониев и ооспор начинается на 9—12-й
день, они обнаруживаются в 75% дисков. В участках листьев:
с ооспорами не наблюдается бесполого спороношения.
Вышеуказанными работами не предусматривались стериль-
ное изолирование паразитов и выращивание их в виде чистых
культур, но разработанные методы оказались пригодными для
исследовательских целей.
Большим научным достижением в области изучения облигат-
ных паразитов было установление возможности выращивания
их в стерильных условиях на изолированных тканях.
Пионером в этом деле является французский ученый Морель (Morel,
1948), который, использовав методы Карреля (A. Carrel) с живот-
ными тканями, Уайта (Ph. R. White) и Готре (R. Gautheret) —
с растительными, занялся разработкой способа культивирования
облигатных паразитов на каллусах.
Перед Морелем, когда он начинал исследования, стояла двой-
ная задача: выращивание ткани растения-хозяина и осуществле-
ние ее инокуляции. Проблема реализации «ассоциированных
культур», как их называет Морель, в то время не была исследована;
имелись лишь работа Уайта по культивированию вируса табач-
ной мозаики на тканях корня томата и работа Сегретэна (G. Segre-
tain) по выращиванию того же вируса на тканях растений табака.
Морель блестяще справился с поставленной задачей. Он вырастил
изолированные ткани целого ряда растений-хозяев тех облигат-
ных паразитов, которых собирался культивировать, разработал
способы асептической изоляции этих грибов, провел инокуляцию
54
культур изолированных тканей и осуществил многочисленные
пассажи грибов-паразитов в чистой культуре. Все исследования
подробно освещены в его трудах, с изложением мельчайших дета-
лей методического характера и полученных результатов (1944—
1948 гг.).
Техника культуры in vitro была применена Морелем для
plasmopara viticola и некоторых облигатных паразитов.
Основной интерес представляет описание метода работы с куль-
турой гриба Р. viticola. Подготовку инокулюма для заражения
изолированных тканей винограда проводят следующим образом.
Листья винограда с «маслянистыми» пятнами без спороношения
или со слабым налетом быстро споласкивают 95%-ным спиртом,
погружают на 10 мин в 7%-ный раствор гипохлорита кальция,
после чего прополаскивают в воде и помещают в стерильные чашки
Петри между двумя слоями влажной фильтровальной бумаги.
Через 24 ч при 20° пятна покрываются конидиеносцами, с которых
можно снять асептически зооспорангии. Таким же способом гриб
был выделен и из кусочков пораженных ветвей после стерилиза-
ции их гипохлоритом кальция, удаления коры и инкубирования;
через 8—10 дней эти срезы ветвей, помещенные на желатиновую
среду, покрываются спороношением. Для эффективного заражения
тканевых культур Морелем были испытаны разные варианты ме-
тодов инокуляции. Наиболее частые случаи бактериального
загрязнения были при помещении зооспорангиев вместе с кусоч-
ками листьев в воду. Автор предпочитает сепарирование зооспо-
рангиев с листьев, оставленных в течение 1.5 ч в воде до образо-
вания зооспор, с последующим нанесением капли взвеси на ткань;
другой предпочитаемый вариант опыта — помещение капли воды
на ткань и внесение в нее зооспорангиев. Для снятия спор с листьев
Морель использует пипетки Пастера с очень тонким отломанным
концом. Каплю стерильной воды с поверхности листа со спороно-
шением оттягивают при помощи пипетки и переносят в стерильную
пробирку. Для инокуляции берется примерно 250 000 зоо-
спорангиев в 1 мл. Через 1.5 ч после выхода зооспор другой пипет-
кой суспензию наносят на ткань. Для того чтобы капля не скаты-
валась, на ткани делают скальпелем небольшую выемку. Ткани
меньше загрязняются посторонними микроорганизмами в тех
случаях, когда зооспорангии вносят в капли стерильной воды,
нанесенные заранее на ткань.
Первое появление спороношения возможно в культурах не
ранее чем через 12 дней, хотя в природных условиях инкубацион-
ный период продолжается всего 5—6 дней. Начало развития споро-
ношения наблюдается всегда на месте инокуляции, но затем кони-
диеносцы появляются на всей поверхности ткани, что происходит
в течение 1—1.5 мес. При инокуляции тканей, пассированных
много раз, инкубационный период становится длиннее, чем при
посеве гриба на первичные каллусы, кроме того, и вероятность
заражения в первом случае меньше (30—4О°/о), чем во втором
55
(60—70%), да и налет в этом случае менее густой. Таким образом,
ткани растения-хозяина, многократно пассированные, более
устойчивы по отношению к паразиту, чем первичные каллусы.
Культура гриба, даже многократно пересеянная, длительно со-
храняет жизнеспособность.
На срезах каллуса, пораженного грибом, обнаруживается
присутствие хорошо развитого мицелия с гифами 8—10 мкм толщ.,
сильно разветвленными, проходящими по межклетникам парен-
химной ткани, с большим количеством гаусториев шаровидной или
грушевидной формы, не отличающихся от гаусториев, образую-
щихся на мицелии в тканях винограда в естественных условиях.
Так как ткань без кутикулы, то гифы часто выходят на ее поверх-
ность в виде боковых ветвей, достигающих 500 мкм дл. Никогда
не наблюдалось, чтобы они вросли в питательную среду. Споран-
гиеносцы возникают только на внутриклеточном мицелии и ни-
когда на воздушном. Они выступают группами по 3—4 между
клетками ткани. Их форма и размеры сильно варьируют. На пер-
вичных каллусах они достигают 1000 мкм дл. и многократно раз-
ветвлены, тогда как на пересеянных каллусах не превышают
250 мкм дл. и мало разветвлены. Половые органы и ооспоры в ино-
кулированных тканях не наблюдались.
Постепенно под влиянием гриба изолированная ткань расте-
ния-хозяина начинает отмирать. Это происходит обычно через
1.5 мес, когда гриб заселяет всю культуру. Чтобы сохранить гриб
в живом состоянии, Морель предлагает следующий способ пере-
сева. Кусочек пораженной ткани срезают и помещают в стериль-
ную чашку Петри; с ее поверхности скальпелем срезают тонкие
(не более 1 мм толщ.) пластинки с конидиеносцами, которые осто-
рожно переносят пинцетом на свежий каллус в каплю воды, зара-
нее на него нанесенную. Мицелий на вырезанном кусочке остается
некоторое время живым и продолжает давать спороношение,
благодаря чему заражение свежего каллуса удается легко. Тех-
ника пересева позволила Морелю поддерживать гриб в культуре
в течение 3 лет.
Метод Мореля с некоторым видоизменением способа инокуля-
ции каллусов использовался Бубальсом (Boubals, 1959) в моно-
графической работе по иммунитету винограда к мильдью. Он при-
менил наряду с другими методами исследования и культуру тка-
ней. Автор исходил из того, что изолированные ткани в культуре
не имеют ни эпидермиса, ни устьиц, что позволяет изучать непо-
средственно реакцию клеток растения-хозяина на внедрение рост-
ков зооспор, развитие мицелия и образование на нем гаусториев.
Различия в устойчивости тканей растений отдельных видов, гиб-
ридов и сортов проявлялись в опытах Бубальса в неодинаковой
быстроте роста мицелия внутри ткани и на ее поверхности, в раз-
ном характере гиф, выходящих наружу, а также в сроках и коли-
честве образующихся спорангиеносцев.
56
Технику Мореля применил и Накамура (Nakamura, 1965) при
культивирования in vitro гриба Peronospora brassicae. Для куль-
туры ткани турнепса он использовал специальную среду:
Ca(NO3)2 - 500 мг, MgSOr7H2O - 125 мг, КН2РО4 - 125 мг,
KNO3 — 125 мг, FeCl3-6H2O — 1 мг, ZnSO4-7H2O — 1 мг,
Н3ВО3 — 1 мг, MnSO4-4H2O — 0.1 мг, CuSO4.5H2O — 0.03 мг,
KI — 0.01 мг, тиамин — 1 мг, а-нафтолуксусная кислота —
0.5—1 мг, глюкоза — 30 г, агар-агар — 9 г, вода дистиллиро-
ванная — 1 л. Из здорового корня турнепса вырезают небольшие
кусочки (размером около 1 см3), которые затем культивируются
в пробирках, содержащих по 1 мл агаровой среды. Каллусы растут
20—30 дней, после чего инокулируются. Для получения иноку-
люма конидии гриба переносят с листьев вначале на ломтики тур-
непса, размещенные в чашках Петри (инокулюм, взятый непо-
средственно с листьев, содержит постороннюю микофлору).
Ломтики с 7-дневным налетом опускают в раствор сулемы (0.1%)
на 15 мин, после чего удаляют верхнюю часть ломтика. На сле-
дующий день гриб снова спорулирует на свежей поверхности лом-
тика, и тогда на нее наносят каплю стерильной воды, которую
затем оттягивают пипеткой и переносят на каллус. Гриб разви-
вается внутри каллуса, образуя мицелий с характерными гаусто-
риями, и на поверхности — спороношение. При помощи такого
способа автор многократно пассировал гриб и поддерживал его
в культуре in vitro.
Гриффин и Кулей-Смит (Griffin, Coley-Smith, 1968) получили
тканевые культуры Humulus lupulus L. и рост на них гриба
Pseudoperonospora humuli. Заражались ткани стебля, выращенные
на твердой D-среде Гильдебрандта. Для приготовления инокулюма
использовались дезинфицированные с поверхности отрезки стебля
или черешков листьев хмеля, диффузно пораженных грибом
Р. humuli; во влажной камере на них обильно возникали конидие-
носцы и воздушные гифы, служившие для инокуляции каллусов.
Внутри каллуса развивался межклеточный мицелий с гаусториями,
а на поверхности — воздушный спороносящий и бесплодный ми-
целий. Наблюдалось также врастание мицелия внутрь питатель-
ной среды в местах ее соприкосновения с каллусом. Попытки
получения культуры гриба вне тканей остались безрезультатными.
В связи с исследованиями по ложной мучнистой росе подсол-
нечника Н. С. Новотельнова в 1965 г. изыскивала методы, поз-
воляющие изучить морфогенез и биологию возбудителя болезни —
гриба Plasmopara heliantht f. heltanthi. Особенностью гриба явля-
ется сильно выраженный полиморфизм с образованием в цикле
развития двух разных типов спороношения: на корнях (|3) и на
листьях (а). На листьях гриб имеет характер, типичный для рода
Plasmopara, тогда как на корнях спороношение обнаруживает
сходство со спороношением рода Phytophthora. Проведенные опыты
с искусственным заражением растений спорами того и другого
типа показали способность их к одновременному развитию в жиз-
57
ненном цикле гриба (см. рис. 25) и принадлежность к одному
и тому же организму. Подтверждение этому получено и в опытах
по культивированию гриба Plasmopara helianthi в стерильных
условиях на проростках подсолнечника. Разработанные приемы
культивирования гриба (Новотельнова, 1965, 1966а, 19696) поз-
волили убедиться в том, что оба спороношения генетически свя-
заны. Инокулюм, изолированный с листьев (а-спороношение)
и нанесенный на выращенные в пробирках в стерильных условиях
проростки подсолнечника, вызывает заражение последних и обра-
зование на 5—6-й день после инокуляции на семядольных листьях,
стебле и корнях спороношение другого типа ([3-спороношение).
Наши данные были позднее подтверждены в исследовании
Р. helianthi (Бе1апоё, Hamant, 1972) при культивировании на кал-
лусе, выращенном из гипокотиля подсолнечника. Гриб развивался
в виде (3-спороношения на каллусе и вокруг него в пределах до
1 см на модифицированной среде Уайта, но не был способен к даль-
нейшему росту при пересевах.
Рассмотрение данных, полученных исследователями при помощи
новых методов, показывает,что лабораторное культивирование обли-
гатных паразитов позволяет более точно, чем допускают исследова-
ния в природных условиях, установить закономерности биологии
этих организмов. Нужна дальнейшая разработка методов культи-
вирования облигатных паразитов, что должно способствовать
совершенствованию наших представлений об онто- и филогенезе
этих грибов, а также служить практическим целям, обеспечивая
возможность изучения механизма многих сложных явлений, таких
как иммунитет, реакций на яды и другие факторы окружающей
среды.
ИСКУССТВЕННОЕ И ЕСТЕСТВЕННОЕ ЗАРАЖЕНИЕ
Инокуляция — искусственное заражение растений — необхо -
димый метод исследования, всегда применяемый при изучении
фитопатогенных грибов, и в частности пероноспоровых, без ко-
торого невозможно установление биологии видов, их онтогенеза,
специализации. Он служит также чисто практическим ^елям —
определению зависимости развития видов от условий среды и
на этой основе их прогнозированию, испытанию защитных меро-
приятий, выявлению устойчивых форм культурных растений.
Инокуляция растений несложный процесс, однако для перо-
носпоровых грибов требуется особый подход. Это вызвано тем,
что Peronosporales, являясь земноводными организмами, в цикле
развития которых имеется водная стадия, представленная зооспо-
рами, особенно чувствительны к условиям влажности. Одновре-
менно они проявляют высокую чувствительность и к температуре,
быстро теряя жизнеспособность при повышении последней.
При инокуляции готовится суспензия спор (конидий, зооспо-
рангиев) в воде. Для получения зооспор суспензию выдерживают
58
предварительно при пониженной температуре. При испытании
различных инфекционных нагрузок проводится точный подсчет
взвешенных спор в жидкости определенного объема. Это осуще-
ствляется в специальных счетных камерах различных типов, при-
меняемых в работах с подсчетом спор любых грибов (Наумов,
1932).
Прежде всего при инокуляции проверяется жизнеспособность
зооспорангиев (конидий). Изучение их жизнеспособности, а также
особенностей прорастания проводится и для диагностики, и как
необходимый этап исследований биологии видов, и с целью испы-
тания подавляющего воздействия на патогены различных физи-
ческих и химических факторов.
Прорастание пропагативных спор изучалось у Phytophthora in-
festans (Новотельнова, 1937), Cystopus candidus (Новотельнова, Ми-
насян, 1970), Peronospora destructor (Каттерфельд, 1926; Талиева,
1968, и др.), Р. tabacina (Пересыпкин, Зражевская, 1967; Бабаян,
Григорян, 1970; Бебия, 1975, и др.), Plasmopara helianthi (Ново-
тельнова, 1966а и др.). С. Ю. Станявичене (1970) изучала про-
растание конидий у 24 видов пероноспоровых грибов. А. К. Вя-
лых и В. Г. Жерягин (1977) провели большое исследование
прорастания зооспорангиев Cystopus tragopogonisc, применением ма-
тематической обработки результатов опытов. Н. С. Новотельнова
(1937, 19666) выявила двувершинную кривую прорастания зооспо-
рангиев Phytophthora infestans, что связано со способом их про-
растания: образованием зооспор или мицелиальных трубок. Тем-
пературные пределы для последних более широкие.
Экспериментальное изучение процесса прорастания пропага-
тивных спор пероноспоровых грибов дается с большим трудом.
Это зависит чаще всего от возраста спор, а иногда от среды про-
растания — воды, температурных условий и т. п. Обычно мы ис-
пользуем только свежесобранный материал из спороношения од-
нодневного возраста, вода — дождевая, речная, водопроводная,
простая или стерильная; при затрудненном прорастании хорошие
результаты дает роса, собранная в утренние часы с листьев рас-
тений. В воду для стимуляции прорастания рекомендуется по-
мещать кусочки тканей специфического хозяина. Суспензия спор
готовится нами в маленьких бюксах с небольшим количеством
воды или в висячих каплях на покровных стеклах, отделенных
от предметных не стеклянными кольцами, прикрепленными вазели-
ном, а несколькими слоями влажной фильтровальной бумаги разме-
ром с площадь покровного стекла и с вырезанным внутри кругом.
В таких камерах исследуется зависимость прорастания спор от
температурных условий, освещенности, возраста спор и других
факторов. Важнейшей задачей является установление кардиналь-
ных для прорастания температур — максимальной, минимальной,
оптимальной, способа прорастания, продолжительности жизнеспо-
собности спор, подвергнутых воздействию обычных природных
Условий.
59
Известно очень много вариантов инокуляции растений приме-
нительно к разным объектам. Ниже даются выборочно некоторые
из них, которые должны послужить основой для последующей
разработки и усовершенствования.
Для инокуляции растений грибами из рода Pythium суще-
ствует несколько методик. В пробирку с культурой гриба, вы-
ращенного на картофельно-глюкозном агаре, вливают 10 куби-
ков воды, сильно взбалтывают и поливают этой суспензией почву
под всходами или субстрат, на котором проращивают семена (1 про-
бирка на 1 горшок). После 2-1— 3-разового полива можно увлаж-
нять почву или субстрат обычной водой. При внесении культуры
гриба непосредственно в почву (лучше стерильную) под семена
или молодые растения инокулюм выращивают в чашках Петри
на тонком слое питательной среды. Вносят гриб из 1 чашки Пе-
три в 2—3 горшка. При достаточном увлажнении субстрата и
оптимальной температуре (для многих патогенных видов рода Py-
thium 15—20° С) питиевой корневой гнилью поражается более
50% растений.
В работах по исследованию видов Phytophthora испытаны и
описаны разные приемы инокуляции. Из них менее известны спо-
собы работы с видами, паразитирующими на древесных растениях.
Для инокуляции растений грибом Ph. cinnamomi (Munro, 1945)
изоляты гриба выращивают на картофельном агаре, на стериль-
ных раздробленных зернах овса или пшеницы. Растения выби-
вают из горшков (по возможности предотвращается повреждение
корней), и инокулюм прикладывают сбоку к земляному кому
в 3—5 местах возле каждого растения, после чего их снова по-
мещают в горшки. В теплицах соблюдается благоприятный для
развития гриба режим температуры и влажности. Успешность
применения метода подтверждается результатами опытов: увядает
более 50% растений, первые симптомы отмечаются через неделюг
полное развитие заболевания — через месяц после иноку-
ляции.
Для изучения патогенных свойств отдельных видов Phyto-
phthorai^QKy^^yiGT клубни картофеля, которые тщательно отмы-
вают от приставших почвенных частиц, после стерилизации спир-
том на их поверхности в трех местах надсекают кожуру. Под
надсеченную кроющую ткань каждого клубня подкладывают ино-
кулюм только одного изолята, причем мицелий помещают вместе
с небольшим кусочком агара (контролем служат неинокулирован-
ные надрезы). Клубни содержат во влажных камерах под стеклян-
ными колпаками при 24°. Обычно через 6 дней клубни в местах
заражения буреют и загнивают. Камерун (Cameron, 1960, 1962а)
предлагает инокулировать изолятами гриба Ph. cinnamomi сеянцы
грушевых деревьев, выращиваемых на аэрируемом питательном
растворе при температуре 18° днем и 13° ночью, что обеспечи-
вает хорошее развитие корневой системы. Количество инокулюма
на сосуд — суспензия мицелия, снятого с питательной среды из
60
двух чашек Петри. Симптомы появляются на 10—28-й день после
инокуляции.
При инокуляции видами Phytophthora древесных и кустарни-
ковых пород черенки или 1—2-летние саженцы (Atkinson, 1965)
помещают в лабораторных условиях при 18—27° в глиняные или
пластмассовые горшки со стерилизованной почвой. Инокулюм го-
товят на гороховом отваре (150 г на 1 л воды) в колбах (объемом
250 мл) и закладывают в почву рядом с корнями каждого расте-
ния; условия повышенной влажности создаются полиэтиленовым
покрытием и помещением горшков в воду.
Разработан метод инокуляции деревьев и ветвей грибом
Ph. cactorum: срезанные ветви толщиной 3—4 см и длиной 15—20 см,
заделанные с концов растопленным парафином, заражают грибом
путем закладки его на агаровых блоках в просверленные в коре
глубокие отверстия, достигающие камбия.
При заражении листьев фенхеля грибом Ph. syringae (Novi-
ello, Snyder, 1962) рекомендуется агаровые диски (5 мм в диам.)
с мицелием, вырезанные с края активно растущей колонии и
наложенные на пластинку листа, покрывать сверху ватой, смо-
ченной стерильной водой для защиты от высыхания. Заражение
удается при предварительном поранении листа стерильным скаль-
пелем. Инокуляция осуществляется также нанесением на листья
суспензии зооспор (принимаются меры предосторожности для пре-
дотвращения высыхания капель в течение 36 ч); опыт проводится
при температуре 15°; симптомы появляются на 3—8-е сут-
ки.
Иванчева-Габровска (1968; Иванчева-Габровска, Димитров, 1968)
для искусственного заражения растений табака грибом Ph. ni-
cotianae рекомендует внесение под растения предварительно сте-
рилизованной почвы, за неделю до этого зараженной 10—15-днев-
ной культурой гриба, выращенной на кукурузно-песчаной среде;
затем производится ежедневный полив для обеспечения необхо-
димой влажности. Один из способов инокуляции в водной куль-
туре саженцев цитрусовых грибом Ph. nicotianae (Klotz, Wolfe,
1958, 1960) и модификация его (Wong, Varchese, 1966) состоят
в выращивании растений в сосудах типа аквариумов с особыми
приспособлениями для равномерного распределения инокулюма,
вносимого в минеральный питательный раствор. Другая техника
инокуляции цитрусовых деревьев грибом Ph. nicotianae (White-
side, 1971) включает изготовление водонепроницаемого воротника
трубчатой формы высотой 30 см, толщиной около 15 мм с внутрен-
ним диаметром 25 см и наложение его на основание ствола. После
закрепления воротника на месте шов и основание ствола зали-
ваются смесью парафина с вазелином (1 : 1) и внутрь вносится
инокулюм в виде 10 мл суспензии, содержащей на 1 мл 0.5-104—
8.0-104 зооспор. Для инокуляции более толстых стволов водо-
непроницаемый воротник наполняется до краев суспензией в коли-
честве 200—400 мл, содержащей 1.0-104 зооспор, или почвой
61
с инокулюмом. Воротник оставляют на стволе в течение 2—3 су-
ток.
Для заражения земляники грибом Ph. fragariae (Hickman,
English, 1951) инокулюмом служат диски, вырезанные из края ак-
тивно растущей колонии гриба на бобовом агаре, которые на-
носят на корни непосредственно под поверхностью почвы. При
регулярном поливе заметное поражение корней наблюдается че-
рез 3 недели. Тест — длина корня. Инокулюм можно готовить
также в виде культуры гриба на овсяно-песчаной смеси, которая
вносится в почву под растения трижды: в начале опыта, на
второй и четвертой неделе. Соблюдаются условия повышенной
влажности и ежедневного освещения при температуре 10—13°. Ви-
доизмененный способ заключается в выдерживании выращенных
в стерильной почве инокулированных растений в течение 3 не-
дель в воде со сменой ее каждые 4 дня; тест — покраснение тка-
ней корня и образование в них оогониев.
При изучении Ph. megasperma var. sojae Кауфман и Гердеман
(Kaufmann, Gerdemann, 1958) установили, что наиболее эффек-
тивны внесение инокулюма на семенах овса и инокуляция про-
ростков сои нанесением мицелия на гипокотили растений. Джон-
сон и Килинг (Johnson, Keeling, 1969) рекомендуют следующие
способы инокуляции: прокол стебля под семядольными листьями
10—14-дневных сеянцев копьевидной иглой, предварительно по-
груженной в полужидкий субстрат с 10—14-дневной культурой
гриба; внесение инокулюма в верхний слой стерилизованной па-
ром почвы (гриб выращивается на стерилизованных семенах овса,
растения инкубируются при 21°).
Инокуляция растений томатов грибом Ph. cryptogea (Pethy-
bridge, Lafferty, 1919) успешно осуществляется при помощи вне-
сения инокулюма, приготовленного из 6-дневной культуры гриба,
в небольшие ранки, сделанные скальпелем в стебле на уровне
почвы, с последующей обвязкой этой части стебля.
Искусственное заражение растений грибами из семейств Рего-
nosporaceae и Cystopaceae проводится с использованием свежего
материала и в оптимальных условиях для прорастания спор от-
дельных видов пероноспоровых грибов.
Много разных вариантов инокуляции предложено в исследо-
ваниях биологии Plasmopara viticola, Р. helianthi, Peronospora ta-
bacina. Cystopus candidus, C. tragopogonis и др. Нами приводятся
отдельные примеры.
Методика искусственного заражения винограда видом Plasmo-
para viticola была детально разработана многими исследователями.
Заражение лучше удается с нижней стороны листа. Материалом
для инфекции служат молодые конидии, которые образовались
в ночь, предшествующую заражению. Старые конидии имеют по-
ниженную жизнеспособность и прорастают хуже. Взвесь конидий
рекомендуется держать только в стеклянной посуде, а не метал-
лической, так как конидии очень чувствительны к солям тяжелых
62
металлов. Суспензию конидий надо распылять как можно тоньше,,
температуру во все время опыта поддерживать в пределах 20—25°.
Для проведения опытов в течение всего года надо иметь по-
стоянно вегетирующие растения для их инокуляции спорами гриба.
Для инокуляции можно также использовать отдельные изолиро-
ванные листья или высечку из них.
Кроме того, инокуляция проводится погружением испытуемого
объекта в суспензию. Приготовляют взвесь из свежих зооспоран-
гиев в дождевой или дистиллированной воде. Из листьев готовят
высечки определенного диаметра с помощью сверла для пробок. Эти
высечки окунают во взвесь и выдерживают в ней 3 ч при темпе-
ратуре около 20°. Предпочтительно погружать во взвесь со сфор-
мированными уже зооспорами, при этом экспозиция сокращается
до 1 ч. Затем высечки листьев извлекают из взвеси и помещают
во влажные камеры (чашки Петри) нижней стороной листа вверх.
В течение инкубационного периода фильтровальную бумагу в чаш-
ках необходимо смачивать.
В природных условиях благоприятны для естественного за-
ражения виноградной лозы продолжительный (до 10 ч) обиль-
ный дождь, смачивающий почву и листья, и температура
у поверхности почвы не ниже 11°. Наиболее восприимчивы к зара-
жению самые молодые листья размером не более 2—3 см2. За-
ражение осуществляется в течение 3—5 ч в ночное и утреннее-
время. Ягоды в раннем возрасте легко заражаются, но потом их
устойчивость повышается.
Различные способы инокуляции растений предложены в рабо-
тах с Р. helianthi,
В работе по созданию устойчивых к ложной мучнистой росе-
гибридов и сортов подсолнечника Г. В. Пустовойт использует
при их оценке высокую инфекционную нагрузку. Применяемый;
инокулюм — 0.5 млн зооспор на одно растение; заражение про-
изводится в фазе «коленца» при 16—19° и влажности воздуха 100%.
Метод заражения, разработанный А. Я. Панченко (1965), поз-
воляет проводить оценку не только в летний, но и в зимний пе-
риод в условиях теплицы. Семена испытуемых растений высе-
ваются в почву, заложенную в ящики размером 65x45x10 см,
рядками на расстоянии 2 см между растениями и 6—8 см между
рядками. В качестве контроля высевается 1 ряд поражаемого
сорта. Для приготовления суспензии в химический стакан ем-
костью 0.5 л наливается на дно вода и помещается 10 пораженных
ложной мучнистой росой растений раннего возраста. Стакан за-
крывается сверху крышкой. После образования на листьях на-
лета (через 12—14 ч) споры снимаются кисточкой в воду в отдель-
ный сосуд, суспензия доводится до объема 2 л и ставится при
13—15° на 1.5—2 ч для проращивания зооспорангиев. Оценивае-
мые растения опрыскиваются суспензией в период выхода под-
семядольного колена проростка на поверхность почвы, что про-
исходит обычно на 4-й день после посева при 25—26°. 2 л взвеси
63
спор достаточно для обработки проростков в числе 80 образцов
(10 ящиков). Опыт проводится при 16—18°. Через 10 дней ящики
покрывают полиэтиленовой пленкой для выявления налета. Метод
позволяет круглогодично проводить строгую браковку селекцион-
ного материала.
Для отбора форм, устойчивых к ложной мучнистой росе под-
солнечника, В. Ф. Кукин (1965) применяет метод заражения
2—3-дневных проростков зооспорами с последующей посадкой их
в грунт. Автор считает, что метод эффективен при проведении
работ по иммунологической оценке подсолнечника.
В. А. Молдаван и В. И. Живило (1965) испытывали 2 способа
заражения подсолнечника ложной мучнистой росой. В первом
случае проросшие семена выдерживались в суспензии зооспор
в течение 12—14 ч при 17—20° и в темноте, после чего их выса-
живали в пикировочные ящики (размером 50x40x15 см) или
в поле. Спустя 6—8 дней проводился подсчет заболевших расте-
ний. Для выявления спороношения растения обильно поливали
водой и на ночь покрывали полиэтиленовой пленкой. Во втором
случае семена подсолнечника высаживали в почву по 100 штук
в пикировочные ящики и появившиеся всходы заражали суспен-
зией зооспор. Оба способа обеспечивали высокий процент зара-
жения растений.
Дальнейшая разработка методов заражения подсолнечника тем
же видом возбудителя представлена в ряде последующих работ
(Goosen, Sackston, 1968; Cohen, Sackston, 1973,1974; Viranyi, 1977).
При изучении специализации Cystopus candidus применяют те
же принципы искусственного заражения, что и в работах с пред-
ставителями сем. Peronosporaceae (Новотельнова, Минасян, 1970).
Инокулюм готовят в виде суспензии спор в дождевой воде. Для
инокуляции растений используют материал, содержащий не ме-
нее 40% проросших зооспорангиев. Инокуляцию проводят в ве-
черние часы, для того чтобы заражение осуществилось ночью,
когда условия температуры и влажности наиболее благоприятны
для сохранения жизнеспособности зооспор.
А. К. Вялых и В. Г. Жерягин (1977) разработали метод ино-
куляции С. tragopogonis. Растения (по 5 штук) выращивали в почве
в двухлитровых сосудах. Суспензия готовилась в водопроводной
воде с содержанием зооспорангиев 104/мл. Растения инокулиро-
вали в фазе семядольных листьев. При инокуляции использовался
распылитель.
Существенное значение в патогенезе болезни имеют бессимптом-
ный период (от момента заражения до появления первых симпто-
мов болезни) и период развития патогена (от прорастания споры
до образования следующей генерации спор). Исследование про-
должительности инкубационного периода в зависимости от метео-
рологических факторов, главным образом от температуры, — не-
обходимое условие для возможности прогнозирования болезней
растений в природе и установления сроков защитных мероприя-
64
тий (Наумова, 1939; Степанов, 1962; Захарова, 1965; Степанов,
Чумаков, 1972, и др.). Эта зависимость представлена различными
исследователями в виде кривых и номограмм. Корреляция между
инкубационным периодом развития мильдью винограда, вызывае-
мого Plasmopara viticola, и температурой выражена в виде кривой,
получившей название «кривая Мюллера» по имени исследователя,
установившего эту зависимость. Позднее Н. П. Олтаржевский
(1933, 1939, 1955) внес коррективы и дополнения к этой кривой
для соответствующего применения ее в разных климатических
зонах СССР. А. А. Шатским (1935) для прогноза мильдью вино-
града рекомендован подсчет суммы эффективных температур.
«Правило эффективных температур» применялось в дальнейшем
и при прогнозировании других болезней (Степанов, 1962; Филип-
пова, 1968; Степанов, Чумаков, 1972, и др.). Н. А. Наумовой
(1939) установлена зависимость продолжительности инкубацион-
ного периода Phytophthora infestans от сочетания минимальной,
максимальной и средней температур воздуха, представленная в виде
широко известной номограммы, используемой при прогнозе фи-
тофторы картофеля.
У большинства пероноспоровых грибов конец инкубационного
периода определяется появлением на листьях «маслянистых» пя-
тен, что зависит преимущественно от температуры, тогда как
появление спороношения — от выпадения росы, дождя. Поэтому
при исследованиях, связанных с инкубационным периодом, не-
обходимо учитывать указанные факторы, а также восприимчи-
вость сорта, пораженные органы самого растения.
При благоприятных условиях инкубационный период у перо-
носпоровых грибов может быть очень коротким, всего 2—
3 дня. У Phytophthora infestans он равен 3 суткам при 22.5°,
у Peronospora tabacina — 3 суткам при среднесуточной темпера-
туре 18е, у Plasmopara helianthi — 2 суткам при развитии гриба
в тепличных условиях. Повышение и понижение температуры вле-
чет за собой увеличение продолжительности инкубационного пе-
риода. Например, известно, что в природных условиях низкие
ночные температуры задерживают развитие Р. viticola и значи-
тельно увеличивают период инкубации (Олтаржевский, 1933).
При изучении вопроса о сроке появления спороношения (обра-
зование налета) важно фиксировать температурные условия и
влажность. Чаще всего спороношение обнаруживается после вы-
падения росы, дождя. Указывается, 'что решающий фактор в по-
явлении спороношения Р. viticola на листьях винограда — затяж-
ные вечерние дожди и теплые влажные ночи с обильными росами
и туманом. Спорангиеносцы выходят из устьиц в темноте после
полуночи. Н. А. Наумова (1965) отмечает, что спороношение Phy-
tophthora infestans возникает на листьях картофеля всегда на рас-
свете после 5—6-часового увлажнения растений.
Вопросы зимовки грибов и сохранения инфекционного начала
являются особенно важными и всегда интересуют исследователей.
5 Н. С. Новотельнова и др.
65
Для их решения наряду с биологическими проводятся анатомо-
морфологические исследования как инокулированных, так и есте-
ственно зараженных растений. Для выявления зимующих форм
пероноспоровых грибов анализируются различные покоящиеся
органы растений: клубни, луковицы, семена. Известны многочис-
ленные исследования мицелия Phytophthora infestans в клубнях
картофеля (Наумова, 1939; Бордукова, 1941; Пшедецкая, Чере-
панова, 1961а, и др.), мицелия Peronospora destructor в луковицах
(Каттерфельд, 1926; Viranyi, 1974, и др.), мицелия Plasmopara
helianthi в репродуктивных органах и семенах подсолнечника
(Новотельнова, 1963в, 1966а).
Сложным, но необходимым этапом исследований является изу-
чение вопросов, связанных с формированием, развитием и про-
растанием ооспор у видов пероноспоровых грибов. Имеются вы-
сокоэффективные исследования в этой области для ряда видов:
Peronospora hyoscyami (Бахтин, 1926), Р. tabacina (McGrath, Mil-
ler, 1959; Пересыпкин, Мархасева, 1964; Бабаян, Шалджян, 1965;
Боровская, 1968а, 19686), Р. brassicae (Куприянова, 1957), Plas-
mopara viticola (Макрушина, 1961, 1962, 1966, 1970; Николаев,
1973, и др.).
Способ проращивания ооспор Р. viticola заключается в том,
что осенью просматривают пораженные листья винограда и
участки с ооспорами вырезают; затем их закладывают во влажный
песок или в мокрую фильтровальную бумагу, которую помещают
в природных условиях в почву на глубину 5—10 см, а весной
выкапывают и в лаборатории исследуют при различной темпера-
туре жизнеспособность ооспор.
Установлено, что формирование ооспор у Р. viticola в Молда-
вии (Макрушина, 1970) начинается с июня (с первых генераций
гриба) и происходит до октября, усиливаясь к осени. Большее
количество ооспор обнаруживается в листьях винограда менее
восприимчивых сортов, в тканях с малой интенсивностью беспо-
лого спороношения и более активно в засушливую погоду. Уста-
новлено также, что продолжительность жизнеспособности ооспор
не превышает 2 лет.
А. А. Бабаян и Л. К. Шалджян (1965) изучали образование
ооспор у Peronospora tabacina в Армении. Пораженные листья
табака закладывались во влажную почву или влажную фильтро-
вальную бумагу и содержались при 18—20°. В лабора-
торных условиях на 8-е сутки в полуразложившихся листьях
отмечались сформировавшиеся в большом количестве ооспоры.
Листья можно закладывать и в полевых условиях в почву на
глубину 3—4 см, многочисленные ооспоры образуются на 8—14-е
сутки.
Крайне трудно изучение прорастания ооспор, однако найдены
методы, дающие положительные результаты. По данным А. Д. Ли-
пецкой (1939), прорастание ооспор Plasmopara viticola удается сти-
мулировать чередующимися замораживанием и оттаиванием, за су-
66
шиванием и увлажнением. Установлено, что после перезимовки
не на поверхности, а под слоем почвы ооспоры остаются жизне-
способными и прорастают. Способствуют прорастанию темпера-
тура около 16°, понижение ее в ночные часы и подъем в дневные.
Необходимо длительное увлажнение ооспор: в условиях Анапы
в феврале 12.5 суток, в марте 7.3, в апреле—мае 5—6, в июне
вообще трудно добиться прорастания. Аналогичные результаты
при изучении прорастания того же вида гриба получены А. Т. Мак-
рушиной (1970) в Молдавии. Автором приводятся прекрасные мик-
рофотографии зародышевых зооспорангиев, образующихся на кон-
цах ростковых трубок ооспор. М. Ф. Боровская (19686) приво-
дит микрофотографии ооспор Peronospora tabacina, прорастающих
только ростковыми трубками.
Весеннее возобновление пероноспорового гриба в природе —
показатель его перезимовки. Необходимы обследования полей
в ранневесенний период. Для изучения рассеивания спор и ана-
лиза заспоренности воздуха используются «спороловушки», чаще
всего в виде предметных стекол, смазанных различными клей-
кими веществами, экспонированных на различной высоте и за-
щищенных от дождя какими-нибудь приспособлениями.
В течение вегетационного периода проводятся обследования по-
севов и учет больных растений.
УЧЕТ В ПОЛЕВЫХ УСЛОВИЯХ1
Для выявления и учета болезней, вызываемых пероноспоро-
выми грибами, необходимо проводить в хозяйствах маршрутное
обследование 2—3 раза за вегетационный период. Площадь обсле-
дования должна составлять не менее 10% от всех посевов (поса-
док) определенной культуры. Отмечаются характер почвы, ее
рельеф, агротехника, климатические условия года и сорт.
Для оценки степени поражения результаты обследований пред-
ставляются в цифровом выражении, в баллах с использованием
разработанных шкал.
Шкала для учета корневой гнили злаков
0 — здоровые растения,
1 — слабое побурение основания стебля,
2 — заметное побурение основания стебля,
3 — сильное побурение основания стебля,
4 — погибшие растения.
Для учета корневой гнили зерновых или овощных культур
берутся пробы, количество которых должно быть больше при
слабом распространении болезни. При распространении болезни
в виде очагов измеряется их площадь, которая приводится в про-
центах к общей площади посева (посадки). Рекомендуется выде-
лять на поле 4 пробных участка площадью 0.1 га (32x32 м) или
0.25 га (50 Х50 м). Площадь очагов устанавливается шагомерно
(шаг — 0.5 м), величины суммируются, сумма делится на число
очагов для получения средней площади очагов. Гибель растений
на каждой пробной площадке вычисляется по формуле
МН . 100
г= р
где Г — гибель растений (%), М — средняя площадь очагов,
Н — число очагов, Р — размер пробной площадки. Процент ги-
бели растений на всем поле вычисляется как среднее арифмети-
ческое из процентов гибели растений па всех пробных площадках.
Развитие болезни отражает усредненную интенсивность пора-
1 Методы учета приводятся по рекомендациям «Указателей возбудите-
лей болезней сельскохозяйственных растений» (1971, 1975), а также по
Н. С. Новотельновой и Е. И. Легенькой (1963), Новотельновой и К. А. Пы-
стиной (1978).
68
жения для одного растения, участка или определенной террито-
рии и вычисляется по формуле
2 (а» б). 100-
“ NK
где Рб — развитие болезни (%), 2 — сумма произведений (а-б),
а — число пораженных растений, б — балл поражения, N — общее
количество учтенных растений (здоровых и больных), К — выс-
ший балл шкалы.
Пример вычисления. На площади 20 га осмотрено 200 расте-
ний, из них балл 0 имеют 120 растений, балл 1 — 35, балл 2 — 17, балл 3 —
13, балл 4 — 8, балл 5 — 7, отсюда
0 + (35 • 1) + (17 • 2) + (13 . 3) + (8 • 4) + (7 • 5)
Рб=-----------------200^5---------------’ 100 = 17-5%-
Для характеристики состояния полей в хозяйствах, районе или
области в целом определяют средневзвешенный процент развития
болезни. Для этого находят сумму произведений площади на со-
ответствующий ей процент развития болезни и полученный ре-
зультат делят на всю обследованную площадь. Например:
Поля хозяйства . .
Площадь, га ...
Развитие болезни, %
12 3 4
5 10 15 20
32 14 53 17
Таким образом, средневзвешенный процент развития болезни
для хозяйства будет равен
(5 • 32) + (10 • 14) + (15 . 53) + (20 • 17) _
5 + Ю+ 15+ 20 — /0.
Результаты обследований заносятся в учетные карточки.
Аналогичным образом производится учет поражения корне-
вой гнилью в теплицах и парниках. В каждом обследуемом пар-
нике за несколько дней до выемки рассады берут 10 проб по
10 растений в каждой. Пробы располагают равномерно: под первой
рамой в углу, под второй — в середине и т. д. Среди учетных
растений выявляют пораженные и по их числу определяют про-
цент поражения рассады.
При учете корневой и прикорневой гнили плодовых культур,
в частности семечковых или косточковых, необходимо ежегодное
трехразовое маршрутное обследование. Площадь учета должна
быть не менее 10% территории сада, количество осматриваемых
деревьев: на участках до 100 га 50 деревьев, на территории до
1000 га на каждые 100 га прибавляют еще по 5 деревьев. В про-
цессе учета у каждого дерева осматривается ствол в нижней его
части и при наличии разрывов и некроза коры отмечаются па-
тологические изменения в кроне.
69
Шкала для учета прикорневой штамбовой гнили
многолетних деревьев
О — здоровое дерево,
1 — небольшие разрывы коры, не превышающие 10 см2, прикрытые
валиком каллуса; крона неповрежденная,
2 — разрывы в основании штамба занимают площадь 20—70 см2,
древесина в пораженных участках побуревшая; в кроне усы-
хание отдельных ветвей,
3 — разрывы в основании штамба глубокие, опоясывают дерево;
в кроне усыхают отдельные ветви, листья мелкие, светлой
окраски,
4 — глубокие опоясывающие раны в основании штамба, дерево
усохшее.
При обнаружении пораженных растений с симптомами, типич
ными для корневой и прикорневой гнили, вызываемой низшими
грибами, необходимо подтверждение диагноза. С этой целью про-
водится изолирование возбудителя из тканей пораженного расте-
ния и дальнейшая его идентификация.
Запись результатов полевых наблюдений
Дата учета Обследован- ная площадь Сорт Число деревьев Количество пораженных деревьев
балл
0 1 2 3 4
Учет проводят по диагонали поля. На площади до 2 га осмат-
ривают 100 растений (10 проб по 10 рядом стоящих растений),
на площади 10 га — 200 растений, на площади 50 га — 500 рас-
тений, на площади свыше 50 га — дополнительно по одной пробе
на каждые 10 га. В зависимости от площади участка расстояния
между местами взятия проб могут быть различными (20, 50, 100 м).
Данные обследований должны отражать распространенность,
или частоту встречаемости болезни, интенсивность поражения
растений и развитие болезни, или степень пораженности растений.
Распространенность болезни показывает процент пораженных
растений от общего количества осмотренных в пробе, на участке,
в поле и определяется по формуле
г| = ^.1ОО,
где г — распространенность болезни (%), а — число пораженных
растений, К — общее количество учтенных растений (здоровых и
больных).
70
Форма учета появления ложной мучнистой росы на возделываемых культурах
Область (крац, республика) . . .
Район ........................
Пункт ОСП.....................
Культура .....................
Пид болезни (гриба) ..........
Хозяй- ство Бригада, отделение Поле, площадь посева, га Сорт Срок посева Предше- ственник, способ посева, удобрение Дата первого (видимого) появления Дата учета Обследо- вана площадь, га Заражена площадь Просмо- трено растений Из них пора- жено Процент поражения растений Средний балл пораже- ния
га э/о
Интенсивность поражения показывает размер пораженной по-
верхности листьев того или другого растения, она определяется
глазомерно по шкале и выражается в баллах.
Шкала для учета других болезней, вызываемых
пероноспоровыми грибами
О — отсутствие поражения,
1 — слабое поражение, поражены единичные листья, не более
10% на растении,
2 — среднее поражение, поражено до 50% всех листьев,
3 — сильное поражение, поражено свыше 50% всех листьев,
4 — поражены не только листья, но и репродуктивные органы,
5 — наиболее сильное поражение, растения недоразвиты или пол-
ностью погибли.
Развитие болезни, или степень пораженности, вычисляется по
вышеприведенной формуле.
Результаты обследований заносятся в учетные карточки.
Аналогичная форма учета используется и для растений за-
крытого грунта с видоизменениями применительно к размерам
обследуемой площади.
ДИАГНОСТИКА
Изучение болезней, вызываемых пероноспоровыми грибами, и
идентификация возбудителей включают многие приведенные выше
этапы исследований. Прежде всего в течение вегетационного периода
осуществляются визуальные наблюдения над растениями, устанав-
ливается их морфологический статус и выявляются патологические
симптомы. Последние часто очень характерны и помогают быстро
диагносцировать болезни в поле. К таковым относятся склероспороз,
белая ржавчина, пероноспороз, ложная мучнистая роса — болезни
с хорошо выраженными характерными симптомами. При корневой
и прикорневой гнили симптомы менее специфичны, так как увя-
дание и усыхание растений вызываются многими возбудителями
и идентификация последних возможна лишь в результате примене-
ния разнообразных методов исследования для выявления морфоло-
гии спороношений.
В настоящем справочнике аннотировано свыше 150 видов и раз-
новидностей пероноспоровых грибов на 172 видах растений, приве-
денных по культурам в виде конспективной симптоматологической
таблицы. В ней представлены следующие культуры: хлебные и
крупяные; бобовые, зернобобовые и крупяные; картофель, овощные
и бахчевые; технические и масличные; плодово-ягодные, виноград
и цитрусовые; декоративные, цветочные, лекарственные; оранже-
рейные.
Собранный обширный материал по пероноспоровым грибам,
вызывающим болезни культурных растений, основан на личных
исследованиях авторов и на литературных данных, в основном
отечественных. Представленный в таблице, где даются симптомы
заболевания, вызываемые тем или иным возбудителем, он дол-
жен облегчить обнаружение болезней при проведении обследова-
ний в полях, садах, оранжереях.
73
Питающее растение
Возбудитель
Симптомы поражения
Сроки обследования
Пшеница.
Рожь.
Ячмень, овес.
Хлебные и крупяные культуры
Pythium arrhenomanes, Р. aristo- На корнях и корешках мягкая В период всходов.
sporum, P. debaryanum, P. gramini- colum, P. irregulare, P. tardicrescens, P. volutum. светло-коричневая гниль. Листья бледной окраски, побеги недораз- виваются. При сильном поражении листья отмирают и растения по- гибают.
Sclerospora macrospora. На листьях пятна в виде темно- бурых точек, окруженных более светлым кольцом отмершей ткани. Колос гипертрофируется, изгиба- ется, вместо него развиваются зе- леные листочки. Колос зеленый в то время, когда здоровые экземпляры желтеют. Внутри тканей ооспоры. В период всходов и в фазу колошения.
Pythium aphanidermatum, P. ari- stosporum, P. arrhenomanes, P. deba- ryanum, P. graminicolum, P. ir- regulare, P. volutum. Симптомы поражения те же, что на пшенице. В период всходов.
Sclerospora secalina. i Пятна на листьях желто-бурые, располагаются вдоль краев листьев полосами или охватывают всю ли- стовую пластинку. Поверхность ли- стьев мелкошероховатая от присут- ствия ооспор. Листья постепенно засыхают. С появлением всхо- дов вплоть до начала колошения.
Pythium aristosporum, P. arrhe- nomanes, P. debaryanum, P. grami- nicolum, P. hypogynum, P. irregu- lare, P. iwayamai, P. papillatum, P. rostratum, P. splendens, P. toru- losum, P. ultimum, P. vexans, P. vo- lutum. Симптомы поражения те же, что на пшенице. В период всходов.
Питающее растение
Возбудитель
Ячмень, овес.
Sclerospora macrospore,.
Рис.
Pythium ar г he по manes, Р. deba-
ryanum, Р. dissotocum, Р. echinocar-
рит, Р. graminicolum, Р. paddicum,
Р. rostratum.
Phytophthora oryzae.
Sclerospora macrospora.
Кукуруза.
Pythium aphanidermatum, P. ar-
rhenomanes, P. debaryanum, P. gra-
minicolum, P. irregulare, P. paroe-
candrum, P. splendens, P. volutum.
Phytophthora nicotianae var. pa-
rasitica.
Sclerospora maydis.
Продолжение
Симптомы поражения
Сроки обследования
Колос гипертрофированный, взду-
тия голубовато-зеленого цвета; ча-
сто остается покрытым верхними
гипертрофированными листьями.
Внутри тканей ооспоры.
Симптомы поражения те же, что
на пшенице.
Корневая гниль.
Поражена метелка. На колосках
вздутия и мясистые голубовато-зе-
леные наросты, зерно не развива-
ется. Внутри тканей ооспоры.
Почернение и утончение корне-
вой шейки. На поверхности об-
разование белого налета мицелия.
Гниль корня и корневой шейки.
Поражение листьев, влагалищ,
мужских соцветий. На них рас-
плывчатые пятна в виде бурых
полос, с нижней стороны слабый
белый налет, размочаливание
листьев. При сильном поражении
отставание роста растений, гибель
всходов.
В фазу колошения.
В период всходов.
То же.
В фазу колошения.
В период всходов.
То же.
В период всходов и
в фазу выметывания
султанов.
Oi
Продолжение
Питающее растение Возбудитель Симптомы поражения Сроки обследования
Просо, могар, чу- миза. Pythium arrhenomanes, Р. debarya- пит, Р. graminicolum, Р. hydnospo- гит. Потемнение и утончение корне- вой шейки, поникание растений, белый налет на пораженной части. В период всходов.
Sclerospora graminicola, S. gra- minicola var. setariae-italicae. На листьях пятна в виде продоль- ных полос с быстро исчезающим бе- ловатым налетом. Внутри тканей ооспоры. При сильном поражении ткани разрушаются, и лист расщеп- ляется продольно, ооспоры высы- паются в виде золотисто-коричне- вого порошка. Соцветия деформи- руются, части цветков ненормально увеличиваются и зеленеют. Семена не развиваются. В фазу колошения.
Гречиха. Phytophthora nicotianae var. para- sitica. На семядольных листьях округ- лые бурые пятна с концентриче- скими кругами, на- нижней сто- роне нежно-белый налет. Гниль сеянцев. В период всходов.
Peronospora fagopyri. Бобовые, зерно-бобовые и кор] Поражение листьев и цветков. На листьях неясно выраженные округлые светло-желтые пятна, С нижней стороны нежный серова- тый налет. Цветки остаются бес- плодными. новые культуры в период цветения.
Горох, пелюшка. Pythium debaryanum, P. hydnospo- rum, P. ultimum, P. vexans. Почернение и загнивание корне- вой шейки, иногда белый паутини- стый налет. в период всходов.
Питающее растение Возбудитель
Горох, пелюшка. Phytophthora erythroseptica, Ph. са- ctorum. Peronospora pist.
Бобы кормовые. Pythium debaryanum.
Peronospora fabae.
Фасоль. Pythium acanthicum, P. debarya- num, P. ultimum. Phytophthora infestans var. pha- seoli, Ph. nicotianae n&t. parasitica.
Соя. Pythium debaryanum. Phytophthora megasperma var. sofae. Peronospora manshurica.
Чечевица. Pythium debaryanum. Peronospora lentis.
Люпин. Pythium debaryanum.
Клевер. Phytophthora cactorum, Ph. me- gasperma var. sofae. Pythium debaryanum.
II родолжение
Симптомы поражения Сроки обследования
Гниль сеянцев. В период всходов.
На листьях бледно-желтые ок- руглые пятна, с нижней стороны серовато-фиолетовый налет. В период цветения.
Признаки поражения те же, что на горохе. В период всходов.
На листьях красновато-бурые пятна, с нижней стороны серовато- фиолетовый налет. В фазу цветения.
Признаки поражения те же, что на горохе. В период всходов.
На листьях бурые расплывчатые Во вторую половину
пятна, с нижней стороны сероватый паутинистый налет. Гниль бобов. вегетации.
Корневая и прикорневая гниль. В период всходов.
То же. То же.
На листьях бурые пятна, с ниж- ней стороны серовато-фиолетовый налет. В период цветения.
• Корневая и прикорневая гниль. В период всходов.
На листьях желтоватые пятна, с нижней стороны серовато-фиоле- товый налет. В период цветения.
Признаки поражения те же, что на горохе. В период всходов.
Прикорневая гниль. То же.
Корневая и прикорневая гниль. » »
Питающее растение Возбудитель
Клевер. Peronospora pratensis, Р. trifol- lii-repentis, Р. trifollii-ar vensis, P. trifollii-alpestris.
Люцерна. Pythium arrhenomanes, P. deba- ryanum, P. helicoides, P. mamilla- tum, P. paroecandrum, P. pulchrum, P. rostratum, P. salpingophorum, P. splendens, P. ultimum, P. vexans. Phytophthora cryptogea, Ph. cac- torum, Ph. megasperma. Peronospora aestivalis.
Вика. Pythium arrhenomanes, P. deba- ryanum, P. graminicolum, P. ir- regulare, P. rostratum, P. splendens. Peronospora viciae, P. viciae-sati- vae, P. majori, P. sepium.
Эспарцет. Peronospora ruegeriae.
Чина. Pythium rostratum, P. ultimum. Peronospora fulva, P. lathyri-pa- lustris, P. orobi.
Донник. Phytophthora cactorum. Peronospora meliloti.
Продолжение
Симптомы поражения Сроки обследования
На листьях желтоватые пятна, От периода всходов.
ограниченные жилками, с нижней стороны серовато-фиолетовый на- лет. Общее угнетение растений. Корневая и прикорневая гниль. В период всходов.
Прикорневая гниль.
На листьях светлые пятна, с ниж-
ней стороны коричневато-фиолето-
вый налет, укорочение междо-
узлий.
Корневая и прикорневая гниль.
На листьях бледные пятна, со
временем засыхающие, с нижней
стороны серовато-фиолетовый налет.
На листьях бледные пятна,
с нижней стороны серый налет.
Корневая и прикорневая гниль.
На листьях желтоватые пятна,
с нижней стороны серовато-фиоле-
товый налет.
Корневая гниль.
На листьях желтоватые пятна,
с нижней стороны коричневато-
фиолетовый налет.
То же.
От периода всходов.
То же.
В период цветения.
То же.
В период всходов.
В период цветения.
В период всходов.
В период цветеция.
II родолжение
Питающее растение Возбудитель Симптомы поражения Сроки обследования
Сераделла. Р. ornithopi. Картофель, овощи На листьях бледно-желтые пятна, с нижней стороны серовато-фиоле- товый налет. ые и бахчевые культуры В период цветения.
Картофель. Pythium acanthicum, Р. aphani- dermatum^ Р. debaryanum, Р. de- liense, P. ultimum, P. vexans. Корневая и прикорневая гниль. В период всходов.
Phytophthora infestans. Ph. erythroseptica. Поражение листьев, стеблей и клубней. На листьях в период цве- тения бурые округлые пятна, по краям которых с нижней стороны в сырую погоду выступает белый налет. На стеблях продольные бу- рые пятна. В жаркую погоду рас- тения засыхают, в сырую гниют. На клубнях слегка вдавленные свинцово-серые пятна твердой кон- систенции, под кожурой ткань ржа- вого цвета. Розовая гниль клубней. Через 10—15 дней после начала массового цветения картофеля и позже. В период клубнеоб- разования.
Томаты. Pythium debaryanum. Phytophthora infestans. Потемнение и перетяжка корне- вой шейки, иногда с сероватым на- летом на поверхности. На листьях бурые пятна, иногда с нижней стороны налет. Мякоть плодов ржаво-бурая, твердой кон- систенции. В сырую погоду плоды загнивают. В период всходов. В фазу зеленого плода и позже.
Питающее растение Возбудитель
Томаты. Phytophthor a nicotianae. Ph. capsici, Ph. cryptogea, Ph. drechsleri.
Баклажаны. Ph. infestans.. Ph. capsici. Peronospora tabacina.
Перец. Pythium aphanidermatum, P. ul- timum. Phytophthora nicotianae var. pa- rasitica, Ph. capsici. Peronospora tabacina.
Капуста кочанная, цветная, брюссель- ская. Pythium aphanidermatum, P. de- baryanum, P. ultimum. Phytophthora megasperma. Peronospora brassicae.
Продолжение
Симптомы поражения Сроки обследования
Корневая гниль, темные пятна На ранних стадиях
с размягченной тканью в любой развития (рассада в теп-
части стебля, гибель растений. лицах).
Заболевание плодов под назва- В фазу зеленого
нием «олений глаз». плода.
На листьях бурые расплывчатые пятна, с нижней стороны белый налет. В период цветения.
На листьях хлоротичные округ- От периода всходов
лые пятна, с нижней стороны се- ровато-фиолетовый налет. В жар- кую погоду пораженные участки листьев засыхают. в парниках до созрева-
ния семян в поле.
Признаки поражения те же, что на баклажанах. В период всходов.
Поражение листьев, плодов и гниль сеянцев. От периода всходов.
Признаки поражения те же, что на баклажанах. В период цветения.
Признаки поражения те же, что на томатах. В период всходов.
Корневая гниль. То же.
Поражение надземных, иногда В парниках в фазу
подземных частей. На листьях хло- двух настоящих листьев.
ротичные, на стеблях и стручках В поле от посадки до
бурые пятна. На пораженных ча- стях в сырую погоду белый рых- лый налет. Иногда гибель рассады в парниках, отмирание растений в поле. уборки.
Питающее растение
и -------------------
о
Возбудитель
Новотельнова и др.
Капуста кочанная,
цветная, брюссель-
ская.
Cystopus candidus.
Репа, брюква, тур-
непс, редька, редис.
Хрен.
Морковь, укроп,
петрушка, пастернак.
Анис.
Фенхель.
Pythium debaryanum,
Phytophthora cryptogea, Ph, nico-
tianae.
Peronospora brassicae.
Cystopus candidus.
Pythium aphanidermatum.
Peronospora cochleariae.
Cystopus candidus.
Pythium aphanidermatum, P. de-
baryanum, P. oligandrum, P. pa-
roecandrum, P, polymastum, P. spi-
nosum, P. ultimum,
Plasmopara dauci, P. pastinacae,
P. petroselini, P. anethi, P. peucedani.
P. pimpinellae.
Phytophthora syringae,
Plasmopara mei-foeniculi.
Продолжение
Симптомы поражения Сроки обследования
Поражение листьев, стеблей, стручков. На пораженных частях небольшие округлые пятна и бело- ватые блестящие пустулы, из кото- рых при созревании высыпаются споры в виде белой порошковидной массы. Пораженные органы гипер- трофируются, стручки на больных цветоносах не образуются. В период цветения.
Признаки поражения те же, что на томатах. Прикорневая гниль. В период всходов. То же.
Признаки поражения те же, что на капусте. В период всходов и позже.
То же. В период цветения.
Корневая и прикорневая гниль. В период всходов.
На листьях расплывчатые пятна, с нижней стороны белый налет. На листьях пустулы. В период всходов и позже. То же.
Корневая и прикорневая гниль рассады. В период всходов.
На листьях неясные бледные пятна, позднее буреющие, с нижней стороны обильный налет. В период цветения.
На листьях пятна, налет. От периода всходов.
Прикорневая гниль. В период всходов.
На листьях пятна, налет. От периода всходов.
Питающее растение Возбудитель
Сельдерей. Phytophthora cryptogea. Plasmopara apii.
Цикорий^ салат эн- дивий. Bremta lactucae var. lactucae.
Эстрагон. Артишок. Шпинат. Peronospora sulfurea. Bremta lactucae, B. scolymi. Pythium anandrum, P. aphanider- matum, P. debaryanum, P. irregulare, P. ultimum. Peronospora spinaciae. Cystopus occidentalis.
Щавель. Pythium mamillatum. Peronospora rumicis.
Ревень. Phytophthora nicotianae var. para- sitica. Peronospora jaapiana.
Салат. Pythium debaryanum. Bremia lactucae var. lactucae.
Продолжение
Симптомы поражения Сроки обследования
Прикорневая гниль. В период всходов.
На листьях пятна, с нижней стороны налет. В период вегетации.
На листьях хлоротичные пятна, позже побурение и усыхание ли- стьев. На листьях пятна, налет. На листьях пятна. От периода всходов. То же. » »
Признаки поражения те же, что на томатах. В период всходов.
На листьях желтые или бурые пятна, с нижней стороны серовато- фиолетовый налет. В период цветения.
На листьях пустулы. От периода всходов.
Признаки поражения те же, что на томатах. В период всходов.
На листьях расплывчатые пятна, с нижней стороны сероватый налет. В период цветения.
Корневая гниль. В период всходов.
На листьях круглые бурые пятна, с нижней стороны белый или серо- вато-фиолетовый налет. В период цветения.
Признаки поражения те же, что на томатах. В период всходов.
Поражение листьев и соцветий. На листьях бледно-желтые углова- тые пятна, с нижней стороны белый налет, на соцветиях" семенников белый слабый налет. В период цветения.
Продолжение
Питающее растение Возбудитель Симптомы поражения Сроки обследования
Огурцы, арбузы, Pythium aphanidermatum, Р. de- Признаки поражения те же, что В период всходов и
дыни. baryanum, Р. irregulare, Р. ultimum. Peronospora (=Pseudoperonospora) cubensis. на томатах. На листьях угловатые желтовато- бурые пятна, с нижней стороны серовато-фиолетовый налет, листья засыхают. плодоношения. В период цветения.
Тыква, кабачки. Pythium aphanidermatum, P. de- baryanum, P. oligandrum, P. ul- timum. Phytophthora capsici. Peronospora (= Pseudo peronospora) cubensis. Признаки поражения те же, что на томатах. Гниль сеянцев и плодов. Признаки поражения те же, что на огурцах. В период всходов и плодоношения. То же. В период цветения.
Лук, чеснок. Pythium debaryanum, P. irregu- lare, P. mamillatum, P. spinosum, P. ultimum. Phytophthora porri. Peronospora destructor. Признаки поражения те же, что на томатах. Корневая и листовая гниль. На листьях бледно-зеленые пятна, серовато-фиолетовый налет. Иногда все листья поражены, растения отмирают. В период всходов. От периода всходов. В период вегетации.
О * Каперцы. Cystopus capparidis. Технические и мае. Пустулы на органах растений, личные культуры То же.
оо w Свекла. Pythium aphanidermatum, P. de- baryanum, P. irregulare, P. ultimum. Побурение и перетяжка подзем- ной части семядольного колена, покоричневение корешков, загнива- ние всходов. Белый войлочный на- лет на пораженных частях рас- тений. В период всходов.
Питающее растение Возбудитель
Свекла. Phytophthora drechsleri, Р. те- gasperma, Peronospora schachtti. Cystopus bliti.
Подсолнечник. Pythium irregulare, P. splendens* Phytophthora drechsleri. Plasmopara helianthi. Cystopus tragopogonis f. helianthi.
Продолжение
Симптомы поражения
Сроки обследования
Корневая гниль, увядание ли-
стьев, полегание сеянцев. На позд-
них стадиях развития — загнива-
ние нижнего конца главного корня
в местах отхождения боковых ко-
решков, потеря тургора, полное
увядание.
На свекле 1-го года поражение
молодых листьев; они хлоротичные,
хрупкие, с закрученными внутрь
краями, с нижней стороны плот-
ный серо-фиолетовый налет. На вы-
садках поражены не только листья,
но и цветоносы. При сильном по-
ражении растения погибают.
На верхней стороне листьев блед-
ные пятна, на нижней белые бле-
стящие пустулы.
Корневая и прикорневая гниль.
Корневая гниль.
На листьях хлоротичные рас-
плывающиеся пятна, с нижней
стороны нежный белый налет, ко-
торый позже становится серова-
тым, густым, войлочным. Карли-
ковость, иногда гибель растений.
На верхней стороне листьев свет-
лые пятна, на нижней белые пу-
стулы.
От периода всходов.
Свекла 1-го года на-
чиная с фазы 4—6-го
листьев и до уборки
урожая. Высадки —
в период вегетации.
В период вегетации.
В период всходов.
То же.
От периода всходов.
То же.
Питающее растение Возбудитель
Лен. Pythium debaryanum, Р. perni- . ciosum. Phytophthora cactorum. Peronospora Uni.
Крамбе. P. crambe.
Хлопчатник. Phytophthora nicotianae var. pa- rasitica.
Табак, махорка. Pythium aphanidermatum, P. de- baryanum, P. perniciosum. Phytophthora nicotianae var. nico- tianae. Peronospora tabacina.
ОС
сл
Продолжение
Симптомы поражения Сроки обследования
Корневая гниль. » » На листьях светло-желтые пятна, с нижней стороны нежный белый налет. На листьях светло-желтые пятна, с нижней стороны налет. Корневая гниль. Утончение основания стебля, увя- дание и гибель всходов. Гниль корней, пятна на стеблях, сгнивание нижних листьев и круп- ные округлые пятна с налетом на листьях, расположенных выше. Больная рассада желтовато-зеле- ная, листья с закрученными кра- ями, с нижней стороны сиреневато- голубоватый паутинистый налет. При сильном развитии болезни растения гибнут. Заболевание про- является сначала на нижних ли- стьях, затем на листьях среднего и верхнего ярусов в виде хло- ротичных маслянистых пятен, с нижней стороны покрытых нале- том. Могут поражаться стебли, па- сынки, бутоны, цветки и семенные коробочки. В период всходов. То же. В период вегетации. То же. В период всходов. То же. От периода всходов. В парниках в фазу «ушек» и перед выбор- кой рассады. В полевых условиях в фазу 5— 6-го листьев, в период бутонизации и позже.
00
05
Питающее растение Возбудитель
Клещевина. Phytophthora nicotianae var. para-^
sitica.
Конопля. Pythium aphanidermatum, P. de-
baryanum.
Peronospora (= Pseudoperonospora)
cannabina.
Кунжут. Phytophthora cactorum.
Ph. nicotianae чах. parasitica.
Сафлор. Ph. drechsleri.
Bremia lactucae чах. carthami.
Горчица. Pythium debaryanum.
П родолжение
Симптомы поражения Сроки обследования
На листьях крупные сливаю- щиеся бурые пятна с концентриче- ской зональностью, в центре более светлые, с нижней стороны белова- тый налет. В период цветения.
Побледнение листьев сеянцев, увядание и отмирание всходов. На пораженных частях обильный вой- лочный налет. В период всходов.
На листьях округлые желтова- тые пятна, часто сливающиеся, с нижней стороны серовато-фиоле- товый налет. При сильном разви- тии болезни листья засыхают и опадают. В период цветения.
На семядольных листьях бурые округлые пятна, на настоящих желтые, с сероватым налетом с ниж- ней стороны. От периода всходов.
Поражение листьев, стеблей и корней. Пятна на стеблях, сгнива- ние нижних листьев и крупные ок- руглые пятна с налетом на выше расположенных листьях. В период цветения.
Корневая гниль. На всех фазах раз- вития.
На листьях большие бурые пятна неясных очертаний, с нижней сто- роны нежный белый налет. В период цветения.
Корневая и прикорневая гниль. В период всходов.
Питающее растение Возбудитель
Горчица. Peronospora brassicae f. brassi- cae-nigrae. Cystopus candidus.
Рыжик. Pythium debaryanum. Peronospora camelinae. Cystopus candidus.
Хмель. Peronospora (=Pseudoperonospora) humuli.
Канатник. Plasmopara skvortzovii.
оо Кориандр. P. nivea.
Продолжение
Симптомы поражения
Сроки обследования
Листья хлоротичные, с нижней
стороны белый рыхлый налет. Бо-
лезнь развивается сначала на ниж-
них, потом на верхних ярусах ли-
стьев.
На листьях бледные пятна и бе-
лые блестящие пустулы.
Побурение, загнивание основа-
ния стебля, белый налет на пора-
женных частях. Всходы увядают
и засыхают.
Поражение листьев, стеблей.
Больные растения сильно отстают
в росте. На нижней стороне листьев
сплошной белый налет. На стебле
такой же налет в виде муфты.
Признаки поражения те же, что
на горчице.
На листьях угловатые желто-
вато-зеленые или бурые пятна,
с нижней стороны едва заметный
грязно-фиолетовый налет. Моло-
дые побеги деформируются и при-
обретают колосо- или шишковид-
ную форму. На «шишках» бурые
штриховидные пятна. Пораженные
листья, «шишки» и стебли засыхают.
На листьях угловатые бледно-зе-
леные, буреющие пятна, с нижней
стороны налет.
На листьях пятна.
В период цветения.
То же.
В период всходов.
В фазу бутонизации.
В период цветения.
То же.
» »
В период вегетации.
П родолжение
Питающее растение Возбудитель Симптомы поражения Сроки обследования
Плодово-ягодные культуры, виноград и цитрусовые
Яблоня.
Pythium debaryanum, Р. torulosum,
Р. ultimum.
Phytophthora cactorum, Ph. sy-
ringae.
Груша, вишня,
черешня.
Pythium aphanidermatum, P. de-
baryanum, P. intermedium, P. me-
galacanthum, P. pulchrum, P. ul-
timum, P. vexans.
Phytophthora cactorum.
Утончение главного корня, потем-
нение ткани, язвы на корневой
шейке сеянцев.
Гниль корневой шейки де-
ревьев в возрасте от 10 лет. На за-
гнивших участках кора растрески-
вается, темнеет и слегка впадает,
место поражения выделяется фиоле-
тово-коричневой окраской. При над-
резах обнаруживаются шоколадно-
коричневые участки коры. Древе-
сина обычно остается неповреж-
денной. В кроне пораженных де-
ревьев листья на концах ветвей
краснеют, скручиваются, а иногда
отмирают.
На плодах округлые коричневые
пятна твердой консистенции с ха-
рактерным блеском. На чечевич-
ках конидиеносцы гриба со спорами.
Признаки поражения те же, что
на яблоне.
Прикорневая гниль.
В период вегетации.
То же.
В период всходов.
То же.
Питающее растение Возбудитель
Земляника. Pythium debaryanum, Р. prolife- rum, Р. ultimum, Р. sylvaticum. Phytophthora cactorum. Ph. fragariae. i
Продолжений
Симптомы поражения
Сроки обследования
Мокрая гниль стеблей.
Поражаются плоды чаще у расте-
ний 3-го года. На плодах перед
созреванием появляются водяни-
стые размягченные участки различ-
ной величины с окраской от бледно-
розовой к лиловой или голубовато-
фиолетовой. Позднее пятна стано-
вятся желто-коричневыми. Мякоть
стекловидная, водянистая и частью
окрашена в коричневый цвет. Иногда
ягоды становятся кожистыми и
твердыми.
Вначале покраснение проводящей
ткани центрального цилиндра
корня. Позднее корни отмирают,
начиная с нижнего конца, и окра-
шиваются в бурый цвет. Появля-
ется симптом «крысиный хвост»
(верхняя часть первичного корня
остается еще светлой, а нижняя
чернеет). Молодые листья тускло-
голубовато-зеленые. У сильно по-
раженных растений листья крас-
неют и увядают.
Растения дают много мелких
плодов. Болезнь может сопровож-
даться карликовостью и гибелью
растений.
Ранней весной на рас-
тениях 1-го года.
В фазу созревания
ягод.
С ранней весны.
CD
О
Питающее растение Возбудитель
Земляника. Peronospora fragariae.
Смородина, кры- жовник. Phytophthora megasperma, Ph. ca- ctorum. Plasmopara ribicola.
Ежевика, малина. Peronospora rubi.
Виноград. Pythium aphanidermatum, P. de- baryanum, P. irregulare, P. mamil- latum, P. rostratum, P. ultimum. Plasmopara viticola.
Лимон, апельсин, мандарин. Pythium aphanidermatum, P. de- baryanum, P. irregulare, P. mega- lacanthum, P. rostratum, P. ulti- mum, P. vexans.
Продолжение
Симптомы поражения Сроки обследования
На листьях пятна, с нижней сто- роны налет. Корневая гниль саженцев. В период вегетации. То же.
На листьях коричневые углова- тые пятна, с нижней стороны обильный налет. На листьях пятна, с нижней сто- роны налет. Корневая и прикорневая гниль. » » На ранних фазах раз- вития.
Поражение всех надземных ор- ганов. На листьях маслянистые желтовато-зеленые пятна, с ниж- ней стороны сероватый налет. Пятна буреют, количество их увеличи- вается, листья отмирают. Пятна и налет появляются и на поражен- ных цветках, бутонах, завязях, ягодах, молодых побегах; ягоды буреют. Корневая и прикорневая гниль. В фазы бутонизации и образования завязей. На ранних фазах раз- вития.
Продолжений
Питающее растение Возбудитель Симптомы поражения Сроки обследования
Лимон, апельсин, мандарин. Phytophthora citrophthora, Ph. cit- ricola, Ph. nicotianae var. parasitica. Ph. syringae. На листьях темно-бурые пятна. На плодах коричневая плотная гниль, постепенно охватывающая весь плод. При поражении сажен- цев отмирают верхушки побегов, ветвей и стволиков. Через трещины выделяется камедь (гоммоз). Са- женцы быстро погибают. В период вегетации.
Мушмула. Ph. cactorum. Увядание и усыхание. То же.
Абрикос. Pythium debaryanum. Корневая и прикорневая гниль. Чаще в период всхо- дов.
Слива. P. debaryanum, P. oligandrum. Лесные i То же. культуры То же.
Хвойные породы: Pythium aphanidermatum, P. de- Поражение молодых проростков Начиная с фазы ви-
сосна, ель, пихта, baryanum, P. intermedium, P. ir- (до одревеснения тканей). Стебель лочки и 1-й пары ли-
кедр, тисс, листвен- ница, кипарис. regulare, P. torulosum, P. pyrilo- bum, P. ultimum. Phytophthora cactorum, Ph. cin- namomi. в области корневой шейки стано- вится вялым и утончается, растение желтеет, вершиной полегает на почву и полностью загнивает. Во влажную погоду на корневой шейке развивается белый мицелий. Корневая и прикорневая гниль сеянцев, дефолиация и гибель де- ревьев. стьев. В период вегетации.
Лиственные по- роды: каштан, грец- кий орех, пробко- вый дуб, лавр, хин- ное дерево, пекан. СО Pythium debaryanum. Корневая и прикорневая гниль сеянцев. На ранних фазах раз- вития.
Продолжение
Питающее растение Возбудитель Симптомы поражения Сроки обследования
Лиственные поро- ды: каштан, грецкий орех, пробковый дуб, лавр, хинное дерево, пекан. Phytophthora cinnamomi, Ph. сат- bivora, Ph. cactorum. Кора и древесина корней и ство- лов приобретают темно-бурую, почти черную окраску, на поражен- ных участках сокоистекающие тре- щины, усыхание корней, изъязвле- ния стволов, «обугливание» коры; симптомы известны под названием «чернильная болезнь» каштана, проб- кового дуба. В период вегетации.
Эвкалипт. Ph. cinnamomi. Корневая и прикорневая гниль, вилт, хлороз и опадение листьев. То же.
Бук. Ph. cactorum. Корневая и прикорневая гниль всходов. В период всходов.
Клен, ильм. » » При хроническом течении бо- лезни мелколистность; усыхание ветвей. В период вегетации.
Свидина. » » Образование ран на стволах. Кора отлущивается, становится ко- ричневой. При опоясывающей гнили отми- рание ветвей и деревьев. То же.
Декоративные, цветочные, лекарственные культуры
Амарантус. Cystopus bliti. На листьях, стеблях, цветках » »
прикрытые пустулы, при нии порошащие. созрева-
Астра. Pythium acanthicum, P. debarya- num, P. me galacant hum, P. ultimum. Загнивание корня и стебля. В период всходов.
Phytophthora cryptogea. Гниль корня. От периода всходов
Питающее растение Возбудитель
Астра. Plasmopara asterea.
Базилик. Peronospora lamii.
Бархатцы. Phytophthora cryptogea.
Белена. Peronospora hyoscyami.
Белладонна. Pythium debaryanum.
Борец. Phytophthora erythroseptica, Plasmopara pygmaea.
Валериана. Phytophthora cactorum, Peronospora Valerianae,
Василек. Premia centaureae.
Гвоздика. Phytophthora cactorum.
Георгин. Ph. cryptogea.
Герань, пеларго- ния. Pythium splendens, Plasmopara pusilia.
Peronospora conglomerata.
Продолжение
Симптомы поражения
Сроки обследования
На листьях пятна, с нижней сто-
роны налет.
На листьях пятна, с нижней сто-
роны рыхлый налет.
Корневая гниль, побурение сте-
блей, поникание и гибель растений.
На листьях, позже буреющих и
засыхающих, пятна. Растения не-
доразвиваются и погибают.
Побурение и загнивание корне-
вой шейки; стебель утончается,
всходы гибнут.
Прикорневая гниль.
На листьях пятна, с нижней
стороны беловатый налет.
Корневая и прикорневая гниль.
На листьях хлорртичные пятна,
с нижней стороны серовато-фиоле-
товый налет.
На листьях отдельные непра-
вильно разбросанные пятна.
Гниль, опоясывающая стебель
у корневой шейки, увядание и ги-
бель растений.
Побурение нижней части стебля,
гниль корня.
Гниль корней, стебля и черенков.
На листьях пятна, с нижней
стороны налет.
То же.
От периода всходов.
То же.
» »
» »
» »
В период всходов.
То же.
От периода всходов.
То же.
» »
» »
» »
» »
» »
» »
» »
Питающее растение
Возбудитель
Гербера. Pythium acanthicum. Phytophthora cryptogea.
Гипсофила. » » Plasmopara gypsophilae.
Гладиолус. Phytophthora cactorum, Ph. cryp- togea.
Девясил. Дельфиниум. Cystopus tragopogonis. Plasmopara pygmaea f. delphini.
Душистый горо- шек. Женьшень. Peronospora fulva. Pythium debaryanum. Phytophthora cactorum. Plasmopara panacis.
Золотарник. P. solidaginis.
Ирис. Phytophthora cactorum, P. cryp-
Кларкия (годеция). togea. Ph. cactorum. Peronospora arthuri.
Продолжение
Симптомы поражения
Сроки обследования
Прикорневая гниль рассады после
пикировки.
Загнивание стебля и корней.
Корневая гниль и увядание рас-
тений.
На листьях пятна, с нижней сто-
роны налет.
Загнивание луковиц и цвето-
носов.
На листьях и стеблях пустулы.
На листьях пятна, с нижней сто-
роны налет.
То же.
Гниль корня.
Корневая гниль, полегание всхо-
дов. На листьях темные пятна,
с нижней стороны слабый налет.
На листьях светлые угловатые
пятна, ограниченные жилками,
с нижней стороны сероватый слабый
налет.
Пятна покрывают всю пластинку
листа. Диффузное поражение всхо-
дов.
Гниль корневища.
Корневая гниль и общее увядание.
На листьях пятна, с нижней сто-
роны налет.
В период всходов.
в период вегетации.
То же.
» »
» »
» »
» »
» »
В период всходов.
в период вегетации.
То же.
От периода всходов.
То же.
» »
» »
Питающее растение Возбудитель
Лилия. Pythium debaryanum, Р. ultimum. Phytophthora cactorum, Ph. nico- tianae var. parasitica.
Львиный зев. Ph. cactorum, Ph. cryptogea. Peronospora antirrhini.
Мак. Pythium debaryanum, P. megala- canthum, P. oligandrum, P. spino- sum, P. ultimum. Phytophthora cactorum, Ph. cryp- togea. Peronospora arborescens.
Мальва. Phytophthora cactorum, Ph. nico- tianae var. parasitica.
Маттиола (левкой). Pythium debaryanum. . Phytophthora cactorum, Ph. cryp- togea. Peronospora matthiolae.
П родолжение
Симптомы поражения Сроки обследования
Поражение луковиц, стеблей, От периода всходов.
реже листьев, цветков. Растения увядают, поникают, гибнут. Гниль луковиц, стеблей, цветков. То же.
» »
Загнивание стебля в области кор- невой шейки. » »
Полегание всходов. На листьях при поражении взрослых растений хлоротичные пятна. » »
Корневая и прикорневая гниль. » »
Полегание, вилт. » »
На листьях, стеблях, коробоч- ках пятна и сероватый рыхлый на- лет. Листья буреют и засыхают, стебли искривляются, коробочки недоразвиваются. » »
Корневая гниль, гниль стеблей и листьев. » »
Прикорневая гниль, загнивание цветоносов и гибель растений. » »
Корневая и прикорневая гниль. » »
На листьях пятна, с нижней сто- роны сероватый налет. » »
о
о
Питающее растение Возбудитель
Маттиола (левкой). Cystopus candidus.
Мята. Peronospora stigmaticola.
Наперстянка. P, digitalidis.
Настурция. Phytophthora cactorum. Peronospora nasturtii-aquatici Cystopus candidus.
Незабудка. Петуния. Пиретрум. Портулак. Peronospora myosotidis. Phytophthora cryptogea, Ph. in- festans. Cystopus tragopogonis. C. portulacae.
Примула. Pythium debaryanum, P. irregulare.
Пион. Phytophthora cactorum. To же.
Резеда. Peronospora crispula.
Cystopus candidus.
Продолжение
Симптомы поражения
Сроки обследования
Листья желтеют, буреют, отми-
рают; стебли, цветки, плоды часто
гипертрофированы, на них блестя-
щие беловатые пустулы.
На листьях пятна, с нижней сто-
роны налет.
На листьях пятна, с нижней сто-
роны серовато-фиолетовый налет.
Гниль корней и корневой шейки.
На листьях пятна.
На листьях и стеблях пустулы;
искривление стебля.
На листьях пятна.
Корневая гниль и увядание.
На листьях и стеблях пустулы.
Ветви и листья видоизменяются,
на них пустулы.
Побурение стеблей и корневая
гниль.
Корневая гниль.
Гниль корней, почернение и от-
мирание побегов, листьев, полега-
ние всего растения.
На листьях пятна, с нижней сто-
роны налет.
На листьях и стеблях белые пу-
стулы.
От периода всходов.
В период вегетации.
То же.
В период всходов.
В период вегетации.
То же.
» »
В период всходов.
В период вегетации.
То же.
В период всходов.
То же.
Весной.
В период вегетации.
То же.
Продолжение
Новотельнова и др.
Питающее растение Возбудитель Симптомы поражения Сроки обследования
Роза. Peronospora (= Pseudoperonospora) sparsa. На листьях, черешках и цветоно- сах пятна, с нижней стороны ли- стьев сероватый налет. В период вегетации.
Ромашка аптечная. P. leptosperma. На листьях, реже на стеблях пятна, с нижней стороны рыхлый обильный налет. То же.
Сирень. Phytophthora cactorum, Ph. sy- ringae. Загнивание корней, ветвей, увя- дание соцветий. » »
Тюльпан. Pythium debaryanum, P. inter- medium, P. sylvaticum, P. ultimum. Phytophthora cactorum, Ph. cryp- to gea. Гниль луковиц, искривление и гниль цветоносов, опадение цветков Гниль луковиц, искривление и гниль цветоносов. Весной.
Флокс. Peronospora phloxidis. На листьях пятна, с нижней сто- роны налет. В период вегетации.
Хризантема. Pythium sp. Корневая гниль, увядание всего растения и полегание. В период всходов.
Шалфей. Peronospora swinglei. Оранжерейньп На листьях пятна, с нижней сто- роны налет. е растения В период вегетации.
Азалия, рододен- дрон. Phytophthora cactorum, Ph. cin- namomi. Корневая гниль; побурение, свер- тывание и поникание листьев; за- сыхание ветвей, почек. У 2—3-летних расте- ний.
Алоэ. Pythium debaryanum. Загнивание корневой шейки и от- мирание всего растения. В период вегетации.
Бегония. P. debaryanum, P. ultimum. Корневая гниль, побурение стеб- лей, гниль черешков листьев. То же.
CD
00
Питающие растение Возбудитель
Каланхое. Phytophthora cactorum, Ph. cryp- togea.
Калла. Ph. capsici, Ph. cryptogea, Ph.eryth- roseptica, Ph. richardiae.
Кактус. Pythium irregulare. Phytophthora cactorum, Ph. ni- cotinae var. parasitica.
Каттлейя. Колеус. Лотос. Синнингия (глок- синия) . Ph. cactorum. Pythium sp. Peronospora lotorum. Pythium debaryanum, Pythium sp. Phytophthora cryptogea.
Фиалка. Pythium sp. Phytophthora cactorum, Ph. cryp- togea. Peronospora violae.
Цинерария. Phytophthora megasperma, Ph. cin- namomi. Bremia lactucae.
П родолжение
Симптомы поражения
Сроки обследования
Поражение основания стебля,
загнивание цветоносов.
Гниль корневища, мякоть ста-
новится бурой или красноватой.
Листья желтеют, увядают. Задер-
живается рост всего растения; фор-
мируются неполноценные цветки.
Мягкая и слизистая гниль корня
и базальной части стебля.
То же.
В период вегетации.
То же.
В разные фазы разви-
тия.
То же.
Побурение и гниль корней. Корневая гниль. На листьях пятна. Корневая гниль. На листьях и черешках водя- нистые пятна, корневая гниль. Увядание и корневая гниль. То же. » » В период всходов. В период вегетации. В разные фазы разви- тия. То же. » » » »
На листьях пятна, с нижней стороны сероватый рыхлый налет. Корневая гниль. » » » »
На листьях светлые пятна, с ниж- ней стороны войлочный налет; листья съеживаются, засыхают. » »
НАИБОЛЕЕ ВРЕДОНОСНЫЕ БОЛЕЗНИ
Грибы порядка Peronosporales вызывают многочисленные опас-
ные болезни культурных растений, которые отличаются характер-
ными симптомами и в зависимости от принадлежности возбуди-
теля к тому или иному роду носят соответствующее название.
Терминологически различают следующие болезни: питиевая
корневая, прикорневая и плодовая гниль, вызываемая видами
Pythium^ фитофторовая корневая, прикорневая и плодовая гниль,
вызываемая видами Phytophthora (фитофтороз — более общее на-
именование; частный случай: фитофтора картофеля и томатов —
название, принятое у нас по давней традиции); склероспороз,
вызываемый видами Sclerospora] пероноспороз, вызываемый ви-
дами Peronospora] ложная мучнистая роса (мильдью) — название,
часто употребляемое для всех болезней, вызываемых пероноспо-
ровыми грибами, но, по нашему мнению, может быть использо-
вано применительно к болезням, вызываемым видами родов Plas-
mopara и Bremia (но не видами рода Peronospora (—Pseudopero-
nospora)] белая ржавчина, вызываемая видами Cystopus.
Болезни, вызываемые пероноспоровыми грибами
ПИТИЕВАЯ КОРНЕВАЯ, ПРИКОРНЕВАЯ
И ПЛОДОВАЯ ГНИЛЬ
Грибы из рода Pythium приурочены к растениям, находящимся
на ранних фазах развития, со слабо развитыми защитными по-
кровами, они вызывают преимущественно гибель сеянцев и моло-
7*
99
дых растений и лишь изредка способны нападать на окрепшие
взрослые экземпляры (например, Р. irregulare, вызывающий со-
судистый некроз свеклы).
Сельскохозяйственные культуры
Питиевая корневая гниль известна для многих сельскохозяй-
ственных растений — зерновых и зернобобовых, овощных и тех-
нических культур.
Корневая гниль пшеницы. Кроме пшеницы поражаются и дру-
гие хлебные злаки, а также травы (Subramaniam, 1928; Vanter-
pool, Truscott, 1932; Drechsler, 1936; Vanterpool, 1938; Vanterpool,
Sprague, 1942; Методические указания. . ., 1977). Вначале болезнь
проявляется в виде светло-коричневых пятен и размягченных уча-
стков на корнях; надземная часть желтеет и недоразвивается. При
сильном поражении в прикорневой части стебля, на соломине,
образуются темно-бурые пятна, переходящие в мягкую гниль.
Возможна гибель всего растения. Корневая гниль пшеницы вы-
зывается многими видами рода Pythium. Наиболее патогенны
Р. aristosporum, Р. arrhenomanes, Р. graminicolum, Р. tardicres-
cens, Р. volutum.
У Р. aristosporum гифы 2.5—6.5 мкм в диам., много аппрес-
сориев, спорангии мешковидные, вздутые, прорастают ростком.
Оогонии конечные или промежуточные, 21—36 мкм в диам.,
оболочка гладкая. Антеридии диклинные или андрогинные, по
2—4 булавовидных антеридия на 1 антеридиальной ветви; 3—6 и
более рядом с оогонием. Ооспора темно-коричневая, аплероти-
ческая, с гладкой оболочкой, иногда 2 ооспоры в 1 оогонии.
У Р. arrhenomanes гифы 2.0—5.5 мкм в диам. Зооспорангии
мешковидные, лопастные, 20 мкм, прорастают 20—50 зооспорами
или ростковой трубкой. Оогонии округлые, конечные, редко про-
межуточные, 17—56 мкм в диам., оболочка гладкая. Антеридии
диклинные, на 1 антеридиальной ветви по 4 антеридия; к оого-
нию примыкает 16—25 крючкообразных антеридиев. Ооспора пле-
ротическая, 15—54 мкм в диам., с гладкой бесцветной оболочкой
1.2—2.0 мкм толщ.
У Р. graminicolum гифы 2.5—7.0 мкм в диам. Зооспорангии
мешковидные, нитевидно вздутые, прорастают зооспорами; циста
8—11 мкм в диам. Конидии округлые, 24—36 мкм в диам., обра-
зуются в старых культурах, прорастают ростком. Оогонии округ-
лые, 16—36 мкм в диам., оболочка гладкая. Антеридии моноклин-
ные, на длинной антеридиальной ветви; 1—6 рядом с оогонием;
антеридиальная клетка булавовидная с плоской вершиной.
Ооспора плеротическая, 15—35 мкм в диам., с гладкой бесцветной
оболочкой 1.2 мкм толщ.
У Р. tardicrescens гифы 2.5—5.0 мкм в диам., с тонкими боко-
выми или шишковидными аппрессориями. Спорангии вздуто
нитевидные, дольчатые, лопастные, прорастают ростком. Оогонии
округлые, конечные 17—30 мкм в диам., оболочка гладкая. Анте-
100
Рис. 16. Гниль плодов кабачка, вы-
званная Pythium ultimum Trow.
ридии булавовидные, моноклин-
ные, редко диклинные; 2—3 ря-
дом с оогонием. Ооспора апле-
ротическая, 16—26 мкм в диам.,
с гладкой бесцветной оболочкой
1.2—2.0 мкм толщ.
У Р. volutum гифы 2.5—
5.5 мкм в диам. Зооспорангии
дольчатые, вздуто нитевидные,
мешковидные; прорастают зоо-
спорами; циста 10—14 мкм
в диам. Антеридии диклинные;
3—10 рядом с оогонием; антери-
диальная клетка крючкооб-
разноизогнута, антеридиальная
ветвь несет по 4 антеридия и
часто закручена вокруг оого-
ниальной. Ооспора аплеротичес-
кая, 14—36 мкм в диам.,
с гладкой бесцветной оболоч-
кой 2 мкм толщ.
Плодовая гниль тыквенных.
Плоды тыквенных (арбуз, дыня,
огурец, тыква) поражаются гри-
бом Р. ultimum на всех фазах
развития, особенно в фазе завязей и активного роста. В местах
соприкосновения с почкой на плодах появляются влажные,
быстро увеличивающиеся пятна. Плоды вначале твердые, по мере
заселения их гнилостными бактериями становятся мягкими.
При повышенной температуре и влажности на них появляется
ватообразный пышный мицелий. Картина болезни у разных пред-
ставителей тыквенных различна. На плодах арбуза появляются
быстро увеличивающиеся коричневые пятна; маленькие плоды
чернеют, прекращают рост и гибнут. Пораженные плоды дынь
и огурцов водянистые, мякоть рыхлая, при надавливании легко
трескаются. На плодах тыкв и кабачков образуются бесцветные
подкожные пятна, вследствие чего внешние признаки болезни
часто незаметны; кожица плода остается твердой, в то время как
мякоть полностью сгнивает (рис. 16). Гниль плодов тыквенных,
вызванная Р. ultimum, часто встречается в парниках.
У Р. ultimum гифы 1.7—6.5 мкм в диам., разветвленные.
Зооспорангии сферические, конечные, реже промежуточные, 12—
25 мкм в диам., прорастающие ростковой трубкой. Оогонии конеч-
ные, промежуточные, 19—23 мкм в диам., оболочка гладкая.
101
Рис. 17. Прорастание зооспорангиев Pythium aphanidermatum (Edson) Fitzp.
Антеридий обычно моноклинный, одиночный, сидячий, без длин-
ной антеридиальной ветви; редко 2 антеридия, тогда 1 диклинный.
Ооспора аплеротическая, 14—18 мкм в диам., оболочка гладкая,
до 3 мкм толщ.
Корневая гниль клевера. Вследствие корневой гнили наблю-
дается изреживание и выпадение этой культуры. Болезнь обна-
руживается в ранний период развития всходов и проявляется
в побурении и утончении корневой шейки и нижней части стебля,
в результате чего растение поникает и гибнет. При повышенной
влажности воздуха и почвы на пораженной части растения обра-
зуется обильный паутинистый налет мицелия гриба. Корневая
гниль клевера вызывается комплексом грибов, относящихся
к родам Fusarium, Rhizoctonia и Pythium (Р. debaryanum).
У Р. debaryanum2 гифы 2.0—7.0 мкм в диам., зооспорангии
прорастающие ростковой трубкой или с образованием зооспор.
Оогонии 10—40 мкм в диам., оболочка гладкая. Антеридии
булавовидные, соприкасаются с оогонием верхушечной частью;
2—5 рядом с оогонием. Ооспора аплеротическая, 8—30 мкм
в диам., с гладкой оболочкой 1—4 мкм толщ.
Корневая гниль свеклы. Известны две болезни свеклы, вызы-
ваемые питиевыми грибами. Первая из них, «корнеед», обуслов-
лена комплексом высших грибов (Phoma betae, Rhizoctonia ader-
holdii, Fusarium oxysporum, Alternaria tenuis) и низших из рода
Pythium (Тверской, 1955). Корнеед поражает свеклу в самой
2 При повторном упоминании вида его описание не приводится.
102
ранней фазе развития и выражается в потемнении, размягчении
и гибели всходов еще до выхода их на поверхность почвы в фазе
«вилочки» и до образования первых настоящих листьев. При
поражении, вызываемом литиевыми грибами, корень и подземная
часть стебля становятся водянистыми, развивается светло-бурая
гниль; надземная часть увядает, желтеет. Пораженные участки
затем засыхают и становятся темно-бурыми, почти черными.
В местах поражения развивается мицелий, распространяющийся
и в почве рядом с больным растением. В комплексе питиевых
грибов чаще всего встречаются Р. debaryanum, Р. ultimum, Р. apha-
nidermatum, Р. paroecandrum, Р. spinosum, Р. conidiophorum.
У Р. aphanidermatum гифы 2.5—7.3 мкм в диам. Зооспорангии
мешковидно вздутые, разветвленные, расположены группами
общим размером 50—1000 мкм дл., 4—20 мкм шир., прорастающие
зооспорами; циста 7.5—12 мкм в диам. (рис. 17). Оогонии конеч-
ные, редко промежуточные, сферические, 16—35 мкм в диам.
Антеридии моноклинные, диклинные, промежуточные, реже ко-
нечные, бочонковидные, цилиндрические, булавовидные, широко
прилегают к оогонию верхушечной частью; 1—2 рядом с оогонием.
Ооспора аплеротическая, 12—28 мкм в диам., с гладкой оболочкой.
У Р. paroecandrum гифы 2.5—8.5 мкм в диам., имеются изогну-
тые булавовидные аппрессории. Зооспорангии округлые, эллипсо-
идальные, промежуточные, реже конечные, 12—33 мкм в диам.,
прорастают зооспорами; циста 9—11 мкм в диам. Оогонии сфе-
рические, конечные, реже промежуточные, включающие часть
поддерживающей гифы, 11—30 мкм в диам., с гладкой оболочкой.
Антеридии моноклинные, диклинные, грушевидные, клювовидно
изогнутые; 1—5 рядом с оогонием. Ооспора аплеротическая,
10—22 мкм в диам., с гладкой оболочкой 1.1—1.5 мкм толщ.
У Р. spinosum гифы 2.4—5.8 мкм в диам. Зооспорангии округ-
лые, веретеновидные, конечные, промежуточные, 14—33 мкм
в диам., прорастают ростковой трубкой, реже зооспорами; циста
9 мкм в диам. Оогонии овальные, конечные или промежуточные,
13—27 мкм в диам., оболочка шиповатая; шипы конические или
ровные с тупой вершиной, 5—8 мкм дл., 1.5—2.0 мкм шир. Анте-
ридии моноклинные, реже диклинные или гипогинные; 1—2 рядом
с оогонием. Ооспора плеротическая, 10—25 мкм в диам., с глад-
кой оболочкой.
У Р. conidiophorum гифы слабо ветвистые, 2.5—6.5 мкм в диам.
Зооспорангии конечные, на длинных прямых спорангиеносцах,
прорастают ростковой трубкой. Оогонии конечные, сферические,
6.5—16 мкм в диам. Антеридии не обнаружены. Ооспора плеро-
тическая.
Другая литиевая корневая гниль свеклы называется «черный
сосудистый некроз». В отличие от корнееда болезнь проявляется
на корнях в более поздний период, на вполне сформировавшихся
корнеплодах. Сосуды корня буреют, при разрезании корня на
поверхности среза в условиях повышенной влажности появляется
.103
мицелий гриба. При хранении корнеплоды загнивают. Известно,
что эта болезнь может быть замечена на растениях, находящихся
еще в фазе 2—4 настоящих листьев. Сначала желтеют отдельные
участки пластинки между жилками, а затем весь лист, который
вскоре засыхает. Больные растения отстают в росте.
Возбудитель — Р. irregulare. Гифы 2—8 мкм в диам. Зоо-
спорангии конечные, промежуточные, округлые, яйцевидные,
грушевидные, 10—30 мкм в диам., прорастают ростковой трубкой,
реже — с образованием зооспор. Оогонии промежуточные, реже
конечные, сферические, 9.5—28 мкм в диам., оболочка неровного
контура с несколькими выростами. Антеридии булавовидные,
клювовидные, соприкасаются с оогонием верхушечной частью;
1—4 рядом с оогонием. Ооспора аплеротическая, 8—25 мкм
в диам., с гладкой оболочкой 1—4 мкм толщ. В СССР заболевание
отмечено в Курской обл. (Легенькая, 1963, 19666).
Корневая гниль гороха. Проявляется в виде побурения и во-
дянистой гнили корней и основания стебля. Вначале наблюдается
обесцвечивание пораженной ткани и коровой части корня, кото-
рая становится мягкой, водянистой. Гниль быстро прогрессирует
и охватывает кору, центральный цилиндр корня и стебель. Ткани
стебля выше корневой шейки разрушаются и ссыхаются. Желтеют
и отмирают нижние листья, поникает верхушка стебля, растение
засыхает. Заражение корневой системы возможно в течение всего
периода вегетации. Болезнь обнаружена в Нечерноземной зоне
РСФСР (Котова, 1968).
Возбудитель — Р. hydnosporum. Гифы 3—7 мкм в диам. Зоо-
спорангии округлые, 12—30 мкм в диам., прорастают ростковой
трубкой. Оогонии сферические, конечные, промежуточные, 19—
30 мкм в диам., оболочка равномерно шиповатая; шипы 3—
6 мкм дл. Антеридии гипогинные или эпигинные. Ооспора апле-
ротическая, 16—24 мкм в диам., с гладкой оболочкой 1—2 мкм
толщ.
Корневая гниль табака. Заболевание становится заметным
сразу при появлении всходов. Семядольные листья вначале сохра-
няют нормальную зеленую окраску, но увядают и распласты-
ваются по поверхности почвы. У всходов корень и стебель раз-
мягчаются и слегка буреют. Затем побурение захватывает все
ткани стебля и распространяется на листья и черешки, которые
становятся темно-зелеными и быстро теряют тургор. При пора-
жении более взрослой рассады в нижней части стебля образуется
перетяжка. В парниках при поражении рассады характерно
образование очагов (Богоявленская, Пыстина, 1974). Возбуди-
телем, вызывающим корневую гниль табачной рассады в СССР,
является, как правило, Р. perniciosum (в 1976—1977 гг. из боль-
ной рассады табака в Краснодарском крае выделен Р. aphani-
dermatum).
У Р. perniciosum гифы разветвленные, бесцветные, 2.0—6.0 мкм
в диам. Зооспорангии нитевидные, внешне неотличимые от веге-
104
тативной гифы, одиночные или группами, 12—60 мкм дл. При про-
растании протоплазма через эвакуационную трубку перетекает
в везикулу, где происходит формирование зооспор. Оогонии
сферические, 18—30 мкм в диам., со слабо бугорчатой угловатой
оболочкой. Антеридии булавовидные, 1, реже 2 рядом с оогонием,
соприкасаются с последним вершиной. Ооспора аплеротическая,
одиночная, с гладкой или угловатой оболочкой желтоватого
цвета; прорастают ооспоры ростком (примерно через 2 мес).
Лесные культуры
Корневая гниль сеянцев сосны и других хвойных деревьев.
Болезнь характеризуется тем, что молодые (до 6 мес) сеянцы
вначале желтеют и увядают, затем полегают. Полегание вызывает
гриб Р. debaryanum совместно с другими почвенными грибами
(Карпова-Бенуа, 1934; Ванин, 1955).
Цветочные культуры
Корневая и стеблевая гниль бегонии. Черная гниль стебля,
вызываемая Р. ultimum, причиняет большой вред растениям
в теплицах и садах. Поражаются корни и корневая шейка, те-
ряется тургор, и надземные органы растения полегают, разви-
вается мягкая гниль. При поражении растений Р. debaryanum
наблюдаются полегание в основном сеянцев и гниль молодых
растений, полученных при разведении из листьев, а также гниль
черешков и пластинок листа. В дальнейшем растения гибнут.
Корневая и прикорневая гниль герани, пеларгонии. Наиболее
сильно болезнь проявляется на молодых растениях, но может
поражать и взрослые. Гниль черенков начинается с основания
и прогрессирует вверх, вызывая увядание и отмирание листьев.
У взрослых растений побурение стебля развивается от основания
к вершине. Нижние участки загнивают, листья опадают, и расте-
ние гибнет.
Возбудитель — Р. splendens. Гифы относительно толстые
(6.5 мкм в диам.), встречаются серповидные аппрессории. Зоо-
спорангии сферические, конечные, с гладкой оболочкой, 22—
49 мкм в диам., прорастают чаще 1—6 ростковыми трубками.
Оогонии сферические, конечные, с гладкой оболочкой, 25—
34 мкм в диам. Антеридии моно- и диклинные, булавовидные,
изогнутые, соприкасаются с оогонием вершинной частью; 1—8 ря-
дом с оогонием. Ооспора не заполняет оогоний, оболочка гладкая,
до 4 мкм толщ.
Корневая гниль дельфиниума. Корневая гниль и гниль осно-
вания стебля приводят к увяданию растения. Возбудитель —
Р. ultimum. Гриб поражает виды Delphinium и их гибриды в теп-
личных и полевых условиях. Болезнь проявляется при повышен-
ной влажности воздуха и пониженной температуре (Dodge, Ri-
ckett, 1948).
8 Н. С. Новотельнова и др.
105
Корневая гниль каттлейи. Заболевание довольно редкое. Вна-
чале появляются овальные или циркулярные темно-коричневые
или черноватые пятна на листьях. Пятна окружены светло-
коричневыми зонами и перемежаются со светло-зелеными участ-
ками здоровой ткани. При увеличении пятен листья буреют и
погибают. Иногда возникают бурые пятна без зон. Возбудитель—
Р. splendens.
Корневая гниль глоксинии. Поражаться могут рассада и взрос-
лые растения. Загнивают корень и прикорневая часть стебля.
В условиях влажной почвы на загнивающих участках заметен
паутинистый налет мицелия гриба. Возбудитель — Р. debaryanum
(Кулибаба, 1970).
Корневая гниль астр. Поражаются в основном молодые расте-
ния. Буреют и загнивают прикорневая часть стебля и корень,
растение вянет и гибнет (Пыстина, 1977).
Возбудитель — Р. acanthicum. Гифы 2.5—3.8 мкм в диам.
Зооспорангии промежуточные, конечные, мешковидные, часто
в комплексах до 60 мкм дл., прорастают с образованием зооспор.
Оогонии сферические, конечные, промежуточные, 15—25 мкм
в диам., оболочка шиповатая; шипы тупоконечные, 4—5 мкм дл. и
2 мкм шир. в основании (рис. 18, см. вкл.). Антеридии моноклинные,
редко диклинные; 1—2 рядом с оогонием; антеридиальная клетка
иногда с 1—2 перетяжками на доли, соприкасается с оогонием
или всей боковой поверхностью, или сильно расширяющейся
вершиной. Ооспора плеротическая, 12—22 мкм в диам., с гладкой
оболочкой 1—1.5 мкм толщ.
Корневая гниль гвоздик. Чаще поражаются гвоздики вегета-
тивного происхождения. Черенки на высоте 1—2 см над поверх-
ностью почвы размягчаются и загнивают. Первые признаки пора-
жения проявляются в увядании и усыхании нижних листьев,
позже увядают и верхние. При повышенной влажности и затенен-
ности почвы на пораженных частях растения можно наблюдать
образование беловатого паутинистого налета — мицелия гриба.
Возбудитель — Р. debaryanum.
ФИТОФТОРОВАЯ КОРНЕВАЯ, ПРИКОРНЕВАЯ
И ПЛОДОВАЯ ГНИЛЬ. ФИТОФТОРОЗ
У фитофторовых грибов возможности преодоления возрастных
барьеров растений-хозяев значительно шире, чем у питиевых.
Большинство видов Phytophthora приурочено чаще к нисходящей
фазе развития питающих растений (соответственно теории имму-
ногенеза М. С. Дунина), например Ph. infestans, нападающий
на растения картофеля обычно после или во время периода цве-
тения.
Фитофторовые грибы причиняют многим растениям заболе-
вания, обозначаемые как корневая и прикорневая гниль. Кроме
того, широко распространены фитофторозы, обусловливающие
106
заболевания и других органов растений. К ним относится пора-
жение подземных органов растений, таких как клубни, луко-
вицы и т. д.
Сельскохозяйственные культуры
Фитофтора картофеля, или «поздняя гниль» . Поражает раз-
личные части растения по мере их развития. В осенний период,
с наступлением дождей и прохладных ночей, заболевание про-
является вначале на листьях. На них возникают одиночные круп-
ные пятна, которые постепенно разрастаются и буреют, листья
вянут, свешиваются вниз и засыхают (рис. 19). Буро-коричневые
пятна образуются и на черешках листьев и стебле в виде отдель-
ных или сплошных удлиненных полос. Пораженная ботва выгля-
дит на поле как побитая морозом. На нижней поверхности еще
зеленых листьев, по краям пятен, при влажной погоде заметен
нежный белый налет, состоящий из спороносцев со спорами,
которые сбиваются ветром или дождем и, просачиваясь через
почву, достигают клубней. Заражение последних осуществляется
через трещины в кожуре, что происходит также и в период уборки
картофеля, особенно при соприкосновении клубней с пораженной
ботвой. Больные клубни легко обнаруживаются по резко очер-
ченным вдавленным пятнам на кожуре серовато-бурого цвета;
при разрезе под ними видны окрашенные в ржаво-коричневый
цвет расплывчатые участки неразмягченной ткани. В поражен-
ные клубни проникают сапрофитные грибы и бактерии, вызы-
вающие гниение, в связи с чем часть урожая картофеля про-
падает во время хранения. При повышенной влажности воздуха
на кожуре клубня из чечевичек иногда выступают спороносцьь
Возбудитель — Ph. infestans. Мицелий зимует в клубнях
и весной проникает в ростки, находящиеся в зоне пораженных
глазков. На поверхности ростков гриб образует спороношение —
источник возобновления болезни. Мицелий гриба разветвленный,
иногда с небольшими вздутиями, образует хламидоспороподобные
тела и гаустории. Спорангиеносцы моноподиального ветвления
или неправильно разветвленные; ветви с четковидными взду-
тиями в местах прикрепления зооспорангиев, способные к возоб-
новлению роста после отчленения спор. Зооспорангии обильные,
лимоновидные, реже овально удлиненные, 29x19 мкм и более
(до 59x31), с небольшим выступающим сосочком и клеткой-
ножкой. Антеридии амфигинные, ооспоры формируются лишь
в парных культурах.
Розовая гниль клубней картофеля. Болезнь вызывает мягкую
гниль клубней, мякоть которых окрашивается в розовый цвет
(позднее она темнеет).
Возбудитель — Ph. erythroseptica. Мицелий разветвленный,
с крупными вздутиями и хламидоспороподобными телами, с гау-
сториями. Спорангиеносцы симподиально разветвленные, зоо-
спорангии овальные, неправильной формы, иногда асимметричные,
8*
107
Рис. 19. Лист картофеля, поражен-
ный Phytophthora infestans (Mont.)
dBy.
с уплощенной вершиной и без
сосочка, иногда с короткой нож-
кой. Антеридии всегда амфигин-
ные. Ооспоры образуются в
одиночных культурах.
Фитофтороз томатов. Пора-
жает только надземные части
растений, преимущественно
плоды, иногда листья. На ли-
стьях образуются бурые пятна,
а на стеблях темно-коричневые
полосы, мякоть незрелых пло-
дов становится ржаво-коричне-
вой и приобретает твердую кон-
систенцию. Возбудитель — Ph.
infestans.
Фитофторовая прикорневая
«южная» гниль томатов. В юж-
ных районах (Черноморское по-
бережье и др.) поражает при-
корневую часть растений в открытом грунте. На рассаде болезнь
сходна с «черной ножкой». Стебли размягчаются, утончаются, расте-
ния гибнут. В северных районах болезнь обнаруживается в теплицах
(отмечена в хозяйствах Московской и Ярославской областей).
Особенно сильно поражается рассада в теплицах для осенне-
зимней культуры при повышенной температуре воздуха. Пора-
жаются и плоды, находящиеся преимущественно вблизи поверх-
ности почвы: образуются пятна водянистой гнили с концентриче-
скими зонами («олений глаз») от серовато- до красновато-корич-
невого цвета и обильным белым мицелием на поверхности.
Болезнь вызывается несколькими видами: Ph. capsid (мицелий
с торулозными вздутиями, не образующий хламидоспор или
гаусториев; спорангиеносцы симподиально разветвленные; зоо-
спорангии овальной или неправильной формы, иногда несимме-
тричные, с 1 или 2—3 хорошо заметными выступающими сосоч-
ками, отрываются с участком гифы; антеридии амфигинные)/,
Ph. cryptogea (мицелий разветвленный, гифы со вздутиями, хла-
мидоспоры и гаустории не образуются; спорангиеносцы симпо-
диально разветвленные; зооспорангии овальные или неправиль-
ной формы, без сосочка; антеридии всегда амфигинные; органы
спороношения образуются лишь при особых условиях культи-
вирования); Ph. nicotianae чэл. parasitica (гифы без вздутий,
образующие хламидоспоры; зооспорангии от широкоовальных
108
до шаровидных с полушаровидным выступающим сосочком;
антеридии всегда амфигинные; органы полового спороношения
образуются лишь в парных культурах); Ph. drechslert (мицелий
со вздутиями, без хламидоспор и гаусториев; спорангиеносцы
симподиально разветвленные; зооспорангии удлиненные, без за-
метного сосочка, уплощенные на вершине; органы спороношения
образуются в небольшом количестве).
Фитофторовая штамбовая гниль яблони. Проявляется в виде
гнили ствола, корней и плодов. Поскольку гниль приурочена
к корневой шейке и часто опоясывает ствол, то ее именуют еще
воротниковой или шейковой гнилью. Заболевание характери-
зуется тем, что поражается часть ствола, находящаяся в непо-
средственной близости к почве и, как правило, близко к месту
прививки. В пораженном участке наблюдаются западание тканей,
фиолетово-коричневая окраска, часто он отделяется от здоровой
части разрывами коры. При надрезах штамба обнаруживаются
обширные пятна на внутренних частях коры, окрашенные в шоко-
ладно-коричневый цвет. Древесина обычно остается здоровой.
Вследствие того что поражение захватывает частично и сосудистую
систему, вскоре в кроне дерева обнаруживаются вторичные симп-
томы болезни — скручивание листьев на концах ветвей, их по-
краснение и сбрасывание.
С возрастом деревьев увеличивается их подверженность бо-
лезни. В природных условиях сильно поражаются экземпляры
в возрасте 12—14 лет, тогда как в возрасте 5—6 лет деревья за-
метно устойчивее. Однако искусственным путем удается осуще-
ствить заражение и молодых деревьев и даже однолетних привоев.
Болезнью поражаются также и плоды, причем не только опав-
шие и соприкасающиеся с почвой, но и находящиеся в кроне
деревьев в процессе созревания. Иногда фитофтороз плодов обнару-
живается в период хранения на складах. Главным диагностическим
признаком является темно-коричневая зона вокруг семенной камеры,
обнаруживаемая при разрезе пораженного яблока. Гниль плодов обычно
начинается с небольшого светло-коричневого пятна, резко очерчен-
ного на поверхности яблока. Она довольно быстро прогрессирует, од-
нако консистенция плода остается твердой, а поверхность блестящей.
Возбудитель — Ph. cactorum. Гриб образует мицелий с вет-
вями, отходящими под прямым углом и здесь часто с перетяжками,
а также с многочисленными клубневидными вздутиями; на гифах
формируются хламидоспоры и гаустории, проникающие в полости
клеток растения. Спорангиеносцы симподиально разветвленные,
несут овальные или лимоновидные зооспорангии с заметным
сосочком и небольшой ножкой, 20—55x17—46 мкм, в среднем
34x27 мкм; обильно образуются и органы полового размножения,
причем антеридии парагинные, реже амфигинные, ооспоры 20—
30 мкм в диам.
Болезнь широко распространена почти во всех странах мира,
принимает иногда эпифитотийный характер. Кроме яблони она
109
поражает грушу, вишню, сливу. Установлено, что чаще всего
гриб проникает в растение в месте прививки и окулировки. Бо-
лезнь развивается и в результате прикопки саженцев в почве,
содержащей отмершие части больных растений, а также при
горизонтальном формировании кроны и соприкосновении ветвей
с поверхностью почвы (Новотельнова, 1969а).
Прикорневая штамбовая гниль яблони вызывается также
Ph. syringae. Заболевание протекает с аналогичными симптомами,
но возбудитель имеет другие характерные особенности. Мицелий
его обычно в виде изогнутых гиф и торулозный, иногда с узлова-
тыми разветвлениями, не образующий хламидоспор, но с ните-
видными гаусториями, проникающими в клетки хозяина. Споран-
гиеносцы симподиально разветвленные, несущие зооспорангии
удлиненной формы с уплощенной вершйной и нерезко выступаю-
щим плоским сосочком, отрывающиеся с длинной ножкой. Анте-
ридии парагинные и амфигинные, ооспоры не всегда удается
обнаружить у изолятов. Этот вид поражает и молодые яблони
в питомниках.
Фитофторовая корневая и прикорневая гниль вишни. До-
вольно распространенное заболевание (в СССР не отмечено),
вызывающее значительные потери урожая в плодовых садах.
Пораженные деревья весной не развиваются или образуют мел-
кие желтоватые вялые листья. Такие деревья быстро погибают,
так же как и деревья, кажущиеся иногда весной здоровыми,
но засыхающие в течение первых жарких летних дней. У них
сильно поражены и разрушены грибом корни и нижняя часть
ствола вблизи от поверхности почвы, где обнаруживаются отми-
рающие участки коры.
Возбудители — разные виды Phytophthora: Ph. cactorum,
Ph. drechslert, Ph. megasperma. К характерным морфологическим
признакам Ph. megasperma относятся следующие: мицелий со
вздутиями и образованиями торулозного типа, образующий хла-
мидоспоры; спорангиеносцы простые или симподиально развет-
вленные, прорастающие нередко через опустевшие зооспорангии
и образующие затем новые овальные или лимоновидные зооспо-
рангии, особенностью которых являются закругленная вершина
и отсутствие заметного сосочка, они отрываются обычно с участком
гифы. Антеридии чаще парагинные, чем амфигинные. Зооспоран-
гии и ооспоры формируются быстро при искусственном культивиро-
вании. Установлено, что фитофторовая корневая и прикорневая
гниль вишни распространена в садах с плохо дренированной почвой.
Фитофторовая гниль корней земляники. Крайне вредоносное
заболевание, называемое «покраснение осевого сосудистого ци-
линдра корня», приводит обычно к прекращению развития боко-
вых корней и ползучих побегов и общему увяданию и отмиранию
растений.
Возбудитель — Ph. fragariae. Мицелий с гаусториями, про-
никающими в клетки растения; характерной особенностью
110
Рис. 20. Кончик корня земляники со спо-
роношением Phytophthora fragariae Hickm.
гриба является образование хлами-
доспор. Спорангиеносцы большей
частью мало дифференцированные,
часто прорастают через опустевшие
зооспорангии (пролификация); зоо-
спорангии удлиненно-овальные, чаще
обратногрушевидные, без сосочка
и ножки, отрывающиеся вместе с до-
вольно длинным участком несущей
гифы, 39—90x22—55 мкм, в сред-
нем 60X 38 мкм, при прорастании об-
разуют по 40—50 зооспор или рост-
ковые трубки; обильно образуются
также органы полового размножения,
причем антеридии и парагинные, и ам-
фигинные, а ооспоры свободно лежащие
в оогонии. Гриб заражает растения осенью или ранней весной
путем проникновения инфекционных ростков зооспор в корневые
волоски и развития внутри корня эндогенного мицелия (рис. 20).
Появлению и развитию болезни способствуют повышенная влаж-
ность почвы и температура 14—18° (Говорова, 1964).
Фитофтороз земляники. Вредоносная болезнь, характеризую-
щаяся поражением надземных (иногда и подземных) частей расте-
ния (рис. 21). Плоды легко загнивают с образованием водянистых
участков с синеватым оттенком; ягоды впоследствии приобретают ко-
ричневатую окраску и становятся кожистыми; поражаются также
черешки листьев; корни буреют, разрушаются; растения увядают,
засыхают (Андреева, 1968, 1977). Возбудитель — Ph. cactorum.
Прикорневая гниль крыжовника. Болезнь проявляется в виде
загнивания корневой шейки растения (Натальина, 1965). Воз-
будитель — Ph. cactorum.
Фитофтороз цитрусовых. Заболевание проявляется в виде
темно-бурых пятен на листьях лимона, апельсина, мандарина.
На плодах коричневая плотная гниль, постепенно охватывающая
весь плод. При поражении саженцев отмирают верхушки побегов,
ветвей и стволиков. Через трещины выделяется камедь. Са-
женцы быстро погибают.
Возбудитель — Ph. citrophthora. Мицелий разветвленный,
иногда образует хламидоспоры. Спорангиеносцы простые или
разветвленные, часто с подспорангиальными вздутиями 5—7 мкм
в диам. Зооспорангии овальные, лимоновидные, обратногруше-
видные или неправильной формы, 25—80x19—40 мкм, с хорошо
заметным сосочком, без ножки, обрывающейся вместе со споро-
носной ветвью. Органы полового размножения не обнаружены.
111
Лекарственные, цветочные
(и оранжерейные) растения
Фитофтороз женьшеня. Вредоносная болезнь, широко распро-
страненная в местах возделывания женьшеня. При поражении
сеянцев болезнь протекает очень быстро, вызывая гибель боль-
шинства всходов. Вначале на листьях в любом месте возникают
темно-зеленые сочные пятна, быстро охватывающие всю поверх-
ность листьев, а затем черешки и стебли. На пораженных частях
растений иногда обнаруживается спороношение гриба в виде
пушистого серовато-белого налета. На пораженных корнях раз-
вивается полумягкая гниль. Болезнь отмечена в Приморском
крае (Бункина, 1960). Здесь она появляется в мае—июне и вызы-
вает гибель растений (рис. 22).
Возбудитель — Ph. cactorum. Он образует спороношение в виде
симподиально разветвленных спорангиеносцев, выступающих на
поверхность пораженной ткани через устьица или прободая
оболочки. Зооспорангии эллипсоидальные или лимоновидные,
с хорошо заметным сосочком 20—55x17—46 мкм (или крупнее).
Зрелые зооспорангии разносятся по плантации ветром и дождем и,
Рис. 21. Земляника, пораженная Phytophthora cactorum (Leb. et Cohn) Schroet.
1 — здоровый, 2 — пораженный плодоносец.
112
Рис. 22. Женьшень, пораженный Phytophthora cactorum (Leb. et Cohn) Schroet.
заражая новые растения, осуществляют развитие новых генера-
ций пропагативных спор за 3—6 дней. Гриб образует также поло-
вые покоящиеся споры — ооспоры (20—30 мкм в диам.), которые,
по данным исследователей, способны сохраняться в почве в жизне-
способном состоянии до 4 лет. Ввиду широкой специализации гриб
способен поражать женьшень переходя на него с других растений.
Фитофторовая гниль корневой шейки кактуса — черная сли-
зистая «воротниковая» гниль прикорневой части растений. Возбу-
дитель — Ph. cactorum. Он сохраняется в почве и отсюда напа-
дает на растения.
Прикорневая гниль каланхое. Заболевание вначале прояв-
ляется у основания стебля в виде побурения. Гниль продвигается
вверх в цветоносы, которые вместе с листьями и цветками увя-
дают. Возбудитель — Ph. cactorum, ооспоры которого обнару-
живаются в пораженных тканях растения.
Корневая гниль каллы. Поражаются корни, часть из них
буреет и сгнивает, оставшиеся становятся водянистыми, размяг-
чаются. Корневище поражается так же. Симптомы болезни на
ИЗ
надземных частях растения проявляются в период начала цвете-
ния. На листьях возникают желтеющие полосы, они увядают,
буреют и отмирают. Цветки недоразвиваются, имеют иногда по-
буревший на вершине лепесток. Эти явления обнаруживаются
последовательно на развивающихся цветоносах. Возбудители —
Ph. richardiae, Ph'. cryptogea, Ph. erythroseptica. Они способны
относительно долго сохраняться в почве оранжерей в жизне-
способном латентном состоянии.
Гниль основания стебля лилии. Стебель поражается непосред-
ственно под поверхностью почвы. Пораженные части сморщива-
ются, растение вянет, поникает и отмирает. Грибы Ph. cactorum
и Ph. nicotianae var. parasitica известны как возбудители бурой
гнили не только стебля, но и луковиц, листьев и цветков.
Корневая и луковичная гниль тюльпана. Поражается основа-
ние луковицы. Возбудитель — Ph. cryptogea. Он проникает в ра-
стение через корни. Развитие болезни сопровождается увяданием
и гнилью цветоносов и засыханием бутонов, пожелтением и увя-
данием, иногда покраснением вершин листьев.
СКЛЕРОСПОРОЗЫ
Сельскохозяйственные культуры
Склероспороз пшеницы. Болезнь поражает большей частью
нижние листья растений, внутри которых образуются скопления
ооспор. Поражение обнаруживается в виде темных пятен, обве-
денных небольшим желтым кольцом, отделяющим их от неповреж-
денной, зеленой поверхности листа. Иногда сильно пораженные
растения остаются стерильными и зелеными в то время, когда
здоровые растения уже желтеют и созревают. Колосья могут
гипертрофироваться и иногда не выходят из влагалища верхнего
листа. Тычинки и пестик дегенерируют и могут принимать форму
зеленых листьев.
Возбудитель — Sclerospora macrospora — не образует беспо-
лого спороношения, а лишь ооспоры, расположенные одиночно и
группами в тканях листовых пластинок. Ооспоры коричневатого
цвета, круглые или неправильно овальные, имеют две оболочки:
внешнюю (темную, неравномерную по толщине, около 6.0 мкм)
и внутреннюю (толщиной 4.0 мкм). Ооспоры 50—55 мкм в диам.
S. macrospora кроме пшеницы найдена и на других хлебных
злаках: овсе, ячмене, кукурузе и на кормовых и дикорастущих
злаках: пырее, лисохвосте, овсянице и др.
Склероспороз ржи. Поражаются весной еще не окрепшие после
зимовки молодые растения. Болезнь сопровождается прекраще-
нием процесса ассимиляции пораженных листьев и приводит
к значительному ослаблению растений. Диагностическим призна-
ком болезни является наличие продольных пятен на листьях
с мелкошероховатой поверхностью в тех местах, где находятся
114
Рис. 23. Просо, пораженное Sclero-
spora graminicola (Sacc.) Schroet.
ооспоры, что заметно даже
невооруженным глазом. Боль-
ные листья никогда не размо-
чаливаются и не растрески-
ваются.
Возбудитель — S. secalina —
образует только половое споро-
ношение (оогонии весьма легко
обнаруживаются в тканях после
кипячения листьев в воде или
после удаления из них воздуха
длительным выдерживанием сре-
зов в воде). Диаметр оогониев
33—38 мкм. Антеридии округ-
лой, эллипсоидальной или гру-
шевидной формы, по 1 около
каждого оогония, 14.7 х 18.0 мкм.
Ооспоры ярко-желтой или зо-
лотисто-бурой окраски, 31 —
46 мкм в диам., оболочка их
плотно прирастает к оболочке
оогония, как у всех представи-
телей этого рода. S. secalina най-
дена только на ржи (Наумов,
1948, 1949).
Склероспороз риса. Поражение сопровождается образованием
на колосках вздутий и мясистых наростов голубовато-зеленого
цвета, семена не развиваются. В тканях вздутий формируются
ооспоры, расположенные одиночно или группами, 70x56 мкм
в диам. Возбудитель — S. macrospora.
Склероспороз кукурузы. Болезнь приводит к появлению бело-
вато-желтых или грязно-зеленых пятен, расположенных на ли-
стьях в виде полос, на которых чаще с нижней стороны выступает
слабый белый налет. Пораженные листья буреют, деформируются
и размочаливаются, волокна оставшихся проводящих пучков
скручиваются. Возбудитель — S. maydis — образует неправильно
разветвленные спорангиеносцы, формирующие овальные или эл-
липсоидальные бесцветные конидии 28—45x15.5—23 мкм.
Склероспороз проса, чумизы, могара. Болезнь приводит к по-
ражению зеленых клеток растений; нетронутыми остаются только
жилки листьев. Пораженные засохшие участки выпадают, и
к концу вегетации вместо листьев остаются только пучки волокон,
из которых при малейшем соприкосновении с ними высыпаются
115
совершенно зрелые ярко-оранжевые или цвета ржавчины ооспоры
(рис. 23). Зараженное растение сильно кустится, и вместо 2—3
образуется большое количество дегенерированных, бесплодных
метелок. На просе развитие болезни иногда ограничивается лишь
появлением на поверхности листьев матового грубого беловатого
нестирающегося налета, а в остальном просо имеет совершенно
нормальный вид.
Возбудитель — S. graminicola. На нижней поверхности пора-
женных листьев из устьиц одиночно или пучками выходят споран-
гиеносцы, образуя в массе сероватый, мало заметный и быстро
исчезающий налет. Спорангиеносцы 10 мкм дл., 10—14 мкм толщ.г
с короткими плотными ветвями, конечные стеригмы заостренные,
короткие. Зооспорангии почти шаровидные, 18—22x15—18 мкм.
Оогонии желтовато-коричневые, округлые, 45—50 мкм в диам.г
с оболочкой 5—18 мкм толщ. Ооспора шаровидная, гладкая,.
32—36 мкм в диам., с тонкой прозрачной оболочкой 2 мкм толщ.
ЛОЖНАЯ МУЧНИСТАЯ РОСА
Сельскохозяйственные культуры
Ложная мучнистая роса подсолнечника. Болезнь проявляется
в нескольких формах. При наиболее сильном поражении расте-
ния чахлые, целиком пронизанные мицелием, все органы покрыты
налетом спороношения (1-я форма). Эти растения погибают.
При 2-й форме пораженные растения развиваются, но резко от-
стают в росте; некоторые проходят все фазы развития вплоть
до образования семян, но остаются карликовыми. На больных
растениях изменяется окраска листьев. По этому признаку
болезнь можно обнаружить в поле рано весной, на растениях,
находящихся еще в фазе семядольных листьев. У них на листьях
заметны слегка пожелтевшие участки ближе к основанию. На
настоящих листьях пятна более резко выражены и выделя-
ются светло-зеленой мозаичной окраской, растекающейся вдоль
жилок на темно-зеленом фоне здоровой части листа от основания
листа к вершине (Новотельнова, 1966а).
На нижней стороне пластинок мозаичных листьев во влажную
погоду выступает налет. Вначале он нежный, белый, но позднее
становится сероватым, густым, войлочным. На нижних листьях
налет образуется чаще всего вдоль жилок у основания листа,
а на более высоко расположенных налетом покрывается вся ниж-
няя поверхность пластинки. Иногда, особенно в период продол-
жительных дождей, налет возникает также и на верхней стороне
листьев, но при продолжительной засухе он может вовсе не про-
явиться даже на листьях карликовых, явно пораженных расте-
ний. В этом случае для уточнения болезни можно искусственно
вызвать налет, помещая больные листья в условия влажной ка-
меры. Иногда в поле встречаются пораженные растения без
116
Рис. 24. Мицелий с гаусториями у
Plasmopara helianthi Novot. в кожуре
-семени подсолнечника.
признаков общего поражения
(как это наблюдается при 1-й
и 2-й формах болезни). С виду та-
кие растения кажутся здоровыми,
но на листьях имеют отдельные
разбросанные угловатые светлые
пятна, ограниченные жилками, на
которых в условиях высокой влаж-
ности воздуха появляется белый
налет спороношения гриба. Эти
пятна являются результатом мест-
ной инфекции (3-я форма), тогда
как 1-я и 2-я формы болезни
представляют собой общее (диф-
фузное) поражение.
Помимо указанных форм отме-
чены еще форма прикорневая
(спороношение образуется только
на корнях и внутри корневой
шейки при наличии в ней полос-
лей) и форма корзиночная (позд-
няя форма поражения растений,
находящихся в фазе созревания
семян) (Тихонов, 1968).
Возбудитель — Plasmopara he-
lianthi f. helianthi (=P. hal-
stedii) — поражает подсолнечник
и однолетние виды рода Helian-
thus L. (Новотельнова, 1961,
19626, 1963а, 19636). Мицелий
гриба неклеточный, бесцветный, толстый, ветвистый, распрост-
раняется внутри растения диффузно, в том числе в семенах
(рис. 24), и проходит по межклетникам, образуя многочисленные
пузыревидные гаустории. Спорангиеносцы 150—750 мкм выс.,
разветвленные, с ветвями, отходящими под прямым углом. Ко-
нечные ветки несут на вершинах по 3—6 стеригм, на которых
образуются зооспорангии гриба, имеющие эллипсоидальную форму
< уплощенными боками и бугорком на вершине (рис. 25, см. вкл.),
величина их 15—30x12—21 мкм. В воде они прорастают с обра-
зованием 7—10 и более зооспор, крупных, 7.5—12x7—9 мкм;
оптимальная температура образования их 16—18°, минималь-
ная 5—6°, максимальная 28°. При благоприятных условиях зоо-
споры прорастают с образованием ростковых трубок, проника-
ющих в ткани подсолнечника, и вызывают заражение. В меж-
117
клетниках тканей пораженных растений в весенне-летний период
закладываются покоящиеся споры. Ооспоры шаровидные, 23 —
32 мкм в диам., со слабо складчатой оболочкой, желтоватые,
встречаются и в листьях, и (чаще) в подземной части больных
растений. В виде ооспор гриб зимует внутри растительных остат-
ков, т. е. последние являются основным источником инфекции.
Возможна также передача болезни семенами (Новотельнова, 1963в).
Первичное заражение подсолнечника в поле, обусловленное
прорастанием ооспор возбудителя, осуществляется ранней вес-
ной вскоре после появления всходов, когда низкие темпера-
туры воздуха сменяются повышенными и выпадают дожди и
росы. В этих условиях стимулируются прорастание ооспор и об-
разование зооспор, являющихся источником первичной инфек-
ции. Начиная с периода образования спороношения на листьях
первично пораженных растений в поле уже имеется источник
вторичной инфекции. Образовавшиеся зооспорангии при наличии
благоприятных условий быстро прорастают и вызывают зараже-
ние новых растений, что обусловливает массовое появление бо-
лезни. Возбудитель ложной мучнистой росы подсолнечника об-
ладает высокой выносливостью и приспособленностью к условиям
среды. Низкие температуры на поверхности почвы зимой
(до —30° С) и высокие температуры в весенне-летний период,
а также засуха не предотвращают развития болезни в зоне ее
распространения.
Ложная мучнистая роса винограда. Поражаются все надзем-
ные органы. На листьях образуются маслянистые желтовато-
зеленые пятна, на нижней стороне которых образуется сероватый
налет. Пятна постепенно буреют, количество их увеличивается,
и листья отмирают. Подобные пятна и налет появляются на пора-
женных цветках, бутонах, завязях, молодых побегах. Созревшие
ягоды буреют и засыхают.
Возбудитель — Р. viticola — образует моноподиально раз-
ветвленные спорангиеносцы до 647.5 мкм выс., с раскидистыми,
ветвями, многократно разветвленными большей частью под пря-
мым углом, имеющими стеригмоподобные окончания. Зооспо-
рангии 14.5—23.2x10.1—15.9 мкм, эллипсоидальные, бесцвет-
ные или слегка желтоватые. Бесполое спороношение возникает
весной, несколько позднее формируются ооспоры, которые,
как показали исследования А. Т. Макрушиной (1966, 1970),
появляются только в листьях, старых и молодых, с июня по
октябрь. Их больше образуется в засушливую, чем в дождливую
пору. Жизнеспособными ооспоры остаются не более двух лет.
Установлена их устойчивость к воздействию низкой температуры
и замораживанию. Они успешно прорастают при благоприятных
условиях начиная с января—февраля, если в природных условиях
находятся в растительных остатках на глубине 10—15 см или
в пониженных влажных местах, под слоем листьев и не подверга-
ются высушиванию.
118
Цветочные, лекарственные культуры
Ложная мучнистая роса астр. Заболевание довольно широко
распространено в Западной Сибири, Новосибирской, Омской и
Кемеровской областях, вызывает вначале пятнистость и некроз
листьев, а впоследствии полное засыхание. Пятна угловатые,
небольшие или расплывчатые, ограниченные жилками (рис. 26,
см. вкл.), с нижней стороны с сероватым; налетом.
Возбудитель — Р. asterea f. callistephi (Новотельнова, 19636).
Спорангиеносцы короткие, 200—250x12—20 мкм, с короткими,
уродливо вздутыми ветвями. Зооспорангии сливовидные, 18—
36x12—24 мкм, вытянутые у основания, с сохранившейся
клеткой-ножкой (рис. 27). Ооспоры золотистые, 39—60 мкм
в диам.
Ложная мучнистая роса астр вызывается и другим пероноспо-
ровым грибом — Basidiophora entospora. Она отмечена на план-
тациях в Техасе, где вызывала поражение, достигавшее 30%.
У нас этот гриб на астрах не обнаружен.
Ложная мучнистая роса василька. Болезнь проявляется в виде
отдельных, неравномерно разбросанных светлых или краснова-
тых, позднее буреющих пятен на листьях растений, особенно на
ранних фазах развития. На нижней стороне листьев выступает
обильный пушистый белый налет.
Возбудитель — Bremia lactucae. Конидиеносцы выступающие,
многократно дихотомически под острым углом разветвленные,
190—400 мкм выс., с конечными, дланевидно расширенными вет-
вями. Конидии почти шаровидные или широкоэллипсоидальные,
17-24 Х10—19 мкм, прорастающие с образованием ростковых
трубок.
Ложная мучнистая роса женьшеня. Заболевание отмечено на
женьшене в 1955 и 1956 гг. в Приморском крае (Бункина, 1960).
На листьях обнаружены угловатой неправильной, часто рас-
плывчатой формы пятна светло-бурого цвета, ограниченные жил-
ками. На нижней стороне листовой пластинки скупо выступает
грязно-белый или сероватый налет спороношения гриба. Пятна
засыхают и крошатся, и листья становятся дырчатыми. Заболе-
вание проявлялось очагами на сравнительно небольшой площади
0.02 га. Развитие болезни было кратковременным. Оно отмечено
в конце июня, максимального развития достигло 10—15 июля,
после чего пораженных растений на плантациях больше не за-
мечено. В следующем году обнаружены лишь единичные больные
экземпляры.
Возбудитель — Plasmopara panacis. Спорангиеносцы выходят
из устьиц по 2—3, у основания желтоватые, на вершине бесцвет-
ные, 145—200 (320) х 6.6—8.0 мкм, в основании вздутые (10—
12 мкм шир.). Зооспорангии бесцветные, округлые, 14—16.5 X
X 11.5—16.5 или 19.8X16.5 мкм. Ооспоры не обнаружены.
119
Рис. 27. Спорангиеносцы и споры Plas-
mopara asterea Novot.
Ложная мучнистая роса золо-
тарника. Болезнь проявляется на
всходах золотарника диффузно,
с образованием крупного пятна,
охватывающего почти весь лист
и расположенного вдоль главной
жилки, с нижней стороны плот-
ный, войлочный, беловатый налет.
В СССР заболевание обнаружено
в Ленинградской обл. и на Коль-
ском полуострове (Новотельнова,
19636).
Возбудитель — Р. solidaginis.
Спорангиеносцы 200—550 х (6) 10—
15 мкм. Зооспорангии 15—27x12—21 мкм, округлые, слегка
вытянутые у основания. Ооспоры не обнаружены.
Ложная мучнистая роса цинерарии. У больных растений
листья со светлыми пятнами на верхней стороне и войлочным
беловатым налетом на нижней. Листья съеживаются и засыхают.
Болезнь особенно сильно поражает всходы.
Возбудитель — Bremta lactucae, Конидиеносцы 180—500 мкм
выс., выступающие группами из устьиц, дихотомически
разветвленные, с дланевидными расширениями на конечных вет-
вях. Конидии шаровидные, широкоэллипсоидальные, 13—20 X
Х12—19 мкм, прорастают с образованием ростковых трубок.
Ооспоры желтоватые, 25—40 мкм в диам., с толстой желтовато-
коричневой гладкой или слегка бородавчатой оболочкой.
ПЕРОНОСПОРОЗ
Сельскохозяйственные культуры
Пероноспороз гречихи. Заболевание проявляется на листьях
и цветках. На листьях пятна небольшие или крупные, округлые
или расплывчатые, желтоватые, просвечивающие, водянистые
(рис. 28), на нижней стороне выступает едва заметный серовато-
фиолетовый рыхлый налет. В соцветии пораженные цветки по-
крываются нежным сероватым налетом, потом приобретают ко-
ричневую окраску и засыхают (Еленев, 1922).
Возбудитель — Peronospora fagopyri. Мицелий гриба эндо-
фитный. Конидиеносцы 380—500 мкм выс., 3—4-кратно ди-
хотомически разветвленные. Конидии яйцевидные, 16—26 X
120
Зак. 891
Рис. 25. Спорангиеносцы Plasmopara helianthi Novot. (а- и З-спороношения).
Рис. 26. Лист астры, пораженный Plasmopara asterea
Novot.
Рис. 28. Лист гречихи, пораженный
Peronospora fagopyri Elenev.
X12—16 мкм. Оогонии округ-
лые, коричневатые, 38—46 мкм
вдиам., с гладкой оболочкой.
Ооспоры шаровидные, коричне-
ватые, гладкие, 22—25 мкм
в диам.
В течение лета заражение
происходит конидиями, для
прорастания которых тре-
буются капельно-жидкая влага
в виде росы или дождя и
сравнительно высокая темпера-
тура.
Пероноспороз сои. Болезнь
вызывает сильное поражение сои
во всех зонах ее выращивания.
Первое появление гриба отмеча-
ется весной на семядольных листьях, позднее на настоящих
листьях, бобах и семенах, т. е, поражается все растение. При по-
вышенной влажности воздуха и умеренной температуре болезнь
быстро распространяется в поле от растения к растению. На верх-
ней стороне листовых пластинок обнаруживаются светло-зеленые
угловатые или расплывчатые «маслянистые» пятна, постепенно
буреющие в связи с наступающим некрозом тканей. При диффуз-
ном поражении гриб сильно воздействует на растение, рост его
прекращается, оно теряет нормальный облик и некоторое время
вегетирует в виде малооблиственного и неплодоносящего карлика,
у которого потом листья засыхают и опадают (на нижней стороне
листьев серый налет).
Возбудитель — Р. manshurica. Мицелий гриба межклеточный,
с нитевидными гаусториями, сплетенными в виде вытянутого
мотка. На пораженных органах возникает бесполое спороношение
в виде многочисленных многократно дихотомически разветвлен-
ных спороносцев 250—500 мкм выс., у которых на вершинах
конечных ветвей формируются широкоэллипсоидальные, реже
шаровидные конидии 14—30x18—24 мкм, прорастающие с об-
разованием ростков. Ооспоры 25—48 мкм в диам., желтоватые,
с оболочкой гладкой или с неправильной сеткой.
Распространение гриба и заражение других растений про-
исходят в течение всего вегетационного периода, чему способ-
ствуют повышенная влажность воздуха и умеренная температура.
Жаркая и сухая погода неблагоприятна для этого паразита.
Сохраняется гриб в виде мицелия в семенах и в виде ооспор,
которые образуются в семядольных и настоящих листьях и в бо-
9 Н. С. Новотельнова и др.
121
бах, причем обнаруживаются также на поверхности семян в виде
пленочек кремового цвета, выходящих наружу при дифферен-
циации плода. Ооспоры сохраняются в жизнеспособном состо-
янии в репродуктивных органах растений около полутора лет,
а в остатках вегетативных частей в природе не более одного года.
Пероноспороз люцерны. Болезнь вызывает поражение отдель-
ных вегетативных или репродуктивных органов (листьев, стеб-
лей, бобов и др.) или поражает диффузно все растение. В послед-
нем случае рост задерживается, растение недоразвивается, отли-
чается более светлой, желтоватой окраской и, как правило, гибнет.
При местной инфекции на листьях возникают разной вели-
чины мозаичные пятна, на нижней поверхности выступает гряз-
новато-белый, позднее серовато-фиолетовый налет. Бобы разви-
ваются иногда деформированные, а семена в них неполноценные.
Возбудитель — Р. aestivalis. Мицелий с гаусториями. Кони-
диеносцы 185—600 мкм выс., 4—8-кратно дихотомически развет-
вленные, с дугообразно изогнутыми ветвями. Конечные ветви
расходятся под прямым, острым, реже тупым углом, с заострен-
ными, реже притупленными концами. Конидии широкооваль-
ные, широкоэллипсоидальные, с заметным сосочком, 16—37 X
Х9—27 мкм, буроватые или серовато-фиолетовые. Ооспоры
шаровидные, 20—30 мкм в диам., с гладкой или бугорчатой зо-
лотистой оболочкой.
Повсеместно встречается в южных регионах, а также отмечен
в Прибалтике.
Пероноспороз шпината. Вредоносное и широко распростра-
ненное заболевание. Если болезнь проявляется рано, в фазу
семядольных листьев, растения погйбают. При более позднем
поражении на настоящих листьях образуются отдельные или сли-
вающиеся светлые пятна, на нижней стороне которых появля-
ется плотный войлочный серовато-фиолетовый налет. Поражен-
ные листья засыхают, сильно пораженные растения гибнут.
Возбудитель — Р. spinaciae. Мицелий межклеточный, с га-
усториями. Конидиеносцы дихотомически разветвленные. Кони-
дии 22—32x15—22 мкм, имеются ооспоры.
Пероноспороз огурцов. Болезнь развивается особенно при
возделывании огурцов в парниках и теплицах, где причиняет
значительный ущерб. Характеризуется образованием угловатых,
сначала желтовато-зеленых, позднее буреющих пятен, увеличи-
вающихся в размере и охватывающих полностью листья, с ниж-
ней стороны которых образуется обильный серовато-фиолетовый
налет. Листья сморщиваются, засыхают и крошатся, а при повы-
шенной влажности воздуха загнивают.
Возбудитель — Р. (=Pseudo peronospora) cubensis. Мицелий
с гаусториями. Конидиеносцы 120—320 мкм выс., дихотомически
разветвленные под острым углом, конечные ветви тупые или слабо
заостренные. Зооспорангии широкоовальные и эллипсоидальные,
20-28 Х16—20 мкм, с сосочком, прорастающие с образованием
122
Рис. 29. Цветонос лука, пораженный Pero-
nospora destructor (Berk.) Casp.
зооспор. Ооспоры шаровидные, 36—
43.5 мкм в диам., с гладкой желтой обо-
лочкой.
Заболевание особенно вредоносно в
Приморском крае в связи с преобладанием
высокой влажности воздуха. Поражение
растений достигает иногда 50%.
Пероноспороз лука. Поражаются виды
рода Allium. Очень вредоносное и ши-
роко распространенное заболевание. По-
раженные листья и цветоносы недоразви-
ваются, искривляются (рис. 29). Пероно-
опороз часто сопровождается гнилью, вы-
зываемой грибом Stemphy Ilium allii Oud.
(Абдуллаев, 1951). Проявляется пероно-
спороз в виде удлиненных, сначала
хлоротичных, потом бурых пятен, распо-
ложенных вдоль листа или цветоноса и
занимающих его значительную часть. На
пятнах выступает светлый, но темнеющий
до дымчато-фиолетового цвета рыхлый
налет. Пораженные органы засыхают,
сжимаются (Шошиашвили, Кирмелаш-
вили, 1950; Blotnicka, 1974).
Возбудитель — Peronospora destructor.
Мицелий около 8 мкм в диам., с суже-
ниями до 1.5 мкм и утолщениями до
37 мкм, с извилисто лентовидными гаус-
ториями, располагается в межклетниках
листьев и наружных и внутренних чешуях
луковиц. Конидиеносцы 240—650 мкм
выс., 4—6-кратно дихотомически, ниж-
ние иногда симподиально разветвленные,
основание конидиеносца обычно взду-
тое. Ветви слегка дугообразно согнутые, на концах притуп-
ленные или заостренные. Конидии яйцевидно-продолговатые,
грушевидные, 19—60x18—29 мкм, желтоватые или серовато-
фиолетовые. Ооспоры шаровидные, 25—35 (40) мкм в диам.,
в разных частях листа, часто рядами вдоль сосудов. При
прорастании ооспор образуются ростки с макроконидиями
на вершинах (McKay, 1957; Стенина, 1974). Мицелий в поражен-
ных луковицах чаще является источником возобновления ин-
фекции из года в год (Каттерфельд, 1926; Viranyi, 1974).
9*
123
Пероноспороз свеклы. Болезнь поражает растения рода Beta,
в том числе свеклы столовой и сахарной на всех стадиях их раз-
вития. Появляется ранней весной (март—апрель) на семенниках
безвысадочной свеклы, где образуются первые очаги болезни.
Возникающие на листьях споры разносятся ветром и воздушными
течениями и попадают на посевы. В посевах болезнь обнаружи-
вается уже на самых ранних стадиях развития свеклы. Поражен-
ные экземпляры отличаются от здоровых значительно более блед-
ной окраской и резко выраженной курчавостью семядольных
и настоящих листьев. При поражении точки роста развитие ра-
стений приостанавливается, в жаркое время дня растения при-
вядают. По мере роста возникают мелкие недоразвитые толстые
и курчавые листья с завернутыми книзу краями, иногда с красно-
ватой пигментацией пластинки. На более старых листьях гриб
вызывает пятна различной величины и более бледной окраски.
Болезнь часто приводит растения к полной гибели.
Возбудитель — Р. schachtii. Мицелий гриба эндофитный, вет-
вистый, с многочисленными нитевидными разветвленными га-
усториями, способен зимовать в верхней части корнеплодов.
Конидиеносцы бесцветные, многократно дихотомически раз-
ветвленные, 250—650 мкм выс. Конидии яйцевидные, дымчатого
цвета, 21—27x16—20 мкм. Ооспоры шаровидные, желтовато-
коричневые, 36—40 мкм в диам., с толстой гладкой оболочкой,
образуются редко.
Интенсивное развитие гриба наблюдается в мае—июне, в период
начала плодоношения — «стрелкования». В летний засушливый
период развитие гриба может элиминироваться, но осенью,
с изменением погодных условий в сторону повышения влажности
и понижения температуры, возобновляется и на вновь образу-
ющихся листьях появляется свежее спороношение. Летнее споро-
ношение развивается обычно на нижней стороне листовых пла-
стинок в виде серовато-бурого с фиолетовым оттенком войлочного
налета. На формирование конидий большое влияние оказывает
температура воздуха. Они чувствительны к сухости воздуха и по-
вышенной температуре и более выносливы при пониженной тем-
пературе. Так, при 25° С зрелое спороношение образуется во
влажных условиях уже через 15 ч, а при 4° для того же про-
цесса требуется более 1.5 сут. В сухом воздухе при 1—4° конидии
сохраняют жизнеспособность до 2 мес. В почве конидии теряют
способность к прорастанию через 3—6 дней под воздействием
почвенных абиотических и биотических факторов, из которых
не последнюю роль играют населяющие почву виды микроскопи-
ческих сапрофитных грибов, бактерий, низших животных.
В течение вегетационного периода гриб образует и половое
спороношение во всех органах растений: листьях, стеблях, цвет-
ках, семенах. Половой процесс наступает довольно рано, но чаще
связан с наступлением жаркого летнего периода, а осенью —
холодного. Роль ооспор в жизненном цикле гриба неясна, так как
124
прорастание их, по наблюдениям исследователей, сомнительно.
По-видимому, гриб резервируется главным образом в виде ми-
целия в растениях безвысадочной свеклы, корнях маточной свеклы
и семенах.
Р. schachtii очень вредоносен, он вызывает преждевременное
усыхание и отмирание в поле большого количества растений,
приводит к значительному снижению урожая корнеплодов и их
сахаристости, уменьшению урожая семян, ухудшению их посев-
ных качеств — уменьшению абсолютного веса, энергии прора-
стания, всхожести и полному бесплодию цветоносов (Синицына,
1965).
Пероноспороз табака. Болезнь обнаруживается в парниках
и в поле на протяжении всего периода вегетации табака. Поражен-
ные в парниках всходы имеют желтовато-зеленые листья с ча-
стично загнутыми вниз краями. С нижней стороны лист покрыва-
ется серовато-фиолетовым налетом; иногда налет появляется и
на верхней стороне листьев. При раннем поражении, как и при
сильном развитии болезни, рассада погибает. При поражении
рассады в более позднем возрасте на листьях появляются хлоро-
тичные «маслянистые» пятна, на жилках и черешках бурые и
вдавленные. Иногда болезнь носит скрытый характер — пора-
женные всходы внешне почти не отличаются от здоровых. Исход
болезни в таком случае зависит от ухода за рассадой.
В полевых условиях пероноспороз табака может поражать
все органы растения: листья (пластинки, жилки, черешки),
стебли, цветки, коробочки и корни. На листьях возникают хлоро-
тичные «маслянистые» пятна разного размера, распространяющи-
еся обычно от черешка к вершине листа либо разбросанные по
всей листовой пластинке. При сильном развитии болезни много-
численные пятна сливаются и занимают значительную часть
листа или полностью весь лист. Плотный серовато-фиолетовый
налет на нижней стороне листьев по мере отмирания листьев
становится грязно-серым или бурым. В сухую жаркую погоду
развитие налета не наблюдается. Пораженные стебли и корни
буреют. При диффузном поражении наблюдается отставание
в росте или гибель растений (Канчавели и др., 1967; Сикан, Вол-
жина, 1974).
Возбудитель — Р. tabacina. Мицелий гриба эндофитный,
сильно разветвленный, с многочисленными лопастными гаусто-
риями. Конидиеносцы 320—500 мкм выс., 4—6-кратно дихото-
мически разветвленные. Конидии бесцветные, овальные с неболь-
шим бугорком и ножкой в месте прикрепления к конидиеносцу,
17-30 Х12—19 мкм. Ооспоры 20—60 мкм в диам., темно-бурые.
Бесполое спороношение развивается в сырую, пасмурную погоду,
в ранние утренние часы. В течение вегетационного периода гриб
образует многие генерации спор, хорошо переносит зимовку
в климатических условиях табаководческой зоны и, ежегодно
возобновляясь, наносит существенный ущерб растениям табака
125
Рис. 30. Хмель с колосовидным побегом, пораженный Peronospora (=Pseudo-
peronospora) humuli Miy. et Tak.
в теплицах и на плантациях. Он вызывает эпифитотии и настолько
вредоносен, что поставил под угрозу табаководство во многих
странах мира. Поражает и другие пасленовые (перец, петуния)
(Легенькая, 1970).
Рассада заражается весной перезимовавшими в растительных
остатках ооспорами (источником инфекции могут быть и семена
с пораженных растений); сильно пораженные растения обычно
гибнут, а оставшиеся, перенесенные в полевые условия, становятся
центрами очагов распространения болезни. При влажной и дожд-
ливой погоде на нижней стороне пораженных листьев появля-
ется спороношение гриба; сначала оно голубоватого цвета (от-
сюда название болезни «голубая плесень»), затем становится серо-
ватым. Гриб поселяется в цветках, поражает образующиеся се-
мена и, таким образом, также наносит ущерб культуре табака
(Боровская, 1968а, 19686, 1970). В ряде случаев форма поражения
и форма существования гриба внутри хозяина не дают прямых
указаний к этиологии и диагностике: и симптомы поражения,
и сам гриб под влиянием хозяина и факторов среды не обусловли-
вают типичной для пероноспоровых грибов картины поражения
(Богоявленская, 1971).
126
Рис. 31. Спорангиеносцы гриба Pe-
ronospora (=Pseudoperonospora) hu-
muli Miy. et Так. с зооспорангиями,
образующими зооспоры.
Пероноспороз конопли. Бо-
лезнь проявляется в пораже-
нии листьев, на которых вна-
чале возникают хлоротичные
участки, имеющие угловатую
форму и ограниченные жил-
ками. С прогрессированием бо-
лезни пятна сливаются, буреют,
листья засыхают. На поражен-
ных листьях, большей частью
с нижней стороны, развивается
серовато-фиолетовый налет.
Возбудитель — Р. (=Pseu-
doperonospora) cannabina — обра-
зует мицелий с гаусториями. Спорангиеносцы 120—280 мкмвыс.,
дихотомически разветвленные, с ветвями, отходящими под острым
углом, с фиолетовым оттенком, конечные ветви с заостренными
окончаниями отходят большей частью под прямым углом. Зо-
оспорангии 18—36x4—24 мкм, с небольшим сосочком, имеют
слабо фиолетовую окраску. В межклеточных тканях формиру-
ются желтоватые шаровидные ооспоры 27—87 мкм в диам., с глад-
кой оболочкой.
Пероноспороз хмеля. Поражаются молодые побеги, которые
становятся колосовидными (рис. 30). Такие побеги имеют укоро-
ченные междоузлия, на них развиваются мелкие скрученные
морщинистые, уменьшающиеся к вершине листья. Уродуются
и главные и вторичные побеги, появляются так называемые
«белые побеги», частично лишенные хлорофилла. Возбудитель
болезни обнаруживается также в листьях, кажущихся внешне
здоровыми, но вскоре на них возникают угловатые пятна светло-
зеленой окраски, листья буреют, засыхают и отмирают. Позднее
возбудитель проходит в цветки, а затем и в «шишки» хмеля, ко-
торые остаются недоразвитыми и бледно-зелеными, постепенно
буреют, засыхают и преждевременно опадают.
Возбудитель — Р. (=Pseudoperonospord) humuli. Мицелий чаще
диффузный, с гаусториями, формирует на пораженных органах
довольно плотный войлочный налет от серовато-фиолетовой до
бурой окраски. Спороносцы многократно дихотомически ветвя-
щиеся, 180—300 мкм выс., с ветвями, отходящими под прямым
углом и дугообразно изогнутыми, с заостренными вильчатыми
конечными отрогами. Зооспорангии овальной и эллипсоидальной
формы, 13—32 Х10—20 мкм, прорастающие с образованием зоо-
127
спор (рис. 31). Внутри пораженных тканей формируются шаро-
видные желтоватые ооспоры 27—87 мкм в диам.
Пероноспороз капусты. Широко распространенное заболева-
ние рассады и взрослых растений. При поражении рассады и се-
менников болезнь наиболее вредоносна, поскольку диффузно
поражается все растение: стебли, цветки, стручки — и ему на-
носится существенный ущерб. При поражении взрослых растений
гриб не столь вредоносен ввиду того, что поселяется преимуще-
ственно на нижних листьях (рис. 32, см. вкл.); однако болезнь способ-
ствует проникновению в растение факультативных паразитов и
приводит к уменьшению лежкости качанов (Анисимов, 1962).
Возбудитель — Р. brasstcae. Мицелий межклеточный, с гаусто-
риями. Конидиеносцы 250—460 мкм выс., разветвленные дихото-
мически, с изогнутыми ветвями, отходящими под острым углом.
Конидии широкоовальные, эллипсоидальные, 18—28x15—23 мкм.
Ооспоры шаровидные, 22—30 мкм в диам., отмечены в листьях
и в кочерыге (Куприянова, 1957); прорастания их не наблюдалось.
Мицелий гриба, обнаруженный внутри и в оболочке семян, со-
храняет жизнеспособность в течение 6 лет и является источником
ежегодного возобновления болезни. В повышении устойчивости
капусты к пероноспорозу большое значение придается регуляции
светового режима (Поляков, Владимирская, 1964).
Цветочные, лекарственные культуры
Пероноспороз аптечной ромашки. Заболевание вызывает на
листьях, реже на стеблях пятна с рыхлым обильным налетом.
Соцветия большей частью остаются непораженными.
Возбудитель — Р. leptosperma. Конидиеносцы 375—533 X
Х10.8—13 мкм, конечные ветви прямые или слабо изогнутые.
Конидии эллипсоидальные, 19—48.7x16.5 мкм, иногда с неболь-
шим бугорком. Ооспоры 30—32 мкм в диам.
Пероноспороз белены. Относительно широко распространенное
заболевание. Вредоносность его проявляется в снижении лекарст-
венной ценности пораженных растений вследствие засыхания
листьев и значительного уменьшения в них содержания алкало-
идов (Енкина, 1970). Встречаются 3 формы болезни, из которых
одна, диффузная, обнаруживается в фазу всходов, при этом
растения недоразвиваются, достигают высоты не более 1—10 см
и погибают; на семядольных листьях выступает конидиальный
налет. При второй форме, тоже диффузной, болезнь обнаружива-
ется в фазе розетки: листья сморщенные вдоль укороченной
жилки, хлоротичные, особенно у основания, на нижней стороне
с обильным плотным серовато-фиолетовым налетом; растения
второго года вегетации тоже поражаются грибом. Третья форма
локальная, встречается в виде отдельных крупных или небольших
светлых пятен па листьях (рис. 33, см. вкл.).
128
Рис. 32. Лист капусты со спороношением Peronospora brassicae
Gaeum.
Эак. 891
Рис. 33. Лист белены, пораженный
Peronospora hyoscyami dBy.
Заболевание ежегодно отмечалось в Новосибирской обл. в пи-
томнике лекарственных растений, особенно в годы с высокой
относительной влажностью воздуха. Развитию болезни спо-
собствовали загущенный посев и повышенная влажность воз-
духа.
Возбудитель — Р. hyoscyami. Мицелий с гаусториями до
9.7 мкм дл. и 2.0 мкм толщ. Конидиеносцы 162—423 (750) х
Хб—8(10) мкм, 2—6-кратно дихотомически разветвленные, ко-
нечные ветви до 45 мкм дл. или короче, отходят под прямым,
реже острым или тупым углом, прямые, иногда слегка согнутые,
заостренные, реже притупленные. Конидии сливовидные,
21.6—27x16—18 мкм. Ооспоры шаровидные, 30—44 мкм в диам,,
с гладкой или складчатой желтовато-коричневой оболочкой.
Пероноспороз мака. Заболевание проявляется на протяжении
всего периода вегетации, начиная от всходов и до созревания
коробочек. Поражаются листья, стебли, цветоносы, цветки и
коробочки. Листья сморщиваются, на них возникают желтоватые
«маслянистые» пятна, покрытые с нижней стороны сероватым
рыхлым налетом; пораженные части листа буреют, листья засы-
хают. Стебли и цветоносы прекращают рост, обычно сильно ис-
кривляются и тоже покрываются налетом. Одни бутоны отмирают,
другие развиваются, но образующиеся цветки и коробочки мень-
ших размеров, чем на здоровых растениях (Зюбин, 1963), наблю-
даются уродливости цветков и коробочек.
Возбудитель — Р. arborescens. Мицелий с многочисленными
разветвленными гаусториями. Конидиеносцы 300—850 мкм выс.,
3—8-кратно дихотомически разветвленные. Конидии яйцевидные,
дымчатые, 19—20x18—19 мкм. Ооспоры многочисленные, шаро-
видные, темно-коричневые, 30—38 мкм в диам., с толстой оболоч-
кой.
При заражении листьев гриб прободает кутикулу и эпидермис,
проникает в стебель, спускается в корень, поднимается вверх
и заходит в цветки, а затем в семена, где и сохраняется из года
в год. Таким образом, растительные остатки и семена являются
источником инфекции. Р. arborescens паразитирует на растениях
сем. маковых. Он поражает виды маков, относящиеся к лекар-
ственным, декоративным и техническим растениям, и распростра-
нен во всех макосеющих районах, особенно в восточной и юго-
восточной Европе. Сильно воздействует на урожай мака лекар-
ственного, вызывая иногда гибель растений, а также снижая
количество алкалоидов у пораженных экземпляров.
Пероноспороз наперстянки. Болезнь проявляется на листьях
вначале в виде беловатого, впоследствии серовато-фиолетового
налета. Листья желтеют, буреют и отмирают.
Возбудитель — Р. dtgitalidis. Конидиеносцы200—400мкмвыс.,
3—8-кратно дихотомически разветвленные, с ветвями, отхо-
дящими под острым углом, 10—30 мкм дл. Конидии эллип-
соидальные, 18—37x11—21 мкм. Ооспоры не обнаружены.
129
Пероноспороз фиалки. Болезнь проявляется в возникновении
на верхней стороне листьев неравномерно разбросанных пятен,
с нижней стороны которых образуется рыхлый сероватый налет.
Нижние листья, соприкасающиеся с почвой, поражаются раньте
и вскоре отмирают.
Возбудитель — Р. violae. Конидиеносцы 200—400 мкм выс.,
3—7-кратно дихотомически разветвленные, с ветвями, отходя-
щими под острым углом, с конечными ветвями прямыми или крюч-
кообразно согнутыми, 10—17 мкм дл. Конидии овальные или эл-
липсоидальные, 18—32x13—21 мкм. Ооспоры шаровидные, 25—
30 мкм в диам., с толстой гладкой светло-желтой оболочкой.
БЕЛАЯ РЖАВЧИНА
Сельскохозяйственные и цветочные культуры
Белая ржавчина подсолнечника. Пораженные растения имеют
характерные симптомы: пустулы на нижней стороне листьев и
светло-зеленые угловатые пятна на верхней стороне. Пустулы
мелкие, белесоватые, часто собранные группами, иногда слива-
ющиеся; найдены только на листовых пластинках, а на черешках,
стебле, корзинке и других органах не обнаружены. На поражен-
ных растениях искривлений и деформаций не наблюдалось.
Пока эта болезнь обнаружена только в Краснодарском крае
(Новотельнова, 1962а, 1962в, 1966) и почти не изучена. В связи
с опасностью, которую она представляет при расширении ареала
и возможном усилении вредоносности, необходима постановка
специальных исследований по изучению болезни и ее возбудителя.
Возбудитель — Cystopus tragopogonis f. helianthi. Под эпидер-
мисом листа развиваются собранные в группы спорангиеносцы
36—48 мкм выс., на которых цепочками образуются шаровидные,
сдавленные с боков зооспорангии 18—20.4x15.6 мкм, имеющие
в экваториальной части кольцевидное утолщение; здесь же раз-
виваются и стерильные (непрорастающие) клетки шаровидной
или неправильной формы, 16.8 мкм в диам., с оболочкой до
2 мкм толщ. (рис. 34). Прорастание зооспорангиев с образованием
зооспор происходит при наличии капельной влаги и благопри-
ятной температуре воздуха. Оптимальна пониженная температура
в пределах 8—16°.
Белая ржавчина капусты. Болезнь проявляется в виде под-
эпидермальных белых выпуклых и блестящих пятен, при разрыве
эпидермиса порошащих. Поражаются листья рассады, а также
стебли и цветоносы.
Возбудитель — С. candidus var. candidus. Мицелий межкле-
точный, с шаровидно-овальными гаусториями. Спорангиеносцы
булавовидные, тесно скученные, 30—40 мкм выс. На них образу-
ются цепочки зооспорангиев 12—18 мкм в диам., округлые,
иногда немного угловатые. Внутри тканей формируются шаро-
130
Рис. 34. Спорангиеносцы и спорангии Cysto-
pus tragopogonis (Pers.) Schroet. f. helianthi
Novot.
видные желтовато-коричневые ооспоры
с мелкобугорчатой оболочкой.
С теми же признаками поражения
известна белая ржавчина других овощ-
ных культур: репы, турнепса, брюквы,
редьки, редиса, хрена, а также гор-
чицы, рыжика, крамбе, относящихся
к масличным культурам.
Белая ржавчина семенников крестоцветных» Одно из наиболее
вредоносных заболеваний семенников крестоцветных. Поража-
ются листья, стебли, цветоносы, цветки и стручки. На листьях
бледно-желтые мелкие пятна 0.3—0.5 мм в диам., единичные
или многочисленные, на нижней стороне образуются беловатые
блестящие подушечки — пустулы гриба. Такие же пустулы
покрывают все пораженные части растений. На стеблях и цвето-
носах они сливаются в виде длинных полос или муфты. При раз-
рушении эпидермиса, прикрывающего подушечки, высыпается
белая порошковидная масса: зооспорангии гриба — возбудителя
болезни. Стебли и цветоносы искривляются. Стручки на поражен-
ных цветоносах, как правило, не развиваются (Легенькая, 1963).
Возбудитель — С. candidus. Мицелий эндофитный, диффуз-
ный, с многочисленными пузыревидными гаусториями. Конидие-
носцы булавовидные, 30—40 мкм выс., образуются слоем в пусту-
лах под эпидермисом. На них возникают цепочками округлые
зооспорангии 12—18 мкм в диам. Ооспоры шаровидные, желто-
вато-коричневые, 50—63 мкм в диам., оболочка мелкобугорчатая.
Зимует гриб в растительных остатках в виде ооспор и в виде
мицелия в семенах. Для его весеннего возобновления необходима
повышенная влажность почвы и воздуха, а для прорастания зоо-
спорангиев — капельно-жидкая влага и температура не выше
15-18°.
Белая ржавчина амарантуса. Болезнь проявляется на листьях,
стеблях, цветках в виде прикрытых вначале эпидермисом белых,
слегка желтоватых пустул, при созревании порошащих. Стебли
и цветки часто искривляются.
Возбудитель — С. bliti. Спорангиеносцы булавовидные,
16.5—46x11.2—13.8 мкм, слоем возникают под эпидермисом,
поднимают его и впоследствии разрывают. На спорангиеносцах
цепочками образуются округлые зооспорангии 12.6—22 х
X 12—18.9 мкм, имеющие по экватору утолщение оболочки и
прорастающие с образованием зооспор. В тканях формируются
шаровидные ооспоры 46—66 мкм в диам. с темно-коричневой
сетчатой оболочкой.
131
Белая ржавчина маттиолы. На листьях, чаще на нижней сто
роне, возникают многочисленные белые блестящие выпуклые
пустулы. Листья желтеют, буреют и отмирают; стебли, цветки,
плоды часто гипертрофируются. Возбудитель — С. candidus.
Белая ржавчина каперцев. Болезнь поражает каперцы с обра-
зованием характерных выпуклых округлых пустул, расположен-
ных на надземных органах.
Возбудитель — С. capparidis. Мицелий с округлыми гаусто-
риями. Спорангиеносцы короткие, булавовидные, располага-
ющиеся тесным слоем. Зооспорангии округлые, 13—20 х
Х12—21 мкм. В межклетниках формируются ооспоры 30—
40 мкм в диам., с бородавчатой оболочкой.
Белая ржавчина портулака. Болезнь, довольно широко распро-
страненная в разных регионах страны. Проявляется в виде бле-
стящих беловато-желтоватых пустул на листьях. Пустулы вна-
чале прикрыты эпидермисом и после его разрыва порошат.
Возбудитель — С. portulacae. Мицелий межклеточный, с яй-
цевидными гаусториями. Спорангиеносцы короткие, булавовид-
ные, 20—40 мкм выс., расположены тесным слоем. Зооспорангии
образуются в цепочках, широкоовальные, иногда угловатые,
с равномерно утолщенной оболочкой 14—22x12—16 мкм, про-
растающие с образованием зооспор. Вершинный зооспорангий
округлый, 20—22 мкм в диам., толстостенный, бесплодный или
прорастает ростком. Ооспоры шаровидные, 35—60 мкм в диам.,
с сетчатой желтовато-коричневой оболочкой.
Болезнь наносит иногда значительный вред в Армении, сни-
жая качество зеленой массы портулака, употребляемого в пищу
(Тетеревникова-Бабаян, 1964).
В технологии сельскохозяйственного производства в борьбе
с болезнями, вызываемыми пероноспоровыми грибами, наиболь-
шее значение имеют предупредительные мероприятия. Среди
них на первом месте стоят карантинные меры по охране нашей
территории от завоза болезней растений из зарубежных стран
и предотвращение распространения новых болезней из отдель-
ных появившихся очагов.
Для предупреждения ложной мучнистой росы и пероноспоро-
зов осуществляют в первую очередь культивирование устойчи-
вых сортов в сочетании с применением биологически обоснован-
ных агротехнических, а в ряде случаев и химических меропри-
ятий.
К наиболее важным агротехническим приемам относится
уничтожение растительных остатков, так как в них сохраняются
зимующие формы грибов. С той же целью необходимо строгое
соблюдение чередования культур и пространственной изоляции
(не менее 1 км) от полей, зараженных в предыдущем году, а также
изоляции посевов семецников от посевов первого года. Необхо-
димо при прорывке пораженных растений уничтожать их; свое-
132
временно проводить удаление сорняков и падалицы; уборку
урожая вести в сжатые сроки.
В связи с тем что многие из перечисленных болезней переда-
ются семенами, требуется, чтобы семеноводческие хозяйства,
расположенные в зоне распространения болезней, применяли
правила защиты посевов от ложной мучнистой росы и пероноспо-
розов и не допускали к посеву и рассылке семена с зараженных
полей.
Совершенствование методов борьбы с болезнями, вызываемыми
пероноспоровыми грибами, и разработка новых эффективных
средств борьбы с ними требуют фундаментального и всесторон-
него изучения этих организмов. Прежде всего должна быть
изучена биология вредных видов. Особое значение имеют иссле-
дования по их онтогенезу и фазам развития, а также установление
зависимости существования видов от факторов внешней среды
(температуры, влажности, освещенности и др.). Необходимо
изучение способов и мест сохранения видов и условий их ежегод-
ного возобновления в природе. Сведения по этим вопросам можно
получить только при проведении специальных работ по культи-
вированию видов в лабораторных условиях и испытанию их
в природных условиях.
Многие пероноспоровые грибы являются облигатными пара-
зитами. При облигатном паразитизме между паразитом и хозя-
ином складываются особые отношения, при которых два орга-
низма длительное время сосуществуют, причем один поглощает
питательные вещества из клеток другого, не вызывая их гибели.
В чем сущность этих взаимоотношений? Каков механизм их осу-
ществления? Для того чтобы ответить на эти вопросы, необходимо
цитологическое изучение пораженных растений.
В фитопатологических исследованиях световая микроскопия
используется очень широко, однако зачастую возможности этого
метода реализуются не полностью. Недостаточно часто применя-
ются фазово-контрастная, темнопольная и люминесцентная микро-
скопия. Очень ценны гистохимические исследования, позволя-
ющие выявить физиологические изменения в мицелии и клетках
гриба и в клетках пораженного растения. Световая микроскопия
незаменима при изучении морфологии видов, при выявлении
распространения мицелия в тканях растений. Однако ее возмож-
ности очень ограниченны, если дело касается внутриклеточных
структур.
В 60-х годах в практику фитопатологических исследований
вошла электронная микроскопия. Этот метод открыл исследова-
телям новый, многообразный мир внутриклеточных структур.
Электронно-микроскопическое изучение патогенных грибов
в тканях растений дает возможность изучить структурные основы
взаимоотношений между патогеном и хозяином, проследить из-
менения в клетках хозяина, лежащие в основе иммунных реакций
растений.
133
При изучении иммунитета растений, особенно в случаях
болезней, вызываемых пероноспоровыми грибами — облигатными
паразитами, необходимо применять электронно-микроскопиче-
ский метод, который должен послужить основой для соответ-
ствующих работ. Ультраструктурные исследования имеют здесь
особую ценность. В комплексе с биохимическими и генетическими
изысканиями они дают наиболее полную и точную информацию
о строении, функциях и характере взаимоотношений между па-
разитом и хозяином.
Дальнейшее изучение пероноспоровых грибов должно раз-
виваться в следующих направлениях.
Изучение морфологии и изменчивости видов, а также внутри-
видовой дифференциации, необходимых для селекционных целей.
Установление жизненного цикла видов и выяснение роли
покоящихся стадий в сохранении и возобновлении инфекции,
а также роли семян в передаче инфекции, необходимых для био-
логического обоснования эффективного комплекса зональных си-
стем защиты растений.
Уточнение симптомов болезней, вызываемых пероноспоровыми
грибами, для разработки методов ранней диагностики болезней.
Изучение ареалов патогенных видов и выявление новых оча-
гов наряду с изучением экологических особенностей видов для
успешного прогнозирования их распространения на территории
СССР и своевременного профилактического блокирования.
Определение физиолого-биохимических особенностей перо-
носпоровых грибов (ферментативной активности, токсичности
и др.), раскрывающих паразитические свойства возбудителей.
Изучение взаимоотношений патогенов с растениями-хозяевами
на микроскопическом и субмикроскопическом уровнях; опреде-
ление факторов устойчивости для дальнейшей рационализации
семеноводства и сортосмены, исключающей или понижающей
приспособительную изменчивость вредных видов.
Изыскание новых сочетаний приемов, подавляющих или сдер-
живающих развитие и распространение вредоносных видов,
с целью создания интегрированных систем зональных меропри-
ятий.
Несомненно, что нужна дальнейшая разработка методов ис-
следований, которые позволят полнее изучить скрытые особен-
ности биологии пероноспоровых грибов для эффективной защиты
культурных растений.
134
ЛИТЕРАТУРА
Абдуллаев С. Г. Пероноспора лука в Азербайджане и меры борьбы
с ней. — Тр. ВИЗР, 1951, вып. 3, с. 49-57.
Андреева Н. Ф. О фитофторе на землянике. — Микол, и фитопатол.,
1968, т. 2, вып. 6, с. 471—474.
Андреева Н. Ф. Фитофтора на землянике. — Защ. раст., 1977, № 4,
с. 47.
Анисимов А. М. Влияние микроэлементов и антибиотиков на устой-
чивость рассады капусты к ложномучнистой росе. — Тез. докл. науч,
конф. Харьк. с.-х. ин-та, 1962, с. 33.
Асатиани Ш. Ш. К вопросу о патогенности Pythium ultimum Trow
в условиях Грузии. — Матер. 3-й закавказ. конф, по спор. раст. АН
ГрузССР, Тбилиси, 1968, с. 112—114.
Бабаян А. А., Григорян Н. Прорастание конидий Peronospora
tabacina Adam на листьях сортов табака с различной устойчивостью к пе-
роноспорозу. — Биол. журн. Армении, 1970, т. 23, вып. 8, с. 21—25.
Бабаян А. А., Шалджян Л. К. Развитие Peronospora tabacina
Adam в период вегетации на листьях табака в Армении. — Матер,
закавказ. конф, по спор, раст., Баку, 1965, с. 140—146.
Бахтин В. С. Об ооспорах Peronospora hyosciami. — Матер, по микол.
и фитопатол., 1926, т. 5, вып. 2, с. 158—160.
Бебия Е.А. О продолжительности жизни конидий Peronospora tabacina. —
Микол, и фитопатол., 1975, т. 9, вып. 1, с. 51—52.
Белозор Н. И. К вопросу о распространении фитофторы на территории
СССР в отдельные годы. Сб. тр. аспирантов и молодых науч, сотруд.
ВИР, Л., 1960, с. 305-310.
Богоявленская Р. А. Необычное проявление пероноспороза та-
бака. — Микол, и фитопатол., 1971, т. 5, вып. 6, с. 506—508.
Богоявленская Р. А., Пыстина К. A. Pythium perniciosum
Serb. — возбудитель рассадной гнили табака. — Микол, и фитопатол.,
1974, т. 8, вып. 4, с. 336—341.
Бордукова М. В. Больные фитофторой клубни являются передатчи-
ками болезни. — Вести, овощев. и картоф., 1941, № 2, с. 87—93.
Боровская М. Ф. Пероноспороз табака в Молдавии. Кишинев, 1968а.
30 с.
Боровская М. Ф. Ооспоры — источник возобновления пероноспороза
табака. — Микол, и фитопатол., 19686, т. 8, вып. 4, с. 311—315.
Боровская М. Ф. Проникновение и сохранение инфекции пероноспо-
роза табака в семенах. — В кн.: Вопросы защ. раст. Т. 1. Кишинев,
1970, с. 146—156.
Б р у и д з а К. Паразитные грибы культивируемых растений Литовской ССР
и некоторые вопросы биологии паразитных грибов. Вильнюс, 1961.302 с.
Бункина И. А. Итоги изучения болезней женьшеня. — Матер, к изуч.
женьшеня и лимонника, 1960, вып. 4, с. 131—162.
Ванин С. И. Лесная фитопатология. М.—Л., 1955, с. 178—180.
Венцлавович Ф. С., Новотельнова Н. С., ГуржиевГ. А.
Нарушение жизнедеятельности подсолнечника при поражении ложной
мучнистой росой. — Тр. по прикл. бот., ген. и селекц., Л., 1970, т. 42,
вып. 1, с. 178—184.
135
Виноградов В. А. Расовый состав Peronospora tabacina Adam и оценка
устойчивости исходного селекционного материала. Автореф. канд. дис.
М., 1977. 16 cJ
Вялых А. К., Ж е р я г и н В. Г. Условия заражения амброзии полынно-
листной (Ambrosia artemisiifolia) возбудителем белой ржавчины Albugo
tragopogonis Schroet. — Микол, и фитопатол., 1977, т. 11, вып. 2, с. 135—
140.
Вялых А. К., Ланецкий В. П. Люминесцентно-микроскопический
метод определения жизнеспособности спор грибов рода Albugo. — Микол,
и фитопатол., 1974, т. 8, вып. 2, с. 155—157.
Говорова Т. Ф. Заболевание земляники, вызываемое грибом Phyto-
phthora fragariae Hickm. — Бюл. Главн. бот. сада, 1964, вып. 54, с. 105—
НО.
Голубева О. Г. О структуре и функциях гаусториев паразитиче-
ских грибов. — Микол, и фитопатол., 1979, т. 13, вып. 1, с. 69—77.
Горленко М. В. Миграция фитопатогенных микроорганизмов. М.,
1975. 107 с.
Дмитриева С. В., Беккер 3. Э. Некоторые данные о природе
волютиновых гранул Penicillium chrysogenum. — Цитология, 1962, т. 4,
вып. 6, с. 691—695.
Дорожкин Н. А. Современное состояние изученности фитофтороза
картофеля. — Докл. науч. конф, по защ. раст., Вильнюс, 1959, с. 239—
251.
Дорожкин Н. А., Шуканов А. С. К вопросу о перезимовке воз-
будителя пероноспороза сахарной свеклы Peronospora schachtii в условиях
Белоруссии. — Докл. АН БССР, 1965, т. 9, вып. 3, с. 208—211.
Дунин М. С. Иммуногенез и его практическое использование. — Тр.
Моск. с.-х. акад., 1946, вып. 40, с. 1—145.
Е л е н е в П. Ф. Болезнь гречихи. — Хозяин, 1922, № 7, с. 6—9.
Енкина Т. В. Патогенные микромицеты на травянистых растениях Но-
восибирской области. Канд. дис. 1970. 232 с. (Бот. сад Зап.-Сиб. отд-ния
АН СССР).
Захарова Т.И. Методические указания по разработке системы прогноза
фитофтороза помидор. Л., 1965, с. 1—10.
3 ю б и н Б. Пероноспороз опийного мака. — Колхозно-совхозное произ-
водство Киргизии, 1963, № 9, с. 22.
Ибрагимов Г. Р. Причины эпифитотий Peronospora tabacina Adam
Азербайджанской ССР. — Микол, и фитопатол., 1968, т. 2, вып. 2, с. 121.
Иванчева-Габровска Т. Имунитетни проучвания при чернилката
по тютюна {Phytophthora parasitica). (Устойчивость сортов табака к по-
чернению стебля). — Науч. тр. н.-и. ин-та защ. раст., София, 1968,
ч. 1, с. 65—103.
Иванчева-Габровска Т., Димитров Д. Г. Проучване влия-
нието на кършенето, филизенето и филизоотглеждането при тютюна
върху устойчивостта му спрямо чернилката {Phytophthora parasitica var.
nicotianae). — Бюлетин по растителна защита, София, 1968, т. 7, кн. 1
и 2, с. 72-91.
Канчавели Л. А., Сакварелидзе Н. А., Кикачеи-
ш в и л и 3. Н., Тогонидзе Е. В. Особенности развития пероно-
спороза табака и основные биологические свойства Peronospora tabacina
Adam. — Тр. Ин-та защ. раст. ГрузССР, 1967, т. 19, с. 114—116.
Каратыгин И. В. Строение патогена и его развитие в тканях растений
хозяев как критерий устойчивости к грибной инфекции. — Микол,
и фитопатол., т. 2, вып. 5, 1968, с. 414—420.
Карпова-Бенуа Е. И. Полегание сеянцев хвойных пород в лесных
питомниках и борьба с этим заболеванием. Л., 1934. 62 с.
Каттерфельд Н. О. К биологии развития Peronospora schleideni Ung. —
Болезни раст., 1926, № 2, с. 71—87.
Корецкий П. М. Ложная мучнистая роса сои на Дальнем Востоке. —
В кн.: Наука — сельскому хозяйству. Хабаровск, 1966, с. 45—49.
136
Коршунова А. Ф., Чумаков А. Е., Щекочихина Р. И.
Защита пшеницы от корневых гнилей. Изд. 2-е. Л., 1976, с. 184.
Котова В. В. Новое заболевание гороха, вызываемое Pythium hydnospo-
rum (Mont.) Schroet. — Микол, и фитопатол., 1968, т. 2, вып. 5, с. 420—
426.
Кузичева В. В. Некоторые физиолого-биохимические особенности
гриба Peronospora schachtii Fuckel. — В кн.: Систематика, экология,
физиология почвенных грибов. Киев, 1975, с. 172—173.
Кукин В. Ф. Иммунитет подсолнечника к инфекционным болезням и прак-
тическое использование его в селекции. — Тез. докл. 4-го Всесоюз. совещ.
по иммунитету с.-х. раст., Кишинев, 1965, с. 177—179.
Кулибаба Ю. Ф. Патогенная микофлора цветочно-декоративных куль-
тур в субтропиках Краснодарского края. — Тр. ВИЗР, 1970, вып. 29,.
ч. 1, с. 62—72.
Куприянова В.К. Особенности биологии Peronospora brassicae Gaum.—
Бот. журн., 1957, т. 42, вып. 5, с. 760—763.
Курсанов Л. И. Микология. М.—Л., 1933. 480 с.
Легенькая Е. И. Распространение и вредоносность болезней сельско-
хозяйственных растений, вызываемых пероноспоровыми грибами в Цен-
трально-черноземной зоне СССР. — Тр. ВИЗР, 1963, вып. 19, с. 72—80.
Легенькая Е. И. Грибы порядка Peronosporales — возбудители болез-
ней сельскохозяйственных растений в Центрально-черноземной зон&
РСФСР. Канд. дис. 1966а. 169 с. (ВИЗР).
Легенькая Е. И. Новый для СССР вид — Pythium irregulare Buis. —
Новости сист. низш. раст., 19666, с. 141—143.
Легенькая Е. И. О специализации Peronospora tabacina Adam в Цен-
трально-черноземной зоне СССР. — Тр. ВИЗР, 1970, вып. 29, ч. 1, с. 91 —
97.
Легенькая Е. И. Болезни сельскохозяйственных растений, вызывае-
мые пероноспоровыми грибами в Центрально-черноземной зоне РСФСР. —
Науч. тр. Курск, с.-х. ин-та, Воронеж, 1971, вып. 4, с. 156—162.
Липецкая А. Д. К биологии зимних спор Plasmopara viticola. — Защ.
раст., 1939, № 18, с. 162-163.
Макрушина А. Т. Методика изучения биологии ооспор, возбуждаю-
щих мильдью. — Виноградарство и садоводство Крыма, 1961, с. 12.
Макрушина А. Т. Методика наблюдений за ооспорами мильдью. —
Виноделие и виноградарство СССР, 1962, № 4, с. 43.
Макрушина А. Т. Биологические особенности развития ооспор миль-
дью и условия, необходимые для первичного заражения виноградников. —
Сб. тр. Молд. фил. ВИЗР, 1966, с. 66.
Макрушина А. Т. Биологические особенности ооспор Plasmopara viti-
cola Berl. et de Toni. — В кн.: Вопросы защ. раст. Т. 1. Кишинев, 1970,
с. 117—127.
Методические указания по диагностике корневых гнилей хлебных
злаков. Л., 1977. 58 с. Авт.: А. А. Бенкен, А. Ф. Коршунова, А. Я. Семе-
нов, М. А. Элбакян, Л. К. Хацкевич.
Молдаван В. А., Живило В. И. Разработка метода искусственного
заражения подсолнечника ложной мучнистой росой. — Тез. докл. 4-го
Всесоюз. совещ. по иммунитету с.-х. раст., Кишинев, 1965, с. 184—186.
Натальина О. Б. Болезни ягодных культур. Доклад-обобщение опуб-
ликованных работ. Л., 1965. 79 с.
Наумов Н. А. Методы микроскопических исследований в фитопатологии.
М.—Л., 1932. 224 с.
Наумов Н. А. Методы микологических и’фитопатологических исследо-
ваний. М.—Л., 1937. 272 с.
Наумов Н.А. Болезнь озимой ржи, вызванная новым видом пероноспоро-
вого гриба Sclerospora secalina N. Naum. — Сб. тр. ВИЗР, 1948, № 1,
с. 51-59.
Наумов Н. А. Новый гриб на озимой ржи. — Бот. матер. Отд. спор,
раст. БИН АН СССР, 1949, т. 6, с. 79-80.
Ю н. С. Новотельнова и др.
137
Наумов Н. А. Современное состояние и основные вопросы микогеогра-
фии. — Тр. ВИЗР, 1972а, вып. 33, с. 5—16.
Наумов Н. А. Основные закономерности географического распределения
болезней сельскохозяйственных растений. — Тр. ВИЗР, 19726, вып. 33,
с. 17—31.
Наумова Н. А. Инфекция картофеля Phytophthora infestans de Вагу
от больных клубней. — Бюл. защ. раст., 1939, т. 1, вып. 20, с. 94—102.
Наумова Н. А. Фитофтора картофеля. Изд. 2-е. Л., 1965. 182 с.
Николаев А. Н. Особенности ооспорообразования у Plasmopara viticola
(Berk, et Curt.) Berl. et de T. — Микол, и фитопатол., 1973, т. 7, вып. 1,
с. 8-11.
Новотельнова Н.С. Влияние температуры и влажности на прораста-
ние конидий Phytophthora infestans (Mont.) de Вагу. — Защ. раст., 1937,
№ 12, с. 79—88.
Новотельнова Н. С. К вопросу о ложной мучнистой росе подсол-
нечника. (Таксономия и специализация возбудителя). — Сб. док л. науч,
конф, по защ. раст. (Таллин—Саку, 4—7 августа 1960 г.), 1961, с. 129—138.
Новотельнова Н. С. Биологические особенности Plasmopara halstedii
(Farl.) Berl. et de Toni на подсолнечнике. — Бот. журн., 1962а, т. 45,
вып. 9, с. 1283—1300.
Новотельнова Н. С. Plasmopara halstedii (Farl.) Berl. et de Toni
как сборный вид. (Обоснование к таксономическому подразделению
рода Plasmopara на сложноцветных). — Бот. журн., 19626, т. 47, вып. 7,
с. 970—981.
Новотельнова Н. С. Белая ржавчина на подсолнечнике. — Защ.
раст., 1962в, № 7, с. 57.
Новотельнова Н. А. Новые виды Plasmopara на Compositae. —
Бот. матер. Отд. спор. раст. БИН АН СССР, 1963а, т. 16, с. 73—83.
Новотельнова Н. С. Обзор видов Plasmopara, паразитирующих
на сложноцветных. — Матер. 2-го симпоз. по вопр. исслед. мико- и ли-
хенофлоры прибалтийских республик, Вильнюс, 19636, с. 111—118.
Новотельнова Н. С. Характер паразитизма возбудителя ложной
мучнистой росы подсолнечника при заражении семян. — Бот. журн.,
1963в, т. 48, вып. 6, с. 845—860.
Новотельнова Н. С. Полиморфизм пероноспорового гриба из рода
Plasmopara. — Бот. журн., 1965, т. 50, вып. 3, с. 301—312.
Новотельнова Н.С. Ложная мучнистая роса подсолнечника. М.—Л.,
«Наука», 1966а. 150 с.
Новотельнова Н. С. Новая специализированная форма Cystopus
tragopogonis Schroet. — Новости сист. низш. раст., 19666, с. 126—128.
Новотельнова Н. С. Фитофтороз яблони. — Сб. обзор, статей М-ва
сельск. хоз-ва СССР, 1969а, с. 9—12.
Новотельнова Н. С. Обзор методов изучения грибов сем. Perono-
sporaceae dBy. в лабораторных условиях. — Микол, и фитопатол., 19696,
т. 3, вып. 6, с. 529—537.
Новотельнова Н. С. Фитофторовые грибы. Л., «Наука», 1974. 150 с.
Новотельнова Н. С., Легенькая Е. И. Болезни сельскохо-
зяйственных растений, вызываемые пероноспоровыми грибами. (Методи-
ческое пособие по диагностике болезней и определению возбудителей).
Л., 1963. 40 с.
Новотельнова Н. С., Минасян М. А. К биологии Cystopus
candidus (Pers.) Lev. и Cystopus tragopogonis Schroet. — Тр. ВИЗР, 1970,
вып. 29, ч. 1, с. 121—129.
Новотельнова Н. С., Пыстина К. А. Корневая и прикорневая
гниль культурных растений, вызываемая низшими грибами. (Диагностика
болезни в условиях Нечерноземья). Л., 1978. 79 с.
Овчинникова А. М. Грибные болезни сои. — В кн.: Болезни и вре-
дители сои на юге Дальнего Востока и меры борьбы с ними. Владивосток,
1971, с. 5—72.
138
Олтаржевский Н.П. Исследование инкубационных периодов миль-
дыо винограда в условиях континентального климата и их значение
в профилактике. — Экспер. ботаника, 1934, вып. 1, с. 223—233.
Олтаржевский Н. П. Определение минимальной критической
температуры проявления мильдью винограда. — Докл. ВАСХНИЛ,.
1939, вып. 6, с. 31—34.
Олтаржевский Н. П. Принципы исследования инкубационного пе-
риода мильдью винограда. — Природа, 1946, № 5, с. 68—69.
Олтаржевский Н.П. Принципы применения инкубационной кривой
мильдью виноградной лозы Plasmopara viticola Berl.et de Toni по климати-
ческим зонам СССР. Автореф. канд. дис. Л., 1955. 15 с.
Панченко А. Я. Ускоренный метод оценки на устойчивость к ложной
мучнистой росе. — Селекция и семеноводство, 1965, № 2, с. 52—54.
Патрикеева М. В. Методика выделения моноспоровых культур Phyto-
phthora infestans. — Микол, и фитопатол., 1970, т. 4, вып. 5, с. 459-460-
Пересыпкин В. Ф., Зражевская Т. Г. Жизнеспособность
конидий Peronospora tabacina Adam. — Микол, и фитопатол., 1967, т. 1,
вып. 3, с. 235—240.
Пересыпкин В. Ф., Мархасева В. А. Некоторые данные об ус-
ловиях формирования ооспор Peronospora tabacina Adam. — В кн.: Перо-
носпороз табака. Кишинев, 1964, с. 19—23.
Плюснина Р. А. Устойчивые к ложной мучнистой росе сортообразцы
огурца. — Защ. раст., 1975, № 11, с. 54—55.
Поляков И. М., Владимирская М. Е. Роль светового режима
в устойчивости капусты к ложной мучнистой росе. — Тр. ВИЗР, 1964„
вып. 21, ч. 2, с. 18—24.
Попова Л. А., Каратыгин И. В. Дифференциация жгутиков зоо-
спор в спорангиях Plasmopara viticola (Berk, et Curt.) Berl. et de T. —
Микол, и фитопатол., 1976, т. 10, вып. 2, с. 139—141.
Попушой И. С., Гринберг Ш. М. Роль агротехнических меро-
приятий в повышении устойчивости подсолнечника к возбудителю ложной
мучнистой росы. — Изв. АН МолдССР, 1967, № 9, с. 81—93.
Прозина А. А. Ботаническая микротехника. М., 1960. 206 с.
Пшедецкая Л. И., Черепанова Н. П. Применение люминес-
центного метода для изучения структуры мицелия Phytophthora infestans:
(Mont.) dBy. в ткани пораженного растения. — Бот. журн., 1961а, т. 46г
вып. И, с. 1655—1662.
Пшедецкая Л. И., Черепанова Н. П. Сохранение Phytophthora
infestans (Mont.) dBy. в культуре. — Вести. ЛГУ, 19616, т. 21, вып. 4Г
с. 23-31.
Пыстина К. А. О распространении грибов рода Sclerospora в СССР. —
Новости сист. низш. раст., 1969, т. 6, с. 92—96.
Пыстина К. А. Грибы рода Pythium Pringsh., обнаруженные в СССР. —
Новости сист. низш. раст., 1972, т. 9, с. 91—94.
Пыстина К. А. Методы выделения грибов рода Pythium Pringsh. из при-
родных местообитаний. — Микол, и фитопатол., 1973, т. 7, вып. 3, с. 249—
252.
Пыстина К. А. Новый для СССР вид рода Pythium — Р. acanthicum.
Drechs. — Новости сист. низш. раст., 1977, т. 14, с. 111—113.
Рокицкий П. Ф. Биологическая статистика. Минск, 1967. 320 с.
Сакварелидзе Н. А. Материалы к изучению фикомицетов, распро-
страненных на бахчевых и огородных культурах в Грузии. — Тр. Ин-та.
защ. раст. АН ГрузССР, 1949, вып. 4, с. 135—157.
Самуцевич М. М. Техника фитопатологических исследований. Л.,
1931. 97 с.
Семихненко П. Г., Белевцев Д. Н. Сроки прорывки подсол-
нечника в районах распространения ложной мучнистой росы. — Вести,
с.-х. науки, 1964, № 6, с. 30—36.
10*
139-
С и к а н Л. 3., Волжина С. 10. Зависимость гибели табака от степени
поражения пероноспорозом. — Микол, ифитопатол., 1974, т. 8, вып. 5,
с. 415-419.
Синицына Н. Защита безвысадочной свеклы от пероноспороза. —
Защ. раст., 1965, № 9, с. 22—23.
Смирнов Е. С. Таксономический анализ. М., 1969. 185 с.
Станявичене С. Ю. Проращивание конидий некоторых видов пероно-
споровых грибов. — Тр. АН ЛитССР, сер. В, вып. 1 (51), с. 25—28.
Стенпна Н. В. Изучение ложной мучнистой росы лука в центральной
зоне Краснодарского края и мер борьбы с ней. Автореф. канд. дис. Красно-
дар, 1974. 21 с.
Степанов К. М. Грибные эпифитотии. Введение в общую эпифитотио ло-
гик» грибных болезней растений. М., 1962. 471 с.
Степанов К. М., Чумаков А. Е. Прогноз болезней сельскохозяй-
ственных растений. Изд. 2-е. Л., 1972. 271 с.
Та л лева М. Н. Физиологические особенности прорастания конидий
Peronospora destructor (Berk.) Casp. — возбудителя ложной мучнистой
росы лука. — В кн,: Физиология иммунитета растений. М., 1968,
с. 120—128.
Тверской Д. Л. Корнеед сахарной свеклы и значение грибов в его
развитии. — Докт. дис. 1955. 668 с. (БИН АН СССР).
Тверской Д. Д., Жукова К. П. Сравнительная агрессивность
возбудителей корнееда сахарной свеклы. — Сахарная пром-сть, 1948,
№ 3, с. 40—44.
Тетеревникова-Бабаян Д. Н. Болезни овоще-бахчевых куль-
тур в Армении и меры борьбы с ними. Т. 2. Ереван, 1964. 359 с.
Тихонов О. И. Новые формы поражения подсолнечника ложной мучни-
стой росой. — Бюл. науч.-техн. информ, по масл. культурам, Краснодар,
май 1968, с. 52—54.
Тупеневич С. М. Защита картофеля от главнейших болезней. Л.,
1973. 144 с.
Уикли Б. Электронная микроскопия для начинающих. М., 1975. 329 с.
Указатель возбудителей болезней сельскохозяйственных растений.
Вып. 1 (по зерновым культурам). Л., 1971. 62 с.
Указатель возбудителей болезней сельскохозяйственных растений.
Вып. 3 (по плодовым культурам). Л., 1975. 70 с.
Филиппова Г. Г. Прогноз продолжительности инкубационного периода
Phytophthora infestans (Mont.) de By. на томатах. — Микол, и фитопатол.,
1968, т. 2, вып. 3, с. 254—255.
Хохряков М. К. Методическое руководство по сбору, пересылке, опре-
делению и составлению коллекций болезней сельскохозяйственных куль-
тур. Л., 1936. 21 с.
Хохряков М. К., Новотельнова Н.С., ПотлайчукВ. И.
К вопросу о возникновении некоторых новых болезней растений в СССР.
Вильнюс, 1963а, с. 21—-27.
Хохряков М. К., Новотельнова Н. С., ПотлайчукВ. И.
Новые грибные болезни культурных растений в СССР. — Тр. ВИЗР,
19636, вып. 17, с. 216—247.
Чумаков А. Е., Минкевич И. И., Власов Ю. И., Гаври-
лова Е. А. Основные методы фитопатологических исследований. —
Науч. тр. ВАСХНИЛ, М., 1974, с. 1-190.
Шатский А. Л. Лечение виноградной лозы от мильдью по инкубацион-
ным периодам. — Защ. раст., 1935, № 6, с. 75—85.
Шишкина А. К. Этиология чернильной болезни пробкового дуба (Quer-
cus suber L.) в Грузинской ССР. — Тез. канд. дис. Тбилиси, 1946, с. 1—6.
Шишкина А. К., Ц а н а в а Н. И. Материалы к изучению болезней
сеянцев и саженцев каштана съедобного (Castanea sativa Mill.) в Грузин-
ской ССР. Сообщ. 2-е. Тр. Ин-та защ. раст. ГрузССР, 1973, вып. 24,
с. 251-260.
140
Шошиашвили И. И., Кирме л ашви л и Н. С. К вопросу о вре-
доносности пероноспоры лука. — Тр. Ин-та защ. раст. ГрузССР, 1950,
вып. 7, с. 179—188.
Э лбакян М. А., Бороздина К. И. Обеззараживание искусствен-
ных субстратов в зимних теплицах (методические указания). Л., 1967.
10 с.
Я ч е в с к и й А. А. Основы микологии. М.—JL, 1933. 1036 с.
Ainsworth С. С., Sparrow F. К., S u ssman A. S. The fungi,
an advanced treatise. Vol. IVB. New York, 1973, p. 160—190.
A r x J. A., von. Pilzkunde. Lehre, 1967. 356 S.
A r x J. A., von. The genera of fungi sporulating in pure culture. Lehre, 1970.
288 S.
Atkinson R. G. Phytophthora species inciting root rot of Ghamaecyparis
lawsoniana and other ornamentals in coastal British Columbia. — Ganad.
Journ. Bot., 1965, vol. 43, N 11, p. 1471—1475.
Banihashemi Z. A new technique for isolation of Phytophthora and
Pythium species from soil. — Pl. Dis. Rep., 1970, vol. 54, p. 261—263.
Blackwell E. Species of Phytophthora as water moulds. — Nature, 1944,
vol. 153, N 3886, p. 496.
Blotnicka K. No we poylady na zjamisko procesu chorobowego maczniaka
rzckomego gebuli (Peronospora destructor (Berk.) Gasp.). — Hodowla roslin,
aklim i nasien, 1974, t. 18, z. 2, s. 131—150.
Boubals D. Contribution a 1’etude des causes de la resistance des Vitacees
au mildiou de la vigne (Plasmopara viticola (B. et G.) Berl. et de T.) et de
leur mode de transmission hereditaire. — Ann. Inst. nat. rech. agron.,
ser. B, Paris, 1959, ann. 9, N 1, p. 1—233.
Burnett J. H. Fundamentals of mycology. London, 1968. 546 p.
Cameron H. R. Infection of pear root with Phytophthora cinnamomi. —
Phytopathology, 1960, vol. 50, N 9, p. 630.
Cameron H. R. Effect of hydrogen ion concentration on growth rates
of Phytophthora sp. — Phytopathology, 1962a, vol. 52, N 8, p. 727.
Cameron H.R. Susceptibility of pear roots to Phytophthora. — Phytopatho-
logy, 1962b, vol. 52, N 12, p. 1295—1297.
Carling D. E., White J. A., Brown M. F. The influence of fixa-
tion procedure on the ultrastructure of the host-endophyte interface of ve-
sicular-arbuscular mycorrhizae. — Canad. Journ. Bot., 1977, vol. 55, N 1,
p. 48-51.
C e j p K. Houby. T. I, II. Praha, 1957. 490, 407 p.
Cohen Y., Sackston W. E. Factors affecting infection of sunflowers
by Plasmopara halstedii. — Canad. Journ. Bot., 1973, vol. 51, N 1, p. 15—
22.
Cohen Y., Sackston W. E. Seed infection and latent infection of sun-
flowers by Plasmopara halstedii. — Canad. Journ. Bot., 1974, vol. 52,
p. 231—238.
Davison E. Cytochemistry and ultrastructure of hyphae and haustoria
of Peronospora parasitica. — Ann. Bot., 1968, vol. 32, N 127, p. 613—621.
De В a г у A. Untersuchungen fiber die Peronosporeen und die Grundlagen
eines natiirlichen Systems der Pilze (Vierte Reihe der Beitrage zur Morpho-
logic und Physiologic der Pilze). — Abh. Senckenberg Naturw. Gesellschaft.,
1881, Bd 12, S. 225.
Delanoe D., H am ante. Contribution a Г etude de Plasmopara heli-
anthi Novot. en milieu semi-artificiel: variabilite biologique et morpholo-
gique de ce parasite. — C. r. Acad, sci., ser. D, 1972, t. 274, N 9, p. 1280—
1281.
Dodge В. O.,, R ick e t t H. W. Diseases and pests of ornamental plants.
New York, 1948. 638 p.
Drechsler Ch. Pythium graminicolum and P. arrhenomanes. — Phytopatho-
logy, 1936, vol. 26, N 4, p. 676-683. F
141
Elsner P. R., Horton J.G., Bowen G. G. Fine structure of sporan-
gia of Phytophthora infestans/during temperature induced differentiation. —4
Amer. Journ. Bot., 1967, vol. 54, p. 649.
Fischer A. Die Pilze Deutschlands, Oesterreichs und der Schweiz. IV Abt.
Phycomycetes. Jena, 1892. 505 S.
Fitzpatrick H. M. The lower fungi. Phycomycetes. New York—London.
1930. 331 p.
Fraymouth J. Haustoria of the Peronosporales. — Trans. Brit. Mycol.
Soc., 1956, vol. 39, N 1, p. 79—107.
F r e z z i M. J. Las especies de Phytophthora en la Argentina. — Rev. in-
vestigac. agric., Buenos-Aires, 1950, vol. 4, N 1, p. 47—133.
Gaumann E. Vergleichende Morphologie der Pilze. Jena, 1926. 626 S.
Goosen P. G., Sackston W. E. Transmission and biology of sunflower
downy mildew. Ganad. Journ. Bot., 1968, vol. 46, N 1, p. 5—10.
Griffin M. J., Coley-S mith J.R. The establishment of hop tissue
cultures and their infection by downy mildew, Pseudoperonospora humuli^
under aseptic conditions. — First Intern. Congress of Plant pathology,.
London, 1968, p. 72.
Guttenberg H., Schmoller H. Kulturversuche mit Peronospora;
brasicae Gaeum. — Arch. Mikrob., 1958, Bd 30, S. 268—279.
Harrison R. W. A method of isolating vesicular-arbuscular endophytes
from roots. — Nature, 1955, vol. 175, N 4453, p. 432.
Hickman G. J., Englisch M. P. Factors influencing the development,
of red core in strawberries. — Trans. Brit. Mycol. Soc., 1951, vol. 34,
N 2, p. 223-236.
Hickman G. J., Goode P. M. A new method of testing the pathogeni-
city of Phytophthora fragariae. — Nature, 1953, vol. 172, N 4370, p. 211 —
212.
Hine R. B. Effect of streptomycin and pimaricin on growth and respiration
of Pythium species. — Mycologia, 1962, vol. 54, N 6, p. 640—646.
Hine R. B., Luna L. V. A technique for isolating Pythium aphanider-
matum from soil. — Phytopathology, 1963, vol. 53, N 6, p. 727—728.
Hohl H. R., Hamamoto S. T. Ultrastructural changes during zoospore
formation in Phytophthora parasitica. — Amer. Journ. Bot., 1967, vol. 54,
N 9, p. 1131—1135.
Hohl H. R., Stossel P. Host-parasite interfaces in a resistent and sus-
ceptible cultivar of Solanum tuberosum inoculated with Phytophthora
infestans. — Ganad. Journ. Bot., 1976, vol. 54, N 9, p. 900—912.
Izard G. Sur une methode de culture de Peronospora tabacina Adam en labo-
ratoire. — G. r. Acad, sci., 1960, t. 251, N 25, p. 3063—3065.
Johnson H. W., Keeling B. L. Pathogenecity of Phytophthora
megasperma isolated from subterranean clover roots. — Phytopathology,
1969, vol. 59, N 9, p. 1279—1283.
Kaufmann M. J., Gerdemann J.W. Root and stem rot of soybean
caused by Phytophthora sofae n. sp. — Phytopathology, 1958, vol. 48,
N 4, p. 201-208.
Klebahn H. Krankheiten des Flieders. Berlin, 1909. 75 S.
Klemmer H. W., Nakano R. Y. Technique in isolation of Pythia-
ceous fungi from soil and diseased pineapple tissue. — Phytopathology,
1962, vol. 52, N 9, p. 955-956.
Klinkowski M., Schmiedeknecht M. Der falsche Mehltau
des Tabaks. Peronospora tabacina Adam, ein fur Deutschland bisher unbe-
. kannte Tabakkrankheit. — Nachrichtenbl. deutsch. Pflanzenschutzdienst,
N. F., 1960, Jg. 14, H. 4, S. 61-73.
Klotz L. J., Wolfe T. A. Techniques for isolating Phytophthora spp.
which attack citrus. — PL Dis. Rep., 1958, vol. 42, p. 675—676.
Klotz L. J.,Wolfe T. A. The production and use of zoospore suspensions
of Phytophthora spp. for investigations on diseases of citrus. — Pl. Dis.
Rep., 1960, vol. 44, p. 572—573.
142
Kreisel H. Grundziige eins natiirlichen Systems der Pilze. Jena, 1969.
240 S.
Lacey J. The infectivity of soils containing Phytophthora infestans, —
Journ. Appl. Biol., 1965, 56, p. 363—380.
Lechoczky J. A szoloperonoszpora (Plasmopara viticola (Berk, et Curt.)
Berl. et de Toni) oospora Kepzesenek vizsgalata. — Novenytermeles Buda-
pest, 1965, t. 14, N 3, p. 261—270.
Leonian L. H. Identification of Phytophthora species. — West. Virg.
Univ. Agric. Exp. Sta. Bull., 1934, N 262, p. 1—36.
Machacek J. A simple method of obtaining Pythium cultures free from
bacteria. — Phytopathology, 1934, vol. 24, N 3, p. 301—304.
Manning W. J., Grossan D. F. Effects of a particular soil bacterium
on sporangial production in Phytophthora cinnamomi in liquid culture. —
Phytopathology, 1966, vol. 56, N 2, p. 235—237.
McGrath H., Miller P. R. Blue mold of tobacco. — Tobacco, 1959,
vol. 148, N 7, p. 11—13; N 8, p. 14—18; N 9, p. 19—23; N 10, p. 24—28.
McKay R. The longevity of the oospores of onion downy mildew Peronospora
destructor (Berk.) Gasp. — Sci. Proc. Roy. Dublin Soc., 1957, vol. 27,
N 12, p. 295-307.
M с К e e n W. E. Electron microscopy studies of a developing Pythium oogo-
nium. — Ganad. Journ. Bot., 1975, vol. 53, N 20, p. 2354—2360.
McLaughlin O. The isolation of Pythium from soil at various seasons
of the year as related to soil temperature and moisture. — Phytopathology,
1947, vol. 37, N 1, p. 15.
Mehrlich F. P. Nonsterile soil leachate stimulating to zoosporangia pro-
duction by Phytophthora sp. — Phytopathology, 1935, vol. 25, N 4, p. 432—
435.
Melhus J. E. Experiments on spore germination and infection in certain
species of Oomycetes. — Wiscon. Agr. Exp. St. Res., 1911, Bull. XV,
p. 25-91.
Meredith G. H. A quick method of isolating certain phycomycetous fungi
from soil. — Phytopathology, 1940, vol. 30, N 12, p. 1055—1056.
Morel G. Recherches sur la culture associee, de parasites obligatoires et de tis-
sus vegetaux. — Ann. Epiphyt., 1948, t. 14, fasc. 3, p. 123—234.
Munro M. G. D.A root rot of Cineraria and a study of the species of Phyto-
phthora concerned. — Trans. Brit. Mycol. Soc., 1945, vol. 28, N 3—4,
p. 115-153.
Nakamura H. The use of tissue culture in the study of obligate parasites. —
Proc. Intern. Conf. Plant tissue cult. Berkeley, 1965, p. 535—539.
Newhook F. J. The association of Phytophthora spp. with mortality of Pi-
nus radiata and other conifers. I. Symptoms and epidemiology in shelter-
belts. — New Zeal. Journ. Agric. Res., 1959, vol. 2, N 4, p. 808—843.
No viello G., Snyder W. G. A Phytophthora disease of fennel. —
Phytopath. Zeitschr., 1962, Bd 46, H. 2, S. 139—163.
Nusbaum G. J., Lucas G. B., Chaplin J. F. Estimating the
inoculum potential of Phytophthora parasitica var. nicotianae in the soil. —
Phytopathology, 1952, vol. 42, N 5, p. 286.
Pethybridge G. H. On the rotting of potato tubers by a new species
of Phytophthora having a method of sexual reproduction hitherto undescri-
bed. — Sci. Proc. Roy. Dublin Soc., n. s., 1918, vol. 13, N 35, p. 529—
564.
Pethybridge G. H., Lafferty H. A. A disease of tomato and
other plants caused by a new species of Phytophthora. — Sci. Proc. Roy.
Dublin Soc., n. s., 1919, vol. 15, N 35, p. 487—505.
Petrlik Z., Stys Z. Metoda laboratorni kultivace peronospory chme-
love — Peronoplasmopara humuli Miy. et Так. na sadovych rizcich. —
Geska mykologie, 1962, Roc. 16, s. 1, p. 56—62.
P i о t h L. Gh. Untersuchungen fiber anatomische und physiologische Eigen-
schaften resistentner und anfalliger Reben in Beziehung zur Entwicklung
143
von Plasmopara viticola. — Zeitschr. Pflanzenziicht., 1957, Bd 37, H. 2r
S. 127-158.
Rands R.D., Dopp E. Variability in Pythium arrhenomanes in relation
to rot of sugarcane and corn. — Journ. Agr. Res., 1934, vol. 49, p. 189—221.
Romero S., Erwin D. G. Variation in pathogenicity among single-
oospore cultures of Phytophthora infestans. — Phytopathology, 1969, vol. 59r
N 8, p. 1310—1317.
Schmitthenner A. F. Prevalence and virulence of Phytophthora,
Aphanomyces, Pythium, Rhizoctonia and Fusarium isolated from diseased
alfalfa seedlings. — Phytopathology, 1964, vol. 54, N 5, p. 1012—1018.
Schroeter J. Peronosporineae. — In: Engler A., PrantlK.
Die natiirlichen Pflanzenfamilien nebst ihren Gattungen und wichtigeren
Arten insbesondere der Nutzpflanzen. 1 Teil, I Abt. Jena, 1897, S. 108—119.
Schultz H. Arbeitsmethoden bei Kultur und Infektionsversuchen mit
Pythium-Arten. — Zentralbl. Bakteriol., Parasitenk. und Infektionskr.,
1942, Bd 105, N 14-16, S. 248.
Schwinn F. J. Der Nachweis von Phytophthora cactorum (Leb. et Cohn)
Schroet. im Boden. — Mitteilungen Biol. Bundesanstalt Land- u. Forst-
wirtschaft, 1961, H. 104, S. 42—44.
Sewell G. W. F., Wilson J. F. Branch, stem and collar rot of apple
caused by Phytophthora species of the cactorum group. — Nature, 1963,
vol. 200, N 4912, p. 1229.
Shepherd G. J., Mandryk M. Effect of metabolites and antimetabo-
lites on the sporulation of Peronospora tabacina Adam. — Austral. Journ.
Biol., 1964, vol. 17, N 4,'p. 878-892.
Shepherd G. J., Stuart F., Mandryk M. The isolation and main-
tenance of single spore lines Peronospora tabacina Adam. — Plant Industry
Ann. Rep. 1962—1963, Australia, Canberra, 1963, p. 49—59.
Sleeth B. Agar medium and technique for isolation Pythium free from
bacteria. — Phytopathology, 1945, vol. 35, N 12, p. 1030—1031.
Srinivasan К. V. A technique for isolating pathogenic cultures of Py-
thium from the soil. — Gurr. Sci., 1958, vol. 27, N 5, p. 180.
Strugger S. Untersuchungen fiber die vitale Fluorochromierung der He-
fezelle. — Flora, 1943, Bd 37, N 1—2, S. 73—94.
Subramaniam L. S. Root rot and sclerotial disease of wheat. — Bull.
Agr. Res. Inst., 1928, N 177, p. 1—7.
Tompkins G. M., Richards B. L., Tucker G. M., Gard-
ner M. W. Phytophthora rot of sugar beet. — Journ. Agr. Res., 1936»
vol. 52, N 3, p. 205—216.
Tompkins G. M., Tucker G. M. Phytophthora rot of honey-dewmelon.
Journ. Agr. Res., 1937, vol. 54, N 12, p. 933—944.
Vaartaja O. Selectivity of fungicidal materials in agar cultures. — Phyto-
pathology, 1960, vol. 50, N 3, p. 870—873.
Vanterpool Z. G. Some species of Pythium parasitic on wheat in Canada
and England. — Ann. Biol., 1938, vol. 25, N 3, p. 528—543.
Vanterpool T. G., Sprague R. Pythium arrhenomanes in the nor-
thern great plains. — Phytopathology, 1942, vol. 32, N 1, p. 327—328.
Vanterpool T. G., Truscott J. H. L. Studies on browning root
rot of cereals. II. Some parasitic species of Pythium and their relation to the
disease. — Canad. Journ. Res., 1932, vol. 6, N 1, p. 68—93.
Viranyi F. Studies on the biology and ecology of onion downy mildew
(Peronospora destructor (Berk.) Fr.) in Hungary. I. Overwintering of the
pathogen in onion bulbs. — Acta Phytopath. Acad. Sci. Hung., 1974,
vol. 9, N 3—4, p. 311—314.
Viranyi F. An improved method for detecting systemic infection of sun-
flower seedlings caused by Plasmopara halstedii. — Acta Phytopath. Acad.
Sci. Hung., 1977, vol. 12, N 3—4, p. 263—267.
Warcup J. H. The soil-plate method for isolation of fungi from soil. —
Nature, 1950, vol. 165, N 4187, p. 117.
144
Whiteside J. 0. Some factors affecting the occurence and development
of foot rot on citrus trees. — Phytopathology, 1971, vol. 61, N 10, p. 1233—
1238.
Wolf F. A., McLean R. A. Sporangial proliferation in Peronospora
tabacina. — Phytopathology, 1940, vol. 30, N 3, p. 264—267.
Wong T. K., Varchese G. Assesing the susceptibility of local citrus
species to Phytophthora root rot in Malaya. — Exp. Agric., 1966, vol. 2,
N 2, p. 305-308.
Zentmyer G. A., Marshall L. A. Factors affecting sporangial pro-
duction by Phytophthora cinnamomi. — Phytopathology, 1959, vol. 49,
N 3, p. 556.
145
УКАЗАТЕЛЬ ВИДОВ
ПЕРОНОСПОРОВЫХ ГРИБОВ *
♦Basidiophora entospora Roze et Cornu
117
Bremia centaureae Syd. 91
— lactucae Regel 14, 80, 96
— — var. carthami Milovtzeva 84
— — var. lactucae 80
— scolymi Jacz. 80
Cystopus bliti (Biv.-Bern) Lev. 82,
90, 129
— candidus (Pers.) Lev. 6, 13, 24, 51,
59, 62, 64, 79, 85, 94, 128—130
— capparidis dBy. 81, 130
— occidentalis Lev. 80
— portulaceae (Dekin et Passy) Lev.
6, 94, 130
— tragopogonis Schroet. 6, 59, 62,
64, 82, 92, 94
— — f. helianthi Novot. 128, 129
Peronospora aestivalis Syd. 76, 120
— antirrhini Schroet. 93
— arborescens (Berk.) dBy. 12, 93,
127
— arthuri Farl. 92
— brassicae Gaeum. 12, 51, 57, 66,
78, 79, 126
— — f. brassicae-nigrae Gaeum. 85
— camelinae Gaeum. 85
— cochleariae Gaeum. 79
— conglomerata Fckl. 91
— crambe Jacz. 83
— crispula Fckl. 94
— destructor (Berk.) Casp. 12, 59,
65, 81, 121
— digitalidis Gaeum. 94, 127
— fabae Jacz. et Serg. 75
— fagopyri Elenev 12, 74, 118, 119
* — fragariae Rose et Cornu 88
— fulva Syd. 76, 92
— hyoscyami dBy. 12, 66, 91, 127
— jaapiana Magn. 80
— lamii Braun 91
— lathyri-palustris Gaeum. 76
— lentis Gaeum. 75
— leptosperma Gaeum. 12, 94, 126
— lini Schroet. 83
— lotorum Syd. 96
— majori Gaeum. 76
— manshurica (N. Naum.) Syd. 12
75, 119
— matthiolae Gaeum. 93
— meliloti Syd. 76
— myosotidis dBy. 94
— nasturtii-aquatici Gaeum. 94
— ornithopi Gaeum. 77
— orobi Gaeum. 76
— parasitica (Pers.) Fr. 14, 40, 51
— phloxidis E. Vassil. 95
— pisi Syd. 75
— pratensis Syd. 76
— rubi Rab. 88
— ruegeriae Gaeum. 76
— rumicis Cda. 80
— schachtii Fckl. 6, 11, 15, 82, 122,
123
— sepium Gaeum. 76
— spinaciae Laub. 80, 120
— stigmaticola Raunkier 94
— sulfurea Gaeum. 80
— swinglei Ell. et Kell. 95
— tabacina Adam 6, 10, 50, 52, 53,
59, 62, 65, 66, 78, 83, 123
— trifolii-alpestris Gaeum. 76
— trifolii-arvensis Syd. 76
— trifolii-hybridi Gaeum. 12
— trifolii-repentis Syd. 12, 76
— Valerianae Fckl. 91
— viciae (Berk.) Gaeum. 15, 76
— viciae-sativae Gaeum. 76
— violae dBy. 96, 128
— (=Pseudoperonospora) cannabina
(Otth) Hoerner 84, 125
— (—) cubensis (Berk, et Curt.) Ro-
stow. 81, 120
— (—) humuli (Miy. et Так.) E. E.
Wils. 52, 57, 85, 124, 125
— (—) sparsa Jacz. 95
Phytophthora cactorum (Leb. et Cohn)
Schroet. 6, 9, 10. 45, 75, 76, 83—
96, 106—112
— cambivora (Petri) Buis. 90
1 Виды, не обнаруженные в СССР, отмечены звездочкой.
146
— capsisi Leonian 47, 78, 81, 96, 106
— cinnamomi Rands 9, 44, 48, 60,
89, 90, 95, 96
*— citricola Saw. 89
— citrophthora (R. E. Sm. et E. H.
Sm.) Leonian 48, 89, 109
— cryptogea Pethyb. et Laff. 47,
62, 76, 78—80, 89—96, 106, 112
— drechsleri Tucker 47, 48, 78, 82,
84, 106-108
— erythroseptica Pethyb. 46, 48, 75,
77, 91, 96, 105, 112
— fragariae Hickman 46, 62, 87, 88,
108, 109
— infestans (Mont.) dBy. 6, 9, 18,
45, 46, 49, 50, 59, 65, 66, 77, 78,
94, Ю4—106
— — var. phaseoli (Thaxt.) Leo-
nian 75
— megasperma Drechs. 76, 82, 88,
96, 108
— — var. sojae (Kaufmann et Ger-
demann) Hildebrand 62, 75
— nicotianae B. de Haan 21, 61, 78, 79
— — var. nicotiana 83
— — var. parasitica (Dast.) Water-
house 48, 73—75, 78, 80, 83, 84,
89, 93, 96, 106, 112
* — oryzae(Itoet Nagai)Waterhouse73
* — porri Foister 81
* — richardiae Buis. 96, 112
— syringae (Kleb.) Kleb. 46, 61,
79, 86, 89, 95, 108
Plasmopara anethi Jermal. 79
— apii Savul. et O. Savul. 80
— asterea Novot. 24, 91, 118
— — f. callistephi Novot. 117
— dauci Savul. et O. Savul. 79
— gypsophilae Jacz. 92
— halstedii (Farl.) Berl. et de T. 115
— helianthi Novot. 6, 11, 14, 24,
58, 59, 62, 63, 65, 82, 115
---f. helianthi 57, 115
— mei-foeniculi Savul. et O. Savul. 79
— nivea (Mart.) Schroet. 14, 85
— panacis (Bunk.) Bond, et Bunk.
92, 117
— pastinaceae Savul. et 0. Savul. 79
— petroselini Savul. et 0. Savul. 79
— peucedani Nannf. 79
— pimpinellae Savul. et 0. Savul. 79
— pusilia (dBy.) Schroet. 91
— pygmaea (Ung.) Schroet. 91
— — f. delphinii Gapon. 92
— ribicola Schroet. 88
— skvortzovii Miura 85
— solidaginis Novot. 92, 118
— viticola (Berk, et Curt.) Berl.
et de T. 6, 10, 14, 24, 50, 55, 62,
66, 88, 116
Pythium acanthicum Drechs. 75, 77,
90, 92, 104
— anandrum Drechs. 80
— aphanidermatum (Edson) Fitzp. 5,
9, 42, 43, 72, 73, 77—81, 83, 84.
86, 88, 89, 100—102
* — aristosporum Vanterpool 72, 98
*— arrhenomanes Drechs. 72—74, 76,
98,
— conidiophorum Jokl 101
— debaryanum Hesse 5, 7, 8, 41, 43,
72-77, 79-81, 83-95, 100, 101,
103, 104
* — deliense Meurs 77
— dissotocum Drechs. 73
— echinocarpum Ito et Tokunaga 73
— graminicolum Subr. 72—74, 76, 98
*— helicoides Drechs. 76
— hydnosporum (Mont.) Schroet. 7,
74, 102
*— hypogynum Middleton 72
— intermedium dBy. 41, 86, 89, 95
— irregulare Buis. 8, 72, 73, 76,
80-82, 88, 89, 94, 96, 98, 102
*— iwayamai Ito 72
— mamillatum Meurs 76, 80, 81, 88
— megalacanthum dBy. 86, 88, 90, 93
— monospermum Pringsh. 41
*— oligandrum Drechs. 79, 81, 89, 93
*— paddicum Hirane 73
— papillatum Matthews 72
— paroecandrum Drechs. 73, 76, 79,101
— perniciosum Serbinow 9, 83, 102
* — polymastum Drechs. 79
— proliferum dBy. 87
— pulchrum Minden 76, 86
* — pyrilobum Vaartaja 89
— rostratum Butler 41, 72, 73, 76, 88
*— salpingo phorum Drechs. 76
— spinosum Saw. 8, 79, 81, 93, 101
— splendens Braun 72, 73, 76, 82,
91, 103, 104
* — sylvaticum Campbell et Hendrix
87, 95
* — tardicrescens Vanterpool 72, 98
— torulosum Coker et Patterson 72,
86, 89
— ultimum Trow 5, 7, 8, 43, 72,
74—81, 86-90, 93, 95, 99, 101, 103
* — vexans dBy. 41,72,74, 76, 77,86, 88
* — volutum Vanterpool et Truscott
72, 73, 98, 99
Pythium sp. 95, 96
Sclerospora graminicola (Sacc.)
Schroet. 6, 74, 113, 114
— — var. setariae-italicae Trav. 74
— macrospora Sacc. 72, 73, 112, 113
— maydis (Rac.) Butler 73, 113
— secalina N. Naum. 72, 113
147
УКАЗАТЕЛЬ ПИТАЮЩИХ РАСТЕНИЙ
Абрикос 89
Азалия 95
Алоэ 95
Амарантус 90, 129
Анис 79
Апельсин 88, 89, 109
Арбуз 81, 99
Артишок 80
Астра 90, 91, 104, 117
Базилик 91
Баклажан 78
Бархатцы 91
Бегония 95, 103
Белена 12, 91, 126
Белладонна 91
Бобы кормовые 75
Борец 91
Брюква 79, 129
Бук 90
Валериана 91
Василек 91, 117
Вика 76
Виноград 6, 10, 54, 56, 62, 63, 65,
66, 88, 116
Вишня 86, 108
Гвоздика 91, 104
Георгин 91
Герань 91, 103
Гербера 92
Гипсофила 92
Гладиолус 92
Глоксиния 96, 104
Годеция 92
Горох 7, 8, 74, 75, 102
Горошек душистый 92
Горчица 84, 85, 129
Гречиха 12, 74, 118, 119
Груша 86, 108
Девясил 92
Дельфиниум 92, 103
Донник 76
Дуб 9
— пробковый 89, 90
Дыня 47, 81, 99
Ежевика 88
Ель 8, 89
Женьшень 10, 92, 110, 111, 117
Земляника 9, 46, 62, 87, 88, 108—
110
Золотарник 92, 118
Ильм 90
Ирис 92
Кабачки 81, 99
Кактус 96, 111
Каланхое 96, 111
Калла 96, 111
Канатник 85
Каперсы 81, 130
Капуста 7, 12, 78, 79, 126, 128
Картофель 9, 46, 65, 66. 77, 97,
104—106
Каттлейя 96, 104
Каштан 9, 89, 90
Кедр 89
Кипарис 89
Кларкия 92
Клевер 7, 8, 12, 75, 76, 100
Клен 84
Клещевина 84
Колеус 96
Конопля 84, 125
Кориандр 85
Крамбе 83, 129
Крыжовник 88, 109
Кукуруза 7, 73, 112, ИЗ
Кунжут 84
Лавр 89, 90
Левкой 93, 94
Лен 78, 83
Лилия 10, 93, 112
Лимон 88, 89, 109
Лисохвост 112
Лиственница 89
Лотос 96
Лук 12, 65, 81, 121
Львиный зев 93
Люпин 7, 75
Люцерна 76, 120
148
Мак 12, 93, 127
Малина 88
Мальва 93
Мандарин 88, 89, 109
Маттиола 93, 94, 130
Махорка 83
Могар 74, 113
Морковь 79
Мушмула 89
Мята 94
Наперстянка 94, 127
Настурция 94
Незабудка 94
Овес 72, 73, 112
Овсяница 112
Огурцы 7, 8, 12, 81, 99, 120
Орех грецкий 9, 89, 90
Пастернак 79
Пекан 89, 90
Пеларгония 91, 103
Пелюшка 74, 75
Перец 78, 124
Петрушка 79
Петуния 94, 124
Пион 94
Пиретрум 94
Пихта 89
Подсолнечник 6, И, 58, 63—65, 82,
И -116, 128
Портулак 94, 130
Примула 94
Просо 74, ИЗ, 114
Пшеница 8, 72, 98, 112
Ревень 80
Редис 13, 79, 129
Редька 79, 129
Резеда 94
Репа 79, 129
Рис 73, ИЗ
Рододендрон 95
Рожь 72, 112
Роза 95
Ромашка аптечная 12, 95, 126
Рыжик 85, 129
Салат 7, 80
— эндивий 80
Сафлор 84
Свекла 6—8, И, 81, 82, 98, 100,
101, 122
Свидина 90
Сельдерей 80
Сераделла 77
Синнингия 96
Сирень 46, 95
Слива 89, 108
Смородина 88
Сосна 8, 89, 103
Соя 12, 62, 75, 119
Табак 6, 8, 11, 52, 61, 66, 83, 102, 121
Тисс 89
Томаты 7, 9, 47, 62, 77, 78, 97, 106
Турнепс 79, 129
Тыква 81, 99
Тюльпан 95, 112
Укроп 79
Фасоль 75
Фенхель 46, 61, 79
Фиалка 96, 128
Флокс 95
Хинное дерево 89, 90
Хлопчатник 83
Хмель 52, 57, 85, 124, 125
Хрен 79, 129
Хризантема 95
Цикорий 80
Цинерария 44, 96, 118
Черешня 86
Чеснок 81
Чечевица 75
Чина 76
Чумиза 74, ИЗ
Шалфей 95
Шпинат 7, 80, 120
Щавель 80
Эвкалипт 90
Эспарцет 76
Эстрагон 80
Яблоня 86, 107, 108
Ячмень 72, 73, 112
149
ПРИНЯТЫЕ СОКРАЩЕНИЯ
ИМЕН АВТОРОВ ВИДОВ
ПЕРОНОСПОРОВЫХ ГРИБОВ
Adam — D. В. Adam
Ch. Benois — Ch. A. Benois
Berk. — M. J. Berkeley
Berl. — A. N. Berlese
Biga — M. L. Biga
Biv.-Bern. — A. de Bivona-Bernardi
Br. — С. E. Broome
Buis. — C. J. Buisman
Butler — E. J. Butler
Byzova — Z. M. Byzova
DC. — A. de Candolle
Carne — W. M. Carne
Casp. — J. X. R. Caspary
Cda. — A. C. J. Corda
Chester — K. G. Chester
Cif. - R. Cifferi
Cke. — M. C. Cooke
Cohn — F. J. Cohn
O. Const. — 0. Constantinescu
Cornu — M. M. Cornu
Curt. — M. A. Curtis
Czerep. — N. P. Czerepanova
Dast. — J. F. Dastur
dBy. — A. de Bary
J. J. Davis — J. J. Davis
Desm. — J. В. H. J. Desmazieres
Drechs. — C. Drechsler
Elenev — P. F. Elenev
Ell. - G. B. Ellis
Farl. — W. G. Farlow
Fckl. — K. W. G. L. Fuckel
Ferr. — T. Ferraris
A. Fisch. — A. Fischer
Fitzp. — H. M. Fitzpatrik
Foister — С. E. Foister
Fr. — E. M. Fries
Gaeum. — E. Gaeumann
Gapon. — N. T. Gaponenko
Grev. — R. K. Greville
Gray — A. Gray
A. Gust. — A. Gustavson
B. de Haan — Breda de J. Haan, van
Halst. — B. D. Halsted
Hartig — R. Hartig
Hesse — R. Hesse
Hickman — C. J. Hickman
Hoerner — G. R. Hoerner
Humph. — J. E. Humphrey
150
Ideta — A. Ideta
Ito — S. Ito
Jacz. — A. A. Jaczewski
Jermal. — J. Jermalaviciute
Jorst. — I. Jorstad
Kalymb. — В. K. Kalymbetow
Kell. — W. A. Kellermann
Kleb. — H. Klebahn
Ktze — К. E. 0. Kuntze
Kuehn — J. G. Kuehn
Laff. - H. A. Lafferty
Lagh. — N. G. Lagerheim
Laub. — K. R. Laubert
Leb. — H. Lebert
Leonian — L. H. Leonian
Lev. — J. H. Leveille
Lib. — M. A. Libert
Lob. — A. I. Lobik
Matthews — V. D. Matthews
McAlp. — D. McApline
Middleton — J. T. Middleton
Miura — M. Miura
Miy. — K. Miyabe
Mont. — C. Montagne
Nannf. — J. A. Nannfeldt
N. Naum. — N. A. Naumov
Novot. — N. S. Novotelnova
Osipian — L. L. Osipian
Otth — G. H. Otth
Pass. — G. Passerini
Pat. — N. T. Patouillard
Pers. — С. H. Persoon
Petersen — H. E. Petersen
Pethyb. — G. H. Pethybridge
Petri — L. Petri
Peyronel — B. Peyronel
Pringsh. — N. Pringsheim
Rab. — C. L. Rabenhorst
Rac. — M. Raciborski
Rands — R. D. Rands
Rayss — T. Rayss
Regel — E. A. von Regel
Rostow. — S. I. Rostowzew
Roum. — C. Roumeguere
Roze — E. Roze
Sacc. — P. A. Saccardo
Sarejanni — J. A. Sarejanni
Savul. — Tr. Savulescu
0. Savul. — О. Savulescu
Saw. — К. Sawada
Schlecht. — D. F. L. von Schlechten-
dahl
Schneider — A. Schneider
Schroet. — J. Schroeter
Shaw — Ch. G. Shaw
Skal. — V. Skalicky
E. H. Sm. - E. H. Smith
R. E. Sm. — R. E. Smith
Sten. — N. P. Stenina
Swing. — D. B. Swingle
Syd.— H. Sydow
de T. — G. B. de Toni
Так. — Y. Takahashi
Thaxt. — E. Thaxter
Tokunaga — J. Tokunaga
Tranz. — W. A. Tranzschel
Trott. — A. Trotter
Tucker — G. M. Tucker
Tul. — E. L. R. Tulasne
Uljanish. — W. I. Uljanishchev
Ung. — F. J. A. N. Unger
Vanterpool — T. C. Vanterpool
Vassyag. — M. P. Vassyagina
Vien.-Bourg. — G. Viennot-Bourgin
Waterhouse — G. M. Waterhouse
Wrob. — A. Wroblewski
151
ОГЛАВЛЕНИЕ
Введение.......................................................... 3
Общие сведения ................................................... 5
Систематическое положение ..................................... 5
Симптомы поражения ............................................ 6
Распространение и вредоносность................................ 6
Особенности строения и развития.................................. 14
Мицелий ...................................................... 14
Репродуктивные органы......................................... 18
Цикл развития ................................................ 25
Методы исследования ............................................. 28
Гербаризация.................................................. 28
Измерение, обработка данных................................... 29
Цитологическое изучение живого и фиксированного материала 30
Изолирование и культивирование Pythiaceae и Phytophthoraceae 40
Изолирование и культивирование Peronosporaceae и Cystopaceae 50
Искусственное и естественное заражение........................ 58
Учет в полевых условиях.......................................... 68
Диагностика ..................................................... 73
Наиболее вредоносные болезни..................................... 99
Литература...................................................... 135
Указатель видов пероноспоровых грибов ......................... 146
Указатель питающих растений..................................... 148
Принятые сокращения имен авторов видов пероноспоровых грибов 150
Наталья Сергеевна Новотельнова,
Кира Анатольевна Пыстина, Ольга Георгиевна Голубева
ПЕРОНОСПОРОВЫЕ ГРИБЫ —
ПАТОГЕНЫ КУЛЬТУРНЫХ РАСТЕНИЙ
В СССР
Справочник по диагностике и методам исследования
Утверждено к печати
Ботаническим институтом им. В. Л. Комарова Академии наук СССР
Редактор издательства Г. Т. Никифорова. Художник И. П. Кремлев
Технический редактор М. Э. Карлайтис. Корректор Г. Н. Атлас
ИБ № 8889
Сдано в набор 6.12.78. Подписано к печати 11.05.79. М-27103. Формат бОхЭО1/^.
Бумага № 2. Гарнитура обыкновенная. Печать высокая. Печ. л. + 4 вкл.
(112 печ. л.) = 10 усл. печ. л. Уч.-изд. л. 10.96. Тираж 1300. Изд. № 7144. Тип. зак. 981.
Цена 60 к.
Ленинградское отделение издательства «Наука»
199164, Ленинград, В-164, Менделеевская лин., 1
Ордена Трудового Красного Знамени Первая типография издательства «Наука»,
199034, Ленинград, В-34, 9 линия, 12
ИСПРАВЛЕНИЕ
В указателях номера страниц начиная со с. 72 следует уве-
личивать на 2.
60 к.
«НАУКА»
Ленинградское
отделение